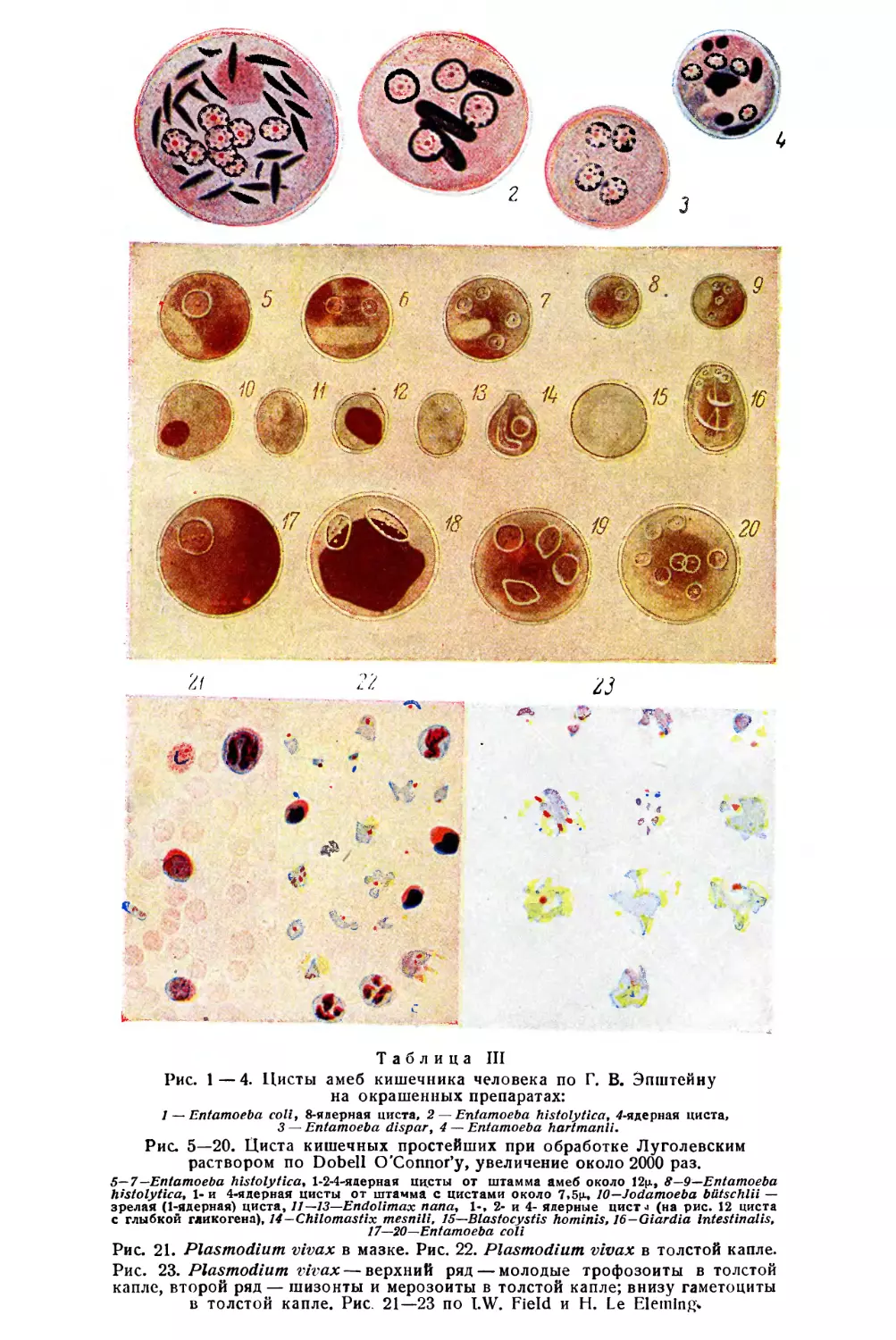
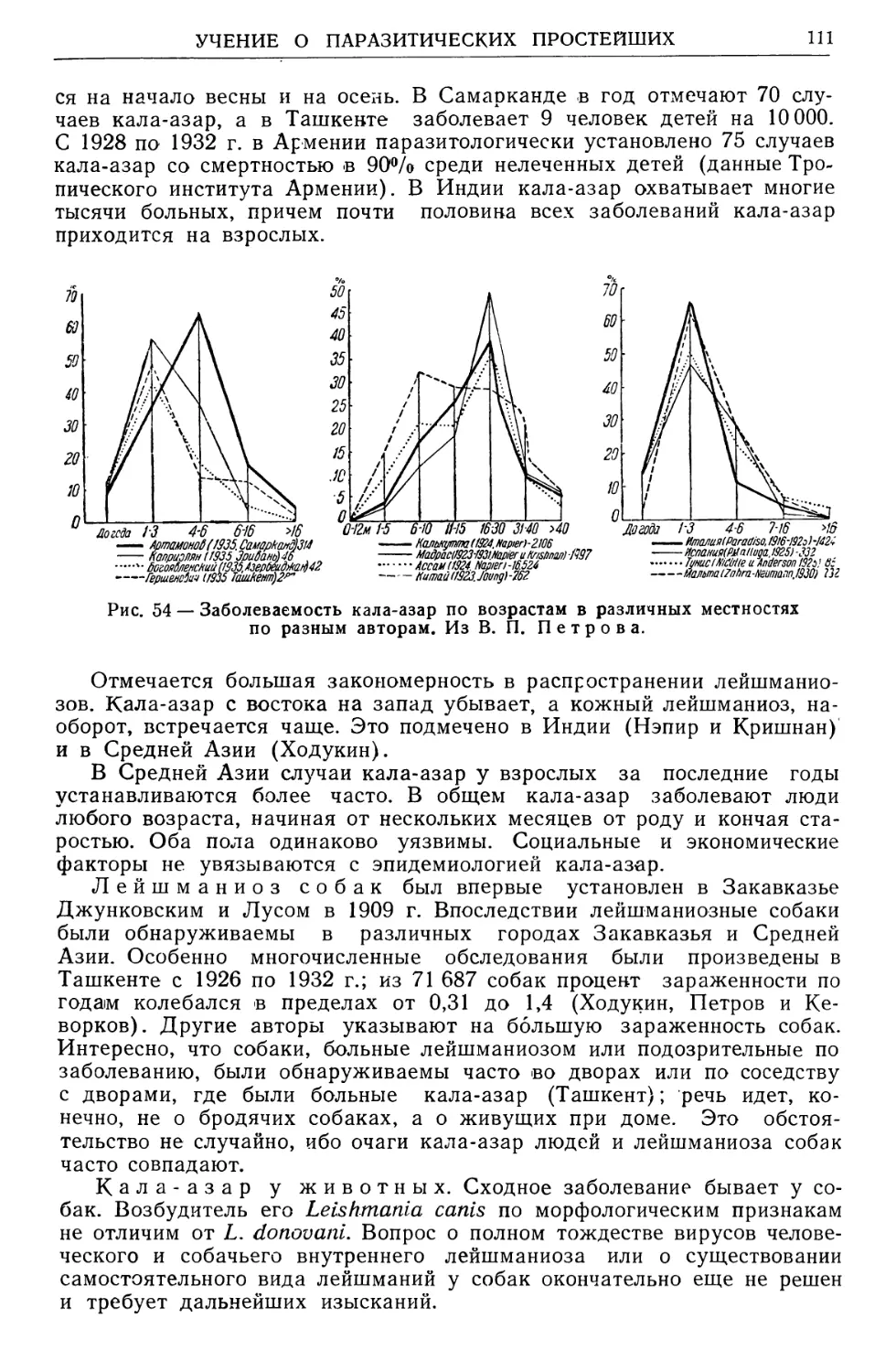
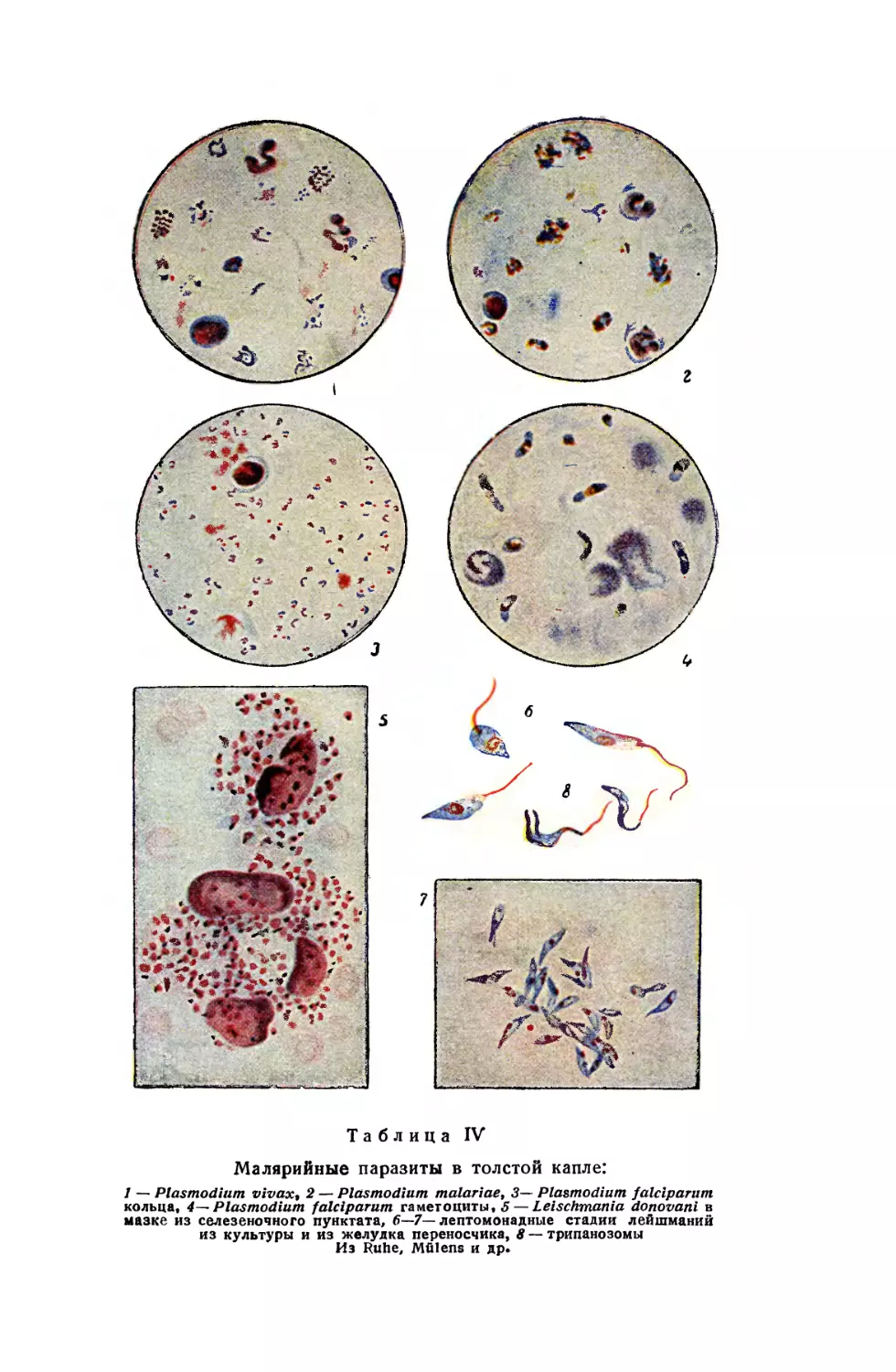
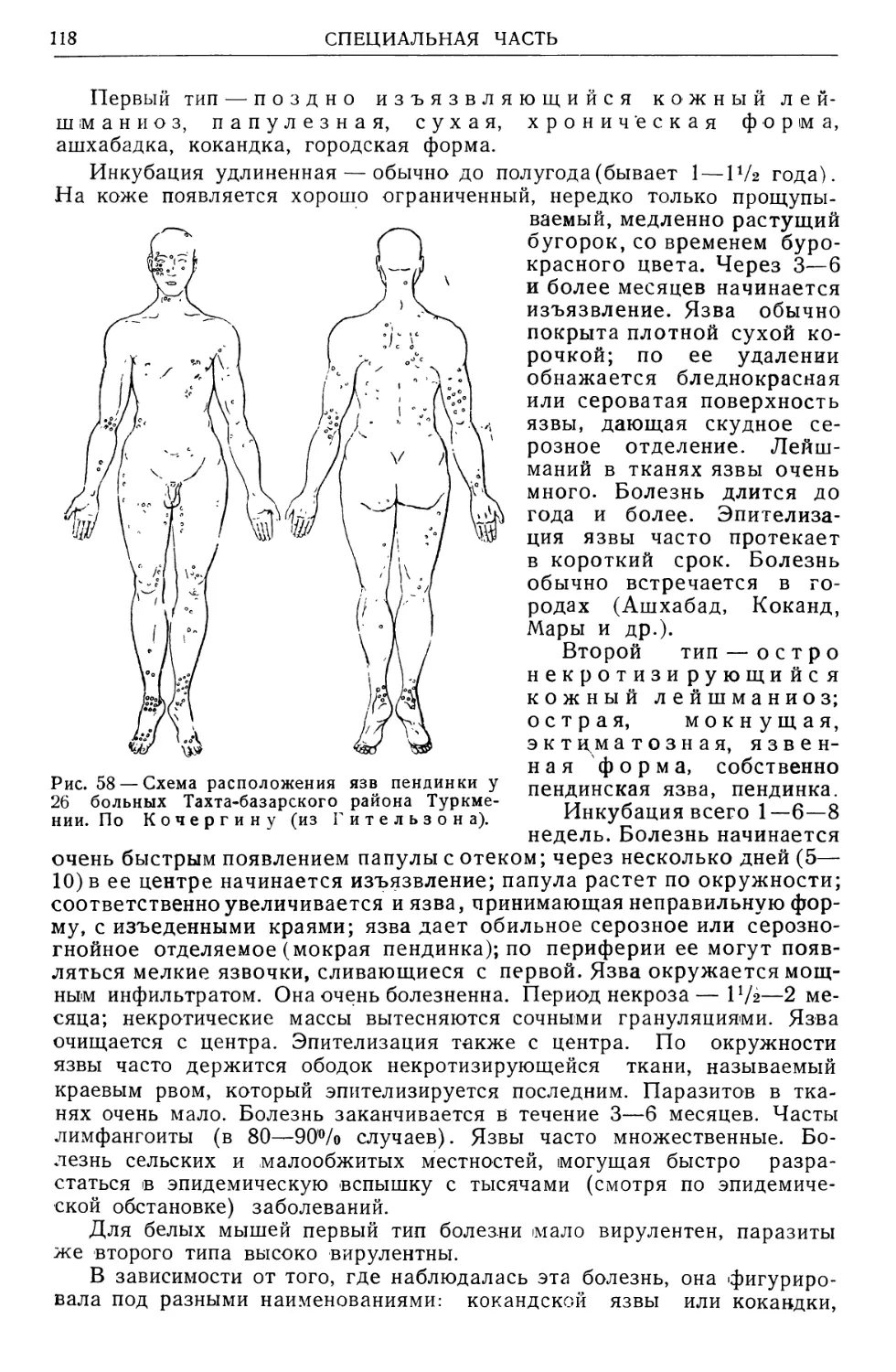
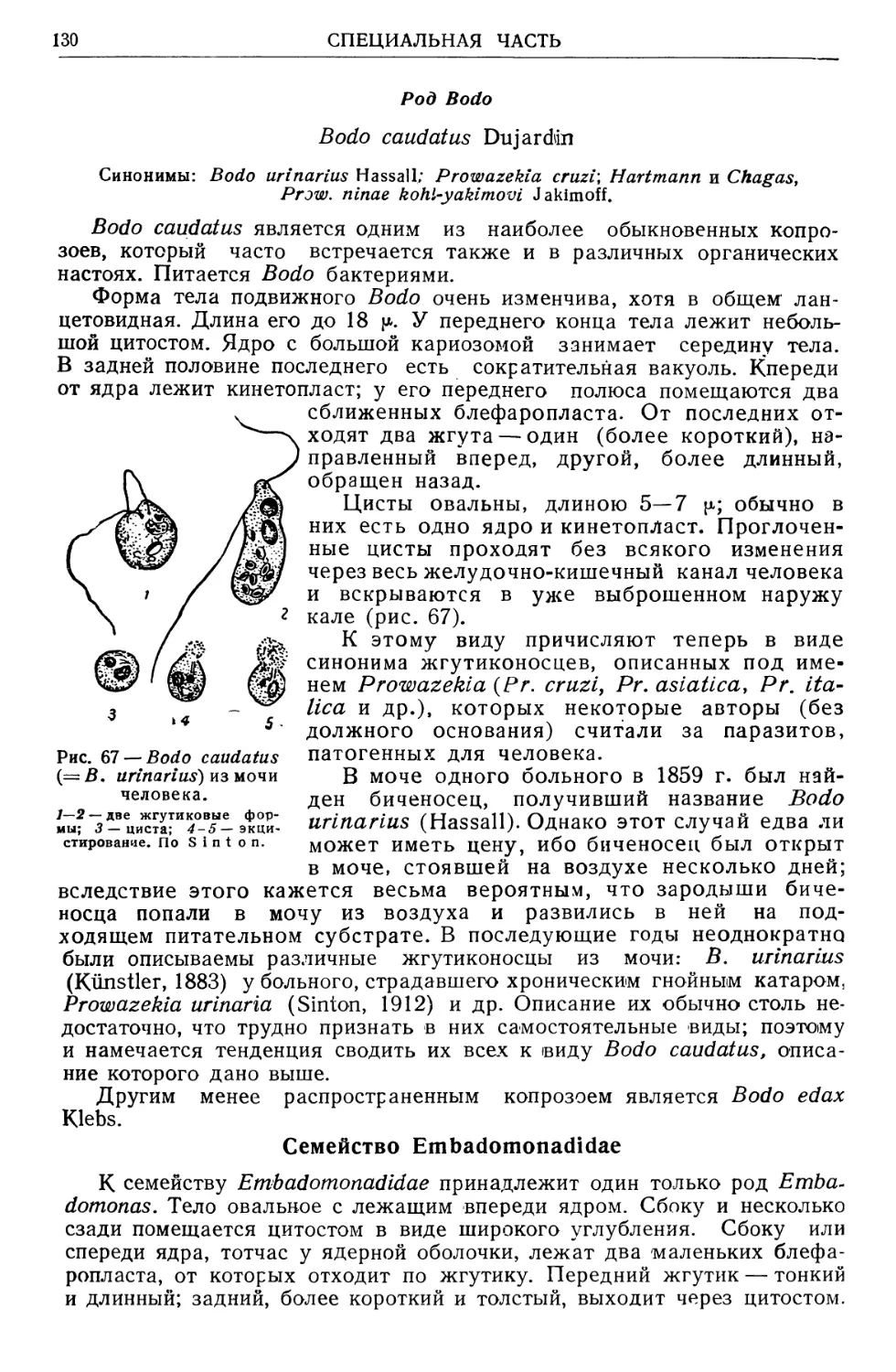
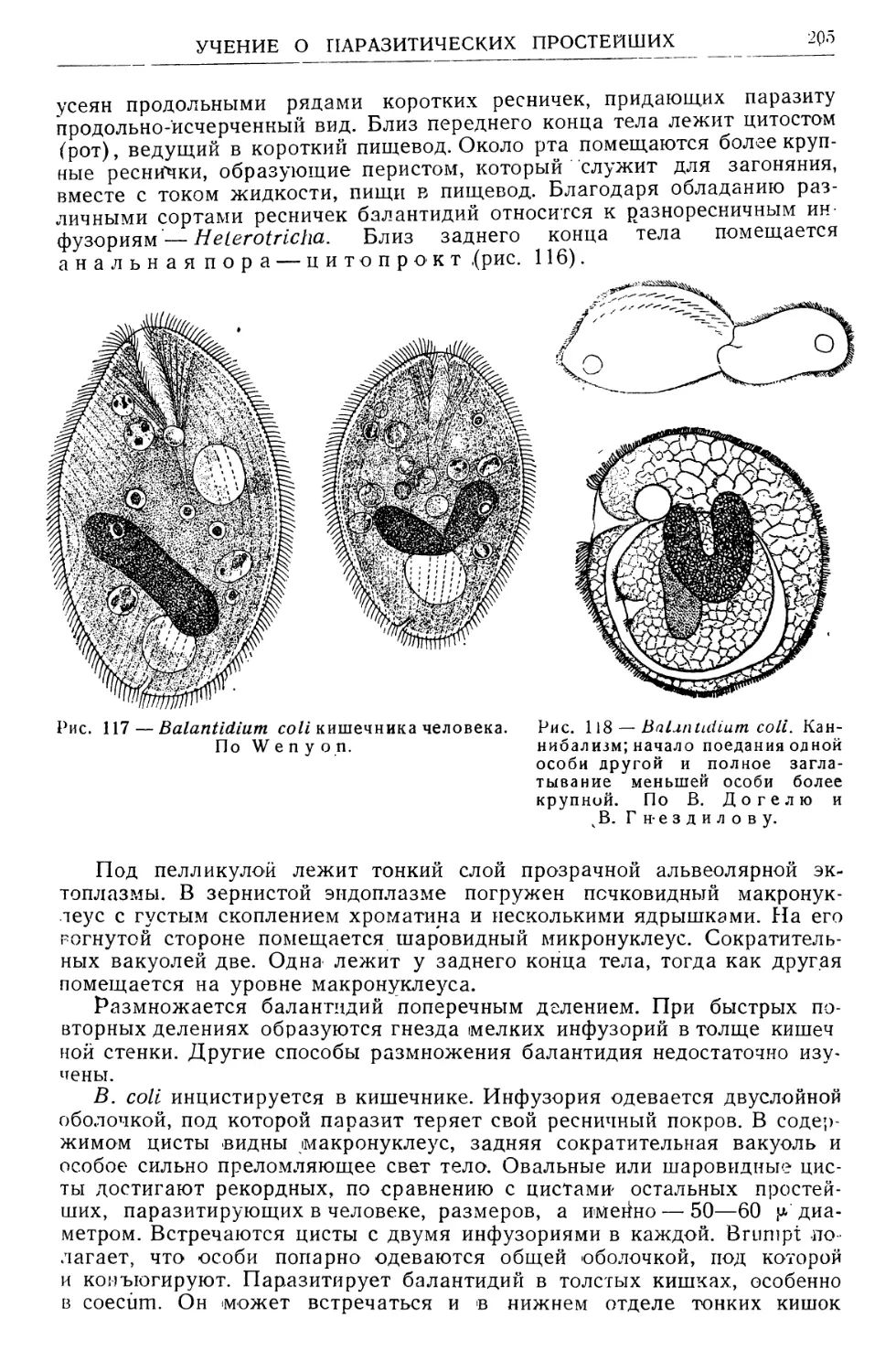
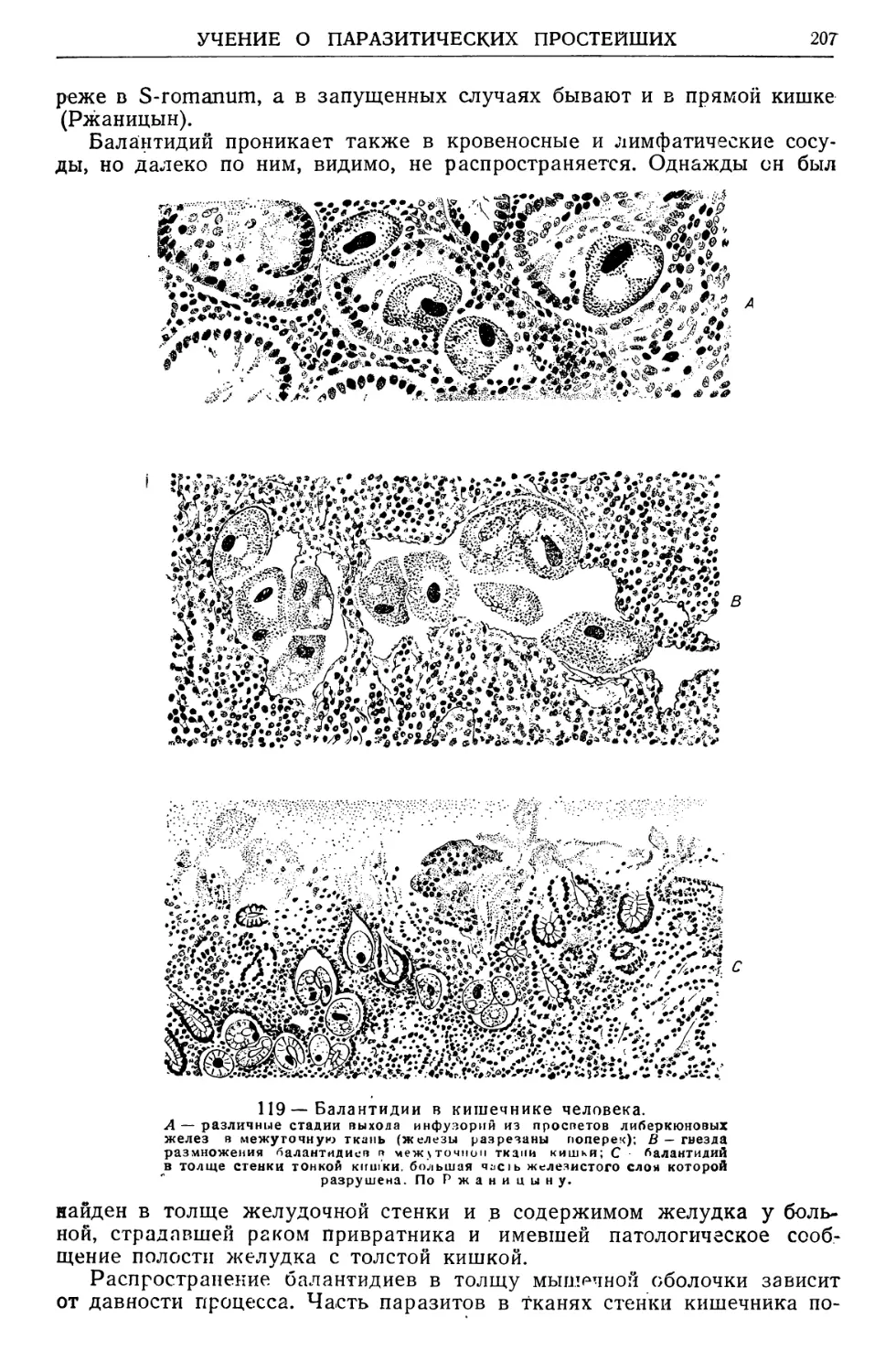
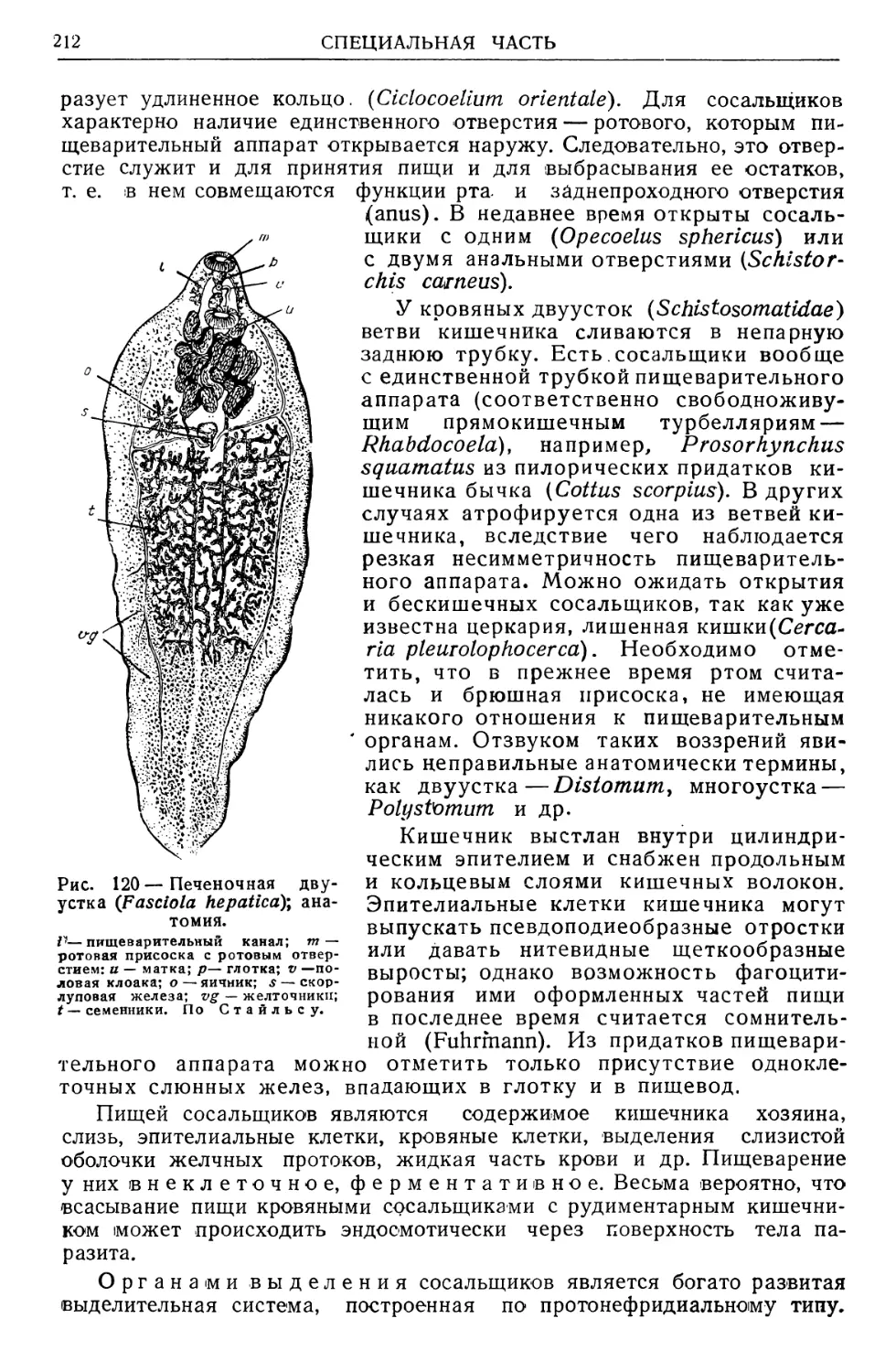
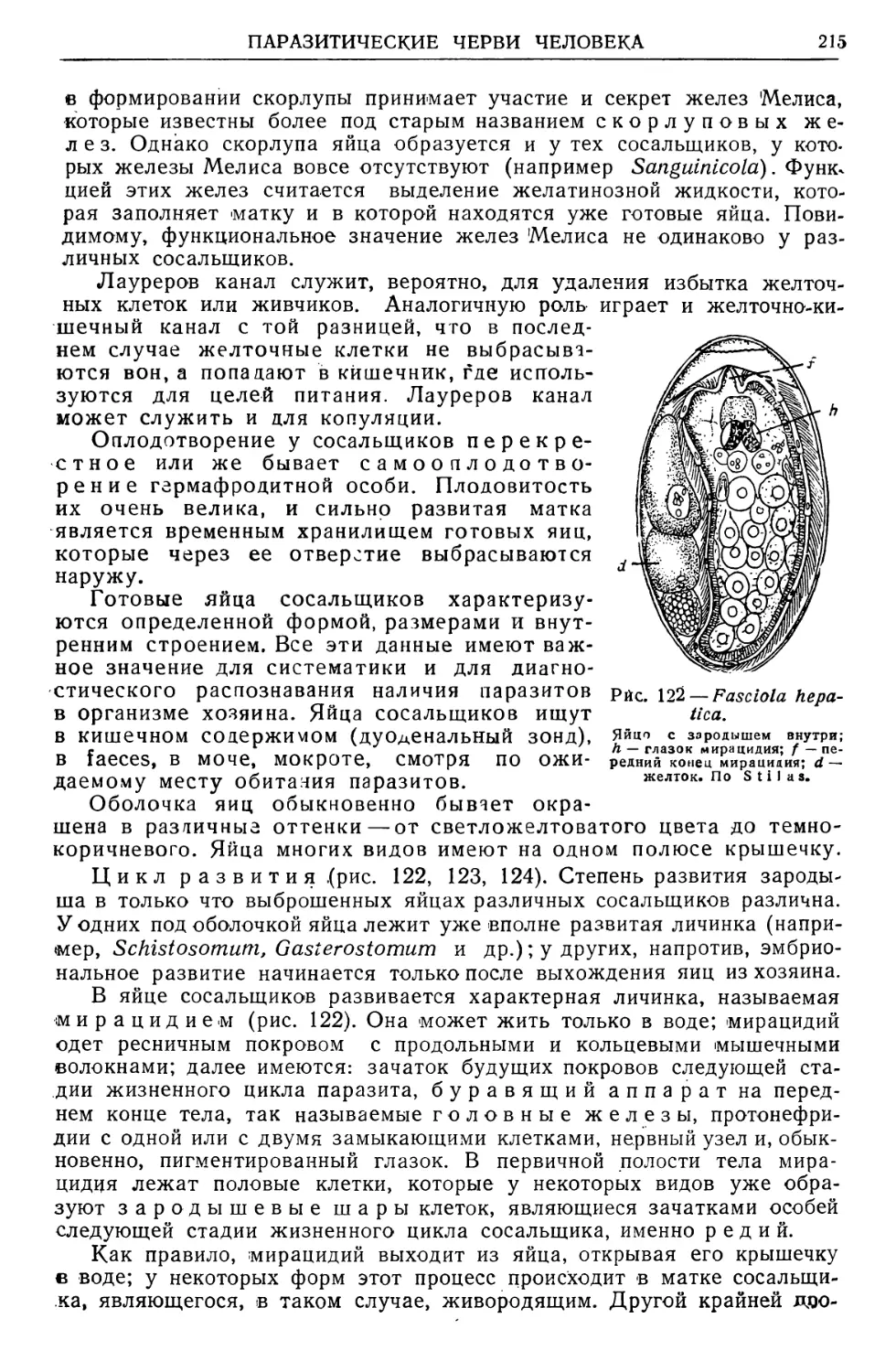
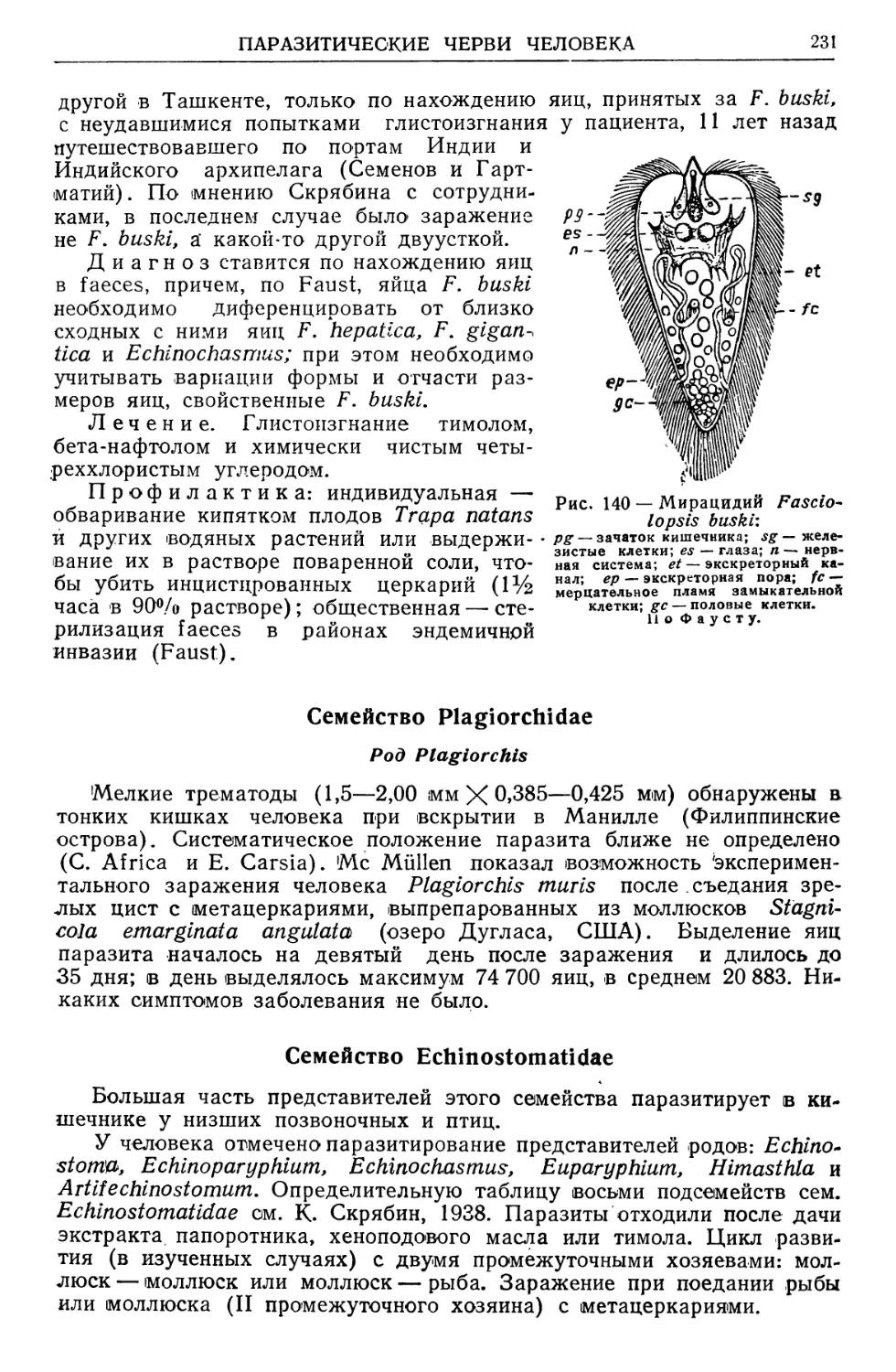
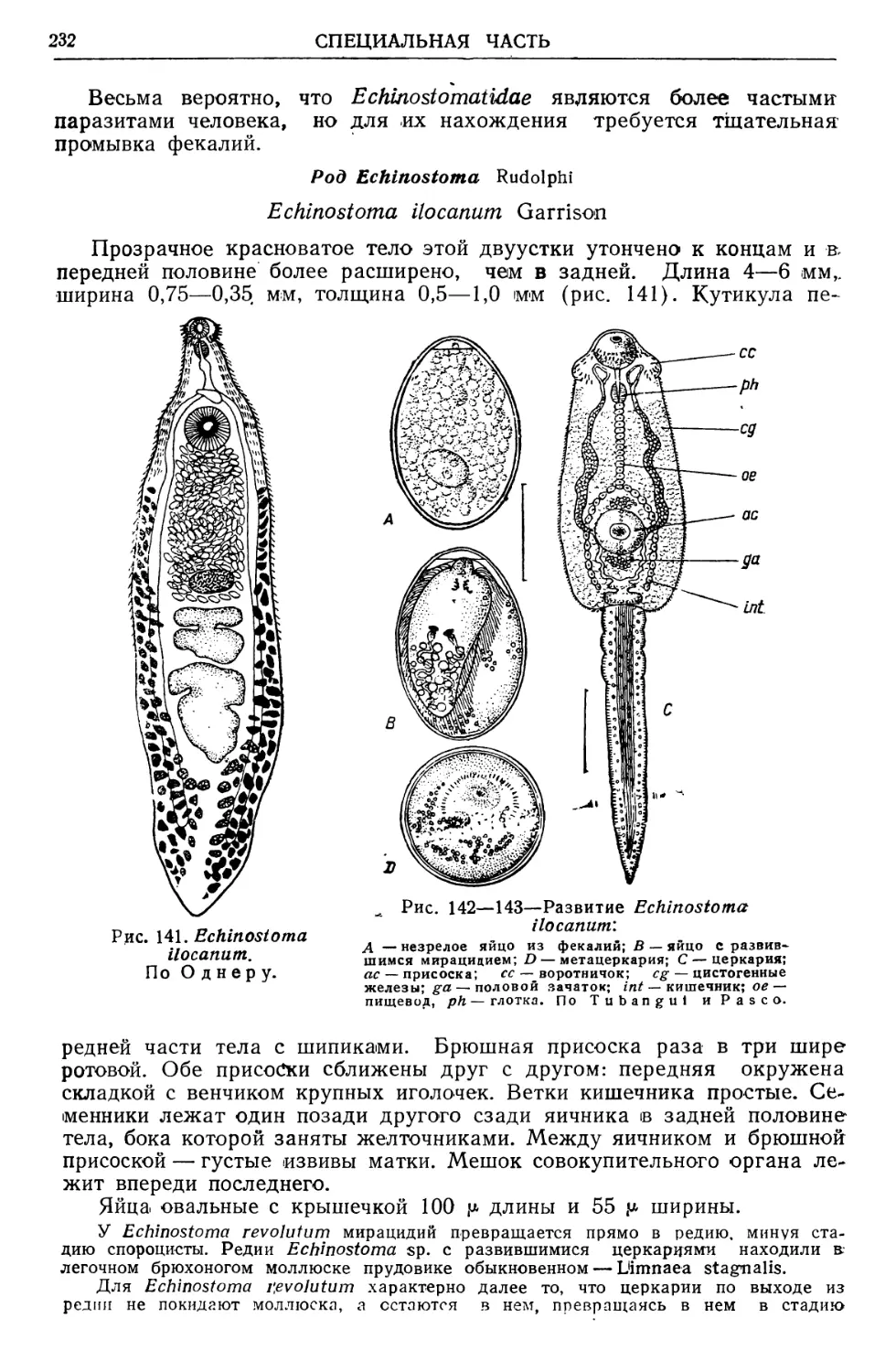
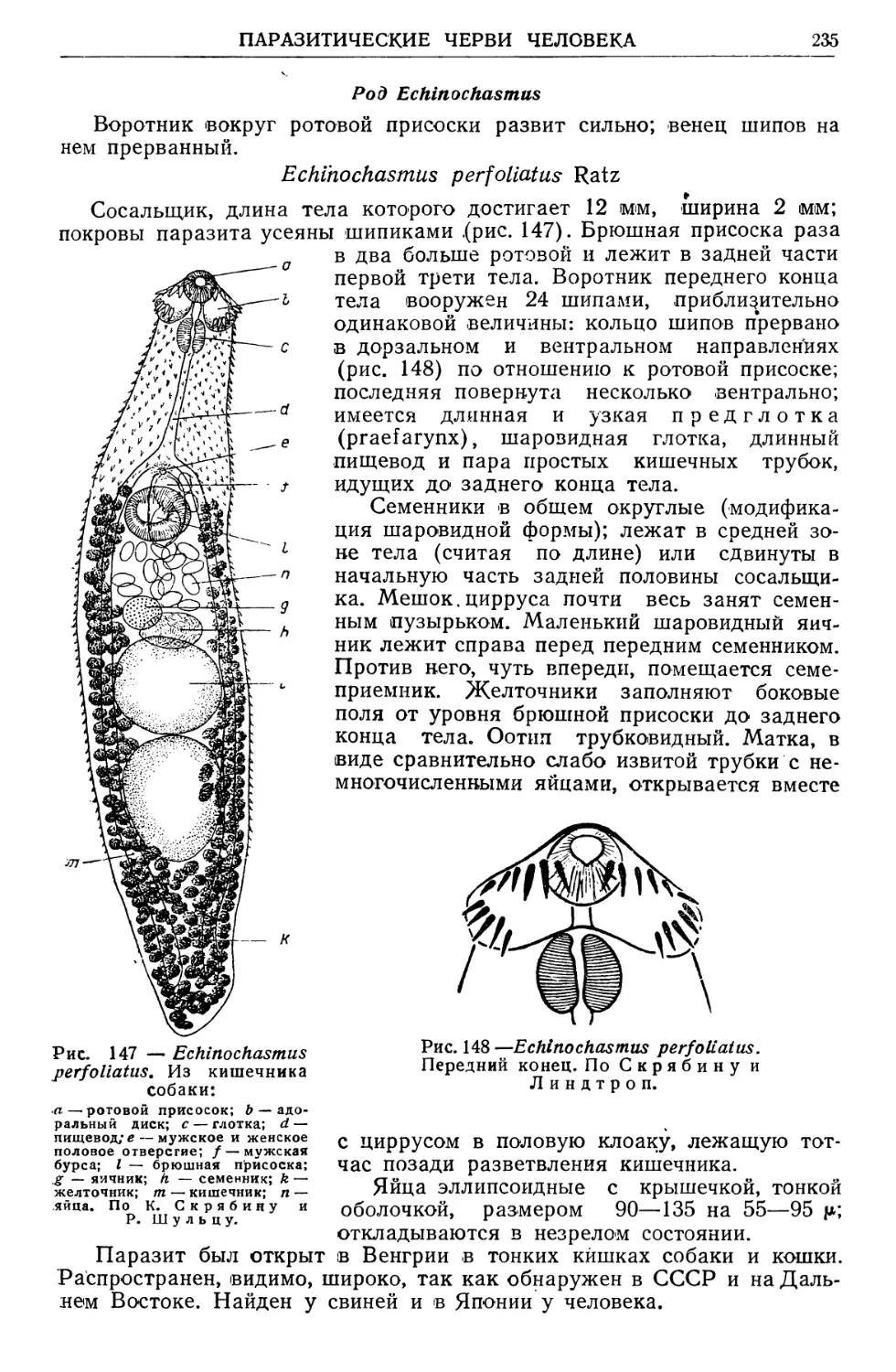
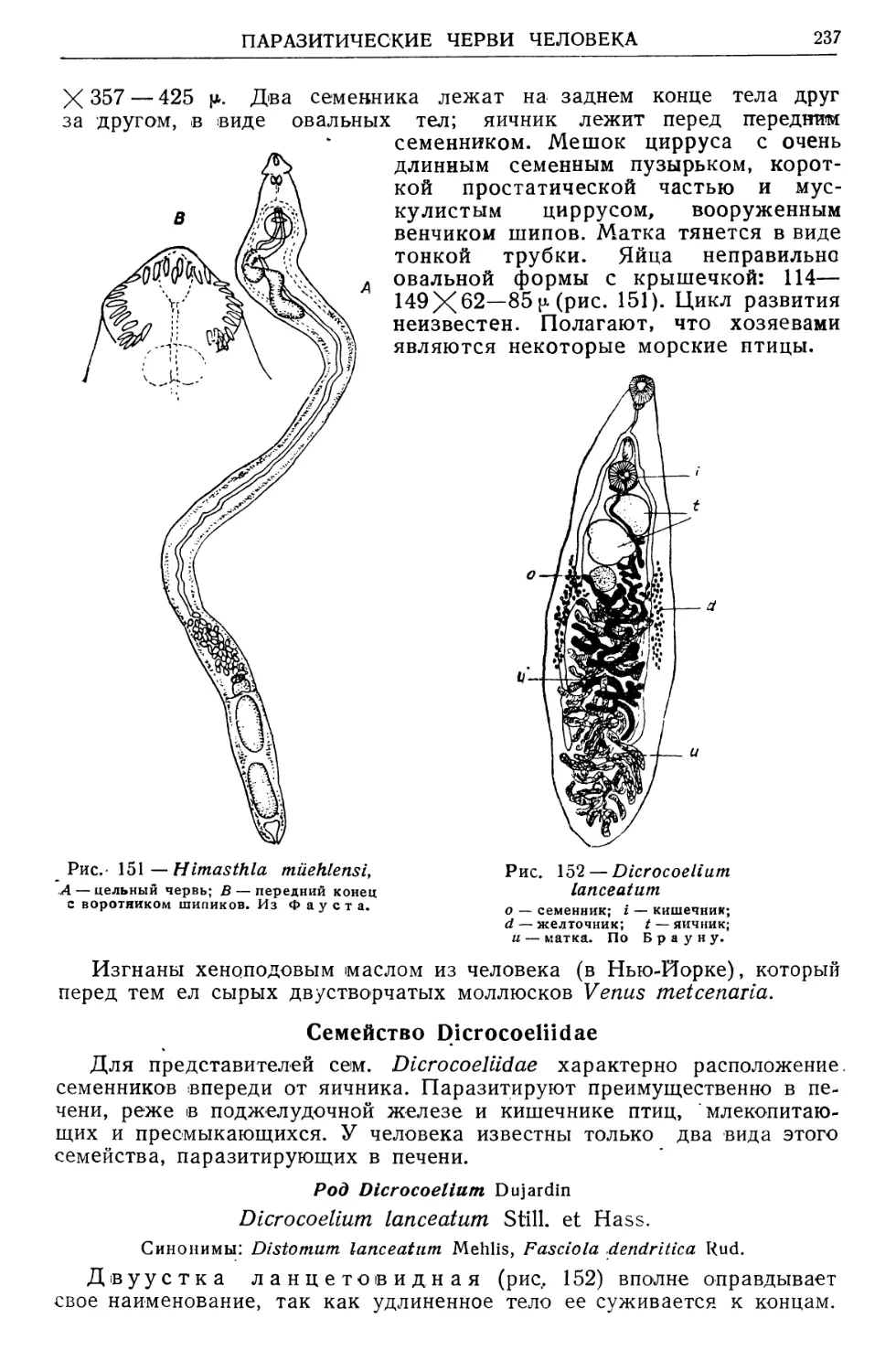
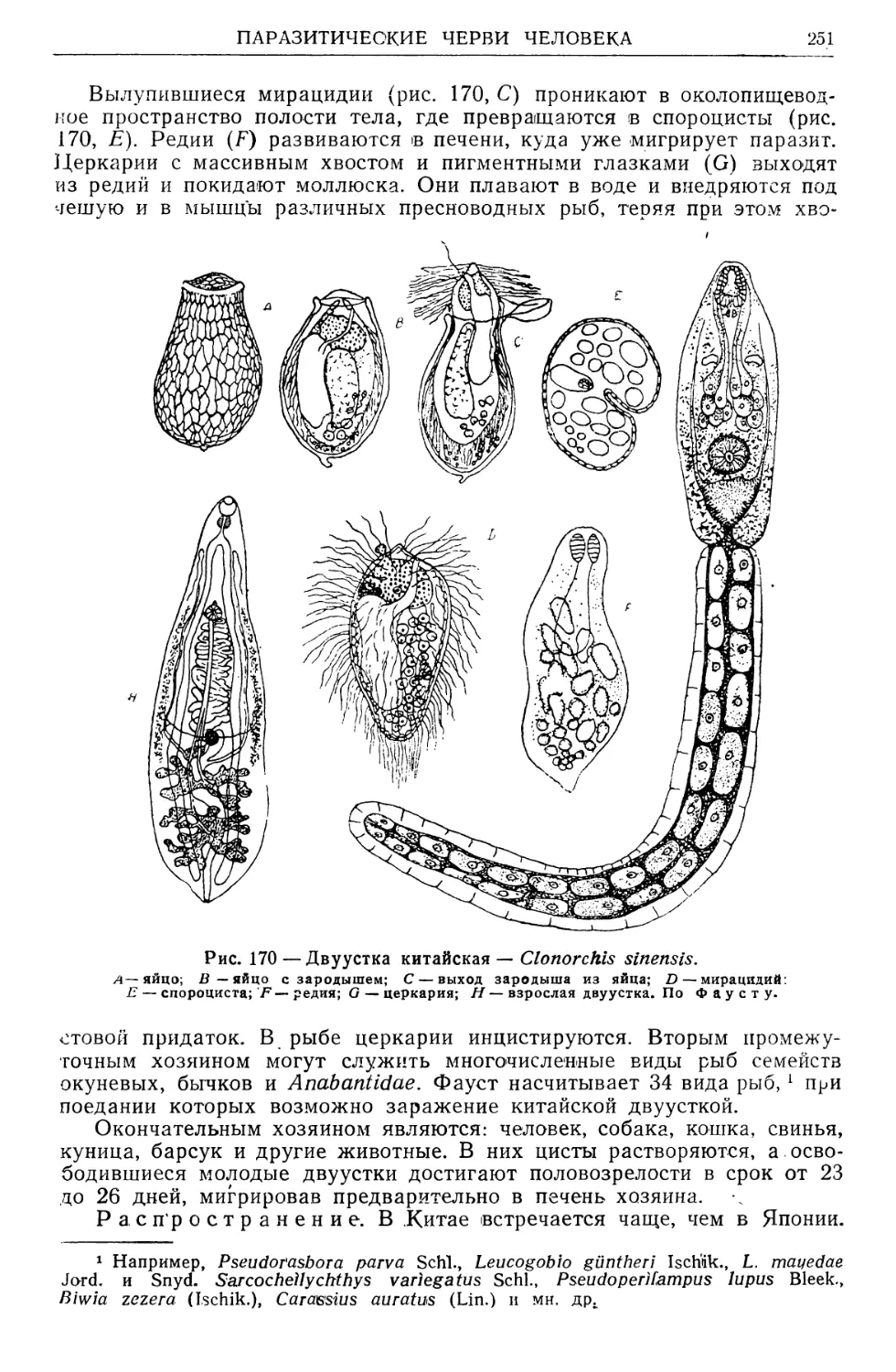
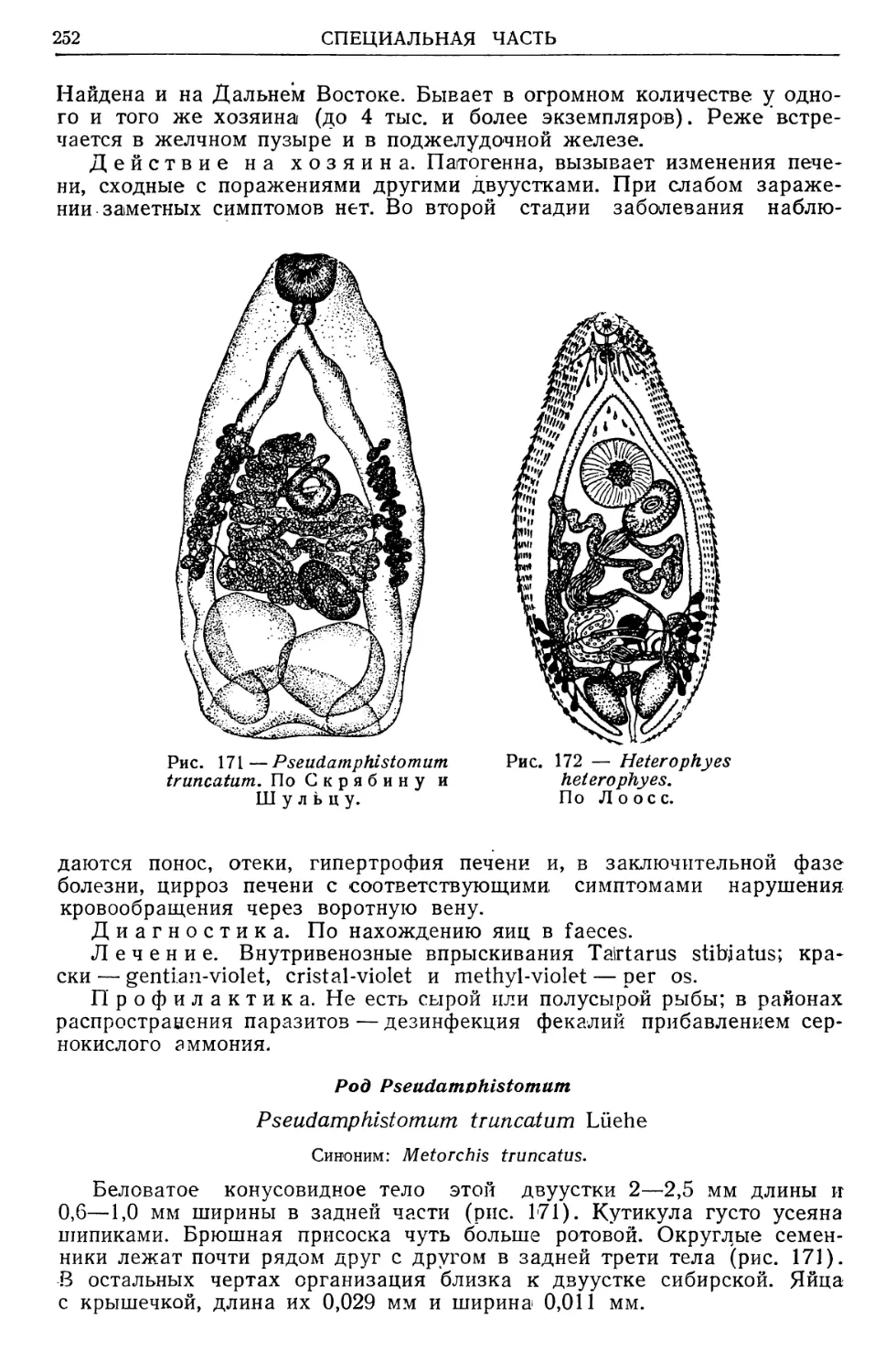
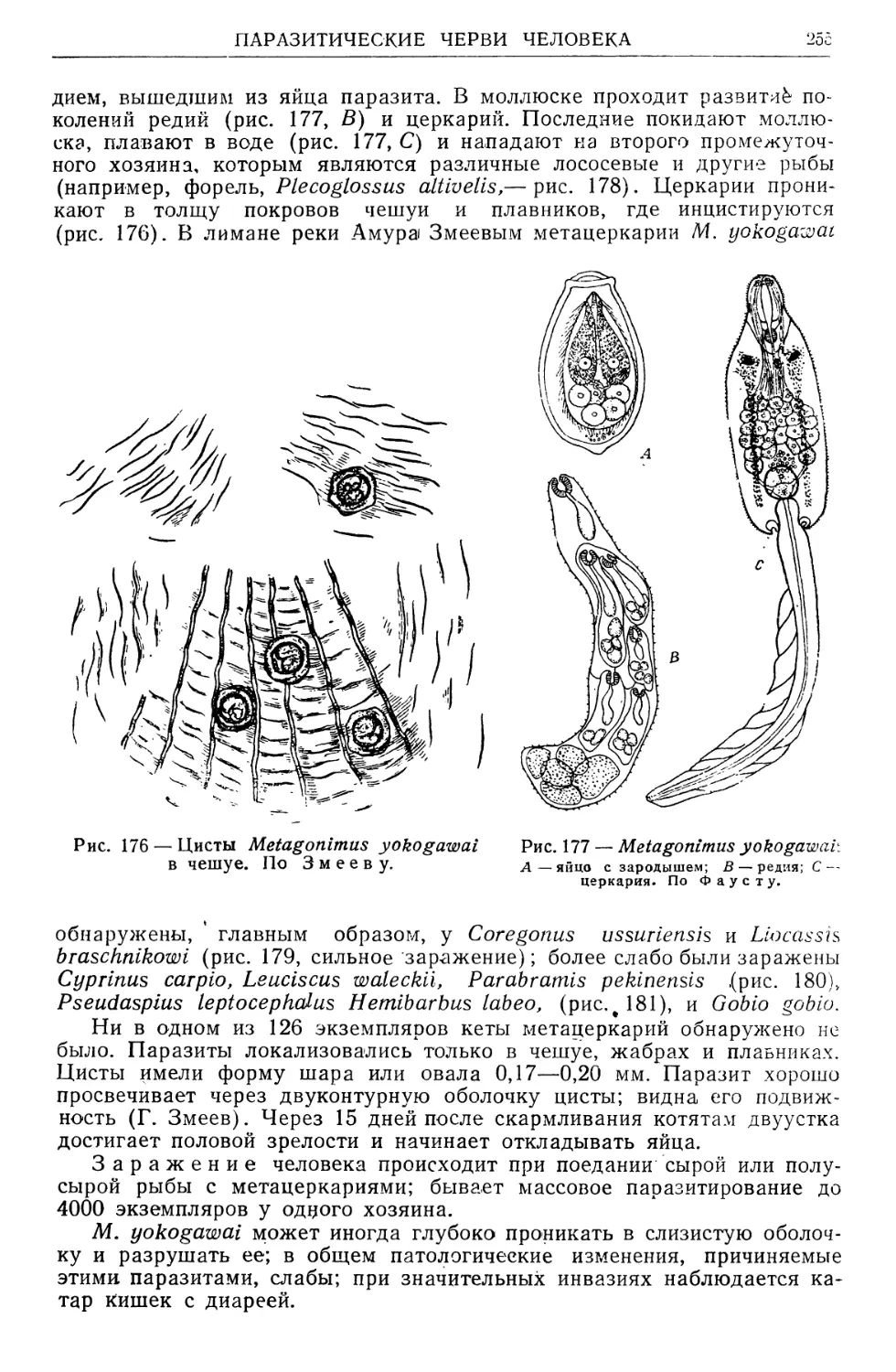
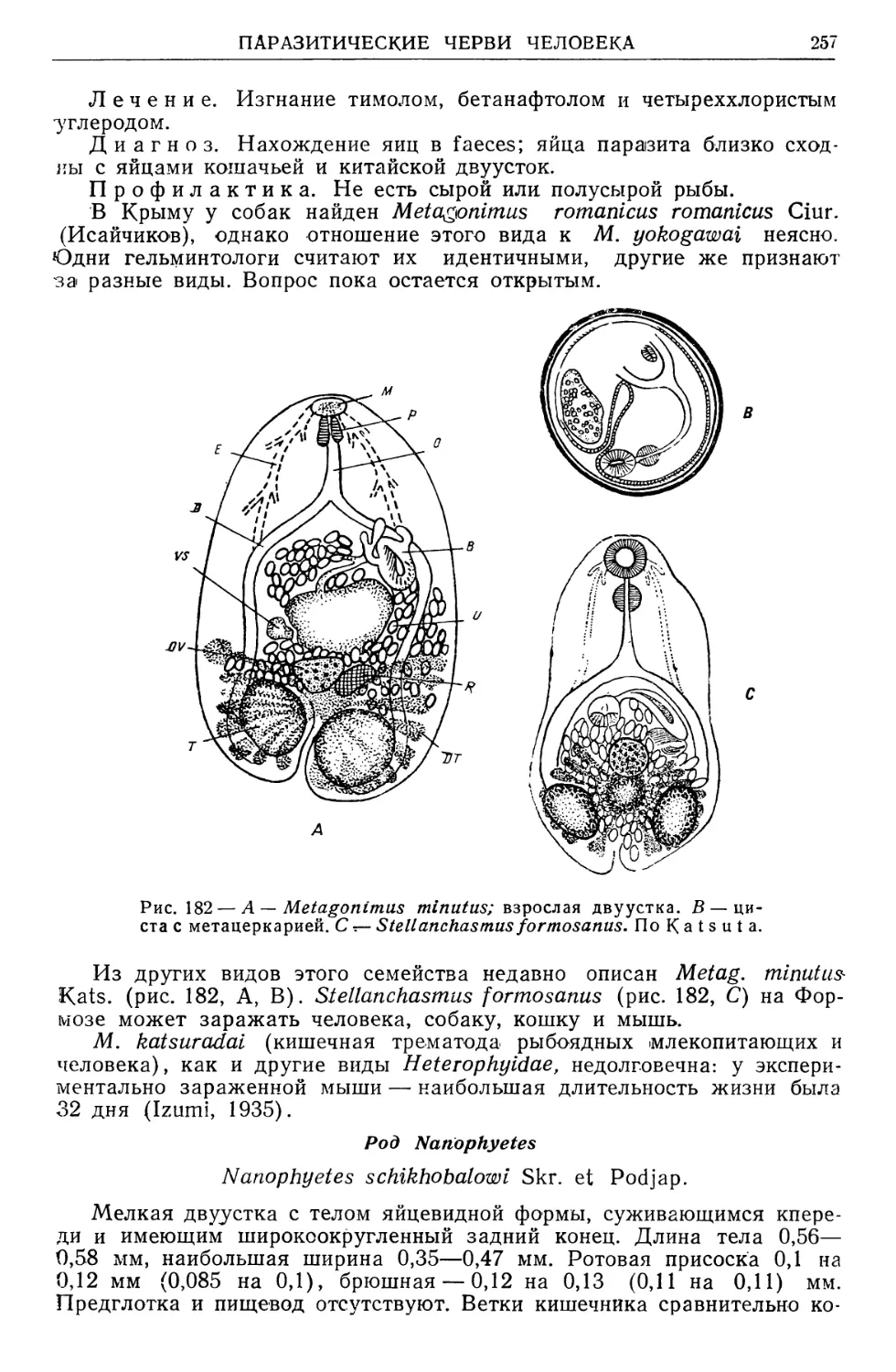
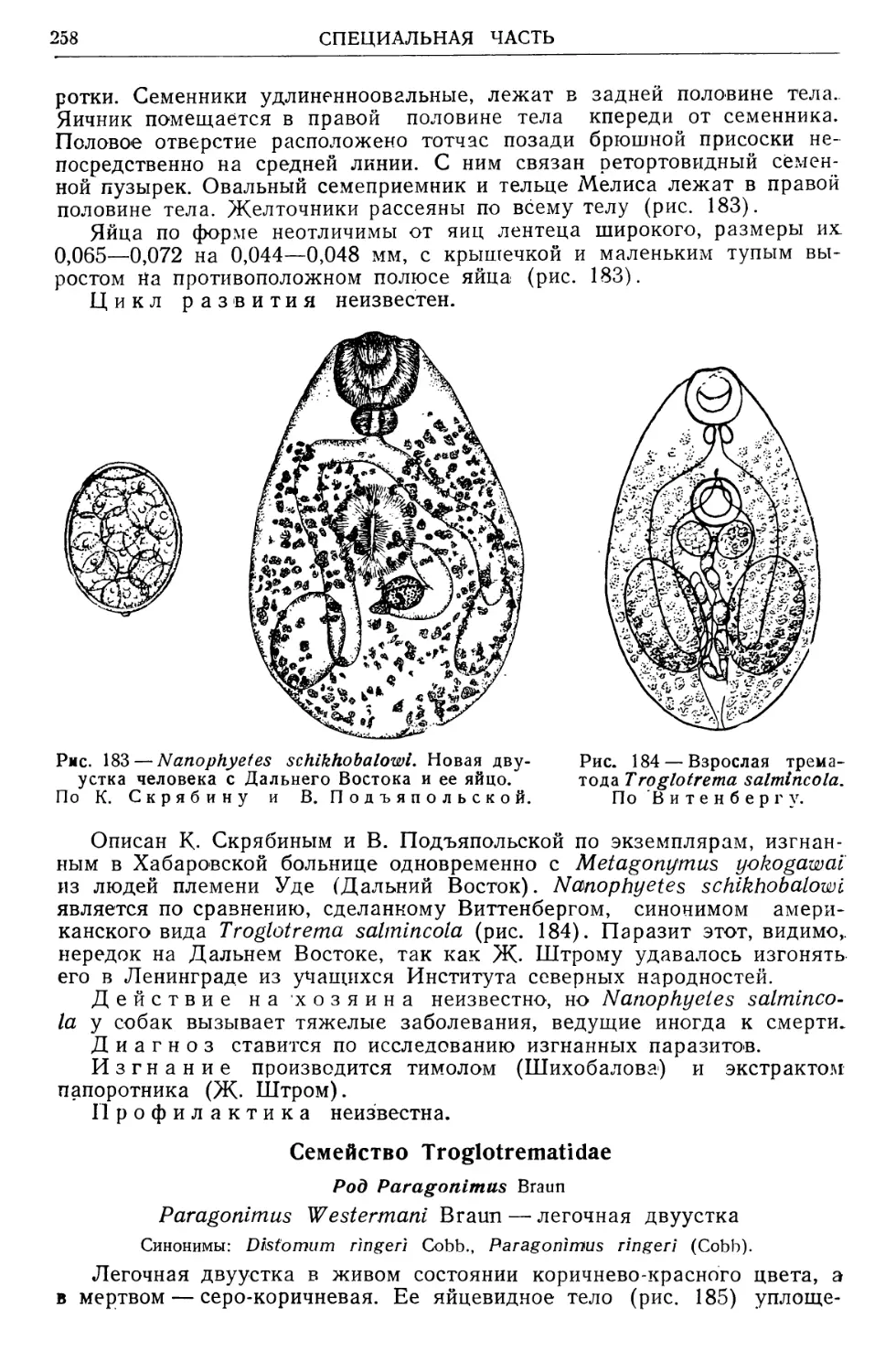
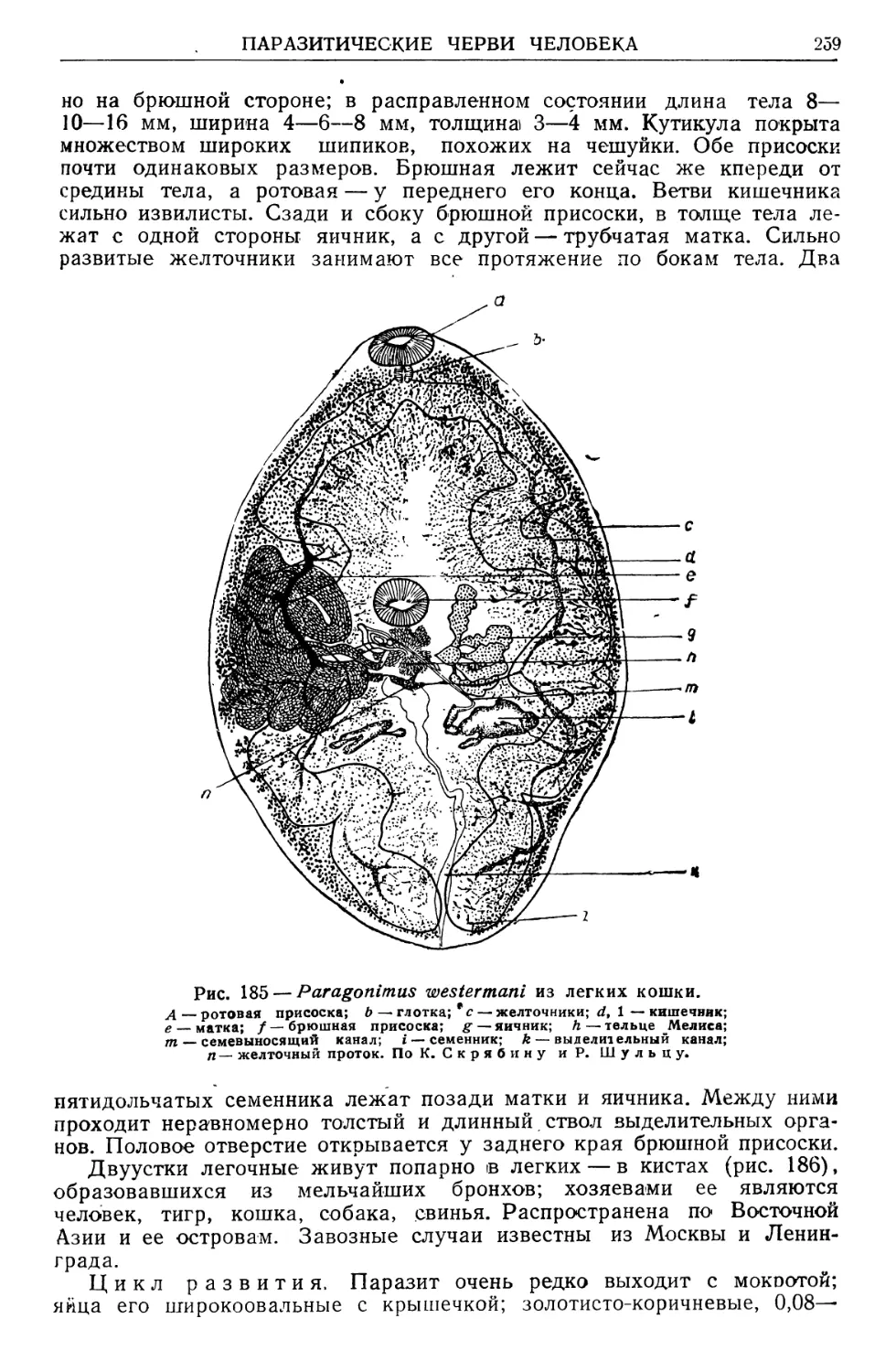
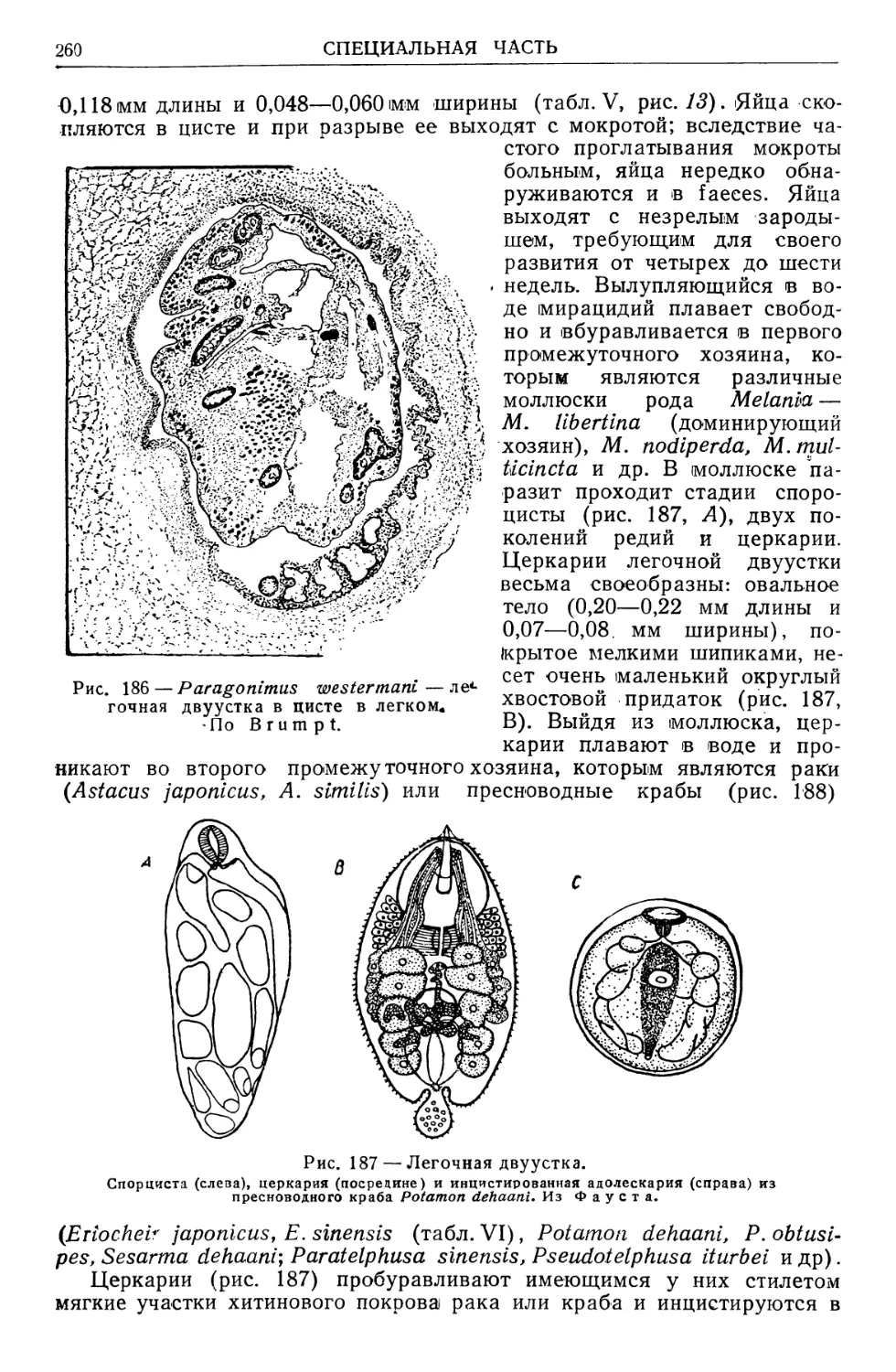
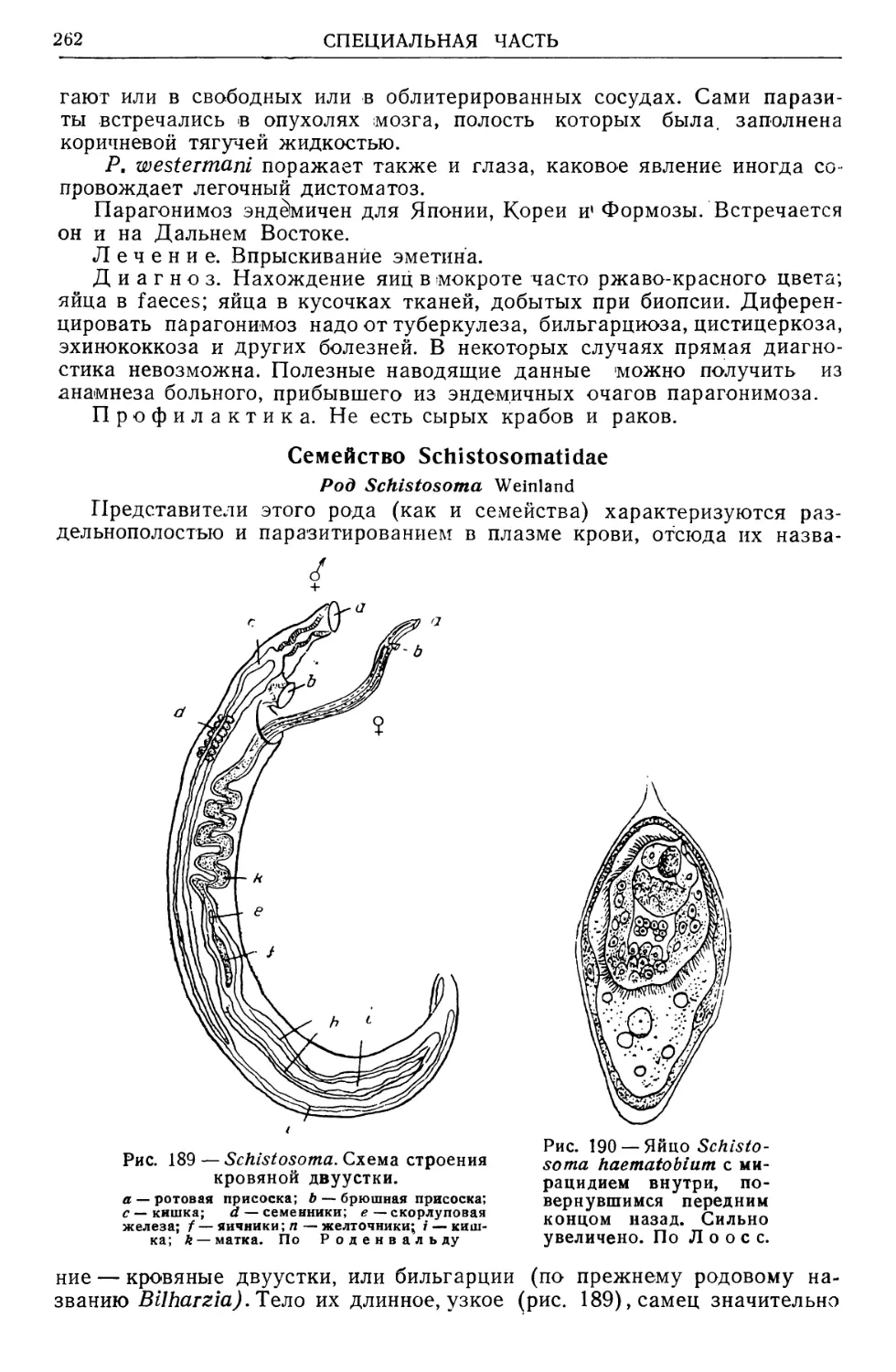
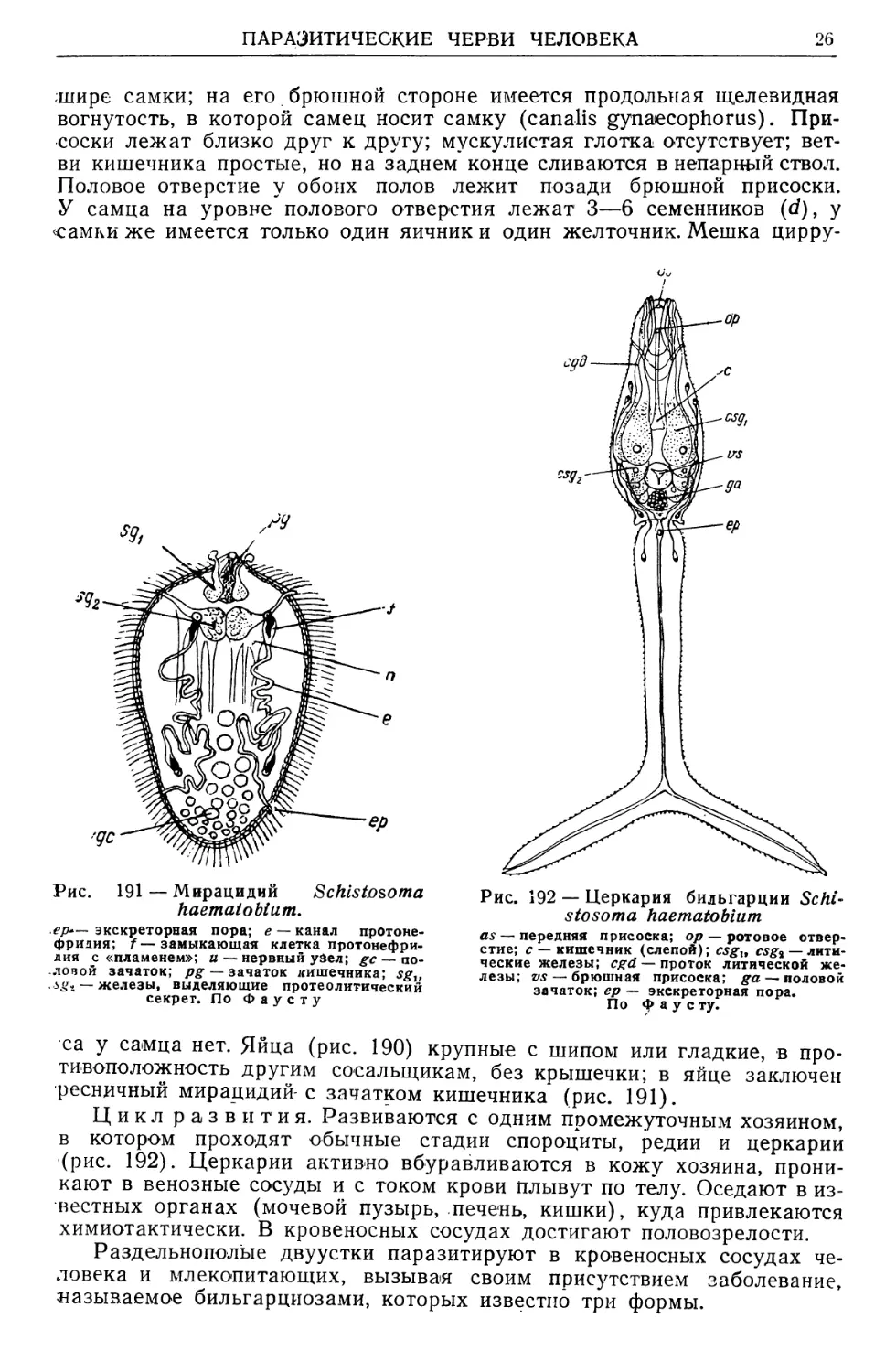
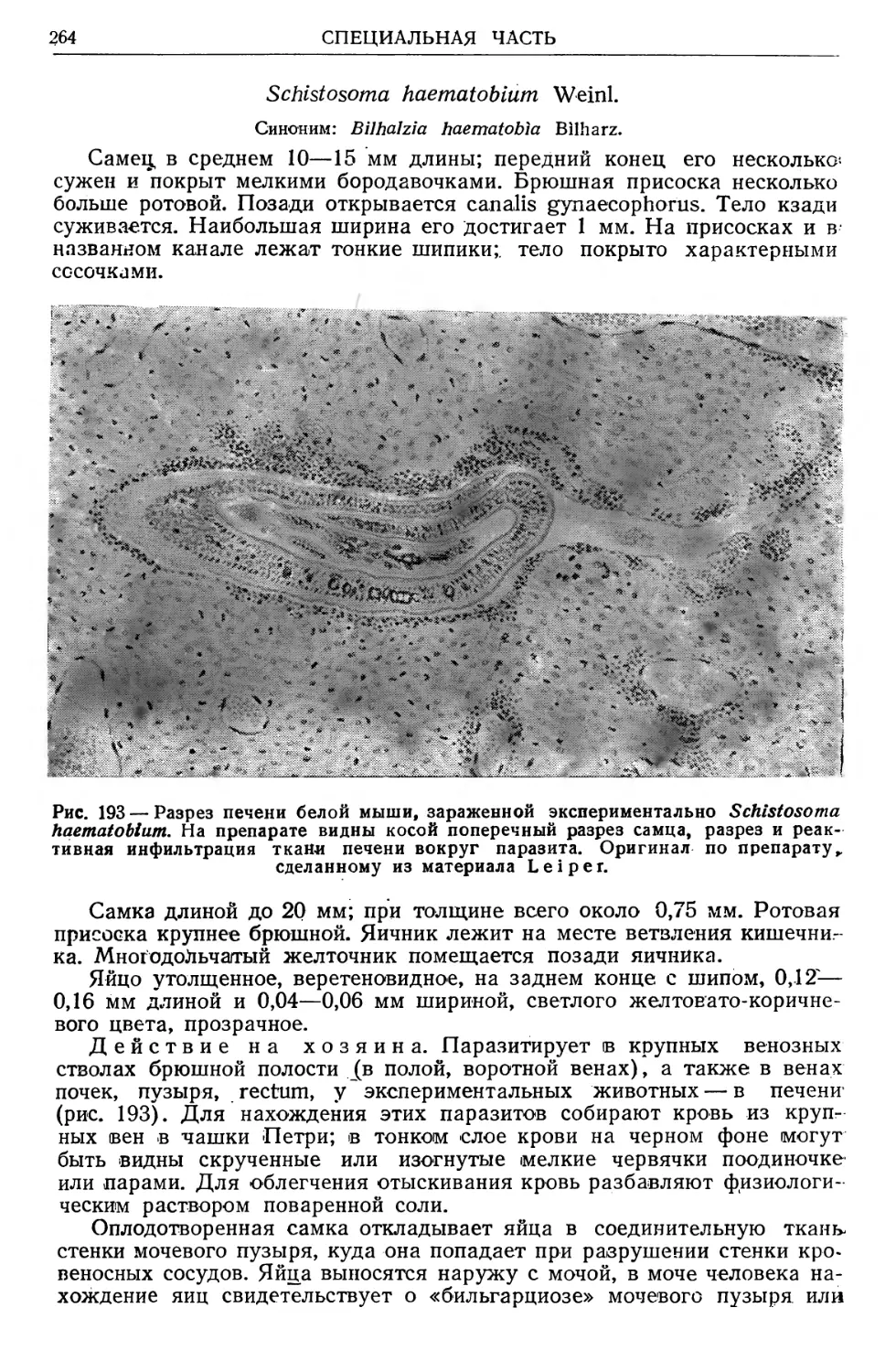
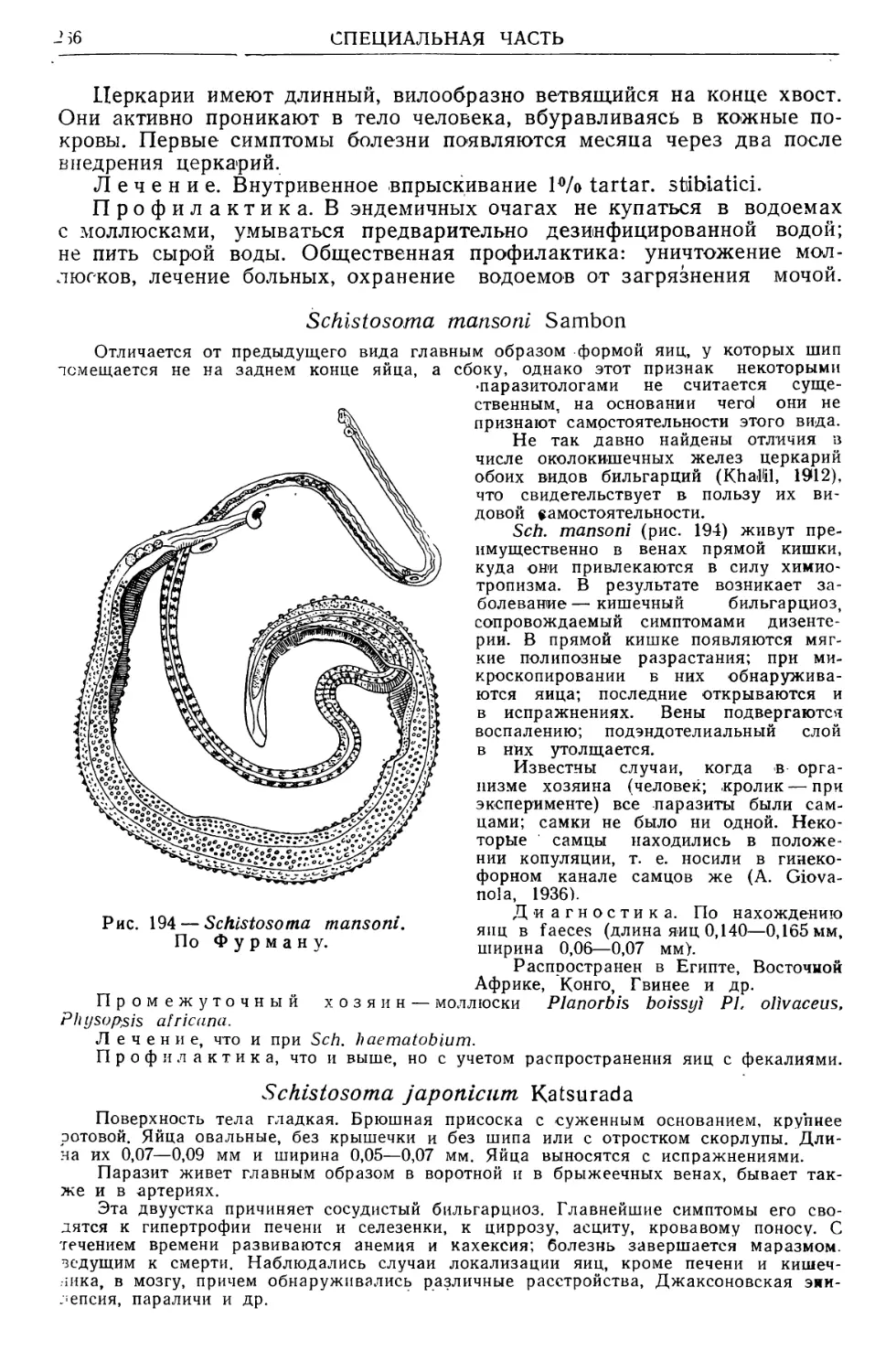
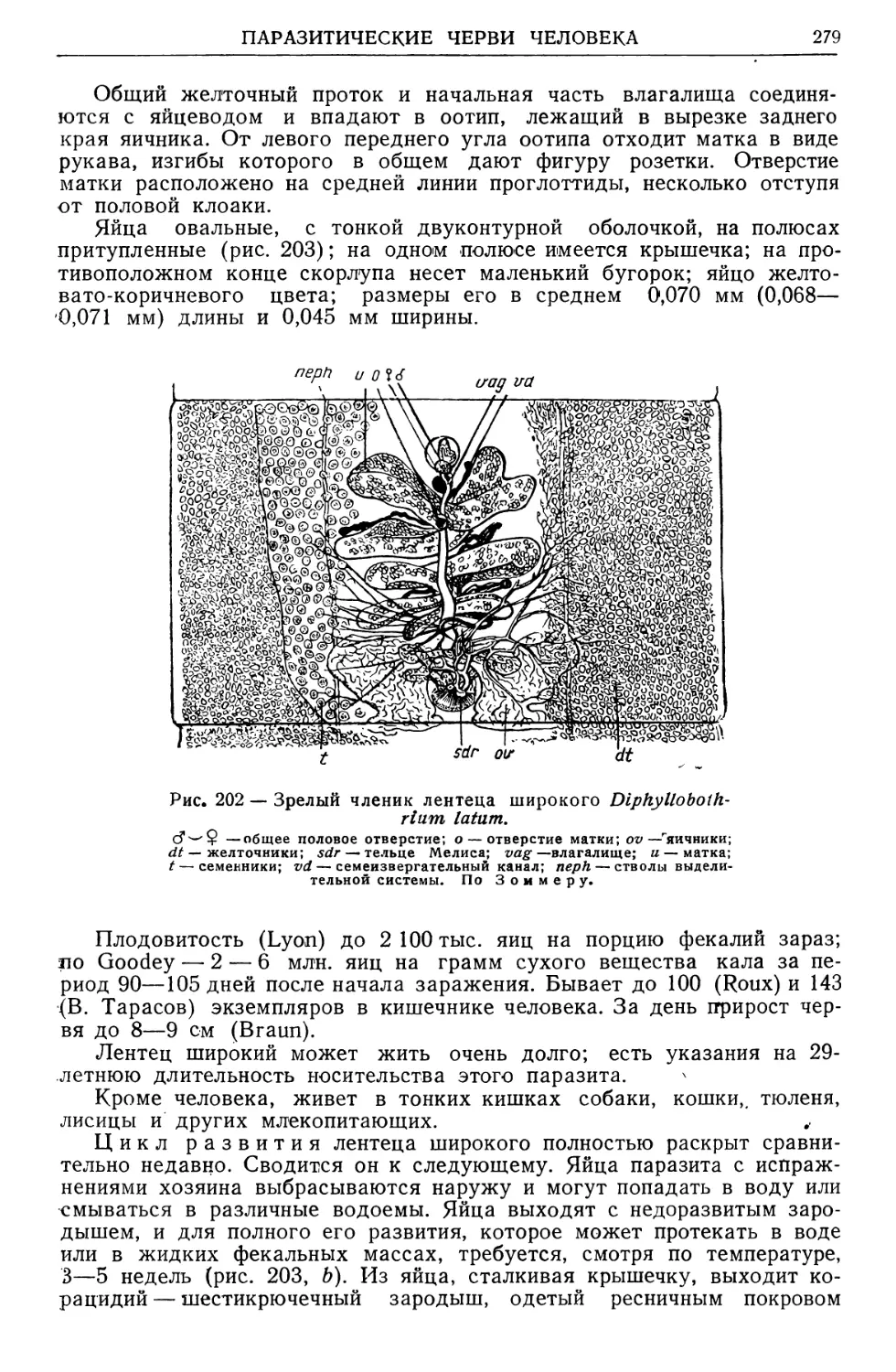
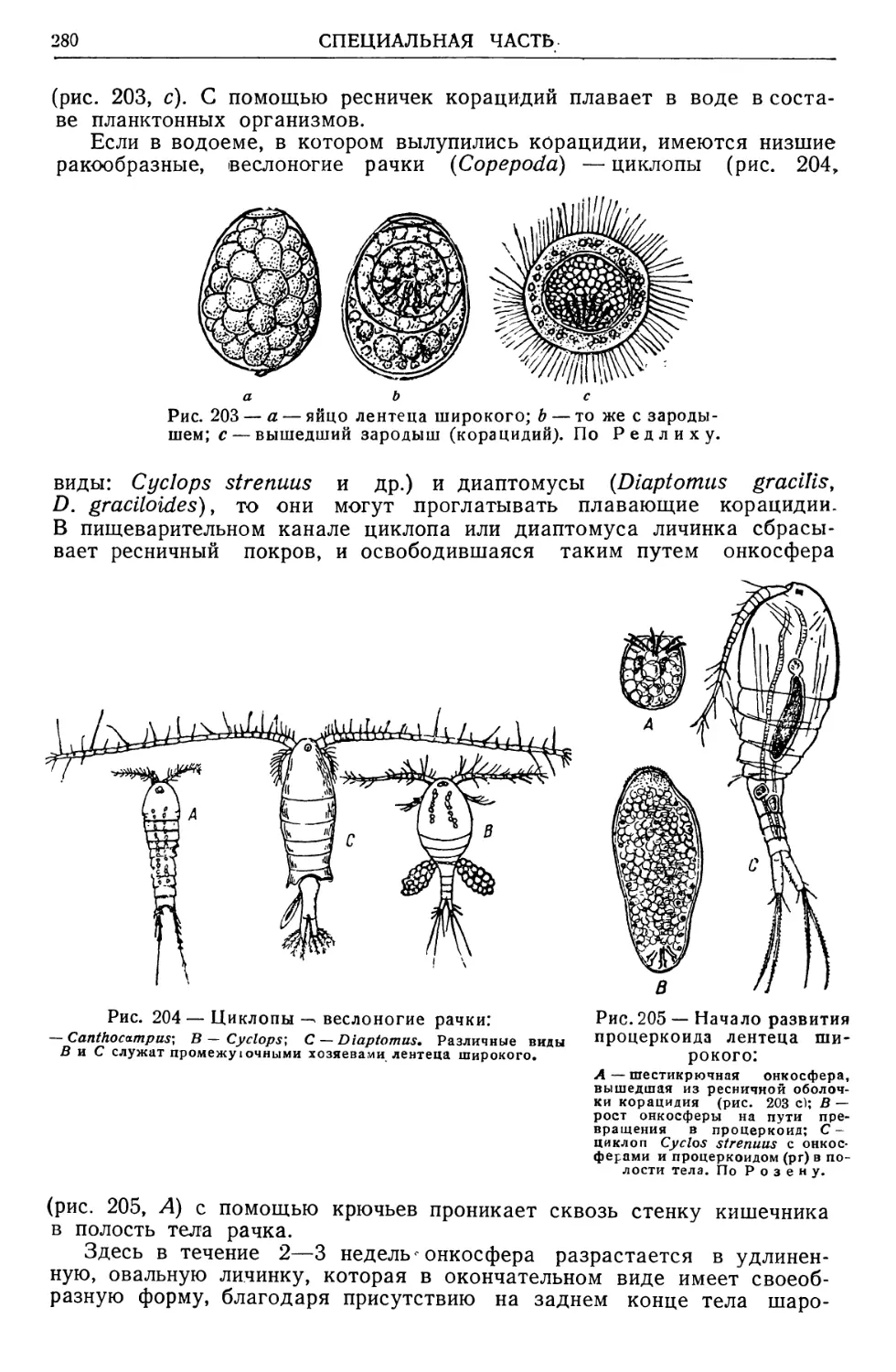
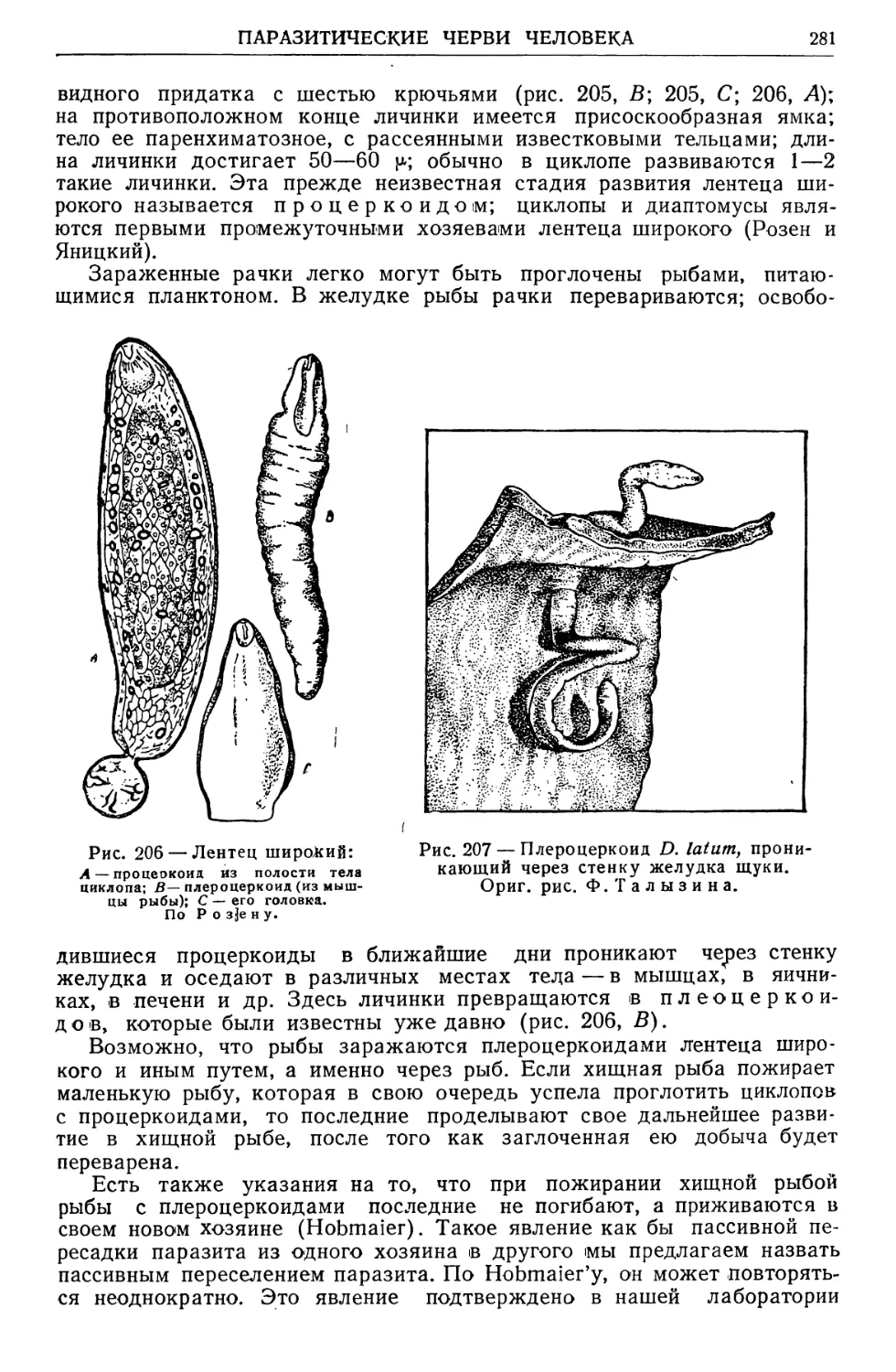
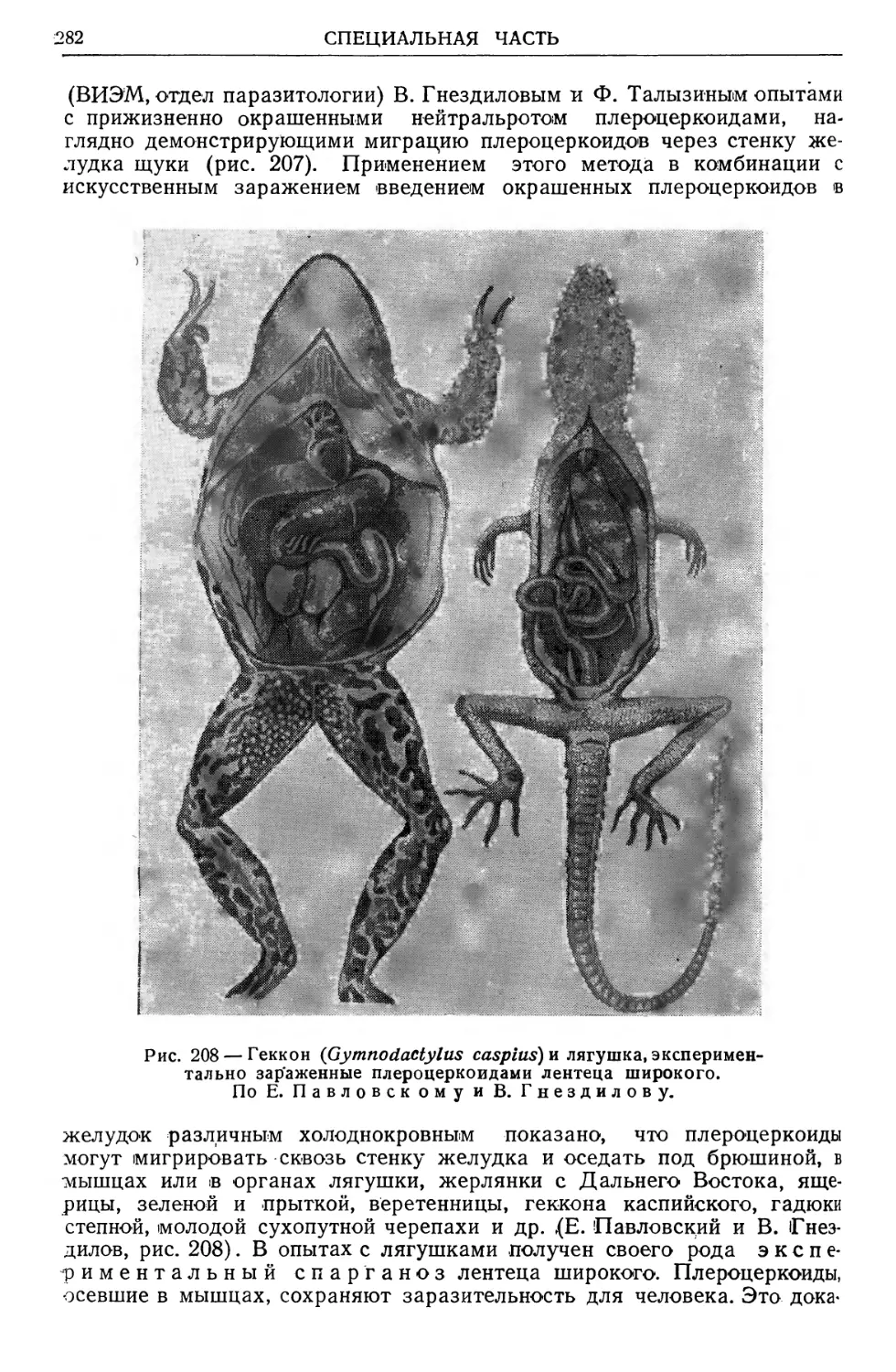
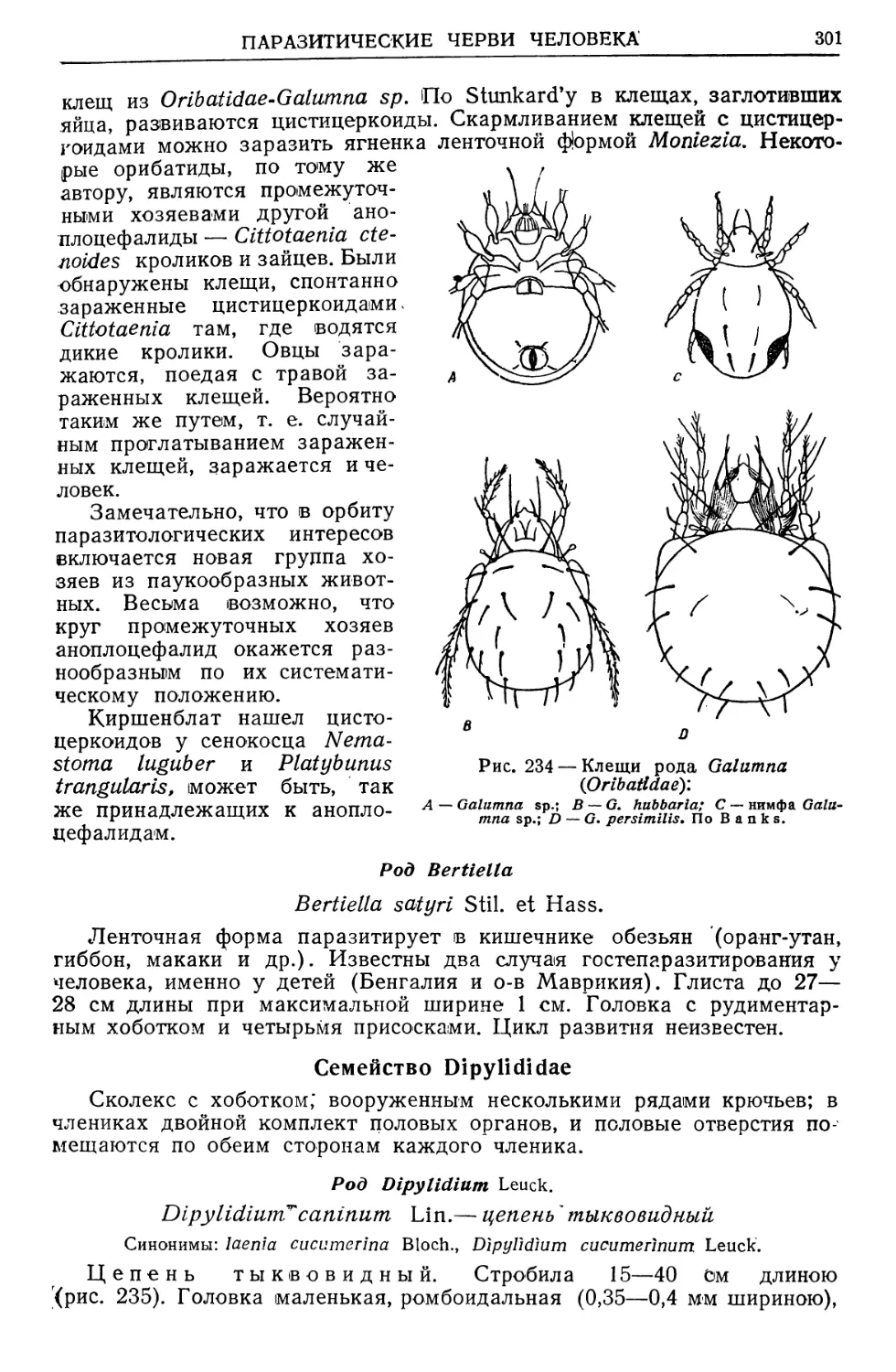
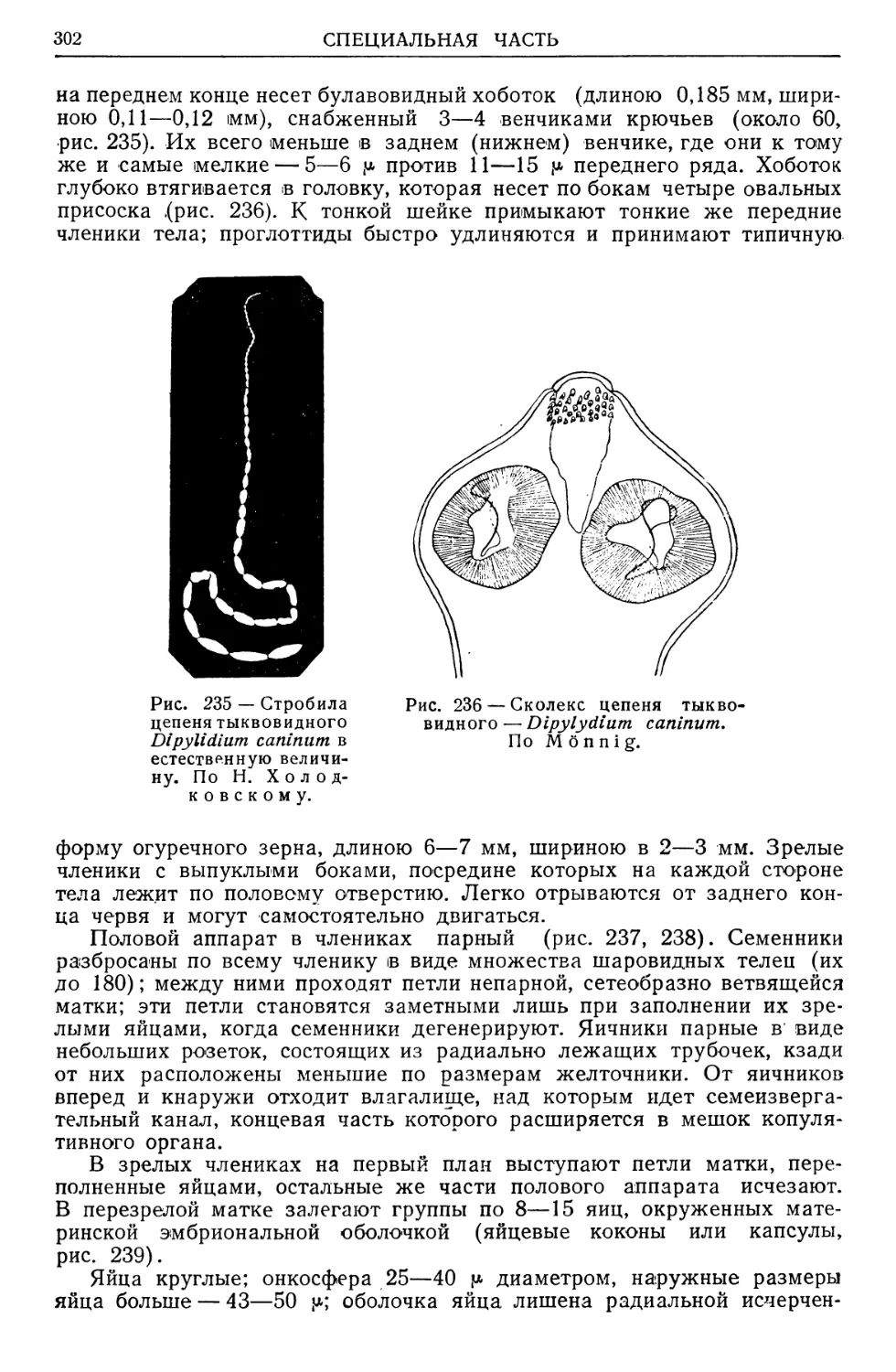
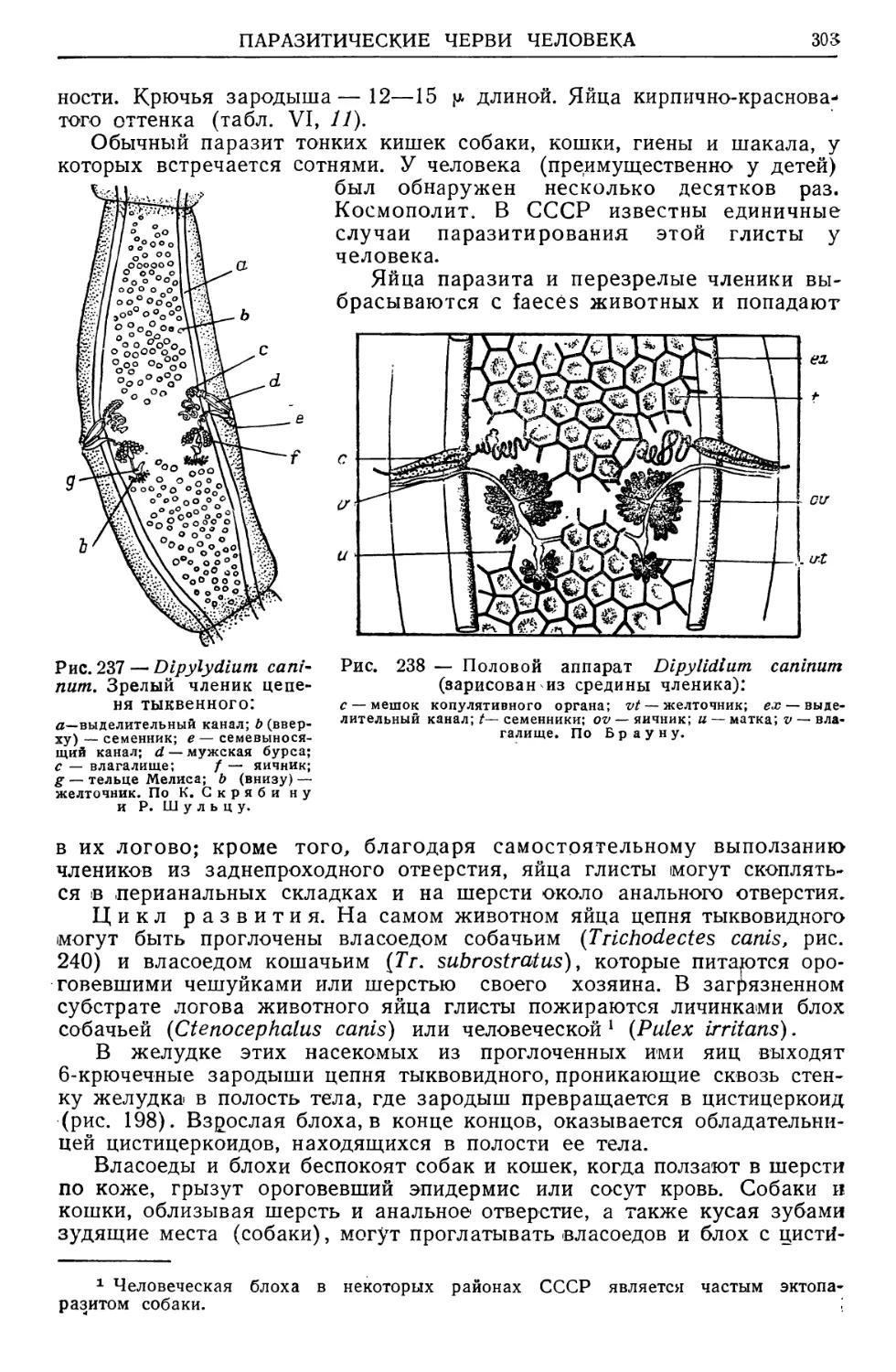
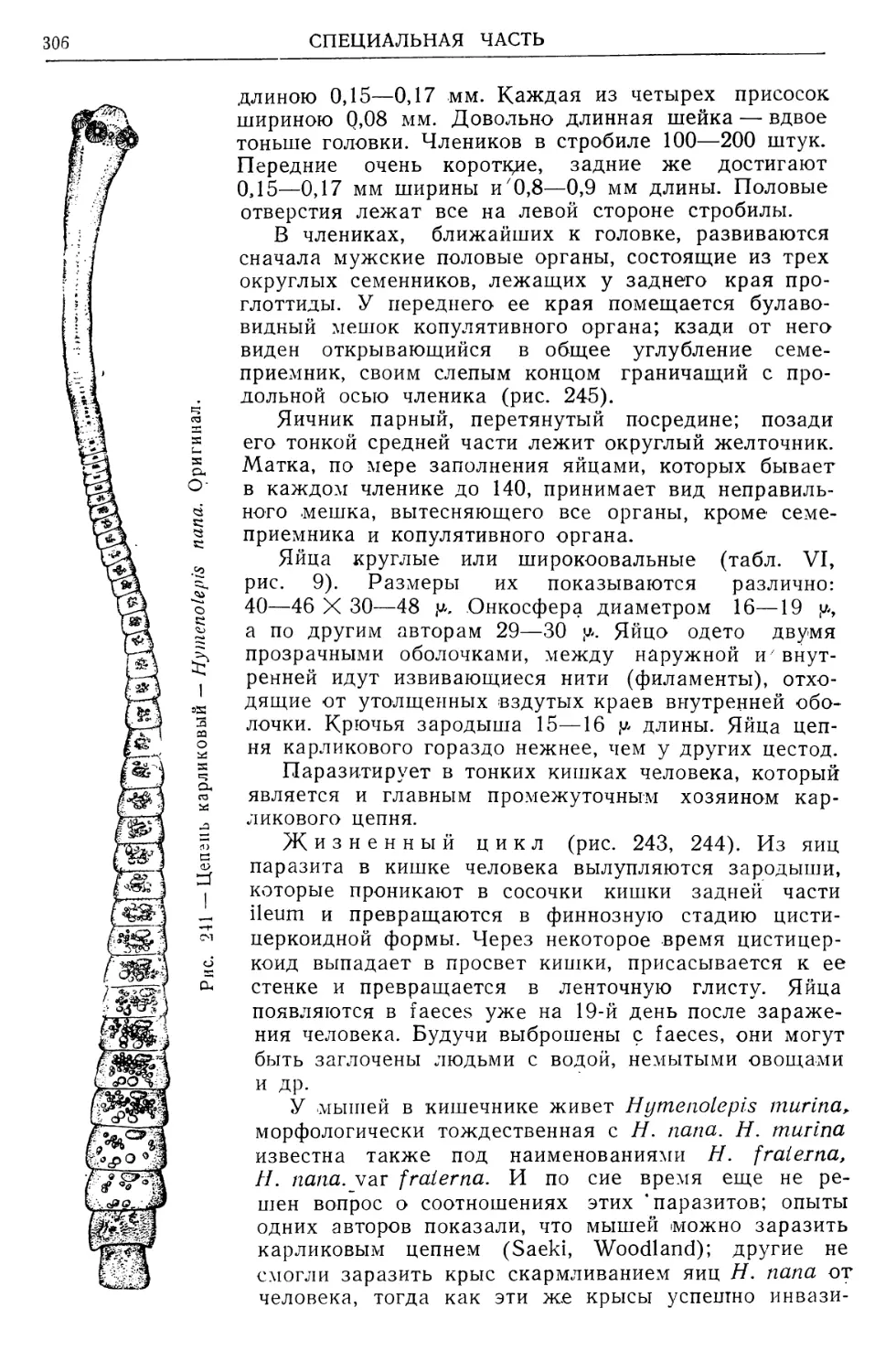
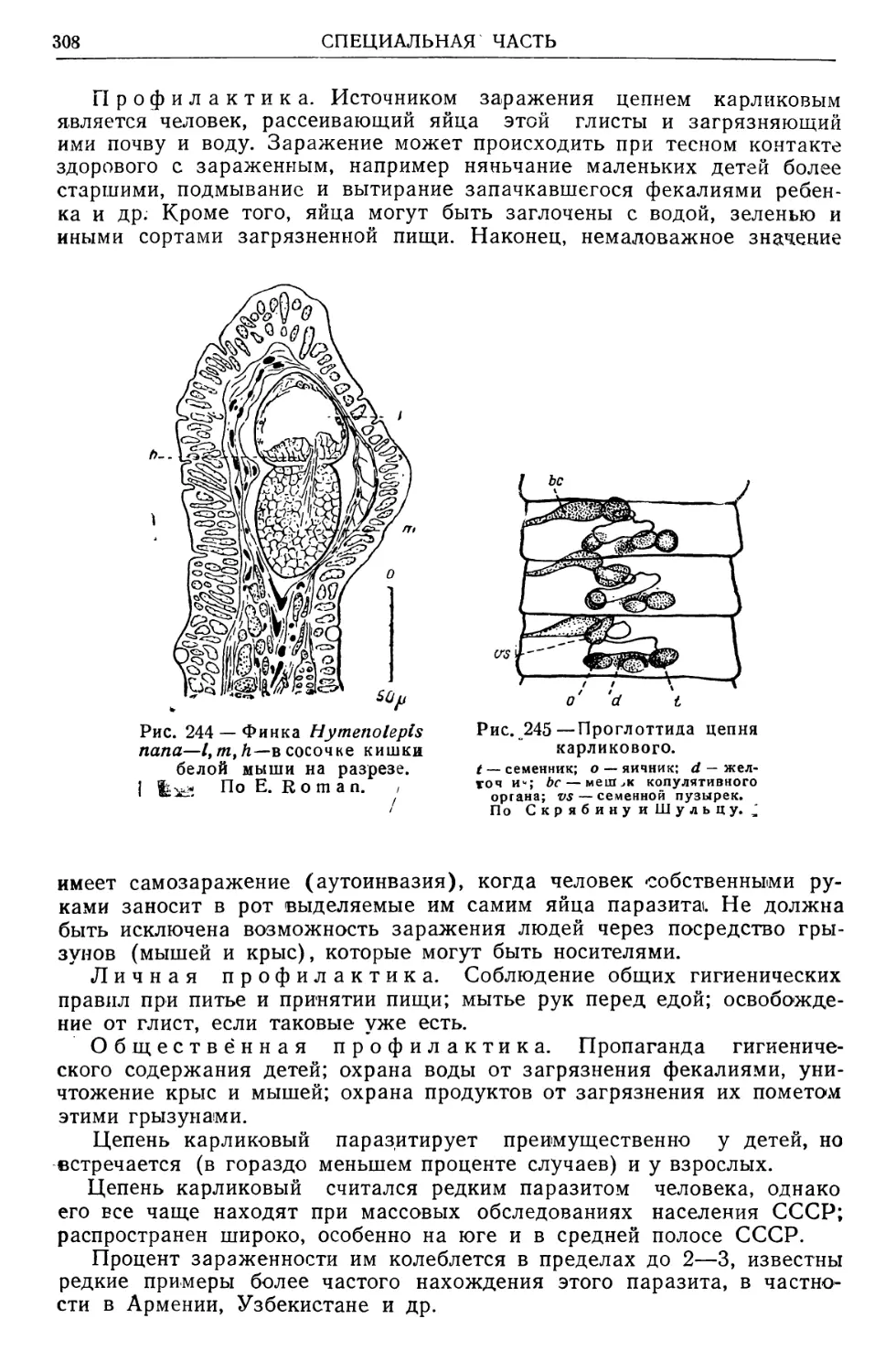
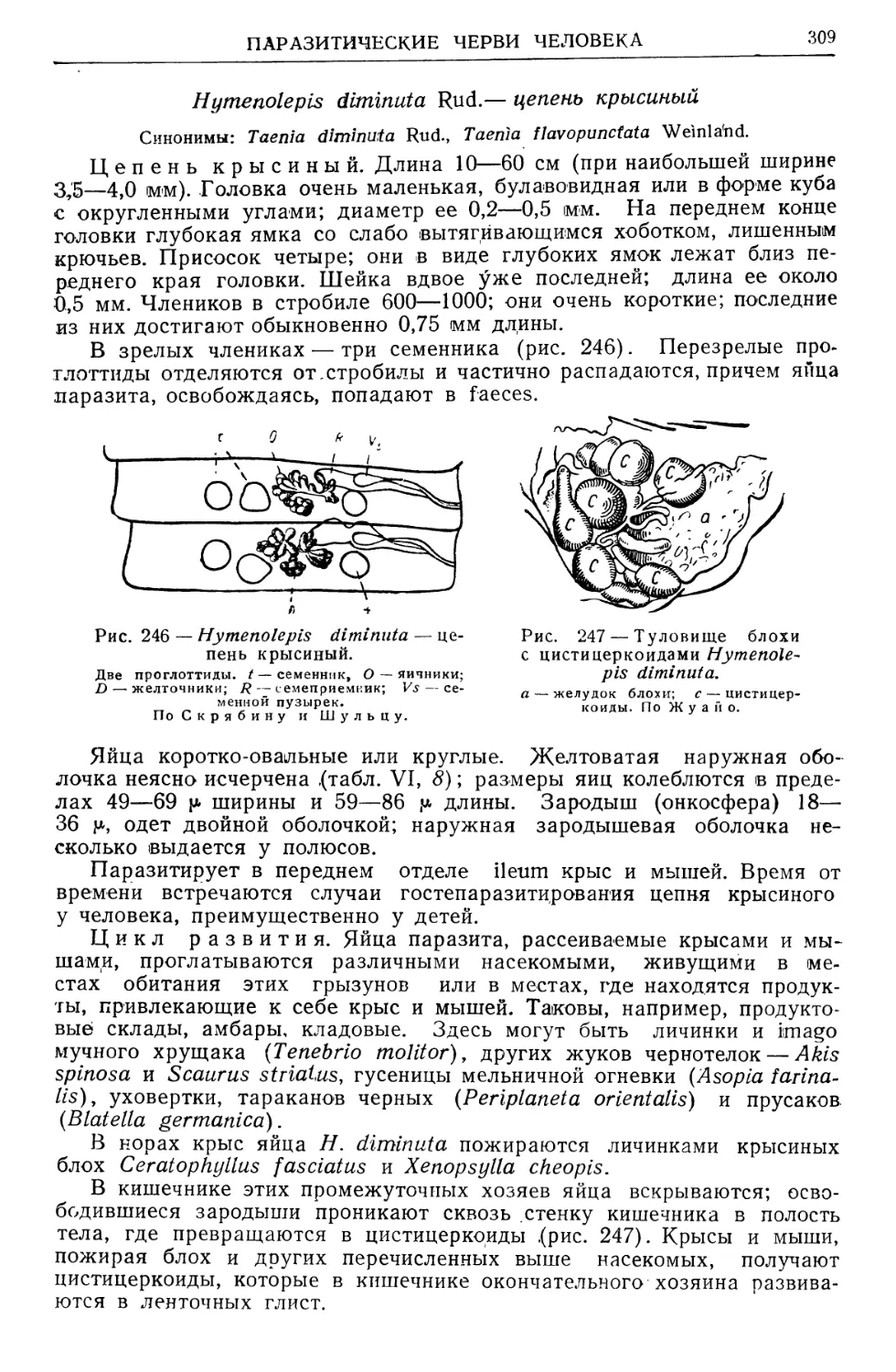
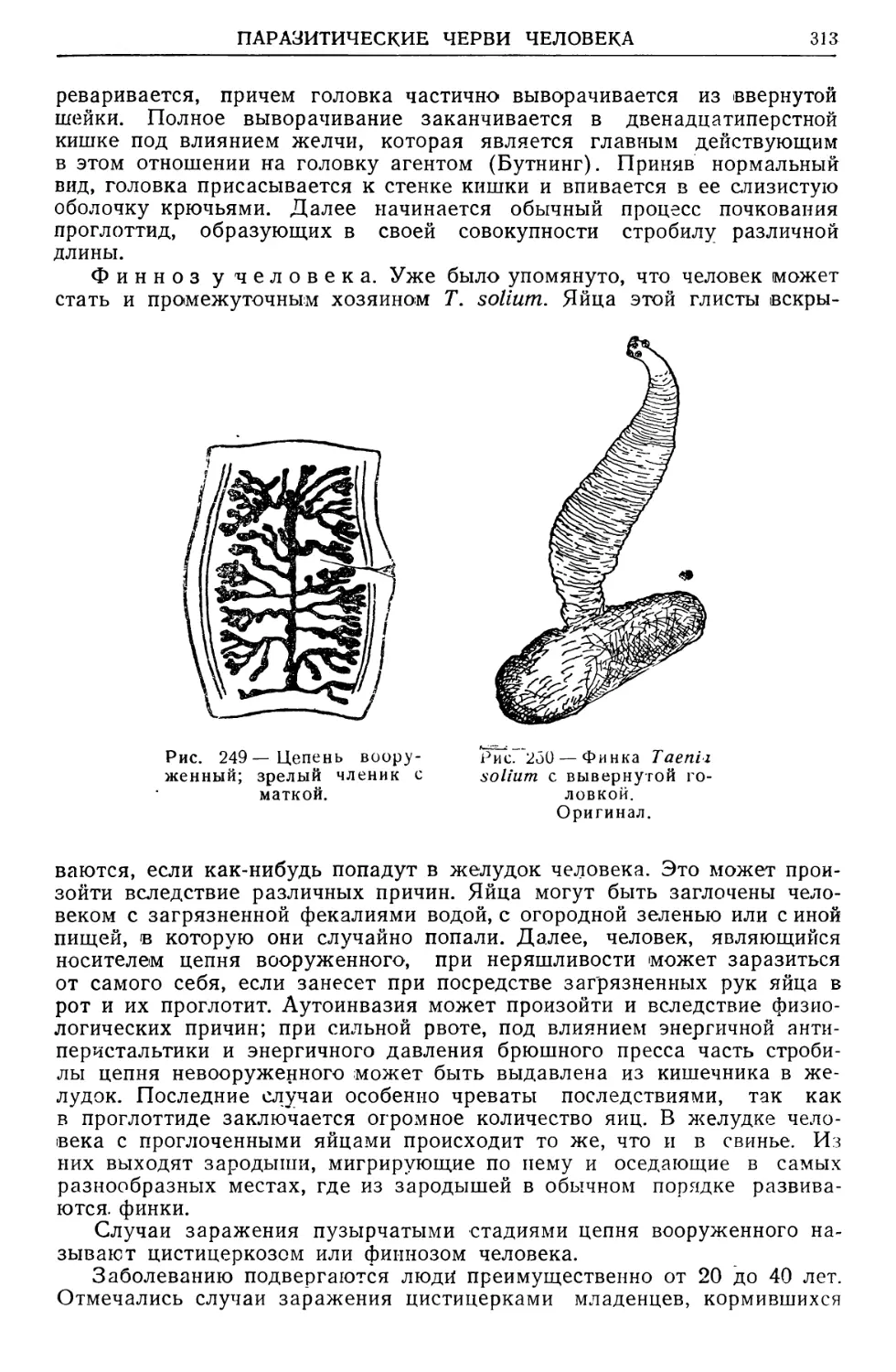
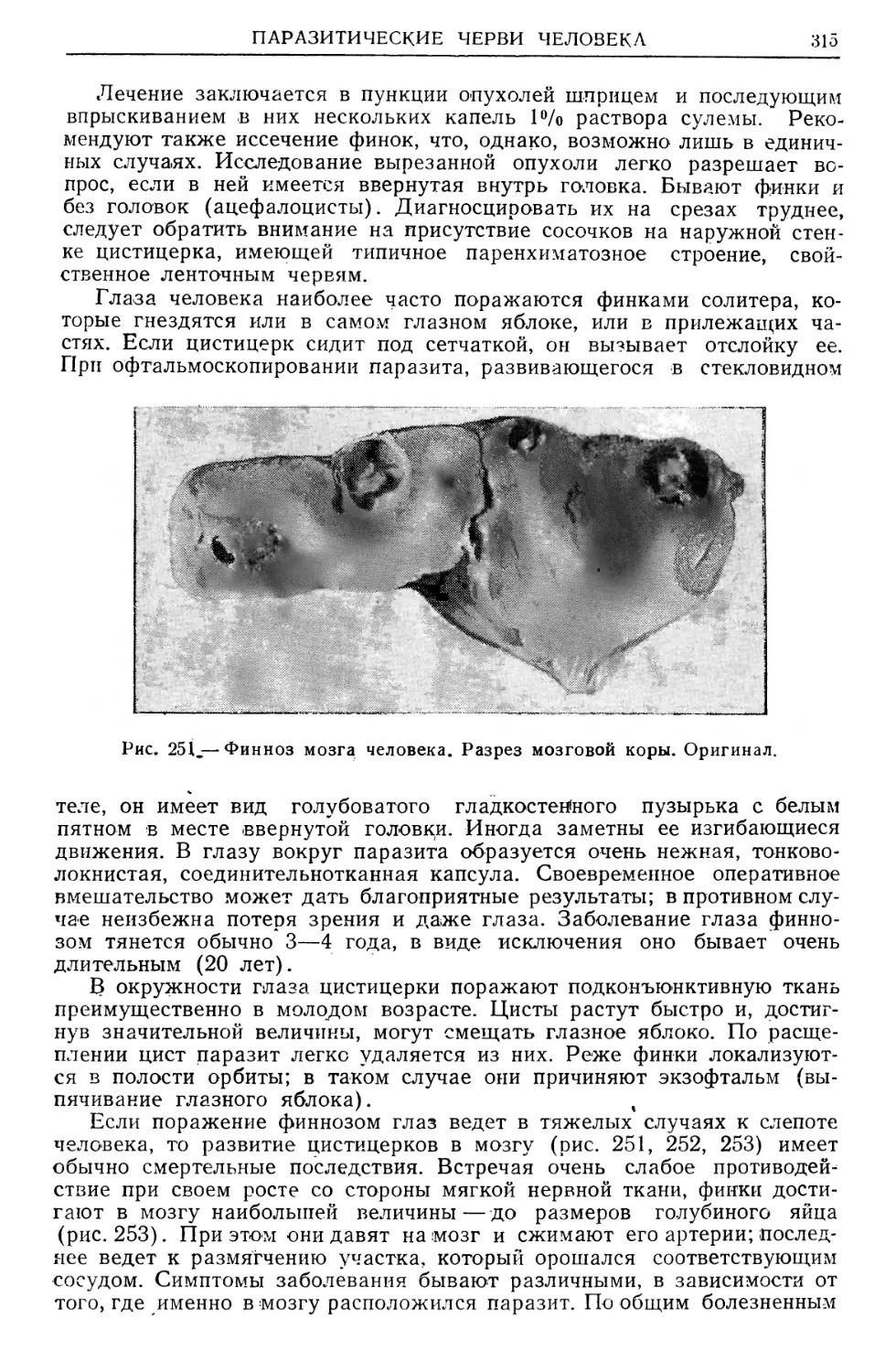
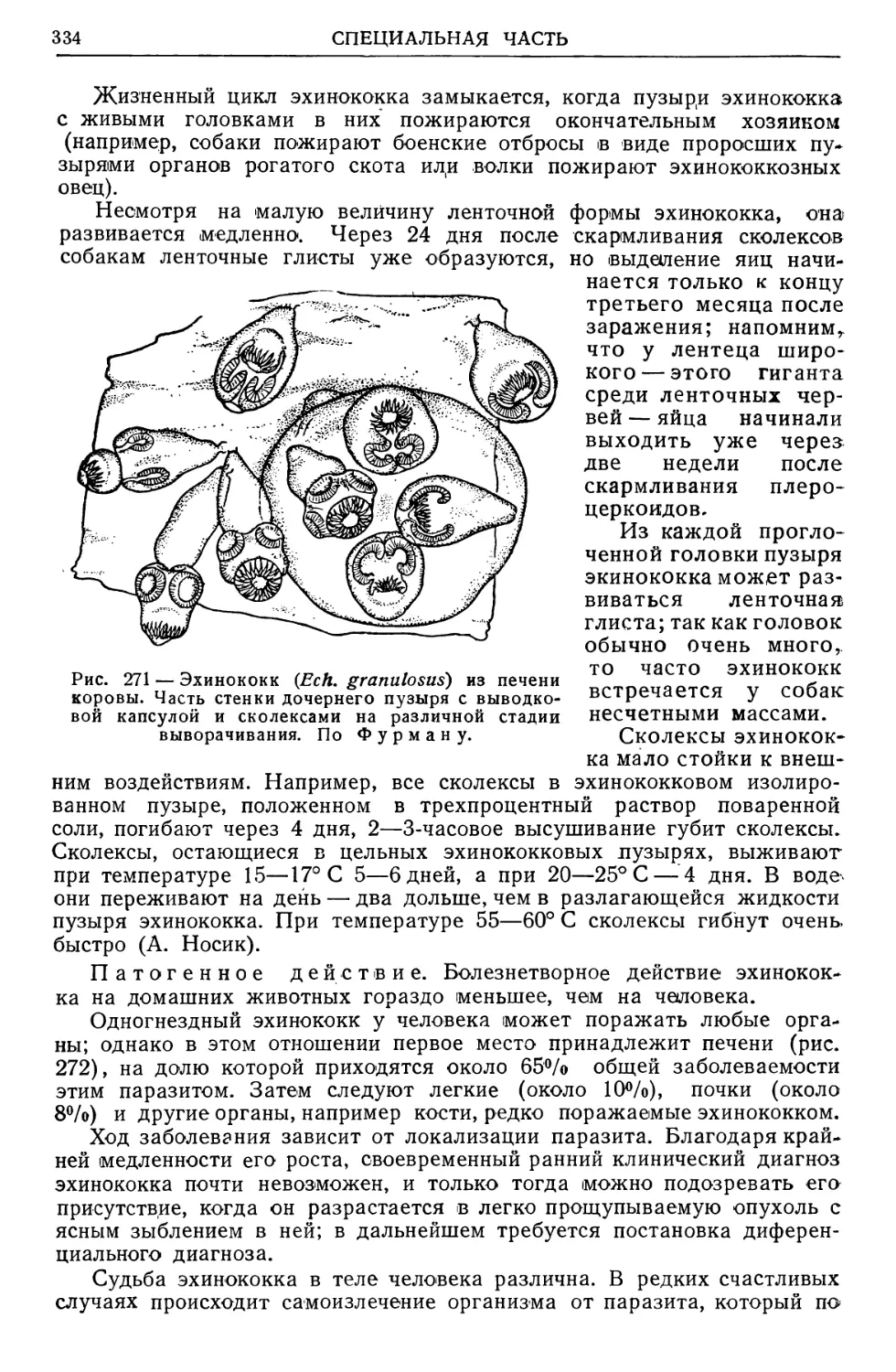
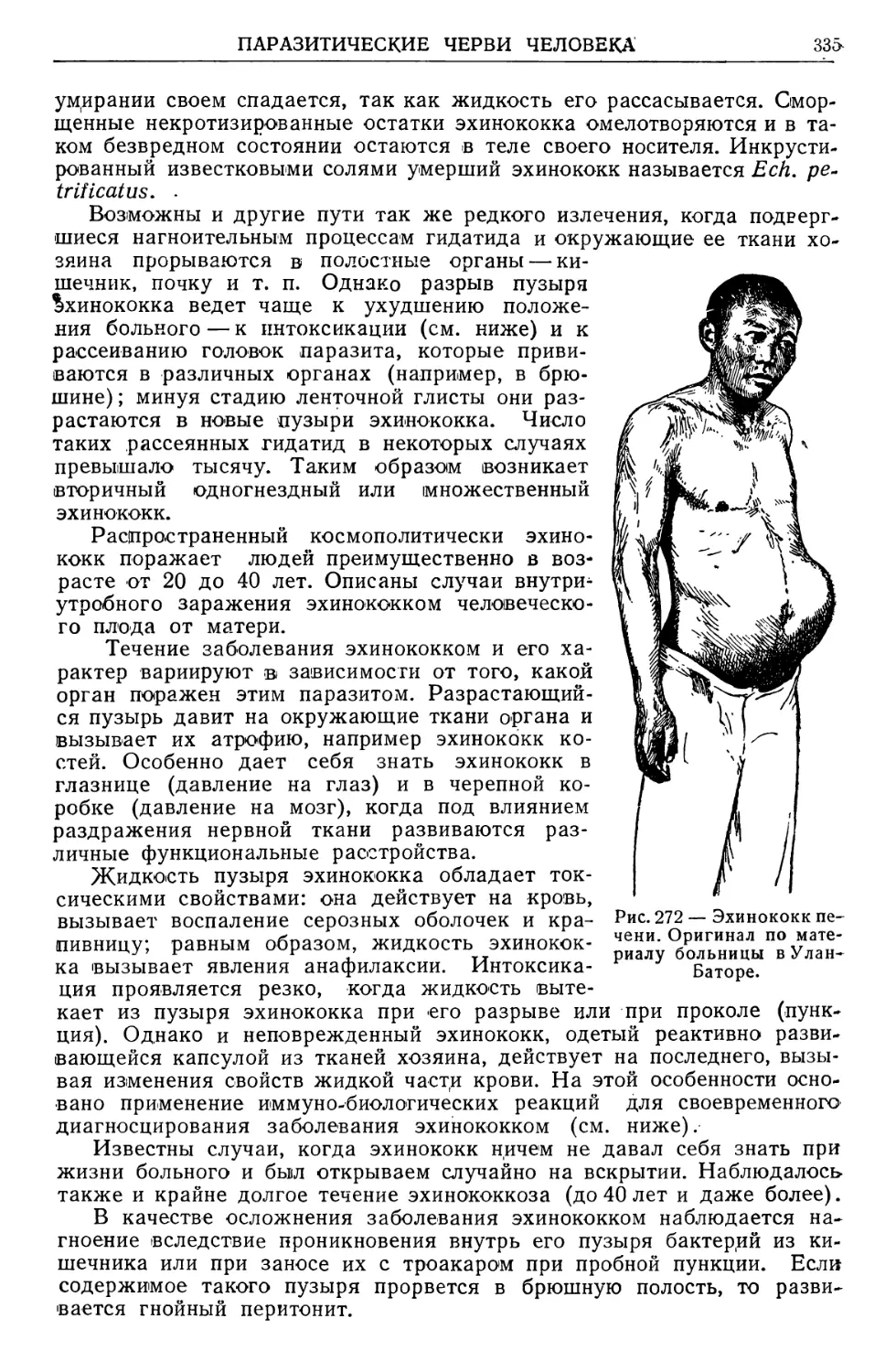
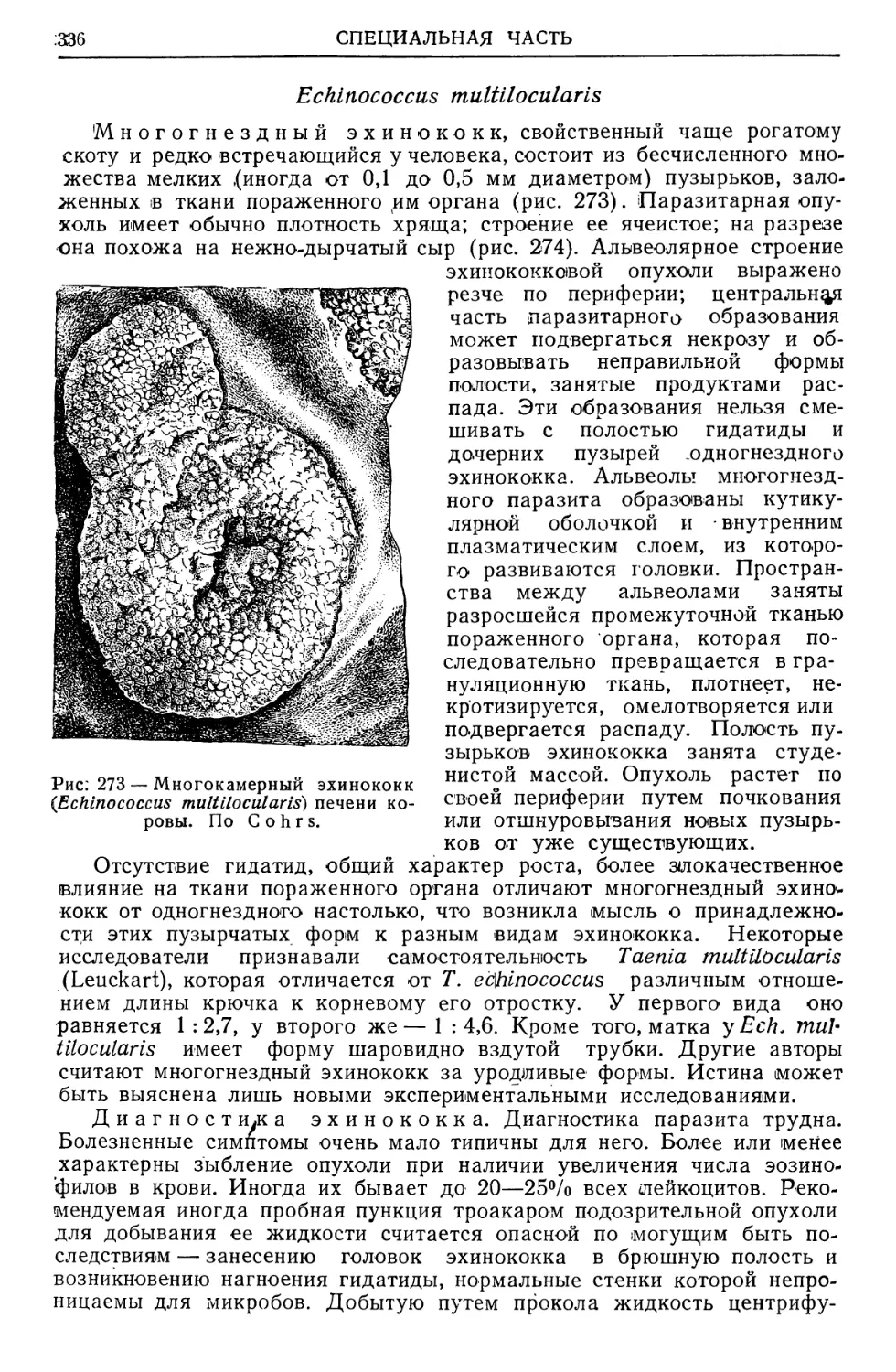
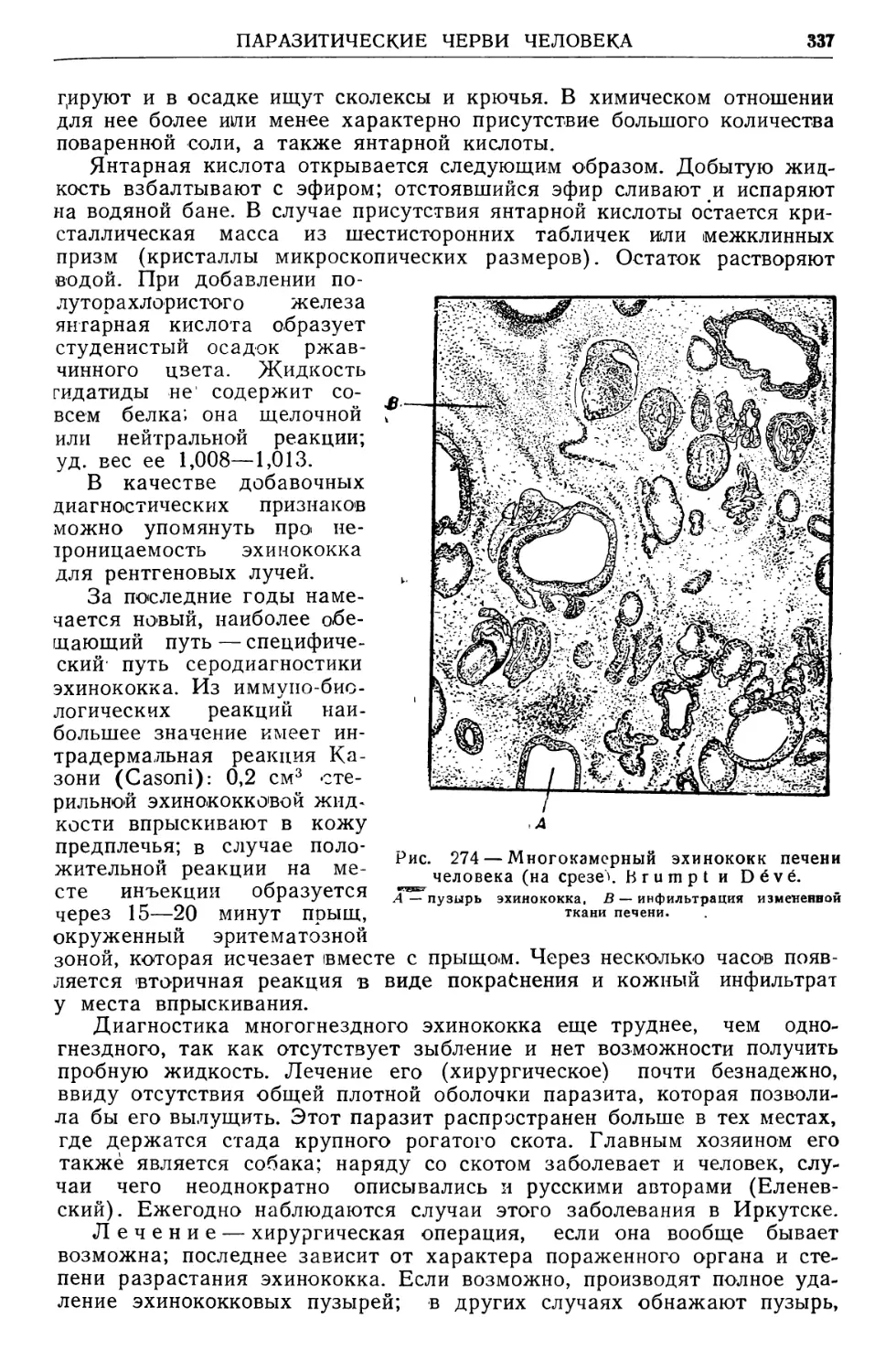
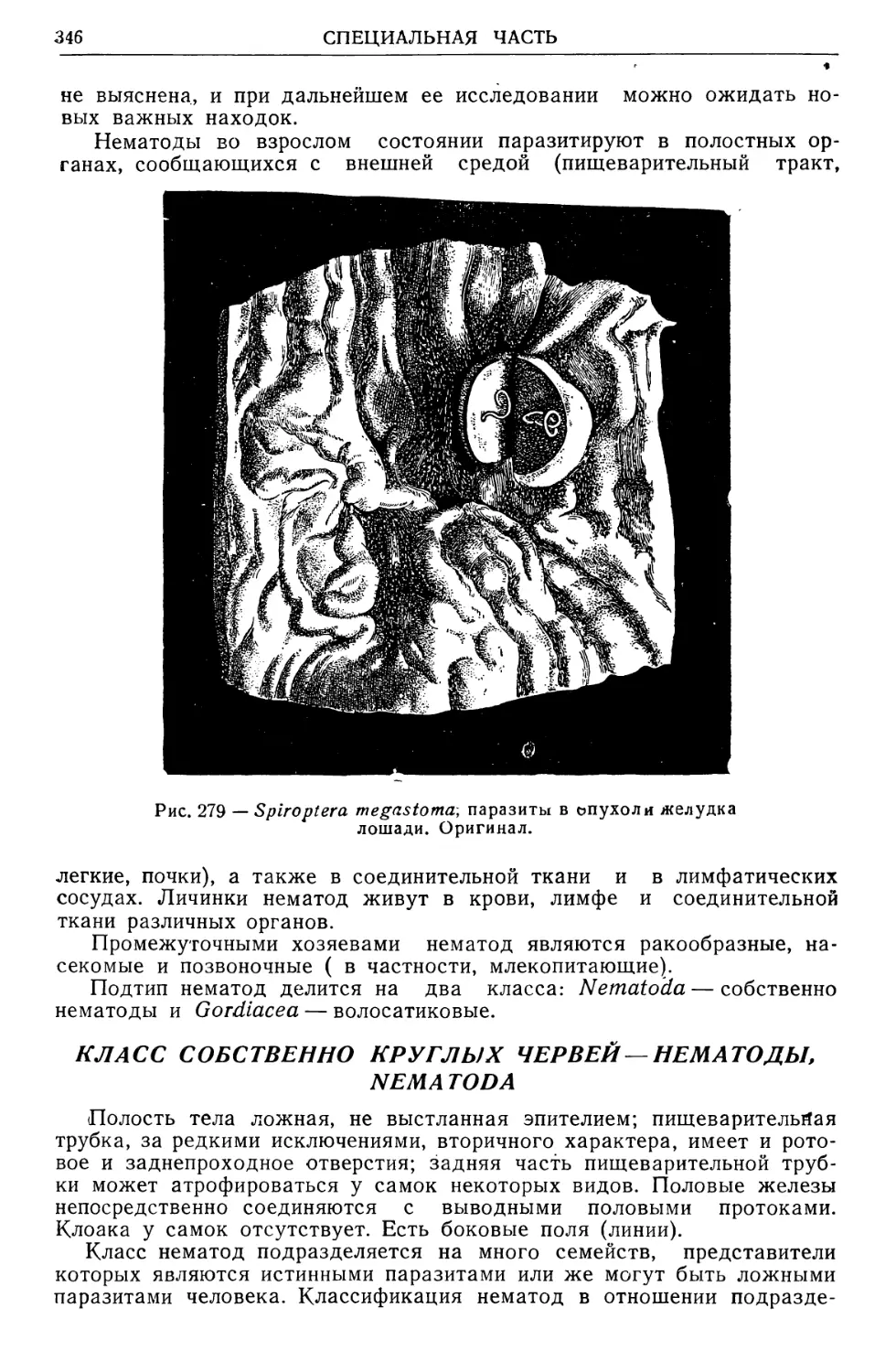
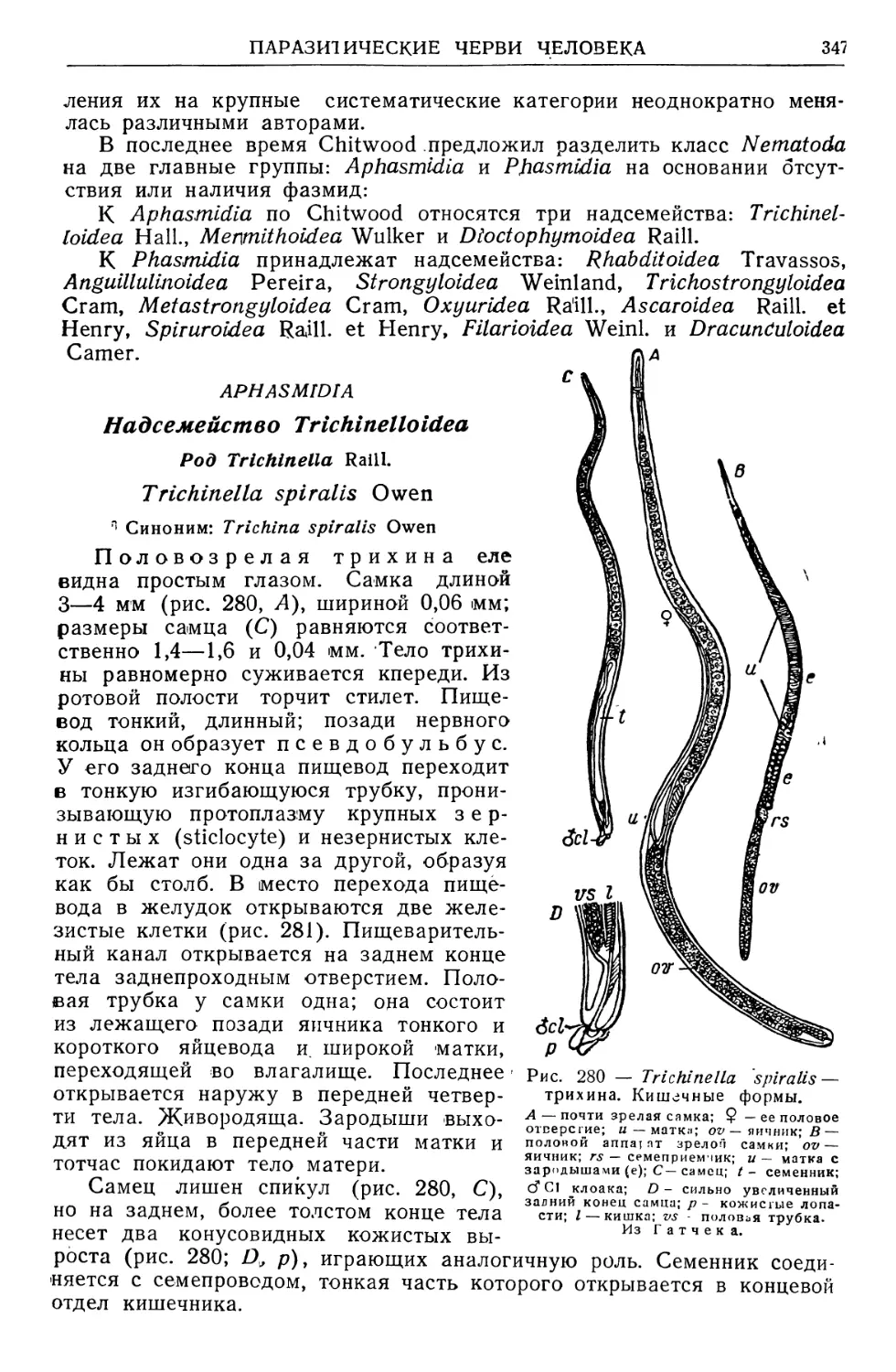
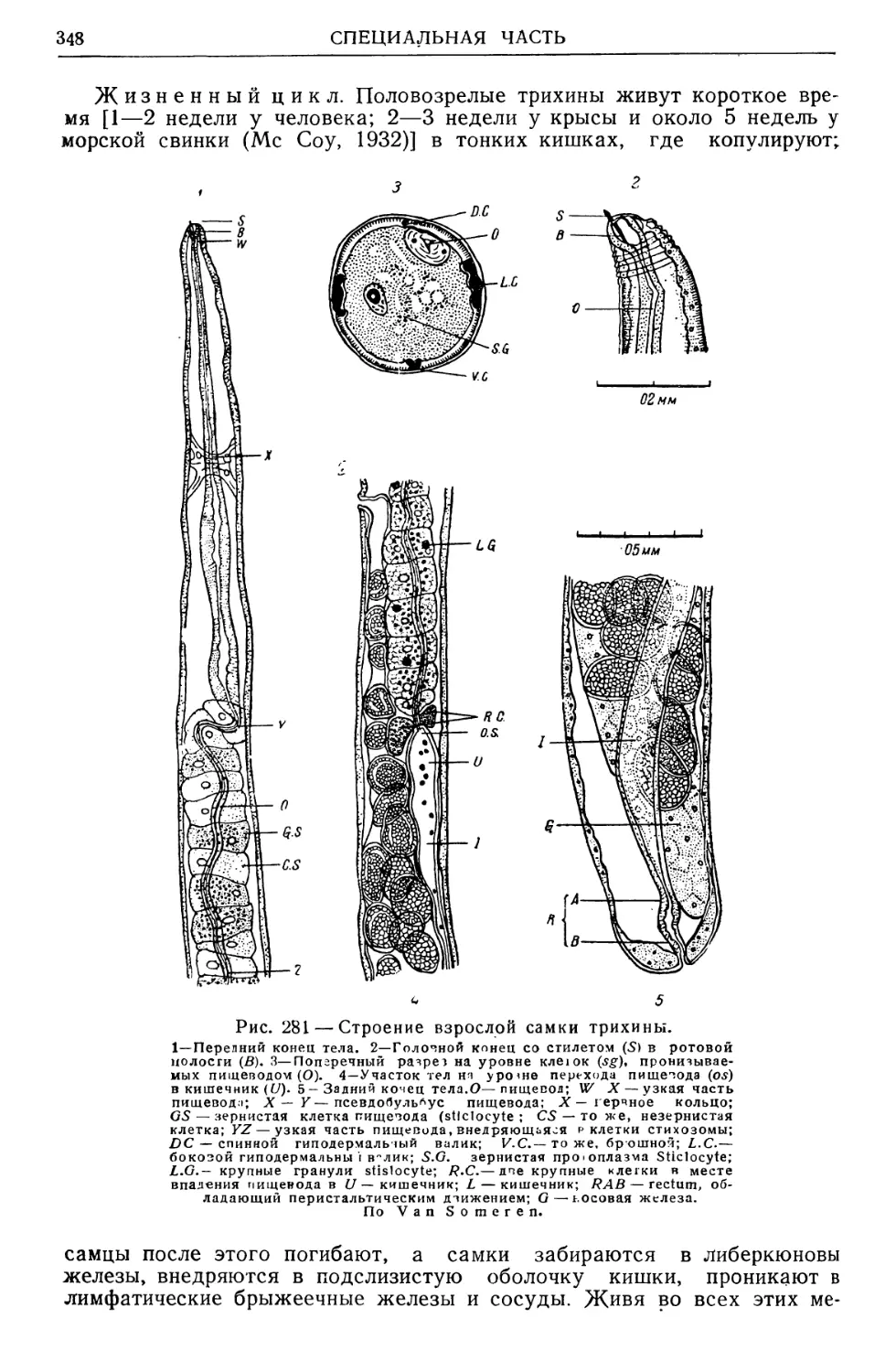
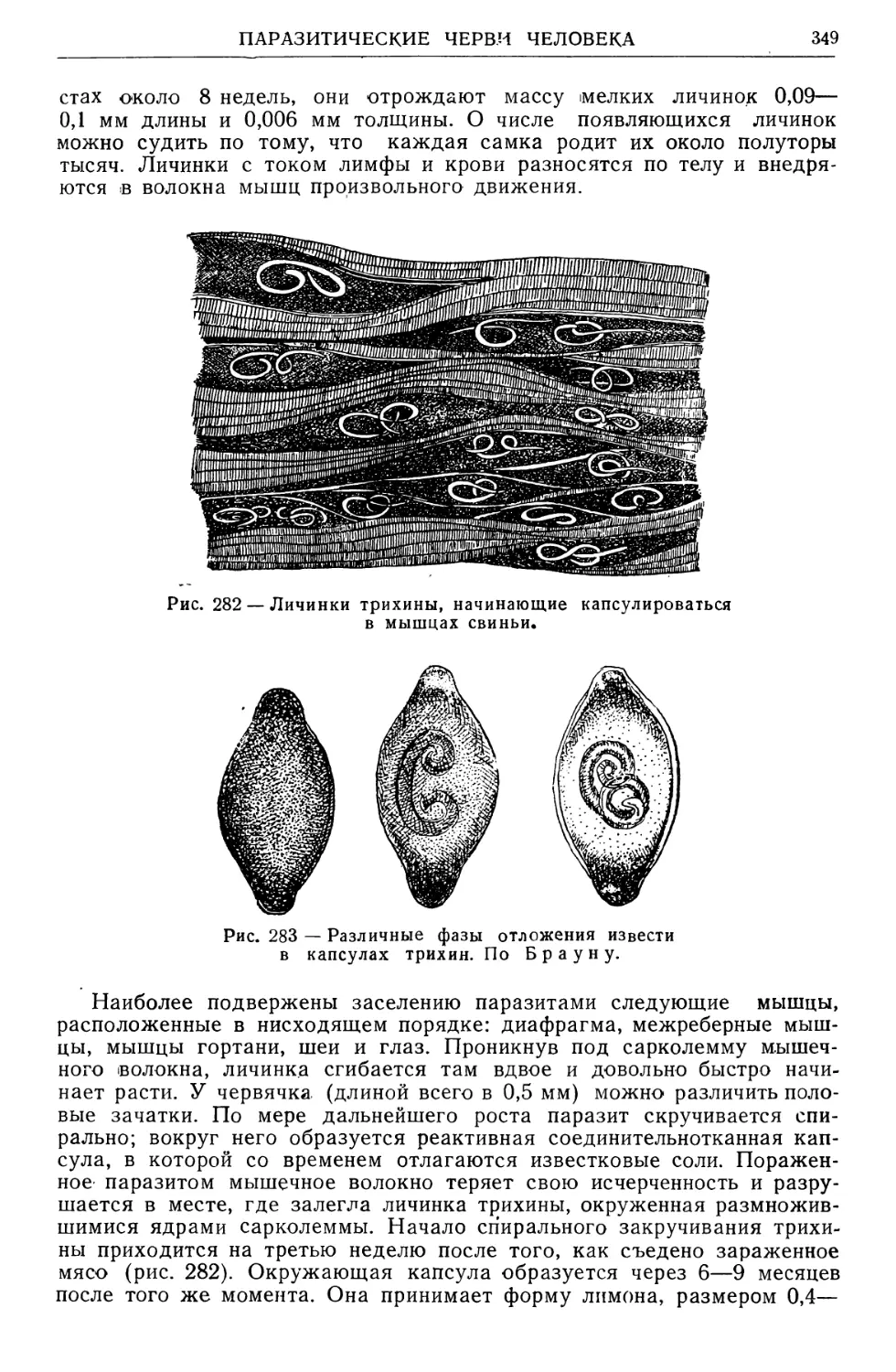
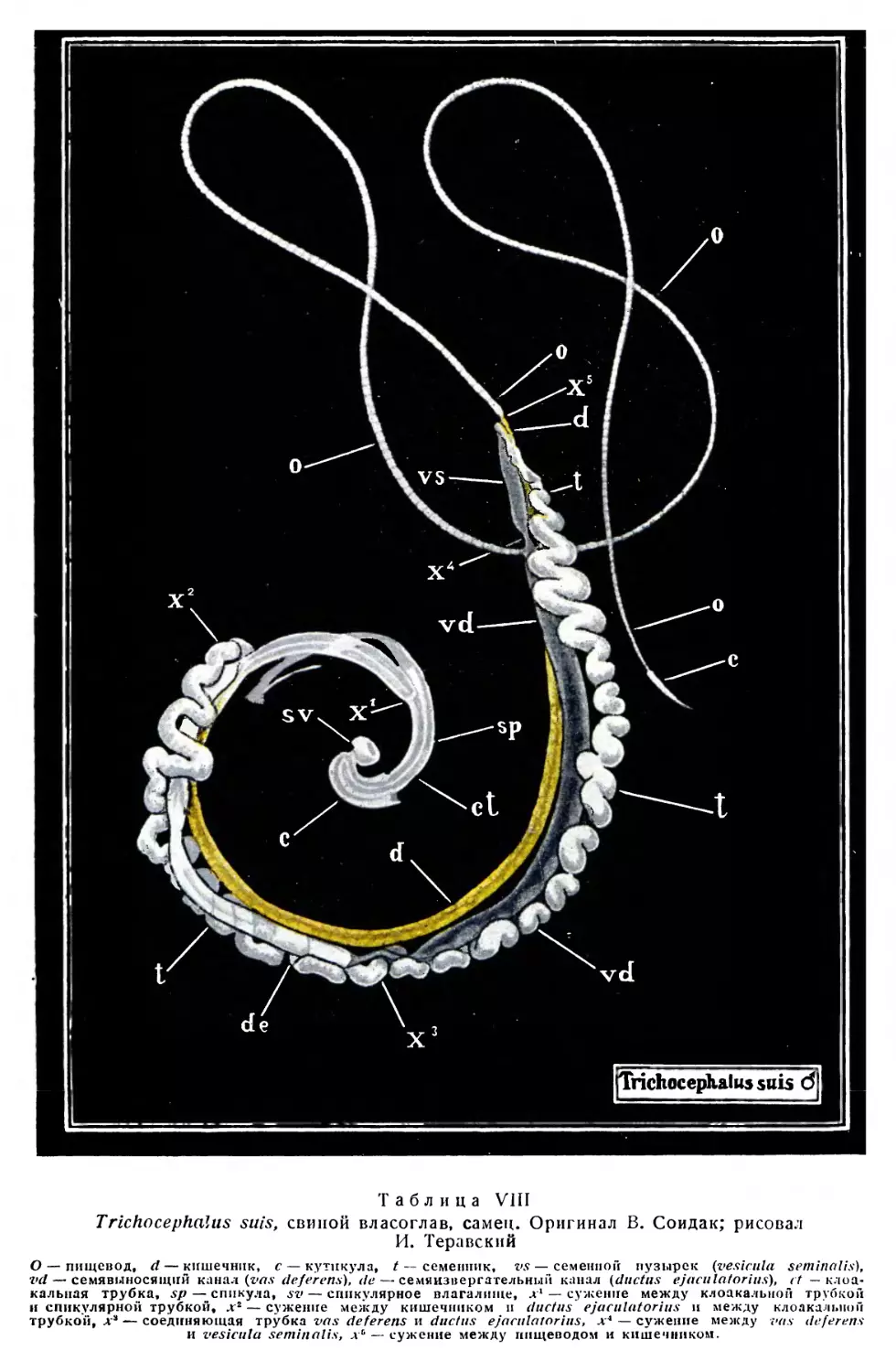
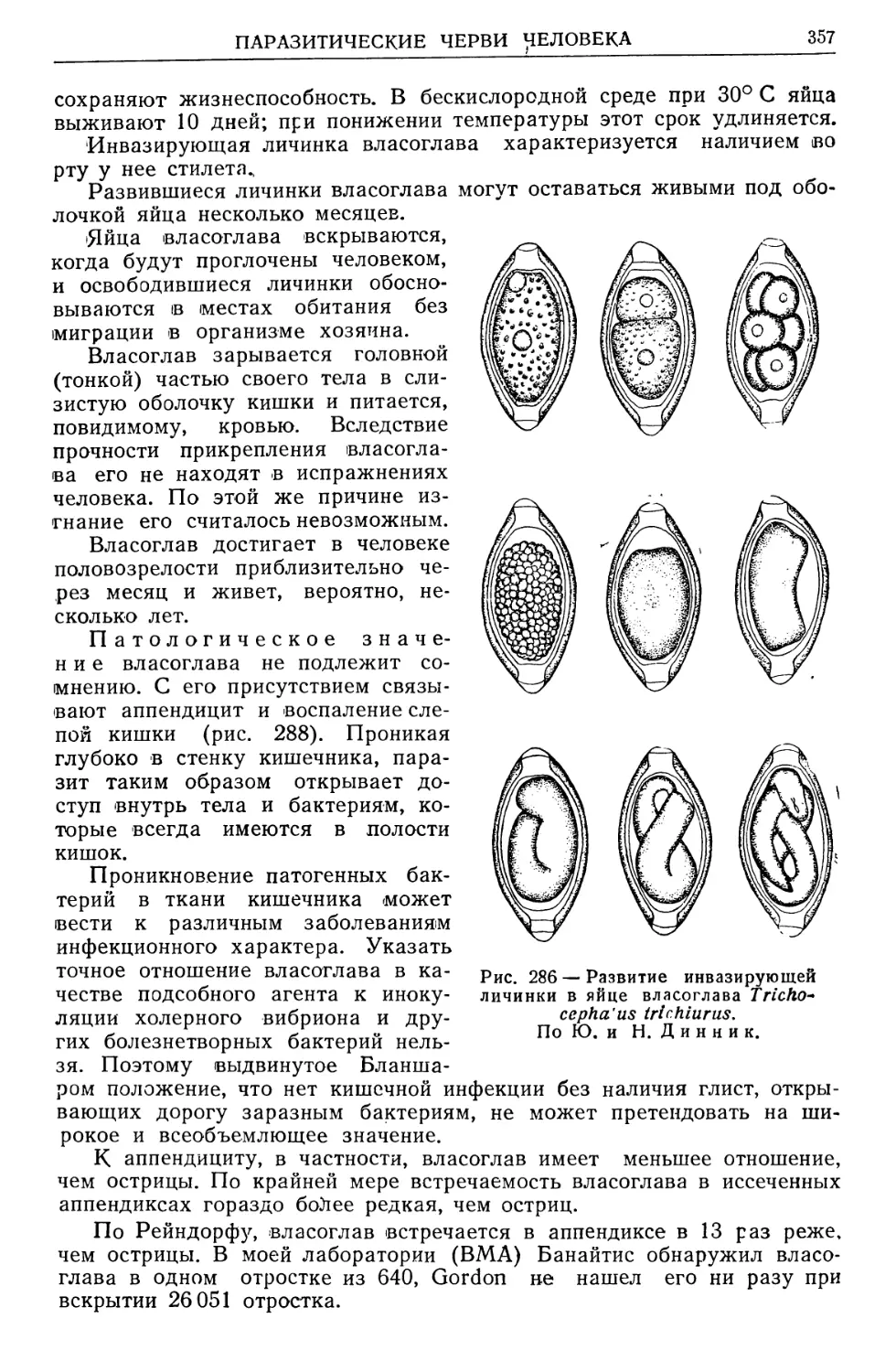
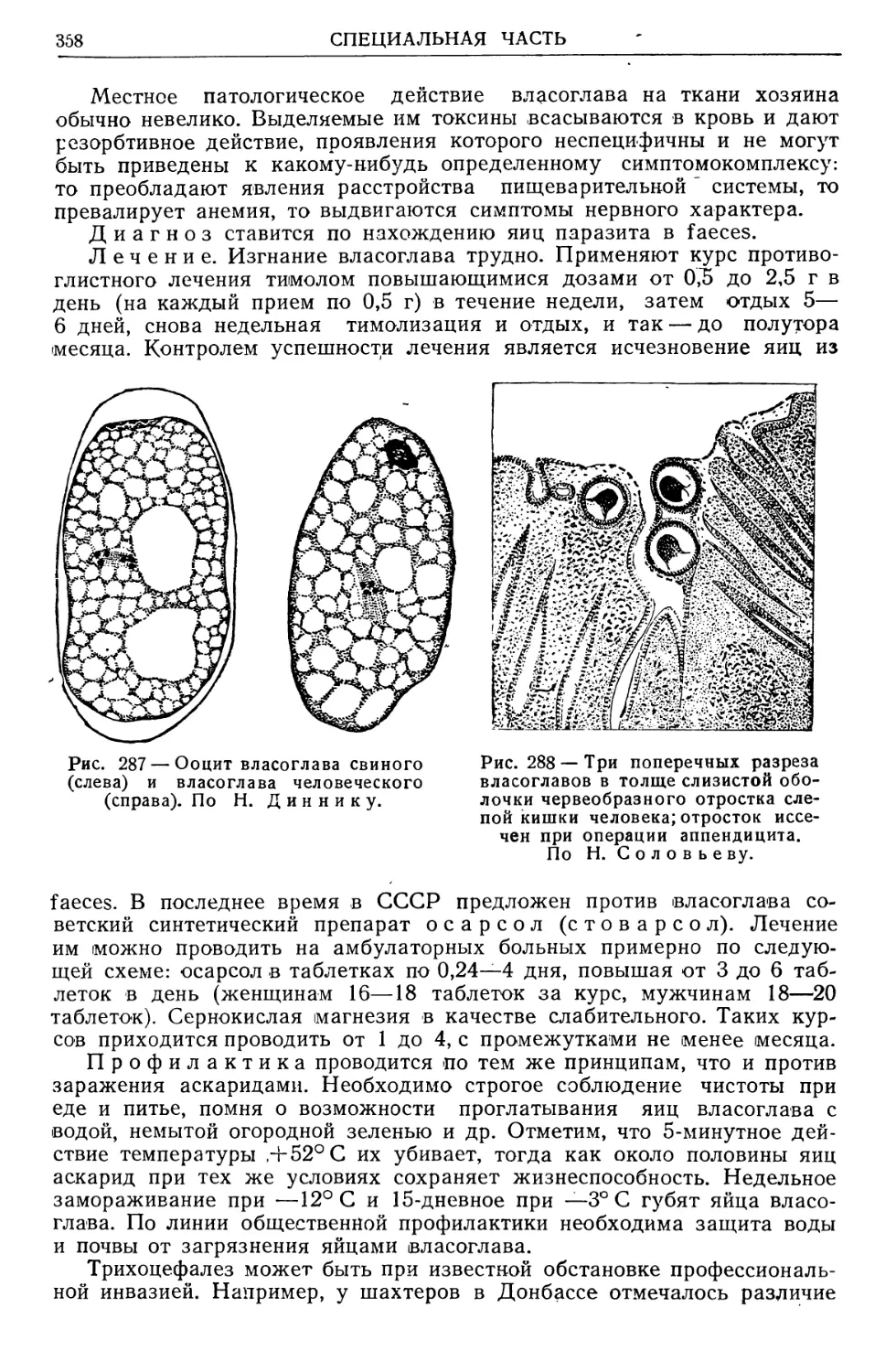
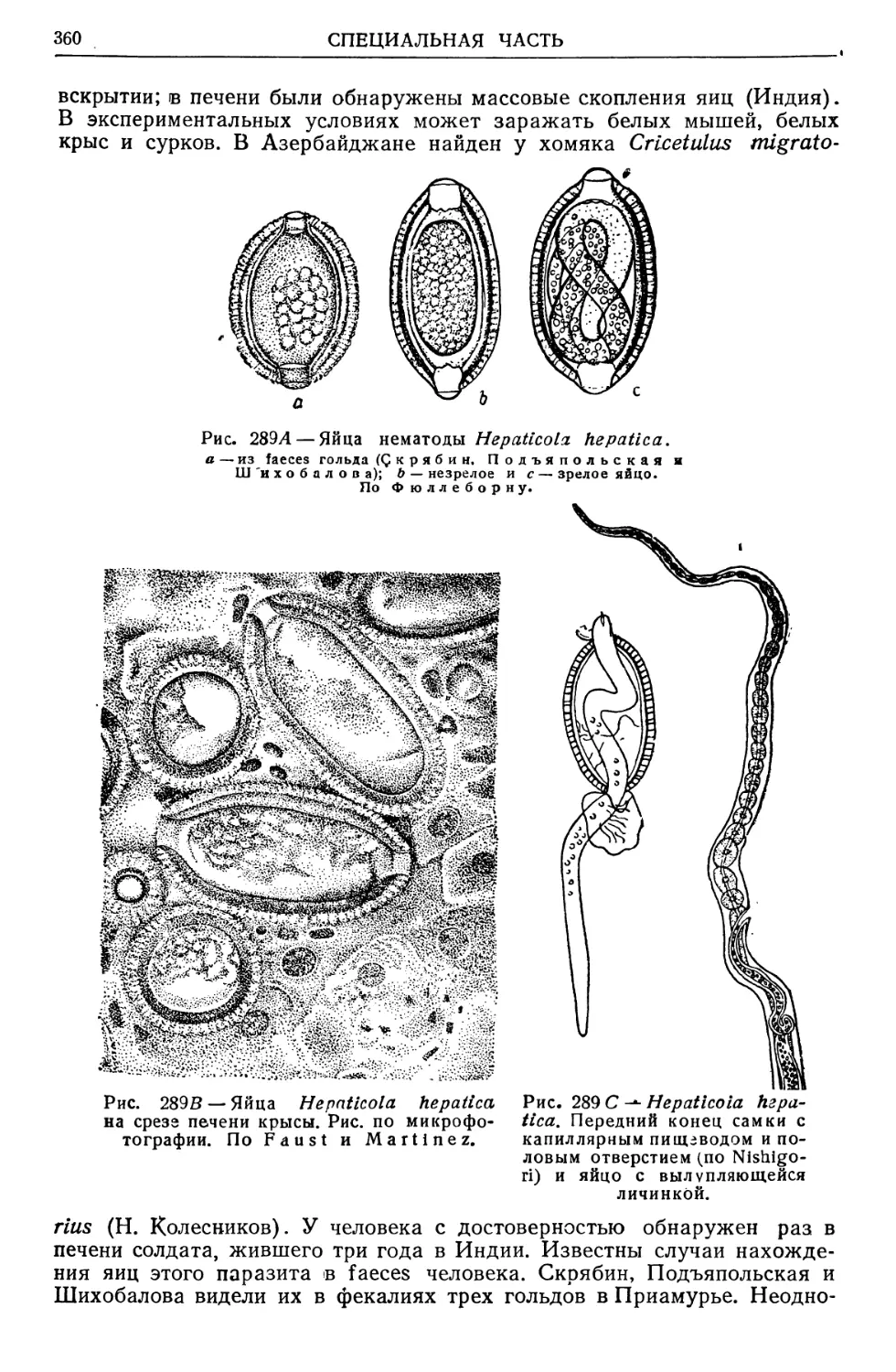
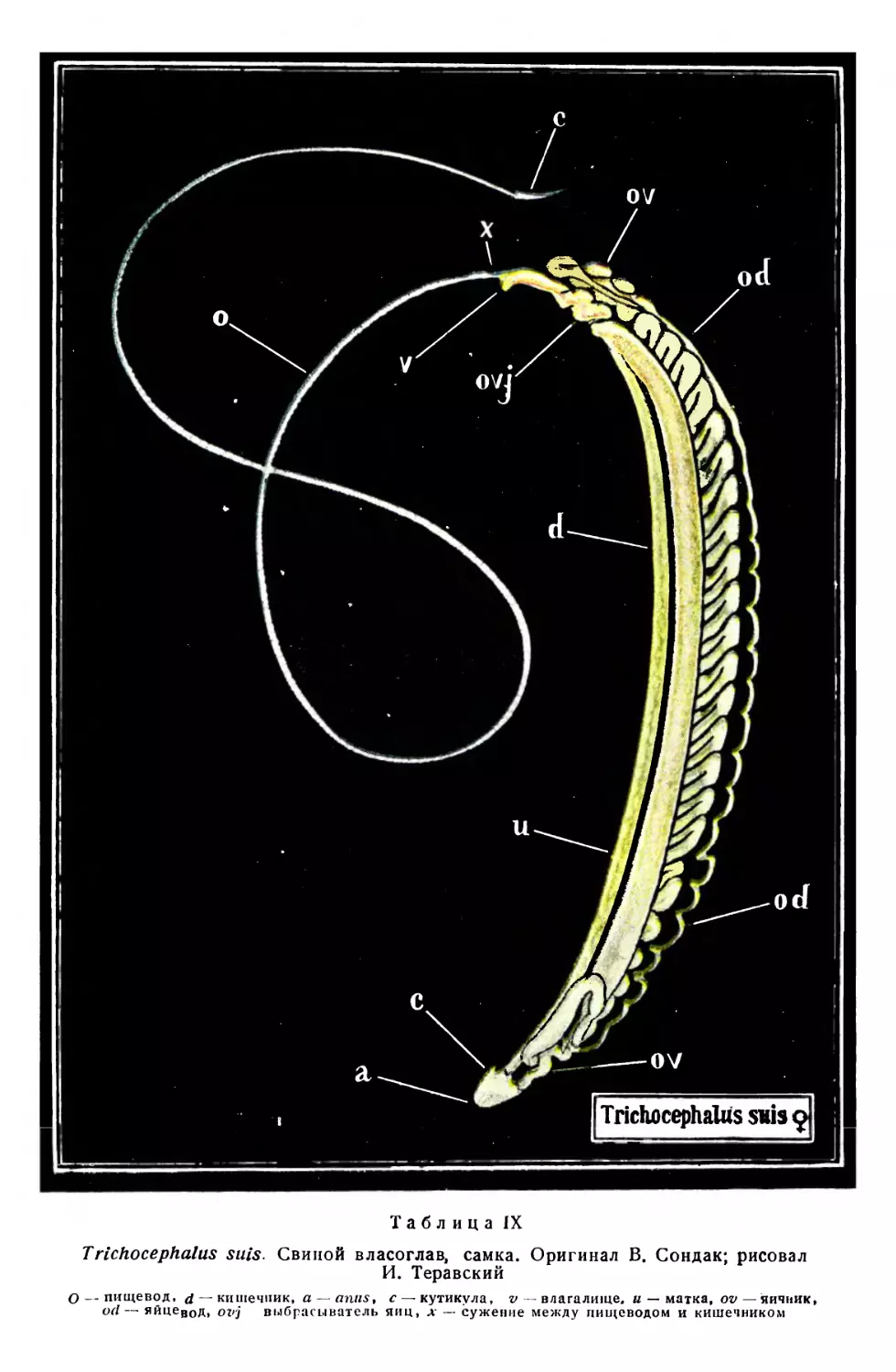
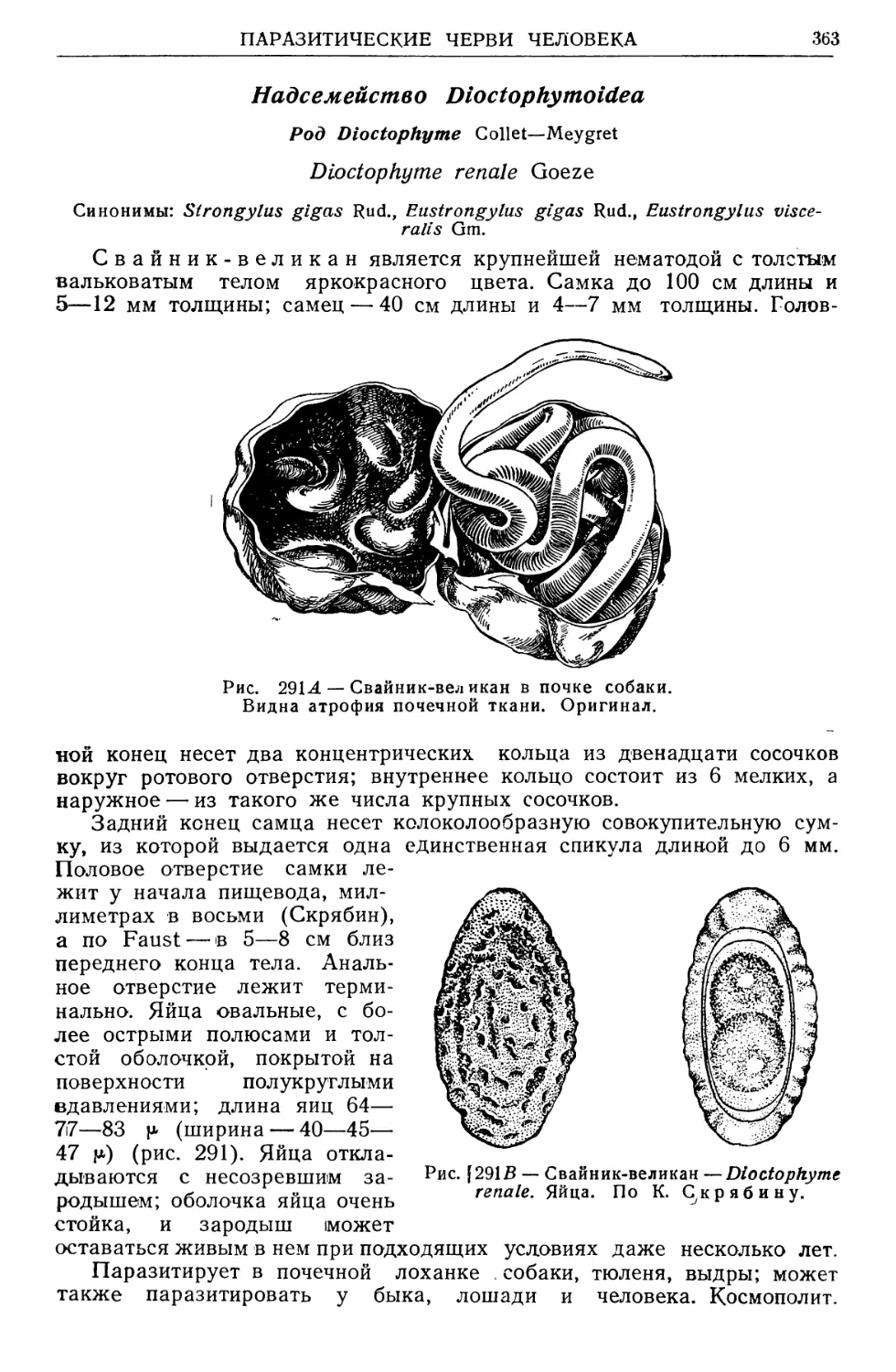
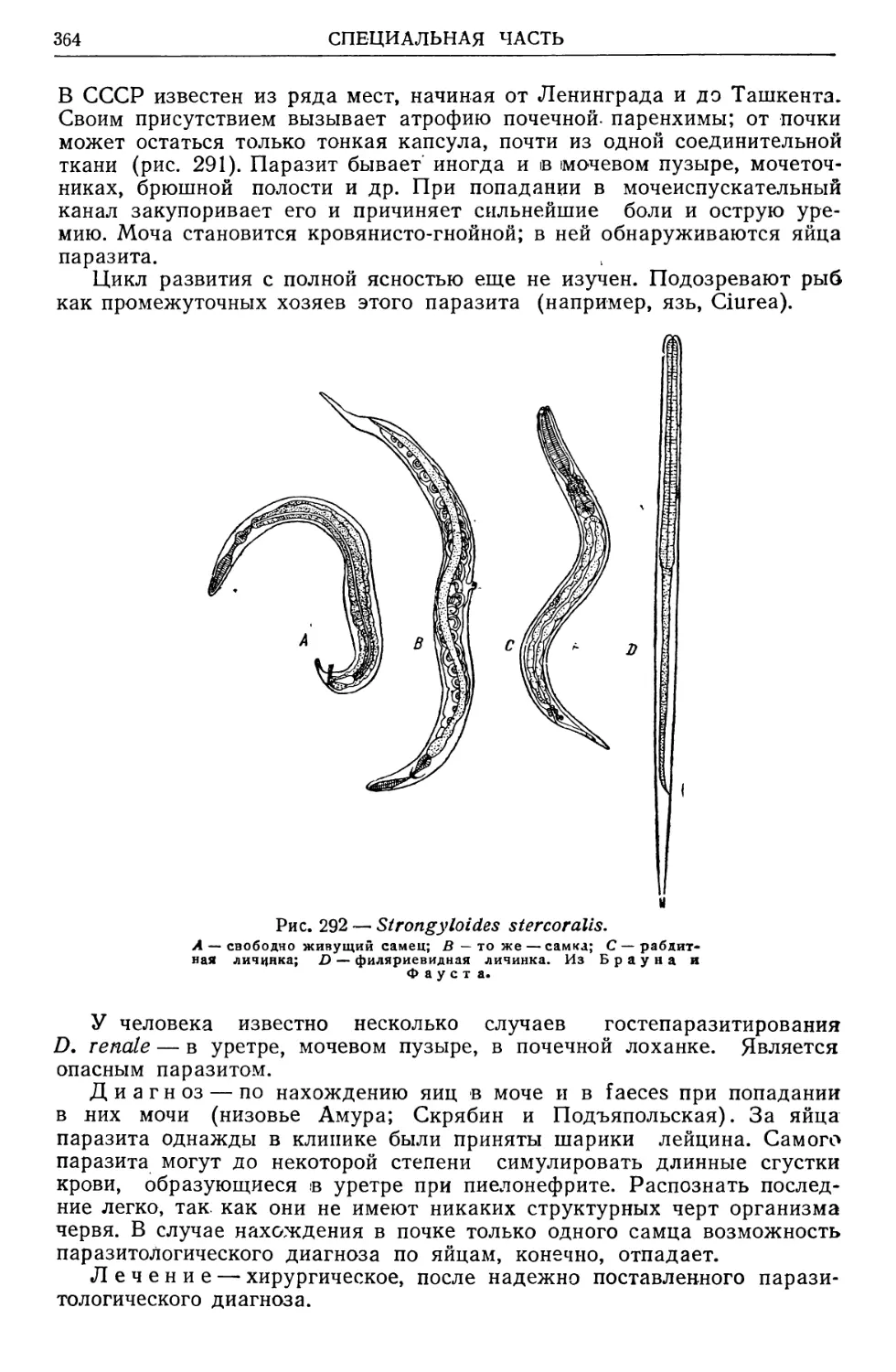
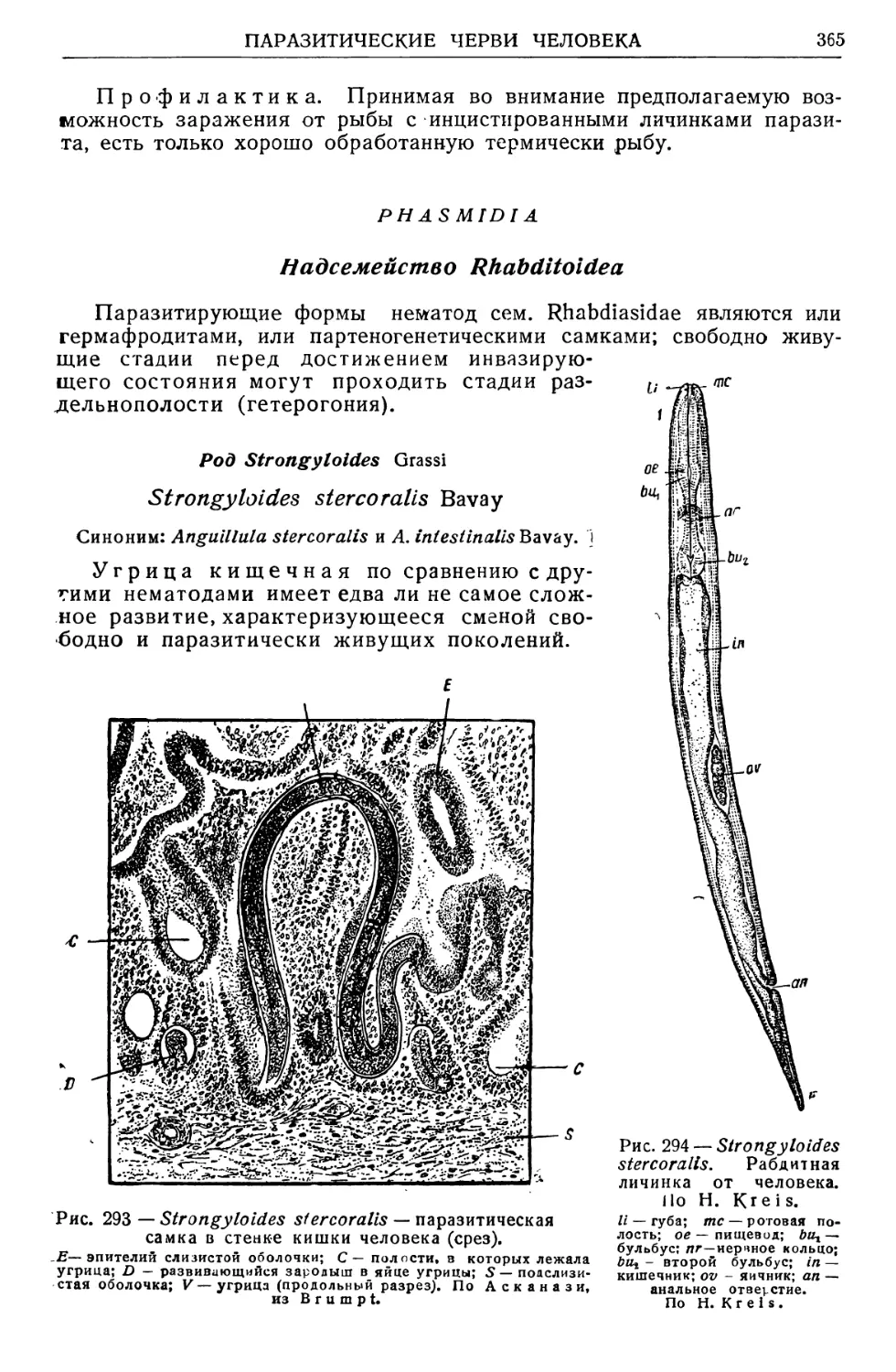
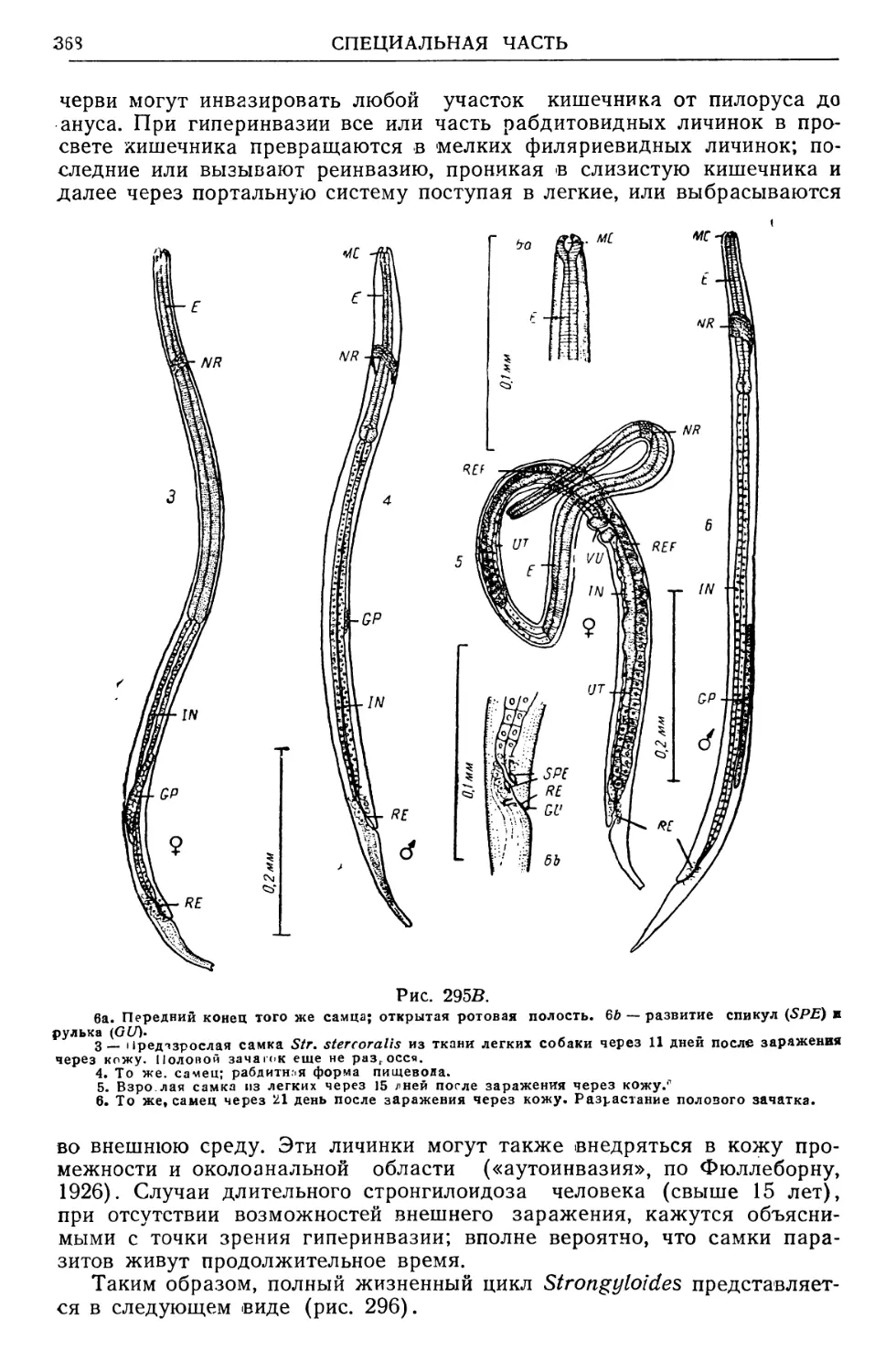
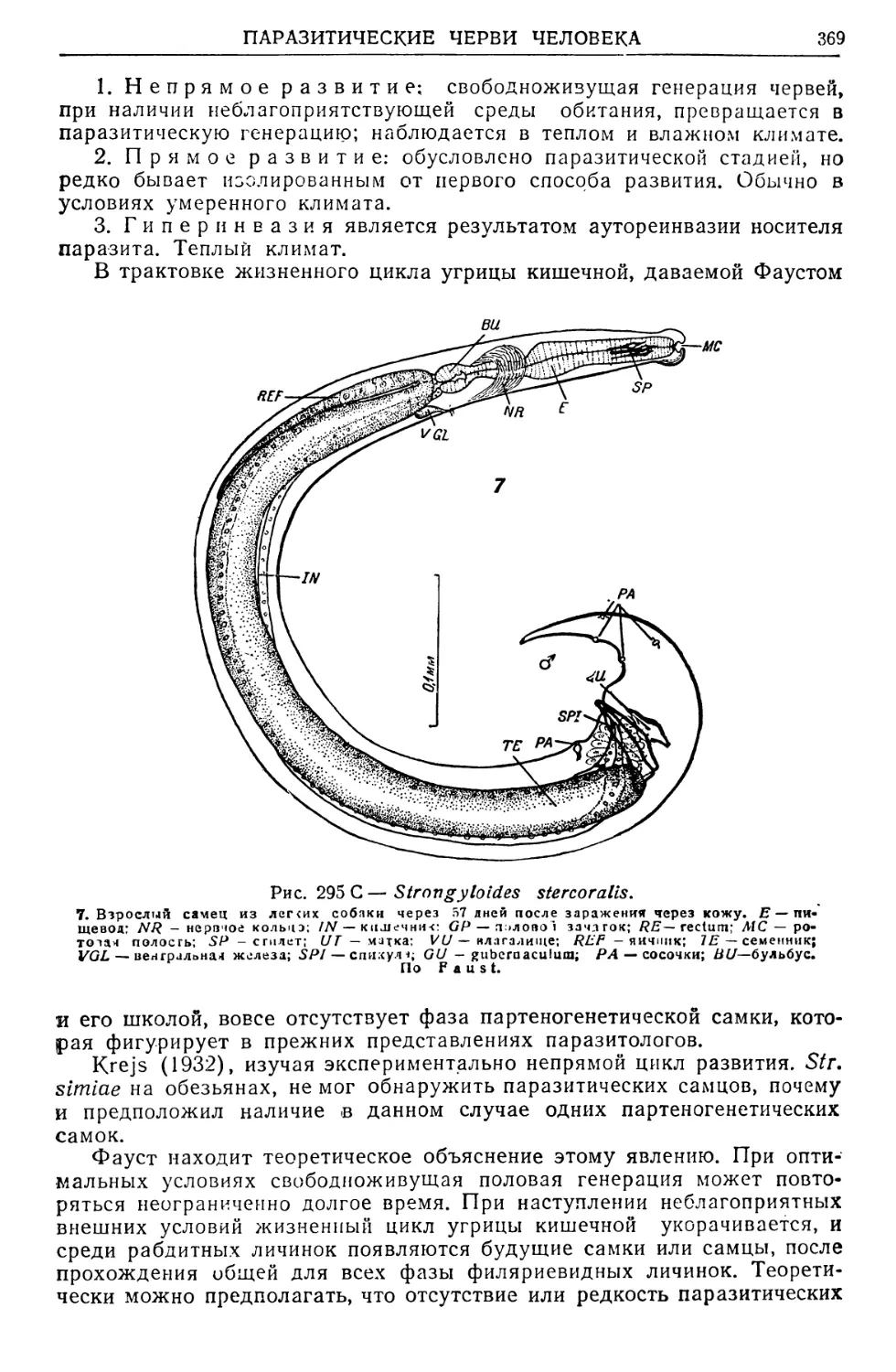
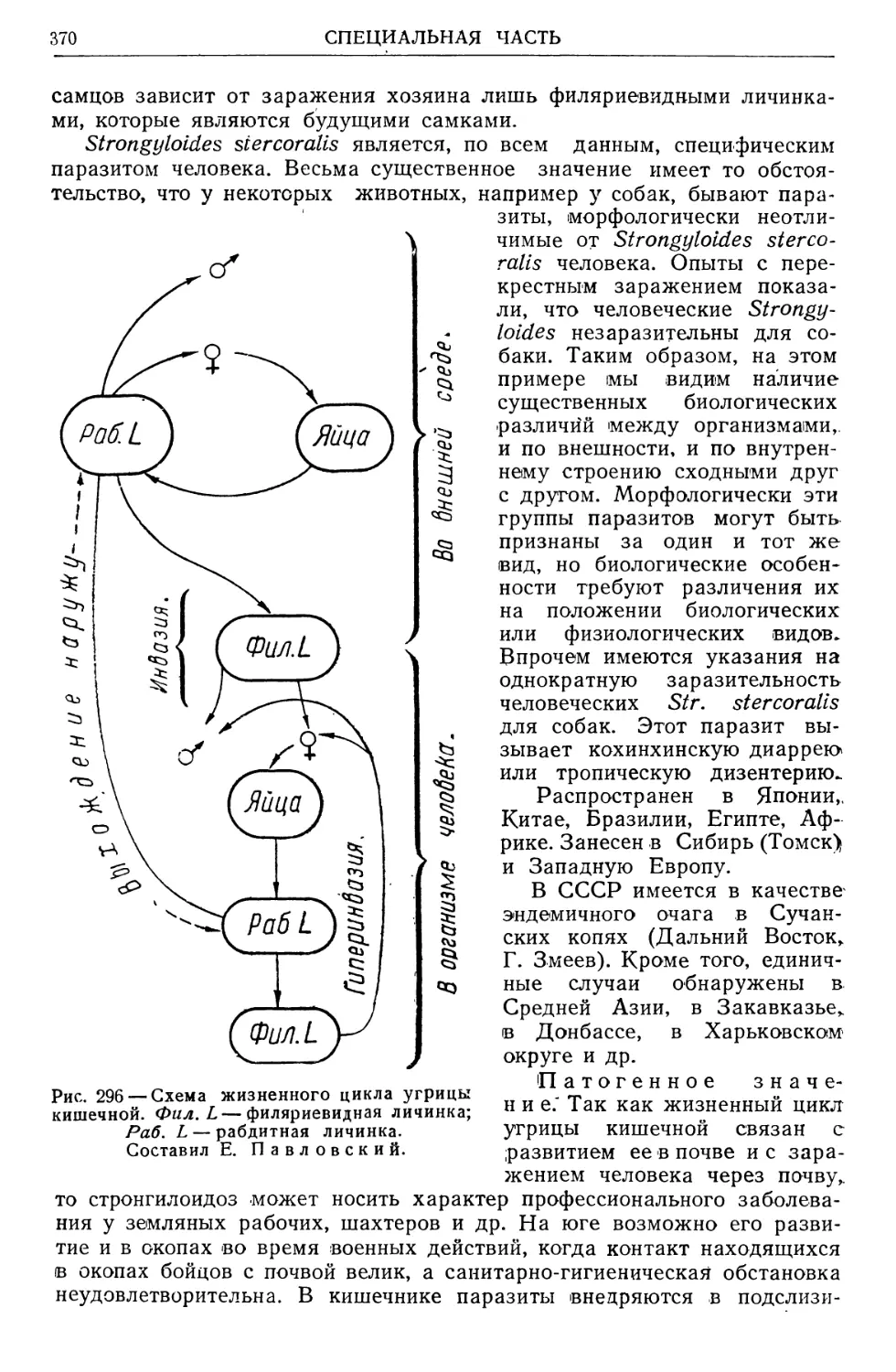
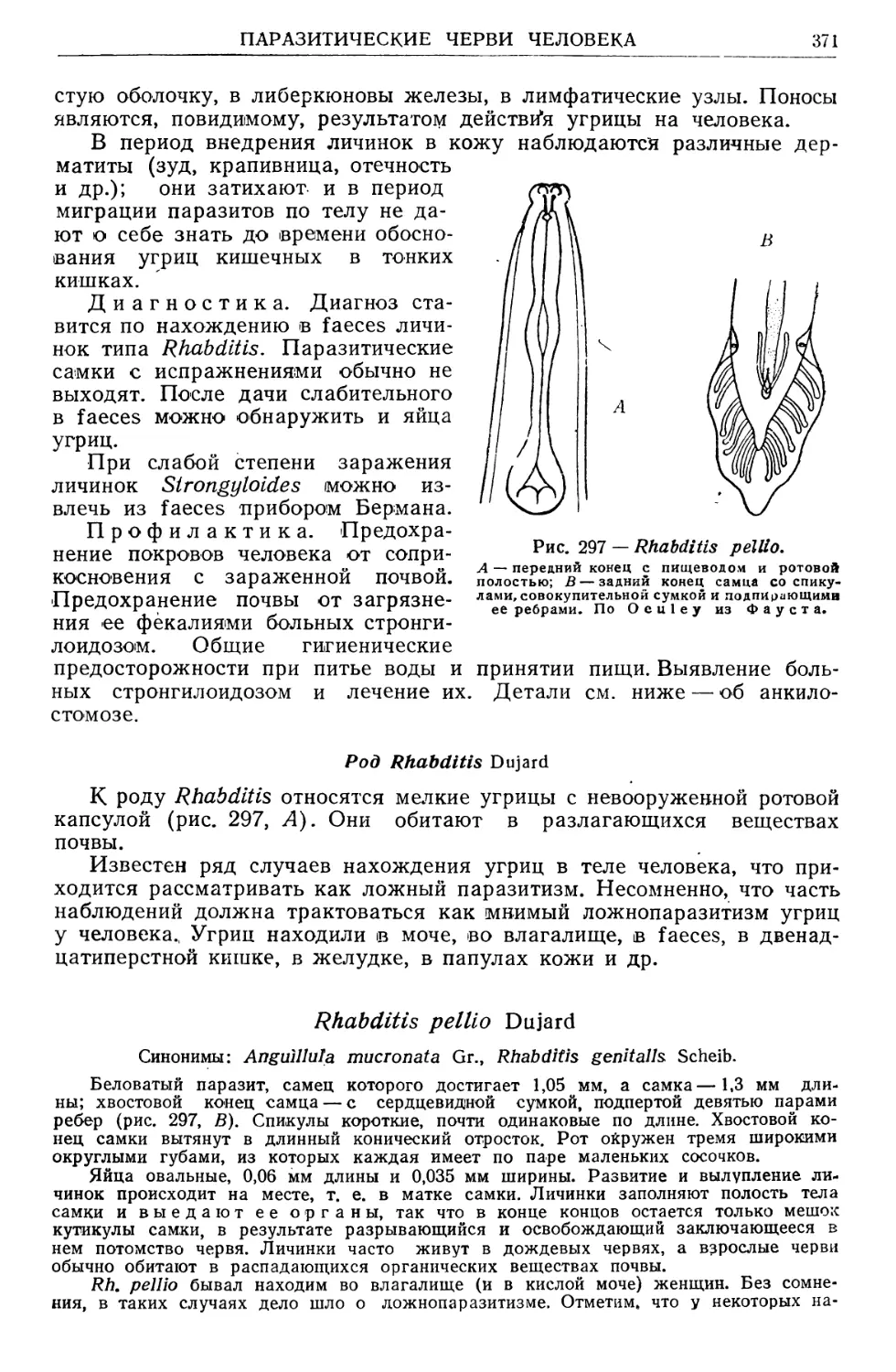
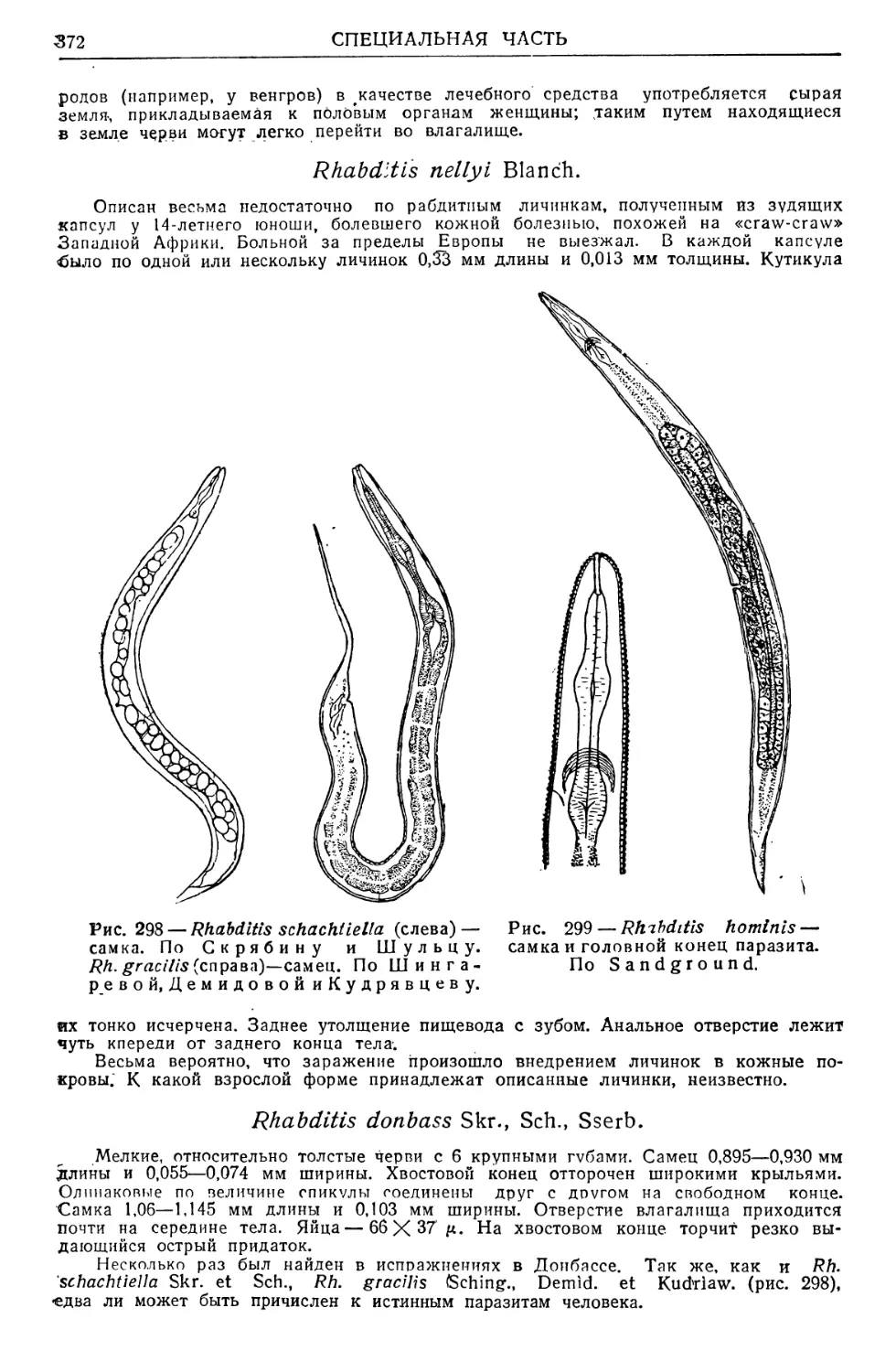
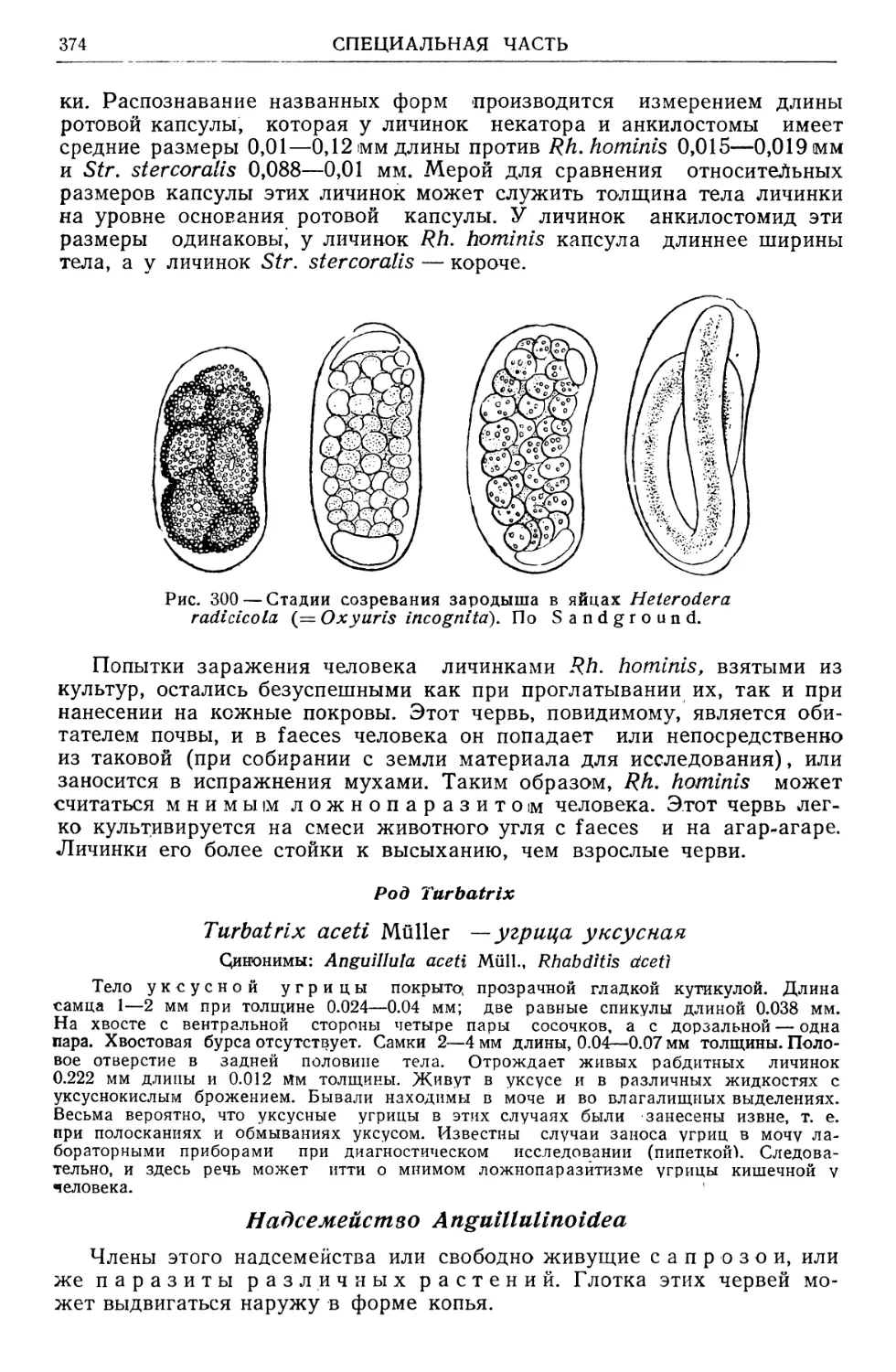
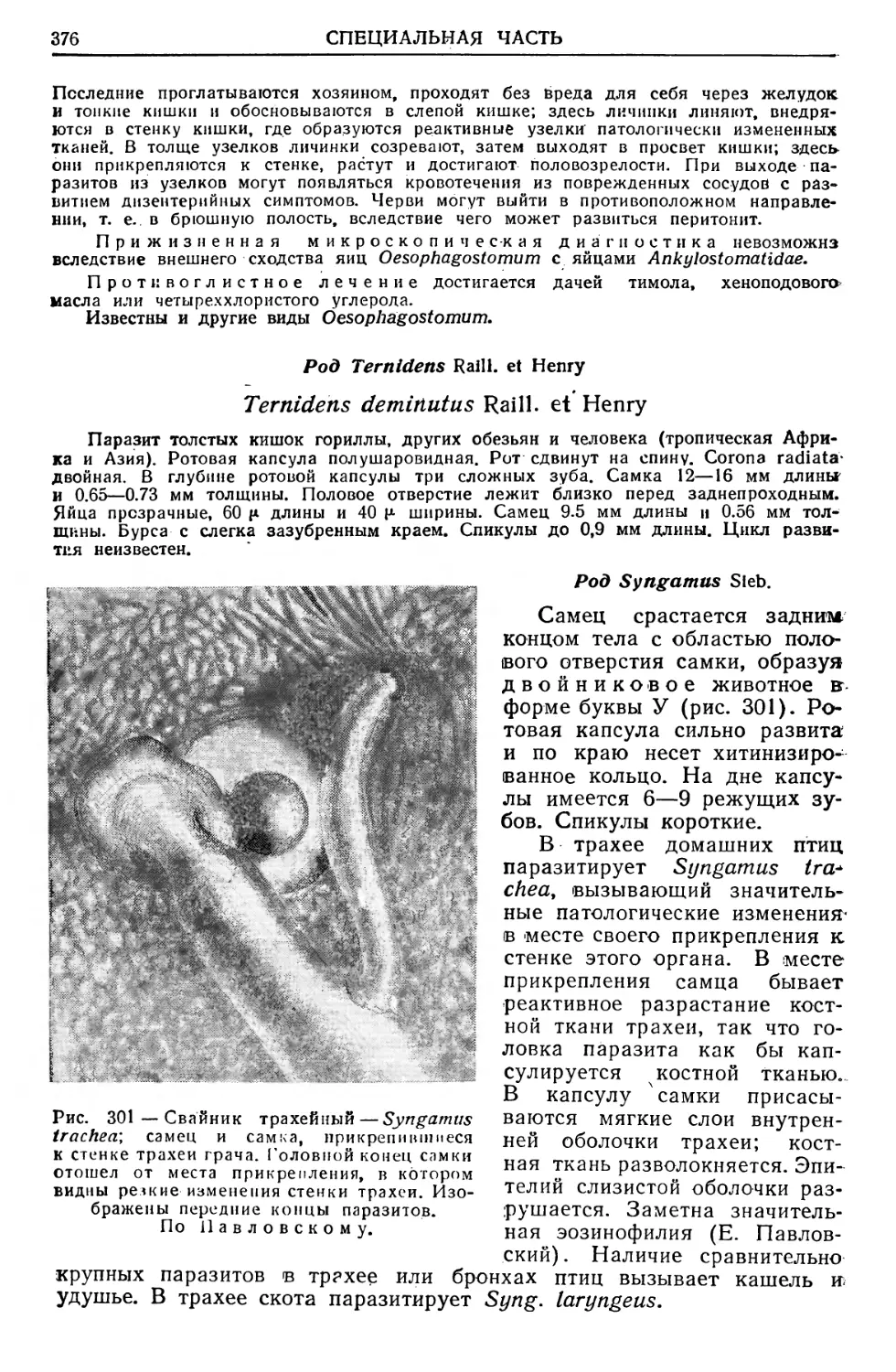
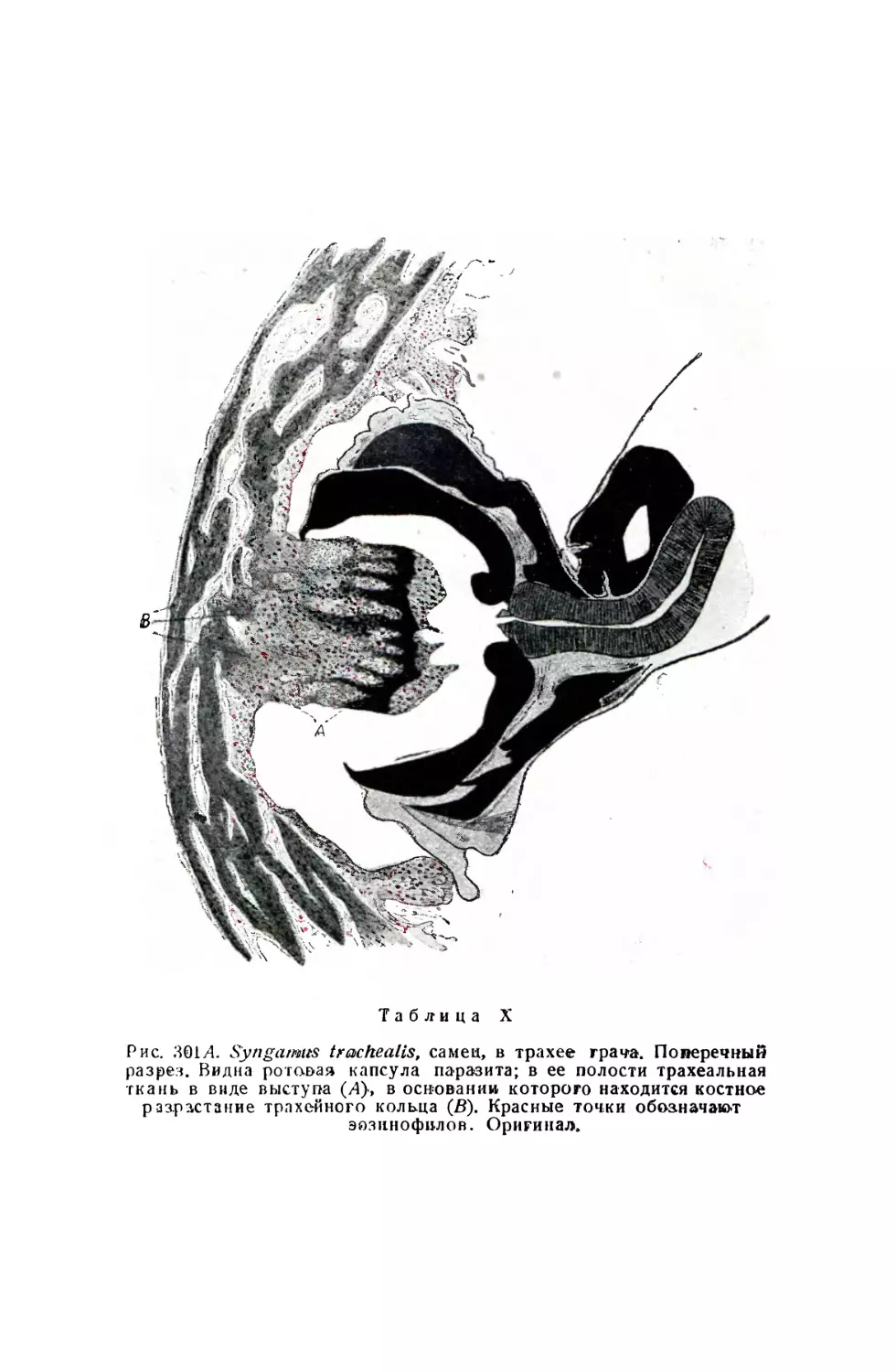
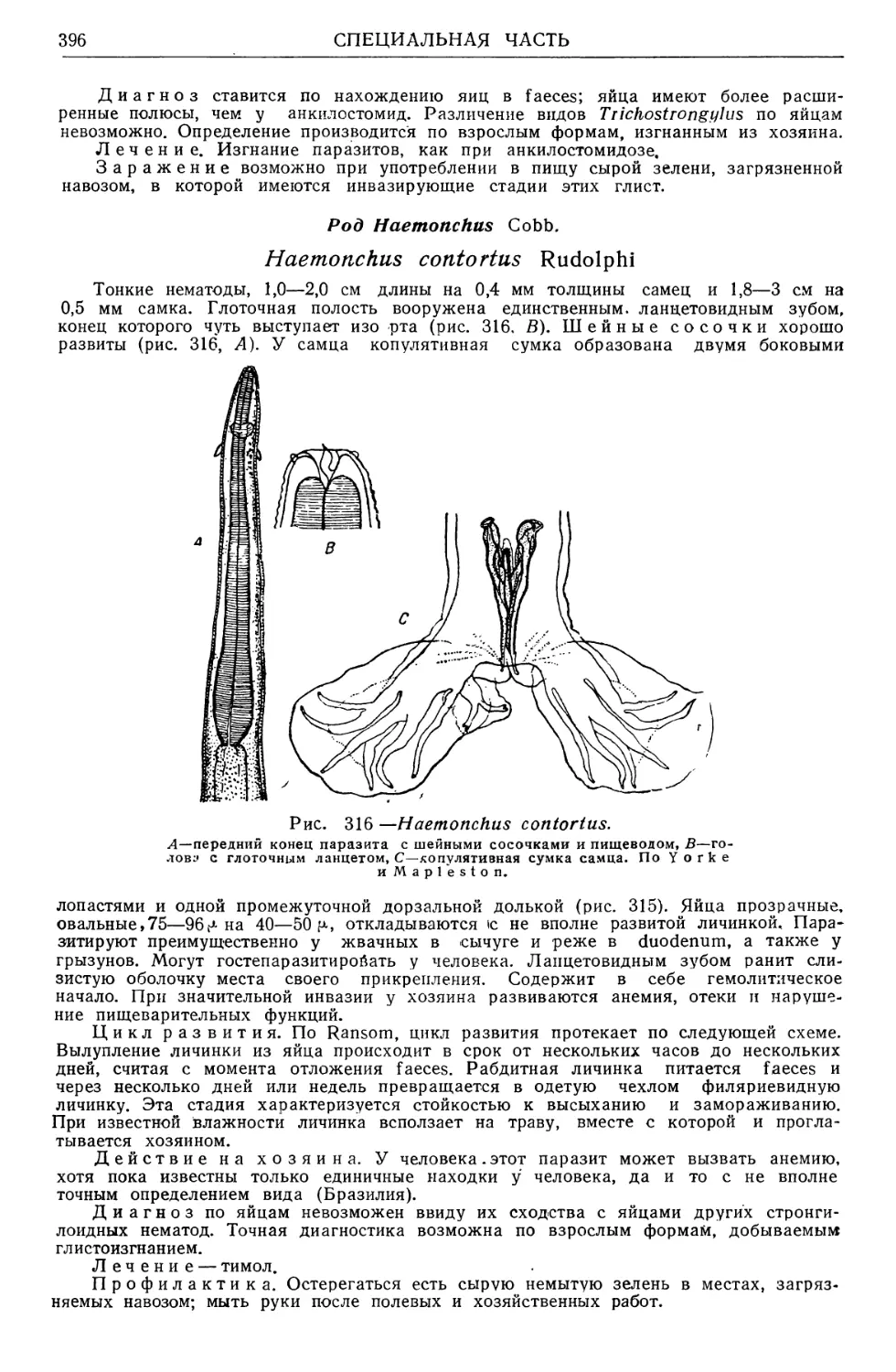
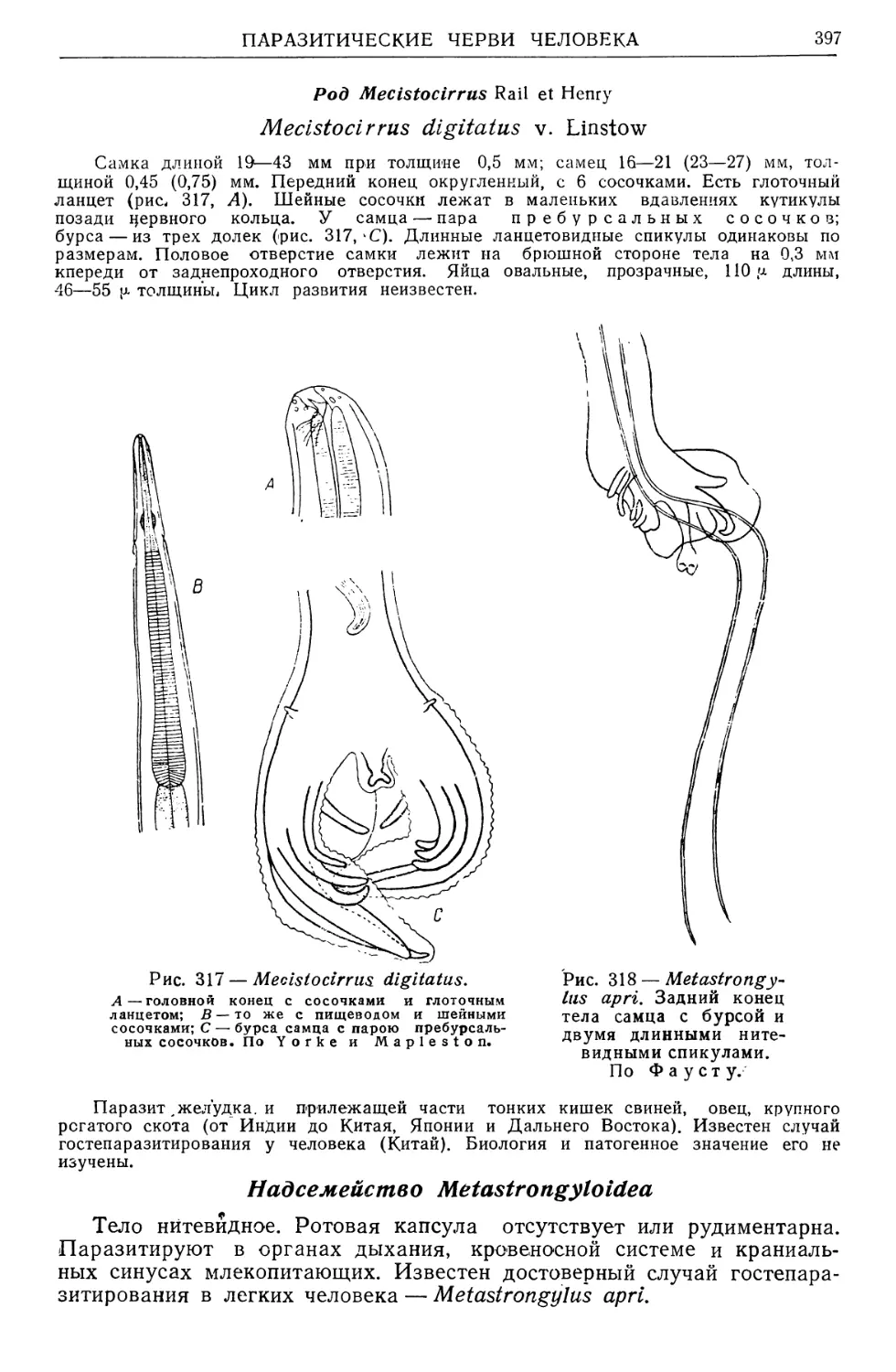
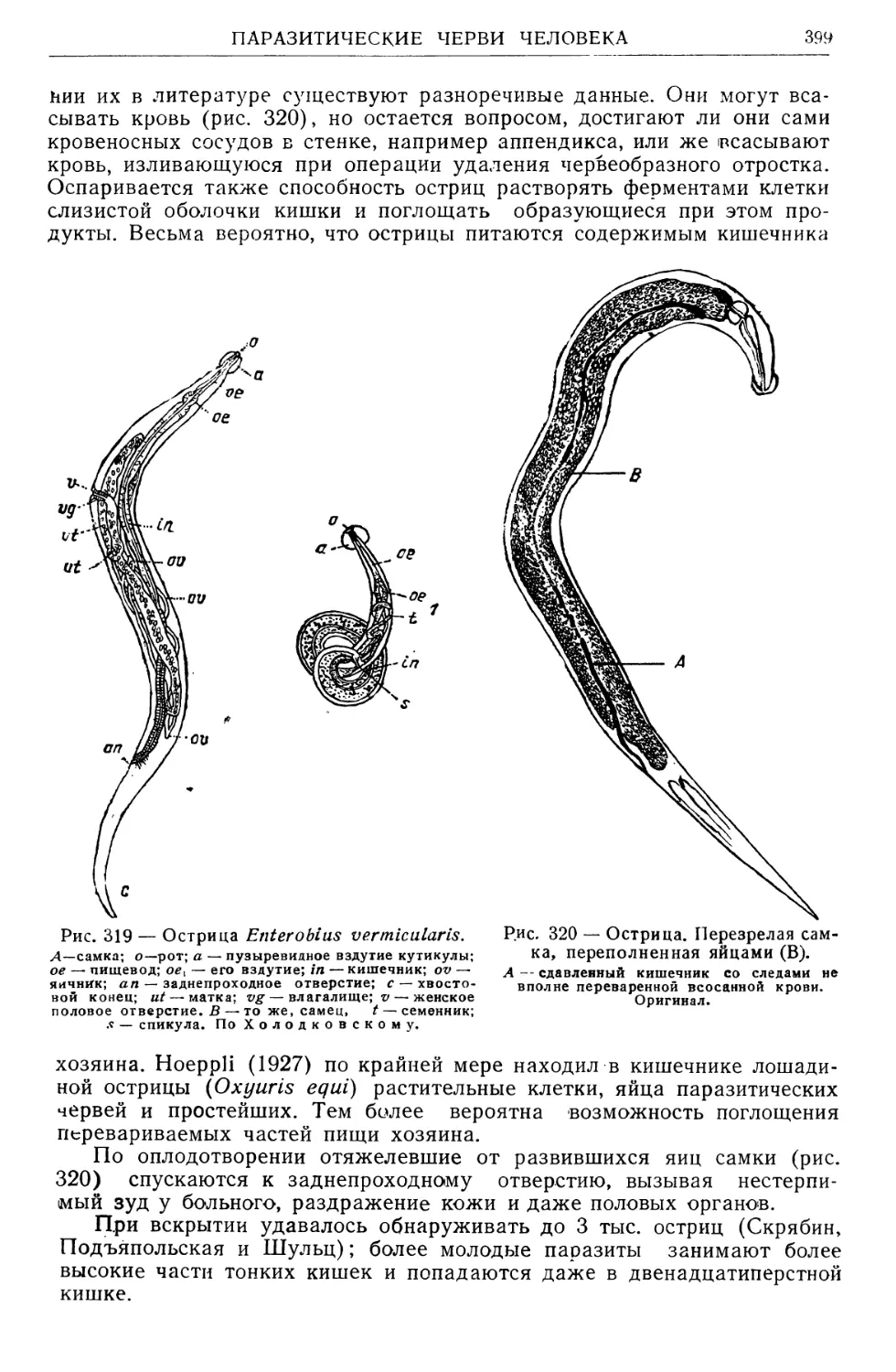
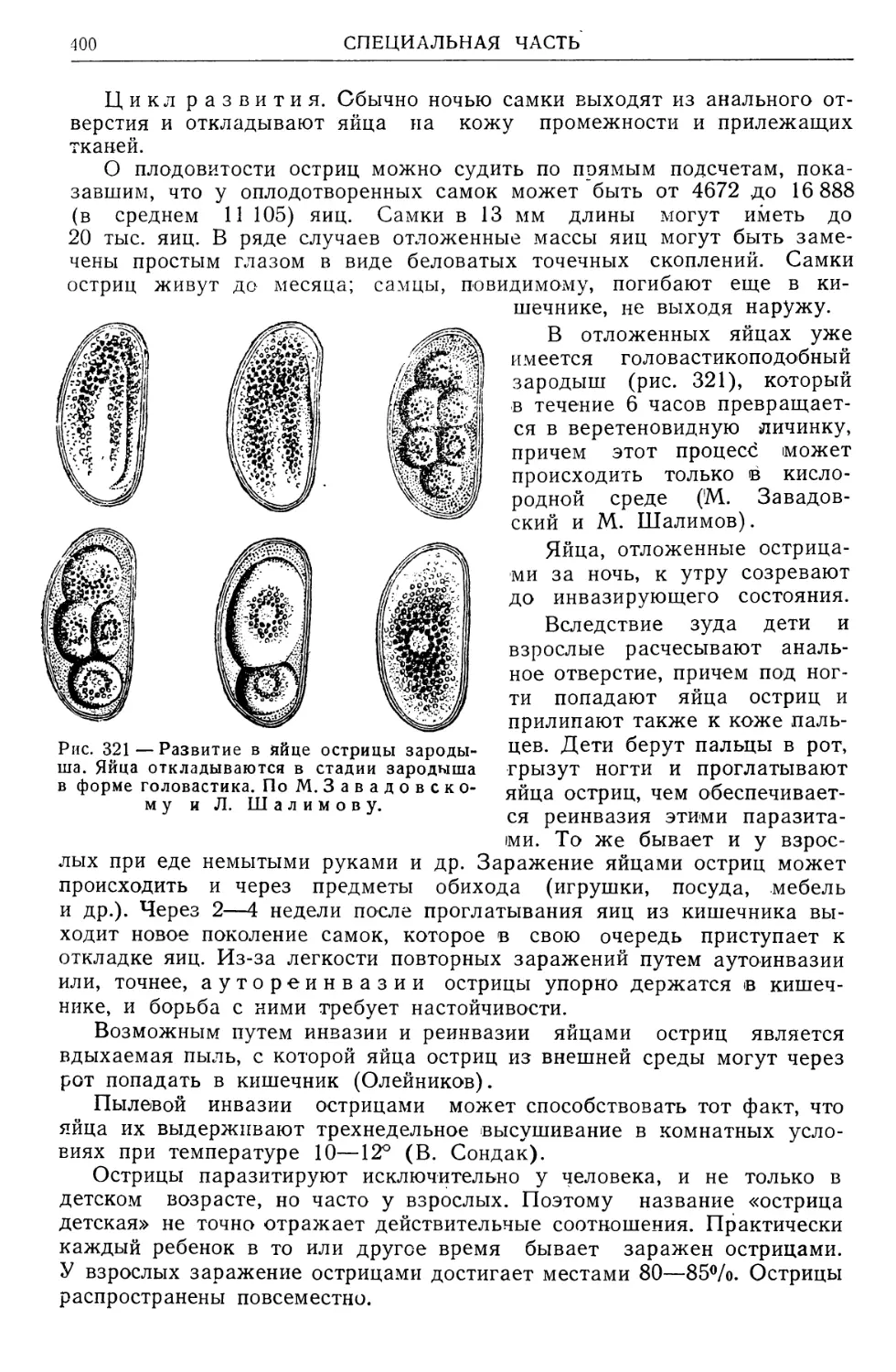
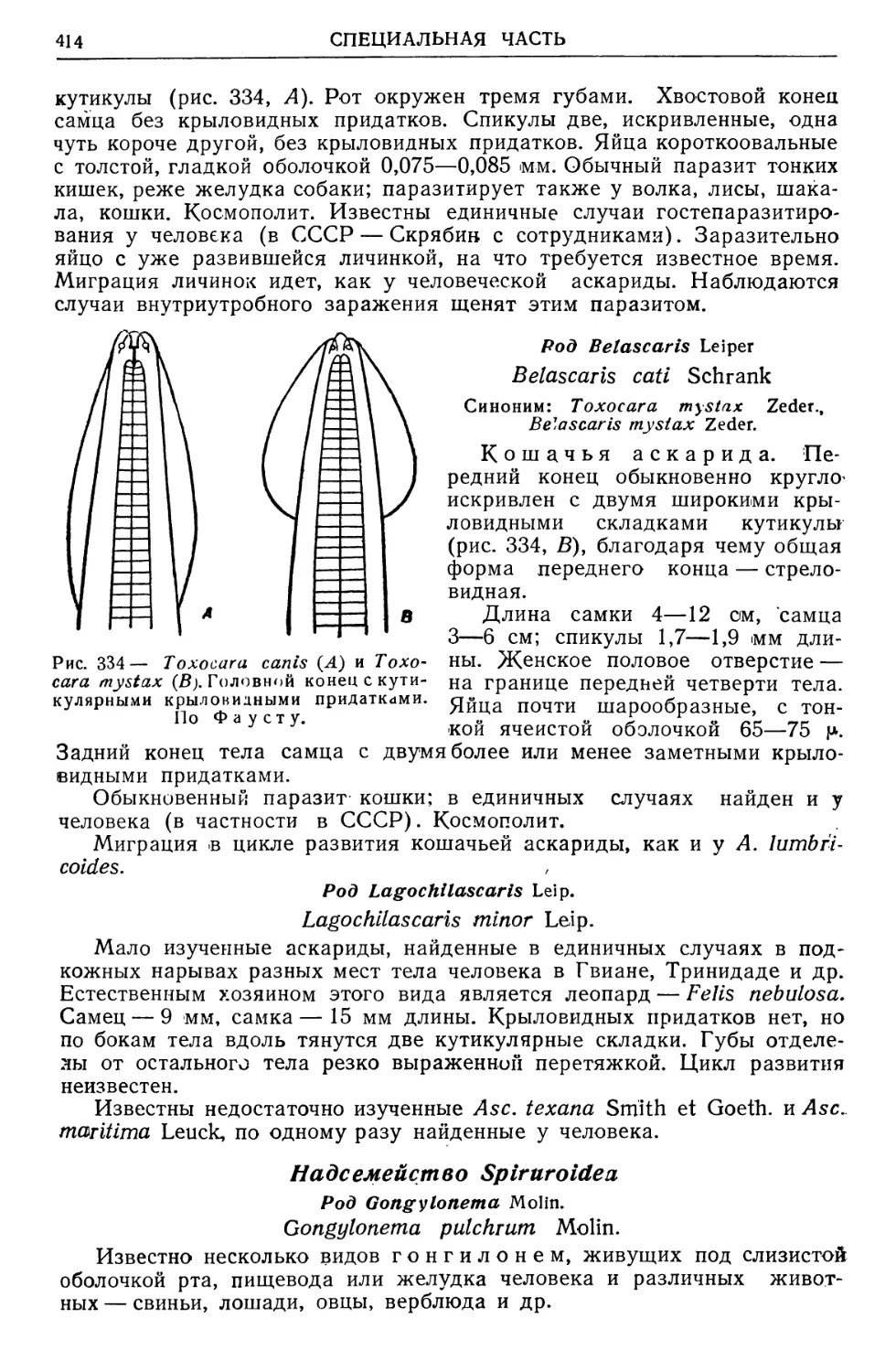
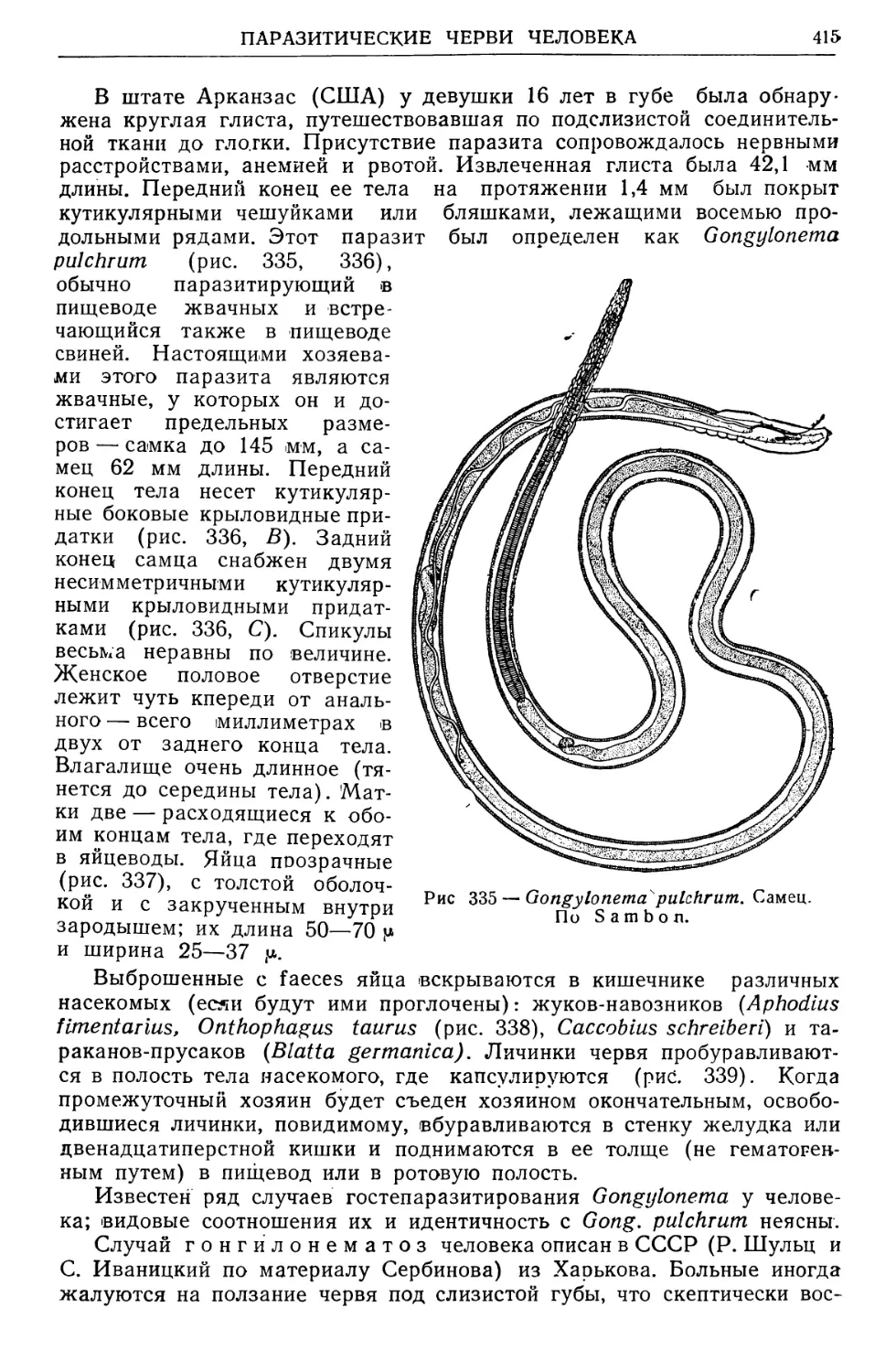
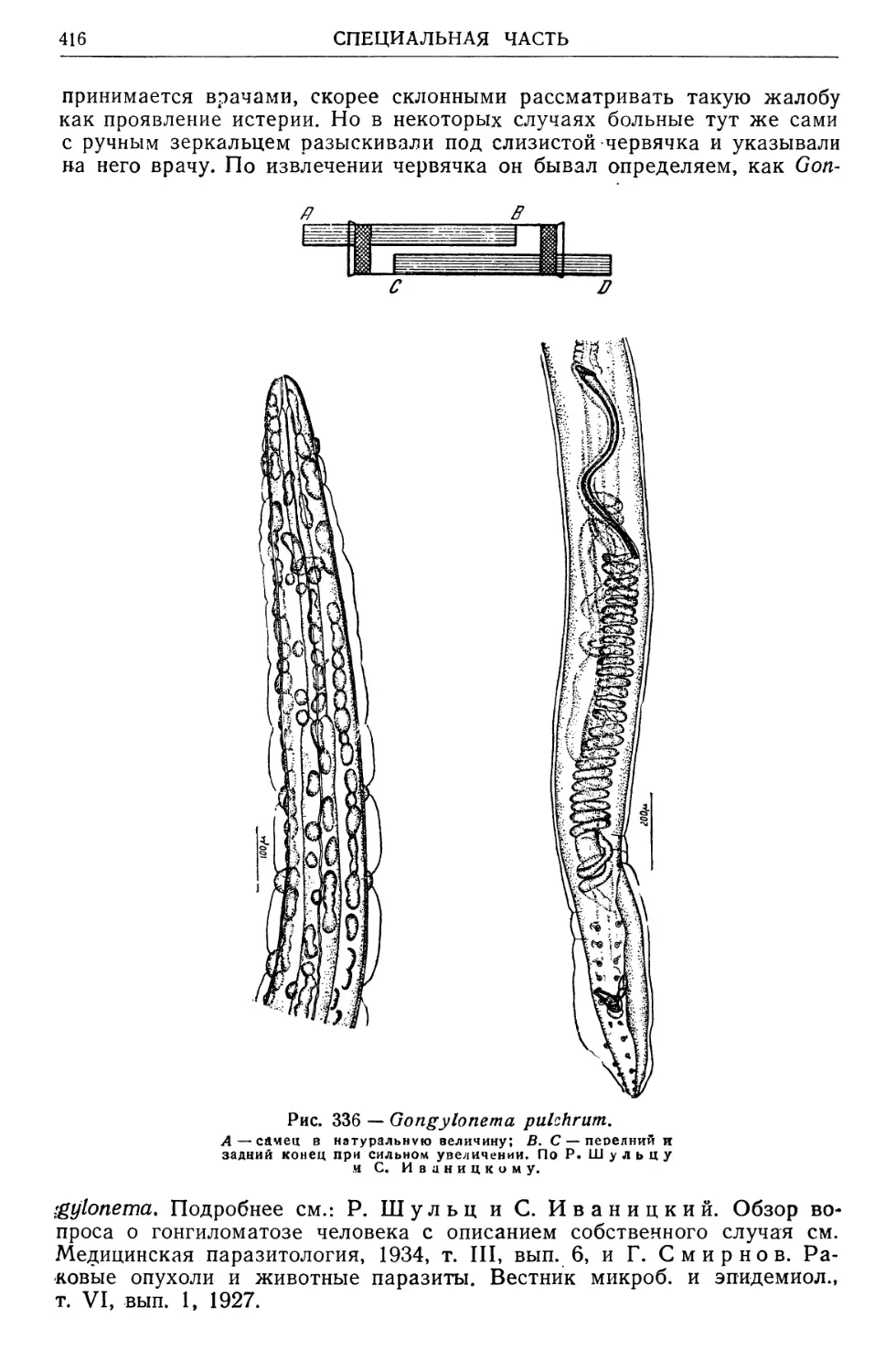
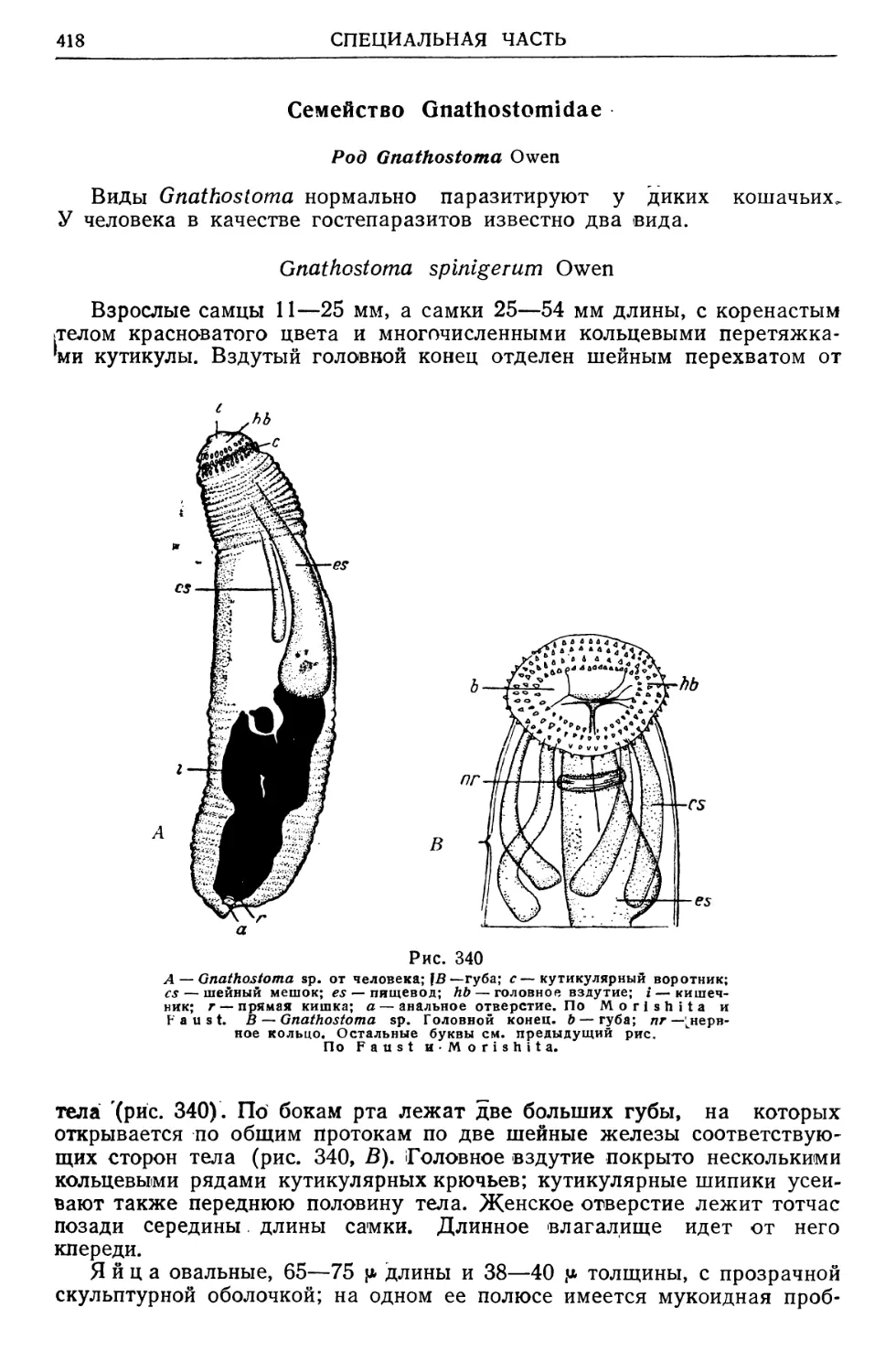
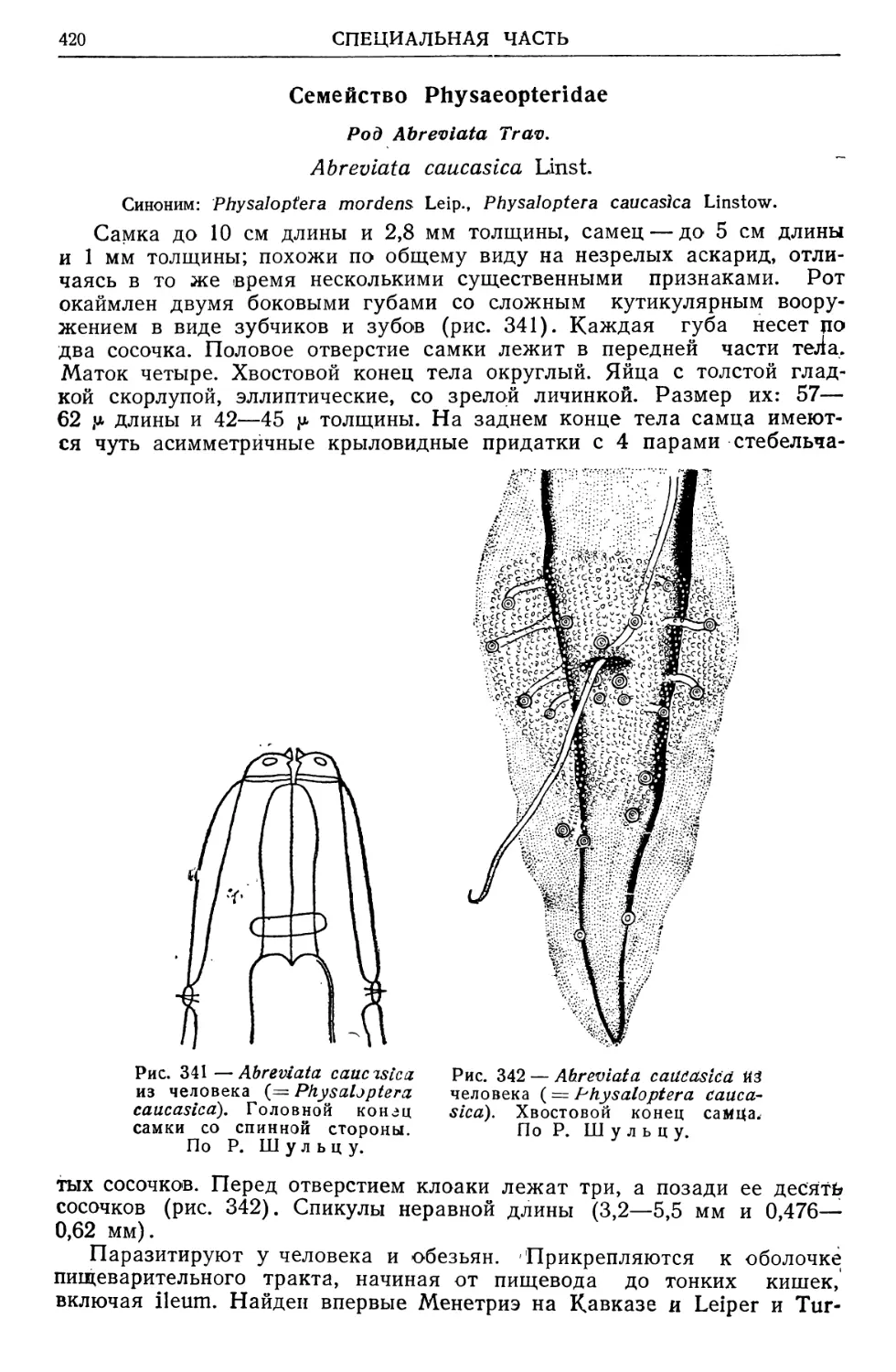
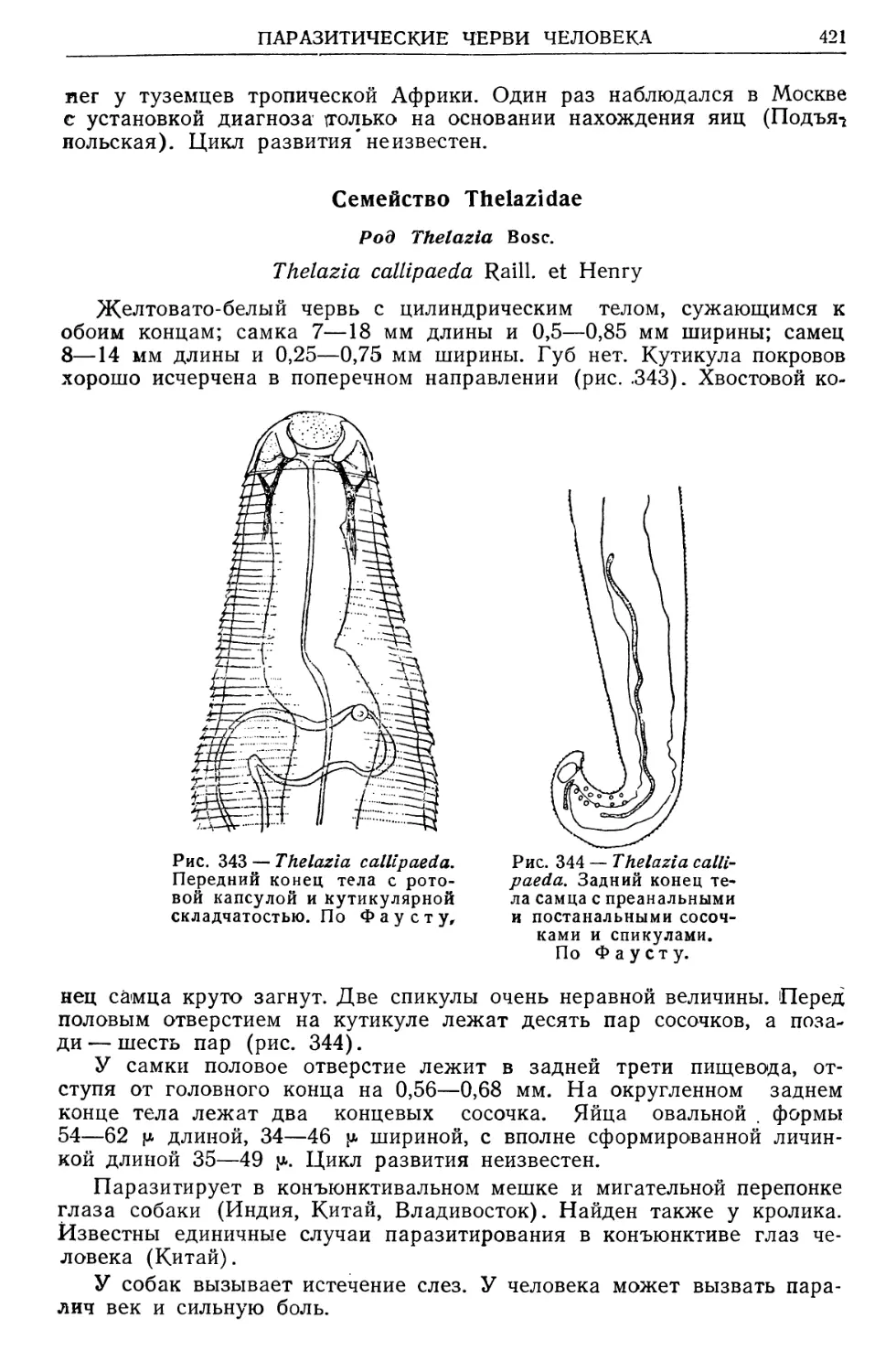
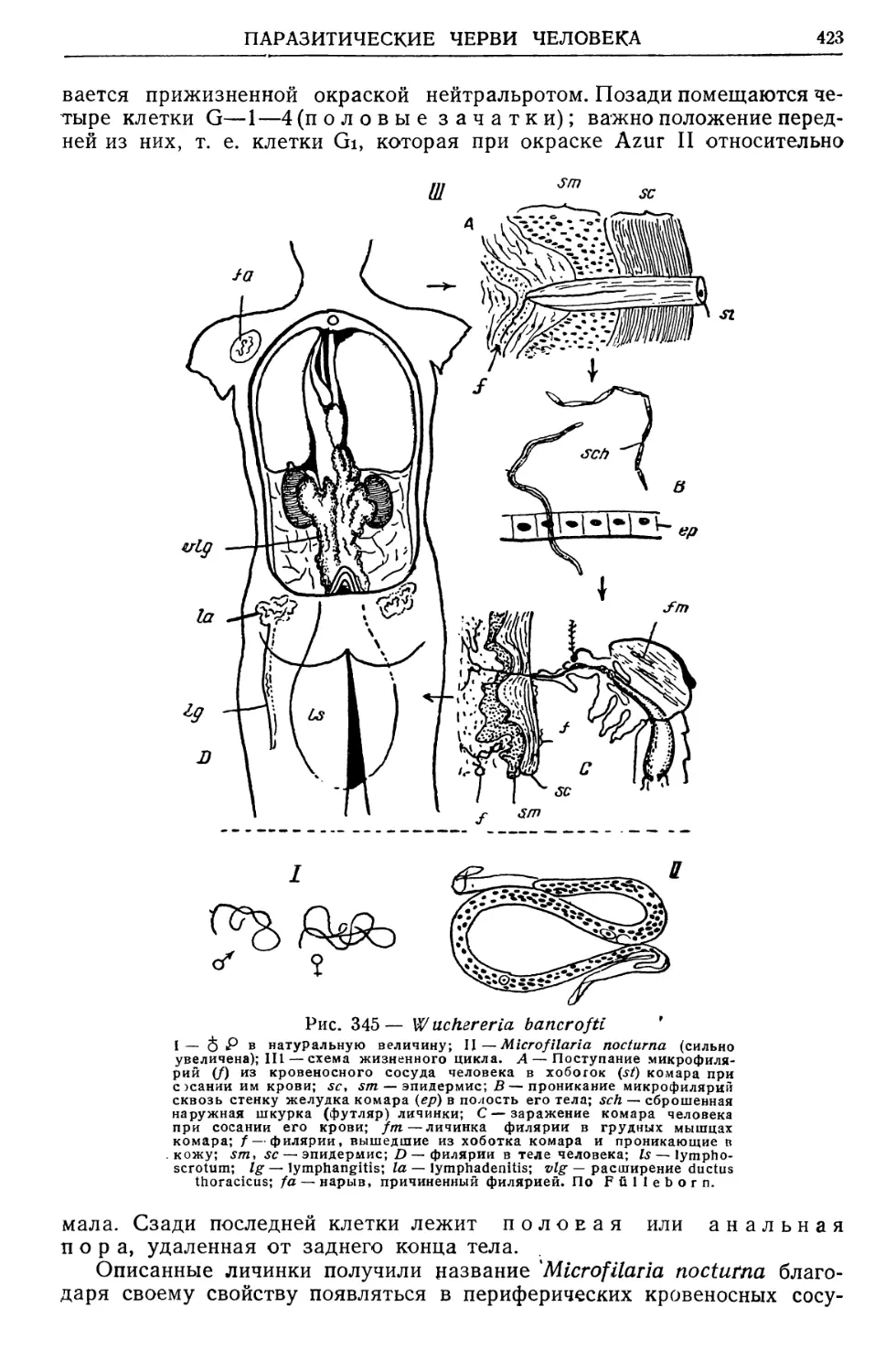
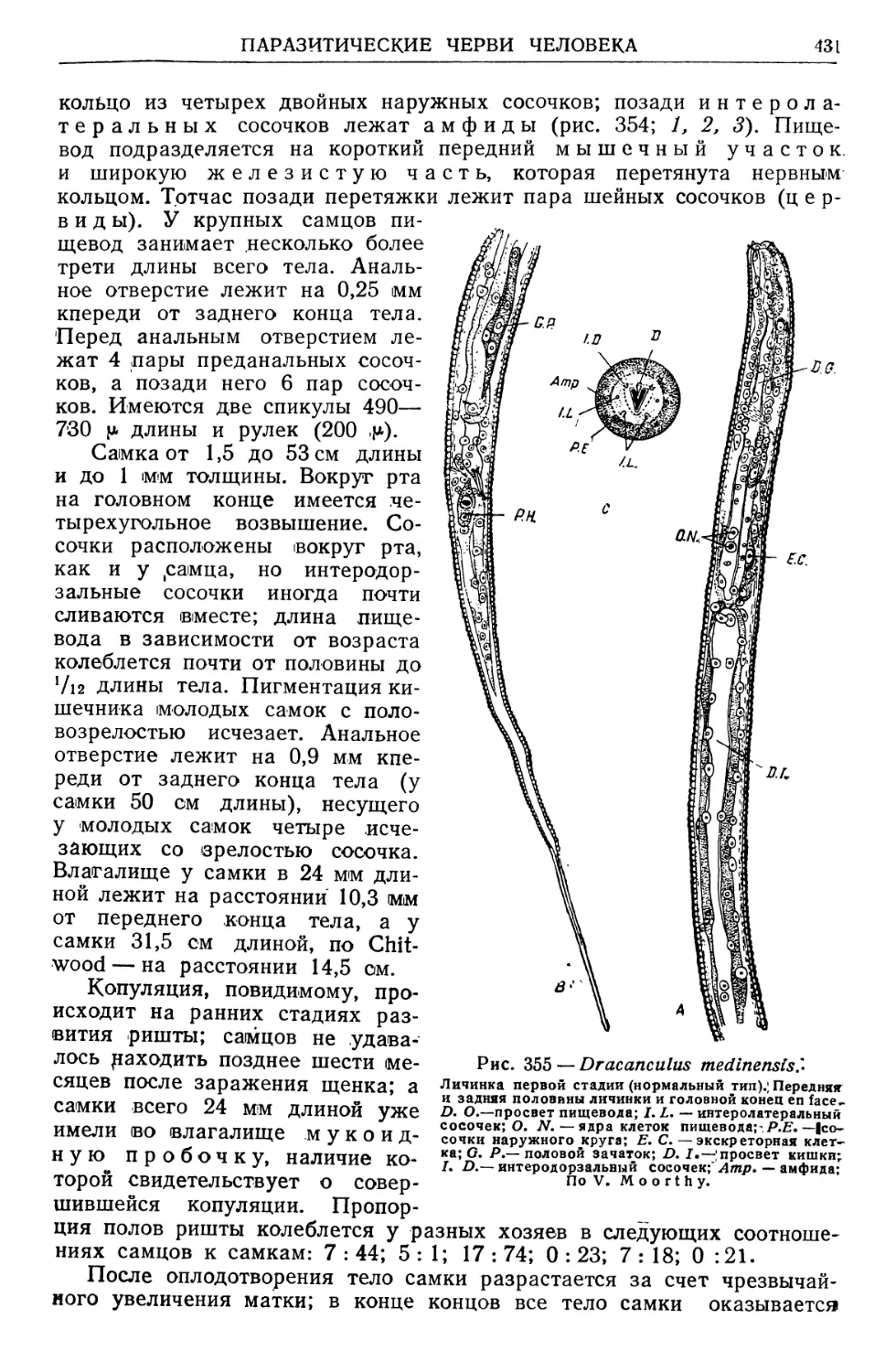
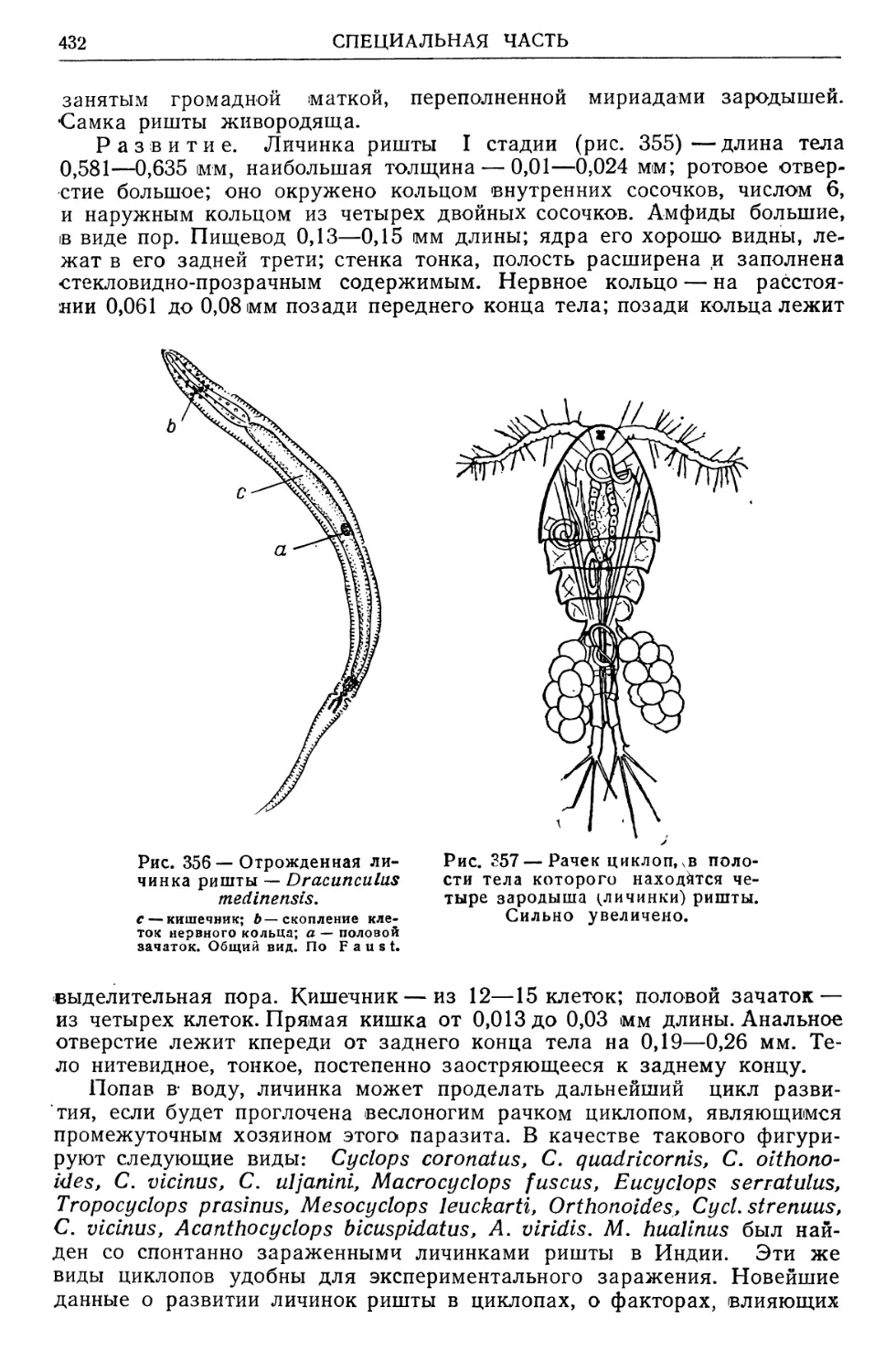
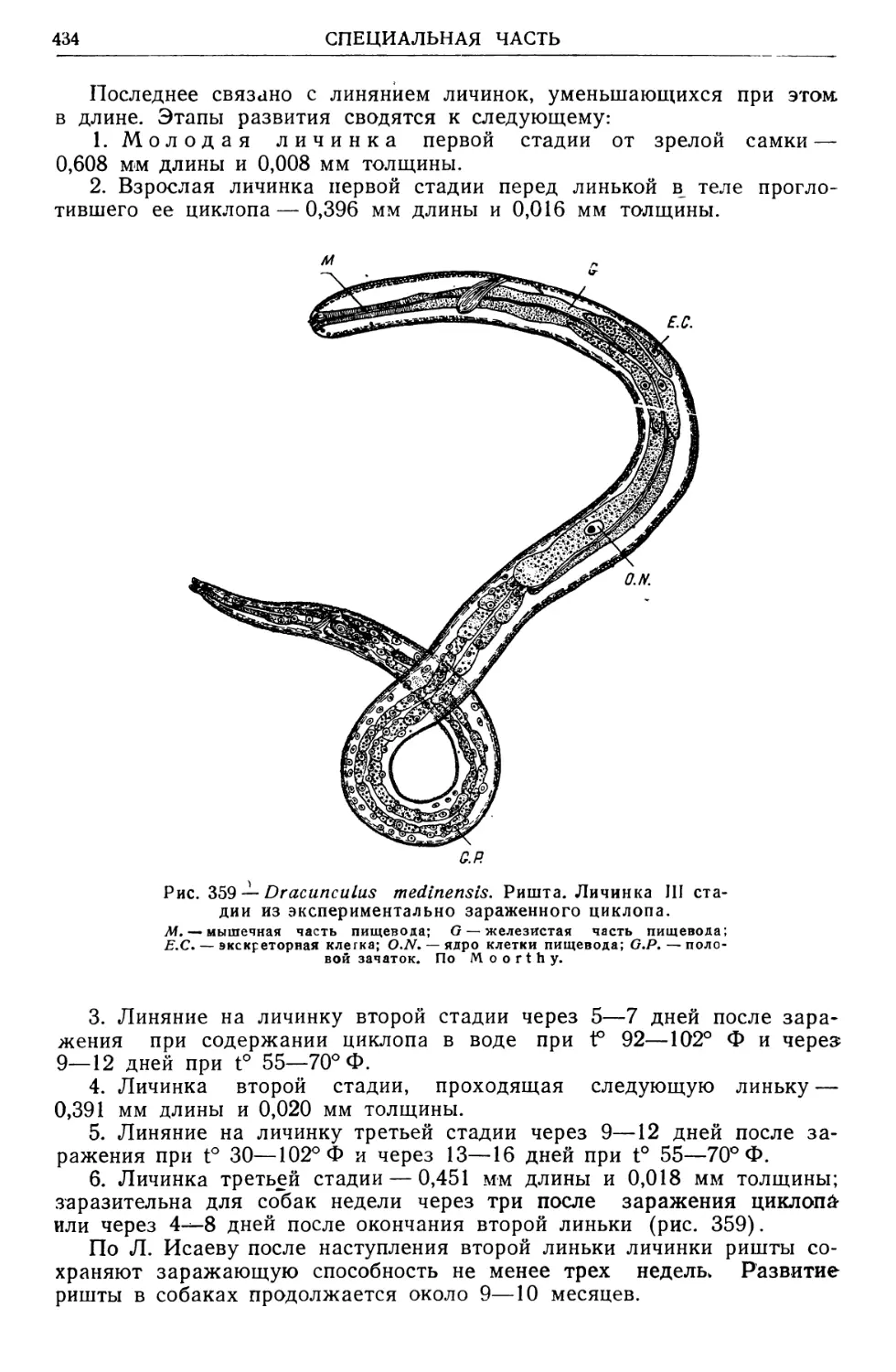
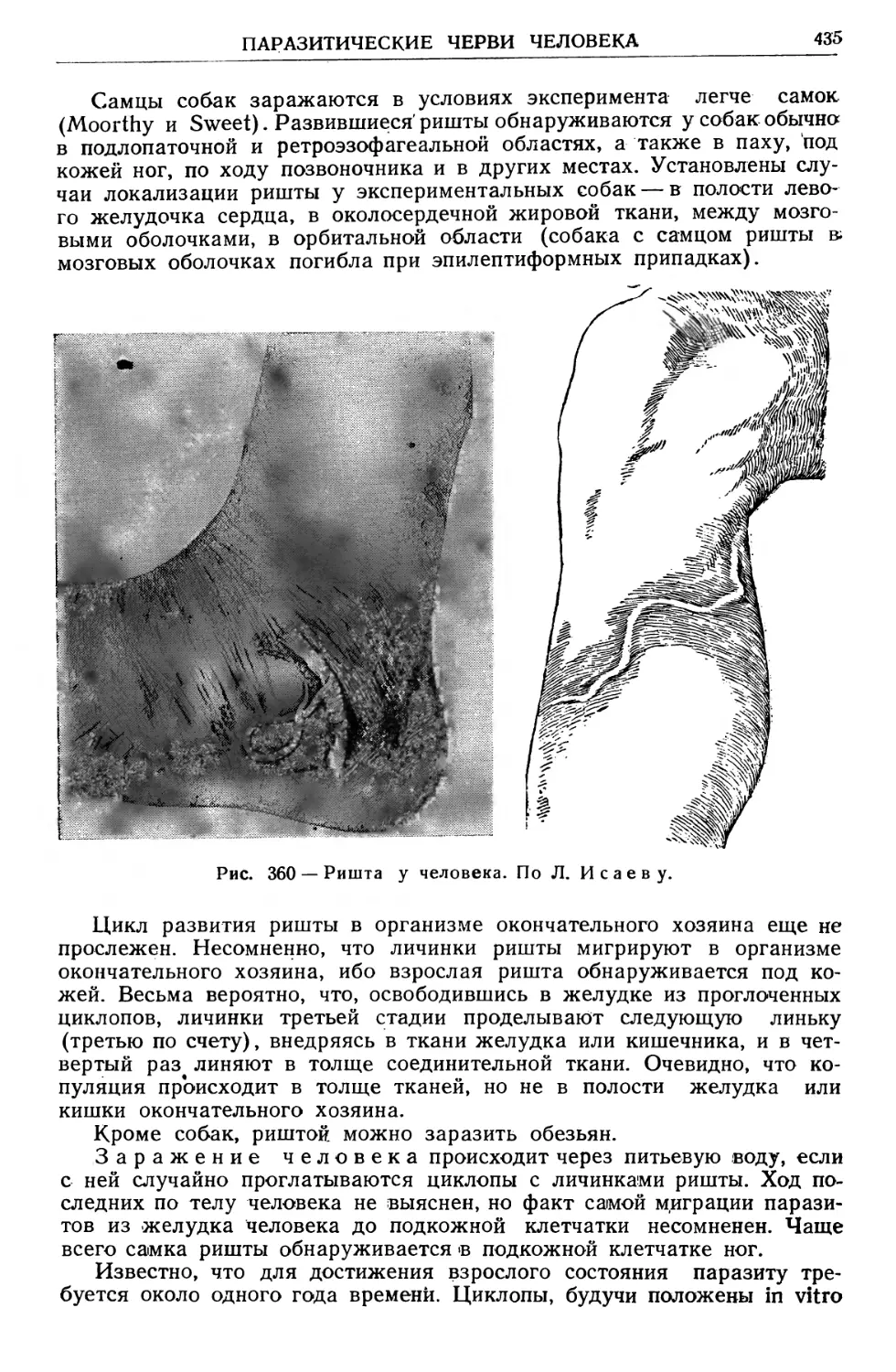
/































