/



































Text
Государственное образовательное учреждение
высшего профессионального образования
«МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ЛЕСА»
С. П. Погиба, Г. А. Курносов, Е. В. Казанцева
МЕТОДЫ КОЛИЧЕСТВЕННОЙ ГЕНЕТИКИ
В ЛЕСНОЙ СЕЛЕКЦИИ
Рекомендовано к изданию Редакционно-издательским
советом университета в качестве учебно-методического пособия
к лабораторным работам для студентов специальностей
250201 Лесное хозяйство и
250203 Садово-парковое и ландшафтное строительство
4-е издание
Москва
Издательство Московского государственного университета леса
2008
П43
Рецензент: профессор М. Д. Мерзленко
Работа подготовлена на кафедре селекции, генетики и дендрологии
Погиба, С. П.
П43 Методы количественной генетики в лесной селекции : учеб.-методич.
пособие / С. П. Погиба, Г. А. Курносов, Е. В. Казанцева. - 4-е изд. -
М.; ГОУ ВПО МГУЛ, 2008. - 32 с.
Учебное издание
Погиба Светлана Петровна
Курносов Геннадий Анатольевич
Казанцева Елена Васильевна
МЕТОДЫ КОЛИЧЕСТВЕННОЙ ГЕНЕТИКИ
В ЛЕСНОЙ СЕЛЕКЦИИ
Редактор Е. Г. Петрова
Компьютерный набор и вёрстка Е. В. Казанцевой
Подписано в печать 28.01.2008. Формат 60x90 1/16. Бумага 80 г/мЕ. 2.
Ризография. Усл. печ. л. 2,0. Доп. тираж 200 экз. Заказ № 96.
Издательство Московского государственного университета леса. 141005, Мытищи-5,
Московская обл., 1-я Институтская, 1, МГУЛ.
E-mail: izdat@mgul.ac.ru
По вопросам приобретения литературы издательства ГОУ ВПО МГУЛ
обращаться в отдел реализации.
Телефон: (498) 687-37-14.
© С. П. Погиба, Г. А. Курносов,
Е. В. Казанцева, 2000
© ГОУ ВПО МГУЛ, 2008
3
Предисловие
Настоящие методические указания разработаны для студентов
дневного, вечернего и заочного обучения по специальностям 2604.00 и
2605.00, изучающих генетику и селекцию.
Явления наследственности и изменчивости на уровне отдельных
признаков организма изучают две ветви генетики - факториальная и
количественная. Термин количественная генетика был введен ДС.
Фальконером в 1960 году. Ее называют математической, биометрической,
статистической, хотя вероятностно-статистические и биометрические
подходы используют все отрасли генетики.
В лесной селекции используют ряд апробированных методов
генетического анализа. Кроме классического гибридологического анализа,
проводится генетический анализ количественных признаков древесных
растений. К количественным признакам относятся приросты, высоты,
диаметры, длина волокна, то есть хозяйственно-ценные признаки. Этот
анализ не ставит целью выявление конкретных генов, число их, типы
взаимодействия, локализацию. Задачей генетического анализа
количественных признаков являются прогнозирование уровня развития
признака в популяциях и опенка наследственна обусловленной
изменчивости. При этом наиболее часто применяемыми показателями
оценки количественных признаков являются арифметическое среднее,
дисперсия и коэффициент вариация.
В селекционной работе часто приходится сравнивать среднее
значение выборочных совокупностей, каждая из которых представляет
различные варианты отбора. Опенка достоверности различия между
средними двух эмпирических совокупностей решается с помощью критерия
Стьюдента. С помощью критерия / проверяется та или иная генетическая
гипотеза. Рассчитав дисперсию, селекционер сможет перейти к следуещему
этапу генетических исследований: выделить из фенотипической дисперсии,
генотипическую, то есть рассчитать коэффициент наследуемости в
широком смысле Н2, а затем рассчитать коэффициент наследуемости в
узком смысле h2, то есть рассчитать долю аддитивной варианты в общей
фенотипической. И, самое главное, рассчитать генетическое улучшение щи
однократном отборе R. Техника вычисления коэффициента наследуемости
может сильно варьировать в зависимости от изучаемого материала и
Особенностей применяемых методов, а также от способа выращивания
материала и учета влияния внешних факторов. В этой работе выбран
методики, нашедшие применение в лесном хозяйстве и озеленении. Для
закрепления материала после каждой темы даны задачи,
составлены по фактическим материалам, выбраным в литературных
источниках, авторефератах и на основании оригинальных воследовапй
авторов.
4
1. МЕТОД ОЦЕНКИ ВАРИАБИЛЬНОСТИ ПРИЗНАКА
При изучении изменчивости лесных древесных растений пользуются
биометрическими методами исследований, входящими в раздел
статистической генетики.
Основным показателем, характеризующим степень изменчивости
изучаемого признака, является коэффициент вариации S%.
В биометрии используют следующие условные обозначения:
1) N - объем генеральной выборки;
2) п - объем выборочной совокупности выборки;
3) АГ-числовое значение признака;
4) X (или М) - средняя арифметическая;
5) Xi - любой член вариационного ряда;
6) Хил - минимальное значение признака данного ряда;
7) Хю - максимальное значение признака в данной выборке;
8) lim - разность между Х^ и Х„1п ;
9) f - количество особей, имеющих одинаковое значение данного
признака;
10) К - класс (группа особей, имеющих одинаковое числовое
значение);
11) i - величина классового интервала;
12) X-знак суммирования;
13) б2 - дисперсия (варианса или варианта) для генеральной
совокупности;
14) б - среднее квадратическое отклонение для генеральной
совокупности;
15) S2 — дисперсия (варианса или варианта) для выборки;
16) S - среднее квадратическое отклонение для выборки;
17) S%(Cv, V) - коэффициент вариации;
18) Sx (mx)- ошибка средней арифметической;
19) t - нормированное отклонение;
20) td - показатель достоверности разницы между средними
арифметическими двух выборок (двух сортов, популяций и
т. п.).
Объем выборной совокупности (выборка п), извлеченной из
генеральной совокупности N должна быть достаточно велика. В лесном
хозяйстве для вычисления средних таксационных показателей в
средневозрастных и приспевающих древостоях он равен 200, в спелых - 100
- 150. В генетике обычно анализируют 25 - 100 и более растений. При этом
анализ выборки начинается с составления вариационного ряда, который,
затем может быть разбит на классы К. На основании показателей
вариационного ряда строят вариационные кривые.
5
Задача. В опытных культурах измерены высоты одной семьи ели
(Picea abies (L.) Karst.} (потомства от свободного опыления - полуснбсы
плюсового дерева) (табл. 1). Рассчитать, как варьирует признак. Можно ли
отобрать перспективные растения на быстроту роста в первой поломвв
жизни (ранняя диагностика). Построить гистограмму или график
распределения. Сравнить с потомством другого плюсового дерева, дать
заключение о том, отличаются ли эти деревья по данному признаку и какое
из них должно быть использовано в дальнейшей работе, если высота
сеянцев второго дерева 20,5 ± 0,4 см.
Таблица 1
_________________________________Высоты одной семьи ели европейской____
_______________________________________________________________________Н, см_
20 18 17 22 19 19 20 21 20 21 20 21 18 19 17 22 21
18 18 20 19 21 21 19 20 18 21 20 19 19 17 20 22 18
24 19 21 20 18 19 18 20 21 21 18 19 21 18 22 20 17
19 20 19 18 21 20 20 19 17 22 19 17 22 19 19 20 18 ;
19 20 20 17 19 21 20 19 20 19 22 20 19 19 20 20 17 ;
19 18 21 21 20 19 19 19 20 17 18 22 18 20 20 19 20 j
В приведенном примере вариационный ряд удобно сгруппировать в
классы, для чего рассчитывается классовый интервал по формуле
X шах-Хит
/_ К '
Число классов К опрыклястсл по табл. 2.
Таблица 2
Число классов
Объем выборки Чнуло классов
25-40 5- 6
40-60 6- 8
60-100 7-10
100 - 200 8-12
200 10-15
Дисперсия S2 рассчитывается по формуле
л-1
6
Среднее квадратичное отклонение вычисляется по формуле
V л-1
Коэффициент вариации S% рассчитывается по формуле
о
5% = 4x100%.
Для расчета основных параметров вариационного ряда удобно
пользоваться таблицей (табл. 3), в которую внесены результаты
вычисления.
