/

































































































































































































































































































Author: Голубев В.Н.
Tags: экология растений этология растение и окружающая среда биология
Year: 1965
Similar




Text
В.Н. ГОЛУБЕВ
ЭКОЛОГО'БИОИОГИЧ ЕС КИЕ
ОСОБЕННОСТИ
ТРАВЯНИСТЫХ РАСТЕНИЙ
И РАСТИТЕЛЬНЫХ СООБЩЕСТВ
ЛЕСОСТЕПИ
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
ВОСТОЧНО-СИБИРСКИЙ БИОЛОГИЧЕСКИЙ ИНСТИТУТ
В.Н. ГОЛУБЕВ
ЭКОЛ ОГО-БИО4ОГИЧ ЕСКИ Е
ОСОБЕННОСТИ
ТРАВЯНИСТЫХ РЛСТЕНИЙ
И РЛСТИТЕЛЬНЫХСООБЩЕСТВ
ЛЕСОСТЕПИ
ИЗДАТЕЛЬСТВО «НАУКА»
Москва 1965
УДК 581.526
Ответственный редактор
доктор биологических наук М. В. КУ Л ЬТ И А С О В
Введение
Советская геоботаническая школа уделяет большое внимание ана-
лизу эколого-биологических особенностей растений 1 как важному сред-
ству познания структуры и динамики растительных сообществ.
Лесостепь Советского Союза в основной своей части является об-
ластью распространения мощных высокогумусных черноземов, что и оп-
ределяет ее интенсивное сельскохозяйственное освоение. Эти плодород-
нейшие черноземы в значительной мере обязаны своим происхождением
природной растительности, которая в настоящее время очень сильно
деградировала под влиянием хозяйственной деятельности и в более или
менее естественном состоянии сохранилась только в специальных запо-
ведниках. Природная флора лесостепи заключает большое количество
видов растений, преимущественно травянистых, отличающихся разнооб-
разными эколого-биологическими признаками. Среди них встречаются
как выходцы из более южных (настоящих) степей, так и виды, распро-
страненные значительно севернее лесостепи — в лесной зоне. Некоторые
типично бореальные виды находят в лесостепи южную границу своего
распространения. Таким образом, в лесостепи концентрируется бога-
тейший комплекс видов с разной историей развития и происхожде-
ния. Поэтому всестороннее изучение эколого-биологических свойств ви-
дов лесостепи, отражающих характер приспособления их к условиям
существования, представляет особенный интерес, который еще более
повышается в связи с тем, что многие из этих видов имеют большое
практическое значение и частично уже введены в культуру.
В настоящей работе предпринята попытка сопряженного изучения
биоморфологии подземных и надземных органов травянистых растений
европейской лесостепи в связи с условиями почвенной и воздушной
среды1 2, определяющей экологическую направленность исследования,
которая составляет фактическую и методическую основу в обсуждении
любого биологического признака и явления.
Чем дальше отделяет нас время от начала данной работы, тем
больше возникает свидетельств в необходимости глубокого познания
1 Наши эколого-биологические исследования касаются той сферы жизнедеятельно-
сти растений, которая проявляется в разнообразных морфологических структурах и их
изменениях. Поэтому в более точном смысле данное исследование ограничивается рам-
ками экологической морфологии растений, идентичной в нашем понимании биоморфо-
логии.
2 Работа является одним из разделов общей темы комплексных исследований «Об-
мен и превращение веществ и энергии в основных биогеоценозах лесостепи Европей-
ской части СССР>, в которых принимали участие Почвенный институт им. В. В. Доку-
чаева, Ботанический институт АН СССР, Институт географии АН СССР, Институт
морфологии животных АН СССР и другие организации. Базой полевых наблюдений,
проводившихся нами в 1959—1962 гг., служил Стрелецкий участок Центрально-Черно-
земного заповедника им. проф. В. В. Алехина (Курская область). Биоморфологии под-
земных органов травянистых растений лесостепи посвящена отдельная книга (В. Н. Го-
лубев, 1962в).
3
биоморфологии растений каждого зонального типа растительности.
Только на этой основе можно успешно развивать физиологические иссле-
дования компонентов природных ценозов в неразрывной связи с усло-
виями существования и вскрывать закономерности взаимоотношения
растений, их фитоценотическую и средообразующую роль. Эти вопро-
сы в конечном итоге получают сейчас признание в качестве основопо-
лагающих, в плане решения которых намечаются пути и перспективы
дальнейшего развития фитоценологии (Сукачев, 1956; Сукачев, Зонн,
1961; Лавренко, 1959; Работнов, 1962; Шенников, 1962, 1963; и др.).
Кардинальная проблема науки о растительных сообществах — это со-
хранение и преобразование естественных и моделирование искусствен-
ных ценозов в режиме максимальной продуктивности в данных эколого-
географических условиях.
В узком смысле биоморфология изучает целостный цикл жизни ра-
стений, проявляющийся в разнообразных морфологических структурах,
закономерно изменяющихся в сезонном, индивидуальном и историче-
ском развитии. Наибольшее внимание в работе уделено вопросам се-
зонного развития: характеру и длительности вегетации, ритмам цве-
тения, перезимовке растений. Там, где это возможно, устанавливается
связь биологии надземных органов с распространением корневых си-
стем, условиями влагообеспеченности и другими факторами экологиче-
ского режима.
Большое место отводится классификации биоморфологических
структур, основанной на историко-генетическом принципе. Развитием
и логическим завершением этого принципа является обсуждение вопро-
сов эволюции биоморфологических структур (гл. III). На конкретных
систематических циклах видов (роды Potentilla, Scrophularia, Planta-
go и др.) показаны вероятные пути и особенности исторического разви-
тия жизненных форм. Для обобщения этих вопросов были привлечены
выводы более ранних наших работ, а также ряд неопубликованных
данных. Классификация отдельных биоморфологических признаков, ис-
пользуемая в различных разделах книги, преследует цель системати-
зации материала и служит первой ступенью в дальнейших поисках
морфогенетического синтеза его и интерпретации.
Но уже сейчас на основании имеющихся данных можно говорить
о существовании глубоких корреляций между различными признаками
и свойствами, а значит и о возможности интегрального понимания био-
морфологических типов растений (жизненных форм).
Подчеркивая фитоценотическую направленность исследования, надо
отметить, что .в сферу изучения был вовлечен весь видовой состав изу-
чавшихся ценозов, насчитывающий свыше 500 видов. Это исключает
случайность в количественной оценке сообществ по различным биоло-
го-морфологическим признакам компонентой, которая еще далеко не
преодолена в геоботанической практике. Наряду с этим выделяются
и основные компоненты, эколого-биологический анализ которых позво-
ляет углубить характеристику сообществ и выделить их типические
черты. !
Придавая большое значение полноте охвата видового состава сооб-
ществ, мы должны предостеречь от переоценки флористических дан-
ных в различного рода геоботанических работах, в том числе в клас-
сификации фитоценозов, что имеет место в работах зарубежных
ботаников школы Цюрих — Монпелье. Исследования показывают, что
биология одних и тех же видов в разных эколого-фитоценотических и
географических условиях изменяется. Например, в условиях Стрелец-
кой степи под Курском мезофильные Rumex acetosa, Stellaria graminea
и другие приобретают среднелетний перерыв вегетации и ведут себя
как эфемероиды. Отсюда ясно, что один лишь флористический состав
4
сообществ, без тщательного анализа эколого-биологических особенно-
стей видов в данных условиях, не может служить полноценной основой
для их классификации. Это обстоятельство в равной мере следует от-
нести и к некоторым работам по изучению природы и происхождения
растительности луговых степей, рассматривающим видовой состав без
какой-либо предварительной биоэкологической оценки видов в данных
условиях.
Для характеристики фитоценозов необходимо перевести биоморфо-
логические особенности компонентов в стройную систему количествен-
ных показателей, обеспечивающих анализ и определение существенных
признаков, связанных с эколого-географическими условиями развития
растительности. Эти вопросы находят отражение в гл. VII, где выявле-
ны количественные закономерности биоморфологических признаков
надземных органов растений, слагающих ценозы луговой степи, тра-
вяной покров дубовых лесов и дна логов. Впервые проведен опыт
сопряженного анализа биоморфологических признаков растений в над-
земной и подземной сферах на основе ярусного расчленения раститель-
ности. Для луговой степи установлено правило количественной коорди-
нации видов по различным признакам в принадлежности к соответ-
ствующим (симметричным) ярусам подземной и надземной частей
травяного покрова.
Логическим следствием изучения эколого-биологических особенно-
стей растений и их состава по этим признакам в ценозах луговых сте-
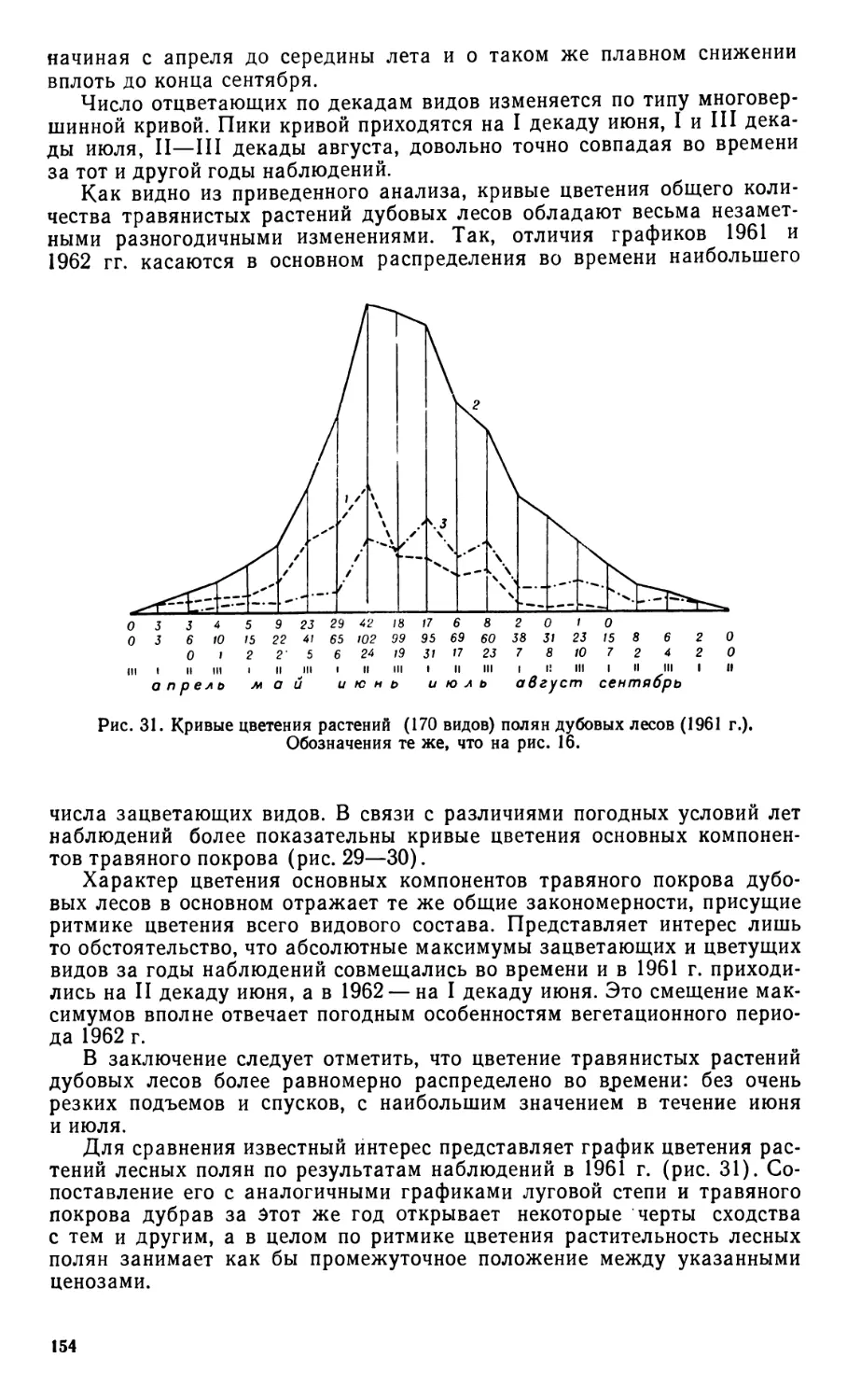
пей, травяном покрове дубовых лесов и дна логов является обсуждение
вопроса о природе и происхождении растительности лесостепи, преи-
мущественно луговых степей. Совокупностью разнообразных данных
обосновывается самобытность и оригинальность этого зонального типа
растительности, аутохтонно развивавшегося под влиянием природных
условий лесостепного ландшафта.
В качестве основного объекта эколого-биологических исследований
луговой степи взят вариант, сохраняющийся при сенокосном режиме.
Вторичное происхождение косимой степи очевидно. Однако эта произ-
водность носит не случайный, а вполне закономерный характер. Ос-
новными факторами формирования растительности являются общие
природные условия лесостепи и покос, действующие систематически,
непрерывно, что и обусловливает определенное постоянство раститель-
ности косимой степи. Последняя находится в тесной генетической связи
с некосимой степью, а точнее — с первобытными луговыми степями
лесостепи. Таким образом, косимая степь представляет закономерный
продукт природных условий лесостепи, длительного исторического раз-
вития растительности и воздействия антропогенного фактора, приобре-
тающего ныне всеобъемлющий космополитический характер.
Глава о практической значимости эколого-биологических исследова-
ний растений лесостепи является завершающей.
Помимо наблюдений в природе, для решения ряда вопросов исполь-
зовался эксперимент. Так, только экспериментальным способом выяс-
нены факультативный характер лесостепных эфемероидов, особенности
зимнего покоя растений.
Все латинские названия растений без указания авторов (за исклю-
чением гл. III) даны по П. Ф. Маевскому (1954). Ссылки в тексте на
особенности почв и их водного режима приводятся без пояснений, так
как соответствующие материалы подробно освещены ранее (Голубев,
1962в).
Некоторые вопросы биоморфологии отражены в настоящей книге
недостаточно или вовсе пропущены. По этому вопросу следует заметить,
что данное исследование нельзя рассматривать как законченный свод
всех или хотя бы главных проблем, положений и фактов по экологии и
5
биологии растений. В век стремительного полета биологической мысли
трудно очертить круг «основных проблем» того или иного научного на-
правления. Но мы и не ставили перед собой целей обзорного по-
рядка. Субъективный отбор материала, как мы уже отметили, неизбе-
жен и в силу объективных обстоятельств. Заранее соглашаясь с воз-
можным упреком в известной неполноте анализа эколого-биологиче-
ских особенностей растений и растительных сообществ лесостепи, мы
тем не менее полагаем, что сообщаемые сведения являются первым и
весьма необходимым элементом познания биоморфологии растений
любого типа растительности.
В работе над книгой благотворную роль сыграли дружеская под-
держка и советы Г. И. Дохман, Т. А. Работнова, И. Г. Серебрякова,
Н. С. Камышева, В. Г. Барышникова, которым я сердечно признателен.
Особую благодарность выражаю своему учителю — Михаилу Василье-
вичу Культиасову, взявшему на себя труд редактирования монографии.
Квалифицированную помощь в проведении фенонаблюдений в при-
роде мне оказала В. С. Жмыхова, которой приношу большую благо-
дарность.
Чувство горячей признательности выражаю проф. Ф. Э. Реймерсу,
проявившему большой интерес к моей работе и благоприятствовавшему
ее окончанию и публикации.
в
Г лава 1
Погодные условия периода наблюдений
Общая характеристика природных условий района исследований
уже дана в нашей работе (Голубев, 1962в). В настоящей главе приво-
дятся некоторые метеорологические и другие данные, преимуществен-
но по температурному режиму, осадкам и водному режиму почв, необхо-
димые для понимания особенностей сезонного развития растений по
годам.
По данным метеостанции «Стрелецкая степь» Центрально-Чернозем-
ного заповедника, температурный режим и осадки за годы наших наб-
людений характеризуются следующими показателями, приведенными
в табл. 1, 2 и на рис. 1—4. Чтобы рельефнее представить экологические
особенности развития растительности в эти годы, принимая во внимание
совокупность метеорологических явлений, приводим развернутую их
характеристику.
1959 год. Среднегодовая температура соответствовала средней мно-
голетней (4,8°), сумма осадков была на 46,1 мм меньше среднегодовой
величины (510 мм).
Зима стояла теплая с частыми оттепелями и дождями. Январская
средняя температура более чем вдвое была выше средней многолетней,
на 2,3° была выше и температура февраля. Оттепели с дождями наблю-
дались с 1 по 5 января, с 9 по 11 и с 23 по 24. На степи появи-
лись проталины, снег сильно осел. Небольшие оттепели отмечались и в
феврале.
Снеготаяние началось с 18 марта, но на первом этапе происходило
медленно. На степи снег полностью растаял 1 апреля. В период 8—
21 апреля заморозки отсутствовали, а затем вновь возобновились. 24,
25 и 30 мая снова отмечены легкие заморозки на почве (от 0 до —1,8°).
Среднемесячные температуры марта и мая были близки к средним
многолетним значениям, а температура апреля превышала многолет-
нюю на 1,6°. Осадки весенних месяцев приближались к средним мно-
голетним величинам.
Весенний запас влаги в почве после таяния снега (в апреле) состав-
лял (в мм) в трехметровой толще на некосимой степи 1018,7, косимой
степи— 1088,8, в лесу — 877,1, на некосимой поляне — 922,8.
Летние месяцы отличались засушливостью. Сумма осадков в июне
и августе была в два раза меньше средних многолетних. Совершенно
бездождными или с малым количеством осадков были III декада июня,
II и III декады июля и августа. Эти сухие периоды сопровождались
очень высокими среднесуточными температурами воздуха. В июле мак-
симальная температура воздуха равнялась 33,5°, в августе — 34,4°.
Сентябрь выдался довольно влажный и относительно прохладный.
Количество осадков вдвое превышало среднюю многолетнюю сумму.
Первый заморозок на почве зарегистрирован 8 сентября (—0,7°), далее
заморозки отмечались 18, 19, 22 и 26 сентября. Октябрь был сухой
и весьма холодный. Два раза наблюдались снегопады, однако снег
1
Таблица 1
Среднемесячная и среднегодовая температуры (в °C) воздуха (метеостанция «Стрелецкая
степь» Центрально-Черноземного госзаповедника, 1959—1962 гг.)
Год Месяц Среднего- довая
I п Ш IV V VI VII VIII IX X XI XII
1959 -4,1 —6,0 -3,8 6,3 12,5 17,0 21,6 17,7 9,3 2,3 -4,6 —10,0 4,8
1960 —6,5 —8,2 -6,8 5,1 13,7 19,4 21,4 16,9 10,3 6,0 —1,0 0,6 5,7
1961 —6,4 —4,0 0,5 6,1 11,8 19,0 19,8 18,0 11,8 6,1 —0,8 —6,1 6,3
1962 —5,0 —8,3 —2,8 8,9 14,4 16,1 16,5 17,3 12,0 7,2 1,4 —6,8 6,0
Средняя многолетняя температура —9,3 -8,7 —4,0 4,7 12,9 16,8 18,7 17,6 11,9 5,3 —1,4 —6,7 4,8
Таблица 2
Среднемесячная и годовая суммы осадков (в мм) (метеостанция «Стрелецкая
степь» Центрально-Черноземного госзаповедника, 1959—1962 гг.)
Год Месяц Годовая
I II Ш IV V VI VII VIII IX X XI XII
1959 56,3 22,9 16,2 30,3 43,1 29,2 52,2 38,4 85,0 27,6 14,4 48,3 463,9
1960 60,6 48,2 7,3 12,8 42,4 22,3 91,3 126,2 31,7 90,7 68,6 54,0 646,1
1961 23,2 19,5 38,9 55,7 165,1 39,0 50,5 39,3 31,2 18,0 38,8 40,2 559,4
1962 38,6 37,6 51,4 23,8 47,9 46,4 76,0 45,2 31,0 14,3 34,1 23,2 469,5
Среднее многолетнее количество осадков 25 19 21 31 55 59 65 77 42 48 36 32 510
быстро таял. Со II декады ноября стояли сильные морозы — до —20,7°.
Почва сильно промерзла, образовались трещины. Снежный покров
установился 25 ноября.
В течение вегетационного периода влага расходовалась преиму-
щественно из гумусового и переходного горизонтов. Влажность
карбонатного горизонта в пределах границы смены наносов и в смежных
с нею слоях сохранялась довольно высокой: на некосимой степи она
равнялась величинам, соответствующим интервалу от полной до наи-
меньшей влагоемкости (Роде, 1952), на косимой степи была равной
наименьшей влагоемкости. Впрочем, на косимой степи иссушение почвы
распространилось довольно глубоко, и верхняя граница наименьшей
влагоемкости проходила на 180 см от.поверхности.
Иной водный режим почвы наблюдался в лесу. Ниже 2 м влаж-
ность почвы в течение всего теплого сезона была в пределах величины
влажности завядания и менее. В почве некосимой поляны на этом же
уровне и несколько выше сохранялась влажность в пределах между наи-
меньшей влагоемкостью и влажностью разрыва капиллярных связей.
На некосимой и косимой степи наибольшее иссушение слоя почвы
от 0 до 50 см наблюдалось в июле с запасом влаги соответственно
в 90,2 и 85 мм, В лесу и на некосимой поляне в этом же слое наимень-
ший запас влаги отмечен в первой половине сентября (111 и 92 мм).
В трехметровой толще минимальный запас влаги в почве некосимой
и косимой степи установлен в августе (784 и 749 мм), а в лесу и в почве
некосимой поляны — в сентябре (516 и 639 мм).
8
•с
30 1 /95? г.
-30 J
месяц
Г I I I I I I I I I I I
I II III IV V VI VII VIII IX X XI XII
Рис. 1. Ход среднесуточных и минимальных температур в 1959—1960 гг., поданным
метеостанции «Стрелецкая степь» Центрально-Черноземного заповедника
1 — среднесуточные температуры; 2 — минимальные температуры
-30 J
Рис. 3. Суточные суммы осадков (в мм) за 1959—1960 ггм по данным метеостанции «Стрелецкая степь»
Рнс. 4. Суточные суммы осадков (в мм) за 1961—1962 ггм по данным метеостанции «Стрелецкая степь»
С конца апреля по октябрь запас влаги в почве в трехметровом слое
уменьшился в лесу на 319,7 мм, некосимой поляне—на 228,5 мм,
некосимой степи — на 191,8 мм, косимой степи — на 283 мм. В начале де-
кабря снеговой покров не превышал 3 см. 16 и 18 декабря прошли
сильные снегопады и высота снега в степи достигла 24 см, в лесу —
39 см. В конце месяца была оттепель, высота снегового покрова резко
упала, на степи местами появились проталины.
1960 год. Среднегодовая температура была на 0,9° выше средней
многолетней. Сумма осадков превышала среднюю величину на
136,1 мм.
Зима стояла теплая. Среднемесячные температуры января и фев-
раля были несколько выше средних за эти месяцы. Снежный покров
в январе на степи достигал 34 см высоты, в лесу — 49 см. I декада
февраля была очень холодная. В середине месяца наступила оттепель.
В результате чередования положительных температур с морозами на
поверхности снега и почвы образовалась ледяная корка.
Среднемесячная температура марта была на 2,8° ниже средней мно-
голетней. Снежный покров увеличился на степи до 45 см, в лесу — до
62 см. Потепление началось только с 27 марта. Вследствие ежеднев-
ных заморозков освобождение почвы от снега затянулось. На степи
снег растаял 7 апреля, в лесу— 13-го. Апрель был сухой и теплый: ко-
личество осадков почти в 2,5 раза было меньше средней многолетней ве-
личины, среднемесячная температура превышала многолетнюю на 0,4°;
II декада характеризовалась очень теплой погодой: осадки мая и тем-
пература были близки к среднемноголетним.
После снеготаяния глубина весеннего промачивания почвы некоси-
мой и косимой степи равнялась 50—60 см, в лесу— 130 см, некосимой
поляны в Дуброшине — 230 см. Весенний запас влаги (в мм) в трех-
метровой толще на некосимой степи составлял 720, косимой — 740,
в лесу — 761, на некосимой поляне — 892.
Лето было жаркое, вначале сухое, затем довольно влажное. Сред-
немесячные температуры июня и июля заметно превышали средние
многолетние (на 2,6 и 2,7°). Июнь отличался засушливостью. В июле
количество осадков превысило среднюю многолетнюю величину в 1,5,
а в августе — в 2 раза. Дожди выпадали неравномерно. С 5 по 7 июля
шли обложные дожди, давшие за 3 дня 71,8 мм осадков. В июле во
II и почти всей III декаде осадки отсутствовали. 6 августа сильный
ливневой дождь дал 45,3 мм осадков.
Температуры осенних месяцев были близки к средним многолет-
ним. Сентябрь характеризовался сухой погодой, тогда как осадки ок-
тября и ноября почти вдвое превышали многолетние величины. Пер-
вый заморозок на почве отмечен 14 сентября. В октябре осадки выпа-
дали почти ежедневно. В течение месяца почва не замерзала. Первый
снег выпал 26 октября, но вскоре стаял. В ноябре положительные тем-
пературы воздуха держались до 14-го. После этого начались морозы.
18 ноября установился снежный покров. В конце месяца наступило по-
тепление и снег сошел почти полностью.
В течение вегетационного периода во всех исследованных биотопах
расход влаги из почвы шел в основном из верхнего метрового слоя.
С конца апреля по ноябрь запас воды в почве в трехметровом слое
уменьшился в лесу на 185,5 мм, на некосимой поляне — на 205,4 мм.
На некосимой и косимой степи расход воды полностью компенсиро-
вался осадками, выпавшими в сентябре и октябре.
Влажность карбонатного горизонта почвы во время всего периода
наблюдений на всех площадках, кроме леса, была высокой, равной наи-
меньшей влагоемкости и величинам, близким к ней. В почве некосимой
и косимой степи влага из карбонатного горизонта практически не рас-
12
ходовалась. В лесу, как и в предыдущие два года, карбонатный гори-
зонт был сухой: за вегетационный период убыль влаги составила
52,4 мм. Значительный расход влаги из карбонатного горизонта с ап-
реля по ноябрь зарегистрирован на некосимой поляне— 180,8 лии.
Дожди, прошедшие в начале июля, увлажнили почву на всех уча-
стках только до 30—40 см. К концу июля верхняя часть гумусового
горизонта луговой степи опять сильно иссушилась. Минимальный за-
пас влаги в трехметровом слое почвы отмечался на некосимой степи
и поляне в августе (611 и 552 мм), на косимой степи — в июне
(647 мм), в лесу — в сентябре (585 мм).
Осадки, выпавшие в конце сентября, октябре, увлажнили почву до
наименьшей влагоемкости и близким к ней величинам в степи и на лес-
ных полянах до глубины 70—80 см, в лесу — до 40 см.
Декабрь был необычно теплым. Среднемесячная температура равня-
лась +0,6°. К 6 декабря снеговой покров целиком разрушился. Весь
месяц почва не замерзала.
1961 год. Среднегодовая температура превышала среднюю много-
летнюю на 1,5°. Сумма осадков за год была выше многолетней вели-
чины на 49,4 мм.
Зима, как и предыдущая, стояла теплая. Среднемесячная темпера-
тура января была на 2,9° выше средней многолетней, февраля — на
4,7°. Отмечались частые оттепели, туманы. До середины января покры-
тая снегом площадь составляла 10—20%. С 16 января установился
снеговой покров мощностью 16—17 см. В феврале он повысился в сте-
пи до 19 см, в лесу — до 23 см.
Весна была теплой и влажной. Средняя температура марта равня-
лась +5° (средняя многолетняя —4°), осадки почти вдвое превышали
многолетнюю величину. В III декаде большинство дней характеризо-
валось положительными среднесуточными температурами. Первые про-
талины наблюдались 10 марта. К 17 марта снег стаял полностью. Осад-
ки апреля также преобладали над средней многолетней суммой почти
в два раза. Заморозки зарегистрированы 2 и 3 апреля с 11 по 15 и с
21 по 25. Май отличался прохладной погодой, среднемесячная тем-
пература была меньше многолетней на 1,1°. Последний заморозок на
почве наблюдался 6 мая. В мае выпало рекордное количество осад-
ков— 165,1 мм, что в три раза превышает среднюю многолетнюю ве-
личину. Особенно сильные дожди прошли 12 мая (18,2 мм), 15
(13,5мм), 19 (22,5мм), 22 (15,5 леи) и 31 (31,5мм).
Весеннее промачивание почвы после снеготаяния было более глу-
боким по сравнению с прошлым годом. На некосимой степи почва
промокла до 250 см глубины, косимой степи — до 270 см, в лесу — до
200 см, на некосимой поляне — до 230 см. Запас влаги на всех участ-
ках превышал прошлогодний в среднем на 100—200 мм. В трехметро-
вом слое в апреле он достигал (в мм) на некосимой степи 958,5, на ко-
симой степи — 918,4, в лесу — 971,7, на некосимой поляне — 964,7.
Майские осадки, выпавшие на влажную почву, значительно увеличили
запас влаги, особенно на косимой степи и в лесу.
Лето было жарким и сухим. Среднемесячные температуры заметно
превышали средние многолетние, а количество осадков в полтора раза
было меньше средних многолетних величин. Зной сохранялся до 20 ав-
густа, а затем температура резко упала.
Осень стояла теплая и сухая. Осадки октября в 2,5 раза были мень-
ше средней многолетней суммы. Первый заморозок отмечен 22 сентяб-
ря. Снеговой покров установился 15 ноября и сохранялся до конца
месяца.
В течение вегетационного периода расход влаги из почвы происхо-
дил в основном из гумусового горизонта. Влажность карбонатного
13
горизонта на всех участках, за исключением третьего метра (200—
300 см) в лесу, в течение всей вегетации была высокой (около наимень*
шей влагоемкости). Минимальный запас влаги в слое 0—50 см наблю-
дался на некосимой степи в июле (80 мм), косимой степи и некосимой
поляне — в августе (89 и 95 мм), в лесу — в сентябре (97 мм). В трех-
метровой толще минимум влаги отмечен на косимой степи — в авгу-
сте (706 мм), некосимой степи и в лесу — в сентябре (667 и 496 мм),
на некосимой поляне — в октябре (589 мм).
С 1 по 10 декабря температура воздуха поднялась выше 0°. 2 декаб-
ря весь снег растаял. Почва оттаяла. Во II декаде наступили умерен-
ные, а в III — сильные морозы. Снеговой покров установился 10 декаб-
ря и достигал в степи 18 см, в лесу — 21 см.
1962 год. Среднегодовая температура превышала многолетнюю
на 1,2°. Сумма осадков за год была ниже средней многолетней на
40,5 мм.
В январе стояла относительно теплая погода. Мощность снегового
покрова на степи равнялась 32 см, в лесу — 47 см. Февраль был холод-
нее января.
Весна была теплая. Среднемесячные температуры заметно превы-
шали средние многолетние. В марте осадки были более чем в два раза
обильнее в сравнении со средней многолетней величиной. Осадки апреля
и мая были несколько меньше средних многолетних. В I декаде марта
наблюдались частые оттепели. Снег почти полностью сошел. Во II де-
каде прошли снегопады, метели. Снежный покров частично восстано-
вился. К концу месяца на степи снег растаял полностью, в лесу и на
северных склонах логов он сохранялся до 4—7 апреля. Последний за-
морозок отмечен 30 апреля.
Весенний запас влаги в трехметровом слое во всех биотопах был
ниже по сравнению с прошлым годом. На некосимой степи он состав-
лял в апреле 729 мм, на косимой степи — 775 мм, в лесу — 735 мм.
Лето было холодное и довольно сухое. Из четырех лет наблюдений
июнь отличался наименее низкой температурой. Осадков выпало мень-
ше средней многолетней величины, но больше, чем в каждые из преды-
дущих лет. Июль также стоял холодный. Хотя количество осадков
(76 мм) несколько превышало среднюю многолетнюю величину, но
большая их половина (44,2 мм) пришлась на один день — 8 июля.
Август был жаркий, особенно первые две декады. Осадков выпало
в полтора раза меньше средней многолетней величины.
Осень характеризовалась теплой и сухой погодой. Среднемесячные
температуры всех осенних месяцев заметно превосходили средние мно-
голетние. Осадки сентября на 11 мм были меньше средней многолет-
ней суммы.
Первый заморозок отмечен 16 сентября. Теплый октябрь отличался
сильной сухостью: за месяц выпало всего 14,3 мм осадков, что более
чем в три раза меньше средней многолетней величины. Ноябрь был
весьма теплый. Отрицательные среднесуточные температуры наблю-
дались только во II декаде. Сумма осадков была близка к многолет-
ней.
В течение вегетационного периода расход влаги, как и в прошлые
годы, шел преимущественно из гумусового горизонта. Запас влаги с
апреля по октябрь в трехметровом слое уменьшился в лесу — на 218 мм,
некосимой степи — на 139 мм, на косимой степи — на 136 мм. Наи-
меньший запас влаги в слое 0—50 см установлен на некосимой степи
в октябре (72 мм), на косимой степи — в июне и октябре (91 мм),
в лесу — в сентябре (106 мм). Минимум влаги в трехметровой толще
отмечался в лесу и некосимой степи в октябре (517 и 590 мм), на коси-
мой степи — в сентябре (637 мм).
14
В первые две декады декабря сильные морозы отсутствовали. К кон-
цу II декады температура воздуха понизилась до —15, —18°. Постоян-
ный снеговой покров установился 19 декабря.
Таким образом, каждый год отличается своими особенностями
в температурном режиме, осадках, водном режиме почв и других явле-
ниях по их значению (величине) и распределению во времени. Это со-
здает чрезвычайно пеструю экологическую обстановку для проявления
многообразных биолого-морфологических признаков растений и в це-
лом для развития растительности в разные годы.
Конечно, разногодичные колебания метеорологических факторов в
количественном выражении характерны для любой зоны. Но среди
смежных (лесной и степной) зон лесостепь выделяется исключительно
высокой амплитудой колебаний метеорологических факторов, что со-
ставляет специфическую черту лесостепной зоны. Конкретные примеры
этих колебаний достаточно отражены в литературе (Мильков, 1950;
Афанасьева, Голубев, 1962, и др.). Так, по данным курской метеостан-
ции в 1938/39 гидрологическом году выпало 379 мм осадков, а в
1952/53 — 800 мм. Большие колебания в разные годы наблюдаются в
температурном режиме, продолжительности снежного покрова, число
ясных и пасмурных дней и т. д.
Указанные климатические особенности лесостепи играют выдающу-
юся роль в формировании ее растительного покрова. Решающее значе-
ние приобретают крайние отклонения, частота их повторяемости, вы-
полняющие функции отбирающего фактора. Таким образом, все суще-
ствующие в лесостепи растения приспособились к резким изменениям
погодных условий в отдельные годы. В развитии растительности осо-
бенно велика роль осадков, значительные отклонения которых от сред-
ней величины вызывают разительные перемены в составе растительно-
сти (преимущественно луговых степей) в различные годы.
Г лава 2
Основные биоморфологические типы
структур надземных органов растений
Надземные органы растений характеризуются разнообразными мор-
фологическими признаками, имеющими определенное эколого-биологи-
ческое значение. В особенностях надземных органов отражается специ-
фика эколого-исторического развития растений, их приспособленность
к тому или иному комплексу окружающей среды. В строении надземных
органов можно, например, различать форму и структуру листьев (ши-
рокие мезоморфные, склероморфные игловидные, чешуйчатые, мяси-
стые суккулентные и т. д.), опушение листьев и стеблей, особенности
пространственного развития побегов (прямостоячие, лазящие, вьющие-
ся и др.), ветвления, их количественные соотношения у особей, разме-
ры в высоту и многое другое. Существенное значение имеют признаки
анатомической структуры стеблей, которые используются в выделении
основных биоморф растений: деревянистых и травянистых. Легко пред-
ставить, какой сложной, громоздкой оказалась бы классификация, в ко-
торой за основу выделения биоморфологических типов принимались бы
перечисленные признаки.
Поэтому в целях классификации вполне оправданным является вы-
членение биоморфологических признаков более общего и частного зна-
чения. В плане биоморфологии и применительно к задачам познания
структуры фитоценозов наибольший интерес представляют лишь неко-
торые, но существенные и четко выраженные признаки, общие для всех
покрытосеменных, допускающие возможность сравнения растений, при-
надлежащих к различным экологическим типам и жизненным формам.
К категории признаков общего значения следует отнести характер
облиствления побегов, способы их нарастания и цикличность развития;
эти признаки свойственны подавляющему большинству автотрофных
покрытосеменных. В этом смысле можно говорить об историко-генети-
ческом характере данных признаков. Такое понимание указанных
структурных особенностей отнюдь не отрицает их экологического зна-
чения. Возникновение и формирование их связано с приспособлением
растений к освоению внешних условий среды обитания. Развитие при-
способлений имело и имеет весьма широкое биологическое значение
в процессе формирования покрытосеменных как нового прогрессивного
типа высших растений. Особенности облиствления, нарастания и цик-
личности развития побегов вполне проявились уже у первых поселен-
цев суши — псилофитов и в дальнейшем совершенствовались и услож-
нялись, чем определялся путь филогенетического развития высших рас-
тений.
В выделении основных биоморфологических типов структур надзем-
ных органов травянистых растений нами используются характер об-
листвления побегов и способы их возобновления. По этим признакам в
составе изучавшихся видов можно различать такие группы растений:
1) безрозеточные симподиальные; 2) розеточные: полурозеточные сим-
16
подиальные, розеточные симподиальные, полурозеточные моноподиаль-
ные, розеточные моноподиальные.
Впервые, по-видимому, эта классификация структурных форм побе-
говой системы была разработана Т. Ирмишем (Irmisch, 1859) на при-
мере клеверов, произрастающих в Германии. В дальнейшем характер
облиствления побегов, способы возобновления и длительность их раз-
вития у травянистых растений умеренных широт были всесторонне
изучены и классифицированы Е. Вармингом. Эти биоморфологические
особенности использовались им при разработке системы жизненных
форм (Warming, 1884, 1908, 1909).
Установленные Т. Ирмишем и Е. Вармингом категории структурных
форм побегов прочно вошли в научную и учебную литературу по мор-
фологии растений (например, Velenovsky, 1907; Goebel, 1928—1933;
Troll, 1937—1942, 1954—1957; Серебряков, 1952 и др.) и широко ис-
пользуются в разнообразных классификациях жизненных форм (Drude,
1913; Raunkiaer, 1905, 1934; Warming, Graebner, 1918 и др.).
Весьма важный структурный признак — длительность развития по-
бегов— специально здесь не рассматривается, так как недавно был
глубоко освещен в работах И. Г. Серебрякова и его учеников. Вместе
с тем следует отметить, что существует известное соответствие между
типом структуры побегов и цикличностью их развития.
Так, побеги безрозеточных симподиальных растений развиваются
по моноциклическому или близкому к нему озимому типу. По существу
моноциклическими являются и генеративные побеги моноподиальных
растений. Среди розеточных (sensu lato) симподиальных растений на-
блюдается все известное разнообразие типов развития побегов, т. е.
моноциклические, озимые, ди-, плейо- и полициклические. Установлен-
ная И. Г. Серебряковым (1959а) высокая изменчивость цикла развития
побегов в пределах одной особи или у разных рядом растущих особей
одного вида свойственна именно розеточным симподиальным расте-
ниям.
Прежде чем перейти к анализу указанных выше основных групп
травянистых растений по биоморфологическим особенностям надзем-
ных органов, несколько слов посвятим терминологии.
Термин «розеточный» побег употребляется очень давно и сейчас
трудно установить, когда он впервые введен в литературу. Что касается
понятия «безрозеточный» побег, то его определение вполне четко фор-
мулируется Вармингом (см. его классификацию жизненных форм, кото-
рая приводится в книге Kirchner, Loew, Schroter, 1908). В русской
ботанической литературе термин «безрозеточные» растения употребляет
А. Г. Воронов (1952), но определения его не дает. К безрозеточным
растениям он, в частности, относит Achillea nobilis, у которой как
главный, так и побеги последующих поколений проходят фазу розетки.
В нашем понимании такие растения относятся к розеточным, у соб-
ственно же безрозеточных растений междоузлия побегов обычно вытя-
нуты и розеточная форма не образуется (Голубев, 19576). И. Г. Се-
ребряков (1952) называет эти растения нерозеточными.
Обратимся теперь к более подробному анализу указанных выше
биоморфологических типов структур надземных органов растений. Из-
вестное значение имеет уточнение определений основных типов, в трак-
товке которых нередко наблюдаются расплывчатость и неясность. Боль-
шой интерес представляет также выявление экологической обусловлен-
ности типов структур. Чрезвычайно важные вопросы эволюционного
развития структурных форм надземных побегов и их морфолого-гене-
тических связей рассматриваются в следующей главе. В обосновании и
разработке затронутых вопросов мы не сочли возможным ограничи-
ваться только фактическими материалами из флоры средней лесостепи.
2 В- Н. Голубев
17
Все эти вопросы носят принципиальный характер, их радикальное
решение доступно только с общих позиций, с использованием обширной
литературы и фактических данных из различных ботанико-географиче-
ских областей.
Всего можно выделить два основных типа структуры надземных по-
бегов растений: безрозеточный (долихоморфный) и розеточный (брахи-
морфный). Различия растений этих типов в конечном итоге сводятся
к способности или, наоборот, неспособности развивать розеточные побе-
ги, т. е. побеги со сближенными узлами, несущими нормально развитые
зеленые листья. При этом розеточный побег может представлять лишь
фазу в развитии более крупной структурной единицы — монокарпиче-
ского побега. В ряде случаев розеточный побег является самостоятель-
ным структурным образованием. Он живет либо неопределенно долгое
время, лишь постепенно нарастая с верхушки, как это бывает у моно-
подиальных растений, либо после более или менее продолжительной
жизнедеятельности отмирает, что отмечено у Carex pilosa (Серебряков,
1952). Наличие розеточных побегов у одного и того же вида не исклю-
чает возможности развития удлиненных безрозеточных побегов.
Безрозеточные растения не способны к развитию розеточ-
ных побегов и обладают только удлиненными побегами с вытянутыми
междоузлиями, вследствие чего листовые розетки не образуются. Ниже
будет показано, что различия растений по структуре побеговой системы
носят качественный характер и имеют определенное морфологическое
и экологическое значение.
Надо отличать настоящие розеточные растения от ложнорозеточных.
У первых, как уже сказано ранее, розеточная форма возникает в ре-
зультате предельного сокращения длины междоузлий при нормальном
развитии в узлах листьев срединной формации. Но розеточная форма
растения (не побега!) может возникать вследствие образования на кор-
невище (или каудексе) многочисленных облиствленных побегов с удли-
ненными междоузлиями, располагающихся приземно и радиально. Та-
кие растения следует называть ложнорозеточными. Соответствующая
этому типу структура описана в литературе у высокогорных рас куль-
турной люцерны (Medicago sativa) А. И. Беловым (1929). Ложнорозе-
точная форма наблюдалась нами неоднократно в позднеосенцее время
у целого ряда растений, образующих в это время зимующие в зеленом
состоянии побеги возобновления (безрозеточной структуры), плотно
прижимающиеся к почве, радиально расходящиеся от корневища, на-
пример у Medicago falcata, Hypericum perforatum, Stellaria graminea.
Cerastium caespitosum и др.
Резкой грани между безрозеточным и розеточным типами структур
не существует. Наблюдаются различные переходы, свидетельствующие
либо о превращении безрозеточного типа в розеточный, как отражение
нормального и наиболее распространенного пути эволюционного преоб-
разования структуры побеговой системы, либо вторичной трансформа-
ции розеточного типа в безрозеточный. Поэтому изучение каждого кон-
кретного случая переходных форм структуры представляет большой
интерес.
Переходная структура в ювенильном возрасте обнаружена у Artemi-
sia vulgaris, Bupleurum falcatum, Hieracium virosum, Linum flavum
(рис. 5). У перечисленных растений первые надсемядольные междоуз-
лия настолько укорочены, что образуется некоторое подобие розеточной
формы. Однако возникающее при этом листорасположение все же ти-
пично розеточным признать нельзя.
Иногда фаза розетки наблюдается только в развитии главного по-
бега, а побеги последующих поколений формируются по безрозеточно-
му типу. В этом можно усматривать явление рекапитуляции, т. е. по-
18
Рис. 5. Переходная структура побегов в ювенильном возрасте:
а — Artemisia vulgaris; б — Bupleurum falcatum; в — Hieracium virosum; г—Linum
flavum
вторение розеточной структуры, которой характеризовались предки
данного вида. Подобные случаи зарегистрированы у Artemisia campest-
ris, Melandrium album, Paeonia tenuifolia, Silene cucubalus, Thalictrum
minus, Th. simplex, Th. aquilegifolium. Как известно, у Trifolium repens
главный побег развивается в форме розетки с моноподиальным нарас-
танием, в пазухах листьев которой образуются безрозеточные ползучие
побеги.
Но бывает явление обратного порядка, когда главный побег разви-
вается по безрозеточному типу, а отдельные побеги последующих поко-
лений в онтогенезе проходят розеточную фазу. Эта особенность мор-
фогенеза установлена у Geranium sanguineum (рис. 6).
Первичным, более древним типом побеговой системы является без-
розеточный. Среди безрозеточных растений в средней лесостепи
2*
19
Рис. 6. Развитие Geranium sanguineum на первом году жизни в питомнике:
a — 25.V, б — 22.VI, в — 15.VIII 1861г. (посев 1.IX 1960 г.)
встречаются многолетние поликарпические: Adonis vernalis, Astragalus
glycyphyllus, Clematis recta, Cucubalus baccifer, Gypsophila paniculata,
Stachys recta и др. (рис. 7), многолетние и двулетние монокарпические
(Melilotus officinalis, М. albus), однолетние озимые (Alyssum calyci-
пит, Arenaria serpyllifolia, Acinos thymoides, Veronica verna, V. arven-
sis и др.) и яровые растения (Bidens tripartitus, Polygonum aviculare,
P. hydropiper и др.).
Особенно отчетливо безрозеточный тип побегов проявляется в ста'
дии проростков тех видов, у которых гипокотиль, эпикотиль и последу'
ющие междоузлия главного побега являются вытянутыми (рис. 7). Но
20
Рис. 7. Безрозеточный тип побегов в ювенильном возрасте растений:
a — St achy s recta; б —Clematis recta; в — Gypsophila paniculata; г — Lathy r us pisiformis;
d — Astragalus glycyphyllus; e — Cucubalus baccifer; ж — Adonis vernalis
и в этой стадии у некоторых видов уже появляются черты переходного
значения, когда, например, недоразвивается гипокотиль или гипокотиль
и эпикотиль.
У ряда поликарпических растений монокарпические побеги начина-
ют формироваться еще в конце лета и осенью, зимуют в зеленом состоя-
нии, а весной (или зимой) частично отмирают, частью продолжают рост
и развитие за счет верхушечного роста (Dianthus deltoides, Asperula
cynanchica, Galium mollugo, Medicago falcata, Polygala comosa, Vero-
nica prostrata и др.). Это растения с озимыми безрозеточными побега-
ми. Озимые растения встречаются и среди безрозеточных монокар-
пиков.
Все безрозеточные растения объединяет четкое единство структур-
ного типа и биологическое расчленение на две подгруппы: с озимыми
21
и яровыми моноциклическими побегами. Их возобновление всегда сим-
подиальное.
Розеточные растения характеризуются симподиальным или
моноподиальным возобновлением побегов. В той и другой группе раз-
личаются растения полурозеточные и собственно розеточные, в зависи-
мости от характера облиствления генеративных побегов.
Полурозеточные симподиальные растения. Их монокарпические по-
беги развиваются по следующей схеме: почка — розеточный побег —
удлиненный облиствленный генеративный побег. Вначале из почки фор-
мируется укороченный вегетативный побег с розеточными листьями. При
переходе в генеративное состояние из терминальной почки розеточного
побега образуется удлиненный цветоносный побег с зелеными листья-
ми. После плодоношения удлиненная часть монокарпического побега
обычно отмирает. На сохраняющейся базальной его части, т. е. стеб-
левой части бывшего розеточного побега, формируются почки возобнов-
ления, а затем новые розеточные побеги, повторяющие цикл развития
материнского.
Длительность розеточной фазы монокарпического побега у разных
видов весьма варьирует. По этому признаку выделяются моноцикличе-
ские, озимые, ди- и полициклические растения. При длительном суще-
ствовании (свыше года) розеточные побеги нарастают моноподиально.
Следует подчеркнуть, что в характеристике полурозеточных симпо-
диальных растений наиболее важными признаками являются наличие
розеточной фазы в развитии монокарпических побегов и облиствленность
формирующегося затем удлиненного генеративного побега. При этом не-
важно, отличаются ли листья генеративного побега от розеточных или
они весьма сходны. Таким образом, наше определение полурозеточных
симподиальных растений несколько шире, чем приводимое И. Г. Сереб-
ряковым (1952, стр. 156), согласно которому к полу розеточным симпо*
диальным относятся лишь растения с резкими различиями в строении
розеточных и стеблевых листьев.
Конечно, не все розеточные побеги переходят в фазу удлиненного
генеративного побега: часть их отмирает в вегетативном состоянии.
Во флоре средней лесостепи полурозеточные симподиальные расте-
ния— самая многочисленная группа поликарпиков. Сюда относятся:
Filipendula hexapetala, Fragaria viridis, Myosotis suaveolens, Ranuncu-
lus polyanthemus, Salvia pratensis, Scabiosa ochroleuca, Scorzonera pur-
purea, Senecio integrifolius, Serratula heterophylla, Veratrum nigrum
и многие другие. На рис. 8 показаны ювенильные особи Cnidium dubi-
um, Potentulla recta, Ranunculus auricomus, R. repens, главный побег
которых имеет типичную розеточную форму.
К полурозеточным симподиальным растениям принадлежит и боль-
шая часть злаков. Действительно, жизненный цикл побегов злаков, по
Т. И. Серебряковой (1961, 1962), складывается из следующих фаз: за-
чаточного побега, развертывания низовых листьев, неветвящегося розе-
точного побега, кущения побега, формирования соцветия и цветонос-
ного стебля, цветения и плодоношения, вторичной деятельности корне-
вища и зоны кущения. Если учесть, что цветоносные побеги многих
злаков несут стеблевые зеленые листья, то все такие виды обладают
признаками, характерными для полурозеточных симподиальных расте-
ний. Во флоре лесостепи можно назвать следующие полурозеточные
злаки: Agropyron intermedium, Agrostis syreistschikovii, Bromus ripari-
us, Calamagrostis epigeios, Helictotrichon pubescens, H. schellianum,
Phleum phleoides и др.
В характеристике структурных особенностей полурозеточных сим-
подиальных растений имеет значение не только последовательность ро-
зеточной и безрозеточной (удлиненного генеративного побега) фаз, но
22
1СМ
Рис. 8. Полурозеточный тип структуры побегов. В ювенильном возрасте
у растений формируются типичные листовые розетки:
а — Ranunculus repens-, б — Potentilla recta-, в — Ranunculus auricomus; г — Cnidium dubium
и их распределение во времени. По этому признаку можно выделить сле <
дующие варианты структурных форм монокарпических побегов:
1. В первый год (или в первые годы) развивается розетка листьев. Весной буду-
щего года из терминальной почки вновь образуется некоторое количество розеточных
листьев и только после этого формируется удлиненный ортотропный генеративный по-
бег. Указанная особенность присуща большинству видов данного типа структуры (Ви-
nias orientalis, Centaurea scabiosa, Crepis pannonica, Hieracium fallax и др.). У многих
относящихся сюда растений генеративные побеги могут развиваться по моноцикличе-
скому типу, т. е. непосредственно из почек, закладывающихся на корневищах в пред-
шествующий год. В этом случае стадия вегетативного розеточного побега, формирую-
щегося в прошлом году, выпадает.
2. В первый год формируется розетка листьев, на зиму отмирающих или частично
зимующих, из верхушечной почки которой на будущий год сразу развивается удлинен-
ный генеративный побег; розеточные весенние листья в основании генеративного побега
отсутствуют (Geranium pratense, Stipa dasyphylla, St. pennata, St. stenophylla, Valeria-
na rossica и др.).
3. Существует и смешанный тип, когда часть особей вида развивается подобно пер-
вому, а другая часть подобно второму варианту.
Кроме того, надо принимать во внимание многообразие побегов у одинаковых и у
разных видов:
а) в год развития генеративных побегов весной в их основании нет розеточных ве-
гетативных побегов (Senecio integrifolius и др );
б) весной и в начале лета генеративные особи обладают как генеративными, так
и розеточными вегетативными побегами (Centaurea jacea и др );
в) переходная группа растений, в которой часть особей развивается по первому
типу, а другая — по второму.
По типу полурозеточных симподиальных растений протекает разви-
тие многих двулетних и многолетних монокарпиков, у которых вначале
формируется вегетативная розетка листьев (рис. 9, а), а в дальнейшем
(на второй год или позднее) облиствленный генеративный побег. После
плодоношения растения нацело отмирают. Таким образом, по сравне-
нию с поликарпическими полурозеточными симподиальными растения-
23
Рис. 9. Ювенильные особи Libanotis intermedia (a)\Succisa pratensis (6); Potentilla alba (в)
ми здесь как бы сохраняется только одно поколение монокарпических
побегов. Выделяя специфику морфогенеза данной группы растений, мы
их называем полурозеточными монокарпиками, примером которых слу-
жат Berteroa incana, Campanula cervicaria, C. patula, C, sibirica, Car-
duus nutans, Cirsium polonicum, Echium rubrum, Erysimum marschal-
lianum, Libanotis intermedia, Tragopogon orientate, Trinia multicaulis,
Turritis glabra, Verbascum lychnitis и др.
Сходную структуру надземных органов имеют некоторые озимые
однолетники, развивающие в конце лета и осенью розетку листьев,
а весной следующего года — облиствленный генеративный побег (Сар-
sella bursa-pastoris, Draba nemorosa, Lithospermum arvense, Myosotis
sparsiflora и др.). Особенно много полурозеточных озимых однолетни-
ков среди сорных растений. Конкретные материалы о таких растениях
приводятся в работах И. В. Попова (1920), А. И. Мальцева (1932),
Э. Корсмо (1933), И. Т. Васильченко (1937), С. А. котта (1944,
1955) и др.
В приведенном ряду полурозеточных растений общий характер
структуры побегов остается неизменным, прогрессивно сокращается
лишь продолжительность жизни растений. Следует отметить, что по
длительности развития побегов полурозеточные монокарпики имеют
наибольшее сходство с полурозеточными поликарпиками с ди- и поли-
циклическими побегами, а озимые однолетники — с поликарпиками,
обладающими озимыми побегами. Впрочем, нет резкой грани и между
монокарпическими двулетниками и озимыми однолетниками (полурозе-
точного типа), так как некоторые двулетние монокарпики способны раз-
виваться в форме озимых однолетников, а отдельные озимые однолет-
24
ники иногда развиваются по типу двулетних монокарпиков, т. е.,
появляясь весной в виде розетки, они пребывают в таком состоянии
весь вегетационный период и только на следующий переходят к цве-
тению.
Нельзя резко разграничить поликарпические растения и монокар-
пики, так как последние изредка после плодоношения сохраняются жи-
выми и способны на другой год к формированию цветоносных побегов
второй генерации, например Cynoglossum officinale и Echium rubrum.
С другой стороны, встречаются поликарпические растения, в условиях
луговой степи развивающиеся по типу двулетних монокарпиков, на-
пример Anthemis tinctoria.
Интересный случай представляют некоторые корнеотпрысковые рас-
тения, генеративные особи которых отмирают подобно монокарпикам.
Но к моменту отмирания цветущие особи успевают образовать вегета-
тивное потомство из придаточных ^очек на’ боковых корнях. Такой
жизненный цикл мы наблюдали у Pieris hieracioides. Сюда же следует
отнести, по-видимому, и Falcaria vulgaris.
Розеточные симподиальные растения. Отличие этой категории тра-
вянистых растений от предыдущей заключается в том, что все ассими-
лирующие листья сосредоточены в прикорневой розетке, а цветоносные
побеги представлены одним вытянутым междоузлием (цветочная стрел-
ка), либо покрыты чешуевидными или рудиментарными листьями и
состоят из нескольких междоузлий. Поскольку возобновление данных
растений симподиальное, цветоносные побеги формируются из терми-
нальных почек розеточных побегов. Развитие монокарпических побегов
можно изобразить такой схемой: почка — розеточный побег — безлист-
ный генеративный побег.
Среди поликарпиков Стрелецкого участка заповедника к розеточ-
ным растениям принадлежат Allium flavescens, Anthericum ramosum,
Salvia nutans, Taraxacum officinale и др. Большое число розеточных
форм отмечено у осок: Carex humilbs, С. spicata, С. verna, С. vulpina
и др. У некоторых из перечисленных видов листья обладают длинными
влагалищами, и листовые пластинки выносятся на стебель. По листо-
расположению они уподобляются, таким образом, полурозеточным рас-
тениям, хотя в морфологическом отношении это типичные розеточные,
цветоносные стебли которых в надземной части лишены узлов, несущих
зеленые листья.
Но среди осок встречаются виды, по структуре побегов занимающие
промежуточное положение между полурозеточными и розеточными рас-
тениями, так как помимо розетки в основании цветоносного побега
имеется несколько сближенных узлов с отходящими зелеными листья-
ми; такова Carex leporina. Из злаков аналогичной переходной структу-
рой характеризуется Molinia coerulea.
Розеточной структурой обладают некоторые озимые однолетники,
например Androsace septentrionalis. Розеточные двулетние монокарпики
не обнаружены.
Полурозеточные мдноподиальные растения обладают постоянно ве-
гетативными моноподиально нарастающими розеточными побегами,
в пазухах листьев которых формируются облиствленные генеративные
побеги. После плодоношения генеративные побеги полностью отмирают
и в образовании корневища не участвуют. Корневище развивается из
осевой части розеточных моноподиальных побегов. Последние могут
ветвиться, вследствие чего разветвляется и корневище.
Полурозеточные моноподиальные растения являются исключительно
многолетними поликарпическими. В флоре средней лесостепи к ним
относятся Alchemilla micans, A. pastor alls, Betonica officinalis, Gentiana
cruciata, Geum rivale, G. urbanum, Polygonum bistorta, Potentilla alba,
P. erecta, P. patula, P. thuringiaca, Sanguisorba officinalis, Succisa pra-
tensis, Trifolium montanum, T. pratense. В ювенильном возрасте форми-
рование структуры идет по типу обычных розеточных (симподиальных)
растений (см. рис. 9, б, в).
Розеточные моноподиальные растения отличаются от предыдущей
группы наличием безлистных генеративных побегов. У них также име-
ются постоянно вегетативные моноподиально нарастающие розеточные
побеги, в пазухах листьев которых формируются безлистные цветоносы.
Сюда принадлежат виды подорожника: Plantago lanceolata, Р. media,
Р. stepposa и др. Как правило, все эти растения обычно являются поли-
карпиками. Но среди видов есть и однолетние формы, по своей струк-
туре аналогичные многолетним поликарпическим формам.
Заканчивая обзор структурных форм розеточных растений (в широ-
ком смысле), следует отметить одно интересное явление, связанное с
пространственным расположением розеточных листьев. По этому при-
знаку все растения можно разделить на две группы: со стабильным
пространственным размещением и переменным. Нами замечено, что у
ряда растений, например у Campanula persicifolia, Hieracium pilosella.
Myosotis suaveolens, Turritis glabra и других, зимующие листья ранней
весной (и осенью) образуют розетку, плотно прижимающуюся к почве,
а молодые листья текущего года развиваются в вертикальном направ-
лении или косо вверх. Эта сезонная периодичность 'пространственного
размещения листьев соответствует изменениям экологического режима
и является приспособительным признаком.
Большое значение имеет изучение эколого-географических законо-
мерностей развития розеточных растений, на что указывал И. Г. Сереб-
ряков (1952, стр. 159). Специальных исследований по этому вопросу
нет, но встречаются отдельные высказывания и приводятся некоторые
факты, лишь частично раскрывающие особенности географической ло-
кализации розеточных растений и экологические условия их развития.
По имеющимся сведениям, розеточные растения (в широком смыс-
ле) особенно характерны для высокогорных местообитаний: нивально-
го; альпийского и субальпийского поясов (Варминг, 1902; Warming,
Graebner, 1918; Schrdter, 1926; Коровин, 1934; Голоскоков, 1949; Але-
хин, 1950, Ь951; Дохман, 1954; и др.). Кроме того, некоторые указания
на приуроченность отдельных розеточных растений к высокогорьям
имеются в работах К. В. Станюковича (1949), И. М. Культиасова
(1955), А. П. Стешенко (1956) и др.
По данным Е. Варминга (1902), розеточные растения встречаются
также в полярных странах, в степях и пустынях, среди эпифитов и оби-
тателей скал в тропических областях. В большом количестве розеточ-
ные растения представлены в тундровой (Городков, 1935) и лесной
зонах. По нашим подсчетам, из 300 изученных видов Подмосковья
160 являются розеточными (всех категорий), что составляет 53% от
общего числа. В луговых степях Северного Казахстана розеточные рас-
тения достигают 55%, а в настоящих степях, в разных ассоциациях,
количество их колеблется от 60 до 82% (Борисова, 1961). Впрочем,
по данным Г. И. Дохман (1954) для европейской территории, число
розеточных растений в южных степях вдвое меньше по сравнению с
луговыми степями. Есть основания предполагать, что в более южных
районах, в области среднеазиатских пустынь, число розеточных расте-
ний еще меньше, чем в настоящих степях. Но далее на юг, в горных
цепях Тянь-Шаня и Памиро-Алая количество розеточных растений в
составе растительности опять резко возрастает. Таким образом, в гео-
графической локализации розеточных растений наблюдается опреде-
ленная закономерность.
Ввиду необычайной широты географического распространения
розеточных растений, биологическое значение розетки должно оцени-
26
ваться по-разному в зависимости от особенностей экологии растений.
Например, по Е. Вармингу (1902), у тропических ананасовых листовая
розетка выполняет функцию резервуара, в котором накапливается и
хранится дождевая влага. У агав же расположение листьев в форме ро-
зетки предохраняет растения от чрезмерного испарения и вредного дей-
ствия солнечных лучей. Что касается альпийских и арктических расте-
ний, то их розеточная форма, приземистое расположение листьев, по
Е. Вармингу, защищают растения от иссушающего действия ветров и
холода. Благодаря тому что листья находятся на самой поверхности поч-
вы, растения оказываются в более благоприятном температурном ре-
жиме.
Замечательные факты влияния условий среды на выработку эозр-
тОчной структуры получены для растений высоких поясов гор. Климати-
ческие условия альпийских высот весьма полно проанализированы Шре-
тером (Schrdter, 1926).
Особенности альпийского климата накладывают неизгладимый от-
печаток на жизнь и весь внешний облик растений. Характерные черты
морфогенеза альпийских растений — укорочение междоузлий и розе-
точная форма роста, малооблиствленные или безлистные короткие цве-
тоносы, ускоренный ритм развития, способность к раннему зацветанию
и другие — все это есть следствие влияния альпийских условий. Как
показывают опыты Бонье (Bonnier, 1890, 1910), при переносе равнин-
ных растений с удлиненными побегами в высокогорье они приобретали
низкорослую розеточную форму. Сходные данные приводит Г. И. Поп-
лавская (1948): в альпийском поясе Крымских гор (Яйла) Cerastium
hiebersteinii развивается в виде невысокого розеточного растения, тогда
как на меньшей высоте, в лесном поясе, ясколка является высокорос-
лым растением и розеточных побегов не образует.
О вероятности непосредственного влияния на образование розеточ-
ных побегов специфического состава света высокогорий свидетельству-
ют опыты с бриофиллум Б. С. Мошкова (1951). На дневном освещении
бриофиллум развивает удлиненные побеги с вытянутыми междоузлия-
ми, а при освещении сине-фиолетовыми лучами рост междоузлий тор-
мозится и возникает розеточная форма побега. Можно предположить,
что обилие в альпийском поясе ультрафиолетовых лучей, близких к
сине-фиолетовым, оказывает еще более сильное тормозящее действие
на рост междоузлий, о чем говорят и данные Г. Люндегорда (1937).
Но не только особая часть спектрального состава солнечного излу-
чения вызывает сокращение прироста междоузлий стебля и образова-
ние розеточной формы побега. В целом все солнечное (излучение, как
известно из физиологии, влияет на сокращение прироста стебля в дли-
ну, а это, в свою очередь, благоприятствует выработке розеточных
форм. Экспериментально доказано, что в темноте некоторые розеточные
растения, например Sempervivum assimile, S. tectorum, Taraxacum of-
ficinale, теряют розеточную форму (Wiesner, 1907).
Увеличение прироста междоузлий и выход из розеточного состояния
побегов мы наблюдали в опытах искусственного окучивания Pimpinella
saxifraga и Knautia arvensis. Эти растения относятся к стержнекорне-
вым симподиальным полурозеточным травам. Опыт заключался в том,
что с осени растения засыпались с поверхности слоем почвы от 10 до
15 см. На следующий год из почек возобновления вместо розеток раз-
вивались подземные ортотропные побеги в насыпанном слое почвы,
представляющие типичные корневища. Но как только верхушка корне-
вища достигала дневной поверхности, так сразу же рост осевой части
прекращался, а из терминальной почки формировалась листовая ро-
зетка. Сходным образом реагирует Trifolium montanum на покрытие
«слоем наилка во время весеннего половодья в условиях поймы. При
27
этом от короткого вертикального корневища (каудекса) формируются
тонкие ортотропные корневища, по длине соответствующие толщине
покрывающего слоя ила. При выходе на поверхность они развивают ли-
стовые розетки.
Необычайно эффектное влияние затенения на выход из розеточной
формы мы наблюдали у среднеазиатских растений. Вегетативные ро-
зетки Rosularia alpestris, R. paniculata, R. turkestanica, заложенные в
гербарий, по прошествии трех месяцев оказались проросшими в удли-
ненные облиствленные вегетативные побеги.
Ряд фактов о влиянии света на форму роста побегов получен нами
в наблюдениях над однолетними розеточными сорняками. Например,
у Matricaria matricarioides, Senecio vulgaris в результате затенения
происходило удлинение междоузлий, что обусловливало замену розе-
точной формы роста на безрозеточную. Рассмотрим это явление более
подробно. ।
Matricaria matricarioides является однолетником, способным развиваться по ози-
мому и яровому типу. При осеннем прорастании семян образуется проросток, главный
побег которого представляет листовую розетку, прилегающую к поверхности почвы.
В открытом поле при весеннем прорастании семян проросток ромашки также пред-
ставляет приземистое розеточное растение. Из терминальной почки розетки позднее
формируется удлиненный облиствленный генеративный побег. В других условиях, при
значительном затенении у ярового проростка ромашки, эпикотиль и последующие
междоузлия хорошо развиты, вследствие чего растение развивается по безрозеточному
типу, минуя стадию розетки. Такие проростки мы в изобилии находили на грядках,
разбитых под пологом яблоневого сада. Аналогичное явление в соответствующих
условиях обнаружено у Senecio vulgaris.
Нечто подобное установлено в опытах выращивания Achillea millefolium. Посев се-
мян произведен осенью в ящики, которые выдерживались под снегом до весны. В конце
апреля, после того как почва освободилась от снега, ящики внесены в жилое помеще-
ние и поставлены против окна, обращенного на запад. У развившихся проростков глав*
ный побег обладал вытянутыми междоузлиями. В то же время проростки, развившиеся
в питомнике при прямом солнечном освещении, имели вид настоящих розеточных расте-
ний с редуцированными междоузлиями главного побега (Голубев, 19586).
В сомкнутом травостое, при боковом затенении, обычно розеточные растения Agri-
monia eupatoria, Campanula glomerata, Sisymbrium polymorphum способны развиваться
по безрозеточному типу.
Вместе с тем в условиях сильного освещения некоторые безрозеточные растения
образуют розеточные побеги, особенно в стадии проростков, что установлено нами
у Artemisia absinthium, A. austriaca, A. vulgaris, Aster amellus, Bupleurum falcatum, La-
vatera thuringiaca и др.
Все эти факты, как нам кажется, говорят о существовании прямого
влияния освещения на форму роста побегов. Затенение розеточных рас-
тений в ряде случаев вызывает увеличение прироста междоузлий в
длину, вследствие чего побеги развиваются по безрозеточному типу,
а усиление освещенности обусловливает иногда формирование розеточ-
ных побегов у безрозеточных растений.
В выработке розеточной структуры большую роль, несомненно,
играл температурный фактор. В этой связи весьма показательно, что
розетку часто рассматривают как орган перезимовки и покоя (Лиси-
цын, 1947; Дохман, 1954; и др.). Несмотря на то, что у многих растений
розеточные листья не доживают до следующего вегетационного перио-
да и отмирают в предыдущий осенне-зимний сезон, даже в этих случа-
ях розеточная форма побега обусловливает базитонное 'размещение
почек возобновления — на самой поверхности почвы. Такое положение
почек и в целом листовых розеток обеспечивает благополучную перези-
мовку в условиях сурового климата.
В Арктике при особо неблагоприятных температурных условиях
многие растения развиваются исключительно в форме розеток прикор-
невых листьев.
О влиянии пониженной температуры на выработку розеточной
структуры говорят, по-видимому, известные в литературе факты сезон-
28
кого диморфизма однолетних растений, когда в развитии их по озимому
типу образуется зимующая розетка, а по яровому — безрозеточное рас-
тение. Подобное явление обнаружено у Thlaspi arvense, Descurainia
sophia, Senecio vulgaris (Мальцев, 1926), Lycopus arvensis (Васильчен-
ко, 1937), Malcolmia grandiflora (Нечаева, Василевская, 1945; Нечаева,
1958), Lepidium filifolium (Келлер, 1948) и др. Об этом же свидетель-
ствуют исследования С. А. Котта (1944), показавшего связь розеточной
формы роста с неблагоприятными температурами и другими условия-
ми, на примере растений Сибири.
Таким образом, можно заключить, что розеточный рост и розеточная
структура побегов являются важной приспособительной особенностью
растений к переживанию холодного зимнего сезона. Являясь в боль-
шинстве гемикриптофитами, розеточные растения хорошо отвечают кли-
матическим условиям умеренно холодных и альпийских областей. Это
обстоятельство, на наш взгляд, служит весьма веским косвенным дока-
зательством влияния температурного фактора на выработку розеточной
структуры побегов у травянистых растений.
Анализ показывает, что не может быть однозначной оценки эколо-
го-биологического значения розеточной формы роста побегов. Свет,
температура, ветер и другие факторы в их разнообразных сочетаниях
играют существенную роль в утверждении розеточной структуры. Но
такая оценка была бы неполной, если в вопросе о выработке розеточно-
го типа побегов исходить только из пассивной роли растений, реагиру-
ющих на внешние условия сокращением роста междоузлий. Розеточная
форма побегов доставляла растениям громадные преимущества в том
отношении, что в ограниченном объеме пространства они получали,
благодаря розеточным побегам, возможность развития максимальной
листовой поверхности. А это неизмеримо повышало способность к про-
дуцированию органического вещества. В прогрессе продуктивности и
следует, вероятно, усматривать основной эволюционно-биологический
смысл возникновения и становления розеточной структуры побегов. На
этой основе, в связи с теми или иными контрастами климатических
факторов, могли уже сформироваться своеобразные приспособительные
типы, у которых розеточная форма роста является ответной реакцией и
конкретным воплощением биоморфологического процесса развития,
определяющегося способностью растения при этой морфологической
структуре достигать наибольшей продуктивности.
Сопоставляя формы структуры подземных и надземных органов изу-
ченных растений в данных условиях существования легко убедиться,
что прямого соответствия между ними нет, о чем уже упоминалось нами
ранее (Голубев, 1962в). Развитие приспособлений и структурных форм
в подземной и надземной сферах совершалось под влиянием различных
факторов, что и определило отсутствие непосредственных структурных
корреляций.
Глава 3
Некоторые закономерности эволюции
биоморфологических структур растений
Вопросы эволюции биоморфологических структур растений чрезвы-
чайно многогранны и широки. В рамках проблемы жизненных форм 1
они были предметом неоднократного обсуждения. Весьма обстоятель-
ные обзоры литературы приводятся в работах А. Л. Тахтаджяна (1948),
М. В. Сеняниновой-Корчагиной (1949), А. П. Шенникова (1950), М. В.
Культиасова (1950), И. Г. Серебрякова (1954а, б, 1955а, 1962) и др.
Ниже мы коснемся лишь самых общих вопросов эволюции биомор-
фологических структур растений, главным образом в том объеме, ко-
торый диктуется содержанием настоящей работы. Основное же внима-
ние уделяется анализу биоморфологических структур и их генетическим
отношениям в пределах определенных систематических категорий, пре-
имущественно родов.
Широкое использование систематико-филогенетических данных для
раскрытия закономерностей исторического развития жизненных форм
ограничивается сравнительно малым количеством монографий, полно-
ценно освещающих филогению тех или иных систематических единиц.
Но даже при наличии таких монографий в большинстве случаев сведе-
ния о структуре вегетативных органов рассматриваемых видов обычно
отсутствуют совершенно или приводятся весьма отрывочно и неполно.
Это и понятно, так как филогения покрытосеменных все еще базируется
в основном на признаках генеративных органов. Поэтому из системати-
ко-филогенетических работ, как правило, можно заимствовать лишь
данные о родственных отношениях видов, структурно-биологические
особенности которых приходится устанавливать специально. В этих це-
лях мы использовали изучение структуры видов в условиях их есте-
ственного произрастания, а также путем выращивания в питомнике из
семян и по гербарным образцам.
Привлечение данных филогенетической систематики для выяснения
особенностей исторического развития жизненных форм весьма перспек-
тивно. Об этом свидетельствуют работы Г. Мейзеля (Meusel, 1951,
1952), Шлитлера (Schlittler, 1955, 1957), В. Г. Хржановского (19586),
К. Ю. Эйхвальда (1959), Г. И. Родионенко (1961) и др. Нельзя, одна-
ко, думать, что филогенетическая систематика способна разрешать все
вопросы соматической эволюции. Напротив, очень часто приходится об-
ращаться к эволюционной морфологии, чтобы определить генетические
отношения тех или иных циклов форм. Наиболее плодотворным являет-
ся комплексный анализ эволюции биоморфологических структур, на
основе координации и синтеза систематико-филогенетических и эволю-
ционно-морфологических представлений.
1 Термин «жизненная форма» здесь и в дальнейшем нами понимается в узком
смысле и соответствует понятию «тип биоморфологической структуры». Эта оговорка
представляется необходимой вследствие разноречивой трактовки жизненных форм
в современной экологической литературе.
30
ОБЩИЕ ПОЛОЖЕНИЯ
Перед тем как приступить к анализу эволюции биоморфологических
структур внутри отдельных родов цветковых растений, было бы инте-
ресно коснуться некоторых особенностей развития всего типа покрыто-
семенных: области их первоначального происхождения, исходной жиз-
ненной формы, направлений трансформации структуры в связи с
определяющими ее факторами и других, раскрывающих содержание
проблемы биоморфологической эволюции.
В вопросе о месте происхождения покрытосеменных существуют две
основные точки зрения, подробно освещенные в работах А. Л. Тахта-
джяна (1957) и В. Н. Васильева (1958). Согласно одной из них, по
времени более ранней, местом развития покрытосеменных были высо-
кие широты северного полушария, его арктические области. Уточняя
район возникновения покрытосеменных, Сюорд и Конвей (Seward and
Conway, 1935) указывают на Гренландию как первичную колыбель
цветковых растений, откуда они расселялись циркумполярно и мигри-
ровали к югу, распространяясь по всему земному шару. Начало этим
представлениям положили палеоботанические исследования О. Геера,
открывшие в Гренландии существование теплолюбивой флоры антофи-
тов. Ряд последующих исследований обнаружил остатки теплолюбивой
флоры на Колыме и в других пунктах Арктики. Эти палеоботанические
находки относили к нижнемеловому времени.
Позднее ископаемые остатки ангиосперм были найдены южнее ука-
занных пунктов, но их возраст считали более молодым. В силу того что
первые сведения о древних покрытосеменных, по воле случая, поступи-
ли из арктических областей, создалось впечатление, что эти области
являются и центром развития антофитов (Васильев, 1958). Современ-
ные исследования ископаемой флоры Комо в Гренландии показывают,
что возраст ее должен датироваться не нижнемеловым, а верхнемело-
вым временем (Вахрамеев, 1952; и др.). Поэтому нет никаких основа-*
ний принимать ископаемые остатки Арктики за наиболее древних пред-
ставителей покрытосеменных.
Весьма веским аргументом в отрицании арктического происхождения
цветковых растений и их миграции к югу являются доводы В. Н. Ва-
сильева (1958) против самой возможности подобной миграции. Автор
приходит к заключению, что каждый флористический комплекс при
распространении встречает сопротивление со стороны сходного с
ним комплекса, находящегося в условиях, не худших, чем первый.
В силу указанных причин смешанная арктическая флора тертьера при
ухудшении условий существования не сдвинулась с места и вымерла
in situ.
В последнее время все большее число сторонников приобретает ги-
потеза тропического и субтропического происхождения покрытосемен-
ных. По подсчетам М. И. Голенкина (1927), большая часть семейств
покрытосеменных сосредоточена в тропиках, из них 70 — исключитель-
но тропические, главное ядро других 90 семейств лежит также в тро-
пиках и только 40 семейств являются собственно бореальными. Флоро-
генетический анализ многих представителей бореальных семейств, про-
веденный В. Н. Васильевым (1958), помог установить, что корни их
происхождения тяготеют к восточной или юго-восточной внетропиче-
ской Азии. Эти данные показывают, как в процессе эволюции тропиче-
ские типы первичных покрытосеменных, переходя в теплоумеренные
условия, постепенно трансформировались в современные бореальные.
А. Л. Тахтаджян (1957), применяя филогенетический метод, приходит
к выводу о том, что наиболее примитивные семейства покрытосемен-
ных приурочены к окраинам и островам Тихого океана. Родиной
31
покрытосеменных он считает древнюю Катазию, куда входили террито-
рии Китая и Индо-Китая.
За тропическое происхождение ангиосперм высказывались: Галлир
(Hallier, 1912), Синнот, Бейли (Sinnott, Bailey, 1914а, б), М. Менз-
бир (1922), Б. М. Козо-Полянский (1922), Бьюс (Bews, 1925),
В. Б. Мак-Дуголл (1935), М. Г. Попов (1940а, 1948), Е. В. Вульф
(1944), Бейли (Bailey, 1949), А. П. Шенников (1950), Е. И. Проскоря-
ков (1950), В. Шафер (1956), П. У. Ричардс (1961), И. Г. Серебряков
(1962) и др. А. А. Яценко-Хмелевский (1958) также склоняется к при-
знанию тропического или субтропического происхождения покрытосе-
менных на основании анатомических данных. Наиболее примитивными
являются гомоксилярные двудольные, большинство которых и сейчас
сконцентрировано в области Новой Каледонии и прилегающих остро-
вов. Гомоксилярные двудольные не способны развиваться в засушли-
вых областях. Поэтому первые покрытосеменные, по всей вероятности,
были приурочены к теплым и влажным биотопам.
Несмотря на значительное число работ, посвященных выяснению
родины покрытосеменных, этот вопрос еще далек от удовлетворитель-
ного разрешения. Выше были освещены только две основные точки
зрения на происхождение ангиосперм. В действительности их суще-
ствует большое количество, причем периодически появляются все новые
и новые гипотезы. Можно, например, отметить гипотезу горного про-
исхождения, ныне приобретающую новых сторонников (Штемпель,
1959; и др.).
Биоморфологическая эволюция покрытосеменных в основном своем
русле протекала от деревьев к травам. По мнению А. Л. Тахтаджяна
(1948), первичными антофитами были «небольшие деревца с очень
слабо разветвленной кроной и относительно немногочисленными тол-
стыми ветвями» (стр. 269). Палеоморфное значение древесных форм
по сравнению с травянистыми обосновывают Е. Джеффри (Jeffrey,
1899), Г. Галлир (Hallier, 1905), И. Бейли (Bailey, 1911), Б. М. Козо-
Полянский (1922, 1945), М. И. Голенкин (1927), М. Г. Попов (1940а,
1948), А. Л. Тахтаджян (1943, 1948, 19546), В. В. Алехин (1936),
А. А. Гроссгейм (1950), Е. И. Проскоряков (1950), М. В. Культиасов
(1950), Г. Ганзен (Hansen, 1956), Е. П. Коровин (1958), В. Г. Хржанов-
ский (1958а, б), К. Ю. Эйхвальд (1959), И. Г. Серебряков (1954а, б,
1955а, 1962) и др. Для доказательства вторичности травянистых форм
ангиосперм используются анатомические, палеоботанические и систе-
матические данные, результаты ботанико-географических и биолого-
морфологических исследований. Литература по этим вопросам обстоя-
тельно проанализирована А. Л. Тахтаджяном (1948) и И.. Г. Серебря-
ковым (1962). Здесь мы коснемся лишь анатомических и других
доказательств, которые одними из первых использованы для выведения
травянистых растений из древесных. В этих целях обратимся вначале
к отличиям деревьев от трав по анатомическим признакам.
Как известно (Василевская, 1954; и др.), основное отличие древес-
ных растений от трав заключается в том, что у первых наблюдается
неопределенно долгое (до конца жизни растения) утолщение стебля,
тогда как утолщение травянистых стеблей ограничено. В прекращении
утолщения весьма велика роль цветения и плодоношения. Если создать
условия для продолжения деятельности камбия, то развитие трав будет
мало чем отличаться от деревьев. Возникнет структура стебля, сходная
с таковой у древесных растений. Подобные опыты удались с подсолнеч-
ником при благоприятных для роста условиях и с клещевиной — при
удалении цветоносных побегов. Следует иметь в виду, что тропические
виды клевещины являются древесными растениями, а виды подсолнеч-
ника в субтропиках и тропиках — полукустарниками.
32
Древесные стебли характеризуются тем, что ксилемные элементы
образуют сплошное кольцо, очень мало места в центральном цилиндре
приходится на узкие сердцевинные лучи, клетки которых в ксилемной
части сходны по строению с некоторыми клетками древесины.
Травянистые стебли отличаются большой раздельностью сосудисто-
волокнистых пучков, сердцевинные лучи состоят из клеток, близких по
строению к клеткам паренхимы первичной коры и сердцевины.
Изучение ранних фаз развития травянистых растений показало, что
нижние участки стеблей вместе с гипокотилем рекапитулируют некото-
рые черты более древней древесной структуры. Так, по исследованиям
В. Г. Александрова и О. Г. Александровой (1928), в гипокотиле подсол-
нечника древесина представлена сплошным цилиндром, состоящим из
перемежающихся участков ксилемы пучков и древесинной паренхимы,
что служит убедительным доказательством происхождения травянисто-
го типа структуры от древесного. Таким образом, в этом и подобных
случаях «нижняя часть травянистого стебля обладает более древними
чертами структуры по сравнению с верхней» (Александров, 1954,
стр. 227).
Ряд ретенций описывает А. Л. Тахтаджян (1948). Например, у тра-
вянистых Zygophyllaceae стебли в базальной части древеснеют и имеют
такое же строение, как у древесных представителей семейства. Сходное
явление наблюдается у травянистых видов рода Sophora, особенно
у S. prodanii Anders.
Травянистый тип структуры выводится из древесного и на основании
анатомического изучения перфорации сосудов. По исследованиям
А. А. Яценко-Хмелевского (1948), исходным и наиболее примитивным
типом перфорации сосуда является лестничная с многочисленными пе-
рекладинами и с более или менее выраженным окаймлением каждой
перфорации. Более специализирован и эволюционно подвинут тип про-
стой перфорации сосуда, возникающий в результате постепенного
уменьшения числа перекладин и исчезновения окаймления. 80 семейств
цветковых растений, состоящих исключительно или в большинстве сво-
ем из травянистых видов, характеризуются простой перфорацией со-
судов, тогда как семействам древесных растений более свойственна
лестничная перфорация. Простая перфорация сосудов имеется у Ranun-
cutaceae, Geraniaceae, Papaveraceae, Cruciferae, Cucurbitaceae, Campa-
nulaceae, Compositae (за немногими исключениями) Solanaceae, Scro-
phulariaceae, Boraginaceae Primulaceae, Umbelliferae и др. Таким об-
разом, коррелятивная связь анатомического строения проводящей си-
стемы с общей структурой свидетельствует в пользу производного
характера травянистых растений.
Вторичность травянистых растений покрытосеменных в сравнении с
древесными вытекает из анализа различных систематических групп.
По данным А. Л. Тахтаджяна (1948), в порядке Ranales наиболее низ-
ко организованные семейства состоят исключительно из древесных
форм. Таковы Magnoliaceae, Winteraceae, Illiciaceae, Schizandraceae,
Degeneriaceae, Hemantandraceae, Anonaceae, Eupteleaceae и др. Более
подвинутые представители раналиевых относятся уже к травянистым
формам. Аналогичная закономерность имеет место во многих других
порядках, семействах и даже родах, «в которых встречаются как дре-
весные, так и травянистые формы и где с известной уверенностью
можно говорить об относительной древности форм, судя по строению
цветка, примитивные типы оказываются древесными, а более новые
травянистыми» (Серебряков, 1962, стр. 132).
Ботанико-географическими исследованиями установлено, что макси-
мальное сосредоточение древесных растений наблюдается в тропиках,
где находится родина покрытосеменных, а в умеренной и холодной
3 В. Н. Голубев .33
зонах преобладают травянистые виды (Sinnott, Bailey, 19146). Эти
данные лишний раз свидетельствуют в пользу первичности древесных
растений. Развитие древесной формы в тропиках определяется климати-
ческими факторами, при которых, по образному выражению М. И. Го-
ленкина (1927), «все растения как бы стремятся сделаться деревьями»
(стр. 38).
Большинство авторов признает три основные ступени биоморфоло-
гической трансформации ангиосперм: дерево — кустарник — трава.
В развернутом же виде схемы преобразования древесных в травянистые
формы усложняются введением промежуточных ступеней и детализа-
цией основных типов. Так, А. Л. Тахтаджян (1948) принимает следую-
щую цепь превращений: деревья — кустарники — полукустарники —
многолетние травы — однолетники. Сходная схема эволюции воспроиз-
водится Г. Галлиром (Hallier, 1905) и М. Г. Поповым (1940а, 1948).
Этот редукционный по морфологии и прогрессивный в эволюционном
отношении ряд превращений, по М. Г. Попову (1948), включает такие
звенья: дерево — лиана (кустарник) — (полукустарник)—многолетняя
трава— (двулетник) —однолетник.
Несмотря на полное единодушие в отношении основного направления
трансформации сомы покрытосеменных, конкретные пути структурно-
морфологической (габитуальной) эволюции разработаны еще недоста-
точно.
Представляется весьма вероятной многоканальность развития тра-
вянистых растений из древесных, возникновение их особыми путями на
основе трансформации разных исходных биоморф в различных филе-
тических линиях. Возможны следующие варианты преобразования де-
ревянистой структуры в травянистую:
1. Деревья — лианы древесные—(обычные травы)—лианы травя-
нистые. Большое число фактов в пользу этого пути эволюции приводит
М. Г. Попов (1948). Например, Campanulaceae — типичное лесное мезо-
фильное семейство из травянистых представителей — имеет в своем
филетическом базисе лиану (Сашрапшпаеа, частично Codonopsis).
Лиановидные предки установлены в родах Rosa (Хржановский, 19586),
Rubus (Эйхвальд, 1959). Путь от деревьев к деревянистым и травяни-
стым лианам обосновывают М. В. Сенянинова-Корчагина (1949),
И. Г. Серебряков (19556). На возможность развития травянистых рас-
тений из древесных в тропических лесах в результате приспособления
к лазящему и вьющемуся образу жизни указывает А. Л. Тахтаджян
(1948, стр. 30).
2. Как полагает А. Л. Тахтаджян (1943, 19546), травянистые расте-
ния могли возникнуть непосредственно из деревьев в результате неоте-
нии. В подтверждение этого взгляда автор приводит следующие сооб-
ражения. Стебли травянистых растений вполне сравнимы с первыми
годичными кольцами соответствующих древесных предков, но с реду-
цированной древесиной. Редукция вторичной древесины вызвана умень-
шением камбиальной активности, что является ведущим фактором раз-
вития травянистых стеблей, сопровождавшегося усиленной их парен-
химатизацией (Sinnott, Bailey, 1914а, б, 1915). Увеличение
относительного объема паренхимной ткани есть результат простого
расширения лучей. «Во всех же существенных своих признаках травя-
нистый стебель похож на первое годичное кольцо родственных древес-
ных форм. Поэтому процесс редукции от древесного типа к травяни-
стому сводится по существу к постепенной или внезапной фиксации
«травянистых» структур, древесного стебля» (Тахтаджян, 19546,
стр. 93).
3. Нормальный ряд трансформации с промежуточными ступенями
как выражение процесса сокращения длительности жизненного цикла
34
надземных скелетных осей (Серебряков, 1955а, 1962). В этом ряду
И. Г. Серебряков помещает следующие основные биоморфы: высоко-
ствольные деревья — немногоствольные деревья — кустарники — кус-
тарнички— травянистые многолетники — травянистые однолетники.
4. Развитие трав через промежуточную стадию полукустарника и
полукустарничка. Промежуточное положение полукустарников между
деревянистыми растениями и травами принимается большинством ис-
следователей, предложивших системы жизненных форм: Гризебахом
(Griesebach, 1872); Друде (Drude, 1913); Вармингом, Гребнером (War-
ming, Craebner, 1918); Браун-Бланке (Braun-Blanquet, 1928); Дю-Рие
(Du-Rietz, 1931); И. К. Пачоским (1921); А. В. Прозоровским (1935,
1940); Б. С. Закржевским, Е. П. Коровиным (1935); М. С. Шалытом
(1955); Р. А. Ротовым (1959) и др. На переходное значение полукустар-
ников указывает В. И. Талиев (1933). В. В. Алехин (1950) отмечает ха-
рактерную особенность полукустарников — перезимовку нижних частей
стеблей и отмирание их верхушек и рассматривает полукустарники
как промежуточную форму между кустарником и травой. А. Л. Тахта-
джян (19546) допускает наряду с внезапным превращением дерева в
траву и постепенное преобразование через стадии кустарника и полу-
кустарника.
Несмотря на то что мнение о промежуточном характере полукустар-
ников в ряду эволюции от деревьев к травам является почти общепри-
нятым, все-таки оно нуждается в существенных коррективах. Во-пер-
вых, по происхождению полукустарники представляют сборную группу,
берущую начало от древесных и травянистых форм (Козлова, 1953;
Серебряков, 19546; Рачковская, 1957; Голубев, 19606). Во-вторых,
в аридных областях, где сосредоточен максимум полукустарников, по-
следние, как правило, являются заключительным звеном биоморфоге-
неза и не дают выхода к развитию других биоморф, в том числе тра-
вянистых (Голубев, 19606). Преобразование полукустарника в траву
может происходить за пределами аридных областей, что имело место,
по всей вероятности, довольно редко.
Еще более категорично высказывается О. Н. Радкевич (1928). Она
пишет: «В противоположность деревьям мц имеем у полукустарников
решительную тенденцию к образованию мягких элементов во вторич-
ных тканях многолетних органов, так что ни в коем случае нельзя их
рассматривать как промежуточное звено между структурой трав и
деревьев, как это делала теория Негели для кустарников» (стр. 435).
5. Развитие травянистых форм протекало и в параллельных главно-
му— побочных направлениях эволюции жизненных форм: в ряду сте-
лющихся подушковидных, суккулентных и корнеотпрысковых форм,
а также среди форм с опадающими ветвями — эремофитов (Серебря-
ков, 1955а).
Эволюция жизненных форм от деревьев к травам совершалась под
направляющим воздействием условий существования. На заре развития
покрытосеменных на земле был, по-видимому, ровный, теплый и влаж-
ный климат, способствовавший формированию древесных растений.
С приближением к концу тертьера (плиоцену) и плейстоцену у полюсов
возникает резко обозначившаяся климатическая периодичность. Усло-
вия различных времен года получают большую выраженность, чем это
было в предыдущую эпоху.
Выработка травянистой структуры является приспособлением к пе-
риодической смене климатических факторов. Происходит интенсивная
паренхиматизация всех тканей, окружающих листовые следы. Бла-
годаря этому усиливается отток пластических веществ и увеличиваются
их запасы. Вместе с тем периодическое наступление времен года с ясно
выраженным неблагоприятным сезоном вызывает угнетение камбиаль-
з*
35
ной деятельности. Вследствие этого вторичные элементы флоэмы и кси*
лемы не образуются, развитие сосудисто-волокнистых пучков останав-
ливается на стадии первичного строения их элементов. В конечном
итоге возникают стебли с ограниченным приростом в толщину, что и
характеризует травянистую структуру (Sinnott, Bailey, 1914а, б; Тах-
таджян, 1948; Александров, 1954; и др.).
Травянистые растения лучше приспособлены к переживанию небла-
гоприятного времени года благодаря наличию приземных или скрытых
в почве почек. Эта линия составила геофильную ветвь эволюции, свя-
занную с зонами умеренно холодного климата в северном и южном
полушариях. Много трав появилось в условиях высокогорного клима-
та. С другой стороны, возникновение трав происходило под воздействи-
ем аридизации климата, но в противоположность холодным странам,
в этих условиях наблюдалось массовое развитие однолетников.
В связи с различиями морфологической структуры побегов древес-
ных растений процесс перехода от деревьев к травам приобретал свои
особенности в разных структурных типах. Как показано в гл. II, по
структуре побегов травянистые растения разделяются на два типа: ро-
зеточный и безрозеточный. Аналогичные типы свойственны и древесным
растениям: деревьям, кустарникам и полукустарникам. Чтобы яснее
представить различия в путях эволюционного развития травянистых
растений из древесных соответственно структурным особенностям побе-
гов, охарактеризуем отмеченные типы — безрозеточный (долихоморф-
ный) и розеточный (брахиморфный)—для древесных растений.
Долихоморфный тип структуры. Сюда относятся древес-
но-кустарниковые растения, у которых подавляющая масса вегетатив-
ных побегов слагается удлиненными побегами — долихобластами, в год
своего возникновения несущими ассимилирующие листья. Междоузлия
этих побегов, хотя и могут значительно изменяться в своих линейных
размерах, обычно достаточно выражены, так что листья располагаются
на некотором расстоянии друг от друга и никогда не образуют розетки.
Длина же годичных побегов, как это показал еще В. Любименко (1900),
может сильно варьировать: от довольно длинных до весьма коротких
с самыми постепенными переходами между этими крайними типами.
Необходимо подчеркнуть, что укороченные побеги здесь возникают как
вследствие некоторого сокращения длины междоузлий, так и вслед-
ствие предельного сокращения их числа. В результате крайнего выра-
жения этого процесса возникает побег с одним листом или двумя ли-
стьями, в случае супротивного листорасположения. Дальнейшее угнете-
ние ростовых процессов приводит к образованию «спящих» почек. На
этом основании можно заключить, что данные короткие побеги пред-
ставляют собой сильно редуцированные побеги долихобластного типа:
сколь бы коротки они не были, листовая розетка обычно не образуется.
Брахиморфный тип структуры. Основу скелета побеговой
системы этих растений, подобно предыдущим, составляют удлиненные
побеги — долихобласты. На удлиненных побегах в качестве боковых
развиваются розеточные вегетативные побеги — брахибласты. Междо-
узлия этих побегов недоразвиваются, в результате чего образуются
короткие побеги, несущие розетку зеленых листьев. В отличие от укоро-
ченных побегов растений долихоморфного типа структуры, рассматри-
ваемые брахибласты имеют более или менее значительное число
узлов, а следовательно и листьев срединной формации. Другую особен-
ность брахибластов составляет то, что они в течение всей своей жизни,
будучи в вегетативном состоянии, нарастают всегда моноподиально.
Определяя брахиморфный (розеточный) тип структуры как совокуп-
ность удлиненных и розеточных вегетативных побегов, мы должны под-
черкнуть необычайную пластичность структуры этих растений, возмож-
,?6
ноСть взаимных переходов и превращении удлиненных побегов в
укороченные и обратно. Эта закономерность, как увидим ниже, имела
огромное значение в трансформации древесных растений в травяни-
стые.
Есть все основания полагать, что безрозеточный и розеточный типы
структур травянистых растений ведут свое происхождение от соответ-
ствующих типов структуры древесных форм.
В рамках безрозеточного типа структуры побеговой системы разви-
тие травянистых растений из древесных достигалось в результате мак-
симального сокращения длительности жизни скелетных осей (до одного
года и менее). Удлиненный, с вытянутыми междоузлиями, тип побегов
сохранялся, но у трав эти побеги стали однолетними, отмирающими на
зиму. В этом ряду проявляется закономерность, открытая И. Г. Серебря-
ковым, охарактеризованная выше.
Возможность данного направления и пути эволюции подтвержда-
ются целым рядом фактов. В роде Sambucus подавляющая масса ви-
дов — деревянистые растения безрозеточного типа, скелетные оси ко-
торых живут несколько лет. Наши исследования Sambucus racemosa
показали, что в первые годы развития из семени побеги этого вида явля-
ются однолетними, отмирающими к зиме до самого основания. Еже-
годное отмирание годичных побегов удерживается до 3—4-летнего воз-
раста, а побеги следующих генераций переходят к многолетнему образу
жизни (Голубев, 1959а). Таким образом, на раннем этапе жизни раз-
витие Sambucus racemosa совершается по типу травянистого растения.
Если бы эта ювенильная черта утвердилась во всем жизненном*цик-
ле, деревянистое растение Sambucus racemosa преобразовалось бы в
травянистое. Анализ видов Sambucus показывает, что в его составе есть
травянистый представитель — S. ebulus L., обладающий безрозеточны-
ми побегами, как и остальные виды рода. Пример этого травянистого
вида наглядно убеждает, что в результате сокращения длительности
жизни скелетных осей до одного года (практически меньше одного
года) возникает травянистая форма.
Сходный пример приводит Б. А. Федченко (1922), описавший един-
ственный травянистый вид в роде Tamarix — Т. spiridonowii В. Fedtsch.
Это, по Федченко, самый молодой представитель рода. Однако он вы-
ражает опасение, что найденный вид, возможно, является юношеской
формой какого-либо кустарникового представителя тамариксов. По
структуре побегов тамариксы относятся к безрозеточному типу.
Очень интересные данные, касающиеся перехода деревянистых расте-
ний в травы, приводит Б. М. Козо-Полянский (1945). В опыте культу-
ры древесно-кустарниковых экзотов в ботаническом саду Воронежского
университета многие из них, вследствие ежегодной потери надземных
побегов под влиянием вымерзания, приобретали черты травянистых
растений. Таковы Pawlownia tomentosa Steud., Diervilla florida S. et. Z.,
Cytisus supinus L., Sorbaria sorbifolia А. Вг. и другие, среди которых от-
мечены летнезеленые деревья и кустарники, вечнозеленые кустарники,
лианы. Весьма поразительные видоизменения приобретала Buddlea
davidii Franch., которая за короткое время перестроилась в траву, со-
хранив способность к семенному размножению. Б. М. Козо-Полянский
заключает, что «доктрина Бэйли — Синнота — Джеффри, имеющая
сравнительно-морфологическое обоснование, согласуется с данными фи-
логенетического метода (Гетчинсон) и отчасти палеостатистического
(Циммерманн): травы произошли от древесных растений. Наши случаи
превращения биоморф или типов Раункиера воочию демонстрируют,
каким путем могла осуществиться переделка деревьев в травы и како-
вы могли быть природные факторы этой переделки» (стр. 49). Сходное
явление наблюдалось у Buddlea lindleyana, Caryopteris incana Miq.,
37
Coronilla etnerus L. Айришем (цит. по Б. М. Козо-Полянскому, 1945)
в США (центральная часть).
В типе розеточных растений процесс преобразования древесной фор-
мы в травянистую протекал несколько иначе. Можно допустить, что
при этом междоузлия главного побега недоразвивались, вследствие
чего возникала розеточная форма, как это и сейчас наблюдается в юве-
нильном состоянии у некоторых видов Berberis (Голубев, 1959а). Но
в розеточном состоянии главный побег и побеги последующих поколе-
ний у трав способны жить несколько лет, а у моноподиальных розеточ-
ных трав — в течение всей жизни растения. Значит, если у розеточных
растений переход к травянистым и сопровождался сокращением об-
щей продолжительности жизни растений, то длительность развития ске-
летных осей часто сохранялась на определенном уровне, измеряемом
иногда несколькими или многими годами. Отсюда ясно, что развитие
трав в этих случаях нельзя оценивать как процесс предельного сокра-
щения длительности жизни скелетных осей — до одного года и менее.
Здесь надо отличать качественное преобразование структуры побегов,
выражавшееся в резком уменьшении прироста стеблевой оси побега,
имевшем своим следствием безболезненную перезимовку в живом со-
стоянии, без потерь через отмирание.
Вероятность эволюционного преобразования структуры розеточного
типа растений с надземными многолетними побегами подтверждают
данные, приводимые Е. П. Коровиным (1958) для родов ферула и до-
рема. Предки их, по-видимому, были представлены в третичных лесах
стеблевыми формами с многолистными розетками на вершине стеблей.
С увеличением засушливости климата или под влиянием похолодания у
предковых форм происходят редукция стебля и выработка розеточных
форм. Интересные сведения о трансформации некоторых розеточных
форм деревьев в розеточные травы приводит Е. Корнер (Corner, 1949,
1954). Безрозеточный тип структуры побегов, несомненно, является более
древним признаком по сравнению с розеточным типом (Голубев, 19576,
1959а, б, 1960г). Схема филогенетических отношений типов побегов дре-
весных и травянистых растений представляется в следующем виде:
Производность розеточного типа древесных растений хорошо моти-
вирована Н. В. Дылисом (1958) на примере лиственницы. У Larix более
специализированными побегами являются брахибласты, первичными —
ауксибласты, что подтверждается, во-первых, вторичностью появления
брахибласт в онтогенезе, во-вторых, коррелятивной связью с появле-
нием укороченных побегов — смены первоначального очередного листо-
расположения более подвинутым пучковым. По мнению Н. В. Дылиса,
предковая форма Larix обладала только удлиненными безрозеточными
побегами (ауксибластами) с одиночно расположенной хвоей. Эта струк-
тура побегов весьма типична для тропиков, где обилие тепла и влаги
стимулирует постоянное и мощное отрастание все новых побегов, обес-
печивающих потребности растений в органическом веществе. Под влия-
нием изменения климата в сторону сокращения тепла, влаги и про-
зе
должительности вегетации, с которым столкнулась предковая форма
лиственницы в известное время при миграции или на месте, исключав-
шего возможность систематического развития сильных приростов, воз-
никает новая структура побегов — розеточные брахибласты. «Образо-
вание укороченных побегов с пучковым расположением хвои сильно
увеличило поверхность скелетных ветвей, способную нести хвою, и этим
резко увеличило густоту кроны и дало возможность растениям быстро
развивать сильный ассимиляционный аппарат с минимальной затратой
запасных питательных веществ» (Дылис, 1958, стр. 140).
Трансформация безрозеточного типа структуры в розеточную имела
место и среди травянистых растений. В качестве примера приведем
некоторые данные о структуре и генетических связях отдельных циклов
форм бобовых. Безроэеточными побегами обладают виды Cicer, Vicia,
Lathyrus.
По М. Г. Попову (1929), эти роды, объединяемые в трибу Vicieae, на-
ходятся в родственных отношениях с трибой Sophoreae, корни проис-
хождения которой восходят к Mimosoideae. Но растения этих триб по
характеру побегов должны быть отнесены к безрозеточному типу. Из
групп близко родственных к этому ряду бобовых можно назвать трибу
Trifolieae, где уже в массе появляются розеточные формы. Trifolieae
выводятся из Sophoreae через группу Dalbergieae. Филогенетические
данные позволяют заключить о производном характере розеточной
структуры травянистых растений, развившейся из безрозеточной. Этот
вывод подтверждается и анализом видов в пределах собственно рода
Trifolium. По данным Е. Г. Боброва (1947), наиболее примитивной в
этом роде является секция Lupinaster, к которой принадлежит и «ли-
дер» секции — Т. lupinaster L., характеризующийся безрозеточной
структурой побегов. Следовательно, палеоморфность безрозеточного
типа здесь хорошо филогенетически мотивирована, так как многие бо-
лее эволюционно подвинутые виды клеверов развиваются уже в форме
розеточных растений. Безрозеточные Т. alpestre L. и Т. medium L. отно-
сятся к секции Trichostoma подрода Lagopus. Эта секция является весь-
ма древней и близко примыкает к исходной секции Stenostoma (Боб-
ров, 1947).
«Едва ли можно сомневаться в том,— пишет А. И. Толмачев
{1957),— что более или менее равномерная облиствленность стеблей
представляет собой первичный признак по сравнению с сосредоточен-
ностью всех листьев в прикорневых розетках, над которыми возвы-
шаются безлистные цветоносы» (стр. 1449). Эти данные основываются
на изучении филогении рода Draba. Автором приводится конкретный
материал, иллюстрирующий все ступени трансформации, начиная от
форм со значительным числом стеблевых листьев, через формы, обла-
дающие далеко отстоящими друг от друга листьями, до растений с без-
листными стрелками. Близкие закономерности выявлены И. Н. Свеш-
никовой (1951а, б) в филогенетическом развитии цветоносных побегов
у Primula. Древние абиссино-аравийские виды (Primula verticillata
Forsk. и др.) сохранили облиственный стебель, в то время как более
подвинутые формы характеризуются безлистными цветоносами. Эволю-
ция последних совершалась от многоярусных, мутовчатых до простых
зонтиковидных и даже головчатых соцветий.
В пределах рода Dianella из семейства лилейных развитие форм,
у которых основная масса листьев сосредоточена в прикорневых розет-
ках, а цветоносы почти безлистные, от растений с облиствленными снизу
доверху маловетвистыми побегами достоверно мотивируется Шлиттле-
ром (Schlittler, 1957).
Наряду с описанными основными направлениями эволюции био-
морфологических структур побеговой системы растений возможны
39
и другие, например возникновение розеточных трав из безрозеточных
деревянистых. Вероятен и обратный путь эволюции — развитие безрозег
точных растений из розеточных (Голубев, 1959а, б). Конкретизация и
раскрытие всех мыслимых путей преобразования структуры побеговой
системы растений в процессе эволюции представляют интереснейшую
область будущих исследований.
Травы, в свою очередь, испытали очень сложную и многообразную
эволюцию, биоморфологическое содержание которой рассматривается
ниже на примерах отдельных родов. Здесь же надо отметить лишь глав-
ные ступени эволюционного преобразования структуры травянистых
видов. Большинство исследователей выделяет три главные ступени из-
менения: многолетние поликарпические, многолетние и двулетние мо-
нокарпические формы и однолетники.
В указанном ряду наиболее молодой биоморфой являются однолет-
ники (Hansen, 1956). Среди однолетников высшую специализацию пре-
терпели эфемеры, максимальное развитие которых связано с аридными
областями [Краснов, 1899; Сполдинг (Spalding, 1909); Штокер (Sto-
cker, 1928); Коровин, 1934; А. В. Прозоровский, 1940; Попов, 1948; Ко-
ган, 1949; Полковниченко, 1958]. Йх жизненный цикл, начинающийся
у озимых осенью, а у яровых — ранней весной, с наступлением жаркой
и сухой погоды быстро завершается формированием зрелых плодов
и полным отмиранием.
Исследования показывают, что некоторые виды эфемеров способны
развиваться как по озимому, так и яровому типу (Васильченко, 1937;
Нечаева, Василевская, 1945; Никитин, 1957; Полковниченко, 1958; Не-
чаева, 1958). По эколого-биологическим особенностям эфемеры пред-
ставляют весьма гетерогенную группу. Одни из них неустойчивы к жару,
и раннее окончание вегетации связано с установлением в природе высо-
ких температур, другие же обладают сравнительно высокой жароустой-
чивостью вегетативных органов, и раннее окончание вегетации в природе
обусловлено нарастающей сухостью почвы (Полковниченко, 1957).
Е. И. Проокоряков (1950) подчеркивает наиболее характерное свойство
эфемеров — способность их семян переносить в течение длительного
времени воздействие сухого жара.
Весьма пластичным оказывается признак продолжительности веге-
тации. В южных районах многие из них, начав вегетацию с осени, про-
должают ее и в зимнее время, конечно при соответствующих благог
приятных условиях (Проскоряков, 1950; Запрометова, 1956; Бурыгин,
1957; Нечаева, 1960; и др.).
Факты большой пластичности эфемеров очень разнообразны (Тара-
канов, 1940; Шенников, Иоффе, 1944; Сергеев, 1945; Полунина, 1950;
Запрометова, 1955; и др.; Волкова, 1958; Бурыгин, Запрометова, 1959).
Можно вполне согласиться с мнением В. В. Никитина, А. Я. Полковни-
ченко (1955) о том, что высокая экологическая и биологическая ла-
бильность эфемеров является следствием молодости этой жизненной
формы и приспособленности к весьма неустойчивым и резко меняю-
щимся факторам внешней среды в аридной зоне.
Генетическая связь однолетников и эфемеров с многолетними и
более длительно вегетирующими формами проявляется в способности
их к длительной вегетации в благоприятные по климатическим усло-
виям годы и возможности развития дополнительных побегов из спящих
почек, подобно кущению у многолетников (Нечаева, 1958). Последнее
явление подробно изучено В. В. Скрипчинским (1958а, б) у однолетних
злаков в сравнении с многолетними. В генетических реконструкциях и
установлении связи однолетников с родственными им многолетниками
дополнительным критерием может служить форма роста побегов
(Meusel, 1955).
40
Как полагает Е. П. Коровин (1958), возникновение эфемеров, по
крайней мере в области Средиземноморья, относится к концу третич-
ного периода. Исходными, по всей вероятности, были растения мезо-
фильного корня, испытавшие преобразование в условиях открытых ра-
стительных формаций. Развитие терофитов достигло высшего уровня
в четвертичном периоде (Hansen, 1956).
Переход многолетних травянистых растений в однолетние можно
вызвать искусственно — воздействием условий культуры. В опытах
А. И. Купцова (1952, 1955) многолетние кок-сагыз, широкий и другие
в культуре превращались в однолетники, т. е. зацветали на первом году
жизни и отмирали. В наших опытах резко сокращал продолжительность
жизни в питомнике заповедника Anthyllis polyphylla W. et К. При осен-
нем посеве и осеннем появлении всходов этот вид на следующий год
достигает мощного развития, цветет, плодоносит и полностью отмирает.
В некоторые годы отдельные особи развивались в форме двулетнего мо-
нокарпика. По типу двулетника проходила жизненный цикл в питомни-
ке (а также в природе на обнажениях) Anthemis tinctaria L. В условиях
хорошего увлажнения и минерального питания Б. А. Келлер (1934)
выращивал многолетний полукустарничек Artemisia pauciflora Web.
В результате растение цвело, плодоносило и закончило свою жизнь как
однолетнее растение.
Е. И. Проскоряков (1950) допускает возможность непосредственного
превращения деревянистых растений в однолетние. Интересные сообра-
жения об эволюции жизненных форм и развитии однолетников высказы-
вает М. Г. Попов (1948). Им устанавливается корреляционная связь
между типом жизненной формы и структурой цветка. Так, в системе
Fumariaceae лианы имеют бисимметрический цветок, а многолетние
травы чаще всего моносимметрический, но возможен и бисимметриче-
ский цветок; однолетники — исключительно моносимметрический зиго-
морфный цветок. При этом от лианы к однолетнику неизменно убывают
размеры цветка. Таким образом, по мнению М. Г. Попова, переход от
одной жизненной формы к другой сопровождался резким изменением
цветка. Это скачкообразное изменение структуры цветка было столь зна-
чительным (родового или секционного значения), что терялась связь с
предковой формой. Несмотря на то, что изложенные соображения не-
достаточно аргументированы фактически, они заслуживают определен-
ного внимания. Весьма возможно, что многие филогенетические гиатусы,
отсутствие связующих звеньев, как раз и обусловлены этой закономер-
ностью эволюции. Во всяком случае факты неотенического развития как
будто бы хорошо согласуются с изложенными принципами.
Выше мотивирована концепция развития биоморфологических типов
(жизненных форм), получившая наибольшее признание и распростра-
нение. Вполне естественно, что некоторые ученые в ряде положений
отстаивают свои воззрения, в той или иной мере не совпадающие с указан-
ными. Так, например, сначала господствовало мнение о том, что первич-
ными были травянистые растения покрытосеменных, из которых, в про-
цессе усложнения организации развились древесные формы (Velenovsky,
1905—1913; Голенкин, 1927; АгЬег, 1928; и др.). Сейчас эта точка зрения
окончательно оставлена и имеет чисто исторический интерес. Но принци-
пиальная основа этой концепции продолжает удерживаться в ряде слу-
чаев применительно к оценке эволюции главных биоморф травянистых
растений. Приверженцы взгляда от «простого» к «сложному» находят,
что исходными формами трав являются однолетники, которые затем дают
начало многолетним травам. В другом варианте этой же гипотезы в ка-
честве первичных рассматриваются многолетние монокарпики. И. К. Па-
чоский (1921) полагает, что поликарпические травы возникли из много-
летних монокарпических, а современные однолетники представляют
41
группу новейшего происхождения. В обоснование своей точки зрения
автор не приводит каких-либо доказательств.
А. Н. Криштофович (1950) на основании палеоботанических данных
склоняется к мысли об одновременном появлении древесных и травя-
нистых растений.
В опыте построения филогенетической системы рода Parietaria
А. В. Ярмоленко (1941) принимает в качестве исходного цикл Manocyma,
заключающий однолетники. Более подвинутая подсекция Руспосугпа со-
стоит из многолетних растений. В подсекции Manocyma всего два вида:
Parietaria erecta и Р. mauritanica. Первый из них, приуроченный к ши-
роколиственным лесам Средней Азии и Юго-Восточной Европы, являет-
ся древним реликтовым типом. Второй вид обитает на каменистых мес-
тах, которые служат уже типичными местообитаниями Parietaria. Кроме
того, Manocyma обладает раскидистыми соцветиями с прицветниками,
представляющими, по мнению А. В. Ярмоленко, первичный признак.
С другой стороны, пример с пажитниками (Trigonella) мог бы слу-
жить основанием к аналогичному заключению: однолетние виды распро-
странены шире многолетних и обитают в Австралии (Г. suavissima
LindL), Тибете (Т. tibetica Vass.), в Капской области (Т. glabra Thunb.),
т. е. на разных обособленных ныне материках. В соответствии с бота-
нико-географическими представлениями такие виды следует считать бо-
лее древними, чем родственные им виды локального распространения.
Однако И. Т. Васильченко (1953) предполагает, что многие однолетние
пажитники являются сорными или полусорными растениями, в их рас-
селении большую роль играет деятельность человека; таким путем не-
которые виды могли попасть в Австралию и Южную Африку и натура-
лизоваться. Таким образом, И. Т. Васильченко находит возможным со-
хранить в силе положение о производном характере однолетников.
В статье, посвященной систематике рода Suaeda, М. М. Ильин (1936)
приходит к выводу о том, что жизненные формы однолетников и кустар-
ничков могут возникать в одном и том же цикле видов независимо и
параллельно из общих предков. Он констатирует, что «нерационально
относить однолетники и кустарнички в разные секции. Мы знаем, что
пути эволюции проходят сквозь эти искусственные перегородки и в одном
цикле могут дать как однолетники, так и кустарники, мало того, в преде-
лах одного и того же вида нередко появляются разные типы» (стр. 42).
Весьма крайние и недостаточно мотивированные взгляды на эволю-
цию жизненных форм травянистых растений отстаивает В. Н. Вороши-
лов (1959), предложивший для биоморфологических типов валерианы
следующую схему эволюции: неизвестный древовидный предок — эпи-
котильный однолетник — розеточный двулетник — замещающий двулет-
ник, корнеклубневой — замещающий двулетник, мочковатый со столо-
нами— то же без столонов — многолетник с настоящим корневищем.
Если канонизация какого-либо из перечисленных положений реши-
тельно неприемлема, то это не означает, что в сложном лабиринте эво-
люции жизненных форм покрытосеменных они совершенно исключают-
ся. Напротив, в каждом конкретном случае, в пределах того или иного
рода, имеются свои отличия эволюции жизненных форм и весьма вероят-
но, что некоторые из описанных путей эволюции находят реальное во-
площение. Задача, очевидно, заключается в том, чтобы, не впадая в шаб-
лон и избегая крайности, раскрыть истинное содержание и пути эволю-
ции биоморфологических структур растений.
