






















































































































































Author: Стороженко В.Г.
Tags: лесопродукты и их использование условия окружающей среды биология леса лесное хозяйство лесохозяйственные науки экология лесные сообщества биогеоценология
ISBN: 978-5-8125-1015-2
Year: 2007
УСТОЙЧИВЫЕ
ЛЕСНЫЕ
СООБЩЕСТВА
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ИНСТИТУТ ЛЕСОВЕДЕНИЯ
ПРОГРАММА ФУНДАМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ
«БИОЛОГИЧЕСКИЕ РЕСУРСЫ РОССИИ:
ФУНДАМЕНТАЛЬНЫЕ ОСНОВЫ РАЦИОНАЛЬНОГО
ИСПОЛЬЗОВАНИЯ »
В.Г. Стороженко
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА
(ТЕОРИЯ И ЭКСПЕРИМЕНТ)
МОСКВА
2007
УДК 630*884.+630*114ю6+630*884/885+630*181.9
ББК43.4
С 82
Устойчивые лесные сообщества. Теория и эксперимент. В.Г. Сторо-
женко. — Тула: Гриф и К. — 192 с.
На основе анализа фундаментальных исследований классиков русского
лесоводства и биогеоценологии, зарубежных экологов, современных
авторов и оригинальных экспериментальных работ обоснованы теоретические
положения устойчивости лесных сообществ.
Рассмотрены фитоценотические критерии коренных устойчивых лесов,
включающие возрастные, породные, возобновительные, горизонтальные
параметры древостоев. Определены санитарные показатели коренных
устойчивых лесов, количественные и качественные параметры древесного
отпада. Изучены микоценотические критерии устойчивых Сообществ.
Обосновано новое в биогеоценологии направление — лесная микоценология,
рассмотрены понятия «больного» и «здорового» лесного сообщества.
Определены количественные и качественные параметры участия грибов дерево-
разрушающего биотрофного комплекса в функционировании устойчивых
коренных лесов. Предложена методика разделения лесов по градациям
устойчивости.
Ответственный редактор:
доктор биологических наук М.Г. Романовский.
Рецензенты:
доктор биологических наук В.И. Крутов,
доктор биологических наук, профессор Л.Г. Гарибова.
Stable forests communities. The theory and experiment. V.G. Sto-
rozhenko.— 192 с
ISBN 978-5-8125-1015-2
© В.Г. Стороженко, 2007
ОГЛАВЛЕНИЕ
Введение 5
1. Предисловие 7
2. Краткая методика исследований 13
3. Содержание понятий «устойчивость» и «устойчивые
лесные сообщества» 16
4. Теоретические положения устойчивости лесных
сообществ 20
5. Понятия «климаксового» и «коренного» лесного
сообщества 39
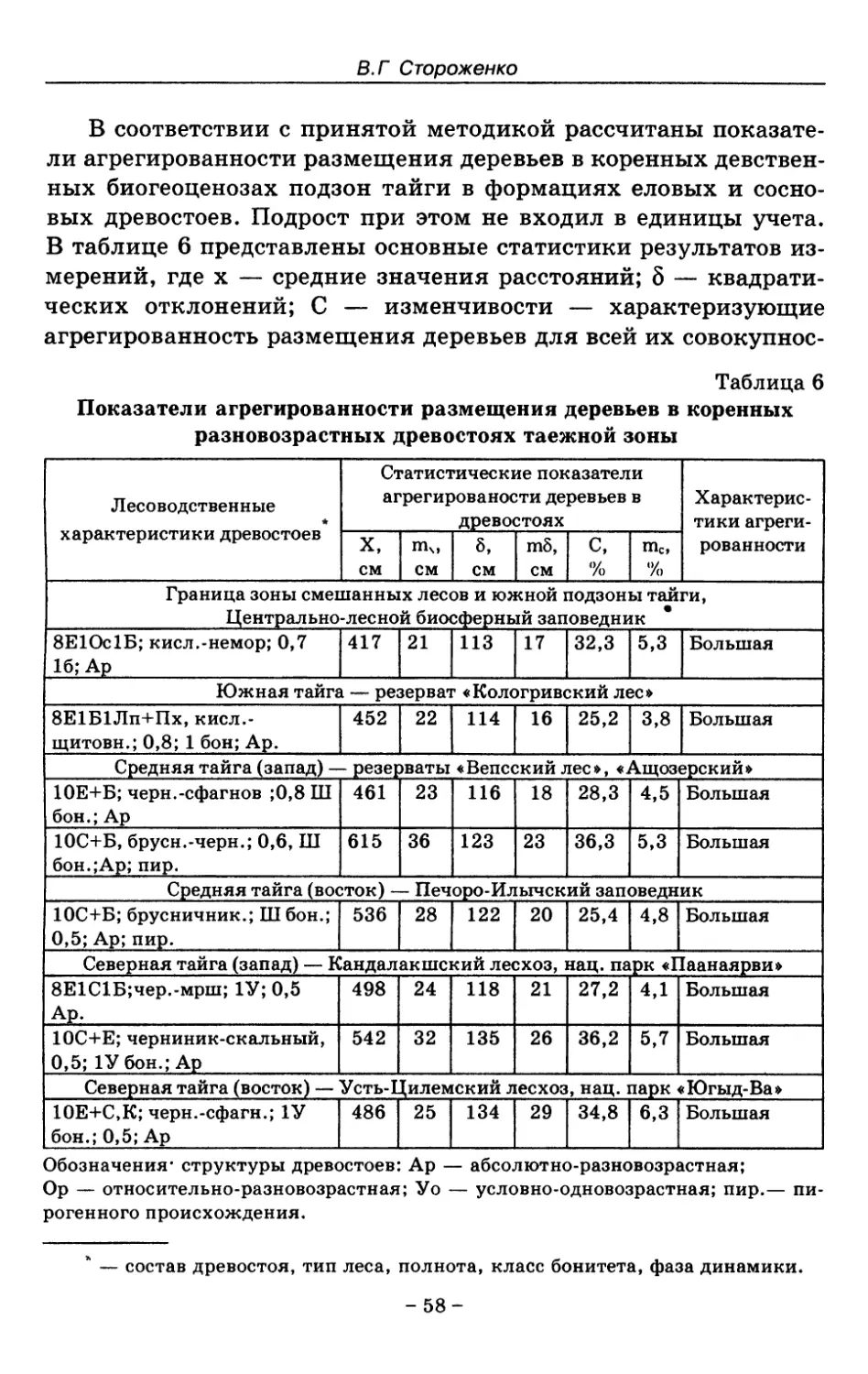
6. Фитоценотические критерии устойчивого лесного
сообщества 37
6.1. Сложность структурного строения лесного
сообщества 41
6.2. Возрастная структура древостоя лесного
сообщества 41
6.3. Породный состав древостоя 47
6.4. Горизонтальная структура древостоя 54
6.5. Возобновительные структуры лесного
сообщества 69
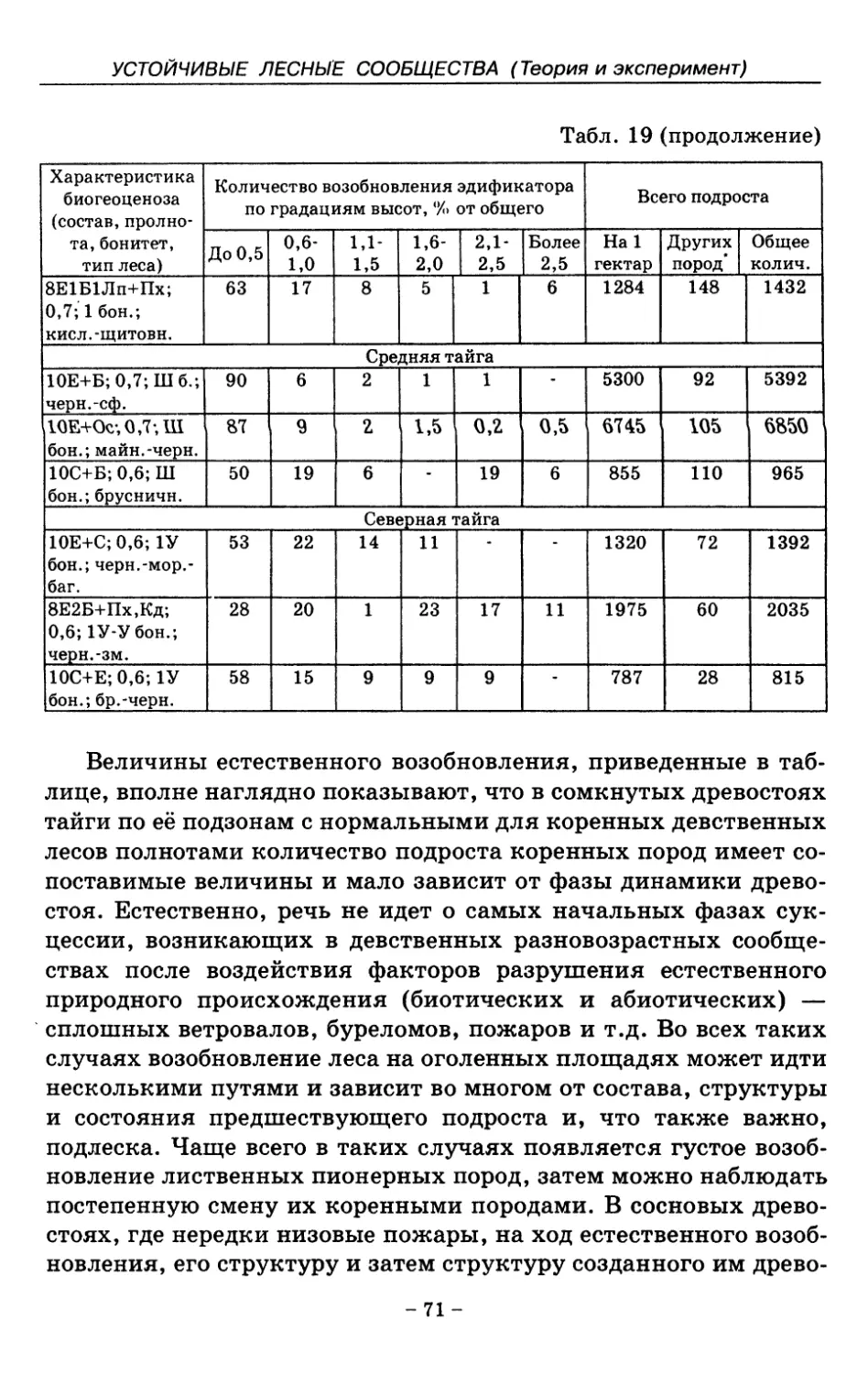
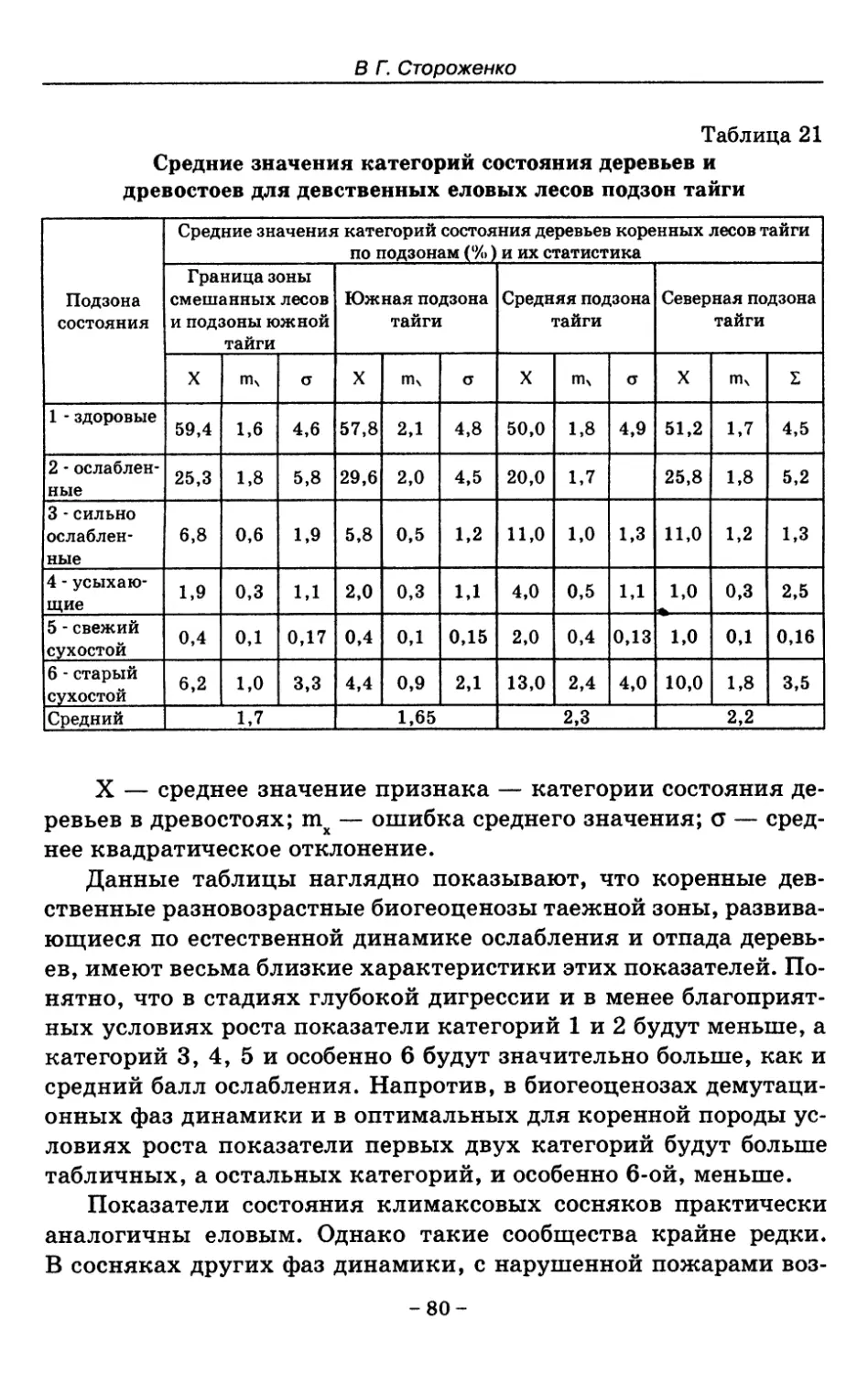
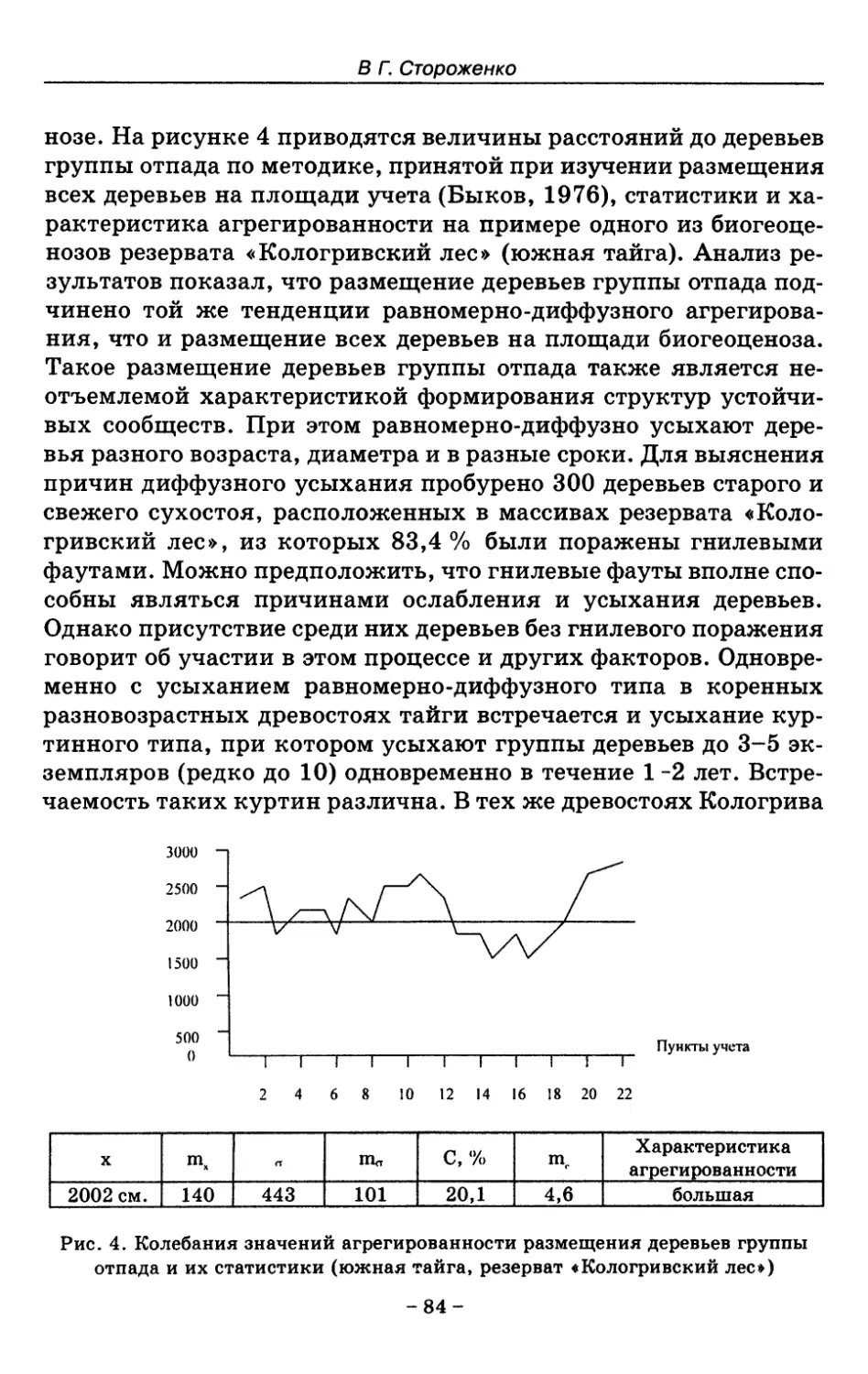
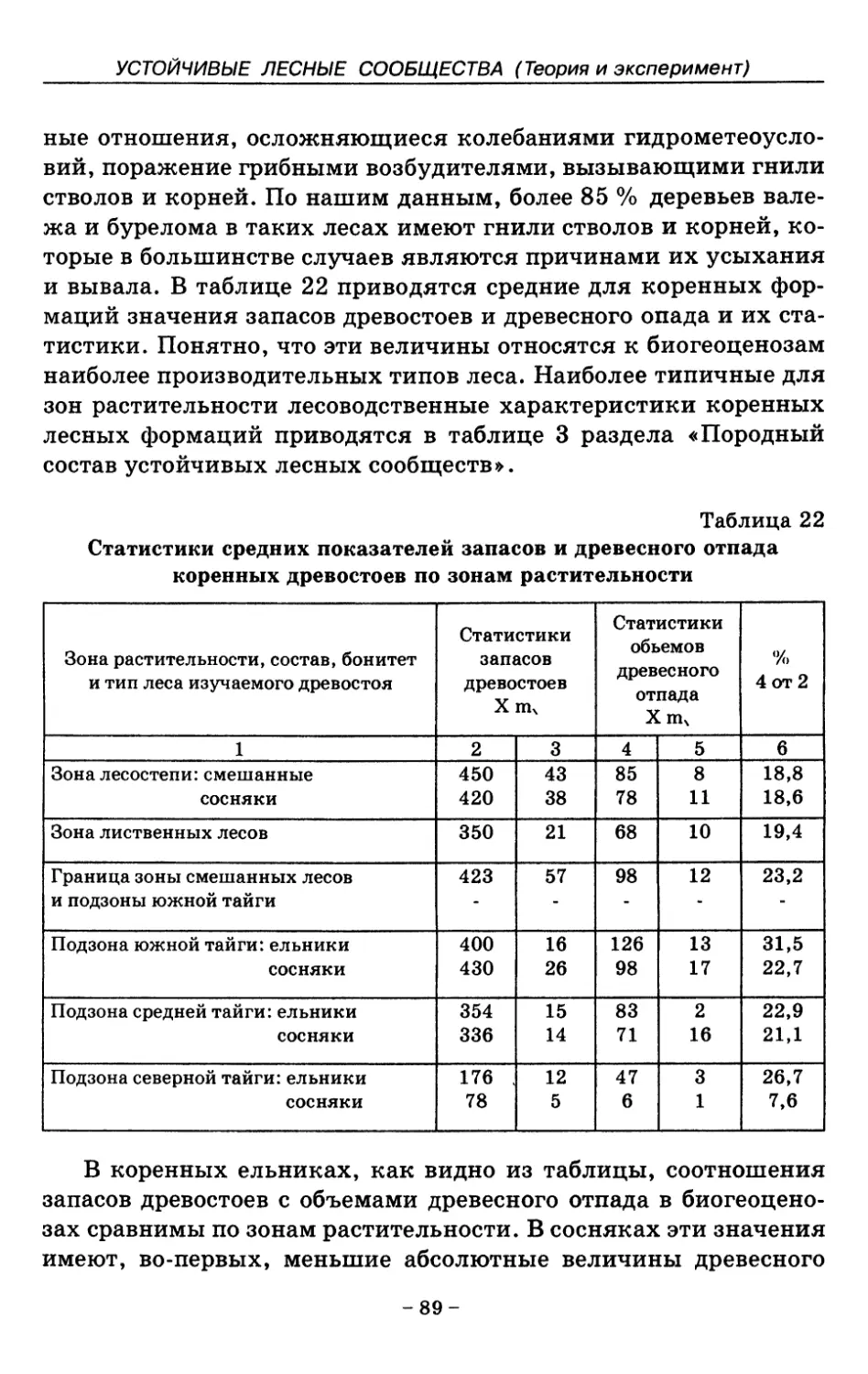
7. Показатели состояния устойчивых лесных сообществ . . 75
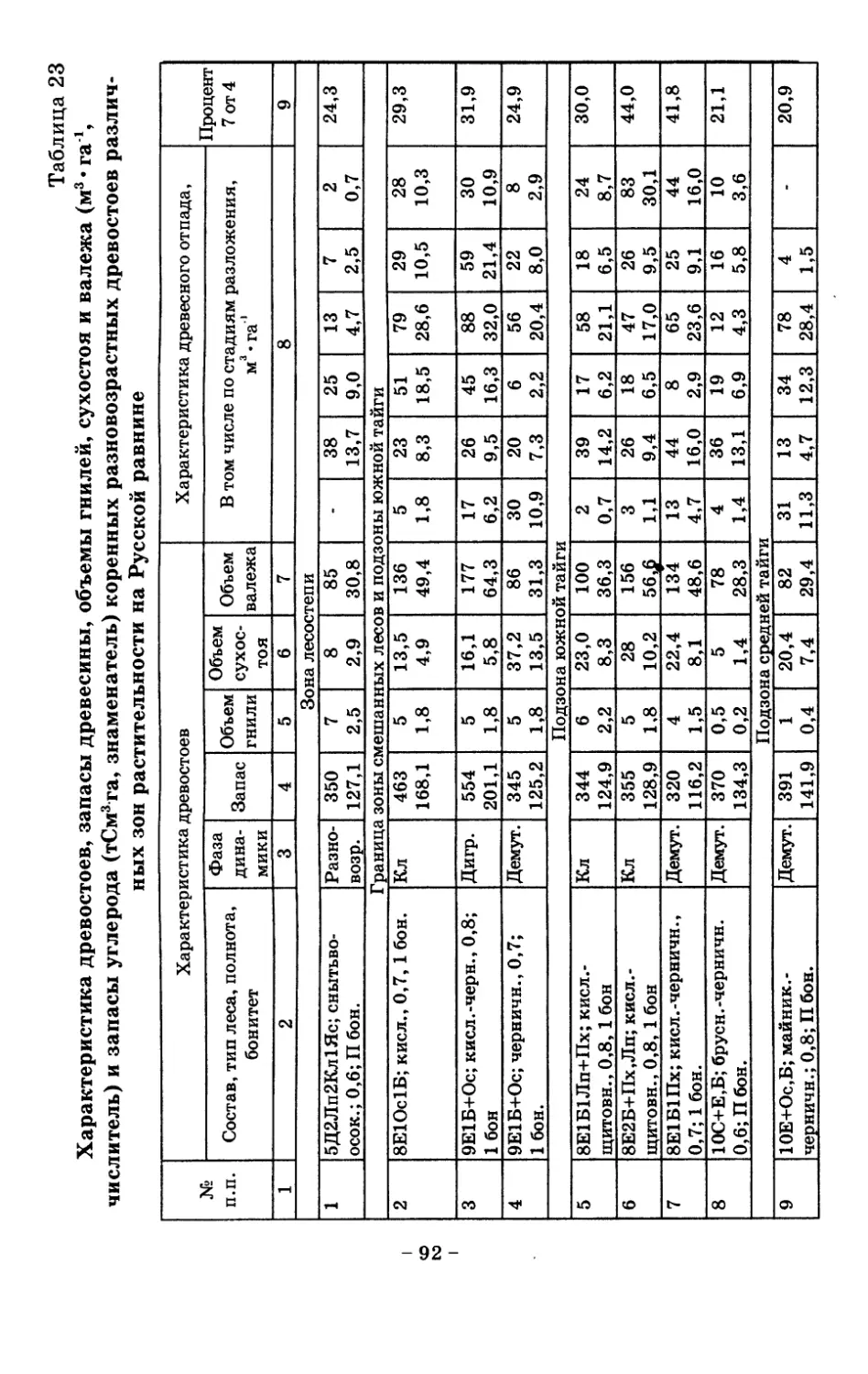
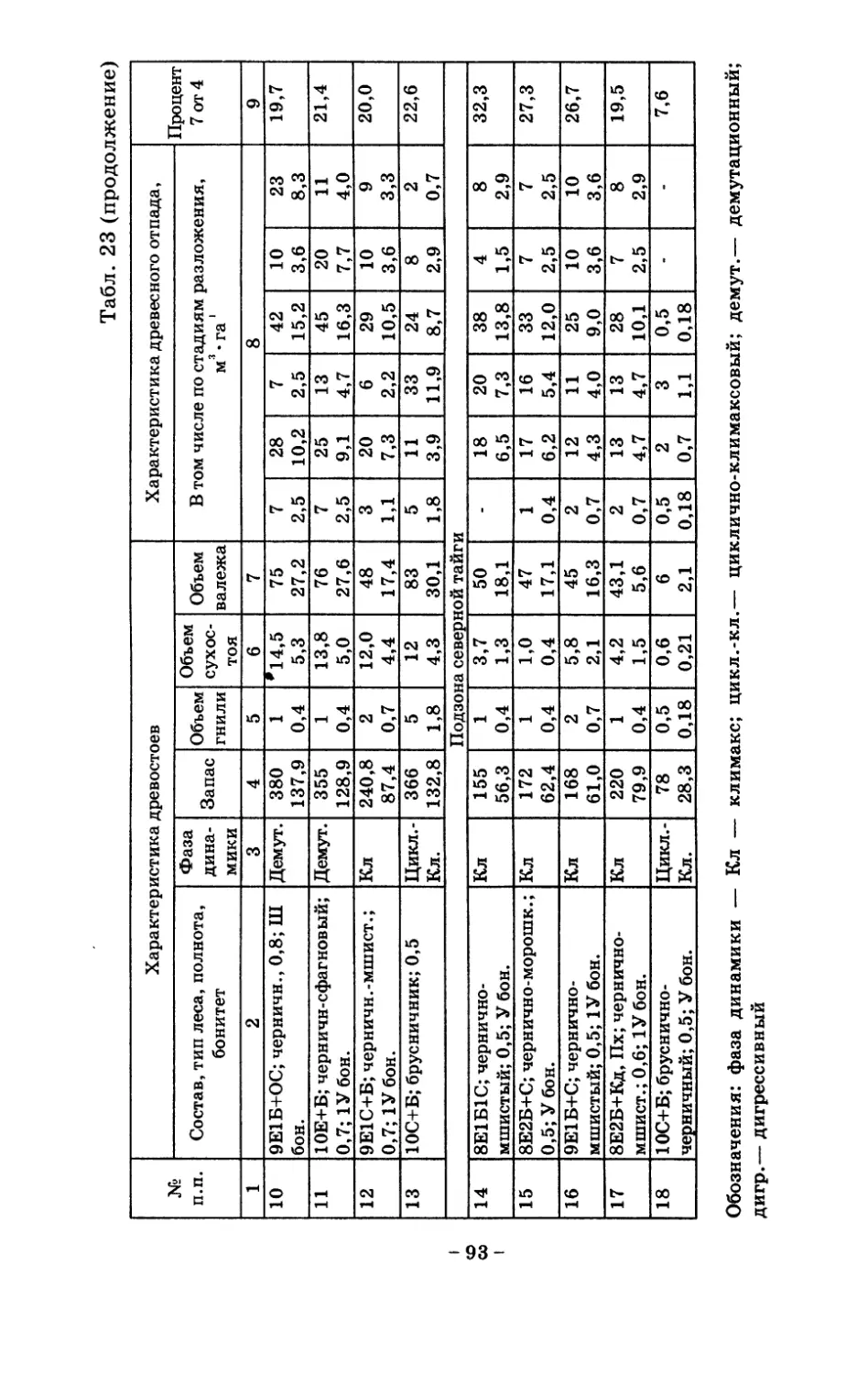
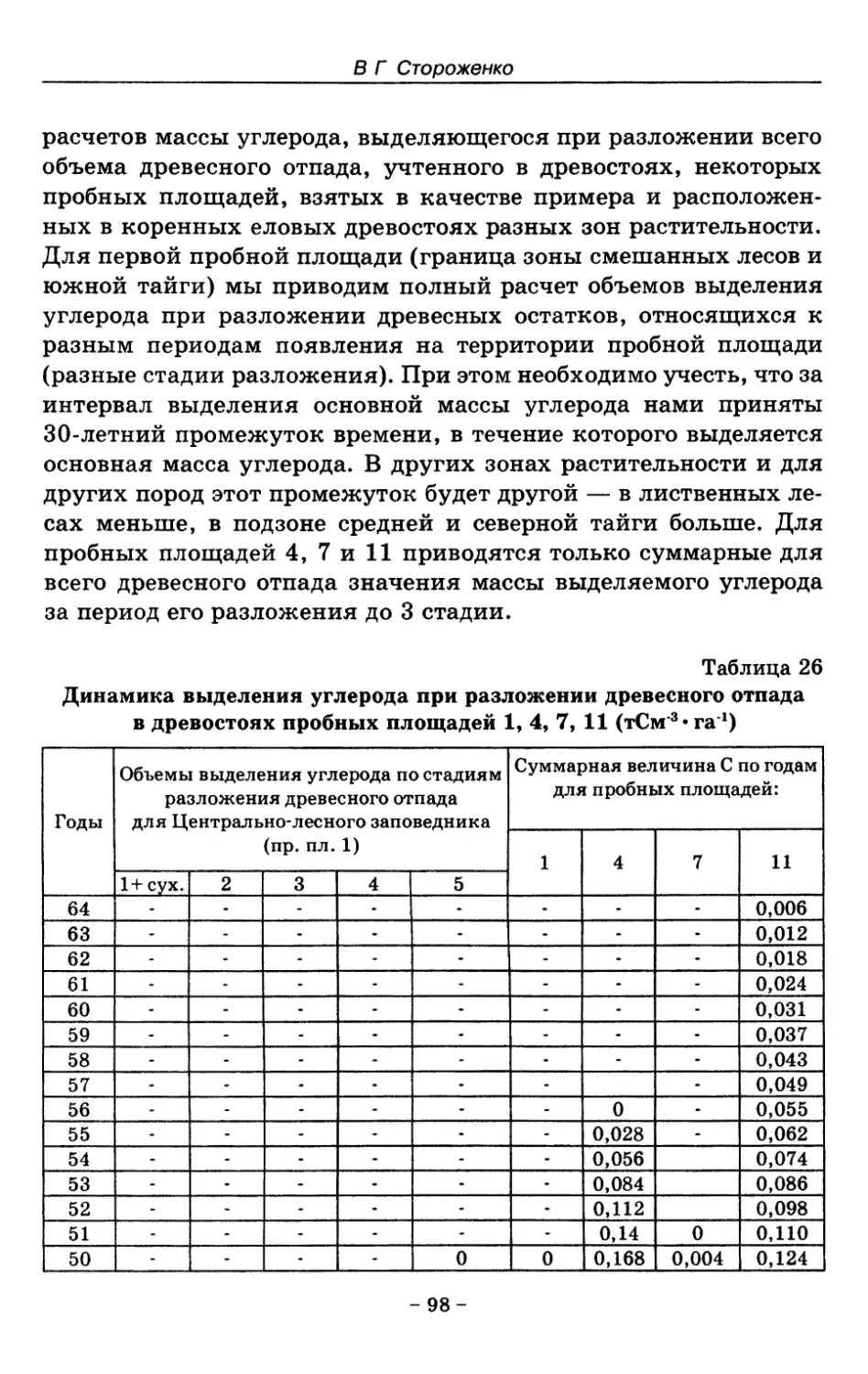
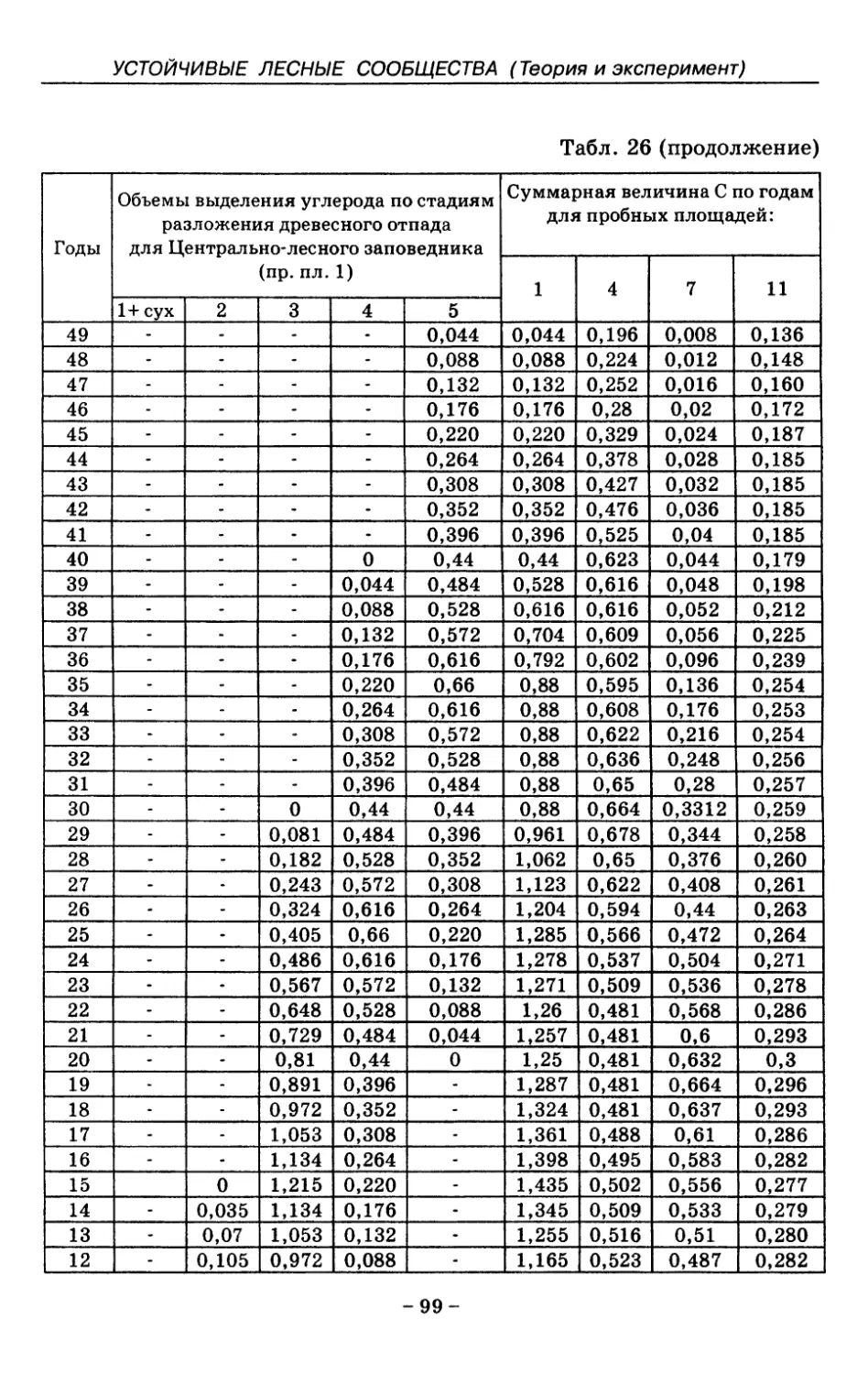
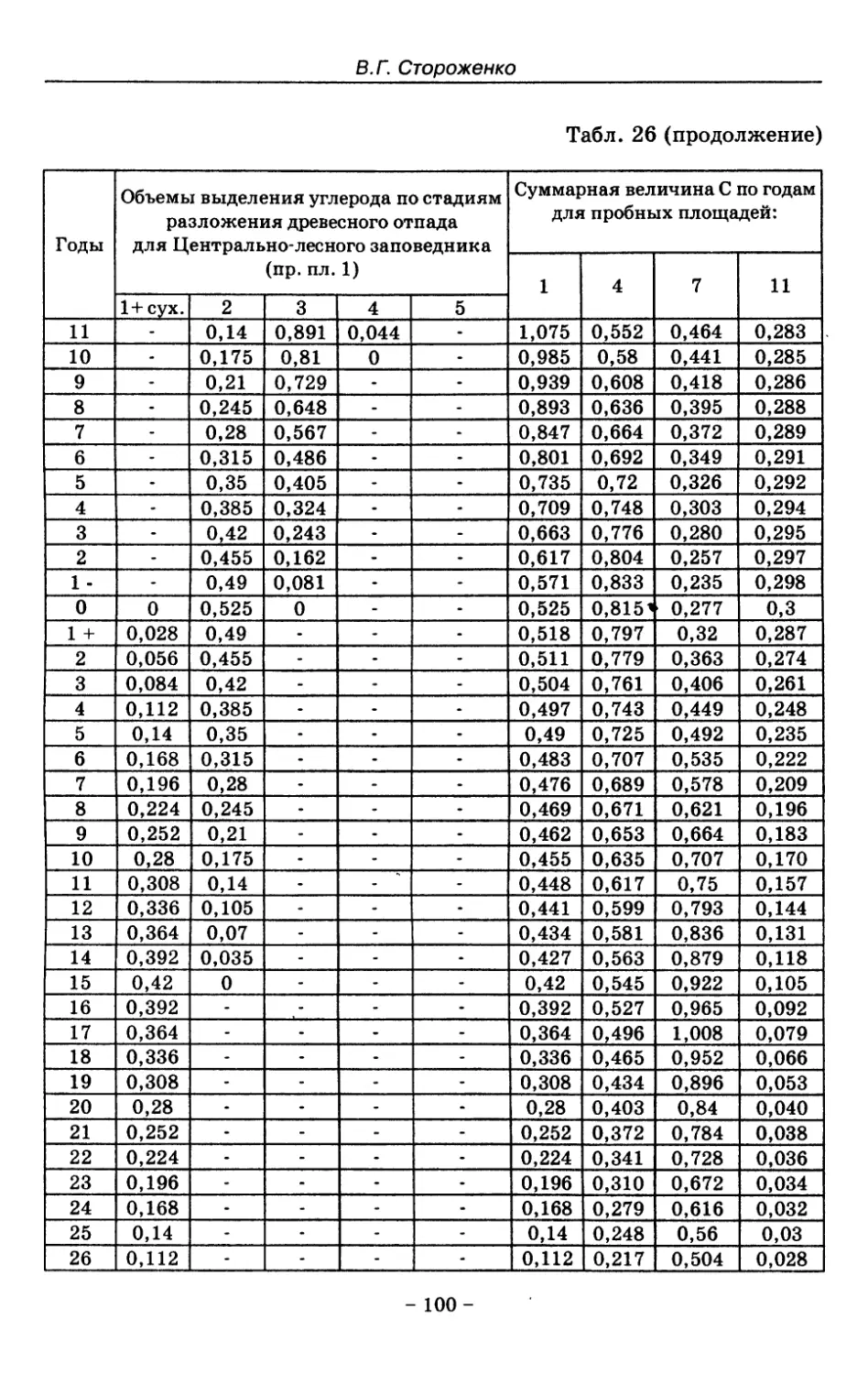
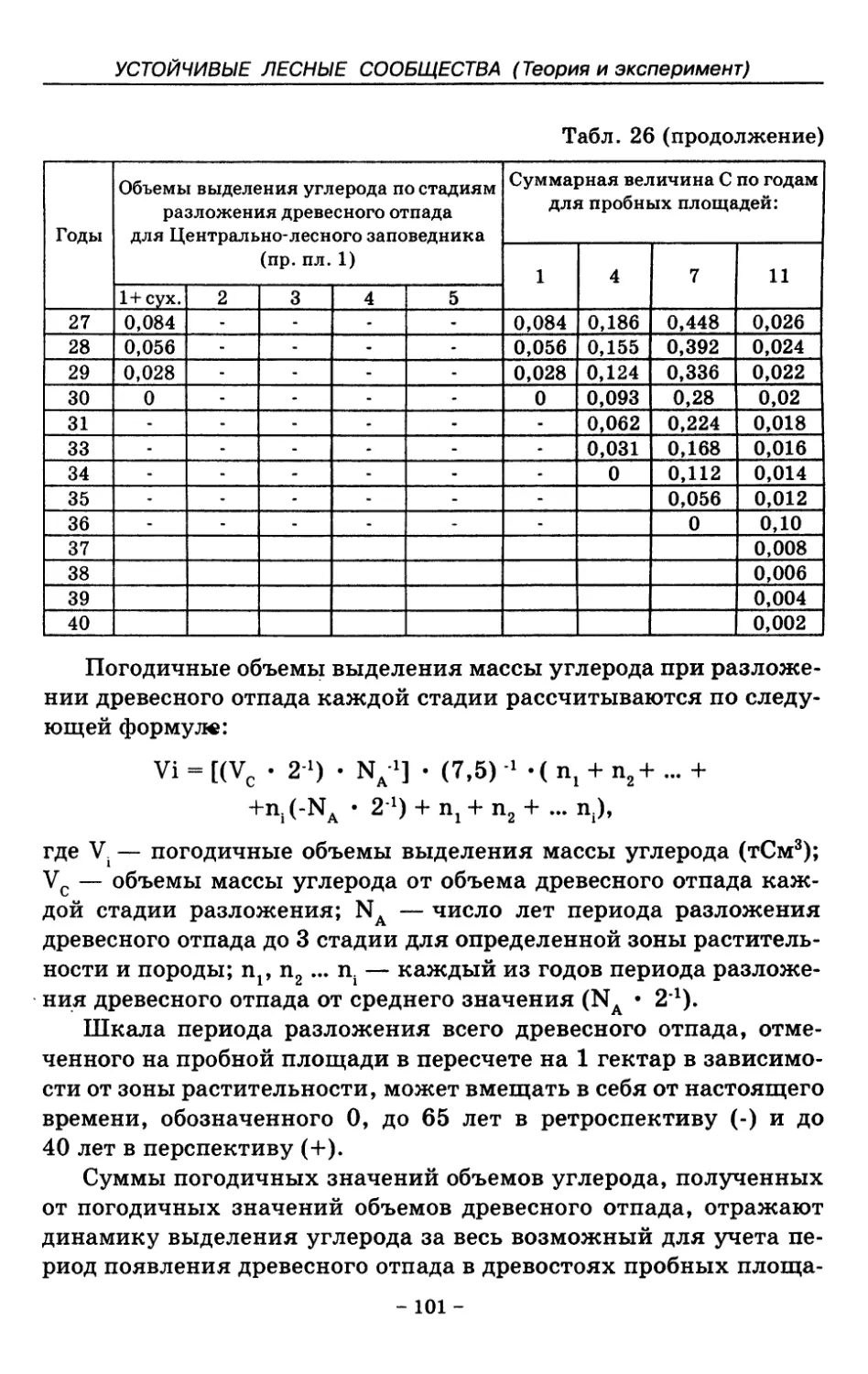
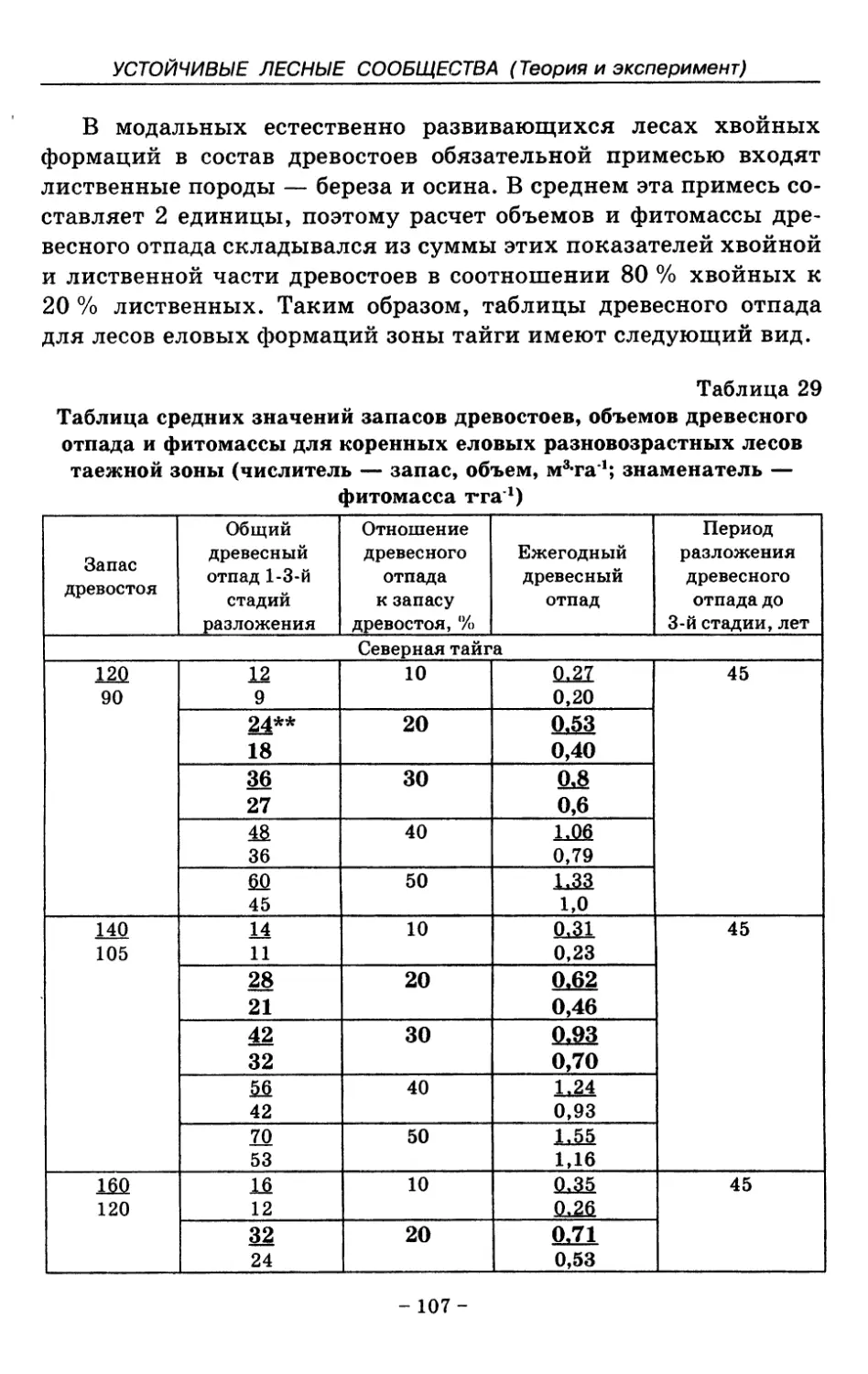
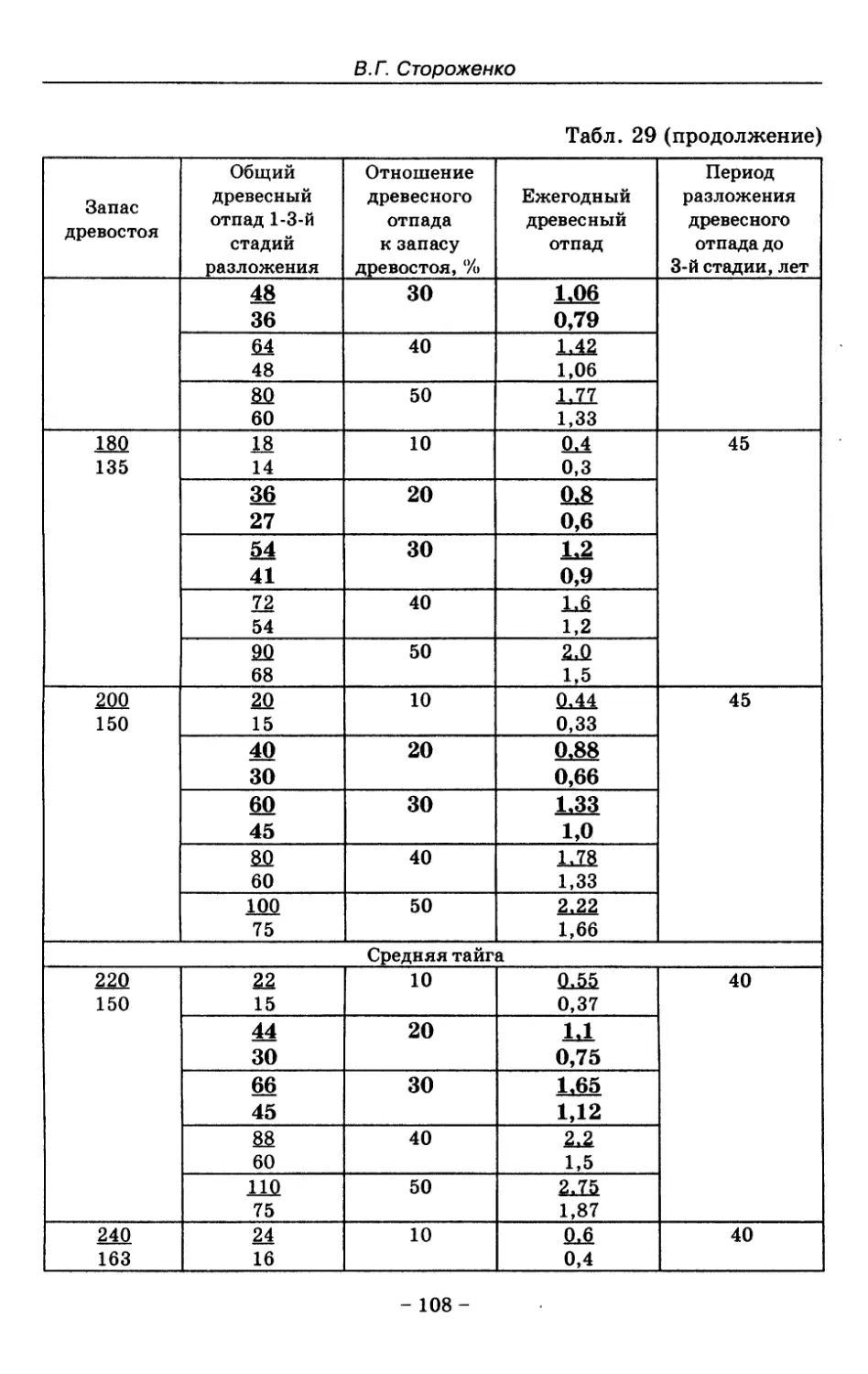
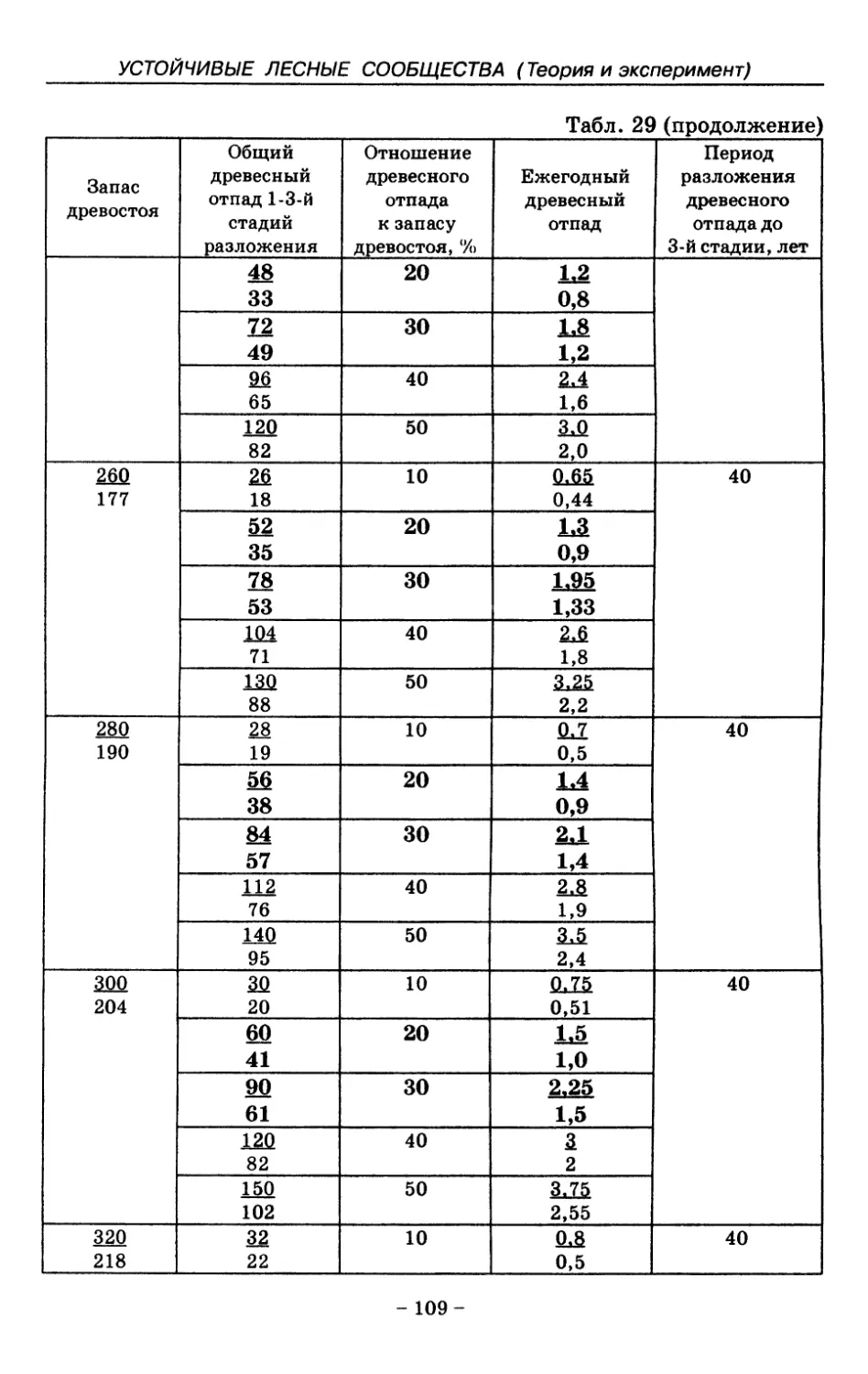
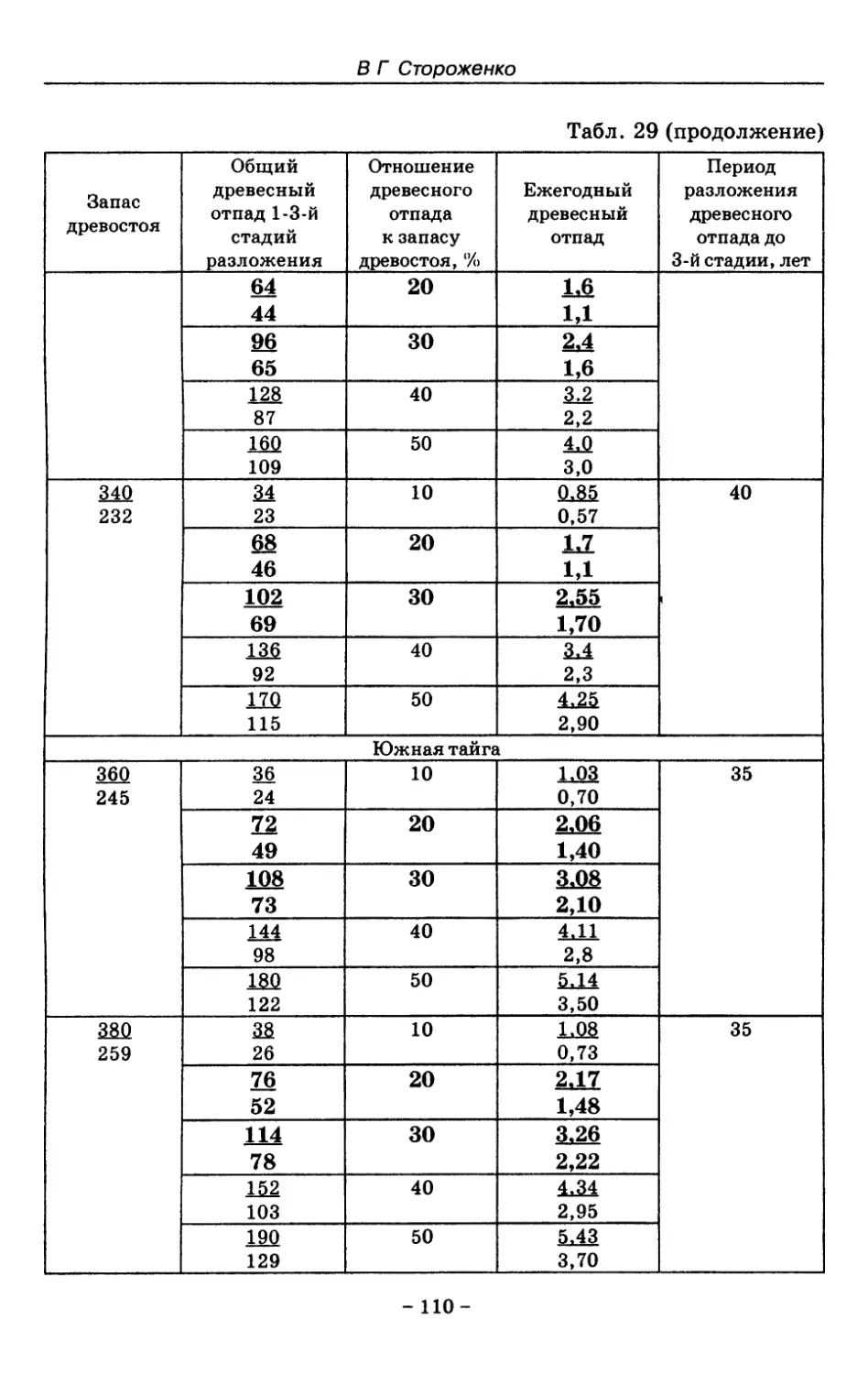
8. Показатели древесного отпада устойчивых лесных
сообществ 86
8.1. Объемные показатели древесного отпада коренных
лесов 87
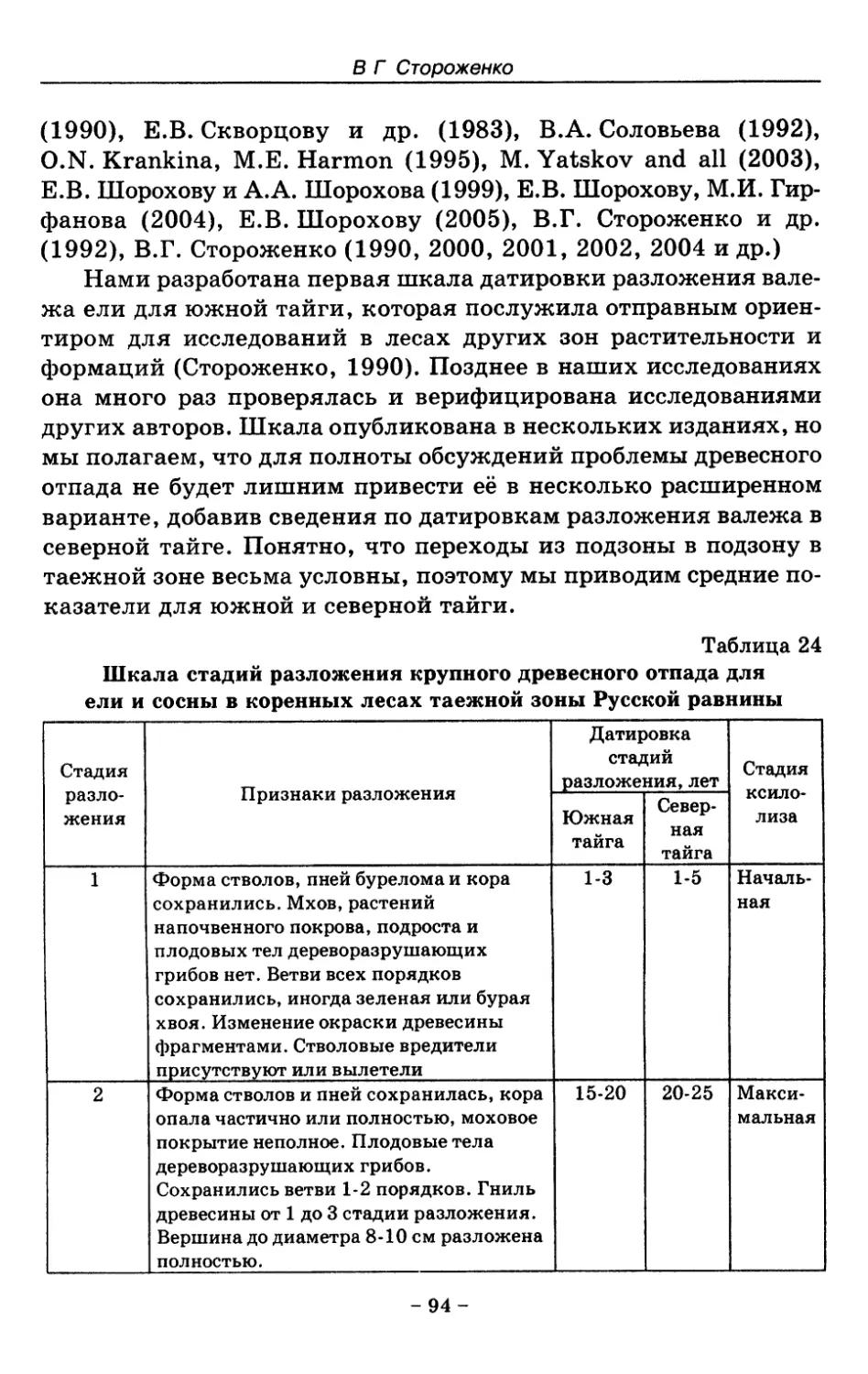
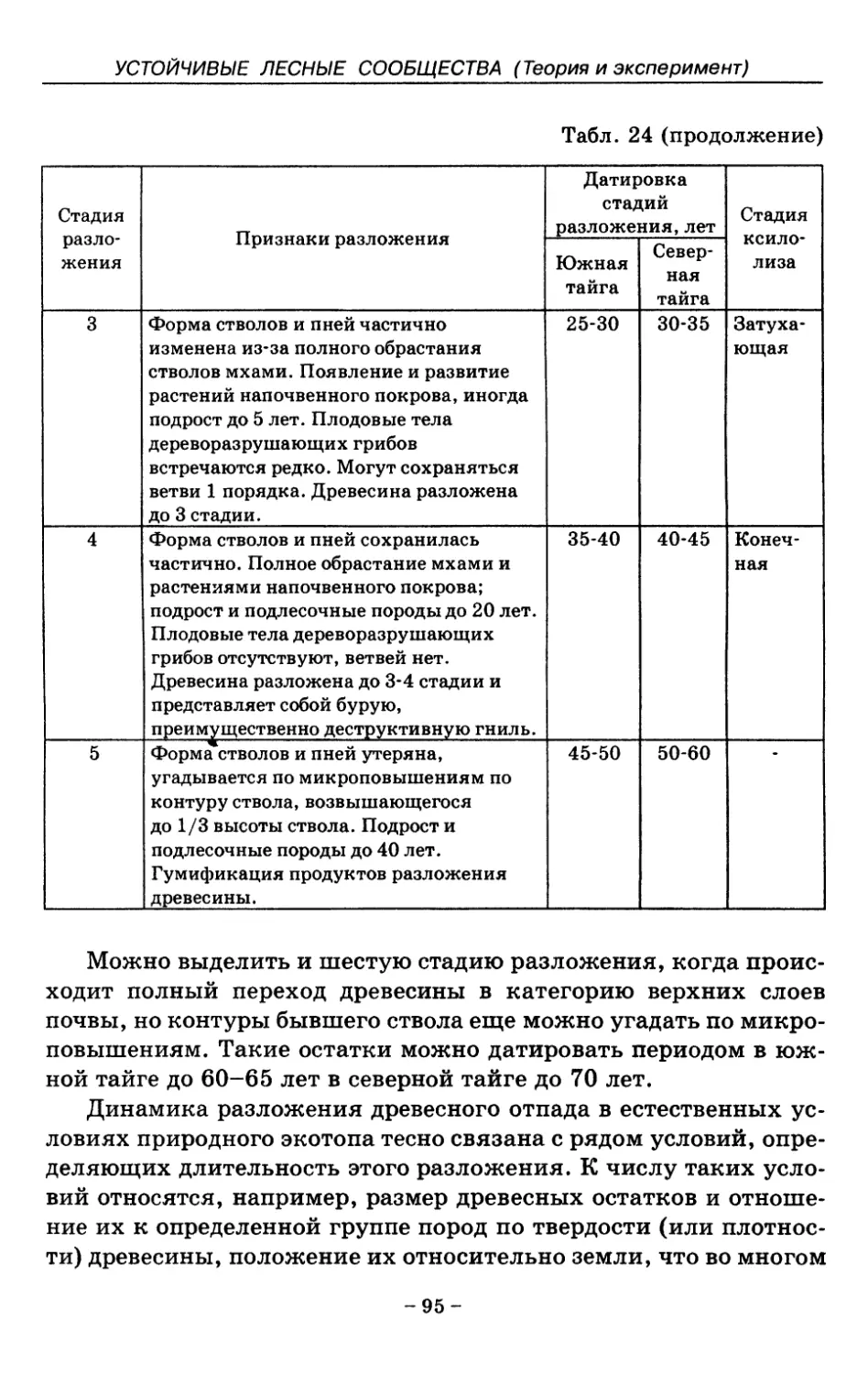
8.2. Процессы разложения древесного отпада 90
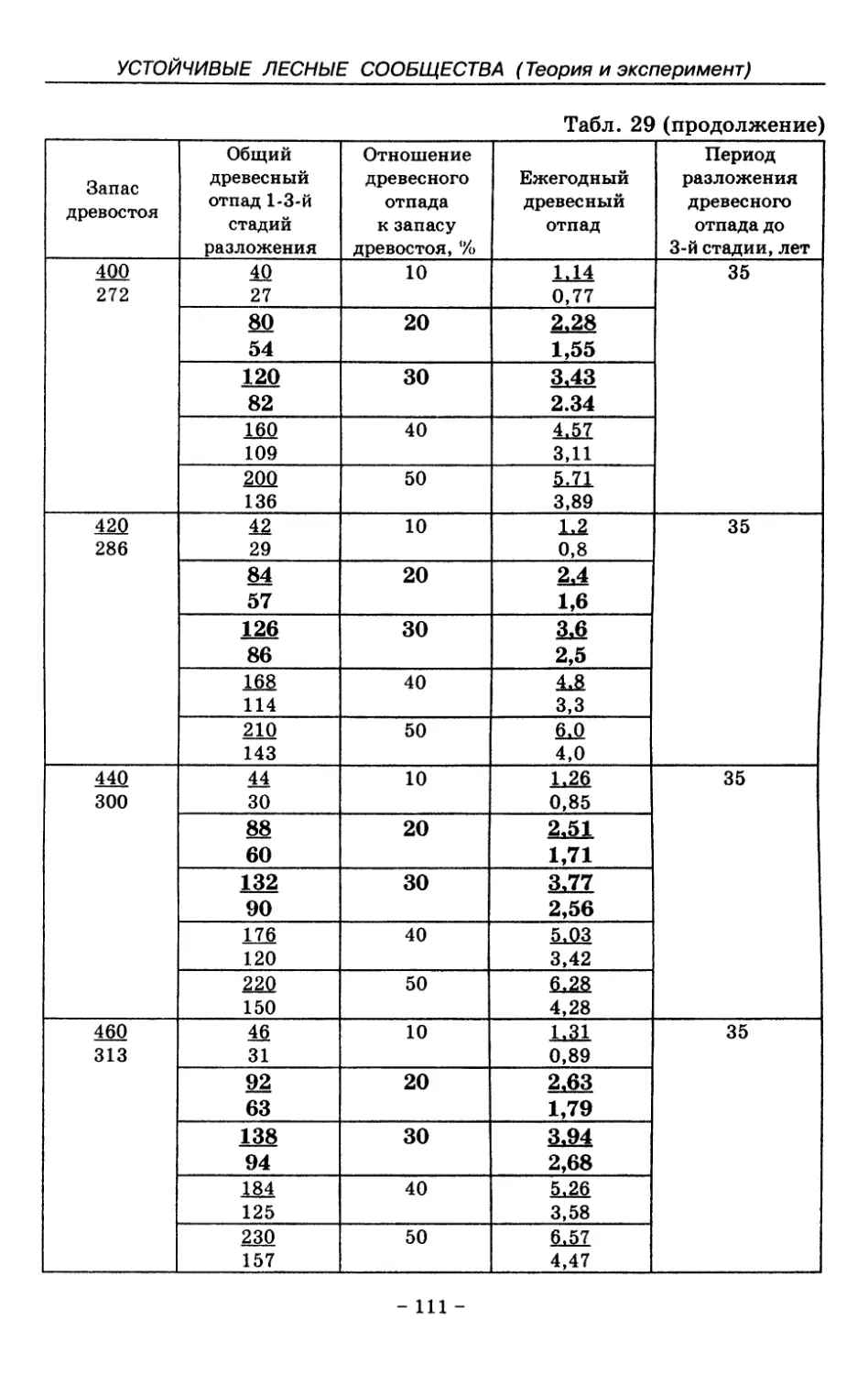
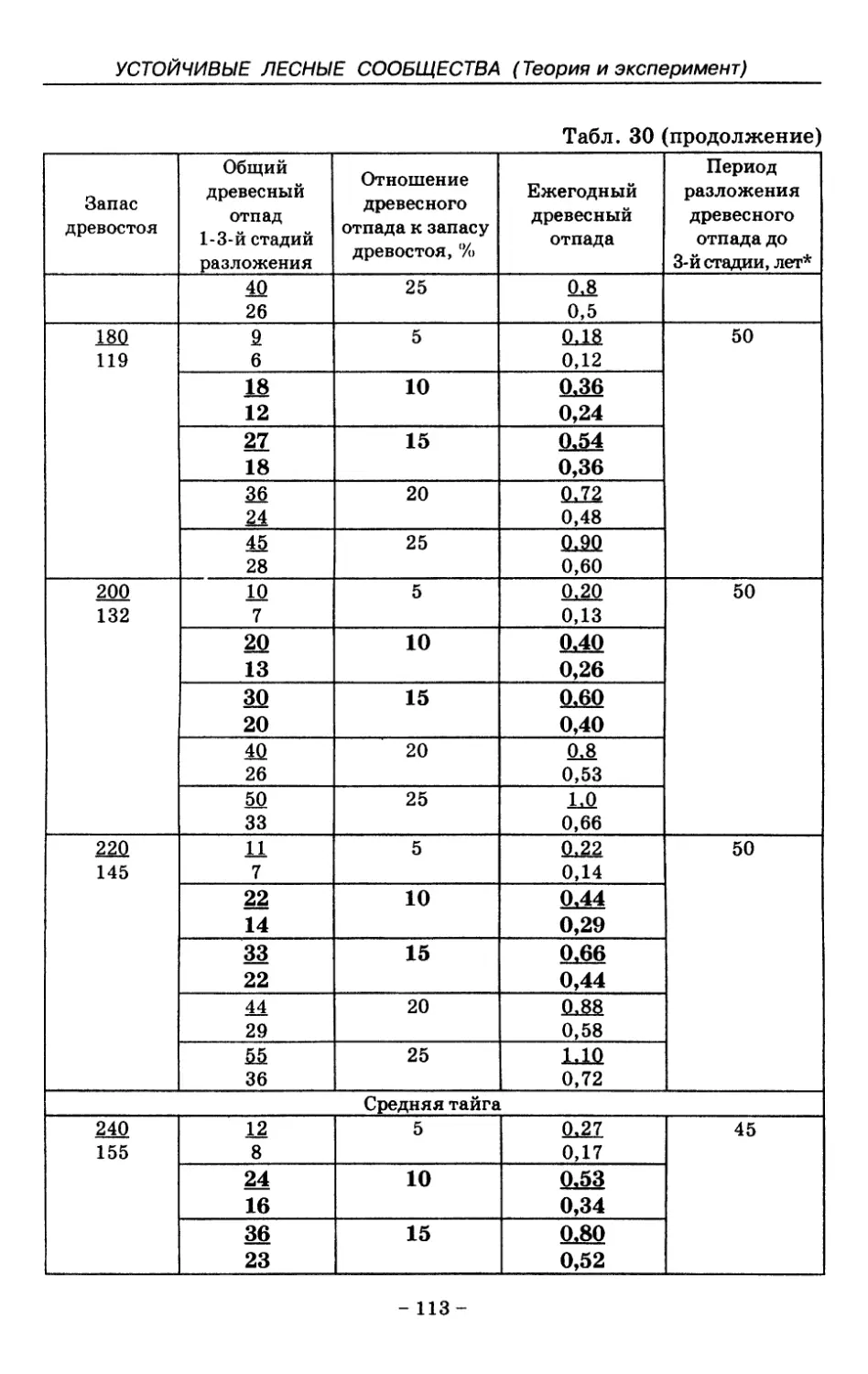
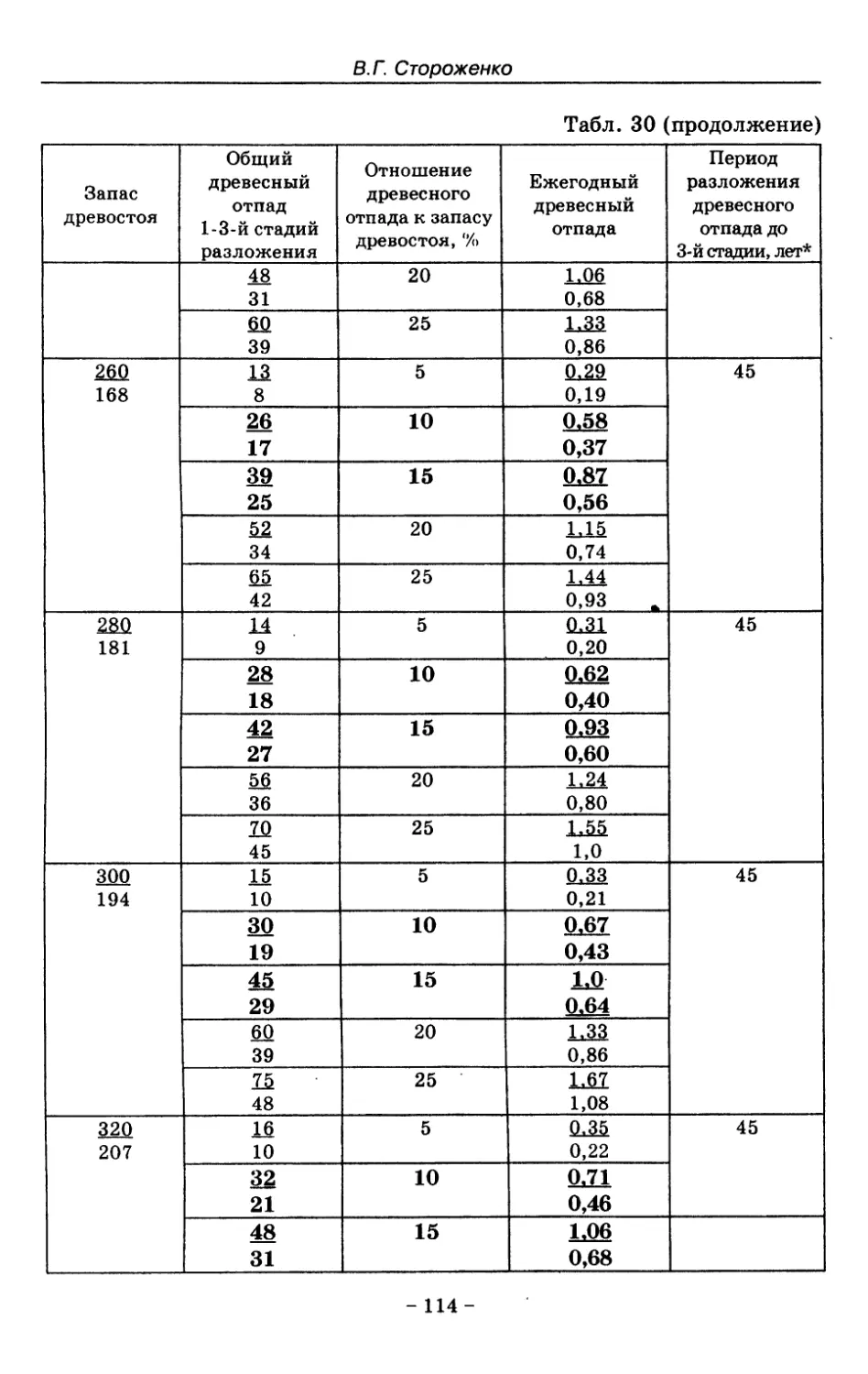
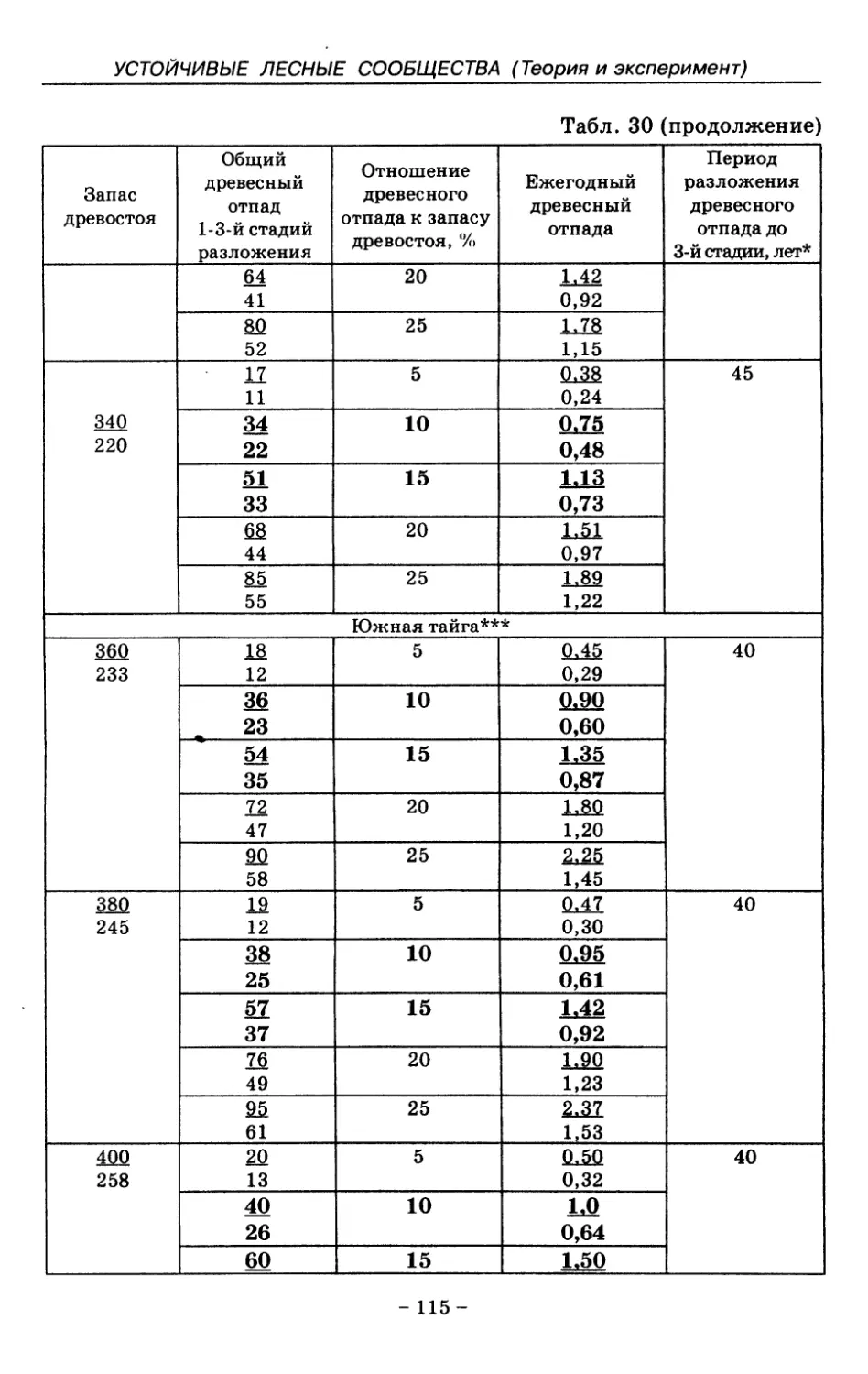
8.3. Таблица древесного отпада для коренных модальных
лесов таежной зоны Русской равнины . . . . 104
9. Микоценотические критерии устойчивых лесных
сообществ 118
9.1. Понятия «больного» и «здорового» лесного
сообщества 118
9.2. Микоценоз и микоценология 126
9.3. Параметры участия грибов дереворазрушающего
биотрофного комплекса в структурах устойчивых
лесов 131
-3-
В. Г. Стороженко
9.3.1. Связь грибного поражения коренных
древостоев с их возрастной структурой . . 131
9.3.2. Связь грибного поражения коренных
древостоев с их горизонтальной
структурой 139
9.3.3. Особенности видового состава дереворазру-
шающих биотрофных грибов коренных
разновозрастных устойчивых лесов . . . 141
10. Критерии устойчивости лесных сообществ 160
11. Градации устойчивости лесных сообществ 165
Заключение 176
Литература 191
ВВЕДЕНИЕ
Осознанная человечеством реальная угроза
самоуничтожения или перерождения в результате изменения им же самим
пространства жизнеобитания приводит, наконец, к поискам путей
разумного регулирования потребностей природопользования и
возобновительных возможностей биосферы.
Изучение возобновительных способностей природных
экосистем, к числу которых относятся и лесные сообщества,
объяснение закономерностей и тенденций в их функционировании,
поиски путей восстановления их структурных и функциональных
возможностей является в настоящее время одной из важнейших
парадигм естественных наук и лесоведения в частности.
Эти тенденции становятся все более отчетливыми и
значительными не только на уровне научных разработок, но и на
уровне государственной политики. Еще в 1990 году на встрече
высших руководителей лесохозяйственных и лесопромышленных
ведомств стран СЭВ, ФРГ и Канады принято обращение к
правительствам стран о необходимости скорейшей переориентации
приоритетов лесных комплексов на неистощительное
лесопользование, сохранение и восстановление лесов с естественно
сформированными структурами. Известны решения конференции
ООН по окружающей среде и развитию в Рио-де-Жанейро (1992)
и т.д.
Лес — это одна из важнейших компонент биосферы,
поддерживающих газовый и водный баланс атмосферы и суши, другие
важные биологические параметры функционирования биосферы
как отдельных регионов, так и в целом планеты. В то же время
это одна из наименее защищенных ее структур, подвергающаяся
разрушающим воздействиям эндогенных природных и
антропогенных абиотических факторов. Но если природные
абиотические воздействия в бесконечной эволюционной динамике
«заставили» лесные сообщества выработать механизмы
восстановления своих структур и функций в кратчайшие сроки с
минимальными для них потерями, то антропогенные, особенно
техногенные, воздействия в корне изменяют природную среду, вызы-
-5-
В Г Стороженко
вая необратимые последствия в структурах лесных экосистем,
нарушая балансовые отношения между их компонентами. Эти
воздействия приобретают огромные масштабы и являются уже
сейчас причиной многих изменений в функционировании
биосферы Земли.
В этой связи перед учеными-лесоводами встает ряд проблем,
важность которых трудно переоценить. К числу таких проблем
можно отнести следующие.
1. Изучение структур и закономерностей функционирования
не затронутых антропогенной деятельностью девственных
сообществ, как эталонов эволюционно развивающихся устойчивых
сообществ, находящихся на разных этапах сукцессионного
развития.
2. Выработка концептуальных положений теории
устойчивости лесных сообществ.
3. Определение параметров устойчивых лесных сообществ.
4. Разработка научно обоснованных рекомендаций по
восстановлению нарушенных лесов и ведению лесного ^хозяйства на
формирование устойчивых лесных сообществ долговременного
функционирования и использования.
5. Разработка концепций и схем оптимальной структуры
лесов по их лесоводственным характеристикам и целевым задачам
использования в рамках границ лесохозяйственных
предприятий и административных территорий.
В предлагаемом издании мы попытались систематизировать
наши разработки для решения трех первых положений. Мы
надеемся, что эти разработки в определенной мере будут
способствовать решению четвертой и пятой, конечной, наиболее
важной практической задачи оптимального сосуществования
