/






























































Text
ЖС16огг)Ны
расп) utt)O л b^bi I, ! JA Cl p Z t)pO(lV1OP0 30M/1CI
ГОСГЕОЛИЗДАТ • 1047
МИНИСТЕРСТВО ГЕОЛОГИИ СОЮЗА ССР
А. Н. ДРУЖИНИН
ЖИВОТНЫЙ и
РАСТИТЕЛЬНЫЙ МИР ПРОШЛОГО ЗЕМЛИ
Под редакцией проф. С. С. Турова Иллюстрации В. А. Ватагина
*
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО ГЕОЛОГИЧЕСКОЙ ЛИТЕРАТУРЫ Москва 1947 Ленинград
ЭТУ КНИГУ ПОСВЯЩАЮ СВЕТЛОЙ ПАМЯТИ ДОРОГОГО УЧИТЕЛЯ ПРОФЕССОРА МАРИИ ВАСИЛЬЕВНЫ ПАВЛОВОЙ, ПЕРВОЙ ЖЕНЩИНЫ-АКАДЕМИКА В СССР, ВПЕРВЫЕ ПОЗНАКОМИВШЕЙ ШИРОКИЙ КРУГ ЧИТАТЕЛЕЙ НАШЕЙ РОДИНЫ С МИРОМ ИСКОПАЕМЫХ ЖИВОТНЫХ.
Автор
ПРЕДИСЛОВИЕ
Прошлое Земли — возникновение и развитие жизни на ней—скрыто от нас давностью миллионов лет, однако пытливый ум человека шаг за шагом разгадывает эти тайны. Теперь мы зпаам мноТое о том, как изменялись очертания мерей и континентов, как одна эпоха истории Земли сменялась другой, как изменялись условия жизни на Земле, в связи с чем многочисленные формы животных и растений исчезали, уступая дорогу новым видам. Сложная картина эволюционного развития живых организмов Говорив о том, что они не оставались постоянными.
В напряженной, непрекращающейся борьбе за существование, проявляющейся Во всех сторонах жизни, выживали только наиболее приспособленные организмы. Так возникали новые виды животных и растений. Смена физико-географических условий и борьба за существование вызывали исчезновение одних и появление других групп животных и растений. Изучение развития животного и растительного мира на протяжении истории Земли представляет одну из самых интересных, захватывающих страниц естествознания.
Дать широкому кругу читателей — ученикам и преподавателям средней школы, студентам педагогических институтов, всем, интересующимся естествознанием, связное представление о развитии наземных животных и растений — такова цель этой книги.
Для того чтобы дать представление о возникновении и развитии жизни на Земле, приходится предварительно знакомить читателя с эволюцией водной фауны и флоры. Понимание всей совокупности грандиозных процессов эволюции
жизни на Земле немыслимо без представления о геологическом времени, об образовании земной коры и горных пород и связанного с этим деления всей истории Земли на периоды и эпохи.
После приобретения этих кратких сведений читатель познакоми!ся с развитием жизни на Земле по отдельным периодам, а начиная с кайнозоя — по отдельным эпохам. Последнее оправдывается тем, что с кайнозоя — эры млекопитающих—эволюция их шла сравнительно быстро, и разница в фауне двух соседних эпох относительно не меньше той, какая существовала между пресмыкающимися двух следующих друг за другом периодов.
Иллюстрации книги — реконструкции фауны отдельных периодов — сделаны художником В. А. Ватагиным при консультации, в то время заведующего филогенетическим отделом Зоологического музея МГУ, А. Н. Дружинина и по плану, разработанному последним при участии проф Б. С. Матвеева.
Все реконструкции животных оригинальны Некоторые животные реконструируются впервые, например, все представители гондванской фауны триасового периода. Работа потребовала большой затраты труда и времени. Только объединенными усилиями, благодаря знаниям специалистов-зоологов и таланту художника удалось воссоздать образы ископаемых чудовищ. Последнему обстоятельству особенно способствовало то, что в лице В. А. Ватагина мы имеем не только художника-анималиста, но и зоолога.
Директор Зоологического музея МГУ
С. С. Туров
В BE Д
ЕНИЕ
емля около 3—5 млрд, лет тому назад отделилась от Солнца! в видё струи раскаленного газа и» приняв шарообразную форму, стала вращаться вокруг него. Сначала Земля была настолько раскалена, что сама, светила подобно маленкому Солнцу. Существующие-в мировом пространстве силы притяжения и отталкивания обусловили дальнейшее формирование Земли. Силы притяжения приводили к сокращению объема Земли, а силы отталкивания создавали лучеиспускание Земли и выбросы из нее вещества в мировое пространство, подобно протуберанцам, наблюдаемым на Солнце. Только по прошествии многих миллионов лет, остывая, Земля покрылась тонкой первичной корой.
Первичная земная кора была образована кристаллическими породами, т. е. породами, которые полностью состоят из различных кристаллов. Примером кристаллической породы может служить гранит. Земная кора была сначала весьма непрочной: она часто трескалась и прорывалась. Огненножидкое содержимое земного ядра изливалось по трещинам на ее поверхность и снова застывало. Температура земной поверхности в те времена была так высока, что на ней не могло еще быть жидкой воды. Вся вода, образующая современные океаны, моря, озера и реки, окружала горячую Землю в виде пара.
Проходили миллионы лет. Земное ядро остывало все больше и больше. Силы притяжения и отталкивания продолжали действовать. Первые приводили к сокращению земной поверхности, что выражалось в сжатии ее коры в складки. Эти складки образовали первичные горы. Силы отталкивания вызывали расширение земной поверхности, а это приводило к выпячиванию ее отдельных участков.
Одновременно с поднятием земной коры при ее растяжении возникали новые прогибы и разломы. Образовавшиеся таким образом возвышенности наметили границы первичных мате-6
риков, а прогибы—вместилища для будущих океанов и морей.
К этому времени охлаждавшиеся водяные пары стали падать на поверхность Земли бесконечными ливнями. Первое время, когда земная поверхность была еще очень горяча, вода снова тотчас же испарялась. По мере дальнейшего охлаждения Земли вода испарялась уже не так быстро и постепенно заполнила прогибы в се коре. Так возникли первичные океаны и моря.
Под действием физических и химических факторов: неравномерного нагревания, растворяющих свойств воды, морского прибоя, ветров, дождей, вулканических взрывов и т. д. — кристаллические горные породы материков начали трескаться и разрушаться. Обломочный материал в виде камней, песчинок и мельчайших частиц уносился потоками ливней с материков в моря и океаны, где осаждался. В результате этого на дне морей и океанов возникли первичные осадочные горные породы: галечники, пески, глины и др. До начала формирования первичных материков и океанов жизнь на Земле не существовала, так как этому препятствовала высокая температура.
О существовании жизни на Земле мы заключаем по тем редким остаткам животных и растений, которые, окаменев, сохранились в осадочных горных породах. Эти организмы когда-то жили в водных бассейнах, а затем были погребены отлагающимся материалом. В некоторых случаях органическое вещество животного постепенно было замещено неорганическим. Так образовались прежде и образуются теперь окаменелости, т. е. минеральные образования, повторяющие форму, а иногда и все внутреннее строение умершего животного или растения. В таком виде обычно сохраняются только твердые части животного: кости, раковины, панцыри. Такие окаменелости могут сохраняться миллионы лет. В пластах, сложенных древнейшими осадочными породами, остатков животных и расти-
тельных организмов не найдено. Эти отложения по времени их образования относятся к архейской (первичной), или азойской (безжизненной), эре истории Земли.
Изучая зародышевое развитие современных животных и растений, ученые пришли к выводу, что первые организмы представляли собою комочки живого вещества, то есть протоплазмы. Лишенные скелета, эти первичные живые существа едва ли могли сохраняться в ископаемом состоянии. Кроме того, под давлением вышележащих слоев земной коры и под действием высокой температуры архейские осадочные породы были, вероятно, превращены в кристаллические, поэтому если в них случайно имелись и сохранившиеся организмы, они были полностью уничтожены в процессе кристаллизации этих пород.
Первые редкие остатки животных и растений были найдены в наиболее молодых осадочных породах из древних отложений. Эти отложения относятся к протерозойской эре, т. е. к эре первичной жизни на Земле.
В протерозойских пластах обнаружены уже не только микроскопические одноклеточные существа, имеющие скелет, например радиолярии^ но также и относительно крупные многоклеточные водоросли и морские животные. Здесь были найдены губки, черви, ракообразные, двустворчатые слизняки (моллюски) и также с двустворчатой раковиной внешне на них похожие плечено гие животные (см. кембрийский период). Все перечисленные существа принадлежали к различным типам беспозвоночных животных. Беспозвоночными эти организмы называются потому, что в их скелете мы не находим характерного для позвоночных животных спинного хребта, или позвоночника, состоящего из отдельных позвонков. У позвоночных животных спинной хребет образует основу их внутреннего скелета, т. е. скелета, лежащего в их теле, при^оытого снаружи мускулами и кожей.
У большинства беспозвоночных животных скелет образуется на поверхности тела, поэтому его называют наружным скелетом. Таким скелетом, например, будет раковина улитки. У беспозвоночных, лишенных раковины, кожа может выделять на своей поверхности особое плотное вещество — хитин. Такой хитин мы находим у насекомых. У многих раков хитин пропитывается известью и образует твердый панцирь.
В Тех же случаях, когда v беспозвоночных возникает внутренний скелет, он никогда не образует позвонков. Иногда этот скелет представлен отдельными известковыми или кремневыми иголочками, как у губок. Эти иголочки мбгут срастаться вместе, и тогда в коже животных мы наблюдаем пластинки или иглы. За подобное строение скелета морских звезд, морских ежей и близких к ним животных называют иглокожими. В редких случаях срастающиеся иголочки образуют внутри тела животного
цельный стержень. Подобный стержень можно видеть в колонии красного коралла, из которого вытачивают бусы, запонки и другие изделия.
Познакомившись со строением внешнего и внутреннего скелета беспозвоночных животных, мы видим, что он легко отличим от скелета позвоночных. Поэтому ученые могли безошибочно определить, что окаменевшие остатки протерозойских животных принадлежали исключительно беспозвоночным животным. Позвоночные, даже наиболее низкоорганизованные, как мы увидим дальше, появились на Земле значительно позднее. К позвоночным животным относятся: круглоротые или миноги, рыбы, земноводные (лягушки и тритоны), пресмыкающиеся (ящерицы, змеи, черепахи и крокодилы), птицы и млекопитающие, т. е. звери, кормящие своих детенышей молоком.
Несмотря на то, что протерозойские организмы были беспозвоночными, они имели уже весьма сложное строение. Чтобы стать такими, какими мы их видим в протерозойское время, им пришлось пройти длинный путь последовательного усложнения своей организации.
Предки протерозойских животных — первые живые существа — появились задолго до этого времени. Жизнь должна была появиться на Земле уже в начале азойской эры.
Во всей толще осадочных пород, отложившихся на протерозойских слоях, остатки животных и растений уже обильны и разнообразны. При этом ученые заметили, что животные, обитавшие на Земле во время отложения нижних слоев, отличались от животных средних слоев. Ископаемые остатки организмов из верхних слоев с очевидностью показывают, что эти организмы не были одинаковы с предыдущими и по своему строению уже значительно приближались к современным.
Время, в течение которого на Земле отлагались осадочные породы, содержащие уже обильные органические остатки, на основании различий в этих остатках, разделяют на три эры: 1) палеозойскую (эру древней жизни), 2) мезозойскую (эру средней жизни) и 3) кайнозойскую (эру новой жизни) (табл< I). В свою очередь, эры разбиваются на меньшие отрезки времени, называемые периодами. Время каждого периода делится на эпохи. Критерием для подобного деления снова служит различие в составе окаменелостей. Между отложениями следующих друг за другом периодов, и тем более эпох, резких границ нет. Животное население одной эпохи постепенно, пойти незаметно, сменяется населением другой. Но, если мы возьмем всех животных, или, как принято говорить, всю фауну любой эпохи или периода в целом, то эта фауна будет заметно отличаться от фауны как предыдущего, так и последую* щего напластования.
В палеозойской эре насчитывают пять периодов. Самый древний из этих периодов назы-
деется кембрийским (кембрий), далее следуют: силурийский (силур), девонский (девон), каменноугольный (карбон) и наиболее молодой в этой эре — пермский (пермь).
Мезозойская эра делится на три периода: триасовый (триас), юрский (юра) и меловой (мел).
Кайнозойская эра слагается из двух периодов: третичного и четвертичного.
Третичный период включает пять эпох: 1) палеоцен — эпоха древней жизни, 2) эоцен — эпоха зари новой жизни, 3) олигоцен — эпоха с малым числом современных животных, 4) миоцен— эпоха с немногими современными животными, 5) плиоцен — эпоха со многими современными животными.
Четвертичный период состоит из двух эпох: 1) плейстоцен — эпоха с большинством современных животных и 2) современная эпоха. Плейстоцен часто называют также л е д н и к о вым периодом, а современную эпоху — послеледниковым.
Чтобы реальнее представить себе развитие жизни на Земле в отдельные геологические периоды, нужно иметь в виду, что для измерения геологического времени непригодны столетия и тысячелетия, которыми пользуются для изучения истории человеческого общества. Даже десятки и сотни тысяч лет оказываются слишком мелкими единицами для определения геологического Времени, которое может измеряться только миллионами лет.
Сперва ученые предполагали, что, зная скорость образования морских осадков в современных условиях и толщину осадочных пород того или другого периода, можно приблизительно оп' ределить время, которое потребовалось для их формирования. Они считали, что таким путем можно определить и возраст самой Земли. Но этот метод оказался неточным, так как скорость образования осадочных пород далеко не всегда остается постоянной и может изменяться от целого ряда причин.
Тогда для определения возраста Земли стали искать иных методов. Так, пробовали использовать общеизвестный факт наличия соли в морской воде. Речная вода называется пресной, но она все же содержит некоторое количество солей, которые непрестанно вымываются в реки из окружающих горных пород. Зная, какое количество солей ежегодно приносят реки в море, и общее количество соли в нем, можно представить себе, сколько лет понадобилось, чтобы данное количество соли успело накопиться в морской воде.
В настоящее время определить возраст Земли пытаются путем изучения процесса распада радиоактивных элементов. Из химии известно, что все огромное разнообразие существующих веществ слагается из различных ком
бинаций небольшого числа элементов. Раньше думали, что эти элементы всегда остаются в неизменном виде. Теперь же известно, что некоторые из них, а именно радиоактивные элементы, с течением времени изменяются и превращаются в другие элементы. Например, металл т о-р и й с течением времени переходит в радий, а радий, в свою очередь, превращается в свинец. Химики установили также время, необходимое для перехода тория в радий и радия в свинец. Изучая минералы, содержащие торий, взятые из различных горных пород, и определяя в них степень распада, т. е. превращения тория в свинец, можно определить число лет, которое прошло с момента образования данной горной породы.
Полученные различными способами цифры, определяющие время образования осадочных пород на Земле, значительно отличаются друг от друга. Приведенные ниже цифры нельзя считать абсолютно точными, но они могут дать относительно правильное представление о продолжительности той или другой эры. того или другого периода.
Продолжительность всего четвертичного периода, с которым связана история появления и развития на Земле человеческого рода, по такому подсчету равна приблизительно 1 млн. лет. Но эта огромная цифра покажется малой по сравнению с продолжительностью всей кайнозойской эры, равной примерно 60 млн. лет. Предшествующая ей мезозойская эра продолжалась на Земле не менее 160 млн. лет, а палеозойская эра — не менее 350 млн. лет.
Протерозойская эра по своей длительности приблизительно равна всем трем вышеперечисленным, более молодым эрам. Она тянулась не менее 500 млн. лет. Еще более длинной была азойская эра, продолжавшаяся около 700 млн. лет. Сложив цифры, показывающие длительность отдельных эр, мы можем определить продолжительность всего» геологического периода истории Земли. Он равен, примерно, 1770 млн. лет.
Даже в течение своей краткой *лизни человек может добиться изменений в породах домашних животных и в сортах культурных растений. Он может наблюдать, как под влиянием морского прибоя, течения рек, дождя и ветра, хотя и незначительно, но все же меняются очертания морских и речных берегов. Землетрясения и извержения вулканов иногда приводят и к более значительным изменениям поверхности Земли.
Если одной человеческой жизни достаточно, чтобы заметить некоторые изменения, происходящие на Земле и в ее органическом мире, то какие же колоссальные перемены в расположении материков и морей, в строении животных и растений должны были произойти на Земле за 1770 млн. лет?! Некоторое представление об этом дает читателю все изложенное в настоящей книге.
ПАЛЕОЗОЙСКАЯ ЭРА
КЕМБРИЙСКИИ ПЕРИОД
ачало первого периода палеозойской эры—кембрия—отдалено от нас при
близительно 570 млн. лет. Продолжительность
этого периода определяется примерно в 65 млн. лет.
Кембрийскому периоду предшествовала эпоха интенсивного горообразования, с которой, возможно, были связаны местные явления оледенения, например, в Австралии и в Китае. Распределение на Земле морей и материков в кембрийское время существенно отлйчалось ’ от Современного^ В южном полушарии расцолагал-с^г ряд КОПтийентальных массивов на территории современной Бразилии, Восточной Африки, ИйИшг тг * Авётра^ййГ с^Ж°Ж'ХШлуЖЙИЙ современная Северная Америка вместе с Гренландией представляли единый континент. Обширная суша занимала территррщо довременной Европейской части СССР. Северные’ материки были отделены от южных широкой полдср>|.., мо« рй^сйЦогочислённьши островами. Это море называют окёанйй^Гетис.
Наземных растений в кембрии еще не было, поэтому материки представляли безмолвную голую пустыню, но в морях уже была богатая разнообразная жизнь.
Из водных растений здесь встречались как очень мелкие (микроскопические), так и крупные водоросли (табл. II, 1), а также разнообразные и многочисленные известковые водоросли (табл. II, 2), которые, подобно булыжникам, устилали морское дно.
В кембрийских морях уже жили представители всех известных нам типов беспозвоночных животных. Позвоночных животных в кембрии еще не было.
В воде плавали одноклеточные организмы, покрытые тонкими раковинами, и медузы (табл. II, 3). На дне моря жили губки (табл. II, 4), различные черви (табл. II, 5 и 6),
многочисленные археоциаты строили подводные рифы (табл. II, 7, 8, 9, 10 и 11). По своему строению археоциаты похожи на губки, но отчасти напоминают и кораллы. Часто на vac поверхности селились мелкие известковые водоросли (табл. II, 12). Почти половину всего морского населения в кембрии составляли плеченогие (табл. II, 13. 14).
Плеченогие, подобно двустворчатым моллюскам, имеют раковину, состоящую из двух половин — створок. У моллюсков эти створки прикрывают правую и левую половины тела животного, а у плеченогих — брюшную и спинную поверхность его. От заднего конца брюшной части тела у плеченогих отходит тоненький стебелек, при помощи которого они прикрепляются ко дну. В переднем конце тела, около рта, находятся спирально закрученные щупальцы, которые обычно называют руками. Они служат животному и для привлечения пищи и для дыхания.
Изредка попадались настоящие двустворчатые и брюхоногие моллюски. Последние имели раковинку в виде колпачка, состоящую всего из одной створки. Жили в то время и первые предки современных осьминогов, каракатиц и корабликов, которые называются головоногими моллюсками. Головоногие жили в трубчатых раковинах, разделенных на отдельные камеры. Одетые в тонкие конические раковины, плавали в воде крылоногие моллюски. Крылоногими их называют потому, что от нижней части тела, называемой ногою, у них в стороны торчат два выроста, напоминающие крылышки.