Основные пути эволюционного изменения подземных органов травя-
нистых растений отчасти рассматривались нами раньше (Голубев, 1962в).
Развитие подземных органов мыслится в такой последовательности:
стержнекорневые — кистекорневые — рыхлокустовые — длиннокорневищ-
ные— столонообразующие растения (Высоцкий, 1915; Казакевич, 1922;
42
Серебряков, 19546; Голубев, 1957а, б). В этом ряду, несомненно, имели
место редукция отдельных звеньев цепи и разнообразные отклонения.
Есть все основания признать в качестве анцестральной стержнекор-
невую структуру травянистых наземных покрытосеменных, что согла-
суется с данными филогенетики и эмбриологии. Известно, что стержне-
корневой системой характеризуются двудольные растения, а мочковатой
{кистекорневой) — однодольные. По современным представлениям,
однодольные являются производными двудольных, составляя более мо-
лодую ветвь антофитов. Это положение не теряет своей силы и в случае
принятия бифилетического происхождения однодольных (Кузнецов, 1936;
Буш, 1940; Гроссгейм, 1945; и др.), так как исходными типами для
устанавливаемых этими авторами двух стволов эволюции однодольных
все же являются двудольные растения. Отсюда вытекает, что стержне-
корневой тип — первичный, а мочковатый — вторичный, более позднего
происхождения.
В эмбриональном состоянии зародыши большинства однодольных об-
ладают только одним главным корнем, придаточные корни появляются
позднее, при прорастании семени и образования ювенильного растения.
Формирование одного главного корня в эмбриональном состоянии у за-
родыша подавляющего числа однодольных с последующей быстрой ут-
ратой его жизнедеятельности в фазе проростка указывает на филогене-
тическую ценность этого признака и является, несомненно, рекапиту-
ляцией анцестральной структуры — стержнекорневого типа двудольных,
что лишний раз подкрепляет мысль о вторичности однодольных. Рекапи-
тулятивный характер зародышевого первичного корня у Nymphaeaceae,
лишенного всякой функции, отмечает Б. М. Козо-Полянский (1937).
Что касается случаев отсутствия главного корня в дефинитивном
возрасте у многих двудольных, то в фазе зародыша <и первых этапов
формирования всхода и проростка у них имеется только один главный
корень. Следовательно, здесь также проявляются черты палеоморфной
структуры, удерживаемые в онтогенезе благодаря наследственности, а,
возможно, и в силу жизненных потребностей растения на данном этапе
развития.
Развитие жизненных форм по линейному типу протекало, очевидно, в
весьма ограниченных масштабах (Серебряков, 1955а). Более вероятна
сопряженная радиальная дивергенция, когда исходная структура пре-
терпевала всевозможные изменения, получала «надстройки» или, наобо-
рот, наблюдалось выпадение отдельных структур. В одно и то же время,
например, стержнекорневая структура могла трансформироваться в кис-
текорневую, рыхлокустовую, длиннокорневищную и иные структуры,
причем каждая из них, в свою очередь, могла стать источником разви-
тия других, более подвинутых (следующих за ней в приведенном ряду)
типов структур. В ныне употребляемых линейных схемах эволюции по
преимуществу выражено абстрагированное объединение целого ряда
радиальных дивергенций, соответствующих развитию жизненных форм
в разнообразных систематических группах (Голубев, 1959а, 1961).
Конкретное воплощение высказанных идей и взглядов найдет место в
специальной части филогенетического анализа биоморфологических
структур растений в избранных нами родах. Здесь только можно,кратко
напомнить о существовании вместе с основным параллельных рядов раз-
вития структур травянистых растений. Во флоре умеренно холодной
области нашей страны хорошо выделяются ряды ползучих, клубне-
вых и корнеотпрысковых растений, намеченные И. Г. Серебряковым
(1955а).
Биолого-морфологический анализ развития структуры ползучих тра-
вянистых растений и данные по филогении отдельных систематических
групп показывают, что ползучие травы возникали независимо и разно-
43
временно в различных направлениях преобразования структуры рас-
тений в качестве боковых ответвлений на разных этапах становления
жизненных форм травянистых растений основного ряда (Голубев,
1961). Этот вывод в полной мере применяем и к формированию
клубневых (Голубев, 1960ж) и корнеотпрысковых растений (Голубев,
1960а).
Решающим фактором возникновения ползучих растений явились
почвенно-климатические условия, благоприятствующие укоренению сте-
лющихся побегов. Весьма правдоподобно следующее предположение: в
развитии самих стелющихся побегов некоторую роль могло сыграть та
обстоятельство, что предками отдельных видов трав были лиановые фор-
мы, как это установлено для Rubus (Эйхвальд, 1959). Образование пол-
зучих побегов обеспечивало широкие возможности расселения растении
по площади часто в таких условиях, где развитие подземных побегов
встречало препятствия, например в сообществах с сильно задернованной
слабо проветриваемой почвой (сырые луга и 1. п.). Обилие ползучих
растений в лесах, особенно хвойных, связано, по всей вероятности, с
трудностью полового воспроизведения и размножения.
Формирование клубневых растений направлялось почвенно-климати-
ческими факторами областей с наличием периодически засушливого се-
зона. Вследствие приспособления растений к данным условиям возника-
ло местное разрастание тканей разных органов (корня, стебля и листа)
в связи с отложением запасных пластических веществ, а часто и влаги,
что и определило развитие клубневых растений.
Другим направлением биоморфогенеза в аридных областях было
развитие многообразных корнеотпрысковых растений. Об этом свиде-
тельствуют статистические дачные, полученные Г. Н. Высоцким (1915,
1926), Л. И. Казакевичем (1922) и др. Однако крайние условия аридно-
сти, большая сухость и жара, например, в пустынях Средней Азии, для
которых лучше всего подходит название «знойные» (Коровин, 1961),
уже менее благоприятны для проявления свойства корнеотпрысковости.
Закладка придаточных почек на корнях находится в связи с отмиранием
боковых корней, которое происходит наиболее интенсивно на одно-дву-
летних корнях, где и наблюдается усиленное их формирование (Васи-
левская и Кондратьева, 1955). Этот процесс, несомненно, возможен лишь
при известном оптимуме тепла и влаги.
Такие условия, с одной стороны обеспечивающие обильное развитие
поверхностных корней и их ветвление, а с другой — массовое отмирание,,
оптимально выражены в степной зоне, где и обнаружено наибольшее
число корнеотпрысковых растений.
Несколько слов о характере нарастания и ветвления у покрытосемен-
ных растений. Первичной формой, по всей вероятности, было моноподи-
альное ветвление (Тахтаджян, 1948, 1956; Серебряков, 1952; Хржанов-
ский, 1958а; и др.), из которого симподиальное ветвление развилось, по-
видимому, в результате образования терминального цветка или соцветия
и путем отмирания верхушки побега в вегетативном состоянии (Серебря-
ков, 1952). Последнее подтверждается и Б. М. Козо-Полянским (1945),
делающим вывод о превращении моноподиального ветвления в симподи-
альное в эволюционной перспективе из факта отмирания верхушек по-
бегов у экзотов под влиянием весенних заморозков и зимних холодов.
По этому поводу он пишет, что «мы как бы присутствуем при этой ме-
таморфозе, некогда имевшей место в морфогенезе многих растений вне
рамок наблюдений и опытов, а также приобретаем возможность причин-
ного понимания этого явления, по крайней мере, для некоторых случаев»
(стр. 46).
Однако, как вытекает из эволюционной концепции развития жизнен-
ных форм древесных растений Е. Корнера (Corner, 1949), терминальное
44
положение соцветий, обусловливающее симподиальное нарастание,
является примитивным признаком.
Что касается эволюционных взаимоотношений моноподиального и
симподиального роста и возобновления у травянистых растений, то этот
вопрос представляется еще слабо разработанным. В распространении
моноподиальных растений установлена определенная географическая за-
кономерность. На равнинной территории максимум моноподиальных
растений приурочен к арктической и лесной зонам (Серебряков, 1952).
Как показали наши исследования, в южных горах наибольшее число
растений моноподиального возобновления обитает в альпийском поясе,
заметное количество их встречается в древесно-кустарниковом поясе и
почти совершенно они отсутствуют в поясе разнотравно-сухой степи и в
эфемеровой горной полупустыне. Получается вполне «симметричная»
картина: максимальное обилие моноподиальных растений сопряжено с
наиболее суровыми климатическими условиями (Арктика и альпика в
горах).
Альпийская флора является сравнительно молодой. Ее развитие
начинается со второй половины третичного периода (Кузнецов, 1909;
Diels,1910; Лавренко, 1938; Вульф, 1944; и др.). Данное обстоятельство
вместе с максимальной концентрацией моноподиальных растений в вы-
сокогорьях может служить доказательством признания вторичности мо-
ноподиального возобновления у альпийских покрытосеменных, развив-
шегося из симподиального (Голубев, 1960в).
Этот вывод обосновывается и с позиций общих эволюционно-мор-
фологических закономерностей и развития травянистых растений из
древесных (Голубев, 1959а).
Существование основного эволюционного ряда, в котором травы яв-
ляются производными древесных форм, не исключает в отдельных слу-
чаях обратного хода соматической эволюции — от трав к полудрёвес-
ным и древесным растениям. Наиболее широко процесс одревеснения
стеблей травянистых растений выражен в аридных областях (Velenov-
sky, 1905—1913; Радкевич, 1926, 1928; Хржановский, 1940; и др.), част-
яое 'проявление его выразилось в развитии жизненной формы полуку-
старников и полукустарничков из травянистых предков. Вероятность об-
разования полукустарников из трав была высказана Н. А. Козловой
(1953), а затем, с помощью систематико-филогенетического анализа,
убедительно доказана И. Г. Серебряковым (19546), Е. И. Рачков-
ской (1957). Нами приводятся дополнительные материалы (Голубев,
49606, г).
Так, в роде Thermopsis исходными формами были мезофильные тра-
вы (Чефранова, 1958), и, следовательно, имеющиеся в этом роде виды с
деревянеющими основаниями побегов являются вторичными, как подоб-
ные же виды Trigonella и Medicago, предками которых были также ме-
зофильные многолетние травы (Васильченко, 1949, 1953).
Но особенно убедительные свидетельства возможности перехода тра-
вянистой структуры стебля в древесную и обратно дают анатомические
исследования, в частности мало цитируемые в современной ботанической
литературе, но весьма содержательные работы О. Н. Радкевич (1926,
1928), а также В. Г. Александрова и О. Г. Александровой (1929).
О. Н. Радкевич установила, что в условиях интенсивной солнечной
инсоляции и сухого жара происходит склерофитизация тканей тра-
вянистого стебля, отождествляющая его со структурой деревянистого
стебля. Автор считает наиболее важными признаками склерофита одре-
веснение сердцевинных и ксилемных лучей. «Этот экологический признак
вновь приближает травянистый стебель к его древесному прототипу ибо,
как известно, у деревьев склероз ксилемных лучей — явление почти уни-
версальное. Да и вообще, по мере разрастания стелярного конца по
45
направлению к своему основанию, стебель склерофита приобретает все
большее сходство с древесным...» (Радкевич, 1928, стр. 430).
В. Г Александров и О. Г. Александрова (1929) нашли, что процесс
раздревеснения стеблей является обратимым и в природе находит ши-
рокое распространение.
По данным А. Л. Тахтаджяна (1948), все древесные представители
Chenopodiaceae имеют вторичное происхождение. Аналогичный вывод де-
лается и в отношении древесных однодольных. Однодольные произошли
от водных неотенических форм травянистых двудольных (Тахтаджян,
1943, 19546). Вторичное утолщение стеблей древесных однодольных про-
исходит за счет образования вторичных сосудисто-волокнистых пучков,
разделенных паренхимной тканью, что обусловлено деятельностью кам-
бия, который залагается в самом внутреннем слое коры или в перицикле.
Этот тип утолщения наблюдается у некоторых пальм, Aloe, Cordyline,
Dasylirion, Sanseviera zeylonica, Veratrum viride, Dracaena, Kniphofia,
Nolina, Jucca, Agave, Dioscorea и немногих других (Тахтаджян, 1948).
Однако, как отмечается многими исследователями (Lindinger, 1909;
и др.), между первичной и вторичной камбиальной активностью резких
отличий нет. Последняя является результатом продолжения клеточных
делений, ограниченных периферической зоной первичного цилиндра.
В. Г. Александров (1954) вскользь сообщает о том, что некоторые виды
пальм, несмотря на твердую консистенцию тканей стволов, в основе сво-
ей структуры являются травянистыми растениями.
Можно было уже убедиться, что решающую роль в построении дока-
зательства и вывода о вторичности или первичности тех или иных струк-
тур играют данные филогении. Но сами эти данные часто основываются
на предположениях и гипотезах, что в значительной мере снижает их
категорическую ценность. Поэтому и в случае использования положений
филогенетической систематики можно говорить лишь о более или менее
вероятных аспектах решения того или иного вопроса.
Другая, и по существу диаметрально противоположная точка зрения,
исходит из того, что древесные однодольные являются первичными, корни
происхождения которых восходят к древесным двудольным растениям.
Палинологические исследования Л. А. Куприяновой (1948, 1954) по-
казывают, что пальмы представляют древнюю группу, имеющую само-
стоятельное происхождение — от гипотетических проангиосперм. В свою
очередь, пальмы являются исходными для развития ветроопыляемой ли-
нии однодольных, в том числе Cyclanthaceae, Pandanaceae, Spargania-
ceae, Thyphaceae, Centrolepidaceae, Restionaceae, Flagellariaceae и Gra-
mineae. В данном ряду первые звенья составляют древесные растения
последующие — травянистые. На этом основании можно утверждать, что
среди однодольных, как и большинства двудольных, намечается анало-
гичный характер эволюции жизненных форм — от древесных к травя-
нистым. Однако были ли предками пальм древесные растения,— в
системе Л. А. Куприяновой остается неясным.
• Этот пробел восполняет система А. А. Гроссгейма (1945), в которой и
порядок Principes (с семействами Palmae и Cyclanthaceae), и порядок
Pandanales (с семействами Pandanaceae, Sparganiaceae, Thyphaceae)
выводятся через какие-то еще неизвестные промежуточные ступени не-
посредственно из Anonales, представленного древесными растениями.
Таким образом, идея сходства общего направления эволюции жизнен-
ных форм от деревьев к травам среди двудольных и однодольных полу-
чает новое подтверждение.
В системе Н. И. Кузнецова (1936) данный ствол однодольных, куда
помимо Palmae, Pandanaceae, Sparganiaceae, Thyphaceae, добавляются
Araceae и Lemnaceae (в системе Л. А. Куприяновой эти два семейства
составляют отдельную линию эволюции однодольных), занимает также
46
обособленное положение. Генетически порядок Spadiciflorae, объединя-
ющий перечисленные семейства, связывается с порядком Piperales, ко-
торый выводится от гипотетических Protogymnospermae. Среди Pipera-
les есть настоящие древесные формы.
Н. А. Бушем (1940) приводятся сходные данные о филогении рас-
сматриваемого ствола однодольных, имеющего своим основанием Pi-
perales. Последние, в отличие от системы Н. И. Кузнецова, выводятся
из Polycarpicae через семейство Lactoridaceae.
Таким образом, данные систем Н. И. Кузнецова и Н. А. Буша также
говорят в пользу ранее сделанного вывода о сходстве соматической эво-
люции двудольных и однодольных. Это заключение разделяет И. Г. Се-
ребряков (1955а).
Приведенные противоречивые данные о происхождении древесных
однодольных ярко свидетельствуют о том, что только синтезирование
данных филогенетической систематики, эволюционной морфологии, ана-
томии и т. д. может обеспечить высокую достоверность представлений
об эволюции жизненных форм.
Заканчивая обзор некоторых фактических данных по биоморфоло-
гической эволюции покрытосеменных, следует (хотя бы весьма конспек-
тивно) коснуться общих вопросов эволюции жизненных форм, составля-
ющих известную основу для их анализа в объеме определенных систе-
матических категорий.
Эволюция жизненных форм совершалась и совершается посредством
развития видов. Вне эволюции видов самостоятельно протекающего про-
цесса исторического развития жизненных форм не существует. Родст-
венные отношения жизненных форм, т. е. происхождение, можно выяс-
нить только на основании анализа генеалогии видов внутри конкретных
систематических единиц.
Вместе с тем на базе исторически определенной ступени организа-
ции (типа, класса) происходит эволюция структуры растений на основе
приспособления к условиям существования в форме идиоадаптивной
радиации (Тахтаджян, 1957; Вилли, 1959; и др.) по принципу классиче-
ской, дарвиновской лучевой (веерной) дивергенции. Она протекает сход-
ным образом во многих систематических группах более мелкого таксоно-
мического ранга — порядка, семейства, рода. На эту особенность сома-
тической эволюции, как всеобщую закономерность параллельной
изменчивости, уже давно обратили внимание многие исследователи.
«Параллельная направленная изменчивость признаков, — пишет В. Г.
Хржановский (1958а),— явление общего порядка, она, несомненно, ши-
роко выходит за пределы рассмотренных родов (роз, малин, лапчаток.—
В, Г.). Перед нами замечательная закономерность формообразователь-
ного процесса, особенно детально изученная Н. И. Вавиловым (1935) и
сформулированная в его работе «Закон гомологических рядов в наслед-
ственной изменчивости» (стр. 307). Параллельную изменчивость морфо-
структур отмечают Е. Л. Любарский (1959, 1961), В. Н. Голубев (1960>
и др.
Утверждая существование тесной связи эволюции жизненных форм
с развитием видов, следует сделать оговорку в том смысле, что здесь
говорится прежде всего о процессе видообразования по типу викаризма,
совершающемся аутохтонно, на основе экологической дифференциации
анцестрального типа в разных частях его ареала. Более подробно эта
сторона проблемы нами мотивирована ранее (Голубев, 1960д). Здесь же
отметим лишь отдельные, наиболее существенные моменты, необходи-
мые в дальнейшем для правильного понимания и оценки филогении взя-
тых для анализа родов.
Сопоставление фактов формирования ареалов и географических за-
кономерностей видообразования дает основание предполагать, что в
47
любой данный исторический момент вид занимает всю возможную для
его расселения территорию. Ареал вида есть показатель всех использо-
ванных экологических и биологических возможностей вида к его распро-
странению. Следует лишь оговориться, что данный исторический момент
нами понимается как сравнительно короткий промежуток времени в не-
сколько столетий, а иногда и того меньше. В геологическом масштабе
такой период представляет весьма малую величину, в сущности мгнове-
ние. На протяжении этого времени при относительно устойчивых клима-
тических условиях вид достигает своих крайних пределов распростране-
ния. В последующей истории существования он испытывает изменения
границ своего ареала исключительно под влиянием окружающих усло-
вий, вызывающих либо подавление жизнедеятельности и сокращение
ареала, либо оказывающих благоприятное влияние в занятии новых
территорий, причем продвижение пропорционально этому влиянию на
распространение. Однако с изменением одних факторов расселения мо-
жет быть связано возникновение или, наоборот, исключение других воз-
можностей к распространению. Словом, характер распространения
всегда находится в сложной функциональной зависимости от целого
ряда взаимосвязанных факторов.
Их постоянные изменения вызывают и вековые смещения границ
ареалов. Однако эти медленные преобразования ареалов следует свя-
зывать с соответствующими изменениями внешних условий, и их нельзя
оценивать как еще незакончившееся расселение видов, не достигших
своих климатических границ.
Вполне обосновано мнение И. К. Пачоского (1910), согласно кото-
рому, новая раса (вид) возникает «на значительной, иногда на всей
площади, занятой расщепляющейся материнской расой. Вследствие
этого раса, еще задолго до выделения ее в нечто самостоятельное, за-
нимает уже известный ареал, причем этот последний является результа-
том изменения на месте, а не миграции» (стр. VI). Таким образом, новые
виды развиваются аутохтонно в рамках ареала материнского типа.
Идея аутохтонного развития видов была высказана еще раньше и
тесно связана с учением о географических расах и видовых сериях
{рядах), фундаментальные основы которого, как известно, заложены
работами К. И. Максимовича (Бобров, 1959), С. И. Коржинского (1893)
и В. Л. Комарова (1901а, б, 1908, 1927, 1934, 1940). К. И. Максимовичем
и С. И. Коржинским было четко сформулировано понятие географиче-
ской расы, а В. Л. Комаровым оно было развито далее и обогащено в но-
вом методе видовых рядов. В его понимании, расы — реально сущест-
вующая генетическая категория, характеризующаяся определенным
ареалом.
Идеи В. Л. Комарова о географических закономерностях видообра-
зования глубоки и четко очерчены. В. Л. Комаров (1901а) писал: «Кли-
матические расы..., соответствующие определенным участкам земной
поверхности..., образуются благодаря постепенной вековой дифференци-
ровке климатов на территориях, некогда в климатическом отношении
однородных. Общему сборному типу соответствуют здесь не только об-
щая территория, но и единые климатические условия... За дифференци-
ровкой общего климата данной единой территории сборного типа на
отдельные подобласти наступает и медленная дифференцировка этого
типа на некоторое число подчиненных рас или видов, равное числу обо-
собляющихся в климатическом отношении областей. Изменение здесь
начинается не с одного или немногих индивидуумов, но распространя-
ется на все поколения, населяющие вновь расщепившиеся территории.
Мы имеем здесь как бы пассивную миграцию отдельных рас в разные
климаты, причем расы эти сами не переселяются, а изменяются медлен-
но и постепенно, но неотразимо... Иначе, процесс образования видов
48
идет путем выработки и дифференцировки климатических рас» (стр. 61).
Еще ранее В. Л. Комаров в этой же работе указывает, что «каждая груп-
па видов связана генетически происхождением от одного древнего ро-
доначальника, занимавшего некогда всю или почти всю территорию
этих видов» (стр. 11).
В. Л. Комаров в ясной форме показал, как в рамках единого ареала
предкового вида аутохтонно развиваются новые виды параллельно эко-
логической дифференциации отдельных частей исходного ареала. Ряд
(серия) викарных видов у систематиков представляет собой группу .род-
ственных видов, происшедших из предкового вида в пределах его древ-
него ареала.
Другие пути видообразования у покрытосеменных — по типу грёга-
ризма (Юзепчук, 1936, 1939, 1958), в результате гибридизации (Попов,
1927; Комаров, 1940; Пояркова, 1953; Толмачев, 1958; Троицкий, 1959;
и др.) — вследствие иной интерпретации и в силу их ограниченного рас-
пространения в природе и слабой изученности, пока не могут быть при-
няты во внимание.
Каждый вид имеет свою первичную область формирования, обуслов-
ленную биоэкологической спецификой предкового вида и констелляцией
условий жизни в пределах исходного ареала. Последующие общеклима-
тические, геоморфологические и другие изменения вызывают видоизме-
нения первичного ареала вида, что усложняет возможности выяснения
центров происхождения видов. Аутохтонное развитие видов отнюдь не
отрицает самых разнообразных миграций как отдельных видов, так и
их комплексов, неоднократно совершавшихся в историческом прошлом.
В процессе приспособления растений к почвенно-климатическим и
ценотическим условиям вырабатывались многообразные биоморфологи-
ческие структуры или жизненные формы. Внешняя структура, габитус
как основной биолого-морфологический показатель жизненной формы,
определяется физиологическими свойствами, биохимическими процесса-
ми, биологией развития и внутренними особенностями строения рас-
тений, тесно связанными с их филогенией и экологией. В оценке жизнен-
ной формы нужно учитывать как надземные, так и подземные органы.
Содержание эволюции жизненных форм составляет преобразование
приспособительной структуры растений под влиянием изменения усло-
вий существования на арене геологической истории земли и в связи с
перемещениями растений по территории. Таким образом, эволюция жиз-
ненных форм должна отражать направления развития биоморфологи-
ческих структур, вскрывать исходные, промежуточные и конечные звенья
трансформации.
Тип биоморфологической структуры определяется как условиями
внешней среды, так и приспособительными особенностями растений в
их историческом развитии. С этой точки зрения эволюционная проработ-
ка жизненных форм должна опираться на данные филогенетической
систематики.
В более узком смысле привлечение систематики совершенно необхо-
димо для филогенетического анализа жизненных форм растений кон-
кретной таксономической категории.
Исходя из целей данной работы, нам нужно было выбрать такой род,
который бы обнимал как можно большее разнообразие биоморфологи-
ческих структур в надземной и подземной частях растений и со стороны
общебиологических особенностей: деревянистые и травянистые растения,
а среди последних — многолетние поликарпические и монокарпические
виды, безрозеточные и розеточные растения с симподиальным и моно-
подиальным возобновлением; растения, обладающие различными мета-
морфозами и структурами подземных органов: стержнекорневые, стерж-
некистекорневые и кистекорневые, разные формы корневищных растений
4 В. Н. Голубев
49
и т. Вместе с тем по такому роду должна быть хорошая монографи-
ческая обработка в систематико-филогенетическом отношении.
Указанным требованиям довольно полно отвечает род Potentilla,
который мы и выбрали в качестве основного объекта исследований.
Фрагментарно рассматриваются особенности развития биоморфологиче-
ских структур в роде Scrophularia и некоторых других.
ЭВОЛЮЦИЯ ЖИЗНЕННЫХ ФОРМ В РОДЕ POTENTILLA
Систематика рода Potentilla
Роду Potentilla посвящена великолепная монография Т. Вольфа
(Wolf, 1908), которая весьма подробно охватывает видовое богатство
рода, морфологические особенности и экологию видов, их географи-
ческое распространение, родственные связи и вероятную историю раз-
вития. Хотя после выхода в свет монографии Вольфа прошло уже более
полустолетия, но многие ее положения сохранили свое значение и по сей
день. В обработке лапчаток СССР С. В Юзепчук (1941) несколько ото-
шел от систематических подразделений Вольфа, однако эти изменения
больше касаются таксономии. Среди них наиболее важным является
вопрос о принадлежности кустарниковых форм лапчаток — следует ли
их принимать за самостоятельный род Dasiphora или же считать видами
Potentilla, как это делает Вольф. Мнение Вольфа в этом вопросе не
одиноко. Кустарниковые формы помещаются среди Potentilla А. А. Грос-
сгеймом (1952), Б. К. Шишкиным (1954) и др.
В настоящей работе мы придерживаемся объема рода Potentilla,
принятого Вольфом, т. е. включая виды Dasiphora, Comarum, Duchesnea.
При этом мы руководствуемся тем, что все эти формы, как и Patentilla
s. str., в своем происхождении ведут начало от одного или немногих
близкородственных типов. Эти систематические категории — виды ли
одного рода, либо цикл близких родов — представляют более или менее
уклонившихся потомков, развившихся на основе дифференциации ан-
цестрального типа по принципу лучевой дивергенции. Одни из современ-
ных форм ближе стоят к предковому типу и мало изменились в течение
длительного периода, другие — претерпели разнообразную трансформа-
цию, сильно уклонились от предкового типа и на протяжении своей
истории в свою очередь вновь и неоднократно могли трансформировать-
ся, давая начало новым циклам форм.
В монографии Вольфа насчитывается 305 пронумерованных видов.
Позже был выделен целый ряд новых видов, главным образом за счет
перевода уже известных разновидностей в ранг видов. Однако какой-
либо кардинальной переработки системы Вольфа рода Potentilla не было.
И если некоторые систематики, например С. В. Юзепчук (1941), выска-
зывают неудовлетворенность системой Вольфа, то это относится преи-
мущественно к объему отдельных полиморфных видов. В целом же
система Вольфа в настоящее время является наиболее разработанной
и мы кладем ее в основу анализа эволюции жизненных форм лапчаток.
Наибольшую филогенетическую ценность в систематике лапчаток
имеют особенности опушения плодиков и форма и характер прикрепле-
ния столбика к завязи. По признаку опушения плодиков род разделяет-
ся на две секции — опушенноплодиковые (Trichocarpae) и голоплодико-
вые (Gymnocarpae). Деление рода на две секции имеет место и в
известной работе Энглера и Прантля (Engler, Prantl, 1894). В зависи-
мости от строения столбика и места его прикрепления к завязи выделя-
ются следующие подсекции: среди Trichocarpae — Rhopalostylae и Nema-
tostylae, в Gymnocarpae—Closterostylae,, Conostylae, Gomphostylae и
Leptostylae. Кроме того, подсекция Nematostylae делится на две серии.
50
Первая из них (Suffruticulosae) включает полукустарниковые формы,
вторая (НегЬасеае) —травянистые формы. На две серии делится и под-
секция Conostylae: Eriotrichae (шерстистоволосые) и Orthotrichae (пря-
моволосые). Основной же классификационной единицей является, по
Вольфу, группа. В роде установлена 31 группа.
Как уже отмечалось выше, в работах советских систематиков Poten-
tilla наблюдается тенденция к укрупнению отдельных таксонов, выделяв-
шихся Вольфом. Так у С. В. Юзепчука (1941) и А. А. Гроссгейма (1952)
группы Вольфа преобразованы в секции. Секционное разделение прини-
мается и в настоящей работе. В системе С. В. Юзепчука (1941) группа
Fruticosae выделена в самостоятельный род Dasiphora, rpynna Bifurcae —
в подрод Schistophyllidium, группа Biflorae — в подрод Micropogon,
группа Palustres—в род Comarum, группа Eriocarpae и серия НегЬасеае—
в подрод Fragariastrum, подсекция Closterostylae с единственной труп-
пой Rupestres — в подрод Closterostyles, подсекция Conostylae — в под-
род Hypargyrium, подсекция Gomphostylae — в подрод Dynamidium,
подсекция Leptostylae — в подрод Chenopotentilla. Таким систематиче-
ским разделением Potentilla подчеркивается обособленность отдельных
циклов форм, далеко зашедший процесс дифференциации. Вместе с тем
это должно говорить и о сравнительно большой древности рода. Впро-
чем, палеонтология мало что может рассказать о прошлой истории и
возрасте рода. Одно несомненно, что некоторые виды лапчаток сущест-
вовали уже в третичном периоде (Криштофович, 1941).
Биоморфология1 и эколого-географические особенности
распространениявидов
В характеристике структурного типа видов Potentilla мы будем ру-
ководствоваться положениями, рассмотренными и мотивированными в
предыдущих наших статьях (Голубев, 1957а б, 1958а, 1959а, б, 19606, г)
и принятыми в настоящей работе.
В состав Potentilla s. 1. входят невысокие кустарники, полукустарни-
ки, травянистые многолетние, двулетние и однолетние растения. По ха-
рактеру структуры побеговой системы деревянистых и травянистых форм
подавляющее большинство видов Potentilla должно быть отнесено к ро-
зеточному типу. Лишь отдельные единичные виды, по-вйдимому древнего
происхождения, характеризуются безрозеточной структурой побегов. Эту
особенность следует рассматривать в качестве анцестрального признака,
сохранившегося у современных видов в силу действия закона эволюци-
онной разновозрастности признаков (Тахтаджян, 19546). Наличие таких
древних черт в строении представляет исключительную ценность и ин-
терес для реконструкции палеоморфного предкового типа.
В своем распространение и происхождении лапчатка является го-
ларктическим родом. Его ареал охватывает почти все северное полуша-
рие, за исключением южной оконечности Северной Америки, полуострова
Флориды, центральной части полуострова Мессопотамии, Моллукского
1 В определении биоморфологических признаков были использованы все имеющие-
ся сборы в гербариях Ботанического института АН СССР, Всесоюзного института аро-
матических и лекарственных растений, Главного ботанического сада АН СССР. Кроме
того, работая в ГБС АН СССР, мы выписали семена по делектусам из всех наиболее
известных ботанических садов мира и выращивали из них растения как на питомнике
ГБС (Москва), так и в Центрально-Черноземном заповеднике в двух вариантах: на
открытом степном участке и в условиях затенения на поляне в дубовом лесу (Дубро-
шина). Важное значение имело ' изучение биоморфологии лапчаток непосредственно
в природных условиях: в Московской и Смоленской областях, Прикарпатье, в лесосте-
пи и южнотаежной зоне Западной Сибири, европейской лесостепи, Западном Тянь-
Шане, Заилийском Алатау, Северном Кавказе, Крыму, лесостепи Прибайкалья. Во всех
этих районах нам довелось работать в последние 10 лет.
4*
51
полуострова и южной оконечности полуострова Индокитая. Лишь не-
многие виды доходят до экватора или заходят за экватор к югу, но в
южном полушарии нет ни одной специфической группы лапчаток. Таким
образом, колыбелью возникновения и развития лапчаток является сс
верное полушарие земли.
Данные о современном распространении видов и групп в сопоставле-
нии с палеогеографическими могут служить некоторым материалом для
оценки их возраста. Так, например, распространение видов на всех трех
материках Голарктиса — в Северной Америке, Евразии и Африке (в
северной части) —свидетельствует о том, что они уже существовали в
ту пору геологической истории земли, когда эти материки соединялись
вместе и имелась возможность обмена флор между тремя континентами.
Существование соединения Северной Америки с Евразией, по Арльдту
(Arldt, 1907), датируется палеогеном. В миоцене связь эта сильно со-
кращается, тогда как Африка широкой полосой соединяется с Европой
посредством Пиренейского полуострова.
Однако узость ареала других видов еще не дает прямых указаний на
их молодость. Вообще к оценке возраста ныне живущих видов надо
подходить осторожно, особенно принимая во внимание закон эволюцион-
ной разновозрастности признаков.
Экология видов лапчаток чрезвычайно разнообразна. Они встреча-
ются во всех зонах северного полушария, начиная от тундры и кончая
полупустынями и пустынями. В условиях высоких гор виды лапчаток
произрастают почти во всех поясах, но особенно разнообразны в видо-
вом и количественном отношении в альпийском и субальпийском поясах.
Столь большое разнообразие условий существования лапчаток явилось
причиной выработки у них многочисленных биоморфологических струк-
тур, к обзору которых мы и переходим.
Деревянистые лапчатки
Первой и, надо полагать, наиболее древней секцией лапчаток явля-
ется группа Fruticosae, объединяющая, по Вольфу, всего два вида —
Р. fruticosa L., Р. davurica Nestl. Центральное место занимает Р. frutico-
sa, которая является весьма полиморфным, и очевидно, сборным видом.
Виды секции 1 — Fruticosae представляют собой невысокие кустарни-
ки розеточного типа структуры до 1,5 м высоты. Крона куста состоит из
долихобластов и розеточных брахибластов. Цветоносные побеги бывают
разных категорий — как розеточными, так и безрозеточными. Первые из
них способны развиваться по ди- и полициклическому типу, в зависимо-
сти от условий, но возможно и моноциклическое развитие их. Безрозе-
точные цветоносные побеги всегда моноциклические. Нарастание побе-
гов осуществляется симподиально и лишь укороченные розеточные веге-
тативные побеги растут моноподиально. В неблагоприятных условиях
розеточные побеги имеют довольно значительный возраст, что соответ-
ствует и наблюдениям Н. В. Дылиса (1958). В подземной части разви-
вается стержнекорневая или стержне-кистекорневая структура.
Ареал секции весьма обширный. Он охватывает горные районы
Северной Америки, Среднюю и приатлантическую Европу, юг СССР,
всю Сибирь, Центральную, Восточную и Среднюю Азию. Обнаружива-
ется тяготение Р. fruticosa к горным странам, где она поднимается до
2—3 тыс. метров, а в Гималаях — до 5 тыс. метров, встречается на
каменистых склонах. Растет она также в низкогорьях и предгорьях, в
северных равнинных странах предпочитает сырые или временно затоп-
ляемые места.
В зависимости от условий структура растений довольно резко варь-
ирует. На подверженных сильным ветрам участках гор в западном
52
Тянь-Шане мы встречали стелющуюся форму лапчатки [Dasiphora раг-
vifolia (Fisch.) Juz.], побеги которой буквально выполняли все неровно-
сти каменных плит. В низкогорном поясе в укрытых от ветра местах
развиваются довольно высокие кустарники с прямостоячими побегами.
Замечательную форму структуры Р. fruticosa s. 1. мы наблюдали у
экземляров, собранных в высокогорьях Гималайских гор (Гербарий
БИНа). Молодая особь состояла из дерновинки розеточных побегов, при-
жатых к почве. У отдельных розеток в пазухах листьев заметны цвето-
ножки с единственным цветком. Следовательно, нарастание побегов
данных экземпляров в высокогорьях осуществляется моноподиаль-
но. Взрослые хорошо развитые особи обладают системой корневищ, при-
жимающихся к почве и слегка засыпаемых щебнем и мелкоземом, окан-
чивающихся дерновинкой розеточных моноподиальных побегов. Эти
корневища представляют осевые части розеточных моноподиально на-
растающих побегов.
Формообразование внутри секции Fruticosae совершалось по типу
викаризма, на основе дифференциации анцестрального типа в различ-
ных частях ареала предкового типа, Это положение хорошо отображено
в системе С. В. Юзепчука (1941). Он выделяет три ряда (серии) в роде
Dasiphora. К первому ряду — Fruticosae — отнесены D. fruticosa (L.)
Rydb. и D. phyllocalyx Juz., ко второму — D. parvifolia и D. dryadanthoi-
des Juz., к третьему — D. davurica (Nestl.) Korn, и D, mandshurica (Ma-
xim.) Juz. Все перечисленные виды характеризуются сходной структурой,
описанной выше.
Секция занимает довольно обособленное положение. Руководящий
систематический признак секции — булавовидный столбик не имеет ана-
логов в других секциях, за исключением секции Bifurcae.
Наличие такой тесной связи между Fruticosae и Bifurcae представ-
ляет большой интерес для выяснения филогении жизненных форм рода,
на чем мы остановимся подробнее ниже. Здесь же укажем, что отдельные
признаки сходства Fruticosae обнаруживаются также со всеми деревя-
нистыми представителями лапчаток — секциями Xylorrhizae, Biflorae,
Palustres, Tridentatae и Eriocarpae.
Изолированное положение в системе Fruticosae, что нашло отраже-
ние в выделении их многими систематиками в самостоятельный род, ши-
рокое географическое распространение и явные признаки географической
дифференциации — все это свидетельствует о большой древности сек-
ции, которая тем не менее еще сохранила некоторые признаки сходства
с другими лапчатками.
Секция 2 — Bifurcae, по Вольфу, монотипная и заключает один вид
Р. bifurca L. Она представляет собой длиннокорневищный полукустар-
ничек с деревянеющим корневищем и ежегодно низко отмирающими по-
бегами. По структуре побегов должна быть отнесена к безрозеточному
типу, возобновление побегов симподиальное. Однако в сборах из Тибета
и Кашмира надземные побеги этой лапчатки представляют укороченные
розетки. Иногда группы скученных розеточных побегов образуют доволь-
но плотные подушечки (высокогорья Кашмира!).
В первые годы развития из семени у лапчатки вильчатой формирует-
ся типичный стержневой корень (Борисова, 1960). Первые плагио-
тропные корневища образуются от основания вертикального главного
побега, находящегося в почве. С возрастом система главного корня
отмирает и растение переходит на питание при помощи придаточных
корней.
Ареал секции охватывает центральные и южные районы Европейской
части СССР и почти всю Азию, включая Армению, Иран, Кавказ, Сред-
нюю Азию, Сибирь, Монголию, Тибет и др. Преобладающими местами
обитания являются степи, сухие глинистые и каменистые склоны, изредка
53
луга, лиственничные и сосновые леса, пустыни, альпийские луга и высо-
когорные пустыни.
Вид следует также признать сборным. С. В. Юзепчук выделяет в этой
секции ряд самостоятельных видов: Л bifurca, Р, orientalis Juz.,P. moor-
croftii Wall.,P. semiglabra Juz., и P. imbricata Kar. et Kir., которые хоро-
шо отличаются экологически. Здесь также отчетливо проступает процесс
дифференциации некогда единого типа. Развитие новых форм протека-
ет аутохтонно в различных частях обширного ареала под влиянием при-
способления к специфическим условиям жизни.
Как указывает Вольф, родственные связи Bifurcae тяготеют к Fruti-
cosae, что проявляется в общих чертах строения столбика. С другой сто-
роны, Bifurcae являются носителями наиболее примитивного признака —
безрозеточного типа структуры побегов. В этой связи можно предпола-
гать, что Bifurcae — отдаленные потомки, сохранившие примитивные
черты морфологической структуры. Это слепая и весьма древняя группа
лапчаток, предки которой в других своих поколениях явились родона-
чальником розеточных форм деревянистых лапчаток, подобных совре-
меннььм Fruticosae.
Из секции 3 — Xylorrhizae наиболее изученным видом является
Р. lignosa Willd., второй ее вид Р. lindenbergii Lehm. Р. lignosa представ-
ляет полукустарничек с очень разветвленным корявым деревянистым
каудексом и мощным стержневым корнем. Структуру побегов следует
отнести к розеточному типу, нарастание побегов моноподиальное.
Розеточные побеги способны вытягивать свои междоузлия. В результа-
те ветвления образуются дерновинки моноподиально нарастающих вет-
вей, осевые части которых, нередко покрытые почвой, уподобляются кор-
невищам. При ортотропном росте розеточные побеги рано отмирают.
Вид довольно редкий, узко локального распространения. Обитает на
скалах в Северном Иране и Курдистане, поднимается до высоты 2 200 м
и выше.
Упомянутый второй вид секции обитает в Гималаях и представляет
полукустарничек, сходный по структуре с первым видом.
Филогенетически, по строению цветков и плодов, секция обнаружи-
вает более тесные связи с Biflorae, чем с Fruticosae. Лишь по форме
листьев имеется близкое сходство с Fruticosae, но они не рассечены. Все
же положение секции довольно обособленное.
Структура деревянистых побегов Xylorrhizae в виде толстых монопо-
диальных корневищ имеет исключительное значение для понимания га-
битуальной эволюции лапчаток.
Секция 4 — Biflorae заключает два вида: Р. biflora Willd. и Р. articu-
lata Franch. Оба вида являются полукустарничками с моноподиально
нарастающими побегами, которые развиваются то в форме приземных
корневищ, то в виде плотных подушек. В последнем случае отми-
рающие листья не опадают, одевая побеги плотным чехлом. Обычно
междоузлия побегов слабо развиты, и их структуру можно принять розе-
точной, хотя типичных розеток не образуется. Подземные органы —
стержневая корневая система.
Ареал Р. biflora занимает Северную Америку, Чукотский полуостров,
Алтай, Джунгарию, Тарбагатай, Тянь-Шань, Саяны, Даурию, Северную
Монголию. Растет на скалах в альпийском поясе гор. Р, articulata встре-
чается в тех же местах обитания, поднимаясь в горы до 4500 м, распро-
странена в Китае в провинции Юннань.
Секция Biflorae генетически ближе всего примыкает к секции Xylor-
rhizae, что основывается на сходстве строения пестиков. Небольшие отли-
чия касаются опушения плодика. Кроме того, у Biflorae длинный и тон-
кий столбик прикрепляется несколько ближе к верхушке завязи, чем у
Xylorrhizae.
54
Секция 5Palustres заключает Р. salesowiana Steph, и Р. palustris
Scop. Первый вид является типичным стержнекорневым полукустарни-
ком. А. П. Стешенко (1956) относит этот вид к кустарничкам, оговари-
ваясь, что рост побегов и цикл их развития протекает у него так же, как
у полукустарничков. В условиях высокогорий Памира, по данным
А. П. Стешенко, скелет куста Р. salesowiana образуют стелющиеся по
поверхности почвы многолетние ветви. Они составлены из ряда разно-
возрастных побегов. У основания стареющих ветвей отрастают молодые
розеточные вегетативные побеги, которые, по А. П. Стешенко, способны
перезимовывать и в течение 4—5 лет продолжать верхушечный (моно-
подиальный) рост. Затем концы таких побегов либо отмирают, либо из
их верхушечной почки развивается цветоносный побег.
Р. palustris представляет длиннокорневищный полукустарничек с де-
ревянеющими корневищами. По структуре побегов этот вид надо отнести
к безрозеточному типу. Возобновление побегов симподиальное.
Как показывают исследования Ирмиша (Irmisch, 1861), главный корень
проростка не получает значительного развития и в дальнейшем от-
мирает.
Р. salesowiana распространена в горах Средней и центральной Азии,
на, Алтае; растет на каменистых и щебнистых склонах, по берегам
речек. Ареал Р. palustris охватывает почти всю Голарктику, исключая
некоторые районы Центральной Азии и южные приморские районы Азии.
Обитает в тундрах, болотах и заболоченных лугах и лесах, по берегам
озер и речек.
Узкая и далеко зашедшая специализация и особенности распростра-
нения свидетельствуют о большой древности секции Palustres. Она об-
наруживает лишь отдаленные связи с деревянистыми лапчатками. С сек-
цией Fruticosae ее сближает форма большой почти пентагональной
чашечки с крупными рецептакулами и другие признаки. По структуре
побеговой системы Р. palustris напоминает Р. bifurca: оба вида относят-
ся к безрозеточному типу. Боковой столбик, прикрепленный посредине
завязи, сближает Palustres с Xylorrhizae.
Секция 6 — Tridentatae заключает три вида (Р. tridentata Sol., Р. ат-
bigua Camb., Р, miyabeiMakino). Р. tridentata, распространенная в севе*
ро-восточной части Северной Америки, представляет длиннокорневищ-
ный полукустарничек розеточного типа структуры, весьма близко напо-
минающий по развитию нащи грушанки. В почве образуются длинные,
довольно тонкие деревянистые корневища, которые, загибаясь вверх,
выходят на поверхность, образуя розетку тройчатосложных листьев.
Позднее из верхушечной почки розетки формируется облиствленный
цветоносный побег. Но в некоторых случаях цветоносный побег с вытя-
нутыми междоузлиями развивается моноциклически, минуя фазу розет-
ки. В этом явлении мы усматриваем рекапитуляцию палеоморфной без-
розеточной структуры. В ювенильном возрасте Р. tridentata обладает
хорошо выраженным стержневым корнем, который затем элиминирует
в связи с развитием системы длинных плагиотропных побегов. Сходны-
ми чертами морфогенеза характеризуется Р. ambigua, побеги которой,
аналогично предыдущему виду, способны развиваться по безрозе-
точному типу. Обитает в альпийском и субальпийском поясах Гима-
лаев. Последний вид секции — Р, miyabei известна лишь из одного места
на вершине потухшего вулкана острова Хоккайдо. Является стержне-
корневым полукустарничком розеточного типа структуры, с симподиаль-
ным возобновлением побегов. С возрастом, однако, как и два других
вида секции, способна развивать длинные корневища.
Секция отличается суббазальным длинным столбиком, сильно опу-
шенными плодиками и тройчатыми листьями с трехзубчатыми листоч-
ками. Близкая связь устанавливается через Р. eriocarpa Wall, с секцией
55
Eriocarpae. С другими секциями деревянистых лапчаток она обнару-
живает мало сходства.
Последняя секция 7 (деревянистых лапчаток) — Eriocarpae исклю-
чительно азиатская и состоит из четырех видов, из которых два вида
(Л dickinsii Franch. et Sav. и P. ancistrifolia Bge.) восточноазиатские, один
вид (P. eriocarpd) обитает в альпийском и субальпийском поясах Гима-
лаев и четвертый вид — Р. elatior Willd. встречается в альпийских рай-
онах Западного Кавказа и Понтийских гор. Все виды в начале развития
являются стержнекорневыми полукустарничками, позднее у Р. eriocarpa
и Р. ancistrifolia образуются дерновинки корневищ с розетками листьев
на верхушке. Необходимо отметить, что у Р. eriocarpa, обитающей в вы-
сокогорьях, побеги нарастают моноподиально, а цветоносные по-
беги слабо облиствленные и укороченные. Нередко корневищеподобные
пооеги образуют плотные подушки. Конечно, это будут факуль-
тативные подушки (Станюкович, 1949), так так в менее суровых местах
обитания плотноподушковидная форма не образуется. У Р. ancistrifolia
при общем симподиальном возобновлении отдельные розеточные побе-
ги длительно остаются вегетативными и превращаются в плагиотропные
корневища, нарастающие моноподиально.
С. В. Юзепчук относит эту секцию уже к травянистым лапчаткам.
Фактические данные показывают, что все секции деревянистых форм
лапчаток обнаруживают известную обособленность, что указывает на их
древнее происхождение. Об этом же свидетельствуют и особенности рас-
пространения ряда видов, ареалы которых охватывают Северную Аме-
рику, Европу и Азию. Большинство видов рассмотренных секций тя-
готеет в своем распространении к горным странам. Отдельные виды в
горах являются специализированными альпийцами. Характер распрост-
ранения секций деревянистых форм лапчаток позволяет с определен-
ностью указать на область Северной Америки, Восточной Азии и Древ-
него Средиземноморья (в понимании Попова, 1927, 1931; Вульфа, 1944)
как на первичную колыбель формирования лапчаток. Деревянистые лап-
чатки, несомненно, являются наиболее древними и исходными предста-
вителями рода.
Анцестральными формами были, по-видимому, безрозеточные стерж-
некорневые кустарники, отдельные черты которых еще и сейчас сохрани-
лись у таких весьма древних форм, как Р. bifurca и Р. palustris. Факты
развития побегов у Р. tridentata и Р. ambigua по безрозеточному типу
подтверждают это. Безрозеточный тип побегов соответствовал благо-
приятным климатическим условиям развития древних лапчаток в до-
третичное и реннетретичное время. Под влиянием горообразовательных
процессов и общих изменений климата тертьера вырабатываются розе-
точные формы лапчаток, подобные современным Fruticosae.
Дошедшие до наших дней деревянистые формы лапчаток представ-
ляют скромные уцелевшие остатки древесных предковых форм. Поэтому
биоморфа лапчаток — полукустарнички — имеют «кустарниковое» про-
исхождение, т. е являются производным древесных предков. Вместе с
тем современные полукустарнички представляют и конечные звенья
трансформации анцестральных форм. Следовательно, форму полукус-
тарничков у лапчаток, по-видимому, нельзя рассматривать в качестве
переходной от древесных к травянистым, что доказывается изолирован-
ным положением их в системе рода, отсутствием близких связей с тра-
вянистыми видами. Формы травянистых растений развились из деревя-
нистых древних лапчаток. Их образование знаменовало новое направ-
ление эволюции, которое в отдельных случаях протекало параллельно
эволюции деревянистых потомков — полукустарничков.
Для понимания общих экологических закономерностей развития био-
морфолсгических структур лапчаток интересны те специфические преоб-
56
разования деревянистых видов, которые без затруднения можно связать
с определенными условиями. Из подобного рода фактов особенную цен-
ность представляют следующие: 1. Переход безрозеточного типа структу-
ры в розеточную в высокогорьях, что наблюдается у Р. bifurca. 2. С ухуд-
шением условий жизни в высоких поясах гор увеличивается продолжи-
тельность жизни вегетативных розеточных побегов, которые в эту фазу
всегда нарастают моноподиально. 3. Зарегистрированы случаи с Р. fruti-
cosa, когда в высокогорьях рост и возобновление обычно симподиальных
побегов изменялись на моноподиальное. Подобная трансформация сим-
подиального роста в моноподиальный наблюдается и у других деревя-
нистых видов в аналогичных условиях. 4. Как при длительном нараста-
нии розеточных побегов, так и при переходе на моноподиальное их во-
зобновление осевые части побегов становятся корневищеподобными
образованиями. 5. В суровых условиях высокогорий происходит нередко
агрегация надземных побегов деревянистых лапчаток в плотные подуш-
кообразные формы. Эти плотные подушки у большинства лапчаток яв-
ляются факультативными и в иных более благоприятных условиях не
образуются. Однако виды секции Biflorae типичны для альпийского поя-
са и в низкогорьях не встречаются, что позволяет считать облигатным
характер подушек ее видов.
Следует особо подчеркнуть, что уже внутри деревянистых лапчаток
ясно намечается преобразование формы роста побегов: из надземных
кустарникового типа они переходят к припочвенному и далее внутрипоч-
венному образу жизни на основе усиления розеточной формы роста,
характеризующей сокращение линейных размеров побегов. Этот пере-
ход вызван суровыми условиями высокогорных районов, с которыми
связано их распространение. В случае моноподиального роста геофиль-
ных побегов их цикл развития продолжается весьма длительное время.
В результате сохранения верхушечного роста активное состояние таких
побегов может совпадать с длительностью жизни особей. Но и в случае
симподиального роста, а также при отмирании верхушек моноподиаль-
ных побегов и формировании замещающих осей, эти структурные кате-
гории— побеги обладают длительным жизненным циклом. Значит, рас-
сматриваемая форма роста побегов геофильных деревянистых лапчаток,
являющаяся прообразом структурных форм травянистых лапчаток, не-
сомненно связанных с первыми и морфологически, и эволюционно-гене-
тически, в своем возникновении определялась не предельным сокраще-
нием жизненного цикла скелетных осей, как это имело место в развитии
некоторых кустарничков (Серебряков, 19546, 1955а) и безрозеточных
трав (Голубев, 19576, 1959а), а изменением направления роста, сокра-
щением размеров годичных побегов и переходом к припочвенному и
внутрипочвенному существованию.
Можно сделать некоторые выводы относительно развития подземных
органов в процессе формообразования. Примитивной структурой под-
земных органов деревянистых лапчаток является стержнекорневой тип.
Стержнекорневая система характерна, по крайней мере в раннем воз-
расте, для всех видов деревянистых лапчаток, за исключением разве
Р. palustris. Многие виды сохраняют стержневую корневую систему в те-
чение всей жизни. Ряд других видов выработал новую приспособитель-
ную особенность — способность к образованию длинных корневищ. Вто-
ричность этого признака мотивирована нами ранее (Голубев, 1957а, 6;
19596). Здесь же интересно отметить, насколько филогенетические дан-
ные подкрепляют это положение. Как можно заметить, виды наиболее
древней секции Fruticosae являются стержнекорневыми растениями.
Однако секция Bifurcae, а также Р. palustris из секции Palustres, не-
сущие признаки анцестральной, безрозеточной структуры, обладают уже
длинными корневищами, а стержневой корень либо совсем не выражен
57
(P. palustris), либо формируется только в ранний период жизни расте-
ния (Р. bifurca). Отсюда мы вправе заключить, что в одном и том же
организме, при относительно примитивном состоянии одних признаков,
другие способны к прогрессивной эволюции. Таким образом, здесь нахо-
дит проявление классический принцип эволюционной разновозрастности
признаков, в данном случае в области вегетативной сферы.
Рыхлокустовая и плотнокустовая структуры побегов деревянистых
лапчаток являются, несомненно, вторичными, возникшими на основе
кустарниковой вследствие преобразования формы роста побегов в ус-
ловиях высокогорий. Стержнекорневая структура подземных органов
осталась без существенных изменений в качестве характерного призна-
ка новых жизненных форм (подушек и рыхлокустовых).
Обобщая все сказанное, можно заключить, что деревянистые лапчат-
ки являются весьма древними представителями рода, значительно обо-
собившимися в результате далеко зашедшей дифференциации, сохранив-
шими в своей организации значительное количество примитивных
признаков.
Современные виды — продукт аутохтонного развития потомков пред-
кового типа, занимавшего широкий ареал в области современных гор
Восточной Азии, Древнего Средиземноморья и Северной Америки. Фор-
мообразование рода совершалось главным образом по принципу луче-
вой дивергенции, но известную роль в видообразовании, вероятно, игра-
ла и гибридизация среди травянистых форм. Родиной лапчаток была, по-
видимому, область древнего Ангарского материка.
Травянистые лапчатки
Секция 8 — Speciosae является восточносредиземноморской. Рас-
пространение секции простирается на южную и юго-восточную Европу,
от Италии к востоку до Малой Азии и Кавказа. Ареалы ее пяти видов
почти нигде не перекрывают друг друга и, следовательно, характеризу-
ются отчетливым викаризмом. Все виды — высокогорные растения, оби-
тающие на высоте от 1500 до 2700 м над ур. моря.
Отличается от других секций травянистых лапчаток наличием густо-
го тонкого войлочного опушения из кудрявых волосков на стеблях и
листьях; опушены и пестики, по крайней мере в незрелом состоянии.
Листья тройчатосложные. Все виды являются растениями с мощным
стержневым корнем, который заметно одревесневает, что служит дока-
зательством близости Speciosae к деревянистым лапчаткам. Вообще
между полукустарниковыми и травянистыми лапчатками по признаку
более или менее выраженного одревеснения нижних частей побегов или
части стержневого корня резкой границы провести нельзя. Укороченные
розеточные побеги, нарастающие моноподиально, превращаются в тол-
стые корневища, которые у Р. oweriniana Rupr., Р. apennina Ten., Р. deo-
rum Boiss. et Heldr. и P. kionaea Halac. образуют над поверхностью поч-
вы довольно плотные дерновинки. У Р. speciosa Willd. толстые корневи-
ща плотных подушек не образуют. Цветоносные побеги у всех видов
занимают боковое положение, развиваясь в пазухах розеточных листьев.
Эти побеги облиствлены слабо или почти безлистные, что связано с аль-
пийским образом жизни.
Секция 9 — Nitidae является в географическом отношении антиподом
предыдущей. Ее следует отнести к западносредиземноморскому геогра-
фическому элементу. Два ее вида — Р. nitida L. и Р. alchimilloides Lap.—
произрастают в высокогорно-альпийских районах и только Р. saxifraga
Ard. спускается на скалы нижнего горного пояса.
Стебли и листья видов Nitidae также опушены густыми блестящими
волосками, однако под покровом простых волосков войлок всегда отсут-
58
ствует. Секция близко примыкает к предыдущей — Speciosae. Виды ха-
рактеризуются хорошо развитой стержневой корневой системой. Над-
земные побеги розеточные, моноподиально нарастающие. У Р. nitida
междоузлия розеточных побегов иногда вытягиваются, вследствие чего
образуются стелющиеся побеги, укореняющиеся придаточными
корнями. Розеточные укороченные побеги Р. nitida часто образуют плот-
ные подушечки. У Р. saxifraga из розеточных побегов возникают мощ-
ные корневища, нередко значительно возвышающиеся над почвой, но к
вегетативному передвижению растение, по-видимому, не способно. Обе
секции, очевидно, имеют общее происхождение. Они развились в области
Средиземноморья в процессе приспособления к различным условиям в
разных частях ареала предкового типа.
Секция 10 — Curvisetae относится к восточноазиатскому географиче-
скому элементу. Р. curviseta Hook. f. и Р. pteropoda Royle. обитают в
высокогорьях Гималаев, а третий вид секции — Р. colletiana Aitch. встре-
чается в Афганистане на высоте свыше 3000 м над ур. моря.
По характеру опушения и некоторым другим признакам секция
Curvisetae близко примыкает к Nitidae. Все виды являются типичными
стержнекорневыми растениями. Укороченные моноподиально нараста-
ющие розеточные побеги образуют плотные подушечки. Цветонос-
ные побеги развиваются в пазухах листьев розеток, короткие и почти
безлистные.
Секция 11 — Crassinerviae является средиземноморской высокогор-
ноальпийской. Ее семь видов распространены от Пиренеев до Кавказа, со-
ставляя почти правильный ряд замещения (Р. valderiana L., Р. haynal-
diana Jka., Р. brachypetala Fisch, et Mey., P. nivalis Lap., P. crassinervia
Viv., P. libanotica Boiss., P. grammopetala Мог.). Стебли и листья не име-
ют войлочного опушения, оттопыренно-волосистые или мохнатые, воло-
систыми являются и плодики.
Все виды представляют стержнекорневые растения. Из укороченных
розеточных моноподиально нарастающих побегов со временем формиру-
ются весьма толстые корневища, которые у Р. grammopetala и Р. crassi-
nervia иногда образуют плотные подушки, у других же видов ча-
ще формируются рыхлые дерновинки, но возможны и плотные поду-
шечки, как у Р. nivalis. Обращает на себя внимание уже отмеченная ра-
нее для Р. nitida способность некоторых розеточных побегов 'превра-
щаться в удлиненные. В результате розеточность теряется, образуется
удлиненный с вытянутыми междоузлиями стелющийся побег, который
вновь способен перейти к розеточному росту (что обусловлено распуска-
нием все новых листьев при недоразвитии междоузлий). Это явление
довольно часто встречается у видов лапчаток из других секций и заслу-
живает особого внимания. В сущности здесь имеет место превращение
укороченных моноподиальных розеточных побегов в моноподиальные
же удлиненные стелющиеся побеги. Такая пластичность биоморфоло-
гической структуры открывает возможность к вегетативному расселению
по площади и размножению, когда расползающиеся побеги переходят
к самостоятельному питанию посредством придаточных корней.
Как и в предыдущих секциях с альпийско-горными видами, можно
отметить у ряда видов Crassinerviae наличие невысоких слабо облист-
вленных цветоносных побегов.
Секция 12 — Caulescens, состоящая из двух видов, должна быть от-
несена к западносредиземноморскому географическому элементу. Р. elu-
sion a Jacq. обитает в высокогорьях западных Балкан, а Р. caulescens L.
распространена от Балкан во всех горных странах южной Европы, в Пи-
ренеях, горах Испании и Алжира.
Виды секции характеризуются нежными тонкими листьями и почти
целиком опушенными плодиками. Может быть секция представляет
59
связующее звено между древними Crassinerviae и сравнительно молоды-
ми Fragariastra.
По структуре вегетативных органов виды сходны с предыдущей сек-
цией. Оба вида являются стержнекорневыми: розеточные моноподиаль-
ные побеги образуют либо плотные подушечки (Р. caulescens)*
либо рыхлые дерновины (Р. clusiana). По особенностям развития кор-
невищ и строению листьев Р. caulescens весьма близко напоминает
Р. alba из следующей секции Fragariastra.
Секция 13 — Fragariastra заключает четыре вида. Они должны быть
отнесены к европейско-горному географическому элементу, тяготея к сре-
диземноморским странам. Распространены в среднегорных, низкогор-
ных и равнинных районах. Обитают на лугах, в светлых лесах, горных
склонах и т. д. ВидыР. montana\^xoi.,P. micrantha^am., Р. sterilis Garke.
являются еще характерными горными растениями, хотя могут встре-
чаться и на равнине (Р. micrantha и Р. sterilis). Лишь Р. alba L. можно
уже считать настоящим равнинным видом, она поднимается вверх не бо-
лее чем на 600 м над ур. моря.
Виды отличаются тем, что плодики опушены только в основании,
а в остальной части голые. Это последняя и самая молодая секция опу-
шенноплодных лапчаток (Potentillae trichocarpae), весьма близко при-
мыкающая к секциям Caulescentes и Crassinerviae.
Все виды, исключая Р. alba, в молодом возрасте являются стержне-
корневыми растениями, особенно Р. micrantha, которая не теряет стерж-
невого корня и во взрослом состоянии. Нарастание и возобновление по-
бегов моноподиальное, цветоносные побеги слабые, почти безлистные и
развиваются в пазухах розеточных листьев. Нарастание розеточных
побегов обусловливает развитие корневищ, укореняющихся придаточ-
ными корнями. Так, Р. alba является уже типично корневищным расте-
нием.
Розеточные побеги всех видов, но в особенности Р. montana и
Р. sterilis, способны преобразовываться в стелющиеся моноподиальные
побеги с вытянутыми междоузлиями.
Описанная группа видов замечательна во многих отношениях. Во-
первых, она прекрасно иллюстрирует пути перехода высокогорных лап-
чаток в низкогорные и равнинные формы. Несмотря на почти полную
утрату связи Р. alba с высокогорными условиями, она стойко сохраняет
характерные биоморфологические признаки высокогорных лапчаток:
моноподиальный рост, слабые почти безлистные цветоносные побеги.
Во-вторых, в секции ясно обозначаются признаки утраты стержневого
корня и переход на питание с помощью придаточных корней. В основе
этого преобразования лежит тенденция к формированию моноподиаль-
ных корневищ, развивающихся за счет надземного нарастания розе-
точных побегов, нередко переходящих в удлиненные стелющиеся побеги
с вытянутыми междоузлиями.
Секцию 14 — Rupestres составляют голоплодные лапчатки. Среди
Gymnocarpae эта секция является весьма древней. Как указывает Вольф
(Wolf, 1908), древний третичный корень Rupestres ближе всего прибли-
жался к современной шире всех распространенной Р. rupestris. По ха-
рактеру ареала секция относится к голарктическому географическому
типу. Часть ее видов исключительно североамериканские (Р. arguta
Pursh., Р. glandulosa Lindl., Р. rhomboidea Rydb., P. cuneifolia Th. W.),
другая часть — евразиатские, в ней Р. polyphylla Wall, и Р. fulgens
Wall.— гималайские, Р. tianschanica Th. W. — западнотяныпанский,
Р. poteriifolia Boiss.— южноиранский вид, Р. geoides М. В.— крымско-
кавказский, Р. calycina Boiss. et Bal.— малоазиатский. Лишь P. rupest-
ris L- является собственно голарктическим видом, произрастая в Север-
ной Америке, Европе и Азии. Большинство видов — горные и высокогор-
ные растения.
60
В систематическом отношении виды секции характеризуются вере-
теновидными почти базальными столбиками, непарноперистыми листья-
ми без войлочного опушения и другими признаками. Специфику био-
морфологической структуры составляют стержнекорневая структура под-
земных органов, часто в онтогенезе преобразующаяся в кистекорневую,
и симподиально нарастающие побеги розеточной структуры. В развитии
монокарпических побегов выделяются фазы почки, розеточного побега
и удлиненного облиствленного цветоносного побега.
Можно предположить, что секция происходит от каких-то анцестраль-
ных деревянистых лапчаток, обладавших розеточным типом структуры
и симподиальным возобновлением. Их развитие, однако, не было связано
с альпийскими условиями высоких гор, а скорее с среднегорьями и низ-
когорьями с засушливым климатом и скалисто-каменистым субстра-
том горных склонов. Древность секции доказывается ее изолированным
положением в системе, что послужило причиной для выделения ее в
ранг подрода Closterostyles.
Следующая секция 15 — Multifidae отличается хорошо выраженным
войлочным опушением вегетативных органов и коротким толстым стол-
биком. Заключает 27 видов, широко распространенных в Голарктике.
Но из них лишь два вида обитают на обоих материках — в Северной
Америке и Евразии (Р. pulchella R. Вг.— в арктической зоне Европы,
Азии и Северной Америки и Р. multifida L.), остальные виды либо соб-
ственно североамериканские (Р. pseudo-sericea Rydb., Р. rubricaulis
Lehm., Р. bipinnatifida Dougl., P. litoralis Rydb., P. pennsylvanica L.,
P. glabrella Rydb.), либо азиатские (P. verticillaris Steph., P. soongorica
Bge., P. sericea L., P. alexeenkoi Lipsky, P. polyschista Boiss., P. discolor
Bge., P. potaninii Th. W., P. lechenaultiana Ser., P. sikkimensis Th. W.,
P. baltistana Th. W., P. approximata Bge., P. fedtschenkoana Siegfr.
P. sischanensis Bge., P. hololeuca Boiss., P. chinensis Ser., P. niponica Th
W., Pi sibtrica Th. W.).
Сюда же следует отнести и Р. ewersmanniana Fisch, являющуюся эьь
демом Южного Урала. Последний вид Р. hispanica Zimm, распространен
в южной Испании, Северной Африке и на Кавказе. В экологическом
отношении секция весьма неоднородна. Два вида произрастают в тунд-
рах Арктики (Р. pulchella, Р. rubricaulis), значительное количество видов
распространено в горах, вплоть до альпийского пояса: Р. alexeenkoi,
Р. potaninii, Р. lechenaultiana, Р. sikkimensis, Р. baltistana, Р. fedtschen-
koana, Р. sischanensis, Р. hololeuca. Большинство остальных видов сек-
ции обитают на равнинах, в степях и пустынях, хотя нередко поднима-
ются и высоко в горы, как и Р. sibirica.
Вольф считает эту секцию древнейшей исходной группой среди Со-
nostylae eriotrichae. Она показывает родство со всеми другими секциями
коническистолбиковых шерстистоволосых лапчаток Graciles, Haema-
tochroae, Niveae, Argenteae, Collinae. Последние развивались из предков
Multifidae через редукцию перистых листьев до тройчатых или пальча-
тых, в результате удлинения столбика и других модификаций.
Все виды в ювенильном возрасте являются сгержнекорневыми расте-
ниями. В дальнейшем, однако, стержнекорневая система заменяется не-
редко кистекорневой или стержне-кистекорневой, например у Р. multi-
fida, Р. soongorica, Р. alexeenkoi, Р. discolor и др. Побеговая система ро-
зеточного типа. Большинство ^идов являются моноподиальными расте-
ниями. Розеточные побеги в процессе нарастания дают начало корневи-
щам, которые образуют либо рыхлую дерновинку (Р. hololeuca), либо
плотные подушечки, как Р. pulchella, Р. sericea (иногда), Р. rub-
ricaulis, Р. polyschista. В форме роста корневищ решающую роль играют
условия внешней среды. Так, в высокогорных условиях Р. pulchella ра-
стет в форме стержнекорневого подушковидного растения, цветоносные
61
побеги совершенно безлистные, едва возвышающиеся над поверхностью
подушки. В более благоприятных условиях существования растение раз-
вивается в форме, близкой к рыхлокустовой, все растение становится
крупнее, листья с длинными черешками.
Наряду с моноподиальными секция включает несколько симподиаль-
ных видов: Р. approximata, Р. bipinnatifida, Р. pensylvanica, Р. sibirica.
У Р. niponica, обнаруживаются оба способа нарастания и возобновле-
ния — симподиальное и моноподиальное. Необходимо отметить, что сим-
подиальные виды — обитатели равнин, степных и других местообитаний.
Симподиальное нарастание и возобновление здесь, по-видимому, разви-
лось из моноподиального. В отличие от Rupestres, где симподиальное
возобновление и нарастание является признаком древним, первичным,
унаследованным от деревянистых предков, в секции Multifidae монопо-
дий следует признать вторичным признаком. Возможность взаимных
переходов моноподиального и симподиального роста видов хорошо под-
крепляется указанным примером с Р. niponica. Таким образом, выска-
занные И. Г. Серебряковым (1952) предположения о генетических
взаимосвязях моноподиального и симподиального возобновления нахо-
дят достаточно убедительную фактическую мотивировку.
Секция 16 — Graciles, заключающая, по Вольфу, 24 вида, являет-
ся целиком североамериканской. Ее представители распространены
главным образом в горах запада Северной Америки, лишь отдель-
ные виды спускаются на равнину, например, Р. crinita Gray, и Р. effusa
Dougl.
Виды секции отличаются очень длинными столбиками, к основанию
конически расширяющимися, раза в два превышающими плодик.
Нами изучено 11 видов из 24 (Р. hippiana Lehm., Р. effusa Dougl.,
Р. osterhoutiana Th. W., P. subjuga Rydb., P. tenerrima Rydb., P. breweri
Wats., P. filicaulis Rydb., P. macounii Rydb., P. rubripes Rydb., P. ambi-
gens Greene., P. crinita Gray., P. lemmonii Greene., P. luteo-sericea Rydb.,
P. pulcherrima Lehm., P. gracilis Dougl., P. blaschkeana Turcz., P. pecti-
nisecta Rydb., P. flabelliformis Lehm., P. brunescens Rydb., P. Candida
Rydb., P. etomentosa Rydb., P. oaxacana Rydb., P. fastigiata Nutt., P. nut-
tallii Lehm.). Все изученные виды обладают стержневой корневой систе-
мой, длительно сохраняющейся в онтогенезе. Лишь у единичных видов
стержнекорневая структура подземных органов заменяется кистекорне-
вой (Р. fastigiata). По способу нарастания побегов большинство видов
являются моноподиальными, хотя, по-видимому, есть и симподиальные
формы (Р. blaschkeana, Р. etomentosa). Весьма интересными биоморфо-
логическими особенностями обладает Р. breweri, у которой в пазухах
розеточных (прикорневых) листьев формируются удлиненные ползучие
побеги, укореняющиеся в узлах. Они гомологичны цветоносным побегам.
Необходимо отметить, что в популяции всегда находятся особи с обыч-
ными, прямостоячими цветоносными побегами. Р. breweri — ползучее
стержнекорневое растение с еще не установившимися чертами биомор-
фологической структуры, показывающей переходы от типичных стерж*
некорневых растений к стержнекорневым ползучим.
Чрезвычайно своеобразна географически секция 17 — Haematochroae,
большая часть видов которой сосредоточена в Мексике, а три вида —
Р. nepalensis Hook., Р. argyrophylla Wall, и Р. atrisanguinea Lodd.— рас-
пространены в Гималаях. Все виды секции являются высокогорно-аль-
пийскими.
Генетически секция близко примыкает к предыдущей, с которой ее
сближает однотипное строение столбика. Однако по красной окраске
цветков Haematochroae отличается от Graciles.
Все виды следует отнести к стержнекорневым, лишь у Р. argyrophylla
стержнекорневая структура нередко преобразуется с возрастом в кисте-
62
корневую через промежуточную стержне-кистекорневую структуру. Во-
зобновление побегов большинства видов — моноподиальное (Р. ehrеfi-
ber giana Schlecht., Р. comaroides N. В., P. lozani Rose et Paint., P. lep-
topetala Lehm., P. argyrophylla Wall., P. atrisanguinea Lodd.). P. horrida
Rydb., P. thurberi Gray., P. atrirubens Rydb. характеризуются симподи-
альным возобновлением. Учитывая филогенетические связи и положение
в системе рода следует признать, что в секциях Multifidae, Graciles, Нае-
matochroae моноподиальное возобновление является первичным призна-
ком, утвердившимся с момента формирования этих секций, а симподи-
альное возобновление отдельных видов возникло из моноподиального.
Один вид секции — Р. nepalensis, в отличие от всех выше рассмотрен-
ных, представляет монокарпическое растение, часто с двулетней продол-
жительностью жизни. Подобно остальным видам секции, Р. nepalensis
является стержнекорневым растением; в первый год у него формируется
розетка прикорневых листьев, а на следующий из терминальной почки
розетки возникает длинный облиствленный генеративный побег. Несом-
ненно, это сравнительно молодой биоморфологический тип лапчаток,
свидетельствующий, каким образом стержнекорневые розеточные поли-
карпики могли трансформироваться в монокарпики сходной биоморфо-
логической структуры.
Основа распространения видов секции 18 — Niveae лежит в Восточ-
ной Азии и Северной Америке. Лишь главный ее представитель — Р, ni-
vea L. занимает всю Голарктику. Секция состоит из аркто-альпийских
видов, из которых одни являются исключительно арктическими (Р. vil-
losa Pall.— северо-восток Азии и Аляска, Р. vahliana Lehm.— Гренлан-
дия, Арктика Северной Америки и о. Врангеля), другие — исключитель-
но альпийскими, составляющими основное ядро секции (Р. saundersiana
Royle., Р. delavayi Franch., Р, pringlei Wats., P. betonicifolia Poir., P. eve-
stita Th. W., P. concinna Rich., P. concinnifortnis Rydb., P. ollanceolata
Rydb., P. bicrenata Rydb.), и, наконец, главный вид Р. nivea является*
аркто-альпийским растением, произрастающим в Арктике Европы, Азии
и Америки с одной стороны, а с другой — в высокогорно-альпийском по-
ясе центрально- и восточноазиатских гор, на Кавказе и в Кордильерах
Северной Америки.
Секция Niveae характеризуется сильно укороченным столбиком, и в
этом отношении она ближе примыкает k Multifidae. В основании стол-
бик ясно конический. В лице Р. nivea Вольф усматривает древний корень,
всей секции.
Исходной структурой подземных органов является стержнекорневой
тип. В ходе онтогенеза стержнекорневая система у отдельных видов от-
мирает, заменяясь системой придаточных корней, развивающихся на
более или менее длинных корневищах, например у Р. villosa, Р. nivea,
Р. evestita. При этом корневища иногда образуют рыхлые дерновины.
Указанные виды с помощью корневищ способны к вегетативному пере-
движению по площади. Корневища Р. saundersiana и Р. vahliana, напро-
тив, образуют плотные подушки, а стержнекорневая система сохраняет-
ся в течение всей жизни растений. Первый из этих видов — альпийское
растение Гималайских гор, Тибета, гор в провинции Юннань, второй
же — типично арктическое растение. Как в тех, так и в других условиях
растения развиваются в форме плотных подушек.
Рост побегов всех видов секции Niveae осуществляется моноподйаль-
но. В объяснении моноподиального роста надо учитывать причины гене-
тического и экологического порядка. Травянистые предки Niveae, близ-
кородственные современным Multifidae и имевшие с ними общие корни,
были, несомненно, моноподиальными растениями, а экологические
особенности местообитаний поддерживали из поколения в поколение
признак моноподиального роста и возобновления.
63
Ареал секции 19 — Argenteae является европейско-передне- и сред-
неазиатским. Видовое разнообразие секции сосредоточено в Централь-
ной и Юго-Восточной Европе. Сюда входят растения засушливых место-
обитаний — степей, сухих горных склонов и др. Совершенно отсутствуют
в альпийских и арктических районах. По строению столбика секция
близко примыкает к Niveae.
Все виды в начале онтогенеза стержнекорневые, в дальнейшем весь-
ма часто превращаются в стержне-кистекорневые и кистекорневые. Рост
и возобновление побегов симподиальные: каждый монокарпический по-
бег проходит фазы почки, розетки и облиствленного удлиненного генера-
тивного побега. Из девяти видов секции нами изучено пять: Р. dealbata
Bge., Р. argentea L., Р. meyeri Boiss., P. canescens Bess., P. pindicola
Hausskn. (остальные — P. tomentosa Ten., P. virgata Lehm., P. molissi-
nia Lehm., P. eremica Th. W. не изучались).
Центральноевропейской, в основном, является и подсекция Collinae,
включающая 16 видов. Лишь некоторые из них распространены в Перед-
ней Азии: Р. svanetica Siegfr., Р. armeniaca Siegfr., Р sommieri Siegfr.
et KelL, P. lazica Boiss. et Bal. Преобладающее количество видов оби-
тает в нижне- и среднегорном поясах, а отдельные виды спускаются на
равнину [Л wibeliana Th. W., Р. johanniniana Goir., P. sordida (Fries.)
Zimm., P. radiati Lehm., P. thyrsiflora (Hills.) Zimm., P. silesiaca Uechtr.,
P. wiemanniana Giinth. et Schumm., P. leucopolitana P. J. Miill., P. prae-
cox F. Schultz., P. rhenana P. J. Miill., P. opizii Dornin.]. Единичные виды
поднимаются в субальпийский пояс гор: Р. lazica, Р. alpicola De la Soie.
Подобно основной секции, виды Collinae характеризуются стержнекор-
невой структурой подземных органов, часто преобразующейся в кисте-
корневую и даже рыхлокустовую. Переход к кистекорневой структуре
наблюдается у Р. sordida, Р. leucopolitana, Р. rhenana у Р. silesiaca, Р.
wiemanniana, Р. ргаесох; формируется структура, близкая к рыхлокусто-
вой. Большинству видов свойственно симподиальное нарастание побе-
гов. Лишь Р. silesiaca отличается неустойчивым нарастанием: большая
часть побегов формируется моноподиально, но иногда цветоносные по-
беги образуются не в пазухах розеточных листьев побегов, а из их тер-
минальной почки, вследствие чего дальнейшее возобновление такого по-
бега осуществляется симподиально. При общем симподиальном нараста-
нии и возобновлении монокарпических побегов у Р. ргаесох отдель-
ные розеточные побеги, длительно остающиеся вегетативными, в тече-
ние известного периода нарастают моноподиально. Все симподиальные
виды подсекции обладают довольно высокими облиствленными побегами.
Секция Argenteae s. 1. занимает среди Conostylae eriotrichae заклю-
чительное место. Как было показано выше, остальные секции конически-
столбиковых шерстистоволосых лапчаток состоят в основном из моно-
подиальных видов. Отсюда следует заключить, что симподиальный рост
видов Argenteae развился из моноподиального. Экологически он опреде-
лялся условиями засушливого климата средне- и низкогорий.
Секцию 20 — Tanacetifoliae составляют коническистолбиковые пря-
моволосые лапчатки (Conostylae orthotrichae). Главной областью рас-
пространения этой секции является азиатский континент. Лишь отдель-
ные виды проникают в Европу до центра Европейской части СССР. Ее
представители совершенно отсутствуют в Америке. Большинство видов
секции обитает в высокогорно-альпийских условиях, например, Р. ban-
nechalensis Camb., Р. gerardiana Lindl., P. griffithii Hook, f., P. clarkei
Hook, f., P. kryloviana Th. W., P. rigidula Th. W., P. coriandrifolia Don.
Только отдельные виды спускаются на равнину — Р. tanacetifolia Th.
W. Р. viscosa Don., P. sanguisorba Willd., P. pimpinelloides L.
€4
Виды секции характеризуются простым нередко железистым опуше-
нием, непарноперистыми листьями, коротким и толстым столбиком, ко-
торый короче зрелого плода или равен ему.
Все виды в начале онтогенеза являются стержнекорневыми. С тече-
нием времени формируется кистекорневая (Р. visiana Рапе), рыхлоку-
стовая (Р. kryloviana, Р. pitnpinelloides), а иногда и плотнокустовая
структуры (Р. astragalifolia Bge., Р. coriandrifolia). Высокогорно-аль-
пийские виды характеризуются моноподиальным нарастанием побегов.
Те виды, которые обита’ют в горах и на равнине (Р. tanacetifolia, Р. vis-
cosa, Р. sanguisorba), отличаются в одних случаях моноподиальным, а в
других—симподиальным ростом. Двум видам — Р. pitnpinelloides и
Р. visiana — свойственно симподиальное нарастание. Как в ряде преды-
дущих примеров у высокогорно-альпийских видов цветоносные побеги
слабо развиты, почти безлистные, низкие. Плотнодерновинное развитие
корневищ, образующих нечто вроде плотной подушки, связано, с одной
стороны, с альпийскими условиями высоких гор (Р. coriandrifolia), а с
другой,— с пустынно-степными условиями (Р. astragalifolia). Указанная
экологическая приуроченность некоторых подушковидных растений от-
мечалась еще Хаури (Hauri, 1912), Хаури, Шретер (Hauri С. Schroter,
1914).
Возможно, что разнообразие способов роста и возобновления побегов
указывает на исходный характер секции, особую пластичность струк-
туры ее представителей, обусловившую впоследствии разные направле-
ния специализации.
Видовое многообразие секции 21 — Rectae сосредоточено в южной Ев-
ропе— на Балканском полуострове и вокруг Черного моря. Отдельные
виды расселялись из этого центра в разных направлениях: к северо-во-
стоку— до Центральной Европы, на восток в Азию — до Ирана и Тура-
на. Виды секции отсутствуют в арктических, субарктических и высоко-
горно-альпийских районах, во всей Восточной Азии и Америке. По Воль-
фу, Rectae представляет молодую, неотипную группу, в которой интен-
сивно идет процесс развития и дифференциации новых форм. Между
отдельными видами и вариациями часто сохраняются формы переходно-
го значения.
Виды секции характеризуются жестким опушением вегетативных над-
земных органов из длинных и коротких волосков. Столбик, как и в пре-
дыдущей секции, толстый и короткий, короче зрелого плодика или рав-
ной с ним длины.
Все виды (Р. recta L., Р. laciniosa W. К., Р. transcaspia Th. W., Р. пи-
rensis Boiss. et Hsskn., P. hirta L., P. gilanica Th. W., P. adriatica Murb.,
P. detommasii Теп., P. taurica Willd.) вначале являются типично стержне-
корневыми растениями, с возрастом же корневая система преобразуется
в кистекорневую, например, у Р. recta, Р. transcaspia, Р. hirta, Р. detom-
masii. Этот переход осуществляется через промежуточную стержне-кис-
текорневую структуру.
Развитие монокарпических побегов проходит по трехфазному циклу:
почка — розеточный вегетативный побег — облиствленный удлиненный
генеративный побег. Следовательно, рост и возобновление побегов сим-
подиальные. Базальные части монокарпических побегов сохраняются и
функционируют в качестве корневищ, на которых образуются придаточ-
ные корни. ,
В дальнейшем формируется дерновинка корневищ, на которых одно-
временно возможно развитие нескольких цветоносных побегов, что осо-
бенно ярко проявляется в условиях культуры. Так, на экспозициях Глав-
ного ботанического сада АН СССР Р. transcaspia образует довольно
крупные рыхлые кусты. Такую структуру можно уже называть рыхло-
кустовой.
5 В. Н. Голубев
65
Секция служит замечательным примером того, как в молодой группе,
сформировавшейся на равнине и низкогорьях Средиземноморской обла-
сти с ксерическими условиями, развивается симподиальное нарастание
побегов, которое накладывает свой отпечаток на биоморфологические
черты. Исходным типом роста и возобновления побегов в этой секции
был, по-видимому, моноподиальный.
Секции 22 — Rivales, по признанию Вольфа, генетически неоднород-
на и представляется искусственной, что подтверждается данными био-
морфологического анализа ее видов. Отдельные ее виды обнаруживают
близость к секциям, далеко отстоящим друг от друга. В большинстве
это проявляется во внешних признаках, особенно в разнообразии расчле-
нения листьев. Тем не менее весьма константным, резко бросающимся
в глаза признаком является длинный и тонкий столбик, четко характе-
ризующий секционную специфику видов.
Распространение совпадает с ареалом всего рода лапчаток, т. е. яв-
ляется циркумполярным, голарктическим. Секция заключает многолет-
ние поликарпики и монокарпические малолетники, среди которых встре-
чаются четырех-, трех-, двух- и однолетние виды. Часто один и тот же
вид в разных условиях развивается то в форме многолетнего, то в форме
двух- и однолетнего монокарпика. Некоторые многолетние поликарпиче-
ские виды обитают в высокогорьях, каковы Р. heterosepala Fritsch.,
Р. dombeyi Nestl., Р. richardii Lehm., P, monanthes Lindl., P. regeliana
Th. W., P. desertorum Bge., P. bungei Boiss., P. kotschyana Fenzl. Все эти
виды по способу нарастания и возобновления побегов следует отнести к
моноподиальным растениям с розеточной формой роста моноподиальных
побегов. Эти виды обладают стержнекорневой структурой подземных
органов, но у отдельных видов в онтогенезе стержнекорневая структура
изменяется, переходя в кистекорневую (Р. monanthes) или рыхлокусто-
вую (Р. bungei). Наряду с моноподиальными имеются среди Rivales и
стержнекорневые симподиальные виды, например Р. rivalis Nutt. Послед-
няя распространена на западе Северной Америки, от Орегона до Мекси-
ки, и предпочитает песчаные места больших рек и озер.
Особенно следует выделить Р. cryptotaeniae Maxim, и Р. centigrana
Maxim., обитающие в Японо-Китайской области в лесах и их опушках,
в кустарниках. У Р. cryptotaeniae, в отличие от большинства травянистых
лапчаток, наблюдается безрозеточный тип развития побегов. Пока труд-
но решить, принимать ли эту особенность за рекапитуляцию палеоморф-
ной структуры предковых форм лапчаток, т. е. расценивать как первич-
ный признак, или же рассматривать ее как вторичное образование,
возникшее из розеточного типа структуры. Мы склоняемся именно к
последнему заключению, полагая при этом, что безрозеточные побеги
развивались из розеточных под влиянием условий подпологового произ-
растания. При обилии влаги и значительном затенении, стимулировав-
ших и благоприятствовавших росту, стадия розетки должна была транс-
формироваться в фазу удлиненного вегетативного побега с растянутыми
междоузлиями. О молодости этого вида можно судить также и по явно
вторично развивавшемуся признаку — кистекорневой структуре подзем-
ных органов. Это одна из немногих лапчаток, у которой главный корень
недоразвивается еще в раннем возрасте и формируется кистекорневая
система придаточных корней мезофильного строения. Корни тонкие, не-
глубокие, многочисленные. Изредка образуются и розеточные вегетатив-
ные побеги.
Р. centigrana обладает другой интересной особенностью — способ-
ностью развивать ползучие столонообразные побеги, которые формиру-
ются из цветоносных побегов. Последнее подтверждается тем, что в от-
дельных случаях ползучие побеги не образуются, и растения обладают
тогда обычными, может быть несколько более нормального, вытянуты-
66
ми цветоносными побегами, занимающими ортотропное положение. Цвет-
ки одиночные, в пазухах листьев. При развитии ползучих побегов про-
исходит лишь удлинение междоузлий, полегание побегов и укоренение в
узлах придаточными корнями. Длинные- цветоножки с единственным
цветком формируются также в пазухах теперь уже розеточных листьев
на ползучих ветвях. Подземные органы Р. centigrana имеют типичную
кистекорневую структуру, что, как и у Р. cryptotaeniae, связано со спе-
цификой эдафических условий обитания: первая произрастает во влаж-
ных лесах, на каменистых, богатых источниками местах; вторая лапчат-
ка— в лесах, кустарниках, на лесных опушках и заброшенных пашнях.
Эти условия обитания, как показывают проведенные исследования (Го-
лубев, 1957а, б; Борисова, 1959; и др.), направляют морфогенез подзем-
ных органов в сторону развития наряду с длиннокорневищной и кисте-
корневой структур.
С Р. centigrana сходна Р. wallichiana Del., у которой формируются
удлиненные столонообразные ползучие побеги. В начале онтогенеза ра-
стение обладает стержневой корневой системой. Возникающие на столо-
нах новые особи имеют часто кистекорневую структуру. Филогенетиче-
ская связь кистекорневой структуры со стержнекорневой проявляется
здесь весьма отчетливо. Также ясны и пути развития растений с ползу-
чими столонообразными побегами. Последние у двух описанных лапча-
ток представляют собой метаморфизированные цветоносные побеги,
формирующиеся в пазухах розеточных листьев побега исходной семен-
ной и вегетативно развившейся особей.
Наконец, среди Rivales имеется специфическая группа монокарпиче-
ских растений ограниченной продолжительности жизни. Цикл развития
этих растений завершается в два-три года, а некоторые виды являют-
ся однолетними. Сюда относятся: Р. supina L., Р. millegrana Engelm.,
Р. biennis Greene, Р. norvegica L., P. pentandra Engelm., P. intermedia L.,
P. asperrima Turcz. Все эти виды являются стержнекорневыми, глав-
ный корень которых проникает на более или менее значительную глуби-
ну. В начале онтогенеза все перечисленные виды развиваются в форме
розеточных растений: главный побег укорочен в виде прикорневой розет-
ки листьев. Лишь у Р. supina розеточная фаза в развитии главного и бо-
ковых побегов выражена неотчетливо.
ОдйоЛетний цикл развития достоверно известен у Р. norvegica. Р. su-
pina. Остальные указанные виды, по-видимому, только в исключитель-
ных случаях характеризуются однолетним развитием, более типично
развитие по типу двулетников, а нередко и трех-четырехлетников. Неко-
торые из них также способны развиваться по типу полицикликов. Таким
образом, эта группа лапчаток доставляет разнообразный и ценный
материал, показывающий, как стержнекорневые розеточные растения по-
лициклического развития преобразовывались в ди- и моноциклические ма-
лолетники и однолетники. Необходимо подчеркнуть, что в процессе транс-
формации основные черты морфологической структуры предкового типа
остались без существенных изменений, преобразовывался цикл развития
на основе сокращения общей продолжительности жизни. Вместе с тем
существенные новообразования отмечаются в структуре цветоносного по-
бега. Так у Р. supina надземный побег превращается по существу в
большое соцветие, функции которого заключаются в максимальной реп-
родукции семян: побег довольно высокий, обильно разветвленный, с мно-
гочисленными листьями, в пазухах которых формируются одиночные
цветки.
Касаясь экологии развития монокарпических малолетних лапчаток,
можно отметить следующие моменты. Все перечисленные виды отсут-
ствуют в Арктике и высокогорных районах. Местами их обитания
являются низкогорья и равнины. Р. supina обитает у берегов рек, на
5*
67
травянистых склонах, пастбищах, на краю дорог, <возле жилья почти по
всей Голарктике, Р. millegrana— в бассейне р. Миссисипи и реже в за-
падных штатах, Р. biennis — в западных штатах Северной Америки, от
Колумбии до Аризоны и Калифорнии, Р. norvegica— по берегам рек,
краяхМ полей и дорог, паровым полям, огородам, пустошам, вырубкам
умеренной и субарктической зоны Старого и Нового Света (в Северной
Америке выделяют особый вид Р. monspeliensis L.)> Р. pentandra — в
Канаде и средней полосе США, Р. intermedia — на лугах, степях, зале-
жах, паровых полях в посевах, садах и по краям дорог средней Евро-
пы, Р. asperrima — по каменистым местам и утесам Даурии, Лено-Ко-
лымского и Зее-Буреинского районов.
Эколого-географические особенности монокарпических видов гово-
рят об их приуроченности к областям мезофильного климата. Лишь от-
дельные виды произрастают в местах недостаточного увлажнения. Раз-
витие их обычно связано с условиями нарушенного почвенного покрова.
Приведенные данные свидетельствуют, по-видимому, о том, что разви-
тие монокарпических лапчаток не было обязано воздействию аридных
условий, а скорее всего определялось мезофильными условиями в ме-
стах с нарушенным почвенным покровом. Чрезвычайно широкое распро-
странение Р. supina и Р. norvegica указывает на то, что возникновение
малолетних форм лапчаток совершилось в сравнительно отдаленное вре-
мя. Вместе с тем бесспорно и то, что современный их ареал значитель-
но шире первичного и завоеван в результате последующей экспансии.
Огромную роль, вероятно, в распространении однолетних лапчаток сыг-
рал человек. Произрастание же Р. supina и Р. norvegica на культурных
землях, без сомнения, способствовало превращению их в однолетние
формы, что находит подтверждение и в данных А. И. Купцова (1925,
1955). Как указывает И. Г. Серебряков (1952), в лесной и степной зонах
СССР число однолетних и двулетних растений достигает максимума на
окультуренных почвах.
Резюмируя, можно сказать, что секция Rivales заключает стержне-
корневые моноподиальные и симподиальные виды, кистекорневые с пол-
зучими столонообразными побегами и стержнекорневые моноцикличе-
ские малолетники. Такое биоморфологическое разнообразие связано с
широкой экологической амплитудой секции. Однако этот факт побуж-
дает к более внимательному систематическому анализу секции, который,
возможно, покажет генетическую гетерогенность и необходимость ее раз-
деления на ряд самостоятельных секций.
В отличие от предыдущей, секция 23 — Persicae является весьма мо-
нолитной. Все ее 19 видов — обитатели альпийского или (реже) субаль-
пийского пояса гор. Большинство видов сосредоточено в горах Ирана
(Р. elvendensis Boiss., Р. argyroloma Boiss. et Hoh., P. persica Boiss. et
Hsskn., P. curdica Boiss. et Hoh., P. pannosa Boiss. et Hsskn., P. mallota
Boiss., P. nuda Boiss., P. flaccida Th. W., P. aucheriana Th. W., P. crypto-
phila Вопил., P. petraea Willd.)w. P. argaea Boiss. et Bal. выходит за пре-
делы Ирана в Закавказье, Армению и Малую Азию. Собственно мало-
азиатские Р. pulvinaris Fenzl. и Р. nudicaulis Boiss. et Bal. Эндемиком
Кавказа является Р. ruprechtii Boiss. Ареал Р. flabellata Reg. et Schmalh.
ограничивается Памиро-Алаем, а Р. komaroviana Th. W. — Памиро-Ала-
ем и Копет-Дагом. Обособленными ареалами характеризуются Р. тайга
Th. W. (северная Африка) и Р. nevadensis Boiss. (горы Сиерра-Невада
в Испании).
Виды Persicae представляют собой невысокие многолетние расте-
ния, лишенные войлочного опушения, с непарноперистыми или пальча-
тыми (но не тройчатыми!) листьями. Столбик очень тонкий, нитевидный,
по длине в 2—3 раза превышает зрелый плодик, в основании конусо-
видно утолщенный.
68
Все виды являются розеточными моноподиальными стержнекорневы-
ми растениями; цветоносные побеги разной степени специализации: сред-
ней величины облиствленные (Р. elvendensis), короткие слабо облист-
вленные (таких большинство — Р. nuda, Р. cryptophila и др.) и, наконец,
от почти безлистных (Р. flaccida) до совершенно безлистных, как у
Р, pulvinaris. Впрочем, в зависимости от условий обитания этот признак
варьирует.
В процессе онтогенеза у отдельных видов стержнекорневая структу-
ра подземных органов преобразуется в стержне-кистекорневую (Р. аг-
gaea, Р. argyroloma, Р. pulvinaris) или рыхлокустовую, как у Р. neva-
densis. Моноподиально нарастающие розеточные побеги часто тесно друг
к другу прижимаются, становятся корневищами и образуют плотные
подушечки. Последние наблюдаются у Р. flabellata, Р. pulvinaris, Р. пи-
da, Р. cryptophila, иногда у Р. nevadenisis.
Вольф относит Persicae к неотипным группам; она возникла в Ар-
мянско-Иранской горной стране.
Распространение видов секции 24— Grandiflorae ограничивается пре-
имущественно горами Европы: Р. delphinensis Gren. et God.— в Альпах.
Р. pyrenaica Ram.— в Пиренеях, Р. grandiflora L.— в Альпах и Пире-
неях, Р. montenegrina Pant.— в западной части Балкан. Несколько обо-
собленно распространена Р. umbrosa Stev. — эндем Крыма. Р. Ьиссоапа
Clem, обитает на Олимпе в Малой Азии. Виды секции являются по пре-
имуществу высокогорными, основное ее ядро составляют растения аль-
пийских маттов.
Для секции характерен довольно толстый столбик, от утолщенного
основания постепенно утончающийся, в полтора раза длиннее зрелого
плодика.
По структуре подземных органов виды секции относятся к стержне-
корневым растениям. Побеги отличаются моноподиальным нарастанием.
Оси розеточных побегов становятся корневищами, образующими часто
рыхлые дерновинки (Р. pyrenaica, Р. umbrosa). В противоположность
Persicae растениям свойственны довольно мощные облиствленные цвето-
носные побеги и тройчатосложные или пятерные листья.
Распространение видов секции 25 — Chrysanthae диффузное. Р. sphe-
nophylla Th. W. и P. orbiculata Th. W. являются эндемиками Кавказа,
P. szovitsii Th. W. и P. adscharica Somm. et Lev. — иранско-кавказскими,
P. lipskyana Th. W. и P. sericata Th. W. — эндемиками Памиро-Алтая,
P. schrenkiana Rgl.— эндемиком Джунгарско-Тарбагатайского района.
P. kashmirica Hook. f. обитает в Кашмире. P. tollii Trautv.— эндемик
Лено-Колымского района. В арктической Сибири распространена Р. sti-
pularis L., спускаясь к югу Обского и Лено-Колымского районов. Евро-
пейской является Р. thuringiaca Bernh. Лишь ареал Р. chrysantha Trev.,
более широкий в сравнении с предыдущими, она распространена в Юго-
славии, Венгрии, Джунгарско-Тарбагатайском районе, на Урале, Алтае,
в Обском и Иртышском районах.
Экологически секция также неоднородна. Часть видов обитает в высо-
когорьях (Р. kashmirica, Р. sericata, Р. lipskyana, Р. adscharica, Р. orbi-
culata, Р. schrenkiana и др.), часть выходит на равнины Европы и Сиби-
ри (Р. chrysantha, Р. thuringiaca, Р. longipes) и, наконец, Р. stipularis
распространена в тундрах Сибири и альпийском поясе сибирских гор.
Chrysanthae характеризуются коротким, толстым столбиком, немно-
го короче зрелого плодика или равным ему, с сосочками. Все виды в пер-
вые годы жизни являются стержнекорневыми растениями, но с возра-
стом многие из них становятся кистекорневыми. Таковы Р. chrysantha,
Р. szovitsii, Р. longipes, Р. tollii. Что касается способа нарастания и во-
зобновления побегов, то в этом отношении многое еще остается неясным.
Верно лишь то, что моноподиальное нарастание более тесно связано с
69
альпийскими условиями обитания видов, тогда как симподиальное чаще
наблюдается у видов, произрастающих на равнине и в ксерических усло-
виях низкогорий.
Как указывает Вольф (Wolf,1908), секция Chrysanthae переживает
эпоху расцвета формообразования. С этой точки зрения и надо решать
биоморфологическое разнообразие вэдов секции, пути и условия разви-
тия их приспособительной структуры.
Секция 26 — Multijugae, содержащая 12 видов, целиком является
американской. Большая часть видов сосредоточена в Калифорнии
(Р. multijuga Lehm., Р. millefolia Rydb., Р. hickmanii Eastw., P. rosulata
Rydb., P. saxosa Greene), другие, обитая в Калифорнии, произрастают и
за ее пределами в Западной и Северной Америке (Р. drummondii Lehm.,
Р. plattensis Nutt., Р. diversifolia Lehm.); в штате Аризона обитает Р. ari-
zonica Greene, Р. monidensis A. Nels.— в Канаде, Р. condicans Numb, et
Bonpl.— в горах центральной Мексики.
Многие виды секции являются высокогорно-альпийскими растения-
ми. Но, произрастая в высокогорьях, они способны спускаться и на рав-
нину (Р. plattensis, Р. diversifolia).
У видов Multijugae столбик в 1,5—2 раза длиннее зрелого плодика,
очень тонкий и лишь в основании коротко и конически утолщается.
Листья непарноперистые. Ближе всего к этой секции стоят Graciles и
Ranunculoides, имеющие между собой тесное родство и общий ареал
формообразования.
Как и большинство других лапчаток, Miltijugae являются стержне-
корневыми растениями. Однако в процессе онтогенеза у части видов
обнаруживается трансформация стержнекорневой структуры в кистекор-
невую (Р. hickmanii, Р. aissecta Pursh.) и даже в рыхлокустовую (Р. di-
versifolia). Развитие рыхлокустовой структуры выражается в том, что
первичная корневая система отмирает, а из розеточных побегов форми-
руется система корневиц, укореняющихся придаточными корнями.
Подавляющее число видов обладает моноподиальным ростом и возобнов-
лением розеточных побегов. Цветоносные побеги, как правило, низкие,
слабые, с небольшим количеством редуцированных листьев, либо почти
безлистные. Но, по-видимому, несколько единичных видов характеризу-
ются симподиальным возобновлением побегов, как у Р. dissecta.
Ареал следующей секции 27— Ranunculoides почти совпадает с аре-
алом Multijugae. Большинство видов распространено в западной части
Северной Америки от полярного Севера до Мексики, т. е. приурочено к
горным системам (Р. brevifolia Nutt., Р. grayi Wats., Р. acuminata Н. М.
Hall., Р. flabellifolia Hook., Р. townsendii Rydb., P. palmeri Th. W., P. sub-
coriacea Rydb., P. rydbergiana Rose., P. ranunculoides H. et B. ex Nestl.,
P. wheeleri Wats., P. subviscosa Greene., P. ramulosa Rydb., P. sierrae-
blancae Woot. et Rydb.). Один вид распространен на восточном побе-
режье Гренландии и северной оконечности Лабрадора (Р. ranunculus
Bge.), один — в Японии (Р. matzumurae Th. W.), а Р. fragiformis Willd.
распространяется от Аляски через Алеутские и Курильские острова в
Северо-Восточную Азию (Северо-Восточный Китай, побережье Охот-
ского и Берингова морей, Камчатка, Джунгария, Алтай, Анадырский край.
Местами обитания подавляющего большинства видов служат высоко-
горно-альпийские районы. Растения поднимаются вверх до 3—3,5 тыс. м
над ур. моря. Лишь Р. ranunculus, да отчасти Р. fragiformis являются
арктическими растениями.
Столбики Ranunculoides в 2—3 раза длиннее зрелого плодика, тон-
кие, при основании коротко конусовидно утолщенные.
Большая часть видов относится к стержнекорневым моноподиально-
розеточным растениям (Р. fragiformis, Р. ranunculus, Р. rydbergiana,
Р. ranunculoides, Р. wheeleri). Но уже среди ряда других видов основ-
70
ным типом структуры во взрослом состоянии становится рыхлокустовая
структура, близкая к длиннокорневищной. Корневища формируются за
счет наземных побегов с вытянутыми междоузлиями. Таковы Р. brevifo-
lia, Р. flabellifolia, Р. matzumurae. Их отдельные розеточные побеги
вытягиваются и превращаются в стелющиеся корневища, постепенно
погружающиеся в грунт. После некоторого периода развития по безрозе-
точному типу они вновь могут превращаться в настоящие розеточные по-
беги с укороченными междоузлиями. Цветоносные побеги средней длины
или короткие слабо облиствленные, либо почти без листьев.
Крупная секция 28 — Aureae, объединяющая 28 видов, имеет чрез-
вычайно широкое географическое распространение: от полярной зоны до
горных цепей Южной Европы и Средней Азии, занимает всю Сибирь и
Восточную Азию. В Северной Америке она представлена лишь тремя
видами: Р. robbinsiana Oakes., Р. emarginata Pursh., Р. alpestris Hall, f.,
из которых собственно американский только первий вид.
Большинство представителей секции являются обитателями высоко-
горного пояса, часто входя в состав альпийских маттов [Р. dubia (Crtz.)
Zimm., Р. kuramensis Th. W., P. frigida Vill., P. robbinsiana, P. elegans
Cham, et Schlecht., P. gelida С. A. Mey, P. tephroleuca Th. W.', P. doubjon-
neana Camb., P. seidlitziana Bien., P. subpalmata Led., P. aurea L., P. ter-
nata C. Koch., P. australis Kras., P. rigoana Th. W., P. cappadocica Boiss.,
P. geranioides Willd., P. gaudinii Crml., P. tommasiniana F. Schultz.,
P. cinerea Chaix., P. velutina Lehm.]. Один вид характерен для высоких
широт Арктики — Р. emarginata Pursh., распространенный циркумполяр-
но на островах Северного Ледовитого океана и частично на побережье.
Р. alpestris Hall. f. распространена как в альпийском и субальпийском
поясе гор Малой Азии, Кавказа, Армении, так и в арктической и суб-
арктической зоне Европы, Азии и Америки. Частично заходит в европей-
скую Арктику и упоминавшаяся ранее Р. gelida. Последнюю, четвертую,
эколого-географическую категорию составляют равнинные лапчатки уме-
ренного/ климата, лишь в некоторых случаях поднимающиеся в горы.
Сюда относятся Р. heptaphylla L., произрастающая на лугах, полянах,
опушках сосновых лесов по всей средней Европе, включая центральные
области Европейской части СССР, Р. humifusa Willd. et Schlecht., рас-
пространенная от центральных областей Европейской части СССР на во-
сток— в Западной и Восточной Сибири, Средней Азии и Предкавказье,
она обитает на типичных и луговых степях, по опушкам сухих лесов и
песчаным местам. Далее сюда необходимо включить Р. verna L. Ее
главными областями распространения служат равнины, холмистые ланд-
шафты и низкогорья Средней и большей части Западной Европы. К этой
же категории относятся Р. patula W. et К., Р. arenaria Borkh., Р. suba-
caulis L. и лишь отчасти — Р. velutina и Р. tommasiniana.
Секция отличается гвоздевидными столбиками без утолщения в ос-
новании, но утолщенными на верхушке под рыльцем, равными зрелому
плодику или немного короче его. Весьма характерна редукция листьев
на цветоносных побегах, последние слабые, короткие, равные или не-
сколько длиннее прикорневых листьев.
Все виды секции в начале онтогенеза обладают стержневым корнем.
Но лишь у немногих видов стержнекорневая структура подземных орга-
нов сохраняется во взрослом состоянии, что имеет место у Р. frigida,
Р. elegans, Р. geranioides. Подавляющая же масса видов с возрастом из-
меняет структуру подземных органов. Здесь можно обнаружить после-
довательные ступени преобразования от стержнекорневой к длиннокор-
невищной структуре через стержне-кистекорневую, кистекорневую и рых-
локустовую структуры, что наблюдается у Р. arenaria, Р. alpestris, Р. ter-
nata, Р. aurea, Р. australis. Значительно чаще конечным звеном преоб-
разования стержнекорневой структуры является рыхлокустовая. Таких
71
видов большинство — Р. kuratnensis, Р. gelida, Р. tephroleuca, Р. subpal-
mata, Р. heptaphylla, Р. humifusa, Р. verna, Р. gaudinii, Р. tommasiniana,
Р. cinerea, Р. velutina, Р. subacaulis. Трансформация структуры в онто-
генезе у части видов останавливается на этапе кистекорневой (Р. dubia,
Р. emarginata, Р. rigoana, Р. patuld). Наконец, у Р. cappadocica форми-
руется типичная стержне-кистекорневая структура.
Для лучшего понимания морфогенеза видов этой секции необходи-
мо подчеркнуть, что все они характеризуются моноподиальным нараста-
нием и возобновлением побегов. Последние представляют собой обычно
розеточные побеги с укороченными междоузлиями, медленно нарастаю-
щие своей верхушкой. У стержнекорневых форм эти розеточные побеги,
как правило, не укореняются придаточными корнями и основной питаю-
щей артерией растения служит стержневая корневая система. У части
видов, однако, от слабо разрастающегося и ветвящегося корневища об-
разуются придаточные корни, вследствие чего возникает стержне-кисте-
корневая структура. При некотором увеличении годичного прироста кор-
невищ осей розеточных побегов, их обильном ветвлении и укоренении
придаточными корнями, вызывающем утрату функционального значения
главного корня, формируется рыхлокустовая структура. Необходимо об-
ратить внимание на тот факт, что в образовании рыхлокустовой, а тем
более длиннокорневищной структуры особое значение имеет отмечав-
шаяся уже способность развития обычно укороченных побегов по безро-
зеточному типу. При этом происходит вытягивание междоузлий, обра-
зуется длинный стелющийся побег, укореняющийся придаточными кор-
нями и становящийся корневищем. Позже эти же побеги могут разви-
ваться в форме розеточных.
У ряда видов укороченные розеточные побеги образуют плотные по-
душечки, как, например, у Р. frigida, Р. elegans, Р. cappadocica.
Таким образом, в условиях альпийского и субальпийского поясов ут-
верждается моноподиальное возобновление. Отдельные виды характе-
ризуются подушковидной формой роста побегов. Моноподиальность,
развившаяся в условиях высокогорий, тоже в Арктике, прочно удержи-
вается в качестве константного признака и у равнинных видов секции,
имеющих, несомненно, альпийское происхождение.
Далее, для секции характерна выработка наиболее прогрессивного и
молодого по возрасту признака — длиннокорневищной структуры. Эти
факты хорошо согласуются с филогенетическими данными. Вольф пока-
зал, что, несмотря на общую филогенетическую древность секции, в ее
недрах в сравнительно недавнее время обнаружился новый прилив нео-
типного формообразования, которое наблюдается и в настоящее время.
Любопытно подчеркнуть, что длиннокорневищные формы относятся в
большинстве своем именно к молодым процветающим циклам секции
Аигеае. Наиболее полиморфные, испытывающие ясные признаки регио-
нальной дифференцировки виды, например Р. aurea, Р. alpestris, харак-
теризуются и четко выраженной способностью к образованию длинных
корневищ. Значит, общая древность группы еще ничего не говорит об
уровне организации ее современных представителей.
Следующая секция 29 — из гвоздевидностолбиковых лапчаток Fraga-
rioides состоит всего из двух видов — Р. fragarioides L., Р. freyniana
Bornm., распространенных в Восточной Азии. Пергвый вид обитает на
равнинах и невысоких горах, сухих лугах, в лесах и кустарниках уме-
ренной и субарктической части Восточной Азии. Второй вид встречает-
ся на влажных и заливных лугах, в сухих разреженных лесах и кустар-
никах Приморского края, Японии, восточной части Северо-Восточного
Китая.
Секция Fragarioides стоит обособленно среди других гвоздевидно-
столбиковых лапчаток. По-видимому, это весьма древняя восточноазиат-
72
ская группа, причем Р. fragarioides, очевидно, ближе стоит к предковому
типу, чем Р. freyniana.
В начале развития у обоих видов обнаруживается первичный корень,
позднее формируется кистекорневая структура. В пазухах розеточных
листьев развиваются длинные столоновидные ползучие побегие. В узлах
побеги укореняются, и из укоренившейся почки образуется новая листо-
вая розетка, в пазухах листьев которой могут формироваться цветонож-
ки с одиночным цветком и новые ползучие побеги. От стеблевой части
розетки возникает пучок придаточных корней. О том, что ползучие
побеги гомологичны цветоносным побегам, свидетельствуют факты
нормального развития ортотропных цветоносных побегов у обоих видов.
Нарастание основных осей побегов осуществляется моноподиально.
Предпоследняя секция 30 — из гвоздевидностолбиковых лапчаток —
Tormentillae состоит из восьми видов [Р. erecta (L.) Натре., Р. ргосит-
bens Sibth., Р., reptans L., P. canadensis L., P. flagellaris Willd., P. simulat-
rix Th., W., P. indica Th. W., P. hemsleyana Th. W.]. Из них один вид амери-
канский— Р. canadensis, один вид европейский —Р. procumbens, два
вида азиатско-европейские — Р. erecta, Р. reptans, три вида азиатские —
Р. hemsleyana (центральный Китай), Р. simulatrix, Р. flagellaris и один
вид азиатско-американский — Р. indica.
Необходимо отметить, во-первых, то обстоятельство, что большинство
видов, за исключением разве Р. indica, являются исключительно обита-
телями равнинных территорий. Во-вторых, виды этой секции приуроче-
ны в своем произрастании к более или менее влажным местообитаниям:
болотам, влажным и тенистым лесам, опушкам, лугам, берегам рек
и озер.
Указанные особенности и определили биоморфологические черты вхо-
дящих сюда видов. Семенные особи еще имеют достаточно выделяющий-
ся главный корень, но у вегетативно развившихся экземпляров форми-
руется типичная кистекорневая структура подземных органов. Цвето-
носные побеги, прямостоячие у Р. erecta, трансформируются в ползучие,
стелющиеся по земле, в узлах которых образуются новые розеточные
кистекорневые особи. Ползучие побеги развиваются из пазух розеточных
листьев и, следовательно, растения являются моноподиальными. Разви-
тие ползучих побегов из цветоносных стимулировалось достаточно влаж-
ными условиями обитания. Кистекорневая структура подземных орга-
нов есть функция тех же эдафических условий жизни. Р. erecta является
короткокорневищным растением, главный корень у нее недоразвивается
еще в раннем возрасте, а короткое и толстое корневище возникает из
гипокотиля и укороченной оси главного и боковых побегов (Голубев,
19566, 19586; Пошкурлат, 1958). Сходной короткокорневищной структу-
рой характеризуется и Р. procumbens, у которой, однако, в начале онто-
генеза стержневой корень выделяется отчетливо. Данное обстоятельства
дополнительно говорит в пользу признания за этим видом гибридоген-
ного происхождения — Р. erecta X Р. reptans, как предполагает Вольф
(Wolf, 1908).
Секция Tormentillae определенно весьма древняя, о чем свидетельст-
вует широкое распространение ее видов. Вместе с тем это естественная
группа, все виды которой связаны близким родством и происходят от
одного корня. Но кто были предками Tormentillae, остается неясным.
Концентрация видов секции в Восточной Азии, несомненно, указывает
на то, что именно здесь совершалось первичное развитие видов секции.
Моноподиальное нарастание видов как филогенетический признак мож-
но истолковать в пользу принятия в качестве предковых каких-то пока
еще не установленных альпийских форм.
Последняя секция 31 —Anserinae, если не учитывать всюду в Голарк.-
тике^распространенную Р. anserina L., является южноазиатской. Ее
73
виды Р. peduncularis Don., Р. tatsienluensis Th. W., P. leuconota Don.,
P. microphylla Don. распространены в Гималаях и провинциях Юннань
и Сычуань. Р. рариапа Focke. Р. parvula Hook. f. встречаются в Новой
Гвинее.
Все виды являются типично высокогорно-альпийскими растениями,
за исключением Р. anserina, произрастающей по берегам рек, озер, пру-
дов, по дорогам, пастбищам и в Европе, Азии и Северной Америке.
Все ее семенные особи обладают хорошо развитой стержневой корне-
вой системой. Позднее стержнекорневая система преобразуется в кисте-
корневую. Кистекорневая структура формируется и у вегетативно
развивающихся особей. Розеточные побеги нарастают моноподиально.
В пазухах листьев розеточных побегов у Р. anserina, Р. leuconota обра-
зуются столонообразные ползучие побеги, укореняющиеся в узлах пуч-
ком придаточных корней. В каждом таком узле формируется новое розе-
точное кистекорневое растение. Цветоножки Р. anserina образуются в
пазухах листьев ползучих побегов, в пазухах розеточных листьев веге-
тативно развившихся и семенных особей. Следовательно, ползучие по-
беги здесь гомологичны цветоносным побегам. Как показали наши иссле-
дования в Центрально-Черноземном заповеднике морфогенеза Р. anse-
rina, в начале жизни розеточного побега на столоне формируется один
глубоко уходящий корень, равносильный стержневому корню семенных
экземпляров. И лишь позднее от стеблевой части розетки возникает еще
несколько придаточных корней.
Р. peduncularis не образует ползучих побегов, и в пазухах ее розеточ-
ных листьев развиваются облиствленные невысокие цветоносные побеги.
Розеточные побеги Р. microphylla образуют плотные подушечки. Этот
вид обитает в Гималаях на высоте 3—5 тыс. м над ур. моря и подуш-
ковидная форма его побегов является ответной реакцией растения на
суровые условия высоких гор. Интересно отметить, что и цветоносные
побеги Р. microphylla почти безлистные и лишь слегка возвышаются
над поверхностью подушки.