человека и биосферы, частью которой он, по забывчивости, себя уже
не считает.
Мы понимаем, что некоторые положения, представленные на
суд читателя в монографии, могут быть спорными, особенно
положения теоретического направления и рекомендации
практического их применения. Тем не менее мы считаем вполне
своевременным поделиться с заинтересованным читателем своими
мыслями, мы уверены в правоте основных идей и
стратегических направлений в изучении проблем устойчивости и будем
признательны за благожелательное обсуждение работы.
-6-
1. ПРЕДИСЛОВИЕ
Современные научные представления о возникновении и
развитии растительности на Земле выстраивают этот процесс в ряд
эволюционных скачков, обусловленных климатическими,
геологическими и связанными с ними гидрологическими
изменениями на планете.
Выход растений на сушу из водной среды, где они
зародились, и далее эволюционные преобразования в строении органов,
в системах их питания и размножения как приспособительные
реакции на геоклиматические колебания на повторяющиеся
периоды потепления и похолодания, приводившие к глобальным
изменениям условий обитания, теория филоценогенеза и
дискретного развития бореальных и неморальных формаций
растительности (теория континиумов) обозначили основные вехи
эволюции растительности на планете и очертили пути
формирования видов и растительных формаций. Движущими факторами
на этих путях эволюции растений и растительных сообществ
признаются геоклиматические. В общем можно говорить, что
представления об ареалах пород, формационной организации
лесов вполне объяснены как теоретически, так и обширным
палеоботаническим материалом (Толмачев, 1954; Покровская, 1937;
Синская, 1933; Сочава, 1961; Сукачев, 1974 и др.).
Одновременно с процессом видообразования, формированием
растительной сферы, распространением растений по территории
суши, формационным построением растительных континиумов
развивались и фитосоциальные отношения внутри растительных
группировок, позволившие в итоге рассматривать эти
группировки как сообщества растений.
Параллельно с автотрофными организмами развивались и
организмы гетеротрофной группы, утилизирующие
наращиваемую автотрофами биомассу. Грибная биота прошла все этапы
эволюционного развития вместе с растительным и животным
миром. Новейшие теории о происхождении грибов рассматрива-
-7-
В. Г. Стороженко
ют их как самостоятельную ветвь развития биоты, начинающую
свое формирование от флагеллят (Flagellata), первичных
бесцветных жгутиковых, через хитридиомицеты (Chytridiomycetes)
к неподвижным грибам. Существуют и другие теории
происхождения грибов, но все они рассматривают эту группу гетеротрофов
как отдельное царство в системе органического мира (Гарибова,
1999; Кусакин, Дроздов, 1994, 1998; Тахтаджан, 1973; Cavalier-
Smith, 1991; Margulis, Schwartz, 1982; Ainsworth, Bisby, 1995
и др.).
Эволюция биогеосоциальных отношений в экосистемах
растительных сообществ, в том числе и лесных, до современного
уровня рассмотрена в теоретических обобщениях Г.Ф. Морозова
(1928, 1970), В.Н.Сукачева (1972), Е.М. Лавренко (1959),
А.А.Корчагина (1976), A.G. Tansley (1920, 1929); G.W. Selleck
(1960); F.E. Clements (1916, 1936), R.H. Whitteker (1975) и др.
В основном эти теоретические обобщения построены на
анализе процессов развития естественно формирующихся лесных
сообществ, изучение которых дает возможность представить и
осветить с наибольшей полнотой и достоверностью тенденции и
закономерности функционирования организмов и сообществ,
составляющих природные комплексы и в их числе лесных
сообществ.
Современная история лесов, их формационный состав,
породная и возрастная структура во многом определяются антропоген-"
ной деятельностью, теми хозяйственными, техногенными,
рекреационными воздействиями, которые приводят к замещению
коренных эволюционно развивающихся лесов и лесных
экосистем на искусственные, производные от них, или
условно-коренные.
Искусственные леса, появившиеся на месте утраченных
коренных формаций, создавались и изучались с позиций их
хозяйственной и биологической продуктивности. Значение
разновозрастных, естественно развивающихся лесов как средообразую-
щего компонента еще в недавнем прошлом, а во многом и в
современных условиях, или занижалось, или умалчивалось, или
вообще отрицалось. Было очевидно, что делалось это
преднамеренно, являлось составной частью государственной политики
поддержания экономики страны за счет варварского
использования лесных ресурсов. Такая затратная сырьевая направленность
-8-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
экономики, связанная с неконтролируемым использованием
ресурсов, характерна для всех стран, прошедших или проходящих
периоды социальных, политических и экономических
катаклизмов или становления.
Вообще, двадцатый век стал для подавляющего числа стран
мира веком экономического скачка, связанного с огромным по
масштабам техническим прогрессом, для некоторых государств,
напротив,— веком политических катаклизмов, связанных с
обрушением экономики. И в том и в другом случае потребности
промышленности мало соизмерялись с возможностями
обеспечения ее природными ресурсами без катастрофических
последствий для последней.
История лесной науки и лесного хозяйства России в XX
столетии иллюстрирует это в гипертрофированных размерах.
Первые два десятка лет XX столетия российская лесная наука и
практика лесоводства были заняты созданием теории строения и
функционирования лесных сообществ и лесохозяйственных
правил эксплуатации лесов, в которых напрямую почти не
рассматривались проблемы устойчивости. Устойчивость растительных
сообществ не рассматривалась как одно из важнейших
стратегических качеств леса. В то же время в трудах ведущих русских и
зарубежных ученых лесоводов, экологов того времени
содержатся основные теоретические предпосылки, объясняющие многие
положения и понятия устойчивости лесов, употребляется
термин «устойчивость».
Невостребованность разработки теории устойчивости лесов в
тот период можно связать еще и с тем, что значительная часть
объемов лесозаготовок велась с использованием технологий,
обеспечивающих сохранение лесной среды, коренных формаций
лесов, учитывающих и использующих самовозобновительные
свойства лесов. Сохранение этого качества в создаваемых лесах
как бы подразумевалось во всей хозяйственной деятельности
лесоводов того времени. Широко применялись узколесосечные и
постепенные рубки с сохранением подроста главных пород, что
способствовало быстрому возобновлению леса без изменения
(или с малым изменением) его формационного состава.
Затем в 30-е годы государство перешло к эпохе строительства
коммунизма, что ознаменовалось резким ростом объемов сплош-
-9-
В. Г. Стороженко
ных рубок на огромных площадях и хищническим истреблением
лесов. Это стало возможным благодаря появлению огромной
массы дешевой рабочей силы, появившейся в результате репрессий,
возведенных в ранг государственной политики. Лесное хозяйство,
как государственная структура, призванная обеспечивать
адекватное по качественным и количественным параметрам
лесовозобновление после лесозаготовок, было практически
отстранено от формирования лесной политики, эту функцию передали
лесной промышленности, в задачи которой и по сей день входит
получение в максимальных количествах деловой крупномерной
древесины. Едва ли не самым трагичным по своим последствиям
для лесного хозяйства и лесов в целом стал упадок культуры
ведения лесного хозяйства, лесоэксплуатации, лесопользования,
лесовосстановления и те моральные потери, которые связаны с
потребительским отношением к лесам и к природе вообще. В
течение последующих десятилетий культивировалось отношение к
лесу как к самовоспроизводимому сырью. Главная цель системы
ведения лесного хозяйства заключалась в «формировании
высокопродуктивных чистых и смешанных насаждений из тех
древесных пород, которые соответствуют данным условиям
произрастания и к моменту главной рубки образуют максимальный
запас высококачественной древесины» (Атрохин, Кузнецов,
1989).