Одну треть всех животных, живших в кембрии, составляли близкие к ракам трилобиты (табл. II, 15). Их тело состоит из большого числа отдельных сегментов — колец. На каждом
ТАБЛИЦА II
КЕМБРИЙСКИЙ ПЕРИОД. ОБИТАТЕЛИ КЕМБРИЙСКОГО МОРЯ
1— водоросли; 2— известковые^водоросли; 3— медузы; 4— губки; 5 и б—кольчатые черви; 7, 8, 9, 10, 11—рьзхжвые археоциаты;
12— водоросли, поселившиеся на археоциатах; 13, 14—плеченогие; 15—трилобиты; 16—морская звезда; 17, 18—ползающие морские кубышки (голотурии); 19— плавающая голотурия; 20—текоидеи; 21— карпоидеи; 22—морские стрелки—сагитты
кольце находится пара ножек. Каждая ножка в свою очередь состоит из отдельных члеников. Передние членики тела трилобита срослись и образовали голову животного. На ней сверху выступают два сложно устроенных глаза, снизу лежат челюсти и одна пара усиков. Среди трилобитов были мелкие формы длиной всего 3—4 см, но встречались и гиганты длиной до 70 см. Кембрийское море уже населяли и настоящие раки. Последние, в отличие от трилобитов, имеют на голове не одну, а две пары усиков. Вместе с ними жили предки современных раков-мечехвостов и ракоскорпионов (см. силурийский период).
Весьма разнообразными были в кембрии животные, которых называют иглокожими. Это название они получили от того, что в коже их имеются известковые пластинки, на которых часто сидят известковые иглы. Иглокожие, имеющие шарообразную форму, получили название «морских ежей», так как они действительно напоминают настоящих ежей, свернувшихся в клубки.
В кембрии морских ежей еще не было, но уже были морские звезды (табл. II, 16), у кото рых, как у большинства иглокожих, хорошо заметно деление тела на пять лучей, так как оно по-слроено по пятилучевой симметрии. Кроме морских звезд, в кембрии жили различные морские
кубышки — голотурии. Одни из них ползали по дну (табл. II, 17, 18), другие вели пхавающий образ жизни (табл. II, 19). Были тут и сидячие — прикрепленные ко дну иглокожие животные. Одни из них, как текоидеи, прикреплялись непосредственно спинной стороной своего тела (табл. II, 20), а у других, как карпоидеи (табл. II, 21), для прикрепления имелся специальный стебелек. Тело карпоидей еще было двусторонне симметричным) а не пятилучевым. Двусторонняя симметрия наблюдается только у свободно двигающихся животных. Когда же животные переходят к сидячему образу жизни, двусторонняя симметрия строения их тела заменяется радиальной или многолучевой. Карпоидеи, недавно перешед шие к сидячему образу жизни, еще сохранили черты строения их свободно двигавшихся предков.
Изучение иглокожих показало, что из всех беспозвоночных животных они являются самыми ближайшими родичами позвоночных. Родственными позвоночным и одновременно иглокожим животным были уже жившие в кембрии морские стрелки—сагитты (табл. II, 22). Этих животных иначе называют щетинкочелюстными червями за особенность строения их челюстей, напоминающих по форме щетинки. Настоящих позвоночных животных в кембрии еще не было, но первичные их предки должны уже были существовать.
СИЛУРИЙСКИЙ ПЕРИОД
коло 500 млн. лет назад кембрийский период сменился силурийским, длившимся примерно 100 млн. лет. Силурийские материки хотя и были.похожи на .кембрийскиел но по своим очертаниям, несколько.... отличались, них. Бблыпая г часты современной Европы и современной Азии , находилась под водой. В на-«?але периода^ материки., были плоскими,, но к концу его, £ р^ультате горообразовательных процессов, создались мшцные горные цепи._Одна из таких мощных горных цепей.возникла на месте современной Норвегии и соединила Европейский матери£^Хде^ Так возник огромный, Северо-Атлантический континент. Вулкану деятельность в течение силурийского"периода Лыла весьма интенсивной.
Поверхность материков еще попрежнему оставалась пустынной и безжизненной, и только в самом конце периода по морским побережьям появились первые наземные растения — псило ф и т ы, которые гораздо больше напоминали морские бурые водоросли, чем обычные наземные растения (рис. 1). У псилофитов, как и у водорослей, тело еще было не расчленено на стебель, корень и лист, а представляло целостное слоевище. Подземная часть растения была похожа на корневище, несущее волоски. От подземной части отходили и наземные части растения. Это были ветвящиеся трубочки, постепенно уменьшавшиеся в диаметре. Вершины этих трубочек часто были закручены. На концах трубочек помещались особые мешочки—с п о р а н-г и и, в которых были заключены многочисленные одноклеточные споры. При помощи этих спор пси-лофиты размножались. Часто основание наземной части растения было покрыто вместо листочков особыми колючками. Кожица псилофитов имела мелкие дыхательные отверстия, называемые устьицами, которые характерны только для
наземных растений. В отличие от водорослей, псилофиты в центре своих трубочек и в слоевище содержали особые группы клегок. По одним из них двигались вода и минеральные вещества из нижней части растения в верхнюю, а по другим опускались органические вещества в нижние его части. Благодаря этой особенности строения псилофиты занимают промежуточное положение между водорослями и настоящими наземньг ми уже не слоевцовыми растениями. Все настоящие наземные растения являются прямыми потомками псилофитов.
Суша силура была почти необитаема, но море уже имело жизнь богатую и разнообразную. В воде были взвешены мельчайшие одноклеточные, покрытые раковинкой, дырочники — фора-миниферы и лучистки — радиолярии. На дне морей обитали многочисленные кремневые губки и различные черви. Вместо вымерших кембрийских археоциат жили колониальные животные, похожие по строению на современных пресноводных гидр,—строматопоры. Они строили целые подводные рифы. Соединенные вместе известковые скелеты особей колонии создавали массивные подводные рифы, наподобие современных коралловых.
В строительстве рифов со строматопорами соперничали близкие к современным восьмищу-пальцевым кораллам колониальные кораллы табуляты. Свое название табуляты получили от латинского названия «табула», что означает «доска» или «днище». И действительно, в трубчатом скелете каждой особи табулят всегда можно найти большое число горизонтальных перегородок, т. е. днищ или табул.
В силуре также жили представители ныне вымершей группы четырехлучевых кораллов. Однако все они еще были одиночными формами. Интересны также жившие в силуре родственные нашим гидрам колониальные граптолиты,
одетые снаружи роговым скелетом. Среди граи-толитов были как свободно плавающие колонии, так и прикрепленные к камням или к водорослям. Иглокожих в силуре было больше, чем в кембрии. Здесь, кроме ранее существовавших, появились похожие на морских звёзд змеехвостки или офиуры, морские ежи, особенно многочисленные сидячие формы: морские лилии и шаро-вики. В конце периода к ним присоединились морские бутоны.
Плеченогих было много, но двустворчатые и брюхоногие моллюски встречались попрежнему редко. Головоногие моллюски были близки к современному кораблику. Раковины их, прямые или изогнутые, а в некоторых случаях совсем закрученные, как у кораблика, имели длину до метра. Знакомые нам по кембрию трилобиты были особенно многочисленны и разнообразны в нижнем силуре.
В опресненных лагунах жили гигантские ракоскорпионы длиной до 1,5 м. Поздние их представители достигали в длину до 3 м. Ракоскорпионы, которых также называют гигантскими раками, были похожи и на раков и на скор-
Рис. 1. Псвлофит
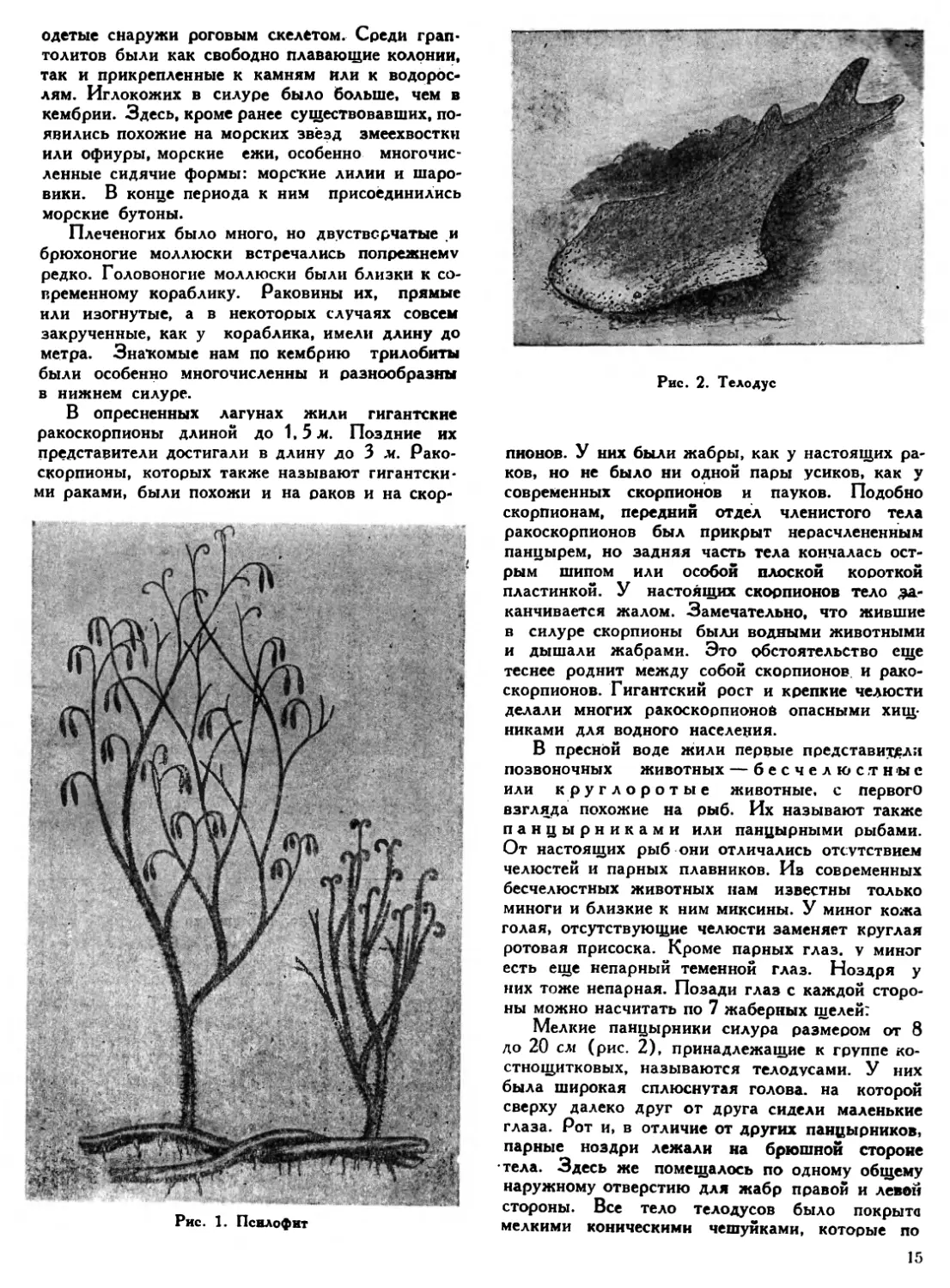
Рис. 2. Телодус
пионов. У них были жабры, как у настоящих раков, но не было ни одной пары усиков, как у современных скорпионов и пауков. Подобно скорпионам, передний отдел членистого тела ракоскорпионов был прикрыт не расчлененным панцырем, но задняя часть тела кончалась острым шипом или особой плоской короткой пластинкой. У настоящих скорпионов тело заканчивается жалом. Замечательно, что жившие в силуре скорпионы были водными животными и дышали жабрами. Это обстоятельство еще теснее роднит между собой скорпионов и ракоскорпионов. Гигантский рост и крепкие челюсти делали многих ракоскорпионов опасными хищ-никами для водного населения.
В пресной воде жили первые представители позвоночных животных — бесчелюстные или круглоротые животные, с первого взгляда похожие на рыб. Их называют также панцырниками или панцырными рыбами. От настоящих рыб они отличались отсутствием челюстей и парных плавников. Из современных бесчелюстных животных нам известны только миноги и близкие к ним миксины. У миног кожа голая, отсутствующие челюсти заменяет круглая ротовая присоска. Кроме парных глаз, у миног есть еще непарный теменной глаз. Ноздря у них тоже непарная. Позади глаз с каждой стороны можно насчитать по 7 жаберных щелей:
Мелкие панцырники силура размером от 8 до 20 см (рис. 2), принадлежащие к группе костнощитковых, называются телодусами. У них была широкая сплюснутая голова, на которой сверху далеко друг от друга сидели маленькие глаза. Рот и, в отличие от других панцырников, парные ноздри лежали на брюшной стороне тела. Здесь же помещалось по одному общему наружному отверстию для жабр правой и левой стороны. Все тело тело ду сов было покрыта мелкими коническими чешуйками, которые по
Рис. 3. Цефаласпкс
своему строению напоминали маленькие зубы. Эти чешуйки состояли из дентина» а сверху были покрыты эмалью.
Более сложное строение имели крупные паи-цырники, образующие группу разнощитковых— цефаласписов (рис. 3). Цефаласписы в длину достигали 50 см. Голова их была закована в сплошной панцырь. От задних краев головы отходили, наподобие плавников, особые выросты, покрытые мелкими чешуйками. Тело цефаласписов было покрыто узкими пластинками, вытянутыми в поперечном направлении. Очень сближенные глаза располагались на верхней стороне черепа, между ними находился третий, теменной, глаз. Впереди него лежала непарная, как у миног,- -ноздря. За глазами и по бокам головы у цефаласписов помещались мощные электрические органы, которые являлись надежной защитой от врагов, главным образом гигантских ра коскорпионов. Несколько жаберных отверстий было расположено на нижней поверхности головы. Из современных рыб электрические органы встречаются у электрических скатов и у южноамериканских электрических угрей. При прико
сновении к электрическим органам этих рыб ощущается ток значительной силы.
В лагунах попадались гораздо более подвижные рыбообразные, бесщитковые панцырники, называемые биркениями, длиной до 10 см (рис. 4). Тело их, сдавленное с боков, было покрыто узкими пластинками и чешуей. Глаза и жаберные щели лежали по бокам головы. Теменной глаз и непарная ноздря располагались на верхней стороне головы. Рот напоминал рыбий, но был еще без челюстей. Таким образом, первые позвоночные на Земле были даже не рыбы, а круглоротые— ближайшие родичи современных миног. Они не могли активно захватывать пищу и питались тем, что случайно попадало в их бесчелюстной рот. Первые настоящие рыбы, называемые хрящевыми, родственные современным акулам, появились в небольшом количестве только в самом конце силура. Они уже имели парные плавники и настоящие челюсти. Хрящевые рыбы хорошо плавали и могли активно захватывать добычу. Для панцырников они были сильными а опасными конкурентами, постепенно приведшими их к гибели.
Рис. 4. Биркения
ДЕВОНСКИЙ ПЕРИОД
Римерно 400 млн. лет отделяют нас от начала девона и 350 млн. лет от его конца. За это время материки и моря успели значительно изменить свои очертания. В начале девона океан Тетис сильно сузился, а северные. и южные материки заметно выросли. Пот их берегам громоздились высокие горы.задерживадщие влажный морской воздух, благодаря чему ;гериках господствовал сухойЛ резко континентальный климат. К концу периода горы сгла^лис^ море "залило большие Участки суши. Климат на Земле стал влажным и теплым. К этому времени Южная Америк^додт^цедц^ в*
морскую пучину. Северо-Атладтия^стхая^ суша существовала^прпрежнем^. В северном полушарии, кроме этой сущцк^уще9тцрвад^донтг1ц1енг в северной, части теперешней, Азии* захватывавший также Чукотку И Аляску.
Еще бедная наземная жизнь девонского пе-рио^сос^^ берегов, число крартений*однако, ^ постепенно увеличивалось. Среди псилофитов, значительно более многочисленных и разнообразных, чем в силуре, попадались и такие растения, которые во многом уже напоминали наши современные папоротники, хвощи (водяные елочки) и плауны, особенно пышно развивавшиеся в следующем, каменноугольном периоде. Это были растения уже с настоящими листьями и стеблями. Все растения девона размножались микроскопическими клетками — спорами, почему и называются споровыми в отличие от с е-м е н н ы х, размножающихся семенами. В середине девона появились и семенные папоротники (рис. 5), занимавшие переходное положение между споровыми и семенными растениями.
Эти замечательные растения, порой достигавшие размеров деревьев, по строению своих листьев и стеблей почти ничем не отличаются от настоящих папоротников, с которыми они
связаны узами кровного родства. На концах их листьев сидели уже не спорангии, набить е одноклеточными спорами, а настоящие крупные одиночные семена. Семена крупных растений достигали размеров лесного ореха. Семенные папоротники были предками всех семенных растений.
В девоне появились первые настоящие сухопутные беспозвоночные: скорпионы, многоножки и бескрылые насекомые. В конце периода появились первые наземные позвоночные животные из класса земноводных — ихтиостеги.
Все земноводные дышат легкими, а вместо парных плавников имеют лапы, но, подобно рыбам, они мечут икру в воду. Их молодь первое время живет в воде и дышит жабрамц. Среди современных земноводных мы знаем бесхвостых жаб и лягушек, хвостатых тритонов и саламандр и безногих змееобразных тропических червяг. Девонские земноводные ихтиостеги не относились ни к одной из известных нам теперь групп. Ихтиостеги — это древнейшие стегоцефалы, т. е. покрытоголовые земноводные. Они называются так потому, что многочисленные толстые кости в виде панцыря одевали их голову. Ихтиостеги — еще настоящие водные животные. На черепных костях у них еще сохранились каналы, в которых помещались особые органы чувств (органы боковой линии), характерные для рыб. Очевидно, ихтиостеги постоянно жили в пресноводных водоемах и болотах и на сушу выходили лишь для того, чтобы переползти в соседний новый водоем в том случае, когда пересыхал старый.
Земноводных в девоне было еще мало, но зато встречалось много разнообразных рыб. Последние обитали главным образом по рекам и опресненным морским лагунам. Среди них было особенно много хрящевых, или иначе, акулообразных рыб. В отличие от современных
акул, у которых скелет действительно хрящевой, большинство девонских акулообразных рыб имело на теле костный панцырь. Такими были небольшие астеролеписы, длиной до 75 см (табл. III, 1), которых относят к рыбам-антиар-хам. Задняя половина их тела с брюшными плавниками и хвост очень напоминали те же части акулы, но вся передняя половина туловища, голова и передние плавники были покрыты костной броней из отдельных щитков. Грудные плавники имели вид членистых валькообразных придатков и использовались рыбами для ползания по дну. Челюсти у астеролеписов были слабые, и потому они писались мелкими беспозвоночными животными и водорослями.
У кокостеуса, размером около 0,75 м (табл. III, 2), хвост кончался длинной тонкой нитью, а голова и передняя половина туловища, как и у астеролеписа, были закованы в прочный пан цырь, состоявший из двух частей. Две половины панцыря—головная и туловищная—соединялись между собой суставом так, что кокостеус мог, н отличие от всех остальных рыб, опускать и поднимать голову. По этому признаку кокостеуса и близких к нему рыб объединяют в группу артро-дира, т. е. суставошейных рыб. От туловищной части панцыря у них отходили боковые выросты — измененные грудные плавники, кроме которых имелись еще парные брюшные плавники и непарные — спинной и анальный. Костные челюсти образовывали особые острые выросты,
заменявшие кокостеусам зубы. В концд девона рыбы, родственные кокостеусу, перешли жить из лагун в открытое море, и многие из них достигли огромных размеров, как, например, хищ* ная динихтис, доходившая в длину до 9 м.