Столбики видов секции тонко- и коротконитевидные, короче зрелого
плодика, прикрепляются к середине плодика сбоку.
Морфологически изолированное положение секции и ее распростра-
нение по всей Голарктике говорит о большой древности. Это, по-види-
мому, палеотипная группа, ближайшие предки которой были высокогор-
ными формами.
♦ ♦ *
Подводя итоги обзора эволюции жизненных форм видов Potentilla,
можно сделать следующие выводы:
1. Ни один из современных видов Potentilla нельзя принять за форму,
признаки которой совпадают с предковым типом. До наших дней дошли
лишь отдельные осколки палеоморфной структуры. Ее признаки рассе-
яны у разных видо<в. Все же в наиболее древних секциях лапчаток на-
блюдается максимум примитивных признаков.
2. Исходной жизненной формой являются деревянистые лапчатки,
в травянистые виды представляют формы производные. Среди травя-
нистых господствующее положение имеют поликарпические растения.
Монокарпические малолетние и однолетние виды единичны и занимают
заключительное место в биоморфологическом ряду трансформации.
Монокарпики появились независимо в разных направлениях эволюции
лапчаток, но они в своем возникновении, по-видимому, не были обязаны
действию аридных условий. Их развитие, по всей вероятности, связано
с интенсификацией обмена веществ, процессов роста и регенераци-
онной способности, обусловивших предельное сокращение длительности
жизни. а
74
3. Переход деревянистых лапчаток в травянистые, на основании име-
ющихся фактов, очевидно, нельзя рассматривать как процесс предель-
ного сокращения длительности жизни надземных скелетных осей. Он
совершался на основе геофилии, при которой побеги растений имеют
возможность длительного приземного или внутрипочвенного роста, осо-
бенно при моноподиальном возобновлении.
4. Жизненная форма подушек формируется как среди древесных, так
и травянистых лапчаток и представляет специализированную биомор-
<J)y, имеющую обособленное положение, без перехода к каким-либо дру-
гим структурам. Это конечное звено процесса узкой специализации,
совершавшейся в своеобразных условиях внешней среды. Подушки обя-
заны своим развитием крайним условиям существования в высокогорьях.
Но в современную эпоху отдельные подушковидные формы характер-
ны и для пустынно-степных местообитаний. Из деревянистых лапчаток
тюдушковидная форма роста побегов формируется у Р. eriocarpa,
Р. bifurca, из травянистых— у Р. nitida, Р. pulchella, Р. frigida и др.
Большинство подушковидных лапчаток являются факультативными.
5. Безрозеточный тип структуры побегов наблюдается, как правило,
у наиболее примитивных представителей рода и его следует признать
исходным по сравнению с розеточным. Наряду с первичной встречается
и вторичная безрозеточная структура, развившаяся из розеточной, на-
пример, у Р. cryptotaeniae.
6. В условиях высокогорий отмечается приземное и внутрипочвенное
развитие побеговой системы лапчаток, что знаменовало возникновение
геофильной линии эволюции. Примитивные черты геофилии отмечаются
уже у деревянистых лапчаток, где прослеживаются все этапы перехода
побегов от надземного к подземному существованию, как, например, у
Р. fruticosa, Р. ancistrifolia, Р. salesowiana и др. Дальнейшее развитие
геофилия получает у травянистых лапчаток. Для лапчаток особенно ха-
рактерно образование подземного побега — корневища, которое часто
называют стержнем. Это довольно толстая стеблевая ось, как правило,
моноподиальная, обычно несущая на верхушке листовую розетку. Весь-
ма замечательно, что у отдельных видов годовой прирост корневищных
побегов может резко варьировать.
7. Первичным способом нарастания и возобновления побегов лапча-
ток был симподиальный (Р. fruticosa и др.). В связи с альпийско-аркти-
ческими условиями утверждается моноподиальное возобновление.
В процессе дальнейшего формообразования вторичные, позднее развив-
шиеся типы лапчаток обладают ростом и возобновлением побегов, кото-
рый следует в каждом случае по-разному оценивать с точки зрения его
происхождения. Установлены моноподиальные и симподиальные формы,
корни происхождения которых находятся среди моноподиальных лап-
чаток. Но есть и аналогичная группа моноподиальных и симподиальных
форм, развившихся из симподиальных предков. Таким образом, в про-
цессе эволюции лапчаток моноподиальное и симподиальное возобнов-
ления побегов могли неоднократно сменять друг друга. Экологически мо-
ноподиальное возобновление тесно связано с суровыми условиями жиз-
ни в высокогорьях и на равнине — в Арктике. У некоторых моноподиаль-
ных видов арктическо-альпийского происхождения этот способ нараста-
ния и возобновления побегов стойко удерживался при изменении усло-
вий распространения и жизни, например, в области степей.
8. Недостаточно ясно представляется положение лапчаток с ползучи-
ми побегами. Если следовать концепции В. Г. Хржановского (1958а, б)
и К. Ю. Эйхвальда (1959) о тоь£, что анцестральные формы лапчаток
были близкими к лиановидным типам Rosa и Rubus, то способность к
развитию ползучих побегов, которые являются гомологами цветоносных,
можно рассматривать в качестве рекапитуляции лиановидной предковой
75
формы. В пользу этого, казалось бы, говорит и то обстоятельство, что
многие лапчатки с ползучими побегами относятся к древним циклам
форм. Но возможна, по-видимому, и иная интерпретация, а именно вто-
ричное развитие ползучих побегов из обыкновенных ортотропных цвето-
носных в результате эколого-биологической специализации.
9. Исходной формой корневой системы лапчаток является стержне-
корневая. У некоторых видов стержневая структура удерживается в те-
чение всей жизни растений. Однако у других в онтогенезе стержнекор-
невая структура заменяется стержне-кистекорневой и далее — кистекор-
невой. Только у единичных видов кистекорневая система становится'
основной формой с самого начала индивидуального развития, как это
обнаружено у Р. cryptotaeniae и Р. centigrana. Редукция главного корня
наблюдается и у многих длиннокорневищных видов. Но и у них в раннем
возрасте система главного корня достаточно выражена, исключая разве
Р. palustris.
НЕКОТОРЫЕ ДАННЫЕ ПО ЭВОЛЮЦИИ ЖИЗНЕННЫХ ФОРМ
В РОДАХ SCROPHULARIA, LATHYRUS И PLANTAGO
В отличие от Potentilla, виды Scrophularia в большинстве являются
безрозеточными растениями. С этой точки зрения интересно сопоставить
особенности эволюции биоморфологических структур норичника с пре-
дыдущим родом.
В определении биоморфологических признаков видов нами использо-
ваны все имеющиеся гербарные материалы по Scrophularia, хранящиеся
в Ботаническом институте АН СССР им. В. Л. Комарова. В ограничен-
ных масштабах были привлечены и материалы непосредственных наблю-
дений в природе. Анализ морфогенеза S. nodosa L. опубликован в спе-
циальной статье (Голубев, 19586).
Филогения рода Scrophularia разработана значительно слабее в срав-
нении с Potentilla. Тем не менее исследование Г. Штифельхагена (Stie-
felhagen, 1910) достаточно подробно освещает видовое богатство рода.
В сводке Штифельхагена учтено 143 вида Scrophlaria. В пределах
СССР, по С. Г. Горшковой (1955), насчитывается 74 вида.
В отличие от Веттштейна (A. Engler, К. Prantl, 1897), выделявшего
три секции внутри рода, оба указанных выше автора различают всего
две секции — Anastomosantes и Tomiophyllum. К Anastomosantes отно-
сятся травы и кустарниковидные растения, характеризующиеся листья-
ми, жилки которых, особенно с нижней стороны, отчетливо анастомози-
руют друг с другом. Tomiophyllum в большинстве представляют полу-
кустарники ксерофильной структуры, бедно облиствленные, листовые
жилки не анастомозируют или анастомозируют только у отдельных ви-
дов неотчетливо.
Анализ распространения показывает, что обе секции преимуществен-
но свойственны странам Древнего Средиземноморья (в понимании Попо-
ва, 1927, 1929, 1931; Вульфа, 1944). Однако среди Anastomosantes мно-
гие виды являются центральноазиатскими. Сейчас трудно еще решить,
являются ли они потомками наиболее древних типов рода или же ви-
дами, заселившими территории Центральной и Восточной Азии, Даль-
него Востока в более позднее время. М. Г. Попов (1927) высказывает
мнение, что Tomiophyllum представляет группу иранского центра.
В целом род Scrophularia является почти исключительно обитателем
северного внетропического или бореального царства, преимущественно*
Старого Света. Только два вида' заходят в пределы Северной
Америки.
Большинство видов рода приурочено к аридным районам и ксериче-
ским местообитаниям. Анализ биоморфологических особенностей видов
76
обнаруживает их соответствие господствующим условиям существова-
ния.
Центрами видового разнообразия Scrophularia у нас являются Кав-
каз и Средняя Азия.
На Кавказе сосредоточено до 32 видов норичника, из них 11 —энде-
миков; в Средней Азии отмечено 23 вида, а число эндемичных видов
достигает 15.
Основными местообитаниями видов норичника на Кавказе и Средней
Азии являются средние и верхние пояса гор. Лишь ограниченное количе-
ство видов спускается к подножьям гор. Щебнисто-каменистые осыпи,
голые скалы — таковы преобладающие разности субстратов, на которых
поселяются виды норичника.
Изученные виды по признакам биоморфологической структуры мож-
но подразделить на следующие группы:
I. Стержнекорневые полукустарники (3. donetzica Kotov., 3. leucoclada Bge., 3. va-
riegata M. B.).
II. Многолетние поликарпические травы
А. Стержнекорневые. Во флоре Европейской части к этой жизненной форме
принадлежат 3. cretacea Fisch., 3. rupestris M. В. ex. Willd., 5. sareptana Kleopov.;
в Средней Азии распространены 3. fedtschenkoi Gorschk., 3. gontscharovii Gorschk.,
5. integrifolia Pavl., o. pamiro-alaica Gorschk., 3. tadshicorum Gontsch., 3. turcomanica
Bornm. et Sint., 3. canescens Bong., 3. czapandaghii B. Fedtsch., 3. frigida Boiss., 3. in-
cisa Weinm., 3. striata Boiss., 3. xanthoglossa Boiss.; на Кавказе произрастают 3. late-
riflora Trautv., 3. thesioides Boiss. et Buhse., 3. amplexicaulis Benth., 3. canina L.,
3. grossheimii B. Schischk., 3. orientalis L., 3. rostrata Boiss. et Buhse., 3. rupestris M. B.
-et Willd.; в Сибири обитают 3. altaica Murr., 3. multicaulis Turcz., 3. incisa Weinm. и др.
Б. Кистекорневые. 3. nodosa L., 3. czernjakowskiana B. Fedtsch., 3. amgunen-
sis F. Schmidt, и др.
В. Корневищные. 3. divaricata Ledb., 3. oldhamii Oliver, и др.
Г. Переходные формы.
1. Растения, обладающие стержневым корнем и способные формировать плагиотроп-
ные симподиальные удлиненные корневища (корневищно-стержнекорневые): 3. zerav-
schanica Gorschk., 3. ruprechtii Boiss., 3. minima M. B., 3. hyrcana Grossh.
2. Растения, развивающие довольно толстое и короткое корневище (типа 3. nodo-
sa) при сохранении стержневого корня: 3. litwinowii В. Fedtsch., 3. heucheriiflora
Schrenk.
3. Обнаружены случаи, когда один и тот же вид в разных условиях относится
к разным жизненным формам (3. rutifolia Boiss., 3. alata Gilib.).
Все перечисленные растения в большинстве являются безрозеточными.
III. Двулетние монокарпики
А. Стержнекорневые: 3. dissecta (В. Fedtsch.) Gorschk., 3. pruinosa Boiss.,
3. sangtodensis B. Fedtsch., 3. atropatana Grossh., 3. chrysantha J. et Sp., 3. kotschyana
Benth., 3. nachitschevanica Grossh., 3. olgae Grossh. и др.
Б. Кистекорневые: 3. scopolii Hoppe et Pers., 3. lunariifolia Boiss. et Buhse.,
3. vernalis L.
IV. Однолетние монокарпики
А. Стержнекорневые: 3. ilvensis C. Koch., 3. olgae Grossh.
Б. Кистекорневые: 3. vernalis L.
Из приведенного видно, что одни и те же виды способны развиваться то в форме
двулетних, то однолетних монокарпиков. Среди монокарпиков зарегистрированы настоя-
щие розеточные формы.
Стержнекорневые виды свойственны ксерическим местообитаниям, причем в особо
засушливых местах, с недостатком влаги в поверхностном слое почвы, развивается глу-
бокая стержневая корневая система. У растений, приуроченных к подпологовому произ-
растанию в лесах, влажным лугам и другим подобным местам, формируется кистекор-
невая система, а корневища иногда становятся удлиненными.
На основании проведенного анализа можно заключить, что основ-
ные биоморфологические типы структур подземных органов видов Scru-
phularia весьма близко совпадают с таковыми у Potentilla. Однако над-
земные побеги в большинстве случаев безрозеточного типа, розеточный
тип появляется у сравнительно молодой группы монокарпиков в единич-
ных случаях. Производный характер розеточных видов здесь представ-
ляется весьма очевидным. Четко определяется экологическая обуслов-
ленность стержневой и кистевой корневых систем, из которых исходной
является первая.
Генетическая связь указанных типов доказывается наличием пере-
ходных форм и пложением видов в филогенетической системе. Напри-
мер, в очаге формирования видового богатства Tomiophyllum — иранском
центре основную массу видов составляют стержнекорневые безрозеточ-
ные растения.
Пример Scrophularia наглядно показывает, что основные структур-
ные типы повторяются в различных (подчас весьма далеко отстоящих
систематически) родах покрытосеменных. Этот параллелизм жизненных
форм свидетельствует о существовании общих экологических и эволюци-
онных закономерностей развития, совершавшегося по принципу адап-
тивной иррадиации. Соматическая дифференциация покрытосеменных
растений подчиняется общей закономерности в соответствии с функци-
ональными потребностями, обусловливающими направленность измене-
ний вегетативных органов. В наземных условиях основные органы авто-
трофных антофитов — корневая система, стебли и листья функциональ-
но необходимы, и этим определяется их устойчивость и сохранение в бес-
численном ряду поколений при возможных значительных метаморфозах
и изменениях в их форме и других признаках.
Экологическая дифференциация сомы сопряженно с констелляцией
условий жизни обогащает и разнообразит основные формы структур ра-
стений, но не выводит их за пределы «функционального кодекса». Пов-
торяемость типов структур у видов в различных систематических подраз-
делениях связана как с явлениями конвергенции, так и с общей направ-
ленностью структурно-морфологических изменений, обусловливающей
известную константность основных соматических типов.
* * *
Близко к Scrophularia разнообразие биоморфологических структур у
Lathyrus. Все виды данного рода относятся к безрозеточным. Среди изу-
ченных во флоре СССР можно отметить стержнекорневые (L. canescens
Grent. et Godr. и др.) и кистекорневые [L. gmelinii (Fisch.) Fritsch.r
L. pisiformis L.]. В результате редукции главного корня и преобладаю-
щего развития корневищ возникли типичные длиннокорневищные виды:
(L. pratensis L., L. palustris L.). Замечательным подтверждением этого1
пути эволюции является наличие видов переходной структуры — корне-
вищно-стержнекорневых растений, у которых одновременно присутству-
ют стержневой корень, иногда весьма мощный, и система длинных пла-
гиотропных корневищ (L. humilis Fisch., L. incurvus Roth, и др.) Наряду
с многолетними имеется много однолетних видов с безрозеточными по-
бегами (L. dominianus Litv., L. hirsutus L., L. inconspicuus L. и др.). Ин-
тересно отметить, что однолетники обладают коротким стержневым кор-
нем. Стержнекорневая структура в ювенильном возрасте обнаружена у
всех кистекорневых и типичных длиннокорневищных видов чины. Все
это убеждает в том, что анцестральной структурой подземных органов,
чины была стержневая корневая система. Несмотря на разнообразную
трансформацию подземных органов и изменения общей продолжитель-
ности жизни, тип структуры надземных побегов оставался весьма ста-
бильным.
* * *
Большой интерес для филогенетического анализа жизненных форм
представляют виды Plantago. В гербарии Ботанического института
АН СССР нами просмотрено 210 видов из различных стран мира и вы-
явлено разнообразие биоморфологических типов, которые в общих чер-
78
тах совпадают co многими родами покрытосеменных. Из основных био-
морф выявлены кустарники и кустарнички (Р. bismarckii Ndl., Р. arbo-
rescens Poir., P. hirsuta, P. macrorhiza Poir., P. princeps Chain., P. subu-
lata), полукустарники и полукустарнички (P. cynops L., P. stricta Scho-
usb., P. sinaica Barn., P. webbii Barn., P. velutina), многолетние поли-
карпические (P. altissima W. К., P. arachnoidea Schrenk., P. decipiens
Barn.) и малолетние монокарпические травы — двулетние и однолетние
(Р. arenaria W. К., Р. obversa Morris, Р. pumila L., Р. pusilia Nutt.,
P. squarrosa Murr. и др.).
По структуре побегов большинство видов относится к розеточным.
Однако среди деревянистых форм встречаются виды, у которых листья
равномерно покрывают стебли, что обнаружено у Р. arborescens, Р. ara-
bica Boiss., Р. asperrima Gand, и др. Все эти виды характеризуются без-
розеточными побегами. Но среди деревянистых видов есть и такие, у ко-
торых листья развиваются пучками на верхушке побегов. Такую струк-
туру можно оценивать уже в качестве розеточной, например, у Р. bis-
marckii, Р. hirsuta и др.
Между указанными типами встречается и промежуточная структура,
как у Р. cynops.
Безрозеточная структура побегов наблюдается у некоторых однолет-
ников, где она, бесспорно, вторичного происхождения [Р. cardiniana
Wall., Р. indica L., Р. ramosa (Gilib.) Aschers. и др.]. Производный ха-
рактер безрозеточной структуры здесь подтверждается тем, что у по-
следних двух видов иногда встречаются особи, развивающиеся в форме
розеточных растений.
Обращает на себя внимание характер роста и возобновления побегов
Plantago, который у большинства видов является моноподиальным,
в том числе у деревянистых форм, травянистых многолетних и даже
однолетних. У последних формируется обычно листовая розетка, оста,
ющаяся вегетативной, генеративные побеги развиваются в пазухах ро-
зеточных листьев. Таким образом, если признать происхождение травя-
нистых форм подорожника из деревянистых (а это, конечно, является
весьма правдоподобным), то будет справедливым заключить, что в дан-
ном роде при переходе от древесных к травянистым растениям тип ро-
ста и возобновления побегов сохранился без изменений. Моноподиаль-
ность Plantago тем более замечательна, что виды его, в отличие от Ро*
tentilia, являются, как правило, растениями аридных условий и обла-
стей. Лишь единичные виды, например Plantago arachnoidea заходят
в альпийский пояс гор. Следствием приспособления к засушливым усло-
виям существования было массовое развитие однолетников. Впрочем,
у некоторых многолетних видов уже возникает симподиальное возобнов-
ление. Так, у Р. albicans L. наряду с моноподиальным нарастанием розе-
точных побегов возможно и симподиальное. Более часты случаи терми-
нальной закладки цветоносных побегов у однолетних форм: Р. canescens
Schrad. (есть особи и с боковыми цветоносами), Р. pumila, Р. squarrosa
и др. Но они все же не характерны.
Своеобразна жизненная форма «подушечек», образуемых плотно
прижатыми друг к другу «колонновидными» розеточными побегами.
В роде Plantago четко прослеживается эволюция корневой системы.
У преобладающего числа видов развита стержнекорневая система, в том
числе у многих деревянистых видов (Р. arborescens, Р. asperrima, Р. me-
dia L., Р. maxima Juss.).
Но уже в онтогенезе ряда видов наблюдается преобразование с воз-
растом стержневой корневой системы в стержне-кистекорневую и кмсте-
корневую (Р. borneoudi Desne., Р. brutia Ten., Р. cordata Lam.). Типич-
ная кистекорневая структура отмечена у Р. brachyphylla Edgew., Р. сог-
nuti Gouan., Р. durvillei Del. и др. Этапы эволюционного преобразования
79
10. Последовательные этапы преобразования стержнекорневой системы в кистекорне-
вую на примере некоторых видов подорожников:
а — Plantago media', б — Р, lanceolata-, в — Р. major (к — корневище, гк — главный корень,
пк — придаточный корень)
структуры корневой системы Plantago хорошо иллюстрирует ряд: Р. me-
dia, Р. lanceolata, Р. major L. (рис. 10).
Таким образом, и в роде Plantago мы находим типы структур, по-
добные ранее рассмотренным. Однако можно заметить, что в каждом
из этих родов существуют свои особенности в морфогенезе и эволюции
биоморфологических структур, пусть даже мелкие и незначительные.
Поэтому параллелизм жизненных форм в различных родах не исключа-
ет индивидуальных отличий в эволюции, в путях и характере приспосо-
бительной изменчивости. За внешним однообразием основных структур-
ных типов скрывается исключительное богатство и разнообразие эволю-
ционного творчества природы. Но даже сходные типы структуры могут
возникать под влиянием разных экологических условий.
Анализ биоморфологических структур видов Gentiana, Oxytropis и
других открыл существование подобных же типов.
* * *
Предпринятое исследование убедительно показало, что в разнообра-
зии и эволюции вегетативного тела автотрофных покрытосеменных име-
ют место общие закономерности. Привлечение систематико-филогенети-
ческих данных лишний раз подтвердило справедливость эволюционных
принципов, развитых во вводной части настоящей главы.
Заканчивая обзор, следует еще раз подчеркнуть, что в оценке габи-
туально-соматической эволюции покрытосеменных надо исходить из ос-
новных вегетативных типов, обеспечивающих жизнедеятельность. Сома
автотрофных антофитов состоит из трех главных органов: корня, стебля
и листа, составляющих энергетическую основу, удерживающихся благо-
даря функциональной необходимости. В зависимости от условий среды
80
эти органы способны видоизменять свою форму и выполняемые функ-
ции. Распространенным повсюду процессом трансформации сомы покры-
тосеменных в их эволюции и возникновении бесчисленного разнообразия
форм является адаптивная радиация, в своем морфологическом значении
не выходящая за ранг девиаций и анаболий.
Частные приспособления захватили как область вегетативной, так и
генеративной сферы. Изменения генеративных органов воспринимаются
как процесс дифференциации, приведший к возникновению разнообра-
зия в видах, родах, семействах и классах. Таким образом, в эволюции
вегетативных и генеративных органов существуют известные различия,
которые, однако, нельзя искусственно возвышать до степени коренных.
Но в силу функционально-экологических закономерностей, отмеченных
выше, основные соматические типы отличаются известной констант-
ностью и в ограниченном разнообразии повторяются в многочисленных
систематических группах покрытосеменных.
В сфере индивидуального и сезонного развития частные приспособ-
ления, необычайно многообразные в своих проявлениях, обусловливают
конкретный облик видов, их эколого-биологические свойства и признаки.
Последние во многом определяют особенности распространения расте-
ний,, количественное развитие в различных условиях среды и характер
и свойства слагаемых ими ценозов. Фитоценотическая роль отдельных
видов в известной степени является функцией эколого-биологических
особенностей растений и экологического режима биотопов. Анализу ос-
новных биологических явлений в сезонном развитии растений, проявля-
ющихся в разнообразных биоморфологических структурах и их измене-
ниях, а также соответствующих особенностей ценозов, компонентами ко-
торых они состоят, и посвящаются следующие главы. Данная глава
является, таким образом, необходимой теоретической предпосылкой и
основанием для такого анализа.
Глава 4
Характер и длительность вегетации
травянистых растений
Вегетация, или формирование и функционирование аппарата зеле-
ных органов, осуществляющих ассимиляцию, представляет наиболее
важный момент в жизнедеятельности растений. Ассимиляция обуслов-
ливает накопление органического вещества в биосфере, в этом заклю-
чается планетарный характер вегетации, ее космическая роль.
В особенностях вегетации растений находят отражение разнообраз-
ные экологические воздействия окружающей среды. Познание этих осо-
бенностей открывает новые критерии для выявления сущности различ-
ных ценозов и их сравнительной оценки. Сопоставление ритма вегетации
и развития одних и тех же видов, произрастающих в разных местооби-
таниях и ценозах, позволяет определить значение и роль экологических
условий в жизни растений и ценозов.
В биоморфологическом отношении вегетация характеризуется не
только формированием побегов, но и их отмиранием и выходом из сфе-
ры ассимиляции. Нарастание и отмирание надземных органов представ-
ляют две стороны одного и того же процесса — вегетации. Поэтому в
анализе особенностей вегетации травянистых растений лесостепи мы ка-
саемся обеих ее сторон, выступающих в обоюдной связи как взаимно
противоположные начала.
Понятие вегетации, в более широком смысле, включает жизнедея-
тельность растения —его подземных и надземных органов как вегетатив-
ных, так и генеративных. На этом основании в настоящей главе рассмат-
риваются и некоторые закономерности образования и отмирания генера-
тивных побегов.
Вегетация, ее ход и особенности — это одна из характерных черт био-
морфы растений. Среди травянистых растений жизненные формы поли-
карпических, многолетних и двулетних монокарпических и однолетних
видов отличаются своими чертами вегетации.
Само собой разумеется, что вегетация идет как слаженный процесс
жизнедеятельности надземных и подземных органов в их функциональ-
ной взаимосвязи. При всякой возможности, в соответствии с имеющими-
ся данными по биоморфологии, мы стремились увязать состояние веге-
тации с характером распространения корневой системы и эдафическими
условиями, главным образом водного режима мощных черноземов.
Касаясь истории вопроса, надо отметить работы Ж. Массарта (Mas-
sart, 1910), которым выделено 12 типов вегетации на примере растений
западноевропейской флоры. В составленной им схеме показаны основ-
ные этапы жизни растений в подземной и надземной сферах, обозначены
период подготовки к цветению и момент цветения (плодоношения), ста-
дия жизни в облиствленном состоянии (короткая и длительная вегета-
ция), наличие или отсутствие зеленых органов в зимний сезон. Для мо-
нокарпических растений указано время, которое проходит между момен-
том созревания семян и их прорастание.
82
Разделение растений на коротко- и длительновегетирующие с дальл
нейшей детализацией прочно вошло в практику геоботанических иссле-
дований (Коровин, 1934, 1961; Алехин, 1935; А. В. Прозоровский, 1940;
Понятовская, 1952; и многие другие).
Следующим этапом по пути углубления биоморфологической основы
познания вегетации являются работы И. Г. Серебрякова и его учеников
по исследованию ритмов развития и побегообразования растений в раз-
личных географических зонах (Серебряков, 1947, 1952, 1959а, 1961, 1962;
Кошкина, 1954; Серебрякова, 1956, 1959, 1960, 1962; Белостоков,’ 1957,
1962; Пономарева, 1959; Денисова, 1960а, б; Борисова, 1960; Борисова-
Гуленкова, 1960; и др.). По методике, предложенной И. Г. Серебряко-
вым, в вегетативной сфере устанавливаются: появление, рост и последу-
ющее отмирание ассимилирующих органов; число генераций листьев в
течение года; продолжительность жизни листьев каждой генерации; на-
личие Или отсутствие зеленых листьев в течение зимнего периода; отно-
сительная площадь листьев, остающихся на зиму; характер почек возоб-
новления; время заложения зеленых листьев в почках возобновления.
В нашей работе изучение вегетации подчинено задачам фитоцено-
тического анализа основных растительных группировок лесостепи и
в целях сравнения с другими ценозами большое внимание уделяется
разработке количественных показателей.
Фитоценотический анализ вегетации был бы неполным, если из его
сферы исключить ярусное распределение органов вегетации в надземной
части фитоценозов. Рассмотрение ярусного распределения надземных
органов растений будет завершением структурного анализа основных тра-
вянистых группировок лесостепи во всей толще среды, занятой растения-
ми, так как ярусное сложение подземной части растительности проанали-
зировано нами раньше (Голубев, 1962в). Вопросу ярусного строения
надземной части травянистых фитоценозов и синузий и некоторым ас-
пектам динамики ярусности посвящается заключительный раздел этой
главы.
ФОРМИРОВАНИЕ НАДЗЕМНЫХ ОРГАНОВ
Начало вегетации
За начало вегетации здесь принимается момент надземного развития
молодых ассимилирующих органов, а также начало ассимиляции пере-
зимовавших зеленых листьев и побегов. Наиболее легко начало вегета-
ции обнаруживается у летнезеленых растений, развивающих весной из
зимующих почек новые надземные органы. Сложнее обстоит с выявле-
нием вегетации у зимнезеленых растений, выходящих из-под снега с зе-
леными листьями. Если последние вскоре после схода снега отмирают,
то это, по-видимому, служит верным признаком неспособности их к фо-
тосинтезу, за начало вегетации в этом случае следует принимать момент
отрастания новых ассимилирующих органов. Когда же перезимовав-
шие зеленые органы сохраняются более или менее продолжительное
время после стаивания снега, появляется основание считать, что фотосин-
тетические реакции в них начинаются одновременно с установлением
благоприятного термического режима. В условиях Стрелецкой степи ве-,
гетация зимнезеленых растений начинается, как правило, спустя декаду
после окончательного разрушения снегового покрова/ как только почва
оттает и слегка прогреется, конечно, при устойчивых положительных
температурах в дневное время. В этот срок обычно наблюдается и замет-
ное отрастание новых органов у большинства травянистых растений.
Особенно ранним началом вегетации характеризуются Primula veris, Viola hirta,
Aegopodium podagraria и другие, приступающие к формированию и росту листьев еще
под снегом и с исчезнованием последнего (почти без перерыва) начинающие вегетиро-
6*
83
вать. Столь ранняя вегетация наблюдается и у некоторых других видов с подснежным
ростом ассимилирующих органов, анализ которого дается в следующей главе.
Наиболее обширную группу растений лесостепи составляют виды, заметная веге-
тация которых начинается во вторую декаду после таяния снега, т. е. обычно в середи-
не апреля. Сюда относятся многие летнезеленые виды и большинство зимнезеленых
растений (Adonis vernalis, Allium flavescens, Anthyllis polyphylla, Asperula tinctoria,
Bromus inermis, Centaurea scabiosa, Delphinium cuneatum, Campanula sibirica, C. persi-
cifolia, Viscaria vulgaris, Leucanthemum vulgare у др.).
В следующую группу включаются растения, начинающие вегетировать в третью де-
каду после схода снега, т. е. обычно в конце апреля (Anthericum ramosum, Asparagus
officinalis, Campanula bononiensis, Convolvulus arvensis, Eryngium planum, Hieracium
umbellatum, Peucedanum oreoselinum, Pieris hieracioides и другие — растения луговой
степи; Actaea spicata, Adenophora liliifolia, Agrimonia eupatoria, Clematis recta, Cucu-
balus baccifer, Scrophularia nodosa, Solidago virgaurea, Rubus saxatilis — растения ду-
бовых лесов; Gladiolus imbricatus, Lythrum virgatum — растения логов). Еще позднее,
в начале мая, приступают к вегетации Aconitum nemorosum, Antitoxicum officinale,
Astragalus cicer, Epipactis latifolia, Gentiana pneumonanthe, Hieracium virosum, Inula
salicina, Lilium pilosiusculum, Molinia coerulea, Orobus niger, Phragmites communis,
Polygonatum officinale, Serratula coronata, Symphytum officinale, Trifolium medium и др.
Все перечисленные растения мы объединяем в одну категорию — с поздним началом ве-
гетации.
Таким образом, признак начала вегетации дифференцирует расте-
ния примерно на три эколого-биологические группы — с ранндм, средним
и поздним началом вегетации. С момента снегосхода до начала веге-
тации самых поздних растений проходит около месяца с некоторыми
колебаниями. Конечно, в разные по метеорологическим условиям годы
наблюдаются известные сдвиги в начале вегетации, однако указанные
различия в сроках начала вегетации растений отмеченных групп принци-
пиально сохраняются.
ТаблицаЗ
Влияние условий местообитания на начало вегетации (наблюдения 1962 г.)
Вид Местообитание
некосимая степь косимая степь дубовый лес
Anthericum ramosum 28. IV 27.IV
A rtemisia vulgaris 24. IV 24. IV 25. IV
В rachypodium pinnatum 26. IV — 25. IV
Clematis recta 27. IV 3.V 29. IV
Iris aphylla % . . . 17.IV 14.IV —
В условиях центральной лесостепи европейской части СССР с ее теп-
лыми почвами, промерзающими на незначительную глубину с поверх-
ности, градации в сроках начала вегетации нельзя связывать с особен-
ностями распространения корневых систем, как это установлено для рай-
онов с холодными почвами (Свешникова, 1952). Так, среди растений
с самым поздним началом вегетации имеются мелкокорневые (Antitoxi-
cum officinale, Epipactis latifolia, Inula salicina, Lilium pilosiusculum
и др.), их большинство; среднекорневые (Hieracium virosum, Serratula co-
ronata и др.); глубококорневые (Astragalus cicer, Peucedanum oreoseli-
num, Pieris hieracioides и т. д.). Очевидно, различия в возобновлении ве-
гетации вызваны эколого-историческими условиями развития данных
видов.
Особенности условий местообитания также сказываются на времени
возобновления вегетации одних и тех же видов, что можно видеть
из табл. 3.
Как правило, режим некошения степи вызывает некоторое запазды-
вание вегетации в сравнении с косимой степью, но бывают и исключе-
ния. Сходное влияние на начало вегетации оказывают и условия дубо-
84
вого леса. Однако реакции растений на условия произрастания строго
индивидуальны и определяются их эколого-биологическими свойствами.
Следует подчеркнуть, что начало вегетации хотя и близко подходит к
понятию начала роста, но все же не совпадает с ним и по существу, и по
срокам. Так, у многих геофитов рост почек в почве начинается еще до
окончательного схода снега или вскоре после этого, но до начала веге-
тации нередко проходит значительное время, в течение которого моло-
дые ростки выйдут на поверхность и образуются зачаточные зеленые
ассимилирующие органы. С другой стороны, ассимиляция многих зим-
незеленых растений начинается до того, как возобновятся ростовые
процессы и станут формироваться новые веегетативные органы.
Коротковегетирующие растения
К коротковегетирующим мы относим растения, вегетация которых
длится не полный вегетационный период, с теми или иными пропуска-
ми— перерывами. Среди коротковегетирующих растений средней лесо-
степи намечается целый ряд биоморфологических типов, которые опре-
деляются, главным образом, продолжительностью вегетации — от самой
краткой до более растянутой.
Весенние геоэфемероиды. Характеризуются очень коротким жизнен-
ным циклом надземной сферы, ограниченным весенним периодом. Рост
и развитие этих видов часто начинается еще под снегом, а с момента
схода снега прогрессивно усиливается, в течение незначительного вре-
мени растения успевают отцвести и принести плоды и семена, после чего
надземные органы отмирают. Вегетация в текущем году обычно не во-
зобновляется. К этой группе принадлежат: Anemone ranunculoides, Fica-
ria verna, Fritillaria meleagris, Fritillaria ruthenica, Gagea lutea, G. mini-
ma, G. erubescens, Ranunculus pedatus, Scilla sibirica и др.
Перечисленные растения обладают разнообразными запасающими
органами: корневищами, клубнями и луковицами. Именно благодаря
запасам пластических веществ они способны к подснежному росту и
энергичному возобновлению надземных органов, которые отличаются
весьма интенсивной ассимиляцией. Почти одновременно с формирова-
нием зеленых побегов за счет их деятельности начинается развитие но-
вых запасающих органов с резервами питательных веществ, а запасы
прошлогодних постепенно истощаются. Как правило, запасающие орга-
ны недолговечны и ежегодно обновляются, но есть и многолетние органы.
Жизнедеятельность весенних геоэфемероидов протекает в то время,
когда поверхностные слои почвы еще увлажнены талыми весенними во-
дами. Все эти растения мелкокорневые, причем корни распространяются
в самом верхнем горизонте. Однако в силу указанного обстоятельства
они находятся в условиях достаточной влагообеспеченности. Раннее
прекращение вегетации и переход в состояние покоя является хорошим
приспособлением, предохраняющим растения от действия засухи.
Максимальное сосредоточение геоэфемероидов наблюдается в облас-
тях с аридным режимом: степях, полупустынях и пустынях, а также в
соответствующих поясах гор (Варминг, 1902; Краснов, 1899; Келлер,
1938а, б; Коровин, 1934; А. В. Прозоровский, 1940; Лавренко, 1940; Пав-
лов, 1948; Troll, 1937 и др.).
Весенне-раннелетние геоэфемероиды, вегетирующие в первую поло-
вину вегетационного периода — до конца июня и более — и не восста-
навливающие в текущем году надземных органов. К данной группе гео-
эфемероидов относятся Hyacinthella leucophaea, Pedicular is kaufmannii,
Ranunculus illyricus, Verbascum phoeniceum и др. Перечисленные расте-
ния все без исключения являются мелкокорневыми, обычно с поверх-
ностным распространением корней.
85
Весенне-раннелетние эфемеры. Это однолетние растения с очень ко-
ротким циклом развития. Наиболее распространены в заповеднике ози-
мые формы, начинающие свое развитие с осени предыдущего года, иног-
да даже раньше — с конца лета, отмирающие обычно в июне. Таковы
Alyssum calycinum, Androsace septentrionalis, Arenaria serpyllifolia, Dra-
ba nemorosa, Linum catharticum и др.
Интересную биоморфу представляет Arenaria serpyllifolia, которая в
середине лета целиком теряет листья и сильно подсыхает, а с наступле-
нием влажной погоды в конце лета на засохших, казалось бы, побегах в
пазухах отмерших листьев формируются короткие веточки с цветками
вторичного цветения. Изредка подобное явление наблюдается у Acinos
thymoides.
Весенне-раннелетние эфемеры характеризуются весьма мелкой кор-
невой системой.
Малолетние монокарпики. Жизненный цикл этих растений складыва-
ется из вегетативной фазы, продолжающейся один год или несколько
лет, и генеративной фазы, после прохождения которой растения пол-
ностью отмирают. В первую фазу растения вегетируют в течение всего
вегетационного периода, сохраняя зеленую листовую поверхность. В год
цветения и плодоношения вегетация генеративных особей продолжается
до середины лета (конец июня —начало июля), а затем, после плодоно-
шения, они обычно засыхают и отмирают. Сюда входят: Barbarea vulga-
ris, Berteroa incana, Campanula patula, C. sibirica, Carduus hamulosus,
Erysimum marschallianum, Jurinea arachnoidea, Tragopogon orientate,
Trinia multicaulis, Turritis glarba и др.
Однако нередко после плодоношения надземные побеги отдельных
видов полностью теряют лишь листья, подсыхают частично и стебли, но
большая их часть сохраняется живой, и в более влажную и прохладную
погоду конца лета и начала осени из пазух отмерших листьев формиру-
ются цветки второй генерации (вторичное цветение). Особенно обильным
из года в год повторяющимся вторичным цветением отличаются Campa-
nula sibirica и Berteroa incana. Вторичное цветение описанного типа наб-
людается и у Campanula patula, Carduus hamulosus, Erysimum mar-
schallianum, Tragopogon orientate, Turritis glabra.
По типу двулетних монокарпиков в условиях заповедника развивает-
ся Anthriscus silvestris, который в лесной зоне является многолетним
поликарпическим растением. Произрастая под пологом дубрав в доволь-
но большом количестве, он после отцветания и плодоношения в середи-
не лета целиком отмирает, включая и подземные органы. Но вегетатив-
ные особи, как и другие вышеназванные монокарпики, вегетируют весь
теплый сезон.
Весьма оригинальным развитием характеризуется Chaerophyllum bul-
bosum, вегетативные особи которого вегетируют лишь в первую полови-
ну лета, а затем теряют надземные органы. Генеративные же особи ве-
гетируют более продолжительное время — до середины августа, когда на-
ступает плодоношение, и растения оканчивают свой жизненный цикл.
Большинство рассматриваемых растений обладает неглубокой кор-
невой системой (до 1 м и менее). Лишь два вида Carduus hamulosus и
Tragopogon orientate в условиях луговой степи являются глубококорне-
выми (свыше 2 м).
Эфемероиды — это растения со среднелетним перерывом вегетации,
совпадающим с наиболее засушливым периодом. В изученных ценозах
они обнаружены только в составе растительности луговой степи. Эфе-
мероиды лесостепи отличаются целым рядом специфических особенно-
стей. Во-первых, все они являются факультативными, так как при уста-
новлении более благоприятных условий увлажнения в середине лета спо-
собны вегетировать в течение всего вегетационного периода. Во-вторых,
86
как следствие вышеуказанного признака, по длительности и характеру
вегетации они являются весьма пластичными растениями, очень чутко
реагирующими на режим увлажнения почвы.
Наши эфемероиды обладают мелкими корневыми системами, не вы-
ходящими за пределы гумусового горизонта мощных черноземов. По-
этому в период максимального иссушения гумусового горизонта, проис-
ходящего обычно в середине лета (конец июня — июль), они до мини-
мума сокращают транспирирующую поверхность вплоть до полного
отмирания всех надземных органов. Но как только улучшается режим
почвенной влаги, эфемероиды отвечают на это возобновлением своей
вегетации. Отмеченная черта проявляется и в ранневесеннее время: с
установлением достаточно теплой погоды эфемероиды одними из пер-
вых начинают вегетировать, образуя молодые листья.
По степени выраженности эфемероидных признаков рассматривае-
мые растения распадаются на ряд подгрупп с весьма постепенными пе-
реходами. Наиболее типичную группу эфемероидов луговой степи Со-
ставляют Arenaria ucrainica, Erigeron acer, Myosotis suaveolens, Senecio
integrifolius, Serratula heterophylla, Valeriana rossica и др. Их генератив-
ные особи в любые по метеорологическим условиям годы в надземной
части целиком отмирают в середине вегетационного периода. Это дава-
ло повод для трактовки некоторых из приведенных видов в качестве об-
лигатных эфемероидов (Голубев, 1962а). Однако экспериментальная
проверка признака перерыва вегетации в зависимости от внешних фак-
торов не подтвердила этого положения. Оба вида Myosotis suaveolens,
Senecio integrifolius были высажены в питомнике и регулярно полива-
лись в периоды засухи. В результате можно было убедиться, что на сме-
ну листьям весенней генерации в середине лета формируются мощные
листовые розетки второй генерации, обеспечивающие беспрерывную асси-
миляцию в течение всего теплого времени года. И тот и другой вид от-
личаются очень мелкой корневой системой, проникающей не глубже
30 см. В условиях луговой степи в этом слое наблюдается особенно силь-
ное иссушение почвы, главным образом за счет интенсивного поглоще-
ния влаги корнями других растений. Можно предположить, что в
поверхностном слое почвы, где распространяются корни крестовника и
незабудки и где все поступающие атмосферные осадки почти тотчас же
усваиваются корнями других растений, эти виды оказываются неспособ-
ными конкурировать и в период среднелетнего дефицита почвенной вла-
ги в верхнем горизонте прекращают свою вегетацию.
Аналогично указанным видам ведут себя в природе и опыте Arenaria
ucrainica, Erigeron acer, Serratula heterophylla, Valeriana rossica.
Следует обратить внимание на то обстоятельство, что у всех перечис-
ленных эфемероидов молодые особи догенеративного возраста, как пра-
вило, сохраняют зеленую поверхность в течение всего летнего сезона.
Это, несомненно, замечательное свидетельство вынужденности переры-
ва вегетации взрослых особей, вызываемого исключительно влиянием
термально-гидрологической обстановки, возникающей в ценозах луговой
степи. Вместе с тем в этой черте лесостепных эфемероидов обнаружива-
ется удивительное сходство с коротковегетирующими (в генеративном
состоянии) монокарпиками, вегетативные особи которых также харак-
теризуются длительной вегетацией.
Особый интерес вызывают факты эфемероидного типа развития в ус-
ловиях луговой степи некоторых мезофильных растений, широко рас-
пространенных севернее, на лугах лесной зоны, где они являются дли-
тельновегетирующими. Из этой категорий растений назовем Cerastium
caespitosum, Rumex acetosa, Stellaria graminea, у которых в середине
лета происходит полное или почти полное отмирание всех надземных ор-
ганов. Однако те же виды, произрастающие на полянах, под пологом
87
леса и в логах заповедника, отличаются длительной вегетацией. Особен'
но показателен Rumex acetosa, у которого в указанных условиях сохра-
няются живыми все лето генеративные побеги с развитыми на них листь-
ями. На степи же все надземные органы полностью засыхают уже в
конце июня — начале июля; лишь в августе, когда спадает жар и уве-
личивается влажность верхнего горизонта почвы, у засохших ранее
особей отрастают розеточные листья. Эта особенность поведения щавеля
кислого легко объясняется, если вспомнить, что его корневая система,
простертая горизонтально, углябляется не более чем на 10—15 см.
Дольше, чем на степи, вегетируют на лесных полянах Arenaria ucrai-
nica, Myosotis suaveolens, Serratula heterophylla и др.
Описанное явление служит доказательством своеобразия экологиче-
ского режима луговой степи в сравнении с лугами лесной зоны. Возмож-
ность глубокого преобразования жизненного цикла мезофильных расте-
ний в условиях луговой степи свидетельствует о том, что по одному лишь
списку видов в тех или иных ценозах, без тщательного анализа их эко-
лого-биологических особенностей в данных условиях, нельзя обоснован-
но судить о природе ценозов и решать вопросы их классификации.
Полуэфемероидные растения в сухой период летнего сезона сокраща-
ют до минимума листовую поверхность, но полностью, однако, не прек-
ращают вегетацию. Эта группа весьма многообразна и богато представ-
лена видами лишь в ценозах луговой степи.
Можно выделить группу злаков, у которых в середине лета отмира-
ют генеративные побеги вместе с листьями в их основании, а у розеточ-
ных вегетативных побегов резко сокращается число живых листьев. Наи-
более жизнеспособными оказываются молодые листья в центре розеток.
Сюда относятся Anthoxanthum odoratum, Briza media, Bromus riparius,
Helictotrichon pubescens, H. schellianutn, Phleum phleoides, Poa angusti-
folia и др. Все эти растения обладают сравнительно неглубокой корневой
системой (в пределах 0,5—1 м).
Оригинальная форма приспособления к сокращению транспираций
наблюдается у генеративных особей Euphorbia subtilis. Все листья на
цветоносных побегах в конце первой половины лета отмирают и опадают,
но стебли сохраняются живыми (в зеленом состоянии) и ассимилируют
до конца вегетационного периода. Вегетативные же особи и побеги этого
вида в большинстве случаев летом сохраняют листву и вегетируют пол-
ный сезон. Таким образом, в условиях луговой степи Euphorbia subtilis
в генеративной фазе характеризуется афиллией, возникающей с конца
июня. Это явление, несомненно, находится в связи с недостаточной влаго-
обеспеченностью растения (начиная с середины лета), обладающего-
мелкой корневой системой, к тому же весьма слабо разветвленной.
Интересно, что у некоторых видов только часть особей в середине ве-
гетационного периода засыхает, в то время как другие являются дли-
тельно вегетирующими, например Orobus pannonicus. Раскопка подзем-
ных органов засыхающих особей показала, что они обладают неглубокой
корневой системой, в пределах 50—70 см. Корни длительновегетирующих
особей углубляются до 1 л и несколько более.
Разнообразие в биоморфологическом составе популяций по продол-
жительности вегетации весьма ярко выражено у ряда изученных видов.
Особенно нагляден пример с Salvia pratensis. А. П. Ильинский (1937)
относит его к понтическому степному элементу и отмечает характерный
признак — окончание жизненного цикла до августа. Однако в условиях
Стрелецкой степи в цикле развития шалфея наблюдаются отклонения
от этого типа. Можно выделить следующие группы особей по ритму раз-
вития:
1. Растения, генеративные побеги которых полностью отмирают в
июле. В их основании еще с начала лета формируются вегетативные ро-
88
зетки <в количестве 1—4, вегетирующие до осени. К этой категории при-
надлежит наибольшее число особей популяции.
2. Растения с отмирающими в середине лета генеративными побега-
ми. Вегетативные побеги не образуются. Такого рода особи встречаются
редко. Они-то и являются коротковегетирующими.
3. Растения, у которых отмирает лишь цветоносная часть генератив-
ного побега, а облиствленная — с двумя-тремя парами листьев сохраняет-
ся живой в течение всего вегетационного периода. Группа также мало-
численна, но представлена несколько обильнее, чем предыдущая.
Шалфей луговой развивает глубокую корневую систему, проникаю-
щую до границы смены наносов, в горизонт постоянного увлажнения.
Поэтому наличие коротковегетирующих особей, по-видимому, является
отголоском былого ритма развития, выработавшегося в условиях понти-
ческих степей. В настоящее время распространение и максимальное
обилие шалфея лугового приурочено к лесостепной зоне, где он являет-
ся характерным компонентом луговых степей Европейской части СССР.
Большое разнообразие форм вегетации наблюдается у глубококорне-
вого растения Nonnea pulla. Часть его особей имеет по одному генера-
тивному побегу ранневесенней генерации, отцветающему в конце июля.
Вегетация длится весь летний сезон. У других особей развивается тоже
по одному побегу, но в поздневесеннее время. Поэтому нарастание вет-
вей побега и формирование новых цветков продолжается дольше, за-
хватывая и август. В третьей группе на особи формируется несколько по-
бегов, отмирающих в июле. В их основании развивается одна или более
розеток зеленых листьев, осуществляющих вегетацию до конца сезона.
У следующей группы особей цветоносные побеги в числе 2—5 образу-
ются в разное время. Самые ранние отмирают в середине лета, другие
продолжают цвести и вегетировать до осени. Наконец, встречаются осо-
би, надземные части которых в июле целиком отмирают и в текущем се-
зоне вегетация не возобновляется.
Полуэфемероидный тип развития на степи свойствен многим дву-
дольным, теряющим значительную часть листовой поверхности лишь в
засушливые годы. Таковы Achyrophorus maculatus, Dianthus andrzejows-
kianus, Draba sibirica, Filipendula hexapetala, Polygala comosa, Primula
veris, Prunella vulgaris, Ranunculus polyanthemus, Trifolium montanum,
Veronica austriaca, V. prostrata, V. chamaedrys и др. Эти же вицы, оби-
тающие в более благоприятных условиях увлажнения, например под по-
логом леса (Primula veris, Veronica chamaedrys), на лесных полянах
(Filipendula hexapetala, Polygala comosa, Prunella vulgaris, Ranunculus
polyanthemus и др.) вегетируют длительно, без какой-либо заметной деп-
рессии в середине лета.
Среди отмеченных опять-таки встречаются настоящие мезофильные
растения, например Prunella vulgaris, Veronica chamaedrys и другие,
ареалы которых заходят далеко к северу, в лесолуговую зону. Именна
эти виды особенно чувствительны к засухе и недостатку почвенной влаги.
У них наблюдается явление завядания листьев в знойные часы и дни
разгара лета, обычно предваряющее отмирание транспирирующих орга-
нов, если режим влажности почвы прогрессивно ухудшается. При еще
достаточной влажности тургор побегов Primula veris, Prunella vulgaris,
Veronica chamaedrys в ночное время восстанавливается. Конечно, при
очень резком иссушении и эти виды теряют свои надземные, органы.
Большинство полуэфемероидных растений относится к группе мелко-
корневых, отчасти — среднекорневых, что и обусловливает характерную
реакцию на недостаток влаги в почве в верхних слоях, возникающий под
луговой степью в середине лета.
Ясно очерченную и своеобразную группу коротковегетирующих пред-
ставляют летне-осенние однолетники-полупаразиты: Euphrasia tatarica,
89
•Odontites serotina. Их вегетация начинается в июне, массовое цветение
наступает в августе, а отмирание и образование покоящихся семян —
в сентябре. Эти виды обильно представлены на косимой степи и хорошо
'Становятся заметными вскоре после сенокоса.
Прямую противоположность по времени вегетации занимают весен-
не-раннелетние однолетники-полупаразиты: Melampyrum cristatum, Rhi-
nanthus major. Жизненный цикл их начинается вскоре после стаивания
снега и заканчивается в конце июня — начале июля.
К коротковегетирующим можно отнести и многолетний полупаразит
Thesium ramosum, надземные побеги которого, несущие помимо цветков
желтовато-зеленые листья, отмирают в июле.
Прежде чем перейти к анализу длительновегетирующих растений,
следовало бы очень кратко упомянуть о некоторой переходной группе со
среднепродолжительной вегетацией. К ней мы относим растения, надзем-
ные органы которых отмирают в конце июля — августе и более в теку-
щем году не возобновляются. Это такие растения как Paeonia tenuifolia,
большинство особей Veratrum lobelianum, некоторые особи Salvia praten-
sis и Adonis vemails.
Длительновегетирующие растения
К этой группе относятся виды, вегетация которых продолжается в те-
чение всего летнего периода и может осуществляться в весенний и осен-
ний периоды (полностью или частично). Наличие длительной вегетации
не означает, что листовой аппарат растения, образовавшийся в начале
вегетации, остается без изменения. Напротив, даже у длительновегети-
рующих видов нередко происходит значительное отмирание листовой по-
верхности, наряду с которым возможно и ее отрастание. Так или иначе,
но в наиболее критические периоды летнего сезона эти растения еще спо-
собны сохранять аппарат ассимиляции и вегетировать без какого-либо
перерыва. Наблюдаются лишь более или менее высокие спады вегетации.
В биоморфологическом отношении среди длительновегетирующих рас-
тений выделяется несколько различных групп, обладающих спецификой
не только в структуре, но и в физиологических свойствах.
1. Длительновегетирующие растения, обладающие глубокой или сред-
ней по глубине корневой системой, не испытывающие в засушливое вре-
мя резкого недостатка в усвояемой влаге. Как правило, для них харак-
терны более или менее крупные, относительно широкие листья, высокие
стебли и в общем мезоморфный облик всех надземных органов. Наиболь-
шее выражение эта группа имеет в условиях луговой степи. Мезоморф-
ность многих относящихся сюда видов часто приводит к ложному
представлению о них как о типичных мезофитах. Но это представление
неверное, ибо всем таким растениям свойственна высокая интенсивность
транспирации, а присутствие их в степи объясняется развитием глубокой
корневой системы, добывающей влагу из горизонта постоянного увлаж-
нения. Сюда относятся: Bromus inermis, Centaurea ruthenica, C. scabiosa,
Coronilla varia, Delphinium cuneatum, Knautia arvensis, Onobrychis are-
naria, Phlomis tuberosa, Salvia pratensis, Scabiosa ochroleuca и др. По
своему распространению эти виды являются преимущественно лесостеп-
ными.
Особую группу длительновегетирующих растений составляют луговые
и опушечные мезофиты, которые в лесолуговой зоне являются мелко-
корневыми, а в условиях луговой степи формируют глубокую корневую
систему, проникающую почти до горизонта постоянной влажности. Это
Achillea millefolium, Campanula persicifolia, Leucanthemum vulgare и др.
К длительновегетирующим мезоморфным растениям относятся глубо-
кокорневые монокарпики, которые и в вегетативном и генеративном со-
90
•стояниях вегетируют полный сезон: Libanotis intermedia, Pastinaca Sil-
vestris, Peucedanum lubimenkoanum, Verbascum lychnitis, V. orientate
и др.
2. Длительновегетирующие растения с узкими листьями и другими ксе-
роморфными признаками (присутствие войлочного опушения, толстой
кутикулы и пр.). При наличии опушения листья могут быть более ши-
рокими. Обладают глубокой или средней по глубине корневой системой.
Из этих растений можно назвать Agropyron intermedium, Asparagus of-
ficinalis, Centaurea marschalliana, Galium verum, Scorzonera purpurea,
Stipa capillata, St. dasyphylla, S. pennata, S. stenophylla и др.
3. Ксероморфные мелкокорневые растения, характеризующиеся неко-
торыми физиологическими свойствами, обусловливающими возможность
развития и длительной вегетации при относительном недостатке почвен-
ной влаги, а также ряд мезоморфных растений со специфическими био-
морфологическими особенностями. Развивают мелкую корневую систе-
му, распространяющуюся исключительно в пределах гумусового гори-
зонта. Произрастание растений ограничено преимущественно луговой
степью. В этой группе можно наметить ряд более мелких подразделений:
а) растения с войлочным опушением листьев: Artemisia austriaca, Linosyris villosa,
Veronica incana и др.;
б) листовые суккуленты: Sedum acre, S. thelephium;
в) растения с относительно широкими листьями суккулентной консистенции, с вы-
сокой водоудерживающей способностью: Anthericum ramosum, Antitoxicum officinale,
Iris aphylla, Polygonatum officinale и др.;
г) мезоморфные розеточные растения нижнего яруса травостоя: Fragaria viridis,
Potentilla alba, P. humifusa, P. patula, Veronica spicata и др.;
д) разеточные растения мезоморфной структуры с прижимающимися к почве листь-
ями и ползучие растения: Hieracium bauhinii, Н. pilosella, Plantago media, Trifolium re-
pens и др.;
e) узколистные растения: Asperula cynanchica, A. tinctoria, Carex humilis, C. michelii,
C. verna, Festuca sulcata, Koeleria gracilis и пр.
Существуют и другие градации растений с менее четкими внешними признаками,
указывающими на характер приспособления к условиям недостаточного водоснаб-
жения.
4. Мезофильные мелкокорневые растения с широкими листьями,
свойственные достаточно увлажненным местообитаниям: дубовым лесам,
дну и северным склонам логов. Некоторые представители этого типа
встречаются и в степи, но в соответствующих микропонижениях и дру-
гих депрессиях рельефа. Таковы: Aegopodium podagraria, Centaurea pseu-
dophrygia, Convallaria majalis, Geranium pratence, G. silvaticum, Pulmo-
naria angustifolia, Ranunculus acer и др. Вегетация этих растений длится
беспрерывно весь сезон. Возможно дробление группы на ряд более мел-
ких категорий на основании различий в формах листьев, отношении к
световому режиму и т. д.
Конечно, с точки зрения продолжительности вегетации разделение
выделенных основных четырех категорий растений на подчиненные не
вызывается особой необходимостью. Проведенное расчленение лишь
обогащает экологическое содержание рассматриваемых растений и со-
вершенно не преследует цели их дифференциальной таксономии.
Рост и возобновление побегов и листьев
в течение вегетационного периода
В анализе вегетации большое значение имеет выяснение особенно-
стей структуры генеративных побегов: наличие или отсутствие листьев в
основании и по длине побегов, а также сроков их формирования вместе
с листовым аппаратом. Кроме того, необходимо учитывать возможность
развития у генеративных особей вегетативных побегов и количество
генераций их листьев. Большой интерес представляет установление
91
характера вегетации побегов вегетативных особей, которые в ценозах,
как правило, составляют подавляющее число от общей суммы побегов.
Иначе говоря, нужен «популяционный» анализ вегетации, когда прини-
маются в расчет побеги всех типов и у всех возрастных групп, исключая
разве молодые всходы и проростки.
Прежде всего надо уточнить, что следует понимать под второй,
третьей (и т. д.) генерациями листьев и побегов. По И. Г. Серебрякову
(1961), у травянистых многолетников Подмосковья вторая генерация
листьев и побегов «образуется за счет выхода на поверхность почвы вер-
хушек корневищ или раскрытия почек возобновления в основании весен-
них побегов» (стр. 87).
Для оценки вегетации не существенно, имеем ли мы дело со второй
генерацией листьев или побегов. Но в морфолого-биологическом отно-
шении эти два явления строго разграничены. Генерация побегов — это
совокупность новых побегов, образовавшихся из почек возобновления
или зачаточных побегов возобновления. Если побеги облиствлены, то с
развитием одной генерации побегов сопряжено и формирование одной ге-
нерации листьев. В дальнейшем, однако, вегетативные розеточные и без-
розеточные побеги нередко снова способны к росту и образованию новой
генерации листьев. У розеточных побегов вторая генерация листьев воз-
никает терминально, над листьями первой генерации, при недоразвитом
состоянии стебля розеточного побега. Возможно, конечно, и формиро-
вание боковых розеток в пазухах листьев материнской. У безрозеточных
растений вторая генерация листьев образуется за счет верхушечного
или пазушного возобновления роста побегов первой, весенне-раннелетней
генерации. При пазушном развитии происходит ветвление исходного
побега, формирование побегов обогащения (Серебрякова, 1952). Из
сказанного видно, что понятия «генерация побегов» и «генерация
листьев» могут быть нетождественными.
Частным случаем развития второй генерации листьев и побегов яв-
ляется уже описанное выше для монокарпиков формирование пазушных
побегов с листьями и цветками на стеблях, потерявших листья в середи-
не лета и слегка подсохших. Указанное явление установлено у Berteroa
incana, Campanula patula, C, sibirica, Sisymbrium loeselii, Turritis glabra
и других растений.
Вторичный верхушечный и пазушный рост побегов весьма часто на-
блюдается у многих безрозеточных растений. Повторно растут как ве-
гетативные, так и генеративные побеги, с листьями или без листьев
(вследствие раннего отмирания). Всякий раз, когда образовавшиеся при
вторичном росте побеги облиствлены, можно говорить о развитии второй
генерации листьев. Формирование второй генерации листьев у безрозе-
точных растений вполне тождественно развитию второй генерации ли-
стьев у вегетативцых розеточных побегов. Разница лишь в том, что у
безрозеточных растений образование второй генерации листьев проис-
ходит одновременно с ростом стеблей, что и определяет безрозеточный
тип побегов.
Ритмам сезонного развития растений Стрелецкой степи посвящена
работа М. А. Борисовой-Гуленковой (1960), близко касающаяся вопро-
сов формирования органов вегетации. Однако возможности использова-
ния в фитоценологических целях опубликованных материалов весьма
ограничены в силу целого ряда погрешностей, имеющих, на наш взгляд,
в такого рода работах принципиальный характер. Поэтому мы позволим
себе более подробно остановиться на анализе недостатков работы, ко-
торые заключаются в следующем. Во-первых, нет принципиального
единства в графическом изображении ритма развития растений. Для
одних видов приводятся данные о развитии как генеративных побегов и
особей, так и вегетативных (Berteroa incana, Salvia nutans, Valeriana
92
rossica и др.), а для других — данные о вегетативных побегах (особях)
либо совсем опускаются (Falcaria vulgaris, Pulsatilla patens, Sanguisor-
ba officinalis, Veratrum nigrum и др.), либо вводятся в график только
для позднелетне-осенней генерации (Bunias orientalis, Draba sibirica,
Erigeron acer). Можно подумать, что у перечисленных видов с весны ве-
гетативных побегов (особей) не существует вовсе. Во-вторых, допущены
многочисленные ошибки. Так, не отмечено формирование второй гене-
рации листьев у розеточных побегов в середине лета у Achyrophorus та-
culatus, Centaurea scabiosa, Cynoglossum officinale, Hieracium pilosella,
Jurinea arachnoidea, Pieris hieracioides, Potentilla argentea, Primula ve-
ris, Pulmonaria angustifolia, Salvia pratensis, Turritis glabra и др. He
точно указано время развития второй генерации листьев у Centaurea
mars chailiana, Leontodon hispidus и других, помеченное осенними ме-
сяцами, тогда как в действительности формирование второй генерации
происходит еще в июле. В-третьих, не находит отражение в графиках
наличие вторичного (дополнительного) роста побегов, в результате вет-
вления или верхушечного прироста весенних побегов, и образование
второй генерации листьев у Galium verum, Stachys recta и других без-
розеточных растений. В-четвертых, исключена экологическая оценка
процессов морфогенеза и возможность их изменения под влиянием фак-
торов внешней среды. Хотя наблюдения проводились автором в течение
двух лет (1952 и 1953 гг.), резко отличавшихся по количеству осадков
и другим метеорологическим показателям, имевшие место различия в
развитии не отмечены в статье, а графики фактически являются обоб-
щенными. При этом остается неясным, каким образом выполнялось это
обобщение.
В нашей работе анализом охвачено развитие листовой поверхности
генеративных и вегетативных особей и побегов. Как уже сказано выше,
вегетативные побеги образуются и на генеративных особях. У безрозе-
точных растений мы различали число генераций побегов и отдельно —
число генераций листьев, которые, как видно из предыдущего, могут не
совпадать.
По количеству генераций листьев за вегетационный период все рас-
тения можно разделить на две основные группы: 1) с одной генерацией,
2) с двумя и более генерациями.
Растения с одной генерацией листьев. Среди расте-
ний этого типа встречаются безрозеточные и розеточные формы, с быс-
трым и растянутым формированием надземных органов, однолетние,
двулетние и многолетние растения.
Виды, обладающие длительным периодом роста и формирования над-
земных побегов, растягивающимся на большую часть вегетационного
периода, занимают промежуточное положение между растениями с од-
ной и двумя генерациями листьев. В сущности виды, у которых
образуются весенне-летняя и летне-осенняя генерация листьев, очень
•близко стоят к видам с растянутым ростом надземных побегов, отли-
чаясь лишь наличием некоторого гиатуса между двумя периодами роста.
Но принимая во внимание, что у последних возникает всего лишь одна
генерация побегов, эта категория растений присоединяется к группе с
одной генерацией листьев.
Безрозеточные растения. Сюда относятся однолетние растения (Alys-
sum calycinum, Linum catharticum, Veronica arvensis V. verna и др.);
двулетние монокарпики (Melilotus officinalis, M. albus) и многолетние
поликарпики (Adonis vernalis, Asparagus officinalis, Clematis recta, Del-
phinium cuneatum, Gypsophila paniculata, Inula ensifolia, I. hirta, I. sali-
cina, Linosyris vulgaris, Lithospermum officinale, Orobus pannonicus^
Paeonia tenuifolia, Polygonatum officinale, Thalictrum minus, Th. simplex.
Veronica spuria, V. teucrium и др.). Среди этой группы встречаются и
93
виды с растянутым ростом побегов, например Delphinium cuneatum, Lb
nosy г is vulgaris, Galatella rossica и др.
Розеточные растения. Одной генерацией листьев в данный сезон ха-
рактеризуются однолетние эфемеры: Androsace septentrionalis, Arabidop-
sis thaliana, Draba nemorosa, Lithospermum arvense и др. Вегетация этих
растений весьма краткая. Продолжительная вегетация свойственна мо-
нокарпикам: Tragopogon orientale (в вегетативном состоянии), Libanotis
intermedia, Falcaria vulgaris, Peucedanum lubimenkoanum и др. Наиболь-
шее число растений с одной генерацией листьев встречается среди поли-
карпиков. Здесь можно выделить виды, листовой аппарат которых
формируется весной, отличающиеся эфемерностью вегетации: Gagea
erubescens, G. lutea, Hyacinthella leucophaea, Pedicularis kaufmannii, Ra-
nunculus illyricus, R. pedatus, Scilla sibirica и др. Есть и длительновегети-
рующие поликарпики: Anthericum ramosum, Brachypodium pinnatum,
Campanula rapunculoides, Iris aphylla, Phlomis tuberosa, Pulsatilla patens,
Salvia nutans, Senecio schwetzowii и др.
Образование одной генерации листьев и побегов в указанных случаях
происходит лишь в условиях естественного развития растений, при от-
сутствии покоса и выпаса. Эти воздействия могут коренным образом
изменять тип развития побегов, и у многих видов отрастает вторая ге-
нерация— отава. Лишь отдельные растения вполне устойчивы в харак-
тере развития надземных органов, и покос или стравливание не вызы-
вает восстановления утраченных листьев и побегов. Таковы Adonis ver-
nalis, Paeonia tenuifolia, Polyginatum officinale, Veratrum nigrum; V. lo-
belianum.
На рубеже между растениями с одной и двумя генерациями листьев
и побегов можно поместить виды, у которых во второй половине лета и
осенью формируются мелкие по размерам побеги возобновления с весь-
ма незначительной ассимилирующей поверхностью, как правило, уходя-
щие под зиму. Образование побегов возобновления представляет, несом-
ненно, явление развития второй генерации ассимилирующих органов.
Однако в силу ограниченной листовой поверхности значение этой генера-
ции побегов в общем энергетическом балансе растений невелико. Основ-
ная роль в накоплении органического вещества принадлежит побегам
первой, весенней генерации. Все такие растения по биологической осо-
бенности ближе стоят к видам с одной генерацией листьев и побегов,
куда они и отнесены. Вполне очевидно, что зачисление их в указанную-
категорию нисколько не затушевывает переходного характера данной
группы между формами с одной и двумя генерациями листьев. Здесь,,
таким образом, мы отступаем от чисто формального, морфологического
принципа, отдавая предпочтение биологической сущности явлений, в
данном случае составляющей наибольший удельный вес. Описанные по-
беги возобновления развиваются у Asperula cynanchica, Brachypodium
pinnatum, Clinopodium vulgare, Galium mollugo, G. verum, Hypericum
hirsutum, H. perforatum, Linum perenne, Origanum vulgare, Poa nemoralis,
Polygala comosa, Veronica prostrata и др. Конечно, иногда наблюдается
сочетание у одних и тех же растений разных типов развития побегов во-
зобновления, например, у Asperula cynanchica, Galium mollugo, G. verum*
Medicago falcata и др.
Растения с двумя и более генерациями листьев.
Надо различать растения с четко выраженной, полной второй генера-
цией листьев, достигающих нормальной для данного вида величины, со
значительной общей активной поверхностью, играющих существенную
роль в вегетации. Наряду с этим существуют растения, у которых вто-
рая генерация листьев неполная, из одного — трех листьев, часто не
достигающих нормальных размеров. Здесь мы имеем переход к видам,
образующим одну генерацию листьев (Stipa pennata, St. stenophylla
94
и др.). Некоторые особи и побеги упомянутых видов формируют одну ге-
нерацию листьев, что подчеркивает их переходный характер.
Растения с двумя генерациями листьев в лесостепи — самые мно-
гочисленные. К ним относятся как безрозеточные виды, так и розе-
точные.
1. Безрозеточные растения. Вторая генерация листьев у них возника-
ет в результате вторичного верхушечного нарастания вегетативных по-
бегов и ветвления вегетативных и генеративных побегов в надземной
части. Возможен и третий способ образования генерации побегов в се-
редине лета — развитие их из почек возобновления. Чаще всего это бы-
вает у стержнекорневых растений, у Которых побеги отрастают от корот-
кого корневища (или от каудекса, корневой шейки), например у Lotus
corniculatus, Onobrychis arenaria, Stachys recta и др.
Развитие второй генерации листьев вследствие верхушечного роста
или ветвления вегетативных и генеративных побегов находит оригиналь-
ное выражение у некоторых однолетников (Arenaria serpyllifolia и др.),
о чем уже сообщалось выше.
Gypsophila muralis, распространенная на достаточно увлажненных
местах, по дну логов фомирует побеги, ветвящиеся в течение почти все-
го летнего сезона, без какого-либо перерыва. Этот тип развития в рав-
ной степени можно отнести как к растениям с двумя и более генерация-
ми листьев, так и к предыдущей группе — с одной генерацией.
Большое разнообразие видов с вторичным ростом побегов, обуслов-
ливающим развитие побегов обогащения, установлено среди безрозеточ-
ных поликарпиков. Сюда относятся: Astragalus cicer, Bupleurum falca-
tum, Convolvulus arvensis, Coronilla varia, Dracocephalum ruyschiana,
Euphorbia semivillosa, Galium verum, Lotus corniculatus, Prunella gran-
diflora, Salvia verticillata, Stachys recta, Tanacetum vulgare, Thymus mar-
schallianus, Trifolium alpestre и др.
Среднелетняя депрессия в вегетации, вызванная засыханием листьев,
как это указано для Arenaria serpyllifolia, наблюдается и у поликарпика
Stellaria graminea, возобновляющего свою вегетацию во второй полови-
не лета (на степи) за счет вторичного роста побегов, когда улучшается
увлажнение 1и становится прохладнее.
2. Розеточные растения. Вторая генерация листьев формируется у
вегетативных побегов вегетативных и генеративных особей. Нам не из-
вестны розеточные растения лесостепи, у которых бы настоящая вторая
генерация листьев развивалась на генеративных побегах. Возможно до-
полнительное ветвление генеративных побегов в области соцветия, но-
возникающие цветоносы, как правило, лишены хорошо развитых листь-
ев как у Centaurea jacea, С. pseudophrygia, Pieris hieracioides, Scabiosa
ochroleuca и др.
Из двулетних монокарпиков с двумя генерациями листьев можно
назвать Anthriscus silverstris, Barbarea vulgaris, Berteroa incana, Cam-
panula patula, C. sibirica, Carduus hamulosus, C. nutans, Centaurea pseu-
domaculosa, Cirsium polonicum, Cynoglossum officinale и др. Разумеется,
дополнительная генерация листьев образуется только у вегетативных
особей перечисленных видов.
Весьма обширна группа поликарпических многолетников с двумя ге-
нерациями листьев: Achillea millefolium, Achyrophorus maculatus, Ajuga
genevensis, Arenaria ucrainica, Campanula persicifolia, Centaurea mar-
schalliana, Filipendula hexapetala, Fragaria viridis, Hieracium fallax,
Leucanthemum vulgare и др. Формирование листьев происходит как у
весенних розеточных побегов, так и за счет развития новых розеточных
побегов на верхушках длинных корневищ (Achillea millefolium, Aegopo-
dium podagraria, Cirsium heterophyllum и др.), надземных столонах
(Fragaria vesca, F. viridis, Potentilla anserina и др.), в виде боковых
95
разветвлений на ползучих побегах (Antennaria dioica), в основании мате-
ринских розеток и побегов у короткокорневищных растений (Leontodon
hispidus, Potentilla argentea, Valeriana rossica и т. д.), в том числе
рыхлокустовых и плотнокустовых злаков и осок (Carex spicata, С. vulpi-
па, Festuca pratensis, Koeleria gracilis, Phleutn phleoides и др.), на корнях
размножения в качестве корневых отпрысков (Ajuga genevensis, Inula
britannica).
Сроки и особенности вторичного роста и развития второй генерации
листьев у растений тесно связаны с метеорологическими условиями и
экологическими чертами местообитаний. Весьма показательны факты
различия в формировании второй Генерации листьев у одних и тех же
видов, произрастающих в разных эколого-ценотических условиях. По
наблюдениям в 1962 г., на степи у розеточных вегетативных побегов
Sanguisorba officinalis, Serratula inermis, Rumex confertus отрастание
листьев второй генерации не обнаружено. Вместе с тем в условиях дна
степных логов, где влажность почвы значительно выше, очень часто про-
исходило формирование листьев второй генерации у Sanguisorba officina-
lis, несколько реже — у Rumex confertus, Serratula inermis. Отмеченные
виды — мезофильные растения, этим и объясняется их отрицательная
реакция в условиях луговой степи. Но большинству ее типичных предста*
вителей свойственно двукратное формирование листового аппарата.
Столь же отчетливы экологические закономерности во вторичном
росте и образовании побегов обогащения у безрозеточных растений.
Вегетативные побеги Trifolium medium на дне лога в июле трогаются
в рост. Образуется новая генерация листьев и побегов за счет верхушеч-
ного прироста. В это же время в условиях некосимой степи вторая гене-
рация листьев отсутствует. То же можно сказать о Dracocephalum гиу-
schiana, который на лесных полянах формирует многочисленные пазуш-
ные вегетативные побеги обогащения, развивающиеся на генеративных
и вегетативных побегах весенне-раннелетней генерации. На косимой
степи способность к образованию летне-осенней генерации побегов не
проявляется. Примеры подобного рода, при желании, можно значитель-
но умножить. Все они убедительно свидетельствуют, что при улучшении
водного и питательного режима количество генераций листьев у расте-
ний увеличивается вплоть до состояния беспрерывного роста и формиро-
вания листьев и побегов в течение всего вегетационного периода. Однако
возможности повышения количества и периодичности образования ли-
стьев при улучшении условий роста у разных видов неодинаковы.
В определенных комплексах экологических факторов в процессе истори-
ческого развития отбирались и закреплялись особые типы возобновле-
ния вегетативной сферы. Вместе с тем характер репродукции вегета-
тивных побегов и листьев у растений того или иного ценоза является
ярким индикатором экологических условий местообитания, в числе ко-
торых важная роль принадлежит водному режиму почв.
Что касается сроков, то формирование второй генерации листьев в
большинстве случаев приходится на середину лета — июль. Очень редко
развитие второй генерации происходит несколько позднее — в августе.
Как правило, третья генерация листьев у растений исследованных био-
топов не развивается. Даже если зимующие побеги возобновления при-
нимать за особую генерацию побегов, то и в этом случае обычно более
двух генераций не бывает, так как закладка их чаще всего падает на
середину лета, т. е. совмещается с формированием побегов обогащения
или второй генерации листьев розеточных побегов. На сроки и интенсив-
ность развития второй генерации побегов и листьев большое влияние
оказывают метеорологические условия, главным образом режим влаж-
ности. Очень сухая погода в середине лета резко тормозит формирова-
ние второй генерации.
96
Способность многих растений к образованию в середине —конце
лета второй генерации листьев и побегов является замечательным свой-
ством, благодаря которому они используют все экологические возмож-
ности биотопов для ассимиляции и накопления максимальной продук-
ции органического вещества. В этом находит проявление общая законо-
мерность развития, в основе которой лежит способность растений к
максимальной продуктивности в данных условиях существования и ее
совершенствование в процессе развития функциональных приспособле-
ний и реакций (М. В. Культиасов, 1953, 1962). Очень велико значение
формирования зачаточных побегов возобновления в конце вегетацион-
ного периода. Из них в следующем году формируется новая генерация
ассимилирующих органов, приступающих к фотосинтезу тем быстрее,
чем полнее и лучше они были развиты в предшествующий сезон.
ОТМИРАНИЕ ЛИСТЬЕВ И ПОБЕГОВ
В ТЕЧЕНИЕ ВЕГЕТАЦИОННОГО ПЕРИОДА
Вопросы отмирания надземных органов уже затрагивались при ана-
лизе различных типов вегетации и в других местах. Но они заслуживают
и самостоятельного рассмотрения в морфологическом и ином отноше-
ниях, чтобы более полно и разносторонне обрисовать характер веге-
тации.
Наряду с нарастанием и развертыванием надземных органов
травянистых растений происходит такое же постоянное закономерное
отмирание их. Большое значение имеет отмирание самых активных
ассимилирующих органов — листьев и полное засыхание надземных
побегов.
Отмирание листьев начинается уже в первые две недели после схода
снега. Это — перезимовавшие и, вероятно, не участвующие в ассимиля-
ции листья. Темпы отмирания усиливаются к концу мая — началу июня.
Они прогрессивно повышаются в июне и начале июля, затем с периоди-
ческими подъемами и спусками продолжаются в течение всего вегета-
ционного периода.
Наиболее просто отмирание надземных органов протекает у весен-
них и весенне-раннелетних эфемероидов. Эфемероиды Anemone ranun-
culoides, Corydalis halleri, Ficaria verna, Fritillaria meleagris, Gagea lu-
tea, Ranunculus pedatus, Scilla sibirica и другие теряют надземные органы
полностью в конце мая — начале июня, включая одновременно вегета-
тивные и генеративные побеги у вегетативных и генеративных особей.
Несколько позднее отмирают вегетативные и генеративные побеги и
особи у Hyacinthella leucophaea, Pedicularis kaufmannii, Ranunculus illy-
ricus.
Разновременное отмирание вегетативных и генеративных побегов и
листьев наблюдается у однолетних и двулетних монокарпиков и поли-
карпических растений. Внутри основных биоморф по характеру отмира-
ния надземных органов выделяются дополнительные категории по про-
должительности жизни органов вегетации.
Однолетние растения. По продолжительности жизни они подразде-
ляются на весенне-раннелетние эфемеры, весенне-среднелетние, весенне-
позднелетние и летне-осенние формы. В этих группах могут встречаться
как озимые, так и яровые виды, исключая летне-осенние, которые все
являются яровыми.
У весенне-раннелетних эфемеров розеточной структуры (Androsace
septentrionalis, Draba nemorosa и др.) в конце мая — начале июня за-
сыхают розеточные листья, а в конце июня растения полностью отмира-
ют. У безрозеточных эфемеров (Allyssum calycinum, Veronica verna и
др.) первыми засыхают листья в основании побегов, а затем отмирание
7 В. Н. Голубев
97
распространяется на остальную часть. Период жизнедеятельности огра-
ничивается теми же сроками, что и у розеточных растений.
К весенне-среднелетним однолетникам относятся растения, отмира-
ющие в конце июля — первых числах августа (Bromus japonicus, В. mol-
lis, Dracocephalum thymiflorum, Melampyrum cristatum, некоторые особи
Arenaria serpyllifolia, Acinos thymoides и др.).
Весенне-позднелетние однолетники отмирают в конце августа, но
возможно продление жизнедеятельности на осенние месяцы. Это будут
Crepis tectorum, Lappula myosotis, Tragopogon majus, некоторые особи
Acinos thymoides, Arenaria serpyllifolia, Viola tricolor, полупаразиты Me-
lampyrum arvense, M. nemorosum и др.
Летне-осенними являются полупаразиты Euphrasia tatarica, Odonti-
tes serotina, начинающие вегетировать в июне, а отмирающие осенью.
У всех рассмотренных групп засыхание листьев происходит от основа-
ния побегов кверху.
Как правило, однолетним растениям с менее продолжительной веге-
тацией свойственна неглубокая, поверхностная корневая система. Такая
же закономерность отмечена и для однолетников более южных районов
(Константинова, 1954; Свешникова, 1955; Нечаева, 1958; и др.).
Двулетние монокарпики розеточного типа структуры. У этих расте-
ний первыми, как правило, отмирают розеточные листья в основании
генеративных побегов. В конце мая — первых числах июня отмирают
розетки генеративных особей у Barbarea vulgaris, Berteroa incana, Cam-
panula patula, C. sibirica, Carduus hamulosus, Centaurea pseudomaculosa,
Cynoglossum officinale, Elisanthe viscosa, Erysimum marschallianum,
Oenothera biennis, Silene parviflora, Tragopogon orientate. Во вторую
половину июня отмирают розеточные листья у Carduus nutans, С arum
carvi, Echium rubrum, Jurinea arachnoidea, Senecio jacobaea, Trinia multi-
caulis\ в июле — у Artemisia scoparia, Cirsium polonicum, Daucus carota,
Falcaria vulgaris, Libanotis intermedia, Pastinaca silvetsris, Verbascum
lychnitis, V. orientate, в августе—у Peucedanum lubimenkoanum, Seseli
annuum.
Вслед за розеточными начинают засыхать и листья на генеративных
побегах в направлении снизу вверх. Полное отмирание генеративных осо
бей происходит либо в первую половину вегетационного периода, как это
имеет место у коротковегетирующих монокарпиков, либо во вторую по-
ловину, частично — в осеннее время (у длительновегетирующих).
У вегетативных особей розеточных монокарпиков листья в розетках
сохраняются в течение всего вегетационного периода, частично за счет
развития второй генерации, в то время как листья первой (весенне-ран-
нелетней) генерации постепенно отмирают. Исключение составляет, как
уже указывалось ранее, Chaerophyllum bulbosum, вегетативные особи
которого характеризуются эфемерным циклом развития, а вегетация ге-
неративных особей продолжается до конца лета.
Отмирание листьев в побегах безрозеточных малолетних монокарпи-
ков совершается в порядке, свойственном вообще для безрозеточных рас-
тений.
Розеточные поликарпические растения. Сроки отмирания розеточных
листьев в основании генеративных побегов весьма сильно варьируют.
Здесь также выделяется группа растений, листья которых отмирают в
конце мая — первых числах июня: Achillea millefolium, Agrostis syreist-
schikovii, Alopecurus pratensis, Arenaria ucrainica, Leucanthemum vulga-
re, Myosotis suaveolens, Rumex acetosa и др.; во вторую половину июня —
Achillea nobilis, Campanula persicifolia, Cirsium pannonicum и др.; в
июле — Artemisia latifolia, Carex praecox, Crepis pannonica, Eryngium
planum и др.; в августе — Peucedanum oreoselinum, Scorzonera purpurea,
Senecio schwetzowii и др.
98
Отмирание, начавшееся в сфере розеточных листьев, затем постепен-
но перемещается и на генеративные побеги. Однако в отмирании стеблей
генеративных побегов существуют свои закономерности. У большинства
растений после отмирания листьев на побеге снизу вверх постепенно за-
сыхает и сам стебель побега вместе с разветвлениями. Но существует и
определенная последовательность, когда усыхает сначала верхушка ге-
неративного побега в области соцветия или цветка, а затем отмирание
захватывает все новые зоны побега (вместе с листьями, если таковые
имеются) в направлении вниз. Эта особенность присуща Achyrophorus
maculatus, Campanula persicifolia, Cirsium pannonicum, Geum rivate,
G. urbanum, Hieracium pratense, Scorzonera purpurea, Silene nutans, Sti-
pa pennata, Thymus marschallianus и др.
Весьма своеобразной чертой многих видов лесостепи, главным обра-
зом, луговой степи, является довольно раннее отмирание генеративных
побегов. Но вегетация растений, как правило, не прекращается, так как
имеется резерв вегетативных побегов.
Растения с отмирающими целиком в середине лета надземными ор-
ганами здесь не рассматриваются, так как относятся к эфемероидам и
другим коротковегетирующим формам, характеристика которых была
дана ранее.
В начале июня отмирают генеративные побеги Carex humilis, С. ver-
па, Potentilla humifusa, Pulsatilla patens, Taraxacum officinale и др.
В конце июня — начале июля совершается отмирание генеративных
побегов Briza media, Centaurea mars chailiana, Crepis praemorsa, Draba
sibirica, Festuca sulcata, Helictotrichon pubescens, Hieracium pilosella,
Koeleria gracilis, Leontodon hispidus, Poa angustifolia, Stipa dasyphylla,
St. pennata и др.
Более подробно количественные данные о составе растений по при-
знаку отмирания листьев на генеративных побегах и самих цветоносных
побегов приводятся в гл. VII.
Метеорологические условия разных лет могут вносить некоторые из-
менения в наступление сроков и характер отмирания листьев и побегов.
Но установленные общие закономерности отмирания надземных орга-
нов растений в определенных эколого-ценотических условиях лесостепи
сохраняют свою силу в любые годы.
Ярусное сложение надземной части
травянистых ценозов и синузий
Некоторые вопросы ярусного сложения фитоценозов уже затраги-
вались раньше (Голубев, 1962в). Однако желательно расширить анализ
явлений ярусности Применительно к надземной части растительных со-
обществ.
Развитие представлений о ярусности растительных сообществ в над-
земной сфере восходит к середине XIX века. ЕщеА. Кернер (Кегпег, 1863)
обратил внимание на неоднородность структуры лесных сообществ в
вертикальном направлении и выделил пять горизонтов, соответствующих
современному пониманию ярусов. Далее можно указать на работу
Р. Хульта (Halt, 1881), различавшего в составе лесных ценозов до семи
ярусов, из которых первые три образуются деревьями и кустарниками,
а остальные — травянистыми растениями.
В отечественной ботанической литературе первые сведения о ярусном
расчленении сообществ связаны с именами С. Коржинского (1888) и
А. Гордягина (1889). Оба упомянутых автора касаются ярусного сложе-
ния лесных фитоценозов. С. И. Коржинский выделял в широколиствен-
ном лесу только основные структурные категории: высокоствольные де-
ревья, кустарники и травянистый покров. По мнению С. И. Коржинского
7*
(1888) , благодаря ярусному сложению ценоза «достигается более полная
утилизация пространства, так что на известном участке может сущест-
вовать большое число индивидуумов» (стр. 100). А. Я. Гордягин же скло-
няется к более дробному пониманию ярусов и намечает подразделение
травяного покрова в лесу по крайней мере на три самостоятельных яруса.
Большое внимание изучению ярусного расчленения леса уделял Г. Ф. Мо-
розов (1949).
О ярусном сложении травостоя луговой степи упоминает Б. А. Кел-
лер (1916), выделявший всего два яруса: нижний — злаковый, в состав
которого входят и листья ковылей, и верхний — из высокостебельного
разнотравья.
Из последующих работ по анализу структуры сообществ особый ин-
терес для нас представляют работы А. В. Кожевникова (1929), В. В. Але-
хина (1934), В. В. Алехина, А. А. Уранова (1933). Первая касается мор-
фологической структуры травяного покрова широколиственного леса, а
работы В. В. Алехина — ярусного сложения травостоя Стрелецкой степи.
В надземной части подпологового травянистого покрова А. В. Кожев-
ников выделял четыре яруса:
1-й (0—6 см)—ползучие побеги Ajuga reptans, Glechoma hederacea, Galeobdolon
luteum и др.
2-й (6—15 см)—Anemone ranunculoides, Asarum eurupaeum, Gagea lutea, Majant-
hemum bifolium, ортотропные побеги Ajuga genevensis, Galeobdolon luteum, низкорос-
лые экземпляры Pulmonaria obscura.
3-й (15—52 cm)—включающий полностью побеги Mercurialis perennis, Orobus ver-
nus, Ranunculus cassubicus, Stellaria holostea, вегетативные части Carex pilosa, прикор-
невые листья Aegopodium podagraria-, низкорослые экземпляры Polygonatum multiflo-
rum, Aegopodium podagraria.
4-й (от 52 см и выше до 120 см)—стебли Aegopodium podagraria, Milium effu-
sum, Thalictrum aquilegifolium, высокорослые экземпляры Polygonatum multiflorum.
Приведенные данные показывают, что 'в основу ярусного расчлене-
ния травостоя А. В. Кожевниковым положено пространственное распре-
деление отдельных органов растений и особей, в каком бы возрастном
состоянии они ни находились. Отдельные части и особи растений могут
входить в состав разных ярусов.
Принцип морфологической и экологической сущности ярусности раз-
рабатывался В. В. Алехиным (1934, 1936). При отнесении растений к
тому или иному ярусу В. В. Алехин отводил решающую роль высоте по-
бегов. На Стрелецкой степи В. В. Алехин выявил 6 ярусов раститель-
ности, градиентами которых служат следующие предельные высоты по-
бегов: 1-й ярус — 80—90 см, 2-й — 50 см, 3-й — 40 см, 4-й — 20 см,
5-й ярус представлен мелким разнотравьем и ползучим клевером и 6-й—
туидиевым мхом.
В. В. Алехин (1936) полагал, что виды одного яруса более или ме-
нее экологически равноценны. Кроме того, ярусы в некоторых пределах
обладают самостоятельностью и в известном смысле могут отождест-
вляться с синузиями. Вопрос о принадлежности к ярусам подроста вы-
сокорослых видов в разном возрасте В. В. Алехиным специально не об-
суждался. Этот последний находит освещение в работах В. Н. Сукачева
(1915, 1928, 1934 и др.), помимо ярусов, различавшего еще пологи Он
писал, что растения верхних ярусов, образующих верхний полог, на
ранних этапах онтогенеза могут располагать побеги на высоте одного из
нижних ярусов. Низкорослое состояние растений первой величины воз-
никает и в результате угнетения. В обоих случаях В. Н. Сукачев не ре-
комендует причислять эти особи к нижним ярусам. Молодые особи от-
носятся к подросту, а отставшие в росте — к угнетенной части того яру-
са, которому растения свойственны в пору полного развития. «Поэтому
одна порода одновременно в одном и том же сообществе (а также и ас-
100
социации) не может входить в два и более ярусов, но может распо-
лагаться в разных пологах» (Сукачев, Зоин, 1961, стр. 64). В этой
связи В. Н. Сукачев различает временное пребывание растения в том
или другом ярусе и постоянную принадлежность к определенному
ярусу.
В. Н. Сукачевым разработана четкая и очень содержательная кон-
цепция ярусного расчленения сообществ. Сущность ярусов заключается
не только в различиях по высоте растений, а главным образом в том,
что растения разных ярусов отличаются по экологии и играют в ценозе
свою роль. Ярусность возникла вследствие длительного исторического
процесса, благодаря чему оказалось возможным полнее использовать
«производительные силы» местообитаний и ослабить конкурентные взаи-
моотношения компонентов сообщества. Таким образом, по В. Н. Сука-
чеву, ярус представляет экологическое понятие, поскольку объединяет
растения одинаковой формы роста и сходной экологии, вместе с тем это
и фитоценотическое понятие, так как между растениями одного яруса
возникают свои особые взаимодействия, отличающиеся от взаимодейст-
вий в других ярусах и между ярусами. Кроме того, ярусы в известной
степени обладают самостоятельностью.
Г. И. Дохман (1960) выступает против условного стратиграфического
принципа выделения ярусов, базирующегося на высоте генеративных
побегов растений, проводя четкую грань между ярусом и пологом. Она
настаивает на том, что единственно правильная трактовка заключается
в понимании яруса как исторически обусловленного сочетания видов,
не входящих в состав других ярусов, и как структурной единицы эколо-
гической и фитоценотической сущности. На этом основании Г. И. Дох-
ман относит к одному ярусу «расположенные на одной высоте активные,
т. е. ассимилирующие части взрослых особей того или иного вида (цве-
тущих или в той или иной стадии бутонизирующих), а также особи дан-
ных видов, хотя и лишенных генеративных стеблей, но своей вегетатив-
ной частью находящиеся на той же высоте, что и полновозрастные осо-
би» (стр. 53). Молодые особи Г. И. Дохман выносит за пределы ярусов
и по аналогии с лесными ценозами причисляет их к пологу. Г. И. Дох-
ман, вслед за В. Н. Сукачевым и В. В. Алехиным, склоняется к призна-
нию известной самостоятельности ярусов, объединяющих экологически
равноценные виды растений, что едва ли правильно.
Вряд ли можно возражать против того, что, например, всходы дре-
весных растений в лесу находятся в составе самых нижних ярусов тра-
вянистой подпологовой растительности. Каждое высокорослое растение
в процессе своего развития переходит из яруса в ярус, пока не займет
окончательного пространственного положения, свойственного взрослому
состоянию. В этом случае следует говорить о временных компонентах
того или иного яруса, в отличие от постоянных, составляющих основу
ярусов. Вполне понятно, что каждый вид в качестве постоянного ком-
понента ярусной структуры сообщества характеризуется вполне опре-
деленным положением.
Точно так же совершенно естественно распределение некоторых частей
растений в разных ярусах. В ряде случаев их уже нельзя считать вре-
менными элементами, а наоборот, нужно принимать органическими, игра-
ющими определенную и закономерную роль в сложении ярусов. Так,
например, у сныти в лесу листья образуют характерный (в дубовых
лесах заповедника по счету 3-й) ярус, в то время как цветоносные побе-
ги выходят в более высокий 2-й ярус.
Значит, в расчленении травянистых сообществ на ярусы необходи-
мо учитывать особенности пространственного размещения ассимилиру-
ющих органов и высоту генеративных побегов. Наблюдения показывают,
что высота цветоносных побегов косвенно, посредством влияния на
।
101
режим опыления с помощью насекомых и ветра, оказывает воздействие
на величину семенной продуктивности и, следовательно, их нельзя
исключать из структурной характеристики травостоя. У большинства
травянистых растений стебли генеративных побегов зеленые, часто с бо-
лее или менее редуцированными листочками, и сами являются ассимили-
рующими органами. Кроме того, они производят известное затенение
ниже расположенной массы листьев, изменяют режим обдувания ветром
транспирирующих органов и т. д. Нельзя также забывать, что высота
генеративных побегов обусловливает возможности разбрасывания и раз-
носа созревающих плодов и семян и в этом отношении имеет определен-
ное биоценотическое значение.
Таким образом, и в области надземной сферы мы находим яркие
свидетельства того, что ярусы представляют экологические ниши фи-
тоценоза и его структурные элементы, образуемые как характерными
эдификаторными и доминантными растениями, так и отдельными частя-
ми побегов растений, принимающих участие в сложении разных ярусов.
Второй случай, конечно, исключает отождествление ярусов с синузиями,
а также трактовку экологической равноценности компонентов определен-
ного яруса. Растения более высоких ярусов в процессе онтогенеза из се-
мени или вегетативных зачатков последовательно размещают надземные
органы во всех последующих ярусах, начиная с самых нижних. Этот
факт, разумеется, не дает никакого повода относить ювенильные и угне-
тенные особи к характерным компонентам того или иного яруса. Они
являются всего лишь временными элементами ярусов. Однако выделение
помимо ярусов новой категории — пологов, т. е. элементов вертикального
расчленения сообществ в связи с возрастным состоянием и линейными
размерами основных эдификаторов ценозов верхних ярусов, как это де-
лают В. Н. Сукачев (1928, 1934), В. Н. Сукачев и С. В. Зонн (1961), вряд
ли оправдано. Пологи — понятие излишнее, по крайней мере для травя-
нистых ценозов, к тому же весьма неопределенное в эколого-фитоцено-
тическом отношении, между тем как развиваемые здесь представления о
ярусах с устойчивыми, руководящими и временными компонентами, на
наш взгляд, хорошо отражают существо явлений, четко передают эколо-
го-фитоценотические особенности вертикальной структуры сообществ и
изменение экологической среды развития в онтогенезе растений, входя-
щих в верхние ярусы.
Несостоятельность представлений об экологической равноценности
компонентов одного яруса подтверждается также тем, что нередко на-
блюдается несовпадение ярусного распределения подземных и надземных
органов видов. Например, низкорослое растение Centaurea marschalliana,
которое в надземной части мы относим к 6-му ярусу растительности лу-
говой степи, обладает корнями, углубляющимися почти до 2 м. A Sedum
acre, принадлежащий к 6-му ярусу в надземной части, имеет мелкие кор-
ни, распространяющиеся 'не глубже 30 см. Наоборот, Rumex acetosa с
такими же поверхностными корнями в травостое входит в состав 2-го
яруса. Отсюда ясно, что понятие об экологической равноценности компо-
нентов отдельных ярусов, в виду хорошо обоснованных исключений, дол-
жно быть оставлено, как не отвечающее действительному положению
вещей.
Что касается соотношения понятий яруса и синузии, то отождествле-
ние этих категорий возможно только в частных случаях и то с извест-
ными натяжками и оговорками.
Конкретный фактический материал по ярусности надземной части
растительности луговой степи, травяного покрова дубовых лесов, дна
и северных склонов логов приводится в гл. VII.
Важнейшей особенностью структуры надземной части травянистых
сообществ и синузий является ее периодичность, т. е. ежегодное разру-
102
шение и возобновление. С этой чертой структуры связан другой ее при-
знак — высокая динамичность. Последняя обусловлена тайже многообра-
зием ритмов развития компонентов, составляющих травянистые ценозы.
Надземная сфера травянистых растений на протяжении вегетацион-
ного периода, как можно было видеть, находится в постоянном измене-
нии: вначале это рост и развитие органов, позже (а часто и одновременно
с нарастанием) совершается противоположный процесс — отмирание
активных органов. Но если структура надземных побегов каждого вида
находится в неуклонном преобразовании, то как следствие этого про-
цесса должна закономерно изменяться и структура надземной части
растительности.
В ^анализе структуры следует исходить из фаз вегетации как опреде-
ленных этапов в структурном развитии растительности. В оценке дина-
мики структуры растительности луговой степи можно различать следу-
ющие этапы:
1. Одноярусная фаза сообщества цветковых растений (мхи не принимаются в рас-
чет, хотя в равной мере являются компонентом одноярусного сообщества) открывается
с момента схода снега и начала вегетации. В первое время вегетации низкорослые
и высокорослые виды образуют приземный ярус преимущественно вегетативных орга-
нов. Несколько позднее или одновременно включаются цветущие весенние растения,
цветоносные побеги которых небольшие по размерам (ранневесенняя фаза)
2. Фаза формирования второго яруса за счет выдвижения вверх более высокорос-
лых побегов (средневесенняя фаза).
3. Возникновение третьего яруса, сопровождающееся отмиранием у многих видов
розеточных листьев, находящихся в самом нижнем ярусе (поздневесенняя фаза).
4. Развитие структуры наивысшей сложности. Ярусы высшего класса формируются
главным образом генеративными побегами, но цветущие растения встречаются и во
всех остальных ярусах, образованных цветковыми растениями. Продолжается отмира-
ние листьев в нижнем и следующем над ним ярусах. Полностью выпадают синузии
весенних и весенне-раннелетних эфемеров, эфемероидов, геоэфемероидов и генератив-
ные особи коротковегетирующих монокарпиков, размещавших надземные органы с пер-
вого (от поверхности почвы) по третий ярусы (раннелетняя фаза).
5. Продолжение формирования самого высокого яруса за счет вдвигания генера-
тивных побегов поздно развивающихся видов (Agropyron intermedium, Delphinium си-
neatum и др.). Происходит отмирание генеративных побегов (полностью или с верхуш-
ки) рано отцветающих растений в верхнем и других ярусах. Усиливается процесс усы-
хания листьев во всех ярусах и возникает вторая генерация листьев (среднелетняя
фаза).
6. Массовое отмирание растений и разрушение структуры надземной части ценоза.
Вторичная дифференциация нижнего яруса как зимующей .в зеленом состоянии струк-
турной категории сообщества (осенняя фаза).
Z
Таким образом, в сезонном развитии структуры надземной части тра-
вяного покрова следует учитывать: а) последовательные фазы усложне-
ния структуры от ранневесенней одноярусной до наивысшей точки;
б) максимальное насыщение пространства ассимилирующими частями,
а также отдельных ярусов; в) обновление составных частей каждого
яруса — исчезновение старых путем отмирания и вхождение новых, позд-
нее развивающихся побегов и растений; г) роль генеративных побегов —
активная в момент цветения и преимущественно пассивная после обсе-
менения и отмирания; д) подрост, т. е. вегетативные особи большинства
видов, тем более всходы и ювенильные растения, располагающие асси-
милирующие органы в нижнем или следующем за первым ярусе, реже —
в других, болеем высоких ярусах, как это имеет место у ковылей с их
длинными листьями; е) разрушение структуры травостоя, включая раз-
ложение и минерализацию отмерших побегов.
Обобщая приведенные данные о ярусном сложении надземной части
травянистых ценозов и принимая во внимание строение их подземной
части (Голубев, 1962в), можно наметить в эколого-биологическом от-
ношении следующие ступени и сферы структурного расчленения со-
обществ:
103
1. Область генеративного процесса, размножения и рассеивания пло-
дов и семян. Пространственно это верхняя сфера надземной части
ценозов, насыщенная генеративными побегами. У низкорослых растений
функции полового размножения и рассеивания плодов совмещаются со
следующей ступенью вертикального расчленения сообществ.
2. Область воздушного питания растений (поглощение света и улав-
ливание углекислого газа), отчасти водообмена между растениями и
воздушной средой. Это область размещения ассимилирующих органов
растений — побегов с листьями. Процесс фотосинтеза частично совер-
шается и в предыдущей сфере ценозов посредством зеленых частей ге-
неративных побегов.
3. Область возобновления и умножения побегов растений, осущест-
вления вегетативного размножения с помощью ползучих побегов и раз-
нообразных побеговых подземных органов. Пространственно эта область
охватывает приземную часть ценозов и поверхностный слой почвы.
В данной части травянистых сообществ сосредоточивается и наиболь-
шее количество запасных пластических веществ в органах запаса: кор-
невищах, клубнях, луковицах, отчасти в корнях.
4. Область почвенного питания и водоснабжения растений. Она ох-
ватывает все пространство распространения корней, которые заходят
и в самый верхний слой почвы.
Глава 5
Перезимовка травянистых растений
Характер перезимовки биологически связан со многими другими яв-
лениями жизнедеятельности растений, особенно с ростом и формирова-
нием побеговых органов в летне-осенний период. Поэтому в анализе
перезимовки надо принимать во внимание особенности всего жизнен-
ного цикла растений и те условия, в которых он протекает. Например,
режим сенокошения оказывает глубокое влияние на ход морфогенеза
в послепокосный период, что сказывается и на характере перезимовки.
В биоморфологическом отношении чрезвычайно важным представ-
ляется изучение состояния растений сразу после снеготаяния, продол-
жительности жизни перезимовавших листьев и побегов, позволяющей
косвенно судить об их жизнедеятельности, состояния растений перед
установлением снежного покрова, характера отмирания листьев в зим-
ний период, подснежного развития, глубины и длительности зимнего
покоя. Все эти моменты и были включены в программу наших исследо-
ваний. Под наблюдение взято большинство травянистых растений лу-
говой степи, дубовых лесов, лесных полян, дна и склонов степных логов.
По возможности мы стремились установить качественные и количест-
венные различия в перезимовке одних и тех же видов в разные по
метеорологическим условиям годы, а также в разных экологических усло-
виях и в связи с режимом сохранения травянистой растительности за-
поведника — в условиях сенокошения и абсолютной зяповедности.
Изучение перезимовки приобретает большую содержательность,
если ее увязывать с закономерностями исторического (разв'ития расте-
ний, с путями выработки тех или иных приспособлений к переживанию
неблагоприятного зимнего сезона. Руководящим является то обстоя-
тельство, что вечнозеленые растения представляют первичную катего-
рию, связанную с третичной растительностью, а листопадность и летне-
зеленость — признаки вторичного, производного характера. Необходи-
мо отметить, что одни и те же внешние формы перезимовки нередко
имеют разное происхождение и обусловлены различными факторами.
Так, свойство вечнозелености может возникать вторично, из летнезеле-
ных типов, в результате приспособления растений к краткому вегетаци-
онному периоду. Это обеспечивает максимальное использование соляр-
но-теплового режима для продуцирования органического вещества. Веч-
нозеленость (зимнезеленость) индуцируется также некоторыми экологи-
ческими условиями. В зоне умеренно холодного климата таким индукто-
ром является повышенное (до известных пределов) увлажнение почвы,
стимулирующее ростовые процессы.
ТИПЫ РАСТЕНИЙ ПО СПОСОБАМ ПЕРЕЗИМОВКИ
Широкой известностью пользуется классификация жизненных форм
растений по способам перезимовки, предложенная Раункиером (Raun-
kiaer, 1905, 1934), в основу которой положен признак высоты почек
105-
возобновления над уровнем почвы в зимний период и степень их защи-
щенности. Эта классификация неоднократно детализировалась разны-
ми авторами, и ее основные пять жизненных форм (фанерофиты, хаме-
фиты, гемикриптофиты, геофиты, терофиты) были разделены на значи-
тельное число более мелких биоморф. Весьма полный обзор этих до-
полненных классификаций жизненных форм по способам перезимовки
можно найти в работах Рюбеля (Rubel, 1930), Б. А. Келлера (1938),
Браун-Бланке (Braun-Blanquet, 1928).
Классификация Раункиера не лишена противоречий и слабых сторон
(Пачоский, 1921, 1926; Кожевников, 19376; Келлер, 1938а; Амелин,
1953; Голубев, 1956а; Зозулин, 19596; и др.). Обнаружено, что одни и
те же виды нередко совмещают признаки разных типов (Лапшина, 1928;
Котт, 1949; Воронов, 1952; и др.). Вместе с тем морфологические разли-
чия отдельных жизненных форм не находят ясного подкрепления в био-
логических свойствах, обусловливающих своеобразие перезимовки
(Амелин, 1953; Голубев, 1956а). Не получил отражения в классифика-
ции Раункиера и признак наличия или отсутствия зеленых зимующих
органов. Этот пробел был восполнен Е. И. Лапшиной (1928), несколько
модернизировавшей систему. Но факты существования растений с зи-
мующими листьями и побегами в области умеренно холодного климата
были известны значительно раньше (Irmisch, 1850; Areschoug, 1895;
Lindforss, 1907; Massart, 1910; Jeswiet, 1914; Linkola, 1922; Riibel, 1925;
и др.).
Из отечественных работ следует особо выделить исследования
Е. И. Лапшиной (1928, 1929), которыми с почти исчерпывающей полно-
той охвачены разнообразные случаи проявления зимнезелености и пере-
живания зелеными органами неблагоприятного времени года. Установ-
ленные Е. И. Лапшиной закономерности вполне применимы не только
к перезимовке растений под Ленинградом, но и в других зонах и об-
ластях. Автором был поставлен также вопрос о продолжительности
жизни перезимовавших зеленых органов, но, к сожалению он не доведен
до стадии систематических полевых наблюдений.
В пределах каждого биологического типа Раункиера Е. И. Лапшина
(1928) выделяет две группы: растения, перезимовывающие почками и
зимующие в зеленом состоянии. Для «еиоторых типов делается попытка
разделить растения по структурному признаку на розеточные и расте-
ния с облиствленными побегами (соответствующие типу безрозеточных
растений). Е. И. Лапшиной описан ряд фактов, свидетельствующих о
возможности взаимного перехода этих двух структурных форм рас-
тений.
Поскольку разнообразие растений по структурному признаку имеет
самостоятельное значение, то включение его в качестве характеристики
зимнего состояния растений оправдано, хотя все же в ограниченном
значении. Надо ясно представить, что структурный тип является кон-
ституционным признаком и находит проявление на протяжении всей
жизни растений, как в зимний, так и летний периоды. И, наоборот, из-
менение морфологического типа побегов в связи с перезимовкой имеет
большой интерес в эколого-биологическом отношении, в понимании
процессов перестройки структуры и приспособления к переживанию не-
благоприятного периода.
Структурные формы зимующих органов растений более четко раз-
делены А. Г. Вороновым (1952), различавшим розеточные и безрозеточ-
ные растения.
Наличие зеленых зимующих органов зарегистрировано у растений
различных ботанико-географических зон. Особенно многочисленные ис-
следования по выявлению зимнего состояния растений в связи с прак-
тическими запросами оленеводства были выполнены в условиях Край-
ни
•него Севера — в тундре и лесотундре (Игошина, 1937; Темноев, 1939;
Глинка, 1939; Александрова, 1940; Андреев, 1940, 1948; Ковакина, 1952;
Кильдюшевский, 1956; Серебряков, 1961, 1963; и др.).’ Факты перези-
мовки травянистых растений в зеленом состоянии в лесолуговой зоне
приводятся в работах Е. И. Лапшиной (1928, 1929), О. В. Саркисовой-
Федоровой (1929), А. В. Кожевникова (19376), И. Г. Серебрякова
(1947, 1952), И. Г. Серебрякова, Т. М. Галицкой (1951), М. М. Шик
(1953), Т. И. Серебряковой (1956), В. Н. Голубева (1956а, б),
А. П. Пошкурлат (1962) и других, в лесостепной и степной зонах —
в работах С. И. Данилова (1937), Н. А. Прозоровского (19406, 1947),
С. А. Котта (1944),.А. Г. Воронова (1952), А. А. Горшковой, О. П. Род-
ченко (1959), А. М. Семеновой-Тян-Шанской, Н. И. Никольской (1960),
А. А. Горшковой (1960а, б), М. А. Борисовой (1960), М. А. Борисовой-
Гуленковой (1960), О. П. Родченко (1961 У, В. Н. Голубева (19626)
и др. Интересное явление зимней вегетации растений в некоторых рай-
онах Средней Азии отмечается М. В. Культиасовым (1946), Н. С. За-
прометовой (1956), В. А. Бурыгиным (1957), Н. Т. Нечаевой (1960),
Л. X. Наабер, И. Л. Захарьянц (1961), Н. С. Запрометовой, И. В. Есипо-
вой (1961) и др.
В работах И. Г. Серебрякова (1947, 1952, 1959а, 1961, 1962) изуче-
ние зимнего состояния растений получает дальнейшее развитие и ме-
тодически новое решение. Помимо регистрации зеленой поверхности
в зимний период, определяется ее относительная площадь, продолжи-
тельность жизни листьев после схода снега и пр.
Исследованиями установлено, что величина зеленой массы в зим-
ний сезон не остается постоянной, а прогрессивно падает, иногда
вплоть до полного отмирания, совершающегося у разных видов в различ-
ные сроки (Ковакина, 1952; Кильдюшевский, 1956; Серебряков, 1947;
и др.).
Скорее всего отмирают те листья, которые сформировались раньше
(в летний период), а позднее развившиеся листья, в позднелетне-осен-
нее время, сохраняются дольше, нередко благополучно перезимовывают
и продолжают ассимилировать. Очень четко эта особенность перези-
мовки листьев проявляется у монокарпических растений: Verbascum
lychnitis, Oenothera biennis, Senecio jacobaea, Carduus nutans, C. hamu-
losus и др. Вегетативные особи этих видов к осени развивают приземи-
стые розетки листьев разного возраста и размеров. На периферии ро-
зетки находятся наиболее крупные ранее остальных сформировавшиеся
листья, а в центре — весьма мелкие, возникшие позднее всех. Крупные
старые листья осенью и в течение зимы почти полностью отмирают, за
исключением разве самых оснований листьев, т. е. черешков с частью
листовой пластинки. Промежуточные по возрасту и величине листья
отмирают либо только с самой верхушки, либо разрушение распростра-
няется дальше—до середины пластинки. Самые молодые мелкие ли-
сточки в центре розеток сохраняются целиком, благополучно перезимо-
вывают и в весеннее время способны не только ассимилировать, но и
расти. Весьма заметный рост зачаточных перезимовавших листьев и
превращение их в довольно крупные пластинки весной установлены
у Verbascum lychnitis, Senecio jacobaea и др. Сходная картина состава
уходящих под зиму розеточных листьев, их отмирания и роста после
перезимовки наблюдается у многих поликарпических растений (Plan-
tago media, Р. lanceolata и др.), в том числе злаков (Bromus riparius,
Helictotrichon pubescens и др.).
Характер перезимовки зависит от фазы развития растений (Котт,
(1949). Последнее, в свою очередь, находится в определенной связи с хо-
зяйственным использованием растительности. Сенокос и выпас — сме-
щающие фазы развития — влияют и на зимнее состояние растений.
107
Существенное влияние на перезимовку, в том числе на состояние
зеленой поверхности, оказывают многочисленные внешние факторы: мощ-
ность, плотность и другие признаки снегового покрова, время его уста-
новления и разрушения, зимние оттепели, температурный режим в
осенно-зимнее время и т. д. Все эти факторы действуют в совокупности,
и подчас очень трудно решить, какой из них имеет главенствующее зна-
чение.
Каузальная сторона перезимовки растений (устойчивость к низким
температурам, зимний покой и пр.) всесторонне и широко освещается
в работах физиологического цикла (Перетолчин, 1904; Максимов, 1952;
Мороз, 1940, 1948; Туманов, 1940, 1951; Генкель, 1948; Генкель и Окни-
на, 1948а, б; Сергеев, 1953; Петровская, 1955; Барановский, 1955; Род-
ченко, 1958; Сергеев, Сергеева, Мельников, 1961; и др.).
Присутствие зимующих зеленых органов у растений большинством
исследователей рассматривается как приспособление к жизни в усло-
виях сокращенного вегетационного периода, так как растения получают
возможность ассимилировать в ранневесеннее и позднеосеннее время
при благоприятном температурном режиме.
Несмотря на кажущуюся очевидность этого положения, специаль-
ные физиологические исследования ассимиляции зеленых зимующих ор-
ганов в предзимнее и ранневесеннее время отсутствуют и приведенное
заключение все еще остается в значительной мере априорным. Но ука-
занное объяснение значения зеленых зимующих органов представляет-
ся недостаточным также в том отношении, что ограничивается рамками
сприспособительной пользы». Между тем многообразие зимнезеленого
состояния, несомненно, связано и с происхождением растений, с эколо-
го-генетическими особенностями их становления и развития.
Биоморфологическое изучение перезимовки травянистых растений
лесостепи на территории Центрально-Черноземного заповедника, пре-
имущественно растений Стрелецкой степи, проводилось Н. А. Прозо-
ровским (1940, 1947), А. М. Семеновой-Тян-Шанской, Н. И. Никольской
(1960), М. А. Борисовой-Гуленковой (19606). Н. А. Прозоровским
(1940) исследовано осеннее и зимнее состояние около 150 видов луго-
вой степи в условиях некосимого режима. Анализировалось положение
зимующих почек относительно поверхности почвы, их количество, раз-
витие зеленых листьев и побегов. Однако наблюдения за состоянием
растений сразу после стаивания снега не были проведены, а они, как по-
лагал сам Н. А. Прозоровский, должны внести ряд поправок к данным,
полученным в результате осенне-зимних обследований. А. М. Семенова-
Тян-Шанская перечисляет несколько видов, зимующих с зелеными
листьями, а также некоторые летнезеленые виды. Работа М. А. Борисо-
вой-Гуленковой лишь в общих чертах характеризует особенности пере-
зимовки растений Стрелецкой степи. Существенным упущением ее яв-
ляется отсутствие указаний, при каком режиме использования (коси-
мом или некосимом) находились взятые под наблюдения растения.
По биоморфологическим особенностям перезимовки все растения
можно подразделить на две основные группы: летнезеленые и зимнезе-
леные. Однако четкой границы между ними провести нельзя. Существо-
вание постепенных переходов приобретает большую наглядность, если
привести более дробное разделение растений, учитывая различия в со-
хранении и отмирании зеленых органов, их новообразовании в зимний
период и другие признаки:
1. Собственно летнезеленые растения. Отмирание зеленых листьев совершается-
осенью, еще до наступления холодной погоды с отрицательными температурами. В зим-
ний период новообразования зеленых органов не происходит, и после схода снега* расте-
ния некоторое время остаются без ассимилирующей поверхности (Adonis vernalis, Anti-
toxicum officinale, Convallaria majalis, Paris quadrifolia, Pulsatilla patens, Thalictrum
lucidum, Veratrum nigrum и др.).