Интересы производства определяли переориентацию лесной
науки на разработку методов и способов быстрого выращивания
наиболее производительных лесов, чем и объясняется
преобладание в лесовосстановлении искусственного лесовыращивания
регулярными посадками. Интенсивно разрабатывались теории и
технологии сплошнолесосечных концентрированных рубок с
применением тяжелой агрегатной техники. При таких рубках
уничтожалось (и до сих пор уничтожается) до 80-100 % живого
напочвенного покрова и верхних гумусированных слоев почвы.
Даже в лесах 1 группы повсеместно оправдывается применение
сплошнолесосечных рубок.
Устойчивость как важнейшее качество лесов в системе
лесного хозяйства не рассматривалось, так как по положениям
принятой государственной лесной политики содержание лесов,
обладающих этим качеством, не соответствовало экономической
потребности строя и государства.
-Ю-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таким образом, к концу прошлого столетия на огромных
площадях страны, включая регионы таежной зоны, естественные
леса были заменены или производными, или искусственными.
Уже к концу столетия сырьевые базы многих леспромхозов
пришли в упадок, истощились, даже в отдаленных регионах Севера
России и Сибири. Закрытие по этой причине лесных
предприятий обострило социальные проблемы регионов, где лесодобыча
являлась, а во многих случаях является и сейчас основным
источником жизни людей.
И все же к концу столетия в среде ученых лесоводов стали
слышны голоса, обосновывающие целесообразность,
экономическую и экологическую необходимость формирования в части
наших лесов, как основного, именно этого качества. Среди них
наиболее активным сторонником этого направления стал
выдающийся русский лесовод А.В. Побединский (1988, 1989).
В последнее десятилетие на рубеже XXI века обострившиеся
экологические проблемы, связанные с глобальным потеплением
климата, нарушениями углеродного, водного, энергетического
баланса экосистем планеты, вырубкой лесов, деградацией почв
и т.д., вынуждают человечество обращать все большее внимание
на необходимость изменения приоритетов потребительского
природопользования на приоритеты учета, сохранения и
восстановления средообразующих функций природных, в том числе
лесных, комплексов.
При этом признается необходимость добиться такого
компромисса, при котором достигается оптимальный баланс между
потребляемым человеком природным ресурсом и сохранением
необходимого для человека и природных комплексов уровня
экологической оптимальности.
Для практического осуществления таких целей научное
осознание этих проблем должно быть подкреплено адекватной
государственной политикой.
Именно сейчас идут поиски наиболее приемлемых моделей
оптимального сосуществования человека и природы, которое
трактуется как устойчивое развитие регионов. Во многих
странах приняты проекты модельных лесов, отвечающие именно
этим задачам. Однако следует признать, что это лишь фрагменты
тех региональных, а в дальнейшем и глобальных стратегий, ко-
-11-
В. Г. Сторожен ко
торые должны составить общепланетарную стратегию жизни на
Земле.
Именно сейчас необходимы и актуальны исследования,
связанные с целым рядом проблем, обеспечивающих это взаимное
оптимальное сосуществование природы и человека,
рациональное природопользование и т.д. В числе наиболее актуальных
проблем в этом контексте следует определить проблему
оптимизации структур лесных экосистем, обладающих свойствами
устойчивости во времени, свойствами саморегуляции и
самовосстановления. Только эти качества лесных сообществ способны
обеспечить долговременное существование лесов и сохранность
лесной среды в оптимальном для нормального функционирования
природы качественном и количественном состоянии. Насущным
является изучение целого ряда проблем, составляющих
целостную разработку, включающую теоретическое обоснование
понятия устойчивости на уровне концептуальных подходов к
содержанию самого понятия устойчивого леса, описание структур
устойчивых лесных сообществ, определение способЬв и методов
формирования устойчивых лесов. Применительно к лесным
сообществам многообразие трактовок понятия устойчивости
показывает незавершенность теоретических исследований в этом
направлении. Существуют расхождения в определении критериев
устойчивого леса, и тем более не сформированы принципы
разделения лесов по устойчивости.
При этом, говоря об устойчивости лесного сообщества,
следует помнить и о сохранении этого качества у других структур
лесных экосистем — почвенного комплекса, зооценоза — поскольку
параметры всех этих структур тесно связаны друг с другом.
В настоящем издании мы постарались систематизировать
накопившиеся за длительный период работ исследования,
проведенные в лесах различного происхождения, состава и структур
для объяснения в первую очередь для себя, а в дальнейшем для
заинтересованного читателя, содержания и фактическое
наполнения понятий «устойчивость» и «устойчивые лесные
сообщества».
-12-
2. КРАТКАЯ МЕТОДИКА ИССЛЕДОВАНИЙ
Методические решения при планировании и проведении
работ определялись целевыми задачами и особенностями
исследований. В число основных задач входило несколько позиций.
1. Определить основные теоретические положения
устойчивости лесных сообществ, обозначить смысловые и
терминологические приоритеты в проводимых исследованиях.
2. Изучить структуры фитоценозов и грибных дереворазру-
шающих комплексов устойчивых лесных сообществ на Русской
равнине.
3. Изучить закономерности коадаптационной динамики
функционирования фитоценозов и грибных дереворазрушающих
комплексов в структурах биогеоценозов — возрастных, породных,
горизонтальных, валежных.
4. Оценить в количественном и качественном выражении
параметры ослабления древостоев, закономерности и динамику
формирования древесного отпада как одну из важнейших
структур лесных биогеоценозов в коренных устойчивых лесах.
5. Определить закономерности формирования комплексов
биотрофных дереворазрушающих грибов в структурах лесных
биогеоценозов.
6. На основе полученных закономерностей сформулировать
комплекс критериев, определяющих устойчивые лесные
сообщества.
7. Разработать шкалу отнесения лесов к градациям
устойчивости.
Исследования проводились в разновозрастных, не
затронутых антропогенным воздействием, в таежной зоне
преимущественно девственных лесных массивах коренных формаций всех
зон растительности — от зоны лесостепи до северной подзоны
тайги. Основными районами сбора информации в коренных
лесах по зонам растительности служили: в зоне лесостепи — древо-
-13-
В. Г. Стороженко
стой с преобладанием дуба в Теллермановском лесничестве
Института лесоведения РАН в Воронежской области; в зоне
лиственных лесов — коренные широколиственные леса из липы,
ясеня, клена, дуба, вяза Тульских засек в Тульской и Калужской
областях; в зоне смешанных лесов и на границе смешанных
лесов и подзоны южной тайги — еловые, сосновые, и смешанные с
лиственными породами леса Московской и Тверской областей и
девственные разновозрастные леса Центрально-лесного
биосферного государственного заповедника (ЦЛБГЗ); в подзоне южной
тайги — девственные древостой резервата Кологривский лес; в
подзоне средней тайги — девственные ельники резервата
«Вепсский лес» и сосняки резервата «Ащозерский» в Ленинградской
области, девственные же сосняки и ельники национального
парка «Водлозерский» в Карелии (южная часть) и Печоро-Илычско-
го заповедника (Коми); в подзоне северной тайги — ельники
Кандалакшского лесхоза Мурманской обл., ельники и сосняки
национального парка «Паанаярви» (Карелия) и Архангельской
области, ельники Усть-Цилемского лесхоза и национального
парка «Югыд-Ва» Коми.
Таким образом, создана сеть объектов, охватывающая
основные коренные формации лесов по зонам растительности. В
каждом пункте наблюдений закладывались серии постоянных и
временных пробных площадей, на которых проводился цикл
исследований, включающий в себя следующий состав работ: перечет
деревьев по категориям состояния с фиксацией всех фаутов
деревьев по шкале, изложенной в «Санитарных правилах в лесах
Российской Федерации» (1992); нумерация деревьев и
картирование их расположения на площади выделенного участка
(пробной площади), величины пробных площадей варьировали:
постоянных — от 0,2 до 0,6 га; временных — от 0,01 до 0,25 га; лесо-
водственно-таксационное описание древостоя; бурение всех
деревьев возрастным буром Пресслера, начиная с диаметра 6 см и
выше, определение диаметров и возрастов деревьев по кернам;
определение средних высот деревьев каждого яруса древостоя по
валежным стволам; описание подроста разного качества и
подсчет его количества на пробной площади; определение наличия и
типа гнилей (коррозионные или деструктивные) по кернам
бурения; определение видов биотрофных дереворазрушающих грибов
возбудителей гнилей, если это было возможно в полевых услови-
-14-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ях, или отбор образцов гнилей для последующего выращивания
чистых культур этих грибов и их идентификации по
определителям чистых культур (Nobles, 1948, 1965; Stalpers, 1978) и
собственным, пока не опубликованным данным; картирование и
описание древесного отпада с разделением его по стадиям
разложения с применением разработанной нами методики (смотри
раздел, посвященный древесному отпаду); определение видов и
встречаемости дереворазрушающих грибов комплекса ксилотро-
фов на валеже. На верхних складах лесосек проводилось
бурение пораженных дереворазрушающими грибами биотрофного
комплекса деревьев и определялись протяженность и
расположение гнилей в стволах для определения объемов гнилей деревьев и
древостоев. Проведенные исследования позволили описать
структуры фитоценозов, динамику их развития, положение в
сукцессионном ряду, видовой состав, встречаемость и структуру
комплексов биотрофов на живых деревьях и ксилотрофов на
древесном отпаде, определить динамику накопления и разложения
валежа и объемы гнилевого поражения, составить таблицы
древесного отпада коренных девственных разновозрастных лесов
тайги.
Более частные методики различных исследований изложены
в разделах работы.
Сопоставление и оценка совместного функционирования
субъектов фитоценоза и микоценоза позволили описать ряд
закономерностей в генезисе коренных лесов, обосновать
целесообразность выделения в рамках общего учения о лесной био-
геоценологии самостоятельного научного направления — мико-
ценологии; определить критерии устойчивых лесных сообществ
и рассмотреть возможность практического применения этих
исследований для разделения лесов по градациям этого качества.
-15-
3. СОДЕРЖАНИЕ ПОНЯТИЙ «УСТОЙЧИВОСТЬ»
И «УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА»
Общечеловеческой, планетарной парадигмой конца
прошлого — начала текущего века и, вероятно, будущих времен можно
признать стремление человечества привести в некий баланс его
разнообразные потребности и ресурсные и восстановительные
возможности окружающей его среды и в целом биосферы Земли.
Этот баланс ограничен прежде всего второй составляющей —
восстановительной способностью биосферы и ее продуцирующей
биомассу автотрофной компонентой. Эволюционно в течение
длительного исторического периода формировавшееся
природное сообщество автотрофных и гетеротрофных организмов, в
которое входил и человек, в определенный период своего развития
стало испытывать с его стороны все большее давление. В
последние два века это давление в результате бурно развивающегося
прогресса возросло настолько, что почти по всей территории
планеты восстановительные способности биосферы не могут
компенсировать потребности человека в ее ресурсах. Деятельность
человека рапространяется на все сферы Земли — от верхних слоев
земли до атмосферы — и выходит в открытый космос. Причем
интенсивность и масштабы этого воздействия непрерывно
возрастают. К наиболее разрушительным воздействиям следует
отнести хозяйственную деятельность человека по вырубке лесов,
изменению состава и свойств почв, гидрологических природных
комплексов, техногенные нагрузки развивающейся
промышленности и транспорта, загрязнение природной среды бытовыми и
промышленными отходами, периодически возникающие в
разных регионах планеты вооруженные конфликты и т.д. Все эти (и
многие другие) воздействия приводят в конечном итоге к общей
токсикации биосферы, экологическим катастрофам, изменению
газового баланса и структуры атмосферы, изменению состава и
структуры растительности и животного мира, нарушению
эволюционно формировавшихся связей и функций компонентов
биосферы. В результате человечество, кроме житейских проблем,
-16-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
обеспечения себя пищей, одеждой, топливом, техническим
оснащением, транспортом и развлечениями, приобрело и глобальную
проблему выживания на собственной планете. Поиски решения
этой проблемы становятся в последние десятилетия для многих
государств, содружеств и объединений государств,
международных организаций разных рангов и направлений одной из
основных задач внутренней и внешней политики. Начиная с
сороковых годов прошлого века, когда была принята первая
Международная конвенция по охране природы и сохранению фауны
западного полушария (Вашингтон, 12 октября 1940 г.) (Страхов и
др., 2001), и вплоть до настоящего времени, многочисленными
международными соглашениями были продекларированы
неотложные необходимости немедленно приступить к реализации
парадигмы устойчивого развития территорий, где потребности
человека ставились бы в подчинение к восстановительным
возможностям природы. Важнейшими из них можно признать
Конференцию ООН по окружающей среде и развитию в
Рио-де-Жанейро (1992), Страсбургскую (1990), Хельсинскую (1993),
Лиссабонскую (1998) конференции министров лесного хозяйства,
Монреальский процесс (1993-1998), Программу действий по лесам
стран «восмерки» (Эдинбург, 1998) и др.
Особое внимание в этих документах и соглашениях уделяется
лесной составляющей биосферы как основной продуцирующей
наземную биомассу структуре и в наибольшей степени
подвергающейся уничтожению и трансформации.
Во многих цивилизованных странах с устоявшейся развитой
экономикой, как мы уже упоминали выше, приняты к действию
и реализованы в практике проекты создания модельных лесов,
задачей которых явилось практическое осуществление идеи
устойчивого развития территорий, пока в небольших масштабах.
Термин «устойчивое развитие» стал одним из самых
распространенных не только при обсуждении вопросов, связанных с
экологическими проблемами, но постоянно употребляется в самых
различных отраслях и масштабах деятельности человека, а
иногда и смыслах. Концепция устойчивого развития впервые была
сформулирована комиссией по проблемам развития и
окружающей природной среде при ООН под председательством премьер-
министра Норвегии Гру Харлем Брункланд и представлена в
решениях Конференции ООН по окружающей среде и развитию
-17-
В. Г. Стороженко
в Рио-де-Жанейро (1992 г.). Её главное содержание
декларировалось как «сочетание экономического роста, связанного с
потреблением природных ресурсов и сохранением природных
экосистем» (Страхов и др., 2001). Более подробные сведения о
глобальных процессах развития идей взаимного сосуществования
цивилизаций и природной среды можно извлечь из книги
процитированных выше авторов.
В этой глобальной парадигме одно из ведущих положений
занимает лес, лесные экосистемы, как важнейший компонент,
обеспечивающий газовый баланс атмосферы, водный и
органический баланс огромных территорий суши, как источник
промышленных и продовольственных ресурсов. Если рассматривать
устойчивое развитие территорий как оптимальное
сосуществование экономического развития и природного, в данном случае
лесного, комплекса, то возникает несколько сценариев этого
сосуществования. Они определяются степенью развития каждой из
составляющих. Территория может успешно устойчиво
развиваться, когда лесные экосистемы по лесистости, составу пород,
структуре древостоев будут удовлетворять экономический рост,
но в то же время не соответствовать коренным природным
параметрам по тем же позициям. Однако в этом случае лесные
экосистемы, отвечая требованиям устойчивого развития территорий в
понимании человека, не будут отвечать устойчивому развитию
биосферы в её «понимании» и не будут выполнять тот
оптимальный объем функций, который сформировал коренное состояние
биосферы и атмосферы. Может существовать некий баланс
между в разной степени измененными природными и в разной
степени развитыми экономическими комплексами. В этом случае
оптимальность структур и функций обеих составляющих
определяется, конечно, человеком и зависит от его осознания
необходимости ограничивать потребности экономического роста, или
оптимизировать этот рост, в зависимости от возможностей
природной среды. С другой стороны оптимальное развитие биосферы
возможно только при полном отсутствии воздействия на неё
человека, что, конечно, невозможно. Исходя из этого необходимый
баланс все равно будет оптимальным только для баланса, но не
для природной среды.
Поэтому, декларируя устойчивое развитие, как важнейшую
парадигму современной цивилизации, необходимо в основные
-18-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
приоритеты ставить все же именно оптимальное
функционирование биосферы, обеспечивающей жизнь на Земле. Развитие
экономики должно быть подчинено этому приоритету, если
человечество хочет выжить и процветать в поколениях. При этом тезис
об «оптимальном функционировании природной среды» может
корректироваться человеком.
Даже этих соображений достаточно, чтобы обосновать
безусловную необходимость сохранения и изучения коренных, не
затронутых антропогенными воздействиями лесных экосистем, как
эталонов природы. Но перед тем как перейти к
непосредственному описанию экспериментальной части работы мы полагаем
необходимым обратить внимание читателя на довольно обширное
поле теоретических поисков, посвященных проблеме
устойчивости, объему и содержанию этого понятия.
-19-
4. ТЕОРЕТИЧЕСКИЕ ПОЛОЖЕНИЯ УСТОЙЧИВОСТИ
ЛЕСНЫХ СООБЩЕСТВ
Не касаясь истоков возникновения и содержания самого
понятия устойчивости, определяющего различные явления и
состояния материи в механике, физике, строительном деле и т.д.,
которые, можно предположить, восходят к глубокой древности,
перейдем непосредственно к обсуждению понятия устойчивости,
применяемого в лесной науке и практике и связанных с этим
понятием терминов.
Прежде всего следует сказать, что в биологии и, в частности,
лесоведении оживленные обсуждения на тему об устойчивости
развернулись с середины 20-го столетия, когда биологические
системы стали пытаться описывать языком кибернетики. По
замечанию Ю. Одума (1986): «Экосистема —■ это кибернетический
организм, так как управляющие функции сосредоточены внутри
ее и дифференцированы». Именно тогда возникла необходимость
определиться с дефиницией этого понятия. В то же время
значительно раньше, в начале двадцатого века, работами Ф. Клемент-
са (Clements, 1916, 1932) было положено начало теории
климакса растительных сообществ, которая рассматривает понятие
устойчивости и объясняет условия, определяющие это качество.
Впоследствии учение климакса расширялось и детализировалось
как самим Клементсом, так и другими авторами. Наиболее
авторитетные разработки в содержании этого понятия принадлежат
В.Н. Сукачеву (1972), Г. Селлеку (Selleck, 1960), Р. Уиттекеру
(Whitteker 1974, 1975). Подробнее с историей развития учения о
климаксе можно ознакомиться в работах В.Д. Александровой
(1964), Б.М. Миркина, Г.С. Розенберга (1978) и др. Более
обстоятельно понятие климакса мы рассматриваем в разделе 5.
Вообще же несмотря на то, что понятие устойчивости
применительно к лесным сообществам оформилось довольно давно,
еще в XIX веке, теоретические исследования, связанные с его
объяснением, находились в глубоком отрыве от практики
лесного дела в России.
-20-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Можно предположить, что понятие устойчивости, как
характеристики лесного сообщества, стало востребовано и начало
употребляться со второй половины девятнадцатого века.
Причиной этому явились зафиксированные немецкими лесоводами во
второй половине XIX века, значительные по площади развалы в
массивах еловых искусственных лесов в Южной Баварии,
созданных в период с середины XVIII по начало XIX века. Именно
тогда обозначилась проблема устойчивости лесов и
необходимость определения характеристик древостоев, обладающих этим
качеством. И неоспоримой заслугой германских лесоводов, и в
частности К. Гайера, явилось целевое формирование именно
этого качества у искусственных, неустойчивых, простых по лесо-
водственным характеристикам лесов путем изменения их
структур на более сложные с помощью группово-выборочных рубок.
С этим понятием связаны работы А. Рудзкого в 70-80-х годах
XIX века, который, сравнивая преимущества сплошных и
выборочных рубок говорил: «Действие крайних явлений в
разновозрастном лесу умеряется, и получаются отношения более
сложные и потому более способные выдержать разрушительный
напор одного какого-то явления» (1881).
Позднее Г.Ф. Морозов (1928, 1970), характеризуя одновоз-
растные и разновозрастные леса, замечал, что «тем сообщество
лесное будет совершеннее, чем оно в большей степени использует
всю географическую обстановку..., чем больше точек
соприкосновения между членами одного сообщества, чем многообразнее
их взаимные отношения, тем устойчивее такое сообщество в
биологическом отношении, тем более обеспечено возобновление
такого леса и тем легче он залечивает раны, причиняемые
человеком или разными стихийными бедствиями, как, например,
пожарами или нападением насекомых..., чем больше
биосоциальных кругов входит в состав сообщества, взаимно влияя друг на
друга, тем лесное сообщество будет более устойчиво».
Основными качествами устойчивого леса Г.Ф. Морозов,
таким образом, определял соответствие фитоценоза условиям его
произрастания, сложное строение леса, включающее все
многообразие присущих экотопу растений, непременное присутствие
достаточного естественного возобновления, наличие всех ценоти-
ческих структур, составляющих лесное сообщество, в том числе
и полночленных грибных сообществ.
-21-
В Г Стороженко
Как видим, на заре российского лесоводства вообще не
давали определения понятию устойчивости, а характеризовали само
лесное сообщество, обладающее этим качеством. Но как термин
он вполне адекватно современным понятиям характеризовал
наиболее совершенные, сложные по структурным параметрам
леса.
Позднее, с развитием лесоводства, понятие устойчивости все
больше формализовывалось и из «качества»
трансформировалось в «способность», что все больше обособляло это понятие от
его материнского корня, то есть от самого лесного сообщества.
Уже некоторое время спустя В.Н. Сукачев (1972) давал
определение устойчивости как «способности» растительного
сообщества «сохранять свой состав и строй в течение продолжительного
времени. Эта устойчивость обусловлена способностью
сообщества к самовозобновлению». Сравнивая трактовки этого
термина, приведенные Г.Ф. Морозовым и В.Н. Сукачевым, нетрудно
заметить существенные различия в определении понятия
устойчивости. Г.Ф. Морозовым приводится описание строения
сообщества, состав которого определяет качество — устойчивость.
В.Н. Сукачевым отмечаются некоторые качества сообщества, из
которых складывется общее свойство — устойчивость, но при
этом не объясняется, каким должен быть этот состав и строй.
В то же время, рассматривая понятие климакса и климаксовых
растительных сообществ, В.Н. Сукачев (1975, т. 3, стр. 255)
приводит 12 позиций, присущих, по его мнению, «выработанным»,
климаксовым сообществам. Уже к этому времени в какой-то
мере можно говорить об обособлении этих двух понятий.
Очевидно, что понятие климакса в экологии и лесоведении,
в частности, окончательно сформировалось, утвердилось и было
интернационализировано, а понятие устойчивости осталось
предметом обсуждений и споров исследователей самых
различных направлений и дисциплин — лесоводов, экологов, биологов
других направлений, кибернетиков, математиков, физиков
и т.д.
Все большее расхождение в смысловом содержании этих
терминов поддерживалось как раз исследованиями
кибернетического направления, формализующими понятия.
Осознавая невозможность, по крайней мере на современном
этапе развития кибернетики, описать математическим языком
-22-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
все многообразие связей в природных, в том числе лесных, даже
самых простых по структуре, сообществах, исследователи этого
направления пытались (и пытаются) описать понятие
устойчивости, используя отдельные свойства или характеристики
сообщества, формализуя тем самым все понятие. Так, А.Н. Горбань и
В.А. Охонин (1986) отмечают, что «переход от качественного
биологического языка к понятиям точных определений
неоднозначен и часто труден. Смысл биологического термина дробится,
возникает набор альтернативных определений, поэтому
выбор адекватного математического аналога понятию
«устойчивость»... до сих пор не принят окончательно». С этим трудно не
согласиться.
Можно привести многочисленные примеры такого
фрагментарного подхода к понятию устойчивости, сопровождающиеся
сложнейшими математическими описаниями, которые однако
кончаются пока выводами, приведенными выше.
Вслед за исследователями этого направления многие
лесоводы и специалисты лесного хозяйства также, не вдаваясь глубоко
в сущность понятия, определяют его элементарно понятными
определениями, такими как «способность биогеоценозов
выдерживать изменения, вызванные извне, или восстанавливаться
после них» (Рожков, Козак, 1989). Более математики, чем
биологи А.И. Горбань и В.А. Охонин (1986) различают
«внутреннюю устойчивость» как способность системы к малым
изменениям численности разновидностей, слагающих биогеоценоз, и
«внешнюю устойчивость» как способность сохранять состав при
малом количестве внедряющихся в биогеоценоз особей.
Устойчивость по Ляпунову — это способность системы
сохранять число видов при её возмущении...
Устойчивость по Холлингу — это сохранение взаимосвязей
при возмущении состояния системы...
Устойчивость по Лагранжу — это сохранение всех видов при
отсутствии равновесного состояния системы...
Устойчивость по Флейшману — это сохранение структуры и
функционирования системы с помощью активного подавления
вредных факторов.
Устойчивость по Ю.М. Свирежеву и Д.О. Логофету (1978) —
это неизменяемость во времени, сохранение числа видов,
постоянство численности популяций, составляющих сообщество.
-23-
В Г Стороженко
А. Д. Арманд (1989) «устойчивой считает систему, которая
любым способом сохраняет инвариант в течение времени,
значительного относительно времени смены инвариант». При этом
«инвариантом может быть состояние системы, или структура,
или способ функционирования, или траектория саморазвития».
Ю. Одум (1986) различал «резистентную» и «упругую»
устойчивость.
Более полную и более приближенную к описанию самого
объекта трактовку понятия устойчивости (стабильности, гомео-
стазиса) приводит В.А. Быков (1983) — «особенность,
заключающаяся в хорошо развитой сложной структуре (большое
количество биогоризонтов), значительной интегрированности ценопо-
пуляций в консорции, высокой степени замкнутости системы,
выровненном отношении вводимой и теряемой энергии,
константном отношении биомассы к мортмассе, большой
информативности».
Ю.П. Демаков (2000), подробно обсуждая проблему
устойчивости, приводит свою дефиницию этого понятия — «это свойство
высокоорганизованных систем поддержания в определенных
рамках значений основных параметров своего состояния в
неустойчивой среде, достигаемое путем эффективного гашения
внешнего возмущающего воздействия во внутренних цепях за счет
различных адаптации и наличия обратных связей между всеми
элементами, выработанное в процессе длительной эволюции
живой материи и направленное на успешное ее продолжение».
И здесь же добавляет, что это понятие «практически
тождественно понятию их жизнеспособности». Обращает на себя внимание
конец этого определения. Выходит, что «это свойство»
вырабатывается системой в процессе её длительного эволюционного
развития, то есть в процессе формирования последних стадий
сукцессии коренных разновозрастных сообществ.
М.Г. Романовский (2002) предлагает «судить об
устойчивости по реакции экосистемы на определенное однотипное
воздействие. Критериями устойчивости могут служить: время,
необходимое для восстановление систем (подсистем) определенного
ранга, скорость наблюдаемых изменений, вероятность
разрушения систем, время их существования (долговечность)».
Как видим, даже довольно близкие к настоящему времени
исследования расссматривают понятие как «способность» или
-24-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
«свойство». Далее автор приводит 14 позиций,
характеризующих необходимые для сохранения устойчивости лесных
сообществ свойства. Однако и в этих описаниях фигурируют понятия
«меры», «способность», «условие» и т.д., но не приводятся
характеристики собственно устойчивого лесного сообщества. Тем
не менее, выстраивая в той же монографии систему
экологической типологии, автор, не акцентируя внимание на этом
термине, приводит некоторые параметры устойчивых коренных
лесов. Правильными и ценными в числе этих позиций нам
представляется отнесение автором к антропоцентричному понятие
«болезни» в естественном, девственном лесу. Это положение
подробно рассматривалось нами ранее (Стороженко и др., 1992) и
обсуждается в настоящей монографии.
Можно привести и совсем недавние публикации, в которых
делается попытка разобраться с термином «устойчивость».
А.А. Рожков (2003), повторяя наши выводы более чем
десятилетней давности (Стороженко и др., 1992), разделяет понятия
«устойчивости» и «состояния» леса, которые в современной
теории и практике зачастую трактуются как синонимы. И это,
безусловно, верно. Однако, пытаясь обозначить критерии
устойчивости лесо», автор, во-первых, не совсем определился с
понятиями «девственных», «коренных» и «естественных» лесов,
различия между которыми необходимо четко представлять,
обсуждая понятие «устойчивость». Во-вторых, одним из основных
критериев устойчивости он определяет возраст древостоев. На
наш взгляд, принципиально неверным является вывод о том, что
«молодняки и жердняки — это возрастной период с
неопределенной устойчивостью», если не привязывать эти древостой к их
происхождению. Тем более, делить жизненный период
естественно возникшего и развивающегося лесного сообщества от
молодняка до спелого или перестойного древостоя на периоды
устойчивости и неустойчивости, по нашему убеждению, неверно
с точки зрения эволюции формирования сообщества. Качество
устойчивости формируется постоянно и постепенно от
возникновения древостоя и до полной его «выработанности». Говорить о
каких-то этапах и тем более устойчивости лесных культур в
течение их жизненного цикла принципиально некорректно.
Лесные культуры изначально являются неустойчивой структурой и
будут таковой всю свою жизнь до периода, пока не появятся при-
-25-
В Г Стороженко
знаки коренного возобновления и последующего формирования
коренного биогеоценоза по пути сукцессионного развития.
Можно констатировать, что в толковании понятия
«устойчивость» существует множество трактовок и это с очевидностью
показывает незавершенность теоретических исследований в этом
направлении.
Таким образом, оценивая описанные выше и другие
характеристики устойчивости, приведенные различными авторами,
можно выделить несколько позиций, объединяющих эти
описания: способность системы возвращаться в исходное состояние
после возмущающих воздействий, сохранение числа видов,
сохранение взаимосвязей, сохранение структуры и
функционирования, неизменяемость во времени....
На наш взгляд, любая из приведенных позиций может
оцениваться критически.
Способностью возвращаться в исходное состояние после
возмущающих воздействий обладает практически люфое лесное
сообщество при условии сохранения коренного экотопа, геоценоза
(условий влагообеспеченности, климатических условий,
структуры подстилающих и покровных пород). Проходя через ряд
сукцессионных преобразований, сообщество через определенный
исторический период может вернуться в исходное состояние, но
шанс этот очень мал. В ходе сукцессии изменятся условия
коренного экотопа и сообщество будет иметь уже другие
характеристики даже в фазе климакса.
Сохранение числа видов при любых возмущающих
воздействиях невозможно. Возвращение к тому же числу видов после
воздействий по сути своей тот же путь сукцессионных смен на
пути к наиболее сбалансированному состоянию лесной системы,
то есть к климаксу. При этом, как правило, не рассматривается,
от какого состояния или сукцессионного этапа идет отсчет числа
видов.
Сохранение взаимосвязей в лесном сообществе невозможно
при возмущающих воздействиях. Возвращение к тем же связям,
как и в предыдущем случае,— путь сукцессионных смен даже от
нулевого состояния (например, после сплошной вырубки) до
фазы климакса, опять же при сохранении основных
характеристик геоценоза.
-26-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Сохранение структуры и функционирования — весьма
неопределенная формулировка, если не учитывать временной
фактор и начальное состояние сообщества. Напрашивается та же
схема вопросов, что и выше: после воздействий и структура и
функционирование не остаются в сохранности, а возвращение
нарушенных структуры и функционирования к тому же
состоянию в биологических системах очень проблематично. Скорее
всего и в этом, и в предыдущих случаях полного возвращения не
произойдет никогда. Вероятнее всего это будет приближенное к
исходному состояние (но не обязательно в худшем качестве)
опять же как один из этапов сукцессионного развития.
Неизменяемость во времени и в структурном и в
функциональном плане несостоятельна. Любое лесное сообщество может
меняться в сукцессионном процессе в значительных пределах,
при этом может меняться и структура, и связи. Важно
определиться с масштабами этих колебаний и соизмерять их с
понятием устойчивости.
В этом контексте можно привести интерпретацию понятия
устойчивости Лагранжа, которую принимают некоторые
исследователи за наиболее правильную — «динамическая система
стремится не к абсолютной стабилизации своего состояния, а к
удержанию его в определенных пределах при непрерывном
колебательном изменении параметров вокруг определенных точек
равновесия, т. е. траектории изменения параметров ее состояния
ограничены в фазовом пространстве как сверху, так и снизу»
(по Ю.П. Демакову, 2000).
На первый взгляд, все логично и такая трактовка может
удовлетворить многих. Мы тоже бы удовлетворились этой
формулировкой, если бы она объясняла какой должна быть
динамическая система в фазе «верха» и «низа».
Внимательно рассматривая все эти теоретические
рассуждения невольно приходишь к мнению, что их авторы говорят об
одном и том же, тщательно избегая конкретного определения
устойчивого лесного сообщества, обособляя и формализуя это
понятие. Ведь употребление терминов способность сохранять, или
возвращаться, или не изменяться по сути дела никак не
определяет само сообщество, его структуру, поэтому зачастую и
возникают противоречия между определением устойчивости и в
приводимых примерах устойчивых лесов.
-27-
В Г. Стороженко
Более приближены к определению свойств и структур
устойчивого лесного сообщества Ю.И. Дробышев и др. (2003), которые
перечисляют ряд характеристик таких лесов. По их мнению,
однако следует применять термин стабильность, «понимаемый
как сумма устойчивостей сообщества к тем или иным факторам
среды». Всякое свойство лесного сообщества, определяющее его
стабильность, представляет один из видов устойчивости. Здесь
имеет место нарушение причинно-следственных понятий. Далее,
тем не менее, авторы перечисляют и обосновывают, на наш
взгляд, вполне логично свойства фитоценозов, способствующие
повышению их устойчивости (гетерогенность популяции, само-
возобновляемость, присутствие деревьев всех классов Крафта).
Рассматривая содержания формулировок устойчивости,
можно отметить, что все они связаны со способностью лесного
сообщества включаться в сукцессионный процесс. Таким
образом, все трактовки верны, если их определять как способность
лесного сообщества эволюционизировать по естественной
динамике развития, при которой и осуществимы все приведенные
выше определения.
Применяя термин «устойчивость», следует иметь в виду, что
в русском языке — это абстрактное существительное,
образованное от прилагательного «устойчивый» и как понятие оно
обозначает качество какого-то объекта, в данном случае — леснрго
сообщества. Любое качество, любого объекта определяется его
содержанием, его структурными характеристиками, его
строением. С этих бесспорных позиций все вышеприведенные (и другие)
определения устойчивости объясняют лишь оттенки или
вариации качества, которым обладает лесное сообщество.
Следовательно, крайне важно определиться с тем содержанием, с теми
особенностями его структурных параметров, которые
характеризуют лесное сообщество как устойчивое.
-28-
5. ПОНЯТИЯ «КЛИМАКСОВОГО» И «КОРЕННОГО»
ЛЕСНОГО СООБЩЕСТВА
Возвращаясь к истокам понятия климакса, мы прежде всего
должны отметить то, что все исследователи, начиная с
Фредерика Клементса (1936), рассматривают его как одну их стадий сук-
цессионного прЬцесса.
В дословном переводе понятие климакса (climax) в
определении Ф. Клементса (1949) звучит как «конечный этап смен
растительности, продолжающийся в определенном ареале до тех пор,
пока климат остается неизменным. Смена растительности,
ведущая к конечному этапу, является приспособлением к
изменяющимся условиям, а сам климакс соответствует условию
относительной стабильности». Кроме того, он различает еще 8
вариантов климакса — disclimax, eoclimax, panclimax, postclimax,
potential climax, preclimax, proclimax, subclimax. Все они
определяют некоторые фазы развития или виды сообществ,
развивающихся в разных условиях, в том числе климатических. Здесь
необходимо также отметить, что некоторые исследователи, весьма
поверхностно знакомые с историей возникновения и развития
этого термина, склонны утверждать, что он имеет много
трактовок, поэтому не существует определенного понятия климакса, а
значит вполне допустимы вольности в его употреблении. Это
мнение ошибочно, дезинформационно и некомпетентно. На
самом деле понятие климакса одно. Оно определено выше. Другие,
приведенные Ф. Клементсом термины, связанные с климаксом,
описывают другие состояния или строения сообществ или
систем. Именно с понятием, приведенным Ф. Клементсом,
дискутировал В.Н. Сукачев (1972), не соглашаясь с определением
климакса, как стабильной системы. Однако в дословном переводе
определения, приведенного Клементсом, мы не находим прямого
указания на то, что климакс — это застывшее состояние
сообщества. Упоминание о связи смен растительности сообщества с
изменением климата и относительной стабильности говорит ско-
-29-
В Г Стороженко
рее о возможности изменения, чем о возможности
долговременной стабильности.
В.Н. Сукачев (1972, том 1, стр. 364-367) приводит довольно
подробный анализ этого понятия и особенно тезиса о «конечной
стадии сукцессионного процесса». Завершая этот анализ, он
резюмирует, что «учение о климаксовой растительности могло
возникнуть лишь при недооценке того большого влияния, которое
оказывают растительный и животный миры, а также и
микроорганизмы на другие компоненты биогеоценоза. А так как это
влияние не может никогда прекратиться..., то биогеоценотический
процесс... никогда не останавливается. Он все время приводит к
смене (сукцессиям) одних биогеоценозов другими, что
исключает возможность существования настоящего климаксового
биогеоценоза. Можно лишь говорить о том, что биогеоценотический
процесс с течением времени замедляется, а растительный покров
и вместе с ним и животный мир могут приобретать некоторую
относительную устойчивость, но и только». Таким образом,
В.Н. Сукачев, отрицая возможность достижения лесным
сообществом состояния длительной стабильности, тем не менее
утверждал неизбежность сукцессионных смен, объясняя её
постоянными динамическими процессами, меняющими структурные и
функциональные характеристики биогеоценозов (Сукачев, 1972,
т. 1, стр. 357-379). Почему же тогда не может быть
правомерным допущение о возможности достижения сообществом фазы
климакса на определенный срок, как одной из фаз его сукцесси-
онной динамики?
Сближая позиции В.Н. Сукачева и Ф. Клементса, можно
говорить о возможности достижения сообществом стадии
климакса как одной из фаз его сукцесиионного развития, в которой, тем
не менее, сообщество не может находиться долгое время.. При
этом структурные и функциональные параметры сообщества,
состав консортов и консортивных связей должны отвечать
условиям полной сбалансированности, потоки вещества и энергии —
оставаться относительно постоянными определенное время.
В нашем понимании Ф. Клементе говорил о конечном этапе
смен растительности именно в теоретическом понимании,
поскольку странно было бы думать, что здравомыслящий человек
может настаивать на долговременной неизменности
геоклиматической обстановки. Здесь же мы берем на себя смелость не-
-30-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
сколько усомниться в правильности тезиса В.Н. Сукачева о
возможности некоторого замедления биогеоценотического
процесса. На наш взгляд, можно говорить о сокращении амплитуд
колебаний биогеоценотического процесса вокруг (или около) того
состояния, которое мы определяем как климаксовое, при
возможности относительной стабильности климатических
процессов длительное время. Но этого не может быть в природе, а
значит можно усомниться в правильности тезиса о замедлении
биогеоценотического процесса. Интенсивность этого процесса,
вероятно, должна определяться физиологическими процессами
обмена вещества и энергии в лесных сообществах, которые в
определенном экотопе протекают с определенной скоростью. Даже
при приближении к стадии климакса или при нахождении в
нем, на наш взгляд, не совсем правильно говорить о замедлении
биогеоценотических процессов. Вернее, по нашему мнению,
говорить о некоем поле флуктуации, которого лесное сообщество
достигает в процессе сукцессии при сохранении интенсивности и
скорости физиологических процессов и в котором оно может
существовать долгое время, находясь в близких к климаксовым
стадиях сукцессии или периодически проходя эти стадии, даже
при значительных колебаниях климата.
С другой стороны, и В.Н. Сукачев, и другие исследователи
признают и утверждают положение о постоянном эволюционном
стремлении лесных сообществ к некоему состоянию баланса.
Ведь в работах В.Н. Сукачева, посвященных динамике лесных
сообществ, постоянно упоминается термин «выработанные
лесные сообщества», который по сути дела и обозначает состояние
наибольшей сбалансированности всех его структур, то есть
состояние климакса. Другое дело, что достижение этого состояния в
полном виде практически невозможно и во многих случаях
может рассматриваться в идеалистическом понимании. Близкие по
совокупности ряда характеристик к состоянию полной вырабо-
танности биогеоценозы, которые мы определяем как климаксо-
вые, можно встретить, как мы уже упоминали выше, довольно
часто. Может быть, относя какое-то сообщество к климаксовому,
мы иногда делаем некоторую натяжку, рассматривая в качестве
основного критерия такого отнесения строение фитоценоза и не
учитывая при этом другие консорты и консортивные связи
сообщества.
-31-
В Г. Стороженко
Принимая позицию В.Н. Сукачева, с одной стороны, о
постоянном движении всех структур биогеоценоза к состоянию
полной выработанности и в связи с этим, с другой стороны,
невозможности достижения состояния некоей стабильности, даже на
короткое время, как безусловную и в то же время признавая
правильным выдвинутый выше тезис о постоянном стремлении
лесного сообщества к состоянию сбалансированности, можно
характеризовать климакс в основных двух параметрах —
временном и структурном. Таким образом, состояние климакса может
быть определено следующим образом.
Состояние климакса — это такое динамическое состояние
лесного сообщества, при котором оно достигает конечных
этапов сукцессии, характеризующихся оптимальным для экотопа
структурным и функциональным содержанием, составляющих
его ценозов, при сохранении баланса вещества и энергии в
течение неопределенного времени, когда амплитуда колебаний
параметров его структур сокращается до минимальных значений
и находится в пределах определенных границ {поле
флуктуации), которые в свою очередь находятся в поле постоянной
эволюционной геоклиматической динамики территорий.
Конечно, редакция этого определения может меняться, но
содержание его нам представляется отвечающим современным
представлениям об одном из важнейших понятий лесной биогео-
ценологии.
При этом немедленно возникает вопрос о том, какими
параметрами можно определять эти границы. Об этом мы будем
говорить ниже, когда будем рассматривать параметры климаксовых
лесных сообществ.
Если принять такую трактовку, то легко примирить позицию
непрерывной динамики В.Н. Сукачева и позицию
последователей Ф. Клементса о конечной стадии сукцессионных процессов.
На наш взгляд, понятие климакса вполне имеет право на
существование и, более того, должно существовать, и в том
понимании, в котором оно обозначает идеальное (или даже
идеалистическое) состояние вполне, по выражению В.Н. Сукачева,
«выработанного» лесного сообщества, и в понимании «флуктуацион-
ного поля». Об этом же упоминал Е.М. Лавренко, когда говорил,
что «в нашей повседневной работе следует использовать
введенные Ф.Е. Клементсом понятия о сукцессионной серии и климак-
-32-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
се, не придавая, конечно, последнему абсолютного значения»
(Лавренко, 1959).
Климатический фактор среди экзогенных абиотических
факторов, влияющих периодически или спонтанно на изменение
структур биогеоценозов,— один из наиболее значительных и
постоянно воздействующих. Именно он в эволюционном развитии
лесов определяет размах отклонений сообщества от конечного
этапа сукцессионных смен, то есть от климакса.
Наши долговременные и обширные наблюдения за
изменениями структур фитоценозов таежных, естественно
развивающихся, не затронутых антропогенной деятельностью, лесов,
показывают, что в достаточно обширных по площади массивах таких
лесов, особенно еловых, присутствуют биогеоценозы самых
различных структурных показателей, среди которых немало древо-
стоев, имеющих близкие к климаксовым или климаксовые
характеристики Чем севернее по зонам растительности, чем более
замедленно идут обменные процессы в биогеоценозах, тем менее
заметно влияние эндогенных факторов на фитоценозы, тем
меньше амплитуды отклонений структур биогеоценозов от климаксо-
вого состояния при воздействии абиотических факторов, тем они
реже и тем чаще можно встретить на этих территориях участки
климаксовых и близких к ним лесных сообществ. Но это не
значит, что биогеоценотические процессы в таких лесах
замедляются. Скорость протекания этих процессов сохраняется
неизменной для соответствующих формаций, типологических разностей
и зон растительности, но сокращается амплитуда колебаний
вокруг фазы климакса и биогеоценозы чаще попадают в эту фазу
или в климаксовое поле флуктуации.
Среди коренных лесов с большим основанием это
утверждение относится к зональным еловым формациям, в значительно
меньшей степени — к азональным сосновым, подверженным на
большей своей площади воздействию пожаров, меняющих
структуру фитоценозов и увеличивающих амплитуды колебаний от
конечных этапов сукцессионных смен до максимальных
отметок. Тем не менее и в сосняках мы довольно часто находили
биогеоценозы по строению фитоценозов, относящихся к
климаксовым фазам динамики.
Утверждая это, мы должны внимательно и подробно
рассмотреть и обосновать характеристики, определяющие климаксовое
-33-
В. Г Стороженко
сообщество, или определить границы, характеризующие кли-
максовое поле флуктуации. Это мы пытаемся сделать в
следующем разделе.
В работах, касающихся изучения структур лесов, нередко
наряду с терминами «устойчивость», «климакс» употребляются
термины «коренной» лес, «коренной тип» леса, «старовозраст-
ный лес».
Употребляя термин «коренной лес», «коренной тип леса»,
необходимо, на наш взгляд, несколько конкретизировать эти
понятия, так как в литературе можно встретить отличные друг от
друга трактовки. Так, М.Г. Романовский трактует коренной лес
и коренной тип леса как равновесное состояние лесной
экосистемы, не возмущенной внешними воздействиями, к которому она
стремится в ходе естественного развития. При этом климаксо-
вый лес рассматривается как предельная точка эволюции
коренного леса. Соглашаясь с последним, мы вынуждены отметить,
что если лесная экосистема достигает равновесного* состояния, к
которому она безусловно стремится, то это состояние, имея в
виду вышеприведенные суждения, и есть климакс. Отсюда
следует, что автор идентифицирует понятия коренного и климаксо-
вого леса, что не всегда верно. Отметим здесь же, что А.И.
Ильинский (1921) почти параллельно с Ф. Клементсом (1916),
рассматривая смены растительности, определяет заключительную
стадию таких смен как «коренную ассоциацию».
Климаксовый лес, понятно, всегда должен быть отнесен к
коренному, так как в течение долгого эволюционного периода
развития на коренных геологических породах фитоценоз и
геоценоз на определенной площади достигли равновесного
состояния (оговоримся, однако, что это состояние может быть очень
кратковременным или вовсе не достижимым в течение очень
длительного периода времени) или попали в климаксовое поле
флуктуации.
Коренной лес не всегда может быть климаксовым. По
трактовке ГОСТа «Лесоводство. Термины и определения» (1998)
«коренной тип леса — тип леса, объединяющий леса,
характеризующиеся естественно сложившимся составом лесной
растительности, соответствующей относительно стабильным лесораститель-
ным условиям определенного типа». И на наш взгляд, это вполне
обоснованная трактовка понятия. Этот состав растительности по
-34-
УСТОЙЧИВЫЕ -ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
своей структуре в разные периоды сукцессионных смен или на
разных этапах развития биогеоценоза может быть различным и
зачастую далек от характеристик климаксового состояния. Тем
не менее такое лесное сообщество относится к коренному. Так,
например, ельники демутационых или дигрессивных фаз
динамики, в сукцессионных рядах довольно далеко находящиеся от
фазы климакса, но произрастающие на коренных для ели
почвах, безусловно, являются лесами коренными.