Пресноводные лагуны населяли и небольшие, длиной в 10—15 см, стройные рыбки из группы акантоди. Одни из них — диплакантусы (табл. III, 3) — на спине имели по два плавника, другие — хейракантусы (табл. III. 4) — всего лишь один спинной плавник. Тело этих маленьких акулообразных рыб было покрыто щитками и чешуей. Череп состоял из настоящих костей. На переднем крае каждого плавника находились массивные острые шипы. В начале девона у некоторых акантодий между шипами брюшного и грудного плавников сидело еще несколько шипов. Эти шипы указывают на то, что v древних акантодий было несколько пар плавников, а не две, как у обычных рыб.
Кроме акулообразных рыб, в лагунах обитали также родственные между собой двоякодышащие и кистеперые рыбы. В настоящее время двоякодышащие рыбы, получившие свое название от способности дышать не только жабрами, но и плавательным пузырем, почти вымерли. Только в реках Австралии, Африки и Южной Америки живет по одному виду этих замечательных рыб. Зубы, срастаясь, образуют у них крупные гребенчатые пластинки. Плавники кистеперых рыб образованы мясистыми лопастями.
ДЕВОНСКИЙ ПЕРИОД. ОПРЕСНЕННАЯ ЛАГУНА ВЕРХНЕДЕВОНСКОГО МОРЯ /—астеролепис; 2—кокостеус; 3— диплакантус; 4—хейракавтус; 5— диптерус; 6— остеолепнс; 7—голоп-тихиус; 8—птеригот
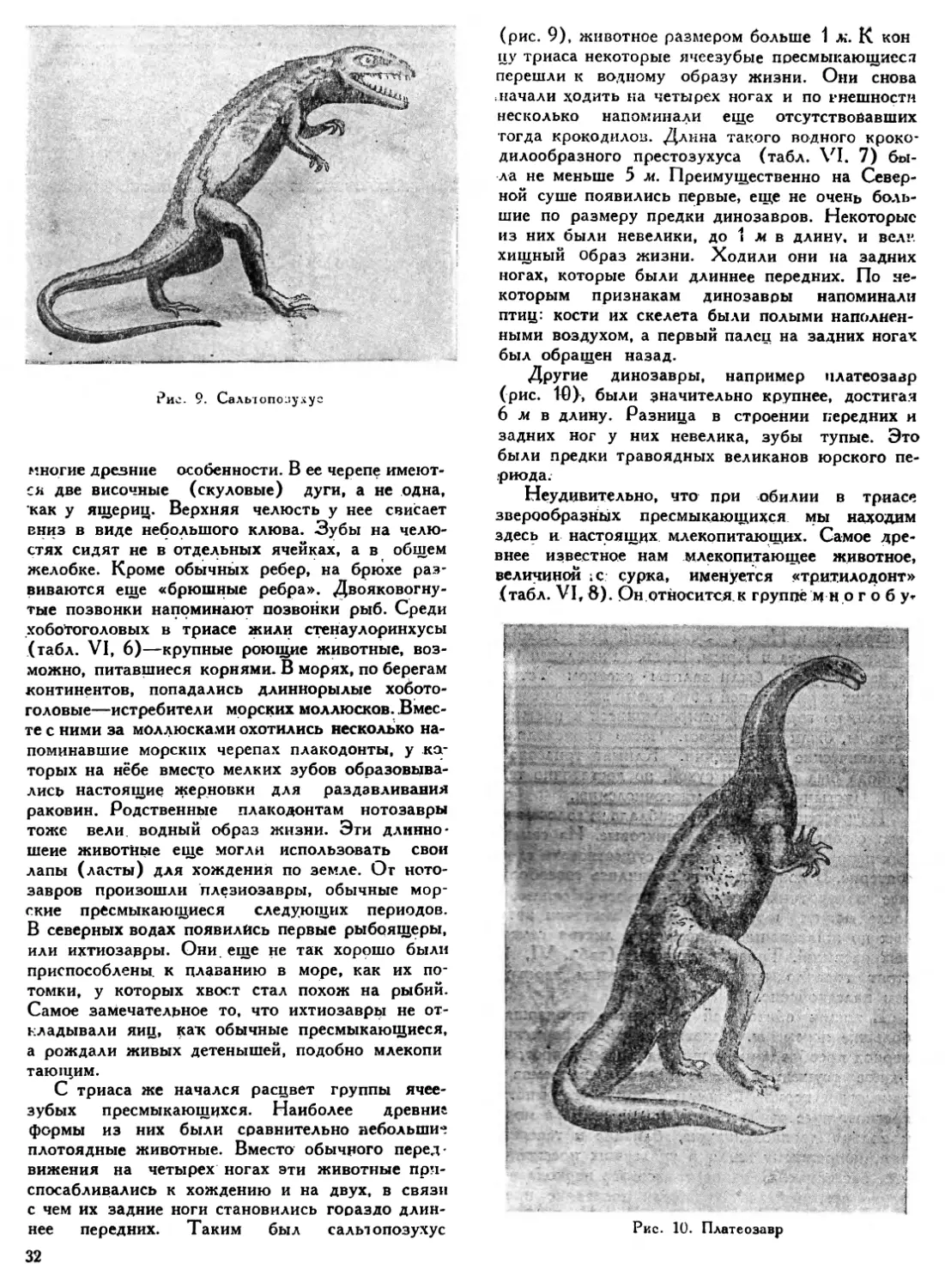
Pile. 6. Аммонит
на которых, как волоски кисти, сидят кожные лучи. За эту особенность строения плавников рыбы получили название кистеперых. Теперь они очень малочисленны. Многопер и другой близкий к нему вид, живущие в реках Центральной Африки, по строению во многом отличаются от своих предков. В последние годы близ берегов Южной Африки была поймана в море латиме-рия — крупная кистеперая рыба длиной больше метра.
Настоящие кистеперые рыбы, как и двоякодышащие, кроме жабр, для дыхания использовали плавательный пузырь, который особым каналом открывался у них на брюшной поверхности пищевода, а не на спинной, как у остальных рыб. У этих рыб имелись внутренние ноздри, соединявшие нос с полостью рта, благодаря чему они могли дышать воздухом прямо через нос, как наземные четвероногие. На спине у них были два плавника. При помощи сходных по строению парных плавников эти рыбы могли ползать по дну. Это приспособление девонских кистеперых и двоякодышащих рыб говорит о том, что они, подобно современным двоякодышащим, жили в пересыха
ющих водоемах или в водоемах, где периодически портилась вода и жаберное дыхание становилось невозможным.
Из двоякодышащих рыб наиболее распространен был диптерус (табл. III, 5), длина которого не превышала 40 см. Особенности строения его черепа и сложных гребенчатых зубов не позволяют видеть в нем прямого предка наземных четвероногих. Зато некоторые кистеперые рыбы по своей организации вполне могли быть прямыми предками земноводных. Девонские кистеперые рыбы очень разнообразны по строению и по величине. Остеолепис (табл. III, 6) не превышал 25 см, аголоптихиус (табл. III, 7), с длинными мясистыми плавниками и крепкими зубами, достигал до метра в длину.
В тех же опресненных бассейнах плавали, правда, еще не очень многочисленные, предки костных рыб (настоящих костистых рыб, таких, как окунь, треска, карп и др., в девоне еще не существовало). Предки костных рыб, жившие в девоне, называются палеонисцидами. В их черепе появились кости, но скелет оставался в основном еще хрящевым. Лопасти хвостового плавника у палеонисцид асимметричны, так же как и у акул.
Многочисленные пресноводные девонские рыбы попрежнему имели страшных врагов — гигантских ракоскорпионов; для защиты от них большинство рыб было одето панцырем. В девоне жили особенно крупные ракоскорпионы. Почти 2-метровый птеригот (табл. III. 8) был вооружен сильными клешнями. Шип на конце его тела был широкий и короткий.
Трилобитов в девонских морях было уже меньше, чем в силуре. Особенно разнообразными являлись плеченогие, из иглокожих процветали морские лилии и бутоны. Среди головоногих моллюсков появились гониатиты — предки будущих аммонитов (рис. 6). Наряду со стро-матопорами и табулятами рифы строили уже и колониальные кораллы из группы четырехлучевых, а также и мшанки. К ранее существовавшим кремневым губкам в девоне прибавились еще губки с известковым скелетом. Среди раков особенно многочисленными были мелкие ракушниковые раки, тело которых одето двустворчатой раковиной.
КАМЕННОУГОЛЬНЫЙ ПЕРИОД
коло 350 млн. лет назад на Земле начался каменноугольный” перирд, (кар-бон). Он'называется так потому, что в течение 100 млн. лет его существования в £олот«рс н озерах из растительных остатков образовывались мощные пласты каменного угля.
За такой длинный промежуток времени суша и море не раз успелу излгенцт^ свои очертания. Материки южного полушария (Южная Америк^^'$рикаГ1%дия и Австралия 1 объединились вобщую сущу,—гГондвану.. Большая часть Южной Азии и северная часть Африки были_,зддиты морем, которое ' рукавом -через Европейскую часть СССР соединялось С полярными ‘ водами.
Материки в северном полушарии по расположению были близки к девонским, но несколько больше залиты, морем.
В начале каменноугольного периода климат был^^аОь^ цр^к" концу он постепенно становился все более сухим и континентальным, вследствие того, что на бь^дщик. плоских.-материках в середине периода сформировались высокие, горы. В конце карбона на Гондване даже скопились ^громШё массы льда; Материки заселялись по берегам морей, рек, озер И болот пышной растительностью.
Уже известные нам из девонского периода споровые растения в камённругольн^ образовывали густые непроходимые заросли. Среди этих зарослей вовидные растения до 2 м в обхвате и до 35 м в вышину. На их стволе в виде ромбов, напоминающих чешую рыб, сохранились следы прикрепления листьев. За эту особенность коры эти растительные гиганты называются лепидодендронами, т. е. чешуйчатыми деревьями (табл. IV, 1). Гиганты-лепидодендроны, как ни странно, ближе всего родственны современным плаунам, длинные тонкие стебли которых стелются
теперь в наших лесах. Папоротники в э^ом периоде достигали размеров настоящих деревьев. Особенно удивительными были, каламиты (табл. IV, 2), которые напоминали современные хвощи, но увеличенные в 20-кратном размере. Высота их достигала 9 м. Наряду со споровыми растениями в каменноугольных лесах произрастали первые семенные растения. Это уже знакомые нам по девону семенные папоротники, а также древовидные кордаиты с большими листьями. Кордаиты являются далекими родичами наших хвойных растений (рис. 7) и, повидимому, происходят от семенных папоротников. Семена кордаитов настолько близки по строению к семенам этих папоротников, что их не всегда можно отличить друг от друга. Кордаиты были высокими деревьями, до 30 м в вышину, и почти до 3 м в обхвате. Их простые продолговатые и часто узкие листья достигали метра в длину. Органы размножения у них были собраны в специальные однополые шишки (мужские и женские). По строению семян кордаиты относятся к голосеменным растениям, семена которых не имеют специальной защитной оболочки. По строению древесины они близки к хвойным, особенно ж араукариям.
Многочисленные насекомые — родичи наших тараканов, кузнечиков и лесных клопов — населяли леса карбона. Здесь же ползали многоножки и скорпионы. Пауки всюду раскидывали свои тенета. В воздухе порхали стрекозы и поденки. Каменноугольные насекомые часто достигали необычайных размеров — до 30 и даже 70 см.
Пресноводные бассейны заболоченных лесов заселяли стегоцефалы, среди них встречались и такие формы, как шести-восьмиметровый эогиринус (табл. IV, 3) с длинным угревидным телом и со слабыми лапами и относительно более короткотелый бафетус (2 м длины), лапы которэ-
го были более развитыми (табл. IV, 4). У всех стегоцефалов, кроме парных глаз, хорошо был развит непарный теменной глаз. Первая жаберная щель рыб, так называемое брызгальце, у стегоцефалов затянулась барабанной перепонкой и образовала отдел среднего уха.
Стегоцефалы были разнообразны по виду; наряду с гигантами жили и мелкие формы, не превышавшие 20 см в длину. Некоторые из них с удлиненным телом и без конечностей напоминали змей. У одних стегоцефалов затылочные углы черепа выпячивались наподобие рогов, у других намечалось исчезновение в черепе
Рис 7. Кордант
костного панцыря. Кроме настоящих мелких стегоцефалов, водоемы карбона были переполнены личинками крупных стегоцефалов. Этих личинок, имевших жабры, называют боанхиозавра-ми (табл. IV, 5).
Стегоцефалы — настоящие хищники. Они питались рыбой, чаще всего многочисленными палеонисцидами. Двоякодышащих рыб было еще много, а кистеперые попадались реже, в особенности к концу периода. Из хрящевых акулообразных рыб вместе с пресноводными акантодиями встречались хищные пл^враканту-сы до 1 м в длину. Их большие мясистые плавники были приспособлены для ползания по дну. На затылочном отделе черепа у плевракантуса находился длинный острый шип.
В каменноугольном периоде от стегоцефалов отделился новый класс наземных животных— пресмыкающиеся. Пресмыкающиеся не мечут икру, а откладывают на суше яйца, подобно современным птицам. Кожа этих животных сухая и ороговевшая, в отличие от земноводных, у которых она влажная, слизистая и мягкая. Благодаря слизистой, легко подсыхающей коже земноводные постоянно были связаны либо с водоемом, либо с очень влажйым лесом, Они могли жить только вблизи воды. Особенности строения тела пресмыкающихся открыли для них неограниченные возможности для за селения суши. Они быстро стали заселять ее и приспособляться к новым различным условиям существования. Первые, относительно.мелкие (до 50 см) верхнекаменноугольные пресмыкающиеся назывались цельночерепными, или котилозаврами. Как и у стегоцефалов, в височном отделе их черепа все кости лежат вплотную друг к другу.
В самом конце каменноугольного периода от котилозавров отделились шлемочерёпйЫе пресмыкающиеся — пеликозавры. Эти животные, не более 1 м в длину, по некоторым особенностям еврей организации напоминали млекопитающих. У пеликозавров в височном отделе черепа кости лежали уже не вплотную друг к другу: между ними образовалось широкое пространство, которое называется височной ямой. Под височной ямой лежала скуловая дуга (скула), сходная с таковой млекопитающих. Зубы этих животных по форме были неодинаковы и напоминали резцы и клыки млекопитающих.
В пресноводных бассейнах Гондваны в конце карбона жили потомки котилозавров — рыбоядные мезозавры, имеющие длину до 1 м, очень рано приспособившиеся к водному образу жизни. Сильный хвост и задние лапы, ставшие ластами, служили мезозаврам для плацдния.
В жизни каменноугольного моря, по сравнению с предыдущим периодом, произошли перемены. Строматопоры и табуляты попрёжнему еще строили рифы, но в этом с ними уже успешно соревновались мшанки. Большинство
КАМЕННОУГОЛЬНЫЙ ПЕРИОД 7—ствол лепидодендрона-, 2—каламиты; 5—рогнривус; 4—бафетус; 5—браихмвватр
четырехлучевых кораллов, ставших колониальными, создавали многочисленные банки. Большие пространства на морском дне были засыпаны ракушками фузулин. Величина последних изменялась от размеров рисового зерна до размеров крупной овсинки. Фузулины, относящиеся к фораминиферам, были донными одноклеточными животными. Их скелеты образовывали известковые пласты толщиной в несколько метров. Повсюду встречались заросли морских лилий. Увеличилось число древних морских ежей и морских бутонов; плеченогих было еще много, но они стали менее разнообразны, чем раньше. Некоторые из них по величине были с голову ребенка.
Предки аммонитов и современных корабликов стали обильными. Среди брюхоногих (одностворчатых) моллюсков появились формы, дышащие не жабрами, а легкими. Все реже встречались ветвистые колонии граптолитов, шарови-ки и трилобиты. Ракоскорпионы почти вымерли; число водных членистоногих заметно сократилось. Зато возросло разнообразие наземных членистоногих. Многочисленные морские рыбы относились к акулообразным, но они еще отличались от современных акул тем, что у большинства из них хотя передние зубы были острые, пригодные для захватывания пищи, зато задние имели плоскую форму и могли быть использо ваны для раздавливания моллюсков — основного корма этих рыб.
ПЕРМСКИЙ ПЕРИОД
gly^Le менее 250 млн. лет назад,нг .З.е^деда-
•ф чался пермский период. Он продолжался около 25 млн. лет. В этот п^ериоХ; осрбё^нр в конце его, суша преобладала над морем. Площадь %Жного материка Гондваны заметцр „ увеличилась; Попреэдему Северр-Атлаэтический>а,.. МД7£-’рик объединял Северную Америку и Европу, включая,, Сорную Африку, ХДОДВОД?
и Южную Азию. Огромные горные j^enH^ поднимались по берегам северных контидантрв, на месте соврёменногб 'Урала и ' Зацаднр-Сибдр-ской низменности.
Гондвана отличалась прохладным климатом и в некоторых ъ местах ^аже испытывала~ оледенение. Климат северных ^материков был резко
На этих ^материках во мнбгих' местах возникали песчавдё" пустыщ!. В Азии в это время происходило мощное угле-^ра^ование.' ^гли" Ътого периода роздали QQHger
На дне пермских морей особенно обильными были закрученные раковины головоногих моллюсков. Края перегородок, разделявших эти раковины на камеры, постепенно принимали извилистую форму, как у аммонитов, самых обычных головоногих моллюсков мезозойской эры. Многие морские беспозвоночные, часто встречавшиеся в предшествующих периодах палеозойской эры, в течение пермского периода окончательно вымерли. К таким животным принадлежали: четырехлучевые кораллы, трилобиты, ракоскорпионы, морские бутоны, древние морские лилии. Все более редкими становились строма-топоры и табуляты. В меньшем количестве попадались плеченогие и древние морские ежи, но многочисленнее стали раковины двустворчатых моллюсков и появились уже настоящие новые
морские ежи, скелет которых, как у. современных ежей, состоял из 20 рядов известковых пластинок.
Наземная жизнь в пермское время была гораздо богаче, чем в какой-либо другой период палеозоя. Она уже не ограничивалась берегами водоемов и все более захватывала сушу благодаря развитию пресмыкающихся. Растительный покров Земли в начале перми мало отличался от растительности каменноугольного периода, но к концу перми он значительно изменился. Папоротникообразные растения постепенно отходили на задний план, а вместо них все большее значение приобретали настоящие семенные растения. Последних часто также называют цветковыми вследствие того, что на них образуются цветы.
Среди цветковых растений различают группу голосеменных и покрытосеменных. К типично голосеменным относятся наши хвойные деревья: ель, сосна, кедр, пихта и др. Голосеменными эти растения называются потому, что их семяпочки, т. е. те образования, из которых возникают будущие семена, лежат в цветке совершенно открыто. Цветок хвойных деревьев называют шишкой. Если взять молодую шишку сосны, ели или кедра и отогнуть ее чешуйки, будет видно, что на каждой чешуйке сидят семяпочки. В зрелой шишке вместо семяпочек сидят развившиеся из них семена.
У покрытосеменных растений семяпочки скрыты в завязи — вздутии в основании пестика цветка. У покрытосеменных растений обычно вокруг развивающихся из семяпочки семян завязь образует плод. Иногда плод вокруг семени образует сухую оболочку, как у ржи или пшеницы. Однако эта оболочка плода может
Рис. 8. Диметр од онт
быть сочной и мясистой, как у яблока, сливы, вишни и т. д. Таким образом, в отличие от голосеменных, у покрытосеменных растений, чтобы добраться до семян, необходимо предварительно очистить защищающие их оболочки.
Все пермские семенные растения принадлежали к голосеменным. В это время попрежнему росли еще широколистые кордаиты, к которым присоединились саговики, или так называемые саговые пальмы, и гиикговые растения, у которых на стволе вместо хвои сидели широкие лопастные листья. Здесь же росли деревья с шиловидными маленькими листочками. Последние, повидимому, являлись родоначальниками современных хвойных.