108
2. Растения с подснежным ростом и формированием листьев и побегов. При бла-
гоприятной погоде у некоторых видов образуются зачаточные зеленые органы еще
в предзимнее время. Решающим условием подснежного роста является наличие талой
почвы в течение всей зимы или в отдельные ее периоды. Поэтому в разные зимы ин-
тенсивность подснежного роста испытывает колебания вплоть до его прекращения
в особо суровые зимы. Большинство видов этой группы — летнезеленые (Lactuca altis-
sima, Milium effusum, Роа nemoralis, Viola hirta и др.), но есть и зимнезеленые (вечно-
зеленые) формы: Роа palustris, Primula veris и др.
3. Летнезеленые растения. Сохраняют зеленые побеги до наступления заморозков,
прекращающих их жизнедеятельность. Но и здесь разные виды обладают неодинаковой
чувствительностью к морозам. У одних надземные органы гибнут при первом даже лег-
ком заморозке (Clematis recta, Nepeta pannonica, Urtica dioica и пр.), зеленая поверх-
ность других сохраняется дольше и изредка уходит под снег, но вскоре, однако, отми-
рает (Orobus vernus, Pulmonaria angustifolia, P. obscura и др.).
4. Растения с зелеными органами, уходящими под зиму и отмирающими в разные
сроки снежного периода (Campanula trachelium, Centaurea pseudophrygia, Cirsium hete-
rophyllum, Potentilla anserina, Solidago virgaurea, Stellaria holostea и др.).
5. Растения с зимующими зелеными органами. Зимующие органы сохраняют зеле-
ную окраску некоторое время после схода снега, но вскоре буреют и отмирают. Весьма
вероятно, что перезимовавшие листья этих растений не способны к ассимиляции (Сеп-
faurea mars chailiana, Jurinea arachnoidea и др.). В неблагоприятные по метеорологи-
ческим условиям весны (яркое солнечное освещение при низких температурах, резкие
смены температур и пр.) по этому типу развиваются некоторые зимнезеленые (вечно-
зеленые) виды (Fragaria viridis, Silene nutans, Thymus marschallianus и др.), вскоре
после схода снега теряющие перезимовавшие зеленые листья.
6. Типичные зимнезеленые (вечнозеленые) растения. Зимуют с зелеными органами,
которые оказываются после схода снега вполне жизнеспособными и ассимилируют бо-
лее или менее продолжительное время весной, иногда летом и даже в осенний период
{Achillea millefolium, Antennaria dioica, Anthyllis polyphylla, Betonica officinalis, Campa-
nula persicifolia, Filipendula hexapetala, Fragaria vesca, F. viridis, Trifolium pratense,
Viola rupestris и др.).
В приведенном ряду биологических типов, внутри которых можно
бы выделить дополнительные подгруппы по количественным признакам,
ла одном полюсе находятся собственно летнезеленые, на другом — зим-
.незеленые и вечнозеленые растения
Нельзя подходить к типизации растений по способам перезимовки
формально, относя к вечно- или зимнезеленым все растения, сохраняю-
щие на зиму зелеными хотя бы зачаточные или единичные листья и по-
беги. Такая классификация не может иметь сколько-нибудь серьезной
научной ценности. Необходимо принимать во внимание количественные
показатели зимующей зеленой поверхности, а также разнообразные
конкретные обстоятельства, обусловливающие ту или иную форму пе-
резимовки.
В дифференциации на летнезеленые и зимнезеленые типы следует
использовать эколого-исторические особенности развития растений,
вскрывая биологическое значение зеленых зимующих органов.
Можно предположить, что вечнозеленость (зимнезеленость) некото-
рых растений является отголоском их вечнозеленой природы в далеком
прошлом, в условиях теплого и влажного климата, и представляет древ-
ний признак. К этой категории ближе всего, вероятно, стоят растения,
у которых перезимовавшие зеленые органы отмирают в первые дни и
недели после разрушения снегового покрова. Вечнозеленость других
видов является молодым прогрессивным, вторично развившимся при-
знаком, возникшим в позднейшее время в результате приспособления
к климату с коротким вегетационным периодом, со снежной и холодной
зимой, при достаточном количестве почвенной влаги. Этот тип вечнозе-
леное™ создает возможность полнее утилизировать для ассимиляции
теплый период года.
1 Растения, перезимовывающие в зеленом состоянии и вегетирующие в течение все-
го теплого времени года, с полным правом могут быть названы вечнозелеными. Если
же в вегетационном периоде бывает перерыв вегетации, то такие растения, зимующие
© зеленом состоянии, относятся к зимнезеленым.
109
Пример «рудиментарной» вечнозелености 1 показывает, как происхо-
дила переработка вечнозеленых растений теплого и влажного климата
в летнезеленые. Процесс этот еще не получил такого развития, как у на-
стоящих летнезеленых растений, листья которых засыхают до наступле-
ния осенне-зимних холодов. Он продолжается и в настоящее время, а на-
личие еще зеленых зимующих, но нежизнеспособных органов является,
по-видимому, свидетельством их вечнозеленой природы в прошлом.
Но существуют и настоящие вечнозеленые травы, генетически связан-
ные с древне-третичными вечнозелеными типами, перезимовывающие
листья которых способны ассимилировать в весеннее время (Asarum
europaeum, Hepatica nobilis, виды Pyrola и других Ericaceae и пр.).
Архаичность вечнозелености некоторых растений хвойных лесов убеди-
тельно мотивирована работами П. Н. Крылова (1898), А. И. Толмачева
(1951, 1954) и др. Большой вклад в решение проблемы вечнозелености
наших травянистых растений могут внести физиологические исследова-
ния, позволяющие вскрывать характер жизнедеятельности перезимовав-
ших листьев. Но эти исследования должны опираться на данные биомор-
фологии, которые уже сейчас позволяют выделять отдельные группы
растений по способам перезимовки, несомненно, отличающиеся и по фи-
зическим свойствам зимующих листьев.
Интересны факты рекапитулятивного характера зимнезелености, про-
являющейся в ювенильном возрасте растений, которые во взрослом со-
стоянии развиваются по типу летнезеленых. Они неоднократно отмеча-
лись в литературе (Тахтаджян, 1948, 19546; Авдошенко, 1949; Шик, 1953;
Серебряков, Чернышева, 1955; Васильченко, 1961; и др.). В наших иссле-
дованиях зимнезеленые ювенильные особи обнаружены у Bupleururn
falcatum, Cytisus ruthenicus, Falcaria vulgaris, Linum flavum, Pyrethrum
corymbosum, Ranunculus polyanthemus и др.
В раскрытии эколого-исторических особенностей развития растений
большое значение имеет выяснение специфики зимнего покоя, которому
ниже посвящается особый раздел.
Принимая во внимание изложенные факты и соображения о прин-
ципах разделения зимнезеленых и летнезеленых растений, мы сочли воз-
можным отнести к последней группе целый ряд видов, обладающих зеле-
ными зимующими органами. Ввиду важности затронутого вопроса,
относящиеся сюда факты заслуживают более подробного рассмотрения.
В качестве летнезеленых нами понимаются виды, сохраняющие зи-
мой малочисленные остатки зеленых листьев, не полностью отмершие,
которые после схода снега живут непродолжительное время или отми-
рают еще в зимний период. Обычно это летние листья, возникшие в со-
ставе определенной генерации позднее остальных. Наличие одного —
двух зеленых зимующих листьев, как правило, подсохших с верхушки
или до середины и почти до основания (но далеко не у каждбй особи,
часто лишь у единичных экземпляров), отмечено нами у Agropyron inter-
medium, Aegopodium podagraria, Brachypodium pinnatum, Calamagrostis
epigeios, C. arundinacea и др. Зеленая поверхность этих растений зимой
настолько мала, а признак дозимнего отмирания большей части надзем-
ных органов так отчетливо выражен, что трактовка данных видов в ка-
честве летнезеленых кажется вполне естественной.
Иную категорию составляют виды, образующие к зиме мелкие зача-
точные листья или побеги, иногда в весьма малом количестве, едва выхо-
дящие на поверхность почвы или скрытые во влагалищах отмерших
листьев целиком, либо на большей части длины и только верхушка мо-
жет быть свободной. Ярким примером последних служат ковыли: Stipa
1 К этой категории относятся растения, листья которых отмирают в разные сроки,
зимнего периода и вскоре после схода снега.
110
capillata, St. dasyphylla, St. pennata, St. stenophylla, у которых все лет-
ние листья на зиму отмирают и лишь внутри влагалища вегетативных
побегов текущего года осенью обнаруживается зачаточный зеленый
лист. Несколько зачаточных розеточных листьев (от 2 до 6), обычно не
превышающих в длину 1 см, формируется осенью у Agropyron caninum,
A. repens, Alchemilla micans, Artemisia compestris, Bunias orientalis, Ca-
rum carvi, Centaurea jacea, Crepis pannonica, Dianthus andrzejowskianus,
Rumex crispus, Scorzonera purpurea, Tragopogon orientate, Trifolium
montanum, Trinia multicaulis и др. Едва заметные над поверхностью поч-
вы безрозеточные побеги формируются у Euphorbia segueriana, Galium
verum, Trifolium alpestre и др. У всех перечисленных растений возмож-
ность ассимиляции зелеными зимующими листьями осенью и ранней
весной близка'к нулю. К тому же на характер развития зачаточных
листьев заметное влияние оказывают метеорологические условия осен-
него периода. В неблагоприятную погоду зачаточные листья совсем не
развиваются. Все подобные растения мы также относим к летнезеленым.
Преимущественно, летнезелеными являются в условиях заповедника
Lathyrus pisiformis, Vicia cracca, V. tenuifolia, у которых зимующие без-
розеточные побеги возникают у немногих особей. Сходны с ними по дан-
ному признаку многие виды разнотравья, как Anemone silvestris, Cen-
taurea marschalliana, Geranium sanguineum, Jurinea arachnoidea, Ranun-
culus polyanthemus и др.
Нечто среднее между двумя вышеприведенными группами составля-
ют Aegopodium podagraria, Anthriscus silvestris, Cnidium dubium и дру-
гие, у которых более регулярно появляются зимующие зеленые листья,
либо весьма позднего развития — зачаточные, либо более ранние по
образованию, уходящие под снег в полуотмершем состоянии. Молодые
зачаточные листья перезимовывают и продолжают рост весной, более
взрослые же листья обычно отмирают зимой. В разные по метеорологи-
ческим условиям годы наблюдаются всевозможные отклонения.
В вегетативных розетках монокарпиков (Cirsium polonicum, Супо-
glossum officinale, Echium rubrum, Verbascum orientate и др.) листья
к зиме обычно отмирают целиком, либо самые молодые центральные
листья, а также основания единичных взрослых сохраняются живыми и
зимуют в зеленом виде. Указанные виды относятся нами к летнезеленым.
От этой группы через ряд переходов намечается уже категория зимнезе-
леных (вечнозеленых) растений, описанная в начале главы, у которых
формируются более крупные зачаточные листья, а остающиеся части
взрослых листьев обладают большей поверхностью.
На основании приведенного анализа следует пояснить, что в разра-
ботанной выше классификации растений по способам перезимовки груп-
пы 1—5, за единичными исключениями, относятся нами к летнезеленым
растениям, а группа 6 — к зимне- (вечно) зеленым. Как показано ранее,
в составе зимнезеленых растений выделяются по меньшей мере две эко-
лого-генетические группы.
По структуре побегов среди зимнезеленых растений наблюдаются
розеточные и безрозеточные формы. Из розеточных можно назвать мно-
гие злаки и осоки: Anthoxanthum odoratum, Arrhenatherum elatius, Briza
media, Bromus riparius, Carex humilis, C. michelii, C. verna, Dactylis
glomerata, Helictotrichon pubescens, H. schellianum и др.; виды разно-
травья: Achillea millefolium, Ajuga genevensis, Antennaria dioica, Campa-
nula persicifolia, Filipendula hexapetala, Fragaria viridis, Leucanthemum
vulgare, Myosotis suaveolens, Potentilla humifusa и др. Безрозеточные
зимующие побеги имеют: Asperula cynanchica, Glechoma hederacea, Hy-
pericum perforatum, Linum perenne, Lysimachia nummularia, Polygala
comosa, Sedum acre, Stellaria graminea, Veronica chamaedrys, V. incana,
V prostrata и др. Как правило, безрозеточные зимующие побеги
ш
располагаются на поверхности почвы или близ нее и в этом отношении
мало отличаются от розеточных растений, зеленые листья которых при-
легают к почве, а почки возобновления или точки роста находятся на
уровне поверхности почвы. Безрозеточные растения часто образуют лож-
ные розетки, например, Medicago falcata. Встречаются безрозеточные
растения, зимующие в форме подушечек (Cerastium caespitosum, Dian-
thus deltoides, Stellaria media, Veronica serpyllifolia и др.), что отмеча-
лось и Е. И. Лапшиной (1928) под Ленинградом. Как справедливо ука-
зывает Лапшина, при подготовке растений к перезимовке изменяется
форма зимующих листьев, их анатомическая структура и другие призна-
ки в сравнении с летними листьями. Будущих исследователей зимовки
растений в этой области, несомненно, ожидают интереснее открытия.
В качестве одной из деталей преобразования зимующих надземных ор-
ганов является изменение их окраски. Листовые розетки некоторых ви-
дов ястребинки Стрелецкой степи в позднеосеннее время так резко
меняют свою окраску, общий габитус и структуру волосяного покрова,
что видовые различия их становятся трудно уловимыми.
Судьба зимующих безрозеточных побегов (побегов возобновления)
разных видов после перезимовки складывается по-разному. У многих
видов они после стаивания снега трогаются в рост верхушкой и из тер-
минальной почки возникают летние побеги. Такой ход развития безрозе-
точных побегов установлен у Acynos thymoides, Asperula aparine, Cera-
stium caespitosum, Dianthus deltoides, Galium mollugo, Hypericum
hirsutum, H. perforatum, Linum perenne, Malachium aquaticum, Moehrin-
gia trinervia, Polygala comosa, Sedum acre, Stellaria graminea, S. palust-
ris, Thymus marschallianus, Veronica chamaedrys, V. incana, V. prostrata,
V. scutellata, V. serpyllifolia и др. Конечно, не все заложенные с осени
побеги возобновления благополучно перезимовывают. Так, у Linum pe-
renne часть побегов отмирает в зимний период и после схода снега. Дру-
гие побеги только теряют все зеленые листья и продолжают рост вер-
хушкой. И лишь некоторое количество перезимовывает без повреждений
и возобновляет нормальный верхушечный рост. Отдельные летние побеги
возникают из почек на корневище.
Иную категорию составляют растения с довольно хорошо развитыми
безрозеточными побегами, уходящими под зиму, благополучно перези-
мовывающими и после более или менее длительной вегетации отмираю-
щими, не дающими перехода к летним побегам. Это Galium vernum,
Lathyrus pisiformis, L. pratensis, Vicia sepium, V. tenuifolia. To же про-
исходит и co взрослыми вегетативными побегами Stellaria holostea, ухо-
дящими зелеными под снег, но в зимний период частично или полностью
отмирающими. Отдельные перезимовавшие побеги хотя и живут непро-
должительное время, но затем также отмирают.
По величине зимующей зеленой поверхности зимнезеленые растения
весьма сильно варьируют. Относительно большая площадь живых листь-
ев наблюдается у Antennaria dioica, Campanula persicifolia, Coronaria
flos cuculi, Elisanthe viscosa, Hieracium pratense, Leucanthemum vulgare,
Silene nutans, Turritis grabra, Viola rupestris, Viscaria vulgaris и др.
Значительно больше число видов, зимующая листовая поверхность
которых невелика: Anthoxanthum odoratum, Briza media, Dracoce-
phalum ruyschiana, Linum perenne, Polygala comosa, Veronica prostrata
и т. д.
Многочисленные факты и наблюдения убеждают что наибольшей
устойчивостью к неблагоприятным условиям и способностью переживать
зимний период в зеленом состоянии характеризуются листья самой по-
следней генерации— позднелетне-осенней. Лучшим подтверждением
этого положения служит то обстоятельство, что если какими-либо воз-
действиями вызвать нарастание новых побегов и листьев в конце вегета-
112
ционного периода у летнезеленых растений, то они могут стать зимнезе-
леными. Надо сказать, чю одним из факторов, нарушающих нормальный
ход роста и вызывающих вторичный рост надземных органов трав,
является покос. Под влиянием покоса многие растения Стрелецкой сте-
пи образуют зеленые зимующие листья и побеги, отсутствующие
в условиях абсолютной заповедности (Geranium sanguineum, Knautia
arvensis, Salvia pratensis и др.). Покос усиливает формирование зеленых
зимующих побегов у Astragalus danicus, Coronilla varia, Onobrychis
arenaria, Vicia tenuifolia и др.
Определенные изменения в перезимовке одних и тех же видов уста-
новлены в связи с погодными условиями разных лет. В «суровые» зимы
листья осенней генерации Primula veris отмирают, тогда как в «теплые»
зимы они сохраняются живыми и весной ассимилируют довольно про-
должительное время. Следует оговориться, что оценки «суровая» и «теп-
лая» зима условны, имеется в виду комплекс факторов, благоприятствую-
щий перезимовке в зеленом виде или, наоборот, затрудняющий ее. Дру-
гим примером может служить Silene nutans, которая в зиму 1959/60 г.
перезимовала в зеленом состоянии на степи удовлетворительно, а зимой
1961/62 г. розетки ее полностью погибли и, вероятно, не столько из-за
низких зимних температур, сколько под влиянием резкого чередования
оттепелей и заморозков в конце зимнего сезона. Аналогичные, но прямо
противоположные отличия в эти же зимы зарегистрированы у Elisanthe
viscosa: ранней весной 1960 г. после схода снега розетки были мертвыми,
а в 1962 г. они вышли из-под снега зелеными, жизнеспособными. В ду-
бовых лесах разногодичные колебания зеленой зимующей поверхности
испытывают Aegopodium podagraria, Anthriscus Silvestris.
Резкие изменения в способах перезимовки одних и тех же видов в раз-
ные по климатическим условиям зимы, например зимовка в зеленом со-
стоянии и без зеленых органов, должны, понвидимому, свидетельствовать
о том, что их зимнезеленость нельзя рассматривать как приспособитель-
ный признак, развившийся в современных условиях среды. Это скорее
всего рекапитуляция свойства вечнозелености предков или же начало
формирования данного признака, возникающего вторично на основе
преобразования летнезеленого типа.
Существенные различия в перезимовке растений возникают под влия-
нием экологических особенностей местообитаний. Так, упоминавшаяся
выше Silene nutans весной 1962 г. в дубовом лесу, в отличие от степи,
имела многолистные зеленые розетки, долгое время сохранявшиеся жи-
выми. Весной 1961 г. Lysimachia nummularia по дну логов вышла из-под
снега с мертвыми листьями, а под пологом дубравы (ур. Дуброшина) —
с зелеными, жизнеспособными. Betonica officinalis на лесных полянах
и под пологом леса является зимнезеленым растением, на некосимой
степи — почти летнезеленым, на косимой — занимает промежуточное
положение.
Весьма неблагоприятны для сохранения зеленых листьев частые зим-
ние оттепели, чередующиеся с резкими холодами. На характер вегетации
перезимовавших зеленых органов большое влияние оказывают погодные
условия ранней весны. Наибольший вред наносит неустойчивость эколо-
гического режима: частые и резкие смены температурного и светового
режимов, осадков и т. д.
Количественные и качественные различия в перезимовке растений
в связи с эколого-ценотическими условиями приводятся в табл. 4. На-
блюдения проводились с 4 по 12 апреля 1960 г. на Стрелецком участке
Центрально-Черноземного заповедника на площадках 0,5 м2. Снег со-
шел 1—2 апреля.
Как видно из таблицы, некосимые участки степи и подпологовая ра-
стительность дубового леса характеризуются бедностью зимнезеленых
8 В. Н. Голуб ев
ИЗ
Таблица 4
Количество перезимовавших в зеленом виде розеток, листьев и побегов
в различных эколого-ценотических условиях
Вид Южный СКЛОН некоей- мого лога • Некоси- ная степь Пастби- ще, пло- щадка № 1 Пастби- ще, площадка № 2 Поляна косимая Поляна некоси- мая Сныте- вая дубра- ва-
Розеточные растения
Achillea millefolium . . 1(1) 1(1) 10(3—4) 3(4-5) — 32(1-2) —
A ego podium podagraria . • — — — — — — 6(1)
Ajuga genevensis . . . . — — — 2(2) 2(1-2) — —
Allium oleraceum . . . — — — — 2(1) — —
Anthriscus silvestris . . — — — — — 2(2)
Arrhenatherum elatius . . — — — — 18(1) 8(1) —
Artemisia absinthium . . 1(1) — — — — — —
Berteroa incana .... 1(10) — — — — — —
Bromus riparius . . . — — — — 56(1—2) — —
Calamagrostis epigeios . . 2(1) — — — — — —
Campanula patula . . . — — — — 2(3) — —
Campanula persicifolia . . — — — — — 6(2-3) —
Carex michelii — — — — 40(2-3) — —
C. humilis — — — 37(4—5) — — —
Centaurea marschalliana . — — — 13(1) — — —
Dactylis glomerata . — — — — 10(2—3) — —
Festuca sulcata .... — — 19(2) 200(1-2) 15(1) 5(1) —
Filipendula hexapetala . . 2(1) 8(1-2) 48(2—3) 35(2—3) 10(2—3) 4(1) —
Fragaria vesca . . . — — — Ю(2) 2(1) 8(2)
F. viridis — 1(2) 10(1—3) 17(1-2) — —
(} urn urbanum .... — — — — 1(2)
Hieracium pilosella . . — 17(3—8) 14(6—8) 2(5)
H. bauhinii — — 1(2) —
Knautia arvensis .... — — 3(2) 2(1)
Leucanthemum vulgare . . — 3(2—3) 2(2—3) 18(2—5) 6(2—3) —
Myosotis suaveolens , . . — 20(3—4) 11(2—3) — __
Plantago lanceolata . . — — — 14(2-3) ___
P. media — -- 19(3—4) 6(1-2) —
Poa angustifolia .... 45(1—2) 35(1—2) — — 14(2)
Potentilla argentea . . . 1(4) 10(5—6) — 12(2)
P. humifusa — 1(2) 47(4) 15(3) —
Primula veris — — 6(2-3) 8(2—3)
Prunella vulgaris . . . — — — 28(4)
Ranunculus polyanthemus — * 1(2) 4(1-2)
Rumex acetosella . . — — 2(1)
Salvia pratensis — — 5(1-2) 5(2-3) 4(1) —
Taraxacum officinale . . — 10(4) 6(3-4) —
Trifolium montanum . . — — 3(1) — — —
T. pratense 1(1) 5(1-2)
Viola hirta 3(1-2)
V. rupestris — 9(2) 43(2—3) 20(3—4) —
Viscaria vulgaris — — 18(9) — —
114
Таблица 4 (продолжение)
Вид Южный склон не- косимого лога • Некоси- мая степь Пастби- ще, площад- ка Кг 1 Пастбище, площадка Кг 2 Поляна косимая Полян- некссиа мая Сныте- вая дубрава
Безрозеточные растения
Galium mollugo .... Genista' tinctoria .... 7 1 3 4(10) 3(4) 4 6 —
Hypericum perforatum . . — — — 6 4 —
Poly gala comosa '. ’. — — 5(5—6) 9^8—10) — — —
Sedumacre . — — 8 4 — — —
Stellaria graminea . . — 1 — — — 22(4—5) —
Thymus marschallianus . — 2 4(6) 2(5-7) — — —
Tri folium repens . . . — 8(3) 4(2) 2 — —
Veronica chamaedrys . . — — — — 18(4-6) 30 —
V. prostrata — — — 5(9) 6 — —
итого: зимнезеленых видов 8 9 20 27 27 12 4
розеточных побегов 53 59 229 427 309 65 17
листьев розеточных побегов . 87 73 772 966 924 99 28
безрозеточных У побе- гов 7 4 28 31 36 62 —
* Цифры вне скобок обозначают количество розеточных или безрозеточных побегов, цифры в
скобках — число листьев на одном побеге.
** При расчете суммы перезимовавших зеленых листьев бралось среднее арифметическое число
листьев, приходящееся на один побег, умноженное на количество розеток.
видов растений в сравнении с пастбищем и косимой лесной поляной. Про-
межуточное положение занимает некосимая лесная поляна.
Наибольшее количество розеточных листьев, перезимовавших зелены-
ми (в расчете на 0,5 л2), наблюдается в фитоценозах пастбища и коси-
мой лесной поляны, весьма малое число листьев подсчитано в сообщест-
вах некосимой степи, некосимого южного степного склона лога и
некосимой поляны. Меньше всего зарегистрировано листьев под пологом
снытевой дубравы. При общей незначительной величине зеленых безро-
зеточных побегов на учетных площадках их количество повышается от
некосимой степи к пастбищу и далее — к косимой и некосимой полянам.
Интересно отметить, что число листьев у розеточных побегов на не-
косимых участках заметно меньше, чем на пастбище и косимых площад-
ках. Это лишний раз подтверждает, что скашивание и стравливание
травостоя имеют своим следствием увеличение количества зеленых зи-
мующих листьев, посредствохм возбуждения ростовых процессов.
ХАРАКТЕР ЗИМНЕГО ПОКОЯ У РАСТЕНИЙ
В осенний период при подготовке травянистых растений к зиме
большая часть надземных органов отмирает, за исключением зимующих
зеленых, и растения переходят в состояние покоя, когда заметные росто-
вые процессы отсутствуют и резко замедляются другие жизненные
функции. Зимний покой является ответственным этапом в общем ритме
сезонного развития растений умеренно холодного климата. Сезонная пе-
риодичность развития выработалась под влиянием климатической пе-
риодичности, и состояние покоя служит замечательным приспособлением,
обеспечивающим переживание растениями неблагоприятного периода.
8*
115
Исследования показывают, что для многих растений воздействие по-
ниженных зимних температур является необходимым условием их даль-
нейшего развития (Diels, 1918; Любименко, Вульф, 1926; Серебряков,
1952, 19596; и др.). Снятие влияния низких температур в осенне-зим-
ний период вызывает у некоторых растений болезненные симптомы:
листья развиваются медленно, не достигают нормальной величины или
совсем не образуются, растения не могут перейти к цветению и в конце
концов погибают (Любименко, Вульф, 1926; Кожевников, 1931, 1939;
Трофимов, 1939; Демидова, 1952; Серебряков, 1959; и др.).
Явление зимнего покоя и связанные с ним зимостойкость и морозо-
устойчивость растений нашли освещение в работах физиологов. Было
установлено, что в зимний период в зимующих органах происходят пре-
вращения запасных веществ. Так, крахмал переходит в сахара и жиры
и благодаря этому растения приобретают устойчивость к морозам (Пе-
ретолчин, 1904; Пояркова, 1924; Петровская, 1955; Родченко, 1958; и др.).
Проблема морозоустойчивости у нас плодотворно разрабатывалась
Н. А. Максимовым (1952), И. И. Тумановым (1940, 1951), а в последнее
время П. А. Генкелем с сотр. (Генкель, 1948; Генкель, Окнина, 1948а, б;
Генкель, Ситникова, 1953; и др.).
П. А. Генкелем установлено, что все клетки растений в период зимне-
го покоя проходят процесс обособления протоплазмы. Плазмодесмы,
соединяющие протопласты клеток в единую систему, втягиваются,
а плазма испытывает процесс обезвоживания, обусловленный накопле-
нием гидрофобных коллоидов — жиров и липоидов. Последние разме-
щаются на поверхности плазмы и со стороны контакта ее с вакуолями
и обеспечивают непроницаемость мезоплазмы как с внешней, так и
с внутренней стороны. Благодаря этому клетки индивидуализируются,
а протоплазма не набухает в воде, значительно медленнее обезвожи-
вается, что и определяет морозустойчивость растений в состоянии покоя.
П. А. Генкель подчеркивает, что индивидуализация клеток возникла
в процессе исторического развития как адаптация к неблагоприятным
условиям окружающей среды. Кроме того, в эмбриональных частях ра-
стений в клетках обнаружено во время покой явление контакта между
протоплазмой и ядром.
Некоторые исследователи полагают, что зимний покой является при-
способлением не только к низким температурам, но и ко всему ком-
плексу неблагоприятных условий в зимний период и выражает собой
признак зимостойкости, а не морозоустойчивости (Васильев, 1953; Несте-
ров, 1957). В южных областях с неустойчивыми зимами, с частыми отте-
пелями приспособление растений шло в направлении выработки более
продолжительного периода покоя. На севере, где зимы устойчивые, как
правило, без оттепелей и, следовательно, нет условий для преждевремен-
ного возобновления вегетации, чаще формируются растения с коротким
периодом покоя (Нестеров, 1957). Я. С. Нестеров обращает внимание
на необходимость разграничения понятий глубины покоя и длительно-
сти его. Глубина покоя определяет собой морозостойкость, а длитель-
ность покоя — зимостойкость растений.
Морфологические приспособления — положение почек возобновления
над уровнем поверхности почвы, наличие почечных покровов и другие,
хотя и имеют известное значение в перезимовывании растений (напри-
мер, Падеревская, 1948, 1963), но не играют при этом решающей роли,
как принято было думать под влиянием работ Раункиера (Амелин, 1953;
Голубев, 1956а; и др.). В этой связи интересно отметить, что многие
растения с подземным расположением почек возобновления (геофиты)
трогаются в рост еще с осени и уходят под зиму с ростками и зачаточ-
ными листьями, иногда выходящими на поверхность, что наблюдалось
у Allium oleraceum, Anemone ranunculoides, Corydalis halleri, Gagea
.116
lutea и др. (Irmisch, 1850; Талиев, 1925; Любименко, Вульф, 1926; Лап-
шина, 1928; и др.). Как показывают наблюдения, продолжительность
покоя часто не стоит в прямой связи с морфологической структурой зи-
мующих органов. Например, одни растения, уходящие под зиму с зеле-
ными листьями, обладают длительным органическим покоем (As arum
еигораеит), другие виды в таком же состоянии характеризуются вы-
нужденным покоем и приступают к вегетации сразу же в любые сроки
осенне-зимнего периода, как только для них создаются нормальные усло-
вия тепла и влаги (Achillea millefolium, Filipendula hexapetala, Leucan-
themum vulgare и др.).
Надо отличать у растений возможность развития листового аппарата
при снятии воздействия низких температур в осенне-зимний период от
способности к генеративному развитию при тех же обстоятельствах (об-
разование членов околоцветника, спорогенез и пр.). Эти процессы могут
не совпадать, и растения, образующие листья в отсутствие охлаждения,
в то же время иногда могут быть не способны к формированию генера-
тивных органов, цветению и плодоношению, даже если зачатки генера-
тивных органов были заложены заранее, в предыдущий вегетационный
период.
Как упомянуто выше, различают покой вынужденный и органический.
Последний, в свою очередь, расчленяется по глубине и продолжительно-
сти. Дильс (Diels, 1918) выделил три категории растений по характеру
и длительности зимнего покоя:
1. Растения с /вынужденным периодом покоя, которые при переносе в оранжерею
до заморозков продолжают беспрепятственно вегетировать — Asperula-тип (Asperula
odor ata, Мег cur tails perennis).
2. Растения, характеризующиеся (при внесении в тепло) замедленным ростом с осе-
ни, постепенно усиливающимся к середине зимы, период покоя относительно короткий
и не очень глубокий (Leucojum-тип): Arum maculatum, Ficaria ranunculoides, Leucojum
vernum, Orchis mascula.
3. Растения, рост которых возобновляется во вторую половину зимы и, значит,
период покоя весьма продолжительный и более глубокий (Polygonatum-тип): Anemone
nemorosa, Convallaria majalis, Polygonatum multiflorum и др.
Из приведенной характеристики видно, что даже у растений третьего
типа органический период покоя несколько короче зимнего периода. Раз-
личия в характере зимнего покоя Дильс связывает с эколого-историче-
скими особенностями развития видов. Растения с вынужденным покоем,
по Дильсу, имеют тропическое происхождение, виды Leucojum-типа
в ритме развития отражают средиземноморское происхождение, а с дли-
тельным органическим покоем соответствуют климату Средней Европы.
Опыты Дильса над выяснением особенностей зимнего покоя у нас
были повторены некоторыми исследователями на различных объектах.
Особое внимание привлекала группа ранневесенних растений (Талиев,
1925; Любименко, Вульф, 1926; Кожевников, 1929, 1931, 1939; Трофимов,
1939; Демидова, 1925; и др). Весьма обстоятельно изучен характер зим-
него покоя у растений Западного Закавказья А. В. Кожевниковым
(1937а), видов западно-сибирской флоры — Г. М. Зозулиным, (1949), ра-
стений подмосковных лесов, лугов и болот — И. Г. Серебряковым
(19596).
Как указывает А. В. Кожевников (1937а), местные экологические
условия могут вызывать заметные отклонения в продолжительности ор-
ганического периода покоя. В этом сказывается селектирующая роль
условий местообитания, что может привести к выработке особых экоти-
пов. Возможность изменения ритма развития растений под влиянием
внешних условий отмечалась еще Ю. Визнером (1892). Весьма эффект-
ные различия в сроках цветения в завйсимости от покрытия побегов сне-
гом у орешника описаны В. А. Штаммом (1953).
117
Анализируя реакцию растений на снятие воздействия низких осенне-
зимних температур при внесении их в оранжерею, И. Г. Серебряков
(19596) выделяет следующие группы:
1. Растения, сохраняющие состояние глубокого покоя в течение всей
зимы и последующих летних месяцев. Они нуждаются в пониженных
зимних температурах не только для генеративного развития, но и возоб-
новления роста. У отдельных видов почки трогаются в рост, но обра-
зуются мелкие, иногда уродливые листья, нарастание которых вскоре
прекращается. Цветочные почки отмирают, не раскрываясь. Сюда отно-
сятся: Asarum europaeum, Epilobium montanum, Hieracium umbellatum,
О rob us vernus, Pyrola rotundifolia, Ramischia secunda, Ranunculus cassu-
bicus, Scrophularia nodosa, Viola rupestris и др.
2. Растения, глубокий покой которых прерывается в январе — февра-
ле, когда почки трогаются в рост, вначале медленно (Filipendula ulmaria,
Solidago virgaurea, Thalictrum aquilegifolium, Viola mirabilis и др.). Эта
группа близка к Polygonatum-типу Дильса.
3. Растения с коротким периодом покоя, распускающиеся спустя
1 —1,5 месяца после пересадки, т. е. в середине ноября — начале декаб-
ря. Однако рост в течение зимы происходит медленно, достигая нормаль-
ной скорости лишь с наступлением весны (A egopodium podagraria, Agro-
ру г on caninum, Antennaria dioica, Calamagrostis arundinacea, Campanu-
la persicifolia, C. trachelium, Chamaenerion angustifolium, Ficaria ranun-
culoides, Geranium silvaticum, Melica nutans, Mercurialis perennis,
Ranunculus auricomus, Succisa pratensis, Viola canina и др.). Группа
близка к Leucojum-типу Дильса.
4. Растения, не обладающие органическим периодом покоя, возоб-
новляющие активный рост сразу после пересадки в тепло. Группа соот-
ветствует Asperula-типу Дильса и заключает наибольшее число предста-
вителей флоры Подмосковья: Achillea millefolium, Alopecurus pratensis,
Anthriscus Silvestris, Anthoxanthum odoratum, Betonica officinalis, Briza
media, Bromus benekenii, Calamagrostis epigeios, Carex pallescens,
C. pilosa, Cirsium heterophyllum, Centaurea jacea, Cerastium caespitosum,
Coronaria flos cuculi, Dactylis glomerata, Deschampsia caespitosa, Festuca
ovina. F gigantea, Fragaria vesca, Galium mollugo, G. uliginosum, Geum
rivale, G. urbanum, Glechoma hederacea, Juncus effusus, Lactuca muralis,
Lathyrus pratensis, Leontodon autumnalis, L. hispidus, Leucanthemum
vulgare, Lysimachia, nummularia, Milium effusum, Phleum pratense, Plan-
tago major, P. lanceolata, Poa pratensis, Potentilla erecta, Pulmonaria
obscura, Ranunculus acer, R. repens, Selinum carvifolia, Stellaria grami-
nea, S. holostea, Trifolium medium, T. pratense, T. repens, Urtica dioica,
Veronica chamaedrys, Vicia sepium и др.
В отличие от генетической концепции Дильсй, И. Г. Серебряков
полагает, что в пределах единой по географическому происхождению
группы встречаются растения, резко уклоняющиеся по глубине и про-
должительности осенне-зимнего покоя. Причину разнокачественное™
И. Г. Серебряков видит в более узких эколого-исторических факторах,
обусловливающих тот или иной тип зимнего покоя. С критическим ана-
лизом воззрений Дильса выступали Г. М. Зозулин (1949), М. М. Шик
(1953).
Наше изучение зимнего периода покоя растений лесостепи проводи-
лось в 1962/63 и 1963/64 гг.1 Было привлечено свыше 250 видов луговой
степи, дубовых лесов, дна и северных склонов логов. В 1962 г. растения
были выкопаны и пересажены в ящики в последней декаде октября,
в 1963 г.— во второй декаде сентября. После пересадки ящики с расте-
ниями устанавливались в лаборатории, где постоянно поддерживалась
1 В этих исследованиях нам оказала большую помощь И. В. Голубева.
118
температура 16—18°. Одновременно с переносом растений в лабораторию
фиксировалось состояние, в котором растения находились в момент пе-
ресадки, а затем проводились регулярные наблюдения до осени следую-
щего года.
Наблюдениями за подопытными растениями было выявлено большое
разнообразие индивидуальных особенностей в характере зимнего покоя
отдельных видов и в их реакциях на измененные условия. Вместе с тем
довольно четко наметилась дифференциация растений на основные груп-
пы по глубине и продолжительности зимнего рокоя, что не исключает
различных переходов от одной группы к другой.
1. Наиболее многочисленны растения первой группы, у которых орга-
нический период покоя отсутствует и возобновление роста листьев и по-
бегов происходит вскоре после внесения растений в лабораторию. Сюда
относятся:
Растения луговой степи
Achillea millsfolium
A. nobilis
Agropyron repens
Agrost is syreistschikowii
Ajuga genevensis
Allium oleraceum
Anemone silvestris
Anthoxanthum odoratum
Anthyllis polyphylla
Arenaria serpyllifolia
A. ucrainica
Artemisia absinthium
A. vulgaris
Asperula cynanchica
Aster amellus
Barbarea vulgaris
Berteroa incana
Betonica officinalis
Bromus inermis
B. riparius
Briza media
Bunias orientalis
Bupleurum falcatum
Campanula bononiensis
C. persicifolia
C. rotundifolia
C. sibirica
Carduus hamulosus
Carex humilis
C. michelii
C. praecox
Centaurea jacea
C. marschalliana
C. scabiosa
Cerastium caespitosum
Cichorium intybus
Coronilla varia
Cy no glossuni officinale
Daucus carota
Draba sibirica
Dracocephalum ruyschiana
Echium rubrum
E. vulgare
Erigeron acer
Erysimum marschallianum
Euphorbia subtilis
Falcaria vulgaris
Festuca sulcata
Filipendula hexapetala
Fragaria viridis
Galium boreale
Galium mollugo
Helichrysum arenarium
Helictotrichon pubescens
H. schellianum
Hieracium bauhinii
H. fallax
H, pilosella
Hypericum perforatum
Jurinea arachnoidea
Knautia arvensis
Roeleria gracilis
Lavatera thuringiaca
Leontodon hispidus
Leucanthemum vulgare
Libanotis intermedia
Linaria vulgaris
Linum perenne
Medicago falcata
M. lupulina
Melandrium album
Myosotis suaveolens
Nonea pulla
Oenothera biennis
Onobrychis arenaria
Pastinaca silvestris
Phleum phleoides
Phlomis tuberosa
Pieris hieracioides
Plantago lanceolata
P. media
P. stepposa
Poa angustifolia
Potentilla alba
P. argentea
P. humifusa
P. patula
Primula veris
Prunella grandiflora
P. vulgaris
Ranunculus polyanthemus
Rumex acetosa
R. thyrsiflorus
Salvia pratensis
Scabiosa ochroleuca
Scorzonera purpurea
Sedum acre
Senecio integrifolius
S. jacobaea
Serratula heterophylla
Silene nutans
Stellaria graminea
119
Tanacetum vulgare Taraxacum officinale Thesium ramosum Thymus marschallianus Tragopogon orientate Trifolium alpestre T. montanum T. repens Trinia multicaulis Turritis glabra Valeriana rossica Verbascum lychnitis V. phoeniceum Veronica austriaca V. chamaedrys V. incana V. prostrata Viola hirta V. г и pest r is Viscaria vulgaris
Растения дубовых лесов
Aegopodium podagraria Agrimonia pilosa Allium oleraceum Anthriscus silvestris Arrhenatherum elatius Campanula trachelium Carex spicata Festuca gigantea Fragaria moschata F. vesca Geranium robertianum G. silvaticum Geum urbanum Glechoma hederacea Moehringia trinervia Nepeta pannonica Origanum vulgare Poa nemoralis Polemonium coeruleum Pul monaria angustifolia Pyrethrum corymbosum Stellaria holostea Veronica chamaedrys V. teucrlum Vicia sepium
Растения дна и северных склонов логов
Alopecurus pratensis
Anthoxanthum odoratum
Anthriscus silvestris
Bromus riparius
Calamagrostis epigeios
Carum carvi
Centaurea pseudophrygia
Cnidium dubium
Coronaria flos cuculi
Dactylis glomerata
Dianthus deltoides
Festuca pratensis
Galium aparine
Geranium palustre
Geum rivale
Leontodon autumnalis
Leucanthemum vulgare
Luzula pallescens
Matricaria inodora
Poa palustris
Polemonium coeruleum
Potentilla alba
P. thuringiaca
Ranunculus acer
Stellaria palustris
Succisa pratensis
Trifolium pratense
Veronica longifolia
V. serpyllifolia
Перечисленные растения биологически неоднородны. Различия каса-
ются интенсивности роста новых надземных органов, продолжительности
их жизни, способности зацветать в условиях оранжереи и других при-
знаков.
Некоторые виды характеризуются очень медленным отрастанием над-
земных органов после их перенесения в помещение. Таковы Agropyron
repens, Bromus inermis, Carex praecox, Dianthus deltoides, Falcaria vul-
garis, Festuca sulcata, Geum rivale, Lavatera thuringiaca, Libanotis inter-
media, Nonea pulla, Phleum phleoides, Salvia pratensis, Verbascum
phoeniceum и др. Весьма возможно, что отдельные виды более отвечают
признакам следующей группы с незначительным периодом покоя. Дру-
гие виды отличаются весьма интенсивным ростом новых органов. Тако-
вы Bromus riparius, Centaurea jacea, Cerastium caespitosum, Daucus caro-
ta, Erigeron acer, Geranium robertianum, Poa nemoralis, Pyrethrum
corymbosum, Rumex thyrsiflorus, Stellaria holostea, Turritis glabra и др.
Среднеинтенсивный рост органов наблюдается у Ajuga genevensis, Car-
duus hamulosus, Helictotrichon schellianum, Jurinea arachnoidea, Tanace-
tum vulgare, Veronica prostrata и др.
У Primula veris формируются мелкие листья, подобные листочкам
осенней генерации, и, таким образом, можно говорить о своеобразном
продлении последней фазы морфогенеза (в истекшем вегетационном
периоде). Очень мелкие листья образуют у Salvia pratensis.
120
Эти факты весьма слабого роста возобновляющихся органов, и их не-
нормальные формы указывают на существование внутренних причин,,
вызывающих торможение роста и нарушение типичного хода морфогене-
за. Значит, одно лишь возобновление роста сразу после внесения расте-
ний в тепло не может служить свидетельством отсутствия периода орга-
нического покоя. На это же указывает и неспособность многих растений,,
тронувшихся в рост, к цветению. Очевидно, в оценке состояния зимнего
покоя нужны физиологические критерии. Однако обсуждение этих во-
просов уже выходит за рамки данного исследования.
Совсем неожиданным оказалось поведение довольйо большого числа
растений, у которых после внесения в оранжерею с осени наступает
активизация роста (разной продолжительности у отдельных видов), сме-
няющаяся затем депрессией и отмиранием сформировавшихся листьев
и побегов. Так, в ноябре — декабре отмерли молодые органы у Achy-
rophorus maculatus, Alopecurus pratensis, Bromus inermis, Coronilla
varia, Rnautia arvensis, Linum perenne, Ranunculus repens, в январе —
у Carduus hamulosus, Cynoglossum officinale, Jurinea arachnoidea, Medi-
cago falcata, M. lupulina, Veronica incana; в феврале — у Arrhenatherum
elatius, Artemisia absinthium, Festuca pratensis, Sedum acre, Taraxacum
officinale, Verbascum phoeniceum; в марте — у Agropyron repens, Ajuga
genevensis, Barbarea vulgaris, Centaurea marschalliana, Falcaria vulga-
ris, Helichrysum arenarium, Helictotrichon schellianum, Hieracium fallax,
Koeleria gracilis, Libanotis intermedia, Thymus marschallianus, Ver-
bascum lychnitis, Veronica austriaca, V. prostrata\ в апреле — у Campa-
nula persicifolia, C. sibirica, Centaurea scabiosa, Helictotrichon pubescens,
Hieracium pilosella; в июне — у Festuca sulcata, Succisa pratensis. У та-
ких видов, как Onobrychis arenaria, Scabiosa ochroleuca, возобновивший-
ся рост продолжается у разных особей различное время, иногда прекра-
щается совсем, а молодые листья отмирают. Остальные перечисленные
выше виды, не обладающие органическим зимним покоем, вегетируют
всю зиму и последующий весенне-летний период.
Представляется вполне правдоподобным, что отмирание части пере-
численных видов обусловлено неблагоприятным режимом для жизнедея-
тельности растений (чрезмерная влажность почвы, ненормальное осве-
щение, температура и пр.). Весьма вероятно, что сказывается и обрезка
корневой системы при пересадке. Но возможны и другие причины фи-
зиологического порядка, связанные с отсутствием зимнего охлаждения.
Из растений с вынужденным периодом покоя зацвели в лаборатории
в ноябре Anemone silvestris, Bupleurum falcatum, Centaurea marschallia-
na, Cerastium caespitosum, Draba sibirica, Matricaria indora, в декабре —
Stellaria holostea, в январе — Carum carvi, Daucus carota, в феврале —
Taraxacum officinale, в марте — Geranium robertianum, Glechoma hede-
racea, Silene nutans, в апреле — Anthyllis polyphylla, Potentilla patula*
в мае — Leucanthemum vulgare, Poa nemoralis, Pyrethrum corymbosumr
Stellaria graminea.
2. Вторую группу составляют растения с коротким периодом покоя
(октябрь — ноябрь). К ним относятся Acinos thymoides, Agropyron inter-
medium, Artemisia austriaca, Galium verum, Lotus corniculatus, Peuceda-
num oreoselinum, Salvia verticillata, Sedum telephium, Senecio schwet-
zowii, Stachys recta, Stipa stenophylla, Vicia tenuifolia — растения луговой
степи; Calamagrostis arundinacea, Campanula rapunculoides, Clinopodium
vulgare, Galium vernum, Ranunculus cassubicus — растения дубовых ле-
сов; Deschampsia caespitosa, Lathyrus pratensis, Lysimachia nummularia,
Molinia coerulea — растения логов.
3. К третьей группе отнесены растения с среднепродолжительным
периодом покоя, возобновляющие рост в декабре — январе. Это: Astra-
galus danicus, Delphinium cuneatum, Inula hirta, J. salicina, Iris aphyl-
12k
la, Polygala comosa, Salvia nutans, Stipa pennata — растения луго-
вой степи; Aconitum lasiostomum, Brachypodium pinnatum, Crepis
sibirica, Lilium pilosiusculum, Solidago virgaurea, Thalictrum aquilegifoli-
um, Viola mirabilis— растения дубовых лесов; Antennaria dioica, Poly-
gonum bistorta, Thalictrum lucidum, Th. simplex — растения логов.
4. Наконец, в четвертую группу можно объединить растения с дли-
тельным периодом покоя. К ним принадлежат следующие растения лу-
говой степи: Adonis vernalis, Anthericum ramosum, Antitoxicum officinale,
Pulsatilla patens, Thalictrum minus, Veratrum nigrum', виды дубовых ле-
сов: Asarum europaeum, Bromus benekenii, Carex montana, Convallaria
majalis, Cucubalus baccifer, Epipactis latifolia, Galium rubioides, Orobus
vernus, Paris quadrifolia, Polygonatum officinale, Ramischia secunda,
Scrophularia nodosa, Serratula inermis, Stachys silvatica\ растения логов:
Ficaria verna, Filipendula ulmaria, Fritillaria meleagris, Ranunculus auri-
comus, Sanguisorba officinalis.
У многих ранневесенних растений, характеризующихся длительным
периодом покоя, с осени возможно некоторое увеличение, почек и даже
их раскрытие. Это явление неоднократно отмечалось в литературе (Та-
лиев, 1925; Любименко, Вульф, 1922; Лапшина, 1928; Кожевников, 1929,
1931; и др.). Локальное возобновление роста осенью наблюдается
у Anemone ranunculoides, Corydalis halleri, Fritillaria meleagris, F. ruthe-
nlca, Scilla sibirica. В зимний период у этих видов в почках возобновле-
ния совершается процесс формирования генеративных органов. Отсюда
следует, что в состоянии органического покоя у растений возможно про-
текание важнейших формообразовательных процессов, т. е. развитие
органов полового воспроизведения при полном отсутствии активной ве-
гетации в условиях оранжереи.
Попутно следует отметить, что у большинства растений с длительным
зимним покоем раньше всех (из длительновегетирующих) кончается ве-
гетация осенью и позднее всех начинается весной, исключая, конечно,
группу ранневесенних эфемероидов.
Среди растений с длительным органическим покоем намечается раз-
деление на две подгруппы. К первой отойдут растения, у которых почки
возобновления не раскрываются совсем и к началу лета или позднее
отмирают. Иногда в конце зимы наблюдается некоторое увеличение по-
чек и даже частичное их раскрытие, но затем они все равно погибают.
Таковы Bromus benekenii, Convallaria majalis, часть особей Galium ru-
bioides, Orobus vernus, Paris quadrifolia, Ramischia setunda, Stachys
silvatica, Veratrum nigrum. Это виды с особо консервативным ритмом
развития, для которых воздействие пониженных температур абсолютно
необходимо для жизнедеятельности.
Виды второй подгруппы уже с середины февраля и позднее трогаются
в рост, образуют нормальные ассимилирующие органы и некоторые спо-
собны даже цвести, например Asarum europaeum, Carex montana.
Своеобразно поведение Pulsatilla patens, приступающей к вегетации
в феврале, но к середине июня целиком теряющей все надземные органы.
Так же ведут себя некоторые особи Serratula heterophylla.
Сравнивая состояние зимнего покоя одних и тех же видов лесостепи
с покоем этих же растений в лесной зоне (Подмосковье, данные И. Г. Се-
ребрякова, 19596), можно заметить следующее:
1. Некоторые виды с глубоким покоем без воздействия пониженных
температур способны возобновлять ростовые процессы и даже перехо-
дить к цветению, что наблюдалось у Asarum еугораеит. Этот вид в лес-
ной зоне в отсутствие зимних холодов не только не цветет, но и не спо-
собен к нормальному росту.
2. Намечается тенденция к сокращению периода покоя у растений
-с неглубоким и малопродолжительным в лесной зоне покоем, вплоть до
122
его полного исчезновения {Aegopodium podagraria, Campanula persicifo-
lia, Geranium silvaticum, Succisa pratensis и др.).
3. Среди растений с длительным или среднепродолжительным покоем
установлено «расслоение», заключающееся в том, что часть особей по
длительности и характеру покоя отвечает особенностям главного типа,
а другая часть резко выделяется сокращенным покоем или его полным
отсутствием. Это явление обнаружено у Inula hirta, Solidago virgaurea,
Viola mirabilis и др.
В целом можно заключить, что условия лесостепи действуют в сторо-
ну некоторого ослабления глубины зимнего покоя и сокращения его про-
должительности, в отличие от условий лесной области. Это влияние
соответствует более мягким климатическим особенностям лесостепной
зоны в сравнении с лесной.
Вместе с тем нужно подчеркнуть, что методика выявления зимнего
покоя требует значительного усовершенствования и перестройки. Было
бы целесообразно не травмировать растения при пересадке, использовав
посев семенами с последующим выращиванием растений в ящиках до
половозрелого состояния под открытым небом. Лишь после этого можно
бы ставить опыт без воздействия зимних холодов. Надо уделять внима-
ние и экологическому режиму в теплицах, при котором осуществляется
эксперимент. Интересно провести наблюдение над растениями, получаю-
щими в зимнее время дополнительное освещение- Чрезвычайно важным
представляется проведение многолетних опытов над одними и теми же
видами, позволяющими вскрыть взаимосвязь между метеорологическими
условиями лет наблюдений и состоянием зимнего покоя. Кроме того,
желательно пронаблюдать состояние зимнего покоя у разновозрастных
особей, в том числе специально у вегетативных и генеративных.
ПОДСНЕЖНЫЙ РОСТ И РАЗВИТИЕ РАСТЕНИЙ
Явление подснежного роста и развития травянистых растений при-
влекает внимание натуралистов, в первую очередь, несомненно, в силу
новизны и оригинальности этого биологического феномена. Подснежное
развитие было обнаружено вначале у альпийских растений в высоко-
горьях Европы и Азии (Сапожников, 1901, Кернер, 1903; Келлер, 1914;
Riibel, 1925; Braun-Blanquet, 1928; и др.). Позже факты подснежного
роста и развития горных растений у нас были значительно умножены
(Семенова, 1939; Семенова-Тян-Шанская, 1948; Запрягаев, Запрягаева,
1948; Наринян, 1948, 1960; Кишковский, 1950; Лакиза, 1953; И. М. Куль-
тиасов, 1955; Яшина, 1961а и др.).
Исследованиями в различных ботанико-географических областях
было установлено, что подснежное развитие растений возможно и в усло-
виях равнины. Оно отмечено у растений Крайнего Севера (Тихомиров,
1950; Ковакина, 1952; Кильдюшевский, 1956; и др.), лесолуговой зоны
(Лапшина, 1928; Кожевников, 1929, 1931; Серебряков, 1947; Серебряков,
Галицкая, 1951; Шик, 1953; Трофимов, 1954; Стржемеская, 1953; Сере-
брякова, 1956; Голубев, 19566; и др.), лесостепной и степной полосы
(Келлер, 1921; Прозина, 1949; Устинова, 1949; Прозоровский, 1947;
и Др.).
В более южных районах с периодическим снежным покровом, когда
зимой он может временами полностью или частично разрушаться, пред-
ставляет интерес уже не само явление зимнего, роста и развития, а воз-
можность зимней вегетации, обусловленной наличием зеленых побегов
и периодическим исчезновением снежного покрова. Зимняя вегетация
растений в среднеазиатских пустынях, полупустынях и других типах ра-
стительности отмечалась неоднократно. Литература по этому вопросу
приводилась ранее.
123
Исследователи указывают, что подснежный рост растений стимули-
руется специфическим экологическим режимом. В альпийском поясе гор,
где выпадает много снега, почва бывает талой в течение всей зимы,
а температура припочвенного слоя удерживается в пределах 0°. Сход-
ные условия наблюдаются и в широколиственных лесах, где подснежное
развитие установлено для многих растений (Кожевников, 1929, 1931;
Серебряков, 1947; и др.). Но ряд других фактов свидетельствует о том,
что подснежный рост и развитие растений возможны и в условиях мерз-
лой почвы (Прозина, 1949; Устинова, 1949; Серебряков, Галицкая, 1951;
Трофимов, 1954; Голубев, 19566; и др.). Впервые зимний рост почек
при отрицательных температурах обнаружен у древесно-кустарниковых
растений (Geleznoff, 1851; Викторов, 1941; и др.).
Возможна ли ассимиляция под снегом молодых распустившихся зи-
мой листьев? Для Окончательного решения этого вопроса еще нет досто-
верных данных. На основании косвенных соображений и фактов можно
с достаточной вероятностью утверждать, что значение подснежной
ассимиляции, если она и существует при особо благоприятных условиях,
в общем энергетическом балансе растений умеренно холодного климата
невелика. Между тем другого рода наблюдения позволяют полагать, что
у некоторых растений под покровом глубокого снега ассимиляция отсут-
ствует совершенно. Выход молодых ростков и листочков из-под снега
в этиолированном виде является лучшим тому свидетельством.
Чем более благоприятен термический режим растений под снегом,
тем вероятнее возможность подснежного роста. Казалось бы, что у рас-
тений с глубоким и длительным органическим зимним покоем подснеж-
ное развитие должно отсутствовать. В действительности у этих растений
часто происходят процессы морфогенеза органов полового воспроизве-
дения при крайне замедленном росте. Здесь наблюдается полная анало-
гия с летним покоем некоторых эфемероидов, в течение которого так-
же совершается , формирование генеративных органов (Трофимов,
1952; и др.).
В условиях лесостепи, на территории заповедника, подснежный рост
листьев и побегов регулярно отмечается у растений дубрав: Anemone
ranunculoides, Corydalis halleri, Fritillaria ruthenica, Gagea lutea, Mer-
curialis perennis, Pulmonaria angustifolia, P. obscura, Scilla sibirica, щ. e.
у ранневесенних растений, а также у ряда других: Aegopodium podagra-
ria, Anthriscus silvestris, Lactuca altissima, Melica nutans, Milium effu-
sum, Poa nemoralis, Stellaria holostea, Taraxacum officinale, Trollius
europaeus, Urtica dioica и др.
Весьма часто подснежный рост наблюдается у растений, произраста-
ющих по влажным днищам логов: Alchemilla micans, Alopecurus praten-
sis, Carum carvi, Ficaria verna, Fritillaria meleagris, Gagea minima, Poly-
gonum bistorta, Ranunculus repens, Rumex confertus, R, crispus и др.
Растения луговой степи проявляют заметный зимний рост почек
возобновления и зачаточных листьев лишь в теплые зимы, когда почва
долгое время не замерзает или периодически оттаивает во время зимних
оттепелей. Особенно интенсивный зимний рост у большого количества
видов Стрелецкой степи установлен в зиму 1960/61 г., когда почва со-
хранялась талой в течение всей зимы {Achillea millefolium, Achyropho-
rus maculatus, Anthoxanthum odoratum, Arenaria ucrainica, Barbarea
vulgaris, Bunias orientalis, Carduus hamulosus, Centaurea jacea, Cichori-
um intybus, Dianthus andrzejowskianus, Filipendula hexapetala, Helic-
totrichon pubescens, Primula veris Poa bulbosa, Pulsatilla patens, Ranun-
culus pedatus, Senecio integrifolius, Trinia multicaulis и др.).
Более ценны, конечно, не простые наблюдения за изменением вели-
чины почек и образованием новых листьев, протекающих в зимний пери*
од, а за структурными изменениями в анатомических элементах вегета-
124
тивных и генеративных органов, процессами спорогенеза и морфогенеза
внутри почек. Подобные исследования выполнены на ряде объектов, и не-
которые закономерности изменения анатомо-морфологических структур
в зимующих органах выявлены с достаточной полнотой (Серебряков,
1947; Устинова, 1949; Прозина, 1949; Душечкин, 1951; Трофимов, 1952,
1954; Яшина, 1961а; и др.). Однако дальнейшее развертывание исследо-
ваний в этом направлении, несомненно, таит в себе широкие возможно-
сти познания новых биологических явлений и закономерностей.
Каково же биологическое значение подснежного роста и развития,
оправдывают ли себя те усилия, которые прилагаются к раскрытию сущ-
ности явления, вносят ли они что-либо новое в познание закономерно-
стей развития растений? Чтобы ответить на эти вопросы, необходимо
принять во внимание различия в подснежном развитии растений. На
основании проведенных исследований можно выделить по крайней мере
две группы видов по характеру зимнего изменения органов:
1. Растения, у которых в зимний период происходит формирование
и рост генеративных органов. По данным И. Г. Серебрякова (1947),
у Stellaria holostea почки возобновления в начале зимы еще не содержат
зачаточных цветков, развивающихся позднее — в период с декабря по
февраль. Сходные процессы морфогенеза зимой выявлены у Milium effu-
sum (Серебряков, 1947), Phleum alpinus (Душечкин, 1951). У Scilla
sibirica и Ornithogalum umbellatum в зимний период наблюдается про-
цесс дифференциации спорогенной ткани в цветках и спорогенез (Усти-
нова, 1949; Прозина, 1949).
Развитие генеративных органов и их отдельных элементов в зимний
период представляет чрезвычайно важный этап онтогенеза (морфогене-
за), часто обусловливающий раннее цветение растений. В этом и заклю-
чается существенная роль подснежного развития, составляющего неотъ-
емлемую часть жизненного цикла, определяющего приспособительные
черты растений.
2. Значительно больше растений, у которых в зимнее время происхо-
дит только увеличение размеров (рост) ранее заложенных органов —
листьев и генеративных побегов. В.этом случае зимний рост не вносит
коренных сдвигов в ритм сезонного развития. В лучшем случае он лишь
несколько ускоряет возобновление надземных частей растений.
Таким образом, явление зимнего роста в узком смысле менее суще-
ственно в биологических отправлениях. Вместе с тем возможность под-
снежного роста указывает на другую особенность растений — на харак-
тер их зимнего покоя. Способность к росту свидетельствует о большой
лабильности приспособительной структуры и об отсутствии органическо-
го длительного покоя у растений.
ДЛИТЕЛЬНОСТЬ ЖИЗНИ ПЕРЕЗИМОВАВШИХ ЛИСТЬЕВ
Вопрос о длительности вегетации перезимовавших листьев травяни-
стых растений впервые у нас был поставлен Е. И. Лапшиной (1928), ко-
торая на основании общего анализа перезимовки и конкретных фактов
пришла к заключению, что перезимовавшие зеленые органы разных ви-
дов обладают различным периодом вегетации, зависящим от метеороло-
гических условий и экологических особенностей местообитаний. Обстоя-
тельные исследования в этом направлении проведены И. Г. Серебряко-
вым (1947, 1952, 1961, 1962, 1963) и его учениками и последователями:
Е. Ф. Кошкиной (1954), Г. М. Денисовой (1960а, б), И. Н. Пономаревой
(Былинкиной) (1959), Г. П. Белостоковым (1962), Т. И. Серебряковой
(1956) и др.
В Стрелецкой степи по методике И. Г. Серебрякова работала
М. А. Борисова-Гуленкова (19606), которая, однако, специальных наблю-
125
дений за продолжительностью жизни перезимовавших листьев не прово-'
дил а.
Нашими исследованиями были охвачены многие зимнезеленые виды
луговой степи при косимом и некосимом режиме, дубовых лесов, дна
и северных склонов логов. Сразу после схода снега производилась мар-
кировка сохранившихся зеленых листьев белой несмывающейся краской.
Листья подснежной генерации не принимались в расчет, отмечались
только развившиеся в предыдущий вегетационный период. Выбирались
вегетативные и генеративные особи, отмечавшиеся пронумерованными
этикетками. Повторность применялась 6—10-кратная. После этикетиро-
вания и маркировки систематически, через 2—3 дня, проводился осмотр
растений и все изменения, в сравнении с состоянием после выхода из-под
снега, заносились в полевой журнал. Временем отмирания отдельного
листа считался момент его полного пожелтения или побурения. Но отме-
чались и промежуточные стадии усыхания: с верхушки листа и до сере-
дины пластинки.
В табл. 5 приводятся сроки окончательного отмирания всех перези-
мовавших листьев у подавляющего числа особей взятых под наблюде-
ние 104 видов в исследованных биотопах. Количественные соотношения
видов с отмирающими листьями, суммированных подекадно, отображе-
ны в табл. 6. Из таблиц видно, что в сроках начала, массового и конца
отмирания листьев растений в разных эколого-ценотических условиях
наблюдаются отчетливые различия. Раньше всего происходит полное
отмирание перезимовавших листьев у отдельных видов в ценозах неко-
симой степи. Так, еще в апреле отмирают листья у Dactylis glomerata,
Festuca pratensis, Thymus marschallianus, в то время как, например,
у последнего вида в условиях северного склона лога отмирание листьев
приходится на середину июня. В объяснении указанных различий надо
иметь в виду, во-первых, что у чабреца на некосимой степи зимуют мощ-
ные длинные побеги, стелющиеся по ковру мертвой растительности,
с листьями весенно-летней генерации (прошлого года).'Правда, в пазу-
хах некоторых листьев находятся боковые укороченные вегетативные
побеги, сформировавшиеся в конце прошлого лета. На склоне же коси-
мого лога у чабреца развиваются невысокие облиствленные, с компакт-
ным расположением, побеги позднелетней генерации. Но раннее отмира-
ние листьев чабреца на некосимой степи нельзя отнести только за счет
различий в характере зимующих листьев, так как и здесь побеги несут
листья второй генерации. Быстрое усыхание этих побегов мы объясняем
тем, что после схода снега почва некосимой степи дольше сохраняется
в мерзлом состоянии и к моменту уже весьма яркого солнечного освеще-
ния еще не успевает прогреться. Поэтому корневая система не обеспечи-
вает достаточным количеством влаги надземные побеги, которые должны
интенсивно транспирировать, пригретые солнечными лучами. Водный ба-
ланс нарушается, и листья. отмирают. На северном склоне экологиче-
ская обстановка совершенно иная: нет яркого освещения, оттаивание
почвы происходит более равномерно по мере стаивания снега, кроме
того, у растений меньше листовая поверхность. Все это и обусловливает
длительное функционирование перезимовавших листьев.
Однако и в более поздние сроки, когда почва становится талой,
у ряда видов некосимой степи листья отмирают раньше, чем на косимой.
Таковы Achillea millefolium, Bromus riparius, Filipendula hexapetala,
Prunella vulgaris, Verbascum lychnitis, Veronica chamaedrys. Причина
этого здесь уже кроется исключительно в различиях самих зимующих
листьев. На косимой степи формируется больше зимующих листьев, при-
чем самые молодые из них возникают позднее, в сравнении с самыми
молодыми листьями на некосимой степи, где, таким образом, листьев
и меньше по количеству и они старше по возрасту. Отмеченная законо-
126
Таблица 5
Сроки окончательного отмирания перезимовавших листьев в разных биотопах
(наблюдения 1962 г.)
Вид Степь косимая Степь некосимая Дно и север* ные склоны логов Дубовый лес
Achillea millefolium 24.V 4.V
Achyrophorus maculatus 15.V —
Acinos thymoides 11. VI
Agropyron intermedium 18.V
A. repens 13.V
Alopecurus pratensis 22.V
Anemone silvestris 14.V
Antennaria dioica 18.VI
Anthemis tinctoria 15.V
Anthoxanthum odoratum ....... 30.V
Anthriscus silvestris 22.V
Anthyllis polyphyIla 5.VI
A rtemisia absinthium .......... 15.V
Asarum europaeum 11.VI
Barbarea vulgaris 15.V 26.V
Berteroa incana 25.V
Betonica officinalis 2.VI 6.VI
Bromus riparius 24.V 19.V
Calamagrostis epigeios ........ 14.V 22.V 30.V
Campanula patula — 30.V —
C. persicifolia 5.VI 3.VII
C. rotundifolia — 30.V
C. sibirica 25.V
Carduus hamulosus 25.V
Carex humilis — 31.V
C. michelii 26.VI 30.VI 29.IX
C. spicata — 22.VI 14.VII 29.IX
C. verna — 11. VI —
Centaurea marschalliana ........ 9.V —
Cerastium caespitosum 28.V —
Chelidonium ma jus — 21.V
Coronaria flos cuculi — 30.V —
Cytisus ruthenicus 30.V — —
Dactylis glomerata — 14.IV 22.V 21.V
Deschampsia caespitosa — — 18.VI 16.VI
Dianthus deltoides — — 13.VI
Dr aba sibirica 26.V
Dracocephcdum ruyschiana — — 30.V
Echium rubrum ll.V
Elisanthe viscosa 28.V —
E rysimum marschallianum 28.V 4.V
Festuca pratensis — “ 21.IV —
F. sulcata — 5.VI —
Filipendula hexapetala 11. VI 29.V —
Fragaria moschata — — 11.VI
F. vesca — — 14.VII
F. viridis 24.V 30.V —
Galium mollugo — 15.V —
G. verum — — 16.VI
Geranium robertianum ........ — — — 6.VI
127
Таблица 5 (продолжение)
Вид Степь косимая Степь некоси- мая Дно и север- ные склоны логов Дубовый лес
Geranium sanguineum 30. V —
Geum rivale — — 15.V —
G, urbanum — — — 6.VI
Glechoma hederacea — — — 31.VIII
Helichrysum arenarium 9.V 19.V — —
Hieracium bauhinii 30.V — —
H. pilosella 26.VI — 1.VII —
H. pratense — — — 26.VI
Hypericum perforatum — 15.V ' —
J uncus effusus ^ ... . — — 26.VII
Jurinea arachnoidea 15.V — —
Knautia arvensis 14.V — — -
Leucanthemum vulgare 26.V — 30.V 21.VII
Linum perenne 18.V — — —
Lysimachia nummularia — — 22.VI —
Myosotis sparsiflora — — . 30.V
M. suaveolens 9.V — .
Nonea pulla 28.V — —
Onobrychis arenaria 18.V —
Phleum phleoides ". — 28.V —
Plantago lanceolata 28.V — — —
P. media 28.V —
Poa angustifolia — ll.VI — —
Polemonium coeruleum — — 26.V
Poly gala comosa 24.V — —
Potentilla alba 24.V —
P. ar gentea 24.V — —.
P. humifusa 24.V — — —
P. thuringiaca — — 15.V —
Primula veris ll.V — —
Prunella vulgaris 16.VI 31.V 6.VI —
Ranunculus auricomus — — 30.V
R. polyanthemus — 26.V — —.
R. repens ' — — 22.V —
Rumex acetosa 24.V — —
Scabiosa ochroleuca 29.V — 30.V
Senecio jacobaea 16.VI — —.
Silene nutans — — 14.VII
Stellaria graminea 24.V 19.V —
S. holostea — — 10.V
Succisa pratensis — — 26.V —
Thymus marschallianus — 14. IV 18.VI —
Tri folium pratense ll.V — — —
T. repens 9.V — — —
Turritis glabra 25.V — — —
Verbascum lychnitis 15.VII 11.VII —
Veronica chamaedrys 2.VI 28. V 30.V U.VI
V. incana 30.VI — —
V. prostrata 26.VI — — —
Vicia sepium — — 14.VII
V. tenuifolia — 30.V — —
Viola hirta 28.V — —
V. rupestris 15.VIII — — —
Viscaria vulgaris — 6. VI 22.V —
128
мерность ясно выступает и в общих количественных соотношениях видов
с отмирающими листьями в подекадном распределении (табл. 6). Во II
и III декадах мая на некосимой степи наблюдается самое интенсивное
отмирание листьев у почти равного количества видов, между тем как на
косимой степи максимум видов с усохшими листьями приходится на
III декаду мая. Подобная же закономерность отмечается и для логов.
Таким образом, в открытых ценозах большая часть видов теряет пере-
зимовавшие листья в III декаду мая. Довольно значительно количество
видов с отмирающими листьями еще во II декаду мая, а также в июне.
Таблица 6
Количественное распределение видов с отмирающими листьями по декадам
вегетационного периода (наблюдения 1962 г.)
Конец июня — это и есть, в основном, предел жизнедеятельности пере-
зимовавших листьев у большинства видов косимой и некосимой степи,
дна и северных склонов логов. Только у единичных видов данных био-
топов перезимовавшие листья сохраняются дольше. На косимой степи
это будет Verbascum lychnitis, Viola rupestris, отмирание листьев кото-
рых соответственно падает на II декаду июля и II декаду августа.
В этот же срок усыхают перезимовавшие листья Verbascum lychnitis
и на некосимой степи.
В логах последними растениями, у которых перезимовавшие листья
дольше всех сохраняют жизнеспособность, являются Carex spicata,
Hieracium pratense и Juncus effusus. Отмирание листьев у этих видов,
с соблюдением той же последовательности, происходит в I, II, III декады
июля.
Резкие отличия в длительности жизни перезимовавших листьев на-
блюдаются у травянистых растений дубовых лесов. Во-первых, макси-
мум отмирания листьев приходится на вторую декаду июня. У других
видов листья сохраняют зеленую окраску в июле (Campanula persicifo-
lia, Fragaria vesca, Leucanthemum vulgare, Silene nutans, Vicia sepium),
в III декаде августа (Glechoma hederacea) и даже в III декаде сентября
(Carex michelii, C. spicata). Поздние сроки отмирания перезимовавших
листьев здесь обусловливаются, несомненно, более умеренным режимом
физических факторов, характеризующихся меньшей интенсивностью и от-
сутствием резких скачков. Не случайно поэтому, что в широколиствен-
ных лесах еще и по сей день сохранились вечнозеленые травы — прямые
потомки вечнозеленых обитателей третичных лесов.
Таким образом, продолжительность жизни перезимовавших листьев
v разных видов и в разных экотопах колеблется в весьма широких преде-
лах: от одной декады, т. е. почти сразу после схода снега, до нескольких
месяцев, чаще всего до двух, что соответствует концу мая — началу
июня. Раннее отмирание определенно свидетельствует о том, что пере-
зимовавшие листья не играют заметной роли в ранневесенней ассими-
ляции и их перезимовку в зеленом виде нельзя рассматривать как
9 Ь. Н. Голубев
129
приспособление к условиям сокращенного вегетационного периода. На-
против, продолжительное пребывание перезимовавших листьев в зеленом
состоянии, без сомнения, указывает на их существенное значение в асси-
миляции.
Нам осталось отметить разногодичные изменения в продолжительно-
сти жизни перезимовавших листьев. Некоторых моментов этого вопроса
мы уже касались при обсуждении характера перезимовки растений
в разные зимы. Вполне естественно, что при резких колебаниях величи-
ны зеленой зимующей поверхности будет соответственно и пропорцио-
нально изменяться длительность жизни перезимовавших листьев. Тем
более очевидны эти колебания в случае внезапной гибели перезимовав-
ших листьев вскоре после схода снега или в период его таяния под влия-
нием неблагоприятных метеорологических условий. Изменения в дли-
тельности жизни могут возникать и в результате тех или иных сдвигов в
ритме развития и морфогенезе растений в прошедшем вегетационном пе-
риоде. Но, как уже подчеркивалось ранее, заметное варьирование по го-
дам в перезимовке и продолжительности жизни перезимовавших
листьев наблюдается у ограниченной группы растений, способ перези-
мовки которых еще строго не стабилизировался.
Значительно больше растений с более или менее устойчивой формой
перезимовки и продолжительностью жизни перезимовавших листьев.
И мы вполне разделяем мнение Е. И. Лапшиной (1928) о том, что в раз-
ные зимы «растения перезимовывают более или менее одинаково»
(стр. 84). В подтверждение этому можно сослаться на совпадение сроков
отмирания перезимовавших зеленых листьев в 1962 и 1961 гг. у следую-
щих растений Стрелецкой степи: Achillea millefolium, Campanula persi-
cifolia, Elisanthe viscosa, Hieracium pilosella, Leucanthemum vulgare,
Plantago lanceolata, Polygala comosa, Potentilla humifusa, Senecio jaco-
baea, Turritis glabra, Veronica chamaedrys, V. incana и др. (режим со-
хранения степи — косимый). Аналогичное совпадение в сроках отмира-
ния перезимовавших листьев установлено за те же годы и для растений
дубовых лесов {Fragaria moschata, Geum urbanum, Veronica chamaedrys
и др.). Обращаясь к перечисленным растениям, можно заметить, что все
они являются типично зимнезелеными с хорошо развитой и довольно
большой зеленой зимующей поверхностью листьев.
Несомненный интерес представляет сопоставление характера перези-
мовки одних и тех же видов в различных ботанико-географических зо-
нах. В этом отношении особенно ценна работа Е. И. Лапшиной (1928),
выполненная в лесолуговой зоне и охватывающая большое количество
видов. Многие из них присущи лесостепным ценозам и, таким образом,
были включены в сферу наших наблюдений. Сравнение открывает уди-
вительное сходство в способах перезимовки одних и тех же видов, про-
израстающих в лесной и лесостепной зонах. Этот факт свидетельствует
о значительной устойчивости признака наличия или отсутствия зеленых
органов в зимний период, что лишний раз говорит в пользу развиваемой
нами мысли о генетической сущности зимнего состояния растений.
Некоторые расхождения в данных сводятся к следующему. Е. И. Лап-
шина к летнезеленым растениям относит Anthoxanthum odoratum, Alope-
curus pratensis, Carex leporina, Epilobium palustre, Melica nutans, Phleum
pratense, Viola canina, которые трактуются нами в качестве зимнезеле-
ных. Впрочем, Е. И. Лапшина в отношении последних трех видов допу-
скает возможность развития зачаточных листьев, зимующих в зеленом
виде. Именно эта особенность и характерна для указанных видов. Что
касается остальных, то несовпадение данных лежит уже в пределах
ошибки и не имеет принципиального значения.
Наоборот, в состав зимнезеленых видов зачислены Agropyron repens,
Роа nemoralis, а также Aegopodium podagraria, Anthriscus Silvestris,.
130
Carutn carvt, Centaurea jacea, Leontodon autumnalis, Pulmonaria obscura,
Ranunculus acer, которые у нас фигурируют как летнезеленые. Из этих
видов Роа nemoralis обладает подснежным развитием, к моменту схода
снега его дерновинки имеют зеленый вид. Видимо, это обстоятельство
и послужило причиной отнесения Роа nemoralis к растениям, зимующим
с зелеными органами. Относительно других перечисленных видов
Е. И. Лапшина сообщает, что их редкие зеленые листья мало устойчивы
к морозам и только при более благоприятных условиях сохраняются зе-
леными и перезимовывают. Но такое свидетельство вполне отвечает
и нашим наблюдениям. Значит, расхождения здесь также непринципи-
альные и касаются лишь таксономии.
Таким образом, следует отметить удивительное тождество в способах
перезимовки многих видов, обитающих под Ленинградом и Курском,
т. е. в очень несходных эколого-ценотических условиях.
Глава 6
Ритмы цветения растении
НОРМАЛЬНОЕ ЦВЕТЕНИЕ
В изучении сезонного развития растений и растительных сообществ
ритмам цветения отводится исключительно важная роль. Цветение каж-
дого вида, как правило, происходит в наиболее благоприятном для дан-
ного вида сочетании факторов внешней среды и вместе с другими эле-
ментами сезонного развития несет на себе печать тех эколого-историче-
ских условий, в которых протекало его формирование.
Исследованиями И. Н. Оловянниковой (1949), А. Н. Пономарева
(1959, 1960а, б, в, г, 1961а, б) и других установлена высокая специфич-
ность ритмов цветения отдельного цветка, особи и популяции растений,
характеризующихся определенной экологией. Ритмы цветения даже си-
стематически близких форм могут заметно отличаться по суточному
ритму и срокам цветения (А. Н. Пономарев, 1959). Наши наблюдения
в Стрелецкой степи показали, что популяционный состав некоторых ви-
дов неоднороден: встречаются ранние и поздние фенологические формы с
весьма значительными сдвигами в ритмах цветения. Числящаяся во фло-
рах Festuca sulcata Hack. (Алехин, 1940; Левицкий, 1957) оказалась
представленной двумя формами — F. valesiaca Koch, и F. pseudovina
Hack., ритмы цветения которых различаются в сроках начала и окон-
чания. Ранние и поздние феноформы (по срокам цветения) свойственны
популяции Роа angustifolia.
Из ритмов цветения отдельных видов слагается ритмика цветения
растительных сообществ. Ее изучение достигается аналитическим и син-
тетическим способами. В аналитическом исследовании важная роль от-
водится выявлению аспектов и составлению фенологических спектров.
Данные о ритмах цветения компонентов сообществ позволяют вы-
вести кривые цветения фитоценозов, которые дают возможность пред-
ставить ритмику цветения обобщенно — в количественном выражении,
что очень важно для сравнительной оценки ценозов и познания особен-
ностей сезонного развития.
Первые исследования аспективности сообществ относятся еще к се-
редине XIX века. Аспективность степных ценозов у нас в числе первых
изучали Г. И. Танфильев (1898), В. В. Алехин (1909, 1925а, б, 1934),
И. К. Пачоский (1910, 1917), М. С. Шалыт (1930), Е. М. Лавренко
(1940) и др. В дальнейшем указание на главные аспекты и их последо-
вательность становится неотъемлемым элементом геоботанического
описания растительности. В Центрально-Черноземной области большой
вклад в изучение аспектов степей и истории их происхождения помимо
В. В. Алехина сделан Н. С. Камышевым (1939, 1946, 1948, 1949, 1951,
1953а, 1956, 1958; и др.).
Особенности аспектов, связанные с цветением обильно развитых ви-
дов, их разнообразие и последовательная смена во времени являются
весьма существенной характеристикой ценозов. Для Стрелецкой степи
132
Рис. 11. Ранневесенний аспект горицвета (Adonis vernalis) на Стрелецкой степи
В. В. Алехиным (1934, 1944) выделено до 11 —12 аспектов. Главные из
них, обусловленные цветением массово представленных растений, сле-
дующие:
1. Pulsatilla patens, Carex humilis (спустя несколько дней после снеготаяния);
2. Adonis vernalis (рис. 11), Hyacinthella leucophaea (конец апреля — начало мая);
3. Primula veris (первая половина мая);
4. Orobus pannonicus, Iris aphylla, Anemone silvestris (середина мая);
5. Myosotis suaveolens, Senecio integrifolius, Ranunculus polyanthemus (конец мая);
6. Salvia pratensis, Tragopogon orientate (первая половина, середина июня); в неко-
торые годы бывает весьма характерный аспект Stipa pennata (рис. 12);
7. Trifolium montanum, Leucanthemum vulgare (рис. 13), Filipenaula hexapetala;
к этим основным аспектабельным видам примешиваются Campanula sibirica, С. persi-
cifolia, Knautia arvensis, Echium rubrum и др. (вторая половина июня);
8. Onobrychis arenaria, Galium verum (рис. 14) (первая половина июля);
9. Delphinium cuneatum (вторая половина июля);
10. Veratrum nigrum (рис. 15) (конец июля, начало августа).
На примере Стрелецкой и других степей лугового типа можно убе-
диться, что они отличаются высокой фенологической насыщенностью.
На лугах к северу от лесостепной зоны и в настоящих степях — к югу
аспективность травянистых сообществ убывает.
Исследованиями установлено, что аспективность Стрелецкой степи
подвержена разногодичным колебаниям. Однако общее количество ас-
пектов и аспектабельных видов в многолетнем ряду изменений остается
весьма постоянным. Изменение аспектов по годам связано, с одной сто-
роны, с колебаниями метеорологических условий, а с другой — с перио-
дичностью цветения, присущей не только древесным, но и травянистым
растениям. Таким образом, разногодичные изменения аспективности
представляют закономерное, а на Стрелецкой степи — и первичное яв-
ление, отнюдь не свидетельствующее о какой-то неустойчивости, сбор-
ности, вторичности ее фитоценозов.
Попытки аналитического изучения ритмов развития компонентов со-
обществ привели к созданию фенологических спектров. Приоритет со-
ставления первого спектра принадлежит Гамсу (Gams, 1918), который
назвал его фено-экологическим. Развитие каждого вида изображается
133
Рис. 12. Аспект ковыля перистого (Stipa pennata) на Стрелецкой степи между первым
отвершком Петрина лога и урочищем «Бабка» в 1963 г. (в начале июня)
Рис. 13. Аспект нивяника (Leucanthetnum vulgare) на Стрелецкой степи
Рис. 14. Среднелетний аспект подмаренника настоящего (Galium verum) на Стрелецкой
степи
Рис. 15. Позднеледаий аспект чемерицы черной (Уeratrum nigrum) на Стрелецкой степи
в виде полосы, наибольшая ширина которой соответствует обилию вида
во время цветения. Гаме допускал раскрашивать полосу так, чтобы от-
разить особенности аспекта, создаваемого данным видом.
Принципы построения спектров по Гамсу были использованы
С. И. Даниловым (1937), И. С. Амелиным (1939) и др.
В нашей стране, независимо от Гамса, графическое изображение рит-
мов развития растений разработал А. П. Шенников (1928). Этот метод
получил широкое распространение как у нас в стране, так и за рубежом.
У А. П. Шенникова график сезонного развития вида также представлен
в виде полосы разной ширины с разной штриховкой и окраской. Он по-
зволяет видеть степень распространения вида, количественные соотно-
шения между генеративными и вегетативными особями, фенологическое
развитие. Вместе с тем фенологический спектр сообщества, состоящий
из графиков развития входящих в него видов, дает представление о всем
ходе фенологического развития сообщества, в том числе и об аспектив-
ности. Для пойменных лугов автором установлено шесть аспектов (ста-
дий развития).
А. П. Шенников писал, что «в интересах фитосоциологии... необхо-
димо фенологические наблюдения над видами внутри сообщества пре-
вратить в средство для фенологического изучения самого сообщества.
Для этого необходимы сопоставление «видовых фенологий» друг с дру-
гом и какое-то их суммирование и обобщение, имеющее целью предста-
вить ход развития сообщества наряду с анализом его отдельных стадий»-
(стр. 5). Метод А. П. Шенникова использовался и используется широким
кругом исследователей.
М. С. Шалыт (1946) предлагает некоторое обобщение методов Гам-
са и Шенникова и изображает спектр в виде полос, ширина которых из-
меняется в соответствии с изменением площади покрытия видов на про-
тяжении вегетационного периода.
В. В. Алехин (19256, 1936) в характеристике фенологического разви-
тия растений и сообществ применил целую систему фенологических знач-
ков, обозначающих фазы, типы фенологического развития и степень уча-
стия видов в аспектах.
Дальнейшим этапом в углублении изучения сезонного развития рас-
тений и его графического изображения является метод, разработанный
И. Г. Серебряковым (1947). Графики сезонного развития, предложенные
И. Г. Серебряковым, позволяют видеть:
I. В вегетативной сфере:
1) появление, рост и последующее отмирание' ассимилирующих органов; 2) число
генераций листьев в течение года; 3) продолжительность жизни листьев каждой гене-
рации; 4) наличие или отсутствие листьев в течение зимнего периода; 5) относительную
площадь листьев, остающихся на зиму; 6) характер почек возобновления; 7) время за-
ложения зеленых листьев в почках возобновления.
II. В-репродуктивной сфере:
1) время заложения соцветий и зачатков цветков в почках возобновления; 2) на-
чало и продолжительность цветения; 3) соотношение цветения и развития листьев;
4) время созревания плодов и обсеменения.
Важнейшим средством обобщения аналитического материала по рит-
мам развития, в том числе и цветения, доставляемого рассмотренными
выше методами, позволяющего в количественной форме представить фе-
нологическое развитие сообщества, является метод фенологических кри-
вых, в частности — кривых цветения.
Первенство в обосновании нового метода принадлежит В. Н. Хитрово
(1910), развернувшему исследования в этом направлении начиная с
1901 г. Однако имя В. Н. Хитрово, к сожалению, было незаслуженно за-
быто в этой области ценофенологии. Еще в 1910 г. на 12-м съезде рус-
136
ских естествоиспытателей и врачей В. Н. Хитрово говорил, что оформ-
ление фенологических наблюдений за ритмами цветения в виде кривых
цветения всех компонентов сообщества является «могущественным мето-
дом биологического анализа элементов формаций и характеристикой по-
следних в разных ботанико-географических зонах» (Хитрово, 1910г
стр. 394).
В. Н. Хитрово лично и вместе с членами своей семьи и сотрудниками
были собраны обширнейшие фенологические материалы по различным
растительным сообществам лесостепи за период почти в 25 лет. Только
самая малая доля материалов увидела свет в нескольких публикациях
А. И. Молозева (1925, 1927).
В 1925 г. В. Н. Хитрово сообщает, что надеется в скором времени
выпустить специальную работу с изображением на графиках и анали-
зом всего хода цветения в сообществах более 600 видов по наблюдениям
с 1901 г. Однако это намерение осталось неосуществленным.
По В. Н. Хитрово (1925), кривая цветения сообщества выводится пу-
тем подсчета числа цветущих видов за каждый день. Сам автор, к со-
жалению, не опубликовал примеров кривых цветения растений и сооб-
ществ. Однако созданный им метод синтетического изучения ритмики
цветения сообществ нашел широкое распространение.
Особенно популярным метод составления кривых цветения становит-
ся после выхода в свет работы А. П. Шенникова (1928), в которой впер-
вые в литературе приведены кривые цветения луговой растительности
на фоне кривых изменения температуры воздуха. Автор указывает, что
кривые цветения можно строить и на фоне кривых изменения влажности
почвы и воздуха, температуры почвы и т. д. Вслед за этим появляется
работа А. В. Кожевникова (1929), где приводится кривая цветения под-
пологовых растений широколиственного леса. В дальнейшем метод по-
строения кривых цветения сообществ прочно входит в ботанические ис-
следования растительности (Семенова, 1939; Серебрякова, 1956; Дени-
сова, 19606; Рещиков, 1961; и др.).
А. М. Семенова (1939) расширяет возможности графического изобра-
жения фенологического развития и приводит для субальпийских лугов.
Юго-Осешии кривые бутонизации, отмирания побегов, созревания пло-
дов. А. П. Балаш (1948, 1955, 1961а, б) при анализе ритмики цветения
приазовских степей использует графики, отображающие изменение ко-
личества зацветающих и цветущих видов по декадам вегетационного пе-
риода. Такой метод составления кривых имеет ряд преимуществ, в осо-
бенности для выявления закономерностей изменения количества зацве-
тающих видов, так как суммирование их за декаду стабилизирует харак-
тер кривой и делает ее более показательной, что нельзя сказать о кривых
зацветающих видов, основанных на ежедневном учете. Более четкую
форму приобретает и кривая общего количества цветущих видов по де-
кадам. Автором этих строк использован метод составления подекадных
графиков изменения числа зацветающих и цветущих видов А. П. Бала-
та с добавлением данных (третьей кривой) о количестве отцветающих,
по декадам видов (Голубев, 19626). Кроме того, ритмика цветения сооб-
ществ характеризуется с помощью указанных графиков двояко: с одной
стороны, кривые цветения выводятся для всего видового состава сооб-
щества, а с другой — для основных компонентов. Этим способом выяв-.
ляется более закономерный характер ритмики цветения ценозов.
Классификация ритмов цветения
Для количественной оценки растительности по ритмам цветения ком-
понентов необходима четко разработанная классификация, допускающая
сопоставимость данных по разным ценозам. Разумеется, для разных
13Г
ботанико-географических областей нельзя принять одну и ту же класси-
фикацию. Системы ритмов должны быть региональными.
В отношении растений умеренной полосы Евразии уже давно приме-
няется классификация ритмов, основанная на распределении цветения
по сезонам вегетационного периода. Так, выделяются ранневесенние,
раннелетние, летние и другие группы растений по ритмам цветения (Ко-
жевников, 19376; Трофимов, 1939). Следует отметить, что в системе
А. В. Кожевникова ритмы цветения характеризуются весьма обобщенно
и расплывчато, а в классификации Т. Т. Трофимова наблюдается неко-
торое несоответствие в названиях групп с календарным делением веге-
тационного периода.
Используя принцип календарного выражения продолжительности и
сроков цветения, мы детализировали отдельные категории ритмов, в свою
очередь объединяя их в соответствующие циклы. Строго календарное
разделение вегетационного периода на общепринятые сезоны года по-
зволяет установить единообразие в определении содержания и таксоно-
мических категорий ритмов, что создает прочную основу для сравнения
ценозов из близких ботанико-географических зон и внутри них по рит-
мам цветения компонентов.
В предлагаемой классификации ритмы цветения растений лесостепи
объединяются в шесть основных циклов в соответствии с периодами теп-
лого времени года I Внутри основных выделяются элементарные циклы.
Наиболее целесообразной классификация растений центральной лесо-
степи по ритмам цветения представляется в следующем виде:
I. Растения весеннего цикла цветения Л
Ранневесенние (1)1 2: Carex humilis, Corydalis halleri, Pulsatilla patens и др.
Средневесенние (2): Fritillaria ruthenica.
Поздневесенние (3): Antennaria dioica, Carex hirta, C. pallescens, C. spicata, Luzula
pallescens и др.
Ранне-средневесенние (1—2): Anemone ranunculoides, Carex verna, Viola mi r a bi-
lls и др.
Средне-поздневесенние (2—3): Anemone silvestris, Barbarea vulgaris, Carex miche-
lii, Orobus pannonicus, Ranunculus pedatus и др.
Весенние (ранне-поздневесенние) (1—3): Adonis vernalis, Draba nemorosa, Primula
veris, Viola rupestris и др.
II. Растения в e с e н н e - л e т н e г о цикла цветения
Ранневесенне-раннелетние (1—4): Glechoma hederacea, Taraxacum officinale.
Средневесенне-раннелетние (2—4): Alyssum calycinum, Cytisus ruthenicus, Myosotis
suaveolens, Thesium ebracteatum и др.
Средневесенне-позднелетние (2—6): Nonea pulla.
Поздневесенне-раннелетние (3—4): Anthoxanthum odoratum, Carum carvi, Euphor-
bia semivillosa, Iris aphylla, Jurinea arachnoidea и др.
Поздневесенне-среднелетние (3—5): Berteroa incana, Stellaria graminea, Poly gala
comosa и др.
Поздневесенне-позднелетние (3—6): Alopecurus geniculatus.
III. Растения летнего цикла цветения
Раннелетние (4): Bromus riparius, Festuca sulcata, Filipendula hexapetala, Tragopo-
gon orientate, Trifolium montanum, Stipa pennata и др.
Среднелетние (5): Agropyron intermedium, Calamagrostis epigeios, Campanula bo-
noniensis, Stipa capillata и др.
1 Здесь принимаются следующие сроки конца периодов вегетационного сезона:
ранневесенний — 5—10 мая, средневесенний—15—20 мая, поздневесенний — 25 —
31 мая, раннеле^ний — 30 июня, среднелетний — 31 июля, позднелетний — 31 августа,
раннеосенний—10—15 сентября, среднеосенний — 25 сентября, позднеосенний — время
наступления устойчивых отрицательных температур.
2 Для удобства подсчетов каждому элементарному циклу присваивается цифровое
обозначение, пропорционально разделению теплого времени года на 9 периодов.
138
Позднелетние (6): Artemisia absinthium, A. latifolia, A. vulgaris, Polygonum hydro-
piper и др.
Ранне-среднелетние (4—5): Antitoxicum officinale. Campanula persicifolia, Leucan-
themum vulgare, Onobrychis arenaria, Phlomis tuberosa и др.
Средне-позднелетние (5—6): Allium flavescens^ Delphinium cuneatum, Helichrysum
arenarium, Veratrum nigrum и др.
Летние (ранне-позднелетние) (4—6): Achillea millefolium, Campanula glomerata,
Medicago falcata, Scabiosa ochroleuca и др.
IV. Растения летне-осеннего цикла цветения
Раннелетне-раннеосенние (4—7): Agrimonia eupatoria, Geranium palustre, Gypsophi-
la muralis (все виды в логу).
Раннелетне-среднеосенние (4—8): Leontodon autumnalis (в логу).
Раннелетне-позднеосенние (4—9): Achillea millefolium (в логу).
Среднелетне-раннеосенние (5—7): Achillea nobilis, Crepis pannonica, Daucus caroia,
Eryngium planum, Sanguisorba officinalis и др.
Среднелетне-среднеосенние (5—8): Inula britannica (в логу), Leontodon autumnalis
(в степи), Serratula inermis (в лесу) и др.
Среднелетне-позднеосенние (5—9): Malachium aquaticum.
Позднелетне-раннеосенние (6—7): Artemisia scoparia, Odontites serotina, Sedum te-
le phium и др.
Позднелетне-среднеосенние (6—8): Aconitum nemorosum, Hieracium umbellatum,
Ji. virosum, Senecio erucifolius, Succisa pratensis и др.
V. Растения осеннего цикла цветения
Ранне-среднеосенние (7—8): Linosyris vulgaris.
VI. Растения весенне-летне-осеннего цикла цветения
Ранневесенме-позднеосенние (1—9): Viola tricolor.
Можно было бы и дальше дифференцировать систему ритмов цве-
тения, выделяя внутри элементарных циклов полные и неполные вариан-
ты. Например, среди раннелетних растений имеются виды, цветущие в
первой и второй половине июня, в середине июня и в течение всего меся-
ца. Таким образом, в элементарном цикле раннелетних растений наме-
чается четыре варианта, из которых первые три — неполные, а четвер-
тый — полный, соответствующий элементарному циклу. Чтобы, однако,
не перегружать количественные характеристики ценозов по ритмам цве-
тения, мы исключаем из учета варианты, оставляя в качестве основных
единиц элементарные циклы, система которых приведена выше.
Сроки цветения растений по годам не остаются постоянными, что вно-
сит известные затруднения в определение ритмов цветения в рамках
приведенной системы. Там, где это было возможным, мы использовали
средние многолетние сроки зацветания и отцветания и только для неко-
торых видов — 3—4-летние наблюдения.
Ввиду обширности материала анализ его дается ниже для всех основ-
ных растительных группировок центральной лесостепи. Этим мы не-
сколько нарушаем принятый порядок, при котором анализ группировок
вынесен в VII главу. Но в целях соблюдения известной «симметрии» в
VII главе оставляется только разбор таблиц, в которых сведен количе-
ственный и процентный состав растений по ритмам цветения. Остальные
же данные помещаются в этой главе.
Обзор фенологического материала
В Центрально-Черноземном заповеднике, организованном в 1935 г.,
накоплены значительные материалы по фенологии степных и лесных тра-
вянистых и древесно-кустарниковых растений. По отдельным видам
имеются фенонаблюдения за 10—17 лет. Этого количества лет наблю-
дений за одними и теми же видами вполне достаточно, чтобы вы-
вести достоверные средние многолетние даты регистрировавшихся
139
фенологических фаз (начала, массового и конца цветения, начала и мас-
сового созревания плодов и семян). Возможно также определить крайние
сроки прохождения фенофаз в различные годы, т. е. амплитуду колеба-
ний фенологических явлений.
Выведение средних многолетних сроков цветения и плодоношения
растений и амплитуды колебаний фенофаз чрезвычайно важно для по-
знания закономерностей сезонного развития растительных сообществ.
Оно имеет и прямое практическое значение: для прогнозирования сроков
сбора плодов и семян древесно-кустарниковых и других полезных расте-
ний, в том числе декоративных, лекарственных, ароматических, для
обоснования сроков сенокошения и т. д. Обобщение многолетних фено-
логических наблюдений и накопление нового фактического материала
составляет основу для изучения ритмов развития растений, что создает
предпосылки для эколого-исторического анализа флоры и раститель-
ности.
Обратимся к краткому обзору литературы. В пределах современной
территории заповедника первые фенологические наблюдения поставлены
на Стрелецкой степи В. В. Алехиным (1909). В 1907 г. он посетил ее
11 раз в различные сроки вегетационного периода и установил последо-
вательную смену красочных аспектов, обусловленных массовым цвете-
нием обильно развитых растений. Яркое художественное описание аспек-
тов Стрелецкой степи, данное В. В. Алехиным, получило широкую изве-
стность и вошло во многие сводки и учебники. Сложная и вместе с тем:
необычайно динамичная смена аспектов стала рассматриваться в каче-
стве одного из важнейших признаков растительности луговых степей.
Поэтому внимание исследователей к изучению аспективности курских:
заповедных степей привлекалось неоднократно (Покровская, 1940; Ка-
ден, 1940; Зозулин, 1955; Голубев, 1960е).
Надо, однако, отметить, что увлечение аспектами наряду с больши-
ми положительными имело и некоторые отрицательные последствия:
работа по кропотливому изучению ритмов развития растений была орга-
низована в недостаточных размерах, подчас нерегулярно. Этим, по-види-
мому, и объясняется то обстоятельство, что в «Летописи природы»
заповедника имеются данные фенонаблюдений лишь по 50 видам степ-
ных растений, 29 видам травянистых растений дубовых лесов и 23 видам
древесно-кустарниковых растений. Конечно, для познания особенностей
фенологического развития растений луговых степей и дубовых лесов та-
кого количества видов явно недостаточно. Исключением является лишь
работа Н. Н. Кадена и Н. В. Гарри (1942), в которой приводятся фено-
материалы по цветению 106 видов Казацкой степи в одном 1936 г.
Интересные сведения по ритмике цветения растений Стрелецкой сте-
пи содержатся в статье И. Н. Оловянниковой (1949). Однако большая
часть полученных материалов осталась неопубликованной. В последнее
время появилась обстоятельная работа о ритмах развития растений
Стрелецкой степи М. А. Борисовой-Гуленковой (19606). Но данные о
продолжительности цветения, изображенные графически, имеют суще-
ственный недостаток в том, что оказывается неизвестным, к какому году
они относятся. Автор отмечает, что наблюдения проводились в 1952 и
1953 гг., а приводятся они обобщенно, без указаний конкретных сроков.
Между тем по погодным условиям эти годы очень резко отличались друг
от друга, что не могло не отразиться на ритмах цветения. Неясно также,
в условиях какого режима сохранения степей проводились наблюдения.
Перечисленным и исчерпывается почти весь круг печатных работ, по-
священных ритмам цветения растений заповедника.
Анализ литературы показывает, что изученность ритмов цветения
растений, а тем более созревания плодов и обсеменения в целом для
заповедника совершенно недостаточна. Дело не меняется и вследствие
140
опубликования статей А. М. Семеновой-Тян-Шанской (1962) и В. В. Ста-
риковой (1963) ; публикаций же по фенологии растений, произрастающих
на соседних с заповедником территориях в пределах Курской области,
нет вообще.
Насколько нам известно, в других областях черноземного центра фе-
нологические наблюдения по срокам опубликованы Н. С. Камышевым
(1954) для Каменной степи; кроме того, уже упоминались выше обшир-
ные наблюдения В. Н. Хитрово в Орловской области за природными
растениями в разных ценозах. Учитывая крайнюю скудость фенологи-
ческих данных о растениях естественных ценозов, мы сочли целесообраз-
ным обобщить весь фактический материал по фенологии растений запо-
ведника, находящийся в «Летописи природы», ряд рукописных материа-
лов и все остальные самостоятельно добытые феноданные.
Наибольшей продолжительностью характеризуются наблюдения над
50 видами Стрелецкой степи, в которых принимали участие Н. Жучков
(1939—1940 гг.), 3. Шкуратенко (1941 г.), И. Н. Оловянникова (1945—
1949 гг.), С. С. Левицкий (1955—1959 гг.), И. В. Голубева (1960 г.),
В. Н. Голубев и В. С. Жмыхова (1961 —1962 гг.). На основании этих
наблюдений нами выведены средние многолетние даты цветения и пло-
доношения (приложения I и II).
Аналогичные фенонаблюдения за этимй же видами проводились в
Казацкой степи в течение семи лет. В них участвовали Н. Гарри
(1936 г.), О. В. Даева (1937 г.) и Г. И. Рыльский (1950—1954 гг.). Об-
общенные средние многолетние данные по результатам указанных на-
блюдений приведены в приложениях III и IV.
Обработаны материалы фенонаблюдений и высчитаны средние мно-
голетние даты начала, массового и конца цветения, начала и массового
созревания плодов и семян 23 видов древесно-кустарниковых и 29 видов
травянистых растений дубовых лесов Стрелецкого и Казацкого участ-
ков заповедника (приложения V, VI, VII и VIII). Сбор исходных данных
осуществлен Н. Жучковым (1940, 1941 гг.), Г. И. Рыльским (1951—•
1954 гг.), С. С. Левицким (1958—1959 гг.), И. В. Голубевой (I960—
1962 гг.).
Приступив к работе в заповеднике в 1959 г., мы поставили целью
значительно расширить фенонаблюдения и провести их над возможно
большей частью видов Стрелецкого участка заповедника, включая все
виды Стрелецкой степи, дубовых лесов, дна и склонов логов. Для выяс-
нения влияния на ритмы развития растений режима заповедности и усло-
вий культуры были организованы фенонаблюдения за одними и теми же
видами на косимых1 и постоянно некосимых участках Стрелецкой степи
и в питомнике. Наибольшей результативности удалось достигнуть в 1961
и 1962 гг. Наблюдения в природных ценозах заповедника за ритмами
развития растений проводились В. С. Жмыховой под нашим руковод-
ством и при непосредственном участии. Фенонаблюдения за развитием
р,астений в питомнике осуществляла И. В. Голубева.
Исчерпывающая полнота видового состава сообществ, над которым
проводились фенонаблюдения, позволила построить фенологические по-
декадные графики, отображающие изменение количества зацветающих,
цветущих и отцветающих видов на протяжении вегетационного периода.
Составление фенологических графиков по ритмам цветения растений
разных ассоциаций в экологических рядах и типов растительности в
разрезе различных зон страны чрезвычайно важно для выявления эко-
лого-географических закономерностей этих явлений и представляет
1 Для завершения полного цикла фенологических наблюдений вегетационного перио-
да на степи с сенокосным режимом сохранения выделялись участки, которые в год на-
блюдений оставлялись нескошенными до конца теплого сезона. Эти участки, во избе-
жание влияния временного некошения, ежегодно менялись.
141
определенную научно-практическую ценность. Весьма важным дополне-
нием к этому является выделение фенологических констант
растительности, основанных на средних многолетних данных по ритмам
цветения главных компонентов. Графическое изображение фенологи-
ческих констант придает им наглядность и создает удобство в географи-
ческих сопоставлениях разных типов растительности. Последнее, однако,
осуществимо лишь при условии проведения длительных стационарных
фенологических наблюдений за основными компонентами изучаемых со-
обществ. В этом отношении заповедники с их разнообразными зональ-
ными типами растительности, не подвергающимися хозяйственному воз-
действию, представляют исключительную возможность, к сожалению
используемую еще далеко недостаточно. Насколько нам известно, в бота-
нической литературе пока нет подобных данных по фенологическим кон-
стантам, которые нами выведены для растительности Стрелецкой степи
на основе обобщения многолетних фенонаблюдений за главными компо-
нентами (137 видами).
Перед тем как перейти к анализу полученных материалов, хотелось
бы несколько слов посвятить вопросам методического характера, касаю-
щимся фенонаблюдений. Последние в 1961 —1962 гг. проводились в наи-
более типичных условиях развития избранных формаций (луговой сте-
пи, дубовых лесов и растительности логов). За начало цветения прини-
малось состояние, когда появлялось несколько цветущих растений, но
не менее 5—10. Массовое цветение, как правило, у большинства видов
охватывает определенный промежуток времени той или иной продолжи-
тельности. Мы фиксировали лишь даты начала вступления растений в
пору массового цветения.
При определении длительности цветения в расчет принималось толь-
ко нормальное цветение. Впрочем, в разграничении нормального и вто-
ричного цветения в ряде случаев встречаются значительные затруднения.
Это касается в первую очередь таких видов, как Achillea millefolium,
Potentilla argentea, Scabiosa ochroleuca, Thymus marschallianus, Trifo-
lium pratense и др.
Ритмы цветения растений
Стрелецкой и Казацкой степей
Сопоставление средних многолетних дат начала, массового и конца
цветения одних и тех же видов, произрастающих на Стрелецкой и Ка-
зацкой степях, открывает следующую картину (см. приложения, I и III).
По подсчетам, около половины изучавшихся видов проходит фенофазы
цветения на Казацкой степи на несколько дней раньше, чем на Стрелец-
кой. Но это ускорение развития весьма незначительное — в пределах
1—3 суток, очень редко до 5 суток или выше.
Выделяя основную особенность фенологического развития растений
Казацкой степи, следует обратить внимание на многочисленные исклю-
чения из общего правила. Последние могут быть подразделены на сле-
дующие категории (в порядке сравнения с ритмами цветения растений
Стрелецкой степи):
1. Все фазы цветения наступают позднее (Centaurea scabiosa, Libanotis intermedia,
Linum perenne, Myosotis suaveolens, Valeriana rossica).
2. Начало и массовое цветение наступают позднее, а конец — раньше (Н elictotrichon
pubescens, Campanula persicifolia)
3. Начало и массовое цветение наступают раньше, а конец — позже (Festuca sul-
cata, Trifolium pratense).
4. Зацветание начинается раньше, массовое цветение совпадает, а заканчивается
позднее (Нyacinthella leucophaea). '
5. Растения с более поздними сроками начала и конца цветения, но с совпадаю-
щими сроками массового цветения (Clematis tecta, Verbascum lychnitis).
142
6. Начало и конец цветения наблюдаются раньше, а массовое — совпадает (Fraga-
ria viridis, Iris aphylla).
7. Совмещается во времени начало цветения, массовое происходит позднее, а ко-
нец цветения наступает раньше (Pulsatilla patens}.
8. Начало и массовое цветение (или только начало) наступают раньше, а конец
цветения совпадает во времени (Carex humilis, Campanula sibirica, Salvia pratensis,
Trifolium montanum).
9. Начало и массовое цветение наступают позднее, а конец цветения совпадает во
времени (Bromus riparius).
В перечисленных отличиях, как и в основном, расхождения в сроках
очень невелики. Можно, конечно, допустить, что отдельные различия в
цветении растений наших степей обусловлены недостаточно продолжи-
тельным рядом лет наблюдений в Казацкой степи. Весьма вероятно, что
с увеличением количества лет некоторые отклонения сгладились бы.
Иначе говоря, случайные расхождения уступили бы место отличиям
закономерного характера. Однако вряд ли справедливо все перечислен-
ные случаи отличия ритмов цветения растений Казацкой и Стрелецкой
степей относить на этот счет. Опыт показывает, что средние многолетние^
полученные за 7—9 лет наблюдений за фазами цветения, весьма мало
изменяются или вовсе не изменяются при добавлении к ряду новых лет
наблюдения и при выведении средних из большего количества лет. Для
Казацкой же степи мы имеем, как правило, 7-летний ряд наблюдений.
На этом основании будет более правдоподобным полагать, что Казац-
кая и Стрелецкая степи являются территориально обусловленными уча-
стками одного и того же типа растительности — луговых степей, что их
особенности в ритмике цветения не выходят за рамки этого типа.
Решающая роль в создании отличий экологического режима рассматри-
ваемых степей, по всей вероятности, принадлежит рельефу. Хорошо
известно, что большая часть территории Стрелецкой степи представляет
уклоны северной экспозиции, тогда как Казацкая степь имеет более
выраженную южную ориентацию. Некоторую роль могут играть и эдафи-
ческие факторы: на Казацкой степи глубже залегает граница смены на-
носов, чаще и ближе к поверхности выходят пески.
Таким образом, анализ фенологических данных позволяет заключить,
что Стрелецкую и Казацкую степи нельзя принимать за различные кате-
гории луговостепной растительности и тем более усматривать принад-
лежность их к разным типам растительности: к остепненным лугам —
с одной стороны, а с другой — к луговым степям.
Чтобы окончательно решить этот вопрос (в плане фенологии) было
бы интересно провести одновременные наблюдения за избранными наи-
более типичными видами на Казацкой степи и участках Стрелецкой се-
верной и южной экспозиции. Последние прекрасно выражены в восточ-
ной части Стрелецкой степи — от гребня водораздела к Хвощеву логу
или между Петриным лесом и Химиной лощиной. Можно не сомневаться,
что ритмика цветения растений этих участков будет приближаться, а воз-
можно, и совпадать с ритмикой цветения растений Казацкой степи.
Ритмы цветения растений Стрелецкой степи
в условиях режима сенокошения
Особую наглядность в выявлении особенностей цветения компонентов
растительности представляют подекадные графики числа зацветающих,
цветущих и отцветающих растений. Подобные графики мы составили как
для всего видового состава Стрелецкой степи (рис. 16—17), так и для
основных компонентов (рис; 18—19) по материалам фенонаблюде-
ний в 1961 и 1962 гг. К их анализу мы возвратимся несколько позже.
Рассмотрим график фенологических констант цветения растений Стре-
лецкой степи (рис. 20). Этот график составлен на основании средних
143:
Рис. 16. Кривые цветения 282 видов Стрелецкой степи в условиях сенокосного режима
(1961 г.)
1— зацветающие; 2 — цветущие и 3 — отцветающие по декадам виды; 1-й ряд чисел — количество
зацветающих видов; 2-й ряд—количество цветущих видов, 3-й ряд—количество отцветающих
видов. Римские цифры — декады
Рис. 17. Кривые цветения 281 вида Стрелецкой степи в условиях сенокосного
режима (1962 г.). Обозначения те же, что на рис. 16
Рис. 18. Кривые цветения основных компонентов (131 вида) растительности
Стрелецкой степи в условиях сенокосного режима (1961 г.).
Обозначения те же, что на рис. 16.
Рис. 19. Кривые цветения основных компонентов (131 вида) растительности
Стрелецкой степи в условиях сенокосного режима (1962 г.).
in
। и м । и hi
август сентябрь
ю л
и
и
и
ю н
и
а
in
ь
in
й
I N Ж
апрель
Рис. 20. Кривые цветения основных компонентов (137 видов) растительности
Стрелецкой степи, по данным средних многолетних сроков прохождения фаз
цветения (константный график). Обозначения те же, что на рис. 16.
многолетних дат начала и конца цветения 137 видов1 — основных цено-
зообразователей Стрелецкой степи, или, как мы их называем, основных
компонентов. Из рис. 20 видно, что кривые зацветающих, цветущих и
отцветающих видов имеют одновершинный характер. Первые цветущие
растения появляются во II декаде апреля, последние — во II декаде сен-
тября. Максимум зацветающих видов приходится на I декаду июня,
цветущих — на II декаду и отцветающих на III декаду июня. Кривая
цветущих видов довольно симметрична, т. е. плавный подъем кривой сме-
няется подобным же спуском. Столь же симметричны левые и правые
части кривых зацветающих и отцветающих видов.
Мы называем этот график константным на том основании, что сред-
ние многолетние сроки начала и конца цветения основных компонен-
тов — величины вполне определенные, а значит и обусловленные ими
формы кривых являются стабильными. С этой точки зрения указанные
фенологические константы отражают характерные черты раститель-
ности, определяемые природными условиями данной ботанико-географи-
ческой зоны. В будущем, когда будут найдены аналогичные константы
для других типов растительности, можно будет более обоснованно судить
о положении курских луговых степей среди смежных типов травянистой
растительности.
Теперь, когда охарактеризованы константные показатели ритмов цве-
тения, следует обратиться к анализу кривых зацветающих, цветущих и
отцветающих видов Стрелецкой степи за 1961 и 1962 гг. (см. рис. 16—17).
Сравнение показывает, что распределение числа зацветающих, цветущих
и отцветающих видов по декадам вегетационного периода в отдельные
годы испытывает известные колебания. Наибольшее сходство с констант-
ными показателями ритмов цветения обнаружили данные наблюдений
1961 г., когда все три максимума, т. е. наибольшие количества зацветаю-
щих, цветущих и отцветающих видов совпали во времени — соответ-
ственно приходились на I, II и III декады июня (см. выше). Аналогич-
ные сведения о распределении во времени зацветающих и цветущих
видов приводятся А. М. Борисовой-Гуленковой (19606). Правда, как уже
отмечено выше, из статьи автора нельзя вывести, за какой год использо-
ваны феноданные при вычерчивании кривых.
Наблюдения 1962 г., характеризовавшегося очень теплой погодой ап-
реля и прохладными июнем и июлем, показали, что максимум зацве-
тающих видов переместился на III декаду мая, а число цветущих (от
всего набора видов)—на III декаду июня. В графике для основных
компонентов произошло смещение максимума зацветающих видов на
III декаду мая и максимума цветущих — на I декаду июня (рис. 19).
Из других особенностей графиков цветения в отдельные годы, со-
ставленных для всего видового богатства Стрелецкой степи, можно отме-
тить следующее. В количестве зацветающих видов помимо абсолютного
(второго) максимума наблюдается менее выраженный (первый) макси-
мум, обычно приходящийся на апрель. Кривая отцветающих видов носит
многовершинный характер. Подъем кривой цветущих видов более крутой
в сравнении со спуском. По годам заметные сдвиги наблюдаются и в
сроках начала и конца цветения, то выходя в пределы марта и октября,
то сужаясь в границы апреля и сентября.
В годы с поздней весной, холодным и влажным летом максимум чис-
ла зацветающих видов будет перемещаться на II—III декады июня,
1 В расчете средних многолетних дат отмеченных видов использованы наши четы-
рехлетние наблюдения (1959—1962 гг.), средние многолетние данные фенонаблюдений
за 50 видами Стрелецкой степи, которые почти все относятся к основным ценозообра-
зовятелям (см. приложение), феноданные за 1955 г. С. С. Левицкого, охватившего
в этот сезон помимо 50 постоянных еще 66 видов, многие из которых являются основ-
ными компонентами, а также четырехлетние наблюдения И. Н. Оловянниковой (1945—
1948 гг.) за 20 видами (не входящими в список основных 50).
146
а максимум цветущих видов — на III декаду июня — I декаду июля.
Именно такими чертами отличаются кривые цветения растений Стре-
лецкой степи по наблюдениям в 1945 г.1 (рис. 21). Из рис. 21 видно,
что максимум зацветающих видов падает на II декаду июня, а цвету-
щих— на I декаду июля. По данным Т. И. Серебряковой (1956), кривые
цветения луговых растений в Дединовской пойме р. Оки (прирусловая
и центральная части) характеризуются точно такими же показателями,
Рис. 21. Кривые цветения некоторых растений (74 видов) Стре-
лецкой степи (1945 г.). Обозначения те же, что на рис. 16.