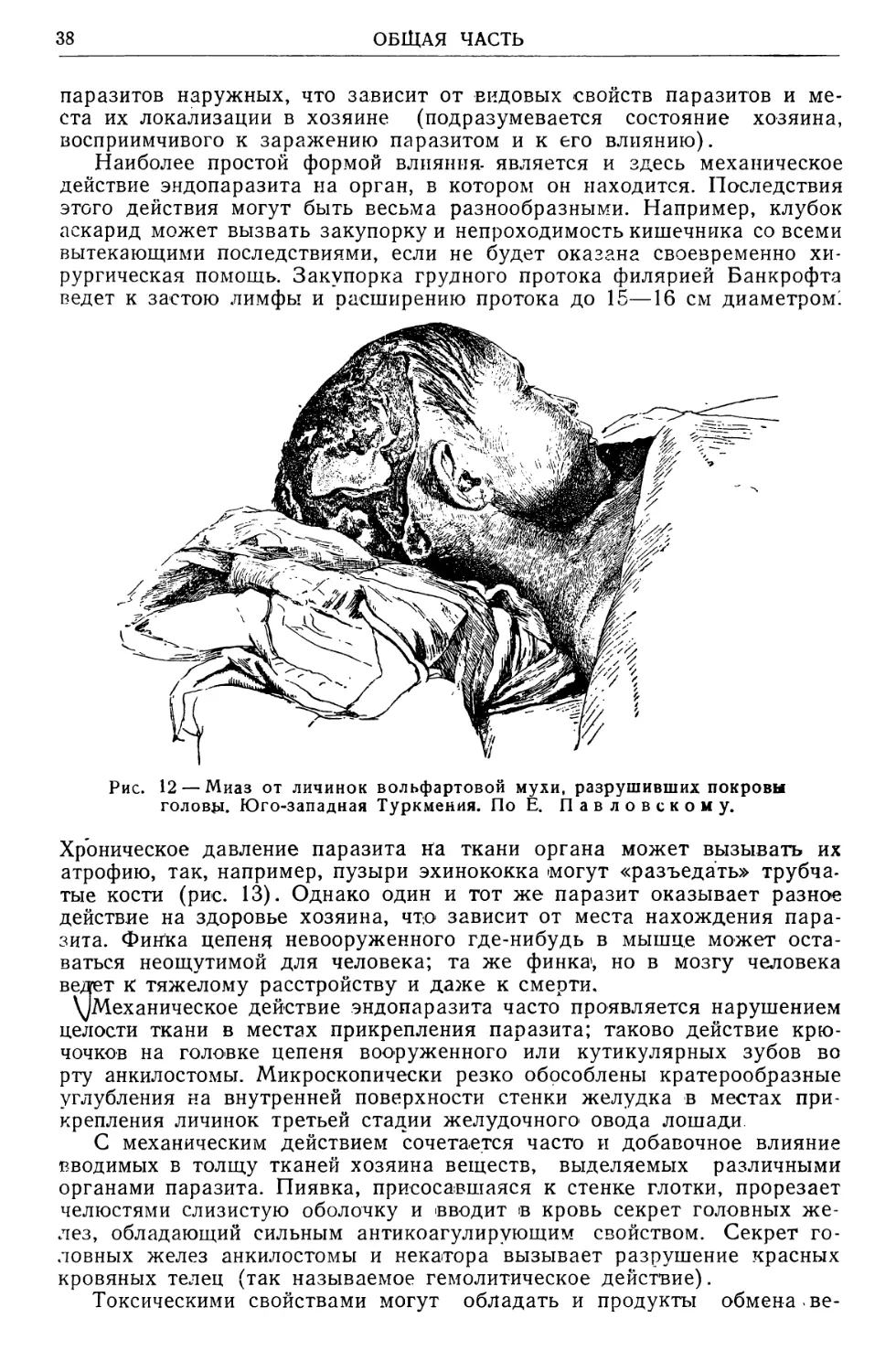








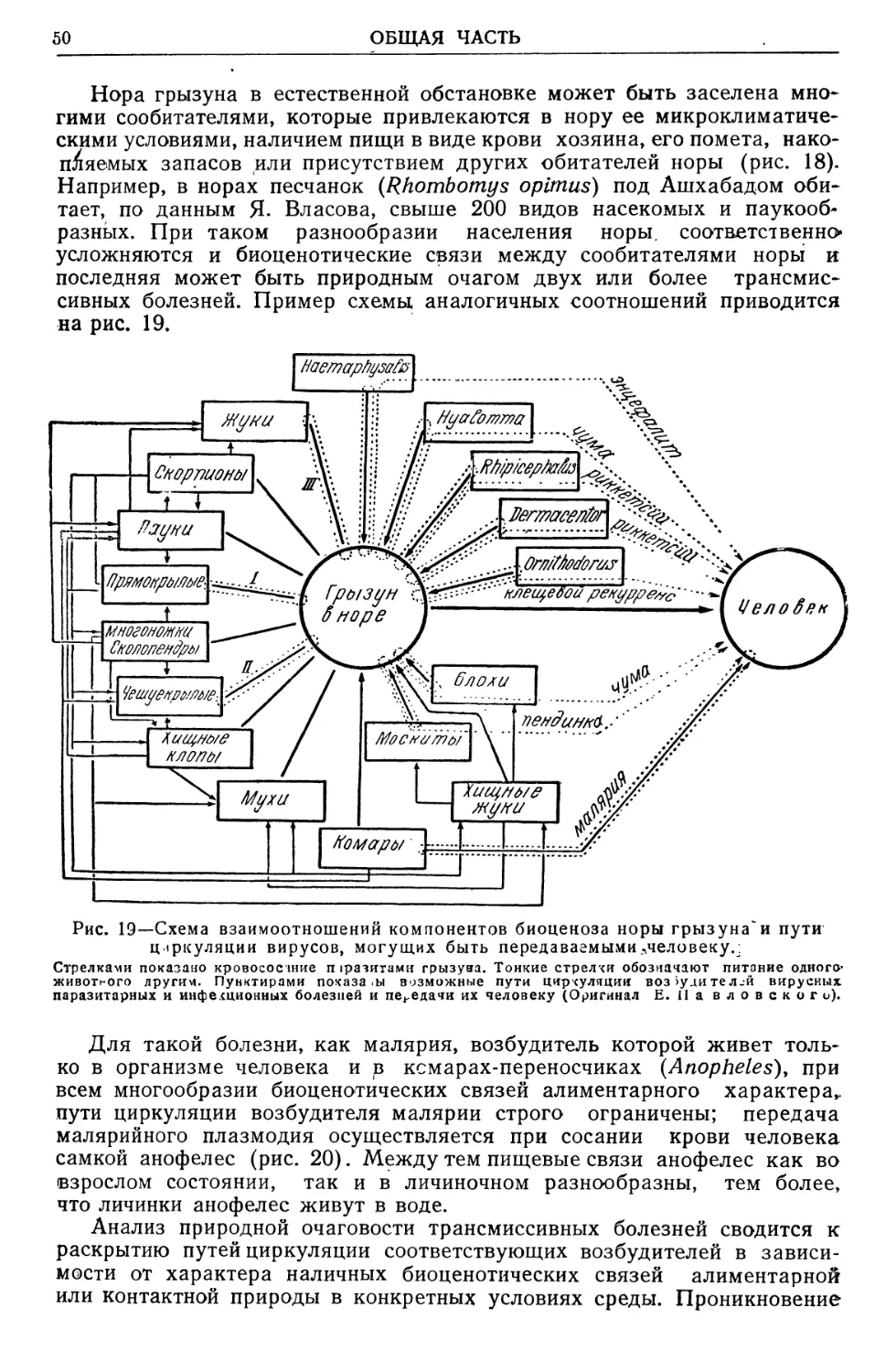
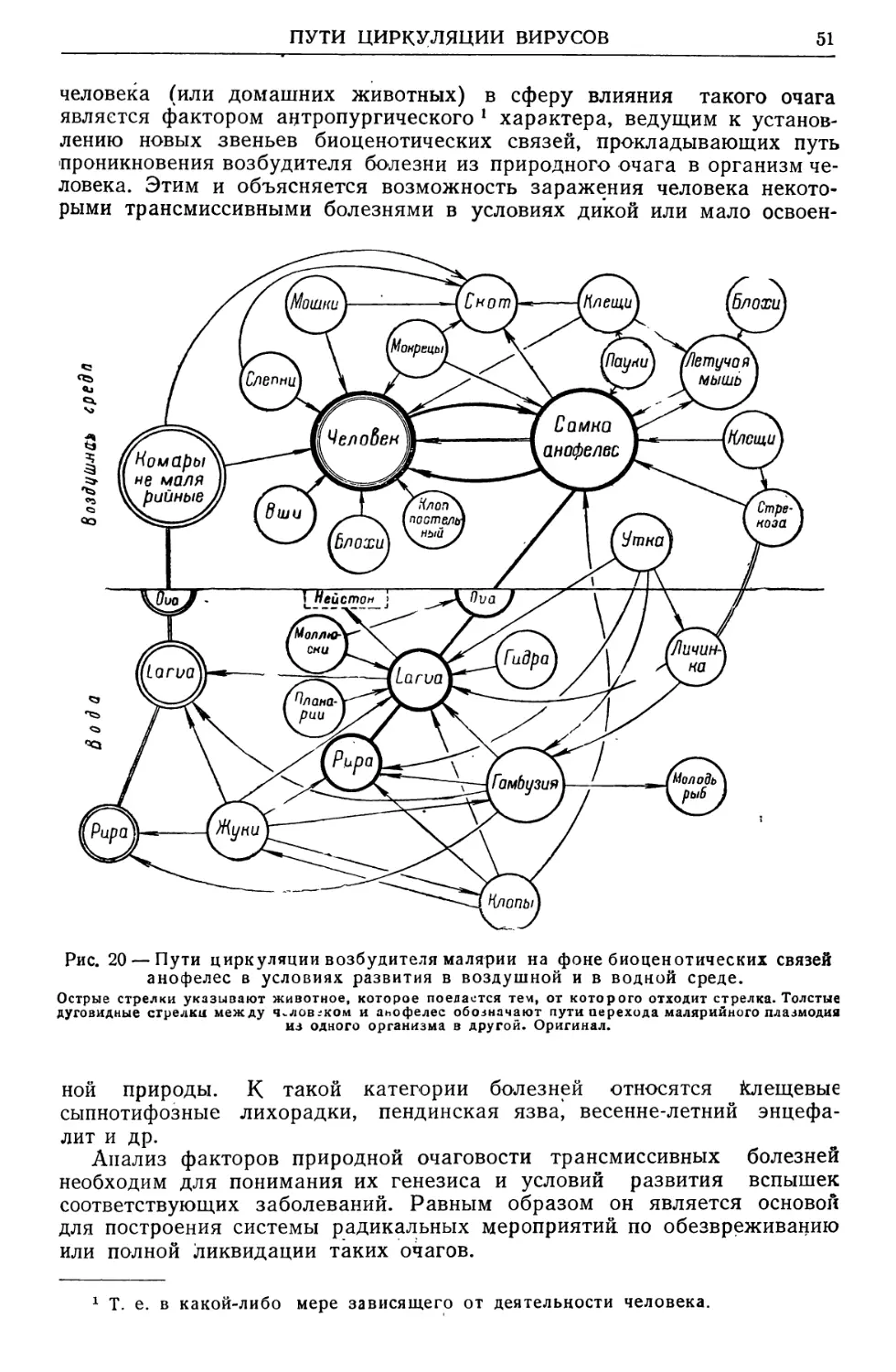


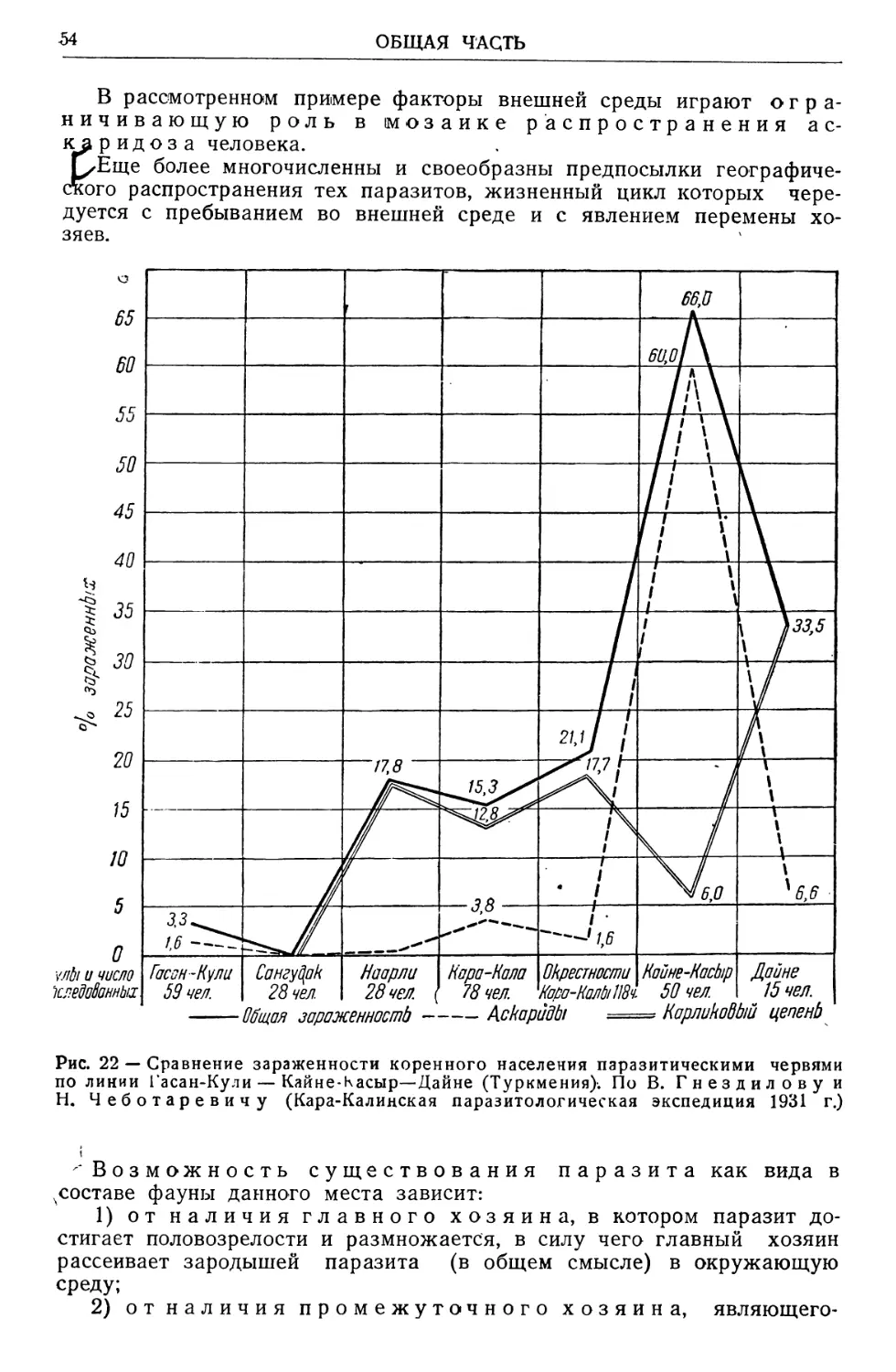








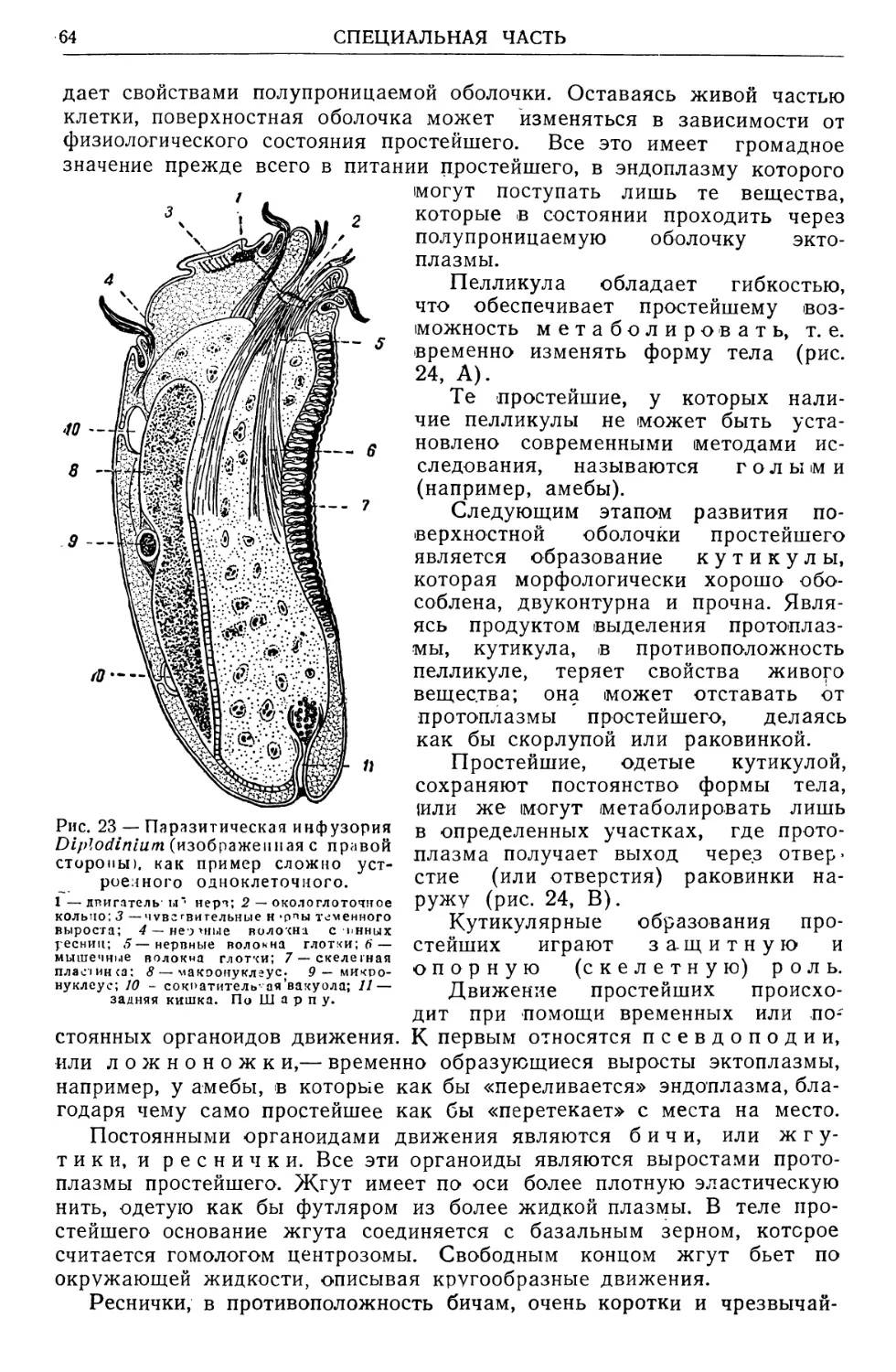
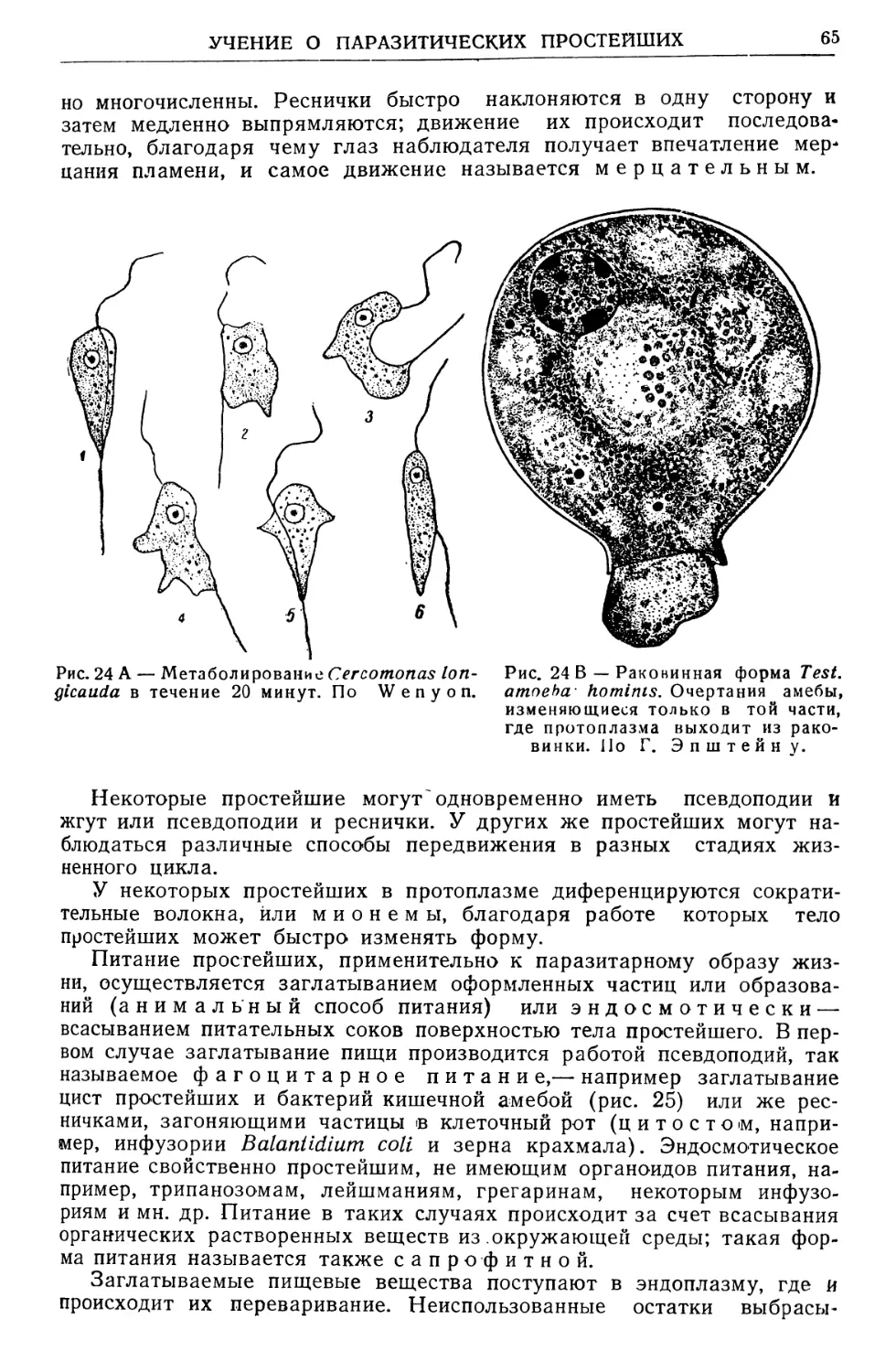
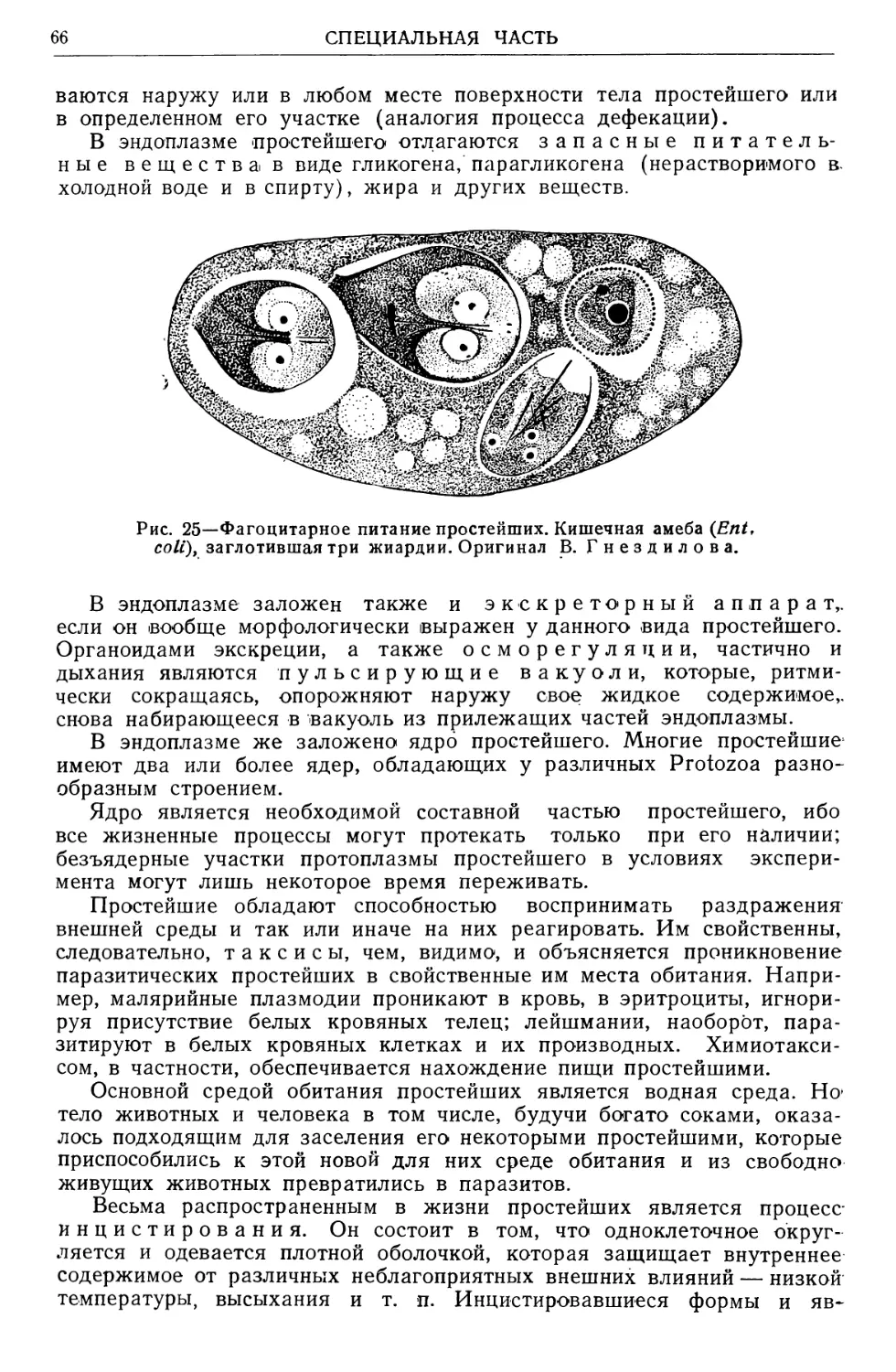
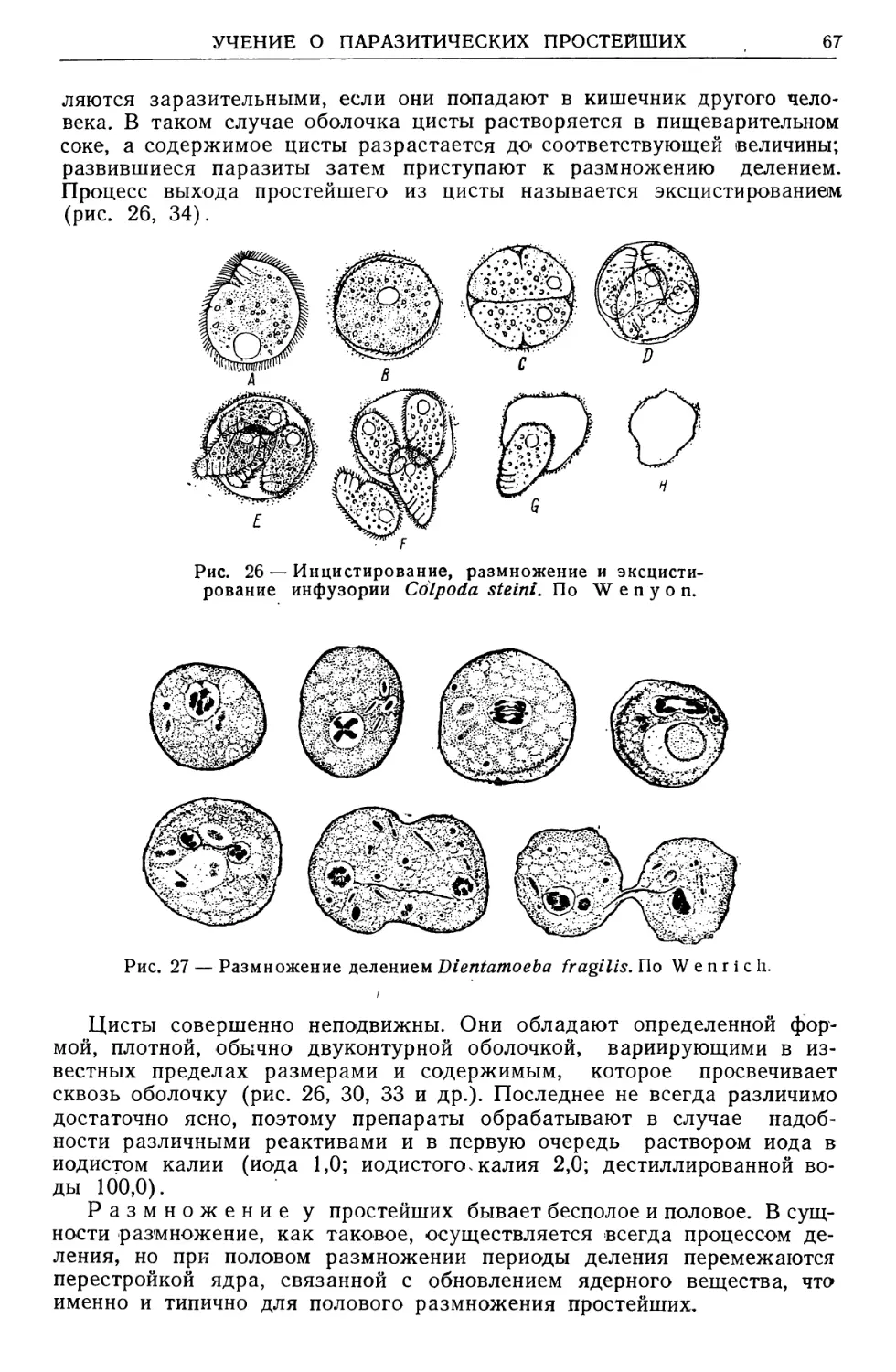

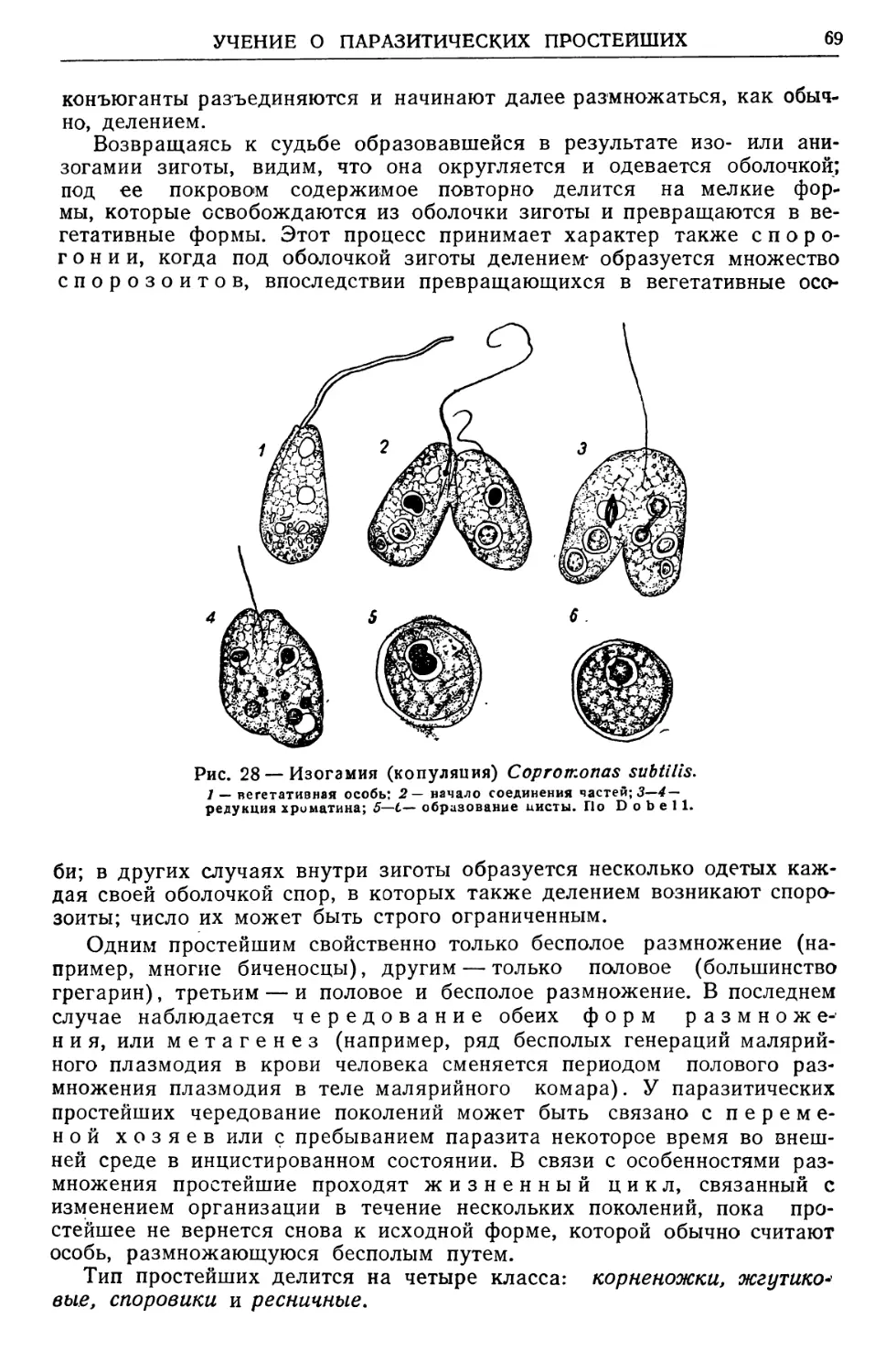


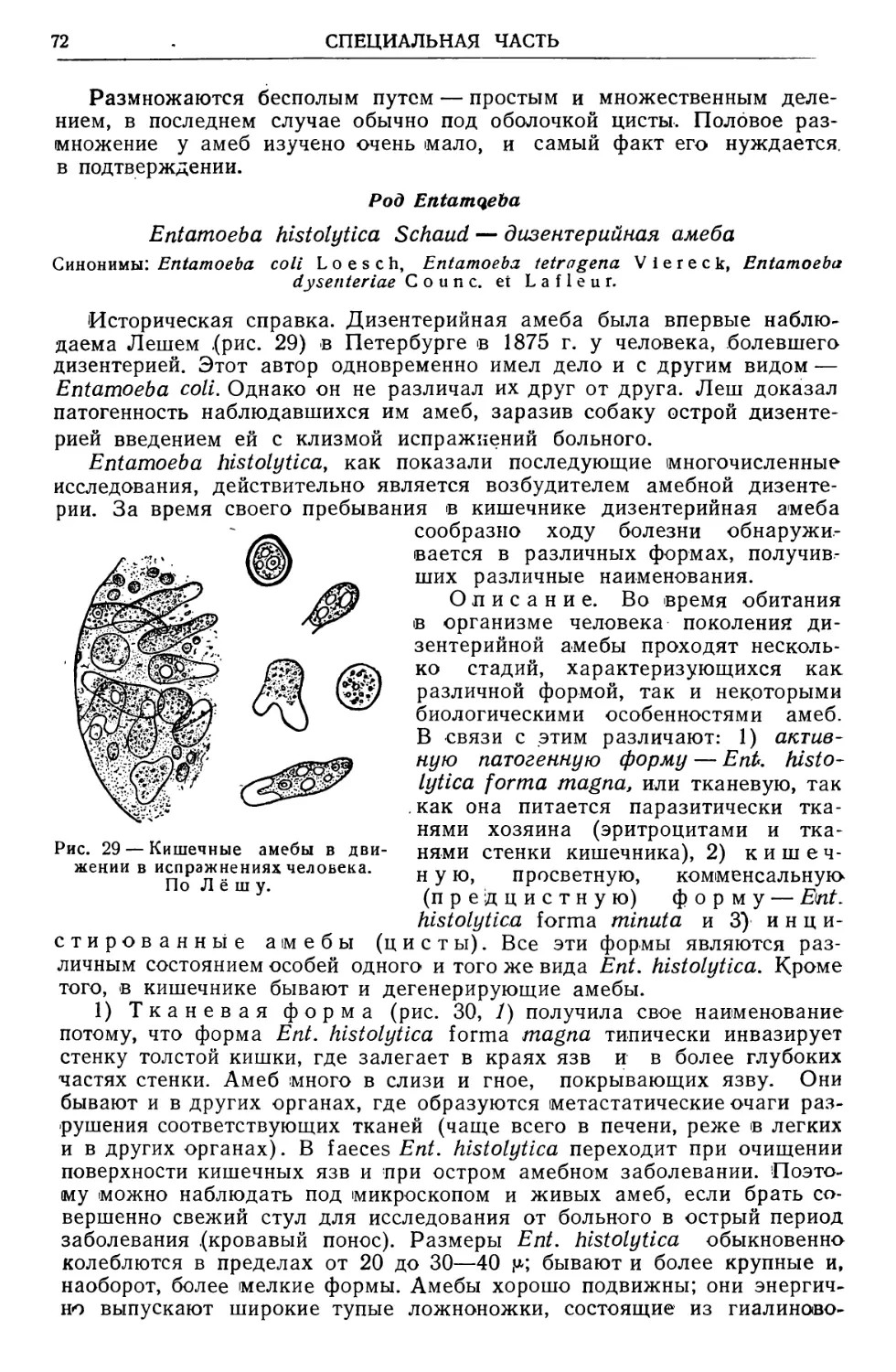
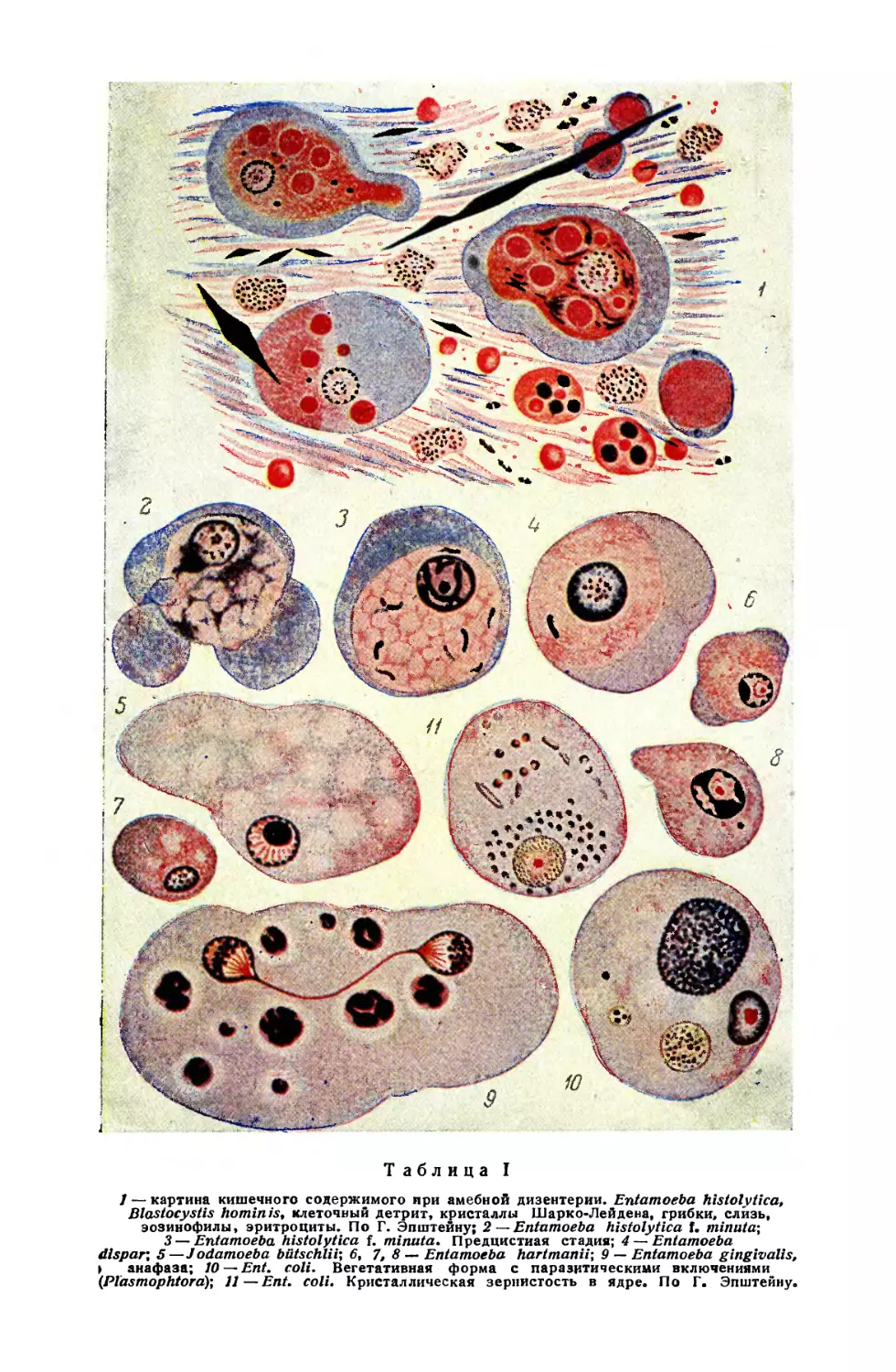
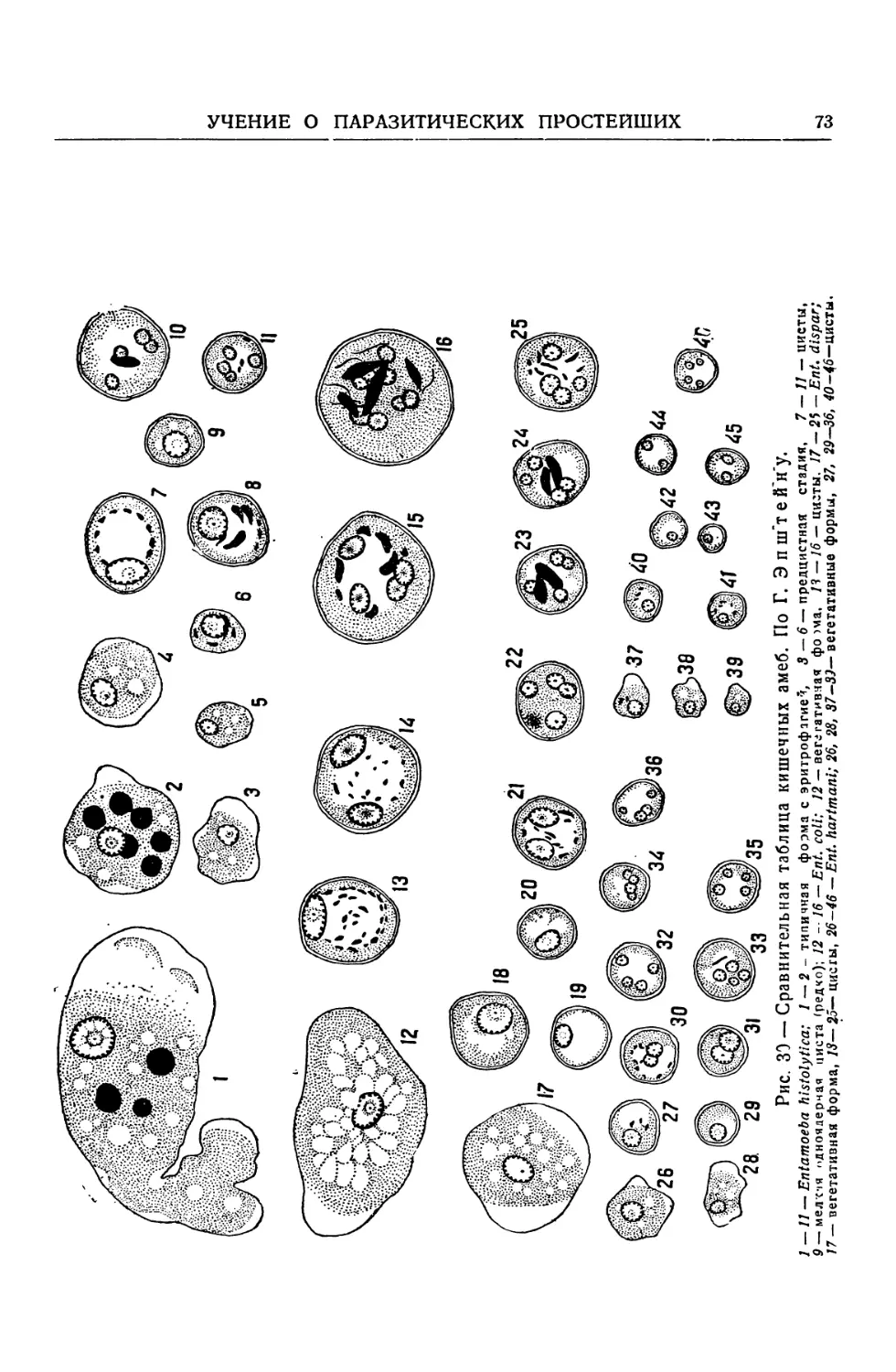




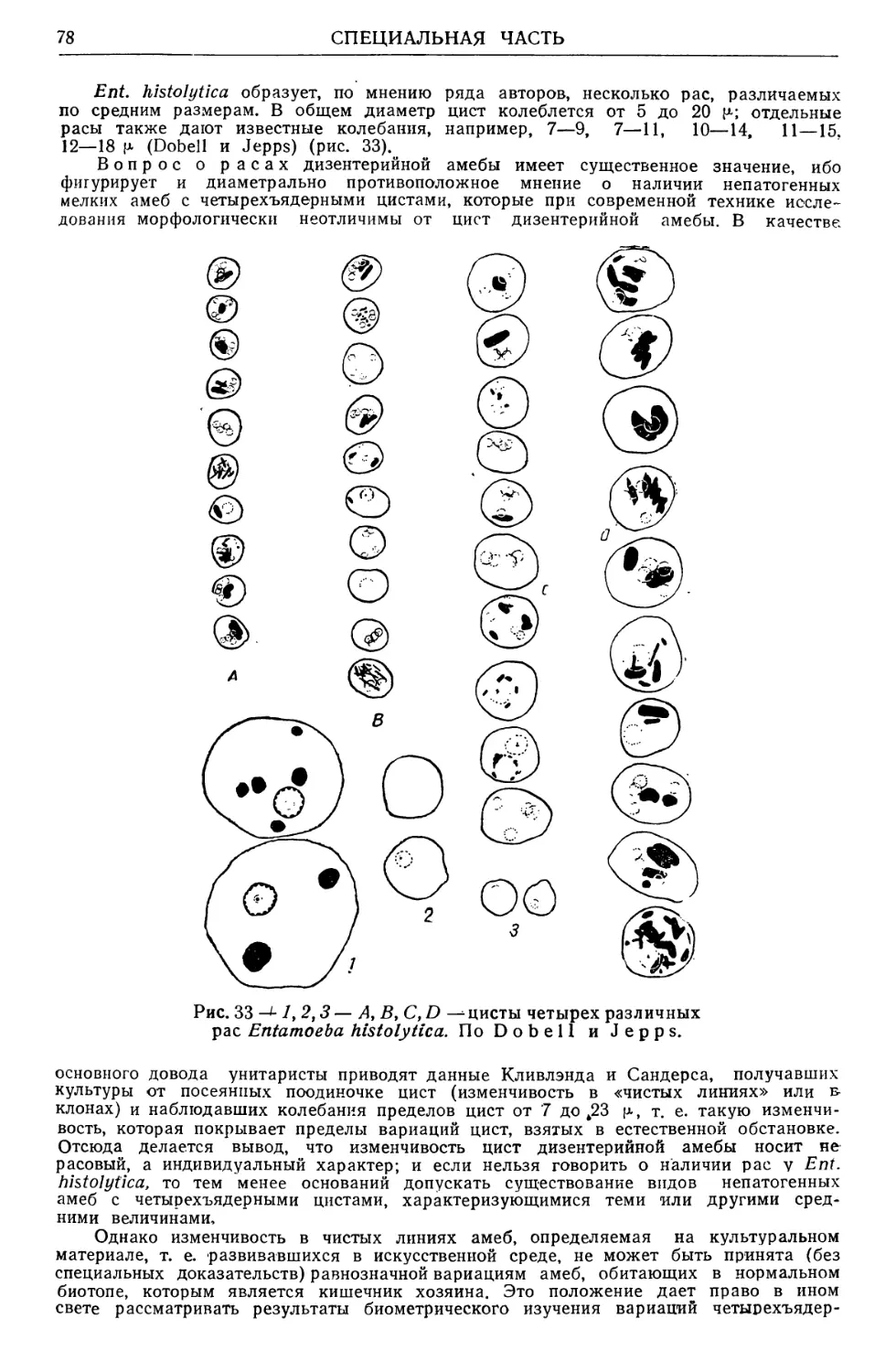


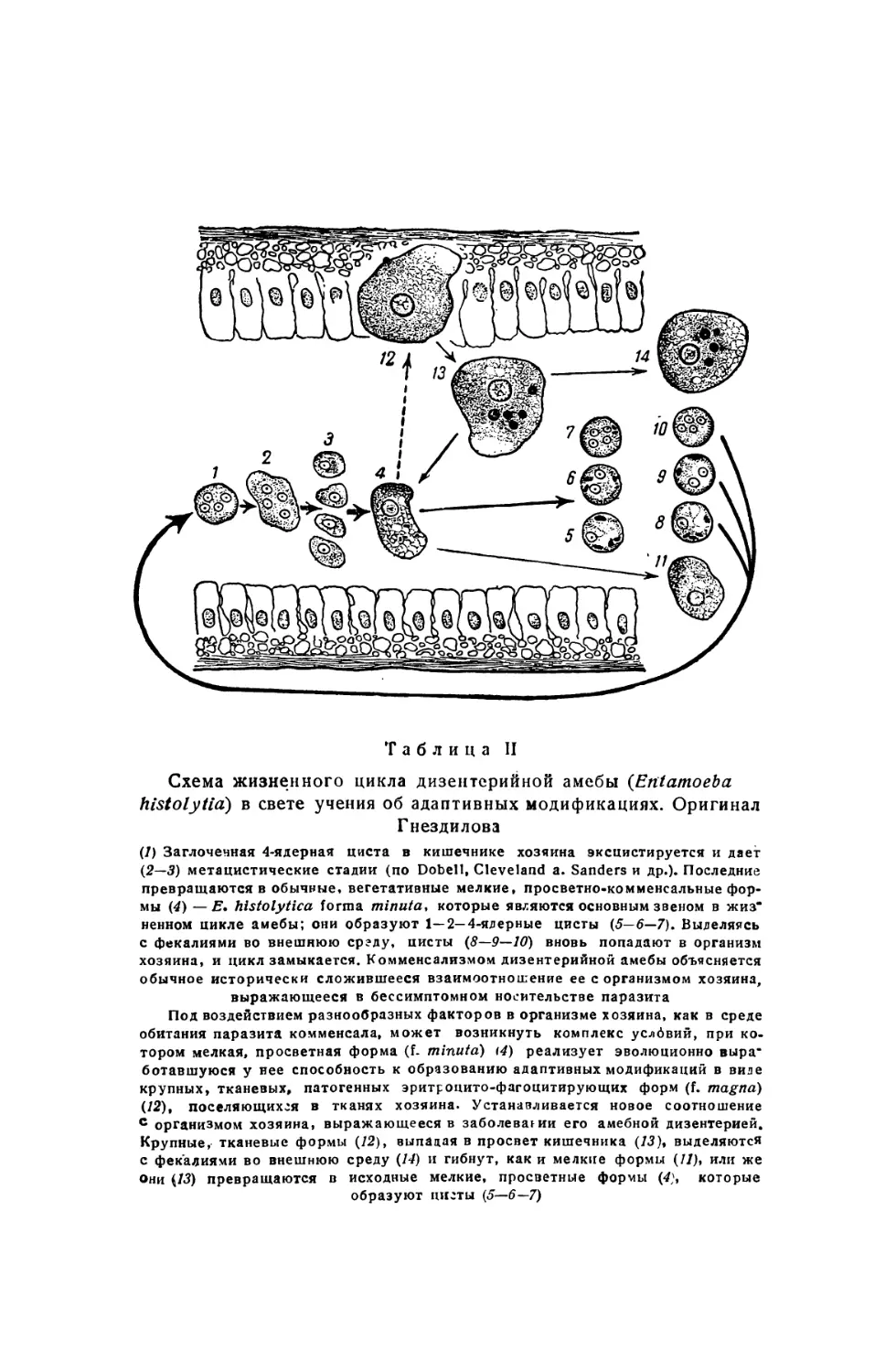
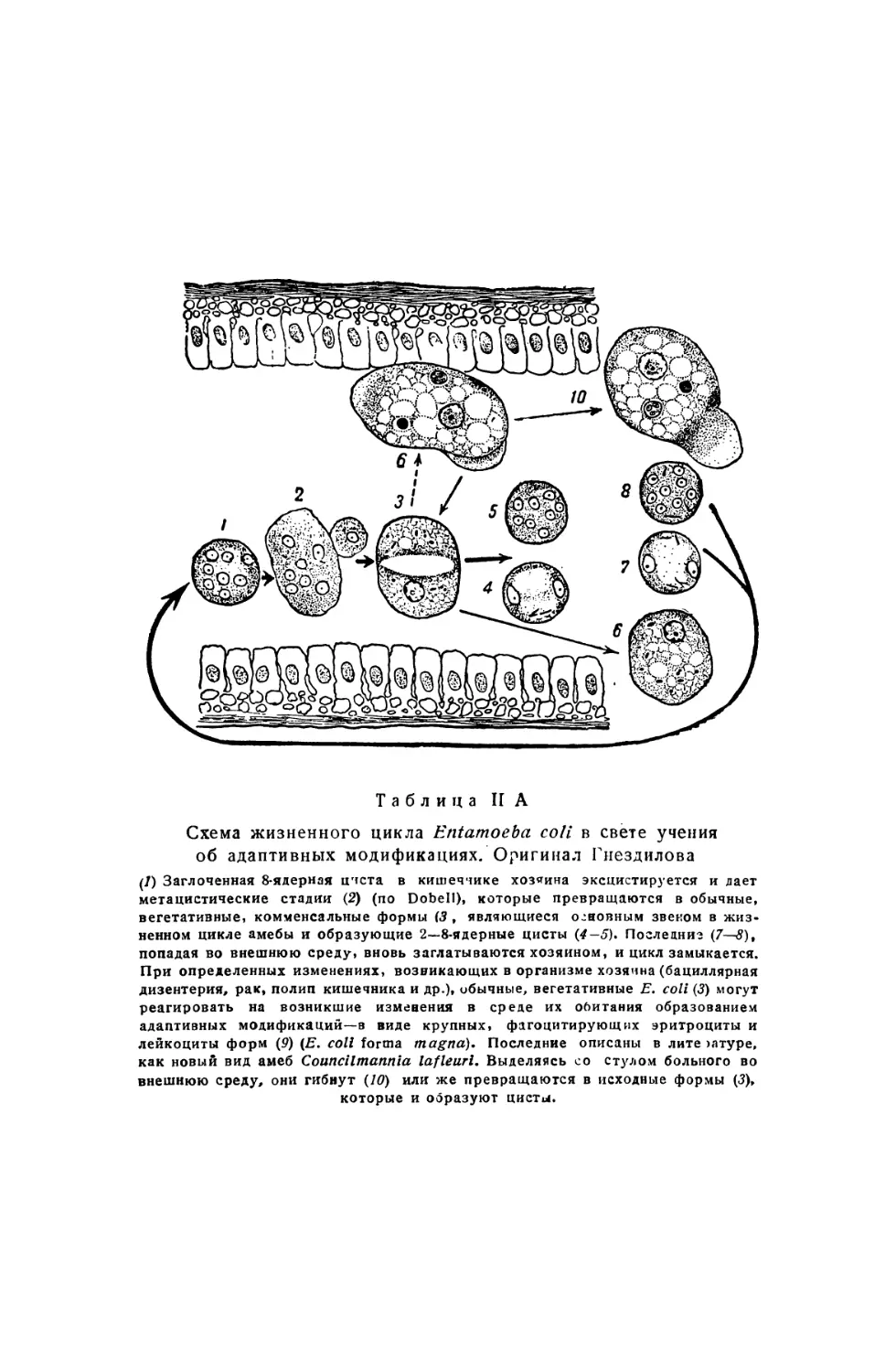

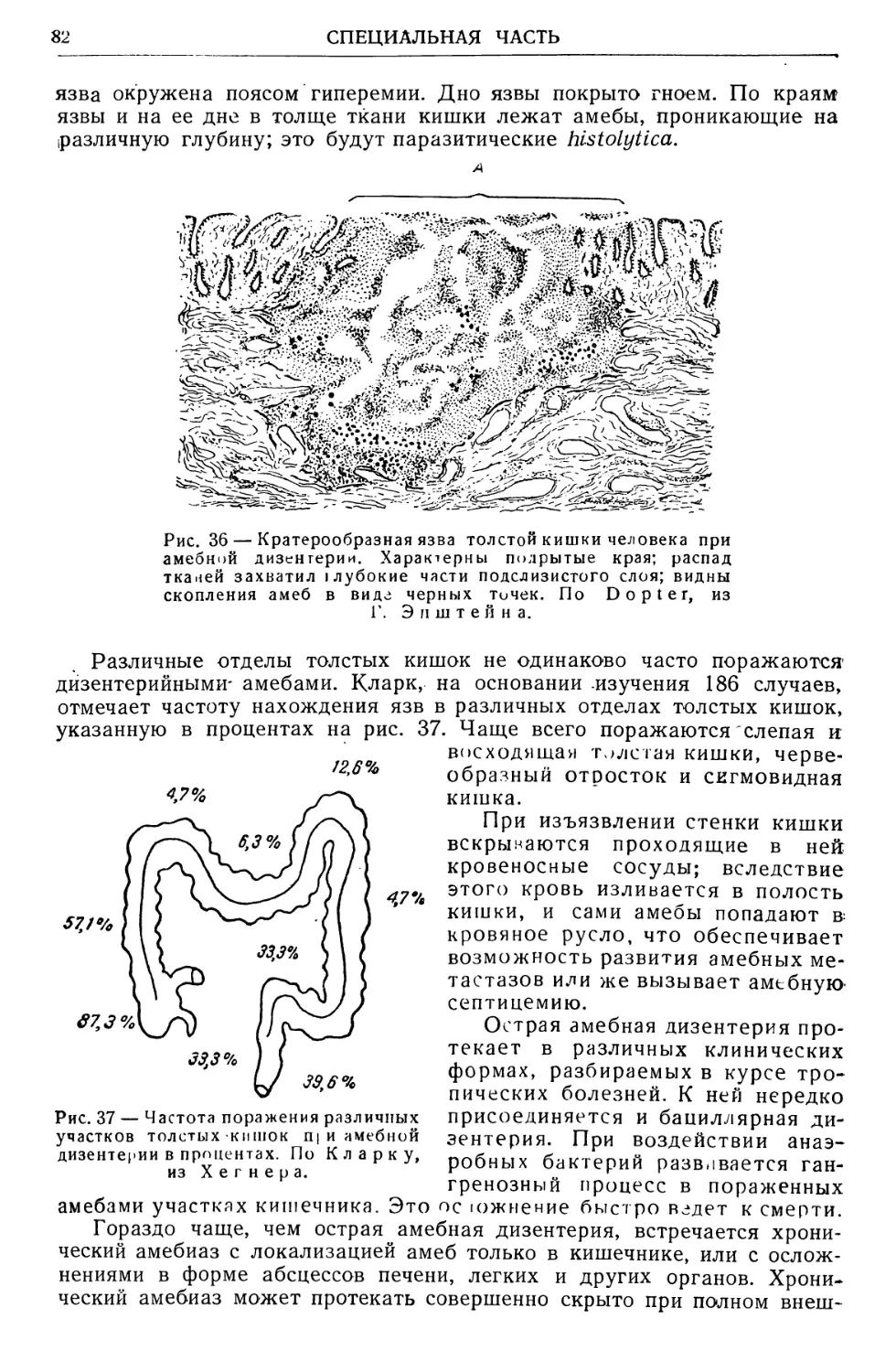

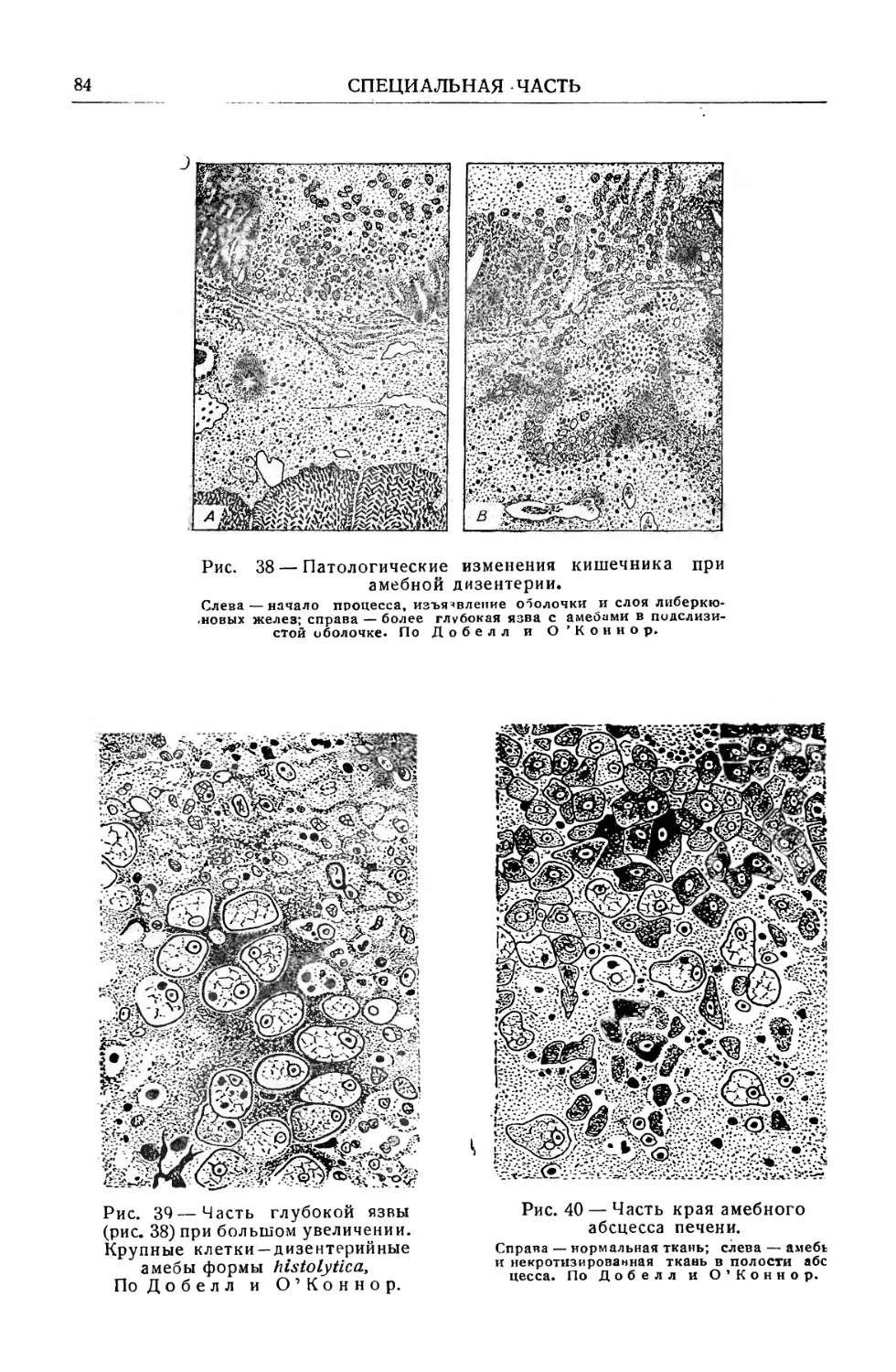
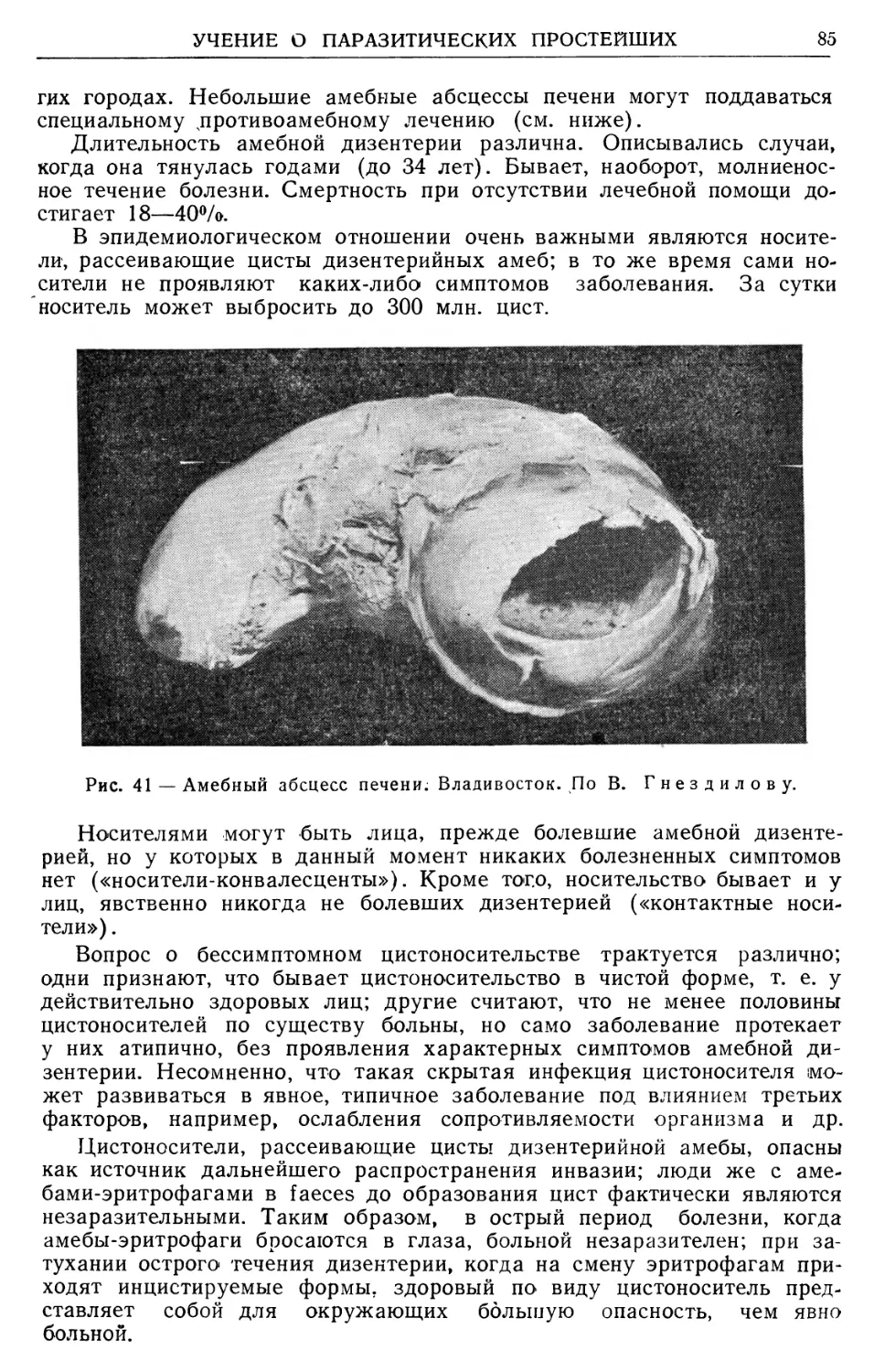
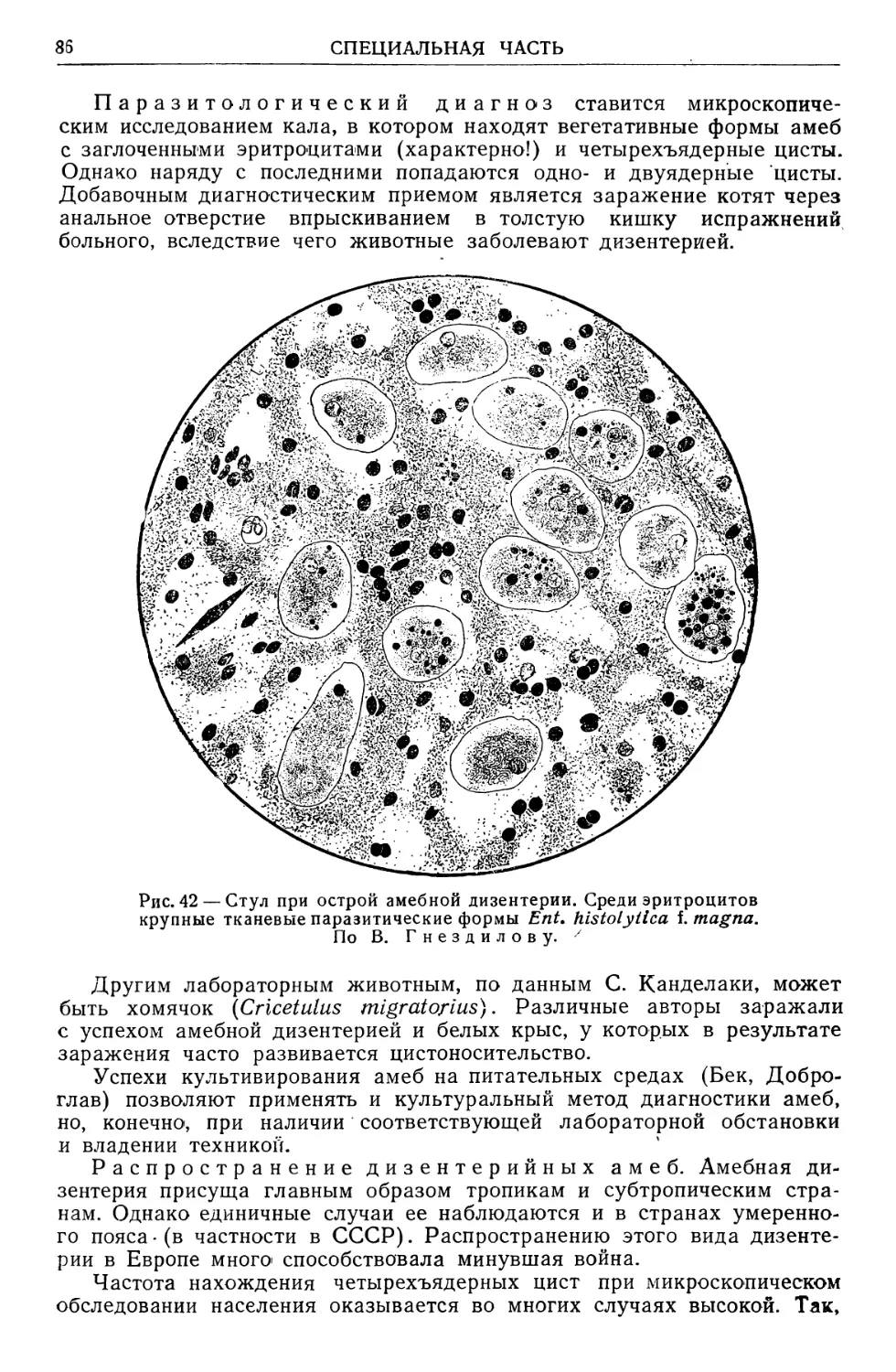





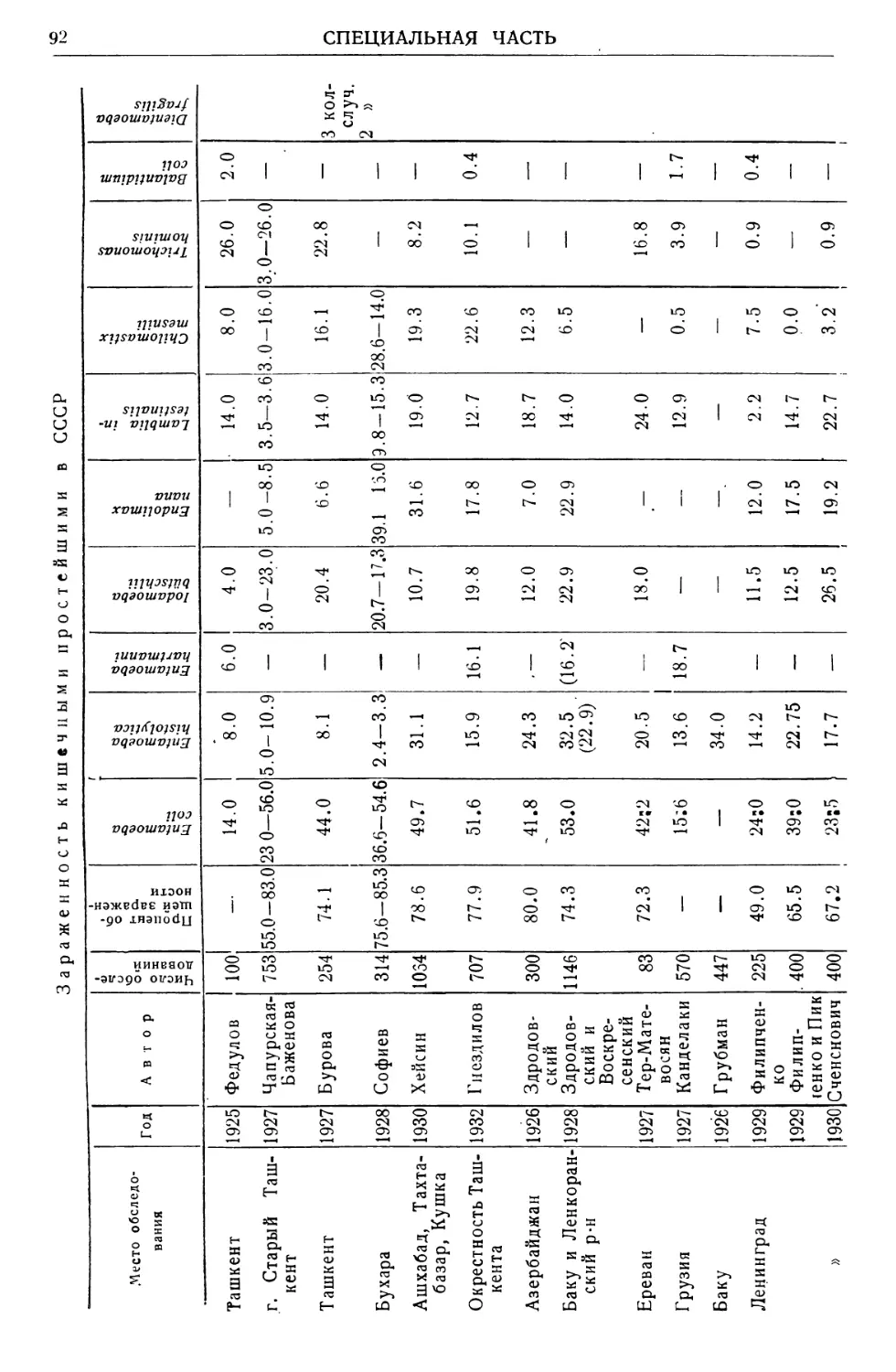
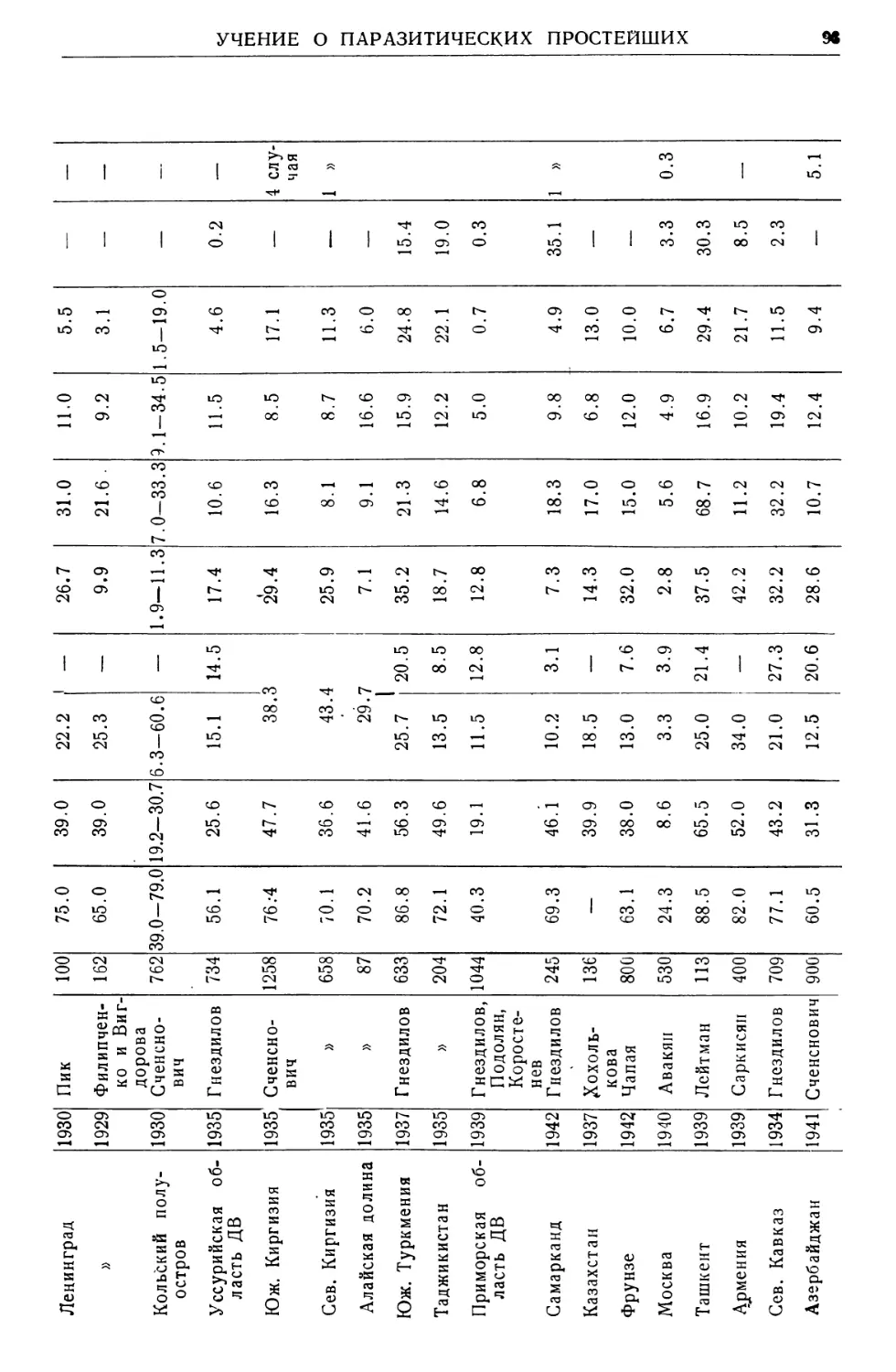
































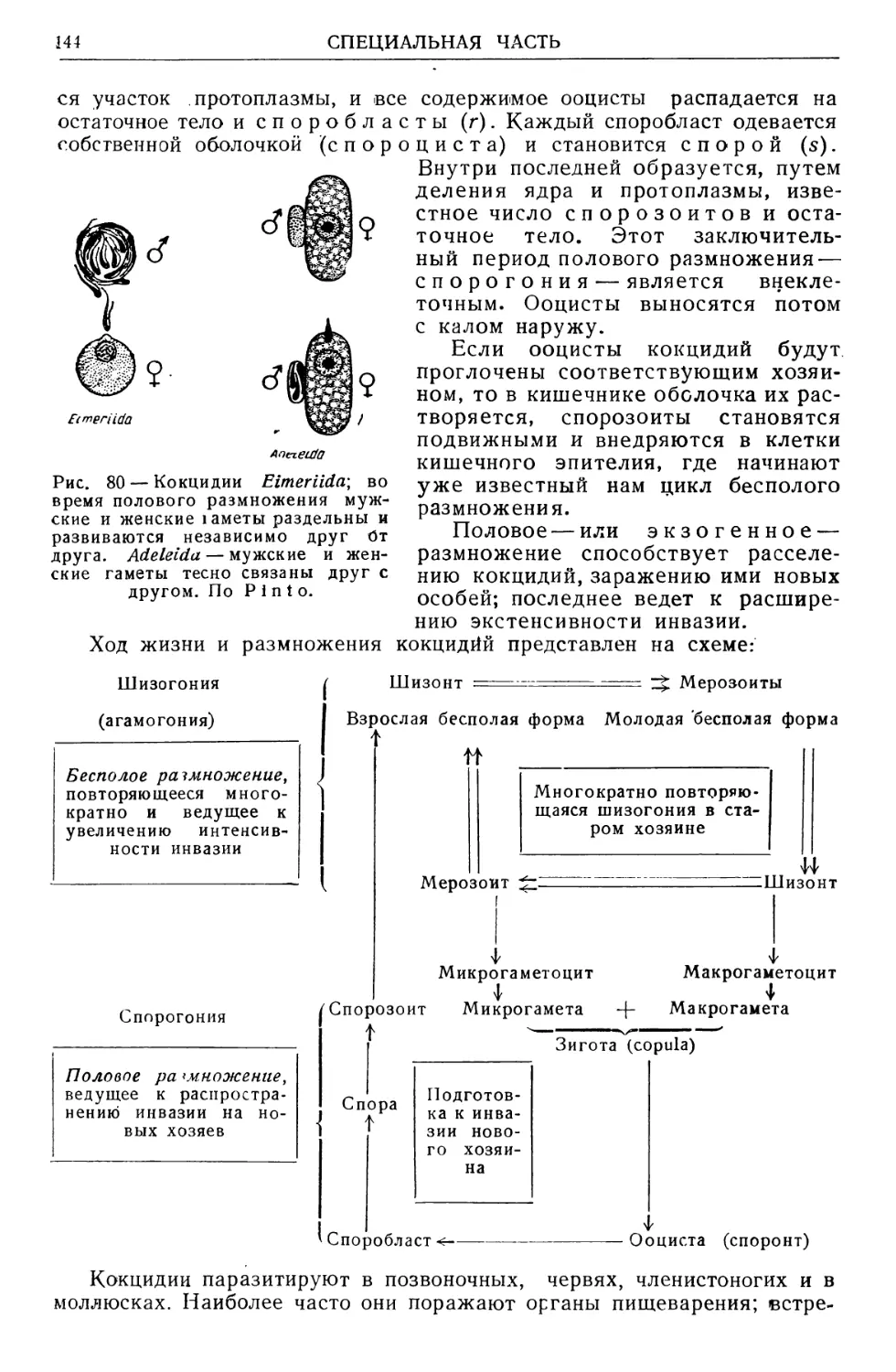


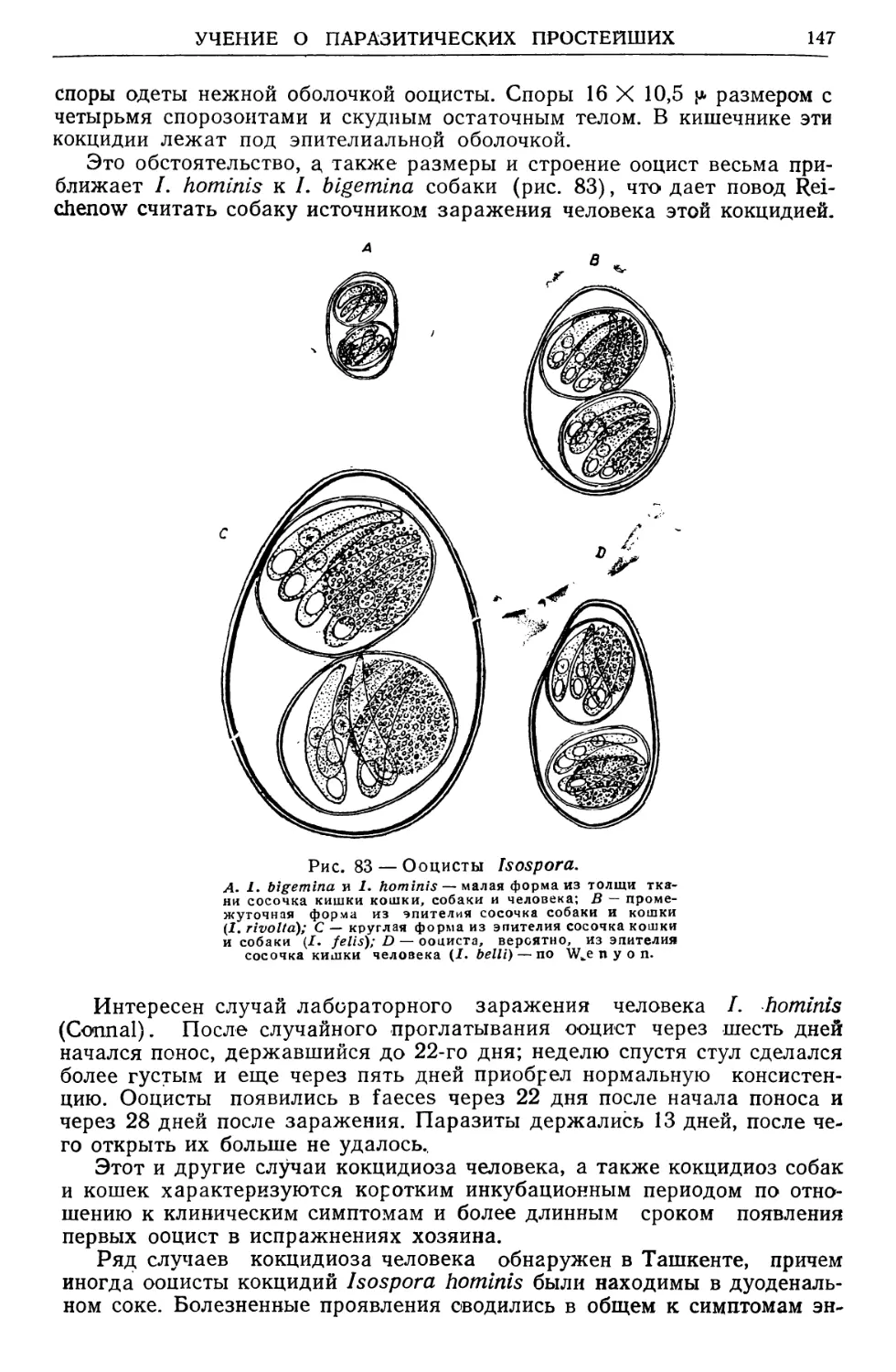

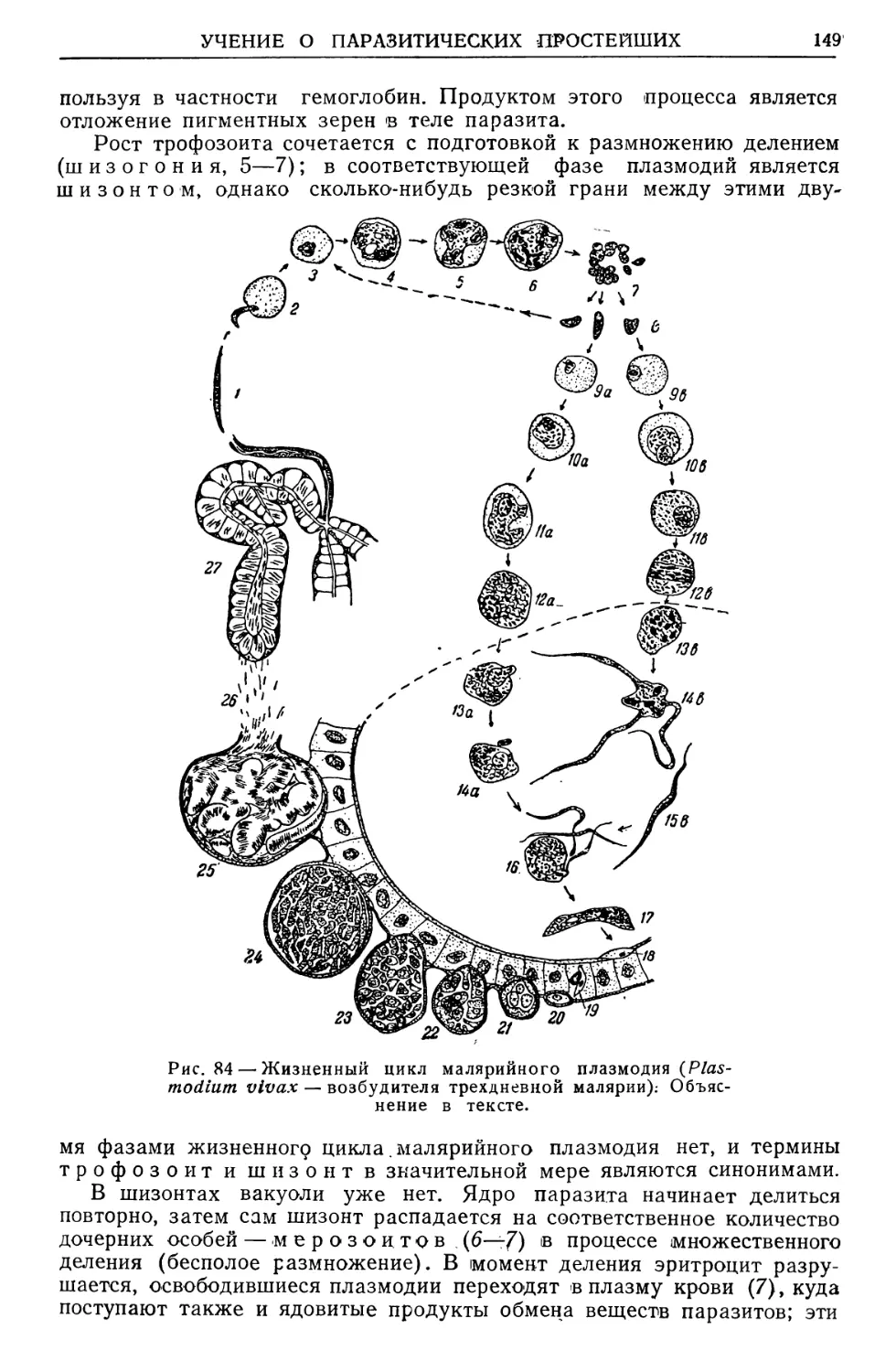


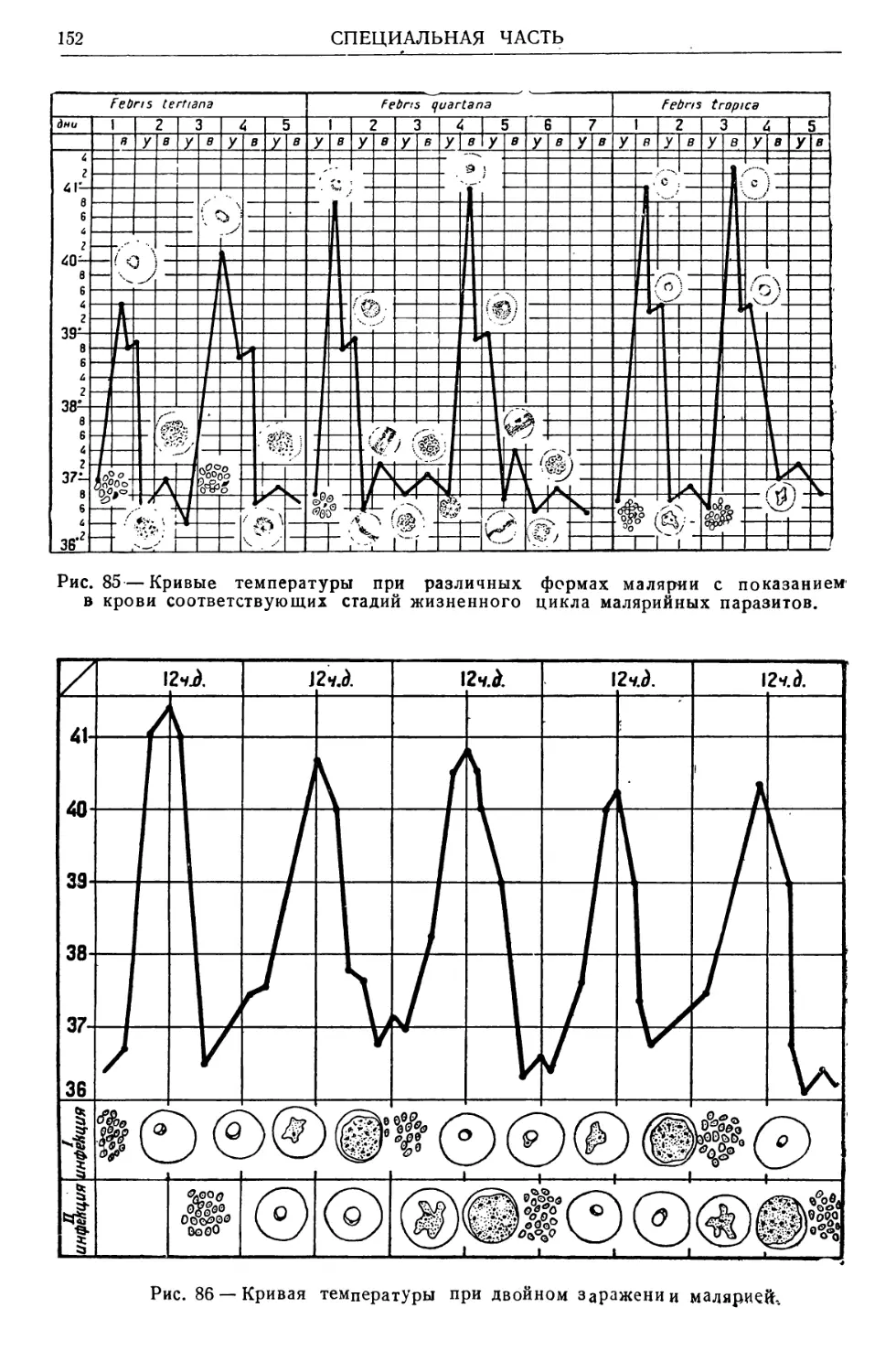
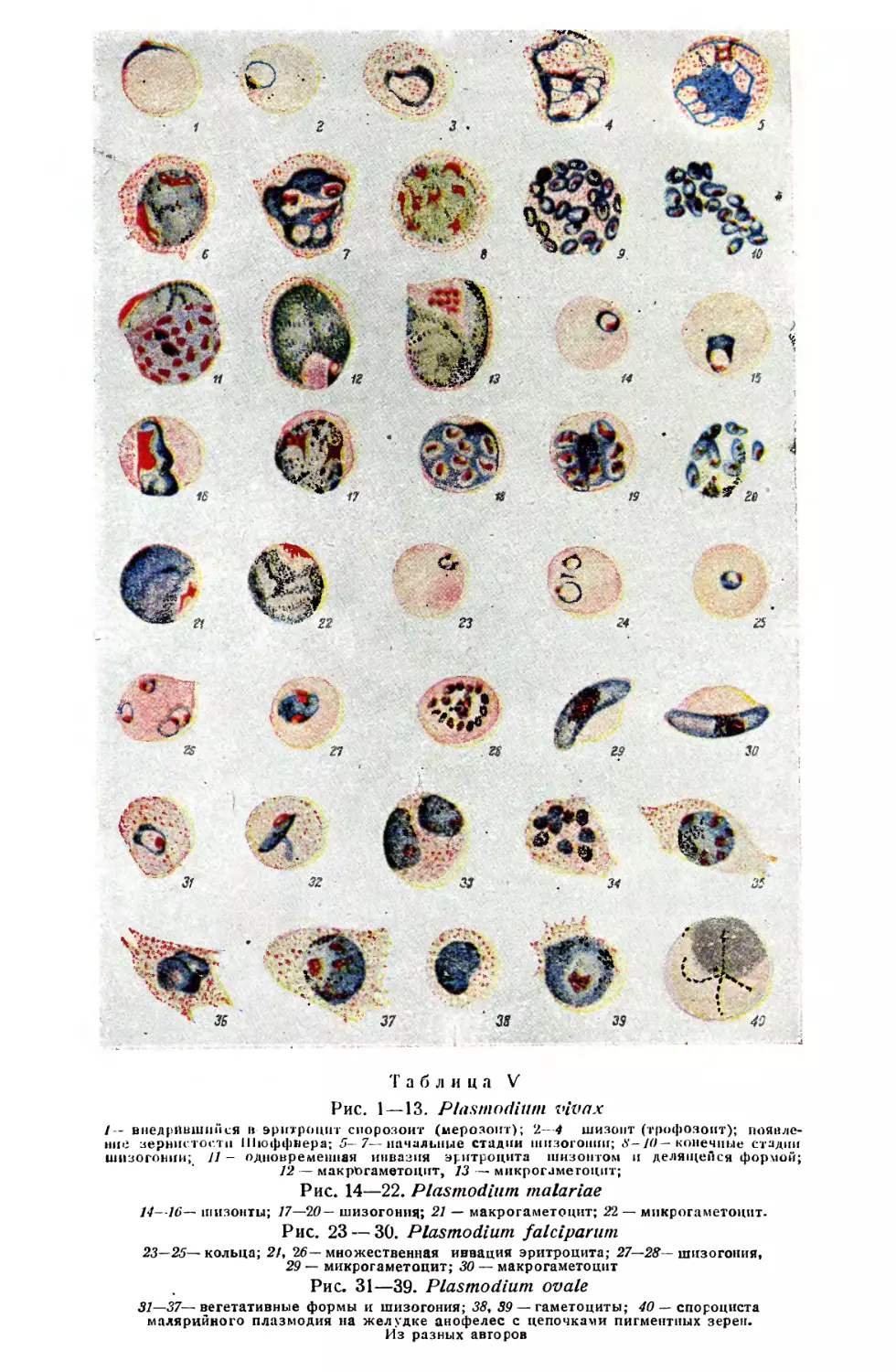

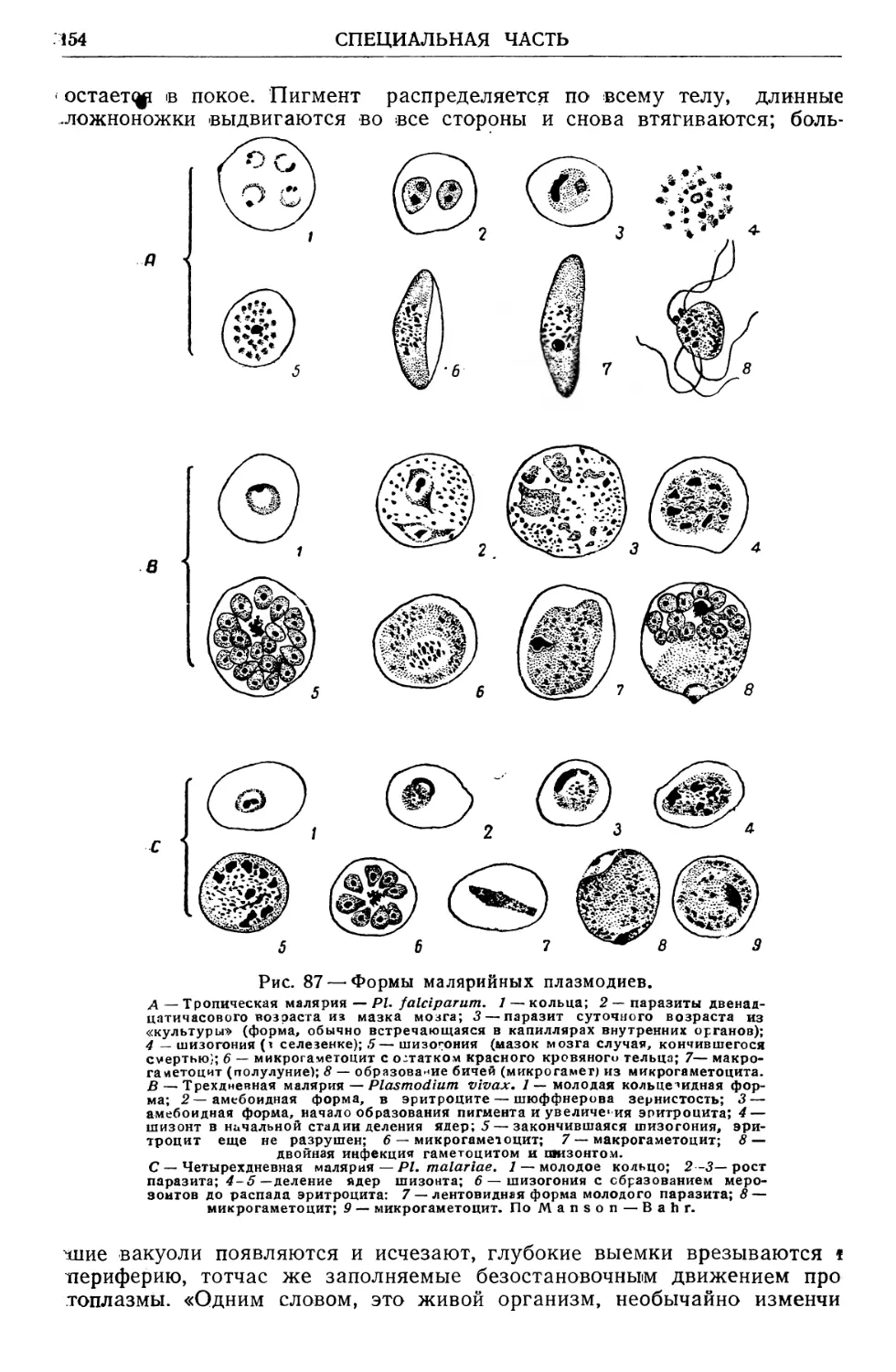






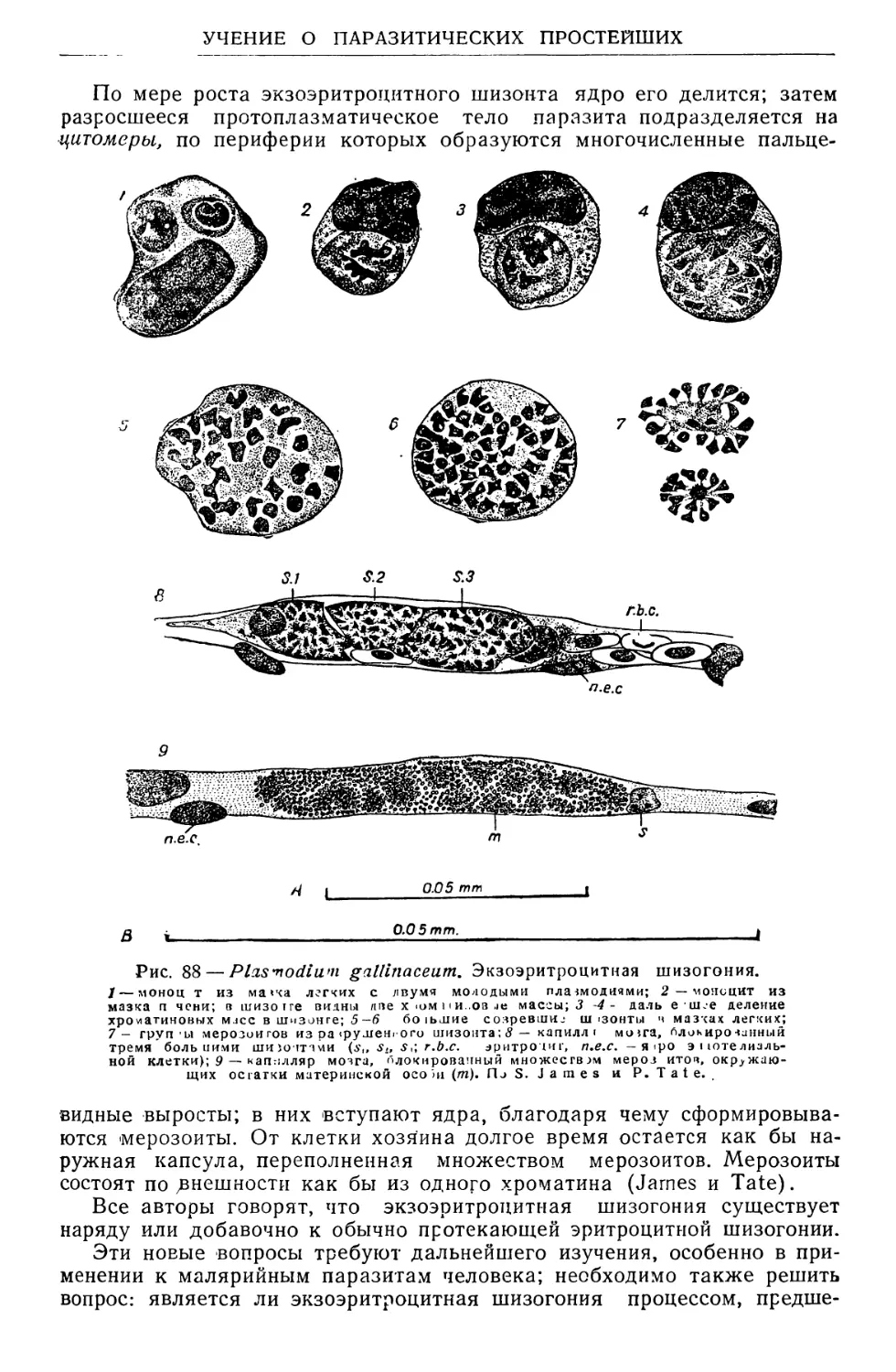




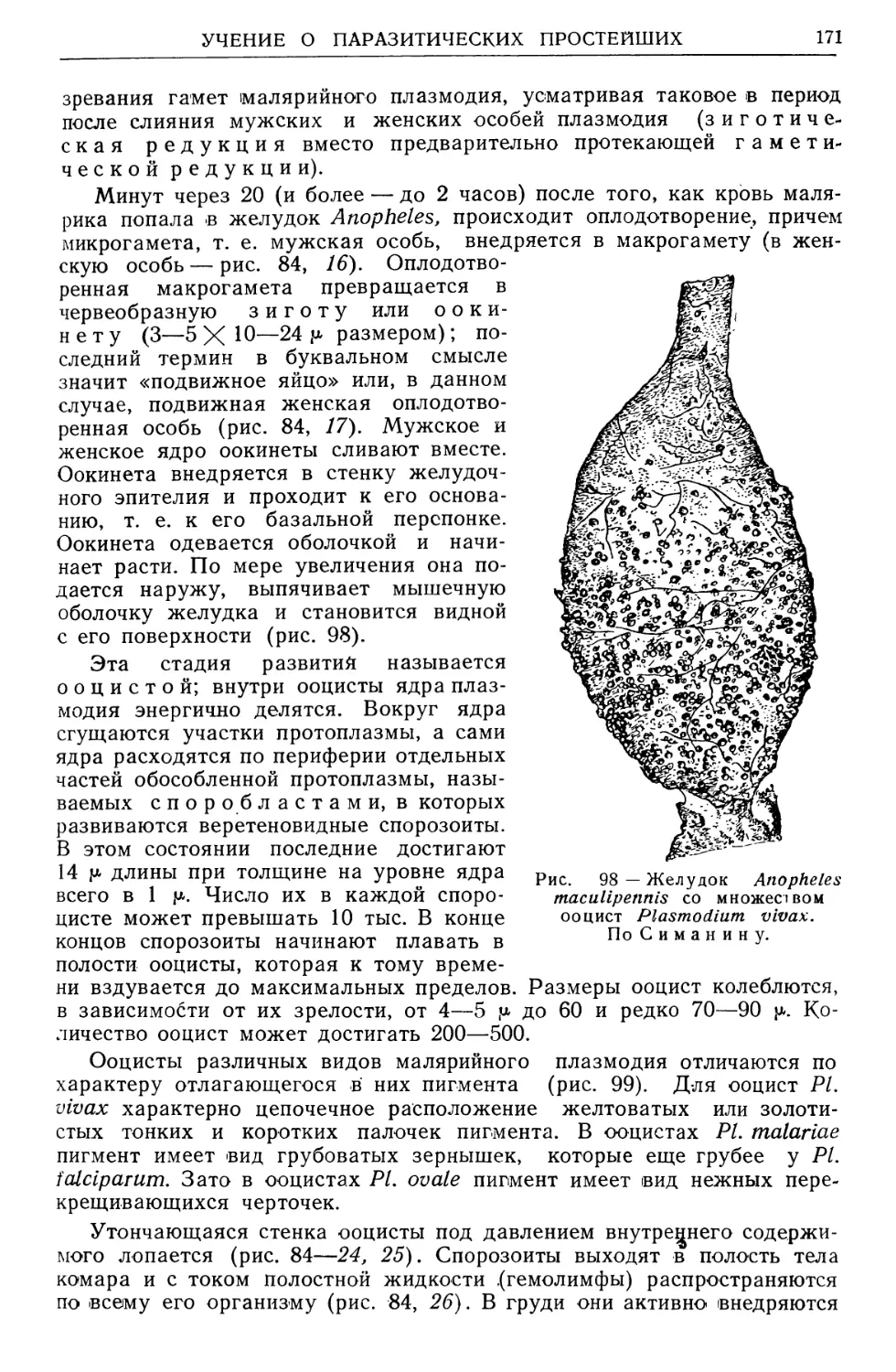

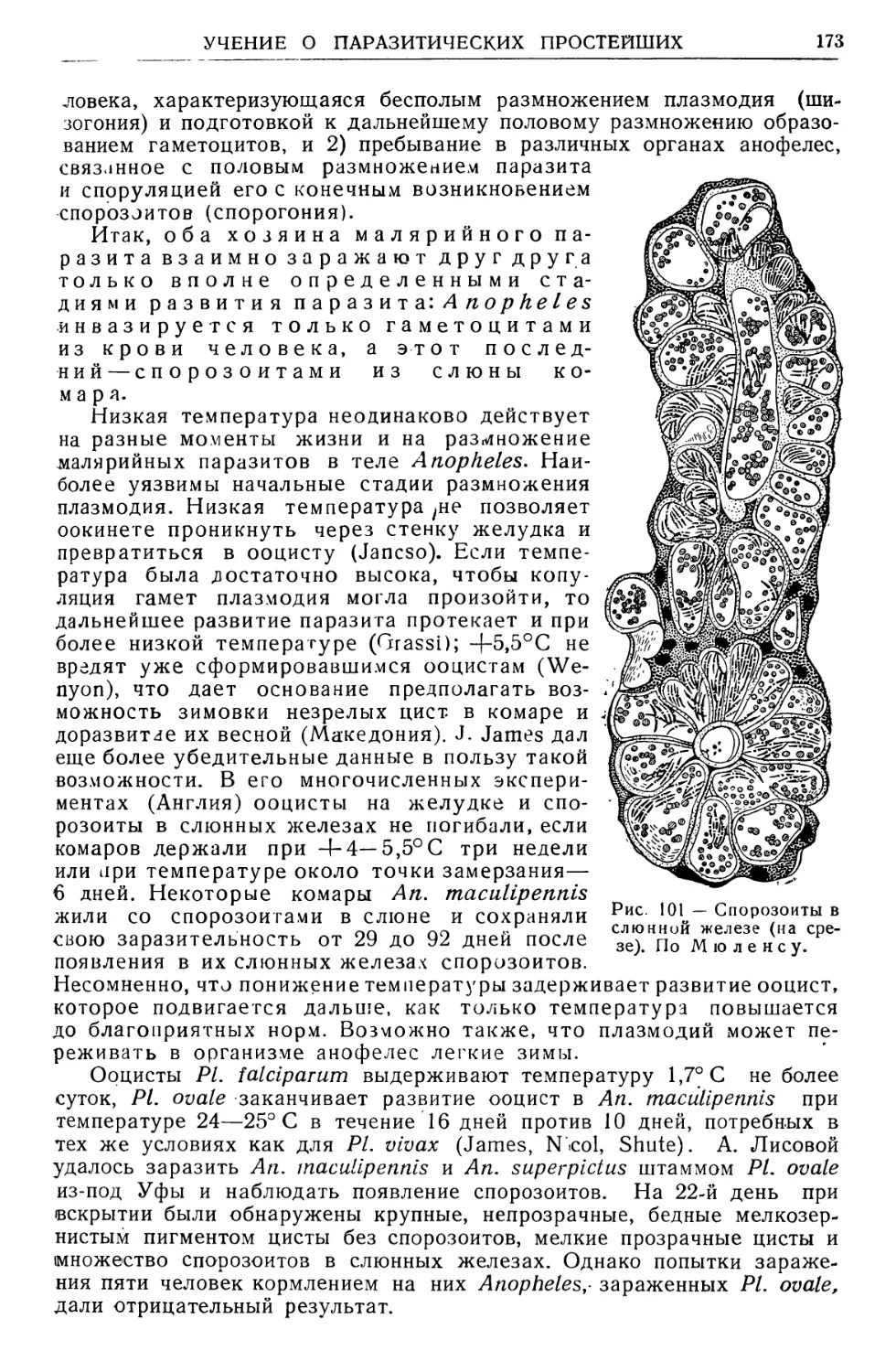



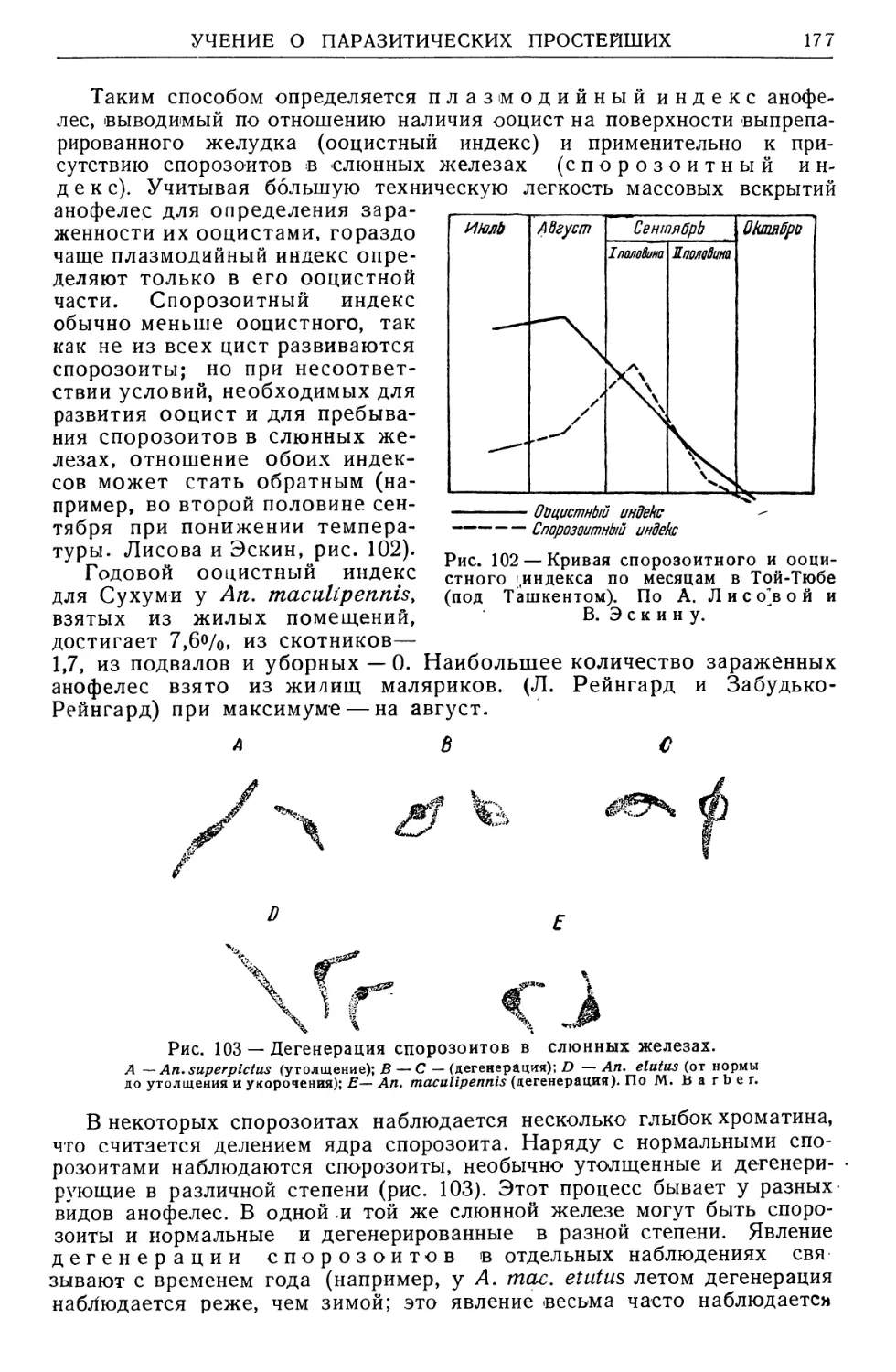





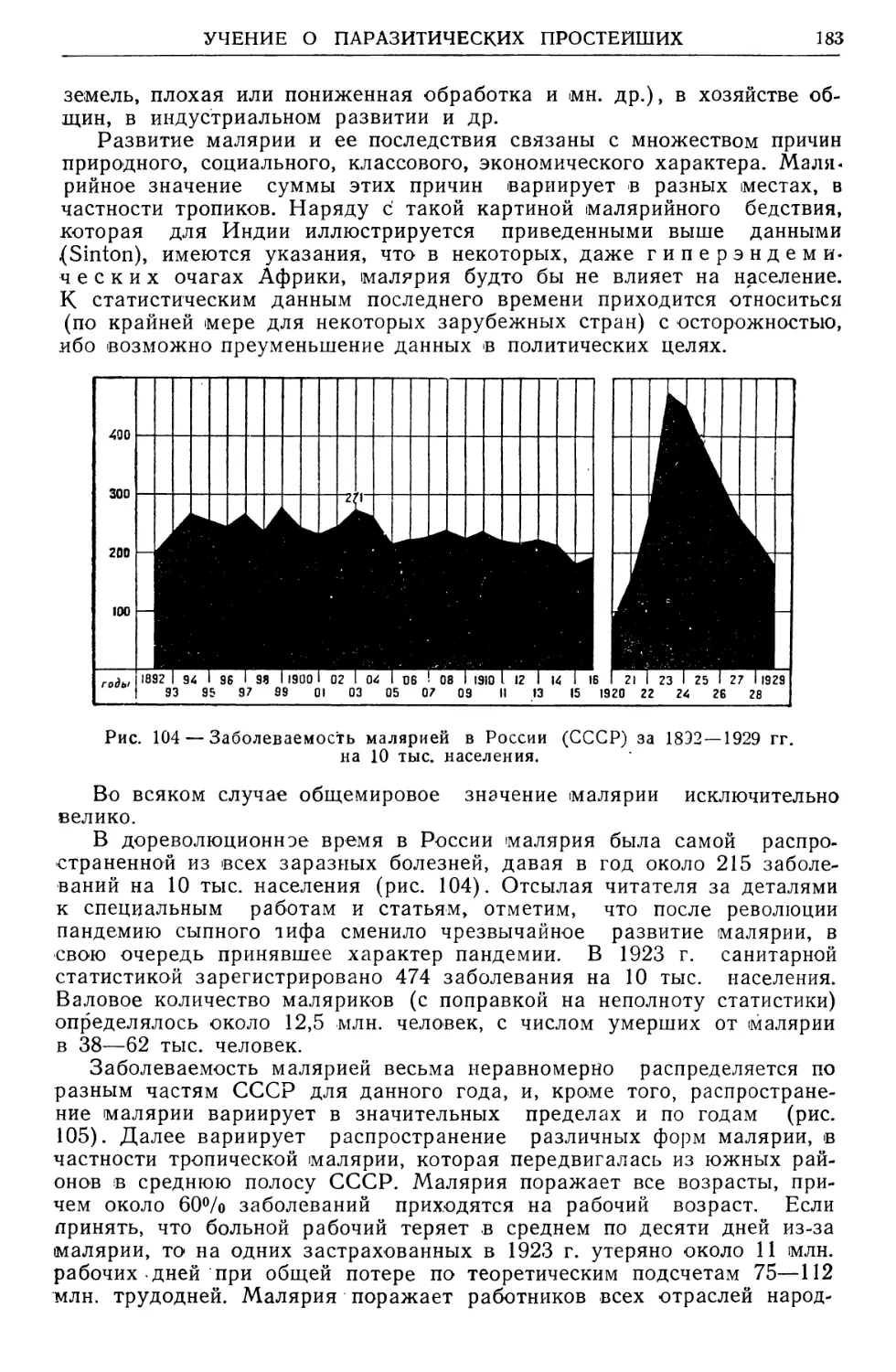
























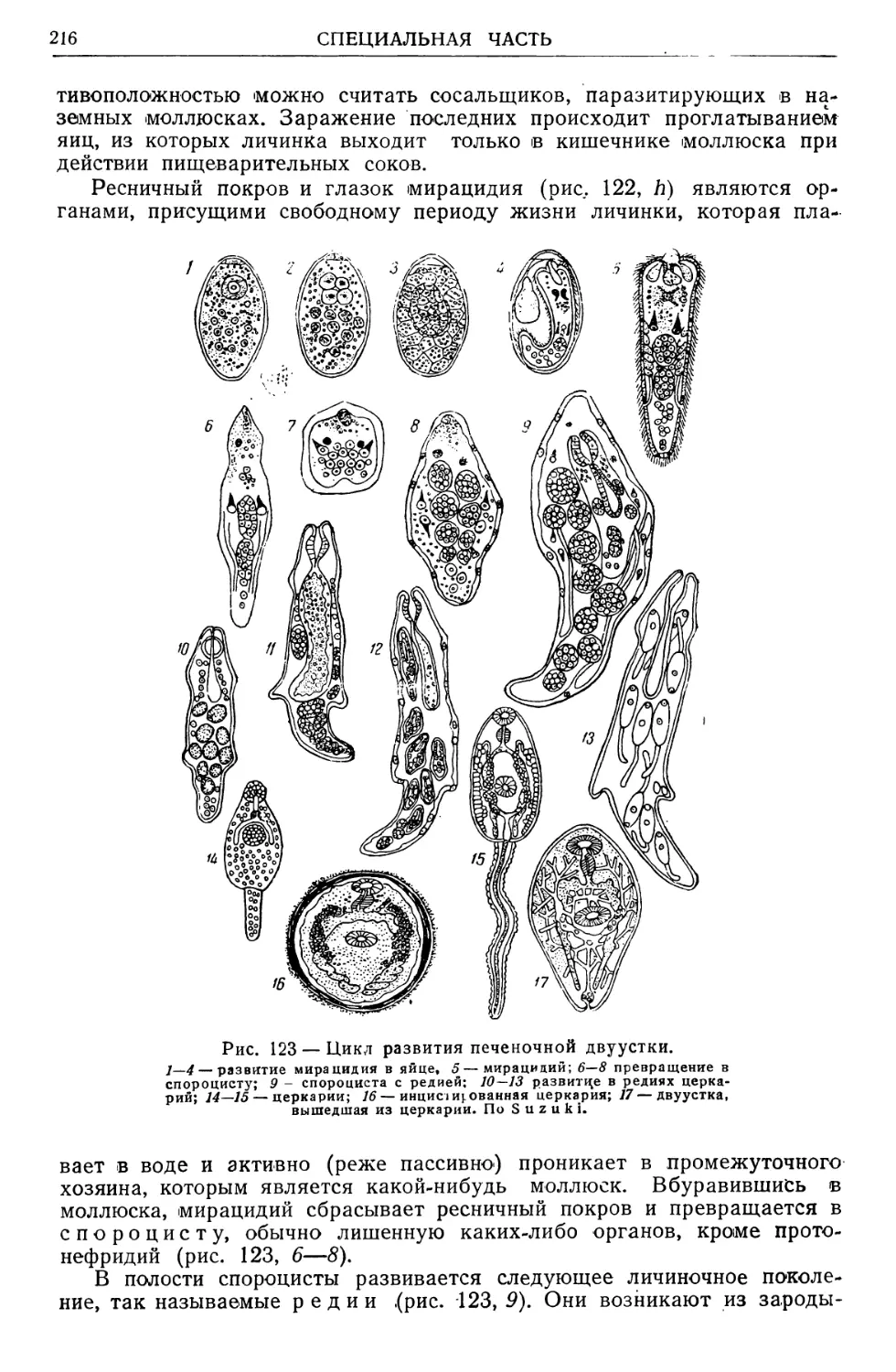



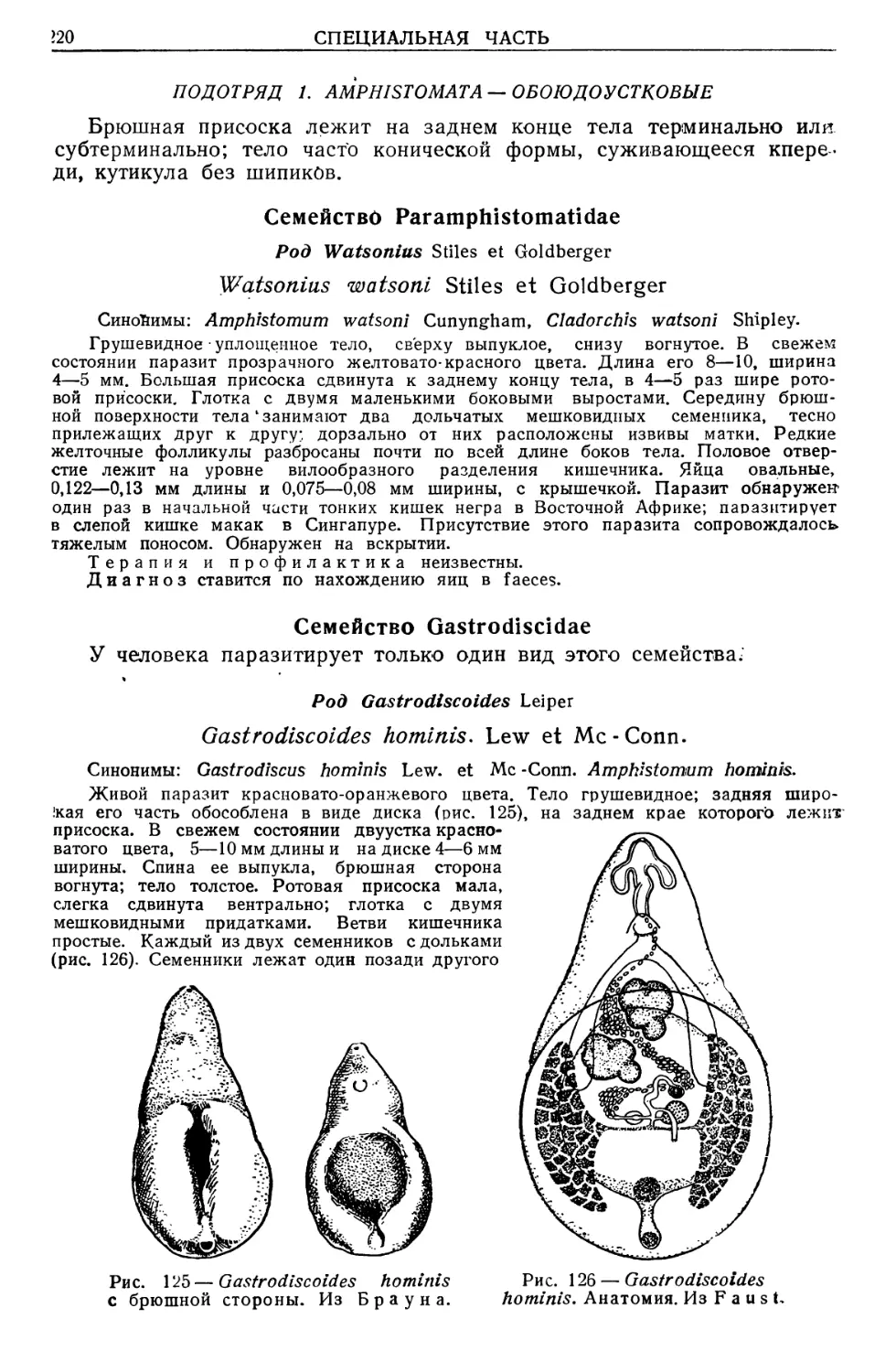
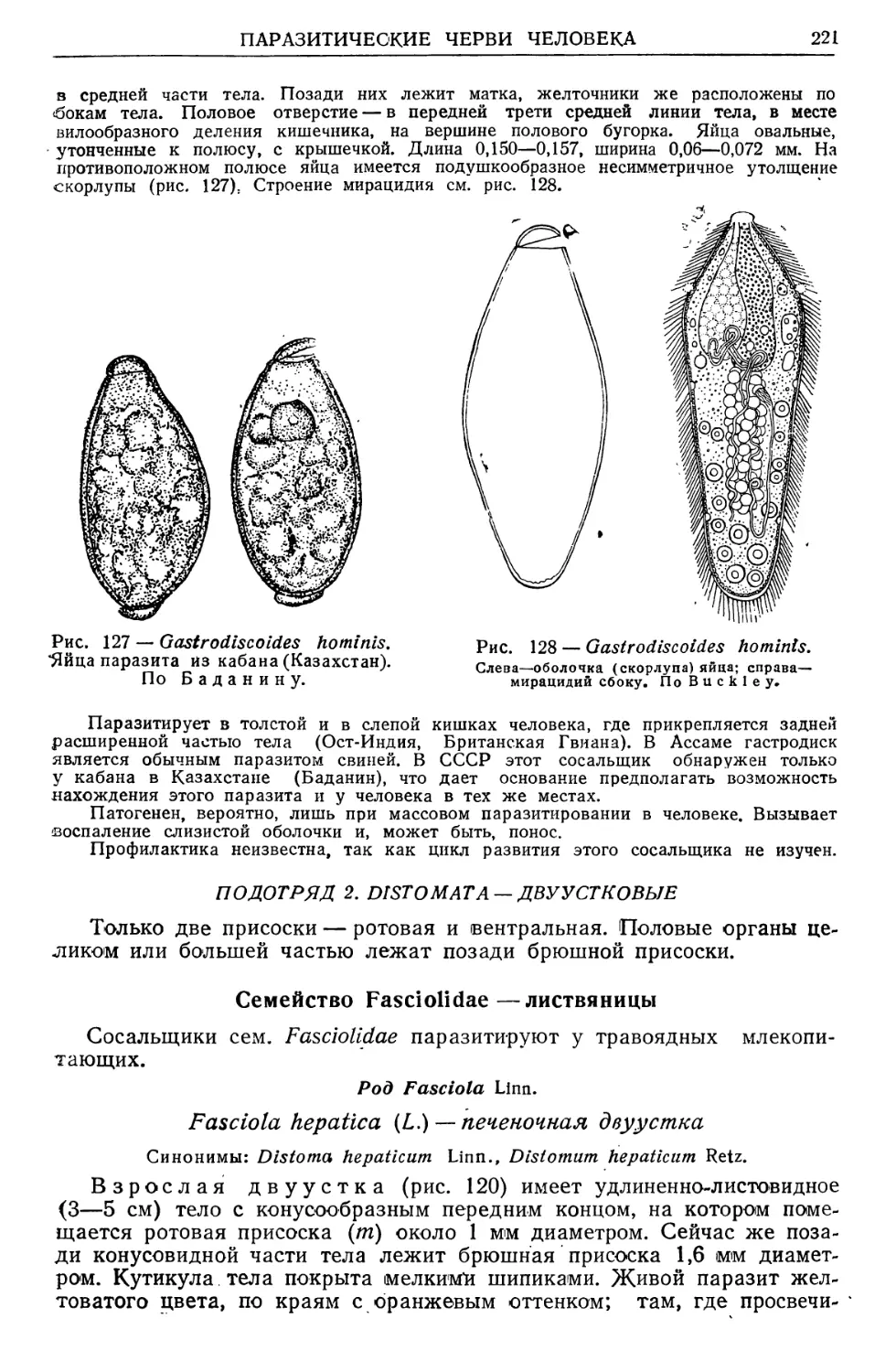
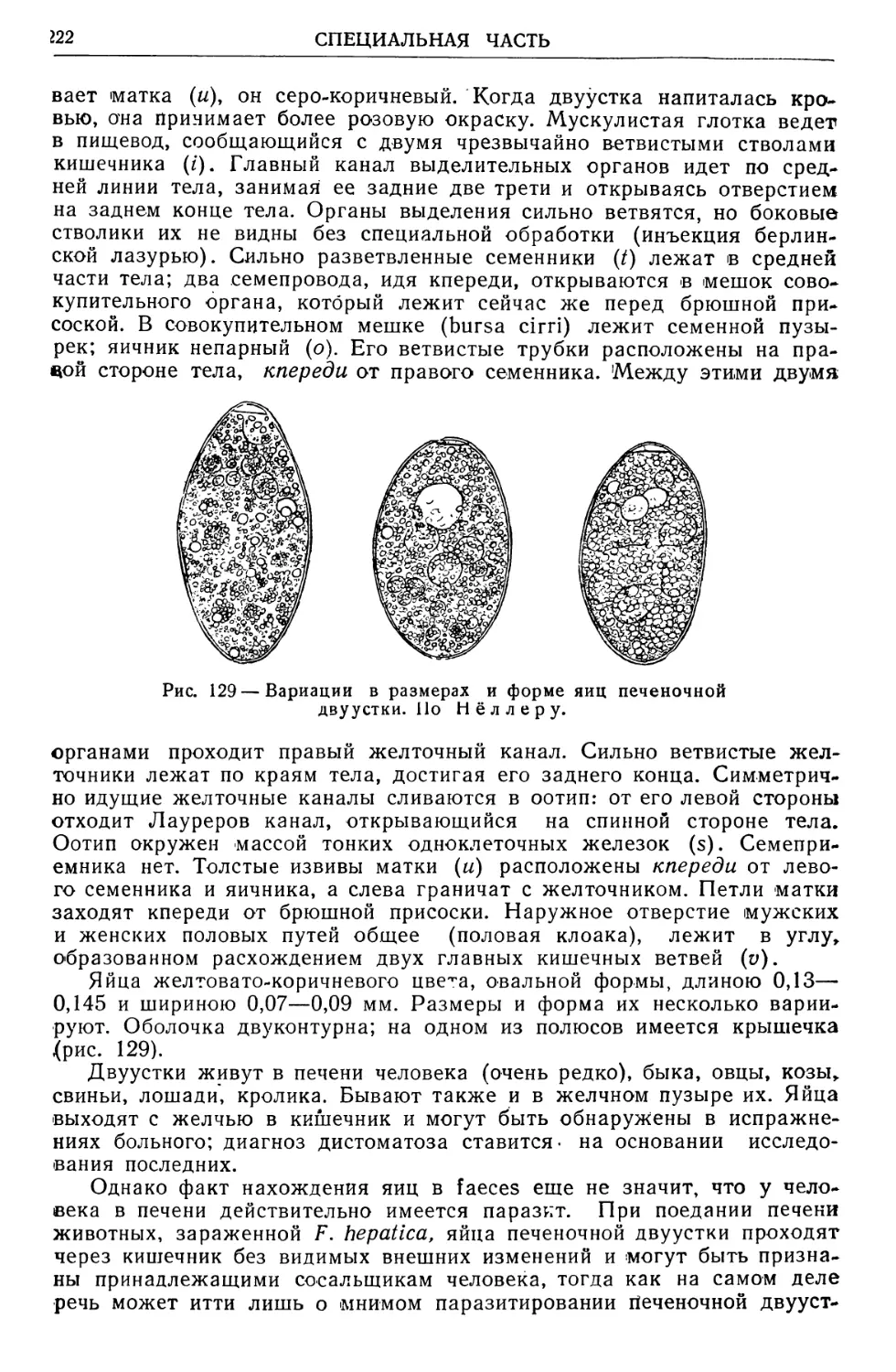































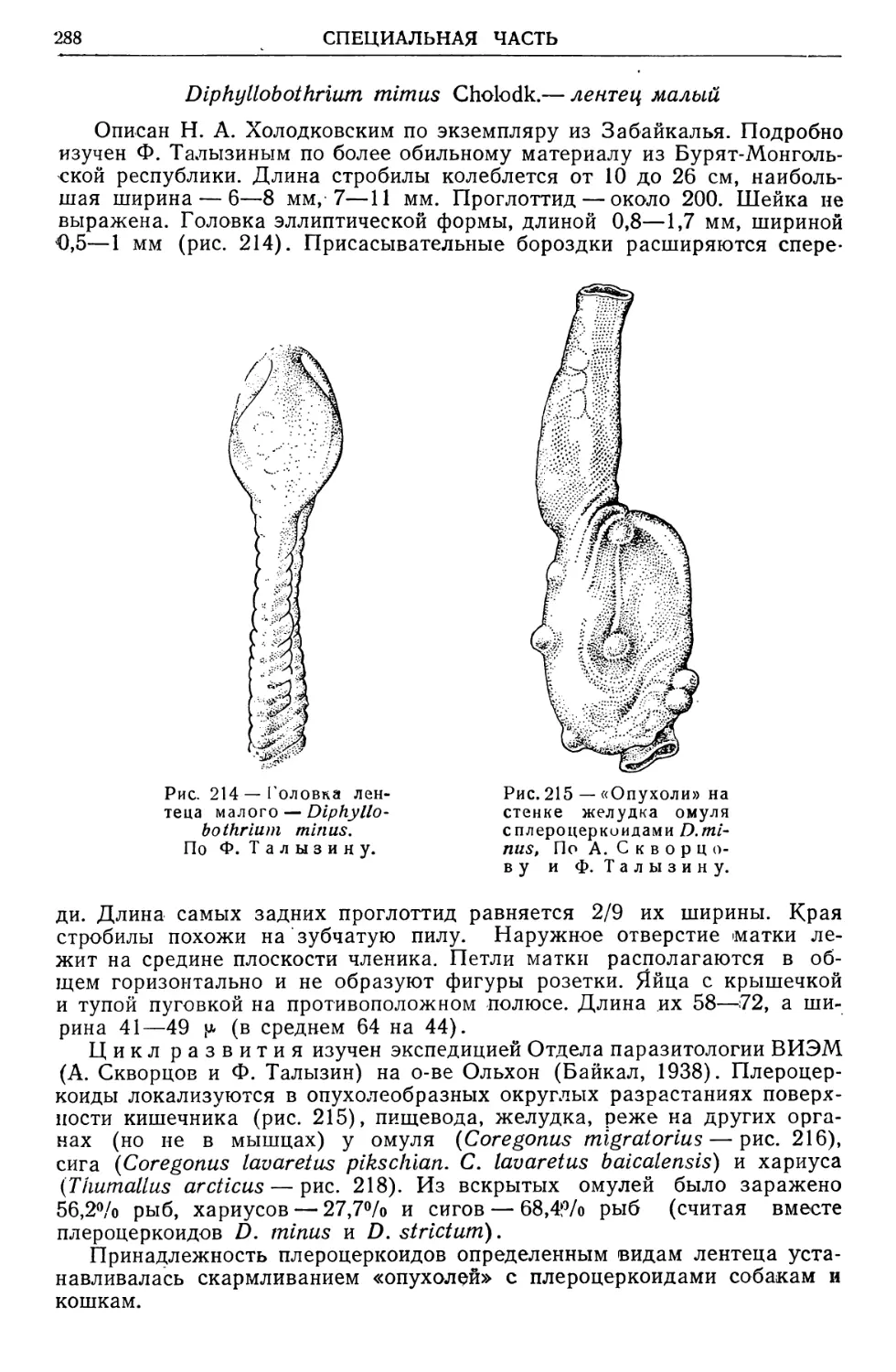
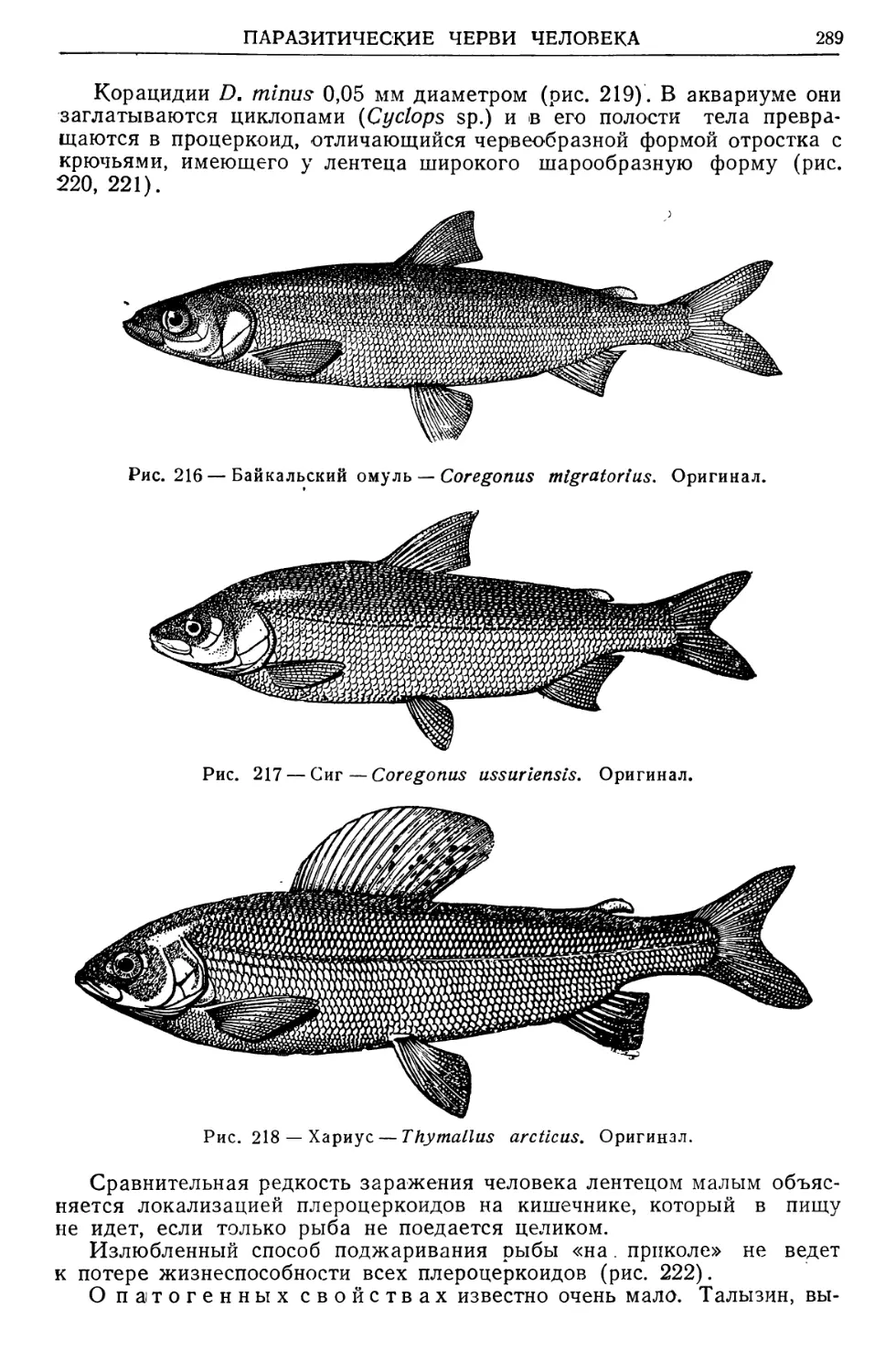
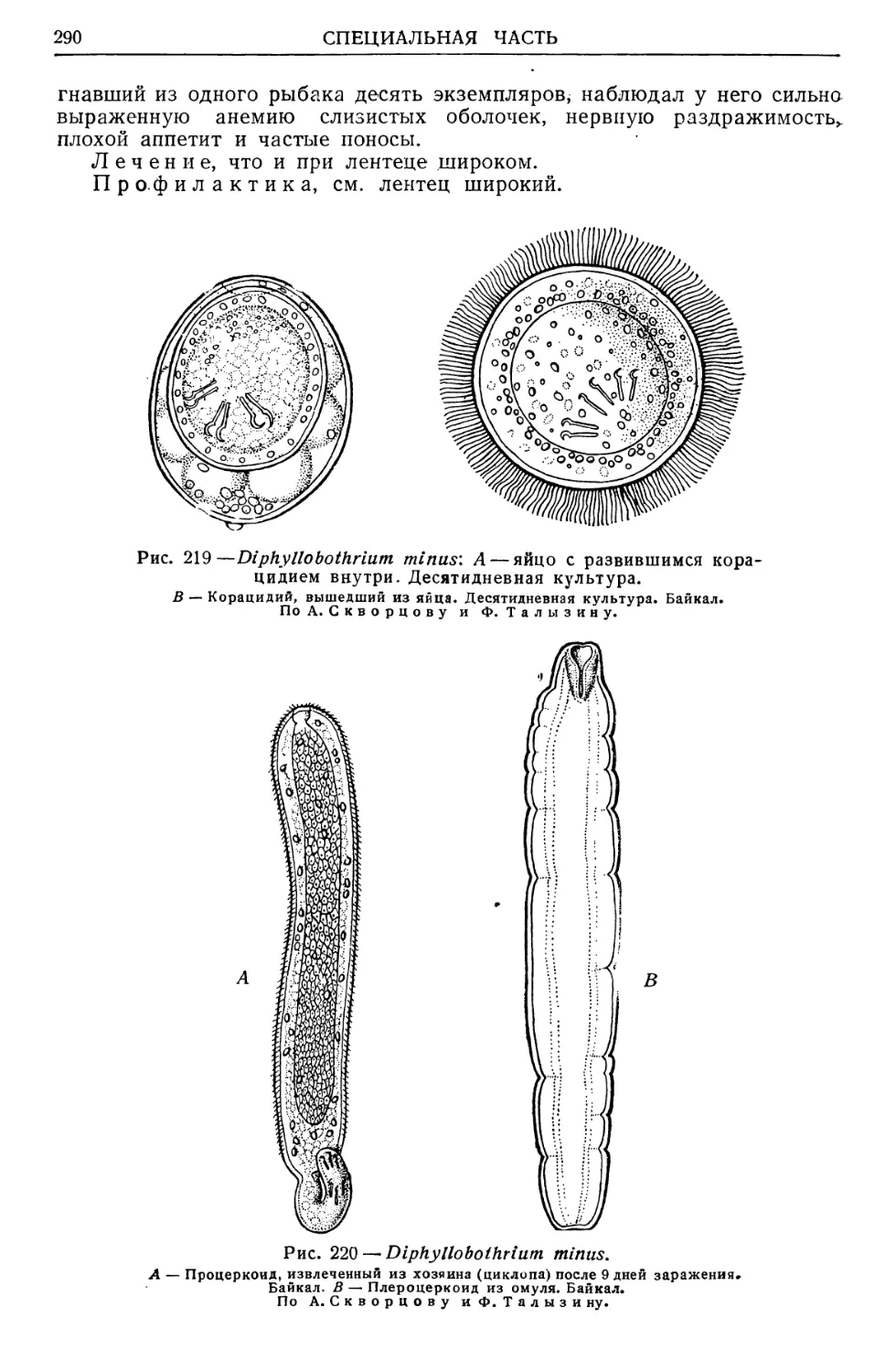
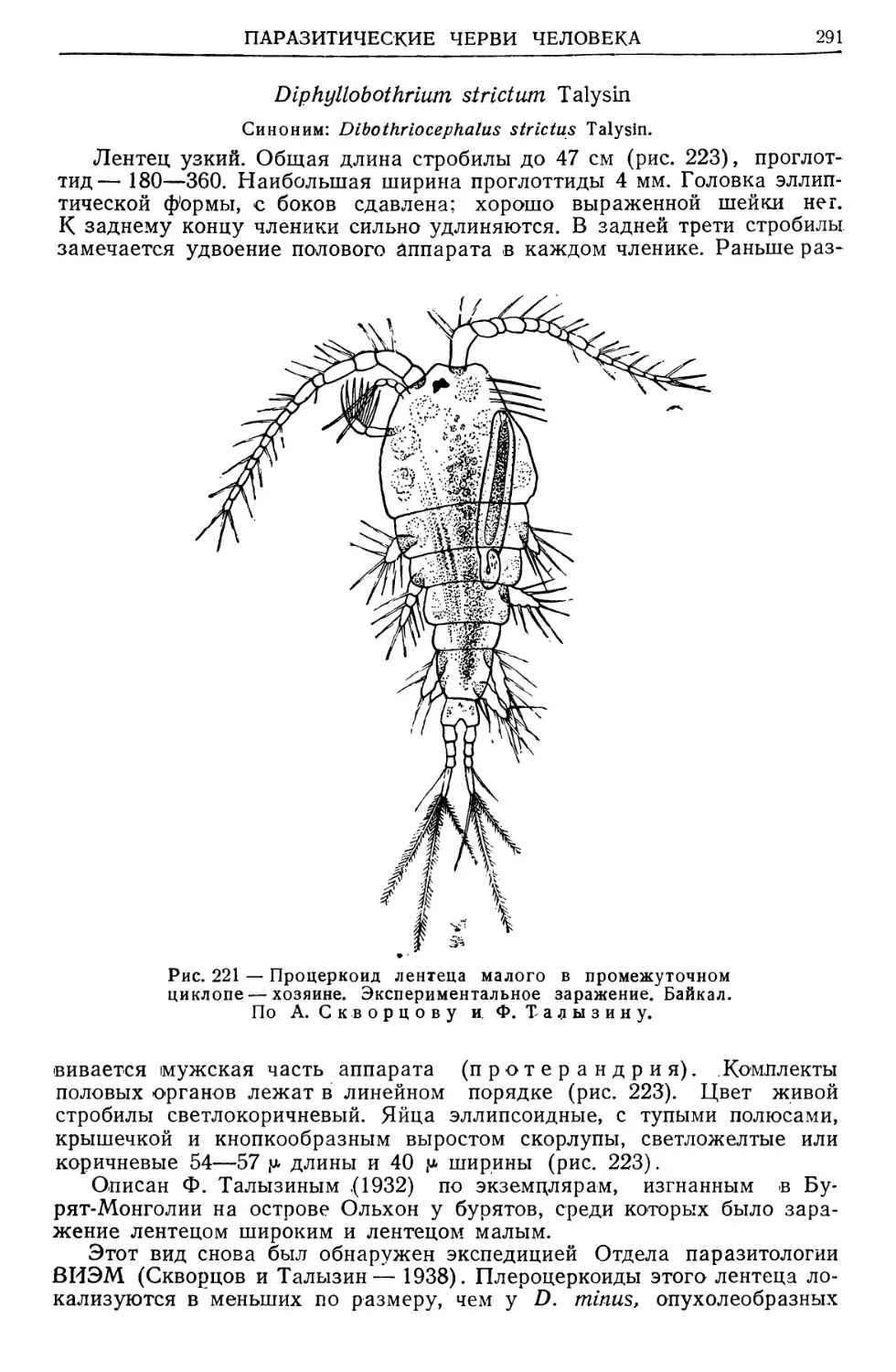












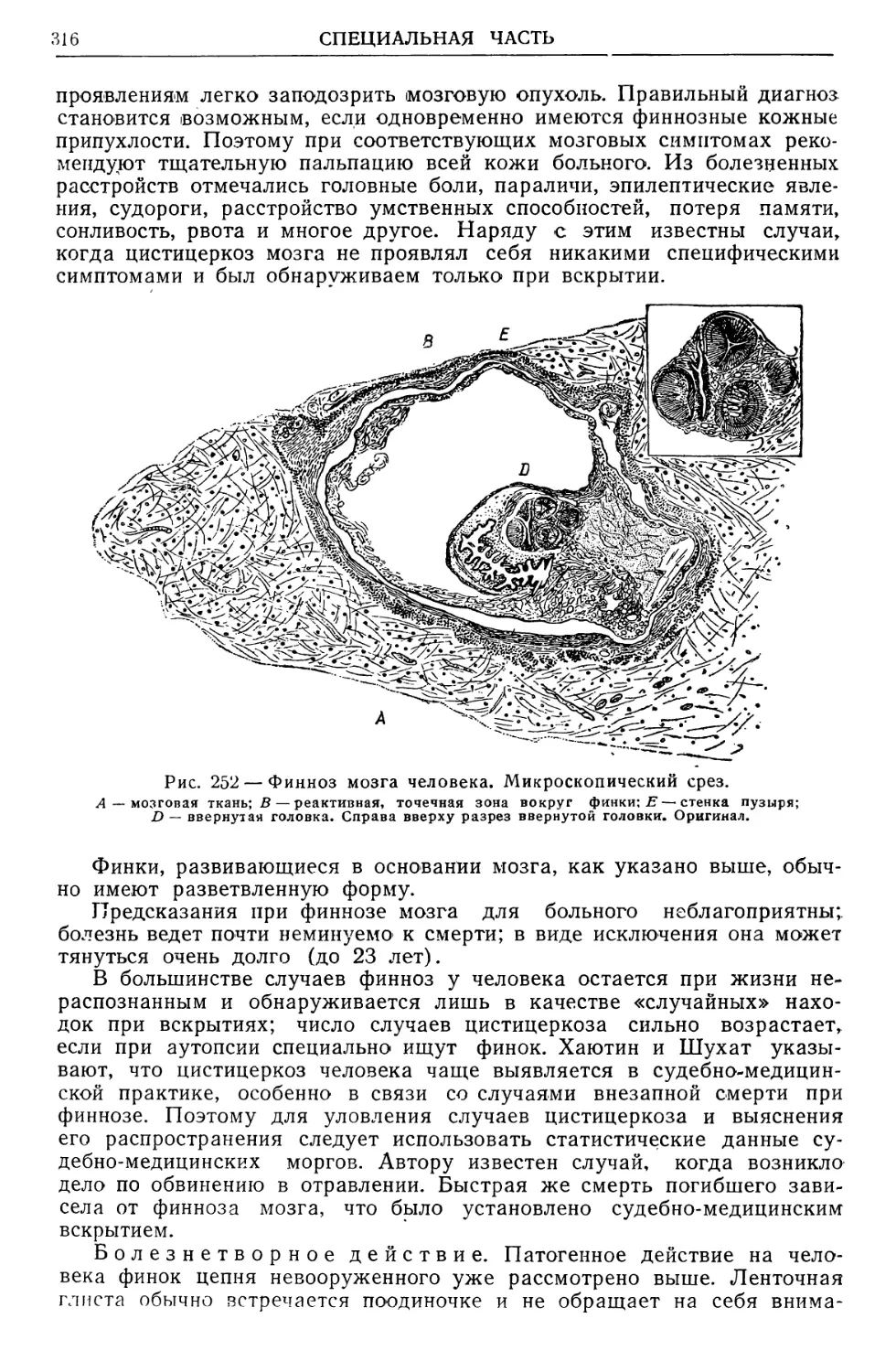
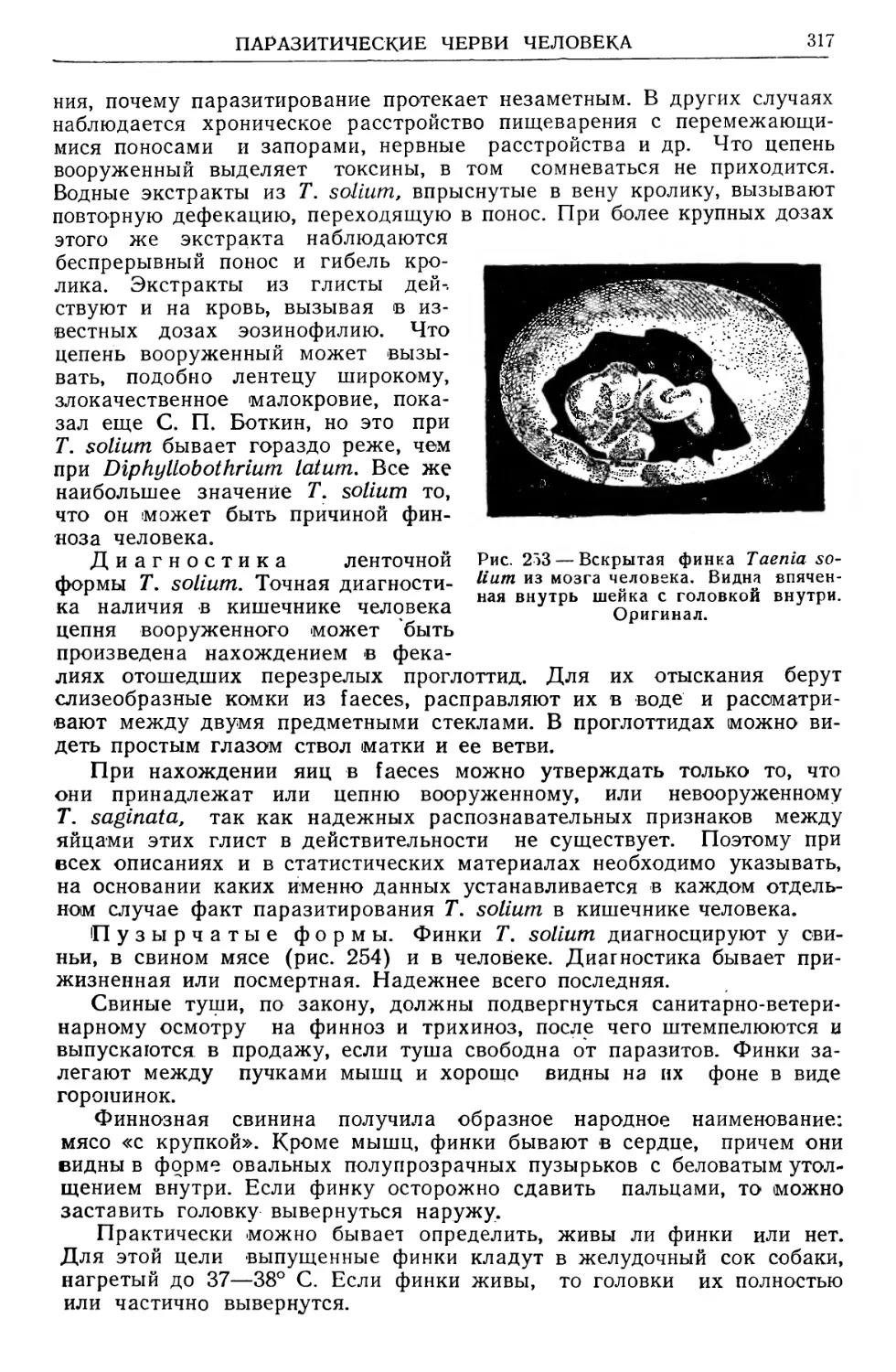
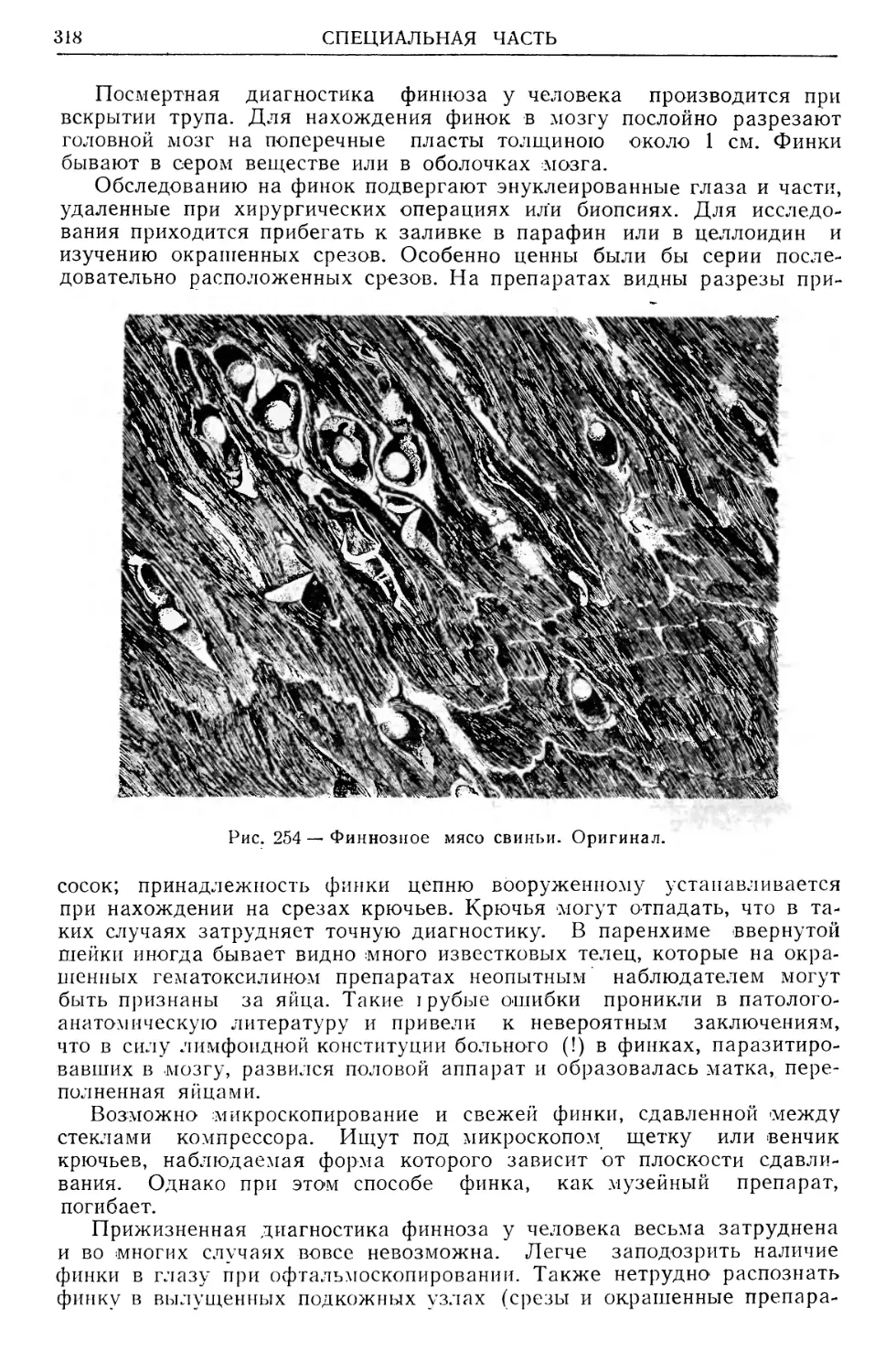


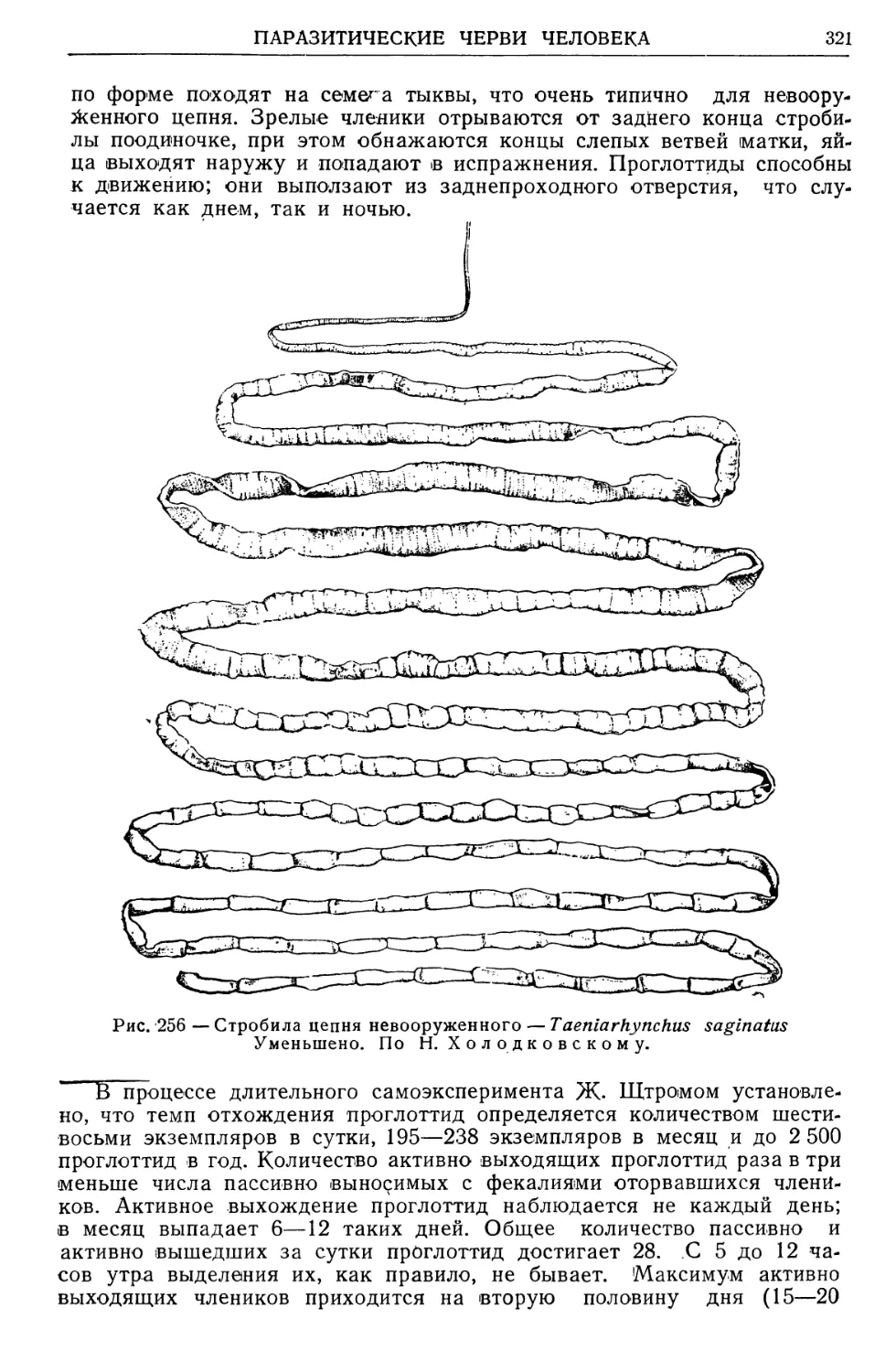
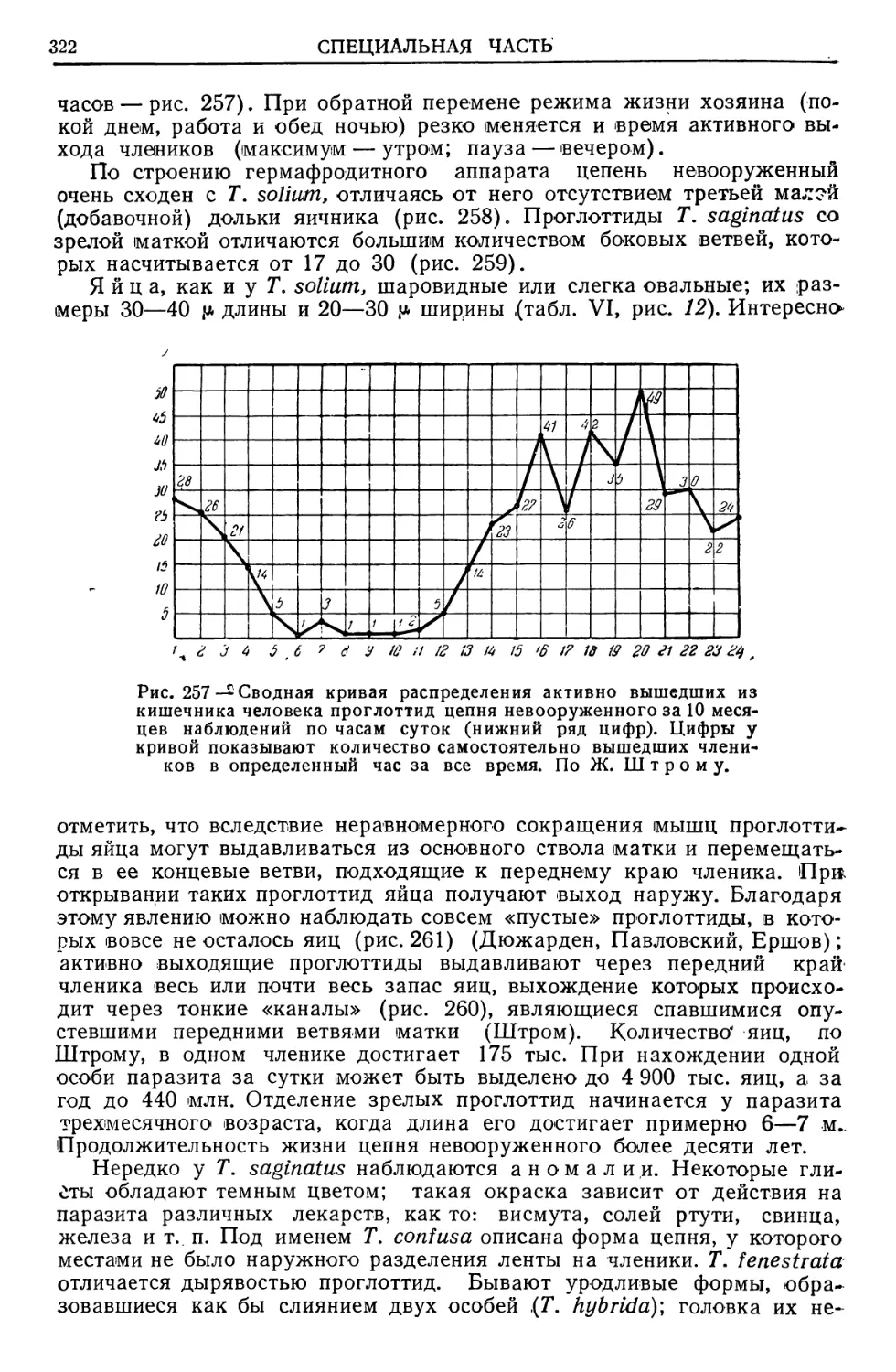
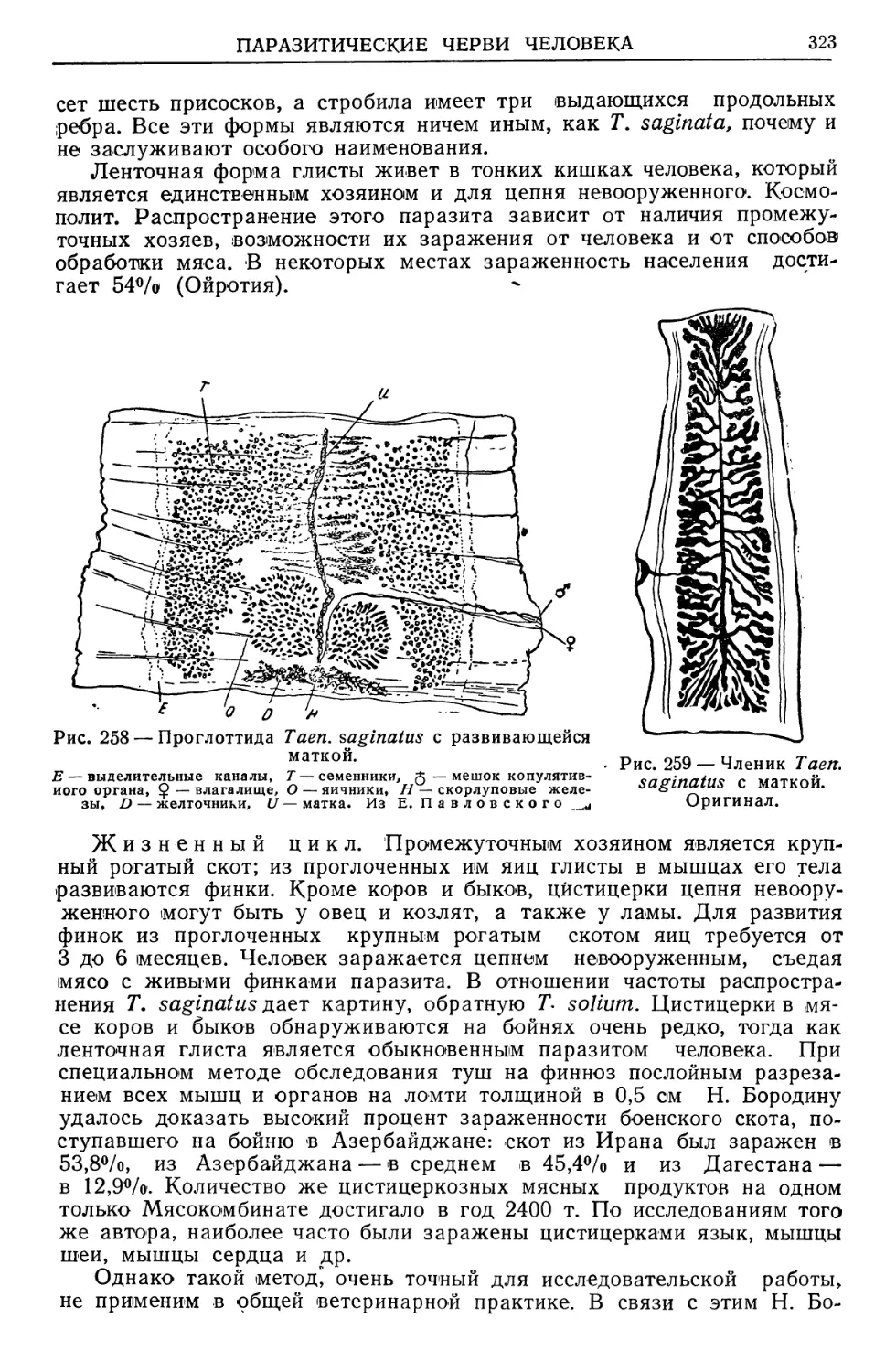
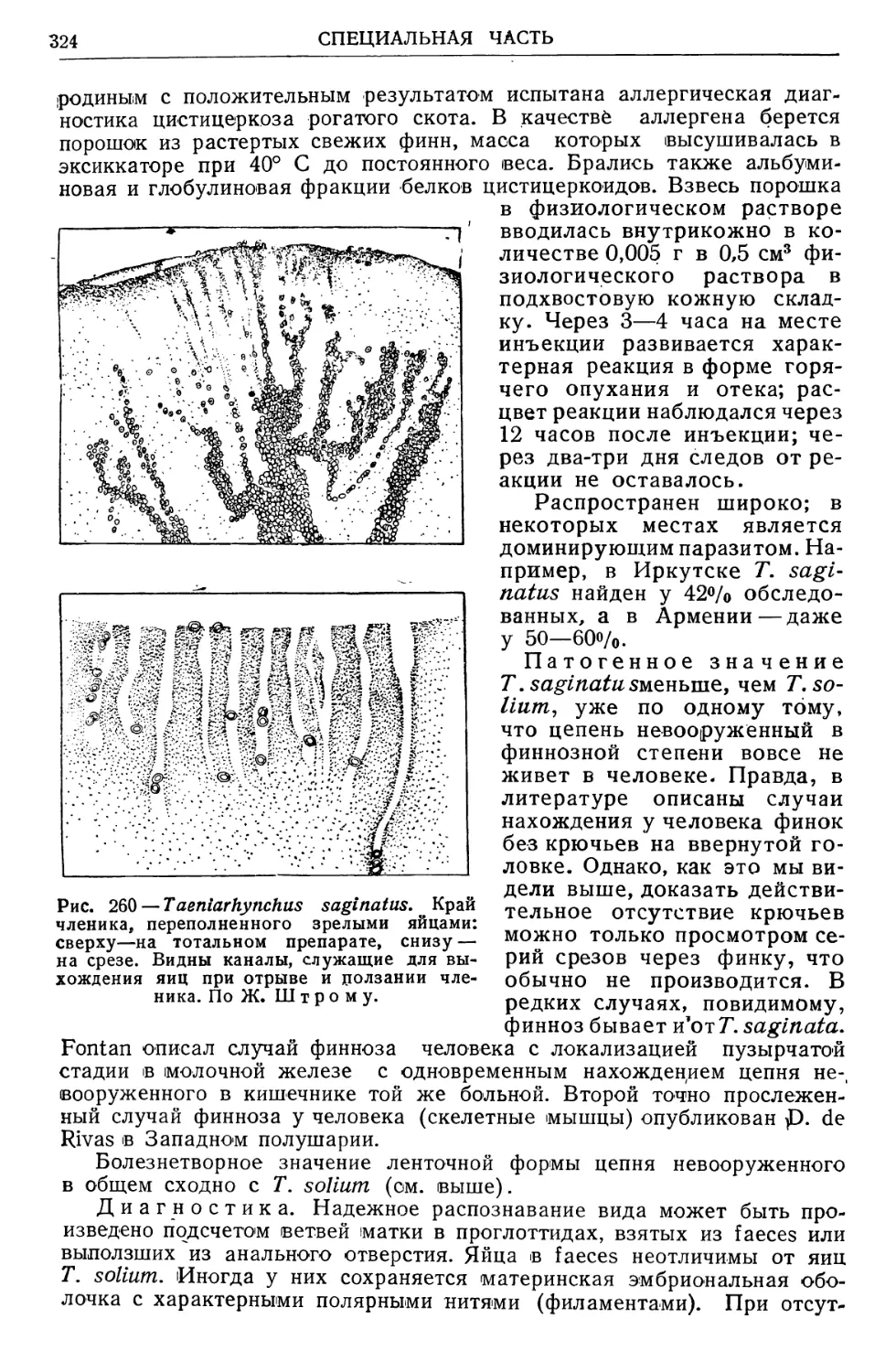
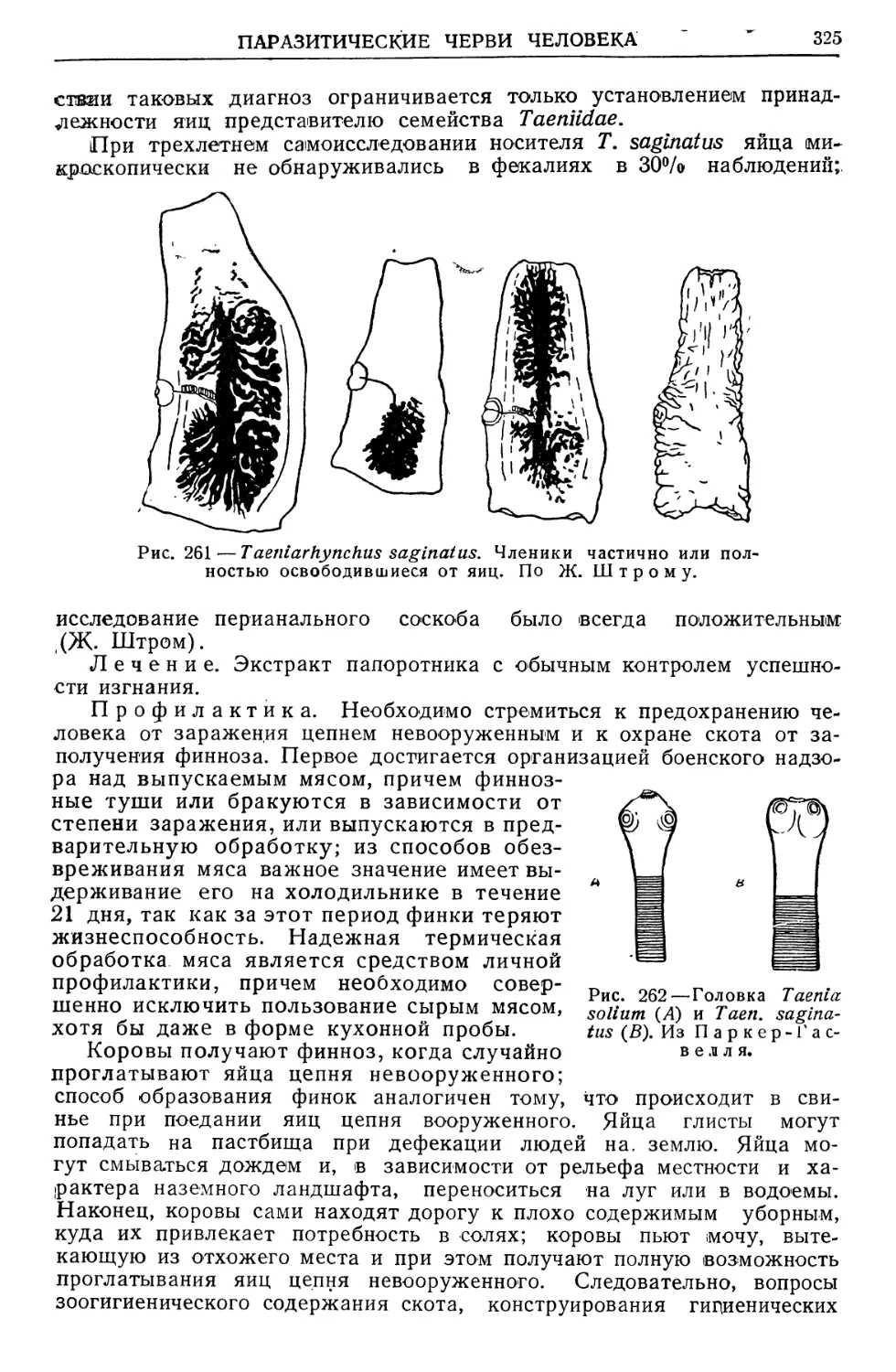














































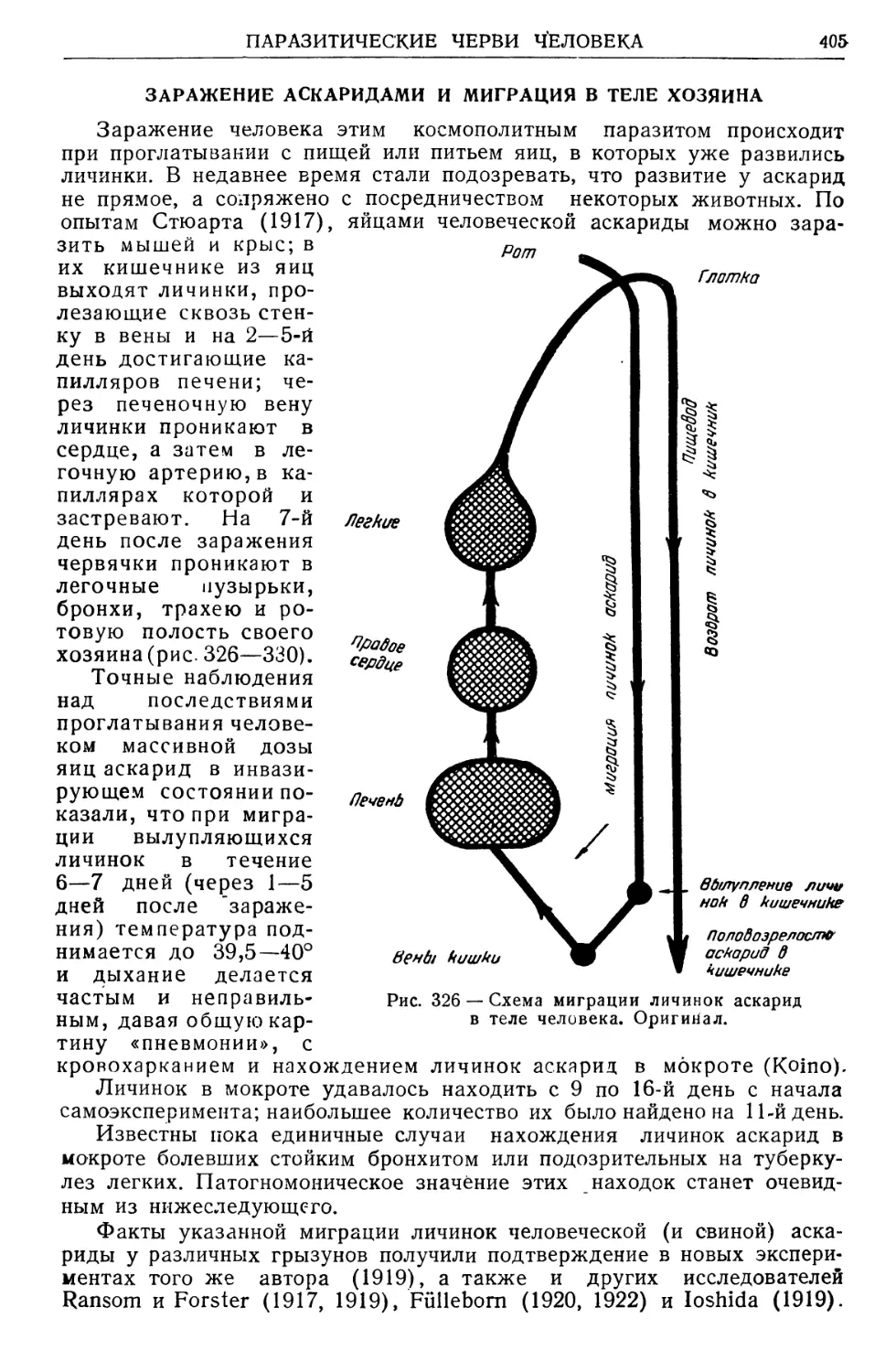
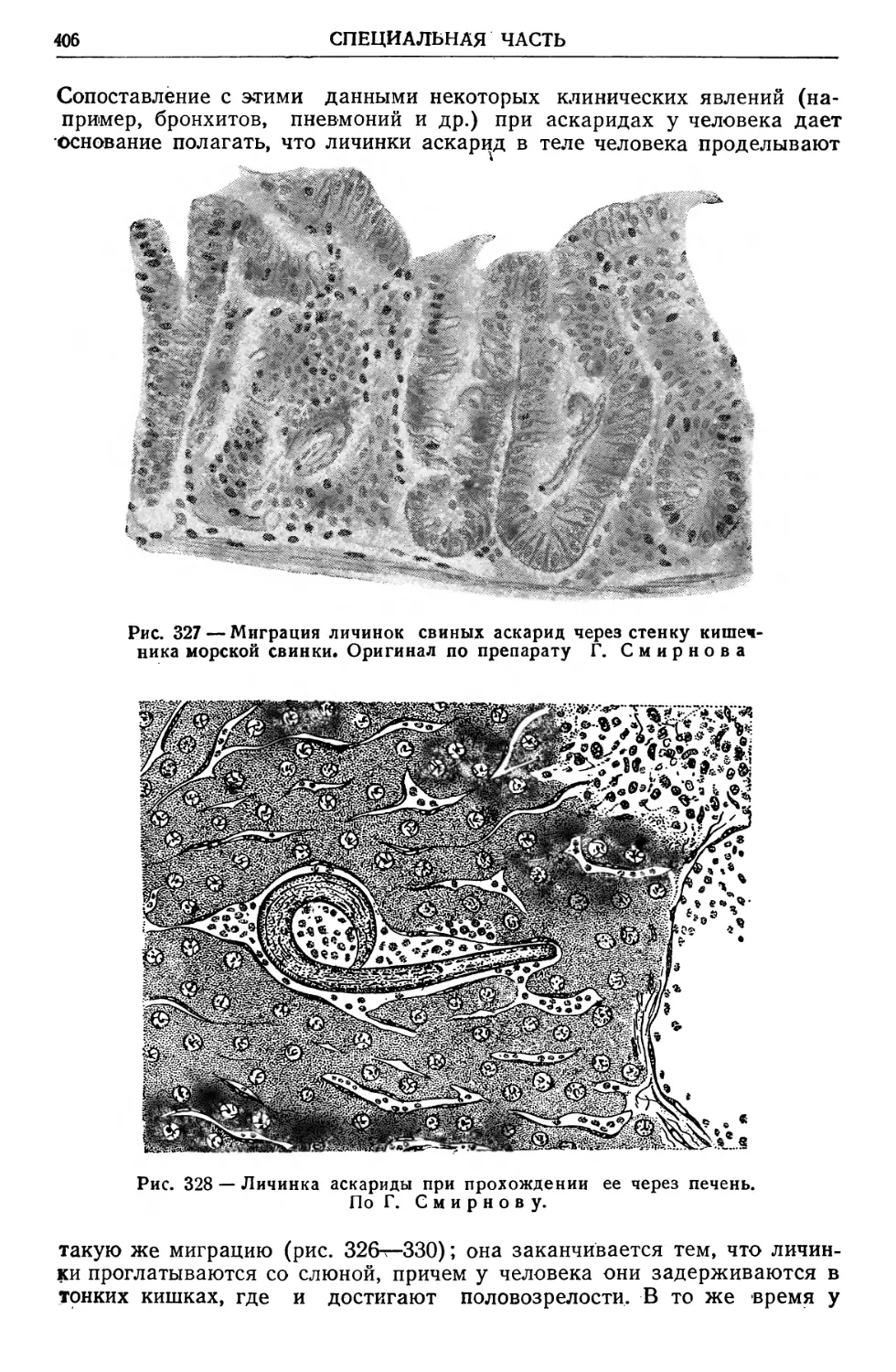
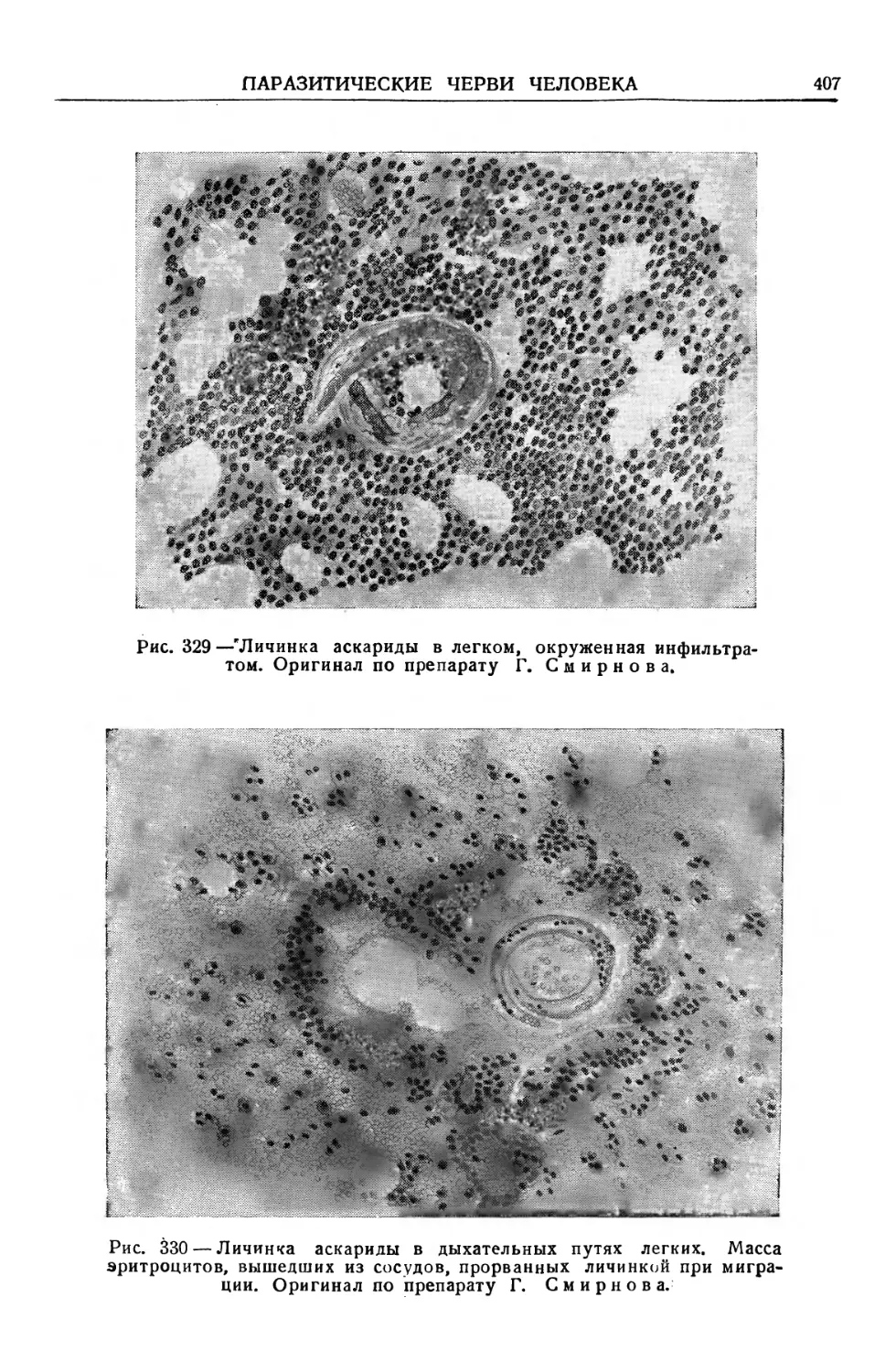