На Северной суше, климат которой был теплее климата Гондваны, все еще росли лепидодендроны (таблица V, 1). Среди растений Гондваны в начале перми особенно часто попадался своеобразный глоссоптерис (табл. V, 2), но и этот папоротник к концу периода переселился на Северную сушу. Через Индию, Персию и Сибирь он достиг мест теперешней Северной Двины.
В начале перми стегоцефалы были еще многочисленны и разнообразны. Среди них выделялись крупные, до 2 м длины, наземные животные. Некоторые, не выдержав конкуренции с пресмыкающимися, вновь возвращались к водному образу жизни, например, двинозавр (1 м длиной), который в Течение всей жизни сохра-26
нял жабры (табл. V, 3). К концу периода число стегоцефалов значительно сократилось.
Быстрое развитие пресмыкающихся к концу пермского периода и одновременное вымирание стегоцефалов можно объяснить изменением климатических условий в связи с преобладанием к этому времени суши над морем. Климат сделался сухим. Многие пресноводные водоемы и болота высохли. Появились многочисленные пустыни. Эти условия благоприятствовали развитию сухолюбивых яйцекладущих пресмыкающихся с ороговевшей кожей и гибельно отра» жалйсь на земноводных, жизнь которых попреж-нему была тесно связана с пресными водоемами. Некоторые из цельночерепных пресмыкающихся, например, небольшая, до 60 см в длину, котлассия (таб. V, 4) и близкая к ней сейму-рия, имели так много общего с земноводными, что трудно даже сказать, кого это животное больше напоминало — земноводное или пресмыкающееся. Котлассию мы называем «переходной формой», потому что строение ее тела с полной очевидностью убеждает в том, что пресмыкающиеся произошли от земноводных.
В свою очередь многие пермские пресмыкающиеся по своему строению напоминают млекопитающих. В это время жили известные уже нам пеликозавры и произошедшие от них зверообразные пресмыкающиеся терапсиды, среди которых нужно искать прямых предков млекопитающих. Из пеликозавров особенно замечателен
ПЕРМСКИЙ ПЕРИОД. ОАЗИС В ПУСТЫНЕ, БЫВШЕЙ НА МЕСТЕ СОВРЕМЕННОЙ СЕВЕРНОЙ ДВИНЫ В ВЕРХНЕПЕРМСКОЕ ВРЕМЯ
7—леяидадеядраи; 4—глоссокгерис; 3—двинозавры; 4—котлассия; 5—дицинодонты; б—иностранцеввя; парейазавр
почти 2-метровый плотоядный диметродонт (рис. 8). У него остистые отростки позвонков почти на 1 м поднимались выше спины и образовывали диковинный гребень.
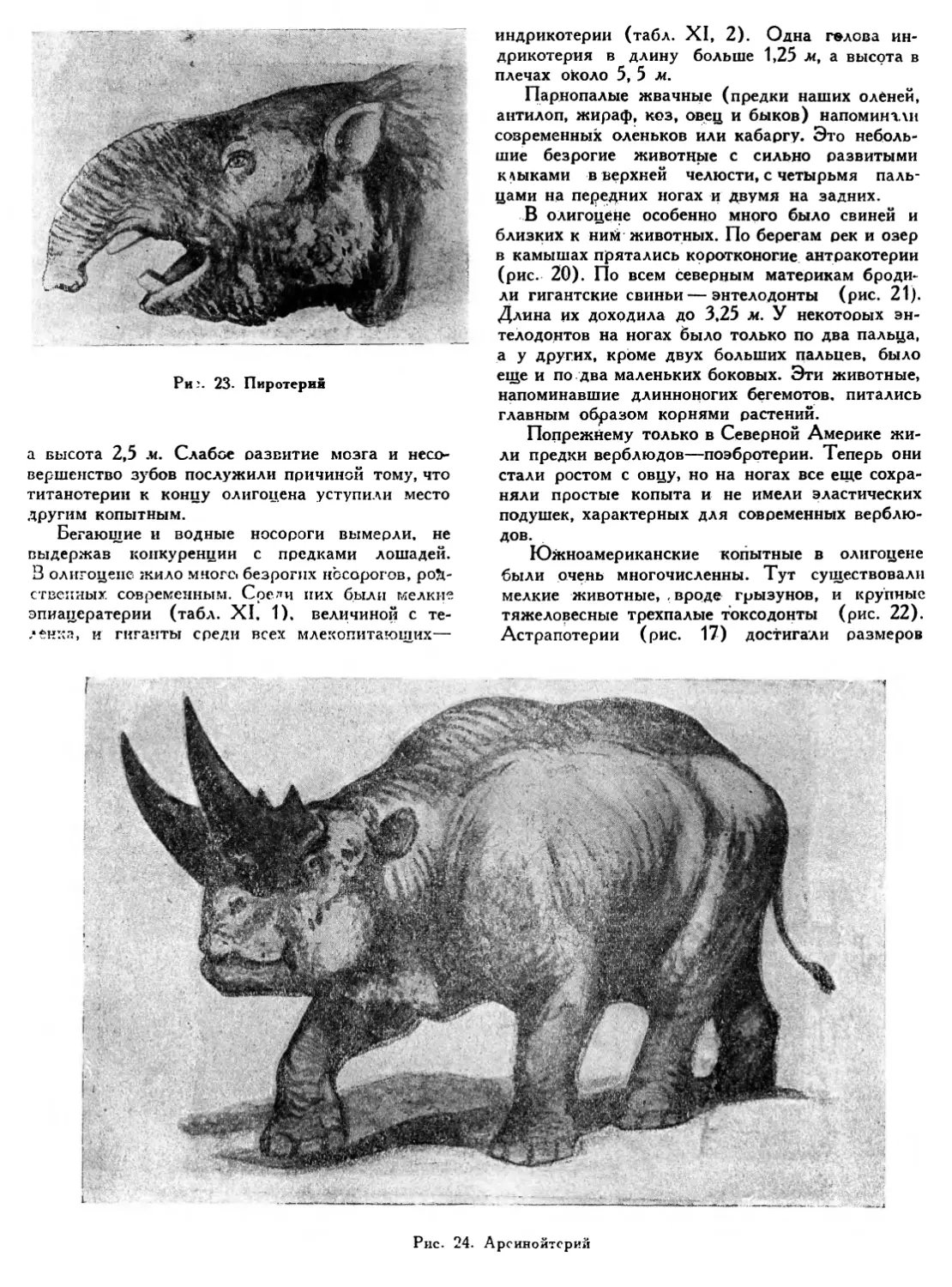
Среди зверообразных пресмыкающихся встречались безобидные травоядные формы, вроде дицинодонта (табл. V, 5), у которого во рту оставалась только одна пара клыкообразных зубов. Очевидно, края челюстей дицинодонта, размеры которого не превышали 1,5 м, были одеты роговым клювом, подобно челюстям черепах.
Большинство зверообразных — хищники. Многие из них, грозные хищники, достигали 4 м в длину. Такова иностранцевия (табл. V, 6>,
вооруженная страшными клыками. Ее излюбленной добычей были крупные 3-метровые парейазавры (табл. V, 7), неуклюжие травоядные цельночерепные пресмыкающиеся.
В самом конце, периода мы находим небольших пресмыкающихся, у которых в височном отделе черепа развивались две височные ямы. Каждая из них была ограничена снизу скуловой дугой. Зубы этих животных сидели в отдельных ячейках, а не в общем желобке, как v большинства пресмыкающихся. И в этих первых ячеезубых пресмыкающихся еще трудно угадать предков будущих крокодилов, летающих драконов и динозавров (ужасных ящеров), с которы ми мы встретимся в последующих периодах.
МЕЗОЗОЙСКАЯ ЭРА
ТРИАСОВЫЙ ПЕРИОД
еиасовый_пе£иод на Земле^ЯМАЖ^ся около 45 млн. лет. С его начала до на-ших: днеи_.шюддал1<ЖОЛИЗИтеАьца3220, млн*, лет. ЕГ триасе суша преобладала над морем. Су шест-вовалодва. крнти^ Северо-Атлантический и Азиатский материки* *^113ОвалиС^
лежала прежняя! ондвана. Азия соединилась с ABCT£aAH|^
“ропа^ ЛЙШ, :Mx^KpJWM.,s J ВШИ жС$-
Бе^ная^ Аф^цка,, были залиты океаном Тетис. Больших горных цепей в это время заново не возникало, но горы, сформировавшиеся в^пр<Е?жние ЬёрйодыГ были еще высоки. Часто происходили вулканические извержения. Клихмат триасового
14зл ра^тгеццц ^заметно ные: саговые, хвойные и гинкговые. Из семенных папррртников^продолжал существовать глос-соптерис. В конце периода появились своеобразные папоротники, особенно многочисленные в
последующем юрском периоде, листочки которых по жилкованию напоминали листья семенных растений. Триасовь^^хво^щи (табл. VI, I) стоят гораздо ближе к современным хвощам, чем палеозойские.
В жизни обитателей материков произошли большие перемены. Начавшееся еще в пермский период преобладание суши над морем и прогрессивное осушение многих пресных водоемов в триасовый период привели к тому, что многие пресноводные рыбы теперь переселились в моря, и только двоякодышащие, близкие к теперешним, попрежнему жили в уцелевших пресноводных бассейнах. В конце триасового периода вымерли стегоцефалы. Это были последние представители лабиринтозубых стегоцефалов, названных так потому, что эмаль на их зубах имела сложное складчатое строение. Все стегоцефалы,
спасаясь от сухого климата и от конкуренции с пресмыкающимися, стали водными, а некоторые даже перешли жить в море. Большинство из них были очень крупными животными. Например, у мастодонтзавра длина черепа достигала 1 м.
В начале триасового периода жили прямые предки современных лягушек. Эти протобатра-хусы (табл. VI, 2) мелкие, длиной в 10 см, животные, по общему складу скорее напоминают жаб, чем настоящих лягушек. Кожа их бугорчатая, задние ноги более приспособлены для плавания, чем для прыжка.
Особенно сильно изменились пресмыкающиеся; цельночерепные окончательно вымирали. Во вторую половину периода появились первые черепахи, у которых в отличие от современных на нёбе еще сохранились зубы, в то время как челюсти были одеты роговым клювом.
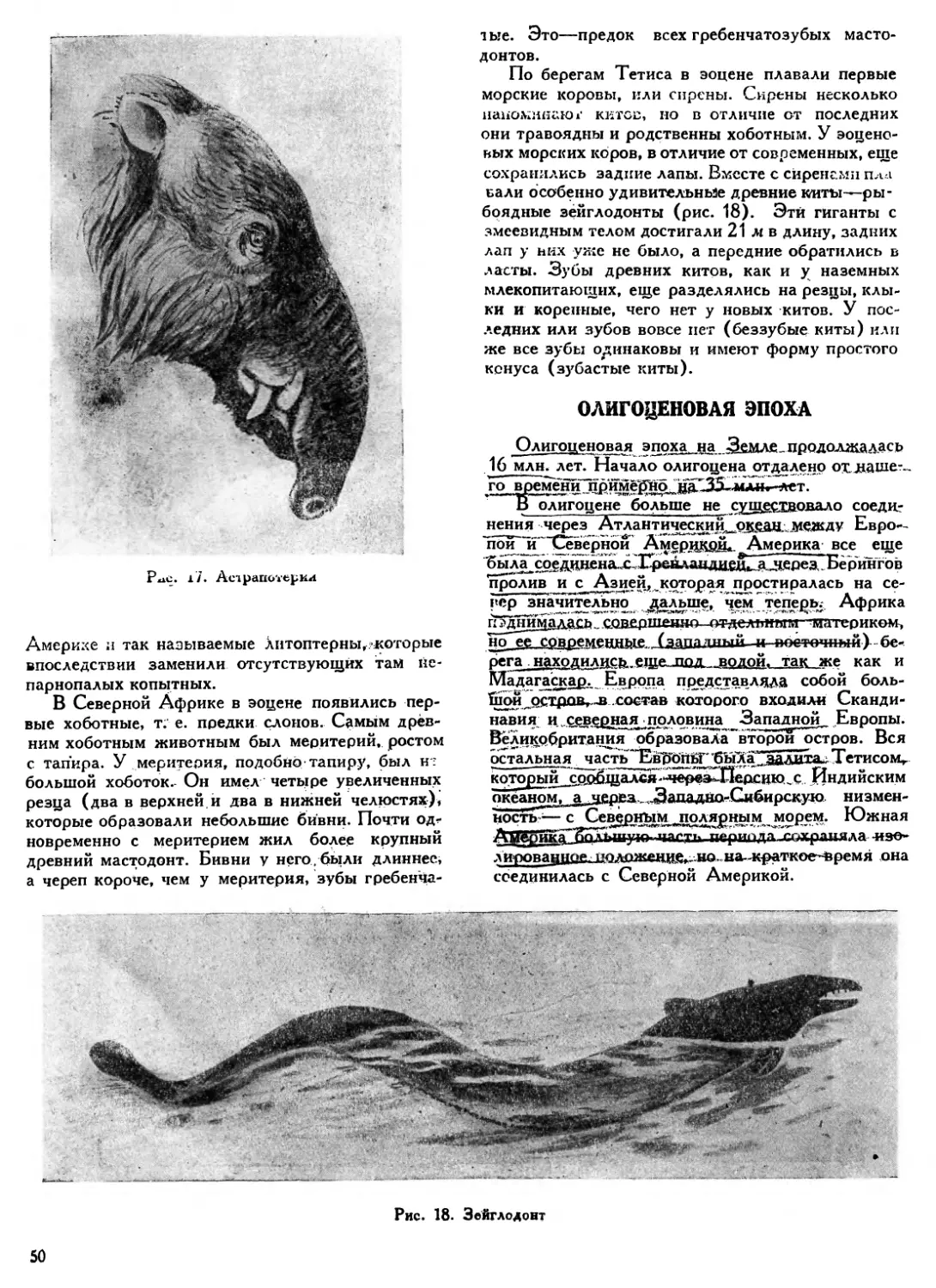
В триасовый период усиленно развивались, но в конце его уже вымерли последние звероподобные пресмыкающиеся. Из них травоядные и уже совершенно беззубые шталекерии (табл. VI, 3) достигали разменов крупного носорога. Меньших размеров был хищный белезодонт (табл. VI, 4) длиной около 1,5 м.
Особенно интересны мелкие зверообразные пресмыкающиеся иктидозавры, близкие к млекопитающим. Так, каромис (табл. VI. 5), животное величиной с крысу, по строению своего черепа уже является настоящим млекопитающим, и только дополнительные кости, имеющиеся в его нижней челюсти, указывают на то, что это животное еще пресмыкающееся.
Из других пресмыкающихся в триасовый период развивались хоботоголовые, ближайшие родичи современной новозеландской гаттерии, которые хотя и похожи на обычных ящериц, но отличаются от них по своему строению. Гаттерия в своем строении до сих пор сохранила
многие древние особенности. В ее черепе имеются две височные (скуловые) дуги, а не одна, как у ящериц. Верхняя челюсть у нее свисает вниз в виде небольшого клюва. Зубы на челюстях сидят не в отдельных ячейках, а в общем желобке. Кроме обычных ребер, на брюхе развиваются еще «брюшные ребра». Двояковогнутые позвонки напоминают позвонки рыб. Среди хоботоголовых в триасе жили стенаулоринхусы (табл. VI, 6)—крупные роющие животные, возможно, питавшиеся корнями. В морях, по берегам континентов, попадались длиннорылые хоботоголовые—истребители морских моллюсков. Вместе с ними за моллюсками охотились несколько напоминавшие морских черепах плакодонты, у которых на нёбе вместо мелких зубов образовывались настоящие жерновки для раздавливания раковин. Родственные плакодонтам нотозавры тоже вели водный образ жизни. Эти длинно-шеие животные еще могли использовать свои лапы (ласты) для хождения по земле. От ното-завров произошли плезиозавры, обычные морские пресмыкающиеся следующих периодов. В северных водах появились первые рыбоящеры, или ихтиозавры. Они. еще не так хорошо были приспособлены, к плаванию в море, как их потомки, у которых хвост стал похож на рыбий. Самое замечательное то, что ихтиозавры не откладывали яиц, как обычные пресмыкающиеся, а рождали живых детенышей, подобно млекопи тающим.
С триаса же начался расцвет группы ячеезубых пресмыкающихся. Наиболее древние формы из них были сравнительно небольшие плотоядные животные. Вместо обычного передвижения на четырех ногах эти животные приспосабливались к хождению и на двух, в связи с чем их задние ноги становились гораздо длиннее передних. Таким был салыопозухус 32
(рис. 9), животное размером больше 1 ж. К кон цу триаса некоторые ячеезубые пресмыкающиеся перешли к водному образу жизни. Они снова ..начали ходить на четырех ногах и по внешности несколько напоминали еще отсутствовавших тогда крокодилов. Длина такого водного крокодилообразного престозухуса (табл. VI. 7) была не меньше 5 м. Преимущественно на Северной суше появились первые, еще не очень большие по размеру предки динозавров. Некоторые из них были невелики, до 1 м в длину, и вели хищный образ жизни. Ходили они на задних ногах, которые были длиннее передних. По некоторым признакам динозавры напоминали птиц: кости их скелета были полыми наполненными воздухом, а первый палец на задних ногах был обращен назад.
Другие динозавры, например цлатеозавр (рис. 10), были значительно крупнее, достигая 6 м в длину. Разница в строении передних и задних ног у них невелика, зубы тупые. Это были предки травоядных великанов юрского периода.
Неудивительно, что при обилии в триасе зверообразных пресмыкающихся мы находим здесь и настоящих млекопитающих. Самое древнее известное нам млекопитающее животное, величиной ;с сурка, именуется «тритилодонт» (табл. VI, 8). Он.относится, к группе м но г о б у*
Рис. 10. Платеозавр
ТРИАСОВЫЙ ПЕРИОД. ФАУНА ЮЖНОГО МАТЕРИКА ГОНДВАНЫ
горчатозубых млекопитающих, называемых ?ак потому, что у них на коренных зубах в два или три ряда были расположены многочисленные бугорки. Клыков у них не было. Одна пара резцов в верхней челюсти и единственная пара в нижней были увеличены. Многобугорча-
тсзубые питались растительной пищей. Вероятно они еще откладывали яйца, а не рождали живых детенышей, так же как и современные австралийские однопроходные млекопитающие: утконос и ехидна. Современные яйцекладущие млекопитающие — беззубы, но зародыши утконоса имеют зачатки зубов многобугорчатого типа. Поэтому многобугорчатозубых считают ближайшими родичами австралийских однопроходных, у которых еще сохраняется много особенностей, характерных для пресмыкающихся.