Таблица3
Результаты вычислений основных параметров вариационного ряда
Границы классов f Xi-X (X-V (X,-X)2><f
17 8 136 -2,4 5,76 46,08
18 17 306 -1,4 1,96 3,02
19 26 494 -0,4 0,16 4,16
20 28 560 0,5 0,36 10,08
21 14 294 1,6 2,56 35,84
22 7 154 2,6 6,76 47,32
I n=Zf=100 X 1944 Z 176,8
Средняя арифметическая взвешенного ряда вычисляется по формуле
Дисперсия
rt 176
п-1 99
Среднее квадратичное отклонение
V п-1
Коэффициент вариации S% применяется для сравнения изменчивости
признаков растений одной семьи (сорта), разных признаков у особей одной
выборки или изменчивости одного и того же признака у разных семей
(сортов):
7
S% = ylOO% = ^-100% = 6,9%.
Оценка коэффициента вариации проводится по табл. 4 (Мамаев,
1970).
Таблица 4
Шкала изменчивости признаков
Уровень изменчивости Очень высокий Высокий Средний Низкий Очень низкий
Коэффициент вариации Более 40 21-40 10-20 I 7-12 I Менее 7 i
То есть у потомства плюсового дерева ели уровень изменчивости по
высоте низкий, выборка однородна по данному признаку, исходя из
значений S%, принятых в лесном хозяйстве.
Ошибка средней вычисляется по формуле:
S-=±-7==±-£= = ±0,13, то есть Х = 19,4±0,03.
' -А Лоо
Расчет нормированного отклонения t тюзяошкх определить, на
сколько минимальная и максимальная величина выборки отличается от
среднего арифметического в вариационном ряду:
Xi-X .
$ ’
1 5 1Д4 5 134
то есть данная выборка укладывается в пределы от -2а до +2ст.
Всюду, где имеют дело с массой случайных явлений, значение
показателя t при нормальном распределении бывает близким к 2 или 3. Это
правило формулируется как правило трех сигм. Все вариации, как бы они
ни различались, укладываются в пределы от X - За до Х+За (16а).
Согласно этому правилу, в пределах X ±1а находится 68,28 % вфнант
выборочной совокупности, в пределах ~Х 12а - 95,4 % , а в гфеделах
Х±3сг-99,73 % •
Установлены следующие три уровня значимости в порядке
возрастания строгости оценки достоверности биометрических показателей:
5 %, 1 %, 0,1 %. Им соответствуют в том же порядке вырастать строгости
оценки следующие доверительные уровни: 95 %, 99 % и 99,9 %.
Выраженные в долях единицы уровни значимости равны соотвегствешо
0,05; 0,01 и 0,001, а доверительные уровни в долях единицы представляется
8
следующими числами: 0,95; 0,99; 0,999. Для биологических исследований
во многих случаях достаточно принимать 5 %-ный уровень значимости
или 95 %-ный доверительный уровень (что одно и то же), при котором
считают достаточным, если подтвердится существенность выводов в 95
случаях из 100. Событие, которое имеет вероятность появления менее 5 %
(р<0,05), менее 1 % (р<0,01) практически не встречаются, поэтому их
относят к числу редко встречающихся и считают возможным пренебречь.
Однако в селекции, когда работают с очень редким признаком, возможно
брать материал и за пределами - Зо и +5<т.
В селекционных работах часто требуется сравнение двух величин.
Так, в нашем примере нам необходимо сравнить два «плюсовых» дерева по
потомству на быстроту роста. Рассчитанный пример дает нам среднее
значение высоты сеянцев плюсового дерева ели (одна семья) х = 19,4 ± о,4см,
высота сеянцев другого плюсового дерева ели (вторая семья)
х = 20,5 ± 0,4см. Достоверно ли отличается потомство этих двух деревьев по
испытуемому признаку?
Коэффициент достоверности рассчитывают по формуле
/ _ 19,4-20,5 1,1 1,1^
i]SF,+Sh А13а+0,42 6,196 0,4
Вычисленный коэффициент сравнивается с коэффициентом
достоверности Стьюдента, который при 5%-ном уровне значимости равен
1,96, при 1%-ном - 2,58, при 0,1%-ном - 3,30. Если полученный
коэффициент достоверности меньше критерия Стьюдента, то различие двух
выборок не достоверно.
В нашем примере G = 2,75, что свидетельствует о высокой степени
достоверности, то есть потомство второго дерева достоверно отличается от
потомства первого дерева по быстроте роста и может быть рекомендовано
для дальнейшей работы по этому признаку.
Задачи к теме 1: Методы оценки вариабельности признака
Задача 1 Средний вес семян пихты сибирской (Abies sibirica Ldb.) (X),
ошибка (S'*), коэффициент вариации признака (S%) представлены в табл. 5.
Рассчитать, достоверно ли отличие деревьев по весу семян? Какие деревья
могут быть отобраны для дальнейшей работы по данному признаку?
Задача 2 На пробной площади измерена высота 98 деревьев ели
европейской (Picea abies (L.) Karst.) (табл. 6). Вычислить коэффициент
вариации S%, построить график. Можно ли рекомендовать популяцию ели,
9
из которой взята данная выборка, для отбора на быстроту роста? Групповой
или индивидуальный отбор вы будете проводить?
Таблица 5
Средний вес семян пихты сибирской и его изменчивость в пределах дерева
Номер дерева X ±S7 5% j
1 7,9810,24 17,4 ।
2 8,8110,18 в,з 1
3 7,1710,10 9,9 ;
4 4,3710,06 8,6
5 5,5510,09 13,1 I
6 5,3610,09 12,9 j
Таблица 6
________________Высоты деревьев ели европейской__________________
____________________________Н, см________________________________।
21 20 18 17 22 19 19 20 21 20 21 20 21 18 19 17 22
18 18 18 20 19 21 21 19 20 18 21 20 19 19 17 20 22
17 21 19 21 20 18 19 18 20 21 21 18 19 21 18 20 22
19 19 20 19 18 21 20 20 17 22 19 17 22 19 20 18 19
17 19 20 20 17 19 21 20 19 20 18 22 20 19 19 20 20
20 19 18 21 21 20 19 19 19 20 17 18 22 18 20 20 19
Задача 3. Тополь советский пирамидашлый (R ’Sowietica pyramidalis)
представляет собой семью гибридов, полученных академком
А.С. Яблоковым от скрещивания тополя белого (Р. alba L.)c тополем Болле
(Р. Bolleana Louche). Ниже приведены средние высоты пирамидальшх и
раскидистых 3-летних гибридов в различных вариантах скрещишпдя.
Пирамидальные- 170, 179, 191, 146, 143, 133, 135, 128, 141, 158, 205,
157,120,114,157,236,175 см.
Раскидистые: 158, 110, 97, 109, 160, 148, 154, 114, 138, 144, 116, 150,
172,138,149 см.
Варьируют ли по высоте пирамидальные и раскидистые гибриды?
Достоверно ли их различие по высоте?
Задача 4. В Щелковском учебно-опытном лесхозе была «упирута»
селекционная инвентаризация и отобраны плюсовые деревья ей
европейской (Picea abies (L.) Karst). Отбор плюсовых деревьев ведется по
фенотипу. Проводится их проверка по потомству на быстроту роста.
Потомства деревьев №3 (семья) и №5 выращивались в идентячшх
условиях. Отличаются ли по данному признаку (быстрота роста) эти
деревья? Высоты одновозрастных саженце», см, представлена в табл. 7.
10
Таблица 7
Высоты одновозрастных саженцев ели европейской______
№ плюсо- вого дерева Высоты саженцев, см
Дерево 3 Т86 190 165 182 182 182 180 173 157 179 164
146 174 144 156 156 165 160 160 161 144 153
1152 151 173
Дерево 5 162 163 190 188 147 146 145 157 162 186 175
147 145 145 155 174 180 148 175 145 144 153
165 141 164
Задача 5. В пределах одной популяции деревья пихты сибирской
(Abies sibirica Ldb.) различаются по величине шишек, которая может быть
использована для их селекционной характеристики. Рассчитать, отличаются
ли два дерева пихты сибирской по длине шишек, если известны X ± Sр, S %
(табл. 8).
Таблица 8
Средняя величина шишек пихты сибирской и уровень ее изменчивости
Год урожая Дерево 1 Дерево 2
X±Sj, см S% У±£*,см S%
1960 6,37±0,15 10,6 6,19±0,08 13,3
1961 6,10±0,20 12,6 - -
1962 6,51±0,10 12,2 5,89±0,20 9,5
1963 8,71±0,05 10,1 6,64±0,06 14,3
1964 6,43±0,07 7,1 6,00±0,08 12,2
1965 6,02±0,16 9,0 4,71±0,07 13,2
1966 6,80±0,06 11,0 6,46±0,07 12,9
1967 - - 5,80±0,19 Н,6
1968 8,05±0,07 13,2 7,16Ю,04 11,1
1969 7,50±0,10 14,2 6,46±0,06 8,7
1970 6,ЗОЮ,09 13,7 5,90±0,06 10,9
Задача 6. Испытательные культуры ели европейской (Picea abies (L.)