Зачастую возникают некоторые разночтения в
соотношении понятий «коренной», «девственный», «разновозрастный» и
«старовозрастный» лес. Все эти понятия иногда
отождествляются, что приводит к непониманию и подмене смысла в трактовке
конкретных лесов.
Понятие «девственный лес» относится к лесному сообществу,
развивающемуся без вмешательства антропогенных факторов
воздействия, естественным эволюционным путем, на какой бы
стадии сукцессии оно ни находилось, какими бы структурными
параметрами оно не характеризовалось — дигрессия, демутация,
условно-одновозрастное, относительно-разновозрастное,
абсолютно-разновозрастное и т.д. Не корректно привязывать
понятие «девственный» лес к понятию абсолютно-разновозрастный,
как это часто делается. Единственный критерий такого
сообщества — это исторически естественное эволюционное развитие
при участии только геоклиматических воздействий.
Понятно, что девственный лес всегда является коренным.
Но в то же время коренной лес не всегда может быть
девственным, так как может быть в некоторой степени изменен, например,
антропогенным или техногенным воздействием. В то же время
девственный лес, как сказано выше, может иметь различную
структуру — от условно-одновозрастной до
абсолютно-разновозрастной, в зависимости от того, на какой фазе сукцессионной
смены он находится после воздействия на него различных
факторов (но не антропогенных). Отсюда следует, что и коренной лес
также может иметь различную возрастную структуру — от
условно-одновозрастной до абсолютно-разновозрастной. Интересно
отметить, что некоторые исследователи склонны в некоторых
случаях идентифицировать понятия девственный и климаксо-
вый лес (Дыренков, 1984, стр. 16). Действительно, такая иденти-
-35-
В Г Стороженко
фикация может иметь место при условии полной выработанное-
ти девственного биогеоценоза.
Понятие «старовозрастный лес», на наш взгляд, никак не
связано с приведенными выше терминами, так как относится
или к элементу леса, или к древостою простой одновозрастной
или условно-одновозрастной структуры. Идентифицировать
понятие «старовозрастный лес» с понятиями «коренного»,
«девственного» или «разновозрастного» леса не корректно. Это
понятие не структурного, а временного порядка.
Таким образом, разделяя перечисленные понятия, можно
видеть, что термины «коренной», «девственный», имеют
структурное содержание, термин «старовозрастный» — временное, а
термин «климаксовый» — динамическое.
Соотнося перечисленные выше понятия с понятием
«устойчивый лес» можно утверждать, что климаксовый и устойчивый
лес — понятия идентичные. Понятия «коренной»,
«девственный» и «разновозрастный», как мы увидим ниже, не всегда
могут отвечать условиям устойчивого лесного сообщества. И
напротив — устойчивый лес не всегда должен быть коренным или
абсолютно-разновозрастным, или девственным, или тем более
старовозрастным.
-36-
6. ФИТОЦЕНОТИЧЕСКИЕ КРИТЕРИИ УСТОЙЧИВОГО
ЛЕСНОГО СООБЩЕСТВА
Переходя к описанию характеристик, присущих климаксо-
вому лесному сообществу, необходимо еще раз акцентировать
внимание читателя на том, что, как было отмечено выше, эти же
структурные и функциональные характеристики описывают и
устойчивое лесное сообщество. Другими словами, лесное
сообщество в фазе климакса максимально отвечает условиям
устойчивого леса. Но, забегая вперед, надо упомянуть и о том, что
устойчивым лесное сообщество может быть и не обладая всеми
чертами климаксового леса. То есть понятие устойчивого лесного
сообщества шире понятия климаксового леса. Здесь мы
попытаемся в общих чертах, не рассматривая многообразие консортов
и консортивных связей, глубокие физико-химические аспекты
функционирования биогеоценозов, представить параметры
устойчивых лепных сообществ, анализируя структуры и
параметры биогеоценозов климаксовых фаз динамики.
Итак, в результате вышеприведенных рассуждений
становится ясно, что все исследователи сходятся в убеждении, что в
конце пути эволюционного естественного развития лесные
сообщества, проходя серию сукцессионых смен, всегда
стремятся достигнуть фазы климакса, полной выработанности
(Сукачев, 1972), «коренной ассоциации» (Ильинский, 1921).
Именно эта фаза является вершиной развития лесных сообществ в
определенном экотопе и именно с этой фазой связывают наивысшее
структурное, функциональное совершенство лесного сообщества
и именно в этом состоянии его определяют как наиболее
устойчивое (Морозов, 1970, т. 1, стр. 317, 515; Сукачев, 1975,
стр. 254; Быков, 1983; Дыренков, 1984; Романовский, 2003, Сто-
роженко, 1992 и др.)
Для того чтобы приблизиться к ответу на интересующий нас
вопрос о том, какими характеристиками должно обладать кли-
максовое лесное сообщество, необходимо обозначить, во-первых,
-37-
В Г Стороженко
условия, при которых возможно достижение им состояния
климакса, во-вторых, условия его функционирования.
Обращаясь к цитированным выше авторам, изучавшим эту
проблему, и высказывая наше убеждение, основанное на
масштабных исследованиях лесов различного формационного состава,
происхождения, состояния, структурных параметров,
расположенных в различных зонах растительности, можно определить
по крайней мере два уровня условий, обязательных для
достижения лесным сообществом фазы климакса или максимальной
структурной и функциональной устойчивости.
Первым, коренным, условием, при котором лесное
сообщество может достигнуть состояния климакса или устойчивости,
является соответствие флористического комплекса и в его числе
фитоценоза геоклиматической основе. Этот тезис выдвигал еще
Карл Гайер в семидесятых годах девятнадцатого века, определяя
условия формирования устойчивых лесов в Южной Баварии
(по Г.Ф. Морозову, 1970, т. 1, стр. 515). Далее можно
перечислять практически всех авторов, касавшихся определений кли-
максовых и устойчивых лесов, начиная с Ф. Клементса (1916),
Г.Ф. Морозова (1930), В.Н. Сукачева (1964), Браун-Бланке
(1964), Р. Уиттекера (1975), С.А. Дыренкова (1984), а также
современных авторов — Ю.П. Демакова (2000) и М.Г.
Романовского (2002) и др. Это соответствие изначально может быть описано
и с точки зрения теории континиумов (рефугиумов), которая
рассматривает расселение растительности после последнего
оледенения из определенных геологических областей и по которой в
определенных геоморфологических регионах формировался
определенный флористический комплекс, в том числе и породный
состав древесного яруса (Величко, 1989; Толмачев, 1954).
По геоклиматическим условиям вся территория России, как
известно, разделена на лесорастительные зоны и провинции,
которые выделены согласно естественным границам
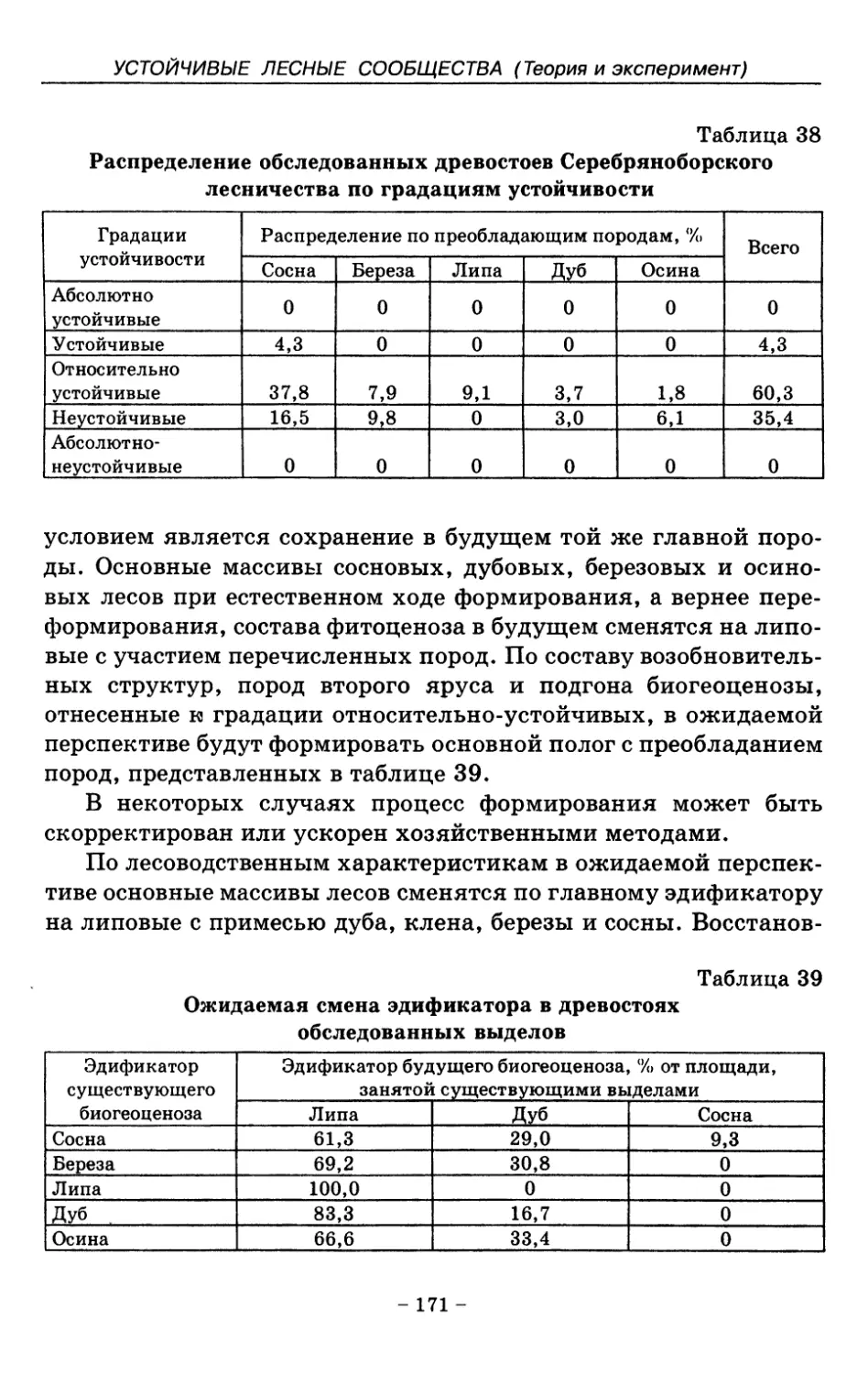
распространения коренных лесных формаций. Наиболее обоснованным и
развернутым нам представляется лесорастительное районирование,
предложенное С.Ф. Курнаевым (1973). Существуют более
мелкие, региональные деления. Несколько упрощенная зонально-
типологическая схема для лесов Русской равнины, в которой
описаны экологические ряды для коренных формаций
европейской части России, разработана А.В. Побединским, А.Я. Орло-
-38-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
вым, доработана и оформлена М.Г. Романовским и приведена в
его монографии (Романовский, 2002). В пределах градаций
геоклиматической основы закреплены коренные лесные формации,
характеризующиеся определенным набором типов леса с
биогеоценозами определенных структур.
Вторым условием достижения лесным сообществом
состояния климакса и нахождения в нем какого-то времени является
неизменность климатических и связанных с ними
гидрологических и почвенных характеристик в определенном экотопе
длительное время. Длительность этого времени может быть описана
скоростью сукцессионных смен или обменных физиологических
и энергетических процессов в биогеоценозе, положением
биогеоценоза в сукцессионном ряду и тем периодом, который
необходим ему для достижения фазы климакса. Этот период может
определяться годами, десятилетиями, а иногда и столетиями.
Во многих случаях в эти эволюционные процессы вносят свои
решающие коррективы разнообразные антропогенные воздействия
(хозяйственные, рекреационные, техногенные), биотические и
абиотические воздействия природного характера.
Понятно, что если первое условие можно рассматривать как
относительно стабильное, мало изменяющееся с течением
времени, то второе условие практически не выполнимо. Колебания
климата, приводящие к разномасштабным изменениям в
структурах биогеоценозов, происходят или спонтанно, или с
определенной периодичностью довольно часто и делают достижение
лесным сообществом фазы климакса весьма проблематичным.
Тем не менее, как мы уже упоминали выше, лесные сообщества,
обладающие, по нашему мнению, характеристиками климаксо-
вых (по крайней*'мере по показателям структур фитоценозов и
микоценозов), в массивах девственных разновозрастных лесов
встречаются довольно часто. Другое дело, в какой полноте эти
характеристики присущи тому или иному сообществу и в какой
области климаксового флуктуационного поля оно находится.
Понятие «область климаксового флуктуационного поля»
вводится нами впервые и может быть определено следующей
дефиницией.
Флуктуационное поле климаксового лесного сообщества —
область вариаций параметров всех его ценотических структур,
консортов и консортивных связей, восстановительных и де-
-39-
В Г Стороженко
структивных процессов баланса потребляемой, сохраняемой
(депонируемой) и выделяемой энергии, при которых величины этих
параметров не выходят за границы показателей наивысших
градаций возрастной динамики древостоев —
абсолютно-разновозрастного древостоя. При этом допускается концентрация
объемов деревьев в одном возрастном поколении не более 40 % от
общего запаса древостоя.
Этот показатель является величиной максимальных
значений объемов деревьев в возрастных поколениях, при которой
биогеоценоз переходит из одной возрастной структуры в другую,
она принята в лесоводстве для характеристик возрастных
структур древостоев (Дыренков, 1984).
Переход сообщества в другую градацию возрастной
структуры означает изменение его положения в сукцессионном ряду и,
следовательно, переход из стадии климакса в стадию демутации
или дигрессии, что влечет за собой изменение и других
параметров ценотических структур биогеоценозов: парцеллярной
структуры продуцентной составляющей, количества и качества
древесного отпада (валежа), породного состава и количества
возобновления, параметров микоценоза и зооценоза.
Условия функционирования лесного сообщества
определяются эволюционной задачей формирования оптимальных структур,
способных в максимальной степени поддерживать баланс
вещества и энергии в лесном сообществе и противостоять воздействию
разрушающих факторов.
Таким образом, если представить, что в определенном экото-
пе соблюдены оба условия и анализируемый биогеоценоз достиг
фазы климакса или попал в область климаксового флуктуацион-
ного поля, то он должен обладать рядом характеристик, которые
с разной степенью подробности обозначены многими авторами.
Нами достаточно подробно они описаны ранее (Стороженко
и др., 1992). Среди этих характеристик наиболее общими для
всех сообществ, по мнению практически всех исследователей
(в том числе и нашему мнению, Стороженко, 1992), можно
определить сложность структурного строения всех ценотических
структур биогеоценоза и его способность к самовоспроизводству,
то есть к поддержанию достаточного для последующих
поколений количества подроста коренных и сопутствующих пород.
Остальные характеристики в определенной мере с ними связаны.
-40-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
6.1. Сложность структурного строения лесного сообщества
Эта характеристика имеет несколько параметров,
описывающих различные показатели ценотических структур
биогеоценоза — фитоценоза, микоценоза, зооценоза. В структуре
фитоценоза, то есть продуционной составляющей биогеоценоза,
рассматриваются показатели возрастной, породной, горизонтальной,
вертикальной и парцеллярной структур. В структуре его редуци-
онной составляющей мы ограничиваемся анализом структур
микоценоза, в котором рассмотрены его количественные и
качественные характеристики — закономерности формирования
видового состава дереворазрушающих грибов, их распространения,
соотношения видов грибов биотрофного комплекса, объемные
показатели поражения и др. Особым и очень важным в
структуре лесного биогеоценоза объектом изучения явился древесный
отпад или валеж, параметры которого также имеют закономерно
изменяющиеся величины в биогеоценозах различного
динамического положения и устойчивости.
6.2. Возрастная структура древостоя лесного сообщества
Характеристика возрастной структуры фитоценоза является
едва ли не самой важной при отнесении биогеоценозов к
определенной категории устойчивости.
Сукцессионные смены, проходящие естественным
эволюционным путем в динамике биогеоценозов при их трансформации от
начальных этапов развития из одновозрастных или чаще услов-
но-одновозрастных древостоев в разновозрастные, описаны
довольно подробно многими авторами (Воропанов, 1946; Баранов,
Григорьев, 1955; Ивашкевич, 1929; Корчагин, 1974; Казимиров,
1971; Комин, Семечкин, 1970, Сукачев, 1972; Стороженко,
1992, 1994; Романовский, 2002 и многие другие). Здесь мы лишь
напомним читателю основные периоды этой трансформации.
Возникновение первого поколения древостоя после
спонтанного исчезновения коренного насаждения (вырубка, пожар,
ветровал, биогенные факторы и т.д.) происходит через разное
количество лет в зависимости от экотопа, лесорастительной зоны и
коренной породы и вида и характеристик фактора воздействия.
-41-
В. Г Стороженко
В среднем, по данным М.Г. Романовского (2002), для ели этот
период равен 2-10 годам, для сосны 1-6 годам. Причем эти сроки
увеличиваются от более сухих типов леса к более влажным и от
южных регионов к северным. Первое поколение уже существует
и обеспечивает воспроизводство коренной породы в тех случаях,
когда в результате ветровалов или буреломов разрушается
древостой, имеющий естественное возобновление коренной породы.
Первое поколение имеет, как правило, условно-одновозрастную
структуру. Заметим, что градации возрастных структур дре-
востоев мы приводим по схеме С.А. Дыренкова (1984),
поскольку считаем её наиболее приближенной к встречающимся в
лесных экосистемах древостоям разного строения. Условно-одно-
возрастные древостой характеризуются 40-летней разбежкой
крайних возрастов деревьев основного яруса древостоя, не
считая подроста.
В условиях коренного произрастания ельников (рамени, су-
рамени), которые при естественном воспроизводстве на
опустевших площадях, как правило, не образуют загущенного
возобновления, на начальных этапах сукцессии формируются смешанные
из ели, березы или осины условно-коренные условно-одновозраст-
ные древостой, имеющие значительную примесь лиственных
пионерных пород. Начало формирования второго поколения
ели очень растянуто, но в основном приходится на период
80-120 лет, когда постепенно выпадает лиственная часть
древостоя. Возрастная структура такого древостоя далеко не полная.
Последующие поколения ели формируются в основном в период,
когда наступает физиологическое старение деревьев первого
поколения ели. Период от возникновения первого
условно-коренного, условно-одновозрастного древостоя до формирования
абсолютно разновозрастного сообщества может составлять
300-400 лет в зависимости от зоны растительности и типа леса.
Далее формирование последующих возрастных поколений идет
по мере вывала или усыхания отдельных деревьев или групп
деревьев предыдущих поколений.
Предельные возраста первого поколения для ели могут
составлять от 300 до 400 лет в зависимости от зоны растительности
и типа леса, но увеличиваются от юга к северу и от восточных
границ Русской равнины к западным. Возрастной ряд древостоя
у ельников, при отсутствии в период формирования разновозраст-
-42-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ной структуры спонтанных факторов разрушения, имеет до
9 возрастных поколений и равномерно убывающую по числу
деревьев в поколениях структуру.
Для сосняков, способных образовывать загущенное
возобновление в условиях боровых экотопов на опустевших площадях,
период интенсивного изреживания и формирования второго
поколения менее растянут, но сконцентрирован, как и для
ельников, в отрезке от 80 до 120 лет. При этом отпад деревьев имеет
чаще диффузный характер, может быть очень интенсивным,
после которого в древостое остается до 400 деревьев. Такое
количество деревьев характерно для разновозрастного девственного
соснового древостоя. Полнота при этом снижается до 0,5-0,6, что
вполне достаточно для появления подроста сосны и
формирования последующих поколений. Разрыв между первым и вторым
поколениями может составлять 60-80 лет. Последующие
поколения возникают по мере постепенного и значительно более
растянутого вывала деревьев первого поколения после 100-120 лет.
Предельные возраста первого поколения у сосны могут
составлять 500 и более лет. В северотаежных сосняках Мурманской
области нам встречались отдельные сосны в возрасте 560 лет.
Возрастной ря/( таких древостоев может включать до 14
возрастных поколений. Предельный возраст деревьев и сосны, и ели на
Русской равнине уменьшается от западных регионов к
восточным и от северных к южным.
Важным признаком, характеризующим степень выработан-
ности, или степень приближения биогеоценоза к состоянию
климакса, является вид распределения деревьев разного возраста в
поколениях. Этот вид может быть показан в числовом
выражении — количество деревьев в поколениях, и в объемном —
объемы деревьев в поколениях. При полной выработанности
древостоя, или климаксе, распределение деревьев в древостое может
быть описано кривыми, представленными на рисунке 1. В
качестве примера возрастных структур ельников, близких к
выработанным сообществам, приводим данные анализа древостоев
постоянных пробных площадей, заложенных в девственных
ельниках Центрально-лесного биосферного заповедника в Тверской
области и резервате Кологривский лес в Костромской области
(южная тайга), в резервате Вепсовский лес (средняя тайга), в
Архангельской области, Карелии и Коми (северная тайга).
-43-
В. Г. Стороженко
X
40
35
30
25 "I
15 Ч
10 -|
5
— 2
1
1 1 1 1 1 1 1 1 1 Г— у
0 40 80 120 160 200 240 280 320 360 и т.д.
X — число деревьев (штук) или их объемы (%);
У — возрастные поколения (0—360 и т.д. лет)
Рис. 1. Теоретически ожидаемое распределение деревьев по их числу (1)
и объемам (2) в возрастных поколениях в климаксовых коренных древостоях
Таблица 1
Соотношение объемов деревьев в возрастных поколениях
еловых древостоев разных фаз динамики по подзонам тайги
Характеристика
древостоя
1 (состав,
тип леса)
Фаза
динамики
Распределение объемов деревьев по возрастным
поколениям, % от общего для древостоя
До
40
Южная тайга — Е
8Е10с1Б;
кисл.-
неморал.
9Е1Б+Ос,
черничник
9Е1Б+Ос,
кисл.-черн.
8Е1Б10с+С,
сфагн,-
чернич.
Кл
Кл
Дг
Дм
0,1
од
0,1
0,1
41-
80
81-
120
121-
160
161-
200
201-
240
241-
280
281-
320
321-
400
ентрально-лесной биосферный заповедник
10,1
1,1
0,7
2,1
31,9
28,1
4,2
65,6
26,4
30,8
30,0
9,8
19,5
15,2
39,2
3,4
21,0
16,2
25,8
11,9
8,5
-
7,1
-
-
-
-
Резерват «Кологривский лес» *
8Е2Б+Пх,
черн.-кисл.-
зеленомошн.
10Е+Лп,Б,
Пх, кисл.-
щитовн.
Кл
Дг
од
од
0,7
1,4
6,3
27,1
16,1
21,3
16,0
26,3
9,3
23,8
51,5
44
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 1 (продолжение)
Характеристика
древостоя
(состав,
тип леса)
8Е1Б1Лп+
Пх, кисл.-
щитовн
9Е1Б+Пх,
сф.-чер.-
морошк
Фаза
динамики
Кл
Кл
Распределение объемов деревьев по возрастным
поколениям, % от общего для древостоя
До
40
0,1
2,7
41-
80
1,0
14,9
81-
120
2,4
20,7
121-
160
20,1
24,0
161-
200
32,5
16,0
201-
240
8,4
4,1
j 241-
280
! 35,4
17,6
281-
320
321-
400
Средняя тайга — резерват «Вепсский лес»
8Е2Б, майн-
кисл.
9Е1Б, майн.-
кисл.
8Е2Б, сф.-
чкрн.
Кл
Дм
Дм
0,1
0,1
0,1
5,0
7,2
1,0
15,4
47,1
20,0
36,5
11,4
51,3
26,0
5,9
16,1
12,2
13,5
6,4
4,8
14,9
5,2
-
-
-
-
-
-
Северная тайга — Архангельская обл., Карелия, Коми |
10Е+Б,С,
брусн.-черн.
9Е1Б, черн.-
морошк
9Е1С+К,б,
долгомошн
! 8Е2Б+С,
черн.-мшист
Кл
Кл
Дм
Кл
Подрост
1990шт/га
Подрост
1370шт/га
Подрост
1975 шт/га
Подрост
834 шт/га
5,2
12,5
47,5
27,0
2,8
25,0
20,0
12,0
12,1
35,9
30,7
32,0
8,1
15,1
1,8
29,0
24,2
9,5
-
-
10,1
2,0
-
-
37,5 1
-
-
-
Представленные в таблице древостой по возрастным
параметрам являются абсолютно-разновозрастными. Большинство из
них относится к климаксовым биогеоценозам.
Чем более неравномерной по объемам деревьев в возрастных
поколениях становится структура древостоя и чем меньшим
числом возрастных поколений в возрастном ряду до предельных
возрастов деревьев представлена структура древостоя, тем меньшей
устойчивостью обладает биогеоценоз.
Практика лесного дела показывает, что наименьшей
устойчивостью обладают абсолютно-одновозрастные древостой,
искусственные насаждения, лесные культуры. Всем известны факты
массового поражения культур ели и сосны возбудителями гниле-
вых болезней, приводящие зачастую к полному их распаду и
нередко к смене биогеоценозов. К числу таких возбудителей отно-
-45-
В Г Стороженко
сятся в первую очередь дереворазрушающие грибы биотрофного
комплекса: корневая губка — Heterobasidion annosum (Ft.)
Bref. и некоторые виды комплекса Armillaria. В большинстве
случаев это A. borealis и A. ostoyae (Звягинцев, 2002; Корхонен,
2004). Здесь сразу необходимо обратить внимание читателя на
то, что дереворазрушающие грибы биотрофного комплекса,
вызывающие поражение и гибель живых деревьев, способные к
массовому распространению, приводящему к расстройству и
распаду древостоев, являются организмами этих же биогеоценозов.
Это не экзогенные факторы воздействия на структуры
биогеоценоза в целом, такие как, например, антропогенные, техногенные
или климатические, к которым применим этот термин, а
эндогенные структуры ценотического уровня, обладающие
свойствами активно влиять на перестройку структур продуцентов с
помощью прямого воздействия на составляющие их организмы или
изменяя между ними консортивные связи. К таким структурам
относятся микоценозы, гетеротрофная редуцентная
составляющая биогеоценозов и, в первую очередь, дереворазрушающие
грибы, одной из функций которых, наряду с деструктивной,
является функция регуляции структур лесного сообщества. Подробнее
функции таких грибов, а также грибов ксилотрофного
комплекса будут рассмотрены ниже в разделе, в котором обсуждаются
понятия «больного» и «здорового» лесного сообщества.
Принимая такую трактовку поведения биотрофных грибов, нельзя
говорить о неустойчивости или устойчивости лесного сообщества к
дереворазрушающим грибам. Можно говорить о неустойчивом
лесном сообществе, рассматривая причиной его неустойчивости,
в числе других факторов, структурное несовершенство его
биогеоценоза. Здесь нельзя забывать о возможном участии в
приобретении такого качества как неустойчивость и базовых
факторов, таких как несоответствие (или неполное соответствие)
породы коренному экотопу.
Даже при искусственном воспроизводстве леса соответствие
породы коренному экотопу при правильной агротехнике посадок
и соблюдении других лесоводственных приемов посадки,
обеспечивающих формирование оптимальных параметров структур
древостоев, отвечающих требованиям экотопа, как правило, не
приводит к массовым эпифитотиям болезней, вызванных
грибами биотрофного комплекса именно потому, что структура таких
древостоев отвечает требованиям устойчивости и биогеоценоз
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
развивается по законам коренных природных сукцессионных
смен с участием комплексов грибов как биотрофной, так и сап-
ротрофной групп. Такой биогеоценоз проходит путь
постепенного усложнения структур и приближения к фазе абсолютной раз-
новозрастности или даже к фазе климакса при отсутствии
воздействия на него эндогенных факторов разрушения. Эти
положения в равной мере относятся и к коренным ельникам, и
коренным соснякам, и к коренным биогеоценозам зоны
смешанных и лиственных лесов.
6.3. Породный состав древостоев
Породный состав древостоя фитоценоза является одним из
обязательных компонентов, определяющих структурное
совершенство лесных биогеоценозов и, соответственно, их
устойчивость. Для более осмысленного понимания современных
представлений о коренных породных ассоциациях лесной зоны мы
сделаем небольшой экскурс в историю возникновения тайги,
основанный на современных палеоботанических и геологических
исследованиях ведущих специалистов в этих областях.
Принимая теорию рефугиумов, возникновения и расселения
растительности (Толмачев 1954, Герасимов, 1990, Величко,
1980, Цвелев, 2005), как основную при естественном
эволюционном формировании флористических комплексов в текущий
геологический период после последнего оледенения, следует
рассматривать современные породные структуры лесов как
производные от этих архаичных коренных породных комплексов,
сформировавшихся, однако, на существующей с тех
исторических периодов геологической основе. После последней фазы
последнего Валдайского оледенения (12-55 тыс. лет назад),
наиболее холодного этапа всего плейстоцена (третий криогенный этап
оледенения) (Герасимов, 1990) и позднее, когда ледниковая
ситуация в Европе перестала существовать (8-9 тыс. лет назад)
(Величко, 1980), на просторах Русской равнины, на
возвышенных местоположениях, автохтонно формировались острова
тайги коренных еловых формаций. Позднее эти острова слились в
единый современный зонально-типологический комплекс
Европейской тайги. В течение длительного исторического периода и
до наших дней породный состав коренных естественно
сформированных флористических комплексов тайги в пределах
макросу-
В Г Стороженко
геоморфологических регионов Русской равнины изменялся
незначительно и только под влиянием долговременных
глобальных колебаний климата. Гораздо более значительные
трансформации породного состава тайги произошли и продолжают
происходить под влиянием антропогенных воздействий. По данным
учета площадей основных лесообразующих пород лесного фонда
России (2005) только в период с 1988 по 2002 годы, то есть всего
за 14 лет, площади хвойных древостоев (сосны, ели,
лиственницы и кедра) уменьшились на 20332,8 тыс. гектаров, а площади
мягколиственных пород (березы и осины) увеличились на
12821,9 тыс. гектаров. При этом площади спелых и перестойных
древостоев хвойных, к которым относятся практически все
коренные девственные леса тайги, уменьшились на 4295,8 тыс. га,
а площади лесов этих же возрастных категорий
мягколиственных увеличились на 6465,6 тыс. га. Эти цифры наглядно
показывают, что даже, казалось бы, в наиболее цивилизованный
период нашей истории происходят огромные по масштабам
трансформации лесных экосистем под влиянием антропогенных
воздействий, когда леса коренных формаций стремительно
сменяются производными древостоями. Что уж говорить о более
ранних периодах государственного опустошения северных
коренных таежных лесов, о чем мы уже упоминали выше.
Таким образом, суммируя все вышесказанное, следует
определить, что коренными породами являются те, которые,
во-первых, по исторической эволюционной памяти соответствуют эко-
топу, в более широкой трактовке — рефугиуму, и в еще более
широкой трактовке — лесорастительной зоне, которая может
включать несколько рефугиумов. Коренные породы образуют
коренной породный состав древостоя, который определяет
коренной тип леса. По ГОСТу «Лесоводство» (1998) «коренной тип
леса — тип леса, объединяющий леса, характеризующиеся
естественно сложившимся составом растительности,
соответствующей относительно стабильным лесорастительными условиями
определенного типа».
Понятно, что в полной мере коренным породным составом
обладают только девственные леса. По тому же ГОСТу
девственный лес — «естественный лес, не испытывающий заметного
хозяйственного и антропогенного воздействия, изменяющийся на
протяжении периода многих поколений лесообразующих
древесных пород только вследствие природных процессов».
-48-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Зависимость коренной породы от коренного экотопа
всеобъемлюща в динамических процессах сукцессионных смен от
первых ступеней формирования до последних климаксовых фаз
динамики коренного фитоценоза. В разных зонах растительности
скорость движения биогеоценоза в сукцессионном процессе
различна, и наиболее наглядно формирование породного состава
коренного древостоя в этом процессе можно проследить в древосто-
ях зоны лиственных лесов, где обменные процессы в
биогеоценозах протекают значительно быстрее и отчетливее, чем в зоне
тайги. Древостой Тульских засек представляют собой насаждения
самого различного происхождения — от очень уже
малочисленных коренных теневых смешанных древостоев до производных с
преобладанием той или иной породы и искусственных чистых по
составу культур. Как в коренных, так и в производных древосто-
ях Тульских засек естественного происхождения с измененным
породным составом и культурах нами в течение 20 лет велись
наблюдения за изменением породного состава насаждений.
В таблице 2 приведены сравнительные характеристики
показателей состава древостоев разного возраста, происхождения и
породного состава. Представленные леса относятся к особо
охраняемым, и руб1Ш в них не ведутся, за исключением выборки
сухостойных деревьев.
Таблица 2
Изменение породного состава древостоев лиственных
пород в древостоях Яснополянского лесничества Тульских засек
Происхождение
древостоев
Естественный,
условно-
одновозрастный
Естественный
разновозрастный
Лесоводственные характеристики древостоев
1973
7Д1Яс1Кл1Лп+Вз,Б;0,9;
снытьевый; 40 лет;
одноярусный
8Д1Яс1Лп; 0,7; снытьевый,
двухъярусный:
1 ярус: 8Д1Яс1Лп, Д,Яс —
более 200 лет; Лп-более 100
лет;
2 ярус — 6Лп2Кл1Яс1Вз —
от 60 до 100 лет.
2005
ЗДЗЛп2Яс2Кл+Вз,Б;0,7;
снытьевый; 60 лет;
одноярусный
5ДЗЛп2Яс+Вз; 08;
снытьевый; трехъярусный:
1 ярус: 5ДЗЛп2Яс, Д,Яс — '
более 200 лет, Лп — более
100 лет;
2 ярус — 6Лп2Кл1Яс1Вз —
от 60 до 100 лет.
-49-
В Г. Стороженко
Табл. 2 (продолжение)
Происхождение
древостоев
Перестойные
культуры
Лесоводственные характеристики древостоев
1973
10Б; 0,5; разнотравный;
одноярусный, подрост —
Лп,Д,Кл,Яс,Вз-густой, 0,9.
2005
10Б; 0,2; разнотравный,
двухъярусный,
1ярус — 10 Б, 110 лет;
2 ярус — Лп,Д,Кл,Яс,Вз—
25-30 лет.
Из данных таблицы наглядно видно, что для условий зоны
лиственных лесов, коренных экотопов, производные древостой
даже в относительно короткий период времени стремительно
изменяют свой породный состав, приближаясь по структурным
параметрам к характеристикам коренного древостоя. Особенно
стоит отметить, что наиболее активно свое место в составе
древостоев этой зоны отвоёвывает липа, восстанавливая быяые
лидирующие позиции. Понятно, что по мере продвижения к северу
сукцессионный цикл, в течение которого происходит смена
производных доминант на коренные породы, увеличивается, и
полное восстановление коренного породного состава древостоя от
начальных этапов может произойти только за 300-400 лет.
Какой же породный состав характерен для коренных
устойчивых древостоев различных зон растительности? Частично в
таблице 1 представлены породные составы коренных
девственных древостоев еловых лесов зоны тайги на конкретных
пробных площадях. В таблице 3 приведены важнейшие формацион-
ные, типологические и лесоводственные параметры коренных
лесов, характерных для наиболее представленных экотопов
зональных почв от регионов лесостепи до северной подзоны тайги.
В таблице использованы данные зонального районирования
С.Ф. Курнаева (1973), экологических рядов А.В. Побединского,
А.Я. Орлова и М.Г. Романовского (2002), а также наши
экспериментальные данные постоянных пробных площадей,
заложенных в коренных древостоях всех зон растительности на Русской
равнине.
-50-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таблица 3
Наиболее распространенные коренные зональные
формации на Русской равнине
Зона,
подзона
растительности
Зона
лесостепи
Зона
лиственных лесов
Зона
смешанных лесов
! Подзона
южной
тайги
Наиболее
распространенные формации
Дубравы
нагорные,
склоновые,
солончаковые,
поименные
Липово-дубово-
ясенево-
кленовые
Сообщества
смешанного
с елью, липой,
дубом, березой,
осиной, кленом,
ольхой,ясенем
состава в разных
соотношениях
в зависимости
от экотопа
Западные и
центральные
регионы —
зональные
еловые
сообщества.
Восточные
регионы —
еловые с
примесью
пихты,
лиственницы, кедра
Наиболее
распространенные
типы леса
Снытьевые, зеленчуковые,
осоковые, пролесниковые,
черемшовые,
папоротниковые, ландышевые, черно-
ольховые, ежевичные, вет-
ляковые, осокорниковые и
переходные между ними
Снытьевые, зеленчуковые,
осоковые, черемшовые,
пролесниковые,
папоротниковые, травяно-
крапивные и переходные
между ними
Кисличные, черничные,
зеленчуковые,
разнотравные, кислично-
снытьевые, чернично-
сфагновые, сфагновые и
переходные между ними
Кисличные, кислично-
папоротниковые,
кислично-черничные,
черничные, зеленомошные,
долгомошные чернично-
сфагновые, сфагновые и
переходные между ними
Характеристики
древостоя
(преобладающие
породы, возрастная
структура)
Дуб, ясень, клен,
липа, вяз + береза,
осина;
разновозрастные
Липа, дуб, ясень,
клен, вяз + береза,
осина,
разновозрастные
Ель, липа, дуб,
береза, осина, клен,
ясень, ольха; от 1а
(суглинистые
дренированные
почвы) до Y бонитета
(оторфованные,
приручьевые),
разновозрастные
Ель, береза, осина,
липа, ольха, сосна;
от 1а (суглинистые
дренированные
почвы) до Y бонитета
(оторфованные,
приручьевые),
разновозрастные
-51-
В. Г. Стороженко
Табл. 3 (продолжение)
Зона,
подзона
растительности
Подзона
средней
тайги
Подзона
северной
тайги
Наиболее
распространенные формации
Западные и
центральные
регионы —
зональные
еловые
сообщества.
Восточные
регионы —
еловые с
примесью пихты,
лиственницы,
кедра.
Западные и
центральные
регионы —
зональные
еловые
сообщества.
Восточные
регионы —
еловые с
примесью пихты,
лиственницы,
кедра.
I Азональные сосновые
сообщества суборевых
и боровых экотопов
Наиболее
распространенные
типы леса
Кисличные, кислично-
папоротниковые,
кислично-черничные,
черничные, зелено-
мошные, долгомошные
чернично-сфагновые,
сфагновые и переходные
между ними.
Черничные, чернично-
брусничные, чернично-
морошковые, чернично-
бруснично-зеленомошные,
чернично-багульниковые,
чернично-сфагновые,
сфагновые, ерниковые и
переходные между ними
Характеристики
древостоя
(преобладающие
породы, возрастная
структура)
Ель (на востоке 1
пихта, лиственница,
кедр), сосна береза,
осина, ольха, от П
(суглинистые
дренированные
почвы) до Y бонитета
(оторфованные,
приручьевые);
разновозрастные
Ель $на востоке 1
пихта, лиственница,
кедр), сосна береза,
осина, ольха,
от Ш (суглинистые
дренированные
почвы) до Ya
бонитета
(оторфованные,
приручьевые);
разновозрастные
В южных областях (лесостепь, лиственные
леса) — сложные сосняки с примесью дуба, липы,
черной ольхи, березы, лещины; чистые сосняки
лишайниковые, лишайниково-мшистые и
переходные между ними.
В таежной зоне — чистые сосняки
лишайниковые, лишайниково-брусничные,
черничные, долгомошные, сфагново-черничные и
сфагново-брусничные, сфагновые, травяно-
сфагновые, кустарничко-сфагновые и переходные
к более богатым раменным условиям с примесью
ели, осины, березы, ольхи черной и др.
От условно-одновозрастных (пирогенных) до
абсолютно-разновозрастных.
-52-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Приведенными выше показателями можно характеризовать
не только девственные климаксовые и близкие к ним по
структурным параметрам сообщества, но и леса условно-коренных
условно-одновозрастных, относительно-разновозрастных
структур, то есть леса, находящиеся на разных этапах сукцессионного
развития, но относящихся к коренным сообществам. Все они при
наличии возобновительных структур, способных обеспечить
воспроизводство коренных пород, относятся к устойчивым
сообществам.
В то же время в лесной практике и лесоведении известны
многочисленные примеры условно-коренных, естественно
сформированных древостоев, как чистых по составу, так и с
примесью различных пород, произрастающих в коренных условиях,
но обреченных на распад и смену доминант. Очень
характерным примером этого утверждения могут служить древостой Се-
ребряноборского лесничества Института лесоведения РАН.
Древостой входят в зону смешанных лесов, значительную их часть
составляют коренные и производные сосняки, липняки,
березняки и смешанные с этими породами древостой, естественного
происхождения, произрастающие на почвах преимущественно
легкого механического состава плакорных местоположений,
террасах р. Москвы, в большинстве своем условно-одновозрастного
строения. Причем чистые сосняки или древостой с
преобладанием сосны составляют более чем 50 % всей площади лесов. В
основном это древостой естественного происхождения, в которых,
как правило, присутствует примесь лиственных пород — береза,
дуб, липа, осина. Большинство выделов имеет очень густой
подлесок из лещины, рябины, черемухи, ольхи, липы, который
препятствует появлению и росту естественного возобновления
сосны. Этому же препятствуют и довольно высокие полноты
древостоев — 0,6-0,8. Обследование древостоев Серебряноборского
лесничества по критериям устойчивости и смены доминант
показало, что основные массивы сосновых, дубовых, березовых и
осиновых лесов по количеству и составу возобновительных
структур, пород второго яруса и подгона в ожидаемой перспективе
будут формировать основной полог с преобладанием пород,
представленных в таблице 4.
-53-
В Г Стороженко
Таблица 4
Ожидаемая смена эдификатора в древостоях
обследованных выделов
Эдификатор
существующего
биогеоценоза
Сосна
Береза
Липа
Дуб
Осина
Эдификатор будущего биогеоценоза, % от существующего
Липа
61,3
69,2
100,0
83,3
66,6
Дуб
29,0
30,8
0
16,7
33,4
Сосна
9,3
0
0
0
0
По лесоводственным характеристикам в ожидаемой
перспективе основные массивы этих лесов сменятся по главному эдифи-
катору на липовые с примесью дуба, клена, березы и сосны.
Восстановление чистых сосняков, как и ельников, не ожидается.
Только 9,3 % существующих сосняков имеют перспективы в
следующем поколении иметь в первом ярусе сосну с примесью
других пород. Причем среди них наиболее представлены сосновые
леса, не имеющие подроста коренных пород. В составе этих лесов
у 22,2 % древостоев зафиксированы очаги усыхания, вызванные
распространением такого биотрофа, как корневая губка
(Heterobasidwn annosum (Fr.) Bref. Все они относятся к
искусственным древостоям. Из данных таблицы можно также
заключить, что все эти древостой, достигнув возраста
физиологического старения войдут в фазу распада и сменятся скорее всего
лиственными породами. Следовательно, сосновые биогеоценозы, как
и большинство выделов других пород (кроме липы), приходится
отнести к неустойчивым сообществам в недалеком будущем со
сменой эдификатора и формации в целом.
Из всего вышеизложенного следует, что одним из
важнейших условий долговременного функционирования устойчивых
лесных сообществ является соответствие состава пород всех
ярусов древостоев, включая подрост, коренному экотопу.
6.4. Горизонтальная структура лесного биогеоценоза
В структуре лесного биогеоценоза одной из основных
характеристик его строения является размещение деревьев по
площади. Взаимное оптимальное расположение растений в простран-
-54-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
стве коренных естественно развивающихся сообществ
вырабатывается в длительном процессе их эволюционной коадаптации.
Характеристики этого расположения входят в состав параметров
устойчивых сообществ наряду с параметрами возрастной и
породной характеристик. В устойчивых коренных древостоях
формируется такое размещение отдельных деревьев или групп
деревьев разного возраста и пород, которое обеспечивает постоянную
смену поколений эдификатора и сопутствующих пород,
постоянную занятость всей площади биогеоценозов, а значит, сложную
структуру сообщества и его пространственную устойчивость.
Характер размещения деревьев по площади коренного
биогеоценоза может определяться многими факторами, среди
которых следует выделить световую и корневую конкуренции,
рельеф местности, особенности гидрологии, размещение валежных
стволов, влияние абиотических факторов (ветровал, снеголом и
др.), биотических воздействий (грибные болезни, энтомовредите-
ли, животные).
Размещение деревьев по площади описывается такими
понятиями как «агрегированность» и «мозаичность».
Понятно, что искусственные древостой, созданные
известными по схемам размещений регулярными посадками, не
нуждаются в каких-либо описаниях.
Термин «мозаичность» употребляется для характеристики
группового размещения деревьев, при котором экземпляры,
обладающие определенными свойствами или признаками,
пространственно обособляются на площади биогеоценоза.
Термин «агрегированность» или, употребляемый как
синоним, термин «контагиозность» применяется в геоботанике и
фитоценологии для обозначения характера размещения объектов
наблюдения по площади учета — в нашем случае это характер
размещения деревьев по площади лесного сообщества.
Структурная пространственная неоднородность лесных
биогеоценозов коренных лесов показана в работах многих авторов.
Предложены принципы выделения структурных элементов их
горизонтального и вертикального строения — синузии,
парцеллы, биогеогоризонты (Бяллович, 1980; Дылис, Уткин,
Успенская, 1964; Морозов, 1928; Сукачев, 1973; Миркин, Розенберг,
1978; Санников, Локосова, 2001 и др.). Эти исследования дают
основу для более глубокого изучения характера и особенностей
-55-
В Г Стороженко
размещения деревьев по площади биогеоценозов как между
всеми особями, составляющими древостой (агрегированность), так
и в связи с их возрастной структурой (мозаичность). При этом
однако следует упомянуть, что исследования предыдущих
авторов в этом направлении напрямую не связывались с понятием
устойчивости древостоев и обозначенные выше два параметра
горизонтальной структуры древостоев не рассматривались как
одни из критериев функционирования устойчивых лесных
сообществ.
Целью наших исследований горизонтальных структур на
постоянных пробных площадях, созданной нами сети
стационарных наблюдений, охватывающей наиболее типичные сообщества
коренных девственных разновозрастных лесов всех зон
растительности, ставилось решение двух задач. Во-первых,
представить степень агрегированности размещения всех деревьев,
слагающих древостой биогеоценоза, на его площади и сравнить эти
величины в коренных девственных устойчивых древостоях всех
подзон тайги для того, чтобы определить правомерность
отнесения (или неотнесения) этого показателя к критериям
устойчивости лесов. Во-вторых, представить характер размещения в тех
же древостоях деревьев, относящихся к определенным
возрастным поколениям, то есть охарактеризовать мозаичность
размещения деревьев разного возраста по площади устойчивых
сообществ или мозаичность горизонтального строения древостоя по
возрастному признаку для тех же целей. Одновременно с этими
исследованиями на тех же постоянных пробных площадях
изучались показатели агрегированности размещения пораженных
деревьев в древостоях, и эти сведения приведены в разделе 9.3.