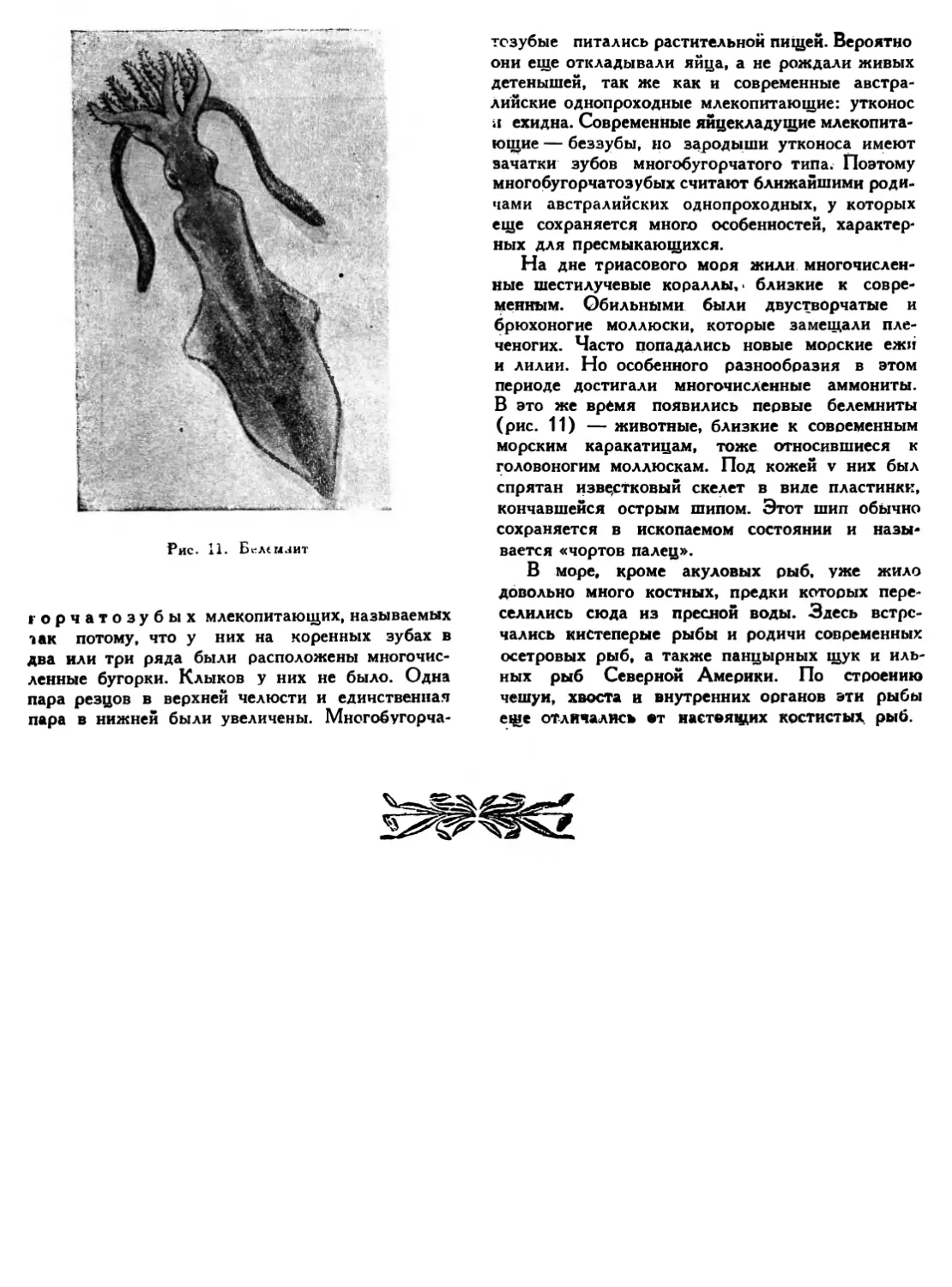
На дне триасового моря жили многочисленные шестилучевые кораллы,* близкие к современным. Обильными были двустворчатые и брюхоногие моллюски, которые замещали пле-ченогих. Часто попадались новые морские ежи и лилии. Но особенного разнообразия в этом периоде достигали многочисленные аммониты. В это же время появились первые белемниты (рис. 11) — животные, близкие к современным морским каракатицам, тоже относившиеся к головоногим моллюскам. Под кожей v них был спрятан известковый скелет в виде пластинки, кончавшейся острым шипом. Этот шип обычно сохраняется в ископаемом состоянии и называется «чортов палец».
В море, кроме акуловых рыб, уже жило довольно много костных, предки которых переселились сюда из пресной воды. Здесь встречались кистеперые рыбы и родичи современных осетровых рыб, а также панцырных щук и ильных рыб Северной Америки. По строению чешуи, хвоста и внутренних органов эти рыбы еще отличались от настоящих костистых рыб.
ЮРСКИЙ ПЕРИОД
родолжительность юрского периода исчисляется приблизительно в 35 млн. лет. С его начала донаШих дней прошло 175 млн. лет. В отличие от предыдущих периодов, в юрский период море преобладало над сушей. Северная суша распалась на части. Азия уже не соединялась с Северной Америкой. Последняя хотя и вХб'дила в состав Севёро;Атлантического матерят ка, но_этот материк - уже ^захватывал Европу. На ее .местебущевКло" море, &ртррре проникло, и в Азию, покрывая собой территорию срвр*> менйых южных горных хребтов._Из всей Евро; пейской суши только Скандинавия и Финлян; дня'поднимались островом. Рельеф юрских, континентов был ровный. Старые горы успел 4 сгладиться/ а формирование новых кряжей только' что нддадось,. Климат юрского перихдл ровный, тзплый и влажный. Он способствовав пышйдйу развитию раётйтёльностй и богатому угдеобразовйм^
Из споровых растений особенно многочисленными были папоротники и хвощи, а из голосеменных — гшнкговые/и ^саговые пальмы^ Из настоящих хвойных в юре появились араукарии и секвой. Среди этих растений порхали первые бабочки.
В теплых водах юрских морей шестилучевые кораллы и известковые губки строили массивные рифы. Многочисленные сидячие двустворчатые моллюски создавали на дне обширные колонии. Часто попадались здесь также заросли морских лилий. Наблюдалось обилие морских ежей. Некоторые из них теряли шаровидную форму. Особенно многочисленными были белемниты и аммониты.
Число морских рыб в юрское время заметно возросло. Здесь уже плавали акулы, близкие к современным. Около дна держались плоские скаты. Многочисленными в юре были костные рыбы. По своему строению эти костные рыбы
занимали среднее положение между осетровыми и настоящими костистыми, первых представителей которых мы встречаем именно в юре. Стегоцефалы исчезли, их заменяли теперь амфибии современного типа, среди которых были уже настоящие лягушки. Суша, воздух и море находились во власти пресмыкающихся.
Вдоль берегов островов и континентов в юрских морях плавали многочисленные и очень разнообразные пресмыкающиеся: удивительные морские крокодилы и длинношеие плезиозавры (табл. VII, 1; рис. 12), достигавшие иногда 15 м в длину. Передние и задние лапы, ставшие плавниками, служили плезиозаврам как хорошие весла. Длинная шея позволяла на мелководье, не ныряя, доставать корм непосредственно со дна. Поодаль от берегов, подобно нынешним дельфинам, плавали многочисленные рыбоящеры — ихтиозавры (рис. 13). Среди них попадались как мелкие, длиной 1.5 — 2 м, так и настоящие гиганты до 8 — 12 м. На суше жили многочисленные, впервые появившиеся в юре крокодилы. Одни из них по размерам близко подходили к современным крокодилам, другие были величиной с ящерицу. У многих из них в коже развивался тяжелый костный панцирь. Встречались здесь уже совершенно беззубые черепахи, изредко попадались первые настоящие ящерицы.
Хоботоголовые пресмыкающиеся — родичч теперешней гаттерии — были довольно обычны. Но особенно поражают совершенно изумительные гиганты — динозавры, доходившие по весу до 50 т. Следовательно, динозавр в 10—12 раз был тяжелее взрослого слона. Длина динозавра от кончика хвоста до конца морды может доходить до 26,5 м. Таков диплодок (табл. VII, 2)— изумительное четвероногое животное с непропорционально маленькой головой, с длинной шеей и таким же хвостом. Это животное,
питавшееся мягкой растительной нищей, большую часть времени проводило в болотистых водоемах, так как в воде ему было легче передвигать свое массивное тело и прятаться от врагов. А врагами были хищные пресмыкающиеся, ходившие на двух ногах мегалозавры, вооруженные страшными зубами. У них, так же как и у птиц, на ногах три пальца смотрели вперед, а первый палец был обращен назад. Передние их лапы были невелики. У некоторых хищников на носу находился небольшой рог.
Кроме диплодока и близких к нему ящеров, в юре жили также менее крупные травоядные динозавры, называемые пти-цетазовыми. Свое название они получили оттого, что их тазовые кости очень напоминали те же кости птиц. Одни из них ходили только на двух зад-
них ногах, а другие — на четырех. Среди таких динозавров особенно интересен 6—8 метровый стегозавр
(табл. VII, 3). Передние лапы его были коротки, а задние длинны. На спине в два ряда* сидели массивные костяные плитки, защищавшие стегозавра от нападения хищника сверху. Хвост стегозавра, несший две пары острых шипов, служил серьезным оружием против врагов, нападав
ших сбоку. Передние части челюстей этого животного были одеты роговым клювом.
Пресмыкающиеся не только бродили по суше, они носились и в воздухе. В начале юры порхали только длиннохвостые имеющие зубы рамфоринхи (табл. VII, 4), позднее появились короткохвостые птеродактили, величиной со среднюю
Рис. 13. Ихтиозавр
птицу, а некоторые — с воробья. Четвертый палец их передних лап постепенно вытягивался. К нему прирастала летательная кожная перепонка, натянутая между передними и задними лапами. Несмотря на то, что по образу жизни и некоторым чертам строения птеродактили напоминали птиц, их все же нельзя назвать близкими родственниками последних.
В конце юры на Земле существовали очень любопытные животные, по своему строению напоминавшие одновременно и пресмыкающихся и птиц. Это были так называемые первоптицы (археоптериксы) (табл. VII, 5; рис. 14).
Археоптериксы—длиннохвостые лазящие животные величиной с голубя. На передних трехпалых лапах и на хвосте у них росли перья, а пальцы передних лап были свободны, как у
ЮРСКИЙ ПЕРИОД. ЖИЗНЬ МОРСКОГО ПОБЕРЕЖЬЯ СЕВЕРО-АТЛАНТИЧЕСКОГО МАТЕРИКА В ВЕРХНЕЮРСКОЕ ВРЕМЯ
7 — плезиозавры; 2— диплсдок; 3—стегозавр; 4—рамфоринхи; 5—археоптерикс; б— амфитсрий
Рис. 14. Археоптерикс
ящерицы, и вооружены когтями. В их челюстях имелись зубы. Археоптериксы были близкими родичами настоящих птиц, но летать они еще не могли и только перепархивали с дерева на дерево.
В юрский период звероподобных пресмыкающихся уже не было. Кроме живших в предыдущем периоде многобугорчатозубых
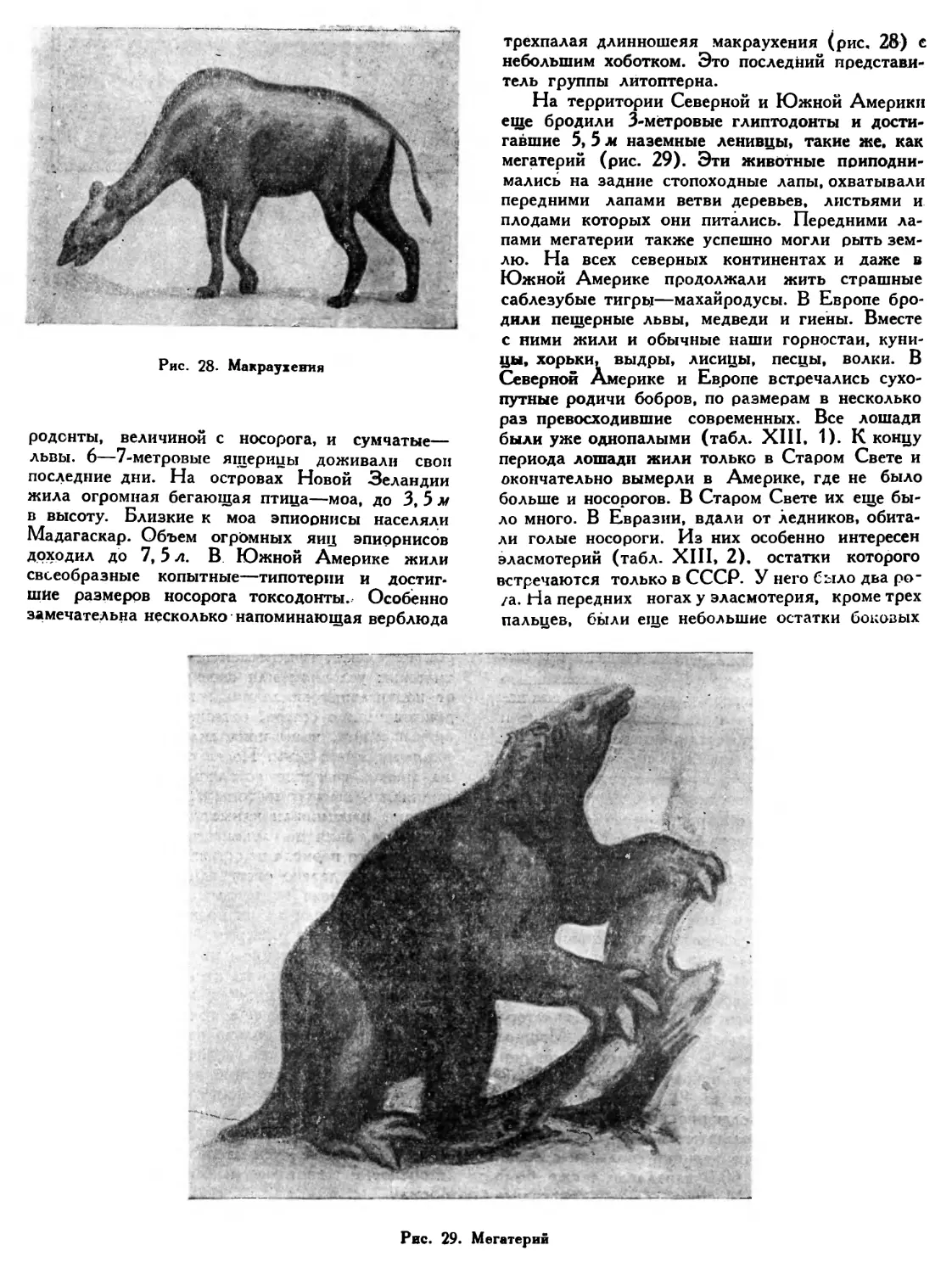
яйцекладущих млекопитающих, в юре пояжилодь мелкие, величиной с мышь или крысу, пакте* терии, каким, например, был амфитерий (табл. VII, 6). Эти древесные, возможно, ночные животные, прятавшиеся днем от страшных пресмыкающихся, были общими предками современных сумчатых и обычных плацентарных (высщих) млекопитающих. Сумчатые животные в наше время живут только в Австралии и Южной Америке. В отличие от обычных плацен-* арных млекопитающих детеныши сумчатых родятся очень маленькими и недоношенными. Мать долгое время носит новорожденных в особой сумке на животе, где они питаются молоком. Обычные, или плацентарные, млекопитающие получили свое название от слова «плацента». Че0ез плаценту, или детское место, устанавливается тесная связь между развивающимся зародышем и материнским организмом. Через нее же совершается питание зародыша. На\ичие плаценты у млекопитающих дает им «возможность рождать уже хорошо развитых детенышей. Поэтому новорожденный жеребенок или теленок, после появления на свет, сразу же способен следовать за матерью. Эта особенность дает детенышу большое преимущество в борьбе за жизнь.
В юре жили и такие млекопитающие, которые не оставили после себя потомков. Это трехконусозубые и трехбугорчатозубые млекопитающие, небольших размеров. Самые крупные из них были не больше кошки.
меловой период
начала мелового периода, продолжавшегося около 80 млн. лет, до наших дней прошло приблизительно 140 млн. лет.
Это время в жизни Земли было богато событиями. Море и суша неоднократно меняли свои очертания. Ближе к концу периода море отвоевало у суши особенно большие пространства, но в самом конце мелового периода оно быстро дт^тупйлрЖМЙЬ .Г2йХваВ?.г.Ум®нь1иилась в размерах. Она объединяла, ^теперь только Африку и Южную Америку. Австралия образовала сРвёрпхенйб изолированный континент. Новая Зеландия поднялась в виде, , острова. Большая часть Азии вместе с Ост-Индским архипелагом образовала материк, которыйчерез БеринговоI море соединялся с высокогорной сушей, тянувшейся по западному берегу Северной Америки. Северо-Атлантический континент, хотя: и меньшийпо площади, все еще продолжал существовать. Скандинавия и Финляндия образовали большой остров. Меньшие острова лежали в Южной Европе. Западная Европа, южная и западная половины Европейской ча-стИ СССРГ Туркестан и центральная часть Азии (место современных Гималайских гор) находились под, врдой.
Грандиозные горные цепи,воздвигались по побережью Тихого океана. Здесь же происходили обильные извержения вулканов. Климат мелового периода, теплый и влажный^ к концу периода стал несколько более прохладным;, резче наметилась разница между отдельными кли-магическими поясами.
В начале мелового периода растительность •©стояла в основном из саговых пальм и хвойных деревьев, но к ним все больше и больше стали присоединяться настоящие покрытосеменные растения. В середине периода они уже размножились настолько, что заметно оттеснили На задний план своих голосеменных предков.
Происхождение покрытосеменных растений от голосеменных не вызывает сомнений, но установить среди последних прародительскую группу пока еще не удалось.
Быстрое размножение покрытосеменцдос растений во второй половине мелового периода одно время пытались связать с их светолюби-востью. Ученые предполагали, что по каким-то причицам_во цторой половине мела значительно возросла сила солнечного света, падающего на поверхность Земли. Это обстоятельство Дала возможность светолюбивым покрытосеменным растениям обогнать в развитии их тенелюбивых предшественников. Нужно думать, что не только изменение в количестве солнечного света на Земле способа процветанию покрытосеменных растений. Их многочисленные, с шире* кой поверхностью листья благоприятствовали образованию и накоплению в растениях крахмала, т. е. необходимого питательного вещества. Их семена, развивающиеся в завязях и покрытые в зрелом состоянии плодовыми оболочками, были значительно лучше защищены от неблаго* приятных воздействий среды, чем у голосеменных растений. Все перечисленные особенности покрытосеменных, несомненно, давали им значительные жизненные преимущества перед дру-гими растениями.
Прежнее мнение, что покрытосеменные появились внезапно только в меловом периоде, оказалось неверным, так как остатки их найдены теперь и в юрских отложениях. Эти древние покрытосеменные растения принадлежали к группе двудольных покрытосеменных. Двудольными они называются потому, что у их зародыша, находящегося в семени, имеются две семядоли два первичных листочка. Эта группа среди покрытосеменных растений значительно более многочисленна, чем группа однодольных, у кеторых в семени имеется только одна семя
доля. Среди двудольных растений имеется большое число древесных и кустарниковых форм, среди однодольных преобладают травянистые растения. Поскольку обширные травянистые покровы на Земле развились значительно позднее лесных, весьма вероятно предположение ученых о том, что однодольные растения произошли от двудольных.
Однодольные травянистые растения характеризуются коротким жизненным циклом плодоносящей части. Это дает им преимущество в переживании неблагоприятного времени года. Зиму они переживают или в виде семян или в виде йодземных корневищ, клубней и луковиц. Поэтому однодольные относительно редки в тропиках и преобладают в умеренном климате. Их усиленное развитие на Земле, как мы увидим дальше, возможно, было связано с постепенным охлаждением климата. Уже в начале мелового периода из двудольных встречались на Земле платаны, лавры, магнолии. Вскоре к ним присоединились дубы, клены, фикусы. Из однодольных в мелу известны различные пальмы, лилейные растения и др. К концу периода Земля впервые покрылась койром из трав и цветов. На ней возникли леса и луга, похожие на современные.
Название мелового периода происходит от слова «мел». Мелом называется осадочная порода, состоящая из скопления раковинок одноклеточных морских животных — фораминифер, смешанных с известняковым илом. В течение мелового периода бесчисленные раковинки отмерших животных, падая на морское дно. скоплялись там целыми многометровыми пластами, которые с течением времени уплотнялись и образовали твердую Торную породу — мел.
Кроме простейших, в меловых морях были очень обильны кораллы, губки и двустворчатые моллюски. Аммониты, вначале многочисленные, в конце периода окончательно вымерли. Среди них появились формы с неправильно раскрученными,раковинами. Белемниты также почти все вымерли. Часто встречались десятиногие раки—родичи нашего речного рака, и в изобилии появились куцые крабы. Настоящие костистые рыбы начали успешно вытеснять оставшихся костных рыб. В, пресных водоемах жили лягушки и хвостатые саламандры. Пресмыкающиеся достигли высшей точки своего развития, но в конце периода началось их массовое вымирание.