ПП1 „ созл3®1 двадцать лет назад. О™ представляют собой
плюсовых деревьев ели различного географического
ЧРОвсхождения. Отобрать по высоте наиболее перспективные семьи и дать
и
оценку материнским деревьям. Высоты потомства плюсовых деревьев
представлены в табл. 9.
Таблица 9
Высоты потомства плюсовых деревьев ели различного
географического происхождения
№ плюсо- вого дерева Высоты потомства, см |
Московская область
Дерево 5 6,9 6,0 7,5 6,3 6,8 6,8 6,9 6,9 6,3 6,8 7,0 6,0 7Д
6,4 6,9 6,8 7,5 7,0 6,9 7,0 6,5 5,9 6,9 5,2 6,8 7,0
7,1 5,3 7,0 5,2 6,3 5,5 6,4 7,0 6,8 6,8 6,8 6,1 5,3
5,5 6,0 6,0 5,3 7,3 6,9 7,3 6,8 6,0 6,9 7,4
Дерево 6 7,9 6,8 7,7 6,9 6,9 5,8 6,6 6,9 8,0 6,8 6,8 6,8 7,8
6,5 7,0 6,6 6,8 7,0 7,9 7,2 6,7 72 5,1 7,1 6,9 7,1
5,2 7,2 6,8 7,5 6,9 7,5 6,5 7,2 4,9 7,2 6,9 5,4 6,0
6,6 6,0 6,8 6,6 6,5 6,6 7,4 7,2 6,0 5,2 6,8
Дерево 7 7,3 6,8 7,4 52 5,8 5,7 5,7 6,5 7,4 52 7,4 6,8 6,8
6,0 5,9 6,5 6,5 6,8 6,8 6,0 7,0 6,5 6,8 5,9 5,9 5,8
6,9 6,0 5,9 52 72 6,8 6,9 5,9 52 5,1 7,0 5,9 7,0
5,7 6,4 7,0 7,0 52 5,7 6,8 5,9 5,5 6,4 52
Белоруссия
Дерево 11 7,6 6,7 4,8 7,9 7,1 5,0 7,6 5,8 7,6 7,6 7,6 6,9 52
6,6 6,9 6,9 6,7 7,0 7,6 32 62 32 4,9 5,8 7,0 6,9
8,2 7,9 6,9 6,8 6,1 6,8 4,7 5,4 7,0 62 6,8 7,1 62
6,3 6,8 6,9 4,7 6,9 7,9 6,4 6,9 6,9 6,8 8,9
Дерево 12 6,4 7,5 7,1 6,9 8,0 8,4 32 7,1 6,4 8,4 7,4 7,8 7,1
6,4 6,8 8,2 8,1 7,8 7,4 32 8,1 6,9 8,2 62 7,4 4,8
6,2 6,9 8,1 8,0 8,3 8,4 6,8 6,3 6,9 4,1 6,8 8,4 6,9
7,9 7,9 8,3 6,5 4,3 7,4 9,0 6,3 8,0 6,2 8,4
Костромская область
Дерево 3 4,6 6,6 5,5 5,9 4,9 5,5 5,5 4,6 42 5,9 62 62 6,7
5,5 7,2 6,9 6,9 6,5 4,0 4,9 6,8 6,7 6,9 5,6 72 42
4,0 6,7 6,2 5,4 7,0 5,5 5,8 5,5 5,4 62 6,0 5,5 42
4,0 6,6 5,5 5,9 5,6 5,5 4,2 42 5,8 4,0 6,4
Дерево 4 5,3 5,4 7,6 6,9 6,9 6,9 7,0 5,4 5,4 5,3 5,4 5,5 7,4
6,1 7,0 5,2 5,4 5,5 5,5 7,0 6,3 5,5 6,8 6,8 7,5 6,6
6,7 6,3 5,2 6,3 5,0 7,3 6,8 6,6 6,9 6,7 6,3 5,0 6,9
5,5 5,3 5,3 6,8 6,8 7,0 6,6 6,8 6,8 6,7 6.8
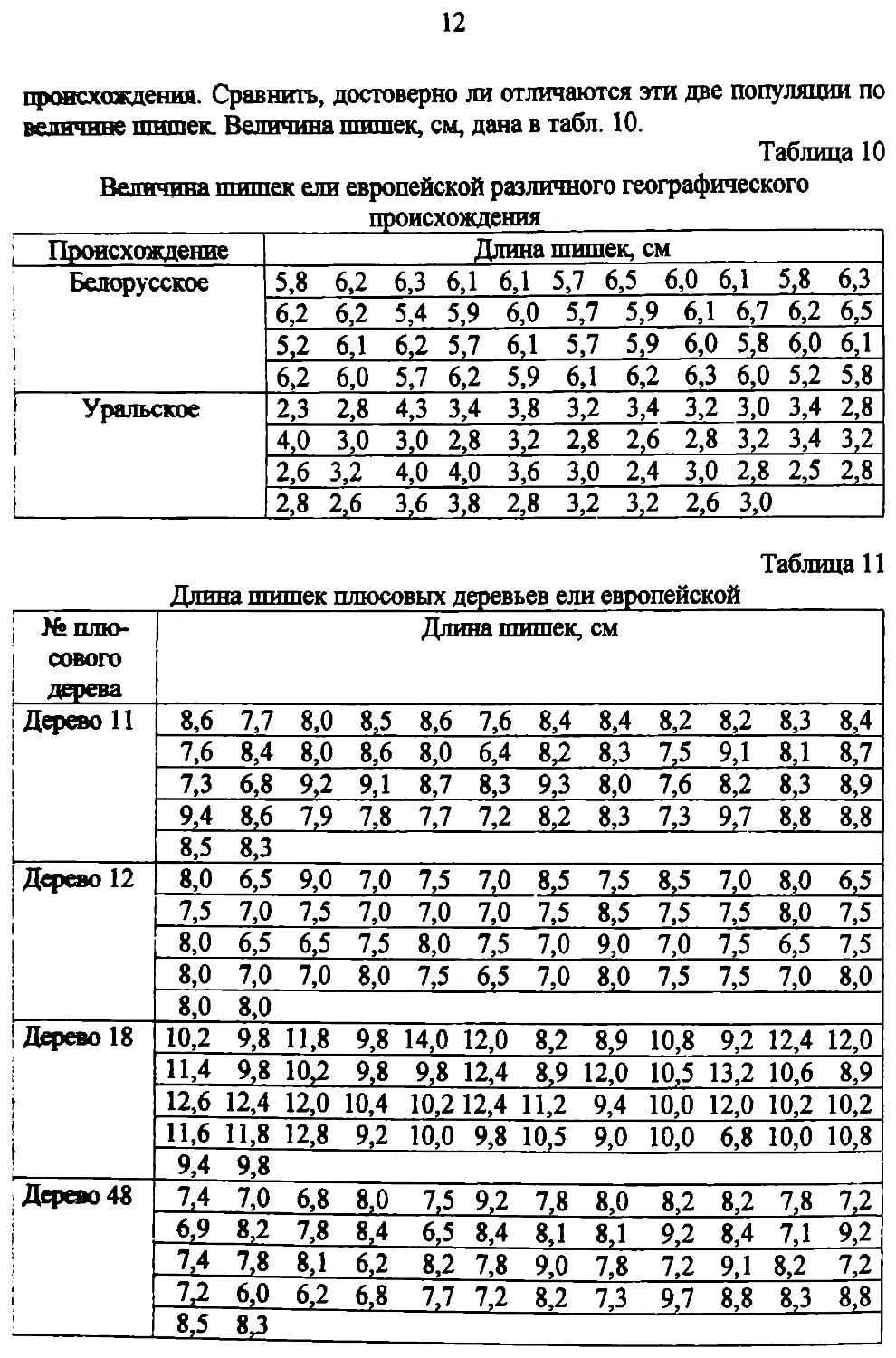
Задача 7. Рассчитать коэффициент вариации длины шишек с—
европейской (Picea abies (L.) Karst) белорусского и уральского
12
происхождения. Сравнить, достоверно ли отличаются эти две популяции по
величине шишек. Величина шишек, см, дана в табл. 10.