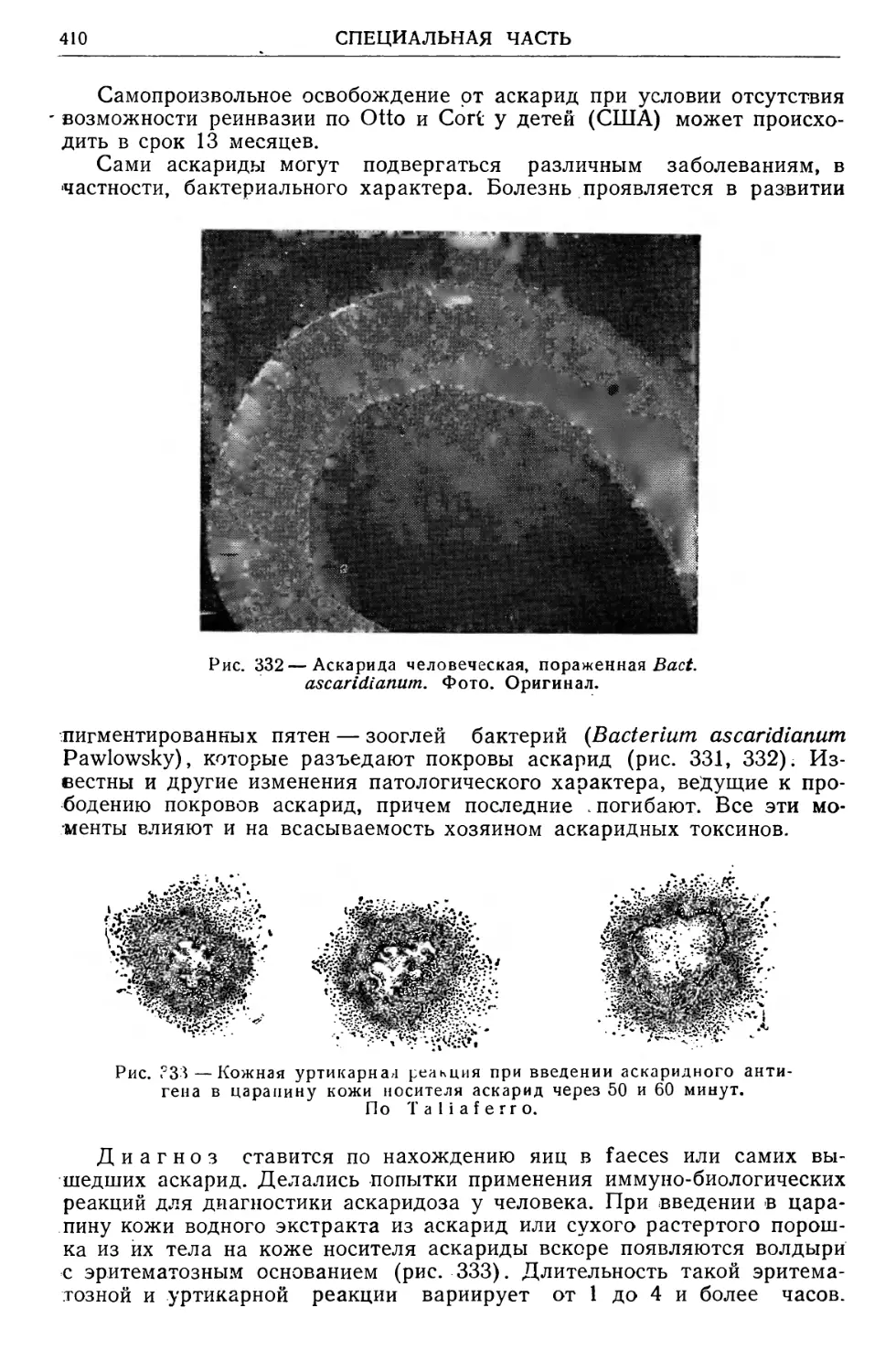






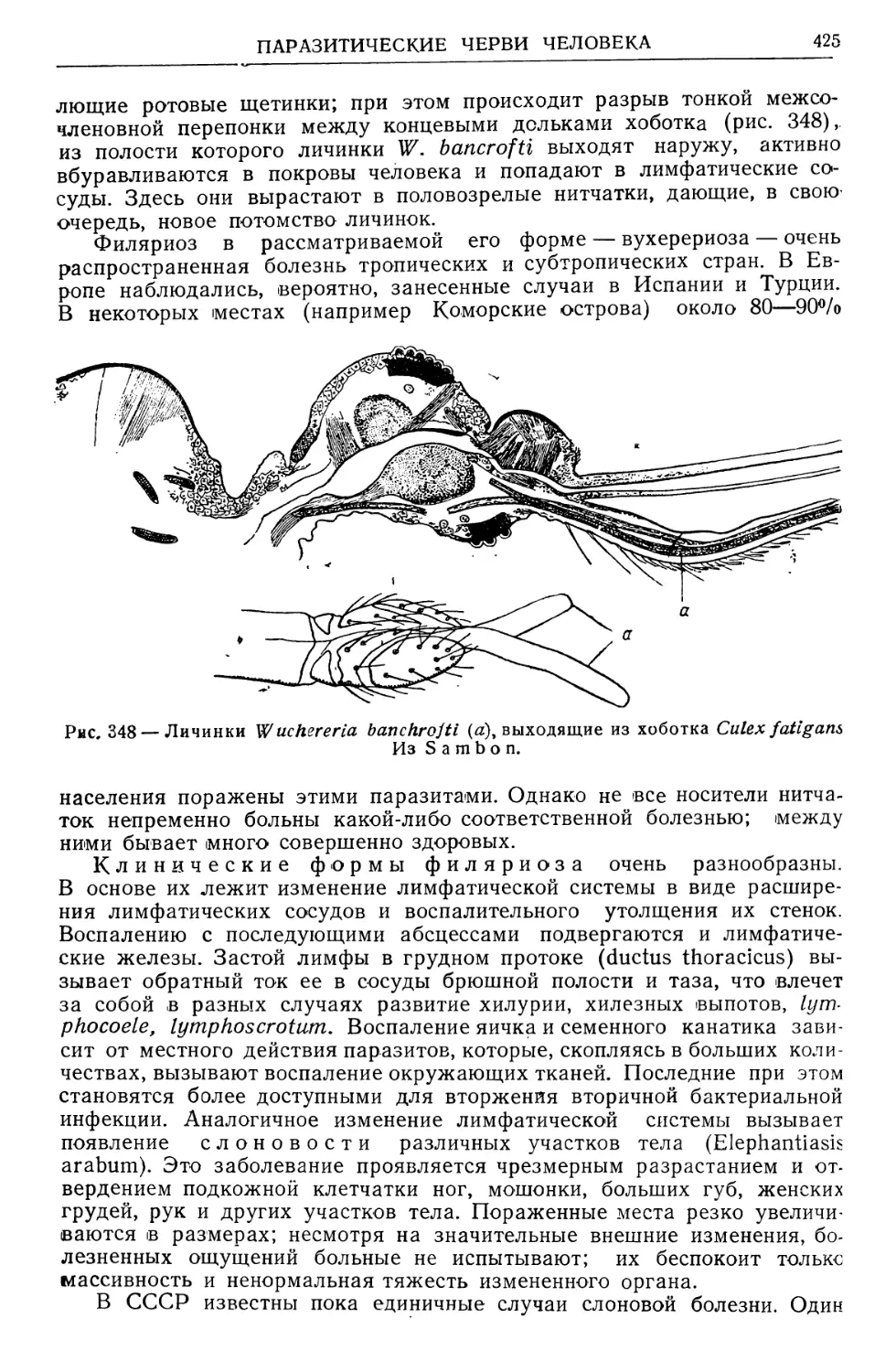

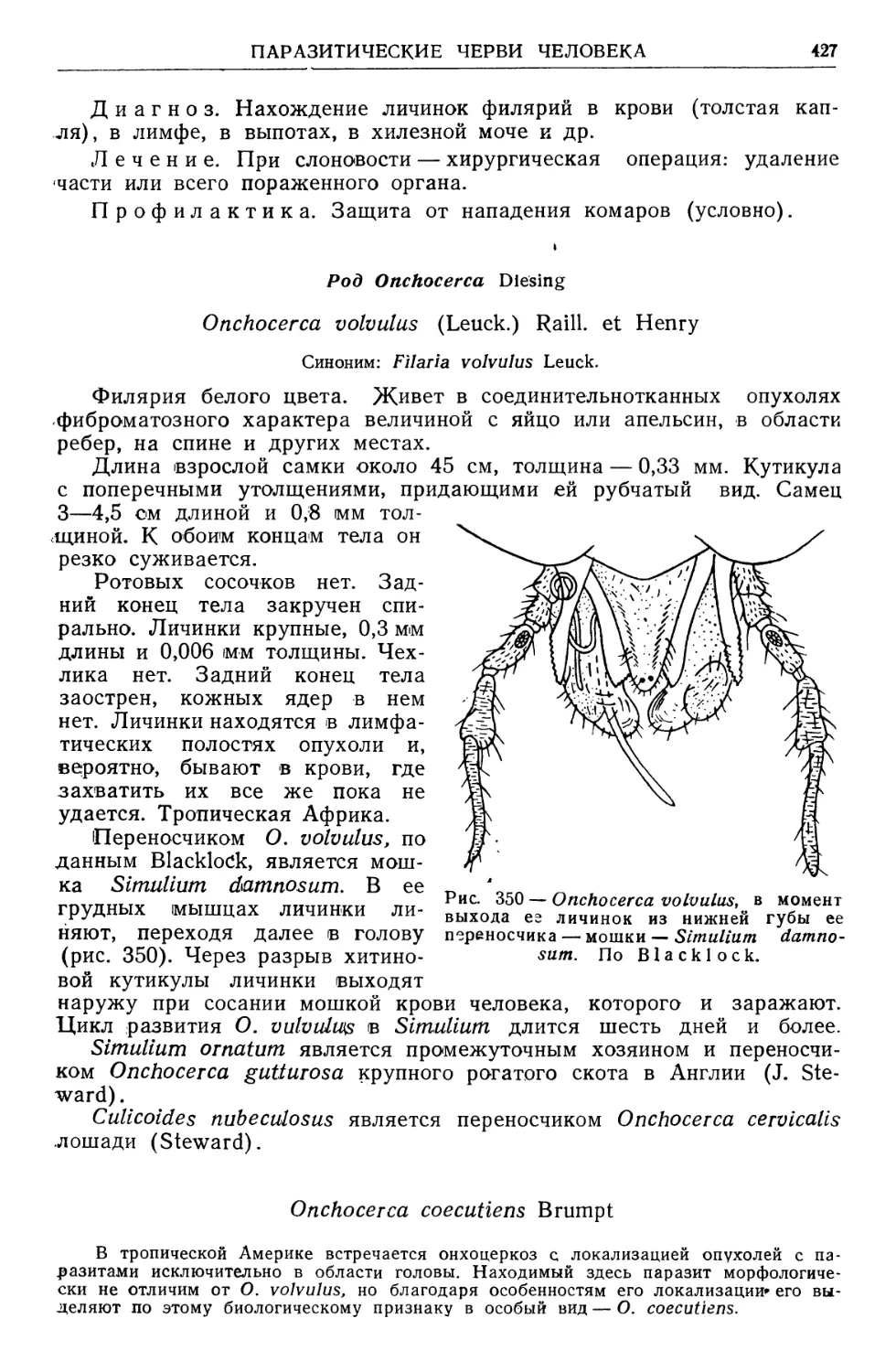

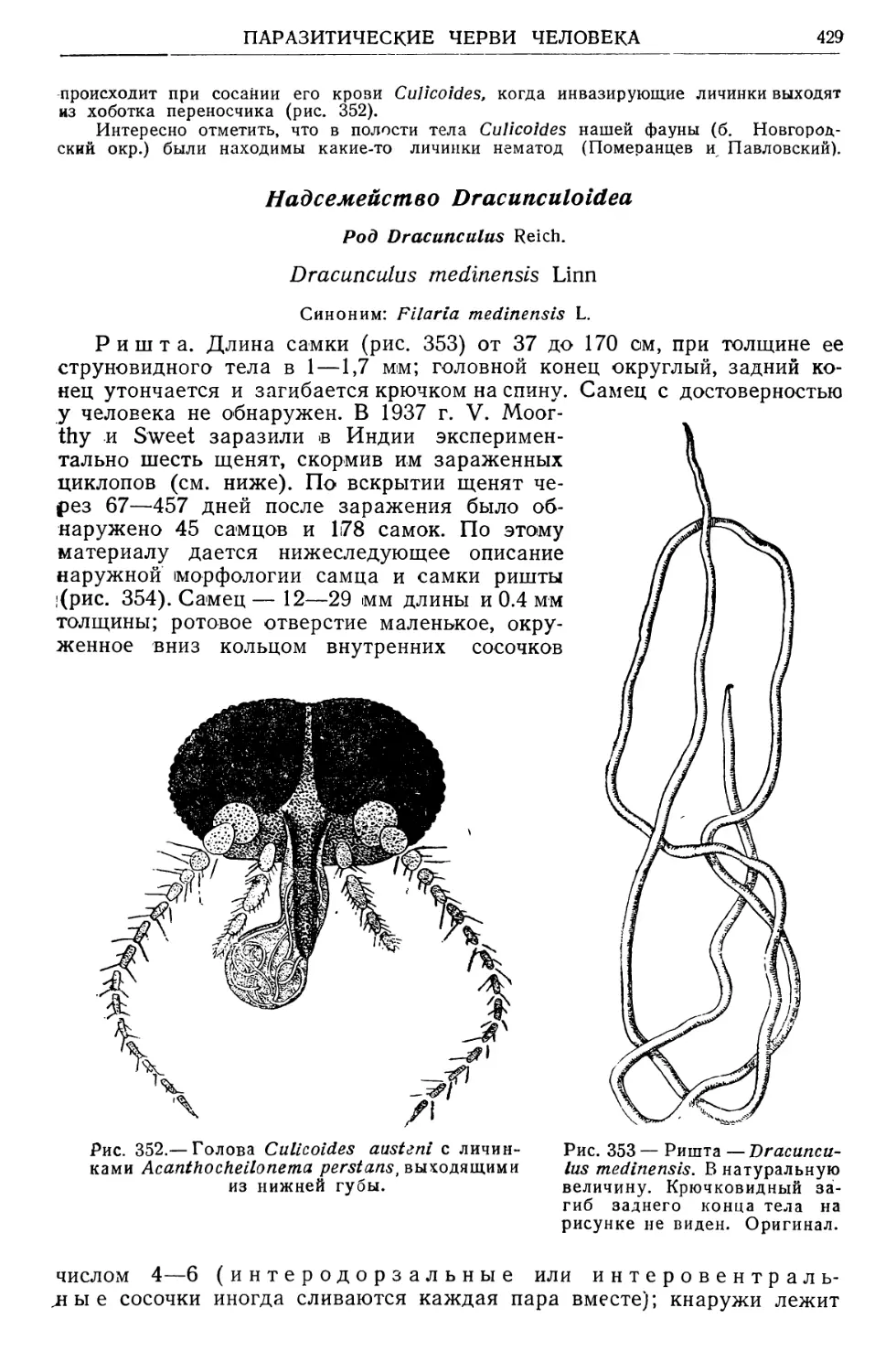
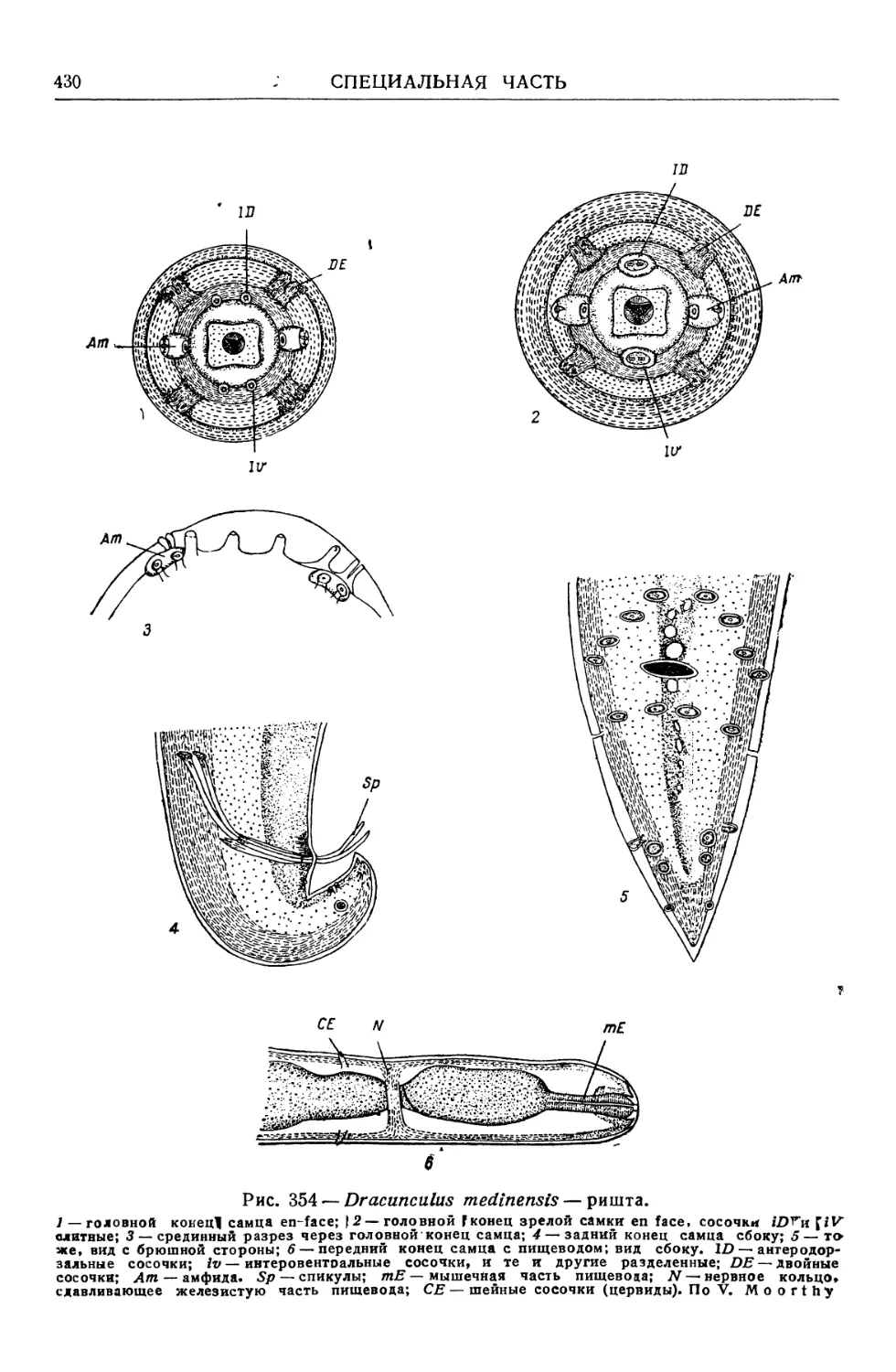


























































