Быстрое вымирание большинства пресмыкающихся в конце мелового периода находится в теснейшей связи с изменением физико-географических условий и с прогрессивным развитием млекопитающих. Как мы говорили, в конце мелового периода начались быстрое отступание моря и подъем континентов, заболоченные пространства вдоль морских берегов стали осушаться, травоядные динозавры лишились на Земле убежищ и корма. Очутившись на сухо
путье, лишенные обычного корма и с трудом-передвигавшиеся, они неминуемо должны были погибнуть. Их гибель повлекла за собой вымиг рание в основном ими питавшихся хищных динозавров. Остальные пресмыкающиеся должны были выдерживать жестокую конкуренцию с млекопитающими, которые к концу мела значительно размножились. Последние обладали' многими преимуществами в своей организации по сравнению с пресмыкающимися и были значительно лучше их приспособлены к новым условиям климата и растительности.
Основные преимущества млекопитающих заключались: в постоянной температуре, тела* и в волосяном покрове, предохраняющем животное от холода, в живорождении и в выкармливании детенышей молоком, в прогрессивном строении головного мозга и органов пищеварения, в частности зубов, которые стали служить млекопитающим не только для захвата корма, но и для его пережевывания.
Немногие пресмыкающиеся, напоминавшие свойх современных представителей, пережили меновой период. Но в начале и середине описываемого периода еще попрежнему морские воды бороздили ихтиозавры, у берегов плавали длинношеие плезиозавры и 13-метоовые морские змееобразные ящерицы мозазавры. Появи-лйсь и мелкие водные ящерицы — долихозав-ры. Наземные ящерицы напоминали современных. Появились первые наземные змеи. Среди < ерепах встречались гиганты до 3,5 в длину. По воздуху, вдоль морских берегов, носились теперь только бесхвостые птеродактили. Из. них совершенно беззубый птеранодонт (табл. VIII, 1) имел в размахе крыльев 8 лс. Затылочная часть его 2-т>тетрового черепа была вытянута назад, как противовес огромному клюву, под которым висел кожаный мешок, так же как у современного пеликана. Этот мешок служил животному подсачком при добывании рыбы, которой он питался.
Травоядных динозавров, родичей диплодока, теперь стало заметно меньше, особенно на северных материках, но хищники процветали. Здесь жил величайший из когда-либо бывших на Земле хищников — тиранозавр (табл. VIII, 2). Длина его тела с хвостом равна 14,5 м. Когда он стоял в обычной для него позе на задних ногах, голова его была поднята над землей почти, на 6 м. Тиранозавр был грозой всего живущего. Он не щадил ни травоядных ящеров, ни своих сородичей. Часто в его пасть попадали похожие на страусов струтиомимусы (табл. VIII, 3), стройные длинношеие и длинноногие бегуны, с сильным хвостом. Они, так же как и птицы, были беззубы и имели роговой клюв. Трехпалые передние лапы служили им для хватания. Струтиомимусы были различной величины—от 1,5 до 6 м. Эти животные питались яйцами, которые похищали из гнезд других пресмыкающихся.
МЕЛОВОЙ ПЕРИОД. ЖИЗНЬ МОРСКОГО ПОБЕРЕЖЬЯ СЕВЕРО-АТЛАНТИЧЕСКОГО КОНТИНЕНТА В ВЕРХНЕМЕЛОВОЕ ВРЕМЯ
7 —птеранодонт; 2—тиранозавр; 3—струтиомимус; #—траходон; 5—трицератопс; 6—нхтяорннс
Быстрота бега давала им возможность ускользнуть от преследования многих ящеров.
Чрезвычайно многочисленны в меловом периоде. были птицетазовые , динозавры . Траходон (табл. VIII, 4), до 9—10 м в длину, жил по берёгам водоемов. Передние расширенные части его челюстей, напоминавшие утиный клюв, были одеты роговым чехлом, дальше в. челюстях во: много рядов сидели мелкие многочисленные зубы. Таких зубову траходона во рту было больше 2000. Он питался, подобно современным уткам, смешанным кормом: растениями и мелкими беспозвоночными, которых ловил на мелководье. В Северной Америке найдены настоящие окаменевшие мумии этого животного, на которых даже сохранились чешуйки, покрывавшие его коЖу. Растительноядный 6-метровый трицера-Tohc (табл. VIII, 5) ходил на четырех ногах. Для защиты от хищников он имел три крепких рога, украшавшие его череп, и огромный костный воротник, как броней закрывавший ему шею. Передние половины челюстей, были вооружены крепким клювом. В задней половине челюсти в один ряд сидели зубы. В Монголии найдены сохранившиеся до сего времени целые гнезда яиц предков трицератопса. Многие яйца были с зародышами.
В меловой период существовали настоящие птицы, но, в отличие от современных, у них еще сохранились зубы. Здесь встречались и мелкие, величиной с голубя, летающие ихтиорнисы (табл. VIII, 6) и крупные, до 1,5 м длины, нелетающие морские гесперорнисы (рис. 15). Они прекрасно плавали и ныряли при помощи задних лап. В процессе эволюции они утратили крылья. От крыльев у них сохранились лишь плечевые кости.
Млекопитающие были еще немногочисленны. Среди них наиболее обычны многобугорчатозубые. Трехконусозубые и трехбугорчатозубые полностью вымерли. Потомки мелких юрских панто-терий образовали теперь две совершенно самостоятельные группы: сумчатых и плацентарных млекопитающих. Меловые сумчатые—небольшие древесные животные, напоминающие нам современных американских сумчатых крыс — опоссумов. Настоящие плацентарные млекопитающие тоже невелики. Одни из них близки к современным ежам, т. е. насекомоядным животным. У других меловых млекопитающих увеличивались клыки, коренные зубы становились более острыми. Эти животные становились похожими на маленьких хищников.
КАЙНОЗОЙСКАЯ ЭРА
ТРЕТИЧНЫЙ ПЕРИОД
ПАЛЕОЦЕНОВАЯ ЭПОХА
/т II
ду? L а палеоценовую эпоху приходится при»
Т’ мерно 7 млн. лет. С ее начала до ца-ших дней прошло 60 млн.лет.
Море, начавшее отступать в конце мелового периода, еще больше сократилось в размерах. Северо-Атлантический континент соединялся теперь с поднявшейся из морской пучины' Европой, но юг современной"’ Европейской части СССР был залит, еще соединилась с Южной Климат палеоцена был ровный, теплый и влажный.^Дддьмы и Древовидные папоротники в большом количестве росли на всех северных .материках. Вечнозеленые субтропические. деревья,...
массу палеоценовых . десрв,. Значительнореже встречались предки наших деревьев, „с .опадающей листвой.
В палеоценовых морях появились первые нуммулиты — наиболее крупные из одноклеточных организмов> Они были настолько многочисленными, что из скопления их раковин образовались пласты мощных нуммулитовых известняков. Палеоценовые кораллы и губки близки к современным. Из иглокожих особенным распространением пользовались морские ежи. Плеченогие и мшанки стали значительно менее разнообразными, чем прежде. Из моллюсков явно преобладали двустворчатые и брюхоногие, заменившие почти исчезнувших головоногих. Членистоногие были близки к современным.
В палеоцене кончилось господство пресмыкающихся. Ни на суше, ни на воде, ни на воздухе не было больше гигантов мелового периода. На Земле остались жить лишь черепахи (табл. IX, 1), ящерицы, змеи (табл. IX, 2) и крокодилы, очень близкие к современным. В
воздухе в это время летали уже настоящие, беззубые птицы (табл. IX, 3). Птицы, имевшие зубы, навсегда исчезли с лица Земли.
Млекопитающие, занимавшие весьма подчиненное положение в природе на протяжении всей мезозойской эры, теперь становились все многочисленнее и разнообразнее. Их быстрому процветанию способствовало вымирание пресмыкающихся. В палеоцене еще жили многобугорчатозубые млекопитающие. Относящийся к последним травоядный птилодус был ростом приблизительно с кошку (табл. IX. 4). Его передние зубы-резцы напоминали резцы крысы. Другие многобугорчатозубые, были крупнее пти-лодуса. Они достигали размеров бобра. Из сумчатых животных особенно часто встречались древесные сумчатые крысы — опоссумы (табл. IX, 5). Эти маленькие пятипалые зверьки, возможно, были предками более молодых сумчатых.
Настоящие плацентарные млекопитающие были уже разнообразны, но по многим особенностям своей организации они еще недалеко ушли от меловых насекомоядных предков. По Земле бегали предки ежей и близких к ним животных. По деревьям карабкались насекомоядные, похожие на современных тупай. Эти животные по образу своей жизни и по строению уже приближались к современным полуобезьянам, или лемурам. Здесь же в листве деревьев скрывались и отдаленные предки настоящих обезьян, по строению напоминающие современного долгопята. Долгопят — это маленькое, большеглазое животное, прыгающее на задних ногах. Прежде его относили к лемурам, но теперь по особенностям строения черепа выделяют в особую группу и сближают с обезьянами.
В палеоцене появились настоящие летучие мыши, предками которых были древесные насекомоядные млекопитающие. От последних, образуя особую ветвь, в палеоцене же отделились растительноядные тениодонты. Эти животные по внешности несколько напоминали теперешних ленивцев, но отличались от них довольно большим хвостом. Многие из них, например псита-котерий, достигали размеров волка (табл. IX, 6). Для своего века это были очень крупные животные.
В палеоцене появились хищники—креодонты. Они еще значительно отличались от современных хищников и имели много общего с насекомоядными. Их мозг был мал. когти на пальцах тупые и похожи на копытца. Палеоценовые креодонты имели острые клыки, но у них среди коренных зубов еще не было отдельных больших режущих так называемых «хищных» зубов. По образу жизни и питанию креодонты сильно различались между собой. Трицентес (табл. IX, 7), величиной с хорька, вооруженный острыми когтями, питался насекомыми и лишь иногда трупами мелких млекопитающих или пресмыкающихся. Длиннохвостый, ростом, с медвежонка^ и.похожий на него кленодонт (табл. IX, 8) хотя и кормился животной пищей, но не отказывался и от растительной, подобно современным медведям. Попадались и настоящие травоядные, копытные животные. Из последних, напоимер, эупротогония (табл. IX, 9), длиной до 80 см, принадлежала к первичным копытным — кондиляртрам. У этих копытных было еще много общего с хищниками. У них мы находим большие клыки и на пальцах копытообразные или даже настоящие когти. На ногах у них по пяти пальцев и самый длинный из них—третий (средний) палец, как у современных непарнопалых (тапиров,
носорогов и лошадей). Весьма вероятно, что эупротогония — далекий предок этих животных. В тени лесов жили довольно крупные, величиной со среднюю собаку, представители группы амблипод—пантоламбды (табл. IX. 10). Эти приземистые пятипалые копытные с массивным хвостом и сильными клыками были отдаленными родственниками слонов (хоботных животных).
ЭОЦЕНОВАЯ ЭПОХА
Продолжительность эоценовой эпохи определяют примерно в VT млн. лет. С ее окончания до довременной эпохи прошло около 35 млн. лет. В эоцене море снова наступало на сушу. Африка и Южная Америка не были связаны больше между .собой. Не существовало соединения и между Северной и Южной Америкой. Африканский континент распался на три острова. Азия попрежне-му соединялась с Северной Америкой, а Америка через Гренландию и Атлантический океан была Спаяна с Великобританией. Западная Европа и Скандинавия образовали самостоятельную сушу, тянувшуюся далеко на север. Южная Европа, почти весь юг Европейской части СССР и большая часть Центральной Азии были залиты морем.
Климат в эоцене был теплый. Вееролистные и финиковые пальмьг^пбпрёжнёму широко росли по северным материкам, которые были покрыты &ещшзелеными лесами.
Жизнь эоценового леса была богата и разнообразна. Среди цветов носились бабочки и другие насекомые. Днем их преследовали многочисленные птицы, а ночью совы (табл. X, 1) и летучие мыши. Многие эоценовые птицы потеряли способность летать и бегали, как современные страусы. Некоторые из них. например североамериканская дйатрима, отличались гигант-:ким ростом (рис: 16), который доходил до 2 м. Похожие на ежей насекомоядные млекопитающие попрежнему жили на земле, некоторые из них вели древесный образ жизни. В лесу жили настоящие полуобезьяны, первые лемуры (табл. X, 2) — маленькие нотарктусы. Появились первые грызуны, по сложению напоминавшие современных белок, а также удивительные тиллотерии (табл. X, 3) — животные величиной с крупного медвежонка, похожие по стооению зубов на грызунов. На самом деле это не грызуны, а своеобразные выродки насекомоядных.
Древние неполнозубые палеанадонты были близки к общим предкам всех неполнозубых. Их представитель метахейромис (табл. X, 4), длиной около 0,5 м, по общему складу больше всего напоминал современного броненосца, хотя панцырь у него был еще зачаточный. Передние лапы метахейромиса были вооружены острыми и длинными когтями, которыми он рыл землю, на задних лапах — когти короткие и тупые. Вместо исчезнувших коренных зубов во рту у
ПАЛЕОЦЕНОВАЯ ЭПОХА. ЖИЗНЬ ЛЕСА СЕВЕРО-АТЛАНТИЧЕСКОГО КОНТИНЕНТА 1—водяная черепаха; 2—змея; 3—кулики; 4—птилодус; 5—опоссум; 6— псятакотернй; 7—трипентес; 8— кленодонт; 9— ауоротогония; 10—пантоламбда
него образовались рогбвыё подушки. Клыки были значительно увеличены.
Прятавшиеся в тени деревьев хищные креодонты становились крупными и сильными животными. Правда, у некоторых из них. например у дромоциона (табл. X, 5). так и не образовалось настоящих хищных зубов, а когти стали тупыми. Этот длинноногий бегун почти достигал размеров медведя. Равный по величине дромоциону патриофелис (табл. X. 6) был плотным, приземистым зверем. Пальцы его широких лап соединялись перепонкой. Он жил около во* ды и плавал, вероятно, не хуже выдры. У пат-риофелиса были хищные зубы, но они не соответствовали зубам современных хищных.
Наряду с названными креодонтами существовали небольшие, похожие на куниц миациды, хищные зубы которых вполне соответствовали, по положению в челюстях тем же зубам современных хищных; последние являются прямыми потомками миацид. Миациды победили креодонтов в жизненной борьбе. Их главное преимущество над креодонтами заключалось в относительно уже хорошо развитом головном мозге. Они были более ловкими, чем креодонты, и с большим успехом могли преследовать травоядных копытных животных, которые к этому времени значительно изменились и усовершенствовались. В начале эоцена все еще продолжали жить первичные копытные — кондиляртры, но число их постепенно уменьшалось, и с середины эоцена они больше уже не существовали. Амблиподы, потомки палеоценовой пантоламбды, достигали огромных размеров. Так, длина уинтатерия (табл. X, 7) больше 4,5 л<, а высота его около 2,5 м. Уинта-терий имел, кроме сильных клыков, ещё и три тары рогов.
Но ни гигантский рост, ни сильное вооружение не спасло амблипод от вымирания. Ничтожный головной мозг и несовершенные зубы не обеспечивали им успеха в борьбе за существование, они уступили место на Земле другим копытным.
Особенно многочисленны в эоценовых лесах были непарнопалые копытные. К ним относятся предки лошадей — эогиппусы. Это мелкие животные, величиной с лисицу. Зубы эогиппусов с низкими коронками указывают на то, что эти животные; питались сочной, мягкой растительностью. Их четырехпалые передние и трехпалые задние ноги были хорошо приспособлены для хождения по мягкой, болотистой почве леса.
Лесные болота служили убежищем тяжелым водным носорогам—аминодонтам. похожим на бегемотов. Длина их доходила до 4 м. По более твердой почве легко двигались стройные бегающие носороги — гиракодонты. Все носороги были совершенно лишены рогов, и это название сохранено за ними только потому, что по строению скелета и зубов они были похожи на современных носорогов. В лесных болотах жили 48
предки тапиров, ёЩе лишенные хоботка, и близкие к тапирам лофиодонты, у которых хоботок уже развился. В лесах современной Западной Европы предки лошадей начали вымирать в конце эоцена, и их место занимали палеотерии, трехпалые животные с небольшим хоботком. В Северной Америке к этому времени появились палеозиопсы—первые представители гигантских и двурогих титанотериев. Безрогий палеозиопс (табл. X, 8) был величиной с тапира. На передних ногах он имел по четыре пальца, на задних— по три. Мозг его был поразительно мал.
Парнопалых копытных, у которых, как и у свиней, оленей и быков, сильнее других пальцев развиваются средний и безымянный (по счету 3-й и 4-й), вначале эоцена жило еще немного, В конце эоцена их стало значительно больше. В Европе жили длиннохвостые водные анаплотерии и напоминающие одновременно зайца и кабаргу скачущие мелкие ценотерии. В Северной Америке были обычны своеобразные четырехпалые ореодонты и совершенно диковинные длиннохвостые агриохерусы. Зубы агриохерусов напоминают зубы коровы, их пятипалые передние и четырехпалые задние лапы вооружены когтями. В Америке встречались первые предки верблюдов и лам, относящиеся к мозоленогим парнопалым. Мозоленогими этих животных называют потому, что у них на подошве развиваются толстые эластические подушки, копытца же имеют вид ногтей. Эти первые верблюды— протилопусы были еще величиной с кролика. На передних лапах у них имелось по четыре пальца, на задних—по два, пальцы заканчивались еще обычными копытами. В Европе вместо предков верблюдов жили близкие к ним двупалые ксифодонты. Предков и родичей свиней в эоцене мы почти не встречаем*
В Южной Америке в описываемое время появились совершенно особые травоядные животные, называемые н о т у н г у л а т а, или южноамериканские копытные. История их проникновения в Южную Америку довольно сложная. Первые представители этих копытных уже в палеоцене обитали в Центральной Азии. Отсюда они перекочевали в Северную Америку и далее перешли в Южную. После их прихода в Южную Америку, связь между Северной и Южной Америкой нарушилась. Копытных и хищников в Южной Америке почти не было, поэтому пришельцы стали благополучно жить на новом материке. Первыми из южноамериканских копытных известны мелкие пятипалые животные с увеличенными резцами, как у грызунов, которых они напоминают по образу жизни. Большой палец на их передних лапах мог противопоставляться остальным так, как это наблюдается у обезьян. Вместе с мелкими южноамериканскими копытными бродили пятипалые астрапотерии (рис. 17), величиной с тапира. Их голова с хоботом и большими клыками напоминала одновременно голову тапира и свиньи. Жили в Южной
ЭОЦЕНОВАЯ ЭПОХА. ЖИЗНЬ ЛЕСА СЕВЕРО-АТЛАНТИЧЕСКОГО КОНТИНЕНТА В СРЕДНЕЭОЦЕНОВОЕ ВРЕМЯ
7—сова; 2—лемур нотарктус; 3— тиллотерии; 4—метахейромис; 5—дромоциои; 6— патрнофелис; 7—уинтатерий; 8— палеозиопс
Америке и так называемые Литоптерны, которые впоследствии заменили отсутствующих там непарнопалых копытных.
В Северной Африке в эоцене появились первые хоботные, т; е. предки слонов. Самым древним хоботным животным был меритерий, ростом с тапира. У меритерия, подобно тапиру, был н: большой хоботок. Он имел четыре увеличенных резца (два в верхней и два в нижней челюстях)» которые образовали небольшие бивни. Почти единовременно с меритерием жил более крупный древний мастодонт. Бивни у него, были длиннее, а череп короче, чем у меритерия, зубы гребенча
тые. Это—предок всех гребенчатозубых мастодонтов.