Таблица 10
Величина шишек ели европейской различного географического
Происхождение Длина шишек, см
Белорусское 5,8 6,2 6,3 6,1 6,1 5,7 6,5 6,0 6,1 5,8 6,3
6,2 6,2 5,4 5,9 6,0 5,7 5,9 6,1 6,7 6,2 6,5
5,2 6,1 6,2 5,7 6,1 5,7 5,9 6,0 5,8 6,0 6,1
6,2 6,0 5,7 6,2 5,9 6,1 6,2 6,3 6,0 5,2 5,8
| Уральское i 2,3 2,8 4,3 3,4 3,8 3,2 3,4 3,2 3,0 3,4 2,8
4,0 3,0 3,0 2,8 3,2 2,8 2,6 2,8 3,2 3,4 3,2
2,6 3,2 4,0 4,0 3,6 3,0 2,4 3,0 2,8 2,5 2,8
2,8 2,6 3,6 3,8 2,8 3,2 3,2 2,6 3,0
Таблица 11
№ плю- сового ! дерева Длина шишек, см
{Дерево 11 8,6 7,7 8,0 8,5 8,6 7,6 8,4 8,4 8,2 8,2 8,3 8,4
1 i 1 7,6 8,4 8,0 8,6 8,0 6,4 8,2 8,3 7,5 9,1 8,1 8,7
7,3 6,8 9,2 9,1 8,7 8,3 9,3 8,0 7,6 8,2 8,3 8,9
9,4 8,6 7,9 7,8 7,7 7,2 8,2 8,3 7,3 9,7 8,8 8,8
8,5 8,3
Дерево 12 8,0 6,5 9,0 7,0 7,5 7,0 8,5 7,5 8,5 7,0 8,0 6,5
7,5 7,0 7,5 7,0 7,0 7,0 7,5 8,5 7,5 7,5 8,0 7,5
8,0 6,5 6,5 7,5 8,0 7,5 7,0 9,0 7,0 7,5 6,5 7,5
8,0 7,0 7,0 8,0 7,5 6,5 7,0 8,0 7,5 7,5 7,0 8,0
8,0 8,0
! Дерево 18 10,2 9,8 11,8 9,8 14,0 12,0 8,2 8,9 10,8 9,2 12,4 12,0
11,4 9,8 10,2 9,8 9,8 12,4 8,9 12,0 10,5 13,2 10,6 8,9
12,6 12,4 12,0 10,4 10,212,4 11,2 9,4 10,0 12,0 10,2 10,2
i 11,6 11,8 12,8 9,2 10,0 9,8 10,5 9,0 10,0 6,8 10,0 10,8
9,4 9,8
'Дерево 48 7,4 7,0 6,8 8,0 7,5 9,2 7,8 8,0 8,2 8,2 7,8 7,2
6,9 8,2 7,8 8,4 6,5 8,4 8,1 8,1 9,2 8,4 7,1 9,2
7,4 7,8 8,1 6,2 8,2 7,8 9,0 7,8 7,2 9,1 8,2 7,2
7,2 6,0 6,2 6,8 7,7 7,2 8,2 7,3 9,7 8,8 8,3 8,8
8,5 83
13
Задача 8 Измерено по пятьдесят шишек с 4 плюсовых деревьев ели
европейской (Picea abies (L.) Karst.) (табл. 11). Рассчитать как варьирует
длина шишек в пределах дерева (эндогенная изменчивость), отличаются ли
деревья по данному признаку, достоверны ли эти отличия. Может ли длина
шишек использоваться как систематический признак?
2. ОЦЕНКА НАСЛЕДОВАНИЯ КАЧЕСТВЕННЫХ ПРИЗНАКОВ
Для того чтобы установить, достоверно или случайно наблюдаемое
явление теоретическому, в биометрии используются особые критерии, к
которым относится критерий Пирсона или критерий Хи-квадрата Он
представляет собой сумму квадратов отклонений эмпирических частот р
от частот теоретических или ожидаемых р', ытххянууъ к теоретическим
частотам р'\
^_Пр~р)2
р'
При этом используется нулевая теория. Предполагают, что
несоответствие эмпирических и теоретических частот случайно, то есть
между этими частотами никакой разницы нет. Если же Х(р -pi* ® равно О,
то может изменяться от 0 до ® , поэтому критерий Пирсона
фактический // сравнивают со стандартным И если то
отклонение носит случайный характер для принятого уровня значимости с
учетом степеней свободы К, что проверяется по табл. 12.
Таблица 12
Значение У при разных степенях свободы (по Фишеру с
сокращениями)________________________________________
Число степеней свободы Вероятность р
0,99 0,95 0,90 0,75 0,50 0,25 0,10 0,05 0,025 0,01
1 — — 0,02 0,1 0,45 1,32 2,71 3,84 5,02 6,63
2 0,02 0,10 0,21 0,58 1,39 2,77 4,61 5,99 7,38 9,21
3 0,11 0,35 0,58 1,21 2,37 4,11 6,25 7,81 9,35 1134
4 0,30 0,71 1,06 1,92 3,36 5,39 7,78 9,49 11,14 13,28
5 0,55 1,15 1,61 2,67 4,35 6,63 9,24 11,07 12,83 15,09
При этом должны соблюдаться следующие условия:
1) вариационный ряд должен включать не менее 50 варианте®;
2) крайние классы не должны быть менее 5, инпе они
объединяются с частотами соседних классов и по вторичному числу классов
устанавливается число степеней свободы;
14
3) при оценке эмпирических распределений по нормальному или
биномиальному распределению число степеней свободы равно: K=N-3\
4) по закону Пуассона K=N-2\
5) в генетических расчетах число степеней свободы в простейших
спутад равно числу классов, уменьшенному на единицу.
Задача, В природе имеются красношишечные и зеленошишечные ели
(Picea abies (L.) Karst.). При скрещивании этих двух форм во втором
поколении F2 было подучено 407 особей с зелеными шишками и 143 - с
красными. Соответствует ли данное расщепление ожидаемому по схеме
моногибридного скрещивания при полном доминировании 3:1 ?
В приведенной задаче имеется два класса: зеленошишечные и
красношишечные формы ели, значит, число степеней свободы /С=2-1=1.
£я при Х=1 и /7=0,05 равен 3,84. Общая численность семян 550, из них э/д
ожидается зеленых, то есть (3x550) / 4= 412,5 и *4 - красных, то есть
14x550=137,5 - это теоретические ожидаемые частоты. Сравним эти
величины с полученными в опыте и рассчитаем критерий / (табл. 13).
Таблица 13
Расчет критерия
Показатели Количество шишек Всего
зеленых красных
Данные опыта р 407 143 550
Ожидаемые р' 412,5 137,5 550
Ряз&жльр-р' 5,5 5,5
(р-рт 30,25 30,25
(р-р’У р' 30,25 412,5 ^ = 0,22 137,5 Л=0,29
йЛг-3,8; значит, расщепление по окраске стробилов
соответствует 3:1, а отклонение носит случайный характер.
Метод ^-квадрата дает возможность сравнивать различные
численные отклонения при разных объемах выборок в одном масштабе, но
он не применим к значениям, выраженным в процентах и относительных
числах.
Задачи к теме 2: Оценка наследования качественных признаков
Задача 9. Дуб черешчатый (Quercus robur L.) имеет две феноформы:
Ра*4™010 ргаесох) и позднюю (f. tardiflord), которые передают это
свойство своему потомству. Поздняя форма устойчива к весенним
заморозкам. В Тульских засеках имеются обе формы. При селекционной
15
инвентаризации на пробной площади из 768 деревьев: 585 - ранняя форма;
183 - поздняя форма. Соответствует ли фактически наблюдаемое
расщепление теоретическому 3:1?
Задача 10, С целью увеличения выхода мужских особей у тополя
белого (Populus alba L.) была проведена обработка семян мутагеном. В
результате было получено 227 мужских и 220 женских особей. Является ли
увеличение мужских особей результатом мутаций или отклонение носит
случайный характер? (Теоретическое расщепление 1:1).
Задача 11. Академиком АС. Яблоковым при получении
пирамидальных тополей были использованы для скрепшияиия тополь белый
(Populus alba L.) и тополь Болеана (Populus Bolleana Lauch.). Первый - с
раскидистой формой кроны, второй - пирамидальный. При этом в 1937 году
был получен 361 саженец, из них пирамидальных 131; в 1938 году - 217
саженцев, из них пирамидальных 76; в 1939 году - 394 саженца, из них
пирамидальных 184. Соответствуют ли полученные данные расщеплению
при моногибридном скрещивании 3:1.
Задача 12. Чернокорые особи Betula pendula Roth, встречаются редко.
При скрещивании белокорой особи с чернокорой из 100 деревьев 57
белокорых и 43 чернокорых. Ожидаемое расщепление 1:1. Соответствует
ли теоретически ожидаемое расщепление практическому?