Text
E.H. t
ПАВЛОВСКИМ
,*>*е
руководство
по
’АРАЗИТОЛОГШ
ЧЕЛОВЕКА
.Академик
Е.Н. ПАВЛОВСКИЙ
руководство
ПО ПАРАЗИТОЛОГИИ
ЧЕЛОВЕКА
ДАТЕ Л ОСТ В«.
иемии илу'
СССР
I
АКАДЕМИЯ НАУК СОЮЗА ССР
Академик Е. Н. ПАВЛОВСКИЙ
Действительный член Академии Медицинских Наук СССР,
Генерал-лейтенант медицинской службы,
Лауреат Сталинской премии
РУКОВОДСТВО
ПО ПАРАЗИТОЛОГИИ
ЧЕЛОВЕКА
С УЧЕНИЕМ О ПЕРЕНОСЧИКАХ
ТРАНСМИССИВНЫХ БОЛЕЗНЕЙ
ТОМ I
Пятое издание, переработанное
и значительно расширенное,
с 374 рис. в тексте
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА-1946 —ЛЕНИНГРАД
ПОСВЯЩАЕТСЯ
моим ученикам и сотрудникам, внес-
шим энергичной, самоотверженной, порой
опасной для жизни работой в экспеди-
циях, в лабораториях и преподаванием
богатый вклад в развитие советской па-
разитологии и в познание природы на-
шей Социалистической Родины, особенно
ее далеких окраин, на пользу охраны
здоровья трудящихся.
ПРЕДИСЛОВИЕ К ПЯТОМУ ИЗДАНИЮ
Пятое издание «Курса паразитологии человека» закончено было
спустя шесть лет после сдачи в производство второго издания. Истек-
ший период знаменуется бурным ростом различных отраслей советской
паразитологии как в теоретическом, так и в прикладном отношениях.
Далеко вперед продвинулось изучение паразитологических особенно-
стей окраинных зон Советского Союза. Больше и больше ширится прак-
тика экспедиционных паразитологических исследований. Растет науч-
ная производительность различных научно-исследовательских парази-
тологических ячеек на периферии, углубляется научно-исследователь-
ская работа центральных паразитологических лабораторий, что находит
себе отражение выпуском» трудов, сборников, монографий. Больше и
больше продвигаются в практику научные достижения последних лет.
Только по одной группе отделов паразитологии в Зоологическом ин-
ституте Академии Наук СССР и в Таджикском филиале АН СССР^ во
Всесоюзном институте экспериментальной медицины имени А. М. Горь-
кого с филиалом в Сухуми, по Кафедре общей биологии и паразитоло-
гии Военно-медицинской академии РККА им. С. М. Кирова и по много-
численным экспедициям, осуществляемым нами по заданиям Нарком-
здрава СССР, Главного военно-санитарного управления наркомздравов
СССР и союзных республик, и в порядке выполнения плановых
работ перечисленных выше паразитологических ячеек, работающих
под общим руководством автора, можно отметить следующие крупные
достижения за истекший шестилетний период.
А. По переносчикам возбудителей вирусных, бактериальных и па-
разитарных болезней: а) открытие трех новых переносчиков клещевого
возвратного тифа и разработка методов клещевой профилактики;
б) установление новой для СССР болезни — марсельской сыпнотифоз-
ной лихорадки с подтверждением роли клеща Rhipicephalus sanguineus,
как переносчика ее возбудителя. Этим открытием впервые в советском
здравоохранении поставлен вопрос о диференциации «сборного» сып-
ного тифа на самостоятельные формы, что подтвердилось в следующий
же год установлением нового клеща-переносчика — Dermacentor nut-
talli для изучаемой еще другой формы сыпнотифозной клещевой лихо-
радки; в дальнейшем был открыт еще новый переносчик клещевого
сыпного тифа в Приморье — Haemaphysalis concinna\ этими открытиями
постулируется необходимость проведения клещевой профилактики соот-
ветствующих форм сыпного тифа; в) открытие факта спонтанной зара-
женности вирусом клещевого (весенне-летнего) энцефалита некоторых
пастбищных клещей, установление роли их, как специфических перенос-
чиков возбудителя этой также новой болезни, выяснение механизма
передачи вируса клещами, раскрытие путей циркуляции вируса в очаге
6
ПРЕДИСЛОВИЕ К ПЯТОМУ ИЗДАНИЮ
инфекции; обоснование природной очаговости клещевого энцефалита и
разработка клещевой профилактики, как основного звена в борьбе с
весенне-летним энцефалитом; г) открытие в СССР комаров—перенос-
чиков вируса японского энцефалита установлением спонтанной зара-
женности их и экспериментом; детальное изучение их экологии и эпи-
демиологического значения; д) детальное изучение роли многих видов
слепней и комаров, мухи-жигалки, различных клещей и других экто-
паразитов в передаче возбудителя туляремии и слепней в распростра-
нении возбудителей сибирской язвы; е) экспериментальное доказатель-
ство трансовариальной передачи вируса лихорадки паппатачи через два
поколения Phlebotomus pappatasii', детальное изучение фауны москитов
и особенно их экологии .(Средняя Азия, Крым) с широким охватом ста-
ций дикой природы и успешные опыты работы по борьбе с лихорадкой
паппатачи методами борьбы с москитами; ж) установление ряда новых
контаминирующих переносчиков возбудителей острых кишечных болез-
ней, обширное изучение синантропных мух в Таджикистане и проведе-
ние опыта борьбы с ними в крупном масштабе; з) работы по фауне,
учету, экологии, эпидемиологическому значению гнуса и методами
защиты от него человека (защитные отпугивающие сетки); и) теорети-
ческое обоснование и детальная разработка проблемы природной очаго-
вости трансмиссивных болезней человека в связи с разработкой крае-
вой паразитологии СССР и др.
Б. По протозойным заболеваниям: а) установление по долине Мурга-
ба (Туркмения) факта массовой зараженности песчанок (Rhombomys
opimus) паразитами пендинской язвы, что дает основание сводить эту
болезнь к зоонозам; блестяще проведенные изучение условий существо-
вания очагов и показательный опыт ликвидации заболеваемости пен-
динкой в природном ее очаге в зоне освояемой пустыни на Мургабе
(Туркмения) методом затравливания нор песчанок хлорпикрином и
другими способами; б) исключительные по объему исследования ано-
фелес на зараженность паразитами малярии в ее злостном очаге и в
очаге анофелизма без малярии; в) обширные сравнительные обследо-
вания населения на кишечных простейших в связи с эпидемиологией
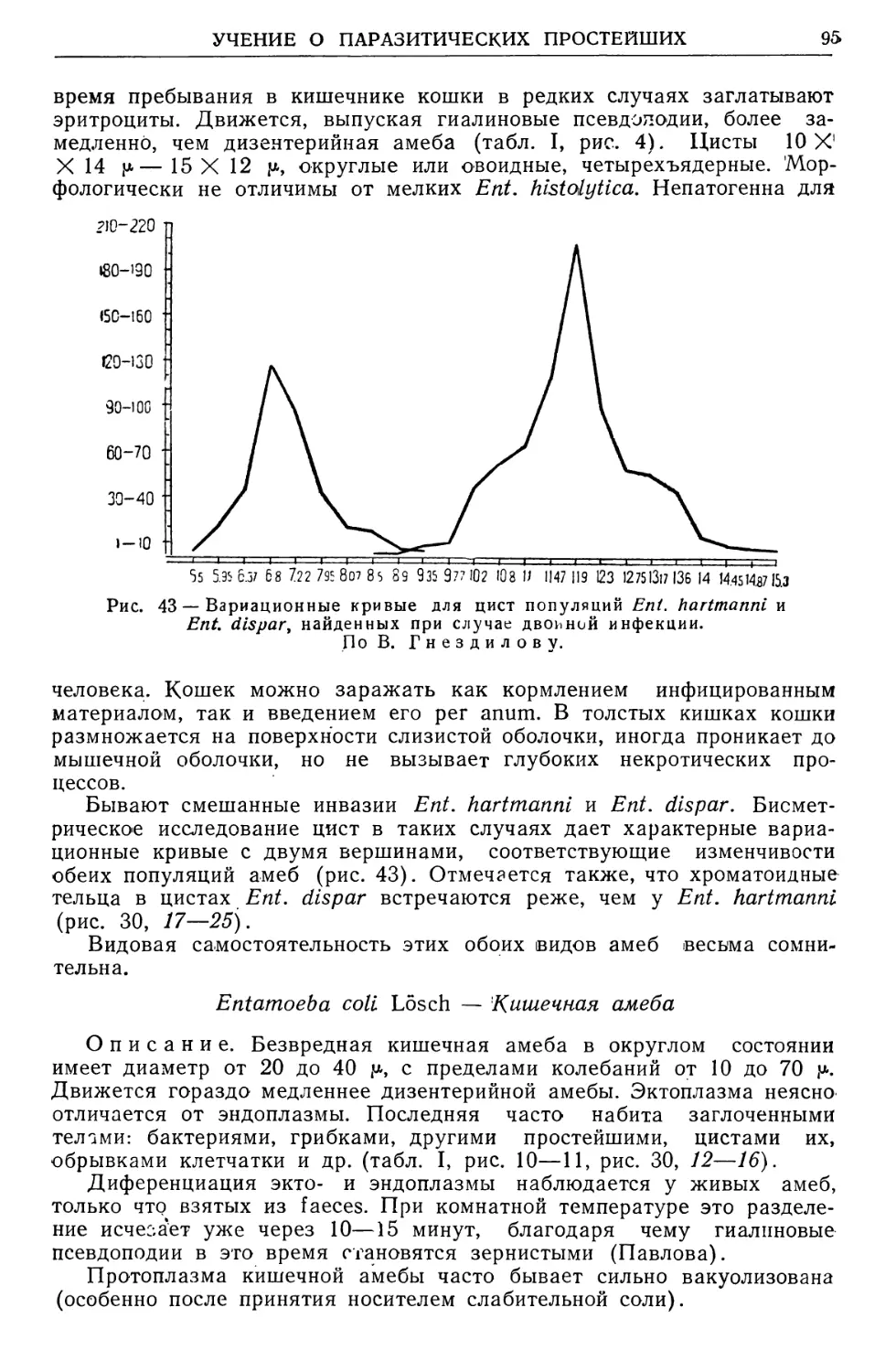
амебной дизентерии и установлением ее действительных очагов.
В. По гельминтологии — ряд экспериментальных исследований: а) по
патогенезу глистных инвазий; б) по токсичности паразитических чер-
вей; в) над факторами становления организма хозяином паразитов;
г) по раскрытию жизненного цикла байкальского лентеца; д) по борь-
бе с анкилостомидозами, тениаринхозом и дифиллоботриозом в соответ-
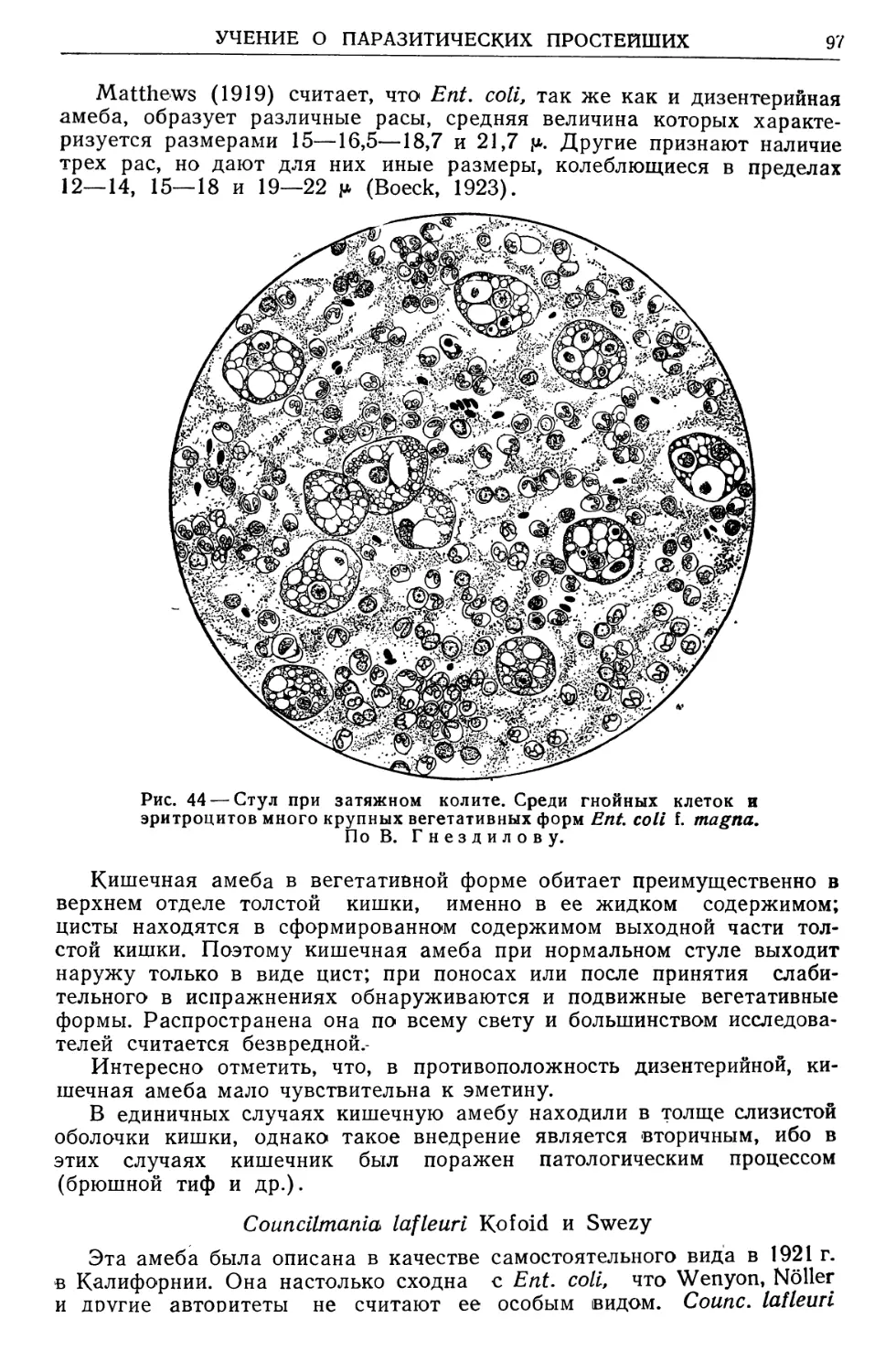
ствующих очагах; е) по биологии цепеня невооруженного; ж) по гео-
графическому распространению паразитических червей человека и др.
По докладу автора 14 апреля 1939 г. на пленуме Ученого медицин-
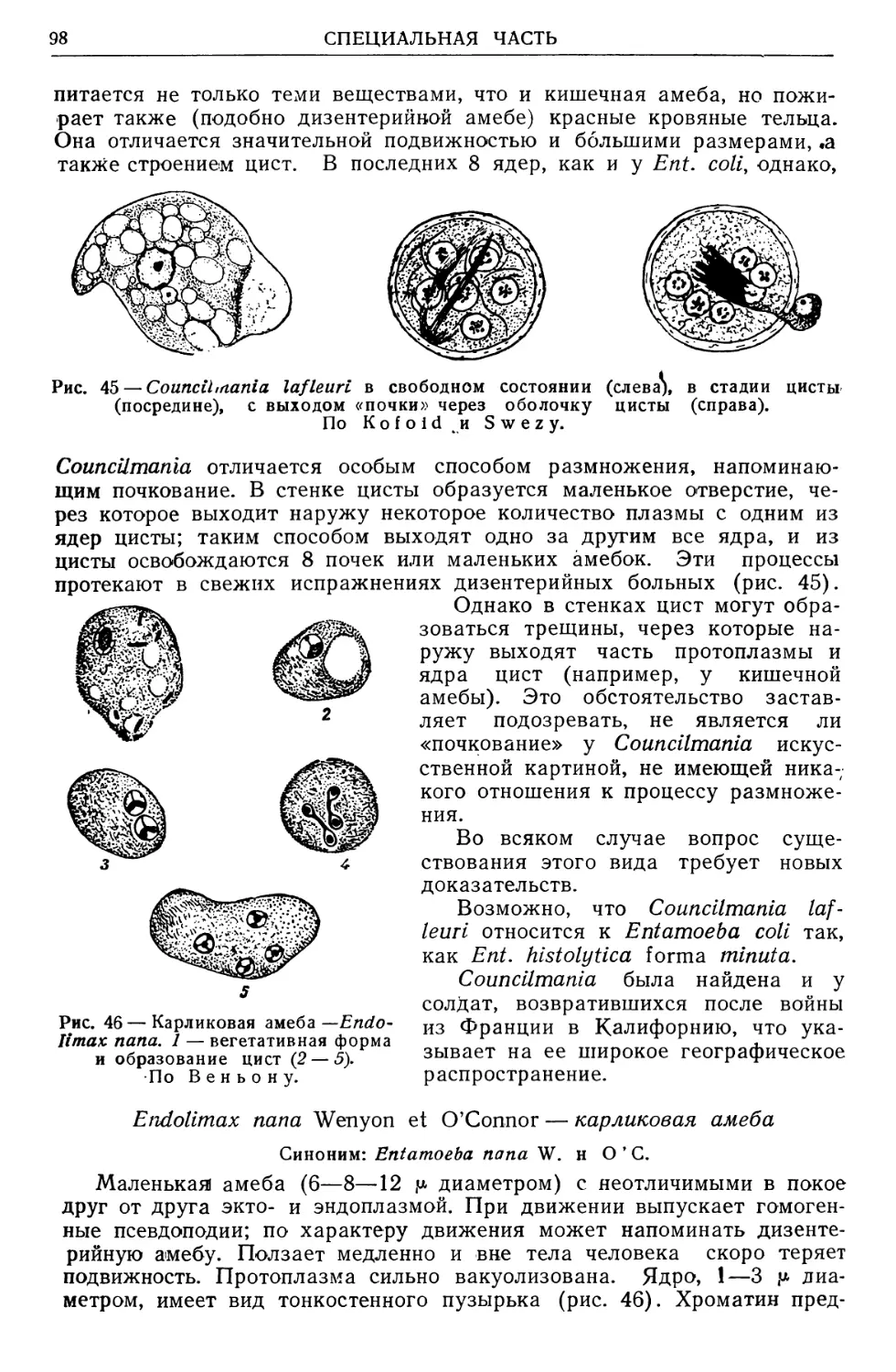
ского совета Наркомздрава СССР о «современном состоянии учения о
переносчиках инфекционных и паразитарных болезней применительно
к интересам здравоохранения в СССР» принято было постановление
об учреждении в мединститутах кафедр или доцентур по паразитоло-
гии и паразитарным болезням, что привело к утверждению Комитетом
по делам высшей школы при Совнаркоме СССР 6 кафедр для медин-
ститутов субтропических зон СССР и введение в курс общей биологии
обязательного раздела по даразитологии.
К этому следует добавить, что при указанной выше группе парази-
тологических отделов и лабораторий за 6 лет организовано около
60 паразитологических экспедиций и издано, включая 1939 г., 33 тома
ПРЕДИСЛОВИЕ К ПЯТОМУ ИЗДАНИЮ
7
трудов, сборников, учебников, пособий и книг по различным вопросам
паразитологии, не считая журнальных статей и участия в коллектив-
ных изданиях.
Все основные новости внесены в пересмотренный и расширенный
текст руководства; использованы также важнейшие опубликованные
данные из остальной советской и иностранной литературы. Общий план
книги оставлен прежним; значительно расширена общая часть ориги-
нальными по идее главами; добавлены новые главы по переносчикам
сыпнотифозных лихорадок, бактерийных инфекций, патогенезу, эпиде-
миологии и профилактике глистных инвазий, а также ссылки на па-
разитологическую литературу по . важнейшим вопросам. Заново пере-
смотрены иллюстрации; заменен ряд старых изображений и прибавле-
но много новых рисунков в тексте и цветные таблицы. Большая часть
новых рисунков оригинальна.
Как и во втором издании, автор ставил своей задачей изложить в
пропорциональном масштабе основные разделы паразито-
логии— протозоологию, гельминтологию и арахноэнтомологию с уче-
нием о переносчиках вирусных, риккетциозных, спирохетозных и бакте-
риальных инфекций и инвазий применительно к интересам советского
здравоохранения вообще и Красной Армии в частности.
Автор продолжает держаться точки зрения, что вопросам клиники
и терапии паразитарных болезней должен быть посвящен специальный
учебник; поэтому в предлагаемом курсе соответствующие материалы
носят подсобный характер.
Для полного изучения медицинской паразитологии необходимо
пользоваться также «практикумом медицинской паразитологии», со-
ставленным коллективом сотрудников автора под его редакцией (Био-
медгиз, 1935).
В окончательном виде книга является новым руководством, делаясь
одновременно пособием справочного характера для врача и для биоло-
га. Как и прежде, автор просит сообщать ему замеченные недостатки,
которые неизбежны в книге, оперирующей весьма разнообразным и
крупным по научной величине и практическому значению материалом.
. . Ленинград, 1.II. 1940
Кафедра общей биологии и пара-
зитологии ныне имени академика
Е. Н. Павловского Военно-меди-
цинской академии
им. С. М. Кирова
Заслуженный деятель науки, академик,
генерал-лейтенант медицинской службы,
лауреат Сталинской премии
Е. Н. Павловский
Руководство, подготовленное к печати к 1 февраля 1940 г., выпу-
скается в свет лишь в 1945 г. (на 27-м году преподавания этой дисци-
плины в Военно-медицинской академии имени Кирова) в первой своей
половине, посвященной общему введению, учению о паразитических
простейших их медицинской гельминтологии. Изменений текста по срав-
нению с рукописью 1940 г. сделать не было возможности за очень не-
многими исключениями. За этот период выпущено 3-е издание в виде
«Краткого курса биологии паразитов человека» (Ленинград, 1941) и
4 е издание — «Учебник паразитологии человека» для медвузов (Мед-
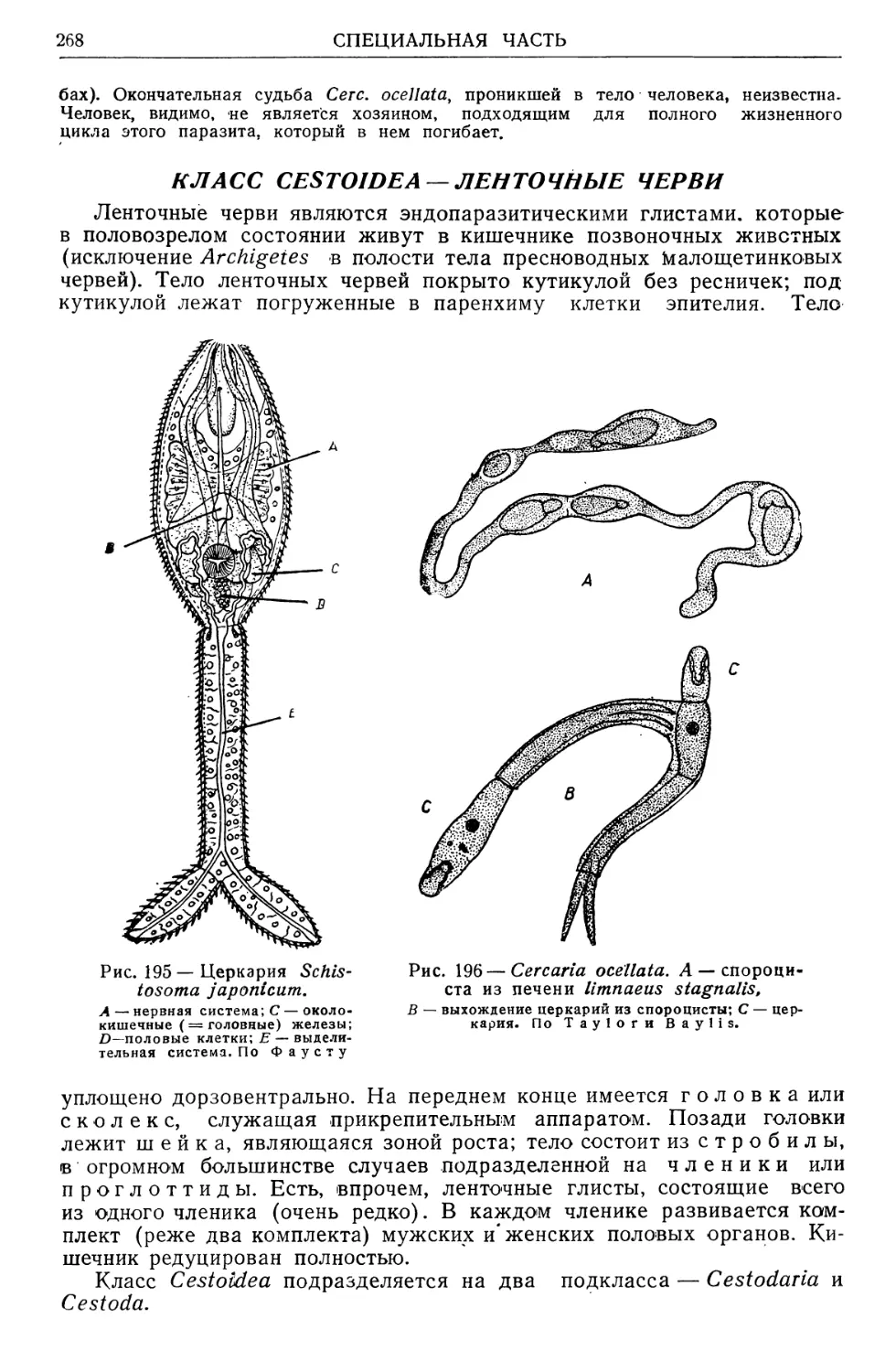
гиз). Часть, посвященная медицинской арахнологии, энтомологии и
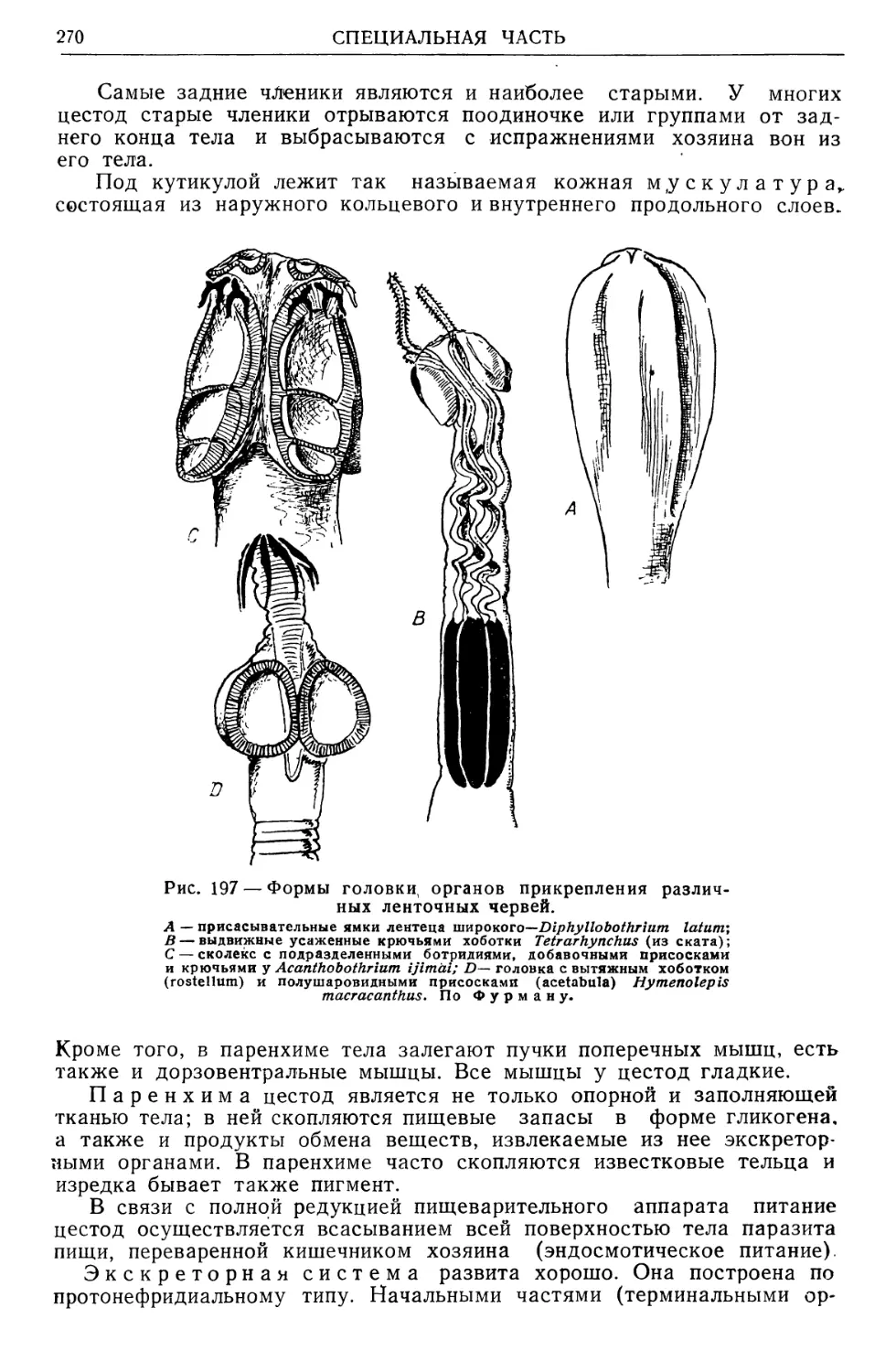
учению о переносчиках трансмиссивных болезней, выйдет вторым то-
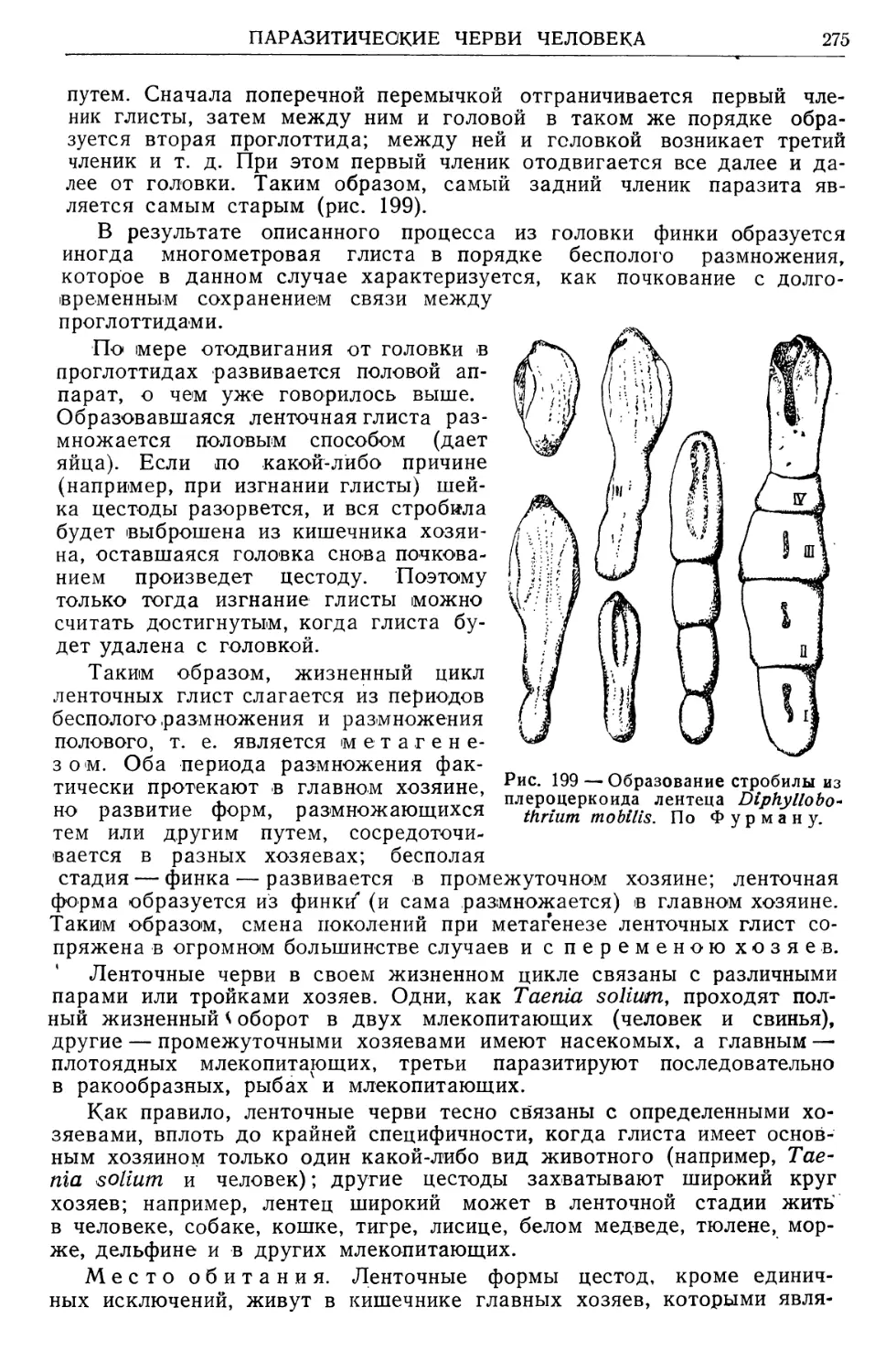
мом руководства.
Акад. Е. Н. Павловский
Москва 20.П. 1944
ИЗ ПРЕДИСЛОВИЯ АВТОРА КО ВТОРОМУ ИЗДАНИЮ
Составленные автором «Руководство к практической паразитологии
человека» (1924) и «Насекомые и заразные болезни человека» (1928)
давно уже разошлись; поэтому перед автором возникла задача пере-
издания этих пособий для Изучения паразитологии человека. За прошед-
шие годы паразитология дала громадный рост в СССР в связи с тре.
бованиями, которые были выдвинуты развитием советского здравоохра-
нения на профилактической базе (борьба с малярией; борьба с тропи-
ческими паразитарными болезнями, глистными инвазиями и др.). Науч-
но-исследовательская, научно-практическая и преподавательская работа
по кадрам требует ряда специальных пособий и, в первую очередь,
основной курс паразитологии человека. Пятнадцать лет практики пре-
подавания паразитологии в Военно-медицинской академии дают автору
основание предложить для широкого использования основной учебник,
обнимающий собою три отрасли паразитологии: протозоологию, гель-
минтологию и арахноэнтомологию с учением о переносчиках инфекций
и инвазий человека. В предлагаемом виде «Курс паразитологии» яв-
ляется совершенно переработанным и в своей большей части заново
составленным по сравнению с первым изданием руководства, 'что де-
лает этот курс фактически новой книгой, отвечающей по своему содер-
жанию теоретическому курсу. ,
ОБЩАЯ ЧАСТЬ
ОПРЕДЕЛЕНИЕ ПАРАЗИТИЗМА; ФОРМЫ ПАРАЗИТОВ
ВО ВРЕМЕНИ И ПРОСТРАНСТВЕ
Паразитами называют животных, которые живут
за счет особей другого вида, будучи биологически и
экологически тесно связаны с ними в своем жизнен-
ном цикле на большем или меньшем его протяжении.
Паразиты питаются соками тела, тканями или переваренной пищей
своих хозяев, причем такой паразитический образ жизни является
специфическим видовым признаком данного пара-
зита, многократно (в противоположность хищни-
кам) пользующегося для питания своим хозяином.
Кроме того, паразиты постоянно или временно используют организм
хозяина, как территорию своего обитания.
Подобные определения необходимы для того, чтобы выявить поня-
тие «л о ж н о п а р а з и т и з м» и отграничить ложнопаразитов от пара-
зитов истинных, а самих паразитов отличить от хищников.
Явление хищничества весьма широко распространено в природе.
Хищник нападает на добычу и ее уничтожает — кошка пожирает мышь,
волк терзает овцу. Добыча, следовательно, служит для однократного
питания хищника, причем используемое им животное погибает.
Паразиты же пользуются заражаемым ими животным или чело-
веком для повторного, многократного кормления, причем вовсе не
убивают или сразу не убивают своего хозяина, хотя последний может
погибнуть через более или менее длительный срок вследствие забо-
левания, вызванного паразитом.
Резкое разграничение явлений паразитизма и хищничества невоз-
можно, потому что в длинном ряде случаев паразитизм исторически
вылился из хищничества, и паразитизм по отношению к свобод-
ной жизни животных должен быть рассматриваем как явление вто-
ричное.
Трактовка того или другого животного как паразита или как хищ-
ника часто является относительной, потому что определение соответству-
ющего качества животного производится по отношению его к другим
животным. Когда пиявка нападает на тритона, высасывает всю его
кровь и тем его убивает, то, пиявку по отношению к тритону мы дол-
жны счесть за хищника; пиявка же, присасывающаяся к лошади, овце
или человеку, является для этих существ паразитом. Различие тут в
величине между тритоном и названным млекопитающим и объеме за-
ключающейся в них крови. Потеря всей крови имеет для тритона, ко-
нечно, иные последствия, чем высасывание нескольких кубических
«сантиметров крови одной пиявкой у человека.
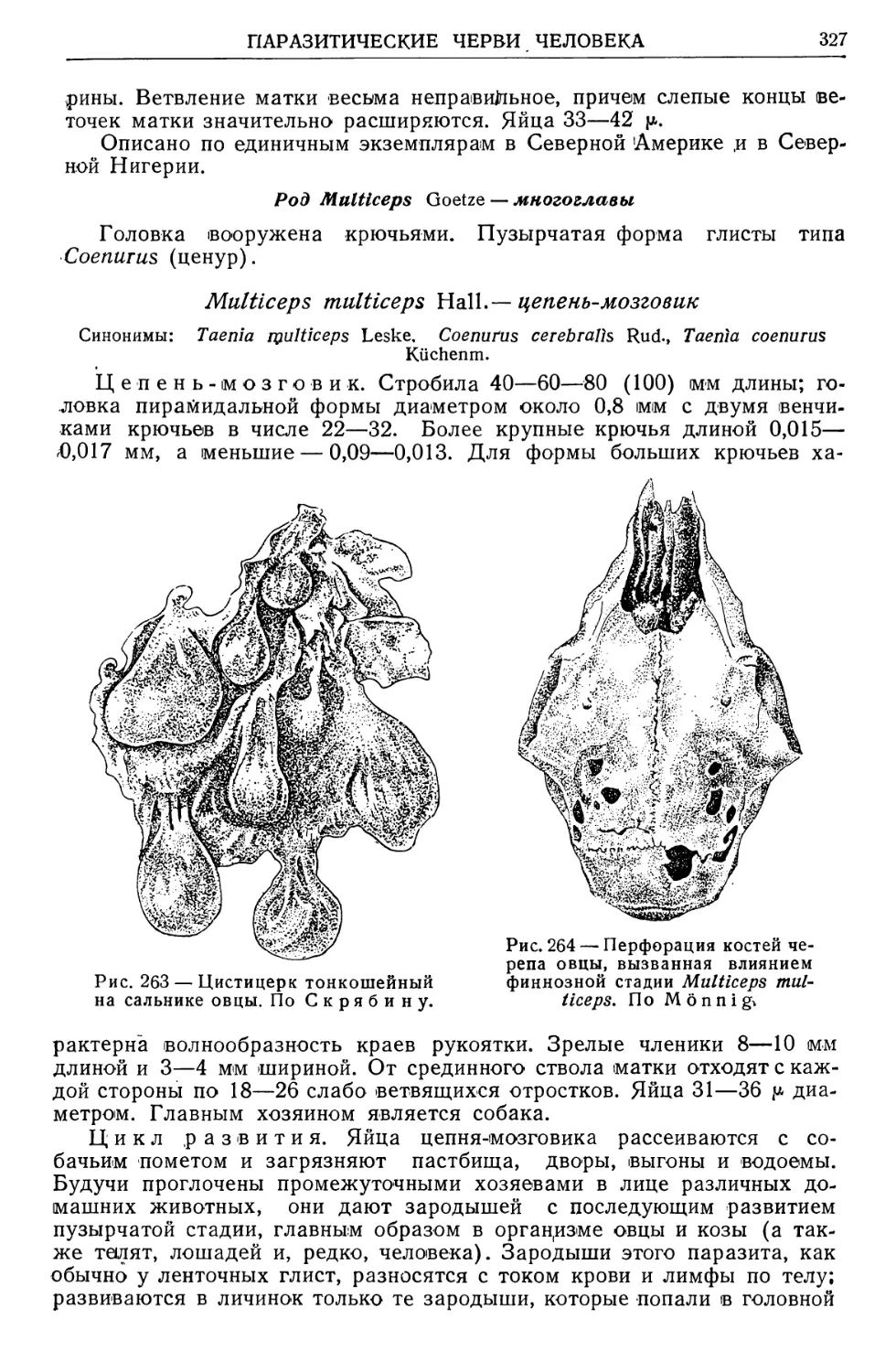
12
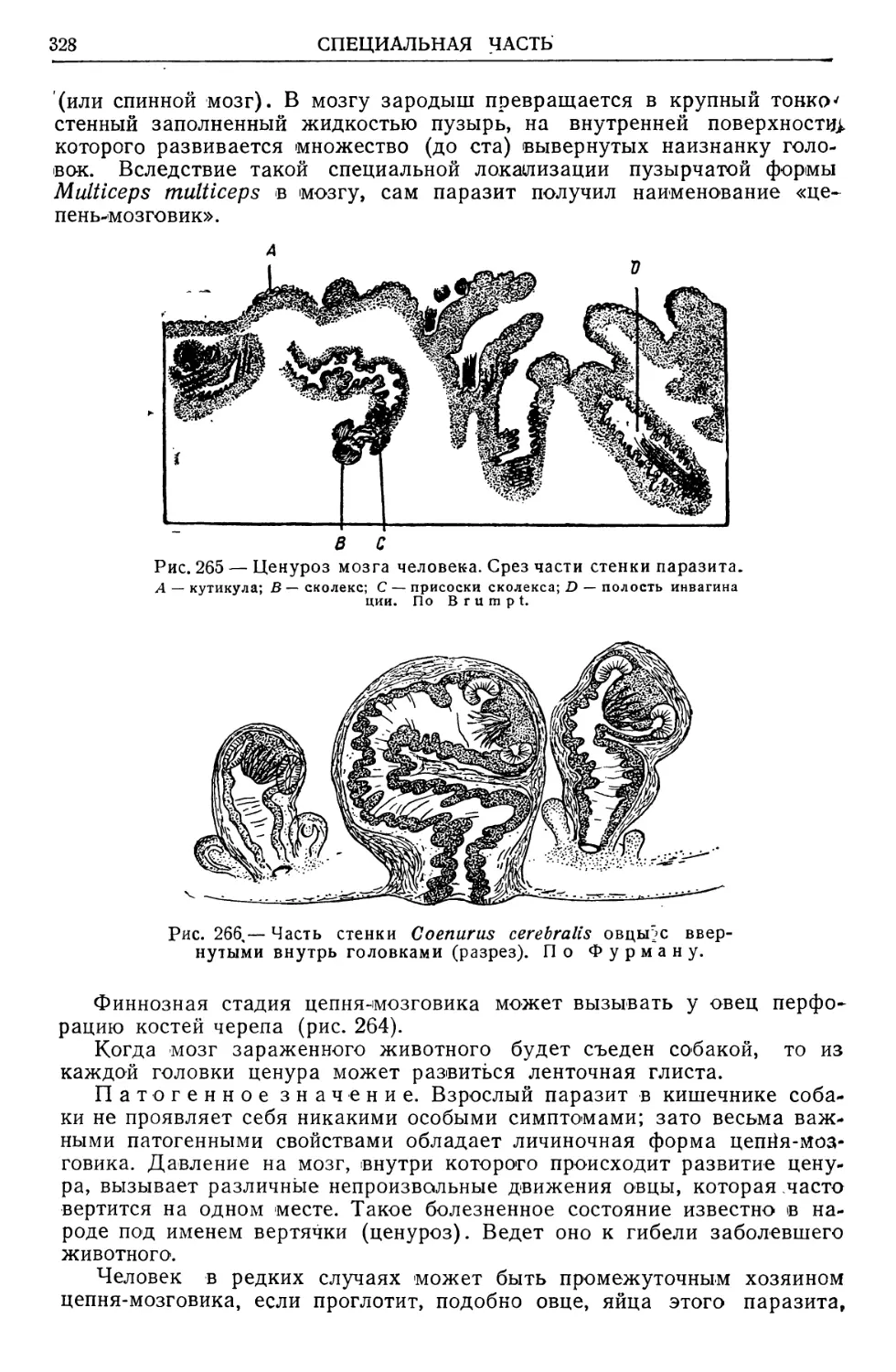
ОБЩАЯ ЧАСТЬ
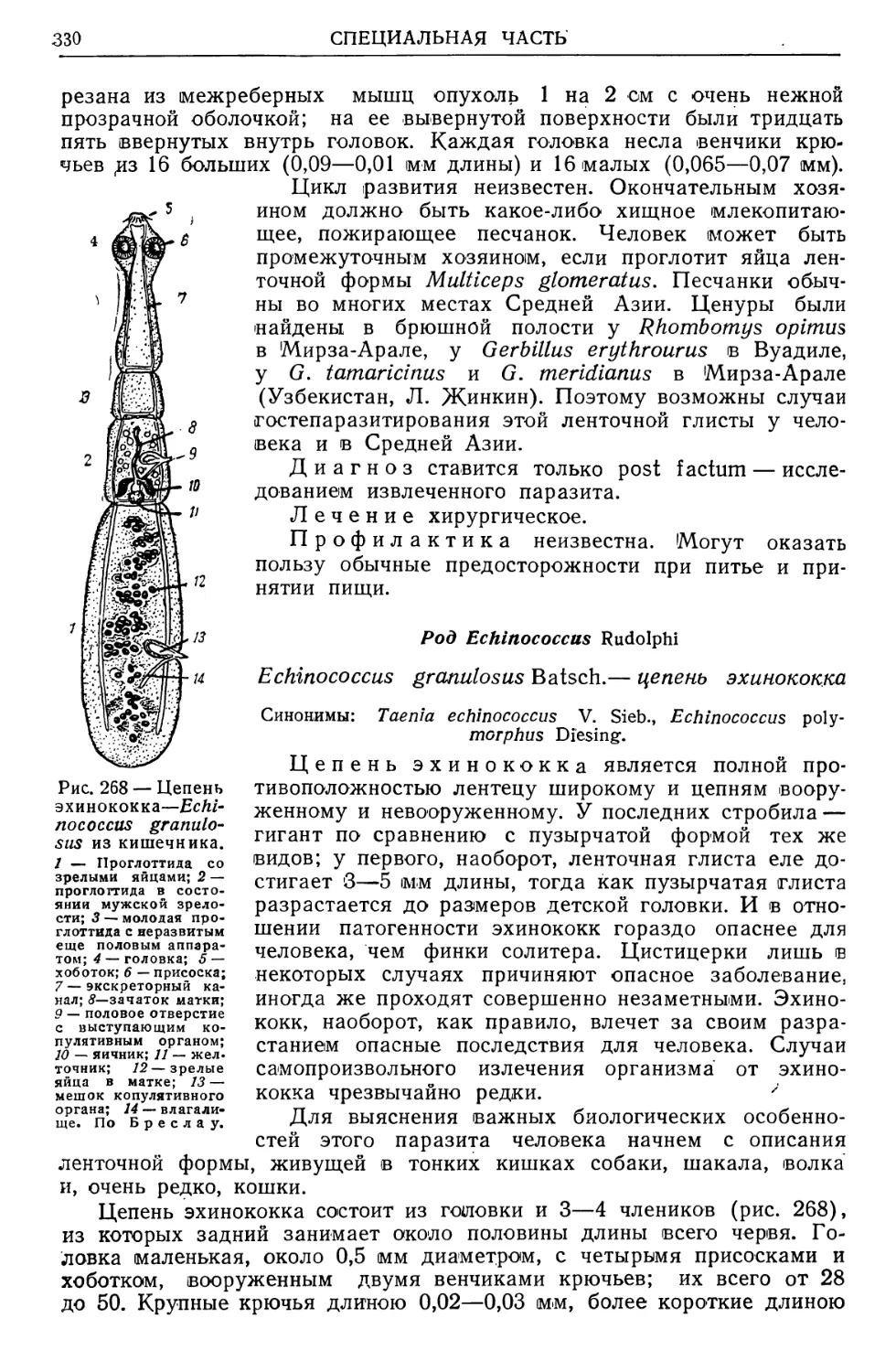
Однако соотношения паразита и хозяина ни в коем случае не ис-
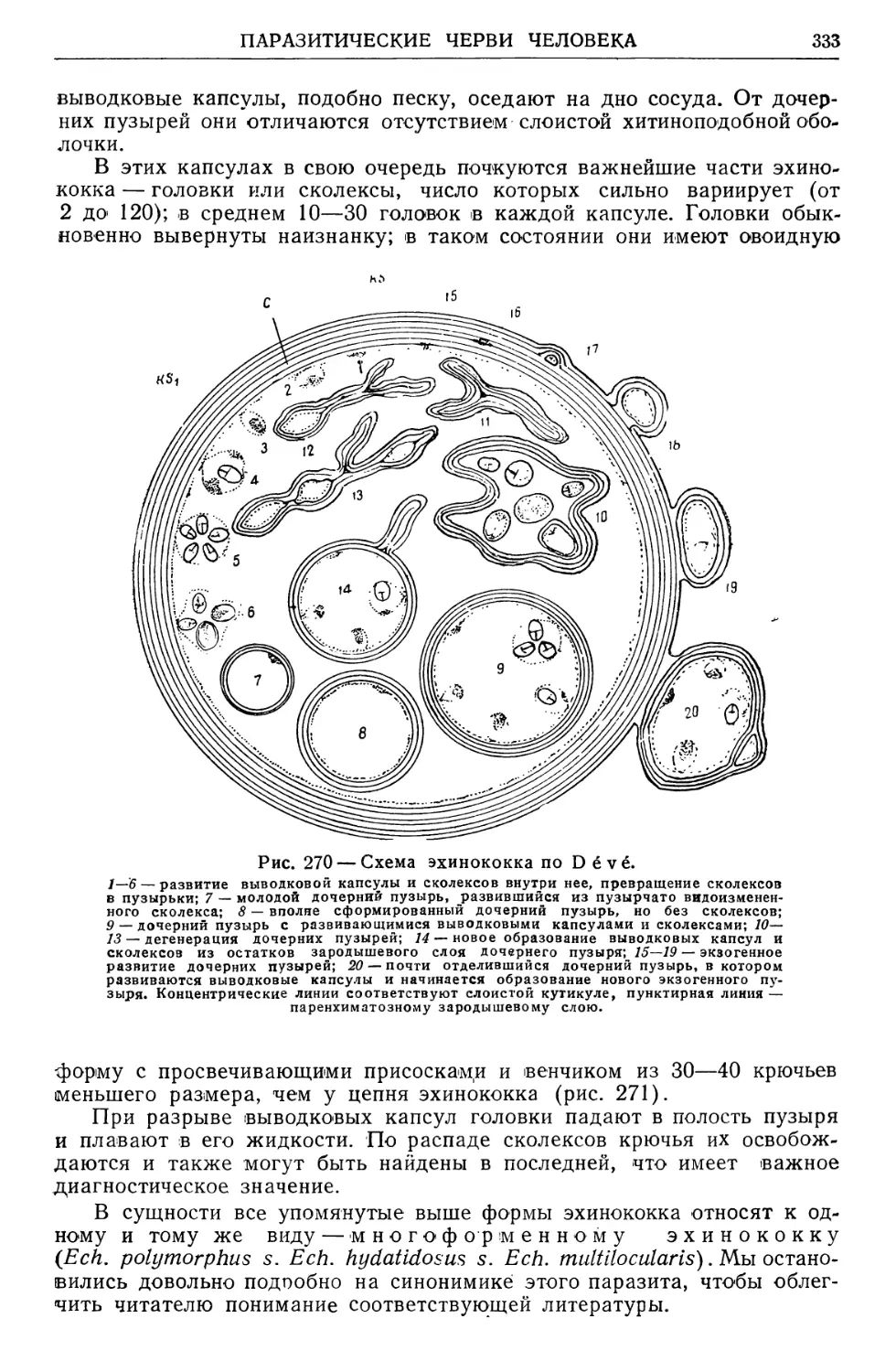
черпываются установлением такой механической пропорции между
величиной тела паразита и хозяина и количеством потребляемых пер-
вым тканей последнего. Паразиты проявляют также специальные воз-
действия на хозяина введением в него своих соков или иных веществ
своего тела (слюна, экскременты, токсические начала и др.). Резуль-
тат действия паразита на хозяина определяется также качеством этих
веществ, количеством вводимой их массы, чувствительностью и инди-
видуальной реакцией данной особи соответствующего вида хозяина к
поступающим из паразита в ее организм веществам.
/-^Переходя к разграничению понятий паразит и ложнопаразит, отме-
тим, что Л-Ожнопа р а зитами являются такие ^свободноживушир
существа, которые в состоянии некоторое время жить внутри тела дру-
гого организма, например в кишечнике, если случайно туда попадут с
пищей, и вредить своему временному и случайному хозяину.
Список ложнопаразитов с каждым годом растет. Ими часто бывают,,
например, личинки различных мух — комнатной мухи — Musca domestic
са (живут в навозе и экскрементах), серой мясной мухи — Sarcophaga
carnaria (живут на падали), иловой мухи — Eristalis tenax (живут в
илу), сырной мухи — Piophila easel (живут в сыре, соленой рыбе) и
мн. др. Личинки мух обладают стойкостью к химическому влиянию
внешней среды и поэтому способны противостоять действию пищевари-
тельных соков кишечника, благодаря чему могут известное время жить
в столь необычайной для себя среде.
Ложнопаразиты могут быть и в других органах, кроме кишечника.
Таковы случаи тканевого миаза, или миазы ран, причиняемые личинка-
ми падальной мухи — Lucilia caesar, а также Sarcophaga carnaria и др.
В то же время нахождение в тканях живого человека личинок
вольфартовой мухи относится к явлениям истинного паразитизма,
потому что они не могут жить нигде, кроме как в живых тканях орга-
низма-- ~
СВ практическом отношении важно помнить, что ложнопаразиты мо-
гут быть истинными и мнимыми. И с т и н н ы е .ложно ща.р а з.и-^-ы\
действительно- бывают в организме и обнаруживаются в нем самом или
в его отбросах. 'Мнимыми ложнопа раз и т а м и являются вся-
кие животные, которые извне 'пПладаЮт'в отбросы тела человека или
животного, представляемые для исследования -врачу или натуралисту.
Если фекалии некоторое время лежат открытыми, то мухи могут отло-
жить на них яйца, из которых в тепле в короткий срок выходят личин-
ки. При запоздалом исследовании врач легко может принять вылупив-
шихся личинок за недавних обитателей кишечника пациента и припи-
сать им причину тех или других болезненных симптомов. Необходимо
также иметь в виду случаи, когда больные (чаще — истерией) умыш-
ленно подбрасывают различных животных в экскреты с целью ввести
врача в заблуждение. Поэтому ко всем случаям ложнопаразитизма г,
с коими приходится сталкиваться в жизни, необходимо относиться кри-
1 От ложнопаразитов следует отличать омеопара зитов, т. е. образования»
похожие на паразитов, но не являющиеся животными организмами. Эти образова-
ния часто симулируют по внешности паразитов и ошибочно принимаются за та-
ковых. Свертки слизи из кишечника могут напоминать по форме аскарид; раз-
личные растительные обрывки также имеют иногда внешнее сходство с червями;
распавшиеся элементы растительной ткани напоминают яйца или личинок глист
и т. д. Для избежания неправильного диагноза определение паразитов должно
производиться по присущим им систематическим признакам.
ОПРЕДЕЛЕНИЕ ПАРАЗИТИЗМА
13
тически. Еще большая строгость оценки находимых фактов нужна при
предполагаемом научном описании соответствующего материала.
То существо, которое кормит паразита, называется его хозяином.
Однйгпаразиты имеют очень широкий круг хозяев — таковы, например,
самки комаров, пьющие кровь различных млекопитающих и птиц,
клещ —! Ixodes ricinus, сосущий кровь не только названных животных,
но и ящериц. Такие паразиты называются полифагами, {э в р и ксе н-
н ы е_л_ар а з и т ы). Их. днтиподами являются паразиты, “живущие за
счет хозяина одного определённого вида или за счет весьма ограничен-
ного круга хозяев; этим свойством характеризуется, например, голов-
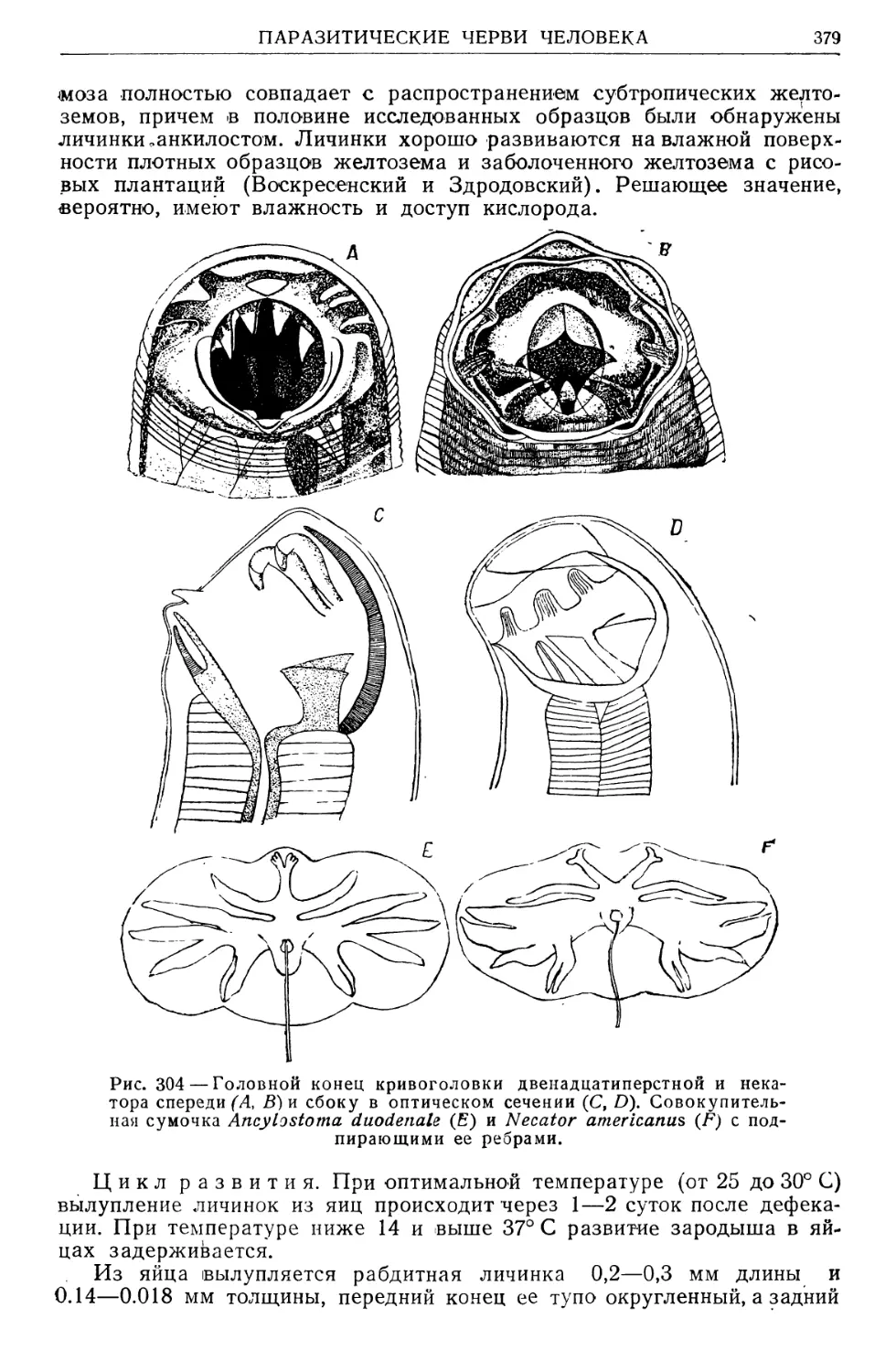
ная вошь — Pediculus capitis, Кривоголовка двенадцатиперстная —
Ancylostoma duodenale живет в двенадцатиперстной кишке только чело-
века; равным образом исключительно человеческим паразитом является
цепень вооруженный — Taenia solium во взрослом состоянии (моно-
кс е н н ы е _ц..а.р.а з и т ы).
' Определение круга хозяев, свойственных данному паразиту, связано
с долгими и многочисленными наблюдениями и, как таковое, может
оказаться^все^же. относительным.
z '^Однако приводимые категории паразитов, использующих хозяев од-
ного или многих видов, не могут быть резко разграничены друг от
друга, так как бывают примеры переходного характера. Например,
можно говорить о стеноксенных п аразитах, которые, имея
определенный вид хозяина, все же могут паразитировать и на других
хозяевах (например, чесоточный клещ лошади и человек). "Наконец
при массовых обследованиях хозяев и их паразитов бывают крайне
редкие или единичные случаи нахождения паразитов у совершенно не-
свойственных им хозяев (например, цестода жвачных животных Л1о-
nie^ia у человека).
/ Паразитов следует характеризовать также, исходя из свойственно-
сти их хозяину. Под таким углом зрения можно говорить о паразитах
специфичных для данного вида хозяина и никогда в настоящее
время не встречающихся у каких-либо других хозяев (например, маля-
рийные плазмодии, человек и анофелес). Паразиты,, часто встречаю-
щиеся у рассматриваемого хозяина, являются для него обычными (на-
пример, обычцым паразитом собаки является собачья блоха). Наконец,
обычные дл^ известных хозяев паразиты могут при случа^дом стече-
нии обстоятельств попасть в несвойственного им хозшшау понятие
«свойство» принимается по отношению к филогенезу паразитов и хо-
зяина, распространяя таковое и на генезис соотношений паразита и
его хозяев в составе биоценозов мест их обитания./ При таком стече-
нии обстоятельств говорят о-е-дула иных, (и в то же время истин-
ных) паразитах, например цепень огуречный (Dipylidium сатпит) —
паразит собаки (и кошки) — бывает и у человека, если последний про-
глотит собачью блоху иди власоеда с личинками (цистицеркоидами)
этого ленточного червя.I
ГВ крайних степенях'-случайности говорят о парадоксальных
паразитах (И. Иофф).
Ь природе часты случаи, когда~паразит проделывает сложный цикл
развития за счет нескольких хозяев (гетероксенные парази-
ты). Это явление бывает в различных формах. Так, клещ Ixodes ricinus
в каждой фазе метаморфоза нападает на какого-либо хозяина. Следо-
вательно, он нуждается в трех хозяевах по числу фаз превращения —
личинки, нимфы и imago. Его поэтому называют треххозяинным кле-
щом. Напротив, Boophilus decoloratus является однохозяинным клещрм,
14
ОБЩАЯ ЧАСТЬ
так как, напав на корову в фазе личинки, он проделывает на ней весь
метаморфоз и отпадает лишь в виде оплодотворенной и упитанной
самки.
у Наряду с этим бывает перемена хозяев и у внутренних паразитов
/эндопаразитов); но такое явление осложняется совершающимся па-
раллельно чередованием поколений паразита. ’Малярийный
плазмодий живет в эритроцитах человека, где размножается бесполым
путем (шизогония); в крови же образуются и половые формы плазмо-
дия. Дальнейший жизненный цикл может иметь место только в самке
комара анофелес, в которой происходит половое размножение плазмо-
дия. В связи с этим человек является промежуточным хозяи-
ном плазмодия, анофелес же — главным хозяином. Каждый
хозяин последовательно заражается паразитом от другого хозяина:
человек -> анофелес -> человек — анофелес и т. д.
f f f f
бесполое половое бесполое половое
поколение поколение поколение поколение
малярийного плазмодия
Приведенный пример характерен тем, что каждое поколение пара-
зита имеет только одного свойственного ему хозяина (человек, род
Anopheles).
/ --.У других паразитов жизненный цикл усложняется еще более благо-
даря наличию, кроме одного главного, двух промежуточных хозяев и
стадий свободной жизни паразита; таковы, например, лентец широкий
(Diphyllobothrium latum), печеночная двуустка (Fasciola hepatica)
и др. (см. стр. 217, 218).
Многие паразиты имеют узко ограниченный круг хозяев, в преде-
лах которого исторически развертывался филогенез данного вида па-
разита. Такие паразиты могут по стечению обстоятельств попасть не в
своего природного хозяина, а совсем вч другой организм, и 4в то же вре-
мя продолжать в нем нормально^житьЛПримером может служить цепень
тыквовидный (Dipylidium camnum), в стадии ленточной глисты нор-
мально живущий в кишечнике собаки или кошки. Его промежуточным
хозяином являются эктопаразиты собаки — блохи и власоеды (Tricho-
dectes), в которых из проглоченных ими яиц червя развиваются фин-
ки — цистицеркоиды (стр. 303). Собака чешет зубами зудящие места ко-
жи и проглатывает зараженных блох или власоедов; в кишечнике собаки
из заглоченных цистицеркоидов развиваются ленточные черви и т. д.
ЕЕтГ^человек случайно проглотит зараженную паразитами блоху,
то и в его кишечнике разовьется нормальная ленточная форма цепеня
тыквовидного. Биологически эта глиста является чуждым для человека
паразитом, и весь цикл ее развития приспособлен к взаимоотношении:
собак, кошек, с одной стороны, и их эктопаразитов — с другой. Однакс
среда кишечника человека практически оказывается пригодной для
развития и жизни Dipylidium caninum, если сюда попадет его финка
Таким образом, наш паразит по отношению к человеку является как
бы гостепаразитом (к.с е н о п а.р а з и т о м) или чужепара-
з и т о м. Другим примером ксенопаразитизма может служить
аскарида собачья, в единичных случаях обнаруженная и у человека
Само собой разумеется, что ксенопаразитизм, являясь раз нови дностьк
истинного паразитизма, должен быть резко 'отличаем от ложнопарази
тизма.
Раз паразиты живут за счет своих хозяев, то подразумевается, чт<
первые вредят последним. Однако патогенные свойства выявлены дале-
ОПРЕДЕЛЕНИЕ ПАРАЗИТИЗМА
ко не для всех паразитов. Наряду с этим имеется ряд фактов, указы-
вающих на безвредность некоторых паразитов или даже на известную
пользу, оказываемую ими своим хозяевам.
Таким образом, резкое разграничение явлений хищничества, пара-
зитизма истинного и паразитизма ложного невозможно так же, как не-
возможно категорическое отграничение форм симбиоза 1 в широком
смысле.
Перейдем к рассмотрению форм паразитизма во времени. Раз-
личают^хя.е.нный и постоянный паразитизм. ~
А. П-аразити зм временный. Градации его весьма разнооб-
разны; их мы рассмотрим, начиная с наиболее примитивных форм.
Паразит живет свободно в открытой природе и нападает на хо-
зяев только для питания и на время питания (сосание крови); таковы
самки комаров (Culicidae), самцы и самки слепней (Tabanidae), самки
москитов (Phlebotomus) и др.; личинки всех поименованных насекомых
вовсе не являются паразитами. Клещи Ixodidae также могут быть при-
мером временных паразитов, число раз принятия пиЩи которых для
разных видов строго'определенно. Питаются кровью и личинки этих
клещей.
Паразит живет близ хозяина — в его жилье, гнезде, норе или
логове; а) на хозяина переходит только взрослая форма для сосания
крови, каковая процедура длится в общем недолго; таковы, например,
блохи (Aphanipterdy, в то же время их личинки питаются непаразити-
ческим путем; б) на хозяине кормятся все стадии развития паразита,
начиная с личинки: клопы сем. Cimicidae (жилье человека, гнезда птиц
и летучих мышей); клещи сем. Argasidae: Argas persicus (курятники),
Orptthodorus papillipes (жилье человека, стойла животных) и др.
Б. Паразитизм постоянный. 1. Паразит проводит на хо-
зяине все время в течение той или другой фазы своей жизни; таковы
овода и некоторые мухи (сем. Larvivoridae); постоянными паразитами
являются личинки этих насекомых, тогда как imago их живут свободно
41вовсе не питаются, (фазовый ..паразитизм).-
2. Паразит всю свою жизнь проводит на или в хозяине; например,
пухоеды (Mallophagd) и вши, приклеивающие яйца к перьям или воло-
сам хозяина; трихина (Trichinella spiralis) и гемо.споридии также ни
одной минуты на современном этапе своей эволюции не проводят вне
своих хозяев (перманентный паразитизм).
менее разнообразны формы паразитизма по локализации пара-
зитов в организме хозяина. Под таким углом зрения различаются:
А. Наружные паразиты, или эктопаразиты, лишь
временно садятся на поверхность тела (покровы) хозяина для приня-
тия пищи,— например, комары, слепни, пиявки; другие эктопаразиты
не только питаются, но и вообще живут на коже хозяина — Mallopha-
ga, вши. Для эктопаразитов, тесно связанных с хозяином в смысле
постоянства их обитания, можно предложить обозначение — сомати-
ческие эктопаразиты (вши и др.).
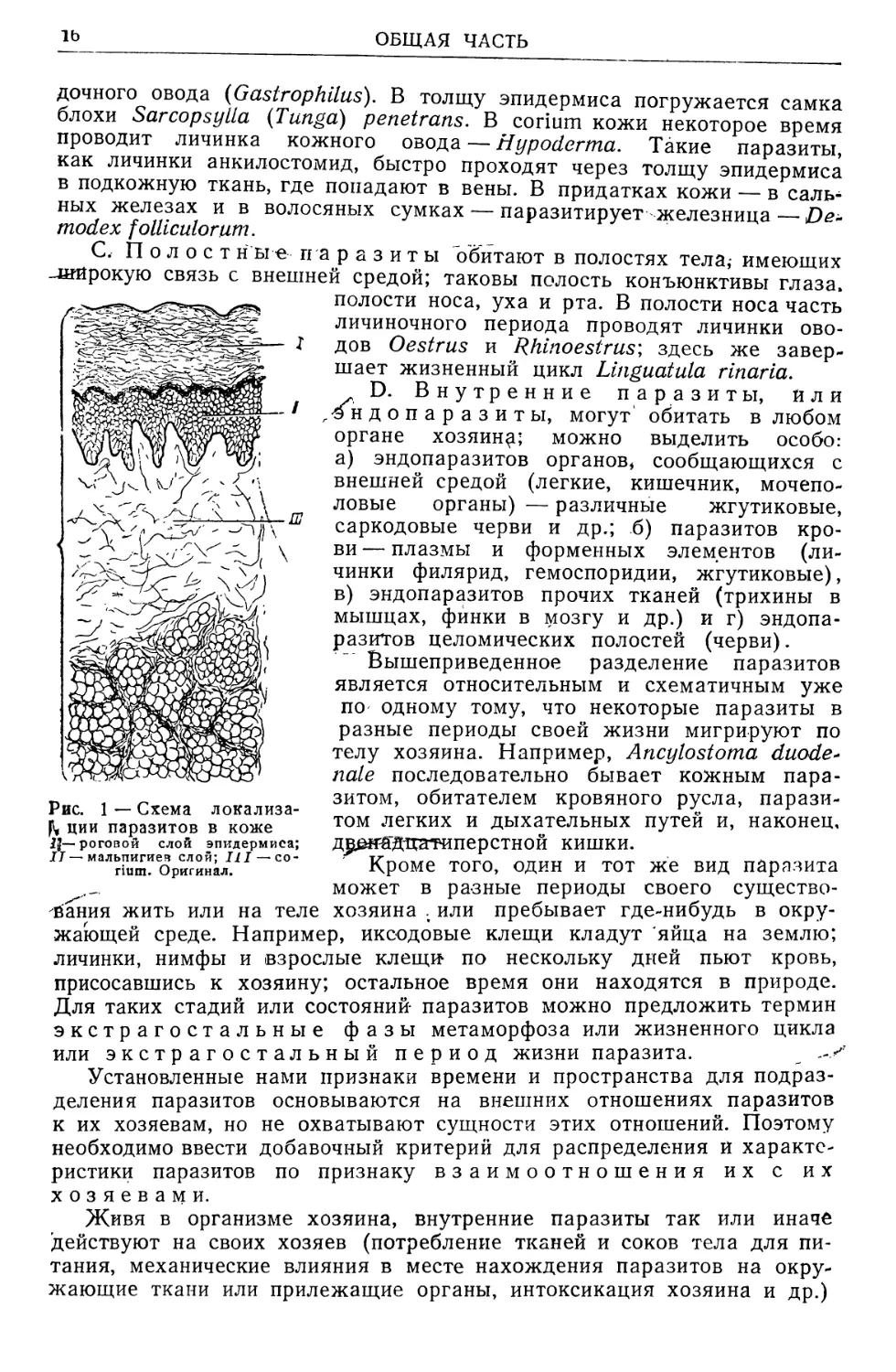
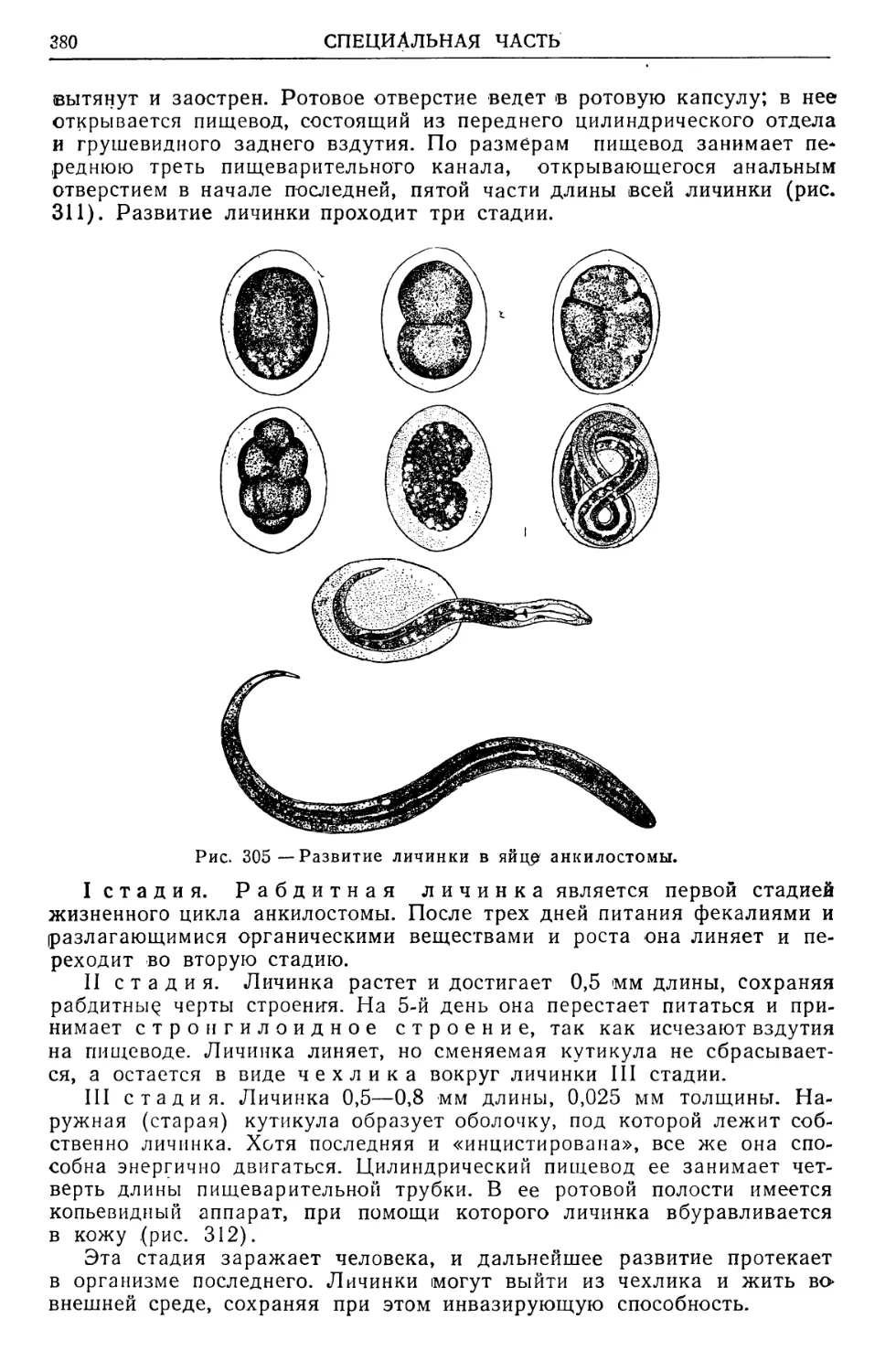
В. В кожные паразиты (рис. 1), живущие в самой толще
кожных покровов; в роговом слое эпидермиса млекопитающих буравит
ходы и живет чесоточный зудень (Sarcoptes scabiei); в мальпигиевом
.слое кожи человека может жить личинка первой фазы развития желу-
1 Под симбиозом в данном случае подразумеваются формы сожительства раз-
личных видов животных, как основанные на взаимно оказываемой пользе (муту-
ализм), так и индиферентные или характеризующиеся односторонней пользой, без
вреда для другого сожителя («нахлебничество» или комменсализм), паразитизм и др.
1b
ОБЩАЯ ЧАСТЬ
Рис. 1 — Схема локализа-
ции паразитов в коже
If—роговой слой эпидермиса;
IT — мальпигиев слой; III — со-
пит. Оригинал.
жить или на теле
дочного овода (G astrop hilus). В толщу эпидермиса погружается самка
блохи Sarcopsylla (Tanga) penetrans. В corium кожи некоторое время
проводит личинка кожного овода — Hypoderma. Такие паразиты,
как личинки анкилостомид, быстро проходят через толщу эпидермиса
в подкожную ткань, где попадают в вены. В придатках кожи — в саль-
ных железах и в волосяных сумках — паразитирует железница—>Ое-
modex folliculorum.
С. П о л о с т н ы е паразиты обитают в полостях тела, имеющих
.дийрокую связь с внешней средой; таковы полость конъюнктивы глаза»
полости носа, уха и рта. В полости носа часть
личиночного периода проводят личинки ово-
дов Oestrus и Rhinoestrus', здесь же завер-
шает жизненный цикл Linguatula rinaria.
D. Внутренние паразиты, или
•эндопаразиты, могут' обитать в любом
органе хозяину; можно выделить особо:
а) эндопаразитов органов, сообщающихся с
внешней средой (легкие, кишечник, мочепо-
ловые органы) — различные жгутиковые,
саркодовые черви и др.; .6) паразитов кро-
ви — плазмы и форменных элементов (ли-
чинки филярид, гемоспоридии, жгутиковые),
в) эндопаразитов прочих тканей (трихины в
мышцах, финки в мозгу и др.) и г) эндопа-
разитов целомических полостей (черви).
Вышеприведенное разделение паразитов
является относительным и схематичным уже
по одному тому, что некоторые паразиты в
разные периоды своей жизни мигрируют по
телу хозяина. Например, Ancylostoma duode-
nale последовательно бывает кожным пара-
зитом, обитателем кровяного русла, парази-
том легких и дыхательных путей и, наконец,
д^жедтщтаперстной кишки.
Кроме того, один и тот же вид паразита
может в разные периоды своего существо-
хозяина . или пребывает где-нибудь в окру-
жающей среде. Например, иксодовые клещи кладут яйца на землю;
личинки, нимфы и взрослые клещи по нескольку дней пьют кровь,
присосавшись к хозяину; остальное время они находятся в природе.
Для таких стадий или состояний паразитов можно предложить термин
экстрагоста л ьные фазы метаморфоза или жизненного цикла
или экстрагостальный период жизни паразита. _
Установленные нами признаки времени и пространства для подраз-
деления паразитов основываются на внешних отношениях паразитов
к их хозяевам, но не охватывают сущности этих отношений. Поэтому
необходимо ввести добавочный критерий для распределения й характе-
ристики паразитов по признаку взаимоотношения их с их
хозяевами.
Живя в организме хозяина, внутренние паразиты так или иначе
действуют на своих хозяев (потребление тканей и соков тела для пи-
тания, механические влияния в месте нахождения паразитов на окру-
жающие ткани или прилежащие органы, интоксикация хозяина и др.)
ОПРЕДЕЛЕНИЕ ПАРАЗИТИЗМА
17
я в то же время сами подвергаются влиянию со стороны организма
хозяина (гуморальные воздействия тканевых соков и крови, защитная
реакция в форме капсуляции вокруг тканевых паразитов с последую-
щим их обизвествлением, депрессивное воздействие организма на па-
разитов в связи с интенсивностью витаминного питания, влияние изме-
няющегося химизма крови, возрастные цСчсугубо индивидуальные со-
стояния хозяина и мн. др.).
Паразит и хозяин являются, следовательно, по-
вседневными партнерами в жизненных соотноше-
ниях; результат взаимодействия их противоречивых интересов опреде-
ляется видовыми свойствами этой биологической системы организмов,
индивидуальными качествами и состояниями данной пары партнеров и
конкретными особенностями среды обитания паразита и хозяина на
фоне их филогенетического развития. Последующая судьба биологиче-
ской системы наших партнеров может вести к одному из трех случаев:
а) паразит и хозяин благополучно сосуществуют без какого-либо
заметного вреда для себя;
б) в паре партнеров хозяин становится страдающим элементом бла-
годаря присутствию в нем жизнедеятельного паразита;
в) в аналогичной паре дефектные условия жизни создаются для па-
разита, который поэтому не может проявить своих вредоносных свойств
и погибает раньше обычного срока.
Такие взаимоотношения ведут к различным последствиям для хо-
зяина:
а) хозяин всегда остается по отношению к находящемуся в нем па-
разиту здоровым;
б) хозяин заболевает благодаря влиянию жизнедеятельности живу-
щего в нем паразита;
в) несмотря на присутствие в своем организме паразита, могущего
вызвать заболевание, хозяин остается здоровым (паразитоноситель-
-ство).
Благодаря повседневным длительным наблюдениям установлено,
что одни паразиты никогда не вызывают болезней у хозяина — их по-
этому называют н е п а т о г е н н ы м и; другие же могут, как правило,
причинять заболевания; их в связи с этим относят к болезнетвор-
ным, или патогенным.
Однако присутствие в теле хозяина патогенного паразита не обяза-
тельно ведет к заболеванию данной особи хозяина, которая может
оставаться совершенно здоровой и быть в то же время источником
распространения болезнетворного паразита. Такое явление (широко
распространенное) носит название паразитоносительства.
В заключение мы должны подчеркнуть, что нет резкой границы ни
между свободным и паразитическим образом жизни, ни между сим-
биозом, паразитизмом и хищничеством, ни между патогенными й не-
патогенными паразитами. Следовательно и соответствующие определе-
ния носят относительный характер; то же имеет место и по отношению
к различным формам паразитизма. Явления этой области природы на-
столько многообразны' как в своем настоящем, так и в историческом
прошлом, что распределить их с пунктуальной точностью по создавае-
мым нашим мышлением рубрикам без натяжки во многих случаях со-
вершенно невозможно.
Но в целях практических, с допущением некоторой условности, па-
разитов распределяют по указанным выше формальным категориям
паразитизма, что имеет под собою научную базу.
18
ОБЩАЯ ЧАСТЬ
ОБЪЕМ И СОДЕРЖАНИЕ ПАРАЗИТОЛОГИИ
Паразитология является наукой, всесторонне изучающей сложный
комплекс явлений паразитизма в свете соотношений паразитов и их
хозяев и, как таковая, по основным объектам и методам исследования,,
относится 'к разрядам биологических и медицинских (resp. ветеринар-
ных) дисциплин. Ведению ее. подлежат паразиты из мира животных
или растений, живущие за счет животных или растений (животная и.
растительная паразитология), и хозяева самих паразитов, поскольку
таковые связаны с последними. ’Мы будем касаться только паразитов
животной природы.
Пути изучения паразитов (resp. паразитизма) многочисленны и
многообразны.
Первой задачей паразитологии является определение вида парази-
та, подлежащего нашему изучению,— этим ведает систематика. Систе-
матическое изучение паразитов тех или других хозяев данной местно-
сти позволяет судить о фауне паразитов; группировка соответ-
ствующих фактов ведется или по хозяевам в целом или по их отдель-
ным органам; так, можно изучать фауну паразитов кур Европы или
исследовать фауну трематод почек птиц Средней Азии. Накопление по-
добных исследований дает материал для выяснения зоогеографии
паразитов, каковые данные необходимо изучать в свете распростра-
нения хозяев паразита.
Изучение строения (анатомии) паразитов, помимо самодовлеющего
интереса, близко касается той же систематики, а также физиологии
паразитов и объясняет нам вместе с нею многое в действии паразитов
на их хозяев. Важной задачей паразитологии является исследование
жизненного цикла, размножения и других особенностей жизни парази-
тов, находящихся в разной -степени зависимости от своих хозяев, как
главных, так и 'промежуточных (собственно биология паразитов).
В отношении изучения хозяев паразитов приходится прибегать к
соответствующим зоологическим специальностям, например к малако-
логии (учение о мягкотелых или моллюсках), энтомологии, ихтиологии^
маммалиологии, или териологии и др.
Далее стоит на очереди определение характера влияния внеш-
ней среды на паразита, причем для эндопаразита тако-
вой средой является прежде всего организм самого
хозяина («организм, как среда обитания»). Таким образом, к эко-
логии паразитов относится и изучение действия хозяина на своих
паразитов, включая сюда, помимо обычного влияния хозяина на пара-
зита, также и специальные воздействия, являющиеся результатом осо-
бых изменений организма хозяина (например, введение лекарственных
веществ, прямо или косвенно влияющих на паразитов, серологические
изменения организма хозяина, приводящие к состоянию стойкости про-
тив заражения паразитами, и др.). Это накладывает на экологию пара-
зитов своеобразный отпечаток, ибо связывает эту отрасль паразитологии
с физиологией и патологией хозяев и с терапией их паразитарных болез-
ней. К сфере экологии паразитов человека относятся также особен-
ности труда и быта людей, ибо бытовые и профессиональные моменты
могут оказать решающее влияние на частоту распространения и даже
на самое распространение тех или иных паразитов человека. В каче-
стве частного примера укажем на зависимость распространения в раз-
личных провинциях Китая китайской двуустки у людей от способа при-
готовления рыбы в пищу (полусырой или хорошо обработанной терми-
ОБЪЕМ И СОДЕРЖАНИЕ ПАРАЗИТОЛОГИИ
19
чески); в связи с этим .географическое распространение китайской
двуустки в Китае (рис. 21), если мы будем судить о нем по нахождению
этих паразитов у людей или у собак, будет не одинаковым (в районах,
где нет этих паразитов у людей, они бывают у собак). Следовательно,
данные этнографии и изучения быта в свете экологических, классовых
и культурных отношений может объяснить многое в паразитологии че-
ловека с точки зрения качественного отличия его, как хозяина парази-
тов, от животных, в свою очередь дающих приют этим последним.
Палеопаразитология, а также филогения стремятся
выяснить пути эволюции или историю развития паразитов, причем эво-
люция паразитов естественным образом связана с эволюцией их хозяев,
почему и должна быть рассматриваема именно в этой связи.
Особняком стоит изучение болезней паразитов и их паразитов
(сверхпаразитов), что, впрочем, в значительной мере может быть от-
несено к экологии паразитов.
Все эти направления паразитологических исследований вращаются
в сфере общей, или теоретической паразитологии.
Паразитизм имеет огромное практическое значение, о каковом на-
гляднее всего можно судить, изучая действие паразитов на хозяев и
экономическое значение паразитов. В связи с прикладной ролью пара-
зитологии обращено особое внимание на выработку методики парази-
тологических исследований и диагностики паразитарных или инвазион-
ных болезней (или паразитозов). Логическим завершением при-
кладной паразитологической работы является изыскание мер борьбы
с паразитами и способов предохранения хозяев от заражения ими (про-
филактика паразитозов). Наконец, ценные сведения дает изучение роли
паразитов и вызываемых ими заболеваний в природе и в истории
культуры.
Намеченные выше пути паразитологических исследований в своей
совокупности дают массу разнообразных сведений, образующих целую
энциклопедию знания, величина и дробность содержания которой ста-
нет еще более очевидной, если мы детальнее рассмотрим хотя бы одно
из направлений интересующего нас комплекса вопросов, например, уче-
ние о влиянии паразитов на своего хозяина.
К освещению этой проблемы подходят с разных сторон, изучая
а) патологические изменения органов хозяина под влиянием данного
паразита (патологическая анатомия и гистопатология), б) функцио-
нальные изменения.организма хозяина (патологическая физиология),
в) токсическое действие паразита (фармакология и токсикология),
г) симптомы и течение заболевания, вызываемого паразитом (клиника
инвазионных болезней), и т. д.
В приведенной нами схеме подразделения паразитологии на части
мы касались паразитов вообще, не выделяя их по хозяевам.
Жизненное значение паразитологии определило также и другой
путь подразделения .нашей дисциплины; критерием такого подразделе-
ния является группировка паразитов по их хозяевам применительно к
значению первых для последних.
'Мы имеем: а) паразитологию человека, или медицинскую паразито-
логию; б) паразитологию ветеринарную, или домашних животных,—
к этой же ветви можно отнести и паразитологию промысловых живот-
ных; в) паразитологию вредных для человека или его хозяйства жи-
вотных; г) паразитологию вообще диких животных; д) паразитологию
са'мйх паразитов и е) паразитологию различных растений.
Распределение материала и по этим ветвям паразитологических
20
ОБЩАЯ ЧАСТЬ
дисциплин является относительным и непостоянным. Например, изуче-
ние трипанозом антилоп получило прямое отношение к паразитологии
человека, когда выяснилось, что эти млекопитающие служат резервуа-
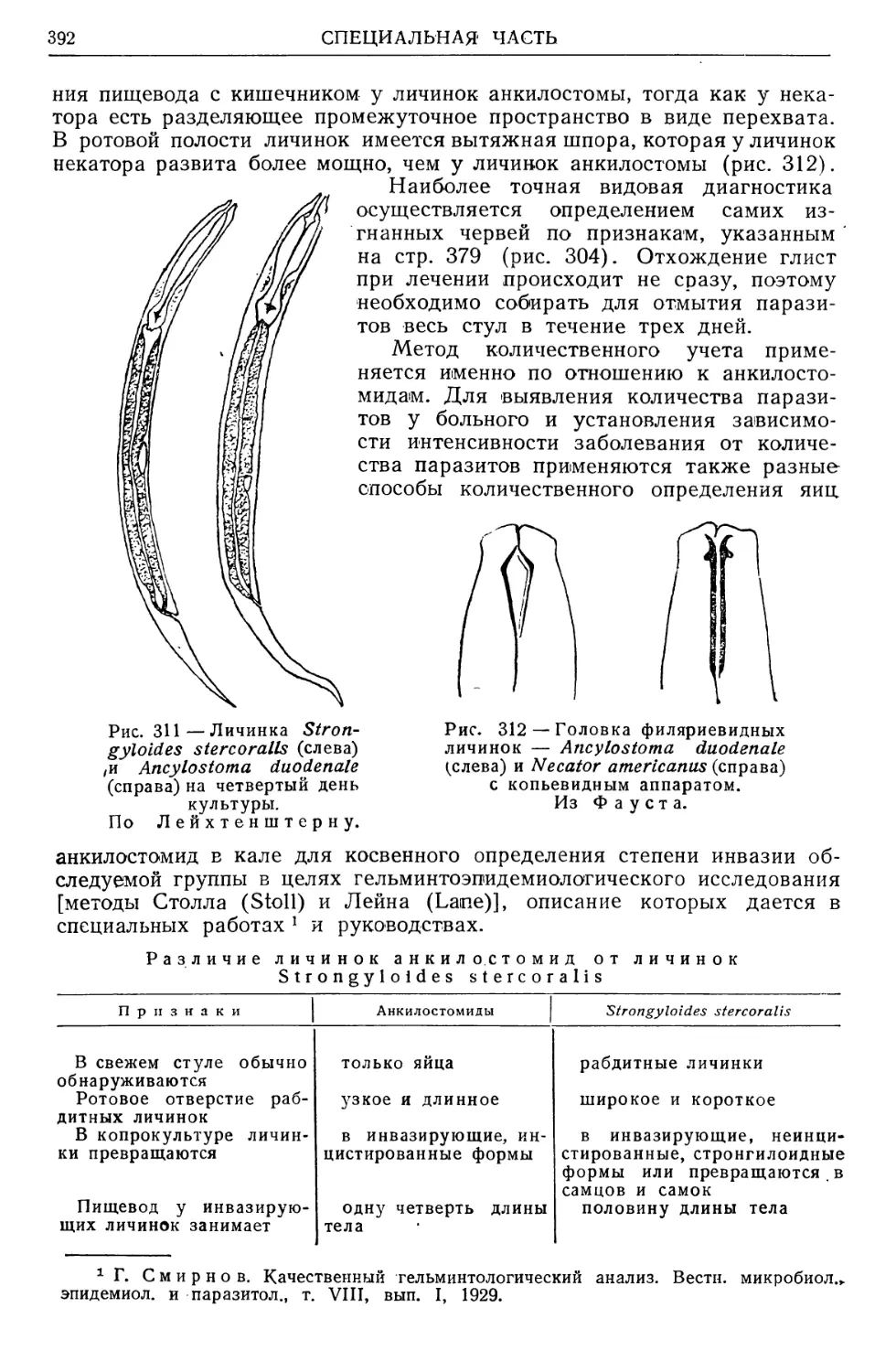
Рис. 2 — Схема отношения паразитологии
к сопредельным дисциплинам. Оригинал.
ром трипанозом сонной болезни, переносимых мухами тсе-тсе от анти-
лоп в организм человека, заболевающего в конце концов сонной бо-
лезнью. С другой стороны, исследование паразитов крыс может иметь
ближайшее отношение и к медицинской и к ветеринарной паразитоло-
гии, ибо крысы могут заражать некоторыми паразитами как человека,
так и свиней.
Маммалиолигия
Орнитология
Герпетология
Ихтиология
Малакология
Энтомология
Гельминтология
Протозоология
Рис. 3—-Содержание паразитологии как
дисциплины, по Hall.
Таким образом, паразитология в целом является огромным по ве-
личине и важности самодовлеющим комплексом данных, добы-
ваемых методами самых разнообразных теоретических и прикладных
дисциплин (биология, медицина, ветеринария, агрономия, зоотехника,
экономика), и имеет в то же время свои прямые задачи и методы ис-
следования. Это делает паразитологическую литературу разбросанной
ОБЪЕМ И СОДЕРЖАНИЕ ПАРАЗИТОЛОГИИ
21
до крайности. В отношении сводок таковой наблюдается чрезвычайная
неравномерность. Наряду с исключительной скудостью книг по общей
паразитологии дело со специальной медицинской и отчасти ветеринар-
ной паразитологией обстоит в мировой литературе более благополучно.
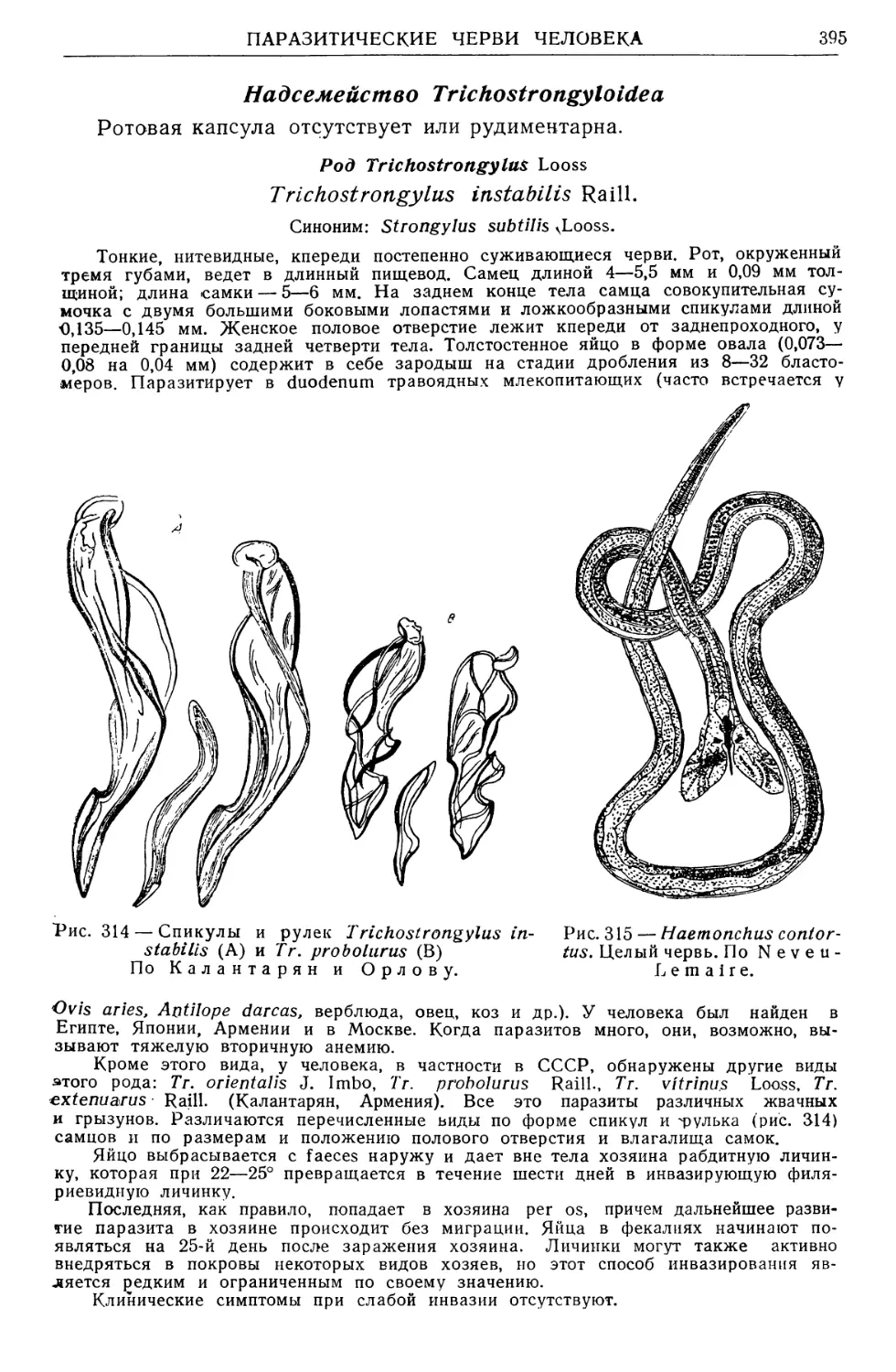
влияние ПАРАЗИТА . I ВЛИЯНИЕ ХОЗЯИНА
НА ХОЗЯИНА I НА ПАРАЗИТА
ВЗАИМОДЕЙСТВИЯ ПАРАЗИТОВ И ИХ ХОЗЯЕВ
_____ОРГАНИЗМ, КАК СРЕДА ОБИТАНИЯ
кдш щи цу ин tug дц^
Рис. 4 — Схема основных исследовательских
направлений в паразитологии.
По Е. Павловскому.
Так как число вновь открываемых паразитов неизменно растет, то
выявилась тенденция к выделению в самостоятельные ветви специаль-
ной паразитологии учений о паразитических простейших, червях и чле-
нистоногих. В литературе поэтому уже давно начали появляться книги,
посвященные медицинской энтомологии, или протозоологии, или гель-
минтологии. Несомненно, что в будущем такая дробность будет еще
более увеличиваться, ибо уже и теперь мы имеем отдельные крупные
монографии, посвященные, например, клещам Ixodoidea, или немато-
дам позвоночных животных, или ленточным червям млекопитающих
и мн. др.
Параллельно идет и ограничение специализации паразитологов одни-
ми группами каких-либо паразитов (например, систематика блох, систе-
матика трематод и т. д.).
В то же время такая диференциация не может и не должна вести
к отчуждению и к замыканию в себе указанных трех ветвей паразито-
логии, ибо в жизненных циклах простейшие и глисты часто бывают не-
разрывно связаны с различными членистоногими; кроме того, сам хо-
зяин бывает местом одновременного обитания самых разнообразных
паразитов, комбинированное влияние которых на организм хозяина
может быть выяснено лишь при учете всех их специфических особен-
ностей. Эта важная часть паразитологии, к сожалению, почти вовсе не
затронута разработкой. Кроме того, именно синтез основных парази-
22
ОБЩАЯ ЧАСТЬ
тологических направлений дает возможность ставить широкие, общие
паразитологические проблемы, не сковывая таковые рамками узкой
специализации.
ПОНЯТИЕ ОБ ИНВАЗИИ
Инвазией называется заражение организма какими-либо паразита-
ми животной природы. В параллель с инфекционными болезнями (бак-
териозы) — инвазионными болезнями или паразитозами называют
такие заболевания, возбудителями которых являются животные пара-
зиты или ложнопаразиты.
Многие паразиты обладают инвазирующими способностями только
в определенной стадии своего жизненного цикла, другие же могут за-
ражать хозяина в течение всей своей жизни.
Вошь может переходить на человека и в личиночной и во взрослой
фазах. Блохи инвазируют своих хозяев только во взрослом состоянии.
Яйца аскарид и власоглава инвазируют человека только тогда, когда
в них развился зародыш в форме червячка, и т. д.
Объектом инвазии являются хозяева соответ-
ствующего паразита или иногда хозяева, обычно не свойствен-
ные нашему паразиту. Объект инвазии может заражаться паразитами
или в любой момент жизни или преимущественно в определенном воз-
расте. Например, малярией человек может заразиться, начиная с вну-
триутробной жизни и кончая старостью.
В отношении календаря инвазия может происходить или в любое
время года, или в определенные сезоны, или даже в определенное вре-
мя суток. Трихинами, например, можно заразиться когда угодно, лишь
бы была трихинозная свинина; глистные инвазии, связанные с почвой,
в умеренном климате зимой имеют мало шансов на дальнейшее рас-
пространение. Вшами можно заразиться в течение всего года, но боль-
шее распространение вшивость приобретает зимой, благодаря скученно-
сти людей. Заражение болезнями, распространяемыми переносчиками,
возможно только тогда, когда есть самые переносчики. Без комара
анофелес малярией заразиться нельзя; поэтому первичные случаи ма-
лярии, как правило, бывают только тогда, когда есть активный пере-
носчик, т. е. в теплое время года. Некоторые анофелес нападают на че-
ловека обычно в сумерки и ночью, поэтому и заражение малярией при-
урочиватся к этому времени суток.
Источником инвазиров ани я являются биотопы
(т. е. типичные места обитания) тех или других ста д и й р а з в и-
тия паразитов. Смотря по месту обитания, биотопами паразитов
являются части тела хозяина, пораженные паразитами,— так называе-
мые гостальные биотопы (например мышцы для трихины), или
же биотопы в обычном понимании этого термина (водоемы, хозяй-
ственные биотопы и др.).
Источником инвазирования может быть организм в целом. Напри-
мер, чесоткой заражаются при контакте с чесоточным. Различные сре-
ды обитания служат или более постоянным местом нахождения пара-
зита (например, обитаемая нора грызуна и блохи), или же временным
этапом пребывания такового (водоем и личинки комаров).
Данная среда может быть специфической для того или другого
паразита (вода и корацидий лентеца широкого) или же паразит попа-
дает в нее случайно, будучи в состоянии жить и в других средах (яйца
глист в водоеме и фекалиях в уборной).
ПОНЯТИЕ ОБ ИНВАЗИИ
23
Весьма важным обстоятельством является тот факт, что сам хо-
зяин может служить источником заражения для
самого себя. Такое явление носит название аутоинвазии; на-
пример, повторное заражение острицами при грызении ногтей, под ко-
торые попадают яйца остриц при расчесывании заднего прохода.
Важным источником инвазий могут служить носители пара-
зитов и животные — резервуары вирусов. Лица, перебо-
левшие амёбной дизентерией, могут рассеивать четырехядерные цисты
и заражать окружающих. Антилопы в тропической Африке бывают за-
ражены безвредными для них трипанозомами сонной болезни; будучи
почерпнуты из крови антилоп мухой тсе-тсе и переданы ею человеку,
трипазономы вызывают у него сонную болезнь.
Способы проникновения инвазирующих начал в
организм бывают активные или пассивные. Паразиты
могут активно проникать в тело хозяина, причем активность прояв-
ляется самим паразитом (проникновение в кожу личинок анкилостом)
или же его самкой, пристраивающей свое потомство на хозяина (вы-
брасывание личинок в глаза человека самкой овечьего овода).
При пассивном проникновении паразиты попадают в хозяина
без каких-либо усилий или действий со своей стороны. Например, чело-
век проглатывает яйца аскарид, поедая в сыром виде немытую ого-
родную зелень. Личинки нематоды Gon&ylonetna развиваются в тара-
кане, когда он проглотит ее яйца.
Пути проникновения инвазирующих начал в хо-
зяина двояки: паразиты проникают через кожные покровы или через
естественные отверстия органов — прежде всего и чаще всего через рот;
паразиты могут внедриться через anus, половую щель, отверстие моче-
испускательного канала, ноздри и др.
При проникновении через покровы — одни паразиты пользуются
уже имеющимися нарушениями целости покровов (экскориации, цара-
пины, раны, язвы и др.), другие же сами пробуравливают кожу.
Замечательны случаи внутриутробной инвазии хозяина пара-
зитами, когда он находится еще в теле матери на положении развива-
ющегося зародыша или даже яйца в яичнике. Примеры такого рода
известны у человека. От больной малярией, сонной болезнью, кала-азар
и некоторыми другими паразитарными болезнями матери развиваю-
щийся в ней плод может получить соответствующих паразитов через
поврежденную плаценту. Этим объясняются случаи так называемой
врожденной малярии (см. ниже).. Некоторые нематоды, при миграции
своей в организме беременной женщины или плодоносящей самки, мо-
гут активно проникать через плаценту в организм плода, который рож-
дается уже с паразитами. Примеры таких отношений видим у Ancylo-
stoma duodenale, собачьих аскарид, Schistosoma и др.
В фазе яйца хозяин заражается паразитом у клещей Ornithodorus,
являющихся переносчиками спирохет клещевого рекурренса.
Из желудка самки клеща, напившейся кровью возвратнотифозного
больного, спирохеты проникают в полость тела и внедряются в яич-
ник, заражая яйца. При откладке таких яиц спирохеты остаются в них
живыми. Когда из яиц выходит следующее поколение клещей, оно со-
держит в себе спирохет и может заражать ими людей (т р а н с о в а-
риальная передача инвазий).
Продолжительность инвазий весьма различна; она опре-
деляется длительностью жизни самого паразита, если он не размно-
жается -в организме хозяина, и влиянием на него хозяина. Например,
24
ОБЩАЯ ЧАСТЬ
гибель трихин в капсулах при омелотворении их, выбрасывание тяже-
лых самок аскарид из кишечника и др. При размножении паразитов
теле хозяина длительность инвазии зависит от интенсивности размно-
жения, свойств паразита и наличной или приобретаемой по мере пре-
бывания паразитов стойкости хозяина (иммунитет при животных пара-
зитах). При последующих размножениях паразитов этот процесс замед-
ляется и даже вовсе прекращается; благодаря этому инвазия из явной
переходит в скрытую .(латентная инвазия). Последняя может
снова переходить в явную без добавочного заражения хозяина новой
партией паразитов (рецидивы инвазии).
ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
Уже было указано, что ни один орган тела хозяина не
застрахован от возможности поселения в нем того
или другого паразита. В качестве примера рассмотрим дан-
ные, касающиеся человека. На поверхности его тела живут вши (Pedi-
cuius), здесь же размножающиеся. В толще рогового слоя эпидермиса
прокладывают ходы чесоточные клещи. Несколько глубже — в мальпи-
гиевом слое эпидермиса буравит ход личинка желудочного ово-
да (Gastrophilus), бывающая гостепаразитом человека. В сальных же-
лезах паразитирует угрица прыщевая.
В подкожной клетчатке залегает и мигрирует по ней личинка кож-
ного овода Hypoderma, также могущая быть гостепаразитом человека.
Личинки вольфартовой мухи, являющиеся настоящими тканевыми
паразитами, могут разрушать кожные покровы и мышцы, «разъедая»
язвы и раны. Изъязвления кожи вызывают кожные лейшмании. В под-
кожной и межмышечной соединительной ткани локализуется у челове-
ка финка цепеня вооруженного, а в саркоплазме самого мышечного
волокна оседают личинки трихин.
Хрящевая поверхность суставов может «разъедаться» дизентерий-
ными амёбами, которые метастатически иногда попадают в -суставы.
Сами трубчатые кости разрушаются при соответствующей локализации
пузырями эхинококка.
Пищеварительный аппарат от входа до выхода бывает заселен раз-
личными паразитами.
В деснах могут паразитировать личинки вольфартовой мухи; в на-
лете зубов бывает Amoeba buccalis. Под слизистой губ и щек залегает
Gongylonema. К стенке носоглотки или к деснам присасываются пиявки.
Желудок является местом врытия капсул трихин и оболочки заро-
дышей цепеня вооруженного, откуда последние начинают миграцию'
через стенку желудка.
В двенадцатиперстной кишке паразитируют лямблии, некатор, ан-
килостома и другие паразиты. В тонкой кишке обитают паразитические
простейшие — амёбы, лямблии и др. Здесь же живут ленточные черви
(цепень невооруженный, цепень вооруженный, лентец широкий и др.).
В слепой и в толстой кишках паразитирует власоглав; в червеобраз-
ном отростке часто встречаются острицы, обычно паразитирующие в
дистальной части тонкой кишки и в слепой кишке. В толстых кишках
живут аскариды, дизентерийные и кишечные амёбы, балянтидий и дру-
гие паразитические простейшие. В прямой кишке, в ее венах, живет
бильгарция {Schistosoma mansoni), вызывающая кишечный бильгар-
циоз. В складках перианальной области откладывают яйца выползаю-
щие из анального отверстия острицы и членики цепеня невооруженного.
ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
2,
В придаточных железах пищеварительного канала паразитируют:
в слюнных железах — эхинококк; в печени — печеночная двуустка
(в желчных ходах), личинки пятиустки и эхинококк в ткани печени;,
бильгарции Schistosoma japonicum — в венах печени и брыжжейки;
в желчном пузыре бывают лямблии и печеночная двуустка; в поджелу-
дочной железе находили эхинококка.
В органах дыхания паразиты бывают, начиная с полости носа (ли-
чинки вольфартовой мухи, пиявки); по трахее мигрируют личинки ан-
килостомид и аскарид. В толще легких локализуются финки цепеня
вооруженного, эхинококк, легочная двуустка и др.
Органы кровообращения и лимфообращения хозяина играют в жиз-
ни паразитов громадную роль в качестве пути (миграции их по телу
хозяина и как основное место обитания.
В стенке сердца бывают эхинококк и финки. (развитие их протекает
в соединительнотканных прослойках этого органа); в толще самих
мышечных элементов сердца паразитируют трипанозомы — возбудители
болезней Чагаса в форме лейшманий.
В эндотелии сосудов кожи локализуются риккетсии сыпного тифа.
В полости вен живут бильгарции; в плазме крови обитают трипанозо-
мы сонной болезни; в эритроцитах живут и размножаются ма-
лярийные плазмодии, а в белых кровяных клетках — лейшмании.
В костном мозгу нападению малярийных паразитов подвергаются уже
молодые эритроциты; белые кровяные тельца костного мозга и лимфа-
тических узлов также поражаются лейшманиями. В полости грудного
протока живет филярия Банкрофта. В селезенке бывает эхинококк;
в лимфатических узлах—личинки пятиустки (Linguatula rhinaria).
’Мужские органы размножения: на glans penis бывает пендинская
язва; в придатке яичка — дизентерийные амёбы.
В яичнике бывает эхинококк. В полость матки и труб через влага-
лище заползают острицы; во влагалище не редок Trichomonas vagi-
nalis.
В лоханке почек паразитирует Dioctophyme renale, в мочевом пузы-
ре— бильгарции.
В головном мозгу бывают финки цепеня вооруженного; то же в
задней камере глаза; в хрусталике находили молодых заблудших дву-
усток. В конъюнктиве глаза бывают личинки вольфартовой мухи, могу-
щие совершенно уничтожить глаз. Паразитируют они и в наружном
слуховом проходе, разрушая барабанную перепонку.
Приведенный перечень паразитов является далеко не исчерпываю-
щим, но и из него видно, что некоторые паразиты не имеют строго
определенного места обитания, другие же, наоборот, живут только в
одном каком-либо органе. Примером первых являются финки и эхино-
кокк, а вторых — цепень невооруженный или лентец широкий, живу-
щие лишь в тонких кишках.
Возможность обитания в разных местах тела связана прежде всего
со способностью паразита проделывать миграцию в организме хозяина,
путь которой начинается от «ворот» внедрения паразита до места его
окончательной локализации. Миграция паразита, в точном понимании
этого явления, сопряжена с проникновением паразита из одной систе-
мы органов в другую и с активным прохождением через ткани различ-
ных органов.
Миграционные пути паразитов для многих из них связаны с местом
внедрения паразита в ткани хозяина. Например, церкарии бильгарций
проникают ц тело человека из воды в толщу кожных покровов; затем,.
26
ОБЩАЯ ЧАСТЬ
попав в вены, задерживаются, смотря по виду паразита, в печени, в мо-
чевом музыре или в венах прямой кишки.
Более кружный путь во время миграции проделывают личинки ас-
кариды, которые через стенку кишки проникают в сосуды, затем через
печень проходят в нижнюю полую вену, далее с током крови забрасы-
ваются в правое сердце и в малый круг кровообращения; из капилляр-
ной сети альвеол легких личинки активно проникают в полость альвеол
и по дыхательным путям поднимаются через трахею к гортани и глот-
ке. Здесь они проглатываются хозяином и, наконец, оседают в кишеч-
нике, где и достигают половозрелости.
Однако некоторые личинки могут пройти через капиллярную сеть
легких и попасть в легочные вены, откуда они вымываются в большой
круг кровообращения. Дальнейшая судьба этих личинок различна. Они
заносятся в разные органы, где и застревают. Фюллеборн наблюдал
оседание таких личинок с последующим реактивным капсулированием
в мозгу собаки (Toxocara cants).
Некоторые паразиты могут проделывать миграцию по разным пу-
тям. Например, личинки Ancylostoma duodenale — кривоголовки двена-
дцатиперстной — мигрируют или через кожу в кишечник (обычный
путь) или из кишечника через печень, сердце, легкие, пищевод, желу-
док — обратно в кишечник.
Феномен миграции встречается у паразитов чаще, чем о том пока
известно в науке.
Интересно, что кратчайший и, казалось бы, наиболее естественный
путь продвижения паразита к месту его окончательного обитания да-
леко не всегда является фактическим маршрутом миграции. Например,
обычно думали, что освободившаяся из проглоченной цисты молодая
печеночная двуустка из желудка переходит в двенадцатиперстную киш-
ку под влиянием химиотаксического привлечения желчи и по желчному
протоку проникает в печень (Лейкарт). Однако есть веские основания
считать, что молодая двуустка проникает в печень через систему во-
ротной вены, что связано, во-первых, с проникновением двуустки через
стенку желудка в сосуды и, во-вторых, с прохождением ее из капил-
ляров воротной вены через паренхиму печени в ее желчные каналы
(Ciurea).
Если паразит проделывает миграцию по путям кровообращения и
тока лимфы, то в случае каких-либо нарушений обычного хода этого
процесса паразит может осесть, сохраняя свою жизнеспособность, в со-
всем необычном для него месте. В таких случаях говорят о «заблудив-
шихся» паразитах; например, молодую двуустку находили в хруста-
лике, аскариды локализовались под кожными покровами и т. п.
Прохождением всего миграционного пути не истощается способ-
ность паразита к миграции. Е. Павловскому и В. Гнездилову удавалось
вызывать повторную миграцию плероцеркоидов лентеца шиоокого,
извлеченных из мышц ерша и введенных искусственно в желудок щуке,
откуда они снова проникали в мышцы.
Замечательны случаи, когда паразиты, передвигающиеся с током
венозной крови по телу, оседают в венах определенных органов. При-
меры этого в отношении различных бильгарций уже упоминались выше.
Очевидно, условия обитания для паразитов в венах печени, мочевого
пузыря и прямой кишки существенно различны, и разные виды биль-
гарций адаптировали каждый к определенным условиям обитания, бу-
дучи привлекаемы к соответствующему органу, вероятно, химиотакси-
ческими влияниями.
ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
27
Различные части организма, понимая под таковыми его органы и
ткани, представляют чрезвычайное разнообразие условий для существо-
вания в них паразитов.
Паразитарное население хозяина или, другими словами, его пара-
зитофауна вариирует весьма разнообразно; установление характерной
для данного вида хозяина паразитофауны может быть обеспечено при-
менением метода полного учета паразитов (Е. Павловский, В. Догель)
каждой особи хозяина. Однако этот метод применяется пока в ограни-
ченных пределах и то чаще для мелких животных.
Хотя человек в отношении его паразитофауны изучен гораздо лучше
животных (но не исчерпывающе!), все же из года в год происходят
открытия новых паразитов. Если в 1911 г. сводка паразитофауны чело-
века и домашних животных уложилась на 256 страницах крупного
шрифта с рисунками (Goedelst), то в 1925—1928 гг. определительные
каталоги насекомых и паукообразных эпидемиологического и санитар-
ного 'значения и паразитов человека вышли четырьмя книжками
в 408 стр. мелкой печати (Stiles и Hassal).
Для изучения проблемы «организм как среда обитания» необходимо
исходить от индивидуального материала, т. е. от каждой отдельной
особи хозяина по возможности во всей полноте ее паразитологического
населения.
Возьмем два-три конкретных примера. Urchs (1925) наблюдал
29-летнего пациента с прогрессивным истощением и анемией, лечивше-
гося от туберкулеза. У него были найдены следующие паразиты: кри-
воголовка (Necator), власоглав, кровяная двуустка (Schtsthosoma
mansbni), микрофилярии нитчатки (Wuchereria bancrofti) в крови;
яйца легочной двуустки (Paragonimus) и мигрирующие личинки, неви-
димому, аскариды в мокроте.
Такое сочетание паразитов в одном «пункте населения», т. е. в
больном, оказывает мощное действие на среду обитания, т. е. на орга-
низм хозяина всех этих паразитов, состояние которого, как мы видели
выше, было тяжелым. У этого человека обследование было произведе-
но только на паразитических червей; следовательно, полная заражен-
ность его паразитами могла оказаться еще большей.
Весьма показательный пример разнообразия гельминтофауны у од-
ной особи хозяина приводит К. Скрябин, обнаруживший у дикой
утки в Новочеркасске 17 видов паразитических червей в следующей
комбинации: в двенадцатиперстной кишке — 1 трематода Metorchis
xantosomus; в тонких кишках — трематоды — 3 Echinostoma revolution,
51 Strigea gracilis; ленточные черви — 56 Fimbriaria fasciolarls, 38 Ну-
те note pis carenula, 24 H, compressa, 2 H. anatina и 3 H. collaris; в сле-
пой кишке — 2 Hymenolepis compressa и 3 нематоды Capillaria anatis;
в прямой кишке — трематоды — Schistogonimus varies и Prostogonimus
sp. (по одной) и 4 скребня Polymorphus minutus\ под кутикулой зоба —
32 нематоды Amidostomum anseris; в трахее — 2 трематоды Т г acheo-
philus sisowi; в опухоли зоба—16 нематод Amidostomum anseris и
7 Tetrameres fissipinus\ в крови — 5 раздельнополых двуусток — Bilhar-
ziella polonica — всего 251 глиста. На других паразитов (наружные
паразиты, простейшие и др.) этот экземпляр утки обследован не был.
У двухлетней лошади Краузе нашел в Белграде 500 аскарид, 190
остриц, несколько миллионов стронгилид, 214 Sclerostomum, 69 Taenia
perfoliata, 287 нитчаток и 6 цистицерков.
Практика паразитических вскрытий показала приуроченность нахо-
ждения паразитов к определенным частям органов хозяина.
28
ОБЩАЯ ЧАСТЬ
Такая приуроченность зависит, очевидно, от своеобразия различных
органов и тканей тела хозяина в отношении биофизических и биохими-
ческих особенностей их, к которым адаптируют те или другие парази-
ты. Несомненно, что условия обитания малярийного плазмодия и его-
питания в эритроците иные, чем условия жизни лейшмании в белом,
кровяном тельце или в клетке ретикуло-эндотелия. Другим же парази-
там свойствен более широкий диапазон адаптаций; например, возбу-
дитель болезни Чагаса (Бразилия) (Trypanosoma cruzi) у человека
может поражать поперечнополосатые мышечные волокна, мышечные
клетки сердца, корковые клетки надпочечника, клетки невроглии, яич-
ника и других органов.
Приуроченность паразитов к обитанию в тканях может принимать
крайне специальный характер, когда паразитами избирается в каче-
стве места обитания лишь определенная часть клетки. Таковы прото-
плазматические паразиты, каковыми являются лейшмании; бывают и
ядерные паразиты, например Nucleophaga в амебе Endolimax williamsi.
В тех случаях, когда паразит по ходу своего жизненного цикла попа-
дает не в обычное для него место обитания, он погибает. Например,,
типичным местом обитания наездника Ageniaspis fuscicollis является
зародыш, развивающийся в яйце Яблоновой моли. Самка наездника,
откладывая свои яйца в яйцо Яблоновой моли, может проникнуть яй-
цекладом в ткань развивающегося зародыша или в желток, оставшийся
от яйца; соответственно этому развиваться будут лишь те яйца наезд-
ника, которые попали в ткани зародыша Яблоновой моли; яйца же, от-
ложенные в желток, дегенерируют и всасываются вместе с ним.
Трактуя организм, как среду обитания, необходимо четко обосновать
понятие «внешняя среда» по отношению к эндопаразитам.
Внешней средой для эндопаразита является
прежде всегото, чтонепосредственно его окружает.
Так, для шизонта малярийного паразита средой является строма оби-
таемого им эритроцита, далее плазма крови, в которой плавает зара-
женный эритроцит, далее тот орган, в котором проходят кровеносные
сосуды, и организм хозяина в целом, вне которого находится «офици-
альная» внешняя среда в обычном понимании этого термина.
Место непосредственного обитания паразита в
организме можно называть эндостацией; так, эндо-
стацией трипанозомы сонной болезни является плазма крови; следую-
щей категорией местообитания является «гостальный» биотоп
(т. е. биотоп, как часть тела хозяина), в рассматриваемом примере—>
кровяное русло.
Эндостация и гостальные биотопы находятся в теснейшей зависимо-
сти друг от друга. Взятые в целом, они образуют высшее единство —
организм хозяина, используемый для обитания рядом чуждых организ-
мов в лице паразитов животной или растительной природы.
Перед нами налицо явление интусусцепции («вложения») одного
организма (или множества организмов) в другой, что ведет к установ-
лению тесных форм взаимодействия эндопаразитов и
среды их обитания в лице организма хозяина.
Нигде резче мы не увидим примеров разнообразных изменений сре-
ды обитания под влиянием эндопаразитов, как результат их влияния на
организм хозяина. Вся патология паразитозов развертывает ряд не-
оспоримых примеров; упомянем хотя бы интоксикацию организма при
анкилостомидозе, инфантилизм при болезни Чагаса, злокачественное
малокровие при паразитировании лентеца широкого и др.
ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
29
Организм не является пассивным по отношению к его заселенцам.
Реакция капсуляции эндопаразитов с последующим омелотвореиием
капсулы ведет к гибели замурованных в них паразитов; выработка про-
тивотел приводит организм в состояние иммунитета по отношению к
повторным инвазиям; фагоцитарные процессы могут губительно влиять
на паразитов и т. д.
Понимая под внешней средой эндопаразитов организм заселенного
ими хозяина, мы можем отнести к сфере экспериментальной экологии и
все искусственные воздействия на эту внешнюю среду, которыми мы
можем ее частично изменить и тем влиять на паразитов. Дачей санто-
нина внутрь изменяется среда организма, и живущие в кишечнике
аскариды легче изгоняются вон. Такой же эффект можно получить и
при впрыскивании сантонина под кожу. В последнем случае также про-
исходит общее изменение среды организма, доходящее до кишечника и
обитающих в нем аскарид.
Описанные процедуры суть не что иное, как терапевтическое вме-
шательство; в таких соотношениях прямая терапия паразитЛз, будучи
ультряслециальной дисциплиной, идеологически должна занять место
среди отраслей прикладной экологии в ряду мероприятий по борьбе (хи-
миотерапия, серотерапия и др.).
В новейшей паразитологии, особенно в США, все яснее и яснее про-
является тенденция рассматривать влияние паразита на хозяина и об-
ратно — хозяина на паразита во взаимной связи; символом этого на-
правления является и установившаяся терминология: host — parasi-
te relations, что мы перевели бы, как соотношения «системы
хозяин — паразит». Под таким заголовком выходят и отдельные
книги, и ряд экспериментальных работ. Аналогичное направление опре-
деляется и в советской паразитологии.
Рассматривая паразитологические проблемы в таком новом свете, мы
должны беречься от ската на метафизические позиции. Уже подчеркива-
лось выше, что надо исходить из сугубо конкретных отправных точек
и памятовать, что помимо видовых и индивидуальных особенностей дан-
ного паразита и данного хозяина существенную роль в соотношениях
«хозяин — паразит» играют и те состояния, в которых находятся
оба партнера биологической системы х-паразит и у-хозяин.
В качестве примеров зависимости эндопаразитов от среды их обита-
ния и взаимных влияний паразитов и их хозяев остановимся на некото-
рых экспериментальных работах над паразитическими червями.
Многие исследования произведены над собачьей анкилостомой
(Ancylostoma caninurn), которая весьма удобна для лабораторного
экспериментирования как в отношении культивирования яиц, так и за-
ражения подопытных животных.
Собачья анкилостома — паразит собаки — может жить и в кишеч-
нике кошки. Один и тот же вид, следовательно, обитает в двух различ-
ных средах обитания, как собака и кошка, относящиеся к различным
семействам хищных млекопитающих. Сказывается ли на паразитах и в
какой мере пребывание этих глист в duodenum собаки или кошки?
Me Соу изучал интенсивность продукции яиц Ancylostoma caninum,
обитавших в кошке; самки этого «кошачьего штамма» давали в
среднем 2350 яиц в сутки каждая, в то же время самки того же вида,
жившие в собаке, откладывали в сутки по 16 тыс. яиц. Разница, как
видим, весьма резкая. Если «собачьим штаммом» анкилостомы заразить
кошку, то самки следующего поколения резко снижают половую про-
дукцию, давая в сутки по 2340 яиц. Противоположная картина наблю-
30
ОБЩАЯ ЧАСТЬ
дается при обратном эксперименте, когда «кошачьим штаммом» зара-
жают собаку: половая продукция самок в таком случае поднимается
до 11 160 яиц в сутки на одну глисту. Следовательно, степень плодови-
тости не является прирожденным расовым признаком изучаемого пара-
зита, а зависит от общего нутритивного действия хозяина на паразита.
A. caninum встречается в различных местах, то в собаках, то в кош-
ках, то и в тех и в других хозяевах; если заражать котят штаммом, по-
лученным от паразитов кошек, то половозрелости достигают 45|О/о ли-
чинок, а в щенках — менее одного процента личинок. При заражении
щенят собачьим штаммом вызревает половина всех личинок, а в котя-
тах— менее чем 5ю/о их. Морфологически оба штамма не отличимы
друг от друга, но можно говорить о некоторых физиологических отли-
чиях, связанных с адаптацией A. caninum к тому или к другому хозяину
(A. Scott).
При экспериментах с заражением хозяина паразитами необходимо
считаться с возможностью наличия приобретенного иммунитета, что мо-
жет смазать все результаты опыта. Приведем пример.
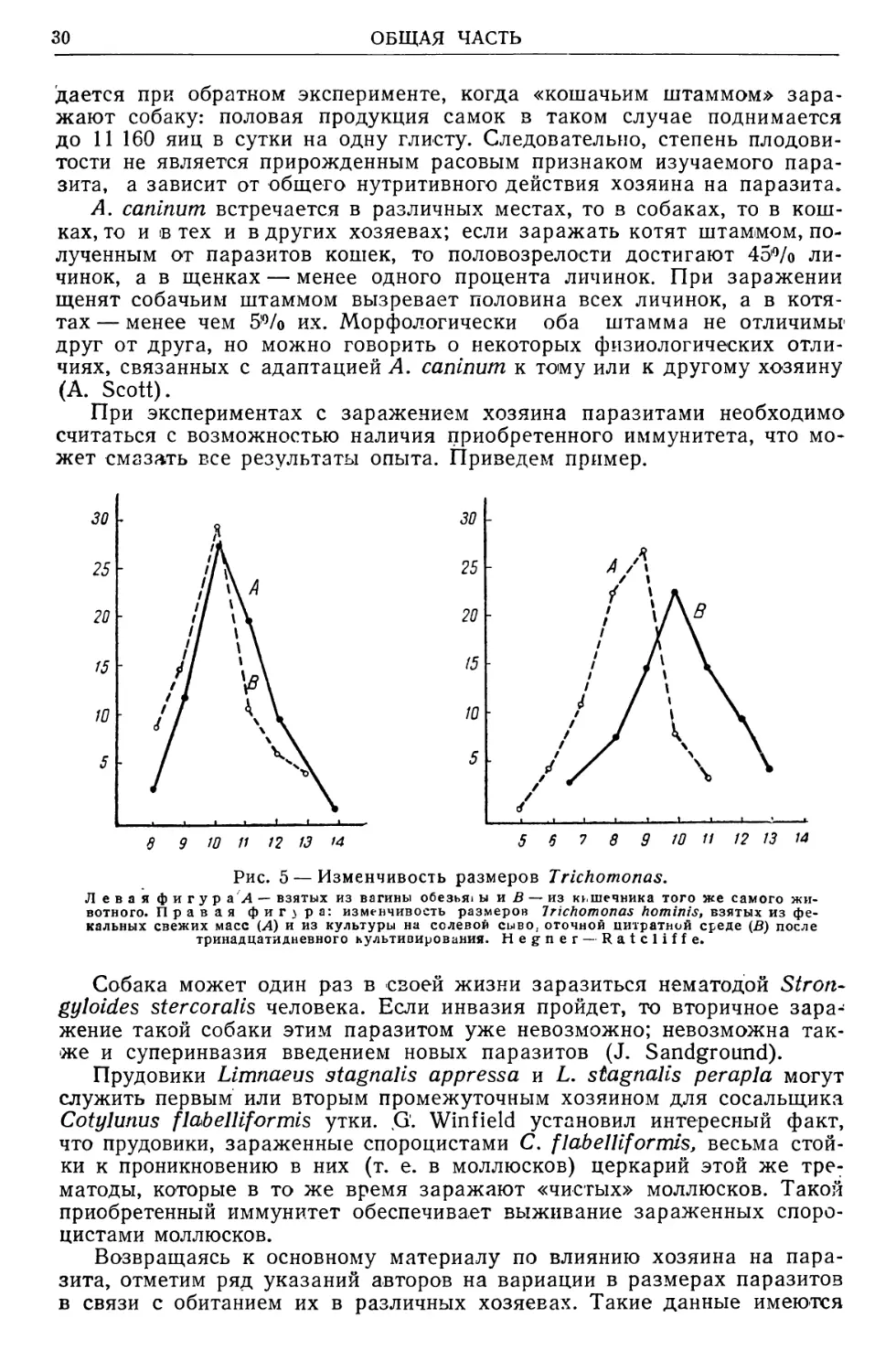
/4
Рис. 5 — Изменчивость размеров Trichomonas.
Л е в а я фигура Л — взятых из вагины обезья» ы и В — из кьшечника того же самого жи-
вотного. Правая ф и г з р а: изменчивость размеров Trichomonas hominis, взятых из фе-
кальных свежих масс (Л) и из культуры на солевой сыво, отечной цитратной среде (В) после
тринадцатидневного культивирования. Hegner — Ratcliffe.
Собака может один раз в своей жизни заразиться нематодой Stron-
gyloides stercoralis человека. Если инвазия пройдет, то вторичное зара-
жение такой собаки этим паразитом уже невозможно; невозможна так-
же и суперинвазия введением новых паразитов (J. Sandground).
Прудовики Limnaeus stagnalis appressa и L. stagnalis perapla могут
служить первым или вторым промежуточным хозяином для сосальщика
Cotylunus flabelliformis утки. G. Winfield установил интересный факт,
что прудовики, зараженные спороцистами С. flabelliformis, весьма стой-
ки к проникновению в них (т. е. в моллюсков) церкарий этой же тре-
матоды, которые в то же время заражают «чистых» моллюсков. Такой
приобретенный иммунитет обеспечивает выживание зараженных споро-
цистами моллюсков.
Возвращаясь к основному материалу по влиянию хозяина на пара-
зита, отметим ряд указаний авторов на вариации в размерах паразитов
в связи с обитанием их в различных хозяевах. Такие данные имеются
ОРГАНИЗМ КАК СРЕДА ОБИТАНИЯ
31
для Clonorchis sinensis, Schistosoma japonicutn — из паразитических
червей, для Trichomonas (рис. 5) из простейших и др.
Анкилостомы также изучались в указанном отношении. Собачьи
анкилостомы вариируют по длине у разных хозяев. Изучение анкило-
стом «кошачьего» и «собачьего» штаммов выяснило, что способность к
росту паразитов в общем у данных хозяев унаследуется паразитами, но
норма роста и окончательная величина червей контролируются организ-
мом хозяина, как средой обитания. Выдвинутый вопрос решается не
гак просто, так как помимо взаимных влияний хозяина и паразита мо-
гут играть большую роль характер и степень заселения органа парази-
тами.
При скоплении множества трематод одного вида в узких желчных
протоках печени размеры выросших паразитов бывают меньшими, чем
при единичном паразитировании. Трематода Psilotrema spiculigerum
достигает больших размеров в кишечнике утки, чему мыши (Mathias).
Яйца, личинки и взрослые Ancylostoma braziliense проявляют тен-
денцию к несколько большему росту в собаке, чем в кошке (М. Sarles).
Пока можно говорить лишь о накоплении частичных наблюдений и
выводов, так как рост паразитов связан со всей жизнедеятельностью
их в хозяине. Со стороны хо-
зяина играют роль факторы юооо -
видового, индивидуального по- 0Q0
рядка и, наконец, те состо-
яния, в которых находится юо.ооо -
данная особь хозяина в то
или другое время. 80.000 -
Одним из факторов влия-
ния на паразита является воз-
раст хозяина; чтобы обеспе- 4о.ооо •
чить точность выводов из
экспериментов, необходимо 20.000 •
исключить влияние приобре-
тенного иммунитета, о значе-
0 5 10 15 20 25 30 35 40 45
нии которого упоминалось
выше. При таких условиях
работы (М. Sarles) выяснено,
что Апс. braziliense достигает
половозрелости в кошках
медленнее, чем в котятах; из
личинок этой анкилостомы
Рис. 6 — Связь интенсивности размножения
бразильской анкилостомы с возрастом оби-
таемого ею хозяина.
На оси абсцисс отмечены дни после заражения хо-
зяина анкилост< мами, а на оси ордцнат — количество
яиц, откладываемых паразитом за сутки. Сплошная
кривая касается анкилостом, паразитирующих в котя-
тах, а прерывистая — в кошках.
только 3,96+ 1,92% вырастают во взрослых червей в кошках и
32,34 + 4,05% — в котятах.
Тот же паразит при обитании в котятах дает гораздо большую про-
дукцию яиц, чем во время жизни в кошках (М. Sarles, рис. 6).
Вся длительность жизни бразильской анкилостомы при паразитиро-
вании во взрослых кошках менее 2 недель, тогда как в молодых кош-
ках паразиты живут до 32 недель!
С другой стороны и возраст паразита отражается на его жизненных
функциях. Выше мы привели пример зависимости интенсивности про-
дукции яиц паразита от пребывания его в том или в другом хозяине.
Однако такой вывод можно делать лишь с учетом возраста паразитов.
Пример: средняя суточная продукция яиц самками бразильской анки-
лостомы, живущими в кошке, увеличивается с возрастом самок от 231
яйца на 21—22-й день жизни паразита до 2448 яиц к 43—45-му дню и
32
ОБЩАЯ ЧАСТЬ
4244 яиц к 50—51-му дню. Приблизительно тот же прогресс продукции
наблюдается и у Л. braziliense, живущих в собаке.
Все эти вопросы весьма сложны, и при анализе причин к ним надо
подходить с разных сторон. В частности, на половую продукцию пара-
зита, кроме факторов вида и возраста хозяина и возраста самого червя,
влияет момент перенаселения гостального биотопа паразитами того же
•самого вида. Для собачьей анкилостомы показано, что половая продук-
ция у этих паразитов уменьшается при большей интенсивности инвазии,
чем при малом количестве паразитов в кишечнике — и обратно (М.
Sarles).
При развитии нескольких особей паразита в хозяине для ряда видов
отмечается явление конкуренции между паразитами одного и того же
вида. Например, при развитии живших 240 дней в искусственно зара-
женной лягушке трех плероцеркоидов лентеца широкого, проглоченных
в процессе самоэксперимента человеком (В. Гнездилов), из кишечника
были изгнаны три ленточных червя 6, 3 и 1,5 м длиной с соответствен-
ной разницей в размерах их проглоттид.
Говоря ,о «перенаселении», или конкуренции, мы имеем в виду лишь
особей того же самого вида паразита. Вопрос усложняется еще более
тогда, когда в хозяевах находим разнообразные по своим составляю-
щим, паразитоассоциации. Таким образом выдвигается вопрос опара-
зитоценозах в хозяевах, причем под такими «сообществами» пони-
мается вся совокупность как животных, так и бактериальных организ-
мов, обитающих в том или в другом гостальном биотопе или в хозяине
в целом (Е. Н. Павловский).
Ряд наблюдений был произведен над влиянием кишечной протозой-
ной фауны и бактериальной флоры друг на друга. Если содержать крыс
на диэте, обеспечивающей развитие грам-отрицательных бактерий, то
количество паразитирующих в кишечнике простейших — Endamoeba
muris и Chilomasiix bettencourti увеличивается. Диэта, благоприятству-
ющая разрастанию протеолитических анаэробов с' уменьшением грам-
положительных и грам-отрицательных бактерий, ведет к уменьшению
количества паразитических простейших кишечника .(Н. Ratcliffe).
Теперь коснемся примеров влияния «состояния» организма хозяина
на живущих в нем паразитов. Если собаке, зараженной активным
штаммом Trypanosoma equiperdum, впрыскивать инсулин, то количе-
ство глюкозы в крови уменьшается и число трипанозом в крови падает
(Savino). Инъекциями инсулина можно влиять и на ход малярийных
приступов у птип (Mac Pougall). При частичном авитаминозе цыплят
(диэта без витамина В) заражение их нематодой' Ascaridia perspicillum
идет успешнее, и глисты в таких цыплятах вырастают крупнее, чем в
контрольных животных. Необходимо отметить, что A. perspicillum мо-
жет оказывать весьма заметное действие на пораженных ею цыплят
(рис. 7). Правда, такие «состояния» организма хозяина вызываются в
условиях эксперимента, но несомненно наличие различных «состояний»
организма и в естественной обстановке; нет ничего невероятного в том,
что при сезонном изменении пищевого режима грызунов, преобладания
по видовому составу и фармакологическим свойствам той или другой
растительной пищи — фауна паразитов их кишечника подвергается
соответственному воздействию.
Итак, валовое влияние хозяина на живущего в нем паразита сла-
гается из множества тесно связанных друг с другом моментов, причем
сам объект влияния — паразит, как член паразитоценоза данного био-
топа, воздействует на среду своего обитания, т. е. на хозяина.
ВЛИЯНИЕ ПАРАЗИТИЧЕСКОГО ОБРАЗА ЖИЗНИ НА ПАРАЗИТОВ 33
Очевидно, что для анализа этого сложного клубка взаимоотношений
должны быть мобилизованы по принципу комплексирования методы
сопредельных наук, имеющих ближайшее касательство к паразитоло-
Рис. 7 — Эффект влияния паразитирования нематод
Ascaridia на рост цыплят.
Сверху — зараженный паразитами цыпленок, снизу— контрольный
(здоровый) такого же возраста. Ackert Herrick.
гии — экспериментальной зоологии, общей патологии, биохимии, физио-
логии, серологии, микробиологии и др.
В этой плоскости, паразитологические проблемы должны получить
широкое общебиологическое и общепатологическое значение.
ВЛИЯНИЕ ПАРАЗИТИЧЕСКОГО ОБРАЗА ЖИЗНИ
НА ПАРАЗИТОВ
Паразитизм является крайне специальной категорией взаимоотноше-
ний между организмами различных видов, которые характеризуются, с
одной стороны, своеобразием питания паразита за счет организма хо-
зяина, с другой — пространственным использованием хозяина паразитом
илщ_в качестве места постоянного, или временного обитания.
л Переход от обычной формы жизни к паразитизму в процессе филоге-
нетического развития данного вида, связанный с борьбой за существо-
И
ОБЩАЯ ЧАСТЬ
?ис. 8—Голова блохи Pettinoctenus pavlovskyi.
Сильно развитые ктенидии на голове и груди.
Оригинал.
вание и естественным отбором, как правило, вел к приспособлению па-
разита к новым условиям существования.
В связи с этим при рассмотрении организации паразита (включая
и физиологические его особенности) необходимо выявить для надлежа-
щей оценки все характерные для него адаптации, т. е. приспособления,
обусловливающие возможность поддержания индивидуального и видово-
го существования в своеобразных условиях частичной или полной зави-
симости от хозяина.
"Адаптации паразитов весьма разнообразны. Для групповой их харак-
теристики следует выделять адаптации прогрессивные и регрессивные;
первые связаны с более сильным развитием или даже с новообразова-
нием некоторых органов па-
разита; вторые ведут к раз-
личной степени редуцирова.
ния или к полному исчез-
новению некоторых частей
телЪ паразита и иногда к
крайнему преобразованию
как внешнего его вида, так
и внутреннего строения.
Рассмотрим некоторые
примеры.
[ Для большинства пара-
зитов крайне важно надеж-
но держаться за своего хо-
зяина, ибо многие паразиты,
почему-либо потерявшие
своего хозяина, ни при ка-
ких условиях в другого хо-
зяина не попадут и поэтому
будут обречены на верную
гибель (например, аскари-
ды, вышедшие из кишеч-
ника) .
Поэтому у многих паразитов развиваются органы прикрепления или
за счет усиления развития и некоторого переформирования уже суще-
ствующих органов (например, ног), или в качестве специальных ново-
приобретенных частей.
Цепкие, сильные, снабженные хорошо развитыми коготками на кон-
цах лапок ноги вшей или густо волосистая нога блох дают представле-
ние об адаптивных применениях конечностей насекомых, перешедших
к наружному паразитированию.
Густые гребенки ктенидий на' голове и груди некоторых видов блох
(рис. 8) облегчают им возможность цепляться за волоски шерсти хо-
зяина.
Иногда колоссально развитые присоски у некоторых сосальщиков,
присоски и крючочки на головке ленточных глист способствуют надеж-
ному прикреплению этих внутренних паразитов к стенкам кишечника
и противодействию изгоняющей- силе его перистальтики.
У кровососущих членистоногих ротовые органы видоизменяются от
типичного грызущего ротового аппарата в колюще- или режуще-сосу-
щие органы (вши, блохи, клопы, слепни), причем последние могут одно-
временно служить и органами прикрепления к покровам хозяина) (иксо-
довые клещи). ^7
ВЛИЯНИЕ ПАРАЗИТИЧЕСКОЮ ОЬРАЗА ЖИЗНИ НА 11АРАЗИ1О15
30
^Пищеварительный канал у некоторых кровососущих паразитов при-
обретает емкие боковые выросты, что позволяет, например пиявкам
и иксодовым клещам выпивать большое количество крови; самки иксо-
довых клещей в насыщенном состоянии могут становиться в сотни раз
более тяжелыми, чем в голодном состоянии. Это обстоятельство и осо-
бенности обмена вещества обеспечивают пиявкам и клещам возможность
долгого голодания. На-
пример, клещи могут го-
лодать до 5—6 лет, а
платяная вошь не более
10 дней. Иногда способ-
ность к голоданию обу-
словливается лишь од-
ними физиологическими
особенностями паразита
(например, постельный
клоп)..^
Физиологической осо-
бенностью . внутренних
паразитических червей
является обладание анти-
ферментными свойства-
ми, определяющими воз-
можность глистам жить
в переваривающей среде;
при полном или частич-
ном умирании ткани па-
разита теряют антифер-
ментную стойкость и не-
медленно переварива-
ются.
"Иё менее характерной
физиологической адапта-
цией является приобре-
тение кишечными пара-
зитами способности жить
почти в полных анаэроб-
ных условиях за счет
интрамолекулярного ды-
Рис. 9 — Пищеварительный аппарат самки клеща
Dermacentor silvarum в отпрепарированном виде.
А — петли желудка, на олнеиные всоса ной кровью; В - слюн-
ные железы; С — трубки выделительных органов. Увеличено.
Оригинал.
хания, но с сохранением
еще прежней способности потреблять кислород атмосферного воздуха
(оплаты Гарнише с Triaenophorus nodulosus).
‘^Важнейшим условием возможности существования для паразитов
является необходимость попасть в организм хозяина, что сопряжено со
множеством случайностей. Неудивительно поэтому, что громадное ко-
личество яиц или личинок паразитов погибает, не попадая в необходи-
мые для них специальные условия существования. В связи с этим Гаран-
тией поддержания существования вида многих паразитов является их
чрезвычайная плодовитость, связанная с соответствующим строением
половых органов и с наличием физиологических предпосылок, обеспечи-
вающих продуцирование больших масс живого вещества и питательных
материалов (желтка) в виде откладывания яиц. О степени плодовито-
сти паразитов можно судить по примеру цепеня невооруженного, могу-
щего выделять ежедневно до 5 млн. яиц на 1 г фекалий (Ж. Штром).
36
ОБЩАЯ ЧАСТЬ
; Адаптирование паразитов к паразитическому образу жизни часто ве-
дет к ослаблению развития или к полной утере некоторых органов,
обычно по принципу редукции от неупотребления.
Так, приобретение специальных органов прикрепления может делать
бесполезными и ненужными ранее ймевшиеся органы передвижения в
виде ног и их мышц. Последние, равно как и органы зрения, начисто
исчезают у усоногих паразитических рачков (например, Sacculina) и
веслоногих рачков, живущих на жабрах рыб (рис. 10—11).
Рис. 10 — Паразитическая редукция весло-
ногого эктопаразитического рачка с жабр
трески. Мурман. Оригинал.
Рис. 11 — Циклоп — свободно жи-
вущий веслоногий рачек Cyclops
strenuus для сравнения его внешнего
вида с паразитическим рачком
рис. 10. Mdnnig.
Иногда наблюдаются редукции, кажущиеся парадоксальными, на-
пример полное исчезновение пищеварительного аппарата, органы кото-
рого, казалось бы, необходимы при лю$ых условиях существования (на-
пример, у ленточных червей).
Дело объясняется тем, что при обитании в среде, где происходит
переваривание пищи ферментами хозяина, ленточные черви перешли к
собственному питанию всасыванием перевариваемой вне их организма
пищи всей поверхностью своего тела (эндосмотическое п и та-
н и е); интересно, что содержащиеся в теле цестод ферменты (протеаза,
амилаза) не выделяются наружу, но служат для переработки эндосмо-
тически всасываемой пищи, подвергающейся предварительному воздей-
ствию пищеварительных ферментов хозяина.
В конечном результате прогрессивные и регрессивные адаптации мо-
гут до неузнаваемости изменить облик и внутреннее строение паразита
(ср. рис. 10 и 11: два веслоногих рачка — свободноживущий и парази-
тический). О систематическом положении последнего можно судить не
ОБЩИЙ ХАРАКТЕР ДЕЙСТВИЯ ПАРАЗИТОВ НА ХОЗЯЕВ
37
по строению взрослой (половозрелой) особи, но по особенностям
эмбрионального развития и превращения.
Так, в частности, принадлежность саккулины и лернеи к ракообраз-
ным устанавливается благодаря формам их личинок и возможности про-
слеживания их превращения.
ОБЩИЙ ХАРАКТЕР ДЕЙСТВИЯ ПАРАЗИТОВ
НА ХОЗЯЕВ
Формы действия паразитов на хозяев чрезвычайно разнообразны.
Рассмотрение этого материала относится к компетенции специальной
паразитологии, поэтому ознакомимся здесь с общим характером влия-
ния паразитов на своих хозяев, помня, что один и тот же паразит мо-
жет оказывать различное действие на хозяев разных видов. Но и на
одного и того же хозяина паразит может влиять не одинаково, что за-
висит от многих причин (количество паразитов в данной особи хозяина,
место их нахождения, состояние паразитов и хозяина и мн. др.).
* Первой формой влияния паразита на хозяина является механи-
ческое раздражение, испытываемое, хозяином при соприкосно-
вении его с эктопаразитами; таково щекотание при ползании эктопа-
разитов по телу, к этому прибавляется местное действие паразита,
когда он вкалывает ротовые части в покровы для сосания крови, одно-
временно впрыскивая в толщу покровов слюну. Примером такого влия-
ния может служить укол комара с болезненными ощущениями зуда и
жжения в месте укола (местное токсическое действие слюны биохими-
ческого характера)^ При массовом нападении комаров эффект отдель-
ных уколов суммируется, благодаря чему общий характер действия
становится для хозяина тягостным и нестерпимым {ср. также невоз-
можность спанья при нападении клопов). К таким моментам физиоло-
гического характера присоединяются и патологические изменения тка-
ней в месте укуса, в форме элементов дерматологического _ характера
(папулы, волдыри и др.) или в виде чисто патологических изменений
тканей с общей внешней реакцией покровов (воспалительная краснота,
отек и др.).
Токсические начала, эктопаразитов, всасываясь в кровь и разносясь
по телу, могут вызывать общую или, как говорят, резорптивную реак-
цию организма; например, при большом завшивлении человека может
наблюдаться токсическая лихорадка, как ответ организма на поступле-
ние множества доз слюны вшей.
Другие паразиты могут производить более или менее грубое разру-
шение тканей, нахождение паразита даже в ороговелых слоях эпидер-
миса вызывает —в случае паразитирования чесоточного зудня — тяго-
стные симптомы чесотки.
Несоразмерно более крупные по величине личинки желудочного ово-
да лошади (Gastrophilus) в первой стадии метаморфоза ьмогут вбурав-
ливаться в мальпигиевы слои эпидермиса человека, где также прокла-
дывают себе ходы. Несмотря на больший механический эффект разру-
шения тканей, болезненные ощущения человека гораздо более слабы и
терпимы, чем при чесотке.
/ Как следующую ступень действия эктопаразитов отметим обширные
разрушения тканей, которые причиняются паразитическими личинками
вольфартовой мухи, могущими «разъедать» раны или, например, пол-
ностью уничтожить глазное яблоко или покровы головы (рис. 12).
Эффект действия внутренних паразитов еще более разнообразен, чем
38
ОБЩАЯ ЧАСТЬ
паразитов наружных, что зависит от видовых свойств паразитов и ме-
ста их локализации в хозяине (подразумевается состояние хозяина,
восприимчивого к заражению паразитом и к его влиянию).
Наиболее простой формой влияния- является и здесь механическое
действие эндопаразита на орган, в котором он находится. Последствия
этого действия могут быть весьма разнообразными. Например, клубок
аскарид может вызвать закупорку и непроходимость кишечника со всеми
вытекающими последствиями, если не будет оказана своевременно хи-
рургическая помощь. Закупорка грудного протока филярией Банкрофта
ведет к застою лимфы и расширению протока до 15—16 см диаметром'.
Рис. 12 — Миаз от личинок вольфартовой мухи, разрушивших покровы
голову!. Юго-западная Туркмения. По Е. Павловскому.
Хроническое давление паразита йа ткани органа может вызывать их
атрофию, так, например, пузыри эхинококка могут «разъедать» трубча-
тые кости (рис. 13). Однако один и тот же паразит оказывает разное
действие на здоровье хозяина, что зависит от места нахождения пара-
зита. Фийка цепеня невооруженного где-нибудь в мышце может оста-
ваться неощутимой для человека; та же финка, но в мозгу человека
ведет к тяжелому расстройству и даже к смерти.
^Механическое действие эндопаразита часто проявляется нарушением
целости ткани в местах прикрепления паразита; таково действие крю-
чочков на головке цепеня вооруженного или кутикулярных зубов во
рту анкилостомы. Микроскопически резко обособлены кратерообразные
углубления на внутренней поверхности стенки желудка в местах при-
крепления личинок третьей стадии желудочного овода лошади
С механическим действием сочетается часто и добавочное влияние
вводимых в толщу тканей хозяина веществ, выделяемых различными
органами паразита. Пиявка, присосавшаяся к стенке глотки, прорезает
челюстями слизистую оболочку и вводит в кровь секрет головных же-
лез, обладающий сильным антикоагулирующим свойством. Секрет го-
ловных желез анкилостомы и некатора вызывает разрушение красных
кровяных телец (так называемое гемолитическое действие).
Токсическими свойствами могут обладать и продукты обмена ве-
ОБЩИЙ ХАРАКТЕР ДЕЙСТВИЯ ПАРАЗИТОВ НА ХОЗЯЕВ
39
ществ эндопаразитов, всасываемые стенкой кишечника, если паразиты
живут в его просвете, или поступающие от тканевых паразитов непо-
средственно в тканевые соки или в кровь. В первом случае токсические
Рис. 13 — Разъедание эхинококком берцовых костей человека.
Фотография поЕ. Павловскому.
Рис. 14—Отек соединительной ткани и воспалительная
инфильтрация с эозинофилией вокруг личинки кожного
овода Hypoderma в стенке пищевода коровы. Микро-
фотография. Ори1инал.
вещества подвергаются действию барьера печени (что может ослаблять
или нейтрализовать их действие); во втором — действие их является
прямым.
40
ОБЩАЯ ЧАСТЬ
Реакция организма может быть местная или общая. Например, во-
круг мигрирующих личинок второй стадии кожного овода коров (Hypo-
derma, рис. 14) в стенке пищевода образуется, сильный отек с обильной
мелкоклеточной инфильтрацией и эозинофилией. В качестве общей ха-
рактерной реакции организма на действие эндопаразитов можно ука-
зать на состояние инфантилизма, наблюдаемое иногда при болезни Ча-
Рис. 15—Инфантилизм
у 19-летней больной бо-
лезнью Ч а г а с a (Try-
panosoma cruzi) в Бра-
зилии. По R. Kraus.
гаса (трипанозомоз в Бразилии). В других
случаях при той же болезни развивается
зоб (рис. 15, 16). Практически трудно дифе.
ренцировать в каждом отдельном случае
влияния токсинов эндопаразитов и их
Рис. 16 — Паразитический зоб при
болезни Ч а г а с а.
По R, Kraus.
экскреторных продуктов, тем более, что некоторые паразиты действуют
в обоих отношениях. К реакциям организма гистопатологического ха-
рактера на тех или других эндопаразитов относятся: цирротические
изменения печени при паразитировании печеночной двуустки; аденома-
тозные разрастания в желчных протоках при кокцидиозе печени кроли-
ка; папилломы в мочевом пузыре человека при бильгарциозе Schistoso-
ma haematobium), принимающие иногда злокачественный характер (рак
мочевого пузыря) и мн. др.
Следует отметить также явления капсуляции некоторых тканевых
паразитов, например, трихин, за счет реактивного преобразования ткани
вокруг личинки паразита, внедрившейся в поперечнополосатое мышеч-
ное волокно и вызвавшей его перерождение в месте своей локализации.
ТД-Таконец, паразиты могут способствовать побочному внедрению в
организм возбудителей инфекции. Например, через места прикрепления
ПОНЯТИЕ О ПАТОГЕННОСТИ ПАРАЗИТОВ
4!
паразитических червей (где происходит нарушение целости слизистой
оболочки, что ведет к нарушению ее барьерной функции) болезнетвор-
ные бактерии из полости кишечника могут проникать в ткани и вызвать
заболевание; так, например, связывают некоторые случаи аппендицита
с нахождением остриц в просвете червеобразного отростка слепой
кшпки.
^Обычно паразиты оказывают на своего хозяина комбинативное дей-
ствие, важнейшие составные элементы которого были нами только что
рассмотрены.
ПОНЯТИЕ О ПАТОГЕННОСТИ ПАРАЗИТОВ,
ПАРАЗИТОНОСИТЕЛЬСТВЕ И О РЕЗЕРВУАРАХ ВИРУСОВ
Наряду с патогенными бактериями своевременно выдвинуть понятие
о патогенных животных. Таковыми являются животные, веду-
щие паразитический образ жизни, переносчики возбудителей вирусных,
бактериальных и паразитарных болезней и ядовитые животные. Однако
не всякий паразит, могущий обитать в теле человека, обладает болезне-
творными свойствами, и не всякий болезнетворный паразит непременно
вызывает заболевание своего хозяина. Иллюстрацией такого положения
служат частые случаи паразитоносительства, аналогами ко-
торых являются случаи бациллоносительства.
ПаразитоносительствОлМ называется такое состояние, когда хозяин
жизнедеятельного паразита не проявляет никаких симптомов заболева-
ния, связанного с заражением паразитом и с последующим его суще-
ствованием. Такое бессимптомное инвазирование хозяина в ряде случа-
ев не ведет к последующим клиническим проявлениям; но, оставаясь
здоровым, паразитоноситель может рассеивать вирулентные паразитар-
ные начала и быть источником распространения соответствующей инва-
зии.
К паразитоносительству относятся явления различного порядка. За-
разившиеся паразитом, но не заболевшие соответствующей болезнью,
являются здоровыми паразитоносителями. Оставаясь здоровы-
ми, такие паразитоносители могут претерпевать важные изменения в
своем организме, например, приобретая невосприимчивость к повторно-
му заражению.
В других ^случаях имеет место скрытый (латентный) паразитизм,
который в течение более или менее длительного срока предшествует
клиническому проявлению паразитарной болезни. Например, при маля-
рии бывают случаи длительной (в течение нескольких месяцев)
инкубации, в течение которой человек является носителем малярий-
ного плазмодия, но без малейших проявлений болезни. Наконец, явле-
ния паразитоносительства устанавливаются нередко уже после оконча-
ния течения инвазионной болезни. Таково, например, га мето носитель-
ство у хронических маляриков.
Бессимптомное паразитоносительство в ряде случаев нелегко распо-
знаваемо от скрытой формы заболевания, когда признаки
последнего могут быть установлены лишь при тщательном патологиче-
ском обследовании человека. Например, носительство лентеца широкого
нередко не сопровождается обращающими на себя внимание симптома-
ми; в частности, злокачественное малокровие развивается лишь у части
носителей. Однако детальными исследованиями крови носителей Н. Ра-
гоза показал, что у них имеются изменения крови, являющиеся как бы
начальной фазой могущего -развиться в будущем злокачественного
малокровия.
n
ОБЩАЯ ЧАСТЬ
В связи с этим естественно рассмотреть вопрос о патогенности, как
о свойстве паразитов. Будучи особенностью качественного порядка, п а-
тогенность паразита распознается не как особен-
ностьпаразита«в себе», а по отношению к организму
хозяина, с которым так или иначе связан оцениваемый паразит.
Один и тот же паразит может быть патогенен для одного хозяина и
безвреден или вовсе не заразителен для другого.
,В главе «Организм, как среда обитания паразитов» показано дей-
ствие организмов хозяина и его паразитов друг на друга. В конечном
счете состояние паразитоносительства хозяина или
его заболевание паразитарной болезнью является
результатом взаимных влияний организмов хозяи-
на и заселяющих его паразитов в конкретных усло-
виях среды.
Отсюда естественно заключение об относительности качества пато-
генности паразита, ибо оно развертывается на живом и изменчивом по
своим состояниям субстрате, коим является организм хозяина. Следует
также учитывать условия влияния также изменчивых факторов внеш-
ней среды и на паразита, и на хозяина во всех моментах их контакта
и взаимодействия; само собой разумеется, что и различные особи или
поколения паразитов в свою очередь изменчивы в отношении своих
свойств, в частности действия на хозяина. Этими причинами и обуслов-
ливается широта диапазона изменчивости состояния хозяина—от бес-
симптомного паразитоносительства до тяжелейших проявлений парази-
тарной болезни.
Известно не мало примеров, когда паразит определенного вида жи-
вет в диком (или в диких) животном и паразитирует также и у челове-
ка. Например, трипанозома Крупа — возбудитель болезни Чагаса у че-
ловека— бывает в крови у броненосца, для которого она, видимо, не-
патогенна. В организме броненосца эта трипанозома проходит нормаль-
ный цикл развития. Броненосец, поэтому, рассматривается как резер-
вуар возбудителя болезни Чагаса. Это млекопитающее обеспечивает
существование болезни Чагаса в природе при наличии насекомых-пере-
носчиков (поцелуйный клоп) этого вируса, которые могут передать воз-
будителя болезни от броненосца человеку.
Животные резервуары вируса1! могут быть в той или другой мере
подвержены его болезнетворному действию, но этим не умаляется их
роль в поддержании очага вируса в природе. Например, по новейшим
исследованиям Н. Латышева, до 60% грызунов (песчанка и др.)
очаге пендинской язвы (Ю. Туркмения) заражены кожными лейшма-
ниями. Эти грызуны и являются резервуаром, откуда москиты-перенос-
чики получают при кровососании паразитов пендинки и передают их
человеку, заболевающему в конечном счете пендинской язвой.
В сходном порядке резервуаром вируса клещевого рекурренса в
Средней Азии являются крысы (Г. Змеев, М. Кешишьян).
Животными резервуарами вируса, по Брумпту, следует считать те
организмы, в которых паразиты могут достигать половозрелое™ и про-
дуцировать яйца или цисты, или же приходить в видимое или невиди-
мое состояние, при котором, в естественных условиях, возможен прямой
или непрямой переход вируса от одного хозяина к другому. Поиски жи-
вотных резервуаров вирусов являются одним из важнейших направле-
ний в исследовании эпидемиологии паразитарных и трансмиссивных бо-
лезней, раскрывающим условия природной очаговости многих из этих
болезней. Указанное направление исследований тесно контактирует с
биологией, экологией и фауной изучаемых животных.
ФАКТОРЫ СТАНОВЛЕНИЯ ОРГАНИЗМА ХОЗЯИНОМ ПАРАЗИТОВ
43
ФАКТОРЫ СТАНОВЛЕНИЯ ОРГАНИЗМА ХОЗЯИНОМ
ПАРАЗИТОВ
Источником заражения хозяина паразитами, за редкими
исключениями внутриутробной инвазии, является внешняя среда, пони-
мая под таковой, когда это необходимо, и других особей того же вида
хозяина. Что касается способа заражения организма паразита-
ми, то это обстоятельство характеризуется прежде всего характером
пути проникновения паразита в тело хозяина.
Все варианты форм заражения можно свести к двум категориям:
а) проникновение паразитов в полостные органы, имеющие сооб-
щение с внешней средой, и
б) внедрение паразитов в организм через покровы с нарушением их
целости.
Если оставить в стороне сравнительно редкие случаи поступления
паразитов через уретру, влагалище, анальное отверстие, а также отвер-
стия носа и уха, то первую категорию способов заражения можно ко-
ротко охарактеризовать как инвазирование per os — с пищей, питьем,
или через предметы, которые так или иначе могут попадать в рот. Вто-
рая категория способов заражения характеризуется как инвазирование
контактного характера, когда поверхность тела соприкасается с непо-
средственным источником инвазии.
Становление особи хозяином паразитов зависит от совокупного дей-
ствия факторов трех категорий, которое рассмотрим применительно к
заражению хозяина паразитами per os.
I. Первым условием возможности заражения организма эндопарази-
тами является наличие такого анатомического строения и таких физио-
логических особенностей хозяина, которые не препятствуют: 1) проник-
новению вирулентного паразита в тело хозяина, 2) продвижению пара-
зита до места обитания в хозяине и 3) существованию паразита са
всеми его жизненными отправлениями в организме хозяина. Условимся
называть все обстоятельства такого рода факторами, предраспола-
гающими организм к заражению и обитанию в нем рассматри-
ваемого паразита.
Обратимся к примерам.
Толстая кутикула желудка взрослой черепахи непроницаема в усло-
виях эксперимента для плероцеркоидов лентеца -широкого, тогда как
последние проникают сквозь нее у молодой черепахи (Е. Павловский
и В. Гнездилов). Молодые белые крысы менее стойки к заражению
цестодами Hymenolepis fraterna и Н. папа, чем крысы взрослые (при-
чинным моментом является совокупность физиологических особенностей
возрастного состояния). С возрастом почти прекращается заражаемость
крупного рогатого скота личинками кожного овода (Hypoderma bovis).
Зарастание жаберных щелей головастика лягушки делает невозможным
дальнейшее обитание в этой части тела личинки Polystomum integerri-
тит, которая продвигается в пищеварительный канал и со временем
пробирается из клоаки в мочевой пузырь лягушки. Строение рта, уса-
женного коническими роговыми зубами, миноги, и способ поедания ею
рыб исключает возможность проглатывания ею плероцеркоидов лентеца
широкого, хотя в условиях эксперимента при искусственном введении
их в желудок миноги они начинают пробуравливаться через стенку пи-
щевода и желудка (Е. Павловский и В. Гнездилов).
И. При наличии в составе членов биоценоза организма, предраспо-
ложенного к заражению (в силу его благоприятного анатомического
14
ОБЩАЯ ЧАСТЬ
строения и физиологического состояния), и вирулентного паразита —
заражение хозяина определяется самою возможностью проникновения
паразита в тело хозяина. Последнее осуществляется тогда, когда пара-
зит и хозяин являются действующими звеньями «цепи» или «цикла»
питания в силу существования между ними биоценотических связей
алиментарного или трофического характера.
Промежуточным хозяином цепеня огуречного (Dipylidium caninum}
является собачий или кошачий власоед, в котором из проглоченных им
яиц глисты развиваются цистицеркоиды. Собака и кошка легко про-
глатывают власоедов при лизании • языком шерсти. Из проглоченных
цистицеркоидных власоедов в кишке окончательного хозяина развива-
ются ленточные черви. В то же время между человеком и власоедами
собак и кошек нет связей алиментарного характера, и человек' свободен
от соответствующих ленточных червей. Однако в редких случаях про-
глатывания при каких-либо особых обстоятельствах указанных проме-
жуточных хозяев цепень огуречный нормально развивается и в кишеч-
нике человека. В рассматриваемом случае анатомические и фи-
зиологические черты организаций человека пред-
располагают его к заражению Dipylidium caninum, но от-
сутствие факторов определяющего порядка в ф^ о р im е
биоценотических связей алиментарного характе-
ра исключает, как правило, возможность паразити-
рования этой цестоды в организме человека.
III. При наличии предрасположенного к заражению хозяина и виру-
лентного паразита (факторы предрасполагающие) и при осуществлении
проникновения паразита в организм хозяина (факторы, определяющие
возможность заражения) последний не всегда будет заражаться рассма-
триваемым паразитом.
Ознакомимся с примерами. Если малярийный комар, насосавшийся
крови малярика с жизнедеятельными гаметоцитами тропической маля-
рии, попадает в температуру ниже 15° С, то он не приобретает заража-
ющих свойств, так как в его желудке не произойдет оплодотворения
плазмодия и последний погибнет. В данном случае фактор внешней
среды — низкая температура — при наличии всех прочих благоприятных
условий пресекает развитие плазмодия и исключает возможность ста-
новления данной самки анофелес хозяином спорозоитов плазмодия.
Различные виды эндопаразитов неодинаково чувствительны к дей-
ствию низкой температуры. Удачно зараженные малярийными плазмо-
диями — Plasmodium vivax и Р. ovale — малярийные комары (Anopheles
maculipennis} при содержании их на леднике удерживают способность
заражать человека малярией в течение нескольких месяцев; аналогич-
ную способность по отношению к паразиту тропической малярии анофе-
лес может сохранять лишь несколько дней (Джемс и Шют).
Такое же значение может иметь и повышение температуры. В усло-
виях опыта можно заразить геккона плероцеркоидами лентеца широко-
го, взяв их из рыбы и введя прямо в желудок геккона при комнатной
температуре; если же зараженного геккона поместить в термостат при
35° С, то миграции плероцеркоидов в нем не совершается и сами пле-
роцеркоиды гибнут уже к ближайшему утру (Е. Павловский и
В. Гнездилов).
В конечном счете при наличии всех благоприятных условий стано-
вление данной особи хозяином вирулентного пара-
зита зависит от факторов осуществляющего поряд-
к а {факторы среды).
ПОНЯТИЕ О ТРАНСМИССИВНЫХ БОЛЕЗНЯХ
45
Резюмируя все приведенные соображения, мы можем утверждать,
что с т а н о в л е н и е данной особи хозяином определен-
ного вирулентного паразита зависит от совокупности влия-
ния факторов трех категорий:
1) когда анатомо-физиологические черты организации предрас-
полагают особь хозяина к заражению вирулентным паразитом;
2) когда биоценотические связи алиментарного характера опреде-
ляют возможность проникновения вирулентного паразита в чувстви-
тельного к заражению хозяина и
3) когда факторы внешней среды, прямо или косвенно действующие
и на восприимчивого хозяина, и на вирулентного паразита, в конечном
счете осуществляют возможность приживания вирулентного па-
разита, проникшего в организм чувствительной особи хозяина (Е. Пав-
ловский и В. Гнездилов).
ПОНЯТИЕ О ТРАНСМИССИВНЫХ БОЛЕЗНЯХ
Болезни человека и животных классифицируются прежде всего но
их этиологии. По этому принципу выделяются группы инфекционных
и паразитарных болезней.
К инфекционным болезням относятся: бактериальные болезни или
бактериозы, возбудителями которых являются различные патоген-
ные бактерии (холера, брюшной тиф, дифтерия и др.), грибковые
заболевания — микозы (например, антиомикоз), болезни, вы-
зываемые фильтрующимися вирусами (желтая лихорадка, сезонные
энцефалиты), спирохетозы (возвратный тиф, лептоспироз, сифи-
лис) и риккетциозы (различные формы сыпнотифозных лихора-
док).
К паразитарным болезням или паразитозам относятся болезни,
вызываемые патогенными простейшими (протозоозы), паразитиче-
скими червями (г е л ь м и н т о з ы), патогенными паукообразными, в
частности клещами (акаринозы), и насекомыми (энтомозы).
Наряду с такой группировкой болезней — по этиологическому прин-
ципу, их классифицируют, далее, смотря по характеру путей их распро-
странения, т. е. по принципу эпидемиологическому.
Капельные инфекции, при которых возбудители болезни вы-
брасываются изо рта с капельками слюны с последующим вдыханием
их здоровыми людьми, едва ли имеют аналогов среди болезней парази-
тарной природы. Быть может, простейшие полости рта иногда переда-
ются таким путем.
В отношении пылевых инфекций, при которых возбудитель
распространяется с пылью в сухом виде, есть предположение, что яйца
остриц могут распространяться именно таким путем. Цисты паразити-
ческих простейших при высыхании фекалий могут попадать в воздух;
но вопрос в том, сохраняют ли они жизнедеятельность и могут ли в
сухом состоянии заражать здоровых людей, когда с вдыхаемой пылью
попадают в рот и проглатываются в пищеварительный канал.
Зато пищевые или алиментарные инвазии весьма
обычны и многообразны. Под пищевыми инвазиями подразумеваются
все те паразитные болезни, возбудители которых'поступают в организм
хозяина per os-, будь то пищевые продукты, получаемые от инвазирован-
ных промежуточных хозяев (например, финнозное мясо), или ,же ин-
вазирующие стадии паразитов (созревшие яйца, цисты и др.), которые
человек проглатывает с загрязненной пищей, водой или через предме-
46
ОБЩАЯ ЧАСТЬ
ты обихода, применяемые при еде или питье, или, наконец, через по-
средство загрязненных рук (большая часть глистных болезней, болезни,
вызываемые паразитическими простейшими кишечника и др.).
Многие паразиты проникают в организм хозяина контактным
путем, т. е. при соприкосновении человека с зараженным больным,,
загрязненными предметами обихода, при пребывании на зараженном
субстрате (почва) или при соприкосновении с зараженной водой. Для
таких контактных инвазий характерна способность некоторых эндопа-
разитов активно проникать через неповрежденные кожные покровы или
слизистые оболочки хозяина.
Контактным путем передается при копуляции Trypanosoma
equiperdum — возбудитель случной болезни лошадей. Вши переходят на
здорового человека при соприкосновении с завшивленным, с его платьем
или постельными принадлежностями. То же характерно и для чесоточ-
ного клеща. Из почвы личинки анкилостомид активно вбуравливаются
в кожу человека, когда он обнаженными покровами соприкасается
с участками зараженной почвы. В аналогичном порядке происходит за-
ражение человека бильгарциями через вбуравливание церкарий в по-
кровы человека при нахождении его в инвазированном водоеме.
К группе контактных инвазий следует причислить также случаи,
когда самка пристраивает свое потомство на тело хозяина или в его
полостные органы. Таковы личинкородящие оводы и мухи, на лету «вы-
стреливающие» отрождаемыми личинками в глаза, в носовую полость,
в ушной проход или в ранки на теле. Аналогичное явление представляет
откладывание яиц некоторыми мухами на покровы ослабленного хозяи-
на, что ведет к последующему развитию миаза (поражение личинками
мух).
К контактным инвазиям относятся в свою очередь случаи, когда хо-
зяин «набирается» голодных пастбищных клещей при соприкосновении
с растительностью, на которой эти клещи сидят.
К этой же категории болезней сопричисляются случаи внутриутроб-
ных инвазий (см. выше), так как развитие плода1 в теле матери можно
рассматривать как взаимный контакт перманентного характера.
Последняя группа паразитарных, равно как и инфекционных болез-
ней, которую следует выделить особо, характеризуется тем, что воз-
будители их передаются через посредничество
переносчиков, каковыми являются главным обра-
зом насекомые и клещи. Такие болезни автором предложено
называть трансмиссивными.
Следует различать трансмиссивные болезни par exellence. На со-
временном этапе существования жизни на земле болезни такого рода
передаются исключительно через переносчиков, но не какими-либо дру-
гими способами; например, малярийный плазмодий и его переносчик —
анофелес. Такие болезни можно назвать облигатно-трансмис-
сивными. Для ряда других болезней передача через посредство пе-
реносчиков является лишь одним из путей их распространения.
Например; туляремия распространяется контактным путем (сдирание
шкурки больного грызуна), через потребление зараженной пищи или
воды или через посредство различных переносчиков (вши грызунов, ко-
мары, слепни, клещи и др.). В таких случаях устанавливают различные
в эпидемиологических отношениях преимущественные формы эпидемий
туляремии, и В том числе эпидемии трансмиссивные, когда болезнь рас-
пространяется именно через переносчиков. В таком случае мы имеем
дело с факультативно-трансмиссивной болезнью.
ПУТИ ЦИРКУЛЯЦИИ ВИРУСОВ
47
Изучение эпидемиологии трансмиссивных болезней должно вестись
комплексно с активным и равноправным участием паразитологов-энто-
мологов; ведущая роль в выяснении путей передачи возбудителя болез-
ни через посредство переносчика принадлежит паразитологии или энто-
мологии. К их компетенции относится выявление всех форм биоценоти-
ческих связей, характером которых определяются как пути циркуляции
вируса болезни, так и методы пресечения таких путей.
ПУТИ ЦИРКУЛЯЦИИ ВИРУСОВ. ПРИРОДНАЯ
И АНТРОПУРГИЧЕСКАЯ ОЧАГОВОСТЬ ТРАНСМИССИВНЫХ
БОЛЕЗНЕЙ
Внутренние и наружные паразиты поддерживают свое существова-
ние, как видов, переходя (активно или пассивно) в процессе размноже-
ния из организма одного хозяина в организм другого хозяина; оба эти
хозяина могут относиться к одному и тому же виду или, наоборот —
принадлежат к далеко отстоящим по зоологической системе существам.
Раскрывая весь круг возможных хозяев для паразита и рассматри-
вая моменты, определяющие переход паразита от хозяина к хозяину
или во внешнюю среду и опять в хозяина, мы получаем представление
о путях циркуляции паразитов в природе.
Рассмотрим подходящие примеры.
Для стеноадаптивных паразитов и особенно для паразитов-монофа-
гов пути циркуляции узко ограничены; например, человеческие вши
переходят с человека на человека при контакте; побочным путем яв-
ляется пребывание вшей в снятой одежде или в постельных принад-
лежностях с последующим переходом на человека при контакте его с
зараженными предметами:
человек -> вошь -> человек -> вошь -> человек -> вошь -> человек
I
платье--------вошь---------> человек
Для паразитов, питающихся за счет нескольких видов хозяев, пути
циркуляции оказываются более многообразными, например:
трихина крыса трихина -> крыса -> трихина
tl II
трихина трихина человек
Н I t
свинья -> трихина -> свинья -> трихина -> человек -> трихина
Более определенны пути циркуляции паразитарных начал, когда за-
вершение жизненного цикла паразита связано с переменой хозяев.
Например, для цепеня невооруженного (Taeniarhynchus saginatusy
собака (—)
собака (—) <- яйца -> корова — финка -> человек — цепень
________--
С фекалиями человека яйца цепеня могут быть проглочены многи-
ми животными, но развиваются далее в финок . они, как правило, в
организме коровы. Финнозное мясо коровы могут поедать различные
хищные животные, но ленточный червь развивается из финок только
ОБЩАЯ ЧАСТЬ
в кишечнике человека. Таким образом, на путях циркуляции парази-
тарное начало может попадать в «тупики», где оно и погибает. В разо-
бранном примере таким тупиком для финки является организм собаки.
Пути циркуляции весьма усложняются при полифагии паразитов,
при наличии двух промежуточных хозяев и при пребывании паразитов
также и во внешней среде.
В аналогичном порядке следует рассматривать пути циркуляции
возбудителей трансмиссивных болезней протозойной, бактериальной
или вирусной природы. Для некоторых болезней такие пути много-
Рмс. 17— Схема путей циркуляции спирохет клещевого возвратного тифа из
организма переносчика в организм чувствительных к заражению животных и
человека.
Стрелками показан источник питания для отмеченных паразитов. Из постельного клона
дальнейшего выхода спирохедам нет. Оригинал.
образны и неспецифичны. Например, бактерии брюшного тифа могут
быть получаемы комнатными мухами при сосании мочи больного или
при соприкосновении с его фекалиями. Далее муха может переносить
их на посуду, пищу, попадать сама в жидкости, выпиваемые челове-
ком, и этими и другими путями передавать возбудителя брюшного тифа
человеку. Рассмотренный пример характеризуется, кроме того, диф-
фузностью путей циркуляции брюшнотифозных бактерий, которые рас-
пространяются во внешней среде и попадают в человека также и неза-
висимо от посредничества мух.
Противоположные отношения представляют трансмиссивные болезни
с явно выраженной очаговостью их локализации. В таких
случаях возбудитель болезни гнездится в определенном, часто резко
ограниченном месте, каковым является, например, биотип (т. е. типич-
ное местообитание) переносчика. В частности, очагом клещевого воз-
вратного тифа, возбудителем которого являются спирохеты, а перенос-
чиками — различные виды клещей рода Ornithodorus, в естественной
ПУТИ ЦИРКУЛЯЦИИ ВИРУСОВ
49
обстановке полупустыни, служит нора грызуна; хозяин норы — носитель
•спирохет — является источником заражения клещей Ornithodorus, жи-
вущих и размножающихся здесь же в норе и питающихся кровью гры-
зуна. В дальнейшем клещи передают спирохеты другим грызунам,
вследствие чего поддерживается очаг клещевого спирохетоза в при-
роде (рис. 17).
Однако могут возникать очаги этой болезни в ближайшем сосед-
стве с человеком. Очагом такого рода является крысиная нора в
Рис. 18 — Характер сооляошений компонентов биоце-
ноза норы грызуна. Иод хозяином подразумевается
строитель норы; он же является и хозяином для мно-
гих паразитов—членов биоценоза норы.
Но Е. Н. Павловскому.
жилье или в помещении для скота, обитаемая, кроме крысы, и клеща-
ми Ornithodorus. Спирохетозный вирус «циркулирует» от крысы к кле-
щу, потом опять к крысе и т. д.; но клещи, выходя из норы в помеще-
ние, могут нападать на человека и сосать его кровь; при этом путь
циркуляции спирохеты захватывает организм человека, который забо-
левает клещевым возвратным тифом. В тех случаях, когда клещи
Ornithodorus поселяются в трещинах стен жилья и питаются кровью
живущих здесь же людей, создается антропургический очаг
клещевого возвратного тифа.
50
ОБЩАЯ ЧАСТЬ
Нора грызуна в естественной обстановке может быть заселена мно-
гими сообитателями, которые привлекаются в нору ее микроклиматиче-
скими условиями, наличием пищи в виде крови хозяина, его помета, нако-
пляемых запасов или присутствием других обитателей норы (рис. 18).
Например, в норах песчанок (Rhombomys opimus) под Ашхабадом оби-
тает, по данным Я. Власова, свыше 200 видов насекомых и паукооб-
разных. При таком разнообразии населения норы, соответственно*
усложняются и биоценотические связи между сообитателями норы и
последняя может быть природным очагом двух или более трансмис-
сивных болезней. Пример схемы аналогичных соотношений приводится
на рис. 19.
Рис. 19—Схема взаимоотношений компонентов биоценоза норы грызуна'и пути
циркуляции вирусов, могущих быть передаваемыми ^человеку.:
Стрелками показано кровосос шие п «разигами грызуна. Тонкие стрелки обозначают питание одного’
животного другим. Пунктирами показа ы возможные пути циркуляции воз>удител_й вирусных
паразитарных и инфекционных болезней и передачи их человеку (Оригинал Е. II а в л о в с к о г и).
Для такой болезни, как малярия, возбудитель которой живет толь-
ко в организме человека и в ксмарах-переносчиках (Anopheles), при
всем многообразии биоценотических связей алиментарного характера,,
пути циркуляции возбудителя малярии строго ограничены; передача
малярийного плазмодия осуществляется при сосании крови человека
самкой анофелес (рис. 20). Между тем пищевые связи анофелес как во
•взрослом состоянии, так и в личиночном разнообразны, тем более,
что личинки анофелес живут в воде.
Анализ природной очаговости трансмиссивных болезней сводится к
раскрытию путей циркуляции соответствующих возбудителей в зависи-
мости от характера наличных биоценотических связей алиментарной
или контактной природы в конкретных условиях среды. Проникновение
ПУТИ ЦИРКУЛЯЦИИ ВИРУСОВ
51
человека (или домашних животных) в сферу влияния такого очага
является фактором ацтропургического 1 характера, ведущим к установ-
лению новых звеньев биоценотических связей, прокладывающих путь
проникновения возбудителя болезни из природного очага в организм че-
ловека. Этим и объясняется возможность заражения человека некото-
рыми трансмиссивными болезнями в условиях дикой или мало освоен-
Рис. 20 — Пути циркуляции возбудителя малярии на фоне биоценотических связей
анофелес в условиях развития в воздушной и в водной среде.
Острые стрелки указывают животное, которое поедается тем, от которого отходит стрелка. Толстые
дуговидные стрелки между ч^лов^ком и анофелес обозначают пути перехода малярийного плазмодия
из одного организма в другой. Оригинал.
ной природы. К такой категории болезней относятся клещевые
сыпнотифозные лихорадки, пендинская язва, весенне-летний энцефа-
лит и др.
Анализ факторов природной очаговости трансмиссивных болезней
необходим для понимания их генезиса и условий развития вспышек
соответствующих заболеваний. Равным образом он является основой
для построения системы радикальных мероприятий по обезвреживанию
или полной ликвидации таких очагов.
T. е. в какой-либо мере зависящего от деятельности человека.
52
ОБЩАЯ ЧАСТЬ
ОСОБЕННОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ
ПАРАЗИТОВ, ПАРАЗИТАРНЫХ И ТРАНСМИССИВНЫХ БОЛЕЗНЕЙ
И ПРИНЦИПЫ ПАРАЗИТОЛОГИЧЕСКОГО РАЙОНИРОВАНИЯ
Географическое распространение паразитарных болезней опреде-
ляется наличием: а) паразита — возбудителя данной болезни, б) орга-
низмов (resp. людей), чувствительных к заражению паразитом, и
в) условий внешней среды, которые способствуют осуществлению за-
ражения паразитом хозяина и заболеванию последнего.
Факторы географического распространения паразитов различны,
будем ли мы изучать паразитозы, исключительно свойственные челове-
ку, или же рассматривать нозогеографию паразитарных болезней, об-
щих некоторым диким животным и человеку. В последнем случае
распространение соответствующего паразитоза может быть неодинако-
вым, смотря по тому, по отношению к какому организму мы его рас-
сматриваем: к человеку или к дикому животному. Например, лентецом
широким и китайской двуусткой человек заражается, поедая сырую
или полусырую рыбу. Стало быть, в конечном результате возможность
нахождения у людей в данной географической точке указанных па-
разитов при наличии всех естественных к тому пред-
посылок решается обстоятельствами бытового и
культурного характера, именно прежде всего способом
приготовления пищи. Если рыбу принято хорошо варить или жарить,
то и зараженная паразитами рыба фактически делается для человека
безвредной. Животные же, всегда пожирающие рыбу сырой, неизмен-
но заражаются рассматриваемыми паразитами так же и там, где у
человека они очень редки или вовсе отсутствуют. Само собой разу-
меется, что мы обсуждаем факты заражаемости паразитами с прин-
ципиальной точки зрения, оставляя в стороне вопросы интенсивности и
экстенсивности соответствующей инвазии.
Конкретный пример, иллюстрирующий указанное положение дела,
дает Фауст. В тех провинциях Китая, где принято есть термически
хорошо ‘обработанную рыбу, заболевания китайской двуусткой редки
по сравнению с провинциями, где рыбу едят сырой или полусырой;
в то же время зараженность этим паразитом собак и тут и там яв-
ляется одинаковой. Следовательно, характер распространения китай-
ской двуустки будет различным, смотря по тому, по каким хозяевам
мы будем ее изучать (рис. 21). Отсюда общий вывод, что для суж-
дения о паразитологическом статусе данной гео-
графической точки необходимо не только обследо-
вание человека в отношении его п а р а з ртт о ф а у н ы,
но и диких, и домашних животных, могущих быть
хозяевами паразитов, общих с человеком.
При оценке географического распространения паразитов примени-
тельно к человеку, как к хозяину, необходимо иметь в виду чрезвы-
чайную его подвижность, особенно при современных скоростных спо-
собах передвижения. Этим обстоятельством определяется возможность
обнаружения людей, зараженных паразитами, в таких местах, которым
находимые в них паразиты вовсе не свойственны. Например, на вра-
чебный прием в Ленинграде попадали больные с анкилостомами или
пораженные легочной двуусткой; между тем Ленинград очень далек
от ближайших границ ареалов названных паразитов. Эти моменты так-
же необходимо учитывать при паразитологическом-обследовании насе-
ления или отдельных больных.
ОСОБЕННОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ ПАРАЗИТОВ 53
Известно, что ряд паразитов строго адаптировал к человеку и
вместе с Homo sapiens широко распространился по земле. Примерами
таких космополитных паразитов являются вши человека, чесоточный
зудень, острица и др. И вши, и зудень, будучи постоянными паразита-
ми человека, живущими на его теле или в толще эпидермиса, при раз-
личных бытовых условиях менее подвержены непосредственному
влиянию факторов внешней среды, что и обеспечивает их весьма ши-
рокое географическое распространение. Они есть там, где есть человек.
Нередко даже и для практически космополитных паразитов отме-
чается мозаичность их распространения, когда на одних
участках паразиты есть, а на других их нет, или же разница касается
ФОРМОЗА
'ХАЙНАН
Зона с высоком зараже-
нием людей
Тоже с позлим зараже-
нием
То же сбысоким^зараже
(наем ре-
То же с низким
зяев
КОРЕЯ
Рис. 21 — Карта распространения Clonorchis sinensis (двуустка китайская) по
констатированию ее у человека и у рззервуарных хозяев в эндемичных очагах
в Китае и в Японии. По Faust и Khaw. <
интенсивности и экстенсивности инвазии. Аскарида человеческая широ-
ко распространена по земле; однако наличие ее в данной точке опреде-
ляется (кроме факторов антропургического характера) качественной
природой субстрата, да который попадают ее нуждающиеся в созрева-
нии зародыша яйца, влажностью, непосредственной инсоляцией и т. д.
Например, в Гассан-Кули на Каспийском море (Туркмения) из 59 об-
следованных аскариды были обнаружены лишь водном случае, а в ауле
Кайне-Касыр из 50 человек — в 60%. Гассан-Кули лежит на плоской
солончаково-песчаной равнине (пользуется привозной питьевой водой),
а Кайне-Касыр — в верховьях долины р. Сумбара, богат оросительной
сетью, садами, заболоченностями; в то же время оба эти пункта рас-
положены в бассейне реки Атрека (и Сумбара) в юго-западном углу
Туркмении (В. Гнездилов и Н. Чеботаревич, рис. 22).
54
ОБЩАЯ ЧАСТЬ
В рассмотренном примере факторы внешней среды играют огра-
ничивающую роль в мозаике распространения ас-
каридоза человека.
L/Еще более многочисленны и своеобразны предпосылки географиче-
ского распространения тех паразитов, жизненный цикл которых чере-
дуется с пребыванием во внешней среде и с явлением перемены хо-
зяев.
Рис. 22 — Сравнение зараженности коренного населения паразитическими червями
по линии Гасан-Кули — Кайне-Касыр—Дайне (Туркмения). По В. Гнездилову и
Н. Чеботаревичу (Кара-Калинская паразитологическая экспедиция 1931 г.)
' Возможность существования паразита как вида в
^составе фауны данного места зависит:
1) от наличия главного хозяина, в котором паразит до-
стигает половозрелости и размножается, в силу чего главный хозяин
рассеивает зародышей паразита (в общем смысле) в окружающую
среду;
2) от наличия промежуточного хозяина, являющего-
ОСОБЕННОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ ПАРАЗИТОВ 55
ся непременным звеном в жизненном цикле паразита (напомним, что
бывают паразиты с двумя промежуточными хозяевами);
3) от наличия факторов внешней среды, благоприят-
ствующей существованию экстрагостальных стадий развития парази-
тов, и
4) от осуществления возможностей заражения и
промежуточного и основного хозяина соответствующими стадиями па-
разита.
При выпадении любого из этих моментов дальнейшее существова-
ние рассматриваемого поколения паразита прекращается. В частности,
в отсутствии промежуточного хозяина дальнейшее существование пара-
зита, как вида, оказывается невозможным. Поэтому бильгарциоз
может быть только там, где в водоемах живут соответствующие виды
моллюсков (например, Bulinus contorus, Blanfordia nosophora).
Однако указываемая зависимость не является абсолютной. Поэтому
необходимо подчеркнуть также и то положение, что ареал географиче-
ского распространения известного паразита (при наличии свойственного
ему основного хозяина) может быть гораздо уже, чем ареал распро-
странения его промежуточного хозяина. Например, ришта распростра-
нена Уже, чем виды циклопов — ее промежуточных хозяев.
Еще более многообразны факторы географического распростране-
ния трансмиссивных болезней паразитарной природы. Так, клещевой
возвратный тиф может быть только там, где распространены его пере-
носчики, какими являются некоторые из видов клещей рода Ornithodo-
rus, Но не непременно в каждой точке ареала распространения клеща-
переносчика будет встречаться клещевой возвратный тиф. Аналогич-
ная зависимость особенно ярко иллюстрируется на примере распростра-
нения весенне-летнего энцефалита, очаги которого занимают простран-
ство меньшее, чем широкий ареал распространения одного из его пере-
носчиков — пастбищного клеща Ixodes persvlcatus.
^Приведенные примеры и множество фактов сходного значения дают
основания ставить вопрос о паразит о логическом райониро-
вании Советского Союза по признакам природных,
хозяйственных и бытовых факторов, обусловливаю-
щих возможность существования (или преимуще-
ственного наличия) определенных паразитов, пара-
зитарных или трансмиссивных болезней человека,
как в известной мере характерных на данный мо-
мент элементов краевой паразитологии (и еще
шире — краевой патологии) изучаемого района. На-
пример, известно, что анкилостомидозы свойственны некоторым частям
республик Закавказья и Средней Азии; .очаги пендинской язвы типичны
для Южной Туркмении. Эндемиками Байкала являются дифиллобо-
ттжозы, вызываемые Diphyllobothrium minor и D. strictum.
^Выявление паразитологических характеристик районов СССР имеет
громадное значение как в отношении обороны страны, так и для куль-
турно-хозяйственного освоения новых районов или для реконструкции
уже давно обжитых мест. Такое выявление должно сочетаться с рабо-
той сходного значения по эпидемиологическому районированию терри-
тории СССР. Необходимо знать, чем именно характеризуется данный
район, с какими заболеваниями вероятно или пока даже неизбежно
могут столкнуться пришлые элементы населения — рабочие, воинские
части и др., равно как и коренное население соответствующего района..
56
ОБЩАЯ ЧАСТЬ
Такие данные будут играть руководящую роль в планировании и на-
правлении лечебной и санитарно-профилактической работы Нарком-
здрава. Однако действительное значение паразитологических принци-
пов районирования еще более широко, ибо они должны были быть
учтены в качестве специфических предпосылок при осуществлении но-
вых задач экономического районирования СССР, предусматривавшихся
в докладе товарища В. М. Молотова на XVIII съезде ВКП(б) третьим
пятилетним планом. Жизнь показала реальное значение проблемы
краевой паразитологии в условиях Отечественной войны, когда при рас-
положении участков фронтов в различных природных условиях созда-
вались непредвиденные возможности развития некоторых трансмиссив-
ных болезней, например клещевого (весенне-летнего) энцефалита.
В условиях как нормального строительства, так и особо в восстано-
вительный период наличие особых паразитарных и трансмиссивных бо-
лезней человека и сельскохозяйственных животных, болезней, прини-
мающих характер вспышек или дающих в определенный сезон большой
рост заболеваемости, может существенно нарушать экономические рас-
четы как в отношении использования живой силы, так и получения
животных продуктов и сырья. Например, на строительствах могут быть
срывы в очагах, где есть лихорадка паппатачи, когда она в сезон рабо-
ты поражает контингенты пришлых рабочих. При всей относительной
легкости этой болезни экономическое значение ее (равно как и оборон-
ное) велико.
Из области ветеринарной паразитологии отметим примеры, когда
породистый скот, ввозимый в зоны, пораженные или неблагополучные
по пироплазмозу, может подвергаться массовым заболеваниям и по-
гибать, чем грубо нарушаются планы завоза скота в целях поднятия
животноводства в дефицитных районах.
В заключение еще раз подчеркнем, что картина паразитологическо-
го (resp. и эпидемиологического) районирования может отражать
положение дела «на сегодня»; по мере прогрессирования оздоровитель-
ных работ соответственно меняется и паразитологический status райо-
нов, что должно своевременно учитываться для руководства в после-
дующей оздоровительной работе. Более или менее детальное паразито-
логическое районирование СССР ввиду чрезвычайного объема этой
задачи — дело будущего; но и теперь все же можно выявить характер-
ные показатели для крупных краев или республик, что стоит в пря-
мой зависимости от степени изученности соответствующей территории.
Огромную роль в этом отношении сыграли и сыграют паразитологиче-
ские экспедиции, бурное развитие которых так характерно для после-
октябрьской эпохи. Весьма важно и то, что мощно растет сеть научно-
исследовательских и научно-практических учреждений на местах (са-
нитарно-бактериоДогические институты, тропические институты с
системой малярийных станций, противочумные станции и Др.). Уже
назрела потребность составления специальных пособий, характеризую-
щих паразитологические особенности районов СССР. Первый опыт
такого рода осуществлен автором и его сотрудниками (А. Гуцевич.
В. Гнездилов, Г. Змеев, П. Петрищева, Б. Виноградов), составившими
печатающуюся книгу «Паразитология Дальнего Востока и Приморья»;
Кассирским и Буровым дана книга «Тропические болезни Средней Азии».
БИОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ БОРЬБЫ С ИНВАЗИЯМИ
57
БИОЛОГИЧЕСКИЕ И ЭКОЛОГО-ПАРАЗИТОЛОГИЧЕСКИЕ
ОБОСНОВАНИЯ БОРЬБЫ С ПАРАЗИТАРНЫМИ
И ТРАНСМИССИВНЫМИ БОЛЕЗНЯМИ
Борьба с инвазиями производится воздействиями на парази-
тов или непосредственно в местах их пребывания в хозяине (например,
оперативное удаление эхинококка и финок, горячие клизмы против не-
которых кишечных паразитов, ртутная мазь при площицах и др.), или
косвенным путем — воздействием различных лекарственных веществ,
вводимых per os., под кожу или в кровь. Большую роль шрает химио-
терапия при различных болезнях, вызываемых кровепаразитами. Приме-
нение иммунизации при инвазионных болезнях находится в самом за-
чаточном состоянии, и то по отношению к отдельным паразитам. При
некоторых инвазиях лечение вовсе невозможно.
Тем большее значение принимает профилактика инвазий
человека, проводимая в порядке мероприятий общественного и инди-
видуального характера.
у т и профилактики определяются следующими направле-
ниями:
1) прямое и косвенное уничтожение паразитов и их зародышей в
открытой природе и в организме хозяев;
2) пресечение возможности заражения человека теми или други-
ми паразитами;
3) внедрение общей санитарной грамотности (санпросвет).
Профилактика инвазии сводится к построению и
проведению в жизнь систем разнообразных пред-
упредительных мероприятий, которые комбинируются,
смотря по характеру инвазии и по обстановке, в которой необходимо
вести с ней борьбу.
Возможность организации профилактических мер против инвазии
может быть обеспечена только точным знанием:
1) фауны болезнетворных паразитов человека;
2) биологии и условий жизни (экология) паразитов и переносчиков
в соотношении их с обстановкой труда и быта человека, как объекта
инвазии;
3) болезнетворного влияния паразитов на хозяина и обратного
влияния хозяина на паразита;
4) податливости паразитов и их хозяев к различным мерам воз-
действия.
Ответ на выдвигаемые требованиями профилактики вопросы дает
паразитология как наука, изучающая паразитов или точнее — биологи-
ческую систему паразита и его хозяина — со всех специальных точек
зрения. Будучи в своей основе, со стороны паразитов, ветвью зооло-
гии, она теснейшим образом смыкается с медициной в лице патологии
и эпидемиологии, и именно в этой связи она и должна быть особенно
энергично изучаема в СССР, где в процессе построения социализма идет
плановая борьба с паразитарными болезнями, этим тяжким наследием
капиталистической России.
В условиях социалистического строительства, Отечественной войны и
восстановительного периода требования, предъявляемые к паразитоло-
гии, столь же велики, как разнообразны и сферы ее приложения.
58
ЛИТЕРАТУРА
ЛИТЕРАТУРА
Большая медицинская энциклопедия. Много статей по паразитологии.
Виноградов Б. С., Павловский Е. Н. и Флеров К. К. Звери
Таджикистана, их жизнь и значение для человека. Труды Таджик. базы АН
СССР, т. I. Зоология и паразитология, изд. АН СССР, 1935.
Графф и Линетов. Паразитизм и паразиты, изд. Брокгауза и Эфрона,
1910.
Догель В. А. Курс общей паразитологии. Учпедгиз. 1941 (Лит.).
Иванов А. В. Морфологические адаптации к паразитическому образу жиз-
ни. «Проблемы общей паразитологии». Лен. гос. университет. Ученые записки
№ 13, 1937.
Кассирский И. А. и Бурова Л. Ф. Тропические болезни Средней
Азии. Издат. УзССР, Ташкент, 1935, изд. 2-е.
Лейкарт Р. Общая естественная история паразитов. СПБ, 1881.
Мордвилко А. К. Происхождение явления промежуточных хозяев у жи-
вотных паразитов. Ежегодник Зоолог, музея АН СССР, т. XIII, 1908.
Павловский Е. Н. Организм, как среда обитания. Природа № 1, 1934.
Павловский Е. Н. Руководство к практической паразитологии человека.
1924.
Павловский Е. Н. Паразитология за двадцать лет послеоктябрьской эпохи.
Математика и естествознание в СССР, изд. АН СССР, 1938.
Павловский Е. Н. Учение о биоценозах в приложении к некоторым
паразитологическим проблемам. Известия АН СССР, Отд. матем. и естест. наук,
№ 4, 1937.
Павловский Е. Н. Итоги работ совещания по паразитологическим про-
блемам, посвященного природной очаговости трансмиссивных болезней. Зоологи-
ческ. журнал, т, XIX, вып, 2, 1940.
Павловский Е. Н. О природной очаговости инфекционных и паразитарных
болезней. Вестник АН СССР, № 10, 1939.
Павловский Е. Н. Методы учета наружных паразитов и переносчиков
возбудителей заразных болезней домашних животных. Сельколхозгиз, 1931.
Павловский Е. Н. Ядовитые животные СССР, Госиздат. Мед. литер. 1931.
Павловский Е. Н. Практикум по зоологии. Учпедгиз. Ленинград, 1938,
изд. 2-е.
Павловский Е. Н. Методы- и принципы преподавания паразитологии в
высшей медицинской школе. Вести, микробиол., эпидем. и паразитологии, № 3, 1928.
Павловский Е. Н. (ред.). Вопросы паразитологии военно-санитарного
значения. Сборник экспедиц. работ кафедры общей биологии и паразитологии.
Тр. Военно-мед. акад. РККА им. С. М. Кирова, т. VIII, 1937.
Павловский Е. Н. (ред.). Паразитологические и биологические исследова-
ния военно-санитарного и теоретического значения. Второй сборник экспедицион-
ных и лабораторных работ кафедры общей биологии и паразитологии. Тр. Военно-
мед. акад. РККА им. С. М. Кирова, т. XVIII, 1939.
Павловский Е. Н. Третий сборник экспедиционных и лабораторных работ
кафедры общей биологии и паразитологии. Тр. Военно-мед. акад. РККА им.
С. М. Кирова, т. XIX, 1939.
Павловский Е. Н. (ред.) Паразитология Дальнего Востока. Медгиз (печа-
тается).
Павловский Е. Н. (ред.). Практикум медицинской паразитологии. Биомед-
гиз, Ленинград, 1935.
Павловский Е. Н. (ред.). Патогенные животные. Тр. Отд. паразитологии
ВИЭМ им. А. М. Горького, т. II, изд. ВИЭМ. 1936.
Павловский Е. Н. (ред.). Вопросы краевой паразитологии. Тр. Отд. пара-
зитологии ВИЭМ им. А. М. Горького, т. III, Медгиз, 1938.
ЛИТЕРАТУРА
59
Павловский Е. Н. (ред.). Животные паразиты и некоторые паразитарные
болезни человека в Таджикистане. Сборник работ Среднеазиатской паразитологиче-
ской экспедиции Зоологического музея АН СССР, Постоянной комиссии по изуче-
нию малярийных комаров при Зоол. ин-те АН, Кафедры зоологии и паразитологии
Военно-мед. акад. РККА, Республиканской б-цы им. Дадабаева в Дюшамбе и дру-
гих учреждений. Изд. Наркомздрава Таджикской ССР, Типогр. АН СССР, 1929.
Павловский Е. Н. (ред.). Материалы по паразитологии и фауне* Южного
Таджикистана. Тр. Таджик, компл. экспедиции 1932 г., вып. 10, изд. АН СССР.
Ленинград, 1935.
Павловский Е. Н. (ред.). Малярия и другие задачи паразитологии Юж-
ного Таджикистана, Тр. Таджик, базы АН СССР, т. VI.— Зоология и паразитология.
Изд. АН СССР. 1936.
Павловский Е. Н. (ред.). Вопросы паразитологии и патологии Западного
Памира и других районов Таджикистана. Тр. Таджик, базы АН СССР, т. XI.— Зоо-
логия и паразитология. Изд. АН СССР, 1940.
Павловск и й Е. Н. (ред.). Мургабская паразитологическая экспедиция
1930 г. АН СССР и Наркомздрава Туркмении. Труды Совета по изучению произво-
дительных сил при АН СССР, серия Туркменская, вып. 2. Изд. АН СССР и Нарком-
здрава Туркмении. 1932.
Павловский Е. Н. (ред.). Труды Каракалинской и Кзыл-Атрекской парази-
тологической экспедиции 1931 г. и материалы по фауне Туркмении. Тр. Совета по
изучению производительных сил при АН СССР, серия Туркменская, вып. 6. Изд.
АН СССР и Наркомздрава Туркмении. Ленинград, 1934.
[Павловский], Берг и Ж. Штром (ред.) Проблемы паразитологии и фау-
ны Туркмении. Тр. Совета по изучению производительных сил АН СССР, серия
Туркменская, вып. 9. Изд. АН СССР, 1937.
Павловский Е. Н. (ред.). Вредители животноводства. Работы по перенос-
чикам пироплазмозов и эктопаразитам. Изд. АН СССР, 1935.
Па вловский Е. Н. -(ред.). Закавказская паразитологическая экспедиция в
Армению 1931 г.— Тр. Совета по изучению производительных сил при АН СССР, серия
Закавказская, вып. 11. 1934.
Па вловский Е. Н. (ред.). Вредители сельскохозяйственных животных и борь-
ба с ними. По материалам паразитологической экспедиции в Северном Казахстане
1932 г. Тр. Совета по изучению природных ресурсов при АН СССР, серия Казахская,
вып. 4, 1935.
Павловский Е. Н. (ред.). Материалы по вредителям животноводства и
фауне преимущественно Южного Казахстана. Тр. Казахстанского филиала АН СССР,
вып. 2, 1937.
Паразиты, переносчики и ядовитые животные.— Сборник работ, посвященный
25-летию научной деятельности профессора Е. Н. Павловского (1909—1934).
Изд. Всесоюзн. инет, эксперимент, медицины (ВИЭМ) им. А. М. Горького. 1935.
Профессору Е. Н. Павловскому к 25-летию научной деятельности (1909—
1934 г.). Тр. Тадж. базы АН СССР, т. V.— Зоология и паразитология. Изд.
АН СССР, Москва. 1935.
Сборник работ к 25-летию научной деятельности акад. Е. Н. Павловского. Тр.
Военно-мед. акад. РККА им. С. М. Кирова, т. XXV.
Скрябин К. И. Симбиоз и паразитизм. 1923.
Издания в СССР с паразитологическим содержанием
Паразитологический сборник Отдела паразитологии и Комиссии по изучению
малярийных комаров Зоологического института АН СС.СР. Изд. АН СССР, т. J.
1930; т. II —1931; т. III— 1932; т. IV— 1934; т. V— 1935; т. VI — 1936; т. VII —
1940; т. VIII—1940; т. IX — печатается (под ред. Е. Павловского).
^Медицинская паразитология. Журн., Москва.
60
ЛИТЕРАТУРА
За социалистическое здравоохранение Узбекистана, Медицинская мысль Узбе-
кистана, журн., Ташкент.
Вес шик микробиологии, эпидемиологии и Ьаразитологии, Саратов.
Лабораторная практика, журн., Москва.
Архив Азербайджанского института микробиологии и гигиены им. Мусабекова,
Баку.
Известия Казахского филиала Академии Наук СССР (некоторые выпуски цели-
ком посвящены паразитологии).
Известия Таджикского филиала Академии Наук СССР (то же).
Труды Тропического института Армении — т. I.— Изд. Наркомздрава Армении.
Москва — Эривань, 1924; т. II—Эривань, 1935.
Труды Тропического института Наркомздрава Абхазской АССР. Вып. I, II и
III. Изд. Наркомздрава Абхаз. АССР, АБГИЗ, Сухуми, 1937.
Труды Тропического института Наркомздрава Азербайджанской ССР, т. П,
Баку, 1936.
Узбекистанский паразитологический сборник под ред. Н. И. Ходукина, т. I, II,
Ташкент.
Труды Узбекского института экспериментальной медицины, т. IV. Проблемы
тропической патологии. Ташкент. 1940.
Труды III съезда по борьбе с малярией и другими тропическими заболеваниями
в ССР Армении. Госиздат ССР Армении, Эривань, 1933.
СПЕЦИАЛЬНАЯ
ЧАСТЬ
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
(медицинская протозоология)
Тип Protozoa — простейшие
К типу простейших принадлежат микроскопически мелкие
животные, тело которых представляет собою клетку, т. е. состоит из
некоторой массы протоплазмы с заключенным в ней ядром. Однако
очень многие простейшие в порядке своего филогенетического разви-
тия отошли от этой примитивной схемы строения. Изменения в орга-
низации коснулись увеличения числа ядер и диференциации прото-
плазмы. Для многих простейших характерна двуядерность, причем их
ядра имеют каждое свое физиологическое значение. Бывают и много-
ядерные простейшие.
Диференцировавшиеся морфологически части протоплазмы про-
стейшего, выполняющие определенную функцию, называются орга-
ноидами или органеллами. Таковы, например, органоиды
движения (жгутики биченосцев, реснички инфузорий и др.), скелетные
органоиды, органоиды выделения и др. Благодаря глубоко зашедшей
морфологической и функциональной диференцировке частей многие
простейшие приобрели сложное строение (рис. 23), сохраняя в то же
время принцип одноклеточное™ своей организации; поэтому понятие
«простейшее» является относительным, ибо в пределах этого типа жи-
вотных существует огромное разнообразие форм организации.
Простейшим, как животным организмам, свойственны все основные
жизненные функции: комплекс процессов обмена веществ с ассимиля-
цией и диссимиляцией, движение, раздражимость, размножение. Им
присуща также способность приспособления к определенным условиям
существования (адаптация).
Протоплазма простейших обычно диференцируется на поверхност-
ный слой — эктоплазму, которая отличается гомогенностью и
большей плотностью, и на внутреннюю массу — эндоплазму, зер-
нистую по структуре и более жидкую по консистенции. В эндоплазме
часто наблюдаются передвижения ее частей — «токи», называемые
циклозом. В эндоплазме имеются различные включения и всегда
содержится ядро (или ядра). Под влиянием действия солей окружаю-
щей среды (простейшие, как правило, живут в жидкой среде — вода,
соки тела и др.) коллоиды поверхности эктоплазмы осаждаются в виде
очень тонкой «поверхностной кожицы», или пелликулы; при мест-
ных нарушениях ее целости она восстанавливается. Состоя из липо-
идов, эта поверхностная кожица в физико-химическом отношении обла-
64
СПЕЦИАЛЬНАЯ ЧАСТЬ
дает свойствами полупроницаемой оболочки. Оставаясь живой частью
клетки, поверхностная оболочка может изменяться в зависимости от
физиологического состояния простейшего. Все это имеет громадное
значение прежде всего в питании простейшего, в эндоплазму которого
могут поступать лишь те вещества,
которые в состоянии проходить через
полупроницаемую оболочку экто-
плазмы.
Пелликула обладает гибкостью,
что обеспечивает простейшему воз-
можность метаболировать, т. е.
временно изменять форму тела (рис.
24, А).
Те простейшие, у которых нали-
чие пелликулы не может быть уста-
новлено современными методами ис-
следования, называются голыми
(например, амебы).
Следующим этапом развития по-
верхностной оболочки простейшего
является образование кутикулы,
которая морфологически хорошо обо-
соблена, двуконтурна и прочна. Явля-
ясь продуктом выделения протоплаз-
мы, кутикула, в противоположность
пелликуле, теряет свойства живого
вещества; она может отставать от
протоплазмы простейшего, делаясь
как бы скорлупой или раковинкой.
Простейшие, одетые кутикулой,
сохраняют постоянство формы тела,
|или же могут метаболировать лишь
в определенных участках, где прото-
плазма получает выход через отвер’
стие (или отверстия) раковинки на-
ружу (рис. 24, В).
Кутикулярные образования про-
стейших играют за-щитную и
опорную (скелетную) роль.
Движение простейших происхо-
дит при помощи временных или по-
К первым относятся псевдоподии,
Рис. 23 — Паразитическая инфузория
Diplodinium (изображенная с правой
стороны), как пример сложно уст-
роенного одноклеточного.
1 — двигатель ы’ нерв; 2 — окологлоточное
кольцо; 3 —чувггвительные н «рвы теменного
выроста; 4 —неэ'ные волокна ситных
ресниц; 5—нервные волокна глотки; 6 —
мышечные волокна глотки; 7 — скелетная
плас1инса; 8 — макоонуклеус. 9 — микро-
нуклеус; 10 - сокоатитель-ая’вакуолд; 11 —
задняя кишка. По Шарпу.
стоянных органоидов движения.
или ложноножк и,— временно образующиеся выросты эктоплазмы,
например, у амебы, в которые как бы «переливается» эндоплазма, бла-
годаря чему само простейшее как бы «перетекает» с места на место.
Постоянными органоидами движения являются бичи, или жгу-
тик и, и реснички. Все эти органоиды являются выростами прото-
плазмы простейшего. Жгут имеет по оси более плотную эластическую
нить, одетую как бы футляром из более жидкой плазмы. В теле про-
стейшего основание жгута соединяется с базальным зерном, которое
считается гомологом центрозомы. Свободным концом жгут бьет по
окружающей жидкости, описывая кругообразные движения.
Реснички, в противоположность бичам, очень коротки и чрезвычай-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
65
но многочисленны. Реснички быстро наклоняются в одну сторону и
затем медленно выпрямляются; движение их происходит последова-
тельно, благодаря чему глаз наблюдателя получает впечатление мер-
цания пламени, и самое движение называется мерцательным.
Рис. 24 А — Метаболирование Cercomonas Ion- Рис. 24 В — Раковинная форма Test,
gicauda в течение 20 минут. По W е п у о п. amoeba homines. Очертания амебы,
изменяющиеся только в той части,
где протоплазма выходит из рако-
винки. По Г. Эпштейну.
Некоторые простейшие могут'одновременно иметь псевдоподии и
жгут или псевдоподии и реснички. У других же простейших могут на-
блюдаться различные способы передвижения в разных стадиях жиз-
ненного цикла.
У некоторых простейших в протоплазме диференцируются сократи-
тельные волокна, или мионемы, благодаря работе которых тело
простейших может быстро изменять форму.
Питание простейших, применительно к паразитарному образу жиз-
ни, осуществляется заглатыванием оформленных частиц или образова-
ний (анимальный способ питания) или эндосмотически —
всасыванием питательных соков поверхностью тела простейшего. В пер-
вом случае заглатывание пищи производится работой псевдоподий, так
называемое фагоцитарное питани е,— например заглатывание
цист простейших и бактерий кишечной амебой (рис. 25) или же рес-
ничками, загоняющими частицы в клеточный рот (цитостом, напри-
мер, инфузории Balantidium coll и зерна крахмала). Эндосмотическое
питание свойственно простейшим, не имеющим органоидов питания, на-
пример, трипанозомам, лейшманиям, грегаринам, некоторым инфузо-
риям и мн. др. Питание в таких случаях происходит за счет всасывания
органических растворенных веществ из .окружающей среды; такая фор-
ма питания называется также сапрофитной.
Заглатываемые пищевые вещества поступают в эндоплазму, где и
происходит их переваривание. Неиспользованные остатки выбрасы-
66
СПЕЦИАЛЬНАЯ ЧАСТЬ
ваются наружу или в любом месте поверхности тела простейшего или
в определенном его участке (аналогия процесса дефекации).
В эндоплазме простейшего отлагаются запасные питатель-
ные вещества в виде гликогена, парагликогена (нерастворимого в.
холодной воде и в спирту), жира и других веществ.
Рис. 25—Фагоцитарное питание простейших. Кишечная амеба {Ent,
colt), заглотившая три жиардии. Оригинал В. Гнездилова.
В эндоплазме заложен также и экскреторный аппарат,,
если он вообще морфологически выражен у данного вида простейшего.
Органоидами экскреции, а также осморегуляции, частично и
дыхания являются пульсирующие вакуоли, которые, ритми-
чески сокращаясь, опорожняют наружу свое жидкое содержимое,,
снова набирающееся в вакуоль из прилежащих частей эндоплазмы.
В эндоплазме же заложено ядро простейшего. Многие простейшие
имеют два или более ядер, обладающих у различных Protozoa разно-
образным строением.
Ядро является необходимой составной частью простейшего, ибо
все жизненные процессы могут протекать только при его наличии;
безъядерные участки протоплазмы простейшего в условиях экспери-
мента могут лишь некоторое время переживать.
Простейшие обладают способностью воспринимать раздражения
внешней среды и так или иначе на них реагировать. Им свойственны,
следовательно, таксисы, чем, видимо, и объясняется проникновение
паразитических простейших в свойственные им места обитания. Напри-
мер, малярийные плазмодии проникают в кровь, в эритроциты, игнори-
руя присутствие белых кровяных телец; лейшмании, наоборот, пара-
зитируют в белых кровяных клетках и их производных. Химиотакси-
сом, в частности, обеспечивается нахождение пищи простейшими.
Основной средой обитания простейших является водная среда. Но*
тело животных и человека в том числе, будучи богато соками, оказа-
лось подходящим для заселения его некоторыми простейшими, которые
приспособились к этой новой для них среде обитания и из свободно
живущих животных превратились в паразитов.
Весьма распространенным в жизни простейших является процесс-
инцистирования. Он состоит в том, что одноклеточное округ-
ляется и одевается плотной оболочкой, которая защищает внутреннее
содержимое от различных неблагоприятных внешних влияний — низкой
температуры, высыхания и т. п. Инцистировавшиеся формы и яв-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
67
ляются заразительными, если они попадают в кишечник другого чело-
века. В таком случае оболочка цисты растворяется в пищеварительном
соке, а содержимое цисты разрастается до соответствующей величины;
развившиеся паразиты затем приступают к размножению делением.
Процесс выхода простейшего из цисты называется эксцистированием
(рис. 26, 34).
Рис. 26 — Инцистирование, размножение и эксцисти-
рование инфузории Cdlpoda steini. По W е п у о п.
Рис. 27 — Размножение делением Dientamoeba fragilis. По W е п г i с h.
Цисты совершенно неподвижны. Они обладают определенной фор-
мой, плотной, обычно двуконтурной оболочкой, вариирующими в из-
вестных пределах размерами и содержимым, которое просвечивает
сквозь оболочку (рис. 26, 30, 33 и др.). Последнее не всегда различимо
достаточно ясно, поэтому препараты обрабатывают в случае надоб-
ности различными реактивами и в первую очередь раствором иода в
иодистом калии (иода 1,0; йодистогокалия 2,0; дестиллированной во-
ды 100,0).
Размножение у простейших бывает бесполое и половое. В сущ-
ности размножение, как таковое, осуществляется всегда процессом де-
ления, но при половом размножении периоды деления перемежаются
перестройкой ядра, связанной с обновлением ядерного вещества, что
именно и типично для полового размножения простейших.
68
СПЕЦИАЛЬНАЯ ЧАСТЬ
Бесполое, или а г а м н о е, размножение сводится к делению
тела простейшего. Сначала делится ядро, а затем протоплазма расще-
пляется на два дочерние тела, получающие каждое по ядру. Деление
совершается в поперечном (инфузории), в продольном (биченосцы)
или в любом направлении; оно обычно бывает равномерным (обычная
форма размножения) или неравномерным (почкование); в последнем
случае от тела простейшего отделяется малая часть протоплазмы
с ядром.
Вариантом бесполого размножения является множественное
деление, или шизогония; ядро простейшего делится последо-
вательно несколько раз, благодаря чему простейшее становится много-
ядерным. Затем протоплазма одновременно разделяется на столько до-
черних частей, сколько образовалось ядер (или меньше); пример —
малярийный плазмодий в крови человека. Само собой разумеется, что
продукты множественного деления будут сильно отличаться по вели-
чине от материнской особи; их называют мерозоитами или ага-
метами. В последующей своей жизни они питаются, растут и пре-
вращаются в свойственную данному виду вегетативную форму, назы-
ваемую общим термином — трофозоит. При бесполом размноже-
нии биченосцев базальное зерно бича делится надвое, причем сам
бич остается в' соединении с одним из дочерних зерен; у другой до-
черней особи жгут образуется заново из базального зерна.
Одним простейшим, как, например, некоторым жгутиковым, свой-
ственно лишь бесполое размножение последовательным делением.
У других простейших, по ходу их размножения, особи целиком
превращаются сначала в незрелые, .потом в зрелые половые фор-
и ы, попарно сливающиеся друг с другом, что является процессом
(опуляции. Половые особи простейших называются гаметами.
3 простейшем случае они морфологически неразличимы друг от друга
(отсутствие полового диморфизма); такие гаметы соединяются друг с
Хругом (изогамия, рис. 28), образуя зиготу (некоторые биченосцы).
В других случаях половые особи простейших отличаются друг от
Хруга по величине, форме и степени подвижности. Более крупные и
иенее подвижные называются макрогаметами и соответствуют
кенскому полу (аналог яиц многоклеточных); менее крупные и более
подвижные называются микрогаметами (мужской пол — аналог
кивчиков многоклеточных). Степень диференциации гамет может
крайне вариировать — от слабо уловимых отличий до резко выражен-
ного полового диморфизма (рис. 28).
Незрелые половые особи называются соответственно макро- и
окрогаметоцитами. Макро- и микрогаметы сливаются попар-
но (анизогамия) и также образуют зиготу, в которой оба половые
ндра сливаются в одно.
Своеобразно протекает половое размножение у инфузорий, которые
нишь на некоторое время попарно соединяются друг с другом (так
называемая конъюгация). Во время конъюгации разрушается
5ольшое ядро каждого конъюганта (макронуклеус), микро-
I у к л е у с повторно делится, образуя в конце концов два генера-
тивных ядра; одно из них переходит в другого конъюганта и обрат-
ю (остальные ядра рассасываются). Таким образом осуществляется
)бмен ядерным веществом между конъюгантами. В каждом конъю-
'анте собственное и пришлое генеративное ядра сливаются вместе, по-
:ле чего делением восстанавливаются макро- и микронуклеус; сами
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
69
конъюганты разъединяются и начинают далее размножаться, как обыч-
но, делением.
Возвращаясь к судьбе образовавшейся в результате изо- или ани-
зогамии зиготы, видим, что она округляется и одевается оболочкой;
под ее покровом содержимое повторно делится на мелкие фор-
мы, которые освобождаются из оболочки зиготы и превращаются в ве-
гетативные формы. Этот процесс принимает характер также споро-
гонии, когда под оболочкой зиготы делением- образуется множество
спорозоитов, впоследствии превращающихся в вегетативные осо
Рис. 28 — Изогамия (копуляция) Copromonas subtilis.
1 — вегетативная особь: 2— начало соединения частей; 3—4 —
редукция хроматина; 5—С— образование цисты. По Dobell.
би; в других случаях внутри зиготы образуется несколько одетых каж-
дая своей оболочкой спор, в которых также делением возникают споро-
зоиты; число их может быть строго ограниченным.
Одним простейшим свойственно только бесполое размножение (на-
пример, многие биченосцы), другим — только половое (большинство
грегарин), третьим — и половое и бесполое размножение. В последнем
случае наблюдается чередование обеих форм размноже-
ния, или метагенез (например, ряд бесполых генераций малярий-
ного плазмодия в крови человека сменяется периодом полового раз-
множения плазмодия в теле малярийного комара). У паразитических
простейших чередование поколений может быть связано с переме-
ной хозяев или с пребыванием паразита некоторое время во внеш-
ней среде в инцистированном состоянии. В связи с особенностями раз-
множения простейшие проходят жизненный цикл, связанный с
изменением организации в течение нескольких поколений, пока про-
стейшее не вернется снова к исходной форме, которой обычно считают
особь, размножающуюся бесполым путем.
Тип простейших делится на четыре класса: корненожки, жгутико-
вые, споровики и ресничные.
70
СПЕЦИАЛЬНАЯ ЧАСТЬ
Из всех простейших инфузории достигают высшей степени органи-
зации, благодаря чему можно говорить о сложности организа-
ции простейших. Эта сложность сохраняется и у многих парази-
тических инфузорий, например офриосколецид (рис. 115).
Паразиты человека имеются среди собственно споровиков, жгути-
ковых, саркодовых и инфузорий.
Паразитические простейшие обитают в самых разнообразных орга-
нах своих хозяев; кожные покровы, подкожная клетчатка, мышцы,
части скелета, различные паренхиматозные органы у различных хо-
зяев могут быть поражены теми или другими простейшими. Особенно
легко подвержены заражению ими пищеварительный тракт и его про-
изводные, куда эти паразиты попадают в вегетативном или в инцисти-
рованном состоянии. Наконец, весьма излюбленной средой обитания па-
разитических простейших является кровь; в кровяное русло простей-
шие попадают или через кожные покровы, будучи вводимы кровосо-
сущими переносчиками, или же активно проникают через слизистые
оболочки, главным образом органов пищеварения.
В теле человека встречаются различные простейшие, которые про-
являют неодинаковое отношение к своему хозяину.
Одни из этих форм являются истинными паразитами, так как пи-
таются частями тела хозяина, производят разрушение его тканей, вьи
зывают интоксикацию своими ядовитыми выделениями и в конце кон-
цов дают известный симптомокомплекс специфического заболевания
(патогенные простейшие). Примером таких паразитов яв-
ляется дизентерийная амеба, питающаяся при известных условиях эри-
троцитами человека и разъедающая стенку кишки, малярийный плазмо-
дий и др. Такие простейшие являются болезнетворными, или
п а т о г е н н ы iM и.
Другие простейшие также типически живут в кишечнике человека,
но питаются бактериями и остатками непереваренных частей пищи.
Такие существа являются нахлебниками (ком м ен с а л и ст а-
ми), не причиняющими своему хозяину никакого вреда (например,
кишечная амеба Entamoeba coli)\ их поэтому называют непатоген-
н ы м и простейшими.
Абсолютной грани между патогенными и непатогенными простей-
шими не существует, так как во многих случаях некоторые болезне-
творные микроорганизмы фактически не вызывают у своего хозяина
явного болезненного состояния (паразитоносительство).
Третья группа простейших может быть отнесена к копрозоям.
Цисты их попадают в кишечник человека, но, в отличие от выше-
указанных паразитических простейших, они проходят без изменения
через весь кишечник и с извержениями выбрасываются наружу. Кал
играет роль питательного субстрата; в нем из цисты выходит ее содер-
жимое — в форме простейшего того или другого вида.
Очевидно, что такие простейшие, цисты которых транзитом прохо-
дят через организм хозяина, не оказывая на него никакого действия,
не могут быть отнесены к какой-либо категории паразитизма, и о них
можно говорить лишь как о мнимых паразитах.
Наконец, необходимо упомянуть о нередко встречающихся случаях,
когда простейшее извне попадает в faeces, мочу или в другие объекты,
служащие для диагностического обследования (например, с водой при
мытье посуды, с пылью и др.). Чтобы дать действительную оценку
таким случаям мнимого паразитизма, необходимо твердое знание тех
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
71
форм простейших, которые являются действительными обитателями ор-
ганизма человека, и знакомство с формами мнимопаразитарного зна-
чения. Так как результаты диференциальной паразитологической диаг-
ностики определяют как характер лечения больного, так и построение
профилактических мероприятйй, то в специальном материале нашей
книги мы даем сведения не только о болезнетворных простейших, но и
о непатогенных их видах, а также и о некоторых формах, могущих
быть мнимыми паразитами.
В пищеварительном аппарате человека встречаются представители
всех четырех классов простейших.
КЛАСС RHIZOPODA—КОРНЕНОЖКИ
(SARCODINA — САРКОДОВЫЕ)
Голые или покрытые раковиной простейшие. Тело
делится на наружную, более плотную и гомогенную протоплазму (эк-
топлазму) и внутреннюю, зернистую (эндоплазму); движутся, выпу-
ская отростки тела, называемые ложноножками или псевдоподиями.
Последние образуются в различных местах поверхности простейшего,
благодаря чему при движении форма одноклеточного постоянно изме-
няется. Органоиды движения в виде бичей встречаются очень редко;
наряду с ними сохраняются и псевдоподии. Пища захватывается лож-
ноножками. Ядро одно или их несколько; оно лежит в эндоплазме.
Размножаются делением в свободном состоянии или под оболочкой
цисты. Тип 1 — амеба.
Паразитические корненожки человека имеются среди представите-
лей АтоеЫпа.
ОТРЯД, AMOEBINA — АМЕБЫ
Тело голое, лишенное раковинки или внутренних скелетных образо-
ваний; движутся при помощи псевдоподий, являющихся временными
органоидами движения. Эндоплазма обычно вакуолизована и содержит
заглоченные пищевые частицы; эктоплазма — стекловидно прозрачна.
Представители этого отряда известны под общим наименованием
амеб. Паразитируют главным образом в кишечнике, реже в других
органах. У паразитических амеб в протоплазме отсутствует пульсирую-
щая вакуоль. Питаются паразитические амебы бактериями, пищевым
детритом кишечника, другими простейшими, иногда себе подобными
(каннибализм, проявляющийся заглатыванием молодых форм или цист
своего вида), эритроцитами человека, переваренными тканями тела
хозяина и др.
1 Нередко случается, что один и тот же вид животного описывается в разное
время разными учеными под различными наименованиями, что объясняется или*
незнакомством с литературой (иногда трудно доступной), или использованием для
описания вариирующих особей, или благодаря применению иным методов иссле-
дования и других критериев для установления вида. В конце концов устанавли-
вается, что несколько научных (латинских) наименований соответствуют в сущ-
ности одному и тому же виду, t. е. по существу однозначны. В таких случаях
виду животного присваивается первое данное ему научное наименование, все же
остальные (позднейшие) наименования обозначаются, как синонимы. При строгом
применении научной номенклатуры после видового обозначения ставится фамилия
автора, описавшего вид, и год опубликования описания. Например, Trypanosoma
spermophili, Laveran, 1911. Необходимо отметить, что при сведёнии синонимики не
все авторы иногда бывают согласны друг с другом. Поэтому один и тот же вид
животного может в некоторых случаях фигурировать в разных работах под раз-
ными научными наименованиями;
72
СПЕЦИАЛЬНАЯ ЧАСТЬ
Размножаются бесполым путем — простым и множественным деле-
нием, в последнем случае обычно под оболочкой цисты. Половое раз-
множение у амеб изучено очень мало, и самый факт его нуждается,
в подтверждении.
Род Entamqeba
Entamoeba histolytica Schaud — дизентерийная амеба
Синонимы: Entamoeba coli Loesch, Entamoeba tetragena V 1 e г e c k, Entamoeba
dysenteriae Counc. et Lafleur.
Рис. 29 — Кишечные амебы в дви-
жении в испражнениях человека.
По Лёшу.
Историческая справка. Дизентерийная амеба была впервые наблю-
даема Лешем .(рис. 29) в Петербурге в 1875 г. у человека, болевшего
дизентерией. Этот автор одновременно имел дело и с другим видом —
Entamoeba coli. Однако он не различал их друг от друга. Леш доказал
патогенность наблюдавшихся им амеб, заразив собаку острой дизенте-
рией введением ей с клизмой испражнений больного.
Entamoeba histolytica, как показали последующие многочисленные
исследования, действительно является возбудителем амебной дизенте-
рии. За время своего пребывания в кишечнике дизентерийная амеба
сообразно ходу болезни обнаружи-
вается в различных формах, получив-
ших различные наименования.
Описание. Во время обитания
в организме человека поколения ди-
зентерийной амебы проходят несколь-
ко стадий, характеризующихся как
различной формой, так и некоторыми
биологическими особенностями амеб.
В связи с этим различают: 1) актив-
ную патогенную форму — Ent. histo-
lytica forma magna, или тканевую, так
как она питается паразитически тка-
нями хозяина (эритроцитами и тка-
нями стенки кишечника), 2) кишеч-
ную, просветную,
(п р е д ц и с т н у ю)
histolytica forma minuta и 3) и н ц и-
стированные амебы (цисты). Все эти формы являются раз-
личным состоянием особей одного и того же вида Ent. histolytica. Кроме
того, в кишечнике бывают и дегенерирующие амебы.
1) Тканевая форма (рис. 30, /) получила свое наименование
потому, что форма Ent. histolytica forma magna типически инвазирует
стенку толстой кишки, где залегает в краях язв и в более глубоких
частях стенки. Амеб много в слизи и гное, покрывающих язву. Они
бывают и в других органах, где образуются метастатические очаги раз-
рушения соответствующих тканей (чаще всего в печени, реже в легких
и в других органах). В faeces Ent. histolytica переходит при очищении
поверхности кишечных язв и при остром амебном заболевании. Поэто-
му можно наблюдать под микроскопом и живых амеб, если брать со-
вершенно свежий стул для исследования от больного в острый период
заболевания .(кровавый понос). Размеры Ent. histolytica обыкновенно
колеблются в пределах от 20 до 30—40 бывают и более крупные и,
наоборот, более мелкие формы. Амебы хорошо подвижны; они энергич-
но выпускают широкие тупые ложноножки, состоящие из гиалиново-
комменсальную
форму — Ent.
Таблица I
1 — картина кишечного содержимого при амебной дизентерии. Entamoeba histolytica,
Blastocystis hominis, клеточный детрит, кристаллы Шарко-Лейдена, грибки, слизь,
эозинофилы, эритроциты. По Г. Эпштейну; 2— Entamoeba histolytica f. minuta\
3 — Entamoeba histolytica f. mi nut а. Предцистиая стадия; 4 — Entamoeba
dispart 5 —Jodamoeba butschlii\ 6, 7, 8 — Entamoeba hartmanii\ 9 — Entamoeba gingivalis,
> анафаза; 10— Ent. coli. Вегетативная форма с паразитическими включениями
(PTasmophtora)', 11—Ent. coli. Кристаллическая зернистость в ядре. По Г. Эпштейну.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
Рис. ЗЭ — Сравнительная таблица кишечных амеб. По Г. Эпш’тейн'у.
1 — 11 — Entamoeba histolytica; 1—2 -типичная форма с эритрофзгие'Ъ 3 — 6 — предцистная стадия, 7 — 11 — цисты,
9— мел счя пдноядерчая циста (редчо); 12 - 16 — Ent. coll; 12 — вегегативная фо >ма, /? — 16 — цисты. 17 — 24 — Ent. dispar;
/7 — вегетативная форма, 13—25—цисгы, 26—46 — Ent.hartmani;26, 28, 37—3}—вегетативные формы, 27, 29—36, 40—46—цисты.
Со
74
СПЕЦИАЛЬНАЯ ЧАСТЬ
прозрачной эктоплазмы, в которую позднее вливается и эндоплазма
(рис. 32, А). Слой эктоплазмы вариирует по толщине.
Стекловидная эктоплазма хорошо диференцирована от эндоплазмы
даже в покойнохМ состоянии амебы. На поверхности эктоплазмы иногда
образуется довольно плотная пелликула.
Движение дизентерийной амебы связано с энергичным цик лозом
эндоплазмы, в которой развиваются токи самого разнообразного на-
правления, принимая часто характер вращательных или вихреобраз-
ных движений; однако не всегда развитие бурных токов эндоплазмы
ведет к передвижению самой амебы (Г. Эпштейн).
Рис. 31 —Дизентерийная амеба.
А — крупнозернистый распад фагоцитированных эритроцитов;
В — фагоцитоз эритроцита, эозинофила", бактерий; С — фагоцитоз
эритроцита, бактерий, кристаллов Шарко — Лейдена.
По Г. Эпштейну. .
При обычном движении где-нибудь на поверхности амебы появ-
ляется вырост эктоплазмы, образуя более или менее широкую псевдо-
подию (лобоподию), которая выпячивается в одном направлении; в
нее как бы переливается эндоплазма, благодаря чему амеба передви-
гается с места на место. Гиалиновые псевдоподии могут образовы-
ваться в нескольких участках поверхности амебы, но в своем дальней-
шем развитии доминирует одна какая-нибудь псевдоподия, которая в
своем движении увлекает эндоплазму; Бывают случаи движения амеб,
у которых нет диференцировки на экто- и эндоплазму; такая форма
движения бывает у экземпляров, только что выделенных из организма
или взятых тотчас из культуры и наблюдаемых при температуре 37°.
Наиболее активное движение наблюдается при температуре тела и при
pH 6,5 (Е. Павлова); при комнатной температуре свежие экземпляры
амеб из фекалий и из культур образуют гиалиновые лобоподии.
Эндоплазма мелкозерниста и часто заключает в себе пищевые ва-
куоли с содержимым. Ядро лежит в эндоплазме. В живой Ent. histoly-
tica оно видно хуже, чем у Ent. coli. Диаметр ядра колеблется от 4 до
7—8 у. Как правило, ядро одно, шарообразной формы. Оно одето тон-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
75
кой одноконтурной оболочкой, под которой лежит тонкий слой хрома-
тиновых зернышек. Почти в самом центре ядра помещается маленькая
хроматиновая кариозома, окруженная светлым ободком. Сеть линина
расположена в общем радиально. Ядро в целом относится к категории
«бедных хроматином (табл. I, рис. 31).
Рис. 32. А — Entamoeba histolytica форма rninuta (предцистная
стадия); В — она же с хроматоидными включениями; С — хрома-
тоидные включения в типичной Entamoeba histolytica.
По Г, Эпштейн у.
Дегенеративные процессы, затрагивающие амебу, касаются прежде
всего тонкой структуры ее ядра. Периферийный слой хроматина рге-
ряет правильность своего расположения и разбивается на неправиль-
ные скопления;1 кариозома отходит ч от центра ядра, увеличивается в
.размерах и др.
При специальной гистологической обработке в ядре Ent. histolytica
можно найти эксцентрично лежащее зерно, которое, вероятно, являет-
ся внутриядерным центральным аппаратом. Кроме всех этих образо-
ваний в ядрах иногда находится по одному эксцентрично лежащему
оксифильному ядрышку.
Деление дизентерийных амеб изучено недостаточно. Описываемые
разнообразные формы амитотического деления на деле; быть может,
являются стадиями дегенерации, а не действительного амитоза.
При кариокинетическом делении из хроматина' ядра формируется
шесть хромозом.
Делящиеся амебы в содержимом кишечника встречаются исключи-
тельно редко. По Dobell, деление ядра у Е. histolytica является про-
межуточным по своему типу между настоящим кариокинезом и ами-
тозом.
Ent. histolytica питается, кроме жидкой пищи (эндосмотическое
питание — см. ниже), прежде всего эритроцитами, затем лейкоцитами
(амеб, заглотивших красные кровяные тельца, называют э р и т р о ф а.-
г a м и).
В среднем амеба заглатывает 6—8 эритроцитов, но бывает их и
несколько десятков. Фагоцитированные эритроциты перевариваются,
совершенно не изменяясь в своей гомогенной структуре, они как бы
«стаивают» с поверхности. Интересно, что в теле дизентерийной амебы
не . удается обнаружить следов железа микрохимическими реакциями;
одним из объяснений этого является предположение, что гемоглобин в
процессе усвоения эритроцитов выделяется из тела амебы вовсе без
разложения или последнее протекает в размерах, неуловимых. совре-
менными методами исследования (Courtelin).
1 Karyamoebina falcata Kofoid и Swezy, невидимому, является Е. histolytica
с дегенеративными процессами в ядре.
76
СПЕЦИАЛЬНАЯ ЧАСТЬ
Способность амеб фагоцитировать эритроциты наблюдалась также и
в культурах Е. histolytica, что было показано Беком и Доброглавом
(Boeck, Dbrohlav, 1925) и другими авторами, (Dobell,. Frey, iMeleney
и др.). Способность амеб к эритрофагоцитозу связана с особенностями
среды. Frey и Meleney показали, что свежая лошадиная сыворотка
задерживает заглатывание эритроцитов амебами; в фильтратах амеб-
ной культуры с лошадиной сывороткой, нагретых до 70° С, эритрофа-
гоцитоз наиболее энергичен; в чистом растворе Рингера он, наоборот,
наиболее слаб. Наиболее выраженный фагоцитоз эритроцитов дизен-
терийной амебой наблюдается при pH от 5,6 до 6,5; сверх pH 7,8 этот
процесс выпадает (Е. Павлова).
В эти же пределы укладываются данные Е. Хейсина.
Бактерии Ent. histolytica заглатывает в виде исключения и то,
вероятно, в состоянии дегенерации. Заглоченные эритроциты в живой
амебе просвечивают в виде округлых желтоватых телец; диаметр их
по мере переваривания уменьшается (рис. 31—В; 30, 1—2). Потреб-
ляются также и другие клетки тела и кристаллы Шарко-Лейдена
(рис. 31, С). Не редкость, когда в эндоплазме амебы совсем нет сле-
дов оформленных пищевых частиц.
Включения указанных выше сортов являются экзогенными, так как
они поступают в амебу извне. Кроме них в протоплазме Ent. histoly-
tica бывают включения, формирующиеся в результате жизнедеятель-
ности амебы; таковыми — эндогенными — включениями являются, на-
пример, окрашивающиеся ядерными красками хроматоидные тела в фор-
ме различных по величине зерен, глыбок, трабекул и др. (рис. 32, С).
2) Ent. histolytica forma minuta живет в просвете, в содержимом
толстой кишки. Размеры амеб колеблются от 7 до 15 р и несколько
выше. Мельчайшие minuta описаны были в качестве особого вида —
Ent. hartmanni Prowazek. Эктоплазма развита слабее, чем у Ent. histo-
lytica forma magna, и скопляется главным образом в ложноножках
(рис. 32 — А, В). Подвижность вариирует в зависимости от подготов-
ленности амеб к инцистированию; во всяком случае, она меньше, чем у
вегетативной формы. Перед инцистированием minuta вовсе теряет по-
движность. Эндоплазма ее бывает сильно вакуолизована; предцистные
амебы не заглатывают ни эритроцитов, ни каких-либо клеток; питание
их комменсальное. В вакуолях иногда видны бактерии, которые, неви-
димому, не являются главной пищей minuta. Перед самым инцистиро-
ванием в эндоплазме появляются и увеличиваются в количестве хрома-
тоидные или сидерофильные (т. е. окрашивающиеся железным
гематоксилином) тельца (рис. 32, В), а также образуется гликогеновая
вакуоля (проба иодом).
Ядро предцистной стадии в общем сходно с формой Ent. histolytica
forma magna, хроматин у первой скопляется неравномерно под оболоч-
кой ядра в более крупные массы. Предцистные формы бывают трудно
отличимы или вовсе не отличимы от соответствующих форм Ent. coli.
Мелкие расы minuta могут быть легко смешаны с Endolimax папа.
Значение встречающихся в подобных случаях амеб может быть
выяснено многократными исследованиями одного и того же больного
в течение хода болезни, что дает возможность проследить смену одной
формы дизентерийной амебы другой ее формой и распознать типичные
цисты.
Мелкие, просветные, предцистные формы дизентерийной амебы в
толще тканей не живут и поэтому обнаруживаются в полужидких ис-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
11
пражнениях; их появление совпадает с ослаблением острого течения
болезни и началом выздоровления. В связи с таким переходным перио-
дом, одновременно с предцистными амебами обычно встречаются веге-
тативные формы и даже цисты. Замена вегетативных форм предцист-
ными происходит энергично при лечении амебной дизентерии эметином
и др. Рецидивы амебиаза сопряжены с повторным преобладанием круп-
ных, паразитических форм над мелкими, просветными, комменсальными.
3) Цисты Ent. histolytica круглые или слегка овоидные (рис. 30,
7—11). Диаметр их колеблется в пределах в общем от 7 до 15,4 у. Ве-
личина цисты зависит от среды и состояния, при котором они измеря-
ются. Истинные размеры получаются при измерениях в физиологиче-
ском растворе; почти такой же индиферентной в этом отношении средой
является люголевский раствор. Цисты, фиксированные и заключенные
в канадский бальзам, являются меньшими по своему размеру процентов
на 10 сравнительно с нормой.
Двухконтурная оболочка цисты тонка и бесцветна. Толщина ее в
цистах среднего размера достигает 0,5 р. В неокрашенных цистах ядра
трудно различимы или вовсе не видны. Строение цисты зависит от сте-
пени ее зрелости. В извержениях обычно встречаются все стадии созре-
вания цист. ’Молодые формы одноядерны (рис. 30, 7—9\ табл. III,
рис. 5, 8), затем ядро делится надвое (двуядерная циста) (табл. III,
рис. 6). Далее следует новое деление с образованием четырехъядерных
цист, каковые и являются типичными для histolytica (рис. 30, 10;
табл. III, рис. 7, 9).
Строение ядра у двуядерных и четырехъядерных цист обычно ха-
рактеризуется «серповидным' скоплением» хроматиновых зерен под
оболочкой ядра, каковая форма ядра типична для Е. minuta.
Кроме ядер, в цистах есть так называемые хроматоидные
тела (рис. 30, 8; табл. III, рис. 2), имеющие вид палочек или стол-
биков с округлыми концами, окрашивающиеся железным гематокси-
лином; в виде сильно преломляющих свет телец, они заметны, и в не-
окрашенных цистах. В долго лежащих цистах они исчезают. Независи-
мо от этого отмечают, что хроматоидные тела гораздо чаще встре-
чаются у одно- и двуядерных цист, чем у четырехъядерных. Вторым
включением в цистах Ent. histolytica является гликогеновая вакуоля
(становится видной при действии люголевского раствора), имеющая
ббльшие размеры в одноядерных цистах (рис. 30, 5).
Со временем гликоген потребляется инцистированной амебой, и
циста лишается вакуоли. Иногда ядра лежат парами у противополож-
ных точек поверхности цисты. Цисты встречаются в оформленном
(твердом) стуле носителей или в периоде внешне, здорового состояния
болевшего.
Встречаются различные виды атипичных цист; таковыми (Г. Эп-
штейн) являются цисты с перетяжками или неправильно контуриро-
ванные, с несоразмерно крупными ядрами, с ядрами, различными по
своим размерам, уродливыми по форме, иногда вовсе безъядерные
и др. Дегенеративные цисты встречаются на всех стадиях зрелости.
Вышедшие с фекалиями наружу цисты продолжают созревать,
о чем можно судить по увеличению количества четырехъядерных цист.
В стуле пятичасовой давности Hegner наблюдал 82% одноядерных
цист, 9% двуядерных и 9% четырехъядерных. Через 22 часа после де-
фекации в том же стуле оказалось 12% четырехъядерных цист и 29%
двуядерных.
78
СПЕЦИАЛЬНАЯ ЧАСТЬ
Ent. histolytica образует, по мнению ряда авторов, несколько рас, различаемых
по средним размерам. В общем диаметр цист колеблется от 5 до 20 и; отдельные
расы также дают известные колебания, например, 7—9, 7—11, 10—14, 11—15.
12—18 р* (Dobell и Jepps) (рис. 33).
Вопрос о расах дизентерийной амебы имеет существенное значение, ибо
фигурирует и диаметрально противоположное мнение о наличии непатогенных
мелких амеб с четырехъядерными цистами, которые при современной технике иссле-
дования морфологически неотличимы от цист дизентерийной амебы. В качестве
Рис. 33 7, 2,3 — А, В, С, D —цисты четырех различных
рас Entamoeba histolytica. По Dobell и Jepps.
основного довода унитаристы приводят данные Кливлэнда и Сандерса, получавших
культуры от посеянных поодиночке цист (изменчивость в «чистых линиях» или и
клонах) и наблюдавших колебания пределов цист от 7 до А23 ц, т. е. такую изменчи-
вость, которая покрывает пределы вариаций цист, взятых в естественной обстановке.
Отсюда делается вывод, что изменчивость цист дизентерийной амебы носит не
расовый, а индивидуальный характер; и если нельзя говорить о наличии рас у Ent.
histolytica, то тем менее оснований допускать существование видов непатогенных
амеб с четырехъядерными цистами, характеризующимися теми или другими сред-
ними величинами,
Однако изменчивость в чистых линиях амеб, определяемая на культуральном
материале, т. е. развивавшихся в искусственной среде, не может быть принята (без
специальных доказательств) равнозначной вариациям амеб, обитающих в нормальном
биотопе, которым является кишечник хозяина. Это положение дает право в ином
свете рассматривать результаты биометрического изучения вариаций четырехъядер-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
79'
ных цист (морфологически относимых оптом к Ent. histolytica), указывающих на
существование групп особей, изменчивость которых колеблется вокруг различных
средних величин.
Конечно, и одна биометрика, взятая сама по себе, не является еще ключом к
практическому распознаванию различных видов амеб, которых одни соединяют в
общий вид Ent. histolytica, а другие описывают в качестве самостоятельных видов
Ent. tenuis, Ent. minutissima, Ent. dispar и др.; но в качестве опорного пункта
для дальнейших исследований данные биометрики имеют бесспорное значение, осо-
бенно при сочетании с другими методами исследования и с результатами эпиде-
миологических обследований (см. ниже).
Инцистирование амеб происходит в просвете кишки; в толще тканей
цисты никогда не образуются.
Оболочка цисты стойка к желудочному соку: растворяется она при
действии трипсина; благодаря этому проглоченные с загрязненной пи-
щей и водой цисты дизентерийной амебы вскрываются лишь в тонких
кишках.
Перед эксцистированием содержимое цисты начинает двигаться;
в стенке цисты образуется маленькое отверстие, через которое амеба
выходит наружу (рис. 34); далее быстро следует деление ядер, после
чего восьмиядерная материнская амеба делится на восемь маленьких
дочерних амеб; амебы переходят в толстые кишки, где и обосновы-
ваются. Повидимому, возможны случаи эксцистирования дизентерийных
амеб в кишечнике того же самого хозяина.
Амебы могут инцистироваться и в культуре; стимулом являются
присутствие в среде некоторых бактерий (например, В. mesentericus)
или особенности состава питательной среды (например, прибавление
рисовой муки). В культуре может происходить непрерывная смена
процессов инцистирования и эксцистирования.
Инкубационный период при опытах Walker и Seilard на людях с
дизентерийными амебами колебался от 20 до 95 дней, тогда как цисты
были обнаружены в испражнениях дней через девять.
Стойкость цист к воздействиям внешних факторов и различных
химикалий различна. Высушивание на воздухе их убивает очень бы-
стро; во влажном кале при комнатной температуре цисты выживают
около месяца; в разведенных водой faeces цисты можно обнаружить до
шестой недели; в то же время хорошо отмытые цисты переживали в
дестиллированной воде при температуре 12—22° С до семи месяцев.
Температуру в 68° С в воде цисты выдерживают пять минут, а темпе-
ратуру ледника — свыше восьми дней.
.Этот срок в опытах Tsuchiga с промытыми цистами удлинялся до
35 дней, тогда как при комнатной температуре тот же автор наблюдал
89
СПЕЦИАЛЬНАЯ ЧАСТЬ
переживания непромытых цист в течение 3—6 дней, а промытых —
в течение 7—9 дней; в фекалиях цисты жили всего 9—11 дней. Такие
различия в результатах опытов зависят, видимо, как от вариаций
свойств различных штаммов амеб, так и от сочетания влияния полно-
стью неучитываемых факторов внешней среды.
По данным, добытым в Ленинграде, цисты в faeces при 37° С при
доступе кислорода не живут больше суток; в тех же условиях при
13—17° С они выдерживают не менее 2 недель, а при 0° сохраняются
больше месяца; при переменных температурах (средняя — ниже 0°,’
а минимальная —21° С) цисты выживают до 100 дней.
Некоторые дезинфицирующие вещества оказывают сильное действие
на цисты. Крезол в разведении 1 : 250 убивает их в течение 5—15 ми-
нут, сулема 1 : 1000 — через 4 часа, 5% формалин не влияет на них в
течение 5 дней. Цисты весьма чувствительны к действию слабых раз-
ведений эметина (даже до 1 : 5 000 000).
Заключение о переживании или гибели цист производится на осно-
вании применения эозиновой пробы; если цисты окрашиваются эозином,
значит, они погибли; неокрашивающиеся считаются живыми. Однако
такое допущение может не всегда отражать действительное положение
дела. Поэтому выживаемость была проверяема посредством посева
бывших под опытом цист на искусственные питательные среды. Эти
опыты дали результаты, в принципе совпадающие с эозиновой пробой.
Следует отметить необходимость проверки сохранения патогенности
амеб, развивающихся из цист, подвергавшихся различным внешним
воздействиям; таковые воздействия могут затронуть также и патоген-
ные свойства амеб при сохранении способности содержимого обрабо-
танных цист к размножению.
4) Дегенерирующие формы Ent. histolytica. В очень ста-
рых случаях амебной'дизентерии, а также у больных, подвергавшихся
лечению,, наблюдаются1 резко дегенерирующие формы Ent. histolytica
(табл. I, рис. 6, 7). Ядро у таких амеб теряет круглую форму: хрома-
тин выходит из него и рассыпается глыбками в протоплазме (карио-
рексис); в других амебах на первый план выступают в ядрах явления
пикноза. В конце-концов ядро как таковое исчезает (рис. 35), а на
поверхности амебы образуются почкообразные выросты, ошибочно при-
нятые Шаудином за цисты.
Дегенерирующие формы амеб часто наблюдаются одновременно с
нормальными предцистными амебами; очевидно, что в таких случаях
дегенерация началась еще во время пребывания амеб в кишечнике.
Дегенеративные процессы в амебах начинаются также вскоре после
выхода их из кишечника. Так как для диагностических целей прихо-
дится брать материал из faeces, то для исследования попадаются не
нормальные амебы, а уже подвергнувшиеся той или другой степени
дегенерации, которая прежде всего отражается на тонком строении
ядра. Некоторыми исследователями такие дегенерирующие амебы были
описаны в качестве новых видов (например, Ent. tetragena Viereck,
Е. africana Hartm. и др.).
Дегенеративные изменения могут носить у амеб временный харак-
тер, и от них амебы могут оправляться. Например, при действии радия
на культуры дизентерийной амебы наблюдаются изменения ядер в
форме распада кариозомы, отхождения периферических зерен оболочки
ядра, вытягивания его, отделение безъядерных участков протоплазмы
Таблица II
Схема жизненного цикла дизентерийной амебы (Erttatnoeba
histolytia) в свете учения об адаптивных модификациях. Оригинал
Гнездилова
(7) Заглоченная 4-ядерная циста в кишечнике хозяина эксцистируется и дает
(2—3) метацистические стадии (по Dobell, Cleveland a. Sanders и др.). Последние
превращаются в обычные, вегетативные мелкие, просветно-комменсальные фор-
мы (4) —F. histolytica forma minuta, которые являются основным звеном в жиз*
ненном цикле амебы; они образуют 1—2—4-ядерные цисты (5—6—7). Выделяясь
с фекалиями во внешнюю среду, цисты (8—9—10) вновь попадают в организм
хозяина, и цикл замыкается. Комменсализмом дизентерийной амебы объясняется
обычное исторически сложившееся взаимоотношение ее с организмом хозяина,
выражающееся в бессимптомном носительстве паразита
Под воздействием разнообразных факторов в организме хозяина, как в среде
обитания паразита комменсала, может возникнуть комплекс услдвий, при ко-
тором мелкая, просветная форма (f. minuta) (4) реализует эволюционно вира*
ботавшуюся у нее способность к образованию адаптивных модификаций в виде
крупных, тканевых, патогенных эритронито-фагоцитирующих форм (f. magna)
(12), поселяющихся в тканях хозяина. Устанавливается новое соотношение
с организмом хозяина, выражающееся в заболевали его амебной дизентерией.
Крупные, тканевые формы (12), выпадая в просвет кишечника (13), выделяются
с фекалиями во внешнюю среду (14) и гибнут, как и мелкие формы (11), или же
они (13) превращаются в исходные мелкие, просветные формы (4), которые
образуют цисты (5—5—7)
Таблица ПА
Схема жизненного цикла Entamoeba coli в свете учения
об адаптивных модификациях. Оригинал Гнездилова
(/) Заглоченная 8-ядерная циста в кишечнике хозяина эксцистируется и дает
метацистические стадии (2) (по Dobell), которые превращаются в обычные,
вегетативные, комменсальные формы (3 , являющиеся основным звеном в жиз-
ненном цикле амебы и образующие 2—8-ядерные цисты (4—5). Последние (7—5),
попадая во внешнюю среду» вновь заглатываются хозяином, и цикл замыкается.
При определенных изменениях, возникающих в организме хозяина (бациллярная
дизентерия, рак, полип кишечника и др.), обычные, вегетативные Е. coli (3) могут
реагировать на возникшие изменения в среде их обитания образованием
адаптивных модификаций—в виде крупных, фагоцитирующих эритроциты и
лейкоциты форм (9) (Е. coli forma magna). Последние описаны в лите >атуре,
как новый вид амеб Councilmannia lafleuri. Выделяясь со стулом больного во
внешнюю среду, они гибнут (10) или же превращаются в исходные формы (3),
которые и образуют цисты.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
81
и др.; после прекращения опыта амебы возвращались в прежнее нор-
мальное состояние (Нессет и Кофоид, 1928).
Цикл развития. Хотя изучению паразитических амеб посвя-
щена огромная литература, все же некоторые вопросы соотношения
форм Ent. histolytica и посейчас в глазах многих авторов остаются
спорными, тем более что полностью жизненный цикл этих амеб еще
не прослежен.
Рис. 35 — Дегенерирующие дизентерийные амебы с разными стадиями
разрушения ядра. По Г. Эпштейну.
Заражение человека происходит проглатыванием цист; по взгляду
одних авторов, выходящие из цист амебы размножаются делением,
оставаясь просветными, комменсальными по питанию формами Ent.
histolytica forma minuta. ‘После ряда поколений образуются предцист-
ные формы, инцистирующиеся здесь же в толстой кишке. Цисты могут
выделяться годами человеком без каких-либо симптомов кишечных за-
болеваний. При даче слабительного с жидким стулом выходит много
Ent. histolytica forma minuta и предцистных форм.
При наличии особых условий Ent. histolytica forma minuta увеличи-
вается в размерах, внедряется в стенку кишки и превращается в пато-
генную Ent. histolytica forma magna, которая размножается в инвазиро-
ванных ею тканях (табл. II). Эти поглощающие эритроциты формы при
недостатке питания делятся на более мелкие амебы и превращаются
таким образом в форму minuta и далее инцистируются. С такой точки
зрения Ent. histolytica forma magna не является непременным звеном
в жизненном цикле амеб формы minuta. По другим взглядам, Ent.
histolytica обусловливает первую фазу инвазии: далее она превращается
в Ent. minuta, которая долго живет в кишке, где инцистируется или
снова превращается в форму histolytica, что сопряжено с появлением
рецидива амебной дизентерии. Первый взгляд автору кажется более
приемлемым.
Патогенность дизентерийных амеб
Ent. histolytica forma magna скопляются и выделяют протеолитиче-
ский фермент, который растворяет клетки слизистой оболочки толстых
кишок. Действительно, экстракт из культуры histolytica обладает цито-
литическим свойством. Разжиженные клетки потребляются амебами,
которые, как указывалось выше, заглатывают также целые клеточные'
элементы (эритроциты). При более или менее длительном действии
амеб на кишечную стенку (рис. 38,39) последняя изъязвляется на раз-
личную глубину до muscularis mucosae, muscularis и даже до брюшины.
При перфорации брюшины развивается перитонит.
Для язвы характерна подрытость ее краев (рис. 36). По периферии
82
СПЕЦИАЛЬНАЯ ЧАСТЬ
язва окружена поясом гиперемии. Дно язвы покрыто гноем. По краям
язвы и на ее дне в толще ткани кишки лежат амебы, проникающие на
различную глубину; это будут паразитические histolytica,
А
Рис. 36 — Кратерообразная язва толстой кишки человека при
амебной дизентерии. Характерны подрытые края; распад
тканей захватил 1лубокие части подслизистого слоя; видны
скопления амеб в виде черных точек. По D о р t е г, из
Г. Эпште й н а.
Различные отделы толстых кишок не одинаково часто поражаются-
дизентерийными' амебами. Кларк, на основании .изучения 186 случаев,
отмечает частоту нахождения язв в различных отделах толстых кишок,
указанную в процентах на рис. 37. Чаще всего поражаются слепая и
Рис. 37 — Частота поражения различных
участков толстых'кишок П| и амебной
дизентерии в процентах. По Кларку,
из X е г н е р а.
амебами участках кишечника. Это ос южнение быстро ведет
восходящая толстая кишки, черве-
образный отросток и сигмовидная
кишка.
При изъязвлении стенки кишки
вскрываются проходящие в ней
кровеносные сосуды; вследствие
этого кровь изливается в полость
кишки, и сами амебы попадают в:
кровяное русло, что обеспечивает
возможность развития амебных ме-
тастазов или же вызывает амебную-
септицемию.
Острая амебная дизентерия про-
текает в различных клинических
формах, разбираемых в курсе тро-
пических болезней. К ней нередко
присоединяется и бациллярная ди-
зентерия. При воздействии анаэ-
робных бактерий развивается ган-
гренозный процесс в пораженных
к смерти.
Гораздо чаще, чем острая амебная дизентерия, встречается хрони-
ческий амебиаз с локализацией амеб только в кишечнике, или с ослож-
нениями в форме абсцессов печени, легких и других органов. Хрони-
ческий амебиаз может протекать совершенно скрыто при полном внеш-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
83
нем здоровье носителей histolytica, у которых в кишечнике, по данным
некоторых авторов (Джемс и др.), имеются очень маленькие язвочки,
вызванные дизентерийными амебами. Хронический амебиаз часто про-
текает с неопределенными симптомами кишечного или нервного ха-
рактера.
Одновременно с такими поражениями кишечника изменяется и ха-
рактер стула, становящегося дизентерийным. Испражнения делаются
жидкими, кровянистыми и слизистыми, нередко походя на земляничное
желе.
Поражение тканей кишечника амебами осложняется вторичной бак-
териальной инфекцией, которая поступает с амебами, проникающими
через слизистую оболочку кишечника в подслизистую ткань. Повиди-
мому, большее значение имеет поступание бактерий в некротизирован-
ные части тканей, измененных вследствие патогенного действия амеб.
В присутствии гемолитического стрептококка и некоторых других
бактерий патогенность Ent. histolytica в опытах с котятами сильно по-
вышается (Spector). Весьма вероятна возможность синергетического
действия и амеб, и бактерий на организм.
Характерным осложнением амебной дизентерии являются метаста-
тические абсцессы печени, вызванные заносом в нее через систему
воротной вены Ent. histolytica. Амебы и здесь действуют на клетки
органа так же, как в кишечнике. Вследствие некроза и растворения
ткани в печени образуется различной величины полость абсцесса, за-
полненная тягучим гноем. В последнем содержатся остатки клеток,
элементы крови, желчь, а также кристаллы гематоидина и холестерина.
Амебы лежат обычно у стенок абсцесса (фиг. 40). Цист в амебных
абсцессах никогда не образуется. В характерных случаях в гное нет
никаких бактерий. Абсцесс печени может вскрыться в легкие, желудок,
двенадцатиперстную кишку или в полость брюшины.
Вследствие своего анатомического положения на пути кровяного
тока из кишечника к сердцу, печень является органом, где наиболее
часто развивается амебный абсцесс (рис. 41). Реже таковой может
быть в легких, селезенке, мозгу, в мышцах. Амеб находят иногда в
коже, в перикарде, аппендиксе, в моче (редко), в семепроводе, се-
менных пузырьках, в семеннике, в суставах и в других органах.
Нахождение дизентерийных амеб в суставах при arthritis deformans
дало повод Кофоиду считать этих паразитов возбудителем данной бо-
лезни, что требует подтверждения.
В редких случаях амебный абсцесс может развиться даже без оче-
видного дизентерийного поражения кишечника. Наблюдались примеры,
когда амебный абсцесс обнаруживался впервые только при вскрытии
умершего, тогда как при жизни никаких симптомов, указывающих на
его присутствие, не отмечалось. Сроки развития амебных абсцессов
весьма разнообразны. Они могут возникнуть во время острого течения
амебной дизентерии, вскоре после нее или спустя' много (до 20 и бо-
лее) лет и обнаруживаться, когда от амеб в кишечнике не остается
и следа.
Частота осложнения амебной дизентерии печеночным абсцессом
различна. Из 186 случаев Кларка абсцесс наблюдался в 51%. Крэг
дает меньшую цифру — 33%. В Баку на 3424 вскрытия за 10 лет най-
дено 124 случая печеночных абсцессов. В двадцатилетней хирургиче-
ской практике Окиншевич (Баку) насчитал двести больных с абсцес-
сом печени. Это осложнение отмечалось в Одессе, Астрахани и в дру-
84
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 38 — Патологические изменения кишечника при
амебной дизентерии.
Слева — начало процесса, изъязвление оболочки и слоя либеркю-
«новых желез; справа — более глубокая язва с амебами в подслизи-
стой оболочке. По Добелл и О’Коннор.
Рис. 39—Часть глубокой язвы
(рис. 38) при большом увеличении.
Крупные клетки —дизентерийные
амебы формы histolytica,
По Добелл и О’Коннор.
Рис. 40 — Часть края амебного
абсцесса печени.
Справа — нормальная ткань; слева — амебь
и некротизированная ткань в полости абс
цесса. По Добелл и О’Коннор.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
85
гих городах. Небольшие амебные абсцессы печени могут поддаваться
специальному дротивоамебному лечению (см. ниже).
Длительность амебной дизентерии различна. Описывались случаи,
когда она тянулась годами (до 34 лет). Бывает, наоборот, молниенос-
ное течение болезни. Смертность при отсутствии лечебной помощи до-
стигает 18—40%.
В эпидемиологическом отношении очень важными являются носите-
ли, рассеивающие цисты дизентерийных амеб; в то же время сами но-
сители не проявляют каких-либо симптомов заболевания. За сутки
носитель может выбросить до 300 млн. цист.
Рис. 41 — Амебный абсцесс печени. Владивосток. По В. Гнездилову.
Носителями могут быть лица, прежде болевшие амебной дизенте-
рией, но у которых в данный момент никаких болезненных симптомов
нет («носители-конвалесценты»). Кроме того, носительство бывает и у
лиц, явственно никогда не болевших дизентерией («контактные носи-
тели») .
Вопрос о бессимптомном цистоносительстве трактуется различно;
одни признают, что бывает цистоносительство в чистой форме, т. е. у
действительно здоровых лиц; другие считают, что не менее половины
цистоносителей по существу больны, но само заболевание протекает
у них атипично, без проявления характерных симптомов амебной ди-
зентерии. Несомненно, что такая скрытая инфекция цистоносителя мо-
жет развиваться в явное, типичное заболевание под влиянием третьих
факторов, например, ослабления сопротивляемости организма и др.
Цистоносители, рассеивающие цисты дизентерийной амебы, опасны
как источник дальнейшего распространения инвазии; люди же с аме-
бами-эритрофагами в faeces до образования цист фактически являются
незаразительными. Таким образом, в острый период болезни, когда
амебы-эритрофаги бросаются в глаза, больной незаразителен; при за-
тухании острого течения дизентерии, когда на смену эритрофагам при-
ходят инцистируемые формы, здоровый по виду цистоноситель пред-
ставляет собой для окружающих большую опасность, чем явно
больной.
86
СПЕЦИАЛЬНАЯ ЧАСТЬ
Паразитологический диагноз ставится микроскопиче-
ским исследованием кала, в котором находят вегетативные формы амеб
с заглоченными эритроцитами (характерно!) и четырехъядерные цисты.
Однако наряду с последними попадаются одно- и двуядерные цисты.
Добавочным диагностическим приемом является заражение котят через
анальное отверстие впрыскиванием в толстую кишку испражнений
больного, вследствие чего животные заболевают дизентерией.
Рис. 42 — Стул при острой амебной дизентерии. Среди эритроцитов
крупные тканевые паразитические формы Ent. histolytica t.magna.
По В. Гнездилову. 7
Другим лабораторным животным, по данным С. Канделаки, может
быть хомячок (Cricetulus migratoriusj. Различные авторы заражали
с успехом амебной дизентерией и белых крыс, у которых в результате
заражения часто развивается цистоносительство.
Успехи культивирования амеб на питательных средах (Бек, Добро-
глав) позволяют применять и культуральный метод диагностики амеб,
но, конечно, при наличии соответствующей лабораторной обстановки
и владении техникой.
Распространение дизентерийных амеб. Амебная ди-
зентерия присуща главным образом тропикам и субтропическим стра-
нам. Однако единичные случаи ее наблюдаются и в странах умеренно-
го пояса-(в частности в СССР). Распространению этого вида дизенте-
рии в Европе много1 способствовала минувшая война.
Частота нахождения четырехъядерных цист при микроскопическом
обследовании населения оказывается во многих случаях высокой. Так,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
87
из 154 обследованных в США студентов университета цистоноситель-
ство было обнаружено у 53,2%. Здоровые европейцы в Тунисе дали
40,4% цистоносительства. В Англии цисты были найдены у 10—11%
выздоравливающих солдат.
В СССР 1 — в Ленинграде цистоносительство при однократном об-
следовании установлено среди разных групп населения у 22,75% и
25,3%, а у детей — в 9,5% и 17,7%. На Кольском полуострове четы-
рехъядерные цисты найдены у 63—60,6% обследованных групп (Счен-
снович). В Мургабской долине носительство определяется цифрой
30—35% (Хейсин). Эти авторы считают всякую четырехъядерную
цисту ,за цисту дизентерийной амебы. Действительная зараженность
окажется еще более высокой, если производить повторные поиски
цист, а не ограничиваться только однократным обследованием. Этому
имеются доказательные примеры. Так, при двенадцатикратном обсле-
довании каждого изх тридцати больных амебной дизентерией Веньон и
О’Коннор впервые нашли амеб при первом обследовании в 10 случаях,
при втором — в 3 случаях/ при третьем — тоже у 3 больных, при
четвертом — у 4, при пятом — у 5, при шестом — у 3 и, наконец, при
восьмом — еще у 2 человек. Поэтому шестикратное обследование боль-
ного со взятием от него материла в разное время считается достаточ-
ным для точного установления факта зараженности простейшими.
Хотя обследование населения СССР на кишечных простейших про-
изводилось лишь спорадически, все же бросается в глаза крайнее несо-
ответствие значительного количества случаев, относимых к нахождению
Ent. histolytica, малому числу заболеваний амебной дизентерией. Первое
объясняется тем, что к Ent. histolytica обычно относят на основании
микроскопического исследования всякую четырехъядерную цисту, отве-
чающую определенным признакам, что, с точки зрения плюралистов,
неверно.
Поэтому возникает вопрос: насколько все четырехъядерные цисты
действительно являются цистами дизентерийной амебы? Brumpt выдви-
нул гипотезу существования наряду с патогенной Ent. histolytica без-
вредной Ent. dispar, а также Ent. hartmanni, которые морфологически
не отличаются от первой. Другие авторитеты стоят на унитарной точ-
ке зрения и полагают, что Ent. histolytica не всегда проявляет пато-
генность в вполне ощутимой форме, что может зависеть от степени
сопротивляемости организма хозяина и от воздействия внешней среды.
Интересны опыты со скармливанием цист здоровым. Исход опытов
бывал весьма разнообразен и не зависел от источника происхождения
цист, т. е. брались ли они от здоровых носителей или от переболевших
дизентерией. Бывали случаи, когда заражение цистами вызывало за-
болевание только при втором или даже третьем пассаже.
Ряд исследований был посвящен изучению патогенных свойств раз-
личных штаммов дизентерийных амеб на котятах, являющихся хоро-
шими для этой цели лабораторными животными. Meleny и Fray в шта-
те Тенесси (США) брали штаммы амеб из местностей, где преобла-
дало здоровое носительство или, наоборот, доминировала эпидемия
амебной дизентерии. Оказалось, что все штаммы амеб, взятых как от
больных, так и от здоровых (носителей), проявляли различную степень
патогенности для котят.
1 Общие данные о валовой зараженности человека кишечными простейшими
в СССР приводятся в таблице на стр. 92—93.
88
СПЕЦИАЛЬНАЯ ЧАСТЬ
В Армении (Т. Матевосян, Саркисян и Цатурян) для аналогичной
цели были использованы культуры, выделенные 1) от больного острой
амебной дизентерией, 2) от хронического не леченного эметином боль-
ного, 3) то же от леченного эметином и 4) от бессимптомного носите-
ля; все эти культуры оказались патогенными для котят и не давали
определенных отличий друг от друга в отношении тяжести заболеваний
котят. Штаммы от хроников и носителя были менее вирулентны, чем от
острого амебного дизентерика.
Пассирование первых через котят поднимало их патогенность. Ав-
торы полагают, что культуры форм histolytica более патогенны, чем
культуры форм minuta.
Случаи амебной дизентерии несомненно встречаются в умеренном
и даже в холодном климате, но несравненно реже, чем в южных, суб-
тропических и тропических широтах. Поскольку они действительно-
автохтонны, следует выяснять особо, так как контактная передача ин-
вазии возможна от носителей, приехавших на север из южных мест,
которым присуща амебная дизентерия (Г. Эпштейн). Во всяком случае
в умеренном климате чрезвычайно велико несоответствие в частоте на-
хождения у человека цист Ent. histolytica и заболеваемости амебной
дизентерией, которая наблюдается лишь в около 3 процентах к числу
носителей .(В. Гнездилов).
Источники заражения дизентерийными амебами
Заражению амебной дизентерией подвержены все возрасты. Резер-
вуаром вируса и, следовательно, основным’ источником существования
и распространения дизентерийных амеб служит человек. Выбрасывае-
мые им с испражнениями цисты загрязняют почву при дефекации на
землю или при пользовании примитивными отхожими местами, особен-
но поглощающего типа. Если ямы-приемники фекалий не имеют непро-
ницаемых стенок (что наблюдается чаще всего), то обеспечивается за-
грязнение подземных вод, а через них и источников питьевой воды (ко-
лодцы и пр.). При антисанитарном быте малокультурных народов и
слоев населения не редкостью является и прямое загрязнение водоемов
затекающими фекалиями.
Вспомним длительность переживания цист в воде (стр. 79), и нам
станет ясным значение загрязнения воды и использования сырой воды
для питья в деле распространения амебной дизентерии. Кроме того,
распространению цист способствуют мухи, муравьи и тараканы, являю-
щиеся механическими переносчиками цист как посредством лапок, ног,
так и благодаря рассеиванию собственных испражнений, в которых
могут быть жизнедеятельные цисты (поедание фекалий больных).
Что эти насекомые действительно могут переносить цисты дизенте-
рийных амеб, доказывается данными обследований мух и тараканов на
спонтанную зараженность их цистами. По наблюдениям Wenyon (Еги-
пет) и Л. Буровой (Ташкент), мухи приблизительно в двух процентах
были заражены цистами. Однако эпидемиологическое значение может
иметь распространение насекомыми-переносчиками вполне жизнедея-
тельных и, следовательно, вирулентных цист. Опыты со скармливанием
цист мухам и тараканам показывают, что цисты без вреда для себя
проходят через кишечник этих насекомых; кал последних оказывается
заразительным для котенка.
Возможность распространения цист с пылью трактуется различно,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
89-
зато несомненны случаи загрязнения пищевых продуктов при посред-
стве упомянутых посредников, а также рук человека (хлеб, молоко,
сахар и др.). Важную роль в заражении амебной дизентерией могут
играть овощи (загрязнение в земле, удобряемой фекалиями), сухие
фрукты и другие продукты. Наконец, бесспорно значение контактной
инвазии, т. е. непосредственного соприкосновения с носителем цист
или с предметами, бывшими в его употреблении (белье, платье, иг-
рушки, посуда; частый контакт нянек и детей и др.).
Особо следует подчеркнуть значение цистоносителей среди работ-
ников пищевой ^промышленности, так как эта категория людей может
играть весьма существенную роль в загрязнении пищевых продуктов
и предметов обихода столовых и через их посредство заражать здоро-
вых потребителей.
Военно-санитарное значение амебной дизенте-
рии бесспорно, так как больные острой формой выходят из строя,
хроники же являются, вследствие ослабленное™, неполноценным бое-
вым элементом. В военных условиях создаются особо благоприятные
условия для развития этой болезни. Отмечено, что пребывание коло-
ниальных войск в Закавказье в период оккупации оставило после себя
следы в виде усиления эпидемии амебиаза.
Передвижения войсковых частей и, в их составе, цистоносителей
способствуют распространению амебной дизентерии, и обратно— пребы-
вание красноармейцев в районах, известных в качестве очагов этой
болезни, облегчает ее заполучение по некоторым причинам общего
характера. Резкая перемена климатических условий может прямо или
косвенно влиять на сопротивляемость организма. Таково может быть,
например, значение летнего перегревания организма в наиболее жар-
ких областях СССР. Изменение пищевого режима, связанное с клима-
том, также воздействует как на организм человека, так и на бактери-
альную флору кишечника, в известном составе необходимую, по мнению
некоторых авторов, для благосостояния дизентерийных амеб в ки-
шечнике человека. Наконец контакт красноармейцев с коренным насе-
лением, а также соприкосновение со средой, примитивной в санитарно-
гигиеническом отношении (например, пользование подозрительными
по загрязненности источниками для питья и др.), без сомнения, благо-
приятствуют заражению бойцов дизентерийными амебами. В условиях
войсковой части и на кораблях большую роль могут сыграть цисто-
носители из поваров, кладовщиков, резчиков хлеба и др. В окопной
жизни, при естественной скученности бойцов и стесненности в сани-
тарно-гигиеническом отношении распространение дизентерийных, амеб
в соответствующих климатических условиях также может быть облег-
чено. Данные о фактической зараженности красноармейцев дизенте-
рийными амебами и другими простейшими скудны. Отметим цифры,
приводимые В. Гнездиловым для одного из среднеазиатских лагерей
(таблица на стр. 92—93).
Следует отметить случаи хронически рецидивирующих кровавых
поносов, при которых больных отправляют для санаторного лечения на
курорты (например, в Ессентуки), причем ни в процессе отбора боль-
ных, ни во время обследования их на курорте не производится про-
тозоологического исследования фекалий. В результате просматривают-
ся 'Случаи хронической амебной дизентерии и лечение, естественно, ока-
зывается безрезультатным. Дело доходит иногда до постановки вопроса
о демобилизации больного из-за невозможности службы. Между тем
50
СПЕЦИАЛЬНАЯ ЧАСТЬ
при правильной протозоологической диагностике и надлежащем лече-
нии, казалось бы, безнадежный больной быстро поправляется и при-
обретает трудоспособность. Больные с протозойными кишечными за-
болеваниями должны подвергаться специфическому лечению на месте
и являются противопоказанными для лечения на курорте.
В эпидемиологическом отношении важен вопрос о существо-
вании животных — резервуаров дизентерийных амеб
и о значении этих носителей для распространения названных парази-
тов. Отмечены случаи спонтанной амебной дизентерии (и даже с пече-
ночным абсцессом) у собак и кошек в тропиках и субтропиках. Мыши
и крысы бывают заражены амебами, которые некоторыми авторами
считаются идентичными Ent. histolytica. Известны амебы у свиней, но
видовое значение их толкуется противоречиво.
Интересно наблюдение Г. Эпштейна и А. Авакяна, обследовавших
случай острого амебиазного колита у |7-летнего ребенка в Москве.
Родственники его не бывали в местах, эндемичных по амебной ди-
зентерии, но некоторые из них являлись здоровыми цистоносителями.
В квартире было много крыс. При протозоологическом обследовании
их были найдены четырехъядерные цисты; они были культивированы;
культурой полученных амеб удалось заразить котят, погибших при
типичных проявлениях амебной дизентерии. Эти наблюдения обосно-
ванно толкуются как пример положительного' значения крыс в рас-
пространении амебной дизентерии, т. е. как носительство крысами.
Ent. histolytica. Впервые этот факт был установлен в США, где обна-
руженные амебы были выделены в вариетет Ent. histolytica var. mu*
rina Chiang. Кроме этого вида, у крыс в кишечнике бывают и соб-
ственные непатогенные амебы — Ent. muris Grassi.
Естественно, что носительство Ent. histolytica наблюдается у наи-
более близких к человеку животных — различных обезьян, как в дикой
природе .(например, у Macacus philippensis, Hegner), так и в зоопар-
ках. В Сухуми, в обезьяннике филиала ВИЭМ, 62% павианов-гамадри-
лов и макак оказались носителями амеб с четырехъядерными цистами
(Бурова, Воскресенский). Обезьянья Ent. histolytica ни в материале
из фекалий, ни в культурах не дает морфологических отличий от дизен-
терийной амебы человека. В пользу их видовой идентичности говорит
также возможность заражения котят цистами амеб от Macacus rhesus
(Kassel), от Macacus nemestrinus и гамадрила (Сухуми, Бурова). Из
лабораторных животных дизентерийными амебами могут быть заражены
морская свинка, кролик, хомяк, обезьяны.
Борьба с амебиазом. Лечение больных амебной дизентерией
производится алкалоидом ипекакуаны — эметином в форме солянокис-
лой соли (Emetinum hydrochloricum). Из новых средств важен ятрен
(иод-оксихинолин-сульфоновая кислота), а также риванол. Последние
два препарата весьма пригодны и для очистки цистоносителей, что
является моментом профилактического воздействия. Для предохране-
ния от заражения при контакте с цистоносителями полезен прием
внутрь мышьяковистых препаратов. Эметин впрыскивается под кожу
или внутримышечно по 0,04—0,08 г в день в течение 4—6—8 дней, по-
вторяя циклы инъекций через различные промежутки времени .(2—4—
5 дней). Сама ипекакуана применяется в форме порошков по 2,0 в день
натощак или через несколько часов после еды. Эметин обладает куму-
лятивным действием, что должно быть учитываемо при лечении амеб-
ной дизентерии.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
91
Эметин весьма токсичен для амеб; он убивает культуральные амебы
в разведении 1 : 100 000; 1 : 10 000 эметина убивает амеб в слизи стула.
Эметин оказывает действие и на культуры Ent. histolytica, характер
которого зависит от реакции среды. Большая щелочность среды спо-
собствует усилению действия эметина; при pH выше 6,8 он убивает
культуры амеб в разведении 1:500 000 (Носина).
Ятрен дается per os по 0,25—0,5—0,75 г повторно в разных комби-
нациях в течение месяца или применяется в форме клизм.
Риванол употребляется в дозах по 0,03—0,05 три раза в день в те-
чение шести дней или в форме клизм (1 : 3000—5000).
Риванол убивает дизентерийных амеб (из культур) в растворе
1 : 1000 в течение суток.
При наличии амебных абсцессов печени рекомендуется предвари-
тельно испробовать лечение ятреном.
Из мышьяковых препаратов применяют стоварсол, особенно при-
годный для лечения хронической амебной дизентерии (по 0,25 г 3—4
раза в день в течение недели); следует отметить, что синтезирован
советский препарат осарсол, используемый также для лечения амебной
дизентерии. Осарсол принимают по 0,25 три раза в день натощак. По-
сле шестидневного лечения столько же дней отдыха. Проводят 3—5 та-
ких курсов лечения.
Новым препаратом является Carbarson (4 — carbamino-phenyl —
arsenil — acid), в котором содержится 28,85% мышьяка; прописывают
по 0,25 два раза в день в желатиновых капсулах десять дней подряд.
Общее количество потребного вещества определяется из расчета
0,075 г на кило веса на десять дней.
Некоторыми авторами .небезуспешно применялся аллизатин (пре-
парат чеснока), в частности при лечении хронической застарелой амеб-
ной дизентерии (Schweitzer, Экваториальная Африка). Весьма вероятно,
что обильное потребление чеснока в пищу может влиять на уменьше-
ние заражаемости людей кишечными простейшими.
* Следует отметить недавние опыты Faust и Kagy (1934), показав-
ших, что кормление собак печенью или экстрактом из печени вело к
быстрому улучшению течения экспериментального амебиаза у них
и к заживлению амебных язв толстых кишок. Вместе с тем бедная ви-
таминами пища (Faust и Scott) и диэта, обильная углеводами (Hegner),
неблагоприятно влияют на течение амебной дизентерии или предраспо-
лагают к заболеванию этим паразитом.
Перемена климата, именно переезд больных из жарких стран в
умеренный или холодный пояс, ведет часто к быстрому улучшению
амебной дизентерии, даже без специального лечения и обратно (Craig,
1934; Snijders, 1932).
Социальная профилактика достигается выявлением при
массовых обследованиях цистоносителей, особенно среди лиц, работаю-
щих по питанию, и недопущением их к занятиям, при которых они
могут заражать пищевые продукты, воду или готовую пищу.
Особенно показателен в этом отношении пример вспышки амебной
дизентерии в Чикаго (1933, Топпеу с сотрудниками), когда было вы-
явлено 185 случаев заболевания амебиазом (с 19 смертями) и 193 но-
сителя цист Ent. histolytica. Все заболевшие или останавливались или
питались в двух гостиницах, среди кухонного персонала которых были
обнаружены носители цист дизентерийной амебы, послужившие источ-
ником заражения пострадавших.
Зараженность кишечными простейшими в СССР
Место обследо- вания Год А в т о р Число обсле- дований Процент об- щей заражен- ности 1 Entamoeba coli Entamoeba histolytica Entamoeba hartmanni lodamoeba biitschlii Endolimax папа Lamblia in- testinalis Chilomastix mesnili Trichomonas ho minis Balantidium coli Dientamoeba fragilis
Ташкент 1925 Федулов 100 — 14.0 8.0 6.0 4.0 — • 14.0 8.0 26.0 2.0
г. Старый Таш- кент 1927 Чапурская- Баженова 753 55.0-83.0 23 0-56.0 5.0-10.9 — 3.0-23.0 5.0-8.5 3.5-3.6 3.0-16.0 3.0-26.0 —
Ташкент 1927 Бурова 254 74.1 44.0 8.1 — 20.4 6.6 14.0 16.1 22.8 — 3 кол- случ. о
Бухара 1928 Софиев 314 75.6-85.3 36.6-54.6 2.4-3.3 — 20.7-17,3 39.1 15.0 9.8-15.3 28.6-14.0 — — 2 »
Ашхабад, Тахта- базар, Кушка 1930 Хейсин 1034 78.6 49.7 31.1 — 10.7 31.6 19.0 19.3 8.2 —
Окрестность Таш- кента 1932 Гнездилов 707 77.9 51.6 15.9 16.1 19.8 17.8 12.7 22.6 10.1 0.4
Азербайджан Баку и Ленкоран- ский р-н Ереван Грузия 1926 1928 1927 1927 Здродов- ский Здродов- ский и Воскре- сенский Тер-Мате- восян Канделаки 300 1146 83 570 80.0 74.3 72.3 41.8 53.0 42;2 15;6 24.3 32.5 (22.9)’ 20.5 13.6 (16.2 18.7 12.0 22.9 18.0 7.0 22.9 18.7 14.0 24.0 12.9 12.3 6.5 0.5 16.8 3.9 1.7
Баку 1926 Грубман 447 — — 34.0 — — — — —
Ленинград 1929 Филипчен- 225 49.0 24:0 14.2 — 11.5 12.0 2.2 7.5 0.9 0.4
» 1929 1930 ко Филип- пенко и Пик Сченснович .400 400 65.5 67.2 39:0 23; 5 22.75 17.7 — 12.5 26.5 17.5 19.2 14.7 22.7 o.o 3.2 0.9 —
СПЕЦИАЛЬНАЯ ЧАСТЬ
Ленинград 1930 1929 Пик Филиппен- 100 162 75.0 65.0 39.0 39.0 22.2 25.3 1 —
Кольский полу- 1930 ко и Виг- дорова Сченсно- 762 39.0-79.0 19.2-30.7 6.3-60.6
остров Уссурийская об- 1935 вич Гнездилов 734 56.1 25.6 15.1 14.5
ласть ДВ Юж. Киргизия 1935 Сченсно- 1258 76 .*4 47.7 38. 3
Сев. Киргизия 1935 вич » 658 70.1 36.6 43.4
Алайская долина 1935 » 87 70.2 41.6 29. 7
Юж. Туркмения 1937 Гнездилов 633 86.8 56.3 25.7 20.5
Таджикистан 1935 » 204 72.1 49.6 13.5 8.5
Приморская об- 1939 Гнездилов, 1044 40.3 19.1 11.5 12.8
ласть ДВ Самарканд 1942 Подолян, Коросте- нев Гнездилов 245 69.3 46.1 10.2 3.1
Казахстан 1937 Хохоль- 136 — 39.9 18.5 —
Фрунзе 1942 кова Чапая 800 63.1 38.0 13.0 7.6
Москва 1940 Авакян 530 24.3 8.6 3.3 3.9
Ташкент 1939 ЛсЙтман 113 88.5 65.5 25.0 21.4
Армения 1939 Саркисян 400 82.0 52.0 34.0 —
Сев. Кавказ 1934 Гнездилов 709 77.1 43.2 21.0 27.3
Азербайджан 1941 Сченснович 900 60.5 31.3 12.5 20.6
26.7 31.0 11.0 5.5 — —
9.9 21.6 • 9.2 3.1 — —
1.9—11.3 7.0-33.3 9.1-34.5 1.5-19.0 — —
17.4 10.6 11.5 4.6 0.2 —
2*9.4 16.3 8.5 17.1 — 4 слу- чая
25.9 8.1 8.7 11.3 — 1 »
7.1 9.1 16.6 6.0 —
35.2 21.3 15.9 24.8 15.4
18.7 14.6 12.2 22.1 19.0
12.8 6.8 5.0 0.7 0.3
7.3 18.3 9.8 4.9 35.1 1 »
14.3 17.0 6.8 13.0 —
32.0 15.0 12.0 10.0 —
2.8 5.6 4.9 6.7 3.3 0.3
37.5 68.7 16.9 29.4 30.3
42.2 11.2 10.2 21.7 8.5 —
32.2 32.2 19.4 11.5 2.3
28.6 10.7 12.4 9.4 — 5.1
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШ:
X
94
СПЕЦИАЛЬНАЯ ЧАСТЬ
Дальнейшие мероприятия сводятся к предупреждению заражения
почвы и воды (конструирование отхожих мест, ограждение водоемов
от заражения), обеззараживанию испражнений с цистами (О,5°/о кре-
зол), охране пищевых продуктов от. загрязнения насекомыми (борьба
с мухами, тараканами и муравьями, живущими в домах); должна быть
учитываема легкость заражения при поедании немытых сырых и су-
шеных фруктов и овощей. Наряду с этим необходима широкая сани-
тарно-просветительная работа, направленная к приобретению населе-
нием санитарно-гигиенических навыков и неукоснительному примене-
нию их в повседневной жизни. Методы протозоологического исследова-
ния больных должны получить соответствующее широкое применение в
больничной обстановке для уточнения диагностики протозойных забо-
леваний и контроля успешности проводимого лечения.
Entamoeba hartmanni Prowazek
Описана Провачеком в Самоа. Вегетативная форма. мелких разме-
ров— 6—8, не более 10 tu. В покоящемся состоянии экто- и эндоплазма
не различаются друг от друга. При движении образуются прозрачные
псевдоподии из гиалиновой эктоплазмы. Ядро 2—3 у, построено сходно
с Ent. histolytica. Встречаются двуядерные особи. В вакуолистой эндо-
плазме находятся заглоченные бактерии; эритроцитов эта амеба не
трогает. Перед инцпстированием в протоплазме около ядра образуется
хроматоидное тело в виде палочки, похожей на бактерию (табл. I,
рис. 6—8). Цисты 6—8—10 (и. В протоплазме встречаются разнообраз-
ной формы хроматоидные тела. Оболочка цист заметна плохо. Ядра
на свежих препаратах не видны. IB зрелых цистах четыре ядра и са-
ми цисты похожи на мелкие цисты дизентерийной амебы (табл. II,
рис. 26—46). Поэтому возникает вопрос о видовой самостоятельности
Ent. hartmanni. Ряд авторов считает ее за мелкую расу Ent. histolytica'
(Веньон, О’Коннор, Добелл и др.), другие же (Нбллер, Бруг, Г. Эп-
штейн и др.) признают ее самостоятельным видом, главным отличием
которого является его безвредность, так как четырехъядерными цистами
менее 10 р диаметром не удается заразить кошек амебной дизенте-
рией. В культурах не удалось видеть переход Ent. hartmanni в Ent.
histolytica. Биометрически обе эти амебы колеблются в размерах, каж-
дая вокруг своей средней величины, что показано и на материале из-
Средней Азии (В. Гнездилов).
При обследовании на патогенных простейших обычно все четырехъ-
ядерные цисты считаются за Ent. histolytica. Таким смешением, ве-
роятно, и объясняются большой процент и широкое распространение-
цистоносительства, огулом относимого к Ent. histolytica.
Пока еще нет способов диференцирования Ent. hartmanni от Ent.
histolytica при массовых микроскопических обследованиях населения на
кишечных простейших.
Другим непатогенным видом амеб с четырехъядерными цистами
Brumpt считает Ent. dispar, отличающуюся несколько более крупными
размерами, чем Ent. hartmanni.
Entamoeba dispar Brumpt
Мелкая амеба, в покойном состоянии 10X20 и, в вытянутом —
25 X 5—6 у. Ядро 2—3,5 у. Протоплазма вакуолизована, в ней бывают
заключены микроорганизмы. Эритроцитов в протоплазме нет; амебы во>
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
95
время пребывания в кишечнике кошки в редких случаях заглатывают
эритроциты. Движется, выпуская гиалиновые псевдоподии, более за-
медленно, чем дизентерийная амеба (табл. I, рис. 4). Цисты 10 X1
X 14 —15 X 12 tu, округлые или овоидные, четырехъядерные. 'Мор-
фологически не отличимы от мелких Ent. histolytica. Непатогенна для
55 5.95 6.37 6 8 7 2 2 7 95 807 8 5 89 935 977 102 Ю8 П 1147 119 123 12751317136 14 14.451487'15.3
Рис. 43 — Вариационные кривые для цист популяций Ent. hartmanni и
Ent. dispar, найденных при случае двойной инфекции.
По В. Гнездилову.
человека. Кошек можно заражать как кормлением инфицированным
материалом, так и введением его per anum. В толстых кишках кошки
размножается на поверхности слизистой оболочки, иногда проникает до
мышечной оболочки, но не вызывает глубоких некротических про-
цессов.
Бывают смешанные инвазии Ent. hartmanni и Ent. dispar. Биомет-
рическое исследование цист в таких случаях дает характерные вариа-
ционные кривые с двумя вершинами, соответствующие изменчивости
обеих популяций амеб (рис. 43). Отмечается также, что хроматоидные
тельца в цистах Ent. dispar встречаются реже, чем у Ent. hartmanni
(рис. 30, 17—25).
Видовая самостоятельность этих обоих видов амеб весьма сомни-
тельна.
Entamoeba coli Losch — Кишечная амеба
Описание. Безвредная кишечная амеба в округлом состоянии
имеет диаметр от 20 до 40 р, с пределами колебаний от 10 до 70 (и.
Движется гораздо медленнее дизентерийной амебы. Эктоплазма неясно
отличается от эндоплазмы. Последняя часто набита заглоченными
телами: бактериями, грибками, другими простейшими, цистами их,
обрывками клетчатки и др. (табл. I, рис. 10—И, рис. 30, 12—16).
Диференциация экто- и эндоплазмы наблюдается у живых амеб,
только что взятых из faeces. При комнатной температуре это разделе-
ние исчеза'ет уже через 10—15 минут, благодаря чему гиалиновые
псевдоподии в это время становятся зернистыми (Павлова).
Протоплазма кишечной амебы часто бывает сильно вакуолизована
(особенно после принятия носителем слабительной соли).
96
СПЕЦИАЛЬНАЯ ЧАСТЬ
Эритроциты заглатываются кишечной амебой в исключительно ред-
ких случаях. Тканей своего хозяина она не трогает.
Возможно, что эритрофагия Ent. coli является результатом опсони-
рования ее какими-либо бактериями, сообитателями амеб в кишечни-
ке. Как и у дизентерийной амебы, в протоплазме Ent. coli бывают эн-
догенные хроматоидные включения.
Ядро Ent. coli 4—8 р диаметром. Оно в общем богаче хромати-
ном, чем у дизентерийной амебы. Под самой оболочкой ядра лежит
более толстый слой базофильных (хроматиновых) зернышек (толщиной
около 1 tu), тесно прилежащих друг к другу. Кариозома с круглым
срединным зерном хроматина лежит несколько эксцентрично, тогда
как у Ent. histolytica она занимает центральное положение. Между ка-
риозомой и периферическим слоем хроматина также бывают разброса-
ны зернышки этого вещества (табл. I, рис. И). В общем, ядро Ent.
coli имеет более грубое строение, чем у Ent. histolytica. В живой ки-
шечной амебе ядро видно хорошо.
Дегенерирующие формы Ent. coli не отличимы от таковых форм
Ent. histolytica. Перед цистообразованием кишечные амебы делятся на
более мелкие предцистные стадии, которые также очень похожи на
соответствующие стадии Ent. histolytica.
Цисты Ent. coli от 10 до 30 и даже до 38 р диаметром. Оболочка
их более толста, чем в цистах дизентерийной амебы. Протоплазма тон-
козерниста и настолько прозрачна, что ядра могут быть сосчитаны в
живой и неокрашенной цисте*. Число ядер зависит от стадии созрева-
ния цисты (рис. 30, 13—16). В только что образующейся цисте одно
большое ядро, в протоплазме — большая вакуоль с гликогеном (табл. 1,
рис. 13). Ядро делится надвое, причем количество гликогена достигает
наибольшей степени .(табл. I, рис. 14); затем следует новое деление
с образованием 4-ядерных цист (табл. I, рис. 15). В результате заклю-
чительного деления возникают 8-ядерные цисты, которые и типичны
для Ent. coli (табл. I, рис. 16; табл. III, рис. 1, 20). Иногда этим дело
не заканчивается, вследствие чего появляются цисты с 12 и даже
16 ядрами. Бывают в редких случаях цисты с 20 и даже с 32 ядрами.
Важно наличие у Ent. coli и 4-ядерных цист, которые, впрочем, попа-
даются нечасто. Это обстоятельство необходимо иметь в виду при учете
цистоносителей; многие авторы все 4-ядерные цисты относят к Ent.
histolytica} этим, вероятно, и объясняется в известной части очень вы-
сокий процент цистоносителей дизентерийной амебы.
Размеры ядер мельчают при увеличении их количества. Типичное
строение ядер 'можно наблюдать уже на четырехъядерных цистах. Хро-
матиновые зернышки располагаются под оболочкой ядра равномерным
слоем, тогда как в цистах дизентерийной амебы они скопляются с од-
ной стороны ядра в виде полулунной массы.
В протоплазме цист бывают хроматоидные тельца в виде глыбок
неправильной формы с заостренными концами; размеры хроматоидных
телец вариируют весьма значительно; эти образования вовсе могут от-
сутствовать в зрелых цистах.
Инцистирование происходит чаще на стадии двух ядер, которые
лежат на периферии большой центральной вакуоли цисты; на перифе-
рии же размещаются и хроматоидные тельца разнообразной формы;
последние в редких случаях могут отсутствовать. Цисты могут быть
поражены грибками, которые иногда наблюдаются и у вегетативных
особей (Г. Эпштейн).
Таблица III
Рис. 1—4. Цисты амеб кишечника человека по Г. В. Эпштейну
на окрашенных препаратах:
1 — Entamoeba coli, 8-ялерная циста, 2 — Entamoeba histolytica, 4-ядерная циста,
3 — Entamoeba dispar, 4 — Entamoeba hartmanli.
Рис. 5—20. Циста кишечных простейших при обработке Луголевским
раствором по Dobell О'Соппог’у, увеличение около 2000 раз.
5—7 —Entamoeba histolytica, 1-2-4-ядерная цисты от штамма амеб около 12р., 8—9—Entamoeba
histolytica, 1- и 4-ядерная цисты от штамма с цистами около 7,5р.. 10—Jodamoeba biitschlii —
зрелая (1-ядерная) циста, ll—13—Endolimax папа, 1-, 2- и 4- ядерные цист»! (на рис. 12 циста
с глыбкой гликогена), 14—Chilomastix mesnili, 15—Blastocystis hominis, 16—Giardia intestinalis,
17—20—Entamoeba coli
Рис. 21. Plasmodium vivax в мазке. Рис. 22. Plasmodium vivax в толстой капле.
Рис. 23. Plasmodium vivax — верхний ряд — молодые трофозоиты в толстой
капле, второй ряд — шизонты и мерозоиты в толстой капле; внизу гаметоциты
в толстой капле. Рис. 21—23 по I.W. Field и Н. Le Fleming-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
97
Matthews (1919) считает, что Ent. coli, так же как и дизентерийная
амеба, образует различные расы, средняя величина которых характе-
ризуется размерами 15—16,5—18,7 и 21,7 ,и. Другие признают наличие
трех рас, но дают для них иные размеры, колеблющиеся в пределах
12—14, 15—18 и 19—22 f* (Boeck, 1923).
Рис. 44 — Стул при затяжном колите. Среди гнойных клеток и
эритроцитов много крупных вегетативных форм Ent. coli f. magna.
По В. Гнездилову.
Кишечная амеба в вегетативной форме обитает преимущественно в
верхнем отделе толстой кишки, именно в ее жидком содержимом;
цисты находятся в сформированном содержимом выходной части тол-
стой кишки. Поэтому кишечная амеба при нормальном стуле выходит
наружу только в виде цист; при поносах или после принятия слаби-
тельного в испражнениях обнаруживаются и подвижные вегетативные
формы. Распространена она по всему свету и большинством исследова-
телей считается безвредной.-
Интересно отметить, что, в противоположность дизентерийной, ки-
шечная амеба мало чувствительна к эметину.
В единичных случаях кишечную амебу находили в толще слизистой
оболочки кишки, однако такое внедрение является вторичным, ибо в
этих случаях кишечник был поражен патологическим процессом
(брюшной тиф и др.).
Councihnania lafleuri Kofoid и Swezy
Эта амеба была описана в качестве самостоятельного вида в 1921 г.
в Калифорнии. Она настолько сходна с Ent. coli, что Wenyon, Ndller
и другие авторитеты не считают ее особым видом. Соипс. lafleuri
98
СПЕЦИАЛЬНАЯ ЧАСТЬ
питается не только теми веществами, что и кишечная амеба, но пожи-
рает также (подобно дизентерийной амебе) красные кровяные тельца.
Она отличается значительной подвижностью и большими размерами, «а
также строением цист. В последних 8 ядер, как и у Ent. coli, однако,
Рис. 45 — Councilmania lafleuri в свободном состоянии
(посредине), с выходом «почки» через оболочку
По К о f о 1 d k и S w е z у.
в стадии цисты
(справа).
цисты
Councilmania отличается особым способом размножения, напоминаю-
щим почкование. В стенке цисты образуется маленькое отверстие, че-
рез которое выходит наружу некоторое количество плазмы с одним из
ядер цисты; таким способом выходят одно за другим все ядра, и из
цисты освобождаются 8 почек или маленьких амебок. Эти процессы
протекают в свежих испражнениях дизентерийных больных (рис. 45).
Однако в стенках цист могут обра-
Рис. 46—Карликовая амеба -Endo-
Итак папа. 1 — вегетативная форма
и образование цист (2 — 5).
По В е н ь о н у.
зоваться трещины, через которые на-
ружу выходят часть протоплазмы и
ядра цист (например, у кишечной
амебы). Это обстоятельство застав-
ляет подозревать, не является ли
«почкование» у Councilmania искус-
ственной картиной, не имеющей ника-
кого отношения к процессу размноже-
ния.
Во всяком случае вопрос суще-
ствования этого вида требует новых
доказательств.
Возможно, что Councilmania laf-
leuri относится к Entamoeba coli так,
как Ent. histolytica forma minuta.
Councilmania была найдена и у
солдат, возвратившихся после войны
из Франции в Калифорнию, что ука-
зывает на ее широкое географическое
распространение.
Endolimax папа Wenyon et O’Connor — карликовая амеба
Синоним: Entamoeba папа W. н О ’ С.
Маленькая! амеба (6—8—12 р диаметром) с неотличимыми в покое
друг от друга экто- и эндоплазмой. При движении выпускает гомоген-
ные псевдоподии; по характеру движения может напоминать дизенте-
рийную амебу. Ползает медленно и вне тела человека скоро теряет
подвижность. Протоплазма сильно вакуолизована. Ядро, 1—3 р диа-
метром, имеет вид тонкостенного пузырька (рис. 46). Хроматин пред-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
99
ставлен только крупной кариозомой; у дегенерирующих особей она
распадается на несколько зерен разнообразного вида.
Питается бактериями. Цисты карликовой амебы овальны, размером
8—10 X 7—8 у; ядер в них 1—2—4 (редко 6—8). Кариозома в ядрах
цист лежит эксцентрично или вплотную прилежит к какому-либо
участку ядерной оболочки. В протоплазме цист содержатся сильно
преломляющие свет зернышки волутина и гликогена (табл. II, рис.
II —13). Цисты карликовой амебы во влажных испражнениях сохра-
няют жизнеспособность до трех недель.
Живет в тонких кишках, непатогенна, встречается часто; геогра-
фическое распространение ее широкое как в умеренных, так и в жар-
ких странах.
Ent. histolytica, Ent. coli и Endolimax папа являются наиболее обык-
новенными обитателями кишечника человека. Ввиду возможности в не-
которых случаях смешения этих форм, приводим таблицу диферен-
циального распознавания цист названных амеб по их признакам:
Цисты Ent. histolytica End. папа Ent. coli
Величина в микронах 5/6—15/20 7 3/5—12/16 11/14—22/35
Форма Шаровидная, иногда асимме- трично округлая Эллипсоидная, у мелких форм — овоидная, у крупных — часто непра- вильная, иногда шаро- видная Шаровидная, иногда эллипсоидная, овоид- ная или неправильная
Jodamoeba biltschlii Dobell
Синонимы: Entamoeba biltschlii Prowazek, Pseudolimax wenyoni Brumpt.
В округлом состоянии 9—17 p диаметром (крайние размеры 5—24—
30 у). У покоящейся амебы экто- и эндоплазма неразличимы. При
передвижении вытягивается червеобразно. В эндоплазме много загло-
ченных бактерий (табл. I, рис. 5-; табл. III, рис. 10).
Ядро — в виде пузырька, диаметром 2—3 у (iDobell) или 3—6 &
(Noller); хроматин представлен только большой центрально лежащей
кариозомой. Между последней и оболочкой ядра — светлая зона с
зернышками, окрашивающимися то плазматическими, то ядерными
красками.
Амеба, готовая к инцистированию, лишена пищеварительных вклю-
чений, плазма ее стекловидная; ядро относительно большое. Оболочка
цисты сравнительно толста, двуконтурна (что характерно), но бесцвет-
на; она ясно видна на фиксированных препаратах. В протоплазме
цисты имеется большая, занимающая иногда весьма значительное про-
странство, постоянная вакуоль с гликогеном. Кроме того, есть и грану-
лы волютина. Ядро одно. Зернышки хроматина скопляются в полулун-
ную по форме кучку, прилежащую с одной стороны к кариозоме. Бы-
вают и двуядерные цисты (по Dobell’y — являющиеся редкостью).
Цисты овальные или круглые, 8—12—20 (и в поперечнике. Бывают
удлиненные, или неправильные по форме, что затрудняет установление
характерных для данного вида размеров.
Jodamoeba biltschlii является распространенной обитательницей ки-
100
СПЕЦИАЛЬНАЯ ЧАСТЬ
шечника человека. При твердом стуле выносятся наружу только ци-
сты. В жидких извержениях находятся и амебы.
Этот вид является непатогенным для человека.
Entamoeba gingivalis Gross — ротовая амеба
Синоним: Entamoeba buccalis (Prowazek)
Является первой паразитической амебой, найденной у человека
(1819, Gross). Штейнберг наблюдал их в Киеве в 1862 г. Они часто
встречаются в кариозных зубах, а также в белом мягком налете, по-
крывающем зубы (табл. I, рис. 9). Известны с полной достоверностью
только вегетативные формы. Размеры амеб колеблются от 6 до 60 н
Рис. 47 — Ротовая амеба—Ent. gingivalis из налета
кариозных зубов. В некоторых амебах заглоченные
эритроциты. По Wenyon.
в диаметре, но обычно величина не превышает 20 р. По виду Ent. gingi-
valis похожа на дизентерийную амебу (рис. 47). Эктоплазма хорошо
различима от эндоплазмы. Двигается при помощи широких псевдопо-
дий из гиалиновой протоплазмы, отличающихся по своему характеру
от ложноножек дизентерийной амебы. Кариозома в ядре лежит цент-
рально или эксцентрично. Она такого же типа, что и у кишечной аме-
бы, но несколько меньших размеров. Ядра обычно видны и в живой
амебе.
Ent. gingivalis питается бактериями и лейкоцитами. Главной пищей
ее являются белые кровяные клетки, однако остается неясным, загла-
тывает ли амеба самые лейкоциты или только голые ядра их (так на-
зываемые слюнные тельца). Последние компактны по структуре и си-
дерофильны при окрашивании железным гоматоксилином; поэтому их
иногда принимали за заглоченных эритроцитов.
Ent. gingivalis, по указанию ряда компетентных авторов, способны
заглатывать красные кровяные тельца. Однако они в несколько минут
исчезали на глазах исследователей. Остается неясным, почему эритро-
циты обнаруживаются в Ent. gingivalis очень редко; причин может быть
две: амебы заглатывают их часто, но они перевариваются очень быстро,
благодаря чему их трудно захватить в самой амебе; или же эри-
трофагоцитоз оказывается исключительно редким явлением.
Наличие цист Ent. gingivalis отрицается огромным большинством ав-
торов. Эпштейн утверждает, что цисты встречаются редко. Находимые
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
101
Цисты Ent. histolytica End. папа Ent. coli
При иссле- довании в свежем со- стоянии Большей частью силь- но преломляют свет, стекловидны, часто неправильно вакуо- лизованы; слегка се- ровато-голубого цве- та; ядра редко види- мы; гликоген неви- дим Менее сильно прелом- ляют свет, но с силь- но преломляющими свет зернышками; се- ро-синеватого цвета; ядра невидимы; гли- коген различим Наиболее слабо пре- ломляют свет, го- могенные, фарфо- ровидные; серо- синеватого цвета; ядра явственно различимы; глико- ген виден также ясно
При окра- шивании иодом и эозином Протоплазма светло- зеленовато-желтая; грубо вакуолизова- на, у мелких рас тонкозерниста. Ядра заметны вследствие сильного лучепре- ломления в них; край их толст; различима центральная гранула. Гликоген диффуз- ный; если он собран в общую массу, то с неопределенными краями. Цисты окра- шиваются в красный цвет скоро Протоплазма зеленова- то-желтая, со множе- ством мелких пре- ломляющих свет ва- куолек. Ядра нераз- личимы, с тонким краем и перифериче- ским хроматиновым телом. Гликоген гу- стой в виде одной или нескольких глы- бок, резко централь- ных. Цисты проти- востоят окраске дольше Протоплазма грубо- однообразно-зер- нистая, темновато- коричневая. Ядра хорошо различи- мы, с тонким зер- нистым краем и центральным хро- матиновым телом. Гликоген лежит центрально; края его неясны. Ци- сты обычно про- тивостоят окраске дольше
Ядра (число) 1 в 30—45о/о; 2 в 13—ЗОо/о; 3 — редко; 4 в 22—55% 1 — весьма обыкновен- но; 2 — менее обычно; 4— обычно для мелких рас; очень редко для крупных 1 — очень редко; 2 — редко; 4 — очень редко; 8 — весьма обыкно- венно; 16 — очень редко
Строение ядра Центральная гранула различима; перифе- рический хроматин распределен равно- мерно маленькими пластинками Центральной гранулы нет; ядерная оболоч- ка неразличима; пе- риферический хрома- тин в виде един- ственной глыбки, прилежащей к ядру Центральная грану- ла различима; пе- риферический хро- матин в виде не- многих больших пластин или те- лец у ядерной оболочки
Хроматоид- ные тела В виде единственного большого бруска или нескольких меньших палочек с округлыми концами. Имеются в общем у 50% цист В виде нескольких ок- руглых или непра- вильных масс. Встре- чаются редко В виде крупных или мелких «заноз» с четырехугол ь н ы- ми краями. Есть в общем у 5- 10% цист
Стенка ци- сты Тонкая, различимая Изменчивой толщины, менее различимая Более толстая, хо- рошо видимая
Величина рас Хорошо различимы: мелкие расы от 7, большие расы от 13—14 jx. Наиболее часты — около 10 р. Хорошо различимы; одни—5—7, другие— 8—Ю, еще большие— 12—15 р. (наиболее часты) Менее различимы; одни около 15, другие около 18 р. и больше
Примечание. Ent. histolytica берется по основному признаку четырехъядер-
ности без диференциации на сомнительные виды.
102
СПЕЦИАЛЬНАЯ ЧАСТЬ
И-М в полости рта одноядерные цисты он приписывает Ent. gingivalis.
Вопрос о патогенном значении этих амеб трактуется различно. На-
хождение больших количеств амеб при альвеолярной пиоррее дало по-
вод считать их причастными к патологии этого заболевания; на этом
основании для лечения амебной альвеолярной пиорреи рекомендовался
эметин. Другие наблюдатели считают Ent. gingivalis безвредными оби-
тателями ротовой полости. Факт нахождения амеб в распаде раковой
опухоли дна ротовой полости не считается доказательством их пато-
генности, так как амебы не являлись тканевыми паразитами, а были
в тканевом детрите.
Jodamoeba schewiakovi G. Epstein
для человека вид —
Г. Эпштейном описан также непатогенный
Jodamoeba schewiakovi.
Рис. 48 — Ent. gingivalis — характерное скопление
амеб в ротовой полости вокруг нити Leptotr'ix.
По Эпштейну.
Довольно большая амеба с крупнокариозомным ядром. Тело в вы-
тянутом состоянии до 55 р длины; диаметр ядра 6—7 р. При движе-
нии эндоплазма редко обособляется от эктоплазмы. Цисты субоваль-
ные 12—20 X 10—17 у, одноядерные с крупной эксцентрически лежа-
щей гликогеновой вакуолью. Кишечник человека. Непатогенна.
Род Dientamoeba
Dientamoeba fragifys Jepps et Dobell
Мелкие амебы (13,5—12 tu; в среднем 9 jx) с хорошо различимыми
экто- и эндоплазмой. Движутся посредством широких, листовидных или:
зубчатых по краям псевдоподий (табл. IV, рис. 26, 27). Почти 4/? осо-
бей этого вида амеб двуядерны. Ядра равны по величине (0,8—2,3 у
в поперечнике; чаще — 2 у). В ядре — кариозома, состоящая из куч-
ки хроматиновых зерен. В периферической зоне ядра хроматина нет.
Живет, вероятно, в ободочной кишке. Очень нежна и может быть
получена только из еще теплого кала. Цист в организме человека не
образует, что дает основание считать ее за гостепаразита.
Найдена в Англии, в Германии, в СССР.
D. fragitis (рис. 27) при ближайшем ее исследовании оказывается
довольно частым паразитом человека; в Голландии Бруг (1936) нашел
ее у 36°/о обследованных обитателей приюта после дачи солевого сла-
бительного. Применение метода культивирования в три раза повысило
число положительных находок. Вегетативные формы этой амебы обыч-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
103
но обнаруживаются в твердом стуле. Культуры можно получить из
твердого стула через 67г часов, а из жидкого—через 137г часов после
дефекации. Хранение культур при комнатной температуре (16—20° С)
вредно влияет, особенно на молодые культуры. Культура D. fragilis
развивается лишь в присутствии обильных бактерий; при их недостатке
культура погибает в один день.
Чувствительность культур к комнатной температуре говорит против
допущения, что D. fragilis является свободно живущей амебой, лишь
случайно попадающей в кишечник человека. Вероятно, вегетативная
форма этой амебы может проходить через желудок человека без осо-
бенного для себя вреда. Из животных эта амеба найдена у обезьян
(Н. Хегнер).
Г. Эпштейн считает, что ему один раз удалось видеть цисту D. fra-
gills. Циста четырехъядерная, в форме эллипса, длиной около 14 р.
Все ядра лежат рядом, ближе к одному из полюсов цисты. Ядра круп-
нокариозомного типа. В цистах включений не наблюдалось.. Родство
этих цист с D. fragilis им устанавливается на основании тождества
строения ядер.
Род Dimastigamoeba
Dimastigamoeba gruberi Schardinger
Синоним: Naegleria gruberi Wilson.
Амеба в округленном состоянии 7—15 р (по Шардингеру 12—24 р)
диаметром. При ползании растягивается до 32—40 р длины. Ядро
3—4 р (4—8 рь) диаметром. В нем большая центральная кариозома и
рассеянные зерна хроматина в ахроматиновой зоне. Есть одна сократи-
тельная вакуоль. Движется при помощи широкой ложноножки из
эктоплазмы. В эндоплазме — вакуоли с заглоченными бактериями.
При некоторых обстоятельствах эта амеба превращается в свободно
плавающую жгутиковую стадию. Амеба округляется в овал, на перед-
нем конце которого залегает ядро. От последнего отходят два длинных
бича. Сократительная вакуоль лежит .на противоположном полюсе ова-
ла. Жгутиковая форма может терять бичи и снова принимать облик
амебы.
Цисты шарообразны, в поперечнике 8—12 р. В них одно ядро и не-
сколько хроматоидных телец. Стенка цисты двойная. Очень характерно
присутствие в стенке 3—8 мелких пор. Через одну из них. амеба поки-
дает цисту.
Dimastigamoeba gruberi встречается в извержениях человека в каче-
стве копрозойной формы. Это' один йз наиболее обыкновенных свобод-
но живущих видов амеб.
Не будем останавливаться на других копрозойных формах (Trima-
stigamoeba philipertsis, Hartmanella hyalina, Craigia (Paramoeba) homi-
nis, Sappinia diploidea и др.) и перейдем к рассмотрению паразитиче-
ских жгутиковых.
КЛАСС MAST1GOPHORA SIVE FLAGELLATA —
ЖГУТИКОВЫЕ ИЛИ БИЧЕНОСЦЫ
Тело покрыто тонкой оболочкой (пелликулой). Для движения слу-
жат особые органоиды в виде бичей или жгутиков, числом от
одного до восьми и более. Ядро в виде пузырька с центрально лежа-
щим телом, называемым кариоз о мой. У основания жгута лежит
104
СПЕЦИАЛЬНАЯ ЧАСТЬ
базальное тельце или блефаропласт; кроме того, с орга-
ноидом движения могут быть у разных биченосцев связаны и другие