По берегам Тетиса в эоцене плавали первые морские коровы, или сирены. Сирены несколько напоминаю г китов, но в отличие от последних они травоядны и родственны хоботным. У эоценовых морских коров, в отличие от современных, еще сохранились задние лапы. Вместе с сиренами пл а вали особенно удивительные древние киты—рыбоядные зейглодонты (рис. 18). Эти гиганты с змеевидным телом достигали 21 м в длину, задних лап у них уже не было, а передние обратились в ласты. Зубы древних китов, как и у наземных млекопитающих, еще разделялись на резцы, клыки и коренные, чего нет у новых китов. У последних или зубов вовсе пет (беззубые киты) или же все зубы одинаковы и имеют форму простого конуса (зубастые киты).
ОЛИГОЦЕИОВАЯ ЭПОКА
Олигоценовая эпох^на продолжалась 16 млн, лет. Начало олигоцена отдалено от. .наше-... го ^е^ш^рийёр^О-
В олигоцене больше не существовало соединения через Атлантическийо^едцм Евро--пои^2* Амердкрй*. Америка все еще
?”***"* -С а черр? Берингпп
пролив и с^ Азией,которая простиралась на север значительно дальше, чем теперь. Африка поднималась.со^ер1пеняо--офдел!>мь1м этаюриком, но ее современные (запанный и нйвтмушый). берега н^рдились^еше^дол „водой, так же как и Мадагаскар. Европа представляла собой боль-йюй остроВт^в состав которого входили Скандинавия иховсЩВЯ^прлови^ Западной^ Европы. Великобритания об разевала "второио ст ров. Вся остальная часть который ср.общался^-череа^Персию^с Индийским океаном, а через, Западно-Сибирскую низменность— с Севернъни дюлярным морем. Южная АД^рикУ^олыную-чагть периода хохрапяла изе-лировацнре цоложение. но, ыа-жраткое-время она соединилась с Северной Америкой.
В конце олигоцена в результате грандиозных горообразовательных процессов поднялись Ги-малайскпе горыг^лимат олигоцена,умеренный и нха^кный? по сравнению с климатом эоцена был Несколько б^С№*1^<№ладным и сухим. Пальмы росли на северных мате^шсах^е Та^Г^оби^ц попрежнему здесь еще госпо' леные леса. Среди них стало больше лвогшье;_.ц лиственных деревьев, периодически сбрасываю*-III ИХ листву.
Главные перемены в жизни олигоценовых морей заключались в уменьшении нуммулитов, которые к концу эпохи вымерли, и в возрастании числа костистых рыб.
На суше стало значительно больше насекомоядных млекопитающих. Появились землеройки, кроты. В воздухе вместе с обычными насекомоядными летучими мышами носились более крупные летучие мыши, питавшиеся плодами. В лесах все еще попадались первичные белкообразные грызуны, но вместе с ними жили и настоящие белки, предки мышей, зайцев, родственных зайцам пищух, а также предки дикобразов. Эти грызуны, попав в Южную Америку, стали здесь разнообразнее и многочисленнее, чем на других континентах. Вымерших древних неполнозубых теперь вЮжнсй Америке заменили новые неполнозубые броненосцы и их близкие родичи глиптодонты. Паицырь броненосцев состоял из отдельных колец, подвижно соединенных между собой, а у глиптодонтов он был цельный, как у
черепах. В лесах попадались пеовы^, еще небольшие наземные ленивцы. На северных материках малочисленные древние хищники—креодонты—доживали последние дни. Их победили в борьбе за существование настоящие хищники— предки собак, куниц», виверр и кошек. Кроме них, жили хищники—медведе-собатси. Медведе-собаки с олигоцене,6ыли невелики. Размер их колебался между размерами лисицы и волка. У .них был длинный хвост, как у собак, и ходили они, как собаки,—опираясь лишь на пл льны ног, а не на вс з стопу, как медведи. Но зубы у этих животных постепенно становились все более тупыми, как у медведей, и служили им для растирания растительной пищи. Олигоценовые кошки уже разделяются на две группы. У одних и верхние и нижние клыки развиты более или менее одинаково, как у наших кошек, у других—саблезубых кошек—страшные верхние клыки по размеру значительно превосходили нижние. Добычи у хищников было много.
В олигоценовых лесах все еще жили настоящие тапиры й вместе с ними трехпалые предки лошадей — мезогиппусы. питавшиеся мягким, сочным кормом. Последние стали крупнее и достигли размеров овчарки, а переселившиеся в Европу анхитерии по росту были уже не меньше пони. Особенно развивались в Северной Америке титанотерии. достигшие гигантских размеров. Большинство из них имело парные рога. Длина мегацерспса. (рис. 19) равна почти 5 м,
Рис. 20. Антракотегн’4
Рис. 21. Энтелодонт
ОЛИГОЦЕНОВАЯ ЭПОХА - чииацератернй; 2— иидрикотерий
Ри 2. 23. Пиротерий
а высота 2,5 м. Слабое развитие мозга и несовершенство зубов послужили причиной тому, что титанотерии к концу олигоиена уступили место другим копытным.
Бегающие и водные носороги вымерли, не выдержав конкуренции с предками лошадей. В олнгоцепе жило много безрогих носорогов, родственных современным. Среди них были мелкие эпиацератерии (табл. XI. 1). величиной с теленка, и гиганты среди всех млекопитающих—
индрикотерии (табл. XI, 2). Одна голова ин-дрикотерия в длину больше 1,25 м, а высота в плечах около 5, 5 м.
Парнопалые жвачные (предки наших оленей, антилоп, жираф, коз, овец и быков) напоминали современных оленьков или кабаргу. Это небольшие безрогие животные с сильно развитыми клыками в верхней челюсти, с четырьмя пальцами на передних ногах и двумя на задних.
В олигоцёне особенно много было свиней и близких к ним животных. По берегам рек и озер в камышах прятались коротконогие антракотерии (рис. 20). По всем северным материкам бродили гигантские свиньи—энтелодонты (рис. 21). Длина их доходила до 3.25 м. У некоторых эн-телодонтов на ногах было только по два пальца, а у других, кроме двух больших пальцев, было еще и по два маленьких боковых. Эти животные, напоминавшие длинноногих бегемотов, питались главным образом корнями растений.
Попрежнему только в Северной Америке жили предки верблюдов—поэбротерии. Теперь они стали ростом с овцу, но на ногах все еще сохраняли простые копыта и не имели эластических подушек, характерных для современных верблюдов.
Южноамериканские копытные в олигоцене были очень многочисленны. Тут существовали мелкие животные, , вроде грызунов, и крупные тяжеловесные трехпалые токсодонты (рис. 22). Астрапотерии (рис. 17) достигали размеров
Носорога. Одни копытные из группы литоптер-на напоминали стройных трехпалых лошадей, а другие были предками плейстоценовых макра-ухений (см. плейстоцен). Заменяя отсутствовавших в Южной Америке хоботных, бродили крупные пиротерии (рис. 23). У них развивался настоящий хобот, а резцы образовывали короткие бивни.
Африканский континент попрежнему населяли настоящие хоботные. Особенно многочисленны были древние мастодонты, предки гребенчатозубых мастодонтов. С ними вместе жили фи-омии. Нижняя челюсть фиомии была уже удлиненной, но бивни и хобот оставались еще короткими. Зубы у фиомии бугорчатые. Фиомии— предки бугорчатозубых мастодонтов. Вместе с предками слонов жили предки настоящих собакообразных обезьян— парапитеки и даже первые Человекообразные обезьяны—проплиопитекн. Из нижнего олигоцена Африки известны своеобразные арсинойтерии—далекие родичи слонов и сирен, пятипалые животные ростом с носорога (рис. 24). На голове у них были две пары рогов.
Олигоценовые сирены, или морские коровы, по своему строению напомичали современных сирен. Древние киты вымерли в олигоцене; к концу периода в море плавали уже короткотелые зубастые киты — сквалодонты и предки беззубых китов. Одни из них (цетотерии) были совершенно беззубые, а другие (патрицетусы) еще полностью сохранили зубы.
МИОЦЕНОВАЯ И ПЛИОЦЕНОВАЯ ЭПОХИ
Миоценовая и плиоценовая эпохи в общей сложности продолжались на Земле около 18 млн. лет. На миоценовое время приходится примерно 12 млн. лет, а на плиоценовое—6. От конца плиоцена до ниших дней прошло около 1 млн. лет.
Начавшееся еще в олигопене отступание моря прогрессивнб^Ъ^одолжалось во ^се. -мио-пли-сденовое вр^.я>м^Аф1ип^нскии мачс^шпе^получил почти современное очертание и соединился мостом с ПЕ5арОПО.й через _срв р^емецщцй_Гибпалтар-ский пррдив^-Европа была соединена-с Азией^и оба материка тянулись на север _ значительно дальше, чем теперь. Великрбритания была полуостровом'Ев^ОД НР му nr та па А я г ь
соединенной с Северной Америкой, а Северная Америка с Гренландией. Австралия занимала изолированное ^положение. Океан Тетис в мио-пене очень сократился^\,разм^^1£2лШ!ЙЕЙким рукавом^сЬедйнялся через Крымско-Кавказскую Область и Персию с Индийским океаном. В плиоцене Тетис распался на три небольших изолированных ~ моря —^Средиземное, Черное и Каспийское.
Г орообразовательиые процессы получили большое развитие и миоцене и еще больше в пли-оценё.ТГ это время поднялись Альпийские. Ал-
Рис. 25.4 Махайродус
тайские, Саянские горы, также Тибет. Памир н другие^кребТы.
~ Климат становился все более холодным. В миоценовое время в Европе уже исче^лй^ пальмы. Их сменили.хвойные ..н. лнетжнные^р^^с, опадающей листвой._ В_ это время росли дубы, клеиьь^^вязы^... ,1ШЮДИ»_ЛШ£Ш^
В связи с похолоданием климата в миоценовую эпоху усиленно .развивались травянистые растения и степи получили весьма широкое распространение. Растительность плиоцена уже почти совсем современная. Морская фауна мио-плиоценового времени была близка к современной. Наземная же фауна млекопитающих за этот период претерпела особенно интересные изменения. Многие из животных этой эпохи по строению были близки к современным* например насекомоядные млекопитающие и летучие мыши. Однако многие животные еще во многом отличались от современных. Грызуны, например» были далеко не такими, как теперь, а бобры—приблизительно в 2 раза, меньше современных. Средн первичных белкообразных грызунов жили диковинные рогатые животные, найденные в миоценовых пластах Северной Америки. Но в то же время в лесах встречались уже настоящие соки, а степи населяли обычные тушканчики. Мышевидные, а также родственные дикобразам грызуны в мио-плиоцене были очень! многочислешш. Родичи дикобразов после соединения обеих Америк в плиоцене широко расселялись по всей Южном и Центральной Америке, а также проникли и на современные Вест-Индские остоова. Многочисленные броненосцы и уже крупные глиптодонты в плиоцене рыли землю своими когтистыми лапами не только в Южной» во к в Северной Америке, где они доходили до широты современного Техаса. Вслед за глиптодонтами в Северную Америку переселились и наземные ленивцы, к концу плиоцена допит ним гигантских разме-
Рис. 26. Гиппарион
Рйс. 27. Моропус
ров. Последние древние хищники—креодонты вымерли в начале миоцена. Все многочисленные хищники северных материков относились уже к настоящим хищникам. В это время от первичных собак отделялись древесные длиннохвостые еноты. Медведе-собаки в мио-плиоценовое время становились все крупнее и крупнее и к концу эпохи догнали по размерам современных медведей.
В конце миоцена жили очень интеоесные хищники, из которых одни по своему Строению занимали переходное положение между собаками и настоящими медведями, а другие — между виверрами и гиенами.
В плиоцене на Евразии уже бегали настоящие гиены и первые медведи. Кошки, особенно саблезубые — махайродусы (рис. 25). во все мио-плиоценовое время были многочисленны, так же как и куницы.
Непарнокопытные в эту эпоху стали менее разнообразны, чем прежде. Исчезли титаноте-рии, лофиодонты, бегающие и водные носороги. Зато умножились предки настоящих носорогов, тапиров и лошадей. Предки лошадей — пара-гиппусы в начале миоцена покинули леса и перешли в степи, где им пришлось бегать по более твердому грунту и питаться более сухими и жесткими кормами, чем прежде. В новых условиях у меригиппусов, трехпалых предков лошадей, средний палец стал гораздо сильнее боковых, которые на бегу уже перестали касаться земли, а коронки зубов постепенно сделались длиннее и выше. От меригиппусов произошел в Севериной Америке гиппарион (рис. 26) — довольно крупная трехпалая лошадь. Ее зубы были устроены сложнее, чему современных лошадей. Гиппарионы в конце миоцена переселились в Евразию, где они особенно размножились в мио-плиоценовое время. От меригиппусов произошли и прямые предки современных лошадей — плио-гиПпусы и протогиппусы. Последние, переселив
шись в Южную Америку, переродились там в горных лошадей—гиппидиумов.
Начиная с миоцена в Северной Америке число носорогов все уменьшалось, а в плиоцене они совершенно исчезли. Зато в Старом Свете носорогов было много. Здесь, наряду с безрогими ацератериями, в мио-плиоценовое время бродили двурогие шлейермахеровы носороги (табл. XII, 1), родственные современным носорогам о-ва Суматры. В нижнем плиоцене жили однорогие предки индийского носорога и двурогие предки африканских носорогов.
Из непарнопалых копытных интересен моропус (рис. 27). Это животное, по росту и по сложению похожее на лошадь, но с более длинной шеей, имело на ногах по три пальца, вооруженных сильными когтями, как у тигра. Такие ноги животное могло использовать для рытья или для того, чтобы, поднявшись на дыбы, держаться когтями пальцев за ствол дерева и наклонять к себе его ветви с сочной листвой. Парнопалые жвачные напоминали по сложению оленей. В начале миоцена они все еще были безрогие, но с большими клыками. В* конце миоцена появились первые настоящие олени с простыми вильчатыми рогами. От оленеобразных предков в Миоценовое же время отделились первые жирафы (табл. XII, 2) и первые антилопы — трагоцеросы (табл. XII. 3). В плиоценовое время существовало большое число антилоп; многие из них по сложению были похожи на быков. В конце плиоцена жили лептобосы, очень близкие по своему строению к настоящим быкам. Начиная с миоцена американские предки верблюдов и лам последовательно становились все крупнее и крупнее. В плиоцене вместо копыт у них на ногах образовались эластические подушки — мозоли. В эту пору верблюды перешли из Северной Америки в Евразию, а ламы — р Южную Америку.
МИО-ПЛИОЦЕНОВАЯ ЭПОХА. ЖИВОТНЫЕ, ОБИТАВШИЕ НА МЕСТЕ СОВРЕМЕННОЙ ЮЖНОЙ ЕВРОПЫ
1—шлейс рмахеров носорог; 2—жирафа; 3—антилопа (трагодерос); 4—длиннорылый мастсдснт: 5—динотерий
Свиньи в мио-плиоценовое время были многочисленны. В начале плиоцена появились первые гиппопотамы, но антракотерии и гигантские свиньи на Земле больше не существовали.
Южноамериканские копытные в миоцене переживали период наивысшего расцвета, и только слонообразные пиротерии не дожили до этого времени. Одни южноамериканские копытные в миоцене все еще оставались мелкими, как кролики, другие перегоняли по размерам медведей. Астрапотерии достигали размеров крупных носорогов, токсодонты по величине приближались к тапирам. Лошадеобразные литоптерны по виду приближались к лошадям, и некоторые из них даже становились однопалыми, но в плиоцене они вымерли, так же как и большинство других южноамериканских копытных.
В начале миоцена из Африки переселились в Евразию и успешно здесь оазвились многочисленные мастодонты, имеющие во рту по четыре бивня (два в верхней челюсти и два в нижней). В конце миоцена они проникли и в Северную Америку. У одних мастодонтов зубы оставались бугорчатыми, как, напримео, у длин-норылого мастодонта (табл. XII, 4), у которого развит уже настоящий длинный хобот; у других мастодонтов зубы становились гребенчатыми.
Среди хоботных плиоцена Индии появился предок настоящих слонов — стегодонт. Его зубы с большим числом гребней по своему строению занимали среднее положение между зубами мастодонтов и настоящих слонов. У современных слонов в каждой половине челюсти находится по одному зубу, который напоминает небольшой жернов и часто весит до 6 кг. Зубы у слона меняются шесть раз в течение его жизни. Зубы у стегодонта менялись так же, как и у слона: новый зуб развивался позади старого и постепенно его вытеснял, так что одновременно в каждой половине челюсти животного было не больше
двух зубов. Верхние бивни у стегодонта большие, а нижние совсем маленькие, исчезающие.
В конце плиоцена по Земле бродили первые, уже настоящие плосколобые слоны. У них во рту была только одна пара верхних бивней, ко ренные зубы состояли из отдельных пластинок, как у современных слонов, а не из гребней. В мио-плиоценовое время вместе с мастодонтами жили еще своеобразные хоботные — динотерии (табл. XII, 5). Некоторые из них были по размерам даже больше слона. Они отличались от всех остальных хоботных тем, что имели одну пару загнутых бивней, которые сидели в нижней челюсти. Коренные зубы динотерия были очень простые, гребенчатые. Питался динотерий сочным мягким кормом.
В миоцене Южной Америки уже жили предки современных широконосых обезьян. В верхнем миоцене в Европе по деревьям карабкались плиопитеки — предки гиббонов, и дриопитеки— общие предки человекообразных обезьян (шимпанзе, гориллы) и человека. В Индии в то же время существовали предки оранг-утанов. В плиоцене человекообразные обезьяны были очень близки к современным своим потомкам, так же как и многочисленные предки собакообразных обезьян уже приближались по строению к современным мартышкам и павианам.
В мио-плиоценовых морях древних китов больше не встречалось. Их заменили новые зубастые и беззубые киты, к плиоцену принявшие современный вид.
Зубастых сквалодонтов постепенно вытесняли обычные кашалоты, нарвалы и дельфины. Дельфины в миоцене были очень длиннорылымн. К плиоцену они приняли современный вид. В конце миоцена из беззубых китов в морях плавали настоящие киты и киты-полосатки. Миоценовые и особенно плиоценовые морские коровы (сирены) по строению приближались к современным.
ЧЕТВЕРТИЧНЫЙ ПЕРИОД
ПЛЕЙСТОЦЕНОВАЯ ЭПОХА
xuKX/l
ffyVzJL иллион лет отделяет современность от начала последней, плейстоценовой эпохи. Для истории Земли это ничтожное время. Однако в начале описываемого периода наша Земля была еще далёко не такая, как теперь. Австралия все еще соединялась с Новой Гвййеёи и Тасманией. Северная.Америка попрёжнему^бы-ла объедин£няи<елодькос_Ю й^Гренландией.ачерез Берингов проливонанег посредственно связывалась, с Азией. Японские и Ост-Индские острова соединялись с Великобритания все ещё была полуостровом^ Европы, так же, как и ^теперешние астровк^Новой Земли. Мадагаскар соединялся с Африкой, последняя соединялась с Европой через-Испанию?-^ дальше на востоке через Аравию и Кавказ. Современное Средиземное море, замкнутое со всех сторон, распалось на два моря-озера.
С началом плейстоцена на Земле было связано заметное похолодание климата. Снега, выпавшие зимой, не успевали полностью растаивать за лето и из года в год накапливались в приполярных странах. Снег постепенно превращался в лед. Так возникли колоссальные ледники, которые ползли в направлении экватора и покрывали огромные пространства. Мощное оледенение развилось в северном полушарии. Огромнейший ледник покрывал Северную Америку приблизительно до 40° северной широты, всю Гренландию, Исландию, Англию, Германию, Скандинавию и большую часть Европейской части СССР. Другие ледники сползали с Альп. Вся Азия севернее 60-й параллели также была покрыта ледником. Мощные ледники спускались в то время и с Гималайских гор. Южные материки, за исключением горных цепей, оставались
свободными от льда. Настоящее оЛедёнёние здесь переживали только Антарктика и южная часть Южной Америки.