что и ритмика цветения растений Стрелецкой степи в 1945 г. Таким обра-
зом, можно заключить, что во влажные холодные годы с поздней весной
ритмика цветения растительности луговых степей приближается к рит-
мике цветения лугов лесной зоны.
Сравнивая приведенные кривые цветения за 1961—1962 гг. с подоб-
ным графиком для степей донского Приазовья, составленным А. П. Ба-
лашом (1961а), легко убедиться, что характер цветения растений Стре-
лецкой степи в 1962 г. напоминает таковой растений южных степей. Так,
максимум зацветающих видов падает на III декаду мая. Смещение же
максимума цветения у нас на III декаду июня объясняется, по-видимо-
му, прохладной погодой этого месяца. В южных степях пик кривой цве-
тущих видов приходится на II декаду июня (Балаш, 1961а). Т. И. Сере-
брякова и А. П. Балаш отмечают, что приведенные ими графики зацве-
тающих и цветущих видов характеризуют типичные черты цветения со-
ответствующих ценозов.
Обобщая сказанное, можно утверждать, что ритмика цветения рас-
тительности Стрелецкой степи по годам испытывает заметные колеба-
ния, в сухие годы с ранней и теплой весной приобретая черты цветения
растительности южных степей, а во влажные и холодные годы с поздней
весной приближается к цветению луговых сообществ лесной зоны. Одна-
ко во всей сложности разногодичных отклонений ритмика цветения рас-
тительности Стрелецкой степи отличается специфическими особенностя-
ми, находящими выражение в фенологических константах цветения.
Эти константы характеризуют зональные особенности курских луговых
степей.
Ритмы цветения растений Стрелецкой степи
в условиях абсолютной заповедности
Несмотря на то, что в ряде работ имеются указания на отличия в ас-
пектах косимых и некосимых участков заповедных степей (Прозоров-
1 В составлении графика мы использовали фенонаблюдения 74 видов 1945 г., заим-
ствованные из рукописи И. Н. Оловянниковой «Экология и ритм цветения растений
Стрелецкой степи», находящейся в научной библиотеке Центрально-Черноземного за-
поведника. По свидетельству И. Н. Оловянниковой, 1945 г. был влажным и холодным,
среднегодовая температура воздуха ниже многолетней на 1,5°, весна была поздняя
и холодная, лето — холодным и дождливым.
10*
147
ский, 1940а; Зозулин, 1955; Камышев, 1956, 1958), конкретные данные
о ритмах цветения растений в этих условиях в литературе отсутствуют.
Между тем такого рода сведения, безусловно, заслуживают серьезного
внимания.
Мы располагаем двухлетними сравнительными наблюдениями за фе-
нологией цветения и плодоношения растений на абсолютно заповедных
и косимых участках Стрелецкой степи. Основная масса видов изучалась
на некосимом участке № 1 между урочищами Дедов Веселый и Дубро-
шиной, по соседству велись наблюдения за этими же видами в условиях
режима сенокошения.
Анализ фенологических данных показывает, что большинству видов
на некосимой степи свойственно некоторое запаздывание сроков про-
хождения фаз цветения, как правило, в пределах 2—7 суток. Только
очень немногие виды отличаются в ритмах цветения на более значитель-
ные сроки. Так, например, цветение Asperula cynanchica на некосимой
степи продолжительнее на 8—15 суток в сравнении с косимой степью,
но, повторяем, таких видов немного, и весьма вероятно, что большие
различия в сроках цветения обусловлены особого рода причинами.
В этой общей закономерности фенологического развития растений не-
косимой степи находят проявление особенности ее экологического режи-
ма, неоднократно отмечавшиеся в литературе (Прозоровский, 1940а;
Афанасьева, 1948; Семенова-Тян-Шанская, 1960а, б). Но в ритмах цве-
тения некоторых видов некосимой степи наблюдаются исключения из об-
щего правила, не столь уже редкие.
Весьма значительно число видов, характеризующихся более или ме-
нее одинаковыми ритмами цветения на косимой и некосимой степи
(Allium oleraceum, Artemisia absinthium, A. vulgaris, Berteroa incana,
Bromus riparius, B. inermis, Euphorbia subtilis, Festuca pratensis, Pri-
mula veris, Pulmonaria angustifolia, Ranunculus illyricus, Scorzonera pur-
purea, Stipa pennata, Thesium ebracteatum, Veratrum nigrum, Vicia tenui-
folia и др.). Интересную группу образуют виды, запаздывающие в начале
цветения, а оканчивающие его одновременно с видами на косимой степи
(Astragalus danicus, Carex verna, Centaurea jacea, C. scabiosa, Inula
hirta, Leontodon hispidus, Orobus pannonicus). Прямую противополож-
ность представляют Barbarea vulgaris, Betonica officinalis, Brachypodium
pinnatum, Campanula persicifolia, Carex praecox, Peucedanum lubimen-
koanum, цветение которых начинается одновременно, а конец запазды-
вает по сравнению с косимой степью. Есть также виды, зацветающие
раньше, а оканчивающие цветение позднее (Bupleurum falcatum, San-
guisorba officinalis) и, наоборот, начинающие цвести позже при более
раннем отцветании (Linum flavum). Встречаются и другие вариации
в ритмах.
Обращает на себя внимание то обстоятельство, что одни и те же виды
в разные годы по-разному реагируют на внешние условия, создавая чрез-
вычайно сложную мозаику индивидуальных вариаций в ритмах цвете-
ния в одном и том же биотопе. Все это свидетельствует о чрезвычайно
многообразных связях ритмов развития с условиями существования,
о своеобразном влиянии определенных комплексов факторов, приходя-
щихся в разные годы на различные отрезки вегетационного периода.
Каждый вид индивидуален в своих реакциях на внешние условия и их
различные комбинации. Долголетние ряды параллельных наблюдений
за ритмами цветения растений косимой и некосимой степи могут доста-
вить ценнейший материал для познания экологических особенностей
растений.
Сопоставим теперь графики цветения растительности некосимой степи
с косимой, вычерченные по фенологическим данным за 1962 г. (рис. 22).
Сравнение показывает, что кривые для всех видов некосимой и косимой
148
степи чрезвычайно близки, если не сказать тождественны. Максимум
зацветающих видов приходится на III декаду мая, цветущих — на III де-
каду июня. Вполне объяснимо смещение первого небольшого подъема
кривой зацветания (в сравнении с косимой степью) на III декаду апреля,
что обусловлено более поздним стаиванием снега, прогреванием почвы
О 6 8 5 17 45 38 26 20 16
О Ь 14 18 JI 73 100 104 107 100
0 1 4 3 II 22 17 23 17
। II Hi । Il Hi ) и ill •
апрель май июнь и
16 9 2 1 0 » О
99 84 68 53 38 21 /5 2 1 0
24 18 16 15 18 6 13 1 1 0
и ill I и ill I il in l и
юль август сентябрь
Рис. 22. Кривые цветения 210 видов Стрелецкой степи в условиях
абсолютной заповедности (1962 г.). Обозначения те же, что на рис. 16.
и началом вегетации. В III декаду июня одинаково с косимой степью
наблюдается увеличение числа отцветающих видов, правда не абсолют-
ное. Если обратиться к графику для основных компонентов некосимой
степи (рис. 23), то и здесь обнаруживается высокая степень сходства.
О 1 2 6 16 8 II 11 14 9 5 5 Ю 2 ' 2 О
I II III । II III I II III i II 111 I II HI । II Hl •
апрель май июнь и юл ь август сентябрь
Рис. 23. Кривые цветения основных компонентов (103 видов) Стрелецкой степи
в условиях абсолютной заповедности (1962 г.).
Обозначения те же, что на рис. 16.
Поэтому следует полагать, что ритмы цветения растений некосимой
и косимой степи имеют весьма незначительные отличия, а количествен-
ные соотношения в ритмике цветения растительности степи в обоих ва-
риантах режима заповедности довольно близкие, позволяющие думать,
что нет принципиальных различий в ритмике цветения, которые бы корен-
ным образом разграничивали степи с сенокосным режимом использова-
ния и абсолютным заповедным.
149
Ритмы цветения луговостепных растений
в условиях культуры
На питомнике Центрально-Черноземного заповедника собрана кол-
лекция, насчитывающая 180 видов, преимущественно луговой степи. Пи-
томник расположен в плакорных условиях Стрелецкого участка. Расте-
ния высажены на делянках в 1 м2 каждая. Наблюдения проводились,
как правило, за хорошо развитыми прижившимися растениями. Отдель-
ные виды в годы наблюдений были представлены молодыми особями,
выращенными из семян, и их фенологическое развитие несколько отли-
чалось от более взрослых особей.
Сравнение ритмов цветения растений в условиях культуры с теми же
видами на косимых участках Стрелецкой степи дало возможность выде-
лить целую серию групп растений, отличающихся определенными осо-
бенностями цветения в питомнике. Группы эти следующие:
1. Ритмы цветения тождественны или весьма близко совпадают (Carex humilis,
Centaurea jacea, Festuca sulcata, Potentilla patula, Rumex thyrsiflorus, Senecio integrifo-
lius, Stipa pennata, Veratrum nigrum).
2. Все фазы цветения — начало, массовое и конец — наступают раньше. Самая мно-
гочисленная группа, насчитывающая до 35 видов (Allium oleraceum, Anthericum ramo-
sum, Bfomus inermis, Carex praecox, Dactylis glomerata, Delphinium cuneatum, Dianthus
deltoides, Dracocephalum ruyschiana, Filipendula hexapetala, Helictotrichon pubescens,
Hieracium pilosella, Hypericum perforatum, Inula hirta, Iris aphylla, Jurinea arachnoidea,
Leucanthemum vulgare, Peucedanum oreoselinum, Phleum phleoides, Polygala comosa, Po-
lygonatum officinale, Potentilla argentea, Pulsatilla patens, Scorzonera purpurea, Sedum
acre, Silene nutans, Verbascum lychnitis, Veronica teucrium, Viola rupestris, Viscaria vul-
garis и др.).
3. Все фазы цветения наступают позже — на 1—2 и большее число суток (Achillea
millefolium, Artemisia austriaca, Campanula glomerata, Centaurea scabiosa, Draba sibi-
rica, Erysimum marschallianum, Galium boreale, G. verum, Genista tinctoria, Phlomis
tuberosa, Potentilla humifusa, Stachys recta, Tragopogon majus, Veronica incana и др.).
4. Сроки начала цветения совпадают, но цветение оканчивается раньше (Clematis
recta, Onobrychis arenaria, Veronica austriaca).
5. Сроки начала цветения совпадают, цветение оканчивается позже (Leontodon his-
pidus, Myosotis suaveolens, Scabiosa ochroleuca, Serratula heterophylla).
6. Цветение начинается раньше, сроки конца цветения совпадают (Arenaria ucrai-
nica, Carex michelii, Libanotis intermedia).
7. Цветение начинается позже, сроки конца цветения совпадают (Achyrophorus
maculatus).
8. Цветение начинается позже, а оканчивается раньше (Acinos thymoides, Adonis
vernalis, Agrostis syreistschikovii, Anemone Silvestris, Centaurea marschalliana, Lotus
corniculatus, Falcaria vulgaris, Primula veris).
9. Цветение начинается раньше, оканчивается позже (Asperula tinctoria, Aster amel-
ias, Campanula sibirica, Dianthus andrzejowskianus, Knautia arvensis, Lavatera thurin-
giaca, Linum nervosum, L. perenne, Medicago falcata, Poa angustifolia, Potentilla alba,
Ranunculus polyanthemus, Sanguisorba officinalis, Veronica spicata).
10. Менее определенные отклонения в ритмах цветения, к тому же в разные годы
проявляющиеся по-разному.
Можно было бы, конечно, с большей или меньшей вероятностью мо-
тивировать причины всех этих отличий в цветении растений в условиях
культуры. Но это слишком бы далеко увело нас от основной темы. Изла-
гая весь этот материал, мы тем самым хотели подчеркнуть огромное
значение эксперимента в распознавании экологической природы расте-
ний, дающего, как можно было убедиться, весьма неожиданные резуль-
таты.
Хорошо известно, сколь гетерогенна по своему происхождению и
экологическим особенностям формирования флора центральной лесосте-
пи вообще и заповедника в частности (Зозулин, 1955; Дохман, I960).
В выяснении конкретных условий исторического развития видов далеко
недостаточно одного лишь ареалогического анализа. Необходимо при-
влечение разнообразных сведений, в числе которых экологическим свой-
ствам должно уделяться самое серьезное внимание.
150
Рис. 24. Кривые цветения луговостепных растений (119 видов) в условиях
культуры (1960 г.). Обозначения те же, что на рис. 16.
I Н III I II III I II III I II III
апрель май июнь июль
। и in i и in i и in i
август сентябрь октябрь
Рис. 25. Кривые цветения луговостепных растений (123 видов) в условиях
культуры (1961г.). Обозначения те же, что на рис. 16.
/5
6
20
5
2
20
О
.11
U
и
14
III
й
4
III
август
। и hi
апрель
о
in । и
сентябрь
/4
а
ь
Рис. 26. Кривые цветения луговостепных растений (160 видов) в условиях
культуры (1962 г.). Обозначения те же, что на рис. 16.
Наши сравнительные наблюдения за ритмами цветения растений в
природных условиях и культуре еще не всегда проводились удовлетво-
рительно. Но цель этого небольшого раздела — преимущественно мето-
дического характера и заключается в том, чтобы показать, как много
может дать эксперимент в познании реакций растений на внешние фак-
торы, их скрытых потенциальных возможностей индивидуальных откло-
нений. Дальнейшее совершенствование методики сравнительных наблю-
дений и накопление многолетних данных по ритмике цветения представ-
ляет нужную и интересную работу в общем комплексе ботанических ис-
следований заповедника.
Коллекции питомника содержат все основные виды луговой степи.
Поэтому мы решили вычертить графики цветения растений в культуре
и сопоставить их с аналогичными кривыми косимой степи. Для состав-
ления графиков использованы фенонаблюдения за 1960—1962 гг.
(рис. 24—26). Как видно из рисунков, цветение растений в 1960 и 1961 гг.
было очень сходно между собой и с характером цветения косимой степи
в 1961 г. (см. рис. 16).
Более внимательного анализа заслуживают кривые цветения расте-
ний в питомнике в 1962 г. (рис. 26). Абсолютный максимум зацветающих
видов приходится на III декаду мая, т. е. как на косимой степи. После
резкого падения кривой зацветания в I декаду июня наблюдается но-
вый— второй подъем во II декаду июня. В этой связи интересно напом-
нить, что точно такую же форму имеет кривая зацветающих видов для
степей донского Приазовья (Балаш, 1961а). На II же декаду июня при-
ходится и максимум общего числа цветущих видов. Таким образом, цве-
тение луговостепных растений в питомнике в 1962 г. наряду с чертами,
свойственными особенностям цветения растений в естественных условиях
косимой степи, приобретает ряд признаков, характеризующих фенологи-
ческое развитие южных степей. Вместе с тем интересно подчеркнуть на-
личие сходного изменения ритмики цветения растений в условиях куль-
туры и косимой степи под влиянием погодных условий.
Ритмы цветения травянистых растений
дубовых лесов
Какие-либо специальные сведения о ритмах цветения травянистых
растений дубовых лесов заповедника в литературе отсутствуют. Нам по-
этому кажется своевременным обобщить фенонаблюдения разных лет и
вывести средние многолетние даты начала, массового и конца цветения,
начала и массового плодоношения растений, за которыми велись наблю-
дения. К сожалению, таких видов в общей сложности оказалось около
30 (см. приложения VII, VIII).
Обратимся к анализу кривых цветения травянистых растений дубо-
вых лесов за 1961 и 1962 гг. (рис. 27—30). Рассмотрим вначале цветение
всего видового состава травянистых растений (рис. 27—28). Кривая
зацветающих видов носит многовершинный характер. Первый весьма
низкий подъем кривой цветения наблюдается в апреле и в зависимости
от погодных условий в разные годы может смещаться в ту или иную сто-
рону. Второй подъем кривой цветения за оба года наблюдений прихо-
дится на III декаду мая. Абсолютный максимум зацветающих растений
отмечается в июне, причем в 1961 г. он зарегистрирован во II декаду,
а в 1962 г. — в III декаду июня. Последние зацветающие виды обнару-
жены в III декаде августа.
Общее количество цветущих видов изменяется в течение вегетацион-
ного периода по типу одновершинной кривой. Максимум за оба года при-
ходится на I декаду июля. Форма кривой довольно симметрична, что
свидетельствует о равномерном увеличении числа цветущих видов,
152
Рис. 27. Кривые цвет ения 104 видов травянистых растений дубовых лесов
(1961 г.). Обозначения те же, что на рис. 16.
апрель мои июнь июль август сентябрь
Рис. 28. Кривые цветения 150 видов травянистых растений дубовых
лесов (1962 г.). Обозначения те же, что на рис. 16.
основных компонентов (45 видов) травяного
г.). Обозначения те же, что на рис. 16.
Рис. 29. Кривые цветения
покрова дубрав (1961
02 5068954
0 2 7 7 12 19 24 22 21
0 1 1 4 7 5 3
I II III I II III I II III
апрель май июнь
3 0 2 0
21 14 15 10 8 5 2 10
7 1 5 2 3 3 1 10
I II III I II III I II III
июль август сентябрь
Рис. 30. Кривые цветения основных компонентов (44 видов) травяного
покрова дубрав (1962 г.). Обозначения те же, что на рис. 16.
начиная с апреля до середины лета и о таком же плавном снижении
вплоть до конца сентября.
Число отцветающих по декадам видов изменяется по типу многовер-
шинной кривой. Пики кривой приходятся на I декаду июня, I и III дека-
ды июля, II—III декады августа, довольно точно совпадая во времени
за тот и другой годы наблюдений.
Как видно из приведенного анализа, кривые цветения общего коли-
чества травянистых растений дубовых лесов обладают весьма незамет-
ными разногодичными изменениями. Так, отличия графиков 1961 и
1962 гг. касаются в основном распределения во времени наибольшего
ОЗ 3 4 5 9 23 29 42 18
0 3 6 10 15 22 4/ 65 102 99
О 1 2 2 5 6 24 19
in I и in । ll Hi । ll HI
а прель май июнь
17 6 8 2 О 10
95 69 60 38 31 23 15 8 6 2 О
31 17 23 7 8 10 7 2 4 2 О
I II III I I! Ill I И III I II
и ю л ь август сентябрь
Рис. 31. Кривые цветения растений (170 видов) полян дубовых лесов (1961 г.).
Обозначения те же, что на рис. 16.
числа зацветающих видов. В связи с различиями погодных условий лет
наблюдений более показательны кривые цветения основных компонен-
тов травяного покрова (рис. 29—30).
Характер цветения основных компонентов травяного покрова дубо-
вых лесов в основном отражает те же общие закономерности, присущие
ритмике цветения всего видового состава. Представляет интерес лишь
то обстоятельство, что абсолютные максимумы зацветающих и цветущих
видов за годы наблюдений совмещались во времени и в 1961 г. приходи-
лись на II декаду июня, а в 1962 — на I декаду июня. Это смещение мак-
симумов вполне отвечает погодным особенностям вегетационного перио-
да 1962 г.
В заключение следует отметить, что цветение травянистых растений
дубовых лесов более равномерно распределено во времени: без очень
резких подъемов и спусков, с наибольшим значением в течение июня
и июля.
Для сравнения известный интерес представляет график цветения рас-
тений лесных полян по результатам наблюдений в 1961 г. (рис. 31). Со-
поставление его с аналогичными графиками луговой степи и травяного
покрова дубрав за Этот же год открывает некоторые черты сходства
с тем и другим, а в целом по ритмике цветения растительность лесных
полян занимает как бы промежуточное положение между указанными
ценозами.
154
Рис. 32. Кривые цветения растений (170 видов) дна и ’северных склонов
степных логов (1961 г.). Обозначения те же, что на рис. 16.
Рис. 33. Кривые цветения растений (214 видов) дна и северных склонов
степных логов (1962 г.). Обозначения те же, что на рис. 16.
апрель л» а й июнь июль август сентябрь
Рис. 34. Кривые цветения основных компонентов (63 видов) растительности
дна и северных склонов степных логов (1961 г.).
Обозначения те же, что на рис. 16.
Ритмы цветения растений дна
и северных склонов логов
На рис. 32—35 изображены кривые цветения растений, входящих в
ценозы дна и северных склонов логов. Количество всех изученных зацве-
тающих видов изменяется по декадам по типу неясно выраженной много-
вершинной кривой (рис. 32, 33). Абсолютный максимум за годы наблю-
дений (1961 — 1962) приходится на первую декаду июня, небольшой подъ-
ем отмечается в апреле, а в 1962 г.— еще в III декаду июня и II декаду
июля. Первые цветущие растения появляются в I—II декадах апреля,
последние — во II—III декадах августа.
Кривая общего числа цветущих видов носит одновершинный харак-
тер с максимумом в III декаде июня. Обе половины кривой довольно
симметричны, но все-таки подъем ее несколько круче, чем спуск.
Рис. 35. Кривые цветения основных компонентов (70 видов) растительности
дна и северных склонов степных логов (1962 г.).
Обозначения те же, что на рис. 16.
С максимумом цветущих видов совпадает наибольшее число отцве-
тающих видов. Отцветание происходит скачками, вследствие чего и кри-
вая имеет несколько вершин. Особенно резкое возрастание числа отцве-
тающих видов помимо первого максимума в III декаде июня происходит
в августе — сентябре, с колебаниями в сроках в отдельные годы. Из ска-
занного видно, что основные показатели цветения — абсолютные макси-
мумы зацветающих, цветущих и отцветающих видов — при учете всего
их обилия, за годы наблюдений не изменяли своего распределения во
времени. Наибольшее «напряжение» периода цветения приходится на
июнь.
Иную картину по годам представляют кривые цветения основных ком-
понентов растительности логов (рис. 34, 35). Так, в 1961 г. все три мак-
симума падали на II декаду июня. В 1962 г. произошло их расхождение
во времени. Максимум зацветающих видов пришелся на III декаду мая,
а цветущих и отцветающих — на III декаду июня. Таким образом, можно
констатировать, что характерный в 1962 г. сдвиг максимума зацветаю-
щих видов, относящихся к основным компонентам, на более ранние сро-
ки имел место и среди растений логов, подобно тому как это наблюда-
лось в луговой степи и травяном покрове дубрав.
Описанные факты, бесспорно, убеждают в том, что общезональные
погодные условия оказывают влияние на все растительные сообщества
данного ландшафта независимо от их топографического распределе-
ния — на плакоре или иных элементах рельефа. Короче го«воря, каждый
фитоценоз в той или иной зоне несет на себе печать зональных особен-
ностей, а поэтому широко развивавшиеся прежде представления об ин-
тразональных явлениях в прямом значении этих слов должны быть остав-
лены как несоответствующие действительным природным закономерно-
стям.
156
Влияние на ритмы цветения растений
эколого-ценотических условий
Имеющиеся в нашем распоряжении данные фенологических наблю-
дений позволяют выяснить характер цветения некоторых видов, произ-
растающих в разных ценозах: на степи, в дубравах и логах. В табл. 7
Таблица?
Характер цветения некоторых видов в разных биотопах в 1962 г.
Вид Начало Массовое Конец
степь коси- мая лес лог степь коси- мая лес лог степь коси- мая лес лог
Achillea millefolium 6.VI 15.VI 6.VI 19.VI 25.VI 18. VI 4.IX 27.IX 2.Х
Agropyrum repens 20.VI 25.VI 25.VI 27.VI 4. VII 29.V1 13.VII 28. VII 20.VII
Allium oleraceum 18.VII 21.VII — 24.VII 27.VII — 16.VIII 28.VIII —
Alopecurus pratensis 22.V 25.V 17.V 27.V 29.V 23.V 31.V 9.VI 8. VI
Antitoxicum offici- nale 25.V 30. V — 3.VI 18.VI — 18.VII 19.VII
Betonica officinalis 25.VI 9.VI1 25.VI 5.VII 17.VII 4.VII 24.VIII 29.VIII 28. VIU
Bromus inermis . 22.VI 23.VI 25. VI 27.VI 29.VI 3.VII 12.VII 9.VII 12.VII
Calamagrostis epi- geios 10.VII 9.VII 13.V1I 14.VII 13.VII 19.VII 23.VII 27.VII 26. VII
Carex montana . . 23. IV 22.IV 23. IV z6.IV 25. IV 26.IV 3.V 5.V 3.V
Clematis recta . . 2.VI 3.VI — 8.VI 14. VI — 4.VII 9.VII
Filipendula hexape- tala 31.V — 3.VI 5.VI — 8.VI 26.VI __ 3.VII
Galium boreale . . 2.VI 7.VI 6.VI 8.VI 14. VI 13.VI 10.VII 10. VII 12.VII
Gtechoma hederacea 23.IV 29.IV 28. IV 3.V 5.V 4.V 8. VI 10.VI 18.VI
Hieracium umbella- tum 31.VII — 2.VIII 13.VIII — 16.VIU 25.IX — 25.IX
Inula hirta .... 5.VI — 7.VI 15. VI — 16.VI 6.VII — 7.VII
Iris aphylla .... 14.V 15.V — 21.V 24.V — 2.VI 6.VI —
Leucanthemum vul- gare 4.V1 6.VI 6.VI 13.VI 20.VI 13. VI 18.VII 27.VII 22.VII
Phlomis tuberosa . 4.VI 7.VI — 15.VI 20.VI — 23.VII 17.VII —
Primula veris . . 23. IV 25. IV 21.IV 1.V 29. IV 26. IV 24.V 29.V 26.V
Veronica chamaedrys 12.V 15.V 14.V 23.V 24. V 23.V 11.VI 14.VI 20.VI
мы выборочно приводим сроки фенофаз, на основании которых можно
судить о степени влияния эколого-ценотических условий на ритмы цве-
тения. Таблица показывает, что эколого-ценотическое окружение ока-
зывает определенное влияние на прохождение фаз цветения одних и тех
же видов. Отличия в сроках бывают весьма различные, в зависимости от
индивидуальных особенностей видов, причем в разных фазах это влия-
ние может сказываться по-разному. Как правило, в условиях луговой
степи растения проходят фазы цветенйя в более ранние сроки, чем под
пологом леса. Однако этого нельзя сказать о логах, где одни виды цветут
раньше, у других — фазы цветения смещаются на более поздние сроки
в сравнении с косимой степью. Все же, за редкими исключениями, отли-
чия в сроках прохождения отдельных фаз цветения в разных ценозах бы-
вают незначительными, в пределах 1—2 — 6—8 суток, но иногда превы-
шают и 10—15 суток.
Проведение параллельных наблюдений в течение многих лет за цве-
тением одних и тех же видов в разных ценозах дает ценнейший мате-
риал к познанию потенциальных экологических свойств и признаков рас-
тений.
157
Состав основных растительных сообществ заповедника
по продолжительности цветения их компонентов
Зная фенологические особенности цветения растений луговых степей,
дубовых лесов, дна и северных склонов логов, легко представить их со-
став по продолжительности цветения. Этот признак имеет существенное
значение в географических сопоставлениях различных типов раститель-
ности.
По продолжительности цветения выделяются длительноцветущие рас-
тения, у которых период цветения продолжается свыше одного месяца,
и короткоцветущие с периодом цветения до одного месяца (Серебряко-
ва, 1956). Как показывают результаты наших фенологических наблюде-
ний, в области лесостепи встречается немало растений, у которых в раз-
ные по погодным условиям годы продолжительность цветения изменяет-
ся настолько, что в одни годы они должны быть отнесены к длительно-
цветущим, а в другие — к короткоцветущим. Таковы Clematis recta, Dra-
ba nemorosa, Myosotis suaveolens, Salvia pratensis, Veronica chamaedrys,
Viola rupestris и др. (в условиях луговой степи). Эти виды можно обозна-
чить как промежуточные. Интересно отметить, что эколого-исторические
причины такого поведения видов могут быть различными. Удлинение
цветения части видов определяется тем, что погодные условия данного
года соответствуют их экологической природе и, наоборот, аналогичная
реакция вызывается при иных экологических свойствах растений.
Накопление наблюдений за разнообразными реакциями растений на
погодные условия, зафиксированными в течение длительного ряда лет,
создает неисчерпаемый источник познания экологии растительных
видов.
В составе растительности Стрелецкой степи насчитывается длитель-
ноцветущих видов 116 (42,3%), короткоцветущих— 123 (45%) и проме-
жуточных— 35 (12,7%). Сходные соотношения сохраняются и среди
основных компонентов: длительноцветущих видов 50 (36,5%), корот-
коцветущих— 68 (50%), промежуточных—19 (13,5%). Таким обра-
зом, господствующими на Стрелецкой степи являются короткоцвету-
щие виды. Это обстоятельство отмечалось еще И. Н. Оловянниковой
(1949) и М. А. Борисовой-Туленковой (19606). По этому признаку расти-
тельность Стрелецкой степи заметно отличается от луговых ценозов, где
длительноцветущие виды насчитывают до половины (от 41,4% до 50,7%)
от всего видового состава (Серебрякова, 1956).
В травяном покрове дубовых лесов зарегистрировано 57 длительно-
цветущих видов (38%), 70 — короткоцветущих (46,6%), 23 — промежу-
точных (15,4%). Но среди основных ценозообразователей преобладают
длительноцветущие — 22 вида (50%), а на короткоцветущие приходится
лишь 42% (19 видов), промежуточные составляют 8% (4 вида). Такие
особенности фенотипического состава растений дубовых лесов объяс-
няются тем, что под полог проникает много луговостепных короткоцвету-
щих видов, которые вместе с подобными лесными становятся наиболее
обильной группой. Но когда остаются основные компоненты, т. е. преиму-
щественно неморальные элементы, то соотношения фенотипических групп
изменяются в пользу господствующего положения длительноцветущих
видов.
Растительность дна и северных склонов логов содержит в своем со-
ставе 108 длительноцветущих видов (50,7%), 85 — короткоцветущих
(40%), 20 — промежуточных (9,3%). Это соотношение остается без из-
менений и среди основных компонентов: длительноцветущих видов 38
(54,3%), короткоцветущих — 27 (38,6%), промежуточных — 5 (7,1%).
Преобладающее значение длительноцветущих видов указывает на изве-
стное сходство этой категории растительности с луговой.
158
ВТОРИЧНОЕ ЦВЕТЕНИЕ
Вторичное цветение растений в области лесостепи выходит из разряда
частных случаев и представляет широко распространенное, из года в год
повторяющееся закономерное явление. Поэтому его изучение имеет су-
щественное значение в эколого-биологической характеристике расти-
тельности. Но прежде чем приступить к его анализу на территории запо-
ведника, необходимо договориться, что следует понимать под вторичным
цветением, и установить разнообразие его биоморфологических типов.
Как показывает само название, вторичное цветение вида заключает-
ся в развитии второй генерации цветоносных побегов и цветков, следую-
щей, как правило, за первой генерацией с некоторым перерывом во вре-
мени. Вид здесь понимается как разновозрастная популяция особей. Вто-
ричное цветение обычно приходится на вторую половину вегетационного
периода или осень. Указание на разрыв между обычным (нормальным)
и вторичным цветением должно разграничивать последнее от явления
длительного беспрерывного цветения за счет многократного ветвления
соцветия и образования все новых и новых веточек с цветками. Однако
и эта оговорка не исключает существования всевозможных пер!еходов,
когда трудно установить границу между нормальным и вторичным цве-
тением.
Вопросы вторичного цветения растений привлекали у нас внимание
многих исследователей (Аненков, 1850; Алехин, 1909; Илличевский, 1925,
1931, 1937; Галахов, 1935, 1937, 1950; Кожевников, 1939; Серебряков,
1947, 1952; Хохлов, 1947; Зарубин, 1949; Середин, 1951; Коровин и Туль-
чиев, 1948; Зворыкина, 1953; Сергеев, 1953, 1955; Немкова, 1954; Каряг-
дыев, Мищенко, 1956; Камышев, 1958; Борисова-Гуленкова, 1959, и др.).
В работах С. О. Илличевского, Н. Н. Галахова, И. Г. Серебрякова доста-
точно полно охвачено почти все разнообразие биоморфологических типов
вторичного цветения, но развернутой их классификации не дано. В этих
и других исследованиях большое место уделялось выяснению причин,
обусловливающих вторичное цветение.
На основании указанных работ и личных наблюдений в природе ниже
предлагается следующая классификация типов вторичного цветения: про-
дленное, настоящее, вторичное и неотеническое. Рассмотрим эти типы.
Продленное цветение
Относящиеся к этому типу случаи вторичного цветения заключаются
в продлении цветения за счет развития новых побегов с цветками или
отдельных цветков на генеративных побегах текущего года. К моменту
вторичного цветения исходные генеративные побеги находятся в различ-
ных стадиях: незрелых плодов, созревания плодов и обсеменения. Воз-
можно и частичное засыхание побегов, уже обсеменившихся, а также
потеря всех листьев, когда живыми сохраняются только стебли.
Этот тип вторичного цветения незаметно сливается с типом растяну-
того (длительного) цветения, характеризующегося появлением все но-
вых ветвей и цветков, в то время как первые уже отцвели и даже обсе-
менились. Примером таких растений служат Hypericum perforatum, Sca-
biosa ochroleuca, Serratula inermis и др. Наличие растянутого цветения
не исключает у тех же самых видов различных форм вторичного цве-
тения.
Среди растений лесостепи продление цветения в рамках вторичного
цветения осуществляется следующими способами:
1. Происходит возобновление верхушечного роста цветоносных побе-
гов и образование цветков вторичного цветения (Campanula rapunculoi-
des, Erysimum marschallianum, Euphrasia tatarica, Galium verum, Lina-
159
ria vulgaris, Phlomis tuberosa, Stachys recta, Thymus marschallianus
и др.).
Следует особо выделить группу моноподиальных растений с удли-
ненными ползучими или розеточными побегами. У Trifolium repens в ус-
ловиях луговой степи бывает остановка роста ползучих побегов, которые
вновь трогаются в рост с наступлением влажной и прохладной погоды.
На вновь возникших участках побегов в пазухах листьев нередко
формируются стрелки с головками цветков. Такой ход морфогенеза дает
полное право на отнесение цветения к вторичному. Между тем в лесолу-
говой зоне и по дну логов заповедника ползучие побеги растут беспрерыв-
но, благодаря чему регулярно появляются и цветоносные побеги. Нечто
подобное наблюдается у Plantago lanceolata, Р. media, Trifolium praten-
se и других, у которых разрыв между первой и последующей генерация-
ми пазушных цветоносов бывает только на степи, тогда как в более
влажных условиях гиатус во времени не выражен. Все эти случаи вто-
ричного цветения моноподиальных растений следует относить за счет во-
зобновления верхушечного роста, но не генеративных, а вегетативных
побегов.
Резче обозначен промежуток между первичным и вторичным цвете-
нием у Centaurea marschalliana, Potentilla alba, P. humifusa, P. patula.
Вторая генерация цветоносных побегов может быть совершенно не вы-
ражена. Здесь возобновление верхушечного роста розеточных монопо-
диальных побегов с образованием цветоносных является ненормальным
и происходит обычно в будущем году.
Этот тип вторичного цветения следует уже рассматривать в качестве
настоящего. Однако в экспериментальных условиях, на делянках питом-
ника при систематическом поливе, удается вызвать почти беспрерывное
цветение в течение вегетационного периода и у этой группы моноподиаль-
ных растений.
2. Формирование новых побегов и цветков протекает в пазухах ли-
стьев генеративного побега этого года, ниже соцветия, а также в соцве-
тии в результате вторичного роста и ветвления. Эта форма вторичного
цветения встречается наиболее часто и обнаружена у Achillea millefo-
lium, Acinos thymoides, Arenaria serpyllifolia, Berteroa incana, Carduus
hamulosus, Campanula patula, C. persicifolia, C. rapunculoides, C. rotun-
difolia, C. sibirica, Crepis tectorum, Centaurea scabiosa, Delphinium
cuneatum, Genista tinctoria, Pieris hieracioides, Potentilla, argentea,
Linum perenne, Lotus corniculatus, Onobrychis arenaria, Senecio jacobaea.
Sisymbrium loeselii, Stachys recta, Tragopogon orientate, Turritis glabra
и др.
Перечисленные типы вторичного цветения, весьма близкие между со-
бой, возникают у растений без какого-либо травмирования, в естествен-
ных условиях развития и являются следствием благоприятного эколо-
гического режима, способствующего продлению ростовых процессов и
генеративному развитию.
3. Развитие цветоносных побегов происходит в основании материн-
ского^ невысоко над поверхностью почвы, но не от корневища. Этот тип
вторичного цветения часто возникает под влиянием сенокошения или
других причин, нарушающих целостность побегов (поедание животны-
ми, вредителями и т. д.). На остающихся пеньках материнских побегов,
в пазухах живых или отмерших листьев возникают новые побеги, неред-
ко переходящие к цветению. В естественных условиях, в отсутствие хо-
зяйственного использования травостоя, развитие цветоносных побегов
указанным способом наблюдается сравнительно редко, но в доагрикуль-
турную эпоху, когда в степях изобиловали дикие травоядные животные,
этот способ вторичного цветения, без сомнения, составлял обычное по-
всеместно распространенное явление. На скашиваемой степи и в других
ио
(Wftyfonax с таким Же рёжямом зарегистрированы следующие растения,
дающие после покоса цветоносные побеги от пеньков скошенных: Anthe-
mis tinctoria, Antitoxicum officinale, Berteroa incana, Campanula persici-
fo'lia, C. rapunculoides, C. sibirica, Carduus hamulosus, Cichorium intybus,
Crefiis teetotum, Dctacus carota, Delphinium consolida, Echium vulgare.
Erigeron acer, Erysimum marschallianum, Hieracium umbellatum, Hype-
ricum perforatum.; Knautia arvensis, Linum perenne, L. flavum, Lotus cor-
niculatus, Matricaria inodora, Medicago falcata, Melampyrum nemorosum,
Pieris hieracioides, Potentilla argentea, R hi nan thus major, Scabiosa ochro-
leuca, Scutellaria galericulata, Sedum telephium, Senecio jacobaea, Sisym-
brium loeselii, Solidago virgaurea, Stachys recta, Trifolium hybridum, Ver-
bascum lychnitis, Veronica spuria, Viola tricolor и др. Интересно отме-
тить, что среди перечисленных имеются как поликарпические, так и мо-
нокарпические малолетние и однолетние растения.
От рассмотренного типа есть все переходы к настоящему вторичному
цветению. Так, под влиянием покоса весьма часто пробуждаются почки
в основании пенька, т. е. фактически уже на корневище. Этот случай вто-
ричного цветения отмечен у Achillea millefolium, Aegopodium podagra-
ria, Centaurea jacea, C. scabiosa, Clematis recta, Delphinium cuneatum,
Echium rubrum, Erigeron acer, Euphorbia esula, Filipendula hexapetala,
Galium boreale, G. verum, Helichrysum arenarium, Lathyrus pratensis,
Linaria vulgaris, Melandrium album, Peucedanum oreoselinum, Polygala
comosa, Salvia verticillata, Selinum carvifolia, Serratula inermis, Silene
cucubalus, Tragopogon orientate и др. Цветоносные побеги при этом раз-
виваются как из почек, так и из розеточных побегов.
Следует подчеркнуть, что все охарактеризованные типы вторичного
цветения имеют внутреннюю связь и, как правило, свойственны одним
и тем же растениям. Так, если данный вид способен к вторичному цве-
тению посредством развития боковых дополнительных побегов в области
соцветия, то вполне вероятно, что при скашивании у него чаще и легче
Может сформироваться цветоносный побег от пенька или в самом осно-
вании пенька. Все это свидетельствует о большой пластичности растений
с продленным вторичным цветением, о выёоких потенциальных возмож-
ностях продуктивности и генеративного развития. Эти же растения от-
личаются неглубоким зимним покоем или его полным отсутствием. Таким
образом, здесь с особенной отчетливостью выступает взаимосвязь при-
знаков в организмах, которые в своей совокупности могут служить на-
дежным средством познания эколого-биологической природы растений и
выделения интегральных типов биоморфологических структур. Исследо-
вания в этой области исключительно перспективны. На этом пути иссле-
дователей ожидает возможность теоретического расчета и предвидения
признаков растений по одному или немногим исходным. Повышается и
роль эксперимента — могучего средства познания потенциальных при-
знаков и свойств растений.
Настоящее вторичное цветение
Сюда относятся все случаи пролептического (преждевременного) раз-
вития генеративных побегов, предназначенных для будущего года. Имен-
но эту категорию и предлагает называть вторичным цветением в соб-
ственном смысле И. Г. Серебряков (1947). Почки и другие вегетативные
зачатки, из которых образуются цветоносные побеги, могут находиться
в разных стадиях развития — от содержания вполне сформированных
генеративных органов до их полного отсутствия. Установлено, что у рас-
тений с почками, в которых уже в конце лета и осенью заложены все
элементы будущего цветоносного побега, включая цветки с микро- и ма-
кроспорангиями, вторичное цветение бывает очень редко (Кожевников,
ИВ Н Голубев
161
1939; Серебряков, 1947, 1952). Этот тип разбивается на ряд более мелких
подразделений.
1. Цветоносные побеги развиваются из почек на корневище в осно-
вании материнского побега, в которых побег будущего года сформиро-
ван полностью, включая соцветие н цветки (Anemone Silvestris, Arenaria
ucrainica, Dianthus andrzejowskianus, Pulsatilla patens и др.).'
2. Генеративные побеги образуются из терминальной почки розеточ-
ных побегов особей, которые в текущем году цвели и плодоносили (Achil-
lea millefolium, Achyrophorus maculatus, Filipendula hexapet ala, Leucan-
themum vulgare, Nonea pulla, Scabiosa ochroleuca и др.).
3. Цветоносные побеги формируются из безрозеточных побегов во-
зобновления (Polygala comosa, Sedum acre, Veronica prostrata и др.).
Неотеническсе цветение
К этому типу принадлежат случаи цветения во второй половине лета
и осенью молодых недостаточно развитых растений и побегов, нормаль-
ное цветение которых наступает в более позднем возрасте. Данная фор-
ма цветения выявлена у озимых однолетних, двулетних и многолетних
поликарпических растений.
Особенно часто у многолетников неотеническое (и по времени — позд-
нее) цветение наблюдается при выращивании растений из семян в пи-
томнике на первом году жизни, например, у Antitoxicum officinale, Вир-
leurum falcatum, Centaurea ruthenica, Delphinium cuneatum, Onobrychis
arenaria, Salvia pratensis, Tanacetum vulgare, Thymus marschallianus,
Trifolium alpestre, T. montanum, Veronica spuria и многих других.
Из малолетних монокарпиков на первом году жизни в природных
условиях осенью иногда неотенически зацветают Campanula patula, Ery-
simum marschallianum, Jurinea arachnoidea.
Позднелетнее развитие цветоносных побегов из молодых розеточных,
нормально переходящих к цветению только в следующий вегетационный
период или еще позже, наблюдается у Achillea millefolium, Campanula
persicifolia, Centaurea scabiosa, Salvia pratensis, Scabiosa ochroleuca
и др. Такая форма неотенического цветения тесно смыкается со второй
и третьей группами настоящего вторичного цветения. Оба варианта, не-
сомненно, весьма широко распространены в природе; при одном и том
же конечном результате, т. е. позднем зацветании, они отличаются лишь
по стадии развития побегов, на которой осуществляется переход к вто-
ричному цветению.
Выяснению причин вторичного цветения, как уже указывалось, уделя-
лось серьезное внимание (Аненков, 1850; Шрейбер, 1908, 1911; Галахов,
1935, 1937; Коровин, Тульчиев, 1948; Зарубин, 1949; Середин, 1951; Зво-
рыкина, 1953, и др.). Были установлены многочисленные реагенты, вы-
зывающие вторичное цветение. Все их разнообразие можно свести к двум
основным группам: искусственным, связанным с деятельностью челове-
ка, куда входит и травмирование растений (надломы и другие поврежде-
ния побегов), и естественным, главным образом климатическим.
К искусственным факторам, индуцирующим вторичное цветение, от-
носятся воздействие газами (сернистым, природным), большими дозами
минеральных удобрений, применением летней обрезки и пр. (Хохлов,
1947; Сергеев, 1955, и др.).
Гораздо большее значение имеют естественные факторы. В их оценке
нет единого.мнения. Одни исследователи склонны выдвигать на первый
план климатические аномалии: сильные и поздние заморозки весной,
сильные засухи летом, ливневые дожди при повышенной температуре
воздуха, обилии прямой солнечной радиации и пр. (Галахов, 1937; Нем-
кова,' 1954; Сергеева, 1955, и др.); другие — основную причину видят в
162
благоприятных для роста климатических условиях (Илличевский, 1925,
1931, 1937; Долгошев, 1954, и др.). Вторичное цветение растений и вы-
зревание плодов чаще и в больших размерах наблюдается на юге и в
западных районах, чем на севере и восточных районах. Это объясняется
большей продолжительностью вегетационного периода на юге и западе.
На ограниченной территории, например в лесах, вторичное цветение чаще
бывает у растений опушек, в условиях лучшего солнечного освещения
(Долгошев, 1954). Близкие высказывания можно найти у С. О. Илличев-
ского (1931), отмечающего связь вторичного цветения с обильными лет-
ними осадками и теплой осенью.
В указанных взглядах нет противоречия, так как преждевременное
раскрытие почек и формирование цветоносных побегов, безусловно, сти-
мулируются как искусственными факторами, так и различного рода рез-
кими, аномальными климатическими явлениями. С другой стороны, все
факторы, благоприятствующие интенсивному росту и большей его про-
должительности (теплая влажная погода летом и осенью), одновременно
способствуют продлению цветения и повторному зацветанию. В южных
районах с более продолжительным вегетационным периодом и более
благоприятным термическим режимом в условиях хорошего увлажнения
вторичное цветение становится массовым явлением. В области лесостепи,
где при сравнительно большой продолжительности вегетационного сезо-
на нередко отмечается весьма оптимальная комбинация тепла и влаги,
вторичное цветение растений наблюдается весьма часто у значительного
количества видов.
Однако вследствие колебаний метеорологических условий по годам,
в лесостепи довольно резких, происходят и разногодичные изменения
вторичного цветения. Варьируют разнообразие видов, количество цве-
тущих особей, интенсивность цветения. Происходит также перераспреде-
ление вторично цветущих видов по местообитаниям: в разных условиях
одни и те же виды по годам цветут неодинаково обильно.
Можно с уверенностью полагать, что особенности прохождения всего
жизненного цикла растениями накладывают свой отпечаток и на харак-
тер вторичного цветения. Нужно учитывать наличие периодичности цве-
тения (и плодоношения) у травянистых растений, особенности возраст-
ного состава популяций в разные годы и т. д.
Разумеется, не все виды одинаково реагируют на внешние условия
возбуждением вторичного цветения. Как указывалось выше, виды с за-
ранее сформированными цветоносными побегами и цветками в почках
возобновления (с конца лета и осенью) очень редко переходят к вторич-
ному цветению. Наоборот, растения, в почках возобновления которых
осенью нет сформированных генеративных органов, цветут вторично
чаще и обильнее. Если для первых в индуцировании вторичного цвете-
ния преобладающую роль играют факторы, выводящие почки возобнов-
ления из состояния покоя, то для вторых решающее значение имеют
условия, стимулйрующие ростовые процессы.
Таким образом, к оценке причин и возможностей вторичного цвете-
ния надо подходить дифференцированно, с учетом эколого-биологиче-
ских особенностей растений.
Особенно сильно вторичное цветение стимулируется покосом. Поэто-
му на косимых степях, лесных полянах и логах заповедника большин-
ство видов, слагающих эти ценозы, способно к вторичному цветению.
Сенокошение обусловливает развитие генеративных побегов, не только
от срезанного пенька, но и пролептическое пробуждение почек возобнов-
ления на корневищах, переход к цветению розеточных побегов, находя-
щихся на' разных стадиях морфогенеза, проявление неотенического цве-
тения ювенильных особей, оказывающихся после уборки сена в лучших
условиях освещения и минерального питания. Столь обильное вторичное
tl*
163
цветение нельзя, конечно, относить за счет влияния одного только поко-
са. Оно возможно лишь при наличии в окружающей среде соответствую-
щих благоприятных условий. Таким образом, возникновение массового
вторичного цветения растений является чутким реагентом на специфи-
ческие природные особенности лесостепи, ее термально-гидрологический
режим.
Не зацветают после скашивания очень немногие виды луговой степи
с весьма консервативным ритмом развития: Adonis vernalis, Iris aphylla,
Paeonia tenuifolia, Polygonatum officinale, Veratrum nigrum. He зареги-
стрировано за четыре года наблюдений вторичное цветение у следующих
весенне-раннелетних геоэфемероидов: Anemone ranunculoides, Corydalis
halleri, Ficaria verna, Fritillaria meleagris, F. ruthenica, Gagea erubes-
cens, G. lutea, G. minima, Hyacinthella leucophaea, Pedicularis kaufmannii,
Ranunculus illyricus, R. pedatus, S cilia sibirica, а также у Carex humilis
C. michelii, C. verna, C. praecox, Salvia nutans, Trinia multicaulis.
Отсутствует вторичное цветение, даже в случае скашивания, у расте-
ний с растянутым развитием и поздними сроками цветения: Agropyron
intermedium, Allium flavescens, Artemisia absinthium, A. austriaca,
A. campestris, A. scoparia, A. vulgaris, Calamagrostis epigeios, Crepis pan-
nonica, Eryngium planum, Gypsophila paniculata, Hieracium virosum, Li-
nosyris vulgaris.
Из травянистых растений дубовых лесов, помимо отмеченных гео-
эфемероидов, вторично не зацветают: Actaea spicata, Astragalus glycyp-
hyllus, Calamagrostis arundinacea, Carex michelii, C. montana, C. spicata,
Convallaria majalis, Epipactis latifolia, Galium rubioides, Lilium pilosius-
culum, Lychnis chalcedonica, Mercurialis perennis, Orobus niger, 0. ver-
nus, Paris quadrifolia, Platanthera bifolia, Polygonatum multiflorum,
P. officinale, Pulmonaria angustifolia, P. obscura, Ranunculus cassubicus,
Rubus saxatilis, Scrophularia nodosa, Thalictrum aquilegifolium, Verbas-
cum nigrum и некоторые другие.
Вторичное цветение не наблюдалось у следующих видов дна и север-
ных склонов логов: Carex caespitosa, С. hirta, С. leporina, С. pallescens,
С. riparia, С. vesicaria, С. vulpina, Crepis praemorsa, Filipendula ulma-
ria, Galatella rossica, Gladiolus imbricatus, Iris sibirica, Juncus effusus,
Molinia coerulea, Serratula coronata, Thalictrum lucidum, Th. simplex, Va-
leriana exaltata, Veratrum lobelianum и некоторых других.
В приведенных списках обращает на себя внимание то обстоятель-
ство, что основная масса указанных видов обладает длительным органи-
ческим покоем. В ритме их развития соблюдается строгая последователь-
ность периода вегетации и покоя как характерная наследственно за-
крепленная особенность. Здесь мы сталкиваемся с известной «нормиро-
ванностью» развития надземных органов, исключающей формирование
дополнительных цветоносных побегов. Лишь у Pulsatilla patens, также
характеризующейся длительным покоем, вторичное цветение происходит
в небольшом количестве экземпляров. Интересно отметить, что большое
число видов, не цветущих вторично, обладает одним периодом роста
надземных органов и одной генерацией листьев. Но есть исключения из
этого правила (Eryngium planum, Pulmonaria angustifolia и др.).
Подавляющее большинство остальных видов, обитающих в луговой
степи, дубовых лесах и логах лесостепи, способно к вторичному цвете-
нию (полные списки видов этих биотопов приводятся в гл. 7).
На степи после сенокоса регулярно происходит вторичное цветение
у длительновегетирующих глубококорневых растений Asparagus offici-
nalis, Centaurea scabiosa, Leucanthemum vulgare, Nonea pulla, Onobry-
chis arenaria, Pieris hieracioides, Salvia pratensis, Scabiosa ochroleuca,
Senecio jacobaea, Stachys recta, Taraxacum officinale, Verbascum lychni-
tis и др. Вторичное цветение у них обильно происходит как в засушли-
164
вые, гак и во влажные годы. Это находит объяснение в устойчивой вла-
гообеспеченности растений, благодаря глубокой корневой системе.
Для обильного цветения мелкокорневых растений в конце лета необ-
ходимо достаточное увлажнение за счет осадков. Наиболее обильно и
устойчиво в разные годы цветет Asperula cynanchica, обладающая интен-
сивной (хотя и неглубокой) корневой системой. Весьма часто вторично
цветут Arenaria ucrainica, Berteroa incana, Campanula sibirica, Draba si-
birica, Erigeron acer, Galium boreale, Leontodon hispidus, Plantago media,
Stellaria graminea, Trifolium pratense и др.
Интенсивное и регулярное вторичное цветение после покоса наблю-
дается у многих растений логов: Bromus inermis, Galium boreale, G. mol-
lugo, Geranium palustre, G. pratense, Leontodon hispidus, Leucanthemum
vulgare, Plantago lanceolata, P. media, Stellaria graminea, Taraxacum
officinale, Trifolium pratense, T. repens и некоторых других.
На некосимых степи, логах, лесных полянах количество вторично цве-
тущих растений заметно сокращается. Основной формой становится про-
дленное цветение за счет ветвления и нарастания исходных генератив-
ных побегов. В некосимой степи продленное вторичное цветение обнару-
жено у Achillea millefolium, Antitoxicum officinale, Arenaria serpyllifo-
lia, Campanula glomerata, C. patula, C. persicifolia, C. rapunculoides,
C. rotundi(olia, C, sibirica, Carduus hamulosus, Centaurea pseudophrygia,
C. scabiosa, Coronilla varia, Delphinium cuneatum, Echium rubrum, Ery-
simum marschallianum, Galium verum, Helichrysum arenarium, Hyperi-
cum perforatum, Knautia arvensis, Linaria vulgaris, Linum perenne, Medi-
cago falcata, Onobrychis arenaria, Pieris hieraciodes, Phlomis tuberosa,
Potentilla argentea, Scabiosa ochroleuca, Senecio jacobaea, Stachys recta,
Stellaria graminea, Thymus marschallianus, Tragopogon orientate, Tur-
ritis glabra и др.
Реже бывают случаи настоящего вторичного цветения — развитие
цветоносных побегов из почек на корневищах или розеток (Achillea mil-
lefolium, Anthericum ramosum, Arrhenatherum elatius, Bromus riparius,
Centaurea marschalliana, Draba sibirica, Knautia arvensis, Leontodon hi-
spidus, Salvia pratensis, Silene nutans, Sisymbrium polymorphum, Stipa
pennata, St. stenophylla, Leucanthemum vulgare и др.). Цветоносные побе-
ги иногда формируются у безрозеточных растений из укороченных побе-
гов возобновления (Polygala comosa, Veronica prostrata) или из почек
на корневищах (Vicia tenuifolia).
Изредка на некосимой степи происходит неотеническое вторичное
цветение в результате преждевременного перехода в состояние половой
зрелости молодых вегетативных особей (Campanula patula, Trifolium
pratense) или побегов (Linaria vulgaris, Veronica chamaedrys).
По характеру вторичного цветения и видовому составу цветущих рас-
тений весьма близко jc некосимой степи стоят временно некосимые уча-
стки степи, где исключается сенокошение только в текущем году.
Сроки вторичного цветения, его фенология, зависят от многих
причин. На скашиваемых участках фенология цветения зависит как от
метеорологических условий, так и от времени покоса. При скашивании
степи и логов в середине июля апогей вторичного цветения приходится
на август, а в теплую погоду обильное цветение продолжается и в пер-
вую декаду сентября.
Начало цветения, как правило, приходится на первые числа августа,
но возможно и в июле. Раньше всех обычно приступают к цветению на
скошенной степи Leontodon hispidus, Nonea pulla, Plantago media, Tri-
folium pratense и др.
Конец цветения резко зависит от температурного режима сентября.
Но в этот первый месяц осени отцветает основная масса видов, а отдель*
ные растения встречаются в цветущем состоянии и в октябре.
16?
В более поздние сроки протекает вторичное цветение на нескашивае-
мых участках.
Однако и здесь наибольший подъем цветения приходится на август и
первые дни сентября.
Как в нормальном, так и вторичном цветении наблюдаются разно-
годичные колебания, связанные, с одной стороны, с особенностями метео-
рологических условий, а с другой — с эколого-биологическими свойства-
ми видов. Общая периодичность цветения, присущая травянистым рас-
тениям в такой же степени, как и древесным, накладывает печать и на
вторичное цветение. О разногодичных изменениях вторичного цветения
свидетельствуют следующие факты. Из четырех лет наблюдений только
в 1960 г. исключительно обильно цвел на косимой степи Galium boreale.
Во второй половине августа этого года западинки в Стрелецкой и Ка-
зацкой степях буквально белели от массового цветения подмаренника.
Напротив, резкая вспышка вторичного цветения Arenaria ucrainica
отмечена в 1959 г., тогда как в последующие она цвела менее
обильно.
В результате вторичного цветения у ряда растений завязываются
вполне жизнеспособные семена, которые частично распространяются еще
осенью или уходят под снег. По наблюдениям в 1960 г. зрелые плоды
с семенами на побегах, сформировавшихся от пеньков скошенных, завя-
зались у Carduus hamulosus, Erigeron acer, Pieris hieracioides, Scabiosa
ochroleuca, Tragopogon orientale и др. В настоящем вторичном цвете-
нии— при развитии побегов из почек и розеток на корневищах — зрелые
плоды обнаружены у Anthericum ramosum, Helichrysum arenarium, Juri-
nea arachnoidea, Leontodon hispidus, Linaria vulgaris, Nonea pulla, Ono-
brychis arenaria, Plantago media, Polygonum bistorta, Ranunculus acer,
Stachys recta, Taraxacum officinale, Trifolium montanum, Viola mirabilis
и др. Вследствие вторичного роста и цветения исходных генеративных
побегов развились зрелые плоды с семенами у Campanula rotundifolia,
Knautia arvensis, Scabiosa ochroleuca и др.
В систематизации знаний о вторичном цветении необходимо в даль-
нейшем более обстоятельное накопление фактических данных о сроках
начала, массового и конца вторичного цветения, возможности образова-
ния зрелых плодов и семян. Большой интерес представляет учет обилия
вторично цветущих растений в различных местообитаниях (косимой и
некосимой степи, логах, на лесных полянах и в лесах) и выявление био-
морфологических типов вторичного цветения. Эти данные позволят бо-
лее объективно оценить место и значение вторичного цветения в общем
ритме сезонного развития растительности. Изучение вторичного цвете-
ния за ряд лет на специально выделенных пробных площадях, достаточ-
но обширных, даст возможность судить о характере разногодичной из-
менчивости вторичного цветения и ее причинной обусловленности.
Изложенным сделана лишь первая попытка наметить круг вопросов
по вторичному цветению, подлежащих изучению, и их частичное разре-
шение на примере растительности центральной лесостепи. Вполне оче-
видно, что затронутые вопросы сохраняют свое значение и в отношении
других растительных формаций. Особого внимания заслуживает изуче-
ние причин вторичного цветения, раскрытие которых необходимо для со-
знательного использования в практических интересах способности расте-
ний к вторичному цветению и плодоношению, в создании более продук-
тивных форм полезных растений.
166
выводы
1. Ритмы цветения растений Стрелецкой и Казацкой степей имеют
незначительные отличия. На последней, как правило, фазы цветения на-
ступают на несколько (1—3—5) дней раньше. Это обстоятельство вместе
с наличием многообразных отклонений в ритмах цветения одних и тех
же видов .на обеих степях позволяет заключить, что,Стрелецкая и Казац-
кая степи в настоящее время являются территориально обособленными
участками одного и того же типа растительности (луговых степей) и осо-
бенности в ритмике цветения не выходят за рамки этого типа.
2. Большинству видов абсолютно заповедных участков Стрелецкой
степи свойственно некоторое запаздывание сроков прохождения фенофаз
цветения, в пределах 1—5 суток с отдельными более резкими отклоне-
ниями. Однако кривые цветения косимой и некосимой степи не имеют
существенных различий. Этот факт, а также наличие разнообразных
отклонений в ритмах цветения дает основание полагать, что нет принци-
пиальных различий в ритмах цветения растений косимых и некосимых
участков степи. Данный вывод ни в коей мере не распространяется на
состояние аспективности тех и других участков.
3. Условия культуры оказывают определенное влияние на ритмы цве-
тения луговостепных растений. Реакции растений на эти условия весьма
индивидуальны, что создает предпосылки к более глубокому познанию
экологической природы растений. Этим же целям могут служить и срав-
нительные наблюдения за цветением растений некосимой степи.
4. Ритмика цветения растительности Стрелецкой степи носит пере-
менный характер. В сухие годы с ранней и теплой весной она прибли-
жается по своему характеру к ритмике цветения южных степей. Во влаж-
ные годы с поздней и холодной весной ритмика цветения уподобляется
той, которая свойственна лугам лесной зоны. Основными показателями
«степного» типа ритмики будет приуроченность максимума зацветаю-
щих видов к III декаде мая, а наибольшего числа цветущих видов —
к I декаде июня. «Луговой» тип ритмики цветения характеризуется мак-
симумом зацветающих видов во II декаде июня, цветущих видов —
в I декаде июля. На основании анализа средних многолетних данных по
ритмам цветения главных компонентов Стрелецкой степи (137 видов)
выведен константный график цветения растительности, в котором макси-
мум зацветающих видов приходится на I декаду июня, цветущих — на
II и отцветающих—на III декаду июня. Таким образом, по указанным
фенологическим константам, растительность Стрелецкой степи занимает
промежуточное положение между южностепным и луговым типами рас-
тительности. Установленная закономерность в фенологическом развитии
Стрелецкой степи аналогична той, которая обоснована в отношении вод-
ного режима мощных черноземов (Афанасьева, 1962), и является, не-
сомненно, частным случаем более общей природной закономерности, ко-
торой подчинены многообразные явления и процессы, протекающие в
лесостепном ландшафте в его центральной полосе.
5. В цветении растений травяного покрова дубовых лесов наиболее
характерной чертой является приуроченность абсолютного максимума
зацветающих видов ко II декаде июня, а цветущих — к I декаде июля.
В годы с резкими погодными аномалиями эти соотношения могут изме-
няться.
6. Абсолютный максимум зацветающих видов дна и северных скло-
нов логов за годы наблюдений приходится на I декаду июня, а цвету-
щих — на III декаду июня.
7. Эколого-ценотические условия вызывают заметные сдвиги в сро-
ках цветения. В ряду сообществ: косимая степь — дубовый лес — расти-
тельность дна и северных склонов логов у одних и тех же видов наиболь-
167
шие отклонения в фазах цветения наблюдаются в дубовом лесу, менее
выражены в степи и логах. Надо подчеркнуть, что указанные сдвиги во
времени на разных фазах цветения нередко имеют различную величину.
8. По продолжительности цветения видов в составе растительности
Стрелецкой степи и травяного покрова дубрав преобладают коротко-
цветущие растения (45 и 46,6%), а на дне и северных склонах логов —
длительноцветущие (50,7%). Во всех исследованных ценозах обнаруже-
ны также виды промежуточного значения, у которых в разные по усло-
виям годы цветение то растягивается во времени (свыше одного месяца) ,
го сокращается и укладывается в период меньше одного месяца.
9. Проведенное обобщение наблюдений по ритмам цветения растений
и растительных сообществ заповедника в средней полосе лесостепи яв-
ляется необходимой основой для дальнейшего анализа причинной обу-
словленности ритмов развития растений в системе многосторонних свя-
зей их с условиями внешней среды и зоо-фитоценотическим окружением.
10. В растительных сообществах лесостепи вторичное цветение на-
блюдается у большого числа видов и представляет закономерное, часто
повторяющееся явление.
11. Все разнообразие форм вторичного цветения объединяется в три
основных типа: продленное, нормальное и неотеническое.
12. Вторичное цветение растений стимулируется искусственными фак-
торами, разного рода резкими аномальными климатическими факторами,
а также экологическими условиями, благоприятствующими росту и раз-
витию растений. Последняя причина играет ведущую роль в возникно-
вении массового вторичного цветения растений лесостепи.
Глава 7
Эколого-биологический состав компонентов
основных растительных группировок
центральной лесостепи
В этой главе проводится анализ эколого-биологического состава ком-
понентов растительности центральной лесостепи по признакам надзем-
ных органов. Сделана также попытка установления коррелятивной связи
в развитии надземных органов с некоторыми биоморфологическими осо-
бенностями подземных органов.
Ниже приводятся почти исчерпывающие списки видов травянистых
растений луговой степи, дубовых лесов и дна логов с указанием для
каждого вида его фитоценотической роли, т. е. принадлежности к ком-
понентам разных степеней, и различных биоморфологических признаков.
Среди них учтены основные типы структур надземных органов, способы
возобновления побегов, принадлежность растений к ярусам надземной
части растительности, продолжительность вегетации, способы перезимов-
ки, количество генераций листьев и побегов за вегетационный период,
ритмы цветения, сроки отмирания листьев в основании генеративных по-
бегов (на всем их протяжении) и полного усыхания побегов, а также их
средняя высота в 1962 г. Биоморфологический состав растительных груп-
пировок лесостепи по указанным признакам приводится для всего видо-
вого богатства и основных компонентов.
В таблицах биоморфологического состава приняты следующие условные обозна-
чения:
По типам структуры надземных органов: бр — безрозеточные, пр — полурозеточные,
р — розеточные растения.
По способам возобновления побегов: м — моноподиальные, с — симподиальные.
По принадлежности к ярусам надземной части растительности: цифрами указаны
номера ярусов, из них 1-й — срмый высокий, далее — в убывающем порядке; дополни-
тельная цифра, заключенная в скобки, обозначает номер яруса, в котором расположена
основная масса листьев растения.
По продолжительности вегетации: д — длительновегетирующие растения; коротко-
вегетирующие подразделяются на полуэфемероиды (пэд), эфемероиды (эд), геоэфеме-
роиды (гэ), растения среднепродолжительнрй вегетации (с), монокарпики (м), озимые
эфемеры (оэ), яровые эфемеры (э), эфемеры летне-осенней вегетации (эло).
По способам перезимовки: л — летнезеленые, з — зимнезеленые, в — вечнозеленые
растения.
По количеству генераций листьев и побегов в вегетационном периоде: 1 — растения
с одной генерацией; 2о — с основной одной и осенней генерацией побегов возобновле-
ния; 1р — с одной генерацией побегов, характеризующихся растянутым ростом; 2 — ра-
стения с двумя генерациями; 2в — с двумя генерациями, причем вторая представлена
побегами обогащения (у безрозеточных растений).
Ритмы цветения обозначены цифрами, соответствующими периодам вегетационного
сезона: 1—ранневесенний (оканчивающийся 5—10 мая); 2 — средневесенний (15—
20 мая|; 3 — поздневесенний (25—31 мая); 4 — раннелетний (30 июня); 5 — среднелет-
ний (31 июля); 6 — позднелетний (31 августа); 7 — раннеосенний (10—15 сентября);
8 — среднеосенний (25 сентября); 9 — позднеосенний, длящийся до наступления отрица-
тельных температур; сроки отмирания листьев в основании генеративных побегов, па
всем их протяжении и полного усыхания побегов приводятся по декадам, исчисляемым
от начала года.
Принципы выделения основных компонентов и расчленения их на три категории
изложены нами ранее (Голубев, 1962в).
W
ЛУГОВАЯ СТЕПЬ
В составе растительности Стрелецкой степи (на плакоре, в западин-
ках и южных склонах логов) учтено 270 видов цветковых растений и
один вид мха (Thuidium abietinum). Биоморфологическая характеристи-
ка антофитов приводится в табл. 8.
Состав растений Стрелецкой степи по основным типам структуры
надземных побегов представлен в табл. 9. Наиболее обильную видами
группу составляют полурозеточные растения (55%). Второе место зани-
мают безрозеточные (37%), третье — розеточные растения (8%). Это
соотношение сохраняется для компонентов в целом и их отдельных кате-
горий. Только в составе компонентов первой категории количество без-
розеточных растений равно числу розеточных.
Подавляющее большинство видов являются симподиальными (93%)
(табл. 10). На долю моноподиальных приходится только 7% от всего
видового разнообразия. Сходная пропорция наблюдается и среди основ-
ных компонентов, а компоненты первой категории все без исключения
возобновляются симподиально.
Распределение видов по ярусам надземной части растительности до-
вольно равномерное. Оно сохраняет свое значение и в отношении компо-
нентов. Но принадлежность к ярусам компонентов первой категории за-
метно отличается. Более всего насыщенным оказывается 3-й ярус, а во
2-м и 4-м ярусах эти компоненты отсутствуют (табл. 11).
В надземной части растительности косимой степи выделяется всего
семь ярусов. Виды луговой степи во взрослом состоянии при косимом
режиме относятся только к одному ярусу, соответствующему линейным
размерам их генеративных побегов. В составе ярусов находятся следу-
ющие наиболее обычные растения.
1-й ярус (свыше 60 см): Agropyron intermedium, Asparagus officinalis, Bromus iner-
mis, B. riparius, Calamagrostis epigeios, Centaurea scabiosa, Delphinium cuneatum и др.
2-й ярус (50—60 см): Achyrophorus maculatus, Knautia arvensis, Linum perenne,
Onobrychis arenaria, Stipa pennata, St. stenophylla, Tragopogon orientate, Valeriana ros-
sica и др.
3-й ярус (40—50 см): Euphorbia semivillosa, Filipendula hexapetala, Galium verum,
Leucanthemum vulgare, Poa angustifolia, Vicia tenuifolia и др.
4-й ярус (30—40 см): Adonis vernalis, Arenaria ucrainica, Asperula cynanchica,
A. tinctoria, Geranium sanguineum, Orobus pannonicus, Ranunculus polyanthemus, Sene-
cio integrifolius, Stellaria graminea и др.
5-й ярус (15—30 см): Carex michelii, Iris aphylla, Myosotis suaveolens, Poly gala
comosa, Primula veris, Pulsatilla patens и др.
6-й ярус (5—15 см): Carex humilis, Centaurea marschalliana, Fragaria viridis, Hya-
cinthella leucophaea, Potentilla alba, P. humifusa, P. patula, Viola rupestris и др.
7-й ярус (0—5 см): моховой покров из Thuidium abietinum.
Цифры, обозначающие предельную высоту того или иного яруса,
нельзя принимать в качестве абсолютных, так как в различные по кли-
матическим условиям годы развитие надземных побегов у одних и тех
же видов может изменяться. Возможны колебания высоты в зависимо-
сти от условий местообитания. Но несмотря на это, соотношения в про-
странственном расположении надземных органов растений бывают
весьма устойчивыми, т. е. указанные 7 ярусов представляют явление
довольно постоянное, закономерное. Несколько беднее структурное по-
строение фитоценозов некосимой степи.
Основная масса листьев растений располагается в разных участках (пятнах) в го-
ризонте от 30 до 60 см высоты, выше которого выдвигается 1-й ярус (свыше 1 м высо-
ты); в него входят побеги Asparagus officinalis, Agropyron intermedium, Bunias orienta-
Us, BrQmus inermis, Centaurea scabiosa, Cichorium intybus, Clematis recta, Dactylis glo-
merata, Delphinium cuneatum, Libanotis intermedia, Peucedanum oreoselinum, Veratrum
tagrum, Verbascum lychnitis и др. В качестве особого подъяруса (до 80 см высоты)
можно выделить генеративные побеги Anthericum ramosum, Centaurea jacea, Falcaria
i/ulgaris, Knautia arvensis, Onobrychis arenaria, Scabiosa ochroleuca, Senecio jacobaea,
Serratula inermis, Stipa pennata, St. stenophylla и др.
iVd
Таблица 8
Биоморфологический состав компонентов растительности Стрелецкой степи
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов• Принад- лежность к ярусам раститель- ности 4 Продол- житель- ность вегетации Способ перези- мовки » Количест- во генера- ций листьев и побегов Ритм цве- тения •• Срок отмирания (декады от начала года) ••• Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
Achillea millefolium K2 ПР С 3 Д В 2 4—6 15 25 27 45
А. nobilis пр С 3 Д в 2 5—7 18 25 29 33
Achyrophorus maculatus .... Кз пр С 2 пэд л 2 4 21 72
Acinos thymoides Кз бр 6 оэ 3 1 4—5 17 20 — 19
Aconitum nemorosum , . бр С 2 Д л 1р 5—8 18 27 29 48
vernalis Кз бр С 4 Д л 1 1-3 19 22 27 13
Agrimonia eupatoria пр С 2 Д л 1 5—6 19 26 28 51
Agropyron intermedium .... К1 пр С 1 Д л 1 5 19 26 28 105
A. repens пр С 1 Д л 1 >4-5< 18 26 28 99
Agrostis syreistschikowii .... Кз пр с 4 Д в 2 4 16 23 26 39
А №a genevensis Кз пр с 6 пэд в 2 3-4 16 1 20 22 23
Allium flavescens р с 5 Д л 1 5-6 24 — 26 25
A. oleraceum Кз р с 4 эд 3 2о >5—6 20 — 24 41
A. rotundum р с 3 с 3 1 >4—5 21 — 25 37
Alopecurus pratensis пр с 3 пэд в 2 3 15 21 22 48
Alyssum calycinum бр 6 оэ 3 1 2—4 16 ’ 18 20 13
A. desertorum бр 6 оэ 3 1 2-4
A nd rosace septentrionalis .... Кз р 6 оэ 3 1 2-3 15 — 19 13
• Пропуски в отметке способов возобновления относятся к монокарпическим растениям — однолетникам и двулетникам.
* • Значок > обозначает, что цветение начинается не с начала соответствующего периода вегетации, отмеченного цифрами, а смещено примерно на f/« периода
в сторону более поздних сроков; значок < обозначает, что цветение оканчивается раньше конца соответствующего периода вегетации, примерно на V* его часть.
• •• Для более легкой ориентировки в сроках отмирания укажем, что декады с 13 по 15 являются соответственно 1, 2 и 3-й декадами мая, декады 16—18 — июня,
19—21 — июля, 22—24 — августа, 25—27 — сентября, 28—30 — октября и т. д. В трех графах «Сроки отмирания» тире (—) обозначает, что данное явление (признак)
отсутствует в силу биологических особенностей вида, а пропуск — об отсутствии у нас соответствующих наблюдений.
a
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Anemone silvestris Кз пр С 4
Antennaria dioica пр С 6
Anthemis tinctoria ....... пр С 4
Anthericum ramosum Кз Р С 2
Anthoxanthum odoraium .... пр С 4
Anthriscus silvestris пр С 1
Anthyllis polyphyIla Кз пр 5
Antitoxicum officinale Кз бр С 4
Arenaria serpyllifolia Кз бр 6
A. ucrainica Кз пр С 4
A rrhenatherum elatius пр С 1
Artemisia absinthium бр С 1
A. austriaca бр С 4
A. campestris бр С 1
Л. latifplia пр С 2
A. scoparia пр 3
A. vulgaris бр С 1
Asparagus officinalis К, бр С 1
Asperula cynanchica Кз бр С 4
A. tinctoria Кз бр с 4
Aster amellus бр с 3
Astragalus cicer бр с 3
A. danicus Кз бр с 6
Таблица 8 (продолжение)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала ^ода) Средня» высота (за 1962 г.) генератив- - ных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
пэд Л 2 2-3 17 __ 22 30
Д В 1 3 20 15
Д В 2 4-5 16 23 26 39
д Л 1 5 26 — 26 65
пэд 3 2 3—4< 15 19 20 30
м 3 2 4 17 20 21 90
с в 2 4 17 21 23 30
д л 1 4-5 19 26 27 32
оэ 3 _ 1 3—4 16 18 19 6
ЭД 3 2 3—4< 15 19 21 38
д в 2 4 16 20 22 90
д в 2в 6 17 27 29 99
д в 2в 6—7 19 27 30 38
Д л 2в 6-7 19 25 30 96
д л 1 6 19 27 28 65
д л' 2 6-7 20 26 , 30 81
д л 2в 6 18 27 29 149
д л 1 4 25 28 89
д в 2о 4—5 18 25 29 23
д л 1 4 18 23 27 40
д л 2в 6—8 19 28 29 52
д л 2в >4-5 18 26 29 44
д в 2 4 18 27 29 20
Вид Основ- ной компо- нент Тнп структуры надземных побегов Способ {возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Barbarea vulgaris пр 4
Berteroa incana к» пр 4
Betonica officinalis пр м 2
В r achy podium pinnatum . . . пр С 2
Briza media Кз пр с 3
Bromus inermis Кз пр с 1
B. japonicus пр 5
B. mollis пр 4
Bromus riparius К1 пр с 1 ‘
Bunias orientalis Кз пр с 1
Bupleurum falcaium бр с 3
Calamagrostis epigeios .... Кз пр с 1
Campanula bononiensis Кз пр с 2
C. glomerata пр с 2
C. patula Кз пр 4
C. persicifolia Кз пр с 2
C. rapunculoides бр с 2
C. rotundifolia пр м 4
C. sibirica Кз пр 4
Carduus hamulosus Кз пр 1
C. nutans пр 2
Carex humilis К1 р с 6
C. michelii Кз р с 5
Таблица 8 (продолжение)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев н побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов в СЖ
листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
М 3 2 2—3 15 18 20 26
м 3 2 3—5 15 19 25 30
А В 2 5-6 26 26 54
А Л 1 >4—5<^ 18 24 ' 24 68
пэд В 2 4 16 19 20 44
А л 1 >4-5< 18 26 29 91
оэ 3 1 4 15 20 21 29
оэ 3 1 4 16 20 21 47
пэд 3 2 4 16 19 23 70
А л 2 4 17 21 24 72
А л 2 5-7 20 27 30 56
А л 1 5 19 26 27 105
Д л 1 5 18 24 26 78
А л 2 4-6 17 26 27 47
м 3 2 4 15 18 21 41
А в 2 >4-5< 17 20 24 64
А л 1 5-6 18 23 26 100
А в 20 4—6 — » 27 29 42
м 3 2 4—5< 16 20 23 43
м 3 2 4—5 16 21 24 78
д в 2 >4—5 18 23 26 112
Д в 1 1 15 — 16 3
А в 1 2-3 19 — 22 15
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Carex praecox Кз ПР С 5
C. verna Кз Р С 6
Carum carvi пр 4
Centaurea jacea пр С 3
C. marschalliana Кз пр м 6
C. pseudomaculosa пр 2
C. ruthenica пр с 1
C. scabiosa Кз пр с 1
Cerastium caespitosum бр 6
Cichorium intybus пр с 1
Cirsium arvense бр с 1
C. pannonicum пр с 2
C. polonicum пр 1
Clematis recta К» бр с 1
Convolvulus arvensis Кз бр с 3
Coronilla varia Кз бр с 3
Crepis pannonica пр с 1
C. praemorsa р с 4
C. tectorum пр 4
Cynoglossum officinale пр 2
Cytisus ruthenicus Кз бр 1
Dactylis gio me rat a пр с 1
Daucus carota пр 3
Таблица 8 (продолжение)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
Д В 2 2—3 19 22 21
Д В 1 1—2 15 — 17 8
м Л 2 3—4 17 20 21 50
д Л 2 5-6 17 25 27 64
д Л 2 3—4< — 18 19 10
д 3 2 5—6 16 24 29 70
д Л 2 4—5 17
д л 2 5 17 27 28 79
оэ 3 2 3-4 16 21 24 18
д л 2 5-6 20 25 27 98
д л 2в 5—6 19 27 28 81
д л 2 >4-5 17 22 26 99
д л 2 6—7 20 26 27 110
д л 1 4 18 26 26 106
д л 2 4—6 18 26 26 49
д в 2о >4-5 18 27 28 41
д л 2 5—7< 21 27 27 99
с л 1 4 19 50
э л 1 4—6< 16 22 24 40
м л 2 4 17 21 22 59
Д л 2в 2-4< 18 29 25
Д в 2 4 16 21 24 73
Д л 2 5—7 19 25 28 56
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Delphinium cuneatum Кз бр С 1
Dianthus andrzejowskianus . . Кз пр С 3
D. deltoides бр с 6
Dr aba nemorosa Кз пр 6
D. sibirica Кз пр с 6
Dracocephalum ruyschiana . . Кз бр с 5
D. thymiflorum бр 6
Echium rubrum Кз пр 2
Elisanthe viscosa пр 2
Erigeron acer Кз пр с 5
Eryngium planum пр с 3
Erysimum marschallianum . . Кз пр 2
Euphorbia segueriana Кз бр с 4
E. semivillosa Кз бр с з •
E. subtilis Кз бр с 4
E. virgata бр с 3
Euphrasia tatarica бр 6
Falcaria vulgaris Кз пр 2
Festuca pratensis Кз пр с 2
F. sulcata Кз пр с 4
Filipendula hexapetala К1 пр с 3
Fragaria viridis Кз р с 6
Gagea erubescens Кз пр с 6
Таблица 8 (продолжение)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генератив- ных побегов (полное усыхание)
Д Л 1 5—6< 18 26 26 НО
ПЭД 3 2 4 16 21 26 62
пэд 3 2 4—5 17 21 24 23
ОЭ 3 1 1—3 15 16 18 10
пэд 3 2 1—3 17 18 18 9
Д Л 2в 4—5< 17 27 27 29
ОЭ 3 1 3-4 16 20 22 23
д Л 2 4 17 27 28 64
м 3 2 4 16 19 21 66
ЭД 3 2 4 17 21 22 23
Д Л 2 5—7 21 25 27 42
м 3 2 4—5< 15 19 24 61
Д Л 2в 4 17 22 27 38
д Л 2в 3-4< 16 25 27 42
пэд Л 2в 3—4<^ 16 19 24 31
Д Л 2в 3—4 17 20 27 48
эло л 1 6-8 22 27 30 И
д л 1 5 -6 19 25 27 66
пэд 3 2 4 16 20 23 48
д в 2о 4 16 18 20 33
д в 2 4 20 22 25 64
д в 2 3—4< 22 10
гэ л 1 1 15 16 16 6
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Galium boreale Ks бр С 4
G. mollugo бр С 3
G. verum K1 бр с 3
Genista tinctoria бр с 3
Gentiana cruciata пр м 4
Geranium pratense пр с 4
G. sanguineum Кз пр с 4
Gypsophila paniculata . ... бр с 2
Helichrysum arenarium Кз пр с 5
Helictotrichon pubescens .... Кз пр с 2
H. schellianum Кз пр с 3
Heracleum sibiricum пр с 1
Hieracium bauhinii К, пр с 2
H. cymosum пр с 2
H. fallax Кз пр с 2
H. florentinum Кз пр с 2
H. pilosella р с 6
H. umbellatum бр с 1
H. virosum бр с 1
Hyacinthella leucophaea .... Кз р с 6
Hypericum elegans бр с 4
H. perforatum Кз бр с 4
Inula ensifolia . бр с 3-
Таблица 8 (продолжение)
Продол- житель- ность . вегетации Способ перези- мовки Количест- во гене- раций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных. побегов, в с.ц
листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
Д Л 2в 4—5< 15 28 28 33
Д В 2в 4—5< 16 22 27 58
Д Л 2в 4—5 16 25 27 54
Д Л 2в 4—5 19 30 — 44
Д л 1 5-6 — 28 29 30
Д л 2 >4-6< 23 26 27 50
Д л 1 4-5 — 26 27 30
Д л 1 5 20 68
Д л 2 5-6 17 25 27 31
пэд 3 2 4< 15 18 20 62
пэд 3 2 4 16 19 21 55
д л 2 >4—5 21 22 23 94
д в 2 4 19 22 23 56
Д л 2 4—5< 72
Д в 2 4 19 22 24 71
Д в 2 4 19 23 24 65
Д в 2 4 — 20 18
Д л 1Р 6—8 20 26 28 92
Д л 1Р 6—8 20; 28 29. 80
гэ л 1 1-3 18 — 17 9,
д в 2о >4-5 20 29 29 35
д в 2о 5-6 18 30 30 51
Д л 1 СП 19 27 27 39
Таблица 8 (продолжение)
12 в. Н. Голубев
Срок отмирания
Принад- лежность к ярусам раститель- ности Количест- во генера- ций листьев и побегов (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Продол- житель- ность вегетации Способ перези- мовки Ритм цве- тения листьев у основа- ния гене- ративных побегов листьев генератив- ных побегов генератив- ных побегов (полное усыхание)
Inula hirta I, salicina Кз бр бр с с
Iris aphylla Кз Р с
Jurinea arachnoidea Кз пр
Knautia arvensis Кз пр с
Koeleria gracilis Кз пр с
Lappula myosotis пр
Lavatera thuringiaca бр с
Leontodon autumnalis пр с
L, hispidus Кз Р с
Leucanthemum vulgare Кз пр с
Libanotis intermedia .... Кз пр
Linaria vulgaris бр с
Linosyris vulgaris бр с
Linum flavum бр с
L. nervosum бр с
L. perenne Кз бр с
Lithospermum arvense пр
L. officinale бр с
Lotus corniculatus Кз бр с
Luzula pallescens Кз пр с
Medicago falcata Кз бр с
M. lupulina бр
4 Д л 1 4—5< 18 27 27 32
4 д л 1 >4—6< 18 27 27 51
5 д л 1 3—4< 25 — 23 21
3 м л 2 >3—4< 17 19 20 49
2 д л 2 4—5 16 24 26 78
3 д в 2о 4 16 19 20 45
4 оэ 3 1 >3-6< 15 21 24 31
1 д л 1 5-6 18 30 30 92
6 д л 2 5-8 — 28 35
4 пэд л 2 4 — 19 46
3 д в 2 4—5< 16 23 26 52
1 д л 1 >4—5 20 22 24 87
3 д л 2в 5-6 18 26 28 52
3 д л 1р 7—8 18 28 29 51
4 д л 2в 4—5 18 30 30 48
3 д л 2в 4 17 26 27 51
2 д в 2 4 18 27 29 68
5 оэ 3 1 2—4 15 19 21 18
3 д л 1 4—5< 17 25 27 49
5 д л 2в >4—6 21 30 30 24
6 эд 3 2 3 16 18 21 21
3 д л Зр 4—6 19 28 29 32
5 оэ 3 1 4—6 19 24 26 11
Таблица 8 (продолжение)
178
Принад- лежность к ярусам раститель- ности Количест- во генера- ций листьев и побегов Срок отмирания (декады от начала года) Средняя
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Продол- житель- ность вегетации Способ перези- мовки I Ритм цве- тения листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генератив- ных побегов (полное усыхание) высота (за 1962 г.) генера- тивных побегов в см
Medicago romanica бр С 3 Д Л *Р 4—6 19 28 28
Melampyrum cristatum . . . Ка бр 5 Э Л 1 4 17 20 20 22
Melandrium album бр С 2 д 3 2в 4—5 18 27 29 60
Melica transsilvanica пр С 2 д в 2о >4 17 19 21 58
Melilotus albas бр 3 д л 1 5-6 19 24 30 57
M. officinalis бр 2 д 3 1 >4—6< 18 23 27 91
Myosotis suaveolens Ki пр С 5 эд 3 2 2—4< 16 18 19 17
Nepeta pannonica бр С 2 д л 1 >4-5 17 24 24 78
Nonea pulla Кз пр С 5 д л 2 2—6 15 27 28 13
Odontites serotina Кз бр 5 эло л 1 6—7 22 27 29 22
Oenothera biennis пр 1 м 3 2 >4-5 16 24 26 85
Onobrychis arenaria Ка бр С 2 Д л 2в 4—5 17 29 29 36
Orobus pannonicus Кз бр С 4 с л 1 2—3 18 24 27 30
Paeonia tenuifolia бр С 5 с л 1 >2—3 19 26 27 25
Pastinaca Silvestris пр 1 д л 1 5-6 19 24 26 88
Pedicularis kaufmannii Кз пр С 4 гэ л 1 4 18 19 20 33
Peucedanum lubimenkoanum . . пр 1 д л 1 5-6 24 26 29 122
P. oreoselinum Кз пр с 1 д л 1 5—6< 24 24 25 103
Phleum phleoides Ка пр с 3 пэд 3 2 >4 15 19 21 48
Ph. pratense пр с 2 Д в 2 >4—5< 17 24 26 61
Phlomis tuberosa Кз пр с 2 Д л 1 4-5 20 23 25 73
Pieris hieracioides Ка пр с 2 Д л 2 5—6 19 23 28 57
Plantago lanceolata Кз р м 5 Д в 2 3—4 — 22 29
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
6ZI
Plantago media Кз р м 5
Р. stepposa р м 4
Роа angustifolia Кз пр с 3
Р. bulbosa , пр с 4
Polygala comosa Кз бр с 5
Polygonatum officinale .... Кз бр с 5
Potentilla alba Кз пр м 6
P. argentea Кз пр с 5
P. humifusa Кз пр м 6
P. patula Кз пр м 6
P. recta пр с 5
Primula veris Ki р с 5
Prunella grandiflora бр с 5
P. vulgaris бр с 6
Pulmonaria angustifplia .... пр с 5
Pulsatilla patens .' . Кз р с 5
Pyrethrum corymbosum . . . . пр с 2
Ranunculus illyricus пр с 4
R. pedatus пр с 5
R. polyanthemus Кз пр с 4
Rhinanthus major Кз бр 5
Rumex acetosa Кз пр с 2
R. acetose/la пр с 4
Таблица 8 (продолжение)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генера- тивных побегов в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыхание)
Д В 2 3-4 - 21 24
Д Л 2 4—5< — 21 54
пэд В 2 4 16 18 19 51
эд 3 1 4< 16 18 19 28
с В 2 3—5< 17 23 27 21
Д Л 1 4 18 26 27 30
д в 2 2-3 — 17 17 7
д в 2 4—5 17 25 29 23
д в 2 1—3 — 17 17 7
д в 2 2—4< — 18 18 10
д в 2 4—5 19 24 26 24
пэд 3 2 1-3 23 — 20 19
Д л 2в >4—6< 20 27 27 31
Д в 2 4—5 20 24 24 12
Д л 2 1—3 16 17 18 12
Д л 1 1 26 — 17 18
д л 1 4-5 22 25 26 72
гэ л 1 4< 16 18 19 36
гэ л 1 2-3 15 16 16 14
пэд л 2 3-4 23 24 35
э л 1 4 17 19 20 ’ 33
эд 3 2 3—4< 16 18 19 68
д 3 2 3—4 16 22 26 35
g
Вид; Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Rumex confertus пр С 1
R. thy r si ftorus пр С 1
Salvia nutans . . Р С 1
S. pratensis К1 пр С 3
S. verticillata бр С 3
Sanguisorba officinalis .... пр м 1
Scabiosa ochroleuca Кз пр с 2
Scorzonera purpurea Кз пр с 3
S. taurica пр с 3
Scutellaria hastifolia .... бр с 6
Sedum acre Кз бр с 6
S. telephium бр с 4
Senecio integrifolius Кз пр с 4
S. jacobaea Кз пр 2
S. schwetzowii пр с 1
S. erucifolius пр с 1
Serratula heterophylla Кз пр с 2
S. inermis пр с 2
Seseli annuum пр 3
Silene parviflora пр 3
S. chlorantha пр 3
S. cucubalus бр с 3
S. nutans Кз пр с 3
Таблица 8 (продолжение)
Продол- житель* кость вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генера- тивных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генератив- ных побегов (полное усыхание)
ПЭД Л 1 4 17 21 23 97
Д Л 2 4 20 25 26 77
Д Л 1 4 25 — 23 71
пэд Л 2 3-4 21 24 24 49
Д Л 2в >4—6< 19 24 26 42
Д Л 1 5-7 26 28 29 83
д В 2 >4-6< 17 24 28 63
д Л 1 4 22 26 27 40
д Л 1 4
д Л 1 4-5 18 29 29 22
пэд 3 2в 4—5< 18 20 21 9
д Л 1Р 6—7 18 27 28 40
ЭД 3 2 3—4< 16 18 19 38
Д 3 2 >4-6< 18 24 27 65
Д Л 1 5—6 24 26 28 112
д Л 1Р 6-8 19 27 30 109
ЭД Л 2 4 16 19 21 83
д Л 1 6-7 21 25 29 69
Д Л 2 5-6 22 26 30 76
м 3 2 4 16 20 24 87
д Л 2 4
с Л 1 4-5 18 23 24 58
пэд в 2 4 16 21 22 47
ВИД Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов
Sisymbrium polymorphum . . . пр С
Solidago virgaurea пр С
Stachys recta К1 бр С
Stellaria graminea Ka бр с
Stipa capillata пр с
St. dasyphylla пр с
St. pennata Ka пр с
St. stenophylla ........ Ka пр с
Tanacetum vulgare бр с
Taraxacum officinale K3 р с
Thalictrum minus Ka бр с
Th. simplex бр с
Thesium ebracteatum бр с
Th. ramosum бр с
Thymus marschallianus .... Ka бр с
Tragopogon ma jus пр
T. orientate Ka пр
Trifolium alpestre K8 бр с
T. medium бр с
T. montanum Ka пр м
T. pratense Кз пр м
T. repens Кз бр м
T, strepens ......... бр
Таблица 8 (продолжение)
Принад- лежность к ярусам раститель- ности Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генератив- ных побегов (полное усыхание)
2 С Л 1 3-4 15 20 25 64
2 Д Л 1 >5-7 24 27 29 70
3 Д Л 2в 4—5< 18 27 29 55
4 ЭД 3 2в 3-5 15 19-21 21—23 20
1 Д Л 1 5 21 24 26 89
2 д л 1 4< 17 18 19 61
2 д л 2 4< 17 18 19 66
2 д л 2 >4 17 19 21 81
1 д л 2в 5—6 18 29 29 84
6 д в 2 1—3 — 17 И
2 д л 1 4-5 21 27 29 76
2 д л 1 5 19 27 28 92
6 д л 1 2—4< 18 21—29 23—30 21
6 пэд л 1 3-4 18 21—28 23-28 19
5 д в 2в 4-5 22 22 23 19
3 оэ л 1 4 18 20 24 77
2 м л 1 4 16 19 23 62
4 д л 2в 4 18 24 26 29
4 д л 1 >4—6 18 28 28 46
3 пэд л 2 4 — 23 23 41
5 пэд 3 2 3-4 — 20 22 21
6 д в 2в 4—5 19 9
5 оэ 3 1 4—6< 18 22 24 26
00 •
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Trinia multicaulis Кз пр 4
Turritis glabra Кз пр 2
Valeriana rossica Кз пр С 2
Veratrum nigrum Кз пр С 1
Verbascum lychnitis Кз пр 1
V. orientate пр 1
V. phoeniceum пр 1
Veronica austriaca Кз бр С 5
V. chamaedrys Кз бр С 5
V. incana Кз бр С 4
V. prostrata Кз бр С 6
Veronica spicata бр С 4
V. spuria бр С 2
V. teucrium бр С 2
V. verna бр 6
Vici a cracca бр С 3
V. tenuifolia Кз бр С 3
Viola hirta Кз р м 6
V. pumila 6
V. rupestris Кз пр м 6
V. tricolor бр 5
Viscaria vulgaris пр с 3
Таблица 8 (окончание)
Продол- житель- ность вегетации Способ перези- мовки Количест- во генера- ций листьев и побегов Ритм цве- тения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, В /7ЛС
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генератив- ных побегов (полное 'усыхание)
М 3 1 3—4< 17 19 22 32
м 3 2 4< 15 18 23 62
ЭД Л 2 >3-4 17 19 20 61
д Л 1 >5—6< 25 — 26 93
д 3 2 >4-5 21 25 27 115
д л 1 5-6 21 25 27 116
с л 1 >3-4 20 — 21 72
д л 1 4 18 24 28 27
пэд 3 2в 3—4< 17 21 24 21
д в 2 5—7 19 28 28 30
пэд 3 2 2-3 16 19-24 24 19
д в 2 >4—6< 21 27 28 27
Д л 1 5-6 18 26 27 45
Д л 1 4-5 18 27 29 44
оэ 3 1 2—4< 15 17 19 5
д л 1 4-6 18 30 30 29
д л 1 4 18 27 27 44
д л 1 1 30 — 23 4
д л 1 2-3
д в 2 1-3 16 30 30 5
оэ 3 1р 1—9 20 30 30 16
д в 2 4 19 22 22 37
Таблица 9
Состав растений Стрелецкой степи по типам структуры надземных побегов
Побег Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Безрозеточный 99 37 43 31 2 22 12 26 29 35
Полурозеточный .... 148 55 78 57 5 56 30 65 43 53
Розеточный 22 8 16 12 2 22 4 9 10 12
Итого. . . . 269 100 137 100 9 100 46 100 82 100
Таблица 10
Состав растений Стрелецкой степи по способам возобновления побегов
Возобновление Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Симподиальное .... Моноподиальное 200 15 93 7 105 10 91,3 8,7 9 100 33 5 86,9 13,1 63 5 92,6 7,4
Итого. . . . 215 100 115 100 9 100 38 100 68 100
Таблица 11
Распределение растений Стрелецкой степи по ярусам в надземной части растительности
Ярус Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество । % коли- чество % коли- чество % коли- чество % коли- чество %
1-й (>60 см) 44 16 15 10,9 2 22,2 5 10,9 8 9,7
2-й (50—60 см) 54 20 26 19,0 — — И 23,9 15 18,3
З-й (40—50 см) 50 19 22 16,1 4 44,5 9 19,5 9 11,0
4-й (30—40 см) 49 18 27 19,7 — — И 23,9 16 19,5
5-й (15—30 см) 35 13 24 17,5 2 22,2 4 8,7 18 22,0
6-й (5—15 см) 7-й (0—5 см). Туидие- вый мох 38 14 23 16,8 1 11,1 6 13,1 16 19,5
Итого.. . . 270 100 137 100 9 J 100 46 100 82 100
183
2-й ярус (до 30—60 см) — лиственный, слагается главным образом листьями ковы-
лей, костров и других злаков, а также разнотравьем, т. е. их вегетативными и генера-
тивными побегами: Achillea millefolium, Asperula tinctoria, Bromus riparius, B. inermis,
Campanula sibirica, Coronilla varia, Galium boreale, G. verum, Geranium sanguineum,
Iris aphylla, Linum perenne, Linar ia vulgaris, Orobus pannonicus, Pedicular is kaufman-
nii, Poa angustifolia, Polygonatum officinale, Salvia pratensis, Stachys recta, Stellaria
graminea, Stipa pennata, St. dasyphylla, St. stenophylla, Trifolium alpestre, T. montanum,
Vicia tenuifolia и др.
3-й ярус (до 20 см) составлен низкорослыми видами: Centaurеа marschalliana, Fra-
garia viridis, Potentilla humifusa, P. patula, Primula ver is, Thymus marschallianus и др.
Ярус мха отсутствует.
Многоярусность растительности луговой степи свидетельствует о
чрезвычайно высокой насыщенности живой органической массой воз-
душной среды обитания, что, в свою очередь, обусловливает высокую
интенсивность круговорота веществ.
Сравнение показывает, что количество ярусов в надземной и под-
земной части растительности совпадает. Таким образом, в известном
смысле получается симметричная картина структурного устройства
ценоза.
Таблица 12
Состав растений Стрелецкой степи по продолжительности вегетации
Тип растения Всего Всего^компонентов категории
видов ‘ компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чест- во % коли- чест- во % коли- чество %
Длительновегетирующие . 178 66,4 82 59,8 5 55,6 29 63,1 48 58,5
Коротковегетирующие: полуэфемероиды . . 27 10,1 23 16,8 3 33,2 7 15,2 13 15,9
эфемероиды .... И 4,1 i 10 7,3 1 11,2 5 10,9 4 4,9
геоэфемероиды . . . 5 1,9 3 2,2 — — — — 3 3,7
среднепродолжитель- ной вегетации:. . . 9 3,4 3 2,2 — — — — 3 3,7
монокарпики .... 16 6,0 9 6,6 — — 3 6,5 6 7,3
озимые эфемеры . . 17 6,3 4 2,9 — — — — 4 4,8
яровые эфемеры . . 3 ' 1,1 2 1,5 — — 2 4,3 — —
эфемеры летне-осен- ней вегетации . . 2 0,7 1 0,7 — — — — 1 1,2
Итого. . . . 268 100 137 100 9 1 100 46 100 82 100
По характеру вегетации преобладающее место занимают длитель-
новегетирующие растения (66,4%), остальные 33,6% приходятся на
коротковегетирующие (табл. 12). Среди коротковегетирующих наибо-
лее обильными являются полуэфемероиды. (10,1%), далее в убываю-
щем порядке идут озимые эфемеры (6,3%), монокарпики (6%), эфе-
мероиды (4,1%), среднепродолжительной вегетации (3,4%), геоэфеме-
роиды (1,9%), яровые эфемеры (1,1%) и летне-осенние эфемеры
(0,7%). Близкие соотношения указанных групп характерны и для
основных компонентов, где только сменились местами эфемероиды,
занимающие третье место (7,3%), монокарпики (6,6%) и озимые
эфемеры (2,9%). Компоненты первой категории относятся к длитель-
новегетирующим (55,6%), полуэфемероидам (33,2%) и эфемерои-
дам (11,2%).
184
В приведенных данных привлекает внимание большое участие в
травостое луговой степи коротковегетирующих растений. По старым
данным В. В. Алехина (1935), и Г. М. Зозулина (1955) они совершенно
отсутствовали.
Массовым по обилию коротковегетирующим факультативным эфе-
мероидом является мох Thuidium abietinum, сплошным напочвенным
ковром покрывающий степень. В ранневесеннее и осеннее время этот
мох, напитанный влагой, пышно разрастается, а в летний сезон сильно
пересыхает и впадает в состояние, близкое к анабиозу. Как известно,
мхи-эфемероиды весьма характерны для аридных областей, в том чис-
ле даже пустынных (Е. П. Коровин, 1958). Туидиевый мох в своем
распространении заходит в более северные от лесостепи районы, а так-
же встречается в южных горах. В этих биотопах он может развиваться
как длительновегетирующее растение. Поэтому наличие у него летней
депрессии в вегетации в условиях луговой степи свидетельствует о спе-
цифичности экологической обстановки последней. Это заметно отли-
чает растительность луговой степи от лугового типа растительности.
Присутствие многочисленных коротковегетирующих растений по-
казывает, что наряду с сохранением живой органической массы в со-
ставе растительности луговой степи происходит также значительное
отмирание надземных побегов и листьев и выпадение их из сферы ас-
симиляции. Поэтому установившееся представление об отсутствии лет-
него перерыва вегетации в луговых степях (Алехин, 1909, 1934, 1936;
Шалыт, 1950; Зозулин, 1955) должно быть заменено дифференциаль-
ным, соответствующим указанным выше особенностям вегетации рас-
тительности. В этом отношении луговые степи заключают в себе неко-
торые признаки луговой и южностепной растительности, а в своей
Т абл ица 13
Состав растений Стрелецкой степи по способам перезимовки
Тип растения Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Летнезеленые 154 57,0 66 48,0 4 44,5 27 58,8 35 42,7
Зимнезеленые .... 58 21,5 33 24,1 3 33,3 7 15,2 23 28,1
Вечнозеленые . . 58 21,5 38 27,9 2 22,2 12 26,0 24 29,2
Итого ... 270 100 137 100 9 100 46 100 82 100
совокупности представляют своеобразное качество, эколого-биологиче-
скую особенность компонентов луговых степей. Следует оговориться,
что распространенное мнение о наличии в настоящих степях перерыва
в вегетации, возникшее под влиянием работ В. Р. Вильямса (1922),
является ошибочным. Исследования, проведенные в южных степях,
убеждают, что и в этом типе растительности развивается значительное
количество видов, вегетирующих беспрерывно в течение всего вегета-
ционного периода (Гурский, 1934, 1945; Балаш, 1961а, б). Конечно,
хорошо выражена и группа растений со среднелетним перерывом веге-
тации. При этом величина отмирающей массы надземных органов в
настоящих степях выше, чем в луговых.
По способам перезимовки выделяются две основные группы: летне-
зеленые, уходящие под снег без зеленых органов, и растения, зимую-
щие в зеленом состоянии. Соотношения этих групп в общем уравно-
185
вешиваются (табл. 13). От всего видового богатства растений летне-
зеленые составляют 57%, остальные 43% приходятся на зимнезеленые
и вечнозеленые, представленные в равном количестве. Среди компо-
нентов число летнезеленых уменьшается до 48%. Еще ниже оно для
компонентов первой категории (44,5%), но заметно возрастает в сле-
дующей группе — компонентов второй категории (58,8%).
Летнезеленые растения более отвечают степному типу климата.
В степях снег зимой подвержен сильному сносу, в результате чего
складываются неблагоприятные условия для перезимовывания зеле-
ных листьев. На сокращение зеленой зимующей листовой поверхности
оказывает влияние и недостаток почвенной влаги, тормозящий процес-
сы роста. В целом зимнее состояние растений луговой степи ярко под-
черкивает ее переходный характер между сообществами лугового типа
в лесной зоне, где процент зимнезеленых растений увеличивается* и
они становятся преобладающей группой, и ценозами настоящих степей,
в которых доминирующее положение занимают летнезеленые виды.
Особенно показательна специфика луговостепной растительности по
количеству генераций листьев и побегов у ее компонентов (табл. 14).
Таблица 14
Состав растений Стрелецкой степи по количеству генераций листьев и побегов
в вегетационном периоде
Всего
Всего компонентов категории
Группа растений видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
101 37,6 42
9 3,4 6
2 22,2 12
С одной генерацией . .
С основной одной и осен-
ней генерациями побе-
гов возобновления . .
С одной генерацией побе-
гов, характеризующих-
ся растянутым ростом
С двумя генерациями
С двумя генерациями (вто-
рая — в виде побегов
обогащения)............
9 3,4
116 43,3
33 12,3 16
Итого ... 268 100 137
30,6
4,4
0,7
52,6
11,7
100
— 1
5 55,6 26
2 22,2 4
9 100 46
6,5 3 3,7
2,2 — —
56,6 41 50,0
8,7 10 12.2
100 82 100
Более половины видов характеризуется развитием второй генерации,
приуроченной к началу второй половины лета (55,6%). Эта цифра еще
больше повышается для компонентов (64,3%), а среди компонентов
первой категории на растения с двумя генерациями листьев и побегов
падает 77,8%. Высокое значение этих растений сохраняется среди ком-
понентов второй и третьей категорий (65,3 и 62,2%).. Таким образом,
в луговой степи резко выражены два импульса новообразования листо-
вой поверхности: весенний и средне-позднелетний. Условия среды не
только позволяют, но и благоприятствуют осуществлению двухтактного
роста. В более северных и влажных местообитаниях с луговой расти-
тельностью, как правило, ростовые процессы совершаются беспрерыв-
но в течение вегетационного периода, за исключением некоторых стро-
го моноциклических ранних растений, например Allium oleraceum, Ve-
ratrum lobelianum и др. (Серебрякова, 1956). Южнее, в настоящих сте-
пях, второй период роста слабо очерчен, весьма низкой интенсивности
и смещен во времени на более поздние сроки (Белостоков, 1962).
186
Акцентируя внимание на растениях с двумя генерациями листьев,
нельзя оставлять в тени и виды с одной генерацией, играющие нема*
лую роль в составе растительности луговой степи (44,4%). Монохрон-
ный тип нарастания ассимилирующей поверхности экологически более
соответствует южностепным местообитаниям. Значит, влияние чисто-
степного (южностепного) элемента в луговых степях проявляется
весьма выразительно.
Таблица 15
Состав растений Стрелецкой степи по ритмам цветения
Тип растения Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Ранневесенние (1) 4 1,5 4 2,9 1 11,1 1 2,2 2 2,5
Поздневесенние (3). Ранне-средневесенние 3 1,1 1 0,7 — — — — 1 1,2
(1-2) 1 0,4 1 0,7 — — — — 1 1,2
Весенние (1—3). . . . Средне-поздневесенние 9 3,3 8 5,8 1 11,1 4 8,7 3 3,7
(2-3) 11 4,1 7 5,2 — — — — 7 8,5
'Средневесенне-раннелет- ние (2—4) Средневесенне-позднелет- 8 3,0 3 2,2 1 11,1 1 2,2 1 1,2
ние (2—6) 1 0,4 1 0,7 — — — — 1 1,2
Поз д невесен не- ран нел ет- ние (3—4) Поз д невесе н не- среднелет- 28 10,3 19 13,9 1 11,1 8 17,4 10 12,2
ние (3—5) 3 1,1 3 2,2 — — 1 2,2 2 2,5
Поз дневесен не- позднелет- ч
ние (3—6) 1 0,4 — — — — — — — —
Раннелетние (4) . . 65 24,1 43 31,4 2 22,2 18 39,1 23 28,1
Среднелетние (5) . . . . 8 3,0 5 3,7 1 11,1 1 2,2 3 3,7
Позднелетние (6) . . . • 3 1,1 —
Ранне-среднелетние (4—5) 53 19,6 26 19,0 2 22,2 7 15,2 17 20,6
Летние (4—6) Средне-позднелетние (5— 20 7,4 6 4,4 — — 2 4,3 4 4,9
6) Средней етне-раннеосенние 28 10,3 8 5,8 — — 3 6,5 5 6,1
(5—7) 8 3,0 1 0,7 — — — — 1 1,2
Среднелетне-среднеосен- ние (5—8) Позднелетне-раннеосен- 2 0,7 — — — — — — — —
ние (6—7) Позднелетне-среднеосен- 7 2,6 1 0,7 — — — — 1 1,2
ние (6—8) Ранне-среднеосенние 5 1,8 — — — — — — —
(7—8) Ранневесенне-позднеосен- 1 0,4 — — — — — — —
ние (1—9) 1 0,4 — — — — — — —
Итого ... 270 100 137 100 9 100 46 100 82 100
В составе луговой степи насчитывается 22 группы растений с раз-
личными ритмами цветения (табл. 15). Наибольшее число растений
цветет в раннелетний период — в течение июня (24,1%). Из других
богато представленных групп можно отметить ранне-среднелетние
187
(19,6%), поздневесенне-раннелетние (10,3%), средне-позднелетние
(10,3%), летние (7,4%), средне-поздневесенние (4,1%). Соотношение
основных компонентов по ритмам цветения очень близко к приведен-
ному для всего видового состава ценозов. Первое место занимают ран-
не-летние растения (31,4%), далее идут ранне-среднелетние (19,0%),
поздневесенне-раннелетние (13,9%), средне-позднелетние (5,8%), весен-
ние (5,8%), средне-поздневесенние (5,2%), летние (4,4%). Таким обра-
зом, в растительности луговой степи преобладают весенне-раннелетние
фенотипы.
Таблица 16
Динамика отмирания листьев в основании генеративных побегов растений Стрелецкой степи
Месяц Порядковый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
V 15 25 10,4 16 13,1 1 11,1 5 12,5 10 13,7
VI 16 44 18,2 27 22,1 3 33,4 10 25,0 14 19,2
17 41 16,9 24 19,5 — — ' 10 25,0 14 19,2
18 50 20,7 23 18,4 1 11,1 5 12,5 17 23,3
VII 19 33 13,6 14 12,3 1 11,1 5 12,5 8 и,о
20 17 7,0 4 3,3 1 11,1 — — 3 4,1
2! 12 5,0 4 3,3 1 11,1 1 2,5 2 2,7
VIII 22 6 2,5 3 2,4 — — 1 2,5 2 2,7
23 2 0,8 1 0,8 1 11,1 — — — —
24 5 2,1 1 0,8 — —у — — 1 1,4
IX 25 3 1,2 2 1,6 — — 2 5,0 — —.
26 3 1,2 2 1,6 — — — — 2 2,7
27 —
X 28
29
30 1 0,4 1 0,8 — — 1 2,5 — —
Итого . . 242 100 122 100 9 100 40 100 73 100
Отмечая основную приуроченность цветения луговой степи, нельзя
совершенно игнорировать летние (средне-позднелетние) и летне-осен-
ние фенотипы, которые в общей массе видов играют заметную роль.
Если учесть широкое распространение вторичного цветения, приходя-
щегося на вторую половину вегетационного периода, то говорить о фе-
нологической ненасыщенности луговой степи как будто бы не прихо-
дится. Эти факты склоняют к тому, чтобы отказаться от мнения
В. В. Алехина (1935) о фенологической ненасыщенности Стрелецкой
степи. Кроме того, нельзя судить о степени фенологической насыщен-
ности только по распределению во времени цветения. Во вторую поло-
вину лета совершаются важнейшие в биологическом отношении про-
цессы созревания и распространения плодов и семян, которые также
являются элементами фенологического (сезонного) развития растений.
Рассматривая динамику отмирания листьев в основании генератив-
ных побегов (табл. 16), следует отметить, что начало отмирания прихо-
188
дится на III декаду мая и заканчивается во II декаде сентября, кроме
одного (вида, листья которого усыхают в последней декаде октября.
Отмирание имеет три максимальных периода. Первый максимум при-
ходится на I декаду второй (абсолютный) на III декаду 'июня.
Незначительное усиление отмирани^ наблюдается еще в III декаду ав-
густа. Наиболее интенсивное усыхание листьев совершается с последней
декады мая по I декаду августа. Таким образом, уже с конца весны
и в течение всего лета происходит процесс отмирания листьев в осно-
вании генеративных побегов.
Таблица 17
Динамика полного отмирания листьев генеративных побегов растений Стрелецкой степи
Месяц Порядковый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
VI 16 3 1,3 2 1,7 2 2,9
17 4 1,7 2 1,7 — — 1 2,4 1 1,5
18 19 8,2 14 12,1 1 14,3 7 17,1 6 8,8
VII 19 23 9,9 19 16,4 1 14,3 10 24,5 8 11,8
20 18 7,7 8 6,9 — — 2 4,9 6 8,8
21 16 6,9 7 6,0 — — 1 2,4 6 8,8
VIII 22 14 6,0 7 6,0 1 14,3 3 7,3 3 4,4
23 14 6,0 9 7,8 — — 6 14,7 3 4,4
24 21 9,0 9 7,8 1 14,3 — — ' 8 11,8
IX 25 20 8,6 9 7,8 1 14,3 3 7,3 5 7,4
26 26 11,2 10 8,6 1 14,3 1 2,4 8 11,8
27 32 13,7 12 10,3 1 14,3 3 7,3 8 11,8
X 28 10 4,3 3 2,6 — — 1 2,4 2 2,9
29 5 2,1 2 1,7 — — 2 4,9 — —
30 8 3,4 3 2,6 — — 1 2,4 2 2,9
Итого . . 233 100 116 100 7 100 41 100 68 100
Отмирание листьев основных компонентов носит более стройный и
выразительный характер. Максимум отмирания приходится на I дека-
ду июня (22,1% видов). Затем число видов, теряющих в основании
генеративных побегов листья, постепенно падает. Лишь в сентябре на-
блюдается слабый подъем.
Первые виды, с целиком усохшими листьями на цветоносных побе-
гах, отмечаются в I декаде июня (табл. 17). Усыхание листьев с пере-
менными подъемами и спусками продолжается до конца октября. Аб-
солютный максимум видов падает на III декаду сентября, когда их
генеративные побеги полностью теряют листья (13,7%). Первый же пик
отмирания отмечается в 1 декаде июля (9,9%). Вообще в первые две
декады июня количество видов с отмершими листьями незначительно,
а в остальное время вегетационного периода число видов, теряющих
листья, распределяется по декадам довольно равномерно. Весьма близ-
кая картина отмирания листьев на цветоносах наблюдается и среди
компонентов.
189
В течение июня генеративные побеги отмирают у небольшого коли-
чества видов (табл. 18). Но уже с I декады июля и вплоть до конца
октября ежедекадно генеративные побеги отмирают у значительного
числа видов. В III декаде сентября происходит отмирание побегов у
наибольшего числа видов (14,7%). Почти тождественные закономер-
ности в отмирании цветоносных побегов наблюдаются среди компонен-
тов.
Таблица 18
Динамика отмирания генеративных побегов растений Стрелецкой степи
Месяц Порядковый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
VI 16 3 1,2 2 1,5 1 11,1 1 1,2
17 6 2,3 6 4,5 — — 1 2,2 5 6,2
18 4 1,5 3 2,2 — — 1 2,2 2 2,5
VII 19 14 5,4 9 6,7 1 11,1 5 11,1 3 3,8
20 15 '5,8 10 7,5 1 11,1 7 15,6 2 2,5
21 20 7,7 И 8,2 — — 5 11,1 6 7,5
VIII 22 15 5,8 10 7,5 — — 1 2,2 9 11,2
23 16 6,2 11 8,2 1 11,1 6 13,4 4 5,0
24 25 9,7 13 9,7 1 11,1 3 6,7 9 11,3
IX 25 6 2,3 4 3,0 1 11,1 — — 3 3,8
26 27 10,4 И 8,2 — — 3 6,7 8 10,0
27 38 14,7 20 14,9 1 11,1 5 11,1 14 17,5
X 28 26 10,0 И 8,2 1 11,1 2 4,4 8 10,0
29 29 11,2 10 7,5 1 11,1 5 11,1 4 5,0
30 15 5,8 3 2,2 — — 1 2,2 2 2,5
Итого . . 259 100 134 100 9 100 45 100 80 100
На примере отмирания отдельных частей генеративных побегов и
полного их усыхания можно убедиться, что отмирание у растений лу-
говой степи сопутствует вегетации на протяжении всего летнего перио-
да. Это касается не только генеративных, но и вегетативных побегов,
последовательно теряющих часть своих листьев на периферии розеток
или в нижней части безрозеточных побегов.