образования в форме парабазального тельца, кинетону-
клеуса и др. Кинетонуклеус и блефаропласт, взятые вместе, обо-
значаются часто общим термином — кинетопласт. У некоторых
биченосцев имеется опорное образование в виде аксостиля. Пища
принимается или в определенном месте тела (называемом клеточным
ртом — цитостом), или же всасывается всей поверхностью тела
(осмотическое питание трипанозом, лямблий). Размножаются
делением, которое происходит всегда в продольном направлении от
переднего конца. Передним считается тот конец тела, который несет
бичи. Некоторые формы инцистируются. Типы: Trichomonas, трипано-
зома.
Паразитические жгутиковые человека принадлежат к отрядам
Protomonadida и Diplomonadida.
О ТРЯД PRO TOMONADIDA
Жгутиков мало, редко более шести; ядро одно (монозойные формы).
Семейство Trypanosomidae — трипанозомовые
Биченосцы с одним жгутом; паразитируют в животных (позвоночных
и беспозвоночных) и в растениях. Тело в общем удлиненное; у типич-
ных форм по его краю идет волнообразная перепонка (ундулирующая
мембрана); жгутик начинается от блефаропласта ризопластом,
каковым именем называется участок жгута, лежащий в протоплазме
биченосца. Позади блефаропласта лежит палочковидный или округлен-
ный кинетонуклеус, красящийся ядерными красками, подобно хромати-
ну. Присутствие кинетонуклеуса типично для всех представителей
сем. Trypanosomidae.
У некоторых трипанозомид имеется ундулирующая или
волнообразная перепонка, представляющая собой вырост
оболочки тела (пелликулы или перипласта); жгут идет по свобод-
ному краю ундулирующей мембраны и на переднем конце тела высту-
пает свободно. Жгутик может отсутствовать, равно как и волнообраз-
ная перепонка. Цитостома нет. Питаются эндосмотически.
Ядро пузырчатое с кариозомой. Размножаются исключительно про-
дольным делением, т. е. бесполым путем.
Размножение начинается делением блефаропласта и кинетонуклеуса
каждого на две части. Жгут делящейся особи остается соединенным
с одним из блефаропластов, тогда как из другого, дочернего блефаро-
пласта, вырастает новый жгут.
Если есть ундулирующая мембрана, то новый жгут продвигается
по ее ребру рядом со старым, и лишь позднее волнообразная перепонка
начинает расщепляться в продольном направлении.
После этого начинается деление ядра, обычно перешнурованием.
Затем происходит продольное деление тела биченосца на две дочерние
особи. У некоторых форм бывает и множественное деление сразу на
несколько особей.
Полный жизненный цикл некоторых Trypanosomidae связан с про-
хождением различных стадий как в основном хозяине, так и в перенос-
чике. Заражение трипанозомовыми бывает прямым, т. е. непосредствен-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
105
ио от больного к здоровым, и непрямым, т. е. при посредничестве
переносчиков, которыми являются различные насекомые и пиявки.
Основными морфологическими формами семейства Trypanosomidae
являются типичные представители родов Leptomonas, Crithidia, Trypa-
nosoma и Leishmania (рис. 49).
Рис. 49 — Характерные формы и классификация некоторых жгутиковых.
По W е п у о п.
Leptomonas имеет удлиненное тело со жгутом, основание которого и
кинетопласт помещаются в переднем отделе тела далеко от ядра;
колеблющаяся перепонка отсутствует.
Crithidia — тело также удлиненное, но кинетопласт лежит непосред-
ственно впереди от ядра; имеется слабо выраженная ундулирующая
мембрана.
Trypanosoma — тело веретеновидное, уплощенное, изогнутое; кинето-
пласт лежит позади ядра у заднего конца тела; благодаря этому уве-
личивается часть жгута, связанная перепонкой с телом трипанозомы;
колеблющаяся перепонка развита хорошо.
Leishmania — овальное или круглое тело очень малых размеров;
жгут отсутствует или есть только его основная внутрипротоплазменная
часть; кинетопласт имеется; лейшманиеподобные формы, одетые плот-
ной оболочкой, называются цистами.
Перечисленные формы являются либо самостоятельными видами
соответствующего рода, либо формами жизненного цикла одного ка-
кого-либо вида.
Так, Leptomonas в своем полном жизненном цикле проходит л е п-
томонадную и лейш маннозную стадии.
Crithidia проходит три формы — лейшмании, лептомонадную и к р и-
тидиальную.
Leishmania бывает в форме Leptomonas и собственно лейшмании.
Trypanosoma equiperdum — возбудитель случной болезни лошадей
имеет только одну типичную трипанозомную форму.
Trypanosoma cruzi — возбудитель болезни Чагаса, помимо трипано-
106
СПЕЦИАЛЬНАЯ ЧАСТЬ
зомной формы в теле человека, принимает лейшманиеподобный вид; в
организме же переносчика («поцелуйный клоп» — Conorrhinus, Triato-
та) проходит стадии критидиальную, лейшманиеподобную и стадию
метацикли ческих трипанозом.
Все названные биченосцы являются, паразитами позвоночных и бес-
позвоночных животных, причем цикл развития этих паразитов проходит
или в одном хозяине, или в двух, относящихся к различным видам.
Первый вариант развития называется моногенетически м, вто-
рой — дигенетически м.
К моногенетическим формам принадлежат роды Leptomonas, Qrithi-
dia и Herpetomonas, паразитирующие в кишечном канале насекомых,
клещей и других беспозвоночных, где протекает весь цикл их развития.
Они имеют цистную стадию.
Дигенетическое развитие связано с переменой хозяев, при-
чем одним хозяином является позвоночное животное, а другим — бес-
позвоночное, питающееся кровью первого (например, рыба и пиявка;
человек и муха тсе-тсе; крупный рогатый скот и слепни).
Таким образом, при дигенетическом развитии происходит смена мест
обитания биченосцев: кровь или ткани позвоночного хозяина и пищева-
рительный аппарат беспозвоночного, которое одновременно является
переносчиком соответствующего биченосца.
К дигенетическим формам принадлежат представители родов Trypa-
nosoma (у некоторых видов выпадает участие переносчика и передача
паразита происходит контактным путем) и Leishmania. Представители
рода Phytomonas живут в соке растений (молочаи) и передаются раз-
личными насекомыми:
Способы заражения хозяина паразитами из семейства Trypanoso-
midae разнообразны. Они сводятся к проглатыванию инвазирующих
форм паразитов и к проникновению их в окончательного хозяина при
сосании его крови переносчиком. Инвазирующие формы в виде лейшма-
ниальных цист рассматриваемых нами биченосцев выбрасываются на-
ружу с калом промежуточного хозяина (т. е. переносчика), в случае
если местом их конечного образования является задняя кишка. Зара-
жение происходит или при проглатывании кала, или при поедании зара-
женного хозяина. Такой способ инвазии называют контаминатив-
н ы м. Интересно отметить, что контаминативным путем проникают
через кишечник даже некоторые трипанозомы, обитающие в крови;
например, овечья — Trypanosoma melophagium попадает в овец при
поедании ими зараженных трипанозомами рунцов (бескрылая кукло-
родная муха, паразитирующая на коже овец — Melophagus ovinus).
Из пищеварительного аппарата овцы метациклические трипанозомы
активно проникают в ее кровь.
Инвазирующие формы других биченосцев образуются в слюнных
железах или в хоботке переносчика, и передача их происходит при со-
сании переносчиком крови окончательного хозяина; паразиты впрыски-
ваются последнему со слюною промежуточного хозяина (он же и пере-
носчик паразита) или вымываются слюною из его хоботка.
Такой и н ок у л я тив н ы й способ заражения осуществляется слеп-
нями, мухою тсе-тсе, пиявками и др.
В тех случаях, когда биченосец проделывает в переносчике хоть ка-
кой-либо цикл развития, говорят о специфическом переносе.
Механическая передача паразитов бывает тогда., когда в переносчике
биченосцы вовсе не размножаются.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
107
Примером последнего может служить передача некоторых трипано-
зом в случаях «прерванного кормления». Если переносчик пьет кровь
зараженного трипанозомами хозяина и будет почему-либо согнан с не-
го до окончательного насыщения, то при ближайшей посадке на новое,
здоровое животное он может занести хоботком трипанозомы, которые
не успели еще погибнуть за время между начальным и окончательным
кормлением.
Среди трипанозомовых есть весьма важные паразиты человека.
Род Leishmania
Биченосцы рода Leishmania, называемые общим именем л е й ш м а-
н и й, являются паразитами позвоночных животных, в которых имеют
вид безжгутиковых внутриклеточных организмов.
Жгутиковую лептомонадную стадию они проходят в
насекомых, которые являются их переносчиками.
Leishmania donovani Laveran et Mesnil
L. donovani является возбудителем общего лейшманиоза, называемо-
го различными наименованиями: кала-азар, тропическая спленомегалия,
болезнь дум-дум, черная лихорадка, висцеральный или внутренно-
стный лейшманиоз, детский кала-азар, детский лейшманиоз, средизем-
номорский лейшманиоз, тропическая кахексия и мн. др. В Азербайджа-
не эту болезнь называют кара-азар.
Описание. L. donovani морфологически не отличима от L. tropi-
ca—возбудителя кожного лейшманиоза (см. ниже). Паразиты имеют
круглую или овальную форму, размером от 2 до 4 р (изредка попада-
ются и вдвое более крупные формы). Протоплазма красится в синий
цвет краской Романовского. В теле лейшмании ближе к центру или
сбоку лежит крупное ядро в виде фиолетово-красного зернышка; кроме
ядра имеется блефаропласт (кинетопласт) в виде короткой палочки
или. округлого тельца (рис. 50). Иногда заметна нить, отходящая от
блефаропласта к периферии паразита. Морфологически она соответ-
ствует основанию жгутика, свободная часть которого у лейшманий от-
сутствует.
Лейшмании, как правило, залегают внутри протоплазмы клеток раз-
личных органов; в нейтрофилах и моноцитах периферической крови, а
также в эндотелии капилляров кровеносной и лимфатической системы,
селезенки, лимфатических желез, костного мозга (миэлоциты), печени,
кожи и др. Часто бывает большое количество лейшманий (100—200),
которые и «забивают» протоплазму зараженных клеток. При разруше-
нии последних лейшмании освобождаются и могут быть обнаружены по
одиночке. В единичных случаях находили в селезенке и костном мозгу
лептомонадную (т. е. жгутиковую) форму превращения лейшманий, ко-
торая свойственна переносчикам этого паразита. Встречаются и стадии
деления лейшманий; сначала делится блефаропласт, затем ядро и, на-
конец, протоплазма (рис. 51).
Течение кала-азар. Инкубационный период при кала-азар
гянется от немногих недель до нескольких (3—6—9) месяцев.
Тропическая спленомегалия сопровождается неправильной и не под-
дающейся хинину упорной лихорадкой: кожа бледная, сухая; часты
изъязвления слизистой оболочки рта и геморрагии, селезенка (а также
и печень) постепенно увеличивается, нередко до громадных размеров
(рис. 52); исхудание вследствие мышечной атрофии и исчезновения
108
СПЕЦИАЛЬНАЯ ЧАСТЬ
подкожного жира. Болезнь идет быстро или принимает хроническое те-
чение (1—3 года). Смертность очень велика.
Диагностика кала-азар., Несомненный диагноз при жизни
ставится нахождением паразитов в клетках селезенки, в печени, в ко-
стном мозгу, лимфатических железах или в периферической крови.
Следует иметь в виду, что иногда
встречаются случаи дегенеративных из-
менений зернистости лейкоцитов (грану-
лоцитов), могущие до известной степе-
ни симулировать паразитов кала-азар
(Аринкин), но не имеющие никакого
положительного диагностического значе-
ния. При установлении диагноза необхо-
дим также подробный анамнез, бывал ли
больной в прошлом (и когда) в местах
бесспорно эндемичных по кала-азар. Из-
вестен, например, случай диагносциро-
вания кала-азар в Иркутске, на осно-
вании которого было сделано утвержде-
ние, что этот город относится к мест-
Рис. — Leishmania donovani в
мононуклеаре из периферической
крови. Из М. М а у е г.
ностям эндемичным по лейшманиозу.
Однако ближайшее рассмотрение анамнеза показало, что больная
приехала в Иркутск из Узбекистана по совету врачей, где она болела
«упорной малярией, не поддающейся хинной терапии»: по правильно-
му толкованию В. Петрова, этот случай должен быть по месту зара-
жения приписан не Иркутску, а Узбекистану.
В мазках крови больше шансов найти паразитов в конце заболева-
ния. Приходится искать их на многих препаратах, иногда в течение
нескольких дней.Бы-
Рис. 51 — Деление Leishmania infantum.
1— парабазальное тело, ядро, 2—аксонема, 3—выступание
жгута из тела, окруженного «шлаковой» массой, 4-7 — деление
паразита в форме ячменного зерна, 8 — двойное деление.
По А. Алексееву и А. Конахову.
ли случаи получения
лейшманий в куль-
турах из крови, в
которой морфологи-
чески паразиты не
определялись.
Более надежные
результаты дает ис-
следование пульпы
селезенки, которую
добывают пункцией
этого органа. Боль-
ного кладут на пра-
вый бок и плотно
прижимают кожу к
средней части селе-
зенки. В придавлен-
ное и растянутое
пальцами место ко-
жи вкладывают на половину длины иглу шприца, затем поднимают
поршень, который и втягивает в канюлю некоторое количество се-
лезеночной пульпы. Иглу быстро извлекают, и содержимое канюли
употребляют для мазков или для посева на NNN-arap. В культурах
развиваются жгутиковые формы лейшманий (рис. 53).
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
109
Аналогичным способом производят пункцию лимфатических желез.
Костный мозг добывают трепанацией головки большой берцовой кости
или из ребра. Проще и практичнее всего добывать костный мозг вкалы-
ванием бировской иглы в грудину до
момента, когда будет ощущаться упор
вошедшей иглы в заднюю стенку гру-
дины (М. Аринкин, Кассирский). Поль-
зуются также предохранительным
щитком, надеваемым на расстоянии
одного сантиметра от конца иглы.
Прокол производят на уровне третьего
межреберного пространства. На иглу
надевают шприц и медленно насасы-
вают некоторое количество костного
мозга. Добытое вещество выдувают
шприцем на предметное стекло для
приготовления мазка, а остающуюся
массу смывают на посев (среда NNN-
агар).
Простой процедурой по сравнению
с добыванием селезеночной пульпы
является пункция лимфатической же-
лезы (например, паховой) для диагно-
стической цели. Производится она в
аналогичном порядке, но применяют
иглу шприца.
Важным фактом является устано-
вление наличия лейшманий в непо-
врежденной коже больных, что на-
блюдалось даже в некоторых случаях,
когда никакими другими методами
диагноза на кала-азар нельзя было
поставить. Исследование произво-
дится на кожных соскобах.
Паразиты кала-азар могут выде-
ляться в вирулентном состоянии с
носовой слизью, а также с испраж-
нениями (человек) и даже с мочой
'хомяк). В трупах лейшмании исче-
зают очень быстро (через 2—5 ча-
сов).
РиС. 52 — Кала-азар у девочки (Сред-
няя Азия). Сильное увеличение пече-
ни и селезенки, водянка живота.
По фотографии Артамонова.
Добавочным методом диагностики кала-азар является реакция
Нэпира (Napier). К 1 см3 сыворотки больного прибавляют две капли
4О°/о формальдегида. Пробирку встряхивают; в положительном случае
сыворотка быстро желатинизируется, твердеет и в ближайшее время
приобретает молочно-перламутровый цвет. Отмечают, что эта реакция
чаще дает положительный результат лишь с третьего месяца болезни.
Распространение кала-азар
Наличие кала-азар в прежней России было установлено венскими
врачами Sluka и Zarfl, диагносцировавшими внутренностный лейшма-
ниоз у девятилетнего ребенка, привезенного из Ташкента в Вену для
но
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 53 — Leishmania donova.nl. Свобод-
ные и аггломерированные формы из
культуры. По Wenyon.
лечения нетипичной лихорадки, принимавшейся ранее за малярию
(1909).
Затем единичные случаи (более или менее достоверные) описыва-
лись и русскими врачами (Никифоров, Петров и др.). Первые сведения
о распространении кала-азар в Средней Азии были добыты экспеди-
цией проф. В. Якимова (1913). За-
тем изучение кала-азар у нас замер-
ло, и лишь в послеоктябрьскую*
эпоху стало энергично развиваться
трудами многих врачей Средней
Азии и Закавказья (Артамонов,
Л. Исаев, С. Канделаки, В. Петров,.
Г. Терехов, Н. Ходукин и мн. др.).
В Средней Азии, по сводке
В. Петрова, кала-азар известен в
Ташкенте, Коканде, Самарканде,
Катта-Кургане, Шахризябсе, Ст.
Бухаре, Чарджоу, Керках, Ашха-
баде, под Кушкой, в Тахта-базаре,
в округах Кашка-дарьинском, Сур-
хан-дарьинском, Андижанском.
Единичые случаи наблюдаются
в Фергане и в Южном Таджики-
стане.
Высказывавшиеся ранее мнения,
что кала-азар в Средней Азии распространен преимущественно по ли-
нии Средне-Азиатской железной дороги, с заходом глубже в страну
вокруг крупных населенных пунктов,— едва ли отражает действи-
тельные границы распространения этой болезни. Глубинные кишлаки
не были еще предметом исследований на кала-азар.
В Закавказских республиках эндемичными по кала-азар являются
Кировабад (Ганджа), Шемаха, Казах, Баку, Нахичевань и др. (Азер-
байджан), Тбилиси, Кахетия (Грузия), Ереван и др. (Армения). Отме-
чен случай и в Дагестане.
В настоящее время мы имеем уже и оригинальную литературу на
русском языке, касающуюся различных сторон изучения этой болезни
(Артамонов, Ходукин, Петров и др.). За пределами СССР кала-азар
распространен вокруг Средиземного моря (средиземноморский лейш-
маниоз) , в Индии, восточной части Китая и в некоторых других мес-
тах. В общем к северу кала-азар поднимается до 42° с. ш.
Благодаря применению более точных паразитологических, серологи-
ческих и других методов диагностики выявляются с каждым годом все
новые и новые случаи кала-азар. Именно этим обстоятельством объяс-
няется «рост» заболеваемости кала-азар за последние годы; например,
в Италии из 2 400 случаев кала-азар 1 60.0 были установлены за деся-
тилетие с 1918 по 1928 г.
В Средней Азии и в Армении наибольший процент заболевающих
кала-азар приходится на детей (87,5—Эв^/о всех заболеваний в Турке-
стане) ; в Индии же более поражается юношеское население. Болезнь
эта у нас не имеет эпидемического характера, но она эндемична и свя-
зывается часто с определенными очагами (районы и кварталы города,
отдельные дома и др.). Наибольшее количество заболеваний приходит-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
111
ся на начало весны и на осень. В Самарканде в год отмечают 70 слу-
чаев кала-азар, а в Ташкенте заболевает 9 человек детей на 10 000.
С 1928 по 1932 г. в Армении паразитологически установлено 75 случаев
кала-азар со смертностью в 90% среди нелеченных детей (данные Тро-
пического института Армении). В Индии кала-азар охватывает многие
тысячи больных, причем почти половина всех заболеваний кала-азар
приходится на взрослых.
Рис. 54 — Заболеваемость кала-азар по возрастам в различных местностях
по разным авторам. Из В. П. Петрова.
Отмечается большая закономерность в распространении лейшманио-
зов. Кала-азар с востока на запад убывает, а кожный лейшманиоз, на-
оборот, встречается чаще. Это подмечено в Индии (Нэпир и Кришнан)
и в Средней Азии (Ходукин).
В Средней Азии случаи кала-азар у взрослых за последние годы
устанавливаются более часто. В общем кала-азар заболевают люди
любого возраста, начиная от нескольких месяцев от роду и кончая ста-
ростью. Оба пола одинаково уязвимы. Социальные и экономические
факторы не увязываются с эпидемиологией кала-азар.
Лейшманиоз собак был впервые установлен в Закавказье
Джунковским и Лусом в 1909 г. Впоследствии лейшманиозные собаки
были обнаруживаемы в различных городах Закавказья и Средней
Азии. Особенно многочисленные обследования были произведены в
Ташкенте с 1926 по 1932 г.; из 71 687 собак процент зараженности по
годам колебался в пределах от 0,31 до 1,4 (Ходукин, Петров и Ке-
ворков). Другие авторы указывают на большую зараженность собак.
Интересно, что собаки, больные лейшманиозом или подозрительные по
заболеванию, были обнаруживаемы часто во дворах или по соседству
с дворами, где были больные кала-азар (Ташкент); речь идет, ко-
нечно, не о бродячих собаках, а о живущих при доме. Это обстоя-
тельство не случайно, ибо очаги кала-азар людей и лейшманиоза собак
часто совпадают.
Кала-азар у животных. Сходное заболевание бывает у со-
бак. Возбудитель его Leishmania canis по морфологическим признакам
не отличим от L. donovani. Вопрос о полном тождестве вирусов челове-
ческого и собачьего внутреннего лейшманиоза или о существовании
самостоятельного вида лейшманий у собак окончательно еще не решен
и требует дальнейших изысканий.
112
СПЕЦИАЛЬНАЯ ЧАСТЬ
Для их диференциации пользовались различными серологическими
реакциями — агглютинации, связывания комплемента, феномена нагруз-
ки и др. В ряде опытов было доказано, что некоторые штаммы со-
бачьих лейшманий идентичны L. donovani (Ходукин, Софиев, Кевор-
ков, Здродовский, Воскресенский).
Положительным моментом, говорящим в пользу идентичности
L. donovani и L. canis, считается совпадение ареалов распространения
кала-азар и собачьего лейшманиоза (Средняя Азия, Италия и др.).
Однако это не является повсеместным; так, в Индии, где кала-азар
распространен, собачьего лейшманиоза нет вовсе, хотя собаки и подда-
ются искусственному заражению
Важен факт заражаемости различных животных лейшманиями. За-
ражению подвержены хомячок Cricetulus migratorius (Канделаки),
являющийся отличным лабораторным животным, и Cr. griseus— в Ки-
тае. Попытки отыскания естественно-зараженных лейшманиозом хомяч-
ков пока давали отрицательные результаты.
Из других животных экспериментально заражаются обезьяны, ша-
калы, белки, суслики, морские свинки, белые мыши. Суслики и хомяки
особенно удобны для опытов с заражением через москитов (Phlebo-
tomus).
Условия заражения. Возможность непосредственного контак-
тного заражения кала-азар новейшими данными отрицается. В частно-
Рис. 55 — Leishmania pulicis из чело-
веческой блохи. Аггломерированные
формы из культуры на среде NNN-arap.
Сильно увеличено. По Weny’on.
сти, не приходится говорить о за-
ражении per os.
Следует, впрочем, вспомнить,
что неоднократно бывали обнару-
живаемы вирулентные лейшмании
в зеве и полости носа больных
кала-азар как в странах Европы,
так и в Азии.
Если и удавались единичные
опыты с заражением обезьян и ки-
тайского хомячка скармливанием
им органов больных лейшманиозом
животных, то в этих случаях нель-
зя исключить возможности про-
никновения вируса через ссадины
слизистой оболочки или иные ее
повреждения. Зато не вызывает
сомнения участие каких-то пере-
носчиков, являющихся посредни-
ками в деле распространения лей-
шманиозной инвазии. В поисках
таковых были перепробованы раз-
личные кровососущие насекомые.
Одно время фигурировала блошиная теория распространения кала-азар (Basile,
Visentini и др.). Авторам удавалось заражать здоровых собак кормлением на них
блох, снятых с лейшманиозных. Подтверждение этой теории ^видели в факте нахож-
дения в желудке различных блох биченосцев лептомонадной формы, которых счи-
тали за стадии развития Leishmania donovani. Однако тщательное изучение обшир-
ных материалов показало, что, несмотря на внешнее сходство (рис. 55), Leptomonas
(Herpetomonas) ctenocephali не имеет ничего общего с возбудителем кала-азар. Да-
лее, биченосцы были находимы в кишечнике блох таких местностей, где кала-азар
не было и не может быть. Что же касается «удачных» опытов заражения собак че-
рез посредство блох, то при этих экспериментах не исключалась возможность
Таблица IV
Малярийные паразиты в толстой капле:
1 — Plasmodium vivax, 2 — Plasmodium malarias, 3— Plasmodium falciparum
кольца, 4— Plasmodium falciparum гаметоциты, 5 — Leischmania donovani в
мазке из селезеночного пунктата, 6—7— лептомонадные стадии лейшманий
из культуры и из желудка переносчика, 8 — трипанозомы
Из Ruhe, Mfilens и др.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
113
побочной естественной инвазии, независимой от воздействий экспериментатора и не
учитываемой им. Таким образом блошиная теория распространения кала-азар не
имеет под собой фактического фундамента.
Не лучше обстоит дело и с оценкой постельных клопов в качестве переносчиков
внутренностного лейшманиоза. При кормлении индийского постельного ‘ клопа Cimex
rotundatus на кала-азарных больных с паразитами в периферической крови удава-
лось заражать это насекомое лейшманиями; проглоченные паразиты принимали
лептомонадную форму и размножались. Точно так же удавалось заражать клопов
кормлением их культурами лейшманий и блошиных биченосцев — Leptomonas pulicis.
Однако заразить какое-либо животное лейшманиозом чрез кормление на нем кло-
пов с паразитами кала-азар не удается. Возможно, что размножение лейшманий в
кишечнике клопа сводится к временной культуре этих микроорганизмов в сравни-
тельно большом количестве крови, выпитой клопом (Wenyon). Находка будто бы
стадий развития лейшманий в слюнных железах клопов (Adie) трактуется как сме-
шение безжгутиковых форм лейшманий с микроспоридиями. Таким образом, клопов
также нельзя признать переносчиками кала-азар, тем более что и эпидемиологи-
ческие соображения говорят против причастности их к передаче L. donovani; в
Индии клопы живут повсюду, а кала-азар имеет ограниченные ареалы распростра-
нения, в которых болезнь эта связана с определенными домами.
«Поцелуйный клоп»—Cononhinus rubrofasciatus, комары — Anopheles maculipen-
nis и другие эктопаразиты человека — вши, комнатная муха и клещ Ornithodorus
savignyi также не являются переносчиками кала-азар.
В последнее время все яснее и яснее определяется истинное отно-
шение москитов к передаче кал а-а з а р. Английские авторы
наблюдали в Индии образование и размножение лептомонадных форм
в кишечнике москитов Phlebotomus argentipes после кормления их на
больных с лейшманиями в периферической крови; мало того, жгутико-
вые формы L. donovani проявляли склонность к занятию передней части
пищеварительного аппарата, в частности глотки москита («передняя
позиция» — рис. 56). В Китае аналогичные факты наблюдались у Phi.
chimensis. Жгутиковые лейшмании были находимы и у «диких» Phi.
argentipes^ но одних морфологических данных мало, чтобы говорить о
естественной зараженности москитов именно возбудителем кала-азар.
Опыты заражения китайского хомяка, белых мышей и добровольцев
кормлением на них инфицированных лейшманиями. Phi. argentipes
(Индия) дали отрицательный результат.
Учитывая положительную роль москитов -в переносе возбудителей
кожного лейшманиоза и сходство развития в москитах L. tropica и L. do-
novani, можно с большой долей вероятия принять положение, что вну-
тренностный и средиземноморский лейшманиоз передается москитами;
в Индии таким переносчиком считается Phi. argentipes, причем проце-
дура сосания крови лишь в редких случаях ведет к заражению объекта
сосания; Adler полагает, что заражение человека происходит при раз-
давливании инфицированных москитов и внедрении лейшманий через
поврежденную кожу. Важно отметить, что ареал распространения Phi.
argentipes в Индии часто совпадает с местами развития кала-азар.
Известны случаи спонтанной зараженности москитов лейшманиями,
когда насекомые были пойманы в жилье больного кала-азар
(Шорт и др.).
Н. Ходукин с сотрудниками опубликовал опыт получения в Ташкен-
те в условиях, близких к естественным, заражейия здоровой собаки от
собаки, больной внутренностным лейшманиозом, при посредстве самок
москитов (для опытов брались Phi. papatasii и Phi. sergenti). Зараже-
ние проявилось в форме образования язвочек на носу, которые были
очень богаты лейшманиями. Необходимо повторение экспериментов в
условиях строгого контроля. Помимо основного указания, интересно то.
что заражение, происшедшее от собаки с внутренностным лейшманио-
114
СПЕЦИАЛЬНАЯ ЧАСТЬ
зом, пока проявилось в форме кожного поражения. Это, с своей сто-
роны, может свидетельствовать в пользу признания идентичности вируса
внутренностного и кожного лейшманиоза собак; серологическая иден-
тичность этих вирусов также доказана. Переносчиком средиземномор-
ского лейшманиоза, по работам в Алжире, считается Phi. major var.
perniciosus, хорошо заражающийся на собаках с кожными поражения-
ми (Parrot и др.).
Р — хоботок; Ph — глотка; Рг — преджелудок; Л1 — желудок; Мр — впадение
мальпигиевых сосудов; Л1г — задняя кишка; Р — ректальный пузырь: I — момент
инвазии (?); II — X — внутриклеточное размножение лейшманий в человеке;
1 — 4 — превращение лейшманий в жгутиковые формы в желудке москита; 5—9—
развитие инвазирующих форм лейшманий в глотке москита.
Ph. chinensis и Ph. sergenti var. mongolensis можно заразить пара-
зитами кала-азар кормлением этих москитов на больном или на хо-
зяине, инфицированном лейшманиями. Заражение от человека удается
е гораздо меньшем числе случаев, чем от хозяина. Развивающиеся
флягеллятные формы продвигаются из желудка в заднюю кишку
москита. Ph. sergenti обычно погибает, когда принятая им кровь пере-
варивается (Wu и Sun)z Повидимому, в С. Китае переносчиком кала-
азар является Ph. chinensis. В этом же виде москита были находимы
флягеллятные формы лейшманий в деревне, где был очаг кала-азар в
С. Китае.
Отмечена способность лейшманий сохраняться в личинках и в ним-
фах Rhipicephalus sanguineus, но действительное значение этого клеща
в передаче кала-азар остается неясным и скорее сомнительным.
Наконец, следует отметить, что в ряде опытов удавалось прямое
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
115
заражение паразитами кала-азар животных per os — крыс, хомяка,
обезьян; но этот путь заражения едва ли играет какую-либо роль в
эпидемиологии кала-азар.
Наблюдались также случаи контактного заражения кала-азар при
продолжительном совместном пребывании здоровых обезьян или хомяка
с больными особями. Но все эти данные не колеблют представления о
главном пути передачи кала-азар через переносчика (москита), ибо в
противном случае осталась бы совершенно необъяснимой ярко выражен-
ная очаговость кала-азар.
Собаки должны быть признаны резервуарами возбудителя кала-азар;
отмечается большая инвазированность лейшманиями кожи собак, неред-
ко при отсутствии каких-либо внешних патологических изменений; это
обстоятельство облегчает москитам возможность заражаться лейшма-
ниями при сосании крови собак.
В о е н н о-с анитарное значение кал а-а з а р, учитывая, что
огромный процент заболеваний падает на ранний детский возраст в
СССР, едва ли может считаться большим. Среди взрослого населения
пока наблюдались лишь единичные случаи кала-азар (В. Петров). Воз-
можно, что при более широком применении паразитологической диагно-
стики, количество кала-азар, открываемого среди взрослых в Средней
Азии, возрастет. Английские войска в Индии в 1869 г. весьма страдали
от кала-азар, и самое открытие возбудителя этой болезни — L. donova-
ni — было сделано Донованом в Лондоне на мазках селезенки умерше-
го от кала-азар в Лондонском военном госпитале (1900) солдата, слу-
жившего перед тем в Индии.
Если в дореволюционное время кала-азар, как и другие тропические
болезни, привлекал к себе внимание лишь единичных исследователей и
врачей, то после революции, в связи с культурным ростом Средней
Азии и Закавказья, вопросам изучения и борьбы с эпидемичными тропи-
ческими болезнями уделяется должное внимание. Этому способствует
наличие сети тропических станций, опирающихся на тропические инсти-
туты, некоторые санитарно-бактериологические институты и другие
учреждения. В частности, широкая обследовательская работа была про-
делана в Ташкенте (см. Н. Ходукин, 1929).
Борьба с кал а-а з а р. По линии лечения больных хорошие ре-
зультаты дает химиотерапия (препараты сурьмы: Stibenyl, Stibosan и
старый препарат Stibio-kalium-tartaricum). Рвотный камень приме-
няется в 2% стерильном растворе, который впрыскивается в вену у
взрослых, начиная с 1 см3 и последовательно доводя до 8—10,0 см3.
Детские дозы регулируются по возрасту и весу ребенка. Лечение длит-
ся долго.
Стибозан, неостибозан и антимозан применяются в 5% растворе, а
стибенил в 1—2% растворе (на физиологическом растворе поваренной
соли) в форме внутривенных инъекций, производимых через день. Дози-
ровка этих препаратов по возрасту приводится на таблице стр. 116 (из
В. Петрова). Несколько лет назад начали производить советский пре-
парат сурьмин (стибозан); он не свободен пока от токсических свойств,
вследствие чего у больных бывают чаще рвота и понос.
Профилактика. Для уничтожения резервуаров вируса — вы-
лавливание и истребление лейшманиозных собак, что в ряде мест велс
к снижению заболеваний кала-азар; борьба с москитами, как с возмож-
ными переносчиками кала-азар (см. том II).
116
СПЕЦИАЛЬНАЯ ЧАСТЬ
Дозировка препаратов сурьмы (из В. Петрова)
Стибенил Стибозан | Неостибозан Антимозан
Я Я к ее
Я Ч Я св к ев Я ев
Возраст 5 ! св я я я = а св Я Я - Я <□
ч з ч 3 =: 3 • Ч S
ев ев М ев ев св И ев ев св аз ев ев ев ш ев
Я СП Я СП Я -СП Я СП О’ « Я СП Я « Я сп
св О св О св о а о ев О ев О св О св О
X « я »=с я ч я ч Я Ч Я Ч я ч я ч
Дозировка по возра-
сту
Цо 3 лет .... 0.03 0.1 0.05 0.1 0.05 0.1 0.05» 0.1
.3—5 лет . .... 0.05 0.12 0.075 0.2 0.075 0.2 0.075
5—9 » 0.06 0.2 0.1 0.25 0.1 0.25 0.1 0.2
'3—15 » 0.075 0.3 0.1 0.3 0.1 0.5 0.1 0.3
Взрослым 0.1 0-4 0.15 0.4 0.15 0.45 0.15> —
Дозировка по весу
До 13 кг 0.04 0.1 0.05 0.1 0.05 0.1 0.05 0.1
13—23 » 0.06 0.12 0.1 0.2 0.1 0.2 0.075 0.2*
^23—27 » . . 0.08 0.25 0.15 0.25 0.15 0.25 ft. 01 (к.2.5
ЧЗвыше 0.15 0.3 0.2 0.3 0.2 0.3—0.45 G-15 0..3
Leishmania tropica (Wright)
L. tropica является возбудителем кожного лейшманиоза, из*
местного в СССР под именем пендинки, в прежней литературе фигури-
ровавшей и под именем сартской язвы. Этот паразит открыт военным
врачом П. Ф. Боровским в Ташкенте в 1882 г.
Морфологически паразиты не отличимы от L. donovani (см. выше):
-локализуются в протоплазме макрофагов и эндотелиальных клеток; при
распаде клеток паразиты лежат свободно в выпоте язвы. Как и
Z. donovani, L. tropica хорошо культивируется на искусственных пита-
тельных средах — в частности, на среде NNN-arap.1 В культурах через
несколько дней развиваются жгутиковые формы типа Leptomonas (ср.
рис. 53, 55).
Кожный лейшманиоз распространен в жарких и теплых странах до
40° сев. широты. Нередко он носит строго эндемичный характер, чем и
объясняется большое число местных наименований этой болезни —
алеппская язва, bouton d’Orient, багдадская язва, пендинская язва
и т. д. В пределах СССР южный лейшманиоз наблюдается на Кавказе:
Ленкорань, Ганджа, Баку, Барда (Азербайджан), Гори (Грузия) и в
•Средней Азии по долинам рек Мургаба, частично Атрека, Теджена,
•частично Аму-Дарьи (Туркмения), частично по Зеравшану, Сыр-Дарье,
Кашка-Дарье и др. (Узбекистан).
В качестве эндемичных местностей по пендинской язве известны-
Тахта-Базар, Мерв, Теджен, Серахс, Ашхабад, Чарджоу, Керки (Турк-
мения) ; в Узбекистане — Фергана, Андижан, Коканд, Самарканд, Кат-
та-Курган, Старая Бухара, Бек-Буди, Хорезм; в Таджикистане (Ленин-
абад) и Айвадж (на Аму-Дарье). Пендинка бывает и в районах, при-
лежащих к упомянутым городам.
Болезнь эта может принимать иногда эпидемический характер, чаще
1 Дестиллированной воды 900.0 г, агар-агара 14 г, поваренной соли 6.0 г; в
отдельные пробирки прибавляют дефибринированной крови кролика в половин-
ном объеме.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
117Г
же она гнездится ограниченными очагами (например, в Ташкенте, Бу-
харе, Джизаке, Мерве, Тахта-Базаре, Серахсе, Теджене, Чарджоу».
Керках, Самарканде, Термезе, Ашхабаде, Тегеране и др.). После из-
вестного инкубационного периода (несколько недель или месяцев) на
коже .лица или открытых частей тела появляются буровато-красные
пятнышки, превращающиеся в узелки. Верхушка последних изъязвляет-
ся :В£дедствие некроза ткани. После этого язва, как говорят,, начинает
Рис. 57 — Больные пендинкой; внизу — пендинка на руке. По Гительзону.
«цвести», что длится от 3 до 18 месяцев. Она увеличивается в размерах
до величины серебряного рубля и даже ладони. Серозно-гнойный выпот
ее и отделения засыхают коркой (рис. 57). Под образовавшимся стру-
пом развиваются грануляции. Язва медленно зарубцовывается от цент-
ра к периферии, причем образующиеся рубцы могут давать сильные
стягивания. Локализация язв показана на рис. 58.
Иногда при кожном лейшманиозе наблюдается припухание близле-
жащих лимфатических желез; в их клетках также обнаруживаются опи-
сываемые паразиты. В некоторых случаях кожного лейшманиоза
L. tropica обнаруживали в периферической крови (Patton).
По современным исследованиям болезнь наблюдается в двух фор-
мах, которые наши исследователи склонны считать двумя различными
болезнями (Латышев, Кожевников и др.).
118
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 58 — Схема расположения язв пендинки у
26 больных Тахта-базарского района Туркме-
нии. По Кочергину (из Г и т е л ь з о н а).
Первый тип — поздно изъязвляющийся кожный лей-
шманиоз, папулезная, сухая, хроническая форма,
ашхабадка, кокандка, городская форма.
Инкубация удлиненная — обычно до полугода (бывает 1 — IV2 года).
На коже появляется хорошо ограниченный, нередко только прощупы-
ваемый, медленно растущий
бугорок, со временем буро-
красного цвета. Через 3—6
и более месяцев начинается
изъязвление. Язва обычно
покрыта плотной сухой ко-
рочкой; по ее удалении
обнажается бледнокрасная
или сероватая поверхность
язвы, дающая скудное се-
розное отделение. Лейш-
маний в тканях язвы очень
много. Болезнь длится до
года и более. Эпителиза-
ция язвы часто протекает
в короткий срок. Болезнь
обычно встречается в го-
родах (Ашхабад, Коканд,
Мары и др.).
Второй тип — остро
некротизирующийся
кожный лейшманиоз;
острая, мокнущая,
э к т иди атозная, язвен-
ная форма, собственно
пендинская язва, пендинка.
Инкубация всего 1—6—8
недель. Болезнь начинается
очень быстрым появлением папулы с отеком; через несколько дней (5—
10) в ее центре начинается изъязвление; папула растет по окружности;
соответственно увеличивается и язва, принимающая неправильную фор-
му, с изъеденными краями; язва дает обильное серозное или серозно-
гнойное отделяемое (мокрая пендинка); по периферии ее могут появ-
ляться мелкие язвочки, сливающиеся с первой. Язва окружается мощ-
ным инфильтратом. Она очень болезненна. Период некроза—Р/г—2 ме-
сяца; некротические массы вытесняются сочными грануляциями. Язва
очищается с центра. Эпителизация также с центра. По окружности
язвы часто держится ободок некротизирующейся ткани, называемый
краевым рвом, который эпителизируется последним. Паразитов в тка-
нях очень мало. Болезнь заканчивается в течение 3—6 месяцев. Часты
лимфангоиты (в 80—9О°/о случаев). Язвы часто множественные. Бо-
лезнь сельских и малообжитых местностей, могущая быстро разра-
статься в эпидемическую вспышку с тысячами (смотря по эпидемиче-
ской обстановке) заболеваний.
Для белых мышей первый тип болезни мало вирулентен, паразиты
же второго типа высоко вирулентны.
В зависимости от того, где наблюдалась эта болезнь, она фигуриро-
вала под разными наименованиями: кокандской язвы или кокавдки,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
119
бухарской или восточной язвы и др. У туркмен пендинка известна под
именем пеше-хурда, что в переводе означает «укус дурной мухи». Так
народная прозорливость подметила связь пендинской язвы с укусами
летающих насекомых.
Замечается своеобразная закономерность обратной пропорционально-
сти интенсивности заражения пендинкой и численности населения по-
ражаемого ею селения. Так, в Ашхабаде заболевает 2—3°/о населения,
а в мелких поселках процент заболеваемости возрастает до 70—80 и
даже до ста (Н. Латышев).
Кожный лейшманиоз у животных. Кожный лейшманиоз в
качестве естественного заболевания бывает у собак (Leishmania canis —
Средняя Азия,- Персия, Индия), обычно сопровождаемого лейшманиозом
внутренних органов. В 1931 г. в Ираке установлены новые случаи спон-
танного заражения домашних кошек кожным лейшманиозом с образова-
нием язв на носу. Выделенные паразиты морфологически и в культурах
не отличимы от L. tropica, полученных от обыкновенной «багдадской
язвы». Патолого-анатомическая картина поражения весьма сходна с
тем, что наблюдается в язвах у людей и у собак (Machattie, Mills и
Chadwick). Вообще, у кошек кожный лейшманиоз встречается гораздо
реже, чем у собак. Описаны случаи поражения L. tropica и верблюдов.
Вопросы соотношения L. tropica, L. donovani и L. canis окончательно
еще не решены. Лейшманиями из спонтанной кожной язвы собаки в
Багдаде удалось заразить человека как культурой, так и через искус-
ственно зараженного москита (Phi. papatasii— Adler и Theodor, 1930).
Серологические реакции обнаруживают весьма большое сходство, если
не полное тождество, лейшманий, вызывающих кожные поражения лю-
дей и собак (в Ираке, 1930 г., Mills, Machattie и Chadwick). Из рус-
ских авторов Ходукин и Софиев считают, что серологически L. dono-
vani идентична L. canis.
В пользу признания идентичности возбудителей кала-азар и пендин-
ки говорит также факт перехода у собак местного заболевания в общее
и обратно (Шевченко), что, видимо, связывается с изменениями виру-
лентности одного и того же возбудителя. Отмечается большая зара-
женность некоторых пород собак в питомниках (например Ташкент),
по сравнению с собаками, живущими в окружении (Радзивиловский,
Шевченко).
Пути заражения. При наличии лейшманиозных язв возможно
механическое распространение их при расчесах, частом соприкоснове-
нии пораженных и здоровых частей тела и др.; бывают также и мета-
стазы возбудителя с образованием новых язв.
Источником первичного заражения пендинской язвой являются
москиты. До установления этого факта переносчиков тщетно искали
среди клопов, человеческой и собачьей блох, вшей, комаров, мух-жига-
лок, комнатных мух, собачьих кровососок (Hyppobosca canind). Однако
в некоторых из этих насекомых L. tropica может развиваться в усло-
виях опыта; это обстоятельство еще не делает таких насекомых пере-
носчиками кожного лейшманиоза.
Необходимо отметить, что в некоторых местах Средней Азии, в
частности Туркмении, москиты часто пьют кровь гекконов — Gymno-
dactylus caspius (Петрищева).
О причастности Phlebotomus к распространению кожного лейшма-
ниоза предположил впервые Wenyon, нашедший в их кишечнике в
г. Алеппо жгутиконосцев типа Leptomonas. Экспериментально к этому
120
СПЕЦИАЛЬНАЯ ЧАСТЬ
вопросу подошли Ed. Sergent, Parrot, Donation и Eteguet (Алжир), за-
разившие человека лейшманиозом втиранием в скарифицированную
кожу растертых Phi. papatasii, привезенных из зараженной местности
(из оазиса Бискра). Язва образовалась через 84 дня после заражения.
Аналогичные результаты получили в Иерусалиме Adler и Theodor с той
разницей, что они пользовались самками Phi. papatasii, естественно
инвазированными, жгутиковыми. Пищеварительный тракт москитов
выпрепаровывался и инокулировался в скарифицированную кожу лю-
дей. В одном случае удалось вызвать появление язвы, клинически
тождественной с восточной язвой и содержавшей в себе лейшманий,
которых удалось также культивировать на питательной среде.
Недавно эти же исследователи произвели удачные опыты с зараже-
нием Phi. papatasii культурой лейшманий. Инфицированные москиты в
части опытов вызвали появление лейшманиозной язвы в месте сосания
кожи человека.
Весьма вероятным переносчиком кожного лейшманиоза в Пенджабе
является Phi. sergenti] в его кишечнике лейшмании принимают жгути-
ковую форму уже на шестой день после кормления москита заразным
материалом (на лейшманийных язвах). Жгутиковые формы лейшманий
из желудка Phi. sergenti, будучи привиты обезьяне макаке, вызывают у
нее заражение и развитие папулы с последующим изъязвлением
(Sinton).
Адлер и Теодор заразили также человека кожным лейшманиозом,
привив ему паразитов из Phi. perniciosus, накормленного на собаке^
больной общим лейшманиозом.
Phi. sergenti также, видимо, имеет отношение к передаче лейшма-
ниоза. Весьма возможно, что такую же роль играет и Phi. caucasicus.
Если факт возможности передачи москитами лейшмании человеку
установлен точно, то нельзя пока сказать того же про самый механизм
передачи и про детали эпидемиологических взаимоотношений.
В частности, возникает вопрос о резервуарах лейшманиозного виру-
са. Является ли источником заражения москитов лейшманиями только
человек, или есть йные пути получения паразитов москитами? Ed. Ser-
gent, Lemaire и Senevet высказали мысль, что резервуаром изучаемого
вируса является алжирский геккон (рептилия) Tarentola mauritanica,
от которого москит и почерпает лейшмании; из органов 15,7% исследо-
ванных гекконов им удалось выкультивировать жгутиковых типа Lep-
tomonas. Wenyon сомневается в высказанной гипотезе, a Laveran не
добился успеха в своих попытках заразить гекконов L. tropica.
Некоторые пресмыкающиеся, например хамелеон, имеют собствен-
ных лейшманиеподобных паразитов (рис. 59).
Совсем иные пути в разрешении проблемы эпидемиологии кожного
лейшманиоза наметил в последующее время R. Strong; он ‘ обнаружил,
что паразитирующие в соке молочайных растений (Euphorbia) жгутй-
ковые в Центральной и Южной Америке очень похожи на L. tropica.
На молочае питается растительный клоп Chariesterus cuspidatus, кото-
рый и всасывает жгутиковые с молочным соком растения. Если этот
клоп делается добычей ящерицы Cnemidophorus lemniscatus, то в ее
кишечнике жгутиковые приобретают патогенные свойства. Если при-
вить в кожу обезьяне содержимое кишечника такой ящерицы со жгути-
ковыми, то на коже развивается язва, содержащая типичные лейшма-
нии. Жгутиковые молочая патогенны для этого растения; пройдя же
через организм насекомого и ящерицы, они приобретают свойства,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
121
патогенные для млекопитающих. Эти данные, конечно, нуждаются в
проверке и подтверждении, чтобы признать за ними, значение ключа
к выяснению эпидемиологии кожного лейшманиоза. Отметим, что за-
ражение молочая жгутиковыми (рис.
Рис. 59 — Leishmania chameleonis из кло-
аки хамелеона Chameleo vulgaris.
По W е п у о п.
60, 61) обнаружено в различных
Рис. 60 — Биченосец Leptomonas
{Phytomonas) davidi из сока ин-
дийского молочайного растения
Euphorbia. По R о W, из W е п у о п.
странах, в том числе недавно и в СССР (Leptomonas davidi в По-
волжье, И. Иофф). Промежуточным хозяином и переносчиком этого
паразита в Португалии является клоп Stenocephalus agilis Scop., <в
в слюнных железах которого раз-
вивается инфицирующая форма
Leptomonas davidi.
Основные факты, на которых
зиждется москитная теория рас-
пространения кожного лейшмани-
оза, по сводке Адлера, заключается
в следующем:
1) L. tropica была обнаружена
в качестве естественного паразита
Phi. papatasii.
2) Лейшманийная стадия L. tro-
pica превращается в лептомонад-
ную жгутиковую форму, которая
затем размножается в желудке Phi.
papatasii и Phi. sergenti.
3) L. tropica в лептомонадной
Рис. 61 — Leptomonas (= Phytomonas)
davidi (4) на разрезе сосудов Euphorbia
segeialis. По Франса.
стадии проявляет тенденцию к за-
нятию в теле москита «передней
позиции» —• в частности скопляется
в хоботке; однако заражение че-
ловека через хоботок случается относительно редко.
4) L. tropica становится заразительной для человека после пребыва-
ния в течение девяти дней в теле Phi. papatasii при температуре между
19 и 23° С и через 6 дней в теле Phi. sergenti при t° около 27° С.
5) Культуры L. tropica, которые стали незаразительными для чело-
века, снова приобретают патогенные свойства после прохождения через
Phi. papatasii.
122
СПЕЦИАЛЬНАЯ ЧАСТЬ
6) Phi. papatasii и Phi. argentipes имеют бактериологически стериль-
ный пищеварительный тракт. Это обстоятельство весьма способствует
пригодности москитов быть хозяевами лейшманий, так как ни L. tropica,
ни L. donovani не могут сколько-нибудь продолжительное время жить
вместе с бактериями.
Уже теперь накопились данные, говорящие в пользу того, что раз-
ные географические штаммы L. tropica приспособились к определенным
видам москитов. Так, в Палестине L. tropica, повидимому, связана с
Phi. papatasii, а в Багдаде — с Phi. sergenti. i
Передача кожного лейшманиоза происходит или при уколе моски-
тами (редко для Phi. papatasii, обычно — для Phi. sergenti), или при
раздавливании зараженных москитов на скарифицированной коже, или
при внесении инвазирующего материала на слизистые оболочки или в
конъюнктиву глаза.
Отделом паразитологии ВИЭМ были организованы продолжительные
экспедиционные исследования очага пендинской язвы в долине Мургаба
в Туркмении (Н. Латышев) в целях выработки системы мероприятий
по ликвидации очагов пендинки в небольших селениях в зоне полупу-
стыни, освояемой в процессе социалистического строительства. Меро-
приятия строились по линии борьбы с москитами (о чем см. ниже, в
главе о москитах). В поисках мест выплода москитов обследовались
норы различных диких животных, в частности грызунов. Исследование
самих хозяев некоторых нор показало, что много песчанок (Rhornbo-
mys opimus) и сусликов (Spermophylopsis leptodactylus) спонтан-
но заражены пендинской язвой. Паразитами от песчанки удалось зара-
зить волонтера, у которого развилась классическая пендинка в месте
введения в кожу инфицирующего материала, и обратно, паразитами от
человека можно было заразить песчанку; эти эксперименты доказывали
тождество лейшманий песчанки и человека (Латышев). Норы этих
грызунов (как указывали Я. Власов и П. Петрищева) являются местом
выплода массы москитов.
Дальнейшее обследование москитов из нор грызунов привело к об-
наружению до 35'% Phlebotomus с флягеллятными формами лейшманий
в желудке и глотке. Сопоставление всех этих фактов приводит Н. Ла-
тышева к выводу, что кожный лейшманиоз является ти-
пичным зоонозом, т. е. болезнью грызунов, очаги ко-
торой поддерживаются в дикой природе независимо
от присутствия человека вследствие сообитания
в норах грызунов и москитов; последние передают
пендинку от одного грызуна к другому.
Когда в зону такого естественного очага пендинки попадает человек,
на него могут нападать для кровососания спонтанно-зараженные моски-
ты, почерпнувшие лейшманий от диких грызунов. Финалом является
заболевание человека пендинской язвой. Следовательно, естественным
рассадником пендинки являются норы диких грызунов. В таком освеще-
нии становятся понятными и заболевания людей пендинкой в безлюд-
ных местностях и высокая заболеваемость кожным лейшманиозом не-
больших поселений во вновь освояемых местах, где контакт человека с
дикой природой тесен. Эти же факты властно диктуют необходимость
борьбы с москитами не только в биотопах антропургического проис-
хождения, но и в дикой природе в известном радиусе от оздоровляе-
мого' селения (Н. Латышев).
Иммунитет. Перенесение заболевания вызывает появление
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
123
иммунитета к последующим заражениям. Однако это состояние дости-
гается лишь тогда, когда язва прошла все нормальное течение до
конца.
Экспериментальный лейшманиоз также вызывает иммунитет (Мар-
циновский и Щуренкова). Иммунитет не абсолютен, так как имеются
примеры (правда, редкие) повторных заражений после перенесенного
уже заболевания. Встречаются также люди, от природы не восприимчи-
вые к заражению пендинкой. Переболевшие песчанки вовсе не при-
обретают иммунитета (А. Крюкова).
Значение кожного лейшманиоза в военн о-с а н и т а р •
ном отношении очень большое. В некоторых гарнизонах Индии
заболевало до 50—7О,{)/о> солдат. Аналогичная картина наблюдалась и в
Средней Азии. В 1885 г. в Мургабском отряде русских войск заболело
около ЗСИ/о, а в 1888 г. некоторые части в районе Мургаба дали пого-
ловную заболеваемость. В 1913 г. в гарнизоне Термеза обнаружено
67,8% болевших пендинкой. Недавно наблюдалась вспышка пендинки
в Серахсе и др. В то же время обращает на себя внимание факт, что
некоторые прежние очаги пендинской язвы теряют свое значение. Так,
в 60—70-х годах XIX столетия почти все туземное население Ташкента
подвергалось в детстве заболеванию пендинкой. Теперь эта болезнь в
Ташкенте является редкостью (если, конечно, исключить приезжих
больных). В Узбекистане в 1931 г. отмечено всего 8049 случаев пен-
динки (Ходукин), а число лиц с активной формой этой болезни в Мер-
ве в 1927 г. равнялось 5,2% всего населения (Сегаль и Гительсон).
Равным образом улучшилось в аналогичном отношении и состояние
Термеза.
Борьба с кожным лейшманиозом. Терапия крайне разно-
образна, что указывает на отсутствие специфических средств. Принци-
пы ее также различны.
Разрушение пораженных тканей с паразитами в них производится
гальванокаутером, диатермокоагуляцией, сурьмяной или мышьяковой
мазями (болезненны), инъекцией хлороформа с эфиром, оперативным
иссечением и пр. В результате могут иногда образоваться рубцы,
большие, чем от самого заболевания. Латентный лейшманиоз усколь-
зает от лечения. Действие на самих’лейшманий пытаются оказать при-
менением риванола, ятрена, протаргола, ксероформа и т. д. Повидимо-
му, наиболее действительна терапия впрыскиванием в бугорок атебри-
на (Фларер). Советский препарат акрихин дает вполне хороший
эффект.
При поздно изъязвляющемся кожном лейшманиозе для обкалы-
вания й имбибиции пораженных тканей употребляется 3% раствор
акрихина в количестве до 10 см3 с одновременным применением
1% риванола, 10% протаргола или 10% ихтиола и других мазей. При
лечении остро некротизирующегося кожного лейшманиоза больные
часто обращаются поздно — в стадии некротических изъязвлений. При
множественных поражениях, которые часто сопровождаются лимфан-
гоитами, рекомендуется общее лечение: белый стрептоцид, акрихин,
сурьма, метиленовая синька; местно производится обкалывание акри-
хином; с успехом применяются 1% ятреновые компрессы; можно
использовать риваноловую мазь, влажные повязки из риванола, марган-
цовокислый калий.
Когда язва очистится, следует применять местное лечение для уско-
рения эпителизации язвы; это достигается применением мазей — рива-
124
СПЕЦИАЛЬНАЯ ЧАСТЬ
половой, протарголовой или ихтиоловой с хлороформом пополам с ра-
стительным маслом, сока маклюры в чистом виде или в смеси с хлоро-
формом.
Из физиотерапевтических методов лечения применяют заморажива-
ние углекислотой, ионтофорез, кварцевую лампу.
Наконец, имеется попытка применения вакцинотерапии для лечения
пендинки.
Совершенно особняком стоят методы лечения пендинки, предложен-
ные акад. Филатовым: пересадка кожи трупа, консервированной на
холоду 7 дней (тканевая терапия); размер трансплантата в 12—15 см2.
Язвы заживают в срок около месяца. В случае необходимости делают
вторичную пересадку кожи (пересадка делается в области ягодицы или
в другие места). Испробованы также, видимо, с благополучным резуль-
татом, подкожные инъекции водного экстракта листьев алоэ, сохраняв-
шихся 15 дней в темноте. Инъекции делались в окружности язв по
2—4 см3. Для полного курса лечения требовалось около 30 инъекций
(рис. 16, А, Б, рис. 17, А, Б).
Обилие методов лечения и пестрота конечных результатов зависят
в каждом отдельном случае от формы болезни, фазы ее течения и
иммунобиологических процессов в организме больного.
Профилактика достигается искусственным заражением чело-
века на закрытых местах тела — способ, применяемый евреями на
Ближнем Востоке к своим детям; вакцинация производится материалом
от язвы.1
Попытки вакцинации людей убитыми культурами L. tropica пока не
дали удовлетворительных результатов.
Учитывая значение москитов в распространении пендинской язвьц
необходимо препятствовать им в сосании пораженных покровов челове-
ка; это достигается закрытием язв повязкой. Общей мерой является
борьба с москитами (см. II том) и уничтожение собак, пораженных
лейшманиозом.
Паразитологическая диагностика. Диагноз ставится
по нахождению паразитов на мазках отделений и грануляций язвы,,
окрашенных по Романовскому. При открытой язве берут пункцией (на-
сасывание шприцем) содержимое из подрываемого участка кожи у края'
язвы; материал идет на мазок и для засева на питательную среду.
Жгутиковые формы L. tropica в культурах не отличимы от лептомо-
надной стадии L. donovani. Паразитов можно найти на окрашенных
срезах иссеченного кусочка края язвы (биопсия). Наконец, хорошим
способом является прививка грануляций белым мышам, у которых не-
редко процесс генерализуется, поражая внутренние органы (при «су-
хой» форме пендинки).
В Южной Америке широко распространен кожный лейшманиоз, отличающийся
от аналогичного заболевания Старого Света глубоким поражением слизистых обо-
лочек полости рта, носа, 'неба. Болезнь течет долго. Возбудителем является Leish-
mania brasillensis; он отличается от L. tropica и L. donovani по серологическим
свойствам, что является, со своей стороны, доказательством видовой самостоятель-
ности L. brasiliensis. Переносчиком его считается Phlebotomus intermedins.
Род Trypanosoma Gruby
К характеристике рода Trypanosoma, данной выше (стр. 105), доба-
вим, что у огромного большинства их жгут продолжаемся свободно от
1 Необходимо предварительно исключить наличие у донора сифилиса и ту-
беркулеза.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
12г)
переднего конца тела; у некоторых же форм свободной части жгута
нет совсем. Трипанозомы являются паразитами позвоночных (от рыб
до человека), проделывающими, как правило, полный жизненный цикл
в двух хозяевах; вторым хозяином является беспозвоночное животное
(пиявки, членистоногие), которое служит переносчиком , трипанозом
позвоночным животным. Редкие случаи, когда трипанозомы передаются
от одного позвоночного другому без какого-либо участия переносчиков,
являются вторичными особенностями соответствующего вида и его
позднейшим свойством (например, трипанозома случной болезни лоша-
дей — Trypanosoma equiperdum).
Наряду с безвредными трипанозомами имеются патогенные виды
первостепенного значения для человека и для домашних животных.
Трипанозом человека в пре-
делах СССР нет и едва ли мо-
жет быть, если не считать при-
езда трипанозомозного боль-
ного, что не имеет никакого
отношения к истинному гео-
графическому распростране-
нию три»анозом. Некоторые
сведения о них дать необхо-
димо, так как трипанозомы
являются классическим объек-
том изучения явлений пере-
дачи кровепаразитов при по-
средстве переносчиков.
У позвоночных трипанозо-
мы паразитируют преимущест-
венно в крови, а также в лимфе,
цереброспинальной жидкости Рис. 62-Агглютинация трипанозом сонной
И др. Ьывают и тканевые фор- болезни в крови человека. Из Т a liaferro.
мы трипанозом. В беспозво-
ночном хозяине (переносчике) трипанозомы обитают в кишечном ка-
нале и в его придатках.
Переносчики передают трипанозом или механически (когда они в
них не изменяются и не размножаются), или же передача, бывает спе-
цифической, когда в переносчике трипанозомы проделывают определен-
ный цикл развития; поэтому говорят также о циклическом способе
передачи этих кровепаразитов. В результате цикла развития трипано-
зомы в переносчике достигают, инвазирующего состояния; такие трипа-
нозомы называются метациклическими.
Трипанозомы попадают в промежуточного хозяина per os. Одни
виды их и не проникают дальше хоботка переносчика, другие проходят
в пищеварительный канал. Выход метациклических трипанозом из тела
переносчика (в процессе передачи трипанозом) может быть двояким.
Одни виды проходят в процессе жизненного цикла через весь кишечник
и в метациклическом состоянии выбрасываются наружу per artum пере-
носчика, передающего в таких случаях трипанозом контаминативно.
В других случаях трипанозомы выводятся из тела переносчика per os,
с той только разницей, что в цикле своего развития они заходят в
слюнные железы (но не через полость тела!), откуда вымываются со
слюной при инокуляции ее в покровы хозяина, кровь которого пьет
переносчик (рис. 64).
12(5
СПЕЦИАЛЬНАЯ ЧАСТЬ
В связи с жизнью в двух хозяевах, весьма отдаленных друг от дру-
га как по систематическому положению, так и по биохимическим и фи-
зиологическим особенностям, трипанозомы проделывают более или ме-
нее сложный цикл развития, который рассмотрим у двух нижеописы-
ваемых видов этих паразитов, относящихся к патогенным видам. Есть
и непатогенные виды, не причиняющие никакого вреда своему позво-
ночному .хозяину, например, Tr. melophagium овец или Tr. lewisi крыс.
Патогенность, как свойство того или иного вида трипанозом, устанав-
ливается по взаимоотношению с определенным видом хозяина; универ-
сально патогенных трипанозом нет; другими словами, трипанозо-
ма, патогенная для одного хозяина, может быть
безвредной для хозяев другого вида, в которых она в
состоянии жить. Такие животные называются резервуарами трипано-
зомного вируса. Считается, что трипанозомы безвредны для их есте-
ственных хозяев; если трипанозомы попадают в новых хозяев и
используют их, то процессы адаптации паразита к хозяину протекают
медленно; пока еще эволюционно не установились определенные
взаимовлияния организмов паразита и хозяина, трипанозомы оказы-
ваются патогенными для своих новых хозяев; в календарном отноше-
нии такой процесс взаимной адаптации длится очень долго (вероятно,
в течение целых геологических эпох).
Trypanosoma gambiense Dutton — трипанозома сонной болезни
Одним из жестоких и смертельных заболеваний человека тропиче-
ской Африки является сонная болезнь, распространение которой совпа-
дает с ареалом обитания мухи тсе-тсе (Glossina palpalis). Возбудите-
лем сонной болезни является жгутиконосец — Trypanosoma gambiense
(рис. 62), который передается при посредничестве мухи тсе-тсе.
Паразит держится в плазме крови человека и в его цереброспи-
нальной жидкости. Трипанозомы могут переходить через плаценту из
крови матери и заражать плод. Известны также случаи передачи три-
панозом ребенку при кормлении его грудью. Не всякий носитель три-
панозом проявляет симптомы сонной болезни.
Тело трипанозом узкое, длинное, ланцетовидное (15—30 X 1,4—2 у).
Оно легко изгибается, и формы его на препаратах весьма разнообраз-
ны. Удлиненно-овальное ядро лежит почти посредине клетки. На зад-
нем конце ее помещается блефаропласт, от которого отходит жгутик;
последний волнообразно изогнут; между ним и телом трипанозомы
натянута прозрачная ундулирующая мембрана. От противоположного
конца трипанозомы жгут продолжается на некоторое протяжение сво-
бодно (рис. 63, А). Трипанозома движется благодаря колебаниям жгу-
та и ундулирующей перепонки, а также вследствие изгибания тела.
При окраске по Романовскому протоплазма трипанозомы красится
в бледносиний цвет, главное ядро, блефаропласт и жгутик — в красный,
ундулирующая мембрана —в бледнорозовый.
Размножаются трипанозомы продольным делением: сначала делится
блефаропласт, и трипанозома становится двужгутиковой (рис. 63, D);
затем делится ядро и, наконец, наступает расщепление протоплазмы,
начиная с переднего конца паразита.
Когда муха тсе-тсе пьет кровь больного трипанозомозом или кровь
здоровых носителей, то судьба проглатываемых трипанозом оказывает-
ся различной:
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
127
и размножаться в кишечнике.
Рис. 63 — Трипанозомы сонной болезни. Тг.
gambiense и их переносчик — муха тсе-тсе (Glos-
sina palpalis).
А — обыкновенные формы трипанозом в крови челорека;
D <— делящиеся трипанозомы из крови человека; Р—
прохождение трипанозом через хоботок тсе-тсе; / —
трипанозомы в желудке мухи; S — они же в слюнных
железах мухи; ^ — длинные формы трипанозом, прохо-
дящие из желудка в слюнные железы через хоботок; В —
критидиальные формы, развивающиеся из длинных три-
панозом в слюнных железах; С — метациклические трипа-
нозомы, развивающиеся из критидий в слюнных железах
(инвазирующая стадия трипанозомы). По Wenyon.
1) Трипанозомы могут разрушиться и исчезнуть в течение двух-трех
суток параллельно с перевариванием крови.
2) Трипанозомы не все исчезают с перевариванием крови; остав-
шиеся погибают при последующем принятии крови.
3) Трипанозомы могут п
несмотря на вторичное со-
сание крови.
4) Они могут пережи-
вать и размножаться в зобу,
причем кишечник может
быть совершенно свободен
от паразитов. Жизнь три-
панозом длится в таком
случае дней двенадцать.
5) Трипанозомы могут
быть в большем или в мень-
шем числе как в зобу, так
и в кишке одной и той же
мухи.
6) Полностью или час-
тично переваренная кровь,
оставшаяся от первого со-
сания, может быть заме-
щена свежей кровью при
последующем принятии пи-
щи без того, чтобы трипа-
нозомы исчезли из желудка.
Именно эта комбинация и
ведет к приобретению пе-
реносчиком стойких зара-
жающих свойств (Wenyon).
Полный цикл развития
трипанозом в Glossina palpalts идет следующими этапами (рис. 63, 64):
а) Трипанозомы с кровью заглатываются в желудок переносчика.
Здесь проходит первая фаза их размножения делением с образованием
веретеновидных трипанозом со свободным жгутиком (рис. 64—3).
б) Трипанозомы передвигаются в преджелудок, где принимают уз-
кую и длинную форму (рис. 64, 2); миграция их продолжается далее
кпереди; они входят в хоботок и через канал гипофаринкса проникают
в слюнные железы (рис. 63, S; 64, 4).
в) В слюнных железах проходит второй период размножения с
образованием веретеновидных критидий (рис. 63, В), которые в свою
очередь превращаются в метациклические трипанозомы (рис. 63, С),
переходящие в гипофаринкс (рис. 64, 5). Эти метациклические трипа-
нозомы и являются заразительными для человека, когда попадут в его
организм при сосании крови мухой тсе-тсе. Таким образом, Glossina
palpalis осуществляет циклический перенос трипанозом инокулятивным
путем, т. е. впрыскиванием трипанозом в тело человека через гипофа-
ринкс.
Весь цикл развития в переносчике тянется около двадцати дней.
Резервуаром трипанозомного вируса являются различные африкан-
ские антилопы, для которых Tr. gambietise совершенно безвредна.
Трипанозомы в крови бывают обыкновенно малочисленными, поэто-
му обнаружить их легче удается на препаратах толстой капли.
128
СПЕЦИАЛЬНАЯ ЧАСТЬ
Необходимо отметить, что «сонная болезнь», наблюдаемая иногда
в Европе, ничего общего не имеет с классической сонной болезнью,
возбудителем которой является Trypanosoma gambiense больного, а
переносчиком — Glossina palpalis.
Рис. 64 — Путь трипанозом сонной болезни в мухе тсе-тсе.
J — полость губы, через которую проходят (трипанозомы; 2 — преджелудок с длин-
ными трипанозомами; 3 —желудок, где трипанозомы размножаются делением и образуют
веретеновидные трипанозомы со свободным жгутом; 4 — слюнные железы, где происхо-
дит вторая фаза размножения с образованием веретеновидных критидий, превращаю-
щихся в инвазирующих (метациклических) трипанозом; 5 — гипофаринкс, через кото-
рый трипанозомы со слюной выходят из тсе-тсе.
Из W еп у о п, по Lloyd и Johnson.
В последнее время определяются весьма благоприятные перспекти-
вы лечения сонной болезни химиотерапией (германии — Bayer 205;
трипарсамид и др.).
Trypanosoma cruzi Chagas
Синоним: S chi z о try pantun cruzi Chagas
Трипанозома Круца является возбудителем болезни Чагаса (Cha-
gas), эндемичной для Ю. Америки (Бразилия). Паразиты при острой
инфекции часто помногу встречаются в периферической крови. Они
имеют вид трипанозом с заостренным задним концом, близ которого
лежит крупный блефаропласт. Длина трипанозомы 14—20 у,.
Замечательно то, что эти трипанозомы не размножаются в перифе-
рической крови. Проникнув в клетки различных органов (сердце, попе-
речнополосатые мышцы, гладкие мышцы кишки, центральная нервная
система и др.), они округляются, принимают лейшманиеподобную фор-
му и энергично размножаются делением, переполняя занятую клетку.
Затем паразиты принимают снова трипанозомную форму и поступают
в русло крови (рис. 65, А, В; рис. 66). Все это повторяется несколь-
ко раз.
Болезнь Чагаса протекает различно; в острой форме она может да-
вать среди детей до одного года смертность до 30%. Хроническое за-
болевание более свойственно старшему возрасту. Проявления ее раз-
личны: опухание щитовидной железы (Thyreoiditis parasitaria —
рис. 16) с явлениями микседемы, сердечные расстройства, параличи,
инфантилизм (рис. 15) и др.; такое преобладание симптомов зависит
от преимущественного скопления паразитов в сердечных мышечных
элементах (рис. 66), в мозгу и др.
Переносчиком паразитов и распространителем болезни является
крупный клоп сем. Reduviidae—Triatoma megista (= Conorhinus
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
12S
megistus); живет он в хижинах и нападает ночью на спящих, высасы-
аает кровь часто из кожных покровов лица; такая локализация уколов
этого переносчика является причиной именования его «поцелуйным
клопом».
Заглоченные с кровью трипанозомы переходят в ректальную часть
кишечника , этого клопа, где размножаются и превращаются в крити-
диальные формы. Спустя некоторый срок образуются метациклические
трипанозомы, выбрасываемые наружу с испражнениями поцелуйного
клопа. Заражение человека происходит при попадании метацикличе-
ских трипанозом в места уколов кожи переносчиком или в расчесанные
места.
Рис. 65 — Trypanosoma cruzi в клопе Тria-
toma и в некоторых тканях человека.
А — лейшманиеобразные формы в мышце сердца;
В — трипанозомные формы зам же, С — тринэЕО-
зсмы в крови; S— трипанозомы в желудке клопа;
D — размножение критидий в ректальном мешке
клопа; /? —ректальная фаза развития; £—мета-
циклические трипанозомные инвазирующие фор-
мы; Р — хоботок клопа. По W е п у о и.
Рис. 66 — Trypanosoma cruzi — лей-
шманийные формы на срезе ткани
сердца человека. По Chagas.
Трипанозомы, занесенные на слизистые оболочки или конъюнктиву
глаза, активно проникают внутрь даже при условии их целости.
Таким образом передача инвазии сводится к циклической контами-
нации. В условиях опыта Trypanosoma cruzi может развиваться в по-
стельных клопах, в клещах Ornithodorus и Rhipicephalus.
Срок цикла развития трипанозом в поцелуйном клопе вариирует от
пяти до пятнадцати дней.
Резервуаром трипанозом Круца являются различные виды броненос-
цев (Dasipus), двуутробок (Didelphis) и др.
Семейство Bodonidae
Мелкие биченосцы с двумя жгутами на переднем конце тела, свя-
занными каждый с базальным тельцем; жгуты неравной длины. К че-
ловеку могут иметь некоторое отношение лишь свободноживущие фор-
мы, являющиеся иногда мнимыми или ложными паразитами.
130
СПЕЦИАЛЬНАЯ ЧАСТЬ
Род Bodo
Bodo caudatus Dujardin
Синонимы: Bodo urinarius Hassall; Prowazekia cruzi\ Hartmann и Chagas,
Prow, ninae kohl-yakimovi Jakimoff.
Рис. 67 — Bodo caudatus
(= В. urinarius) из мочи
человека.
1—2 — две жгутиковые фор-
мы; 3 — циста; 4-5— экци-
стирование. По Sinton.
Bodo caudatus является одним из наиболее обыкновенных копро-
зоев, который часто встречается также и в различных органических
настоях. Питается Bodo бактериями.
Форма тела подвижного Bodo очень изменчива, хотя в общем лан-
цетовидная. Длина его до 18 у. У переднего конца тела лежит неболь-
шой цитостом. Ядро с большой кариозомой занимает середину тела.
В задней половине последнего есть сократительная вакуоль. Кпереди
от ядра лежит кинетопласт; у его переднего полюса помещаются два
сближенных блефаропласта. От последних от-
ходят два жгута — один (более короткий), на-
правленный вперед, другой, более длинный,
обращен назад.
Цисты овальны, длиною 5—7 рь; обычно в
них есть одно ядро и кинетопласт. Проглочен-
ные цисты проходят без всякого изменения
через весь желудочно-кишечный канал человека
и вскрываются в уже выброшенном наружу
кале (рис. 67).
К этому виду причисляют теперь в виде
синонима жгутиконосцев, описанных под име-
нем Prowazekia (Pr. cruzi, Pr. asiatica, Pr. ita-
Иса и др.), которых некоторые авторы (без
должного основания) считали за паразитов,
патогенных для человека.
В моче одного больного в 1859 г. был най-
ден биченосец, получивший название Bodo
urinarius (Hassall). Однако этот случай едва ли
может иметь цену, ибо биченосец был открыт
в моче, стоявшей на воздухе несколько дней;
вследствие этого кажется весьма вероятным, что зародыши биче-
носца попали в мочу из воздуха и развились в ней на под-
ходящем питательном субстрате. В последующие годы неоднократно
были описываемы различные жгутиконосцы из мочи: В. urinarius
(Kiinstler, 1883) у больного, страдавшего хроническим гнойным катаром,
Prowazekia urinaria (Sinton, 1912) и др. Описание их обычно столь не-
достаточно, что трудно признать в них самостоятельные виды; поэтому
и намечается тенденция сводить их всех к виду Bodo caudatus, описа-
ние которого дано выше.
Другим менее распространенным копрозоем является Bodo edax
Klebs.
Семейство Embadomonadidae
К семейству Embadomonadidae принадлежит один только род Emba-
domonas. Тело овальное с лежащим впереди ядром. Сбоку и несколько
сзади помещается цитостом в виде широкого углубления. Сбоку или
спереди ядра, тотчас у ядерной оболочки, лежат два маленьких блефа-
ропласта, от которых отходит по жгутику. Передний жгутик — тонкий
и длинный; задний, более короткий и толстый, выходит через цитостом.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
131
Род Embadomottas
Embadomonas intestinalis Wenyon et O’Connor
Очень маленький биченосец, овальной или сердцевидной формы (4—9 X 3—4 р,).
Питается бактериями. Размножается делением. Грушевидные цисты очень малы —
4,5—6,4 /л длины; они похожи на цисты Chilomastix (см. ниже). В окрашенных цистах
наблюдается характерная особенность строения: по длине цисты видны две дуго-
образно изогнутые линии, между которыми помещается ядро. В каком участке ки-
шечника живет Embadomonas, с точностью еще не установлено.
Паразит найден в Египте (Wenyon и O’Connor) в дизентерийном стуле. Биче-
носцы в свежем материале оживленно подвижны, причем орудием движения является
передний жгут; задний же играет направляющую роль. Е. intestinalis найден и в
других местах: в Лондоне у человека, прибывшего из Китая, в Калифорнии и др.
Его можно культивировать.
Хотя паразит и встречается в диаррейном стуле, но это не говорит еще в пользу
признания его патогенным для человека. Пока является редким биченосцем человека.
Семейство Chilomastigidae
Сем. Chilomastigidae характеризуется наличием трех передних жгу-
тиков и одного направленного назад и лежащего в ротовой щели. Ти-
пичные грушевидные цисты с одним ядром и различимой ротовой
щелью. Единственный род — Chilomastix.
Род Chilomastix
Chilomastix mesnili Wenyon
Синонимы: Cercomonas hominis (Davaine), Trichomonas intestinalis (Leuckart)
(pro parte).
Тело грушевидное, асимметричное, как бы перекрученное с хвостовым придатком
различной длины. Длина Chilomastix колеблется в крайних пределах от 4 до 21 р-,
а в средних — от 8 до 15 р- (рис. 68). Паразит одет пелликулой; протоплазма его
густа.
Ядро (овальное или шаровидное) лежит у переднего конца тела. В нем одна
или несколько кариозом и хроматиновые зернышки. Сбоку ядра лежит вдоль по
длине тела ротовая щель, отличающаяся большими размерами — она достигает трети
или половины длины Chilomastix. Рот окаймлен двумя неравными губами.
Между ядром и поверхностью переднего конца паразита лежит кучка мелких
блефаропластов {Dobell, 1920). От них отходят: а) три передних свободных жгута,
одинаковых по длине и толщине, б) две фибриллы, из которых каждая подпирает
соответствующую губу рта, и в) задний жгут, лежащий вдоль ротовой щели. Аксо-
стиль и ундулирующая мембрана отсутствуют.
Chilomastix питается бактериями кишечника.
Цисты этого паразита. имеют форму, близкую к очертаниям лимона или груши,
длиною 7,5—8,5—10 р-. Оболочка цисты топка, бесцветна и слегка утолщена на
более остром конце. В живой цисте не видно ничего, кроме гомогенного содержи-
мого с зернышками волютина. В фиксированных и окрашенных цистах можно най-
ти ядро, блефаропласты, фибриллы, окаймляющие губы, и задний бич, идущий
вдоль рта (табл. III, рис. 14).
Задний волочащийся жгутик связан с поверхностью тела биченосца.
Кроме него, есть один или более передних жгутиков.
Chilomastix живет в толстых кишках. Он не патогенен и нередко встречается
одновременно с другими обитателями кишечника, как то: лямблиями, кишечными
амёбами и др. В больном кишечнике Chilomastix наводит для себя, вероятно, более
благоприятные условия существования. ЦисТы на вскрытиях обнаруживаются в со-
держимом прямой • кишки.
Распространен этот паразит по всему свету. Обнаруживается в диаррейном
стуле как в свободном состоянии, так и в инцистированном виде. В оформленных
испражнениях бывают только цисты.
В Ленинграде обнаружен в 5—6ю/о обследованных взрослых; на Кольском полу-
острове до 10% обследованных были с этими паразитами.
132
СПЕЦИАЛЬНАЯ ЧАСТЬ
Семейство Cercomonadidae
Род Cercomonas
Gercomonas longicauda Dujardin
Тело Cercomonas longicauda амебоидной формы (5—10 p- длиной). Ядро лежит
в его переднем конце. От блефаропластов отходят два жгута: передний — длинный
(раза в три длиннее тела) и задний — короткий. Питается бактериями, которых за-
хватывает ложноножками. Пульсирующей вакуоли нет. Размножается продольным
делением.
Рис. 68 — Chilomastix mesnili
(полусхематично).
1—3 — передние жгутики; 4—6—блефаро-
пласты; 7 — внутриядерный ризопласт; 8 —
яд^о; 9 — кариозома; 10 — вараоиль; 12 —
цитосюм; 13— ротовой жгутик или ун-
дулирующая мембрана; 14 — спиральная
борозда; 15—парабазальное тело.
По Кофоид и Свизи.
Рис. 69 — Trichomonas angusta,
схема строения.
1 —передний жгут; 2— ундулирующая
мембрана; 3- блефарс пласты; 4 — jh-
зопласг, соеяиняющий ядро с блефзро-
пласт( м; 5 — цитостом; 6— ядро; 7 —
парабазальное тело; 8 — вакуоли; 9—
аксостилярные хромидии; 10 — краевая
фибрилла; 11 — аксос i иль; 12 — цито-
пл: этатические хромидии; 13 — задние
аксостильные тельца; 14 — задний жгу-
тик. По Кофоид и Свизи.
Цисты шаровидные с одним яд{5ом и множеством сильно преломляющих свет
грапул. Они стойки к высыханию. Диаметр их 4—5 р.
Изолирован из faeces овец и коз. Повидимому, может встречаться в качестве
копрозоя у человека.
Род Tricercomonas
Tricercomonas intestinalis Wenyon et O’Connor
Синоним: Enteromonas hominis da Fonseca.
Маленький овальный или округлый биченосец, в живом состоянии 4—10 р- длины,
3—6 р-ширины; пузырчатое ядро с вентральной кариозомой лежит у переднего края
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
123
паразита. У переднего полюса ядра помещаются два маленьких блефаропласта, от
которых отходят четыре жгутика. Три из них свободны, четвертый идет назад, тес-
но прилегая к телу и от заднего его конца отходит свободно. Рта нет; ундулирую-
щая мембрана также отсутствует. В протоплазме — вакуоли с заглоченными бакте-
риями.
Цисты овальные, 6—8X3—4^ . В них — одно, два или четыре ядра, что ти-
пично именно для Enteromonas. В протоплазме бывают также и хроматоидные тела.
В точности неизвестно, в какой части кишечника живет этот жгутиконосец. В пользу
его патогенности достаточных данных не имеется.
Enteromonas найден в Македонии, Бразилии, Египте, Индии и во французской
Гвиане.
Семейство Trichomonadidae
Жгутиков несколько, цитостом есть (рис. 69, 5; 70, е), от блефаро-
пласта отходит аксостиль (рис. 69, 70, а), который пронизывает все
тело и выходит, несколько наружу на заднем конце биченосца. Задний
жгут может быть связан ундулирующей мембраной с телом биченосца.
Род Trichomonas
Trichomonas hominis Davaine
Рис. 70—Схема стро-
ения Trichomonas.
а — осевой опорный Стер-
жень (аксостиль); b — ос-
новная фибрилла; с—волно-
образная перепонка; d —
основное тельце; е — цито-
c. о и; п — ядро; о — бичи
в их различных положе-
ниях. По R odenwald t.
Синоним: Trichomonas intestinalis Leuck. (pro parte).
Маленький биченосец овальной формы с заостренным шиповид-
ным выростом па заднем конце тела. Форма тела может изменяться и
становиться то шарообразной, то веретеновидной, то вовсе неправиль-
ной. В зависимости от этого и размеры его колеблются от 5 до 15 у.
Фиксировать его трудно; при фиксации тело
Trichomonas округляется, а детали строения ста-
новятся плохо различимыми.
Пузыревидное ядро лежит у самого переднего
конца паразита. В нем заключена маленькая карио-
зома и несколько зерен хроматина. Ядерная обо-
лочка выражена хорошо. Между ядром и передним
краем помещается несколько мелких скученных
вместе блефаропластов (рис. 69, 3). От них отхо-
дят три или четыре передних свободных и оди-
наковых по длине жгута и ундулирующая пере-
понка (рис. 69, 2), подпертая по свободному изо-
гнутому краю нитью, которая отходит от заднего
конца тела в виде короткого жгутика; место соеди-
нения волнообразной перепонки с телом Tricho-
monas также обозначено основной фибриллой
(рис. 70,’&). По продольной оси паразита прохо-
дит аксостиль — опорный (слабо красящийся) стер-
жень, конец которого образует упомянутый выше
шип заднего конца тела.
Пища в виде' бактерий заглатывается через
клеточный рот (рис. 69, 5), лежащий в виде щели
вдоль ядра. Ундулирующая мембрана занимает
противоположную поверхность тела (рис. 69,2). Trichomonas питается,
вероятно, и эндосмотически, всасывая жидкие ^пищевые вещества
поверхностью тела.
Размножается Trichomonas продольным делением.
Относительно йнцистирования Trichomonas hominis в литературе со-
гласных данных нет. Цисты его с достоверностью неизвестны.
134
СПЕЦИАЛЬНАЯ ЧАСТЬ
Живут эти паразиты в толстых кишках (Dobell и O’Connor).
Trichomonas могут терять жгуты и принимать амебоидную форму с
остатками ундулирующей мембраны.
Заражение Tr. hominis происходит при проглатывании этих биченос-
цев с загрязненной пищей или водой. Опыты на животных говорят за
способность Tr. hominis проходить через желудок без вреда для себя.
Рис. 71 — Биченосцы:
1 — Trichomonas vaginalis; 2 — Pentatrichomonas ardindeiteili;
3 — Tr, buccalis\ 4 — Ir. hominis; 5 — Tr. fecalis. По разным авторам, из W. Powell.
В опытах Hegner здоровые крысы получали трихомонадную инвазию
слепой кишки через четыре дня после ©ведения Tr. muris ib желудок.
Tr. hominis отличаются вообще значительной стойкостью; так, при 0°
эти биченосцы погибали через два с половиной часа, при 50° С—черёр
45 минут, при 65° С — через 7 минут. В фекальных массах они живут
долго (Pentatrichomonas ardindeiteili, по Кофоиду и Свизи, до 24 дней).
Живые биченосцы были находимы в отрыгиваемой мухами жидкости
через пять минут после кормления мух зараженными faeces. Этот про-
межуток времени является достаточным для того, чтобы муха успела
перенести биченосцев на значительное расстояние от места заражения
(Веньон и О’Коннор).
Вопрос о вредоносности Tr. hominis для человека неясен. В некото-
рых случаях в них находили заглоченные эритроциты, a Wenyon (1920)
нашел этих простейших в подслизистой оболочке стенки толстой киш-
ки (рис. 72). Однако эти факты нуждаются в комментировании. Отсут-
ствие тканевой реакции вокруг внедрившихся паразитов заставляет ду-
мать, что Trichomonas проникли вглубь тканей или перед смертью чело-
века, или тотчас после смерти, при начавшемся изменении эпителия
слизистой оболочки. Таким образом, эти факты не говорят в пользу
признания Trichomonas патогенным простейшим.
Заглатывание Trichomonas эритроцитов было наблюдаемо и на куль-
турах, особенно когда в них преобладали пятижгутиковые формы (Pen-
tatrichomonas). Trichomonas несомненно способен заглатывать свобод-
ные эритроциты, поступившие под влиянием сторонних причин в полость
кишки (или примешанные к культуре). Помимо заглатывания эритро-
цитов наблюдается и обратное выталкивание их из Trichomonas
(«рвота», В. Гнездилов).
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
135
Есть указание, что Trichomonas может находиться в желудке в
случаях резкого понижения или полного отсутствия кислотности желу-
дочного сока. В связи с этим его находили при раковых опухолях же-
лудка, но никакого специфического соотношения между этими фактами
нет.
Рис. 72—Часть разреза толстой кишки человека с
Trichomonas hominis, проникшими в толщу ткани (обо-
значены стрелками). По Wen у on.
Нередко Trichomonas встречается у людей, страдающих поносами.
В таких случаях можно говорить, что Trichomonas вызывает дизенте-
рийные явления, иди наоборот — заболевания кишечника создают бла-
гоприятные условия для жизни Trichomonas.
Из доводов противоположного характера отметим, что котят можно
заразить Tr. hominis, выделенными из faeces человека. У них развивал-
ся энтерит с некротическим поражением выстилки толстых кишок.
В одном из городов Перу наблюдалась вспышка острой дизентерии;
у всех болевших (около 150 человек) был найден Tr. hominis, который
был обнаружен также в цистерне с питьевой водой, которой пользо-
вались все заболевшие. После очистки цистерны дальнейшие заболева-
ния прекратились.
Однако подобные случаи, которые, казалось бы, убедительно говорят
в пользу признания Tr. hominis болезнетворными для человека, отмеча-
ются редко по сравнению с частотой нахождения этих биченосцев у че-
ловека. Необходимо заметить, что только тогда статистика нахождения
этого простейшего даст убедительную картину, когда наряду с больны-
ми будут обследованы и массы здоровых людей с целью выяснения сте-
пени зараженности их Trichomonas и другими кишечными биченосцами.
Сравнимых цифр частоты нахождения Trichomonas у больных и у
здоровых пока очень мало. Между тем, техническая трудность микро-
136
СПЕЦИАЛЬНАЯ ЧАСТЬ
скопического открытия Trichomonas в оформленном стуле очевидна;
присутствие в нем паразитов может быть легче обнаружено посевом
кусочков faeces на питательные среды для культивирования простейших.
Несомненна способность Trichomonas жить в иных условиях, чем в ки-
шечнике, так как этих паразитов обнаруживали в гное амёбного абсцес-
са печени и в крови.
Интересно отметить, что сравнительный анализ faeces, мочи, крови,
желудочного сока, кровяного давления'у носителей Trichomonas не дал
никаких материалов для признания этого паразита патогенным.
Tr. hominis принадлежит к широко распространенным рбитателям
толстых кишок человека.
Trichomonas elongata Steinberg
Штейнберг нашел во рту человека несколько видов рода Trichomonas, а именно
Tr. elongata (рис. 73), Tr. caudata и Tr. flagellata; Prowazek отнес обнаруженных
им простейших к виду Tr. hominis. Весьма вероятно, что все названные биченосцы,
вместе с Tr. buccalis Goodey и Wellings (рис. 74), который, видимо, отличается от
типичного Tr. hominis и Тг. vaginalis отсутствием свободного жгутика на заднем
конце ундулирующей мембраны, относятся к виду ТГ. elongata.
Описывались и другие Hagellata изо рта человека, относящиеся к родам Monas,
Bodo, Cercomonas, но описание их (Штейнберг, 1862) настолько недостаточно, что
при отсутствии проверочных исследований новейшими протозоологическими метода-
ми нельзя установить их точное систематическое положение.
Trichomonas был обнаруживаем в гное из фолликулов миндаликов, в экссудате
при плевритах, в откашливаемом материале при гангрене легких и др. Повидимому,
соответствующие паразиты по своему происхождению были связаны с Trichomonas
полости рта. ,
Trichomonas vaginalis Donnee
Tr. vaginalis — паразит влагалища; крупнее Tr. hominis, имеет в
длину 15—25 и даже 30 р (рис. 71, /, 75). Тело широкоовальное или
грушевидное, при движении весьма пластичное. От переднего конца
отходят четыре жгутика, связанные с блефаропластом, образованным
кучкой пяти тесно лежащих зерен. От блефаропласта к овальному
ядру паразита отходит короткий стержневидный ризопласт. С блефаро-
пластом соединены также 1) парабазальная фибрилла, которая идет
к концу тела паразита, огибая ею ядро; 2) подпирающая и базальная
фибриллы короткой ундулирующей мембраны, доходящей до половины
или двух третей длины тела и 3) аксостиль. Последний на свежем
объекте не виден, кроме выдающегося заднего конца, обладающего
ригидностью. Протоплазма на препаратах тонкозернистая. В ней раз1
бросаны зерна, окрашивающиеся ядерными красками. Эти хрома-
тиновые (?) гранулы лежат также по сторонам аксостиля. Заглочен-
ные бактерии редко встречаются в протоплазме этого биченосца,
взятого из влагалища; в культурах же заглатывание бактерий —
дело обычное; наблюдается также заглатывание крахмальных зерен и
эритроцитов. Клеточный рот с достоверностью не обнаружен. Ядро
удлиненноовальное, лежит в передней трети паразита. Размножается
делением (рис. 76). Блефаропласт делится надвое; с каждой до-
черней половиной остаются в соединении по два жгута; аксостиль реду-
цируется; в ядре начинается формирование четырех хромозом. Между
расходящимися дочерними блефаропластами остается связующая
нить — парадесмоз. К одному из блефаропластов отходят старая унду-
лирующая мембрана и парабазальная фибрилла. Ядро в это время про-
ходит стадию профазы кариокинетического деления. От второго дочер-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
137
него блефаропласта начинает расти ундулирующая мембрана с базаль-
ной фибриллой. В стадии экваториальной пластинки кариокинеза от
обоих блефаропластов образуются новые аксостили. В анафазе хорошо-
Рис. 73 — Trichomonas elongata.
По Wenyon.
Рис. 74—Trichomonas
buccalis. По К о ф о и д у.
различимы по четыре дочерних хромозомы. Блефаропласты далеко от-
ходят Друг от друга; соответственно удлиняется и парадесмоз. Он со-
храняется еще и в фазе деления, когда формируются дочерние ядра; в
это время от каждого блефаропласта отрастают по два жгута, благо-
даря чему образуется как бы двойной организм (W. Powell).
Рис. 75 — Trichomonas vaginalis. По Brumpt.
Встречаются дегенерирующие формы, иногда похожие на Blasio-
cystis.
Tr. vaginalis встречается у женщин всех возрастов в слизи влагали-
ща, выделения которого имеют кислую реакцию при вагините и белях.
Его находили также в мочевом пузыре при катаре и цистите. -Частота
обнаружения Tr. vaginalis различна—10% у обследованных женщин во
138
СПЕЦИАЛЬНАЯ ЧАСТЬ
Франции (Brumpt), обыкновенен в Англии (Wenyon). В Москве Поно-
жина нашла влагалищного биченосца у 16 женщин с гнойными белями
из числа 29 человек с этим симптомом и из общего числа 55 взрослых
женщин, тогда как 22 девочки в возрасте от 2 до 14 лет были свобод-
ны от заражения. Tr. vaginalis может переходить и приживается в вос-
паленной уретре мужчин. Его находили также*в центрифугатном осад-
ке мочи и в секрете предстательной железы.
Особый интерес представляет факт нахождения Tr. vaginalis в крови
людей, что удавалось установить культивированием этих биченосцев по-
севами крови.
Мнения по вопросу о патогенности влагалищного биченосца расхо-
дятся. Одни считают, что он является возбудителем трудно излечимых
катаров половых путей и мочевого пузыря; другие видят в нем безвред-
Рис. 76 — Trichomonas vaginalis, деление,
По W. Powell.
ного паразита, который находит себе благоприятные условия жизни в
уже патологически измененных от других причин органах, причем он
может поддерживать сильное воспаление. Третьи усматривают мало
оснований приписывать какое-либо патогенное значение этому пара-
зиту.
Заражение происходит при половых сношениях и при пользовании
общими предметами, например полотенцем, общей постелью, наконец,
общей уборной.
При орошении влагалища щелочными растворами или просто холод-
ной водой (ниже ,4-15° С) паразит погибает. Однако излечение не
удается просто и скоро, что зависит, вероятно, от длительности и стой-
кости основного заболевания пораженного органа. Так, двухмесячное
спринцевание влагалища Kai. hypermanganicum вело к исчезновению
паразитов, которые снова появились после прекращения лечения. Реко-
мендуют также спринцевание щелочным раствором хины (чайная ложка
Ntr. bicarbonicum, 0,3 Chin, muriat.).
Wenyon высказывает весьма интересное соображение о видовых со-
отношениях Tr. vaginalis, Tr. intestinalis и Tr. gingivalis, предполагая,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
139
что эти биченосцы являются в сущности одним видом, вариации разме-
ров которого зависят от различия в питании при паразитировании их в
разных органах (рис. 71).
Однако попытка пересадки кишечных Trichomonas hominis человека
во влагалище не дали результатов (Kassel и Gafford); тот же паразит,
введенный во влагалище обезьяны (Macacus rhesus), жил там до два-
дцати дней, но не размножался (Hegner). В культурах Trichomonas
vaginalis, Tr. buccalis и Tr. hominis (рис. 71) сохраняют свои морфоло-
гические отличия и не переходят один в другой, что говорит за видовую
самостоятельность этих паразитов (W. Powell).
ОТРЯД DIPLOMONADIDA
Представители этого отряда имеют парные ядра и другие органоиды,
расположенные симметрично, поэтому тело таких простейших построе-
но по двубоковой симметрии. Из трех родов этого отряда — Hexamita,
Giardia и Trepomonas у человека паразитирует только Giardia.
Род Giardia (Lamblia)
Giardia intestinalis Lambl
Синоним: Lamblia intestinalis.
Рис. 77 — Giardia {Lamblia) intestinalis; схема
строения.
Вид сбоку и наискось. 1 — хвостовая пара жгутиков:
2 — средняя пара жгутиков; 3 — аксостили; 4— передняя
пара жгутиков; 5—парабазальное тело; 6 — фибрилла,
окружающая присоску; 7 — ядро; 8 — блефаропласты; 9 —
краевая фибрилла; 10 — вентральная пара жгутиков.
По Роденвальд.
Двусторонне симметрична. Тело co спинной стороны имеет прибли-
зительно грушевидную форму. Передний конец паразита расширен и
округлен, задний заострен. На вентральной стороне расширенной поло-
вины тела имеется вогнутость в месте так называемого присасыва-
тельного д и с|к а. Длина
паразита колеблется от 10
до 18 рь. Тело его одето
пелликулой.
У ламблии два овальных
ядра (рис. 77). лежащие в
широкой части ’"тела дор-
зально от присоска. В ядре
крупная удлиненная карио-
зома.
Между ядрами, вдоль по
длине тела, проходят два
параллельно лежащие аксо-
стиля или аксонемы, иду-
щие до заднего конца тела.
Жиардия имеет четыре па-
ры жгутиков, симметрично
отходящих от блефаропла-
стов, расположение кото-
рых и число различно описывается разными наблюдателями. Пара
жгутиков отходит от заднего конца тела, остальные отходят снизу
и с боков.
Приблизительно в середине тела ламблии помещаются так называе-
мые парабаза л ьные те л а. в. виде запятых или изогнутых пало-
чек. Значение их неизвестно.
Для принятия пищи никаких органоидов нет; пища всасывается всей
поверхностью тела паразита, благодаря чему протоплазма жиардии сво-
бодна от каких-либо включений.
140
СПЕЦИАЛЬНАЯ ЧАСТЬ
перстной. Отмечены случаи обитания
Рис. 78—Giardia (Lamb1, id) intestinalis
в стадиях деления. По Добелл и
О’Коннор.
Паразиты размножаются продольным делением, процесс которого
протекает сложно. Сначала делятся ядра, потом удвояются присасыва-
тельный диск, блефаропласты, осевые стержни и бичи. Затем начинает-
ся расщепление тела с его широкого конца. Некоторые фигуры деле-
ния жиардии изображены на рис. 78. Жиардии . могут делиться как в
свободноподвижном состоянии, так и в инцистированном виде. Стадии
деления подвижных жиардий могут быть находимы в стуле в исклю-
чительно редких случаях.
Жиардии живут в тонких кишках, главным образом в двенадцати-
их в желчном пузыре. Своей при-
соской они садятся на свобод-
ную поверхность эпителиаль-
ных клеток слизистой обо-
лочки кишечника. Полагают,
I что этим нарушается секре-
ция кишечного сока.
В нижнем отделе кишеч-
ника жиардии инцистируются.
Тело паразита округляется,
принимает яйцевидную форму
и одевается довольно толстой
оболочкой. В' цисте паразит
готовится к продольному делению, причем его органоиды удвояются.
Ядер в цисте четыре; аксостили, парабазальные тела также расще-
пляются; в конце концов внутренняя структура цист делается слож-
ной (табл. II, рис. 16). Длина цист 10 — 14 у-, ширина 7,5 — 9 рь.
В стуле нормальной консистенции встречаются только инцистирован-
ные жиардии; вегетативные формы можно обнаружить при поносах или
после дачи обильного слабительного. При исследовании дуоденальным
зондом в добываемом соке легко можно обнаружить жиардии, если они
есть у пациента. Цисты погибают при высыхании, но во влажном кале
сохраняют жизнеспособность недели две. Воеск наблюдал их сохране-
ние вне организма в течение нескольких месяцев.
В опытах на крысах инфицированию жиардиями благоприятствует
жировая диэта. Молочная, жировая, растительная, лактозная и сме-
шанные диэты благоприятствуют заражению жиардиями; значение же
мясной и дрожжевой диэт обратно противоположное, что может иметь
и лечебное, и профилактическое значение (Б. Глуховцев). Неблаго-
приятное влияние на жиардиоз протеиновой диэты констатирует Тсучиа
(Tsuchia).
Жиардии широко распространены. С уточнением клинической диагно-
стики они чаще и чаще обнаруживаются у больных.
Вопрос о болезнетворном значении жиардии трактуется противоре-
чиво. Несомненно, что эти паразиты бывают у здоровых людей. Присут-
ствие цист в испражнениях носителей непостоянно: они то исчезают на
различный срок, то снова появляются. В некоторых случаях жиардиоза
бывают периодические поносы с выхождением масс прозрачной слизи,
весьма богатой паразитами. Возможно, что выделение масс слизи яв-
ляется результатом раздражения слизистой оболочки кишечника множе-
ством паразитов.
Wenyon допускает, что некоторые люди более чувствительны к жи-
ардиям, чем другие, и что периоды поноса соответствуют времени ин-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
141
тенсивного размножения паразитов. Несомненна способность жиардий
пробираться через печеночный проток в желчный пузырь, где их находи-
ли при холецистите; однако считать их причиной соответствующего за-
болевания достаточных данных пока нет. Хотя и нельзя вынести окон-
чательного решения о патогенном значении жиардий, все же мнение о
их безвредности поколеблено.
Экстенсивность инвазии жиардиями определяется различно. Из
20 тыс. обследованных с 1916 по 1919 гг. паразиты были обнаружены у
12% (Hegner и Payne). Другие авторы дают иные цифры: 6,5% и даже
2,1%. Интересно отметить, что дети заражены в большем количестве,
чем взрослые. Очевидно, с возрастом приобретается большая стойкость
к заражению этим паразитом. Из русских данных отметим материалы
по Ленинграду 9,2—14,7% заражения взрослых и 20% заражения в
Туркмении (см. стр. 92—93).
Заражение жиардиями происходит при проглатывании цист, рассеи-
вание которых во внешней среде происходит по тем же путям, что и
других кишечных простейших (Ent. histolytica, Tr. hominis и др.). Поэто-
му и меры профилактики против жиардиоза в общем совпадают с про-
филактикой амёбной дизентерии (см. выше, стр. 91).
Лечение производится осарсолом — 3 раза в день по 0,25 г, 3—4 дня
подряд. Таких курсов лечения проводят 3—4.
КЛАСС SPOROZOA - СПОРОВИКИ
К классу споровиков относятся исключительно паразитические про-
стейшие. Во взрослом состоянии органоидов движения нет. Биченосные
формы появляются только в период полового размножения. Питаются
осмотически. Пульсирующая вакуоль отсутствует. Характерный способ
размножения путем спорообразования. После предшествовавшего поло-
вого процесса (копуляции) слившиеся вместе мужская и женская особи
одеваются оболочкой и повторно делятся, вследствие чего образуются
так называемые спорозоиты.
Формы, делящиеся бесполым путем, похожи на амёб.
В последнее время класс споровиков .некоторыми протозоологами
разбит на два класса — книдоспоридии (Cnidosporidia) и собственно
споровики (Sporozod). К первым относятся — микс^споридии, микро-
споридии. саркоспоридии и гаплоспоридии; ко вторым — грегарины,
кокцидии и кровеспоровики.
ПОДКЛАСС TELOSPORIDIA
Телоспоридии часть своей жизни проводят внутриклеточно; во
взрослом состоянии они одноядерны; образование спор приурочивается к
определенному периоду жизни. Из трех отрядов телоспоридий — Grega-
rinida, Coccidia и Haemosporidia — важнейшие для человека паразиты
имеются среди кровеспоровиков (Haemosporidia); грегарины вовсе не
паразитируют у человека, а кокцидии имеют далеко недостаточно выяс-
ненное значение для его патологии; но ознакомление с последними важ-
но для понимания жизненного цикла у кровеспоровиков. Кокцидиоз же
сельскохозяйственных животных и домашней птицы имеет большое эко-
номическое значение.
142
СПЕЦИАЛЬНАЯ ЧАСТЬ
ОТРЯД I. COCCIDI А —КОКЦИДИИ
Кокцидии являются типичными паразитами эпителиальных клеток,
причем бесполый и половой периоды размножения протекают в одном и
том же хозяине. 1
Впервые кокцидия проникает в эпителиальную клетку (например,
печени или кишечника) в стадии спорозоита (рис. 79 A, f), вышед-
шего из проглоченной хозяином ооцисты (/). Спорозоит округляет-
ся и становится маленькой клеткой, лежащей в протоплазме клетки
хозяина (а). Кокцидия растет, питаясь за счет клетки, в которой она
находится (на стадии роста паразит называется трофозоитом);
Рис. 79 А — цикл развития кокцидии. Схема (Добелл и
О’Коннор). Объяснение в тексте.
ядро трофозоита делится повторно, причем образуется многоядерная
стадия паразита — шизонт (с); последний распадается делением на
столько дочерних особей, называемых мерозоитами, сколько в
шизонте было ядер (rf). ’Мерозоиты являются бесполыми особ~я-
м и; процесс их образования — шизогония — есть не что иное, как
множественное деление (бесполое размножение).
Мерозоиты выходят из эпителиальной клетки, где развивался шизонт
(е), и попадают в полость органа. Будучи активно подвижными, они
внедряются в незаряженные эпителиальные клетки (7), где снова начи-
нается цикл шизогонии, и так много раз подряд. Этот бесполый пери-
од жизни кокцидий — эндогенное размножение — ведет к
усилению заражения того хозяина, в которого попали ооцисты кокци-
1 Некоторые имеют двух хозяев, например, Karyolysis lacertae — ящерицу
(Lacerta muralis) и клеща (Liponyssus saurarum), Aggregata— краба и головоногого
моллюска.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
143
дии. Интенсивность зараженности хозяина зависит от количества по-
павших в него ооцист и энергии их размножения (конечно, при соот-
ветствующей податливости организма хозяина).
В конце концов шизогония затихает, и у кокцидий наступает п о-
ловое размножение. Мерозоиты одного из бесполых поколений,
внедрившись в клетки эпителия, больше уже не делятся бесполым пу-
тем, но диференцируются в половые особи; незрелые мужские особи
Рис. 79 В — Isospora felts в эпителии. /. bigemina под эпителием.
Разрез через сосочек кишки кошки; mig — микрогаметоцит I. felis\ mag —
макрогаметоцит I. fells; ос — ооцисты I. bigemina. По Wenyon.
называются микрогаметоцитами, а женские — макрогаме-
тоцитами. Макрогаметоцит разрастается в клетке хозяина до зна-
чительных размеров; часть хроматина при этом выбрасывается из ядра
(что соответствует процессу редукции хроматина при созревании поло-
вых клеток многоклеточных организмов)*; и макрогаметоцит превра-
щается в зрелую женскую особь, называемую макрогаметой
(/, т, и, о).
В то же время в микрогаметоците (h) ядро делится несколько раз:
около ядер обособляются участки протоплазмы, вследствие чего каждый
микрогаметоцит дает много зрелых, веретеновидных, хвостатых, актив-
но подвижных особей, называемых микрогаметами (А); кроме
зрелых микрогамет, микрогаметоцит дает еще остаточное тело.
Микрогаметы выходят из клетки (/?), в которой они образовались, и
активно проникают в макрогаметы (о); этот процесс является оплодо-
творением.
Так обстоит дело у Eimeriida. У Adeleida соединение происходит
между макрогаметой и микрогаметоцитом, который лишь после этого
дает небольшое число микрогамет. Ядра (пронуклеусы) обеих особей
сливаются, в результате чего образуется одноядерная клетка, называе-
мая зиготой. У кишечных кокцидий зигота выпадает в просвет киш-
ки, одевается оболочкой и в этой стадии называется уже ооци-
стой (р). Ядро ооцисты делится' и дает несколько дочерних ядер
(<7, на рисунке их четыре). Вокруг каждого дочернего ядра обособляет-
144
СПЕЦИАЛЬНАЯ ЧАСТЬ
ся участок протоплазмы, и все содержимое ооцисты распадается на
остаточное тело и спороблас
собственной оболочкой (споро
Anneufa
Рис. 80 — Кокцидии Elmer iida\ во
время полового размножения муж-
ские и женские 1аметы раздельны и
развиваются независимо друг бт
друга. Adeleida — мужские и жен-
ские гаметы тесно связаны друг с
другом. По Pinto.
т ы (г). Каждый споробласт одевается
циста) и становится спорой (s).
Внутри последней образуется, путем
деления ядра и протоплазмы, изве-
стное число спорозоитов и оста-
точное тело. Этот заключитель-
ный период полового размножения —
спорогония — является внекле-
точным. Ооцисты выносятся потом
с калом наружу.
Если ооцисты кокцидий будут,
проглочены соответствующим хозяи-
ном, то в кишечнике оболочка их рас-
творяется, спорозоиты становятся
подвижными и внедряются в клетки
кишечного эпителия, где начинают
уже известный нам цикл бесполого
размножения.
Половое — или экзогенное —
размножение способствует расселе-
нию кокцидий, заражению ими новых
особей; последнее ведет к расшире-
нию экстенсивности инвазии.
Ход жизни и размножения
кокцидйй представлен на схеме:
Шизогония
Шизонт -------- -----
Мерозоиты
(агамогония)
Взрослая бесполая форма Молодая бесполая форма
Бесполое размножение,
повторяющееся много-
кратно и ведущее к
увеличению интенсив-
ности инвазии
Многократно повторяю-
щаяся шизогония в ста-
ром хозяине
Спорогония
Половое ра умножение,
ведущее к распростра-
нению инвазии на но-
вых хозяев
Мерозоит
Микрогаметоцит
Ф
Спорозоит Микрогамета
Спора
Подготов-
ка к инва-
зии ново-
го хозяи-
на
Шизонт
Макрогаметоцит
4
Макрогамета
Зигота (copula)
Споробласт <---------------—
Ооциста (споронт)
Кокцидии паразитируют в позвоночных, червях, членистоногих и в
моллюсках. Наиболее часто они поражают органы пищеварения; ветре-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
145
чаются также в половых органах, почках, кровеносной системе, жировом
теле и др. Некоторые кокцидии являются паразитами ядер клеток (на-
пример, Cyclospora caryolytica у крота).
Рис. 81 — Ооцисты кокцидий.
а. ах, b — Isospora belli; с — е - Eimeria sardinae=E. oxyspora = Е.
snijdersi— транзитные ложнопаразиты человека; d — Eimeria
clupearum сельди ~Е. wenyoni— транзитный ложнопаразиг че-
ловека. По Добелл и О’Коннор.
Макрогаметы и микрогаметы кокцидий развиваются в различных
клетках. Взрослые микрогаметоциты не меньше или даже крупнее, чем
макрогаметоциты; из первых образуется множество микрогамет.
Рассмотрим некоторых представителей подотряда Eimeriida.
Семейство Eimeriidae
Кокцидии, находимые у человека, относятся к сем. Eimeriidae. Они
являются наименее изученными паразитами человека.
Многих представителей этого семейства (род Eimeria и др.) в насто-
ящее время необходимо рассматривать как ложнопаразитов человека,
попадающих в кишечник вместе с зараженной ими пищей (рыбы) и об-
наруживаемых впоследствии в испражнениях.
Род Eimeria
Eimeria clupearum Theldhan
Синоним: Eimeria wenyoni' Dobell.
Обнаружены в испражнениях человека на стадии ооцисты. Ооциста шаровидная
(рис. 81), диаметром около 20 р.. Одета двуслойной оболочкой: внутренний слой тон-
кий, наружный толстый, с поверхности шероховатый. В ооцисте четыре овальных
146
СПЕЦИАЛЬНАЯ ЧАСТЬ
споры размером 10 X 7 р- ; в спорах по два червовидных спорозоита и по одному
или по два остаточных тела (рис. 81, d). Найдена у больного, приехавшего из Гал-
липоли; другие случаи были из Салоник (Wenyon). Thomson и Robertson показали,
что ооцисты Eimeria wenyon! тождественны ооцистам Eim. clupearum, которые бы-
вают в печени сельди, салаки и макрели. Таким образом Е. wenyon! не может счи-
таться паразитом человека. В действительности бывают случаи мнимого паразитиро-
вания Е. clupearum, когда люди едят рыбу, зараженную этой кокцидией.
Eimeria sardinae Thelo’han
Синонимы: Eim. oxyspora Dobell; Eim. snijdersi Dobell.
Рис. 82 — Ооциста кокцидии
Eimeria sardinae (Томсон и
Робертсон).
E. oxyspora была обнаружена в испражнениях человека в форме шарообраз-
ных ооцист, с четырьмя спорами в каждой и с остаточным телом; диаметр ооци-
сты около 36 jx. Она одета двойной оболочкой; наружная оболочка имеет шерохо-
ватый вид благодаря прилипанию частиц кала (рис. 81, с).
Споры в форме оселка 30—32 X 7,5 р- . Оболочка спор также двойная; состоит
из внутренней оболочки (эндоспор) и тонкой наружной оболочки (э п и с п о р).
Внутри заключены две тонкие/ длинные и слегка
перекрученные своими концами споры. Кроме
них, есть еще остаточное тело в виде несколь-
ких зерен. Все это видно на свежем объекте.
Найдена у больного, бывавшего в Ю. Америке,
в Индии, на Цейлоне.
Eimeria snijdersi (рис. 81, с), найденная од-
нажды на Суматре в испражнениях больных,
имеет ооцисты несколько более крупные, чем
Eim. oxyspora (диаметр их около /40—48 ?л)-
Споры несколько отличаются по величине (рис.
82). Thomson и Robertson доказали тождество
Е. oxyspora и Е. snijdersi, с одной стороны, и
рыбьей кокцидии Е. sardinae — с другой.
Е. sardinae (рис. 82) паразитирует в се-
менниках сельди, салаки и сардинок. Желтова-
тые ооцисты этого вида колеблются в пределах
размеров 35—45 р-. Длина спор 30—35 р<. Упо-
мянутые выше авторы после поедания жарены .к
молок сельди и принятия слабительного нахо-
дили ооцисты у себя в стуле. Reichenow внес
существенное дополнение к этим опытам, пока-
зав, что при поедании сырых молок и
при нормальной деятельности желудочно-кишечного аппарата ооцисты в испражне-
ниях не открываются.
Несомненно, что и в случаях с. Е. oxyspora и Е. snijdersi мы имеем дело с
мнимым паразитизмом кокцидий, которые были съедены с рыбой и слу-
чайно обнаружены при микроскопировании кала больных.
Род Isospora
Isospora belli Wenyon
Синоним: Isospora hominis Dobell.
В свежем кале человека Isospora belli встречается в виде удлиненных ооцист
25—33X 13,5—16 р< с неразделившимся еще содержимым в виде шаровидной зер-
нистой массы (рис. 81, а). Один полюс ооцисты тупой,* другой—более тонкий со
слабой перетяжкой перед ним. Через два-три дня в спороцисте развиваются две
споры 12—14 X 7—9 р- , а в каждой споре образуется по четыре спорозоита и по
одному большому остаточному телу. Стадии развития неизвестны. Обнаруживались
в районе восточной части Средиземного моря (Греция, Галлиполи, Месопотамия,
Египет). Прямых доказательств патогенности I. belli для человека нет. Попытки за-
ражения различных животных (мышей, щенят и др.) ее зрелыми спорами не дали
положительных результатов. В случае кокцидиоза человека было бы очень важно
исследовать, если к тому представляется возможность, стенку тонкой кишки на
срезах для изучения совершенно неизвестного еще цикла развития /. belli.
Isospora hominis Railliet et Lucet
Кокцидия, которую нашел у человека Kjellberg в 1860 г. Ооцисты
выходят с испражнениями в совершенно зрелом состоянии, причем обе
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
147
споры одеты нежной оболочкой ооцисты. Споры 16 X 10,5 размером с
четырьмя спорозоитами и скудным остаточным телом. В кишечнике эти
кокцидии лежат под эпителиальной оболочкой.
Это обстоятельство, д также размеры и строение ооцист весьма при-
ближает /. hominis к /. bigemina собаки (рис. 83), что дает повод Rei-
chenow считать собаку источником заражения человека этой кокцидией.
Рис. 83 — Ооцисты Isospora.
А. 2. bigemina и 1. hominis — малая форма из толщи тка-
ни сосочка кишки кошки, собаки и человека; В — проме-
жуточная форма из эпителия сосочка собаки и кошки
(I. rivolta); С — круглая форма из эпителия сосочка кошки
и собаки (Z. fells); D — ооциста, вероятно, из эпителия
сосочка кишки человека (1. belli) — по Wte п у о п.
Интересен случай лабораторного заражения человека I. hominis
(Connal). После случайного проглатывания ооцист через шесть дней
начался понос, державшийся до 22-го дня; неделю спустя стул сделался
более густым и еще через пять дней приобрел нормальную консистен-
цию. Ооцисты появились в faeces через 22 дня после начала поноса и
через 28 дней после заражения. Паразиты держались 13 дней, после че-
го открыть их больше не удалось.
Этот и другие случаи кокцидиоза человека, а также кокцидиоз собак
и кошек характеризуются коротким инкубационным периодом по отно-
шению к клиническим симптомам и более длинным сроком появления
первых ооцист в испражнениях хозяина.
Ряд случаев кокцидиоза человека обнаружен в Ташкенте, причем
иногда ооцисты кокцидий Isospora hominis были находимы в дуоденаль-
ном соке. Болезненные проявления сводились в общем к симптомам эн-
[48
СПЕЦИАЛЬНАЯ ЧАСТЬ
тероколитов. Лечение осарсолом давало благоприятный результат (Ча-
пурская-Баженова и Лейтман). Кокцидий человека находили также в
Закавказье (Грузия и Азербайджан). В Абхазии Зиминым выявлено
25 случаев кокцидиоза человека (/. hominis) нахождением ооцист в фе-
калиях. У большинства имелись субъективные жалобы и кишечные рас-
стройства. У некоторых больных ооцисты при повторных анализах обна-
руживались в течение четырех месяцев. Размеры незрелых ооцист в
фекалиях вариировали в пределах от 25—33 X 12—16 до 12—14 X
X 7—9 р.
Диагностика кокцидиоза производится микроскопированием faeces,
разбавленных водой или физиологическим раствором, а также дуоде-
нального сока. В свежем препарате ищут ооцисты кокцидий. При хра-
нении проб faeces при комнатной температуре в течение нескольких
дней может происходить дозревание ооцист с окончательным формиро-
ванием спорозоитов.
Заражение происходит при проглатывании зрелых ооцист со споро-
зоитами. Следовательно, профилактика кокцидиоза проводится по тем
же принципам, что и по отношению к другим паразитическим простей-
шим кишечника человека. Кокцидии печени кролика Eimeria stiedae в
человеке паразитировать не могут.
ОТРЯД II. HAEMOSPORIDIA—КРОВЕСПОРОВИКИ
Кровеспоровики являются паразитами кровяных телец различных
позвоночных. Размножаются, подобно кокцидиям, бесполым и половым
путем; но в отличие от большинства этих споровиков, гемоспоридии ме-
няют хозяев, причем цикл бесполого размножения проделывают в по-
звоночном хозяине, а половое размножение — в насекомом или в дру-
гом членистоногом животном. Последние являются, следовательно,
переносчиками кровеспоровиков по отношению к позвоночным хозяевам.
Для человека из гемоспоридий имеют значение только некоторые пред-
ставители семейства Plasmodidae.
Семейство Plasmodidae
Стадии бесполого размножения паразитов и гаметоциты локализуют-
ся в красных кровяных тельцах хозяина. К семейству Plasmodidae при-
надлежат возбудители птичьей малярии (Proteosoma ргаесох и другие
виды) и паразиты малярии человека (Plasmodium vivax, PL malariae
и PL falciparum).
Возбудитель малярии, открытый в 1880 г. Лавераном, проделывает
в крови человека лишь часть своего жизненного цикла, а именно период
бесполого размножения (рис. 84).
Цикл развития малярийного паразита в теле человека начинается с
внедрения в эритроцит (1, 2); паразит действительно проникает внутрь
эритроцита, но не остается на его поверхности, как неверно утвержда-
лось в некоторых работах. Плазмодий в эритроците растет—фаза тро-
фозоита (2), и из перстневидной формы кольца с центральной ва-
куолей и утолщением, с ядром где-либо в толще периферически распо-
ложенной протоплазмы (5) превращается в амебовидную; последняя
характеризуется не только вариирующей формой, но и действительной
подвижностью, если наблюдать под микроскопом живой объект (4, 5).
Растущий трофозоит питается за счет содержимого эритроцита, ис-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
149
пользуя в частности гемоглобин. Продуктом этого процесса является
отложение пигментных зерен в теле паразита.
Рост трофозоита сочетается с подготовкой к размножению делением
(шизогония, 5—7); в соответствующей фазе плазмодий является
шизонтом, однако сколько-нибудь резкой грани между этими дву-
Рис. 84 — Жизненный цикл малярийного плазмодия {Plas-
modium vivax — возбудителя трехдневной малярии).- Объяс-
нение в тексте.
мя фазами жизненного цикла малярийного плазмодия нет, и термины
трофозоит и шизонт в значительной мере являются синонимами.
В шизонтах вакуоли уже нет. Ядро паразита начинает делиться
повторно, затем сам шизонт распадается на соответственное количество
дочерних особей — м е р о з о и то в (6—/) в процессе множественного
деления (бесполое размножение). В момент деления эритроцит разру-
шается, освободившиеся плазмодии переходят в плазму крови (7), куда
поступают также и ядовитые продукты обмена веществ паразитов; эти
150
СПЕЦИАЛЬНАЯ ЧАСТЬ
продукты всасываются и своим влиянием на организм человека вызы-
вают ощущение потрясающего озноба и затем приступа лихорадки.
Затем мерозоиты внедряются в здоровые эритроциты * (3) и в них про-
делывают такой же цикл бесполого размножения (шизогония).
Наряду с дальнейшим течением шизогонии наблюдается превраще-
ние некоторых мерозоитов в незрелые половые формы, называемые
гаметоцитами; различают мужские незрелые особи — м и к р о га-
метоциты (9Ь—12b) и женские — макрогаметоциты (9а—
12а). Появление первых гаметоцитов наблюдается даже во время пер-
вого припадка малярии, но, как правило, число их возрастает по мере
дальнейшего течения малярии.
Эту стадию развития плазмодия можно охарактеризовать как подго-
товку к половому размножению; оно протекает в теле основного хозяи-
на малярийного плазмодия, т. е. комара рода Anopheles, если самка
комара насосется крови больного с половыми формами паразита и если
условия внешней среды (в частности, температура) будут благоприят-
ствовать размножению плазмодия в организме переносчика.
В желудке Anopheles (рис. 84) гаметоциты созревают (см. ниже
стр. 171) и превращаются в макро- (13а—14а) и микрогаметы
(13b—15b), которые соединяются попарно и образуют оокине-
ты (17). Последние проникают через стенку желудка (18) на его на-
ружную поверхность (19) и здесь превращаются в ооцисты (20),
содержимое которых делится много раз подряд.и образует множество
спорозоитов веретеновидной формы (21—24). Параллельно про-
исходит увеличение ооцисты, которая в конце концов лопается (25) и
освобождает спорозоиты (26). Последние с током гемолимфы разно-
сятся по телу анофелес, внедряются в его слюнные железы и прони-
кают в слюнной канал железы (27). При сосании таким комаром крови
человека спорозоиты (1) впрыскиваются со слюной анофелес в кровь
человека, чем и определяется заражение последнего малярийными
паразитами и, как правило, последующее заболевание малярией.
Схема размножения кокцидий и малярийных паразитов
и их отношений к своим хозяевам
Название паразитов
Кокцидии
Малярийные паразиты
Название хозяев
Млекопитающее (яв- г Человек (первый Анофелес (второй
ляется единственным хозяин) хозяин)
хозяином)
•Формы размножения
я последовательность
заражения хозяев
бесполое->половое
__________Ф
4
бесполое-»половое
бесполое — -»половое
бесполое-»половое
и т. д.
бесполое --»половое
________________Ф
Ф
бесполое— -»половое
Ф
и т. д.
Общая схема жизненного цикла малярийных паразитов сходна с
тем, что мы видели раньше у кокцидий. Однако имеется и существенное
отличие. Рассмотренные нами кокцидии семейства Eimeriidae пределы-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
151
вают бесполое и половое размножение п одном и том же органе одного
и того же хозяина,1 тогда как перемена поколений малярийных парази-
тов связана с переменой хозяев, причем в том хозяине, где происходит
половое размножение, паразиты мигрируют из одних органов в другие,
достигая в конце концов места естественного выхода наружу, в част-
ности слюнных желез комара.
В настоящее время роль малярийного плазмодия как возбудителя
перемежающейся лихорадки не подлежит никакому сомнению, и иссле-
дование крови больных, подозрительных в отношении малярии, сдела-
лось обычным методом диагностики.
Перемежающаяся лихорадка характеризуется правильно чередую*
щимися приступами высокой температуры, отделенными друг от друга
безлихорадочными днями. В свя^зи с длительностью последних разли-
чают три формы малярии:
1) Ежедневная лихорадка (febris quotidiana) — когда при-
ступы лихорадки приходятся на одни и те же часы каждого дня; такая
форма болезни является сложной и зависит от комбинированного зара-
жения малярией (рис. 86),
2) Трехдневная лихорадка, когда приступы озноба и лихо-
радки повторяются через день (febris tertiana, рис. 85, Д); через день
же бывают и более неправильные приступы трехдневной летне-осенней
лихорадки, характеризующейся злокачественным течением (febris
tertiana maligna, f. tropica — тропическая малярия).
3) Четырехдневная лихорадка (febris quartana simp-
lex) — когда за каждым лихорадочным днем следуют два безлихора-
дочных (рис. 85). •
Естественная классификация форм малярии основывается на опре-
делении вида того кровепаразита, который причиняет соответствующее
заболевание. Зоологически различают 4 вида малярийного плазмодия
(рис. 85, 86):
1) Plasmodium vlvax и Pl. ovale, дающие трехдневную лихорадку —
febris tertiana — или терциану.
2) Plasmodium malariae — возбудитель четырехдневной лихорадки
f. quartana — или квартаны.
3) Plasmodium falciparum, вызывающий тропическую малярию —
f. tertiana maligna — или тропику.
Клинический диагноз осложняется нередким комбинированием раз-
личных форм малярии у данного больного, который может заразиться
несколькими генерациями одного и того же вида паразита или же за-
получает плазмодиев, относящихся к различным видам.
В связи с этим ежедневная лихорадка может вызываться различны-
ми причинами, а именно:
a) febris quotidiana — ежедневная летне-осенняя лихорадка
зависит от последовательного заражения больного двумя генерациями
Pl. falciparum;
б) febris tertiana duplex вызывается аналогичным зараже-
нием генерациями Plasmodium vivaxA
Схематически ход приступов можно изобразить так: первая инвазия,
например, 15, 16, 17, 18, 19 июня и вторая инвазия—16, 17, 18, 19 и
1 Представители некоторых других семейств кокцидий проделывают смену
хозяев в своем жизненном цикле.
152
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 85—Кривые температуры при различных формах малярии с показанием
в крови соответствующих стадий жизненного цикла малярийных паразитов.
Рис. 86 —Кривая температуры при двойном заражении малярией.
Таблица V
Рис. 1 — 13. Plasmodium vivax
/— внедрившийся в-эритроцит спорозоит (мерозоит); 2—4 шизонт (трофозоит); появле-
ние зернистости 111 юффнера; 5--7— начальные стадии шизогонии; 6’-10— конечные стадии
шизогонии; 11 - одновременная инвазия эритроцита шизонтом и делящейся формой;
12 — макрогаметоцит, 13 — микрогамегоцит;
Рис. 14—22. Plasmodium malarias
14-16— шизонты; 17—20— шизогония; 21 — макрогаметоцит; 22 — микрогаметоцит.
Рис. 23 — 30. Plasmodium falciparum
23—25—кольца; 2/, 26—множественная новация эритроцита; 27—28— шизогония,
29 — микрогаметоцит; 30 — макрогаметоцит
Рис. 31—39. Plasmodium ovale
31—37—вегетативные формы и шизогония; 38, 39 — гаметоциты; 40 — спороциста
малярийного плазмодия на желудке анофелес с цепочками пигментных зерен.
Из разных авторов
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
153
20 июня и т. д. (лихорадочные дни жирным шрифтом); в результате
двух последовательных инвазий лихорадка бывает каждый день;
в) febris quartana triplex — тоже ежедневная лихорадка,
но зависящая от последовательного заражения тремя генерациями
Pl. malariae,
В таком случае схема распределения лихорадочных дней принимает
другой вид, а именно:
первая инвазия 123
вторая инвазия 2 3
третья инвазия 3
4 5 6 7 мая и т. д.,
4 5 6 7 8 мая и т. д.,
4 5 6 7 8 9 мая и т. д.
Бывают также и неправильные формы малярии, например febris
quartana duplex, когда за двумя лихорадочными днями следует один
безлихорадочный день; дело объясняется заражением двумя последова-
тельными генерациями Pl. malariae, т. е.
первая инвазия 1 2 3 4 5 6 7 мая и т. д.,
вторая инвазия 2 3 4 5 6 7 8 мая и т. д.
Приведенными примерами не исчерпывается все возможное разно-
образие комбинированных заболеваний малярией. Разобраться в них
с точностью можно лишь при тщательном исследовании кровепарази-
тов, причем для успеха диагноза в запутанных случаях приходится
брать кровь больного через каждые два часа, чтобы таким образом
проследить последовательность развития малярийных плазмодиев.
Род Plasmodium
Plasmodium vivax Grass! et Felletti
Начальной стадией развития паразита трехдневной малярии в кро-
ви человека является мерозоит (или спорозоит), внедрившийся в эри-
троцит. После того как паразит малярии проник в красное кровяное
тельце, он начинает стадию развития, называемую шизонтом.
В течение первых четырех часов паразит, величиной 1,5—3 у, имеет
округлую или амебоидную форму, объясняемую подвижностью плазмо-
дия в живом состоянии. В протоплазме шизонта образуется централь-
ная вакуоль; он принимает поэтому кольцевидную или перстневидную'
форму; ядро, окруженное светлым ободком, лежит у края паразита-
(рис. 87, В, 1—2), В протоплазме последнего начинает отлагаться пиг-
мент в виде мелких зернышек, являющихся продуктом потребления:
гемоглобина эритроцита паразитирующим в нем малярийным плазмо-
дием. Зараженный эритроцит начинает увеличиваться в размерах, и в
его строме появляется характерная шюффнерова зернистость
или крапчатость (табл. V, рис. 3—5); она обнаруживается при силь-
ной окраске мазка крови краской Романовского в виде кирпичнокрас-
ных зернышек. Пораженный плазмодием эритроцит обесцвечивается*
что можно наблюдать в живой крови. В дальнейшем шизонт растет*
сохраняя кольцевидную форму, и достигает размера в половину диа-
метра эритроцита, который в свою очередь продолжает увеличиваться.
Протоплазма шизонта в это время окрашивается более густо. Коли-
чество отлагаемого пигмента возрастает. В начале вторых суток ваку-
оль постепенно исчезает: шизонт в живом состоянии энергично подви-
жен. Этот период высшего развития вегетативной деятельности шизон-
та характеризуется значительным усилением амебоидного движения.
Он принимает самые необыкновенные формы и ни на минуту не
154
СПЕЦИАЛЬНАЯ ЧАСТЬ
- остает(|р в покое. Пигмент распределяется по всему телу, длинные
.ложноножки выдвигаются во все стороны и снова втягиваются; боль-
Рис. 87 — Формы малярийных плазмодиев.
д — Тропическая малярия — Pl. falciparum. 1 — кольца; 2 — паразиты двенад-
цатичасового возраста из мазка мозга; 3— паразит суточного возраста из
«культуры» (форма, обычно встречающаяся в капиллярах внутренних органов);
4 — шизогония селезенке); 5— шизогония (мазок мозга случая, кончившегося
смертью); 6 — микрогаметоцит с остатком красного кровяного тельца; 7— макро-
гаметоцит (полулуние); 8 — образование бичей (микрогамет) из микрогаметоцита.
В — Трехдневная малярия — Plasmodium vivax. 1 — молодая кольцевидная фор-
ма; 2— амебоидная форма, в эритроците — шюффнерова зернистость; 3—
амебоидная форма, начало образования пигмента и увеличения эритроцита; 4 —
шизонт в начальной стадии деления ядер; 5—закончившаяся шизогония, эри-
троцит еще не разрушен; 6 — микрогаметоцит; 7 — макрогаметоцит; 8—
двойная инфекция гаметоцитом и шизонтом.
С — Четырехдневная малярия — Pl. malariae. 1 — молодое кольцо; 2-3— рост
паразита; 4-5— деление ядер шизонта; 6 — шизогония с образованием меро-
зоитов до распада эритроцита: 7 — лентовидная форма молодого паразита; 8 —
микрогаметоцит; 9 — микрогаметоцит. По Manson — Bahr.
чаше вакуоли появляются и исчезают, глубокие выемки врезываются 1
периферию, тотчас же заполняемые безостановочным движением про
топлазмы. «Одним словом, это живой организм, необычайно изменчи
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
155
вое и приковывающее к себе внимание наблюдателя зрелище и остав-
ляет впечатление, что он заслуживает свое имя «vivax» (Шаудинн).
Протоплазма шизонта окрашивается' краской Романовского в сине-
ватый цвет, а ядро—в красный (табл. V, рис. 1 —11).
Ядро его несколько увеличивается, размеры эритроцита продолжают
возрастать до Р/з—Р/2 нормальной величины (рис. 87, В, 3). Во вторую
половину вторых суток шизонт, теряющий подвижность, заполняет до
двух третей диаметр эритроцита. Приблизительно через 40 часов жиз-
ни шизонта ядро его начинает повторно делиться с образованием
нескольких дочерних ядер (рис. 87, В, 4). Пигмент скучивается в
центре паразита, который к концу вторых суток разделяется на 12—24
дочерних мерозоита (шизогония, рис. 87, В, 5). Одновременно происхо-
дит разрушение зараженного эритроцита с освобождением в плазму
крови образовавшегося поколения мерозоитов и кучки пигментных зе-
рен. Пигмент впоследствии фагоцитируется эндотелиальными клетками
и моноцитами хозяина. Итак, весь цикл развития Pl. vivax от одного
деления до другого длится 48 часов.
Plasmodium malariae Lave ran
Молодые шизонты паразита четырехдневкой лихорадки сходны с
'соответствующими стадиями развития Pl. vivax, но мало подвижны.
В течение первых суток развития различия между ними нет (рис. 87,
G, /). Позднее указанного срока паразит Pl. malariae вытягивается в
длину и часто в форме ленты как бы опоясывает эритроцит, теряя свою
слабую подвижность (рис. 87, С, 7). Первоначально узкая лента в те-
чение следующих суток развития утолщается в два или три раза и пиг-
ментируется, занимая собой почти 3Л кровяного тельца. Это последнее
в размерах не увеличивается и не бледнеет.
В последние сутки развития шизонт теряет лентообразную форму, и
по краям его образуются вырезки, причем разрушенный пигмент эри-
троцита скопляется в центральную кучку. За двенадцать часов до деле-
ния паразита шизонт заполняет собой весь эритроцит (рис. 87, С, 4—5).
Шизонт делится на 8 (реже 6—12) мерозоитов (рис. 87, С, 6), которые
располагаются вокруг центральной кучки пигмента с известной пра-
вильностью, благодаря чему эту стадию развития сравнивают с марга-
риткой.
Полный цикл развития Р/. malariae завершается в крови человека в
72 часа.
Plasmodium falciparum Welch
Некоторыми протозоологами паразиты тропики выделяются в род
Laverania и относятся к виду Laverania malariae Grassi й Felletti. Дру-
гим синонимом является наименование Pl. immaculatum Grassi и
Felletti.
Плазмодий тропической малярии обладает важной биологической
особенностью: он только часть своего развития проделывает в крови
периферических сосудов, так как самая шизогония в громадном боль-
шинстве случаев происходит в капиллярах таких внутренних органов,
как печень, селезенка.
Типичной формой паразита, находимого на обычных мазках крови,
являются мелкие тонкоконтурные и резко очерченные кольца- с зерни-
156
СПЕЦИАЛЬНАЯ ЧАСТЬ
стым утолщением (ядром), сбоку (рис. 87, А, 1). Эти мелкие кольца
плазмодия достигают всего только 1/в диаметра эритроцита. Их можно
найти на высоте лихорадочного приступа. С течением времени кольца
растут и увеличиваются почти вдвое; нередко они бывают двуядерными
и серповидными.
К началу безлихорадочного периода кольца плазмодия разрастаются
до 1/з диаметра эритроцита, сохраняющего все время свои нормальные
размеры и окраску. Эти «крупные кольца» Pl, falciparum, будучи взяты
сами по себе, не отличимы от колец других плазмодиев малярии.
К концу безлихорадочного периода они исчезают из, периферической
крови.
В окрашенных по Романовскому пораженных паразитами эритроци-
тах отмечается яркая мауреровская пятнистость, образуемая нескольки-
ми крупными красными пятнами. Согласно данным 'Мошковского, ее
можно трактовать как артефакт, связанный с процедурой приготовле-
ния препарата.
Дальнейшее развитие Pl. falciparum сосредоточивается в глубине
тела. Готовящиеся к делению паразиты имеют вид неправильных дисков
(рис. 87, А, 2, 5) с комочками грубого темнокоричневого пигмента. По-
следний скопляется в центре плазмодия, распадающегося на 10—20
мерозоитов, более мелких, чем у других малярийных паразитов (рис. 87,
А, 4, 5). Собирающийся делиться шизонт Pl. falciparum почти вдвое
меньше аналогичной стадии развития Pl. malariae,
В редких случаях при тяжелой тропической малярии шизогония мо-
жет происходить в периферических сосудах, и тогда стадии деления
обнаруживаются; и на мазках крови периферических сосудов.
Полный цикл развития Pl, falciparum длится от 24 до 48 часов.
Сравнительная таблица малярийных плазмодиев
Plasmodium vivax Plasmodium malariae Pl, falciparum
Топогра- фия па- разита В периферической крови все стадии беспо- лого размножения В периферической кро- ви только кольцевые формы. Шизогония — в капиллярах мозга, пе- чени, селезенке
Шизого- ния 1 Молодые шизонты в виде кольца, равного по величине г/3 диаметра эритроцита,оживленно подвижны —>- Кольца очень малы — ’/б-’Л Диаметра эритро- цита, оживленно под- вижны. Только они и встречаются (из форм, шизогонии) в перифе- рической крови
Взрослые шизонты амебоидно подвижны, неправильной округлой формы. Пигмент в ви- де одной или несколь- ких кучек. Шизонт до- стигает величины Р/г нормального эритро- цита Шизонты неподвиж- ны, округлой формы, не достигают величины нормального эритро- цита -> Встречаются в пери- ферической крови лишь в редких случаях; в об- щем, более или менее сходны с Pl. malariae
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
157
Plasmodium vivax Plasmodium malariae falciparum
Эритро- цит Шизонт делится на 12—24 мерозоита не- правильной формы Мерозоитов чаще 8, реже 12, деление более правильное Шизогония обычно в капиллярах внутрен- них органов, редко в периферической крови; мерозоитов 10—20, го- раздо меньших по ве- личине, чем у других малярийных паразитов. Деление неправильное
Пораженный парази- том эритроцит увели- чивается раза в 1^6 и обесцвечивается Пораженные паразиго чиваются и не обесцвеч м эритроциты не увели- иваются
Длитель- ность цикла развития 48 часов 72 часа 24—48 часов
Гаметы Взрослые гаметы крупнее эритроцита, не- правильной дисковид- ной формы. Пигмент коричневый, тонкозер- нистый Величиной с эритро- цит, неправильной дис- ковидной формы. Пиг- мент темнокоричневый, более грубозернистый Гаметы в виде прямых или изогнутых «полу- луний». Мужская гаме- та с рассеянным пигмен- том, женская с пигмен- том посредине
Малярийные плазмодии можно культивировать по методу Басса и
Джонса. К крови, взятой от малярика, прибавляют 50% раствор дек-
строзы, смесь дефибринисуют и, разлив по пробиркам, ставят в термо-
стат при 40° С. В таких культурах наблюдается шизогония. Проникают
ли мерозоиты в незараженные эритроциты — этот вопрос остается
спорным.
Plasmodium ovale Stephens
Pl. ovale, описанный в 1922 г. у больного из Восточной Африки
(Stephens), имеет более основательные данные считаться самостоятель-
ным видом малярийного плазмодия из числа всех новых видов или раз-
новидностей малярийных паразитов, описания которых опубликованы
различными исследователями. Он обладает как бы синтетическими при-
знаками по отношению к основным видам малярийных паразитов и не-
которыми свойственными ему особенностями.
По общему впечатлению на мазках крови, окрашенных по Романов-
скому, Pl. ovale напоминает крупные экземпляры Pl. malariae в эритро-
цитах, имеющих изменения, сходные с теми, что наблюдаются при
Pl. vivax.
Кольцевидные паразиты лежат часто в увеличенных эритроцитах с
зернистостью, похожей на зернистость Шюффнера (зернистость
Джемса). Пораженные эритроциты бледнеют.
В дальнейшем шизонт принимает неправильную форму, теряет ва-
куоль и удлиняется, часто подобно лентовидной форме Pl. malariae.
Пигмент обильный, черно-бурого цвета. Эритроциты с Pl. ovale часто
принимают овальную, угловатую или фестончатую форму.
Pl. ovale в толстой капле крови не различим от Pl. vivax.
Деление богатого хроматином ядра . паразита начинается, когда
158
СПЕЦИАЛЬНАЯ ЧАСТЬ
плазмодий занимает еще не более половины пораженного эритроцита.
Пигмент скудный. Близкий к разделению на уже готовые мерозоиты,
шизонт занимает часто весь эритроцит с желтоватым или желтовато-
бурым пигментом в центре. Мерозоитов образуется 8—10, реже—12.
Ядра их крупные и сами мерозоиты раза в два больше, чем у Pl. vivax.
Цикл шизогонии, как и у Pl. vivax, длится 48 часов.
Гаметоциты трудно дифференцируются от аналогичных стадий
Pl. vivax и Pl. malariae.
Указанные особённости Pl. ovale сохраняет при пассажах на про-
грессивных паралитиках.
Наблюдавшийся в различных местах тропической Африки, на Филип-
пинских островах и в других местах, Pl. ovale отмечен и в СССР
(Эскин) у больного из-под Уфы (смешанная инфекция с Pl. vivax).
Проведено 5 пассажей через прогрессивных паралитиков. Приступы
купировались акрихином, у 7 больных наблюдалось отсутствие рециди-
вов в течение 6—9 месяцев после лечения L
Переносчиком Pl. ovale в условиях эксперимента может быть
An. maculipennis (см. II том).
Половые формы малярийного плазмодия
в крови человека
После нескольких приступов малярии энергия бесполого размноже-
ния плазмодия начинает падать, и наряду с обычными шизонтами в
крови обнаруживаются новые формы развития паразита, а именно — его
половые особи. В крови человека развиваются незрелые половые осо-
би — гаметоциты. Они образуются из мерозоитов, различных по счету
поколений малярийного плазмодия. Гаметоциты неспособны к амебовид-
ному движению. В сформированном состоянии онщ крупнее шизонтов,
готовящихся к размножению. Зерна пигмента в гаметоцитах имеют
палочкообразную форму, они крупнее и грубее, чем пигмент шизонтов.
Пигмент более равномерно рассеян по протоплазме гаметоцита, чем у
шизонта.
Гаметоциты характеризуются большей стойкостью к хинину, чем
прочие стадии малярийного плазмодия в крови человека. Зато они хо-
рошо поддаются действию плазмохина и его советского' аналога плаз-
моцида, обладающих гаметоцидными свойствами. Отмечается большая
чувствительность к плазмохину гамет паразитов тропической малярии.
Plasmodium vivax
Начальные стадии развития гаметоцитов довольно трудно отличимы
от мерозоитов, зато взрослые гаметоциты отличаются в крови хорошо.
Они характеризуются более правильной округлой формой, без псевдо-
подий, и большим количеством более грубого пигмента.
Макрогаметоциты — величиной с эритроцит или еще крупнее (до
12 X 16 р). Протоплазма красится довольно густо. Ядро маленькое, бо-
лее или менее компактное. Темнокоричневый пигмент в форме палочек
рассеян по всей клетке, включая довольно ясно очерченный бледный
участок протоплазмы вблизи ядра, который встречается, однако, не
всегда (рис. 87, В, 7). Пигмента в макрогаметоците скопляется: больше,
чем в шизонтах, и его зернышки гораздо крупнее.
1 В. Эскин. Случай естественного заражения человека Pl. ovale в, СССР.
«Медицинская паразитология и паразитарные болезни», т. VII, вып. 1. 1938.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
15&-
Микрогаметоцит несколько меньше по величине. Плазма его красит-
ся бледнее, чем у макрогаметоцита. Ядро более рыхлое и диффузное.
Пигмент светлокоричневого цвета. Лишенное пигмента пятно не резко
ограничено. Оно лежит в средине паразита, занимая иногда до поло-
вины всей его поверхности. Зрелые гаметоциты не сохраняют следов
эритроцита, в котором они развились (рис. 87, В, 6).
Первые единичные гаметоциты при терциане появляются при четвер-
том или пятом цикле бесполого размножения, что соответствует девято-
му или десятому дню заражения.
Plasmodium malariae
Гаметоциты PL malariae очень похожи на половые особи PL vivax.
Единственное отличие — это разница в величине, именно, первые мень-
ше последних (6.8—8,5 аи против 12—16 ?), но позДнее и это различие
сглаживается (рис. 87, С, 8, 9). Пигмент их более груб.
Plasmodium falciparum
Первые гаметоциты PL falciparum появляются не ранее, как через
пять-семь дней после первого приступа тропической малярии, обыкно-
венно на 10—12-й день после начала болезни.
Гаметы плазмодия тропической малярии очень характерны в полу-
взрослом и во взрослом состояниях. Формирующаяся гамета имеет вна-
чале вид клина, лежащего у края эритроцита. Взрослая гамета полу-
лунной или, точнее, сосискообразной формы. По мере роста гамета
удлиняется, причем эритроцит становится овальным (рис. 87, А, 6).
Длина ее раза в полтора превышает диаметр нормального красного-
кровяного тельца. Пигмент скопляется в средней части гаметы, всякие
остатки эритроцита исчезают, и гамета в виде «полулуния» плавает
свободно в крови (рис. 87, А, 7).
Мужские и женские особи PL falciparum довольно трудно различимы
друг от друга. Можно отметить, что макрогаметоцит более длинен, узок
и богат пигментом, а микрогаметоцит — относительно короток, широк и
беден пигментом. Ядро первого более компактно, причем пигмент со-
средоточивается вокруг него, тогда как ядро микрогаметоцита более
рыхлое, и пигмент распределяется по протоплазме паразита.
Протоплазма микрогаметоцита красится по Романовскому—Гимза в
бледноголубой или зеленоватый цвет, а макрогаметоцита — в синий
цвет.
Длина гаметоцитов 9—14, а ширина 2—4 у.
Остатки эритроцита бывают видны в форме оболочки, имеющей в
физическом профиле как бы зазубренный вид.
Полулуния Pl. falciparum, равно как и шизонты, в стадии малых ко-
лец являются опорными формами при постановке диагноза тропической
малярии исследованием крови.
ОБЩЕЕ ТЕЧЕНИЕ МАЛЯРИИ И РЕЦИДИВЫ
Спорозоиты, попав со слюной малярийного комара в кровь человека,
превращаются в шизонты, которые размножаются последовательными
поколениями.
Заражение человека малярией в экспериментальных условиях удает-
ся при впрыскивании растертых в сыворотке слюнных желез анофелес
150
СПЕЦИАЛЬНАЯ ЧАСТЬ
со спорозоитами в кровь человека (James и др.). Аналогичный резуль-
тат получается при втирании спорозоитов паразитов птичьей малярии в
кожу птиц.
При инокуляции крови с паразитами тропической малярии кровь
привитого становится вирулентной в отношении последующего пассиро-
вания через 20 часов после инъекции, что приблизительно соответствует
длительности периода шизогонии Pl. falciparum. Если же инъицировать
внутривенно эмульсию слюнных желез анофелес с спорозоитами тро-
пики, то паразитов можно видеть в толстой капле, начиная с восьмого
дня после инъекции, клинические же проявления болезни наступают на
10-й день. В то же время кровь привитого становится заразительной
для человека при ее инъекции через 6 дней после инъекции споро-
зоитов. Очевидно, что шестидневный срок необходим для развития спо-
розоита до трофозоита (Чука, Баллиф и др.). С этими данными гармо-
нируют опыты с Pl. relictum птиц. Если на канарейке накормить экспе-
риментально инфицированных комаров, то ее кровь остается незарази-
тельной пои инъекциях последней в течение 65 часов; заражение
удается лишь после 3 дней инкубации; это дало основание Рафаэле
выдвинуть гипотезу о .прохождении спорозоитами внутриклеточной ста-
дии в ретикуло-эндотелии, после чего уже паразиты проникают в эри-
троциты, где и начинают уже известный цикл развития.
Не было недостатка и в морфологических исследованиях. Рафаэле
проследил процесс шизогонии Pl. elongatum в клетках ретикуло-эндоте-
лия канарейки, протекающий в дополнение к уже известной шизогонии
в эритроцитах. Аналогичные явления наблюдались также у кур при
заражении их Pl. gallinaceum (James и Tate, Brumpt и др.). Было уста-
новлено, что эндотелиальные стадии шизогонии часто наблюдаются в
мозгу.
Для всех стадий шизогонии, протекающих вне и независимо от
эритроцитов птицы, предложен индиферентный термин — экзоэри-
троцитная шизогония.
Положение дела с этой новой проблемой можно резюмировать выво-
дами из работы James и Tate (1938).
Экзоэритроцитные шизонты никогда не имеют в себе пигмента; это
и понятно, так как они паразитируют в клетках, лишенных гемоглоби-
на (моноциты, клетки ретикуло-эндотелия); эти стадии паразита отли-
чаются крупной величиной и образованием 50—60 и даже более меро-
зоитов; повидимому, в этом отношении играет роль и большая величи-
на клеток, являющихся местом обитания экзоэритроцитных шизонтов.
Последние же встречаются, как правило, в лейкоцитах русла крови, но
развиваются в эндотелии различных органов: мозга, легких, печени,
селезенки.
Мозг является особенно важным местом развития экзоэритроцитных
шизонтов; в нем экзоэритроцитная шизогония развертывается в первую
очередь, но не ранее появления обычных паразитов в периферической
крови. Чаще всего экзоэритроцитные стадии обосновываются в капил-
лярах мозга, что в ряде случаев ведет к их закупорке. Благодаря этому
у кур наблюдаются параличи и смерть в течение нескольких дней.
Интересно, что такой исход бывает также и у кур, вылеченных хинином
от «периферической инфекции» малярии. Этот факт указывает на то, что
экзоэритроцитные паразиты недоступны для влияния хозяина: они раз-
виваются у кур одинаково как при инъекции зараженной Pl. gallina-
ceum крови, так и при введении спорозоитов.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
По мере роста экзоэритроцитного шизонта ядро его делится; затем
разросшееся протоплазматическое тело паразита подразделяется на
цитомеры, по периферии которых образуются многочисленные пальце-
। 0.05 mm ।
0.05 mm.
Рис. 88 — Plasmodium gallinaceum. Экзоэритроцитная шизогония.
]— моноц т из мата легких с двумя молодыми плазмодиями; 2 — моноцит из
мазка и чени; в шизотге видны две х юм i1 и..ов je массы; 3 -4- даль е *шее деление
хроматиновых мисс в шизонге; 5—6 бо 1ьлие созревши.* ш 1зонты ч мазках легких;
7— труп'Ы мерозоитов из ра «руленгого шизонта; 8 — капилл < мочга, бликирочинный
тремя боль ними шиючтами (slf s,, s с, г.Ь.с. зритрохиг, п.е.с. — я <ро энотелиаль-
ной клетки}; 9— капилляр мозга, блокированный множествам мероз иточ, окружаю-
щих остатки материнской осо >и (т). Пэ S. James и P.Tate. .
видные выросты; в них вступают ядра, благодаря чему сформировыва-
ются мерозоиты. От клетки хозяина долгое время остается как бы на-
ружная капсула, переполненная множеством мерозоитов. Мерозоиты
состоят по .внешности как бы из одного хроматина (James и Tate).
Все авторы говорят, что экзоэритроцитная шизогония существует
наряду или добавочно к обычно протекающей эритроцитной шизогонии.
Эти новые вопросы требуют дальнейшего изучения, особенно в при-
менении к малярийным паразитахМ человека; необходимо также решить
вопрос: является ли экзоэритроцитная шизогония процессом, предше-
162
СПЕЦИАЛЬНАЯ ЧАСТЬ
ствующим обнаружению шизогонии в эритроцитах, или же эти процес-
сы идут независимо друг от друга, или в какой-либо корреляции один
за другим. Этот вопрос нуждается в дальнейшей разработке. Интересны
Рис. 89 — Plasmodium gallinaceum.
Внизу — часть среза селезенки с большим
шизонтом S в эндотелиальной клетке ве-
нозного синуса; т — скопление мерозоитов
вокруг остатков материнской особи; nhc—
дегенерирующее ядро эндотелия; р — маля-
рийный пигмент; spe — клетка селезенки. По
S. James и Р. Tate. Вверху — разрез
легкого. slt s4, s3 — 5g—большие шизонты
в клетках, прилежащих к альвеоле; nhc—
ядро клетки, пораженной плазмодием. По
S. J a m е s и Р. Т at е.
попытки обнаружения спорозоитов в
тканях человека. Boyd и Kitchen (1939)
на срезах биопсированных паховых
лимфатических желез, ближайших к
месту кормления спорозоитных ано-
фелес на бедре человека, видели
спорозоитов, которые лежали или в
соединительной ткани трабекул узла
или в околоузловой соединительной
ткани (рис. 90). Такая участь пости-
гает лишь часть спорозоитов, кото-
рые не попали прямо в кровь. Если,
нет каких-либо препятствующих об-
стоятельств (повышенная сопротив-
ляемость организма хозяина, наличие
в крови лекарственных веществ и др.),
то через известный период у чело-
века ^появляется первый приступ ма-
лярии. Длина такого срока инкуба-
ции вариирует для каждого вида
малярийного плазмодия. ПриР/. vivax
(терциана) инкубационный период
колеблется от 14 до 18 дней (более
редкими крайними сроками являются,
7—20 дней); при Р/. malariae (квар-
тана)— 18 — 21 день (15—25 дней) и
при Pl falciparum (тропика) —9 —
12 — 19 дней.
За инкубацией следует продро-
мальный период, длящийся неопреде-
ленное время от нескольких часов до
3 дней с нетипичными болезненными
проявлениями в форме чувства общей
разбитости, головной боли и др.
За ним следуют периодические
приступы малярии.
Затем наступают лихорадочные
приступы. Длительность приступа
лихорадки, включая и ее падение, ко-
леблется от 8 до 16 часов.
При тропической малярии приступ лихорадки удлйнен, и температура
падает не сразу и не равномерно. Поэтому настоящих безлихорадочных
периодов может вовсе не быть.
Однако накопилось большое количество случаев, которые можно
объяснить принятием длинного срока инкубации до нескольких месяцев
и даже до одного года. О таком длительном латентном периоде гово-
рят при случаях первичной малярии, падающих на раннюю весну или
на зиму, когда исключается ближайшая возможность естественного за-
ражения через анофелес. Подобный вывод можно сделать, только
исключив раннее нетипичное (или забытое больным) заболевание маля-
рией; в противном случае весеннее заболевание является уже не ре-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
163
зультатом первичной инвазии, а рецидивом. Необходимо также иметь в
виду возможность зимования спорозоитов малярийного плазмодия в
организме анофелес, благодаря чему первичное заражение может про-
исходить от таких комаров при благоприятной обстановке и зимой и
ранней весной.
Рис. 90 — Спорозоиты малярийного плазмодия в соеди-
нительной ткани около лимфатического узла человека.
По М. Boyd и S. Kitchen.
Длительную инкубацию называют иначе первичным
латентным периодом, он сменяется затишьем или вторичным
латентным периодом.
Размножение паразитов в, крови идет прогрессивно, но не все па-
разиты выживают, в противном случае кровь в короткий срок была бы
заполнена плазмодиями. Известно^ количество паразитов фагоцитирует-
ся белыми кровяными клетками; другая часть находится под влиянием
или погибает от гуморальных воздействий. Необходимо отметить, что
механическая пропорция между количеством паразитов и интенсивно-
стью заболевания не имеет общей силы потому, что вирулентность
самих плазмодиев и состояние организма малярика подвержены коле-
баниям. Этим и объясняются такие парадоксальные случаи, когда ма-
лярия протекает очень тяжело при скудном количестве паразитов в
периферическом русле крови и обратно: бывали примеры легкого те-
чения болезни при большом количестве паразитов.
Повидимому, малярийным плазмодиям свойственны некоторые се-
зонные изменения их особенностей. При заражении людей Pl. vivax в
процессе маляриотерапии через , уколы анофелес в зимнем квартале
Boyd, Kitchen и Muench наблюдали меньшее количество заражений с
более длительной инкубацией и более коротким клиническим течением;
рецидивы появляются в первые восемь недель после прекращения пер-
вичных приступов. В (летнем же квартале было много успешных зара-
164
СПЕЦИАЛЬНАЯ ЧАСТЬ
жений при наименьшей инкубации и Han6chneeJ длинном течении болез-
ни; наибольшее количество рецидивов наблюдалось через 24 недели
после первых приступов.
Неодинакова и индивидуальная восприимчивость людей к зара-
жению- малярией.
Вторичный латентный период связан с прекращением
шизогонального размножения, причем паразиты или вовсе исчезают из
периферической крови, или находятся в ней в виде гаметоцитов. Если
впрыскивать людям кровь маляриков, содержащую в себе только одни
гаметоциты, заражения малярией не произойдет; обратный эффект по-
лучится при впрыскивании крови с шизонтами.
Отсюда делается заключение, что гаметоциты не заразительны для
человека и поэтому не являются сами по себе болезнетворными
для него. Если характеризовать значение всех стадий развития маля-
рийного плазмодия в крови человека, то можно сказать, что возбуди-
телями экспериментальной малярии являются шизонты и мерозоиты, но
не гаметоциты.
Длительность вторичного латентного периода может быть весьма
значительна и исчисляться годами (2—4—6—10 лет). Человек может
забыть, что у него когда-то была малярия и вместе с тем может вне-
запно снова заболеть этой болезнью не за счет нового заражения (так
называемой реинвазии), а в порядке возврата ранее бывшего за-
болевания, т. е. рецидива.
По данным Шаудинна, рецидивы объясняли партеногенозом макро-
гаметоцитов, которые долгое время остаются в недеятельном состоя-
нии, а затем под влиянием различных инсультов (переохлаждение ор-
ганизма, циэтетическое воздействие, травмы, кровотечения, воздей-
ствие ультрафиолетовых лучей солнца и мн. др.) начинают размно-
жаться делением без предшествующего процесса слияния с микрога-
метоцитами. Возникающие таким путем дочерние особи внедряются в
эритроциты и в дальнейшем ведут себя как нормальные шизонты,
вызывая приступы малярии.
Картина такого девственного размножения, когда макрогаметоци-
ты будто бы дают некоторое количество мерозоитов и остаточное те-
ло, морфологически не отличима от так называемых двойных инва-
зий. Последние характеризуются внедрением в один и тот же эритро-
цит двух паразитов на разных стадиях их жизни, причем каждый па-
разит ведет независимо от другого «свою линию жизни» — например,
делится, когда другой находится на положении гаметоцита или ра-
стущего шизонта (табл. V, рис. И; рис. 87, В, S). Поэтому гипотеза дев-
ственного размножения макрогаметоцитов не имеет под собой базы, а
многие авторы объясняют рецидивы тем, что в организме в скрытом
состоянии сохранились шизонты, которые при подходящих к тому ус-
ловиях начинают снова размножаться со всеми вытекающими отсюда
последствиями.
Малярия может прекратиться у некоторых людей в естественном
порядке, т. е. без специального лечения. Такое выздоровление называет-
ся спонтанным. Абсолютным выздоровлением надо считать полное
исчезновение малярийных паразитов из организма человека. Нормаль-
ными сроками этого процесса считают 3—5 лет.
Рецидивы могут быть вызваны искусственно — рентгенизацией
селезенки, впрыскиванием адреналина и др. (способ провокации
малярии) для последующего уничтожения паразитов лечением.
От рецидивов следует отличать реинфекцию (точнее — р е и н-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
165
Рис. 91 —Увеличение
селезенки у малярика.
По Широкого-
рову.
вазию), или повторное (т. е. новое) заражение человека, который,
в силу тех или других причин вовсе освободился от первичной инфек-
ции или у которого она затихла; однако такое разделение на практике
не всегда возможно провести в смысле установления диференциальной
диагностики у больного.
Наконец, суперинфекцией (суперинвазией) называют
добавочное или последовательное заражение малярийным плазмодием
того же самого или другого вида паразита (например, суперинфекция
Pl. vivax на Pl. falciparum).
Одним из характерных проявлений малярии является увеличение
селезенки, край которой в норме не выходит за границы левых ребер.
Увеличение селезенки при хронической малярии
свидетельствует не только о зараженности чело-
века малярией, но и о сопротивляемости органи-
зма этой инфекции. Показательны примеры тяже-
лых и смертельных случаев малярии, при которых
селезенка сохраняла свои нормальные размеры.
Известны случаи увеличения селезенки до
6,4 кг весом.
При тропической малярии селезенка увеличи-
вается не в такой степени, как при терциане.
При общем учете распространения малярии
отмечается ее сезонность, проявляющаяся резким
поднятием заболеваемости в определенные сезо-
ны года. Случаи заболевания малярией могут
время От время наблюдаться круглый год, но все
же для малярии типична сезонность, связанная с
теплым периодом года. О ходе заболеваемости
малярии судят по обработке материалов учета
заболеваемости. Обычно учитывается валовая за-
болеваемость. Статистические данные оформляют
в виде месячной кривой, отражающей календар-
ное течение заболеваемости. Рассмотрение боль-
ших материалов из разных в широтном располо-
жении местностей СССР позволяет сгруппировать
малярийные кривые в две категории. Для ряда
местностей типична кривая, дающая максималь-
ный подъем в первую половину лета (рис, 92);
такой же ход заболеваемости характерен для се-
верной половины территории. В южной половине
кривая малярии дает второй — большой осенний
подъем (рис. 93). Наблюдаются и различные от-
клонения этих кривых. Весенний подъем заболе-
ваемости малярией обусловливается как рециди-
вами, так и первичной заболеваемостью с дли-
тельной инкубацией терцианой. Осенний подъем
зависит от свежих случаев терцианы, от реци-
дивов и первичных заболеваний тро шческой малярией. При раз-
дельном учете первичных заболеваний и рецидивов терцианы в южных
(местностях кривая заболеваемости малярией характеризуется также
двувершинностью. При учете заболеваемости необходимо отмечать
срок нахождения или время прибытия малярика в соответствующее
место, где он подвергается учету. Валовые статистические данные име-
ет часто относительное значение для малярийной характеристики места
166
СПЕЦИАЛЬНАЯ ЧАСТЬ
учета, отражая лишь фактическое наличие маляриков, часть которых
могла приехать в точку учета, заразившись малярией совсем в другом
месте (подробнее см. стр. 180, 181).
При регистрации маляриков необходимо точно различать первич-
ное заболевание и первичное посещение. Первич-
ны м im а л я р и ко м является человек, впервые^заболевший маля-
рией. Первично обратившимся считается всякий малярик,
Рис. 92 — Северный тип кривой маля-
рии (по заболеваемости в Ленинград-
ской области в 1934—1937 гг., в процен-
тах к годовой). По А. Якушевой.
Рис. 93 — Южный тип кривой малярии
(по заболеваемости в Дагестанской
АССР в 1931, 1933 гг., в процентах к
годовой). По А. Якушевой.
который впервые обращается в учетный год в лечебное учреждение
из-за малярии. Первично обратившимися могут быть первичные маляри-
ки, малярики-рецидивисты и малярики с реинфекцией. Рецидивирующие
малярики являются недолеченными хрониками; наличие их в данном
месте является показателем необходимости противорецидивной профи-
лактики (см. стр. 196). Точное установление первичных для данной мест-
ности заболеваний малярией говорит о том, что эпидемическая вспыш-
ка малярии не изжита, что диктует применение специальных мероприя-
тий. Сравнение малярийных кривых по годам дает представление о росте
или затухании эпидемичности малярии в месте ее учета (рис. 94, 95).
Кривые малярии строятся по абсолютному числу маляриков, или по
расчету на 10 тыс. населения, или по индексам.
Для характеризования зараженности населения и малярийности
местности определяют некоторые индексы, а' именно:
1. Паразитный индекс — процент паразитоносителей среди
населения, обследованного микроскопированием окрашенных толстых
капель; особое значение имеет определение процента гаметоносителей.
ибо только эта категория маляриков заражает анофелес и, следова-
тельно, является источником дальнейшего распространения малярии
в районе.
2. Селезеночный индекс — процент лиц с увеличенной се-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
167
Рис. 94 — Двувершинная кривая заболе-
ваемости малярией ^Куйбышевская обл.
1932—1934 гг., в процентах к годовой).
По А. Якушевой.
лезенкой (рис. 96) по отношению к осмотренному населению; этот ин-
декс является показателем распространения или экстенсивности маля-
рии в обследуемом районе.
Так как дети прежде всего заболевают малярией и реагируют
на эту болезнь резче, чем взрослые, у которых может вырабатываться
известная степень относитель-
ной невосприимчивости (стр. 180),
то определение селезеночного
индекса по возрастам имеет су-
щественное значение; количе-
ственные показатели селезеноч-
ного индекса днот более точную
картину, если исходить из опре-
деления его у детского населе-
ния; маляриологи в Индии счи-
тают, что соответствующий ин-
декс менее 1Оо/о характеризует
местность как здоровую; 10 —
25% соответствуют умеренной
малярийности; 25 — 50% свиде-
тельствуют о сильной малярий-
ности и, наконец, индекс выше
50% говорит о весьма тяжелой
малярийности.
3. Индекс Росса иллю-
стрирует среднюю степень увеличения селезенки у обследованного
населения, или интенсивность малярии. При осмотре всех обследуемых
делят на четыре категории:
а — с нормальной по размерам селезенкой;
Ь — с селезенкой, выступающей из-под края ребер на 3 пальца;
с — с селезенкой, выступающей из-под края ребер на 4—5 пальцев;
d —с селезенкой, выступающей из-под края ребер более чем на
5 пальцев.
Индекс вычисляется по формуле:
а -I- 3/? —6с —I— 9^
“ а b с d
Под а, Ь, с, d подставляют численное обозначение соответствую-
щей группы обследованных; при отсутствии лиц с увеличенной селе-
зенкой индекс Росса равняется единице; повышение его до 5 и более
свидетельствует о весьма тяжелом поражении населения малярией.
Для определения увеличения селезенки удобно пользоваться способом
С. Канделаки.
Индекс Росса, выраженный в механистической форме, не всегда то-
чен, ибо селезенка при тропике увеличивается в меньшей степени, чем
три терциане, что в известной мере уменьшает значение этого вида.
Для наибольшего уточнения характеристики малярийности опреде-
ляют эндемический индекс, под которым подразумевается
общий процент лиц с объективными признаками малярии (наличие
только паразитов в крови; наличие только увеличения селезенки и од-
новременное наличие обоих признаков).
Паразитный и селезеночный индексы, каждый в отдельности, не
168
СПЕЦИАЛЬНАЯ ЧАСТЬ
гарантируют уловления всех маляриков среди обследуемых, так как
Рис. 95 — Движение малярии по Волхово-Ильменю за 1932 — 1934 гг.
По Г. Шубу и В. Федорову.
бывают люди с увеличенной селезенкой, но без паразитов в крови в
момент исследования. Необходимо также помнить, что увеличение
селезенки бывает и не у малярийных больных (например, кала-
азар и др.).
W ® S3 О
W 2.7 3.V17 В Ш. 1907
Рис. 96 — Колебания в величине селезенки. (Пастеровский
институт в Алжире).
Необходимо отметить, что «одномоментное» обследование населения
на малярийность не дает исчерпывающе точной картины, так как
малярийные паразиты могут временами (или на более или менее дол-
гий срок) исчезать из крови; селезенка также может значительно
колебаться в размерах у одного и того же человека (влияние лечения
и другие причины; рис. 96).
Наконец необходимо помнить, что зараженность обследованного на-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
169
селения малярией не всегда стоит в полном соответствии с малярийно-
стью местности, так как могут быть приезжие малярики, которые за-
разились малярией не в месте их обследования. Поэтому важно дифе~
ренцировать коренное население от пришлого и среди коренного насе-
ления обследовать особенно тщательно детей, как элемент наиболее
оседлого характера, могущий, следовательно, наиболее точно харак-
теризовать малярийность пункта работ.
Рис. 97 — Падение индексов обследования на малярию, как показатель оздо-
ровления малярийного очага (с. Хамидие Кабардинской АССР).
1 — эндемический индекс; 2 — селезеночный индекс; 3 — паразитарный индекс.
По М. Рашиной и А. Хованской.
Падение индексов при повторных обследованиях на малярию одной
и той же точки или района является показателем оздоровления маля-
рийного очага (рис. 97).
ИСТОЧНИКИ ЗАРАЖЕНИЯ МАЛЯРИЕЙ
Заражение от человека. В естественных условиях человек
может заразиться малярией от человека в порядке внутриутроб-
ной инвазии: от матери, больной малярией, плазмодии могут прой-
ти сквозь плаценту и через сосуды пупочного канатика попасть «в плод;
такое прохождение зараженных эритроцитов от матери в плод мысли-
мо при различных поражениях плаценты (например, сифилис), которая
теряет вследствие этого свои барьерные свойства. Еще легче может
произойти смешение зараженной крови матери с кровью плода при от-
торжении плаценты. Наконец, мыслимо заражение во время самих
родов при попадании крови , матери в перерезанную пуповину или сса-
дины кожи плода.
Указанные способы заражения малярией называются к о н г е ни-
т а л ь н ы м и, а самая форма малярии врожденной (ложная на-
следственность).
Искусственное заражение людей малярией легко достигается впры-
скиванием крови малярика с шизонтами человеку, причем для успеха
дела бывает достаточно ничтожного ее количества (капля или даже
меньше). Искусственная или экспериментальная маля-
рия широко применяется для лечения прогрессивного паралича (Wag-
ner v. Jaurreg) (пиретотерапия). Бывают случайные заражения
малярией при инъекциях (например, анестезия при экстракции зубов.
170
СПЕЦИАЛЬНАЯ ЧАСТЬ
шприцем, которым ранее производилась инъекция в десны малярика и
‘без достаточной дезинфекции шприц был использован для здорового
от малярии человека и т. п.) При лечебной практике накопился побоч-
ный биологический материал, говорящий в пользу некоторого различия
течения и особенностей естественной и экспериментальной малярии.
Последняя, в частности, дает гораздо меньше рецидивов и с чрезвычай-
ной успешностью поддается лечению хинином. Инкубационный период
экспериментальной малярии при прививке крови малярика значитель-
но короче, чем при заражении крови через укол анофелес; для возбу-
дителя терцианы он в среднем равняется 10—12 дням против 17 дней
(при колебаниях от 3 до 31 дня); для паразита квартаны— 14 дней —
против 21 -(при колебаниях от 11 до 50 дней) и для тропики — 5 дней
против 10—12 (при колебаниях от 2 до 14 дней).
Для экспериментального заражения малярией необходима кровь с
шизонтами плазмодия; кровь только с гаметоцитами вовсе не зарази-
тельна. Количество вводимой крови не имеет существенного значения;
достаточно бывает даже следов крови в игле шприца, но практически
инъицируют 1—2 см3 дефибринированной крови.
Возможны случаи заражения малярией при переливании крови даже
тогда, когда у донора микроскопически паразиты не обнаруживаются
и когда в его анамнезе не значится явно перенесенной малярии.
Заражение через переносчика. В настоящее время точ-
но установленным положением является признание специфическим
переносчиком возбудителей различных форм малярии комаров рода
Anopheles, что экспериментально установлено Россом и Грасси. Заме-
чательно, что дикари Восточной Африки интуитивно связали представ-
ление о малярии с комарами, ибо и малярия, и комары ими называют-
ся одним и тем же словом «мбу» (Кох).
Кроме анофелес никаких других переносчиков малярийных парази-
тов не известно.
Когда Anopheles напивается крови малярика, то всасывает в себя
различные стадии развития паразита. Мерозоиты и другие стадии бес-
полого размножения в желудке комара погибают, тогда как гаметы
приступают к созреванию; из макрогаметоцита выталкивается часть
ядерного вещества с протоплазмой (редукционное тельце,
рис. 84, 14 а), причем он созревает в макрогамету или женскую особь
паразита. Микрогаметоциты подвергаются резко бросающимся в глазч
изменениям. Ядра их распадаются на несколько частей, подходящих к
поверхности плазмодия, который начинает выпускать как бы длинные
и тонкие псевдоподии или бичи .(рис. 84, 14 Ь; рис. 87, А, 8). Эти отро-
стки протоплазмы с ядром внутри отделяются от остаточного тела
микрогаметоцита в качестве зрелых мужских особей или микрогамет.
Они имеют вид бичей длиной 20—25 при толщине всего в 1 Из
каждого микрогаметоцита образуется от 4 до 8 подвижных змееобраз-
ных микрогамет и остаточное тело.
Интересно отметить, что микрогаметы могут образовываться из
микрогаметоцитов вне тела анофелес и вне русла крови человека; их
образование можно видеть под микроскопом в капле свежей крови под
влиянием ее охлаждения.
Их-то и заметил впервые Лаверан, при микроскопировании капли
крови малярика; по их колебательным движениям Лаверан дал откры-
тому им возбудителю малярии название OscUlaria malariae. Следует
отметить, что в связи с указанными выше особенностями полового цик-
ла кокцидий (стр. 144) в сходном порядке начинают толковать ход со-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
171
Рис. 98 — Желудок Anopheles
maculipennis со множеством
ооцист Plasmodium vivax.
ПоСиманину.
Размеры ооцист колеблются,
зревания гамет малярийного плазмодия, усматривая таковое в период
после слияния мужских и женских особей плазмодия (зиготиче-
ская редукция вместо предварительно протекающей г а м е т и-
ческой редукции).
Минут через 20 (и более — до 2 часов) после того, как кровь маля-
рика попала -в желудок Anopheles, происходит оплодотворение, причем
микрогамета, т. е. мужская особь, внедряется в макрогамету (в жен-
скую особь — рис. 84, 16). Оплодотво-
ренная макрогамета превращается в
зиготу или ооки-
нету (3—5 X 1°—24 размером); по-
следний термин в буквальном смысле
значит «подвижное яйцо» или, в данном
случае, подвижная женская оплодотво-
ренная особь (рис. 84, 17). Мужское и
женское ядро оокинеты сливают вместе.
Оокинета внедряется в стенку желудоч-
ного эпителия и проходит к его основа-
нию, т. е. к его базальной перепонке.
Оокинета одевается оболочкой и начи-
нает расти. По мере увеличения она по-
дается наружу, выпячивает мышечную
оболочку желудка и становится видной
с его поверхности (рис. 98).
Эта стадия развитий называется
ооцистой; внутри ооцисты ядра плаз-
модия энергично делятся. Вокруг ядра
сгущаются участки протоплазмы, а сами
ядра расходятся по периферии отдельных
частей обособленной протоплазмы, назы-
ваемых спор об ластами, в которых
развиваются веретеновидные спорозоиты.
В этом состоянии последние достигают
14 у* длины при толщине на уровне ядра
всего в 1 у-. Число их в каждой споро-
цисте может превышать 10 тыс. В конце
концов спорозоиты начинают плавать в
полости ооцисты, которая к тому време-
ни вздувается до максимальных пределов
в зависимости от их зрелости, от 4—5 р до 60 и редко 70—90 jx. Ко-
личество ооцист может достигать 200—500.
Ооцисты различных видов малярийного плазмодия отличаются по
характеру отлагающегося в них пигмента (рис. 99). Для ооцист Р1.
vivax характерно цепочечное расположение желтоватых или золоти-
стых тонких и коротких палочек пигмента. В ооцистах Pl. malariae
пигмент имеет вид грубоватых зернышек, которые еще грубее у Р1.
falciparum. Зато в ооцистах Pl. ovale пигмент имеет вид нежных пере-
крещивающихся черточек.
Утончающаяся стенка ооцисты под давлением внутреннего содержи-
мого лопается (рис. 84—24, 25). Спорозоиты выходят в полость тела
комара и с током полостной жидкости .(гемолимфы) распространяются
по всему его организму (рис. 84, 26). В груди они активно внедряются
172
СПЕЦИАЛЬНАЯ ЧАСТЬ
в слюнные железы, вероятно, привлекаемые химиотаксически, прони-
кают в слюнные клетки и сквозь них проходят в полость железы и в ее
выводной проток (рис. 100, 101).
А 8 С П
Рис. 99—Различие пигмента в ооцистах
малярийных плазмодиев.
А—тропической малярии; В — четирехднеаной малярии:
С — трехдневной малярии; D — птичьей малярии (Daniels).
Если теперь Anopheles со спорозоитами в слюне станет сосать
кровь здорового человека, то он впрыскивает ему возбудителей маля-
рии, т. е. спорозоиты. Последние, попав в кровь человека, внедряют-
ся в эритроциты, теряют в них веретеновидную форму и становятся
Рис. 100 — Продольный разрез спинки анофелес: в полости тела
в гемолимфе видно огромное количество веретеновидных споро-
зоитов малярийного плазмодия. По Мюленсу.
молодыми шизонтами, которые растут и переходят к бесполому раз-
множению. С этой частью их жизненного цикла мы уже ознакомились
выше.
Человек явственно заболевает перемежающейся лихорадкой после
инкубационного периода, , требующегося для превращения спорозоитов
в шизонты и £ля размножения этих последних, результаты чего нам
уже известны.
Таким образом замыкается полный жизненный цикл малярийного
плазмодия — цикл, слагающийся из двух стадий: 1) жизнь в крови че-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
173
ловека, характеризующаяся бесполым размножением плазмодия (ши-
зогония) и подготовкой к дальнейшему половому размножению образо-
ванием гаметоцитов, и 2) пребывание в различных органах анофелес,
связанное с половым размножением паразита
и споруляцией его с конечным возникновением
спорозоитов (спорогония).
Итак, оба хозяина малярийного па-
разита взаимно заражают друг друга
только вполне определенными ста-
диями развития паразита: Anopheles
инвазируется только гаметоцитами
из крови человека, а этот послед-
ний — спорозоитами из слюны ко-
мара.
Низкая температура неодинаково действует
на разные моменты жизни и на размножение
малярийных паразитов в теле Anopheles, Наи-
более уязвимы начальные стадии размножения
плазмодия. Низкая температура ^не позволяет
оокинете проникнуть через стенку желудка и
превратиться в ооцисту (Jancso). Если темпе-
ратура была достаточно высока, чтобы копу-
ляция гамет плазмодия могла произойти, то
дальнейшее развитие паразита протекает и при
более низкой температуре (Grass!); -f-5,5°C не
вредят уже сформировавшимся ооцистам (We-
nyon), что дает основание предполагать воз-
можность зимовки незрелых цист в комаре и
доразвитле их весной (Македония). J. James дал
еще более убедительные данные в пользу такой
возможности. В его многочисленных экспери-
ментах (Англия) ооцисты на желудке и спо-
розоиты в слюнных железах не погибали, если
комаров держали при -4-4—5,5°С три недели
или при температуре около точки замерзания—
6 дней. Некоторые комары An, maculipennis
жили со спорозоитами в слюне и сохраняли
свою заразительность от 29 до 92 дней после
появления в их слюнных железах спорозоитов.
Рис. 101 — Спорозоиты в
слюнной железе (на сре-
зе). По М ю л е н с у.
Несомненно, что понижение температуры задерживает развитие ооцист,
которое подвигается дальше, как только температура повышается
до благоприятных норм. Возможно также, что плазмодий может пе-
реживать в организме анофелес легкие зимы.
Ооцисты Pl. falciparum выдерживают температуру 1,7° С не более
суток, Pl. ovale заканчивает развитие ооцист в Art. maculipennis при
температуре 24—25° С в течение 16 дней против 10 дней, потребных в
тех же условиях как для Pl. vivax (James, Ncol, Shute). А. Лисовой
удалось заразить An. maculipennis и An. superpictus штаммом Pl. ovale
из-под Уфы и наблюдать появление спорозоитов. На 22-й день при
вскрытии были обнаружены крупные, непрозрачные, бедные мелкозер-
нистым пигментом цисты без спорозоитов, мелкие прозрачные цисты и
(множество спорозоитов в слюнных железах. Однако попытки зараже-
ния пяти человек кормлением на них Anopheles,- зараженных Pl. ovale,
дали отрицательный результат.
174
СПЕЦИАЛЬНАЯ ЧАСТЬ
Сроки- развития малярийных плазмодиев в комаре
Anopheles (в сутках)
1. Anopheles maculipennis
Название паразитов Температура °C Авторы
15—17 17—20 18-25 20-22 22—24 25 28—30 39
15 11 Roubaud.
Plasmodium f 53 2 16 1 Wenyon Jancso
vivax 15-16 10 или более 9 7 Grassi James
19 20 7 Grassi
Pl, falciparum { 20 14 Roubaud.
2. Anopheles superpictus
2: 1.5—23.7° 21—29.7°
PI. vivax Pl. falciparum 17 . 15 Wenyon.
Более детальные данные по длительности спорогонии
у Anoph. maculipennis (добыты Б. Николаевым)
Температура °C
Вид паразита 16-17 17-18 18-19 19-20 о -4 21-22 22-23 СЯ in 04 CD 04 04 00 04 о
Pl. vivax 45 32 26 22 19 16 13 11 10 9 8 7—6 5 5.5
Pl. falciparum . . — — — 26 — 20 16 14 12 11 10 9 8 8
Pl. malariae .... — — — — — — — 18 16 — — — — —
Ходукин с Лисовой и Симанин экспериментально доказали, что
An. elutus (Ташкент) и An. maculipennis (Барнаул), взятые с зимовок,
могут в тепле зимой заражаться малярийными плазмодиями. Поэтому
в известных условиях вполне возможно и зимнее первичное заражение
человека малярией.
Присутствие в слюнных железах комара подвижных спорозоитов не
обусловливает обязательности заражения малярией каждым таким ко-
маром любого • человека .(Bruce Mayne), в то же время Mitzmain
наблюдал заражение одним и тем же экземпляром Anopheles девяти
человек из двенадцати в 14 дней. Для заражения малярией может
быть достаточно только одного укола инфицированного анофелес.
Другие авторы указывают, что в большинстве случаев необходимо
повторное сосание крови человека анофелес и что для заражения че-
ловека требуется от 13 до 31 укола (Hanschell). ♦
Спорозоиты паразитов терцианы и тропики могут жить в слюнных
железах анофелес до 4 месяцев, но заразительность комаров сохра-
няется лишь в течение 2—3 месяцев; снижаться же она начинает уже 1 2
1 За это время при тех же условиях ооцисты Pl. falciparum еще недостаточно
созрели.
2 Эта температура, по Jancko, является нижним пределом развития плазмодия
тропической малярии в комаре.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
175
через месяц для Pl, vivax и, начиная с 20-го дня,— для PL falciparum,
считая этот срок со времени проникновения спорозоитов в слюнные
железы комара.
Влажность имеет меньшее значение для развития плазмодия, неже-
ли температура. Из других факторов некоторые побочные или случай-
ные обстоятельства также могут исключить возможность развития
плазмодия в желудке комара. Такое действие оказывают различные
кислые растительные соки, всасываемые самкой комара. Они созда-
ют неблагоприятную для плазмодия реакцию среды в желудке анофе-
лес, и еще Celli заметил, что в тех местностях Италии, где культиви-
руются томаты, малярия относительно редка. Питание анофелес
кровью животных и птиц может также делать их невосприимчивыми к
заражению плазмодием.
James находил не восприимчивых к заражению комаров в природе.
Их необходимо было выдерживать некоторый срок исключительно на
питании человеческой кровью, чтобы потом можно было бы с успехом
инвазировать плазмодием.
На заражаемость комара, кроме уже указанных моментов, влияют
количество выпитой им крови, относительное богатство ее гаметоцита-
ми, достаточное количество микрогаметоцитов, которых вообще разви-
вается меньше, чем макрогаметоцитов, и факт предварительного лече-
ния малярика хинином или синтетическими препаратами. Хинин губит
гаметоциты Pl. vivax и не действует на Pl. falciparum, поэтому анофе-
лес могут быть заражены плазмодиями тропической малярии на боль-
ных, принимавших хинин, тогда как при аналогичных условиях Pl. vi-
vax развивается в комаре редко (Wenyon).
Важным обстоятельством является факт потери микрогаметоцитами
способности созревать, т. е. эксфлягеллировать после приема
маляриком 0,03 синтетического хинолинового препарата плазмохина
(М. Bahr), что подтверждено многочисленными наблюдениями и опы-
тами. Такое гамостатическое действие препарата, пресекающее под-
готовку к половому размножению, ведет к тому, что анофелес, пью-
щие кровь таких маляриков, сами не заражаются малярийными пара-
зитами; это же в конечном результате при достаточно экстенсивном
применении плазмохина способствует значительному снижению заболе-
ваемости малярией в данной точке.
Плазмоцид (советский аналог плазмохина) обладает аналогичными
свойствами. Например, в Дагестане при лечении плазмоцидом 975 че-
ловек отмечено снижение индекса зараженности анофелес по сравне-
нию с участком, где производилась хининйзация, и за весь сезон гаме-
тоциты тропической малярии были найдены лишь у 6 человек (Пикуль,
Сергиев и Тибурская).
Однако указанное гамотропное действие плазмоцида имеет предел.
При дозе 0,12 в день в отношении Pl. vivax плазмоцид действует в те-
чение 27г суток после приема последней дозы. При дозе 0,09 действие
ее прекращается ко вторым суткам. При тропической малярии, лечен-
ной плазмоцидом (0,09 в течение одного дня) и акрихином (по 0,3 в
течение пяти дней), больной остается незаразительным для комаров
не менее шести дней после последнего приема плазмоцида (Тибурская).
Эти сроки важны для установления рациональных норм приема
лечебных препаратов (см. ниже инструкцию по применению синтетиче-
ских препаратов).
Возвращаясь к вопросу о связи богатства 'крови малярика гаме-
тами и заражаемости анофелес, пьющего его кровь, отметим, что
176
СПЕЦИАЛЬНАЯ ЧАСТЬ
An. maculipennis может быть заражен Pl. falciparum даже при на-
личии всего 10 гамет в 1 мм3 крови (Вещезеров). Число развиваю-
щихся ооцист не прямо пропорционально количеству проглатываемых
гамет, а меньше. Mitzmain наблюдал, что большее число их (Pl. fal-
ciparum) проходит через кишечник и выбрасывается с испражнениями
комара, и лишь малый процент развивается дальше в его желудке.
Наряду с вышеприведенными результатами Вещезерову вовсе не уда-
валось заразить Anovheles кровью человека с 372 гаметами Pl. mala-
riae в 1 мм3. Имеет, повидимому, значение и возраст гаметоцитов; по
Крэгу (Craig), они должны быть не моложе 12 дней. Повторность
кормления данного анофелес на данном гаметоносителе увеличивает
шансы заражения комара малярийными паразитами.
Паразиты квартаны труднее передаются через анофелес, что
повело к неоправдавшемуся предположению Маршу, будто бы этот
вид малярийного плазмодия имеет какого-то другого переносчика, а
не анофелес. Опыты передачи Pl. malariae давали положительный
результат с A. quadrimaculatus (США, М. Broyd и Stratman — Tho-
mas) и A. maculipennis airoparvus (de Buck).
Плазмодиями четырехдневной малярии удачно заражала в Сухуми
An. maculipennis, An. bifurcatus и An. nigripes Гендельман.
Попытки некоторых исследователей заразить Anopheles на боль-
ных, привитых кровью малярика, кончались неудачно. W. York и
W. Wright были счастливее в этом отношении. Они показали что 53—
54 пассажа Pl. vivax через организм людей в течение 31А лет ни-
сколько не ослабили способности плазмодия к гаметообразованию
и инфицирующей силы гаметоцитов. Анофелес заражался на послед-
нем больном и своим уколом заражал людей, никогда не болевших
малярией.
Заражение анофелес плазмодиями малярии, видимо, не влияет на
среднюю продолжительность его жизни. Однако, по показаниям дру-
гих авторов, Anopheles maculipennis, зараженные малярийными па-
разитами, летом быстро погибают; таким образом, до проявления
потребности повторного сосания крови человека доживает лишь не-
большое число самок, что уменьшает . шансы заражения людей
малярией (Голландия, de-Buck и A. Swellengrebel). Очевидно, что
длительность жизни зараженных анофелес в различных наблюдениях
зависела не только от самой зараженности их плазмодиями малярии,
но и от других факторов, совокупное действие которых ведет к раз-
ным результатам в отношении продолжительности жизни анофелес.
Не все виды анофелес являются в данном месте переносчиками
любой формы' малярийного паразита. Возможно, что разные виды
малярийных комаров переносят преимущественно разные виды
плазмодия (Wenyon). Главным носителем Pl. vivax в Македонии
Wenyon считает An. maculipennis, а для Pl. falciparum — An. super-
pictus. По Kinoshita,— Pl. falciparum на Формозе переносится Ап.
listoni и, в противоположность Pl. vivax, вовсе не развивается в Ап.
hyrcanus.
Эпидемиологическое значение разных видов анофелес устанав-
ливается как вышеизложенными опытами с искусственным зараже-
нием их плазмодиями малярии, так и обследованием естественной
зараженности плазмодиями в данной местности; для обнаружения
последней вскрывают множество самок, ищут в них ооцисты и спо-
розоиты (последнее — дело более трудное, чем первое) и определя-
ют процент зараженности комаров.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
177
Таким способом определяется плазмодийный индекс анофе-
лес, выводимый по отношению наличия ооцист на поверхности выпрепа-
рированного желудка (ооцистный индекс) и применительно к при-
сутствию спорозоитов в слюнных железах (спорозоитный ин-
декс). Учитывая большую техническую легкость массовых вскрытий
анофелес для определения зара-
женности их ооцистами, гораздо
чаще плазмодийный индекс опре-
деляют только в его ооцистной
части. Спорозоитный индекс
обычно меньше ооцистного, так
как не из всех цист развиваются
спорозоиты; но при несоответ-
ствии условий, необходимых для
развития ооцист и для пребыва-
ния спорозоитов в слюнных же-
лезах, отношение обоих индек-
сов может стать обратным (на-
пример, во второй половине сен-
тября при понижении темпера-
туры. Лисова и Эскин, рис. 102).
Годовой ооцистный индекс
для Сухуми у An. maculipennis,
взятых из жилых помещений,
достигает 7,6°/о, из скотников—
1,7, из подвалов и уборных —0.
анофелес взято из жилищ маляриков.
Рейнгард) при максимуме — на август.
А в
--------Спорозоитный индекс
Рис. 102 — Кривая спорозоитного и ооци-
стного ^индекса по месяцам в Той-Тюбе
(под Ташкентом). По А. Лисо'вой и
В. Эскину.
Наибольшее количество зараженных
(Л. Рейнгард и Забудько-
D
С
Рис. 103 — Дегенерация спорозоитов в слюнных железах.
А — An. superpictus (утолщение); В — С — (дегенерация); D — An. elutus (от нормы
до утолщения и укорочения); Е— An. maculipennis (дегенерация). По М. Barber.
В некоторых спорозоитах наблюдается несколько- глыбок хроматина,
что считается делением ядра спорозоита. Наряду с нормальными спо-
розоитами наблюдаются спорозоиты, необычно- утолщенные и дегенери-
рующие в различной степени (рис. 103). Этот процесс бывает у разных
видов анофелес. В одной .и той же слюнной железе могут быть споро-
зоиты и нормальные и дегенерированные в разной степени. Явление
дегенерации спорозоитов в отдельных наблюдениях свя
зывают с временем года (например, у A. mac. etutus летом дегенерация
наблюдается реже, чем зимой; это явление весьма часто наблюдается
178
СПЕЦИАЛЬНАЯ ЧАСТЬ
на Кипре), с возрастом спорозоитов и с другими причинами. Несомнен-
но, что дегенерирующие спорозоиты теряют заражающую способность;
это может вести при соответствующих условиях к ослаблению передачи
малярии в данной местности или к уменьшению •эпидемиологического'
значения вида анофелес, которому в сильной степени присущ рассма-
триваемый феномен .(Barber).
Swellengrebel с сотрудниками наблюдал дегенерацию спорозоитов
у A. maculipennis в Голландии во время зимования зараженных самок,,
начиная с ноября; в январе уже не оставалось нормальных спорозои-
тов. Ооцисты также дегенерируют, что начинается раньше дегенера-
ции спорозоитов.
Экспериментальное заражение разных видов анофелес плазмодиями
малярии в СССР производилось рядом исследователей. Весьма удачны
были опыты Вещезерова в районе Адлера с заражением An. maculi-
pennis и An. bifurcatus. Оба вида были резко восприимчивы к Pl. fal-
ciparum. Удавалось также заражение их и Pl. vivax. В первом случае
развитие плазмодиев было прослежено до внедрения спорозоитов в
слюнные железы анофелес. Ходукину в единичных случаях удалось
найти цисты на желудке An. pulcherrimus (Голодная степь) после за-
ражения комаров PI. vivax и Pl. malariae. Авербух удалось заразить,
до 40% этого вида комара, но естественное заражение его малярий-
ными паразитами в Туркестане пока обнаружено только нахождением
спорозоитов в слюнных железах комаров, взятых в августе из жилых
помещений одного из поселков Голодной степи (Завадский).
Симанин не мог заразить среднеазиатских An. hyrcanus плазмоди-
ем тропической малярии. 136 подопытных комаров дали отрицатель-
ный результат, тогда как из 41 комара видов An. maculipennis var.
sacharovi, An. pulcherrimus и An. superpictus заразились 34 комара,,
соответственно в следующих процентах: 90, 84 и 77.
Лисовой также не удалось заразить An. hyrcanus паразитами тро-
пической малярии; в другой серии опытов с Pl. vivax из 17 комаров
заразились три (Ташкент). В Биробиджане Гуцевичу удалось заразить
двух комаров из 26, также Pl. vivax, но на Дальнем Востоке и в Би-
робиджане, где имеется единственный вид A. hyrcanus, роль его в эпи-
демиологии малярии при настоящих условиях мала. Такое же мнение
высказывают о соответствующей роли этого вида анофелес в эпидемио-
логии малярии в Приазовских плавнях (Данилова и Будышко).
An. nigripes в условиях эксперимента заражается всеми тремя ви-
дами малярийного плазмодия (Гендельман).
Недавно найденный в фауне СССР An. algeriensis подвержен зара-
жению плазмодиями тропической малярии (А. Лисова).
Естественная зараженность анофелес плазмодиями — плазмодийный
индекс,— превышающий 10%, считается исключительной редкостью
(Wenyon). Зараженность анофелес колеблется по месяцам года, что
видно из нижеприводимых данных (см. табл, на стр. 179).
В Украинском протозойном институте из 560 вскрытых комаров
зараженных было два. М. Парцванидзе в Мингрелии нашел только
одного зараженного анофелес на тысячу. Топорков и Ефремов в
Астрахани нашли ооцисты у 7 An. maculipennis из 187 вскрытых са-
мок (август, сентябрь). Из 4490 An. maculipennis, вскрытых в течение
четырех лет в Днепропетровске (В. Горицкая), малярийными плазмо-
диями были заражены 33 комара. Важно подчеркнуть, что в жилых
строениях, хозяйственных постройках и в помещениях для скота наи-
больший процент зараженных анофелес был обнаружен в домах маля-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
179
риков (например, 4 зараженных анофелес на 52, и далее: 4 из 40, 4:
из 16). Первый зараженный анофелес был обнаружен 29 мая со зре-
лыми цистами; наиболее позднее нахождение комара с цистами и спо-
розоитами пришлось на 14 ноября. Отмечено также, что процент зара-
женных комаров, при прочих равных условиях, увеличивается в поме-
щениях со скученным населением, живущим в антисанитарной обста-
новке и не охраняемом от нападения комаров (Лисова и Эскин).
В конечном счете количество инфицирующих анофелес зависит в
каждом частном случае от количества комаров, пьющих кровь челове-
ка, количества гаметоносителей и количества анофелес, выживающих
до проникновения спорозоитов в слюнные железы анофелес. Пример
расчета количества зараженных анофелес возьмем из данных Данило-
ва и Будышко (плавни Приазовья). В станице Тривенской анофелес,
пьющих кровь человека, 22‘°/о, гаметоносителей — 3% и 16% заражен-
ных анофелес доживают до появления спорозоитов. Общий процент
зараженных комаров — 0,01 от всей комарийной популяции, т. е. 1
на 1000, что близко соответствует данным вскрытия анофелес.
Хотя зараженные плазмодиями комары и являются в общем ред-
костью, но каждый из них может повторно заразить несколько чело-
век. Даже после сосания 30—40 человек в слюнных железах анофелес
остаются многочисленные спорозоиты (James). Очевидно, эти органы
время от времени пополняются спорозоитами, выходящими из последо-
вательно созревающих, и лопающихся цист.
В общем эпидемиологическое значение различных анофелес в дан-
ной местности зависит, помимо наличия маляриков, от длинного ряда
факторов: количества гаметоносителей, известной степени антропофи-
лии комаров (см. главу о комарах), обстоятельств, благоприятствую-
щих половому размножению плазмодиев (температура, влажность, ре-
акция среды и химический состав содержимого пищеварительной
трубки анофелес), склонности данного вида анофелес заражаться
той или иной формой малярийного паразита, плотности анофелес, от
длительности жизни насосавшегося малярийной кровью комара (обсто-
ятельства, имеющего особо важное значение), от восприимчивости лю-
дей'к заражению малярией. Под индексом плотности анофе-
лес понимается среднее количество анофелес на сбор в той или дру-
гой постройке (хлев, дом, погреб и др.). Передача малярии анофелес
до известной степени пропорциональна плотности анофелес, конечно,
при осуществлении основного условия — литания анофелес кровью че-
ловека. Имеет также значение вид анофелес. Например, в Биробиджане,,
в Дальневосточном крае и в. Приморье единственным известным пока:
видом является An. hyrcanus. Местная заболеваемость малярией неве-
180
СПЕЦИАЛЬНАЯ ЧАСТЬ
лика, что зависит по линии переносчика от комбинации следующих об-
стоятельств: 1) слабая восприимчивость An. hyrcanus к заражению
плазмодиями малярии, 2) малое количество- перезимовывающих особей
и позднее начало их размножения, 3) краткость периода активной дея-
тельности большинства комаров, 4) относительно слабый контакт с че-
ловеком (А. Гуцевич).
Трудно сказать, существуют ли люди, абсолютно не восприимчивые
к заражению малярией, но несомненно, что некоторые весьма стойки в
указанном отношении. Несомненно также, что с возрастом коренное на-
селение малярийной местности становится более выносливым к малярии,
и хроническая инфекция протекает гораздо легче, чем у недавно при-
бывших людей. "Повидимому, вырабатывается относительный несте-
рильный (т. е. не связанный с полным исчезновением паразитов из
организма) иммунитет. Однако среди, казалось бы, невосприимчи-
вого коренного населения бывают сильные обострения малярии, когда
в данную местность вливаются незараженные пришельцы. Последние
заражаются малярией; вирулентность местных штаммов паразитов, ве-
роятно, усиливается при циркуляции их через свежих людей; благодаря
этому паразиты оказывают повышенное влияние' на коренное население,
которое добавочно заражается через комаров анофелес. Аналогичное
значение имеет приезд гаметоносителей из других местностей; при этом
происходит внесение нового штамма малярийных плазмодиев с боль-
шей вирулентностью для коренного населения, у которого уже вырабо-
талась известная невосприимчивость к местной малярии.
Наибольшее значение в распространении малярии в СССР играет
An. maculipennis} в Средней Азии и на Кавказе такую же роль играет
An. superpictus. Эпидемиологическое значение An. hyrcanus и An. pul-
cherrimus, невидимому, гораздо меньше. An. bifurcatus и An. plumbens,
в силу своих экологических особенностей, едва ли играют особо замет-
ную роль в распространении малярии, но в районном отношении значе-
ние их может быть вполне актуальным (санатории, стройки и др.). В на-
стоящее время изучается эпидемиологическое значение недавно обна-
руженного в СССР An. algeriensis (Средняя Азия, Кавказ), который, по
Sergent, является переносчиком малярии.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ МАЛЯРИИ
Географическое распространение малярии очень ши-
роко. При его выяснении необходимо условиться, что считать достаточ-
ным критерием для суждений о захвате малярией новых местностей.
Если мы будем судить лишь по появлению в таких местах больных с
«первичной малярией», то одного этого факта еще недостаточно для
утвердительного решения поставленной задачи. «Первичная» для боль-
ного малярия может быть не местного происхождения. Если такой
больной раньше жил в малярийной местности, то у него в прошлом
могли быть незаметно прошедшие приступы болезни, которая при пере-
езде больного на новое место дала новый рецидив; такой рецедив мо-
жет появиться спустя много лет после первого заболевания. Далее,
инкубация при малярии далеко не всегда ограничивается шаблонными
12—14 днями — период ее может затянуться с осени до весны. Поэто-
му при обнаружении весенней первичной малярии следует всегда точно
выяснить, когда и откуда приехал данный больной.
В обоих случаях может оказаться, что у обследуемых больных ма-
лярия не местная, т. е. она не вызвана уколами местных анофелес, а
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ рРОСТЕИШИХ Д81
привезена больными в самих себе из другого пункта. Следовательно,
при таких обстоятельствах мы можем говорить лишь о распростране-
нии больных малярией в новой для последней местности, являющейся
лишь субстратом для проявления в сущности уже готовой болезни.
Теоретически допустима возможность заражения и никогда не вы-
езжавшего из данной местности человека малярией не местного про-
исхождения. Пассивный транспорт анофелес может происходить на
большие расстояния, вследствие этого анофелес (со спорозоитами в
слюнных железах) в вагонах прямого сообщения может быть привезен
с Кавказа или из Туркестана, например, в Ленинград и здесь заразить
на вокзале людей, никогда не выезжавших из города. Такие случаи
первичной малярии тоже не могут быть поставлены на счет Ленин-
града.
Реальное распространение малярии по новьш для нее пунктам мо-
жет быть признано тогда, когда выплодившийся на местах анофелес
здесь же напивается крови малярика (хотя бы и приезжего), когда
экологические условия вполне благоприятствуют развитию спорозоитов
и появлению их в слюнных железах комара и когда, наконец, послед-
ний инвазирует плазмодием местное население (аутохтонная ма-
ляр и я). Только такие случаи малярии являются доподлинно первич-
ными для данной местности, и только они определяют ареал распро-
странения малярии в отношении наличия источников заражения этой
болезнью (Е. Павловский).
В эпидемиологическом отношении далеко не одно и то же такие
факты, когда у приезжих больных, уже с малярийными плазмодиями,
впервые появляется малярия в данной местности (без связи с мест-
ными анофелес) или когда больной действительно получает малярию в
той же местности от зараженного плазмодиями местного анофелес.
Эти соображения должны побуждать врачей к уточнению регистра-
ции «первичных» случаев малярии. При обнаружении таковых должна
быть принята в расчет и выяснена экологическая возможность появле-
ния того или иного случая малярии в данный срок применительно к
месту пребывания больного и к природным условиям за время, пред-
шествовавшее его заболеванию.
При таком подходе к определению границ ареала распространения
малярии оказывается, что севернее 60° с. ш. нет оснований предпола-
гать возможности полного созревания плазмодиев в анофелес, а следо-
вательно, и допускать наличие «первичной» малярии. В прилегающих к
60° с. ш. местностях б. Ленинградской, Вологодской, Вятской и Перм-
ской губ. заражение людей от первого поколения весеннего анофелес
теоретически мыслимо в благоприятные годы не ранее 1Q—20 августа,
а в неблагоприятные — около !—20 сентября, и то если допустить од-
новременное наличие всех оптимальных условий, благоприятствующих
как развитию плазмодия, так и самому заражению комара и человека.
В реальной обстановке природы и жизни северной полосы Европейской
части СССР таковые комбинации могут иметь место лишь в исключи-
тельно редких случаях, поэтому ко всем случаям появления «первич-
ной» малярии здесь следует относиться критически (Колодизнер).
Прямые наблюдения и эксперименты в Сольвычегодске (Архангель-
ской области), являющемся наиболее малярийной точкой северной по-
лосы РСФСР, показали, что анофелес, вылетающие из зимовок, не
играют сколько-нибудь существенной роли в передаче малярии. В пере-
зимовавших и зараженных кровью малярика An. maculipennis messeae
в условиях свойственного их дневкам микроклимата развития парЦзи-
132
СПЕЦИАЛЬНАЯ ЧАСТЬ
тов малярии не наблюдается; но при содержании комаров при +18—
25° С спорогония течет до конца. Анофелес первой генерации, зара-
женные 22—23 июня, дают спорозоиты к 4 июля. Заражение комаров
может происходить и до середины августа, но спорогония из-за похо-
лодания не успевает закончиться. Следовательно, период возможного
заражения людей от анофелес ограничивается одним месяцем (В. Ре-
менникова).
Впрочем, следует учитывать, что анофелес более или менее долгое
время проводят в жилье человека, где температура, конечно, выше
температуры определяемой на метеорологических станциях. Это обсто-
ятельство может способствовать развитию малярийного плазмодия в
теле анофелес и при неблагоприятной для этого процесса «внешней»
температуре.
В отношении продвижения малярии на восток отмечается своеоб-
разная закономерность, обусловливаемая сменой одного вида анофелес
другим. Там, где распространен только A. hyrcanus (Приморская, Ус-
сурийская, Хабаровская области и, вероятно, также Биробиджан), мест-
ная малярия слаба, что зависит от слабой восприимчивости A. hyrca-
nus к заражению плазмодиями, малым числом перезимовывающих осо-
бей и поздним началом размножения, непродолжительностью активной
деятельности большого числа A. hyrcanus и слабым контактом его с
человеком в силу экзофильного образа жизни (А. Гуцевич).
Восточной границей распространения малярии практически является
зона в районе Благовещенска, где оканчивается распространение Ап.
maculipennis и отмечается присутствие An. hyrcanus. При изменении
указанных выше условий роль A. hyrcanus в передаче малярии на
восток может возрасти.
В Сибири к северу распространение малярии уже, чем анофелес;
решающим фактором здесь являются климатические условия, неблаго-
приятствующие спорогонии плазмодия в анофелес.
СОЦИАЛЬНОЕ И ЭКОНОМИЧЕСКОЕ ЗНАЧЕНИЕ МАЛЯРИИ
Малярия является весьма распространенной болезнью, особенно
жаркого, теплого и прилежащих к ним стран умеренного пояса.
Ее 'значение для тропических стран ярко иллюстрируется данными
для Индии (Sinton, 1936). Заболеваемость малярией в этой стране
определяется колоссальной цифрой в 100 млн. человек; к данной цифре
необходимо добавить еще 25—75 млн. других заболеваний, развиваю-
щихся на почве малярии. Ежегодная смертность непосредственно от
малярии и от косвенных с ней связей поражает по крайней мере 2 млн.
человек. Дефицит рождаемости в одной только Бенгалии исчисляется
800 тыс. человек в год; на долю малярии следует отнести 37.8% абор-
тов. Половое бессилие, уменьшение числа зачатий, увеличение количе-
ства мертворождений также связываются с действием малярийной
инвазии. Всеми этими факторами определяется замедление прироста
населения. Малярия оказывает тяжелое влияние на физическое разви-
' тие детей и взрослых («б олотное худосочи е»). Потери за год —
1) от непосредственной смертности определяются суммой 67.5 млн. ф. ст.;
2) от понижения трудоспособности после малярии 22—55 млн. ф. ст.;
3) в заработной плате в связи с малярией—18 млн. ф. ст.; 4) на по-
гребальные расходы — 0.5 мл. ф. ст.; 5) стоимость медицинской помо-
щи — 15 млн. ф. ст.; 6) остаются также не поддающиеся сколько-ни-
будь, определенному учету потери в сельском хозяйстве (заброшенность
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
183
земель, плохая или пониженная обработка и мн. др.), в хозяйстве об-
щин, в индустриальном развитии и др.
Развитие малярии и ее последствия связаны с множеством причин
природного, социального, классового, экономического характера. Маля-
рийное значение суммы этих причин вариирует в разных местах, в
частности тропиков. Наряду с такой картиной малярийного бедствия,
которая для Индии иллюстрируется приведенными выше данными
{Sinton), имеются указания, что в некоторых, даже гиперэндеми-
ческих очагах Африки, малярия будто бы не влияет на население.
К статистическим данным последнего времени приходится относиться
(по крайней мере для некоторых зарубежных стран) с осторожностью,
ибо возможно преуменьшение данных в политических целях.
400
300
200
100
годы
1892 I 94 I 96 । 98 I 1900 I 02 I 04 I 06 08 I 1910 I 12 I U I 16 I 21 I 23 I 25 I 27
93 95 97 99 01 03 05 07 09 II 13 15 1920 22 24 26 28
Рис. 104 — Заболеваемость малярией в России (СССР) за 1892—1929 гг.
на 10 тыс. населения.
Во всяком случае общемировое значение малярии исключительно
велико.
В дореволюционное время в России малярия была самой распро-
страненной из всех заразных болезней, давая в год около 215 заболе-
ваний на 10 тыс. населения (рис. 104). Отсылая читателя за деталями
к специальным работам и статьям, отметим, что после революции
пандемию сыпного тифа сменило чрезвычайное развитие малярии, в
свою очередь принявшее характер пандемии. В 1923 г. санитарной
статистикой зарегистрировано 474 заболевания на 10 тыс. населения.
Валовое количество маляриков (с поправкой на неполноту статистики)
определялось около 12,5 млн. человек, с числом умерших от малярии
в 38—62 тыс. человек.
Заболеваемость малярией весьма неравномерно распределяется по
разным частям СССР для данного года, и, кроме того, распростране-
ние малярии вариирует в значительных пределах и по годам (рис.
105). Далее вариирует распространение различных форм малярии, в
частности тропической малярии, которая передвигалась из южных рай-
онов в среднюю полосу СССР. Малярия поражает все возрасты, при-
чем около 60% заболеваний приходятся на рабочий возраст. Если
принять, что больной рабочий теряет в среднем по десяти дней из-за
малярии, то на одних застрахованных в 1923 г. утеряно около 11 млн.
рабочих дней при общей потере по теоретическим подсчетам 75—112
млн. трудодней. Малярия поражает работников всех отраслей народ-
184
СПЕЦИАЛЬНАЯ ЧАСТЬ
ного хозяйства: сельское хозяйство, лесоразработки, торфоразработки,
рыбные промыслы, стройки железных и шоссейных дорог, гидротехни-
ческие работы, работы по созиданию электрических станций, железно-
дорожный и водный транспорт и мн. др. Ущерб от малярии слагается
из недопроизводства рабочими и населением ценностей за период бо-
Рис. 105 — Малярийные районы в СССР в 1913, 1923,
1926 и 1928 гг. с заболеваемостью от 400% и выше.
Из Эпштейна.
лезни, пониженной работоспособности после болезни или в промежут-
ках между рецидивами, потерь страхкассы по оплате временной нетру-
доспособности. К этому прибавляются возможные срывы производства
в тех или других его звеньях вследствие поголовного заболевания ма-
лярией или текучести рабочей силы, вызываемой той же причиной; на-
конец, содержание специальной противомалярийной организации, до-
полнительная зарплата и дополнительные отпуска в малярийных мест-
ностях являются новыми расходами, вызванными развитием малярии.
Военно-санитарное значение малярии весьма велико. Войны, как
правило, сопровождались сильнейшим развитием малярии в войсках.
В американскую гражданскую войну (1861 —1865) в некоторых
штатах по реке 'Миссисипи наблюдалась поголовная заболеваемость
войск. Во время русско-турецкой войны в русской армии заболело
610 591 человек, т. е. 247 на тысячу, и умерло 0,5 на тысячу, причем
малярия составляла до 7/8 всех заболеваний. При продвижении в Турк-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
185
мению (1880—1881 гг.) в русских войсках заболело от малярии 8411 че-
ловек. Еще более тягостная заболеваемость от малярии наблюдалась
при завоевании колоний и при колониальных войнах англичан, францу-
зов и др. в Африке и в Азии.
Мировая война, в свою очередь, способствовала значительному
росту малярии.
Рис. 106 — Заболеваемость малярией в царской и Красной армии
на 1000 человек.
В германской армии заболеваемость малярией на 100 тыс. человек
среднего состава составляла на 1-й год войны 35, на 2-й—132, на
3-й — 620 и на 4-й— 1370 человек; на балканском фронте заболевае-
мость эта достигала на 3-й год войны 9260 и на 4-й — 23 240 на 100
тыс. Еще большей рост заболеваемости был отмечен на турецком
фронте. I !
• Однако малярия имеет для армии столь существенное значение не
только в военное время, но и в мирное. Нередко различным частям
войск — гарнизоны крепостей, пограничные заставы и др.— приходится
стоять в крайне неблагоприятных по малярии пунктах.
Неудивительно поэтому, что некоторые места расквартирования
войск стяжали себе печальную славу гиблых именно потому, что
являлись очагами злостной малярии. В частности, фигурировавший в
народных песнях эпитет «погибельный» Кавказ объяснялся крайней
малярийностью многих его районов; в ту пору, когда малярия была
непобедима, правительству приходилось упразднять некоторые укреплен-
ные пункты, так как гарнизон в них регулярно вымирал от малярии.
К таким неблагополучным стоянкам относился, например, ныне пре-
красный курорт Гагры, где в прежнее время за три года гарнизон це-
ликом выходил из строя.
Заболеваемость в царской и Красной армиях малярией давала зна-
чительные колебания, представленные на рис. 106. В 1923 г. переболе-
ло 14.2% всего состава (142,0 на 1000),, что стояло в связи с общей
заболеваемостью населения и той специальной опасностью, которой
186 СПЕЦИАЛЬНАЯ ЧАСТЬ
подвергалась Красная армия по роду своей службы (необходимость
пребывания и операций в малярийных местностях).
В дальнейшем наблюдается большое снижение — до 28,9 на 1000 в
1929 г. Совершенствование медицинского и санитарно-гигиенического
обслуживания Красной армии уже теперь обеспечивает меньшую за-
болеваемость малярией красноармейцев, чем гражданского населения.
Инструкция по борьбе с малярией в Красной армии издана приложе-
нием к приказу начальника ВСУ РККА № 67 от 6 апреля 1931 г.
В дальнейшем она повторялась с необходимыми усовершенствованиями.
Огромная заболеваемость малярией после сыпнотифозной эпидемии
в 1920 г. вызвала в СССР появление сети противомалярийных учреж-
дений в форме малярийных или тропических станций, опирающихся на
республиканские тропические институты. г В 1920 т. был открыт Тропи-
ческий институт в ’Москве, а в следующем году возникли первые
семь малярийных станций; число их в 1923 г. достигало 71, а в 1929 г.
их было более 200. В настоящее время сеть малярийных станций ра-
стет еще шире.
Являясь основными ячейками в деле борьбы с малярией, станции
повели как лечебную, так и профилактическую работу. Результаты
противомалярийной борьбы сказались понижением в распространении
и в интенсивности малярии, что видно, например, из кривой на рис. 104,
которая уже к 1929 г. давала крутое падение заболеваемости с 474 до
183 на 10 тыс. населения.
По данным русских и иностранных эпидемиологов, развитию маля-
рийной эпидемии в СССР благоприятствовала совокупность следующих
обстоятельств:
1) небывалое передвижение людских масс в связи с блокадой, граж-
данской войной, беженством и мешечничеством, сопровождавшееся
заносом малярии через гаметоносителей в местности, до того благо-
получные по малярии;
2) особенное потепление в 1921 —1922 гг., благоприятствовавшее
размножению анофелес;
3) заболачивание огромных пространств вследствие разрушения
оросительных систем (Средняя Азия, Кавказ) и вследствие прекраще-
ния обработки почвы во многих местах;
4) большое уменьшение поголовья скота, отвлекающего, в качестве
источника питания, анофелес от человека;
5) полное отсутствие хинина во время гражданской войны вслед-
ствие блокады и необычайно возросшей потребности в нем, в связи с
накоплением огромного количества гаметоносителей;
6) усиление восприимчивости населения, у которого голод ослабил
иммунитет к малярии.
Однако достигнутые в отношении снижения малярии результаты не
были везде прочно закреплены, и в некоторых районах СССР в 1931 и
1932 гг. отмечен значительный подъем малярии, особенно высокий в
осенние месяцы и характеризующийся к тому же усилением тяжести
течения болезни.
Причинами такого подъема малярии являются:
1) осеннее развитие малярии 1931 г. и связанная с ним большая
волна рецидивов весной 1932 г.;
2) обилие осадков и последующее жаркое лето, что весьма благо-
приятствовало размножению анофелес;
3) большая передвижка рабочей силы без достаточного малярийно-
го обслуживания;
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
187
4) недооценка со стороны ряда здравотделов угрозы малярии и не-
достаточная настойчивость в выполнении противомалярийных меро-
приятий. 1
Бурный рост социалистического строительства и задачи улучшения
культурно-бытовых условий жизни трудящихся требуют и сами создают
необходимые предпосылки для решительной борьбы с малярией (раз-
витие химической и нефтяной промышленности, машиностроения, авиа-
строение, коллективизация сельского хозяйства, землепользование,
мелиорация, расширение и укрепление медицинской сети, улучшение
качества медицинского обслуживания трудящихся, ликвидация негра-
мотности и мн. др.).
В царской России организованной борьбы с малярией, как с бо-
лезнью общего первостепенного значения, не велось; лишь Пироговское
общество врачей давало почин организации борьбы с малярией в от-
дельных местах, сильно пораженных этой болезнью. По инициативе
этого общества была открыта в Батуми первая и единственная в то
время малярийная станция. Создание в Тбилиси института по борьбе
с малярией не было разрешено.
В 1920 г. после окончания гражданской войны в Москве по ини-
циативе проф. Е. Марциновского создается Тропический институт (ныне
Центральный институт малярии), на который ложилась обязанность
научно-практического обоснования борьбы с малярией и подготовка
кадров противомалярийных работников. В следующем году были орга-
низованы первые восемь малярийных станций. Сеть их начала быстро
расти. Всего (кроме Москвы) было до тринадцати тропических инсти-
тутов в южной зоне СССР; на 1945 г. числилось 1200 малярийных стан-
ции (включая отделения санитарно-бактериологических институтов и
тропические диспансеры) и 1488 малярийных пунктов. Кроме этих
учреждений в деле борьбы с малярией играют существенную роль вре-
менные противомалярийные отряды (250) и хинизаторско-бонификаци-
онные бригады (в 1945 г. их было 5000) при общих медицинских уча-
стках или связанные с хозяйственными организациями и колхозами.
Существенную роль в стимулировании научно-исследовательской
работы в энтомологической части малярийной проблемы сыграла орга-
низованная в 1924 г. Е. Павловским и А. Штакельбергом при Зоологи-
ческом институте АН СССР Постоянная комиссия по изучению маля-
рийных комаров (и других переносчиков) —помощью периферийным,
работникам научными консультациями, проведением научных заседа-
ний, редактированием и печатанием работ, составлением и изданием
пособий для исследовательских и практических работ, предоставлением
рабочих мест, рассылкой энтомологической литературы, изданием «Па-
разитологического сборника», организацией паразитологических экспе-
диций и др. Комиссия эта вросла в структуру .Зоологического институ-
та АН СССР на положении Отдела паразитологии.
Борьба с малярией возглавляется Отделом по борьбе с малярией
Противоэпидемического управления Наркомздрава СССР. В нарком-
здравах союзных республик имеются малярийные отделы или группы.
На обязанности этих центральных ячеек лежит разработка с институ-
тами планов противомалярийной борьбы, финансирование, снабжение
лечебными средствами, профилактическими материалами, аппаратурой,
•обеспечение транспортом и др. ’Малярийные станции организованы
двух родов: областные (краевые) и районные. Последние организуются
1 М. Рашина. Тези-сы доклада на совещании директоров тропинов, 1932.
188
СПЕЦИАЛЬНАЯ ЧАСТЬ
там, где заболевает малярией более 3% населения или где возможны
вспышки малярии в силу специальных условий (например, торфоразра-
ботки, ирригационные культуры и др.).
Районная малярийная станция осуществляет всю практическую ра-
боту по борьбе с малярией на подлежащей ее ведению, территории че-
рез посредство малярийных пунктов, отрядов, бригад и др. Малярий-
ные пункты организуются в точках, где заболеваемость более 10!%. Они
имеют в своем ведении одно или несколько близких друг к другу се-
лений. Временные малярийные отряды организуются на малярийный се-
зон, особенно на срок развертывания сельскохозяйственных работ, и
действуют они, естественно, в сельских местностях. При этих отрядах
или при больницах на период малярийных вспышек развертываются
сезонные койки для тяжелых случаев малярии; в 1937 г. было 3 932 кой-
ки. Наконец, низовой ячейкой являются бригады хинизаторов и бони-
фикаторов (см. ниже раздел 6 малярийных комарах) при сельских вра-
чебных участках или же содержимые за счет средств хозяйственных
организаций и колхозов. Для курсового систематического лечения широ-
ко привлекается в помощь медицинскому персоналу широкая колхозная
общественность (среди колхозников вербовались лица для раздачи ле-
карств; их с 1936—1937 гг. насчитывалось около 50 тыс. человек).
ЛЕЧЕНИЕ И ЛЕКАРСТВЕННАЯ ПРОФИЛАКТИКА МАЛЯРИИ
Борьба с малярией сводится к проведению комплексной системы
мероприятий для воздействия на возбудителя малярии в человеке и
для влияния на переносчика — комаров анофелес; в сферу противома-
лярийных работ включаются также воздействия в широком смысле
слова на среду обитания анофелес и человека.
Борьба с паразитами малярии в теле человека сводится к лечебным
процедурам, среди которых первое место занимает лечение хинином; в
последнее время выдвигают плазмохин, • являющийся синтетическим
продуктом типа хинолина; применяют также химиотерапевтические пре-
параты групп мышьяка (например, сальварсан) и органические краски
(метиленовая синька и др.).
Беспорядочное лечение малярии является вредным, так как ведет
к приобретению малярийными паразитами такого больного хинино-
устойчивых свойств. Поэтому лечение малярии должно быть
строго индивидуальным, тем более, что на характер влияния лекарства
на малярийного паразита оказывает действие и организм больного. Из-
вестны случаи, когда один и тот же штамм малярийного плазмодия,
привитый двум человекам, у одного оказался хининоустойчивым, а у
другого поддавался хинному лечению. Хинин влияет сильнее на меро-
зоитов и на молодые шизонты; половые формы паразита стойки и к
этому веществу.
Лечению подвергаются не только явные больные малярией, но и га-
метоносители, в целях их стерилизации и обезвреживания, как источ-
ников заражения анофелес. Недавно предложенный синтетический
препарат плазмохин обладает хорошими гаметоцидными свой-
ствами при тропической малярии.
Все малярийные больные, обращающиеся за помощью в лечебные
учреждения, должны быть обеспечены систематическим курсовым ле-
чением в поликлиниках, амбулаториях малярийных станций или пунк-
тов. Схемы такого лечения разработаны в деталях (см. ниже инструк-
цию). В 1936 г. было пользовано 4 935 100 человек.- '
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
189
Однако ограничиваться ожиданием, что (малярик сам придет за по-
мощью, не приходится. Поэтому в практику противомалярийной рабо-
ты вошли массовые обследования населения на малярию в начале или
•в конце года методами взятия крови (толстая капля с последующей
окраской по Романовскому) или анамнеза. Например, в 1936 г. было
обследовано 32 898 800 человек. Выявленные активные малярики и па-
разитоносители направляются на систематическое курсовое лечение.
Переболевшие в текущем году малярией, если у них во время об-
следования нет клинических проявлений болезни, подвергаются по
спискам, противорецидивному лечению (в 1936 г. было обслужено
4 440 200 человек).
Наконец, в наиболее пораженных малярией местах или к особо
уязвимым по малярии группам населения применяется массовая ле-
карственная профилактика, охватившая в 1936 г. 744 500 человек.
Рис. 107 — Изменение Pl. vivax в эритроцитах под влиянием подкожного или
внутривенного введения акрихина. Ядро не окрашивается.
Пигмент собирается в кучки и выталкивается из паразита, который как бы распадается.
По Ф. Зельмановой.
В СССР быстро развернулись удачные работы по синтезированию
некоторых противомалярийных препаратов. Выяснена формула нового
заграничного препарата атебрина; он же получен синтетически под
именем акрихина. Добыт плазмоцид и идут дальнейшие работы в
том же направлении. Производится проверка терапевтического эффек-
та этих препаратов, продвигаемых в заводское производство. Совет-
ские синтетические препараты хинолинового ряда обладают определен-
ным паразитоцидным и купирующим малярийную лихорадку действием
при различной степени токсичности этих препаратов (Е. Тареев с со-
трудниками). Акрихин обладает шизонтотропными свойства-
м и; под его действием протоплазма плазмодия как бы разрывается, а
ядро теряет способность окрашиваться ядерными красками, пигмент
собирается в кучки. Эти изменения обнаруживаются уже через два
часа после внутривенного или подкожного введения 0,2 г акрихина
(рис. 107). У Pl. vivax они соответствуют тому, что наблюдал Джемс
после введения 0,6 г атебрина per os (Ф. Зельманова). К этому следует
добавить, что шизонты терцианы быстро исчезают на периферической
крови; резкое уменьшение их количества обнаруживается уже через
четыре часа, а через 16—18 часов после инъекции акрихина они про-
падают. Гаметоциты исчезают позднее, через 2—4 и более дня
(Н. Демина).
В отношении влияния хинина и атебрина на спорозоиты плазмодия
тропической малярии Чука с сотрудниками показал, что при прямом
19а
СПЕЦИАЛЬНАЯ ЧАСТЬ
действии указанных лекарственных веществ в растворе 1 : 2 500 на.'
взвесь спорозоитов, добытых из слюнных желез анофелес, спорозоиты &
ряде случаев сохраняют свою инфицирующую способность.
В настоящее время работает завод «Акрихин», построенный по лич-
ному указанию т. Сталина. Выпуск акрихина покрывает необходимую-
потребность страны.
Вопросы борьбы с малярией по линии воздействия на переносчика
и на окружающую среду рассматриваются ниже, в главе о комарах;,
там же освещаются основные наметки мероприятий по ликвидации ма-
лярии.
В результате широко поставленной борьбы с малярией в СССР в:
первую половину1936 г. заболеваемость малярией снизилась по срав-
нению с таким же периодом 1935 г. на 35%, а смертность упала на
40% (Сергиев).
Результаты противомалярийной работы за 9 месяцев в 1939 г. при-
вели к уменьшению первичной обращаемости по малярии по всему
Советскому Союзу в целом на 35,7% сравнительно с такими же меся-
цами 1938 г. Значительно уменьшились малярийные индексы при мас-
совых обследованиях населения. На первом месте по снижению маля-
рии на 1939 г. стоит Грузинская ССР (П. Сергиев).
Частные примеры результатов противомалярийной борьбы можно
видеть на рис. 97.
Малярия остается все еще первой после гриппа болезнью по своей
распространенности.
За последние годы накопился большой опыт применения советских
синтетических препаратов в борьбе с малярией, основанный на ряде
экспериментальных работ. Учет результатов научных и научно-практи-
ческих исследований и опытного применения хинина и синтетических
препаратов в различных эпидемиологических условиях отражен в ин-
струкции, составленной Центральным тропическим институтом, приня-
той совещанием директоров Тропических институтов и утвержденной
президиумом Ученого медицинского совета Наркомздрава СССР
26 марта 1928 г. Новым изданием этой переработанной на основе ши-
рокого опыта (1944 г.) инструкции мы и закончим главу о малярийных
паразитах.
Утвержцена президиумом
Ученого медицинского совета
Наркомздрава CCCPj
16/III 1944 г.
ИНСТРУКЦИЯ
ПО МЕДИКАМЕНТОЗНОЙ ТЕРАПИИ И ПРОФИЛАКТИКЕ
МАЛЯРИИ
Медикаментозную терапию и профилактику малярии следует проводить на
основе особенностей течения малярийной инфекции и свойств отдельных противо-
малярийных препаратов.
ОБЩЕЕ ТЕЧЕНИЕ МАЛЯРИЙНОЙ ИНФЕКЦИИ
Наиболее распространенной формой малярии в СССР является трехдневная
малярия. Значительно реже даже в южных районах Советского Союза встречается
тропическая малярия. Четырехдневная форма малярии встречается в большинстве
районов СССР в виде отдельных случаев или редких очаговых заболеваний.
В климатических условиях СССР заражение малярией происходит, как правило,,
только в течение III квартала года.
Длительность инкубационного периода трехдневной малярии различна.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
191
В районах северной и средней полосы СССР преобладает форма с длительным
инкубационным периодом — от 7 до 11 месяцев с момента заражения; значительно
реже встречается форма с коротким инкубационным периодом — от 14 до 20 дней
с момента заражения.
В южных районах, наоборот, преобладает форма с короткой инкубацией и
много реже наблюдаются случаи с длительной инкубацией.
При трехдневной малярии с короткой инкубацией после окончания первой се-
рии приступов в ближайшие 3—5 недель наблюдаются так называемые ближайшие
рецидивы, за которыми следует, если в результате лечения не наступило полного
излечения, длительный период, охватывающий зимние месяцы, в течение которого
острых проявлений малярии у больного не отмечается.
В течение весенних месяцев отмечается волна так называемых отдаленных ре-
цидивов, которая, в зависимости от климатических условий, начинается в марте —
апреле и достигает максимума в мае — июне.
Наблюдения показали, что отдаленные рецидивы наблюдаются в 30—50%
случаев при трехдневной малярии с короткой инкубацией. В значительно меньшем
проценте рецидивы бывают осенью, т. е. через год после заражения, и еще реже —
весной второго года болезни. При трехдневной малярии с длительной инкубацией
первые приступы лихорадки начинаются весной — в апреле — июне. После купиро-
вания этой серии приступов в течение 3—5 недель имеют место ближайшие реци-
дивы, которые могут наблюдаться в течение всего III квартала.
Весной следующего года, т. е. через 1% года после заражения, рецидивы1
имеют место только в 7—10% случаев.
Помимо рецидивов с явными клиническими приступами, у многих больных в те
же сроки наблюдается паразитрносительство, т. е. появление паразитов в перифе-
рической крови без наличия приступов лихорадки.
Е громадном большинстве случаев больные трехдневной формой малярии вы-
здоравливают через 18—20 месяцев с момента заражения.
Менее изученная четырехдневная малярия может давать рецидивы в более дли-
тельный срок — свыше 3 лет.
Тропическая малярия протекает всегда с короткой инкубацией и про-
является лихорадочными приступами во вторую половину лета, рецидивируя до
поздних осенних месяцев и следующей весной, или нередко проявляется весной толь-
ко в виде эпидемиологически опасного паразитоносительства. Общая длительность
тропической малярии при однократном заражении меньше года.
Ввиду того что однократно перенесенная малярийная инфекция не дает иммуни-
тета, в очагах часто наблюдаются повторные заражения и заболевания малярией в
течение нескольких лет подряд. Это дало повод старым авторам говорить о дли-
тельно, в течение многих лет, продолжающейся малярии, а также породило мнение
о неизлечимости малярии.
При повторных заражениях нередко происходит самокупирование приступов с
последующей длительной циркуляцией паразитов в крови.
Диагноз малярии
Диагноз малярии ставится по клинической картине обязательно с подтвержде-
нием присутствия плазмодиев анализом крови. Лечение можно начать немедленно,
не дожидаясь результатов анализа крови.
В целях наиболее раннего выявления больных малярией во время эпидемиче-
ского сезона необходимо брать кровь у всех лихорадящих и у всех подозрительных
на малярию для обнаружения паразитоносителей.
При первичном отрицательном анализе крови исследование необходимо повторить.
Общие принципы лечения малярии
Исходя из данных о течении малярийной инфекции, основными принципами ле-
чения должны быть следующие:
1. Лечение должно быть начато немедленно по установлении диагноза клини-
чески или паразитологически для быстрейшего купирования приступов и возвраще-
ния больного к труду.
2. Быстрейшее купирование приступов достигается назначением полной, соот-
ветствующей возрасту больного, дозы медикаментов, указанной в нижеприводимых
схемах лечения. Как правило, лечение должно проводиться пероральным . приемом
медикаментов, и только в тяжелых коматозных случаях, а также при рвоте и про-
фузных поносах следует применять инъекции.
3. Для предотвращения ближайших рецидивов каждый больной и паразитоно-
ситель должен провести полный курс систематического лечения по одной из ниже-
указанных схем.
192
СПЕЦИАЛЬНАЯ ЧАСТЬ
4. Каждый больной малярией должен находиться под наблюдением медицинской
организации не менее Р/г лет.
5. В течение этого срока каждый больной должен получить не менее одного
курса противорецидивного лечения.
6. В местностях с большим количеством комаров во время эпидемического се-
зона для' предотвращения распространения малярии лечение больных и паразито-
носителей должно проводиться комбинированным назначением акрихина или хинина
с плазмоцидом.
СВОЙСТВА ПРОТИВОМАЛЯРИЙНЫХ ПРЕПАРАТОВ
а) ОБЩАЯ ХАРАКТЕРИСТИКА
Основными лечебными и профилактическими противомалярийными препаратами
являются акрихин и хинин, быстро разрушающие бесполые формы плазмодиев —
шизонты, бесполое размножение которых собственно и вызывает лихорадочные при-
ступы и другие клинические проявления малярии. Плазмодии трехдневной, четырех-
дневной и тропической малярии (кроме гаметоцитов тропической малярии) исчезают
в крови после 3—5 дней акрихинизации и хинизации. Температура снижается до
нормы при трех- и четырехдневной малярии после Р/г—3 дней лечения, при тро-
пической малярии — после 2—4 дней лечения и скорее при лечении большими доза-
ми и инъекциями. На гаметоцитов тропической малярии ни акрихин, ни хинин не
действуют. Так же слабо они действуют и на гаметоцитов трехдневной и четырех-
лневной малярии.
В случае отсутствия основных противомалярийных препаратов для купирования
приступов трехдневной малярии может быть применен осарсол.
Нередко применяемый в практике лечения малярии новарсенол употреблять не
следует, во-первых, потому, что он не только не действует на тропическую форму
малярии, но нередко провоцирует тяжелые приступы ее,. а во-вторых, потому, что
у малярийных больных он может вызвать тяжелое заболевание энцефалитом.
Иными свойствами обладает плазмоцид. Значительно слабее действуя на бес-
полые формы плазмодиев, чем акрихин и хинин, и потому непригодный для купи-
рования приступов, он гораздо сильнее действует на половые формы (гаметоциты)
всех видов малярийных плазмодиев (включая полулуния тропической малярии). Уже
спустя 1—2 суток после приема плазмоцида гаметоциты теряют способность зара-
жать комара, а спустя 4—5 суток полностью исчезают из крови. Кроме того, в
комбинации с акрихином (и хинином) плазмоцид, вследствие несколько отличного ха-
рактера своего воздействия на плазмодиев, сильнее снижает процент рецидивов.
б) АКРИХИН
Акрихин, синтетический противомалярийный препарат советского производства,
имеет в основе акридиновое ядро и представляет собой двусолянокислую соль 2-мет-
окси-6-хлор-9-8-диэтиламино-а-метилбутиламиноакридина, или, по другому обозна-
чению, 7-метокси-3-хлор-9-8-диэтиламино а- метил-бутиламиноакридина.
Молекулярный вес 508,5. Мелкий кристаллический порошок интенсивно желтого
(канареечного) цвета, очень горький на вкус. Растворяется в комнатной воде до 3%.
при подогревании — до 4—5% и выше.
Выпускается акрихин в виде:
1) желтых таблеток с содержанием в каждой по 0,1 акрихина (для взрослых)
или по 0,05 акрихина (для детей);
2) зеленых таблеток (подкрашенных метиленовой) синыо с содержанием в каж-
дой по 0,1 акрихина и по 0,02 плазмоцида (для взрослых) или по 0,05 акрихина и
по 0,01 плазмоцида (для детей);
3) акрихина для инъекций чистого порошка для приготовления растворов.
Приготовление растворов акрихина для инъекции
4,0 акрихина в порошке раствбряют в 100 мл (см3) дестиллированной воды,
фильтруют и стерилизуют текучим паром на водяной бане в течение 30 минут. Ра-
створ при правильной стерилизации остается прозрачным и годен для применения
в течение 2 недель.
При охлаждении раствора акрихин частично выпадает в осадок, который при
легком подогревании снова легко растворяется.
При отсутствии акрихина в порошке растЬор для инъекции можно приготовить
мз таблеток (желтых). 40 таблеток по 0,1 акрихина каждая измельчают в ступке,
добавляют 100 мл дестиллированной воды комнатной температуры (но не выше
20°) и растворяют в течение 3 часов. Раствор тщательно отфильтровывают от крах-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ 193
-----------------ц------------------------------------------------------------
мала и других нерастворимых в воде при комнатной температуре составных частей
таблеток и стерилизуют, как сказано выше.
Стерильный 4% раствор акрихина вводят подкожно или внутримышечно по
5—10 мл (т. е. по 0,2—0,4 акрихина);, раствор хорошо переносится больными и не
дает инфильтратов. При болезненности можно прикладывать грелку на место
инъекции. В редких случаях акрихин вводят в вену в количестве 2,5 мл 4% раствора
(т. е. 0,1 акрихина). Вводить надо медленно во избежание коллапса.
Акрихин быстро поступает в кровь после приема внутрь и еще быстрее после
подкожного или внутримышечного и особенно внутривенного введения. Выводится
акрихин из организма мочой медленно — в основного в течение ближайших 2 недель.
Для определения выделения акрихина берут в делительную воронку или в
цилиндр 50 мл нефильтрованной мочи, прибавляют 5 мл 40% раствора едкого натра
и 10 мл серного эфира. Смесь осторожно перемешивают (без взбалтывания) и
оставляют стоять на 24 часа.
При наличии акрихина (эфирная вытяжка) верхний слой жидкости дает зелено-
вато-желтую флюоресценцию, интенсивность которой соответствует концентрации ак-
рихина в моче.
Акрихин хорошо переносится больными, мало токсичен.
Побочное действие акрихина:
а) Прокрашивание покровов в своеобразный желтый цвет совершенно безвредно
и пе зависит от поражения печени. Оно мож«т держаться во все время лечения и
профилактики акрихином и еще несколько недель спустя.
б) Описанные ранее в связи с приемом акрихина диспептические явления (тош-
нота, рвота) и головная боль вряд ли зависят от действия акрихина; они являются
следствием самого малярийного' приступа.
При передозировке акрихин может вызвать нервные явления в виде:
а) скоро проходящего «акрихинового опьянения», которое выражается в двига-
тельном и речевом возбуждении, известной потере ориентировки длительностью от
одного до нескольких часов;
б) «акрихинового психоза» с галлюцинациями, возбуждением или депрессией и
другими синдромами, длительностью до недели и редко дольше, также бесследно
проходящего.
В виде редкого исключения нервные явления могут наступить и после обычных
доз акрихина, особенно при парэнтеральн- его введении у лиц с неустойчивой пси-
хикой. При появлении нервных, осложнении лечение акрихином прекращают, вводят
большие количества жидкости для быстрейшего выведения из организма акрихина,
дают глюкозу, успокаивающие средства, назначают ванны.
Акрихин противопоказан у лиц с психическими заболеваниями, а также у «лиц,
дававших и раньше нервные, явления под влиянием акрихинизации. Относительно
акрихин противопоказан у лиц с лябильными психическими реакциями, при резком
нарушении выделительной функции почек (гипостенурия, анурия, резкая азотемия),
при холемии, наличии чрезвычайно резкого прокрашивания покровов от длитель-
ного применения акрихина, при установлении отсутствия или задержки выделения
его мочой.
в) X И Н И Н
Хинин, естественный алкалоид хинного дерева, имеет в основе хинолиновое ядро
и' представляет метоксихинолин-винилхинуклидинкарбинол. Молекулярный вес
378,26. Применяется чаще в виде следующих солей:
1. Солянокислый хинин — тонкие белые шелковистые игольчатые кристаллы
чрезвычайно горького вкуса. Содержат около 82% хинина-основания с приведенной
выше формулой. Растворим в 30 частях холодной и в 1 части кипящей воды.
2. Двусолянокислый хинин — бесцветные кристаллы сильно горького вкуса. Со-
держит около 73% хинина-основания. Растворим в 0,7 части воды.
3. Сернокислый хинин — белые игольчатые кристаллы чрезвычайно горького
вкуса. Содержит свыше 72%. хинина-основания. Растворим в 800 частях холодной и
в 25 частях кипящей воды.
Солянокислый и сернокислый хинин применяются внутрь. в порошках, в таб-
летках, в облатках, в капсулах; солянокислый хинин применяется также в растворе.
Долго лежавшие таблетки следует проверять на распадаемость в воде.
Для инъекций применяются следующие растворы:
1. Двусолянокислый хинин по 1—2 мл 25—50% раствора (т. е. по 0,5 двусоляно-
кислого хинина).
2. Солянокислый хинин: а) 10% раствор солянокислого хинина на физиологи-
ческом растворе (вводится в подогретом до 35° виде); б) хинопирии — солянокислого
хинина 3,0, антипирина 2,0, дестиллированной воды 6,0. Каждый миллилитр рас-
твора содержит 0,3 хинина. Эти растворы легко приготовить в простой обстановке.
194
СПЕЦИАЛЬНАЯ ЧАСТЬ
Растворы хинина стерилизуются обычным путем, обладают сильно раздражаю-
щим местным действием (особенно кислые растворы двусолянокислого хинина)..
Растворы хинина следует вводить глубоко в подкожную клетчатку, лучше в боль-
шом разведении (например, с солевым раствором), шприцем с сухой иглой, хотя и
таким путем не всегда удается избежать некроза тканей.
При внутривенном введении хинин может вызвать внезапную смерть, особенно
при тяжелом общем состоянии больных и у лиц с идиосинкразией к хинину.
Введение хинина через прямую кишку или путем ионтофореза для лечения ма-
лярии не должно применяться, так как не обеспечивает поступления необходимого
количества хинина в кровь.
В последние годы под названием совхинета (советского хинета) применяется
смесь сернокислого хинина, цинхонидина, цинхонина и других алсалоидов однолет-
них культур хинного дерева, выращенных в Абхазии и Аджарии. Совхинет при на-
значении внутрь в дозах сернокислого хинина хорошо переносится и обладает пол-
ным действием хинина. л
Хинин быстро всасывается и выводится из организма в основном в течение 2
дней. Присутствие хинина в моче, свободной от белка, обнаруживают по белому
кольцу при переслаивании мочи с сулемово-иодисто-калиевым раствором.
Хинин нередко вызывает побочные действия — шум в ушах, головокружение,
рвоту, сердцебиение, дрожание рук, бессонницу, маточные кровотечения. Лучше пере-
носится при прибавлении кофеина, бромистого кальция, аспирина. Длительное
применение терапевтических доз хинина на здоровое сердце неблагоприятного влия-
ния не оказывает.
От больших доз хинина наблюдались токсические явления — слепота, глухота,
резкая головная боль и другие нервные явления, и даже смерть при явлениях кол-
лапса (от 6,0 и более хинина). Против амавроза рекомендуют нитриты, папаверин,
кофеин, при коллапсе — возбуждающие средства.
При идиосинкразии к хинину уже минимальные дозы могут вызвать дерматит
(крапивницу, эрйтему), лихорадку, распад крови, маточное кровотечене. Против
явлений идиосинкразии применяют эфедрин, адреналин и питуитрин, хлористый каль-
ций.
Хинин противопоказан: при идиосинкоазии (особенно опасно внутривенное вли-
вание хинина), у больных с гемоглобинуригной лихорадкой, при заболеваниях сред-
него и внутреннего уха; относительно пр а^вопоказан при декомпенсации сердца и
в поздние месяцы беременности.
г) ОСАРСОЛ
Осарсол — синтетический, содержащий мышьяк препарат, представляет 4-окси-
3-ацетоаминофениларсиновую кислоту. Выпускается в таблетках по 0,25 для приема
внутрь.
Побочные токсические действия: дерматиты, повышение температуры, геморра-
гический энцефалит, поражение печени, раздражение почек.
Проведения лечения малярии осарсолом следует избегать при тяжелых заболе-
ваниях печени и почек.
д) ПЛАЗМОЦИД
Плазмоцид, советский синтетический препарат, представляет метиленбиссали-
циловокислую соль 6-метокси-8-диэтиламинопропил-аминохинолина. Молекулярный
вес 863,41. Оранжево-желтый кристаллический порошок, нерастворимый в воде, слабо
горького вкуса. Выпускается только вместе с акрихином в зеленых таблетках (под-
крашенных метиленовой синью) с содержанием в каждой по 0,02 плазмоцида и по
0,1 акрихина для взрослых или по 0,01 плазмоцида и по 0,05 акрихина для детей.
Максимальная суточная доза плазмоцида для взрослых 0,06, т. е. 3 зеленые
таблетки; детям дают соответственно меньше (см. далее). Ни при каких условиях
эту суточную дозу нельзя превышать ввиду токсичности плазмоцида.
Плазмоцид в принятой в последние годы дозе в 0,06 за сутки хорошо перено-
сится и не приводит к каким-либо побочным или токсическим действиям; общепри-
нятая комбинация с акрихином, повидимому, улучшает переносимость плазмоцида.
При передозировке плазмоцида наблюдаются побочные явления в виде болей в
голове и в подложечной области, парестезии и невралгические боли в области
тройничного нерва, а при значительной передозировке — также в виде мозжечковой
атаксии, полиневрита и атрофии зрительного нерва.
При появлении вышеназванных симптомов со стороны нервной системы необхо-
димо прекратить дальнейший прием плазмоцида, промыть желудок, вводить боль-
ному больше жидкости, назначать щелочи, кофеин и проводить ретробульбарные'
инъекции О,1°/о раствора атропина; избегать, кроме настоятельных показаний, нарко-
тиков. Целесообразно применение витамина Bi, никотиновой кислоты, препаратов;
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
195
печени. Все случаи плазмоцидовой интоксикации подлежат регистрации и извеще-
нию вышестоящих органов здравоохранения.
Плазмоцид противопоказан:
1. При заболеваниях глазного дна и зрительного нерва.
2. У лиц, страдающих энцефалитом или менинго-энцефалитом или перенесших
эти заболевания раньше.
3. У лиц, реагировавших ранее на лечение плазмоцидом токсическими явле-
ниями.
4. У грудных детей.
Следует особенно подчеркнуть и разъяснять населению, что плазмоцид, принятый
даже в больших ведущих к смерти дозах, не вызывает у беременных абортов или
преждевременных родов.
Схемы лечения и профилактики малярии
Систематический курс лечения
Всем остролихорадочным больным малярией как свежей, так и рецидивной сле-
дует проводить систематический курс лечения длительностью около месяца, состоя-
щий из трех циклов акрихинизации (в эпидемический сезон вместе с плазмоцидом!
по одному' из трех нижеперечисленных вариантов, или из четырех циклов хинизации.
Систематический курс лечения акрихином с плазмоцидом
Первый вариант
1 Циклы и перерывы Продолжи- тельность в днях Суточная лоза в г Примечание
акрихин плазмоцид
Первый цикл 5 0.3 0.06 Суточная доза может
Перерыв *. 7 10 — — быть дана в один-два
Второй цикл . .... 3 0.3 0.06 приема
Перерыв . . . .... 7—10 — —
Третий цикл 3 0.3 0.06
При тяжелых случаях малярии для быстрейшего купирования приступов назна-
чается удвоенная доза акрихина в первый день лечения. Это обеспечивает более .
быстрое достижение необходимой для терапевтического эффекта концентрации акри-
хина в крови больного (второй и третий вариант).
Второй вариант
Цикл и перерывы Продолжи- тельность в днях Суточная доза п г Порядок приема медикаментов
акрихин плазмоцид
Первый цикл ( 1 0.6 0.06 За один прием 0.3 ак- рихина и 0.04 плазмо- цида. Через 6 часов
Перерыв 1 U 7—10 0.3 0.06 прием 0.3 акрихина и 0.92 плазмоцида В 1 -2 приема
Второй цикл 3 0.3 0.06 В 1—2 приема
Перерыв Третий цикл . . 7—10 3 0.3 0.06 В 1—2 приема
196
СПЕЦИАЛЬНАЯ ЧАСТЬ
Третий вариант
Циклы и перерывы Продолжи- тельность в днях Суточная доза в г Порядок приема медикаментов
акрихин ] 1ЛЗЗМ0 1ИД
1 Подкожно или внутри- мышечно вводят 7.5 мл
Первый цикл 1 0.6 0.04 4% раствора акрихина и одновременно внутрь дают 0.3 акрихина и 0.04 плазмоцида
1 3 0.3 0:04 В 1—2 приема
Перерыв . . , 7—10 — — —
Второй цикл . 3 0.3 0.06 В 1—2 приема
Перерыв . . 7—10 — — —
Третий цикл ' 3 0.3 0.06 В 1—2 приема
Примечание. При тяжелом течении малярии или при пепрекращающихся
приступах, например, при тропической малярии на юге, первый цикл акрихини-
зации может быть продолжен до 7 дней. Однако, если лихорадка не прекра-
щается на 3—4-й день лечения, необходимо всегда пересмотреть диагноз маля-
рии и исключить другие заболевания как причину лихорадки.
Лечение акрихином без плазмоцида проводится по тем же схемам, только жел-
тыми таблетками.
Систематический курс лечения хинином
Циклы и перерывы Щ одолжи- тельность в днях Суточная доза в г Число приемов в день
Первый цикл . . • 3—4 1.0—1.2 2
Перерыв .... .... . ... 4 — —
Второй цикл .... 3 1.0-1-2 2
Перерыв 4 • — —
Третий цикл . . 3 1.0-1.2 2
Перерыв . . 4 — —
Четвертый цикл ... .... 3 1.0—1.1 2
В тяжелых случаях трехдневной малярии и при тропической малярии дают
внутрь по 1,5 солянокислого хинина (или по 2,0 сернокислого) в сутки в два-три
приема или при показаниях вводят хинин под кожу по 0,5—1,0 два раза в день.
Лечение осарсолом
Применяется для купирования приступов трехдневной малярии (при тропической
малярии противопоказано).
Осарсол назначают 3 дня подряд по 0,75 в сутки в три приема по одной таблет-
ке в 0,25, с повторением такого же- цикла после четырехдневного промежутка.
В дальнейшем проводят два трехдневных цикла хинином или акрихином.
Противорецидивное лечение
Проявление ближайших рецидивов малярии, как празило, предотвращается пол-
ным месячным курсом систематического лечения. Если приступы малярии у больного
были в начале эпидемического сезона (весной и лете м), то через 1—IV2 месяца по
окончании проведенного курса лечения рекомендуется провести для предотвращения
могущих быть рецидивов противомалярийное лечение по той же схеме, как и систе-
матический курс лечения. Но суточная доза дается меньше на одну треть (две таб-
летки вместо трех).
Всем больным, имевшим острое проявление малярии в предыдущем году, для
предотвращения отдаленных рецидивов в весенние месяцы текущего года назначают
противорецидивное лечение по нижеуказанной схеме.
Сроки проведения противорецидивного лечения для каждой республики, края,
области устанавливаются в зависимости от сроков наступления в данной местности
весенней волны рецидивов трехдневной малярии.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
197
Если массовое проявление рецидивов наблюдают в середине мая, то массовое
противорецидивное лечение необходимо ничинать с 1 мая; если в начале мая, то с
15 апреля ит. д.
Лицам, болевшим в предыдущем году тропической малярией,- противорецидивное
лечение назначают таблетками акрихина с плазмоцидом с момента наступления ак-
тивности малярийных комаров.
Правильно и своевременно проведенное противорецидивное лечение снижает
в 2—3 раза процент рецидивов у лиц, больных малярией, по сравнению с боль-
ными, не подвергавшимися противорецидивному лечению.
Всякий остролихорадочный рецидив малярии и выявленный паразитоноситель
требуют повторения систематического курса лечения акрихином или хинином в пол-
ной дозе.
'Схема противорецидивного лечения акрихином. Первый
цикл — 5 дней, второй и третий циклы — по 3 дня с десятидневными промежутками
между циклами. Каждый день противорецидивного лечения дают внутрь по 0,2
акрихина (две желтые таблетки) в один прием.
Схема противорецидивного лечения хинином: четыре цикла по 3 дня каждый
с четырехдневными промежутками. Каждый день противорецидивного лечения дают
внутрь по 0,6 солянокислого хинина (или по 0,75 сернокислого) в сутки в одид-два
приема.
Общественная химиопрофилактика
В местностях с большим количеством комаров и большой заболоченностью, где
невозможно или нецелесообразно проводить противокомариные мероприятия, для
предотвращения распространения малярии рекомендуется проводить общественную
химиопрофилактику. Она состоит в том. что всем лицам, болевшим малярией в пре-
дыдущем году после проведения противорецидивного курса лечения, а такЖе всем
болевшим острыми проявлениями малярии в настоящем году и выявленным парази-
тоносителям после проведения курса систематического лечения назначают, начиная с
15 июня—1 июля, т. е. с момента _ вылета первой генерации комаров, регулярные
приемы акрихина с плазмоцидом по две таблетки в день — 2 дня подряд с четырех-
пятидневным перерывом или 1 день с 2 днями перерыва — в течение всего эпидеми-
ческого сезона.
Сроки проведения общественной химиопрофилактики устанавливаются: в север-
ных районах до 15 августа, в районах средней полосы до 1 сентября и на юге до
15 сентября—1 октября, в зависимости от климатических условий.
Такой регулярный прием акрихина с плазмоцидом оказывает гибельное действие
на половые формы малярийных паразитов, циркулирующих в крови маляриков, и
лишает их способности заражать комаров анофелес.
При недостатке медикаментов и невозможности охватить общественной химио-
профилактикой всех болевших малярией в предыдущем году, в обязательном порядке
должны быть охвачены все болевшие тропической формой малярии.
Правильное и аккуратное проведение общественной химиопрофилактики резко
сокращает количество зараженных комаров и тем самым предотвращает появление
свежих случаев заболеваний малярией среди населения, проживающего в данной
местности.
Проведение общественной химиопрофилактики в первую очепедь показано в
районах рисосеяния, на рыбных промыслах и на торфоразработках, в местах
стоянки гарнизонов, а также в организованных коллективах, живущих вдали от на-
селенных пунктов, но среди большой заболоченности (например, воинские части, по-
селки новостроек и т. п.). В этих условиях все болевшие малярией в предыдущем
году и паразитоносители, а также имевшие острые проявления малярии в текущем
году члены коллектива должны быть охвачены общественной химиопрофилактикой.
Химиопрофилактика здоровых от малярии
Акрихин и хинин, систематически принимаемые здоровыми людьми в дозах, не-
сколько сниженных против лечебных, предотвращают в большом проценте случаев
наступление остролихорадочных проявлений малярии после происшедшего зараже-
ния. В этих дозах акрихин и хинин хорошо переносятся даже при многомесячном
применении.
Профилактику акрихином или хинином следует проводить у не болевших маля-
рией и прибывших в сильно пораженную малярией местность. Начинают химиопро-
филактику со времени массового появления комаров, но не ранее 1 июля (для при-
бывших в очаг после 1 июля — со дня прибытия), и продолжают весь эпидемический
?езон, а в случае выезда из малярийного очага еще в течение месяца по оставлении
очага.
198
СПЕЦИАЛЬНАЯ ЧАСТЬ
В сильно пораженных малярией местностях, где невозможно проводить противо-
комариные мероприятия, химиопрофилактику акрихином (или хинином) следует
проводить поголовно всему населению. Перед началом химиопрофилактики всем
остролихорадочным малярикам и паразитоносителям проводят пятидневный лечебный
цикл акрихином (по 0,3 акрихина в сутки) или хинином (по 1,0 в сутки).
Для быстрейшего уничтожения гаметоцитов, особенно при тропической малярии,
химиопрофилактику в эпидемический сезон проводят акрихином с плазмоцидом.
Схема химиопрофилактики акрихином
Каждые 2 дня подряд дают по 0,2 акрихина в один прием с трех-пятидневными
перерывами.
Длительность перерывов между приемами медикаментов устанавливается в за-
висимости от эпидемиологических условий (чем риск заражения больше, тем меньше
должен быть перерыв).
Схема химиопрофилактики хинином
Первый вариант—ежедневно по 0,4 хинина в один прием, второй вариант —
каждую неделю дают 3 дня подряд внутрь по 0,5 хинина в один прием.
Приложения
а) Лечение малярийной комы
Лечение следует начинать возможно раньше в виДе инъекций больших доз акри-
хина или хинина.
Акрихин вводят в первый день в дозе 0,4—0,6 за сутки внутримышечно: или по
0,2 два-три раза с промежутками в 6—8 часов, или первую дозу в 0,4. Можно сна-
чала ввести 0,1 акрихина внутривенно (2,5 мл 4°/о раствора, лучше вместе с 20—40
мл 40% раствора глюкозы; вводить надо медленно, в течение 3—5 минут), и тотчас
же 0,2 акрихина внутримышечно, а остальное количество акрихина ввести спустя
8—12 часов.
Хинин вводят в первый день в дозе 2,0—2,5 подкожно в два-три приема. Начать
лечение можно с внутривенного введения-0,25 хинина (лучше на растворе глюкозы
или на физиологическом растворе; вводить обязательно медленно, в течение не ме-
нее 3—5 минут; необходимо предварительно удостовериться, что больной переносил
хинин раньше хорошо, иначе — при идиосинкразии — может наступить внезапная
смерть), вслед за этим ввести 0,5—0,75 хинина под кожу. Остальное количество
хинина вводят в течение суток.
При явлениях сосудистой слабости (частый Малый пульс, пустые вены) одно-
временно вводят под кожу физиологический раствор и тонизирующие средства:
стрихнин по 2—3 мг сразу, кофеин, камфору, эфедрин, адреналин, коразол и др.
Показано также внутривенное введение 2,0 уротропина или 1,0 хлористого каль-
ция в растворе.
Второй и третий день лечения проводят инъекциями акрихина или хинина в дозе
0,4 акрихина или 2,0 хинина в сутки. По возвращении сознания и при отсутствии
поносов переходят на обычное лечение путем приема препаратов внутрь через рот.
По тем же правилам лечат алгидную малярию (обязательно с введением боль-
ших количеств солевого раствора), молниеносную трехдневную малярию и другие
тяжелые осложнения малярии. Инъекционный метод лечения приходится применять
и при упорных рвотах, и при профузных поносах.
б) Лечение гемоглобинурийной лихорадки
Переливание одногруппной крови в количестве 300—400 мл, назначение щелочей,
вливание солевого раствора, тонизирующих средств, хлористого кальция и раствора
глюкозы. При анурии — общее согревание тела, диатермия в области почек, около-
почечная новокаиновая блокада. При продолжающихся лихорадочных приступах с
плазмодиями в крови — акрихин в обычных дозах (хинин противопоказан!)
в) Лечение малярии у детей
В большинстве случаев малярия протекает у детей тяжелее, чем у взрослых.
Дети чаще дают рецидивы и длительно являются паразитоносителями. В районах
средней полосы Союза ССР у детей наблюдаются тяжелые, молниеносно протекаю-
щие формы трехдневной малярии, часто заканчивающиеся летально. Поэтому на ле-
чение детей, больных малярией, должно быть обращено особенное внимание, и они,
как правило, должны находиться под тщательным наблюдением персонала педиат-
рических учреждений, Лечение малярии у детей проводится по вышеуказанным схе-
мам, но в соответствующих возрасту дозах.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
199
Повозрастные дозы акрихина и плазмоцида
Возрастная группа Суточная доза акри- хина при приеме внутрь Доза акрихина при внутримышечном ле- чении в мл 4% рас- твора акрихина Суточная доза плаз- моцида
До 1 года От 1 » до 2 лет . . . » 2 >> » 4 » ... » 4 » » 6 » . . » 6 » » 8 » ... » -»8 » » 12 » ... » 12 » » 16 » ... Старше 16 лет 0.025 0.05 0.075 0.1 0.15 0.15—0.2 0.25 0.3 0.5—0.1 1.0-1.25 1.5-2.0 2.0-3.0 3.0—4.0 4.0—5.0 5.0—6.0 6.0—7.5 Не назначают 0.01 0.015 0.02 0.03 0.03-0,04 0.05 0.06
Детям до 4—5 лет акрихин дают внутрь в растворе, суточную дозу дают в два
приема. Раствор выписывают, назначая 10 разовых доз в рецепте. Например, ре-
бенку Р/г лет раствор из 0,25 акрихина на 50,0 воды — по одной чайной ложке
<т. е. по 5 мл два раза в день).
Детям акрихин можно давать также с джемом, предварительно измельчив таб-
летки, или в форме драже акрихина.
В тяжелых случаях малярии лечение надо начинать с внутримышечных инъек-
ций раствора акрихина, который хорошо переносят даже грудные дети.
Детям до 1 года хинин дают в сутки из расчета по 0,01 на месяц жизни, но
не более 0,1; детям от 1 года до 10 лет — по 0,1 на год жизни; от 10 до 15 лет —
по 1,0 в сутки; детям свыше 15 лет назначают дозу взрослого.
Инъекций хинина детям следует избегать ввиду частого образования абсцессов.
Эйхинин (этилугольный эфир .хинина), почти безвкусный белый легкий порошок с
содержанием около 80% хинина-основания, дается в дозах, в полтора раза боль-
ших, чем хинин; в тяжелых случаях малярии препарат недостаточно активен.
Детям до 1 года плазмоцид противопоказан.
г) Лечение малярии у беременных женщин
У беременных лечение малярии следует проводить по общим правилам и осо-
бенно настойчиво ввиду вредного влияния малярии на организм матери и на разви-
вающийся плод. Для лечения рекомендуется применять акрихин, так как он не ока-
зывает действия на матку.
Хинин, ввиду свойства его раздражать матку, следует применять только в дроб-
ной дезировке, например, по 0,2 пять раз в день. В тяжелых' случаях малярии
лечение проводят инъекциями.
Следует особенно подчеркнуть, что ни акрихин, ни плазмоцид даже в больших
дозах (плазмоцид даже при. смертельном отравлении) не вызывает аборта или
преждевременных родов.
Кормящие женщины могут проводить лечение акрихином или хинином в обыч-
ных дозах. Средства эти переходят в молоко лишь в ничтожных количествах, не
нарушающих кормления грудью, но совершенно недостаточных для получения лечеб-
ного эффекта у грудных детей, больных малярией.
д) Общий режим и симптоматическое лечение
Постельный режим необходим в тяжелых случаях малярии. Особой диэты при
малярии не требуется; в частности, разрешаются яйца, рыба, фрукты и сладости,
прием в пищу которых старыми авторами не рекомендовался ввиду их свойства
якобы провоцировать рецидивы малярии.
В ближайшие недели * после лихорадочных приступов рецидивы могут наступать
после резкого охлаждения или, наоборот, перегревания тела, промачивания ног,
физического переутомления, горячей ванны, а также во. время гриппа, ангины или
после вакцинации и т. д. Чтобы предупредить рецидивы, больным делают ванны,
вакцинацию и т. д. в дни очередного цикла лечения или, например при гриппе и
других случайных заболеваниях, назначают предварительно очередной цикл лечения.
При всяком лихорадочном заболевании или плохом самочувствии у малярика тре-
буется исследование толстой капли крови для выявления возможной активЯй ма-
лярии или паразитоносительства.
200
СПЕЦИАЛЬНАЯ ЧАСТЬ
Выздоравливающим после острого приступа полезны мышьяк, железо, иод, пре-
параты печени, витамины.
Осложнения малярии геморрагическим колитом и долевой пневмонией вызы-
ваются, как правило, дизентерийной палочкой и соответствующим пневмококком и
должны лечиться сульфидином и аналогичными препаратами.
Метиленовая синь, иод, сульфидин, стрептоцид оказывают недостаточное или
спорное действие на плазмодиев и для купирования малярийного приступа не долж-
ны применяться. Совершенно не действуют на плазмодиев квасцы, рвотной камень,
подсолнечник, эвкалипт, сирень, полынь и многие другие предлагаемые народной
медициной растительные средства.
ПОДКЛАСС NEOSPORIDIA
Споры образуются в течение всей жизни споровика. Из отрядов рас-
смотрим миксоспоридий и саркоспоридий.
ОТРЯД 1. MYXOSPORIDIA — МИКСОСПОРИДИИ
Миксоспоридии являются в большинстве случаев паразитами рыб;
отдельные виды живут в амфибиях и пресмыкающихся. Паразиты могут
жить во всех системах органов хозяина; по месту обитания взрослых
Рис. 108 — Myxobolus в коже карпа; споровики образуют
в коже бедые цисты. Из Плен.
Рис. 109 — Голова судака с кожей и жабрамн, поражен-
ными Myxobolus sp.
Споровик образует на теле судака шаровидные молочно-белые
цисты. По В. Д о г е л ю.
форм в хозяине различают миксоспоридий полостных (например, в мо-
чевом или желчном пузыре) или тканевых. . В тканях они лежат или
внутйжлеточно (редко), или заключены в цистах. Мы рассмотрим при-
меры только последних. У карповых рыб в листочках жабр бывают
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
201
узелки или вздутия благодаря присутствию Myxobolus miilleri Biitschli;
М. pfeifferi (Thelohan) может поражать различные органы усача (Bar-
bus fluviatilis) — почки, печень, селезенку, яичник и главным образом
мышцы. Вследствие развития больших капсул паразита на теле рыбы
видны припухлости; сами опухолеобразные разрастания паразита до-
стигают V2—2 см диаметром и даже более — доходя до размеров кури-
ного яйца (рис. 108—109).
Миксоспоридии многоядерны. В процессе полового размножения яд-
ра копулируют попарно; вокруг слившейся пары ядер обособляется уча-
сток протоплазмы паразита, называемый пан-
споробластом. Последний превращается в
двё многоядерные споры с амебоидным' заро-
дышем внутри. В теле миксоспоридии разви-
вается несметное количество спор с двумя
полярными капсулами каждая.
Споры весьма своеобразны и разнообразны
по форме. Они одеты двустворчатой оболоч-
кой; в капсуле, внутри, спрятана свернутая
спирально нить, благодаря чему спора получа-
ет некоторое конвергентное сходство со стрека-
тельной капсулой кишечнополостных животных.
При гниении погибшей рыбы споры осво-
бождаются и поступают в воду. С ней они
заглатываются здоровыми рыбами. В пищева-
рительном канале из спор выбрасываются ни-
ти полярных капсул, и спора таким образом
укрепляется на слизистой оболочке. Затем из
споры, по открытии ее оболочек, выходит
амебообразный зародыш, который проникает
через стенку кишечника и с током крови или
лимфы попадает на место паразитирования.
Миксоспоридии бывают причиной заболева-
ний и гибели рыб. Об этих паразитах прихо-
дится упоминать в медицинской паразитоло-
гии по косвенным причинам. Миксоспори-
дии часто встречаются в съедобных рыбах,
Рис. 110—Споры.
А —Myxobolus pfeifferi; В -
Hennegyia acerinae; С — Неп-
negyia zschokkei. Из К у до.
например, крупные капсулы Hennegyia zschokkei (рис. НО, С) в.
мышцах сигов, Myxobolus в коже карпа (рис. 108). Нахождение в мыш-
цах рыб заметных или крупных капсул может служить поводом к бра-
ковке рыб. Однако эти капсулы могут быть легко удалены из мяса ры-
бы, которое остается вполне пригодным для еды. При микроскопирова-
нии содержимого таких капсул хорошо видны характерные споры пара-
зита (рис. НО). Миксоспоридии ни при каких условиях не могут пара-
зитировать у человека, млекопитающих или птиц. 1
ОТРЯД 2. SARCOSPORIDIA — САРКОСПОРИДИИ
Саркоспоридии, известные прежде под названием мясных споро-
с п е р м и й, являются паразитами мышц млекопитающих, а также птиц
1 Подробнее о миксоспоридиях см. в книге проф. В. А. Догеля «Паразитарные
заболевания рыб». Ленинград, 1932. Для определения можно пользоваться опре-
делителем того же автора. «Пресноводные Myxosporidia СССР», Ленснабтехиздат,
1932 (вып. IV Определителей организмов пресных вод СССР. Всес. инет, озерно
речного рыб. хозяйства).
202
СПЕЦИАЛЬНАЯ ЧАСТЬ
и пресмыкающихся. Реже саркоспоридии паразитируют в соединитель-
ной ткани. Паразиты имеют вид удлиненных или округлых тел, от микро-
скопических размеров до величины нескольких сантиметров. Они одеты
оболочкой, состоящей из нескольких слоев (рис. 111). Более крупные
Рис. 111 — Продольный разрез мишерова мешка Sarcocystis
в мышце быка. По Wenyon.
саркоспоридии видны простым глазом, отличаясь сероватым или серо-
вато-белым цветом от основного фона мышечной, ткани. Обызвествлен-
cl £ tc
к **
Рис. 112 —Продольный разрез
через пристеночную часть ми-
шерова мешка овцы (Sarcocystis
tenella).
т — мускул; tc — соединительная ткань;
Zf— волокнистая зога; Е — оболочка;
cl — внутренние перегородки; spb — ка-
меры со споробластами; ch, sp — ка-
меры со спорами. По Алексееву.
Рис. 113 — Заражение саркоспоридиями.
Спора, внедрившаяся в эпителий кишечни-
ка подопытного животного. По Гарт-
ману и Пашеру.
ные паразиты еще резче бросаются в глаза. У взрослых паразитов под
наружной оболочкой образуется множество перегородок, в простран-
ствах между которыми лежат серповидные тела — споры (рис. 112)..
В отличие от спор книдоспоридий они не имеют полярных капсул, хотя
у спор некоторых саркоспоридий бывают на одном из полюсов нитевид-
ные отростки. Цикл развития саркоспоридий изучен недостаточно.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
203
При переваривании проглоченных спор из их оболочки освобождает-
ся амебовидный паразит, который проникает в слизистую оболочку ки-
шечника (рис. 114); здесь в эпителиальных клетках паразиты растут
и размножаются множественным делением. Далее, мерозоиты, невиди-
мому, мигрируют в мышцы, где оседают в мышечных волокнах. По ме-
ре роста образуется многоядерный паразит, который/ размножается
плазмотомией (разделение масс протоплазмы на части с несколь-
кими ядрами каждая). Паразиты превращаются в «м и ш е р о в ы
мешки», заполненные бесчисленным количеством спор длиной 10—
15 р. Так протекает развитие мышиной саркоспоридии Sarcocystis mu-
sts. У других саркоспоридий неизвестны стадии их жизни от заглаты-
вания спор до появления молодых паразитов в мышечных волокнах.
Рис. 114 — Sarcocystis Undemanni из мышечных
волокон гортани человека.
А —продольный разрез; В — поперечный разрез поражен-
ного мышечного пучка; С — изолированная спора. Из
В г и. m р t.
Паразиты проходят стадию споробластов, дающих споры. Заражение
происходит через поедание мышечных волокон со спорами паразитов.
Из примеров саркоспоридий приведем следующие: Sarcocystis
misheriana Kuhn — часта в мышцах свиней, tenella Railliet в форме
гороховидных утолщений наружной поверхносли пищевода овец, S. ma-
rts Blanch в мышцах тела мышей и пр. Известно несколько случаев
нахождения саркоспоридий у человека (рис. 114) в мышцах гортани
(S. Undemanni), сердца, плеча и др. Судить о действительной патоген-
ности их для человека данных нет. У саркоспоридий животных найдено
токсическое начало — саркоспоридиотоксин .(саркоци-
стин, саркоспоридии), ядовитый при опытах с лабораторными
животными. Весьма вероятно, что токсичность саркоспоридий влияет и
на хозяев этих паразитов, что можно ожидать лишь от сильных пора-
жений саркоспоридиями. При наличии саркоспоридий в тушах (мише-
204
СПЕЦИАЛЬНАЯ ЧАСТЬ
ровы мешочки) туши бракуются, если мясо сильно водянисто или
сильно обесцвечено; сало или жир в таких случаях выпускаются без
препятствия. Пищевод, пораженный саркоспоридиями (В alb lane а), на-
правляется на утилизацию.
КЛАСС С1ЫАТА РЕСНИЧНЫЕ, ИЛИ ИНФУЗОРИИ
Тело покрыто пелликулой, несущей множество ресничек, являющих-
ся органоидами движения. Реснички или равномерны по длине (Holo-
tricha — равноресничные), или различны в этом отношении (Hete-
Рис. 115—116 — Balantidium coli.
Слева — в живом состоянии (внизу виден цитопрокт, над
ним большая ©колоректальная вакуола); справа—фиксиро-
ванный и окрашенный железным гематоксилином, Ориг. рис.
Алексеева.
rotricha — разноресничные). Цитофаринкс обычно есть. Органои-
дами выделения служат пульсирующие вакуоли. В эндоплазме два
ядра — макронуклеус и микронуклеус. Первое играет роль в вегетатив
ной жизни инфузорий, второе выступает на первый план в период поло-
вого размножения (конъюгация). Размножаются делением, которому в
известные периоды может предшествовать конъюгация. Инфузории
могут инцистироваться. Тип — инфузория-туфелька. 1
Род Balantidium
s Balantidium coli — балантидий
Единственной достоверной паразитической инфузорией человека яв-
ляется балантидий — Balantidium coli, самый крупный из паразитиче-
ских простейших человека. Размеры тела балантидия колеблются в пре-
делах 30—200 на 20—70 р . Обычные средние размеры его равняются
50—70 р длины и 40—70 у ширины. Паразитизм очейь мало отразился
на организации этой инфузории. Тело ее несимметрично. Передний ко-
нец заострен более, чем задний. Инфузория одета пелликулой, допус-
кающей некоторые изменения формы тела при движении. Балантидий
1 Термин «тип» применяется к трем различным понятиям: 1) тип в смысле
высшей систематической категории (тип простейших, червей и др.); 2) тип — в
смысле характерного для данной группы животного (как в данном случае) и
3) тип — тот именно экземпляр животного, по которому описывается новый вид
животного.
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
усеян продольными рядами коротких ресничек, придающих паразиту
продольно-исчерченный вид. Близ переднего конца тела лежит цитостом
(рот), ведущий в короткий пищевод. Около рта помещаются более круп-
ные реснички, образующие перистом, который служит для загоняния,
вместе с током жидкости, пищи в пищевод. Благодаря обладанию раз-
личными сортами ресничек балантидий относится к разноресничным ин-
фузориям'— Heterotricha. Близ заднего конца тела помещается
анальная пора — цитопрокт .(рис. 116).
Рис. 117 — Balantidium coli кишечника человека.
По Wenyon.
Рис. 118 — BaLintidium coll. Кан-
нибал изм; начало поедания од ной
особи другой и полное загла-
тывание меньшей особи более
крупной. По В. Догелю и
кВ. Г н-е з д и л о в у.
Под пелликулой лежит тонкий слой прозрачной альвеолярной эк-
топлазмы. В зернистой эндоплазме погружен почковидный макронук-
леус с густым скоплением хроматина и несколькими ядрышками. На его
гогнутой стороне помещается шаровидный микронуклеус. Сократитель-
ных вакуолей две. Одна лежит у заднего конца тела, тогда как другая
помещается на уровне макронуклеуса.
Размножается балантидий поперечным делением. При быстрых по-
вторных делениях образуются гнезда мелких инфузорий в толще кишеч
ной стенки. Другие способы размножения балантидия недостаточно изу-
чены.
В. coll инцистируется в кишечнике. Инфузория одевается двуслойной
оболочкой, под которой паразит теряет свой ресничный покров. В содер-
жимом цисты видны макронуклеус, задняя сократительная вакуоль и
особое сильно преломляющее свет тело. Овальные или шаровидные цис-
ты достигают рекордных, по сравнению с цистами остальных простей-
ших, паразитирующих в человеке, размеров, а имейно — 50—60 у диа-
метром. Встречаются цисты с двумя инфузориями в каждой. Brumpt по-
лагает, что особи попарно одеваются общей оболочкой, под которой
и конъюгируют. Паразитирует балантидий! в толстых кишках, особенно
в coecum. Он может встречаться и в нижнем отделе тонких кишок
206
СПЕЦИАЛЬНАЯ ЧАСТЬ
(Ржаницын, 1923). Питается балантидий углеводами (крахмалом) и
различными остатками в каловых массах. Заглатывает также красные
и белые кровяные тельца и обрывки тканей. В культурах балантидий,
бедных питательными веществами, нередко наблюдается каннибализм,
когда крупные инфузории целиком заглатывают меньших по размеру
(рис. 118, Догель и Гнездилов).
Вне хозяина подвижные балантидии могут жить лишь несколько ча-
сов. Балантидии из свиней в анаэробных условиях при комнатной темпе-
ратуре живут до 10 дней (Rees). Цисты балантидия гораздо более стой-
ки. При комнатной температуре во влажной среде они живут около 2-х
месяцев; в сухом состоянии в тени выдерживают 1—2 недели, а на
прямых солнечных лучах погибают через 3 часа. В желчи выдерживают
15 дней, в моче— 10 дней, в желудочнбхМ соке— 12 часов, в 5% рас-
творе карболовой кислоты — 3 часа, в 10% формалине — 4 часа (Ohi,
1924).
Заражение происходит через заглатывание цист; не исключается
возможность инвазирования и подвижными балантидиями; по крайней
мере у некоторых подопытных животных инфузории безнаказанно про-
ходили через желудок, кишечник и открывались живыми в слепой киш-
ке (морская свинка в опытах Hegner). Где и при каких условиях про-
исходит вскрытие заглоченных цист — еще неизвестно.
Балантидии могут жить и размножаться в кишечнике в некоторых
случаях без всякого патогенного влияния на его ткани. Однако В. colt
встречается не только в содержимом толстых кишок, но и в толще их
стенок. На препаратах они могут быть обнаружены только при раннем
вскрытии тела (не-позднее 3—5—-6 часов после смерти).
Новые авторы отмечают возможность обнаружения балантидиев при
вскрытии' более чем через сутки после смерти.
Причину внедрения балантидиев из просвета кишки в ее стенку, что,
является началом проявления патогенного влияния балантидиев на
организм, Догель и Гнездилов видят в обеднении кишечного содержи-
мого углеводами и прежде всего крахмалом. «Гликогеновый голод»,,
вероятно, и толкает балантидиев к миграции в ткани кишечника.
Инфузории пробираются в либеркюновы железы, протискиваются
между клетками слизистой оболочки, повидимому, растворяют ее фер-
ментами, благодаря чему образуются язвочки, и проникают в подсли-
зистую ткань (рис. 119, Д); здесь балантидии делятся и образуют це-
лые гнезда, проникая глубже (рис. 119, В) до muscularis. В. coli некро-
тизирует ткани, вследствие чего образуются язвы, несколько отличаю-
щиеся от язв при амебной дизентерии своим гангренозным характером.
В зависимости от длительности поражения язвы при балантидиозе-
бывают разной величины — от макового зерна до серебряного рубля.
Разбросаны они обыкновенно на здоровой по внешнему виду слизистой
оболочке, которая лишь в некоторых местах и в слабой степени бывает
гиперемирована.
Балантидиозная язва имеет валикообразные подрытые края с хороша
выраженным яркокрасным пояском по периферии, вследствие капилляр-
ных кровоизлияний. Дно язвы или зеленовато-желтого цвета, если она
покрыто гноем, или же, при наличии некроза, грязнобурого или черно-
го цвета.
Одиночные язвы чаще овальной формы, своей длинной осью лежат
перпендикулярно к продольной оси кишечника. Язвы, образовавшиеся
слиянием нескольких меньших язв, имеют неправильные очертания: они
помещаются обычно в слепой кишке, боковых изгибах толстой кишки,
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
207
реже в S-romanum, а в запущенных случаях бывают и в прямой кишке
(Ржаницын).
Балантидий проникает также в кровеносные и лимфатические сосу-
ды, но далеко по ним, видимо, не распространяется. Однажды он был
119—Балантидии в кишечнике человека.
А — различные стадии выхода инфузорий из просветов либеркюновых
желез в межуточную ткань (железы разрезаны поперек); В — гнезда
размножения балантидиев в меж>точноп ткани кишки; С балантидий
в толще стенки тонкой кишки, большая ч^с)ь железистого слоя которой
разрушена. По Ржаницын у.
найден в толще желудочной стенки и в содержимом желудка у боль-
ной, страдавшей раком привратника и имевшей патологическое сооб-
щение полости желудка с толстой кишкой.
Распространение балантидиев в толщу мышечной оболочки зависит
от давности процесса. Часть паразитов в тканях стенки кишечника по-
208
СПЕЦИАЛЬНАЯ ЧАСТЬ
гибает, рассасываясь или подвергаясь фагоцитированию. При достиже-
нии балантидиями субсерозной оболочки кишечника может развиться
перитонит (Н. Жук).
Болезненные проявления при балантидиозе вариируют от поноса до
дизентерии, сходной с амебной. На почве первичных поражений кишеч-
ника, причиняемых балантидием, может развиться хроническая дизенте-
рия или колит. Смертность при балантидиозе довольно большая: по
статистике Strong — 29% (1904).
Диагноз ставится по нахождению живых инфузорий в испражнениях;
паразиты оживленно плавают в свежем жидком кале и легко видимы
вследствие своих больших размеров.. Отыскание цист не имеет практи-
ческого значения, так как у человека они образуются редко.
Уже давно замечено, что у различных животных в кишках также
бывают балантидии; в частности балантидиями свиней удалось заразить
обезьян. Для свиней балантидии совершенно непатогенны, и у этих хо-
зяев паразиты легко инцистируются. Весьма вероятно, что балантидии
свиньи, человека и обезьян относятся к одному и тому же виду
В. coli; хотя опыты Grass! и Callandruccio не дали результатов, их нель-
зя считать окончательно решающими вопрос. Весьма вероятно, что
именно свиньи являются источником заражения человека балантидио-
зом через проглатывание цист балантидиев от свиней.
Например, под Ташкентом в свиноводческом совхоз^ среди рабочих
наблюдалось большое количество носителей балантидий, тогда как в
самом Ташкенте балантидиоз человека исключительно редок (Чапур-
ская, Баженова, Лейтман, Фиалковская).
Аналогичное значение, видимо, имеют и кабаны (Средняя Азия,
В. Гнездилов). В анамнезах многих больных значился контакт со свинь-
ями (уборка животных, хлева и др.). Кроме того, возможно зараже-
ние балантидиями при колбасном производстве или заготовке кишеч-
ного сырья (промывка свиных кишок, надувание .кишок ртом для их
калибровки и др.).
Балантидии широко распространены по свету.
К 1935 г. был описан 201 случай балантидиоза человека, и только
14 из них были исследованы микроскопически.
Лечение балантидиоза трудно, так как нет специальных лекарств.
Применяются клизмы из азотнокислого хинина (0,75 г : 300 см3 воды),
азотнокислого серебра (1,0 : 3000,0) и др.
ДРУГИЕ ПРЕДПОЛАГАЕМЫЕ ПАРАЗИТИЧЕСКИЕ ИНФУЗОРИИ
КИШЕЧНИКА ЧЕЛОВЕКА
У одного и того же больного Шаудинн обнаружил две инфузории и
описал их в качестве новых видов под наименованием Balantidium mi-
nut ит и Nyctotherus faba. Повторные нахождения .этих паразитов бы-
ли сомнительны. В качестве обитателей кишечника описывались и дру-
гие различные виды инфузорий: Chilodon uncinatus, Uronema caudatum,
Colpoda cucullus, C. steini.
Colpoda steini бывает ложнопаразитом у различных животных, в ча-
стности у слизняка Agriolimax agrestis, обитая в его кишечнике, ман-
тийной полости, гермафродитной железе, белковой железе и в других
органах. На слизняков действует неблагоприятно, видимо обусловливая
их частичную гибель (Reynold).
При большом развитии обследований faeces человека на паразити-
ческих простейших ни одна из инфузорий, кроме В. coli, не оказалась
признанной бесспорным паразитом человека. Для некоторых из них да-
УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
209
же сомнительно, чтобы они действительно были в кишечнике обследо-
ванного больного. Возможно, что они были занесены с водой извне в
исследуемые испражнения, или уже в инцистированном состоянии про-
шли через желудочно-кишечный канал человека и вышли из цист в
faeces на положение копрозойных форм. Следовательно, здесь возмож-
ны случаи мнимого ложнопаразитизма.
ЛИТЕРАТУРА
Гительсон И. И. Кожный, лейшманиоз (пендинская язва). Туркменгосиздат, Аш-
хабад, 1933.
Догель В. А. Подцарство Protozoa, тип простейших. Руководство по зоологи^
т. 1, под ред. Л. Зенкевича, Москва, 1937.
Ирен М. Г. Инфузорная дизентерия (балантидиоз). Гос. изд. научно-техн, и соц.-
экон. литературы УзССР, Ташкент, 1939.
Куш ев Н. Е. Лекции по малярии. Саратов, 1925.
Латышев Н. И. Малярия и борьба с ней. Гос. изд. Москва, 1923.
Н о х т. Амебная дизентерия. Перев. Тер-Джаняна, Тифлис.
Павловский Е. Н. Важнейшие достижения советской паразитологии по про-
тозоологии и гельминтологии за 20 лет. Природа, № 11, 1937.
Павловский Е. Н. (ред.) Практикум медицинской паразитологии. А. Гуцевич.
Малярийные плазмодии; В. Гнездилов. Паразитические простейшие. Биомедгиз,
Ленинград, 1935.
Петров В. П. Кала-азар (лейшманиоз внутренних органов). Изд. Комитета науки
УзССР, Ташкент, 1938.
Т а р е е в. Клиника малярии.
Топорков Ф. М. Малярия. Материалы к учению об эпидемиологии и пато7
генезе. Астраханский гос. мед. ин-т, Сталинградское книгоиздат., 1939.
Фавр В. В. Опыт изучения малярии в России в санитарном отношении. Харьков,
1903.
Ходукин Н. И. Основные проблемы эпидемиологии кала-азар в связи с эпи-
демиологией собачьего лейшманиоза в Средней Азии. Ташкент, 1929.
Шахматов А. П. Руководство по малярии, ч. 1 (паразитология, клиника, диа-
гностика, лабораторная методика и лечение малярии). Изд. «Красное знамя»,
Томск, 193’9.
Штакельберг Н. С. Библиография малярии (эпидемиология, переносчики,
борьба с малярией, организация противомалярийного дела в СССР). 1771—1935.
Под ред. акад. Е. Н. Павловского. Изд. АН СССР.
Шуб Г. М. и Федоров В. Г. К эндемии малярии бассейна оз. Ильмень и план
борьбы. Изд. Леноблздравотдела и института им. Пастера, 1935.
Щупак Н. Б. Ламблиоз. Госмедиздат УССР, 1939.
Эпштейн Г. В. Патогенные простейшие, спирохеты и грибки. Москва, 1931.
Эпштейн Г. В. Материалы по эпидемиологии амебиаза. Медицинская паразито-
логия и паразитарные болезни, т. I, вып. 5—6, 1932, и ряд статей разных ав-
торов по протозоологии в том же журнале.
Эпштейн Г. В. Паразитические амёбы. 1941.
Эпштейн Г. В. Техника микроскопического исследования простейших. 1940.
Якимов В. Л. Лейшманиозы. Труды экспедиции по изучению тропических болез-
ней людей и животных Туркестанского края в 1913 г., т. I, Петроград, 1915.
Якимов В. Л. Болезни домашних животных, вызываемые простейшими. 1931.
Архив Русского протистологического общества.
Большая медицинская энциклопедия. Статьи: Амёбы, Лейшманиоз, Малярия
и др.
Медицинская паразитология и паразитарные болезни. Журнал..
Труды малярийных съездов, экспедиций, тропических институтов.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
(медицинская гельминтология)
Тип Vermes—Черви
Среди червей имеется большое количество (около 175 видов) пара-
зитических форм, которые принадлежат к подтипам плоских ч е р-
в е й (Platodes), скребней (Acanthocephali), круглых червей
(Nematodes); меньшую роль играют пиявки (Hlrudinea) и в о л о са-
тиновые (Gordiacea). Паразитические черви, кроме пиявок и Gor-
diacea, издавна известны под научным именем собирательного, но не
систематического значения Helminthes или Helminthozoa. В связи с
этим отрасль паразитологии, изучающая их, получила название гель-
минтологии. С термином Helminthes по значению совпадает слово
глисты, происходящее от латышского корня glidet, что значит'
скользкий.
В специальном материале даются дальше лишь общие сведения об
экстенсивности глистных инвазий, ибо в одном и том же пункте про-
цент зараженности данным паразитическим червем может весьма ва-
риировать у различных групп населения.
Подтип Плоские черви — Platodes
Все черви являются многоклеточными организмами:
(Metazoa), построенными из производных трех зародышевых листков —
эктодермы, энтодермы и мезодермы. Тело их двухсторонне-сим-
метричное, листовидное или лентовидное; в огромном большинстве
случаев плоские черви гермафродитны; пищеварительный канал,
по основным особенностям организации, соответствует гастровас-
кулярной пищеварительной полости кишечнопо-
лостных животных; вторичной полости тела (целома) у плоских
червей нет; в связи "с этим нет и кровеносной системы. Органов дыха-
ния также нет; органы выделения построены по протонефри-
диальному типу, т. е. состоят из внутренней замыкающей
клетки с подвижным жгутом, свисающим в полость отходящего от
клетки канала; такие каналы сливаются в более крупные и в конце
концов открываются наружу. Нервная система узловая; она образована
из головных ганглиев и отходящих от них нервных с т в о-
лов, соединяющихся перемычками друг с другом. Из трех классов
плоских червей — турбеллярий (Turbellaria), сосальщиков
(Trematoda) и ленточных червей (Cestoidea) — паразиты чело-
века имеются среди последних двух групп. Турбеллярии, за редким»
исключениями, свободноживущи.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
211
КЛАСС СОСАЛЬЩИКИ—TREMATODES
Сосальщики имеют несегментированное тело листовидной или
овальной формы; являясь потомками турбелляриеподобных предков,
сосальщики сохраняют ресничный покров только в личиночной стадии.
Пищеварительный канал есть; большей частью гермафродитны; раз-
множаются с превращением или со сменой поколений; все сосальщи-
ки — паразиты (наружные или внутренние); почти все во взрослом
состоянии являются паразитами позвоночных.
Строение сосальщиков. Все последующие данные касаются
эндопаразитических сосальщиков (Digenea), так как только их пред-
ставители паразитируют 'у человека .(рис. 120).
Форма тела уплощенная, языкообразная, листовидная, удлинен-
ная, коническая, цилиндрическая и др. Размеры тела чаще всего ко-
леблются в пределах от 1 мм до 2—3 см. Тело одето кутикулой, кото-
рая у некоторых видов покрыта шипиками. Органами прикрепления к
хозяину являются присоски — мускулистые полые диски с шипиками,
крючьями или без них. Количество присосок и местоположение их
вариируют у разных видов. Одна — ротовая присоска окружает
ротовое отверстие, другая или остальные присоски, если их несколько,
оканчиваются слепо и не имеют никакого отношения к пищеваритель-
ной системе; лежат они на брюшной стороне тела. Близ переднего
края брюшной присоски Открывается половое отверстие. Выделительная
пора (экскреторное отверстие) у эндопаразитов помещается обыкно-
венно на заднем конце тела.
Кутикула сосальщиков выделяется эпителиальными клетками,
которые погружены в паренхиму паразита и не образуют сплош-
ного эпителиального покрова. С поверхностью кутикулы они соединяют-
ся тонкими отростками. Между этими клетками иногда располагаются
одноклеточные железы. Под кутикулой лежат мышцы, которые по на-
правлению своих волокон диференцируются на три слоя: наружный —
кольцевой, средний — диагональный и внутренний — продольный. Вме-
сте с кожными покровами эти слои образуют кожно-мускульный
мешок, заполненный внутренними органами, дорзовентральными
мышцами и особой сетеобразной соединительной тканью, лежащей
везде в промежутках между органами; ее называют паренхимой. При-
соски снабжены экваториальными, радиальными и меридиональными
мышечными волокнами и являются специальными мышечными орга-
нами. Мышцы у сосальщиков гладкие.
Пищеварительная система начинается ротовым отвер-
стием, лежащим на переднем конце тела (т. е. терминально) или
несколько отступя кзади на брюшной поверхности (т. е. с у б т е р ми-
н а л ь н о) на дне ротовой присоски. Ротовая полость (cavum
ors) ведет в мускулистую глотку (pharynx) обычно шаровидной
формы. Далее кзади отходит короткий пищевод (oesophagus), впа-
дающий в кишечник (intestinum). Все эти части, кроме кишечник:,
называют передней кишкой, которая эмбриологически развивается из
наружного зародышевого листка и выстлана изнутри кутикулой. Соб-
ственно кишечник, как правило, образован двумя кишечными трубками,
оканчивающимися слепо у заднего конца тела (рис: 137). Обыкновенно
они простые, но у некоторых сосальщиков более или менее богато вет-
вятся (Fasciola hepatica, рис. 136). Своими задними концами кишечные
трубки могут соединяться друг с другом, благодаря чему кишечник об-
212
СПЕЦИАЛЬНАЯ ЧАСТЬ
т. е. в нем совмещаются
Рис. 120—Печеночная дву-
устка {Fasciola hepatica)\ ана-
томия.
Г— пищеварительный канал; тп —
ротовая присоска с ротовым отвер-
стием: и — матка; р— глотка; v —по-
ловая клоака; о — яичник; 5 — скор-
луповая железа; vg — желточникп;
/—семенники. По Стайльсу.
разует удлиненное кольцо. (Ciclocoelium orientate). Для сосальщиков
характерно наличие единственного отверстия — ротового, которым пи-
щеварительный аппарат открывается наружу. Следовательно, это отвер-
стие служит и для принятия пищи и для выбрасывания ее остатков,
функции рта и заднепроходного отверстия
(anus). В недавнее время открыты сосаль-
щики с одним {Opecoelus sphericus) или
с двумя анальными отверстиями {Schistor-
chis carneus).
У кровяных двуусток {Schistosomatidae)
ветви кишечника сливаются в непарную
заднюю трубку. Есть.сосальщики вообще
с единственной трубкой пищеварительного
аппарата (соответственно свободноживу-
щим прямокишечным турбелляриям —
Rhabdocoela), например, Prosorhynchus
squamatus из пилорических придатков ки-
шечника бычка {Cottas scorpius). В других
случаях атрофируется одна из ветвей ки-
шечника, вследствие чего наблюдается
резкая несимметричность пищеваритель-
ного аппарата. Можно ожидать открытия
и бескишечных сосальщиков, так как уже
известна церкария, лишенная кишки(Сегса-
ria pleurolophocerca). Необходимо отме-
тить, что в прежнее время ртом счита-
лась и брюшная присоска, не имеющая
никакого отношения к пищеварительным
' органам. Отзвуком таких воззрений яви-
лись неправильные анатомически термины,
как двуустка —Distomum, многоустка —
Potystomum и др.
Кишечник выстлан внутри цилиндри-
ческим эпителием и снабжен продольным
и кольцевым слоями кишечных волокон.
Эпителиальные клетки кишечника могут
выпускать псевдоподиеобразные отростки
или давать нитевидные щеткообразные
выросты; однако возможность фагоцити-
рования ими оформленных частей пищи
в последнее время считается сомнитель-
ной (Fuhrmann). Из придатков пищевари-
тельного аппарата можно отметить только присутствие однокле-
точных слюнных желез, впадающих в глотку и в пищевод.
Пищей сосальщиков являются содержимое кишечника хозяина,
слизь, эпителиальные клетки, кровяные клетки, выделения слизистой
оболочки желчных протоков, жидкая часть крови и др. Пищеварение
у них внеклеточное, ферментативное. Весьма вероятно, что
всасывание пищи кровяными сосальщиками с рудиментарным кишечни-
ком может происходить эндосмотически через поверхность тела па-
разита.
Органами выделения сосальщиков является богато развитая
выделительная система, построенная по протонефридиальному типу.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА 213
Замыкающие внутренние клетки дифференцируются на собственно клеточное тело,
воронку1 и начальный капилляр. Все это вместе образует «концевой орган»; распо-
ложение их, число и другие детали строения выделительной системы имеют боль-
шое значение в систематике сосальщиков. От тела замыкающей клетки в воронку
свисают длинные реснички; ресничная выстилка и пучки длинных жгутов бывают и
в более крупных стволах экскреторной системы, с которыми соединяются концевые
органы. Главные стволы и экскреторный пузырь являются образованиями синци-
тиального характера. Экскреторный пузырь открывается пброй, снабжен-
ной кольцевыми замыкательными мышцами (sphincter,). Выделительная пора ле-
жит терминально или субтерминально у заднего конца тела на вентральной или
дорзальной поверхности. У сосальщиков Monogenea выделительная система пол-
ностью парная. Собственно экскреторной частью выделительной системы являются
выделительные каналы, плазматическая стенка которых выбрасывает в просвет
канала продукты обмена веществ. Концевые органы служат для регулирования
тканевого осмотического давления, что достигается выделением в полость воронки
избытков воды, всасываемой в организм. Прогоняемая действием ресничек и би-
чей вода вымывает собственно экскреторные продукты из выделительных каналов
и уносит их (т. е. продукты) с собой.
Кровеносной системы 'у сосальщиков нет; следовательно,
распространение всасываемой переваренной пищи происходит имби-
бицией («пропитыванием»).
Нервная система образована парой мозговых узлов, лежа-
щих по бокам глотки и соединяющихся друг с другом дорзальной ко-
миссурой. ' От ганглиев идут три пары передних и три пары задних
нервных стволов (пара брюшных, пара спинных и пара боковых), со-
единяющихся перемычками друг с другом. Органы чувств (напри-
мер глаза) у взрослых эндопаразитических сосальщиков отсутствуют;
личинки их (м и р а ц и д и и), а также церкарии имеют органы
зрения, что связано со свободным пребыванием в воде этих стадий жиз-
ненного цикла сосальщиков. В редчайших случаях и половозрелые
эндопаразитические сосальщики имеют рудиментарные органы зрения в
виде пигментных пятен (Acanthopsolus).
Органы размножения. Огромное большинство сосальщиков
является гермафродитами и имеет полные комплекты мужских и жен-
ских половых органов. Кровяные сосальщики (Schistosomatidae) раз-
дельнополы. Органы размножения развиты весьма сильно, причем пре-
обладают над прочими органами тела.
Мужские половые органы (рис. 120) состоят обыкновенно
из пары семенников (testes), выводных путей для половых продук-
тов и копулятивных органов. Семенники имеют разную форму — округ-
лых или лопастных тел, трубчатых ветвистых органов и др. Лежат они
обыкновенно в задней половине тела паразита и в его срединной (ме-
дианной) части. От каждого семенника кпереди отходит семепровод
(vas deferens) в форме тонкой трубки; оба семепровода сливаются в
непарный семеизвергательный канал (ductus ejaculatorius\
ведущий в циррусовую сумку (bursa cirri). Упомянутый канал
расширяется здесь в семенной пузырек (vesicula seminalis) или
образует несколько изгибов; в него, или в дистальную часть, соединяю-
щуюся с циррусом, впадают одноклеточные железки, образующие пакет
простатических желез (gl. prostaticae), далее половые пути
переходят в концевой мышечный отдел в виде трубки.— копулятив-
ный орган или cirrus. Последний может выворачиваться наружу
через мужское половое отверстие.
Женские половые органы (рис. 120) имеют следующие
1 Не смешивать с концевой ресничной воронкой метанефридий кольчатых
червей.
214
СПЕЦИАЛЬНАЯ ЧАСТЬ
составные части: единственный яичник (ovarium), яйцевод (ovi-
duct), пара желточников (vitellogenes), желточные про-
токи, семеприе мн и к (receptaculum seminis). Все эти органы со-
общаются с особой камерой, называемой оотип. В нее открывается
множество одноклеточных железок (железы Мелиса) и из оотипа
выходит начальная часть матки .(uterus), имеющей трубчатое строе-
ние. Кроме того, от оотипа отходит на спинную сторону тела лауре-
ров канал, открывающийся самостоятельным отверстием (canalis
Laureri). Выходная часть матки, играющая роль влагалища, от-
крывается рядом с отверстием цирруса в половую клоаку или
непосредственно на поверхность тела. Лауреров канал имеется у мно-
гих Digenea; у других сосальщиков бываёт желточно-ки ш е ч н ы й
канал, соединяющий место слияния яйцевода и желточных каналов
с кишечником (ductus vitello-intestinalis).
Рис. 121 —Fasciola hepatica. Процесс образования скорлупы яйца.
1 — вителлогенная клетка из дольки желточника; 2 - вителлогенные каетки
в начальной час:и матки; у клгтос выделяются гранули «е цестза, >бразую-
шего на поверхности группы клето< оболочку яйца слиянием гр»нул; 3 — жел-
точн ..е клетки зретого яйца; 4 клетей те <ь Мелиса; 5 - яйцеелегка из кон-
цевой части яйцевода; 6 — яйце «летка в «яйцо, проде [ываютая кариокине-
тическое деление. Ио К о u г i и N a v s s.
Яичник вырабатывает яйцевые клетки, которые через яйцевод по-
ступают в оотип. Желточники дают клетки, богатые желтком, которые,
в сущности, являются недоразвитыми и измененными (иначе говоря —
абортивными) яйцами. При копуляции семя от копулирующего партне-
ра поступает через извивы матки в оотип и в семеприемник (если по-
следний не отсутствует).
Желточные клетки тоже выходят со своей стороны в оотип. Здесь
(или в яйцеводе) происходит оплодотворение яйцевой клетки, которая
с некоторым количеством желточных клеток одевается прочной оболоч-
кой или скорлупой в качестве сформировавшегося яйца поступает
в матку. Скорлупа образуется за счет капелек желтоватого вещества,
выделяемого желточными клетками (рис. 121); весьма возможно, что
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
215
в формировании скорлупы принимает участие и секрет желез ’Мелиса,
которые известны более под старым названием скорлуповых же-
лез. Однако скорлупа яйца образуется и у тех сосальщиков, у кото-
рых железы Мелиса вовсе отсутствуют (например Sanguinicold). Функ-
цией этих желез считается выделение желатинозной жидкости, кото-
рая заполняет матку и в которой находятся уже готовые яйца. Пови-
димому, функциональное значение желез Мелиса не одинаково у раз-
личных сосальщиков.
Лауреров канал служит, вероятно, для удаления избытка желточ-
ных клеток или живчиков. Аналогичную роль
шечный канал с той разницей, что в послед-
нем случае желточные клетки не выбрасыва-
ются вон, а попадают в кишечник, где исполь-
зуются для целей питания. Лауреров канал
может служить и для копуляции.
Оплодотворение у сосальщиков перекре-
стное или же бывает самооплодотво-
рение гермафродитной особи. Плодовитость
их очень велика, и сильно развитая матка
является временным хранилищем готовых яиц,
которые через ее отверстие выбрасываются
наружу.
Готовые яйца сосальщиков характеризу-
ются определенной формой, размерами и внут-
ренним строением. Все эти данные имеют важ-
ное значение для систематики и для диагно-
играет и желточно-ки-
Рйс. 122 — Fasciola hepa-
tica.
Яйцо с зародышем внутри;
h — глазок мирацидия; f — пе-
редний конец мирацидия; d —
желток. По Stilus.
стического распознавания наличия паразитов
в организме хозяина. Яйца сосальщиков ищут
в кишечном содержимом (дуоденальный зонд),
в faeces, в моче, мокроте, смотря по ожи-
даемому месту обитания паразитов.
Оболочка яиц обыкновенно бывает окра-
шена в различные оттенки — от светложелтоватого цвета до темно-
коричневого. Яйца многих видов имеют на одном полюсе крышечку.
Цикл развития .(рис. 122, 123, 124). Степень развития зароды-
ша в только что выброшенных яйцах различных сосальщиков различна.
У одних под оболочкой яйца лежит уже вполне развитая личинка (напри-
мер, Schistosomiun, Gasterostomum и др.); у других, напротив, эмбрио-
нальное развитие начинается только после выхождения яиц из хозяина.
В яйце сосальщиков развивается характерная личинка, называемая
•мирацидие-м (рис. 122). Она может жить только в воде; мирацидий
одет ресничным покровом с продольными и кольцевыми мышечными
волокнами; далее имеются: зачаток будущих покровов следующей ста-
дии жизненного цикла паразита, буравящий аппарат на перед-
нем конце тела, так называемые головные железы, протонефри-
дии с одной или с двумя замыкающими клетками, нервный узел и, обык-
новенно, пигментированный глазок. В первичной полости тела мира-
цидця лежат половые клетки, которые у некоторых видов уже обра-
зуют зародышевые шары клеток, являющиеся зачатками особей
следующей стадии жизненного цикла сосальщика, именно редий.
Как правило, мирацидий выходит из яйца, открывая его крышечку
в воде; у некоторых форм этот процесс происходит в матке сосальщи-
ка, являющегося, в таком случае, живородящим. Другой крайней одо-
216
СПЕЦИАЛЬНАЯ ЧАСТЬ
тивоположностью можно считать сосальщиков, паразитирующих в на-
земных моллюсках. Заражение последних происходит проглатыванием
яиц, из которых личинка выходит только в кишечнике моллюска при
действии пищеварительных соков.
Ресничный покров и глазок мирацидия (рис. 122, й) являются ор-
ганами, присущими свободному периоду жизни личинки, которая пла-
Рис. 123 — Цикл развития печеночной двуустки.
1—4 — развитие мирацидия в яйце, 5—мирацидий; 6—8 превращение в
спороцисту; 9 - спороциста с редией; 10—13 развитее в редиях церка-
рий; 14—15 — церкарии; 16 — инцисчи].ованная церкария; 17 — двуустка,
вышедшая из церкарии. По Suzuki.
вает в воде и активно (реже пассивно) проникает в промежуточного
хозяина, которым является какой-нибудь моллюск. ВбуравившиСь <в
моллюска, мирацидий сбрасывает ресничный покров и превращается в
спороцисту, обычно лишенную каких-либо органов, кроме прото-
нефридий (рис. 123, 6—S).
В полости спороцисты развивается следующее личиночное поколе-
ние, так называемые редии .(рис. 123, Р). Они возникают из зароды-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
217
Рис. 124 — Адолескарий.
По Скворцову.
шевых клеток без оплодотворения таковых, следовательно, в цорядке
девственного размножения или партеногенеза. Редии прорывают стенку
спороцисты и освобождаются, оставаясь в том же хозяине. Строение
редий более сложно. На переднем конце их удлиненного тела поме-
щается ротовое отверстие, ведущее в глотку и в непарную пищевари-
тельную трубку. Парная выделительная система развита хорошо. В по-
лости тела редии из зародышевых клеток развиваются хвостатые ц ер-
кар и и, являющиеся следующей личиночной формой сосальщика
(рис. 123, 10—13). У редий некоторых видов сосальщиков церкарии
отрождаются через особое отверстие; церкарии выходят из редий и
активно покидают тело моллюска.
Строение церкарий (рис. 123, 14 и 15) разнообразно, но при всем
разнообразии можно отметить некоторые общие черты. Округлое или
удлиненное тело церкарии несет на
заднем конце хвост, являющийся
органом плавания в воде. Церка-
рия имеет пару присосок—рото-
вую и брюшную; первая ведет в
глотку, пищевод и вилообразно
ветвящийся кишечник. Отлично
развита выделительная система.
Кишечник может быть представлен
мешкообразным впячиванием или
вовсе отсутствовать. Из внутрен-
них органов отметим мозговые
ганглии, с которыми связаны глаз-
ки, зачатки полового аппарата и
некоторые органы специально ли-
чиночного значения, как головные
железы, называемые иначе слюн-
ными, кожные железы, выделяю-
щие материал для оболочки при
инцистировании, и буравящий ап-
парат для облегчения внедрения
в тело хозяина.
Свободно плавающая церкария
одних сосальщиков одевается ци-
стой, которая образуется из секрета цистной железы. Инцистирование
происходит или на поверхности воды, или на погруженных в воду ра-
стениях. При инцистировании церкария округляется, и циста прини-
мает шарообразную форму (рис. 123, 16). У других сосальщиков цер-
кария активно вбуравливается в тело окончательного или предпослед-
него хозяина, пользуясь буравящим стилетом и действуя на покровы
хозяина секретом головных желез, обладающим способностью раство-
рять ткани тела. В предпоследнем хозяине церкарии инцистируются и
в этом состоянии называются метацеркариями. Инцистированные
в свободном состоянии церкарии известны под именем адолескарий
{рис. 124).
Попав активно или пассивно в окончательного хозяина, церкарии
превращаются в половозрелых сосальщиков, причем проделывают ино-
гда весьма длинную миграцию, пока попадут к месту постоянного оби-
тания в теле хозяина.
Таким образом полный жизненный цикл сосальщиков сложен. Изу-
218
СПЕЦИАЛЬНАЯ ЧАСТЬ
чен он полностью сравнительно у немногих видов. Схематически его
можно изобразить в следующем виде:
* главный хозяин половозрелый сосальщик 1 яйца
внешняя среда вылупление из яиц мирацидиев, плавание мирацидиев в виде, разыскивание промежуточного хозяина и вне- дрение в него
первый промежу- точный хозяин V превращение мирацидия в спороцисту, партеногенетиче- ское развитие в спороцисте редий 4 развитие (партеногенетическое) в редиях нового поколения редий или церкарий 4 выход церкарий из редий и из промежуточного хозяина в воду
внешняя среда вода второй промежу точный хозяин плавание в воде 4 4 проникновение во второго проме- инцистирование в жуточного хозяина и инцистиро- воде (адолеска- вание — стадия метацеркарйи рия)
главный хозяин 4 „ 4 поедание метацеркарии со вторым проглатывание промежуточным хозяином глав- адолескарий ным главным хозяи- ном освобождение молодого сосальщика из цисты метацер- карии, или адолескарии, рост, миграция по телу и половозрелость в окончательном месте обитания. По- ловое размножение (обоеполое), откладка яиц и т. д., как выше, в начале столбца
Таким образом, у сосальщиков имеется чередование поколений, со-
пряженное с переменой хозяев, причем, по преобладающим воззре-
ниям, паразиты на всех стадиях жизненного цикла размножаются
половым путем, но в различных его формах, а именно однополое
(девственное — партеногенетическое) размножение редий
и церкарий сменяется обоеполым размножением половозрелых
гермафродитных особей. Такой цикл размножения называется
гетерогонией.
В последнее время выдвинута иная точка зрения на цикл развития
сосальщиков. В половых клетках спороцист и редий не обнаружено
процессов созревания, нет и яичников, так как яйца лежат непосред-
ственно в полости тела молодых спороцист и редий и дают прямой за-
родышевый путь через все стадии жизненного цикла сосальщика. По-
коления редий и церкарий возникают в порядке полиэмбрионии
(деления зародышей, т. е. их бесполого размножения). При такой
трактовке не приходится говорить ни о гетерогении, ни о метагенезе,
педогенезе или метаморфозе сосальщиков, у которых налицо прохож-
дение прямого зародышевого полового пути через добавочные ‘ личи-
ночные стадии, размножающиеся полиэмбрионией (Brooks, 1930).
Первым промежуточным хозяином сосальщиков бы-
вают различные моллюски; вторым хозяином служат моллюски,
личинки насекомых, рыбы, земноводные, реже другие животные —
вплоть до млекопитающих. Второго хозяина может и не быть вовсе.
Окончательным хозяином служат различные позвоночные.
Жизненный цикл различных сосальщиков дает много разнообразных
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
219
отклонений от рассмотренной выше схемы. Особенности, касающиеся
сосальщиков, паразитирующих у человека, будут указаны далее в спе-
циальной части.
Взрослые сосальщики паразитируют в самых разнообразных орга-
нах животных, начиная с пищеварительного канала, печени, легких, мо-
чевых органов и кончая такими исключительными местами обитания,
как евстахиева труба или конъюнктивальный мешок.
Сосальщики, в общем, проявляют меньшую-специфичность по отно-
шению к промежуточному и основному хозяину, чем другие паразити-
ческие черви.
КЛАССИФИКАЦИЯ СОСАЛЬЩИКОВ
Класс Trematoda, характеристика которого дана на стр. 211, делится
на два подкласса — Monogenea и Digenea.
ПОДКЛАСС MONOGENEA — ЭКТОПАРАЗИТИЧЕСКИЕ
СОСАЛЬЩИКИ
Сосальщики, являющиеся наружными паразитами, или обитающие
в полостных органах, имеющих широкое сообщение с внешней средой
(например мочевой пузырь бесхвостых амфибий); присосок не менее
трех, часто больше; на задней — прикрепительном органе — обыкно-
венно имеются хорошо развитые крючья или аналогичные придатки
иной формы; органы прикрепления вообще развиты очень сильно. Мат-
ка короткая, содержит обычно только одно яйцо. Развитие простое,
без перемены хозяев (прямое). Яйца часто своеобразной формы —
с длинными нитевидными отростками.
Паразиты кожи, жабр различных рыб, мочевого пузыря, кожи,
глотки земноводных и др.; у человека эти сосальщики никогда не
встречаются.
ПОДКЛАСС digenea—эндопаразитические
СОСАЛЬЩИКИ
Сосальщики, являющиеся внутренними паразитами; органы прикре-
пления их развиты гораздо слабее, чем у эктопаразитических сосаль-
щиков; присосок две, реже одна; вокруг половой клоаки иногда бывает
присосковидное образование, матка длинная, содержащая очень боль-
шое количество яиц; развитие с превращением и переменой хозяев. Из
двух отрядов этого подкласса — Gasterostomata 1 и Prosostomata —
паразиты человека относятся к последнему. _
ОТРЯД, PR3S3ST3MATA — ПЕРЕ ЦНЕРЭТЫЕ СОСАЛЫЦПКП
Ротовое отверстие на переднем конце тела или близ него и, как
правило, открывается в ротовую присоску. Брюшная присоска (если
она не отсутствует) лежит позади ротовой на брюшной стороне или
па заднем конце тела. Паразиты человека имеются только среди пред-
ставителей двух подотрядов.
1 Передняя присоска редуцирована. Рот открывается непосредственно на
брюшной стороне тела, кишечник в виде простого мешка; паразиты рыб (Bucepha-
lus pusillus).
>20
СПЕЦИАЛЬНАЯ ЧАСТЬ
ПОДОТРЯД 1. amphistomata — обоюдоустковые
Брюшная присоска лежит на заднем конце тела терминально или
субтерминально; тело часто конической формы, суживающееся кпере-
ди, кутикула без шипикОв.
Семейство Paramphistomatidae
Род Watsonius Stiles et Goldberger
Watsonius 'watsoni Stiles et Goldberger
Синойимы: Amphistomum watsoni Cunyngham, Cladorchis watsoni Shipley.
Грушевидное • уплощенное тело, сверху выпуклое, снизу вогнутое. В свежем
состоянии паразит прозрачного желтовато-красного цвета. Длина его 8—10, ширина
4—5 мм. Большая присоска сдвинута к заднему концу тела, в 4—5 раз шире рото-
вой присоски. Глотка с двумя маленькими боковыми выростами. Середину брюш-
ной поверхности тела‘занимают два дольчатых мешковидных семенника, тесно
прилежащих друг к другу; дорзально от них расположены извивы матки. Редкие
желточные фолликулы разбросаны почти по всей длине боков тела. Половое отвер-
стие лежит на уровне вилообразного разделения кишечника. Яйца овальные,
0,122—0,13 мм длины и 0,075—0,08 мм ширины, с крышечкой. Паразит обнаружен*
один раз в начальной части тонких кишек негра в Восточной Африке; паразитирует
в слепой кишке макак в Сингапуре. Присутствие этого паразита сопровождалось
тяжелым поносом. Обнаружен на вскрытии.
Терапия и профилактика неизвестны.
Диа гноз ставится по нахождению яиц в faeces.
Семейство Gastrodiscidae
У человека паразитирует только один вид этого семейства;
Род Gastrodiscoides Lei per
Gastrodiscoides hominis. Lew et Me-Conn.
Синонимы: Gastrodiscus hominis Lew. et Me -Conn. Amphistomum hominis.
Живой паразит красновато-оранжевого цвета. Тело грушевидное; задняя широ-
!кая его часть обособлена в виде диска (рис. 125), на заднем крае которого лежит
присоска. В свежем состоянии двуустка красно-
ватого цвета, 5—10 мм длины и на диске 4—6 мм
ширины. Спина ее выпукла, брюшная сторона
вогнута; тело толстое. Ротовая присоска мала,
слегка сдвинута вентрально; глотка с двумя
мешковидными придатками. Ветви кишечника
простые. Каждый из двух семенников с дольками
(рис. 126). Семенники лежат один позади другого
Рис. 125—Gastrodiscoides hominis
с брюшной стороны. Из Брауна.
Рис. 126 — Gastrodiscoides
hominis. Анатомия. Из F a u s L
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
221
в средней части тела. Позади них лежит матка, желточники же расположены по
«бокам тела. Половое отверстие — в передней трети средней линии тела, в месте
вилообразного деления кишечника, на вершине полового бугорка. Яйца овальные,
утонченные к полюсу, с крышечкой. Длина 0,150—0,157, ширина 0,06—0,072 мм. На
противоположном полюсе яйца имеется подушкообразное несимметричное утолщение
скорлупы (рис. 127), Строение мирацидия см. рис. 128.
Рис. 127 — Gastrodiscoides hominis.
Яйца паразита из кабана (Казахстан).
По Баданину.
Рис. 128 — Gastrodiscoides hominis.
Слева—оболочка (скорлупа) яйца; справа—
мирацидий сбоку. По Buckley.
Паразитирует в толстой и в слепой кишках человека, где прикрепляется задней
расширенной частью тела (Ост-Индия, Британская Гвиана). В Ассаме гастродиск
является обычным паразитом свиней. В СССР этот сосальщик обнаружен только
у кабана в Казахстане (Баданин), что дает основание предполагать возможность
нахождения этого паразита и у человека в тех же местах.
Патогенен, вероятно, лишь при массовом паразитировании в человеке. Вызывает
воспаление слизистой оболочки и, может быть, понос.
Профилактика неизвестна, так как цикл развития этого сосальщика не изучен.
ПОДОТРЯД 2. DISTOMATA — ДВУУСТКОВЫЕ
Только две присоски — ротовая и вентральная. Половые органы це-
ликом или большей частью лежат позади брюшной присоски.
Семейство Fasciolidae —листвяницы
Сосальщики сем. Fasciolidae паразитируют у травоядных млекопи-
тающих.
Pod Fasciola Linn.
Fasciola hepatica (L.) — печеночная двуустка
Синонимы: Distoma hepaticum Linn., Distomum hepaticum Retz.
Взрослая двуустка (рис. 120) имеет удлиненно-листовидное
(3—5 см) тело с конусообразным передним концом, на котором поме-
щается ротовая присоска (т) около 1 мм диаметром. Сейчас же поза-
ди конусовидной части тела лежит брюшная присоска 1,6 мм диамет-
ром. Кутикула, тела покрыта мелкими шипиками. Живой паразит жел-
товатого цвета, по краям с оранжевым оттенком; там, где просвечи-
122
СПЕЦИАЛЬНАЯ ЧАСТЬ
вает матка (и), он серо-коричневый. Когда двуустка напиталась кро-
вью, она принимает более розовую окраску. Мускулистая глотка ведет
в пищевод, сообщающийся с двумя чрезвычайно ветвистыми стволами
кишечника (/). Главный канал выделительных органов идет по сред-
ней линии тела, занимая ее задние две трети и открываясь отверстием
на заднем конце тела. Органы выделения сильно ветвятся, но боковые
стволики их не видны без специальной обработки (инъекция берлин-
ской лазурью). Сильно разветвленные семенники (/) лежат в средней
части тела; два семепровода, идя кпереди, открываются в мешок сово-
купительного органа, который лежит сейчас же перед брюшной при-
соской. В совокупительном мешке (bursa cirri) лежит семенной пузы-
рек; яичник непарный (о). Его ветвистые трубки расположены на пра-
вой стороне тела, кпереди от правого семенника. Между этими двумя
Рис. 129—Вариации в размерах и форме яиц печеночной
двуустки. Но Н ё л л е р у.
органами проходит правый желточный канал. Сильно ветвистые жел-
точники лежат по краям тела, достигая его заднего конца. Симметрич-
но идущие желточные каналы сливаются в оотип: от его левой стороны
отходит Лауреров канал, открывающийся на спинной стороне тела.
Оотип окружен массой тонких одноклеточных железок (s). Семепри-
емника нет. Толстые извивы матки (и) расположены кпереди от лево-
го семенника и яичника, а слева граничат с желточником. Петли матки
заходят кпереди от брюшной присоски. Наружное отверстие мужских
и женских половых путей общее (половая клоака), лежит в углу»
образованном расхождением двух главных кишечных ветвей (у).
Яйца желтовато-коричневого цве^а, овальной формы, длиною 0,13—
0,145 и шириною 0,07—0,09 мм. Размеры и форма их несколько варии-
руют. Оболочка двуконтурна; на одном из полюсов имеется крышечка
{рис. 129).
Двуустки живут в печени человека (очень редко), быка, овцы, козы»
свиньи, лошади, кролика. Бывают также и в желчном пузыре их. Яйца
выходят с желчью в кишечник и могут быть обнаружены в испражне-
ниях больного; диагноз дистоматоза ставится- на основании исследо-
вания последних.
Однако факт нахождения яиц в faeces еще не значит, что у чело-
века в печени действительно имеется паразит. При поедании печени
животных, зараженной F. hepatica, яйца печеночной двуустки проходят
через кишечник без видимых внешних изменений и могут быть призна-
ны принадлежащими сосальщикам человека, тогда как на самом деле
речь может итти лишь о мнимом паразитировании Печеночной двууст-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
223
ки (транзитные яйца). О необходимых предосторожностях диагностики
см. ниже в абзаце о Dicrocoeliurri lanceatutn.
Цикл развития печеночной двуустки слагается из
следующих стадий: яйцо выходит с желчью в кишечник и с испражне-
ниями выбрасывается наружу. Развитие зародыша в яйце начинается
после его выхода из тела хозяина. В воде из яйца выходит мирацидий.
Оптимальная температура для развития зародыша в яйце печеноч-
ной двуустки колеблется в пределах от 25 до 30° С. При +35° С яйца
погибают после 5 дней; 4-40—45° С убивают яйца в 1 час. При
i+50°C — яйца погибают еще скорее. При t° 22—24° С созревшие за-
родыши (мирацидии) начинают выходить из яиц через 20—22 дня.
При t° 10—15° С развитие яиц идет, но замедлено. При t° 0—10° С
Рис. 130 — Яйцо Fas-
ciola hepatica, пора-
женное грибком Са-
tenaria ang illulae.
Но Д и н н и к у.
Рис. 131
А — Прудовик малый — Limnaeus
minutus. В — Обычкиве) ный прудо-
вик Linina. stagnalis. С болотный
прудоник- Linin. / alustris. Из Р и м-
ского-Корсакова и Рай-
кова.
эмбриональное развитие останавливается, но яйца сохраняют жизне-
способность, что обнаруживается при перенесении их в оптимальные
условия развития. Ниже —6° С яйца не выживают.
Если яйца находятся в темноте, то развившиеся в них мирацидии
могут оставаться в яйце живыми иногда даже свыше двух месяцев.
При перенесении таких яиц на свет (даже электрический) мирацидии
начинают выходить из яиц через 15—30 минут на естественном свету
и часа через 13/4 —т-при электричестве.
Перед выходом мирацидия крышечка яйца открывается; в его от-
верстие прорывается эмбриональная оболочка и мирацидий активными
движениями выбирается из яйца, что -происходит в одну — полторы
минуты.
Яйца F. hepatica чувствительны к высыханию. Если их оставить на
стекле при t° 20—22° С и 72°/о отн. влажности на несколько минут,
пока испарится вода, то яйца погибают. В шариках помета овцы они
могут оставаться живыми 4—7—12 и, вероятно, более дней, смотря по
температуре и влажности окружающей среды. В бескислородной влаж-
ной среде они могут жить до 5 дней. Развитие яиц может происходить
иногда и в загнившей воде.
.224
СПЕЦИАЛЬНАЯ ЧАСТЬ
При прочих равных условиях яйца развиваются быстрее в болотной
воде, чем в водопроводной или дестиллированной.
Иногда яйца фасциолы поражаются грибками (рис. 130); такие
яйца, конечно, зародышей не дают.
из яиц печеночной двуустки происходит
Выхождение мирацидиев
.при pH = 4,2—7,2 (оптимум 6,5; А.
Рис. 132 — Litnnaea truncatula. Оригинал.
Скворцов и сотрудники). В водо-
емах, в которых живут Ыт-
naeus truncatula, зараженные
F. hepatica (см. ниже), pH ко-
леблется от 6,7 до 7,3 (те же
авторы).
Вышедшие мирацидии в де-
стиллированной воде живут
около суток, а в болотной 36 —
40ч часов. В физиологическом
растворе они погибают через
2 часа, а в рингеровской жид-
кости живут 16—20 часов.
Оптимальная реакция среды
колеблется в пределах значе-
ния pH от 6,5 до 7,5, т. е. от
слабой щелочной до слабо ки-
слой. Вышедший мирацидий вбуравливается в промежуточного хозяина,
которым является брюхоногий моллюск — прудовик малый (Лей-
карт); по Noller и Spren (1926), в Европе аналогичное зна-
чение играют и другие виды: L. stagnalis и L. palustris (рис.
131). В лимфатических пространствах моллюска мирацидий пре-
вращается в спороцисту (рис. 123), которая перебирается в лимфатиче-
Рис. 133 — Physa fontinalis (увеличено). Оригинал.
екие пространства печени; здесь в спороцисте партеногенетически разви-
вается дочернее поколение редий; последние также внутри себя и также
партеногенетически дают или редий, или церкарий. Последние выходят
из редий, покидают моллюска, плавают в воде и через некоторое время
инцистируются на погруженной в воду траве или на поверхности воды.
Срок жизненного цикла по Скворцову/ спороциста — на 3 день после за-
ражения; спороциста с редиями I поколения — на 21 день; редии без
•спороцист (?)—на 34 день; на 48—49 день — редии с редиями II по-
коления или редии I с церкариями; на 60 день — редии II поколения
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
225
или с зародышами церкарий; на 78—80 день — редии II с готовыми
церкариями и свободные церкарии. От мирацидия до церкария 50—80
дней, а от яйца до церкария 70—100 дней. В экспериментальных усло-
виях у кролика живет более трех лет (Montgomery, 1931), а у скота и
овец — пять лет и больше (Sandground). Мирацидии могут заражать
(проникать в) Planorbis planorbis, Physa fontinalis (рис. 133) и разные
виды Lininaea, но до церкарий развитие идет только в L. truncatula
(рис. 132; Лейкарт, Синицын, Скворцов). Человек и животные прогла-
тывают цисты с водой или с травой. В двенадцатиперстной кишке цисты
вскрываются, и молодые двуустки через печеночный проток проползают
в желчные ходы печени, где и достигают половозрелое™.
Другие авторы считают, что молодые паразиты попадают в печень
гематогенным путем, т. е. внедряются в стенки пищеварительного ка-
нала и через кишечные вены попадают в воротную вену, откуда пара-
Рис. 134 — Двуустковый пирроз (cirrhosis disto-
malosa) левой доли печени и викарная гипертро-
фия остальных долей. Печень овцы. По С о hrs.
зитам прямой ход в печень. Наконец, есть третье мнение, согласно
которому молодые двуустки непосредственно из кишечника пробира-
ются в печень, прободая стенку кишки и вбуравливаясь в прилежащую
часть печени, проникая через ее капсулу.
Что молодые двуустки действительно попадают в кровь, в том со-
мнений нет, так как только таким путем можно объяснить случаи из-
вращенной локализации этих сосальщиков, когда их находили в крови,
в подкожных нарывах, в легких, в желудочках мозга, в хрусталике
глаза и др. Возможно также внутриутробное заражение плода от ма-
тери.
Паразиты глаза были описаны в качестве самостоятельных видов —
Oistornum ophthadmobium, Monosfromtun lentis. Изучены они крайне
недостаточно. Случаи «блуждания» F. hepatica бывают у животных, где
их находили под брюшиной, в соединительной ткани, между мышцами,
в селезенке и в других местах. Паразиты обычно окружены цистой, в
которой они чаще всего атрофируются.
Действие на хозяина. При нормальной локализации печеноч-
ные двуустки пробираются в желчные протоки и достигают перифери-
ческих желчных капилляров, где становятся половозрелыми; взрослые
крупные паразиты возвращаются в желчные протоки большего диамет-
ра. В местах нахождения двуусток желчные пути образуют дивертику-
лы, эпителий протоков дает папилломатозные разрастания и в некото-
рых случаях вовсе разрушается; стенки протоков склерозируются;
226
СПЕЦИАЛЬНАЯ ЧАСТЬ
$
® них наблюдается .инфильтрация лейкоцитами с эозинофилами. В ме-
стах разрушения слизистой выстилки протоков двуустки проникают не-
посредственно в ткань печени (рис. 135), где застревают и откладывае-
мые ими яйца. Вследствие этого образуются очаги воспаления (иногда
в форме абсцессов), где печеночная ткань в конце концов замещается
тканью соединительной (цирротический процесс). Печень опухает и
становится бугристой (рис. 134).' Двуустки закупоривают желчные
протоки, обусловливают застой желчи и развитие желтухи у больного.
Больной ощущает боли в печени (печеночные камни), у него развива-
Рис. 135 — Ходы в печени овцы, проделанные
молодыми двуустками.
Слева вверху свежий ход (с двуусткой) еще без реактив-
ного воспаления; остальные ходы старые, с ограниченной
воспалительной реакцией. По С о h г s.
ются малокровие и асцит, смотря по степени цирротических изменений
печени ;и сдавливания воротной вены. Общее действие паразитов объ-
ясняется интоксикацией хозяина продуктами жизнедеятельности F. he-
patica. Инвазия осложняется побочными инфекциями 1вследствие про-
никновения бактерий в ткань печени через места с разрушенным эпи-
телием желчных путей и, может быть, при самой миграции паразита,
если она идет гематогенным путем.
Полный эффект действия обнаруживается при более или менее
сильных инвазиях человека печеночными двуустками. Нахождение одно-
го-двух экземпляров не обусловливает всей картины поражения печени,
но при извращенной локализации и один паразит, в зависимости от
места его нахождения, может дать тяжелый эффект.
Заражение человека происходит при питье сырой или не-
фильтрованной воды или при поедании сырой зелени, поливаемой во-
дой из водоемов, где есть инцистированные церкарии сосальщика. При
еде сырой козьей или бараньей печени с двуустками последние приса-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
227
Сываются к слизистой оболочке глотки, которую и раздражают. В дан-
ном случае налицо ненормальная локализация печеночного паразита,
зависящая от случайных обстоятельств. Действие паразита на своего
хозяина, в зависимости от перемены места обитания, совершенно
иное. Двуустки проявляют в печени токсическое действие на организм
человека и резко изменяют ее нормальное строение; в глотке же они
причиняют местное раздражение и очень легко удаляются из нее.
Глоточным дистоматозом называют болезнь, когда дву-
устки присасываются к слизистой оболочке глотки. Своим присутствием
они вызывают отечное воспаление глотки, гортани, носовой полости,
рта, евстахиевых труб и других окружающих частей. Побочными симп-
томами являются одышка, потеря голоса и даже удушье, которое в тя-
желых случаях может быть причиною смерти.
Болезнь эта описана A. Khouri у жителей Сирии (Liban), где она
известна под именем Halzoun. Диагноз ставится непосредственным
исследованием глотки. Излечивается болезнь рвотными средствами и
алкогольными полосканиями.
Случаи ф а с ц и о л е з а человека все чаще и чаще обнару-
живаются в СССР. Впервые у нас F. hepatica найдена 12 октября
1923 г. при операции женщины в Баку, у которой из желчного протока
вышла печеночная двуустка (хирург Финкельштейн; определение пара-
зита произведено И. И. Широкогоровым и подтверждено автором по
препарату, поступившему в музей патогенных животных кафедры об-
щей биологии и паразитологии Военно^медицинской академии; заметка
об этом случае была дана в первом издании нашего учебника в 1924 г.).
В январе 1924 г. Калантарян нашла печеночных двуусток на вскрытии
трупов мужчины и женщины в Армении. Аналогичные находки были в
Средней Азии (Ташкент).
Была сделана попытка отсева сомнительных случаев фасциолеза
людей post factum. Из описанных свыше сотни случаев фасциолеза
Кеворков и Сыркина считают фасциолез истинным в сорока случаях.
Известно уже несколько десятков случаев обнаружения F. hepatica
у человека или ее яиц в faeces людей. Однако не все эти указания от-
носятся к случаям явного фасциолеза. Наряду с бесспорным фасциоле-
зом, несомненно, фигурируют и случаи фасциолеза мнимого.
Пр едсказание исхода истинного фасциолеза зависит от
степени инвазии и от локализации паразитов. При тяжелой инвазии
оно неблагоприятно. Возможно ли самопроизвольное излечение при
самостоятельном выходе сосальщиков из печени человека, сказать
трудно. Если такое допущение и выдвигается в качестве возможного,
то единственно на основании нахождения вышедших паразитов в ки-
шечнике у скота. Однако проведение решительного заключения только
по аналогии едва ли возможно.
Лечение. Экстракт корневища папоротника (Extr. filic- mar.
aether.) находит применение в ветеринарной практике. У человека с
успехом применяется эметин (Emetinum hydrochloricum) в форме вну-
тривенных вливаний 1% раствора или в виде внутримышечных впрыс-
киваний. Об успехе судят по исчезновению яиц двуустки из faeces
больного.
Профилактика: а) индивидуальная — не пить сырой или не-
профильтрованной воды из водоемов, заселенных моллюсками; не
есть сырой огородной зелени, поливаемой водой из таких же водоемов;
не есть сырой козьей, бараньей или бычьей печени; б) общественная —
228
СПЕЦИАЛЬНАЯ ЧА’СТЬ
уничтожение брюхоногих .моллюсков в водоемах; очистка водоемов от
моллюсков. Мероприятия по ветеринарной, линии — см. специальные
руководства.
Рис. 136 — Дву-
устка гигантская
с зарисованным
кишечником.
По Коббольду.
Fasciola gigantica Cobb.— двуустка гигантская
Сийоним: Distomifm gigant icum Dies! ,n g.
Двуустка гкга11Тская» отличающаяся от предыдущего вида большими размерами
{25 75 мм длины и 3—12^ мм ширины), коротким головным конусом, большей
величиной брюшной присоски и более выдвинутым вперед поло-
жением семенника, не всеми исследователями признается за само-
стоятельный вид. Тело ее более узкое, почти с параллельными
боками (рис. 136). Яйца 150—190р- длиной, 70—90 р- шириной.
В Туркестане обнаружена в печени коров; в Африке паразитирует
у буйволов, овец, коз и жирафф. Вызывает в печени сходные
патологические изменения, что и F. hepatica. Один раз обнару-
жена и у человека (в Рио-де-Жанейро): после сильного припадка
кашля с кровянистой мокротой вышла из больного двуустка в
272 см длины. Едва ли можно сомневаться в том, что это был
«заблудившийся» паразит.
Описанный Пигулевским случай ' нахождения F. gigantica у
человека в Ташкенте по переисследованию Н. Кеворкова не под-
твердился; но Кеворковым и Сыркиной обнаружен на трупном
материале F. gigantica в одном из 56 вскрытых трупов (Ташкент).
Диагностика: по нахождению яиц паразита в faeces.
Профилактика: как и при F. hepatica (см. выше).
Род Fasciolopsis Looss
Fasciolopsis buski Lank
Синонимы: Distomum cr'assum Busk., Distomum rathouisi,
Fasciolopsis fiillebofni Rodenw.
В новейшей паразитологической литературе прояв-
ляется тенденция к признанию пяти описанных видов
Fasciolopsis (F. buski, F. rathouisi, F. fiilleborni, F. god-
dardi и F. spinifera) за один вид F, buski.
Тело паразита листовидное или удлиненное (без
переднего конусовидного сужения), достигает 7,5 см
длины и 1,5—2,0 см ширины (рис. 137). Таким обра-
зом, этот сосальщик является самой крупной дву-
усткой человека; средние размеры ее составляют
около 30X12 мм. Величина паразитов вариирует
в зависимости от хозяев и от степени инвазии.
При множественной инвазии (паразит встречается иногда сотнями;
максимальная зараженность достигала 3 721 экземпляра) сосальщики
меньшего размера; единичные обитатели хозяина достигают наиболь-
шей величины.
Цвет живого паразита похож на окраску мяса или крови. Брюш-
ная присоска лежит очень близко от субтерминально расположенной
ротовой присоски и в четыре-шесть раз больше последней (2—3 мм
против 0,6 и меньше). Кутикула покрыта легко отпадающими шипи-
ками. Пищеварительный аппарат состоит из очень короткой предглот-
ки, шаровидной глотки, еле выраженного пищевода и двух длинных
неветвящихся трубок, достигающих почти самого заднего конца тела
(рис. 137). . „
Семенники развиты очень сильно. Их два: они образованы системой
сложно ветвящихся толстых трубок и лежат друг за другом в среднем
поле задней половины тела. Семевыносящие каналы впадают в боль-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
229
шой семенной пузырек, лежащий в мешке цирруса, который тянется
почти на четверть длины всего тела. Разветвленный яичник мал, ле-
(жит рядом с немного большей по размерам скорлуповой железой. Изви-
вы матки помещаются между передним
кишечника и брюшной присоской, на
краю которой и открывается половая
клоака оотипом, куда впадает множество
одноклеточных железок, одетых снаружи
соединительнотканной капсулой. Яйце-
вод отдает от себя Лауреров канал и
перед впадением в оотип соединяется с
общим желточным протоком. Семепри-
емник отсутствует. Сильно развитые
желточники занимают боковые края па-
разита.
Светлокоричневые яйца вариируют по
форме (эллипсоидальные или овальные)
и величине: 130—140 у длины, 80—
85 у ширины (Faust); 0,179—0,211 у
длины и 72—91 у ширины (Скрябин с
сотрудниками). Скорлупа яйца 'тонкая;
крышечка маленькая, уплощенная, на
противополо.жном полюсе или несколько
сбоку имеется небольшое ограниченное
утолщение скорлупы яйца (рис. 138).
Паразиты обитают в тонких кишках
и, в виде исключения, в желудке чело-
века и свиньи. Плодовитость их велика;
зрелая самка дает в день около1 25 тыс.
яиц. Мирацидий развивается в яйце,
когда оно находится в воде. При темпе-
ратуре 80—90° F на это требуется от 3
до 7 недель. Вылупившиеся мирацидии
(рис. 140) активно плавают в воде,
смотря по температуре, от 6 др 52 часов.
Промежуточным хозяином является один
из моллюсков Planorbis coenostis, Seg-
mentina nitidella, S. schmakeri и др. Ми-
рацидий (рис. 140) внедряется в моллю-
ска и превращается в спороцисту, кото-
рая отличается наличием кишечника как
у редий, но без глотки. Через 3—4 .дня
в спороцисте начинают развиваться ре-
дии, выходящие из нее на 9—10-й день.
В редиях развивается новое поколение
редий, внутри которых уже образуются
церкарии. Последние покидают моллюска
краем семенников, ветвями
Рис. 137 — Fasciolopsis buski. По
К. Скрябину, Подъяполь-
ской и Статировой.
и плавают в воде. Осев на
различных водяных растениях, например водяном орехе (Trapa natans)
или Eliocharis tuberosa, церкарии инцистируются, образуя шары диа-
метром 216—187 у.
3 а р а ж е н.и е происходит при обдирании зубами оболочки сырых
плодов водяного ореха, употребляемых в пищу китайцами. На одном
орехе насчитывали до двухсот ‘инцистированных церкарий.
230
СПЕЦИАЛЬНАЯ ЧАСТЬ
F. buski является паразитом человека и свиней (Центральный и
Южный Китай, Индо-Китай, Сиам и прилежащие острова, начиная с
Формозы).
Патогенность паразита для человека несомненна.
Клинический эффект зависит от степени инвазии, хотя прямая пропор-
Рис. 138 — Яйца Fasciolopsis buski. По Скрябину, Подъяполь-
ской и Статировой.
циональность между количеством паразитов и тяжестью заболевания
обнаруживается не всегда; бывают случаи, когда люди с меньшим чис-
лом паразитов страдают гораздо сильнее, чем более обильно заражен-
ные пациенты.
Проявления болезни наступают месяца через три после заражения.
При слабой инвазии симптомы болезни слабы и неопределенны .(напри-
Рис. 139 — Вылупление мирацидия Fasciolopsis buski из яйца.
По Barlow.
мер боль подложечкой); более сильное заражение вызывает симптомы,
которые могут повести к предположению о наличии у больного язвы
желудка. Понос сменяется периодами запора. Позднее понос делается
постоянным, развиваются отеки лица, ног, водянка живота и другие
симптомы. Интоксикация от действия паразитов может повести к смер-
ти больного.
В СССР известны два случая завозного фасциолопсидоза — один,
установленный изгнанием за два приема 190 экземпляров паразита в
Москве у семилетней китаянки (Скрябин, Подъяпольская и Статирова),
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
231
другой в Ташкенте, только по нахождению яиц, принятых за F. buski,
с неудавшимися попытками глистоизгнания у пациента, 11 лет назад
путешествовавшего по портам Индии и
Индийского архипелага (Семенов и Гарт-
матий). По мнению Скрябина с сотрудни-
ками, в последнем случае было* заражение
не F. buski, а какой-то другой двуусткой.
Диагноз ставится по нахождению яиц
в faeces, причем, по Faust, яйца F. buski
необходимо диференцировать от близко
сходных с ними яиц F. hepatica, F. gigan-,
tica и Echinochasmus; при этом необходимо
учитывать вариации формы и отчасти раз-
меров яиц, свойственные F. buski.
Лечение. Глистоизгнание тимолом,
бета-нафтолом и химически чистым четы-
реххлористым углеродом.
Профилактика: индивидуальная —
обваривание кипятком плодов Trapa natans
и других водяных растений или выдержи-
вание их в растворе поваренной соли, что-
бы убить инцистцрованных церкарий (1%
часа в 90°/о растворе); общественная — сте-
рилизация faeces в районах эндемичной
Рис. 140 — Мирацидий Fascio-
lopsis buski'.
pg— зачаток кишечника; sg — желе-
зистые клетки; es — глаза; п— нерв-
ная система; г/— экскреторный ка-
нал; ер — экскреторная пора; fc —
мерцательное пламя замыкательной
клетки; gc — половые клетки.
11 о Фаусту.
инвазии (Faust).
Семейство Plagiorchidae
Род Plagiorchis
'Мелкие трематоды (1,5—2,00 мм X 0,385—0,425 мм) обнаружены в
тонких кишках человека при вскрытии в Манилле (Филиппинские
острова). Систематическое положение паразита ближе не определено
(С. Africa и Е. Carsia). Me Mullen показал возможность 'эксперимен-
тального заражения человека Plagiorchis muris после съедания зре-
лых цист с метацеркариями, выпрепарованных из моллюсков Stagni-
cola emarginata angulata (озеро Дугласа, США). Выделение яиц
паразита началось на девятый день после заражения и длилось до
35 дня; в день выделялось максимум 74 700 яиц, в среднем 20 883. Ни-
каких симптомов заболевания не было.
Семейство Echinostomatidae
Большая часть представителей этого семейства паразитирует в ки-
шечнике у низших позвоночных и птиц.
У человека отмечено паразитирование представителей родов: Е chino-
stoma, Echinoparyphium, Echinochasmus, Euparyphium, Himasthla и
Artife chino st отит. Определительную таблицу восьми подсемейств сем.
Echinostomatidae ом. К. Скрябин, 1938. Паразиты отходили после дачи
экстракта папоротника, хеноподового масла или тимола. Цикл разви-
тия (в изученных случаях) с двумя промежуточными хозяевами: мол-
люск— моллюск или моллюск—рыба. Заражение при поедании рыбы
или моллюска (II промежуточного хозяина) с метацеркариями.
232
СПЕЦИАЛЬНАЯ ЧАСТЬ
Весьма вероятно, что Echinostomaiidae являются более частыми
паразитами человека, но для их нахождения требуется тщательная
промывка фекалий.
Род Echinostoma Rudolphi
Echinostoma ilocanum Garrison
Прозрачное красноватое тело этой двуустки утончено к концам и в.
передней половине более расширено, чем в задней. Длина 4—6 мм,,
ширина 0,75—0,35 мм, толщина 0,5—1,0 мм (рис. 141). Кутикула не-
Рис. 141. Echinostoma
ilocanum.
По О д н е р у.
Рис. 142—143—Развитие Echinostoma
ilocanum:
А — незрелое яйцо из фекалий; В — яйцо с развив-
шимся мирацицием; D — метацеркария; С — церкария;
ас — присоска; сс — воротничок; eg — цистогенные
железы; ga— половой зачаток; int — кишечник; ое —
пищевод, ph — глотка. По Tubangui и Pasco.
редней части тела с шипиками. Брюшная присоска раза в три шире
ротовой. Обе присоски сближены друг с другом: передняя окружена
складкой с венчиком крупных иголочек. Ветки кишечника простые. Се-
менники лежат один позади другого сзади яичника в задней половине
тела, бока которой заняты желточниками. Между яичником и брюшной
присоской — густые извивы матки. Мешок совокупительного органа ле-
жит впереди последнего.
Яйца овальные с крышечкой 100 у длины и 55 у ширины.
У Echinostoma revolution мирацидий превращается прямо в редию, минуя ста-
дию спороцисты. Редии Echinostoma sp. с развившимися церкариями находили в
легочном брюхоногом моллюске прудовике обыкновенном — Limnaea stagnalis.
Для Echinostoma r.evolutum характерно далее то, что церкарии по выходе из
редии не покидают моллюска, а остаются в нем, превращаясь в нем в стадию
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
23&
Agamodistomum, которую можно считать эквивалентной метацеркарии. Заражение
окончательного хозяина происходит при поедании моллюсков с Agamodistomum.
Паразитирует в кишечнике человека на Филиппинских о-вах (изгнан
тимолом).
Цикл развития Ech. ilocanum (рис. 142, 143 по Тубангуи и Паско).
Первый промежуточный хозяин — моллюск (Guraulus prachadi)\ церка-
рии, по выходе из него, проникают в мантию других пресноводных
моллюсков, являющихся вторым промежуточным хозяином, где пре-
вращаются в метацеркарии, например, в Pila luzonica, употребляемого
сырым в пищу населением Филиппинского архипелага. Эксперимен-
тально можно заразить белых мышей, кошку и обезьяну.
Еchinostoma suprartyfex Lane
Синоним: Echinostoma malayanum (Leiper).
Два раза был обнаружен в тонких кишках человека около Малак-
ки. Вокруг ротовой присоски воротниковый вырост с плотным
венцом шипов.
Локализация паразита в теле человека неизвестна. Не выяснен
цикл развития, равно как и патогенное значение.
Симптомы заражения Ech. suprartyfex сходны с фасциолопсидозом..
Изгнание удалось применением экстракта корневища папоротника.
Echinostoma jassyense Leon et Ciurea
Синоним: Euparyphiutn jassyense.
Красноватый сосальщик 5,4—7,6 мм длины и 1 —1,3 мм наибольшей
ширины (рис. 144). Околоротовой воротниковый вырост вооружен
27 шипами двух размеров. Ротовая присоска около 22 |л диаметром;
брюшная гораздо крупнее (73 ^). Яйца с крышечкой, овальные, 132 —
154 р длиной и 79—85 шириной. Цикл развития и патогенное зна-
чение неизвестны. Изгнан румынскими гельминтологами (Leon и
Ciurea) из кишечника перса, страдавшего поносами и леченного
тимолом.
Echinostoma paraulum pietz
Плоское удлиненное тело, несколько сужающееся кпереди и закруг-
ленное на заднем конце; длина 7,7 мм, наибольшая ширина—1,4 мм;
кутикула усеяна мелкими шипиками, особенно густо лежащими на вен-
тральной стороне сосальщика. Адоральный диск —воротник — в
форме почки 0,48 мм ширины; он несет 37 одинаковых по размерам
шипов (0,055 мм длины); расположение их см. на рис. 146, I.
Ротовая присоска (0,48 мм ширины) лежит субвентрально. Она в
полтора раза меньше вентральной присоски, лежащей на границе пер-
вой и второй четвертей тела сосальщика, считая по длине. Сравнитель-
но длинный пищевод ветвится тотчас перед половыми отверстиями.
Слепые ветви кишечника доходят почти до заднего конца тела.
Семенники двулопастевидные, округлые, лежат один за другим на
границе средней и задней трети длины тела. Половая бурса овальная:
она лежит между развилком кишечника и брюшной присоской, не-
сколько заходя за передний край последней.
Яичник круглый; лежит асимметрично, позади средины длины тела;
между ним и последним семенником помещается тельце ’Мелиса. Жел-
точники в виде сравнительно крупных фолликулов густо лежат по бо-
!34
СПЕЦИАЛЬНАЯ ЧАСТЬ
кам тела, располагаясь от заднего конца тела почти до уровня заднего
края брюшной присоски. Извивы матки, лежащей между яичником и
брюшной присоской, не заходят за ветви кишечника. Яйца 100 X 60 ц.
Ech. paraulum является паразитом кишечника утки, гагары, голубя.
Известен из Германии и СССР. В единственном экземпляре обнару-
Рис. 144 — Echino-
stoma jassyense.
По Леону.
Рис. 145 — Echino-
stoma paroulum.
Цельный экзем-
пляр. По К. С к р я-
б и н у.
Рис. 146 — Echinostoma paraulum.
I— Адоральный диск с 37 шипами от человека. II—
Схема строения выводных половых путей (рис.
Ж. Штрома). А — брюшная присоска; J — кишечник;
•vs — семенной пузырек; $ — женское половое отвер-
стие; <3 — мужское половое отверстие; О—пищевод.
По К. Скрябину.
жен у человека в Просницком районе Кировской области (В. Подъя-
польская) попутно при промывке фекалий после изгнания цепеня не-
вооруженного.
Патогенное значение для человека неизвестно. Ech. paraulum пато-
генен для голубей и при массовой инвазии может вызывать их гибель.
Подробнее ом. К- Скрябин. Echinostoma paraulum — новый паразит че-
ловека. Медицинская паразитология и паразитические болезни. 1938,
т. VII, вып. I.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
235
Род Echinochasmus
Воротник ©округ ротовой присоски развит сильно; венец шипов на
нем прерванный.
Echinochasmus perfoliatus Ratz
Сосальщик, длина тела которого достигает 12 мм, ширина 2 мм;
покровы паразита усеяны шипиками .(рис. 147). Брюшная присоска раза
в два больше ротовой и лежит в задней части
первой трети тела. Воротник переднего конца
тела вооружен 24 шипами, приблизительно
одинаковой величины: кольцо шипов прервано
в дорзальном и вентральном направлениях
(рис. 148) по отношению к ротовой присоске;
последняя повернута несколько' вентрально;
имеется длинная и узкая предглотка
(praefarynx), шаровидная глотка, длинный
пищевод и пара простых кишечных трубок,
идущих до заднего конца тела.
Семенники в общем округлые (модифика-
ция шаровидной формы); лежат в средней зо-
не тела (считая по длине) или сдвинуты в
начальную часть задней половины сосальщи-
ка. Мешок.цирруса почти весь занят семен-
ным пузырьком. Маленький шаровидный яич-
ник лежит справа перед передним семенником.
Против него, чуть впереди, помещается семе-
приемник. Желточники заполняют боковые
поля от уровня брюшной присоски до заднего
конца тела. Оотип трубковидный. Матка, в
виде сравнительно слабо извитой трубки с не-
многочисленными яйцами, открывается вместе
к
147 — Echinochasmus
Рис.
perfoliatus. Из кишечника
собаки:
а—-ротовой присосок; Ь — адо-
ральный диск; с — глотка; d —
Рис. 148—Echinochasmus perfoliatus.
Передний конец. По Скрябину и
Линдтроп.
пищевод; е — мужское и женское
половое отверстие; f — мужская
бурса; I — брюшная присоска;
,g — яичник; /г — семенник; k —
желточник; т — кишечник; п —
.яйца. По К. Скрябину и
Р. Шульцу.
с циррусом в половую клоаку, лежащую тот-
час позади разветвления кишечника.
Яйца эллипсоидные с крышечкой, тонкой
оболочкой, размером 90—135 на 55—95 /л;
откладываются в незрелом состоянии.
Паразит был открыт в Венгрии в тонких кишках собаки и кошки.
Распространен, видимо, широко, так как обнаружен в СССР и на Даль-
нем Востоке. Найден у свиней и в Японии у человека.
236
СПЕЦИАЛЬНАЯ ЧАСТЬ
Цикл развития изучен недостаточно. Первым промежуточным
хозяином считается моллюск Parafossarulus striatus (и другие виды
того же рода). Вторым хозяином служат различные рыбы. В них были
находимы инцистированные метацеркарии, которые по скармливании
их кошкам, собакам и людям давали соответствующих сосальщиков.
Семнадцать видов рыб значатся среди возможных вторых хозяев на*
Рис. 149 — Blicca bjoerkna — густера — второй промежуточ-
ный хозяин Echinochasmus perfoliatus и Opisthorchis feli-
neus (Neveu-Lemaire и Pellegrin). Оригинал.
Рис. 150 — Pseudorasbora parva. Оригинал.
шего паразита, кроме таких обычных видов, как щука, язь, линь, лещ,
густера (Blicca bjorkna— рис. 149), среди вторых хозяев значатся Gna-
thopogon, elongatus, Pseudorasbora parva (рис. 150), Opsarichtys unciro-
stris и др.
Паразитирует в тонких кишках. Значительной патогенностью, види-
мо, не обладает. Для человека является, вероятно, гостепаразитом.
Диагностика по нахождению яиц в faeces.
Лечение, как при Fasciolopsis buski.
Профилактика. Не есть рыбы иначе, как хорошо проваренной
или прожаренной.
Род Himasthla
Himasthta miichlensi Vogel
Узкие длинные сосальщики—11 —17,7 мм длины, 0,41—0,67 мм
ширины. Циркуморальный диск в форме почки, состоит из 32 шипов. Ро-
товая присоска 118—145 X 94 — 123 у; брюшная присоска 358—410 X
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
237
X 357 — 425 р. Два семенника лежат на заднем конце тела друг
за другом, в виде овальных
Рис.- 151—Himasthla miiehlensi,
Л —цельный червь; В — передний конец
с воротником шипиков. Из Фауста.
тел; яичник лежит перед передним
семенником. Мешок цирруса с очень
длинным семенным пузырьком, корот-
кой простатической частью и мус-
кулистым циррусом, вооруженным
венчиком шипов. Матка тянется в виде
тонкой трубки. Яйца неправильно
овальной формы с крышечкой: 114—
149X62—85р. (рис. 151). Цикл развития
неизвестен. Полагают, что хозяевами
являются некоторые морские птицы.
Рис. 152 — Dicrocoelium
lanceatum
о — семенник; i — кишечник;
d — желточник; t — яичник;
и — матка. По Брауну.
Изгнаны хенрподовым маслом из человека (в Нью-Йорке), который
перед тем ел сырых двустворчатых моллюсков Venus metcenaria.
Семейство Dicrocoeliidae
Для представителей сем. Dicrocoeliidae характерно расположение,
семенников впереди от яичника. Паразитируют преимущественно в пе-
чени, реже в поджелудочной железе и кишечнике птиц, млекопитаю-
щих и пресмыкающихся. У человека известны только два вида этого
семейства, паразитирующих в печени.
Род Dicrocoelium Dujardin
Dicrocoelium lanceatum Still, et Hass.
Синонимы: Distomum lanceatum Mehlis, Fasciola .dendritica Rud.
Двуустка ланцетовидная (рис. 152) вполне оправдывает
свое наименование, так как удлиненное тело ее суживается к концам.
238
СПЕЦИАЛЬНАЯ ЧАСТЬ
Задний конец тела тупокруглый, передний — более заострен. Длина
5—15 мм, ширина 1,5—2,5 мм. На полупрозрачном беловатом теле
двуустки довольно ясно темными полосками просвечивают трубочки
матки (рис. 152, п). Кутикула лишена шипиков. Присоски лежат до-
вольно близко друг около друга. Ротовая лежит терминально, 0,5 мм
диаметром, а брюшная — 0,6 мм. Ветви кишечника простые (/), до зад-
него конца тела они не доходят. Выделительная система состоит из
длинного трубчатого экскреторного пузыря, открывающегося на заднем
конце тела, а спереди принимающего в себя две поперечные ветви; по-
следние отходят от двух боковых стволов, дающих боковые ответвле-
ния. Два неправильно округлых или слабо дольчатых семенника (о>
лежат сейчас же за брюшной присоской — один позади другого. Все
пространство, начиная от семенников и до заднего конца тела, занято
темнобурыми трубками матки. Яичник (округлой формы) один (t);
лежит позади заднего семенника. Желточники занимают бока средней
трети тела (d), прикрывая собою ветви кишечника.
Яйца овальные, с толстой скорлупой, с маленькой крышечкой на
одном из полюсов, уплощенные с одной стороны. Длина их 38—45
ширина 22—30 В незрелом состоянии они желтоватые, позднее же
делаются темнокоричневыми.
Паразитирует в печени овцы, быка, коз, зайцев, свиней. У человека
обнаружена только в единичных случаях (Европа, Азия, Африка и
Америка).
Яйца откладываются с уже развитым зародышем.
Цикл развития протекает следующим образом (Mattes и Neu-
haus) (рис. 153).
Яйца откладываются с вполне развитым мирацидием, покрытым
редкими ресничками и имеющим буравящий стилет. Пигментного глаз-
ка нет; его редукция объясняется, видимо, тем, что мирацидий (III) не
выходит во внешнюю среду, но вылупляется лишь иногда, когда яйцо^
будет съедено одним из наземных моллюсков {рис. 154), являющимся
промежуточным хозяином ланцетовидной двуустки (Helicella erfcetorum,
Н. candidula, Zebrina detrita, Torquilla frumentum, Theba cartusiana и>
Euomphalia strigella). Мирацидий вылупляется из яйца в пищеводе
моллюска (ZV), проходит в желудок и в протоки печени, затем актив-
но проникает в междолечную соединительную ткань печени, где и пре-
вращается в разветвленную спороцисту первого порядка (V). Внутри
ее образуется множество спороцист второго порядка. Оболочка мате-
ринской спороцисты распадается. В дочерних спороцистах развиваются
церкарии с буравящим стилетом и длинным хвостом (V7, VII) у
описанные ранее под именем Cercaria vitrina. Церкарии выходят из
спороцист и активно проникают через ткань печени в венозные сосуды
и попадают в капилляры легкого. Здесь церкарии мигрируют в полость
легкого, где и инцистируются; инцистированные церкарии склеиваются
поверхностной слизью вместе по нескольку экземпляров, образуя
«сборные цист ы» (VIII). Последние при дыхательных движениях
моллюска выдавливаются через отверстие мантии наружу и со слизью
моллюска попадают на поверхность растений, по которым ползает мол-
люск (IX). Когда овцы поедают корм с жизнедеятельными сборными
цистами, то последние вскрываются в двенадцатиперстной кишке и ге-
матогенным путем через портальную вену мигрируют в печень, где и
превращаются в половозрелых двуусток.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
239»
Очевидно, что заражение человека может произойти при случайном
проглатывании «сборных цист».
По А. Скворцову, промежуточным хозяином этой трематоды для
Московской области является Helicella candidula (1934); в Азовско-
Рис. 153 —Цикл развития двуустки ланцетовидной. По Н е й г а у з у.
Черноморском крае таковым является наземный моллюск Theba frutici-
cola и Th. carthusiana (рис. 154). В теле моллюска личиночные формы
D. lanceatum достигают инвазирующей стадии в течение 82—134 дней
(в условиях лаборатории). В окончательном же хозяине паразит начи-
нает выделять яйца через 70—81 день после съедания зараженного
моллюска овцой или кроликом (В. Пухов, Е. Кривошта и Л. Величкин).
240
СПЕЦИАЛЬНАЯ ЧАСТЬ
Действие на хозяина. Паразит несомненно обладает пато-
генными свойствами. Желчные пути печени в местах нахождения па-
разитов расширяются, эпителий гипертрофируется с. образованием аде-
номоподобных разрастаний; окружающая соединительная ткань ин-
фильтрируется различными формами лейкоцитов, разрастается и при-
нимает склеротический характер. Вовлекается в патологический про-
£
Рис. 154— Моллюски — промежуточ-
ные хозяева двуустки ланцетовид-
ной:
А, В — Hdicella unifasciata Poir (— Н. can-
didula Stud.), (3. Европа); C,D — Theba
cartas, i.ia Mill!. (Крым, Кавказ); E — Zeb-
rina [Buliminus) detrita Mull. (Кавказ).
Оригинал.
Рис. 155 — Церкария Dicro-
coelium dendriticutn с брюш-
ной стороны с экскреторной
системой (а) и литическими
железами (by, отдельно изо-
бражен стилет, лежащий
в передне-спинном отделе
ротовой присоски. Из Ф о-
гель и Фауста.
цесс и эпителиальная ткань печени с атрофией ее клеток и замеще-
нием их соединительной тканью. В общем, патологические процессы
при дикроцелирзе слабее, чем при фасциолезе. Меньшую вредоносность
двуустки ланцетовидной авторы ставят в связь с отсутствием шипи-
ков, с меньшими размерами паразита, «быть может с меньшей токсич-
ностью», причем меньшую токсэмию объясняют малыми размерами
червя. Едва ли такие моменты механического характера могут быть
выдвинуты в качестве решающего объяснения. Двуустка кошачья не-
многим больше двуустки ланцетовидной, а патологический эффект от
нее неизмеримо больший. С другой стороны, токсичность паразита свя-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
241
зывается не с размерами его тела, а со свойствами ядовитых начал;
крупные паразиты могут быть и бывают слабо токсичными, а мелкие —
сильно ядовитыми. Поэтому характеристику паразита шожно давать
только на основании совокупности его свойств.
Dicr. lanceatum распространен очень широко. У животных он встре-
чается иногда одновременно с печеночной двуусткой. В иностранной
литературе описано около двух десятков случаев дикроцелиоза челове-
ка. В СССР было насчитано свыше 125 случаев дикроцелиоза. Дей-
ствительно, D. lanceatum в печени человека паразитировать может;
Калантарян нашла его дважды при вскрытии трупов. Однако огромное
большинство случаев дикроцелиоза (в частности, в Донбассе) было
установлено банальным способом исследования faeces на яйца глист.
Между тем, как показал опытами Ж. Штром, яйца ланцетовидной и
печеночной двуусток выделяются с faeces у лиц, съевших вареную
или жареную баранью печень с этими паразитами, в течение трех —
пяти дней. Переисследование им faeces ряда лиц, в которых были пре-
жде обнаружены яйца £>. lanceatum, дало отрицательные результаты.
Равным образом наблюдались случаи семейного дикроцелиоза, когда
у всех трех членов некоей семьи были найдены яйца D. lanceatum,
позднее также исчезнувшие. Vogel (1919) и Brumpt (1927) высказали
предположение, что яйца D. lanceatum, могут быть съедены с заражен-
ной печенью и, следовательно, не иметь никакого отношения к зара-
женности самого человека двуустками. Штром, независимо от назван-
ных авторов, пришел к заключению, что наряду с действительным за-
ражением человека печеночной и ланцетовидной двуустками бывают
случаи ложного (по нашей терминологии — мнимого) фасциолеза и
дикроцелиоза, когда при микроскопировании faeces попадаются яйца
паразитов, лишь проходящих через кишечник («т ранзитные»
яйца).
В свете этих данных теряют силу те случаи «дикроцелиоза», кото-
рые были установлены на основании обычного микроскопирования
faeces. Во втором издании «Трематодозы печени человека» (1928)
Скрябин и Шульц принимают в расчет соображения работ Vogel (1919),
Brumpt (1927) и Штрома (1927) и считают, что часть опубликованных
случаев дикроцелиоза и фасциолеза человека должна быть отнесена к
псевдотрематодозам (мнимым инвазиям двуустками). К сожа-
лению, нет возможности в большинстве случаев установить по опубли-
кованным голым фактам нахождения яиц в faeces, к какой именно ка-
тегории инвазии — явной или мнимой — должен быть отнесен каждый
такой случай.
В 1927 г. Скрябин и Шульц настаивали на необходимости полного
игнорирования всех статистических данных по проценту заражения лю-
дей острицами, когда соответствующая диагностика проводилась обыч-
ными методами микроскопирования faeces — методами, оказавшимися
в отношении точности данных «скомпрометированными». Аналогичное
положение создалось и для старых случаев дикроцелиоза и фасциоле-
за, если не было приведено особых доказательств их действительности.
Все эти обстоятельства дали толчок к уточнению диагностики ди-
кроцелиоза и аналогичных глистных инвазий. Faeces исследуется повтор-
но, чтобы убедиться в постоянстве присутствия яиц. Обследуемых пе-
реводят на растительно-молочную диэту или исключают из пищевого
рациона печень; выясняют анамнестически, что ел обследуемый в бли-
жайшие дни; добывается дуоденальным зондом содержимое двенадцати-
перстной кишки и исследуется на яйца. Скрупулезность обследования
242
СПЕЦИАЛЬНАЯ ЧАСТЬ
необходима также и ввиду того, что сырая печень употребляется в ка-
честве лекарственного вещества в клиниках; в нашей практике был слу-
чай, когда принесли от больной с тяжелым анемическим состоянием и
измененной печенью faeces с яйцами D. lanceoium\ при разговоре с
врачом выяснилось, что больная каждый день получает сырую печень; по
предложению автора, печень была исследована и оказалась зараженной
сосальщиками.
Необходимо подчеркнуть реальность существования дикроцелиозов
человека. Поэтому особенное значение приобретают работы, в которых
дается диференциация дикроцелиозов,
открываемых у людей — на истинные и
мнимые. Наконец, и в клинике начинают
отыскивать симптомы, ро которым мож-
но судить о действительном наличии ин-
вазии. Так, при ‘наличии яиц ланцето-
видной двуустки у больных дикроцелио-
зом "отмечают постоянные и усиливаю-
щиеся по ночам боли в правом подре-
берье, частые головные боли, голово-
кружение, горечь и сухость во рту,
частые тошноты, иногда рвоту,. субикте-
ричность склер, увеличение печени, плот-
ность и болезненность ее при пальпации
и простукивании реберной дуги (Егоров,
Маликов). Были сделаны попытки
установления микроскопических отли-
чий содержимого яиц (т. е. зароды-
шей) D. lanceatum, подвергавшихся ку-
линарной обработке при приготовлении
пищи, от яиц свежих (Блажин), но
практическая приложимость этих данных
весьма ограничена, так как изменения
Рис. 156 — Eurytrema pancreaticum в структуре*^ародыша, наступающие при
а — ротовая присоска; ь - брюшная при- высокой температуре, не всегда ЕЫра-
соска; с — семенник; v — желточник; о— r J к г
яичник; и — матка; i — кишечник; go — ЖеНЫ ДОСТЗТОЧНО реЗКО (наПрИМер, В
мужское и женское половое отверстие в толстых кусках печени), И В ТвМНОКО-
по к.скрябину и р. шульцу. ричневых яйцах не могут быть рассмо-
трены (Тарасов).
Диагностика. По нахождению яиц, с исключением возможности
алиментарного их происхождения.
Лечение. Вливание эметина.
Профилактика Профилактические меры могут носить предпо-
ложительный характер, так как неизвестен способ попадания церкарий
паразита в окончательного хозяина; следует остерегаться есть сырые
растительные продукты.
Род Eurytrema Looss
Eurytrema pancreaticum Looss
Синонимы: Distomum pancreaticum Janson^, \Dicrocoelium pancreaticum
Rail, и Marotel.
Тело. более короткое и широкое, чем у Dicrocoelium lanceatum, со слегка гоф-
рированными краями (рис. 156). Ротовая присоска очень большая. Семенники лежат
по бокам задней половины брюшной присоски. Яичник расположен сбоку от обще-
го желточного канала. Желточники лежат в третьей четверти тела. Матка в задней
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
243
половине тела заходит кпереди от правого семенника. Яйца неразличимы по ве-
личине и окраске от яиц Dicr. lajiceatum.
Паразитирует в протоке поджелудочной железы свиней, в желчных ходах ро-
гатого скота (Гонконг, Северный Китай).
В СССР этот паразит обнаружен у овец на Дальнем Востоке (К. Скрябин и
Р. Шульц), в Казахстане (Баданин), Южной Киргизии (Ф. Талызин). Один раз най-
ден у человека в Гонконге.
Изменения в поджелудочной железе сходны с патологическими поражениями при
фасциолезе.
У овец эуритрематоз протекает в виде хронического интерстициального панкреа-
тита. При сильных инвазиях наблюдается острый некротизирующий панкреатит и
некротизирующее воспаление протоков поджелудочной железы. Какую-то роль в
этих процессах играет и вторичное: внедрение микрофлоры из двенадцатиперстной
кишки (В. Всеволодов).
Диагноз по яйцам может быть поставлен только и тех местностях, где отсут-
ствует Dicr. lanceatum. В противном случае различительная диагностика невозможна.
Цикл развития паразита неизвестен, профилактика—также.
Семейство Opisthorchidae
Род Opisthorchis Blanch.
Opisthorchis felineus Riv.— кошачья двуустка
Двуустка кошачья, или сибирская (рис. 157), имеет
удлиненное тело, спереди приостренное, сзади округленное. В свежем
виде она желто-оранжевого или желтоватого цве-
та, почти совсем прозрачная, исключая среднюю
часть тела, где лежит матка. Длина — 8—13 мм,
ширина—1—2—2—3 мм. Паразиты из печени чело-
века крупнее, чем из печени кошки (6,1 X 7 мм
против 4,5 ХМ мм. Н. Плотников). Шипиков на
кутикуле нет. Ротовая присоска 0,25 мм диаме- 9
тром; задняя немного больше. Ветви кишечника (z)~
простые, доходят до заднего конца тела, гранича
с выходной частью толстого извитого выделитель-
ного канала, идущего посредине задней трети те-
ла. По бокам его изгибов лежат, наискось один
позади другого, два розетковидных семенника (Z).
Средняя треть тела, в пространстве между ветвя-
ми кишечника, занята изгибами трубчатой матки,
которые могут закрывать собою лежащие у ее
заднего конца яичник и семенной пузырек. Кна-
ружи от ветвей кишечника в средней трети тела
с каждой стороны помещаются желточники.
Цикл развйтия. Желтоватые овальные
яйца суживаются к переднему полюсу (табл. V,
рис. 7), где помещается крышечка. Размеры их
значительно вариируют: в среднем они 0,026—
0,03 мм длины и 0,01—0,015 мм ширины. Парази-
тирует в печени (встречается также и в кишечни-
ке) собаки, кошки, россомахи, лисицы, песца, со-
боля, свиньи, человека в Европе и в Азии. Из ла-
бораторных животных ею заражаются кролик и
морская свинка. Паразитирует также в поджелу-
дочной железе (редко). Первым промежуточным,
хозяином, по Vogel (1932), является моллюск —
Bithynium leachii (рис. 158); вторым промежуточ-
ным хозяином, по Ciurea, служат рыбы— плотва
Рис. 157 — Кошачья
двуустка — Opisthor-
chis felineus.
р — ротовая присоска; Z—
кишечник; С? $—половые
отверстия; и — матка; d—
желточники; dd— желточ-
ные протоки; rs—семепри-
емник; о — оотип; t —се-
менники; с — выделитель-
ный канал; ре — экскре-
торная пора. Оригинал
Е. Павловского.
244
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 158 — Bythynium leachil. Оригинал.
Рис. 159— Scardinius erythrophthalmus — красноперка. Оригинал.
Рис. 160 — Opisthorchis felineus.
Мирацидий. По Фогелю.
Рис. 161 —Opislhorchis feli-
neus. Яйцо с мирацидием.
По Фогелю.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
245
(Leucis cus rutilus) и язь (Idus melanotus), а также линь (Tinea vul-
garis), лещ (Abramis Ьгата) и красноперка (Scardinius erythrophthal-
mus,— рис. 159) густера — Blicca bjorkna (рис. 149), в разных внутрен-
них органах которых попадаются инцистированные молодые двуустки.
Мирацидии (рис. 160) не вылупляются, как обычно, в воде из по-
павших туда яиц (рис. 161) паразита. Моллюски проглатывают яйца,
которые потом вскрываются под действием пищеварительных соков
(Vogel), что происходит в задней кишке моллюска. Вышедший мираци-
дий проникает в полость тела и здесь превращается в спороцисту
Рис. 162 — Opisthorchis felineus.
Спороциста. По Фогелю.
Рис. 163 — Opisthors-
his felineus. Редия.
По Фогелю.
(рис. 162), в которой образуются редии (рис. 163); в освободившихся
редиях развиваются церкарии (рис. 164), активно покидающие мол-
люска и некоторое время плавающие в воде. При наличии подходящего
вида рыбы церкарии вбуравливаются в кожные покровы и в мышцы и
превращаются здесь в метацеркарии, которые капсулируются двойной
оболочкой. Когда рыба с метацеркариями съедается окончательным хо-
зяином, то в его желудке перевариваются мышцы рыбы и наружная тол-
стая оболочка капсулы метацеркарии; личинка остается одетой тонкой
внутренней оболочкой, которая, по Н. Плотникову, переваривается в
дуоденальном соке. Освободившиеся из капсул молодые двуустки про-
двигаются из полости тонкой кишки в желчный пузырь и в печень по
желчному протоку (Фогель, Плотников); здесь двуустки растут дней
двенадцать и через месяц начинают выделять яйца. Длительность жиз-
ни О, felineus у кошки свыше трех лет (Плотников).
Заражение человека и животных происходит через поедание сырой
или вяленой рыбы.
По работам Плотникова и Зерчанинова в Тобольске и Обдорске,
промежуточными хозяевами кошачьей двуустки являются язь, чебак
4G
СПЕЦИАЛЬНАЯ ЧАСТЬ
(Rutilus rutilus lacustris—’рис. 168) и сибирский елец (Leuciscus
leuciscus bfyicalensis,— рис. 169). Одетые толстой соединительнотканной
оболочкой и гиалиновой капсулой цисты с метацеркариями О. felineus
залегают в мускулатуре и в подкожном жировом слое этих рыб числом
эт нескольких штук до 1500 экземпляров. Из обследованных там же
эечных (Обь и Иртыш) чебаков зараженными оказались 10% рыбы, из
Рис. 164 а и b — Opisthorchis felineus. Церкарии. По Фогелю.
я зем — 47,6% и из ельцов —- 55,8%. Рыбы тех же видов и того же
возраста из заречных озер Тобольска были вовсе свободны от цист ме-
га церкарий. Рыбы начинают заражаться этим паразитом со стадии
малька; интенсивность и экстенсивность инвазии увеличиваются с воз-
растом рыбы.
Быпрепарованные из мышц рыбы метацеркарии могут жить в физио-
логическом растворе поваренной соли до четырех месяцев, сохраняя
инвазирующую способность в течение месяца. В солевых растворах они
гибнут в 10 дней (5—10% NaCl) и в три дня (15—20% NaCl). При
засоле рыбы натиранием ее солью в разрезанной полости брюха или
через разрезанную спинку язя в 1 кг весом все личинки в цистах поги-
бают через 10 дней.
Копчение холодное и горячее, которому предшествует обработка
рыбы солью, как правило, убивает паразитов. Замораживание не сразу
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
247
Рис. 165 —Схема миграционного пути кошачьей двуустки
во время развития ее в организме моллюска Bithynla,
а'—рот; Ь—,пищевод; g — желудок; /— печень; d — анальное отверстие;
*с — жаберные пластинки; R — рении; гг — церкарии. По Фогелю.
Рис. 166 — Инкапсулированная личинка
кошачьей двуустки в мыщцах линя (слева)
и личинка или молодая двуустка, выну-
тая в свежем состоянии из цисты (справа).
По Ч и у р е а.
РисЛ1б7 — Opisthorchis
felineus — молодая дву-
устка.
Cj— выделительные каналы,
i \ По Фоге лю.
248
СПЕЦИАЛЬНАЯ ЧАСТЬ
убивает личинок в цистах; например, цисты могут выживать в мясе ры-
бы при t° —2 —12° С до 3 суток. Сильно высушенная рыба1 становится
безвредной.
Патологическое значение паразита несомненно. С ним
ознакомимся по описанию К. Виноградова, открывшего этого сосальщи-
ка в печени человека в Сибири {Томск).
Рис. 168 — Чебак — Rutilus rutilus lacustris. Оригинал.
Печень уменьшена в объеме, бугриста; капсула оморщена, ткань дря-
бла и окрашена в темнозеленый цвет в виде гнезд различной величины
(от лесного ореха до кулака и более), между которыми выделяются участ-
ки ткани бледной с буроватым оттенком и мускатной. Зеленые гнезда
представляют собою печеночную ткань, набухшую, сочную, со сглажен-
ными дольками, выдающуюся на поверхности разреза. Желчные прото-
ки по всей печени наполнены густой темной желчью, смешанной со
Рис. 169 — Сибирский елец — Leuciscus leuciscus baikalensis. Оригинал.
слизью. Желчный пузырь растянут до размеров гусиного яйца и напол-
нен желтовато-зеленоватой слизистой жидкостью, в которой находится
несколько рыхлых, легко раздавливаемых конкрементов черного цвета-
ст едва заметной величины до конопляного зерна.
В значительном проценте случаев (около 36%) описторхоз человека
протекает без каких-либо клинических проявлений, что зависит от сте-
пени интенсивности инвазии и от соотношений «паразит — хозяин». По-
ражению может подвергнуться и поджелудочная железа, в которой
Таблица VI
Г — Финноз сердца свиньи (Тaenia solium), Оригиналы
2 — яйцо Fasciola hepatica; 3 — nnno^Paragonimus Westerman! ringeri; 4 — яйцо Dicrocoelium
lanceatum; 5 — яйцо Metagonimus "yokogawai; 6 — яйцо Clonorchis sinesis; 7 — яйцо Opis-
thorchis felineus; 8— яйцо Hymenolepis diminuta; 9 — яйцо Hymenolepis папа; 10 — яйцо
Diphyllobothrium latum; 11 — яйцо Dipylidium caninum; 12 — яйцо Taeniarhynchus sagin at us
с эмбриональной оболочкой и с филаментами; 13 — яйцо Ascaris lumbricoides с поверх-
ностной структурой; 14 — Ascaris lumbricoides, яйцо в оптическом разрезе; 15, 16 — яйцо
Trichicephalus trichiurus; 17— яйцо Hepaticola hepatica; 18 — яйцо Enterobius vermicular is,
несколько полежавшее; 19 — яйцо Enterobius, свежее яйцо. По К. Скрябину и Р. С. Шульцу.
Яйца, изображенные на рисунках 5—7, имеют на нижнем полюсе не видные на рисунках
штифтиковидное утолщение оболочки.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
249
Плотниковым обнаружены О. felineus у 36% носителей этого паразита.
-Патолого-анатомические изменения в развитых случаях ведут к меж-
дольковому и внутридольковому циррозу и к некрозу паренхимы под-
желудочной железы.
Микроскопические изменения печени заключались в следующем.
Кроме воспаления стенок желчных протоков, имелся цирроз, ограничен-
ный участками различной величины, чередовавшимися с нормальной
тканью. По большей части паренхима печени находилась в состоянии
бурой атрофии. Глубокий цирроз наблюдался на участках печени, окра-
шенных желчью в темнозеленый цвет. В этих местах печеночные доль-
ки нередко во всю толщу прорезаны были грануляционной тканью, пе-
ченочные же клетки являлись рассеянными среди этой ткани отдельно1
или небольшими группами.
Такие изменения печени могут считаться результатом паразитирова-
ния двуусток, которые, закупоривая желчные каналы, вызывают жел-
туху и, раздражая стенки желчных протоков, обусловливают своим при-
сутствием развитие воспалительных процессов, переходящих и на пече-
ночную ткань. Цирротические изменения печени могут влиять и на
воротную вену, сдавливать ее просвет и причинять тем расстройство
брюшного кровообращения. Упомянутые выше конкременты в желчном
пузыре имели каждый внутри себя яйца двуустки, на которые отлага-
лись кристаллы холестерина с желчным пигментом. При длительности
такого процесса мелкие конкременты могут превратиться в желчные
камни. Клинически описторхоз протекает при явлениях гепатита, холе-
цистита, ангиохолита или панкреатита, однако без каких-либо специфи-
ческих особенностей, которые можно было бы связать с описторхозом.
Реактивные воспалительные процессы в печени, вызванные парази-
тированием в ней двуустки, могут иметь и благоприятное для организма
течение. Кулябко наблюдал случай, когда 18 кошачьих двуусток плава-
ли в совершенно замкнутой соединительнотканной капсуле печени кош-
ки. Такая киста образовалась из желчного протока, стенки которого
срослись вследствие слипчивого воспаления. Инкапсулированные пара-
зиты были совершенно изолированы, и выбраться из капсулы, без на-
сильственного разрыва ее стенок, не могли. Благодаря такой изоляции
они не могли более причинять вреда своему хозяину.
Ярко выраженный описторхоз кошек и пушных зверей может вести
к циррозу печени и к смерти зараженных животных. Иммунитета к су-
перинвазиям или к реинвазиям при описторхозе не развивается; этим
и объясняется накопление в печени иногда1 колоссального количества
кошачьих двуусток (см. ниже).
Распространение. Двуустка кошачья довольно широко рас-
пространена в Европейской части СССР; у человека особенно часто
этот паразит встречается в Сибири, в частности в Западной, а. также
в Уральской области. На вскрытиях в Томске двуустка кошачья была
обнаружена в 7%, а в Тобольске в 60% случаев. В низовьях Оби опи-
сторхоз установлен у 85,2% всех обследованных, при общей зараженно-
сти глистами в 89% (вогулы, остяки и меньше зыряне и самоеды);
дети до 4-летнего возраста инвазированы уже наполовину вследствие
принятого обычая кормить их рыбой, начиная с годовалого возраста
(В. Подъяпольская).
Описторхоз несомненно доказан и на Украине (в Никопольском рай-
оне, М. Миронова).
Из животных собаки несколько менее заражены, чем кошки; в То-
больске все обследованные кошки имели описторхоз с количеством па-
>50
СПЕЦИАЛЬНАЯ ЧАСТЬ
разитов до 5 тыс. В других случаях отмечалось присутствие лишь еди-
ничных паразитов.
У человека находили в печени до 25 321 (!) экземпляров этой -дву-
устки (Плотников и Зерчанинов).
Диагноз ставится по нахождению яиц (табл. V, рис. 7) в faeces
и в дуоденальном соке. Возможно паразитирование О. felineus и у сви-
ней, что следует учитывать при диагностике человеческих случаев, хотя
в общем возможность нахождения яиц этого паразита алиментарного
происхождения гораздо более редка, чем по отношению к двуустке
ланцетовидной или двуустке печеночной.
Лечение — Н. Плотниковым рекомендован гексахлорэтан.
Профилактика общественная пока трудно осуществима, так
как она связана с охраной водоемов от загрязнения фекалиями зара-
женных собак, кошек и людей. Ветеринарный надзор за рыбой в целях
изъятия илй недопущения на рынок зараженной рыбы практически не-
осуществим. Остается более надежной профилактика личная: не есть
сырой или провяленой рыбы. Массовое обезвреживание рыб может
быть осуществлено выдерживанием их на холоду при t° 2—12° С в те-
чение 5—6 дней.
Opisthorchis noverca Braun
Двуустка индийская относится к тому же роду, что и двуустка сибирская, так
как имеет два семенника (слабодольчатые, не розеточные) в задней трети тела.
Длина тела 9—12 мм, ширина 2 мм. Кутикула покрыта мелкими шипиками. Ротовая
Ърисоска гораздо больше брюшной. Ветви кишечника простые, не доходят до зад-
него конца тела. Между семенниками проходит тонкий извилистый выделительный
канал, ветвящийся на переднем конце. Бока средней части тела заняты желточника-
ми, прерванными посредине. Матка образует меньшее количество извивов, чем у
О. felineus, лежащих в передней половине тела.
Яйца овальные, с крышечкой, длиною 0,034 мм, шириною 0,021 мм. Паразити-
рует в печени человека (Индия) и собаки. Развитие неизвестно.
Возможно заражение человека и другими видами трематод рода Opisthorchis,
например, О, viverrinus (двуустка виверры).
Род Clonorchis Looss
Clonorchis sinensis Looss — двуустка китайская
Синоним: Clonorchis endemicus Looss.
Красновая полулрозрачность двуустки с удлиненным телом, сужи-
вающимся к переднему концу. Длина 10—25 мм, ширина 3—5 мм. Ку-
тикула без шипиков. Передняя присоска 0,6 мм диаметром, брюшная
около 0,470 мм. Ветви кишечника простые. Типичны два Ветвистых се-
менника, лежащих друг за другом в задней части тела. Матка обра-
зует небольшое число изгибов в средней трети тела. Желточники тянут-
ся от брюшной присоски до переднего семенника. В паренхиме тела
пигментные зернышки.
Яйца 0,028—0,035 мм длины и 0,012—0,019 мм ширины (в среднем
0,029 на 0,016). Они почти черного цвета (табл. V, рис. 6). Задний
полюс их тупокруглый, к переднему полюсу, занятому крышечкой, яйцо
суживается.
Цикл развития. В отложенном яйце уже имеется сформиро^
ванный мирацидий (рис. 170). Яйца вскрываются (рис. 170,С) в пище-
воде моллюсков, когда будут ими проглочены. Первыми промежуточны-
ми хозяевами являются моллюски — Parafossarulus striatulus, Bythinia
fuchsiana. В. longicornis.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
251
Вылупившиеся мирацидии (рис. 170, С) проникают в околопищевод-
ное пространство полости тела, где превращаются в спороцисты (рис.
170, Е). Редии (F) развиваются в печени, куда уже мигрирует паразит.
Церкарии с массивным хвостом и пигментными глазками (G) выходят
из редий и покидают моллюска. Они плавают в воде и внедряются под
чешую и в мышцы различных пресноводных рыб, теряя при этом хвэ-
Рис. 170 — Двуустка китайская — Clonorchis sinensis.
л—яйцо; В — яйцо с зародышем; С — выход зародыша из яйца; D — мирацидий:
Е — спороциста; F — редия; G — церкария; Н — взрослая двуустка. По Фаусту.
стовой придаток. В рыбе церкарии инцистируются. Вторым промежу-
точным хозяином могут служить многочисленные виды рыб семейств
окуневых, бычков и Anabantidae. Фауст насчитывает 34 вида рыб, 1 при
поедании которых возможно заражение китайской двуусткой.
Окончательным хозяином являются: человек, собака, кошка, свинья,
куница, барсук и другие животные. В них цисты растворяются, а . осво-
бодившиеся молодые двуустки достигают половозрелое™ в срок от 23
до 26 дней, мигрировав предварительно в печень хозяина.
Распространение. В Китае встречается чаще, чем в Японии.
1 Например, Pseudor'asbora parva Schl., Leucogobio giintheri Isch'ik., L. mayedae
Jord, и Snyd. Sarcochellychthys variegatus Schl., Pseudopen fampus lupus Bleek.,
Biwia zezera (I.schik.), Carassius auratus (Lin.) и мн, др^
252
СПЕЦИАЛЬНАЯ ЧАСТЬ
Найдена и на Дальнем Востоке. Бывает в огромном количестве у одно-
го и того же хозяина (до 4 тыс. и более экземпляров). Реже встре-
чается в желчном пузыре и в поджелудочной железе.
Действие на хозяина. Патогенна, вызывает изменения пече-
ни, сходные с поражениями другими двуустками. При слабом зараже-
нии заметных симптомов нет. Во второй стадии заболевания наблю-
Рис. 171—Pseudamphistomum
truncatum. По Скрябину и
Шульцу.
Рис. 172 — Heterophyes
heterophyes.
По Л о ос с.
даются понос, отеки, гипертрофия печени и, в заключительной фазе
болезни, цирроз печени с соответствующими симптомами нарушения
кровообращения через воротную вену.
Диагностика. По нахождению яиц в faeces.
Лечение. Внутривенозные впрыскивания TaJrtarus stibjatus; кра-
ски — gentian-violet, cristal-violet и methyl-violet — per os.
Профилактика. He есть сырой или полусырой рыбы; в районах
распространения паразитов — дезинфекция фекалий прибавлением сер-
нокислого аммония.
Род Pseudamphistomum
Pseudamphistomum truncatum Liiehe
Синоним: M etorch is truncatus.
Беловатое конусовидное тело этой двуустки 2—2,5 мм длины и
0,6—1,0 мм ширины в задней части (рис. 171). Кутикула густо усеяна
шипиками. Брюшная присоска чуть больше ротовой. Округлые семен-
ники лежат почти рядом друг с другом в задней трети тела (рис. 171).
•В остальных чертах организация близка к двуустке сибирской. Яйца
с крышечкой, длина их 0,029 мм и ширина 0,011 мм.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
253
Паразитирует в печени тюленей, лисиц и кошек. Возможный пара-
зит человека; последний может заражаться им от рыб.
Повидимому, была найдена в единственном экземпляре среди коша-
чьих двуусток в печени человека в Томске Виноградовым. Обнаружена
у кошек в различных местах Европейской части СССР (Харьков,
б. Самарская губ. и др.).
Цикл развития и характер патогенности для человека неизвестны.
Семейство Heterophyidae
Род Heterophyes Cobb.
Heterophyes heterophyes Still, et Hass.— двуустка египетская
Очень мелкий паразит — 1—2 мм длины и 0,4—0,5 мм ширины (рис. 172). Удли-
ненное тело разделяется на более узкую переднюю и более широкую заднюю части.
Кутикула густо усеяна шипиками. По середине плоского тела лежит крупная брюш-
ная присоска, раза в три бблыпая, чем ротовая. Длинный пищевод делится на две
простые кишечные ветви. Два овальные семенника лежат между задними концами
кишечных трубок и выделительным каналом. Кпереди от них на средней линии те-
ла помещается яичник. Желточники развиты слабо. Они представлены каждый при-
близительно 14 фолликулами, лежащими нд кишечных трубках на уровне яичника.
Семеприемник велик. Изгибы матки занимают свободные места задней половины
тела и не заходят кпереди за брюшную присоску. Большой, лежащий вне мешка
совокупительного органа, семенной пузырек и влагалище открываются в половую
клоаку. Отверстие последней окружено широким присоскообразным валиком с вен-
чиком разветвленных щетинок внутри него. Кольцевидный валик этот лежит сбоку
и кзади от брюшной присоски. Светлобурые овальные яйца с толстой скорлупой и
крышечкой 0,02—0,03 мм длины и 0,015-—0,017 мм ширины.
Яйца очень сходны с яйцами других представителей сем. Heterophyidae,
В Японии имеется близкий вид Н. katsuradai (человек, собака).
Цикл развития. Яйца откладываются с вполне развитым мирацидием; однако он
во внешней среде не вылупляется. Последнее происходит лишь тогда, когда яйцо
будет проглочено пресноводным моллюском Pironelia conica (Египет, по Khalil.). В
первом промежуточном хозяине паразит проходит стадию спороцисты, затем дает
одно или два поколения редий. Выходящие из тела моллюска церкарии инцистируют-
ся в кефали (Mugil cephalus — рис. 173) и в некоторых других рыбах. Человек за-
ражается, употребляя в пищу полусырую или. сырую рыбу.
Паразиты могут вызывать раздражение кишечника, колики, понос со слизисты-
ми испражнениями, поверхностные некрозы слизистой оболочки (Khalil).
Яйца могут проникать через стенку кишки, оседать в мезентериальных железах,
попадать даже в сердечные заслонки, в миокард и в мозг. В зависимости от их
локализации могут развиваться симптомы сердечной недостаточности или мозговых
кровоизлияний, ведущих к смерти (Africa).
Паразитирует в тонких кишках человека (Египет), собаки, кошки, лисицы и
птицы — Milvus parasiticus.
Профилактика. Не есть сырой или полусырой рыбы.
Род Metagonimus Katsur.
Metagonimus yokogawai Katsurada
'Мелкая двуустка с грушевидно контурированным телом (рис. 174),
длиной 1—2,5 мм, шириной 0,4—0,8 мм. Кутикула густо покрыта шипи-
ками, лучше развитыми в передней половине тела. Ротовая присоска
0,07—0,09 мм в поперечнике; брюшная соединена с половой присоской
в одно целое размером 0,12—0,145 мм длины и 0,075—0,1 мм ширины;
все это образование сдвинуто в правую сторону, считая от продольной
средней линии тела (рис. 174),. Пищевод длинный. Ветви кишечника
оканчиваются у заднего конца тела. • Есть предглотка и шаровидная
глотка. Круглые или овальные семенники помещаются у заднего конца
тела; между ними лежит экскреторный пузырек. Кпереди от семенников
лежат семеприемник и яичник. Желточники занимают боковые поля в
251
СПЕЦИАЛЬНАЯ ' ЧАСТЬ
задней половине тела в виде двух групп по 10—12 фолликулов в каж-
дой. Извивы матки помещаются в задней половине тела.
Яйца светлого желтовато-коричневого цвета 0,026—0,032 мм длины
и 0,015—0,017 мм ширины, с крышечкой на более узком полюсе; на
Рис. 173—Кефаль {MugИ cephalus)— второй промежуточ-
ный хозяин Heterophyes heterophyes. По Neveu-Lemaire
и Pellegrin.
противоположном округ’лом полюсе имеется маленькое пуговчатое утол-
щение скорлупы (рис. 177, Л).
Паразитирует в слизистой оболочке кишечника! человека, собаки,
кошки и пеликана на! Дальнем Востоке и в Японии. В пределах СССР
Рис. 174 — Metagonimus yokogawai
из кишечника собаки:
а—.ротовая присоска; b — глотка; с — пи-
щевод; d — кишечник; е— половая присоска;
f — семепровод; у — яичник; h — семеприем-
ник; i — желточник; k — семенник
По К. Скрябину и Р. Шульцу.
Рис. 175 — Melania атигеп-
sis. Оригинал.
обнаружена у 13,б°/о обследованных гиляков, гольдов, ульчей и удехей-
цев (по низовьям Амура и Уссури; К. Скрябин с сотрудниками).
Цикл развития. Первым промежуточным хозяином являются
моллюски Melania libertina, М. ebenina, Blanfordia noso-phora, Piradus
cin&ulatus и др., которые заражаются вбуравливающимся в них мираци-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
25с
днем, вышедшим из яйца паразита. В моллюске проходит развитий по-
колений редий (рис. 177, В) и церкарий. Последние покидают моллю-
ска, плавают в воде (рис. 177, С) и нападают на второго промежуточ-
ного хозяина, которым являются различные лососевые и другие рыбы
(например, форель, Plecoglossus altivelis,— рис. 178). Церкарии прони-
кают в толщу покровов чешуи и плавников, где инцистируются
(рис. 176). В лимане реки Амура Змеевым метацеркарии М. yokogawai
Рис. 176 — Цисты Metagonimus yokogawai
в чешуе. По Змееву.
Рис. 177 — Metagonimus yokogawai
А — яйцо с зародышем; В — редия; С —
церкария. По Фаусту.
обнаружены, ' главным образом, у Coregonus ussuriensis и Liocassis
braschnikowi (рис. 179, сильное заражение); более слабо были заражены
Cyprinus carpio, Leuciscus waleckii, Parabramis pekinensis .(рис. 180),
Pseudaspius leptocephalus Hemibarbus labeo, (рис. 1181), и Gobio gob io.
Ни в одном из 126 экземпляров кеты метацеркарий обнаружено нс-
было. Паразиты локализовались только в чешуе, жабрах и плавниках.
Цисты имели форму шара или овала 0,17—0,20 мм. Паразит хорошо
просвечивает через двуконтурную оболочку цисты; видна его подвиж-
ность (Г. Змеев). Через 15 дней после скармливания котятам двуустка
достигает половой зрелости и начинает откладывать яйца.
Заражение человека происходит при поедании сырой или полу-
сырой рыбы с метацеркариями; бывает массовое паразитирование до
4000 экземпляров у одного хозяина.
7И. yokogawai может иногда глубоко проникать в слизистую оболоч-
ку и разрушать ее; в общем патологические изменения, причиняемые
этими паразитами, слабы; при значительных инвазиях наблюдается ка-
тар кишек с диареей.
256
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 181 — Hemibarbus labeo. Оригинал.
Т а б л и ц а VII
Пресноводный краб — Eriochelr sinensis. Оригинал.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
257
Лечение. Изгнание тимолом, бетанафтолом и четыреххлористым
углеродом.
Диагноз. Нахождение яиц в faeces; яйца паразита близко сход-
ны с яйцами кошачьей и китайской двуусток.
Профилактика. Не есть сырой или полусырой рыбы.
В Крыму у собак найден Metagpnimus romanicus romanicus Ciur.
(Исайчиков), однако отношение этого вида к М. yokogawai неясно,
©дни гельминтологи считают их идентичными, другие же признают
за разные виды. Вопрос пока остается открытым.
Рис. 182 — А — Metagonimus minutus; взрослая двуустка. В — ци-
ста с метацеркарией. С ?— Stellanchasmus formosanus. По К a t s u t a.
Из других видов этого семейства недавно описан Metag. minutus
Kats. (рис. 182, А, В). Stellanchasmus formosanus (рис. 182, С) на Фор-
мозе может заражать человека, собаку, кошку и мышь.
М. katsuradai (кишечная трематода рыбоядных млекопитающих и
человека), как и другие виды Heterophyidae, недолговечна: у экспери-
ментально зараженной мыши — наибольшая длительность жизни была
32 дня (Izumi, 1935).
Род Nanophyetes
Nanophyetes schikhobalowi Skr. et Podjap.
Мелкая двуустка с телом яйцевидной формы, суживающимся кпере-
ди и имеющим широкоокругленный задний конец. Длина тела 0,56—
0,58 мм, наибольшая ширина 0,35—0,47 мм. Ротовая присоска 0,1 на
0,12 мм (0,085 на 0,1), брюшная — 0,12 на 0,13 (0,11 на 0,11) мм.
Предглотка и пищевод отсутствуют. Ветки кишечника сравнительно ко-
258
СПЕЦИАЛЬНАЯ ЧАСТЬ
ротки. Семенники удлиненноовальные, лежат в задней половине тела..
Яичник помещается в правой половине тела кпереди от семенника.
Половое отверстие расположено тотчас позади брюшной присоски не-
посредственно на средней линии. С ним связан ретортовидный семен-
ной пузырек. Овальный семеприемник и тельце Мелиса лежат в правой
половине тела. Желточники рассеяны по всему телу (рис. 183).
Яйца по форме неотличимы от яиц лентеца широкого, размеры их
0,065—0,072 на 0,044—0,048 мм, с крышечкой и маленьким тупым вы-
ростом на противоположном полюсе яйца (рис. 183).
Цикл развития неизвестен.
Рис. 183 — Nanophyetes schikhobalowi. Новая дву-
устка человека с Дальнего Востока и ее яйцо.
По К. Скрябину и В. Подъяпольской.
Рис. 184 — Взрослая трема-
тода Т го giotrema salmincola.
По 'Витенберг у.
Описан К. Скрябиным и В. Подъяпольской по экземплярам, изгнан-
ным в Хабаровской больнице одновременно с Metagonymus yokogawai
из людей племени Уде (Дальний Восток). Nanophyetes schikhobalowi
является по сравнению, сделанному Виттенбергом, синонимом амери-
канского- вида Troglotrema salinincola (рис. 184). Паразит этот, видимо,,
нередок на Дальнем Востоке, так как Ж. Штрому удавалось изгонять
его в Ленинграде из учащихся Института северных народностей.
Действие на хозяина неизвестно, но Nanophyetes salminco-
la у собак вызывает тяжелые заболевания, ведущие иногда к смерти.
Диагноз ставится по исследованию изгнанных паразитов.
Изгнание производится тимолом (Шихобалова) и экстрактом
папоротника (Ж. Штром).
Профилактика неизвестна.
Семейство Troglotrematidae
Род Paragonimus Braun
Paragonimus Westermani Braun — легочная двуустка
Синонимы: Distomum ringeri Cobb., Paragonimus ringeri (Cobb).
Легочная двуустка в живом состоянии коричнево-красного цвета, а
в мертвом — серо-коричневая. Ее яйцевидное тело (рис. 185) уплоще-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
259
но на брюшной стороне; в расправленном состоянии длина тела 8—
10—16 мм, ширина 4—6—8 мм, толщина 3—4 мм. Кутикула покрыта
множеством широких шипиков, похожих на чешуйки. Обе присоски
почти одинаковых размеров. Брюшная лежит сейчас же кпереди от
средины тела, а ротовая — у переднего его конца. Ветви кишечника
сильно извилисты. Сзади и сбоку брюшной присоски, в толще тела ле-
жат с одной стороны яичник, а с другой — трубчатая матка. Сильно
развитые желточники занимают все протяжение по бокам тела. Два
Рис. 185 — Paragonimus westermani из легких кошки.
Л — ротовая присоска; b — глотка; 9 с — желточники; dt 1 — кишечник;
е — матка; / — брюшная присоска; g — яичник; h — тельце Мелиса;
т — семевыносящий канал; i — семенник; k — выдели!ельный канал;
п— желточный проток. По К. Скрябину иР. Шульцу.
иятидольчатых семенника лежат позади матки и яичника. Между ними
проходит неравномерно толстый и длинный ствол выделительных орга-
нов. Половое отверстие открывается у заднего края брюшной присоски.
Двуустки легочные живут попарно в легких — в кистах (рис. 186),
образовавшихся из мельчайших бронхов; хозяевами ее являются
человек, тигр, кошка, собака, свинья. Распространена по Восточной
Азии и ее островам. Завозные случаи известны из Москвы и Ленин-
града.
Цикл развития. Паразит очень редко выходит с мокоотой;
яйца его широкоовальные с крышечкой; золотисто-коричневые, 0,08—
260
СПЕЦИАЛЬНАЯ ЧАСТЬ
0,118 мм длины и 0,048—0,060 мм ширины (табл. V, рис. 13). Яйца ско-
пляются в цисте и при разрыве ее выходят с мокротой; вследствие ча-
Рис. 186 — Paragonimus westermani—ле<
точная двуустка в цисте в легком^
•По Brumpt.
стого проглатывания мокроты
больным, яйца нередко обна-
руживаются и в faeces. Яйца
выходят с незрелым зароды-
шем, требующим для своего
развития от четырех до шести
недель. Вылупляющийся в во-
де мирацидий плавает свобод-
но и вбуравливается в первого
промежуточного хозяина, ко-
торым являются различные
моллюски рода Melania —
М. libertina (доминирующий
хозяин), М. nodiperda, М. mul-
ticincta и др. В моллюске па-
разит проходит стадии споро-
цисты (рис. 187, Л), двух по-
колений редий и церкарии.
Церкарии легочной двуустки
весьма своеобразны: овальное
тело (0,20—0,22 мм длины и
0,07—0,08. мм ширины), по-
крытое мелкими шипиками, не-
сет очень маленький округлый
хвостовой придаток (рис. 187,
В). Выйдя из моллюска, цер-
карии плавают в воде и про-
никают во второго промежуточного хозяина, которым являются раки
(Astacus japonicus, A. Simins') или пресноводные крабы (рис. 188)
Рис. 187 — Легочная двуустка.
Спорциста (слева), церкария (посредине) и инцистированная адолескария (справа) из
пресноводного краба Potamon dehaani. Из Фауста.
(Eriocheir japonicus, Е. sinensis (табл. VI), Potamon dehaani, Р. obtusi-
pes, Sesarma dehaani-, Paratelphusa sinensis, Pseudotelphusa iturbei и др).
Церкарии (рис. 187) пробуравливают имеющимся у них стилетом
мягкие участки хитинового покрова рака или краба и инцистируются в
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
261
По данным Jokogawa, молодые
кишечника в брюшную полость, миг-
188 — Пресноводные крабы.
Рис.
1 — Potamon dehaani; 2 — Sesarma dehaani; 3 — Eri-
ocheir japonicus — вторые промежуточные хозяева
легочной двуустки.
По Nakagawa, из Brumpt.
в бронхиолах.
множество лей-
фиброзная ткань и
киста, вмещающая в
двух или нескольких
которые окружены
его мышцах, печени и т. п. Цисты шарообразные, жемчужно-белого
цвета (рис. 187, С).
При поедании зараженных крабов или раков сырыми цисты освобо-
ждаются в желудке окончательного хозяина и вскрываются в его тон-
ких кишках.
Действие на хозяина,
двуустки проникают через стенку
рируют сквозь диафрагму и пле-
вры в легкие (рис. 186), где и
обосновываются
Вокруг осевших паразитов в тка-
нях скопляется
коцитов и среди них эозинофилы.
Развивается
образуется
себе одного,
паразитов,
гнойной красноватой, жидкостью.
Киста выдается на поверхности
легких бугорками величиной с
большую горошину. Кисты могут
соединяться друг с другом и об-
разовывать обширные полости,
дающие объективные симптомы
расширения бронхов. Поражение
легких ведет к развитию диффуз-
ного цирроза.
Легочный дистоматоз (или па-
рагонимоз) сказывается хрониче-
ским кашлем, более сильным
утром при вставании. Припадки
кашля оканчиваются отхожде-
нием мокроты ржавчинного цве-
та. К ней может примешиваться
и кровь, если при развитии па-
разита резорвутся кровеносные
сосуды. В тяжелых случаях при-
соединяются явления плеврита и
гемоптое. Болезнь может симу-
лировать туберкулез легких.
Местоположение паразита в
легких очень благоприятно для
метастаза яиц и, может быть,
самих паразитов. Действительно,
при вскрытии яйца были находи-
мы в брюшине, брыжейке, сальнике и т. п. Легочные двуустки могут
поражать печень, селезенку, поджелудочную железу, предстательную
железу, стенку кишечника и др. Большое значение для больного имеют
случаи локализации яиц в мозгу. Эта болезнь в Японии известна да-
леко не в единичных случаях. Протекает она при самых различных
явлениях — Джаксоновской эпилепсии, общих судорогах, половинных
параличах и т. п. Симптомы неспецифичны для самого паразита, а за-
висят от того, какой участок мозга им поражен. Яйца двуустки зале-
262
СПЕЦИАЛЬНАЯ ЧАСТЬ
гают или в свободных или в облитерированных сосудах. Сами парази-
ты встречались в опухолях мозга, полость которых была, заполнена
коричневой тягучей жидкостью.
A westermani поражает также и глаза, каковое явление иногда со-
провождает легочный дистоматоз.
Парагонимоз эндемичен для Японии, Кореи и' Формозы. Встречается
он и на Дальнем Востоке.
Лечение. Впрыскивание эметина.
Диагноз. Нахождение яиц в мокроте часто ржаво-красного цвета;
яйца в faeces; яйца в кусочках тканей, добытых при биопсии. Диферен-
цировать парагонимоз надо от туберкулеза, бильгарциоза, цистицеркоза,
эхинококкоза и других болезней. В некоторых случаях прямая диагно-
стика невозможна. Полезные наводящие данные можно получить из
анамнеза больного, прибывшего из эндемичных очагов парагонимоза.
Профилактика. Не есть сырых крабов и раков.
Семейство Schistosomatidae
Род Schistosoma Weinland
Представители этого рода (как и семейства) характеризуются раз-
дельнополостью и паразитированием в плазме крови, отсюда их назва-
+
Рис. 189 — Schistosoma. Схема строения
кровяной двуустки.
а — ротовая присоска; b — брюшная присоска;
с — кишка; d — семенники; е — скорлуповая
железа; f — яичники; п — желточники; г — киш-
ка; k — матка. По Роденвальду
Рис. 190 — Яйцо Schisto-
soma haematobium с ми-
рацидием внутри, по-
вернувшимся передним
концом назад. Сильно
увеличено. По Л о о с с.
ние — кровяные двуустки, или бильгарции (по прежнему родовому на-
званию Bilharzia). Тело их длинное, узкое (рис. 189), самец значительно
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
26
шире самки; на его брюшной стороне имеется продольная щелевидная
вогнутость, в которой самец носит самку (canalis gynaiecophorus). При-
соски лежат близко друг к другу; мускулистая глотка отсутствует; вет-
ви кишечника простые, но на заднем конце сливаются в непарный ствол.
Половое отверстие у обоих полов лежит позади брюшной присоски.
У самца на уровне полового отверстия лежат 3—6 семенников (<7), у
самки же имеется только один яичник и один желточник. Мешка цирру-
Рис. 191—Мирацидий Schistosoma
haematobium.
ер— экскреторная пора; е — канал протоне-
фридия; f—замыкающая клетка протонефри-
дия с «пламенем»; и — нервный уЗел; gc — по-
ловой зачаток; pg — зачаток кишечника; sglf
*51 —железы, выделяющие протеолитический
секрет. По Фаусту
Рис. 192 — Церкарии бильгарции Schb
stosoma haematobium
as — передняя присоска; op — ротовое отвер-
стие; с — кишечник (слепой); csg^ — лити-
ческие железы; cgd — проток литической же-
лезы; vs — брюшная присоска; ga — половой
зачаток; ер — экскреторная пора.
По Ф а у с ту.
са у самца нет. Яйца (рис. 190) крупные с шипом или гладкие, в про-
тивоположность другим сосальщикам, без крышечки; в яйце заключен
ресничный мирацидий-с зачатком кишечника (рис. 191).
Цикл развития. Развиваются с одним промежуточным хозяином,
в котором проходят обычные стадии спороциты, редии и церкарии
(рис. 192). Церкарии активно вбуравливаются в кожу хозяина, прони-
кают в венозные сосуды и с током крови плывут по телу. Оседают в из-
вестных органах (мочевой пузырь, печень, кишки), куда привлекаются
химиотактически. В кровеносных сосудах достигают половозрелости.
Раздельнополые двуустки паразитируют в кровеносных сосудах че-
ловека и млекопитающих, вызывая своим присутствием заболевание,
называемое бильгарциозами, которых известно три формы.
264
СПЕЦИАЛЬНАЯ ЧАСТЬ
Schistosoma haematobium Weinl.
Синоним: Bilhalzia haematobia Bilharz.
Самец, в среднем 10—15 мм длины; передний конец его несколько-
сужен и покрыт мелкими бородавочками. Брюшная присоска несколько
больше ротовой. Позади открывается canalis gynaecophorus. Тело кзади
суживается. Наибольшая ширина его достигает 1 мм. На присосках и в
названном канале лежат тонкие шипики;. тело покрыто характерными
сосочками.
Рис. 193 — Разрез печени белой мыши, зараженной экспериментально Schistosoma
haematobium. На препарате видны косой поперечный разрез самца, разрез и реак-
тивная инфильтрация ткани печени вокруг паразита. Оригинал по препаратуF
сделанному из материала L е i р е г.
Самка длиной до 20 мм; при толщине всего около 0,75 мм. Ротовая
присоска крупнее брюшной. Яичник лежит на месте ветвления кишечни-
ка. МногодоЛьчаггый желточник помещается позади яичника.
Яйцо утолщенное, веретеновидное, на заднем конце, с шипом, 0,12—
0,16 мм длиной и 0,04—0,06 мм шириной, светлого желтовато-коричне-
вого цвета, прозрачное.
Действие на хозяина. Паразитирует в крупных венозных
стволах брюшной полости (в полой, воротной венах), а также в венах
почек, пузыря, rectum, у экспериментальных животных — в печени
(рис. 193). Для нахождения этих паразитов собирают кровь из круп-
ных вен в чашки Петри; в тонком слое крови на черном фоне могут
быть видны скрученные или изогнутые мелкие червячки поодиночке-
или парами. Для облегчения отыскивания кровь разбавляют физиологи-
ческим раствором поваренной соли.
Оплодотворенная самка откладывает яйца в соединительную ткань
стенки мочевого пузыря, куда она попадает при разрушении стенки кро-
веносных сосудов. Яйца выносятся наружу с мочой, в моче человека на-
хождение яиц свидетельствует о «бильгарциозе» мочевого пузыря или
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
265
о пузырном бильгарциозе. Болезнь проявляется отхождением несколь-
ких капель или даже кубиков крови в конце мочеиспускания. В более
тяжелых случаях моча окрашена вся в красный цвет (гематурия). Яй-
ца отыскивают в слизистых или кровянистых хлопьях мочи. Сам пара-
зит гнездится в венах пузыря и на свободу не выходит.
При эндоскопировании пораженного мочевого пузыря обнаруживает-
ся гиперемия слизистой оболочки с мелкими кровоизлияниями. Послед-
ствиями местного воспалительного процесса является гипертрофия во-
локнистой соединительной ткани; такие участки имеют характерный вид
округлых неправильной формы площадок, точно посыпанных песком.
Под ножом они хрустят. В других случаях гипертрофируются слизистая
и подслизистая оболочки и образуют полипозные разрастания, сидящие
на ножках или похожие на цветную капусту. Они очень богаты крове-
носными сосудами и чрезвычайно нежны. Во всех этих образованиях
находится множество яиц Sch. haematobium.
Рано или поздно, описанные выше патологические элементы пузыря некроти-
зуются и распадаются, вызывая образование язв. Продукты их распада — кровь, об-
рывки тканей яйца бильгарции попадают при этом в мочу. Полипы при своем раз-
растании в некоторых случаях заполняют полость пузыря и закрывают отверстие
мочеточников, вызывая этим впоследствии гидро- и пионефроз.
Вокруг яиц могут отлагаться фосфорнокислые соли, вследствие чего возникают
пузырные камни паразитарного происхождения. Иногда такие же камни образуются
в мочеточниках и почках, вызывая гидронефроз.
К побочным явлениям бильгарциоза относятся и мочевые фистулы, которые на-
блюдались в некоторых случаях во множестве, до 50 штук у одного больного. В
стенках их также множество яиц паразита; когда они пропитываются известковыми
солями, ткань, окружающая их, становится твердой и хрустит под ножом. В стенке
фистул встречаются и взрослые паразиты, которые могут быть открыты на срезах
мочевого пузыря в венах и лимфатических сосудах.
Бильгарциоз у женщин, помимо гематурии, проявляется иногда подострым вос-
палением влагалища. Слизистая оболочка его утолщается, делается как бы перга-
ментной и легко растрескивается по разным направлениям. На внутренней стенке
влагалища нередко образуются разрастания, похожие на полипов или на головки
цветной капусты. Иногда они заполняют собой все влагалище и заходят на наруж-
ные половые органы и в шейку матки. Эти папилломатозные разрастания легко мо-
гут быть смешаны с эпителиомами. Отличить их можно, если исследовать кусочек подо-
зрительного образования на яйца Sch. baendatobium. Для этой цели кусочек или раз-
рушают ЗО°/о раствором едкого кали, или искусственно переваривают пепсином.
В обоих случаях жидкости дают отстояться и в осадке ищут под микроскопом яйца
паразита: (рис. 190).
Длительность жизни паразитов, определяемая по выхождению яиц,
достигает 27—28 лет, что повидимому не является пределом
(Christopherson, 1924).
Паразит распространен в Африке, Египте, Аравии, Месопотамии,
Кипре, Португалии, Индии и других странах.
В Армении описан единственный случай пузырного бильгарциоза
у больного, бывшего ранее в Персии и в Багдаде (Месопотамия); не-
сомненно, что случай этот является завозным (Кечек и Нанасян).
Цикл развития паразита связан с водой. Если выброшенные
наружу яйца остаются в неразведенной моче, то зародышей из них не
вылупляется. Разведенная вчетверо водой моча способствует вылупле-
нию мирацидия.
Мирацидий одет ресничками, с двумя парами железистых клеток,
нервным узлом, протонефридиями, зачатком кишечника и с зачаточны-
ми клетками.
Промежуточным хозяином являются моллюски: Bulinus dtjbowskii,
В. contortus, В. innesi — в Египте, Planorbis metidjensbs var dufouri —
в Португалии и др.
2 56
СПЕЦИАЛЬНАЯ ЧАСТЬ
Церкарии имеют длинный, вилообразно ветвящийся на конце хвост.
Они активно проникают в тело человека, вбуравливаясь в кожные по-
кровы. Первые симптомы болезни появляются месяца через два после
внедрения церкарий.
Лечение. Внутривенное впрыскивание lVo tartar, stibiatici.
Профилактика. В эндемичных очагах не купаться в водоемах
с моллюсками, умываться предварительно дезинфицированной водой;
не пить сырой воды. Общественная профилактика: уничтожение мол-
люсков, лечение больных, охранение водоемов от загрязнения мочой.
Schistosoma mansoni Sambon
Отличается от предыдущего вида главным образом формой яиц, у которых шип
помещается не на заднем конце яйца, а сбоку, однако этот признак некоторыми
Рис. 194 — Schistosoma mansoni.
По Фурману.
•паразитологами не считается суще-
ственным, на основании чего1 они не
признают самостоятельности этого вида.
Не так давно найдены отличия в
числе околокишечных желез церкарий
обоих видов бильгарций (КЬаШ, 1912),
что свидетельствует в пользу их ви-
довой самостоятельности.
Sch. mansoni (рис. 194) живут пре-
имущественно в венах прямой кишки,
куда они привлекаются в силу химио-
тропизма. В результате возникает за-
болевание— кишечный бильгарциоз,
сопровождаемый симптомами дизенте-
рии. В прямой кишке появляются мяг-
кие полипозные разрастания; при ми-
кроскопировании в них обнаружива-
ются яйца; последние открываются и
в испражнениях. Вены подвергаются
воспалению; подэндотелиальный слой
в них утолщается.
Известны случаи, когда в орга-
низме хозяина (человек; кролик — при
эксперименте) все паразиты были сам-
цами; самки не было ни одной. Неко-
торые самцы находились в положе-
нии копуляции, т. е. носили в гинеко-
форном канале самцов же (A. Giova-
nola, 1936).
Диагностика. По нахождению
яиц в faeces (длина яиц 0,140—0,165 мм,
ширина 0,06—0,07 мм).
Распространен в Египте, Восточной
Африке, Конго, Гвинее и др.
Промежуточный хозяин — моллюски Planorbis boissyi PL olivaceus,
Physopsis africana.
Лечение, что и при Sch. haematobium.
Профилактика, что и выше, но с учетом распространения яиц с фекалиями.
Schistosoma japonicum Katsurada
Поверхность тела гладкая. Брюшная присоска с суженным основанием, крупнее
ротовой. Яйца овальные, без крышечки и без шипа или с отростком скорлупы. Дли-
на их 0,07—0,09 мм и ширина 0,05—0,07 мм. Яйца выносятся с испражнениями.
Паразит живет главным образом в воротной и в брыжеечных венах, бывает так-
же и в артериях.
Эта двуустка причиняет сосудистый бильгарциоз. Главнейшие симптомы его сво-
дятся к гипертрофии печени и селезенки, к циррозу, асциту, кровавому поносу. С
течением времени развиваются анемия и кахексия; болезнь завершается маразмом,
ведущим к смерти. Наблюдались случаи локализации яиц, кроме печени и кишеч-
ника, в мозгу, причем обнаруживались различные расстройства, Джаксоновская эяи-
пепсия, параличи и др.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
267
Распространен в Японии и в Китае. Известны завозные случаи в СССР.
Промежуточным хозяином служат моллюски — Blanfordia nosophora,
Bl. formosana, Hemibia hupensfa.
Основным хозяином, кроме человека, бывают собака, кошка, свинья, овца, коза,
лошадь и др. Предполагается, что Sch. japonicum паразитирует у овец на Дальнем
.Востоке.
Диагноз — по нахождению яиц в faeces.
Терапия и профилактика те же, что и для других бильгарций.
Сравнительная таблица признаков человеческих бильгарций
Sch. haematobium Sih. mansoni Sch. japonicum
Взрослый самец 12—14 мм длины Кутикула с тонкими бугорками Соединенная часть ки- шечных трубок ко- роткая Семенников 4 12 мм длины Кутикула с более крупными бугорка- ми Соединенная часть ки- шечных трубок длин- ная Семенников 8 9—22 мм длины Кутикула без бугорков Общая часть кишечных трубок равняется ’/s—’/в длины тела Семенников 7—8
Взрослая самка 2Э мм длины Матка длинная Яичник в задней по- ловине тела 14—15 мм длины Матка короткая с 1—3 яйцами Яичник в передней половине тела 12—26 мм длины Матка с 50—300 яйца- ми Яичник почти на сере- дине тела
Яйца С концевым шипом 0.150—0.060 мм; выходят с мочой С боковым шипом 0.15—0.06 мм; выходят с faeces С укороченным шипом 0.08—0.065 мм; выходят с faece»
Schistosoma turkestanicum Skrjab.
Самец имеет около 80 семенников. Яйца на обоих полюсах с различными по
форме шипами. Длина яиц 0.072—0.174 мм, ширина 0.022—0.026 мм.
Описана по материалу от крупного рогатого скота в Аулиэ-ата (К. Скрябин) из
крозеносных сосудов печени. Найдена на Аральском море у кошки (Н. Попов)
Обнаружена также в Ираке, где оказалась опасным вредителем скота. Парази-
тирует у овец, коз, буйволов, верблюдов, лошадей, мулов и др. животных. Яйца,
скопляясь в оболочках кишечника, ведут к образованию узлов, обесценивающих цен-
ность кишок, как сырья. Промежуточным хозяином является Lymnaea tenera euphra-
tica. Человек, домашние и дикие птицы, видимо невосприимчивы к заражению церка-
риями Sch. turkestanicum (С. Machattie). Цикл развития этой трематоды в СССР
остается еще неизученным.
Trichobilharzia ocellata. La Vallete Saint-Georges
Паразит птиц (Anas circia); обитает в сосудах слизистой оболочки кишечника и
в брыжейке. Промежуточный хозяин — пресноводные моллюски — Limnaea palustris,
Planorbis rotundatus и др.
Церкария этого* паразита описана под именем Cercaria ocellata (рис. 196). Эта
церкария вбуравливается в кожные покровы купающихся людей, в результате чего
на коже появляется сыпь, сопровождаемая резкой зудящей болью. Субъективные
ощущения достигают наибольшей силы на 2—3-fij день. Болезненные проявления
заканчиваются к 10—18 дню, когда с места внедрения церкарии в кожу спадает
струпик (Фогель). Развитие сыпи после купанья В некоторых озерах Геомании vna-
лцалось настолько, что жители вовсе прекращали купанье в некоторых из них. Ин-
тересно, что аналогичные явления наблюдались в СССР, в частности в Белом озере
»(Косино, под Москвой), но специальных исследований там не производилось (Декс-
268
СПЕЦИАЛЬНАЯ ЧАСТЬ
бах). Окончательная судьба Cere, ocellata, проникшей в тело человека, неизвестна.
Человек, видимо, не является хозяином, подходящим для полного жизненного
цикла этого паразита, который в нем погибает.
КЛАСС CESTOIDEA —ЛЕНТОЧНЫЕ ЧЕРВИ
Ленточные черви являются эндопаразитическими глистами, которые
в половозрелом состоянии живут в кишечнике позвоночных животных
(исключение Archigetes в полости тела пресноводных Малощетинковых
червей). Тело ленточных червей покрыто кутикулой без ресничек; под
кутикулой лежат погруженные в паренхиму клетки эпителия. Тело
Рис. 195— Церкария Schis-
tosoma japonicum.
А — нервная система;С—около-
кишечные ( = головные) железы;
О—половые клетки; Е — выдели-
тельная система. По Фаусту
Рис. 196—Cercaria ocellata. А — спороци-
ста из печени limnaeus stagnalis,
В — выхождение церкарий из спороцисты; С — цер-
кария. По T ay 1 о г и Baylis.
уплощено дорзовентрально. На переднем конце имеется головка или
сколекс, служащая прикрепительным аппаратом. Позади головки
лежит шейка, являющаяся зоной роста; тело состоит из стробилы,
в огромном большинстве случаев подразделенной на членики или
проглоттиды. Есть, впрочем, ленточные глисты, состоящие всего
из одного членика (очень редко). В каждом членике развивается ком-
плект (реже два комплекта) мужских и* женских половых органов. Ки-
шечник редуцирован полностью.
Класс Cestoidea подразделяется на два подкласса — Cestodaria и
Cestoda.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
269
ПОДКЛАСС CESTODARIA — НЕРАСЧЛЕНЕННЫЕ
ЛЕНТОЧНЫЕ ЧЕРВИ
Одночленистые ленточные черви с одной или двумя присасыва-
тельными ямками на переднем конце тела. Имеется один комплект
половых органов. Зародыш с 10 крючьями. Примеры — АлгрШНпа fo-
Насеа в полости тела осетровых рыб. Нерасчлененные ленточные чер-
ви не имеют никакого отношения к человеку.
ПОДКЛАСС CESTODA — РАСЧЛЕНЕННЫЕ ЛЕНТОЧНЫЕ ЧЕРВИ
Тело огромного большинства цестод подразделено на членики.
Если паразит снаружи не расчленен, то у него наблюдается метамерная
повторность половых органов по длине тела. В единичных случаях
тело паразита состоит из одного членика с одним комплектом органов
размножения. Таким образом, признак расчлененности не является все-
объемлющим для. подкласса Cestoda. Зато наличие шести крючьев в за-
родыше, выходящем из яйца (против 10 крючьев ликофоры заро-
дыша Cestodaria), характеризует всех представителей Cestoda, К этому
подклассу принадлежат многие важные эндопаразиты человека.
ОРГАНИЗАЦИЯ CESTODA
У типичных цестод передний конец тела суживается и несет на се-
бе головку, являющуюся органом прикрепления к хозяину. Головка
снабжена или присасывательными ямками (ботрии, рис. 197, Л),
ограниченными каждая двумя губами, или настоящими полушаровид-
ными присосками (acetabula, рис. 197, D), лежащими крест-накрест.
В центре свободной поверхности головки между присосками бывает
также более или менее развитой втяжной хоботок (rostellum), не-
сущий на себе различно развитое вооружение из кутикулярных крючь-
ев микроскопического размера. Форма, величина и количество крючьев
имеют значение в систематике цестод. Указанные органы прикрепления
присущи ленточным червям, паразитирующим в человеке. У цестод,
живущих в различных животных, бывает большое разнообразие в строе-
нии головки, ее присосок, крючьев и т. д. Бывают, например, четырех-
хоботные ленточные глисты, сильно складчатые ботрии с добавочными
присосками и др. (рис. 197, В и С).
Ленточные черви во взрослом состоянии обитают в кишечнике че-
ловека. Ботриями паразит защемляет ворсинки кишки; присосками он
прикрепляется к ее стенке; крючьями впивается в слизистую оболочку,
как бы становясь таким образом «на якорь».
За головкой идет узкая шейка, являющаяся зоной роста и почкова-
ния червя. На некотором расстоянии, кзади от головки, лентовидное
тело цестоды подразделяется поперечными насечками на членики
(проглоттиды — рис. 200); количество члеников у разных видов
вариирует весьма значительно; у некоторых цестод члеников всего
три; их антиподами являются виды с 3—4 тысячами члеников; это от-
ражается и на общих размерах паразитов, длина которых колеблется
от нескольких миллиметров до 10—12 м.
Форма проглоттид является вариантами прямоугольника или трапе-
ции; позади шейки лежат короткие членики; длина их более кзади
увеличивается, они могут становиться квадратными; при дальнейшем
продвижении кзади они делаются относительно узкими и длинными.
Углы члеников бывают закруглены, что сглаживает геометричность
их формы; в некоторых случаях членики делаются более овальными.
270
СПЕЦИАЛЬНАЯ ЧАСТЬ
Самые задние членики являются и наиболее старыми. У многих
цестод старые членики отрываются поодиночке или группами от зад-
него конца тела и выбрасываются с испражнениями хозяина вон из
его тела.
Под кутикулой лежит так называемая кожная мускулатура,
состоящая из наружного кольцевого и внутреннего продольного слоев.
Рис. 197 — Формы головки, органов прикрепления различ-
ных ленточных червей.
А — присасывательные ямки лентеца широкого—Diphyllobothrium latum*,
В — выдвижные усаженные крючьями хоботки Tetrarhynchus (из ската);
С — сколекс с подразделенными ботридиями, добавочными присосками
и крючьями у Acanthobothrium ijimai; D— головка с вытяжным хоботком
(rostellum) и полушаровидными присосками (acetabula) Hymenolepis
macracanthus. По Фурману.
Кроме того, в паренхиме тела залегают пучки поперечных мышц, есть
также и дорзовентральные мышцы. Все мышцы у цестод гладкие.
Паренхима цестод является не только опорной и заполняющей
тканью тела; в ней скопляются пищевые запасы в форме гликогена,
а также и продукты обмена веществ, извлекаемые из нее экскретор-
ными органами. В паренхиме часто скопляются известковые тельца и
изредка бывает также пигмент.
В связи с полной редукцией пищеварительного аппарата питание
цестод осуществляется всасыванием всей поверхностью тела паразита
пищи, переваренной кишечником хозяина (эндосмотическое питание).
Экскреторная система развита хорошо. Она построена по
протонефридиальному типу. Начальными частями (терминальными ор-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
271
ганами) ее являются клетки с внутриклеточной воронкой и колеблю-
щимся жгутом. Эти клетки замыкают капиллярные трубочки, впадаю-
щие в главные стволы, которых типически по два с каждой стороны
тела; они идут продольно по бокам стробилы, выделительные
стволы пронизывают всего паразита и, если у него еще цела первая
по времени образования ,(т. е. самая старая) проглоттида, то в ней все
выделительные стволы открываются наружу одним общим отверстием,
предварительно впадая в маленький концевой экскреторный пу-
зырек. При отрыве члеников протонефридиальная система естественно
открывается двумя или четырьмя самостоятельными отверстиями, при-
надлежащими каждое главному стволу соответствующей стороны тела.
Продольные стволы по месту своего залегания являются спинными
и брюшными; последние обычно бывают развиты сильнее первых, ко-
торые могут вовсе исчезать.
В головке цестоды главные стволы соединяются друг с другом
ответвлениями, образующими более или менее сложное сплетение ка-
нальцев. Кроме того, в каждом членике стробилы у его заднего края
имеется по поперечному соединительному каналу (обычно между
брюшными каналами), вследствие чего вся система принимает лест-
ничный характер.
.Нервная система состоит из поперечно расположенных ган-
глиев, дающих вперед и назад нервные стволы.
Нервные узлы соединяются в нервное кольцо, которых бывает
два. Назад, т. е. в стробилу, отходят два главные нервные с т в о-
л а; они могут иметь тонкие сопровождающие нервные тяжи; кроме
того, субмедианно отходят две пары тонких нервных стволиков; всего
может быть до десяти продольных нервных стволов. По заднему краю
проглоттид нервные стволы соединяются поперечными перемычками.
Все это в совокупности образует центральную нервную систему, от
которой отходят нервы, иннервирующие мышцы и различные органы.
Анатомически развитых органов чувств у цестод. нет, но имеются
веретеновидные чувствительные клетки, лежащие в подкутикулярном
слое и своими периферическими отростками пронизывающие насквозь
кутикулу тела. Органов дыхания и кровеносной системы у лентецов,
как и у других плоских червей, нет.
Все цестоды, кроме рода Dioicocoestus, гермафродитны. Зрелые чле-
ники имеют два полных комплекта органов, т. е. мужские и женские
органы размножения. Развиваются они постепенно в молодых члени-
ках; проглоттиды позади шейки еще не имеют закладок половых ор-
ганов; более кзади появляются части мужского полового аппарата,
еще далее — женского, и в перезрелых члениках некоторых групп це-
стод набитая яйцами матка вытесняет все прочие части как женского,
так и мужского полового аппарата. В описанных случаях можно гово-
рить о протерандрическом гермафродитизме (т. е. о
более раннем развитии мужской части гермафродитных органов).
Мужские органы размножения (рис. 202) у цестод пред-
ставлены семенниками в виде обыкновенно весьма многочисленных
фолликулов; есть виды с очень малым (3—2—1) количеством семенни-
ков. Если рассматривать поперечные срезы через проглоттиду, то мож-
но видеть, что семенники лежат ближе к одной стороне тела; ее на-
зывают спинной. От семенников отходят тонкие семевыносящие ка-
нальцы (vasa efferentia), которые соединяются друг с другом и обра-
зуют vas deferens, идущий к копулятивному органу; последний назы-
вается циррусом (cirrus); лежит он в циррусовой сумке, являющейся
1272
СПЕЦИАЛЬНАЯ ЧАСТЬ
мышечным мешком. В этой сумке помещаются концевые извивы Vas
deferens и семеизвергательный канал (ductus ejaculatofius), образую-
щий расширения, лежащие или внутри, или вне циррусовой сумки:
расширение называется семенным пузырьком внутренним или внешним.
Сумка цирруса впадает рядом с женским половым отверстием в
половую клоаку, открывающуюся или на брюшной поверхности члени-
ка, или на краю проглоттиды на небольшом возвышении, называемом
половым сосочком.
Женский половой аппарат (рис. 202) устроен сложнее.
Основной его частью является яичник обыкновенно двойной или пар-
ный; лежит он ближе к брюшной стороне членика; форма его дольча-
тая, трубчатая, реже сетчатая. От яичника отходит мускулистый «я ft-
цех в ат» (ovicapt), служащий для продвижения зрелых яиц в яйце-
вод. В последний открывается влагалище, образующее у внутреннего
конца расширение — семеприемник — receptaculum seminis; яйцевод
идет далее к тельцу ’Мелиса, известному ранее под именем скорлуповой
железы; в него впадает желточный канал, служащий для проведения
желточных клеток, вырабатываемых желточниками. Последние имеют
вид или фолликулов, или компактного удлиненного тела, или другой
формы.
Полость, принимающая в себе отверстие яичников, желточников и
протоки железок тельца Мелиса, называется оотипом; из ооти-
па выходят влагалище и трубчатая матка, которая или открывается
самостоятельным отверстием, или остается замкнутой. Форма зрелой
(т. е. совершенно развитой матки, наполненной яйцами) характерна:
у одних цестод она имеет вид розетковидно сложенной трубки, у дру-
гих построена в форме продольного ствола с боковыми ветвящимися
отростками, у третьих — в виде поперечно вытянутого мешка и т. д.
Развитая матка может распадаться, благодаря чему яйца паразита
попадают в самую паренхиму проглоттиды; последняя, видоизменяясь,
образует вокруг яиц или групп их капсулы. У некоторых цестод около
матки образуется мощная перенхиматозная капсула, называемая о к о-
ломаточным органом, в который переходят зрелые яйца из
матки.
Влагалище открывается рядом с мужским половым отверстием в
половую клоаку или на брюшной поверхности членика, или на его бо-
ку, причем половые отверстия неправильно чередуются между члени-
ками по обеим сторонам или же все лежат на одной стороне тела.
При копуляции возможны различные случаи. Если рядом лежат
две глисты, то циррус какой-либо проглоттиды одного паразита может
войти во влагалище другой глисты; при наличии всего только одной
цестоды циррус одной проглоттиды вводится в женское половое от-
верстие другого членика. Наконец, бывает и самооплодотворение про-
глоттиды, когда циррус, круто загибаясь, входит в рядом лежащее
женское половое отверстие.
При копуляции живчики проникают во влагалище и скопляются в
семеприемнике. Оплодотворенная яйцевая клетка вместе с одной или
несколькими желточными клетками одевается плотной скорлуповой
оболочкой; последняя является производным желточных клеток, кото-
рые выделяют капельки вещества, образующего оболочку яйца; в ее
формировании, повидимому, принимают некоторое участие и железы
тельца Мелиса, которым ранее приписывалась исключительная роль в
деле образования скорлупы (скорлуповая железа).
Форма яиц у цестод разнообразная — круглая, овальная
{рис. 219), или иных своеобразных очертаний. То она сходна с яйцами
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
273
трематод [яйца с тонкостенной скорлуповой оболочкой и крышечкой
(Pseudophy llidea)], то своеобразна, например, яйца с толстой радиаль-
но исчерченной скорлупой (Taeniidae и др.). У Diphyllobothrium под
скорлупой образуется из бластомер развивающегося зародыша тонкая
эмбриональная оболочка, а у Cyclophyllidea покровная оболочка оде-
вает снаружи радиально исчерченную скорлуповую оболочку и вслед-
ствие своей нежности легко разрушается.
Сформировавшиеся яйца выходят через отверстие матки, если та-
ковое имеется у паразита. При замкнутой матке яйца освобождаются
при отрывании члеников или групп их и при распаде выброшенных на-
ружу проглоттид.
В яйце уже имеется зародыш в виде кучки мелких бластомер;
у Pseudophyllidea зародыш часто бывает одет оболочкой из слоя
ресничного эпителия; такой тип личинки называется корацидием.
Собственно личинка цестод носит название онкосферы; она во-
оружена шестью крючьями.
Жизненный цикл цестод изучен сравнительно у немногих
видов. У большинства их имеется смена основного хозяина одним
промежуточным, более редки случаи развития с двумя промежуточ-
ными хозяевами, и еще реже наблюдается цикл с одним хозяином.
В окончательном хозяине живет глиста, размножающаяся поло-
вым путем; ее половые продукты — яйца — выбрасываются с испраж-
нениями хозяина наружу. В промежуточного хозяина яйца попадают
пассивно, когда заглатываются им с пищей или с водой; онкосфера,
вышедшая из вскрывшегося яйца, превращается в промежуточном
хозяине в личиночную стадию, носящую общее наименование финки
или пузырчатой глисты.
Формы финок разнообразны (рис. 198). Различают следующие
формы их:
1) Цистицерк, или типичная пузырчатая глиста; она состоит из
гонкостенного овоидного или шаровидного пузырька, заполненного
кидкостью; в одной из точек окружности стенка финки вворачивает-
ся внутрь, причем в этой части можно видеть как бы вывернутую на
внутреннюю сторону головку глисты с присосками; при выворачива-
нии наружу головка принимает нормальный вид, характерный для
данного паразита; в вывернутом состоянии головка и шейка гораздо
меньше пузыря; последний является как бы хвостом финки, откуда и
название Cysticercus — цистицерк или в переводе — пузырехвост.
2) Цистицеркоид характеризуется подразделением финки на
переднюю вздутую часть с ввернутой головкой и хвостообразный зад-
ний компактный придаток с эмбриональными крючьями на нем (на-
пример, Dipylidium caninum),
3) Ц е н у р имеет на тонкостенном пузыре много ввернутых внутрь
головок.
4) Эхинококк характеризуется крупными размерами (до вели-
чины детской головы); он переполнен жидкостью, в которой находятся
дочерние тонкостенные пузыри; на их внутренней поверхности
развиваются выводковые капсулы с ввернутыми внутрь голов-
ками; различные уклонения в строении пузырчатой формы эхинококка
см. ниже..
Все эти формы личиночных стадий ленточных глист присущи отря-
ду Cyclophyllidea. Заражение окончательного хозяина происходит при
поедании промежуточного хозяина с финками. Из проглоченных фи-
274
СПЕЦИАЛЬНАЯ ЧАСТЬ
нок в кишечнике хозяина развивается бесполым размножением лен-
точная глиста. !
Цикл с двумя промежуточными хозяевами свойственен Pseudo-
phyllidea; финнозная стадия у этих глист своеобразна в том отноше-
нии, что никакой полости, заполненной жидкостью, у них нет; все
тело личинки паренхиматозное — отсюда и название плероцеркоид
для второй личиночной стадии и процеркоид для первой (рис. 206, А).
Плероцеркоид (рис. 206, В) имеет присасывательные бороздки
(bothriae) на переднем конце тела,
Рис. 198 —Схема строения личиночных форм (финок)
ленточных червей:
а — цйстицерк; Ъ — ценур; с — эхинококк с выводковыми капсу-
лами (под толстой оболочкой) в виде дочерних и внучатых пу-
зырей со сколексами; d — цистицеркоид Dipylidium caninum;
е — плероцеркоид. [Из Ф и б и г е р а.
а процеркоид несет там же присо-
сообразное углубление
и на противоположном
конце — шарообразный
придаток с крючьями..
Пузырчатые глисты'
были известны давно;,
прежде их считали са-
мостоятельными жи-
вотными, хотя и усмат-
ривали большое сход-
ство в строении голов-
ки финок и ленточных
глист. Наличие пере-
полненного жидкостью
пузыря давало также
повод признавать их
за водяночно перерож-
денных глист, и только
точными эксперимен-
тами Кюхенмейстера
(1851) была выяснена
истинная природа этих
форм паразитических
червей, оказавшихся
личиночными стадиями
ленточных глист.
Замечательно, что ленточные глисты и их финнозные формы не
только живут, как правило, в разных хозяевах, но и в разных орга-
нах тела. Ленточные глисты являются вообще обитателями кишечни-
ка, тогда как пузырчатые глисты паразитируют в тканях органов
(соединительная ткань мышц, кожа, печень, мозг, глаз, сердце,’ кости
и мн. др.).
Финки образуются на месте из зародышей, освободившихся при
растворении скорлупы заглоченных яиц в пищеварительном канале.
Снабженная крючьями онкосфера пробирается через стенку кишки и
попадает в кровеносные пути или в полость тела. С кровью онкосфе-
ра плывет и оседает где-либо внутри тела, превращаясь в финку.
Финки могут размножаться бесполым путем в своем хозяине; именно-
этим способом образуются такие многоголовые формы, как эхино-
кокк. Некоторыми учеными самый процесс образования головки счи-
тается бесполым размножением.
Когда финка будет проглочена окончательным хозяином, в его
кишечнике головка и шейка финки выворачиваются обратно и прини-
мают нормальный вид. Пузырь переваривается, а оставшаяся парен-
химатозная часть финки растет и начинает размножаться бесполым.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
275
путем. Сначала поперечной перемычкой отграничивается первый чле-
ник глисты, затем между ним и головой в таком же порядке обра-
зуется вторая проглоттида; между ней и головкой возникает третий
членик и т. д. При этом первый членик отодвигается все далее и да-
лее от головки. Таким образом, самый задний членик паразита яв-
ляется самым старым (рис. 199).
Рис. 199 — Образование стробилы из
плероцеркоида лентеца Diphyllobo-
thrium mobilis. По Фурману.
В результате описанного процесса из головки финки образуется
иногда многометровая глиста в порядке бесполого размножения,
которое в данном случае характеризуется, как почкование с долго-
временным сохранением связи между
проглоттидами.
По мере отодвигания от головки в
проглоттидах развивается половой ап-
парат, о чем уже говорилось выше.
Образовавшаяся ленточная глиста раз-
множается половым способом (дает
яйца). Если по какой-либо причине
(например, при изгнании глисты) шей-
ка цестоды разорвется, и вся стробила
будет выброшена из кишечника хозяи-
на, оставшаяся головка снова почкова-
нием произведет цестоду. Поэтому
только тогда изгнание глисты можно
считать достигнутым, когда глиста бу-
дет удалена с головкой.
Таким образом, жизненный цикл
ленточных глист слагается из периодов
бесполого размножения и размножения
полового, т. е. является метагене-
зом. Оба периода размножения фак-
тически протекают в главном хозяине,
но развитие форм, размножающихся
тем или другим путем, сосредоточи-
вается в разных хозяевах; бесполая
стадия — финка — развивается в промежуточном хозяине; ленточная
форма образуется из финки (и сама размножается) в главном хозяине.
Таким образом, смена поколений при метагенезе ленточных глист со-
пряжена в огромном большинстве случаев и с переменою хозяев.
Ленточные черви в своем жизненном цикле связаны с различными
парами или тройками хозяев. Одни, как Taenia solium, проходят пол-
ный жизненный * оборот в двух млекопитающих (человек и свинья),
другие — промежуточными хозяевами имеют насекомых, а главным —
плотоядных млекопитающих, третьи паразитируют последовательно
в ракообразных, рыбах и млекопитающих.
Как правило, ленточные черви тесно связаны с определенными хо-
зяевами, вплоть до крайней специфичности, когда глиста имеет основ-
ным хозяином только один какой-либо вид животного (например, Tae-
nia solium и человек); другие цестоды захватывают широкий круг
хозяев; например, лентец широкий может в ленточной стадии жить
в человеке, собаке, кошке, тигре, лисице, белом медведе, тюлене, мор-
же, дельфине и в других млекопитающих.
Место обитания. Ленточные формы цестод, кроме единич-
ных исключений, живут в кишечнике главных хозяев, которыми явля-
276
СПЕЦИАЛЬНАЯ ЧАСТЬ
юуся позвоночные. Редким местом обитания цестод служат печеноч-
ные каналы, и то не у человека.
Из кишок пристанищем этих паразитов является тонкая кишка;
здесь цестоды буквально купаются в пище, перевариваемой хозяином.
Среда обитания в виде полости тонких кишок характерна в том отно-
шении, что здесь имеются пищеварительные соки, богатые фермента-
ми, которые легко переварили бы любую ленточную глисту, если бы
у цестод не было специфичной приспособительной особенности, имен-
но стойкости к действию на них ферментов.
Необходимо отметить, что таким важным свойством обладают
только здоровые глисты; если они будут чем-либо повреждены, то
соответствующая часть стробилы, или весь паразит, перевариваются.
В условиях^Ьпыта обработка цестоды электрическим током убивает
паразита, и последний переваривается панкреатическим соком. Дока-
зано, что некоторые глисты в норме выделяют в течение своей жиз-
ни антикиназу, которая нейтрализует панкреатический сок и тем
определяет возможность для глисты жить в опасной для нее об-
становке.
Следует отметить, что противоферментные свойства
присущи не только цестодам, но и другим глистам, обитающим в соот-
ветствующей среде.
Действие ленточных глист на хозяев зависит * от
видовых особенностей паразитов, места нахождения их в хозяине, ко-
личества их и состояния хозяина.
Когда крупные цестоды живут в кишечнике, то потребление ими
в кишечнике пищи за счет хозяина может быть для него очень ощу-
тительным, особенно если учесть постоянное нарастание новых члени-
ков взамен оторвавшихся проглоттид и обильную половую продукцию
глисты.
Механическое действие цестод сказывается в повреждении цело-
сти слизистой оболочки кишечника вследствие внедрения в нее ско-
лекса глисты; присоски некоторых видов цестод могут раздвигать, или
уничтожать эпителий слизистой оболочки в местах прикрепления це-
стоды к стенке кишечника. В обоих случаях повреждение целости сли-
зистой оболочки нарушает ее барьерные функции, чем открывается
доступ в ткани тела патогенных бактерий, если таковые имеются
в кишечнике хозяина. У некоторых видов под влиянием действия
сколекса на стенку кишки образуются настоящие дивертикулы,, в ко-
торые уходит не только головка, но и часть стробилы паразита. Меха-
ническое действие в форме закупорки кишечника клубком перепутанных
цестод бывает очень редко.
Наконец, цестоды выделяют различные токсины, которые всасы-
ваются в организм хозяина и при долгом обитании глисты в теле
вызывают различные патологические явления в форме анемии, раз-
нообразных нервных расстройств и других болезненных симптомов,
зависящих от резорбтивного действия токсинов.
Случаи извращенной локализации ленточных форм цестод в теле
хозяина крайне редки, зато место нахождения пузырчатых форм этих
паразитов весьма вариирует.
Финки, эхинококк и другие личиночные формы цестод могут раз-
виться в самых разнообразных органах, поэтому и значение этих па-
разитов для хозяев будет весьма различно. Финки под кожей практи-
чески безвредны; финки же в мозгу ведут к смерти.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
277
Организм хозяина не представляет собой неизменяющейся среды
обитания для того или другого паразита; изменения в организме
хозяина со своей стороны оказывают различные воздействия на це-
стод; например, характер пищи, возрастные состояния и др. При ста-
рении финки, одетые реактивной капсулой тканей хозяина, подверга-
ются обызвествлению и погибают.
Диагностика ленточных форм цестод производится по на-
хождению яиц или проглоттид, или кусков стробилы в faeces; по
яйцам не всегда может быть дано решающее заключение, так как
яйца некоторых глист не отличимы друг от друга ни по величине,
ни по форме, ни по строению иногда не только среди самих цестод,
но даже между цестодами и трематодами. Примеры первого случая
дают цестоды собак Taenia pisiformis (= Т. serrata), Т. hydatigena
(= Т. marginata) и Multiceps serialis (T. serialis).
Второй пример видим при сравнении яиц лентеца широкого и тре-
матоды Nanophyetes salminicola.
Диагностика пузырчат&х глист затруднена тем, что они являются
тканевыми паразитами. Поэтому об их присутствии приходится судить
по косвенным данным (иммуно-биологические реакции), применять
рентгеноскопию, в ограниченных случаях прибегать к биопсии, произ-
водить пункцию зыблящих образований и др.
КЛАССИФИКАЦИЯ CESTODA
Cestoda подразделяются на четыре отряда:
1. Pseudophyllidea— сколекс с двумя присасывательными ямками
или бороздками, или с одной только ботрией на переднем конце
(рис. 197, А). Матка имеет собственное отверстие, открывающееся на
поверхности проглоттиды; матка розетковидная или мешковидная.
Яйца с крышечкой.
2. Trypanorhyncha9, головка с четырьмя хоботками, усеянными
крючьями (рис. 197, В).
3. Тetraphyllidea-, головка с четырьмя ботридиями (рис. 197, С),
желточников два — в боковых зонах члеников или же фолликулы
желточников рассеяны по всей проглоттиде.
4. Cyclophyllidea-, головка с четырьмя присосками (рис. 197, D);
матка не имеет выводного отверстия; перезрелые проглоттиды отде-
ляются от стробилы; яйца без крышечки.
Паразиты человека имеются только среди представителей отрядов
Pseudophyllldea и Cyclophyllidea.
ОТРЯД PSEUDOPHYLLIDEA
Из семейства отряда Pseudophyllldea у человека паразитируют
представители только одного сем. Diphyllobothriidae.
Семейство Diphyllobothriidae
К сем. Diphyllobothriidae принадлежат виды, паразитирующие
у рыб, птиц и млекопитающих. У человека бывают цестоды только
двух родов — Diphyllobothrium и Diplogonoporus.
278
СПЕЦИАЛЬНАЯ ЧАСТЬ
Род Diphyllobothriam Cobbold
DiphyUobothriutn latum L.— лентец широкий
Синоним: Bothriocephalus latus Brems., Dibothriocephalus latus Liihe.
Известен как паразит человека с глубокой древности.
Стробила состоит из 4000 и более члеников, достигая 2—9 м дли-
ны при ширине до 1,5 ом. В исключительных случаях бывали лентецы
15—20 м длины. Сероватого или желтоватого цвета.
Рис. 200 — Стробила
лентецаширокого/)/-
phy llo bothrium latum.
Головка от 1—5 мм длины удлиненно овальной
формы; на спинной и брюшной узких ее сторонах
находится по одной глубокой присасывательной бо-
розде (рис. 197, Л). За тонкой бейкой следует
тонкая передняя часть стробилы, в которой отдель-
ные членики плохо различимы. Далее кзади лежат
проглоттиды, ширина которых более их длины
(рис. 201). Полного развития половой аппарат до-
стигает приблизительно в 60-м членике, считая спе-
эеди. Средину и заднюю половину стробилы зани-
мают совершенно зрелые членики, в центре поверх-
ности которых просвечивает темное пятно — матка,
заполненная оплодотворенными яйцами. Членики,
освободившиеся от яиц, уменьшаются в объеме,
могут стать почти квадратными и легко отрываются
от заднего конца стробилы.
Проглоттиды с вполне сформированными орга-
нами размножения имеют следующее строение
(рис. 202). ‘Многочисленные семенники в виде мел-
ких фолликулов занимают боковые поля членика,
ближе к его дорзальной
Рис. 201—Части строби-
лы лентенца широкого; в
середине члеников про-
свечивает розетка матки.
стороне. От каждого фол-
ликула отходит тонкий
vas efferens; несколько та-
ких канальцев сливаются
в более крупные стволы,
образующие в конце кон-
цов семеизвергательный
канал, извивающийся ме-
жду складками матки в
срединной зоне членика;
он направляется кпереди,
образует перед концом
семенной пузырек и окан-
чивается мускулистым со-
вокупительным органом (cirrus); последний открывается в передней
части половой клоаки, которая расположена близ переднего края про-
глоттиды.
Яичник в виде переметной сумы лежит у заднего края членика;
. желточников масса, их фолликулы лежат также в боковых полях
проглоттид, но вентрально от семенников. Правый и левый желточные
каналы сливаются в общий желточный проток. По средней линии чле-
ника идет, слегка волнообразно изгибаясь, влагалище; задний конец
его расширяется в семеприемник. Наружное отверстие влагалища ле-
жит позади отверстия мужского копулятивного органа в половой
клоаке.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
279
Общий желточный проток и начальная часть влагалища соединя-
ются с яйцеводом и впадают в оотип, лежащий в вырезке заднего
края яичника. От левого переднего угла оотипа отходит матка в виде
рукава, изгибы которого в общем дают фигуру розетки. Отверстие
матки расположено на средней линии проглоттиды, несколько отступя
от половой клоаки.
Яйца овальные, с тонкой двуконтурной оболочкой, на полюсах
притупленные (рис. 203); на одном полюсе имеется крышечка; на про-
тивоположном конце скорлупа несет маленький бугорок; яйцо желто-
вато-коричневого цвета; размеры его в среднем 0,070 мм (0,068—
0,071 мм) длины и 0,045 мм ширины.
Рис. 202 — Зрелый членик лентеца широкого Diphylloboth-
rium latum.
—общее половое отверстие; о — отверстие матки; ov —гяичники;
dt — желточники; sdr — тельце Мелиса; vag —влагалище; и — матка;
t — семенники; vd — семеизвергательный канал; neph — стволы выдели-
тельной системы. По Зоммеру.
Плодовитость (Lyon) до 2 100 тыс. яиц на порцию фекалий зараз;
no Goodey — 2 — 6 млн. яиц на грамм сухого вещества кала за пе-
риод 90—105 дней после начала заражения. Бывает до 100 (Roux) и 143
(В. Тарасов) экземпляров в кишечнике человека. За день прирост чер-
вя до 8—9 см (Braun).
Лентец широкий может жить очень долго; есть указания на 29-
летнюю длительность носительства этого паразита.
Кроме человека, живет в тонких кишках собаки, кошки,, тюленя,
лисицы и других млекопитающих.
Цикл развития лентеца широкого полностью раскрыт сравни-
тельно недавно. Сводится он к следующему. Яйца паразита с испраж-
нениями хозяина выбрасываются наружу и могут попадать в воду или
смываться в различные водоемы. Яйца выходят с недоразвитым заро-
дышем, и для полного его развития, которое может протекать в воде
или в жидких фекальных массах, требуется, смотря по температуре,
3—5 недель (рис. 203, 6). Из яйца, сталкивая крышечку, выходит ко-
рацидий — шестикрючечный зародыш, одетый ресничным покровом
280
СПЕЦИАЛЬНАЯ ЧАСТЬ
(рис. 203, с). С помощью ресничек корацидий плавает в воде в соста-
ве планктонных организмов.
Если в водоеме, в котором вылупились корацидии, имеются низшие
ракообразные, веслоногие рачки (Copepodd) — циклопы (рис. 204,
а Ь с
Рис. 203 — а — яйцо лентеца широкого; Ь — то же с зароды-
шем; с — вышедший зародыш (корацидий). По Редлиху.
виды: Cyclops strenuus и др.) и диаптомусы (Diaptomus gracilis,
D. graciloides), то они могут проглатывать плавающие корацидии.
В пищеварительном канале циклопа или диаптомуса личинка сбрасы-
вает ресничный покров, и освободившаяся таким путем онкосфера
Рис. 204— Циклопы — веслоногие рачки:
— Canthoca.mpus\ В — Cyclops} С — Diaptomus. Различные виды
В и С служат промежуючными хозяевами лентеца широкого.
Рис. 205 — Начало развития
процеркоида лентеца ши-
рокого:
А — шестикрючная онкосфера,
вышедшая из ресничной оболоч-
ки корацидия (рис. 203 с); В —
рост онкосферы на пути пре-
вращения в процеркоид; С-
циклоп Cycles strenuus с онкос-
ферами и процеркоидом (рг) в по-
лости тела. По Розену.
(рис. 205, А) с помощью крючьев проникает сквозь стенку кишечника
в полость тела рачка.
Здесь в течение 2—3 недель « онкосфера разрастается в удлинен-
ную, овальную личинку, которая в окончательном виде имеет своеоб-
разную форму, благодаря присутствию на заднем конце тела шаро-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
281
видного придатка с шестью крючьями (рис. 205, В; 205, С; 206, Д);
на противоположном конце личинки имеется присоскообразная ямка;
тело ее паренхиматозное, с рассеянными известковыми тельцами; дли-
на личинки достигает 50—60 обычно в циклопе развиваются 1—2
такие личинки. Эта прежде неизвестная стадия развития лентеца ши-
рокого называется процеркоидом; циклопы и диаптомусы явля-
ются первыми промежуточными хозяевами лентеца широкого (Розен и
Яницкий).
Зараженные рачки легко могут быть проглочены рыбами, питаю-
щимися планктоном. В желудке рыбы рачки перевариваются; освобо-
Рис. 206 — Лентец широкий:
А — процеокоид из полости тела
циклопа; В—плероцеркоид (из мыш-
цы рыбы); С — его головка.
По Р о з^е н у.
Рис. 207 — Плероцеркоид D. latum, прони-
кающий через стенку желудка щуки.
Ориг. рис. Ф. Талызина.
лившиеся процеркоиды в ближайшие дни проникают через стенку
желудка и оседают в различных местах тела — в мышцах, в яични-
ках, в печени и др. Здесь личинки превращаются в плеоцеркои-
дов, которые были известны уже давно (рис. 206, В).
Возможно, что рыбы заражаются плероцеркоидами лентеца широ-
кого и иным путем, а именно через рыб. Если хищная рыба пожирает
маленькую рыбу, которая в свою очередь успела проглотить циклопов
с процеркоидами, то последние проделывают свое дальнейшее разви-
тие в хищной рыбе, после того как заглоченная ею добыча будет
переварена.
Есть также указания на то, что при пожирании хищной рыбой
рыбы с плероцеркоидами последние не погибают, а приживаются в
своем новом хозяине (Hobmaier). Такое явление как бы пассивной пе-
ресадки паразита из одного хозяина в другого мы предлагаем назвать
пассивным переселением паразита. По Hobmaier’y, он может повторять-
ся неоднократно. Это явление подтверждено в нашей лаборатории
282
СПЕЦИАЛЬНАЯ ЧАСТЬ
(ВИЭМ, отдел паразитологии) В. Гнездиловым и Ф. Талызиным опытами
с прижизненно окрашенными нейтральротом плероцеркоидами, на-
глядно демонстрирующими миграцию плероцеркоидов через стенку же-
лудка щуки (рис. 207). Применением этого метода в комбинации с
искусственным заражением введением окрашенных плероцеркоидов в
Рис. 208 — Геккон (Gy mno dactylus caspius)n лягушка, эксперимен-
тально зараженные плероцеркоидами лентеца широкого.
По Е. Павловск ому и В. Гнездилову.
желудок различным холоднокровным показано, что плероцеркоиды
могут мигрировать сквозь стенку желудка и оседать под брюшиной, в
мышцах или в органах лягушки, жерлянки с Дальнего Востока, яще-
рицы, зеленой и прыткой, веретенницы, геккона каспийского, гадюки
степной, молодой сухопутной черепахи и др. ,(Е. Павловский и В. Гнез-
дилов, рис. 208). В опытах с лягушками получен своего рода экспе-
риментальный спарганоз лентеца широкого. Плероцеркоиды,
осевшие в мышцах, сохраняют заразительность для человека. Это дока-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
283
зано Гнездиловым самоэкспериментом с проглатыванием плероцерков,
проживших 244 дня в мышцах лягушки.
Плероцеркоид имеет вид белого стерженька длиною около 6 мм
и более (рис. 209). Он хорошо подвижен, и поэтому его размеры мо-
гут колебаться. На переднем конце он несет две присасывательные
бороздки. При движении в воде in vitro передняя часть плероцеркоида
Рис. 209 — Плероцеркоид лентеца широкого в мышцах ерша.
По Е. Павловскому.
раздувается на подобие того, как у очковой змеи. По данным Фурмана,
плероцеркоиды могут размножаться делением, чем он объясняет иногда
весьма значительную зараженность рыб этими паразитами.
Возможность размножения плероцеркоидов делением в настоящее
время решительно опровергается; плероцеркоиды могут подвергаться
распаду на несколько частей, благодаря местному отмира-
нию участков тела (рис. 210); такое явление фрагментации не
имеет ничего общего с размножением (Е. Павловский и В. Гнездилов).
Рис. 210 — Фрагментация плероцеркоида лентеца широкого.
По Е. Павловскому и В. Гнездилову.
Рыбы, следовательно, являются вторыми промежуточными хозяе-
вами лентеца широкого.
Если они будут съедены человеком или различными млекопитаю-
щими в сыром, полусыром, вяленом или другом состоянии, но с пле-
роцеркоидами, сохранившими жизнеспособность, то в тонких кишках
освободившиеся плероцеркоиды прикрепляются к стенке кишечника и
почкованием развиваются во взрослую глисту. Человек, следователь-
но, является окончательным хозяином лентеца широкого.
Известен широкий круг рыб, могущих быть хозяевами этого пара-
зита: щука (Esox lucius), окунь (Perea fluviatilis), ерш (Acerina cer-
nua, рис. 213), налим (Lota vulgaris, рис. 114), форель (Trutta lacust-
284
СПЕЦИАЛЬНАЯ ЧАСТЬ
ris), лосось (Trutta vulgaris), нериус (Salmo umbla), хариус (Thymallus
vulgaris), сиг (Coregonus lavaretus), ряпушка (Coregonus albula), сне-
ток (Osmerus eperlanus), судак (Lucioperca lucioperca), бычок (Cottus
gobio), колюшка (Pygosteus pungitius) и др. В Африке и в С. Америке
есть свои хозяева лентеца широкого среди рыб местной фауны.
Рис. 211 — Налим — Lota vulgaris. По Neveu-Lemaire и Pellegrin.
Рис. 213 — Ерш — Acerina cernua. Оригинал.
За последние 3—4 года перед Отечественной войной в СССР, преи-
мущественно в окрестностях Ленинграда и в Карелии (Конч-озеро и
прилежащие озера), были произведены обширные обследования рыб на
зараженность их плероцеркоидами лентеца широкого. Особенно сильно
были заражены рыбы Невской губы, что понятно, так как сюда сбра-
сываются сточные воды Ленинграда, в котором, конечно, имеется не
мало лиц, зараженных лентецом широким.
По данным В. Г. Гнездилова, щуки в Невской губе были заражены
поголовно, ерши — в 82%, окуни и налимы — в 50%; в районе Петер*
гофа наиболее были заражены налимы (91,6%), затем щуки (88,8%)>
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
285
<ерши (98%) и окуни (53,3) (Петрушевский). В Карелии в районе Конч-
озера на первом месте по заражению стоят щуки (100%), затем нали-
мы и окуни (Г. Петрушевский и И. Павловская).
В эпидемиологическом отношении важно выяснить распределение
плероцеркоидов по частям тела рыб. У щук они встречаются в брюш-
ной мускулатуре в два раза чаще, чем в спинной; у ершей и окуней
отношения получаются обратные. Туловищный отдел рыб в 4—10 раз
чаще заражен плероцеркоидами, чем хвостовой. Из внутренних орга-
нов часто бывает заражен яичник, в котором плероцеркоиды держат-
ся между икринками.
Эти данные важны, так как свидетельствуют о наибольшей засе-
ленности плероцеркоидами как раз тех частей рыб, которые идут в
пищу. Интенсивность инвазии достигает у рыб значительной степени.
Например, у щурят было обнаруживаемо до 211 плероцеркоидов. На
100 г веса рыбы из плероцеркоидов, найденных в мышцах (т. е. в
съедобной части рыб) у мелких ершей, приходилось круглым числом
по 22, а у мелких щук — по 10 экземпляров. Интересно отметить, что
крупные экземпляры рыб заражены относительно меньше, чем мелкие
(Гнездилов).
Итак, распространение лентеца широкого зависит от наличия про-
межуточных хозяев и от способов питания рыбой. Поэтому у человека
в СССР этот паразит распространен в водных районах (Ленинград-
ская область, Карелия, б. Архангельская губ., б. Тобольская губ.,
Якутия и др.), и обратно: рыбы оказываются заражены там, где более
многочисленное население живет по берегам рек и озер. Необходимо,
впрочем, иметь в виду, что в природе лентец широкий может сущест-
вовать и независимо от человека, которого на положении окончатель-
ных хозяев замещают лиса, тюлень, дельфин и другие животные (при-
родная очаговость этого паразита). Зараженность населения этим пара-
зитом может достигать местами 80%.
Лентец широкий эндемичен для прибрежных стран Балтийского
(моря. С эмигрантами из Прибалтики лентец, повидимому, был завезен
в США, где распространился в ряде озер.
Патогенное действие. Лентец широкий бывает причиной
злокачественного глистного малокровия — anaemia perniciosa dibothrio-
cephalica, однако, такое заболевание развивается далеко не при каж-
дом случае паразитирования лентеца широкого в человеке. Количе-
ственные отношения прямой роли не играют. Так, были известны случаи
паразитирования нескольких десятков лентецов широких (до 100) без
каких-либо симптомов заболевания их хозяина, и, наоборот, один един-
ственный экземпляр этого паразита может вызвать развитие злокаче-
ственного малокровия. Продолжительность жизни лентеца широкого в
кишечнике также не является непременным условием заболевания ане
мией.
Глистное малокровие от лентеца широкого развивается вследствие
всасывания токсинов, выделяемых этим паразитом в кишечнике челове-
ка; согласно менее распространенному взгляду, токсическими свойства-
ми обладают вещества, освобождающиеся при переваривании дегенери-
рующих или мертвых частей глисты. Итак, при наличии глисты в ки-
шечнике неминуемо должно происходить всасывание выделяемых ею
токсинов и последовательное развитие симптомов хронического отравле-
ния.
Однако, как это уже указывалось выше, внешние признаки заболе-
вания проявляются далеко не всегда. Очевидно, организм человека об-
286
СПЕЦИАЛЬНАЯ ЧАСТЬ
ладает известной стойкостью в отношении влияния на него глистных
токсинов. Может быть, здесь играет роль степень проницаемости кишеч-
ной стенки данного индвидуума для токсинов. Несомненно, существуют
также моменты, которые изменяют в ту или другую сторону реакцию
организма на поступающие в его кровь токсины. Бесспорное значение
имеет работа кроветворных органов, возмещающих ущерб, причиняемый
глистами в форме разрушения элементов крови' и нарушения пропор-
ционального соотношения сортов ее форменных элементов.
В ярко развивавшихся случаях злокачественного малокровия число
эритроцитов падает до 1—1,5—2 млн.; появляются ядерные эритроци-
ты; наблюдаются пойкилоцитоз и уменьшение количества лейкоцитов;
эозинофилия более или менее выражена; процент гемоглобина падает
до 25—30; восковая бледность лица, чревычайная слабость.
Однако детальное изучение крови у глистоносителей без каких-либо
заметных внешних признаков болезни привело Н. Рагозу к следующим
заключениям, которые касаются не только лентеца широкого, но и дру-
гих ленточных глист человека.
«Постоянное всасывание глистных токсинов ведет, как правило, к по-
стоянному и определенному изменению крови! число эритроцитов па-
дает, появляются дегенеративные и регенеративные формы их, в том
числе макроциты и мегалобласты. Менее стойкие эритроциты раство-
ряются; наряду с этим вырабатываются и более стойкие красные кро-
вяные клетки. Белые кровяные клетки также подвергаются измене-
ниям: при общей лейкопении уменьшается число нейтрофилов, базофи-
лов и больших одноядерных лейкоцитов. Эозинофилия, как правило,
имеется налицо. Случаи ее отсутствия, вероятно, зависят от преждевре-
менного истощения деятельности костного мозга. Антитриптическая си-
ла кровяной сыворотки глистоносителей и больных ясно и постоянно’
повышена».
Таким образом, состояние крови обладателей лентеца широкого ха-
рактеризуется разрушением красных и белых кровяных телец с одно-
временным извращением деятельности костного мозга.
Весьма важно, что гематологическая формула обладателей ленточ-
ных глист очень близка к формуле злокачественного малокровия в своей
как бы неопределившейся или скрытой формой (Н. Рагоза). Следова-
тельно, здоровое состояние глистоносителей является относительным,
ибо у них в крови происходят изменения, могущие при дальнейшем
развитии привести к проявлению злокачественной анемии.
Не было недостатка в попытках выделить из лентеца широкого ток-
сическое начало. В члениках этой глисты были обнаружены протеолити-
ческий фермент, агглютинин (для некоторых сортов крови) и гемоли-
зин. Последний является липоидом. Гемолизин не является секретом
паразита, но заключен в его тканях, откуда освобождается при распа-
дении или мацерации лентеца. В связи с этим в нормальных условиях
гемолизин не поступает в кишечник хозяина и. не влияет поэтому на, его-
кровь. При полном или частичном распаде глисты гемолизин освобож-
дается, всасывается в кровь и вызывает развитие злокачественной ане-
мии .(Tallquist).
Интересно отметить, что экстракты из лентеца широкого, взятого от
неанемичного больного, при впрыскивании их кошкам вызывали резкое
малокровие. Природа глистных токсинов пока все еще остается неясной.
Токсические свойства приписывали липоидам и масляной кислоте глист
и в них видели причину, обусловливающую развитие злокачественного
малокровия; но эта* гипотеза очень мало обоснована.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
287
Недавняя попытка изучения влияния на кровь кошек при желудочном
подкожном и внутримышечном введении экстрактов и эмульсий из цель-
ных обезжиренных лентецов и их липоидов не дали определенных ре-
зультатов; авторы причину неуспеха предположительно видят в том,
что кошки Карелии (работа велась в Петрозаводске) в районе эндеми-
ческого очага дифиллоботриоза приобретают стойкость к токсическим
веществам лентеца широкого (Н. Шихобалова и Н. Попова).
Нераспознанные случаи злокачественной анемии от лентеца широко-
го ведут к смерти. Своевременное установление действительной причи-
ны заболевания и изгнание глисты спасают больного от гибели.
Диагноз ставится по нахождению яиц паразита в faeces. Необхо-
димо иметь в виду, что яйца лентеца! широкого ни ПО' форме, ни по ве-
личине, ни по структуре не отличаются от яиц недавно найденной у че-
ловека в СССР трематоды Nanophyetes salminlcola (Скрябин, Шихоба-
лова, Подъяпольская). Так как jr лентеца широкого матка открывается
самостоятельным отверстием, то яйца обычно обнаруживаются в faeces
в большом количестве.
Лечение. Изгнание глисты после дачи экстракта папоротника.
Контроль успешности глистоизгнания проводится обследованием вы-
гнанной глисты. Если головка ее вышла, то цель достигнута полностью.
Необходимо отметить, что головку иногда трудно распознать, если она
сокращена, и губы присасывательных щелей тесно сомкнуты. При нена-
хождении головки, необходимо недели через 3—4 снова исследовать
faeces пациента на яйца лентеца широкого; опытами ‘установлено, что
уже через две недели после заражения человека и собаки плероцеркои-
дами лентеца широкого в испражнениях можно найти яйца этого
паразита.
Профилактика. Употребление в пищу рыбы только в хорошо
проваренном или прожаренном состоянии. Рыба может быть обезвре-
жена выдерживанием ее на холоду. 172-часовое действие из плероцер-
коидов температуры в 9° ниже нуля и 4-часовое воздействие температу-
ры в 6° ниже точки замерзания не лишало плероцеркоидов подвижности
и способности развиться в ленточную глисту при скармливании соба-
ке.— 10° С убивают плероцеркоидов по истечении трех часов, равно
как и 10-минутное нагревание при температуре Ч7 45—46° С. Поэтому
мерой общественной профилактики является выдерживание рыбы на
холодильнике не менее как в течение двух недель. Соление и маринова-
ние рыбы также быстро убивают плероцеркоидов.
Большую эпидемиологическую роль играет в крупных городах кана-
лизация, если сточные воды отводятся в водные бассейны. При неиз-
бежном массовом попадании в воду яиц лентеца широкого обеспечи-
вается прохождение его жизненного цикла и заражение рыб. Такое же
значение имеют и уборные на судах и барках со сбрасыванием фекалий
прямо в воду. Канализационные отбросы необходимо обезвреживать хло-
ром или фильтровать и лишь после этого спускать в воду. Достоверно
установлено, что при упорядочении и совершенствовании работы систем
канализации зараженность рыб в соответствующих местах плероцеркои-
дами лентеца широкого уменьшается (Т. Magath).
Далее, необходима широкая пропаганда гельминтологических сведе-
ний по линии служащих Нарпита, особенно поваров. Наконец, мерой
общественной профилактики является предохранение водоемов от попа-
дания в них faeces человека.
288
СПЕЦИАЛЬНАЯ ЧАСТЬ
Diphyllobothrium mimus Cholodk.— лентец малый
Описан Н. А. Холодковским по экземпляру из Забайкалья. Подробно
изучен Ф. Талызиным по более обильному материалу из Бурят-Монголь-
ской республики. Длина стробилы колеблется от 10 до 26 см, наиболь-
шая ширина — 6—8 мм, 7—11 мм. Проглоттид — около 200. Шейка не
выражена. Головка эллиптической формы, длиной 0,8—1,7 мм, шириной
0,5—1 мм (рис. 214). Присасывательные бороздки расширяются спере-
Рис. 214 —Головка лен-
теца малого — Diphyllo-
bothrium minus.
По Ф. Т а л ы з и и у.
Рис. 215 — «Опухоли» на
стенке желудка омуля
с плероцеркоидами D. mi-
nus, По А. С к в о р ц о-
ву и Ф. Талызину.
ди. Длина самых задних проглоттид равняется 2/9 их ширины. Края
стробилы похожи на зубчатую пилу. Наружное отверстие матки ле-
жит на средине плоскости членика. Петли матки располагаются в об-
щем горизонтально и не образуют фигуры розетки. Яйца с крышечкой
и тупой пуговкой на противоположном полюсе. Длина их 58—72, а ши-
рина 41—49 у (в среднем 64 на 44).
Цикл развития изучен экспедицией Отдела паразитологии ВИЭМ
(А. Скворцов и Ф. Талызин) на о-ве Ольхон (Байкал, 1938). Плероцер-
коиды локализуются в опухолеобразных округлых разрастаниях поверх-
ности кишечника (рис. 215), пищевода, желудка, реже на других орга-
нах (но не в мышцах) у омуля (Coregonus migratorius— рис. 216),
сига (Coregonus lavaretus pikschlan. С. lavaretus baicalensis) и хариуса
(Thumallus arcticus — рис. 218). Из вскрытых омулей было заражено
56,2% рыб, хариусов — 27,7% и сигов — 68,4% рыб (считая вместе
плероцеркоидов D. minus и D. strictum).
Принадлежность плероцеркоидов определенным видам лентеца уста-
навливалась скармливанием «опухолей» с плероцеркоидами собакам и
кошкам.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
289
Корацидии D. minus 0,05 мм диаметром (рис. 219) . В аквариуме они
заглатываются циклопами (Cyclops sp.) и в его полости тела превра-
щаются в процеркоид, отличающийся червеобразной формой отростка с
крючьями, имеющего у лентеца широкого шарообразную форму (рис.
220, 221).
Рис. 216 — Байкальский омуль — Coregonus migratorius. Оригинал.
Сравнительная редкость заражения человека лентецом малым объяс-
няется локализацией плероцеркоидов на кишечнике, который в пищу
не идет, если только рыба не поедается целиком.
Излюбленный способ поджаривания рыбы «на. приколе» не ведет
к потере жизнеспособности всех плероцеркоидов (рис. 222).
О патогенных свойствах известно очень мало. Талызин, вы-
290
СПЕЦИАЛЬНАЯ ЧАСТЬ
гнавший из одного рыбака десять экземпляров, наблюдал у него сильно
выраженную анемию слизистых оболочек, нервную раздражимость^
плохой аппетит и частые поносы.
Лечение, что и при лентеце широком.
Профилактика, см. лентец широкий.
Рис. 219—Di phyllo bothrium minus'. A— яйцо с развившимся кора-
цидием внутри. Десятидневная культура.
В — Корацидий, вышедший из яйца. Десятидневная культура. Байкал.
По А. Скворцову и Ф. Талызину.
Рис. 220 — Diphyllobothrium minus.
А — Процеркоид, извлеченный из хозяина (циклопа) после 9 дней заражения.
Байкал. В — Плероцеркоид из омуля. Байкал.
По А. Скворцову и Ф. Талызину.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
291
Diphyllobothrium strictum Talysin
Синоним: Dibothriocephalus strictus Talysin.
Лентец узкий. Общая длина стробилы до 47 см (рис. 223), проглот-
тид— 180—360. Наибольшая ширина проглоттиды 4 мм. Головка эллип-
тической ф'ормы, с боков сдавлена: хорошо выраженной шейки нег.
К заднему концу членики сильно удлиняются. В задней трети стробилы
замечается удвоение полового аппарата в каждом членике. Раньше раз-
Рис. 221 — Процеркоид лентеца малого в промежуточном
циклопе—хозяине. Экспериментальное заражение. Байкал.
По А. С к в о р ц о в у и, Ф. Т а л ы з и н у.
вивается мужская часть аппарата (протерандрия). Комплекты
половых органов лежат в линейном порядке (рис. 223). Цвет живой
стробилы светлокоричневый. Яйца эллипсоидные, с тупыми полюсами,
крышечкой и кнопкообразным выростом скорлупы, светложелтые или
коричневые 54—57 длины и 40 у ширины (рис. 223).
Описан Ф. Талызиным .(1932) по экземцлярам, изгнанным в Бу-
рят-Монгол ии на острове Ольхон у бурятов, среди которых было зара-
жение лентецом широким и лентецом малым.
Этот вид снова был обнаружен экспедицией Отдела паразитологии
ВИЭМ (Скворцов и Талызин— 1938). Плероцеркоиды этого лентеца ло-
кализуются в меньших по размеру, чем у D. minus, опухолеобразных
292
СПЕЦИАЛЬНАЯ ЧАСТЬ
выростах поверхности кишечника или желудка хариуса, омуля или си-
га (Скворцов и Талызин).
Патогенное значение неизвестно.
Лечение и профилактика те же, что при лентеце широком.
Diphyllobothrium tungussicum Pod jap. et Gned.
Лентец тунгусский, длина 56—63 см, наибольшая ширина 3,56 мм.
Головка булавовидно утолщается; ее наибольшая ширина 0,68 мм. Го-
ловка нередко отделяется от шейки. Первые членики начинаются на
Рис. 222 — Приготовление рыбы «на рожне» (внутрен-
ности не подвергаются действию высокой температуры).
О-в Ольхон. Байкал. Оригинал Ф. Талызина.
расстоянии 60 мм от головки. Члеников до 502 (рис. 224). Задние про-
глоттиды шире передних. Начало развития полового аппарата заме-
чается с 118-го членика.
Яйца 72—75 длины и 48—51 у ширины, одеты оболочкой с тонкой
альвеолярной структурой.
Описан В. Подъяпольской и М. Гнединой (1932) по экземплярам из
района Нижней Тунгузки и Лены. Паразитирует у якутов и тунгузов
(в Туруханске — у 23,6% обследованных). Одновременно бывает зара-
жение и лентецом широким. У одного хозяина наблюдалось свыше 477
экземпляров лентеца тунгусского.
Патогенное значение и цикл развития — неизвестны.
П р о ф и л al к т и к а: не есть полусырой рыбы.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
293
Diphyllobothrium skrjabini Plotnikoff
Длина до 260 см, ширина на уровне шейки 1,0 мм, в средине длины
стробилы 1,3 см; у заднего конца 2,0 см. Проглоттид до 2700. Членики
й
Рис. 223 — Лентец узкий Diphyllobothrium strictum:
А — стробила, В—головка/ С—членик, D — членик с удвоенным половым аппаратом;
Е—яйцо; ои— яичник, ut — матка; Г—семенник, CR— мешок цирруса, VD— семепро-
вод, V — влагалище, ЕХ — выделительный канал. По Ф. Талызину.
очень коротки; ширина резко преобладает над их длиной. Форма матки
вариирует от бобовидной до розетковидной. Яичник двулопастный гру-
шевидный с слабофестончатыми краями. Яйцо 62—67X43—48 }*
294
СПЕЦИАЛЬНАЯ ЧАСТЬ
(М. Петров). Хозяева — человек (низовье Печоры, Ненецкий националь-
ный округ), собака (Свердловск).
Цикл развития неизвестен.
Diphyllobothrium nenzi М. Petrov
Длина 107 см. Ширина не более 5,5 мм. На протяжении первых
32 см стробилы проглоттиды очень мелки и границ между ними не вид-
но. Бурса цирруса лежит ближе к переднему краю членика. Матка спль-
Рис. 224 — Лентец тунгусский Diphyllo bothrium tun-
gussicum. Зрелая проглоттида. По В. По дъяпол ь-
ской и М. Гнединой.
Рис. 225 — Diphyllobothrium skrjabini. Гермафродитный членик.
По М. Петрову.
но развита; ее петли могут обнимать мешок цирруса. Яичник четырех-
угольный с округленными углами. Семенники и желточники из боковых
полей заходят к переднему краю членика.
Яйцо 59—68 X 425—45 у с крупной крышечкой.
Кишечник человека (низовье Печоры).
Цикл развития неизвестен.
1 Diphyllobothrium giljacicum Rutk.
Длина до 263 см; проглоттид до 1830. Сколекс толщиною макси-
мально 1,062 мм. Ширина всех проглоттид превышает длину — зрелые
проглоттиды 5,22—5,31 мм длины, 7,42—8,7 мм ширины. Половые от-
верстия открываются на брюшной поверхности членика. Матка в виде
узкого петлистого ствола дает до семи петель с каждой стороны. Матка
с зрелыми яйцами имеется в члениках, начиная с 98 см оъ сколекса.
Яичник нежного сетевидного строения в виде двух треугольных крыльев
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
295
с комиссурой. Яйца овальные, 51—57 X 41—44 }л. Изгнан из нескольких
гиляков на западном побережье о-ва Сахалина). Одновременно парази-
тировал и D. latum.
DiphyUobothrium luxi Rutk.
Длина 150 см; члеников до 1 662. Наибольший диаметр сколекса (по
ширине) 0,34 мм. Ширина всех проглоттид больше их длины. Зрелые
членики 2,3 мм длиной и 7,94 мм шириной. Половой аппарат заклады-
вается, начиная почти с 300 членика. Матка дает до 4 (8) петель с каж-
дой стороны. Членики с зрелой маткой начинаются в 48 см от сколекса.
Яичник сетевидный в виде двух равнобедренных треугольников или
(в задних члениках) грушевидной формы. Яйца 60—63 X 46—48 у. Из-
гнан на Сахалине из эвенков и якутов на восточном побережье.
Diphyllobothrium cordaium R. Leuck.
Синонимы: Bothriocephalus cordatus R. Leuck., Dibothriocephalus corda-
tus R. Leuck.
Длина стробилы 80—115—130 см, наибольшая ширина — около 8 мм. Членики
коротки. Головка сердцевидная 2 на 1 мм. Присасывательные борозды идут по
спинной и брюшной сторонам. Шейка не выражена. Матка розеткой с 6—8 ветвями.
Яйца с крышечкой, овальные 75 X 50 р-.
Цикл развития неизвестен. Обыкновенный паразит тюленей, моржей и со-
бак в Гренландии, Исландии и Японии. Возможно паразитирование и у человека.
Вторым промежуточным хозяином этого паразита без сомнения является рыба.
Диагноз, лечение и профилактика, как и при лентеце широком.
Diphyllobothrium mansoni Cobbold.
Синоним: Sparganum mansoni Cobb., Dibothriocephalus mansdni Cobb.
Стробила длиной 60—100—250 см. Головка 1 —1,5 мм длиной и
0,4—0,8 мм шириной, в поперечном сечении почти четырехугольная. Се-
менников 300—540 в членике. Матка образует 3—5 петель. Яйца
52—63,5 у длины и 32—43,5 ширины, к концам более сжаты.
Взрослый паразит обитает в средней зоне тонких кишок собаки,
волка, лисицы, кошки, леопарда, тигра. Паразит распространен в Юж-
ном Китае, в Японии, в Индо-Китае и в Панаме.
Первым промежуточным хозяином являются — циклопы: Cyclops
leuckarli, в полости тела которого может развиться процеркоид, а так-
же Macrocyclops albidus, М. fuscus, Eucyclops serratulus, Mesocyclops
leuckarti, M. oithnoides, Cyclop? vicinus, A canthocy clops, viridis
A. bicuspidatus.
Вторым промежуточным хозяином могут быть лягушки, кошки,
свиньи, крысы, мыши, домашние птицы, змеи и человек. Плероцеркоид-
ная стадия паразита была известна раньше ленточной и была описана
под именем Sparganum mansoni (рис. 227). Их находили у человека
в подкожной клетчатке, брюшной и плевральной полостях, в шейной
мускулатуре, в глазах (японцы и китайцы). Тело Sparganum удлинен-
ное, червеобразное, длиною обычно 20—30 см (иногда до 60. см) при
ширине в 3 мм. На переднем конце имеется головка в виде колбо-
образного утолщения со щелевидными продольными углублениями. Па-
разит лежит в соединительной ткани, окруженный капсулой. Sparganum,
взятый от человека, скармливали собакам и кошкам, причем у них в
кишечнике развивалась ленточная глиста, получившая название Di-
296
СПЕЦИАЛЬНАЯ ЧАСТЬ
phyllobothrium mansoni (Jamata и Joshida). В одном из опытов за 2 ме-
сяца. в кишечнике собаки выросла глиста 2,5 м длины. У собаки на-
блюдалась естественная зараженность сходным паразитом. Личинками,
выведенными из яиц D. mansoni, удавалось заражать Cyclops leuckarti,
который в то же время был стоек к заражению лентецом широким.
При скармливании циклопов с процеркоидами D. mansoni мышам и ля-
гушкам удавалось вызвать в них развитие Sparganum. Таким образом
удалось замкнуть изучение жизненного цикла D. mansoni.
Рис. 226 — А — Diphyllo bothrium nenzi. Гермафродитный
членик. По М. Петрову. В — Diphyllobothrium nenzi.
Яйцо. По М. Петрову.
В некоторых местах Японии до 30—4О?/о лягушек заражены Sparga-
пит, залегающими преимущественно в мышцах.
Человек заражается проглатыванием циклопов с процеркоидами
(питье воды), причем из процеркоида в естественном порядке разви-
вается плероцеркоид. Другим источником заражения являются цыплята
и лягушки. При поедании сырых цыплят или лягушек с плероцеркоида-
ми последние приживаются в теле человека (явление «переселения» па-
разитов, аналогичное повторному заражению щук плероцеркоидами лен-
теца широкого, стр. 282).
Поражение человека плероцеркоидами. D. mansoni известно под
наименованием спарганоза. В местах его нахождения бывают воспали-
тельные припухания и образования абсцессов. Если паразит залегает
в орбите, то наблюдаются отек век, птоз, слезотечение и ощущение
значительной боли.
Так как Sparganum является тканевым паразитом человека, то при-
жизненная диагностика его возможна только в случае извлечения пара-
зита.
Профилактика. Пить прокипяченную или профильтрованную
воду, не есть сырых и полусырых цыплят, лягушек и других позвоноч-
ных, у которых бывает паразит.
В пределах СССР этот паразит у человека не обнаружен. Возможно*
нахождение его на Дальнем Востоке.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
297
В Японии и Флориде был найден другой вид паразита, названный
Sparganum proliferum (рис. 228); он обитает в кожных, легко вылущи-
вающихся кистах; бывает массами в подкожной соединительной ткани»
Рис. 227— Sparganum]
mansoni. По I j i m a
и M u г a t a.
в брыжейке, почках, сердце, легких, в мозгу
и др. Повидимому, более патогенен, чем
D. mansoni. Форма паразита многообразна,,
на нем бывают отростки, которые отпочко-
вываются и дают новых личинок.
Цикл развития неизвестен.
Род Diplogonoporus Loennenberg
Половые органы в каждой проглоттиде
удвоены, в остальном паразиты сходны с
Diphyllobothrium.
Рис. 228 — Куски Sparganum proli-
ferum, увеличенные в четыре раза.
По М. Б р а у н у.
Diplogonoporus grandis R. Blanch.
Головка! неизвестна. Стробила около 10 м длины, при ширине 2,5 см.
Членики очень коротки и широки. На брюшной стороне червя два про-
дольных ряда половых отверстий, лежащих в желобоватых углублени-
Рис. 229—Кусок стробилы Diplongonoporus grandis.
а — а — два ряда половых отверстий. Увеличено.
По Куримото.
ях. Ряды маток отстоят на некоторое расстояние друг от друга, так.
что белесоватая стробила разбивается ими на три пояса — средний
и два боковых (рис. 229).
Обычно паразитирует в кишках китообразных и тюленей. Известны
два случая нахождения этой глисты у человека в Японии.
298
СПЕЦИАЛЬНАЯ ЧАСТЬ
Яйца с крышечкой, широкоовальные, 63—68 р длины и 50 у- шири-
ны, темнокоричневые.
Цикл развития неизвестен. Весьма вероятно, что вторым про-
межуточным хозяином являются рыбы.
Патогенное действие. У зараженных этим паразитом бы-
вают колики, развивается анемия, перемежающиеся поносы и запоры.
Диагноз. По нахождению яиц в faeces или оторвавшихся члени-
ков после дачи слабительного.
Терапия. Экстракт папоротника.
Профилактика неизвестна; провизорно остерегаться есть сы-
рую или полусырую рыбу.
Diplogonoporus Brauni Leon.
Длина светлосерой толстой стробилы около 29 см; головка шири-
ною в 1 мм лишена шейки; несет две широкие ботрии — спинную и
брюшную? Проглоттиды очень коротки при ширине до 6 мм (maximum).
Задние, опорожнившиеся от яиц, членики удлинены. На каждой поверх-
ности червя идет по два желобка; брюшные желобки глубже спинных
и несут в себе половые отверстия.
Известны два экземпляра паразита, выгнанные из человека в Яссах
'(Румыния). В обоих случаях у больных был одновременно и лентец
широкий.
Терапия. Экстракт папоротника.
Профилактика, см. выше.
Род В г aunt a Leon.
Braunia jassyensis Leon.
Дважды найден в Румынии у человека. Длина нерасчлененной лен-
ты паразита 18—20 см при ширине 8—12 мм; на переднем конце ма-
ленькая треугольная головка с двумя рудиментарными присасыватель-
ными бороздками. Самостоятельность этого рода и вида вполне основа-
тельно подвержена сомнению (К. Скрябин и Р. Шульц). Вероятно, эти
паразиты являются какими-нибудь ремнецами (Ligula), нормально па-
разитирующими в личиночной стадии у рыб. Паразиты могли попасть
в кишечник человека при поедании сырой зараженной рыбы; и остаться
там известное время жить (гостепаразитизм). Изгнаны экстрактом па-
поротника.
ОТРЯД CYCLOPHYLLIDEA
Ленточные черви отряда Cyclophyllidea характеризуются наличием
головки с четырьмя полушарообразными присосками, лежащими крест-
накрест; скоде^с может иметь хоботок с одним, двумя или многими
венчиками крючьев. Отверстие половой клоаки помещается на чбоку
членика, матка обычно не имеет выводного отверстия. Яйца лишены
крышечки. Развитие зародыша в яйцах протекает еще в матке; заро-
дыш в яйце не имеет ресничной оболочки, но вооружен шестью крючь-
ями.
Семейство Anoplocephalidae
Богатое видами семейство широко распространенных по свету це-
стод млекопитающих и птиц; особенно свойственны жвачным.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
299
Род Monlezia Blanch
Известны лишь обрывки стробилы. Гермафродитные членики до
1 см ширины и 0,6—1 мм длины. Половой аппарат двусторонний; по-
Рис. 230 — Гермафродитные членики Monlezia sp. из чело-
века. По Е. Ершову и С. Малыгину.
ловые отверстия открываются по бокам члеников. Се-
менники в виде множества мелких фолликулов лежат
в пространстве между обоими выделительными канала-
ми. Семепровод волнообразно изогнут. Циррус с шеро-
ховатой поверхностью (рис. 230). Характерные межпро-
глоттидные железы не обнаружены. Яичник в виде двух
гроздевидных желез, лежащих позади семеприемника,
который развит хорошо (длина его до 0,3 мм). Влага-
лище извитое, довольно длинное. ’Матка в зрелом со-
стоянии мешковидная со множеством яиц.
Яйца весьма своеобразные с онкосферой, имеющей
особый «грушевидный аппарат» (рис. 233). Размеры
яиц — 63 — 66 X 70 у.
Виды рода Monlezia паразитируют у жвачных копыт-
ных (напр. М. expansa, М. benedeni, М. lenticulata и др.
у овец и телят). Обнаруженный у человека случай на-
блюдался в местности, где ежегодные эпизоотии моние-
зиоза обычны.
Единственный случай паразитирования у человека
Monlezia описан В. Ершовым и С. Малыгиным у жен-
щины в Сунском районе Кировской области. Больная
была анемичная, истощенная; жаловалась на боль в
области живота; наблюдались периодические приступы
болезни, особенно после жирной пищи. Часто были
головокружения и отсутствие аппетита; иногда бывали
позывы к рвоте. 8,0 г экстракта папоротника обеспечи-
ли глистоизгнание. После изгнания паразита дней че-
рез 15 прекратились болезненные симптомы, беспокоив-
Рис. 231 — Мо-
niezia expansa.
Стробила.
шие больную: болезненность в области пупка и выше часто периоди-
ческого характера, тошнота, потеря аппетита, головокружения, общая
слабость. Сильная анемичность и истощение больной были причиной
направления ее в больницу. Больная до лечения отмечала выхождение
члеников глисты или кусков стробилы.
Цикл развития Monlezia до последнего времени не поддавался рас-
крытию. Обзор разноречивых точек зрения на развитие этих цестод и,
следовательно, на способы заражения ими дают К. Скрябин и Р. Шульц
(«Гельминтозы крупного рогатого скота и его молодняка», 1937).
В последнее время показано, что развитие совершается с,промежуточ-
ным хозяином, каковым является свободноживущий непаразитический
300
СПЕЦИАЛЬНАЯ ЧАСТЬ
А
В
Рис. [232 — А — Moniezia expansa. Членик с вполне развитой половой
системой:
а — межчлениковая железа; b — семепровод; g— половой сосочек с половой клоакой; е —
яичник; с — желточник (кпереди от него тельце Мелиса); d — семенники; ’ ех— выделитель-
ный канал. По Е. Матевосян.
В — Moniezia expansa. Зрелый членик, переполненный яйцами. По Е. Матевосян*.
Рис. 233 — Moniezia expansa. Яйцо.
А — онкосфера с полным грушевидным аппаратом. В — вполне зрелое яйцо-
По Е. Матевосян
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
301
клещ из Oribatidae-Galumna sp. По Stunkard’y в клещах, заглотивших
яйца, развиваются цистицеркоиды. Скармливанием клещей с цистицер-
гоидами можно заразить ягненка ленточной формой Moniezia. Некото-
рые орибатиды, по тому же
автору, являются промежуточ-
ными хозяевами другой ано-
плоцефалиды — Cittotaenia cte-
itoides кроликов и зайцев. Были
обнаружены клещи, спонтанно
зараженные цистицеркоидами
Cittotaenia там, где водятся
дикие кролики. Овцы зара-
жаются, поедая с травой за-
раженных клещей. Вероятно
таким же путем, т. е. случай-
ным проглатыванием заражен-
ных клещей, заражается и че-
ловек.
Замечательно, что в орбиту
паразитологических интересов
включается новая группа хо-
зяев из паукообразных живот-
ных. Весьма возможно, что
круг промежуточных хозяев
аноплоцефалид окажется раз-
нообразным по их системати-
ческому положению.
Киршенблат нашел цисто-
церкоидов у сенокосца Nema-
stoma luguber и Platybunus
trangularis, может быть, так
же принадлежащих к анопло-
цефалидам.
Рис. 234 — Клещи рода Galumna
(Oribatldaey.
— Galumna sp.; В — G. hubbaria; С — нимфа Galu-
mna sp.; D — G. persimilis. По Banks.
Pod Bertiella
Bertiella satyri Stil. et Hass.
Ленточная форма паразитирует в кишечнике обезьян (оранг-утан,
гиббон, макаки и др.). Известны два случая гостепаразитирования у
человека, именно у детей (Бенгалия и о-в Маврикия). Глиста до 27—
28 см длины при максимальной ширине 1 см. Головка с рудиментар-
ным хоботком и четырьмя присосками. Цикл развития неизвестен.
Семейство Dipylididae
Сколекс с хоботком,' вооруженным несколькими рядами крючьев; в
члениках двойной комплект половых органов, и половые отверстия по-
мещаются по обеим сторонам каждого членика.
Род Dipylidium Leuck.
D ipy lidiuin’'caninum Lin.— цепень ’ тык в о видный
Синонимы: laenia cucumerina Bloch., Dipylidium cucumerinum Leuck.
Цепень тыквовидный. Стробила 15—40 Ом длиною
(рис. 235). Головка маленькая, ромбоидальная (0,35—0,4 мм шириною),
302
СПЕЦИАЛЬНАЯ ЧАСТЬ
на переднем конце несет булавовидный хоботок (длиною 0,185 мм, шири-
ною 0,11—0,12 мм), снабженный 3—4 венчиками крючьев (около 60,
•рис. 235). Их всего меньше в заднем (нижнем) венчике, где они к тому
же и самые мелкие — 5—6 р против И—15 р переднего ряда. Хоботок
глубоко втягивается в головку, которая несет по бокам четыре овальных
присоска {рис. 236). К тонкой шейке примыкают тонкие же передние
членики тела; проглоттиды быстро удлиняются и принимают типичную
Рис. 235 — Стробила
цепеня тыквовидного
Dipylidium caninum в
естественную величи-
ну. По Н. Хол од-
ко в с к о м у.
Рис. 236 — Сколекс цепеня тыкво-
видного — Dipylydium caninum.
По М о и n i g.
форму огуречного зерна, длиною 6—7 мм, шириною в 2—3 мм. Зрелые
членики с выпуклыми боками, посредине которых на каждой стороне
тела лежит по половому отверстию. Легко отрываются от заднего кон-
ца червя и могут самостоятельно двигаться.
Половой аппарат в члениках парный (рис. 237, 238). Семенники
разбросаны по всему членику в виде множества шаровидных телец (их
до 180); между ними проходят петли непарной, сетеобразно ветвящейся
матки; эти петли становятся заметными лишь при заполнении их зре-
лыми яйцами, когда семенники дегенерируют. Яичники парные в‘ виде
небольших розеток, состоящих из радиально лежащих трубочек, кзади
от них расположены меныпие по размерам желточники. От яичников
вперед и кнаружи отходит влагалище, над которым идет семеизверга-
тельный канал, концевая часть которого расширяется в мешок копуля-
тивного органа.
В зрелых члениках на первый план выступают петли матки, пере-
полненные яйцами, остальные же части полового аппарата исчезают.
В перезрелой матке залегают группы по 8—15 яиц, окруженных мате-
ринской эмбриональной оболочкой (яйцевые коконы или капсулы,
рис. 239).
Яйца круглые; онкосфера 25—40 диаметром, наружные размеры
яйца больше — 43—50 у; оболочка яйца лишена радиальной исчерчен-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
303
ности. Крючья зародыша—12—15 р длиной. Яйца кирпично-краснова-
того оттенка (табл. VI, 11).
Обычный паразит тонких кишек собаки, кошки, гиены и шакала, у
которых встречается сотнями. У человека (преимущественно у детей)
Рис. 237 — Dipylydium canh
пит. Зрелый членик цепе-
ня тыквенного:
а—выделительный канал; b (ввер-
ху) — семенник; е — семевынося-
щий канал; d — мужская бурса;
с — влагалище; / — яичник;
g — тельце Мелиса; b (внизу) —
желточник. По К. С к р я б и ну
и Р. Шульцу.
был обнаружен несколько десятков раз.
Космополит. В СССР известны единичные
случаи паразитирования этой глисты у
человека.
Яйца паразита и перезрелые членики вы-
брасываются с faeces животных и попадают
Рис. 238 — Половой аппарат Dipylidium caninum
(зарисован из средины членика):
с — мешок копулятивного органа; vt— желточник; ех — выде-
лительный канал; t— семенники; ov — яичник; и — матка; v — вла-
галище. По Брауну.
в их логово; кроме того, благодаря самостоятельному выползанию
члеников из заднепроходного отверстия, яйца глисты могут скоплять-
ся © перианальных складках и на шерсти около анального отверстия.
Цикл развития. На самом животном яйца цепня тыквовидного
могут быть проглочены власоедом собачьим (Trichodectes cards, рис.
240) и власоедом кошачьим (Tr. subrostratus), которые питаются оро-
говевшими чешуйками или шерстью своего хозяина. В загрязненном
субстрате логова животного яйца глисты пожираются личинками блох
собачьей (Ctenocephalus cants) или человеческой 1 (Pulex irritans).
В желудке этих насекомых из проглоченных ими яиц выходят
6-крючечные зародыши цепня тыквовидного, проникающие сквозь стен-
ку желудка в полость тела, где зародыш превращается в цистицеркоид
(рис. 198). Взрослая блоха, в конце концов, оказывается обладательни-
цей цистицеркоидов, находящихся в полости ее тела.
Власоеды и блохи беспокоят собак и кошек, когда ползают в шерсти
по коже, грызут ороговевший эпидермис или сосут кровь. Собаки и
кошки, облизывая шерсть и анальное отверстие, а также кусая зубами
зудящие места (собаки), могут проглатывать власоедов и блох с цистй-
1 Человеческая блоха в некоторых районах СССР является частым эктопа-
разитом собаки.
304
СПЕЦИАЛЬНАЯ ЧАСТЬ
перкоидами. Последние в пищеварительном канале окончательного хо-
зяина освобождаются и через 12 дней превращаются во вполне разви-
тых глист.
Легкость заражения основного хозяина от промежуточного и обрат-
но— в данном случае очевидна. Этим и объясняется обычно множе-
ственное заражение плотоядных животных цепнем тыквовидным. В то
же время у человека он встречается обычно в единичных экземплярах.
Рис. 239 — Кокон с яйцами Di-
oylldlum caninum. По Скря-
бину и Шульцу.
Рис. 240 — Промежуточный
хозяин цепня тыквовидного—
власоед собачий — Trichodec-
tes canis. Оригинал.
Заражение человека происходит при проглатывании блох или власо-
едов собак или кошек. Это чаще всего случается с детьми, играющими
с этими животными, когда они едят на полу и собака или кошка по-
лучают доступ к тарелке. При отталкивании животных от тарелки их на-
ружные паразиты могут упасть в пищу и быть съеденными ребенком.
Возможно случайное проглатывание промежуточных хозяев цепня
тыквовидного и грудными детьми (известен случай паразитирования
этой глисты у 12-дневного ребенка), вероятно, при пребывании кошки
в люльке ребенка. <
Факт нахождения D. caninum у.человека является показателем са-
нитарно-гигиенического неблагополучия в отношении содержания ре-
бенка или совершенно случайным стечением обстоятельств.
Патогенное значение D. caninum, повидимому, невелико;
у собак и кошек массовое паразитирование не сопровождается какими-
либо заметными болезненными проявлениями; в других случаях отме-
чают истощение хозяев. У человека паразитирование цепня тыквовид-
нсто протекает или без всяких симптомов, или же появляются нерв-
ные и кишечные расстройства, проходящие после изгнания этих глист.
Диагностика. Нахождение в faeces коконов яиц (под микро-
скопом) или выползших члеников (диференцировать от обрывков слизи).
Лечение. Легко поддаются действию экстракта папоротника.
Профилактика. Держать собак и кошек в чистоте (вычесы-
вать эктопаразитов, чистить подстилку и др.), не допускать., тесного
контакта детей с собаками и кошками.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
305
Семейство Davaineidae
Род Davainea Blanch.
Для представителей сем. Davaineidae характерно присутствие мел-
ких крючьев, расположенных по краям присосков. Паразиты птиц и
млекопитающих.
Davainea madagascariensis Blanch.
Длина 25—30 см, ширина до 2,6 мм. Головка около 1 мм диамет-
ром несет толстый втяжной хоботок с двойным венчиком из 90 крючьев,
длиною 0,018 мм. Присоски крупные, округлые. Члеников 500—700;
большая часть их коротка: зрелые проглоттиды длиною около 2 мм,
при ширине в 1,4 мм. Все половые отверстия по одной стороне строби-
лы. Семенников в каждом членике около 50. Матка из перепутанных
трубочек распадается на капсулы с яйцами внутри. Последние круг-
лы—-40 у диаметром; оболочка яйца стекловидно-прозрачна; внутрен-
ний диаметр яйца—15 рч наружная оболочка с двумя нитевидными
отростками. Найден в кишечнике детей туземцев на Мадагаскаре, в
Бангкоке и Гвинее.
Жизненный цикл неизвестен. Промежуточным хозяином пред-
положительно считаются тараканы рода Periplaneta.
Патогенность неизвестна.
Диагноз по нахождению в faeces яиц или члеников с характер-
ной маткой.
Профилактика неизвестна.
Род Raillietina Fuhrm.
Raillietina (?) asiatica Linst.
Синонимы: Taenia asiatica Linst., Davainea asiatica Braun.
Известна по единственному безголовому экземпляру, добытому Ан-
гером в Ашхабаде.
Стробила из почти 750 члеников длиною 298 мм при наиболь-
шей ширине проглоттид — в 1,78 мм. Половые отверстия расположены
все по одной стороне ленты паразита. Матка распадается на 60—70
больших капсул с яйцами. Описание Линстова недостаточно. Необхо-
димо новое изучение этого вида, который вторично, к сожалению, ни-
кому больше не попадался.
Семейство Hymenolepididae
Паразиты кишечника млекопитающих и птиц, дающие сборное се-
мейство, изобилующее видами. Матка мешковидной формы.
Род Hymenolzpis Weinl.
Hymenolepis папа V. Sreb.— цепень карликовый
Цепень карликовый. Как мышь мала по сравнению с сло-
ном, так и цепень карликовый ничтожен по своим размерам против
лентеца широкого. Длина первого, считая всю стробилу, колеблется от
I до 4,5 см (рис. 241) при наибольшей ширине 0,7—0,9 мм. Шарооб-
разная или несколько удлиненная головка (шириною 0,25—0,32 мм)
несет короткий втяжной хоботок с одним венчиком из 20—24 крючьев,
306
СПЕЦИАЛЬНАЯ ЧАСТЬ
длиною 0,15—0,17 мм. Каждая из четырех присосок
шириною 0,08 мм. Довольно длинная шейка — вдвое
тоньше головки. Члеников в стробиле 100—200 штук.
Передние очень короткие, задние же достигают
0,15—0,17 мм ширины их0,8—0,9 мм длины. Половые
отверстия лежат все на левой стороне стробилы.
В члениках, ближайших к головке, развиваются
сначала мужские половые органы, состоящие из трех
округлых семенников, лежащих у заднего края про-
глоттиды. У переднего ее края помещается булаво-
видный мешок копулятивного органа; кзади от него
виден открывающийся в общее углубление семе-
приемник, своим слепым концом граничащий с про-
дольной осью членика (рис. 245).
Яичник парный, перетянутый посредине; позади
его тонкой средней части лежит округлый желточник.
Матка, по мере заполнения яйцами, которых бывает
в каждом членике до 140, принимает вид неправиль-
ного мешка, вытесняющего все органы, кроме семе-
приемника и копулятивного органа.
Яйца круглые или широкоовальные (табл. VI,
рис. 9). Размеры их показываются различно:
40—46 X 30—48 у, Онкосфера диаметром 16—19 у,
а по другим авторам 29—30 у. Яйцо одето двумя
прозрачными оболочками, между наружной и7 внут-
ренней идут извивающиеся нити (филаменты), отхо-
дящие от утолщенных вздутых краев внутренней обо-
лочки. Крючья зародыша 15—16 р длины. Яйца цеп-
ня карликового гораздо нежнее, чем у других цестод.
Паразитирует в тонких кишках человека, который
является и главным промежуточным хозяином кар-
ликового цепня.
Жизненный цикл (рис. 243, 244). Из яиц
паразита в кишке человека вылупляются зародыши,
которые проникают в сосочки кишки задней части
ileum и превращаются в финнозную стадию цисти-
церкоидной формы. Через некоторое время цистицер-
коид выпадает в просвет кишки, присасывается к ее
стенке и превращается в ленточную глисту. Яйца
появляются в faeces уже на 19-й день после зараже-
ния человека. Будучи выброшены с faeces, они могут
быть заглочены людьми с водой, немытыми овощами
и др.
У мышей в кишечнике живет Hymenolepis murina,.
морфологически тождественная с Н. папа. Н. murina
известна также под наименованиями 77. fraierna,
Н. папа, var fraierna. И по сие время еще не ре-
шен вопрос о соотношениях этих ‘паразитов; опыты
одних авторов показали, что мышей можно заразить
карликовым цепнем (Saeki, Woodland); другие не
смогли заразить крыс скармливанием яиц Н. папа от
человека, тогда как эти же крысы успешно инвази-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
307
ровались яйцами Hymenolepis murina, взятыми от мышей. Таким обра-
зом вопрос о видовом тождестве Н. папа и Н. marina требует дальней-
шей разработки.
Яйца цепня карликового, скормленные белым мышам, лишь в
0,8% дают цистицеркоидов, которые располагаются в задней половине
тонкой кишки. На отмытой от слизи вскрытой кишке цистицеркоиды
просвечивают на слизистой в виде белых точек, видимых простым гла-
зом (А. Бадалян).
Белые крысы в опытах с заражением их Н. murina давали 80%
заражения в возрасте от IV2 до 3 месяцев и лишь 12,1% в возрасте
Рис. 242 — Яйцо цепня
карликового.
По Фауст у.
Рис. 243— Цикл развития цепня карлико-
вого (Hymenolepis папа) в кишке мыши.
1 — взрослая глиста; 2—3 -отрывающиеся членики;
1—яйца; 7—финка; 6—8— развитие ленточной
глисты. По Фаусту.
свыше трех месяцев. При аналогичных условиях белые крысы заража-
лись Н. папа лишь в возрасте до одного месяца (Данскер и Пик);
малая заражаемость взрослых крыс объясняется ф-актором возрастной
резистентности. Весьма возможно, что Н. папа и Н, murina окажутся
биологическими видами.
Патогенность цепня карликового в ряде случаев бывает вы-
ражена резко: симптомы интоксикации, нервные расстройства (кон-
вульсии, эпилептические припадки, бессонница), боли в животе, иногда
поносы или запоры, исхудание и др. Эозинофилия бывает часто выра-
жена. Цепень карликовый обычно паразитирует во множестве (извест-
ны случаи нахождения около полутора тысяч экземпляров у больного),
но, как и у других глист, интенсивность заболевания не стоит в прямой
пропорции к количеству глист, паразитирующих у данного больного.
Диагноз ставится по нахождению яиц в faeces.
Лечение. Экстракт папоротника, тимол; Необходимо иметь в ви-
ду, что лечебными процедурами затрагиваются ленточные стадии цепня
карликового, которые в дальнейшем изгоняются; в то же время ци-
стицеркоиды, лежащие в толще ворсинок, остаются нетронутыми; из
них развиваются новые партии ленточных глист, которые должны стать
объектом повторной терапии. Следовательно, недели через 1%—2 лече-
ние надо повторить и последующим микроскопическим обследованием
убедиться в окончательном достижении положительных результатов.
308
СПЕЦИАЛЬНАЯ ЧАСТЬ
Профилактика. Источником заражения цепнем карликовым
является человек, рассеивающий яйца этой глисты и загрязняющий
ими почву и воду. Заражение может происходить при тесном контакте
здорового с зараженным, например няньчание маленьких детей более
старшими, подмывание и вытирание запачкавшегося фекалиями ребен-
ка и др. Кроме того, яйца могут быть заглочены с водой, зеленью и
иными сортами загрязненной пищи. Наконец, немаловажное значение
Рис. 244 — Финка Hymenolepis
папа—I, m,h—B сосочке кишки
белой мыши на разрезе.
I По Е. Roman. ,
Рис. 245 —Проглоттида цепня
.карликового.
t — семенник; о — яичник; d — жел-
точ и*; Ьс — мешjK копулятивного
органа; vs— семенной пузырек.
По Скрябину иШульцу. ;
имеет самозаражение (аутоинвазия), когда человек собственными ру-
ками заносит в рот выделяемые им самим яйца паразита. Не должна
быть исключена возможность заражения людей через посредство гры-
зунов (мышей и крыс), которые могут быть носителями.
Личная профилактика. Соблюдение общих гигиенических
правил при питье и принятии пищи; мытье рук перед едой; освобожде-
ние от глист, если таковые уже есть.
Общественная профилактика. Пропаганда гигиениче-
ского содержания детей; охрана воды от загрязнения фекалиями, уни-
чтожение крыс и мышей; охрана продуктов от загрязнения их пометом
этими грызунами.
Цепень карликовый паразитирует преимущественно у детей, но
встречается (в гораздо меньшем проценте случаев) и у взрослых.
Цепень карликовый считался редким паразитом человека, однако
его все чаще находят при массовых обследованиях населения СССР;
распространен широко, особенно на юге и в средней полосе СССР.
Процент зараженности им колеблется в пределах до 2—3, известны
редкие примеры более частого нахождения этого паразита, в частно-
сти в Армении, Узбекистане и др.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
309
Hymenolepis diminuta Rud.— цепень крысиный
Синонимы: Taenia diminuta Rud., Taenia flavopuncfata W einland.
Цепень крысиный. Длина 10—60 см (при наибольшей ширине
3;5—4,0 мм). Головка очень маленькая, булавовидная или в форме куба
с округленными углами; диаметр ее 0,2—0,5 мм. На переднем конце
головки глубокая ямка со слабо вытягивающимся хоботком, лишенным
крючьев. Присосок четыре; они в виде глубоких ямок лежат близ пе-
реднего края головки. Шейка вдвое уже последней; длина ее около
0,5 мм. Члеников в стробиле 600—1000; они очень короткие; последние
из них достигают обыкновенно 0,75 мм длины.
В зрелых члениках — три семенника (рис. 246). Перезрелые про-
глоттиды отделяются от.стробилы и частично распадаются, причем яйца
паразита, освобождаясь, попадают в faeces.
Рис. 246 — Hymenolepis diminuta — це-
пень крысиный.
Две проглоттиды, / — семенник, О — яичники;
D — желточники; R— семеприемник; Rs — се-
менной пузырек.
По Скрябину и Шульцу.
Рис. 247 — Туловище блохи
с цистицеркоидами Hymenole-
pis diminuta.
а — желудок блохи; с — цистицер-
коиды. По Ж у а й о.
Яйца коротко-овальные или круглые. Желтоватая наружная обо-
лочка неясно исчерчена ,(табл. VI, 8); размеры яиц колеблются в преде-
лах 49—69 уь ширины и 59—86 уь длины. Зародыш (онкосфера) 18—
36 р, одет двойной оболочкой; наружная зародышевая оболочка не-
сколько выдается у полюсов.
Паразитирует в переднем отделе ileum крыс и мышей. Время от
времени встречаются случаи гостепаразитирования цепня крысиного
у человека, преимущественно у детей.
Цикл развития. Яйца паразита, рассеиваемые крысами и мы-
шами, проглатываются различными насекомыми, живущими в ме-
стах обитания этих грызунов или в местах, где находятся продук-
ты, привлекающие к себе крыс и мышей. Таковы, например, продукто-
вые склады, амбары, кладовые. Здесь могут быть личинки и imago
мучного хрущака (Tenebrio molitor), других жуков чернотелок — Akis
spinosa и Scaurus striatus, гусеницы мельничной огневки (Asopia farina-
lis), уховертки, тараканов черных (Periplaneta orientalis) и прусаков.
(Blatella germanica).
В норах крыс яйца Н. diminuta пожираются личинками крысиных
блох Ceratophyllus fasciatus и Xenopsylla cheopis.
В кишечнике этих промежуточных хозяев яйца вскрываются; осво-
бодившиеся зародыши проникают сквозь стенку кишечника в полость
тела, где превращаются в цистицеркоиды ,(рис. 247). Крысы и мыши,
пожирая блох и других перечисленных выше насекомых, получают
цистицеркоиды, которые в кишечнике окончательного хозяина развива-
ются в ленточных глист.
31G СПЕЦИАЛЬНАЯ ЧАСТЬ
Редкие случаи паразитирования Н. diminuta у человека также яв-
ляются следствием проглатывания промежуточных хозяев (вероятнее
всего блох) с цистицеркоидами цепня крысиного.
Патогенное действие для человека изучено очень мало в
виду редкости случаев, подвергавшихся обследованию. Отмечают неко-
торое истощение, поносы, боли в животе, нервные расстройства. В теле
цейня крысиного открыты гемолизины, могущие влиять на кровь хозяев
этой глисты. Диагноз ставится по нахождению яиц паразита в faeces.
Лечение — экстракт папоротника. Отмечают также случаи произ-
вольного выхождения из кишечника или выхождения после дачи слаби-
тельного без противоглистных средств.
Профилактика. Человек заражается при поедании таких ве-
ществ, в которых могут оказаться живые цистицеркоиды цепня крыси-
ного в их промежуточных хозяевах, например дурно испеченный хлеб
из муки, пораженной амбарными вредителями. Однако такой источник
заражения практически играет ничтожную роль. Плохо испечен-
ный хлеб, в котором остаются живые цистицеркоиды, должен быть
настолько сырым, что едва ли он будет съеден, если исключить крайние
случаи голода, истерию или питание сырыми продуктами у полудиких
пародов. В наших условиях, принимая во внимание заражаемость этим
паразитом даже грудных детей (наше неопубликованное наблюдение),
источник заражения следует искать среди блох. Кошка, пожирая крыс
и мышей, может заполучить от них крысиных блох, с цистицеркоидами
Н. diminuta и в свою очередь может передать их человеку. В этом слу-
чае пути заражения детей цепнем крысиным будут аналогичны опи-
санным выше у Dipylidium caninum (стр. 304).
Меры профилактики те же, что и для Dipylidium caninum; кроме
того, надзор за продуктами (охрана от амбарных вредителей, борьба с
домовыми грызунами, термическая обработка пищи).
Семейство Taeniidae
Большей частью крупные ленточные черви, живущие в кишечном ка-
нале человека и домашних животных (плотоядных и всеядных). Личи-
ночные формы, называемые пузырчатыми глистами (цистицерк, цпсти-
церкоид, ценур и эхинококк), являются тканевыми паразитами млеко-
питающих животных (травоядных и всеядных) и человека.
Головка с четырьмя присосками, невооруженная или имеющая
крючья, в виде венчиков. Семенников много, матка — в форме продоль-
ного ствола с боковыми ветвями.
Род Taenia L.
Taenia solium L.' — цепень вооруженный
Стробила 2—3 м длины, в исключительных случаях удлинена до
8 м. Головка 1 мм диаметром при 2—3 мм длины. Ее нередко пигмен-
1 Solium происходит от сирийского слова schuschl, арабского susl или sosl, что
означает — цепочка; в латинизированном написании sosl превратилось в solium. В
литературе solium часто трактуется как производное латинского слова solus, что
означает—единственный. Отсюда установление неправильной связи между таким
названием и якобы типичным паразитированием цепня вооруженного по одному
экземпляру в кишечнике человека. Между тем, по существу дела, solium должно
означать «цепочечный», что вполне соответствует форме цепня, состоящего из
множества проглоттид (Лейкарт).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
311
тированный хоботок несет два ряда крючьев (22—28—32 штуки); пе-
редний ряд их длиною 160—180 задний короче— НО—140 р. Позади
и кнаружи от хоботка лежат четыре полушаровидных присоски, каж-
дая диаметром около 0,5 мм. Шейка длинная до 1 см. Члеников в
стробиле около 900. Ширина передних члеников больше их длины; на
расстоянии приблизительно метра от головки членики квадратной фор-
мы; в этой части глисты происходит оплодотворение проглоттид. Шири-
на зрелых члеников раза в два меньше их длины. Неправильно череду-
ющиеся половые отверстия лежат по бокам члеников. Старые задние
проглоттиды отрываются кусками по 5—6 штук; активным движением
они не обладают.
Рис. 248 — Цепень вооруженный Taenia solium.
Зрелый членик.
ov — яичник; ov. ас—добавочная долька яичника; М—место тельца
Мелиса; U — матка; gl.v -желточник; Вс —сумка цпрруса; Vd —
семепровод. По Скрябину и Р. Шульцу.
Полное развитие полового аппарата наблюдается в члениках, распо-
ложенных, начиная, приблизительно, на расстоянии одного метра от
головки (рис. 248).
Мужской половой аппарат состоит из нескольких сот
округлых семенников, разбросанных в виде зернышек по всему членику
в пространстве между боковыми стволами выделительной системы; от
каждого семенника идет тонкий семепровод; семепроводы соединяются
в общий семеизвергательный канал. Последний идет в виде волнооб-
разно извилистого тяжа поперек членика в той его половине, на боку
которой имеется половое отверстие; семеизвергательный канал перехо-
дит в копулятивный орган, лежащий в особом мешечке. Cirrus выво-
рачивается наружу через отверстие, лежащее кпереди от отверстия
влагалища.
Женский половой аппарат образован двумя яичниками
(ои), состоящими каждый из пучка ветвистых трубок, лежащих з зад-
ней трети проглоттиды; яичники лежат по бокам начальной части мат-
ки и идущего параллельно с ней конца влагалища. Для цепня нево-
оруженного характерной отличительной чертой является наличие
третьей добавочной дольки яичника,в виде небольшого пучка веточек,
помещающегося впереди между стволом матки и влагалищем, т. е.
в той половине проглоттиды, которая несет половые отверстия.
Позади главных долек яичника в поперечном направлении лежит
узкий, вытянутый желточник; между ними и срединной частью яичников
видно шаровидное тельце Мелиса.
Кпереди от последнего отходит неветвистая трубка матки, идущая
312
СПЕЦИАЛЬНАЯ ЧАСТЬ
по средней линии членика. По мере заполнения матки яйцами ее ствол
утолщается и дает боковые ответвления, которые в свою очередь вет-
вятся. По мере развития матки и истощения деятельности продуцирую-
щих частей полового аппарата все его органы редуцируются, и в зре-
лой проглоттиде имеется только матка, переполненная яйцами (рис.
249).
. Вторым характерным признаком цепня вооруженного является на-
личие 7—12 боковых веток зрелой матки (рис. 249), считая число ве-
ток по корням их отхождения от продольного ствола матки. Последняя
лишена самостоятельного отверстия. Женское половое отверстие, т. е.
отверстие влагалища, лежит на дне бокового вдавления (половая клоа-
ка). Влагалище идет позади семеизвергательного канала и у своего
внутреннего конца расширяется в семеприемник, открываясь далее в
тельце Мелиса.
Яйца цепня вооруженного шаровидные, с толстой радиально-ис-
черченной скорлупой коричневого цвета (табл. VI, рис. 12); она являет-
ся собственно внутренней — эмбриональной оболочкой яйца, тогда как
наружная нежная оболочка разрушается. Диаметр яйца — 31—38 у,
под его оболочкой просвечивает онкосфера с шестью крючьями; диа-
метр ее — 20 р. Сходны с яйцами цепня невооруженного.
Цепень вооруженный живет только в тонких кишках человека, имен-
но в передней их половине. Космополит. Встречается часто там, где
едят сырую или полусырую свинину, что будет понятно из рассмотре-
ния никла развития этого паразита.
Жизненный цикл. Яйца цепня вооруженного, рассеиваемые с
испражнениями человека, пожираются случайно самыми различными жи-
вотными, но развиваются они в желудке свиньи. Там скорлупа их рас-
творяется, шестикрючная онкосфера освобождается и с помощью своего
вооружения проникает сквозь стенку желудка или кишек в лимфати-
ческие пути или кровеносные сосуды. С током крови или лимфы заро-
дыши проходят через правое предсердие и желудочек, минуют малый
круг кровообращения и через левую половину сердца попадают в ар-
терии большого круга, по которым и разносятся в различные места те-
ла. Осев в мышцах или в соединительной ткани, зародыш сбрасывает
свои крючья и превращается в пузырчатую глисту или финку. Типиче-
ски он имеет вид овоидного пузырька 15 мм длины и 7—8 мм толщины;
пузырек заполнен жидкостью; одна из длинных сторон его впячена
внутрь и оканчивается вывернутой наизнанку головкой, которая снаб-
жена 4 присосками и крючьями.
На все развитие финки цепня вооруженного (рис. 250) требуется
2!/2—4 мес. Цистицерки Г. solium являются тканевыми паразитами и
как таковые не имеют выхода из тела промежуточного хозяина. Кроме
свиньи, играющей первостепенное значение, финки цепня вооруженного
могут развиваться также в диком кабане, в собаке, кошке и в самом
человеке. Окончательная участь финки может быть двоякая. Если про-
межуточный хозяин погибает естественной смертью, то в нем погибают
и финки. Возможно умирание финки в живом еще хозяине, причем та-
кие финки оказываются или творожисто перерожденными (так называе-
мое казеозное перерождение), или же омелотворенными (т. е. пропита-
ны отложившимися солями извести). Мало-мальски точные сроки пере-
живания финок в свинье неизвестны.
Если сырая или полусырая свинина с живыми финками в ней будет
съедена человеком, то пузыри финок могут повреждаться зубами. Если
же финки проглатываются неповрежденными, то в желудке пузырь пе-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
313
реваривается, причем головка частично выворачивается из ввернутой
шейки. Полное выворачивание заканчивается в двенадцатиперстной
кишке под влиянием желчи, которая является главным действующим
в этом отношении на головку агентом (Бутнинг). Приняв нормальный
вид, головка присасывается к стенке кишки и впивается в ее слизистую
оболочку крючьями. Далее начинается обычный процесс почкования
проглоттид, образующих в своей совокупности стробилу различной