Задача 13. При скрещивании гладкокорой березы повислой (Betula
pendula Roth.) с грубокорой березой пушистой (Betula pubescens Ehrh.) во
втором поколении F2 произошло расщепление на 315 гладкокорых
плакучих, 108 гладкокорых раскидисты^ 101 грубокорых плакучих и 2
грубокорых раскидистых. Установить, соответствует ли это расщепление
дигибридному при полном доминировании 9:3 :3 :1 ?
Задача 14. При свободном ппыпении кяреттьской березы (Betula
pendula Roth var. carelica (Merci.) Hamet-Ahti) и? 720 выращенных растений
320 имели узорчатую текстуру, остальные - безузорчатую. Соответствует
ли это расщепление теоретически ожидаемому - 50 % узорчатых, 50 %
безузорчатых, то есть 1:1?
Задача 15. Клен остролистный ‘Швеллера’ (Acer platanoides
‘Schwedleri) имеет красную окраску7 листьев в первой половине лета. По
данным исследователей, этот признак наследуется до 75 %. Из 6798
растений, выращенных из семян, собранных с клена ‘Швеллера’, 1700
имеют зеленые листья. Соответствует ли это расщепление теоретически
ожидаемому при моногибридном скрещивании при полном доминяроватаи
3:1?
Задача 16. Проведя анализ шишек сосны обыкновенной (Pinas
sylvestris L.) по апофизу студенты установили, что в Кировской области
соотношение их следующее: вдавленный апофиз (f plana) - 29 штук;
выпуклый апофиз (f gibba) - 52 штуки; крючковатый (f. reflexa) - 31 штука,
в Карелии соответственно 31 : 54 : 10; в Московской области 42 : 86 : 39; в
16
Крыму - 5 : 48 : 34. Соответствует ли это расщепление расщеплению при
неполном доминировании 1:2:1?
Чятача 17. При селекционной инвентаризации осинников Латвии
было выделено три формы осины (Popuhis tremula L.) по цвету коры:
зеленые, светло-серые и темно-серые в соотношении 144:290:120.
Соответствует ли это соотношение расщеплению при неполном
доминировании (1:2 :1)?
Задача 18. Ель европейская (Picea abies (L.) Karst.) в Литве
представлена тремя фенологическими формами: рано распускающимися
пг^мми промежуточными и позднораспускающимися в соотношении 298 :
506 : 196. Соответствует ли это соотношении расщеплению при неполном
доминировании (1:2:1)?
Задача 19. Аллели генов Аа контролируют окраску листьев, Вв -
форму кроны. В F2 дигибридного скрещивания получено расщепление по
фенотипу 82АВ : 114 в : ЗЗаВ : 8ав. Проверить с помощью х2 его
соответствие ожидаемому отношению 9 : 3 : 3 :1.
Задача 20. Статистические показатели урожайности деревьев пихты
сибирской (Abies sibirica Ldb.) отличаются в разных типах леса. Так, число
шишек на одном дереве в пихтаче крупнотравном 38,3; в пихтаче
кустарниково-разнотравном - 40,6; в пихтаче папоротниковом - 65,3.
Требуется проверить методом х2 достоверность различной урожайности
пихты сибирской от типа леса.
Задача 21. В кроне сосны обыкновенной (Pinus Silvestris L.) были
подсчитаны мужские и женские стробилы в количестве 205 и 206
соответственно. Соответствует ли эти численности теоретически
ожидаемому соотношению 1:1, и если нет, то к какому типу сексуализации
относится данное дерево?
Задача 22. По данным Калининградского областного ботанического
сада при свободном опылении клена ложноплатанового пурпурно-
листного (Acer pseudoplatanus 'Purpurascens1) в потомстве F] краснолистных
растений насчитывалось 49,7 %, в то время как по данным Лесосеменной
(житной станции Липецкой области этот признак наследовался до 30 %.
Составьте задачи и с помощью критерия Пирсона проверьте, какому типу
скрещивания соответствует расщепление в потомстве моногибридному 3 :
1, анализирующему 1:1?
Зуяча 23 Растения с измененной окраской листьев (красные,
желтые, пурпуровые) можно отбирать по окраске семядольных листьев. У
Лионе Нсдавецкого (Matus rtiedzwetzkyana Dieck.) в семенном потомстве
цри свободном опылении красные листья наследуются в 46,3 %.
Соответствует ли это расщепление теоретически ожидаемому 1:1?
Зздича 24. Клен остролистный, имеющий темно - красную окраску
лвспев в конце лета, известен под названием клена Рейгенбаха (Acer
piatanoides Lf. Retienbachiihort). В Лесостепной опытной станции
17
Липецкой области при размножении его семенами от свободного
опыления на 690 зеленолистных растений приходилось 312 растений с
красными листьями. Проверьте с помощью критерия Пирсона,
соответствует ли это расщепление расщеплению при моногибридном
скрещивании 3 :1?
Задача 25. У туи западной [Thuja occidentalis L.) имеется около 83
форм и культиваров, которые широко используются в озеленении. При
семенном размножении Th. о. ‘Globoза’, имеющей шаровидную форму
кроны из 150 растений 75 растений имели шаровидную, 37 растений -
шаровидно-карликовую и 38 растений - пирамидальную формы. К какому
типу наследования относится это расщепление? Докажите с помощью
критерия Пирсона.
Задача 26. Туя западная ‘Белокончиковая’ (Thyia occidentalis ‘Albo-
Spicatar) представляет собой растение пирамидальной формы с белыми
концами побегов. При семенном размножении 48,5 % растений имели белые
концы побегов и 49,5 % - темно-зеленые. Соответствует ли это
расщепление расщеплению при анализирующем скрещивании?
3. ОЦЕНКА НАСЛЕДОВАНИЯ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ
1) Методы оценки количественных признаков и расчет коэффициента
наследуемости в широком смысле Н2
Лесным селекционерам очень важно знать, наследуются ли признаки, с
которыми ему предстоит работать, или нет. При этом чаще всего лесоводы
работают с признаками, которые можно измерить: высота и диаметры,
приросты, масса плодов и семян, величина их, масляничность и содержание
белков у орехоплодовых, длина волокна у ели, пихты, осины. Признаки,
которые можно измерить, называются количественными ( в отличие от
качественных - окраска, цвет, опушенность и т. д.). Количественные
признаки контролируются чаще всего неаллельными, аддитивными
(суммарными) генами, и они контролируют практически все хозяйственно
ценные признаки. Для того чтобы определить, наследуется ли данный
признак или нет, пользуются методами математической статистики, в
основе которой лежит теория вероятностей. При этом чаще всего
пользуются методом дисперсионного анализа, определяя долю
наследственных (генотипических) и ненаследственных (паратипических,
экологических) факторов в изменчивости признака. Исследуемые признаки
проявляются через фенотипы, поэтому можно сказать, что фенотипическая
дисперсия S2^ равна сумме дисперсий генотипической Sfg и экологической
s2.-.
3^=3^+Л
18
Дисперсия рассчитывается по формуле
л-1
где S2 - варнанса для выборки;
х - среднее значение признака;
х, - любой член вариационного ряда;
л - объем выборочной совокупности выборки (или число измерений).
Наиболее просто наследуемость того или иного количественного
прганака может быть определена в популяции, состоящий из нескольких
различающихся в генотипическом отношении клонов. На одном участке
выращивается несколько клонов (черенкованных или привитых), а затем
сравнивают изменчивость между клонами и внутри них. Вследствие того,
что внутри отдельного клона генотипическая изменчивость отсутствует, то
S*g= 0, исходя из формулы (1) будем иметь 5^= S2^ то есть внутри клона
фенотипическая дисперсия признака равна экологической. Внутри
популяции, то есть между клонами, фенотипическая варнанса определяется
по всем растениям. Средняя же изменчивость признака внутри клонов дает
экологическую дисперсию. Тогда доля генотипической дисперсии в общей
фенотипической равна
s^-s2,.
Она обозначается через 1г (heritability - наследуемость) и называется
коэффициентом наследуемости в широком смысле, то есть рассчитав ее, мы
узнаем, наследуется ли данный признак или его изменения зависят от
экологических условий.
Задача. Определить коэффициент наследуемости высоты растений в
экспериментальной популяции, состоящей из трех клонов тополей (по
Петрову,1973) (табл. 14).
Таблица 14
Расчет экологической дисперсии
Клон 1 Клон 2 Клон 3
Высота росте^й Х.-Х Высота растений Xj -X Высота растений \-х fo-*)2
10 -1 1 12 -1 1 13 1 1
13 2 4 14 1 1 И -1 1
11 0 0 13 0 0 10 -2 4
12 1 1 13 0 0 13 1 1
L 10 . -1 1 14 1 1 12 0 0
! Ю -1 1 12 -1 1 13 1 1
^=8 £=4 £=8
19
— 10 + 13+11+12+10+10
Х'= 6
— 12+14+13+13+14+12
*2~ 6
— 13 + 11+10 + 13 + 12 + 13
_2 _ 1,6+0,8+1,6
----------------- .