Агрегированность размещения деревьев по площади
коренных сообществ
Изучение пространственного размещения деревьев по
площади биогеоценоза осуществляется с помощью различных методик.
В большинстве случаев используются в разных вариантах
линейные и площадные параметры, характеризующие размещение
деревьев (Бузыкин и др., 1983; Быков, 1976; Дутина, 1984; Ипа-
тов,1975; Марков, 1985, Маслов, 1985; Плотников, 1968 и др.).
В некоторых работах предлагаются способы изучения мозаично-
-56-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
сти сообществ с помощью математических методов (Грабарий,
Комарова, 1967; Kent, Dress, 1980 и др.)- Нами для анализа
горизонтальных структур коренных устойчивых
разновозрастных древостоев еловых формаций тайги применена методика
Б.А. Быкова (1976), несколько трансформированная
применительно к целям наших исследований. Краткая методика этих
работ заключалась в следующем.
На плане пробной площади с закартированными и
размещенными на нём деревьями в определенном масштабе, проводится
разбивка всей площади на участки 10x10 м. Из центров пунктов
учета замерялись расстояния до ближайших 6-ти деревьев.
Затем вычислялись средние значения этих расстояний, которые
и являлись линейными характеристиками агрегированности
размещения деревьев на участке всей пробной площади. Частота
расположения центров пунктов учета (густота сети разбивки
участка) зависит от плотности размещения деревьев на площади
сообщества (числа стволов на 1 га) и должна обеспечивать охват
всего количества деревьев на участке. Количество деревьев,
равное 6, берущееся для расчета средней величины расстояний для
каждой точки учета, обоснованное в работах Б.А. Быкова (1976),
А.И. Бузыкина и др. (1983); P. Vincent and all (1976),
обеспечивает оптимальную для подобных исследований точность
получаемых результатов, равную 95 % для пуассоновских
распределений, к которым относятся и распределения деревьев по площади
разновозрастных коренных лесов.
Для определения степени агрегированности деревьев нам
предложено использовать показатель изменчивости признаков
по градациям, употребляемым М.Л. Дворецким (1971).
Значения величин вероятности и агрегированности приведены в
таблице 5.
Таблица 5
Шкала степеней агрегированности размещения деревьев,
определяемых по величине коэффициента изменчивости
(по М.Л. Дворецкому)
Изменчивость
признак, С,%
Степень
агрегированности
До 5
Слабая
6-10
Умеренная
11-20
Значительная
21-50
Большая
Более 50
Очень
большая
-57-
В. Г Стороженко
В соответствии с принятой методикой рассчитаны
показатели агрегированности размещения деревьев в коренных
девственных биогеоценозах подзон тайги в формациях еловых и
сосновых древостоев. Подрост при этом не входил в единицы учета.
В таблице 6 представлены основные статистики результатов
измерений, где х — средние значения расстояний; 5 — квадрати-
ческих отклонений; С — изменчивости — характеризующие
агрегированность размещения деревьев для всей их совокупнос-
Таблица 6
Показатели агрегированности размещения деревьев в коренных
разновозрастных древостоях таежной зоны
Лесоводственные
*
характеристики древостоев
Статистические показатели
агрегированости деревьев в
древостоях
X,
см
тч,
см
6,
см
т§,
см
с,
%
тс,
%
Характеристики
агрегированности
1 Граница зоны смешанных лесов и южной подзоны тайги, 1
Центрально-лесной биосферный заповедник
8Е10с1Б; кисл.-немор; 0,7
16; Ар
417
21
113
17
32,3
5,3
Большая
Южная тайга — резерват «Кологривский лес»
8Е1Б1Лп+Пх, кисл.-
щитовн.; 0,8; 1 бон; Ар.
452
22
114
16
25,2
3,8
Большая
Средняя тайга (запад) — резерваты «Вепсский лес», «Ащозерский» |
10Е+Б; черн.-сфагнов ;0,8 Ш
бон.; Ар
10С+Б, брусн.-черн.; 0,6, Ш
бон.;Ар; пир.
461
615
23
36
116
123
18
23
28,3
36,3
4,5
5,3
Большая
Большая
1 Средняя тайга (восток) — Печоро-Илычский заповедник |
[ 10С+Б; брусничник.; Ш бон.;
0,5; Ар; пир.
536
28
122
20
25,4
4,8
Большая
Северная тайга (запад) — Кандалакшский лесхоз, нац. парк «Паанаярви»
8Е1С1Б;чер.-мрш; 1У; 0,5
Ар.
10С+Е; черниник-скальный,
0,5; 1Убон.; Ар
498
542
24
32
118
135
21
26
27,2
36,2
4,1
5,7
Большая
Большая
Северная тайга (восток) — Усть-Цилемский лесхоз, нац. парк «Югыд-Ва»
10Е+С,К; черн.-сфагн.; 1У
бон.; 0,5; Ар
486
25
134
29
34,8
6,3
Большая
Обозначения* структуры древостоев: Ар — абсолютно-разновозрастная;
Ор — относительно-разновозрастная; Уо — условно-одновозрастная; пир.— пи-
рогенного происхождения.
* — состав древостоя, тип леса, полнота, класс бонитета, фаза динамики.
-58-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ти. Для примера нами приведены показатели агрегированности
размещения деревьев в коренных древостоях ели и сосны по
одному для каждой формации для всех подзон тайги, кроме
сосновых древостев южной тайги. Сосновые древостой климаксовой
структуры рассмотрены только в средней и северной тайге.
В подзоне южной тайги абсолютно-разновозрастных сосняков не
обнаружено, да и в условно-одновозрастных пирогенных
сосняках выделять мозаики нет большого смысла, так как структура
таких древостоев формируется в большинстве случаев не
эволюционным путем, а под огневым воздействием.
Анализ представленных в таблице 6 данных позволяет
констатировать, что в разновозрастных коренных лесах тайги
еловых и сосновых формаций эволюционные законы построения
горизонтальных структур фитоценозов, несмотря на различие в их
климатических, геоморфологических, лесоводственных
характеристиках, одинаковы и направлены на формирование
устойчивых сообществ. В структуре горизонтального строения таких
древостоев можно выделить следующие критерии:
1) размещение деревьев по площади устойчивых коренных
разновозрастных древостоев неравномерно, но эта
неравномерность носит системный, постоянный для всего сообщества
характер. Такое размещение можно рассматривать как
равномерно-агрегированное;
2) равномерно-агрегированное размещение деревьев в таких
сообществах обеспечивает полную занятость площади
биогеоценозов коренных девственных сообществ, непрерывность
горизонтальной структуры и тем самым непрерывность существования
сообществ, то есть является одним из важнейших качеств
постоянно функционирующих устойчивых лесов.
Внимательно анализируя полученные выводы и соглашаясь с
их безусловностью, можно, однако, выдвинуть весьма веское
возражение всеобъемлющего значения факта пространственного
равномерного размещения деревьев по площади биогеоценозов
для характеристики и отнесения сообщества к категории
устойчивых. Можно возразить, что и одновозрастные искусственные
древостой после периода дифференциации приобретают
некоторую неравномерность в горизонтальном строении, однако они
относятся нами к категории неустойчивых. И это было бы
справедливое замечание. Ответом на него послужат следующие
соображения. В естественно эволюционно развивающихся сообще-
-59-
В Г. Стороженко
ствах в структуре горизонтального строения наряду с качеством
неравномерной агрегированности формируется и качество
разновозрастное™. Причем разновозрастность формируется по тому
же закону неравномерной агрегированности, который
выражается в мозаичности размещения групп деревьев по площади
биогеоценоза, относящихся к определенным возрастным поколениям.
Мозаичностпь размещения деревьев в горизонтальной
структуре биогеоценозов
Анализ размещения деревьев по площади коренных
девственных сообществ показал, что вся территория сообществ при
равномерно-агрегированном размещении занята далеко не
равномерно деревьями, относящимися к определенным возрастным
поколениям. Даже визуально в разновозрастном лесу легко
отметить неравномерность в размещение деревьев, относящихся к
различным возрастным поколениям. В цикл наших
исследований на постоянных пробных площадях входило сплошное
бурение деревьев у шейки корня, которое дало возможность
определять возраста деревьев, присутствие и некоторые
характеристики гнилей, о чем мы будем говорить ниже. Проводилось также
сплошное картирование деревьев и группировка их в мозаики
возрастных поколений. Измерение возрастов деревьев и
картирование их размещения по возрастному признаку относительно
друг друга позволило представить характер их размещения в
горизонтальной проекции и описать некоторые особенности
строения и динамики возрастной структуры биогеоценозов.
Нанесенные на план деревья, относящиеся к разным возрастным
поколениям, легко обособляются в отдельные мозаики, то есть группы
деревьев, относящихся к определенному возрастному
поколению. М.А. Проскуряков (1982) такие однородные по возрастному
признаку группы деревьев называл «элементарными группами».
Мозаики правомерно относить к элементарной единице
горизонтального строения фитоценоза, так как они имеют достаточно
четко выраженные границы, отделяющие их друг от друга, как
по площадному признаку, так и по качественным
характеристикам. В нашем случае по возрастному признаку.
Мозаики, как структурные единицы горизонтального
строения фитоценоза, имеют два основным параметра измерения —
-60-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
количественный и качественный. Количественный параметр
характеризует их величину и выражается в м2 площади
биогеоценоза. Качественный параметр характеризует степень
однородности мозаик, которая определяется в долях единицы. Абсолютно
однородные мозаики представляют собой группы деревьев,
состоящие из экземпляров одного возрастного поколения и
обозначающиеся соответственно качественным параметром с
индексом 1. Примесь в составе мозаик деревьев других возрастных
поколений снижает индекс однородности, который может
доходить до 0,0. Мозаики, характеризующиеся индексом
однородности менее 0,5 и имеющие в своем составе деревья разного
возраста в близких по числу деревьев соотношениях, не относятся к
определенному возрастному поколению, обособляются как
мозаики со средним возрастом, составляющих их деревьев.
Можно также отметить, что в отдельных случаях выделить
мозаики по возрастным поколения бывает чрезвычайно трудно.
Это может случиться тогда, когда коренной древостой имеет ус-
ловно-одновозрастную структуру и находится в начальных
стадиях сукцессионного процесса. В таких древостоях все деревья
входят, как правило, во временной интервал одного возрастного
поколения, равного 40 годам. Еще реже встречаются
биогеоценозы, в которых деревья, относящиеся к различным возрастным
поколениям равномерно размещены на больших площадях. Как
правило, в девственных коренных древостоях чистые по
возрастному признаку мозаики не бывают слишком большого размера.
В качестве иллюстраций, доказывающих все вышесказанное, мы
приводим серию таблиц, в которых представлены суммарные
площади выделенных мозаик и степени однородности этих
мозаик. Для древостоев каждой подзоны тайги и Центрального
лесного биосферного заповедника (граница северной подзоны
смешанных лесов и южной тайги) приведены по две пробные
площади еловых древостоев. Сосновые древостой различных
динамических структур рассмотрены только в средней и северной тайге.
В подзоне южной тайги разновозрастные сосняки крайне редки,
да и в условно-одновозрастных пирогенных сосняках выделять
мозаики нет большого смысла, так как структура таких
древостоев формируется в большинстве случаев не эволюционным
путем, а под огневым воздействием.
-61-
В. Г. Стороженко
Таблица 7
Горизонтальное строение разновозрастных еловых древостоев
Центрально-лесного биосферного заповедника ( А,Б,В; табл. 7-9)
А) ПП — 1. 8Е2Б; кисличник; 0,8; 1 бон.;
абсолютно-разновозрастный, климаксовый
Измеряемые
величины
Площадь
мозаик, м1
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
Не
выделена
21,0
2
430
8,9
19,5
3
710
14,7
26,4
4
70
31,9
5
6,4
10,1
6
Не
выделена
0,1
Степень однородности
мозаик
1,0-
0,85
2Ш
53,7
0,85-
0,7
34Q
7,0
0,69-
0,5
1Ш
39,3
Менее
0,5 J
Не
выделены
В биогеоценозе не во всех возрастных поколениях можно
обособить мозаики с чистотой более 0,5. Наибольшие площади
мозаик выделяются в четвертом возрастном поколении, и
наибольшие площади занимают чистые мозаики. Из всего этого можно
сделать выводы о том, что: 1) несмотря на значительные объемы
деревьев старших поколений в древостое, обособить мозаики этих
поколений невозможно, и это значит, что деревья старше 200 лет
довольно равномерно рассредоточены по площади биогеоценоза
и попадают в мозаики разных возрастных поколений; 2) в то же
время вся площадь сообщества сложена мозаиками других
возрастных поколений и групп деревьев, не выделяющихся в
мозаики, нет.
Б) ПП — 2; 9Е1Б+Ос; кислично-черничный; 0,8; 1 бон.;
климаксовый до дигрессивного.
Таблица 8
Измеряемые
величины
Площадь
мозаик, м!
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
700
17,1
25,8
2
115Q
28,1
39,2
3
1Ш
44,6
30,0
4
4,2
5
420
10,2
0,7
6
0,1
Степень однородности
мозаик
1,0-
0,85
2270
55,4
0,85-
0,7
0,69-
0,5
1Ш
44,6
Менее
0,5
В биогеоценозе не выделяются два возрастных поколения —
с 80 до 120 лет и поколение подроста до 40 лет, деревья которых
-62-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
рассредоточены в мозаиках других поколений. Выделенные
мозаики по степени однородности попадают в две градации, причем
больше половины площадей занимают чистые по однородности
мозаики.
В) ПП — 3; 8Е1Б10с+С; чернично-сфагновый; 0,8; П бон.;
к лимаксово-демутационный.
Таблица 9
Измеряемые
величины
Площадь
мозаик, м1
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
18Q
4,3
7,1
2
1М
3,6
3,4
3
2140
51,2
11,9
4
57Q
13,6
9,8
5
21,3
65,6
6
250
6,0
2,1
7
0,1
Степень однородности
мозаик 1
1,0-
0,85
15М
37,5
0,85-
0,7
220
5,5
0,69-
0,5
150
3,7
Менее!
0,5
21М
53,3
Сообщество имеет деревья всех поколений и во всех
поколениях, кроме подроста, можно обособить мозаики. Наибольшие
площади приходятся на мозаики третьего возрастного
поколения, однако запасы деревьев наибольшие в пятом возрастном
поколении. Довольно большие площади (более половины)
занимают однородные по составу мозаики.
Горизонтальное строение разновозрастных еловых дре-
востоев резервата «Кологривский лес» — южная тайга (А,Б;
табл. 10-11)
А) ПП — 1. 8Е1Б1Лп+Пх; кислично-щитовниковый; 0,8;
1 бон.; климаксовый.
Таблица 10
Измеряемые
величины
Площадь
мозаик, ы_
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
230
4,6
35,4
2
Нет
8,4
3
МО
7,0
32,5
4
421fi
84,2
20,1
5
нет
2,4
6
210
4,2
1,0
7
нет
0,1
Степень однородности
мозаик
1,0-
0,85
6,0
0,85-
0,7
16,2
0,69-
0,5
150
3,0
Менее
0,5
Д74Ц
74,8
-63-
В Г. Стороженко
На большей части площади, выделенного под пробную
площадь древостоя, мозаики не обособляются, межмозаичное
пространство имеет очень низкий индекс однородности и занято
деревьями разных возрастных поколений со средним возрастом
близким к возрасту четвертого поколения. Сообщество имеет
характеристики климаксового, но в значительной степени
продвинуто в сукцессионном развитии к дигрессивной стадии.
Б) ПП — 2. 8Е2Б+Пх; чернично-кислично-зеленомошный;
0,8; 1 бон.; климаксовый.
Таблица 11
Измеряемые
величины
Площадь
мозаик, мг
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
Д10
6,2
23,8
2
700
14,0
26,3
3
6S0
13,6
21,3
4
Щ5£
61,0
27,1
5
Щ)
5,2
1,4
6
нет
0,1
7
Степень однородности
мозаик
1,0-
0,85
2Ш
42,4
0,85-
0,7
1Ш
37,8
0,69-
0,5
490
9,8
Менее
0,5
500
10,0
Горизонтальное строение разновозрастных еловых древосто-
ев резервата «Вепсский лес», сосняков резервата «Ащозерский»
и Печоро-Илычского заповедника — подзона средней тайги (А-Г;
табл. 12-15).
А) ПП — 1. 10Е+С, Б; майниково-кислично-черничный; 0,8;
П бон.; климаксовый.
Таблица 12
Измеряемые
величины
Площадь
мозаик, м
%
Объемы
деревьев, %
Распределение деревьев по возрастным
поколениям
1
ш
21,8
5,2
2
нет
16,3
3
7М
45,6
36,7
4
42Q
26,2
24,5
5
ш
6,0
12,2
6
нет
4,1
7
нет
1,0
Степень однородности
мозаик
1,0-
0,85
нет
0,85-
0,7
1Ш
67,5
0,69-
0,5
330
20,6
Менее
0,5
ш
11,9
Древостой имеет один выраженный, но с относительно
пологими крыльями возрастной эксцесс. Тем не менее его можно
относить к разновозрастным древостоям, вошедшим во флуктуаци-
-64-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
онное поле климаксового сообщества. Около половины площади
выделенного сообщества составляют деревья от 200 до 240 лет,
вошедшие в одну мозаику с однородностью выше 0,7. Вообще,
можно отметить, что мозаичность на участке выражена довольно
заметно.
Б) ПП — 2; 10Е +Б; чернично-сфагновый; 0,8 Ш бон.;
относительно-разновозрастный .
Таблица 13
Измеряемые
величины
Площадь
мозаик, м!
%
Объемы
деревьев, %
Распределение деревьев по возрастным
поколениям
1
нет
12,4
2
80
4
21,3
3
Ш
1
58,7
4
ш
16
6,9
5
1320
66
0,7
6
ш
13
7
подрост
Степень однородности
мозаик
1,0-
0,85
m
3,5
0,85-
0,7
0,69-
0,5
1Ш
96,5
Менее
0,5
Древостой довольно однородный по строению, выделяются
только четыре мозаики, на остальной площади деревья разного
возраста рассредоточены относительно равномерно, однородных
по возрасту зугЪзаик всего две. Деревья первого поколения хоть и
присутствуют в древостое, но не выделяются в мозаику.
В) ПП — 3. ЮС; брусничник; 0,5; Ш бон.;
абсолютно-разновозрастный.
Таблица 14
Измеряемые
величины
Площадь
мозаик, м_1
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
1
2
3
3
•
4
6
5
£7fi
23
19
6
ш
15
12
7
30
24
8
32
35
Степень однородности
мозаик
1,0-
0,85
950
38
0,85-
0,7
0,69-
0,5
750
30
Менее
0,5
800
32
Древостой по структуре фитоценоза разновозрастный, однако
с большим эксцессом в область демутации. Причиной этому
можно назвать последствия ранее прошедших пожаров,
значительно изредивших в основном деревья первых поколений. На
-65-
В. Г. Стороженко
оставшихся деревьях этих поколений заметны следы давних
пожаров. Молодые поколения влияния пожаров еще не испытали.
Мозаичность выражена относительно заметно, и довольно
большая площадь занята чистыми по однородности мозаиками.
Г) ПП-4. ЮС + Лц, Е; лишайниково-брусничный; 0,6; Ш
бон.; разновозрастный, дигрессивный.
Таблица 15
Измеряемые
величины
Площадь
мозаик, м1
1%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
1
2
2Ш
46,6
23
3
1££
3,9
37
4
22М
45,3
16
5
3
6
1
7
2
8
-
9
1
10
210
16
Степень однородности
мозаик
1,0-
0,85
8,1
0,85-
0,7
2Ш
46,6
0,69-
0,5
2Ш
45,3
<0,5
Древостой на протяжении своей жизни, судя по строению
каллюсов вокруг ран от подгаров, испытал три пожара и
структура его изменена. Тем не менее его дигрессивный характер ясно
выражен и мозаики сложены в основном деревьями старших
поколений. Однако чистые по однородности мозаики занимают
незначительные площади. Довольно большая площадь учета
содержит всего четыре мозаики.
Горизонтальное строение разновозрастных еловых древосто-
ев Кандалакшского лесхоза (Мурманская обл.), Архангельской
области (центр севера Русской равнины) и национального парка
Югыд-Ва (восток Русской равнины, Предуралье) — подзона
северной тайги (А-В; табл. 16-18).
А) ПП-1. 8Е1С1Б; чернично-морошково-багульниковый; 0,7;
Y бон.; абсолютно-разновозрастный, климаксовый
(Кандалакшский лесхоз).
Таблица 16
Измеряемые
величины
Площадь
мозаик, м*'
%
Объемы
деревьев,%
Распределение деревьев по возрастным
поколениям
1
2,0
2
1280
9,5
3
15,1
4
3440
31,3
5
1660
25,0
6
1620
12,5
7
4,6
Степень однородности 1
мозаик 1
1,0-
0,85
М&1
49,4
0,85-
0,7
1420
17,7
0,69-
0,5
32Q
4,0
<0,5
28,9
-66-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Древостой имеет ясно выраженный эксцесс в четвертом
поколении (160-200 лет). Мозаичность размещения деревьев
довольно отчетливо выражена, причем почти половина площади
учтенного древостоя занято чистыми по составу мозаиками.
Б) ПП-2. 10Е + С, Б; чернично-брусничный; 0,6; 1Y бон.;
абсолютно-разновозрастный.
Таблица 17
Измеряемые
величины
Площадь
мозаик, м!
%
Объемы
деревьев, %
Распределение деревьев по возрастным
поколениям
1
Ж
18
1,0
2
8,6
3
6,6
4
12,2
5
1330
67
17,3
6
130
6
7,1
7
40
2
17,3
8
140
7
9,6
9
15,2
10
5,1
Степень
однородности мозаик
1,0-
0,85
360
18
0,85-
0,7
ш
16
0,69
-0,5
4£
2
<0,5
1290
64
Древостой абсолютно-разновозрастный, климаксовый.
Эксцессы количества деревьев в отдельных поколениях не
выражены. Мозаичность по возрастным поколениям также не
выражена, и большая часть площади сообщества относится к мозаикам
малой величцны однородности.
В) ПП-3. 8Е2Б+К,Пх; чернично-зеленомошный; 0,6; 1Y бон.;
абсолютно разновозрастный, климаксовый с некоторой
динамикой в область дигрессии.
Таблица 18
Измеряемые
величины
Площадь
мозаик, м1
%
| Объемы
(деревьев, %
Распределение деревьев по
возрастным поколениям
1
Ж
1,3
37,0
2
62£
39,1
26,7
3
6,2
29,1
4
85£
53,4
5,4
5
1,8
6
Подрост
Степень однородности мозаик
1,0-
0,85
520
32,5
0,85-
0,7
855
53,4
0,69-
0,5
4,7
<0,5
150
9,4 1
Биогеоценоз находится во флуктуационном поле климакса с
некоторым уклоном в область дигрессии и вплотную
приблизился к нему. Мозаичность размещения деревьев по возрастному
признаку выражена довольно отчетливо двумя поколениями —
-67-
В Г Стороженко
вторым и четвертым. Эти же мозаики обладают высшими
баллами однородности.
Для примера приводится рисунок одной из пробных
площадей ЦЛГЗ, на котором в масштабе 1 : 1000 см изображено
размещение мозаик по возрастным поколениям.
Представленные экспериментальные данные об особенностях
структурного строения коренных девственных разновозрастных
древостоев позволяют сформулировать некоторые важные
положения, которые необходимо учитывать при анализе и
характеристиках устойчивых лесных сообществ.
1. Лесные сообщества, развивающиеся естественным путем, в
ходе эволюционных адаптационных процессов формирования
структур фитоценозов вырабатывают такое размещение особей
по площади биогеоценозов, которое может характеризоваться
двумя видами параметров: а) параметрами мозаичного
размещения групп деревьев, относящихся к одному возрастному
поколению, в результате чего вся площадь биогеоценоза разбивается на
мозаики из групп деревьев разных возрастных поколений; б)
параметрами диффузно размещенных деревьев, относящихся к
различным возрастным поколениям, более или менее
равномерно размещенных относительно друг друга по площади
биогеоценоза. В первом случае, как правило, смена поколений проходит
по схеме группового усыхания деревьев, что нередко можно
встретить в разновозрастных лесах.
Во втором случае смена поколений проходит по схеме
диффузного равномерного усыхания отдельных деревьев старшего
возраста. Это не значит, что из других поколений деревья не
могут выпадать в ходе поражения их грибными болезнями, энто-
мовредителями или в результате напряженных конкурентных
отношений.
2. Мозаичное по возрастному признаку размещение деревьев,
как и диффузное, равномерное, обеспечивает такое же по
характеристикам размещения воспроизводство поколений в процессе
сукцессионных смен.
3. Мозаики возрастных поколений могут иметь различную
степень однородности (чистоты), что также обеспечивает
различную по времени смену поколений на площади сообщества.
4. В целом сложная структура горизонтального размещения
деревьев различных возрастных поколений обеспечивает
лесному сообществу постоянную занятость пространства, непрерыв-
-68-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Рис. 2. Размещение мозаик возрастных поколений в древостое пробной
площади 1 Центрально-лесного биосферного заповедника 1..5 — возрастные
поколения; 0,5... 1,0 — однородность мозаик
ность восстановительных процессов и тем самым является одним
из важных факторов устойчивости и одной из основных
характеристик устойчивых лесных сообществ.
6.5. Возобновительные структуры устойчивых
лесных сообществ
Способность к самовозобновлению, самовоспроизводству,
регенерации при стрессовых нарушениях фитоценозов является
одним из двух основополагающих, краеугольных свойств
устойчивых лесных сообществ. Еще одним свойством такого же
уровня является соответствие породного состава фитоценоза экотопу.
При этом возобновительные структуры в древостоях разного
состава и происхождения могут иметь различные характеристики.
Соотношение показателей породного состава древостоя и
возобновления во многом определяют степень устойчивости лесного
сообщества, перспективы его дальнейшего развития и динамики
формирования его структур. Для примера мы рассмотрим
несколько вариантов таких соотношений и в каждом случае
обсудим перспективы развития биогеоценозов. Но прежде всего при-
-69-
В Г. Стороженко
ведем характеристики возобновления в коренных девственных
климаксовых и близких к ним по структурам лесах, как
эталонов эволюционно развивающихся устойчивых лесных
сообществ, в которых структуры возобновления имеют оптимальные
параметры. В таблице 19 представлены характеристики
возобновления в коренных разновозрастных лесах таежной зоны.
Отметим, что в учеты подроста не вошли всходы, число которых
может достигать 10-20 и более тысяч штук на 1 гектар. К
сожалению, в зонах лесостепи, лиственных и смешанных лесов
естественный ход возобновления изменен из-за изменения структур
биогеоценозов и девственных лесных сообществ, по всей
видимости, не осталось. Здесь мы вынуждены ограничиться лишь
отдельными примерами возобновительных структур в древостоях
разного происхождения и структур.
В таблице представлены результаты учетов естественного
возобновления на конкретных постоянных пробных площадях в
коренных лесах абсолютно-разновозрастных структур по зонам
растительности.
Таблица 19
Характеристики возобновления коренных девственных
лесов таёжной зоны Русской равнины
Характеристика
биогеоценоза
(состав,
полнота, бонитет,
тип леса)
Количество возобновления эдификатора
по градациям высот, % от общего
До 0,5
0,6-
1,0
1,1-
1,5
1,6-
2,0
1 2,1-
1 2,5
Более
2,5
Всего подроста
Hal
гектар
Других I
пород* |
Общее
колич.
Граница зоны смешанных лесов и южной тайги j
7Е1Бос1Лп+Кл,
0,8;1, кисл.-
черн.
8Е 1Б10с+Лп,
0,7; 16 , кисл.
8Е1Б1Ос+С,0,7,
Пб.# сф.-черн.
53
79
74
13
7
9
9
6
7
12
3
4
10 1
3
3
3
2
3
6573
8759
5993
132
138
87
6705
8893
6080
1 Южная тайга
8Е2Б+Ш, 0,8, 1
бон ; черн.-кисл.-
зеленомошн
10Е+Лп,Б,Пх;
0,7; 1 бон. кисл.-
|щитовн.
72
68
14
18
7
8
5
3
1
2
1
1
5880
4230
175
205
6055
4435
* примесь хвойных или лиственных пород — березы, осины, кедра, ивы,
лиственницы и др.
-70-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 19 (продолжение)
Характеристика
биогеоценоза
(состав, пролно-
та, бонитет,
тип леса)
8Е1Б1Лп+Пх;
0,7; 1 бон.;
кисл.-щитовн.
Количество возобновления эдификатора
по градациям высот, % от общего
До 0,5
63
0,6-
1,0
17
1,1-
1,5
8
1,6-
2,0
5
1 2,1-
1 2,5
1
Более
2,5
6
Всего подроста
Hal
гектар
1284
Других 1
пород* 1
148
Общее 1
колич.
1432
1 Средняя тайга |
10Е+Б;0,7;Шб.;
черн.-сф.
10Е+Ос-,0,7-,Ш
бон.; майн.-черн.
10С+Б;0,6;Ш
бон.; брусничн.
90
87
50
6
9
19
2
2
6
1
1,5
-
1
0,2
19
-
0,5
6
5300
6745
855
92
105
ПО
5392 1
6850
965
1 Северная тайга
10Е+С; 0,6; 1У
бон.; черн.-мор,-
баг.
8Е2Б+Пх,Кд;
0,6;1У-Убон.;
черн.-зм.
10С+Е;0,6;1У
{бон.; бр.-черн.
53
28
58
22
20
15
14
1
9
11
23
9
17
9
11
-
1320
1975
787
72
60
28
1392
2035
815
Величины естественного возобновления, приведенные в
таблице, вполне наглядно показывают, что в сомкнутых древостоях
тайги по её подзонам с нормальными для коренных девственных
лесов полнотами количество подроста коренных пород имеет
сопоставимые величины и мало зависит от фазы динамики
древостоя. Естественно, речь не идет о самых начальных фазах
сукцессии, возникающих в девственных разновозрастных
сообществах после воздействия факторов разрушения естественного
природного происхождения (биотических и абиотических) —
сплошных ветровалов, буреломов, пожаров и т.д. Во всех таких
случаях возобновление леса на оголенных площадях может идти
несколькими путями и зависит во многом от состава, структуры
и состояния предшествующего подроста и, что также важно,
подлеска. Чаще всего в таких случаях появляется густое
возобновление лиственных пионерных пород, затем можно наблюдать
постепенную смену их коренными породами. В сосновых
древостоях, где нередки низовые пожары, на ход естественного
возобновления, его структуру и затем структуру созданного им древо-
-71-
В Г. Стороженко
стоя заметно влияет огневое воздействие, в результате которого
уничтожается одно, иногда два поколения подроста, и древостой
впоследствии приобретает прерывистую возрастную структуру.
Данные таблицы также показывают, что количество
естественного возобновления по мере его роста стремительного
сокращается. Сообщество как бы «перестраховывается», заранее
«рассчитывая» определенные величины отпада молодого
поколения, создавая значительный запас прочности своего будущего
через большие объемы текущего возобновления. Причем,
интересно отметить, что, начиная с подроста высотой 2 и более метра,
то есть с возраста в древостоях южной тайги около 30 лет
(до 49 лет при высоте 3-3,5 метров), его количество соизмеримо с
количеством деревьев этого древостоя на одном гектаре. Такое
количество подроста как бы достаточно биогеоценозу для
создания последующих поколений древостоя даже при полной его
гибели.
В условиях северной тайги, когда древостой разрежены и их
полноты составляют не более 0,5-0,6 единиц, казалось бы,
создаются условия для появления и развития большего количества
естественного возобновления. Тем не менее его величины
подчинены той же закономерности количественной оптимальности,
что и в условиях более благоприятных для роста зональных
коренных пород.
Понятно, что выше рассмотрены только общие, но типичные
для девственных разновозрастных коренных лесов примеры
естественного хода возобновления. В зонах лиственных лесов и
лесостепи такие исследования не проводились по причине
отсутствия девственных лесов. Однако стремление коренного экотопа
восстановить на своей площади соответствующий его природе
фитоценоз путем формирования естественного возобновления,
состоящего из коренных пород, можно наблюдать повсеместно.
Приведем несколько примеров такой смены.
В течение длительного времени (более 20 лет) по просьбе
администрации музея-усадьбы Л.Н. Толстого «Ясная Поляна»
нам пришлось наблюдать за состоянием и формированием древо-
стоев различного происхождения — мемориальных посадок
березы, естественно возникших после рубок смешанных древосто-
ев разного возраста. Эти наблюдения позволили наглядно про-
-72-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
следить за изменениями состава древостоев при их естественном
развитии, возникновением и формированием естественного
возобновления. В древостоях музея-усадьбы, которые входят своей
значительной частью в Тульские засеки, не проводится никаких
рубок за исключением выборки сухостойных деревьев. Участки
старовозрастной березы (Старая Абрамовская посадка,
Срезанная посадка, Митрофановская посадка), высаженные еще при
жизни писателя, вплоть до 60-х годов освобождались от
естественного возобновления сначала ежегодными прокосами между
рядами берез (схема 3x3 м), затем его вырубкой. В последние
30 лет естественное возобновление формировалось естественным
путем. По причине предельного возраста береза в последние
несколько десятков лет (около 40) интенсивно отпадает, сухостой
выбирается и полнота древостоя снизилась до 0,2-0,3 единиц.
За этот период из подроста сформировался густой подгон,
выходящий во второй ярус, под которым практически нет травяного
покрова. Состав подгона сложен типичными коренными для
условий зоны смешанных лесов породами — липой, кленом,
ясенем, вязом, дубом. Березы в его составе нет. Понятно, что при
окончательном распаде абсолютно неустойчивого мемориального
березового полога древостой трансформируется в коренное
устойчивое сообщество зонального типа.
Необходимо также отметить, что в некоторых случаях даже в
коренных экотопах ход естественного возобновления коренных
пород, соответствующих этому экотопу, его структура и даже
вообще присутствие, может лимитироваться некоторыми
факторами. Общеизвестны примеры, когда густой подлесок из таких
подлесочных пород как лещина, рябина, бузина, липа,
черемуха, малина и др. может препятствовать появлению подроста
коренных пород. Например, в еловых культурах К.Ф. Тюрмера,
созданных на еловых же почвах, то есть в коренном экотопе, в
кв. 109 Порецкого лесничества Бородинского лесхоза густой
подлесок из лещины, экземпляры которой в диаметре достигают
10-12 см, полностью исключил появление возобновления ели,
несмотря на то, что полнота древостоя невелика — до 0,5
единиц. В других кварталах, в которых нами заложены постоянные
пробные площади, естественное возобновление ели составляет
значительные величины (от 1 до 3 тыс./га), вполне обеспечивает
-73-
В Г Стороженко
сохранность коренной породы после естественного распада
древостоя. Сукцессия в таких сообществах пойдет по пути
восстановления коренного древостоя без каких-либо задержек с
начальных этапов развития.
Не менее известны и подробно изучены состав и структура
естественного возобновления в лесах различных зон
растительности, процессы смены березняков и осинников коренными
ельниками. Здесь мы отсылаем читателя к обширной литературе,
через некоторые работы Н.В. Федорчука и др. (1998), В.И.
Алексеева (1978), А.Н.Мартынова (1992), А.Я.Орлова (1983),
А.Ф. Чмыря (1977), А.Д. Волкова и С.А. Дыренкова (1971) и
многих др.
Вообще же следует заметить, что наши исследования не
преследуют цель анализа этих работ, а только ограничиваются
экспериментальным подтверждением тезиса о краеугольной роли
естественного возобновления в формировании структур
устойчивых лесных сообществ.
В лесах всех зон растительности сопутствующими коренной
породе видами, способными входить в основной полог, являются
так называемые «пионерные» породы — береза, осина, реже ива.
Эти породы, как известно, первыми занимают освободившиеся
из-под леса площади и в ходе сукцессии сменяются на коренные
эдификаторы. В зоне тайги эти породы составляют примесь в
коренных древостоях до 2-3 единиц в зависимости от
динамического положения биогеоценоза и зоны растительности, в
разреженных ельниках северной тайги примесь березы в составе
древостоев достигает 4 единиц. Соответственно в составе
естественного возобновления всегда присутствуют и эти породы.
В определенном смысле их также можно причислить к
коренным азональным породам.
Количество, качество и структура естественного
возобновления в коренных девственных лесах могут лимитироваться
несколькими факторами: видовым составом и густотой подлеска,
подгона и присутствием других ярусов под основным пологом;
полнотой самого древостоя; видом и плотностью напочвенного
покрова; характером мозаичного строения фитоценоза; количеством
и качеством (стадии его разложения) древесного отпада;
воздействием зоогенных факторов (погрызы дикими животными), энто-
могенным и микогенным факторами и, наконец, антропогенными
факторами (рекреационными, техногенными, хозяйственными).
-74-
7. ПОКАЗАТЕЛИ СОСТОЯНИЯ УСТОЙЧИВЫХ ЛЕСНЫХ
СООБЩЕСТВ
Естественно развивающиеся лесные сообщества, не
испытывающие никакого антропогенного влияния, разновозрастные
девственные леса имеют вполне определенную динамику
развития, которая обуславливает параметры различных компонентов
их ценозов. Некоторые характеристики этих компонентов
используются для анализа состояния биогеоценозов несмотря на
то, что сам термин «состояние» имеет весьма неопределенное
содержание и может варьироваться в разных случаях с разным
значением. Вообще в лесозащите, где он употребляется наиболее
часто и по прямому значению, нет сколько-нибудь убедительного
терминологического объяснения этому слову, что не мешает тем
не менее его широкому употреблению практически во всех
лесных дисциплинах и особенно широко в лесной фитопатологии,
энтомологии, экологии и т.д.
Пожалуй, наиболее подробные исследования по разработке
критериев, определяющих состояние древостоев, проведены
Е.Г. Мозолевской (1979, 1973, 1984 и др.). Ею предложена
шкала оценки состояния насаждений, основными показателями
которой явились характеристики текущего отпада, общий размер
усыхания древостоя и пораженность болезнями и вредителями
(1984). Соглашаясь в общем с возможностью применения этих
показателей, мы, однако, не можем согласиться с однозначной
их трактовкой для всех древостоев. Принятая автором величина
естественного отпада, от которой ведется отсчет состояния
сообщества, не может быть величиной одинаково объясняющей
состояние древостоев, например, культур и девственного леса.
Параметры этой величины по-разному характеризуют древостой
различного происхождения и динамических характеристик.
Нельзя согласиться с отождествлением понятий «категория
состояния» и «классы биологической устойчивости», то есть
«состояние» и «устойчивость». Эти понятия, по нашему мнению,
качественно различаются, о чем уже шла речь выше. Тем не ме-
-75-
В Г Стороженко
нее, для определенных целей такая шкала вполне может быть
применима в практике и широко используется в лесозащите.
Наряду с этим существует большое количество
всевозможных подходов к определению состояния древостоев и
содержанию термина. Можно говорить о состоянии по пораженности
древостоев грибными возбудителями, энтомовредителями,
абиотическими факторами и т.д. Существует такой термин, как рубки
по состоянию, который используется при назначении
санитарных рубок. Некоторые исследователи применяют интегральные
показатели состояния, в которые входят ряд характеристик
деревьев (Расин и др., 1988). Вполне правомерно применение этого
термина при отнесении лесов к определенным этапам их сукцес-
сионного положения или оценке перспективы их формирования
и развития. В этом случае используются не патологические, а
лесоводственные критерии оценки биогеоценозов. Но
практически всегда при определении этого показателя применяется
шкала категорий состояния деревьев, изложенная в
«Санитарных правилах в лесах России» (1992). Основой её служит
визуальная оценка характеристик деревьев, их крон и стволов,
которые в совокупности дают возможность отнести их к
определённой категории — от здорового до старого сухостоя. Шкала
подвергается критике со стороны некоторых исследователей за
неизбежный субъективизм при оценке характеристик деревьев и
возможную при этом ошибку, такое положение, конечно, имеет
место, хотя степень неточности или величина ошибки зависит от
опытности и квалификации исследователя. В то же время
приходится признать, что пока не существует более точных методов
оценки состояния деревьев, а тем более насаждений, включая
инструментальные методы (измерение биоэлектрического
потенциала, биохимические методы и т.д.). Даже
компьютеризированная международная программа мониторинга состояния лесов
в основе своей имеет те же визуальные оценки характеристик
деревьев.
Мы не разделяем суждений о том, что визуальная оценка не
отражает истинного состояния деревьев. Допуская возможность
ошибки, особенно в древостоях, пораженных скрытыми
сердцевинными гнилями, мы, тем не менее, считаем, что она может
быть использована в очень многих случаях. При этом
необходимо помнить, что оценивается только внешнее состояние деревьев
-76-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
на текущий период времени, которое нельзя пролонгировать на
длительный срок, а иногда даже на недельный, месячный.
По методике «Санитарных правил...» выделяется 6 категорий
состояния, которые мы приводим ниже.
1 — без признаков ослабления — хвоя зеленая блестящая,
крона густая, прирост текущего года нормальный для данной
породы, возраста условий произрастания и времени года;
2 — ослабленные — хвоя часто светлее обычного, крона
слабоажурная, прирост уменьшен не более чем наполовину по
сравнению с нормальным;
3 — сильно ослабленные — хвоя светло-зеленая или
серовато-матовая, крона ажурная, прирост уменьшен более чем
наполовину по сравнению с нормальным;
4 — усыхающие — хвоя серая, желтоватая или
желто-зеленая, крона заметно изрежена, прирост текущего года еще
заметен или отсутствует;
5 — сухостой текущего года (свежий) — хвоя текущего года
серая, желтая или бурая, крона сильно изрежена, мелкие
веточки сохраняются, кора сохранена или осыпалась лишь частично;
6 — сухостой прошлых лет (старый) — хвоя осыпалась или
сохранилась лишь частично, мелкие веточки, как правило,
обломались, кора осыпалась.
Во многих случаях, особенно при детальных тщательных
обследованиях ценных древостоев общепринятая методика
слишком груба и опытному исследователю она недостаточно подробно
описывает отличия в состоянии отдельных деревьев. В таких
случаях мы используем более детальную шкалу, в которую
входит 10 категорий состояния деревьев.
1 — внешне здоровые — без признаков изреживания крон и
поражения грибными болезнями и вредителями;
1,5 — с признаками ослабления — кроны изрежены на
10-15 % и (или) отмечаются явно выраженные фауты стволов
патологического происхождения: раковые образования,
признаки гнилей, механические повреждения, смолотечения и т.д.;
2,0 — слабо ослабленные — кроны изрежены до 25 % за счёт
сокращения объёма хвои (листвы) и усыхания отдельных
побегов или ветвей, сокращение величины побегов текущего
прироста, явные признаки гнилей или развитые раковые или некроз-
ные образования на стволах, механические повреждения с при-
-77-
В Г Стороженко
знаками раневых гнилей, плодовые тела трутовых грибов,
дефолиация от объедания энтомовредителями;
2,5 — ослабленные — кроны изрежены до 40 %.
Перечисленные выше фауты могут присутствовать в более выраженной
форме. Некоторая дихромация хвои. Начало формирования кисточ-
ности побегов у сосны;
3,0 — сильно ослабленные — кроны изрежены до 55-60 %.
Перечисленные выше фауты могут присутствовать в более
выраженной форме. Явная дихромация хвои и формирование кисточ-
ности побегов за счет опадения хвои прошлых лет и
значительного сокращения приростов нескольких последних лет;
3,5 — очень сильно ослабленные — кроны изрежены до
75-80 %. Хвоя бледно-зеленого цвета с серым оттенком. Могут
присутствовать желтая или рыжая хвоя или отдельные ветви.
Выраженная кисточность побегов, усыхание отдельных ветвей
или побегов. Могут появляться попытки поселения стволовых
вредителей, дупла в месте выхода гнилей, плодовцр тела дерево-
разрушающих грибов;
4 — усыхающие — кроны изрежены более чем на 85 %,
остатки хвои бледно-зеленого цвета, часть хвои может быть
рыжего цвета. Попытки или поселения стволовых вредителей, часть
коры может опасть;
5 — свежий сухостой — остатки рыжей хвои; поселения
стволовых вредителей, плодовые тела грибов, дупла с признаками
гнилей, кора может частично опасть;
6 — старый сухостой — стволовые вредители вылетели, кора
частично или полностью опала.
Свежий сухостой может иметь две градации: сухостой
1-го года и сухостой 2-го года в зависимости от присутствия в
нем вредителей определенного вида или генерации.
В сосновых древостоях любого происхождения — от
искусственных до девственных — одним из распространенных фаутов,
определяющих состояние деревьев начиная с возраста 40-50 лет,
является поражение стволов раком-серянкой, образующей
раковые раны в кроне деревьев на разной её высоте или под кроной,
которые приводят к отмиранию части или всей кроны и гибели
или ослаблению деревьев. Нами составлена шкала определения
степени развития рака-серянки по состоянию раковых ран и
угрозе гибели деревьев.
-78-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таблица 20
Шкала определения степени развития рака-серянки
на стволах сосны
Стадия
развития
раковой
раны
1
2
3
4
Признаки отнесения раковой раны
к определенной стадии развития
Рана занимает до 1/3 окружности ствола и
расположена не ниже 1/3 длины кроны
от вершины дерева
Раковая рана занимает 2/3 окружности
ствола и расположена до 1/2 длины кроны
от вершины
Раковая рана занимает более
2/3 окружности ствола и расположена
до 2/3 длины кроны от вершины
Раковая рана занимает более
2/3 окружности ствола и расположена
под кроной
Степень влияния
раны на состояние
дерева
Ослабление дерева
без угрозы его
гибели — 2
категория состояния
Ослабление дерева
с малой угрозой
его гибели — 3
категория состояния
Ослабление дерева и
большая вероятность
его гибели —
3,5-4 категории
состояния
Ослабление дерева
и 100 % вероятность
его гибели —
4-5 категории
состояния 1
Для более детального ознакомления с болезнью и
особенностями ее распространения и биологии отсылаем читателя к
последним работам белорусских исследователей (Федоров, Ярмо-
лович, 2004).
В цикл исследований на постоянных и временных пробных
площадях в лесах всех зон растительности входило определение
состояния древостоев по принятой нами шкале категорий
состояния. В таблице 21 приведены средние для еловых
биогеоценозов подзон тайги величины этих показателей. Для зон
лиственных лесов и лесостепи, где, как мы уже не раз отмечали, нет
сохранившихся девственных лесов и состояние отдельных пород,
кроме эндогенных факторов функционирования сообществ,
определяется во многом другими, часто неизвестными лесной
науке и практике факторами, мы ограничимся лишь некоторыми
примерами состояния пород и древостоев.