Характер настоящей работы позволяет сопоставить биоморфологи-
ческие особенности надземных органов растений с некоторыми призна-
ками подземных органов. Для сопряженного анализа из последних мы
выбираем глубину распространения корней, определенные градиенты
которой служат основой ярусного расчленения подземной части расти-
тельности. С другой стороны, представляется возможным дать количе-
ственную характеристику по некоторым биоморфологическим призна-
кам в связи с ярусной принадлежностью растений в надземной части 1.
Результаты этого анализа приведены в табл. 19 а — г.
1 Первый опыт анализа биоморфологического состава отдельных ярусов в надзехМ-
ной части растительности Стрелецкой степи проведен Г. И. Дохман (1960).
190
Таблица 19а
Сопряженный количественный анализ видов Стрелецкой степи по биоморфологическим особенностям надземных органов в связи с ярусной
принадлежностью растений в подземной и надземной частях (в скобках лроцент от общего числа растений)
Ярус Тип надземных побегов • -L- ... Способ нарастания и возобновления побегов
безрозеточный полурозеточный розеточный симпоидальный моноподиальный
всего основные компоненты - всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты
В подземной части: 6-й (до 30 см) 19(7,0) 9(6,7) 22(8,2) - 14(10,3) 5(2,0) 3(2,2) 27 (12,6) 16(13,8) 1 (0,5) 1 (0,9)
5-й (до 60 см) 19(7,0) 8(5,8) 25 (9,2) 19(13,9) 8(3,0) 6(4,4) 33(15,4) 23(19,8) 5(2,3) 5(4,3)
4-й (до 1 м) 14(5,2) 6(4,4) 26 (9,8) 9(6,7) 5(2,0) 5(3,6) 36(16,8) 18(15,5) 3(1,4) 2(1,7)
3-й (до 1,5 м) 12(4,4) 1 (0,7) 19(7,0) 6(4,2) 2 (0,8) 1(0,7) 26 (12,0) 5(4,3) 4(1,9) 2(1,7)
2-й (до 2 м) 6 (2,2) 4(2,8) 25 (9,2) 15(11,1) — — 25(11,5) 16(13,8) 3(1,4) 1 (0,9)
1-й (глубже 2 м) 29(10,7) 15(11,1) 31 (11,5) 15(11,1) 2(0,8) 1(0,7) 52 (24,2) 27 (23,3) — —
В надземной части: 1-й (свыше 60 см) 12(4,4) 4(2,9) 31 (11,5) И (8,0) 1 (0,4) — 35 (16,3) 12(10,3) 1 (0,5) —
2-й (50—60 см) 12(4,4) 3(2,2) 41(15,4) 23(16,8) 1 (0,4) 1 (0,7) 41 (19,0) 21 (18,1) 1 (0,5) —
3-й (40—50 см) 23 (8,5) 7(5,1) 26 (9,7) 14(10,2) 1 (0,4) — 42(19,5) 19(16,4) 1 (0,5) 1 (0,9)
4-й (30—40 см) 20(7,4) 13 (9,5) 25 (9,2) 12(8,7) 4(1,5) 2(1,5) 37 (17,1) 23 (19,8) 3(1,4) —
5-й (15—30 см) 15(5,6) 10(7,3) 13(4,8) 8(5,8) 7(2,6) 6(4,4) 24(11,2) 18(15,5) 3(1,4) 3(2,6)
6-й (5—15 см) 17(6,4) 6(4,4) 12(4,4) 10(7,3) 8(3,0) 7(5,1) 20 (9,3) 12(10,4) 7(3,3) 7(6,0)
8
Таблица 196
Сопряженный количественный анализ видов Стрелецкой степи по биоморфологическим особенностям надземных органов в связи с ярусной
принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Длительность вегетации Способ перезимовки Количество генераций листьев и побегов
длительно- вегетирующие коротковегетирующие летнезеленые вечнозеленые зимнезеленые с 1 генерацией с 2 генерациями
всего основные компо- ненты всего основные компо- ненты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты
В подзем-
ной ча- сти:
6-й 9(3,3) 3(2,2) 36 (13,3) 22 (15,3) 17(6,3) 7(5,1) 5(1,9) 3(2,2) 24(8,9) 16(11,6) 31 (11,5) 13(9,5) 15(5,6) 13(9,5)
5-й 26 (9,7) 17(12,6) 26 (9,7) 17(12,6) 16(6,0) 10(7,3) 18(6,7) 15(11,1) 18(6,7) 8(5,8) 19(7,0) 7(5,1) 33 (12,2) 26(19,0)
4-й 34 (12,5) 14(10,2) И (4,1) 6(4,4) 24 (8,9) 8(5,8) 15(5,5) 8(5,8) 6(2,2) 4(2,9) 16(6,0) 8(5,8) 29(10,7) 12(8,7)
З-й 27 (10,0) 5(3,7) 6(2,2) 3(2,2) 18(6,7) 3(2,2) 7(2,6) 2(1,5) 8(3,0) 3(2,2) 12(4,5) 3(2,2) 21 (7,7) 5(3,7)
2-й 28 (10,3) 16(11,7) 4(1,5) 3(2,2) 25 (9,3) 13(9,5) 6(2,2) 5(3,7) 1 (0,4) 1 (0,7) 16(6,0) 6(4,4) 16(6,0) 13(9,5)
1-й 56 (20,7) 28 (20,7) 7(2,6) 3(2,2) 54 (20,0) 25 (18,2) 7(2,6) 5(3,7) 1 (0,4) 1 (0,7) 26 (9,7) 12(8,7) 36 (13,3) 19(13,9)
В над- земной
части:
1-й 38 (14,0) 13(9,5) 6(2,2) 2(1,5) 36 (13,3) 12(8,7) 3(1,1) — 5(1,9) 3(2,2) 23 (8,5) 9(6,6) 21 (7,7) 6(4,4)
2-й 43(15,9) 19(13,9)' И (4,0) 8(5,8) 34(12,5) 15(11,0) 10(3,7) 6(4,4) 10(3,7) 6(4,4) 23 (8,5) 6(4,4) 31(11,5) 21 (15,4)
З-й 36 (13,3) 12(8,7) 14(5,2) 9(6,6) 33 (12,2) 10(7,4) 12(4,4) 8(5,8) 5(1,9) 3(2,2) 15(5,5) 5(3,7) 35 (13,0) 16(11,6)
4-й 26 (9,7) 13(9,5) 23(8,5) 14(10,2) 25 (9,3) 14(10,2) Ю (3,7) 5(3,7) 14(5,2) 8(5,8) 24(8,9) 12(8,7) 25 (9,3) 15(11,0)
5-й 18(6,7) 14(10,2) 17(6,3) 10(7,4) 16(6,0) И (8,0) 9(3,3) 8(5,8) 10(3,7) 5(3,7) 16(6,0) 8(5,8) 19(7,0) 16(11,6)
6-й 19(7,0) 12(8,7) 19(7,0) И (8,0) 10(3,7) 4(2,9) 14(5,2) И (8,0) 14(5,2) 8(5,8) 19(7,0) 9(6,6) 19(7,0) 14(10,2)
Голубев ;193
□ Таблица 19в
Сопряженный количественный анализ видов Стрелецкой степи по биоморфологическим особенностям надземных органов в связи с ярусной
ро принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Ритм цветения
весенние весенне-летние летние летне-осенние осенние весенне-летне-осенние
всего основные компонен- ты всего основные компонен- ты всего основные компоненты всего основные компонен- ты всего основные компонен- ты всего основные компоненты
В подземной части: 6-й ю(3,7) 8(5,8) 15 (5,6) 9(6,6) 18(6,7) 8(5,8) 2(0,7) 1 (0,7) 1 (0,4)
5-й 9(3,3) 7(5,1) 10(3,7) 7(5,1) 31(11,5) 19 (13,9) 3(1,1) i (0/0 — — — —.
4-й 6(2,2) 3(2,2) 6(2,2) 4(2,9) 30 (11,1) 13 (9,5) 2(0,7) (10,4) — — —>
3-й 1(0,4) 1 (0,7) 3(1,1) 1(0,7) 23 (8,5) 6(4,4) 6(2,2) — — — — —
2-й — 2(0,7) 1(0,7) 25 (9,3) 17(12,4) 4(1,5) — — — — —
1-й 2(0,7) 2(1,5) 5(1,9) 4(2,9) 50 (18,5) 25 (18,2) 5(1,9) — — — — —
В надземной части: 1-й — 2(0,7) 1 (0,7) 35(13,0) 14(10,2) 7(2,6)
2-й — — 3(1,1) 2(1,5) 48 (17,8) 25(18,2) 3(1,1) — — — — —
3-й 1 (0,4) — 4(1,5) 3(2,2) 38 (14,0) 18(13,2) 6(2,2) — 1 (0,4) — 1 (0,4) —
4-й 4(1,5) 3(2,2) 11 (4,0) 7(5,1) 31 (11,5) 16(11,7) 3(1,1) 1(0,7) — — — —
5-й 7(2,6) 4(2,9) 9(3,3) 8(5,8) 17(6,3) И (8,0) 1 (0,4) 1 (0/0 — • — — —
6-й 16(5,9) 14 (10,2) 12(4,5) 5(3,7) 8(3,0) 4(2,9) 2(0,7) — — — —
Таблица 19г
Сопряженность распределения видов по ярусам в надземной и подземной частях растительности Стрелецкой степи
(в скобках процент от общего числа растений)
Ярус в подзем-. ной части Ярус в надземной части растительности
1-й 2-й 3-й 4-й 5-й 6-й
всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты
6-Й 1 (0,4) 1 (0,7) 1 (0,4) 12(4,5) 7(5,1) И (4,0) 6(4,4) 21 (7,7) 12(8,7)
5-Й — — 10(3,7) 7(5,1) 8(3,0) 5(3,7) 14(5,1) 9(6,6) Ю (3,7) 7(5,1) 10(3,7) 5(3,7)
4-Й 5(1,9) 2(1^5) 11 (4,0) 4(2,9) Ю (3,7) 2(1,5) 9(3,3) 6(4,4) 6(2,2) 3(2,2) 4(1^5) 3(2,2)
3-Й 4(1,5) 8(3,0) 1(0,7) Ю(3,7) 2(1,5) 8(3,0) 2(1,5) 3(1,1) 3(2,2) —
2-Й 9(3,3) 4(2,9) 9(3,3) 5(3,7) 5(1,9) 4(2,9) 5(1,9) 2(1,5) 3(1,1) 3(2,2) 1(0,4) 1 (0,7)
1-й ,25 (9,3) 8(5,8) 15(5,6) 9(6,6) 17(6,3) 9(6,6) 1 (0,4) 1 (0,7) 2(0,7) 2(1,5) 2(0,7) 2(1,5)
Приступая к обзору данных таблиц, напомним, что в подземной ча-
сти растительности луговой степи в зависимости от глубины проникно-
вения корней растений выделяются следующие ярусы: 6-й — до 30 см
от поверхности, 5-й — до 60 см, 4-й — до 1 м, 3-й — до 1,5 м, 2-й — до
2 м, 1-й — глубже 2 м. В отличие от ранее принятого ярусного расчле-
нения подземной части растительности луговой степи (Голубев, 1962в),
здесь, во-первых, изменяется нумерация ярусов на обратную, т. е. самые
глубококорневые растения относятся к 1-му ярусу, а наиболее мелко-
корневые — к последнему, 6-му ярусу. Во-вторых, глубококорневые
растения (до 250 см и глубже) объединяются в одну группу, слагаю-
щую 1-й подземный ярус, на гом основании, что и те и другие исполь-
зуют влагу горизонта постоянной влажности.
Обратимся к конкретному материалу. Наибольшее число безрозеточ-
ных растений относится к 1-му подземному ярусу, т. е. глубококорне-
вым (10,7%), но их много и среди видов 6, 5 и 4-го ярусов. Такое рас-
пределение безрозеточных растений по ярусам, установленное для всего
видового состава, свойственно и основным компонентам. Полурозе-
точных бо'лее всего также среди растений 1-го яруса (11,5%), но их
много и в 4, 2, 5 и 6-м ярусах. Розеточные растения более характерны
для верхних ярусов подземной части, т. е. 5, 6 и 4-му. Лишь единичные
розеточные растения обладают глубокой корневой системой (табл. 19а).
Моноподиальные растения приурочены к нижним ярусам, особенно
к 5, 4 и 3-му. В первом ярусе подземной части безраздельно господ-
ствуют симподиальные растения (табл. 19а).
Длительновегетирующие растения особенно характерны для 1-го
яруса (20,7%), далее — для 4-го (12,5%), 2-го (10,3%), 3-го (10%) и
5-го (9,7%). Наоборот, максимум коротковегетирующих растений рас-
пространяет корни в 6-м (13,3%), 5-м (9,7%), 4-м (4,1%) ярусах,
далее число видов с короткой вегетацией становится незначительным
(табл. 196). Таким образом, связь продолжительности вегетации рас-
тений луговой степи с глубиной проникновения корневых систем здесь
получает четкую количественную характеристику.
Определенная правильность в ярусном распределении растений
отмечается и по способам перезимовки (табл. 196). Наибольшее
число летнезеленых являются растениями 1-го (20%) и 2-го (9,3%)
ярусов, а зимующих в зеленом состоянии (вечнозеленые и зимнезеле-
ные)— относится к мелкокорневым: 6, 5 и 4-му ярусов (10,8%,
15,4%, 7,7%).
Диаметрально противоположно распределение максимального коли-
чества растений с одной и двумя генерациями листьев и побегов: пер-
вые концентрируются в 6-м (11,5%), а вторые — в 1-м ярусе (13,3%).
Эта пропорция вполне отвечает особенностям экологии нарастания над-
земных органов.
По ритмам цветения главную роль в сложении растительности лу-
говой степи играют весенние, весенне-летние и летние фенотипы, в рас-
пределении которых по ярусам подземной части наблюдается извест-
ная закономерность, чего нельзя сказать о летне-осенних фенотипах
(табл. 19в). Подавляющая масса видов весеннего цикла цветения рас-
полагает свои корни в верхних ярусах — 6, 5 и 4-м. Эта же черта ярус-
ной принадлежности характерна и весенне-летним фенотипам. Однако
в этой группе уже намечается заметное увеличение глубококорневых
видов. В следующей группе растений летнего цветения абсолютный
максимум относится к 1-му ярусу (18,5%), но довольно значительно и
количество видов 5-го (11,5%), 4-го (11,1 %), 2-го (9,3%), 3-го (8,5%)
и 6-го (6,7%) ярусов. В распределении по ярусам растений летне-осен-
него цикла цветения особая приуроченность отсутствует. Это, по всей
вероятности, объясняется тем, что в пору их цветения нет резко напря-
194
женной экологической зависимости развития растений от увлажнения
почвы, так как обычно в это время острый дефицит влажности сглажи-
вается за счет поверхностного промачивания почвы, умеряются и тем-
пературы воздуха.
Остается теперь сопоставить распределение видов по ярусам в под-
земной и надземной частях растительности (табл. 19г). Анализ табли-
цы убеждает, что в общем существует прямое соответствие в распреде-
лении растений по ярусам в подземной и надземной сферах раститель-
ности. Это положение поясняется цифровыми данными. Наибольшее
число глубококорневых растений (1-го яруса подземной части)
относится к 1-му ярусу надземной части (9,3%). Но значительное их
число располагает побеги во 2-м (5,6%) и 3-м (6,3%) ярусах. Виды
2-го яруса подземной части ценозов в наибольшем количестве приуро-
чены к 1-му (3,3%) 'И 2-му (3,3%) ярусам надземной части. Растения
с корневыми системами, входящими в З-й ярус, больше всего представ-
лены в 3-м ярусе надземной части (3,7%), заметное их число встре-
чается во 2-м (3,0%) и 4-м (3,0%) ярусах. Растения, принадлежащие
к 4-му ярусу корневых систем, в наибольшем количестве отмечаются
среди 2-го (4%), 3-го (3,7%), 4-го (3,3%) ярусов надземной части.
Мелкокорневые растения 5-го яруса (в подземной части) довольно рав-
номерно представлены во всех ярусах травостоя, исключая 1-й, но бо-
лее всего их в 4-м ярусе (5,1%). Максимум мелкокорневых (6-го яру-
са) соответствует максимуму низкорослых растений (6-го яруса)
травостоя (7,7%). Таким образом, в количественной характеристике
ярусной принадлежности растений мы находим убедительные свиде-
тельства прямого соответствия ярусного расчленения травянистых це-
нозов в подземной и надземной частях. Этот важный во многих отно-
шениях вывод, разумеется, не отрицает возможности исключений, при-
меры которых приводились в гл. IV, и имеют место для каждого яруса
в подземной и надземной сферах сообществ.
Переходим к количественной характеристике биоморфологических
особенностей надземных органов в связи с ярусным расчленением тра-
востоя в надземной сфере (табл. 19а). Как видно из таблицы, наи-
большее число безрозеточных растений относится к 3 и 4-му ярусам
(8,5 и 7,4%). В остальных ярусах присутствует меньшее и.довольно
равномерное количество. Максимум полурозеточных видов падает на
2-й (15,4%), 1-й (11,5%), 4-й (9,2%) и З-й (9,7%) ярусы. Основная
масса розеточных растений приурочена к 6-му ярусу (3,0%) и 5-му
(2,6%) ярусам. Обнаруживается закономерное убывание количества
симподиальных растений от верхних к нижним ярусам. Максимум мо-
ноподиальных растений приурочен к самому нижнему — 6-му ярусу
(3,3%).
По продолжительности вегетации (табл. 196) в распределении рас-
тений по ярусам можно заметить следующую особенность: число дли-
тельновегетирующих растений последовательно снижается от верхних
ярусов к нижним. Однако среди основных компонентов эта последова-
тельность нарушается и обнаруживаются два максимума в верхних и
нижних ярусах. Такое соотношение довольно точно отражает биоморфо-
логический состав длительновегетирующих растений: часть из них с круп-
ными побегами и глубокой корневой системой, другие — низкорослые
мелкоукореняющиеся. В абсолютном выражении число коротковегети-
рующих растений в нижних трех ярусах (21,8%) значительно превос-
ходит количество в трех верхних (11,4%). Общее число летнезеленых
растений сокращается от верхних ярусов к нижним. Близкая картина
наблюдается и среди основных компонентов. Зимующие в зеленом
состоянии растения (зимне- и вечнозеленые) преобладают в нижних
ярусах, а в верхних число их заметно убывает.
13*
195
По количеству генераций листьев и побегов растений обнаружить
закономерное распределение по ярусам не удается. Это обстоятельство,
по-видимому, указывает на то, что признак количества генераций ли-
стьев и побегов является показателем общих эколого-ценотических
условий и более характеризует зональные особенности растительности,
нежели ее внутреннее структурное расчленение.
Растения весеннего цикла цветения приурочены (табл. 19в) исклю-
чительно к трем нижним ярусам, причем наибольшее количество сосре-
доточено в 6-м ярусе (5,9%). Максимум весенне-летних растений падает
на нижние три яруса, в остальных ярусах они весьма малочисленны.
Основная масса видов летнего цикла цветения относится к верхним
ярусам. Более всего их содержится во 2-м ярусе—17,8% (от всего
видового состава) и 18,2% (от числа основных компонентов). В распре-
делении летне-осенних растений по ярусам отсутствует строгая после-
довательность: хотя в общем составе видов большая их часть находит-
ся в верхних ярусах, но основные компоненты здесь отсутствуют
совершенно.
О координации распределения видов по ярусам в надземной и под-
земной сферах мы уже говорили раньше.
В вышеприведенной характеристике мы наметили лишь основные
ступени и закономерности сопряженности признаков и количественного
состава видов в структурном расчленении растительности. Многие част-
ные моменты сознательно опущены, нет и достаточной детализации
материалов, уже сведенных в таблицы. Но поскольку в работе поме-
щаются все исходные фактические материалы, то при желании эти
пропуски можно восполнить самостоятельно.
Изложенные данные показывают, что ярусное расчленение в над-
земной и подземной частях растительности четко разделяет растения
и по их эколого-биологическим особенностям. Таким образом, здесь
проявляется ясная эколого-фитоценотическая специфика ярусов, по-
зволяющая сравнивать их с экологическими нишами. Однако такая
трактовка ярусов все-таки исключает представление об экологической
равноценности их компонентов. Еще раз привлекая внимание к установ-
ленной соотносительной координации ярусного расчленения ценоза в
надземной и подземной частях и подчеркивая экологическую и фито-
ценбтическую оригинальность, специфику ярусов, мы таким образом
находим новые свидетельства и критерии в пользу концепции о моно-
литности растительного сообщества, развивавшейся у нас В. Н. Сука-
чевым (1915, 1928 и др.). Он писал, что «растения, входящие в состав
сообщества, образуют как бы одно целое, имеющее определенный строй
и структуру» (Сукачев, 1928, стр. 56). Понимание самостоятельной сущ-
ности ценоза вместе с тем предполагает эколого-ценотическую индиви-
дуальность компонентов, степень которой определяется характером
взаимного приспособления видов, их фитоценотической и средообразу-
ющей ролью.
Сравнение биоморфологических признаков растений в двух приве-
денных рядах ярусной принадлежности для подземной и надземной
сфер растительности показывает большое сходство и соответствие меж-
ду ними. Например, длительновегетирующие растения в наибольшем
обилии являются компонентами 1-го яруса корневых систем и верхних
же ярусов надземной части. Полное тождество наблюдается и в ярус-
ном распределении растений по способам перезимовки, ритмам цве-
тения и другим признакам. О соответствии максимального числа видов
по ярусам в надземной и подземной частях уже говорилось выше. Та-
ким образом, рассматриваемое явление выходит из разряда частных
и выражает собой объективную закономерность сложения растительных
сообществ луговой степи, которую можно- определить следующим обра-
1дв
зом: в строении надземной и подземной частей наблюдается соответ-
ствие, преломляющееся, в частности, в одинаковом количестве ярусов.
Если в качестве исходной взять плоскость поверхности земли, то ярус-
ное сложение сообщества приобретает известные черты симметрии в
надземной и подземной сферах относительно этой плоскости. Как пра-
вило, максимальное число видов, обладающих каким-либо биоморфоло-
гическим признаком, свойственное определенному ярусу надземной ча-
сти сообщества, соответствует наибольшему количеству видов по этому
же признаку, входящих в состав аналогичного (симметричного) яруса
подземной части ценоза, и наоборот. Последнее, собственно говоря,
и характеризует установленное для растительности луговой степи пра-
вило количественной координации видов по биоморфологическим осо-
бенностям в связи с ярусным расчленением ценозов в надземной и
подземной частях.
ДУБОВЫЕ ЛЕСА
Из состава травянистых растений дубовых лесов изучено 114 видов,
биоморфологические особенности которых отображены в табл. 20.
Подпологовые травы представлены почти в равном количестве без-
розеточными (43,9%) и полурозеточными (44,7%) видами, 10,5% со-
ставляют розеточные растения. Сходные пропорции сохраняются среди
основных компонентов (табл. 21). По сравнению с луговой степью в со-
ставе травянистой растительности дубовых лесов заметно возрастает
роль растений с безрозеточными побегами. Такое изменение можно
объяснить спецификой светового режима подпологовой растительности,
способствующего развитию побегов по безрозеточному типу.
Господствующим способом возобновления побегов растений дубовых
лесов является симподиальный (94,2% всего видового состава и
91%—компонентов). Роль моноподиальных растений незначительна
(табл. 22).
В вертикальном расчленении травостоя можно выделить четыре яру-
са (табл. 23).
Ьй ярус образуют высокорослые травы, генеративные побеги которых возвышают-
ся свыше 90 см. Сюда относятся Adenophora liliifolia, Anthriscus silvestris, Calamagro-
stis arundinacea, Campanula rapunculoides, Clematis recta, Lilium pilosiusculum, Pyre-
thrum corymbosum и др. От всего видового богатства растения 1-го яруса составляют
27,2 и 22%—от числа основных компонентов.
2-й ярус слагается генеративными побегами растений средней высоты — 60—90 см
(Achillea millefolium, Aegopodium podagraria, Brachypodium pinnatum, Geranium silvati-
cum, Geum urbanum, Inula salicina, Lathyrus pisiformis, Serratula inermis и др.). Эти
виды, от всего набора составляют 33,3 и 41,3% —от числа основных компонентов.
З-й ярус (40—60 см) является основным, в котором концентрируется подавляющая
масса листьев растений с более высокими цветоносными побегами. В снытевой дубра-
ве главную роль в сложении 3-го яруса играют листья Aegopodium podagraria, в дуб-
раве с господством коротконожки перистой — листья Brachypodium pinnatum, в вейни-
ковой ассоциации — листья Calamagrostis arundinacea. Помимо указанного в составе
3-го яруса целиком размещают свои побеги такие растения, как Clinopodium vulgare,
Convallaria majalis, Iris aphylla, Melampyrum nemorosum, Melica nutans, Orobus vernus,
Polygonatum officinale, Ranunculus auricomus, Stellaria holostea и др. Они составляют
19,3% -всех видов и столько же среди основных компонентов.
Низкорослые виды с ползучими побегами образуют 4-й ярус (0—25 см). К ним при-
надлежат Asarum auropaeum, Fragaria vesca, Glechoma hederacea, Primula veris, Pul-
monaria angustifolia, Rubus saxatilis и др.
В травянистом покрове дубовых лесов заповедника господствуют
длительновегетирующие виды (табл. 24). В составе всей учтенной фло-
ры травянистых растений они занимают 92,1%, а среди основных ком-
понентов— 97,8%. Обращает на себя внимание исключительная бед-
ность лесов весенними геоэфемероидами, что уже раньше отмечалось
В. В. Алехиным (1909). Правда, в списке отмечены Anemone ranunculo-
des, Corydalis halleri, Gagea lutea, Scilla sibirica и можно еще добавить
197
Таблица 20
Биоморфологический состав травянистых растений дубовых лесов *
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам расти- тельности Тип по продолжи- тельности вегетации Способ перези- мовки Количе- ство генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота генера- тивных побегов (за 1962 г.] в см
листьев у основания генера- тивных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыхание)
Achillea millefolium Кз пр С 2 Д В 2 4-8 16 29 30 64
Aconitum lasiostomum . . . пр С 1 Д л 1 >4-5 22 25 27 135
A. nemorosum бр С 2 Д л 1р >6—8 15 28 29 93
Actaea spicata бр С 3 Д л 1 >3—4< 22 27 27 50
Adenophora liliifolia бр С 1 Д л 1р >5—6 19 27 28 111
A egopodium podagraria .... Ki пр С 2(3) Д л 2 4—5< 19 24 25 90
Agrimonia eupatoria пр С 2 Д л 1 5—7 18 27 29 84
A. pilosa пр С 2 Д л 1 >4—6 18 27 29 88
Agropyron caninutn пр С 2 Д л 1 5 16 26 30 99
Allium oleraceum р С 3 С 3 2о >5—6 23 25 50
Anemone ranunculoides бр С 4 гэ л 1 1—2 15 15 15 16
Angelica silvesiris пр 1 д л 1 >4-5 22 24 26 132
Anthriscus silvestris Кг пр 1 м л 2 4 18 20 22 116
A rrhenatherum elatius пр С 1 д в 2 4 16 21 23 ИЗ
A rtemisia vulgaris бр ' С 1 д л 2в >5—6 20 29 30 158
Asarum europaeum бр С 4 д в 1 1 18 4
Astragalus glycyphyllus .... бр С 3 д л 2в 5 20 29 30 47
Betonica officinalis К» пр м 2(3) д в 2 5—6 28 29 88
В rachypodium pinnaium . . . Кг пр с 2(3) д л 1 >4-5< 20 27 28 85
Bromus benekenii пр с 1 д в 2о >4-5< 16 23 28 137
Calamagrostis arundinacea . . Кг пр с 1(3) д л 1 5 18 23 29 128
См. сноски к табл. 8, стр.. 171
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Campanula glomerata . . . пр С 2
C. persicifplia Кз пр С 2
C. rapunculoides Кз бр С 1
C. t г ас helium пр С 1
Carex michelii Р. С 4
C. montana Кз Р С 3(4)
C. spicaia Кз Р С 3
Centaurea pseudophrygia .... пр С 2
Chamaenerium angustifplium . . бр С 1
Chelidonium majus пр С 3
Clematis recta Кз бр С 1
CUnopodium vulgare Кз бр С 3
Convallaria majalis Кз р М 3
Corydalis halleri бр С 4
Crepis sibirica пр с 1
Cucubalus baccifer бр с 2
Dactylis glomerata Кз пр с 1
Epilobium montanum бр с 2
Epipactis latiffilia бр с 2
Festuca gigantea пр с 2
Fragaria moschata пр с 4
F. vesca К. пр с 4
Fritillaria ruthenica пр с 3
£
Таблица 20 (продолжение)
Тип по продол- житель* ности ве- гетации Способ перези- мовки Количе- ство гене- раций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота генера- тивных побегов (за 1962 г.), в см
листьев у основания генера- тивных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыхание)
Д Л 2 4—6
Д В 2 >4—5 18 22 26 76
Д Л 1 5-6 18 24 26 96
Д л 1 5—6< 18 28 28 107
Д в 1 2—3 — 23 19
д л 1 1 15 — 16 30
д в 2 3 28 — 29 33
д л 2 >5—6 18 26 28 96
д л 1 5—6< 18 26 27 125
д в 2 3—4 18 25 27 56
д л 1 4—5< 18 26 26 140
д в 2о 5—7 18 29 30 55
д л 1 3-4< 27 — 28 29
гэ л 1 1 15 15 15 15
д л 1 >4-6< 121
д л 2в 5—6 18 26 29 85
д в 2 4 16 21 23 100
д л 2о >4—5 18 25 29 67
д л 1 >5-6< 21 28 29 78
д в 2о 5 19 23 29 92
д в 2 4 25 28
д в 2 3—4 22 13
гэ л 1 2 16 16 16 38
ГО g
Вид Осно- вной ком- понент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам расти- тельности
Gagea lutea пр С 4
Galium boreale Кз бр С 2
G. rubioides К, бр С 2
Geranium silvaticum Кз пр С 2
Geum urbanum ........ Кг пр м 2
Glechoma hederacea Кз бр м 4
Heracleum sibiricum пр 1
Hieracium pratense пр с 3(4)
Hypericum hirsutum бр с 2
H. maculatum бр с 2
Inula hirta К» бр с 2
I. salicina Кз бр с 2
Iris aphylla Кз р с 3
Lactuca altissima пр 1
Lapsana communis пр 1
Laserpitium pruthenicum .... пр - 1
Lathyrus pisiformis Кз бр с 2
Leucanthemum vulgare К» пр с 2
Li Hum pilosiusculum пр с 1
Lychnis chalcedonica бр с 1
Melampyrum nemorosum . . . . Кз бр 3
Melica nutans бр с 3
Mercurialis perennis бр с 3
Таблица 20 (продолжение)
Тип по про- должи- тельности вегетации Способ перези- мовки Количе- ство гене- раций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота генера- тивных побегов (за 1962 г.), в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыхание)
ГЭ Л 1 1 15 15 15 13
Д Л 1 4—5 17 29 29 68
д Л 2в 5 18 29 29 82
д Л 2 3—4 20 25 25 82
д в 2о 4-5 — 24 29 67
д в 2в 2-4 — 25 17
д л 1 >4-5 21 23 24 131
д в 2 4—5< 20 23 29 50
д в 2о 5 18 24 26 75
д л 1 4-5 61
д л 1 4—5 20 29 29
д л 1 5—6< 20 29 29 73
д л 1 3—4< 21 26 42
м л 1 5—6 17 27 29 137
м 3 2 4 19 21 24
д л 1 6-7
д л 1 4 20 27 30 81
д в 2 4-5 17 27 29 74
д л 1 5 20 26 28 98
д л 1 5 19 29 29 98
д л 2в 4-6 18 27 27 39
д в 2о 3—4< 18 29 29 37
д л 1 1 — 29 29 23
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам расти- тельности
Milium effusum пр С 1
Moehringia trinervia бр 4
Myosotis spar si flora пр 4
Neottia nidus-avis бр С 4
Nepeta pannonica Кз бр С 1
Origanum vulgare Кз бр С 2
Orobus niger бр С 2
0. vernus Кз бр с 3
Paris quadri folia бр м 3
Platanthera bifplia р с 3(4)
Poa nemoralis Кз пр с 2
Polemonium coeruleum .... пр с 1
Polygonatum multiflorum Кз бр с 2
P. officinale Кз' бр с 3
Primula veris Кз р с 4
Pulmonaria angu sti folia .... К, пр с 4
P. obscura пр с 4
Pyrethrum corymbosum .... Кз пр с 1
Pyrola minor р с 4
P. rotundifolia р с 4
Ramischia secunda ...... бр с 4
Ranunculus auricomus пр с 3
R. cassubicus пр с 3
Таблица 20 (продолжение)
Тип по про- должи- тельности вегетации Способ перези- мовки Количе- ство гене- раций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота генера- тивных побегов (за 1962 г.), в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыха- ние)
Д В 2о 4 15 22 28 123
Д 3 2в 3—4 19 29 29 19
Д 3 2 3—5 15 23 25 23
Л 1 >3-4 — — 22 25
Д Л 1 >4-5 20 27 27 111
Д В 2о 5—6 18 29 29 80
Д Л 1 4 18 29 29 76
Д Л 1 >2—3 18 31 31 38
Д Л 1 2—3 — 27 27 30
Д Л 1 3—4 22 — 22 48
Д л 1 4 15 21 28 94
Д в 2 4—5 17 27 28 108
Д л 1 3—4< 16 27 27 83
Д л 1 >3—4< 16 27 27 61
Д в 2о 1-3 26 — 21 24
Д л 2 1—3 15 19 19 16
Д л 1 1—3 15 17 18 16
Д л 1 4—5 20 23 27 112
Д в 1 4
Д в 1 4
Д в 1 >4—5< 12
Д л 1 2—4< 18 21 21 29
Д л 1 2—3 21 20 22 36
202
Вид Основ- ной компо- нент Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Rub us saxatilis K8 бр С 4
Sanguisorba officinalis .... пр М 1
Scilla sibirica р С 4
Scrophularia nodosa к» бр с 1
Serratula inermis Кз пр с 2
Solanum dulcamara бр с 2
Solidago virgaurea Кз пр с 1
Stachys silvaiica бр с 2
Stellaria holostea бр с 3
Taraxacum officinale Кз Р с 4
Thalictrum aquilegifplium .... пр с 1
T. simplex бр с 1
Torilis japonic a ........ Tri folium medium пр бр с 2 2
Trisetum sibiricum пр с 2
Urtica dioica Кз бр с 2
Verbascum nigrum пр с 1
Veratrum nigrum Кз пр с 1(3)
Veronica chamaedrys Кз бр с 4
V. teucrium .......... К. бр с 2
Vida sepium ........... Кз бр с 3
V. silvaiica .......... бр с 2
Viola canina .......... бр с 4
V. mirabilis ......... Кз Р-пр м 4
Таблица 20 (продолжение)
Тип по про- должи- тельности вегетации Способ перези- мовки Количе- ство гене- раций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота генера- тивных побегов (за 1962 г.), в см
листьев у основа- ния гене- ративных побегов листьев генера- тивных побегов генера- тивных побегов (полное усыха- ние)
Д Л 1 4 26 28 28 28
Д Л 1 >5—8 28 28 126
ГЭ Л 1 1 15 15 15 9
Д Л 1 4 20 27 29 112
Д Л 1 >5-8 21 28 30 97
Д Л 1 4-6 20 27 — 44
д Л 2 6-7 25 28 29 ИЗ
д Л 1 >4-5 18 28 28 78
д в 1 2—4< 16 29 29 32
д в 2 2-3 — 18 15
д Л 1 4 21 27 27 128
д Л 1 5 19 26 27
д Л 1 5—6 19 26 28 61
д Л 1 5—6 18 28 28 67
д в 2 >4—5< 16 22 25 87
д Л 2о 4—5< 18 28 29 95
д Л 1 5—6 22 26 28 122
д Л 1 6 27 28 128
д в 2в 3—4 18 24
д Л 1 >4—5< 18 27 28 81
д в 1 3-4 17 29 29 47
д в 1 4
д в 1 3-4 22 23
д Л 1 1—2 28 29 8
Таблица 21
Состав травянистых растений дубовых лесов по типам структуры побегов
Побег Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли* чество % коли- чество % коли- чество % коли- чество % коли- чество %
Безрозеточный . . 50 43,9 21 45,6 — — 3 30 18 51,5
Полурозеточный .... 51 44,7 18 39,2 1 100 5 50 12 34,3
Розеточный 12 10,5 6 13,0 — — 2 20 4 11,4
Розеточно-пол у розеточ- ный 1 0,9 1 2,2 — — — — 1 2,8
Итого. . . . 114 100 46 100 1 100 10 100 35 100
Таблица 22
Состав травянистых растений дубовых лесов по способам возобновления побегов
Возобновление Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Симподиальное Моноподиальное .... 98 6 94,2 5,8 40 4 91,0 9,0 1 100 7 2 77,8 22,2 32 2 94,2 5,8
Итого.. . . 104 100 44 100 1 100 9 100 34 100
Таблица 23
Распределение травянистых растений дубовых лесов по ярусам в надземной части
Ярус Всего Всего компонентов категория
видов компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
1-й (свыше 90 см) . . . 31 27,2 10 22,0 — 3 30,0 7 20,0
2-й (60—90 см) .... 38 33,3 19 41,3 1 100 3 30,0 15 42,8
З-й (40—60 см) .... 22 19,3 9 19,5 — — 2 20,0 7 20,0
4-й (0—25 см) 23 20,2 8 17,2 — — 2 20,0 6 17,2
Итого . . . | 114 | 100 46 100 1 100 10 100 35 100
Gagea minima и Ficaria verna, но все эти виды произрастают по лесным
логам, где улучшается режим влажности. Этот факт является убеди-
тельным свидетельством того, что не молодость лесов Стрелецкого
участка заповедника обусловливает отсутствие геоэфемероидов, а осо-
бые гидрологические условия, неблагоприятные для резко выраженных
мезофитов, какими и являются перечисленные выше виды. В недостатке
почвенной влаги кроется и причина отсутствия кустарникового под-
леска в лесах из Corylus avellana.
203
Таблица 24
Состав травянистых растений дубовых лесов по продолжительности вегетации
Тип растения Всего Всего компонентов категории
всего компонентов !-й 2-й 3-й
коли- чество % коли- чества % коли- чество % коли- чество % коли- чество %
Длительновегетирующие
Кор откове гети р у ющие:
геоэфемероиды . . .
монокарпики . . . .
среднепродолжитель-
ной вегетации.........
Итого. . .
104
5
3
1
ИЗ 100 46
100 1 100 10 100 35 100
Таким образом, здесь мы находим новые доказательства в пользу
высказанной ранее мысли (Голубев, 1962в) о том, что своеобразный
комплекс луговой степи и одноярусных дубрав паркового типа Стрелец-
кого участка, выходящих на плато, только в логах приобретающих
сомкнутый характер, представляет явление первичное, древнее, обуслов-
ленное геоморфологическими и геологическими особенностями местно-
сти, ухудшающими водный режим .корнеобитаемого слоя почво-грунта.
Этим объясняется и бедность неморальными элементами леса вообще,
и обилие в лесу степных форм, и большая древность Стрелецкой степи,
которая, несмотря на более широкое в прошлом распространение леса,
не покрывалась им. Имеется в виду, конечно, сравнительно недалекое
прошлое нашего участка, в пределах 1—2 тысяч лет.
Подчеркивая скудость многолетних мезофильных эфемеров в дубо-
вых лесах, нельзя забывать о присутствии коротковегетирующего
Anthriscus silvestris, характеризующегося к тому же в лесах заповед-
ника монокарпностью. Как уже отмечено выше, в лесах лесной зоны
это растение является поликарпическим и длительновегетирующим.
Этот факт еще более рельефно выделяет экологическую особенность ле-
состепных дубрав и проявление общезональных условий на все расти-
тельные формации, свойственные лесостепной зоне.
Своеобразие дубовых лесов выделяется и составом травянистых рас-
тений по способам перезимовки (табл. 25). Ведущая роль принадлежит
летнезеленым видам (66,7 и 65,2%). Весьма значительно число вечно-
зеленых растений (29,8 и 34,8%) в сравнении с зимнезелеными (3,5 и
0%). Вечнозеленость некоторых растений здесь представляет явление
первичное, как наследие вечнозеленых предков третичной эпохи.
В отличие от луговой степи, большинство травянистых растений
дубрав лесостепи (табл. 26) характеризуются одной генерацией листь-
ев и побегов — 73,8% (от всего видового разнообразия) и 91,4% (от чис-
ла основных компонентов). Отсутствие второй генерации можно объяс-
нить недостатком света и неблагоприятным для роста водным режимом
почвы. Под пологом преобладают мелкокорневые растения, доволь-
ствующиеся небольшим запасом влаги в поверхностном слое почвы и
поступающими осадками, значительная часть которых задерживается
кронами деревьев.
По ритмам цветения преобладающей группой травянистых растений
дубовых лесов являются виды ранне-среднелетнего цикла цветения, со-
ставляющие 20,2% от всего набора видов и 26%—от числа основных
204
Таблица 25
Состав травянистых растений дубовых лесов по способам перезимовки
Тип растения Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Летнезеленые 76 66,7 30 65,2 1 100 7 70 22 62,8
Зимнезеленые 4 3,5 — — — — — — — —
Вечнозеленые 34 29,8 16 34,8 — — 3 30 13 37,2
Итого. . . . 114 100 46 100 1 100 10 100 35 100
Таблица 26
Состав травянистых растений дубовых лесов по количеству генерации листьев и побегов
Группа растений Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
С одной генерацией . . . 69 60,5 24 52,2 — — 6 60 18 51,5
С основной одной и осен- ней генерацией побе- гов возобновления . . 12 11,6 5 10,9 2 20 3 8,6
С одной генерацией по- бегов, характеризую- щихся растянутым ростом 2 1,7
С двумя генерациями . 23 20,2 13 28,3 1 100 1 10 И 31,3
С двумя генерациями (вторая — в виде побе- гов обогащения) . . . 8 7,0 4 8,7 — — 1 10 3 8,6
Итого. . . . 114 100 46 100 1 100 10 100 35 100
компонентов (табл. 27). Заметную роль играют средне-позднелетние
(14% и 8,7%), раннелетние (13,2 и 13,0%), поздневесенне-раннелетние
(13,2 и 17,4%) и среднелетние (7,9 и 4,3%) растения. Всего насчиты-
вается 20 категорий различных ритмов цветения, большинство которых
представлено малым количеством видов. По сравнению с луговой сте-
пью под пологом леса преобладают более поздние фенотипы, что объ-
ясняется, конечно, режимом затенения.
Наибольшее число видов, у которых в основании генеративных по-
бегов в этом время полностью отмирают листья, отмечается в III дека-
ду июня (29,1 и 32,5%), т. е. в тот же период, что и у растений луговой
степи (табл. 28). При этом максимумы от всего видового богатства и ос-
новных компонентов совпадают. С рядом подъемов и спусков отмира-
ние листьев в основании цветоносов у разных видов продолжается до
II декады октября. Наибольшая интенсивность отмирания (по количе-
ству видов) наблюдается в период с III декады мая по I декаду августа.
Эта закономерность свойственна и растениям луговой степи.
Первый небольшой подъем отмирания листьев на цветоносных
побегах отмечается в последней декаде мая (4,5%) (табл. 29) f Это
2Ц5
Таблица 27
Состав травянистых растении дубовых лесов по ритмам цветения
Тип растения Всего Всего компонентов категории
ВИДОВ компонентов 1-й 2-й З-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
Ранневесенние (1) ... 6 5,3 1 2,2 1 2,9
Средневесенние (2) 1 0,9 — — — — — —
Поздневесенние (3) . . 1 0,9 1 2,2 — — — — 1 2,9
Ранне-средневесенние (1-2) 2 1,7 1 2,2 1 2,9
Средне-поздневесенние (2-3) 5 4,4 2 4,3 1 10,0 1 2,9
Весенние (1—3) 3 2,6 2 4,3 — — 1 10,0 1 2,9
Средневесенне-раннелет- ние (2—4) 3 2,6 1 2,2 1 2,9
Позд невесенне- р а н нелет- ние (3—4) 15 13,2 8 17,4 2 20,0 6 17,0
Поздневесенне-среднелет- ние (3—5) 1 0,9
Раннелетние (4) .... 15 13,2 6 13,0 — — 2 20,0 4 11,3
Среднелетние (5) . . . . 9 7,9 2 4,3 — — 1 10,0 1 2,9
Позднелетние (6) .... 1 0,9 1 2,2 — — — — 1 2,9
Ранне-среднелетние (4—5 23 20,2 12 26,0 1 100,0 2 20,0 9 25,5
Средне-позднелетние (5-6) 16 14,0 4 8,7 — — 1 10,0 3 8,5
Летние (4—6) 5 4,4 1 2,2 — — — — 1 2,9
Раннелетне- среднеосенние (4-8) 1 0,9 1 2,2 1 2,9
Среднелетне-раннеосенние (5-7) 2 1,7 1 2,2 1 2,9
Среднелетне-среднеосен- ние (5—8) 2 1,7 1 2,2 — — 1 2,9
Позднелетне-раннеосенние (6—7) 2 1,7 1 2,2 — 1 2,9
Позднелетне-среднеосен- ние (6—8) 1 0,9 —
Итого . . . | 114 | 100 I 46 | 100 I 1 1 100 1 10 1 100 I 35 100
геоэфемероиды, которые не входят в состав основных компонентов. Абсо-
лютный максимум видов с отмирающими в это время листьями на
цветоносных побегах падает на III декаду сентября (20,2 и 25%). Весьма
значительное количество видов по данному признаку наблюдается еще
во II декаду октября, а в III декаду этого же месяца листья отмирают
почти у всех видов. Заметные подъемы числа видов зарегистрированы
также в III декаду июля и II декаду августа, т. е. в более поздние сро-
ки, чем на луговой степи, хотя время абсолютного максимума в обоих
биотопах совпадает.
Отмирание генеративных побегов у отдельных видов травянистых
растений дубовых лесов обнаруживается в последнюю декаду мая и
завершается в I декаде ноября (табл. 30). Максимум видов приходится
на II декаду октября (29,1%—от всего количества видов и 33,3% —
от основных компонентов). Весьма значительное число (видов с отмира-
ющими в это время цветоносными побегами наблюдается в I декаду
октября (17,5 и 13,4%) и в II декаду сентября (11,7 и 11,2%).
Смещение максимума отмирания генеративных побегов растений в
206
Таблица 28
Динамика отмирания генеративных побегов травянистых растений дубовых лесов
Месяц Порядковый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли* чество % коли* чество % коли- чество % коли- чество % коли- чество %
V 15 И 11,8 3 7,5 — — 1 11,1 2 6,7
16 10 10,7 4 10,0 — — — — 4 13,3
VI 17 5 5,4 3 7,5 — — — — 3 10,0
18 27 29,1 13 32,5 5 55,6 8 26,7
19 8 8,6 1 2,5 1 100 — — — —
VII 20 13 14,0 8 20,0 — — 1 11,1 7 23,3
21 6 6,4 2 5,0 — — — — 2 6,7
22 6 6,4 — — — — —
VIII 23 1 1,1 —
24 —
25 1 1,1 1 2,5 — — — — 1 3,3
IX 26 2 2,2 2 5,0 — — 1 11,1 1 3,3
27 2 2,2 2 5,0 — — 1 11,1 1 3,3
X 28 1 1,1 1 2,5 — — — — 1 3,3
| Итого | 93 100 40 | 100 1 1 100 9 100 30 100
дубовых лесах по сравнению с луговой степью на более позднее время
можно объяснить более благоприятным термическим режимом под по-
логом леса, отсутствием резких скачков температур.
Обратимся теперь к сопряженному анализу биоморфологических
признаков на основе ярусной принадлежности растений в подземной и
надземной частях (табл. 31а, б, в). Вначале напомним, что в подземной
части травяного покрова дубовых лесов выделяется четыре яруса с глу-
бинами в 30, 60, 100 и 150 см. Более глубокоукореняющиеся виды еди-
ничны, в число компонентов не входят, а потому и не являются строите-
лями соответствующих ярусов. Таким образом, и здесь число ярусов в
надземной и подземной частях травяного покрова совпадает.
Проанализируем распределение видов с определенными признаками
по ярусам подземной части. Наибольшее число безрозеточных растений
(табл. 31а) приурочено к 4-му ярусу (17,6% всего видового состава и
15,2% от числа основных компонентов). В следующих ярусах (в на-
правлении вглубь почвы) число безрозеточных видов уменьшается.
Полурозеточные более всего представлены в 3-м ярусе (18,4%), но мак-
симум основных компонентов приурочен ко 2-му ярусу (17,4%). Розе-
точные растения преобладают в 4-м ярусе (7 и 6,6%), в следующих
глубже — 3 и 2-м ярусах — число их сокращается. По способам возоб-
новления побегов стоит лишь отметить приуроченность моноподиальных
растений к 4-му ярусу.
Поскольку большинство растений дубовых лесов является длитель-
новегетирующими, а коротковегетирующие — не типичными, то усмат-
ривать связь длительности вегетации с той или иной глубиной проник-
новения корневых систем не приходится.
207
Таблица 29
Динамика полного отмирания листьев генеративных побегов травянистых растений
дубовых лесов
Месяц Порядковый номер декады (от начала t года) Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
V 15 4 4,5 — — — — — — — —
16 1 1,1 — _ —
VI 17 1 1,1 — — — — — — — —
18 —
19 1 1,1 1 2,8 __ — — — 1 3,6
VII 20 2 2,3 1 2,8 — — 1 14,3 — —
21 5 5,6 2 5,5 — — 1 14,3 1 3,6
22 3 3,4 1 2,8 — — — — 1 3,6
VIII 23 7 7,9 2 5,5 — — 1 14,3 1 3,6
24 5 5,6 3 8,3 1 100 2 28,5 — —
25 4 4,5 1 2,8 — — — — 1 3,6
IX 26 9 10,1 1 2,8 — — — — 1 3,6
27 18 20,2 9 25,0 — — 1 14,3 8 28,6
28 12 13,5 6 16,7 — — — — 6 21,2
X 29 16 18,0 8 22,2 — — — — 8 28,6
30 —
XI 31 1 1,1 1 2,8 — — 1 14,3 — , —
Итого 89 | 100 36 100 1 100 7 100 28 100
Максимум видов с одной генерацией листьев (табл. 316) падает на
4-й ярус (26,3%), далее число их постепенно снижается. Однако среди
компонентов 'наибольшее количество их приходится на 3-й ярус. Рас-
тения с двумя генерациями листьев в значительном числе представле-
ны в 3 и 2-м ярусах, а также в 4-м ярусе. Больше всего летнезеленых
видов насчитывается в 3-м ярусе (23,8 и 26,0%). Вечнозеленые и зим-
незеленые растения достигают наибольшего обилия в 4 ярусе, в после-
дующих ярусах количество их убывает.
Цветущие весной растения (табл. 31в) в наибольшем количестве
приурочены к 4 и 3-му ярусам, весенне-летние — к 4-му ярусу, летние —
к 3 и 2-му, летне-осенние — к 1-му ярусу.
Четкого соответствия в распределении видов по ярусам в подземной
и надземной частях травяного покрова не наблюдается. Можно лишь
отметить, что наибольшее число растений 4-го яруса надземной сферы
приходится на 4-й же ярус в подземной сфере. Подобной координации
в других ярусах обнаружить не удается (табл. 31в). Таким образом,
здесь наблюдается противойоложная тенденция по сравнению с луговой
степью.
Кратко остановимся на количественной характеристике видов по
биоморфологическим признакам в связи с ярусным распределением
растений в надземной сфере травяного покрова. Максимум безрозёточ-
ных растений падает на 2-й ярус (17,6 и 19,5%), полурозеточных — на
208
Таблица 30
Динамика отмирания генеративных побегов травянистых растений дубовых лесов
Месяц Порядковый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
коли- чество % коли- чество % коли- чество % коли- чество % коли- чество %
V 15 4 3,9 —
16 2 1,9 1 2,2 • — 1 2,9
VI 17 —
18 3 2,9 1 2,2 — — — — 1 2,9
19 1 1,0 1 2,2 — — Т 1 2,9
VII 20 —
21 2 1,9 1 2,2 — — 1 11,1 — —
22 5 4,9 2 4,4 — — 1 11,1 1 2,9
VIII 23 3 2,9 1 2,2 — — — — 1 2,9
24 2 1,9 —
25 7 6,8 3 6,7 1 100 — — 2 5,7
IX 26 6 5,8 4 8,9 — — 1 11,1 3 8,5
27 12 11,7 5 11,2 — — — — 5 14,3
28 18 17,5 6 13,4 — — 3 33,4 3 8,5
X 29 30 29,1 15 33,3 — — 2 22,2 13 37,1
30 7 6,8 4 8,9 — — — — 4 11,4
XI 31 1 1,0 1 2,2 — — 1 11,1 — —
Итого 103 100 45 100 1 100 9 100 35 | | 100
1 и 2-й ярусы (19,3 и 15,9%), розеточных — на 4 и 3-й (6,1 и 5,2%)
(табл. 31а).
В распределении растений по ярусам с разным способом возобнов-
ления особой правильности не отмечается. Нельзя установить приуро-
ченность видов к тем или инь!м ярусам и по длительности вегетации,
так как большинство является длительновегетирующим.
Наибольшее количество видов с одной и двумя генерациями листьев
входит в состав 2-го яруса (табл. 316). К этому же ярусу относится и
максимум летнезеленых растений (24,5 и 28,2%). Зимующие в зеленом
состоянии виды доминируют в 4-м ярусе (12,1%). Цветущие весной
растения преобладают в 4-м ярусе (9,6 и 8,7%), весенне-летние — в
3-м (8,8 и 8,7%), летние — во 2-м (28,1 и 32,5%), летне-осенние —
также во 2-м ярусе (3,5 и 4,4%) (табл. 31 в).
Поярусный анализ количественного состава видов по биоморфологи -
ческим признакам в связи с ярусным расчленением надземной и под-
земной частей травяного покрова дубрав не вскрывает той пропорцио-
нальной координации, которая установлена для растительности луговой
степи. Эта особенность структуры травяного покрова дубовых лесов,
в сравнении с луговой степью, определяется, конечно, меньшей диффе-
ренциацией условий среды в пределах занимаемого растениями про-
странства. Относительной выравненное™ экологического режима впол-
не отвечает и мезофитная природа подпологовых растений.
14 в. Н. Голубев 209
Таблица 31а
Сопряженный количественный анализ травянистых растений дубовых лесов по биоморфологическим особенностям надземных органов в связи
с ярусной принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Тип структуры надземных побегов Способ нарастания и возобновления Длительность вегетации
безрозеточный полурозеточный / розеточный симподиальный моноподиальный длительновегети- рующие коротковегети- рующие
всего основные компо* ненты всего основные компо- ненты всего основные компо ненты всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты
В подземной части: 4-й (до 30 см) 20 (17,6) 7(15,2) 10(8,8) 2(4,3) 8(7,0) 3(6,6) 31 (29,8) 8(18,2) 4(3,8) 3(6,8) 32 (28,3) 12(26,1) 5(4,4)
З-й (до 60 см) 13 (11,4) 6(13,0) 21 (18,4) 5(10,8) 4(3,5) 3(6,6) 33 (31,7) 13(29,5) 1 (1,0) 1(2,3) 36 (31,8) 15(32,6) 3(2,7) —>
2-й (до 1 м) 7(6,1) 4(8,7) 13(11,4) 8(17,4) 1 (0,9) 1 (2,2) 18(17,3) И (25,0) 1 (1,0) 1 (2,3) 19 (16,8) И (23,9) 1 (0,9) 1 (2,2)
1-й (глубже 1 м) 10 (8,8) 4(8,7) 7(6,1)) 3(6,5) — 15(14,4) 7(15,9) 1 (1,0) 17(15,1) 7(15,2) —
В надземной части: 1-й (свыше 90 ел) 9(7,9) 4(8,7) 22 (19,3) 6(13,0) — — 24 (23,1) 9(20,4) 1(1,0) 28 (24,7) 9(19,6) 3(2,7) 1(2,2)
2-й (60—90 см) 20(17,6) 9(19,5) 18(15,9) 10(21,7) — — 35 (33,6) 17(38,6) 2(1,9) 2(4,6) 38 (33,5) 19(41,2) —-
З-й (40—60 см) И(9,6) 5(10,8) 5(4,4) — 6(5,2) 4(8,7) 19(18,3) 7(15,9) 2(1,9) 1 (2,3) 20 (17,7) 9(19,6) 2(1,8) —.
4-й (0—25 см) 10 (8,8) 5(6,6) 6(5,2) 2(4,4) 7(6,1) 3(6,6) 19(18,3) 6(13,6) 2(1,9) 2(4,6) 18(16,0) 8(17,4) 4(3,7) —
Таблица 316
Сопряженный количественный анализ травянистых растений дубовых лесов по биоморфологическим особенностям надземных органов
в связи с ярусной принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Количество генераций листьев и побегов Способ перезимовки
с одной генерацией с двумя генерациями летнезеленые вечнозеленые зимнезеленые
всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты всего основные компоненты
В подземной части: 4-й 30 (26,3) 7(15,2) 8 (7,0) 5(10,9) 18(15,8) 5(10,9) 18 (15,8) 7(15,2) 2(1,7)
З-й 29 (25,5) 10(21,7) 10 (8,8) 5(10,9) 27 (23,8) 12(26,0) 9(7,9) 2(4,4) 2(1,7) —
2-й 9(7,9) 6(13,0) И (9,6) 6(13,0) 14 (12,3) 7 (15,2) 7(6,1) 6(13,0) —
1-й 15 (13,2) 6(13,0) 2(1,7) 1(2,2) 17(14,9) 7(15,2) — — —
В надземной части: 1-й 24 (21,0) 7(15,2) 7(6,1) 3(6,6) 25 (22,0) 9(19,5) 5(4,4) 1(2,2) 1 (0,9)
а . 2-й 27 (23,8) 12 (26,0) Н (9,6) 7(15,2) 28 (24,5) 13(28,2) 10 (8,8) 6(13,0) — —
и З-й 17(14,9) 7(15,2) 5(4,4) 2(4,4) 14 (12,4) 6 (13,0) 7(6,1) 3(6,6) 1 (0,9) —
U 4-й 15 (13,2) 3(6,6) 8(7,0) 5(10,9) 9(7,9) 3 (6,6) 12(10,4) 5(10,9) 2(1,7)
Таблица 31в
Сопряженный количественный анализ травянистых растений дубовых лесов по биоморфологическим особенностям надземных органов в связи
с ярусной принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Ритм цветения Ярус в надземной части травяного покрова
весенние весенне-летние летние летне-осенние 1-й 2-й 3-й 4-й
всего основные компо- ненты всего в) з . S 2 з о Е н ЙВ 38 X о о <и о * я всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты всего основные компо- ненты
В подзем- ной части: 4-й 12 (10,4) 2 (4,4) 14(12,4) 6(13,0) И (9,6) 3(6,6) 1 (0,9) 1(2,2) 2(1,7) 5(4,4) 2(4,4) 13(11,4) 5(10,8) 18(15,8) 5 (10,8)
3-й 5(4,4) 4(8,7) 4(3,5) 3(6,6) 27(23,7) 7(15,2) 2(1,7) — 12 (10,4) 2(4,4) 16(14,1) 7(15,2) 7(6,1) 4(8,7) 3(2,6) 1(2,2)
2-й 1 (0,9) 1 (2,2) 1(0,9) — 18 (15,8) 11 (23,8) 1 (0,9) 1(2,2) 8(7,0) 4(8,7) И (9,6) 8(17,4) 1 (0,9) — 1 (0,9) 1(2,2)
1-й — — — — 13(11,4) 5 (10,8) 4(3,5) 2 (4,4) 9(7,9) 4(8,7) 6(5,2) 2(4,4) 1 (0,9) — 1 (0,9) 1 (2,2)
В надзем- ной части: 1-й 28 (24,6) 9(19,5) 3(2,6) 1 (2,2)
2-й — — 2(1,7) 2(4,4) 32 (28,1) 15 (32,5) 4(3,5) 2(4,4) — — — — — — — —
3-й 7(6,1) 3(6,5) 10(8,8) 4(8,7) 4(3,5) 1(2,2) 1 (0,9) 1(2,2) — — — — — — — —
4-й 11(9,6) 4(8,7) 7(6,1) 3(6,5) 5(4,4) 1(2,2) — — — — — — — — — —
РАСТИТЕЛЬНОСТЬ ДНА ЛОГОВ
В составе растительности дна степных логов проанализировано
144 вида, биоморфологические признаки которых приводятся в
табл. 32.
По структурным особенностям побегов в составе травостоя преоб-
ладают полурозеточные растения (табл. 33). От всего видового разно-
образия они составляют 58,3 и 70%—от числа основных компонентов.
Второе место занимают безрозеточные растения (34,1 и 18,6%),
третье — розеточные (7,6 и 11,4%). Такое соотношение видов по типам
побегов весьма близко напоминает состав луговой степи.
Как и в остальных типах растительности лесостепи, на дне логов
господствуют симподиальные растения (89,3 и 83,3%; табл. 34). На
моноподиальные соответственно приходится 10,7 и 16,7% (от всей фло-
ры и основных компонентов). В вертикальном строении травостоя дна
логов можно выделить 5 ярусов:
1-й ярус (свыше 1 ж) слагают генеративные побеги Angelica silvestris, Bromus
inermis, Heracleum sibiricum, Sanguisorba officinalis, Succisa pratensis, Thalictrum luci-
dum, Veratrum lobelianum и др.
2-й ярус (60—100 см) образуют побеги Alopecurus pratensis, Betonica officinalis,
Campanula glomerata, Centaurea pseudophrygia, Cnidium dubium, Deschampsia caespi-
tosa, Galium mollugo, Poa pratensis, Polygonum bistorta, Selinum carvifolia, Veronica
longifolia и др.
3-й ярус (30—60 см) составляют: Briza media, Galium boreale, Geranium pratense,
Geum rivale, Leucanthemum vulgare, Plantago lanceolata, Stellaria palustris, Trifolium
hybridum и др.
4-й ярус (15—30 см) ограничивается побегами Alchemilla micans, Dianthus deltoi-
des, Luzula pallescens, Potentilla erecta, Ranunculus repens и др.
5-й ярус (0—15 см) образуется местами, если развиваются такие низкорослые ра-
стения, как Glechoma hederacea, (.ysimachia nummularia, Potentilla anserina, Veronica
serpyllifolia, Viola canina и др.
Наибольшее число видов (табл. 35) входит в состав 3-го яруса (33,3
и 32,9%). Велико еще количество видов 2-го яруса (31,9 и 31,4%),
далее идут 4-й ярус (15,3 и 17,1%), 1-й ярус (13,9 и 15,7%) и самое
последнее место по количеству видов занимает 5-й.
Решающую роль в сложении травостоя играют длительновегетиру-
ющие растения, что вполне соответствует благоприятным условиям вла-
гообеспеченности. Из всего видового состава на их долю приходится
94,1 и 91,4%—от числа основных компонентов (табл. 36). Коротко-
вегетирующие растения занимают весьма скромное место. Однако их
заметно больше, чем в дубовых лесах.
По способам перезимовки более всего представлены летнезеленые
виды (55,5%—от состава всей флоры и 52,9%—ют числа основных
компонентов) (табл. 37). В этом отношении растительность дна логов
сближается с луговой степью. В отличие от последней, в логах выделя-
ется группа вечнозеленых растений (41 и 45,7%), наоборот, количество
зимнезеленых растений сведено к крайнему минимуму (3,5 и 1,4%).
Растения с двумя генерациями листьев и побегов (табл. 38) зани-
мают ведущее место в составе растительности дна логов (72,2%—от
всего видового богатства и 81,4%—от числа основных компонентов).
Растения с одной генерацией имеют подчиненное значение (27,8 и
18,6%). Эти цифровые показатели при сопоставлении с соответствую-
щими данными для луговой степи и травяного покрова дубрав красно-
речиво свидетельствуют о том, что экологической причиной развития
второй генерации являются условия влагообеспеченности растений,
стимулирующие ростовые процессы. Благоприятным водным режимом
почвы дна логов объясняется и высокий процент вечнозеленых расте-
ний, перезимовывающих в зеленом состоянии и вегетирующих в тече-
ние всего летнего периода.
212
Таблица 32
ю
Биоморфологнческий состав компонентов растительности дна логов
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
Achillea millefolium Кз пр С 3 Д В 2 4—9 15 29 29 37
Aconitum lasiostomum пр С 2 Д Л 1 >4—5 20 26 27 82
Agrimonia eupatoria пр С 2 Д л 2в >4-7 19 27 29 106
Agrostis alba пр с 2 Д в 2 5 18 24 28 74
A. carUna пр с 2 Д в 2 >4—5< 16 23 27 71
Alchemilla micans пр м 4 Д л 2 3—4< 26 27 19
Alopecurus pratensis Кз пр с 2 Д в 2 3—4 15 20 23 90
Angelica silvestris К, пр 1 Д л 2 >4-5 20 22 24 117
Anthoxanthum oduratum . . . К, пр с 3 Д в 2 3—4< 16 20 22 46
Anthriscus silvestris К, пр с 1 СМ л 2 4 18 20 21 111
Betonica officinalis К, пр м 2 Д в 2 5—6 26 27 61
Bidens tripartitus бр 3 д л 1р 6—7 19 26 28 58
Briza media Кз пр с 3 д в 2 4 18 20 21 51
Bromus inermis Кз пр с 1 д л 2в >4—5< 17 25 28 112
Bunias orientalis пр с 2 д л 2 3-4 17 22 25 81
Calamagrostis arundinacea . . Кз пр с 1 д л 1 5 16 26 27 117
C. epigeios пр с 1 д л 1 5 19 26 28 107
C. lanceolata пр с 2 д л 1 5 18 26 27
Campanula glomerata Кз пр с 2 д л 2 4-5 18 25 27 63
C. patula Кз пр 3 см 3 2 4—5< 16 20 21 58
C. rapunculoides бр с 1 д л 1 5-6 18 25 26 134
Carex caespitosa Кз р с 3 д л 1 >1-3 22 20
* См. сноски к табл. 8. стр. 171
214
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Carex hirta пр С 3
C. leporina Кз пр С 4
C. pallescens .... .... Кз пр с 4
C. riparia .... пр с 2
C. vesicaria пр с 3.
C. vulpina Кз р с 3
Carum carvi Кз пр 3
Centaurea jacea Кз пр с 2
C. pseudophrygia Ki пр с 2
Chaerophyllum bulbosum пр 1
Cirsium arvense Кз пр с 2
C. heterophyllum пр с 2
Cnidium dubium . . . Кз пр с 2
Coronaria flos cuculi . . пр с 3
Creptз praemorsa ... р с 3
Dactylis glomerata Кз пр с 2
Deschampsia caespitosa .... Кз пр с 2
Dianthus deltoides бр с 4
D. stenocalyx бр с 2
Digraphis arundinacea . . пр с 1
Epilobium palustre бр с 2
Euphorbia virgata бр с 2
Festuca pratensis К, пр с 2
F. rubra Кз пр с 2
Таблица 32 (продолжение)
Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения „ i Срок отмирания i . _ (декады от-начала.года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
Д В 2 3 21 22 22 40
Д в 2 3—4< 21 22 22 24
Д л 1 3 22 24 33
Д в 2 3—4< 22 22 22 73
Д в 2 2-3 22 22 22 45
Д в 2 2—3 24 52
СМ л 2 3—4 17 19 21 53
Д л 2 5-6 15 26 26 74
Д л 2 5-6 15 26 26 85
см л 1 >4-5 16 21 24
Д л 2в 5-6 18 26 29 95
Д л 2 4—5< 16 21 24 86
Д л 2 6-7 20 27 30 61
д в 2 4 15 18 21 61
д л 1 3—4 19 47
д в 2 4 15 22 24 81
д в 2 >4 18 21 24 82
д в 2 4-5 16 21 23 26
д в 2в 6—8 22 28 30 67
д л 2в >4 17 23 27 167
д в 2в 5-6 18 24 25 88
д л 2в 3-4 16 . 21 27 69
д в 2 4 16 21 23 66
д в 2 4 16 21 23 61
215
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Ficaria verna Кз пр С 4
Filipendula ulmaria Кз пр С 1
Fritillaria meleagris Кз пр с 4
Galatella rossica бр с 1
Galium boreale Кз бр с 3
G. mollugo Кз бр с 2
G. uligi nosum бр с 3
Gentiana Ungulata * пр 4
G. pneumonanthe бр с 2
Geranium palustre Кз пр с 3
G. pratense Кз пр с 3
G. silvaticum Кз пр с 3
Geum rivale Кз пр м 3
Gladiolus imbricatus ... ... . . пр с 3
Glechoma hederacea ....... Кз бр м 5
Gypsophila muralis .... бр 5
Heracleum sibiricum К, пр 1
Herniaria glabra бр с 5
Hieracium pratense пр с 3
H. umbellatum бр с 2
Inula britannica пр с 3
Iris sibirica пр с 2
Juncus articulatus пр с 3
J. effusus р с 2
Таблица 32 (продолжение)
Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота(за 1962 г.) генератив- ных побегов, в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
ГЭ Л 1 1 15 16 16 10
Д л 2 >4—5 17 27 27 107
ГЭ л 1 1—2 16 16 16 19
Д л 1р 6—7 18 28 29 115
Д л 2в 4—5 16 28 28 42
Д в 2в 4-5 16 24 27 47
Д в 2в 4—5 18 24 29 37
Д 3 2 4—5 18 23 25
Д л 1р 6—8 20 27 30 71
д л 2 4—7 21 28 29 57
д л 2 4—6 21 24 25 40
д л 2 3—4< 20 22 23 48
д в 2 >2—4< 19 21 47
с л 1 4 21 22 23 56
д в 2в 1—4 12
д л 1р 5-6 19 22 26 8
д л 2 4-5 22 23 24 102
д в 2в 4 ‘ 16 18 24 2
д в 2 4—5< 20 23 23 40
д л 2в 6—8 20 27 28 96
д л 2 5—8 19 25 27 33
д л 1 4 — — 26 81
д в 1 4 16 43
д в 1 >4 78
to
2
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ, возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Koeleria delavignei Кз пр С 3
Lathyrus pratensis Кз бр С 3
Leontodon autumnalis К, пр с 4
L. hispidus Кз Р с 3
Leucanthemum vulgare К. пр с 3
Lotus corniculatus бр с 4
Luzula pallescens К» пр с 4
Lycopus exaltatus ....... бр с 2
Lysimachia nummularia .... Ка бр м 5
Lysimachia vulgaris бр с 2
Lythrum virgatum бр с 2
Malachium aquaticum бр с 3
Medic ago f alcata бр с 3
Melampyrum arvense бр 3
Mentha arvensis бр с 3
Molinia coerulea к. р с 2
Myosotis caespitosa пр 4
Phleum pratense К. пр с 2
Plantago lanceolata Ка р м 3
P. media Кз р м 4
Poa palustris пр с 2
P. pratensis • . . К, пр с 2
P. trivialis пр с 2
Polygonum amphibium бр с 3
Таблица 32 (продолжение)
Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов» в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
Д в 2 4 16 20 22 56
Д л 2в 4-5 18 28 28 67
Д ' л 2 4-8 30 - 29 29 30
Д л 2 4 20 52
Д в 2 4—5 16 22 26 50
Д л 2в 5-7 19 26 30 24
Д в 2 3—4< 16 19 21 31
Д л 2в 5-7 18 28 30 92
Д в 2в 4-5 15
Д л 1 5—6< 18 25 27 87
Д л 2в 5-6 19 28 29 75
Д в 2в 5-9 18 24 30 82
Д в 1Р 4—7 19 24 29 54
Д л 1 4—6 18 23 24 35
Д л 2в 5-7 18 29 30 36
Д л 1 >5—6< 23 27 92
Д 3 2 4-5 17 20 24 50
Д в 2 5 17 26 27 91
Д в 2 4 22
Д в 2 3—4 21 31
Д в 2 4 15 19 21 76
Д в 2 4 17 19 21 62
д в 2 4 17 20 22 67
д л 1 18 26 27
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ- возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Polygonum bistorta .......
P. hydropiper...................
Potentilla anserina.............
P. erecta.......................
P. thuringiaca..................
P. supina.......................
Primula veris .........
Prunella vulgaris...............
Ptarmica vulgaris...............
Pulmonaria angustifolia . . . .
Ranunculus acer.................
R. auricomus....................
R. repens..........:............
Rorippa austriaca...............
R. islandica....................
Rumex acetosa...................
R. confertus....................
R. crispus......................
Sanguisorba officinalis . . . .
Scutellaria galericulata . . . .
Sedum purpureum.................
Selinum carvifolia..............
Serratula coronata..............
M S. inermis........................
3 Solidago virgaurea................
к» пр м 2
бр 3
р м 5
пр м 4
Кз пр м 3
пр с 4
Кз р с 4
Кз бр с 4
бр с 2
пр с 3
Кз пр с 2
пр с 3
К, пр с 4
пр с 3
пр с 3
К, пр с 2
Кз пр с 2
пр с 1
Кз пр м 1
бр с 3
бр с 3
Кз бр с 2
пр с 1
Кз пр с 2
пр с 2
Таблица 32 (продолжение)
Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) Средняя высота (за 1962 г.) генератив- ных побегов, в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
Д Л 1 3—4 19 20 89
Д Л 1р 6 18 26 28 72
Д Л 2 4 30 — 24 7
Д Л 2 3—5 — 21 27 23
Д В 2 >3-4 — 21 24 35
Д Л 2 4-6 17 24 27
Д В 2о 1—3 — 21 16
Д в 2 4-5 19 24 26 20
Д л 1Р 5—7 18 26 27 76
Д л 2 1—2 15 16 17 18
Д л 2 3-5 17 20 22 73
Д в 1 2—3 16 17 20 31
Д в 2 3—5 — 20 24 31
Д 3 1 5-6 16 21 26
Д в 2 >4—5 17 24 28 60
Д в 2 3 -4 17 18 20 69
Д л 1 4 17 21 23 99
Д л 1 4 17 21 24 116
Д л 2 5-8 — 29 30
Д л 2в 5-6 18 27 28 58
Д л 1р 6—7 18 23 29 61
Д л 2 5-6 23 26 27 96
Д л 2 >5-6 19 26 27 140
Д л 2 >5-8 21 27 29 85
Д л 2 6—7 26 28 30 74
Вид Основ- ные компо- ненты Тип структуры надземных побегов Способ возобнов- ления побегов Принад- лежность к ярусам раститель- ности
Stachys palustris бр С 2
Stellaria graminea бр С 3
S. palustris к» бр С 3
Succisa pratensis Кз пр М 1
Symphytum officinale пр С 2
Tanacetum vulgare Кз бр с 1
Taraxacum officinale Кз Р/ с 4
Thalictrum lucidum Кз бр с 1
T. simplex бр с 1
Tri folium hybridum бр с 3
T. medium Кз бр с 3
T. pratense Кз пр м 3
T. repens . Кз бр м 4
T. spadiceum бр 4
Trollius europaeus Кз пр с 3
Valeriana exaltata Пр с 1
Veratrum lobelianum Кз пр с 1
Veronica chamaedrys .... бр с 4
V, longifolia Кз бр с 2
V. scutellata бр с 4
V. serpyllifolia бр с 5
Vicia cracca Кз бр с 3
V. sepium бр с 3
Viola canina бр с 5
V. stagnina бр с 5
Таблица 32 (окончание)
Тип по продол- житель- ности ве- гетации Способ перези- мовки Количест- во генераций листьев и побегов Ритм цветения Срок отмирания (декады от начала года) ^Средняя высота(за 1962 г.) генератив- ных побегов, в см
листьев у основания генератив- ных побегов листьев генератив- ных побегов генера- тивных побегов (полное усыхание)
Д Л 2в 4—6 19 24 25 79
Д В 2в 3—5 15 21 24 30
Д В 2в 3-5 15 20 24 30
Д В 2о 6-8 — 27 29 109
Д Л 2 4—6 16 23 25 65
Д Л 2в 5-6 17 25 27 ИЗ
Д В 2 1—4< — — 18 10
Д Л 1 4-5 19 26 27 112
Д Л 1 5
Д В 2 4-6 16 21 27 26
Д л 2в >4-6 18 27 28 34
Д в 2 4—5 20 23 25
Д в 2в 4-6 17
Д 3 1 4 16 20 22 21
Д л 1 2-3 17 19 20 53
Д л 2 5-6 19 27 27 118
С л 1 4—5< 23 24 24 122
Д в 2о 2-4 17 25 26 23
Д л 2в 5-6 18 25 27 85
Д в 2в 4-6 17 31 31 27
Д в 2 >2—4< 17 19 21 8
д л 2в 4-6 18 31 31 47
д в 2в 4 18 27 27 50
д в 2в 2-4 18 29 29 7
д л 2в 2—4 18 29 29 15
Таблица 33
Состав растений дна логов по типам структуры надземных побегов
Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
Побег количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
Безрозеточный 49 34,1 13 18,6 4 26,6 9 16,6
Полурозеточный .... 84 58,3 49 70,0 1 100 10 66,7 38 70,4
Розеточный И 7,6 8 11,4 — — 1 6,7 7 13,0
Итого 144 100 70 100 1 100 15 100 54 100
Таблица 34
Состав растений дна логов по способам возобновления побегов
Возобновление. Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
Симподиальное 118 89,3 55 83,3 1 100 12 80,0 42 84,0
Моноподиальное . . . 14 10,7 И 16,7 — — 3 20,0 8 16,0
Итого 132 100 66 100 1 100 15 100 50 100
Таблица 35
Распределение растений дна логод по ярусам в надземной части растительности
Ярус Всего Всего компонентов категории j
видов компонентов 1-й 2-й 3-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
1 (свыше 1 л) 2 (60—100 см) 3 (30—60 см) 4 (15—30 см) 5 (0—15 см) 20 46 48 22 8 13,9 31,9 33,3 15,3 5,6 И 22 23 12 2 15,7 31,4 32,9 17,1 2,9 1 100 1 8 ' 4 1 . 4 6,7 53,3 26,6 6,7 6,7 10 13 19 И 1 18,0 24,7 35,2 20,3 1,8
Итого.... | 144 | 100 70 100 I 1 100 15 | | 100 54 | 100
219
Таблица 36
Состав растений дна логов по продолжительности вегетации
Гип растения Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
О) ег 5 о ч CQ % количе- ство % количе- ство % количе- ство % количе- ство %
Длительновегетирующие 128 94,1 64 91,4 1 100 13 86,6 50 92,6
Коротковегетирующие: монокарпики .... 4 2,9 3 4,6 — — — — 3 5,6
геоэфемероиды . . . 2 1,5 2 2,9 — — 1 6,7 1 1,8
среднепродолжитель- ной вегетации .... 2 1,5 1 1,4 — •— 1 6,7 — —
Итого .... 136 I 100 70 1 :100 1 1 1 1 100 15 1 100 54 | 100
Таблица 37
Состав растений дна логов по способам перезимовки
Тип растения Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
Летнезеленые 80 55,5 37 52,9 1 100 9 60,0 27 50,0
Зимнезеленые 5 3,5 1 1,4 — — — — 1 1,9
Вечнозеленые 59 41,0 32 45,7 — — 6 40,0 26 48,1
Итого . . . 144 | f 100 70 1 100 1 100 15 | 100 54 | 100
Таблица 38
Состав растений дна логов по количеству генераций листьев и побегов
Группа растений Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
С одной генерацией . . С основной одной и осен- 29 20,1 И 15,7 — — 4 26,6 7 13,0
ней генерацией побегов возобновления .... 3 2,1 2 2,9 — 2 3,7
С одной генерацией побе-
гов, характеризующих- ся растянутым ростом 8 5,6
С двумя генерациями . . С двумя генерациями (вто- 71 49,3 44 62,8 1 100 7 46,8 36 66,7
рая — в виде побегов обогащения) 33 22,9 13 18,6 • — — 4 26,6 9 16,6
Итого . . . 144 100 70 100 1 100 15 100 | 54 100
220
Растения дна логов по ритмам цветения разделяются на 23 катего-
рии, соответствующие количеству элементарных ритмов цветения.
Главная роль принадлежит ранне-среднелетним растениям (17,5 и
21,4%) (табл. 39). Далее в убывающем порядке идут раннелетние
(16,8 и 15,8%), поздневесенне-раннелетние (10,5 и 14,3%), средне-
позднелетние (11,9 и 11,4%), летние (7 и 5,7%), среднелетние (4,2 и
2,9%). Остальные категории ритмов представлены незначительным
количеством видов. Таким образом, в составе растительности дна ло-
гов развиты более поздние фенотипы по сравнению с луговой степью
Таблица 39
Состав растений дна логов по ритмам цветения
Всего Всего компонентов категории
вадов компонентов 1-й 2-й З-й
Тип растения
в? “2 % 2 g О % ВТ §2 % Is % |g %
2 Я St О См « Б
Ранневесенние (1) . . 1 0,7 1 1,4 1 1,8
Поздневесенние (3) . * Ранне-средневесенние (1— 2 1,4 1 1,4 — — — — 1 1,8
2) Средне-поздневесенние 2 1,4 1 1,4 — — 1 6,7 — —
(2-3) 4 2,8 2 2,9 — — — — 2 3,7
Весенние (1—3) .... Ранневесенне- раннелетние 2 1,4 2 2,9 — — — — 2 3,7
(1-4) 2 1,4 2 2,9 — — — — 2 3,7
Средневесенне-раннелет- ние (2—4) Поздневесенне-раннелет- 5 3,5 1 1,4 — — — — 1 1,8
ние (3—4) Поздневесенне-среднелет- 15 10,5 10 14,3 — — 2 13,3 8 15,0
ние (3—5) 5 3,5 3 4,3 — — 2 13,3 1 1,8
Раннелетние (4) . . . . • 24 16,8 4,2 11 15,8 2,9 з 20,0 8 15,0 3,7
Среднелетние (5) ... 6 2 — — 2
Позднелетние (6) ... 1 0,7 —
Ранне-среднелетние (4—5) 25 17,5 15 21,4 — — 4 26,6 11 20,4
Средне-позднелетние (5-6) 17 11,9 8 11,4 1 100 1 6,7 6 11,2
Летние (4—6) Раннелетне-раннеосенние 10 7,0 4 5,7 — — 1 6,7 3 5,6
(4-7) Раннелетне- среднеосенние 3 2,1 1 1,4 — — — — 1 1,8
(4-8) • 1 0,7 1 1,4 — — — — 1 1,8
Раннелетне- позднеосенние (4-9) Среднелетне- раннеосенние 1 0,7 1 1,4 — — — — 1 1,8
(5-7) Среднелетне-среднеосен- 4 2,8 — — — — — — — —
ние (5—8) 3 2,1 2 2,9 — — 1 6,7 1 1,8
Среднелетне-позднеосен- ние (5—9) Позднелетне-раннеосенние 1 0,7 — — — — — — — —
(6-7) ......... Позднелетне-среднеосен- 5 3,5 1 1,4 — — — — 1 1,8
ние (6—8) 4 2,8 1 1,4 — — — — 1 1,8
Итого . . . | 143 1 100 I 70 100 I 1 1 1 100 15 100 54 100
221
и более ранние — в отличие от травяного покрова дубрав, что хо-
рошо согласуется с особенностями экологического режима данного
биотопа.
Отмирание листьев в основании генеративных побегов начинается
с последней декады мая (табл. 40), достигает первого максимума в
I декаду июня—19,2% (от всего видового состава) и 21,2% (от числа
основных компонентов), второго максимума—III декаду июня (24,2%
от всего состава видов) и в III декаду июля (5,8% от числа основных
компонентов), а в августе заканчивается у большинства видов, за еди-
ничными исключениями. Сравнивая отмирание листьев в основании
генеративных побегов у растений дна логов с растениями луговой сте-
пи, можно обнаружить весьма близкое соответствие, особенно в сроках
абсолютных максимумов видов от всей флоры и основных компонентов,
которые совпадают. Некоторое сходство отмирания наблюдается и с
растениями травяного покрова дубовых лесов. Однако максимум видов
среди основных компонентов в дубовых лесах наступает на две декады
позже и совмещается с максимумом от всего состава видов, т. е. с
III декадой июня.
Таблица 40
Динамика отмирания листьев в основании генеративных побегов травянистых
растений дна логов
Месяц Порядко- вый номер декады (от начала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й 3-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
V 15 И 9,2 7 13,5 1 100 3 25,0 3 7,7
16 23 19,2 11 21,2 — — 3 25,0 8 20,5
VI 17 20 16,6 10 19,2 — — 2 16,7 8 20,5
18 29 24,2 9 17,3 — — 1 8,3 8 20,5
19 13 10,8 2 3,8 2 5,1
VII 20 7 5,8 3 5,8 — — — — 3 7,7 f
21 6 5,0 4 7,7 — — 2 16,7 2 5,1
22 5 4,2 2 3,8 2 5,1
VIII 23 3 2,5 3 5,8 — — 1 8,3 2 5,1
24 —
25 •
IX 26 1 0,8 — — — — — — — —
27 —
28
X 29 —
30 2 1,7 1 1,9 — — — — 1 2,6
1 Итого | 120 100 52 | 100 | 1 1 100 | 12 100 39 | 100
Отмирание листьев на генеративных побегах начинается с I декады
июня и с рядом подъемов и спусков продолжается до I декады ноября
(табл. 41). От всего видового разнообразия максимумы видов отмеча-
ются в III декаду июля (11,1%), в III декаду августа (9,5%) и во II де-
222
каду сентября (14,3%). Среди основных компонентов первый максимум
видов падает на I декаду июня (3,5%), второй — на II декаду июля
(17,2%), третий —на II декаду сентября (13,8%).
Таблица 41
Динамика полного отмирания листьев генеративных побегов травянистых растений дна
логов
Месяц Порядко- вый номер декады (от нача- ла года) Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
16 3 2,4 2 3,5 1 7,7 1 2,3
VI 17 1 0,8 — — — — — — — —
18 3 2,4 1 1,7 — — — — 1 2,3
19 8 6,3 6 10,4 — 1 7,7 5 11,4
VII 20 13 10,3 10 17,2 — — 3 23,1 7 15,9
21 14 11,1 5 8,6 — — 2 15,3 3 6,8
22 11 8,7 5 8,6 5 11,4
VIII 23 8 6,4 1 1,7 — — — — 1 2,3
24 12 9,5 4 6,9 — — 3 23,1 1 2,3
25 8 6,4 4 6,9 — — — — 4 9,0
IX 26 18 14,3 8 13,8 1 100 1 7,7 6 13,7
27 И 8,7 5 8,6 — — 1 7,7 4 9,0
28 8 6,4 3 5,2 — 1 7,7 2 4,5
X 29 6 4,7 3 5,2 — — — — 3 6,8
30 —
XI 31 2 1,6 1 1,7 — — — — 1 2,3
Итого | 126 | 100 I 58 | 100 | 1 100 13 100 44 | 100
Сопоставляя отмирание листьев растений дна логов с растениями
луговой степи (от всего состава флоры), можно видеть, что первый мак-
симум видов дна логов возникает на две декады позже, второй — со-
впадает, а третий — отмечается на одну декаду раньше. Абсолютный
максимум основных компонентов дна логов наблюдается на декаду
позднее в сравнении с луговой степью. Таким образом, влияние экологи-
ческого режима дна логов находит ясное отражение и в сроках
отмирания листьев на генеративных побегах. Абсолютный макси-
мум видов дубовых лесов приходится на более поздние сроки, чем у
растений дна логов, что является закономерным следствием эколо-
гических особенностей развития растений под пологом лесостепных
дубрав.
Отмирание генеративных побегов начинается с I декады июня и
оканчивается в I декаду ноября (табл. 42). Среди полного набора ви-
дов первый максимум отмирания наблюдается в I декаду июня (1,5%),
второй максимум — в III декаду июля (8,7%), третий — в III декаду
августа (13%), четвертый, самый высокий,— в III декаду сентября
(19,6%), пятый — во II декаду октября (10,1%). Среди основных ком-
понентов наиболее крупные подъемы отмирания (в количестве видов)
223
Таблица 42
Динамика отмирания генеративных побегов травянистых растений дна логов
Месяц Порядко- вый номер декады (от на- чала года) Всего Всего компонентов категории
видов компонентов 1-й 2-й З-й
количе- ство % количе- ство % количе- ство % количе- ство % количе- ство %
16 2 1,5 2 3,0 1 7,1 1 1,9
VI 17 1 0,7 — — — — — — — —
18 1 0,7 1 1,5 — — — — 1 1,9
19 1 0,7
VII 20 5 3,6 4 6,0 — — 1 7,1 3 5,8
21 12 8,7 9 13,4 — — — — 9 17,3
22 И 8,0 6 9,0 2 14,5 4 7,7
VIII 23 9 6,5 6 9,0 — — 2 14,5 4 7,7
24 18 13,0 10 14,9 — — 3 21,3 7 13,5
25 6 4,4 1 1,5 — — 1 7,1 — —
IX 26 9 6,5 4 6,0 1 100 1 7,1 2 3,9
27 27 19,6 И 16,4 — — 1 7,1 10 19,2
28 11 8,0 5 7,4 — — 1 7,1 4 7,7
X 29 14 10,1 5 7,4 — — 1 7,1 4 7,7
30 9 6,5 2 3,0 — — — — 2 3,8
XI 31 2 1,5 1 1,5 — — — — 1 1,9
|И то г о 138 100 67 100 1 100 14 100 52 100
отмечаются в III декаду июля (13,4%), III декаду августа (14,9%) и
в III декаду сентября (16,4%), т. е. совпадают с данными для всего ви-
дового состава дна логов.
Характер отмирания генеративных побегов растений дна логов весь-
ма сходен с таковым у растений луговой степи: сроки подъемов отми-
рания от всего видового состава и основных компонентов одни и те же.
Здесь, очевидно, проявляются общие закономерности, свойственные от-
крытым травянистым ценозам.
Переходя к сопряженному анализу биоморфологических особенно-
стей растений на основе вертикального расчленения ценозов в надзем-
ной и подземной частях, следует отметить, что в числовом отношении
надземная и подземная ярусность совпадают. Наиболее четко в под-
земной части выделяются три верхних яруса (до 30, 60 и 100 см). До-
вольно редким по наполнению пространства корнями является ярус до
150 см глубины. Условно можно выделить еще один ярус, глубина кор-
ней которого превышает 150 см. Это и будет 1-й ярус. Итак, в подзем-
ной части растительности дна логов вычленяется пять ярусов корней:
1-й ярус — глубже 150 см, 2-й — до 150 см, З-й — до 100 см, 4-й —
до 60 см, 5-й — до 30 см. О ярусах в надземной части растительности
указывалось выше.
Проанализируем принадлежность растений по определенным био-
морфологическим признакам к ярусам подземной части растительно-
сти. Максимум безрозеточных растений (табл. 43а) сосредоточен в
5-м ярусе (21,2 и 8,6%), в глубже расположенных ярусах число их
224
5 В. Н. Голубев
Таблица 43а
Сопряженный количественный анализ травянистых растений дна логов по биоморфологическим особенностям надземных органов в связи с ярусной
принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Тип структуры надземных побегов Способ возобновления Длительность вегетации
безрозеточный полурозеточный розеточный симподиальный моноподиальный длительновегетиру- ющие коротковегети- рующив
всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты
В подземной части: 5-й (до 30 см) 30(21,2) 6(8,6) 18 (12,6) 11 (15,8) 3(2,1) 2(2,8) 37 (28,0) 14 (21,2) 5(3,9) 4(6,0) 47(33,1) 16(22,9) 4(2,8) 3(4,3)
4-й (до 60 см) 9(6,3) 4(5,7) 32 (22,7) 18(25,8) 3(2,1) 2(2,8) 39 (30,1) 20(30,3) 5(3,9) 4(6,0) 43(30,3) 24(34,3) 1(0,7)
3-й ( до 1 м) 6(4,2) 1(1,4) 21 (14,8) 15(21,5) 5 (3,5) 4(5,7) 26 (20,1) 15 (22,8) 4(3,1) 3(4,6) 31(21,8) 19 (27,2) 1(0,7) 1 (1,4)
2-й ( до 1,5 м) 3(2,1) 1(1,4) 9(6,3) 4(5,7) — 10(7,8) 3(4,6) 1(0,8) 1(1,5) Ю(7,1) 3(4,3) 2(1,4) 2(2,8)
1-й (свыше 1,5 м) 1 (0,7) 1(1,4) 2(1,4) 1(1,4) — — 3 (2,3) 2 (3,0) 3(2,1) 2(2,8)
В надземной части: 1-й (свыше 1 м) 5(3,5) 2(2,8) 14 (9,9) 8(11,5) — 14 (10,7) 7(10,6) 2(1,5) 2(3,0) 16(11,2) 8(11,5) 3(2,1) 2(2,8)
2-й (60—100 см) 12 (8,4) 2(2,8) 32 (22,7) 20(28,6) 2(1,4) 1(1,4) 44(33,9) 20 (30,3) 2(1,5) 2(3,0) 46(32,4) 23 (32,9)
3-й (30—60 см) 18 (12,6) 5(7,2) 24(17,0) 14(20,1) 5(3,5) 4(5,7) 38 (29,2) 17(25,8) 4(3,1) 4(6,1) 44(31,0) 21 (30,1) 3(2,1) 2(2,8)
4-й (15—30 см) 7(4,9) 2(2,8) 12 (8,4) 7 (10,0) 3(2,1) 3(4,3) 15 (11,6) 10(15,2) 4(3,1) 2(3,0) 20(14,1) 10(14,3) 2(1,4) 2(2,8)
5-й (0—15 см) 7(4,9) 2(2,8) — — 1 (0,7) — 4(3,1) — 3(2,3) 2(3,0) 8(5,6) 2(2,8) —
Таблица 436
Сопряженный количественный анализ травянистых растений дна логов по биоморфологическим особенностям надземных органов в связи с ярусной
принадлежностью растений в подземной и надземной частях (в скобках процент от общего числа растений)
Ярус Количество генераций листьев и побегов Способ перезимовки
с одной генерацией с двумя генерациями летнезеленые вечнозеленые зимнезеленые
всего основные ком- поненты всего основные ком- поненты всего основные компоненты всего основные компоненты всего основные компоненты
В подземной части: 5-й 16(11,3) 4(5,7) 35 (24,7) 15(21,5) 21 (14,8) 5(7,1) 27(18,9) 13(18,6) 4(2,8) 1 (1,4)
4-й 11(7,8) 4(5,7) 33(23,2) 20(28,6) 24(17,0) 13(18,6) 19 (13,3) 11(15,7)
3-й 7(4,9) 3(4,3) 25 (17,6) 17(24,3) 23 (16,3) 12 (17,1) 9(6,3) 8(11,5)
2-й 4(2,8) 1(1,4) 8(5,6) 1 (0,‘7) 4(5,7) 10(7,1) 5(7,1) 2(1,4) -
1-й 2(1,4) 1 (1,4) 1(1,4) 2(1,4) 2(2,8) —. 1(0,7)
В надземной части: 1-й 10(7,0) 4(5,7) 10(7,0) 7(10,0) 19 (13,4) 10 (14,3) 1 (0,7) 1(1,4)
2-й 10(7,0) 3(4,3) 35(24,7) 19 (27,1) 28 (19,8) 12(17,1) 17(11,9) 10(14,3) .
й 3-й 13(9,2) 2(2,8) 34(24,0) 21 (30,1) 22 (15,5) 11(15 7) 23(16,1) 18 (15,7) 2(1,4) 1(1,4)
°' 4-й 6(4,2) 4(5,7) 16(11,3) 8(11,5) 8(5,6) 4 (5,7) И (7,7) 8(11,5) 3(2,1)
5-й 1(0,7) 7(4,9) 2(2,8) 3(2,1) — 5(3,5) 2(2,8) —.
Таблица 43в
Сопряженный количественный анализ травянистых растений дна логов по биоморфологическим особенностям надземных органов в связи с ярусной
принадлежностью растений в подземной и надземной частях (в скобках—процент от общего числа растений)
Ярус Ритм цветения Ярус в надземной части растительности
весенние весенне-лет- ние летние летне-осенние 1-й 2-й 3-й 4-й 5-й
всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты всего основные компонен- ты
В подземной части: 5-й 5(3,5) 4(5,7) 14(10,0) 8(11,5) 26(18,5) 7(10,0) 5(3,5) 2(1,4) 1 (1,4) 8 (5,7) 2(2,9) 19(13,4) 4(5,7) 17(11,9) 10(14,3) 5(3,5) 2 (2,9)
4-й 2(1,4) 1(1,4) 6(4,3) 4(5,7) 29(20,6) 14(20,0) 7 (5,0) 5(7,2) 6 (4,2) 2(2,9) 18(12,7) 10(14,3) 16(11,3) 11(15,7) 3(2,1) 1 (1,4) 1 (0,7) —
3-й 2(1,4) 2(2,9) 4(2,8) 3(4,3) 20 (14,2) 14(20,0) 6(4,3) 1(1,4) 7(5,0) 5(7,2) 14(9,9) 8(11,5) 7(4,9) 6(8,6) 2^1,4) 1 (1,4) 2(М) —
2-й 1(0,7) — 2(1,4) 1 (1,4) 5(3,5) 3(4,3) 4(2,8) 1 (1,4) 5(3,5) 3(4,3) 4(2,8) 1(1,4) 3(2,1) 1(1,4) — —
1-й — —. 3(2,1) 2(2,9) — —— — — 1(0,7) 1(1,4) 2(1,4) 1 (1,4) — — — —
В надземной части: 1-й 17(12,0) 9(12,8) 3(2,1) 2(2,9)
2-й — — 6(4,3) 4(5,7) 30 (21,3) 16 (22,8) 9(6,4) 2(2,9) — — — — — — —. — — —
3-й 6(4,3) 3(4,3) 8(5,7) 6(8,6) 24(17,0) 12(17,1) 8(5,7) 2(2,9) — — — — — — — — — —
4-й 4(2,8) 4(5,7) 8(5,7) 5(7,2) 8(5,7) 2(2,9) 2(1,4) 1(1,4)
5-й — 4(2,8) 1(1,4) 4(2,8) 1(1,4)
постепенно убывает. Наибольшее количество полурозеточных растений
отмечается в 4-м ярусе (22,7 и 25,8%). Розеточные растения более всего
представлены в 3-м ярусе (3,5 и 5,7%). Максимумы симподиальных и
моноподиальных растений приходятся на один и тот же 4-й ярус.
Длительновегетирующие растения от верхнего яруса к нижнему (от
5-го к 1-му) последовательно убывают в числе, отражая общую количе-
ственную закономерность распределения растений по ярусам. Это озна-
чает, что сопряженной связи между глубиной проникновения корней
растений дна логов с длительностью их вегетации нет.
То же, в основном, можно сказать и о ярусном распределении рас-
тений по количеству генераций листьев, способам перезимовки
(табл. 436), ритмам цветения (табл. 43в). Отдельные отклонения от
общей направленности изменения в количестве видов — от наибольше-
го в поверхностных до наименьшего в глубинных ярусах — лишний раз
подчеркивают отсутствие четкой экологической зависимости между
биоморфологическими особенностями побеговых органов и поярусными
условиями развития корневой системы. В любом горизонте почвы дна
логов растения находят достаточное количество влаги и элементов ми-
нерального питания. Это касается в первую очередь поверхностного
яруса подземной части растительности, где даже мелкокорневые рас-
тения не испытывают затруднений в поглощении влаги. *
Сходная картина наблюдается в количественном распределении ви-
дов по ярусам в надземной части ценозов дна логов (табл. 43, а, б, в).
Почти по всем учтенным биоморфологическим признакам максимум
видов приходится на второй или третий ярусы. Это, конечно, свидетель-
ствует о том, что и в надземной части ценозов дна логов расслоение
растений по ярусам не связано с резкой биоморфологической диффе-
ренцировкой. Вместе с тем в экологических (биоморфологических)
свойствах растений, входящих в разные ярусы, наблюдается близкое
сходство. Ярусное расчленение травостоя в надземной и подземной
сферах лишь указывает на характер дифференциации в освоении рас-
тительностью внешней среды. Ярусное строение этих ценозов и насы-
щение пространства обеспечивает максимальное использование усло-
вий внешней среды и наивысшее продуцирование растительной массы.
В силу достаточной обеспеченности влагой в логах развивается мезо-
фильный тип растений с ограниченными защитными приспособлениями
и поэтому с весьма высокими показателями продуктивности (Культиа-
сов, 1956, 1962), нивелирующими специфику по разнообразным биомор-
фологическим свойствам.
15е
Г лава 8
Проблема природы и происхождения
растительности луговых степей
Выяснение природы и происхождения луговых степей тесно связано
с изучением развития всего лесостепного комплекса растительности.
Однако принимая во внимание, что, во-первых, основными объектами
нашего исследования являются травянистые растения и ценозы и, во-
вторых,— проблема луговых степей заключает ряд специфических, при-
сущих только ей одной, особенностей, мы выделяем анализ ее в каче-
стве самостоятельного раздела. Это, конечно, не исключает из сферы
обсуждения и тех вопросов, которые касаются формирования расти-
тельности лесостепи вообще, т. е. и других зональных подразделений.
Мы почти не рассматриваем здесь вопрос об источниках формирования
флоры и растительности луговых степей, о путях заселения и миграций
флористических элементов на фоне геологической истории соответству-
ющих территорий, так как это непосредственно не соприкасается с ха-
рактером настоящей работы и может отвлечь от основной темы.
ОБЩИЕ ПОЛОЖЕНИЯ
В русской ботанической географии интерес к европейской лесостепи
возник еще в прошлом столетии и дал большую когорту блестящих
имен ее 'исследователей. Течение мысли при этом концентрировалось
преимущественно вокруг «степного вопроса», «проблемы взаимоотно-
шений леса и степи», «степного лесоразведения», «причин безлесия рус-
ских степей». В плане этих вопросов косвенно и затрагивалась пробле-
ма природы лесостепной растительности. Лишь значительно позднее
отдельные исследователи выдвинули задачу ландшафтного изучения
лесостепи как специфического зонального образования с присущими
ему оригинальными чертами растительности, почв, климата, геологиче-
ского строения, истории развития и т. д. В самостоятельную проблему
оформляется и вопрос о природе и происхождении растительности лу-
говых степей.
Большим тормозом в углублении знаний о растительности лесостепи
является представление о ее переходном характере, еще недавно со-
хранявшее свою силу (Буш, 1936; Берг, 1936, 1955; и др.), согласно
которому в лесостепи происходит пестрое смешение лесной и степной
растительности. Это лишь полоса контакта двух разнородных типов
растительности, между которыми идет постоянная борьба. Несостоя-
тельность этого взгляда очевидна уже потому, что любая ландшафтная
зона по своему положению является переходной между смежными зо-
нами. Между тем промежуточный характер зон не может затушевывать
присущие им особенности.
Вопрос о природе и происхождении луговых степей, их геоботани-
ческой классификации и эколого-биологических признаках в настоящее
время является чрезвычайно запутанным. Причины такого положения
228
кроются отчасти -в объективных показателях, отчасти же носят субъек-
тивный характер. В числе объективных причин важнейшим является
то обстоятельство, что не только современники, но и исследователи лу-
говых степей конца XIX и начала XX века наблюдали уже сильно нару-
шенные хозяйственной деятельностью, а потому и значительно изменен-
ные остатки природной растительности, весьма отрывочные, очень не-
полно представляющие растительный покров лесостепи. В начале же
ее колонизации русскими поселенцами геоботаники как науки еще не
существовало, и за редчайшими исключениями мы теперь не имеем
достоверных документов о состоянии растительности первобытных лу-
говых степей. Различный состав растительности сохранившихся от рас-
пашки участков природы лесостепи, являющийся следствием разной
степени ее нарушенности и изменения, нередко порождает многообра-
зие классификационных схем для одних и тех же зональных категорий
растительности.
В настоящей главе мы не претендуем на сколько-нибудь исчерпы-
вающее обсуждение затронутой проблемы. Наша задача — наметить
хотя бы в самых общих чертах ее наиболее существенные аспекты, вы-
явить их содержание и на основе анализа литературы и выполненных
нами исследований в зоне лесостепи сформулировать рациональную
точку зрения на природу и происхождение растительности луговых
степей.
Вначале несколько слов посвятим истории названия «луговые сте-
пи». Название «луговые степи» впервые встречается в работе А. Н. Бе-
кетова (1874), который употребляет его в качестве синонима травяных
(в контексте — травных) степей, распространенных на юге Русской рав-
нины, между турецкой границей и Доном и в других районах. Помимо
луговой А. Н. Бекетов выделяет голодную, песчаную, каменистую и
солончаковую степи. Отсюда можно заключить, что понятие луговой
степи еще точно не определено. Оно в равной степени применимо и к
северным разнотравным, и южным ковыльным степям.
Четкая формулировка луговых степей дается в классических рабо-
тах С. И. Коржинского (1888, 1891, 1901), предложившего разверну-
тую их характеристику в географическом, фитоценологическом и эколо-
го-биологическом отношениях, которая достаточно известна. Дальней-
шее углубление и расширение характеристики растительности дается
в работах И. К. Пачоского (1917), В. В. Алехина (1915а, б, 1934,
1936), Б. А. Келлера (1916, 1923, 1931)), А. Я. Гордягина (1922),
И. И. Спрыгина (1915, 1917, 1925), И. В. Новопокровского (1927, 1931,
1937), И. М. Крашенинникова (19376), Е. М. Лавренко (1940),
Г. И. Дохман (1956), В. Н. Голубева (1962а, б, в) и др.
Черноземно-степная область разделяется С. И. Коржинским (1901)
на две полосы: типичную степную или ковыльно-типчаковую и лугово-
степную или лесостепную. Таким образом, распространение луговых
степей на равнине С. И. Коржинский связывает исключительно с лесо-
степной зоной. Это положение, по существу, принимается затем боль-
шинством исследователей: А. Я. Гордягиным (1901, 1922), И. К. Пачо-
ским (1910), В. В. Алехиным (1915а, б, 1934), П. Н. Крыловым (1915),
Б. А. Келлером (1916, 1923), И. И. Спрыгиным (1922, 1925), И. В. Но-
вопокровским (1922, 1931, 1937), Н. А. Ивановой, И. М. Крашенинни-
ковым (1934), М. В. Марковым (1935), И. М. Крашенинниковым (1939,
1941) и др.
Следует отметить, что С. И. Коржинский (1888, 1891), А. Я. Гордя-
гин (1901) и другие применяют в качестве синонима луговой степи
термин «степной луг», а И. К. Пачоский (1910)—«луговидная степь».
Еще более свободно варьирует названиями П. Н. Крылов (1916), ко-
торый часто говорит о лугах степной области, степных лугах и лугах
229
лесных, т. е. понятие «луг» использует в народном смысле (Хитрово,
1925). Однако, несмотря на разнобой в терминологии, все перечисленные
авторы были единодушны относительно трактовки природы травянистых
группировок, именуемых луговыми степями, которые ими зачислялись
в категорию степного типа растительности. Чтобы оттенить принадлеж-
ность луговых степей к степям И. К. Пачоским (1917), И. И. Спрыги-
ным (1925), В. В. Алехиным (1925а, 1926) предложен для луговых
степей термин «северные степи».
Эти авторы предостерегали от произвольного толкования понятия
«луговой» степи. Так, И. И. Спрыгин (1925) возражал против названия
«луговые» степи на том основании, что, во-первых, различные исследо-
ватели вкладывают в это понятие разное содержание и, во-вторых, лу-
говые ассоциации настолько резко отличаются от травяно-степных по
составу и характеру травостоя, «что совсем неуместно прилагать на-
звание «луговые» к степным ассоциациям» (стр. 235).
Какими же признаками характеризуются луговые степи, в трактовке
их наиболее компетентными исследователями? По С. И. Коржинскому
(1891, 1901), в составе растительности луговых степей главная роль
принадлежит двудольным растениям. Всюду встречаются и степные
злаки, в том числе ковыли (Stipa pennata s. 1., St. capillata), но они
играют подчиненную роль. Местами ковыли могут совершенно отсут-
ствовать. Существенное участие в ценозах принимают степные кустар-
ники. В луговых степях почва почти сплошь задернована злаками и
основаниями побегов других растений. В весенне-летний период луго-
вая степь отличается пышным цветением трав. Высота надземных побе-
гов растений значительно превышает таковую в настоящих степях.
С. И. Коржинский приводит список видов луговой степи 85 названий,
из которых наиболее показательными для тучного чернозема он считает
Adonis vernalis, Anemone silvestris Arenaria longifolia, A. ucrainica,
Astragalus danicus, Asperula tinctoria, Helictotrichon pubescens, Salvia
dumetorum, Scorzonera purpurea. Впрочем, как указывает С. И. Кор^
жинский (1891), в различных частях широтного протяжения луговых
степей состав их флоры может заметно изменяться.
А. Я. Гордягин (1922), соглашаясь в основном с характеристикой
луговых степей С. И. Коржинского и поддерживая мысль, что в их
составе ксерофитные злаки не играют преобладающей роли, как в бо-
лее южных степях, все-таки указывает (по исследованиям в Татарской
республике) на значительное участие типчака (Festuca sulcata). В лу-
говых степях типчак маскируется обильно развивающимися между его
дерновинками двудольными травами.
В составе пензенских луговых степей И. И. Спрыгин (1925) также
устанавливает большую роль типчака. В большем или меньшем коли-
честве к нему примешиваются Stipa dasyphylla, St. pennata, St. steno-
phylla и Helictotrichon pubescens, которые, однако, могут совершенно
отсутствовать. Обильно представлен Bromus riparius. Обычен покров
из Thuidium abietinum на участках, где выпас исключен. Не растут
более южные виды, например Stipa capillata в плакорных условиях.
Развиваются некоторые влаголюбивые виды, количество которых воз-
растает к северу. Всюду обильны двудольные растения, и степи носят
разнотравный характер.
И. К. Пачоский (1917), анализируя изменение растительности сте-
пей б. Херсонской губернии в направлении к лесостепной зоне, указы-
вает, что при этом увеличивается количество видов, входящих в ценозы,
особенно состав двудольных растений. Это связано с улучшением усло-
вий увлажнения. Северные (т. е. луговые) и южные степи в злаковой
основе довольно сходны, но для первых характерны Phleum phleoides,
Stipa dasyphylla, St. pennata, St. pulcherrima, а виды Bromus inermis^
230
В. riparius, Festuca sulcata, Роа bulbosa, Stipa lessingiana, St. steno-
phy,lla, распространены как в южных, так и северных степях. Более
•существенные отличия северных степей от южных автор видит в составе
разнотравья, значительное число видов которого свойственно только
северным степям, например, Adonis vernalis, Campanula sibirica, Echiutn
rubrum, Filipendula hexapetala, Orobus pannonicus, Plantago media, Po-
lygala comosa, Potentilla humifusa, Ranunculus polyanthemus, Salvia
pratensis, Trifolium alpestre, T. montanum и др. И. К. Пачоский обра-
щает внимание на то обстоятельство, что с уплотнением и усложнением
травостоя в луговидных >(т. е. северных) степях происходит ярусная
дифференцировка подземной части растительности, при которой верх-
ние горизонты почво-грунта осваиваются корнями злаков, а корни дву-
дольных проникают в более глубокие слои. Развитие травостоя в север-
ных степях более равномерное в течение вегетационного периода, чем
в южных. В направлении к северным степям увеличивается гумусность
почвы и количество осадков.
Для луговых степей б. Тамбовской губернии В. В. Алехин (1915а)
указывает повсеместное присутствие ковылей и других ксерофильных
злаков: Festuca sulcata, Koeleria gracilis. Автор отмечает, что распро-
странение этих видов не позволяет использовать их для разделения
растительности на подзоны и тем более для расчленения северных и
южных степей. В качестве северной границы луговых степей В. В. Але-
хиным принимается северная граница распространения Amygdalus
лапа и Spiraea crenata, в качестве южной — южная граница распро-
странения Leucanthemum vulgare, Myosotis suaveolens, Veronica cha-
maedrys.
В другой своей работе В. В. Алехин (19156) более подробно моти-
вирует эти положения, подчеркивая, что перистый ковыль, как и другие
ксерофильные злаки (Festuca sulcata, Koeleria gracilis), распространены
в южных и северных степях, правда в первых более обильно. Поэтому
они никоим образом не могут указывать на тот или иной зональный
тип степей. Более показательными являются многие растения северно-
го типа, пределы распространения которых к югу четко зависят от кли-
матических условий местности.
В более поздних работах, в результате анализа курских и вообще
северных степей, В. В. Алехин (1935, 1936, 1951) приводит следующие
главнейшие их признаки:
1. Чередование многочисленных красочных аспектов, сменяющихся
с весны до осени.
2. Господство красочных разнотравных видов, среди которых имеет-
ся довольно много влаголюбивых.
3. Преобладание среди злаков широколистных видов, а также гос-
подство рыхлокустовых и корневищных типов (не плотнокустовых).
4. Из ковылей характерно присутствие двух видов — Stipa pennata
L. и St. stenophylla.
5. Вообще господство растений многолетних.
6. Присутствие сплошного покрова из туидиевого мха.
7. Почти полное отсутствие весенних эфемеров и эфемероидов (из
последних лишь Hyacinthella leucophaea и Gagea erubescens).
8. Незначительное число растений «перекати-поле» (из них лишь
Falcaria vulgaris, Trinia multicaulis, Asparagus officinalis, очень редко
Cypsophila paniculata).
На основании исследований в б. Саратовской губернии и Централь-
но-Черноземной области Б. А. Келлер (1916, 1923, 1931) расчленяет
зону луговых степей на два подтипа: разнотравно-луговые и дерновин-
яо- (ковыльно- и типчаково-) луговые степи, которые обладают своими
•особенностями, главным образом в соотношении разнотравья и плотно-
231
кустовых злаков. Под разнотравно-луговыми Б. А. Келлер подразуме-
вает такие степи, в составе которых типчак и ковыли встречаются в
рассеянном состоянии или отсутствуют, главную роль играют двудоль-
ные и корневищные злаки. В дерновинно-луговых степях роль плотно-
кустовых злаков повышается, по-прежнему очень значительное место
принадлежит разнотравью. Луговые степи отличаются высокой видовой
насыщенностью: на 100 м2 насчитывается до 100 видов, на 1 ж2 —до
40—45 видов. В разнотравно-луговых степях встречаются растения от-
носительно влажных лугов и светлых лесов, которые проникают и в
дерновинно-луговые степи. /
Луговые степи Южного Урала, по И. В. Новопокровскому (1931)г
характеризуются преобладающим развитием луговостепного разно-
травья, обязательным присутствием Stipa pennata и St. stenophylla,
произрастанием небольшого количества луговых видов (Calamagrostis
epigeios, Festuca elatior, Stellaria graminea и др.), светолюбивых лес-
ных (Hieracium umbellatum, Pimpinella saxifraga, Potentilla thuringia-
ca), сравнительно малым количеством степных форм южных степей
(Arenaria longifolia, Centaurea scabiosa, Galium verum, Phlomis tuberosa
и др.), богатством и разнообразием видового состава, преобладанием
разнотравья по массе, густым и высоким травостоем.
Е. М. Лавренко (1940, 1956) вводит в характеристику луговых сте-
пей состав экологических типов по отношению к влаге. Единый у всех
перечисленных и других авторов тип луговых степей Е. М. Лавренко^
следуя некоторым принципам А. П. Шенникова (1935, 1938), расчленя-
ет на остепненные луга и луговые степи, принадлежащие к различным
типам растительности — луговому и степному. «Географически остепнен-
ные луга» Е. М. Лавренко в общем соответствуют «разнотравно-луговым
степям» Б. А. Келлера. Остепненные луга, по Е. М. Лавренко, связаны
в своем распространении на плакоре с выщелоченными черноземами.
Для них характерно преобладание ксеромезофильных видов с примесью
эумезофильных и мезоксерофильных, по преимуществу из группы раз-
нотравья и корневищных злаков. Обычна примесь ксерофильных плот-
нокустовых злаков — ковылей, тонконога, типчака. Луговые же степи
определяются преобладанием мезоксерофильных и эвриксерофильных
видов с примесью ксеромезофильных и эуксерофильных. Следует заме-
тить, что эти экологические подразделения экспериментально (физио-
логически), по крайней мере для растений луговых степей не обоснова-
ны. В их выделении нередко руководствуются чисто субъективными
соображениями. Ясно, что такой подход к классификации травянистой
растительности в какой-то степени обесценивается.
Мы привели основные работы, связанные с оригинальными исследо-
ваниями в различных частях лесостепной зоны, непосредственно касаю-
щиеся определения луговых степей. Из обзора видно, что трактовки
луговых степей подавляющим большинством исследователей не проти-
воречат, а взаимно дополняют друг друга, вскрывая некоторые флори-
стические, морфологические и эколого-биологические особенности це-
нозов. Отдельные частные расхождения в определениях, несомненно,
следует отнести на счет географических различий луговых степей, где
проводились исследования разных авторов, а также объяснить сте-
пенью их изменения под влиянием хозяйственной деятельности.