Таблица 15
Растет фенотипической дисдерсии
Высота растений в популяциях X, - X (X; -X)2
10 -2 4
13 1 1
11 1 1
12 0 0
10 -2 4
10 -2 4
12 0 0
14 2 4
13 1 1
13 1 1
14 2 4
12 0 0
13 -1 1
11 -1 1
10 -2 4
13 1 1
12 0 0
13 1 1
Z216 Z32
20
rJ _ 32 i оо • ij2 = q 292
s>irr1’“’ № ~w—°292'
Вычисляемое значение коэффициента наследуемости свидетельствует
о тем, что изменчивость высоты растений в изучаемой экспериментальной
популяции в значительной большей степени (70,8 %) обусловлена
условиями внешней среды, чем наследственными различиями клонов,
входящих в состав этой популяции (29,2 %).
Считается, что если it > 0,5 (50 %), то признак обусловлен
генетически.
Роне В. М. (1980) предложила более простой способ расчета Н2 для
вегетативно размноженных растений. Генетическая составляющая при этом
была обозначена как Н2, а экологическая - е2. Если 5^=1, тогда 5^=
/f+e2, или 1= 1?+е\ то Н2=1-е2. Если е2=0, то Н2=1, то есть проявление
признака не зависит от экологических условий. При е2 не равно 0,
выраженность признака в клоне меняется, и сопоставление интервала
изменчивости признака в клоне и популяции можно выразить, так:
Lim d = Xmax - Xmin; Limp=Xmax-Xmin,
- где max и min - значения признака в пределах клона и популяции.
е
тг—, тогда Я =1-е
11ТП
lim '
Этот способ не требует нормального распределения признаков -
явления, редкого в природных популяциях.
Рассчитаем предложенную задачу через лимиты:
max -min
13-10 , Л__
max min м
max - min
14-12
14-10
= 1-0,5 = 0,5;
13-10
14-10
= 1-0,75 = 0,25
21
При расчете коэффициента наследуемости в широком смысле через
лимиты Я2<0,5, следовательно признак не является генетически
обусловленным и в большей степени зависит от экологических факторов.
2) Методика расчета коэффициента наследуемости в узком смысле Н2
Узнав, наследуется ли данный признак, селекционер должен знать,
какие гены контролируют его. Генотипическая дисперсия Sfg представляет
собой сумму дисперсий аддитивных генов S2^, эпистатических 5^ и
доминантных :
S2g = S2add + S2e + S2d.
Райт С. разработал теоретическу модель, выражающую связь
фенотипов с генотипами между родственными особями При этом
рассчитывается коэффициент наследуемости в узком смысле А2. Он
показывает долю адлидиштых генов в генотипической составляющей,
контролирующих количественные признаки. При этом исходят из
следующих допущений:
1) изучаемая популяция находится в состоянии генотипического
равновесия;
2) количественные признаки определяются аддидивным действием
генов;
3) связь между фенотипом родителей и потомков может быть
выражена с помощью коэффициента корреляции, то есть эта
связь является прямолинейной.
При этом h2 = 2г^ - для сибсов; А2 = 4г^ - для полусибсов,
где г - коэффициент корреляции родитель - потомок; х - потомок; у -
родитель. Коэффициент корреляции рассчитывается по формуле
Задача. Как известно, карельская береза образует несколько
габигуальных форм: высокоствольные, короткоствольные, кустарниковые и
кустовидные. С каждой из этих форм собраны семена, и выращены в
однородных условиях отдельно по семьям сеянцы в количестве 12000 штук.
Высоты материнских деревьев у, см и двухлетних сеянцев х, см измерены и
средние значения этих показателей представлены в табл. 16.
22
Таблица 16.
Расчет коэффициента наследуемости в узком смысле h_
у, см У\'У (У,-У? X, см х,-х
700 -216,67 46945,89 11,86 -3,78 14,29 819,01
750 -166,67 27778,89 15,74 0,1 0,01 -16,67
! 1700 783,33 613605,89 17,76 2,12 4,49 1660,66
1 1100 183,33 33609,89 19,24 3,6 12,96 659,99
1 700 -216,67 46945,89 20,81 5,17 26,73 1 -1120,18
600 -316,67 100279,89 15,61 ' -0,03 0,0009 9,50
550 -366,67 134446,89 13,81 -1,83 3,35 671,01
750 п -166,67 27778,89 13,26 -2,38 5,66 396,67
1400 483,33 233607,89 12,70 -2,94 8,64 -1420,99
58250 £1265000,01 £140,79 5276,13 £1659
1659______________1659 1659
^1265000,01x76,13 ' ^96304450,76 ~ 9813,48
При свободном опылении (полусибсов) полученный коэффициент
корреляции умножается на 4.
Таким образом, й2 =4^=4*0,169=0,676.
Надежность вычисления коэффициента корреляции зависит от числа
саженца N >100.
Высокий показатель коэффициента наследуемости в узком смысле
(к2=0,676) говорит о том, что высоту карельской березы контролируют
аддитивные (суммарные) гены, так как й2 > 0,25 (Погиба, 1988). Если й2 <
0,25, то признак контролируется другими генами (Чудный, 1985).
3) Методика расчета генетического улучшения
Определив, наследуется ли изучаемый признак Н2, какие гены
млнротируют его фенотипическое проявление й2, лесной селекционер
переходит к следующему этапу работы - отбору.
Дм того чтобы определить генетическое улучшение при отборе по
изучаемому признаку, его рассчитывают для генетически однородного
материала (чистых линий, клонов) через коэффициент наследуемости в
широком смысле Н3 по формуле
R=S*rf,
где R—генетическое улучшение;
23
S- селекционный дифференциал;
Н2 - коэффициент наследуемости.
При семенном размножении генетическое улучшение рассчитывается
через коэффициент наследуемости в узком смысле А7, при этом
учитывается, от контролируемого или от свободного опыления получены
семена.
Для контролируемого опыления
R=S*tf.
Для свободного опыления
Селекционный дифференциал S представляет собой разность между
максимальным значением признака по которому ведут отбор, и его
средним значением в популяции X.
Задача. В популяции ели европейской подеревно измерена величина
шишек, она колеблется от 6,5 до 11,2 см. Средняя длина шишки 8,1см,
коэффициент наследуемости длины тпиттпси if = 0,80. Рассчитать
эффективность отбора или генетическое улучшение по данному признаку,
если в селекционных целях отбираются особи с наиболее крупными
шишками.
Я7=0,80.
<5=11,2-8,1=3,1.
Я= 3,10x0,80=2,48.
Если принять среднюю величину шишки за 100 %, тогда х
рассчитывается при помощи пропорции
8,1 см-100 %
2,48 см - х
Таким образом, в результате однократного отбора генетическое
улучшение равно 2,48 см, или 30,6 %. Величина шишки коррелирует с
количеством семян, а значит, на этот признак можно вести селекцию.
24
к меме 3; Оценка наследования количественных признаков
Ъшта 27. На питомнике МГУЛа имеется коллекция привитых форм
вфелыхой березы (Betula pendula Roth var. Carelica (Merci.) Hamet-Ahti)
трех клонов. Определить, наследуется ли высота карельской березы Н2 и
геяетнчесхое улучшение R, если вести селекцию на быстрорастущие
формы. Коэффициент наследуемости рассчитать через дисперсию и
Высота 1 клона, м.. 9,00 8,50 8,00 8,75 8,25.
Высота 2 клона, м.: 7,00 7,50 8,00 8,00 7,00.
Высота 3 клона, м.: 3,80 3,60 4,00 3,20 3,40.
Чятача 28. В озеленении широко используются виды клематиса
(СТеяюПт L.). Они служат исходным материалом при создании сортов,
сежстм которых ведется на величину цветка (диаметр), окраску,
махровость, сроки цветения, величину кустов. Рассчитайте через лимиты,
наследуется ли величина цветка Н2, с какими из перечисленных видов
предпочтительнее работать по этому признаку и какой результат можно
ожидать при однократном отборе R, если у ниже перечисленных видов
диаметр цветков следующий:
клематис цельнолистный (С. integrifolia L.) - 30 - 50 мм;
клематис кустарниковый лопастной (С. fruticosa Turcz. f. lobata
Maxim.) - 40 - 45 мм;
клематис Питчера (С. pitcheri Torz et Gray) -18 - 20 мм;
клематис фиолетовый (С viticella L) - 30 - 50 мм;
клематис Жакмана (С. х jackmanii Th. Moore) - 90 -140 мм;
клематис Арманда (С. Armandii Franch.) - 50 - 70 мм.