-79-
В Г. Стороженко
Таблица 21
Средние значения категорий состояния деревьев и
древостоев для девственных еловых лесов подзон тайги
Подзона
состояния
1 - здоровые
|2
-ослабленные
3 - сильно
ослабленные
4 -
усыхающие
5 - свежий
сухостой
6 - старый
сухостой
[Средний
Средние значения категорий состояния деревьев коренных лесов тайги
по подзонам (%) и их статистика j
Граница зоны
смешанных лесов
и подзоны южной
тайги
X
59,4
25,3
6,8
1,9
0,4
6,2
ГПх
1,6
1,8
0,6
0,3
0,1
1,0
а
4,6
5,8
1,9
1Д
0,17
3,3
1,7
Южная подзона
тайги
X
57,8
29,6
5,8
2,0
0,4
4,4
тх
2,1
2,0
0,5
0,3
0,1
0,9
а
4,8
4,5
1,2
1,1
0,15
2,1
1,65
Средняя подзона
тайги
X
50,0
20,0
11,0
4,0
2,0
13,0
Шх
1,8
1,7
1,0
0,5
0,4
2,4
а
4,9
1,3
1,1
0,13
4,0
2,3
Северная подзона
тайги
X
51,2
25,8
11,0
1,0
1,0
10,0
тч
1,7
1,8
1,2
0,3
0,1
1,8
I
4,5
5,2
1,3
2,5
0,16
3,5
2,2
X — среднее значение признака — категории состояния
деревьев в древостоях; тх — ошибка среднего значения; с —
среднее квадратическое отклонение.
Данные таблицы наглядно показывают, что коренные
девственные разновозрастные биогеоценозы таежной зоны,
развивающиеся по естественной динамике ослабления и отпада
деревьев, имеют весьма близкие характеристики этих показателей.
Понятно, что в стадиях глубокой дигрессии и в менее
благоприятных условиях роста показатели категорий 1 и 2 будут меньше, а
категорий 3, 4, 5 и особенно 6 будут значительно больше, как и
средний балл ослабления. Напротив, в биогеоценозах демутаци-
онных фаз динамики и в оптимальных для коренной породы
условиях роста показатели первых двух категорий будут больше
табличных, а остальных категорий, и особенно 6-ой, меньше.
Показатели состояния климаксовых сосняков практически
аналогичны еловым. Однако такие сообщества крайне редки.
В сосняках других фаз динамики, с нарушенной пожарами воз-
-80-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
растной структурой действуют те же законы соотношений
деревьев разных категорий состояния при наступлении демутации
или дигрессии, что и для ели.
Вообще, следует заметить, что анализ состояния девственных
коренных лесов ранее не проводился и представленные данные
тем более имеют интерес. Из данных анализа состояния древо-
стоев тайги следует еще один важный вывод — в состав
критериев устойчивости разновозрастных коренных лесных сообществ
важной составляющей входит количественное постоянство
процессов ослабления и отпада в структурах древостоев, которое
связано в том числе с определенными величинами поражения их
грибами биотрофного комплекса, о чём будет сказано ниже. Это
постоянство является также одним из показателей
энергетического баланса и баланса вещества в лесных коренных
девственных сообществах.
В зоне лиственных лесов и лесостепи, в смешанных древосто-
ях с измененными хозяйственной деятельностью структурами,
состояние древостоев определяется состоянием отдельных пород,
которое во многих случаях далеко не равнозначно. Например,
исследования состояния древостоев Тульских засек в нескольких
лесхозах (Прй^пский, Щекинский, Тульский, Одоевский) и
старовозрастных древостоев Теллермановского лесничества
показало, что в наиболее ослабленном состоянии находится дуб —
средние баллы состояния этой породы колеблются в пределах 2,8 —
3,5 балла, в перестойных дубравах эта величина еще ниже.
Другие породы в среднем имеют значительно более высокие баллы
состояния: липа в пределах 1,3-1,8; клен — 1,1-1,5; ясень —
1,5-2,0. В Серебряноборском лесничестве Института
лесоведения РАН (зона смешанных лесов) учет состояния деревьев
разных пород в смешанных с разным соотношением пород древосто-
ях показал, что, действительно, средние значения состояний,
вычисленных для древостоев в целом, в смешанных сообществах
неинформативны, так как в большинстве случаев расхождения в
состоянии пород, слагающих древостой, слишком велики,
чтобы величина средней давала объективную информацию. На
рисунке 3 приводятся величины этих показателей в средних
значениях для деревьев основных пород, слагающих смешанные
древостой.
-81-
В. Г. Стороженко
балл
4
3
2
1
0
1
3,5 1
К4
II
1 ^
1 1
1 г
1 1
4
1,9
2,2
1 ■
■ i
11
С Д Лп Б Ос
породы
Рис. 3. Средние показатели состояния основных
лесообразующих пород Серебряноборского лесничества
Института лесоведения РАН
Дуб, как видно из рисунка, находится в очень ослабленном
состоянии. Количество усохших деревьев в некоторых древосто-
ях достигает 30 % и более. Причинами ослабления и усыхания
дуба на всей территории Русской равнины могут быть названы
несколько факторов, среди которых объедание листвы в течение
ряда лет комплексом листоверток, ранневесенние заморозки,
грибные возбудители различных болезней. Все эти причины,
конечно, имеют место быть и, безусловно, связаны с ослаблением
дуба, но, по всей вероятности, есть еще другие причины, более
общего, может быть, глобального характера, которые пока не
определены, не выявлены. В пользу этих доводов говорит
масштабность явления ослабления и усыхания этой породы на большей
части территории Евразии.
Сравнивая две шкалы категорий состояния деревьев, можно
видеть, что исследователь с любым опытом и квалификацией
безошибочно определит лишь три категории: усыхающие,
свежий и старый сухостой. В определении остальных категорий,
при малом опыте таких наблюдений, возможны большие
субъективные ошибки, которые при расчете средних значений
состояния древостоя в целом еще более усугубятся. Исходя из этого при
расчетах состояния биогеоценозов в целом желательно
учитывать дополнительные показатели, которые помогли бы более
объективно оценивать состояние лесного сообщества. Для
разновозрастных, тем более девственных, лесов, в которых, как
показано выше, средние величины категорий состояния деревьев в
82-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
древостоях практически всех зон растительности близки,
такими дополнительными характеристиками могут являться
относительные величины количества деревьев категорий усыхающих,
свежего и старого сухостоя, пораженности древостоев грибами
дереворазрушающего биотрофного комплекса и объемов валежа
последних 5 лет. Эти показатели определяются или
инструментальным путем (бурение деревьев), или учетом древесного
отпада. Если гипотетически допустить существование сообщества
абсолютно ничем не пораженного, состоящего только из
здоровых деревьев, то интегральный показатель состояния (ИПС)
биогеоценоза будет равен 0. Напротив, если теоретически
предположить существование древостоев, состоящих только из
сухостойных деревьев, на 100 % пораженных грибными болезнями,
имеющих 100 % валежа, то величина ИПС будет находиться в
пределах 0 < ИПС < 3. Таким образом, состояние лесного
биогеоценоза может быть оценено по следующей формуле:
ИПС = С +П + В, где
ИПС — интегральный показатель состояния биогеоценоза,
С — относительная величина количества деревьев группы отпада
(усыхающие, свежий и старый сухостой) — от 0 до 1; П — пора-
женность древостоя дереворазрушающими грибами,
выраженная в относительных еденицах числа пораженных деревьев от
общего их количества на площади учета — от 0 до 1; В —
относительная величина объемов валежа не старше 5 лет от запаса
древостоя на 1 га — от 0 до 1. Для абсолютно-разновозрастных
коренных древостоев, близких к климаксовым структурам, ИПС
находится в пределах от 0,250 до 0,400 единиц. В демутацион-
ных древостоях эта величина уменьшается, в дигрессивных —
увеличивается.
Усыхание деревьев по горизонтальной структуре
биогеоценоза также является одной из характеристик состояния
устойчивых лесных сообществ, которая должна быть описана в ряду
общих показателей усыхания и отпада деревьев.
Из предыдущих исследований определено, что все деревья в
девственных разновозрастных коренных лесах в их
горизонтальных структурах распределены равномерно-агрегированно,
занимая все пространство биогеоценоза. Отличное от общей
тенденции размещение деревьев группы отпада по площади сообщества
может характеризовать динамику процесса отпада в биогеоце-
-83-
В Г. Стороженко
нозе. На рисунке 4 приводятся величины расстояний до деревьев
группы отпада по методике, принятой при изучении размещения
всех деревьев на площади учета (Быков, 1976), статистики и
характеристика агрегированности на примере одного из
биогеоценозов резервата «Кологривский лес» (южная тайга). Анализ
результатов показал, что размещение деревьев группы отпада
подчинено той же тенденции равномерно-диффузного
агрегирования, что и размещение всех деревьев на площади биогеоценоза.
Такое размещение деревьев группы отпада также является
неотъемлемой характеристикой формирования структур
устойчивых сообществ. При этом равномерно-диффузно усыхают
деревья разного возраста, диаметра и в разные сроки. Для выяснения
причин диффузного усыхания пробурено 300 деревьев старого и
свежего сухостоя, расположенных в массивах резервата
«Кологривский лес», из которых 83,4 % были поражены гни левыми
фаутами. Можно предположить, что гни левые фауты вполне
способны являться причинами ослабления и усыхания деревьев.
Однако присутствие среди них деревьев без гнилевого поражения
говорит об участии в этом процессе и других факторов.
Одновременно с усыханием равномерно-диффузного типа в коренных
разновозрастных древостоях тайги встречается и усыхание
куртинного типа, при котором усыхают группы деревьев до 3-5
экземпляров (редко до 10) одновременно в течение 1 -2 лет.
Встречаемость таких куртин различна. В тех же древостоях Кологрива
1 1 1 1 1 1 1 1 ! Г
4 6 8 10 12 14 16 18 20 22
Пункты учета
X
2002 см.
тх
140
«
443
т<т
101
С, %
20,1
тс
4,6
Характеристика
агрегированности
большая
Рис. 4. Колебания значений агрегированности размещения деревьев группы
отпада и их статистики (южная тайга, резерват «Кологривский лес»)
-84-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
было проанализировано 30 таких куртин. Бурение деревьев
показало, что усыхание куртинного типа не связано с развитием
гнилевых фаутов. Средняя величина поражения деревьев в
куртинах составляет 32,0 % (± 5,2 %). Эти величины не превышают
общих уровней поражения биогеоценозов коренных
разновозрастных ельников. Факторы, вызывающие этот вид усыхания
остаются невыясненными. Можно предположить, что
размещение этих куртин каким-то образом связано с парцеллярным
строение биогеоценозов, с их возрастной мозаикой, так как деревья в
куртинах усыхания относятся, как правило, к одному
возрастному поколению. Такой же тип усыхания отмечен в коренных
разновозрастных ельниках других подзон тайги с мозаичной
структурой фитоценозов. В древостоях простых структур как еловых,
так и других формаций куртинное усыхание, как правило,
связано с грибным происхождением очагового поражения деревьев
каким-либо одним видом возбудителя.
Аналогичные усыхания отмечены многими
исследователями. А.Д. Масловым (1972) сделана попытка обобщения этих
сведений и объяснения причин куртинного усыхания ельников.
Основной причиной усыхания такого типа он считает ослабляющее
действие засухи и последующее заселение деревьев стволовыми
вредителями. V этими выводами можно согласиться, но в ряде
случаев заселение усыхающих деревьев стволовыми
вредителями мы не наблюдали.
Подводя итог по разделу, следует отметить, что
распределение деревьев по категориям состояния, характерное для
коренных разновозрастных древостоев и отражающее их динамику
развития, нельзя рассматривать как нормальное для древостоев
другого происхождения, так как динамика их развития и отпада
деревьев формируется по другим законам и зачастую с участием
антропогенных факторов ослабления. К тому же анализируется
состояние древостоев с участием деревьев больших средних
возрастов и значительной частью деревьев предельного возраста, в
то время как в условно-одновозрастных и искусственных одно-
возрастных древостоях возраст основного полога имеет гораздо
более низкие предельные до периода распада показатели. В
искусственных насаждениях никогда нам не встречались даже
отдельные деревья, достигшие возраста 300 лет и более для ели
или 400-500 лет для сосны, в то время как в девственных лесах
это обычное явление.
-85-
8. ПОКАЗАТЕЛИ ДРЕВЕСНОГО ОТПАДА УСТОЙЧИВЫХ
ЛЕСНЫХ СООБЩЕСТВ
Древесный отпад как структура лесных сообществ в
последнее время привлекает все больше внимания специалистов
разных направлений исследований. Прежде всего следует отметить
исследования по изучению процессов накопления и разложения
древесного отпада, которые, надо признать, не слишком
обширны. Особенно это касается лесов, не затронутых антропогенным
воздействием или какими-либо катастрофическими
абиотическими или биотическими факторами разрушения, лесов,
развивающихся эволюционным путем, девственных, находящихся на
разных стадиях естественного формирования. В определенном
объеме эти сведения можно почерпнуть из наших работ (Сторо-
женко и др., 1992; Стороженко, 1990, 1992, 2000а, 20006, 2001,
2004 и др.), работ других авторов (Шорохова, Соловьев, 2002;
Тарасов, 1999; Кузнецов, 1998 и др.), таблиц хода роста древо-
стоев (Третьяков и др.,1952), таблиц древесного отпада (Воропа-
нов, 1973; Швиденко и др., 1997). Последние две группы
источников, однако, могут быть применимы только к лесам простых
структур, нормальным, одновозрастным или, в крайнем случае,
к условно-одновозрастным. В разновозрастных лесах динамика
накопления и разложения древесного отпада в значительной
мере отличается от таковой в нормальных лесах.
Между тем такие исследования крайне важны, что
подтверждается все возрастающим интересом к этой проблеме ученых
различных стран и направлений исследований. Важны они по
нескольким причинам, среди которых наиболее значительными
являются: 1) стремительное сокращение площадей девственных
коренных лесов и трансформация их в производные с
измененными функциональными характеристиками. Мы лишаемся
эталонов эволюционно развивающихся лесных сообществ и в целом
лесных экосистем; 2) необходимость изучения структур и
закономерностей их развития для задач восстановления планетарной
функции лесных экосистем в условиях нарастающих антропо-
-86-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
генных и техногенных воздействий; 3) решения задач изучения
пулов и стоков углерода в лесных экосистемах для расчета
глобальных величин углеродного баланса в природных
экосистемах; 4) далеко не полная изученность количественных,
качественных и функциональных характеристик составляющих
такие экосистемы организмов, групп организмов, ценотических
структур, консортивных взаимоотношений разных уровней,
баланса вещества и энергии и т.д.
В то же время исследований процессов накопления и
разложения древесного отпада, охватывающих обширные регионы
таежной зоны Русской равнины, вообще нет. До сих пор неясно
какие объемы древесного отпада характерны для модальных
коренных девственных лесов на просторах лесной зоны
европейской России.
8.1. Объемные показатели древесного отпада коренных лесов
Многолетние обширные по площади исследования древесного
отпада проводились нами на тех же постоянных и временных
пробных площадях, что и другие исследования по другим
направлениям. Напомним, что основными объектами изучения
явились леса Теллермановского лесничества в Воронежской области
(зона лесостепи), Тульские засеки в Тульской области (зона
теневых лиственных лесов), Московской, Рязанской, Тверской
областей (зона смешанных лесов и граница зоны смешанных лесов и
подзоны южной тайги), Костромской области (подзона южной
тайги), Ленинградской области и Печоро-Илычского
заповедника (подзона средней тайги), Мурманской области, Карелии,
Архангельской области и Коми (подзона северной тайги). На всех
пробных площадях определялись структуры фитоценозов,
состояние и величины текущего отпада, объемы валежа (ветровала и
бурелома) по стадиям разложения по оригинальной,
разработанной нами методике (смотри ниже), параметры поражения
деревьев и древостоев дереворазрушающими грибами биотрофного
комплекса с определением характеристик и возбудителей
гнилей, в том числе объемов гнилей. По линейным параметрам
деревьев вычислялись запасы стволовой древесины в пересчете на
1 гектар, которые значительно отличались от официальных так-
-87-
В. Г Стороженко
сационных показателей. Масса депонированного углерода
вычислялась по конверсионным коэффициентам для древесных
пород (стволы + ветви + пни и корни 4- хвоя), исходя из объемных
запасов стволовой древесины. Для хвойных пород средняя
величина принята равной 0,363 ± 0,015 тС (Уткин и др., 2001). Для
определения объемов и динамики стоков С02 рассчитывали
биомассу древесных остатков и сухостоя для разных периодов их
образования и динамику общего стока за длительный
ретроспективный период. Все расчеты велись для древостоев конкретных
пробных площадей, которые, тем не менее, отражают средние
показатели структур и древесного отпада коренных
разновозрастных древостоев, имеющих естественные величины объемов ва-
лежа. Вычислены некоторые статистики средних величин
запасов древостоев и древесного отпада 1-5 стадий разложения лесов
упомянутых регионов при малой выборке. При этом следует
представлять себе, что динамика накопления древесного отпада
в древостоях различных динамических характеристик и разной
степени деструкции в результате воздействия различных
факторов в одном регионе может быть настолько различной, что
величины средних значений и их ошибки не будут информативными,
хотя минимальные значения могут соответствовать средним
достоверным величинам. Очень показательны в этом отношении
ельники Центрально-лесного биосферного заповедника в
Тверской области (граница зоны смешанных лесов и подзоны южной
тайги), в которых периодически проходят ветровалы и буреломы
различной интенсивности и разброс значений объемов
древесного отпада в выделах в зависимости от степени их сохранности
огромен. Поэтому в настоящем сообщении мы не можем
распространять полученные выводы на все коренные древостой, но,
повторимся, рассматриваем древостой средних характеристик
естественных величин древесного отпада. Одновременно отметим,
что в настоящем разделе характеристики коренных лесов
отвечают всем перечисленным выше условиям структурного
строения и функционирования устойчивых лесных сообществ.
Общим для биогеоценозов всех изученных регионов является
характер усыхания деревьев и обеспеченность естественным
возобновлением. Факторами отпада в таких древостоях
являются естественное изреживание в результате отмирания деревьев,
относящихся к различным возрастным поколениям, конкурент-
-88-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ные отношения, осложняющиеся колебаниями
гидрометеоусловий, поражение грибными возбудителями, вызывающими гнили
стволов и корней. По нашим данным, более 85 % деревьев вале-
жа и бурелома в таких лесах имеют гнили стволов и корней,
которые в большинстве случаев являются причинами их усыхания
и вывала. В таблице 22 приводятся средние для коренных
формаций значения запасов древостоев и древесного опада и их
статистики. Понятно, что эти величины относятся к биогеоценозам
наиболее производительных типов леса. Наиболее типичные для
зон растительности лесоводственные характеристики коренных
лесных формаций приводятся в таблице 3 раздела «Породный
состав устойчивых лесных сообществ».
Таблица 22
Статистики средних показателей запасов и древесного отпада
коренных древостоев по зонам растительности
Зона растительности, состав, бонитет
и тип леса изучаемого древостоя
1
Зона лесостепи: смешанные
сосняки
Зона лиственных лесов
Граница зоны смешанных лесов
и подзоны южной тайги
Подзона южной тайги: ельники
сосняки
Подзона средней тайги: ельники
сосняки
Подзона северной тайги: ельники
сосняки
Статистики
запасов
древостоев
Хтч
2
450
420
350
423
400
430
354
336
176 .
78
3
43
38
21
57
16
26
15
14
12
5
Статистики
объемов
древесного
отпада
Хтч
4
85
78
68
98
126
98
83
71
47
6
5
8
11
10
12
13
17
2
16
3
1
%
4от2
6
18,8
18,6
19,4 !
23,2
31,5
22,7
22,9
21,1
26,7
7,6
В коренных ельниках, как видно из таблицы, соотношения
запасов древостоев с объемами древесного отпада в
биогеоценозах сравнимы по зонам растительности. В сосняках эти значения
имеют, во-первых, меньшие абсолютные величины древесного
-89-
В. Г Стороженко
отпада, во-вторых, гораздо более значительные расхождения в
величинах соотношений. Это связано с двумя причинами: 1)
почти обязательным воздействием пожаров, в результате которых
выгорает валеж, 2) неравномерностью процесса усыхания
деревьев и появления в этой связи древесного отпада. Например, в
резервате «Ащозерский» Ленинградской области (средняя тайга)
нами заложена пробная площадь в естественном послепожарном
условно-одновозрастном сосняке бруснично-черничном, средний
возраст которого составляет 70 лет. Древесный отпад в
результате естественного изреживания в нем составляет 52,8 % от числа
деревьев на пробе, причем средний диаметр деревьев отпада
составляет 89 % от живых деревьев, то есть характер усыхания
мало связан с отмиранием только угнетенных деревьев.
Добавим, что никакой патологии в древостое нет и с этим фактором
усыхание так же не связано. Оставшееся число деревьев
соответствует среднему количеству стволов на 1 гектар, характерному
для разновозрастных девственных сосновых биогеоценозов этой
зоны — около 350 деревьев. При достижении фазы динамики
древостоев, близкой к климаксу, процесс отпада деревьев
выравнивается и приближается к табличным.
8.2. Процессы разложения древесного отпада
Древесный отпад складывается из текущего отпада, то есть
стоящих в древостое деревьев усыхающих, свежего и старого
сухостоя и отпада прошлых лет, то есть стволов вал ежа (ветровала
и бурелома), лежащих в разном положении по отношению к
земле на площади биогеоценоза. Это положение, а также ряд других
факторов определяют скорость разложения древесного отпада, а
значит скорость выделения продуктов ксилолиза. Соотношение
этих величин в биогеоценозах различных зон растительности и
динамических характеристик представляет значительный
интерес для понимания процессов накопления и разложения
древесного отпада в устойчивых лесах. В таблице 23 приведены
характеристики древостоев, запасы сырорастущей древесины, объемы
гнилей, сухостоя и вал ежа и вычисленные запасы углерода в
конкретных древостоях части пробных площадей, заложенных в
-90-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
коренных разновозрастных сообществах основных районов
исследований на Русской равнине.
Запасы углерода, рассчитанные по конверсионным
коэффициентам, как это принято, определяются для древесной
биомассы фитоценоза, не затронутой деструктивными процессами, то
есть для запасов древостоя, исключая запасы сухостоя и объемы
гнилей растущих деревьев. Живая древесина содержит весь
объем биомассы и весь объем углерода, который и
рассчитывается по конверсионным коэффициентам, та часть биомассы
фитоценоза, которая по разным причинам отмерла, сразу вовлекается
в процессы разложения, сопровождающиеся высвобождением из
нее продуктов ксилолиза, в том числе С02 и Н20. Поэтому любой
фрагмент древесного отпада, вовлеченный в процессы
разложения, будет содержать углерода меньше той величины, которая
рассчитывается по конверсионным коэффициентам. В этом
случае важно представить динамику разложения древесного отпада
и динамику выделения углерода на протяжении всего периода
разложения вал ежа. Те величины запасов углерода древесного
отпада, которые приведены в таблице, соответствуют только
факту и количеству древесного отпада, но отнюдь не описывают
динамику процесса его разложения. Вопросы, касающиеся
изучения процессов разложения древесного отпада, в последнее
время являются объектами повышенного внимания в связи с
актуализацией проблем углеродного баланса планеты, углеродных
потоков, глобального потепления.
Некоторые сведения о величине древесного отпада в
коренных лесах приводятся в работах L. Karjalainen и Т. Kuulivainen
(1999), Е.В. Шороховой и А.А. Шорохова (1999), М.Е. Тарасова
(1999) и некоторых других. Наши исследования по изучению
древесного отпада в коренных лесах начаты в 1988 году и
впервые опубликованы в 1990 году (Стороженко, 1990).
Процессы разложения валежа и связанные с ними процессы
эмиссии углерода невозможно изучать без оценки их датировок.
И этой проблеме уделяется пристальное внимание в ряде
научных центров как в России, так и за рубежом. В России первые
исследования в этом направлении проведены П.Ф. Еленевым
(1923), затем многими авторами, среди которых можно назвать
СИ. Ванина (1931), В.Я. Частухина (1948), Н.Т. Картавенко и
Б.П. Колесникова (1962), И.М.Вересову (1968), В.А.Мухина
-91-
со
eg
ев
I s
се
К
ф
ч
л
м
ф
ч
Я
X
и
3
s
ф
\о
о
sr
я
ч
»
ев
РЗ
ф
о
о
о
м
ф
з
н
ев
ft
ее Ф
2 м
О д
5 «
X ф
ft£
8 л
№ Ф
о се
м ф
**■ X
3 w -
о * се
се 2 а
5 « £
J S о
о се £
н ч: к
2 °
|s
§ а
g се
See
ft «
Ф S
Н _
ft ф
се н
^ ч
о
я
5Г
ф -*
Проц
7 от
ада
с
н
ОГОО
№
8
ер
ф
стика др
fr»
арак
X
зтоев
о
ей
ристика др
ф
Харак
ния
ф
азло-
s
«т
Я ecj
5*
о
X
ч
к
TOM^
PQ
Объем
валежа
| 8 к
£ х о
8 1-
2 Я
ф Ч
i* S
» X
о
СО
X
8
Фаза
динамики
ей
олнот
став, тип леса, п
бонитет
'*З
к
Ф
оо
t>
со
ю
Tf
со
СМ
^
есостепи
Зона л
со
-*
см
г>
uad
CM
w ^
*H <*
ю о
<M Oi
oof-
со со
'
« 00
00 £
CO
^ O)
00 см
Ю
t- -
CM
,_,
о *
ю t-
co см
Разно-
возр.
Д2Лп2Кл1Яс; снытьво-
сок.; 0,6;Пбон.
ю о
гН
5!
и
>х
ей
жнойт
анных лесов и подзоны ю
смеш
зоны
Граница
со
аз
см
on".
см о
OS *
СМ О
СО
ел ~г
t- оо
см
ю оо
со с*э
см оо
~ оо
тН
136
49,4
«О»
2*
00
ю -
1-4
СО ~Г
«о оо
т* «О
*■*
£
, 1 бон
0с1Б; кисл., 0,7
1-Н
Ы
00
см
СЛ
гН
СО
^ч в>
со О
СЛ *
о
оо *
00 СМ
СО
~ со
"Э .-Г
т* СО
СО «1
<М СЛ
177
64,3
too
2«
00
ю
1-4
1-1
rt< -
iO ^
ю о
см
Дигр.
...
8*0 -н
Б+Ос; кисл.-чер
н
Ы ю
С35 1-4
СО
о>
^
СМ
*5
<м оо
Tt*
со -
*я S
см
co°i
CSI
о w
см ^
°5
со о
см ю
1> СО
СО 1-н
00
ю *
^ см
ю •
Tf Л
со см
Демут.
0,7;
Б+Ос; черничн.,
н.
И «
СЛ »н
TJ»
>жной тайги
дзона к
о
С
о
о
СО
^ ^
^ 00
оо "1
т* СО
гЧ
00 -
iO i-l
см
t- см
гН сО
Л см
СЛ _j
СО ^
сч >-г
о
100
36,3
Я со
00 00
см °°
~ см
СО ^»
см
— СИ
^ *
^ тс
со см
i-i
й
8Е1Б1Лп+Пх; кисл.
щитовн., 0,8,1 бон
ю
о
^«
^
гЧ
СО ►
00 Й
со
со ^
о
*> г*
TJ4 1>
r-t
00 »
*-< СО
со ^
СМ д.
iH
со^
156
56^
m°i
см О
00
ю »
1-»
О)
х* S
ХО 00
со см
т-4
S
Б+Пх,Лп; кисл.
ювн.,0,8,1 бон
с^> 5
00 3
СО
00
тН
«^
^.я
тР СО
г-4
«■ о»
СО
кО Л^
СО СО
СМ
»rf
^^
^ 2
со "^
134
48,6
ЧгН
041 Oft
см °°
_^ хз
Tf -
1-4
-. см
о -г
см со
со »-*
Демут.
ничн.
БШх; кисл.-чер
1 бон.
00 О
t-
f-l
CM
о to
*-< со
со оо
см со
*-* т#
СЛ OJ
тН со
1-(
СО •»
со со
1-Н
oow*
ь- оо
см
.Л ч*
ю -
ю см
о о
-. со
о •
t- Tf<
со со
тН
Демут.
ничн.
+Е,Б; брусн.-чер
Пбон.
8 ч
гН О
00
чзона средней тайги
о
G
СЛ
О
см
^
оо Jr
»> оо
CM
со см
1rH
со «>
ТН ^44
со »н
1-Н
«3
00 см
ч^
8-1
н
OI
О)
СЛ Й
со ^
i-H|
Демут.
+Ос,Б; майник.-
дичн.; 0,8; Пбон
О ф|
»н }»•
о> J
-92-
Ен
tr g
a t»
И
ада,
Ен
О
и
о
к
ф1
и
0)
а
со
К
рист
акте
а
X
ф
о
&
ю
ф
а
Характеристика д
ч
к
ф
о
ч
го
а
2
&5т
S со
Ен«
О
Ф
ч
S
8
О
Ен
PQ
J3 ф
о 3
2 8 «
О о
£ Я
* 3
8£
СО
К
й
ell
«Г
S
в
§
С
-Г Ен
со о>
g
о
a
Oi
00
t>
o>
Ю
Tj*
CO
CM
rH
b-
o>
1—I
со w
ем oo
о «о
»-• со
CM
(M «
tF »Л
,н
. л
см
00 S
см О
1-Н
ю
fc- о
CM
см
ю„со
2«
ч*
о
о w *
00 £;
со с*э
а
со
о
X
К
я
£
&■
о
о
+
W
н S
Oi ХО
о
ч*
тН
см
о «>
CM fr.
со
ю ►
^ <£>
тН
» ^
тН ^
Ю «"1
<N О)
ю
t- -
см
со -
см
°°©
2ю
Tt<
rH •>
о
ю ft
со см
«
«
о
и
и
$
и
к
я .
а я
+ *ч
§*■:
гн о
гН
о
о
см
05 со
2 е-
тН со
ю
05 «
см О
*-'
rn°i
см
о w
CM ^
1-1
со *
Т-)
со />
^ |>
о*
rH ^
, ь-
см -
о
*£ тР
2 t^
см<*>
й
if
я
я
s
я
^ g
о ^
и *:
o> о
CM
to
Cvj
CM
-si
00 9
CM
^ ^
CM oo
CO •>
CO rH
»ч CO"
^ CO
Ю -
w3
00 s
CO
см w
J^r
oo
Ю *•
rH
«^
^ CM
CO W
о
s
«
я
a
w
+
о
тН
CO
s
2
JtSS
о
8
a
If
Ф
о
cO
X
о
CO
tl
о
со
CM
со
oo 9
w см
— ю
1-4
со
л *
со со
1-Н
ъ™
СМ |>
oo ift
г>П
ю оо
1> СО
СО гН
т)*
о
о w
ю о
i-l ю
^
н
2 *°
* *
о я
со S
h*1
со
ь-
см
ь» •>
см
. л
см
о
W лг
со <n
г"'
«о "*
гН Л
1-Н ^
т*«
о
t- *
^ ^
о ^
гН О
-*
о
см ^
•> см
1—4 с^
0
а
о
2
о
к
а
ф
и1 и
Сн ^
и*
со о
ХО
ь-
<о
см
о Ч
1-1 СО
о»
»-* со
ХО О
«» Oi
ГН О
ГН ^
см со
1-1 TF
л, «>
см •
о
ю°Я
Tf JO
00 тН
ю см
^, t*
см -
о
оо о
<0 гН
1-Н с©
а
О 2
+ в
w ^
Oi S
СЛ
ю
о>
iH
оо 9
см
^ ю
см
гН
со «
см О
^
СОЬ;
rH ^
гН ^
|>
см ~
о
*1<о
3»
см ю
^f 1-1
•^
rH ^
о
о о>
CM oi
см £:
ч
6
а о
s«
^°г
а!
х S
t*
to
'
( 1
хл 00
*9» гН
°°1
со ^J
^
«а
ю00
°6
тН
50 см
«я
°°1
« 2
°d
00 "
i J
и о
а з
О ФП
rH »•
со
93-
В Г Стороженко
(1990), Е.В. Скворцову и др. (1983), В.А.Соловьева (1992),
O.N. Krankina, M.E. Harmon (1995), М. Yatskov and all (2003),
Е.В. Шорохову и А.А. Шорохова (1999), Е.В. Шорохову, М.И. Гир-
фанова (2004), Е.В. Шорохову (2005), В.Г. Стороженко и др.
(1992), В.Г. Стороженко (1990, 2000, 2001, 2002, 2004 и др.)
Нами разработана первая шкала датировки разложения вале-
жа ели для южной тайги, которая послужила отправным
ориентиром для исследований в лесах других зон растительности и
формаций (Стороженко, 1990). Позднее в наших исследованиях
она много раз проверялась и верифицирована исследованиями
других авторов. Шкала опубликована в нескольких изданиях, но
мы полагаем, что для полноты обсуждений проблемы древесного
отпада не будет лишним привести её в несколько расширенном
варианте, добавив сведения по датировкам разложения вал ежа в
северной тайге. Понятно, что переходы из подзоны в подзону в
таежной зоне весьма условны, поэтому мы приводим средние
показатели для южной и северной тайги.
Таблица 24
Шкала стадий разложения крупного древесного отпада для
ели и сосны в коренных лесах таежной зоны Русской равнины
Стадия
разложения
1
2
Признаки разложения
Форма стволов, пней бурелома и кора
сохранились. Мхов, растений
напочвенного покрова, подроста и
плодовых тел дереворазрушающих
грибов нет. Ветви всех порядков
сохранились, иногда зеленая или бурая
хвоя. Изменение окраски древесины
фрагментами. Стволовые вредители
присутствуют или вылетели
Форма стволов и пней сохранилась, кора
опала частично или полностью, моховое
покрытие неполное. Плодовые тела
дереворазрушающих грибов.
Сохранились ветви 1-2 порядков. Гниль
древесины от 1 до 3 стадии разложения.
Вершина до диаметра 8-10 см разложена
полностью.
Датировка
стадий
разложения, лет
Южная
тайга
1-3
15-20
Северная
тайга
1-5
20-25
Стадия
ксило-
лиза
Начальная
Максимальная
-94-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 24 (продолжение)
Стадия
разложения
3
4
5
Признаки разложения
Форма стволов и пней частично
изменена из-за полного обрастания
стволов мхами. Появление и развитие
растений напочвенного покрова, иногда
подрост до 5 лет. Плодовые тела
дереворазрушающих грибов
встречаются редко. Могут сохраняться
ветви 1 порядка. Древесина разложена
до 3 стадии.
Форма стволов и пней сохранилась
частично. Полное обрастание мхами и
растениями напочвенного покрова;
подрост и подлесочные породы до 20 лет.
Плодовые тела дереворазрушающих
грибов отсутствуют, ветвей нет.
Древесина разложена до 3-4 стадии и
представляет собой бурую,
преимущественно деструктивную гниль.
Форма стволов и пней утеряна,
угадывается по микроповышениям по
контуру ствола, возвышающегося
до 1/3 высоты ствола. Подрост и
подлесочные породы до 40 лет.
Гумификация продуктов разложения
древесины.
Датировка
стадий
разложения, лет
Южная
тайга
25-30
35-40
45-50
Северная
тайга
30-35
40-45
50-60
Стадия
ксило-
лиза
Затухающая
Конечная
Можно выделить и шестую стадию разложения, когда
происходит полный переход древесины в категорию верхних слоев
почвы, но контуры бывшего ствола еще можно угадать по
микроповышениям. Такие остатки можно датировать периодом в
южной тайге до 60-65 лет в северной тайге до 70 лет.
Динамика разложения древесного отпада в естественных
условиях природного экотопа тесно связана с рядом условий,
определяющих длительность этого разложения. К числу таких
условий относятся, например, размер древесных остатков и
отношение их к определенной группе пород по твердости (или
плотности) древесины, положение их относительно земли, что во многом
-95-
ВТ. Стороженко
определяет температурный и влажностный режимы процесса
разложения, зона растительности, лесоводственные
характеристики древостоя, особенности экотопа и др.
Стволы деревьев старого сухостоя проходят первую, а иногда
и вторую, стадию разложения в стоячем виде.
Представленная выше шкала создана для коренных древосто-
ев таёжной зоны. Для древостоев зоны лесостепи и лиственных
лесов, в которых леса имеют в основном простые условно-одно-
возрастные или одновозрастные структуры, шкала датировок
разложения древесного отпада имеет несколько иной вид,
привязанный более к возрастным группам лесов и делению пород на
твердо лиственные и мягко лиственные, что связано с
длительностью периода их разложения. Впрочем, визуальные признаки
разложения практически совпадают. В таблице 25 представлены
величины интервалов длительности разложения валежа
лиственных пород по зонам растительности и плотности их
древесины.
Естественно, приведенные данные отражают средние
значения и вариации их в различных экотопах могут быть
значительными. Пни твердолиственных пород, особенно дуба, разлагаются
Таблица 25
Длительность периода разложения древесного отпада по
фракциям деревьев для двух групп пород по возрастным
группам в двух природных зонах, годы
Возрастная
группа
Молодняки
Средневозрастные
Приспевающие,
спелые и
перестойные
Фракция
деревьев
Ствол
Ветви
и корни
Ствол
Ветви
и корни
Ствол
Ветви
и корни
Зона растительности
Лесостепь
Лиственные леса
Твердолиственные
10-15
15-20
8-10
До15
25-30
35-40
10-15
15-20
30-40
50-60
8-12
15-20
8-10
10-15
5-8
до 10
15-20
25-30
10-12
15-20
20-25
35-40
8-10
10-15
Лесостепь
Лиственные леса
Мягколиственные
8-10
10-15
5-8
8-10
15-20
20-25
8-10
10-12
20-25
25-30
8-10
10-15
5-8
10-12
3-5
5-8
8-12
15-20
5-8
8-10
15-18
15-20
5-8
10-12
-96-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
с некоторыми особенностями. Пни большого диаметра
разлагаются неравномерно по диаметру из-за разной влажности
древесины внутренних и наружных слоев. Древесина периферической
части пня более сухая, чем внутренняя, и деятельность дерево-
разрушающих грибов в ней замедлена, высохшие остатки
периферической части пней дуба могут сохраняться до 70-80 лет,
тогда как внутренняя часть пней совершенно разрушается и
древесина мацерируется. Интересно также отметить, что в зоне
широколиственных лесов, то есть на 300-400 км севернее, чем
древостой лесостепи, период разложения древесных остатков тех же
пород на 10-15 лет короче. Это связано, по всей вероятности, с
большей сухостью климата, верхних слоев почвы, подстилки и
соответственно разрушаемых грибами древесных остатков в зоне
лесостепи и с менее благоприятными условиями
функционирования дереворазрушающих грибов.
Период разложения древесины до 3 стадии является
заключительным в процессе выделения продуктов ксилолиза, но
стволы древесного отпада в своем объеме разлагаются неравномерно,
форма ствола валежа даже сгнившего до третьей стадии может
поддерживаться за счет сохраняющейся «арматуры» еще не
сгнивших внутренних сучьев, в связи с чем весь период
разложения ствола растягивается на более длительный срок. И сам
процесс разложения неравномерен по времени. В естественных
условиях характер протекания этого процесс описан весьма скудно.
По некоторым данным его можно аппроксимировать кривой
нормального распределения (Кузнецов, 1998; Тарасов, 1999). В
последнее время обширные исследования в этом направлении
предприняты Е.В. Шороховой (2004, 2005). Интенсивность процесса
разложения стволов валежа нарастает постепенно, по мере
освоения дереворазрушающими грибами всего объема древесины и
также постепенно затухает по мере использования грибами
субстрата.
Имея распределение древесного отпада по стадиям
разложения, годам его образования и период разложения по среднему
модельному дереву, можно рассчитать динамику поступления
углерода за весь период разложения всего объема древесного
отпада, который можно зафиксировать на период его учета в
любом участке леса. В таблице 26 приведены итоговые данные
-97-
В Г Стороженко
расчетов массы углерода, выделяющегося при разложении всего
объема древесного отпада, учтенного в древостоях, некоторых
пробных площадей, взятых в качестве примера и
расположенных в коренных еловых древостоях разных зон растительности.
Для первой пробной площади (граница зоны смешанных лесов и
южной тайги) мы приводим полный расчет объемов выделения
углерода при разложении древесных остатков, относящихся к
разным периодам появления на территории пробной площади
(разные стадии разложения). При этом необходимо учесть, что за
интервал выделения основной массы углерода нами приняты
30-летний промежуток времени, в течение которого выделяется
основная масса углерода. В других зонах растительности и для
других пород этот промежуток будет другой — в лиственных
лесах меньше, в подзоне средней и северной тайги больше. Для
пробных площадей 4, 7 и 11 приводятся только суммарные для
всего древесного отпада значения массы выделяемого углерода
за период его разложения до 3 стадии.
Таблица 26
Динамика выделения углерода при разложении древесного отпада
в древостоях пробных площадей 1, 4, 7, 11 (тСм'3« га1)
Годы
64
63
62
61
60
59
58
57
56
55
54
53
52
51
| 50
Объемы выделения углерода по стадиям
разложения древесного отпада
для Центрально-лесного заповедника
(пр. пл. 1)
1+сух.
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
2
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
3
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
4
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
5
-
-
-
-
-
-
-
-
-
-
-
-
-
-
0
Суммарная величина С по годам
для пробных площадей:
1
-
-
-
-
-
-
-
-
-
-
j
-
-
-
0
4
-
-
-
-
-
-
-
0
0,028
0,056
0,084
0,112
0,14
0,168
7
-
-
-
-
-
-
-
-
-
-
0
0,004
11
0,006
0,012
0,018 |
0,024
0,031
0,037
0,043
0,049
0,055
0,062
0,074
0,086
0,098
0,110
0,124
-98-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 26 (продолжение)
Годы
49
48
i 47
46
45
44
43
42
41
40
39
38
37
36
35
34
33
32
31
30
29
28
27
26
25
24
23
22
21
20
19
18
17
16
15
14
13
1 12
Объемы выделения углерода по стадиям
разложения древесного отпада
для Це* трал ьно-лесного заповедника
(пр. пл. 1)
1+сух
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
2
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
0
0,035
0,07
0,105
L 3
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
0
0,081
0,182
0,243
0,324
0,405
0,486
0,567
0,648
0,729
0,81
0,891
0,972
1,053
1,134
1,215
1,134
1,053
0,972
4
-
-
-
-
-
-
-
-
-
0
0,044
0,088
0,132
0,176
0,220
0,264
0,308
0,352
0,396
0,44
0,484
0,528
0,572
0,616
0,66
0,616
0,572
0,528
0,484
0,44
0,396
0,352
0,308
0,264
0,220
0,176
0,132
0,088
5
0,044
0,088
0,132
0,176
0,220
0,264
0,308 1
0,352
0,396
0,44 !
0,484
0,528
0,572
0,616
0,66
0,616
0,572
0,528
0,484
0,44
0,396
0,352
0,308
0,264
0,220
0,176
0,132
0,088
0,044
0
-
-
-
-
-
-
-
Суммарная величина С по годам
для пробных площадей:
1
0,044
0,088
0,132
0,176
0,220
0,264
0,308
0,352
0,396
0,44
0,528
0,616
0,704
0,792
0,88
0,88 !
0,88
0,88
0,88
0,88
0,961
1,062
1,123
1,204
1,285
1,278
1,271
1,26
1,257
1,25
1,287
1,324
1,361
1,398
1,435
1,345
1,255
! 1,165
4
0,196
0,224
0,252
0,28
0,329
0,378
0,427
0,476
0,525
0,623
0,616
0,616
0,609
0,602
0,595
0,608
0,622
0,636
0,65
0,664
0,678
0,65
0,622
0,594
0,566
0,537
0,509
0,481
0,481
0,481
0,481
0,481
0,488
0,495
0,502
0,509
0,516
0,523
7
0,008
0,012
0,016
0,02
0,024
0,028
0,032
0,036
0,04
0,044
0,048
0,052
0,056
0,096
0,136
0,176
0,216
0,248
0,28
0,3312
j 0,344
0,376
0,408
0,44
1 0,472
0,504
0,536
0,568
0,6
0,632
0,664
0,637
0,61
0,583
0,556
0,533
0,51
0,487
11
0,136
0,148
0,160
0,172
0,187
0,185
0,185
0,185
0,185
0,179
0,198
0,212
0,225
0,239
0,254
0,253
0,254
0,256
0,257
0,259
0,258
0,260
0,261
0,263
0,264
0,271
0,278
0,286
0,293
0,3
0,296
0,293
0,286
0,282
0,277
0,279
0,280
| 0,282
-99-
В. Г. Стороженко
Табл. 26 (продолжение)
Годы
11
10
9
8
7
6
5
4
3
2
1-
0
1 +
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
1 26
Объемы выделения углерода по стадиям
разложения древесного отпада
для Центрально-лесного заповедника
(пр. пл. 1)
1+сух.
-
-
-
-
-
-
-
-
-
-
-
0
0,028
0,056
0,084
0,112
0,14
0,168
0,196
0,224
0,252
0,28
0,308
0,336
0,364
0,392
0,42
0,392
0,364
0,336
0,308
0,28
0,252
0,224
0,196
0,168
0,14
0,112
2
0,14
0,175
0,21
0,245
0,28
0,315
0,35
0,385
0,42
0,455
0,49
0,525
0,49
0,455
0,42
0,385
0,35
0,315
0,28
0,245
0,21
0,175
0,14
0,105
0,07
0,035
0
-
-
-
-
-
-
-
-
-
-
-
3
0,891 !
0,81
0,729
0,648
0,567
0,486
0,405
0,324
0,243
0,162
0,081
0
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
4
0,044
0
-
j
]
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
5
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Суммарная величина С по годам
для пробных площадей:
1
1,075
0,985
0,939
0,893
0,847
0,801
0,735
0,709
0,663
0,617
0,571
0,525
0,518
0,511
0,504
0,497
0,49
0,483
0,476
0,469
0,462
0,455
0,448
0,441
0,434
0,427
0,42
0,392
0,364
0,336
0,308
0,28
0,252
0,224
0,196
0,168
0,14
0,112
4
0,552
0,58
0,608
0,636
0,664
0,692
0,72
0,748
0,776
0,804
0,833
0,815i
0,797
0,779
0,761
0,743
0,725
0,707
0,689
0,671
0,653
0,635
0,617
0,599
0,581
0,563
0,545
0,527
0,496
0,465
0,434
0,403
0,372
0,341
0,310
0,279
0,248
0,217
7
0,464
0,441
0,418
0,395
0,372
0,349
0,326
0,303
0,280
0,257
0,235
0,277
0,32
0,363
0,406
0,449
0,492
0,535
0,578
0,621
0,664
0,707
0,75
0,793
0,836
0,879
0,922
0,965
1,008
0,952
0,896
0,84
0,784
0,728
0,672
0,616
0,56
0,504
11
0,283
0,285
0,286
0,288
0,289
0,291
0,292
0,294
0,295
0,297
0,298
0,3
0,287
0,274
0,261
0,248
0,235
0,222
0,209
0,196
0,183
0,170
0,157
0,144
0,131
0,118
0,105
0,092
0,079
0,066
0,053
0,040
0,038
0,036
0,034
0,032
0,03
0,028
-100-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 26 (продолжение)
Годы
27
28
29
30
31
33
34
35
36
37
38
39
40
Объемы выделения углерода по стадиям
разложения древесного отпада
для Центрально-лесного заповедника
(пр. пл. 1)
1+сух.