Исследователи единодушны в понимании луговых степей как степно-
го типа растительности. Исключением являются воззрения Е. М. Лаврен-
ко, расчленяющего луговые степи на два различных типа — луговой
(остепненные луга) и степной (собственно луговые степи).
Необходимо отметить, что в приведенных характеристиках отсут-
ствуют количественные (в цифровом выражении) оценки признаков.
Это в значительной мере сужает возможности сравнения травянистых
232
ценозов лесостепи и определения их места в геоботанической класси-
фикации растительного покрова.
Уже в самом начале геоботанического изучения луговых степей по
мере накопления фактических данных возникали различные точки зре-
ния относительно их природы. Один из пионеров изучения и наиболее
выдающихся исследователей луговых степей С. И. Коржинский (1888»
1891) полагал, что луговые степи представляют северную окраину
степной области, совмещающуюся пространственно с полосой лесосте-
пи. Вместе с тем С. И. Коржинский развивал концепцию наступления
леса на степь, в процессе которого и возникает лесостепь. Открытые
степные участки в лесостепи сохранились только потому, что лес еще
не успел целиком занять свои позиции. Лесная формация, по
С. И. Коржинскому, является более мощной в сравнении с травянисты-
ми. Взаимное распространение леса и степи в лесостепи он объяснял
борьбой за существование между этими формациями, историческими
особенностями борьбы и совершенно исключал во взаимоотношениях
леса и степи роль климата, топографического характера местности, при-
знаков субстрата, с чем сейчас, конечно, нельзя согласиться.
Если последовательно развивать высказанную мысль С. И. Коржин-
ского, то с логической необходимостью мы должны прийти к выводу,
что при естественных взаимоотношениях между лесом и степью с тече-
нием времени луговая степь будет поглощена лесом. С другой стороны»
С. И. Коржинский (1891) утверждал, что «луговая степь является наи-
более постоянной и наиболее сформированной формацией, не изменя-
ющейся на очень больших протяжениях» (стр. 38). Как видно, в этих
высказываниях сквозит очевидная противоречивость, так как невоз-
можно в одно и то же время говорить об устойчивости луговых степей
при их явной ассимиляции со стороны леса.
Гипотеза наступления леса на степь с различными модификациями
поддерживалась в дальнейшем очень многими исследователями. Среди
ее сторонников можно назвать П. А. Костычева (1890), Г. И. Танфилье-
ва (1894, 1901), А. Н. Краснова (1893, 1894), Л. С. Берга (1922, 1947),
И. И. Спрыгина (1922), Д. Г. Виленского (1924), А. И. Куренцова
(1929), С. А. Захарова (1935), В. 3. Гулисашвили (1948), М. Г. Попо-
ва (1953) и др. Следует подчеркнуть, что во многих из перечисленных
работ не разграничиваются вопросы восстановления леса на ранее по-
крытой лесом площади, а также смещения лесостепи к югу под влия-
нием общего изменения климата в сторону увеличения влажности.
Остается неясной и роль циклических колебаний климата в периодиче-
ских наступлениях и отступлениях леса. Между тем с точки зрения
изучения природы луговых степей эти вопросы, в свете гипотезы на-
двигания леса на степь, являются принципиальными. Но поскольку
здесь они не нашли положительного решения, постольку и само значе-
ние данных работ в выяснении природы луговых степей остается от-
крытым. Одно несомненно, что все указанные выше авторы рассматри-
вали травянистые группировки, контактирующие в лесостепи с лесной
формацией, в качестве степного типа растительности.
Другие сторонники степной природы луговых степей уже более
определенно высказывались в пользу их первичности. Равноправность,
зональность степных и лесных сообществ лесостепи принимались
И. К. Пачоским (1910), В. В. Алехиным (1915а, 1934), П. Н. Крыловым
(1915), И. В. Новопокровским (1927, 1937), Н. А. Ивановой, И. М. Кра-
шенинниковым (1934), М. В. Марковым (1935) и другими, а в настоя-
щее время Н. А. Прозоровским (1949, 1951), Г. И. Дохман (1956, 1960),
Д. И. Сакало (1961) и др.
Противоположную точку зрения отстаивали А. Я. Гордягин (1901),
В. И. Талиев (1901, 1913), Б. А. Келлер (1916, 1923), а также В. Н. Су-
23»
качев (1903), П. А. Баранов (1960), считавшие луговые степи вторич-
ными образованиями, развившимися на месте сведенных лесов. Увле-
каясь полемикой, А. Я. Гордягин (1901) в одном месте своей работы
категорически заявляет, что «луговая степь всегда представляет расти-
тельность производную» (стр. 440). Впрочем, это остается априорным,
так как не подкрепляется какими-либо конкретными фактическими дан-
ными. Анализируя работы перечисленных авторов, можно с уверен-
ностью заключить, что ни один из них в принципе не отрицал возмож-
ность существования первичных луговых степей. В. И. Талиевым,
А. Я. Гордягиным, Б. А. Келлером лишь акцентировался случай форми-
рования луговых степей на месте уничтоженных человеком лесов в об-
ласти лесостепи. Против этого вряд ли возникнут серьезные возра-
жения.
Но в связи с вопросом о природе луговых степей привлекают внима-
ние высказывания А. Я- Гордягина (1901), ставшие затем знаменем
целого направления. Он писал, что «характер флоры луговостепных
участков представляется в высшей степени неустойчивым и вообще
мало типичным» (стр. 441). И далее он подчеркивал, что луговые степи
местами постепенно переходят в лесные луга. Логическое завершение
этих идей мы находим в концепции А. П. Шенникова (1938), согласно
которой в лесостепи зональным типом травянистой растительности яв-
ляются суходольные луга, которые к югу все более «остепняясь», по-
степенно замещаются ассоциациями степных ксерофитов. Расширяя
содержание понятия лугового типа растительности и принимая в ка-
честве такового ассоциации травянистых многолетних мезофитов,
А. П. Шенников недостаточно мотивированно исключал из числа харак-
терных такие признаки луга, как почти сплошное задернение почвы,
высокое проективное покрытие (полнота) травостоя, грунтовое увлаж-
нение (не всегда, но довольно часто), господство мезофильных злаков
и зимнезеленых розеточных гемикриптофитов, отсутствие летнего пе-
рерыва вегетации и т. д. Вполне понятно, что с этих позиций нетрудно
было «зачислить в луга» не только настоящие луговые степи, но и суб-
альпийские высокотравья, луговинные тундры, эфемеровые подгорные
пустыни Средней Азии и пр. С другой стороны, из состава лугов
А. П. Шенников выводит ценозы на пойменных гривах с господством
типчака, кобреэиевые сообщества в горах, белоусники в высокогорьях
и лесной области, замоховелые луга и т. д.
Нет необходимости доказывать, что основания для столь крупной
реформы в геоботанической классификации весьма шатуне и мало
оправданные. Объединение под очень общим приятием «мезофитов»
самых различных экологических типов и жизненных форм растений,
к тому же в то время совершенно не изученных или слабо исследо-
ванных в эколого-биологическом отношении, представляется не только
нелогичным, но и порочным.
Развивая идею зональности лугового типа, А. П. Шенников пред-
лагает переименовать лесостепную зону в лесолуговую. Им же была
выделена и новая категория луговой растительности — «остепненные
луга» (А. П. Шенников, 1935), которая после работы Е. М. Лавренко
(1940) стала широко использоваться ботаниками для обозначения тра-
вянистых группировок, по нашему мнению ближе стоящих к катего-
рии луговых степей, чем луговому типу.
Е. М. Лавренко (1940, 1956) несколько ограничил область распрост-
ранения луговой растительности. Он признал самостоятельное сущест-
вование луговых степей в средней и южной частях лесостепи. Но в трак-
товке травянистых группировок севера лесостепной зоны Е. М. Лаврен-
ко делает уступку концепции А. П. Шенникова, относя их к категории
•остепненных лугов. Эта точка зрения воспринята некоторыми современ-
234
ными геоботаниками, в том числе и работавшими на Стрелецкой степи —
—М. С. Шалытом (1950), Г. М. Зозулиным (1950, 1955, 1959а, б),
А. М. Семеновой-Тян-Шанской (1960а, б).
Из перечисленных авторов наибольшей категоричностью выделяются
суждения Г. М. Зозулина. Какие же факты и положения выдвинуты им
для обоснования луговой природы Стрелецкой степи и отнесения ее к лу-
говому типу растительности? Основным критерием в пользу этого заклю-
чения Г. М. Зозулин считает генетический состав флоры, в котором как
по общему количеству, так и константным видам преобладают растения
луговой исторической свиты.
Чтобы в должной мере оценить основательность подобных утвержде-
ний, интересно привести некоторые высказывания самого автора «исто-
рических свит растительности». В статье, посвященной обзору научной
работы заповедника, Г. М. Зозулин (1959в, стр. 49) сообщает, что им в
течение одного 1952 г. был произведен исторический анализ флоры запо-
ведника в количестве 744 видов. А в более ранней работе автор поясня-
ет, что «по техническим причинам мы не можем останавливаться здесь
на тех соображениях, которыми мы руководствовались при отнесении
каждого вида к той или иной свите...» (Зозулин, 1955, стр. 135). Таким
образом, за один год проводится исторический анализ 744 видов (1), апо
техническим причинам исключается само существо исследования. По-
зволительно спросить, какова же ценность такого исследования? В столь
ответственных выводах, связанных с историческим анализом, нельзя
удовольствоваться одними априорными утверждениями. Нужны веские
факты, мотивированные положения, объективная аргументация.
Ближайшее знакомство с распределением Г. М. Зозулиным видов по
историческим свитам открывает в нем много субъективного. Тдк,
Г. И. Дохман (1956) нашла, что «в реестр луговых растений Зозулиным
совершенно необоснованно вносятся типичные степные виды — компо-
ненты разнотравно-ковыльных степей, например Thalictrum minus L.,
Phlomis tuberosa L., Medicago falcata L., Arenaria ucrainica Spreng., Fi-
lipendula hexapetala Gilib., Salvia stepposa Schost. и т. п. В луговой сви-
те имеются виды, неизменно связанные с северной степью и не встре-
чающиеся на лугах, например, Adonis vernalis L., Tragopogon orientate
L. и др.» (стр. 203—204).
Из этих и других данных становится очевидным, что ссылка на гене-
тический состав флоры в работах Г. М. Зозулина является наименее со-
стоятельной, чтобы использовать ее в качестве аргумента для обоснова-
ния луговой природы Стрёлецкой степи.
Из других положений, выдвигаемых Г. М. Зозулиным, можно отме-
тить следующие: а) отсутствие летнего перерыва вегетации на целинах
заповедника, связанного с физической сухостью почв; б) в составе рас-
тительности луговых степей почти полностью выпадают эфемеры, эфеме-
роиды и «перекати-поле»; в) структура травостоя целин отличается зна-
чительной сомкнутостью и высоким проективным покрытием (90—100%).
Обращаясь к приведенной ранее характеристике луговых степей,
можно заключить, что классики русской ботанической географии и гео-
ботаники (С. И. Коржинский, В. В. Алехин, И. И. Спрыгин и др.) рас-
сматривали данные положения в качестве признаков луговых степей.
Поэтому даже с этой точки зрения использование их Г. М. Зозулиным
для отождествления Стрелецкой степи с луговым типом растительности
представляется неубедительным. Оно несостоятельно и с других пози-
ций. Только чистым недоразумением можно объяснить сохраняющийся
в настоящее время в ботанической литературе тезис' о среднелетнем
перерыве вегетации в южных степях и его отсутствии в луговых.
Весьма авторитетными исследованиями А. В. Турского (1934, 1945),
А. П. Балаша (1961а, б) и других установлено, что в южных степях
235
всегда присутствуют длительновегетирующие растения, имеющие зеленые
листья и побеги в течение всего летнего периода наряду с коротковеге-
тирующими видами. Последние в большом количестве обнаружены и в
луговых степях, о чем подробно говорилось в соответствующей главе этой
книги.
Жизненная форма «перекати-поле» представлена на Стрелецкой сте-
пи такими обильно развитыми видами, как Asparagus officinalis, Falca-
ria vulgaris, Trinia multicaulis и редкими растениями — Peucedanum lu-
bimenkoanum, Gypsophila paniculata, Phlomis pungens.
Важное место в раскрытии природы и происхождения растительного1
покрова лесостепи занимает вопрос о первобытной растительности луго-
вых степей, ибо его выяснение не только расширило бы наши представ-
ления относительно сущности лесостепной растительности, но и позволи-
ло бы судить о происшедших изменениях ее под влиянием культуры.
Единственным пока средством «натурального» познания доагрикультур-
ного растительного покрова является изучение и оценка восстановления
растительности при отсутствии искусственного вмешательства (в запо-
ведниках).
Исследование Г. М. Зозулиным изменения растительности степных
целин под влиянием некошения в условиях абсолютной заповедности
дало повод утверждать, что при этом резко сокращается обилие степного
разнотравья, а некоторые виды совершенно выпадают. Плотнокустовые
ксерофильные злаки — типчак, тонконог, ковыли, а также осока низкая
испытывают сильное угнетение и постепенно исчезают из состава траво-
стоя, причем дольше всех противоборствуют угнетению ковыли. Наи-
большее развитие получают длиннокорневищные луговые злаки — костер
безостый и вейник наземный. Наблюдается мезофитизация (олугове-
ние) травостоя за счет появления некоторых рыхлокустовых широколи-
стных злаков и других луговых мезофитов. В конечном итоге степь «бук-
вально перерождается» и заменяется «зарослями мезофитных корневищ-
ных злаков, среди которых ютятся угнетенные остатки степняков»
(Г. М. Зозулин, 1955, стр. 219). Г. М. Зозулин неоднократно повторяет
высказанное еще А. Я. Гордягиным (1901) мнение о нестойкости флори-
стического и фитоценотического комплекса, определяющего современ-
ную луговую степь, о его производной, вторичной природе. Коренным
типом является луговая растительность.
А. М. Семенова-Тян-Шанская (1960а, б) безоговорочно принимает
тезисы Г. М. Зозулина и вслед за Е. М. Лавренко подчеркивает веду-
щую роль среди видового разнообразия Стрелецкой степи ксеромезо-
фильных и мезоксерофильных растений, особенно корневищных и рыхло-
кустовых злаков, будто бы характеризующих растительность остепнен-
ных лугов.
Некоторые новые положения вводятся М. С. Шалытом (1950), уста-
новившим отсутствие ясной ярусности корневых систем в подземной ча-
сти растительности Стрелецкой степи. Он обращает внимание на сомкну-
тость и равномерность корнестоя, наличие плотной дернины, обилие кор-
невищ, набор жизненных форм и соотношение господствующих видов
растений. Все это, по М. С. Шалыту, служит свидетельством луговой при-
роды нашей степи. Критический разбор указанных положений нами сде-
лан раньше (В. Н. Голубев, 1962в).
Таков в общих чертах арсенал доказательств луговой сущности
Стрелецкой степи и принадлежности ее к луговому типу растительности.
К обсуждению части вышеприведенных выводов мы еще вернемся не-
сколько позднее, в связи с анализом результатов исследования биомор-
фологии компонентов луговой степи. Здесь уместно сослаться на работу
Г. И. Дохман (1956), в которой дается развернутая критика большинст-
ва затронутых вопросов. Г. И. Дохман справедливо отмечает, что на не-
236
скашиваемых участках степи ковыли не только не испытывают угнете-
ния, а получают более мощное развитие. Мы не разделяем мнения
Г. И. Дохман о резком подавлении длиннокорневищных злаков на неко-
симых участках под влиянием прогрессивного развития ковылей или
вследствие их самоугнетения. Но мы решительно отвергаем всякие по-
пытки представить смену растительности Стрелецкой степи при режиме
некошения таким образом, что в ходе ее ковыли постепенно деградиру-
ют. Это утверждение противоречит фактам. Оно расходится и с новей-
шими (еще не опубликованными) исследованиями И. В. Голубевой
возрастного состава популяции Stipa pennata на некосимых участках
Стрелецкой степи. Изучение показало удивительную равномерность ко-
личественного состава всех возрастных групп. Это обстоятельство сви-
детельствует о полной устойчивости популяции, о нормальном семенном
возобновлении ковыля перистого в условиях абсолютной заповедности
•степи.
В подтверждение развиваемой мысли можно сослаться и на мнение
А. Я. Гордягина (1901, 1922), Б. А. Келлера (1903), И. К. Пачоского
(1910, 1917), В. В. Алехина (1925а). Так, А. Я. Гордягин полагал, что
современная разнотравно-луговая степь в процессе восстановления, при
исключении хозяйственного воздействия, преобразовывается в типчако-
во-луговую или ковыльно-луговую степь, в которой ксерофитные злаки
приобретают больший удельный вес, не подавляя, однако, развитие и
других растений. Подобный же взгляд был высказан И. К. Пачоским,
считавшим, что в коренном (первобытном) растительном покрове луго-
вых степей ковыли были более обильны, но никогда не доминировали
над прочей растительностью, как в настоящих ковыльных степях.
Что касается утверждений о выпадении на нескашиваемой степи дру-
гих плотнокустовых злаков и осоки низкой, то они, очевидно, объясня-
ются крайне поверхностным изучением видового состава. Тщательный
осмотр старых некосимых участков выявил присутствие прекрасно раз-
витых дерновин типчака, не испытывающих никаких признаков угне-
тения. Найдены были и осока низкая, и тонконог. Значит, плотнокусто-
вые ксерофитные злаки и осока низкая не исчезают из травостоя под
влиянием режима заповедности, а становятся лишь менее обильными.
Выявление флористического состава некосимых целин заповедника
на большой площади показывает присутствие абсолютно всех видов, ха-
рактерных для косимой степи. Можно даже утверждать (как раз обрат-
ное Г. М. Зозулину), что под влиянием покоса часть видов степи выпа-
дает, например Artemisia latifolia, Aster amellus Lithospermum officinale,
Peucedanum lubimenkoanum, Sedum telephium, Senecio erucifolius и др.
Слабо обоснованной представляется и идея о мезофитизации (олу-
говении) луговой степи под влиянием режима некошения. Костер без-
остый, произрастающий на целинах заповедника, в том числе некоси-
мых участках, относится к степному экотипу (Левицкий, 1955), и его
развитие в этих условиях нисколько не помогает сторонникам луговой
гипотезы улучшить свои позиции. Можно согласиться, что в результате
накопления толстого слоя степного войлока на некосимой степи созда-
ются благоприятные условия для жизнедеятельности длиннокорневищ-
ных растений. Рыхлый войлок — прекрасная среда для обильного раз-
растания корневищ. Но положительное влияние этой среды равнозначно
как в отношении мезоморфных, так и ксероморфных длиннокорневищ-
ных растений. Последние на некосимых целинах заповедника значитель-
но преобладают над первыми и местами достигают высокого обилия. Та-
ковы следующие степные длиннокорневищные виды: Bromus inermis
(степной экотип), Agropyron intermedium, Asperula tinctoria, Galium ve-
rum и др. И вряд ли можно сравнивать по обилию с этими степными
видами какие-либо длиннокорневищные луговые виды некосимой степи.
237
Вейник наземный нельзя принимать в расчет, так как это не луговой вид^
Кроме того, исследования показали наличие у него исключительно глу-
бокой корневой системы, проникающей в условиях луговой степи глубже
3,5 м. Все-таки вейниковые заросли обычно связаны с депрессиями рель-
ефа и различными другими особенностями почво-грунта, улучшающими
водный режим местообитания. В менее благоприятных условиях (пла-
кор, южные экспозиции и пр.) вейник не достигает высокой плотности
развития или же совсем отсутствует. Мало подходит для отождествле-
ния с луговыми растениями и Роа angustifolia, который скорее является
типичным лесостепным ксеромезофитом.
Нельзя, конечно, отрицать, что при некосимом режиме степи несколь-
ко увеличивается обилие рыхлокустовых мезофильных злаков Festuca
pratensis и Dactylis glomerata. Но обилие их нигде не достигает той ве-
личины, по которой их можно было бы отнести к доминантным или суб-
доминантным видам.
Таким образом, факты увеличения обилия мезофильных растений на
некосимой степи не дают каких-либо веских аргументов для отрицания
существования в прошлом в лесостепи луговых степей как зонального
образования. Напротив, скопление огромного количества неразложив-
шихся мертвых растительных остатков на заповедных участках луговых
степей — до 100 ц/га и более (Семенова-Тян-Шанская и Никольская^
1960) —свидетельствует лишь о ксерической обстановке, в условиях ко-
торой процессы минерализации отмирающих надземных органов расте-
ний сильно затруднены. На лугах лесной зоны подобного накопления ве-
тоши и опада не наблюдается, так как значительная часть растительных
остатков в условиях достаточной влажности быстро минерализуется.
Было бы, однако, мало оправданным проводить полную аналогию в
особенностях растительности длительно некосимых участков луговых
степей заповедника и первобытных луговых степей. Режим абсолютной
заповедности в настоящее время представляется в значительной мере
искусственным. Раньше несравненно большую роль играл зоогенный
фактор. Особо велика была роль копытных животных, влияние которых
в настоящее время совершенно исключено. Поедание копытными зеле-
ной массы сокращало количество отмирающих частей растений, а зна-
чит и накопление степного войлока. Кроме того, выпас копытных спо-
собствовал разрыхлению войлока и его удалению путем выдувания вет-
ром (Пачоский, 1917; Лавренко, 1940; Дохман, 1956). Далее, мертвые
растительные остатки в массе поедались мышевидными грызунами, чис-
ленность которых, несомненно, была весьма высокой. Сейчас даже труд-
но представить, сколь велики были последствия зоогенного фактора на
степную растительность. В этом отношении весьма показателен пример
1962 г. Отдельные участки некосимой степи в зимний период были глад-
ко «подстрижены» мышевидными грызунами, неожиданно достигшими
большой численности.
Учитывая все эти обстоятельства, можно предположить, что в соста-
ве растительности первобытных луговых степей удельный вес длинно-
корневищных растений был меньше, чем сейчас на постоянно некосимых
участках. Сокращение мощности рыхлого войлока имело своим следст-
вием угнетение длиннокорневищных растений. По этой же причине бо-
лее обильно быЛо представлено разнотравье, так как оно получало боль-
ше возможностей для семенного возобновления. Господствующее поло-
жение в первобытных луговых степях, несомненно, занимали ковыли.
Последние, как показывает современный пастбищный режим на специ-
ально отведенной площади Стрелецкой степи, мало страдали от весьма
умеренного выпаса, а поедание мышевидными грызунами мертвых побе-
гов в зимнее время лишь благоприятствовало отрастанию надземной ча-
сти в летний период. Вполне понятно, конечно, что луговым степям в
238
прошлом, как и в современную эпоху, были свойственны свои виды ко-
вылей, отличающихся от ковылей зоны типичных (южных) степей.
Таким образом, сопоставление состояния растительности луговых сте-
пей в режиме сенокошения, абсолютной заповедности и умеренного вы-
паса создает объективные предпосылки для оценки характера развития
растительности первобытных степей. Есть все основания предполагать,
что доагрикультурная травянистая растительность лесостепи в своей
основе была ковыльной степью со значительным участием в травостое
разнотравья и корневищных злаков. Более подробные данные о влиянии
сенокошения, выпаса и режима заповедности на степную растительность,
в том числе луговостепную, можно почерпнуть в работах Б. А. Келлера
(1903, 1916, 1923), Г. Н. Высоцкого (1912, 1915), К. Владимирова (1914),
И. К. Пачоского (1917), А. И. Мальцева (1924), В. В. Алехина (1925а,
1934), Н. А. Аврорина (1934), Е. М. Лавренко (1940), Н. А. Прозоров-
ского (1940а), Н. Ф. Комарова (1951), Д. Ф. Федюнкина (1953),
Н. С. Камышева (19536, 1956), Г. И. Дохман (1956), А. М. Семеновой-
Тян-Шанской (1960а, б, в) и др.
В связи с обсуждаемыми вопросами заслуживают внимания выводы
Н. С. Камышева (1956) об изменении растительности Каменной степи
под влиянием режима абсолютной заповедности. Автор указывает, что
заповедные участки представляют явление совершенно особенное, не
похожее ни .на косимую степь, ни на естественную степь, подвергающую-
ся нормальному выпасу. «И развитие растительного покрова их идет по
своим особым закономерностям. К эволюции растительности этих зале-
жей не применима ни одна из схем развития залежей, которые до сих
пор были известны науке» (стр. 61).
Таким образом, даже если в других условиях, например на некото-
рых заповедных участках Стрелецкой степи, смена растительности про-
текает несколько иначе, чем в наблюдениях Н. С. Камышева, то и в
этом случае приведенные выводы не теряют своего исключительного
значения для решения проблемы природы и происхождения луговых сте-
пей. Они недвусмысленно свидетельствуют о том, что и факты мезофити-
зации травяного покрова при установлении режима некошения степей,
и увеличение при этом длиннокорневищных злаков (костра безостого,
вейника наземного, пырея) и даже сорных корнеотпрысковых видов
(осота розового) нельзя использовать в качестве доказательства луго-
вой природы луговых степей и отнесения их к луговому типу раститель-
ности, как это имеет место в работах Г. М. Зозулина. Очень важным
доводом в пользу высказанного заключения является то обстоятельство,
что сходные признаки «перерождения» описаны и для южных степей
(Пачоский, 1917). Но на этом основании вряд ли кто станет утверж-
дать, что на месте южных степей в доагрикультурную эпоху были раз-
виты луговые ценозы.
Объективные свидетельства былой природы луговых степей можно
получить, как уже сказано выше, только путем сопоставления состояния
и развития растительности при сенокосном режиме, в условиях умерен-
ного выпаса и абсолютной заповедности.
Из вышеизложенного становится очевидным, сколь многообразны,
сложны и запутанны вопросы, сопряженные с «проблемой природы и про-
исхождения растительности луговых степей. Наряду с этим выявляется,
что подобные расхождения взглядов обусловлены отсутствием глубоко
разработанной теории луговых степей в ботанико-географическом и фи-
тоценотическом отношениях. Такая теория определила бы сам подход
к изучаемым явлениям, вскрыла бы их основное содержание, объедини*
ла бы громадное количество разрозненных фактов и явлений в одну
стройную систему.
23»
ЛУГОВЫЕ СТЕПИ КАК ЗОНАЛЬНЫЙ ЭЛЕМЕНТ
РАСТИТЕЛЬНОГО ПОКРОВА ЛЕСОСТЕПИ
В травянистых сообществах средообразующая роль отдельных видов
не идет ни в какое сравнение с видами древесных растений. Компоненты
травянистых ценозов характеризуются большей самостоятельностью,
меньшей взаимозависимостью и относительной легкостью изменения сво-
его состава, обилия, жизненности и цругих признаков под влиянием
условий внешней среды. Непреложным следствием этой закономерности
является аутохтонность развития травянистых сообществ в определен-
ных физико-географических районах и областях с присущими им особен-
ностями климата, геоморфологическими, геологическими, гидрологиче-
скими, почвенными и другими условиями. Древесные ценозы также
подчинены данной закономерности, но в силу значительного средообра-
зующего потенциала они способны в некоторых пределах нейтрализовать
воздействие внешних условий, нивелируя их специфику в разных место-
обитаниях. Конечно, нельзя переоценивать последствия этого свойства
древесных растений, поскольку его действие ограничивается именно из-
вестными пределами.
Аутохтонное развитие растительности — всеобщая закономерность,
которой подчинено формирование всех растительных ассоциаций. Зо-
нально-провинциальное расчленение растительности является ярким
тому свидетельством. Отдельные мысли и понятия о развитии фитоце-
нозов на месте под влиянием конкретных условий существования рассе-
яны в работах А. Н. Краснова (1888), А. Л. Тахтаджяна (1946),
В. Н. Васильева (1946, 1958), В. Б. Сочавы (1946), Ан. А. Федорова
(1958) и др.
В разработке теоретических предпосылок к обоснованию аутохтонно-
го развития растительных сообществ велика заслуга М. Г. Попова. Эта
«сторона значения научного наследия выдающегося ученого еще не на-
шла надлежащего акцента в литературе. В своей замечательной работе
«Флора пестроцветных толщ Бухары» М. Г. Попов (1923) установил, что
эта флора в систематическом и эколого-историческом отношениях пред-
ставляет весьма цельный и своеобразный ботанико-географический
комплекс. М. Г. Попов подчеркивал, что чем экологически резче выде-
ляется тот или иной регион, тем более заметными становятся морфоло-
гические отличия видов, живущих в этом районе, по сравнению с близ-
кими формами соседних территорий. Как видно, здесь четко формулиру-
ются идеи, которые в дальнейшем легли в основу учения о филоценоге-
незе В. Н. Сукачева (1944). Вместе с тем в данной работе показано
аутохтонное формирование флоры и растительности на основе местных
литологических, климатических и других природных особенностей стра-
ны. В ряде последующих работ это замечательное положение находит
дальнейшее развитие (Попов, 1929, 1940а, б, 1949). В «Очерке раститель-
ности и флоры Карпат» М. Г. Попов (1949) делает интереснейший вы-
вод о том, что геоботаническое и флористическое районирование земли
должно быть единым. Таким образом, понятие о флористико-ценологи-
ческой самобытности регионов, отличающихся экологически, здесь полу-
чает окончательное завершение.
Отмечая значение работ М. Г. Попова для обоснования теории ауто-
хтонного развития растительного покрова, необходимо оговориться в
том смысле, что сам М. Г. Попов был страстным приверженцем мигра-
ционной гипотезы, которая в определенном значении противоположна
теории аутохтонного развития. Внешние расхождения этих двух точек
зрения особенно обострились после выхода в свет работ М. М. Ильина
(1958), Ан. А. Федорова (1942, 1952, 1958) и В. Н. Васильева (1958).
Анализируя фактические материалы, положенные в основу рассматри-
ваемых концепций, мы пришли к выводу о том, что они нуждаются в
240
расчленении, по крайней мере, на три категории: 1) географические за-
кономерности видообразования, 2) флорогенез в собственном смысле,
3) формирование растительных сообществ. Добытые наукой данные
убеждают, что если видообразование и формирование растительных со-
обществ совершается аутохтонно, в определенных природных условиях
тех или иных районов, то во флорогенезе имеют значение как элементы
аутохтонного развития, так и в еще большей степени разнообразные
миграции видов и флор (Голубев, 1960д). Только смешение этих трех
самостоятельных природных явлений и процессов может порождать про-
тиворечия между миграционной теорией и теорией аутохтонного разви-
тия видов и ценозов.
Мысль об историко-генетической и фитоценотической самобытности
лесостепного ландшафта в наиболее четком ее выражении одним из пер-
вых была высказана В. В. Докучаевым еще в 1892 г. «...Своеобразный
характер самой лесостепи,— писал творец «Русского чернозема»,— есть
явление вполне естественное, от века существующее, а не случайное и
временное, почему и причины его могут и должны корениться только в
постоянно действующих физических особенностях страны и ее геологи-
ческом прошлом...» (Докучаев, 1953, стр. 111). И. К. Пачоский (1910)
рассматривал лесостепь в качестве основного, наиболее древнего типа
растительности по сравнению со степным (к югу) и лесным (к северу)
типами. В пользу этого положения И. К. Пачоский приводит флорогене-
тические доказательства — факт наличия в подольской лесостепи много-
численных реликтов, не встречающихся поблизости, по разбросанных
спорадически в лесостепных местностях более древнего геологического
возраста — Средне-Русской возвышенности, Польше, Добрудже, Венг-
рии и др. Подольская лесостепь имеет возраст не моложе миоценового.
Со времени миоцена, по мнению автора, лесостепь неоднократно меняла
свой экологический облик.
По существу, самостоятельность растительности лесостепи вытекает
из признания ее бизонального характера — равноправности в лесостепи
и лесных и травянистых луговостепных группировок (Алехин, 1915а;
Крылов, 1915; Новопокровский, 1927, 1937; Сакало, 1961; и др.). Этот
вывод весьма наглядно подтверждается высказываниями В. В. Алехина
(1915а), характеризующего лесостепь следующим образом: «Здесь мы
находим небольшие перемежающиеся лески, между которыми раскину-
лись открытые места, покрытые травянистой растительностью. Таким
образом, нужно смотреть на лесостепь не как на случайное соединение
в местах контакта различных растительных форм, наоборот, здесь мы
имеем комплекс растений, вполне приспособленных к определенным
климатическим условиям, к определенным условиям существования.
Это такая же формация, как степная или чисто лесная, но тогда как в
степи, например, сумма внешних условий дает возможность существо-
вать лишь формам травянистым, а в лесах доминируют древесные по-
роды — лесостепь представляет еще более сложную комбинацию различ-
ных -форм (травы и деревья), находящихся в тесной зависимости друг
от друга и от внешних условий» (стр. 13).
Следует особо подчеркнуть, что природные условия лесостепи, ори-
гинальность и своеобразие которых достаточно глубоко и разносторонне
освещены Ф. Н. Мильковым (1950) 1 и другими, накладывают глубокий
1 Несмотря на последовательную мотивировку оригинальности природных условий
лесостепи, автор некритически принимает воззрения Е. М. Лавренко о развитии в лесо-
степи двух различных экологически типов травянистой растительности — лугового
(остепненные луга) и степного (луговые степи). Между тем в лесостепной зоне форми-
руется лишь один тип травянистой растительности — луговые степи, которые нельзя
отождествлять ни со степным, ни с луговым типами. Единству ландшафта здесь соот-
ветствует и единство травянистых сообществ, которые таксономически должны объеди-
няться в самостоятельный тип растительности — луговые степи
16 В. Н. Голубев
241
отпечаток на все ее растительные образования, будь то травянистые или
древесные сообщества. Так, лесные ценозы лесостепи претерпевают оп-
ределенную трансформацию, например, в составе травяного покрова, где
в изобилии появляются луговостепные формы. Факт наличия в лесах ле-
состепи луговостепных травянистых растений в литературе отмечался
неоднократно (Коржинский, 1888, 1891; Алехин, 1909; Крылов, 1916;
Новопокровский. 1927, 1931; Крашенинников, 1937а, 1941; Гордягин,
1901; Пономарев, 1952, и др.).
Особенно заметное влияние степных элементов в степных и тем бо-
лее пристепных лесах оказывается в лесостепи по направлению к восто-
ку. Это обстоятельство было замечено еще С. И. Коржинским (1891).
Ученый, категорически отрицавший наличие группировок, переходных
между лесом и степью, при обследовании восточных губерний Европей-
ской части — Пермской и Уфимской — вынужден был признать,— что
здесь, в лесостепи, возникают такие сочетания растений, «которые при-
дают всей стране, действительно, средний, лесостепной характер, не по-
зволяющий приравнивать ее ни к лесной, ни к степной области...»
(стр. 141).
В наших исследованиях европейской лесостепи oбнaipyжeн целый ряд
видов, которые одинаково хорошо себя чувствуют и в дубравах и луго-
вой степи. Это будут Campanula persicifolia, Carex michelii, Iris aphylla;
Phlomis tuberosa, Primula veris, Serratula heterophylla, Veratrum nigrum,
Veronica chamaedrys и др. Такие виды с полным правом и в прямом
смысле можно назвать лесостепными. Этот факт свидетельствует об из-
вестной выравненное™ экологических условий в лесу и степи, о близких,,
а может быть и равных возможностях развития в лесостепи лесных и
травянистых сообществ, о взаимном проникновении степных и лесных
элементов, об относительно гомогенном лесостепном растительном комп-
лексе в эколого-фитоценотическом смысле.
В Восточной Сибири явление внедрения степных элементов под по-
лог березово-сосново-лиственничных лесов достигает наибольшей выра-
зительности. Однако это обстоятельство не привлекает должного внима-
ния исследователей. Этот процесс расценивается еще со старых пози-
ций — как проявление «остепнения» или свидетельство более широкого
в прошлом распространения к северу степей, подвергающихся ныне
облесению. Между тем наступила пора пересмотреть прежние взгляды
и оценить подобные сочетания растений разных жизненных форм как
нормальные и естественные, издавна существующие, гармонически и е
необходимостью отвечающие природным условиям лесостепного ланд-
шафта.
Ярким защитником теории флористико-фитоценотической самобыт-
ности и самостоятельности лесостепной растительности был И. М. Кра-
шенинников (1937а, б; 1939). Он писал: «Лесостепь в нашем толковании
есть не просто переходная полоса от лесной к степной зоне, а вполне са-
мостоятельная область, которая имела особую историю образования и
развития своей флоры и растительности...» (1939, стр. 96). Автор пола-
гал, что лесостепной флористический комплекс сформировался в плей-
стоцене. Наиболее близко к исходным типам холодной плейстоценовой
степи стоят современные березово-сосно^о-кедрово-лиственничные леса,,
распространенные в районе гор южной Сибири и северной Монголии (об-
ласти южной, средней и восточной Сибири). В плейстоцене на указан-
ной территории лиственничные леса с примесью березы, сосны, кедра
играли очень большую роль и включали разнообразную травянистую и
кустарниковую растительность, виды которой имели центром своего раз-
вития Сибирь, в особенности ее восточную часть. Многие виды плейсто-
ценового флористического комплекса были широко представлены в са-
мых различных ассоциациях сосново-лиственнично-березовой лесостепи.
242
Причины этого И. М. Крашенинников видел в слабой диффенцирован-
ности общих экологических условий, сочетавшейся с суровостью клима-
та в целом. Физико-географический режим плейстоценовой лесостепи
весьма близко напоминал режим современных субальпийских районов
и горной лесостепи средних широт Азии. В качестве примера автор при-
водит области горной Сибири и Хангая, где и сейчас формы, населяю-
щие лесные, лесостепные и субальпийские пояса, очень близки в систе-
матическом отношении и трудно разграничимы так же, как и раститель-
ные сообщества на отдельные типы растительности.
Анализируя разнообразные данные (климатологии, палеоботаники,
палеозоологии, флорогенетики, археологии и др.), Ф. Н. Мильков (1947,
1948, 1950) приходит к выводу о большой древности лесостепного ланд-
шафта, сформировавшегося аутохтонно, не позднее других смежных
ландшафтных зон, вероятно, в плиоцене. С этого времени он претерпел
сложную историю развития, на протяжении которой происходили как
изменения границ и очертаний лесостепной зоны, так и сущность само-
го ландшафта. В настоящее время составными зональными категориями
лесостепного ландшафта Русской равнины являются широколиственные
леса и разнотравно-луговые степи.
Большой вклад в познание специфики растительности лесостепи вно-
сят флорогенетические исследования А. Н. Пономарева (1948, 1949,
1952), которыми установлен целый ряд видов, совмещающихся в совре-
менном распространении преимущественно с лесостепной зоной. Глав-
ной особенностью этих видов является то, что наиболее обильное их
развитие наблюдается в ценозах луговых степей и довольно широкое
распространение в лесостепной зоне вообще. Эта закономерность по
признаку встречаемости в количественном выражении фактически былс
установлена еще П. Н. Крыловым (1916). У этих видов происходит сов-
мещение экологического оптимума с оптимумом физиологическим, бла-
годаря чему они достигают в области лесостепи максимального обилия,
к северу же и к югу от лесостепной зоны их обилие резко идет на убыль
(Голубев, 1962а). Наличие специфических лесостепных видов как из
группы низших беспозвоночных, так и высших животных выявлено и
зоологами.
Среди лесостепных видов А. Н. Пономарев выделяет несколько гео-
графо-генетических элементов. Наиболее универсальным (географиче-
ски) является евразиатский элемент лесостепного комплекса (Achyro-
phorus macutatus, Adonis vernalis, Asparagus officinalis, Campanula sibi-
rica, Filipendula hexapetala, Fragaria viridis, Poa angustifolia, Potentilla
humifusa, Senecio integrifolius, Stipa pennata и др.). Значительный удель-
ный вес занимают виды восточно-континентального элемента, подразде-
ляющегося на южносибирские виды (Artemisia frigida, A. commutata,
Potentilla viscosa и др.), сибирские с дизъюнктивным ареалом (Aster
amellus) и восточно-евразиатские виды, продвинувшиеся сплошными
ареалами за Урал в область Восточной Европы (Artemisia armeniaca,
A, latifolia, Gypsophila altissima, Helictotrichon schellianum, Trifolium
lupinaster и др.). Известную роль играют виды западно-атлантического
элемента, континентальные евразиатские лесостепные виды и др. Несом-
ненный интерес представляют суббореальные луговолесные виды (Вга-
chypodium pinnatum, Dracocephalum ruyschiana, Lathyrus pisiformis, Po-
lygonatum officinale и др.), являющиеся, в сущности, бетулетальными, од-
нако приуроченными в распространении главным образом к лесостеп-
ным зональным ценозам. Оригинальную группу образуют горностепные
виды, которые можно также оценивать в качестве элементов субаркти-
ческих степей (Aster alpinus, Bupleurum multinerve, Helictotrichon de-
sertorum и др.). Трактовку этой последней группы видов в качестве
лесостепных можно принять лишь с рядом оговорок.
16*
243
Большого внимания заслуживают выводы А. Н. Пономарева о широ-
ком экологическом диапазоне многих лесостепных видов, особенно суб-
бореально-бетулетальных и горностепных, способных развиваться как в
луговых степях, так и лесах лесостепи. Вслед за Б. Н. Городковым
(1939) автор видит причину «лесостепной» природы рассматриваемых
видов в особенностях их эколого-исторического прошлого, в вероятности
формирования в светлых хвойных (сосново-лиственничных) и березовых
лесах Сибири: «Благодаря этому,— пишет А. Н. Пономарев (1952,
стр. 324),— одни и те же виды произрастают и в светлых лесах, и в се-
верных луговых степях. Однако признание принадлежности этих видов
к флороценотическому комплексу светлых сибирских лесов отнюдь не
должно заслонять их преимущественного распространения в лесостепи
и их связи с зональными ценозами последней. Это не исключающие,
а взаимно дополняющие стороны одного явления». Автор подчеркивает,
что флорогенетическое понимание лесостепных видов вполне оправдыва-
ется и в ценологическом отношении (Пономарев, 1949). Таким образом,
эти воззрения близко примыкают к взглядам И. М. Крашенинникова.
Из изложенного становится ясным, какую исключительную научную
ценность представляют углубленные эколого-биологические и фитоцено-
тические исследования светлохвойных и березовых лесов лесостепи Во-
сточной Сибири для решения кардинальных проблем лесостепной рас-
тительности на всем ее широтном протяжении.
Итак, приведенные данные согласованно свидетельствуют о том, что
лесостепная растительность представляет собой единство лесного и лу-
говостепного типов растительности, развившихся аутохтонно под влия-
нием природных условий лесостепного ландшафта. Луговые степи как
зональный элемент растительного покрова лесостепи, характеризуются
флористико-фитоценотической самобытностью и самостоятельностью,
которая четко разграничивает их с другими типами травянистой расти-
тельности, в том числе луговыми.
Зональность травянистых и лесных сообществ в лесостепи, а также
развитие лесостепных видов (в прямом смысле) позволяют заключить,
что с уничтожением лесов в зоне лесостепи на их месте неминуемо долж-
ны образоваться травянистые ценозы, по своим признакам совершенно
подобные зональным травянистым сообществам первичного характера,
т. е. должны возникнуть луговые степи, ничем существенно не отличаю-
щиеся от первичных луговых степей. Конечно, восстановление «нормаль-
ной» растительности луговой степи представляет процесс, завершение
которого требует определенного времени. Кроме того, восстановление
растительности в направлении формирования травянистых ценозов воз-
можно только при известных условиях, а именно в отсутствие неумерен-
ного выпаса или в условиях сенокошения, когда развитие древесных
растений подавляется.
В подтверждение сказанного можно сослаться на многочисленные
указания в литературе о близком сходстве первичных и вторичных луго-
востепных сообществ и большой трудности их разграничения. Напри-
мер, А. Я. Гордягин (1922) утверждал, что вторичные луговые степи,
развившиеся на черноземе, мало измененном предыдущим влиянием
леса, «едва ли отличимы от нормальных степей, т. е. не подвергавшихся
облесению» (стр. 199). «До сих пор мы полностью не выяснили,— пишет
М. В. Марков (1935),— чем -вторичные травянистые группировки, воз-
никшие на месте уничтоженного леса в области лесостепи, будут отли-
чаться от «коренных» степных группировок» (стр. 71). Аналогичные со-
ображения и факты приводятся С. И. Коржинским (1891), Г. М. Зозу-
линым (1955) и другими исследователями. На Стрелецком участке Цент-
рально-Черноземного заповедника, по свидетельству Г. М. Зозулина
(1955, стр. 193), «небольшой лесок Бабки в недалеком прошлом соеди-
244
нялся по водоразделу с леском Селиховы Кусты, и после сплошной вы-
рубки пни на водоразделе усиленно выкорчевывались на топливо. Сей-
час здесь «степь», ничем не отличающаяся по растительному покрову
и почвам от основной территории Стрелецкой степи».
Таким образом, тождественность первичных и вторичных луговых
степей, при достаточной длительности развития последних, следует при-
знать установленным фактом; но этот факт еще не находил удовлетвори-
тельного объяснения. С позиций зональности леса и луговой степи в ле-
состепной зоне их взаимозаменяемость представляет явление естествен-
ное и, по-видимому, единственно возможное. А наличие в лесах и луго-
вых степях общих элементов травянистой флоры уже непосредственно
обусловливает развитие на месте сведенных лесов травянистых ценозов,
сходных с коренными. Это обстоятельство было прозорливо подмечено
еще С. И. Коржинским (1891) в отношении березовых лесов в пермской
лесостепи. Аналогичные данные сообщаются и А. Я. Гордягиным (1901).
Развиваемая концепция, как можно было убедиться, опровергает ги-
потезу остепненных лугов, распространенных на северном пределе лу-
говых степей и возникших на месте сведенных лесов в полосе лесостепи,
которая, за отсутствием каких-либо серьезных фактических и теорети-
ческих обоснований, представляется несостоятельной и должна быть ос-
тавлена.
На этом можно было бы и покончить с анализом различных точек
зрения на развитие вторичных травянистых сообществ в лесостепи. Од-
нако здесь возникает один весьма интересный вопрос — какие травяни-
стые ценозы формируются на месте уничтоженных лесов в пограничной
с лесостепью лесной полосе. Отвечая на этот вопрос, надо принять во
внимание ряд обстоятельств. Во-первых, примыкающая к лесостепи лес-
ная полоса неравноценна расположенной севернее. Во-вторых, луговые
сообщества лесной зоны все вторичны, за исключением разве неболь-
ших участков пойменных лугов. Их развитие происходило за счет флори-
стических элементов окружающих лесов и луговых степей, наиболее ме-
зофильных элементов последних. В условиях лесной зоны они испытали
дальнейшую мезофитизацию, выразившуюся в образовании особых луго-
вых экотипов. В-третьих, основным флористическим источником форми-
рования травянистых группировок в пограничной с лесостепью полосе,
конечно, были луговые степи. Эти ценозы, естественно, будут производ-
ными луговых степей, в значительной мере мезофитизированными. Весь-
ма возможно, что такие группировки занимают промежуточное поло-
жение между лугами лесной зоны и луговыми степями. Но даже в этом
случае их нельзя называть остепненными, так как понятие о последних
предполагает изначальное существование лугов, претерпевших затем
остепнение, т. е. внедрение степных элементов. Генетически этот процесс
не мог иметь места в действительности. Проводящийся здесь взгляд на
природу вторичных травянистых группировок, возникших на месте све-
денных лесов, в южной части лесной зоны, весьма близко совпадает с
представлением Д. И. Сакало (1961) о дексерофитизации луговостепных
ценозов в направлении с юга на север, признаваемой им, однако, лишь
для травянистых сообществ в пределах лесостепи.
Единство луговостепного типа растительности в лесостепной зоне не
исключает его меридиональной дифференциации, связанной с соответст-
вующими изменениями климатических, эдафических и других факторов.
Это обстоятельство уже давно отмечено исследователями, что нашло от-
ражение в геоботанической классификации. Ряд авторов выделяет, по
существу, три варианта луговых степей: северные, средние и южные (Па-
чоский, 1910, 1917; Клеопов, Лавренко, 1933; Алехин, 1934, 1936) !;
’ У каждого из перечисленных авторов есть свои отличия в названиях: у И. К. Пъ
чоского — это три полосы лесостепи: северная, средняя и южная; у Ю. Д. Клео-
245
другие (или те же авторы, но в других работах) сокращают число ши-
ротных вариантов луговых степей (и лесостепи) до двух: северного и
южного (Алехин, 1915; Келлер, 1916, 1923; Залесский, 1918; Лавренко,
1940; Крашенинников, 1932) * 2.
Принимая это деление, следует отметить, что четкие границы между
выделенными подразделениями отсутствуют, соединяясь рядом посте-
пенных переходов. Однако нельзя согласиться с мнением Е. М. Лаврен-
ко (1940) о существовании последовательного перехода растительности
луговой степи в луговую (остепненные луга) в северном направлении.
В естественных условиях на северном пределе распространения луговые
степи непосредственно граничили с лесами: в Европейской части Сою-
за — с подзоной дубовых лесов (на что указывает и Е. М. Лавренко,
1940), в Азиатской части — с березовыми, лиственничными, сосновыми
лесами. Наличие переходов от луговой степи к лугам ограничено лишь
пределами лесостепи, в различного рода депрессиях рельефа, где бла-
годаря лучшим условиям водоснабжения развивается растительность,
близкая к луговой и даже болотной. Естественно, что от плакорных уча-
стков в сторону понижения рельефа возможна смена луговостепных це-
нозов близкими к луговым. Особенно многочисленные факты такого
рода описаны Н. А. Ивановой и И. М. Крашенинниковым (1934) для
западносибирской лесостепи, где это явление обусловлено слабым раз-
витием эрозионной сети и наличием замкнутых бессточных впадин и кот-
ловин.
На юге, т. е. на южной границе лесостепи, луговые степи через про-
межуточные ковыльно-разнотравные переходят в настоящие (типчако-
во-ковыльные) степи, что было подробно мотивировано еще И. К. Па-
чоским (1910), В. В. Алехиным (19156), Б. А. Келлером (1916) и др.
Во флористическом составе, структуре, доминирующих группах и ви-
дах, экологических и других особенностях травянистых растительных
сообществ севера и юга лесостепной зоны нельзя найти кардинальных
отличий, на основании которых эти сообщества следовало бы расчле-
нять на два принципиально различных типа — луговой и степной.
Весьма показательно, что многие широко распространенные виды
представлены в лесостепи особыми экотипами, отличающимися по росту,
консистенции ассимилирующих органов и морфолого-систематическим
признакам. Это обстоятельство было отмечено еще П. Н. Крыловым
(1916). Особый лесостепной экотип установлен у Bromus inermis (Смир-
нов, 1945; Коликов, 1950). В качестве самостоятельного вида выделена
Agrostis syreistschikowii из цикла .4. canina (Смирнов, 1938). По словам
П. А. Смирнова (1958), это «характерный элемент полосы луговых сте-
пей Русской равнины» (стр. 52). К сожалению, исследования экотипиче-
ского состава растений лесостепи немногочисленны. Но, судя по имею-
щимся данным, эколого-систематические исследования внутривидовой
дифференцировки эврптопных видов, несомненно, доставили бы богатей-
шие фактические материалы, удостоверяющие эколого-флористическую
оригинальность растительности луговых степей.
В этой связи я позволю себе процитировать одно место в работе
Г. М. Зозулина (1958), как известно категорически отклонявшего само-
пова и Е. М. Лавренко — три варианта луговидной или влажной степи: гигротический, ме-
зотический и ксеротический; у В. В. Алехина (1904) —три полосы северных разнотрав-
ных степей; северная, средняя и южная или три варианта; северный гигрофильный ос-
новной мезофильный и южный ксерофильный (Алехин, 1936). Указанное деление лесо-
степи вообще и луговых степей в том числе принимается и современными исследовате-
лями (Борисова, Исаченко, Рачковская, 1957; и др).
2 У В. В. Алехина луговые степи подразделяются на северный и южный варианты,
у Б. А. Келлера — разнотравно-луговые и дерновинно- (ковыльно- и типчаково-) луго-
вые степи, у К. М. Залесского — красочные (без- или малозлаковые) луговые степи
и луговые степи со Stipa pennata, у Е. М. Лавренко — остепненные луга и луговые
степи, у И. М. Крашенинникова и других — северный и основной варианты.
246
«Тоятельность луговых степей как зонального образования. «Нельзя от-
рицать,— пишет Г. М. Зозулин,— некоторую самобытность эколого-фи-
тоценотических особенностей лугового типа (? — В. Г.) растительности
в лесостепной зоне. Ряд луговых видов присущ преимущественно лесо-
степн...(Anthyllis, polyphylla, Arenaria ucrainica, Filipendula hexapetala-
Phlomis tuberosa, Poa angustifolia, Salvia pratensis, Tragopogon orienta-
te и др.). He исключена возможность, что мы являемся свидетелями
формирования новой исторической свиты растительности, в которой про-
цессы филоценогенеза приведут к формированию новых комплексов
группировок из исходного видового материала, главным образом из лу-
говой (южный вариант) и ковыльниковой (средиземно-голарктический
вариант) исторических свит растительности. Подытоживая сказанное,
можно отметить, что «северные степи» не могут быть фитоценотически
отнесены ни к степям, ни к лугам»... (стр. 821). Эта же мысль проводит-
ся и в более ранней работе Г. М. Зозулина (1955, стр. 189). Какие-либо
комментарии здесь излишни. Но тем более парадоксальным кажется
вывод, следующий за цитированными строками, что луговые степи
следует включать... в луговой тип растительности!
Флористико-фитоценотический анализ растительности луговых сте-
пей со всей очевидностью показывает, что они представляют довольно
монолитный зональный тип травянистой растительности. Поэтому надо
признать, что травянистые растительные группировки в пределах лесо-
степи имеют единое происхождение. «Искусственно разрывать их на ос-
тепненные луга и луговые степи... нет оснований» (Сакало, 1961,
стр. 972).
В зональности луговых степей и самобытности лесостепной расти-
тельности содержится ясный ответ и еще на один вопрос, занимавший
многих исследователей,— почему в лесостепи участки травянистой рас-
тительности, несмотря на сравнительно благоприятные условия, не были
поглощены лесом. С. И. Коржинский (1888, 1891), как было сказано
выше, утверждал, что леса как наиболее мощная и древняя формация
еще не успели занять все открытые пространства в лесостепи и не будь
человека, резко нарушившего процесс естественных взаимоотношений
леса и степи, они неминуемо бы вытеснили травянистые ценозы.
А. Я. Гордягин (1901, 1922), В. И. Талиев (1913), Б. А. Келлер (1916,
1923), Н. Ф. Комаров (1951) и другие выдвинули в качестве причины,
препятствовавшей распространению леса на степные участки, степные
пожары. Но с позиции признания самостоятельности лесостепной расти-
тельности и признания зонального характера лесов и луговых степей
затронутый выше вопрос оказывается беспредметным. Наличие в лесо-
степи открытых травянистых сообществ наряду с островными лесами
представляет явление закономерное, обусловленное природными усло-
виями лесостепной зоны. Теряет смысл и тезис о мощности лесной фор-
мации. В лесостепи, по крайней мере в ее центральной части, как лес,
так и степь обладают одинаковой «мощностью», 'более или менее рав-
ными возможностями в своих взаимоотношениях. Логическим след-
ствием этих положений является то обстоятельство, что наблюдающий-
•ся в современную эпоху процесс наступления леса на степь, отмечаемый
многими авторами (Новопокровский, 1927; Куренцов, 1929; Алехин,
1934; Захаров, 1935; Сочава, 1945; Попов, 1953; Гулисашвили, 1948;
Зозулин, Кусмарцева, 1951; В. Н. Виппер, П. Б. Виппер, 1953; Исачен-
ко, 1954; и др) обусловливается исключительно внешними, главным
образом климатическими факторами. В ряде случаев имеет место про-
стое восстановление леса на территориях, еще недавно находившихся
под лесом. Мы вполне разделяем утверждение Д. И. Сакало (1961)
•о том, что «при зональном наступлении лесной растительности ле*
«состепная зона не создается заново, не является более молодым
247
образованием и не исчезает, а только перемещается вместе с другими
зонами, наступая, в свою очередь, на степь» (стр. 971). Вслед за
Д. И. Сакало мы категорически отклоняем воззрения С. И. Коржинского
(1888, 1891), согласно которым лесостепь возникла вследствие надвига-
ния леса на степь.
В понимании указанной зональной закономерности необходимо при-
нимать во внимание, что леса лесостепной зоны в плакорных условиях —
это не те леса, что развиваются в лесной зоне. В их составе, как уже
говорилось раньше, большую роль играют лесостепные травянистые эле-
менты; измененными являются структура лесных сообществ, их синузи-
альное сложение, полнота и другие признаки и свойства. Бесспорно,
лесостепные леса — это совершенно особая категория лесов, в значи-
тельной мере преобразованных за счет луговостепных элементов и,
соответственно, по своему экологическому режиму: гидрологическим,
термическим, световым и другим условиям. Как правило, лесостепные
леса на плакоре становятся осветленными, паркового характера, обла-
дающими целым рядом фитоценотических особенностей. К сожалению,
за единичными исключениями, обстоятельных работ по изучению изме-
нения лесов в меридиональном направлении, т. е. в поперечном сечении
лесостепной зоны, нет ни по европейской, ни по западносибирской, ни
по восточносибирской лесостепи. В подобном исследовании, основываю-
щемся на теоретических положениях об аутохтонном формировании
растительных сообществ и зональности лесостепной растительности,
можно было бы, между прочим, найти фундаментальные критерии, свя-
занные с решением нашего традиционного ботанико-географического
вопроса о взаимоотношениях леса и степи. Нет сомнения в том, что это
исследование является ценным также в геоботаническом и практичес-
ком отношениях.
БИОМОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ КОМПОНЕНТОВ
ЛУГОВЫХ СТЕПЕЙ КАК КРИТЕРИЙ ИХ САМОБЫТНОСТИ
Своеобразие луговых степей проявляется в многочисленных биомор-
фологических признаках компонентов. Некоторые приспособительные
черты растений Стрелецкой степи в подземной части рассматривались
в нашей более ранней работе (Голубев, 1962в). Эколого-биологические
свойства надземных органов растений луговой степи были подвергнуты
анализу на предыдущих страницах этой книги. Представляется необхо-
димым суммировать некоторые наиболее показательные факты и выво-
ды из области биоморфологии, свидетельствующие о неповторимой са-
мобытности луговых степей как ботанико-географического явления.
Прежде всего укажем на сложную ярусную структуру травостоя ко-
симой степи, в которой выделяется до шести ярусов цветковых растений
и один — моховой. По данным В. Н. Сукачева (1928), на пойменных лу-
гах высокого уровня (в б. Псковской губернии) выделяются всего два
яруса антофитов и один ярус мха. При аналогичном с нашим подходе
к расчленению травостоя на ярусы В. Н. Макаревичем (1963) установ-
лено в обыкновеннополевицевых и душистоколосковых лугах Карель-
ского перешейка (опытная база БИНа «Отрадное») наличие пяти яру-
сов, из которых один— моховой. Таким образом, по сравнению с лугами
лесной зоны ярусное сложение растительности луговой степи значитель-
но сложнее. Оно богаче, разнообразнее и в сравнении со структурным
построением травостоя южных степей.
Исследованиями характера и длительности вегетации выявлено при-
сутствие в составе луговой степи большого количества коротковегети-
рующих растений (33,6% — от всего видового состава и 41,2% — от чис-
ла основных компонентов). Среди них особенно характерна группа
248
полуэфемероидных растений, очень пластичных в своих реакциях на
периодические колебания почвенной влажности. В годы с резким дефи-
цитом влаги эти растения в середине лета теряют значительную часть
листьев и побегов, тогда как во влажные годы они способны пышно раз-
растаться и вегетировать в течение всего вегетационного периода. Па
наличию большого количества коротковегетирующих растений, в том
числе полуэфемероидных, наряду с длительновегетирующими, луговые
степи четко отличаются как от лугов лесной зоны, так и южных степей.
Обращает на себя внимание тот факт, что целый ряд видов, известных в
качестве типичных луговых мезофитов, например Rumex acetosa, Stella-
ria graminea и другие, а также лесолуговых видов—Prunella vulgaris,
Veronica chamaedrys и т. п.— в условиях луговой степи ведут себя как
настоящие эфемероиды или полуэфемероиды. Значение этого факта иск-
лючительно большое. Во-первых, он подчеркивает экологическую специ-
фику луговой степи. Во-вторых, он обязывает по-новому оценивать,
флористический состав ценозов в их геоботанической интерпретации и
классификации. Без надлежащего анализа биоэкологии видов в данных
условиях по одному лишь списку видов нельзя положительно решать
вопросы природы ценозов и их таксономии.
Весьма показательным представляется и состав растений Стрелецкой
степи по способам перезимовки, в котором 57% всех видов приходится на-
летнезеленые, а остальные 43% —на зимне- и вечнозеленые. Среди же
основных компонентов число летнезеленых и растений, зимующих в зеле-
ном состоянии, почти одинаково. По данным В. Н. Макаревича (1963),
для лугов Карельского перешейка количество видов, зимующих в зеле-
ном состоянии (розетками и побегами), составляет 66,7% от всего видо-
вого состава и 87,6% — от общего веса травостоя с единицы площади.
Остальная часть приходится на летнезеленые, зимующие почками и се-
менами. Таким образом, различия луговых степей и лугов по способам
перезимовки их компонентов выступают достаточно рельефно. Нет сом-
нения, что подобные же отличия луговой степи (но качественно противо-
положные) существуют и при сравнении с южными степями. К сожале-
нию, полноценных данных, с учетом всего видового состава и основных
компонентов, по способам перезимовки растений южных степей по^а>
еще нет. Отсюда, между прочим, видно, насколько важно )исследованйе-
биоморфологии компонентов различных типов растительности с охва-
том всего видового состава, что и учтено в настоящей работе.
Оригинальность луговостепной растительности выявляется по харак-
теру нарастания листовой поверхности растений. Для видов луговой сте-
пи свойственно двукратное формирование листьев — в весенний период
и средне-позднелетний. Виды с двумя генерациями листьев и побегов
занимают 55,6% от всего видового состава и 64,3%—от числа основ-
ных компонентов. Большинство же растений лугов характеризуется бес-
прерывным ростом в течение всего теплого времени года (Серебрякова,
1956). У растений настоящих степей второй период роста выражен не-
четко, слабой интенсивности и смещен на более поздние сроки (Белосто-
ков, 1962). Любопытно отметить, что у некоторых мезофильных расте-
ний, например у Trifolium repens, в условиях луговой степи в середине
лета происходит остановка ростовых процессов, в то время как на лугах
лесной зоны они растут беспрерывно весь летний сезон.
Ритмика цветения растительности, оформленная в виде константных
кривых количества зацветающих, цветущих и отцветающих видов по де-
кадам вегетационного периода, основанных на средних многолетних
данных фенологических наблюдений, четко выделяет луговые степи сре-
ди смежных лугового и южностепного типов травянистой растительно-
сти. В константном графике цветения растительности луговой степи мак-
симум зацветающих видов приходится на I декаду июня, цветущих — на
249
II и отцветающих — на III декаду июня. «Луговой» же тип ритмики цве-
тения характеризуется приуроченностью наибольшего числа зацветаю-
щих видов ко II декаде июня, а максимума цветущих — к I декаде
июля (Серебрякова, 1956; Шенников, 1928). В южных степях максимум
зацветающих видов совмещается с III декадой мая, а цветущих с I де-
кадой июня (Балаш, 1961а).
Луговая степь отличается и более разнообразными, ярче выраженны-
ми, более интенсивными красочными аспектами, связанными с массовым
цветением обильно развитых видов. В. В. Алехиным (1934, 1944) выде-
лено для Стрелецкой степи до 10—11 аспектов. В луговых сообществах
(Шенников, 1928; Кожевников, 1936) и южных степях (Танфильев, 1898;
Лавренко, Дохман, 1933; Горшкова, 1954; Балаш, 1961а, б; и др.) коли-
чество аспектов значительно меньше.
В составе растительности Стрелецкой степи преобладают весенне-
раннелетние фенотипы, цветение которых падает на весенний (по конец
мая) и раннелетний (июнь) периоды. На лугах же преобладающей
труппой являются среднелетние фенотипы, цветущие в июле (Макаре-
вич, 1963).
По длительности цветения господство среди растений луговой степи
принадлежит короткоцветущим (45% — от числа всех видов и 50% — от
суммы основных компонентов). Но и длительноцветущие виды играют
очень существенную роль. В составе луговой растительности основная
масса видов относится к числу длительноцветущих растений (Серебря-
кова, 1956). Из-за отсутствия в литературе соответствующих данных со-
поставить луговые степи по двум последним признакам с южными сте-
пями не представляется возможным. Однако уже на основании имею-
щихся публикаций (Балаш, 1961а, б; Белостоков, 1962) существование
различий по составу фенотипов (распределению цветения во времени)
и длительности цветения компонентов между луговыми и настоящими
степями является бесспорным.
Заканчивая этот краткий сравнительный обзор, мы можем сделать
заключение о том, что и в сфере биоморфологических особенностей над-
земных органов растений имеются яркие свидетельства самобытности и
оригинальности луговых степей, по которым их нельзя сблизить ни с лу-
говым, ни с южностепным типами растительности. Значит, луговые сте-
пи, производные лесостепного ландшафта, являются равноправными
членами в системе зональных типов травянистой растительности. В от-
личие от современных лугов лесной зоны, возникших вторично, на месте
сведенных лесов, луговые степи в области естественной лесостепи явля-
ются коренными, первичными образованиями. Конечно, они всюду мог-
ли возникать и вторично на освобождавшихся от леса территориях в
зоне лесостепи, и на примыкающей к ней южной полосе лиственных
лесов. Независимо от своей природы — первичного или вторичного про-
исхождения — луговые степи являются устойчивым зональным типом
растительности, характеризующимся целым рядом специфических при-
знаков флористического, фитоценотического и эколого-биологического
значения. Следует оговориться, что устойчивость зонального типа вовсе
не предполагает слабое проявление динамических процессов. Напротив,
лугово-степным сообществам свойственна исключительная подвижность
структуры, ярко выраженная сезонная динамика и разногодичные изме-
нения, наряду с развитием сукцессий общего значения. Кроме того, впол-
не закономерны изменения взаимного расположения леса и степи в лесо-
степи, а также периодические смещения границ лесостепной зоны в се-
верном и южном направлениях под влиянием циклических колебаний
космических факторов, связанных главным образом с ритмикой солнеч-
ной активности (Шнитников, 1957; Эйгенсон, 1957; Боровский, 1961;
« др.).
550
Убедительные доказательства самобытности лесостепного ландшафта
можно почерпнуть в почвенных исследованиях. Но хорошо известно, что
в лесостепи Европейской части СССР зональными являются серые лес-
ные почвы и выщелоченные черноземы. Под лугами и хвойными лесами
лесной зоны развиты подзолистые почвы, а под южными степями —
обыкновенные черноземы.
Высокой специфичностью характеризуется водный режим почв лесо-
степи. По данным А. А. Роде (1956), обобщившего исследования водно-
го режима почв всех основных зон нашей страны, водный режим почв
лесостепи складывается по периодически промывному типу, в то время
как в зоне лесов господствует промывной тип, а в зоне степей — непро-
мывной.
Как растительный и почвенный покров лесостепи являются показате-
лем ее ландшафтного своеобразия, так и другие природные условия,
особенно климатические, выделяют лесостепь среди смежных ландшафт-
ных зон. Таким образом, мы приходим к выводу о зональности лесостеп-
ного ландшафта вообще и его растительного покрова в частности. Ос-
новные компоненты последнего — леса и луговые степи — экологически
и фитоценотически являются преобразованными типами, в сравнении с
соответствующими формациями лесной зоны и степной области. В этом
выражаются аутохтонные черты растительности лесостепи, по которым
лесостепные леса нельзя отождествлять с лесами лесной зоны, а луговые
степи — с южными степями. По тем же причинам следует проявлять осто-
рожность, подчеркивая антагонизм между лесной и степной раститель-
ностью лесостепи, так как в определенных условиях лес включает в себя
луговостепные элементы и приобретает совершенно новые эколого-фито-
ценотические свойства так же, как травянистые элементы леса входят в
состав луговой степи иногда в качестве весьма устойчивых компонентов,
придающих ее ценозам своеобразные черты. Нет сомнения, что подобный
подход к анализу растительных группировок лесостепи создает предпо-
сылки в поисках принципиально новых решений проблемы взаимоотно-
шений леса и степи — узловой проблемы отечественной геоботаники.
Глава 9
Вопросы практического использования
эколого-биологических исследований
природных растений лесостепи
Многосторонний анализ и обобщение эколого-биологических особен-
ностей растений лесостепи создает предпосылки к познанию механизма-
освоения растениями природных условий этой зоны, путей возникнове-
ния, характера и сущности приспособлений, благодаря которым расте-
ния утверждаются в жизни в данных условиях и способны накапливать
органическое вещество, умножать его запасы в биосфере. В таком аспек-
те эколого-биологические исследования являются необходимой научной
основой для практического использования и реконструкции раститель-
ных ресурсов лесостепи. Более того, знание приспособительных свойств
диких растений, особенно в условиях естественных заповедников, чрез-
вычайно важно и для работы с культурными растениями в их селекции
и агротехнике возделывания высоких урожаев. Об этом очень ярко гово-
рил в свое время Б. А. Келлер (1931). Он писал, что «среди диких степ-
ных трав мы имеем возможность изучать большое богатство средств и
путей, какими эти травы справляются с неблагоприятными свойствами
степного климата... Природа сохранила в наших степях только расте>-
ния, наиболее приспособленные к местным природным условиям. Мы
далеко еще не использовали эту богатую сокровищницу в интересах
сельского хозяйства и должны постоянно иметь ее у себя поблизости в
естественных суровых условиях нашей природы» (стр. 5).
Изучение эколого-биологических признаков и свойств диких растений
представляет несомненный интерес в целях моделирования искусствен-
ных высокопродуктивных ценозов. Совершенно очевидно, что подбирая
растения с разной глубиной проникновения и интенсивностью корневых
систем, с различной структурой надземных органов и разными ритмами
развития, можно достигнуть всестороннего использования экологических
факторов (по принципу естественных сообществ) и получения макси-
мальной продуктивности. Основой такого прогрессивного моделирования
является учет экологических свойств растений в дополнительно заме-
щающих рядах, которые бы соответствовали особенностям экологиче-
ских ниш искусственного биогеоценоза. Практически возможно конст-
руировать весьма произвольные сочетания экологических ниш за счет
как факторов неорганической среды, так и в результате подбора видов
с определенными экологическими и эдификаторными свойствами. Нам
отчетливо представляются те замечательные перспективы, которые от-
крываются на этом пути развития экологии растений и эксперименталь-
ной геоботаники, особенно с внедрением методов и технических возмож-
ностей кибернетики. Подготовка исходного материала для обработки в
счетно-вычислительных машинах специального назначения представляет
необходимый этап и условие в указанном направлении развития. Всесто-
ронний анализ эколого-биологических свойств растений после известной
обработки, несомненно, является одним из существенных звеньев в бу-
252
дущих поисках программного моделирования искусственных ценозов с
произиольно устанавливаемыми качествами.
Рассмотрим конкретные возможности практического использования
.проведенных нами эколого-биологических исследований в текущее
время.
1. Данные о биоморфологических особенностях растений в подземной
и надземной сферах, частично опубликованные раньше (Голубев, 1962в),
являются, в известном смысле, эталонными, характеризующими расте-
ния в условиях естественного произрастания, в отсутствие отрицатель-
ного влияния последствий нерациональной эксплуатации. Многие изу-
чавшиеся нами виды входят в состав травостоев природных сенокосов
•и пастбищ, находящихся в ведении колхозов и совхозов. Сопоставление
состояния растений на лугах и пастбищах лесостепной зоны Европей-
ской части СССР и в условиях заповедника позволит выявить характер
изменения биоморфологических особенностей растений под влиянием
хозяйственной деятельности, степень угнетения и на этой основе изы-
скать методы использования и преобразования кормовых угодий в це-
лях повышения их продуктивности. Этим определяется прикладное зна-
чение обобщений и собранных фактических материалов.
2. Проведенное исследование позволяет выявить наиболее устойчи-
вые формы полезных природных растений, гармонирующих в своих при-
способительных чертах с экологическими условиями центральной лесо-
степи. К ним можно отнести кормовые растения: эспарцет песчаный
(Onobrychis arenaria), костер безостый (Bromus inermis), горошек тон-
колистный (Vicia tenuifolia), пырей средний (Agropyron intermedium).
Все эти виды обладают глубокой интенсивной .корневой системой, про-
никающей вглубь до 2 м и более, т. е. до горизонта постоянного увлаж-
нения. Благодаря этому данные растения в любые по метеорологиче-
ским условиям годы способны давать высокие урожаи.
Среднеглубокие корневые системы развивают на мощных черноземах
в условиях культуры овсяница луговая (Festuca pratensis) и райграс
высокий (Arrhenatherum elatius). Эти виды образуют большую массу
надземных органов, отличающихся высокой питательностью, и довольно
устойчивы к значительным колебаниям выпадающих по годам осадков.
Особенности распределения влаги в мощных черноземах Средне-Рус-
ской возвышенности и ее динамики в совокупности с данными по строе-
нию подземных частей растений и длительности вегетации позволяют
заключить, что если возделываемые культуры в европейской лесостепи
будут обладать глубокой корневой системой, достигающей горизонта по-
стоянного увлажнения (на глубине 2—2,5 м), то тем самым откроется
возможность получать высокие и устойчивые урожаи. Поэтому селек-
ция культурных растений по признакам корневой системы представляет
весьма важную практическую задачу.
В борьбе за высокие урожаи сельскохозяйственных культур в зоне
лесостепи существенную помощь может оказать направленная интенси-
фикации развития корневой системы, особенно на ранних стадиях онто-
генеза культурных растений. В этом процессе формирования мощной
корневой системы в период до наступления засушливой погоды велика
роль минеральных удобрений, обеспечивающих быстрый рост всходов
и укрепление молодых растений и представляющих большие резервы,
реализация которых позволит получать дополнительную прибавку уро-
жая.
3. В природных ценозах заповедника заключено большое число по-
лезных растений, которые либо культивируются, либо еще ждут хозяй-
ственного освоения. В работе по вовлечению природных растений в
культуру обязательно требуются обстоятельные познания их биомор-
фологии. Особенно это касается лекарственных растений. Возьмем,
253
к примеру, адонис или горицвет (Adonis vernalis). Это ценнейшее ле-
карственное растение, катастрофически исчезающее в природных усло-
виях из-за неправильных приемов заготовки лекарственного сырья. Вста-
ет вопрос о его искусственном размножении и выращивании. Однако
опыты с посевами адониса под Москвой терпят неудачу. Есть основания
полагать, что причина неуспеха в выращивании адониса кроется в кон-
серватизме его биоморфологических особенностей. Растение развивает
глубокую (до 2,5 м) корневую систему и требует богатой карбонатами
глубокой хорошо аэрируемой и теплой почвы с достаточным количеством
влаги, хотя бы в нижнем горизонте. Именно такие условия растение на-
ходит в лесостепи, где и следует создавать его плантации. Данные о-
строении корневой системы позволяют рассчитать наиболее приемлемые
размеры междурядий в посевах адониса.
Материалы по корневым системам и биологии надземных органов
могут быть использованы при освоении в культуре и других лекарствен-
ных растений лесостепи: чабреца (Thymus marschallianus), шалфея
(Salvia pratensis), валерианы (Valeriana rossica) и многих других.
Данные о времени цветения и плодоношения, помещенные в прило-
жениях очень важны для определения конкретных сроков сбора лекарст-
венного сырья в лесостепной зоне по отдельным видам: ландыша (Соп-
vallaria majalis), горчака перечного (Polygonum hydropiper), горицвета»
малины (Rubus'idaeus), земляники (Fragaria vesca), зверобоя (Hype-
ricum perforatum), чабреца, шалфея, валерианы, первоцвета (Primula
veris), чернокореня (Cynoglossum officinale) и др. Указанные фенодан-
ные представляют интерес для планирования .времени заготовки плодов
таких растений, как груша (Pyrus communis), вишня (Cerasus frutico-
sa), яблоня (Malus ргаесох), шиповник (Rosa canina), терн (Prunuz
spinosa), рябина (Sorbus aucuparia), черемуха (Padus racemosa), степ-
ная клубника (Fragaria viridis) и др.
4. Результаты ризологических исследований с успехом можно ис-
пользовать в правильном подборе растений для борьбы с эрозией почв»
залужения склонов действующих оврагов. Задача сводится к тому, что-
бы выделить растения, обладающие разветвленной и глубокой корневой
системой, способные на большую глубину укреплять почву, и растения
с относительно мелкой, но интенсивной подземной частью, развивающие
густой поверхностный дерн. Интересно сочетать указанные признаки
с хорошей поедаемостью надземных частей этих растений. В результате
будет достигнуто прекращение эрозии и превращение неудобной земли
в продуктивное угодие. В нашей работе можно найти большой выбор
подобных растений, удовлетворяющих задачам залужения различных
эродированных земель лесостепи (различных по экологическим особенно-
стям), которые необходимо учитывать при подборе тех или иных видов.
Для залужения южных склонов можно рекомендовать следующие
растения: а) Эспарцет песчаный (Onobrychis arenaria) —растение с глу-
бокой сильно разветвленной стержневой корневой системой, вегетатив-
но неподвижное. В надземной части дает высокий урожай питательного
белкового корма; б) Вязель разноцветный (Coronilla varia) — глубоко-
корневое растение, способное быстро разрастаться с помощью корневых
отпрысков. Сено с примесью вязеля поедается всеми видами сельско-
хозяйственных животных; в) Хатьма тюрингенская (Lavatera thuringia-
са) — высокорослое растение с исключительно разветвленной мощной
и глубокой корневой системой; кормовой ценности не имеет; г) Костер
безостый (Bromus inermis)—длиннокорневищное глубококорневое рас-
тение, пронизывающее почву, помимо основных, многочисленными ветви-
стыми боковыми корнями; дает обильный урожай высококачественного
сена. Из мелкокорневых лучше всего подойдут типчак (Festuca sulca-
ta), мятлик узколистный (Роа angustifolia) и др.
254
Перечисленные виды хорошо возобновляются семенами и вполне при-
годны для практического использования без какого-либо предвари-
тельного улучшения породных качеств.
Bromus inermis, Coronilla varia, Onobrychis arenaria, Poa angustifo-
lia прекрасно чувствуют себя и на склонах северной экспозиции.
5. Обобщение ритмики цветения сообществ в виде кривых цветения
с параллельным изучением динамики урожайности ценозов (Голубев,
1963) позволило установить, что сроки максимума цветущих видов сов-
падают с временем наивысшего урожая надземной массы. На основа-
нии графика фенологических констант цветения растительности луговой
степи (см. гл. VI) максимум цветущих видов приходится в среднем на
вторую декаду июня. Этот срок и является временем наибольшей уро-
жайности луговой степи. Таким образом, мы имеем теперь весьма объ-
ективный критерий для определения рациональных сроков сенокошения
травянистых угодий в любой по метеорологическим условиям год, внося
соответствующие коррективы на основе фенологических данных во
влажные и сухие годы. Формирование максимального урожая других ти-
пов сенокосов, отличающихся от луговой степи, происходит, вероятно,
в другие сроки, которые легко установить путем организации фенологи-
ческих наблюдений и выведения кривых цветения.
Мы далеки от мысли, что установленная закономерность исключает
потребность дальнейшего обоснования наиболее экономически выгодных
сроков сеноуборки. Напротив, представляется весьма желательным про-
ведение многолетних опытов со скашиванием в момент максимального
урожая и изучением его влияния на продуктивность угодий в последую-
щие годы, а также анализов питательной ценности травостоя в разные
сроки.
6. Большой, почти неиспользованный резерв природной флоры лесо-
степи заключается в широких возможностях применения многих расте-
ний для декоративного оформления садов, парков, скверов, газонов го-
родов и поселков. Опыт участия Центрально-Черноземного заповедни-
ка в выставках цветов и плодов в г. Курске в 1961 и 1962 г. показывает,
что дикие растения лесостепи пользуются у посетителей неизменным
успехом.
Высокими декоративными качествами выделяются такие растения,
как незабудка степная {Myosotis suaveolens), девясил шершавый {Inula
hirta). нивяник обыкновенный {Leucanthemum vulgare). лен желтый
{Linum flavum). лен украинский {Linum ucrainicum). проломник Козо-
Полянского {Androsace kozo-poljanskii), пиретрум щитковый {Pyrethrum
corymbosum). живокость клиновидная {Delphinium cuneatum). дрок
красильный {Genista tinctoria). крестовник крупнолистный {Senecio
schwetzowii). качим метельчатый {Gypsophila paniculata). типчак (Fes-
tuca sulcata) и др.
Перечисленным, конечно, далеко не исчерпываются возможности ис-
пользования природных растений лесостепи в декоративных целях.
Наша задача — обратить внимание практических работников зелентре-
стов и любителей-садоводов на этот животворный источник оригиналь-
ных интересных растений. Следует отметить, что в культуре многие из
указанных видов заметно увеличивают размеры цветков, отчего их деко-
ративность еще более повышается.
7. Изучение биоморфологии растений, характера вегетации, отмира-
ния органов, способов перезимовки и длительности жизни перезимовав-
ших листьев дало возможность разработать новый метод определения
абсолютной продуктивности травянистых цензов и синузий в надземной
части (Голубев, 1963). Абсолютная продуктивность травянистого расте-
ния измеряется той растительной массой, которая продуцируется в тече-
ние всего вегетационного периода. Для ее определения надо учесть массу
265
всех развившихся у растения листьев, стеблей, цветоносных побегов,
цветков и плодов с семенами. Абсолютная продуктивность надземной
части травянистых ценозов и синузий представляет сумму абсолютной
продуктивности особей всех видов, входящих в их состав, отнесенную
к единице площади.
В основе предложенного метода лежит учет естественно отмирающих
•органов травянистых растений, т. е. листьев, стеблей, венчиков цветков
и т. д., а также созревающих и отделяющихся оорганов — плодов, семян,
вивипарных луковичек и пр. Сбор и учет этих органов производится на
пробных площадках в течение всего вегетационного периода, начиная
с момента схода снега и кончая временем установления устойчивых отри-
цательных температур, когда ростовые процессы заведомо прекращают-
ся. Пробные площадки закладываются осенью предыдущего года, на них
перед установлением снежного покрова собираются и удаляются все
отмершие листья и побеги, но так, чтобы не повредить слой полуразло-
жившейся подстилки и основания почек перезимовки, особенно у дерно-
винных злаков. В ранневесенний учет поэтому попадают только органы,
отмершие у зимнезеленых растений в морозный период. Все перезимо-
вавшие в зеленом виде органы отмирают, как правило, в первую полови-
ну вегетационного периода, очень редко отдельные растения сохраняют
их несколько дольше, но не позднее, чем установится новый снеговой
покров. Эта органическая масса компенсирует ее недобор в репродукции
текущего года, измеряемый массой органов, уходящих под зиму в зеле-
ном состоянии. Таким образом, мерилом абсолютной продуктивности яв-
ляется вся отмирающая за вегетационный период масса надземных,
а также созревающих и естественно отделяющихся органов (плодов, се-
мян и т. д.), которая в сумме соответствует всей формирующейся в дан-
ном году массе живых надземных частей.
Чем чаще производится сбор отмирающих органов на учетных пло-
щадках, тем точнее будут результаты, так как исключается убыль орга-
нического вещества вследствие его разложения. Кроме того, оказывается
возможным учесть массу всех созревающих плодов и семян.
Использование метода основано на том, что сбор на пробных пло-
щадках отмирающих органов не нарушает Существенно ход нарастания
надземной массы на протяжении всего теплого времени года, обусловли-
вающего создание полного урожая. Метод допускает изучение абсолют-
ной продуктивности травянистых цензов и синузий (луговых, болотных,
луговостепных и подобных им сообществ, травянистого покрова в лесах)
и непригоден для ее выявления в ценозах с господством нетравянистых
жизненных форм: полукустарничков, кустарников и т. п.
Определения, проведенные данным методом показывают, что абсо-
лютная продуктивность надземной части луговой степи заметно превос-
ходит наибольшее значение растительной массы в серии последователь-
ных укосов в течение вегетационного периода. Можно с достаточной
вероятностью принять, что абсолютная продуктивность надземной
части Стрелецкой степи колеблется в разные годы от 5 до 10 т воздуш-
но-сухой массы на га.
Абсолютная продуктивность ценозов представляет биологическую
основу создания хозяйственной продуктивности и с этой точки зрения
определение ее в разных сообществах имеет непосредственное практиче-
ское значение. Правильная эксплуатация природных растительных бо^
гатств (в луговодстве, пастбищеводстве, лесоводстве) должна основы-
ваться на знании абсолютной продуктивности растительных угодий.
Следует отметить, что до последнего времени методы определения
абсолютной продуктивности надземной части травяного покрова отсут-
ствовали совершенно. Лишь недавно опубликована работа В. Б. Сочавы,
В. В. Липатовой, А. А. Горшковой (1962), в ней предложен метод «моде-
256
лей» для определения полной продуктивности, анализ и оценка которого
даны нами раньше (Голубев, 1963).
8. Коренной вопрос о природе и происхождении луговых степей и ра-
стительности лесостепного ландшафта вообще имеет прямое отношение
к исключительно важной экономической проблеме природно-сельскохо-
зяйственного районирования территории нашей страны, заключающей
природно-сельскохозяйственную оценку земель, определение экономиче-
ского профиля хозяйств и географическое районирование сельскохозяй-
ственных растений и животных. Для ведения сельского хозяйства в лесо-
степной зоне далеко не безразлично, принимать ли луговые степи за
луговой тип растительности или же рассматривать их в качестве ориги-
нального, самобытного, аутохтонно развившегося типа, весьма полно
гармонирующего с многообразными природными особенностями лесо-
степного ландшафта, в конечном итоге являющегося их производным.
Значительная, а вероятно, и большая часть пахотных земель в лесостепи
расположена на месте первобытных луговых степей. Поэтому весь строй
изучения эколого-биологических и исторических закономерностей разви-
тия растительности луговых степей теснейшим образом переплетается
с вопросами практического значения — экологического подбора сельско-
хозяйственных культур, производства максимального урожая и факто-
ров, его обусловливающих. Некоторые затронутые вопросы уже рассмат-
ривались выше. Здесь же следует подчеркнуть, что представления о
самобытности растительности лесостепи и определяющих ее внешних
условий призваны акцентировать внимание практических работников на
поиски конкретных путей обеспечения наивысшей экономической произ-
водительности земель лесостепи, в соответствии с местными природными
условиями, изгоняя шаблон, а возможно, и заменяя и модернизируя
приемы и методы выращивания сельскохозяйственных культур, апроби-
рованные и разработанные в условиях лесной зоны.
Сообщаемые в работе сведения о природных особенностях лесостепи,
эколого-биологических признаках и свойствах растительности, ее приро-
де и происхождении, на наш взгляд, могут оказать существенную по-
мощь практическим работникам в решении указанных экономических
задач сельского хозяйства в зоне лесостепи.
Заканчивая обзор возможных аспектов практического применения ре-
зультатов биоморфологических исследований в средней лесостепи, сле-
дует отметить, что они создают предпосылки к раскрытию фитоценотиче-
ской роли видов и их средообразующего значения. В данных условиях
внешней среды эколого-биологические особенности растений во многом
определяют их взаимоотношения в сообществе. Здесь мы вступаем в об-
ласть, характеризующую основную проблему современной фитоценоло-
гии и перспективы ее развития, прикладное значение которых трудно
переоценить.
17 В. Н. Голубев
Ритмы цветения некоторых растений Стрелецкой степи Центрально-Черноземного
Вид Начало цветения Массовое
самое раннее самое позднее сумма дней * и количество лет наблюде- ний средняя многолет- няя самое раннее
Achillea millefolium . . . 25.V-39 7.VII—41 277 (6) 15.VI 10.VI—39
Adonis vernalis 8. IV—61 26.V—46 295 (13) 23. IV 24. IV—61
A grору ron intermedium 25.VI—46 10.VII—45 159(5) 2.VII 30.VI—46
Anemone silvestris .... 8.V—50 23.V—58 205 (13) 15.V 12.V—41
Anthericum ramosum . . 15.VI—48 10.VII—45 308(11) 28.VI 20.VI—48
Anthyllis polyphyIla . . 4.VI—61 20.VI—45 53 (5) 11.VI 6.VI—46
Bromus inermis 5.VI—39 5.VII—45 288(12) 24. VI 20.VI—49
B. riparius 26.V—49 27.VI—41,55 632 (15) 11.VI 6.VI—60
Campanula persicifolia 7.VI—50 26.VI—45 179 (11) 16.VI 17.VI—59
C. sibirica 27. V—50,57 14.VI—45 406(H) 5.VI 5.VI—39
Carex humilis 15.IV—50,61 3.V—58 183 (8) 23. IV 20. IV—50
Centaurea marschalliana. . 3.V—39 26.V—58 234(14) 16.V 13.V—50
C. scabiosa l.VI—39 17.VII—41 282 (Н) 25.VI 25.VI—48
Clematis recta 6.VI-50 20.VI—45 72(6) 12.VI 14.VI— 50
Delphinium cuneatum . . 26.VI—39 15.VII—45 476 (13) 6.VII 9. VII—61
Echium rub rum . . 26.V—50 14.VI—45 422 (12) 4.VI 3.VI—50
Festuca sulcata 25.V—39 15.VI—47 459 (12) 7.VI 5.VI—39
Filipendula hexapetala . . 25.V—39 20.VI—45 587 (15) 8.VI 5.VI—48
Fragaria viridis .... 8.V—50 27.V—56 199(H) 18.V 18.V—57
Galium verum 25.V—39 7.VII—41 733 (15) 17.VI 18.VI—46
Helictotrichon pubescens 23.V—57 10.VI—45 285 (9) 1.VI 30.V—57
Hyacinthella leucophaea 24.IV—50,61 14.V—45 297 (9) 3» V 30. IV—50
Iris aphylla 8. V—50,59 29.V—45 223 (12) 18.V 18.V—57
Koeleria gracilis .... 30.V—50 16.VI—58 353 (9) 8.VI 5.VI—49
Leucanthemum vulgare . 25.V—39 27.VI—41 546 (14) 8. VI 11.VI—57
Libanotis intermedia . . 12.VI—57 l.VII—55 196 (9) 22.VI 23.VI—56
Linum perenne .... 19.V—57 6.VI—55 364(13) 28.V 25. V—47
Medicago falcata . . . 8.VI—50 И.VII—40 258 (12) 21.VI 21.VI—50
Myosotis suaveolens . . . 5.V—57 24.V—45 142(12) 12.V 13.V—57
Onobrychis arenaria . . 20.V—48 16.VI—58 501 (15) 2.VI 6.VI 46
Orobus pannonicus . . . 28. IV—50 19.V—59 422 (Ю) 12.V 9.V—57
Phleum phleoides 25.V—39 22.VI—40 421 (9) 15.VI 1O.VI—39
Ph. pratense — — 26(1) 26.VI —
Phlomis tuberosa .... 25.V—39 29.VI—55 476 (И) 12.VI 8.VI—56
Poa angustifolia .... 23.V—57 14.VI—45 245 (7) 4.VI 20.V—50
Primula veris 14. IV—61 15.V—56 474 (15) 1.V 28. IV—61
Pulsatilla patens .... 8. IV—61 30. IV—58 220(11) 20. IV 18. IV—61
Salvia nutans 25.V—50 17.VI— 41 483 (13) 6.VI 6.VI—46
S. pratensis 15.V—48 10.VI—46 368 (15) 24.V 25.V—46
Senecio integrifolius . . 10.V—50 l.VI—55 233 (12) 19.V 17.V—50
Stipa pennata 24.V—45 27.VI—41 365 (И) 2.VI 30.V—48
Thymus marschallianus 27.V—50 26.VI—45 439(11) 9.VI 11.VI—57
Tragopogon orientate . . 25.V—46 17.VI— 41 515(16) 1.VI 30.V—46
Tri folium montanum . . 23.V—57 14.VI—45 499 (15) 2.VI 30.V—57
T. pratense 12.V—50 17.VI—41 242 (9) 26.V 31.V—50
Valeriana rossica 16.V—57 31.V—56 295 (12) 25.V 23.V—57
Veratrum nigrum .... l.VI I—60 26. VI I—45 179 (И) 16.VII 15. VII—60
Verbascum lychnitis . . 18.VI—60 4.VII—49 188 (8) 23.VI 29.VI—60
Veronica chamaedrys . . 16.V—61 25.V—39 61 (3) 20.V 24.V—61
V. prost rata 7.V—57 22.V—58 74(5) 15.V 16.V—57
• Помещенные здесь и в следующих приложениях суммы дней по срокам начала, массового и
сяцу за все годы наблюдений, количество которых указано в скобках. Они могут быть использованы
вых материалов фенологических наблюдений.
258
Приложение I
заповедника в многолетнем ряду (по 1961 г. включительно)
цветение Конец цветения
самое позднее сумма дней • и количест- во лет на- блюдений средняя многолет- няя самое раннее самое позднее сумма дней • и количест- во лет на- блюдений средняя многолет- няя
1.VII—50 161(6) 27.VI 4.VII—39 1.IX—61 94 (3) 31.VII
12.V—58 446 (14) 2.V 12.V—60 6.VI—41 365 (14) 24. V
15.VII—45 183 (5) 7.VII 14.VII—49 20.VII—45 65(4) 16.VII
30. V—58,60 308 (14) 22.V 25.V—48 16.VI—56,58 433 (12) 5.VI
25.VII—58 413(11) 8.VII 15.VII—48 8.VIII—58 208 (7) 30. VII
26.VI—45 98 (6) 16.VI 15.VI—60 5. VII— 45 143 (5) 29.VI
12.VII—41 398 (13) 30.VI 26.VI—39 23.VII—59 436 (11) 10.VII
3.VII—50 244 (15) 16.VI 12.VI—49 7.VII—41,50 307 (12) 26.VI
2.VII—45 257 (И) 23.VI 25.VI—46 18.VII—59,61 246 (6) 11.VII
20.VI— 45 154(11) 14.VI 21.VI—56 27.VII—40 403 (11) 6.VII
6.V—58 198(7) 28. IV 23. IV—50 11.V—58 216 (7) 1.V
9.VI—59 374 (15) 25.V 19.V—50 20. VI—61 630 (14) 14.VI
15.VII—45 339 (9) 8.VII 20.VII— 48 10.VIII—45 230 (8) 27.VII
29.VI—55 136 (7) 19. VI 21.VI—50 15.VII—55 195 (6) 3.VII
21.VII—49 206 (13) 16.VII 25. VII— 48 31. VIII—58 294 (8) 6.VIII
20.VI—45,55 159 (13) 12.VI 18.VI— 46 11.VII—57 284 (10) 28.VI
20.VI—47 152 (И) 14.VI 10.VI—49 23.VI—61 79 (5) 16.VI
26.VI— 45 187 (13) 14.VI 15.VI—48 6.VII—58 374 (14) 27.VI
5.VI—45 375 (14) 27.V 1.VI—50 21.VI—58 129 (11) 12.VI
5. VII—45,46 346 (13) 27.VI 4.VII—39 30. VII—57 177 (И) 16.VII
14.VI—45 307 (8) 7.VI 9.VI—49 26.VI—45 68 (5) 14.VI
19.V—45 428(11) 9.V 10.V—50 29.V—56 204 (10) 20.V
5.VI—45 331 (13) 25.V 26.V—50 10. VI—45,61 469(13) 5.VI
20.VI—45 110 (9) 12.VI 15.VI—49 2.VII—45 134 (6) 22.VI
5. VI1—45 237 (13) 18.VI 18.VI—46 26.VII—61 523 (13) 11VII
10.VII—45 344(11) 31.VI 14.VII—49 1.VIII—45 125 (6) 21.VII
19 VI—58 473 (13) 5.VI 13.VI—46 18. VII—61 276 (10) 28.VI
20. VII— 40 426 (13) 2.VII 25.VII—39 5.VIII—45 162 (5) 1.VIII
25.V—56 285 (13) 22.V 25.V—49 20.VI—61 441 (И) 9.VI
29.VI—58 236(15) 16.VI 26.VI—39 26. VII—45 339 (9) 8.VII
24.V—58 205 (И) 19.V 16.V—50 1.VII—55 364 (И) 2.VI
1.VII—55 178 (8) 22.VI 25.VI—61 6.VII-55 180 (6) 30.VI
61(2) 31.VI — — 10(1) 10.VII
4.VII—49 239 (10) 24.VI 4.VII—39 10.VIII—45 76 (5) 15.VII
29.VI—57 399 (10) 9.VI 9.VI—49 26.VI—45 90 (5) 18.VI
19. V—41,56 569 (14) 11.V 20. V—39,55 8.VI—56 393 (15) 26.V
6.V—58 368 (14) 26.IV 28. IV—50 22.V—41,56 564 (14) 10.V
20.VI—45 178 (13) 14.VI 18.VI—46 11.VII—57 371 (13) 31.VI
15.VI—41 519 (15) 4.VI 9.VI—49 2.VII—41 270 (13) 21.VI
5.VI—45 313 (12) 26.V 30.V—46 16.VI—56 450 (12) 7.VI
17.VI—55 332 (9) 6.VI 5.VI—39,48 22.VI—55 113(9) 13.VI
2.VII—45 182 (10) 18.VI 18.VI—60 20. VII—57 360 (9) 10.VII
22.VI—41 543 (14) 8.VI 10.VI—48 5.VII—61 293 (14) 21.VI
28.VI—41 692 (16) 12.VI 20.VI—48,49 17.VII—41 482 (15) 2.VII
16.VI—58 285(7) 10.VI 28.VI—49 5.VII—59 96 (3) 1 .VII
7.VI—56 422 (13) 1.VI 6.VI—46 24.VI—57 177 (12) 15.VI
1.VIII—45,60 270(11) 25.VII 28. VII—60 23.VIII—61 322 (8) 9.VIII
13.VII—58 279 (8) 5.VII 14.VII—49 19.VIII—58 166 (5) 2.VIII
14.VI—60 156 (5) 31 .V 10.VI—39 20.VI-61 75 (5) 15.VI
26.V—58 112(5) 22.V 30.V—57 12.VI—56 179 (5) 5.VI
конца цветения получены в результате приведения конкретных дат к одному соответствующему ме-
для уточнения средних многолетних показателей ритма цветения по мере накопления в будущем но-
17*
259
Ритмы цветения некоторых растений Казацкой степи Центрально-Черноземного
Вид Начало цветения Массовое
самое раннее самое позднее сумма дней и количество лет наблюде- ний средняя многолет- няя самое раннее
Achillea millefolium • . 16.V—37 22.VI—52 270 (7) 8.VI 12.VI— 36
Adonis vernalis 9. IV—51 27. IV—52 96 (5) 19. IV 27. IV—50
Agropyron intermedium . 23. VI—53 29.VI—51 105 (4) 26.VI 28.VI—53
Alopecurus pratensis . 6.V—37 30.V—52 101 (5) 20.V 16.V—37
Anemone silvestris . . . 6.V—37 26.V—52 92 (7) 13.V 11.V—37
Anthericum ramosum . . 23.VI—51 29.VI—53 77(3) 26.VI 30. VI—54
Bromus inermis .... 12.VI— 36 28.VI—52 206 (4> 20.VI 24.VI—36
B. riparius 6.VI—53 27.VI—50 61 (4) 15.VI 10.VI—53
Campanula persicifolia . . 7.VI—50 26.VI—52 85 (5) 17.VI 20.VI—53
C. sibirica 18.V—36 12.VI— 52 205 (6) 3.VI 6.VI—36
Carex humilis 15. IV—50 27.IV-54 62 (3) 21.VI 20. IV—51
Centaurea marschalliana . 2.V—37 27 V—52 82 (7) 12.V 12.V—50
C. scabiosa 12.VI—36 18.VII—52 150 (5) 30.VI 4.VII—54
Clematis recta . . 6.VI-50 2.VII—52 90(5) 18.VI 14.VI—50
Delphinium cuneatum 20.VI—37 И. VI1—52 180(6) 30.VI 4.VII— 54
Echium rub rum . 21.V-37 13. VI-52 229(7) 2.VI 31.V—37
Festuca sulcata . . 16.V-37 14.VI—52 239 (7) 3.VI 31.V—37
Filipendula hexapetala . . 16.V—37 14.VI— 52 209 (6) 4.VI 31.V—37
Fragaria viridis . 7.V—51 25.V—52 96 (6) 16.V 16.V—37
Galium verum . . . . ’ 31.V—37 29.VI—52 279 (6) 15.VI 18. VI—36
Helictotrichon pubescens 25.V—50 10.VI-54 176(5) 4.VI 7.VI—51
Hyacinthella leucophaea . 22. IV—51 6.V—53 148(5) 30. IV 30. IV—50
Iris aphylla 8.V—51 28.V—52 120 (7) 17.VI 17.V-50
Koeleria gracilis .... 30.V-50 12.VI—52 191 (5) 7.VI 10.VI—50
Leucanthemum vulgare . . 28.V—50 9.VI—52 216 (6) 6.VI 12.VI—36
Libanotis intermedia . . 22.VI—51 7.VI—52 119(5) 24.VI 30.VI—54
Linum perenne 21.V—51 8.VI—52 144 (5) 29.V 25.V—51
Medicago falcata 5.VI—37 22.VI—54 92 (7) 13.VI 18.VI—36
Myosotis suaveolens . 6.V—50 29.V—52 81 (4) 20.V 10.V—51
Onobrychis arenaria 16.V-37 13.VI—52 203 (7) 29.V 26.V-37
Orobus pannonicus . . . 28. IV—50 19.V—53 190 (5) 8.V 9.V—50
Phleum phleoides 26.V—37 22.VI—52 263 (6) 13.VI 10.VI—37
Ph. pratense 17.IV—54 7. VII—52 101 (4) 25.VI 23.VI—54
Phlomis tuberosa * 16.V—37 20.VI—52 224 (6) 6.VI 6.VI—36
Poa angustifolia 3.VI-51 10.VI—52 19(3) 6.VI 20.V-50
Primula veris 7. IV—51 9.V—54 170 (6) 28.IV 30. IV—50
Pulsatilla patens 9. IV—51 30. IV—52 101 (5) 20. IV 22.IV—51
Salvia nutans 16.V-37 13.VI—52 227 (7) 1.VI 31.V—37
S. pratensis 15.V—50 ЗОЛ—52 152 (7) 22.V 21.V—37
Senecio integrifolius 2.V—37 29.V—52 105 (7) 15.V 15.V—50
Stipa pennata 18.V—36 10.VI—50 207 (7) 29.V 23.V—37
Thymus marschallianus . 27.V—50 14.VI—51 189’(5) 7.VI 15.VI—54
Tragopogon orientate . . 16.V—37 8.VI—52 208 (7) 29.V 30.V—36
Tri folium montanum . . 16.V—37 13.VI—52 196 (7) 28.V 26.V—37
T. pratense 12.V—50 7.VI-54 153 (7) 22.V 24.V—36
Valeriana rossica 19.V—50 3.VI—52 133(5) 26.V 29.V—50
Veratrum nigrum 20.VI—37 26. VII—52 235 (6) 9.VII 18. VII— 36
Verbascum lychnitis . . 18. VI—36 1.VII—52 172 (7) 25.VI 25.VI—37
Veronica chamaedrys. . . 11. V—37 •22.V—52 45(3) 15.V 16.V-37
260
Приложение if
заповедника в многолетнем ряду (по наблюдениям в 1936—1937 и 1950—1954 гг.)
цветение Конец цветения
самое позднее сумма дней и количество лет наблюде- ний средняя многолетняя самое раннее самое позднее сумма дней и количество лет наблюде- ний средняя многолет- няя
12.VII—52 171 (7) 24.VI 24. VII—36 27.Х—53 210 (4) 21.IX
15.V—53 245 (7) 5.V 11.V—37 4.VI—52 134(7) 19.V
6.VII—52 128 (4) 2.VII 8.VII—54 15.VII—53 46 (4) 12.VII
6.VI—52 89 (3) • 29.V 25.V—50 18.VI—53 193(5) 8. VI
4.VI—52 150 (7) 21. V 26.V—37 16.VI—54 238 (7) 3.VI
6.VII—51 66 (2) 3.VII 30.VII—51 8.VIII—54 103(3) 3.VIII
5.VII—52 137 (5) 27.VI 29.VI—53 10. VII—52 168(5) 4.VII
3.VII—50 83(4) 21.V1 19.VI—53 7.VII—50 105 (4) 26.VI
3.VII—52 98(4) 25.VI 30.V1—54 18. VII—52 148 (4) 7.VII
20.VI—52 93(6) 15.VI 24.VI-36 16.VI—52 215(6) 6.VII
2.V—54 130(5) 26. IV 23. IV—50 6.V—36 186 (6) 1.V
8.VI—52 159 (7) 23.V 5.V1—37 27.VI—52 105 (7) 15.VI
27.VII—52 50(5) 10.VI1 18.VII—36 12.VIII— 53 117(4) 29.VII
23.VI—53 56(3) 19. VI 21. VI—50 15. VII—52 169 (5) 4.VII
25. VII—52 64(6) 11.VI1 24. VII—36 18. VIII—52 208 (6) 5.VIII
23.VI—52 285 (7) 10.VI / 14.VI—51 7.VII—54 189(7) 27.VI
17.VI—52 292 (7) 11.VI 12.VI—36 20.VI—52 117(7) 17.VI
20.V1—52 263 (6) 13.VI 18.VI—36 5.VII—52 149(6) 25.VI
5.V1—52 162 (6) 27.V 30.V—36 23.VI—52 288 (7) 10.VI
15.VII—52 167 (6) 28.VI 30. VI—36 25.VII—52 267 (6) 15.VII
16.VI—54 47(4) 12.VI 2.VI—50 20.V1—54 62 (5) 12.VI
20.V—52 195 (5) 9.V 10.V—50 31.V—52 104 (5) 21.V
3.VI—52 173 (7) 25.V 26.V-50 16.VI—52 241 (7) 3.VI
18.VI—52 68(5) 14.VI 16.VI—50 27.VI—52 100 (5) 20.VI
23.VI—52 99(6) 16.VI 24.VI—36 20.VI1—52 225 (6) 6. VII
25.VII—52 189 (5) 8.VII 23. VII—51 18. VIII—50 188 (5) 7.VIII
13.VI—52 187 (5) 6.VI 20.VI—53 7.VI1—51 149 (5) 30.VI
6. VII—52 187 (7) 27.VI 24.VI—36 28. IX—52 322 (4) 19.VIII
4.VI—52 92(4) 23.V 27.V—50 18.VI—53 205 (5) 10.VI
1.VII—52 315 (7) 14.VI 30.VI—37 26. IX—53 308 (4) 16.VII
27.V—53 89 (5) 18.V 16.V—50 10. VI—54 142(5) 28.V
30. VI—52 121 (6) 20.VI 22.VI—53 4.VII—52 160(6) 27.VI
14.VII—52 129 (4) 2.VII 28.VI—53 22. VII—52 155 (4) 8.VII
3. VII—52 109 (6) 18.VI 24.VI—36 20. VII—52 244 (6) 11.V1I
15.VI—52 193 (5) 7.VI 14.VI—51 22.VI—52 71 (4) 18.VI
20.V—52 292 (7) 12.V 11.V—37 1.VI—52 157 (7) 22.V
10.V—52 113(4) 28. IV 28. IV—50 15.V—53 246 (7) 5.V
20.VI—62 299 (7) 12.VI 12.VI—36 12.VII—52 196(7) 28.VI
15.VI—52 236 (7) 3.VI 6.VI-36 2.VII—51 144 (7) 21. VI
2.VI—52 170 (7) 24.V 27.V—51 10.VI—52 229 (7) 2.VI
8.VI—52 121 (4) 30.V 30.V—36 12.VI—53 232 (6) 8.VI
22.VI—51 74(4) 19.VI 20.VI—54 20. VII—52 146 (4) 6.VII
12.VI—52 219 (6) 5.VI 10.VI—51 24.VI—52 105 (7) . 15.VI
20.VI—52 285 (7) 9.VI 20.VI—37 25. VII—52 225 (7) 2.VII
15. VI—52 256 (7) 6.VI 12.VI—36 З.Х—52 162 (3) 24.VII
12.VI—52 185 (5) 6.VI 14 .VI—50 18.VI—53 78 (5) 16.VI
30.VII—54 97(4) 24.VII 30.VII—36 18.VIII—52 189 (5) 8.VIII
13.VII—52 249 (7) 5.VII 25.VII—37 15. VIII—50 260 (7) 6.VIIL
1.VI-52 66(3) 22.V 26.V—37 10.VI—52 97 (3) 1.VI
26J
Приложение III
Средние многолетние даты начала и полного созревания плодов и семян некоторых растений
Стрелецкой степи Центрально-Черноземного заповедника (по 1962 г. включительно)
Вид Плодоношение
начало массовое
сумма дней и коли- чество лет наблюде- ний средняя многолетняя сумма дней и количество лет наблюде- ний средняя многолетняя
Achillea millefolium 146 (4) 5.VII 50 (2) 25.VIII
Adonis vernalis 174 (10) 17.VI 302 (И) 27.VI
Agropyron intermedium 196 (5) 8.VIII 73 (3) 24.VIII
Alopecurus pratensis 121 (4) 30.VI 110(3) 7.VII
Anemone silvestris 234 (11) 21.VI 367 (И) 3.VII
Anthericum ramosum 134 (6) 22.VIII 58 (2) 29.VIII
Anthyllis polyphyIla 245 (6) 11.VII 97 (5) 19.VII
Bromus inermis 294 (И) 27.VII 173 (5) 4.VIII
B. riparius 478 (12) 10.VII 132 (8) 16.VII
Campanula persicifolia 160 (6) 27.VII 55 (2) 27.VII
C. sibirica 205 (8) 26. VII 109 (4) 27.VII
Carex humilis 69 (3) 23.V 87 (4) 29.V
Centaurea marschalliana 188 (9) 21.VI 231 (9) 26.VI
Centaurea scabiosa 286 (8) 5.VIII 42 (4) 10.VIII
Clematis recta 84(5) 17.VIII 51 (2) 25.VIII
Delphinium cuneatum 270 (7) 7.VIII 29 (2) 14.VIII
Echium rubrum 436 (9) 18.VII 95 (3) 1.VIII
Festuca sulcata 303 (9) 4.VII 241 (6) 10.VII
Filipendula hexapetala 270 (15) 18.VII 259 (9) 29.VII
Fragaria viridis * 304 (9) 4.VII 284 (7) 11.VII
Galium verum 296 (8) 6.VIII 50 (3) 17.VIII
Helictotrichon pubescens 143 (6) 24.VI 245 (8) 1.VH
Hyacinthella leucophaea 296 (7) 11.VI 157 (7) 22.VI
Iris aphylla 468 (10) 17.VII 223 (7) 1.VIII
Koeleria gracilis 243 (7) 5.VII 47 (4) 12.VII
Leucanthemum vulgare 240 (И) 22.VII 210 (7) 30. VII
Libanotis intermedia 251 (7) 5.VIII 67 (5) 13.VIII
Linum perenne 411(10) 11.VII 193 (9) 21.VII
Medicago falcata 37 (5) 7.VIII 45 (2) 22.VIII
Myosotis suaveolens 182 (10) 18.VI 203 (8) 25.VI
Onobrychis arenaria 240 (И) 22.VII . 224 (7) 1.VIII
Orobus pannonicus 243 (9) 27.VI 217(6) 6.VII
Phleum phleoides 81 (4) 20. VII 123 (5) 24.VII
Ph. pratense 88 (3) 29. VII 30(2) 15.VIII
Phlomis tuberosa 172 (4) 12.VIII 37 (2) 18.VIII
Poa angustifolia 288 (9) 2.VII 289 (7) 11.VII
Primula veris 837 (12) 9.VII 120 (9) 13.VII
Pulsatilla patens 348 (И) 1.VI 390 (10) 8.VI
Salvia nutans 472 (12) 9.VII 149 (8) 19.VII
S. pratensis 444 (15) 30.VI 342 (9) 8.VII
Senecio integrifolius 132 (10) 13.VI 231 (9) 26.VI
Stipa pennata 368(15) 25.VI 253 (9) 28.VI
Thymus marschallianus 167 (6) 28.VII 91 (3) 30.VII
Tragopogon orientate \ 412 (14) 29.VI 306 (9) 4.VII
Trifolium montanum 252 (13) 19.VII 204 (8) 25.VII
T. pratense 206 (5) И.VII 44 (3) 15.VII
Valeriana rossica 306 (13) 23.VI 417(12) 5.VII
Veratrum nigrum 20 (1) 20.VIII — —
Verbascum lychnitis 243 (6) 9.VIII 45 (2) 22.VIII
Veronica chamaedrys 54 (2) 27.VI 118(3) 9.VII
V. prostrata 46 (2) 23.VI —
262
Пр и л о ж с н и в IV
Средние многолетние даты начала и полного созревания плодов и семян некоторых
растений Казацкой степи Центрально-Черноземного заповедника
(по наблюдениям в 1936—1937 и 1950—1954 гг.)