длины.
Финноз у человека. Уже было упомянуто, что человек может
стать и промежуточным хозяином Т. solium. Яйца этой глисты вскры-
Рис. 249—Цепень воору-
женный; зрелый членик с
маткой.
Рис. 250 — Финка Taenii
solium с вывернутой го-
ловкой.
Оригинал.
ваются, если как-нибудь попадут в желудок человека. Это может прои-
зойти вследствие различных причин. Яйца могут быть заглочены чело-
веком с загрязненной фекалиями водой, с огородной зеленью или с иной
пищей, в которую они случайно попали. Далее, человек, являющийся
носителем цепня вооруженного, при неряшливости может заразиться
от самого себя, если занесет при посредстве загрязненных рук яйца в
рот и их проглотит. Аутоинвазия может произойти и вследствие физио-
логических причин; при сильной рвоте, под влиянием энергичной анти-
перистальтики и энергичного давления брюшного пресса часть строби-
лы цепня невооруженного может быть выдавлена из кишечника в же-
лудок. Последние случаи особенно чреваты последствиями, так как
в проглоттиде заключается огромное количество яиц. В желудке чело-
века с проглоченными яйцами происходит то же, что и в свинье. Из
них выходят зародыши, мигрирующие по нему и оседающие в самых
разнообразных местах, где из зародышей в обычном порядке развива-
ются. финки.
Случаи заражения пузырчатыми стадиями цепня вооруженного на-
зывают цистицеркозом или финнозом человека.
Заболеванию подвергаются людй преимущественно от 20 до 40 лет.
Отмечались случаи заражения цистицерками младенцев, кормившихся
314
СПЕЦИАЛЬНАЯ ЧАСТЬ
молоком матери. Очевидно, паразиты проникали в них из материнского
организма через плаценту в пупочный канатик еще в эмбриональном
состоянии.
Онкосфера, осев в каком-либо месте тела, превращается в цисти-
церк, требующий для своего окончательного роста 2—3 месяца. Форма
и величина развивающейся пузырчатой глисты зависят от противодей-
ствия, оказываемого окружающими тканями тела. В паренхиматозных
органах финки солитера величиною с горошину, в мышцах они удлинен-
но овальные, в мягких тканях вырастают до лесного ореха, а в мозгу
и его желудочках раздуваются с голубиное яйцо. В некоторых местах
тела, в мозговых оболочках и под кожей ф цистицерки принимают при-
чудливую ветвистую форму (Cysticercus racemosus).
О локализации финок солитера в теле человека можно судить по
данным Vosgien (1911), проследившего около 800 случаев. Из них
372 случая приходится на глаза (120 в сетчатке, 112 в стекловидном
теле, 84 в конъюнктиве, остальные в передней камере, роговице, радуж-
ной оболочке и др.), 230 случаев на нервную систему, 279 в головном
мозгу и мозговых оболочках, 51 в коже и паренхиматозных органах, 28
в мускулатуре.
Остальные случаи приходятся на сердце, легкие, кости и т. д.
У свиней цистицерки также наиболее часто поражают мозг. Мееро-
вич нашел финки в мозгу почти у 40% цистицеркозных свиней.
Общий процент финноза у свиней сильно колеблется, что зависит
и от действительных условий заражения и от высоты санитарно-ветери-
нарного надзора на мясокомбинатах и бойнях.
При своем росте цистицерки вызывают реакцию со стороны прилежа-
щих тканей хозяина, проявляющуюся образованием соединительноткан-
ной капсулы, инфильтрированной эозинофилами. Этой капсулы не бы-
вает, когдау финка развивается в стекловидном теле, желудочках мозга
и мягкой мозговой оболочке. Воспалительные процессы перекидываются
иногда и на сопредельные ткани, вызывая побочные болезненные эф-
фекты, например, хороидит или воспаление радужной оболочки глаза,
гипертрофию миокарда, менингит и т. п. Однако дело может окончиться
и смертью цистицерка, подвергающегося вместе со своей капсулой обыз-
вествлению.
Для описания всех возможных поражений, причиняемых финками
солитера, потребовалось бы очень много.места. Поэтому мы ограничим-
ся лишь общим освещением главнейших возможностей, начав с цисти-
церкоза подкожной клетчатки как наиболее доступного для диагноза
и лечения.
Обыкновенно^ финки во множестве сидят под кожей и в мышцах,
редко встречаясь одиночными экземплярами. Присутствие паразитов ска-
зывается наличием множества плотных, хрящевой консистенции опухо-
лей, величиною от горошины до небольшого ореха. Они легко подвижны
относительно кожи и более связаны с глубже лежащими мышцами. Ве-
личина всех их обычно почти одинакова, форма шарообразная или
овоидная. Опухоли располагаются чаще симметрично на сгибательных
поверхностях конечностей. Они сами по себе не болезненны, и кожа
над ними не изменена.
Болезненные явления при таких заболеваниях разнообразны. Боль-
ной может ощущать боли по ходу нервов конечностей; последние иног-
да даже парализуются, или пальцы на них скрючиваются. Такие симп-
томы обусловливаются механическим действием финок, расположив-
шихся по соседству с крупными нервными стволами.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
315
Лечение заключается в пункции опухолей шприцем и последующим
впрыскиванием в них нескольких капель 1% раствора сулемы. Реко-
мендуют также иссечение финок, что, однако, возможно лишь в единич-
ных случаях. Исследование вырезанной опухоли легко разрешает во-
прос, если в ней имеется ввернутая внутрь головка. Бывают финки и
без головок (ацефалоцисты). Диагносцировать их на срезах труднее,
следует обратить внимание на присутствие сосочков на наружной стен-
ке цистицерка, имеющей типичное паренхиматозное строение, свой-
ственное ленточным червям.
Глаза человека наиболее часто поражаются финками солитера, ко-
торые гнездятся или в самом глазном яблоке, или в прилежащих ча-
стях. Если цистицерк сидит под сетчаткой, он вызывает отслойку ее.
При офтальмоскопировании паразита, развивающегося в стекловидном
Рис. 251,—Финноз мозга человека. Разрез мозговой коры. Оригинал.
теле, он имеет вид голубоватого гладкостейного пузырька с белым
пятном в месте ввернутой головки. Иногда заметны ее изгибающиеся
движения. В глазу вокруг паразита образуется очень нежная, тонково-
локнистая, соединительнотканная капсула. Своевременное оперативное
вмешательство может дать благоприятные результаты; в противном слу-
чае неизбежна потеря зрения и даже глаза. Заболевание глаза финно-
зом тянется обычно 3—4 года, в виде исключения оно бывает очень
длительным (20 лет).
В окружности глаза цистицерки поражают подконъюнктивную ткань
преимущественно в молодом возрасте. Цисты растут быстро и, достиг-
нув значительной величины, могут смещать глазное яблоко. По расще-
плении цист паразит легко удаляется из них. Реже финки локализуют-
ся в полости орбиты; в таком случае они причиняют экзофтальм (вы-
пячивание глазного яблока). ч
Если поражение финнозом глаз ведет в тяжелых случаях к слепоте
человека, то развитие цистицерков в мозгу (рис. 251, 252, 253) имеет
обычно смертельные последствия. Встречая очень слабое противодей-
ствие при своем росте со стороны мягкой нервной ткани, финки дости-
гают в мозгу наибольшей величины — до размеров голубиного яйца
(рис. 253). При этом они давят на мозг и сжимают его артерии; послед-
нее ведет к размягчению участка, который орошался соответствующим
сосудом. Симптомы заболевания бывают различными, в зависимости от
того, где именно в мозгу расположился паразит. По общим болезненным
316
СПЕЦИАЛЬНАЯ ЧАСТЬ
проявлениям легко заподозрить мозговую опухоль. Правильный диагноз
становится возможным, если одновременно имеются финнозные кожные
припухлости. Поэтому при соответствующих мозговых симптомах реко-
мендуют тщательную пальпацию всей кожи больного. Из болезненных
расстройств отмечались головные боли, параличи, эпилептические явле-
ния, судороги, расстройство умственных способностей, потеря памяти,
сонливость, рвота и многое другое. Наряду с этим известны случаи,
когда цистицеркоз мозга не проявлял себя никакими специфическими
симптомами и был обнаруживаем только при вскрытии.
Рис. 252 — Финноз мозга человека. Микроскопический срез.
А — мозговая ткань; В — реактивная, точечная зона вокруг финки; Е — стенка пузыря;
D — ввернутая головка. Справа вверху разрез ввернутой головки. Оригинал.
Финки, развивающиеся в основании мозга, как указано выше, обыч-
но имеют разветвленную форму.
Предсказания при финнозе мозга для больного неблагоприятны;
болезнь ведет почти неминуемо к смерти; в виде исключения она может
тянуться очень долго (до 23 лет).
В большинстве случаев финноз у человека остается при жизни не-
распознанным и обнаруживается лишь в качестве «случайных» нахо-
док при вскрытиях; число случаев цистицеркоза сильно возрастает,
если при аутопсии специально ищут финок. Хаютин и Шухат указы-
вают, что цистицеркоз человека чаще выявляется в судебно-медицин-
ской практике, особенно в связи со случаями внезапной смерти при
финнозе. Поэтому для уловления случаев цистицеркоза и выяснения
его распространения следует использовать статистические данные су-
дебно-медицинских моргов. Автору известен случай, когда возникло
дело по обвинению в отравлении. Быстрая же смерть погибшего зави-
села от финноза мозга, что было установлено судебно-медицинским
вскрытием.
Болезнетворное действие. Патогенное действие на чело-
века финок цепня невооруженного уже рассмотрено выше. Ленточная
глиста обычно встречается поодиночке и не обращает на себя внима-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
317
Рис. 253 — Вскрытая финка Taenia so-
lium из мозга человека. Видна впячен-
ная внутрь шейка с головкой внутри.
Оригинал.
Для их отыскания берут
ния, почему паразитирование протекает незаметным. В других случаях
наблюдается хроническое расстройство пищеварения с перемежающи-
мися поносами и запорами, нервные расстройства и др. Что цепень
вооруженный выделяет токсины, в том сомневаться не приходится.
Водные экстракты из Т. solium, впрыснутые в вену кролику, вызывают
повторную дефекацию, переходящую в понос. При более крупных дозах
этого же экстракта наблюдаются
беспрерывный понос и гибель кро-
лика. Экстракты из глисты дей-
ствуют и на кровь, вызывая в из-
вестных дозах эозинофилию. Что
цепень вооруженный может вызы-
вать, подобно лентецу широкому,
злокачественное малокровие, пока-
зал еще С. П. Боткин, но это при
Т. solium бывает гораздо реже, чем
при Diphyllobothrium latum. Все же
наибольшее значение Т. solium то,
что он может быть причиной фин-
ноза человека.
Диагностика ленточной
формы 7\ solium. Точная диагности-
ка наличия в кишечнике человека
цепня вооруженного может быть
произведена нахождением в фека-
лиях отошедших перезрелых прог;
слизеобразные комки из faeces, расправляют их в воде и рассматри-
вают между двумя предметными стеклами. В проглоттидах можно ви-
деть простым глазом ствол матки и ее ветви.
При нахождении яиц в faeces можно утверждать только то, что
они принадлежат или цепню вооруженному, или невооруженному
Т. saginata, так как надежных распознавательных признаков между
яйцами этих глист в действительности не существует. Поэтому при
всех описаниях и в статистических материалах необходимо указывать,
на основании каких именно данных устанавливается в каждом отдель-
ном случае факт паразитирования Т. solium в кишечнике человека.
Пузырчатые формы. Финки Т. solium диагносцируют у сви-
ньи, в свином мясе (рис. 254) и в человеке. Диагностика бывает при-
жизненная или посмертная. Надежнее всего последняя.
Свиные туши, по закону, должны подвергнуться санитарно-ветери-
нарному осмотру на финноз и трихиноз, после чего штемпелюются и
выпускаются в продажу, если туша свободна от паразитов. Финки за-
легают между пучками мышц и хорошо видны на их фоне в виде
горошинок.
Финнозная свинина получила образное народное наименование:
мясо «с крупкой». Кроме мышц, финки бывают в сердце, причем они
видны в форме овальных полупрозрачных пузырьков с беловатым утол-
щением внутри. Если финку осторожно сдавить пальцами, то можно
заставить головку вывернуться наружу.
Практически можно бывает определить, живы ли финки или нет.
Для этой цели выпущенные финки кладут в желудочный сок собаки,
нагретый до 37—38° С. Если финки живы, то головки их полностью
или частично вывернутся.
318
СПЕЦИАЛЬНАЯ ЧАСТЬ
Посмертная диагностика финноза у человека производится при
вскрытии трупа. Для нахождения финок в мозгу послойно разрезают
головной мозг на поперечные пласты толщиною около 1 см. Финки
бывают в сером веществе или в оболочках мо-зга.
Обследованию на финок подвергают энуклеированные глаза и части,
удаленные при хирургических операциях или биопсиях. Для исследо-
вания приходится прибегать к заливке в парафин или в целлоидин и
изучению окрашенных срезов. Особенно ценны были бы серии после-
довательно расположенных срезов. На препаратах видны разрезы при-
Рис. 254 — Финнозное мясо свиньи. Оригинал.
сосок; принадлежность финки цепню вооруженному устанавливается
при нахождении на срезах крючьев. Крючья могут отпадать, что в та-
ких случаях затрудняет точную диагностику. В паренхиме ввернутой
шейки иногда бывает видно много известковых телец, которые на окра-
шенных гематоксилином препаратах неопытным наблюдателем могут
быть признаны за яйца. Такие трубые ошибки проникли в патолого-
анатомическую литературу и привели к невероятным заключениям,
что в силу лимфоидной конституции больного (!) в финках, паразитиро-
вавших в мозгу, развился половой аппарат и образовалась матка, пере-
полненная яйцами.
Возможно микроскопирование и свежей финки, сдавленной -между
стеклами компрессора. Ищут под микроскопом щетку или венчик
крючьев, наблюдаемая форма которого зависит от плоскости сдавли-
вания. Однако при этом способе финка, как музейный препарат,
погибает.
Прижизненная диагностика финноза у человека весьма затруднена
и во многих случаях вовсе невозможна. Легче заподозрить наличие
финки в глазу при офтальмоскопировании. Также нетрудно- распознать
финку в вылущенных подкожных узлах (срезы и окрашенные препара-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
319
ты). Финноз внутренних органов (например, легких) может быть с
большим или меньшим вероятием установлен при рентгенофотографи-
ровании и рентгеноскопии. Финноз мозга распознают по косвенным
данным наличия цепня вооруженного в кишечнике, финок — под ко-
жей и др.
Рис. 255 — Деревенское отхожее место, доступное для
посещения свиньям, пожирающим faeces. Оригинал.
У свиней финноз можно установить осмотром век и прощупыванием
языка. Цистицеркоз же мышц и внутренних органов при жизни остает-
ся нераспознанным.
Попытки применения для прижизненной диагностики финноза им-
муно-биологических реакций пока не дали еще практически приложи-
мых результатов.
Лечение. Изгнание ленточной формы производится экстрактом
папоротника. Контролировать успешность изгнания отысканием голов-
ки изгнанной глисты или повторным обследованием faeces на яйца
глисты через 3—4 недели после изгнания.
120
СПЕЦИАЛЬНАЯ ЧАСТЬ
Цельные глисты с головкой представляют музейную ценность; их
надо беречь; порванные же стробилы ценны, как материал для практи-
ческих занятий. Поэтому изгоняемых Т. solium надо консервировать
75% спиртом и сохранять.
Профилактика. Личная: общие санитарно-гигиенические пра-
вила принятия пищи и питья. Не есть сырой или полусырой свинины
(например, проба сырого фарша); не употреблять в пищу свинину,
приобретенную из-под полы и не прошедшую через ветеринарно-сани-
тарный контроль.
Общественная: конструирование уборных так, чтобы свиньи
не могли иметь доступа к faeces и поедать их (рис. 255),
Содержание свиней в переносных загородках для недопущения
бродяжничества их и пожирания ими faeces, разбросанных по земле.
Организация строгого ветеринарно-санитарного надзора над выпу-
ском свиных туш с боен и широкая пропаганда по социалистическому
сектору сельского хозяйства и другим кругам населения сведений об
опасности цепня вооруженного для человека, его экономической вре-
доносности и инструктирование в осмотре и браковке свиного мяса.
В крайнем случае допускается использование мало зараженной свини-
ны после надежной термической обработки или после длительного вы-
держивания в холодильниках или на льду.
Непременное уничтожение (например, сжиганием) изгоняемых глист,
если их не приходится почему-либо сохранять (см. выше).
Обязательное глистоизгнание у всех лиц, зараженных цепнем во-
оруженным.
Для профилактики аутоинвазии и предупреждения развития фин-
ноза непременно обследовать faeces на яйца Т. solium или Г. saginata
и выгнать глист перед хирургической операцией с хлороформным нар-
козом, так как при рвоте легко может произойти аутоинвазия. Равным
образом то же необходимо делать с персоналом морской и авиацион-
ной службы (морская болезнь). Является практически редким парази-
том человека космополитного значения.
Приспособление Г. solium и Г. saginata к человеку происходило во
время и в связи с переходом предков человека от растительной к мяс-
ной пище (Joyeux и Baer).
Taeniarhynchus saginatus. Taenia saginata Goeze.— цепень
невооруженный или бычий
Синонимы: Taenia mediocanellata Kuchenm., Taeniarhynchus mediocanellata
Weinland.
Цепень невооруженный или бычий. Стробила 4—10 м
длины. Грушевидная головка 1—2 мм диаметром. Крючьев на ней нет
(рис. 262, В); вместо хоботка лежит рудиментарный присасывательный
диск. Присосок четыре; форма их эллипсоидная при 0,8 мм длины;
они почти всегда пигментированы. Шейка очень растяжима; в вытяну-
том состоянии тонка; в сокращенном — по ширине мало отличима от
головки. Члеников в стробиле свыше тысячи. Половой аппарат начи-
нает закладываться приблизительно с двухсотой проглоттиды. Матка
намечается в члениках, занимающих среднюю часть стробилы. Поло-
вые отверстия неправильно чередуются по бокам члеников. Дли-
на зрелых проглоттид 16—20—30 мм, ширина 5—7 мм. Матка несет
17—35 боковых ветвей. При сокращении они становятся округлыми и
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
321
по форме походят на семема тыквы, что очень типично для невоору-
женного цепня. Зрелые членики отрываются от заднего конца строби-
лы поодиночке, при этом обнажаются концы слепых ветвей матки, яй-
ца выходят наружу и попадают в испражнения. Проглоттиды способны
к движению; они выползают из заднепроходного отверстия, что слу-
чается как днем, так и ночью.
Рис. 256 — Стробила цепня невооруженного — Тaeniarhynchus saginatas
Уменьшено. По Н. X о л о д к о в с к о м у.
’ В процессе длительного самоэксперимента Ж. Щтромом установле-
но, что темп отхождения проглоттид определяется количеством шести-
восьми экземпляров в сутки, 195—238 экземпляров в месяц и до 2 500
проглоттид в год. Количество активно выходящих проглоттид раза в три
меньше числа пассивно выносимых с фекалиями оторвавшихся члени-
ков. Активное выхождение проглоттид наблюдается не каждый день;
в месяц выпадает 6—12 таких дней. Общее количество пассивно и
активно вышедших за сутки проглоттид достигает 28. С 5 до 12 ча-
сов утра выделения их, как правило, не бывает. 'Максимум активно
выходящих члеников приходится на вторую половину дня (15—20
322
СПЕЦИАЛЬНАЯ ЧАСТЬ
часов — рис. 257). При обратной перемене режима жизни хозяина (по-
кой днем, работа и обед ночью) резко меняется и время активного вы-
хода члеников (максимум — утром; пауза — вечером).
По строению гермафродитного аппарата цепень невооруженный
очень сходен с Т. solium, отличаясь от него отсутствием третьей мал?й
(добавочной) дольки яичника (рис. 258). Проглоттиды Т. saginatus со
зрелой маткой отличаются большим количеством боковых ветвей, кото-
рых насчитывается от 17 до 30 (рис. 259).
Яйца, как и у Г. solium, шаровидные или слегка овальные; их раз-
меры 30—40 р длины и 20—30 ширины (табл. VI, рис. 12). Интересно
Рис. 257-2 Сводная кривая распределения активно вышедших из
кишечника человека проглоттид цепня невооруженного за 10 меся-
цев наблюдений по часам суток (нижний ряд цифр). Цифры у
кривой показывают количество самостоятельно вышедших члени-
ков в определенный час за все время. По Ж. Ш т р о м у.
отметить, что вследствие неравномерного сокращения мышц проглотти-
ды яйца могут выдавливаться из основного ствола матки и перемещать-
ся в ее концевые ветви, подходящие к переднему краю членика. Пр»
открывании таких проглоттид яйца получают выход наружу. Благодаря
этому явлению можно наблюдать совсем «пустые» проглоттиды, в кото-
рых вовсе не осталось яиц (рис. 261) (Дюжарден, Павловский, Ершов);
активно выходящие проглоттиды выдавливают через передний край1
членика весь или почти весь запас яиц, выхождение которых происхо-
дит через тонкие «каналы» (рис. 260), являющиеся спавшимися опу-
стевшими передними ветвями матки (Штром). Количество* яиц, по
Штрому, в одном членике достигает 175 тыс. При нахождении одной
особи паразита за сутки может быть выделено до 4 900 тыс. яиц, а за
год до 440 млн. Отделение зрелых проглоттид начинается у паразита
трехмесячного возраста, когда длина его достигает примерно 6—7 м..
Продолжительность жизни цепня невооруженного более десяти лет.
Нередко у Т. saginatus наблюдаются аномалии. Некоторые гли-
сты обладают темным цветом; такая окраска зависит от действия на
паразита различных лекарств, как то: висмута, солей ртути, свинца,
железа и т._ п. Под именем Т. confusa описана форма цепня, у которого
местами не было наружного разделения ленты на членики. Т. fenestrata
отличается дырявостью проглоттид. Бывают уродливые формы, обра-
зовавшиеся как бы слиянием двух особей (Т. hybrida)\ головка их не-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
323
сет шесть присосков, а стробила имеет три выдающихся продольных
ребра. Все эти формы являются ничем иным, как Т. saginata, почему и
не заслуживают особого наименования.
Ленточная форма глисты живет в тонких кишках человека, который
является единственным хозяином и для цепня невооруженного. Космо-
полит. Распространение этого паразита зависит от наличия промежу-
точных хозяев, возможности их заражения от человека и от способов’
обработки мяса. В некоторых местах зараженность населения дости-
гает 54% (Ойротия).
Рис. 258 — Проглоттида Таеп. saginatus с развивающейся
маткой.
Е — выделительные каналы, Т — семенники, — мешок копулятив-
ного органа, влагалище, О — яичники, Н — скорлуповые желе-
зы, D — желточники, U — матка. Из Е. Павловского^
Рис. 259 — Членик Таеп.
saginatus с маткой.
Оригинал.
Жизненный цикл. Промежуточным хозяином является круп-
ный рогатый скот; из проглоченных им яиц глисты в мышцах его тела
развиваются финки. Кроме коров и быков, цистицерки цепня невоору-
женного могут быть у овец и козлят, а также у ламы. Для развития
финок из проглоченных крупным рогатым скотом яиц требуется от
3 до 6 месяцев. Человек заражается цепнем невооруженным, съедая
мясо с живыми финками паразита. В отношении частоты распростра-
нения Т. saginatus дает картину, обратную Т- solium. Цистицерки в мя-
се коров и быков обнаруживаются на бойнях очень редко, тогда как
ленточная глиста является обыкновенным паразитом человека. При
специальном методе обследования туш на финноз послойным разреза-
нием всех мышц и органов на ломти толщиной в 0,5 см Н. Бородину
удалось доказать высокий процент зараженности боенского скота, по-
ступавшего на бойню в Азербайджане: скот из Ирана был заражен в
53,8%, из Азербайджана — в среднем в 45,4% и из Дагестана —
в 12,9%. Количество же цистицеркозных мясных продуктов на одном
только Мясокомбинате достигало в год 2400 т. По исследованиям того
же автора, наиболее часто были заражены цистицерками язык, мышцы
шеи, мышцы сердца и др.
Однако такой метод, очень точный для исследовательской работы,
не применим в общей ветеринарной практике. В связи с этим Н. Бо-
324
СПЕЦИАЛЬНАЯ ЧАСТЬ
родиным с положительным результатом испытана аллергическая диаг-
ностика цистицеркоза рогатого скота. В качества аллергена берется
порошок из растертых свежих финн, масса которых высушивалась в
эксиккаторе при 40° С до постоянного веса. Брались также альбуми-
новая и глобулиновая фракции белков цистицеркоидов. Взвесь порошка
в физиологическом растворе
Рис. 260 — Тaeniarhynchus saginatus. ~ Край
членика, переполненного зрелыми яйцами:
сверху—на тотальном препарате, снизу —
на срезе. Видны каналы, служащие для вы-
хождения яиц при отрыве и ползании чле-
ника. По Ж. Ш т р о м у.
вводилась внутрикожно в ко-
личестве 0,005 г в 0,5 см3 фи-
зиологического раствора в
подхвостовую кожную склад-
ку. Через 3—4 часа на месте
инъекции развивается харак-
терная реакция в форме горя-
чего опухания и отека; рас-
цвет реакции наблюдался через
12 часов после инъекции; че-
рез два-три дня следов от ре-
акции не оставалось.
Распространен широко; в
некоторых местах является
доминирующим паразитом. На-
пример, в Иркутске Т. sagi-
natus найден у 42% обследо-
ванных, а в Армении—даже
у 50—60%.
Патогенное значение
Т. saginatu SMenbuie, чем Т. so-
lium, уже по одному тому,
что цепень невооруженный в
финнозной степени вовсе не
живет в человеке- Правда, в
литературе описаны случаи
нахождения у человека финок
без крючьев на ввернутой го-
ловке. Однако, как это мы ви-
дели выше, доказать действи-
тельное отсутствие крючьев
можно только просмотром се-
рий срезов через финку, что
обычно не производится. В
редких случаях, повидимому,
финноз бывает и’отТ. saginata.
Fontan описал случай финноза человека с локализацией пузырчатой
стадии в молочной железе с одновременным нахождением цепня не-,
вооруженного в кишечнике той же больной. Второй точно прослежен-
ный случай финноза у человека (скелетные мышцы) опубликован )D. de
Rivas в Западном полушарии.
Болезнетворное значение ленточной формы цепня невооруженного
в общем сходно с Т. solium (см. выше).
Диагностика. Надежное распознавание вида может быть про-
изведено подсчетом ветвей матки в проглоттидах, взятых из faeces или
выползших из анального отверстия. Яйца в faeces неотличимы от яиц
Т. solium. Иногда у них сохраняется материнская эмбриональная обо-
лочка с характерными полярными нитями (филаментами). При отсут-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
325
стейш таковых диагноз ограничивается только установлением принад-
лежности яиц представителю семейства Taeniidae.
При трехлетнем самоисследовании носителя Т. saginatus яйца ми-
кроскопически не обнаруживались в фекалиях в 30% наблюдений;
Рис. 261—Тaeniarhynchus saginatus. Членики частично или пол-
ностью освободившиеся от яиц. По Ж. Ш т р о м у.
исследование перианального соскоба было всегда положительным
(Ж. Штром).
Лечение. Экстракт папоротника с обычным контролем успешно-
сти изгнания.
Профилактика. Необходимо стремиться к предохранению че-
ловека от заражения цепнем невооруженным и к охране скота от за-
получения финноза. Первое достигается организацией боенского надзо-
ра над выпускаемым мясом, причем финноз-
ные туши или бракуются в зависимости от
степени заражения, или выпускаются в пред-
варительную обработку; из способов обез-
вреживания мяса важное значение имеет вы-
держивание его на холодильнике в течение
21 дня, так как за этот период финки теряют
жизнеспособность. Надежная термическая
обработка мяса является средством личной
профилактики, причем необходимо совер-
шенно исключить пользование сырым мясом,
хотя бы даже в форме кухонной пробы.
Коровы получают финноз, когда случайно
проглатывают яйца цепня невооруженного;
способ образования финок аналогичен тому,
Рис. 262—Головка Taenia
solium (А) и Таеп. sagina-
tus (В). Из П а р к е р - Гае-
ве л л я.
что происходит в сви-
нье при поедании яиц цепня вооруженного. Яйца глисты могут
попадать на пастбища при дефекации людей на, землю. Яйца мо-
гут смываться дождем и, в зависимости от рельефа местности и ха-
рактера наземного ландшафта, переноситься на луг или в водоемы.
Наконец, коровы сами находят дорогу к плохо содержимым уборным,
куда их привлекает потребность в солях; коровы пьют мочу, выте-
кающую из отхожего места и при этом получают полную возможность
проглатывания яиц цепня невооруженного. Следовательно, вопросы
зоогигиенического содержания скота, конструирования гигиенических
326
СПЕЦИАЛЬНАЯ ЧАСТЬ
отхожих мест, правильное пользование ими ,и аккуратное содержание
их имеют и в профилактике заражений цепнем невооруженным пер-
венствующее значение.
Taenia infantis Bacigalupo
Описан в Аргентине. У пятилетней девочки изгнана глиста длиной
30 см, состоявшая из 208 члеников с неправильно чередующимися по-
ловым|и отверстиями; головка 3 мм диаметром с двойным венчиком из
35—40 крючьев; самые крупные из них 0,041 мм, а самые малые
0,026 мм. Яйца 35—40 (и.
Хотя эта глиста и очень похожа на Taenia crassicollis кошки, Baci-
galupo считает ее новым видом.
Taenia crassicollis Rud.
Является обычным паразитом кишечника кошки. Пузырчатая ста-
дия, известная под именем Cysticercus fasciolaris, обитает в печенр
мышей и крыс, являющихся промежуточными хозяевами этого парази-
та. Некоторые паразитологи считают возможным паразитирование
Т. crassicollis и у человека на том основании, что сырая крысиная и
мышиная печень употребляется в качестве народного средства при за-
держке мочи (Ютландия), но прямых подтверждений этого предполо-
жения пока еще нет. Интересно отметить, что при экспериментальном
заражении крыс онкосферами Т. crassicollis в ряде опытов удалось по-
лучить саркому печени, разраставшуюся от капсулы, окружающей ци-
стицерков в печени.
Отмечено также увеличение количества миэлоидных клеток в селе-
зенке мышей и крыс как естественно зараженных Cysticercus fasciola-
ris, так и после инъекции им эмульсии из таких же цистицерков: в
аналогичных условиях опыты с хомяком дали отрицательные результа-
ты (Hoeppli и Feng). Космополит.
Taenia hydatigena Pallas
Ленточная глиста живет в кишечнике собаки. Пузырчатая стадия,
известная под именем тонкошейного цистицерка Cysticercus taenuicollis,
встречается в брюшной полости на брыжейке или сальнике овец, реже
свиней, коров, оленей. У человека не бывает ни в какой стадии разви-
тия, но упоминание об этом паразите необходимо для уточнения дифе-
ренциальной диагностики между Cysticercus taenuicollis и эхинококком
при осмотре мяса перед выпуском его в дело. Пузыри С. taenuicollis
(рис. 263) тонкостенны, в них просвечивает по одному выпячивающе-
муся внутрь бугорку, заключающему в себе впяченную внутрь головку.
Если пузырь сдавить, то из него выворачивается тонкая длинная шей-
ка с головкой. Пузырь эхинококка или не имеет никаких хорошо замет-
ных бугорчатых утоищений, иди заключает в себе еще другие дочерние
пузыри. Эти различия важно иметь в виду при осмотре туш для бра-
ковки зараженного мяса.
Taenia confusa Ward.
Стробила 5—8 м длины, состоит из 500—800 члеников, длина боль-
шей части которых, особенно задних, больше ширины. Головка 1,5—1 мм.
Задние членики с развитой маткой 25—33 мм длины и 3,5—9 мм ши-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ, ЧЕЛОВЕКА
327
рины. Ветвление матки весьма неправильное, причем слепые концы (ве-
точек матки значительно* расширяются. Яйца 33—42 р.
Описано по единичным экземплярам в Северной Америке ,и в Север-
ной Нигерии.
Род Multlceps Goetze — многоглавы
Головка вооружена крючьями. Пузырчатая форма глисты типа
Coenurus (ценур).
Multlceps multlceps Hall.— цепень-мозговик
Синонимы: Taenia npulticeps Leske. Coenurus cerebralis Rud., Taenia coenurus
Kiichenm.
Цепень-мозговик. Стробила 40—60—80 (100) мм длины; го
ловка пирамидальной формы диаметром около 0,8 мм с двумя венчи
ками крючьев в числе 22—32. Более крупные крючья длиной 0,015—
0,017 мм, а меньшие — 0,09—0,013. Для формы больших крючьев ха
Рис. 263 — Цистицерк тонкошейный
на сальнике овцы. По Скрябину.
Рис. 264 — Перфорация костей че-
репа овцы, вызванная влиянием
финнозной стадии Multiceps mul-
ticeps. По Monnig^
рактерна волнообразность краев рукоятки. Зрелые членики 8—10 мм
длиной и 3—4 мм шириной. От срединного ствола матки отходят с каж-
дой стороны по 18—26 слабо ветвящихся отростков. Яйца 31—36 р диа-
метром. Главным хозяином является собака.
Цикл развития. Яйца цепня-мозговика рассеиваются с со-
бачьим пометом и загрязняют пастбища, дворы, выгоны и водоемы.
Будучи проглочены промежуточными хозяевами в лице различных до-
машних животных, они дают зародышей с последующим развитием
пузырчатой стадии, главным образом в организме овцы и козы (а так-
же тешят, лошадей и, редко, человека). Зародыши этого паразита, как
обычно у ленточных глист, разносятся с током крови и лимфы по телу;
развиваются в личинок только те зародыши, которые попали в головной
328
СПЕЦИАЛЬНАЯ ЧАСТЬ
(или спинной мозг). В мозгу зародыш превращается в крупный тонко-'
стенный заполненный жидкостью пузырь, на внутренней поверхности^
которого развивается множество (до ста) вывернутых наизнанку голо-
вок. Вследствие такой специальной локализации пузырчатой формы
Multiceps multiceps в мозгу, сам паразит получил наименование «це-
пень-мозговик».
Рис. 265 — Ценуроз мозга человека. Срез части стенки паразита.
А — кутикула; В — сколекс; С — присоски сколекса; D — полость инвагина
ции. По В г u m р t.
Рис. 266,— Часть стенки Coenurus cerebralis овцы^с ввер-
нутыми внутрь головками (разрез). По Фурману.
Финнозная стадия цепня-мозговика может вызывать у овец перфо-
рацию костей черепа (рис. 264).
Когда мозг зараженного животного будет съеден собакой, то из
каждой головки ценура может развиться ленточная глиста.
Патогенное значение. Взрослый паразит в кишечнике соба-
ки не проявляет себя никакими особыми симптомами; зато весьма важ-
ными патогенными свойствами обладает личиночная форма цепйя-моз-
говика. Давление на мозг, внутри которого происходит развитие цену-
ра, вызывает различные непроизвольные движения овцы, которая часто
вертится на одном месте. Такое болезненное состояние известно в на-
роде под именем вертячки (ценуроз). Ведет оно к гибели заболевшего
животного.
Человек в редких случаях может быть промежуточным хозяином
цепня-мозговика, если проглотит, подобно овце, яйца этого паразита,
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
329
рассеиваемые собаками. Известен пока один надежно исследованный
случай ценуроза у человека в Париже. Болезнь началась внезапно на-
ступившей афазией (больной потерял уменье читать и писать). Затем
присоединились и другие симптомы: судороги, бред, коматозное состоя-
ние; приблизительно через год больной умер. В боковом желудочке
мозга при вскрытии был обнаружен ценур с семьюдесятью пятью го-
ловками, заходивший и в самое вещество мозга (рис. 265, 266).
Диагностика у человека пока невозможна. Вертячка овец дает
достаточно ясные признаки, по которым можно заподозрить наличие це-
нур а в мозгу. Окончательный диагноз ставится при вскрытии мозга и
нахождении в нем пузыря паразита. Истинную вертячку надо диферен-
Рис. 267 — Песчанка Rhombomys opimus. Из Е. Павловского.
цировать от ложной, причиняемой личинками овечьего (носового) ово-
да — Rhinoestrus purpureus, паразитирующими в носовой полости и в
сопредельных пазухах овец (ом. во II части главу об оводах). Поэтому
надо вскрывать и носовую полость овец для отыскания в ней личинок
овечьего овода.
Лечение ценуроза человека невозможно.
Профилактика. Личная профилактика сводится к общегигие-
ническим правилам принятия пищи и питья. Избегать тесного соприкос-
новения с собаками.
Общественная профилактика выливается в форму борьбы с ценуро-
зом овец. Необходимо широкое распространен,ие сведений о причинах
и способах заражения этой болезнью среди социалистического сектора
деревни, совхозов, колхозов и прочего' населения, имеющего дело с ов-
цами. При падеже овец от вертячки их головы непременно должны
быть сжигаемы, чтобы предупредить заражение собак при пожирании
ими инвазированного трупа. Сторожевые собаки стад должны быть два
.раза в год очищаемы от глистов.
Multiceps glbmeratus Raill. et Henry
Синонимы: Coenurus glome'ratus Turner et Leiper, Taenia glomerata Brumpt.
Паразит известен пока только в форме пузырчатой глисты типа
ценура, описанной у песчанки. Возможно гостепаразитирование этого
паразита и у человека. В Северной Нигерии у одного туземца была вы-
330
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 268 — Цепень
эхинококка—Echi-
nococcus granulo-
sus из кишечника.
1 — Проглоттида со
зрелыми яйцами; 2 —
проглоттида в состо-
янии мужской зрело-
сти; 3 — молодая про-
глоттида с неразвитым
еще половым аппара-
том; 4 — головка; 5 —
хоботок; 6 — присоска;
7 — экскреторный ка-
нал; 8— зачаток матки;
9 — половое отверстие
с выступающим ко-
пулятивным органом;
10 — яичник; 11 — жел.
точник; 12—зрелые
яйца в матке; 13 —
мешок копулятивного
органа; 14 — влагали-
ще. По Бреслау.
резана из межреберных мышц опухоль 1 на 2 см с очень нежной
прозрачной оболочкой; на ее вывернутой поверхности были тридцать
пять ввернутых внутрь головок. Каждая головка несла венчики крю-
чьев дз 16 больших (0,09—0,01 мм длины) и 16 малых (0,065—0,07 мм).
Цикл развития неизвестен. Окончательным хозя-
ином должно быть какое-либо хищное млекопитаю-
щее, пожирающее песчанок. Человек может быть
промежуточным хозяином, если проглотит яйца лен-
точной формы Multiceps glomeratus. Песчанки обыч-
ны во многих местах Средней Азии. Ценуры были
найдены в брюшной полости у Rhombomys opimus
в ’Мирза-Арале, у Gerbillus erythrourus в Вуадиле,
у G. tamaricinus и G. meridianus в ’Мирза-Арале
(Узбекистан, Л. Жинкин). Поэтому возможны случаи
гостепаразитирования этой ленточной глисты у чело-
века и в Средней Азии.
Диагноз ставится только post factum — иссле-
дованием извлеченного паразита.
Лечение хирургическое.
Профилактика неизвестна. Могут оказать
пользу обычные предосторожности при питье и при-
нятии пищи.
Род Echinococcus Rudolphi
Echinococcus granulosus Batsch.— цепень эхинококка
Синонимы: Taenia echinococcus V. Sieb., Echinococcus poly-
morphus Diesing.
Цепень эхинококка является полной про-
тивоположностью лентецу широкому и цепням воору-
женному и невооруженному. У последних стробила —
гигант по сравнению с пузырчатой формой тех же
видов; у первого, наоборот, ленточная глиста еле до-
стигает 3—5 мм длины, тогда как пузырчатая глиста
разрастается до размеров детской головки. И в отно-
шении патогенности эхинококк гораздо опаснее для
человека, чем финки солитера. Цистицерки лишь в
некоторых случаях причиняют опасное заболевание,
иногда же проходят совершенно незаметными. Эхино-
кокк, наоборот, как правило, влечет за своим разра-
станием опасные последствия для человека. Случаи
самопроизвольного излечения организма от эхино-
кокка чрезвычайно редки. 2
Для выяснения важных биологических особенно-
стей этого паразита человека начнем с описания
ленточной формы, живущей в тонких кишках собаки, шакала, волка
и, очень редко, кошки.
Цепень эхинококка состоит из головки и 3—4 члеников (рис. 268),
из которых задний занимает около половины длины всего червя. Го-
ловка маленькая, около 0,5 мм диаметром, с четырьмя присосками и
хоботком, вооруженным двумя венчиками крючьев; их всего от 28
до 50. Крупные крючья длиною 0,02—0,03 мм, более короткие длиною
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
331
Рнс. 269 — Онкосферы (яйца) эхинококка —
.Ech. granulosus. По Раевской.
0,018—0,022 мм. Половые отверстия в члениках чередуются неправиль-
но. В заднем членике, шир(ина которого доходит до 0,6 мм, лежит
матка в виде продольного тяжа с боковыми дивертикулами. В ней за-
ключается от 400 до 800 яиц, внутри которых заложены зародыши
0,025 мм длины. Яйца коротко-овальные, длина их 32—36 ширина
21—30 у. Оболочка яйца радиально исчерчена (рис. 269).
Жизненный цикл. Промежуточными хозяевами эхинококка
являются: человек, крупный рогатый скот, овцы, козы, верблюды, оле-
ни, антилопы, кролики, мыши, белки, медведи, сами собаки, кошки, ло-
шади, ослы, свиньи и другие животные.
Яйца выходят с испражнениями собаки наружу и прилипают к ее
шерсти. Они могут быть занесены в рот человека и проглочены им,
если гладить рукою заражен-
ную собаку и есть потом ка-
кую-либо пищу немытыми ру-
ками. То же может про-
изойти при поцелуях собаки,
лизавшей перед тем свой
anus. Близкое сожительство
с собаками неминуемо отра-
жается на заболеваемости лю-
дей эхинококком. В север-
ных странах, где собаки
являются ближайшими для
человека домашними живот-
ными, эхинококки у людей
встречаются часто.
Общий ход развития проглоченного яйца совершается, как у Т. so-
Нит. В желудке человека из оболочки яйца выходит зародыш, который
проходит сквозь стенку желудка или кишок и попадает в лимфати-
ческую систему или в кровеносные сосуды. С током крови зародыш за-
носится в различные органы,чаще всего в печень, где и разрастается
в громадную пузырчатую глисту.
К счастью, эхинококк является не столь частым паразитом челове-
ка, зато он довольно обыкновенен у других промежуточных хозяев —
коров, овец, свиней. Распространен широко. В некоторых местах энде-
мичен. Максимальная частота нахождения у человека на секционном
материале обнаружена в Томске за 26 лет — у 0,54% вскрытых тру-
пов, а за 34 года вскрытий в Военно-медицинской академии найден
только у 0,35%. Частота заражения эхинококком крупного рогатого
скота вариировала по разным бойням от 27,9% (Тула) до 83,1% (б. Ку-
банская обл.).
Пузырчатые формы эхинококка разнообразны по своему строению,
что наталкивает на мысль о существовании двух самостоятельных ви-
дов этого паразита — Echinococcus unilocularis и Е. multilocularls
(alveolaris), однако затронутый вопрос остается пока не разрешенным.
Выяснить его можно только путем строго обставленных опытов.
Echinococcus unilocularis
Одногнездный эхинококк. Развитие эхинококка происхо-
дит очень медленно. По опытам Deve, уже через 3 часа после кормления
свиньи яйцами эхинококка в печени ее можно найти молодые эхинокок-
ковые личинки, окруженные скоплением лейкоцитов. Через 27г дня ли-
332
СПЕЦИАЛЬНАЯ ЧАСТЬ .
чинки превращаются в небольшой, комочек протоплазматической массы.
На 7-й день она принимает характерный вид пузырька 0,060—0,070 мм
диаметром. Через месяц пузырьки достигают величины 1 мм и только
через 5 месяцев разрастаются до размеров 1 см. Так, шаг за шагом
растет гидатида 10, 20, даже 30 лет, в конце концов чрезвычайно уве-
личиваясь и вмещая в себя до 16 кг жидкости и более.
О степени развития эхинококка можно судить по случаю, когда в
печени коровы выросли пузыри паразита весом в 64 кг, оставшаяся же
паренхима органа весила всего 1 кг с небольшим: пузыри содержали
в себе около 3V2 ведер жидкости.
По мере роста эхинококка в нем обособляются различные части,
степень и характер развития которых влияют на общий habitus
паразита.
Из личинки первоначально возникает одиночный пузырь, стенка ко-
торого образована из двух частей — из наружной слоистой хитинопо-
добной капсулы (кутикула в свежем состоянии молочно-белого цвета
или слегка желтовата) и из внутренней зародышевой паренхиматозной
оболочки. Последняя образована двумя слоями клеток: одним, приле-
жащим к кутикуле (он состоит из более крупных клеток с отложения-
ми в них (известковых телец), и другим, синцитиального характера,—
граничащим с полостью пузыря. В зародышевой оболочке содержится
гликоген; толщина ее колеблется от 0,010 до 0,025 мм.
Полость пузыря эхинококка заполнена желтоватой жидкостью
удельного веса 1007—1015; в ней содержатся поваренная соль, сахар,
инозит, янтарная кислота и лейкомаины.
Паразита, остающегося на стадии одиночного пузыря, называют
Echinococcus simplex. У человека такая форма наблюдалась под брю-
шиной, в костях и других органах.
В дальнейшем развитии эхинококка можно наблюдать образование
дочерних пузырей (рис. 150); они отпочковываются от материнского
(первичного) пузыря или во внутреннюю его полость (рис. 270,2—6) —
эндогенное образование пузырей, или же отщепляются
кнаружи, внедряясь в окружающие ткани хозяина (рис. 270,15—20),—
экзогенное образование дочерних пузырей.
В первом случае образуется Echinococcus endogenes (Е. hydatidosus,
Е. altricipariens), по преимуществу свойственный человеку, во втором
же возникает Ech. hydatidosus exogenes (Е. granulosus, Е. scolicipa-
riens), встречающийся наиболее часто у животных, в частности у свиней.
Если эхинококк остается на стадии материнского и дочерних дву-
слойных пузырей, в которых не образуется больше никаких новых ча-
стей, то такую форму паразита называют стерильным эхинококком
(Ech. cysticus sterilie s. Acephalocyste). Он отличается недоразвито-
стью головок (сколексов). При отсутствии головок пузырчатая форма
эхинококка неспособна заражать восприимчивых к этому паразиту
животных.
Этой форме паразита противопоставляется Ech. cysticus fertilis,
характеризующийся дальнейшем образованием зародышевых или вы-
водковых камер, почкующихся из внутреннего паренхиматозного слоя
гидатиды или дочернего пузыря (рис. 270). Возникшие камеры малы —
около 0,25—0,5 мм диаметром, но все заметны простым глазом. Они
точно крупинки или песчинки насыпаны на внутренней поверхности
гидатиды. Если выпущенной жидкости эхинококка дать отстояться, то
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
333
выводковые капсулы, подобно песку, оседают на дно сосуда. От дочер-
них пузырей они отличаются отсутствием слоистой хитиноподобной обо-
лочки.
В этих капсулах в свою очередь почкуются важнейшие части эхино-
кокка — головки или сколексы, число которых сильно вариирует (от
2 до1 120); в среднем 10—30 головок в каждой капсуле. Головки обык-
новенно вывернуты наизнанку; в таком состоянии они имеют овоидную
Рис. 270 — Схема эхинококка по D£v£.
1—6— развитие выводковой капсулы и сколексов внутри нее, превращение сколексов
в пузырьки; 7 — молодой дочерний пузырь, развившийся из пузырчато видоизменен-
ного сколекса; 8 — вполне сформированный дочерний пузырь, но без сколексов;
9 — дочерний пузырь с развивающимися выводковыми капсулами и сколексами; 10—
13 — дегенерация дочерних пузырей; 14 — новое образование выводковых капсул и
сколексов из остатков зародышевого слоя дочернего пузыря; 15—19 — экзогенное
развитие дочерних пузырей; 20 — почти отделившийся дочерний пузырь, в котором
развиваются выводковые капсулы и начинается образование нового экзогенного пу-
зыря. Концентрические линии соответствуют слоистой кутикуле, пунктирная линия —
паренхиматозному зародышевому слою.
форму с просвечивающими присоскам,и и венчиком из 30—40 крючьев
меньшего размера, чем у цепня эхинококка (рис. 271).
При разрыве выводковых капсул головки падают в полость пузыря
и плавают в его жидкости. По распаде сколексов крючья их освобож-
даются и также могут быть найдены в последней, что имеет важное
диагностическое значение.
В сущности все упомянутые выше формы эхинококка относят к од-
ному и тому же виду—многоформенному эхинококку
{Ech, polymorphic s. Ech, hydatidosus s. Ech. multilocularis). Мы остано-
вились довольно подробно на синонимике этого паразита, чтобы облег-
чить читателю понимание соответствующей литературы.
334
СПЕЦИАЛЬНАЯ ЧАСТЬ
Жизненный цикл эхинококка замыкается, когда пузыри эхинококка
с живыми головками в них пожираются окончательным хозяином
(например, собаки пожирают боенские отбросы в виде проросших пу-
зырями органов рогатого скота ил,и волки пожирают эхинококкозных
овец).
Несмотря на малую величину ленточной формы эхинококка, она
развивается медленно. Через 24 дня после скармливания сколексов
собакам ленточные глисты уже образуются, но выделение яиц начи-
Рис. 271 — Эхинококк (Ech. granulosus) из печени
коровы. Часть стенки дочернего пузыря с выводко-
вой капсулой и сколексами на различной стадии
выворачивания. По Фурману.
ним воздействиям. Например, все сколексы в
нается только к концу
третьего месяца после
заражения; напомним,,
что у лентеца широ-
кого — этого гиганта
среди ленточных чер-
вей— яйца начинали
выходить уже через
две недели после
скармливания плеро-
церкоидов.
Из каждой прогло-
ченной головки пузыря
экинококка может раз-
виваться ленточная
глиста; так как головок
обычно очень много,
то часто эхинококк
встречается у собак
несчетными массами.
Сколексы эхинокок-
ка мало стойки к внеш-
эхинококковом изолиро-
ванном пузыре, положенном в трехпроцентный раствор поваренной
соли, погибают через 4 дня, 2—3-часовое высушивание губит сколексы.
Сколексы, остающиеся в цельных эхинококковых лузырях, выживают
при температуре 15—17° С 5—6 дней, а при 20—25° С — 4 дня. В воде>
они переживают на день — два дольше, чем в разлагающейся жидкости
пузыря эхинококка. При температуре 55—60° С сколексы гибнут очень-
быстро (А. Носик).
Патогенное действие. Болезнетворное действие эхинокок-
ка на домашних животных гораздо меньшее, чем на человека.
Одногнездный эхинококк у человека может поражать любые орга-
ны; однако в этом отношении первое место принадлежит печени (рис.
272), на долю которой приходятся около 65% общей заболеваемости
этим паразитом. Затем следуют легкие (около 1О°/о), почки (около
8%) и другие органы, например кости, редко поражаемые эхинококком.
Ход заболевания зависит от локализации паразита. Благодаря край-
ней медленности его роста, своевременный ранний клинический диагноз
эхинококка почти невозможен, и только тогда можно подозревать его
присутствие, когда он разрастается в легко прощупываемую опухоль с
ясным зыблением в ней; в дальнейшем требуется постановка диферен-
циального диагноза.
Судьба эхинококка в теле человека различна. В редких счастливых
случаях происходит самоизлечение организма от паразита, который по
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
33>
Рис. 272 — Эхинококк пе-
чени. Оригинал по мате-
риалу больницы в Улан-
Баторе.
умирании своем спадается, так как жидкость его рассасывается. Смор-
щенные некротизированные остатки эхинококка омелотворяются и в та-
ком безвредном состоянии остаются в теле своего носителя. Инкрусти-
рованный известковыми солями умерший эхинококк называется Ech. ре-
trificatus. -
Возможны и другие пути так же редкого излечения, когда подверг-
шиеся нагноительным процессам гидатида и окружающие ее ткани хо-
зяина прорываются в полостные органы — ки-
шечник, почку и т. п. Однако разрыв пузыря
Эхинококка ведет чаще к ухудшению положе-
ния больного — к интоксикации (см. ниже) и к
ра-ссеиванию головок паразита, которые приви-
ваются в различных органах (например, в брю-
шине) ; минуя стадию ленточной глисты они раз-
растаются в новые пузыри эхинококка. Число
таких рассеянных гидатид в некоторых случаях
превышало тысячу. Таким образом возникает
вторичный одногнездный или множественный
эхинококк.
Распространенный космополитически эхино-
кокк поражает людей преимущественно в воз-
расте от 20 до 40 лет. Описаны случаи внутри-
утробного заражения эхинококком человеческо-
го плода от матери.
Течение заболевания эхинококком и его ха-
рактер вариируют в зависимости от того, какой
орган поражен этим паразитом. Разрастающий-
ся пузырь давит на окружающие ткани органа и
вызывает их атрофию, например эхинококк ко-
стей. Особенно дает себя знать эхинококк в
глазнице (давление на глаз) и в черепной ко-
робке (давление на мозг), когда под влиянием
раздражения нервной ткани развиваются раз-
личные функциональные расстройства.
Жидкость пузыря эхинококка обладает ток-
сическими свойствами: она действует на кровь,
вызывает воспаление серозных оболочек и кра-
пивницу; равным образом, жидкость эхинокок-
ка вызывает явления анафилаксии. Интоксика-
ция проявляется резко, когда жидкость выте-
кает из пузыря эхинококка при его разрыве или при проколе (пунк-
ция). Однако и неповрежденный эхинококк, одетый реактивно разви-
вающейся капсулой из тканей хозяина, действует на последнего, вызы-
вая изменения свойств жидкой части крови. На этой особенности осно-
вано применение иммуно-биологических реакций для своевременного
диагносцирования заболевания эхинококком (см. ниже).
Известны случаи, когда эхинококк ничем не давал себя знать при
жизни больного и был открываем случайно на вскрытии. Наблюдалось
также и крайне долгое течение эхинококкоза (до 40 лет и даже более).
В качестве осложнения заболевания эхинококком наблюдается на-
гноение вследствие проникновения внутрь его пузыря бактерцй из ки-
шечника или при заносе их с троакаром при пробной пункции. Если
содержимое такого пузыря прорвется в брюшную полость, то разви-
вается гнойный перитонит.
:336
СПЕЦИАЛЬНАЯ ЧАСТЬ
Echinococcus multilocularis
'Многогнездный эхинококк, свойственный чаще рогатому
скоту и редко встречающийся у человека, состоит из бесчисленного мно-
жества мелких (иногда от 0,1 до 0,5 мм диаметром) пузырьков, зало-
женных в ткани пораженного (им органа (рис. 273). Паразитарная опу-
холь имеет обычно плотность хряща; строение ее ячеистое; на разрезе
юна похожа на нежно-дырчатый сыр (рис. 274). Альвеолярное строение
эхинококковой опухоли выражено
Рис: 273 — Многокамерный эхинококк
{Echinococcus multilocularis') печени ко-
ровы. По С о hrs.
резче по периферии; центральная
часть паразитарного образования
может подвергаться некрозу и об-
разовывать неправильной формы
полости, занятые продуктами рас-
пада. Эти образования нельзя сме-
шивать с полостью гидатиды и
дочерних пузырей юдногнездного
эхинококка. Альвеолы многогнезд-
ного паразита образованы кутику-
лярной оболочкой и внутренним
плазматическим слоем, из которо-
го развиваются головки. Простран-
ства между альвеолами заняты
разросшейся промежуточной тканью
пораженного органа, которая по-
следовательно превращается в гра-
нуляционную ткань, плотнеет, не-
кротизируется, омелотворяется или
подвергается распаду. Полость пу-
зырьков эхинококка занята студе-
нистой массой. Опухоль растет по
своей периферии путем почкования
или отшнуровывания новых пузырь-
ков от уже существующих.
Отсутствие гидатид, общий характер роста, более злокачественное
влияние на ткани пораженного органа отличают многогнездный эхино-
кокк от одногнездного настолько, что возникла мысль о принадлежно-
сти этих пузырчатых форм к разным видам эхинококка. Некоторые
исследователи признавали самостоятельность Taenia multilocularis
(Leuckart), которая отличается от Т. echinococcus различным отноше-
нием длины крючка к корневому его отростку. У первого1 вида оно
равняется 1 :2,7, у второго же— 1 : 4,6. Кроме того, матка у Ech. mul-
tilocularis имеет форму шаровидно вздутой трубки. Другие авторы
считают многогнездный эхинококк за уродливые формы. Истина может
быть выяснена лишь новыми экспериментальными исследованиями.
Диагност и.к а эхинококка. Диагностика паразита трудна.
Болезненные симптомы очень мало типичны для него. Более или менее
характерны зыбление опухоли при наличии увеличения числа эозино-
филов в крови. Иногда их бывает до 20—25% всех лейкоцитов. Реко-
мендуемая иногда пробная пункция троакаром подозрительной опухоли
для добывания ее жидкости считается опасной по могущим быть по-
следствиям — занесению головок эхинококка в брюшную полость и
возникновению нагноения гидатиды, нормальные стенки которой непро-
ницаемы для микробов. Добытую путем прокола жидкость центрифу-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
337
г,ируют и в осадке ищут сколексы и крючья. В химическом отношении
для нее более или менее характерно присутствие большого количества
поваренной соли, а также янтарной кислоты.
Янтарная кислота открывается следующим образом. Добытую жид-
кость взбалтывают с эфиром; отстоявшийся эфир сливают и испаряют
на водяной бане. В случае присутствия янтарной кислоты остается кри-
сталлическая масса из шестисторонних табличек или межклинных
призм (кристаллы микроскопических размеров). Остаток растворяют
водой. При добавлении по-
луторахлористого железа
янтарная кислота образует
студенистый осадок ржав-
чинного цвета. Жидкость
гидатиды не1 содержит со-
всем белка; она щелочной
или нейтральной реакции;
уд. вес ее 1,008—1,013.
В качестве добавочных
диагностических признаков
можно упомянуть про- не-
зроницаемость эхинококка
для рентгеновых лучей.
За последние годы наме-
чается новый, наиболее обе-
щающий путь — специфиче-
ский- путь серодиагностики
эхинококка. Из иммупо-бис-
логических реакций наи-
большее значение имеет ин-
традермальная реакция Ка-
зони (Casoni): 0,2 см3 сте-
рильной эхинококковой жид*
кости впрыскивают в кожу
4
Рис. 274 — Многокамерный эхинококк печени
человека (на срезе\ Brumpt и D6ve.
А — пузырь эхинококка, В — инфильтрация измененной
ткани печени.
предплечья; в случае поло-
жительной реакции на ме-
сте инъекции образуется
через 15—20 минут прыщ,
окруженный эритематозной
зоной, которая исчезает вместе с прыщом. Через несколько часов появ-
ляется вторичная реакция в виде покраснения и кожный инфильтрат
у места впрыскивания.
Диагностика многогнездного эхинококка еще труднее, чем одно-
гнездного, так как отсутствует зыбление и нет возможности получить
пробную жидкость. Лечение его (хирургическое) почти безнадежно,
ввиду отсутствия общей плотной оболочки паразита, которая позволи-
ла бы его вылущить. Этот паразит распространен больше в тех местах,
где держатся стада крупного рогатого скота. Главным хозяином его
также является собака; наряду со скотом заболевает и человек, слу-
чаи чего неоднократно описывались и русскими авторами (Еленев-
ский). Ежегодно наблюдаются случаи этого заболевания в Иркутске.
Лечение — хирургическая операция, если она вообще бывает
возможна; последнее зависит от характера пораженного органа и сте-
пени разрастания эхинококка. Если возможно, производят полное уда-
ление эхинококковых пузырей; в других случаях обнажают пузырь,
338
СПЕЦИАЛЬНАЯ ЧАСТЬ
выпускают его содержимое через троакар, вскрывают стенку пузыря,
удаляют внутреннюю оболочку эхинококка и протирают его изнутри
тампонами, смоченными формалино-глицериновой смесью для убивания
головок паразита.
Профилактика. Предохранение человека от заражения эхино-
кокком достигается мерами, касающимися как самого человека, так
и собак.
Личная профилактика. Остерегаться собак и иметь с ними
наименьший контакт, не целовать их и не позволять детям играть с
ними или спать с ними на кровати; соблюдать общегигиенические
предосторожности при еде и питье; не позволять собакам касаться про-
дуктов питания.
Общественная профилактика. Борьба с бродячими со-
баками и уничтожение их. Организация боенского надзора над выпус-
Рис. 275— Печень овцы с пузырями однокамерного
эхинококка.
каемыми с бойни органами животных (рис. 275). Ни в каком случае
не выпускать в продажу проросшие пузырями органы (мясо для кошек
и собак). Пораженные органы использовать на утилизационных заво-
дах, стерилизовать (например, кипячением) или сжигать. Не допус-
кать собак в боенские помещения и не допускать пожирание ими эхи-
нококкозных органов. Широко распространять среди населения сведе-
ния об опасности эхинококка и о мерах борьбы с ним, чтобы население
само не поддерживало культуру эхинококка скармливанием поражен-
ных органов убойного скота собакам.
Производить регулярное глистоизгнание у собак со сжиганием их
экскрементов. К сожалению, микроскопированием помета собак нельзя
установить зараженность их эхинококком, так как яйца эхинококка пс
форме и размерам очень близки к яйцам других собачьих ленточных
глист: Taenia pisiformis, Т. hydatigena и Multlceps serialis.R&TKHo отме-
тить, что яйца эхинококка выживают 11 дней при высушивании про-
глоттид глисты, взятых из помета собаки. В воде яйца сохраняют
жизнеспособность 16 дней и более.
Отмечено, что в некоторых эндемичных местах женщины заболева-
ют эхинококком гораздо чаще, чем мужчины, что связывают с контак-
том женщин с овцами. Такое положение дела подало повод испанско-
му хирургу Lozano высказать грубо ошибочное утверждение, будто бы
люди заражаются эхинококками не от собак, как это принято «в клас-
сической медицине», а от овец... через их экскременты и мясо. В эпи-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
339
демиологии эхинококка овцы, могут играть некоторую роль, но чисто
механических и пассивных переносчиков яиц эхинококка. Дело об-
стоит так: зараженные эхинококками собаки загрязняют яйцами эхи-
нококка пастбища; овцы, лежа на таком пастбище, пачкают шерсть, и
к ней могут приставать яйца этих глист. Женщины при доении или
стрижке овец имеют с ними более тесный контакт, чем мужчины, по-
этому получают больше шансов проглотить яйца эхинококка, которые
могут быть занесены грязными руками в рот. Если овцеводческие
функции ложатся на мужчин, то они в свою очередь будут заражаться
эхинококками чаще, чем женщины.
Строгое проведение системы профилактических мероприятий про-
тив эхинококка обеспечивает эффективность дела. Пример тому по-
черпнем из практики Исландии, которая являлась одним из классиче-
ских очагов эхинококкоза. 60 лет назад больных эхинококкозом было
там по отношению ко всему населению 2%. В 1890 г. был проведен за-
коЬ о собаках — налог на собак, обязательное лечение их, сжигание
пораженных эхинококком органов. Число собак на острове уменьшилось
в 3—5 раз, и больных людей, по новейшей статистике, насчитывалось
уже менее одного на 1500 человек населения, т. е. заболевание снизи-
лось более чем в 30 раз.
Учитывая рост поголовья скота в связи с реконструкцией живот-
новодства в СССР, необходимо теперь же выдвигать для практическо-
го использования весь комплекс противоэхинококковых мероприятий
как по отношению к пастухам и обслуживающему животных персона-
лу,. так и применительно к пастушьим собакам. Точно так же необходи-
мо следить за эхинококком и в частях Красной Армии, в частности на
пограничных заставах, где контакт человека с собакой может оказать-
ся более или менее тесным, в совхозах, колхозах и др.
Подтип Nematodes — круглые черви
Круглые черви, или нематоды, характеризуются удлинен-
ным, округлым на поперечном разрезе .(откуда и название их) телом,
которое лишено каких бы то ни было следов наружной или внутренней
членистости. Размеры нематод вариируют от микроскопических до дли-
ны метра или несколько более. У некоторых видов передний или задний
конец тела постоянно и характерно изогнут или даже закручен.
Снаружи тело покрыто развитой кутикулой; ресничный покров
всегда отсутствует. При осмотре кутикулы снаружи бывают видны
просвечивающие продольные линии, идущие или по средней
линии спинной и брюшной стороны, или по бокам (это стволы нервной
системы и выделительные каналы). Кутикула может образовывать
различные придатки— крылообразные или воротничковые выросты, со-
сочки разной величины, губы и др. Кутикула с поверхности бывает
гладкая или исчерченная в поперечном, продольном или косом направ-
лениях. Характер исчерченности бывает типичен для некоторых видов
нематод, пример чего дается на рис. 276.
Под кутикулой лежит слой гиподермы, который у взрослых
нематод превращается в зернистую или волокнистую ткань, потеряв-
шую следы клеточного строения или превратившуюся в синцитий. Ку-
тикула у личинок нематод неоднократно сбрасывается, каковой про-
цесс является линькой. Изнутри к гиподерме прирастают мышечные
клетки, продольными ребрышками вдающиеся в полость тела (рис.
343
СПЕЦИАЛЬНАЯ ЧАСТЬ
|-1-!-1-1-1-1-i-1-1-1 1
О Юм 100
ИИШИТО 2
fwmmmwTii -
гтттгттгттптгп *
птптт тгптп «
>
ШШШШШШП) ’
д iqOm. , , , .... юоо д
tzrzozo •
Рис. 276 — размеры и характер попе-
речной исчерченности некоторых не-
матод, паразитирующих или могущих
паразитировать у человека.
1 — масштаб к фиг. 2—8 в микронах; 2 — сам-
ка острицы 2.5-2,6 р.; 3 - самец — 1 р.; 4—
Ancylostoma duodenale — оба пола — 5 р.; 5 —
Necator americanus — оба пола — 6 р.; 6 — вла-
соглав — оба п ла в переднем конце — 6 —
7 р, в заднем конце - 5р.; 7 - Toxocara canis
оба пола — 12—13 р.: 8 — Toxocara mystax —
4,8 — 5#о «л; 9- 10—аскарида челогеческая, оба
пола, 180 - 203 р.. Bondeau du Moyer.
277). Число мышечных клеток на поперечном разрезе червя у разных
паразитов неодинаково. У аскариды их много; таких нематод называют
Polymyaria (рис. 277, 7); если мышечных клеток мало (8 у острицы —
рис. 277, II), то соответствующих паразитов относят к группе Мего-
myaria. И в том и в другом случае мышечные клетки распределены по
квадрантам поперечного сечения паразита, ограниченным продольными
валикообразными выростами гиподермы в полость тела, образующими
так называемые боковые линии
(рис. 277, L) или боковые поля.
Если такие выросты отсутствуют и
мышечные клетки лежат непрерывным
слоем, то соответствующих червей
(Trichotrachelidae) называют Holo-
myaria (рис. 277, III). Кожно-мышеч-
ный мешок ограничивает собой «по-
лость тела», в которой и лежат орга-
ны нематоды.
'Мышечные клетки круглых
червей (рис. 277, ст) устроены свое-
образно. В общем они вытянуты -в
длину и имеют изнутри придаток, в
котором сосредоточены протоплазма
(саркоплазма) клетки, ядро и
опорные фибриллы. В наружной
веретеновидной части клетки дифе-
ренцировано сократительное вещество
фибриллярной структуры (м и о ф и б-
р и л л ы). Мышцы нематод должны
быть отнесены к категории гладких
мышечных волокон. Движения тела
нематод могут быть весьма энергичны
и быстры (например, движение личи-
нок). Замечательно, что даже круп-
ные нематоды ’ могут пролезать в
очень узкие отверстия; известны слу-
чаи, когда аскариды пролезали в
отверстия пуговиц или бусинок, случайно проглоченных человеком.
Мышечные клетки прочно сращены с гиподермой, так как опорные
фибриллы сократительного отдела клетки переходят в слой ги-
подермы.
Мышечные клетки, как правило, лежат одним слоем. Замечателен
факт строгого постоянства числа этих клеток у некоторых нематод
(Martini).
Размеры мышечных клеток могут быть очень велики: у лошадиной
аскариды до 3 мм Длины, а у свайника-великана — до 4 мм.
Между кожно-мускульным мешком и внутренними органами лежит
заполненная жидкостью полостьтела, которая является псевдо-
целем, т. е. ложной полостью тела, лишенной целомической
клеточной выстилки. У некоторых изученных в соответствующем отно-
шении нематод, собственно говоря, нет и псевдоцеля, так как простран-
ство между органами занято изоляционной тканью, состоящей
из перегородок, являющихся производными нескольких соединительно-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
341
тканных клеток. Лакуны между ними заполнены жидкостью (например,
у аскарид). У других нематод соединительная ткань псевдоцеля обра-
зует тяжи (своего рода брыжейку), подвешивающие пищевод или дру-
гие органы. Например, у власоглава между ними лежат синусы ложной
полости теша.
Рис. 278 — Строение самки лошади-
ной аскариды.
1 — губы; 2 — окологл! точное нервное коль-
цо; 3 — глотка; 4 — выделительная система;
5 — фагоцитарные кл лк..; 6 — брюшной л jp-
вный С1вол; 7 — киш чн к; 8 — влагал ице;
9 — матка; 10— яйцевод; 11 — яичник (левой
стороны тела). По Л. В. Ш и м и к е в и ч.
Рис. 277—Поперечные разрезы
через тело крупных глист.
I тип Polymyaria, II тип Meromyaria.
Ill тип Holomyaria; С—кутикула;
Н — гиподерма; ст - мышечные клет-
ки; М. — спинная и брюшная (в рис.
II) линии; L — боковая линия; Chm —
мышечный слой. По В ru ш pt.
Пищеварительный аппарат у нематод, несмотря на их
паразитический образ жизни, сохранился. Он начинается ротовым от-
верстием и, как правило, открывается заднепроходным отвер-
стием (anus). Пищеварительная трубка вытянута в длину (рис.,278)
и в зависимости от развития из разных зародышевых листков делится
на три отдела: переднюю кишку (stomadaeum), эктодер-
мальную среднюю кишку и заднюю кишку .(proctodaeumk
Первая и последняя эмбриологически развиваются из наружного заро-
дышевого листка.
342
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рот круглых червей лежит терминально, т. е. на переднем конце
тела (рис. 278), который у некоторых нематод изогнут вентрально или,
наоборот, дорзально (рис. 304). Строение ротового конца весьма раз-
нообразно и имеет большое значение в систематике нематод.
У одних рот является простым отверстием, ведущим в переднюю
кишку. У других он окружен кожными выростами, называемыми г у б а-
м и (рис. 278, /); у третьих рот вооружен венчиком различных кутику-
лярных придатков, обрамляющих ротовую капсулу. Эти образо-
вания снабжены особыми чувствительными нервными окончаниями.
У некоторых круглых червей, лишенных губ, рот ведет в расширен-
ную полость, называемую ротовой капсулой (рис. 304), воору-
женную различными кутикулярными режущими или колющими придат-
ками (так называемые «з у б ы» нематод в форме разнообразных пла-
стинок, крючьев, зубцов и др.). Эти придатки служат для ранения
тканей органов, к которым нематода «присасывается» ротовой капсу-
лой. Последняя не может быть сравниваема с уже известными нам
присосками плоских червей, так как имеет совершенно другое строение
и, в частности, вовсе лишена собственных мышц.
Рот или ротовая капсула ведут обычно прямо в п,ищевод. Послед-
ний состоит из трубки, выстланной кутикулой; стенка пищевода по-
строена из радиально лежащих мышечных волокон, между которыми
бывают заложены железистые клетки. В спавшемся состоянии
пищевод на разрезе имеет форму трех правильно расположенных ра-
диальных щелей. Длина пищевода различна: он или одинакового диа-
метра на всем протяжении, или же имеет одно или два глоточных
вздутия. Последняя форма пищевода носит специальное обозначение
рабдитной — по аналогии со строением пищевода угриц (рис. 294).
В других случаях пищевод подразделяется на передний — мышеч-
ный — и задний — железистый отделы. От пищевода могут
отходить слепые придатки.
Все части передней кишки, выстланные кутикулой, принимают уча-
стие в процессе линьки, сбрасывая старую выстилку и замещая ее новой.
Средняя кишка, или собственно кишечник, представлена пря-
мой трубкой, построенной из эпителия различной формы, чаще цилин-
дрического, лежащего на базальной перепонке соединительно-
тканной природы. Свободная поверхность клеток превращена в каем-
чатый палочковидно исчерченный слой (который не имеет никакого от-
ношения к ресничкам). В протоплазме эпителиальных клеток имеются
гранулы, включения гликогена, жира, экскреторных продуктов, предсу-
ществующие формы секрета и т. д. У некоторых аскаридовых кишеч-
ник образует направленный кпереди трубчатый вырост, отходящий от
места соединения пищевода со средней кишкой.
Задняя кишка у нематод очень коротка^ она, так же как и
передняя, выстлана «хитиновой» кутикулой и открывается у самок са-
мостоятельным отверстием, а у самцов вместе с половым отверстием
впадает в клоаку. Anus у самок лежит обычно на брюшной стороне,
чуть отступя кпереди от заднего конца тела. У самцов некоторых не-
матод (аскариды, острицы) задняя кишка имеет ректальные же-
лезы. У самок единичных видов нематод задняя кишка, как и аналь-
ное отверстие, отсутствует (например, ришта).
Органы выделения (экскреторная система) нематод весьма
своеобразны. Они состоят из продольных трубок, заложенных в полях
боковых линий (рис. 277, L). Передние концы трубок открываются
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
343
общей порой сзади нервного кольца, у других нематод — две пары ка-
налов: два задних длинных и два передних коротких; все они соеди-
няются короткой поперечной ветвью, образуя фигуру буквы Н. Общий
канал их открывается наружу по средней линии брюшной стороны
тела, ближе к переднему концу. Иногда наблюдается асимметрия в
развитии боковых стволов или полное отсутствие их (власоглав, свай-
ник-великан). При всей своей длине боковые каналы выделительной
системы представляют собой одну разросшуюся клетку, пронизанную
внутриклеточным протоком. Выводная часть (porus excretorius) по-
строена из двух гиподермальных клеток. Никаких следов бичей или
ресничек в выделительных каналах нет. Экскреторная система нематод
считается производным железистых образований покровов.
Функции внутриклеточного выделения присущи особым крупным
•отростчатым фагоцитарным клеткам, которые лежат в изоля-
ционной ткани на боковых полях. Таких клеток бывает 4, 6 или более.
Помещаются они в передней части тела или же разбросаны по всей его
длине. При впрыскивании в тело живой аскариды взвеси зернышек
кармина можно наблюдать захватывание их протоплазмой этих клеток,
что позволяет говорить об их фагоцитарной способности.
Нервная система состоит из окологлоточного к о л ь-
ц а, образованного сплетением нервных волокон, среди которых лежат
дающие их нервные клетки, скопляющиеся иногда в ганглиозные
группы. От нервного кольца кпереди и кзади отходят нервные
стволы, лежащие в гиподермальном слое покровов. Самый мощ-
ный— задний вентральный ствол; его антиподом является дорзальный
ствол; есть также боковые стволы. Все эти части соединяются попереч-
ными комиссурами и по своему ходу имеют местами ганглиозные
скопления нервных клеток (например, анальный ганглий). От
глоточного кольца кпереди отходят стволы к органам чувств (сосочки
и др.).
Органы чувств развиты слабо; можно видеть только различ-
ные сосочки и нервные окончания в покровах, распознаваемые только
на срезах.
Чувствительными сосочками является, видимо, пара так называе-
мых амфид — или боковых органов, лежащих в головной или шей-
ной части (рис. 354; /, 2) нематод; у некоторых нематод имеется пара
или более постанальных ф а з м и д; последние свойственны видам, не
имеющим хвостовых желез..
Органы дыхания и кровообращения у нематод отсут-
ствуют.
Органы размножения. Круглые черви обыкновенно раздель-
нополы. Самцы меньше самок и отличаются от них присутствием ко-
пулятивных органов в форме кутикулярных спикул, направляющего
их движение рулька (gubernaculum) и копулятивной сумки
(bursa copulatrix); отдельные части этого комплекса совокупительного
аппарата у различных видов нематод отсутствуют. Копулятивная сумка
состоит из выроста кутикулы, подпертой также кутикулярными ребра-
ми (рис. 297, В). Форма и расположение всех этих частей копулятив-
ного аппарата важны для систематики нематод. Половое отверстие, как
уже указывалось выше, открывается в клоаку, которая в свою оче-
редь открывается на зацнем конце тела. В ее окружности на покровах
нематоды имеются половые сосочки.
Строение внутренних органов размножения самцов нематод весь-
344
СПЕЦИАЛЬНАЯ ЧАСТЬ
ма просто. Семенник представлен обычно одной тонкой длинной
трубкой, которая расширяется в семепров од, в свою очередь под-
разделяющийся на семенной пузырек и семеизвергатель-
н ы й канал; последний открывается вентрально от средней кишки в
начало задней кишки, которая таким образом, становится половой
клоакой. Живчики нематод своеобразны по форме и лишены бичевид-
ного придатка.
Женские органы размножения парны (рис. 278), но
также имеют трубчатую форму. Я и ч н и к в виде тонких нитей перехо-
дит в более толстые яйцеводы; эти ib свою очередь продолжаются в
толстые стволы матки, которые сливаются в короткое влагалище.
Женское половое отверстие совершенно не связано с пищеварительным
каналом и лежит самостоятельно на вентральной поверхности немато-
ды. У некоторых крупных червей имеется непарная матка (EnterobiusY
есть виды и с непарным половым аппаратом (власоглав).
Яйца нематод одеты тонкой двуконтурной оболочкой (рис. 290);
у некоторых видов оболочка толстая. Есть круглые черви, размножаю-
щиеся живорождением (отрождают личинок).
При копуляции самец охватывает самку копулятивной сумкой (ес-
ли она у него есть) и вводит во влагалище спикулы. У некоторых
нематод копулирующие пары срастаются в таком положении, образуя
фигуру буквы У (например, Syngamus, рис. 301).
Жизненный цикл нематод. Жизненный цикл круглых чер-
вей начинается эмбриональным развитием зародыша под оболочкой
яйца. Откладываемые самкой яйца или содержат еще недоразвитого
зародыша, нуждающегося в дальнейшем развитии, протекающем во
внешней среде, или же обладают вполне сформированной личинкой,
способной заражать соответствующего хозяина. У некоторых видов ли-
чинки развиваются во время пребывания яйца в организме самки и
отрождаются в подвижном состоянии.
Постэмбриональное развитие протекает различно в зависимости от
того, имеются ли промежуточные хозяева у данного вида нематоды
или нет; далее играют роль необходимость смены среды обитания по
ходу метаморфоза и способ заражения хозяина, свойственный тем или
другим круглым червям.
Многие нематоды развиваются прямым -путем, без участия
промежуточных хозяев. У нематод можно различать следующие гра-
дации жизненного цикла.
1. Самка дает яйца, уже способные заражать хозяина, если они
будут им проглочены.
2. Яйца, отложенные самкой, нуждаются в доразвитии зародыша;
последнее зависит от того, куда, в какую среду и в какие •экологиче-
ские условия попадет яйцо, вынесенное с faeces из тела хозяина. При
благоприятных условиях (температура, влажность и др.) в яйце через
известный срок развивается личинка, которая уже может заразить
хозяина, если яйцо будет им проглочено.
3. У некоторых нематод созревшая личинка выходит из яйца и
живет в почве, если тому благоприятствует обстановка. В почве ли-
чинка линяет и достигает инвазирующего состояния, причем в своего
хозяина она попадает пассивно — с пищей или питьем, или внедряется
в него активно через кожу (ср. выше Schistosorrium).
4. Личинки нематод во внешней среде могут проделывать метамор-
фоз, принимая после линьки различный вид и превращаясь в конце кон-
цов в половые стадии. Размножение таких видов характеризуется как
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
345
гетерогония, т. е. смена поколений паразита, размножающегося
разными способами полового размножения, например партеногенети-
ческое, однополое размножение, от которого появляются личинки, со
временем превращающиеся в самцов и самок, размножающихся далее
обоеполым путем.
Жизненный цикл многих нематод усложняется приспособлением к
распространению их при содействии промежуточных хозяев.
Все разнообразие соотношений можно разбить на две категории
случаев:
а) заражение окончательного хозяина нематодами происходит при
поедании им промежуточного хозяина с соответствующими стадиями
развития круглой глисты;
б) промежуточный хозяин фактически является переносчиком
нематод, и заражение окончательного хозяина не связано с гибелью
промежуточного хозяина.
Наконец, необходимо отметить, что нематоды никогда не размно-
жаются в промежуточных хозяевах.
ДЕЙСТВИЕ ПАРАЗИТИЧЕСКИХ НЕМАТОД НА СВОИХ ХОЗЯЕВ
Паразитические круглые черви питаются остатками пищевой каши-
цы кишечника, слизью, эпителиальными клетками, кровью или же со-
ками тканей, которые всасывают всей поверхностью тела.
На хозяев нематоды действуют многообразно. Механическое дей-
ствие проявляется закупоркой кишечника, желчного протока, лимфати-
ческих сосудов, давлением на органы, разрывом органов, атрофией
тканей от давления паразита, прорывом стенки кишечника и др.
Моменты интоксикации играют важную роль, особенно при парази-
тах, выделяющих вещества, разрушающие кровь (например, секрет
шейных желез кривоголовки); но ядовитыми оказываются и экскре-
торные продукты нёматод, живущих в почти анаэробной среде ки-
шечника.
В местах прикрепления к телу хозяина нематоды могут нарушать
целость слизистых оболочек и тем открывать ворота для вторжения
инфекции.
Бесспорное патологическое значение имеет феномен мигра-
ции личинок некоторых нематод в теле хозяина, причем таковое
передвижение личинок совершается по определенным путям.
Наконец, общее действие некоторых нематод ведет к своеобразным
патологическим реакциям местного или общего характера, примеры
чего будут рассмотрены ниже при описании соответствующих парази-
тов. Наличие некоторых нематод в органах ведет к образованию опухо-
лей .(рис. 279).
Паразитические нематоды—лишь часть собственно круглых червей,
среди которых имеется множество свободно живущих видов. Инте-
ресно отметить, что паразитический образ жизни сравнительно мало
отразился- на организации нематод. Круглые черви паразитируют за
счет растений и животных; хозяином их является и человек; в нем
могут встречаться также и ложные паразиты, т. е. свободно живущие
нематоды, случайно попавшие в организм; бывают случаи гостепарази-
тирования круглых червей у человека, т. е. обитание в нем таких па-
разитов, которые свойственны другим хозяевам.
Фауна паразитических нематод человека в СССР полностью еще
346
СПЕЦИАЛЬНАЯ ЧАСТЬ
не выяснена, и при дальнейшем ее исследовании можно ожидать но-
вых важных находок.
Нематоды во взрослом состоянии паразитируют в полостных ор-
ганах, сообщающихся с внешней средой (пищеварительный тракт,
Рис. 279 — Spiroptera megastoma, паразиты в опухоли желудка
лошади. Оригинал.
легкие, почки), а также в соединительной ткани и в лимфатических
сосудах. Личинки нематод живут в крови, лимфе и соединительной
ткани различных органов.
Промежуточными хозяевами нематод являются ракообразные, на-
секомые и позвоночные ( в частности, млекопитающие).
Подтип нематод делится на два класса: Nematoda — собственно
нематоды и Gordiacea — волосатиковые.
КЛАСС СОБСТВЕННО КРУГЛЫХ ЧЕРВЕЙ — НЕМАТОДЫ,
NEMATODA
Полость тела ложная, не выстланная эпителием; пищеварительная
трубка, за редкими исключениями, вторичного характера, имеет и рото-
вое и заднепроходное отверстия; задняя часть пищеварительной труб-
ки может атрофироваться у самок некоторых видов. Половые железы
непосредственно соединяются с выводными половыми протоками.
Клоака у самок отсутствует. Есть боковые поля (линии).
Класс нематод подразделяется на много семейств, представители
которых являются истинными паразитами или же могут быть ложными
паразитами человека. Классификация нематод в отношении подразде-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
347
ления их на крупные систематические категории неоднократно меня-
лась различными авторами.
В последнее время Chitwood предложил разделить класс Nematoda
на две главные группы: Aphasmidia и Pfiasmidia на основании отсут-
ствия или наличия фазмид:
К Aphasmidia по Chitwood относятся три надсемейства: Trichinel-
loidea Hall., Menmithoidea Wulker и Dioctophymoidea Raill.
К Phasmidia принадлежат надсемейства: Rhabditoidea Travassos,
Anguillulinoidea Pereira, Strongyloidea Weinland, Trichostrongyloidea
Cram, Metastrongyloidea Cram, Oxyuridea Ra'ill., Ascaroidea Raill. et
Henry, Spiruroidea Raill. et Henry, Filarioidea Weinl. и
Camer.
DracunCuloidea
О
APHASMIDIA
Надсемейство Trichinelloidea
Род Trichinella Raill.
Trichinella spiralis Owen
п Синоним: Trichina spiralis Owen
Половозрелая трихина еле
видна простым глазом. Самка длиной
3—4 мм (рис. 280, Л), шириной 0,06 мм;
размеры самца (С) равняются соответ-
ственно 1,4—1,6 и 0,04 мм. Тело трихи-
ны равномерно суживается кпереди. Из
ротовой полости торчит стилет. Пище-
вод тонкий, длинный; позади нервного
кольца он образует псевдобульбу с.
У его заднего конца пищевод переходит
в тонкую изгибающуюся трубку, прони-
зывающую протоплазму крупных зер-
нистых (sticlocyte) и незернистых кле-
ток. Лежат они одна за другой, образуя
как бы столб. В место перехода пище-
вода в желудок открываются две желе-
зистые клетки (рис. 281). Пищеваритель-
ный канал открывается на заднем конце
тела заднепроходным отверстием. Поло-
вая трубка у самки одна; она состоит
из лежащего позади яичника тонкого и
короткого яйцевода и, широкой матки,
переходящей во влагалище. Последнее'
открывается наружу в передней четвер-
ти тела. Живородяща. Зародыши выхо-
дят из яйца в передней части матки и
тотчас покидают тело матери.
Самец лишен спикул (рис. 280, С),
но на заднем, более толстом конце тела
несет два конусовидных кожистых вы-
роста (рис. 280; D, р), играющих аналогичную роль. Семенник соеди-
няется с семепроводом, тонкая часть которого открывается в концевой
отдел кишечника.
Р
Рис.
280 — Trichinella spiralis —
трихина. Кишечные формы.
А — почти зрелая самка; $ — ее половое
отверстие; и — матки; ov — яичник; В —
половой аппа;ат зрелой самки; ov—
яичник; rs — семеприем чик; и — матка с
зародышами (е); С—самец; t- семенник;
С? С1 ^клоака; D - сильно увеличенный
задний конец самца; р - кожистые лопа-
сти; I — кишка; vs - половая трубка.
Из Г а т ч е к а.
348
СПЕЦИАЛЬНАЯ ЧАСТЬ
Жизненный цикл. Половозрелые трихины живут короткое вре-
мя [1—2 недели у человека; 2—3 недели у крысы и около 5 недель у
морской свинки (Me Соу, 1932)] в тонких кишках, где копулируют;
Рис. 281 — Строение взрослой самки трихины.
1—Передний конец тела. 2—Голодной конец со стилетом (S) в ротовой
цолосги (В). 3—Поперечный разрез на уровне клеюк (sg), пронизывае-
мых пищеводом (О). 4—Участок тел нз уровне перехода пищевода (os)
в кишечник (Z7). 5 — Задний конец тела.О—пищевод; W X — узкая часть
пищевода; X— У—псевдобуль^ус пищевода; X — герчное кольцо;
GS— зернистая клетка пищевода (stlclocyte ; CS — то же, незернистая
клетка; YZ — узкая часть пищевода, внедряющаяся р клетки стихозомы;
DC — спинной гиподермальчый валик; КС.—то же, брюшной; L.C.—
боковой гиподермальны I в~лик; S.G. зернистая про!оплазма Sticlocyte;
L.G.— крупные гранули stislocyte; /?.С.— две крупные клетки в месте
впадения пищевода в U — кишечник; L — кишечник; RAB — rectum, об-
ладающий перистальтическим движением; G — ь.осовая железа.
По Van Someren.
самцы после этого погибают, а самки забираются в либеркюновы
железы, внедряются в подслизистую оболочку кишки, проникают в
лимфатические брыжеечные железы и сосуды. Живя во всех этих ме-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
349
стах около 8 недель, они отрождают массу мелких личинок 0,09—
0,1 мм длины и 0,006 мм толщины. О числе появляющихся личинок
можно судить по тому, что каждая самка родит их около полуторы
тысяч. Личинки с током лимфы и крови разносятся по телу и внедря-
ются в волокна мышц произвольного движения.
Рис. 282 —Личинки
трихины, начинающие
капсулироваться
в мышцах свиньи.
Рис. 283 — Различные фазы отложения извести
в капсулах трихин. По Брауну.
Наиболее подвержены заселению паразитами следующие мышцы,
расположенные в нисходящем порядке: диафрагма, межреберные мыш-
цы, мышцы гортани, шеи и глаз. Проникнув под сарколемму мышеч-
ного волокна, личинка сгибается там вдвое и довольно быстро начи-
нает расти. У червячка, (длиной всего в 0,5 мм) можно различить поло-
вые зачатки. По мере дальнейшего роста паразит скручивается спи-
рально; вокруг него образуется реактивная соединительнотканная кап-
сула, в которой со временем отлагаются известковые соли. Поражен-
ное паразитом мышечное волокно теряет свою исчерченность и разру-
шается в месте, где залегла личинка трихины, окруженная размножив-
шимися ядрами сарколеммы. Начало спирального закручивания трихи-
ны приходится на третью неделю после того, как съедено зараженное
мясо (рис. 282). Окружающая капсула образуется через 6—9 месяцев
после того же момента. Она принимает форму лимона, размером 0,4—
350
СПЕЦИАЛЬНАЯ ЧАСТЬ
0,25 мм. Обызвествление ее заканчивается к 15—16 месяцам (рис.
283). Этот процесс, начавшись с капсулы, може.т перейти на паразита
и погубить его. Обычно инкапсулированные мьцпечные трихины годами
живут в мышцах. У человека ‘ известен случай паразитирования их в
течение 31 года. Они стойки к вредным внешним воздействиям и даже
в гниющем мясе выживают 2—3 месяца.
Заражение трихинами происходит при съедании трихиноз-
ного мяса с живыми личинками трихин. В желудке’под влиянием пище-
варительного сока капсулы трихин растворяются и личинки освобож-
даются. Они переходят в тонкие кишки, где быстро растут, превра-
щаясь в половозрелые формы. Последние приступают к размножению
уже на 6—7-й день после заражения данного хозяина, т. е. после
поедания им трихинозного мяса. Таким образом, для замыкания пол-
ного жизненного цикла трихины требуются два хозяина, которые могут
принадлежать к одному и тому же виду.
Вопрос о локализации взрослых трихин в кишечнике, видимо, под-
лежит уточнению. Н. Roth в опытах на морских свинках, обильно за-
ражаемых личинками трихин, обнаружил, что большая часть поло-
возрелых червей скопляется в заднем отделе тонкой кишки, в сле-
пой кишке и в начале толстой кишки. Взрослые трихины живут
в кишечнике морской свинки гораздо дольше,, чем у крыс, мышей,
кроликов, собак и других животных. В тонкой кишке свинки трихи-
ны жили от 30 до 37 дней, в слепой — от 47 до 55 дней. Соответ-
ственно длительности жизни удлиняется и период отрождения личи-
нок (22—30 дней).
Трихины могут развиваться и в совершенно необычной для них
среде. Если стерильные живые личинки ввести в зародышевые обо-
лочки куриного зародыша или в амнион крысиного зародыша, то
некоторое количество их достигает половозрелости. Развитие трихин
возможно также в матке беременной крысы; в нормальной же матке
личинки погибают очень быстро (Me Соу).
Следует отметить резкое различие в локализации взрослых и ли-
чиночных форм трихин — тонкие кишки и поперечно-полосатые мыш-
цы — в одном и том же хозяине.
Иногда личинки проскакивают через капилляры мышц и попадают
в другие органы, где и оседают. Этим объясняются случаи атипич-
ной локализации «заблудившихся» личинок в серозных оболочках
брюшной и грудной полости, в перикардии, в цереброспинальной
жидкости, в легких, мозгу, в сетчатке глаза и даже в сердечной
мышце, которая никогда не бывает местом первичной локализации
личинок трихин. Образование капсулы вокруг личинки трихины не
является обстоятельством, абсолютно необходимым для последую-
щего благополучного превращения личинки в половозрелую форму.
Опыты со скармливанием мяса с личинками трихин, образовав-
шимися в результате заражения 17-дневной давности, давали в ряде
случаев положительный результат (Щупаков). Личинки указанного
возраста не только не имели вокруг себя капсулы, но сами еще не
были закручены. В таком состоянии они могут пройти через желудок
и доразвиться в половозрелые формы.
Распространение и способ заражения. Трихины —
космополиты и у людей встречаются чаще в тех местностях, где при
отсутствии ветеринарного контроля широко употребляется в пищу
сырая или полусырая свинина. Свиньи в свою очередь заражаются
трихинами, поедая больных этими паразитами крыс или уничтожая
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
351
остатки зараженных животных на бойнях. Крысы же поедают друг
друга и тем передают одна другой паразитов.
Сравнительно легко заражаются человек, свинья, крысы, мыши,
морские свинки и кролики. Трихины могут встречаться также у собак,
лисицы, барсука, куницы, кошки и медведя. Схемы соотношения хо-
зяев при возможных случаях распространения трихиноза среди жи-
вотных и человека даны на рис. 284.
Патогенное действие. Инкубационный период при трихи-
нозе длится от 15 до 24 дней (чаще 20—24 дня, Л. Поллер).
Рис. 284 — Схемы, рисующиэ различные взгляды на вопрос об
основном хозяине трихины.
I — Лейкарт, II — Ценкер — Сгейбели, III—Зеленский и Щупаков
a, bt с и т. д.— различные виды диких животных. Ио Зеленскому.
Болезненные явления проявляются начиная с внедрения самок три-
хин в стенку кишечника и отрождения ими личинок, проникающих
в общий круговорот крови и лимфы. Паразиты выделяют какие-то
токсины, которые вызывают эозинофилию крови. Впрочем при очень
сильной инвазии этого изменения крови не наблюдается. Эозинофи-
лия становится заметной начиная с 9-го дня, когда в кровяное русло
проникают первые личинки трихин.
Болезнь начинается желудочно-кишечными расстройствами и оте-
ком лица, преимущественно век, который держится около 3—7 дней.
Вскоре после заболевания после озноба повышается температура, до-
ходящая иногда до 40° и выше; она послабляющего типа, причем
падение температуры сопровождается потами. Головная боль, ломота
в теле, разбитость и иногда бред дополняют картину болезни. При
прощупывании различных мышц, например межреберных мышц (VI—
VIII межреберных промежутков кнаружи от сосковой линии), грудной,
дельтовидной, прямой мышцы и др., больной ощущает различной
силы болезненность. Позднее последняя наблюдается и при активных
движениях — жевании, глотании, повороте глаз и т. п. Голос стано-
вится беззвучным и хриплым; язык обложен; общее состояние боль-
ного похоже на брюшйотифозное заболевание. Действительно, три-
хиноз нередко был принимаем врачами за брюшной тиф, тем более
что заболевание трихинозом иногда является семейным или охваты-
352
СПЕЦИАЛЬНАЯ ЧАСТЬ
вающим в короткий срок ряд семей (благодаря общности источника
заражения в виде свиной трихинозной туши, проскочившей в оборот
для питания). В моче обычно резкая диазореакция. В легких случаях
человек выздоравливает недели через три; в тяжелых случаях забо-
левание длится месяца полтора и более. Смертность различна, дети
легче взрослых переносят трихиноз.
Количество паразитирующих личинок весьма вариирует в разных
случаях трихиноза. Интенсивность инвазии определяют средним чис-
лом личинок на 1 г мышц. Очень легкой инвазией считаются случаи,
когда на 1 г мышц приходится менее одной личинки; легкая инва-
зия— 1 —10 личинок; умеренная — при И—50 личинках; средняя —
51—100 личинок; сильная инвазия— 101—500; тяжелая — 501 —1000
личинок. Наблюдалась и еще более интенсивная инвазия. Так, в слу-
чае Coupland на 1 г приходилось 2700 личинок; во всем же теле можно
было предполагать наличие 75 млн. личинок.
Что количество личинок находится в корреляции с тяжестью за-
болевания, это несомненно, но едва ли дело сводится к простой ариф-
метической пропорции, примеры чего увидим ниже в разделе об анки-
лостомидозах.
Тяжесть заболевания лишь в известной мере связана с количе-
ством проглоченных капсул трихин; по Маршу, тяжелое заболевание
бывает, когда проглочено не менее 100 тыс. капсул (иногда это соот-
ветствует 10—20 г свинины).
Косвенным моментом опасного действия трихин является возмож-
ность занесения ими (при внедрении в стенку кишки) различных бак-
терий кишечной флоры (кишечная палочка, бактерии паратифа и др.),
что было устанавливаемо -экспериментально. Диагноз заболевания до-
вольно труден.
При вскрытии трупов для исследования на трихиноз вырезывают
кусочки диафрагмы ближе к ее сухожильной части. Кусочки измель-
чают ножницами и микроскопируют в компрессоре.
Кусочки диафрагмы в общей сложности весом 5 и более граммов
обрабатывают методом переваривания — Бермана. Переваривание
обеспечивается действием свеже приготовленного пепсинного раство-
ра (5 г пепсина, 7 см3 концентрированной соляной кислоты, 1 л воды)
при 37° С. После окончания переваривания лишнюю жидкость отса-
сывают; придонную жидкость с осадком смешивают с водой 37—
45° С и переводят смесь в аппарат Бермана, доливая горячую воду
до сетки воронки. Через час жидкость выпускают в коническую мен-
зурку, со дна которой через два часа выбирают пипеткой осадок
для микроскопирования и подсчета в нем личинок трихин.
Точной статистики трихиноза у человека в СССР и в прежней
России нет. С 1904 по 1907 г. в России отмечено 1940 заболеваний
трихинозом (Мари), каковая цифра, конечно, ниже действительной,
так как во многих случаях трихиноз не распознается врачами как
таковой, а идет под диагнозом брюшного тифа и инфлюэнцы. На сек-
ционном материале в Харькове (1902—1909) установлено наличие
трихин у одно^ четверти вскрытых трупов (Крылов).
При обследовании 1000 диафрагм умерших Nolan и Bozicevich
(США) трихиноз обнаружили в 17,4%.
Несколько лет назад стали говорить о наличии какой-то новой
болезнр (на Северном Кавказе и в Белоруссии), которая получила
название «одутловатки» из-за развития характерных отеков лица.
Была сделана попытка отнесения ее к инфекционным болезням; но
применение, по предложению автора, к этим случаям паразитологиче-
Таблица VIII
Trichocephalus suis, свиной власоглав, самец. Оригинал В. Соидак; рисовал
И. Теравский
О — пищевод, d — кишечник, с — кутикула, / — семенник, vs — семенной пузырек (vesicula seminalis),
vd — семявыносящий канал {vas deferens), de — семяизвергательный канал {ductus ejaculatorius), ct — клоа-
кальная трубка, sp — спикула, sv — сппкулярное влагалище, а1 — сужение между клоакальной трубкой
и спикулярной трубкой, д2 — сужение между кишечником и ductus ejaculatorius и между клоакальной
трубкой, а* — соединяющая трубка vas deferens и ductus ejaculatorius, а4 — сужение между vas deferens
и vesicula seminalis, дь — сужение между пищеводом и кишечником.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
353
ских методов диагноза с несомненностью* показало, что «одутловатка»
есть не что иное, как трихиноз (Алисов).
Диагностика. Паразитологическими способами диагностики
являются:
1) Исследование вырезанных или вырванных специальным гарпу-
ном кусочков дельтовидной или двуглавой мышц (ближе к месту пе-
рехода их в сухожилия) или межреберных мышц. Кусочки сдавливают
между стеклами компрессора и при слабом увеличении микроскопа
ищут в них трихинозные капсулы. Личинки трихин внедряются в мус-
кулы около 10-го дня заболевания; капсулы же образуются в конце
3-й недели болезни.
2) Способ Staubli. У больного берут 0,3—0,4 см3 артериальной
крови и смешивают ее с десятерным количеством 3°/о раствора уксус-
ной кислоты, которая разрушает эритроциты. Смесь центрифугируют
и в осадке ищут под микроскопом убитых кислотой личинок трихин,
которых можно окрасить гематоксилином и другими красками.
Исследование самих половозрелых паразитов в испражнениях
больного едва ли имеет практическое значение, так как трихин чрез-
вычайно трудно искать вследствие их ничтожной тонины.
Подсобным признаком побочного характера является сильная
эозинофилия.
Лечение симптоматическое.
Профилактика. Частота случаев трихиноза является прямо
пропорциональным показателем недостаточности боенского надзора
над выпускаемой в продажу свининой или зависит от употребления
в пищу свинины от животных, убитых не на бойне и, следовательно,
миновавших санитарно-гигиенический контроль.
Так как источником заражения человека трихинами является сви-
ное мясо, то особо важное значение приобретает организация обяза-
тельного ветеринарного контроля над выпускаемой в продажу свини-
ной. Именно трихинозом с его опасностью для жизни человека, в пер-
вую очередь, вызвано издание специальных законоположений по ве-
теринарно-санитарному осмотру мяса и мясных продуктов. Всякая
свиная туша должна подвергаться микроскопическому обследованию
при бойне или в специальных лабораториях.
Аналогичному обследованию подвергаются и копченые продукты,
вроде окороков, грудинки и др. Для трихиноскопии вырезают малень-
кие куски из ножек, диафрагмы, из языка, брюшных, межреберных,
гортанных, ножных мышц. Их сдавливают между двумя пластинками
зеркального стекла (компрессор) и микроскопируют при увеличении
в 50—100 раз. Если в капсуле отложена известь, то ее удаляют дейст-
вием крепкой соляной кислоты. Мясо, прошедшее контроль, штемпе-
люется, а грудинка и окорока снабжаются пломбами.
Трихинозное мясо подлежит обязательному уничтожению, сало же
отправляют в техническую переработку.
В Ленинграде с 1923 г., согласно обязательному постановлению,
живые свиньи, поступающие для убоя и последующего потребления
населением, контролируются на городской бойне; экспортный товар —
на бэконном заводе; парная свинина и окорока проходят через Цен-
тральную микроскопическую станцию, а мороженая свинина контроли-
руется на холодильнике.
Такой повторный контроль выявляет время от времени трихиноз
некоторого количества туш, зараженность которых ускользает от пер-
вого микроскопического обследования.
354
СПЕЦИАЛЬНАЯ ЧАСТЬ
Важность точного и своевременного контроля весьма велика, так
как при доминировании общественного питания попавшая в использо-
вание трихинозная свиная туша может причинить много бед; в Ра-
домно (б. Западная Пруссия) от одной пропущенной надзором трихиноз-
ной свиной туши заболело 40 человек, из которых умерло 15. Органи-
зация в СССР крупных мясных комбинатов обеспечивает совершен-
ство паразитологического контроля над выпускаемыми продуктами.
Наряду с этим необходимо расширение сети ветеринарно-санитарных
учреждений с обязательным микроскопическим контролем свинины в
пределах, покрывающих соответствующую потребность.
Весьма неблагоприятным в профилактическом отношении является
тот факт, что капсулированные трихины весьма стойки к действию'
холода. Замораживание туши до —15° С убивает в ней личинки три-
хин не менее чем в 12-дневный срок .{Ransom); в холодильниках же
температура держится обычно на уровне—10° С, что не влияет на
трихин. Единственным действительным средством считается кипячение
трихинозного мяса в течение получаса по расчету на каждый англий-
ский фунт (Фауст). Соление и копчение не влияют на капсулированных
трихин.
Источником заражения трихинозом свиней являются крысы и боен-
ские отбросы. Поэтому профилактика трихиноза человека должна
быть обеспечена и с этих сторон. Зараженность крыс трихинами раз-
лична и носит гнездный характер. Так, например, в Ленинграде из
шести обследованных рынков трихинозные крысы были обнаружены
на б. Сенном рынке (14%). Большая зараженность крыс обнаружена в
Добрушах Гомельского округа. Боенское дело должно быть везде
поставлено так, чтобы бойня не служила местом заражения свиней,
собак, кошек, крыс трихинами. Другими словами, боенские отбросы
должны быть уничтожены или же быть используемы для целей
утиля, причем к ним не должны иметь доступа перечисленные только
что животные. В отношении свиней необходимо также обеспечить
зоогигиеническое их содержание. Наконец, уничтожение крыс допол-
няет комплекс противотрихинозных мероприятий.
Учитывая большое развитие свиноводства в СССР как в специаль-
ных животноводческих совхозах, так и в колхозах, частях Красной
Армии, пограничных заставах, а также при столовых, фабриках, ко-
оперативах и в домашнем хозяйстве, необходимо широко обеспечить
знание населением всех бед, которые могут быть причинены человеку
свиньями © отношении заражения паразитарными болезнями, в част-
ности трихинозом.
Успешность профилактических мер в прошлом может быть иллю-
стрирована огромным снижением трихиноза свиней в Германии.
В 1879 г. одна зараженная свинья приходилась на 1632 здоровых, а
в 1913 г.— уже на 24 503 здоровых свиньи; но также в Германии мы
почерпаем убедительные доводы в пользу необходимости неустанного
контроля трихиноза. За пять лет перед войной (1910—1914) в Германии
было 184 заболевания людей трихинозом с 10 смертями; в следующую
пятилетку (1915—1919), когда из-за условий военного времени был
ослаблен санитарный надзор и ухудшился уход за свиньями, трихино-
зом заболело уже 366 человек, из которых умерло 32. Не менее по-
казателен пример Ганновера, где после ряда лет полного благополучия
в отношении трихиноза свиней был отменен обязательный осмотр мяса;
в результате вспыхнула эпидемия трихиноза, поразившая 400 человек
с 40 умершими.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
355
Отсюда мы вправе сделать вывод, который действителен не только
по отношению к трихинозу, но который должен быть распространен
вообще на инвазионные болезни: профилактика паразитозов
должна носить перманентный характер и в и дай з.м е-
няться в деталях, смотря по обстоятельствам, но не
прекращаться, когда ею уже достигнуты якобы
«окончательные» результаты.1
Семейство Trichocephalidae
Род Trichocephatus Schrank
Trichocephalus trichiurus L.— власоглав
Синонимы: Trichocephalus dispar Rud., Trichuris trichiura S til.
Хлыстовик, власоглав человеческий — беловатая гли-
ста, местами с красноватым оттенком. Передний конец тела вытянут
нитевидно (откуда и название — власоглав, рис. 285); в нем прохо-
дит только пищевод. Задний конец самца спирально закручен на спин-
ную сторону. Длина самца — 30—40 мм,
самки же 35—50 мм. В толстой части
тела лежат кишки и половые органы,
образованные у самки волнообразно из-
гибающимся трубчатым яичником, кото-
рый переходит в более толстую матку.
Последняя открывается влагалищем близ
переднего конца утолщенной части тела.
Семенник у самца также представлен
волнообразно изогнутой трубкой. У ме-
ста сообщения пищевода с кишкой он
перегибается и переходит в утолщенный
семевыносящий канал, который в задней
трети тела суженным участком соеди-
няется с семеизвергательным каналом.
Спикула у самца одна (2,5 мм длины):
она лежит в выворачивающемся мешоч-
ке, стенки которого усажены мелкими
шипиками. Яйца очень типичные, в
виде боченочков 50—54 & длины и 23
ширины (табл. VI, 15\ рис. 286). Кож-
ночмышечный мешок типа Holomyaria
(рис. 277, III). Живут в слепой кишке
человека, заходят и в червеобразный
отросток; гораздо реже встречаются в
Рис. 285 — Власоглав — Trichoce-
phalus trichiurus.
А~—'самец, В;—самка. Оригинал.
толстых кишках и совсем редко в тон-
кой кишке. Кроме человека, паразитируют также в обезьянах и полу-
обезьянах.
По Schwartz, власоглавы свиньи и человека идентичны; однако
морфологическое сходство должно быть подтверждено и биологически
(взаимной заражаемостью свиньи и человека).
Подробнее о трихинозе см. В. Д. Зеленский. Некоторые вопросы био-
логии и этнологии трихиноза. Вести, микробиологии, эпидемиологии и папазитологии.
т. VIII, вып. 4. 1929. S. Е. Gduld. Trichinosis. 1945, р. 356.
356
СПЕЦИАЛЬНАЯ ЧАСТЬ
Опыт самозаряжения человека инвазирующими яйцами власоглава
свиного дал отрицательный разультат (Ж. Штром и В. Сондак). Не
удалось также с достоверностью заразить поросят инвазирующими
яйцами человеческого власоглава. Все это говорит в пользу видовой
самостоятельности Tr. trichiurus и Tr. suis.
В последнее время Динник добыл новые данные в пользу различия че-
ловеческого и свиного власоглавов. Яйца первого несколько мельче, чем
у второго: средняя величина длины 57,75 + 0,21 против 62,60 + 0,2 ц
и ширины — 28,83 + 0,07 против 29,55 + 0,22 у».
Еще определеннее различия в сроках развития яиц, которое проте-
кает у человеческого власоглава гораздо быстрее, чем у свиного
(см. ниже).
Сроки развития яиц власоглавов человеческого и сви-
ного, по Н. Диннику, различны.
Стадия развития яиц Срок и развитие в сутках и часах
при t° 25о с при t° 30° С
власоглач человече- ский власоглав свиной власоглач человече- ский власоглав свиной
Слияние половых ядер . 1 сут. 4 сут. 13 часов 42 часа
Два бластомера 2 6 1 3
Овальный зародыш .... 12 29 8 16
Цилиндрический зародыш . . 18 37 11 29
Инвазионная личинка 21 55 17 28
Отмечается также большая средняя длина инвазионной личинки у
свиного, власоглава по сравнению с человеческим (158 у против 138).
Свиной власоглав обозначается как вид Trich. suis Schrank.
Наконец, диплоидное число хромозом у них также различно: четыре
у человеческого власоглава и шесть у свиного (рис. 287).
Бросается в глаза параллелизм этих отличий с тем, что установле-
но Для человеческой и свиной аскарид (см. ниже).
Таким образом, видовая самостоятельность рассматриваемых власо-
главов получает новые подтверждения. Космополит. Процент заражен-
ности различных групп населения в различных местах СССР колеблет-
ся в пределах от нескольких процентов до поголовной зараженности
(Азербайджан).
Цикл развития власоглава прямой — без промежуточного хо-
зяина. С faeces выбрасываются яйца, в которых имеются еще не слив-
шиеся в синкарион мужское и женское половые ядра. Слияние ядер
при температуре 28—30° С (в воде) происходит часов через 14—18
после выхода яиц из кишечника чёловека. Затем следует период покоя
(13—18 часов), после чего начинается эмбриональное развитие
(Н. Динник, рис. 286).
Во внешней среде при температуре 28—30° С в яйцах власоглава
развиваются рабдитные личинки, и яйцо становится заразительным для
человека через 28 дней. В воде при 22° С к 41-му дню наблюдения
созревают лишь 19% яиц власоглава, а при 30° — 64%.
Ю. и Н. Динник уточнили сроки развития яиц власоглава: при 15° С
развитие длится 120 дней; при 20° — 57 дней, при 25° — 29 дней, при
30° — 17,5 дней и прц 35°—11 дней. При 40° яйца дробятся непра-
вильно и погибают дня через 3—4. При 0° яйца не развиваются, но
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
357
сохраняют жизнеспособность. В бескислородной среде при 30° С яйца
выживают 10 дней; при понижении температуры этот срок удлиняется.
Инвазирующая личинка власоглава характеризуется наличием во
рту у нее стилета.,
Развившиеся личинки власоглава
лочкой яйца несколько месяцев.
Яйца власоглава вскрываются,
когда будут проглочены человеком,
и освободившиеся личинки обосно-
вываются в местах обитания без
миграции в организме хозяина.
Власоглав зарывается головной
(тонкой) частью своего тела в сли-
зистую оболочку кишки и питается,
повидимому, кровью. Вследствие
прочности прикрепления власогла-
ва его не находят в испражнениях
человека. По этой же причине из-
гнание его считалось невозможным.
Власоглав достигает в человеке
половозрелости приблизительно че-
рез месяц и живет, вероятно, не-
сколько лет.
Патологическое значе-
ние власоглава не подлежит со-
мнению. С его присутствием связы-
вают аппендицит и воспаление сле-
пой кишки (рис. 288). Проникая
глубоко в стенку кишечника, пара-
зит таким образом открывает до-
ступ внутрь тела и бактериям, ко-
торые всегда имеются в полости
кишок.
Проникновение патогенных бак-
терий в ткани кишечника может
вести к различным заболеваниям
инфекционного характера. Указать
могут оставаться живыми под обо-
Рис. 286 — Развитие инвазирующей
личинки в яйце власоглава Tricho-
серка'us trichiurus.
По Ю. и Н. Д и н н и к.
точное отношение власоглава в ка-
честве подсобного агента к иноку-
ляции холерного вибриона и дру-
гих болезнетворных бактерий нель-
зя. Поэтому выдвинутое Бланша-
ром положение, что нет кишечной инфекции без наличия глист, откры-
вающих дорогу заразным бактериям, не может претендовать на ши-
рокое и всеобъемлющее значение.
К аппендициту, в частности, власоглав имеет меньшее отношение,
чем острицы. По крайней мере встречаемость власоглава в иссеченных
аппендиксах гораздо бо)тее редкая, чем остриц.
По Рейндорфу, власоглав встречается в аппендиксе в 13 раз реже,
чем острицы. В моей лаборатории (ВМА) Банайтис обнаружил власо-
глава в одном отростке из 640, Gordon не нашел его ни разу при
вскрытии 26 051 отростка.
358
СПЕЦИАЛЬНАЯ ЧАСТЬ
Местное патологическое действие власоглава на ткани хозяина
обычно невелико. Выделяемые им токсины всасываются в кровь и дают
резорбтивное действие, проявления которого неспецифичны и не могут
быть приведены к какому-нибудь определенному симптомокомплексу:
то преобладают явления расстройства пищеварительной ’ системы, то
превалирует анемия, то выдвигаются симптомы нервного характера.
Диагноз ставится по нахождению яиц паразита в faeces.
Лечение. Изгнание власоглава трудно. Применяют курс противо-
глистного лечения тимолом повышающимися дозами от 0,5 до 2,5 г в
день (на каждый прием по 0,5 г) в течение недели, затем отдых 5—
6 дней, снова недельная тимолизация и отдых, и так — до полутора
месяца. Контролем успешности лечения является исчезновение яиц из
Рис. 287 — Ооцит власоглава свиного
(слева) и власоглава человеческого
(справа). По Н. Д и н н и к у.
Рис. 288 — Три поперечных разреза
власоглавов в толще слизистой обо-
лочки червеобразного отростка сле-
пой кишки человека; отросток иссе-
чен при операции аппендицита.
По Н. Соловье ву.
faeces. В последнее время в СССР предложен против власоглава со-
ветский синтетический препарат осарсол (стоварсол). Лечение
им можно проводить на амбулаторных больных примерно по следую-
щей схеме: осарсол в таблетках по 0,24—4 дня, повышая от 3 до 6 таб-
леток в день (женщинам 16—18 таблеток за курс, мужчинам 18—20
таблеток). Сернокислая магнезия в качестве слабительного. Таких кур-
сов приходится проводить от 1 до 4, с промежутками не менее месяца.
Профилактика проводится по тем же принципам, что и против
заражения аскаридами. Необходимо строгое соблюдение чистоты при
еде и питье, помня о возможности проглатывания яиц власоглава с
водой, немытой огородной зеленью и др. Отметим, что 5-минутное дей-
ствие температуры ,+ 52° С их убивает, тогда как около половины яиц
аскарид при тех же условиях сохраняет жизнеспособность. Недельное
замораживание при —12° С и 15-дневное при —3° С губят яйца власо-
глава. По линии общественной профилактики необходима защита воды
и почвы от загрязнения яйцами власоглава.
Трихоцефалез может быть при известной обстановке профессиональ-
ной инвазией. Например, у шахтеров в Донбассе отмечалось различие
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
359
в проценте заражения власоглавом у наземных и подземных рабочих, с
преобладанием в сторону последних (Скрябин). Отмечают также боль-
шую пораженность власоглавами рабочих полей орошения и ассениза-
ционных обозов. Устройство отхожих мест, предохраняющих почву и
воду от загрязнения фекалиями (и, следовательно, яйцами глист), ве-
дет к понижению зараженности власоглавом. Подземные рабочие более
благоустроенных шахт оказались менее зараженными этими паразита-
ми, чем в шахтах, где дефекация производилась прямо на землю
(Скрябин).
Трихоцефалез по сравнению с аскаридозом распространен менее
экстенсивно. 'Меньшее распространение его в США Cort и Otto объяс-
няют меньшей стойкостью яиц власоглава, по сравнению с яйцами
аскариды, к высыханию. От этого и зависит то, что очаги трихоцефале-
за приурочиваются к тем местам, где близ жилья имеются сырые, за-
тененные биотопы, загрязняемые экскрементами человека.
Trichocephalus muris — мышиный власоглав. У человека не
паразитирует. В экспериментальных условиях легко культивируется на
белых мышах; процент заражающихся мышей по мере дальнейшего
пассирования культуры повышается. С фекалиями выходят яйца на ста-
дии двух неслившихся половых ядер (как и у Tr. trichiurus). До стадии
инвазирующей личинки яйца доходят при содержании их в 1°/о растворз
соляной кислоты при 30° С в течение 18—20 дней. Средний срок до-
стижения половозрелое™ мышиного власоглава в белых мышах —
37,7 дня. Период размножения (кладка яиц) в среднем длится 45,6 дня_
После окончания кладки яиц самки могут еще жить от 2 до 8 дней.
Общая средняя длительность паразитирования Tr. marts в белых мы-
шах— около 88 дней; за это время Tr. muris вырастают — самки
•от 0,165 До 23 мм, а самцы до 19 мм длины, линяя несколько
раз .(предположительно четыре раза). Половая продукция зависит от
числа самок, паразитирующих в мыши; чем больше самок, тем меньше
каждая дает яиц (например, колебания числа яиц на 1 г фекалий в
сутки при одной самке —до 413 яиц, при 55 самках — до 381 яйца на
самку). Мышиный власоглав представляет интерес, как легко культиви
руемый в лаборатории вид власоглава, который может быть использо-
ван в качестве «модели» для предварительного испытания химиотера-
певтических веществ для борьбы с трихоцефалезом (данные Н. Шихо-
баловой). :р
Род Hepaticola Hall
Hepatlcola hepatica Bancroft
Длина самцов и самок от 40 до 120 мм, толщина 0,1—0,12 мм. Раз-
ница между утонченной передней и более толстой задней частью тела
невелика. Спикулы у самца отсутствуют. Половое отверстие самки ле-
жит в 6—7 мм от переднего конца тела.
Яйца трихоцефального типа, но отличаются скульптурой оболочки
в виде множества мелких вдавлений; в оптическом сечении оболочка
яйца производит впечатление радиальной исчерченности; размеры
яиц — 55 на 30 при крайних вариациях длины — 67—49 р и толщины
33—27 р (рис. 289; табл. VI, рис. 17).
Паразитируют у крыс (черной, александрийской), пасюка, мыши,
хомяка, зайца, собак, пекари, шимпанзе, Ateles geoffroyi, Cebus
capucinus— у всех в печени, где откладывают яйца (рис. 289, С). Воз-
можно паразитирование у человека. Первый случай установлен на
360
СПЕЦИАЛЬНАЯ ЧАСТЬ
вскрытии; в печени были обнаружены массовые скопления яиц (Индия).
В экспериментальных условиях может заражать белых мышей, белых
крыс и сурков. В Азербайджане найден у хомяка Cricetulus migrato-
Рис. 289А — Яйца нематоды Hepaticola. hepatica.
а — из faeces гольда (Q к р я б и н, Подъяпольская и
Ш'ихоб ал ов а); b — незрелое и с—зрелое яйцо.
По Фюллеборну.
Рис. 289 С Hepaticola hepa-
tica. Передний конец самки с
капиллярным пищеводом и по-
ловым отверстием (по Nishigo-
ri) и яйцо с вылупляющейся
личинкой.
Рис. 289В — Яйца Hepaticola hepatica
на срезе печени крысы. Рис. по микрофо-
тографии. По Faust и Martinez,
rius (Н. Колесников). У человека с достоверностью обнаружен раз. в
печени солдата, жившего три года в Индии. Известны случаи нахожде-
ния яиц этого паразита в faeces человека. Скрябин, Подъяпольская и
Шихобалова видели их в фекалиях трех гольдов в Приамурье. Неодно-
Таблица IX
Тrichocephalus suis. Свиной власоглав, самка. Оригинал В. Сондак; рисовал
И. Теравский
О — пищевод, d — кишечник, а — anus, с — кутикула, v — влагалище, и — матка, ov — яичник,
od— яйцевод, ovj выбрасыватель яиц, х — сужение между пищеводом и кишечником
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
361
кратно обнаруживались в аналогичных условиях в Панаме и в других
местах. Нахождение яиц в фекалиях человека не может считаться
доказательством его действительной зараженности рассматриваемым
паразитом. Яйца могут быть «транзитными», если человек ранее ел
печень животного, пораженную С. hepatica (см. выше данные Штрома,
стр. 241). A. Foster и С. Johnson установили наличие С. hepatica у пе-
кари (дикая свинья) и у некоторых обезьян, печень которых употреб-
ляется в Панаме в пищу. В фекалиях туземцев деревень и обнаружи-
вались яйца С. hepatica. В таких случаях речь идет о мнимом госте-
паразитировании С. hepatica у человека. Наряду с этим возможны не-
распознаваемые пока при жизни случаи действительного паразитирова-
ния С. hepatica у человека, когда яйца этого паразита не попадают в
кишечник из пораженной печени.
Жизненный цикл в некоторых деталях описывается противо-
речиво. По данным Nichigori, паразит откладывает яйца в ткань
печени, причем они никогда не выходят из нее в кишечник, а остаются
как бы замурованными. Здесь же в них развиваются личинки, и яйца
делаются инвазирующими. При пожирании здоровым животным живот-
ного с указанной формой яиц паразита из проглоченных яиц вылупля-
ются в кишечнике личинки .(рис. 289, В), которые вбуравливаются в
стенку кишки и по портальной системе попадают в печень, где стано-
вятся половозрелыми.
Другие авторы утверждают, что яйца не могут достичь в печени
инвазирующего состояния, что дальнейшее развитие их протекает во
внешней среде. Эту точку зрения поддерживают Скрябин, Подъяполь-
ская, Шихобалова. Очевидно, необходимы дальнейшие исследования
для решения спорных вопросов биологии этой нематоды.
Этот паразит вызывает цирротические процессы в печени.
Диагноз по яйцам в faeces или на секционном материале на со-
скобе из разреза печени.
Лечение неизвестно.
Семейство Capillariidae
Род Thominx
Thominx аёгорЫШз (Creplin) Skr.
Синоним: Capillaria аёгophila.
Тонкая нитевидная нематода (рис. 290) —самка 18,2—18,9 мм дли-
ны при наибольшей толщине до 0,1 мм. Самец — около 17 мм длины
и до 0,06 мм толщины.
Ротовое отверстие лежит на переднем конце. Пищевод самки немно-
го менее $4 длины всего тела.
Анальное отверстие лежит субтерминально. Задний конец тела
округлен. Отверстие влагалища лежит позади уровня заднего конца
пищевода.,
Яйца боченковидные, чуть асимметричные, 65—^1 Р длины и
29—31 р ширины, с «пробочками» на концах. Поверхность оболочки
изборождена извилистыми вдавлениями.
У самца имеются спикула и длинное, с плотными стенками трубко-
видное спикулярное влагалище, усеянное по поверхности
мелкими шипиками.
362
СПЕЦИАЛЬНАЯ ЧАСТЬ
Паразитирует в трахее, легких и в носовой полости различных хищ-
ных (кошка, собака, волк, лиса, соболь, уссурийский енот и др.).
Описанный Н. Корниловичем случай нахождения яиц Trichocepha-
lus dispar на срезах трахеи кошки в б. Юрьеве является, видимо, пер-
вым нераспознанным обнаружением Th. aerophilus в России. Затем
многократно был находим К. Скрябиным (1918 г.), А. Петровым и др.
у кошек; единичные находки известны у собак и волка.
Рис. 290 — Thominx aerophilus, самка. Яйца в двух положениях.
Структура скорлупы яйца (при иммерзионном увеличении).
По К. Скрябину.
Заражение происходит проглатыванием яиц, созревших до инвази-
рующего состояния; соответствующее развитие яиц достигается при
оптимальных условиях в сорокадневный срок (Christenson). Яйца
вскрываются в кишечнике, личинки гематогенным путем мигрируют в
легкие; на все это идет около 7—10 дней. В легких паразиты становят-
ся половозрелыми дней через 40. Живут паразиты не более года.
Известен единственный случай паразитирования Th. aerophilus у
человека, страдавшего тяжелой формой трахео-бронхита; обнаружен-
ные в его мокроте Альперович яйца были отнесены К. Скрябиным
(1939) к Th. aerophilus (рис. 290).
Больной выздоровел приблизительно через год без каких-либо спе-
циальных мероприятий по отношению к паразитам.
Налицо — случай гостепаразитизма типичного паразита хищных у
случайного хозяина, каким оказался человек.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
363
Надсемейство Dioctophymoidea
Род Dioctophyme Collet—-Meygret
Dioctophyme renale Goeze
Синонимы: Strongylus gigas Rud., Eustrongylus gigas Rud., Eustrongylus visce-
ralis Gm.
Свайник-великан является крупнейшей нематодой с толстым
вальковатым телом яркокрасного цвета. Самка до 100 см длины и
5—12 мм толщины; самец — 40 см длины и 4—7 мм толщины. Голов-
Рис. 291Л — Свайник-вел икан в почке собаки.
Видна атрофия почечной ткани. Оригинал.
ной конец несет два концентрических кольца из двенадцати сосочков
вокруг ротового отверстия; внутреннее кольцо состоит из 6 мелких, а
наружное — из такого же числа крупных сосочков.
Задний конец самца несет колоколообразную совокупительную сум-
ку, из которой выдается одна единственная спикула длиной до 6 мм.
Половое отверстие самки ле-
жит у начала пищевода, мил-
лиметрах в восьми (Скрябин),
а по Faust — в 5—8 см близ
переднего конца тела. Аналь-
ное отверстие лежит терми-
нально. Яйца овальные, с бо-
лее острыми полюсами и тол-
стой оболочкой, покрытой на
поверхности полукруглыми
сдавлениями; длина яиц 64—
77—83 р (ширина — 40—45—
47 р) (рис. 291). Яйца откла-
дываются с несозревшим за-
родышем; оболочка яйца очень
стойка, и зародыш может
Рис. [291В — Свайник-великан —Dioctophyme
renale. Яйца. По К. Скрябину.
оставаться живым в нем при подходящих условиях даже несколько лет.
Паразитирует в почечной лоханке . собаки, тюленя, выдры; может
также паразитировать у быка, лошади и человека. Космополит.
364
СПЕЦИАЛЬНАЯ ЧАСТЬ
В СССР известен из ряда мест, начиная от Ленинграда и до Ташкента.
Своим присутствием вызывает атрофию почечной- паренхимы; от почки
может остаться только тонкая капсула, почти из одной соединительной
ткани (рис. 291). Паразит бывает иногда и в мочевом пузыре, мочеточ-
никах, брюшной полости и др. При попадании в мочеиспускательный
канал закупоривает его и причиняет сильнейшие боли и острую уре-
мию. Моча становится кровянисто-гнойной; в ней обнаруживаются яйца
паразита.
Цикл развития с полной ясностью еще не изучен. Подозревают рыб
как промежуточных хозяев этого паразита (например, язь, Ciurea).
II
н
Рис. 292 — Strongyloides stercoralis.
А — свободно живущий самец; В — то же — самка; С — рабдит-
ная личцнка; D — филяриевидная личинка. Из Брауна и
Фауст а.
У человека известно несколько случаев гостепаразитирования
D. renale — в уретре, мочевом пузыре, в почечной лоханке. Является
опасным паразитом.
Диагноз — по нахождению яиц в моче и в faeces при попадании
в них мочи (низовье Амура; Скрябин и Подъяпольская). За яйца
паразита однажды в клинике были приняты шарики лейцина. Самого
паразита могут до некоторой степени симулировать длинные сгустки
крови, образующиеся в уретре при пиелонефрите. Распознать послед-
ние легко, так. как они не имеют никаких структурных черт организма
червя. В случае нахождения в почке только одного самца возможность
паразитологического диагноза по яйцам, конечно, отпадает.
Лечение — хирургическое, после надежно поставленного парази-
тологического диагноза.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
365
Профилактика. Принимая во внимание предполагаемую воз-
можность заражения от рыбы с инцистированными личинками парази-
та, есть только хорошо обработанную термически рыбу.
PH AS MIDI А
Надсемейство Rhabditoidea
Паразитирующие формы нематод сем. Rhabdiasidae являются или
гермафродитами, или партеногенетическими самками; свободно живу-
щие стадии перед достижением инвазирую-
щего состояния могут проходить стадии раз-
дельнополости (гетерогония).
Род Strongyloides Grassi
Strongyloides stercoralis Bavay
Синоним: Anguillula stercoralis и A. intestinalis Bavay. j
У гр и ц а кищечная по сравнению с дру-
гими нематодами имеет едва ли не самое слож-
ное развитие, характеризующееся сменой сво-
бодно и паразитически живущих поколений.
Рис. 293 — Strongyloides stercoralis — паразитическая
самка в стенке кишки человека (срез).
-Е— эпителий слизистой оболочки; С — полости, в которых лежала
угрица; D — развивающийся зародыш в яйце угрицы; 5 — подслизи-
стая оболочка; V—угрица (продольный разрез). По Асканази,
из В г и ш р t.
Рис. 294 — Strongyloides
stercoralis. Рабдит ная
личинка от человека.
Но Н. Kreis.
И — губа; тс — ротовая по-
лость; ое — пищевод; Ьи^—
бульбус; пг—нервное кольцо;
Ьиг - второй бульбус; in —
кишечник; ov - яичник; ап —
анальное отверстие.
По Н. К г е i s .
366
СПЕЦИАЛЬНАЯ ЧАСТЬ
СВОБОДНОЖИВУЩАЯ ГЕНЕРАЦИЯ1
Развитие этой стадии происходит в почве. Эта фаза жизненного
цикла паразита почти постоянно наблюдается в теплом климате, где
влажность и наличие питательных веществ в почве обеспечивают суще-
ствование свободноживущей стадии (рис. 292). Веретеновидный самец
имеет в длину 0,7 мм и 0,04—0,06 мм в плоскости наибольшего диа-
метра. Хвостовые крылья отсутствуют, но имеется губерна-
к у л ю м (рис. 295, 7. Gu) и две спикулы. Задний конец тела заострен и
загнут вентрально. Самка такой же толщины (1 мм X 0,05—0,07 мм);
имеет двойную матку и короткую вульву, которая открывается близ сере-
дины вентральной стороны тела. Развивающиеся яйца наполняют мат-
ку, и тогда последняя занимает большую часть тела червя. Оплодотворен-
ная самка несет частью зрелые яйца, которые в несколько часов закан-
чивают свое развитие, после чего происходит вылупление личинок. По-
явившаяся первая стадия — рабдит о видная личинка (рис. 292^
294) — имеет форму тела и структуру пищевода, типичные для рода; за
булавовидной передней частью пищевода идет позади его середины
суженный отдел и сзади имеется бульбус. Ротовая полость короткая и
малого диаметра. Личинка прожорливо питается за счет органических
веществ почвы, однократно линяет, снова переходит к питанию и быст-
ро вырастает в свободноживущего взрослого червя. При оптимальных
условиях свободноживущая фаза может повторяться неопределенно
долгое время. При неблагоприятных условиях развития рабдитовидная
личинка после питания превращается в активную стадию — филярие-
видную личинку (рис. 292, D), которая имеет длинное нежное
тело и, за исключением выемки на конце тела, напоминает личинку
анкилостомид. Эта личинка является инвазионной для чело-
века почти непосредственно после линьки, но может жить в почве в
течение нескольких недель.
ПАРАЗИТИЧЕСКАЯ ГЕНЕРАЦИЯ
При контакте с кожей и, реже, со слизистой ротовой полости чело-
века филяриевндные личинки проникают в мелкие кровеносные сосуды;
большая часть их достигает с током крови правого сердца и поступает
в легкие. Здесь они разрывают легочные капилляры, выходят в альвео-
лы (рис. 295, /), где после двух последовательных стадий превращают-
ся во взрослых червей (самок и самцов, рис. 295, 3—5). Оплодотворе-
ние самок может происходить в бронхах, трахее, и позднее — в кишеч-
нике, после того как черви спускаются в последний.
Копуляция имеет место раньше проникновения самок в эпителий этих
органов, так как самцы не являются паразитами тканей и после корот-
кого пребывания в организме хозяина выходят с калом. Некоторые
самки иногда внедряются в. эпителий воздухоносных путей и присту-
пают к кладке яиц, но большая часть их поднимается к надгортанни-
ку, проглатывается и затем поступает в кишечник; реже они задержи-
ваются в пилорической части желудка. В дальнейшем личинки мигри-
руют в слизистую, достигают половой зрелости и приступают к кладке
яиц в ткани (рис. 293). Инкубационный период у человека длится око-
ло 28 дней.
1 Цитата из Ch. Craig и Е. Faust. Clinical parasiitology, 1937, стр. 244—246.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
367
Паразитические самцы рабдитовидного типа (рис.
295, 6), почти подобные самцам свободноживущей генерации (рис. 295, 7),
имеют слабо выраженную ротовую полость. Самки — нежные нитевид-
ные черви (2,2 мм X 20—50 рр). Цилиндрический пищевод тянется че-
Рис. 2954.
' 1. Филяриевидная личинка Strongyloides intestinalis из ткани легких собаки чеоез [три дня
после заражения чеоез кожу.
2. Следующая личиночная стадия из ткани легких: NR — нервное кольцо; Е — пищевод; GPS-
половой зачаток; JN — кишка; AN — анальное отверстие; РА — сосочки. По Faust.
рез переднюю треть ‘ или 2/5 тела паразита. Задний конец заострен,
и анальное отверстие занимает средне-вентральное положение, на ко-
ротком расстоянии от конца тела. Вульва открывается близ или немно-
го кзади середины червя. Парная матка (яйцеводы и яичники) тянется
под прямым углом от короткой вульвы, одна из них направляется кпе-
реди, другая кзади.
После полового созревания самка откладывает несколько дюжин
яиц, часть из которых уже заключает в себе личинку. Яйца имеют тон-
кую оболочку, прозрачны, овоидной формы (50—58 1^X30—34 уь );
вскрываются они обычно в тканях. Вышедшие личинки мигрируют в
просвет кишечника и выбрасываются с калом. Наиболее частая лока-
лизация паразитов — двенадцатиперстная кишка (рис. 293) и верхний
отдел тонких кишок; впрочем, за исключением червеобразного отростка
368
СПЕЦИАЛЬНАЯ ЧАСТЬ
черви могут инвазировать любой участок кишечника от пилоруса до
ануса. При гиперинвазии все или часть рабдитовидных личинок в про-
свете кишечника превращаются в мелких филяриевидных личинок; по-
следние или вызывают реинвазию, проникая в слизистую кишечника и
далее через портальную систему поступая в легкие, или выбрасываются
Рис. 295В.
ба. Передний конец того же самца; открытая ротовая полость. 6Ь — развитие спикул (SPE) л
рулька (GU)-
3— 11редтзрослая самка Str. stercoralis из ткани легких собаки через 11 дней после заражения
через кожу. Половой зачаток еще не разгосся.
4. То же. самец; рабдитнпя форма пищевода.
5. Взро.лая самка из легких через J5 лней после заражения через кожу/
6. То же, самец через 21 день после заражения через кожу. Разрастание полового зачатка.
во внешнюю среду. Эти личинки могут также внедряться в кожу про-
межности и околоанальной области («аутоинвазия», по Фюллеборну,
1926). Случаи длительного стронгилоидоза человека (свыше 15 лет),
при отсутствии возможностей внешнего заражения, кажутся объясни-
мыми с точки зрения гиперинвазии; вполне вероятно, что самки пара-
зитов живут продолжительное время.
Таким образом, полный жизненный цикл Strongyloides представляет-
ся в следующем виде (рис. 296).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
369
1. Непрямое развитие; свободноживущая генерация червей,
при наличии неблагоприятствующей среды обитания, превращается в
паразитическую генерацию; наблюдается в теплом и влажном климате.
2. Прямое развитие: обусловлено паразитической стадией, но
редко бывает изолированным от первого способа развития. Обычно в
условиях умеренного климата.
3. Гипер инвазия является результатом аутореинвазии носителя
паразита. Теплый климат.
В трактовке жизненного цикла угрицы кишечной, даваемой Фаустом
Рис. 295 С—Strongyloides stercoralis.
7. Взрослый самец из лег<их собаки через 57 дней после заражения через кожу. Е — пи-
щевод: NR — нервчое колыи; IN — кишечник: GP — лилово i зачаток; RE~ rectum; МС — ро-
тозач полость; SP - стилет; UT — матка; VU — влагалище; REF - яичник; 1Е — семенник;
VGL — вентральная железа; SPI — спикулу GU - guberuaculum; РА — сосочки; BU— бульбус.
По Faust.
и его школой, вовсе отсутствует фаза партеногенетической самки, кото-
рая фигурирует в прежних представлениях паразитологов.
Krejs (1932), изучая экспериментально непрямой цикл развития. Str.
simiae на обезьянах, не мог обнаружить паразитических самцов, почему
и предположил наличие в данном случае одних партеногенетических
самок.
Фауст находит теоретическое объяснение этому явлению. При опти-
мальных условиях свободноживущая половая генерация может повто-
ряться неограниченно долгое время. При наступлении неблагоприятных
внешних условий жизненный цикл угрицы кишечной укорачивается, и
среди рабдитных личинок появляются будущие самки или самцы, после
прохождения общей для всех фазы филяриевидных личинок. Теорети-
чески можно предполагать, что отсутствие или редкость паразитических
370
СПЕЦИАЛЬНАЯ ЧАСТЬ
самцов зависит от заражения хозяина лишь филяриевидными личинка-
ми, которые являются будущими самками.
Strongyloides st er cor alts является, по всем данным, специфическим
паразитом человека. Весьма существенное значение имеет то обстоя-
тельство, что у некоторых животных,
Рис. 296 — Схема жизненного цикла угрицы
кишечной. Фил. L — филяриевидная личинка;
Раб. Z, — рабдитная личинка.
Составил Е. Павловский.
например у собак, бывают пара-
зиты, морфологически неотли-
чимые от Strongyloides stereo-
ralis человека. Опыты с пере-
крестным заражением показа-
ли, что человеческие Strongy-
loides незаразительны для со-
баки. Таким образом, на этом
примере мы видим наличие
существенных биологических
различий между организмами,,
и по внешности, и по внутрен-
нему строению сходными друг
с другом. Морфологически эти
группы паразитов могут быть
признаны за один и тот же
вид, но биологические особен-
ности требуют различения их
на положении биологических
или физиологических видов.
Впрочем имеются указания на
однократную заразительность
человеческих Str. stercoralis
для собак. Этот паразит вы-
зывает кохинхинскую диаррею
или тропическую дизентерию.
Распространен в Японии,,
Китае, Бразилии, Египте, Аф-
рике. Занесен в Сибирь (Томск)
и Западную Европу.
В СССР имеется в качестве
эндемичного очага в Сучан-
ских копях (Дальний Восток,
Г. Змеев). Кроме того, единич-
ные случаи обнаружены в
Средней Азии, в Закавказье,
в Донбассе, в Харьковском
округе и др.
Патогенное значе-
н и е.“ Так как жизненный цикл
угрицы кишечной связан с
развитием ее в почве и с зара-
жением человека через почву,,
то стронгилоидоз может носить характер профессионального заболева-
ния у земляных рабочих, шахтеров и др. На юге возможно его разви-
тие и в окопах во время военных действий, когда контакт находящихся
в окопах бойцов с почвой велик, а санитарно-гигиеническая обстановка
неудовлетворительна. В кишечнике паразиты внедряются в подслизи-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
371
стую оболочку, в либеркюновы железы, в лимфатические узлы. Поносы
являются, невидимому, результатом действия угрицы на человека.
В период внедрения личинок в кожу наблюдаются различные дер-
матиты (зуд, крапивница, отечность
и др.); они затихают и в период
миграции паразитов по телу не да-
ют о себе знать до времени обосно-
вания угриц кишечных в тонких
кишках.
Диагностика. Диагноз ста-
вится по нахождению в faeces личи-
нок типа Rhabditis. Паразитические
самки с испражнениями обычно не
выходят. После дачи слабительного
в faeces можно обнаружить и яйца
угриц.
При слабой степени заражения
личинок Sirongyloides можно из-
влечь из faeces прибором Бермана.
Профилактика. Предохра-
нение покровов человека от сопри-
косновения с зараженной почвой.
Предохранение почвы от загрязне-
ния ее фекалиями больных стронги-
лоидозом. Общие гигиенические
предосторожности при питье воды и
ных стронгилоидозом и лечение их
стомозе.
Рис. 297 — Rhabditis pellio.
А — передний конец с пищеводом и ротовой
полостью; В — задний конец самца со спику-
лами, совокупительной сумкой и подпирающими
ее ребрами. По Oeuley из Фауста.
принятии пищи. Выявление боль-
Детали см. ниже — об анкило-
Род Rhabditis Dujard
К роду Rhabditis относятся мелкие угрицы с невооруженной ротовой
капсулой (рис. 297, Л). Они обитают в разлагающихся веществах
почвы.
Известен ряд случаев нахождения угриц в теле человека, что при-
ходится рассматривать как ложный паразитизм. Несомненно, что часть
наблюдений должна трактоваться как мнимый ложнопаразитизм угриц
у человека., Угриц находили в моче, во влагалище, в faeces, в двенад-
цатиперстной кишке, в желудке, в папулах кожи и др.
Rhabditis pellio Dujard
Синонимы: Anguillula mucronata Gr., Rhabditis genitalis Scheib.
Беловатый паразит, самец которого достигает 1,05 мм, а самка—1,3 мм дли-
ны; хвостовой конец самца — с сердцевидной сумкой, подпертой девятью парами
ребер (рис. 297, В). Спикулы короткие, почти одинаковые по длине. Хвостовой ко-
нец самки вытянут в длинный конический отросток. Рот окружен тремя широкими
округлыми губами, из которых каждая имеет по паре маленьких сосочков.
Яйца овальные, 0,06 мм длины и 0,035 мм ширины. Развитие и вылупление ли-
чинок происходит на месте, т. е. в матке самки. Личинки заполняют полость тела
самки и выедают ее органы, так что в конце концов остается только мешок
кутикулы самки, в результате разрывающийся и освобождающий заключающееся в
нем потомство червя. Личинки часто живут в дождевых червях, а взрослые черви
обычно обитают в распадающихся органических веществах почвы.
Rh. pellio бывал находим во влагалище (и в кислой моче) женщин. Без сомне-
ния, в таких случаях дело шло о ложнопаразитизме. Отметим, что у некоторых на-
372
СПЕЦИАЛЬНАЯ ЧАСТЬ
родов (например, у венгров) в качестве лечебного средства употребляется сырая
земля, прикладываемая к половым органам женщины; таким путем находящиеся
в земле черви могут легко перейти во влагалище.
Rhabditis nellyi Blanch.
Описан весьма недостаточно по рабдитным личинкам, полученным из зудящих
капсул у 14-летнего юноши, болевшего кожной болезнью, похожей на «craw-craw»
Западной Африки. Больной за пределы Европы не выезжал. В каждой капсуле
~ [ длины и 0,013 мм толщины. Кутикула
<было по одной или нескольку личинок 0,33 мм
Рис. 299 — Rh'ibditis hominis —
самка и головной конец паразита.
По Sandground.
Рис. 298 — Rhabditis schachtiella (слева) —
самка. По Скрябину и Шульцу.
Rh. gracilis (справа)—самец. По Ш и н га-
ревой, Демидовой иКудрявцеву.
их тонко исчерчена. Заднее утолщение пищевода с зубом. Анальное отверстие лежит
чуть кпереди от заднего конца тела;
Весьма вероятно, что заражение произошло внедрением личинок в кожные по-
кровы; К какой взрослой форме принадлежат описанные личинки, неизвестно.
Rhabditis donbass Skr., Sch., Sserb.
Мелкие, относительно толстые черви с 6 крупными губами. Самец 0,895—0,930 мм
длины и 0,055—0,074 мм ширины. Хвостовой конец отторочен широкими крыльями.
Одинаковые по величине спикулы соединены друг с другом на свободном конце.
Самка 1,06—1,145 мм длины и 0,103 мм ширины. Отверстие влагалища приходится
почти на середине тела. Яйца—66X37 у.. На хвостовом конце торчит резко вы-
дающийся острый придаток.
Несколько раз был найден в испражнениях в Донбассе. Так же, как и Rh.
schachtiella Skr. et Sch., Rh. gracilis (Sching., Demid, et Kudrlaw. (рис. 298),
•едва ли может быть причислен к истинным паразитам человека.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
37$
Rhabditls hominis Kobayashi
Тело цилиндрическое, утончающееся к переднему и заднему концам.
Кутикула тонко исчерчена в поперечном направлении. Рот с четырьмя
губами. Пищевод распадается на четыре отдела: два трубчатых участ-
ка и два шаровидных вздутия (рис. 299). Длина его от 0,17 до 0,2 мм.
Самец 0,9—1,2 мм длины и 0,03—0,05 мм толщины. На заднем конце
тела имеются кутикулярные крыловидные придатки, подпертые на каж*
дой стороне шестью короткими ребрышками. Две одинаковые повели»»,
чине спикулы имеют шишкообразную головку.
Самка — 1,5—2,0 мм длины и 0,12 мм толщины (рис. 299). Задний
конец вытянут в острый отросток. Половое отверстие помещается на
середине длины тела. У молодых самок в каждом из двух стволов.
матки помещается по 10—50 яиц эллипсоидной формы, 44X28 р;
У старых самок рабдитные личинки вылупляются из яиц внутри их те-
ла. Личинки, выходящие из самок, 0,24—0,3 мм длины и 0,012 мм тол-
щины. Развиваются они во взрослых червей в faeces или на разлагаю-
щемся органическом субстрате.
Личинки Rh. hominis могут быть смешаны с личинками Strongyloi^
des stercoralis. 1
Sandground дает следующую таблицу для их различения:
Дифференциальная диагностика Rh. hominis ис в о б о д н^о живу щих.
Strongyloides stercoralis
Длина .... .......
Ширина .......................
Ротовая капсула ..............
Совокупительная сумочка — есть, хотя
0.9—1.3 мм
0.03 -0.05 мм
0.02 мм длины]
часто незаметна
Rh. hominis" Str. stercoralis]
С а-м цГы 3
'0.7—0.9 мм*
0 035—0.04 мм
0.013 мм длины
отсутствует
С а^м^кли
Длина ....................... 1.4—2 мм 1—1.2 мм
Ширина....................... 0.12 мм 0.05 мм
Ротовая капсула ... ... как у самца как у самца
Размножение . ....................... яйцеживорождение обычно откладка£яиц
Яйца 24 — 44 X 32 — 28 ь...........обычно лежат в каждом
стволе матки в два
ряда — от 20 до 50 обычно лежат в один
штук ряд — от 16 до 18 штук
Личинка (молодаг, рабдитная)
Длина .......... 0.024—0 03 .мм 0.2—0 25 мм
Ширина .... 0.012—0.03 \лм 0.0;6 мм
Ротовая полость 0Л 15 — 0.019 мм 0.008—0.01 мм
Половой зачаток 0.022—0.024 мм длины 0.034—0.0^6 мм длины
Личинка . . обычно развивается в Личинка или развивает*
половозрелую рабдит- ся в половую проме-
ную форму жуточную рабдитную
генерацию, пли пре-
вращает ся в филярие-
видную личинку
Rh. hominis был неоднократно обнаруживаем .у японских школьни-
ков; позднее этот червь был найден и в С. Америке; во всех случаях
диагноз ставился по исследованию faeces. Личинки этого червя могут
быть смешаны с личинками Str. stercoralis и анкилостомид, особенно
если исследуются не свежие испражнения, а постоявшие более суток.
За это время из яиц анкилостомид могут уже выйти рабдитные личин-
374
СПЕЦИАЛЬНАЯ ЧАСТЬ
ки. Распознавание названных форм производится измерением длины
ротовой капсулы, которая у личинок некатора и анкилостомы имеет
средние размеры 0,01—0,12 мм длины против Rh. hominis 0,015—0,019 мм
и Str. stercoralis 0,088—0,01 мм. Мерой для сравнения относительных
размеров капсулы этих личинок может служить толщина тела личинки
на уровне основания ротовой капсулы. У личинок анкилостомид эти
размеры одинаковы, у личинок Rh. hominis капсула длиннее ширины
тела, а у личинок Str. stercoralis — короче.
Рис. 300 — Стадии созревания зародыша в яйцах Heterodera
radicicola (= Oxyuris incognita). По Sandground.
Попытки заражения человека личинками Rh. hominis, взятыми из
культур, остались безуспешными как при проглатывании их, так и при
нанесении на кожные покровы. Этот червь, повидимому, является оби-
тателем почвы, и в faeces человека он попадает или непосредственно
из таковой (при собирании с земли материала для исследования), или
заносится в испражнения мухами. Таким образом, Rh. hominis может
считаться мнимым ложнопаразитом человека. Этот червь лег-
ко культивируется на смеси животного угля с faeces и на агар-агаре.
Личинки его более стойки к высыханию, чем взрослые черви.
Род Turbatrix
Turbatrix aceti Muller — угрица уксусная
Qhhohhmh: Anguillula aceti Mull., Rhabditis dceti
Тело уксусной угрицы покрыто* прозрачной гладкой кутикулой. Длина
самца 1—2 мм при толщине 0.024—0.04 мм; две равные спикулы длиной 0.038 мм.
На хвосте с вентральной стороны четыре пары сосочков, а с дорзальной — одна
пара. Хвостовая бурса отсутствует. Самки 2—4 мм длины, 0.04—0.07 мм толщины. Поло-
вое отверстие в задней половине тела. Отрождает живых рабдитных личинок
0.222 мм длины и 0.012 Мм толщины. Живут в уксусе и в различных жидкостях с
уксуснокислым брожением. Бывали находимы в моче и во влагалищных выделениях.
Весьма вероятно, что уксусные угрицы в этих случаях были занесены извне, т. е.
при полосканиях и обмываниях уксусом. Известны случаи заноса угриц в мочу ла-
бораторными приборами при диагностическом исследовании (пипеткой). Следова-
тельно, и здесь речь может итти о мнимом ложнопаразйтизме угрицы кишечной у
человека.
Надсемейство Anguillulinoidea
Члены этого надсемейства или свободно живущие с а п р о з о и, или
же паразиты различных растений. Глотка этих червей мо-
жет выдвигаться наружу в форме копья.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
375
Anguillulina putrefaciens Kuhn
Синоним: Trichina contorta Botkin.
Этот червь паразитирует на луке. Боткин описал случай нахождения этого ви-
да в рвотных массах больного. Нематоды были, без сомнения, съедены с луком и
в качестве ложнопаразитов обнаружены у данного субъекта.
Heterodera radicicola Greef
\
Синоним: Oxyuris incognita Kofoid et White.
Heterodera radicicola является паразитом редиса, картофеля, брюквы, сельдерея,
моркови и других овощей. При поедании овощей, зараженных этой нематодой, яйца
ее обнаруживаются в faeces. При обследовании на яйца глист в Техасе 140 тыс.
солдат Kofoid и White обнаружили у 429 человек яйца 82—120р- длины и 24—43 р-
ширины, удлиненно-овальной формы, с округлыми полюсами и одной более утолщен-
ной стороной. Оболочка двойная, контурная; на каждом полюсе яйца зеленовато-
синее сплюснутое овоидное образование (рис. 300). Зародыш в стадии то морулы,
то червячка. Сам паразит не был ни разу обнаружен; найденные яйца были отнесе-
ны к новому виду Oxyuris incognita. Опытным путем Sandground доказал, что та-
кие яйца принадлежат Heterodera radicicola. Полярные образования отсутствуют в
свежих яйцах этой нематоды, но появляются вследствие дегенерации содержимого
яйца при изготовлении пищи из зараженных нематодами овощей и при прохождении
яиц через кишечник человека, а также при лежании их в разведенном спирте.
В таких случаях налицо транзит сторонних яиц через кишечник
человека и явление мнимого ложнопаразитизма Heterodera radicicola у человека.
Никакого патогенного значения для человека эта нематода не имеет, но транзит ее
яиц может ввести в заблуждение наблюдателя при диагностическом исследовании
faeces. 1 _
Надсемейство Strongiloidea
Ротовая капсула без зубов или режущих кутикулярных пластинок;
ротовое отверстие, как правило, окружено венцом хитинизированных
придатков (corona radiata). Женское половое отверстие лежит в
задней половине тела. Самцы с парой одинаковых спикул и копулятив-
ной бурсой. Взрослые черви обычно паразитируют в пищеварительном
канале.
Род Oesophagostomum Molin
Паразиты этого рода присущи человекообразным обезьянам, но бы-
вают и у человека, что считается одним из паразитологиче-
ских доказательств родства человека с человеко-
образными обезьянами. В СССР могут быть обнаружены
лишь в качестве завозных паразитов у лиц, заразившихся ранее в тро-
пиках.
Oesophagostomum apiostomum Willach
Синоним: Oesophagostomum brumpti Raill. et Henry.
Может быть назван узелковым червем вследствие локализации его
.личинок в узелках тканей кишечника. Является обыкновенным паразитом толстых
кишок гориллы, оранга и других обезьян Западной Африки, Филиппинских островов
и Китая.
Червь покрыт поперечно-исчерченной кутикулой. Рот окружен шестью околоро-
товыми сосочками; внутри от них расположена corona radiata, состоящая из 12 пи-
рамидальных кутикулярных выростов. В ротовой полости лежат три маленьких зу-
ба. Половое отверстие самки лежит тотчас перед анальным. Яйца очень похожи на
анкилостомные; их длина 60—83 р-, а ширина 27—40 Р- Самка 8,5 мм длины и
0,295—0,325 мм толщины. Самец 8—10 мм длины и 0,3—0,35 мм толщины. На зад-
нем конце его тела имеется колоколовидная бурса, подпертая ребрышками. Спикулы
длинны и искривлены.
Цикл развития. Яйца, выброшенные с faeces, дают рабдитных личинок,
которые живут в почве, дважды линяя и достигая состояния инвазирующей стадии.
376
СПЕЦИАЛЬНАЯ ЧАСТЬ
Последние проглатываются хозяином, проходят без бреда для себя через желудок
и тонкие кишки и обосновываются в слепой кишке; здесь личинки линяют, внедря-
ются в стенку кишки, где образуются реактивные узелки патологически измененных
тканей. В толще узелков личинки созревают, затем выходят в просвет кишки; здесь
они прикрепляются к стенке, растут и достигают половозрелое™. При выходе па-
разитов из узелков могут появляться кровотечения из поврежденных сосудов с раз-
витием дизентерийных симптомов. Черви могут выйти в противоположном направле-
нии, т. е. в брюшную полость, вследствие чего может развиться перитонит.
Прижизненная микроскопическая диагностика невозможна
вследствие внешнего сходства яиц Oesophagostomum с яйцами Ankylostomatidae.
Противоглистное лечение достигается дачей тимола, хеноподового
масла или четыреххлористого углерода.
Известны и другие виды Oesophagostomum.
Род Ternidens Raill. et Henry
Ternidens deminutus Raill. et Henry
Паразит толстых кишок гориллы, других обезьян и человека (тропическая Афри-
ка и Азия). Ротовая капсула полушаровидная. Рот сдвинут на спину. Corona radiata-
двойная. В глубине ротовой капсулы три сложных зуба. Самка 12—16 мм длины
и 0.65—0.73 мм толщины. Половое отверстие лежит близко перед заднепроходным.
Яйца прозрачные, 60 р- длины и 40 р* ширины. Самец 9.5 мм длины и 0.56 мм тол-
щины. Бурса с слегка зазубренным краем. Спикулы до 0,9 мм длины. Цикл разви-
тия неизвестен.
Рис. 301—Свайник трахейный—Syngamus
trachea} самец и самка, при креп шин неся
к стенке трахеи грача. Головной конец самки
отошел от места прикрепления, в котором
видны резкие изменения стенки трахеи. Изо-
бражены передние концы паразитов.
По Павловскому.
крупных паразитов в трахее или
удушье. В
Род Syngamus Sleb.
Самец срастается задним
концом тела с областью поло-
вого отверстия самки, образуя
двойниковое животное в
форме буквы У (рис. 301). Ро-
товая капсула сильно развита
и по краю несет хитинизиро-
ванное кольцо. На дне капсу-
лы имеется 6—9 режущих зу-
бов. Спикулы короткие.
В трахее домашних птиц
паразитирует Syngamus tra*
chea, вызывающий значитель-
ные патологические изменения*
в месте своего прикрепления к
стенке этого органа. В месте
прикрепления самца бывает
реактивное разрастание кост-
ной ткани трахеи, так что го-
ловка паразита как бы кап-
сул ируется костной тканью,,
В капсулу самки присасы-
ваются мягкие слои внутрен-
ней оболочки трахеи; кост-
ная ткань разволокняется. Эпи-
телий слизистой оболочки раз-
рушается. Заметна значитель-
ная эозинофилия (Е. Павлов-
ский). Наличие сравнительно
бронхах птиц вызывает кашель и
трахее скота паразитирует Syng. laryngeus.
Таблица X
Рис. 301 A. Syngatmts trachealis, самец, в трахее грача. Поперечным
разрез. Видна ротовая капсула паразита; в ее полости трахеальная
ткань в виде выступа (Л), в основании которого находится костное
разрастание трахейного кольца (В). Красные точки обозначают
эозинофилов. Оригинал»
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
377
Яйца Syngamus заглатываются хозяином с отделениями дыхатель-
ных путей и выбрасываются с faeces. В сырой земле происходит их
развитие. Развившийся заро-
дыш выходит из яйца. Зарази-
тельными являются яйца со
сформировашимися зародыша-
ми или уже вышедшие личин-
ки. Будучи заглочены хозяи-
ном, личинки мигрируют в лег-
кие, достигают бронхов или
трахеи, где и обосновывают-
ся, превращаясь во взрослые
формы. Известны единичные
случаи гостепаразитирования
Syngamus у человека.
Syngamus kingi Leiper
Этот вид описан по экземпляру,
выкашлянному женщиной на Ан-
тильских островах. После выхода
паразита приступы хронического
кашля у больной прекратились. Весь-
ма вероятно, что S. kingi явтяется
паразитом некоторых кошачьих (пу-
ма) и идентичен S. dispar. Фауст
(1937) сводит его в синонимы к
S. laryngacus Raill.
Рис. 302—A. Necator americanus, В. 'Ancy-
lostoma duodenale; увеличено.
Семейство Ancylostomatidae1
Ротовая капсула развита хорошо; на ее вентральной стороне име-
ются режущие приспособления в форме зубообразных выростов или
полулунных пластинок. Паразитируют в тонких кишках.
Род Ancylostoma Dubini — кривоголовка
Ancylostoma duodenale Dubini — кривоголовка двенадцатиперстная
Головная часть паразита искривлена на спинную сторону, отсюда
и название — кривоголовка.
Красноватые червячки после смерти — белесоватого или серо-желтого
цвета. Самка длиною 10—13 мм, толщиною 0,6 мм, самец соответственно
8—10 мм и 0,4—0,5 мм (рис. 302, 303). Суженный передний конец
искривлен на спинную сторону; он заключает в себе большую ротовую
капсулу, вооруженную несколькими кутикулярными зубами (пара
брюшных передних, два боковых глубоких и- два спинных передних зуб-
ца — рис. 304, А, В). У основания п е р е д н е б р ю ш н ы х зубцов
открываются две одноклеточные головные железы, задним
концом доходящие до середины тела. В ротовую капсулу открывается и
спинная пищеводная железа. Пищевод занимает приблизи-
1 Раздел об апкилостомидах дан в более подробной разработке для показа-
ния основных точек зрения, которые должны быть учитываемы при паразитоло-
гическом изучении вопроса. Эта возможность обеспечивалась наличием щромпой
литературы и сводок по анкилостомидам и анкилостомидозам. К такой детализа-
ции необходимо приближаться в соответствующих вариантах и при изучении дру-
гих паразитических червей первостепенного патологического значения.
378
СПЕЦИАЛЬНАЯ ЧАСТЬ
а ь
Рис. 303 — A. Necator america-
nus,a—копулирующий, в нат.
велич.; b — то же, кривоголов-
ка двенадцатиперстная Ancylo-
stoma duodenale.
тельно Vs длины тела; он впадает в одетую толстой мышечной оболоч-
кой кишку, которая у самки открывается чуть отступя кпереди от зад-
него конца тела, а у самца вместе с половым отверстием впадает в
концевую совокупительную сумочку. Семенники оплетают своим изви-
вами часть кишки и впадают в семенной пузырек, открывающийся в се-
меизвергательный канал. Последний ведет в совокупительную сумочку,
которая Цринимает в себя также и отверстия двух цементных
железок, выделяющих клейкий секрет для прикрепления самца к
самке при копуляции (рис. 303). Спикул две; они равной длины
(до 1,9—2,0 мм). Совокупительная сумочка расширена колоколообразно
и состоит из двух боковых крупных лопастей и срединной маленькой
дольки; стенки бурсы подперты 11 ребрыш-
ками (рис. 304, Е, F).
Кишечник самки также окружен изви-
вами половых трубок двух яичников и двух
маток; стволы матки расходятся один кпе-
реди, другой кзади. Яичники извиты пре-
имущественно в поперечном направлении.
Половое отверстие лежит в начале задней
части тела. На заднем конце тела — ма-
ленький втяжной штифтик.
Яйца одеты тонкой прозрачной оболоч-
кой (рис. 305), зародыш в свежеотложен-
ных яйцах находится на стадии 2—4 (8)
бластомер. Яйца овальные с тупоокруглен-
их вариируют; средняя длина — 60 р, а ши-
ными полосами; размеры
рина 40 р. ’Могут попадаться и несегментированные яйца; при запорах
в задерживаемом кале развитие зародыша в яйцах может достигнуть
стадии рабдитной личинки.
Количество откладываемых самкой яиц вариирует в зависимости от
многих причин: возраста анкилостомы, количества паразитов у данного
хозяина, возраста и характера пищи последнего и др. За сутки самка
откладывает 5—10 тыс. яиц и более. Количество яиц в грамме кала
колеблется в зависимости от количества самок анкилостом у данного
хозяина и от причин, влияющих на половую продукцию этого паразита.
По разным авторам, на одну анкилостому приходится в грамме оформ-
ленного стула 47, 22, 30, 18, 52 яйца.
В самом отложенном кале яйца развиваются очень медленно, хотя
и сохраняют жизнеспособность до двух месяцев. В моче яйца погибают
через Р/г—2 недели. При смешении кала с землей и при попадании
яиц во влажную почву развитие зародыша идет быстро, если налицо
благоприятная температура (около 25° С), порозность и, следовательно,
хорошая аэрация почвы, отсутствие прямого влияния солнечных лучей
(затененность почвы), избытка влажности, кислотности и др. (pH —
= 4,8—5 неблагоприятно для развития личинок).
Физико-химические свойства почвы имеют существенное значение
для развития яиц анкилостомы. Засоленность почвы, обилие в ней
сульфатов и плотность грунта не благоприятствуют развитию зароды-
шей в яйцах, что в конце концов отражается в соответствующем месте
на степени зараженности населения анкилостомами. Наоборот, пороз-
ные — песчаные и песчано-глинистые почвы, богатые гумусом, являют-
ся хорошим субстратом для развития личинок.
В Азербайджане (Ленкоранский уезд) распространение анкилосто-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
379
моза полностью совпадает с распространением субтропических желто-
земов, причем в половине исследованных образцов были обнаружены
личинки .анкилостом. Личинки хорошо развиваются на влажной поверх-
ности плотных образцов желтозема и заболоченного желтозема с рисо-
вых плантаций (Воскресенский и Здродовский). Решающее значение,
вероятно, имеют влажность и доступ кислорода.
Рис. 304 — Головной конец кривоголовки двенадцатиперстной и нека-
тора спереди (А, В) и сбоку в оптическом сечении (С, Z)). Совокупитель-
ная сумочка Ancylostoma duodenale (Е) и Necator americanus (F) с под-
пирающими ее ребрами.
Цикл развития. При оптимальной температуре (от 25 до 30° С)
вылупление личинок из яиц происходит через 1—2 суток после дефека-
ции. При температуре ниже 14 и выше 37° С развитие зародыша в яй-
цах задерживается.
Из яйца вылупляется рабдитная личинка 0,2—0,3 мм длины и
0.14—0.018 мм толщины, передний конец ее тупо округленный, а задний
380
СПЕЦИАЛЬНАЯ ЧАСТЬ
вытянут и заострен. Ротовое отверстие ведет в ротовую капсулу; в нее
открывается пищевод, состоящий из переднего цилиндрического отдела
и грушевидного заднего вздутия. По размерам пищевод занимает пе-
реднюю треть пищеварительного канала, открывающегося анальным
отверстием в начале последней, пятой части длины всей личинки (рис.
311). Развитие личинки проходит три стадии.
Рис. 305 —Развитие личинки в яйцр анкилостомы.
I стадия. Рабдитная личинка является первой стадией
жизненного цикла анкилостомы. После трех дней питания фекалиями и
разлагающимися органическими веществами и роста она линяет и пе-
реходит во вторую стадию.
II стадия. Личинка растет и достигает 0,5 мм длины, сохраняя
рабдитны^ черты строения. На 5-й день она перестает питаться и при-
нимает строп гилоидное строение, так как исчезают вздутия
на пищеводе. Личинка линяет, но сменяемая кутикула не сбрасывает-
ся, а остается в виде чехлика вокруг личинки III стадии.
III стадия. Личинка 0,5—0,8 мм длины, 0,025 мм толщины. На-
ружная (старая) кутикула образует оболочку, под которой лежит соб-
ственно личинка. Хотя последняя и «инцистирована», все же она спо-
собна энергично двигаться. Цилиндрический пищевод ее занимает чет-
верть длины пищеварительной трубки. В ее ротовой полости имеется
копьевидный аппарат, при помощи которого личинка вбуравливается
в кожу .(рис. 312).
Эта стадия заражает человека, и дальнейшее развитие протекает
в организме последнего. Личинки могут выйти из чехлика и жить во»
внешней среде, сохраняя при этом инвазирующую способность.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
281
Стойкость личинок и их поведение. Под водой инвазирую-
щие личинки могут жить до 18 месяцев, но к этому сроку теряют спо-
собность заражения человека; во влажной почве личинки живут более
двух лет. Температура 50—55° С убивает личинок, равно как и одно-
кратное замораживание. Они могут переносить температуру в преде-
лах от 10 до 45° С. Личинки сравнительно стойки к высыханию; без
кислорода могут оставаться несколько дней. Углекислота и гнилостные
газы их быстро убивают. i
Личинки совершают в почве миграции. Передвижение их в гори-
зонтальном направлении весьма ограничено и не распространяется
обычно далее 10 см от места их выплода. В вертикальном направлении
личинки могут пробираться через слой почвы до 40—46 см и более;
впрочем, способность передвижения по вертикали ограничивается свой-
ствами почвы. Глинистые почвы гораздо труднее проницаемы для ли-
чинок, чем почвы с примесью песка. Чем большую миграцию приходит-
ся проделывать личинкам, тем меньшее количество их достигает по-
верхности почвы. На своем пути они подвергаются травматическим воз-
действиям частиц почвы и часто теряют свою наружную оболочку, что
делает их менее стойкими по отношению к вредным воздействиям
внешней среды.
Инвазирующая личинка пищи не принимает и живет за счет ранее
накопленных запасов в своих тканях. Естественная продолжительность
жизни их в почве гораздо короче, чем сроки, устанавливаемые в усло-
виях эксперимента. Почва в тропическом климате очищается от данной
партии личинок через 3—6 недель, если они не подвергаются каким-
либо прямым вредным воздействиям. При более низкой температуре
личинки живут в почве дольше. Грибки и бактерии почвы способствуют
их ранней гибели.
Хотя личинки в эндемичных очагах анкилостомоза сравнительно
скоро погибают в почве, последняя остается зараженной вследствие
последовательного загрязнения ее яйцами анкилостом при дефекации.
Дальнейшая судьба личинок, попавших в тело человека, а также
значение анкилостомоза для человека будут рассмотрены ниже, в отде-
ле «Анкилостомоз».
Ancylostoma braziliense De Faria
Синоним: A. ceylanicum Looss.
Самки длиной 9.1 мм, шириной 0.375 мм; самцы — 7.75—8.5 мм длины и 0.35 мм
ширины. Ротовое отверстие меньше, чем у A. duodenale. Каждая зубная пластинка
несет маленький искривленный передний зуб и два больших задних. Яйца такие же,
как у A. duodenale.
Живет в двенадцатиперстной кишке человека, собаки и кошки в восточной
области тропической Азии, включая Филиппины и Южный Китай, в Панаме, Бри-
танской Гвиане, в Сиерра-Леоне (у леопарда). Цикл развития — как у кривоголовки
двенадцатиперстной.
Ancylostoma ^caninum Ercolani — кривоголовка собачья
Самец 10 мм длины и 0,45 мм ширины; самка—14 мм длины и
0,6 мм толщины. Парные вентральные зубные пластинки ротовой кап-
сулы несут на свободном краю по три зуба. Яйца, как у A. duodenale,
но несколько крупней: 74—84 на 48—54 р.
Довольно обычный паразит собак, кошек, лисиц и некоторых других
хищных. Личинки могут проходить через тело человека, но в нем не
задерживаются и, следовательно, не достигают половозрелости.
Широко распространенный паразит, в частности по СССР.
382
СПЕЦИАЛЬНАЯ ЧАСТЬ
Род Necator Stiles
Necator americanus — некатор
Синонимы: Uncinaria americana Stiles, Ancylostoma americanum Stiles.
Тело серовато-желтого цвета с сильно искривленным дорзально го-
ловным концом (рис. 303). Самец 5,2—10 мм длиной и 0,18—0,24 мм
толщиной; соответственные размеры самки 7,7—13,5 мм и 0,38—0,45 мм.
Ротовая капсула меньше, чем у анкилостом, и лишена вентральных
зубов, вместо которых имеются две острые режущие пластинки. С дор-
зальной стороны против них торчат две пары зубцов. Кутикулярная
выстилка пищевода выступает в полость ротовой капсулы свободно, об-
разуя три зубца. На дорзальной стенке капсулы имеется конический
выступ с отверстием пищеводной железы. Копулятивная сумка самца
уже, чем у анкилостомы. Непарная, спинная лопасть сумки подразде-
лена на две дольки, из которых каждая подперта ребром. Спикулы
длинные (0,943—1,066 мм), с крючком на конце. Половое отверстие
самки лежит кпереди от середины длины тела. На заднем конце самки
кутикулярный штифт отсутствует.
Яйцо, как у анкилостомы, 64—72 р длины и 36—40 толщины.
Паразитирует в тонких кишках (дуоденум и прилежащая часть тон-
кой кишки) человека и гориллы. Встречается и одновременно с A. duo-
de nale.
Распространение: юг Северной Америки, Бразилия, Индия^
Цейлон, Восточная Африка и др. В СССР найден в Закавказье и на
Дальнем Востоке (Сычуанские шахты).
Биологический цикл сходен с циклом жизни A. duodenale.
Длительность жизни анкилостомид человека не превышает макси-
мума в 4—5 лет.
Necator suillus Ackert et Payne
Некатор свиной. Видовое отношение к М americanus не вполне
выяснено. 'Морфологических отличительных признаков видового зна-
чения не обнаружено. Опыты с заражением поросят инвазионными ли-
чинками Necator americanus перкутанно давали отрицательный ,или не-
ясный результат. Человека удалось заразить личинками свиного нека-
тора (Buckley); яйца появились в фекалиях через 54 дня после зара-
жения. В Абхазии у свиней анкилостомозной экспедицией Отдела пара-
зитологии ВИЭМ были наблюдаемы в фекалиях свиней яйца анкило-
стомид, видовое отношение которых осталось невыясненным.
Той же экспедицией (Штром, Смирнов, Сондак, Андреева) под-
тверждена способность яиц некатора человека проходить сквозь кишеч-
ник свиньи с сохранением их жизнеспособности. Свиньи и собаки слу-
жат как бы аккумулятором личинок некатора человека: последние мо-
гут вбуравливаться в их кожу, но жизненного цикла в ней не закан-
чивают, выходя таким образом «из строя» (Brumpt; положительный
гистиотропизм личинок).
ЗАБОЛЕВАНИЯ, ПРИЧИНЯЕМЫЕ АНКИЛОСТОМОЙ И НЕКАТОРОМ
(АНКИЛОСТОМОЗ И НЕКАТОРОЗ)
Взрослые анкилостома и некатор присасываются к слизистой оболоч-
ке тонких кишок и режущим аппаратом ротовой капсулы (зубы, режу-
щие пластинки) нарушают ее целость. Ткань кишечной стенки втяги-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
383
вается в ротовую капсулу и целиком ее заполняет. Нарушение целости
слизистой оболочки ведет к образованию кровоточащих изъязвлений,
которые могут быть множественными не только вследствие присутствия
большого количества паразитов, но и от повторной перемены места
присасывания одной и той же глисты. Благодаря развивающимся ге-
моррагиям в стуле больных может появляться кровь. Повреждение сли-
зистой оболочки кишечника открывает доступ бактериальной инфекции,
что в свою очередь может усугублять основное заболевание хозяина
анкилостомы.
Вопрос о питании анкилостомид1 все еще вызывает
споры. Одни авторы считают их гематофагами, с чем и связывают бо-
лезнетворное действие этих нематод, в частности развитие малокровия.
Другие исследователи указывают, что анкилостомиды питаются за счет
слизистой оболочки, причем случайно могут заглатывать и кровь.
Принимая последнюю точку зрения, причину патогенного действия
анкилостомид естественнее искать в действии их секретов на кровь и
на организм в целом. В экстрактах анкилостомид экспериментально об-
наружены гемотоксины, т. е. вещества, повреждающие эритроциты и
способствующие их разрушению. Эти начала всасываются поврежден-
ной слизистой оболочкой в кровь, где проявляют свои патогенные свой-
ства in vivo. В анкилостомах есть и другие действующие начала, в ча-
стности препятствующие свертыванию крови (антикоагулины).
Общее болезнетворное действие взрослых анкилосто-
мид слагается из влияния различных моментов: потери крови, интокси-
кации и возможной вторичной инфекции. Кроме того, должны быть
учтены и явления миграции личинок анкилостомид по организму чело-
века (см. ниже).
Сила заболевания человека определенного возраста анкилостомозом
и некаторозом зависит прежде всего (но не исключительно) от количе-
ства этих нематод, паразитирующих у данного субъекта.
Smillie и Augustine дают такие соотношения интересующих нас мо-
ментов с указанием соответствующего гемоглобинного индекса (НЬ):
При наличии 1—25 паразитов — очень легкая инвазия 4
» » 26 -100 » — легкая инвазия НЬ = 75.2%
» » 101- -500 » — умеренная инвазия НЬ = 7з.-.%
» 501 -1 000 » — тяжелая инвазия НЬ = 66.8%
» » 1 001 —3 000 » — очень тяжелая инвазия НЬ = 57.6%
Дети и взрослые различно реагируют на паразитирующее в них ко-
личество анкилостомид, что объясняется возрастными отличиями орга-
низма. Дети более чувствительны к анкилостомидным токсинам, чем
женщины, которые в свою очередь менее стойки в указанном отно-
шении, нежели мужчины. Стойкость взрослых старше 40 лет также по-
нижается. Следовательно, степень развития анемии зависит не только
от количества паразитов, но и от возраста больного.
При наличии паразитов менее ста, а у более стойких индивидов и
при паразитировании 200—300 анкилостом, дефекты крови, причиняе-
мые паразитами, компенсируются кроветворением. Поэтому у них ане-
мия не развивается, и обладатели этих нематод практически являются
здоровыми носителями.
При нескольких сотнях анкилостом процессы кроветворения не по-
крывают полностью ущерба, наносимого паразитами: гемоглобина в
1 Анкилостомиды — общее название для анкилостомы и некатора.
384
СПЕЦИАЛЬНАЯ ЧАСТЬ
крови на 10—40% меньше сравнительно с нормой. Такие случаи забо-
левания можно рассматривать, как нарушение компенсации потери ге-
моглобина от гемотоксического действия паразитов.
Более глубокая инвазия с 500—1000 и более анкилостом расстраи-
вает компенсацию потерь гемоглобина, количество которого падает до
половины против нормы.
Количество паразитов и возраст больного, конеч-
но, не являются единственными причинами, обуслов-
ливаю щ и ми степень развития анемии. Бесспорную
роль играет и состояние организма: характер пита-
Рис. 306 — Внедрение личинки анкилостомы в кож-
ные покровы человека (момент заражения человека
анкилостомозом).
\
ния, условия труда и быта, одновременное наличие
других заболеваний, приобретение относительного
и м му нитета и др. Поэтому величины гемоглобинного
индекса у разных авторов весьма вариируют и
имеют, следовательно, относительное значение.
Вопрос о сравнительной токсичности анкилостом и некаторов едино-
гласно не решен. Одни считают первых гораздо более злокачественны-
ми; другие утверждают противное; есть также данные, по которым
один и тот же вид паразита (например, некатор) в разных местностях
вызывает то легкие, то тяжелые заболевания. В частности, тяжелые
случаи некатороза наблюдались в Азербайджане (Здродовский и
Воскресенский).
Заражение анкилостомами и некатором происходит главным об-
разом активным внедрением инвазирующих личинок
из загрязненной почвы в кожные покровы человека
(перкутанная инвазия). Инцистированная личинка при сопри-
косновении голого тела с зараженной почвой привлекается теплом тела
и быстро в него вбуравливается (рис. 306—308), оставляя снаружи сбра-
сываемый чехлик, что является третьей по счету линькой. Вбуравли-
ваться в покровы могут также личинки, уже вышедшие из чехлика.
Попав в лимфатические или кровеносные сосуды, личинки с tokoim
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
385
крови заносятся в правое сердце и через малый круг кровообращения
попадают в капилляры, оплетающие легочные альвеолы (рис. 308):
Мигрирующая личинка является четвертой стадией жизненного цикла
анкилостомы.
Застряв в капиллярной сети легких, личинки активно прорывают
стенку капилляра, окружающие ткани и выстилку легочных альвеол,
причем проникают в полость последних. Из альвеол они ползут в брон-
хиолы и бронхи; при содействии мерцательного эпителия личинки под-
нимаются в трахею, достигают гортани и проходят в глотку. Отсюда
они проглатываются со слюной, пищей или питьем и обосновываются в
тонких кишках.
На прохождение всего этого пути требуется до 7—10 дней. В две-
надцатиперстной кишке личинка растет и анатомически изменяется
Рис. 307 — Личинка анкилостомы, проникшая под роговой слой
эпидермиса хомяка через два часа после заражения.
По Г..Смирнову и Н. Камалову.
вследствие образования у нее ротовой капсулы. На 13-й день она ли-
няет в четвертый раз и превращается в пятую стадию; к этому време-
ни личинка достигает 2,5 мм длины; для превращения ее в половозре-
лую нематоду требуется еще 3—4 недели. В общем, на все развитие в
теле человека, считая с момента внедрения личинки в кожу и до раз-
вития в duodenum самцов и самок, затрачивается 1 — Р/г месяца.
При прорывании стенки альвеол легких некоторые личинки могут
проникнуть в ветви легочной вены и попасть в большой круг кровооб-
ращения; при этом они могут быть занесены с током крови в различ-
ные органы.
Перкутанное проникновение личинок анкилостом и некатора являет-
ся главным способом заражения человека анкилостомидами, по сравне-
нию с которым инвазия через рот отступает на второй план.
При заглатывании человеком инвазирующих личинок анкилостомид
с водой большая часть их превращается в половозрелые формы без ми-
грации, но некоторая часть личинок все* же проделывает миграцию, как
и при кожном заражении.
Личинки различных анкилостомид нередко попадают и не в своих
природных хозяев; например, человек проглатывает личинки кривого-
ловки собачьей. В таком случае личинки проделывают полную мигра-
цию, но все же в конечном счете не задерживаются в кишечнике и вы-
386
СПЕЦИАЛЬНАЯ ЧАСТЬ
брасываются вон. Стимулом для миграции, по Yokogawa, являются не-
благоприятные свойства среды кишечника у необычного хозяина по от-
ношению к личинкам. Последние покидают неподходящую для них
среду обитания, проникают в толщу стенки пищеварительного канала и
таким образом попадают в русло миграционного пути.
Симптомы анкилостомоза и некатороза могут сильнсг
вариировать, в зависимости от интенсивности инвазии, степени стойко-
Рис. 308 — Схема миграции личинок анкило-
стом в теле человека (по идее музея Велком-
ского бюро научных исследований в Лон-
доне).
сти организма и некоторых
привходящих причин.
Симптомы заболевания про-
являются прежде всего по вре-
мени и по пути заражения че-
ловека анкилостомидами. Ли-
чинки их вбуравливаются в
волосяные фолликулы и в ще-
ли отслаивающихся чешуек
эпидермиса. Различные места
кожи данного субъекта неоди-
наково проницаемы для пара-
зитов. Ригидная кожа стари-
ков может быть вовсе недо-
ступной для личинок анкило-
стомид. Особи с более толсты-
ми кожными покровами менее
поддаются заражению, чем
обладатели более тонкой кожи.
Чтобы произошло заражение,,
необходима достаточно высо-
кая температура внешней сре-
ды, так как холод подавляет
подвижность личинок и, следо-
вательно, парализует их актив-
ность.
Личинки кривоголовки две-
надцатиперстной и некатора
оказывают одинаковое дей-
ствие на кожу человека в ме-
сте их внедрения. В опытах с
проникновением малого коли-
чества личинок ощущается
легкий зуд с появлением на месте их вбуравливания маленькой крас-
ной точки, которая через несколько минут преобразуется в волдырек;
через 2—3 часа волдырь пропадает, и на его месте обнаруживается
маленькое красное пятнышко. Когда кожные покровы поражаются мно-
жеством личинок, больной ощущает весьма сильный зуд и жжение,.
сопровождаемые развитием сильного дерматита, симптомы которого
держатся до трех недель. Такие явления наблюдаются при внедрении
личинок, относительно мало загрязненных сторонними бактериями.
Часто цитируются опыты 'Malvoz и Lambinet, которые намазали ко-
жу морской свинки мокротой, богатой туберкулезными палочками, и
положили на мокроту много личинок кривоголовки собачьей. Свинка
погибла от туберкулеза. На основании этого опыта делается заключе-
ние» что личинки анкилостомид могут при внедрении своем в кожу че-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
387
ловека вносить сторонние болезнетворные бактерии и тем осложнять
основную инвазию. Однако, по справедливому замечанию Fiillebom,
нельзя без оговорок переносить данные опыта с тонкокожими животны-
ми на человека. Бактерии имеются лишь на поверхностном чехлике ин-
вазирующей личинки, тогда как кутикула собственно личинки свободна
от них. При внедрении в кожу чехлик разрывается у переднего конца,
из него выходит личинка, погружающаяся в кожные покровы, на по-
верхности которых остается сброшенная и загрязненная бактериями на-
ружная оболочка личинки. Личинки с уже сброшенными чехликами при
гвоем внедрении в кожу, в опытах Augustin, вызывали не большее раз-
дражение кожи, чем инцистированные личинки.
Известны в разных местах кожные поражения, связываемые с пер-
кутанной инвазией личинками анкилостомид; таковы — экзема на ногах,
«Ground itch» и др. Бесспорно, что в их этиологии играет роль вторич-
ная инфекция; последняя связывается с расчесами вследствие зуда.
У рабочих горной промышленности (работа в шахтах, тоннелях
и др<) на ногах и руках иногда образуются зудящие папулы, которые
позднее могут переходить в пустулы и изъязвляться. Эти явления свя-
зывают с проникновением личинок анкилостомид через кожу и вторич-
ными осложнениями последствий этого процесса.
При внедрении в покров человека личинок Arie. braziliensis, Uncina-
На sienocephala — транзитных гостепаразитов человека —
появляется очень маленький волдырек или красное пятнышко, которое
быстро исчезает. Через 1—3 дня в месте внедрения образуются зудя-
щие круглые папулы 2—5 мм диаметром. От них могут отходить ходы,
как при creeping disease. Кожные изменения могут вариировать с об-
разованием пузырчатых ходов и подвергаться осложнениям при рас-
чесах.
Клиническая картина при внедрении личинок A. caninum носит иной
характер; в ней преобладают явления местного блуждающего отека на
руках или ногах с эритемой при отсутствии лихорадки.
Следующим моментом, когда мигрирующие по телу личинки анки-
лостомид могут себя проявить, является время проникновения их из ка-
пилляров малого круга кровообращения в дыхательные полости легких.
Благодаря разрыву капилляров образуются геморрагии, величина ко-
торых зависит от степени инвазии. Бывают также осложнения в форме
бронхитов, и совсем редко случается отек легких.
Анкилостомиды, окончательно обосновавшиеся в кишечнике, вызы-
вают общие симптомы болезни. Со стороны пищеварительного аппара-
та отмечается боль подложечкой, усиливающаяся при давлении. Эти
болезненные ощущения ослабевают, когда едят такие вещества, как
земля или мел. У анкилостоматиков наблюдается часто извращение
вкуса, они инстинктивно ощущают потребность есть землю (геофа-
гия), что можно поставить в связь с умеряющим действием земли на
боли подложечкой, с потребностью наполнения желудка и с извраще-
нием вкуса, как таковым. Аппетит часто повышен; бывают также пери-
оды его ослабления или отсутствия, бывает рвота водянистыми, сли-
зистыми или желчными массами. Понос наблюдается чаще запоров.
В стуле может быть кровь. Бывает прерывистая лихорадка с подня-
тием температуры до 38—39° С.
После некоторого периода нарастают явления анемии с общей сла-
бостью, головокружениями, побледнением покровов и видимых слизи-
стых оболочек. Со стороны кровеносной системы присоединяются серд-
388
СПЕЦИАЛЬНАЯ ЧАСТЬ
цебиения, учащение пульса, одышка. При усилении болезни появляется
отек лица и ног. В качестве дальнейшего осложнения наблюдается во-
дянка живота.
Нервная система подвержена действию анкилостомных токсинов: го-
ловные боли, неврастения, апатия, ослабление памяти, спутанность мыс-
лей, общее понижение умственных способностей, задержка в психиче-
ском развитии —все это в разной степени наблюдается у анкилостом-
ных больных.
Дальнейшее отягчение болезни ведет к общему истощению и даже
смерти.
Распространение. Считается, что почти всего населения
земного шара заражена анкилостомидами, свойственными главным об-
разом жарким и теплым странам. В СССР анкилостомоз встречается
на юге: в Грузии в ее западной части—Абхазия, Аджаристан, Мин*
грелия, Имеретия, Гурия. Процент заражения колеблется в разных ме-
стах Грузии, по разным авторам, от 20 до 73.
В Грузии С. Канделаки и Н. Камалов насчитывают (включая Аб-
хазию и Аджаристан)! около полумиллиона анкилостомидозных; из них
9,1% тяжело больны, 44,6°/о имеют умеренную инвазию и остальные
46,3% являются носителями без каких-либо клинических явлений. Наи-
более пораженной анкилостомидами является предгорная, удаленная
от черноморского побережья полоса, с почвой, богатой красноземом.
Каменноугольные копи (Тквибули) и марганцовые рудники (Чиатуры)
свободны от анкилостомидоза, что Канделаки и Камалов объясняют
низкой температурой.
В Армении, повидимому, анкилостомоз является завозным; про-
цент зараженности обследованных был ничтожен (0,3% по Каланта-
рян). Азербайджан дает наибольшую зараженность населения в Ленко-
ранском районе, там же личинки анкилостомид были находимы в поч-
ве (рис. 309). Закатальский район также является злостным очагом ан-
килостомоза. В Средней Азии имеются лишь единичные случаи этого
гельминтоза, более приуроченного к Туркмении, где он намечался по
линии железной дороги (Кушка, Мерв, Ашхабад, Геоктепе). Известны
случаи сильного и даже поголовного заражения анкилостомидами турк-
менов-призывников из некоторых аулов. В отношении зараженности
анкилостомозом Средняя Азия уступает Закавказью. Завозные случаи
известны и в других местах Средней Азии.
Анкилостомоз есть и на Дальнем Востоке, в частности в Сычуанских
копях. Заносился он сюда главным образом китайцами-рабочими, но
единичные случаи местного заражения наблюдаются и у русских, нико-
гда или долго не покидавших рудника. Тяжелых картин анкилостомоза
на Сычуанском руднике обнаружено не было (Г. Змеев). Завозные слу-
чаи анкилостомоза обнаруживались в Москве, Ленинграде, Одессе и
других городах, что не имеет никаких эпидемиологических последствий.
Пути заражения. Заражению подвергаются чаще люди, сопри-
касающиеся голым телом с зараженной почвой. Это обстоятельство де-
лает анкилостомоз в иных случаях профессиональной бо-
ле з н ью. Земледельцы, рабочие, на земляных работах, горнорабочие в
шахтах, землекопы в тоннелях могут легко заразиться анкилостомозом
в зоне распространения этой болезни. С другой стороны, эти больные
или носители сами заражают почву при дефекации и расширяют воз-
можность распространения этой болезни. Заражение начинается с дет-
ского возраста. В одних случаях экстенсивность инвазии повышается с
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
38<
возрастом, в других (реже) — наблюдается обратная картина, что, мо-
жет быть, объясняется приобретением иммунитета. Резкое поднятие за-
раженности детей совпадает с началом участия их в полевых работах.
Различия в зараженности женщин и мужчин анкилостомозом объяс-
няются неодинаковым участием их в полевых работах, различиями в де-
талях быта и пр.
Социальное значение анкилостомидозов. Зараже-
ние анкилостомидами происходит при наличии благоприятных экологи-
Рис. 309 — Ландшафт в Азербайджане, где личинки анкилостомид были найдены
в почве. По фотографии Воскресенского.
ческих и бытовых условий очень легко, поэтому распространенность
этой инвазии велика. Легкость заражения в связи с тяжестью действия
паразитов на человека делает анкилостомоз и некатороз болезнями
огромного социального значения. В иных случаях поголовно заражаются
семьи, целые селения и даже районы. Дети, пораженные анкилостоми-
дами, сильно задерживаются как в физическом, так и в умственном
развитии. 20-летний юноша порой выглядит 12—13-летним ребенком.
Вследствие влияния паразитов на эндокринные органы задерживается
половое созревание. В массе дети, пораженные анкилостомидами, про-
являют понижение умственных способностей на 30—33% против здо-
ровых.
Равным образом подвергается депрессии и работоспособность взрос-
лых, понижающаяся, смотря по месту, времени, силе заражения и др.,
на 10—80%. Излечение рабочих от анкилостомоза и некатороза ведет к
явному повышению их работоспособности. Вредоносность анкилостомид
усугубляется еще тем, что в их присутствии ослабляется стойкость орга-
низма к другим болезням, как, например, пневмонии.
Кроме того, социальный вред анкилостомидозов отражается на сель-
скохозяйственных работах, в дорожном строительстве, в горном деле и
на обороноспособности страны. Все эти моменты особенно актуальны
390
СПЕЦИАЛЬНАЯ ЧАСТЬ
для периода социалистического строительства южных республик Совет-
ского Союза, а через их посредство и Союза в целом.
Заслуживает особого внимания возможное значение анкилостоми-
дозов для Красной Армии. В буржуазных странах аналогичные соотно-
шения уже подмечены; практически стал на очередь этот вопрос в им-
периалистическую войну, когда пополнения для западного фронта стали
прибывать из стран, пораженных анкилостомозом. Выявилась необходи-
мость предварительного извлечения из состава пополнений всех носи-
Рис. 310 — Дом с земляным полом, без окон; рядом кукурузник, используемый
вместо отхожего места. Очаг анкилостомидоза. Оригинал Г. Смирнова.
телей этих паразитов для полного освобождения их от этих глист и по-
следующей отправки на фронт.
Лечение. Кардинальным моментом лечения является глистоиз-
гнание. В качестве противоглистных средств при анкилостомозах приме-
няются: тимол, в форме очень мелкого порошка в капсулах или в папи-
росной бумаге; дозировка по возрасту (Ashford и King):
До 5 лет............ 0.5 г От 15 до 20 лет ... 3 г
От 5 до 10 лет . 1.0 » От 20 до 60 лет . . 4 »
От 10 до 15 лет 2.0 » Выше 60 лет . . 2—3 »
Лекарство дается в два-три приема с часовыми промежутками. На-
кануне лечения дается солевое слабительное. Утром натощак прини-
мается, как указано выше, тимол; через час-два после приема послед-
ней дозы тимола дается солевое слабительное. Для полного изгнания
паразитов требуется повторное лечение, которое начинают не ранее ис-
течения недели.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
391
Хеноподовое масло >(О1. chenopodii), для взрослых — максимальная
доза — 3,0 см3, разделенные на два-три приема с часовыми промежутка-
ми. Другие авторы дают 2 см3. Препарат обладает токсичностью, осо-
бенно для детей. Через два часа солевое слабительное.
Четыреххлористый углерод (Carboneum tetrachloricum); доза для
взрослых — 3—4 см3, принимаемые сразу. Для детей доза рассчитывает-
ся по 0,13 см3 на каждый год возраста. Предварительной дачи слаби-
тельного не требуется. Препарат дается через час после еды. Дача
солевого слабительного не обязательна.
Сравнительная оценка действия противоглистных
средств на анкилостомид. Тимол действует на некатора силь-
нее, чем на анкилостом. Хеноподовое масло влияет сильнее тимола, осо-
бенно на анкилостом. Четыреххлористый углерод сильно действует на
некатора и слабее на анкилостом. В практической обстановке массового
глистоизгнания четыреххлористый углерод находит себе широкое приме-
нение.
Диагностика. Диагноз ставится по нахождению в faeces яиц
.анкилостом и некатора. Яйца овальные, с тонкой прозрачной оболочкой
и с зародышем на ранней стадии дробления (табл. VI). Размеры
яиц анкилостомы и некатора очень близки; для первой — 53—64 jx
на 34—38 ц, для второго 64—72 р на 36—40 у. Поэтому при микроско-
пировании практически яйца анкилостомы не распознаются от яиц
некатора и диагносцируется в общей форме наличие яиц анкилостомид.
Далее, яйца анкилостомид необходимо дифференцировать от яиц Tri-
chostrongylidae, которые, несколько крупнее яиц некатора и обычно
имеют зародыш на более поздней стадии дробления (16 бластомер и
больше). Наилучшим способом диагностики является метод с
всплыванием яиц в солевом растворе (Кофоид — Фюллеборн).
Важным подсобным способом диагностики может служить культи-
вирование личинок анкилостомид (копрокультура). Кал с яйцами
паразита накладывают слоем на стерильный песок в фильтре из черного
газа, зубчатые края которого выдаются над поверхностью воронки. Во-
ронка на кольце помещается в стеклянный цилиндр, который ставится
в широкий стеклянный сосуд с притертой пластинкой или пробкой; на
дно этого сосуда наливается крепкая серная кислота слоем в 2—3 см.
Песок увлажняется, и закрытый сосуд остается стоять в темноте при
25—30° С.
Вылупившиеся личинки всползают на свободную кайму газового
фильтра, причем хорошо видимы на его черном фоне. Их снимают пла-
тиновой петлей. Упавшие личинки погибают в серной кислоте.
Более простой способ культивирования заключается в смешивании
кала с угольным порошком или со стерильным песком в пропорции
1 часть кала на 3—4 части индиферентного вещества. Смесь кладут
ровным слоем в чашку Коха и оставляют в темноте при 25—30° С. Для
выбирания личинок на культуру наливают теплую воду, в которую в
силу термотропизма и переходят личинки.
Личинок анкилостомид приходится дифференцировать от личинок
Strongyloides stercoralis (рис. 311). Отличия приводятся в таблице на
стр. 392.
Дальнейший дифференциальный диагноз проводится по инвазирую-
щим личинкам анкилостомид между видами Ancylostoma duodenale yl
Necator americanus. При сильном увеличении микроскопа можно ви-
деть прямое соприкосновение внутреннего луковицеобразного расшире-
392
СПЕЦИАЛЬНАЯ ЧАСТЬ
ния пищевода с кишечником у личинок анкилостомы, тогда как у нека-
тора есть разделяющее промежуточное пространство в виде перехвата.
В ротовой полости личинок имеется вытяжная шпора, которая у личинок
некатора развита более мощно, чем у личинок анкилостомы (рис. 312).
Наиболее точная видовая диагностика
осуществляется определением самих из-
гнанных червей по признакам, указанным
на стр. 379 (рис. 304). Отхождение глист
при лечении происходит не сразу, поэтому
необходимо собирать для отмытая парази-
тов весь стул в течение трех дней.
Метод количественного учета приме-
няется именно по отношению к анкилосто-
мидам. Для выявления количества парази-
тов у больного и установления зависимо-
сти интенсивности заболевания от количе-
ства паразитов применяются также разные
способы количественного определения яиц
Рис. 312 —Головка филяриевидных
личинок — Ancylostoma duodenale
(слева) и Necator americanus (справа)
с копьевидным аппаратом.
Из Фауста.
Рис. 311—Личинка Stron-
gyieldes stercoralis (слева)
/И Ancylostoma duodenale
(справа) на четвертый день
культуры.
По Лейхтенштерну.
анкилостомид в кале для косвенного определения степени инвазии об-
следуемой группы в целях гельминтоэпидемиологического исследования
[методы Столла (Stoll) и Лейна (Lane)], описание которых дается в
специальных работах 1 и руководствах.
Различие личинок анкилостомид от личинок
Stг оngу 1 оidеs stercoralis
Признаки | Анкилостомиды | Strongyloides stercoralis
В свежем стуле обычно обнаруживаются Ротовое отверстие раб- дитных личинок В копрокультуре личин- ки превращаются Пищевод у инвазирую- щих личинок занимает только яйца узкое и длинное в инвазирующие, ин- цистированные формы одну четверть длины тела рабдитные личинки широкое и короткое в инвазирующие, неинци- стированные, стронгилоидные формы или превращаются . в самцов и самок половину длины тела
1Г. Смирнов. Качественный гельминтологический анализ. Вести, микробиол.>
эпидемиол. и паразитол., т. VIII, вып. I, 1929.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
393
Профилактика а н к и л о с т о м и д о з ов и борьба с ними,
Профилактика анкилостомидозов основывается на характерных особен-
ностях биологического цикла анкилостомы и некатора с учетом путей
заражения этими паразитами и того факта, что источником заражения
внешней среды является сам человек.
Основные этапы эпидемиологии анкилостомидозов сводятся к сле-
дующим моментам.
1) Хозяином анкилостомы и некатора является человек.
2) При дефекации выбрасываются наружу яйца анкилостомид.
3) Попав на землю, яйца дают при благоприятных условиях личи-
нок, превращающихся в инвазирующие (инцистированные) формы.
Рис. 313 — Неблагоустроенная и удовлетворительно содержимая уборные.
Рис. по фото Тропич. института Грузии.
4) При соприкосновении обнаженных покровов человека с заражен-
ной почвой личинки анкилостомид активно вбуравливаются в кожу и
мигрируют уже известным нам образом по телу, пока не обоснуются в
двенадцатиперстной кишке.
5) Заражение per os имеет гораздо меньшее значение, чем перку-
танная инвазия.
Борьба с анкилостомидозом и профилактика его сводятся к разрыву
указанных эпидемиологических звеньев.
Некоторые животные, пожирающие фекалии человека, могут про-
глатывать яйца анкилостомид. Первую роль в этом отношении играют
свиньи; через их кишечник яйца анкилостомид проходят без вреда для
себя (рис. 313). Аналогичное значение имеют собаки и крысы.
Эти животные содействуют рассеиванию яиц анкилостомид во внеш-
ней среде; результаты зависят от характера места, куда с фекалиями
будут выброшены заглоченные яйца. Последние погибают, если попа-
394
СПЕЦИАЛЬНАЯ ЧАСТЬ
дут на сухие места, особенно подверженные прямому освещению
солнечными лучами. Крысами и собаками яйца могут быть занесены в
подходящие для развития биотопы, но сами биотопы могут быть пра-
ктически недоступными для человека, благодаря чему достигшие инва-
зирующего состояния личинки некатора или анкилостомы не получают
возможности проникнуть в тело человека.
Резервуаром анкилостомидной инвазии и ее источником являются
носители анкилостомид и больные анкилостомозом и некаторозом. Ши-
рокая дегельминтизация ведет к уничтожению или ослаблению перво-
источников инвазии, что может быть обеспечено применением хорошо
действующих противоглистных средств и полным охватом пораженных
групп населения. Для массовой дегельминтизации вполне пригоден
четыреххлористый углерод, обусловливающий изгнание при однократ-
ном применении до 80—98%. Поголовная абсолютная очистка населе-
ния от анкилостомид достижима в местах гнездного его распростране-
ния. В районах пандемий ставится задача наибольшего снижения анки-
лостомидозов, что вполне благоприятно отражается на состоянии
очищаемых людей. Для оценки эффективности снижения инвазии
недостаточно опираться только на сравнение процента зараженных
анкилостомидами до и после глистоизгнания. Правильное представле-
ние можно получить лишь сравнением интенсивности инвазии. Сравне-
ния такого рода делались; падение общего процента зараженности после
глистоизгнания бывало ничтожным, например всего на 2,2%; в то же
время среднее количество анкилостомид на одного человека упало
с 500 до 14. Эта последняя цифра и является решающей для положи-
тельной оценки достигнутых результатов (Smille, Бразилия).
Вместе с тем одна дегельминтизация сама по себе дает весьма
кратковременный эффект, если освобожденные от инвазии возвращают-
ся в прежнюю неизмененную обстановку,— в короткий срок они снова
заражаются анкилостомидами. Поэтому борьба должна вестись ком-
плексно. Наряду с очисткой населения необходимо воздействие на
внешнюю среду для обезвреживания ее как источника заражения
человека анкилостомидами. Задача состоит в прекращении загрязнения
почвы яйцами паразитов. Это может быть достигнуто прекращением
использования почвы для дефекации и введением в обиход пользования
отхожими местами, которые должны быть специально сконструированы.
Такая как будто простая задача чрезвычайно трудно разрешима,
особенно среди населения с недостаточным культурным уровнем раз-
вития. Берега рек и других водоемов, как правило, служат для отправ-
ления естественных надобностей. Применяемые на Востоке омовения,
совершаемые, естественно, около воды, также ведут к загрязнению
почвы. Дурно содержимые отхожие места, подолгу или вовсе не очи-
щаемые, в свою очередь загрязняют воду потоками фекалий. Наконец,
может происходить загрязнение и толщи почвенного слоя из неизоли-
рованных ям — приемников извержений. Близ воды достаточная влаж-
ность благоприятствует развитию личинок анкилостомид в почве.
Вследствие этого такие места, часто посещаемые населением, делаются
важными рассадниками анкилостомидоза.
Для дегельминтизации внешней среды (участки загрязненной почвы,
порции фекалий и пр.) можно применять 4—5% карболовую кислоту,
4% серно-карболовую смесь, 3% сулемо-карболовый раствор, 3% ли-
зол; надежное действие этих веществ по отношению к анкилостомидам
собак показано 3. Раевской.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
395
Надсемейство Trichostrongyloidea
Ротовая капсула отсутствует или рудиментарна.
Род TrichostrongyluS Looss
Trichostrongylus instabllis Raill.
Синоним: Strongylus subtilis xLooss.
Тонкие, нитевидные, кпереди постепенно суживающиеся черви. Рот, окруженный
тремя губами, ведет в длинный пищевод. Самец длиной 4—5,5 мм и 0,09 мм тол-
щиной; длина самки — 5—6 мм. На заднем конце тела самца совокупительная су-
мочка с двумя большими боковыми лопастями и ложкообразными спикулами длиной
0,135—0,145 мм. Женское половое отверстие лежит кпереди от заднепроходного, у
передней границы задней четверти тела. Толстостенное яйцо в форме овала (0,073—
0,08 на 0,04 мм) содержит в себе зародыш на стадии дробления из 8—32 бласто-
меров. Паразитирует в duodenum травоядных млекопитающих (часто встречается у
Рис. 314 — Спикулы и рулек Trichostrongylus in-
stabitis (А) и Tr. probolurus (В)
По Калантарян и Орлову.
Рис. 315 — Haemonchus contor-
tus. Целый червь. По Neveti-
Lemaire.
'Ovis aries, Antilope darcas, верблюда, овец, коз и др.). У человека был найден в
Египте, Японии, Армении и в Москве. Когда паразитов много, они, возможно, вы-
зывают тяжелую вторичную анемию.
Кроме этого вида, у человека, в частности в СССР, обнаружены другие виды
атого рода: Tr. orientalis J. Imbo, Tr. probolurus Raill., Tr. vitrinus Looss, Tr.
extenuarus Raill. (Калантарян, Армения). Все это паразиты различных жвачных
и грызунов. Различаются перечисленные виды по форме спикул и 'рулька (рис. 314)
самцов и по размерам и положению полового отверстия и влагалища самок.
Яйцо выбрасывается с faeces наружу и дает вне тела хозяина рабдитную личин-
ку, которая при 22—25° превращается в течение шести дней в инвазирующую филя-
риевидную личинку.
Последняя, как правило, попадает в хозяина per os, причем дальнейшее разви-
тие паразита в хозяине происходит без миграции. Яйца в фекалиях начинают по-
являться на 25-й день после заражения хозяина. Личинки могут также активно
внедряться в покровы некоторых видов хозяев, но этот способ инвазирования яв-
ляется редким и ограниченным по своему значению.
Клинические симптомы при слабой инвазии отсутствуют.
396
СПЕЦИАЛЬНАЯ ЧАСТЬ
Диагноз ставится по нахождению яиц в faeces; яйца имеют более расши-
ренные полюсы, чем у анкилостомид. Различение видов Trichostrongylus по яйцам
невозможно. Определение производится по взрослым формам, изгнанным из хозяина.
Лечение. Изгнание паразитов, как при анкилостомидозе.
Заражение возможно при употреблении в пищу сырой зелени, загрязненной
навозом, в которой имеются инвазирующие стадии этих глист.
Род Haemonchus Cobb.
Haemonchus contortus Rudolphi
Тонкие нематоды, 1,0—2,0 см длины на 0,4 мм толщины самец и 1,8—3 см на
0,5 мм самка. Глоточная полость вооружена единственным, ланцетовидным зубом,
конец которого чуть выступает изо рта (рис. 316. В). Шейные сосочки хорошо
развиты (рис. 316, А). У самца копулятивная сумка образована двумя боковыми
Рис. 316 —Haemonchus contortus.
Л—передний конец паразита с шейными сосочками и пищеводом, В—го-
лову с глоточным ланцетом, С—копулятивная сумка самца. По Yorke
и М а р 1 е s t о п.
лопастями и одной промежуточной дорзальной долькой (рис. 315). Яйца прозрачные,
овальные,75—96р-на 40—50 р-, откладываются ic не вполне развитой личинкой. Пара-
зитируют преимущественно у жвачных в сычуге и реже в duodenum, а также у
грызунов. Могут гостепаразитиройать у человека. Ланцетовидным зубом ранит сли-
зистую оболочку места своего прикрепления. Содержит в себе гемолитическое
начало. При значительной инвазии у хозяина развиваются анемия, отеки и наруше-
ние пищеварительных функций.
Цикл развития. По Ransom, цикл развития протекает по следующей схеме.
Вылупление личинки из яйца происходит в срок от нескольких часов до нескольких
дней, считая с момента отложения faeces. Рабдитная личинка питается faeces и
через несколько дней или недель превращается в одетую чехлом филяриевидную
личинку. Эта стадия характеризуется стойкостью к высыханию и замораживанию.
При известной влажности личинка всползает на траву, вместе с которой и прогла-
тывается хозяином.
Действие на хозяина. У человека. этот паразит может вызвать анемию,
хотя пока известны только единичные находки у человека, да и то с не вполне
точным определением вида (Бразилия).
Диагноз по яйцам невозможен ввиду их сходства с яйцами других стронги-
лоидных нематод. Точная диагностика возможна по взрослым формам, добываемым
глистоизгнанием.
Лечение — тимол.
Профилактика. Остерегаться есть сырую немытую зелень в местах, загряз-
няемых навозом; мыть руки после полевых и хозяйственных работ.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
397
Род Mecistocirrus Rail et Henry
Mectstocirrus digitatus v. Linstow
Самка длиной 19—43 мм при толщине 0,5 мм; самец 16—21 (23—27) мм, тол-
щиной 0,45 (0,75) мм. Передний конец округленный, с 6 сосочками. Есть глоточный
ланцет (рис, 317, А). Шейные сосочки лежат в маленьких вдавлениях кутикулы
позади цервного кольца. У самца — пара пр ебу р сальных сосочков;
бурса — из трех долек (рис. 317, 1 С). Длинные ланцетовидные спикулы одинаковы по
размерам. Половое отверстие самки лежит на брюшной стороне тела на 0,3 мм
кпереди от заднепроходного отверстия. Яйца овальные, прозрачные, 110 р. длины,
46—55 р толщины. Цикл развития неизвестен.
Рис. 317 — Mecistocirrus digitatus.
А—головной конец с сосочками и глоточным
ланцетом; В — то же с пищеводом и шейными
сосочками; С — бурса самца с парою пребурсаль-
ных сосочков. По Yorke и Maple st оп.
Рис. 318 — Metastrong у-
Lus apri. Задний конец
тела самца с бурсой и
двумя длинными ните-
видными спикулами.
По Фаусту.
Паразит,желудка, и прилежащей части тонких кишек свиней, овец, крупного
рогатого скота (от Индии до Китая, Японии и Дальнего Востока). Известен случай
гостепаразитирования у человека (Китай). Биология и патогенное значение его не
изучены.
Надсемейство Metastrongytoidea
Тело нитевидное. Ротовая капсула отсутствует или рудиментарна.
Паразитируют в органах дыхания, кровеносной системе и краниаль-
ных синусах млекопитающих. Известен достоверный случай гостепара-
зитирования в легких человека — Metastrongylus apri.
393
СПЕЦИАЛЬНАЯ ЧАСТЬ
Род Metastrongylus Mol in
Metastrongylus apri Gmelin
Синоним: Strongylus elongatus Duj.
Нитевидная нематода желтоватого или коричневатого цвета. Около рта две
боковые трехдольчатые губы. Пищевод длинный. Самец 12—25 мм длины, 0.16—
0.22 мм толщины; на его заднем конце бурса с двумя длинными спикулами (рис.
318), самка 20—25 мм длины, 0.4—0.5 мм толщины.
Яйца эллипсоидные 57—100 р- длины и 39—72 р- ширины, с хорошо развитой ли-
чинкой, которая вылупляется и значительный срок может сохранять' жизнеспособ-
ность в земле. Дальнейшее развитие происходит тогда, когда. они будут проглочены
дождевым червем (Lumbicus terrestris). Окончательный хозяин (свинья)! заражается,
пожирая дождевых червей с личинками Metastrongylus. В кишечнике свиньи личинки
линяют, вбуравливаются через его стенки в лимфатические железы брыжейки, здесь
еще раз линяют и через лимфатические пути пробираются в легкие, в бронхах ко-
торых достигают половозрелости. Эти паразиты могут быть причиной гибели свиней.
Заражение человека может происходить таким же путем при съедании овощей
с зараженными дождевыми червями.
Диагноз ставится на нахождение яиц в мокроте; яйца могут быть обнару-
жены и в faeces при проглатывании мокроты.
Терапия неизвестна.
Профилактика — аккуратное приготовление огородной зелени для еды.
Надсемейство Oxyuroidea
Род Enterobius Leach
Enterobius vermicularis L.— острица
\
Синоним: Oxyuris vermicularis L.
Острица. Белого цвета червячки (рис. 319); самка около 1 см,
самец 2—5 мм длины; задний конец последнего закручен спирально, у
самки же он втянут и шиловидно заострен (с).
Ротовое отверстие отгружено тремя втягивающимися губами, покры-
тыми по краям более плотной кутикулой. По бокам тела кутикула
выдается двумя острыми рантами (рис. 323), идущими почти до аналь-
ного отверстия. На переднем конце тела кутикула образует короткие
овоидные симметричные парные расширения, дающие так называемую
головную везикулу. Пищевод кзади утолщается и переходит в
шаровидный бульбус, за которым следует кишечник.
В бульбусе имеется так называемый зубной аппарат, образо-
ванный тремя кутикулярными пластинками, приводимыми в движение
сильными мышцами, прикрепляющимися к основанию «зубов». Бульбус
работает, видимо, как всасывающий насос, и в связи с этим способ-
ствует прикреплению острицы к слизистой оболочке (присасывание).
Острицы питаются, вероятно, содержимым кишечника и могут быть
факультативными гематофагами.
Заднепроходное отверстие самки лежит миллиметра на два кпереди
от заднего конца тела (ап). Яичники и матка парные (ov); отверстие
влагалища расположено в начале второй трети тела (у). Семенник (I)
у самца один, равно как и спикула (s).
Яйца острицы (рис. 321) имеют форму несимметричных овалов с
одной уплощенной стороной (50—60 на 20—32 у); довольно толстая
оболочка их бесцветна, почему яйца довольно легко просматриваются
в испражнениях (табл. VI, рис. 18, 19).
Острицы живут в нижнем отделе тонких кишек, включая слепую
кишку и червеобразный отросток человека, чаще всего у детей. О пита-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
399
Нии их в литературе существуют разноречивые данные. Они могут вса-
сывать кровь (рис. 320), но остается вопросом, достигают ли они сами
кровеносных сосудов в стенке, например аппендикса, или же всасывают
кровь, изливающуюся при операции удаления червеобразного отростка.
Оспаривается также способность остриц растворять ферментами клетки
слизистой оболочки кишки и поглощать образующиеся при этом про-
дукты. Весьма вероятно, что острицы питаются содержимым кишечника
Рис. 320 — Острица. Перезрелая сам-
ка, переполненная яйцами (В).
А — сдавленный кишечник со следами не
вполне переваренной всосанной крови.
Оригинал.
Рис. 319 — Острица Enter obius vermicular is.
Л—самка; о—рот; а — пузыревидное вздутие кутикулы;
ое — пищевод; оех — его вздутие; in — кишечник; ov —
яичник; ап — заднепроходное отверстие; с — хвосто-
вой конец; ut — матка: vg— влагалище; -и — женское
половое отверстие. В — то же, самец, t — семенник;
л — спикула. По Холодковскому.
хозяина. Hoeppli (1927) по крайней мере находил в кишечнике лошади-
ной острицы (Oxyuris equi) растительные клетки, яйца паразитических
червей и простейших. Тем более вероятна возможность поглощения
перевариваемых частей пищи хозяина.
По оплодотворении отяжелевшие от развившихся яиц самки (рис.
320) спускаются к заднепроходному отверстию, вызывая нестерпи-
мый зуд у больного, раздражение кожи и даже половых органов.
При вскрытии удавалось обнаруживать до 3 тыс. остриц (Скрябин,
Подъяпольская и Шульц); более молодые паразиты занимают более
высокие части тонких кишек и попадаются даже в двенадцатиперстной
кишке.
400
СПЕЦИАЛЬНАЯ ЧАСТЬ
Цикл развития. Обычно ночью самки выходят из анального от-
верстия и откладывают яйца на кожу промежности и прилежащих
тканей.
О плодовитости остриц можно судить по прямым подсчетам, пока-
завшим, что у оплодотворенных самок может быть от 4672 до 16 888
(в среднем 11 105) яиц. Самки в 13 мм длины могут иметь до
20 тыс. яиц. В ряде случаев отложенные массы яиц могут быть заме-
чены простым глазом в виде беловатых точечных скоплений. Самки
остриц живут до месяца; самцы, повидимому, погибают еще в ки-
шечнике, не выходя наружу.
В отложенных яйцах уже
имеется головастикоподобный
зародыш (рис. 321), который
в течение 6 часов превращает-
ся в веретеновидную личинку,
причем этот процесс может
происходить только в кисло-
родной среде (’М. Завидов-
ский и М. Шалимов).
Яйца, отложенные острица-
ми за ночь, к утру созревают
до инвазирующего состояния.
Вследствие зуда дети и
взрослые расчесывают аналь-
ное отверстие, причем под ног-
ти попадают яйца остриц и
прилипают также к коже паль-
цев. Дети берут пальцы в рот,
грызут ногти и проглатывают
яйца остриц, чем обеспечивает-
ся реинвазия этими паразита-
ми. То же бывает и у взрос-
лых при еде немытыми руками и др. Заражение яйцами остриц может
Рис. 321—Развитие в яйце острицы зароды-
ша. Яйца откладываются в стадии зародыша
в форме головастика. По М.Завадовско-
му и Л. Шалимову.
происходить и через предметы обихода (игрушки, посуда, мебель
и др.). Через 2—4 недели после проглатывания яиц из кишечника вы-
ходит новое поколение самок, которое в свою очередь приступает к
откладке яиц. Из-за легкости повторных заражений путем аутоинвазии
или, точнее, а у т о ре инвазии острицы упорно держатся в кишеч-
нике, и борьба с ними требует настойчивости.
Возможным путем инвазии и реинвазии яйцами остриц является
вдыхаемая пыль, с которой яйца остриц из внешней среды могут через
рот попадать в кишечник (Олейников).
Пылевой инвазии острицами может способствовать тот факт, что
яйца их выдерживают трехнедельное высушивание в комнатных усло-
виях при температуре 10—12° (В. Сондак).
Острицы паразитируют исключительно у человека, и не только в
детском возрасте, но часто у взрослых. Поэтому название «острица
детская» не точно отражает действительные соотношения. Практически
каждый ребенок в то или другое время бывает заражен острицами.
У взрослых заражение острицами достигает местами 80—85%. Острицы
распространены повсеместно.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
401
Действие на хозяина. В месте своего нахождения в кишеч-
нике острицы присасываются к слизистой оболочке кишки, каковые
участки благодаря действию паразита воспаляются. В слепой кишке и
в аппендиксе наблюдаются нарушения целости слизистой оболочки
вследствие давления головного конца острицы. Дефекты слизистой обо-
лочки (рис. 322) могут возникать и под действием токсических ве-
ществ, выделяемых острицами. В некоторых случаях происходит некро-
тизация тканей кишечника. Острицы могут проникать в подслизистую
оболочку и в патологически измененные ткани или в новообра-
зования кишечника (рис. 323,
С. П. Алфеева). Отмечались слу-
чаи погружения остриц в ткани
кишечника до % длины их тела,
проникновение остриц до наруж-
ного мышечного слоя и даже
до серозного покрова (Frohlich,
1907).
Дефекты слизистой оболочкй
являются воротами вторжения
вторичной инфекции. Неудиви-
тельно поэтому, что паразитиро-
вание остриц в аппендиксе ста-
вят в связь с аппендицитом. По-
казательна цифра в общем
50-процентного нахождения ост-
риц в аппендиксах, иссеченных
при операциях аппендицита
(Н. И. Соловьев). Количество
обнаруживаемых в аппендиксах
остриц вариирует у разных авто-
ров весьма сильно. Наблюдались
также примеры нахождения мно-
жества остриц в аппендиксе без
какой-либо патологической реак-
Рис. 322 — Дефект слизистой оболочки
червовидного придатка, вызванный пара-
зитированием остриц (две острицы пере-
резаны поперек). Придаток иссечен при
операции аппендицита.
По Н. С о л о в ь е в у.
ции (Maclean, 1909). Конечно можно говорить только о косвенном от-
ношении остриц к этиологии аппендицита, тем более, что во многих
случаях аппендицита острицы могут отсутствовать. Показательный
пример получил Gordon, исследовавший 26 051 аппендикс, удаленный
оперативным путем, и нашедший остриц всего только в 311 отростках,
причем в 82% острицы залегали в просвете отростка при отсутствии
каких бы то ни было поражений со стороны тканей. Только в двена-
дцати отростках были обнаружены реактивные явления, связанные с
острицами (точечные геморрагии и др.).
Другие авторы вовсе отрицают значение остриц в этиологии аппен-
дицита. Третьи, как Ашоф, допускают, что острицы, при большом ско-
плении в аппендиксе, могут вызывать аппендицитоподобные
симптомы, симулирующие воспаление отростка (appendicopathia oxyu-
rica).
В качестве основного момента действия остриц на хозяина отмечает-
ся чрезвычайное раздражение анальной зоны, доходящее в некоторых
случаях (от действия токсинов остриц) до нервных расстройств. Вслед-
ствие легкости заползания остриц в половую щель их находили в гени-
402
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 323 — Миома тонкой кишки человека с внед-
рившейся в мышечную ткань острицей, попав-
шей на поперечный разрез.
А — кутикуля} ные крыловидные придатки головы пара-
зита; В-- отечная жидкость между тканью опухоли и
паразитом; С — воспалительная клеточная инфильтрация.
По С. Алфеевой.
однократная дача 4,0 см3 взрослому с
кислой магнезии вела к удалению до
талиях детей и женщин; возможно и более} глубокое проникновение
остриц в их половые пути.
Энтеробиоз, особенно в детском возрасте, является одной из важных
причин вульвовагинита (Шульман и Порецкая, по наблюдениям в
Харькове).
В качестве атипичной локализации остриц отмечают нахождение их
в кожных опухолях и в брюшной полости.
Диагноз ставится по нахождению яиц в соскобе с покровом пери-
анальной области, производимом деревянным шпателем; в faeces яйца
остриц бывают лишь в редких
случаях. Находят их также в
носовой слизи и в грязи под
ногтями (в Германии однажды
при обследовании учащихся
яйца остриц были обнаруже-
ны под ногтями у 60% детей).
Лечение комбинируют
двояко: per os дают тимол
(1,0 г по 3 раза в день два
дня подряд взрослому, детям
5—6 лет — 0,3 г), нафталин
(0,7—0,8 г по 3 раза в день
взрослому, 0,2—0,7 г детям от
4 до 16 лет; процедура длится
два-три дня), сантонин (по
0,1 г три раза в день взросло-
му, детям по 0,005 на каждый
год жизни — два дня подряд),
чередуя с клизмами для уда-
ления самок остриц, спустив-
шихся в прямую кишку; необ-
ходимо регулярное обмывание
анальной области после дефе-
кации и тщательное мытье рук.
В Азербайджане испробо-
ван для борьбы с острица-
ми четыреххлористый углерод;
последующим принятием серно-
900 экземпляров остриц. Детям
дается по 0,1 см3 на г.од возраста (Туаев).
Наилучшим средством против остриц считается, по данным Москов-
ского тропического института, экстракт папоротника, 2,0—2,5 г в тече-^
ние одного-двух дней для взрослых. Детям меньше по соответствующе-
му расчету (В. Подъяпольская). Безвредным для человека и вместе с
тем хорошо действующим веществом является Bism. carbonicum —
40,0 г в два приема в день два раза подряд. Детям дается от 5,0 до-
35,0 г.
Профилактика. Соблюдение общих санитарно-гигиенических
навыков и привитие их детям; надзор за продуктами питания; гельмин-
тологический осмотр пищевиков с последующей выгонкой глист, в част-
ности остриц.
Важным моментом профилактики энтеробиоза является борьба с
микрофекализацией внешней среды в ближайшем окружении
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
403
человека в самом жилище. Яйца остриц находили на партах, на полу,
на ручках дверей, на столиках, игрушках, постелях, постельном белье,
гардинах, посуде, столах и других предметах обихода в квартирах,
школах, столовых, в театре, на военном корабле и в других местах.
Такое рассеивание яиц остриц центром распространения имеет убор-
ные, которыми пользуются носители остриц. Дело идет именно о микро-
фекализации, наличие которой устанавливается микроскопическим
исследованием смоченных глицерином акварельных кисточек, которыми
«подметают» обследуемые объекты. Для уничтожения яиц остриц во
внешней среде пользуются 5% раствором карболовой кислоты или
10% лизолом (В. Сондак). Яйца остриц вообще довольно стойки к
внешним воздействиям.
Род Syphacia Seurat.
Syphacia obvelata Rudolphi
Рот окружен тремя губами; в шейной
части пара сравнительно малых крыло-
видных придатков кутикулы (рис. 324).
Задний конец тела вытянут и заострен
(рис. 324 Л). Самец 1 —1.6 мм длины и
00,1—0,12 мм толщины. На брюшной
стороне хвоста единственная спикула
длинна и слегка изогнута. Рулек корот-
кий. Самка 3,5—3,7 мм длины, 0,3—
0,5 мм толщины; хвост ее длинный; по-
ловое отверстие лежит в 2 мм расстоя-
ния от переднего конца тела; матка
непарная, длинная; имеются два узких
семеприемника. Яйцекладущи. Яйца по-
хожи на яйца остриц, но несколько
крупнее (125 у на 40 р) и более вере-
теновидной формы, с одной стороны
сильно уплощены (рис. 324).
Рис. 324 — Syphacia obvelata
А — самец; В — передний конец тела
сам си; С — яйца. По Р. Шульц.
Паразитирует в кишечнике мышей
и крыс (в СССР мыши заражены в 33%). Известен единственный слу-
чай нахождения этого паразита в faeces человека (ребенка с Филип-
пинских островов).
Яйца Syphacia были находимы в подногтевых пространствах детей
вместе с яйцами остриц (Wilhelmi и Quast), но осталось невыяснен-
ным, паразитировали ли сами Syphacia у этих детей или же яйца были
получены в порядке внешнего загрязнения.
Надсемейст зо Ascaroidea
Род Ascaris L.
A scar is limbricoides L.
Аскарида человеческая (рис. 325). Крупный паразит ве-
ретеновидной формы, красновато-желтого, а после смерти беловатого
цвета. Самка 20—40 см длины, 6 мм толщины; самец 15—25 см длины,
при 3 мм толщины. Его задний конец изогнут на ^брюшную сторону
или даже закручен. Ротовое отверстие окружено тремя губами—одной
дорзальной и двумя вентральными. Первая несет пару чувствительных
404
СПЕЦИАЛЬНАЯ ЧАСТЬ
лежащую половую клоаку, из
Рис. 325 — Аскарида человеческая—
Ascaris lumbricoides. Слева — самец,
справа — самка; о — ротовой, с — хво-
стовой конец, а — половое отверстие
у самца, совмещающееся с заднепроход-
ным. Уменьшено. По Н. Холодно в-
с к о м у.
сосочков, последние же по одному такому же сосочку. Тело одето тон-
ким, сильно напряженным кожно-мускульным мешком. Пищевод кзади
нерезко утолщается, не образуя резко выраженных типичных вздутий.
Кишка оканчивается заднепроходным отверстием, которое у самки ле-
жит сейчас же перед задним концом тела; у самца кишечник откры-
вается вместе с выводным протоком половых путей в терминально
рой высовываются две одинаковые
спикулы, каждая 1,5—2 мм длины.
Анальное отверстие самца окруже-
но 70—75 сосочками. Семенник один,
в виде тонкой трубки, переходящей
в более широкий семенной мешок,
который и открывается в клоаку.
Женские половые органы —
трубчатые, парные (яичник, яйце-
вод и матка), кроме влагалища, от-
крывающегося наружу на брюшной
стороне в начале второй трети
тела.
Выделительные каналы имеют
общее отверстие, помещающееся
близ переднего конца тела на ниж-
ней его стороне.
Яйца паразита (табл. VI, рис.
13, 14) одеты толстой бугристой
(белковой) оболочкой, зародыш на-
ходится в нем в стадии оплодотво-
ренного яйца, еще не начавшего
дробиться. В испражнениях бывают и неоплодотворенные яйца, отли-
чающиеся несколько большей длиной (80 против 50—75) и большим
содержанием желтка, который заполняет собой все яйцо, тогда как
содержимое нормального яйца шаровидно и до полюсов не доходит.
Бывают яйца и без бугристой скорлупы; нередки ненормальные
яйца с грубобугорчатой оболочкой и зернистым содержимым (табл. VI).
Наличие белковой оболочки позволяет яйцам свиной аскариды
потреблять в пять раз меньшее количество кислорода, чем при искус-
ственном удалении антиформином этой оболочки: 4,07 куб. мм 1 000 яиц
с оболочкой и 22,86 куб. мм без оболочки при 30° С (Huff).
Развитие яиц. По выходе из кишечника развиваются в сырой
земле или в воде. При температуре +33° С зародыш развивается в
яйце в течение 9—13 дней. При температуре + 24° С развитие длится
от 10 до 15 дней. При 4-17--|-15°С оно затягивается до 40—45 дней.
При температурах ниже 0° яйца выдерживают разные сроки, что обес-
печивает им возможность перезимования, наблюдавшегося на полях
фильтрации под Москвой (Подъяпольская), на полях биофильтрации
сточных вод под Тулой (Василькова) и в других местах. В комнатных
условиях яйца аскарид развиваются в 40—45 дней, в бане в 15 дней
(Данскер). При благоприятных условиях яйца аскарид могут оставаться
В загрязненной почве живыми 5—6 лет.
Аскарида живет у человека,' вероятно, один-два года. Количество
аскарид, паразитирующих у человека, вариирует от единичных экзем-
пляров (обычная инвазия) до десятков и сотен их. Максимальное коли-
чество. наблюдавшееся у 10-летней девочки Форстером, равнялось 1100.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
405-
ЗАРАЖЕНИЕ АСКАРИДАМИ И МИГРАЦИЯ В ТЕЛЕ ХОЗЯИНА
аскариды можно зара-
Легкие
ЛеченЬ
венЬ
Ррадое
с&рдце
человеческой
Рот
кишки
Рис. 326 — Схема миграции
вЬглупленив личу
нок в кишечнике
Половоэрелосте
аскарид д
кишечнике
личинок аскарид
в теле человека. Оригинал.
Глотка
Заражение человека этим космополитным паразитом происходит
при проглатывании с пищей или питьем яиц, в которых уже развились
личинки. В недавнее время стали подозревать, что развитие у аскарид
не прямое, а сопряжено с посредничеством некоторых животных. По
опытам Стюарта (1917), яйцами
зить мышей и крыс; в
их кишечнике из яиц
выходят личинки, про-
лезающие сквозь стен-
ку в вены и на 2—5-й
день достигающие ка-
пилляров печени; че-
рез печеночную вену
личинки проникают в
сердце, а затем в ле-
гочную артерию, в ка-
пиллярах которой и
застревают. На 7-й
день после заражения
червячки проникают в
легочные пузырьки,
бронхи, трахею и ро-
товую полость своего
хозяина (рис. 326—330).
Точные наблюдения
над последствиями
проглатывания челове-
ком массивной дозы
яиц аскарид в инвази-
рующем состоянии по-
казали, что при мигра-
ции вылупляющихся
личинок в течение
6—7 дней (через 1—5
дней после ’зараже-
ния) температура под-
нимается до 39,5—40°
и дыхание делается
частым и неправиль-
ным, давая общую кар-
тину «пневмонии», с
кровохарканием и нахождением личинок аскарид в мокроте (Koino),
Личинок в мокроте удавалось находить с 9 по 16-й день с начала
самоэксперимента; наибольшее количество их было найдено на 11-й день.
Известны пока единичные случаи нахождения личинок аскарид в
мокроте болевших стойким бронхитом или подозрительных на туберку-
лез легких. Патогномоническое значение этих находок станет очевид-
ным из нижеследующего.
Факты указанной миграции личинок человеческой (и свиной) аска-
риды у различных грызунов получили подтверждение в новых экспери-
ментах того же автора (1919), а также и других исследователей
Ransom и Forster (1917, 1919), Fiillebom (1920, 1922) и loshida (1919).
406
СПЕЦИАЛЬНАЯ ЧАСТЬ
Сопоставление с этими данными некоторых клинических явлений (на-
пример, бронхитов, пневмоний и др.) при аскаридах у человека дает
основание полагать, что личинки аскарид в теле человека проделывают
Рис. 327 — Миграция личинок свиных аскарид через стенку кишеч-
ника морской свинки. Оригинал по препарату Г. Смирнова
Рис. 328 — Личинка аскариды при прохождении ее через печень.
По Г. Смирнову.
такую же миграцию (рис. 326^-330); она заканчивается тем, что личин-
ки проглатываются со слюной, причем у человека они задерживаются в
тонких кишках, где и достигают половозрелости. В то же время у
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
407
Рис. 329—'Личинка аскариды в легком, окруженная инфильтра-
том. Оригинал по препарату Г. Смирнова.
Рис. 330 — Личинка аскариды в дыхательных путях легких. Масса
эритроцитов, вышедших из сосудов, прорванных личинкой при мигра-
ции. Оригинал по препарату Г. Смирнова.
408
СПЕЦИАЛЬНАЯ ЧАСТЬ
подопытных животных проглатываемые личинки в кишечнике не задер-
живаются, вследствие чего и не могут стать там половозрелыми. Уста
новленные факты миграции личинок в корне меняют взгляд на аскарид,
как на сравнительно безобидных паразитов.
Подвергнутые опытам морские свинки (Ftilleborn) погибали на 5-й
день от отека легких вследствие воздействия на этот орган личинок
аскарид. Интересно, что личинки аскарид, проскочившие через капил-
лярную сеть легких, попадали в левое сердце, разносились по боль-
шому кругу кровообращения и застревали в капиллярах мозга и дру-
гих органов.
Joshida проглотил легкие морской свинки с личинками аскарид и че
рез 76 дней нашел в своих извержениях множество яиц аскарид. Сле-
довательно, для достижения половой зрелости аскариде требуется не
менее 89 дней, считая с момента выхода личинок из яйца. Личинки
аскарид получают способность задерживаться в кишечнике лишь после
миграции по телу. Окончательное заражение происходит при проглаты-
вании со слюной мигрировавших в глотку личинок аскарид.
Asada же (1921) утверждает, будто бы личинки аскарид вылупля-
ются во внешней среде и, подобно Strongyloides, активно вбуравливают-
ся в неповрежденную кожу человека, что сомнительно.
По опытам Г. Смирнова, нет достаточных оснований говорить об^
аскаридной пневмонии при миграции личинок аскарид по телу. Вслед-
ствие разрыва капилляров при выхождении из них личинок, проникаю-
щих далее в легочные альвеолы и дыхательные пути, происходят
местные кровоизлияния (рис. 329). При массовой миграции личинок
легкие принимают мраморный вид или приобретают сплошную темно-
вишневую окраску, напоминая печень. Воспалительных явлений обычно
не наблюдается, и всю реакцию органа на прохождение через нега
личинок аскарид можно в. патолого-анатомическом отношении скорее
трактовать• как аскаридную экстравазацию, чем аскариднукь
пневмонию; весь же симптомокомплекс, связанный с миграцией личи-
нок, называется мигроаскаридозом (Смирнов).
Личинки аскарид в процессе миграции имеют своеобразный пищевой
режим. В первой фазе миграции (1—2 суток) личинки питаются, види-
мо, лишь плазмой крови; при прохождении через легкие подросшие
личинки заглатывают красные кровяные тельца; в тканях таких личи-
нок может быть обнаружено железо реакцией Pearls — Nishimara
(Г. Смирнов).
Аскаридная инвазия вызывает некоторые закономерные морфологи-
ческие изменения крови. В отношении понижения процента гемоглоби-
на и количества эритроцитов изменения крови могут быть* считаемы за
анемию, развивающуюся вследствие обширных кровоизлияний в легких
при миграции личинок аскарид. Наличие эозинофилии свидетельствует
о токсическом воздействии мигрирующих личинок аскарид на организм
хозяина (Г. Смирнов и М. Глазунов).
Аскаридная миграция может способствовать вторичным осложне-
ниям или параллельно протекающим патологическим процессам. При-
мером могут служить наблюдения над течением мышиного тифа
(В. Breslau) при одновременно идущей миграции личинок свиных и
человеческих аскарид. Мышиный тиф в таких случаях протекает более
быстро и бурно, что объясняется массовым вторжением бактерий при
нарушении мигрирующими личинками целости кишечного барьера
(В. Подъяпольская и Н. Дедова).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
40<>
БОЛЕЗНЕТВОРНОЕ ДЕЙСТВИЕ ВЗРОСЛЫХ АСКАРИД
Взрослые аскариды живут в тонких кишках, причем иногда встреча-
ются в громадных количествах. При паразитировании в кишечнике
одной-двух аскарид может не быть никаких симптомов; большое коли-
чество аскарид, перевившихся клубком, в некоторых случаях обуслов-
ливало механическую закупорку кишечника, каковое заболевание лечат
хирургическим путем. Однако закупорка кишечника не всегда вызы-
вается только лишь механическим действием клубка аскарид. Известен
случай непроходимости кишечника, вызванный всего только одной аска-
Рис. 331 — Bacterium ascaridianum (А) на кутикуле (С)
аскариды человеческой. М — мышечные клетки. Оригинал.
ридой (Козырев), что объяснялось токсическим влиянием ее, вызвавшим
спазм кишечника. Аналогичное явление наблюдалось и при лентеце
широком, когда непроходимость кишечника вызвала потребность опера-
ции (неопубликованный случай). Бывали наблюдения над миграцией
аскариды в желудок и выбрасыванием ее наружу при рвоте. Внедрение
аскариды в патологически измененную стенку кишки ведет к перфора-
ции кишечника и последующему развитию перитонита. Проникание
аскарид в желудочный проток способствует развитию желтухи. Бывают
и другие случаи ненормальной локализации аскарид в евстахиевой тру-
бе, в черепных пазухах, в мочевых путях, в кожных абсцессах, в желч-
ных путях (с последующим разрывом печени) и мн. др.
Соответственно изложенному, симптомы болезни при аскаридозе
вариируют от неопределенных и малозначительных проявлений (рас-
стройство желудочно-кишечного аппарата, некоторые нервные симпто-
мы и др.) до угрожающих жизни расстройств (прободение кишек,
аппендицит при внедрении аскарид в червеобразный отросток и др.).
Нередки случаи «самопроизвольного» выхода аскарид без примене-
ния каких-либо противоглистных средств. Это явление в ряде случаев,
связывают с лихорадочным состоянием хозяина.
410
СПЕЦИАЛЬНАЯ ЧАСТЬ
Самопроизвольное освобождение от аскарид при условии отсутствия
' возможности реинвазии по Otto и Cort у детей (США) может происхо-
дить в срок 13 месяцев.
Сами аскариды могут подвергаться различным заболеваниям, в
•частности, бактериального характера. Болезнь проявляется в развитии
Рис. 332 — Аскарида человеческая, пораженная Bad.
ascaridianum. Фото. Оригинал.
пигментированных пятен — зооглей бактерий (Bacterium ascaridianum
Pawlowsky), которые разъедают покровы аскарид (рис. 331, 332). Из-
вестны и другие изменения патологического характера, ведущие к про-
бодению покровов аскарид, причем последние . погибают. Все эти мо-
менты влияют и на всасываемость хозяином аскаридных токсинов.
Рис. ?33 — Кожная уртикарна/i реакция при введении аскаридного анти-
гена в царапину кожи носителя аскарид через 50 и 60 минут.
По Taliaferro.
Диагноз ставится по нахождению яиц в faeces или самих вы-
шедших аскарид. Делались попытки применения иммуно-биологических
реакций для диагностики аскаридоза у человека. При введении в цара-
пину кожи водного экстракта из аскарид или сухого растертого порош-
ка из их тела на коже носителя аскариды вскоре появляются волдыри
с эритематозным основанием (рис. 333). Длительность такой эритема-
тозной и уртикарной реакции вариирует от 1 до 4 и более часов.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
411
Однако практического применения эти реакции не получили ни при
диагностике аскаридоза, ни при других гельминтозах, кроме эхинокок-
коза (стр. 333).
Лечение. Сантонин, который, однако, не предотвращает мигра-
ции личинок аскарид (Г. Смирнов), хеноподовое масло.,
В Японии широко применяется для изгнания аскарид макнин, извле-
каемый из травы Digenea simplex; он рекламируется как препарат,
наиболее сильно действующий на аскарид. Однако проверка эффектив-
ности его действия при аскаридозе человека, произведенная Р. Шуль-
цем и Н. Шихобаловой, показала, что макнин обладает гораздо более
слабым действием на аскарид, чем сантонин, давая в среднем 38,3%
изгнаний.
Хорошим средством является гексилрезорцин, который дают после
вечерней клизмы утром и до еды взрослому в количестве 1,5 г, детям
до 12-летнего возраста от 0,4 до 1,0 г. После приема лекарства не есть
пять часов. Затем солевое слабительное.
Успех противоглистной борьбы может быть обеспечен лишь приве-
дением в действие системы потребных мероприятий. Одностороннее
действие их дает кратковременный и сходящий на-нет успех. Так, ре-
зультат одного только лечения при аскаридозе нивеллируется уже че-
рез восемь месяцев, т. ve. освободившееся от аскарид население снова
их приобретает, если одновременно не проводятся общие профилакти-
ческие мероприятия (Otto, Cort).
Профилактика. Соблюдение основных санитарно-гигиенических
требований; обработка фекалий, употребляемых для удобрения, горячей
водой для уничтожения яиц аскарид; введение в практику пользования
уборных, предохраняющих почву и воду от попадания в них яиц аска-
рид; охрана водоемов от фекальных загрязнений; борьба с мухами,
тараканами и другими механическими переносчиками яиц аскарид; чис-
тое содержание жилья и служб; широкая дегельминтизация различных
групп населения с сжиганием выходящих глист; широкое распростра-
нение санитарных сведений среди населения.
Одним из важных звеньев профилактики аскаридоза является уни-
чтожение яиц аскарид во внешней среде («дегельминтизация»
внешней среды). Стойкость яиц человеческой аскариды к внешним воз-
действиям иллюстрируется следующими показателями.
Минутное нагревание яиц при 54° С убивает три четверти яиш
Трехминутное нагревание при 54° С или мгновенное действие темпера-
туры 56° С губит все яйца (Nolf).
К низким температурам яйца аскариды устойчивы настолько, что
перезимовывание их во внешней среде может быть вполне обеспечено,
но —30° С убивают яйца аскарид (Bakker). Более высокие температуры
легче переносятся незрелыми яйцами, чем яйцами с развившимся заро-
дышем. Сорокадневное хранение незрелых яиц и яиц в стадии дробле-
ния в холодильнике при —18,9° С до —26,7° С не препятствует их
дальнейшему развитию (при +24° С оно длится две недели).
Зрелые яйца в тех же условиях погибали через 20 дней хранения на
холоду. При более коротких сроках в яйцах оставались живые личинки,
но заражение животных такими яйцами осталось безрезультатным
(Cram).
Возможность зимования яиц аскарид в условиях Ленинграда была
показана Г. Смирновым.
Для развития аскарид необходима влажность. Однако этот фактор
41?
СПЕЦИАЛЬНАЯ ЧАСТЬ
труднее учитывается количественно, ибо его необходимо определять
применительно к микроклимату биотопов, в .которых находятся яйца
аскарид.
В условиях комнатной температуры + 20— +27° яйца аскариды (из
фекалий) погибали на 13-й день (Коровицкий и Артеменко), причем к
исходу пятых суток уже погибало 60% яиц.
При 30° С и насыщенной влажности почти три четверти всех яиц
созревают к 11-му дню; при той же влажности и температуре 22° ‘С к
16-му дню созрело лишь 39% яиц, а при 77% влажности яйца через
15 дней погибли (Sindler).
Роль влажности косвенно подтверждается увеличением интенсивно-
сти инвазии аскаридами к концу дождливого сезона (Панама, Cort с
сотрудниками). Показателем является увеличение вдвое среднего коли-
чества аскаридных яиц на 1 см3 фекалий (7 910 против 15 810 яиц).
Однако о значении фактора влажности нельзя судить только по дан-
ным метеорологии; интенсивность аскаридоза, например, сходна в Па-
наме (2 540 мм осадков в год) и в Северном Китае (508—762 мм осад-
ков, Cort и Stoll). Очевидно, решающее значение имеет влажность (в
комбинации с другими факторами) тех биотопов, куда попадают фека-
лии с яйцами аскарид.
Прямые солнечные лучи убивают яйца аскарид с 6 по 69 часов дей-
ствия (при + 25—+30° С воздуха и +42° С поверхности почвы,—
Коровицкий и Артеменко); все личинки погибали уже через 27 часов.
Для развития яиц аскарид важен и фактор субстрата (почвенный
или эдафический фактор), значение которого определяется
комбинациями с уже рассмотренными выше факторами внешней среды.
Наиболее неблагоприятна для развития яиц аскарид песчаная, прогре-
ваемая солнцем почва. В черноземе развитие идет несколько замед-
ленно.
При культивировании в тени яйца свиной аскариды (Otto) развива-
ются на песчаной, глинистой и глинисто-зольной почве; на солнце же-
погибли почти все, кроме части яиц (около одной четверти) на глини-
стой и песчаной почве.
Особое значение приобретает использование городского мусора и по-
лей орошения в огородном хозяйстве. На полях орошения (Тула) яйца
аскарид и власоглава были находимы в пробах земли с участков, зали-
тых за полгода до производства анализа. При соблюдении норм залива
участков фекалиями, правильного чередования участков при заливке,
запахивания в первые сутки после заливки и при соблюдении правиль-
ной смены культур поля орошения могут быть безопасными в отноше-
нии распространения глистных инвазий через взращиваемую на них
огородную зелень и корнеплоды (3. Василькова). Широко применяемая
для дезинфекции уборных хлорная известь мало действует на яйца
аскарид. Прибавление к фекалиям даже половинного количества хлор-
ной извести не убивает все яйца аскарид (Затуренская и Вишневская).
Поэтому в отношении профилактики аскаридоза важнее регулярная
очистка пола и сидений уборных от загрязнения фекалиями с мытьем
их 5—10% раствором карболовой кислоты. Действительным средством,
убивающим яйца глист, является негашеная известь в момент гашения.
Распространение аскарид велико, и экстенсивность инва-
зии ими различна. В некоторых районах СССР процент зараженного
аскаридами населения ничтожен; 11,7% отмечается в некоторых местах
Средней Азии; в других аскариды поражают больший процент населе-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
413
ния (Армения, Белоруссия). Общая картина распространения аскарид
в СССР очень пестра; в одном и том же пункте процент зараженности
аскаридами весьма вариирует в зависимости от культурного уровня об-
следуемых групп населения. Это соображение, конечно, имеет общее
значение и для оценки распространения других паразитических червей,
заражение, которыми происходит без посредничества промежуточных
хозяев.
Роль свиней в возможном распространении аскаридоза человека не
вполне ясна. Свиные аскариды морфологически идентичны человече-
ским, хотя, по имеющимся экспериментальным данным, яйца свиной
аскариды — Ascaris suilla — не заражают человека; но миграция
личинок, вышедших из проглоченных яиц, происходит. Следовательно,
у человека могут наблюдаться все симптомы, связанные с феноменом
миграции личинок свиных аскарид, хотя половозрелых аскарид в его
кишечнике не развивается, так как личинки не задерживаются в киш-
ках, а выбрасываются с faeces наружу. Таким образом, можно говорить
о вредоносном действии свиных аскарид в течение периода миграции
их личинок (мигроаскаридоз).
Вопрос о видовых соотношениях человеческой и свиной аскарид
имеет довольно длинную и разностороннюю историю. Ее уместно рас-
смотреть для получения представления о разнообразии путей поисков
видовых критериев паразитических организмов, морфологически неот-
личимых друг от друга.
Число хромозом может быть одним из видовых признаков; в яйцах
аскариды человеческой Койдзуни, Хираиши и Койно засчитывают 12,
а у свиной аскариды— 13 хромозом.
При вариационно-статистическом изучении яиц этих аскарид
И. Хаустовым обнаружены отличия в объеме и в форме яиц; средний
об^ем яиц человеческой аскариды М = 76,960+ 219 у-3, а у свиней —
М = 66,335+ 134 +; среднее отношение длинной оси яйца к попереч-
ной у человеческой аскариды М = 1,2839 + 0,0027, а у свиней
М= 1,3652 + 0,0029.
Скорость развития яиц также неодинакова. При 21 и 24° яйца сви-
ной аскариды развиваются скорее, чем человеческой (455 : 500;
315:360 часов). Величина влияния температуры на скорость развития
(Qio формулы Ван’т Гофф) при 21 и 30—31° у свиной аскариды рав-
няется 2,11, а у человеческой — 2,62 (И. Хаустов). Опыты с перекрест-
ным заражением человека яйцами свиной аскариды и поросят—яйца-
ми человеческой аскариды дают отрицательный результат.
Что же касается эпидемиологических соотношений аскаридоза чело-
века и свиней, то в различных точках наблюдаются пестрые соотноше-
ния, которые не дают возможности делать общие решающие выводы.
По совокупности имеющихся данных есть основание считать человече-
скую и свиную аскариды различными видами, несмотря на отсутствие
признаков морфологического значения.
Род Toxocara Stiles
Toxocara canis Werner
Синоним: Toxascaris limbate Raill. et Hen., Toxascaris marginata Leip,
Toxascaris leonina Linst
Аскарида собачья — самка 6,5—10 см, самец 4—6 см длины.
Передний конец тела искривлен и несет узкие крыловидные придатки
414
СПЕЦИАЛЬНАЯ ЧАСТЬ
случаи внутриутрооного заражения
Рис. 334— Toxocara cards (Л) и Toxo-
cara mystax (В). Головной конец с кути-
кулярными крыловидными придатками.
По Фаусту.
кутикулы (рис. 334, Л). Рот окружен тремя губами. Хвостовой конец
самца без крыловидных придатков. Спикулы две, искривленные, одна
чуть короче другой, без крыловидных придатков. Яйца короткоовальные
с толстой, гладкой оболочкой 0,075—0,085 мм. Обычный паразит тонких
кишек, реже желудка собаки; паразитирует также у волка, лисы, шака-
ла, кошки. Космополит. Известны единичные случаи гостепаразитиро-
вания у человека (в СССР — Скрябин с сотрудниками). Заразительно
яйцо с уже развившейся личинкой, на что требуется известное время.
Миграция личинок идет, как у человеческой аскариды. Наблюдаются
щенят этим паразитом.
Род Belascaris Lei per
Belascaris cati Schrank
Синоним: Toxocara mystax Zeder.,
Belascaris mystax Zeder.
Кошачья аскарида. Пе-
редний конец обыкновенно кругло^
искривлен с двумя широкими кры-
ловидными складками кутикулы
(рис. 334, В), благодаря чему общая
форма переднего конца — стрело-
видная.
Длина самки 4—12 ом, самца
3—6 см; спикулы 1,7—1,9 мм дли-
ны. Женское половое отверстие —
на границе передней четверти тела.
Яйца почти шарообразные, с тон-
кой ячеистой оболочкой 65—75 р.
Задний конец тела самца с двумя более или менее заметными крыло-
видными придатками.
Обыкновенный паразит- кошки; в единичных случаях найден и у
человека (в частности в СССР). Космополит.
Миграция в цикле развития кошачьей аскариды, как и у A. lumbfi-
coides.
Род Lagochilascaris Leip.
Lagochilascaris minor Leip.
Мало изученные аскариды, найденные в единичных случаях в под-
кожных нарывах разных мест тела человека в Гвиане, Тринидаде и др.
Естественным хозяином этого вида является леопард — Felis nebulosa.
Самец — 9 мм, самка—15 мм длины. Крыловидных придатков нет, но
по бокам тела вдоль тянутся две кутикулярные складки. Губы отделе-
лы от остального тела резко выраженной перетяжкой. Цикл развития
неизвестен.
Известны недостаточно изученные Asc. texana Smith et Goeth. и
maritime. Leuck, по одному разу найденные у человека.
Надсемейство Spiruroidea
Род Gongylonema Molln.
Gongylonema pulchrum Molin.
Известно несколько видов гонгилонем, живущих под слизистой
оболочкой рта, пищевода или желудка человека и различных живот-
ных — свиньи, лошади, овцы, верблюда и др.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
415
В штате Арканзас (США) у девушки 16 лет в губе была обнару-
жена круглая глиста, путешествовавшая по подслизистой соединитель-
ной ткани до глотки. Присутствие паразита сопровождалось нервными
расстройствами, анемией и рвотой. Извлеченная глиста была 42,1 мм
длины. Передний конец ее тела на протяжении 1,4 мм был покрыт
кутикулярными чешуйками или бляшками, лежащими восемью про-
дольными рядами. Этот паразит был определен как Gongylonema
pulchrum (рис. 335, 336),
обычно паразитирующий в
пищеводе жвачных и встре-
чающийся также в пищеводе
свиней. Настоящими хозяева-
ми этого паразита являются
жвачные, у которых он и до-
стигает предельных разме-
ров— самка до 145 мм, а са-
мец 62 мм длины. Передний
конец тела несет кутикуляр-
ные боковые крыловидные при-
датки (рис. 336, В). Задний
конец самца снабжен двумя
несимметричными кутикуляр-
ными крыловидными придат-
ками (рис. 336, С). Спикулы
весьма неравны по величине.
Женское половое отверстие
лежит чуть кпереди от аналь-
ного — всего миллиметрах в
двух от заднего конца тела.
Влагалище очень длинное (тя-
нется до середины тела). 'Мат-
ки две — расходящиеся к обо-
им концам тела, где переходят
в яйцеводы. Яйца прозрачные
(рис. 337), с толстой оболоч-
кой и с закрученным внутри
зародышем; их длина 50—70 у
и ширина 25—37 у.
Рис 335 — Gongylonema pulchrum. Самец.
По Sam bon.
Выброшенные с faeces яйца вскрываются в кишечнике различных
насекомых (если будут ими проглочены): жуков-навозников (Aphodius
fimentarlus, Onthophagus taurus (рис. 338), Caccobius schreiberi) и та-
раканов-прусаков (Blatta germanica). Личинки червя пробуравливают-
ся в полость тела насекомого, где капсулируются (рис. 339). Когда
промежуточный хозяин будет съеден хозяином окончательным, освобо-
дившиеся личинки, повидимому, вбуравливаются в стенку желудка или
двенадцатиперстной кишки и поднимаются в ее толще (не гематоген-
ным путем) в пищевод или в ротовую полость.
Известен ряд случ-аев гостепаразитирования Gongylonema у челове-
ка; видовые соотношения их и идентичность с Gong, pulchrum неясны.
Случай гонгилонематоз человека описан в СССР (Р. Шульц и
С. Иваницкий по материалу Сербинова) из Харькова. Больные иногда
жалуются на ползание червя под слизистой губы, что скептически вое-
416
СПЕЦИАЛЬНАЯ ЧАСТЬ
принимается врачами, скорее склонными рассматривать такую жалобу
как проявление истерии. Но в некоторых случаях больные тут же сами
с ручным зеркальцем разыскивали под слизистой червячка и указывали
на него врачу. По извлечении червячка он бывал определяем, как Gon-
Рис. 336 — Gongylonema pulshrum.
А — самец в натуральную величину; В. С — передний и
задний конец при сильном увеличении. По Р. Шульцу
и С. Иваницкому.
gylonema. Подробнее см.: Р. Шульц и С. Иваницкий. Обзор во-
проса о гонгиломатозе человека с описанием собственного случая см.
Медицинская паразитология, 1934, т. III, вып. 6, и Г. Смирнов. Ра-
ковые опухоли и животные паразиты. Вестник микроб, и эпидемиол.,
т. VI, вып. 1, 1927.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
417
Болезненные симптомы при гонгилонематозе различны и непостоян-
ны (фарингит, нервные расстройства и др.). G.neoplasticum, повиди-
Рис. 337 — Яйца Gongy-
lonema из фекалий чело-
века. Панама. По
Faust и Martinez.
Увеличено в 1000 раз.
Рис. 338 — Onttophagus tauras— самка,
промежуточный хозяин Gongylonema.
мому, способствует разрастанию опухолей слизистой оболочки пищева-
рительного канала, чем и объясняется данное ей название.
Диагноз ставится по определению извлеченного паразита.
Рис. 339 — Капсулированная личинка' Gongylonema
из жука Onthiphagus taurus.
Лечение. Извлечение глисты из-под слизистой оболочки прошив-
кой иглой тела паразита. Известен случай, когда тимоловым полоска-
нием эта глиста была стимулирована к выходу и удалена пальцами
(Фауст).
Профилактика. Остерегаться возможного проглатывания тара-
канов-прусаков,
418
СПЕЦИАЛЬНАЯ ЧАСТЬ
Семейство Gnathostomidae
Род Gnathostoma Owen
Виды Gnathostoma нормально паразитируют у диких кошачьих.
У человека в качестве гостепаразитов известно два вида.
Gnathostoma spinigerum Owen
Взрослые самцы 11—25 мм, а самки 25—54 мм длины, с коренастым
телом красноватого цвета и многочисленными кольцевыми перетяжка-
ми кутикулы. Вздутый головной конец отделен шейным перехватом от
Рис. 340
А — Gnathostoma sp. от человека; губа; с — кутикулярный воротник;
cs — шейный мешок; es — пищевод; hb — головное вздутие; i—кишеч-
ник; г — прямая кишка; а — анальное отверстие. По Morishita и
Faust. В — Gnathostoma sp. Головной конец. Ъ — губа; пг —^нерв-
ное кольцо. Остальные буквы см. предыдущий рис.
По Faust и • М о г i s hi t a.
тела (рис. 340) . По бокам рта лежат две больших губы, на которых
открывается по общим протокам по две шейные железы соответствую-
щих сторон тела (рис. 340, В). Головное вздутие покрыто несколькими
кольцевыми рядами кутикулярных крючьев; кутикулярные шипики усеи-
вают также переднюю половину тела. Женское отверстие лежит тотчас
позади середины. длины самки. Длинное влагалище идет от него
кпереди.
Яйца овальные, 65—75 у длины и 38—40 jx толщины, с прозрачной
скульптурной оболочкой; на одном ее полюсе имеется мукоидная проб-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
419
ка; зародыш находится в стадии морулы или гаструлы. На заднем кон-
це самца несколько пар крупных и мелких сосочков вокруг клоаки.
Спикулы неравной длины (1,2 и 0,4 мм).
Действие на хозяина. Паразитируют в опухолеобразных уз-
лах желудка тигра, леопарда, дикой кошки, собаки и др. От Индии до*
Китая, в Европе и в Америке. В узлах паразиты лежат по нескольку
штук, плотно свернутыми, будучи окружены гноем; через отверстие,
которым паразитарный узел сообщается с полостью желудка, яйца
паразита выходят в полость желудка и в конце концов выбрасываются!
наружу. Жизненный цикл в точности неизвестен.
У человека установлено около 50 случаев гостепаразитирования
Gn. spinigerum в абсцессах кожи, в узлообразных разрастаниях покро-
вов и в состоянии блуждания под кожей, причем место передвижения
паразита отекает; отек перемещается с места на место в зависимости
от нахождения паразитов (oedeme ambulant siamois). В других случаях
кожные изменения дают картину линейно ползущего дерматоза—der-
matitis linearis migrans, который надо отличать от «larva migrans» или
«creeping disease» — поражения кожи, обусловливаемого передвиже-
нием в ней личинок желудочного овода — Gastrophilus. И у нормаль-
ных хозяев Gn. spinigerum бывают случаи кожного гнатостомоза с на-
хождением паразитов или в опухолеобразных узлах кожи, или в ходах,
прокладываемых между эпидермисом и соединительной тканью покро-
вов. Не исключается возможность нахождения этого паразита и в стен-
ке желудка человека.
В недавнее время (1933, 1936) выяснено, что из яиц Gn. spinige-
rum через 10—12 дней пребывания в воде при комнатной температуре
выходят очень подвижные личинки; будучи скормлены циклопам, они
проходят в полость их тела и там превращаются в стадию, имеющую
сходство с взрослым червем. Скармливание таких циклопов с 7—30-
дневными личинками кошкам оставалось безрезультатным. Очевидно,
должен быть какой-то второй хозяин. Одним из таковых является пре-
сноводная рыба Clarias batrachus L.; в ее мышцах обнаруживались ин-
капсулированные личинки после скармливания рыбе циклопов с личин-
ками (Chalerm Prommas и Daengsvang, 1936, Сиам). Эксперименталь-
но доказано, что вторым промежуточным хозяином может быть и дру-
гая рыба Ophicephalus striatus.
Диагноз — по определению извлеченного паразита.
Лечение. Извлечение узла с паразитом в нем.
Профилактика. Не есть полусырой рыбы.
Gnathostomum hispidum (Fedtsch.)
Паразитирует, прикрепляясь к стенке желудка домашней свиньи,
кабана; бывает также у коров и один раз добыт из покровов человека.
Более обыкновенен, чем Gn. spinigerum. Распространен в Европе и в
Азии; известен из Средней Азии и других мест СССР. Отличается от
предыдущего вида присутствием 10—12 рядов головных крючьев (про-
тив 8 рядов Gn. spinigerum); кутикулярные щитки, покрывающие перед-
нюю часть тела, имеют по несколько зубчиков. Яйца с выростом на од-
ном из молюсов 74 у длины и 41 толщины. Цикл развития неизвестен.
420
СПЕЦИАЛЬНАЯ ЧАСТЬ
Семейство Physaeopteridae
Род Abreviata Trav.
Abreviata caucasica Linst.
Синоним: Physaloptera mordens Leip., Physaloptera caucasica Linstow.
Самка до 10 см длины и 2,8 мм толщины, самец — до 5 см длины
и 1 мм толщины; похожи по общему виду на незрелых аскарид, отли-
чаясь в то же время несколькими существенными признаками. Рот
окаймлен двумя боковыми губами со сложным кутикулярным воору-
жением в виде зубчиков и зубов (рис. 341). Каждая губа несет до
два сосочка. Половое отверстие самки лежит в передней части теЯа.
Маток четыре. Хвостовой конец тела округлый. Яйца с толстой глад-
кой скорлупой, эллиптические, со зрелой личинкой. Размер их: 57—
62 (и длины и 42—45 tu толщины. На заднем конце тела самца имеют-
ся чуть асимметричные крыловидные придатки с 4 парами стебельча-
Рис. 341 —Abreviata caucisica
из человека Physaloptera
caucasica). Головной конец
самки со спинной стороны.
По Р. Шульцу.
Рис. 342 — Abreviata caticastcd ИЗ
человека ( = Physaloptera cauca-
sica). Хвостовой конец самца.
ПоР. Шульцу.
тых сосочков. Перед отверстием клоаки лежат три, а позади ее десять
сосочков (рис. 342). Спикулы неравной длины (3,2—5,5 мм и 0,476—
0,62 мм).