Задача 29. В промышленном садоводстве все клематисы (Clematis L.)
подразделяются на мелко- и крупноцветковые, кустовые и вьющиеся.
Сортовые клематисы относят к определенным группам (Жакмана, Патенс и
др.); учитывая происхождение по материнской линии, сортовые клематисы
размножаются тфививкой или черенкованием. Рассчитайте, наследуется ли
величина цветка (его диаметр) в группах, относящихся к крупноцветковым
вьющимся. Рассчитайте для каждой группы коэффициента наследуемости
Н3 чсРез дисперсию и лимиты. Рассчитайте через лимиты коэффициент
наследуемости Н3 для всех крупноцветковых сортов. Можно ли проводить
oi6op R по этому признаку внутри группы и между ними. Объясните
полученные результаты.
Сорта грушил Жакмана, диаметр цветков, см:
Бчжвинка - 12,0 15,2 16,0 14,4 13,4 13,9 15,2 15,4 16,0 14,0
Дымчатый - 14,2 16,0 18,0 15,1 16,9 14,0 17,0 18,1 16,4 15,8
Салют Победы - 14,0 12,0 16,0 14,8 13,2 12,5 15,5 12 4 14 6 14 5
Лютер Бербанк - 18,0 16,0 20,0 17,4 18,6 16,9 19,1 20 0 16,0 18,0
25
Сорта группы Патенс, диаметр цветков, см:
БарбараДибм - 17,8 15,5 17,4 15,8 18,0 17,0 16,0 18,0 17,9 16,4
Нелли Мозер - 16,2 18,0 20,0 19,4 17,6 18,0 19,2 19,8 16,8 20,0
Надежда - 14,8 15,4 16,0 15,8 14,9 14,0 15,0 16,2 15,3 14,9
Президент - 16,4 14,2 15,0 18,0 18,0 17,8 15,6 14,8 18,0 17,9
Сорта группы Вигцела, диаметр цветков, см:
Александрит - 12,0 14,0 12,5 13,5 12,7 14,0 13,2 12,0 13,5 12,4
Лесная опера - 11,9 12,0 13,1 14,0 14,5 12,5 13,7 12,3 14,0 12,2
Никитский розовый - 10,0 12,0 14,0 10,5 13,5 12,5 11,5 11,7 12,25 12,0
Рассвет - 14,5 15,6 16,5 17,4 20,0 21,0 17,8 19,2 19,0 18,5
Задача 30. Почти все сорта сирени получены в результате селекции
исходного дикорастущего вида сирени обыкновенной (Syringa vulgaris L.).
Основные признаки характерные для сорта - это цветки и соцветия, их
окраска и величина. Рассчитайте коэффициент наследуемости Н3 и
генетическое улучшение R для нескольких сортов сирени обыкновенной по
диаметру цветка и длине соцветия (табл. 17). Сорта размножены
черенкованием. Коэффициент наследуемости рассчитайте через лимиты.
Таблица 17
Статистические показатели соцветий различных сортов сирени___
Название сорта Диаметр цветка, см Длина соцветия, см
Академик Бурденко 2,8-3,0 22,5 - 23,0
Вивиан-Морель 1,5-1,8 23,5-25,0
Лебедушка 2,5-3,0 27,5-28,0
Маршал Жуков 2,5-3,0 24,0 - 25,0
Память о Вавилове 2,0-2,7 19,5-20,0
Школьница 2,0-2,2 24,0 - 25,0
Экселлент 2,5-3,0 27,0-28,0
Задача 31. Сорта роз группы флорибунда имеют различную величину
цветка и количество лепестков (табл. 18). Наследуют ли эти признаки Н2,
рассчитайте генетическое улучшение R при отборе растений по выше
указанным признакам. Сорта размножаются вегетативно. Каждый сорт
представлен 7 растениями.
Задача 32. При селекции каштана посевного (Castanea saliva Mill) на
урожайность были отобраны 10 материнских деревьев, отличающихся
обильным плодоношением. В дальнейшем была определена урожайность их
потомства. В табл. 19 даны средние значения урожайности каждого
материнского дерева У, кг, и его потомства X, кг. Определил» долю
аддитивных генов в генетической сотавляющей h2 и эффективность
однократного отбора по этому признаку.
26
Таблица 18
Величина цветка различных сортов роз
Сорт Величина цветка, см
Количество лепестков, шт.
Август Зеебауер 8,2 40 9,0 44 8,5 42 8,0 41 8,8 43 9,0 40 8,0 44
Воуч 7,0 7,2 8,0 7,9 7,4 8,0 7,0
20 22 21 23 20 24 24
: Волга-Дон 9,0 10,0 9,2 9,5 9,8 10,0 9,0
28 30 29 29 28 30 28
Волшебница 6,0 7,0 6,5 6,2 6,8 7,0 6,0
27 25 26 25 26 27 26
Капри 7,0 7,1 7,8 8,0 7,6 7,0 8,0
35 40 36 37 38 40 36
Таблица 19
Урожайность каштана посевного _____________
№ дерева 1 2 3 4 5 6 7 8 9 10
У, кг 25 26 27 28 23 24 24 25 26 23
X, кг 28 24 26 26 27 26 26 28 28 24
Задача 33. Известно, что плотность древесины - генетически
обусловленный фактор. Является ли этот признак результатом действия
аддитивных генов у дуба черешчатого (Quercus robur L.)? Плотность
древесины родителей У, кг/м3, и потомства X, кг/м3 даны в табл. 20.
Таблица 20
Плотность древесины у дуба черешчатого
№ д ерева 1 2 3 4 5 6 7 8 9 10
У, кг/м3 709 760 713 717 731 712 714 708 720 742
Х,кг/м3 740 737 717 712 713 717 718 711 755 722
Задача 34. По данным А.В. Чудного (1985) смолопродуктивность сосны
обыкновенной (Pinus sylvestris L.) является генетически обусловленным
признаком, коррелирующим с диаметром дерева. За сезон с одного дерева
можно получить от 0,12 до 4,9 кг живицы. В результате многолетних
исследований были отобраны материнские деревья с максимальным
выходом живицы, и получено их потомство (полусибсы). Выход живицы у
материнских д^мвьев У, кг, и потомства X, кг, представлены в табл. 21.
Какие гены могут оказывать влияние на передачу этого признака? Можно
27
ли в результате однократного отбора улучшить популяцию по признаку
«смолопродуктивность»?
Таблица 21
Выход живицы у сосны обыкновенной
№ дерева 1 2 3 4 5 6 7 8 9 10
У, кг 4,3 4,8 4,5 4,4 4,7 4,6 4,9 4,3 4,4 4,1
X, кг 4,6 4,5 4,7 4,5 4,9 4,4 4,5 4,6 4,7 4,4 I
Задача 35. В результате селекционной инвентаризации было отобрано
10 деревьев сосны кедровой сибирской (Pirrus sibirica (Rupr.) Mayr.},
отличающихся обильным плодоношением Затем проведено
контролируемое опыление, и у выращенного потомства (сибсов) определена
урожайность. Значения урожайности родителей У, кг, и потомства X,
кг, представлены в табл. 22. Можно ли считать, что признак «урожайность»
контролируется аддитивными генами? Можно ли получить генетическое
улучшение при однократном отборе особи с максимальным значением?
Таблица 77
Урожайность сосны кедровой сибирской
№ дерева 1 2 3 4 5 6 7 8 9 10
У, кг 7,5 8,0 6,5 6,0 9,0 7,0 8,5 10 9Л 11
X, кг 8,5 8,5 9,5 8,0 8,5 8,5 8,0 юл 9,0 9,0
Задача 36. Известно,что высота дерева - генетически обусловленный
признак. Докажите при помощи методов математической статистики,
контролируется или не контролируется аддитивными генами высота ели
европейской (Picea abies (L) Karst}. В табл. 23 дана высота материнских
деревьев У, см, и средняя высота потомства X, см, полученного от
свободного опыления в 8-летнем возрасте.
Таблица 23
Высоты ели европейской
№ дерева 1 2 3 4 5 6 7 8 9 10
У, см 2600 2800 2550 2700 2600 2650 2750 2800 2600 2700
X, см 75 85 60 78 65 82 89 62 76 90
Задача 37, Методом поликросса (свободное опыление) было получено
первое поколение F; плюсовых деревьев лиственницы сибирской (Zzrzr
sibirica Ldb.}. Высота материнских деревьев У, см, и средние высоты их
потомства в 9-летних культурах X, см, даны в табл. 24. Является ли признак
«высота» результатом действия аддитивных генов? Возможно ли получить
генетическое улучшение при однократном отборе по признаку «высота»?