0,084
0,056
0,028
0
-
-
-
-
-
2
-
-
-
-
-
-
-
-
-
3
-
-
-
-
-
-
-
-
-
4
-
-
-
-
-
-
-
-
-
5
-
-
-
-
-
-
-
-
-
Суммарная величина С по годам
для пробных площадей:
1
0,084
0,056
0,028
0
-
-
-
-
-
4
0,186
0,155
0,124
0,093
0,062
0,031
0
7
0,448
0,392
0,336
0,28
0,224
0,168
0,112
0,056
0
11
0,026
0,024
0,022
0,02
0,018
0,016
0,014
0,012
0,10
0,008
0,006
0,004
0,002
Погодичные объемы выделения массы углерода при
разложении древесного отпада каждой стадии рассчитываются по
следующей формуле:
Vi = [(Vc • 21) • NA4 • (7,5) -1 •( пг + n2+ ... +
+n.(-NA • 2-1) + n1+n2 + ... п.),
где V. — погодичные объемы выделения массы углерода (тСм3);
Vc — объемы массы углерода от объема древесного отпада
каждой стадии разложения; NA — число лет периода разложения
древесного отпада до 3 стадии для определенной зоны
растительности и породы; пх, п2 ... п. — каждый из годов периода
разложения древесного отпада от среднего значения (NA • 21).
Шкала периода разложения всего древесного отпада,
отмеченного на пробной площади в пересчете на 1 гектар в
зависимости от зоны растительности, может вмещать в себя от настоящего
времени, обозначенного 0, до 65 лет в ретроспективу (-) и до
40 лет в перспективу (+).
Суммы погодичных значений объемов углерода, полученных
от погодичных значений объемов древесного отпада, отражают
динамику выделения углерода за весь возможный для учета
период появления древесного отпада в древостоях пробных площа-
-101-
В. Г. Стороженко
дей. Одновременно с этим следует отметить, что с углублением в
ретроспективу ошибка реальных значений объемов выделения
углерода будет увеличиваться в область их уменьшения,
поскольку из учетов будут уходить разложившиеся стволы
древесного отпада все более крупных размеров. В качестве
иллюстрации можно привести данные учета древесного отпада на одной
из пробных площадей, заложенной в
абсолютно-разновозрастном древостое в Центрально-лесном биосферном заповеднике
(ЦЛБГЗ) (граница зоны смешанных лесов и южной тайги), где в
1969 году, то есть ко дню учета около 35 лет назад, прошел
сплошной вывал деревьев. На день учета от всего количества
деревьев разных диаметров сохранились стволы диаметром от 24
до 46 см. Все они от третьей до пятой стадий разложения и
прослеживаются от 2/3 до 1/2 длины ствола, верхние части стволов
уже не фиксируются. Плодовых тел дереворазрушающих грибов
нет. Древесина разложена до 3 стадии, процесс ксилолиза
находится в заключительной стадии. На месте вывала сформировался
условно-одновозрастный еловый древостой.
В качестве примера состава древесного отпада в устойчивых
сообществах приводим распределение валежа по диаметрам и
стадиям разложения в древостое означенной нами выше пробной
площади (ЦЛБГЗ).
Таблица 27
Распределение стволов древесного отпада по стадиям разложения
и диаметрам в древостое пробной площади 1 (м3га_1/% )
Стадия
разложения
1
2
3
4
5
1 Всего,
M'W/'X)
Группы диаметров стволов древесного
отпада, см
До
16 см
-
1,2
0.9
0,7
0.9
0,7
0.8
0,6
4^
3,2
17-2
-
1x1
1,3
6.2
4,6
2.2
1,6
Ы
1,2
11.0
8,7
25-32
-
8.8
6,4
8.8
6,5
Ях2
2,3
3,7
24.9
19,1
33-40
-
5.8
4,3
20.6
15,2
18.9
13,9
9.3
6,8
54.6
40,2
41-48
4.7
3,6
4,7
3,6
1Л
5,7
2,9
-
21.2
15,6
49-56
-
6.5
4,8
-
11.5
8,4
-
18.0
13,2
Объемы
валежа
4.7
3,6
29.2
21,6
44.4
32,7
40.7
29,8
16.9
12,3
136,Q
100
-102-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Как видим, в любой стадии разложения присутствуют стволы
любых диаметров. Здесь же следует оговориться, что при
измерении стволов древесного отпада начиная с 3 стадии разложения и
особенно 4 и 5 стадий неизбежна некоторая ошибка из-за
частичной потери формы стволов в результате обрастания их
поверхности растениями напочвенного покрова и деструкции древесины.
Возвращаясь к таблице 26, можно видеть, что в каждой
колонке рассчитанных суммарных величин объемов выделенного
углерода для всех объектов (древостой принятых для примера
пробных площадей) выделяется временной период, когда эти
значения имеют колебания вокруг средней величины (при
нормальном распределении процесса разложения), не опускаясь
ниже определенного предела. Так как существуют конечные и
начальные периоды отсчета значений величины, то существуют
и области их возрастания и убывания до нулевых отметок.
В таблице 28 приводятся значения средних погодичных
объемов углерода и их статистики за период, когда фактические
значения массы выделяемого углерода выходят из зон возрастания
и убывания от нулевых отметок.
Таблица 28
Средние величины погодичной массы углерода, выделяющегося
при разложении древесного отпада в древостоях пробных площадей,
и их статистики (тСм^а1)
№ пробной
площади
1
4
7
11
Средние
погодичные
значения массы
С, х
0,983
0,604
0,542
0,291
Период расчета
средних
значений,лет
41
50
43
32
<т
0,272
0,083
0,192
0,016
мх
0,042
0,012
0,029
0,003
Погодичные колебания значений массы углерода здесь
связаны с различной суммарной массой древесного отпада разных
стадий разложения. При этом надо учитывать, что фактические
погодичные значения массы углерода будут отличаться от
вычисленных в большую сторону, поскольку, как упоминалось
выше, в учеты не включена часть разложившегося древесного
отпада, имевшего диаметры меньше, чем стволы, вошедшие
в четвертую и пятую стадии.
- 103-
В. Г Стороженко
Наглядно видно, что от зоны смешанных лесов к подзоне
северной тайги средние величины массы углерода, выделяемого
при разложении древесного отпада, уменьшаются. Понятно, что
это происходит из-за общего снижения объемов древесного
отпада по мере снижения производительности древостоев и
абсолютных величин запаса вал ежа в них. Относительные величины
объемов древесного отпада, представляющие собой отношения
объемов древесного отпада к запасам древостоев и
соответственно выделяемого при его разложении углерода, соизмеримы во
всех зонах растительности. Колебания этих величин зависят
только от фазы динамики биогеоценозов. Этот тезис вполне
согласуется с обозначенными ранее положениями о соизмеримых
величинах показателей состояния и объемах древесного отпада в
коренных лесах на Русской равнине.
Таким образом, можно предположить, что в коренных лесах
естественного происхождения, развивающихся эволюционным
путем и не подверженных каким-либо стрессовым ситуациям,
влияющим на резкое увеличение объемов древесного отпада, по-
годичные величины массы выделяемого при его разложении
углерода будут близки к приведенным в таблице. Эти значения
были бы более точными, если бы удалось восстановить объемы
разложившегося древесного отпада в ретроспективе от 10 и более
лет.
Если принять во внимание то, что значительная часть лесов
таежной зоны Русской равнины может удовлетворять этим
условиям или иметь близкие характеристики, можно рассчитать
(с определённой ошибкой) и массовые значения выделяемого
углерода по зонам растительности. Основная трудность при этом
заключается в определении площадей таких лесов.
8.3. Таблицы древесного отпада для коренных модальных
лесов таежной зоны Русской равнины
До недавнего времени одной из нерешенных задач
практического использования исследований в области изучения процессов
накопления и разложения древесного отпада являлось
составление таблиц древесного отпада для модальных лесов.
-104-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Для нормальных лесов основных лесообразующих пород, то
есть, по сути дела, гипотетических культур, известны таблицы
древесного отпада П.В. Воропанова (1973), включающие
результаты региональных исследований ряда авторов. Несколько более
приближены к естественным коренным лесам таблицы А.З.Шви-
денко (1997). И те и другие рассчитаны на основе таблиц хода
роста нормальных древостоев и разнятся в конечных цифрах
объемов древесного отпада по нашим данным не более чем на
9 %, что в пределах таксационной ошибки. Для
разновозрастных, а тем более для девственных, лесов никаких таблиц не
существует. В то же время такие сведения крайне важны по
нескольким позициям. Во-первых, важно представлять в каком
балансе с объемами древостоев находится древесный отпад в
естественно развивающихся, незатронутых антропогенными
воздействиями, девственных лесах, то есть определить природную
естественную динамику древесного отпада. Эти сведения
являются базовыми для оценки динамики развития лесов других
характеристик. Во-вторых, древесный отпад является величиной,
обладающей огромной динамикой формирования и разложения,
что определяет необходимость изучения баланса скорости
разложения с накоплением древесного отпада и приростами
древостоев. В-третьих, важным является обозначить уровень величин
выделяемых продуктов ксилолиза при разложении древесины
древесного отпада в естественно развивающихся сбалансированных
лесных экосистемах. Но прежде всего для составления таблиц
древесного отпада для модальных лесов следует определить два
основных параметра: запасы древесного отпада для таких
древостоев и скорость его разложения для изучаемых пород. Наши
исследования объемов валежа и скорости его разложения в
течение последних 20 лет проводились в девственных лесах еловых и
сосновых формаций всех подзон таежной зоны. Выводы по этим
исследованиям представлены в соответствующих разделах
выше. На более чем 500 пробных площадях определялись
возрастные структуры древостоев, их запасы, запас валежа по
стадиям разложения согласно разработанной нами шкале датировки
стадий разложения древесного отпада (см. таблицу выше).
Период разложения древесного отпада ели и сосны грибами
ксилотрофного комплекса для стволов 30-40 см в диаметре до
третьей стадии разложения, когда выделяется основной объем
-105-
В Г Стороженко
С02 и воды и процессы ксилолиза древесных остатков затухают,
от подзоны южной до подзоны северной тайги увеличивается в
среднем от 35 до 45 лет для ели и от 40 до 50 лет для сосны. Эти
показатели позднее подтверждены исследованиями других
авторов (Шорохова, Шорохов, 1999; Морозов, 1994 и др.).
Для расчетов объемов выделяющихся продуктов ксилолиза
при разложении древесного отпада необходимо переводить
объемные показатели запасов древесного отпада в его фитомассу.
При этом для расчета фитомассы мы используем переводные
конверсионные коэффициенты, разработанные другими
авторами (Замолодчиков, Уткин, Честных, 2003). Для расчета объемов
фитомассы ежегодного древесного отпада нами предлагается
следующая формула:
= [Удр(х+л)>Р]:Пр
100
где: Ve о — объем ежегодного древесного отпада, м3та1;
Удр(х+л) — запас древостоя (хвойные + лиственные породы),
м3та1; О — отношение древесного отпада к запасу древостоя
(от 1 до 100); Пр — средняя величина периода разложения
древесного отпада до третьей стадии, лет.
Фитомасса рассчитывается через умножение запасов древо-
стоев и объемов древесного отпада на конверсионные
коэффициенты, определенные для отдельных пород, широтных полос,
групп возраста (Замолодчиков, Уткин, Честных, 2003) и могут
закладываться в вышеприведенную формулу.
Таежные леса по версии этих авторов входят в две широтные
полосы. В первую широтную полосу входят леса северной
подзоны тайги, во вторую широтную полосу входят леса средней и
южной подзон тайги. При расчетах показателей древесного
отпада модальных лесов следует пользоваться конверсионными
коэффициентами, относящимися к группе возраста спелые и
перестойные. Для подзоны северной тайги (1-я широтная полоса)
конверсионные коэффициенты составляют для ели — 0,750±0,039;
для сосны — 0,661±0,024; для березы — 0,845±0,089; для
осины— 0,730±0,113. Для подзоны средней и южной тайги (2-я
широтная полоса) конверсионные коэффициенты составляют
для ели — 0,681±0,030; для сосны — 0,646±0,028; для березы —
0,738±0,044; для осины — 0,730±0,113.
- 106-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
В модальных естественно развивающихся лесах хвойных
формаций в состав древостоев обязательной примесью входят
лиственные породы — береза и осина. В среднем эта примесь
составляет 2 единицы, поэтому расчет объемов и фитомассы
древесного отпада складывался из суммы этих показателей хвойной
и лиственной части древостоев в соотношении 80 % хвойных к
20 % лиственных. Таким образом, таблицы древесного отпада
для лесов еловых формаций зоны тайги имеют следующий вид.
Таблица 29
Таблица средних значений запасов древостоев, объемов древесного
отпада и фитомассы для коренных еловых разновозрастных лесов
таежной зоны (числитель — запас, объем, м^га*1; знаменатель —
фитомасса тта"1)
Запас
древостоя
Общий
древесный
отпад 1-3-й
стадий
разложения
Отношение
древесного
отпада
к запасу
древостоя,%
Ежегодный
древесный
отпад
Период
разложения
древесного
отпада до
3-й стадии, лет |
1 Северная тайга |
120
90
140
105
160
120
12
9
24**
18
36
27
4£
36
№
45
14
11
28
21
42
32
42
70
53
16
12
32
24
10
20
30
40
50
10
20
30
40
50
10
20
0.27
0,20
0.53
0,40
0J
0,6
1.06
0,79
1.33
1,0
0.31
0,23
0*62
0,46
0.93
0,70
1.24
0,93
1.55
1,16
0.35
0.26
0.71
0,53
45 1
45
45
-107-
В. Г. Стороженко
Табл. 29 (продолжение)
Запас
древостоя
180
135
■ 200
150
Общий
древесный
отпад 1-3-й
стадии
разложения
48
36
48
Ш
60
18
14
36
27
54
41
72
54
68
20
15
40
30
60
45
Ш
60
100
75
Отношение
древесного
отпада
к запасу
древостоя,%
30
40
50
10
20
30
40
50
10
20
30
40
50
Ежегодный
древесный
отпад
1.06
0,79
1.42
1,06
Ы1
1,33
ол
0,3
0,6
1,2
0,9
1*6
1,2
2*0
1,5
0.44
0,33
0.88
0,66
1.33
1,0
1.78
1,33
2.22
1,66
Период
разложения
древесного
отпада до
3-й стадии, лет
45
45
Средняя тайга |
220
150
240
163
22
15
44
30
66
45
60
110
75
24
1 16
10
20
30
40
50
10
0.55
0,37
и,
0,75
1.65
1,12
2.2
1,5
2.75
1,87
ол
0,4
40
40
-108-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 29 (продолжение)
Запас
древостоя
260
I 177
280
190
300
204
320
218
Общий
древесный
отпад 1-3-й
стадий
разложения
48
33
12
49
96
65
120
82
23.
18
52
35
18
53
i ш
71
130
88
28
19
56
38
84
57
112
76
140
95
20
60
41
90
61
120
82
150
102
32
22
Отношение
древесного
отпада
к запасу
древостоя,%
20
30
40
50
10
20
30
40
50
10
20
30
40
50
10
20
30
40
50
10
Ежегодный
древесный
отпад
0,8
1,2
2,4
1,6
3.0
2,0
0.65
0,44
1Л
0,9
1.95
1,33
1,8
3.25
2,2
0,7
0,5
14
0,9
2л1
1,4
2.8
1,9
3.5
2,4
0.75
0,51
1.5
1,0
2.25
1,5
3.
2
3.75
2,55
0,5
Период
! разложения
древесного
отпада до
3-й стадии, лет
40 |
40
40
40
- 109-
В Г Стороженко
Табл. 29 (продолжение)
Запас
древостоя
340
232
Общий
древесный
отпад 1-3-й
стадий
разложения
64
44
96
65
128
87
160
109
23
68
46
102
69
ш
92
170
115
Отношение
древесного
отпада
к запасу
древостоя,%
20
30
40
50
10
20
30
40
50
Ежегодный
древесный
отпад
1Д
2Л
1,6
3,2
2,2
3,0
0.85
0,57
1,1
2,55
1,70
2А
2,3
4.25
2,90
Период
разложения
древесного
отпада до
3-й стадии, лет
40
Южная тайга
360
245
380
259
36
24
72
49
108
73
144
98
180
122
Ш
26
76
52
114
78
152
103
190
129
10
20
30
40
50
10
20
30
40
50
1.03
0,70
2.06
1,40
3.08
2,10
4JJL
2,8
5.14
3,50
1.08
0,73
2.17
1,48
3,26
2,22
4.34
2,95
5.43
3,70
35
35
-ПО-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 29 (продолжение)
Запас
древостоя
400
272
420
286
440
1 300
460
313
Общий
древесный
отпад 1-3-й
стадий
разложения
40
27
80
54
120
82
160
109
200
136
4£
29
84
57
126
86
168
114
210
143
44
30
§8
60
132
90
176
120
220
150
46
31
92
63
138
94
184
125
230
157
Отношение
древесного
отпада
к запасу
древостоя, %
10
20
30
40
50
10
20
30
40
50
10
20
30
40
50
10
20
30
40
50
Ежегодный
древесный
отпад
1.14
0,77
2.28
1,55
3.43
2.34
4.57
3,11
5.71
3,89
0,8
2,4
1,6
М 1
2,5
L& 1
3,3
6.0
4,0
1.26
0,85 1
2,51
1,71
3.77
2,56
5.03
3,42
6.28
4,28
1.31
0,89
2Ш
1,79
3.94
2,68
5.26
3,58 !
6.57
4,47
Период
разложения
древесного
отпада до
3-й стадии, лет
35
35
35
35
-111-
В Г Стороженко
Табл. 29 (продолжение)
Запас
древостоя
480
327
Общий
древесный
отпад 1-3-й
стадий
разложения
4£
33
96
65
144
98
192
131
240
163
Отношение
древесного
отпада
к запасу
древостоя,%
10
20
30
40
50
Ежегодный
древесный
отпад
1.37
0,93
2.74
1,86
Ml
2,80
5.48
3,73
6.86
4,67
Период
разложения
древесного
отпада до
3-й стадии, лет
35
* Приведены величины периода разложения стволов древесного отпада
диаметром в среднем 30 см.
** Выделенный шрифт — показатели климаксовых и близких к ним
сообществ.
Для лесов сосновых формаций таблица древесного отпада
имеет следующих вид.
Таблица 30
Таблица средних значений запасов древостоев, объемов древесного
отпада и фитомассы для коренных сосновых разновозрастных лесов
таёжной зоны (числитель — запас, объем, м^га1;
знаменатель — фитомасса, тта"1)
Запас
древостоя
i Общий
древесный
отпад
1-3-й стадий
разложения
Отношение
древесного
отпада к запасу
древостоя,%
Ежегодный
древесный
отпада
Период
разложения
древесного
отпада до
3-й стадии, лет*
Северная тайга
160
106
5
16**
11
24
16
32
21
5
10
15
20
QJ3.
0,11
0*32
0,21
&48
0,32
0.64
0,42
50
-112-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 30 (продолжение)
Запас
древостоя
180
119
200
132
220
145
Общий
древесный
отпад
1-3-й стадий
разложения
40
26
2
6
18
12
27
18
Ш
24
45
28
10
7
20
13
30
20
40
26
50
33
11
7
22
14
33
22
44
29
36
Отношение
древесного
отпада к запасу
древостоя,%
25
5
10
15
20
25
5
10
15
20
25
5
10
15
20
25
Ежегодный
древесный
отпада
0,8
0,5
0.18
0,12
0,36
0,24
0.54
0,36
0Л2
0,48
0.90
0,60
0.20
0,13
0.40
0,26
0.60
0,40
0.8
0,53
L0
0,66
0.22
0,14
0.44
0,29
0.66
0,44
0.88
0,58
1.10
0,72
Период 1
разложения
древесного
отпада до
3-й стадии, лет*
50
50
50
1 Средняя тайга
240
155
12
8
24
16
36
23
5
10
15
0.27
0,17
0.53
0,34
0.80
0,52
45
-113-
В. Г. Стороженко
Табл. 30 (продолжение)
Запас
древостоя
260
168
280
181
300
194
320
207
Общий
древесный
отпад
1-3-й стадий
разложения !
48
31
00
39
13
8
26
17
39
25
£2
34
£5
42
14
9
28
18
42
27
56
36
m
45
15
10
30
19
45
29
60
39
15
48
16
10
I 32
1 21
48
31
Отношение
древесного
отпада к запасу
древостоя,%
20
25
5
10
15
20
25
5
10
15
20
25
5
10
15
20
25
5
10
15
Ежегодный
древесный
отпада
1.06
0,68
0,86
0.29
0,19
0.58
0,37
0.87
0,56
1.15
0,74
1.44
0,93 .
0.31
0,20
0,62
0,40
0.93
0,60
0,80
1.55
1,0
0.33
0,21
0.67
0,43
laO
0,64
1.33
0,86
1.67
1,08
0.35
0,22
0.71
0,46
1.06
1 0,68
Период
разложения
древесного
отпада до
3-й стадии, лет*
45
45
45
45 !
- 114-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 30 (продолжение)
Запас
древостоя
340
220
Общий
древесный
отпад
1-3-й стадий
разложения
64
41
80
52
11
11
34
22
51
33
44
85
55
Отношение
древесного
отпада к запасу
древостоя,%
20
25
5
10
15
20
25
Ежегодный
древесный
отпада
1.42
0,92
1.78
1,15
0.38
0,24
0,75
0,48
L13
0,73
1.51
0,97
1.89
1,22
Период
разложения
древесного
отпада до
3-й стадии, лет*
45
Южная тайга*** |
360
233
380
245
400
258
18
12
36
23
54
35
72
47
на
58
19
12
38
25
57
37
16
49
61
Ж
13
40
26
60
5
10
15
20
25
5
10
15
20
25
5
10
15
0.45
0,29
0,90
0,60
1.35
0,87
1.80
1,20
2.25
1,45
QA1
0,30
0.95
0,61
1л42
0,92
1.90
1,23
2*£7
1,53
0.50
0,32
L0
0,64
1.50
40
40
40
-115-
В. Г. Стороженко
Табл. 30 (продолжение)
Запас
| древостоя
420
271
440
284
460
297
Общий
древесный
отпад
1-3-й стадий
разложения
80
52
100
66
21
14
42
27
63
41
84
54
105
68
22
14
44
28
66
43
88
57
1Ш
71
Ш
15
46
30
69
45
59
115
74
Отношение
древесного
отпада к запасу
древостоя,%
20
25
5
10
15
20
25
5
10
15
20
25
5
10
15
20
25
Ежегодный
древесный
отпада
Ш
1,3
2.50
1,61
0.52
0,34
1.05
0,68
1.57
1,01
2.10
1,35
2.63
1,70
0.55
0,35
1.10
0,71
1.65
1,07
2.20
1,42
2.75
1,78
0.57
0,37
1.15
0,74
1.72
1Д1
2£
| 1,49
2.87
1 1,85
Период 1
разложения
древесного
отпада до
3-й стадии, лет*
40
40
40
* Приведены величины периода разложения стволов древесного отпада
диаметром в среднем 30 см.
** Выделенный шрифт — показатели климаксовых и близких к ним
сообществ.
*** Приведенные для сосняков южной тайги данные следует рассматривать
как гипотетические, так как девственных разновозрастных сосняков в этой зоне
не встречено.
-116-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Приведенные данные представляют собой первый опыт
составления таблиц, в которых наряду с числовыми
количественными показателями используются качественные показатели
исследуемых величин. Полезной особенностью их является то, что,
изменяя переменные величины (с названиями столбцов «запасов
древостоев», «общего древесного отпада и «периода его
разложения»), можно находить искомые величины при любых
параметрах динамической структуры древостоев.
-117-
9. МИКОЦЕНОТИЧЕСКИЕ КРИТЕРИИ УСТОЙЧИВЫХ
ЛЕСНЫХ СООБЩЕСТВ
Грибная биота коренных разновозрастных девственных
лесов, как уже было определено выше, является неотъемлемой
составляющей структурного строения биогеоценоза. Микоценоз
формируется и развивается по своим эволюционным законам
оптимального строения и функционирования, которые
согласуются с динамическими процессами развития фитоценоза и
трактуются как закономерности. Длительные согласованные
исследования структур и функций фитоценозов и микоценозов
девственных эволюционно развивающихся лесных сообществ,
проведенные в массивах лесов разных зон растительности и
формационного состава, позволили выявить эти закономерности
и обозначить другие позиции и подходы к пониманию сущности
и значения грибной биоты в функционировании лесов
различного происхождения, структурных характеристик и
динамического положения. Но прежде чем перейти к изложению наших
экспериментальных исследований по изучению структур
микоценозов лесов, мы считаем необходимым изложить наши позиции по
содержанию некоторых терминов, применяемых в лесоводстве,
лесоведении, фитопатологии, микологии, напрямую связанных
с содержанием понятий устойчивости.
9.1. Понятия «больного» и «здорового» лесного сообщества
Понятия «больного» или «здорового» леса связывается с
деятельностью группы грибов, поражающих живые деревья,
вызывающих болезни отдельных деревьев или участвующих в
поражении древостоев, к которым относятся грибы биотрофы, по
типу питания входящие в группу облигатных паразитов,
факультативных сапротрофов или факультативных паразитов.
Наиболее наглядно наши позиции можно обозначить на
примерах деятельности грибов-дереворазрушителей биотрофной груп-
-118-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
пы, которые составляют устойчивые консортивные связи с
высшими растениями. Несколько особняком стоит группа грибов
облигатных паразитов, о которых будет идти речь ниже, в
разделе, в котором обосновывается выделение «микоценологии» как
самостоятельного направления в биогеоценологии.
Пораженность лесов дереворазрушающими грибами, как
одна из эндогенных характеристик качественного состояния
биогеоценозов, может оцениваться с разных позиций
по-разному. Это во многих случаях определяется неправильной оценкой
действительного, определенного природой, положения грибов
биотрофной группы в лесных экосистемах и эволюционных
задач самого лесного сообщества. Зачастую оценка их
деятельности рассматривается с антропоцентрических позиций, не
имеющих ничего общего с эволюционными задачами естественно
развивающихся лесов. Как ни странно, это во многом можно
объяснить с исторической и даже с политической точек зрения.
Можно предположить, что идеи великих русских лесоводов
Г.Ф. Морозова, А. Рудзкого, М.Е. Ткаченко, М.М. Орлова, В.Н.
Сукачева и др., не произойди в истории России социальных
катаклизмов, приведших к планомерному, целенаправленному
уничтожению лесов и повернувших лесную науку на служение
экономическим интересам страны, направили бы русское лесоводство
и лесное хозяйство в несколько другое русло. В значительно
большей степени при хозяйственном использовании лесов
учитывались бы свойства природных сообществ, закономерности их
эволюционного развития, их многогранное, не в последнюю
очередь экосистемное значение для жизни и благополучия
человека. К сожалению, только теперь, с отставанием почти в век,
когда структура лесного покрова изменена на огромных площадях,
когда произошли изменения в глобальных природных циклах,
человечество начинает привыкать к мысли о необходимости
согласовывать свои потребности с восстановительными
возможностями природных экосистем. В нашей стране потребности
промышленного использования природных ресурсов до сих пор во
многих случаях, а в лесном деле почти сплошь, перекрывают
необходимость принимать во внимание и считаться с
эволюционными задачами и возможностями природных экосистем. Именно
антропоцентрическое отношение к природе выработало по
отношению к ней и, в частности, к лесным экосистемам и биогеоцено-
- 119-
В. Г. Стороженко
зам, взятые из обихода человека, термины «болезнь», «больные
древостой», «здоровые древостой», «пораженность» и т.д. В
определенном понимании эти термины применять правомерно, но
в некоторых смыслах — нет. Правомерно, когда
рассматривается внедрение грибного возбудителя в отдельное растение, дерево.
В этом случае, действительно, грибной возбудитель вызывает его
болезнь, гниль древесины, в результате которой дерево
ослабляется и зачастую гибнет. Факт поражения или непоражения
определяется в этом случае таким качеством организма, как
иммунитет (Горленко, 1962). Когда же рассматривается лесное
сообщество как одна из структур природной экосистемы, как
социум, в котором социальные отношения регламентируются
взаимозависимыми связями огромного числа консортов, понятия
«больного» и «здорового» лесного сообщества с
антропоцентрическим наполнением совершенно не соответствует биологии
лесов, их эволюционным, динамическим задачам.
Для объяснения этих положений можно привести несколько
примеров, описывающих различные варианты участия дерево-
разрушающих грибов в генезисе лесных сообществ.
Предварительно следует сделать акцент на изложение одного
из основополагающих положений структурного строения лесных
сообществ. Оно заключается в том, что дереворазрушающие
грибы, как и вся грибная биота лесных биогеоценозов, является
одной из важнейших структур лесов, участвующая в
многогранных консортивных взаимоотношениях со всеми живыми и
неживыми членами биогеоценозов, сопровождающая их
формирование и развитие непрерывно на протяжении всей их исторической
жизни. Нет необходимости подробно объяснять важность их
функций в генезисе лесов. Мы это делали ранее и будем говорить
об этом ниже. Повторим только, что их значение для лесных
экосистем сравнимо разве только со значением для природных
экосистем фитоценоза.
Пример 1. Искусственные леса, культуры хвойных пород,
высаженные регулярным способом на площадях с
инфекционным фоном некоторых биотрофных дереворазрушающих грибов
из порядков Aphyllophorales (Heterobasidion annosum (Fr.)
Bref.— коревая губка) или порядка Agaricales (некоторые виды
рода Armillaria spp.).
-120-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Из практики лесозащиты хорошо известно, что такие
культуры очень часто подвержены очаговому поражению корневой
губкой или опенком, начиная с самого раннего возраста (Алексеев,
1974; Федоров,1984, Василяускас, 1989, Стороженко, 2002,
Rischbeth, 1963, Heterobasidion ..., 1998 и др.). Огромное
количество исследований посвящено разработке мер борьбы с этими
возбудителями. По нашим последним обследованиям, 50 %
культур Серебряноборского лесничества Института лесоведения
РАН имеют очаговое поражение корневой губкой, в то время как
в лесах естественного происхождения условно-одновозрастных
структур этого же лесничества очаговое поражение слабой
степени отмечено только в одном выделе. Причем выделы культур и
естественные сосняков примыкают друг к другу. В «Инструкции
по борьбе с корневой губкой сосны, ели и пихты в лесах СССР»
(1979) отмечено, что пораженность культур этим возбудителем
от 10 % до 40 % и выше характеризуется от слабой до сильной
степени и древостой отводятся в выборочную санитарную рубку,
а при уровнях поражения свыше 40 % — в сплошную
санитарную рубку. Судьба таких древостоев не определена. Во многих
случаях грибное поражение приводит к полной смене всего
биогеоценоза. В некоторых случаях, особенно когда культуры
высажены с учетом характеристик, присущих коренному экотопу,
древостой благополучно доживают до возраста спелости, иногда
перестоя, но всегда распадаются, и последующие поколения в
лучшем случае формируются по схеме начальных этапов
сукцессии коренных древостоев, в худшем случае со сменой эдификато-
ра. Такие древостой вступают в фазу разрушения фитоценоза, их
будущее неопределенно, и их поражение можно определять как
болезнь, а древостой — как больные.
Пример 2. Древостой хвойных пород естественного
происхождения, возникшие даже на площадях сельхозугодий или
аналогичных с культурами почвах, как правило, имеют
естественное возобновление коренных пород и не обязательно эдифи-
катора, редко подвержены очаговому поражению
перечисленными видами возбудителей в течение всего жизненного цикла. В то
же время к возрасту спелости, а тем более перестоя, в таких дре-
востоях накапливается поражение другими видами биотрофных
грибов. Зачастую общее поражение древостоя, складывающееся
-121-
В. Г. Стороженко
как сумма поражений отдельными видами дереворазрушающих
грибов, может превышать поражение древостоев при очаговом
распространении грибов и составлять от 20 до 40 % по числу
деревьев (Стороженко, 2002). По возрастной структуре такие
древостой относятся, как правило, к условно-одновозрастным, по
эволюционному развитию, или динамическому положению они
стоят на низшей ступени сукцессионной лестницы. Но их
перспективы вполне ясны. Отпад деревьев в таких древостоях более
растянут по времени, к возрасту распада основного полога они
обеспечены возобновлением коренной породы. Уже в этом случае
поражение нельзя относить к разряду болезни, а
характеризовать как механизм формирования более сложных структур
фитоценоза и в целом биогеоценоза. Такое сообщество вступило в
фазу формирования качества устойчивости.
Пример 3. Таежные коренные разновозрастные девственные
леса разных фаз динамики. Нашими долговременными
исследованиями лесов таежной зоны на Русской равнине определено,
что все они, каких бы динамических структур не были,
обязательно имеют в составе деревья, пораженные дереворазрушаю-
щими грибами биотрофного комплекса. Поражение древостоев в
таких лесах в фазах дигрессии может достигать 30-40 (45) % от
числа всех деревьев (Стороженко, 1998, 2002 и др.)» Никогда в
таких лесах не бывает очагового поражения каким-либо видом
дереворазрушающих грибов. Состав биотрофных видов,
поражающих живые деревья, не слишком обширен, но всегда включает
несколько видов, представленных в определенных пропорциях в
зависимости от сукцессионного положения лесного сообщества.
Всегда такие леса имеют возобновительные структуры,
способные обеспечить смену поколений. Благодаря всем этим
характеристикам поражение отдельных деревьев в сумме нельзя
определять как болезнь, и они в полной мере обладают качеством
устойчивости.
Сопоставляя приведенные примеры, нетрудно заметить, что
уровни поражения древостоев во всех трех случаях могут быть
близкими. Тем не менее во всех трех случаях характеристики
поражения и лесоводственные параметры древостоев в корне
отличаются друг от друга. Наглядно этот тезис подтверждается
данными таблицы.
-122-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таблица 31
Характеристики лесных сообществ различного происхождения
Происхождение
лесного
сообщества
Лесные культуры
хвойных пород
Условно-коренные, условно-
одновозрастные
леса
Коренные
разновозрастные
девственные леса
различных фаз
динамики
Характеристики
поражения
и возбудителя
Очаговое поражение
куртинного или
диффузного вида,
вызванное одним или
в редких случаях
двумя видами
биотрофных грибов
Очаговое поражение
встречается редко.
Диффузное
неочаговое поражения в
пределах 30-40 %
к возрасту спелости
вызывается
несколькими видами
биотрофов
Очаги биотрофного
поражения
отсутствуют. Диффузное
размещение
деревьев,
пораженных
различными видами дерево-
разрушающих
грибов
Лесоводственные характеристики
лесных сообществ.
Одновозрастные регулярные
культуры, высаженные на земли
с обедненным составом комплекса
почвенной микофлоры или
с инфекционным фоном
возбудителей очагового
распространения, без
возобновительных структур,
в некоренных для эдификатора
условиях роста
В коренных условиях роста
формируются естественные условно-одно-
возрастные леса смешанного состава
с естественным возобновлением
коренных и сопутствующих пород,
структурами валежа. Более богатый
почвенный фон микофлоры.
Характеристики, близкие к начальным
фазам сукцессионного развития
Коренные разновозрастные леса на
разных стадиях динамики,
мозаичной смены поколений,
оптимальные структуры подроста,
валежа и микоценоза
Приведенные характеристики обозначают только
схематичный облик различных по показателям поражения лесов.
Анализируя данные таблицы, следующим образом можно описать
понятия «больного» и «здорового» леса.
В обоих вариантах употребляется термин «поражение».
Однако значение этого термина в зависимости от его
употребления в вариантах также будет различно. Он вполне, если
относится к понятию «больного» сообщества. Один возбудитель,
поражающий и отдельные деревья и способный к очаговому
распространению по площади биогеоценоза, как бы нивелирует эти два
«субъекта» (отдельное дерево и биогеоценоз). В случае «здорово-
-123-
В. Г. Стороженко
го» леса биогеоценоз выступает как субъект социального
устройства, в котором возбудители болезней контролируются консор-
тивными связями, составляющих биогеоценоз консортов. Во
втором случае термин «поражение» в полной мере оправдан только
в одном применении — если он употребляется по отношению к
конкретному дереву, не способному сопротивляться внедрению
патогена, не обладающему достаточным индивидуальным
иммунитетом против возбудителя болезни.
Таким образом, можно различать два типа «поражения».
При первом типе поражения дереворазрушающие грибы в
социальной структуре сообщества выполняют функцию регулятора
структур фитоценоза, и такое поражение можно определить как
болезнь сообщества только в том случае, когда его
характеристики выходят за рамки необходимых для биогеоценоза параметров
и перспективы развития его становятся неопределенными. При
оптимальных для биогеоценоза характеристиках такое
поражение нельзя определить как болезнь. Второй тип «определяется
как поражение отдельного дерева в этом социуме, при котором
дерево действительно болеет. В этом случае, индивидуальное
поражение происходит по принципу отбора из сообщества
наименее устойчивых, обладающих наименьшим иммунитетом к
грибным возбудителям, экземпляров в рамках допустимого общего
для сообщества поражения, которое соответствует динамике,
структуре и сукцессионному положению сообщества.
В таком понимании термины «больной» и «здоровый»
биогеоценозы тесно связаны с понятиями «устойчивый» и
«неустойчивый» биогеоценоз.
Лесное сообщество, определенное как устойчивое, относится
к категории здоровых, так как грибной биотрофный комплекс
является в нем неотъемлемой и необходимой составляющей це-
нотической структурой социального субъекта, биогеоценоза, и
выполняет не разрушающую, а формирующую функцию в сук-
цессионном процессе.
Лесное сообщество, определенное как неустойчивое,
относится к категории больных, если возбудитель, в благоприятных для
воспитания высокой патогенности и агрессивности условиях,
выходит из-под контроля консортивных связей биогеоценоза и
выступает как разрушитель неустойчивого сообщества с
неопределенной перспективой формирования коренного древостоя.
-124-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таким образом, консортивные связи в лесном биогеоценозе
являются тем «механизмом», который определяет формирование в
лесном сообществе качества устойчивости. В качестве консортов
выступает огромное количество объектов фитоценоза, валежа,
геоценоза и зооценоза. В таблице 32 в дополнение к таблице 31
приводятся в кратком изложении внешние признаки больного и
здорового лесного сообщества
Таблица 32
Признаки больного и здорового лесного сообщества
Признаки больного сообщества
Поражение древостоев вызвано одним или
реже двумя видами возбудителей
Поражение деревьев одним видом гриба
носит куртинный или мелкогрупповой
характер
Поражение древостоев одним видом
возбудителя может составлять
от единичных до 100 % деревьев и не
связано с сукцессионным положением
сообщества
Как правило, искусственные монокультуры
или, значительно реже, естественные
условно-одновозрастные леса, возникшие
в измененых коренных экотопах, или в
некоренных условиях роста
Признаки здорового сообщества
Поражение вызвано несколькими
видами грибов, вызывающих
гнили разного качества
Пораженные различными видами 1
грибов деревья относительно
равномерно размещены по
площади биогеоценоза
Поражение древостоя составляет
определенную величину, но не
более 30-40 %, соответствующую
сукцесионному положению
сообщества
Разновозрастные, относительно- 1
разновозрастные или условно-
одновозрастные леса коренных
условий роста, стоящие в ряду
сукцессионных смен |
Повторим, что в качестве агентов поражения здесь имеется в
виду группа дереворазрушающих грибов биотрофного комплекса
микоценоза. Совсем другое поведение и взаимоотношения с авто-
трофами у группы грибов, обладающих свойствами облигатного
паразитизма, таких как ржавчинные грибы, вызывающие,
например, болезни хвои и листьев, некоторые раковые болезни и
некрозы.
Определившись, как мы считаем, с понятиями больного и
здорового лесного сообщества, мы считаем необходимым и
важным обозначить наши позиции в трактовке понятия «микоце-
ноз» и в обосновании нового направления в биогеоценологии —
лесной микоценологии.
- 125-
В. Г. Стороженко
9.2. Микоценоз и микоценология
До недавнего времени в лесоведении, лесоводстве и тем более
в лесном хозяйстве грибы и в том числе грибы дереворазрушаю-
щего комплекса не рассматривались как самостоятельная
структура лесного биогеоценоза. Этот тезис до сих пор
оспаривается, и грибную биоту предлагают считать одним из консортов,
входящих в фитоценоз. Эту точку зрения отстаивал один из
ведущих геоботаников МГУ — Т.А. Работнов, и вместе с ним
Т.Г. Мирчинк, А.П. Шенников, Д.Г. Звягинцев, В.В. Астапенко
и др. Т.А. Работнов дважды: в 1965, а затем в 1985 годах —
категорически выступал против понятия микоценоз. В то же время
каждый, кто проводил исследования в области биогеоценологии
и знаком с трудами основоположника этого учения В.Н.
Сукачева помнит содержание схемы построения биогеоценоза, которую
он привел в своей работе «Основы лесной биогеоценологии»
(1961). Изучая эту схему, можно видеть, какое большое значение
В.Н. Сукачев придавал именно такому устройству биогеоценоза,
в котором «микроценоз» занимает равноправное место с
фитоценозом и зооценозом и такому значению грибов в его структурах.
Если не принимать это устройство лесного биогеоценоза, а
именно так обстоит дело, когда отрицается структура микоценоза, то
надо предлагать что-то взамен, что объединяло бы продуценты и
редуценты в одной структуре. Взамен ничего не предлагалось, и
все возражения вылились в беспочвенную полемику,
затормозившую развитие лесной фитопатологии и микологии. В то же
время в лагере В.Н. Сукачева оставались многие ведущие
микологи и фитопатологи. К их числу можно отнести В.В. Мазинга,
К.А. Каламеэса, Л.Г. Бурову, Н.В. Черемисинова, В.А. Мухина,
Л.В. Гарибову, М.А. Бондарцеву, В.И. Крутова и многих других.
Мы останавливаемся на том, что микробоценоз, как
определил гетеротрофную составляющую биогеоценоза В.Н. Сукачев, а
в нашем понимании микоценоз, входит в состав биоты
биогеоценоза как самостоятельная структура, обладающая
таксономической обособленностью, морфологическим, экологическим и
функциональным строением, наравне с фитоценозом и зооценозом.
Понятие фитоценоза давно принято и устоялось в
лесоведении, широко обсуждается, структура его определена и является
-126-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
предметом изучения фундаментальных наук и практического
использования.
Таксономическая обособленность грибов обоснована макро-
таксономическим делением биоты Земли (Кусакин, Дроздов,
1994, 1998; Cavalier-Smith, 1995; Margulis, Schwartz, 1982 и
др.). Нелишне напомнить принятое в современной биологии
положение о происхождении и месте грибов в филеме
органического мира (Рис. 5, по Л.В. Гарибовой и С.Н. Лекомцевой).
Рис. 5. Положение грибов в системе живого мира
Сейчас сперы о том, являются ли грибы в лесном
биогеоценозе ценотической структурой или не являются, представляются
по меньшей мере некомпетентными.
Морфологическое, экологическое и функциональное
строение микоценоза обосновано нами в предыдущих работах (Сторо-
женко и др.,1992). Здесь мы в кратком изложении напомним
содержание этих исследований.
Морфологическое строение микоценоза складывается, так же
как и фитоценоза, во-первых, из микогоризонтов по высоте
расположения в фитоценозе — микогоризонты подстилочного слоя
(симбионты и разлагающие подстилку грибы), корневого и
комлевого слоя (дереворазрушающие грибы, поражающие корни и
комлевую части деревьев), стволового слоя (грибы, поражающие
стволовую часть деревьев), ассимиляционного слоя (грибы,
поражающие хвою и листву деревьев), древесного отпада (ксилотроф-
ный комплекс грибов). Каждому слою соответствует свой набор
видов, обладающих определенными специфическими
свойствами. Вторым легко фиксируемым в натурных наблюдениях,
свойством, доказывающим присутствие в биоте грибов морфоло-
-127-
В. Г. Стороженко
гического строения, является возможность расчленения его на
микромикоценозы, то есть обособленные друг от друга в
горизонтальной проекции группы грибов, связанные общим субстратом,
но отличающиеся от таких же групп экобиологическими
свойствами. При этом используются те же критерии, что и при
выделении микроценозов при расчленении фитоценоза на более
мелкие структуры.
Экологическое строение микоценоза складывается из мико-
синузий, так же как и фитоценоза. Микосинузии, как и
растительные синузии, можно классифицировать по различным
признакам, как пространственным, так и временным. В
вертикальном строении выделяются микосинузии микогоризонтов —
почвы, подстилки, корней, комлей, стволов, хвои (листвы).
В горизонтальном строении микоценоза выделяются
микосинузии микромикоценозов — отдельных деревьев, валежных
стволов, пней, и т.д. По типу питания можно различить
микосинузии паразитной группы грибов (поражают листья или хвою),
факультативной группы (поражают живые деревья), сапротрофной
группы (разлагают древесный отпад), симбиотрофной группы
(микоризные грибы). По времени функционирования можно
различить микосинузии кратковременные (например, действующие
в течение одного вегетационного периода), длительного
функционирования (например, микосинузии гнилей стволов,
разложения валежа) и т.д.
Функциональное строение микоценоза выстраивается по той
же схеме, что и фитоценоза, в котором за элементарную единицу
функционального строения выделяется консорция. Впервые это
понятие введено В.Н. Беклемишевым (1951), позднее многими
авторами оно детализовано и дополнено. Но наиболее ясно и
понятно оно сформулировано Е.М. Лавренко (1959) — «сочетание
популяций вида высшего растения в данном растительном
сообществе... со связанными с ним популяциями низших растений и
животных». По В.В. Мазингу растительная консорция
разделяется на два концентра, причем первый концентр составляют
организмы, питающиеся за счет автотрофов, второй —
организмы, питающиеся за счет представителей первого концентра.
По этой схеме к первому концентру микоценоза следует отнести
грибы-симбиотрофы, паразиты и грибы факультативной
группы, ко второму концентру — грибы некротрофы.
-128-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Обширнейшая группа грибов, разлагающих древесный
отпад, относится к редуцентам и её можно выделить, по нашему
представлению, в отдельную группу консорции мортмассы.
Таким образом, доказав присутствие у всего состава грибной
биоты биогеоценоза черт, характеризующих это сообщество как
самостоятельную структуру, обладающую морфологическим,
экологическим и функциональным строением, и, принимая во
внимание её макротаксономическую обособленность, можно
утверждать, что грибное сообщество представляет собой структуру
ценотического уровня в иерархии биогеоценоза.
Устойчивость растительного сообщества непосредственно
связана со структурным строением биогеоценоза и микоценоза.