Паразитируют у человека и обезьян. Прикрепляются к оболочке
пищеварительного тракта, начиная от пищевода до тонких кишек,
включая ileum. Найден впервые Менетриэ на Кавказе и Leiper и Tur-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
421
пег у туземцев тропической Африки. Один раз наблюдался в Москве
с установкой диагноза гголько на основании нахождения яиц (Подъя-i
польская). Цикл развития неизвестен.
Семейство Thelazidae
Род Thelazia Bose.
Thelazia callipaeda Raill. et Henry
Желтовато-белый червь с цилиндрическим телом, сужающимся к
обоим концам; самка 7—18 мм длины и 0,5—0,85 мм ширины; самец
8—14 мм длины и 0,25—0,75 мм ширины. Губ нет. Кутикула покровов
хорошо исчерчена в поперечном направлении (рис. .343). Хвостовой ко-
Рис. 343 — Thelazia callipaeda.
Передний конец тела с рото-
вой капсулой и кутикулярной
складчатостью. По Фаусту,
Рис. 344 — Thelazia calli-
paeda. Задний конец те-
ла самца с преанальными
и постанальными сосоч-
ками и спикулами.
По Фаусту.
нец самца круто загнут. Две спикулы очень неравной величины. Перед
половым отверстием на кутикуле лежат десять пар сосочков, а поза-
ди— шесть пар (рис. 344).
У самки половое отверстие лежит в задней трети пищевода, от-
ступя от головного конца на 0,56—0,68 мм. На округленном заднем
конце тела лежат два концевых сосочка. Яйца овальной . формы
54—62 рь длиной, 34—46 шириной, с вполне сформированной личин-
кой длиной 35—49 j*. Цикл развития неизвестен.
Паразитирует в конъюнктивальном мешке и мигательной перепонке
глаза собаки (Индия, Китай, Владивосток). Найден также у кролика.
Известны единичные случаи паразитирования в конъюнктиве глаз че-
ловека (Китай).
У собак вызывает истечение слез. У человека может вызвать пара-
лич век и сильную боль.
422
СПЕЦИАЛЬНАЯ ЧАСТЬ
Диагноз. Определение паразита, извлеченного из глаза.
Лечение. 2% раствор кокаина в конъюнктивальный мешок и
удаление глисты пинцетом.
Профилактика неизвестна.
Надсемейство Filarioidea
Род Wuchereria da Silva Araujo
Wuchereria bancrofti Cobboid
Взрослая нитчатка похожа на конский волос беловатого или ко-
ричневатого цвета. Самка 70—90 мм длины и 0,24—0,28 мм ширины;
соответственные размеры самца — 40—50 мм и 0,1 мм (рис. 345, I).
Задний конец тела округлый; передний приострен и несет кнопкообраз-
ное утолщение. Близ рта лежат два ряда сосочков. Самка имеет две
трубчатые матки, в которых развиваются зародыши (из яиц). Муску-
листое влагалище открывается на брюшной стороне тела на расстоя-
нии 1,2—1,3 мм от его переднего конца. Задний конец тела самца
несет восемь околоанальных и два постанальных сосочка. Из отверстия
половой клоаки торчат две неравных спикулы (0,2 и 0,6 мм длины).
Клоака открывается наружу на расстоянии 0,13 мм кпереди от задне-
го конца тела. Яйца длиной 40 шириной 25 у.
Взрослые нитчатки чаще всего встречаются в припухших лимфати-
ческих железах, семеннике, лимфатических сосудах и т. п. (рис. 345,
III). Скопляясь в больших количествах, они могут закупоривать ductus
thoracicus и вызывать застой лимфы в брюшной полости. Следствием
этого является варикозное расширение лимфатических сосудов, пре-
имущественно мошонки (lymphoscrotum) и брюшной стенки. Некото-
рые варикозные узлы достигают до 30 см диаметром, они наполнены
мутной красновато-белой жидкостью. Растянутые лимфатические сосу-
ды (lymphangitis) могут разрываться. Если это случается в почечной
лоханке, то моча принимает молочный вид (хилурия), и в ней могут
быть открыты как личинки, так и взрослые нитчатки. Паразиты эти бы-
вают также в хилезном асците. Во всех указанных местах встречаются
оба пола нитчаток.
Самки отрождают громадное количество личинок, поступающих в
кровяное русло, где они известны под именем Microfilaria nocturna
(рис. 346, 347).
Процесс отрождения личинок (т. е. жизни самок в человеке) может
длиться многие годы, по Knabe (1932) —более 17 лет.
Длина личинок колеблется от 0,127 до 0,3 мм, а толщина 0,008—
0,01 мм. Они окружены тонкой оболочкой, заметно отстоящей чехлом
от переднего и заднего концов тела. Непосредственно под кутикулой
лежит слой хорошо красящихся гематоксилином ядер, принадлежащих,
вероятно, мускульным клеткам (рис. 346). В хвостовой части этих личи-
нок упомянутых ядер нет, что имеет диагностическое значение, так
как у Microfilaria diurna ядра доходят до заднего конца тела. Кроме
этих клеток, личинки нитчаток имеют и другие, значение которых тол-
куется пока предположительно. Сзади за нервным кольцом (рис. 346)
лежит экскреторная пора и за ней экскреторная клет-
ка (видны при окраске по Романовскому, Гимза). В начале задней по-
ловины личинки помещается «внутреннее тел о»; оно обнаружи-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
423
вается прижизненной окраской нейтральротом. Позади помещаются че-
тыре клетки G—1—4 (половые зачатки); важно положение перед-
ней из них, т. е. клетки Gi, которая при окраске Azur II относительно
Рис. 345 — U7 uchereria barterofti *
I— JPb натуральную величину; II—Microfilaria nocturna (сильно
увеличена); III — схема жизненного цикла. А — Поступание микрофиля-
рий (f) из кровеносного сосуда человека в хобогок (st) комара при
осании им крови; sc, sm — эпидермис; В—проникание микрофилярий
сквозь стенку желудка комара (ер) в полость его тела; sch — сброшенная
наружная шкурка (футляр) личинки; С—заражение комара человека
при сосании его крови; fm—личинка филярии в грудных мышцах
комара; f — филярии, вышедшие из хоботка комара и проникающие в
кожу; sm, sc — эпидермис; D — филярии в теле человека; Is — lympho-
scrotum; lg— lymphangitis; la — lymphadenitis; vlg — расширение ductus
thoracicus; fa — нарыв, причиненный филярией. По F й 1 1 е b о г n.
мала. Сзади последней клетки лежит половая или анальная
пора, удаленная от заднего конца тела.
Описанные личинки получили название 'Microfilaria nocturna благо-
даря своему свойству появляться в периферических кровеносных сосу-
424
СПЕЦИАЛЬНАЯ ЧАСТЬ
дах только ночью; днем же они уходят в глубь тела и держатся в ле-
гочных сосудах, сердце, аорте, почках. В печени, селезенке и костном
мозгу их не бывает. Такая пёриодичность в поведении паразитов свя-
зана с режимом сна, больного. Если
хозяин микрофилярий будет спать
днем, а бодрствовать ночью, то ми-
крофилярии появляются в перифе-
рических сосудах днем, ночью же
их не найти. В последнее время ука-
зывают, что микрофилярии привле-
Рис. 346— Схема строения Microfilaria
nocturna по Rodenwaldt.
J — внутреннее тело; Gx — G4 — половые клетки;
Ер — выделительная пора; N — зачаток нервной
системы; Ez — выделительная клетка; Gp —
•анальная пора. Сильно увеличено.
каются в русло периферической
крови химиотактически действием
слюны комаров, сосущих человека.
Человек заражается нитчаткой
от укола ротовыми органами
различных кровососущих комаров,
(рис. 348). В- качестве промежуточ-
ных хозяев Wuchereria bancroftl
установлены следующие виды их:
Culex fatigans, С. pipiens, С. аппи-
lirostris, Aedes variegatus, Taenio-
rhynchus africanus, Anopheles ros-
si, Л. costalis, A. algeriensis и др.
Для некоторых мест определена естественная зараженность кома-
ров личинками W. bancrofti. На Барбадосе 23% исследованных Culex
fatigans было заражено при
наличии 12—13% населения с
Microfilaria nocturna.
Когда перечисленные насе-
комые сосут вечером или но-
чью кровь носителей вухере-
рий, то плавающие в крови
личинки попадают в желудок
комара. Там они через час —
полтора теряют свою в л а г а-
лищную оболочку и про-
никают сквозь стенку желудка
в полость тела (рис. 345, В).
Залегая затем между мышца-
ми груди, где их можно видеть
уже через сутки, личинки про-
делывают дальнейшее разви-
тие, в течение которого они
приобретают размеры — 1,7 мм
длины и 0,03 мм толщины;
у них появляются кишечник и
другие органы. Затем они рас-
Рис. 347 — Микрофилярии Wuchereria banc-
rofti (слева) и Acanthocheilonema perstans
(справа). По Baylis.
ползаются по телу, и часть
личинок скопляется у основания ротовых органов, а также в самом
хоботке комара. Развитие нитчаток в насекомом длится от 8 до 35 дней,
смотря по температуре.
Когда комар сосет здорового человека, то вонзает в его кожу ко-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
425
лющие ротовые щетинки; при этом происходит разрыв тонкой межсо-
членовной перепонки между концевыми дольками хоботка (рис. 348),
из полости которого личинки W. bancrofti выходят наружу, активно
вбуравливаются в покровы человека и попадают в лимфатические со-
суды. Здесь они вырастают в половозрелые нитчатки, дающие, в свою
очередь, новое потомство личинок.
Филяриоз в рассматриваемой его форме — вухерериоза — очень
распространенная болезнь тропических и субтропических стран. В Ев-
ропе наблюдались, вероятно, занесенные случаи в Испании и Турции.
В некоторых местах (например Коморские острова) около 80—90%
Рис. 348 — Личинки Wuchererta banchrojti (л), выходящие из хоботка Culex fatigans
Из S a m b о п.
населения поражены этими паразитами. Однако не все носители нитча-
ток непременно больны какой-либо соответственной болезнью; между
ними бывает много совершенно здоровых.
Клинические формы филяриоза очень разнообразны.
В основе их лежит изменение лимфатической системы в виде расшире-
ния лимфатических сосудов и воспалительного утолщения их стенок.
Воспалению с последующими абсцессами подвергаются и лимфатиче-
ские железы. Застой лимфы в грудном протоке (ductus thoracicus) вы-
зывает обратный ток ее в сосуды брюшной полости и таза, что влечет
за собой в разных случаях развитие хилурии, хилезных выпотов, 1ут-
phocoele, lymphoscrotum. Воспаление яичка и семенного канатика зави-
сит от местного действия паразитов, которые, скопляясь в больших коли-
чествах, вызывают воспаление окружающих тканей. Последние при этом
становятся более доступными для вторжения вторичной бактериальной
инфекции. Аналогичное изменение лимфатической системы вызывает
появление слоновости различных участков тела (Elephantiasis
arabum). Это заболевание проявляется чрезмерным разрастанием и от-
вердением подкожной клетчатки ног, мошонки, больших губ, женских
грудей, рук и других участков тела. Пораженные места резко увеличи-
ваются в размерах; несмотря на значительные внешние изменения, бо-
лезненных ощущений больные не испытывают; их беспокоит толькс
массивность и ненормальная тяжесть измененного органа.
В СССР известны пока единичные случаи слоновой болезни. Один
-426
СПЕЦИАЛЬНАЯ ЧАСТЬ
из них в классической форме разрастания scrotum (окружность —
IV2 м) и ноги был наблюдаем, у таджика в горах Таджикистана (на
р. Кафирниган, Е. Павловский; рис. 349). Микрофилярий в крови у
этого больного найти не удалось. Это соответствует факту сравнитель-
но редкого нахождения названных паразитов у больных слоновостью,
тогда как носительство Microfilaria nocturna бывает иногда у значи-
тельного количества здоровых.
Рис.349 — Слоновая болезнь (случай из Таджикистана).
По Е. Павловскому.
В противоположность теории признания нитчатки Банкрофта возбу-
дителем слоновой болезни некоторые исследования считают, что «тео-
рия застоя лимфы» должна быть оставлена и что инвазия W. bancrofti
"Сама по себе не дает никаких симптомов; все связанные с филяриозом
патологические проявления зависят от вторичной бактериальной инфек-
ции. Пока остается в силе мнение Fiillebom, что мы все еще не знаем,
в какой мере при слоновости или при ведущем к ней бактериальном
лимфангите причинную роль играет действительно W. bancrofti, или
местная 'бактерийная флора, или какие-то другие еще неизвестные
факторы.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
427
Диагноз. Нахождение личинок филярий в крови (толстая кап-
ля), в лимфе, в выпотах, в хилезной моче и др.
Лечение. При слоновости — хирургическая операция: удаление
-части или всего пораженного органа.
Профилактика. Защита от нападения комаров (условно).
Род Onchocerca Diesing
Onchocerca volvulus (Leuck.) Raill. et Henry
Синоним: Filaria volvulus Leuck.
Рис. 350 — Onchocerca volvulus, в момент
выхода ее личинок из нижней губы ее
переносчика — мошки — Simulium damno-
sum. По Blacklock.
Филярия белого цвета. Живет в соединительнотканных опухолях
-фиброматозного характера величиной с яйцо или апельсин, в области
ребер, на спине и других местах.
Длина взрослой самки около 45 см, толщина — 0,33 мм. Кутикула
с поперечными утолщениями, придающими ей рубчатый вид. Самец
3—4,5 см длиной и 0,8 мм тол-
щиной. К обоим концам тела он
резко суживается.
Ротовых сосочков нет. Зад-
ний конец тела закручен спи-
рально. Личинки крупные, 0,3 мм
длины и 0,006 мм толщины. Чех-
лика нет. Задний конец тела
заострен, кожных ядер в нем
нет. Личинки находятся в лимфа-
тических полостях опухоли и,
вероятно, бывают в крови, где
захватить их все же пока не
удается. Тропическая Африка.
Переносчиком О. volvulus, по
данным Blacklodk, является мош-
ка Simulium damnosum. В ее
грудных мышцах личинки ли-
няют, переходя далее в голову
(рис. 350). Через разрыв хитино-
вой кутикулы личинки выходят
наружу при сосании мошкой крови человека, которого и заражают.
Цикл развития О. volvulus в Simulium длится шесть дней и более.
Simulium ornatum является промежуточным хозяином и переносчи-
ком Onchocerca gutturosa крупного рогатого скота в Англии (J. Ste-
ward).
Culicoides nubeculosus является переносчиком Onchocerca cervicalis
.лошади (Steward).
Onchocerca coecutiens Brumpt
В тропической Америке встречается онхоцеркоз с локализацией опухолей с па-
разитами исключительно в области головы. Находимый здесь паразит морфологиче-
ски не отличим от О. volvulus, но благодаря особенностям его локализации* его вы-
деляют по этому биологическому признаку в особый вид — О. coecutiens.
428
СПЕЦИАЛЬНАЯ ЧАСТЬ
Род Loa Stiles
Loa loa Cobbold
Синонимы: Filaria loa Gyot et Leuck.
Взрослая нитчатка белого цвета, толщиной с тонкую струну. Для паразита ха-
рактерны кснический передний конец тела и бородавчатые бугорки на кутикуле.
Мускульные клетки развиты хорошо, вследствие чего червь на свободе оживленно
движется, в хозяине же передвигается по соединительной ткани со скоростью
до 1 см в минуту. Головной конец несет шишкообразный вырост^ с воронковидным
ротовым отверстием и шесть плоских сосочков. Пищевод короткий—1.0 мм длины.
Г Г ____ ___„ Q. И л.Ж ТТ ГГГ1ТТТТ ЧТ ЛТГ/ЛТТГ»
Рис. 351 — Конец ротовых органов
слепня Chrysops silacea, являющего-
ся перенрсчиком нитчатки Loa loa.
Видно выхождение личинок Loa loa
из концевых долек (labellulae) хо-
ботка. По А. и S. С о n n а 1.
день после сосания крови слепнем. М.
Самка около 6,0 см длины и около
0,5 мм толщины. Наиболее толстая часть
тела позади головы; задний же конец
его шириною всего 0,17 мм. Бородавки
особенно многочисленны на кутикуле близ
хвостовой части паразита. Заднепроходное
отверстие лежит на расстоянии 0,17 мм
от изогнутого на брюшную сторону зад-
него конца тела.
Личинки несколько мельче, чем 7И/с-
roL nocturna (длина 0,25—0,275 мм); от
последних отличаются тем, что ядра до-
ходят до самого заднего конца тела.
В периферической крови встречаются днем,
почему и называются также М. diurna.
Чехлики у них есть.
Взрослые филярии живут в соедини-
тельной ткани различных мест тела, чаще
в конъюнктиве глаза; они могут перепол-
зать из одной глазницы в другую. Причи-
няют местное опухание, называемое «ка-
лабарским отеком».
Личинки продолжают появляться в
крови человека более чем через 15 лет
после выезда его из места заражения
(Cautelen, 1935).
Переносчиком лооза является слепень
Chrysops dimidiatus (Connal). В первый
diurna теряет в его желудке оболочку,
проходит сквозь стенку желудка и залегает в жировом теле или в мышцах. На тре-
тий день у личинки уже сформировывается пищеварительный канал. Личинка с даль-
нейшим ростом делается очень подвижной; на 10—12-й день паразиты собираются в
голове насекомого и выходят наружу при сосании слепнем крови человека (рис. 351)
в его покровы личинки вбуравливаются активно. Тропическая Западная Африка; за-
несена ' и в Южную Америку и Восточную Индию.
Род Acanthocheilonema Cobbold
Acanthocheilonema perstans Manson
Длиной c W. banorofti, но раза в два тоньше. На заднем конце и самцов иг
самок имеется маленькое раздвоение. У самца две очень неравные спикулы, 4 па-
ры преклоакальпых и пара постклоакальных очень маленьких со-
сочков.
Живут в соединительной ткани брюшины, брыжейки, почек, в перикардии и др.
Личинки в периферической крови бывают безразлично и днем и ночью (рис. 347).
Длина их около 0.2 мм. Чехлика нет; задний конец тела с ядрами, тупой.
Распространена в тропической-Африке и в Британской Гвиане. Патогенное зна-
чение этого паразита неясно. Одни считают его безвредным (Brumpt), другие связы-
вают с его присутствием появление отеков, припухание лимфатических желез и
других симптомов.
Переносчиком является мелкое кровососущее двукрылое из длинноусых
Culicoides austeni, в котором проглоченные им с кровью личинки Acanthocheilonema
perstans проделывают цикл развития в течение 7—10 дней. Заражение человека
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
429
происходит при сосании его крови Culicoides, когда инвазирующие личинки выходят
из хоботка переносчика (рис. 352).
Интересно отметить, что в полости тела Culicoides нашей фауны (б. Новгород-
ский окр.) были находимы какие-то личинки нематод (Померанцев и Павловский).
Надсемейство Dracunculoidea
Род Dracunculus Reich.
Dracunculus medinensis Linn
Синоним: Filaria medinensis L.
Ришта. Длина самки (рис. 353) от 37 до 170 см, при толщине ее
струновидного тела в 1—1,7 мм; головной конец округлый, задний ко-
нец утончается и загибается крючком на спину. Самец с достоверностью
у человека не обнаружен. В 1937 г. V. Моог-
thy и Sweet заразили в Индии эксперимен-
тально шесть щенят, скормив им зараженных
циклопов (см. ниже). По вскрытии щенят че-
рез 67—457 дней после заражения было об-
наружено 45 самцов и 178 самок. По этому
материалу дается нижеследующее описание
наружной морфологии самца и самки ришты
1(рис. 354). Самец — 12—29 мм длины и 0.4 мм
толщины; ротовое отверстие маленькое, окру-
женное вниз кольцом
внутренних сосочков
Рис. 352.— Голова Culicoides austeni с личин-
ками Acanthocheilonema perstans, выходящими
из нижней губы.
Рис. 353— Ришта —Dracuncu-
lus medinensis. В натуральную
величину. Крючковидный за-
гиб заднего конца тела на
рисунке не виден. Оригинал.
числом 4—6 (интеродорзальные или интеровентраль-
дые сосочки иногда сливаются каждая пара вместе); кнаружи лежит
430
СПЕЦИАЛЬНАЯ ЧАСТЬ
Рис. 354 — Dracunculus medinensis — ришта.
1 — головной конец! самца en-face; |2—головной Гконец зрелой самки en face, сосочки iD^n [iK
слитные; 3 — срединный разрез через головной конец самца; 4 — задний конец самца сбоку; 5— то
же, вид с брюшной стороны; 6 — передний конец самца с пищеводом; вид сбоку. 1D — а нт ер о дор-
зальные сосочки; iv — интеровентоальные сосочки, и те и другие разделенные; DE — двойные
сосочки; Ат — амфида. Sp — спикулы; тЕ — мышечная часть пищевода; N — нервное кольцо»
сдавливающее железистую часть пищевода; СЕ—шейные сосочки (цервиды). По V. Moorthy
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
431
кольцо из четырех двойных наружных сосочков; позади интерола-
теральных сосочков лежат а м ф и д ы (рис. 354; 1, 2, 5). Пище-
вод подразделяется на короткий передний мышечный участок,
и широкую железистую часть, которая перетянута нервным
кольцом. Тотчас позади перетяжки лежит пара шейных сосочков (ц е р-
в и д ы). У крупных самцов пи-
щевод занимает несколько более
трети длины всего тела. Аналь-
ное отверстие лежит на 0,25 мм
кпереди от заднего конца тела.
Перед анальным отверстием ле-
жат 4 пары преданальных сосоч-
ков, а позади него 6 пар сосоч-
ков. Имеются две спикулы 490—
730 р- длины и рулек (200 н).
Самка от 1,5 до 53 см длины
и до 1 мм толщины. Вокруг рта
на головном конце имеется че-
тырехугольное возвышение. Со-
сочки расположены вокруг рта,
как и у .самца, но интеродор-
зальные сосочки иногда почти
сливаются вместе; длина пище-
вода в зависимости от возраста
колеблется почти от половины до
712 длины тела. Пигментация ки-
шечника молодых самок с поло-
возрелостью исчезает. Анальное
отверстие лежит на 0,9 мм кпе-
реди от заднего конца тела (у
самки 50 см длины), несущего
у молодых самок четыре исче-
зающих со зрелостью сосочка.
Влагалище у самки в 24 мм дли-
ной лежит на расстоянии 10,3 мм
от переднего конца тела, а у
самки 31,5 см длиной, по Chit-
wood— на расстоянии 14,5 ом.
Копуляция, невидимому, про-
исходит на ранних стадиях раз-
вития ришты; самцов не удава-
лось раходить позднее шести ме-
Рис. 355 — Dracanculus medinensis.'-
Личинка первой стадии (нормальный тип).; Передня*
и задняя половины личинки и головной конец en face,.
D. О.—просвет пищевода; I. L. — интеролатеральный
сосочек; О. N.—ядра клеток пищевода;-,Р.Е. —(со-
сочки наружного круга; Е. С. — экскреторная клет-
ка; G. Р.— половой зачаток; D. l.— просвет кишки;
/. D.— интеродорзальный сосочек; Атр. — амфида;
По V. М о о rt h у.
сяцев после заражения щенка; а
самки всего 24 мм длиной уже
имели во влагалище мукоид-
ную пробочку, наличие ко-
торой свидетельствует о совер-
шившейся копуляции. Пропор-
ция полов ришты колеблется у разных хозяев в следующих соотноше-
ниях самцов к самкам: 7 : 44; 5 : 1; 17 : 74; 0 : 23; 7 : 18; 0 :21.
После оплодотворения тело самки разрастается за счет чрезвычай-
ного увеличения матки; в конце концов все тело самки оказывается
432
СПЕЦИАЛЬНАЯ ЧАСТЬ
занятым громадной маткой, переполненной мириадами зародышей.
Самка ришты живородяща.
Развитие. Личинка ришты I стадии (рис. 355) —длина тела
0,581—0,635 мм, наибольшая толщина — 0,01—0,024 мм; ротовое отвер-
стие большое; оно окружено кольцом внутренних сосочков, числом 6,
и наружным кольцом из четырех двойных сосочков. Амфиды большие,
в виде пор. Пищевод 0,13—0,15 мм длины; ядра его хорошо видны, ле-
жат в его задней трети; стенка тонка, полость расширена и заполнена
•стекловидно-прозрачным содержимым. Нервное кольцо — на расстоя-
нии 0,061 до 0,08 мм позади переднего конца тела; позади кольца лежит
Рис. 356 — Отрожденная ли-
чинка ришты — Dracunculus
medinensis.
с — кишечник; Ь— скопление кле-
ток нервного кольца; а — половой
зачаток. Общий вид. По Faust.
Рис. 357 — Рачек циклоп,кВ поло-
сти тела которого находится че-
тыре зародыша (личинки) ришты.
Сильно увеличено.
выделительная пора. Кишечник — из 12—15 клеток; половой зачаток —
из четырех клеток. Прямая кишка от 0,013 до 0,03 мм длины. Анальное
отверстие лежит кпереди от заднего конца тела на 0,19—0,26 мм. Те-
ло нитевидное, тонкое, постепенно заостряющееся к заднему концу.
Попав в- воду, личинка может проделать дальнейший цикл разви-
тия, если будет проглочена веслоногим рачком циклопом, являющимся
промежуточным хозяином этого паразита. В качестве такового фигури-
руют следующие виды: Cyclops coronatus, С. quadricornis, С. oithono-
ides, С. vicinus, С. uljanini, Macrocyclops fuscus, Eucyclops serratulus,
Tropocyclops prasinus, Mesocyclops leuckarti, Orthonoides, Cycl. strenuus,
C. vicinus, Асаnthocyclops bicuspidatus, A. viridis. M. hualinus был най-
ден co спонтанно зараженными личинками ришты в Индии. Эти же
виды циклопов удобны для экспериментального заражения. Новейшие
данные о развитии личинок ришты в циклопах, о факторах, влияющих
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
433
на этот процесс, исчерпаны в новейших работах американских авторов
(1938). х
Федченко, впервые раскрывший развитие ришты во время своего пу-
тешествия по Туркестану (1868), доказал, что промежуточным хозяи-
ном этого паразита является циклоп (рис. 357). По мнению Федченко,
личинка ришты вбуравливается активно в покровы циклопа и прони-
кает в полость его тела. Однако наблюдения других авторов (Шаттон,
Л. Исаев) показали, что циклоп проглатывает личинку ришты
(рис. 358). В дальнейшем личинки проникают через стенку желудка
циклопа в полость тела, что происходит через 1—6 часов после загла-
тывания личинки. -Незрелые формы циклопов заражаются легче, чем
Рис. 358 — А - личинки ришты в полости кишечника
циклопа; В — циклоп, заглатывающий личинку ришты.
Сильно увеличено. По Исаеву.
взрослые, и самки — легче самцов. Паразитирование личинок ришты в
циклопах вредно влияет на них, что ведет к большой смертности (до
7О°/о!) приблизительно к сороковому дню инвазии; большинство же за-
раженных циклопов в лаборатории живут 20—25 дней.
В естественных условиях в циклопах пока находили лишь по одной
личинке ришты, тогда как экспериментально удавалось заражать ци-
клопов даже 15—20 личинками. Однако полное последующее развитие
личинки могут проделать в циклопе лишь в том случае, если их будет
менее пяти. Равным образом не вполне жизнеспособными оказываются
при дальнейшем развитии те личинки ришты, у хозяина которой было
септическое нагноение раны, откуда выходил конец тела самки ришты.
При суперинфекции личинками ришты циклопов, ранее зараженных
личинками Camallanus sweeti, первые проделывают дальнейшее разви-
тие, но в замедленном темпе. Личинки ришты, повидимому, подавляют
развитие яичников зараженного ими циклопа, и последний не отклады-
вает яиц.
Циклопы являются обычной пищей многих рыб. Если молодые гам-
бузии пожирают зараженных циклопов, то с фекалиями названных рыб
выходят живые и активно подвижные личинки. В условиях эксперимен-
та ими можно заразить циклопа, в котором они могут проделать пол-
ное развитие.
434
СПЕЦИАЛЬНАЯ ЧАСТЬ
Последнее связано с линянием личинок, уменьшающихся при этом,
в длине. Этапы развития сводятся к следующему:
1. Молодая личинка первой стадии от зрелой самки —
0,608 мм длины и 0,008 мм толщины.
2. Взрослая личинка первой стадии перед линькой в теле прогло-
тившего ее циклопа — 0,396 мм длины и 0,016 мм толщины.
м
G.P.
Рис. 359 — Dracunculus medinensis. Ришта. Личинка J1I ста-
дии из экспериментально зараженного циклопа.
ЛЬ — мышечная часть пищевода; G— железистая часть пищевода;
Е.С.— экскреторная клегка; O.N. — ядро клетки пищевода; G.P,— поло-
вой зачаток. По М о о г t h у.
3. Линяние на личинку второй стадии через 5—7 дней после зара-
жения при содержании циклопа в воде при t° 92—102° Ф и через
9—12 дней при t° 55—70° Ф.
4. Личинка второй стадии, проходящая следующую линьку —
0,391 мм длины и 0,020 мм толщины.
5. Линяние на личинку третьей стадии через 9—12 дней после за-
ражения при t° 30—102° Ф и через 13—16 дней при t° 55—70° Ф.
6. Личинка третьей стадии — 0,451 мм длины и 0,018 мм толщины;
заразительна для собак недели через три после заражения циклопа
или через 4-^-8 дней после окончания второй линьки (рис. 359).
По Л. Исаеву после наступления второй линьки личинки ришты со-
храняют заражающую способность не менее трех недель. Развитие
ришты в собаках продолжается около 9—10 месяцев.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
435
Самцы собак заражаются в условиях эксперимента легче самок
(Moorthy и Sweet). Развившиеся'ришты обнаруживаются у собак обычна
в подлопаточной и ретроэзофагеальной областях, а также в паху, под
кожей ног, по ходу позвоночника и в других местах. Установлены слу-
чаи локализации ришты у экспериментальных собак — в полости лево-
го желудочка сердца, в околосердечной жировой ткани, между мозго-
выми оболочками, в орбитальной области (собака с самцом ришты &
мозговых оболочках погибла при эпилептиформных припадках).
Рис. 360 — Ришта у человека. По Л. Исаеву.
Цикл развития ришты в организме окончательного хозяина еще не
прослежен. Несомненно, что личинки ришты мигрируют в организме
окончательного хозяина, ибо взрослая ришта обнаруживается под ко-
жей. Весьма вероятно, что, освободившись в желудке из проглоченных
циклопов, личинки третьей стадии проделывают следующую линьку
(третью по счету), внедряясь в ткани желудка или кишечника, и в чет-
вертый раз« линяют в толще соединительной ткани. Очевидно, что ко-
пуляция происходит в толще тканей, но не в полости желудка или
кишки окончательного хозяина.
Кроме собак, риштой можно заразить обезьян.
Заражение человека происходит через питьевую воду, если
с ней случайно проглатываются циклопы с личинками ришты. Ход по-
следних по телу человека не выяснен, но факт самой миграции парази-
тов из желудка человека до подкожной клетчатки несомненен. Чаще
всего самка ришты обнаруживается «в подкожной клетчатке ног.
Известно, что для достижения взрослого состояния паразиту тре-
буется около одного года времени. Циклопы, будучи положены in vitro
436
СПЕЦИАЛЬНАЯ ЧАСТЬ
в 2°/о раствор соляной кислоты, гибнут, а вышедшие из них микрофи-
лярии плавают без вреда для себя в кислой среде.
Ришта бывает у человека в количестве одного, реже нескольких
экземпляров (рис. 360). У мужчин она встречается чаще, чем у жен-
щин. Она причиняет болезнь, называемую дракункулезом, кото-
рая эндемична для Старой Бухары, Персии, Индии, тропической Афри-
ки и других жарких стран.
Болезнь сказывается ощущением чаще всего в ногах зуда в опре-
деленном участке, на котором в подкожной клетчатке прощупывается
затвердение.
Позднее эпидермис изъязвляется, и из глубины вскрывшегося нары-
ва высовывается петля или конец свернутой в клубок ришты, обусло-
вившей своим присутствием воспалительное состояние кожи. Вскоре
после вскрытия нарыва ришта отрождает множество личинок, выходя-
щих струей из передней части ее тела. Если спустя некоторое время
язву обмыть свежей водой, ришта снова выпускает из себя другую
порцию зародышей. Попав в воду при мытье ног больного, личинки мо-
гут заглатываться циклопами, которые, как описано выше, становятся
заразительными для людей и собак.
Затем эпидермис образует крупный пузырь — pemphigus. При лока-
лизации ришты близ суставов больной не может ходить. Заболева-
ние риштой нередко осложняется вторичной инфекцией с развитием
флегмон, синовита, анкилоза и других осложнений.
Диагноз. Образование под кожей сплошного извилистого валика
благодаря продвижению ришты к поверхности тела; нахождение личи-
нок в отделениях вскрывшейся язвы; наличие в язве петель ришты.
Лечение. Туземцы извлекают ришту из вскрывшегося нарыва
медленным способом, наматывая конец червя на палочку (рис. 360).
За день таким путем можно вытянуть 3—4 см паразита; полный же
курс удаления его требует около двух недель. Операция эта произво-
дится с большой осторожностью, так как в случае разрыва паразита
его содержимое выливается в язву, причиняет сильнейшую боль, вы-
зывает флегмону, за которой может наступить и неподвижность су-
става.
Извлечение паразита значительно облегчается, если его усыпить
прикладыванием тампона ваты с хлороформом к отверстию язвы. Ана-
логичные результаты достигаются и при впрыскивании в червя раство-
ра сулемы 1 : 1000. Если диагноз поставлен правильно до вскрытия
язвы, то- червя можно удалить и хирургическим путем.
Для лечения применяют внутривенное впрыскивание Tartarus sti-
biatus — 0,03 г в физиологическом растворе с прибавлением следов
карболовой кислоты. Каждые два дня впрыскивают 0,06 г; всего на
курс лечения идет 0,36 г. Иногда требуется повторное лечение.
Профилактика. Учет всех больных и лечение их. Запрещение
купаться и мыть ноги в хаузах. Регулярная очистка и осушение хау-
зов. Продажу воды для питья в разнос допускать с непременной
фильтрацией воды через плотное полотно (завязывать носик чайника
или жбана). Не пить сырой и непрофильтрованной воды. Учитывая
наличие ришты у собак (Исаев), уничтожать и этот резервуар парази-
тов. Последовательное применение комплекса мер борьбы с риштой
привело почти к полному искоренению этого паразита в исконном эн-
демическом очаге его — в Старой Бухаре (Л. Исаев, Тропин, УзбССР).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
437
Другие недостаточно изученные нитчатки.
В 1938 г. С. Desportes установил зараженность почти половины
водяных ужей (Tropidonotus natrix persa) филярией, очень близкой
к Dr. medinensis и относящейся к виду Dr. oesophagens (Polonio).
В условиях эксперимента ее личинками легко заражается Macrocyclops
fuscus.
Filaria romanorum orientalis Jarcani. Найдены однажды в крови
женщины (Румыния) в личиночной стадии (личинки 1 мм длиной,
0,03 мм толщиной; задний конец тела заострен).
Filaria conjunctivae (Addario). Длина самки 16—20 см, толщина
0,5 (рис. 361), беловатого или буроватого цвета. Кутикула поперечно
исчерчена. Ротовых сосочков нет. Неоднократно была находима в со-
Рис. 361 — Filaria соп ипМиае; передний ко-
нец самки (слева) с кишечником и маткой,
справа задний конец тела с экскреторной
порой (е) и половым отверстием (tr).
По Б р а у н у, из Ward.
Рис. 362 — Filaria (Loa?}
ext г ао с и laris:
А — передний, В - задний
конец; п — нервное кольцо;
v — отверстие влагалища;
а — заднепроходное отвер-
стие; о — яичник.
По К. Скрябину.
единительной ткани брюшной полости, в конъюнктиве и под кожей
человека в Венгрии, Италии, Сицилии и Македонии. В 1919 г. Forbes
открыл самца этой филярии.
Повидимому, идентичные филярии паразитируют также у лошадей и
ослов, что дало повод некоторым исследователям считать этот вид за
синоним Filaria inermis (Grassi).
Переносчиком F. conjunctivae предположительно считается слепень-
златоглазик (Chrysops coecutiens).
Filaria (Loa?) extraocularis Skrjabin. Описана только неполовозре-
лая самка 14,3 см длиною при толщине до 0,612 мм; кутикула попе-
речно исчерчена (рис. 362). Женское половое отверстие лежит на рас-
стоянии 2,04 мм от переднего конца тела (а у F. conjunctivae почти
у самого переднего конца). Сзади анального отверстия два боковых
маленьких сосочка; у переднего конца тела 2 боковых, 2 спинных и
2 брюшных сосочка. Добыта д-ром Владыченским из припухлости во
внутреннем углу глаза крестьянки »в Екатеринодаре. Переносчик (неиз-
вестен. Описаны и другие случаи.
438
СПЕЦИАЛЬНАЯ ЧАСТЬ
Filaria sp. Проф. Холодковскому была доставлена в сухом (!) ви
де филярия, длиною около 10 см, полученная из нарыва на Пальце
крестьянина (Осташковский у. Тверской губ.). По словам доставив-
шего ее студента, у крестьян часто бывают на пальцах нарывы, из
которых и 'выходят подобные черви.
Подтип Acanthocephala—колючеголовые черви,
или скребни
Колючеголовые черви являются возможными паразитами
человека, хотя в естественных случаях в человеке они пока еще не
были обнаружены. Поэтому мы коснемся скребней в самых общих
чертах.
Тело их, округлое на разрезе, несет на переднем конце втяжной
хобот с несколькими рядами крючьев (отсюда и название — колючего-
ловые). Задний конец тела обычно сужен. Кожным покровом является
кожно-мускульный мешок. Под поверхностной кутикулой
лежит так называемая гиподерма (слой видоизмененных кле-
ток), с которой соединены мышечные клетки. От основания хобота
внутрь тела отходят два лопастных выроста гиподермы, называемые
лемнисками. Пищеварительной, кровеносной и дыхательной систем
органов нет. Скребни раздельнополы. Половые органы их тесно связа-
ны с лигаментом (рис. 363) — полым мышечным тяжом, соединяю-
щим дно влагалища хобота с задним концом тела. У самцов на этом
лигаменте лежат два семенника. От них отходят два семепровода, со-
единяющиеся в семеизвергательный канал. Последний открывается на
дно совокупительной сумочки, лежащей на заднем конце тела. Яичники
только у молодых самок, заключенные в их лигаменте; у зрелых особей
яичники распадаются на яйца, выходящие в полость тела при разрыве
лигамента или остающиеся в его полости, когда он не лопается. Нару-
жу они выходят через маточный колокол, яйцеводы и матку.
Оплодотворение у скребней внутреннее. Яйца (рис. 364) выходят из са-
мок и с испражнениями выносятся наружу. Дальнейшее развитие они
проделывают лишь тогда, когда попадают в промежуточного хозяина —
насекомое или ракообразное. В организме последнего развивается ли-
чинка скребня, которая превращается в половозрелую особь, если про-
межуточный хозяин будет съеден хозяином основным.
Скребни паразитируют в кишечнике рыб, птиц, реже в остальных
позвоночных.
Человеческими паразитами могут подозреваться виды рода Масга-
canthorynchus (М. hirudinaceus Pall.) — скребень-великан и скребень
четковидный — Moniliformis moniliformis Brems.
М. hirudinaceus вполне оправдывает свое название — скребень-великан, так как
является самым крупным колючеголовым червем. Самка его 20—35 см длины и
4—10 мм толщины (рис. 365). Соответственные размеры самца 5—10 см и 35 мм.
Его молочнобелое тело сплющено и несет на своей поверхности иногда поперечные
складки, почему мало сведущие наблюдатели могут принять его за Цепня. Складки
после смерти червя еще больше увеличиваются. Тело его сзади несколько сужено.
Булавовидный на своем наружном конце хобот вооружен 5—6 продольными рядами
загнутых назад крючьев. Лигамент у самки не лопается, и яйца остаются в его
полости. Яйца, как и у всех скребней, одеты тремя оболочками; наружная снабже-
на ромбовидными углублениями. Эллипсоидное по своей форме яйцо (0.8—0.1 мм
длины) заключает в себе перегнутый зародыш, на переднем конце которого видны
крючья.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
439
Паразитирует в тонких кишках домашней и дикой свиньи и у пекари Dicotyles
torqufitus.
Впиваясь своим хоботом в стенку кишки, этот скребень выпячивает холмиком
наружу брюшную ее поверхность; благодаря этому о присутствии паразита можно
•судить без вскрытия кишек, по одним только ее выпуклостям. То же наблюдается и
у других скребней, например, Filicollis anatis утки. Промежуточными хозяевами
являются личинки пластинчатоусых жуков — майского жука (Meloiontha
Рис. 364 — Macracanthorhynchus’hirudinaceus.
Яйца. По Озерской, 1930.
Рис. 365—а — Macracanthorynchus hirudina-
ceus скребень - великан и в — Moniliformis
moniliformis — скребень четковидный в на-
туральную величину. По Бремзеру.
Рис. 363 — Схема
строения самца
A canthocephal us.
ex'— влагалище хоботка;
л—лемниски; г—ганглий;
с — семенники; цж — це-
ментные железы.
По Догелю.
vulgaris), бронзовки (Cetonia) и др. Обитая в земле, они проглатывают яйца скреб-
ней, выходящие с калом свиней наружу. В полости тела личинок жуков разви-
ваются личинки скребня. Свиньи же получают этих паразитов, пожирая заражен-
ных личинок бронзовки или майского жука.
Вполне достоверных случаев паразитирования скребня-великана в человеке не
наблюдалось. Оно возможно лишь при условии проглатывания личинок пластинчато-
усых жуков, что едва ли кому придет в голову. Утверждение Линдемана, будто бы
по берегам Волги этот паразит часто бывает у жителей, которые заражаются им,
употребляя в пищу сырую воблу, явно неосновательно, так как вобла не является
промежуточным хозяином скребней.
Moniliformis moniliformis (рис. 365, В). Белое тело скребня четковидного к обоим
концам суживается. Обычно оно изогнуто спирально и всегда несет кольцевые пере-
тяжки, придающие этому паразиту по внешности членистый вид. Самка 7—10 см
440
СПЕЦИАЛЬНАЯ ЧАСТЬ
длины, а самец 4—5 см. Булавовидный или цилиндрический хобот (0,425—0,450 мм
длины и 0,16—0,190 мм ширины) вооружен 14 продольными рядами тонких и силь-
но искривленных крючьев, длиной 0,026 мм. Эллипсоидные яйца, длиной 85 И и
шириной 45 р-.
Паразитирует в кишке различных грызунов (крыс, мышей, хомяков и др.) в
Европе, ^Африке и Бразилии. Промежуточными хозяевами в Европе являются жуки
из семейства чернотелок (Bl?ps mucronata) и американский таракан (Periplaneta
Рис. 366 — Кишечник кряковой утки с скребнем Filicolis antis. На
наружной поверхности кишки видны шаровидные выросты — выпячи-
вания, содержащие в себе головку скребня. Оригинал.
Americana) в Алжире. Каландруччио доказал возможность паразитирования четко-
видного скребня и в человеке, проглотив вынутые из жуков личинки этого червя.
Спустя недели три у наблюдателя стали ощущаться сильные боли в животе, сла-
бость, появился также и понос. Приняв крупную дозу экстракта папоротника, Ка-
ландруччио выгнал из себя 53 скребня.
В естественных условиях заражение человека четковидным скребнем едва ли
может случиться, но самую возможность врачу следует иметь в виду, так как
истеричные больные проглатывают самые неожидаемые объекты и могут не оста-
новиться и перед съеданием тараканов.
КЛАСС КОЛЬЧАТЫЕ ЧЕРВИ — ANNELIDA
ПОДКЛАСС ПИЯВКИ— HI RUDINЕА
Пиявки — кольчатые черви с вторичной мелкой
ложной кольчатостью тела, лишенного параподий. Перед-
ний конец тела утончается, несет ротовую присоску; задний ко-
нец оканчивается более крупным присасывательнымдиск ом.
Весьма подвижны вследствие очень большой растяжимости тела. Пи-
таются кровью различных позвоночных животных. Живут в воде. На-
падают и на водных позвоночных и на животных, погружающихся в
воду. Ротовое отверстие вооружено или хоботом (хоботные
пиявки — Rhynchobdellidae), или зазубренными пильчатыми
челюстями (челюстные пиявки — Gnathobdellidae). Выпивают
зараз много крови вследствие емкости желудка и его слепых выростов..
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
441
Поэтому пиявки издавна применялись в качестве народного средства,
а также и в медицине для оттягивания крови при различных болезнях,
тем более, что сосание крови пиявкой является безболезненным.
Примеры. Пиявка медицинская, Hirudo medicinalis, живущая в прудах,
болотах, озерах; широко распространена на материке Европы.
Египетская пиявка — Limnatis nilotica — побережье Средиземного моря.
Сухопутная пиявка — Haemadipsa ceylanica — Цейлон и мн. др.
Случаи паразитирования пиявок у человека время от времени обна-
руживаются при исследовании носовой полости или носоглотки, причем
первоначально пиявка принимается часто за полип. Истинное положение
дела определяется подвижностью паразита и последующим его извле*
Рис. 367 — Limnatis turkestanlca.
Оригинал.
Рис. 368 — Limnaiis turkestanica, присо-
савшаяся к десне. Наблюдение М. По-
спеловой — Штромв Средней Азии
Рис. по фотографии.
чением. Хотя материалов по паразитированию пиявок у человека в
СССР собрано в общем мало, но все же можно отметить, что таковые
случаи чаще встречаются на юге СССР, в частности в Средней Азии.
Из фауны пиявок на 1926 г. в качестве паразитов человека отмече-
ны были следующие виды:
Hirudo aegyptiaca Blainv. Серьезное поражение у солдат, которые
пили воду из стоячих водоемов. Египет.
Н. medicinalis Lin. Может паразитировать в естественных полостях.
Европа. Северная Африка. Завезена в США.
Н. nipponica Whitimn. Отмечено паразитирование в конъюнкти-
вальном мешке. Япония.
Limnatis granulosa Sav. Паразитирует в носовых ходах. Индия.
L. nilotica Sav. Паразитирует в глотке, гортани, пищеводе, трахее,
442
СПЕЦИАЛЬНАЯ ЧАСТЬ
носовых полостях, в вагине, конъюнктиве. Африка, Канарские острова,
Южная Европа, 'Малая Азия.
Haemopis sanguisuga Linn. Носовая полость. Северная Африка-
77. paludum Tenneut. Носовая полость, гортань. Цейлон.
Haeniadipsa ceylanica Blainv. Эктопаразит человека.
Н. japonica taiwana Oka. Серьезный паразит человека.
* Trachybdella bistriata Pinto. Нападает на человека. Бразилия.
Limnatis turkestanica Plotn. (рис. 367). В Зоологическом музее
имеются экземпляры этого вида из Ашхабада, окрестностей Самаркан-
да, Ташкента, горы. Каратау, района между Курган-Тюбе и Бальджуа-
ном, б. Шир-Абадского бекства и из некоторых мест Восточной Персии.
Заражение человека и животных пиявками происходит при питье во-
ды из водоемов, обитаемых этими паразитами. При питье днем из гор-
сти или из какой-либо посуды пиявка будет замечена, и человек остере-
жется ее проглотить. Питье воды ночью из естественных источников и
питье ничком как ночью, так и днем является моментом, способствую-
щим заражению пиявками. В случаях М. Березняка и К. Лещенко не-
которые пострадавшие связывали заболевание с питьем воды из арыков,
причем один из пациентов почувствовал, что проглотил что-то скольз-
кое, засевшее в пищеводе.
Характерным симптомом нахождения пиявки в носоглотке или в
полости носа является «ничем необъяснимое» кровотечение изо рта и
носа, вполне понятное, если вспомнить, что пиявка выделяет мощный
антикоагулин — гирудин, лишающий кровь способности свертывать-
ся. Гирудин является секретом шейных или головных желез
пиявки; он не ядовит при впрыскивании животным, но влияет на
кровь в смысле понижения способности свертываться. В зависимости
от срока пребывания пиявки в глотке или в полости носа потеря крови
принимает более или менее хронический характер, что в конце концов
может вести к анемии. Окрашивание содержимого желудка в коричне-
вый цвет, a faeces в темный — из-за* примеси крови — наблюдается уже
в первые дни после попадания' пиявки в тело человека.
Существенное значение имеет локализация паразита. Весьма опас-
ным является присасывание пиявки близ голосовой щ,ели или попада-
ние в трахею. По мере насасывания кровью и увеличения в размерах
пиявка закрывает трахею и вызывает асфиксию. Спасение больного
возможно только при немедленном удалении пиявки хирургическим пу-
тем. Для удаления пиявок применяли полоскание крепким раствором
поваренной соли.-
Пиявки, как известно, издавна применялись в медицине для оттяги-
вания крови при различных воспалительных процессах. Потом они бы-
ли вытеснены кровососными банками с насечками и «искусственными»
пиявками. Однако в последние годы врачи чаще и чаще используют
медицинских пиявок при глазных заболеваниях с увеличенным давле-
нием.
С лечебными целями в различных частях света применяются следу-
ющие виды пиявок: Hirudo medicinalis Lin., Европа; Н. nipponica
Whitm., Япония; Н. troctina Johnton, Европа, С. Африка; Limmatis gra-
nulosa Sav., Индия; L. mesomelas Serr. et Vir., Сенегамбия; Haemen-
taria officinalis Fil., Мексика, Северная Америка; Placabdella cateni-
gera Moq. Tand., Крым.
В связи с возможностью использования в медицинской практике
пиявок, особенно на юге, где подходящие виды нередки, следует оста-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
443
ловиться на важном вопросе — сохраняемости различных болезне-
творных начал во всосанной пиявками крови.
Этот момент интересен в двух отношениях. В случае более или ме-
нее долгой сохраняемости микроорганизмов в крови, всосанной пияв-
кой, последняя может быть использована как хранилище для содержа-
ния и пересылки вирусов. С другой стороны, при аналогичных усло-
виях пиявка может сыграть роль переносчика патогенных бактерий.
В первом отношении Пастернацкий показал, что спирохеты возврат-
ного тифа при сохранении пиявок на льду жили до десяти дней. Каг-
wacki и Szokalski наблюдали пребывание живых спирохет в пиявках
до ста дней и указали на возможность размножения спирохет в их же-
лудке.
Судьба бактерий и паразитических простейших, попавших в желудок
пиявок, различна. Весьма вероятно, что различные роды и виды пиявок
неодинаково действуют на находящихся в них бактерий и простейших;
но более или менее, детальные опыты были производимы пока только
с медицинской пиявкой.
К. Steffenhagen и П. Андреев (1910) показали, что брюшнотифоз-
ные бактерии могут быть обнаружены посевом крови из желудка пияв-
ки в срок до 30 дней от момента заражения пиявки; бацилла паратифа В
в аналогичных условиях обнаруживается в течение 3 месяцев; сиби-
реязвенные палочки (заражением свинок кровью) —до 14—17 дней;
туберкулезные палочки человеческого туберкулеза — до 28 дней, а типа
bovinus — до 60 дней; бактерии септицемии свиней — до 38 дней; спи-
рохеты кур — до 7 дней; Trypanosoma equiperdum (заражением крыс) —
до 4 дней; Trypanosoma brucei — до 3 дней; Trypanosoma lewisii по-
движная — до 5 дней; вирус оспы кур — до 20 дней; вирус чумы сви-
ней — 20 дней.
Кроме того, известно, что вирус сыпного тифа сохраняется в пияв-
ках 2 дня.
Однако факт нахождения в явном или скрытом состоянии жизне-
деятельных микроорганизмов в желудке пиявки еще не означает, что
пиявка передаст эти микроорганизмы хозяину, кровь которого она бу-
дет сосать. Во всяком случае, эту способность необходимо доказать
экспериментально параллельно с установлением формы состояния мик-
роорганизмов в крови желудка пиявки, тем более, что указанная спо-
собность противоречиво трактуется в литературе. Одни авторы, как
Leuckart и Pacinotti, считают возможность передачи пиявками патоген-
ных микроорганизмов реальной, тогда как другие, например, Miihling,
полагают/ что роль пиявок в указанном отношении нулевая или почти
нулевая.
Ввиду таких противоречий интересно привести данные уже упомя-
нутых выше Steffenhagen и Андреева об опытах с передачей пиявками
различных патогенных микроорганизмов. Эксперименты с туберкулез-
ными палочками типа bovinus, а также с возбудителями септицемии кро-
ликов и с патогенным для мышей штаммом стафилококка дали отрица-
тельный результат. В то же время опыты с повторным прерванным
кормлением пиявок сначала на зараженном животном, а тотчас после
перерыва — на здоровом были положительны для палочки сибирской
язвы, спирохет кур, вируса оспы кур, Trypanosoma equlperdum и Тг
brucei. Но уже суточный перерыв между двумя кормлениями в указан-
ных опытах давал отрицательный результат. На основании изложенных
данных Steffenhagen и Андреев считают, что передача пиявками воз-
будителей заразных болезней от человека к человеку может происхо-
444
СПЕЦИАЛЬНАЯ ЧАСТЬ
дить только вследствие совпадения исключительно неблагоприятных,
обстоятельств.
Когда обсуждается вопрос о переносчиках возбудителей инфекций
и инвазий, необходимо выявлять форму самого переноса. В освещен-
ных выше соотношениях пиявки могут быть трактованы, как механиче-
ские (не специфические) переносчики бактерий, вирусов и трипанозом,
осуществляющие передачу инфекций в условиях немедленного повтор-
ного сосания пиявками здорового животного после перерыва кормления
их на животном зараженном. Вместе с тем известны виды пиявок, яв-
ляющиеся специфическими переносчиками паразитических простейших,
проделывающих в пиявках часть своего жизненного цикла. Таковы,
например, Hemiclepsis marginata и некоторые Ichthyobdellidae — пере-
носящие трипанозом рыб.
Для полноты характеристики свойств пиявок отметим сохранение
заглоченной ими кровью заключающихся в ней иммунных тел. В част-
ности, в пиявках долго сохраняются агглютинины. Сыворотка парати-
фа В после 78 дней нахождения в желудке пиявки сохранила почти
тот же титр агглютинации, что у исходной сыворотки кролика, исполь-
зованного для кормления пиявки. В аналогичных условиях агглютини?
рующая холерная сыворотка сохраняла свое действие 80 дней; агглю-
тинирующая сыворотка Шига — 40 дней и др. (Steffenhagen и Ан-
дреев).
ОБЩИЙ ХАРАКТЕР ВЛИЯНИЯ ПАРАЗИТИЧЕСКИХ ЧЕРВЕЙ НА ОРГАНИЗМ
В деле оценки значения паразитических червей для их хозяев и, в
частности, для человека наблюдались значительные колебания от пол-
ного игнорирования их присутствия в кишечнике человека до поголов-
ной трактовки всех глист человека, как возбудителей глистных болез-
ней. Теоретически нет основания не допускать возможности существо^
вания патогенных и непатогенных паразитических червей, подобно то-
му, как существуют болезнетворные и безвредные бактерии и простей-
шие
Накопившаяся огромная казуистическая литература дает множество-
примеров наличия разнообразных и нехарактерных симптомов, связан^
ных с присутствием тех или иных паразитических червей в кишечнике
человека. Замечательно то, что таковая связь часто устанавливается
лишь post factum: после изгнания глист те или другие болезненные
симптомы исчезают, откуда делается вывод о предполагаемой зависи-
мости исчезнувших симптомов от паразитирования изгнанного паразита.
Такие обстоятельства лишают возможности дать определенную клини-
ческую характеристику некоторым гельмиктозам, вызываемым самыми
обыкновенными и распространенными глистами человека.
Трудность положения усугубляется еще весьма широким распро-
странением явлений паразитоносительства, когда глисты годами живут
в своем хозяине, ничем не выдавая своего присутствия. Правда, такое
благополучие здоровья хозяина в ряде случаев бывает кажущимся, ибо
при детальном анализе состояния организма обнаруживаются внешне
не проявляющиеся расстройства. Например, при паразитировании лен-
теца широкого у человека без проявлений симптомов злокачественного
малокровия кровь по характеру своей формулы представляет собой
как бы «стертую форму» anaemia perniciosa diphyllobothriocephalica
(Н. Рагоза).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
445
Паразит и хозяин представляют биологическую
систему организмов, живущих один в другом и
взаимно влияющих друг на друга. Организм хозяи-
на является средой обитания паразита, откуда по-
следний черпает все необходимое для существова-
ния и куда он выбрасывает продукты своей жизне-
деятельности.
Степень здоровья хозяина определяется: а) первичными патогенны-
ми свойствами паразита, б) стойкостью хозяина и в) состояниями (на
данный момент) как хозяина, так и паразита (например, характер ди-
эты, принятие лекарств, реакция организма на факторы внешней сре-
ды, болезни паразита и др.).
Рассмотрим общий характер влияния глист на организм хозяина
(resp. человека).
Все разнообразия форм их влияния можно свести к следующим ка-
тегориям:
1) местное механическое и комбинативное действие;
2) общее (резорптивное) влияние (интоксикация);
3) косвенное патогенное действие;
4) миграционное действие.
Указанные категории характеризуют лишь доминирующие направ-
ления действия, которые, как правило, практически сочетаются в раз-
личных комбинациях друг с другом.
1. Местное — механическое и комбинативное дей-
ствие.
Многие паразитические черви имеют специальные органы прикреп-
ления (присоски, крючья, ротовую капсулу и др.)л что особенно свой-
ственно паразитам, обитающим в кишечнике.
Органами прикрепления механически повреждается целость слизи-
стой оболочки кишечника, что ведет к нарушению ее барьерной
функции в отношении находящихся в кишечнике различных бакте-
рий (см. ниже). В месте нарушения слизистой в толщу тканей посту-
пают различные секреты, выделяемые паразитом, что вызывает местную
патологическую реакцию тканей и может вести к общим последствиям
при разрушительном действии этих секретов на кровь (например, соче-
тание действия кутикулярных зубов ротовой капсулы анкилостомы и
секрета ее головных и шейных желез).
Местная реакция проявляется воспалительными процессами, гемор-
рагией, изъязвлением тканей и др.
Чистой формой механического действия является закупорка глиста-
ми просвета обитаемого ими органа, например непроходимость кишеч-
ника при клубке аскарид, требующая для спасения жизни экстренной
операции; закупорка дыхательного горла попавшей в неге аскаридой
с последующей смертью от асфиксии; разрыв печени при росте ано-
мально локализованных в ней аскарид; прорыв дряблой при брюшном
тифе стенки кишечника аскаридой с последующим воспалением брю-
шины.
Однако закупорка кишечника не связана непременно с механиче-
ским действием клубка червей; бывают случай спастической непроходи-
мости как реакции организма на присутствие всего лишь одной аска-
риды или такого «мягкого» паразита, как лентец широкий. Очевидно,
в таких случаях доминирующим моментом является токсическое влия-
ние паразита при особых условиях взаимодействия с организмом хо-
зяина.
446
СПЕЦИАЛЬНАЯ ЧАСТЬ
Соматические паразитические черви (иначе — тка-
невые п а р а з и ты) в процессе роста в толще хозяина неминуемо
оказывают на его органы давление, эффект которого зависит в первую
очередь от «нежности» пораженного органа. Разрастание финки в
мышце может оставаться при жизни незамеченным, тогда как расту-
щая в глазу финка вызывает отслойку сетчатки и в совокупности со
всеми формами своего влияния ведет к потере глаза.
Разнообразные симптомы наблюдаются при развитии финки в тка-
нях мозга, что зависит от локализации финок по отношению к раз-
личным мозговым центрам.
Эхинококк в процессе роста вызывает агрофию «вытесняемых» им
тканей пораженного органа («разъедание» костей, замещение тканей
печени и др.).
Несомненно, что в таких случаях момент механического действия
(давления) сочетается с влиянием поступающих в окружающие ткани
продуктов жизнедеятельности паразита в виде экскреторных или иных
веществ. В результате местных взаимовлияний вокруг паразита обра-
зуется реактивная капсула (например, у эхинококка, мышеч-
ных трихин и др.); наблюдается также перерождение тканей в месте
залегания паразита (например, в поперечнополосатом мышечном во-
локне, пораженном личинкой трихины); соматический паразит воспри-
нимает потребные ему пищевые вещества из окружающих его тканей:
органа, что в свою очередь является обстоятельством патологического
порядка. Интересно отметить, что в стенке формирующихся капсул три-
хин особо разрастается сеть кровеносных сосудов, улучшающая
кровоснабжение капсулы.
Из сказанного ясно, что механическое действие пара-
зита неотрывно сочетается с иными формами влия-
ния его на организм хозяина; поэтому правильнее говорить
о комбинативном влиянии паразита на почве его механического дей-
ствия на организм хозяина.
ОБЩЕЕ (РЕЗОРБТИВНОЕ) ВЛИЯНИЕ (ГЛИСТНЫЕ ИНТОКСИКАЦИИ)
Внешней средой для кишечных паразитических
червей является полость кишечника с ее содержи-
мым, как часть организма человека. Из этой внешней
среды паразит почерпает всасыванием пищевые вещества, перевари-
ваемые ферментами хозяина; в эту же среду паразит выбрасывает про-
дукты обмена вещества, всасываемые кишечником хозяина. Этими
взаимосвязями и определяется в каждом отдельном случае эффект па-
разитирования глисты — от - бессимптомного паразитоносительства до
явного состояния болезни.
Материалы, иллюстрирующие рассматриваемую сторону дела, весь-
ма отрывочны и разнообразны по своему характеру, ибо разные виды
глист в разных хозяевах проявляют, конечно, различные свойства.
Пестрота материалов и далеко недостаточная изученность вопроса
затрудняют суммирование выводов формулированием тех или других
закономерностей. Все же возможно .наметить основные пути изучения
принципиальной по своей важности токсичности паразитических червей.
Из тела червя в полость кишечника поступают следующие естествен-
ные продукты жизнедеятельности паразита:
1) Секреты специальных желез (например, шейные железы анкило-
стомид) .
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
447
2) Экскреты, выбрасываемые выделительной системой червей, и
остатки, выделяемые из кишечника (если таковой имеется у глисты).
Все эти вещества можно отнести к экзотоксинам.
3) Для некоторых кишечных и соматических паразитов естествен-
ным продуктом является полостная жидкость (аскариды, пузыри эхи-
нококка, финки и др.). Нормально она не поступает, как таковая, в
организм хозяина, но может изливаться при травме (разрыв кисты эхи-
нококка) .
4) К естественным продуктам следует также отнести продукты пе-
реваривания больных или погибших глист; эту категорию веществ
условно относят к эндотоксинам. Скорое всасывание больших ко
личеств их ведет к острому отравлению.
Поступание тех или других продуктов жизнедеятельности паразита
в течение его жизни в организм хозяина ведет к хронической интокси-
кации, проявление которой крайне разнообразно и не всегда постоянно
для одного и того же паразита в хозяевах также одного вида.
В результате клинических наблюдений и экспериментальных патоло-
гических и фармакологических исследований добыт ряд фактов, харак-
теризующих токсические свойства паразитических червей. Соответству-
ющие материалы разберем на нескольких важнейших объектах. 1
Что ленточные глисты бывают причиной злокачественного малокро-
вия, это уже давно было установлено клиницистами, давшими такой
форме болезни особое название anaemia perniciosa bothriocephalica,
т. е. анемия, вызываемая лентецом широким—Diphyllobothrium (= Di-
bothriocephalus latus). В гораздо более редких случаях злокачествен-
ную анемию вызывает и солитер — Taenia solium, как то впервые было
установлено С. П. Боткиным (см. также Ortel, T)ervis).
Общие изменения крови у людей, инвазированных лентецом широ-
ким, сказываются уменьшением числа эритроцитов, появлением де- и
регенеративных форм их (в том числе макроцитов и мегалобластов);
к этому присоединяются общая лейкопения, уменьшение числа нейтро-
филов, базофилов и больших мононуклеаров. Эозинофилия, как прави-
ло, имеется. Антитриптическая сила кровяной сыворотки таких больных
ясно повышена (Рагоза).
Зависимость злокачественной анемии от кишечных паразитов не
всегда легко объяснима. Во всяком случае анемия не связана прямо
пропорционально с количеством глист в кишечнике, так как бывали
случаи паразитирования 78—100 ленточных червей у одного и того же
больного без проявления симптомов злокачественного малокровия. Рав-
ным образом и продолжительность пребывания паразита в кишечнике
(лентец широкий может жить до 35 лет) не является непременным
условием возникновения анемии.
Стремясь выяснить причины этой тяжелой болезни, клиницисты вы-
сказали несколько предположений, из которых остановимся на двух
наиболее вероятных. Глистное малокровие зависит от отравления то-
ксинами, свойственными живому и здоровому паразиту, при всасывании
их тканями тела хозяина. По другому взгляду заболевание происходит
лишь тогда, когда сама глиста, умирая и задерживаясь в кишечнике,
подвергается разложению, продукты которого всасываются и отрав-
ляют носителя паразита.
1 Последующий материал заимствован из моей книги «Ядовитые животные
СССР», 1931 (а также в издании «Gifttiere und ihre Gifitgkeit», 1927), с некоторыми;
дополнениями.
448
СПЕЦИАЛЬНАЯ ЧАСТЬ
На общую картину отравления влияют и факторы, касающиеся хо-
зяина, например, проницаемость кишечной стенки для токсинов и нали-
чие моментов, изменяющих реакцию организма на' поступающие в его
кровь гельминтотоксины.
Подтверждение той или иной гипотезы можно получить только спе-
циальными опытами, которые велись в двух направлениях: 1) испыты-
вали действие на организм самих глист или их естественных продуктов
и 2) изучали влияние вытяжек из паразитов. Schauman и Tallquist
(1898) скармливали собакам 30—50 г измельченных или предваритель-
но обработанных трипсином члеников лентеца, а также впрыскивали
им под кожу солевую вытяжку из того же паразита. В обоих случаях
наблюдалось падение почти вдвое числа эритроцитов. При вскрытии
животных было обнаружено отложение кровяного пигмента (сидероз)
в печени и селезенке, что в свою очередь указывает на разрушение
красных кровяных клеток. Таким образом результаты анемии были на-
лицо. Эритроциты собаки (но не кролика) растворялись в вытяжках
лентеца и вне тела. Для этих опытов лентецы брались как от анемич-
ных, так и от неанемичных больных.
Позднее Tallquist нашел в члениках лентеца протеолитический фер-
мент, агглютинин (для кровяных клеток некоторых сортов крови) и
гемолизин. Последний является липоидом, стойким к нагреванию и не
поддающимся действию протеолитических ферментов. Гемолизин не
является секретом, выделяемым паразитом, но содержится в его тканях,
откуда освобождается при распадении или мацерации лентеца. В нор-
мальных условиях гемолизин не поступает в кишечник хозяина и не
оказывает поэтому токсического влияния на кровь. Если же глиста
подвергается каким-либо изменениям, теряет свою жизнеспособность и
распадается аутолитически (чему могут способствовать и ферменты хо-
зяина)*, гемолизин освобождается, всасывается стенкой кишечника и,
поступая в кровь, вызывает развитие злокачественного малокровия.
Tallquist утверждает, что тении не содержат в себе гемолизинов.
Schmaucli впрыскивал кошкам экстракты из лентецов широких, взя-
тых от неанемичных больных, и наблюдал развитие резкого малокро-
вия, причем, наряду с усиленным распадом эритроцитов происходило
и энергичное их новообразование. Гемолитические начала нашел в лен-
теце широком и Schwartz.
Многие исследователи приписывают первичную роль в этиологии
пернициозной анемии липоидам Dibothriocephalus и других глист. Ли-
поиды, действительно, растворяют in vitro эритроциты; однако при
впрыскивании их животным они не вызывают злокачественного мало-
кровия, но могут вести к значительной вторичной анемии.
Действующим началом паразитов другие авторы считают «яд», in
vitro не растворяющий эритроцитов, но при повторных впрыскиваниях
могущий повести к тяжелой первичной анемии с возникновением более
или менее обширных очагов кроветворения в печени и селезенке. Эти
«истинные кровяные яды» и являются, по Seyderhelm,' причиной перни-
циозной анемии.
Труды ряда исследователей показали, что водные, глицериновые и
ширтовые вытяжки Taenia solium, Т. saginatus, Т. expansa, Dipylidium
caninum и других глист, впрыснутые животным, вызывают у них сла-
бость, судороги, сведение задних ног, понижение температуры, а
иногда и смерть.
Ленточная глиста неодинаково ядовита во всех своих частях: отдел
Г. saginatus, прилежащий к головке, менее ядовит, нежели задние чле-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА.
449
ники глисты, набитые яйцами (Parisot и Simonin). Экстракты из этого
паразита при впрыскивании в вены кролику вызывают расстройство
кровообращения и дыхания. В одном из типичных опытов начальное
давление ib a. carotis = 8 см. После впрыскивания оно падает и через
2 минуты = 4 см. Работа сердца замедляется до 40 ударов в минуту;
сила же его значительно возрастает, благодаря чему давление вырав-
нивается и даже повышается до 10—13 см. Далее оно снова падает
и на 5-й минуте = 3 см при 180—200 ударах сердца в минуту. На 7—
8-й минуте — новая фаза замедления сердца; на 10-й минуте — давле-
ние 2 см, фибриллярное сокращение сердца, и на 12-й минуте — смерть
кролика. В первое время опыта дыхание сначала ускоряется; затем
происходит его замедление, ритм же становится неправильным.
Min.g’azzini считает, что тениотоксины ленточных червей плотояд-
ных гораздо ядовитее, чем таких же паразитов травоядных. С таким
взглядом несогласен Сао, полагавший, что токсичность вытяжек из
глист зависит от побочных причин, например, от загрязнения бактерия-
ми. Le pantec, работая с вытяжками из Т, inermis, не мог убедиться
в их ядовитости.
Опыты Calamida, Pomella, Messineo, Bedson и др. устранили сомне-
ние как в самом факте токсичности вытяжек из глист, так и в причи-
нах их ядовитости. Свежие вытяжки, пропущенные через фильтр Шам-
берлена, обладают несомненным гемолитическим действием (для мор-
ской свинки и кролика); по отношению к этим экстрактам лейкоциты
проявили положительный химиотаксис. Впрыскиванием в вену растер-
тых в физиологическом растворе члеников Т. saginatus Proscher
вызвал у кролика явную эозинофилию, более же сильные дозы вытяж-
ки убивали животное. Тениотоксин Т. saginatus состоит из трех дей-
ствующих начал — нейротоксического, гемолитического и эозинофило-
токсического. 'Мельников, применяя солевую вытяжку Т. solium. полу-
чал у животных не только эозинофилию в русле крови, но и увеличе-
ние числа эозинофилов в костном мозгу, что, по мнению автора, свиде-
тельствует о возбуждении костного мозга тениотоксином. Только сла-
бые дозы последнего стимулируют кроветворные органы к выработке
эозинофилов, тогда как большие дозы его в этом отношении недеятель-
ны (Вейнберг).
Parisot и Simonin особо остановились на влиянии впрыскивания экс-
трактов из Т. solium на кишечник. Через 1—2 минуты после введения
в вену слабой дозы экстракта у кролика начинается повторная дефе-
кация, переходящая в понос. Через час животное приходит в норму.
Пятерная доза экстракта вызывает беспрерывный понос и гибель кро-
лика. Слизистая оболочка кишечника у него слущивается, вследствие
чего обнажается mucosa.
Проводившееся в Отделе паразитологии ВИЭМ фармакологическое
изучение токсинов паразитических червей в экстрактах из тела послед-
них показало, что экстракты из лентеца широкого в разведениях даже до
предела 1 : 100 000 000 вызывают в подавляющем большинстве случаев
угнетение работы изолированного сердца через посредство перифериче-
ского аппарата вагуса. Кроме того, ведущий узел и мышца сердца
вследствие непосредственного влияния на них токсина в большинстве
случаев возбуждаются. Аналогичные исследования, произведенные над
экстрактами из цепня невооруженного (1:1000—1:100 000 000), вы-
явили двухфазное действие на работу изолированного сердца: скачала
кратковременное возбуждение, затем длительное угнетение. Возбужда-
ющая фаза выражается увеличением амплитуды и учащением ритма,
450
СПЕЦИАЛЬНАЯ ЧАСТЬ
а угнетение сказывается понижением силы сокращений и урежением
ритма. Действие экстракта при первых пропусканиях обнаруживается
гораздо быстрее и сильнее, чем при последующих. Неправильно рабо-
тающее сердце выравнивает сокращение при действии экстракта цепня
невооруженного. Токсическое начало рассмотренных ленточных червей
следует признать мышечным и ваготропным (Е. Павловский и 3, Ду-
наева).
На кишечник (изолированный и in toto) экстракт лентеца широкого
действует угнетающе на его моторную функцию, но фазы угнетения
быстро сменяются восстановлением нормальных сокращений после про-
мывания рингер-локковской жидкостью (Ф. Талызин).
Pomella, работая с экстрактами Т. plicata и perfoliata, получал не-
редко смертельное отравление морских свинок, а также и выкидыши у
беременных самок. R. Kamada доказал присутствие в теле Hymenole-
pis diminuta гемолизина.
Все эти данные достаточно красноречиво говорят в пользу первич-
ной ядовитости глист, которые содержат В/своем организме
различные действующие начала.
Все же пока остается не совсем ясным способ освобождения тенио-
токсинов. Представляют ли они собой секреты (или экскреты?) пара-
зитов или же являются эндоксинами, которые выходят наружу лишь
при распаде глисты?
Как бы то ни было, тениотоксины поступают в кишечник, где и вса-
сываются его слизистой оболочкой. Разносясь с кровью по телу, они
вызывают различные патологоанатомические изменения, как то: гемор-
рагические поражения почек, печени, стенок кишок, жировое перерож-
дение печени, миелоид селезенки и лимфатических желез.. Из губчатых
слоев надпочечников липоиды исчезают почти совсем и в тяжелых слу-
чаях отравления вовсе не восстанавливаются. Распад эритроцитов обу-
словливается освобождением кровяного пигмента, который и отлагает-
ся в печени, селезенке и в других органах; Различные изменения про-
исходят и в остальных железах внутренней секреции (S. Bedson).
Однако анемия не всегда развивается в присутствии распадающейся
глисгы, когда тениотоксины должны бы всасываться в кровь и вызы-
вать типичную картину заболевания. Эти факты не противоречат ядо-
витости глист вообще, так как и вирулентные патогенные бактерии ни-
чем не проявляют своего присутствия у бациллоносителей (Lazarus).
После тщательных исследований у глистных больных Рагоза (1913)
пришел к заключению, что паразит начинает вредить сво-
ему хозяину уже с первых месяцев своего нахожде-
ния в кишечнике. Гемолитическая формула глист-
ной крови очень близка к формуле пернициозной
анемии; кровь при дифиллоботриозе представляет собой «forme fru-
ste» злокачественного малокровия и обратно — anaemia perniciosa
bothriocephalica в гематологическом отношении тождественна со скры-
той формой данного заболевания. Степень анемии в каждом отдельном
случае является результатом динамического равновесия между виру-
лентностью глисты и компенсаторной деятельностью костного мозга
(Рагоза).
При экспериментальном заражении людей лентецом широким (Le
Bas, 1924) не удалось получить сколько-нибудь серьезных симптомов
отравления. Степень анемии в каждом случае была мала, но самый
характер изменений крови был сходен с тем, что наблюдается при
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
451
пернициозной анемии (Passey и Braine). Такое заключение совпадает
с только что приведенным выше взглядом.
Изучение ядовитых свойств жидкости пузырчатых форм (кист эхи-
нококка, цистицерков и т. nj дало разноречивые результаты. Выделен-
ный из жидкости эхинококка птомаин (Mourson и Schlagdenhauffen)
оказался для лягушки безвредным; равным образом недеятельной была
и чистая жидкость той же глисты в опытах Joest, Лаверана, Мениля,
Шсффарда и других исследователей. Boidin и Laroche заметили, что
не всякий экземпляр эхинококка содержит в своих пузырях ядовитую
жидкость. Можно было наблюдать токсичность последней, если впры-
скивать 510 см3 жидкости, сгустив ее в вакууме до 7ю первоначально-
го объема.
При инъекции 20—100 см3 жидкости бараньего эхинококка в вены
у собак наступали тяжелые симптомы шока. В разведении 1:10 000
эта жидкость вызывает сокращение нерожавшей матки морской свинки.
Промывание переживающего сердца лягушки или жабы жидкостью
эхинококка большей частью поддерживает работу сердца (Giusti и
Hug, 1923).
Количество лейкоцитов у нормального кролика после внутривенного
впрыскивания 2 см® эхинококковой жидкости в опытах Bacigalupo и
Grosso в течение 10 минут падало с 9 800 до 4 800. У животных же с
эхинококком в тот же срок оно, наоборот, увеличивалось (в одном слу-
чае— с 13 000 до 28 000).
При подкожном впрыскивании 1 см3 жидкости человеческого эхино-
кокка, нагревавшейся в течение 1 часа до 60° С, людям, больным эхи-
нококком, наблюдалась местная реакция с отеком, покраснением, а
иногда и болью. Эти явления развивались в течение суток и проходили
через 1—2 дня. У здоровых людей реакции не было (Bac,i|galupo и
Grosso).
В случаях самопроизвольного разрыва пузырей эхинококка, а ино-
гда и при пункции (с целью диагностики) развивается картина гида-
тидной интоксикации, которая сказывается воспалением серозных обо-
лочек и крапивницей.
При повторных впрыскиваниях слабых доз эхинококковой жидкости
легко вызвать явления анафилаксии, причем Вейнбергу удалось полу-
чить все формы анафилаксии, которые бывают и у человека. Токсиче-
ское действие гидатидной жидкости Richet сравнивает с влиянием та-
лассина из мидий (Му tills).
Экспериментальная эхинококковая анафилаксия изучалась и други-
ми исследователями. Нормальная морская свинка (300—500 г весом)
в опытах Deluca погибала от внутривенного впрыскивания 8—10 см3
гидатидной жидкости. В то же время у предварительно сенсибилизиро-
ванной свинки тяжелый анафилактический шок вызывается инъекцией
всего только 2—3 см3 той же жидкости. Анафилактизирующая потен-
ция стенки пузыря эхинококка гораздо выше, чем жидкости тех же
самых цист.
Содержимое Cysticercus cellulosae очень ядовито (Robin, Fiessinger);
таким же свойством отличается и содержимое Coenurus serialis кролика
(Henry, Ciuca).
5 см3 жидкости Cysticercus pisiformis способны убить кролика весом
в 2 кг в 10 минут. После внутривенного впрыскивания жидкости кровя-
ное давление падает на 2—3 см. Ритм сердца замедляется, систолы
усиливаются, и кровяное давление выравнивается. Позднее оно может
внезапно упасть при ускорении сердцебиений до 150—180 ударов в ми-
452
СПЕЦИАЛЬНАЯ ЧАСТЬ
нуту. При больших дозах дыхание сильно нарушается. Временами оно
вовсе останавливается: делаясь поверхностным, дыхание прерывается
неравномерными паузами. Все эти явления имеют место и у тех кроли-
ков, которые не заражены Cysticercus pisifortnis; следовательно, экспе-
риментальное отравление не имеет ничего общего с анафилаксией
(Parisot и Simonin).
При внутрибрюшинном впрыскивании жидкости Cysticercus tenuicol-
lis (Taenia marginata) у морских свинок наблюдается проходящая лей-
копения и затем лейкоцитоз. У них также вырабатываются преципити-
иы по отношению к жидкости этого цистицерка. Реакция преципитации
получается с жидкостью Cysticercus tenuicollis, взятой от разных видов
(собаки, свиньи, овцы); это говорит в пользу того, что содержимое
цистицерка свойственно ему самому и не зависит от того, в каком хо-
зяине он образуется (Beckwith и Scott). В то же время гидатидная жид-
кость эхинококков от различных хозяев не абсолютно одинакова в ко-
личественном отношении по химическому составу, что показал Mazzocco:
В 100 см3 жидкости кисты эхино- кокка от человека В 100 см3 крови чело- века В 100 см3 жидкости кисты бычьего эхинококка В 100 см3 крови быка
Глюкозы . . . 0.090 0.110 0.035 0.070
NaCl 0.587 0.428 0.675 0.653
Мочекислых соедине-
ний . ...... 0.015 0.018 0.014 0.017
Химическая природа гидатидного яда неизвестна. В самой жидко-
сти содержатся янтарная кислота и сахар. Brieger, действуя платиной,
изолировал из этой жидкости вещество, быстро убивающее мышей.
Находясь в печени, эхинококк вызывает уменьшение в ней гликогена
с 61% до 39%; антитоксическая сила экстракта аутолизованной печени,
пораженной эхинококком, по отношению к гемолитическому токсину
этого последнего ослабевает, если такое свойство сравнить с нормаль-
ной печенью (Sparapani).
Круглые черви также были предметом разнообразных иссле-
дований на их. токсичность. Одним из наиболее изучавшихся объектов
являются аскариды человеческая, свиная и -лошадиная. Аскариды выде-
ляют из себя летучее вещество неприятного перечного запаха, которое
раздражает слизистые оболочки и вызывает конъюнктивит. Gold-
schmidt, опросив зоологов, часто имевших дело с аскаридами при лабо-
раторных занятиях, выяснил, что чувствительность людей к токсинам
аскарид различна. Одни вовсе не испытывают никаких неприятных яв-
лений, другие подвержены действию аскарид одного только вида, третьи
испытывают довольно тяжелую форму одышки, кашля и раздражения
слизистых оболочек. Эти данные подтверждают мысль о ядовитости ас-
карид. Какова же причина этой ядовитости? Исследуя токсичность по-
лостной жидкости аскарид при впрыскивании ее различным животным,
некоторые исследователи не получали положительных результатов или
объясняли их вторичной инфекцией (Boycott). Bussano и Flury удалось
вызвать экспериментальный конъюнктивит у морских свинок, кроликов
и собак, действуя на них тем же веществом.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
453
Полостная жидкость аскариды обладает несомненной токсичностью.
Weinberg и Julien наблюдали реакцию лошади на введение в ее глаз
нескольких капель жидкости A. megalocephala. «Уже через 5—10 минут
глаза краснеют, появляется сильное слезотечение, веки сильно опухают,
часто настолько, что совсем закрывают глаза. У 41% лошадей явления
принимают более тяжелый характер: развивается сильная одышка, по-
нос и подчас такой обильный пот, что под лошадью образуется целая
лужа пота. Иногда эти осложнения принимали столь тяжелый харак-
тер, что можно1 было опасаться смертельного исхода» (Вейнберг), од-
нако спустя час или два эти явления обычно проходили. В некоторых
случаях аскаридная жидкость была столь токсична, что 1 см3 раствора
4 : 1 000 вызывал глазную реакцию лошади. Раздражение глаз наблюда-
лось неоднократно и у лиц, вскрывавших аскарид (Raillet, Goldschmidt
и др.).
Несомненное ядовитое (резорптивное) действие проявляет та же
жидкость и при впрыскивании под кожу. У собак наблюдались уста-
лость и рвота, понижение кровяного давления, одышка, парез задних
ног, потеря чувствительности и даже смерть при введении 8—10 см3
полостной жидкости аскариды по расчету на 1 кг веса животного. Ас-
каридотоксин вызывает также и выкидыш у беременных морских сви-
нок, если впрыскивается им под кожу. Токсичность свойственна самой
аскаридной жидкости, так как исследователям удавалось получать по-
следнюю для своих опытов в стерильном состоянии.
Детальное фармакологическое исследование аскариды лошадиной
произведено в отделе паразитологии и фармакологии ВИЭМ В. Шва-
бауэром, который, работая с полостной жидкостью и с водными экстрак-
тами из цельных высушенных аскарид, пришел к следующим выводам.
Полостная жидкость при внутривенном введении (0,5 см3 на 1 кг
веса кошки) вызывает резкое падение кровяного давления при кратко-
временном начальном вагус-пульсе и учащенном пульсе на максималь-
но-низком уровне кровяного давления. При меньших дозах (0,2—
0,5 см3) наблюдается двухфазный эффект: небольшая начальная де-
прессия сменяется продолжительным подъемом давления выше нормы.
Падение кровяного давления зависит от влияния полостной жидкости
на бульбарные центры и на сердечно-сосудистый аппарат непосредствен-
но. При повторных введениях жидкости реакция подопытных животных
(кошки и кролики) ослабевает в значительной степени.
При внутривенном введении экстрактов из тела аскарид (1 : 100—
1 : 500) в количестве 1 см3 на 1 кг веса кровяное давление падает
вследствие прямого действия вещества на сердечно-сосудистую систему.
Изменения работы аппарата кровообращения при внутривенном введе-
нии токсина вызывают отклонения со стороны дыхательного аппарата
в виде начального угнетения и последующего возбуждения. Под влия-
нием полостной жидкости аскарид объем внутренних органов животно-
го резко уменьшается, а объем конечностей увеличивается, что зависит
от центрального влияния испытуемого вещества. Экстракты у лошади-
ной аскариды действуют на сердце теплокровных положительно ино- и
хронотропно. И полостная жидкость и экстракты действуют возбуж-
дающе на утомленную и адинамичную мышцу сердца; действие этих
веществ направлено непосредственно на сердечную мышцу. Повидимо-
му, мышечная часть стенок сосудов также является точкой воздействия
полостной жидкости и экстракта из аскарид (сокращение при более
крепких концентрациях и расширение — при слабых). Фармакологиче-
ская активность полостной жидкости самок аскарид выражена резче,
454
СПЕЦИАЛЬНАЯ ЧАСТЬ
чем у самцов. Наиболее активные действующие начала включает в се-
бя полостная жидкость аскарид. Действие и ее и экстрактов (в част-
ности их белковой элементарной фракции) направлено непосредственно
на сердечно-сосудистую систему, причем точкой приложения являются
мышца сердца и мышечная ткань сосудов (В. Швабауэр).
Жидкость тела A. lumbricoides в умеренной степени задерживает
свертывание крови.
Экстракт из стенки тела (мышцы) и полостная жидкость аскарид
задерживают рост фибробластов в культуре тканей (Hoeppli). Не-
сомненно влияние аскарид, особенно при их гибели (переваривание),
на секреторную функцию желудка. Отмечается, например, понижение
кислотности желудочного сока даже вплоть до ахилии при аскаридозе
и повышение кислотности после изгнания глист (Водонос). Наблюда-
лась также передача действующих начал аскарид с молоком матери ре-
бенку, у которого' развилась диспепсия.
Однако можно сомневаться, насколько ядовитость этой жидкости
может проявляться в «нормальных» условиях пребывания глисты в ки-
шечнике своего хозяина. Дело в том, что жидкость в полости тела аска-
риды замкнута наглухо; она может выступать наружу лишь при нару-
шении целости стенки тела паразита. Обычно же из глисты выходят ее
экскреторные продукты. Они также обладают токсичностью. Если дер-
жать некоторое время живых аскарид в бульоне, в чистом физиологи-
ческом растворе или же смешанном с глюкозой, пептоном и двуугле-
кислой содой, то все эти жидкости становятся ядовитыми (Cafiero, Flu-
гу и др.). В этих случаях остается не вполне ясным, чему именно при-
суща токсичность — секретам или выделениям аскариды. Кожных же-
лез у нее нет, и если кожа в целом не обладает отделительной способ-
ностью, то остается думать только об экскретах глисты, выходящих из
трубок ее боковых линий наружу через общее переднее отверстие.
Впрочем, при заболеваниях аскарид (вроде описанной выше бактери-
альной кожной болезни) возможна перфорация стенки тела аскарид и
изливание полостной жидкости в просвет кишечника с последующим
ее всасыванием-
В общем, понятие «аскаридный токсин» остается очень расплывча-
тым. Повидимому, он является соединением нескольких действующих
начал, одного — растворимого в воде и действующего на нервную си-
стему, другого—растворимого и в алкоголе и сходного по влиянию
с кураре (Vaullegard). Аскаридный токсин проходит без изменения
сквозь фильтр Шамберлена и не теряет способности вызывать глазную
реакцию у лошадей даже после двадцатиминутного нагревания его при
120°. Свернувшиеся от жара белки по обработке их водою так же
токсичны по отношению к глазу, как спиртовые, эфирные вытяжки
и продукты перегонки аскаридного токсина (Вейнберг). По данным
Вейнберга и Seguin, одни части последнего действуют хемотоксически
на лейкоцитов, вызывая эозинофилию, другие же влияют на стенку со-
судов, способствуя отеканию соответствующего участка тела. Bedson
показал, что аскаридные токсины действуют на органы внутренней се-
креции, в первую очередь на кору надпочки, а затем на щитовидную
железу, что в свою очередь вызывает дальнейшие коррелятивные изме-
нения в организме.
Аскаридотоксин влияет также и на нервную систему, что показали
Messineo и Рахманов. В нервных клетках наблюдаются хроматолиз раз-
ной степени и появление большого числа тонких каналов в протоплаз-
ме. Клетки невроглии приобретают амебоидный характер. Фибриллы
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
455
белого вещества мозга изменяются и неправильно вздуваются. Дей-
ствие аскаридотоксина на кровь непостоянно; в одних случаях она не
изменяет своих обычных свойств, в других же остается не свернувшей-
ся. Сильное гемолитическое действие экстракта кишечника аскарид на-
блюдал Schwartz, считающий, что анемия у свиней и лошадей часто
бывает связана с присутствием аскарид в их кишечнике. При аскари-
дозе у детей также наблюдались явления анемии и эозинофилия
(10—22%).
При опытах со впрыскиванием экстракта из аскарид Morenas уда-
лось получить явления анафилаксии, которыми он объясняет некоторые
симптомы аскаридоза.
Flury признает наличие в теле аскарид нескольких фармакологиче-
ских, различно действующих начал, которые в разных случаях, смотря
по обстоятельствам, вызывают весьма разнообразные симптомы в орга-
низме хозяина.
Наконец необходимо остановиться на новейших исследованиях при
моей кафедре (Военно-медицинская академия — С. П. Алфеева) над
чисто патологическими изменениями у морских свинок под влиянием
хронической интоксикации вследствие продолжительного кормления их
свежими или высушенными аскаридами (действие со стороны желу-
дочно-кишечного канала). Преже всего бросается в глаза дистрофиче-
ское ожирение подкожной клетчатки передней части туловища, явля-
ющееся следствием атрофии и ожирения щитовидной железы, функция
которой явно понижается. Происходит также сильное ожирение лимфа-
тических узлов щитовидной железы. В ретикулярном слое коры надпоч-
ки появляется зернистость с содержанием железа, что указывает на
усиленный распад клеток вследствие интоксикации; последняя ведет
также к прогрессирующей атрофии селезенки, при обильном отложе-
нии пигмента в ее пульпе. Из изменений крови отмечаются, уменьше-
ние количества эритроцитов, зернистых клеток, пластинок и увеличение
лимфоцитов. Ядра гладких мышц желудка принимают под влиянием
аскаридного токсина спиралевидную форму, чем можно объяснить на-
рушение моторной функции желудка и колики, наблюдаемые нередко
при аскаридозе.
Гематоэнцефалический барьер также повреждается под влиянием
действия аскаридного токсина (появление в сосудистом сплетении тре-
тьего желудочка головного мозга отека, экссудата, кровоизлияний, уве-
личение бурой зернистости хориоидального эпителия и др.), следствием
чего является развитие менингеальных явлений. Сердце, легкие, пе-
чень и почка не изменяются. В целом же повреждения кроветворной
и эндокринной системы при хроническом отравлении нарушают физио-
логическую корреляцию органов и могут вести к трудно расшифровы-
ваемым заболеваниям (С. П. Алфеева). Другие нематоды также обла-
дают токсическими свойствами, но нет возможности рассматривать
здесь дальнейшие материалы.
В заключение можно сделать вывод, что токсичность исследованных
глист несомненна, но формы проявления ее крайне вариируют от бес-
симптомного паразитоносительства до самых разнообразных и пестрых
по характеру заболеваний, смазывающих «лицо» глистных болезней.
Подобные вариации зависят от того, что состояние зараженно-
го глистами организмахозяинанаданный момент
является результатом взаимного влияния друг на
друга обоих компонентов биологической систе-
мы—хозяин и паразит, из которых каждый изменчив
456
СПЕЦИАЛЬНАЯ ЧАСТЬ
в своих физиологических состояниях. Этого мало: р е-
зультаты взаимного влияния находятся также под
известным модифицирующим воздействием других
обитателей организма хозяина (синергетическое
действие паразитоценоза хозяина на его организ м)„
равно как и под контролем факторов внешней
среды. Необходима даьнейшая научно-исследовательская работа, ко-
торую следовало бы направить на изучение токсичности действующих
начал паразитических червей в той именно форме, в которой эти ве-
щества поступают, проходя через печеночный барьер, в общий круго-
ворот крови. Может быть, по этой линии исследований окажется при-
менимым метод ангиостомии Лондона, которым обеспечено изучение
тонкостей внутреннего обмена веществ.
Косвенное влияние паразитических червей сказывается в отношении
содействия их передаче инфекции вследствие грубого нарушения барь-
ерной функции кишечника; миграционное влияние оказывается слож-
ным и различным на разных этапах его прохождения, что рассмотрено
выше на отдельных объектах.
ПУТИ ЗАРАЖЕНИЯ ЧЕЛОВЕКА ГЛИСТАМИ
Источником заражения человека глистами яв-
ляется внешняя среда — понятие, взятое в широком
смысле по отношению к каждому индивидууму.
К внешней среде, следовательно, относятся субстрат (почва), вода, бли-
жайшее окружение человека (жилье, предметы обихода), пища, раз-
личные животные и растения и, наконец, другие люди (по отношению
к рассматриваемой особи). Первым условием возможности заражения
человека глистами является наличие паразитических червей в инвази-
рующем состоянии (личинки, яйца и др.) во внешней среде (роль
других факторов см. в общей части книги, стр. 44); в дальнейшем бу-
дем их называть просто «паразиты».
Все паразитирующие черви человека могут быть разбиты на три
группы, в связи с тем, как они проникают в его организм. Само собой
понятно, что такая группировка является экологической, но не систе-
матической. Паразиты могут проникать в тело человека: 1) пассив-
но, 2) активно и 3) при посредстве переносчиков.
1. Паразитические черви, проникающие в тело человека пассивно.
Для соответствующей группы глист характерно, что сами паразиты
не предпринимают и не могут предпринимать каких-либо действий, что-
бы попасть в организм хозяина. В этом отношении они являются пас-
сивными или совершенно инертными. Зато активен хозяин, который
обычно в процессе принятия пищи или питья бессознательно и неосве-
домленно для себя проглатывает паразитических червей в инвазирую-
щих фазах их жизненного цикла.
Соответствующая категория паразитов подразделяется на группы в,
зависимости от деталей процесса инвазирования.
А. Пассивное поступи н и е паразитических червей,
естественно заражающих животных, служащих
пищей человека
К соответствующей группе относятся паразитические черви, проме-
жуточные хозяева которых идут в качестве обычной пищи человека.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
45 /
I. Мясо крупного рогатого скота, пораженное финками Т. saginatus,.
служит источником заражения человека цепнем невооруженным, если
употребляется в пищу в сыром виде или при недостаточной термине
ской обработке (проглатывание жизнеспособных и не убитых жаром
финок).
Своего рода профессиональное заражение бывает при пробе сырого
мясного фарша на соленость поварами или хозяйками.
Меры профилактики:
1) ветеринарно-боенский контроль (на финок) за выпуском мяса;
2) обеспечение достаточной термической обработки пищи;
3) не есть сырого мяса.
Рис. 369 — Пути заражения скота финнозом и людей острицами, аскаридами,
власоглавом (через немытую зелень и воду). Оригинал.
По линии уменьшения инвазирующего запаса паразита:
1) плановая дегельминтизация носителей цепня невооруженного с
уничтожением изгоняемых паразитов;
2) предохранение скота от заражения финками:
а) пастьба под надзором;
б) устройство и содержание уборных, не доступных для скота в
отношении питья мочи;
в) не загрязнять почву фекалиями и пользоваться уборными.
II. Свиное мясо 1) с финками Т, solium и 2) с капсулами трихин.
При употреблении в пищу сырой или полусырой финнозной свинины
человек заражется цепнем вооруженным.
Таким же путем, а также при использовании копченых продуктов,
(ветчина, колбасы, иногда сало) обеспечивается заражение трихинами.
Меры профилактики аналогичны указанным выше. Кроме того,
борьба с крысами, поеданием которых свиньи могут заражаться три-
хинами; уничтожение боенских конфискатов; не скармливать боенсюк
отбросы и конфискаты свиньям.
458
СПЕЦИАЛЬНАЯ ЧАСТЬ
III. Мясо рыб, могущих быть промежуточными хозяевами лентеца
широкого (и других видов) и кошачьей двуустки (Opisthorchis felineus),
китайской двуустки (Metagonimus) и близких нематод.
В мышцах рыбы, под кожей, в яичнике и в других органах залега-
ют плероцеркоиды лентеца широкого (щука, налим, окунь, ерш и др.),
лентеца малого (Diph. minus, на Байкале — омуль, хариус, сиг). Не-
сомненно, что и другие виды лентецов, описанные в СССР (см. выше),
имеют вторым хозяином какие-то виды рыб. Человек заражается, съе-
дая сырую или полусырую рыбу.
Рис. 370—Пути заражения лентецом широким. Оригинал.
Дифиллоботриозы характеризуются очаговостью распространения
(применительно к местам первичного заражения человека: Восточная
Сибирь, Карелия, Ленинградская область, Прибалтика и др.), связан-
ного с паразитологическими соотношениями рыб, циклопов и человека,
а также частично собак и свиней.
Рыбы, передающие в качестве промежуточных хозяев различные
трематодозы, имеют в мышцах или в чешуе инцистированные метацер-
карии соответствующих трематод.
Географическое распространение и очаговость трематодозов опреде-
ляется паразитологическими соотношениями рыб, моллюсков и человека
(также и плотоядных).
Описторхоз не заходит восточнее бассейна Оби. В Европейской ча-
сти Союза он обнаружен в бассейне Днепра. Метагонимоз характерен
для бассейна Амура.
Описторхоз связан с потреблением в пищу рыб: язя, чебака (Ruth
lus rutilus). сибирского ельца (Leucis)cus leuciscus baicalensis), леща
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
459
{Abramis), красноперки и других видов. Вторыми хозяевами Metagoni-
mus являются на Амуре корюшка, Hemibarbus labeo, Gobio gobio
(пескарь), Parabramis pekinensis (лещ), Pseudobagrus fulvidraco, Liocas-
sis ussuriensis, L. brazhnikowi и другие виды рыб.
Профилактика:
1) Не есть сырой или полусырой рыбы.
2) Обеспечение надежной термической обработки рыбы и ее очистки
от чешуи и внутренностей.
Меры общего порядка:
1) Уменьшение запаса инвазирующих паразитов дегельминтизацией
с уничтожением изгоняемых глист.
2) Охрана водоемов от попадания фекалий с живыми яйцами глист;
спуск сточных вод в водоемы после отстаивания или химического обез-
вреживания.
3) Выдерживание рыбы в холодильниках, пока не погибнут плеро-
церкоиды.
4) Не скармливать сырых рыбных отбросов или рыб свиньям и со-
бакам.
Для всех рассмотренных примеров характерна та общая черта, что
инвазионность сырых пищевых продуктов является результатом специ-
фического течения жизненного цикла лентецов или сосальщиков.
Б. Пассивное поступалие паразитов с случайно
загрязняемой пищей
Пищевые продукты и готовая пища могут быть загрязняемы инвази-
рующими яйцами глист при различных обстоятельствах, зависящих ча-
сто от самого человека.
При удобрении огородов и поливке их фекалиями и при использова-
нии фекалий для удобрения в парниковом и оранжерейном хозяйстве
земля и произрастающие на ней овощи могут загрязняться яйцами
аскарид, власоглава, остриц трихонстронгилид, анкилостомид, цепня
карликового, цепня вооруженного и др. Все поименованные нематоды
имеют прямое развитие; яйца их нуждаются в довольно долгом (аска-
рида и власоглав) созревании, пока в них не разовьются личинки.
При поедании немытой огородной зелени и овощей могут проглаты-
ваться и яйца указанных выше паразитических червей.
Профилактика:
1) Есть огородную зелень и овощи хорошо обмытыми чистой (кипя-
ченой) водой; не есть прямо вынутые из земли и лишь обтертые
овощи.
2) Обработка фекалий кипятком перед использованием их для удо-
брения парников.
3) Мытье рук перед едой.
4) Дегельминтизация групп населения, как мера уменьшения инва-
зирующего запаса паразитических червей.
В. Поступи ние паразитов с питьевой вод oil
Вода может быть естественным или случайным посредником в зара-
жении человека глистами при пассивном поступании их per os.
I. На поверхности водоемов или на поверхности листьев и стеблей
различных водяных растений инцистируются церкарии печеночной дву-
устки, образуя стадию, называемую адолескарией.
460
СПЕЦИАЛЬНАЯ ЧАСТЬ
При проглатывании с водой адолескарий или при поедании сырой
зелени, которая поливалась водой с адолескариями, человек заражает-
ся фасциолезом.
Меры профилактики:
1. Не пить сырой (или в крайности не профильтрованной через плот-
ный холст) болотной, прудовой, озерной или речной воды. Водопровод-
ная, колодезная и ключевая вода в рассматриваемом отношении без-
вредна.
2. В отношении огородной зелени см. выше.
3. Прямая и косвенная борьба с моллюсками L. minutus — проме-
жуточным хозяином печеночной двуустки.
Рис. 371 — Пути заражения риштой. Оригинал.
4. Массовая дегельминтизация овец и коров четыреххлористым угле-
родом.
II. С водой хаузов (прудов) в очагах ришты (Dracunculus persarum}
проглатываются циклопы, зараженные ее личинками, что ведет к зара-
жению человека и собаки риштой (бывшие очаги в Старой Бухаре).
Профилактика:
1. Не пить сырой или непрофильтрованной воды хаузов.
2. Осушение и периодическая очистка хаузов.
3. Лечение больных и запрещение купаться или мыть ноги в
хаузах.
Ш. В воду в порядке вторичного загрязнения могут попадать до-
стигшие инвазирующего состояния яйца аскарид, власоглава или он-
косферы цепней. Попав с питьевой водой в кишечник человека, они
обеспечивают его заражение соответствующими паразитами.
Профилактика:
1) Не пить сырой воды из подозрительных на загрязнение водоемов.
2) Охранять водоемы от попадания или сброса фекалий.
3) Снабжать шахтеров индивидуальными фляжками с питьевой ки-
пяченой водой.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
461
Г. Пассивное поступание паразитов через посред-
ство загрязненных рук (контактная инвазия)
При несоблюдении правил личной гигиены или при отсутствии ми-
нимально необходимых санитарно-гигиенических навыков легко осу-
ществляется заражение человека некоторыми весьма опасными для
него паразитическими червями.
I. При расчесывании промежности, зудящей вследствие выползания
самок остриц и откладки их яиц, яйца легко забиваются под ногти.
Рис. 372 —Уборные, устроенные непосредственно над водой. Оригинал
по фотографии Г. Смирнова.
При еде пищи грязными руками или скусывании ногтей яйца остриц
проглатываются и способствуют суперинвазии.
Через руки, загрязненные в указанных условиях, яйца остриц могут
быть переносимы на игрушки, ручки дверей, различные предметы оби-
хода и в дальнейшем также через посредство рук проглочены здо-
ровыми детьми или взрослыми, что ведет к первичному энтеробиозу.
Профилактика:
1. Мытье рук перед едой и соблюдение правил личной гигиены.
2. Дегельминтизация носителей остриц с уничтожением последних.
3. Дегельминтизация предметов обихода и борьба с микрофека-
лизацией жилья.
II. При личной нечистоплотности загрязненными руками после де-
фекации носитель цепня вооруженного может занести яйца в рот и
их проглотить. Это ведет к развитию финноза или цистицеркоза, по-
следствия которого зависят от локализации финок.
462
СПЕЦИАЛЬНАЯ ЧАСТЬ
Профилактика:
1. Соблюдение правил личной гигиены.
2. Дегельминтизация носителей цепня вооруженного с уничтожени-
ем паразитов.
3. Борьба с финнозом свиней (см. выше).
III. При близком контакте с собакой, зараженной эхинококком, и
несоблюдении правил личной гигиены яйца эхинококка, приставшие к
шерсти собаки, могут быть загрязненными руками занесены в рот и
проглочены. Заражение эхинококком опасно, так как требуёт хирур-
гической операции, которая, смотря по месту нахождения паразита, не
всегда возможна.
Меры профилактики:
1. Соблюдение правил личной гигиены.
2. Повторная дегельминтизация собак с уничтожением фекалий.
3. Не скармливать собакам проросших эхинококком органов убой-
ного скота.
4. Утилизация пораженных эхинококком органов животных на
бойнях.
5. Уничтожение бродячих собак.
6. Осторожность в обращении с овцами при доении, стрижке шер-
сти, к которой могут пристать яйца эхинококка из фекалий собак на
пастбище (см. пункт 1).
Кроме того, см. выше стр. 481.
Д. Пассивное поступление паразитов в желудок
человека при особых обстоятельствах
I. При сильной рвоте членики и яйца цепня вооруженного могут из
кишечника носителя, вследствие влияния антиперистальтики, попасть
«обратным ходом» в его желудок, результатом чего является развитие
финноза различных органов иногда множественного характера.
Профилактика:
1. Своевременная диагностика и изгнание обнаруженного паразита.
2. Обязательное диагностическое обследование faeces больных с
последующим глистоизгнанием перед наркозом, могущим вызвать
рвоту, и категорий служащих, которые в первое время службы осо-
бенно подвержены рвоте в силу профессиональных условий (летчики,
моряки).
II. При интимной близости с собаками (поцелуи, игры детей и пр.)
возможно проглатывание яиц эхинококка.
Профилактика см. выше.
3. При проглатывании власоедов или блох собак и кошек с цисти-
церкоидами цепня тыквовПдного {Dip. caninum) у человека в кишеч-
нике развивается не свойственная ему, как естественному хозяину,
ленточная глиста.
Профилактика:
1. Не кормить детей на полу и не позволять им есть с одной та-
релки с собакой или кошкой.
2. Не позволять целовать собак или кошек.
3. Дегельминтизация собак и кошек.
4. Борьба с наружными паразитами собак и кошек.
5. Кошки могут косвенно способствовать заражению человека (де-
тей) цепнем крысиным (Hymenolepis diminuta). На кошку могут перей-
ти блохи с пойманной ею крысы; блохи могут быть заражены цисти-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
463-
церкоидами глисты. При проглатывании таких блох человеком, послед-
ний заражается цепнем крысиным.
Профилактика:
1. Не класть кошек в кровать или в люльку к детям.
2. Другие меры см. в предыдущем абзаце.
3. Проглатывание других промежуточных хозяев цепня крысиного
в плохо пропеченном хлебе из муки, пораженной амбарными вредите-
лями (мучным хрущаком — Tenebrio molitor, гусеницами мельничной
огневки — Asopia farinalis) или домовыми обитателями (тараканы).
Рис. 373 — Заражение скота эхинококком на пастбище. Оригинал.
Профилактика.
1. Борьба с амбарными вредителями и домовыми обитателями
(крысы и др.).
2. Высокое качество выпускаемого в продажу хлеба и муки.
II, Паразитические черви, проникающие в организм человека
активно V
Активное проникновение паразитических червей в организм чело-
века осуществляется вбуравливанием (внедрением) их инвазирующих
личинок из зараженной внешней среды в кожные покровы с последую-
щим проникновением в кровеносные сосуды с миграцией по телу
вплоть до места окончательного обитания, каковым, смотря по виду
глисты, являются кишечник или такие органы, как печень или моче-
вой пузырь.
Зараженным субстратом, из которого поступают личинки в тело
человека, бывает почва или вода.
454
СПЕЦИАЛЬНАЯ ЧАСТЬ
А. Внедрение личинок из влажной почвы в тело
человека
I. Инвазирующие личинки анкилостомы, некатора и угрицы кишеч-
ной вбуравливаются в кожу человека при контакте его голыми покро-
вами тела с зараженным субстратом, что осуществляется в условиях
сельскохозяйственных, земляных и шахтных работ, а также в траншей-
ной жизни. Распространение этих паразитических червей приурочено
к югу. У
Рис. 374 — Заражение 'людей и собак эхинококком. Оригинал.
Профилактика.
1) выявление больных и носителей и дегельминтизация их с унич-
тожением изгоняемых паразитов;
2) устройство рационально сконструированных уборных, предохра-
няющих почву от заражения, и искоренение привычки пользоваться для
дефекации огородами и вообще почвой;
3) работа в подозрительных по заражению местах в обуви;
4) не ложиться голым телом на землю в аналогичных местах;
5) запрещение дефекации в отсеках шахт и устройство выносных
очищаемых клозетов.
Б. Активное внедрение паразитов в тело
человека из воды
I. Церкарии бильгарций (шистозом), выходящие из промежуточных
хозяев — моллюсков Bullinus contortus, Planorbis boissyi, Blanfodria
nosophora и др. в воду и плавающие в соответствующем водоеме,
вбуравливаются в кожу человека при купании, хождении в воде,
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
46с
стирке и др. Результат — заболевание соответствующей формой биль-
гарциоза. В СССР бильгарциоз известен лишь в качестве завозных
случаев.
Профилактика:
1) не иметь контакта с водой в подозрительных по заражению во-
доемах;
2) уничтожение в водоемах моллюсков, промежуточных хозяев;
3) уничтожение в водоемах церкарий бильгарций;
4) предохранение водоемов от загрязнения их мочой и фекалиями
человека;
5) лечение больных бильгарциозом как мера уменьшения запаса
паразитов.
III. Передача паразитических червей через посредство
насекомых-переносчиков.
Следует различать насекомых — промежуточных хозяев
глист, от насекомых — переносчиков паразитических
червей, хотя в ряде случаев таковое значение насекомых совпадает.
Насекомые — промежуточные хозяева глист в собственном смыс-
ле — передают соответствующую инвазию, если будут съедены (про-
глочены) окончательным хозяином (см. выше блохи, власоеды, жуки,
тараканы и др.).
Насекомые не входят в пищевой рацион человека, и понятно, что
заражение человека через их посредство является случайным и ред-
ким и самые случаи заражения человека соответствующими глистами
относятся к явлениям гостепаразитизма.
Насекомые — переносчики в собственном смысле этого понятия —
являются кровососущими двукрылыми. В процессе сосания крови они
проглатывают личинок различных нитчаток. Личинки мигрируют по те-
лу насекомого, проделывая часть жизненного цикла, и в инвазирую-
щем состоянии скопляются в хоботке (нижняя губа); прорывая его
тонкую кожицу, личинки при сосании насекомым крови здорового чело-
века выходят наружу и через покровы (быть может, и через ранку)
активно проникают в тело человека. Однако доминирующим является
обстоятельство передачи весьма подвижным двукрылым личинок нит-
чаток человеку, причем переносчик остается живым и может быть
способным к дальнейшей передаче личинок, если таковые имеются в
его теле разных возрастов. Во всех таких случаях дело касается спе-
цифической передачи инвазии.
А. Специфическая передача личинок
паразитических червей насекомыми-переносчиками
1. Личинки («микрофилярии») нитчатки Банкрофта (Wuchereria ban-
crofti) передаются через посредство различных комаров: Culex fati-
gans, An. hyrcanus и др.
В СССР случаи слоновой болезни с микрофиляриями в крови неиз-
вестны.
Профилактика:
1) защита носителей личинок филярий в крови от нападения ко-
маров;
2) защита здоровых от нападений комаров. Борьба с комарами.
2. Личинки филярии калабрского отека (Loa loa) передаются через
слепней Chrysops silaceus (Тропическая Африка).
466
СПЕЦИАЛЬНАЯ ЧАСТЬ
сосущих двукрылых нашей
Рис. 375 — Нога Musca vicina
с приставшими яйцами тениид
Куляб. По Г. Змееву.
В СССР известны единичные случаи филяриоза конъюнктивы гла-
за человека (см. выше Loa extraocularis, стр. 457), но переносчик остает-
ся неоткрытым. Какая-то нитчатка обнаружена в нарывах пальцев
крестьян (б. Тверская губ.), но она не изучена, неизвестен и перенос-
чик ее.
3. Нитчатка Onchocerca volvulus (соединительнотканные опухоли
под кожей) в Тропической Африке передается мошкой Simulium
damnosomum.
4. В Тропической Африке, в Британской Гвиане нитчатка Acan-
thocheilonema perstans передается через посредство мокреца Culicoi-
des austehL
Если в отношении паразитов человека еще неизвестная роль крово-
фауны оказывается небольшой, то обрат-
ное определится при экспериментальном
изучении роли кровососущих двукрылых в
распространении различных филяриозов.
домащних животных, каковая область рабо-
ты еще не затронута.
Б. Механическая (не специфиче-
ская передача яиц и личинок
паразитических червей насеко-
мым и-пере .носчиками
К рассматриваемой категории явлений
относятся все случаи распространения яиц
или личинок глист вследствие загрязнения
ими хоботка, ротовых органов, поверхно-
сти тела и ног различных насекомых при
ползании их по фекалиям или субстрату с
яйцами или личинками глист. При попада-
нии этих насекомых в воду или при полза-
нии их по пищевым продуктам или посуде
они оставляют яйца глист, впоследствии
проглатываемые человеком с водой или с
питьем.
Другим вариантом передачи яляется
заглатывание насекомыми фекалий с яйца-
ми глист и выбрасывание не потерявших
жизнедеятельности транзитных яиц при
пищевые продукты или посуду.
Во всех случаях дело сводится к рассеянию яиц паразитических
червей механической контаминацией, причем в насекомом-переносчике
или на нем паразит не проделывает какой-либо части своего жизнен-
ного цикла, но некоторое время на, нем или в нем переживает.
1. Универсальную роль в механической передаче яиц паразитиче-
ских червей играет комнатная муха, переносящая яйца на хоботке,
лапках, ногах и через кишечник (острица, аскарида, власоглав, цепни,
анкилостомиды и др.).
2. Несколько более ограничена пространственно аналогичная роль
общественных ос — Polistes gallicus (Е. Павловский и В. Сондак) и
шершня восточного (Vespa orientalis), передающих не только яйца
глист, но и личинок анкилостомид.
переносчика на
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
467
3. Из прямокрылых неоднократно устанавливалась спонтанная за-
раженность яйцами аскарид и власоглавов у тараканов (в их кишеч-
нике, Сондак), равно как и в экскрементах.
4. У египетского таракана-черепашки (Polyphaga saussurei) быва-
ли обнаруживаемы в кишечнике яйца TaenUdae, Hymenolepidae и
других глист, не являющихся собственными паразитами этого насеко-
мого (Ю. Таджикистан, Г. Змеев).
Несомненно, что список насекомых — механических переносчиков
яиц глист при дальнейших исследованиях значительно расширится.
Профилактика:
1) борьба с мухами;
2) защита пищи от мух, ос, полистов и «домовых обитателей».
Общим условием профилактики всех гельминтозов человека, ка-
кими бы путями они ни распространялись, является широкая осведом-
ленность врачей и медицинских работников, освоение ими знаний о па-
тогенном значении глистного фактора и о методах борьбы с парази-
тическими червями как в самом организме человека, так и в его окру-
жении.
Второе условие — широкая пропаганда в среде населения сведений
о значении паразитических червей в отношении здоровья человека и о
способах личной и общественной защиты от них.
ОРГАНИЗАЦИЯ ПЛАНОВОЙ БОРЬБЫ С ГЕЛЬМИНТОЗАМИ В СССР
1939 год явился поворотным в деле организации плановой борь-
бы с глистными заболеваниями и глистоносительством в СССР.
В результате работ II Всесоюзного совещания по борьбе с гелъминто-
зами в СССР в III пятилетке выработано и утверждено Наркомздра-
вом СССР положение о гельминтологическом пункте, являющемся
основной ячейкой, ведущей лечебно-профилактическую противоглист-
ную работу.
Плановые массовые противоглистные мероприятия, обязательные
для всего Союза, касаются аскарид и цепней вооруженного и нево-
оруженного; к ним добавляются и другие объекты, смотря по типич-
ным гельминтологическим показателям республик или районов (напри-
мер, анкилостомиды, лентец широкий и др.).
Для ознакомления с организацией противоглистной борьбы при-
ведем, с некоторыми сокращениями, положение о гельминтологических
пунктах, система которых находится в составе Противоэпидемического
управления Наркомздрава СССР, в отделе борьбы с малярией.
1. Организация гельминтологических пунктов
1. Гельминтологические пункты являются лечебно-профилактически-
ми учреждениями, организующими, проводящими и контролирующими
противоглистные мероприятия в обслуживаемых ими районах (обсле-
дование населения, лечебно-профилактические мероприятия, учет дина-
мики глистных заболеваний).
2. Гельминтологические пункты организуются или как самостоя-
тельные учреждения, или при детских поликлиниках, диспансерах, ам-
булаториях, объединенных поликлиниках, тропических и малярийных
станциях и других медицинских учреждениях.
3. В тех районах, где население поражено гельминтозами в 25.%,
468
СПЕЦИАЛЬНАЯ ЧАСТЬ
организуются гельминтологические пункты по одному на район по
спискам, утвержденным Наркомздравами союзных республик.
4. В сельских районах, где распространены в 5% и выше анкило-
стомы, тенииды или карликовый цепень, организуются гельминтологи-
ческие полустационары при районных амбулаториях.
5. Помимо полустационаров при районных амбулаториях органи-
зуются полустационары при гельминтологических пунктах в тех райо-
нах, население которых поражено в 10% и выше тениидозами, анки-
лостомидозами или гименолепидозом, из расчета 1 койка на 6 тыс. насе-
ления (при 10% поражения) и 1 койка на 3 тыс. населения (при 20%
поражения) и т. д.
6. Гельминтологический полустационар может работать:
а) постоянно, если для него отведено специальное помещение
(см. ниже); или б) временно — при отсутствии специального помеще-
ния. В таком случае полустационар развертывается по выходным дням
в амбулаториях, поликлиниках и других медицинских учреждениях.
Число коек во временных полустационарах должно быть соответ-
ственно увеличено, так как койка полустационара, работающего только
по выходным дням, пропустит в год не 300 больных, а только 55—60.
7. Гельминтологический пункт может быть с полустационаром или
без полустационара.
Гельминтологический пункт без полустационара состоит из:
а) кабинета врача;
б) лаборатории;
в) ожидальной комнаты.
При наличии лаборатории и ожидальни в том медицинском учреж-
дении, при котором организован гельминтологический пункт, эти поме-
щения используются и для гельминтологического пункта.
8. Гельминтологический полустационар (временный или постоян-
ный) состоит из:
а) одной или двух палат;
б) ожидальной и раздевальной комнат;
в) душевой (с 2 душами);
г) помещения для дефекаций с промывочной.
9. При планировании помещения полустационара необходимо при-
нять следующий порядок.
Больной из раздевальни проходит в душевую (при наличии тако-
вой), где сдает свою одежду и получает чистое белье, а затем прохо-
дит в палату. При отсутствии душевой больной переодевается в палате.
Из палаты больной должен пройти в близко расположенную комнату
для дефекации. Последняя должна иметь хорошо вентилируемую про-
мывочную, в которую переносятся для анализа фекалии больных.
В промывочной желательно устройство герметически закрытого куба
или бака для кипячения выделенных паразитов.
10. В тех случаях, когда лаборант гельминтологического пункта
может работать в лаборатории того медицинского учреждения, при ко-
тором организован пункт, отдельная лаборатория при пункте не обя-
зательна.
Массовая дегельминтизация при аскаридозе проводится по установ-
ленной инструкции на базе учреждений (школы, детсады, детдома,
производства и пр.).
Массовая дегельминтизация при тениидозах и анкилостомидозах
проводится по установленным инструкциям в полустационаре гельмин-
тологического пункта, а также и во временных полустационарах, орга-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
469
низуемых пунктом на базе других учреждений: детдомов, амбулаторий,
детсадов, поликлиник и др., при ликвидации тениидозных и анкило-
стомидозных очагов.
Амбулаторный прием глистных больных проводится 1—2 раза в
пятидневку, по направлению врачей общей медицинской сети (консуль-
тативный7 прием).
2. Подготовка кадров
1. Заведующий гельминтологическим пунктом ведет работу по по-
вышению гельминтологической квалификации врачей всех специально-
стей в обслуживаемых им районах (лекции, семинары, конференции),
а также проводит подготовку средних и младших гельминтологических
кадров по установленным программам.
2. Заведующий гельминтологическим пунктом оказывает консуль-
тативную помощь всей лечебной сети по диагностике, лечению и про-
филактике гельминтозов на обслуживаемой им территории.
3. Санитарно-профилактическая работа
1. Гельминтологический пункт организует проведение санитарно-
профилактических мероприятий по борьбе с гельминтозами.
2. Контролирует совместно с ГСИ проведение в жизнь санитарных
правил использования нечистот в сепьском хозяйстве, утвержденных
ВГСИ от 26 апреля 1938 г.
3. Заведующий гельминтологическим пунктом проводит санитарно-
просветительную работу как с больным, обращающимся на гельминто-
логический пункт, так и среди населения обслуживаемого района.
Пр имечание. 1. Гельминтологический пункт без полустационара проводит
ту же работу (см. а, б, в), за исключением дегельминтизации больных в полу-
стационаре.
2. Гельминтологический полустационар проводит: а) дегельминтизацию больных
в полустационарах (тениидозных, анкилостомидозных и гименолепидозных), направ-
ляемых лечебными учреждениями; б) учет эффективности дегельминтизации; в) сан-
просветработу с больными в полустационаре; г) повышение гельминтологической
квалификации средних медицинских кадров.
ОБЩИЕ ОСНОВЫ МЕДИКАМЕНТОЗНОЙ БОРЬБЫ С ГЛИСТНЫМИ
ИНВАЗИЯМИ 1
Объектом лекарственного воздействия являются черви в период
нахождения их в организме человека, прежде всего в пищеваритель-
ном канале.
Прежнее деление лекарственных веществ на vermi;cida (убивающие
глист) и vermifuga (собственно глистогонные) не удержалось в фар-
макологии, потому что характер действия лекарств на глист опреде-
ляется иными формами. Поэтому уместно говорить о противоглистных
средствах, давая характеристику их применительно к влиянию каж-
дого вещества на различные виды глист (поскольку требуемые дета-
ли известны в научной литературе).
1 При составлении этой главы использованы материалы из инструкции Нар-
комздрапа СССР по борьбе с аекяоидозом и тениидозами.
В 1943 г. Наркомздравом СССР выпущен сборник организационно-методических
материалов по борьбе с малярией и гельминтозами, 200 стр., 'Медгиз. В нем даны
разнообразные организационные и практические материалы.
Дозировка противоглистных средств, приведенных в тексте Руководства, сверена
со схемой лечения гельминтозных больных указанного сборника.
470
СПЕЦИАЛЬНАЯ ЧАСТЬ
Противоглистные средства могут убивать одних паразитов, пони-
жать тонус («оглушать») других, ослаблять силу мышц третьих и др.
Приведенные в состояние депрессии паразитические черви легко под-
даются изгоняющему влиянию перистальтики кишечника, усиливаемой
слабительными или клизмами.
Значение дегельминтизации двоякое: 1) лечебное — по отношению
к индивидууму и 2) профилактическое — по отношению к коллективу.
Профилактическое значение дегельминтизации связано с уменьшением
общего запаса паразитных начал в людях. Однако указанная цель
достигается лишь при массовой дегельминтизации различных групп
населения, с непременным уничтожением изгоняемых паразитических
червей, чтобы этим предотвратить дальнейшее рассеивание их яиц во
внешней среде, со всеми последующими этапами их жизненного цикла.
Установлено предпочтительное влияние различных лекарственных
веществ на определенные группы паразитических червей.
Обычно противоглистные средства вводят per os, но некоторые из
них могут действовать на кишечных червей и при инъекциях под кожу
или в мышцы.
Для успеха действия обычно бывает необходима подготовка к
дегельминтизации: переведение за 1—% дня на легкую диэту, не даю-
щую объемистых масс содержимого кишечника; предварительное очи-
щение кишечника клизмой или солевым слабительным с вечера; при-
нятие лекарства натощак; выбор и принятие подходящего слабитель-
ного.
Эффект мероприятий определяется детальным исследованием
изгнанных глист промыванием фекалий (отыскание головки ленточно-
го червя) и последующим микроскопическим контролем фекалий на
яйца глист через 1—2—3 месяца.
Из растущего арсенала противоглистных средств отметим наибо-
лее распространенные и принятые в широкой практике.
Детальные данные см. в VIII главе книги «Глистные заболевания
человека» (В. Подъяпольская и В. Капустин, 1937), в «Anthelmintics
апД their uses» (Chopra a. Chandler, London, 1928) и в «А Handbook
of Tropical therapeutics» (Chopra, Calcutta, 1936).
1. Экстракт корневища папоротника — Extraction fi-
llets marls aethereum — токсичен для человека; растворяется
в маслах; поэтому в качестве слабительного касторовое масло при нем
совершенно исключается. Должен быть применяем из свежего материа-
ла, так как при хранении теряет свои полезные свойства. Является
наилучшим средством для изгнания ленточных червей—лентецов и
цепней. Вполне пригоден для изгнания кишечных трематод. Дневная
доза 4,0—7,0.
Rp. Extr. filicis mar,is aeth. 6,0 (s. 7,0).
Div. in part, aequal. Nr. 10.
D. in caps, gelatinos.
S. Принять в течение часа натощак; через 1 — IV2 ч.— солевое
слабительное.
Вследствие раздражения слизистой желудка после принятия па-
поротника бывает рвота, ослабляющая действие этого лекарства.
Для предотвращения ее рекомендуют выпить стакан 1% содового
раствора за полчаса до приема папоротника и запивать проглатывае-
мые капсулы таким же раствором. Папоротник можно прописывать
в пилюлях или вводить самый экстракт дуоденальным зондом в две-
надцатиперстную кишку.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
471
1. Подготовка к лечению
Перед дегельминтизацией проверить анализ мочи.
За 1—2 дня до лечения назначается питательная, но легко усвоя-
емая пища, бедная жирами (белый хлеб, сухари, бульон, крупяные
супы, супы-пюре, молоко, кефир, простокваша, творог, жидкие молоч-
ные каши, мясные котлеты, отварная свежая рыба, кисели, чай, кофе).
Вечером накануне лечения ужин заменяется стаканом чая или кофе
с сухарем и назначается слабительное. Взрослым и детям, начиная с
5-летнего возраста, дается глауберова или английская соль:
В о з р а
5 лет ....
6—7 лет . .
8—9 » ..
10—11 »
12-14 » ...
15 —18 » . .
Взрослые . .
Доза
(в г)
81—102
10—12
12—15
15—20
20-25
25—30
30—40
Венского питья ребенку 2—3 лет дают 1 дессертную ложку; 4—7
лет — 1 столовую ложку (при отсутствии венского питья давать соле-
вое слабительное — английскую или глауберову соль в соответствую-
щих дозах).
Примечание. Детям старше 5 лет можно давать венское питье в следую-
щих дозах: 8—12 лет—2 столовые ложки, 13—15 лет — 3 столовые ложки, взрос-
лым— 4 столовые ложки.
2. Дача противоглистного
В день лечения утром поставить клизму. Натощак в течение полу-
часа маленькими глотками дать выпить V2—1 стакан 1% прохладного
раствора двууглекислой соды. По окончании приема соды дать экстракт
мужского папоротника в капсулах:
Возраст Доза (в г)
5—А лет .... 2.5
7—9 » 3
10- 12 » 3.5
13- 15 » 4
Взрослые 4—6
Капсулы принимаются в течение 30 минут, запиваются 5/2— 1 стака-
ном 1% раствора соды.
Примечание. Лицам, плохо переносящим раствор соды, можно давать
экстракт папоротника без соды. В таких случаях доза папоротника увеличивается:
В о з р а
5—6 лет ..........
7—9 » . . .
10-12 » ....
13-15 »
Взрослые . . .
с т Доза (в г
........... 2—2.5
.............. 2-4
. . . 4-5
. . 5-6
5-7
1 Доза (в г) глауберовой соли.
* Доза (в г) английской соли.
472
СПЕЦИАЛЬНАЯ ЧАСТЬ
Детям до 5 лет экстракт дается с медом или повидлом в следую-
щих дозах:
Возраст Доза (в г)
1 ГОД 0.5
2 года 1
3 » ... 1.5
4 » 2
5—6 лет 2.5
7—9 » 3.0
10—12 » 4.0
13-15 » 4.0—5.0
Экстракт дается двумя полупорциями с перерывом в 30 минут.
3. Выведение паразитов из кишечника
Через 30 минут после последней капсулы (при приеме экстракта
с содовым раствором) дается солевое слабительное в дозах, указан-
ных выше (см. подготовку к лечению). Лицам, принимавшим экстракт
без содового раствора, слабительное (солевое или венское питье)
дается через 1—V2 часа после последней порции . экстракта. Через
Р/гчаса после приема слабительного дается легкий завтрак независимо
от того, подействовало слабительное или не подействовало.
Если через 2 часа после приема слабительного стула нет, нужно
поставить клизму из теплой воды (32—34°) — взрослому из 5 стаканов,
детям соответственно возрасту.
Если паразит вышел без головки, нужно поставить еще 1—3
клизмы.
Примечание. Если у больного во время приема папоротника появилась
тошнота или рвота, рекомендуется полный покой, грелка на живот, мятные капли,
глотание кусочков льда и пр. Если часть папоротника вышла со рвотой, продол-
жить прием его минут через 15—20, но не увеличивать назначенной дозы за счет
удаленного со рвотой.
Противопоказания к лечению экстрактом муж-
ского папоротника: туберкулез легких в активной форме, поро-
ки сердца в состоянии суб- и декомпенсации, язвы желудка и двенад-
цатиперстной кишки с кровотечениями, болезни печени и почек, период
кормления, беременность, менструации. Соблюдать осторожность у лиц
истощенных и у стариков.
Для лечения гименолепидоза в Тропическом институте (Москва)
выработана следующая схема:
1-й День — легкая полужидкая диэта;
2-й день — то же; на ночь солевое слабительное;
3—4-й день — взрослому 2,5—3,0 г экстракта папоротника в два
приема, с перерывом 15—20 мин. Через 1—2 часа солевое слабитель-
ное. Через 1—2 часа легкий завтрак. После стула — обед (полужид-
кая диэта).
5—'13-й день — обычный стол.
14-й день — легкая полужидкая диэта.
15-й день—то же; на ночь слабительное и далее второй курс ле-
чения, как указано выше (В. Подъяпольская и В. Капустин).
Детям дают соответственно пониженные дозы папоротника: 1—3
года — 0,3 г; 3—4 года — 0,6 г; 5—6 лет — 0,9 г; 7—8 лет— 1,2 г;
9—10 лет—1,5 г; 11 —12 лет—1,8 г; 13—14 лет — 2,2 г: 15—16
лет — 2,5 г.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
473
Экстракт корневища папоротника вполне пригоден для изгнания
кишечных трематод Metagonymus и Nanophyetes schichobalovi [-Troglot-
rema (Nanophyetes) "salmincola] (Ж. Штром).
Из препаратов папоротника приготовляют филицилен (Rilicilen).
Доза в порошке для взрослого 3—4 г. В масляном растворе — 6—8 г
(советский препарат). Заграничным препаратом является фильмарон
(Fi-lmaron); доза — 0,85—1,0 г.
2. Четырех хлористый углерод Carboneum tetrachloratum
(CCU) — прозрачная бесцветная жидкость, по запаху приближающая-
ся к хлороформу. Должен применяться лишь в химически чистом
виде.
Является лучшим средством при анкилостомозе, некаторозе, реко-
мендуется при острицах и аскаридах.
R. Carb, tetrachlorati 1,0 куб. см
D. t. dos. in caps, gelat. Nr. 3
принять натощак в течение получаса; через час солевое слабительное.
Доза для детей 0,13 куб. см на каждый год жизни до 15 лет.
Производство в СССР начато в Азербайджане.
3. Тетрахлорэтилен (Tetrachloraethylenum — С2С14) менее
токсичен, чем четыреххлористый углерод, но зато более нестоек при
хранении. Дозировка та же; употребление то же.
4. Гексилрезорцин — Hexyl resorcinol.
5. Хеноподовое масло — 01. Chenopodii из травы — Chenopo-
dium anthelminticum. В опытном порядке хеноподовое масло получено
в Сухумском ботаническом саду. Является отличным средством при
аскаридозе и хорошим при анкилостомидозах.
Дозировка взрослым:
Rp. О1. chenopodii 3 куб. см.
Div. in part, aequal.
D. in caps, gelatin. Nr. 3
S. Принимать через час по одной капсуле; через час после конца
приема — солевое слабительное.
Детям; 1—2 года 3 раза по 2 капли
3—5 лет — по 3—5 капель
6— 10 » — по 6—8 »
11 —16 » — по 8—12 »
Взрослым до 50 лет — по 15 »
» свыше 50 лет — по 8-- 12 (Brumpt)
Обладает токсичностью для человека.
Действующим началом является аскаридол, добываемый в жидком
виде: дается в три раза меньшем количестве, чем Ol. chenopodii.
6. Сантонин — Santoninum — действующее начало цитварного
семени (цвета цитварной полыни Artemisia cinae). Кристаллический
порошок; на свету желтеет.
Под влиянием сантонина аскариды теряют способность удержи-
ваться в кишечнике, благодаря чему легко изгоняются принятием со-
левого слабительного.
Rp. Santonini 0,05
Calomelan. 0,03
Sacch. albi 0,2
Mf. pulv. d. t. d. Nr. 10
474
СПЕЦИАЛЬНАЯ ЧАСТЬ
S. Принимать по 3 порошка в течение трех дней.
Rp. Santonini 0,1
Sacch. albi 0,2
Mf. pulv. d. t. d. Nr. 3
S. Принимать через час по порошку.
При обеих прописях через час после последнего приема — солевое
слабительное.
Детям дается по 0,005 на прием на год жизни и в день до 0,015.
А. Дозировка сантонина
Разовая дача сантонина (в г)
Возраст Доза Возраст Доза
На 1-м году жизни . . 1 год 2—3 года 0.005 0.005—0.01 0.01-0.02 10—12 лет . 0.05
4—5 лет 0.025—0.03 12-14 » . . . 0.05
6—7 » 0.03 - 0.035 15-17 » 0.07-0.08
8—9 » 0.04-0.045 ‘ Взрослые . 0.1
Сантонин назначается 2 раза в день в таблетках или порошках в
течение двух дней подряд по одному из следующих методов.
Первый метод. Первый прием сантонина утром через V2—1 час
после легкого завтрака (чай с небольшим куском хлеба).
Второй прием сантонина через IV2—2 часа после первого, не позже
чем за 1 час перед следующей едой.
Второй метод. Если невозможно дать сантонин в утренние ча-
сы, следует давать первый прием сантонина через 2—3 (лучше 3 ча-
са) после еды, второй прием через IV2—2 часа после первого, не
позже чем за 1 час перед следующей едой.
Для удаления глист из кишечника назначают слабительное.
Виды слабительных, которые могут быть назначены:
а) Слабительная соль — глауберова или англий-
ская.
Дается в, дни приема сантонина через час после второго порошка
в растворенном и хорошо остуженном виде.
Возраст Доза (в г) Возраст Доза (в г)
2—3 года .... 41 61 2 10—11 лет . . . 15 1 20 2
4—5 лет 8 10 12-14 » 20 25
6—7 » . . 10 12 15-18 » 25 30
8—9 12 15 Взрослые 30 40
Б. Венское питье
Дается оба дня лечения по одному разу в день через
1 час после
последнего приема сантонина.
Возраст
.2—3 года
4—7 лет .
8—12 » .
13-15 » .
Взрослые
Доза
1 дессертная ложка
1 столовая ложка
2 столовые ложки
3 » »
4 » »
1 Доза глауберовой соли.
2 Доза английской соли.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
475
В. Ревень (в порошке)
Дается одновременно с сантонином (в таблетках или порошком)
в дозе, превышающей в 10 раз дозу сантонина. На второй день лече-
ния через Р/г—2 часа после второго порошка сантонина с ревенем —
солевое слабительное.
Г. Сложнолакричный порошок
Дается детям в течение двух дней по одному разу в день тотчас
за вторым приемом сантонина.
Возраст Доза
2—3 года................ Vi чайной ложки
4—6 лет............... 1 учайная ложка вровень с краями
8—12 »......... P/g чайные ложки » •» »
Взрослые......... 4 » » » » »
Характерным симптомом отравления сантонином является ксантоп-
сия — видение предметов в желтом свете.
7. Тимол. Добывается из тимьяна и других трав. Является хоро-
шим средством при некаторозе, а также при анкилостомозе,
8. Нафталин — Naphthalinum ригит имеет ограниченное приме-
нение, так как по противоглистным свойствам уступает другим веще-
ствам. Иногда применяется при энтеробиозе.
Rp. Naphthalin! pur. 0,3
Santonin! 0,05
Calomelan. 0,03
Sacchar. lact. 0,02
D. t. d. obi. Nr. 10
S. По одной облатке три раза в день между едой. Избегать жирной
пищи. На ночь водяная или мыльная клизма. Повторить курс лечения
не ранее как через 10 дней. Соблюдать правила личной гигиены.
9. Осарсол — советский стоварсол (Osarsol) — мышьяковый пре-
парат, изготовляемый в СССР с 1923 г. При испытании его при анки-
лостомозе Медведевой — Кобринец и Подъяпольской удалось обнару-
жить в фекалиях больного вышедших самок власоглава, что дало им
повод исследовать противоглистные свойства осарсола при трихоцефа-
лезе.
Осарсол выпускается в таблетках по 0,25 г’.
Дозировка взрослым — разовая — 0.25; дневная — 0.75—1.0, на
курс 4.9—5.0. Детям столько таблеток на курс, сколько ребенку лет;
таблетки по 0.25 для разовых доз делить на 2—4 части.
Порядок дачи осарсола:
I метод.- 1-й и 2-й дни — по 0.25 3 раза в день за час до еды,
3—4—5-й дни — по 0.25 4 раза в день; на 6-й день — солевое слаби-
тельное.
II метод: перед приемом осарсола по I методу 3 дня метиленовая
синька 0.1 3 раза в день после еды; детям — по 0.005 на год возраста.
III метод: 1—2-й день по 0.25 3 раза в день до еды; 3-й день —
3—4 таблетки; 4-й день — слабительное, с 5-го по 9-й день — перерывы.
С 10-го по 13-й день, как 1—4-й день.
IV метод: перед каждым циклом приема осарсола по III методу в
476
СПЕЦИАЛЬНАЯ ЧАСТЬ
течение 3 дней принимать метиленовую синьку, как указано во II ме-
тоде.
10. Эметин — действующее начало ипекакуаны (Бразилия, Psycho-
tria ipecacuanhae); применяется в виде солянокислой соли Emetinurrf
hydrochloricum для лечения соматических паразитов — пененочной
двуустки (Fasc. hepatica), Dicrocoelium lanceatum.
at Rp. Sol. Emetini hydrochi. 2%— 20,0
Внутримышечно no 1,0 куб. см через день. Через 20 дней недельный
перерыв. В случае необходимости курс лечения повторяется.
Rp. Sol. Emetini hydrochi. 1%—20,0
Внутривенно; ежедневно, начиная с 2 куб. см и доходя до 4. Через
10 вливаний недельный перерыв.
И. Сера. Против остриц. Дозы взрослым — разовая 1.0; дневная —
3.0; на цикл—15.0; на курс — 45.0—75.0. Детям на год жизни — 0.05.
Слабительное не назначается. Серу дают 3 раза в день во время еды
5 дней подряд; затем 4 дня перерыв. Таких циклов проводить 3—5.
В дни перерыва — ежедневно клизма на ночь из воды или 1°/о) соды
или поваренной соли. Обмывать задний проход утром и вечером и сма-
зывать его белой, желтой или серой ртутной мазью; при резком зуде —
3—5% анестезиновой мазью.
ИНСТРУКЦИЯ ПО МАССОВОЙ БОРЬБЕ С НЕКОТОРЫМИ
ГЕЛЬМЙНТОЗАМИ
В связи с организацией плановой и массовой борьбы с гельминто-
зами приведем текст инструкции по борьбе с гельминтозами Нарком-
здрава СССР, утвержденной ГСИ, в частности по борьбе с аскаридозом
и тениидозами.
ИНСТРУКЦИЯ ПО БОРЬБЕ С АСКАРИДОЗОМ
Борьба с аскаридозом проводится в массовом масштабе среди орга-
низованных групп взрослого и детского населения (школы, детсады;
производства и пр.) и состоит из следующих этапов.
I. Предварительное обследование.
IJ. Массовая дегельминтизация.
III. Профилактические мероприятия.
ПРЕДВАРИТЕЛЬНОЕ ОБСЛЕДОВАНИЕ
1. В однородном коллективе (дети детсада, школьники приблизи-
тельно одного возраста, рабочие одного производства и др.), включаю-
щем менее 200 человек, обследованию подвергаются все лица.
2. В однородном коллективе, включающем более 200 человек, об-
следованию подвергается «пробная группа» в 100—150 человек. В слу-
чае обнаружения аскаридоза более чем у 50% лиц «пробной группы»
дальнейшего персонального обследования можно не проводить и счи-
тать выявленный процент заражения общим для всего коллектива.
В случае обнаружения аскаридоза менее чем у 50 лиц «пробной
группы» обследованию подлежат все члены коллектива.
3. Методика обследования. Обследование производится путем ко-
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
477
прологического анализа одновременно: 1) по методу Фюллеборна, до-
полненному просмотром осадка со дна, и 2) по методу нативного
мазка.
МАССОВАЯ ДЕГЕЛЬМИНТИЗАЦИЯ
1. Лица, подлежащие дегельминтизации:
а) В случае обнаружения аскаридоза менее чем у 50% обследован-
ного коллектива лечению подвергаются лица с положительным копро-
логическим анализом.
б) В случае обнаружения в «пробной группе» аскаридоза у 50%
лиц и более лечению подвергаются поголовно все лица коллектива.
Противопоказаниями к дегельминтизации являются: острые
и хронические нефрозо-нефриты, острые гастроэнтериты, острое лихо-
радочное состояние.
При общем физическом недоразвитии и резком общем истощении
отдельных членов коллектива лечение проводится под особым наблю-
дением врача.
2. Сроки плановой дегельминтизации:
Ввиду того, что заражение аскаридами чаще происходит летом и
осенью, дегельминтизация в одном и том же коллективе проводится
два раза в год: поздней осенью (октябрь — ноябрь) и в конце зимы
(февраль — март).
Примечание. В местностях с короткой и теплой зимой, в которых зара-
жение происходит почти круглый год, дегельминтизация проводится- 3 раза в год-
1) поздней осенью (октябрь — ноябрь), 2) зимой (февраль — март) и 3) весной
(май — июнь).
3. Этапы дегельминтизации:
1) подготовка к лечению,
2) дача противоглистного,
3) удаление паразитов,
4) обезвреживание выделенных фекалий и паразитов,
5) учет результатов дегельминтизаиии.
А. Подготовка к лечению
Накануне и в дни приема противоглистного предлагается воздер-
живаться от жирной пищи.
Б. Дача противоглистного
Лечение проводится сантонином по предварительно заготовленным
именным спискам, подписанным врачом, с обозначением индивидуаль-
ных доз. Раздача сантонина производится врачом, сестрой-дегельмин-
тизатором в строго индивидуальном порядке с регистрацией каждого
приема. Детали см. в предыдущей главе.
В. Обезвреживание выделенных фекалий и паразитов
а) Ые допускать рассеивания фекалий и паразитов (отсутствие
благоустроенных уборных) вблизи населенных мест, жилых и про-
мышленных зданий и железных дорог.
б) Канализованные уборные (унитаз, пол и стены) в течение 5 дней
после начала дегельминтизации обливать крутым кипятком на близком
478
СПЕЦИАЛЬНАЯ ЧАСТЬ
расстоянии (во избежание остывания воды). Неканализованные убор-
ные (стульчак, пол, стены вблизи стульчака), выгреба и почву вокруг
выгребов заливать 50% раствором хлорной извести.
Г. Учет результатов дегельминтизации
производится
а) По данным опроса о количестве отошедших паразитов в течение
первых 5 дней после начала дегельминтизации при условии, если
имеется индивидуальная посуда для сбора фекалий; количество ото-
шедших аскарид отмечается в дегельминтизационных списках.
б) По данным лабораторного контроля дегельминтизации (повтор-
ное обследование), который производится в период от 10 дней до
1 месяца после окончания дегельминтизации.
Все лица, не освобожденные от аскарид полностью, дегельминтизи-
руются повторно не ранее чем через 15 дней после предыдущего курса.
ПРОФИЛАКТИЧЕСКИЕ МЕРОПРИЯТИЯ
А. Меры общественной профилактики
1. Охрана внешней среды от загрязнения экскрементами:
а) Концентрирование фекалий: устройство в местностях, где отсут-
ствует канализационная сеть, при каждом дворе типовых уборных,
пригодных для пользования взрослыми и детьми.
б) Усиление контроля за содержанием уборных с периодической
дезинфекцией их.
в) Своевременный вывоз мусора из мусорных ящиков и фекалий из
выгребных ям в места, указанные местным саннадзором.
г) Регулярное тщательное подметание дворов и улиц после предва-
рительной их поливки во избежание- рассеивания яиц аскарид с под-
нимающейся пылью.
д) Рациональное использование фекалий в виде удобрения, что до-
стигается одним из следующих методов:
1) внесением фекалий в почву огородных культур овощей, упо-
требляемых в сыром виде, не раньше чем за полный календарный год
до посева;
2) предварительным компостированием фекалий в течение трех
летних месяцев с конским навозом, домовым мусором, торфом.
2. ' Наблюдение за источниками водоснабжения населения. В ме-
стах, где нет водопровода, необходимо следить, чтобы в питьевую во-
ду (колодцы, реки и др.) не могли попасть экскременты. '
3. Борьба с мухами.
4. Наблюдение за хранением продуктов и приготовлением пищи в
общественных столовых. Инструктирование и контроль за тщательным
мытьем рук обслуживающего персонала.
5. Широкая санитарно-просветительная работа среди населения.
Б. Меры личной профилактики.
Профилактика личная может быть сведена к следующим мероприя-
тиям:
а) Не пить сырой воды (исключение может составлять водопровод-
ная обезвреженная вода).
б) Хорошо мыть овощи, фрукты, употребляемые в сыром виде.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
479
в) Соблюдать правила личной гигиены (мыть руки перед едой*
стричь ногти, употреблять в уборных бумагу и пр.).
г) Закрывать пищу от мух. Уничтожать мух.
ИНСТРУКЦИЯ ПО БОРЬБЕ С ТЕНИИДОЗАМИ
Плановая борьба .с тениидозами проводится в массовом и индиви-
дуальном порядке по всей территории СССР.
Борьба с тениидозами слагается из следующих этапов:
I — выявление носителей тениид;
II — дегельминтизация;
III — профилактические мероприятия.
Выявление носителей тениид
Выявление носителей тениид среди организованного населения про-
изводится путем поголовного обследования отдельных групп взрослого
и детского населения (детсады, предприятия и пр.), а среди неоргани-
зованного путем выявления их лечучреждениями и отдельными врачами.
Выявление носителей тениид (бычьего и свиного цепня) произво-
дится:
1) путем опроса об отхождении члеников (обязательно представле-
ние члеников для определения);
2) у лиц, не представивших членик для диагноза, путем одного из
следующих лабораторных методов: а) соскобом с перианальных скла-
док (или ректальным), б) копрологическими анализами (метод мазка,
метод Фюллеборна, дополненный просмотром осадка с дна).
Дегельминтизация
Лечению подвергаются все лица, зараженные тениидами, кроме тех,
кому оно противопоказано по состоянию здоровья (см. ниже).
Лечение тениидозов производится эфирным экстрактом мужского
папоротника в обстановке стационара или полустационара.
Процедура лечения тениидозов состоит из следующих этапов:
1) подготовка к лечению,
2) дача противоглистного,
3) удаление паразитов из кишечника,
4) определение и регистрация выделенных паразитов,
5) обезвреживание фекалий и выделенных паразитов.
Пункты 1—3 см. в предыдущей главе.
Определение и регистрация вышедших паразитов
Все порции фекалий, отходящих после приема папоротника, соби-
рать в отдельные сосуды и промывать водой.
Для этого в сосуд с фекалиями наливают воду, перемешивают,
оставляют стоять до полного осаждения, после чего жидкость сливают,
не затронув осадка, и в сосуд снова наливают воду. Промывку повто-
ряют до тех пор, пока жидкость над осадком не станет прозрачной.
Осадок слить в сосуд, лучше, в черную кюветку, выделить паразитов
и их отдельные куски. Если паразит лишен головки, тщательно искать
ее в осадке, разбавляя его водой и просматривая маленькими порциями.
480
СПЕЦИАЛЬНАЯ ЧАСТЬ
Обезвреживание фекалий и выделенных паразитов
Выделенные во время дегельминтизации фекалии, а также паразиты
и их куски должны быть подвергнуты обезвреживанию. Для этого сле-
дует:
а) выделенных паразитов подвергать термической обработке: кипя-
тить в течение 30 минут в закрытых баках и только после этого спу-
скать в канализацию или выливать в уборные;
б) фекалии и вода после промывки фекалий там, где нет канализа-
ции, должны выливаться в ямы, к которым не должен иметь доступ
крупный рогатый скот и свиньи.
Использование фекалий из выгребных ям для удобрения огородов
под культуры овощей, потребляемых в сыром виде, возможно только
при внесении фекалий в почву за полный календарный.год до посева
овощей или после предварительного компостирования их в течение
трех летних месяцев с конским навозом, домовым мусором, торфом.
Профилактические мероприятия
Профилактические мероприятия в борьбе с тениидозами слагаются
из мер общественной профилактики и мер личной профилактики.
А. Меры общественной профилактики
1. Дегельминтизация лиц, зараженных свиным и бычьим цепнем.
2. Широкая санитарно-просветительная работа среди населения.
3. Для реализации перечисленных ниже мер общественной профи-
лактики тениидозов необходимо войти в контакт с медицинскими
и ветеринарными санорганизациями.
а) Устройство уборных и содержание их в таком виде, чтобы жи-
вотные не могли иметь контакта с испражнениями.
б) Борьба с рассеиванием фекалий на задних дворах, хлевах, ого-
родах, улицах и других местах, где может находиться или проходить
домашний скот.
в) Усиление саннадзора за мясом, употребляемым в общественных
столовых, и за его приготовлением.
г) Усиление саннадзора за мясом, продаваемым на рынках.
д) Наблюдение за тем, чтобы крупный рогатый скот и свиньи не
могли бродить по местам, где имеются скопления человеческих фе-
калий.
Б. Меры личной профилактики
а) Не есть сырого, плохо проваренного или плохо прожаренного
мяса (бычьего и свиного). Не пробовать сырого фарша.
б) Не покупать мяса, не прошедшего ветсаннадзора.
в) В целях предохранения от возможности заражения цистицерко-
зом соблюдать все правила личной гигиены (изгнание ленточной ста-
дии паразита, частое мытье рук, мытье овощей, употребляемых в пищу
в сыром виде, борьба с мухами и пр.).
Примечание. Лечение дифиллоботриоза проводится согласно наставлениям,
изложенным в разделе «Дегельминтизация» (п. II).
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
481
МЕТОДЫ ОБЕЗЗАРАЖИВАНИЯ ФИННОЗНОГО МЯСА
При обнаружении финноза крупного рогатого скота и свиней в ко-
личестве не более 3 финн в обычных для их нахождения мышцах на
большинстве разрядов площадью 40 кв. см мясные туши подлежат
стерилизации, а свыше 3 финн — утилизации.
Обезвреживание условно годных мясопродуктов согласно действую-
щему ветеринарному законодательству производится следующими спо-
собами:
1) Проваривание — финнозное мясо должно вариться не менее
2V2 часов от начала кипения, причем куски должны быть не толще
15 см; трихинозное мясо — то же, но куски не толще 10 см.
2) Стерилизация мяса паром.
3) Посолка мяса кусками не более 27г кг весом; мясо пересыпает-
ся солью и в бочках заливается 25% солевым раствором по расчету
количества рассола к мясу в пропорции 40 : 100. Финнозное мясо счи-
тается обезвреженным через три недели. Однако перед выпуском фин-
нозной солонины финны проверяются на жизнедеятельность.
4) Замораживание мяса ведет к гибели финн, если туши замора-
живают в морозилках при температуре —12—15° С и хранят в ка-
мерах хранения не менее 10 дней; свиные туши замораживаются при
температуре не выше —12° С и выдерживаются в камерах не менее
15 дней.
По опытам Н. Бородина, изолированные финки крупного рогатого
скота погибают после часового замораживания при —8, —10—12—
—14° С, после восьми часов выдерживания при —6° С и через сутки —
при охлаждении до —4° С. Эти опыты, а также обширные эксперимен-
ты над замораживанием финнозных туш позволили укоротить сроки
замораживания и выдерживания на холодильнике финнозного мяса
крупного рогатого скота в целях его полного обезвреживания:
а) замораживание мясных туш до температуры —12°—14° С в тол-
ще мышц;
б) то же при —6° С с односуточным выдерживанием заморожен-
ного мяса в камере хранения при температуре —9°—9,5° С;
в) замораживание до —3° С с последующим выдерживанием мяса
в камере хранения при температуре —5° С в течение недели. Эти
нормы утверждены Главным ветеринарным управлением Наркомзема
СССР.
Перед выпуском и соленое и замороженное мясо проверяются на
жизнедеятельность финн. Приготовляют смесь 80 г бычьей желчи и
20 г физиологического раствора поваренной соли и подогревают ее до
40° С; в нее кладут не менее десятка финн, вынутых из мяса, освобож-
денных от оболочек и отмытых предварительно в теплой воде. Если
финны живы, они вытягивают шейку и производят различные движе-
ния. Наблюдать лучше под лупой.
При трихинозе свиней при наличии единичных трихин в 24 срезах
(кусочках, раздавленных в компрессоре) и отсутствии изменения
мышц — мясо считается условно годным, допускаемым в пищу людям
после стерилизации. В некоторых местах оно абсолютно бракуется.
При нахождении в мышцах туш свиней множественных и разбро-
санных по нескольким частям туш эхинококков, если их нельзя уда-
лить, туши направляются в стерилизацию.
482
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ ЧЕЛОВЕКА
При наличии эхинококков только в окороках или корейках или пле-
чевых мышцах необходимо пораженные места зачистить, а тушу вы-
пускать в пищу.
ЛИТЕРАТУРА
Даниляк И. Д.— Материалы к изучению эхинококковой болезни в Азербайджан-
ской ССР. Изд. Тропического института Наркомздрава АССР, Баку, 1935.
Деятельность двадцати восьми гельминтологических экспедиций в СССР
(1919—1925 г.). Под редакцией К. И. Скрябина, Москва, 1927.
Павловский Е. Н.— Дополнение к классу Turbellaria, отряд Temnocephalida.
Класс сосальщиков (Trematoda); класс ленточных червей (Cestoidea); класс
волосатиковых (Nematomorpha); класс скребней и др. Руководство по зоологии,
т. I. Под ред. Л. Зенкевича, Москва — Ленинград, 1937.
Па вловский Е. Н.— Важнейшие достижения советской паразитологии • по про-
тозоологии и гельминтологии за 20 лет. Журн. Природа № 11, 1937.
Павловский Е. Н. (ред.) — Практикум медицинской паразитологии (Главы по
глистам Н. Костылева и Г. Смирнова.) Биомедгиз, Ленинград, 1935.
Петров А. М.— Глистные инвазии собак и их санитарное и экономическое значе-
ние. Сельколхозгиз. Ленинград — Москва, 1931:
Подъяпольская В. П. и Капустин В. Ф.— Глистные заболевания человека.
Биомедгиз, Москва — Ленинград, 1937.
Работа 25-й союзной гельминтологической экспедиции в Артемовском округе Дон-
басса.— Под ред. К. И. Скрябина и P.-Эд. С. Шульц, Москва, 1926.
Работа 35-й союзной гельминтологической экспедиции в Среднюю Азию, организо-
ванной Дорздравотделом Средне-Азиатской железной дороги.— Под ред.
К. И. Скрябина и P.-Эд. С. Шульц, Москва, 1928.
Работы по гельминтологии.— Сборник, посвященный 30-летию научной деятельности
акад. К. И. Скрябина и 15-летию Всесоюзного института гельминтологии.
Под ред. P.-Эд. С. Шульц и М. П. Гнединой. Изд. Всес. акад. с.-х.
наук им. Ленина, Москва, 1937.
Ра го за Н. И.— Об изменениях крови при ленточных глистах. СПБ, 1913.
Сборник работ по гельминтологии, посвященный проф. К- И. Скрябин у.—
Москва, 1927.
Скрябин К. И. (ред.) Библиотека по гельминтологии (ряд книг многих авторов
по различным группам паразитических червей и по хозяевам, по большей части
ветеринарною значения).
Скрябин К. И. и Шульц Р.<— Основы общей гельминтологии. Сельхозгиз, Мос-
ква, 1940.
Скрябин К. И. и Шульц P.-Эд. С.— Фасциолезы животных и меры борьбы с
ними. Москва, 1935.
Скрябин К. И. и Шульц P.-Эд. С.— Гельминтология. Ветеринарная паразито-
логия и инвазионные болезни домашних животных. Часть 1. Сельхозгиз, Москва.
1937.
Скрябин К. И. и Шульц P.-Эд. С.— Гельминтозы человека, т. I, II, .Москва,
1929, 1931.
Холодковский Н, А. — leones helminthum hominis. Атлас человеческих глист,
вып. 3. »
Труды Бородинской биологической станции в Карелии, т. VI, вып. 2, Ленинград,
1933.
Труды Бородинской биологической станции в Карелии, т. VIII, вып. 1, Ленинград,
1935. Оба сбооника посвящены работам по лер,тепу широкому.
Труды Всесоюзного Института гельминтологии, т. I. Под ред. К. И. Скрябина.
Серия XIV — Ветеринария, Гельминтология, вып. 3. Изд. Всес. акад. с.-х. наук
им. Ленина, Москва, 1935. *
Большая мечипчиская энциклопедия.
См. также журналы, труды экспедиций и съездов (стр. 58—60 настоящей книги).
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ ЖИВОТНЫХ
А
Амеба дизентерийная 24, 72, 74 78,
82, 84
Амеба карликовая 98, 99
Амеба кишечная 24, 66, 70, 95
Амеба ротовая 100, 102
Амебы 71
Амфибии 200
Амфибии бесхвостые 219
Анкилостома 2, 16, 24, 385, 391, 393,
394, 445
Анкилостома собачья 29, 31
Анкилостомиды 2, 16, 383, 387, 388,
391, 392, 466, 467
Аноплоцефалиды 301
Анофелес 14, 150, 170, 174, 175, 179
Антилопы 20, 22, 127, 331
Аскарида кошачья 414
Аскарида лошадиная 341, 452, 453, 455
Аскарида свиная 408, 412, 413, 452, 455
Аскарида собачья 14, 23, 413
Аскарида человеческая 53, 403, 404,
408, 409, 413, 452
Б
Балянтидий 204, 206, 207, 208
Балянтидий обезьян 208
Балянтидий свиньи 208
Балянтидий человека 208
Барсук 251, 351
Белки 112, 331
Бильгарций церкария 464
Бильгарция 24
Биченосцы 103
Блоха кошачья 462
Блоха собачья 13, 303, 462
Блоха человеческая 112. 303
Блохи 14, 15, 22, 34, 112, 303, 304, 309,
310, 462, 465
Блохи крысиные 309, 462
Броненосцы 42, 129
Бронзовка 439
Буйвол 228, 267
Бык 202, 222, 238, 323, 363
Бычки 212, 251, 284
В
Верблюды 119, 267, 331/ 395, 414
Веретенница зеленая 282
Веретенница прыткая 282
Власоглав 24, 27, 341, 343, 344, 355,
457, 459, 460, 466
Власоглав мышиный 359
Власоглав свиньи 355, 356
Власоглав человеческий 355, 356
Власоед кошачий 44, 303
Власоед собачий 44, 303, 304
Власоеды 13, 14, 304, 462, 465
Вобла 439
Волки 295, 334, 362
Волосатиковые 210, 346
Вошь головная 13
Всеядные 310
Вши 15, 24, 34, 46, 52, 113, 119
Выдра 363
Г
Гагара 234
Гадюка степная 282
Гамбузия 433
Гаплоспоридии 141
Геккон 44, 119, 120
Геккон каспийский 282
Гемоспоридии 15, 148
Гиббон 301
Гиены 303
Глисты 210, 444
Голец 360
Голубь 234
Гонгилонема 414
Гориллы 375, 376
Грегарины 65, 141
Грызуны 308, 309, 440
Густера 236, 245
д
Двуустка гигантская 228
Двуустка египетская 253
Двуустка индийская 250
Двуустка китайская 18, 52, 250, 251,
458
Двуустка кошачья 243, 253, 458
Двуустка кровяная 27
Двуустка ланцетовидная 237
Двуустка легочная 27, 52, 258, 261
Двуустка печеночная 14, 26, 221, 477
Двуустка сибирская 243
Двуустки молодые 226
Двуустковые 221
Двуутробки 129
Дельфин 275
Дикроцелиоз 241, 242
Дождевой червь 398
484
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИИ ЖИВОТНЫХ
Е
Елец сибирский 246, 248, 458
Енот уссурийский 362
Ерш 283, 284, 285, 458
«ЦТ»
Жвачные 299, 415
Жвачные копытные 299
Жгутиковые 69, 103, 121
Жиардия 66; 139, 140, 141
Жираффа 228
Желез ница 16
Жерлянка 282
Жук майский 439
Жук чернотелка 440
Жуки 440, 465
Жуки-навозники 415
Жуки пластинчатоусые 439
3
Заяц 238, 301, 359
Земноводные 218
Змеи 295
Зудень 52
Зудень чесоточный 15
И
Иксодовые клещи 34, 35
Инфузории 65, 204
Инфузории паразитические 208
К
Кабан 208, 221, 312, 419
Карп 201
Кета 255
Кефаль 253, 254
Клещ чесоточный 13, 24
Клещи 15, ИЗ, 142, 301
Клоп постельный 35, ИЗ, 119
Клоп поцелуйный 106, 113, 129
Клопы 15, 34
Книдоспоридии 141, 202
Коза 222, 228, 238, 267, 327, 331, 395
Кокцидии 142, 150
Кольчатые черви 440
Колючеголовые черви 438
Колюшка 284
Комары 15, ИЗ, 119, 424, 427, 465
Комар малярийный 44
Корненожки 69, 71
Корова 228, 323, 325, 326, 332, 336,
419
Корюшка 459
Кошачьи 377
Кошка 13, 29, 31, 90, 95, 119, 147, 233,
235, 243, 245, 249, 250, 251, 253, 257,
267, 275, 287, 295, 303, 304, 310, 312,
326, 331, 351, 362, 381, 404, 419, 462
Кошка дикая 419
Краб 142
Крабы пресноводные 260, 261
Красноперка 244, 245, 459
Кривоголовка двенадцатиперстная 13,
377, 378, 379
Кривоголовка собачья 381, 385, 386
Кровеспоровики 141, 148
Кровососка собачья 119
Кровососущие двукрылые 465
Кролик 90, 148, 222, 225. 243, 266,
301, 331, 350, 351, 421, 449, 451
Крот 145
Круглые черви 210, 339, 452
Круглые черви собственно 346
Крупный рогатый скот 43, 267, 323,
331, 397, 427
Крыса 20, 42, 90, 115, 306, 307, 308,
309, 326, 348, 350, 351, 354, 359, 393,
394, 440, 443, 462
Крыса александрийская 359
Крыса белая 43, 86
Крыса черная 359
Куница 251, 351
Куры 160
Л
Ламблии 24, 139
Лейшманий 65, 107
Лейшманий песчанки 122
Лейшманий человека 122
Лейшмания кожная 42
Лентец 275, 448, 470
Лентец байкальский 6
Лентец малый 288, 458
Лентец тунгусский 292, 294
Лентец узкий 291
Лентец широкий 14, 24, 275, 280, 281,
287, 289, 292, 334, 444, 448, 450, 458,
467
Ленточные черви 34, 211, 447
Леопард 295, 381, 419
Летучие мыши 15
Лещ 236, 245
Линь 236, 245, 247
Лисица 243, 253, 275, 295, 362, 381
Личинки анкилостомы 464
Лосось 284
Лошадь 13, 222, 267, 327, 331, 363, 414
Лягушка 282, 283, 295, 296
М
Макака 90, 220, 301
Маклюра 124
Макрель 146
Медведи 331, 351
Медведь белый 275
Мидии 451
Микроспоридии 141, 200
Микроспоридии полостные 200
Микроспоридии тканевые 200
Микрофилярии 27, 465
Млекопитающие 150, 218, 395
Млекопитающие травоядные 221
Многоглавы 327
Многоустка 212
Моллюски 145, 218, 231, 260, 266, 267,
458, 464
Моллюски головоногие 142
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
485
Моль яблоневая 28
Морж 275, 295
Морские свинки 90, 112, 243, 348, 351,
408
Москиты 15, ИЗ
Мошки 427, 460
Мул 267
Муравьи 88
Муха 48, 88, 374, 478, 480
Муха вольфартова 24, 38
Муха-жигалка 119
Муха иловая 12
Муха комнатная 12, ИЗ, 119, 466
Муха падальная 12
Муха серая мясная 12
Муха сырная 12
Муха тсе-тсе 20, 106, 127
Мыши 31, 90, 203, 257, 295, 306. 308,
326, 331, 350, 351, 359, 440, 443
Мышь белая 112, ИЗ, 118, 233, 264,
360
Н
Наездник 28
Налим 283, 284, 285, 458
Насекомое 415
Некатор 379, 393, 394
Некатор свиной 382
Нематоды 339, 340, 341, 342, 344, 346,
429
Нериус 284
Нитчатка Банкрофта 426
Нитчатка Банкрофта, личинки 465
Нитчатки 422, 465
О
Обезьяна 90, 103, 115, 120, 301, 355;
361, 369, 375, 376, 420, 435
Обоюдоустковые 220
Овода 15, 329
Овод желудочный 15, 24, 419
Овод кожный 24, 40
Овод носовой 329
Овод овечий 329
Овца 203, 222, 225, 226, 228, 238, 243,
267, 299, 323, 326, 327, 329, 331, 334,
338, 395, 397, 414, 452, 462
Огневка мельничная 309
Окунь 283, 284, 285, 458
Окуневые (сем.) 251
Олени 326, 331
Омуль 288, 289, 290, 292, 458
Оранг-утанг 301, 375
Ослы 331
Острица детская 400
Острицы 24, 27, 241, 340, 342, 357, 398,
399, 403, 457, 461, 466, 473, 476
Осы 466
Офриосколециды 70
П
Павианы-гамадрилы 90
Пасюк 359
Пекари 359, 361, 439
Песец 243
Пескарь 459
Песчанка 6, 42, 49, 122, 329, 330
Пиявка египетская 441, 442
Пиявка медицинская 441
Пиявка сухопутная 441 ~~
Пиявки 15, 24, 38, 106, 125, 210, 440,
444
Пиявки челюстные 440
Пиявки хоботные 440
Плоские черви 210
Плотва 243
Плотоядные 310, 458
Позвоночные 145, 218, 438
Полуобезьяны 355
Пресмыкающиеся 200, 202
Простейшие 63
Простейшие голые 71
Простейшие паразитические 444
Прудовик болотный 223
Прудовик обыкновенный 223
Прудовик малый 223, 224
Прудовики 30
Прусак 309
Птицы 15, 201, 253, 267, 438
Птицы дикие 267
Птицы домашние 267, 295, 376
Пума 377
Пухоеды 15
Пятиустка 25
Р
Равноресничные 204
Разноресничные 204
Раки 260
Рачки веслоногие 36, 280
Рачки паразитические 36
Ремнецы 298
Ресничные 69
Риккетсии 25
Ришта 342, 429, 430, 432, 434, 435, 436,
460
Ришта личинка 432, 433
Рогатый скот 243
Россомаха 243
Рунец 106
Рыбы 125, 200, 218, 220, 231, 236, 243,
245, 253, 281, 284, 295, 298, 438, 458
Рыбы карповые 200
Ряпушка 284
С
Салака 146
Сардинка 146
Саркоспоридии 141, 200, 201, 204
Саркоспоридии мышиные 203
Свайник великан 340, 343, 363
Свайник трахейный 376
Свинья 90, 203, 206, 208, 222, 229, 235,
238, 243, 250, 251, 267, 295, 312, 314,
319, 326, 331, 350, 351, 382, 393, 397,
398, 414, 415, 419, 452, 458, 481
Свинья дикая 361, 439
Сельдь 146
Сенокосцы 301
486
УК АЗ АТЕ Л Ь РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Сиг 201, 284, 288, 289, 292, 458
Скот 225, 325
Скребни 210, 438
Скребень великан 438, 439
Скребень четковидный 438, 439
Слепни 15, 34, 428, 465
Снеток 284
Собака 13, 19, 29, 31, 90, 111, 114, 115,
147, 235, 243, 249, 250, 251, 253, 257,
258, 267, 275, 294, 295, 296, 303, 304,
312, 326, 328, 329, 331, 334, 338, 339,
350, 351, 359, 362. 363, 370, 381, 382,
393, 394, 419, 421, 435, 452, 458, 460,
462
Соболь 243, 362
Солитер 447
Сосальщики 34, 210, 211, 225
Сосальщики переднеротые 219
Сосальщики эктопаразитические 219
Сосальщики эндопаразитические 211,
219
Спирохеты 443
Споровики 69, 141
Стронгилиды 27
Судак 284
Сурки 360
Суслики 112, 122
Т
Таракан американский 440
Таракан — черепашка египетская 467
Тараканы 88, 305, 463, 465, 467
Тараканы черные 309
Телоспоридии 141
Телята 299, 327
Тении 448, 466
Тигр 275, 295, 419
Травоядные 310
Трематоды кишечные 470, 473
Трипанозома 42
Трипанозома овечья 106
Трипанозома сонной болезни 125, 126
Трипанозомы 20, 65
Трипанозомы рыб 444
Трихина 15, 24, 347, 348
Трихины мышечные 446
Турбеллярии 210
Турбеллярии прямокишечные 212
Тюлени 253, 275, 295, 363
У
Угрица кишечная 365, 370
Угрица прыщевая 24
Угрица уксусная 374
Угрицы 371
Уж водяной 437
Усач 201
Усоногие 36
Утка 31, 234, 439
Утка кряковая 440
Утки дикие 27
Уховертки 309
Ф
Филярия 27
Финка 446, 447
Форель 255, 283
«X
Хамелеон 120
Хариус 284, 288, 289, 292, 458
Хищные 381
Хлыстовик 355
Хомяки 90, 109, 115, 326, 359, 360,
385, 440
Хомяк китайский 113
Хомячок 86, 111
Хрущак мучной 309, 463
ц
Цепень 470
Цепень бычий 320, 47<>
Цепень вооруженный 13, 24, 310, 311,
313, 317, 467
Цепень карликовый 305, 306, 307, 308
Цепень крысиный 308, 310, 463
Цепень мозговик 327, 328
Цепень невооруженный 6, 24, 35, 47,
317, 320, 321, 324, 457
Цепень огуречный 13, 44,. 302
Цепень свиной 479
Цепень тыквовидный 14, 301, 302, 303,
462
Цепень эхинококка 330
Циклопы 36, 280, 289, 290, 291, 295,
419, 432, 433, 434, 435, 458
Цистицерк тонкошейный 326, 327
Ч
Чебак 245, 248, 458
Человек 13, 19, 109, 125, 146, 150, 159,
172, 173, 201, 203, 204, 210, 219, 222,
225, 228, 229, 231, 233, 238, 243, 251,
253, 257, 266, 275, 278, 285, 294, 295,
296, 297, 299, 301, 303, 304, 306, 308,
310, 313, 323, 324, 326, 327, 328, 329,
330, 331, 332, 336, 340, 348, 350, 351,
355, 359, 360, 362, 363, 364, 370, 371,
375, 376, 377, 381, 386, 394, 395, 396,
398, 414, 417, 418, 419, 420, 424, 425,
435, 436, 440, 444, 445, 458, 460, 462,
464, 469
Черви 145, 210
Черви паразитические 210
Червь узелковый 375
Черепаха 43
Черепаха сухопутная 282
Чернотелка 309
Членистоногие 145
Ш
Шакалы 303
Шершень восточный 468
Шимпанзе 357
Шистозомы 464
щ
Щука 236, 281, 282, 283, 284, 285, 458
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ 487
Эхинококк 24, 39/ 310, 326, 331, 334,
446, 462, 463, 464, 481
Эхинококк мног г гнездный 336
Эхинококк многокамерный 336
Эхмнококк одногнездный 331
Эхинококк собак 464
Эхинококка пузыри 447
Я
Язь 236, 245, 364, 458
Ящерица 282
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
А
Abramis brama"245, 459
Abreviata 420
Abreviata caucasica 420
Acanthobotrium ijimai 270
Acanthocephala 210, 438
Acanthocephalus 439
Acanthocheilonema 428
Acanthocheilonema perstans 424, 428,
429, 466
Acanthocyclops bicuspidatus 295, 432
Acanthocyclops viridis 295, 432
Acephalocyste 332
Acerina cernua 283, 284
Adeleidae 143, 144
Aedes variegatus 424
Agamodistomum 233
Ageniaspis fuscicollis 28
Aggregata 142
Agriolimax agrestis 208
Akis spinosa 309
Amidostomum anseris 27
Amoeba buccalis 24
Amoebina 71
Amphilina foliacea 269
Amphistomata 220
Amphistomum hominis 220
Amphistomum watsoni 220
Anabatidae 251
Anas 267
Anguillula aceti 374
Anguillula intestinalis 365
Anguillula mucronata 371
Anguillula putrefaciens 375
Anguillula stercoralis 365
Anguillulinoidea 347, 377
Ancylostoma 377
Ancylostoma americanum 382
Ancylostoma braziliensis 31, 32, 381,
387
Ancylostoma caninum 29, 30, 381, 387
Ancylostoma ceylonicum 381
Ancylostoma duodenale 13, 16, 23, 26,
340, 377, 378, 379, 381, 382, 391, 392
Ancylostomatidae 376, 377
Annelida 440
Anopheles 150, 170, 171, 172, 173, 174
Anopheles algeriensis 178, 180, 424
Anopheles bifurcatus 176, 178, 180
Anopheles costalis 424
Anopheles elutus = Anopheles maculi-
pennis, var. sacharovi 174, 177, 178
Anopheles hyrcanus 176, 178, 179, 180,
182, 465
Anopheles listoni 176 1
Anopheles maculipennis 44, <13, 158
173, 174, 176, 177, 178, 180, 182
Anopheles maculipennis atroparvus 176
Anopheles maculipennis messeae 181
Anopheles nigripes (= An. plumbeus)
176, 178, 180
Anopheles pulcherrimus 178, 180
Anopheles quadrimaculatus 176
Anopheles rossi 424
Anopheles superpictus 173, 174, 176,
177, 178, 180
Anoplocephalidae 298
Antilope darcas 395
Aphaniptera 15
Aphasmidia 347
Aphodius fimentarius 415
Archigetes 268
Argas persicus 15
Argasidae 15
Artifechinostomum 231
Ascaridia perspicillum 32
Ascaris 403
Ascaris lumbricoides 403, 404, 4(4, 454
Ascaris megalocephala 453
Ascaris suilla 413
Ascaroidea 347, 403
Asopia farinalis 309, 463
Astacus japonicus 260
Astacus similis 260
Ateles geoffroyi 359
В
Bacterium ascaridianum 409, 410
Bacterium mesentericus 79
Balantidium coli 65, 92, 204, 206
Balantidum minutum 208
Balbianea 204
Barbus fluviatilis 201
Belascaris 414
Belascaris cati 414
Belascaris mystax 414
Bertiella 301
Bertiella satyri 301
Bilharzia 262
Bilharzia haematobia 264
Bilharziella polonica 27
Bithynia 247
Bithynium leachii 243, 244
Biwia zezera 251
Bythinia fuchsiana 250
Bythinia longicornis 250
Blanfordia formosana 267
Blanfordia nosophora 254, 267, 464
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
489»
Blaps mucronata 440
Blastocystis 137
Blatella germanica 309
Blicca bjorkna 236, 245
Bodo 130, 136
Bodo caudatus 130
Bodo edax 130
Bodo urinarius 130
Bodonidae 129
Boophilus decoloratus 13
Bothriocephalus cordatus 295
Bothriocephalus latus 278
Braunia 298
Braunia jassyensis 298
Bucephalus pusillus 219
Bulinus contortus 55, 265, 464
Bulinus dybowskii 265
Bulinus innesi 265
C
Caccobius schreiberi 415
Camallanus sweeti 433
Capillaria aerophila 361
Capillaria anatis 27
Capillariidae 36
Carassius auratus 251
Catenaria anguillulae 223
Cebus capucinus 359
Ceratophyllus fasciatus 309
Cercaria ocellata 267, 268
Cercaria pleurolophocerca 212
Cercaria vitrina 238
Cercomonadidae 132
Cercomonas 131, 136
Cercomonas hominis 132
Cercomonas longicauda 65, 132
Cestoda 268, 269, 277
Cestodaria 268, 269
Cestoidea 211, 268
Cetonia 439
Chameleo vulgaris 121
Chariesterus cuspidatus 120
Chilodon uncinatus 208
Chilomastiginae 131
Chilomastix 131
Chilomastix bettencoarti 32
Chilomastix mesnili 92, 131, 132
Chrysops coecutiens 437
Chrysops dimidiatus 428
Chrysops silacea 428, 465
Ciclocoelium orientale 212
Ciliata 204
Cimex rotundus 113
Cimicidae 15
Cirrhosis distomatosa 225
Cittotaenia ctenoides 301
Cladorchis watsoni 220
Clarias batrachus 419
Clonorchis 250
Clonorchis endemicus 250
Clonorchis sinensis 31, 53, 250
Clupea 243
Cnemidophorus lemniscatus 120
Cnidosporidia 141
Coccidia 141, 142
Coenurus 327
Coenurus cerebralis 327/328
Coenurus glomeratus 329
Colpoda cucullus 208
Colpoda steini 67, 208
Conorhinus 106
Conorhinus megistus 129
Conorhinus rubrofasciatus 113
Copepoda 280
Copromonas subtilis 69
Coregonus albula 284
Coregonus lavaretus 284
Coregonus lavaretus baikalensis 288:
Coregonus lavaretus pikschian 288
Coregonus migratorius 288, 289
Coregonus ussuriensis 255, 289
Cottus gobio 284
Cottus scorpius 212
Cotylurus flabelliformis 30
Councilmania lafleri 97, 98
Craigia hominus 103
Cricetulus griseus 112
Cricetulus migratorius 86, 360
Crithidia 105, 106
Ctenocephalus canis 303
Culex annulirostris 424
Culex fatigans 424, 465
Culex pipiens 424
Culicidae 15
Culicoides 429
Culicoides austeni 428, 429, 466-
Culicoides nubeculosus 427
Cyclophyllidea 273, 277, 298
Cyclops 289
Cyclops coronatus 432
Cyclops leuckarti 295, 296
Cyclops oithonoides 432
Cyclops quadricornis 432
Cyclops strenuus 36, 280, 432
Cyclops uljanini 432
Cyclops vicinus 295, 432
Cyprinus caprio 255
Cysticercus 273
Cysticercus cellulosae 451
Cysticercus fasciolaris 326
Cysticercus pisiformis 451, 452
Cysticercus ramicosus 314
Cysticercus tenuicollis 326, 452
D
Dasipus 129
Davainea 305
Davainea asiatica 305
Davainea madagascariensis 305
Davaineidae 305
Demodex folliculorum 16
Dermacentor nuttalli 5
Dermacentor silvarum 35
DiaptOmus graciloides 280
Dibothriocephalus 448
Dibothriocephalus cordatus 295
Dibothriocephalus latus 270, 278
Dibothriocephalus mansoni 295
Dibothriocephalus strictus 291
Dicotyles torquatus 439
Dicrocoeliidae 237
490
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Dicrocoelium dendriticum 240
Dicrocoelium lanceatum 223, 237, 239,
241, 242, 243, 476
Dicrocoelium pancreaticum 242
Didelphis 129
Dientamoeba 102
Dientamoeba fragilis 67, 92, 102, 103
Digenea 211, 214, 219
Digenea simplex 411
Dimastigamoeba gruberi 103
Dioctophyme 363, 364
Dioctophyme renale 25, 363
Dioctophymoidea 347, 363
Dioicocoestus 271
Diphyllobothriidae 277
Diphyllobothrium 273, 277, 278, 297,
447
Di phyllobothrium cordatum 295
Diphyllobothrium giljacicum 294
Diphyllobotrium latum 14, 278, 279,
295, 317, 447
Di phyllobothrium luxi 295
Di phyllobothrium mansoni 295, 296,
297
Diphyllobothrium minor 55
Diphyllobothrium minus 288, 289, 291,
458
Di phyllobothrium mobilis 275
Di phyllobothrium nenzi 294, 295
Di phyllobothrium skrjabini 293, 294
Diphyllobothrium strictum 55, 288, 291,
293
Di phyllobothrium tungussicum 292, 294
Diplodinium 64
Diplogonoporus 277, 297
Diplogonoporus brauni 298
Diplogonoporus grandis 297
Diplomonadida 104, 139
Dipylidiidae 301
Dipylidium 301
Dipylidium caninum 13, 14, 44, 273,
301, 302, 303, 304, 310, 448, 462
Dipylidium cucumerinum 301
Distomata 221
Distomum 212
Distomum crassum 228
Distomum giganticum 228
Distomum hepaticum 221
Distomum lanceatum 237
Distomum ophthalmobium 225
Distomum pancreaticum 242
Distomum rathouisi 228
Distomum ringeri 258
Dracunculoidea 347, 429
Dracunculus 429
Dracunculus medinensis 429, 430, 431,
432, 434, 437
Dracunculus oesophagens 437
Dracunculus persarum 460
E
Echinochasmus 231, 235
Echinochasmus perfoliatus 235
Echinococcus 330
Echinococcus altricipariens 332
Echinococcus alveolaris 331
Echinococcus cysticus sterilis 332
Echinococcus cysticus fertilis 332
Echinococcus endogenes 332
Echinococcus granulosus 330, 331, 332,
334
Echinococcus hydatidosus 332, 333
Echinococcus hydatidosus exogenes 332
Echinococcus multilocularis 331, 333,
336
Echinococcus petrificatus 335
Echinococcus polymorphus 330, 333
Echinococcus scolicipariens 332
Echinococcus simplex 332
Echinococcus unilocularis 331
Echinoparyphium 231
Echinostoma 231, 232
Echinostoma ilocanum 232, 233
Echinostoma jassyense 233
Echinostoma malayanum 233
Echinostoma paraulum 233, 234
Echinostoma revolutum 27, 232
Echinostoma superartyfex 233
Echinostomatidae 231
Echinostomidae 231
Eimeria 145
Eimeria clupearum 145, 146
Eimeria oxyspora Д45, 146
Eimeria sardinae 145, 146
Eimeria snijdersi 145, 146
Eimeria stiedae 148
Eimeria wenyoni 145, 146
Eimeridae 145, 150
Eimeriida 143, 144, 145
Embadomonas 130, 131
Embadomonas intestinalis 131
Endolimax nana 76, 93, 98, 99, 101
Endolimax williamsi 28
Entamoeba 72
Entamoeba africana 80
Entamoeba buccalis 100
Entamoeba butschlii 99, 102
Entamoeba coli 66, 70, 72, 73, 74, 76,
92, 95, 96, 97, 98, 99, 101
Entamoeba dispar 73, 79, 87, 94, 95
Entamoeba dysenteriae 72
Entamoeba gingivalis 100
Entamoeba hartmanni 73, 76, 92, 94,
95
Entamoeba histolytica 72, 73, 74, 75,
76, 77, 78, 79, 80, 81, 87, 90, 91, 92,
94, 95, 96, 99, 101, 141
Entamoeba histolytica forma magna 72,
76, 81, 97
Entamoeba histolytica forma minuta
72, 75, 76, 77, 81
Entamoeba minutissima 79
Entamoeba muris 32, 90
Entamoeba nana 98
Entamoeba tetragena 72, 80
Enterobius 344, 398
Enterobius vermicularis 398, 39,9
Enteromonas hominis 132
Enteromonas intestinalis 132
Eriocheir japonicus 260, 261
Eriocheir sinensis 260
Eristalis tenax 12
Esox lucius 283, 284
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ ЖИВОТНЫХ
491
Eucyclops serratulus 295, 432
Euomphalia strigella 238
Euparyphium 331
Euphorbia 120
Eurytrema 242
Eurytrema pancreaticum 242
Eustrongylus gig as 3G3
Eustrongylus visceralis 363
F
Fasciola 221
Fasciola dendritica 237
Fasciola gigantica 228
Fasciola hepatica 211, 214, 221, 476
Fasciolidae 221
Fasciolopsis 228
Fasciolopsis buski 228, 230, 231
Fasciolopsis fiilleborni 228
Fasciolopsis goddardi 228
Fasciolopsis spinifera 228
Filaria conjunctivae 437
Filaria conjunctivae orientalis 437
Filaria (Loa) extraocularis 437
Filaria inermis 437
Filaria loa 428
Filaria medinensis 429
Filaria sp. 438
Filaria volvulus 427
Filarioidea 347, 422
Filicollis anatis 439, 440
Fimbriaria fasciolaris 27
Flagellata 103, 136
Funga 15
G
Galumna 301
Gasterostomata 219
Gasterostomum 215
Gastrodiscidae 220
Gastrodiscoides 220
Gastrodiscoides hominis 220, 221
Gastrodiscus hominis 220
Gastrophilus 16, 24, 37, 419
Gerbillus erythrourus 330
Gerbillus meridianus 330
Gerbillus tamaricinus 330
Giardia 139, 140, 141
Giardia intestinalis 139, 140
Glossina palpalis 126, 127, 128
Gnathobdellidae 440
Gnathopogon elongatus 236
Gnathostoma 418
Gnathostoma spinigerum 418, 419
Gnathostomidae 418
Gnathostomum hispidum 419
Gobio gobio 255, 459
Gongylonema 23, 24, 414, 417
Gongylonema, личинка 417
Gongylonema neoplasticum 417
Gongylonema pulchrum 414, 415, 416
Gordiacea 210, 346
Gregarinida 141
Guraulus prachadi 233
Gymnodactylus caspius 119
H
Haemadipsa ceylonica 441, 442
Haemadipsa japonica taiwana 442
Haemaphysalis concina 5
Haementaria officinalis 442
Haemonchus 396
Haemonchus contortus 395, 396
Haemopis paludinum 442
Haemopis sanguisuga 442
Haemosporidia 141, 148
Haepaticola 359
Haepaticola hepatica 359, 360, 361
Hartmanella hyalina 103
Helicella candidula 239, 240
Helicella ericetorum 238
Helicella unifasciata 240
Helminthes 210
Helmintozoa 210
Hemibarbus labeo 255, 256, 459
Hemiclepsis marginata 444
Hemibia hupensis 267
Hennegyia zschokkei 201
Herpetomonas 106
Herpetomonas ctenocephali 112
Heterodera radicicola 374, 375
Heterophyes 253
Heterophyes heterophyes 252, 253, 254
Heterophyes katsuradai 253
Heterophyidae 253, 257
Heterotricha 204
Hexamita 139
Himasthla 231, 236
Himasthla miihlensi 236, 237
Hirudinea 210, 440
Hirudo aegyptica 441
Hirudo medicinalis 441, 442
Hirudo nipponica 441, 442
Hirudo troctina 442
Holomyaria 340, 341, 355
Holotricha 204
Homo sapiens 53
Hymenolepididae 305, 467
Hymenolepis 305
Hymenolepis anatina 27
Hymenolepis carenula 27
Hymenolepis collaris 27
Hymenolepis compressa 27
Hymenolepis diminuta 309, 310, 450,
462
Hymenolepis fraterna 43, 306
Hymenolepis macrocanthus 270
Hymenolepis murina 306, 307
Hymenolepis nana 43, 305, 306, 307,
308
Hypoderma 16, 24, 39, 40
Hypoderma bovis 43
Hypobosca canina 119
1
Ichthyobdellidae 444
Idus melanotus 245
Isospora 146
Isospora belli 145, 146
Isospora bigemina 147
Isospora felis 143, 147
Isospora hominis 139, 146, 147, 148
Isospora rivolta 147
Ixodes persulcatus 55
Ixodes ricinus 13
Ixodidae 15
492
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
J
Jodamoeba biitschli 92, 99
Jodamoeba schewiakowi 102
К
Karyamoebina falcata 75
Karyolysi lacertae 142
Lacerta muralis 142
Lamblia 139, 140
Lamblia intestinalis 92
Larvivoridae 15
Laverania 155
Leishmania 105, 106, 107
Leishmania brasiliensis 124
Leishmania canis 111, 112, 119
Leishmania chameleonis 124
Leishmania donovani 107, 108, 111, 112,
113, 115, 116, 119, 122, 124
Leishmania pulicis 112
Leishmania tropica 107, 116, 117, 119,
120, 121, 122, 124
Leptomonas 105, 106, 116, 119, 120
Leptomonas ctenocephali 112
Leptomonas davidi 121
Leptomonas pulicis 113
Leptotrix 102
Leuciscus leuciscus baicalensis 246, 248,
458
Leuciscus waleckii 255
Leucogobio guntheri 251
Leucogobio mayedae 251
Ligula 298
Limnatis granulosa 441, 442
Limnatis mesomelas 442
Limnatis nilotica 441
Limnatis turkestanica 441, 442
Linguatula rinaria 16, 25
Liocassis braschnikowi 255. 256, 459
Liocassis ussuriensis 459
Liponyssus saurarum 142
Loa extraocularis 466
Loa loa 428, 465
Lota vulgaris 283, 284
Lucilia caesar 12
Lucioperca lucioperca 284
Lumbricus terrestris 398
Limnaea 225
Lymnaea terebra euphratica 267
Limnaeus minutus 223, 224
Limnaeus palustris 223, 224, 267
Limnaeus stagnalis 223, 224, 268
Limnaeus stagnalis appressa 30
Limnaeus stagnalis perapla 30
Limnaeus truncatula 224, 225
M
Macacus nemestrinus 90
Macacus philippensis 90
Macacus rhesus 139
Macracanthorynchus hirudinaceus 438,
439
Macrocyclops albidus 295
Macrocyclops fuscus 295, 432, 437
Mallophaga 15
Mastigophora 103
Mecistocirrus 397
Mecistocirrus apri 397
Mecistocirrus digitalis 397
Melania 260
Melania amurensis 254
Melania ebenina 254
Melania libertina 254, 260
Melania multicincta 260
Melania nodiperda 260
Melolontha vulgaris 439
Melophagus ovinus 106
Mermithoidea 3£7
Meromyaria 340, 341
Mesocyclops hyalinus 432
Mesocyclops leuckarti 295, 432
Mesocyclops oithnoides 295
Metagonimus 458, 459, 473
Metagonimus katsuradai 253, 257
Metagonimus minutus 257
Metagonimus romanicus romanicus 257
Metagonimus yokagawai 253, 254, 255,
257, 258
Metastrongyloidea 397
Metastrongylus 398
Metastrongylus apri 397, 398
Metazoa 210
Metorchis truncatus 252
Metorchis xantosomus 27
Microfilaria diurna 422, 428
Microfilaria nocturna 422, 423, 426, 428
Milvus parasiticus 253
Monas 136
Moniezia 13, 299, 301
Moniezia benedeni 299
Moniezia expansa 299, 300
Moniezia lenticulata 299
Monogenea 213, 219
Monostomum lentis 225
Mugil cephalus 253
Multiceps 327
Multiceps glomeratus 329, 330
Multiceps multiceps 327, 328
Multiceps serialis 12
Musca domestica 12
Musca vicina 468
Mytilis 451
Myxobolus 201
Myxobolus mulleri 201
Myxobolus pfeifferi 200
Myxosporidia 200
N
Naegleria gruberi 103
Nanophyetes salmincola 258, 277, 287
Nanophy etes schikhobalowi 257, 258,
473
Necator 27, 382
Necator amerancusi 340
Necator americanus 378, 379, 382, 391,
392
Necator kuillus 382
Nemastoma luguber 301
Nematodes 210, 339, 346, 347
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
493
Neosporidia 200
Nucleophaga 28
Nyctotherus faba 208
О
Oesophagostomum 375, 376
Oesophagostomum apiostomum 375
Oesophagostomum brumpti 375
Oestrus 16
Onchocerca 427
Onchocerca cervicalis 427
Onchocerca coecutiens 427
Onchocerca gutturosa 427
Onchocerca volvulus 427, 466
Onthophagus taurus 415, 417
Opecoelus sphericus 212
Opjiicephalus striatum 419
Opistorchidae 243
Opistorchis 243
Opistorchis felineus 243, 244, 245, 246,
247, 249,’ 250, 458
Opisthorchis noverca 250
Oribatidae 301
Ornithodorus 23, 48, 49, 55
Ornithodorus papillipes 15
Ornithodorus savignyi 113
Orthonoides 432
Oscillaria malariae 170
Osmerus eperlanus 284
Ovis aries 395
Oxyuridae 347
Oxyuris equi 399
Oxyuris incognita 375
Oxyuris vermicularis 398
Oxyuroidea 398
P
Parabramis pekinesis 255, 256, 459
Parafossarulus striatulus 236, 250
Paragonimus 27, 258
Paragonimus ringeri 258
Paragonimus westermani 258, 259, 262
Paramoeba hominis 103
Paramphistomatidae 220
Paratelphusa sinensis 260
Pectinoctenus pavlovskyi 34 '
Pediculus capitis 13
Pentatrichomonas ardinenteili 134
Perea fluviatilis 283
Periplaneta 305
Periplaneta americana 309
Periplaneta orientalis 309
Phasmidia 347, 365
Phlebothomus 15, 112, 122
Phlebothomus argentipes 113, 122
Phlebothomus caucasicus 120
Phlebothomus chinensis 113, 114
Phlebothomus intermedius 124
Phlebothomus major var. perniciosus
114, 120
Phlebothomus papatasii 6, 113, 119,
120, 121, 122
Phlebothomus sergenti 113, 114, 120,
121, 122
Phlebothomus sergenti var. mongolen-
sis 114
Physa fontinalis 225
Physaloptera caucasica 420
Physaloptera mordens 420
Physalopteridae 420
Physopsis africana 266
Phytomonas davidi 121
Placobdella catenigera 442
Plagiorchidae 231
Plagiorchis 231
Plagiorchis muris 231
Planorbis boissyi 266, 464
Planorbis coenosus 229
Planorbis metidje‘nsis var. dufouri 265
Planorbis olivaceus 266
Planorbis planorbis 225
Planorbis rotundatus 267
Plasmodiidae 148
Plasmodium elongatum 160
Plasmodium falciparum 148, 151, 154,
155, 156, 157, 159, 160, 162, 165,
173, 174, 175, 176, 178
Plasmodium gallinaceum 160, 161
Plasmodium immaculatum 155
Plasmodium malariae 148, 151, 153,
154, 155, 156, 157, 158, 159, 162,
174, 176, 178
Plasmodium ovale 44, 151, 157, 158,
173
Plasmodium relictum 160
Plasmodium vivax 44, 148, 149, 151,
153, 154, 155, 156, 157, 158, 162,
163, 165, 171, 173, 174, 175, 176,
178, 189
Platodes 210
Platybunus triangularis 301
Plecoglosus altivelis 255, 256
Polistes gallicus 466
Polymorphus minutus 27
Polymyaria 340, 341
Polyphaga saussurei 467
Polystomum 212
Polystomum integerrimum 43
Potamon dehaani 260, 261
Potamon obtusipes 260
Prosorhynchus squamatus 212
Prosostomata 219
Prostogonimus 27
Proteosoma praecox 148
Protomonadida 104
Protozoa 63
Prowazeckia 130
Prowazeckia asiatica 130
Prowazeckia cruzi 130
Prowazeckia italica 130
Prowazeckia ninae kohl-yakimovi 130
Prowazeckia urinaria 130
Pseudamphystomum 252
Pseudamphistomum truncatum 252
Pseudaspius leptocephalus 255
Pseudobagrus fulvidraco 459
Pseudolimax wenyoni 99
Pseudoperilampus lupus 251
Pseudophyllidea 273, 274, 277
Pseudorasbora parva 236, 251
Pseudotelphusa iturbei 260
Psilotrema spiculigerum 31
494
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Pulex irritans 303
Pygosteus pyngitius 284
В
Raillietina asiatica 305
Reduviidae 128
Rhabditis 371
Rhabditis aceti 374
Rhabditis donbass 372
Rhabditis genitalis 371
Rhabditis gracilis 372
Rhabditis hominis 372, 373, 374
Rhabditis nellyi 372
Rhabditis pellio 371
Rhabditis schachtiella 372
Rhabditoidea 347, 365
Rhabdocoela 212
Rhinoestrus 16
Rhinoestrus purpureus 329
Rhipicephalus sanguineus 5, 114
Rhizopoda 71
Rhombomys opimus 6, 50, 122,
329, 330
Rhynchobdellidae 440
Rutilus rutilus 45
Rutilus rutilus lacustiris 246, 248
S
Sacculina 36
Sacocheilychthis variegatus 251
Salmo umbla 284
Sanguinicola 215
Sappinia diploidea 103
Sarcocystis 202
Sarcocystis mischeriana 203
Sarcocystis muris 203
Sarcocystis tenella 203
Sarcophaga carnaria 12
Sarcopsylla penetrans 16
Sarcoptes scabiei 15
Sarcosporidia 201
Scardinius erythrophthalmus 244,
245
Scaurus striatus 309
Schistogonimus varies 27
Schistosoma 23, 262
Schistosoma haematobium 4, 262, 263,
264, 265, 267
Schistosoma japonicum 25, 31, 266,
267, 268
Schistosoma mansoni 24, 27, 266, 267
Schistosoma turkestanicum 267
Schistosomatidae 212, 262
Schistosomum 344
Schistosomum carneus 212
Schizotrypanum cruzi 128
Sclerostomum 27
Segmentina nitidella 229
Sagmentina schmakeri 229
Sesarma dehaani 260, 261
Simulium damnosum 427, 466
Simulium ornatum 427
Sparganum 296
Sparganum mansoni 295
Sparganum proliferum 297
Spermophilopsis leptodactylus 122
Spiroptera megastoma 346
Spiruroidea 414
Sporozoa 141
Stagnicola emarginata angulata 231
Stellanchasmus formosanus 257
Stenocephalus agilis 121
Strigea gracilis 27
Strongyloides 347, 365, 371, 375, 408
Strongyloides intestinalis 368
Strongyloides simile 369
Strongyloides stercoralis 364, 365, 370r
373, 374, 391, 392
Strongylus elongatus 398
Strongylus gigas 363
Strongylus subtilis 395
Syngamus 344, 376, 377
Syngamus dispar 377
Syngamus kingi 377
Syngamus laryngaeus 376, 377
Syngamus trachea 376
Syphacia 403
Syphacia abvelata 408
T
Tabanidae 15
Taenia 310
Taenia asiatica 305
Taenia coenurus 327
Taenia confusa 322, 326
Taenia crassicollis 326
Taenia cucumerina 301
Taenia diminuta 309
Taenia echinococcus 330, 336
Taenia expansa 448
Taenia fenestrata 322
Taenia flavopunctata 309
Taenia glomerata 329
Taenia hybrida 322
Taenia hydatigena 277, 326, 338
Taenia inermis 449
Taenia infantis 326
Taenia marginata 277
Taenia mediocanellata 320
Taenia multiceps 327
Taenia multilocularis 336
Taenia perfoliata 27, 450
Taenia pisiformis 277, 338
Taenia plicata 450
Taenia saginata 317, 320, 448, 449, 457
Taenia serialis 277
Taenia serrata 277
Taenia solium 13, 275, 310, 311, 312,
313, 317, 320, 322, 323, 324, 325,
331, 447, 448, 457
Taeniarhynchus africanus 424
Taeniarhynchus mediocanellata 320
Taeniarhynchus saginatus 47, 320, 321,
322, 323, 324, 325
Taeniidae 273, 310, 325, 467
Tarentola mauritanica 120
Tartarus stibiatus 252, 266
Telosporidia 141 *
Tenebrio molitor 309, 463
Ternidens 376
Ternidens deminutus 376
Tetrameres fissispinus 27
Tetrahpyllidae 277
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
495
Tetrarhynchus 270
Theba cartusiana 238, 239, 240
Theba fruticicola 239
Thelazia 421
Thelazia callipaeda 421
Thelaziidae 421
Thominx 361
Thominx aerophilus 361, 362
Thymallus arcticus 288, 289
Thymallus vulgaris 284
Tinea vulgaris 245
Toxascaris leonina 413
Toxascaris limbata 413
Toxascaris marginata 413
Toxocara 413
Toxocara canis 26, 340, 413, 414
Toxocara mystax 340, 414
Tracheophilus sisowi 27
Trachybdella bistriata 442
Trematodes 211
Trepomonas 139
Triaenophorus nodulosus 35
Triatoma 106, 129
Triatoma megista 128
Tricercomonas 132
Tricercomonas intestinalis 132
Trichina contorta 375
Trichina spiralis 347
Trichinella 347
Trichinella spiralis 15, 347
Trichinelloidea 347
Trichobilharzia ocellata 267
Trichocephalidae 355
Trichocephalus 355
Trichocephalus dispar 355, 362
Trichocephalus muris 359
Trichocephalus suis 356
Trichocephalus trichiurus 355, 356, 357,
359
Trichodectes 14
Trichodectes canis 303, 304
Trichodectes subrostratus 303
Trichomonas 31, 104, 133, 134, 135, 136
Trichomonas angusta 132
Trichomonas buccalis 136, 137, 139
Trichomonas caudata 136
Trichomonas elongata 136, 137
Trichomonas fecalis 134
Trichomonas flagelata 136
Trichomonas gingivalis 138
Trichomonas hominis 133, 134, 135,
136, 139, 141
Trichomonas intestinalis 131, 133, 138
Trichomonas vaginalis 25, 134, 136,
137, 138, 139
Trichostrongylidae 391
Trichostrongyloidea 347, 395
Trichostrongylus 395, 396
Trichostrongylus externatus 395
Trichostrongylus instablis 395
Trichostrongylus orientalis 395
Trichostrongylus probulurus 395
Trichostrongylus vitrinus 395
Trichotrochelidae 340
Trichuris trichiura 355
Trimastigamoeba philippensis 103
Troglotrema (Nanophyetes) salmi ncola
473
Troglotrematidae 258
Tropidonotus natrix persa 437
Tropocyclops prasinus 432
Trutta lacustris 283
Trutta vulgaris 284
Trypanorhyncha 277
Trypanosoma 105, 106, 124
Trypanosoma brucei 443
Trypanosoma cruzi 28, 40, 105, 129
Trypanosoma equiperdum 32, 46, 105,
125, 443
Trypanosoma gambiense 126, 127
Trypanosoma lewisi 126, 443
Trypanosoma melophagium 106, 126
Trypanosoma spermophili 71
Trypanosomidae 104, 105, 106
Turbatrix 374
Turbatrix aceti 374
Turbellaria 210
U
Uncinaria americana 382
Uncinaria stenocephala 387
Uronema caudatum 208
V
Venus metcenaria 237
Vermes 210
Vespa orientalis 466
W
Watsonius 220
Watsonius watsoni 220
Wuchercria 422
Wuchereria bacrofti 27, 422, 423, 424,
425, 428, 465
X
Xenopsylla cheopis 309
Z
Zebrina detrita 240
ИМЕННОЙ УКАЗАТЕЛЬ РУССКИЙ
Авакян 90, 93
Авербух 178
Адлер 120, 121
Алисов 353
Алфеева, С. П. 401, 455
Альперович 362
Ангер 305
Андреев, П. 443, 444
Андреева 382
Аринкин 108, 109
Артамонов 110
Артеменко 412
Ашоф 401
Б
Бадалян 307
Баданин 221, 243
Баженова 208
Баллиф 160
Банайтис 357
Басс 157
Бек 76, 86
Березняк 442
Блажин 242
Бланшар 357
Боровский 116
Бородин 323, 481
Боткин 317, 447
Брицкая 178
Бруг 94, 102
Брумпт 42
Будымко 178, 179
Буров 56, 57
Бурова 58, 88, 90, 92
Бутнинг 313
В
Василькова 404, 412, 414, 422
Вейнберг 449, 450, 454
Величкин 239
Венион 87, 94, 134
Вещезеров 176, 178, 179
Вигдорова 93
Виноградов 56, 58, 248, 253
Витенберг 258
Вишневская 412
Владыченский 437
Власов 50, 122
Власова 49.
Водонос 454
Воскресенский 90, 92, 112, 379, 384
Всеволодов 243
Г
Ганновер 354
Гартматтий 231
Гаршин 35
Гендельман 176, 178
Гительсон 123, 209
Глуховцев 140
Гнедина 292, 482
Гнездилов 26, 32, 43, 44, 45, 53, 54,
56, 66, 88, 89, 92, 93, 94, 95, 134,
206, 208, 281, 282, 283, 284, 285
Графф 58
Грубман 92
Гуцевич 178,^180, 182
д
Данилова 178, 179
Даниляк 482
Данскер 307,^404
Дедова 408
Дексбах 267
Демина 189
Джемс 44, 83, 189
Джонс 157
Джунковский 111
Динник Н. 356
Динник Ю. 356
Добелл 94>
Доброглав 76, 86
Догель В. А. 27, 58, 201, 206, 209
Донован 115
Дунаева 3, 450
Дюжарден 322
Е
Егоров 242
Еленевский 337
Ершов 299, 322
Ефремов 178
2К
Жинкин 330
Жук Н. 208
3
Забудько-Рейнгардт 177
Завадский 178, 400
Затуренская 412
Здродовский 92, 112, 379,« 384
Зеленский 355
Зельманов 189
Зерчанинов 245, 250
Зимин 148
Змеев 42, 56, 255, 370, 388, 466
ИМЕННОЙ УКАЗАТЕЛЬ РУССКИЙ
497
И
Иваницкий 415, 416
Иванов 58
Иофф 13, 121, 310
Ирен 209
Исаев 110, 433, 434, 4 36
Иеайчиков 257
К —
Каландруччио 440
Калантарян 227, 241, 388, 395
Камалов 388
Канделаки 86, 92, 110, 112, 167, 388
Капустин, В. 470, 472
Кассирский 56, 58, 109
Кеворков 111, 112, 227, 228
Кечек 265
Кешишьян 42
Киршенблат 301
Кларк 82, 83
Кливлэнд 78
Кобринец 475
Кожевников 117
Койдзуни 413
Койно 413
Колесников 360
Колодизнер 181
Корнилович 362
Коровицкий 412
Коростелев 93
Костылев 482
Кофоид 81, 83, 134
Кофоид-Фюллеборн 391
Кох 170
Краузе 27
Кривошта 239
Кришнан 111
Крылов 352
Крэг 83
Крюкова 123
Кулябко 249
Кушев 209
Кюхенмейстер 274
Л
Лаверан 148, 170, 451
Латышев 42, 117, 119, 122, 209
Лейкарт 26, 58, 225
Лейна метод 392
Лейтман 93, 148, 208
Леш 72
Лещенко 442
Линетов 58, 305
Лисова 173, 174, 177, 179
Лондон 456
Лус 111
М
Макнин 411
Маликов 242
Малыгин 299
Манасян 265
Мари 352
Марциновский 123, 187
Маршу 176, 352
МатевоСян 88, 92
Медведева 475
Меерович 314
Мельников 449
Менетриэ 420
Менилл 451
Миронова 249
Мордвилко 58
Н
Непир 111
Нессет 81
Никифоров 110
Николаев 174
Ноллер 94
Носиц 334
Носина 91
Нохт 209
О
Окиншевич 83
О’Коннор 87, 94, 136
Олейников 400
П
Павлова 74, 76, 95
Павловская 285
Павловский 26, 27, 32, 38, 39, 43,
44, 45, 49, 50, 58, 59, 181, 187, 209,
282, 283, 288, 322, 376, 426, 429,
450, 466, 482
Парцванидзе 178
Паско 233
Пастернацкий 443
Петрищева 56, 119, 122
Петров 108, 110, 111, 115, 209, 294,
362, 482
Петрушевский 285
Пигулевский 228
Пик 92, 93, 307
Пикуль 175
Плотников 243, 245, 248, 250
Подолян 93
Подъяпольская 230, 234, 249, 258,
287, 360, 361, 364, 404, 408, 409,
411, 470, 472, 475, 482
Поллер Л. 351
Померанцев 429
Поножина 138
Попов 267
Попова 287
Порецкая 402
Поспелова-Штром 441
Провачек 94
Пухов В. 239
Р
Рагоза, Н. 41, 286, 444, 447, 450, 482
Радзициловский 119
Раевская 394
Рафаэле 160
Рахманов 454
Рейнгардт 177, 179
Рейндорф 357
Ременникова 182
Ржаницын 206, 207
Розен 281
Росс 170
С
Сандерс 78
Саркисян 88, 93
498
х ИМЕННОЙ УКАЗАТЕЛЬ РУССКИЙ
Свиэи 134
Сегаль 123
Семенов 231
Сербинов 415
Сергиев 175, 190
Симанин 174, 178
Синицын 225
Скворцов 224, 225, 239, 288, 291, 292
Скрябин К. 27, 59, 230, 231, 234, 241,
243, 254, 258, 267, 287, 298, 299,
350, 360, 361, 362, *363, 364, 399,
482
Смирнов 382, 392, 408, 411, 416, 482
Соловьев 401
Сондак 356, 382, 400, 403, 466
Софиев 92, 112, 119
Статирова 230
Столла метод 392
Стюарт 405
Сченснович 92, 93
Сыркин 227, 228
Т
Талызин 243, 281, 288, 289,. 291, 293,
450
Тарасов 242, 279
Тареев 189
Теодор 120
Терехов 110
Тибурская 175
Топорков 178, 209
Тсучиа 140
Туаев 402
Тубангун 233
Ф
Фавр 209
Фауст 52, 251, 3-54, 368, 377, 417
Федоров 209
Федченко 433
Фиалковская 208
Филатов 124
Филипченко 92, 93
Финкельштейн 227
Фларер 123
Флеров 58
Фогель 245, 267
Форстер 404
Фюллеборн 367, 479
X
Хаустов 413
Хаютин 316
Хегнер 103
Хейсин 76, 92, 141
Хираиши 423
Ходукин 110, 111, 112, 113, 115, 119^
123, 174, 178, 209
Холодковский 288, 438, 482
Хохолькова 93
ц
Цатурян 88
Ч
Чапая 93
Чапурская 208
Чапурская-Баженова 92, 148
Чеботаревич 53, 54
Чука 160, 189
Ш
Шалимов 400
Шамберлен 454
Шардингер 103
Шарпу 64
Шаттон 433
Шаудинн 80, 155, 164, 208
Шахматов 209
Швабауэр 453, 454
Шевченко 119
Шиг 444
Широкогоров 227
Шихобалова 258, 287, 359, 360, 411
Шорт 113
Шоффард 451
Штакельберг 187, 209
Штейнберг 100, 136
Штром 35, 241, 258, 321, 322, 325,.
356, 361, 382, 473
Шуб 209
Шульман 402
Шульц 241, 243, 298, 299, 399, 4Г15,
415, 416, 482
Шу хат 316
Шют 44
щ
Щупак 209
Щупаков 350
Щуренкова 123
Э
Эпштейн 65, 73, 74, 75, 77, 81, 88„
90, 94, 96, 102, 103, 209
Эскин 158, 177, 179
Я
Якимов 110, 209
Яницкий 281
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А
Аборт 132
Авитаминоз 32
Агаметы 68
Arap-NNN 109, 116
Адаптации 34
Адолескарий 217
Адоральный диск 233
Акаринозы 45
Акрихин 123, 190, 192, 193
Акрихиновое опьянение 193
Акрихиновый психоз 193
Аксонемы 139
Аксостиль 104, 136
Актиномикоз 45
Алкалоид ипекакуаны 90
Аллергическая диагностика 324
Аллизатин 91
Алоэ 124
Амавроз 194
Амбарные вредители 463
Амбулаторный прием глистных боль-
ных 469
Амебиаз 77, 82, 90
Амилаза 36
Амфиды 343, 431
Анафилаксия 451, 455
Анафилаксия эхинококковая 451
Английская соль 471, 474
Анемия 450
Анемия злокачественная 447
Анемия пернициозная 450
Анизогамия 68
Анкилостоматики 387
Анкилостомидоз 6, 55, 389, 390
Анкилостомоз 382, 386, 389, 394, 473,
475
Аномалии 322
Апофелизм 6
Антикиназа 276
Антикоагулины 383
Антимозан 115, 116
Антиперистальтика 462
Антиферментные свойства 35
Антиформин 404
Антропургический очаг 51
Аппарат грушевидный 299, 300
Аппендицит 357, 401, 409
Аппендицитоподобные симптомы 401
Ареал распространения малярии 181
Аскаридная экстравазация 408
Аскаридный токсин 454, 455
Аскаридоз 359, 411, 455
Аскаридоток0йн 454, 455
Атаксия мозжечковая 194
Атебрин 123, 189
Атрофия зрительного нерва 194
Аутоинвазия 23, 308, 313, 320, 367
Аутореинвазия 400
Аутохтонная малярия 181
Ацефалоцисты 315
Ашхабадка 118
Б
Базальное зерно 64
Базальное тельце 104
Бактерии паратифа 352
Бактерии септицемии свиней 443-
Бактериозы 22, 45
Балантидиоз 208
Барьерная функция 445
Бациллоносительство 41
Бета-нафтол 231, 257
Бильгарциоз 264
Биологические виды 307, 3701
Биотоп госта льный 22, 28
Биотопы 22, 48
Биофильтрация 404
Биоценоз нор 49
Биоценотические связи 44
Бичи 64, 103, 170
Блефаропласт 104
Блуждание Fasciola hepatica 226-
Боенские отбросы 354
Боковые линии 340, 342
Боковые поля 340
Болезни инвазионные 22
Болезни инфекционные 22, 45
Болезнь дум-дум 107
Болезнь случная 105, 125
Болезнь Чагаса 25, 28, 40, 42, 105г
128
«Болотное худосочие» 182
Борьба с амебиазом 90
Борьба с аскаридозом 476
Борьба с гельминтозами массовая 476-
Борьба с гельминтозами плановая 467
Борьба с глистными инвазиями меди-
каментозная 469
Борьба с инвазиями 56
Борьба с малярией 187, 190
Борьба с москитами 115
Борьба с паразитами малярии 188
Борьба с тениидозами 479
Ботрии 269
Бригады хинизаторско-бонификацион-
ные 187, 188
300
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Брюква 375
Бульбус 398
Буравящий аппарат 215
Бурса 378
В
Вакуола гликогеновая 76, 77
Вариационно-статистическое изучение
яиц 413
Вегетативные особи 69
Венское питье 472, 474
Вертячка 328
Вертячка истинная 329
Вертячка ложная 329
Весенний подъем заболеваемости маля-
рией 165
Ветеринарно-боенский контроль 457
Ветеринарно-санитарный осмотр 353
Ветеринарный контроль 353
Вирулентность штаммов паразитов ма-
лярии 180
Вирус оспы кур 443 /
Вирус сыпного тифа 443
Вирус чумы свиней 443
Влагалище 214, 272, 344
«Вложения» 28
Внедрение личинки анкилостомы 384
Внедрение паразитов в тело 464
Внешняя среда 28, 456
Внутриутробная инвазия 23
Внутриутробное заражение эхинокок-
ком 335
Война Русско-турецкая 184
Волнообразная перепонка 104
Воронка 213
Воротниковый вырост 233
Воспаление слепой кишки 357
Вредители амбарные 310
Всесоюзный институт эксперименталь-
ной медицины 5
Вухерериоз 425
Выведение паразитов из кишечника 472
Выводковые капсулы 273, 333
Выделительная пора 211
Выделительные каналы 213
Выделительные стволы 271
Выздоровление спонтанное 164
Вылупление мирацидия 230
Вытяжная шпора 392
Г
Гальванокаутер 123
Гаметоцидные свойства 158, 188
Гаметоцит 150, 158, 164
Ганглии головные 210
Ганглиозные группы 343
Гастроваскулярная полость 210
Гексилрезорцин 411, 473
Гексахлорэтан 250
Гельминтозы 45
Гельминтологический полустационар
469
Гельминтологический пункт 467, 469
Гельминтология 6
Гельминтология медицинская 210
Гельминтотоксин 448
Гематоидин 83
Гематофаги 382
Гемолизин 448
Гемолимфы 172
Гемолитическое начало 396
Географическое распространение 52
Географическое распространение маля-
рии 180
Геофилия 387
Германии 128
Гермафродиты 210
Гермафродитизм протерандрический
Гетерогония 218, 345, 365
Гидатида 335
Гидатидный яд 452
Гидатиды внутренний паренхиматозный
слой 332
Гименолепидоз 472
Гиперинвазия 368
Гипердндемические очаги малярии 183
Гиподерма 339, 438
Гирудин 442
Гистиотропизм личинок 382
Глазок мирацидия 216
Глауберова соль 471, 474
Гликоген 66
Глист первичная ядовитость 450
Глистное малокровие 447
Глистогонные 469
Глотание льда 472
Глотка 211
Глоточный ланцет 396
Гнус 6
Головка 270, 333
Головная везикула 398
Гонгилонематоз 415, 417
Гостепаразитизм 298, 362, 364, 465
Гостепаразитирование 377, 396, 414,
419
Гостепаразитирование мнимое 361
Гостепаразиты 14, 418
Грибки 96, 223, 224
Губернакулюм 366
д
Дача противоглистного 477
Движение малярии 168
Дегельминтизация 394, 476, 479
Дегельминтизация внешней среды 394,
402 •
Дегельминтизация лечебная 470
Дегельминтизация предметов обияода
461
Дегельминтизация массовая при аска-
ридозе 468
Дегельминтизация массовая при тени-
идозах и анкилостомидозах 468
Дегельминтизация профилактическая
470
Дегельминтизация собак 462
Дегенерация спорозоитов 177
Действие ленточных глистов на хозяев
276
Деление множественное 68, 149
Дефицит рождаемости 182
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
SOI
Диагноз при малярии 191
Диагностика кал^-азар 108
Диагностика финноза 318
Диагностика цестод 277
Диазореакция 352
Диаптомус 280
Диатермокоагуляция 123
Дизентерия амебная 23, 84, 86, 88
Дизентерия тропическая 370
Диск присасывательный 139
Дистоматоз глоточный 227
Дифиллоботриоз 6, 458
Дифтерия 45
Длительная инкубация 41
Домовые обитатели 463, 467
Дракункулез 436
Ж
Жгут 104
Жгутики 64, 103
Жгутики биченосцев 63
Железа межчлениковая 300
Железа цистная 217
Железки цементные 378
Железы головные 38, 215, 377, 442
Железы межпрог лоттидные 299
Железы Мелиса 214
Железы одноклеточные 211
Железы простатические 213
Железы ректальные 342
Железы слюнные одноклеточные 212
Железы хвостовые 343
Железы шейные 345, 418, 442
Железы шейные анкилостомид 446
Желтая лихорадка 45
Желтозем 379
Желточник 213, 214, 272
Желточно-кишечный канал 214, 215
Желточные протоки 213
Желточный канал 272
Жиардиоз 140
Живорождение 344
Живчики нематод 344
Жидкость полости пузыря 332
Жизненный цикл цестод 273
Жилье человека 15
Жир 66
3
Заболеваемость малярией 182, 183, 185
Заболеваемость малярией в царской
и Красной Армии 185.
Заболевание первичное 166
Заболевания аскарид 454
Заболевания грибковые 45
Завод «Акрихин» 190
Закупорка кишечника 469
Замораживание 124, 481
Замыкающие клетки 210, 213
Заражение конгенитальное 169
Заражение профессиональное 457
Заражение человека .225
Заражение через переносчика 170
Зародышевая паренхиматозная оболоч-
ка 332
Зародышевые шары 215
Зернистость Джемса 157
Зернистость Шюффнера 153, 157
Зернистые клетки 347
Зигота 68., 143, 171
Зимование зараженных комаров 178
Зимовка цист плазмодия 173
Зоогеография паразитов 18
Зоонозы 6, 122
Зубной аппарат 398
Зубы 398
Зубы кариозные 100
Зубы нематод 342
И
Идиосинкразия к хинину 194
Изогамия 68, 69
Изоляционная ткань 340
Имбибиция 123, 213
Иммунитет нестерильный 180
Иммунитет приобретенный 30
Иммуно-биологическая реакция 410
Инвазии алиментарные 45
Инвазии пищевые 45
Инвазирующая личинка 381
Инвазия 22
Инвазия внутриутробная 46, 169
Инвазия латентная 24
Инвазия множественная 228
Инвазия перкутанная 384
Индекс паразитный 166, 167
Индекс плазмодийный 177, 178
Индекс плотности анофелес 179
Индекс Росса 167
Индекс селезеночный 166, 167
Индекс спорозоитный 177
Индекс эндемический 167
Индексы малярийные 166
Инкубационный период 79
Инкубационный период эксперименталь-
ной малярии 170
Инкубация 162, 191
Инкубация длительная 163
Инозит 332
Иноку лятивный способ инвазии 106
Институт зоологический 5, 187
Институт малярии центральный 187
Институты тропические 186
Инсулин 32
Интусусцепция 28
Инфантилизм 28, 40, 128
Инфекции пылевые 45
Инфекция двойная 152
«Инфекция периферическая» 160
Инцистирование 66, 67, 79, 96, 217, 224
Иицистированная личинка 380
Инъекция растворов хинина 193
Иод 200
Иод-оксихннамин-сульфо новая кисло-
та 90
Ионтофорез 124
Ипекакуана 90, 476
Источник заражения 43
К
Казеозное перерождение 312
Кала-азар 23, 107, 108, 109, 110, 111,
112; 113, 114, 115
502
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Калабарский отек 428, 465
Календарь инвазии 22
Кала-азар 107
Калий марганцево-кислый 123
Каменноугольные копи 388
Каннибализм 71, 206
Капельные инфекции 45
Капсулы трихин 457
Капсулы хитиноподобные 332
Карболовая кислота 394, 403, 412
Кариозодоа 103
Кафедра общей биологии и паразито-
логии 5
Квартана 151
Кварцевая лампа 124
Квасцы 200
Кинетонуклеус 104
Кинетопласт 104
Кислота янтарная 332, 337
Киста эхинококка 451
Кишечная палочка 352
Кишечник 211
Кишечное сырье 208
Клетка выделительная 414
Клетки желточные 214
Клетки мускульные 422
Клетки мышечные 340
Клетки половые 424
Клетки экскреторные 422
Клеточное тело 213
Кожистые выросты 347
Кожная уртикарная реакция 410
Кожно-мускульный мешок 211, 340, 438
Кокаин 422
Кокандка 119
Кокцидиоз 146
Кокцидиоз печени кролика 40
Колбасное производство 208
Количественный учет 392
Кольца крупные 156
Колюще-сосущие органы 34
Комиссия постоянная по изучению
малярийных комаров 187
Комиссура дорзальная 213
Комменсализм 15
Комменсалисты 70
Компрессор 318
Компрессы ятреновые 123
Контактный путь 46
Контаминативный способ инвазии 106
Концевые дольки 428
Конъюганты 68
Конъюгация 68, 204
Копрозои 70
Копрокультура 391
Копулятивная сумка 343
Копулятивный орган 213
Копуляция 68, 69
Корацидии 22, 273, 280, 289, 290
Коха чашка 391
Кохинхинская диаррея 370
Краевая паразитология 55
Краски органические 188
Красная Армия 390
Крезол 80 '
Кривая двувершинная (заболеваемость
малярией) 167
Критидиальная стадия 105, 106
Крыловидные складки кутикулы 414
Ксенопаразит 14
Ксероформ 123
Ктенидий 34
Кукурузник 390
Культура D. fragilis 103
Культура Ent. histolytica 76
Купирование приступов малярии 191
Курятники 15
Кутикула 64, 211, 332, 339
Кутикулярные зубы ротовой капсулы
445
Кутикулярные ребра 343
Кутикулярные щитки 419
Л
Латентный период вторичный (маля-
рии) 163, 164
Латентный период первичный (маля-
рии) 163
Лауреров канал 214, 215, 2'22
Лейкомаины 332
Лейшманиоз 107
Лейшманиоз висцеральный 107
Лейшманиоз внутренний собачий 111,
114
Лейшманиоз внутренностный 107
Лейшманиоз кожный 107, 114, 116,
118, 119, 122
Лейшманиоз кожный остро некротизи-
рующийся 118
Лейшманиоз кожный поздно изъяз-
вляющийся 118
Лейшманиоз кожный у животных 119
Лейшманиоз кожный форма городская
118
Лейшманиоз кожный форма мокнущая
118
Лейшманиоз кожный форма острая 118
Лейшманиоз кожный форма папулез-
ная 118
Лейшманиоз кожный форма сухая 118
Лейшманиоз кожный форма хроничес-
кая 118
Лейшманиоз кожный форма эктиматоз-
ная 118
Лейшманиоз кожный форма язвенная
118
Лейшманиоз латентный 123
Лейшманиоз собак 111, 114
Лейшманиоз средиземноморский 107,
ИЗ
Лейшманиозная , стадия 105, 106
Лекарственная профилактика малярии
188, 189
Лемниски 438
Ленточные черви 268, 270, 274
Ленточные черви нерасчлененные 269
Ленточные черви расчлененные 269
Лептомонадная стадия 105
Лептоспироз 45
Лечение гемоглобинурийной лихорад-
ки 198
Лечение курсовое 189
Лечение малярии 188, 192, 195
Лечение малярии акрихином с плазмо-
цидом 195
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
503
Лечение малярии осарсолом 196
Лечение малярии противорецидивное
196, 197
Лечение малярии симптоматическое 199
-Лечение малярии у беременных жен-
щин 199
Лечение малярии, схема 195
Лечение малярии хинином 194, 196
Лечение малярийной комы 198
Лечение противорецидивное 189
-Лигаменты 438
Лизол 403
Ликофора 269
Линька 339
Липоиды 448
Лихорадка ежедневная 151
Лихорадка марсельская сыпнотифоз-
ная 5
Лихорадка паппатачи 56
Лихорадка перемежающаяся 151
Лихорадка сыпнотифозная 45
Лихорадка трехдневная летне-осенняя
151
Лихорадка черная 107
Личинка анкилостомиды 46
Личинка власоглава 356
..Личинка рабдитная 365, 379
Личинка рабдитовидная 366
Личинки заблудившиеся 350
Личинки насекомых 218
Личинки трихины 349, 350
Личцночные стадии добавочные 218
Лобоподии 74
Ложная кольчатость 440
Ложная полость тела 340
Ложноножки 64, 71
.Ложнопаразитизм 11
Ложнопаразитизм мнимый 371, 374
Ложнопаразиты 12, 145, 375
Ложнопаразиты истинные 12
Ложнопаразиты мнимые 12
Ложный паразитизм 371
Локализация извращенная 225
Лооз 428
Л гоголевский раствор 74
М
Мазь анестезиновая 476
Мазь мышьяковая 123
Мазь протарголовая 124
Мазь риваноловая 123, 124
Мазь ртутная белая 476
Мазь ртутная желтая 476
Мазь ртутная серая 476
Мазь сурьмяная 123
-Макрогамета 68, 150
Макрогаметоцит 68, 143, 150
Макронуклеус 68, 204
Малярийность местности 169
Малярийные кривые 166
Малярик первичный 166
Малярик рецидивирующий 166
.Маляриотерапия 163
Малярия 23, 150, 152, 175, 188
Малярия врожденная 169
Малярия птиц 148, 160
Малярия трехдневная 149, 151, 153,
190, 191, 192
Малярия тропическая 151, 155, 178.
190, 191, 192
Малярия человека 148
Малярия четырехдневная 151, 155, 199,
191, 192
Малярия экспериментальная (искус-
ственная) 169
Малярия, южный тип кривой 166
Марганцовые рудники 388
Масло хеноподовое 473
Массовая дегельминтизация 477
Матка 214, 242, 344
Маточный колокол 438
«Мбу» 170
Медикаментозная профилактика маля
рии (химиопрофилактика) 190, 195.
197, 198
Медикаментозная терапия малярии 19С
Мембрана ундулирующая 104
Мерозоит 68, 142, 149, 161
Мерцательное движение 65
Местное механическое действие 445
Метаболирование 64, 65
Метагенез 69, 218, 275
Метагонимоз 458 f
Метаморфоз 344
Метаморфоз сосальщиков 218
Метацеркарии 217
Метациклические трипанозомы 125
Метиленовая синька 123, 188, 200
Метод переваривания Бермана 352
Метод с всплыванием яиц в солевом
растворе 391
Механические переносчики трипанозом
444
Миаз 38, 46
Миаз тканевой 12
Миазы ран 12
Миграции в почве 381
Миграция 463
Миграция личинок 345
Миграция личинок анкилостом 386
Миграция личинок аскарид 405, 406,
413
Миграция паразита 25
Миграция плероцеркоида 282
Мигрирующая личинка 395
Мигроаскаридоз 408
Микозы 45
Микрогамета 150
Микрогаметоцит 68, 143, 150
Микронуклеус 68, 204
Микрофекалнзация 402
Микседема 128
Миома 402
Мионемы 65
Миофибриллы 340
Мирацидий 213, 215, 224
Мишеровы мешки 202, 203
Мозаичность распространения 52
Мозговые узлы 213
Монозойные формы 104
Морковь 375
Мукоидная пробочка 431
.504
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Мускулатура (червей) 270
Мутуализм 15
Мышиный тиф 408
Мясокомбинат 323, 354"
Мясные спороспермии 201
Мясо рыб 458
Мятные капли 472
Н
Нафталин 402, 475
Нахлебники 70
Нахлебничество 15
Начальный капилляр 213
Негашёная известь 412
Некатороз 382, 384, 386, 389, 394, 473,
475
Неостибозан 115, 116
Нервные стволы 211, 213, 271, 343
Новарсенол 192
Нозогеография 52
Нора грызуна 49
Носите ли-Конвалесценты 85
Носители контактные 85
Носители паразитов 23
Носители тениид 479
О
Обезвреживание выделенных паразитов
480
Обезвреживание фекалий 480
Обезвреживание фекалий и паразитов
477
Обезвреживание финозного мяса 481
Обследование одномоментное на маля-
рийность 168
Обследования массовые (на малярию)
189
Общая реакция 37
Овощи 375
Огородная зелень 459
Одутловатка 352
Оздоровление малярийного очага 169
Околоматочный орган 272
Окружающая капсула 349
Омеопаразиты 12
Онкосфера 273, 302, 460
Онкосфера эхинококка 331
Оокинета 150, 171, 173
Оотип 214, 272
Ооциста 142, 150, 171, 172, 173, 174
Описторхоз 458
Оплодотворение перекрестное 215
Опорные фибриллы 340
Органеллы 63
Организм как среда обитания 18, 24
Органоиды 63
Органоиды движения 64
Органы выделения 342
Органы зрения рудиментарные 213
Органы прикрепления 34
Осарсол 91, 141, 148, 192, 194, 358, 475
Осенний подъем заболеваемости маля-
рией 165
Осмотическое давление тканевое 213
Отдел паразитологии Зоологического
института 187
Отложение извести в капсулах 349
Отпугивающие сетки 6
Отряды противомалярийные 187, 188
Очаг пендинки 122
П
Палеопаразитология 19
Палочки сибирской язвы 443
Палочки туберкулезные 443
Панспоробласт 201
Парабазальное тельце 104
Парагликеген 66
Парадесмоз 136
Паразитарные болезни 45
Паразитизм 11, 33
Паразитизм временный 15
Паразитизм латентный 41
Паразитизм мнимый 146
Паразитизм перманентный 15
Паразитизм постоянный 15
Паразитизм скрытый 41
Паразитирование пиявок v человека?
441
Паразитическая генерация 366
Паразитический зоб 40
Паразитологическое районирование 55„
56
«Паразитологический сборник» 187
Паразитологические экспедиции 56
Паразитология 18
Паразитология ветеринарная 19
Царазитология медицинская 19, 63
Паразитология растений 19
Паразитоносители 41
Паразитоносительство 17, 41, 70, 444
Паразитоносительство при малярии 191
Паразитофауна 27
Паразитоценозы 32
Паразиты 11, 17, 19, 22, 45
Паразиты вкожные 15
Паразиты внутренние 16
Паразиты гетероксенные 13
Паразиты кровососущие 34
Паразиты малярийные 150
Паразиты мнимые 70
Паразиты моноксенные 13
Паразиты-монофаги 47
Паразиты наружные 15
Паразиты облигатно-трансмиссивные 46»
Паразиты парадоксальные 13
Паразиты полостные 16
Паразиты протоплазматические 28
Паразиты растений 374
Паразиты случайные 13
Паразиты специфичные 13
Паразиты стеноадаптивные 47
Паразиты стеноксенные 13
Паразиты тканевые 446
Паразиты факультативно-трансмиссив-
ные 46
Паразиты эвриксенные 13
Паразиты ядовитые 28
Паренхима 211, 270
Партеногенез 218
Партеногенез макрогаметоцитов 164
Патогенные животные 41
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
505-'
Патогенез 6
Патогенность Fasciolopsis buski для
человека 230
Патогенность паразита 42
Патогенные простейшие 70
Педогенез 218
Пелликула 63, 204
Пендинка 116, 117, 118, 119
Пепсинный раствор 352
Первичная малярия 180
Первично обратившийся 166
Передача инвазии трансовариальная 23
Передача механическая 106
Передача механическая паразитических
червей 466
Передача механическая яиц 466
Передняя позиция 113, 121
Перемена хозяев 69, 106, 218, 275
«Переселение» 32
Перенос специфический 106
Переносчик 46
Переносчики нематод 345
Переносчики паразитических червей 466
Пересадка кожи 124
Перипласт 104
Перкутанное проникновение личинок
385
Перфорация костей 328
Печени абсцесс 83, 84, 85
Печень баранья 226
Печень козья 226
Пеше-хурда 119
Пигмент 153, 171, 172
Пиретотерапия 169
Пироговское общество врачей 187
Пироплазмоз 56
Питание алиментарное 44
Питание анимальное 65
Питание осмотическое 104
Питание сапрофитное 66
Питание фагоцитарное 65, 66
Питание эндосмотическое 36, 65
Пищеварение внеклеточное 212
Пищеварительная полость 210
Пищеварительный канал Ц5
Пищевод 211
Пиявки искусственные 442
Плазмодии малярийные 156, 163, 172,
174, 192
Плазмотомия 203
Плазмохин 158, 175, 188
Плазмоцид 158, 175, 192, 194
Плероцеркоид 26, 32, 43, 44, 274, 281,
283, 285, 291, 295, 296
Плероцеркоид лентеца малого 291
Поваренная соль 332
Поверхностная кожица 63
Повозрастные дозы акрихина и плаз-
моцида 199
Подготовка-к дегельминтизации 470
Подготовка к лечению 471, 477
Подсолнечник 200
Полиневрит 194
Полифаги 13
Полиэмбриония 218
Половая клоака 214, 222, 272
Половое отверстие мужское 213
Половые 'зачатки 423
Половые органы женские^ 213
Половые органы мужские^213
Половые сосочки 343
Половые формы 68
Половые формы малярийного плазмо-
дия 158
Полостная жидкость аскариды 453, 454
«Полулуния» 159, 192
Полустационар 468
Полынь 200
Поля орошения 412
Поля фильтрации 404
Полярные капсулы 201
Полярные нити 324
Пора анальная 423, 424
Пора выделительная 424
Пора половая 423
Пора экскреторная 422
Посещение первичное 166
Постоянство числа клеток 340
Почкование 68, 98
Пребурсальные сосочки 397
Предглотка 235
Пред цистные формы 76
Препараты хинолинового ряда синтети-
ческие 189
Прерванное кормление 107
Приазовские плавни 178, 179
Природная очаговость 6
Природная очаговость трансмиссивных
болезней 50
Присасывательные бороздки 274
Присасывательный диск 440
Присоска ротовая 211, 440
Присоски 34, 211, 269
Приступы малярии 162
Прободение кишек 409
Проверка жизнедеятельности финн 48&
Провокация малярии 164
Проглоттиды 268, 269
Проглоттиды «пустые» 322
Продромальный период малярии 162
Промежуточные хозяева 345
Промежуточные хозяева глист 465
Протеаза 36
Протерандрия 291 ~
Противоглистные средства 470
Противопоказания экстракта папорот-
ника 472
Противоферментные свойства (глист) 276
Протозоозы 45
Протонефридии 210
Профессиональная болезнь 388
Профилактика инвазий 57
Профилактика личная 478, 480
Профилактика (медикаментозная) ма-
лярии, схема 195
Профилактика общественная 478
Профилактика фасциолеза 227
Профилактика фасциолопсидоза 231
Профилактические мероприятия 478
Процеркоид 281, 290
Процессы дегенеративные 75
Псевдоцелом 340
Псеудобульбус 347
Птоманн 451
.506
ПРЕДМЕТНЫЙ- УКАЗАТЕЛЬ
.Пузырехвост 273
Пузыри дочерние 273, 332
Пузырчатая глиста 273
Пузырь материнский 332
Пузырь первичный 332
Пузырь эхинококка 326
Пункты малярийные 187
Пункция 124, 315
Пункция грудины 109
Пункция лимфатических желез 109
Пункция селезенки 108
Пути заражения анкилостомидами 466
Пути заражения глистами 456
Пути циркуляции 47, 48
Пути циркуляции возбудителя 50
Пылевая инвазия 400
Р
Развитие дигенетическое 106
Развитие моногенетическое 106
Размножение агамное 67, 144
^Размножение бесполое 68, 69, 142,
149
Размножение девственное (партеногене-
тическое) 218, 345
Размножение обоеполое 218
Размножение однополое 345
-Размножение половое 143
Размножение экзогенное 144
Размножение эндогенное 142
Разъедание костей 446
Раствор сулемы 315
Расы 78, 97, 101
Рвотный камень 115, 200
Реактивная капсула 446
Реакция Казони 337
Реакция P&arls—‘Nishimura 408
Ревень 475
Регистрация первичных случаев маля-
рии 181
Редии, дочернее поколение 224
Редис 375
Редия 215, 216, 217
Редукция 36
Редукция гаметическая 171
Редукция зиготическая 171
Режим при малярии 199
Режуще-сосущие органы 3
Резервуар возбудителя 42
Резервуары вирусов 23
Резорптивная реакция 37
Реинвазии 164
Реинфекция 164
Реснички 64
Реснички инфузорий 63
Ресничный покров мирацидия 216
Рецидивы инвазии 24
Рецидивы малярии 159, 163, 164, 191,
192
Риванол 90, 91, 123
Риккетсиозы 45
Рот круглых червей 342
Ротовая капсула 342
Ротовая полость 211
Ротовые органы 34
Рулек 395
С
Сальварсан 188
Самозаражение 308
Самоизлечение организма 334
Самооплодотворение 215, 272
Самопроизвольный выход аскарид 409
Самцы рабдитовидного типа 367
Санитарно-просветительная работа 469
Сантонин 29, 402, 411, 473, 475
Саркома печени 326
Саркоплазма 340
Саркоспоридии 203
Саркоспоридиотоксин 203
Саркоцистин 203
Сахар 332
Сверхпаразиты 19
Свободноживущая генерация 366
Свойства шизонтотропные 189
Северный тип кривой малярии 166
Сезонность малярии 165
Сезонные койки 188
Секрет половых и шейных желез 445
Селезенка 108
Сельдерей 375
Семевыносящие канальцы 271
Семеизвергательный канал 213, 272, 344
Семенники 213, 344
Семенной пузырек 213, 344
Семеприемник 214, 272
Семепровод 213, 344
Сера 476
Серебро азотнокислое 208
Серно-карболовая смесь 394
Серокислая магнезия 358
Серповидные тела 202
Симбиоз 15
Синергетическое действие паразитоце-
ноза хозяина 456
Синонимика 71
Синцитий 213, 339
Сирень 200
Сифилис 45
Сколекс 268, 332, 333
Скорлупа яйца 214
Скорлуповые железы 215, 272
Скрытая форма заболевания 41
Слепые придатки 342
Сложнолакричный порошок 475
Слоновая болезнь 465
Слоновость 425
Случная болезнь 46
Смена среды обитания 344
Смертность от малярии 182
Совокупительная сумочка 378
Совхинет (советский хинин) 194
Содовый раствор 470
Солевое слабительное 390, 471, 473,
475
Соль солянокислая 476
Соматические паразитические черви 446
Сонная болезнь 20, 23, 128
Соотношение «хозяин — паразит» 29
Сосальщики живородящие 215
Сосочки интеровентральные 429
Сосочки интеродорзальные 429
Сосочки интеролатеральные 431
Сосочки постклоакальные 428
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
507
Сосочки преклоакальные 428 '
Социальная профилактика 91
Социальное значение малярии 182
Спарганоз 296
Спарганоз экспериментальный 282
Спикулы 343, 395
Спикулярное влагалище 361
Спинная пищеводная железа 378
Спирохетозы 45
Спирохетч кур 443
Споробласт 144, 171
Спорогония 69, 144, 174, 182
Спорозоит 69, 141, 142, 144, 150, 163,
171, 172, 175
Спорообразование 141
Спороциста 144, 216, 224
Споры 144, 201, 202
Способ заражения 43
Стадии эндотелиальные (шизогонии) 160
Стадия цистная 106
Станции малярийные 186, 187, 188
Стертые формы 444
Стибенил 115, >116
Стибозан 115, 116
Стоварсол 91, 358, 477
Стойла животных 15
Стрептоцид 123, 200
Стробила 268
Сулема 80, 436
Сулемо-карболовый раствор 394
Сульфидин 200
Суперинвазия 165
Суперинфекция 165, 433
Сурьма 123
Сурьмин 115
Т
Таджикский филиал 5
Таксисы 66
а л ассин 451
Тела парабазальные 139
Тела хроматоидные 76, 77, 95
Тело остаточное 143,ч170
Тельца сидерофильные 76
Тельце Мелиса 272
Тельце редукционное 170
Тениаринхоз 6
Тениотоксины 449, 450
Теория блошиная распространения ка-
ла-азар 112
Терапия тканевая 124
Термотропизм 391
Терциана 151, 189
Тетрахлорэтилен 474
Течение малярии 159, 190
Тимол 231, 233, 257, 307, 358, 376, 400,
406, 412, 475
Тимоловое полоскание 417
Тимьян 475
«Тип» 204
Тиф брюшной 45, 48
Тиф возвратный 45, 48
Тиф клещевой возвратный 5
Токсичность паразитических червей 6
Транзитные гостепаразиты 387
Транзитные яйца 241
Трансмиссивная болезнь 46
Трематодозы 458
Трипанозомоз 40
Трипарсамит 128
Трихиноз 352
Трихинозная капсула 353
Трихоцефалоз 358
Тропическая кахексия 107
Тропическая спленомегалия 107
Трофозоит 68, 142, 148
Туберкулезная палочка типа bovinus
У
Уборная 325, 464, 480
Увеличение селезенки 165
Ф
Фагоцитарная клетка 343
Фазмиды 343
Фасциолоз мнимый 277
Фасциолоз человека 227
Фасциолопсидоз завозный 230
Фибрилла парабазальная 136
Физиологические виды 370
Филаменты 324
Филицилен 473
Филогения 19
Фильмарон 473
Филяриевидные личинки 366, 392
Филяриоз 425
Филяриоз коньюнктивы 466
Финка солитера 314
Финки 38, 273, 274, 308, 313, 315, 316,
317
Финки омелотворенные 312
Финноз 313, 315, 319, 323, 461, 481
Финноз мозга 316
Финноз свиней 314
Финноз скота 457
Финноз человека 318
Финнозное мясо 45, 47, 318
Формалин 80
Формалино-глицериновая смесь 338
Формула Ван-Гоффа 413
X
Хауз .460
Хеноподовое масло 231, 237, 376, 391
Хилурия 422
Химиопрофилактика малярии обще-
ственная 197
Химиотаксис 66
Хинин 175, 192, 193
Хинин азотнокислый 208
Хинин двусолянокислый 193, 194
Хинин сернокислый 193, 194
Хинин солянокислый 193 •
Хининоустойчивое свойство малярий-
ных паразитов 188
Хинное дерево 194
Хинный щелочной раствор 198
Хищничество 11
Хлорная известь 412
508
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Хлороформ 436
Хозяин 13, 17, 22
Хозяин главный 14, 53
Хозяин окончательный 218
Хозяин промежуточный 14, 54
Хозяин промежуточный второй 218
Хозяин промежуточный первый 218
Холера 45
Холестерин 83
ц
Ценур 273, 310, 327, 330
Ценуроз 328
Ценуроз мозга 328
Ценуроз овец 329
Цервиды 431
Церкарии 213, 217, . 224
Церкарии печеночной двуустки 459
Цикл развития 215
Цикл развития двуустки ланцетовид-
ной 239
Цикл развития лейшманий 114
Цикл развития печеночной двуустки
216, 232
Цикл развития сосальщиков 218
Циклический перенос трипанозом 127
Циклозы 63, 74
Цинхонидин 194
Цинхонин 194
Циркуляция паразитов 47
Цирроз двуустковый 225
Цирротический процесс 226
Циррус 271
Цистицерк 27, 273, 310, 451
Цистицеркоз 313, 461
Цистицеркоид 273, 309, 310
Цисты сборные 238, 239
Цитварная полынь 473
Цитварное семя 473
Цитомеры 161
Цитостом 65, 104
Ч
Чередование поколений 14, 218
Чередование форм размножения 69
Чеснок 91
Чесотка 22
Четыреххлористый углерод 231, 257,
376, 391, 473
Членики 268
Чуженаразиты 14
Ш
Шизогония 68, 142, 144, 149
Шизогония экзоэритроцитная 160, 161
Шизонт 149
Шизонты экзоэритроцитные 161
Э
Эдафический фактор 412
Эвкалипт 200
Экзотоксины 447
Экология паразитов 18
Экономическое значение малярии 182
Экспериментальное заражение анофе-
лес 178
Экстрагостальная фаза 16
Экстракт корневища папоротника 227,
231, 233, 287, 299, 307, 310, 319, 325г
440, 470, 471, 472
Экстракт лентеца широкого 449
Эксфлагелляция, 175
Эксцистирование 67, 79
Эктопаразиты 15
Эктопаразиты соматические 15
Эметин 80, 90, 91, 97, 227, 242, 262, 47&
Эндемичность малярии 166
Эндопаразиты 16
Эндоспоры 146
Эндостация 28
Эндотоксины 447
Энтомозы 45
Энцефалит 192
Энцефалиты сезонные 45
Эозиновая проба 80 t
Эпидемиологическое значение разных
видов анофелес 176, 178, 179
Эпидемиологическое районирование 56
Эписпоры 146
Эритрофаги 75
Эритрофагоцитоз 76
Эхинококк 273
Эхинококк вторичный одногнезднып 335
Эхинококк многогнездный 336
Эхинококк многокамерный 336, 337
Эхинококк многоформенный 333
Эхинококк множественный 335
Эхинококк одногнездный 334
Эхинококк печени 335, 336
Эхинококк стерильный 332
Эхинококковая опухоль 336
Я
I
Ядра генеративные 68
Язва 124
Язва алеппская 116
Язва багдадская 116, 119
Язва бухарская 119
Язва восточная 119
Язва кокандская 119
Язва кратерообразная 82, 84
Язва пендинская 6, 42, 116, 117, 118,
Язва сартская 116
ГЕОГРАФИЧЕСКИЙ И ЭТНОГРАФИЧЕСКИЙ УКАЗАТЕЛЬ
А
Абхазия 148, 194, .388
Аджаристан 388
Аджария 194
Адлер (район Адлера.) 178, 179
Азербайджан 9,2, 93, 107, 110, 116, 148,
323, 356, 360, 378, 384, 388, 389/402,
473
Азия 185, 238, 243, 376, 381, 449
Азия Восточная 259
Азия Средняя 6, 42, 55, 5б, 94, 109,
110, 111, 112, 116, 119, 122, 180, 186,
208, 227, 330, 370, 388, 412, 419, 441
Азовско-Черноморский край 239
Айв ад ж 116
Алайская долина 93
Алеппо 119
Алжир 114, 120, 440
Америка 238, 419
Америка Северная 284, 327, 373, 375,
382, 442
Америка тропическая 427
Америка Центральная 120
Америка Южная 120, 124, 128, 146, 428
Амстердам 179
Аму-Дарья 116
Амур 254, 255, 364, 458
Англия 87, 102, 138, 173, 427
Андижанский округ 110
Антильские острова 377
Аравия 265
Аральское море 267
Аргентина 326
Арканзас 415
Армения 88, 93, 110, 111, 227, 265, 308,
324, 388, 395, 413
Архангельская губ. 285
Архангельская обл. 181
Ассам 221
Астрахань 83, 178
Атрек 53, 116
Аулие-ата 267
Африка 23, 126, 158, 183, 185, 228, 238,
370, 440, 442
Африка Восточная 170, 220, 266, 382
Африка Западная 372
Африка Северная 442
Африка тропическая 376, 421, 427, 428,
436, 465, 466
Африка экваториальная 91
Ашхабад 50, 92, 110, 116, 117, 118,
119, 305, 388, 440, 442
Б
Багдад 119, 122, 265
Байкал 55, 288, 290, 291, 292
Баку 83, 92, 110, 116, 227
Бальджуан 442
Бангкок 305
Барбадос 424
Барда 116
Барнаул 174
Батуми 187
Бек-Буди 116
Белград 27
Белое озеро 267
Белоруссия 352, 413
Бенгалия 182, 301
Биробиджан 178, 179, 182
Бискра 120
Бразилия 28, 40, 133, 370, 382, 394,
396, 440, 442, 476
Бурят-Монгольская республика 288, 291
Буряты 291
Бухара 92, 110, 116
Бухара Старая 436
В
Вена 109
Венгрия 235, 437
Венгры 372
Владивосток 421
Вогулы 249
Волга 439
Вологодская губ. 181
Восток 394
Вуадил 330
Вятская губ. 181
Г
Гагры 185
Галлиполи 146
Гассан-Кули 53, 54
Гвиана Британская 221, 381, 428, 466
Гвиана Французская 133
Гвинея 266, 305
Геок-тепе 388
Германия 102, 234, 267, 354, 402
Гиляки 254
Голландия 103, 176, 178
Голодная степь 178
Гольды 254
Гомельская обл. 354
Гонконг 243
510 ГЕОГРАФИЧЕСКИЙ И ЭТНОГРАФИЧЕСКИЙ УКАЗАТЕЛЬ
Гори 116
Гренландия 295
Греция, 146
Гривенская (станица) 179
Грузия (Грузинская ССР) 92, 110, 116,
148, 190, 388, 393
Гурия 388
д
Дагестанская АССР 166, 175, 323
Дайна 54
Дальний Восток 56, 178, 179, 235, 243,
252, 254, 258, 262, 267, 282, 370, 372,
388, 397
Джизак 117
Днепр 458
Днепропетровск 178, 179
Добруши 354
Донбасс 241, 358, 370, 372
Е
Европа 128, 238, 243, 372, 419, 425, 440,
442
Европа Западная 370
Европа Южная 442
Европейская часть СССР 181, 253, 458
Египет 88, 131, 146, 253, 265, 266, 370,
395, 441
Екатеринодар 437
Ессентуки 89
3
Забайкалье 288
Закавказье 55, 89, ио, 111, 112, 148.
370, 382
Закатальский район 388
Западное полушарие 324
Зеравшан 116
Зыряне 249
И
Иерусалим 120
Имеретия 388
Индийский архипелаг 231
Индия 110, 111, 112, 113, 115, 119, 123,
133, 146, 167, 182, 183, 221, 231, 250,
265, 360, 382, 397, 419, 421, 429, 432,
436, 441, 442
Индо-Китай 230, 295
Ирак 119, 267
Иран 323
Иркутск 108, 324, 337
Иртыш 246
Исландия 295, 339
Испания 425
Италия ПО, 112, 175, 437
К
Кабардино-Балкарская АССР 169
Кавказ 116, 180, 181, 185, 186, 240, 420
Кавказ Северный 93, 352
Казах ПО
Казахстан 93, 221, 243
Кайне-Касыр 53, 54
Калифорния 97, 98, 131
Канарские о-ва 442
Каратау 442
Карелия 284, 285, 287, 458
Каспийское море 53
Катта-Курган ПО, 116
Кафирниган 426
Кахетия ПО
Кашка-Дарьинский округ ПО
Кашка-Дарья 116
Керки ПО, 116, 117
Киев 100
Кипр 178, 265
Киргизия северная 93
Киргизия южная 93, 243
Кировабад ПО
Кировская обл. 299
Китай 18, 52, ПО, 112, 113, 114, 13К
251, 267, 370; 375, 397, 419, 421
Китай Северный 243, 412
Китай Центральный 230
Китай Южный 230, 295, 381
Китайцы 229, 295
Соканд ПО, 116, л
Кольский полуостров 87, 93, 131
Коморские о-ва 425 .
Конго 266
Конч-озеро 284
Корея 262
Косино (под Москвой) 267
Крым 6, 257
Кубанская обл. 331
Куйбышевская обл. 167
Куляб 466
Курган-Тюбе 442
Кушка 92, ПО, 388
Л
Лена 292
Ленинабад 116
Ленинград 52, 80, 87, 92, 93, 131, 141,
258, 259, 354, 364, 388, 411
Ленинградская область (губ.) 166, 181,,
285, 458
Ленкоранский район 92, 378, 388
Ленкорань 116
Лондон 115
М
Маврикия о-ва 301
Мадагаскару 305
Македония 133, 173, 176, 179, 437
Малакка 233
Малая Азия 442
Манилла 231
Мары 118
Мексика! 44 2
Мерв 388
Месопотамия 146, 265
Мингрелия 178, 388
Мирза-Арал 330
Миссисипи 184
ГЕОГРАФИЧЕСКИЙ И ЭТНОГРАФИЧЕСКИЙ УКАЗАТЕЛЬ
51^
Москва 90, 93, 138, 186, 187, 230, 259,
388, 395, 404, 421
Московская обл. 239
Мургаб 6
Мургабская долина 87
Н
Нахичевань 110
Невская губа 284
Негры 220
Ненецкий национ. окр. 294
Никопольский район 249
Нижняя Тунгуска 292
Новочеркасск 27
Новгородский округ 429
Нью-Йорк 237
О
Обдорск 245
Обь 246, 249, 458
Одесса 83
Озеро Дугласа 231
Ойротия 323
Ольхон (о-в) 288, 291, 292
Осташковский уезд 438
Остяки 249
П
Палестина 122
Панама 295, 361, 381, 412, 417
Париж 329
Пенджаб 120
Пермская губ. 181
Персия 119, 265, 436
Персия Восточная 442
Персы 233
Перу 135
Петербург 72
Петергоф 284
Петрозаводск 287
Печоры низовье 294
Поволжье 121
Португалия 121, 265
Приамурье 360
Прибалтика 285, 458
Приморская область Д. В. 93
Приморье 5, 56, 179, 182
Просницкий район (Кировск, обл.) 234
Пруссия Западная 354
Р
Радомно 354
Рио-де-Жанейро 228
Россия 183, 352, 362
РСФСР 181
Румыния 298, 437
С
Салоники 146
Самарканд 93, 110, 111, 116, 117, 442
Самарская губ. 253
Самоа 94
Самоеды 249
Сахалин о-в 295
Свердловск 294
Северная Нигерия 327, 329
Сенегамбия 442
Серахс 116, 117, 123
Сиам 230, 419
Сибирь 182, 248, 249, 370, 458
Сибирь Восточная 458
Сиерра-Леоне' 381
Сингапур 220 •
Сирия 227
Сицилия 437
Сольвычегодск 181
Средиземное море 110, 441
СССР 103, 121, 125, 158, 165, 178, 1834
190, 201, 221, 227, 230, 234, 235, 241..
243, 249, 254, 267, 285, 296, 303, 308L
339, 352, 354, 356, 358, 364, 370, 375.
381, 382, 388, 395, 412, 414, 419, 441.
458, 465, 466, 467
Суматра 146
Сумбар 53
Сунский р-н 299
Сурхан-Дарьинский округ 110
Сухуми 5, 90, 176, 177
Сухумский ботанический сад 473
Сучанские копи 370
США 87, 90, 176, 231, 285, 352, 359г.
410, 415, 441
Сыр-Дарья 116
Сычуанские копи 382, 388
Т
Таджикистан 6, 426
Таджикистан Южный 110, 467
Тахта-Базар 92, 110, 116, 117
Тахта-Базарский район 118
Ташкент 88, 92, 93, 111, 113, 115, 116,
117, 119, 123, 147, 174, 178
Тбилиси 179, 208, 227, 228, 231, 354
442
Тверская губ. 438, 466
Тегеран 117
Теджен 116, 117
Тенесси 87
Термез 117, 123
Техас 375
Ткнибули 388
Тобольск 245, 246, 249
Тобольская губ. 285
Томск 248, 249, 253, 331, 370
Тула 331, 404, 412
Тунгусы 292
Тунис 87
Туркестан 110, 178, 181, 228, 433
Туркмения 6, 38, 53, 54, 116, 119, 122,
141, 388
Туркмения Южная 42, 55, 93
Туруханск 292
Турция 425
У
Уде 258
Удехейцы 254
512
ГЕОГРАФИЧЕСКИЙ И ЭТНОГРАФИЧЕСКИЙ УКАЗАТЕЛЬ
Узбекистан 108, 116, 123, 308, 330
Улан-Батор 335
Ульчи 254
Украина 249
Уссури 254
Уссурийская обл. 93, 182, 249
Уфа 158
Ф
Фергана 110, 116
Филиппинские о-ва 158, 231, 233, 375,
403
Филиппины 381
Флорида 297
Формоза 176, 230, 257, 262
Франция 98, 138
Фрунзе 93
X
Хабаровск 258
.Хабаровская обл. 182
Харьков 253, 352, 402, 415
Харьковский округ 370
Хорезм 116
ц
Цейлон 146, 382, 441, 442
Ч
Чарджоу 110, 116, 117
Чнатуры 388
Чикаго 91
Ш
Шахрисябс 110
Шемаха 110
Шир-Абад 442
Э
Эвенки 295
Эривань 92, 110
Ю
Юрьев 362
Ютландия 326
Я
Япония 235, 251, 253, 254, 262, 267,
295, 296, 297, 370, 395, 397, 441, 442
Якутия 285
Якуты 292, 295
Японцы 295
Яссы 298
УКАЗАТЕЛЬ ЛАТИНСКИХ ТЕРМИНОВ
А Febris tertiana maligna (f. tropica) 151. Febris tropica 151
Acetabula 269 Anaemia perniciosa bothriocephalica 447, 450 Anaemia perniciosa dibothriocephalica 285, 444 Anus 341 Appendicopathia oxyurica 401 Arthemisia cinae 473 Arthritis deformans 83 Filicilen 473 Filmaron 473 G Gentian-violet 252 Glandulae prostaticae 213 Grounditch 387 Gubernaculum 343
В H
Bismut, carbonicum 402 Bothriae 274 ’Bursa cirri 213 Bursa copulatrix 343 Halzoun 227 Hexyl resorcinol 473 1
C Intestinum 211
Canalis gynaecopteprus 263, 264 Canalis laureri 214 Carbamino-phenol-arsenil-acid 91 Carboneum tetrachloricum 391, 473 Cavum oris 211 Cirrus 213, 271 Corona radiata 375 Creeping disease 387, 419 Chenopodium anthelminticum 473 Cristal-violet 252 К Kalium hypermanganicum 133 L Larva migrans 419 Lymphangitis 422 Lymphocoele 425 Lymphoscrotum 422
D M
Dermatitis linearis migrans 419 Ductus ejaculatorius 213, 272 Ductus vitello-intestinalis 214 Methyl-violet 252 N
E Naphtalinum purum 475
Elephantiasis arabum 425 Eliocharis tuberosa 229 Emetinum hydrochloricum 90 Extractum filicis maris aethereum 470 0 Oedeme ambulant siamois 419, Oleum chenopodii 391, 473 Osarsol 475
F Ovarium 213
Febris quartana 151 Febris quartana duplex 153 Febris quartana simplex 151 Febris quotidiana 151 Febris quartana triplex 153 Febris tertiana 151 Febris tertiana duplex 151 Ovicapt 272 Oviduct 213 P Pharynx 211 Porus excretorius 343 Praepharynx 235
514 УКАЗАТЕЛЬ ЛАТИНСКИХ ТЕРМИНОВ
Proctodaeum 341 Psychotria ipecacuanhae 476 т Tartarus stibiatus 436 Testes 213
R Tetrachloraethylenum 473 Thyreoiditis parasitaria 128
Receptaculum seminis 214, 272 Rostellum 269 Trapa natans 229, 231 u
S Uterus 214 V
Santoninum 473 Stibenyl 115 Stibio-kalium-tartaricum 115 Stibosan 115 Sticlocyte 347 Sphincter 213 Stomadaeum 341 V Vas deferens 271, 272 Vasa efferentia 271 Vermicida 469 Vermifuga 46.9 Vesicula seminalis 213 Vitellogenes 213
ИМЕННОЙ УКАЗАТЕЛЬ ИНОСТРАННЫЙ
А
Ackert 33
Adie 113
Adler 113, 119, 120
Asada 40$
Ashford 390
Augustin 383, 387
Connal 147, 428
О‘Connor 131, 134
Cort 359, 410, 411, 412
Coupland 352
Courtelin 75
Craig 91, 176, 366
Cram 411
В
Bacigalupo 326, 451
Baeckwith 452
Baer 320
Bahr M. 175
Bakker 411
Baraber 178
Basile 112
Bayer 128
Bedson 449, 450, 454
Beguet 120
Blacklock 427
Boeck 76, 140
Boidin 451
Boycott 452
Boyd 162, 163
Bozicevich 352
Braine 451
Braun 279
Breslau 408
Brieger 452
Brooks 218
Broyd M. 176
Brumpt 87, 138, 160, 205, 241, 382,
428
de Buck 476
Buckley 382
Bussano 452
C
Cafiero 454
Calamida 449
Callandruccio 208
Cantelen 428
Cao 449
Casoni 337
Celli 175
Chadwick 119
Chagas 128
Chalerm Pronunas 419
Chandler 470
Chitwood 347, 431
Chopra 470
Christenson 362
Christopherson 265
Ciuca 451
Ciurea 26, 233, 364
D
Daengsvang 419
Le Dantec 449
Deluca 451
Dervis 447
Desportes Cf 437
Deve 331
Dobell 69, 75, 76, 78, 99, 131, 13
Donatien 120
Dobrohlav 76
F
Faust 53, 91, 231, 366
Feng 326
Fiessinger 451
Flury 452, 454, 455
Forbes 437
Forster 405
Foster 361
Frey 76, 87
Frohlich 401
Fuhrmann 212
Fiilleborn 405, 408, 426
G
Giavanola 266
Giusti 451
Goedelst 27
Goldschmidt 452, 453
Goodey 279
Gordon 357, 401
Grassi 173, 174, 208
Gross 100, 451
H
Hall 20
Hanschell 174
Hassal 27, 130
Hegner 74, 90, 91, 134, 141, 206
Henry 451
Herrick 33
Hobmaier 281
Hoeppli 326, 399, 454
516
ИМЕННОЙ УКАЗАТЕЛЬ ИНОСТРАННЫЙ
Huff 404
Hug 451
I
Izumf 257
J
James 160, 161. 173, 174, 175, 179
Jancko 173, 174
Jepps 78
Joest 451
Johnson 361
Joyeux 320
Julien 453
К
Kagy 91
Kamada R. 450
Karwacki 443
Kassel 90
Khalil 253
Khaw 53
Khouri 227
King 390
Kinoshita 176
Kitchen 152, 163
Kjellberg 146
Knabe 422
Kofoid 375
Koino 405
Kraus 40
Kreis 369 7
Kiinstler 130 #
L
Lambinet 386
Lane 392
M
Malvoz 386
Martini 340
Mattes 238
Matthews 97
Mayne (Bruce) 174
Mazzocco 452
Meleney 76, 87
Messineo 449, 454
Mills 119
Mingazzini 449
Mitzmain 174, 176
Monnig 36
Montgomery 225
Moorthy 429, 435
Morenas 455
Morson 451
Muhling 443
N
Neuhaus 238
Nichigori J361
Nicolle 1.73
Nolan 352
Nolf 411
N611er 97, 99
О
Ohi 206
Ortel 447
Otto 359, 410, 411
P
Pacinotti 443
Parrot 114, 120
Passey 451
Patton 117
Payne 141
Polonio 437
Pomella 449, 450
Powell 137
Proscher 449
Q
Quast 403
R
Raillet 453
Ransom 354, 396, 405
Ratcliffe 32
Rees 206
Reichenow 146
Reynold 208
Richet 451
Rivas 324
Robertson 146
Robin 451
Roth 350
Roubaud 174
S
Saeki 306
Sandground 30, 225, 373, 375
Sarles 31, 32
Savino 32
Schauman 448
Schlangdenhauff 451
Schmanch 448
Schwartz 355, 448, 455
Schweitzer 91
Scott 30, 91, 452
Seguin 454
Seilard 79
Senevet 120
Sergent 120
Seyderhelm 448
Shute 173
Simonin 449, 452
Sindler 412
Sinton 120, 130, 182, 183
Sluka 109
Smille 394
Smillie 383
Snijders 91
Sparapani 452
ИМЕННОЙ УКАЗАТЕЛЬ ИНОСТРАННЫЙ
517
Staubli 353 Steffenhagen 443, 444 Stephens 157 Steward 427 Stiles 27 Stoll 392, 412 Stratman-Thomas 176 Strong 120, 208 Stunkard 301 Sun 114 Sweet 429, 435 Swellengrebel 176, 178, 179 Szokalski 443 T Tallquist 286, 448 Tate 160, 161 Theodor 119, 120 Thomson 146 Tonney 91 Tsuchia 140 Turner 420 U Urchs 27 Vaullegard 454 Visentini 112 Vogel 241, 243, 245 Vosgien 314 W Walker 79 Weinberg 453 Wenrich 67 Wenyon 65, 67, 88, 97, 113, 119, 120, 127, 131, 134, 138, 140, 146, 173, 174, 175, 176, 178, 179 White 375 Wilhelmi 403 Winfield 30 Woodland 306 Wright W. 176 Wu 114 Y Yokogawa 261, 386 York W. 176 Yoshida 405, 408 Z Zarfl 109
ОГЛАВЛЕНИЕ
Стр.
Предисловие к пятому изданию........................................... 5
Из предисловия автора ко второму изданию .............................. 8
ОБЩАЯ ЧАСТЬ
Определение паразитизма; формы паразитов во* времени и пространстве . . 11
Объем и содержание паразитологии...................................... 18
Понятие об инвазии..........................................• . . . . 22
Организм как среда обитания .......................................... 24
Влияние паразитического образа жизни на паразитов..................... 33
Общий характер действия паразитов на хозяев........................... 37
Понятие о патогенности паразитов, паразитоносительстве и о резервуарах
вирусов...........................................•................ 41
Факторы становления организма хозяином паразитов...................... 43
Понятие о трансмиссивных болезнях..................................... 45
Пути циркуляции вирусов. Природная и антропургическая очаговость транс-
миссивных болезней ................................................ 47
Особенности географического распространения паразитов, паразитарных и
трансмиссивных болезней и принципы паразитологического райониро-
вания ............................................................. 52
Биологические и эколого-паразитологические обоснования борьбы с парази-
тарными и трансмиссивными болезнями................................ 57
Литература............................................................ 58
СПЕЦИАЛЬНАЯ ЧАСТЬ
Учение о паразитических простейших7 (медицинская протозоология)
Тип Protozoa,— простейшие
Класс Rhizopoda—корненожки (Sarcodina — саркодовые)............ 71
Отряд Amoebina — амебы.............................................. 71
Род Entamoeba................................................ 72
Род Dientamoeba............................................. 102
Род Dimastigamoeba.......................................... 103
Класс Mastigophora sive Flagellata — жгутиковые, или биченосцы . . . .
Отряд Protomonadida................................................ . .
Семейство Trypanosomidae — трипанозомовые.........................
Род Leishmania..................................................
Род Trypanozoma Gruby...........................................
Семейство Bodonidae...............................................
Род Bodo .......................................................
Семейство Embadomonadidae ........................................
Род Embadomonas...............................................
Семейство Chilomastigidae.........................................
Род Chilomastix.................................................
Семейство Cercomonadidae..........................................
Род Cercomonas..................................................
103
104
104
107
124
129
130
130
131
131
131
132
132
ОГЛАВЛЕНИЕ 519
Стр,
Род Tricercomonas............................................... 132
Семейство Trichomonadidae....................................... 133
Род Trichomonas............................................... 133
Отряд Diplomonadida..................................................... 139
Род Giardla (Lamblia) . . . •................................ 139
Класс Sporozoa — споровики.............................................. 141
Подкласс Telosporidia.................................................. 141
Отряд I. Coccidi а —кокцидии............................................ 142
Семейство Eimerildae.............................................. 145
Род Eimeria.................................................... 145
Род Isospora.................................................. 146
Отряд II. Haemosporidia — кровеспоровики ............................. 148
Семейство Plasmodidae............................................. 148
Род Plasmodium................................. *............. 153
Общее течение малярии и рецидивы...................... •............... 158
Источники заражения малярией..............'............................. 169
Заражение от человека..........................<........................ 169
Географическое распространение малярии.................................. 180
Социальное и экономическое значение малярии............................. 182
Лечение и лекарственная профилактика малярии..............•............. 188
Инструкция по медикаментозной терапии и профилактике малярии............ 190
Подкласс Neosporidia . . . . ...................................... 200
Отряд 1. Myxosporidia — миксоспоридии................................... 200
ОтРяД 2. Sarcosporidia —саркоспоридии................................... 201
Класс Ciliata— ресничные, или инфузории................................. 204
Род Balantidium................................................ 204
Другие предполагаемые паразитические инфузории кишечника человека . . 208
Литература ........................................................... 209
Паразитические черви человека (медицинская гельминтология)
Тип Vermes — черви
Подтип Platodes — плоские черви ....................................... 210
Класс Trematodes — сосальщики..................•.....................,. 211
Классификация сосальщиков .............................................. 219
Подкласс Mono gene а — Эктопаразитические сосальщики.................... 219
Подкласс Digenea — Эндопаразитические сосальщики ....................... 219
Отряд Pros о stomata — Переднеротые сосальщики.......................... 219
Подотряд 1. Amphistomata — Обоюдоустковые . . . •...................... 220
Семейство Paramphistomatidae...................................... 220
Род Watsonius S t i 1 е s etGoldberger......................... 220
Семейство Gastrodiscidae ......................................... 220
Род Gastrodiscoides Leiper..................................... 220
Подотряд 2. Distomata — Двуустковые..............>...................... 221
Семейство Fasciolidae — Листвяницы ............................... 221
Род Fasciola Linn.............................................. 221
Род Fasciolopsis Looss...........................•............. 228
Семейство Plagiorchidae......................................• . . 231
Семейство Echinostomatidae........................................ 231
Род Echinostoma Rudolphi ...................................... 232
Род Echinochasmus...........................•................. 235
Род Himasthla ................................................. 236
Семейство Dicrocoeliidae.......................................... 237
Род Dicrocoelium Dujardin...................................... 237
Род Eurytrema Looss............................................ 242
Семейство Opisthorchidae.......................................... 243
Род Opisthorchis Blanch........................................ 243
Род Clonorchis Looss........................................... 250
Род Pseudamphistomum Liihe..................................... 252
Семейство Heterophyidae........................................... 253
Род Heterophyes Cobb.........>................................. 253
Род Metagonimus Katsur......................................... 253
520
ОГЛАВЛЕНИЕ
Стр.
Род Nanophyetes..................................................... 257
Семейство Troglotrematidae.............................................. 258
Род Paragonimus Braun............................................... 258
Семейство Schrstosomatidae.............................................. 262
Род Schistosoma Weinland............................................. 262
Класс Cestoidea — ленточные черви............................................ 268
Подкласс Cestodaria—нерасчлененные ленточные черви.................. 269
Подкласс Cestoda—расчлененные ленточные черви............................... 269
Организация Cestoda................................................... . 269
Классификация Cestoda ....................................................... 277
Отряд Pseudophyllidea........................................................ 271
Семейство Diphyllobothriidae........f................................... 277
Род Diphyllobothrium Cobbold......................................... 278
Род Diplogonoporus L&nnenberg........................................ 297
Род Braunia Ldon.................................................... 298
Отряд Cyclophyllidea......................................................... 298
Семейство Anoplocephalidae............................................ 298
Род Moniezia Blanch............................................. . 299
Род Bertiella........................................................ 301
Семейство Dipylidiidae ... •.......................................... 301
Род Dipylidium Leuck................................................. 301
Семейство Davaineidae . . . .......................................... 305
Род Davainea Blanch..........................•............../ . . . • 305
Род Raillietina Fuhrm............................................... 305
Семейство Hymenolepididae.........................*..................... 305
Род Hymenolepis Weinl................................................ 305
Семейство Taeniidae............. . ................................. 310
Род Taenia L........................................................ 310
Род Echinococcus Rudolphi...................................• ... 330
Подтип Nematodes — круглые черви........................................ . 339
Действие паразитических нематод на своих хозяев............................. 345
Класс собственно круглых червей — нематоды. Nematoda ........................ 346
Aphasmidia . ............................................................... 347
Надсемейство Trichinelloidea.............................................. 347
Семейство Trichocephalidae.............................................. 355
Род Trichocephalus Schrank........................................... 355
Род Hepaticola Hall................................................. 359
Семейство Capillariidae...........................•..................... 361
Род Thominx.......................................................... 361
Надсемейство Dioctophymoidea.............................................. 363
Род Dioctophyme Coll.— Meygret ...................................... 363
Phasmidia................................................................ 365
Надсемейство Rhabditoldea................................................. 365
Род Strongyloides Gfassi............................................. 365
Свободноживущая генерация.................................................... 366
Паразитическая генерация.................................................. . . 366
Род Rhabditis Dujard................................................. 371
Род Turbatrix........................................................ 374
Надсемейство Anguillulinoidea ............................................ 374
Надсемейство Strongyloidea ............................................... 374
Род Oesophagostomum Molln............................................ 375
Род Ternidens Raill. et Henry........................................ 376
Род Syngamus Sieb.................................................... 376
Семейство Ancylostomatidae...................................... • ... 377
Род Ancylostoma Dubini — кривоголовка................................ 377
Род Necator Stiles................................................... 382
Заболевания, причиняемые анкилостомой и некатором (анкилостомоз и не-
катороз)................................................................. 382
Надсемейство Trichostrongyloidea.......................................... 395
Род Trlchostrongylus Looss........................................... 395
Род Haemonchus Cobb.................................................. 396
ОГЛАВЛЕНИЕ 521
Стр.
Род Mecistocirrus Rail. etHehry................................. 397
Надсемейство Meiastrongyloidea................................. . . 397
Род Metastrongylus Molin........................................ 398
Надсемейство Oxyuroidea............................................... 398
Род Enterobius Leach............................................ 398
Род Syphacia Seurat.................•... •...................... 403
Надсемейство Ascaroidea..........................................• . . 403
Род Ascaris L.................................................. '403
Заражение эскаридами и миграция в теле хозяина...........•............... 405
Болезнетворное действие взрослых аскарид................................. 409
Род Toxocara Stiles............................................. 413
Род Belascaris Leiper........................................... 414
Надсемейство Spiruroidea............................ . ....... 414
Род Gongylonema Molin........................................... 414
Семейство Gnathostomidae.................♦......................... 418
Род Gnathostoma Owen . . .................................... 418
Семейство Physalopteridae........................................ 420
Род Abreviata Tr a v.......................................... 420
Семейство Thelaziidae............................................ 421
Род Thelazia Bose............................................. 421
Надсемейство Filarioidea.............................................. 422
Род Wuchereria da Si It a Araujo. . ............................ 422
Род Onchocerca Diesing.......................................... 427
Род Acanthocheilonema Cobbold................................... 428
Надсемейство Dracunculoidea........................................... 429
Род Dracunculus Reich......................................... 429
Подтип Acanthocephala— колючеголовые черви, или скребни ...... 438
Класс Annelida — кольчатые черви....................................... 440
Подкласс Hirudinea — пиявки ............................................. 440
Общий характер влияния паразитических червей на организм................. 444
Общее (резорптивноё) влияние (глистные интоксикации)..................... 446
Пути заражения человека глистами......................................... 456
Организация плановой борьбы с гельминтозами в СССР ...................... 467
1. Организация гельминтологических пунктов......................... 467
2. Подготовка кадров .............................................. 469
3. Санитарно-профилактическая работа............................... 469
Общие основы медикаментозной борьбы с глистными инвазиями ............... 469
1. Подготовка к лечению............................................ 471
2. Дача противоглистного........................................... 471
3. Выведение паразитов из кишечника . ........................... 472
Инструкция по массовой борьбе с некоторыми гельминтозами................. 476
Инструкция по борьбе с аскаридозом....................................... 476
1. Предварительное обследование.................................... 476
2. Массовая дегельминтизация....................................... 477
Инструкция по борьбе с тениидозами....................................... 479
Определение и регистрация вышедших паразитов............................. 479
Методы обеззараживания финнозного мяса.................................. 481
Литература.......................................•....................... 481
Указатель русских названий животных ..................................... 483
Указатель латинских названий животных.................................... 488
Именной указатель русский................................................ 496
Предметный указатель .................................................... 499
Географический и этнографический указатель................................509
Указатель латинских терминов............................................ 513
Именной указатель иностранный............................................ 515
Е. Н. ПАВЛОВСКИ И.- РУКОВОДСТВО
ПО ПАРАЗИТОЛОГИИ .ЧЕЛОВЕКА
Печатается по постановлению
Редакционно-издательского совета
Академии Наук СССР
♦
Редактор Издательства Н. Й. Латышев
Технический редактор Е, Н. Симкина
Обложка художника
Д. А. Бажанова
РИСО АН СССР № 2174. А-22411. Изд. № 1091.
Тип. заказ № 468. Подп. к печ. 1/XII 1945 г. Формат
бум. 70X108. Печ. л. 328/< 4-11 вклеек. Уч.-изд. л. 50.
Тираж 8000
2-я типография Издательства Академии Наук СССР.
Москва, Шубинский пер., 10
ОПЕЧАТКИ И ИСПРАВЛЕНИЯ
Стра- ницы Строка Напечатано Должно быть
25 18 св. болезней болезни
55 15 » pontorus contortus
96 27, 29, 30,32 св. (тдбл. 1, рис... .) рис. 31
99 8 св. табл. II табл. III
100 11 сн. гематоксилином гематоксилином
105 16 СЙ. хорошо. хорошо (табл. IV, рис. 8)
107 15 св. переносчиками. переносчиками (табл. IV,
108 5 сн. вкладывают рис. 5, 6, 7). вкалывают
154 4 сн. микрогаметоцит макрогаметоцит
157 После таблицы Сравнительные изображе-
160 3 сн. хозяина ния паразитов малярии в толстой капле крови, часто применяемой при диагно- стике малярии, см. табл. III, рис. 22, 23 и табл. 10, рис. 1, 2, 3, 4. хинина
234 подпись к рис. 145 paroulum paraulum
236 4 сн. miichlensi \
237 подпись у muehlensi
243 к рис. 151 13 сн. miiehlensi J табл. V табл. VI
250 6 св. табл. V табл. VI
260 8 сн. ) 2 св. (табл. V, рис. 13) (табл. VI, рис. 3)
260 4 сн. табл. VI табл. VII
265 19 св. тканей яйца тканей, яйца
266 6 св. stibiatici stibiati
281 14 и 15 сн. плеоцеркоидов плероцеркоидов
283 1 сн. рис. 114 рис. 211
288 1 св. mimus minus
295 14 сн. Acanthocyclops, viridis Acanthocyclops viridis
301 3 сн. taenia Тaenia
332 23 сн. рис. 150 рис. 270
352 1 353 f табл. VIII Соидак; Сондак;
428 14 сн. banorofti bancrofti
439 8 св. Meloiontha Meloiontha
442 И св. горы. Каратау гор Каратау
442 6 сн. Limmatis Limnatis
450 22 св. эндоксинами эндотоксинами
Акад. Павловский.
Руковод. по паразитолог, человека