Обоснуйте ваш ответ.
28
Таблица 24
Высоты деревьев лиственницы сибирской
№ дерева 1 2 3 4 5 6 7 8 9 10
' У, см 3100 2900 2700 3000 2800 2750 2900 2850 2950 2900
i X, см 90 92 84 101 93 91 78 94 86 97
Задача 38. Культуры сосны обыкновенной {Pinus sybestris L.)
заложены из семян, полученых от контролируемого опыления плюсовых
дерстьев (сибсы). Даны средние высоты материнских и отцовских деревьев
У, м, и средняя высота потомства в 30-летнем возрасте X, м (табл. 25).
Какие гены контролируют передачу этого признака? Какое
генетическое улучшение получится при однократеом отборе особи с
максимальным значением признака?
Таблица 25
Высоты деревьев сосны обыкновенной
№ дерева 1 2 3 4 5 6 7 8 9 10
У, м 13,2 13,5 13,8 14,2 14,5 14,8 15,3 15,6 15,8 16,5
1 Х,м 14,0 13,0 14,5 13,2 15,5 15,0 14,8 13,2 14,5 15,5
Задача 39. 40-летние культуры ели европейской (Picea abies (L.) Karst.)
заложены семенами плюсовых деревьев (полусибсы). Определить h2, если
известна высота материнских деревьев У, м, и средняя высота
полусибсового потомства X, м (табл. 26.). Можно ли получить генетическое
улучшение при однократном отборе особи с максимальным значением
признака «высота»?
Таблица 26
Высоты деревьев ели европейской
№ дерева 1 2 3 4 5 6 7 8 9 10
У, м 16,0 16,5 16,8 16,8 17,3 17,7 18,5 18,7 19?5 22,0
Х,м 15,0 16,9 16,5 15,0 18,0 16,0 17,0 15,5 16,0 19,01
Задача 40. 50-летние культуры сосны кедровой сибирской {Pinus
sibirica (Rupr.) Мауг.) заложены семенами плюсовых деревьев (полусибсы).
Расчигать /г2 для признака «высота дерева». Средние высоты родителей У,
м, и потомков X, см, даны в табл. 27. Расчитайте генетическое улучшение.
Задача 41. Культуры сосны кедровой сибирской {Pinus sibirica (Rupr.)
Маут.) заложены семенами, собранный с материнских деревьев,
разичающихся по признаку «масса 1000 семян» У, г. 1>ассчш^Т*’
коэффициент наследуемости для этого признака, если известна масса 1000
штук семян у полусибсового потомства X, г (табл. 28).
29
Таблица 27
Высоты деревьев сосны кедровой сибирской
№ дерева _L_| 2 3 4 5 6 7 8 9 Lioi
У,м 17,0 18,2 18,6 18,8 19,6 19,8 20,1 20,6 21,5 [2X5j
Х,м 22,0 1 19,0 17,0 20,2 22,0 19,0 24,0 18,2 17,0 120,2;
Таблица 28
Масса 1000 штук семян сосны кедровой сибирской
№ дерева 1 2 3 4 5 6 7 8 9 10
У, г п 180 190 200 210 220 230 240 250 260 270
Х.г 200 180 170 260 190 180 170 250 300 280
Задача 42. Культуры сосны кедровой корейской (Pinus koraiensis Sieb.
et Zucc.) заложены семенами, собраными с материнских деревьев,
различающихся по признаку «масса 1000 семян» У, г. Рассчитать
коэффициент наследуемости для этого признака, если известна масса 1000
штук семян у полусибсового потомства X, г (табл. 29).
Таблица 29
Масса 1000 ппук семян сосны кедровой корейской
№ дерева 1 2 3 4 5 6 7 8 9
У, гр 350 360 370 380 390 420 450 480 500 550
Х,гр 400 320 300 350 400 300 380 400 550 400 1
30
Литература
1. Бороеинч С. Принципы и методы селекции растений. - М: Колос, 1984. - 142 с.
1 Ватта КВ , Тихомирова ММ Руководство к практическим занятиям по генетике. -
М.: Просвещение, 1979. - С. 35 - 68; 134 -149; 157 - 158.
3 Вейр Б. Анатаз генетических данных. - М: Мир, 1995. - 400 с.
4. Гулиев ГВ. Геветика. - М: Колос, 1971. - 340 с.
5. Гулиев Г В. Задачник по генетике. - М: Колос, 1980. - 78 с.
6. Гулиев Г В, Мальченко В.В. Словарь терминов по генетике, цитологии, селекции,
совноводству и семеноведению. - М.: Россельхозиздат, 1983. - 240 с.
7. Зайцев Г В. Математическая статистика в экспериментальной ботанике. - М: Наука,
1984.-С. 22 - 23; 142-145.
8. Зжчик МТ. Статистика для лесных специалистов. - М.-Л.: Гослесбумиздат, 1952. 225
с.
9. Котов ММ, Лебедева Э.П. Применение биометрических методов в лесной селекции. -
Горький, 1977 - С. 48 - 60.
10. Лакин Г.Ф. Биометрия. - М: Высшая школа, 1980. - 293 с.
11. Лобашов МВ., Ватта КВ., Тихомирова ММ Генетика с основами селекции. Изд. 2-
е. - М: Просвещение, 1979. - 304 с.
12. Любавская АЛ. Лесная селекция и генетика. - М: лесная промышленность, 1982. -
288 с.
13. Луценко А.И., Луценко Э.К., Ляхова НФ. Введение в биометрию. - РТУ, 1986. - 96 с.
14. Магомедмирзаев ММ Введение в количественную морфогенетику - М: Наука,
1990.-232 с.
15. Мазер К, Джинкс Дж. Биометрическая генетика. - М: Мир, 1985. - 463 с.
16. Мюнтцинг А. Генетика. Общая и приглядная - М: Мир, 1967. - 610 с.
17. Орлова ИН. Генетический анализ. - М: МГУ, 1991. - 318 с.
18. Петров С А. Методы определения и практического использования коэффициента
наследуемости в лесоводстве. - М, 1973. - С. 2 - 7; 12 - 22; 26 - 38.
19. Погиба СП, Зуихина С.П, Шапкин ОМ Генетическая (щенка плюсовых деревьев и
подбор родительских пар для плантаций П и Ш порядков. Методические разработки.
-МгМЛТИ, 1991.-40 с.
20. Погиба СП, Шапкин О.М Методические указания к выполнению контрольных
работ по селекции. - М: МЛТИ, 1991. - 60 с.
21. Райт Д.В. Введение в лесную генетику. - М: Лесная промышленность, 1978. - 470 с.
31
22 Рокицкий ПФ. Биологическая статистика. - Минск Высшая школа, 1967. - 328 с.
23. Ромедер Э. Шенбах Г. Генетика и селекция лесных порол - М: Сельхождат, 1962. -
267 с.
24. Роне В.М Генетический анализ лесных популяций. - И: Наука, 1980. -158 с.
25. Свалов Н.Н. Вариаиоиная статистика. Пособие для студентов лесохозяйственного
факультета. - М: МЛТИ, 1975. - С. 3 - 20.
26. Свиржев Ю.М., Пасеков В.П. Основы математической генетики. - М: Наука., 1982. -
512 с.
27. Смиряев А.В., Гохмая МВ. Биометрические методы в селекции растений. - М.
Агропромиздат, 1985. - 214 с.
28. Смиряев А.В., Мартынов С.П., Кильчевский А.В. Биометрия в генетики и селекции
растений. - М: МСХА, 1992. - 269 с.
29. Урбах В.Ю. Биометрические методы. - М: Наука, 1964. - 415 с.
30. Хайрулин Р.Х. Математические методы в генетике. - Изд-во Казанского ун-та, 1988.
-185 с.
31. Falconer D.S. Introdactions to Quantitative genetics. - Edinburg; London: Oliver and Boyd,
I960.-365 p.
32
Содержание
Предисловие.................................................3
1. Метод оценки вариабильности признака.....................4
Задачи к теме 1: Методы оценки вариабельности признака...8
2. Оценка наследования качественных признаков..............13
Задачи к теме 2: Оценка наследования качественных
признаков...............................................14
3. Оценка наследования количественных признаков............17
1) Методы оценки количественных признаков и
расчет коэффициента наследуемости в широком
смысле Н2..........................................17
2) Методика расчета коэффициента наследуемости в
узком смысле h2....................................21
3) Методика расчета генетического улучшения R.......22
Задачи к теме 3: Оценка наследования количественных
признаков...............................................И
Литература.................................................