Своеобразные климатические условия плейстоцена резко повлияли на животный и растительный мир Земли. Животные и растения, которые не смогли приспособиться к новым климатическим условиям или своевременно отступить от надвигавшегося ледника, погибли. Ледники, двигавшиеся с севера, оттеснили северных животных к югу, и им приходилось жить вместе с южными животными. После отступления ледника многие северяне остались жить на юге, но поднялись высоко в горы, где климатические условия напоминали климат ледниковой эпохи. Площади, занятые ледниками, в течение всего ледникового периода не оставались неизменными. Ледники то далеко отступали от своих прежних границ, то снова надвигались на них или даже широко выходили за их прежние пределы. Наступание ледника принято называть оледенением, а отступание или полное его исчезновение — межледниковой эпохой. Современная эпоха называется послеледниковой. До сих пор еще точно не установлено, было ли на Земле только одно оледенение, которое периодически то увеличивалось, то уменьшалось, или же было три, даже четыре отдельных оледенения, отделенных друг от друга теплыми межледниковыми эпохами. Животный мир в разное время плейстоцена заметно изменялся. Он был богаче, чем теперь, и многие плейстоценовые животные навсегда исчезли с лица Земли.
В Австралии в плейстоцене жили гигантские формы сумчатых. Здесь были травоядные дип-
Рис. 28. Макраухения
родснты, величиной с носорога, и сумчатые— львы. 6—7-метровые ящерицы доживали свои последние дни. На островах Новой Зеландии жила огромная бегающая птица—моа, до 3, 5 jh в высоту. Близкие к моа эпиорнисы населяли Мадагаскар. Объем огромных яиц эпиорнисов доходил до 7,5 л. В. Южной Америке жили своеобразные копытные—типотерии и достигшие размеров носорога токсодонты. Особенно замечательна несколько напоминающая верблюда
трехпалая длинношеяя макраухения (рис, 28) с небольшим хоботком. Это последний представитель группы литоптерна.
На территории Северной и Южной Америки еще бродили 3-мётровые глиптодонты и достигавшие 5, 5 м наземные ленивцы, такие же. как мегатерий (рис. 29). Эти животные приподнимались на задние стопоходные лапы, охватывали передними лапами ветви деревьев, листьями и плодами которых они питались. Передними лапами мегатерии также успешно могли рыть землю. На всех северных континентах и даже в Южной Америке продолжали жить страшные саблезубые тигры—махайродусы. В Европе бродили пещерные львы, медведи и гиены. Вместе с ними жили и обычные наши горностаи, куницы. хорьки, выдры, лисицы, песцы, волки. В Северной Америке и Европе встречались сухопутные родичи бобров, по размерам в несколько раз превосходившие современных. Все лошади были уже однопалыми (табл. XIII, 1). К концу периода лошади жили только в Старом Свете и окончательно вымерли в Америке, где не было больше и носорогов. В Старом Свете их еще было много. В Евразии, вдали от ледников, обитали голые носороги. Из них особенно интересен эласмотерий (табл. XIII, 2). остатки которого встречаются только в СССР. У него было два ро-/а. На передних ногах у эласмотерия, кроме трех пальцев, были еще небольшие остатки боковых
ЧЕТВЕРТИЧНЫЙ ПЕРИОД. ПЛЕЙСТОЦЕНОВАЯ ЭПОХА. ЖИВОТНЫЕ СЕРЕДИНЫ ПЛЕЙСТОЦЕНА, ЖИВУЩИЕ ВДАЛИ ОТ ЛЕДНИКА (ФАУНА ВТОРОЙ МЕЖЛЕДНИКОВОЙ ЭПОХИ)
J—дикая лошадь; 2—эласмотерий; 3—гигантский олень; 4—слон трогонтернй
Рис. 30. Первобытные
люди на охоте. С картины В. Васнецова
пальцев. Его сложно устроенные зубы говорят о том, что это животное питалось твердыми корневищами водных растений. Носорогов с голой кожей во вторую половину плейстоцена сменил носорог, покрытый шерстью (табл. XIV, 1), вооруженный огромным рогом. Прекрасно защищенный от холода, он спокойно мог жить вблизи самого ледника.
Настоящие быки, зубры, бизоны, а также живущий теперь в Гренландии холодолюбивый овцебык жили по материкам северного полушария. Многочисленны и разнообразны в плейстоцене были олени. Вдали от ледников встречались косули, лани и различные олени, называемые благородными. По краю ледника бродили северные олени (табл. XIV, 3). Особенно замечательным был торфяной олень, живший только в ледниковую эпоху, в течение которой он неоднократно изменялся (табл. XIII, 3). Расстояние между концами огромных рогов самцов торфяного оленя было больше 3 м. Стада кабанов бродили по камышам и зарослям вдоль рек. Многочисленные бегемоты населяли весь район вокруг Средиземного моря. Верблюды, размножившиеся в Старом Свете, в Северной Америке вымерли. Хоботных животных в плейстоцене было очень много. Мастодонтов, за исключением американского, на Земле больше не существовало, но было очень много разнообразных слонов. В Европе в начале ледниковой эпохи жили южные слоны и родичи современных африканских слонов—д ревние слоны с прямыми бивнями. В Америке в это время бродил огромнейший слон император. Позднее в Старом Свете появились слоны, родственные нынешнему индийскому виду. Сперва это был
слон трогонтерий (табл. XIII, 4), а за ним появился и настоящий волосатый мамонт (табл. XIV, 2). Целые трупы мамонтов с волосами и мясом не раз были найдены в вечномерзлой почве Сибири.
Самым замечательным событием в плейстоцене было появление на Земле человека. В верхнем плиоцене Старого Света жило несколько видов крупных человекообразных обезьян. Некоторые из этих обезьян по своему строению стояли значительно ближе к человеку, чем современные человекообразные обезьяны. Одни из этих обезьян дожили до начала четвертичного периода (австралопитеки), другие—до его середины (плезиантропы, парантропы). Причиной их гибели, весьма вероятно, было охлаждение климата на Земле, повлекшее за собой оледенение материков. Изменение климата способствовало вымерзанию многих плодовых деревьев, а следовательно, и значительному ухудшению пищевого режима человекообразных обезьян. До наших дней дожили только те обезьяны, которые успе* ли уйти в тропический пояс и усовершенствовать свои руки для лазания по деревьям.
Одна из обезьян, близкая по строению к австралопитеку, понуждаемая голодом, стала спускаться на землю, куда ее манили ягоды, грибы, семена злаков, сочные корни и корневища многих растений, насекомые, ящерицы и т. д. В поисках пищи эта обезьяна сначала случайно, а потом закономерно стала польз<'^ать''ч простейшими природными орудиями. Бросая палку, она пробовала сшибить высоко висящие орехи. Той же палкой она пробовала выкапывать корни. Камнем она разбивала твердую скорлупу ореха. Так, шаг за шагом, обезьяна все более
ст.
ТАБЛИЦА XIV
ЧЕТВЕРТИЧНЫЙ ПЕРИОД. ПЛЕЙСТОЦЕНОВАЯ ЭПОХА. ЖИВОТНЫЕ КОНЦА ПЛЕЙСТОЦЕНА, ЖИВШИЕ У КРАЯ ЛЕДНИКА (ЛЕДНИКОВАЯ ФАУНА) /—волосатый носорог; 2—мамонт; 3— северный олень
совершенствовалась в употреблении природных орудий. Таким образом обезьяна начала трудиться.
Во время трудового процесса при добывании пищи обезьяна пользовалась исключительно передними лапами, т. е. руками. Она все чаще и чаще стала подниматься на дыбы и ходить исключительно на задних лапах, т. е. на ногах. При таком хождении руки, полностью освобожденные от передвижения тела, могли уже всецело быть использованы для хватания различных орудий и для труда. Значительно развившиеся и усовершенствовавшиеся в труде руки, а также органы чувств повлекли за собой усиленное развитие головного мозга. Обезьяна под воздействием труда стала превращаться в человека.
В начале плейстоцена Южную Азию уже заселяли первые обезьяны-люди, или питекантропы,-^настоящие переходные формы между обезьяной и человеком. Первые остатки питекантропа были найдены в 90-х годах прошлого столетия на острове Яве. Недавно там же были обнаружены новые части его скелета (1937 г.). Питекантропы, очевидно, умели пользоваться уже камнями и палками в качестве орудий и оружия, но они еще сохраняли обезьяньи признаки: в черепе у них был надбровный валик, отсут
ствовал подбородок, клыки заметно выдавались и т. д. Их мозг, по объему равный 940 ел?, занимал среднее положение между мозгом человекообразной обезьяны (600 слЛ) и мозгом современного человека (1500 см3).
Немного позже питекантропов Южную Азию населяли близкие к ним синантропы, которые по объему мозга еще более приближаются к современному человеку. Они уже знали применение огня и умели делать грубые каменные орудия. Наконец, в середине плейстоцена на Земле жил «первобытный человек» — охотник, умевший делать уже весьма различные каменные орудия. Этот первобытный человек отличался от современного несколько согнутыми коленями, значительно более длинными руками, покатым лбом, все еще выступающим над глазами надбровным валиком и отсутствием развитого подбородка.
В конце ледникового периода первобытных людей сменили кроманьонцы. По анатомическим особенностям они существенно не отличались от современного «разумного человека». Кроманьонцы были одновременно и отличными охотниками и хорошими художниками. Они оставили нам на стенах пещер прекрасные изображения первобытных быков, носорогов и мамонтов.
ЗАКЛЮЧЕНИЕ
ы проследили, как последовательно изменялись животные и растения, начи
ная с древнейших периодов жизни Земли и
кончая современной эпохой, и убедились, что
во всем их многообразии они появились не сра-
зу, а развивались постепенно.
В докембрийское и кембрийское время на Земле существовали лишь морские беспозвоночные животные и морские водоросли. Начиная с силурийского периода, начали встречаться водные позвоночные, родственные миногам бесче
люстные панцырники и первые настоящие рыбы—родичи современных хрящевых рыб—акул. В то же время появились и первые наземные растения — псилофиты, еще очень близкие к бурым морским водорослям.
В девонское время появляются первые наземные беспозвоночные. Рыбы уже стали разнообразными. В конце девона появились первые земноводные животные и настоящие споровые растения.
Силурийский и девонский периоды можно назвать царством рыб: в это время на Земле рыбы были высшими животными.
В конце каменноугольного периода, называемого периодом земноводных, на Земле появились первые пресмыкающиеся. Кроме многочисленных споровых, в это время уже существовали и голосеменные растения: семенные папоротники и кордаиты.
В пермском периоде, в противоположность уменьшающимся в числе земноводным, все более многочисленными и разнообразными становились пресмыкающиеся. Среди них особенно интересны звероподобные ящеры, напоминавшие по своему строению млекопитающих. В это время вместе со споровыми растениями и древними голосеменными попадались и настоящие голосеменные, близкие к современным саговым пальмам, гинкговым и хвойным деревьям.
Мезозойскую эру с ее тремя периодами называют эрой пресмыкающихся, которые в это время являлись полными хозяевами суши, воды и даже воздуха. Эту эру также можно назвать эрой голосеменных растений.
В триасовый период, когда пресмыкающиеся были еще не очень разнообразны, как и в последующие два периода, появились первые многобугорчатозубые млекопитающие и первые лягушки.
В юрский период, кроме многобугорчатозу* бых млекопитающих, уже жили паитотерии— предки сумчатых и настоящих живородящих плацентарных млекопитающих. Появились пер* вые птицы—археоптериксы.
В меловой период жили хотя и имеющие зубы, но уже настоящие птицы. От паитотериев произошли сумчатые и настоящие плацентарные млекопитающие, близкие к современным насекомоядным. В конце периода покрытосеменные растения были уже близки к нашим травам, лиственным кустарникам и деревьям.
Всю кайнозойскую эру мы называем эрой млекопитающих и покрытосеменных растений. Млекопитающие, подобно мезозойским пресмыкающимся, населяли сушу, воду и даже летали, С самого начала кайнозойской эры удает^ ся последовательно проследить родословную как ближайших родичей человека—обезьян, так и его самого. В палеоцене, кроме обычных насекомоядных млекопитающих, жили еще насекомоядные, напоминающие лемуров, а также родичей современного долгопята,—это были первые предки обезьян. В эоцене существовали настоящие лемуры и те же предки обезьян. В олигоцене известны уже настоящие обезьяны—прямые предки собакообразных и человекообразных обезьян.
В мио-плиоцене жили предки современных человекообразных обезьян и переходные формы
6т человекообразных обезьян к человекоподобным существам. В начале плейстоцена известны питекантропы. В середине плейстоцена появился «первобытный человек». К концу ледникового периода его сменил человек современного типа. Плейстоцен можно назвать веком человека.
Проследив историю развития жизни на Земле, мы убеждаемся не только в том, что жизнь развивалась на Земле постепенно, но и в том, что чем позднее появляется та или другая группа животных или растений, тем выше она организована.
Между высшими и породившими их низшими группами животных и растений часто удается найти переходные группы или, иначе, «переходные формы». Эти формы по своему строению занимают промежуточное положение между обеими группами. Например, девонские кистеперые рыбы занимали среднее положение между рыбами и происшедшими от них земноводными. Они имели не только легкие и плавники, напоминавшие пятипалые лапы, но и череп их был очень похож на череп стегоцефалов. Исходные для всей группы пресмыкающиеся—цельночерепные, существовавшие в каменноугольный и пермский периоды, по многим признакам настолько напоминали породивших их земноводных стегоцефалов, что отличить их от последних часто нелегко даже специалисту. Пермские и особенно триасовые звероподобные пресмыкающиеся, предки млекопитающих, были так похожи1 на последних, что также с трудом могут быть отличимы от них. Юрский археоптерикс — предшественник птиц, занимает переходное положение между настоящими птицами и пресмыкающимися. Среди растений псилофиты являются промежуточными между водорослями и сухопутными растениями, семенные папоротники—между споровыми и семенными растениями и т. д.
Последовательное изменение организмов, в течение миллионов лет идущее одновременно с изменениями лица Земли, объясняется тем, что организмы в связи с постоянно меняющимися внешними условиями должны были вырабатывать все новые и новые приспособления, отвечающие новым условиям среды. Все организмы вообще способны изменяться в самых различных направлениях. Большинство из этих изменений будут бесполезными или даже вредными для организмов, но некоторые случайно могут совпасть с требованиями меняющейся среды. Такой удачно изменившийся организм способен выжить в новых условиях.
Вместе с новой группой животных или растений, которой суждено впоследствии пышно развиться от тех же предков, обычно возникают многочисленные и близкие к ней боковые группы. Они не имеют столь выгодного при данных условиях строения, каким отличается основная группа, и поэтому осуждены на вымирание. Их существование обычно крат-66
повременно. Таким образом, из большого числа попыток природы изменить строение организмов соответственно новым условиям среды только единичные оказываются удачными. Например, когда в девонский период пресноводные рыбы вынуждены были жить в периодически пересыхающих водоемах, выжили только те из них, которые выработали легочное дыхание. Это были сравнительно немногочисленные счастливцы; огромное число их сородичей, не имевших этого дыхания, погибли. Но и из тех рыб. которые выработали легочное дыхание, только одна какая-то группа, благодаря наличию у ней случайно благоприятных для наземной жизни особенностей строения, сумела подняться на высшую ступень организации, т. е. переродиться в земноводных. Это перерождение не для всех рыб прошло удачно и было куплено дорогой ценой— гибелью многих неудачников.
То же самое происходило и при возникновении от земноводных пресмыкающихся, когда возникли яйцекладущие животные с ороговевшей кожей. Только те животные, у которых случайно оба эти признака — яйценоскость и ороговение кожи оказались соединенными в одном организме, смогли выжить, а затем и широко заселить континенты вдали от водоемов. Многочисленные же их яйцекладущие родичи, не имевшие ороговевшей кожи, бесследно исчезли с лица Земли, не оставив потомства.
Изменение организмов всегда стоит в тесной зависимости от изменения жизненных условий, в которых они находятся. Для пояснения этой мысли вспомним еще раз историю развития лошади. Мелкие лесные предки лошадей, бродившие по мягкой болотистой почве, постепенно переселялись #з лесов в места открытых ландшафтов с твердым грунтом и переходили от питания сочными кормами к питанию жесткими, сухими травами. В связи с переходом из леса в степь у лошадей постепенно изменялись ноги и зубы: четырехпалая нога преобразовалась в однопалую, а низкие коронки зубов сделались высокими и' покрылись сложным рисунком эмали и т. д.
Но под понятием меняющихся «условий среды» или «жизненных условий» не следует понимать только изменения неживой природы, как-то: климата, почвы и т. д. Нельзя понимать также всякое изменение организма как приспособление его к изменяющимся условиям неживой природы, как выражение его борьбы с этой природой. Под «жизненными условиями» нужно понимать не только все отношения данного организма к неживой природе, но и его отношения со всеми окружающими организмами, с которыми ему приходится так или иначе сталкиваться.
Организму приходится часто вырабатывать новые приспособления специально для борьбы с другими организмами, чтобы не быть
^^^^^Я<ным ими в жизненной борьбе. Костный трицератопса, покрытая костными же плитками спина стегозавра и особенно его мощный хвост, вооруженный острыми стилетами, — все это специальные приспособления для борьбы с хищными динозаврами. Примеров таких приспособлений можно привести сколько угодно.
Часто окружающие условия неживой природы остаются неизмененными, но среди старых организмов появляется новый конкурент. Тогда этим старым организмам остается или перестроиться для борьбы с новичком или, если они не способны к такому изменению, погибнуть. Так, процветавшие в силурийское и раннедевонское время бесчелюстные панцырники должны были уступить свое место более совершенным конкурентам — рыбам. В пермское время более приспособленные к наземной жизни пресмыкающиеся усиленно вытесняют стегоцефалов. Псило-фитам пришлось отступить под натиском папо-ротников, хвощей и плаунов. Споровые растения сдали позиции голосеменным растениям. Голо
семенные отступили под натиском покрытосеменных и т. д.
Обобщая, можно сказать, что для того, чтобы та или другая порода животных или растений могла существовать на Земле в течение долгого периода времени, она должна быть способной вырабатывать все новые и новые приспособления. Если же животное или растение не может выработать новых приспособлений, отвечающих сменяющимся условиям существования, оно обречено на гибель.
«Переживание» в природе наиболее приспособленных организмов Чарльз Дарвин назвал естественным отбором. За время развития жизни на Земле мы видели, какое колоссальное число самых различных видов животных и растений было уничтожено естественным отбором. Взамен их тем же естественным отбором были созданы новые виды. Представители современной фауны и флоры — это редкие счастливцы, которые, случайно вырабатывая «целесообразные» приспособления к постоянно меняющимся условиям среды, смогли избежать вымирания н сохраниться до наших дней.
СОДЕРЖАНИЕ
Стр~
Предисловие 5
Введение 6-
Палеозойская эра 9
Кембрийский период 11
Силурийский период 14
Девонский период 17
Каменноугольный период 21
Пермский период 25
Мезозойская эра 29
Триасовый период 31
Юрский период 35
Меловой период 39
Кайнозойская эра 43
Третичный период 45
Палеоценовая эпоха 45
Эоценовая эпоха 46
Олигоценовая эпоха 50
Миоценовая и плиоценовая эпохи 55
Четвертичный период 59
Плейстоценовая эпоха 59
Заключение 65
Редактор М. В. Семенова Техредактор А. И. Красновская
______________________________________Обложка художника Е. А, Ганнушкина Сдано в набор 29IW 1947 г.____________Подписано в печать 29/XI 1947 г.
А—11096 Формат 60x92%. У. над. л. 6,5 30.000 ан. в 1 п. л. Объем 4% п. л.
Зак? 505 Тираж 25.000 Цена 5 руб.
1-я типография Госгеолнвдата, 2-я Мещанская, д. 97