Плодоношение
Вид начало массовое
сумма дней и коли* чество лет наблюде* НИЙ средняя многолетняя сумма дней н количество лет наблюде- ний средняя многолетняя
Achillea millefolium 302 (7) . 13.VIII 115 (3) 7.IX
Adonis vernalis 85 (6) 14.VI 223 (7) 2.VII
Agropyron intermedium 38(3) 13.IX 80(3) 27.IX
Alopecurus pratensis 158 (5) 1.VII 86(5) 17.VII
Anemone silvestris 127 (5) 25.VI 233 (6) 9.VI
Anthericum ramosum ........ 46(3) 6.VIII 41 (2) 20.1 X
Bromus inermis 225 (6) 7.VIII 130 (4) 1.IX
B. riparius 71 (5) 14.VII 126 (4) 1.VIII
Campanula persicifolia 127 (5) 25.VII 59(4) 15.VIII
C. sibirica 129 (5) 26.VII 231 (5) 15.VIII
Carex humilis 127 (5) 25.V 169 (5) 3.VI
Centaurea marschalliana 64 (4) 16.VI 130 (5) 26.VI
C. scabiosa 117 (4) 29.VII 84(4) 21.VIII
Clematis recta 155 (4) 8.VIII 122 (4) 30.VIII
Delphinium cuneatum 262 (6) 13.VIII 156 (5) 31.VIII
Echium rub rum 320 (7) 16.VII 272 (6) 14. VIII
Festuca sulcata 223 (7) 2.VII 131 (7) 19.VII
Filipendula hexapetala 133 (5) 26.VII 226 (6) 6.VIII
Fragaria viridis 221 (6) 7.VII 137 (7) 19.VII
Galium verum 192 (5) 8.VIII 233 (5) 15.IX
Helictotrichon pubescens 86 (4) 21.VI 173 (5) 4.VII
Hyacinthella leucophaea 133 (5) 27.V 143(5) 29.VI
Iris aphylla 116(5) 23.VII 184 (6) 30.VII
Koeleria gracilis 183 (5) 6.VII 35 (4) 9.VIII
Leucanthemum vulgare 86 (5) 17.VII 132 (5) 26.VII
Libanotis intermedia 196 (5) 8.VIII 148 (4) 6.IX
Linum perenne 89 (5) 18.VII 158 (5) 31.VII
Medic ago falcata 251 (7) 5.VIII 381 (5) 15.IX
Myosotis suaveolens 149 (5) 30.VI 65(2) 12.VII
Onobrychis arenaria 206 (6) 3.VIII 39(4) 9.IX
Orobus pannonicus 99 (4) 25.VI 88(3) 29.VI
Phleum phleoides 95 (6) 16.VII 289 (6) 17.VIII
Ph. pratense 20 (4) 5.VIII 129 (4) 1.1X
Phlomis tuberosa 166 (6) 28. VII 253 (6) И.VIII
Poa angustifolia 60 (5) 12.VII 196 (5) 8.VIII
Primula veris 94 (5) 19.VII 179 (5) 29.VII
Pulsatilla patens 198 (7) 28.V 99(6) 16.VI
Salvia nutans 211(6) 5.VII 214 (6) 5.VIII
S. pratensis 214 (7) 1.VII 148 (7) 21.VII
Senecio integrifolius ........... 85 (6) 155 (7) 14.Vb 22.VI 105 (5) 224 (7) 21.VI
Stipa pennata 2.VII
Thymus marschallianus 210(5) 12.VII 188 (5) 7.VIII
Tragopogon orientale 128 (6) 21.VI 168 (6) 28.VI
Tri folium montanum 122 (6) 20.VII 97(4) 24.VIII
T. pratense 83 (6) 14.VII 75 (3) 25.VIII
Valeriana rossica 107 (5) 21.VI 162 (5) 2.VII
Verbascum lychnitis 137 (5) 27.VII 59 (3) 19.IX
Veronica chamaedrys 134 (3) 13.VII 93 (3) 31.VII
Приложение V
Ритмы цветения древесно-кустарниковых пород Стрелецкого участка Центрально-Черноземного заповедника в многолетнем ряду
(по 1961 г. включительно)
Вид Начало цветения Массовое цветение Конец цветения
самое раннее самое позднее сумма дней и ко- личество лет наблюдений средняя много- летняя самое раннее самое позднее сумма дней и ко- личество лет наблюдений средняя много- летняя самое раннее самое позднее сумма дней и ко- личество лет наблюдений средняя много- летняя
Acer platanoides . . . 24. IV—50 12.V—53 190 (6) 2.V 27. IV—50 10.V—54 166 (5) 3.V 30.IV—50 22.V—53 203 (5) 11.V
А. tataricum 28.V—53 7.VI—51 128 (4) 1.VI 6.VI—53 8.VI—52 14(2) 7.VI 12.VI—54 16. VI—51 - 56 (4) 14.VI
Amygdalus папа . . . 28. IV—51 17.V—47 200 (6) 3.V 4.V—51 25.V—47 55 (4) 14.V 8.V—51 28.V—61 74(4) 19.V
Cerasus fruticosa . . . 3.V—51 4.VI—41 205 (14) 15.V 9.V—50 8.VI—41 174 (9) 19.V 11.V—50 16.VI—41 363 (14) 26.V
Corylus avellana .... 4. IV—51 20. IV—54 101 (8) 13.IV И. IV—48 23. IV—54 126 (7) 18.IV 9. IV—51 4.V-47 213 (9) 24. IV
Crataegus curvisepala . . 20.V—60 2.VI—56 214 (8) 27.V 17.V—57 6.VI—54 191 (6) 1.VI 30.V—57 12.VI—61 309 (8) 8.VI
Euonymus europaea . . 9.V—51 14.VI—41 379 (15) 25.V 19.V—57 25.VI—41 463 (14) 2.VI 3.VI—50 30.VI—41 152 (12) 13.VI
E. verrucosa 8.V—50 14,VI— 41 351 (15) 23.V 22.V—48 25.VI—41 404 (13) 31.V 5.VI—48 30.VI—55 225 (12) 19.VI
Frangula alnus 20.V—47 7.VI-55 155 (5) 31 .V 8.VI—53 14.VI—55 22 (2) 11.VI 20.VI—53 30.VI—55 100(4) 25.VI
Mai us praecox l.V—50 5.VI-41 194 (15) 13.V 5.V—50 10.VI—41 228 (12) 19.V 9.V—50 14.VI—41 384 (15) 26.V
Padus racemosa .... 28. IV—40 26.V—41 550(14) 9.V 1.V—40 30.V—41 169 (14) 12.V 7.V—50 5.VI—41 333 (16) 21.V
Populus tremula . . . 3.IV—61 30. IV—54 154 (8) 19.IV 13. IV—51 4.V—52 179 (7) 25. IV 18. IV—51 8.V—53 262 (8) 3.V
Prunus spinosa . . . . 27. IV—50 27.V—41 597 (15) 10.V 2.V—50 30.V—41 197 (15) 13.V 9.V—50 14.VI-41 388 (17) 23.V
Pyrus communis 29. IV—50 24.V—61 620 (15) 11.V 2.V—50 28.V—61 224 (15) 15.V 6.V—50 12.VI—41 375 (17) 22.V
Rhamnus cathartica . . 12.V—51 30.V—53 143 (6) 24.V 23.V—51 16.VI—41 225 (7) 1.VI 1.VI—51 25.VI—41 82 (7) 12.V1
Rosa canina 24.V—48 18.VI—58 342(9) 7.VI 1.VI—50 28.VI—55' 100(7) 14.VI 10.VI—49 4. VII—55 198 (9) 22.VI
Rubus idaeus 24.V—49 30.VI—41 496 (14) 5.VI 27.V—49 25.VI—61 330 (8) 10.VI 3.VI—49 12. VII—48 223 (10) 22.VI
Quercus robur var. praecox l.V—50 4.VI—41 151 (10) 15.V 4.V—50 8.VI—41 139 (7) 20.V 6.V—50 16.VI—41 206(9) 23.V
Salix capraea 2. IV—41 30. IV—54 259 (14) 18.IV 10. IV—41 5.V—52 320 (13) 25.IV 19.IV—51 15.V—41 371 (И) 4.V
Sambucus nigra . . . . 5.VI—61 21. VI—55 26 (2) 13.VI 10.VI—61 25.VI—55 35 (2) 17.VI 1.VII—61 14.VII—55 15(2) 7.VII
S. racemosa 1 l.V—60 14.V—61 25 (2) 12.V 18.V—61 24.V—55 42 (2) 21 .V 26.V—61 28.V—60 82 (3) 27.V
Sorbus aucuparia . . 13.V—57 27.V—53 251(11) 23.V 15.V-57 2.VI—55 244 (9) 27.V 21.V—57 U.VI—55 323 (10) 1.VI
Viburnum opulus . . . . 22.V—50 3.VI-52 289 (10) 29.V 27.V—49 21.VI—55 298 (8) 6.VI 3.VI—49 24.VI—58 . 132 (10) 13.VJ
П риложекае VI
Средние многолетние даты начала и полного созревания плодов древесно-кустарниковых
пород Стрелецкого и Казацкого участков Центрально-Черноземного заповедника
(по 1962 г. включительно)
Вид Плодоношение
начало массовое
сумма дней и ко- личество лет наблюдений средняя многолетняя сумма дней и ко- личество лет наблюдений средняя многолетняя
Acer platanoides 70 (2) 4.IX 7(2) з.х
А. tataricum . . 78 (2) 8-VIII 46(2) 23-IX
Amygdalus папа 105 (3) 4.VIII 27(2) 13.VIII
Cerasus fruticosa 225 (14) 16.VII 141 (6) 23.VII
Corylus avellana 99 (5) 20-VIII 99(3) 2.IX
Crataegus curvisepala 198(6) 2.IX 79(3) 26-IX
Euonymus europaea 567 (И) 20.VIII 226 (5) 14.IX
E, verrucosa 512(11) 16.VIII 112(4) 28.VIII
Frangula alnus 112(4) 28.VII 53 (2) 26.VIII
Matus praecox . . 141 (6) 28.VIII 181 (5) 5.IX
Padus racemosa 446(11) 10.VII 117(6) 20-VII
Populus tremula 135 (6) 22.V 71 (3) 24.V
Prunus spinosa 572 (12) 17.VIII 271 (6) 14.IX
Pyrus communis 213(7) 30.VIII 114(3) 7.IX
Rhamnus cathartica 163 (4) 10.VIII 51 (2) 25. IX
Rosa canina 212(9) 24.VIII 202 (4) 19-IX
Rubus idaeus 464 (13) 6.VII 140 (8) 17.VII
Quercus robur var. prae-
cox 410 (6) 6.IX 79 (3) 26. IX
Salix capraea . . 182 (10) 18.V 181 (7) 26- V
Sambucus nigra .... 30(2) 15.VIII 2(1) 2.IX
S. racemosa 141(4) 4-VII 20(1) 20-VII
Sorbus aucuparia 186 (6) 31.VII 71 (2) 4.IX
Viburnum opulus 232 (8) 29.VIII 67 (4) 17.IX
Ритмы цветения некоторых травянистых растений дубовых лесов Стрелецкого и Казацкого
включи
Начало цветения Массовое
Вид самое раннее самое позднее сумма дней и количество лет наблюде- ний средняя мно- голетняя самое раннее
Aconitum lasiostomum , . 19.VI—62 25.VI—54 54 (3) 18.VI 28.VI—61
Actaea spicata 20.V—62. 30.V—52 132 (5) 26.V 25.V—62
Aegopodium podagraria. . 6.VI—62 25.VI—41 125 (9) 14.VI 17.VI—53
Anemone ranunculoides . 15. IV—61 7.V—54 131 (5) 26. IV 21. IV—61
Anthriscus silvestris . . . 29.V—61,62 14.VI— 41 143(4) 6.VI 6.VI—62
Asarum europaeum . 14. IV—61 3.V—54 105(5) 21. IV 20. IV—62
Asperula odorata * . . . 8.V—51 26.V—53 79 (4) 19.V 18.V—51
Brachypodium pinnatum . 17.V—53 4.VII— 51 155(6) 26.VI 20.VI—53
Calamagrostis arundinacea 23.VI—51 10.VII—58 218 (7) 1.VII 27.VI-51
Carex montana . . 18. IV—61 3.V—41,54 134(5) 27.IV 22.IV-61
Clematis recta .... 3.VI—62 12.VI—61 36 (5) 7.VI 10.VI—60
Convallaria majalis . . 7.V—51 10.VI—41 202 (10) 20.V 17.V—62
Fragaria vesca . . . • 14.V—62 20.V—41 50 (3) 17.V 19.V—62
Geranium silvaticum . . 18.V—62 10.VI—41 103(4) 26.V 29. V—60,62
Glechoma hederacea 28. IV—61 27.V—41 114(3) 8.V 5.V—62
Hypopitys monotropa . . 15.VII—53 30.VII—51 99 (4) 25.VII 28.VII—53
Lilium pilosiusculum . . 26.VI—62 7.VII—51 161 (5) 2.VII 29.VI—62
Mercurialis perennis ♦ . 9.VI—61 13.V—54 115(5) 23.IV 13. IV—61
Orobus vernus 29. IV—62 25.V—41 236 (6) 9.V 5.V—62
0. vernus * 29. IV—51 12.V—54 112(3) 7.V 6.V—51
Polemonium coeruleum . . 6.VI—62 16.VI—58 41 (4) 11.VI 12.VI—59
Polygonatum officinale . . 27.V—62 29.V—61 112(4) 28. V 29.V—62
Pulmonaria angustifolia . 7. IV—51 3.V—54 157 (8) 19. IV 17.IV—61
Ranunculus auricomus . . 2.V—61 25.V—41 37 (3) 12.V 13.V—61
Stet lari a holostea ♦. 6.V—51 17.V—53 59 (5) 12.V 15.V—62
Taraxacum officinale . . 29. IV—61 19.V—41 108 (3) 6.V 8.V—61
Trolttus europaeus . . . 14.V—62 20.V—41 50(3) 17.V 18.V—62
U rtica dioica .... 6.VI—62 18.VI—61 24 (2) 12.VI 14.VI—62
Viola hirta ........ 6. IV—61 18.V—60 74(3) 25. IV 17.IV—61
V. mirabilis ....... 21. IV—51 4.V—53 190 (7) 26.IV 25. IV—62
• Наблюдения за данными видами проводились только в Казацком лесу.
266
Приложение VII
участков Центрально-Черноземного заповедника в многолетнем ряду (по 1962 г.
тельно)
цветение Конец цветения
самое позднее сумма дней и количество лет наблю- дений средняя многолет- няя самое раннее самое позднее сумма дней и количество лет наблюде- ний средняя многолетняя
2. VII— 54 89 (3) ' 27.VI 20. VII—54 31. VI1-62 75(3) 25.VII
8.VI—52 131 (4) 2.VI 29.V—62 17.VI—52 191 (5) 7.VI
30.VI—41 211 (9) 23.VI 3.VII-54 27.VII—58 90(8) 11.VII
16. V- 54 169(5) 4.V 2.V—61 24.V—54 86(5) 17.V
30.VI—41 62(4) 15.VI 26. VI—62 * 2. VI1—61 58 (2) 29.VI
9.V—54 139 (5) 28. IV 2.V—61 12. V—53,54 37(5) 7.V
3.VI—53 81 (3) 27.V 2.VI—51 12.VI—52 30(4) 7.VI
6.VII—58 108 (4) 27.VI 25.VI—53 18.VII—51,58 197(5) 9.VII
12.VII—58 245 (7) 5.VII 7.VII—51 25. VII—58 ИО (7) 16.VII
15. V—41 191 (6) 2.V 2f.IV—51 25.V—41 238 (6) 10.V
18.VI—61 73(5) 15.VI 26. VI—60 20. VII— 58 192 (5) 8.VII
30.V—53 221 (9) 24.V 29. V—60 25.VI—41 392 (Ю) 8.VI
30.V—41 78(3) 26.V 14.VI—62 22.VI—61 36(2) 18.VI
14.VI—41 137 (4) 4.VI 11.VI—60 25.VI-41 66(4) 16. VI
30.V—41 43(3) 14.V 10. VI—41,62 2.VII—61 52 (3) 17.VI
7. VIII—54 66 (2) 3.VIII 8.VIII—51 25.VIII—54 68(4) 17.VIII
9.VII—61 142 (4) 5.VII 9.VII—62 20.VII—51 71 (5) 14.VII
17.V—54 146 (5) 29. IV 18. IV—61 24.V—54 176 (5) 5.V
30.V—41 84(6) 14.V 20.V—59 30 V—60 132 (5) 26.V
22.V—54 48 (3) 16.V 17.V—51 5.VI—54 72 (3) 24.V
22.VI—58 66 (4) 16.VI 17.VII—61 11.VIII—58 116 (4) 29.VII
7.VI—54 101 (3) 2.VI 2.VI—62 20.VI—54 48(4) 12.VI
25.V—41 332 (9) 7.V 12.V—59 25.VI—41 255 (9) 28.V
30.V—41 58 (3) 19.V 29.V—62 10.VI—41 1°9 (3) 5.VI
30.V—52 110(5) 22.V 22.V—51 18.VI—52 180 (5) 5.VI
30.V—41 48(3) 16.V 29.V—62 5.VI—41 96 (3) 1.VI
30.V—41 70 (3) 23.V 2.VI—62 12.VI—61 14 (2) 7.VI
30.VI—41 72(3) 24.VI 5.VII—62 13.VII—61 18 (2) 9.VII
18.V—60 134(4) 3.V 8.V—61 5.VI—41 55 (3) 18.V
18.V—53 292 (8) 6.V 6.V-51 22.V—54 85 (6) 14.V
267
Приложение VIIг
Средние многолетние даты начала и полного созревания плодов и семян некоторых
травянистых растений дубовых лесов Стрелецкого и Казацкого участков
Центрально**Черноземного заповедника (по 1962 г. включительно)
Вид Плодоношение
начало массовое
:умма дней и количе- ство лет наблюде- ний средняя многолетняя сумма дней и количество лет наблюде- ний средняя многолетняя
Aconitum lasiostotnum 70(2) 4.VIII 35 (2) 18.VIII
Actaea spicata 73 (3) 24.VII 153 (3) 30-VIII
Aegopodium podagraria 189(6) 31.VII 74 (6) 12.VIII
Anemone ranunculoides 79 (3) 26.V 101 (3) 3.VI
Anthriscus silvestris 32(3) 11.VII 64 (3) 21-VII
Asarum europaeum 56(4) 14.VI 79(4) 20-VI
Asperula cdorata 60(2) 30.VIII 81 (2) 9-IX
Brachypodium pinnatum 80(4) 20. VIII 112(4) 28-VIII
Calamagrostis arundinacea 177 (5) 5.VIII 87 (6) 15-VIII
Carex montana 144 (4) 5.VI 51 (4) 13.VI
Clematis recta 158 (5) 1.VIII 97(5) 20 VIII
Convallaria majalis 55(4) 14. VIII 116(4) 29 VIII
Fragaria vesca 31 (2) 15.VI 52 (2) 26-VI
Geranium silvaticum 39 (2) 20.VI 94(3) 1.V1I
Glechoma hederacea 54(2) 27.V1 66 (2) 3.VII
Hypopitys monotropa 33 (2) 16.IX 28(1) 28-X
Lilium pilosiusculum 153 (4) 7.IX 119(5) 24. IX
Mercurialis perennis 61 (3) 20.VI 88(3) 29.VI
Orobus vernus 138(5) 28.VI — —
0. vernus (в Казацком лесу) .... 70(3) 23.VI 95 (3) 2.VII
Polemonium coeruleum 59 (2) 30.VII 178 (4) 13.VIIT
Polygonatum officinale 64(2) 1.IX 29(2) 14.IX
Pulmonaria angustifolia 152 (5) 30. V 231 (5) 15.VI
Ranunculus auriccmus 33 (2) 16.VI 62(2) 1.VII
Stellaria holostea 70(5) 14.VI 100(5) 20.VI
Taraxacum officinale 87 (3) 29.V 63(2) ' 1.VI
Trollius europaeus 33 (2) 16.VI 48(2) 24.VI
Urtica dioica 59 (2) 30. VIII 74(2) 6.IX
Viola hirta 53 (2) 26.VI 23 (2) 11.VII
V. mirabilis 118(5) 24.VI 193 (6) 2 VII
Л ИТЕРАТУРА
Авдошенко А. К. 1949. Биология северных брусничных.— Уч. зап. Ленингр. гос.
пед. ин-та им. А. И. Герцена, т. 82.
Аврорин Н. А. 1984. Растительность разновозрастных залежей Каменной степи.—
Труды Бот. ин-та АН СССР, серия 3 (геоботаника), вып. 1. Л., Изд-во АН СССР.
Аврорин Н. А. 1956. Переселение растений на полярный север. Эколого-географиче-
ский анализ. М.— Л., Изд-во АН СССР.
Александров В. Г. 1954. Анатомия растений. Изд. 3. М., Изд-во «Сов. наука».
Александров В. Г., Александрова О. Г. 1928. О сосудисто-волокнистых пуч-
ках стебля подсолнечника как объекте экспериментальной анатомии, ч. 1.— Журя.
Русск. бот. об-ва, т. 13, № 3—4.
Александров В. Г., Александрова О. Г. 1929. Является ли одревеснение об-
ратимым или необратимым процессом.— Изв. Главн. бот. сада СССР, т. 27,
вып. 5—6.
Александрова В. Д. 1940. Кормовая характеристика растений Крайнего Севера.—
Труды Ин-та полярн. землед., животноводства и промысл, хозяйства, серия «Олене-
водство», № 11. Л.
Алехин В. В. 1909. Очерк растительности и ее последовательной смены на участке
Стрелецкая степь под Курском.— Труды СПб. об-ва естествоиспыт., отд. ботаники,
т. 40, ч. 4, вып. 1.
Алехин В. В. 1915а. Введение во флору Тамбовской губернии. (Ботанический очерк).
Изд. Тамбовск. губ. земства. М.
Алехин В. В. 19156. Типы русских степей.— Изв. Бот. сада Петра Великого, т. 15,
№ 3—4.
Алехин В. В. 1925а. Растительный покров степей ЦЧО. ’Изд. союза обществ и орга-
низаций по изучению ЦЧО. Воронеж.
Алехин В. В. 19256. Фитосоциология (учение о растительных сообществах) и ее
последние успехи у нас и на Западе.— В кн.: В. В. Алехин, Вл. С. Дохтуров-
ский, А. Е. Ж а д о в с к и й, А. П. Ильинский. Методика геоботанических ис-
следований. Л.— М., Изд-во «Пучина».
Алехин В. В. 1926. Растительность Курской губернии.— Труды Курск, губ. планового
совещ., вып. 4. Курск, Изд-во «Сов. деревня».
Алехин В. В. 1934. Центрально-Черноземные степи. Воронеж, Изд-во «Коммуна».
Алехин В. В. 1935. Проблема фитоценоза и некоторые новые фактические данные.—
Уч. зап. Моск. гос. ун-та, вып. 4. (Сб. трудов Научно-исслед. ин-та ботаники,
№ 1). М.
Алехин В. В. 1936. Растительность СССР в основных зонах.— В кн.: Г. Вальтер,
В. Алехин. Основы ботанической географии. М.— Л., Изд-во биэл. и мед. лит-ры.
Алехин В. В. 1940. Флора Центрально-Черноземного заповедника.— Труды Центр.-
Черноземн. гос. заповедника, вып. 1. М.
Алехин В. В. 1944. География растений. (Основы фитогеографии, экологии и фито-
ценологии). М., Изд-во «Сов. наука».
Алехин В. В. 1950. География растений. (Основы фитогеографии, экологии и геобо-
таники). Изд. 3. М., Учпедгиз.
Алехин В. В. 1951. Растительность СССР в основных зонах. Изд. 2. М., Изд-во
«Сов. наука».
Алехин В. В., У рано в А. А. <1933. Методика исследования степей.— Сов. ботаника,
№2.
Амелин И. С. 1939. Производительность в связи с фенологией некоторых типов кара-
кулеводческих пастбищ.— В сб.: «Освоение пустынь, полупустынь и высокогорий».
М., Изд-во ВАСХНИЛ.
Амелин И. С. 1953. Об эколого-биологической системе растений Раункиера.— Бот.
журн., т. 38, № 4.
Андреев В. Н. 1940. Пастбища и пастбищеобороты в оленеводстве.— В сб.: «Вопро-
269
сы оленеводства Крайнего Севера». Всесоюзн. акад. с.-х. наук. Комисе. Крайнего
Севера. М.
Андреев В. Н. 1948. Корма и пастбища северных оленей.— Северное оленеводство.
М.» Сельхозгиз.
Аненков Н. 1850. Наблюдения над развитием дикорастущих растений Московской
губернии.— Журн. сельск. хозяйства и овцеводства, № 6.
Афанасьева Е. А. 1948. Сезонные и годичные передвижения углесолей в мощных
черноземах Стрелецкой степи.— Труды Центр.-Черноземн. гос. заповедника^
вып. 2. М.
Афанасьева Е. А. 1962. Влияние жизнедеятельности растений на почвенные про-
цессы в черноземах.— Сообщ. лабор. лесоведения АН СССР, вып. 6. М.
Афанасьева Е. А., В. Н. Голубев. 1962. Почвенно-ботанический очерк Стрелец-
кой степи. Курск.
Балаш А. П. 1948. Смена аспектов и фенологический спектр Аксайской степи.— Уч.
зап. фак-та естествозн. Ростовск. пед. ин-та, вып. 1.
Б а л а ш А. П. 1955. Приазовские степи Дона.— Уч. зап. Ростовск. пед ин-та, Юби-
лейный сборник. Ростов-на-Дону.
Балаш А. П. ‘1961а. Степи донского Приазовья.— Бот. журн., т. 46, № 8.
Балаш А. П. 19616. Приазовские степи правого берега Дона. Ростов-на-Дону, Изд-ва
Ростовск. ун-та.
Баранов П. А. 1960. Владимир Николаевич Сукачев. (К 80-летию со дня рожде-
ния).— Изв. АН СССР, серия биол., № 6.
Барановский П. М. 1955. Биологическое значение покоя у растений.— Изв. АН Каз.
ССР, серия биол., вып. 10.
Бейдеман И. Н. 1954. Методика фенологических наблюдений при геоботанических
исследованиях. М.— Л., Изд-во АН СССР.
Бекетов А. Н. 1874. Примечания переводчика к первому тому, стр. 546—575.— В кн.:
А. Гризебах. Растительность земного шара согласно климатическому ее рас-
пределению. Очерк сравнительной географии растений. Ч. 1. СПб.
Белкин Н. И. 1961. Зимостойкость растений (факторы зимостойкости). Кишинев,
Изд-во «Штиинца» Молдавского фил. АН СССР.
Белов А. И. 1929. Особенности культурных люцерн разных стран вида Medicago sati*
va L.— Труды Всесоюзн. съезда по генетике, селекции, семеноводству и племенному
животноводству в Ленинграде 10—16 января ‘1929 г., т. 3. Л.
Белостоков Г. П. 1957. К побегообразованию плотнокустовых злаков.— Бот. журн.,
т. 42, № 8.
Белостоков Г. П. 1962. Ритм сезонного развития растений полупустыни.— Бюлл.
Моск, об-ва испыт. природы, отд. биологии, т. 67, вып. 6.
Белянина Н. Б. 1961. О перезимовке растений можжевеловых и сосновых лесои.
южного берега Крыма.— Труды Гос. Никитск. бот. сада, т. 35, материалы по флоре
и растительности Крыма.*
Б е р г Л. С. 1922. Климат и жизнь. М., Госиздат.
Берг Л. С. 1936. Физико-географические (ландшафтные) зоны СССР, ч. 1. Изд-во
Ленингр. ун-та.
Берг Л. С. 1947. Географические зоны СССР, т. 1. М., Географгиз.
Б е р г Л. С. 1955. Природа СССР. М., Географгиз.
Бобров Е. Г. 1947. Виды клеверов СССР.— Труды Бот. ин-та им. В. Л. Комарова
АН СССР, серия Ч (флора и систематика высших растений), вып. 6. М.— Л., Изд-во
АН СССР.
Бобров Е. Г. 1959. Развитие учения о расе и метод видовых рядов в русской систе-
матической школе.— Бот. журн., т. 44, № 11.
Борисова И. В. 1959. К биологии Potentilla humifusa Willd.— Бот. журн., т. 44, № 4.
Борисова И. В. 1960. Основные жизненные формы двудольных многолетних травя-
нистых растений степных фитоценозов Северного Казахстана.— Бот. журн, т. 45,
№ 1.
Борисова И. В. 1961. Биология и основные жизненные формы двудольных много-
летних травянистых растений степных фитоценозов Северного Казахстана.— Труды
Бот. ин-та им. В. Л. Комарова АН СССР, серия 3 (геоботаника), вып. 13.
Борисова И. В., Исаченко Т. И., Рачковская Е. И. 1957. О лесостепи в Се-
верном Казахстане.— Бот. журн., т. 42, № 5.
Борисова М. А. 1960. Побегообразование у некоторых видов злаков Стрелецкой сте-
пи Курской области.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, т. 57,.
каф. ботаники, вып. 4. «Вопросы биологии растений». М.
Борисова-Гуленкова М. А. 1959. Способы отрастания побегов после скашива-
ния.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, т. 100, каф. ботаники,
вып. 5. «Вопросы биологии растений». М.
Борисова-Гуленкова М. А. 1960. Ритм сезонного развития растений луговой
степи.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 65, вып. 6.
Боровский В. М. 1961. О солеобмене между морем и сушей и многолетней динами-
ке почвенных процессов.— Почвоведение, № 3.
270
Бурыгин В. А. 1957. О зимней вегетации растений в южном Кызыл Куме.— Изв. АН
Узб. ССР, серия биол. наук, № 1.
Бурыгин В. А., Запрометова Н. С. 1959. О сроках весеннего окончания вегета-
ции у некоторых эфемеров и эфемероидов Южных Кызылкумов.— Узб. биол. журя.,
№4.
Буш Н. А. 1936. Ботанико-географический очерк Европейской части СССР и Кавказа.
М.— Л., Изд-во АН СССР.
Буш Н. А. 1940. Курс систематики высших растений. Л.
Вавилов Н. И. 1935. Закон гомологических рядов в наследственной изменчивости.
Изд. 2. М.— Л., Сельхозгиз.
В армии г Е. 1902. Распределение растений в зависимости от внешних условий (эко-
логическая география растений). СПб.
Василевская В. К. 1954. Стебель.— В кн.: В. Г. Александров. Анатомия расте-
ний. М., Изд-во «Сов. наука».
Василевская В. К., Кондратьева Е. А. 1955. Образование почек на корнях
древесно-кустарниковых растений.— Докл. АН СССР, т. 101, № 5.
Васильев В. Н. 1946. Закономерности процесса смен растительности.— Материалы
по истории флоры и растит. СССР, вып. 2. М.— Л., Изд-во АН СССР.
Васильев В. Н. 1958. Происхождение флоры и растительности Дальнего Востока
и Восточной Сибири — Материалы по истории флоры и растит. СССР, вып. 3. М.—
Л., Изд-во АН СССР.
Васильев И. М. 1953. Зимостойкость растений. М., Изд-во АН СССР.
Васильченко И. Т. 1937. Определитель всходов сорных растений СССР. М.— Л.,
Сельхозгиз.
Васильченко И. Т. 1949. Люцерна — лучшее кормовое растение.— Труды Бог.
ин-та им. В. Л. Комарова АН СССР, серия 1 (флора и систематика высших расте-
ний), вып. 8.
Васильченко И. Т. 1953. Обзор видов рода Trigonella.— Труды Бот. ин-та им.
В. Л. Комарова АН СССР, серия 1 (флора и систематика высших растений), вып. 10.
Васильченко И. Т. 1961. О соотношении онтогенеза и филогенеза у высших рас-
тений.— Бот. журн., т. 46, № 12.
Вахрамеев В. А. 1952. Стратиграфия и ископаемая флора меловых отложений За-
падного Казахстана.— Региональная стратиграфия СССР, т. 1. М., Изд-во АН СССР.
В и з н е р Ю. 1892. Биология растений. СПб.
Викторов С. 1941. Зимний рост деревьев и кустарников.— Усп. соврем, биол., т. 14,
вып. 3.
Виленский Д. Г. 1924. Засоленные почвы, их происхождение, состав и способы
улучшения. М., Изд-во «Новая деревня».
В и л л и К- 1959. Биология. М., ИЛ.
Вильямс В. Р. 1922. Естественнонаучные основы луговодства или луговедение.—
Общее земледелие, ч. 2. М., Изд-во «Новая деревня».
Виппер В. Н., Виппер П. Б. 1953. Динамика и взаимоотношения древесной и
травянистой растительности в полезащитных насаждениях Заволжья.— Бюлл.
Моск, об-ва испыт. природы, отд. биологии, т. 58. вып. 1.
Владимиров К. 1914. Залежная и степная растительность в Бобровском у. Воро-
нежской губ.— Труды Бюро по прикл. ботанике, т. 7.
Волкова Е. Н. 1958. Особенности развития пустынных эфемеров в условиях Моск-
вы.— Труды Ин-та ботаники АН Туркм. ССР, т. 4.
Воронов А. Г. 1952. О перезимовывании травянистых растений.— Бот. журн., т. 37,
№ 2.
В о р о ш и л о в В. Н. 1959. Лекарственная валериана. М., Изд-во АН СССР.
Вульф Е. В. 1944. Историческая география растений. История флор земного шара.
М.—Л., Изд-во АН СССР.
Высоцкий Г. Н. 1912. Лесные культуры степных опытных лесничеств с 1893 по
1907 г.— Труды по лесному опытному делу в России, вып. 41.
Высоцкий Г. Н. 1915. Ергеня. Культурно-фитологический очерк.— Труды по прикл.
ботанике, т. 8, № 10—11.
Высоцкий Г. Н. 1926. О некоторых формах корнеотпрыскового возобновления.—
Труды с!льско-господ. боташки, т. 1, вып. 2.
Галахов Н. Н. 1935. Возвраты лета осенью («бабье лето») и вторичное цветение
растений.— Метеорол. вест., № 7—8.
Галахов Н. Н. 1937. Вторичное цветение растений.— Природа, № 1.
Галахов Н. Н. 1950. Вторичное цветение растений в 1949 г.— Природа, № 7.
Генкель П. А. 1948. Состояние покоя у растений как процесс обособления прото-
плазмы.— Вест. АН СССР, № 8.
Генкель П. А., О к н и н а Е. 3. 1948а. О состоянии покоя у растений.— Докл. АН
СССР, т. 57, № 3.
Генкель П. А., О к н и н а Е. 3. 19486. Состояние покоя у растений как процесс обо-
собления протоплазмы клеток.— Труды Ин-та физиол. раст. им. К. А. Тимирязева,
т. 6, вып. 1.
Генкель П. А., Ситникова О. А. 1953. Состояние покоя у растений и морозоус-
тойчивость.—Труды Ин-та физиол. раст. им. К. А. Тимирязева, т. 8.
271
Глинка Д. М. 1939. Пастбищные сезоны в оленеводстве и условия зимнего питания
оленей в ненецком округе —Труды Научно-исслед. ин-та полярн. землед., живот-
новодства и промысл, хозяйства, серия «Оленеводство», вып. 4. Л.
Глумов Г. А., Красовский П. Н. 1961. Залежная растительность южной лесо-
степи Зауралья и ее классификация.— Труды Ин-та биологии Уральского фил. АН
СССР, вып. 27 «Вопросы классификации растительности». Свердловск.
Голенкин М. И. 1927. Победители в борьбе за существоваиие. Изд. ассоциации На-
учно-исслед. ин-та при физ.-матем. фак-те 1 Моск. гос. ун-та.
Голоскоков В. П. 1949. Флора и растительность высокогорных поясов Заилийского
Алатау. Алма-Ата, Изд-во АН Каз. ССР.
Голубев В. Н. 1956а. О биологическом значении геофилии у травянистых расте-
ний.— Бот. журн., т. 41, № 2.
Голубев В. Н. 19566. К биоморфологии природных растений Подмосковья с запаса-
ющими органами побегового происхождения.— Уч. зап. Моск; обл. пед. ин-та, т. 41,
труды каф. ботаники, вып. 1.
Голубев В. Н. 1957а. Материалы к эколого-морфологической и генетической харак-
теристике жизненных форм травянистых растений.— Бот. журн., т. 42, № 7.
Голубев В. Н. 19576. О морфогенезе и эволюции жизненных форм травянистых рас-
тений лесолуговой зоны.— Бюлл. Моск, об-ва испыт. природы, отд. биологии,
т. 62, вып. 6.
Голубев В. Н. 1958а. О морфогенезе жизненных форм травянистых растений Запад-
ной Сибири.— Докл. АН СССР, т. 120, № ‘1.
Голубев В. Н. 19586. О короткокорневищных растениях.— Бюлл. Моск, об-ва испыт.
природы, отд. биологии, т. 63, вып. 3.
Голубев В. Н. 1959а. О морфогенезе деревянистых растений и путях морфологиче-
ской эволюции от деревьев к травам.— Бюлл. Моск, об-ва испыт. природы, отд.
биологии, т. 64, вып. 5.
Голубев В. Н. 19596. О некоторых особенностях морфогенеза жизненных форм
травянистых растений лесолуговой зоны в связи с их эволюцией.— Бот. журн.,
т. 44, № 12.
Голубев В. Н. 1960а. К эколого-морфологической характеристике жизненных форм
травянистых растений лесостепи Западной Сибири.— Бот. журн., т. 45, № 7.
Голубев В. Н. 19606. О месте полукустарников в ряду эволюции жизненных форм
от деревьев к травам.— Бюлл. Главк, бот. сада АН СССР, вып. 36. М.
Голубев В. Н. 1960в. О высотном распределении моноподиальных растений в За-
падном Тянь-Шане.— Бюлл. Главн. бот. сада АН СССР, вып. 39. М.
Голубев В. Н. 1960 г. К вопросу о классификации жизненных форм.— Труды Центр.-
Черноземн. гос. заповедника, вып. 6. Курск.
Голубев В. Н. 1960д. О некоторых ботанико-географических вопросах видообразо-
вания и флорогенеза.— Труды Центр.-Черноземн. гос. заповедника, вып. 6. Курск.
Голубев В. Н. 1960е. Аспективные картины растительности лесостепи Западной Си-
бири.— Труды Центр.-Черноземн. гос. заповедника, вып. 6. Курск.
Голубев В. Н. 1960ж. К морфолого-генетической характеристике клубневых расте-
ний.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, .т. 65, вып. 5.
Голубев В. Н. 1961. К морфолого-генетической характеристике ползучих растений.—
Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 66, вып. 4.
Голубев В. Н. '1962а. К эколого-биологическому познанию растительности луговых
степей.— Бот. журн., т. 47, № 1.
Голубев В. Н. 19626. Растительность. В кн.: Е. А. Афанасьева, В. Н. Голу-
бев. Почвенно-ботанический очерк Стрелецкой степи. Курск.
Голубев В. Н. 1962в. Основы биоморфологии травянистых растений центральной ле-
состепи. Ч. 1. Биоморфология подземных органов.— Труды Центр.-Черноземн. гос.
заповедника, вып. 7. Изд-во Воронежск. ун-та.
Голубев В. Н. 1963. К методике определения абсолютной продуктивности надземной
части травяного покрова луговой степи.— Бот. журн., т. 48, № 9.
Гордягин А. Я. 1889. Естественноисторическое описание Казанской губернии, часть
ботаническая. 1. Ботанико-географические исследования в . Казанском и Лаишев-
ском уездах.— Труды Об-ва естествоиспыт. при Казанск. ун-те, т. 22, вып. 2.
Гордягин А. Я. 1901. Материалы для познания почв и растительности Западной
Сибири.— Труды Об-ва естествоиспыт. при Казанск. ун-те, т. 35, вып. 2.
Гордягин А. Я. 1922. Растительность Татарской республики.— В кн.: «Географиче-
ское описание Татарской республики». Ч. 1. Природа края. Казань.
Городков Б. Н. 1935. Растительность тундровой зоны СССР. М.— Л., Изд-во АН
СССР.
Городков Б. Н. 1939. Есть ли родство между растительностью степей и тундр? —
Сов. ботаника, № 6—7.
Горшкова А. А. 1954. Материалы к изучению степных пастбищ Ворошиловградской
области в связи с их улучшением.— Труды Бот. ин-та им. В. Л. Комарова АН
СССР, серия 3 (геоботаника), вып. 9.
Горшкова А. А. '1960а. Перезимовка растений в Иркутско-балаганской лесостепи.
Научные чтения памяти М. Г. Попова. 3-е чтение. Восточно-Сиб. фял. СО АН СССР,
Иркутск.
272
Горшкова А. А. 19606. Сезонное развитие травянистых растений иркутско-балаган-
ской лесостепи. Вопросы региональн. фенологии и биогеографии.— Бюлл. Восточ-
но-Сиб. фенол, комиссии, № 1. Иркутск.
Горшкова А. А., Родченко О. П. 1959. О перезимовывании некоторых многолет-
них растений в иркутско-балаганской лесостепи. — Бот. журн., т. 44, № И.
Горшкова С. Г. 1955. Норичник — Scrophularia L.—В кн.: «Флора СССР>, т. 22.
М.— Л., Изд-во АН СССР.
Григорьев Ю. С., Паузнер Л. Е. 1963. Материалы по экологии видов Aegilops L.
(К вопросу об экологической природе эфемеров-однолетников) .—Бот. журн., т. 48,
№ 5.
Гроссгейм А. А. 1945. К вопросу о графическом изображении системы цветковых
растений.— Сов. ботаника, т. 13, № 3.
Гроссгейм А. А. 1950. Теория ксероморфогенеза и некоторые вопросы истории
флоры.— Пробл. ботаники, вып. Г. М.— Л., Изд-во АН СССР.
Гроссгейм А. А. 1952. Rosaceae — Leguminosae.— В кн.: «Флора Кавказа», т. 5.
М.—Л., Изд-во АН СССР.
Гроссет Г. Э. 1930. Лес и степь в их взаимоотношениях в пределах лесостепной по-
лосы Восточной Европы. Воронеж.
Гроссет Г. Э. 1961. Колебания границ между лесом и степью в голоцене в свете
учения о смещении зон.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 66,
вып. 2.
Гулисашвили В. 3. 1948. Лесостепь и степи Восточного Закавказья и взаимоотно-
шения между древесной и травянистой растительностью в них.— Изв. Всесоюзн.
геогр. об-ва, т. 80, вып. 2.
Гурский А. В. 1934. Экологические черты лёссовых полупустынь Западного Копет-
Дага.— Сов. субтропики, № 3.
Гурский А. В. 1945. Несколько слов о степях, полупустынях и пустынях. — Бот.
журн., т. 30, № 6.
Данилов С. И. 1937. Характеристика кормового достоинства травостоя зимних паст-
бищ Бурят-Монголии.— Вести, дальневосточного фил. АН СССР, № 25.
Демидова Л. С. 1952. О влиянии осеннего охлаждения на ритм развития некото-
рых травянистых растений.— Бюлл. Моск, об-ва испыт. природы, отд. биологии,
т. 57, вып. 3.
Денисова Г. М. 1960а. Побегообразование и сезонное развитие некоторых злаков
на поемных лугах низовий Северной Двины.— Уч. зап. Моск. гор. пед. ин-та им.
В. П. Потемкина, т. 57, каф. ботаники, вып. 4. «Вопросы биологии растений». М.
Денисова Г. М. 19606. Ритм сезонного развития луговых растений низовий Север-
ной Двины.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, т. 57, каф. бота-
ники, вып. 4. «Вопросы биологии растений». М.
Докучаев В. В. 1953. Наши степи прежде и теперь. М., Сельхозгиз.
Долгошев В. И. 1954. Повторное плодоношение растений.— Природа, № 1.
Дохман Г. И. 1937. Вопросы классификации степей.— Землеведение, т. 39, вып. 4—5.
Дохман Г. И. 1954. Материалы к изучению экологических типов растений северных
и южных степей.— Докл. на совещ. по стационарн. геобот. исслед. (22—25 декабря
19Э1 г.). М.— Л., Изд-во АН СССР. -
Дохман Г. И. 1956. Опыт фитоценологической трактовки генезиса северных степей.—
В сб.: «Академику В. Н. Сукачеву к 75-летию со дня рождения». М. — Л., Изд-во
АН СССР.
Дохман Г. И. 1960. К аналитическому изучению структуры фитоценозов северной
(луговой) степи.-- Труды Центр.-Черноземн. гос. заповедника, вып. 6. Курск.
Душечкин В. И. 1951. В каком состоянии зимуют побеги многолетних трав.— Докл.
АН СССР, новая серия, т. 76, № 6.
Д ы л и с Н. В. 1958. О происхождении брахибласт у Larix. Докл. АН СССР, т. 122. № 1.
Закржевский Б. С., Коровин Е. П. 1935. Экологические особенности главнейших
растений Бетпак-Дала.— Труды Среднеаз. гос. ун-та, серия 8-6 (ботаника), вып. 23.
Залесский К. М. 1918. Материалы к познанию растительности донских степей. Изд.
сенного отд. Донского обл. продов. комитета. Ростов-на-Дону.
Запрометова Н. С. 1955. Удлинение срока вегетации у некоторых эфемеров-эфе-
мероидов.— Докл. АН Узб. ССР, № 6.
Запрометова Н. С. 1956. О зимней вегетации некоторых растений пустынных низ-
когорий.— Докл. АН Узб. ССР, № 2.
Запрометова Н. С., Есипова И. В. 1961. О зимней вегетации некоторых расте-
ний Южных Кызылкумов.— В сб.: «Пастбища Узбекистана». Ташкент, Изд-во АН
Узб. ССР.
Запрягаев Ф. Л., Запрягаева В. И. 1948. Особенности сезонного развития кри-
офильной растительности Гиссарского хребта.— Сообщ. Тадж. фил. АН СССР,
вып. 6.
Зарубин А. Ф. 1949. К вопросу о вторичном цветении грецкого ореха. — Природа,
№ 10.
Захаров С. А. 1935. Борьба леса и степи на Кавказе (по данным почвенных иссле-
дований).— Почвоведение, № 4.
18 В. Н. Голубев
273
Зворыкина К. В. 1953. Фенология и подземные части основных растительных ас-
социаций Ергеней.— Труды компл. научн. экспедиции по вопр. полезащитн. лесо-
разведения АН СССР, т. 2, вып. 5. i
Зозулин Г. М. 1949. Характер зимнего покоя некоторых многолетников западноси-
бирской флоры.— Научно-метод. зап. главк, управления по заповедникам Совета
Министров РСФСР, вып. 12. М.
Зозулин Г. М. I960. Стрелецкая степь. Центрально-Черноземный госзаповедник им.
проф. В. В. Алехина. Курск.
Зозулин Г. М. 1955. Взаимоотношения лесной и травянистой растительности в Цен-
трально-Черноземном гос. заповеднике.— Труды Центр.-Черноземы, гос. заповедни-
ка, вып. 3. Курск.
Зозулин Г. М. 1958. О некоторых вопросах фитоценологии в связи с проблемой про-
исхождения растительности северных степей.— Бот. журн., т. 43, № 6.
Зозулин Г. М. 1959а. Вопросы взаимоотношений леса и степи в центральной части
лесостепной зоны.—Зап. Курск, обл. краеведы, музея, вып. 1.
Зозулин Г. М. 19596. Подземные части основных видов растений и ассоциаций пла-
коров Средне-Русской лесостепи в связи с вопросами формирования растительного
покрова.— Труды Центр.-Черноземн. гос. заповедника, вып. 5. Курск.
Зозулин Г. М. !1959в. Научная работа в Центрально-Черноземном заповеднике.—
Охрана природы и озеленение, вып. «1. М.
Зозулин Г. М., Кусмарцева Н. М. 1951. Изменение границ древесно-кустарнико-
вой и травянистой растительности на опытных участках Центрально-Черноземного
заповедника за 5 лет.— Бот. журн., т. 36, № 3.
Иванов В. В. 1958. Степи Западного Казахстана в связи с динамикой их покрова.—
Зап. Геогр. об-ва СССР, т. 17, новая серия. М.— Л., Изд-во АН СССР.
Иванова Н. А., Кр ашенинников И. М. 1934. К истории развития раститель-
ных ландшафтов Западной Сибири. — Землеведение, т. 36, вып. 1.
Игошина К. Н. 1937. Пастбищные корма и кормовые сезоны в оленеводстве При-
уралья.— Сов. оленеводство. № 10.
Илличевский С. О. 1925. Второе цветение, его механизм и причины в связи с ус-
ловиями цветения вообще.— Журн. Русск. бот. об-ва, т. 10, № 1—2.
Илличевский С. О. 1931. Две аномалии в цветении растений.— Журн. Русск. бот.
об-ва, т. 46, № 5—6.
Илличевский С. О. 4937. О вторичном цветении растений.— Сов. ботаника, № 4.
Ильин М. М. 1936. К систематике рода Suaeda Forsk. и трибы Suaedeae Rchnb.—
Сов. ботаника, № 5.
Ильин М. М. 1958. Флора пустынь Центральной Азии, ее происхождение и этапы раз-
вития.— Материалы по истории флоры и растит. СССР, вып. 3. М.— Л., Изд-во
АН СССР.
Ильинский А. П. 1937. Растительность земного шара. М.— Л., Изд-во АН СССР.
Исаченко Т. И. 1954. Травяной покров в лесных посадках Каменной степи Воронеж-
ской области.— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 3 (геобо-
таника), вып. 9.
Каден Н. Н.. 1940. Очерк растительности Казацкой степи под Курском.— Труды
Центр.^Черноземн. гос. заповедника, вып. 1. М,
Каден Н. Н., Гарри Н. В. 1942. Фенология степных растений и закон Минио-
Илличевского.— Бот. журн., т. 27, № б.
Казакевич Л. И. 1922. Материалы к биологии растений Юго-Востока России.—
Изв. Саратовск. обл. с.-х. олытн. станции, т. 3, вып. 3—4.
Камышев Н. С. 1939. О происхождении степных аспектов. Юб. сб. XX лет Воронеж-
ского ун-та.— Труды Воронежск. гос. ун-та, т. 1 Г, отд. ботаники, вып. 1—2.
Камышев Н. С. '1946. Аспекты степи и прогноз сроков сенокошения.— Труды Воро-
нежск. гос. ун-та, т. 14, вып. 1.
Камышев Н. С. 1948. Динамика растительного покрова степной залежи в условиях
Каменной степи.— Труды Воронежск. гос. ун-та, т. 16, вып. 1.
Камышев Н. С. 1949. Аспекты Хреновской степи и их происхождение.— Труды Во-
ронежск. гос. ун-та, т. 15.
Камышев Н. С. 1961. Аспекты Хреновской степи и прогноз сроков сенокошения.—
Бюлл. об-ва естествоиспыт. при Воронежск. гос. ун-те, т. 7.
Камышев Н. С. 1953а. Фазы развития Каменной степи и проблема происхождения
аспектов дерновинно-луговых степей.— Труды Воронекск. гос. ун-та/ г. 28.
Камышев Н. С. 19536. Роль выпаса в изменении естественных пастбищ Каменной
степи.— Труды Воронежск. гос. ун-та, т. 28.
Камышев Н. С. 1954. Календарь сезонного развития растений в Каменной степи
и использование его в сельском хозяйстве.— Труды Воронежск. гос. ун-та, т. 26.
Камышев Н. С. 1956. Закономерности развития залежной растительности Каменной
степи.— Бот. журн., т. 41, № 1. -
Камышев Н. С. 11958. Фенология Каменной степи и проблема происхождения степ-
ных аспектов.— Труды Воронежск. гос. ун-та, т. 45, вып. 3 (ботаника). Изд-во Во-
ронежск. ун-та.
Карягдыев Н. Н., Мищенко А. С. 1956. Случаи вторичного цветения древесно-
кустарниковых растений в Туркмении.— Изв. АН Туркм. ССР, № 3.
274
Келлер Б. А. 1903. Из области черноземно-ковыльмых степей. Ботанико-географиче-
ские исследования в Сердобском уезде Саратовской губернии.— Труды Об-ва есте-
ствоислыт. при Казанск. ун-те, т. 37, вып. 1.
Келлер Б. А. 1914. По долинам и горам Алтая. Ботанико-географические исследова-
ния. Т. 1.— Труды Об-ва естествоиспыт. при Казанск. ун-те, т. 46, вып. 1.
Келлер Б. А. 1916. К вопросу о классификации русских степей. (По поводу новых
работ В. В. Алехина и П. Н. Крылова). — Русский почвовед, № 16—18.
К е л л е р Б. А. 1921. Растительность Воронежской губернии. Воронеж.
Келлер Б. А. 1923. Растительный мир русских степей, полупустынь и пустынь. Очер-
ки экологические и фитосоциологические. Воронеж.
Келлер Б. А. 1931. Степи Центрально-Черноземной области.— В кн.: «Степи ЦЧО.
(Степные сенокосы и пастбища)». М.— Л., Сельхозгиз.
Келлер Б. А. 1933. Об изучении жизненных форм при геоботанических исследовани-
ях.— Сов. ботаника, № 2.
Келлер Б. А. 1934. Доклад на дискуссии: «Основные установки и пути развития со-
ветской экологии».— Сов. ботаника, № 3.
Келлер Б. А. 1938а. Растение и среда. Экологические типы и жизненные формы.—
В кн.: «Растительность СССР», т. 1. М.— Л., Изд-во АН СССР.
Келлер Б. А. 19386. Главные типы и основные закономерности в растительности
СССР.— В кн.: «Растительность СССР», т. 1. М.— Л., Изд-во АН СССР.
К е л л е р Б. А. 1948. Основы эволюции растений. М., Изд-во АН СССР.
К е р н е р А. 1903. Жизнь растений, т. 2. СПб.
Кильдюшевский И. Д. 1956. Подснежное развитие некоторых видов флоры Ма-
лого Ямала.— Бот. журн., т. 41, № *111.
Кишковский Т. Н. 1950. Условия обитания растений у верхнего предела их рас-
пространения на Восточном Памире.— Бот. журн., т. 35, № 6.
Кишковский Т. Н., Артюшенко 3. Т. 1951. К биологии высокогорных растений
Памира.— Бот. журн., т. 36, № 5.
Клеопов Ю. Д., Лавренко Е. М. 193*3. Сучасний стан клясифжацП украТнських
стешв.— Журн.-6io бот. циклу ВУАН, № 5—6, Ки1в.
Ко в а кин а. В. А. 1952. О перезимовывании некоторых злаков на Крайнем Севере.—
Бот. журн., т. 37, № 5.
Коган Ш. И. 1949. Эфемеры южного Усть-Урта.— Изв. Туркм. фил. АН СССР, № 2.
Кожевников А. В. 1929. К фитосоциологической характеристике липовой часги
38-го квартала Погонно-поемного острова.— Труды по лесному опыт. делу. Цент-
ральная лесная опытн. станция, вып. 6. М., Сельхозгиз.
Кожевников А. В. 1931. О перезимовке и ритме развития весенних растений ли-
пового леса.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 40, вып. 1—2.
Кожевников А. В. 1936. Материалы по динамике ярусности лугового травостоя.—
Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 45, вып. 3.
Кожевников А. В. 1937а. Исследования продолжительности зимнего периода по-
коя у некоторых растений Западного Закавказья.— Труды Бот. сада Моск. гос.
ун-та, вып. il.
Кожевников А. В. 19376. Некоторые закономерности сезонного развития раститель-
ных ассоциаций.— Уч. зап. Моск. гос. ун-та, вып. 11 (биология).
Кожевников А. В. 1939. Весна и осень в жизни растений. М.— Л., Медгиз.
Козлова Н. А. 1953. Анатомо-экологическая характеристика полукустарников Во-
сточного Крыма.— Бот. журн., т. 38, № 4.
Козо-Полянский Б. М. 1922. Введение в филогенетическую систематику высших
растений. Воронеж.
Козо-Полянский Б. М. 1937. Основной биогенетический закон с ботанической
точки зрения. Воронеж.
Козо-Полянский Б. М. 1945. Случаи превращения биоморф культурных растений
и их значение.— Труды Воронежск. гос. ун-та, т. 13, научн. сообщ. и автореф., вып. 1.
Коликов М. С. 1950. Костер. Руководство по апробации с.-х. культур, т. 4 (кормо-
вые растения). М.— Л., Сельхозгиз.
Комаров В. Л. 1901а. Хвойные деревья Манчжурии.— Труды СПб. об-ва естествоис-
пыт., т. 32, вып. 1.
Комаров В. Л. 19016. Флора Манчжурии, 1.— Труды Петербургск. бот. сада, т. 20.
Комаров В. Л. 1908. Введение к флорам Китая и Монголии. Монография рода
Caragana. — Труды Петербургск. бот. сада, т. 29, вып. 2.
Комаров В. Л. 1927. Флора полуострова Камчатки, т. 1. Л., Изд-во АН СССР.
Комаров В. Л. 1934. Предисловие. Флора СССР, т. 1. М.— Л., Изд-во АН СССР.
Комаров В. Л. 1940. Учение о виде у растений. М.— Л., Изд-во АН СССР.
Комаров Н. Ф. 1951. Этапы и факторы эволюции растительного покрова чернозем-
ных степей.— Зап. Всесоюзн. геогр. об-ва, новая серия, т. 13. М., Географгиз.
Константинова А. Г. 1954. Корневые системы некоторых растений эфемеровой
пустыни предгорий Копет-Дага и их распределение в связи с водным режимом поч-
вы.— Труды Ин-та биологии АН Туркм. ССР, серия ботаники, т. 1.
Коржинский С. И. 1888. Северная граница черноземно-степной области восточной
полосы Европейской России в ботанико-географическом и почвенном отношении.
18* 275
I. Введение. Ботанико-географический очерк Казанской губернии.— Труды Об-ва
естествоиспыт. при Казанск. ун-те, т. 18, вып. 5.
Коржинский С. И. 1891. Северная граница черноземно-степной области восточной
полосы Европейской России в ботанико-географическом и почвенном отношении.
2. Фитотопографические исследования в губерниях Симбирской, Самарской, Уфим-
ской, Пермской и отчасти Вятской. — Труды Об-ва естествоиспыт. при Казанск.
ун-те, т. 22, вып. 6.
Коржинский С. И. 1893. Флора Востока Европейской России в ее систематическом
и географическом отношениях.— Изв. Томск, ун-та, кн. V.
Коржинский С. И. 1899. Растительность России.— Энциклоп. словарь Брокгауза
и Ефрона, т. 54. СПб.
Коржинский С. И. 1901. Степи.— Энциклопед. словарь Брокгауза и Ефрона, т. 31А
(62). СПб.
Коровин Е. П. 1934. Растительность Средней Азии и южного Казахстана. М.— Таш-
кент, ОГИЗ, среднеаз. отд.
Коровин Е. П. 1958. Биологические формы и потребность в воде растительных видов
аридной зоны.— Труды Среднеаз. гос. ун-та им. В. И. Ленина, новая серия, вып. 136,
биол. науки, кн. 32 (ботаника).
Коровин Е. П. 1961, 1962. Растительность Средней Азии и южного Казахстана.
Изд. 2, кн. 1—‘1961, кн. 2—4962. Ташкент, Изд-во АН Узб. ССР.
Коровин Е. П., Тульчиев М. 194.8. О весеннем возобновлении и вторичном цве-
тении грецкого ореха в Средней Азии.— Бот. журн., т. 33, № 3.
Короткова Е. И. 1957. Динамика развития типчаково^ковыльной заповедной степи
Аскании-Нова в связи с погодными условиями. — Бот. журн., т. 42, № 6.
К о р с м о Э. 1933. Сорные растения современного земледелия. М. — Л.» Сельхозгиз.
Корчагин А. А. 1956. К вопросу о характере взаимоотношений растений в сооб-
ществе.— В сб.: «Академику В. Н. Сукачеву к 75-летию со дня рождения». М.— Л.,
Изд-во АН СССР.
Костычев П. А. 1890. Связь между почвами и некоторыми растительными форма-
циями.— Труды 8 съезда русск. естествоиспыт. и врачей в 1889 г., т. 1, отд. 5 (бота-
ника). СПб.
Котт С. А. 1944. О некоторых особенностях развития сорных растений Сибири. — Сов.
ботаника, К® 4—5.
Котт С. А. 1949. Перезимовка многолетних сорных растений в посевах кормовых
трав.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 54, вып. 4.
Котт С. А. 1955. Сорные растения и меры борьбы с ними. М., Сельхозгиз.
Кошкина Е. Ф. 1954. Ритм развития луговой растительности низовий Дона.— Уч. зап.
Моск. гор. пед. ин-та им. В. П. Потемкина, т. 37, каф. ботаники, вып. 2.
Краснов А. Н. 1888. Опыт истории развития флоры южной части Восточного Тянь-
Шаня.— Зап. Русск. геогр. об-ва, т. 19. Изд. Академии наук.
Краснов А. Н. 1893. Рельеф, растительность и почвы Харьковской губернии. Докл.
Харьковск. об-ва сельск. хозяйства.
Краснов А. Н. 1894. Травяные степи северного полушария. — Изв. об-ва любителей
естествозн., антропол. и этнографии при Моск, ун-те, т. 83, труды геогр. отд.,г
вып. 1. М.
Краснов А. Н. 1899. География растений.— Основы земледелия, вып. 4, ч. 1. Харьков.
Красовский П. Н., Глумов Г. А. 1946. Основные черты зацелинения залежей
Троицкого лесостепного заповедника.— Уч. зап. Пермского гос. ун-та им. А. М. Горь-
кого, т. 4, вып. 2.
Крашенинников И. М. 1932. Растительность Общего Сырта и бассейна р. Боль-
шого Ика в пределах Башкирской АССР.— Труды Совета по изуч. производств,
сил, серия башкирская, вып. 2. Л.
Крашенинников И. М. 1937а. Анализ реликтовой флоры Южного Урала в связи
с историей растительности и палеографией плейстоцена.— Сов. ботаника, № 4.
Крашенинников И. М. 19376. Взаимоотношения леса и степи на южной окраине
Уральской возвышенности.— Землеведение, т. 39, вып. 6. М.— Л.
Крашенинников И. М. 1939. Основные пути развития растительности Южного
Урала в связи с палеогеографией северной Евразии в плейстоцене и голоцене.—
Сов. ботаника, № 6—7.
Крашенинников И. М. 1941. Растительность Башкирской АССР. Природные ре-
сурсы Башкирской АССР, т. 1. Совет по изуч. производит, сил. М.— Л., ИзД-во
АН СССР.
Криштофович А. Н. 1941. Указания на палеоботанические нахоцки видов Poten-
tilla.—В кн.: «Флора СССР», т. 10. М,— Л., Изд-во АН СССР, стр. 78.
Криштофович А. Н. 1950. Эволюция растений по данным палеоботаники.— Пробл.
ботаники, вып. 1. М.—Л., Изд-во АН СССР.
Крылов П. Н. 1898. Тайга с естественноисторической точки зрения. Научные очерки
Томского края. Томск.
Крылов П. Н. 1915. К вопросу о колебании границы между лесной и степной обла-
стями.— Труды музея Академии наук, т. 14. СПб.
Крылов П. Н. 1916. Степи западной части Томской губернии. Ботанико-географиче-
276
ский обзор. — Труды почвенно-Оот. экспедиций по исслед. колониз. районов Азиатск.
России. Ч. 2. Бот. исслед. 1913 г., вып. 1. Минист. земледелия, Переселенч.
управл. Пг.
Кузнецов Н. И. 1909. Принципы деления Кавказа на ботанико-географические про-
винции.— Зап. Академии наук, серия 8, т. 24, № 1. СПб.
Кузнецов Н. И. 1936. Введение в систематику цветковых растений. Л.
Культиасов И. М. 1955. Особенности экологии высокогорных растений Западного
Тянь-Шаня. М., Изд-во АН СССР.
Культиасов М. В. 1938. Tav-сагыз и экологические основы введения его в культуру.
М.— Л., Изд-во АН СССР.'
Культиасов М. В. 1946. Этюды по формированию растительного покрова жарких
пустынь и степей Средней Азии.— Материалы по истории флоры и растит. СССР,
вып. 2. М.— Л., Изд-во АН СССР.
Культиасов М. В. 1950. Проблема становления жизненных форм у растений.—
Пробл. ботаники, вып. 1. М.— Л., Изд-во АН СССР.
Культиасов М. В. Эколого-исторический метод в интродукции растений.— Бюлл.
Главн. бот. сада АН СССР, вып. 15.
Культиасов М. В. 1956. И. В. Мичурин — основоположник исторического метода
в интродукции растений.— Бюлл. Главн. бот. сада АН СССР, выл. 25.
Культиасов М. В. 1962. Экологические основы интродукции растений природной
флоры.— Труды Главн. бот. сада АН СССР, т. 9. Экология и интродукция растем
ний. М.
Куприянова Л. А. 1948. Морфология пыльцы однодольных растений. — Труды Бот.
ин-та им. В. Л. Комарова АН СССР, серия 1 (флора и систематика высших расте-
ний), вып. 7.
Куприянова Л. А. 1954. Палинологические данные о филогении класса однодоль-
ных растений.— Вопр. ботаники, вып. 1. М.— Л., Изд-во АН СССР.
Купцов А. И. 1952. Превращение диких многолетних растений в культурные мало-
летники.— Докл АН СССР, т. 86, № 5.
Ку пцов А. И. 1955. Переход диких и сорных растений в культурные.— Природа, № 12.
Куренцов А. И. ’1929. Степи в бассейне верхней Оки в б. Орловской губернии.—
Изв. Северо-Черноземн. (б. Шатиловской) обл., с.-х. опытн. станции, т. 3, вып. 2.
Из работ Муратовской бот. базы. Орел.
Лавренко Е. М. 1938. История флоры и растительности СССР по данным совре-
менного распространения растений.— В кн.: «Растительность СССР», т. 1. М.— Л.,
Изд-во АН СССР.
Лавренко Е. М. 1940. Степи СССР. В кн.: «Растительность СССР», т. 2. М.— Л.,
Изд-во АН СССР.
Лавренко Е. М. 1956. Степи и сельскохозяйственные земли на месте степей.— В кн.:
«Растительный покров СССР». Пояснит, текст к геобот. карте СССР. М.— Л.,
Изд-во АН СССР.
Лавренко Е. М. 1959. Основные закономерности растительных сообществ и пути их
изучения.— В кн.: «Полевая геоботаника», т. 1. М.— Л., Изд-во АН СССР.
Лавренко Е., Дохман Г. 1933. Рослиннють Старобмьских стетв.— Журн. Bio-
бот. циклу ВУАН, № 5—6. Ки!в.
Л а к и з а Е. Н. 1953. Весенние растения флоры Закарпатья.— Бюлл. Главн. бот. сада
АН СССР, вып. 16. М.
Лапшина Е. И. 1928. О перезимовывании высших растений, по наблюдениям в окрест-
ностях Петергофа.— Труды Петергофск. естеств.-научн. ин-та, № 5.
Лапшина Е. И. 1929. Перезимовывание высших растений. — Природа. № 2.
Левина Р. Е. 1956. О способах расселения растений в степях.— Бот. журн., т. 41,
№ 5.
Левина Р. Е. 1957. Способы распространения плодов и семян. Изд-во Моск, ун-та.
Левицкий С. С. '1955. Перспективные хозяйственно-ценные формы дикорастущих
кормовых трав.— Труды Центр.-Черноземн. гос. заповедника, вып. 3. Курск.
Левицкий С. С. 1957. Список сосудистых растений Центрально-Чернозёмного госу-
дарственного заповедника.— Труды Центр.-Черноземн. гос. заповедника, вып. 4.
Курск.
Лисицын П. И. 1947. Вопросы биологии красного клевера. М., Сельхозгиз.
Любарский Е. Л. 1959. К вопросу о построении биоморфологических рядов травя-
нистых многолетников.— Бот. журн., т. 44, № 12.
Любарский Е. Л. 1961. Об эволюции вегетативного возобновления и размножения
травянистых поликарпиков.— Бот. журн., т. 46, № 7.
Любимей ко В. Н. 1900. О спящих почках. — Труды СПб. об-ва естествоиспыт., т. 30,
отд. ботаники, вып. 3.
Л ю б и м е н к о В. Н., Вульф Е. В.. 1926. Ранние весенние растения. М.— Л., Госиэ
дат.
Люндегорд Г. 1937. Влияние климата и почвы на жизнь растений. М., Сельхозгиз.
Маевский П. Ф. 1954. Флора средней полосы Европейской части СССР. Изд. 8
М.— Л., Сельхозгиз.
Макаревич В. Н. 1963. Влияние на луговой травостой способов его использова-
277
ния. — Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 3 (геоботаника),
вып. 114. Комплексные экспериментальные геоботанические исследования.
М а к - Д у г о л л В. Б. 1935. Экология растений. М., Учпедгиз.
Максимов Н. А. 1952. Избранные работы по засухоустойчивости и зимостойкости
растений, т. 2. Зимостойкость растений. М., Изд-во АН СССР.
Мальцев А. И. *1924. Фитосоциологические исследования в Каменной степи.— Труды
Бюро по прикл. ботанике и селекции, т. 13, вып. 2.
Мальцев А. И. 1926. О биологических типах сорных растений и мерах борьбы
с ними.— Изв. Гос. ин-та опытн. агрономии, т. 4, № 1—2. Л.
Мальцев А. И. 1932. Сорная растительность СССР. М.—Л., Сельколхозгиз.
Марков М. В. 1935. Лес и степь в условиях Закамья. — Уч. зап. Казанск. гос. ун-та,
т. 95, кн. 7 (ботаника), вып. 2.
Марков М. В. 1958. О доминантах фитоценоза по работам советских геоботаников. —
Бот. журн., т. 43, № 4.
Мензбир М. А. 1922. Тайна Великого океана. М.
Мильков Ф. Н. 1947. О возрасте лесостепного ландшафта и его природе. — Вопр.
географии, сб. 4. М., Географгиз.
Мильков Ф. Н. 1948. Докучаев и некоторые вопросы генезиса лесостепного ланд-
шафта.— Изв. Чкаловск, отд. геогр. об-ва, вып. 2.
Мильков Ф. Н. 1950. Лесостепь Русской равнины. Опыт ландшафтной характеристик
ки. М., Изд-во АН СССР.
Молозев А. И. 1925. Календарь цветения.— В кн.: «Природа Орловского края».
Орловск. губ. планов, комис.
Молозев А. И. 4927. О предсказании с весны сроков зацветания диких и культур-
ных растений.— Труды Шатиловской обл. с.-х. опытн. станции, № 20, серия X
(Бот. база при селе Муратове), вып. 1. Орел.
Мороз Е. С. 1940. Влияние пониженных температур на рост и развитие древесных
растений.— Сов. ботаника, № 5.
Мороз Е. С. 1948. Экспериментально-экологические исследования периода покоя
у древесных растений.— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 4
(эксперим. ботаника), вып. 6. М.— Л.
М о р о з о в Г. Ф. 1949. Учение о лесе. Изд. 7. М.— Л., Гослесбумиздат.
Мошков Б. С. 1951. Влияние спектрального состава лучистой энергии на рост зем-
ляники и бриофиллума.— Докл. АН СССР, т. 76, № 4.
Наабер Л. X., Захарьянц И. Л. 1961. Фотосинтетическая способность растений
эфемеров в южных Кызылкумах. — Бот. журн., т. 46, № 8.
Наринян С. Г. 1948. О сезонной динамике альпийских ковров Арагаца.— Докл. АН
Арм. ССР, т. 9, № 4.
Наринян С. Г. 1960. К экологии и фенологии альпийских ковров на горе Арагац
(Армянская ССР).— Пробл. ботаники, т. 5. М.— Л., Изд-во АН СССР.
Немкова В. К. 1954. Вторичное цветение конского каштана (Aesculus hippocasta-
num L.) в Симферополе в 1953 году.— Изв. Крымского отд. Геогр. об-ва СССР,
вып. 3.
Нестеров Я. С. 1957. Период покоя и зимостойкость плодовых культур.— Докл. АН
СССР, т. 1117, № 3.
Нечаева Н. Т. 1958. Динамика пастбищной растительности Каракумов под влияни-
ем метеорологических условий. Ашхабад, Изд-во АН Туркм. ССР.
Нечаева Н. Т. 1960. О зимней вегетации эфемеровой растительности в Каракумах. —
Труды Туркм. научно-исслед. ин-та животноводства и ветеринарии, т. 2.
Нечаева Н. Т., Василевская В. К- 1945. Особенности развития однолетников
в Каракумах.— Изв. Туркм. фил. АН СССР, № 3—4.
Никитин В. В. 1957. Сорная растительность Туркмении. Ашхабад, Изд-во АН
Туркм. ССР.
Никитин В. В., Полковниченко А. Я. 1955. Об экологической лабильности
эфемеров.— Изв. АН Туркм. ССР, № 5.
Новопокровский И. В. 1922. Естественноисторические районы Юго-Востока
(Донская область, Северный Кавказ, Черноморская губерния).— В сб.: «Юго-Во-
сток», № 2. Орган Крайэкономсовета. Ростов-на-Дону.
Новопокровский И. В. 1927. Некоторые данные о растительности степей и
речных долин Южного Предуралья.— Труды совещаний геоботаников-луговедов
17—22 марта 1927 года при Гос. луговом ин-те. Труды совещаний по вопросам
луговедения и опытн. луговодства, вып. 1. Дмитров.
Новопокровский И. В. 1931. Материалы для познания растительности Южного
Урала. Стерлитамакский и Уфимский кантоны.— Труды Гос. почв, ин-та, вып. 8.
М., Сельхозгиз.
Новопокровскмй И. В. 1937. Зональные типы степей Европейской части СССР.
(Опыт классификации русских степей).— Землеведение, т. 39, вып. 3. М.— Л.
Одишария К. Ю. 1961. Особенности роста и развития главнейших вечнозеленых по-
крытосеменных растений черноморского побережья Кавказа.— Тбилиси, Изд-во АН
Груз. ССР.
Оловянникова И. Н. 1949. Экология и ритм цветения растений Стрелецкой степи.
278
Научно-метод. зап. Главк, упр. по заповедникам Совета Министров РСФСР,
вып. 12. М.
Павлов Н. В. 1948. Ботаническая география СССР. Алма-Ата, Изд-во АН Каз. ССР.
Падеревская М. И. 1958. К вопросу о значении почечных чешуи.— Уч. зап. Кур-
ского гос. пед. ин-та, вып. 7, естеств.-геогр. цикл.
Падеревская М. И. 4963. Об особенностях почек некоторых растений Стрелецкой
степи.— Бот. журн., т. 48, № 3.
Пачоский И. К. 1910. Основные черты развития флоры Юго-Западной России.—
Зап. Новороссийского об-ва естествоиспыт., приложение к 34 т. Херсон.
Пачоский И. К. 1917. Описание растительности Херсонской губернии. II. Степи.
Естеств .-мстор. музей Херсонского губ. земства.
Пачоский И. К. 1921. Основы фитосоциологии. Студенч. комитет с.-х. техникума,
Херсон.
Пачоский И. К. 1926. По поводу статьи проф. В. В. Алехина «Фитосоциология и ее
последние успехи у нас и на Западе».— Журн. Русск. бот. об-ва, т. 11, № <1—2.
Перетолчин К. 1904. Изменения запасных веществ наших деревьев в период зим-
него покоя.— Изв. Имп. лесного ин-та. СПб.
Петровская Т. П. 1955. Состояние покоя цветочных почек древесно-кустарниковых
растений.— Труды Ин-та физиол. раст. им. К. А. Тимирязева, т. 9. М., Изд-во
АН СССР.
Покровская В. М. 1940. Стрелецкая степь в аспективных картинах. — Труды
Центр.-Черноземн. гос. заповедника, вып. 1. М.
Полковниченко А. Я. 4957. К вопросу о биологической природе эфемеров.— Тру-
ды Туркм. с.-х. ин-та им. М. И. Калинина, т. 9. Ашхабад.
Полковниченко А. Я. 1958. Биологические особенности эфемеров Южной Турк-
мении. Автореф. канд. дисс. Л.
Полунина Н. Н. 1950. Средне-Азиатские эфемеры в условиях Москвы. — Бюлл.
Главн. бот. сада АН СССР, вып. 7. М.
Пономарев А. Н. 1948. Лесостепной комплекс северной окраины кунгурской лесо-
степи.— Изв. Естеств.-научи, ин-та при Пермском гос. ун-те им. А. М. Горького,
т. 12, вып. 6.
Пономарев А. Н. 1949. О лесостепном флористическом комплексе Северного и се'
верной части Среднего Урала.— Бот. журн., т. 34, № 4.
Пономарев А. Н. 1952. О лесостепном комплексе и сибирских влияниях во флоре
севера Европейской части СССР.— Изв. Естеств. научн. ин-та при Пермском гос.
ун-те им. А. М. Горького, т. 13, вып. 4—5.
Пономарев А. Н. 1959. О биологической изоляции Festuca sulcata Hack, и Festuca
pseudovina Hack.— Докл. АН СССР, т. 127, № 3.
Пономарев А. Н. 1960а. Экология цветения и опыления злаков.— Научн. докл.
высш. шк. биол. науки, №<1.
Пономарев А. Н. 19606. О протерандрии у зонтичных.— Докл. АН СССР, т. 135,
№ 3.
Пономарев А. Н^ 1960в. Клейстогамия у маревых.— Докл. АН СССР, т. 135, № 5.
Пономарев А. Н. 1960г. Суточный ритм и экология цветения и опыления ко-
стра безостого (Bromus inermis Leyss.).— Научн. докл. высш. шк. биол. науки,
№ 3.
Пономарев А. Н. 1961а. Клейстогамия у ковылей.— Бот. журн., т. 46, № 9.
Пономарев А. Н. 19616. О протерандрии у зонтичных.— Уч. зап. Пермского ун-та,
т. 18, вып. 3.
Пономарева И. Н. (Былинкина). 1959. Ритм сезонного развития растений
иссык-кульской котловины.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина,
т. 100, каф. ботаники, вып. 5. «Вопросы биологии растений». М.
Понятовская В. М. 1952. Стационарное изучение травянистых сообществ лесного
пояса южной Киргизии.— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 3
(геоботаника), вып. 8.
Поплавская Г. И 1948. Экология растений. М., Изд-во «Сов. наука».
Попов И. В. <1920. Из наблюдений над биологией сорно-полевой растительности Во-
ронежской опытной станции. Материалы по опытн. делу Воронежск. губ., вып. ,12.—
Труды Воронежск. с.-х. опытн. станции, № 5.
Попов М. Г. 1923. Флора пестроцветных толщ (краснопесчаниковых низкогорий)
Бухары. (Фрагмент к истории флоры Туркестана).— Труды Туркестанского научн.
об-ва, т. 1.
Попов М. Г. 1927. Основные черты истории развития флоры Средней Азии.— Бюлл.
Среднеаз. гос. ун-та, вып. 15. Ташкент.
Попов М. Г. 4929. Род Cicer и его виды. К проблеме происхождения средиземномор-
ской флоры (опыт морфологической и - географической монографии).— Труды по
прикл. ботанике, генетике и селекции, т. 21 (1928—1929 гг.), № 4. Л.
Попов М. Г. 1931. Между Монголией и Ираном.— Труды по прикл. ботанике, гене-
тике и селекции, т. 26, вып. 3. Л.
Попов М. Г. 1940а. Опыт монографии рода Eremostachys Bge. Новые мемуары Моск,
об-ва испыт. природы, т. 19. М.
279
Попов М. Г. 19406. Растительный покров Казахстана. — Труды Казахск. фил. АН
СССР, т. 48. М.—Л.
Попов М. Г. 1948. Род Erigeron в горах Средней Азии.—Труды Бот. ин-та
им. В. Л. Комарова АН СССР, серия 1 (флора и систематика высших растений),
вып. 7.
Попов М Г. 1949. Очерк растительности и флоры Карпат.— Материалы к познанию
фауны и флоры СССР, новая серия, отд. ботаники, вып. 5 (ХШ). М.
Попов М. Г. 1953. О взаимоотношении леса (тайги) и степи в Средней Сибири.—
Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 58, вып. 6.
Пошкурлат А. П. 1958. Изменение морфологических признаков с возрастом у лап-
чатки прямостоячей.— Potentilla erecta (L.) Натре.—Бюлл. Моск, об-ва испыт.
природы, отд. биологии, т. 63, вып. 3.
Пошкурлат А. П. 4962. Анализ ритма развития первоцвета весеннего.— Бот. журн.,
т. 47, № 2.
Пояркова А. И. 1924. О соотношении между глубиной зимнего покоя, превращением
запасных веществ и холодостойкостью у древесных растений.— Труды Ленингр.
об-ва естествоиспыт., т. 54.
Пояркова А. И. 1953. Sorbocotoneaster Poiark — новый естественный межродовой
гибрид.— Бот. материалы Гербария Бот. ин-та АН СССР, вып. 15.
Прозина М. Н. 1949. Зимний рост у птицемлечника (Ornithogalutn utnbellatum L.).—
Докл. АН СССР, новая серия, т. 64, № 6.
Прозоровский А. В. 1935. О биологических типах растений пустыни.— Бот. журн,
т. 21, №5.
Прозоровский А. В. 1940. Полупустыни и пустыни СССР.— В кн.: «Растительность
СССР>, т. 2. М.—Л., Изд-во АН СССР.
Прозоровский Н. А. 4940а. Изменение растительности Стрелецкой степи при от-
сутствии пастьбы скота и сенокошения.— Труды Центр.-Черноземн. гос. заповед-
ника, вып. >1. М.
Прозоровский Н. А. 19406. Наблюдения над осенним и зимним состоянием степ-
ных растений.— Труды Центр.-Черноземн. гос. заповедника, вып. 1. М.
Прозоровский Н. А. 1947. Данные о зимнем состоянии баранчика в лесостепной
зоне.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 52, вып. 4.
Прозоровский Н. А. 1949. Очерк растительного покрова центрально-черноземных
областей (Воронежской, Орловской и Тамбовской).— Вопр. географии, т. 13. М.,
Географгиз.
Прозоровский Н. А. 11951. Центрально-Черноземный заповедник им. проф.
В. В. Алехина.— В кн.: «Заповедники СССР>, т. 1. М., Географгиз.
Проскоряков Е. И. 1950. О происхождении однолетних эфемеров.— Труды У?
бекск. гос. ун-та, новая серия, № 43.
Работнов Т. А. 1950. Жизненный цикл многолетних травянистых растений в луго-
вых ценозах.— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 3 (геобота
ника), вып. 6.
Работнов Т. А. 1960. Новое руководство по методике геоботанических исследова-
ний.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 65, вып. 4.
Работнов Т. Jk. 1962. Некоторые вопросы изучения эдификаторов в луговых цено
зах.— Пробл. ботаники, вып. 6. М.— Л., Изд-во АН СССР.
Радкевич О. Н. 1926. Развитие склеренхимы в осевых органах среднеазиатских
гелиофитов.— Бюлл. Среднеазиатск. гос. ун-та, вып. 44.
Радкевич О. Н. 4928. Соотношение мягких и твердых тканей у травянистых и полу-
кустарниковых гелиофитов Средней Азии.— Изв. Главн. бет. сада СССР, т. 27,
вып. 4.
Рачковская Е. Н. 1957. К биологии пустынных полукустарничков.— Труды Бот.
ин-та им. В. Л. Комарова АН СССР, серия 3 (геоботаника), вып. 11.
Рещиков М. А. 1961. Степи западного Забайкалья.— Труды Восточно-Сиб. фил. СО
АН СССР, серия биол., вып. 34. М. Изд-во АН СССР.
Рихтер Г. Д. 1948. Роль снежного покрова в физико-географическом процессе.—
Труды Ин-та географии АН СССР, вып. 40. М.— Л.
Ричардс П. У. 1961. Тропический дождевой лес. М., ИЛ.
Р о д е А. А. 1952. Почвенная влага. М.— Л., Изд-во АН СССР.
Роде А. А. 1956. Водный режим почв и его типы.— Почвоведение, № 4.
Родионе н ко Г. И. 1961. Род ирис—Iris L. (Вопросы морфологии, биологии, эво-
люции и систематики). М.— Л., Изд-во АН СССР.
Родченко О. П. 1958. Динамика запасных веществ и изменения в состоянии прото-
плазмы растений люцерны и клевера при перезимовке.— Изв. АН СССР, серия
биол., № 5.
Родченко О. П. '1961. Жизнеспособность некоторых видов растений иркутско-бала-
ганской лесостепи, зимующих с зелеными листьями.— Материалы к конференции
молодых научн. сотрудн. Восточно-Сиб. фил. СО АН СССР, серия биол., вып. 2.
Благовещенск.
Ротов Р. А. 1959. Биолого-морфологические особенности древесно-кустарниковых
жизненных форм растений песчаной пустыни Каракум.— Уч. зап. моек. гор. пед.
280
ин-та им. В. П. Потемкина, т. 100, каф. ботаники, вып. 5. «Вопросы биологии расте-
ний». М.
Сакало Д. И. 1961. Лесостепной ландшафт Европейской части СССР и его расти-
тельность.— Бот. журн., Т. .46, № 7.
Сапожников В. В. 1901. Катунь и ее истоки.— Изв. Томского ун-та, кн. 18.
Саркисова-Федорова О. 1929. К биологии травяного покрова еловых лесов.
Очерки по фитосоциологии и фитогеографии. М., Изд-во «Новая Деревня».
Свешникова В. М. 1952. Корневая система растений Памира.— Труды Ин-та бота-
ники АН Тадж. ССР, т. 4. Душанбе, Изд-во АН Тадж. ССР.
Свешникова В. М. 1955. О корневых системах эфемеров и эфемероидов в Караку-
мах.— Труды Репетекской песчано-пустынной станции АН Туркм. ССР, т. 3.
Свешникова И. Н. 1951а. К морфологии соцветия рода Primula L.— Бот. журн.,
т. 36, № 2.
Свешникова И. Н. 19516. Морфологические исследования некоторых представите-
лей семейства первоцветных (Primulaceae). Автореф. канд. дисс. Л.
Семенова А. М. 4939. Фенологическое развитие субальпийских лугов Юго-Осетии.—
Сов. ботаника, № 4.
Семенова-Тян-Шанская А. М. 1948. Корневые системы растений субальпий-
ских лугов Юго-Осетии.— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 3
(геоботаника), вып. 5.
Семенова-Тян-Шанская А. М. 4960а. Взаимоотношения между живой зеленой
массой и мертвыми растительными остатками в луговых и степных сообществах.—
Журн. общ. биол. АН СССР, т. 21, № 2.
Семенова-Тян-Шанская А. М. 19606. Динамика накопления и разложения
мертвых растительных остатков в луговостепных и луговых ценозах.— Бот. журн.,
т. 45, № 9.
Семенова-Тян-Шанская А. М. 1962. Изменение ритмики развития степных
и луговых сообществ в зависимости от разных режимов их использования.— Пробл.
ботаники, вып. 6. М.— Л., Изд-во АН СССР.
Семенова-Тян-Шанская А. М., Никольская Н. И. 1960. Состав и ди-
намика надземной массы травостоев в степях и на лесных полях Центрально-
черноземного заповедника.— Труды Центр.-Черноземн. гос. заповедника, вып. 6.
Курск.
Сенянинова-Корчагина М. В. 4949. К вопросу о классификации жизненных
форм.— Уч. зап. Ленингр. гос. ун-та, № 107, серия геогр. наук, вып. 5.
Сергеев Л. И. 1945. Растения — лилипуты как показатель ухудшения условий роста
и развития.— Изв. Туркм. фил. АН СССР, .№ 5—6.
Сергеев Л. И. 1953. Выносливость растений. М., Изд-во «Сов. наука».
Сергеев Л. И. 1955. Вторичное цветение растений.— Природа, № 3.
Сергеев Л. И., Сергеева К. А., Мельников В. К. 4961. Морфофизиологическая
периодичность и зимостойкость древесных растений. Уфа, Изд-во АН СССР.
Серебряков И. Г. 4947. О ритме сезонного развития растений подмосковных ле-
сов.— Вести. Моск, ун-та, № 6.
Серебряков И. Г. 1952. Морфология вегетативных органов высших растений. М.,
Изд-во «Сов. наука».
Серебряков И. Г. 1954а. О морфогенезе жизненной формы дерева у лесных пород
средней полосы Европейской части СССР.— Бюлл. Моск, об-ва испыт. природы,
отд. биологии, т. 59, вып. 1.
Серебряков И. Г. 19546. Биолого-морфологический и филогенетический анализ жиз-
ненных форм покрытосеменных.— Уч. зап. Моск, гор. пед. ин-та им. В. П. Потем-
кина, т. 37, каф. ботаники, вып. 2. М.
Серебряков И. Г. 1955а. Основные направления эволюции жизненных форм у по-
крытосеменных растений.— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 60,
вып. 3.
Серебряков И. Г. 19556. О работах VIII международного конгресса ботаников. Эк-
скурсии по Франции и Африке.— Бюлл. Моск, об-ва испыт. природы, отд. биологии,
т. 60, вып. 1.
Серебряков И. Г. 1959а. Типы развития побегов у травянистых многолетников
и факторы их формирования.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина,
т. 100, каф. ботаники, вып. 5. «Вопросы биологии растений». М.
Серебряков И. Г. 19596. Период покоя у некоторых травянистых и древесных ра-
стений Подмосковья.— Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, т. 100,
каф. ботаники, вып. 5. «Вопросы биологии растений». М.
Серебряков И. Г. 1961. Ритм сезонного развития растений Хибинских тундр.—
Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 66, вып. 5.
Серебряков И. Г. 1962. Ритм сезонного развития растений Приполярного Урала.—
Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 67, вып. 3.
Серебряков И. Г. 1963. Некоторые данные о ритме сезонного развития растений
долины реки Пясииы в ее нижнем течении.— Бюлл. Моск, об-ва испыт. природы,
отд. биологии, т. 67, вып. 2.
281
«Серебряков И. Г. Галицкая Т. М. 1951. К биологии сезонного развития болот •
ных растений Подмосковья в связи с условиями их жизни и происхождением.—
Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина, т. 19, каф. ботаники, вып. 1.
Серебряков И. Г., Чернышева М. Б. 1955. О морфогенезе жизненной формы
кустарничка у черники, брусники и некоторых болотных Ericaceae.— Бюлл. Моск,
об-ва испыт. природы, отд. биологии, т. 60, вып. 2.
Серебрякова Т. И. 1956. Побегообразование и ритм сезонного развития растений
заливных лугов средней Оки.— Уч. зап. Моск. гос. пед. ин-та им. В. И. Ленина,
т. 97, каф. ботаники, вып. 3.
Серебрякова Т. И. 1959. О некоторых морфологических закономерностях побего-
образования у овсяницы луговой (Festuca pratensis Huds.).— Бюлл. Моск, об-ва
испыт. природы, отд. биологии, т. 64, вып. 3.
Серебрякова Т. И. 1960. О двух формах кущения у костра безостого (Bromus
inermis Leyss.) в первый год жизни.— Бюлл. Моск, об-ва испыт. природы, отд.
биологии, т. 65, вып. 5.
Серебрякова Т. И. 1961. О формировании куста у овсяницы луговой в первый год
жизни.— В кн.: «Морфогенез растений», т. 1. Изд-во Моск, ун-та.
Серебрякова Т. И. 1962. К вопросу о побегообразовании и кущении у злаков.—
Бот. жури., т. 47, № 3.
Середин Р. М. 1951. О вторичном цветении конского каштана на кавказских мине-
ральных водах.— Природа, № 8.
Скрипчинский В. В. ’1958а. Некоторые закономерности формирования куста у мно-
голетних рыхлокустовых злаков в первый и последующие годы жизни в сравнении
с родственными им однолетними злаками.— Докл. АН СССР, т. 118, № 4.
Скрипчинский В. В. 19586. Зависимость функционирования органов вегетативного
воспроизведения многолетних и однолетних злаков от процесса образования се-
мян.— Докл. АН СССР, т. <118, № 5.
Смирнов В. А. 1945. Костер безостый. Канд. дисс. Пушкин.
Смирнов В. А. 1938. О некоторых видах рода Agrostis L.— Бюлл. Моск, об-ва испыт.
природы, отд. биологии, т. 47, вып. 4.
Смирнов П. А. 4958. Флора Приокско-Террасного государственного заповедника.—
Труды Приокск.-Террасн. гос. заповедника, вып. 2. М.
Сочава В. Б. 1945. Фрагменты горной степи на Среднем Урале.— Сов. ботаника,
т. 13, № 3.
Сочава В. Б. 1946. Вопросы флорогенеза и филоценогенеза манчжурского смешанно-
го леса.— Материалы по истории флсры и растит. СССР, вып. 2. М.— Л., Изд-во
АН СССР.
Сочава В. Б., Липатова В. В., Горшкова А. А. 1962. Опыт учета полной про-
дуктивности надземной части травяного покрова.— Бот. журн., т. 47, № 4.
Спрыгни И. И. 1915. Новая работа из области северных степей (Алехин В. В.
Введение во флору Тамбовской губ. Ботанический очерк).— Труды Пензенск. об-ва
любит, естествозн., т. 2.
Спрыгни И. И. 4917. Ответ Алехину.— Труды Пензенск. об-ва любит, естествозн., т. 3.
Спрыгни И. И. 1922. Борьба леса со степью в Пензенской губернии. Пенза.
Спрыгни И. И. 1925. Из области пензенской лесостепи. 1. Травяные степи Пензен-
ской губернии.— Труды по изучению заповедников. Отд. охраны природы главнауки
НКП, вып. 4. М.
Станюкович К. В. 1949. Растительный покров Восточного Памира.— Зап. Всесоюзн.
геогр. об-ва, новая серия, т. 10. М., Географгиз.
Старикова В. В. 1963. Методика изучения семенной продуктивности растений на
примере эспарцета Onobrychis arenaria.— Бот. журн., т. 48, № 5.
Стешенко А. П. 1956. Формирование структуры полукустарничков в условиях высо-
когорий Памира.— Труды Памирск. биол. станции Бот. ин-та АН Тадж. ССР, т. 50.
Душанбе, Изд-во АН Тадж. ССР.
СтржемескаяА. И. 1953. К биологии мать-и-мачехи.— Уч. зап. Моск. гос. пед. ин-та
им. В. И. Ленина, т. 73, каф. ботаники, вып. 2.
Сукачев В. Н. 1903. Очерк растительности юго-восточной части Курской губернии.—
Изв. Лесного ин-та, вып. 9, неофиц. отд.
Сукачев В. Н. 1915. Введение в учение о растительных сообществах. Пг.
Сукачев В. Н. 1928. Растительные сообщества. (Введение в фитосоциологию).
Л.— М., Изд-во «Книга».
Сукачев В. Н. 4934. Дендрология с основами лесной геоботаники. Л., Гослестехиз-
дат.
Сукачев В. Н. 1944. О принципах генетической классификации в фитоценологии.—
Журн. общ. биологии АН СССР, т. 5, № 4.
Сукачев В. Н. 1956. О некоторых современных проблемах изучения растительного
покрова.— Бот. журн., т. 41, № 4.
Сукачев В. Н., Зоин С. В. 1961. Методические указания к изучению типов леса*
М., Изд-во АН СССР.
Талиев В. И. 1901. Контакт леса и степи в Валкском уезде Харьковской губернии.
Харьков.
282
Т а л и е в В. И. 1904. Нерешенная проблема русской ботанической географии.— Лесной
журн., № 3—4.
Та л ие в В. И. 1913. Введение в ботаническое исследование Харьковской губернии. Изд.
Харьковск. губ. земства.
Т а л и е в В. И. 1925. Биология наших растений. М.— Л., Госиздат.
Т а л и е в В. И. 4933. Основы ботаники в эволюционном изложении. Изд. 7. М.— Л.,
Сельхозгиз.
Т а н ф и л ь е в Г. И. 1894. Пределы лесов на юге России.— Труды экспедиции, снаря-
женной Лесным департаментом под руководством проф. Докучаева. СПб.
Танфильев Г. И. 1896. Доисторические степи Европейской России.— Землеведение,
№2.
Танфильев Г. И. 4898. Ботанико-географические исследования в степной полосе.—
Труды Особой экспедиции Лесного департамента. СПб.
Танфильев Г. И. 1901. К вопросу о безлесии степей. (Ответ В. И. Талиеву).—
Естествозн и география, № 5.
Тараканов К. Н. 1940. Изменение Bromus danthonie Trin. в различных условиях
выращивания.— Докл. АН СССР, т. 29, № 7.
Тахтаджян А. Л. 1943. Соотношения онтогенеза и филогенеза у высших растений
(этюды по эволюционной морфологии).— Научн. труды Ереванск. гос. ун-та, т. 22.
Тахтаджян А. Л. 1946. К истории развития растительности Армении.— Труды Бот.
ин-та АН Арм. ССР, т. 4.
Тахтаджян А. Л. 1948. Морфологическая эволюция покрытосеменных. М.
Тахтаджян А. Л. 1954а. Происхождение покрытосеменных растений. М., Изд-во
«Сов. наука».
Тахтаджян А. Л. 19546. Вопросы эволюционной морфологии растений. Изд-во
Ленингр. ун-та.
Тахтаджян А. Л. 1956. Высшие растения. I. От псилофитовых до хвойных. М.— Л.,
Изд-во АН СССР.
Тахтаджян А. Л. 1957. К вопросу о происхождении умеренной флоры Евразии.—
Бот. журн., т. 42, № 11.
Темноев Н. И. 1939. Перезимовывание надземных органов у некоторых травянистых
растений на Крайнем Севере.— Труды Научно-ислед. ин-та полярного землед., жи-
вотное. и промысл, хозяйства, серия «Оленеводство», вып. 4. Л.
Тихомиров Б. А. 1950. К биологии растений Крайнего Севера.— Бюлл. Моск, об-ва
испыт. природы, отд. биологии, т. 55, вып. 4.
Толмачев А. И. 1951. О некоторых архаичных чертах растений тайги, их экологиче-
ской и исторической обусловленности.— Труды Томск, гос. ун-та, т. 116 (Сб. памя-
ти П. Н. Крылова).
Толмачев А. И. 1954. К истории возникновения и развития темнохвойной тайги.
М.— Л., Изд-во АН СССР.
Толмачев А. И. 1957. К истории развития и географического распространения рода
Draba L.— Бот. журн., т. 42, № 9.
Толмачев А. И. 1958. О происхождении некоторых основных элементов высокогор-
ных флор Северного полушария.— Материалы по истории флоры и растит. СССР,
вып. 3. М.— Л., Изд-во АН СССР.
Троицкий Н. А. 1959. О гибридогенном возникновении видов у растений.— Бот.
журн., т. 44, № 3.
Трофимов Т. Т. 1939. К вопросу о ритме развития ранневесенних растений.—
Научно-метод. зап. Главн. управл. по заповедникам, вып. 5.
Трофимов Т. Т.*1952. Вегетативное размножение хохлатки Галлера (плотной)
(Corydalis halleri Willd.).— Бюлл. Моск, об-ва испыт. природы, отд. биологии, т. 57,
вып. 3.
Трофимов Т. Т. 1954. О подснежном росте и развитии ранневесенних растений.—
Природа, № .1.
Туманов И. И. 1940. Физиологические основы зимостойкости культурных растений.
М., Сельхозгиз.
Туманов И. И. 1951. Основные достижения советской науки в изучении морозостой-
кости растений. 11-е Тимирязевское чтение. М.— Л., Изд-во АН СССР.
Устинова Е. И. 1949. Ритм развития конуса нарастания в луковицах пролески
(Scilla sibirica) в осенне-зимнее время.— Докл. АН СССР, новая серия, т. 64, № 6.
Федоров Ан. А. 1942. Альпийские ковры Кавказа и их происхождение.— Изв. Арм.
фцл. АН СССР, № 9—10.
Федоров Ан. А. 1952. История высокогорной флоры Кавказа в четвертичное время
как пример аутохтонного развития третичной флористической основы. Материа-
лы по четвертичному периоду СССР, вып. 3. М.— Л., Изд-во АН СССР.
Федоров Ан. А. 1958. О флористических связях Восточной Азии с Кавказом (на
примере изучения рода Pyrus L. s. str.). Материалы по истории флоры и растит.
СССР, вып. 3. М — Л., Изд-во* АН СССР.
Федченко Б. А. 1922. О новом однолетнем виде Tamarix из Туркестана.—Бот. ма-
териалы Гербария Главн. бот. сада РСФСР, т. 3, вып. 44—46.
Федюнкин Д. Ф. 1953. Влияние мертвых растительных остатков и степных палов
283
на развитие растительности лесостепного Зауралья.— Изв. естеств.-научн. ин-та при
Пермском гос. ун-те им. А. М. Горького, т. 13, вып. 7.
Хитрово В. Н. 1910. Рациональная постановка фенологии, как метода ботанической
географии. Дневник XII съезда русск. естествоиспыт. и врачей в Москве с 28 де-
кабря 1909 г. по 6 января 1910 г. Отд. 2. Программы и протоколы общих, соеди-
ненных и секционных заседаний. М.
Хитрово В. Н. 4925. Растительность.—В кн.: «Природа Орловского края». Орловск,
губ. планов, комиссии.
Хохлов С. С. 1947. Вторичное цветение плодовых и другие особенности поведения
растений в районе газоносной скважины в окрестностях Саратова.—Сов. ботани-
ка, № 1.
Хржановский В. Г. 1940. Историческое развитие учения о виде.—Уч. зап. Казахск.
гос. ун-та им. С. М. Кирова, т. 3 (биология). Приложение 1. Алма-Ата.
Хржановский В. Г. 1958а. К вопросу о соматической эволюции у покрытосемен-
ных.— Докл. ТСХА, вып. 36. М.
Хржановский В. Г. ,19586. Розы. М., Изд-во «Сов. наука».
Чефранова 3. В. 1958. Материалы к монографии рода термопсис (Thermop-
sis R. Вг.).— Труды Бот. ин-та им. В. Л. Комарова АН СССР, серия 1 (флора и си-
стематика высших растений), вып. 12.
Шалыт М. С. 4930. Геоботанический очерк государственного степного заповедника
«Чапли» (б. Аскания-Нова).— Бюлл. фитотехн. станции Аскания-Нова, т. 1. Мели-
тополь.
Ш а л ы т М. С. 1946. О фитофенологических спектрах.— Сов. ботаника, т. 14, № 4.
Ш а л ы т М. С. 4950. Подземная часть некоторых луговых, степных и пустынных расте-
ний и фитоценозов. Ч. 1. Травянистые и полукустарничковые растения и фитоцено-
зы лесной (луга) и степной зон.— Труды Бот. ин-та им. В. Л. Комарова АН СССРв'
серия 3 (геоботаника), вып. 6.
Шалыт М. С. 1955. Система жизненных форм степных растений.— Уч. зап. Тадж. гос
ун-та, т. 6, Труды фак-та естеств. наук, вып. 1.
III а ф е р В. 1956. Основы общей географии растений. М., ИЛ.
Шенников А. П. 1928. Фенологические спектры растительных сообществ.— Труды
Вологодск. обл. с.-х. опытн. станции, вып. 2.
Шенников А. П. 1935. Принципы ботанической классификации лугов.— Сов. ботани-
ка, № 5.
Шенников А. П. 1938. Луговая растительность СССР.— В кн.: «Растительность
СССР», т. 1. М.—Л., Изд-во АН СССР.
Шенников А. П. 1950. Экология растений. Изд-во «Сов. наука». М.
Шенников А. П. 1962. К созданию единой естественной классификации раститель-
ности.— Пробл. ботаники, вып. 6. М.— Л., Изд-во АН СССР.
Шенников А. П. 1963. Задачи лаборатории экспериментальной геоботаники Бота-
нического института Академии наук СССР.— Труды Бот. пн-та им. В. Л. Комарова
АН СССР, серия 3 (геоботаника), вып. 14. «Комплексные экспериментальные гео-
ботанические исследования».
Шенников А. П., И о ф ф е А. Ф. 1944. К биологии пустынных злаков.— Бот. журн,
СССР, т. 29, № 1.
Шик М. М. ,1953. Сезонное развитие травяного покрова дубравы.— Уч. зап. Моск, гос,
пед. ин-та им. В. И. Ленина, т. 73, каф. ботаники, вып. 2. М.
Шишкин Б. К. 1954. К вопросу о видообразовании.— Бот. журн., т. 39, № 2.
Шнитников А. В. 1957. Изменчивость общей увлажненности материков северного
полушария.— Зап. геогр. об-ва СССР, т. 16.
Шрейбер А. Ф. 1908. Причины вторичного цветения растений.— Изв. Восточно-Сиб.
отд. Русск. геогр. об-ва, т. 39.
Шрейбер А. Ф. 19111. Причины вторичного (осеннего) цветения растений.— Вести, са-
доводства, плодоводства и огородничества, №11.
Штамм В. А. 1953. О причинах разновременного зацветания лещины.— Бюлл. Главн.
бот. сада АН СССР, вып. 16. М., Изд-во АН СССР.
Штемпель Б. М. 1959. К вопросу о местообитании первых покрытосеменных расте-
ний.— Бот. журн., т. 44, № 7.
Шумилова Л. В. 1962. Ботаническая география Сибири. Изд-во Томск, ун-та.
Эйгенсон М. С. 1957. Очерки физико-географических проявлений солнечной активно-
сти. Изд. Львовск. гос. ун-та.
Эйхвальд К. Ю. 1959. Подрод ежевик Cylactis Rafin. Исследование филогенеза од-
ной бореальной растительной группы.— Уч. зап. Тартусск. гос. ун-та, вып. 81.
Юзе п ч у к С. В. 1936. Новые данные по систематике среднеазиатских видов рода
Cousinia Cass.— Труды Бот. ин-та АН СССР, серия 1, вып. 3.
Юзепчук С. В. 1939. Проблема вида в свете учения Дарвина. Сов. ботаника, № 6—7.
Юзепчук С. В. 1941. Лапчатка — Potentilla L.— Флора СССР, т. 10. М.— Л., Изд-во
АН СССР.
Юзепчук С. В. 1958. Комаровская концепция вида, ее историческое развитие и отра-
жение во «Флоре СССР».— В сб.: «Проблема вида в ботанике», т. 1. М.— Л., Изд-во
АН СССР.
284
Ю н о в и до в А. П. 1963. О взаимоотношениях леса и степи в северной части Казах-
станского мелкосопочника.— Бот. журн., т. 48, № 2.
Ярмоленко А. В. 1941. Опыт построения системы рода Parietaria и родственных ему
форм. (Материалы к монографии).— Труды Бот. ин-та им. В. Л. Комарова АН
СССР, серия 1 (флора и систематика высших растений), вып. 5.
Яценко-Хмелевский А. А. 1948. Принципы систематики древесины.— Труды Бот.
ин-та АН Арм. ССР, т. 5.
Яценко-Хмелевский А. А. 1958. Происхождение покрытосеменных, по данным
внутренней морфологии их вегетативных органов.— Бот. журн., т. 43, № 3.
Яшина А. В. 4961а. Подснежное развитие растений.— В сб.: «Роль снежного покрова
в природных процессах». К 60-летию со дня рождения Г. Д. Рихтера. *. М., Изд-во
АН СССР.
Яшина А. В. 19616. О проникновении солнечной радиации в толщу снежного покро-
ва.— В сб.: «Роль снежного покрова в природных процессах». К 60-летию со дня
рождения Г. Д. Рихтера. М., Изд-во АН СССР.
А г b е г А. 1928. The tree-habit tn angiosperms: its origin and meaning.— New Phytolo-
gist, v. 27, N 1.
Areschoug F. W. 1895. Beitrage zur Biologie der geophilen Pflanzen.— Acta Univ.
Lundensis, Bd. 31.
A r 1 d t Th. 1907. Die Entwicklung der Kontinente und ihrer Lebewelt. Leipzig.
Bews J.W. 1925. Plant forms and their evolution in south Africa. London.
Bews J. W. 1927. Studies in the ecological evolution of the angiosperms.— New Phyto-
logist, v. 26, Reprint. London.
Beiley I. W. 1911. The relation of the leaf-trace to the formation of compound rays in
the lower dicotyledons.— Ann. Bot., v. 25.
В a i ley I. W. 1949. Origin of the angiosperms: need for a broadened outlook.— J. Arnold
Arboretum, v. 30.
Bonnier G. 1890. Cultures experimentales dans les Alpes et les Pyrdnees.— Rev. gen.
bot., v. 2.
Bonnier G. 1895. Recherches experimentales sur 1’adaptation des plantes au climat
alpin.— Ann. sci. natur. Bot., t. 20, ann. 19, ser. 7.
В о n n i e r G. 1910. Le monde vegetal. Paris.
Braun-Blanquet D. J. 1928. Pflanzensoziologie. Grundzuge der Vegetationskunde.
Wien.
Corner E. S. 1949. The durian-theory of the origin of the modern tree.— Ann. Bot.,
N. S., v. 13, N 52.
Co r n er E. S. 1954. The durian-theory.— Phyto morphology, v. 3.
Diels L. 1910. Genetische Elemente in der Flora der Alpen.— Engler’s Bot. Jahrb.,
Bd. 44, S. 102.
Diels L. 1918. Das Verhaltnis von Rhytmik und Verbreitung bei der Perennen des euro-
paischen Sommerwaldes — Ber. Deutsch, bot. Ges., Bd. 36, H. 6.
Driide O. 1913. Die Okologie der Pflanzen.— Die Wissenschaft. Einzelndarstellungen
aus der Natur-Wissenschaft und der Technik. Bd. 50.
Du Rietz G. 1931. Life forms of terrestrial flowering plants. I.— Acta phytogeogr.
suecica, v. 3.
Engler A. und Prantl K. 1894, 1897. Die naturlichen Pflanzenfamilien. III. und IV.
Theil. Leipzig. 4
Gams H. 1918. Pnnzipienfragen der Vegetationsforschung.— Vierteljahrschr. Naturforsch.
Ges. Ziirich, Bd. 63.
Geleznoff N. 1851. Observations sur la development des bourgeons pendant 1’hiver.—
Bull. Soc. imp. natur. Moscou, t. 3.
Goebel K. 1928—1933. Organographie der Pflanzen, insbesondeie der Archegoniaten
und Samenpflanzen. 3 Aufl. Theil 1—4. Jena.
Grisebach. A. 1872. Die Vegetation der Erde. Leipzig.
Hal Her H. 1905. Ein zweiter Entwurf des naturlichen (phylogenetischen) System der
Blutenpflanzen.— Ber. Deutsch, bot Ges., Bd. 23.
Hallier H. 1912. Uber Where Landbrucken, Pflanzen- und Volkerwanderungen
zwischen Australasien und Amerika.— Meded. van’s Rijks herbarium, Bd. 8—14.
Hansen M. 1956. Life forms as age indicators. Ringkjobing.
Hauri H. 1912. Anabasis aretioides Mog. et Coss., eine Polsterpflanze der algerichen
Sahara.— Bot. bl., Beih., Bd. 28, Abt. 1, H. 3.
Hauri H. und Schrdter C. 1914. Versuch einiger Ubersicht der Siphonogamen Pol-
sterpflanzen.— Bot. Jahrb., Bd. 50, Suppl.
Hult R. 1881. Forsok till analytisk behandling af vaxtformationerna.— Meddelanden Soc.
pro Fauna et Flora Fennica, v. 8.
Irmisch Th. 1850. Zur Morphologie der monokotylischen Knollen- und Zwiebelgewachse.
Berlin.
Irmisch Th. 1859. Uber Lathyrus tuberosus und einige andere Papillionaceen.— Bot.
Zeit., Jg. 17, N 7—9.
285
I rm is ch Th. 1861. Ober Polygonum amphibium, Lysimachia vulgaris, Comarum palust-
re und Mehyanthes trifoliata.— Bot. Zeit, Jg. 19, N 17—19.
Jeffrey E. C. 1899. The morphology of the central cylinder in the angiosperms.—
Trans. Canad. Inst., v. 6.
Jeswiet J. 1914. Eine Einteilung der Pflanzen der niederlandischen Kustendunen in
okologische Gruppen.— z. Bot. bl., Beih., Bd. 31, Abt 2, H. 2.
К e г n e r A. 1863. Das Pflanzenleben der Donaulander. Jena.
Kirchner O., Loew E., Schrdter C. 1908. Lebensgeschichte der Blutenpflanzen
Mitteleuropas. Bd. 1, Abt. 1. Stuttgart.
Lindforss B. 1907. Die wintergrune Flora. Lunds Univ. Arsskr., N. F., Afd. 2, Bd. 2.
Lindinger L. 1909. Jahresringe bei den Monokotylen der Drachenbaumform.— Natur-
wiss. Wochenschr., N. F., Bd. 8.
Linkola K. 1922. Zur Kenntnis der Oberwinterung der Unkrauter und Ruderalpflanzen
in der Gegend von Helsingfors.—Ann. Soc. zool.-bot. fennicae, t. 1, N 7.
Massart J. 1910. Esquisse de la geographic botanique de la Belgique.—Recueil Inst,
bot. Leo Errera, Bruxells, Tome supplementaire, 7 bis.
Meusel H. 1951. Die Bedeutung der Wuchsform fur die Entwicklung des naturlichen
Systems der Pflanzen.— Feddes Repertorium spec. nov. regn. veget., Berlin, Bd. 54.
Meusel H. 1952. Uber Wuchsformen, Verbreitung und Phylogenie einiger mediterran-
mitteleuropaischer Angiospermen-Gattungen.— Flora, oder Allgem. bot. Zg, Bd. 139,
H. 3.
Meusel H. 1955. Uber Wuchsform und Wuchsdauer mediterraner Einjahrspflanzen.—
Wiss. Z. Martin-Luther-Univ., Halle — Wittenberg. Math.-naturwiss. Reihe, Bd. 4, N 3.
Raunkiaer C. 1905. Types biologiques pour la geographic botanique.— Kgl. Dansk.
vidensk. selskabs. forhandl., Bd. 5.
Raunkiaer C. 1934. The life forms of plants and statistical plant geography. Oxford.
Rubel E. 1925. Alpenmatten-Uberwinterungstadien.— Festschrift Karl Schroter. Verof-
fentl. Geobot. Inst. Rubel in Zurich, H. 3.
Rubel E. 1930. Pflanzengesellschaften der Erde. Bern-Berlin.
Schlittler J. 1955. Vorlaufige Mitteilung uber die organphylogenetischen Zusammen-
hande der wichtigsten grundgestalten bei den Monokotyledonen, mit spezieller Be-
riicksichtigung der Liliaceen und einigen Bemerkungen zu deren Systematik.—Vier-
teljahrsschr. Naturforsch. Ges. in Zurich, Jg, 100, N 3.
Schlittler J. 1957. Die Verbreitung der Liliaceengattung Dianella Lam. im Zusammen-
hang mit der Organdifferenzierung und der Arealbildung.— Vierteljahrsschr. natur-
forsch. Ges. in Zurich, Jg. 102, N 1.
Schmidt E. 1957. Ein Vergleich der Wuchsformen im illyrischen Buchen- und Laubmi-
schwald.— Ber. uber das Geobot. Forschungsinst. Rubel in Zurich f. d. Jahr. 1956.
Zurich.
Schroter C. 1926. Das Pflanzenleben der Alpen. Eine Schilderung der Hochgebirgsflora.
2 Aufl. Zurich.
Seward A. C. and Conway V. M. 1935. Additional cretaceous plants from Western
Greenland.—Kgl. Svenska vetensk. akad. handl. v. 15, N. 3.
Sinnott E. W. and В a i 1 e у I. W. 1914a. Investigations on the phylogeny of the agios-
perms. 3. Nodal anatomy and the morphology of stipules.— J. Amer. bot. Soc., v. 1.
Sinnott E. W. and Bailey I. W. 1914b. The origin and dispersal of herbaceous angi-
osperms.— Ann. Bot., v. 28.
Sinnott E. W. and Bailey I. W. 1915. Changes in the fruit type of angiosperms co-
incident with the development of the herbaceous habit.— Rep. Sci. Bot. Soc. Amer.,
v. 41.
S p a 1 d i n g V. 1909. Distribution and movements of desert plants.— Wash., Carnegie Inst.
Stiefelhagen H. 1910. Sistematische und pflanzengeographische Studien zur Kennt-
nis der Gattung Scrophularia.— Engler’s Bot. Jahrb., Bd. 44.
Stocker O. 1928. Halophytenproblem.— Ergebn. Biol., Bd. 3.
Troll W. 1937—1942. Vergleichende Morphologic der hoheren Pflanzen.— Bd. 1—2,
Berlin.
Troll W. 1954—1957. Praktische Einfuhrung in die Pflanzenmorphologie. 1. Teil: Der
vegetative Aufbau; 2 Teil: Die bluhende Pflanze. Jena.
Velenovsky J. 1905—1913. Vergleichende Morphologic der Pflanzen. Teil 1—4 (T.
1 — 1905, T. 2— 1907, T. 3— 1910, T. 4— 1913). Prag.
Warming E. 1884. Uber Sprofibau, Uberwinterung und Verjflngung.— Engler’s Bot
Jahrb., Bd. 5.
Warming E. 1908. Om planterigets lievsformer. Kobenhavn.
Warming E. 1909. Ecology of plants. Oxford.
Warming E. und Graebner J. 1918. Lehrbuch der okologischen Pflanzengeographie.
3 Aufl. Berlin.
Wiesner! 1907. Der Lichtgenufi der Pflanzen. Leipzig.
Wolf Th. 1908. Monographie der Gattung Potentilla, Bibliotheca bot,. Stuttgart, Bd. 16,
H. 71.
286
ОГЛАВЛЕНИЕ
Введение .................................................................... 3
Глава 1. Погодные условия периода наблюдений................................. 7
Глава 2. Основные биоморфологические типы структур надземных органов
растений.................................................................. 16
Глава 3. Некоторые закономерности эволюции биоморфологических структур
растений.................................................................. 30
Глава 4. Характер и длительность вегетации травянистых растений .... 82
Формирование надземных органов......................................... 83
Отмирание листьев и побегов в течение вегетационного периода .... 97
Глав а 5. Перезимовка травянистых растений.................................105-
Глава 6. Ритмы цветения растений.............................................132
Нормальное цветение.......................................................132
Вторичное -цветение.......................................................159
Глава 7. Эколого-биологический состав компонентов основных растительных
группировок центральной лесостепи........................................169-
Глава 8. Проблема природы и происхождения растительности луговых степей 228
Глава 9. Вопросы практического использования эколого-биологических ис-
следований природных растений лесостепи...................................252
приложения
I. Ритмы цветения некоторых растений Стрелецкой степи Центрально-Черно-
земного заповедника в многолетнем ряду (по 1961 г. включительно) . . . 258
II. Ритмы цветения некоторых растений Казацкой степи Центрально-Чернозем-
ного заповедника в многолетнем ряду (по наблюдениям в 1936—1937 и
*1950—1954 гг.).......................................................260
III. Средние многолетние даты начала и полного созревания плодов и семян не-
которых растений Стрелецкой стеци Центрально-Черноземного заповедника 952
(по 1962 г. включительно)............................................
IV. Средние многолетние даты начала и полного созревания плодов и семян 263
некоторых растений Казацкой степи Центрально-Черноземного заповедника
(по наблюдениям в 1936—1937 и 1950—-.1954 гг.).......................
V. Ритмы цветения древесно-кустарниковых пород Стрелецкого участка Цен-
трально-Черноземного заповедника в многолетнем ряду (по 1961 г. включи-
тельно) ................................................................
VI. Средние многолетние даты начала и полного созревания плодов древесно-
кустарниковых пород Стрелецкого и Казацкого участков Центрально-Черно-
земного заповедника (по 1962 г. включительно).............................. 265
VII. Ритмы цветения некоторых травянистых растений дубовых лесов Стрелец-
кого и Казацкого участков Центрально-Черноземного заповедника в много-
летнем ряду (по 1962 г. включительно).......................................266
VIII. Средние многолетние даты начала и полного созревания плодов и семян
некоторых травянистых растений дубовых лесов Стрелецкого и Казацкого
участков Центрально-Черноземного заповедника (по 1962 г. включительно) 268
Литература..................................................................269
Виталий Николаевич Голубев
Эколого-биологические особенности растений
и растительных сообществ лесостепи
Утверждено к печати
Восточно-Сибирским биологическим институтом Сибирского отделения
Академии наук СССР
Редактор ИА, М. Культиасов
Технический редактор Я. И. Дорохина
Художник ЛН, И. Растащенов
Сдано в набор 29/Х 1964 г. Подписано к печати 18/11 1965 г.
Формат 70х108*/ц. 18 печ. л/Усл. печ. л. 24,66. Уч.-изд. л. 26,1. Тираж 1400 экз.
Т-00333. Изд. № 3612. Тип. зак. № 1395. Темплан 1965 г. № 559
Цена 2 р. 03 к.
Издательство «Наука». Москва, К-62, Подсосенский пер., 21
2-я типография издательства «Наука». Москва, Г-99, Шубинский пер., 10
ОПЕЧАТКИ И ИСПРАВЛЕНИЯ
Страница Строка Напечатано Должно быть
41 9 СВ. широкий цикорий
92 27 св. Серебрякова Серебряков
110 19—20 св. физическим физилогическим
185 7 св. степень степь
207 Затоловок отмирания листьев в
табл. 28 отмирания основании
241 24 св. по но
282 20 св. Смирнов В. А. Смирнов П. А.
В. Н. , Голубев