Чем сложнее структурное строение биогеоценоза, чем
многообразнее его функциональное строение, тем сложнее и
разнообразнее (в пределах оптимальности) по видовому и функциональному
составу микоценоз и тем устойчивее это растительное
сообщество.
Таким образом, рассматривая микоценоз как равноправную с
фитоценозом структуру биогеоценоза вполне правомерно
допустить и вычленить из биогеоценологии, как научного
направления, отдельное самостоятельное научное направление — микоце-
нологию. Такая правомерность, однако, должна основываться на
доказательстве существования в предлагаемом научном
направлении определяющего этот термин содержания — всякое
научное направление должно основываться на закономерностях
поведения или функционирования объекта изучения. Объектом
нашего изучения явились грибы, осуществляющие разложение,
накапливаемой автотрофами биомассы — комплекс дереворазру-
шающих грибов биотрофной и сапротрофной групп, на примере
которых мы постараемся обосновать правомерность выделения
микоценологии как самостоятельного направления в
биогеоценологии. Закономерности поведения этой группы грибов
доказаны исследованиями, представленными ниже.
В любом лесном биогеоценозе биотрофный комплекс грибов
микоценоза складывается из групп грибов разных микогоризон-
тов и микосинузий. По функциональным характеристикам эти
группы грибов также различны и об этом мы говорили выше.
Довольно устойчивая связь отдельных видов и групп грибов с
морфологическим, экологическим и функциональным строени-
-129-
В. Г. Стороженко
ем биогеоценоза определяет степень свободы функционирования
и передвижения их в пространстве экосистем. Например, грибы
симбиотрофной группы жестко привязаны к биогоризонту
прикорневого слоя, грибы сапротрофной группы — к древесному
отпаду и отмершим частям растений, подстилки и корневому
опаду, грибы факультативной группы — к различным частям
стволов и ветвей автотрофов и также ограничены в передвижении в
пространстве. Особую группу грибов биотрофного комплекса в
этом смысле составляют грибы, обладающие свойствами обли-
гатного паразитизма. Как правило, в морфологическом строении
микоценоза эта группа грибов занимает микогоризонт
ассимиляционного слоя, поражая листву, хвою. В пространстве экосистем
эти грибы имеют широкую степень свободы и в наименьшей
степени контролируются консортивными связями как с высшими
растениями биогеоценозов, так и внутри микоценозов. Именно
поэтому такие грибы образуют пандемии. К их числу, как
известно, можно отнести мучнистую росу дуба {Microsphaera
alphitoides), ржавчину хвои ели (Chrysomyxa ledi), многие
другие мучнисто-росяные и ржавчинные грибы. Их
распространение определяется в основном наличием промежуточных хозяев и
климатическими условиями.
Группа грибов факультативных паразитов и факультативных
сапротрофов, к которым относятся рассматриваемые нами виды
дереворазрушающих грибов, довольно жестко связана
консортивными связями со всеми консортами биогеоценоза и за счет
этих связей имеет вполне определенные количественные и
качественные параметры, а в функциональном плане
взаимозависимое влияние с растениями фитоценоза. Это значит, что как
фитоценоз влияет на структуру и деятельность этой группы грибов,
так и грибы влияют на структуру и функционирование
фитоценоза. Из этих связей и складываются закономерности
функционирования грибов.
Изучение общих закономерностей функционирования грибов
в лесных биогеоценозах правильнее осуществлять в
незатронутых антропогеном, эволюционно развивающиеся сообществах,
девственных лесах, находящиеся на разных этапах сукцессион-
ного развития. Понятно, что прежде всего необходимо хорошо
ориентироваться в динамике самих биогеоценозов от начальных
-130-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
этапов до климакса, в чем мы пытались убедить читателя
исследованиями, представленными выше.
Понятно также, что изучение этих закономерностей
невозможно без изучения среды их обитания, то есть структур
фитоценоза, сукцессионных процессов развития биогеоценозов, ва-
лежных структур, геоморфологии территорий, климатических
факторов, определяющих зональность и типологию лесов. Эти
условия мы выполнили, представив результаты
вышеприведенных исследований. Одновременно с этим мы постараемся
выполнить главную задачу этого раздела работы — определения пара-'
метров участия дереворазрушающих грибов в
функционировании устойчивых лесных сообществ.
9.3. Параметры участия грибов дереворазрушающего
биотрофного комплекса в структурах устойчивых лесов
Представленными выше исследованиями выявлен ряд
закономерностей функционирования устойчивых лесных сообществ,
которые непосредственно связаны с характеристиками грибного
поражения коренных устойчивых биогеоценозов. К числу таких
характеристик относятся показатели структурного строения фи-
тоценозов (возрастные и горизонтальные структуры) и валежные
комплексы.
9.3.1. Связь грибного поражения коренных древостоев
с их возрастной структурой
Если соглашаться с коэволюционной динамикой
функционирования консортов в биогеоценозах, участие грибов
дереворазрушающего биотрофного комплекса в возрастных структурах
коренных разновозрастных древостоев должно быть подчинено
динамическим процессам формирования фитоценозов. В
разновозрастном сообществе деревья различных возрастных поколений
можно представить как слои. Каждый слой можно
рассматривать как ступень сукцессионного развития данного
биогеоценоза, где первый слой — это подрост, второй слой — подгон (жерд-
-131-
В. Г. Стороженко
няк), третий слой — приспевающие и спелые, четвертый слой
спелые и перестойные, далее — перестойные. Каждому слою
соответствует своя динамика отпада и причины ему
способствующие.
В возрасте жердняка, наиболее значимой причиной отпада
деревьев являются конкурентная борьба за свет и корневое
питание. В устойчивом сообществе в этот период деятельность грибов
биотрофного комплекса минимальна. После периода активной
дифференциации, когда все отношения между автотрофами в
основном выяснены и биогеоценоз накопил максимальную
биомассу, возникает необходимость в его омоложении и обеспечении
непрерывности воспроизводства. Это становится возможным при
постепенном высвобождении пространства, на котором
возникает новые поколения. Механизмом, способствующим
переживанию древостоя в этом возрастном периоде вплоть до предельного
перестоя, в устойчивом биогеоценозе служит комплекс биотроф-
ных дереворазрушающих грибов. В количественном выражении
этот процесс вполне объясняется данными наших исследований.
В таблице 33 приведены величины поражения коренных
биогеоценозов дереворазрушающими грибами биотрофного комплекса
по зонам растительности, возрастным группам и формациям по
данным конкретных пробных площадей, древостой которых
отражают наиболее представительные в формациях типы леса и
динамические разности биогеоценозов. Анализ таблицы
позволяет сделать ряд важных выводов, характеризующих
закономерности участия грибов биотрофной группы в
функционировании коренных разновозрастных лесов на Русской равнине.
1. В лиственных сообществах зоны лесостепи и смешанных
лесов наибольшие уровни поражения дереворазрушающими
грибами биотрофного комплекса коренных биогеоценозов
естественного происхождения отмечаются в дубовых и сосновых древосто-
ях глубоко перестойного возраста. Причем предельные уровни
поражения составляют не более 50 % по числу деревьев и
приходятся в основном на деревья первого яруса.
2. В сообществах хвойных пород коренных экотопов,
начиная от зоны смешанных лесов и кончая северной подзоной тайги,
средние значения пораженности как еловых, так и сосновых
лесов, во-первых, имеют сопоставимые величины, во-вторых, ле-
-132-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
жат в пределах 30 %. Более занчительные величины отмечены в
древостоях дигрессивных фаз динамики, где в первых
возрастных поколениях сосредоточено большое количество
перестойных деревьев. Ни в одной из более чем пятисот пробных
площадей, заложенных в коренных ельниках и сосняках тайги, не
зафиксированы более высокие уровни поражения грибами био-
трофного комплекса, что говорит о закономерных и достаточных
для коренных биогеоценозов уровнях распространения биотроф-
ных грибов, участвующих в процессах ослабления и отмирания
вполне определенного количества автотрофов, переходящих из
состава продуцентов в состав мортмассы.
3. В этих же сообществах прослеживается вполне
объяснимая и оговоренная выше закономерность нарастания пораженно-
сти по мере увеличения возраста деревьев, как следствие
активизации деятельности биотрофов в перестойной части древостоев.
Эти выводы обосновывают положение об оптимальном и
достаточном для устойчивых коренных биогеоценозов участии
грибов биотрофного комплекса в функционировании естественно
развивающихся лесов.
Не менее важной характеристикой комплекса биотрофных
дереворазрушающих грибов, распределяющихся в возрастных
структурах коренных лесов, являются их качественные
показатели, которые выражаются в соотношении деревьев в лесном
биогеоцеозе, пораженных возбудителями, вызывающими гнили
разных типов — коррозионные и деструктивные. Признаки
отнесения гнилей к определённому типу подробно описаны
(Ванин, Полубояринов, Соловьев, 1969). Здесь мы ограничимся
краткой справкой.
Гнили коррозионного типа вызываются грибами,
ферментативная система которых ориентирована на разложение лигнина
стенок клеток древесины. В результате их воздействия у
лиственных пород образуется гниль светлых окрасок волокнистого
типа. У хвойных пород гниль этого типа имеет более темные
тона — светло-охряные или охряные разной интенсивности с
выцветами белого или темного цвета живого или отмершего
мицелия гриба.
Гнили деструктивного типа вызываются грибами,
разлагающими целлюлозу древесины. Поражение сопровождается утра-
-133-
В. Г. Стороженко
той структуры древесины, гниль приобретает коричневую или
бурую окраску, растрескивается в диаметральном и продольном
направлениях, иногда приобретает порошкообразную структуру.
Деструктивные гнили носят общее название «бурые гнили».
Деление дереворазрушающих грибов биотрофного комплекса
по типу вызываемых ими гнилей имеет весьма важное значение
как качественная характеристика биотрофного поражения
лесов. Возбудители, вызывающие коррозионные гнили, относятся
по типу питания к факультативным сапротрофам, способным к
массовым очаговым распространениям. Это грибы с активной
ферментативной системой, более агрессивные по отношению к
автотрофам нежели грибы, вызывающие деструктивные гнили.
Среди дереворазрушителей-биотрофов эти виды входят в число
агентов, осуществляющих наиболее оперативное разрушение
неустойчивых по структурным параметрам древостоев.
Биотрофные дереворазрушители возбудители деструктивных
гнилей также относятся к факультативным сапротрофам, однако
они не имею способности к эпифитотическому распространению
и могут образовывать в отдельных случаях небольшие по
площади очаги из нескольких пораженных деревьев. Кстати сказать, в
современной фитопатологии и микологии явно прослеживается
путаница или неуверенность в употреблении терминов «биотро-
фы» и «ксилотрофы». Подробное объяснение нашего понимания
смысла этих терминов приведено в одной из наших монографий
(Стороженко, 2002, стр. 16-18). Здесь мы кратко обозначим, что
к биотрофам относятся грибы, обладающие в разной мере
паразитическими свойствами (облигатными или факультативными)
и поражающие живые растения. В нашем случае — это дерево-
разрушающие грибы, поражающие деревья. К ксилотрофам
относятся грибы, проявляющие только сапрофитические свойства,
и выполняющие функцию деструкции отмершей древесины.
Таким образом, разделяя общее поражение древостоев по
типу вызываемых грибами биотрофами гнилей, мы тем самым
даем качественную оценку этим комплексам по активности дере-
воразрушающего биотрофного поля биогеоценоза.
Соотношение видов биотрофов в устойчивых коренных лесах
выявлялось по результатам бурения стволов и определения типа
гнилей. В стационарных условиях из образцов гнилей выделя-
-134-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
со
со
сб
Я"
S
ч
ю
Ен
н
о
к
А
ч
£ к
S =
s §
н о
w S
в х
5 2
о) а
ч »
2 «
Й *
6 в
* ев
I!
к
ф
cd
a
о
И
8 а
0) О)
а а
гН О
со ^
СО м
si
т? 00
гЧ О
о ^
eg <м
гЧ О
со о
А8
тН О
Tf 00
о о
СО
о
к
У
S к ™
М нн О
Н Я Ф
а в я
I «*§
S §
Ч «J
си
«3
«
a
X И
со \о ►*>
,-1 О ft
в « н
5 оо
во
3 *
ев
PQ
о о
« VO
тН О
гЧ <Р
К Я" д
si*
5
2° «
Г? •« ев
*
п
2
3
н
Id
а в
S о
- 135
В. Г. Стороженко
/**ч
а;
жени
Е*
О
tt
О
в
со
со
ч
ю
об
и
яяпо
стою,
X О $^
£ 5Р
ф ф
ft ft
U fcC
^
S
деления
о
и
нх
о возрас
1 с
н
о
ораженн
U!
££
т* S
^ 2
«о 2
СО ^
£§
со «
281-
320
241-
280
201-
240
161-
200
121-
160
• о
00 ,-н
^ 9
т* 00
оо
Лесоводственная
характеристика лесного
сообщества
Tt*
,
•
'
О
СО
^
СМ
см
СО
О
1-4
О
+Ос; черничник;
бон.; Ар; Кл
« ^
9Е1
0,7;
00
°о
1
'
•
iOi
!гЧ
CM
16,5
см
ю
«
+Ос; кисл.-черн.;
бон.; Ар; Дг.
Ю 1"ц
9Е1
0,8;
о>
:rt«
,
'
•
о
СМ
Irt*
i-i
Т*
т-t
О
О
Б10с+С;сфагн.-
яич.; 0,7; П бон.; Ар;
00 V Е{
о
т-1
со
тН
'
'
'
О
ю
СО
Oi
О
О
+Е,Б; бруснично-
истый; 0,6; П бон.; Ор;
14
ига
S
№
1
^
СМ
•
'
О
ю
см
см
о
см
со
00
со
о
о
1Лп+Пх; кислично-
в.;0,8;1бон.;Ар;
8Е1Б
щито
Кл.
см
тЧ
СО
(
•
•
см
со
ХЛ
1Л
со
со
00
см
см
о
+Пх; черн.-кисл.-
ом.;0,8; 1 бон.; Ар;
8Е2Б
зелен
Кл.
со
т""1
1>
1-Н
,
•
•
СМ
со
со
1-4
СМ
Oi
|>
о
о
+С, Б; майник.-кисл.-
яич.; 0,8; П бон.; Ар;
Э5н
О ф Я
-*
тН
СО
,
•
•
00
гН
о
О
СО
о
о
о
Б;чернич.-
ювый; 0,8; Ш бон.;
м
+ S4
10Е
сфа
Ор;
ю
тН
1-1
1-4
,
•
'
'
О
оо
СМ
о
о
В; брусн.-
йн.;0,6;Шбон.;Ор;
10С+
лиша
со
^
ироген.
в
Дм;
эй тайги
X
i
о
Подзона
о
i
•
•
*-*
со
см
о
см
со
^
см
тН
о
10Е+Ос,Б; майник.-
черничн.; 0,8; П бон.; Р;
Дм,
*-
тН
136-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
о -
и 2
* 8
д о ^
fct m
ф ф
ft a
О к
-^
£
«
в
олен
ым пок
[HJ.C
возра
ность по
ажен
а
И
§;
со ч*
»Н О
со w
гН О
<м со
*н О
О ^
тН О
СО ©
Si
• о
00 гн
Дц О
^ 00
эсоводственная
теристика лесного
сообщества
ЧЕ
*
2 *
к
^
СМ
•
'
■
О
ю
о
тН
t>
СМ
СМ
00
О
О
со
о
Д
8*
+ д
W о
гЧ Ю
со
тЧ
1-Н
•
'
•
О
ь-
гН
со
см
со
■ Д
1>
1-4
о
В
о
ю
>>
Гг5
гЛ О
Т И
Ы еб
© .& i?
гН О О
Оз
^
СМ
СО
CM
00
<§
о
ftfl.
Tl - В
+ о о
О \о а
2Эё
О
CM
2
ей
Eh
аз
В
$
5
0>
•
'
•
О
uo
Tt<
CO
о
<N
l>
^
<N
CO
О
о
1
в
Si-
£ч Е->
И 3 2
см
CS1
^
•
со
со
СО
о
*л
о
ь-
со
rj<
OS
со
СО
со
i-Ч
о
о
а
<
• д
О о
д ю
о »
+ 3
CQ о
со 2 К
СМ
СМ
t>
см
■
•
•
о
ю
СМ
СМ
см
О
СО
о
о
§1
в т
в я
С со
3%
И я ч
оо 2 К
со
см
СО
т-Ч
со
о
со
о
СО
со
со
СЧ
см
со
см
о
1-1
о
о
i
3
la.
а т
?^
ii О
rj<
CSJ
СО
тН
•
'
-
О
»л
СО
СО
ю
са
00
о
о
о
1
1И
2 о
О ей
1Л
СМ
cd W s
И $ S
s g s
ей О аЗ
«ИМ
2 8 s
ьГ ^» ей
g Be
й 3
^ S л
ой Я О
О, Ф 0Q
о g о
О С, №
И ft 3
| * 6
в S
ей
Ь*
О
в
§
Н ей
* Я
о
в
о „
ft &*
н о
. ft
со О
я ..
»s
3
в
Е*
ей
В
Ф
5
CQ
В
ft
в
ф ОЙ
£ в
X В
ой fee
х 3
« -ft
ф .
S s
ft в
И 2
В
о о
SS
^в
о ч
Si
0 ..
ей к
1 Л
< s
ей
£ S
-137
В. Г. Стороженко
лись чистые культуры грибов и определялся вид возбудителя.
Общей для всех лесов закономерностью явилось стремление
искомого соотношения к равным количествам деревьев с
коррозионными и деструктивными гнилями, то есть к 1, в наиболее
сбалансированных климаксовых сообществах. С продвижением в
область демутации или дигрессии этот баланс нарушается в
сторону преобладания количества деревьев, пораженных одним или
несколькими видами грибов, вызывающих коррозионные или
деструктивные гнили. При увеличении доли деревьев с
коррозионными гнилями в фазах демутации видовой состав
возбудителей, вызывающих эти гнили, как правило, состоит из
нескольких видов с преобладанием наиболее активных. С углублением в
фазу дигрессии возрастает не только количество деревьев с
присутствием деструктивных гнилей, но и общий видовой состав
различных биотрофных дереворазрушающих грибов, во многих
случаях с преобладанием возбудителей, вызывающих
деструктивные гнили. Этим закономерностям есть вполне логичное
объяснение. В фазе демутации возрастной состав древостоя
омолаживается, приобретает более однородную структуру, деревья
имеют высокую физиологическую активность и доступны для
внедрения очень активных видов биотрофных дереворазрушите-
лей, вызывающих коррозионные гнили. В фазах дигрессии в
древостое накапливается большое количество деревьев
предельных для условий экотопа возрастов, физиологическая
активность которых значительно снижена и такие деревья доступны
для внедрения менее активных по отношению к живым автотро-
фам возбудителей. К тому же к этому возрасту в древостое
накапливаются механические повреждения стволов, которые
облегчают внедрение в деревья патогенов с меньшей активностью.
Таким образом, динамическая структура биогеоценоза в
определенном смысле «диктует» микоценозу его состав и активность, а
микоценоз, в свою очередь «диктует» фитоценозу интенсивность
процесса отпада деревьев из древостоя. В общем виде
полученную закономерность можно представить следующим рисунком.
-138-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Возбудители, вызывающие
коррозионные гнили
Возбудители, вызывающие
деструктивные гнили
Фаза демутации
Клтгжс
соотношение видов
стремится к !
Фаза дигрессии
Увеличение доли одного вида
гриба и гнилей одного типа в
общем поражении
Некоторое увеличение количества
разных видов грибов и доли гнилей
одного типа в общем поражении
Рис. 6. Схематическое изображение закономерностей участия возбудителей
разных видов, вызывающих гнили разных типов в общем поражении
коренных устойчивых лесов
9.3.2. Связь грибного поражения древостоев
с их горизонтальной структурой
Одной неважных характеристик поражения грибами био-
трофного комплекса устойчивых коренных разновозрастных
лесов, то есть характеристик микоценозов, является размещение
пораженных деревьев по площади биогеоценоза. Исследования
такого рода в лесной фитопатологии вообще не проводились. В то
же время, по нашему мнению, они являются важными для
изучения структурного строения микоценозов разновозрастных
лесов и могут в определенной мере ответить на вопрос о степени
влияния грибов биотрофного комплекса на формирование
горизонтальных структур фитоценозов, на тенденции
горизонтального размещения древесного отпада, связанного с поражением
деревьев дереворазрушающими грибами.
Анализ размещения деревьев, пораженных биотрофами,
осуществлялся по той же методике Б.А. Быкова (1976), которая
применялась и при анализе горизонтальных структур
фитоценозов (раздел 5.4.). Для примера приведем расчеты колебаний
значений агрегированности размещения деревьев, пораженных
грибами биотрофного комплекса в ельниках Центрально-лесного
биосферного заповедника и резервата «Кологривский лес». Пред-
-139-
ВТ. Стороженко
ставлены только табличные данные, в которых, кроме
распределения пораженных деревьев, независимо от типа гнилей,
приведены также значения агрегированности деревьев, пораженных
отдельно гнилями коррозионного и деструктивного типов.
Таблица 34
Показатели агрегированности размещения деревьев, пораженных
грибами биотрофного комплекса в устойчивых коренных
разновозрастных лесах (на примере ельников ЦЛБГЗ и Кологрива),
х — расстояние между пораженными деревьями на площади
биогеоценоза
№
п.п.
Тип гнили
Показатели и их статистики
см
m;.
а, СМ
СМ
С,%
тс
Характеристика
агрегированности
Резерват «Кологривский лес» |
1
2
3
Все гнили
Деструктивные
Коррозионные
Все гнили
Деструктивные
Коррозионные
Все гнили
Деструктивные
Коррозионные
895
1498
1253
1000
2106
1146
736
1236
936
49
115
100
46
206
64
39
74
100
245
366
318
233
651
203
215
234
316
34
83
72
32
148
46
27
53
72
27,3
24,4
25,3
23,3
30,9
17,7
29,3
18,9
33,7
4,1 4 Большая
5,4
5,9
3,0
6,8
3,9
4,2
4,1
7,2
Большая
Большая
Значительная
Большая
Значительная
Большая
Большая
Большая
Центрально-лесной биосферный заповедник !
1
2
3
Все гнили
Деструктивные
Коррозионные
Все гнили
Деструктивные
Коррозионные
Все гнили
| Деструктивные
| Коррозионные
1440
1961
2253
1205
2184
1483
1246
1582
11988
54
285
156
43
307
103
49
76
177
320
901
494
222
972
326
237
241
559
41
205
112
31
221
74
35
58
127
29
46
22
23
45
22
31
15
28
5,0
10,1
4,8
5,5
9,8
4,8
4,8
3,3
6,8
Большая
Большая
Большая
Большая
Большая
Большая
Большая
Значительная
Большая
Сравнивая колебания значений агрегированности
размещения всех деревьев фитоценоза и только имеющих гнили разного
типа, можно видеть, что в их пространственном распределении
по площади сообщества наблюдается идентичная тенденция
равномерно-диффузного размещения. Разница заключается только
в значительно больших расстояниях между деревьями, так как
-140-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
пораженных деревьев меньше и расстояния между ними,
естественно, тоже больше. Такие же закономерности присущи всем
исследованным нами разновозрастным устойчивым лесам
независимо от зон растительности и лесных формаций.
Из всего вышеизложенного следует вывод о том, что в
коренных разновозрастных лесах деревья, пораженные дереворазру-
шающими биотрофными грибами независимо от величины
общего поражения древостоя относительно равномерно
распределяются по площади сообщества, поражая отдельные деревья.
Очагового поражения деревьев и активизации распространения
какого-то одного вида биотрофа в таких лесах нет. Такое
распределение биотрофных грибов в горизонтальных структурах лесов
относится к разряду закономерностей их функционирования и
является одним из критериев устойчивости лесных сообществ.
Такая же тенденция характерна и для лесов более простых
структур и даже для лесных культур, созданных в коренных для
эдификатора условиях роста с соблюдением всех возможных
условий, присущих коренному лесу. Примером могут служить
культуры К.Ф. Тюрмера в Порецком лесничестве Бородинского
лесхоза Московской области. До недавнего времени очагов
распространения дереворазрушающих грибов в них практически не
было, несмотря на предельный во многих случаях возраст древо-
стоев для этих условий. Справедливости ради следует добавить,
что статус культур, то есть неустойчивых сообществ, все же
привел эти древостой сначала к локальным вывалам и буреломам, а
затем к массовым усыханиям в результате распространения
короеда-типографа и, как итог, к сплошным рубкам древостоев.
9.3.3. Особенности видового состава комплекса
дереворазрушающих биотрофных грибов коренных
разновозрастных устойчивых лесов
Видовой состав грибного биотрофного дереворазрушающего
комплекса изучен далеко не полно. Известны только основные
виды дереворазрушителей, вызывающих гнили коррозионного
или деструктивного типа. Во многих случаях виды возбудителей
гнилей живых деревьев возможно определить только путем
выращивания чистых культур и идентификации возбудителя по
-141-
В. Г. Стороженко
определителям чистых культур, которых в отечественной
литературе практически нет. Определение по весьма редким
.зарубежным изданиям (Nobles, 1948, 1965; Stalpers, 1978) очень
затруднено из-за сложности методик и долговременности процесса
идентификации. Значительно более полно изучен видовой состав
сапротрофного комплекса (Бондарцев, 1953; Бондарцева, 1986,
1992; Martin, Gilbertson, 1977,1978,1980; Nordic Macromycetes,
1977 и др.)
Наши исследования не ставили целью выявления всего
состава биотрофных и сапротрофных дереворазрушающих грибов на
всех породах лесной зоны Русской равнины. Однако наиболее
часто встречающиеся виды приведены нами ниже.
Таблица 35
Наиболее часто встречающиеся виды дереворазрушающих грибов,
проявляющие факультативные и сапрофитические свойства
на основных древесных породах Русской равнины
№
п.п.
Вид гриба
Порода
Встречаемость
1 Дереворазрушающие грибы, обладающие факультативными свойствами
(биотрофы)
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
Antrodia albida (Fr.) Donk.
Armillaria spp.
Bjerkandera adust a (Fr.) Karst.
B. fumosa (Fr.) Karst.
Climacocystis borealis (Fr.) Kotl. et
Pouz.
Collybia velutipes (Fr.) Quel.
Daedalea quercina Fr.
Diplomitoporus flavescens (Bress.)
Dom.
Fistulina hepatica Fr
Forties fomentarius (L.:Fr.) Fr.
Fomitopsis cajandery (Karst.) Kotl. et
Pouz.
F.pinicola (Sw.: Fr.) Karst.
F. rosea (Alb.et Schw.) Karst.
Ganoderma lipsience (Batsch.) Atk.
G. resinaceum Bourd et Pat.
Hapalopilus croceus (Pers.: Fr.) Donk.
Heterobasidion annosum (Fr.) Bref.
Hymenochaete tabacina (Sow.: Fr.)
Lev.
Л
Х,Л
Х,Л
Л
Х(Л)
л
Д,Кл,Яс
С(Х)
д
л
Е,Пх,С
Х,Л
X
Л(Х)
Л
л
Х(Л)
Д(Л)
Редко
Часто, Оч
Регулярно
Редко
Регулярно
Регулярно
Часто
Редко
Регулярно
Часто
Редко
Часто
Часто
Часто
Регулярно
Редко
Часто, Оч
Регулярно
-142-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 35 (продолжение)
п.п.
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
Вид гриба
Inonotus cuticularis (Bull.iFr.) Karst.
/. dryadeus (Pers.: Fr.) Murr.
/. dryophilus i Berk.)Murr.
/. hispidus (Fr.) Karst.
/. nidus-pici Pil.
/. obliguus (Pers.: Fr.) Pil.
/. obliguus — form, sterllis (Vanin)
Nicol.
/. rheades (Pers.)Bond. et Sing.
Ischnoderma bensolnum (Wahl.:Fr.)
Karst.
/. reslnosum (Fr.) Karst.
Laetiporus sulphureus (Bull.: Fr.)
Murr.
Onnia trlqueter (Lentz.: Fr.) Imaz.
Oxyporus popullnus (Schum.: Fr.)
Donk.
Perenniporia fraxinea (Fr.) Ryv.
Phaeolus schweinitzii (Fr.) Pat.
Phelllnus alni (Bond.) Parm.
Ph. chrysoToma (Fr.) Donk.
Ph. conchatus (Pers.: Fr.) Quel.
Ph. hartlgil (Allesch. Et Schn.) Bond.
Ph. ignlarlus (L.: Fr.) Quel.
Ph. pinl (Thore.: Fr.) Pil.
Ph. robustus (Karst.) Bourd.et Galz.
Ph. tremulae (Bond.) Bond, et Boris.
Pholiota adlposa Fr.
Ph. aurlvella (Fr.) Kumm.
Ph. squarrosa Karst.
Pleurotus ostreatus Jacq.
Polyporus alveolarius (DC.:Fr.) Bond
et Sing.
Pol. squamosus Huds. :Fr
Spongipellis spumeus (Sow.:Fr.) Pat.
Stereum gausapatum (Fr.) Fr.
St. hirsutum (Fr.) Gray.
St. sanguinolentum (Fr.) Fr.
St. frustilosum Fr
Trametes gibbosa (Pers.) Fr. 1
Tyromyces fissilis (Berk, et Curt.)
Donk. |
Порода
Л
!Д(Кл,Вз,Яс)
д
л
д
Б,Ол,Кл,Яс,Вз
Б,Ол
Д,Б,Ос,Ив,Ол
Х(Л)
Л
Л(Х)
Е,Пх,С
Кл,Лп
Яс,Д,0с,Вз
X
Ол
Е,Пх,Лц,С
Л
Пх,Е,С
Л
С
Д(Л) 1
Ос
х,л
л
л
л
л
Л(Х)
л
Д(Л)
л,х
X
д,л
л
Л(Х)
Встречаемость
Регулярно
Часто
Редко
Редко
Редко !
Часто
Часто
Регулярно
Редко 1
Редко
Часто 1
Часто, Оч
Часто |
Редко 1
Часто
Регулярно 1
Часто, Оч 1
Регулярно
Редко 1
Часто 1
Часто, Оч
Часто
Часто, Оч
Регулярно 1
Редко
Регул я но
Регулярно
Редко
Регулярно
Редко
Регулярно
Регулярно
Часто
Редко
Редко 1
Редко
-143-
В. Г. Стороженко
Табл. 35 (продолжение)
п.п.
Вид гриба
Дереворазрушающие грибы сап
| 55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
' 83
84
85
86
87
88
Antrodia albida (Fr.) Donk.
Antrodia crassa (Karst.) Ryv.
A. mollis (Somm.ex Fr.)
A. seriales (Fr.) Donk.
A. xantha (Fr.) Ryv.
A. sinuosa (Fr.) Karst.
Antrodiella semisupina (Berk, et
Curt.) Ryv.
Ar miliaria spp.
Bjerkandera adusta (Fr.) Karst.
B. fumosa (Fr.) Karst.
Ceriporiopsis aneirina (Somm.) Donk.
C. pannocineta (Rom.) Gilb.et Ryv.
1 Cerrena unicolor (Bull.: Fr.) Murr.
Climacocystis borealis (Fr.) Kotl. et
Pouz.
Coriolopsis trogii (Berk.) Dom.
Coriolus pubescens (Schum.:Fr) Quel.
Daedalea quercina Fr.
Daedaleopsis confragosa (Bolt.: Fr.)
Schr.
D. tricolor (Bull.: Fr.) Bond. etSing.
Datronia spereoides (Fr.:Fr.) Ryv.
Diplomitoporus lenis (Karst.) Gilb. et
Ryv.
Fibuloporia donkii Donk.
Fomitopsis cajandery (Karst.) Kotl. et
Pouz
F. pinicola (Sw.: Fr.) Karst.
F. rosea (Alb.et Schw.) Karst.
F. subrosea (Weir.) Bond, et Sing
Fomes fomentarius (L.:Fr.) Fr.
Ganoderma lipsience (Batsch.) Atk.
G. lucidum (Fr.) Karst.
G. resinaceum Bond et Pat.
Gloeophyllum trabeum (Pers.: Fr.)
Murr.
Gl. abietinum (Fr.) Karst.
Gl. odoratum (Wulf.:Fr.) Imaz.
Gl. sepiarium (Fr.) Karst.
Порода
Встречаемость
ротрофы (ксилотрофы) 1
Ос
X
Б,Д,Лп,Ос
Е,С
X
Е,С
Яс,Д,Лп
Х,Л
Л
Яс,Кл,Вз
Л
Л
Л
Е,С,Пх
Л
Ос
Д
л
Ив,Б,Ос
К,Б,Д,Ос
С,Е
С
Е.С
Л,Х
Е,С,Пх
X
Л
Л
Кл,Б,Яс,Ос
Д,Яс,Кл
Д,0с,Вз
Е,С
Е,С
Е,С
Редко
Регулярно
Часто 1
Редко
Регулярно 1
Редко 1
Регулярно
Регулярно
Редко 1
Редко 1
Редко 1
Регулярно
Регулярно
Редко
Регулярно
Часто 1
Регулярно
Регулярно
Редко
Редко
Редко |
Редко
Часто 1
Часто 1
Редко 1
Часто 1
Часто
Регулярно
Регулярно
Редко
Регулярно
Регулярно
Регулярно
-144-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 35 (продолжение)
№
п.п.
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
.113
114
115
116
117
118
119
120
121
122
Вид гриба
Gloeoporus taxicola (Pers.:Fr.) Gil. et
Ryv.
Gloeoporus dichrous (Fr.) Bress.
Hapalopilus nidulans (Fr.) Karst.
Hapalopilus salmonicolor
(Berk.etCurt.)Pouz.
Haploporus odorus (Somm.iFr.) Sing.
Hirschioporus pergameus (Fr.) Bond,
et Sing.
Hymenochaete rubiginosa (Dick.:Fr.)
Lev.
Hymenochaete tabacina (Sow.: Fr.)
Lev.
Inonotus obliguus (Pers.: Fr.) Pil.
/. obliguus — form, sterilis (Vanin)
Nicol.
/. radlatus (Sow.: Fr.) Karst.
J. rheades (Pers.)Bond. et Sing.
J. weirii (Murr.)Kotl. et Pouz.
I rpex lacteus Fr.
Ischnoderma bensoinum (Wahl.iFr.)
Karst.
/. resinosum (Fr.) Karst.
Junghuhnia collabens (Fr.) Ryv.
J. luteo-alba (Karst.) Ryv.
J. nitida (Fr.) Ryv.
J. separabelina (Pouz.) Ryv.
Laetiporus sulphureus (Bull.: Fr.)
Murr.
Lenzites betulina (Fr.) Fr.
Merulius molluscus Fr.
Oligoporus fragilis (Fr.) Gilb.et Ryv. 1
01. floriformis (Quel.) Gilb.et Ryv.
O. caesius (Schrad.:Fr.) Gilb.et Ryv.
O. stipticus (Pers.:Fr.) Gilb.et Ryv.
O. tephroleucus (Fr.) Gilb.et Ryv.
Onnia leporina (Fr.) Jahn.
Onnia triqueter (Lentz.: Fr.) Imaz.
Oxyporus corticola (Fr.) Parm.
O. populinus (Schum.: Fr.) Donk.
Phaeolus schweinitzii (Fr.) Pat.
Phellinus chrysoloma (Fr.) Donk.
Порода
C(X)
Б,Ос,Д
Б,Ос,Д
С
Кл,Вз,Ив
Л
Д,Кл
Л
л
Б,Ол
Л
Ос,Б,Д
Е,С
Д,Б,Ос,Яс
Е
Е,С
Е
Е,С 1
Д,Б,Кл,Яс
Е
Л
Б
Л(Е,С)
Е,С
Е
Е
Е,С
Е
Е
Е
Яс,Б,Ос
Л
С,Е
Е |
Встречаемость
Редко
Редко
Редко 1
Редко 1
Редко 1
Регулярно
Регулярно 1
Регулярно 1
Регулярно 1
Регулярно 1
Регулярно
Регулярно
Регулярно 1
Редко 1
Редко
Редко 1
Редко
Редко 1
Редко
Часто 1
Регулярно 1
Редко
Редко 1
Редко 1
Редко 1
Регулярно 1
Регулярно 1
Редко
Часто 1
Редко 1
Регулярно 1
Часто 1
Часто |
-145-
В. Г. Стороженко
Табл. 35 (продолжение)
№
п.п.
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
! 149
150
151
152
153
154
155
156
157
158
160
161
Вид гриба
Ph. conchatus (Pers.: Fr.) Quel.
Ph. ferruginosus (Schrad.: Fr.) Pat.
Ph. ferrugineofuscus (Karst.)
Bourd.et Galz.
Ph. igniarius (L.: Fr.) Quel.
Ph. Lsabelinus (Fr.) Bourd. et Galz.
Ph. punctatus (Fr.) Pil.
Ph. microporus (Pil.) Parm.
Ph. nigrolimitatus (Rom.) Bourd. et
Galz.
Ph pint (Thore: Fr) Ames.
Ph. viticola (Schw.: Fr.) Donk.
Pholiota adiposa Fr.
Ph. squarrosa Karst.
Phlebiopsis gigantea
Phlebia libida (Fr.) Bres.
Piptoporus betulinus (Bull.: Fr.) Karst.
Pleurotus ostreatus Jacq.
Pleurotus cornicopiae (Pers.) Roll.
Polyporus arcularius Batsch.:Fr.
P. brumalis Pers.:Fr.
P. squamosus Huds.:Fr
P. varius Fr.
Sceletocutis amorpha (Fr.) Kotl. et
Pouz.
Sceletocutis nivea (Jungh.) Kell.
Sceletocutis stellae (Pil.) Kell.
Spongipellis spumeus (Sow.iFr.) Pat.
Stereum hirsutum (Fr.) Gray.
St. sanguinolentum (Fr.) Fr.
Trametes gibbosa (Pers.) Fr.
Tr. ceroina (Schw.) Bres.
Tr. hirsuta (Wulf .:Fr.) Pil.
Tr. pubescens (Schum.rFr.) Pil.
Tr. versicolor ( L.:Fr) Pil.
Trichaptum abietinum (Dicks.: Fr.)
Ryv.
Tr. beforme (Fr.:Klot) Ryv.
Tr. fusco-violaceum (Ehren.: Fr.) Ryv.
Tr. laricinus (Karst.) Ryv.
Tyromyces byssinus (Pers.) Bond.
Tyromyces chioneus (Fr.) Karst.
Порода
0с,Вз,Кл,Яс
Д,Кл,Яс,Лп
E
Л
X
Д,Б,Яс,Лп
Е,С
Е
С
Е,С
Яс,Д,Лп,Кл
Л
Х,Л
X
Б
Л
Л
л
л
л
л
Е,С,Пх
Л
Е
Л
Л
Е,С
Б,Ос
С
Л
л
л
X
Б,Ос
X
Е,Лц
Л
Л(Х)
Встречаемость
Регулярно |
Регулярно
Редко |
Часто 1
Редко 1
Регуляно
Регулярно 1
Регулярно 1
Часто |
Регулярно 1
Регулярно 1
Регулярно
Регулярно 1
Редко
Часто
Часто
Редко
Редко
Редко
Регулярно
Редко
Регулярно
Регулярно
Редко
Редко 1
Регулярно
Часто
Редко
Редко
Регулярно
Регулярно
Регулярно
Часто
Редко
Редко
Редко
Редко
Редко
-146-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Примечание. Виды, вошедшие в список биотрофов и сапро-
трофов одновременно, способны поселяться как на живых,
деревьях, так и на древесном отпаде. Биотроф, переходящий вместе с
валежом на мертвый субстрат, становится ксилотрофом.
Обозначения. Встречаемость: часто — в регионе
обследования (лесничество, лесхоз, район) многократно на каждом пункте
учета (пробная площадь, трансекта и т.д.); регулярно — в
регионе обследования на каждом пункте учета единично; редко — в
регионе обследования не на каждом пункте учета; Оч —
возбудитель способен к очаговому распространению.
Породы: X — хвойные породы; Л — лиственные породы;
Е — ель; С — сосна, Лц — лиственница; Пх — пихта; Д — дуб;
Б — береза; Яс — ясень; Лп — липа; Ос — осина; Кл — клен;
Ол — ольха.
Понятно, что в представленный список вошли далеко не все
виды дереворазрушающих грибов. Большинство из
перечисленных возбудителей хранится в нашей коллекции плодовых тел
дереворазрушающих грибов, которая регулярно пополняется.
Биотрофные виды дереворазрушающих грибов как объект
нашего особого внимания в структурах микоценозов коренных
устойчивых лесов имеют определенные особенности в экологии и
распространенности отдельных видов в деревьях, древостоях и в
массивах коренных формаций по зонам растительности.
В составе приведенного списка можно выделить группу
видов, причиняющих наибольший вред отдельным породам.
Краткие описания этих видов и некоторых особенностей их биологии
в лесах коренных формаций на Русской равнине приведены
ниже.
На лиственных породах во всех зонах растительности, где
они произрастают, наиболее представительны следующие виды.
Fomes fomentarius (L.: Fr.) Fr.— настоящий трутовик
поражает преимущественно среднюю, самую ценную часть ствола
лиственных пород. Гниль белая центральная, коррозионно-де-
структивная трухляво-волокнистая с черными линиями,
отделяющими гниль от здоровой древесины. Гниль может занимать
почти всю длину ствола, способствуя ослаблению его механических
свойств и бурелому. Как правило, плодовые тела образуются в
местах обломов сучьев, глубоких механических повреждений,
достающих до гнили. Они же могут являться и воротами для
-147-
В. Г. Стороженко
проникновения спор гриба и заражения деревьев. Пораженность
древостоев березы, осины, липы в возрасте перестоя может
достигать 50 % и более. Пищевая специализация — от
факультативного паразитизма до сапрофитизма.
Phellinus igniarius (L.:Fr.) Quel — ложный трутовик имеет
близкие с настоящим трутовиком характеристики гнилей>
расположения гнили в стволах деревьев и её объемов. Пути
проникновения и заражения деревьев также одинаковы. Встречаемость
его несколько меньше, чем настоящего трутовика, но, как
правило, поражая древостой лиственных пород вместе с настоящим
трутовиком, значительно увеличивает величину общего
поражения. Пищевая специализация — та же.
Laetiporus sulphureus (Bull.: Fr.) Murr.— серно-желтый
трутовик поражает многие лиственные породы и значительно реже
встречается на некоторых хвойных: сосне, лиственнице, ели.
Гриб вызывает бурую деструктиную гниль центрального или
эксцентрического расположения, обычно локализованную на
2-4 м вверх и вниз от расположения плодового теЛа, но при
значительном развитии болезни достигающую 10 м по длине ствола.
Гниль располагается преимущественно в средней и нижней
части ствола, но может достагать толстых ветвей и опускаться в
корни. Гриб характерен для спелых и перестойных древостоев и
пораженность им может достигать 20 %, а в перестойных
дубравах — до 50 %. Нередко является причиной образования дупел.
Пищевая специализация — от факультативного сапрофитизма
до сапрофитизма.
Комплекс рода Armillaria sp. К этому роду относится
несколько видов, имеющих морфокультуральные различия и
обладающих различной активностью по отношению к древесным
породам — Armillaria mellea (Vahl: Fr.) Kumm.; A. ostoyae (Rom.)
Herink; A. borealis Marxm. & Korhonen; A gallica Marxm. &
Romagn.; A. cepistipes Velen.; A. tabescens (Scop.) EmeL; A ectypa
(Fr.) Lam. (авторы приведены по Korhonen, 2004). В России
живые деревья поражают три вида этого комплекса: A borealis,
A. ostoyae и A. mellea. Причем в северных районах наиболее
распространен А. borealis, в средней полосе — A ostoyae, в более
южных районах A mellea (Звягинцев, 2002; Korhonen, 2004).
Пищевая специализация этих видов от факультативного
сапрофитизма до сапрофитизма. Остальные виды относятся к сапрофи-
-148-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
там. Возбудители вызывают преимущественно периферическую
трухляво-волокнистую гниль, но на хвойных породах, может
встречаться и гниль центрального расположения (Стороженко,
1994). Гниль поражает корни и комлевую часть деревьев, не
поднимаясь выше 1-2 м по стволу. Гриб может поражать деревья
любых возрастов. В искусственных и неустойчивых по
структурным характеристикам древостоях может вызывать очаговое
поражение, образовывая очаги усыхания вплоть до распада дре-
востоев (Соколов, 1964; Федоров, 1984; Стороженко, 2002).
Ареалы распространения описаны Кари Корхоненом (2004). Совсем
недавно в темнохвойных условно-одновозрастных лесах юга
Красноярского края отмечены очаги этого возбудителя. В
коренных разновозрастных лесах очагов никогда не образует и как
биотроф в таких лесах встречается в единичных случаях.
Polyporus squamosus Huds.: Fr.— полипорус чешуйчатый, пе-
стрец поражает стволовую часть деревьев многих лиственных и
реже некоторых хвойных пород. Вызывает центральную белую
волокнистую гниль. Признается как раневый возбудитель,
поэтому особенно опасен в зонах рекреационного влияния, в
перестойных древостоях, где механические повреждения
способствуют его распространению. Пищевая специализация — от
факультативного сапрофитизма до сапрофитизма.
Pleurotus ostreatus Jacq.— вешенка. Широко известный
гриб, плодовые тела которого съедобны и с успехом
культивируются в искусственных условиях. Поражает многие лиственные
древесные и кустарниковые породы. Вызывает светлую
волокнисто-коррозионную гниль стволов и корней сильно ослабленных,
усыхающих и усохших деревьев многих лиственных пород.
Поселяется на границе мертвой и живой древесины, часто в
местах поранений, ошмыгов, увеличивая зону поражения, приводя
к образованию дупел, способствуя облому стволов. Пищевая
специализация от факультативного паразитизма до сапрофитизма.
Phellinus robustus (Karst.) Bourd.et Galz.— ложный дубовый
трутовик вызывает стволовую волокнисто-коррозионную гниль
деревьев лиственных пород, преимущественно дуба.
Расположение гнили в стволе по диаметру центральное или
эксцентрическое с выходом в обломанные сучки, по высоте ствола — от
комлевой части до кроны, включая толстые ветви. Поражая камбий
-149-
В. Г. Стороженко
и луб, гриб может вызывать некротические раны. На стволе
может быть несколько выходов гнили и несколько плодовых тел по
высоте ствола, в результате чего гниль может занимать до 50 %
деловой части ствола. Пораженность дубовых древостоев в
возрасте перестоя может достигать 15-20 % (Вакин, 1954; Черемиси-
нов и др., 1970; Чураков, 2000; Стороженко, 2002 и др.).
Пищевая специализация — от факультативного сапрофитизма до
факультативного паразитизма.
Inonotus dryophilus (Berk.) Murr.— древолюбивый трутовик
поражает стволовую часть деревьев дуба, вызывая светлую
коррозионную гниль стволов. Гриб, как и ложный дубовый
трутовик, проникает в ствол через обломанные сучья и механические
повреждения, что делает его опасным в лесах рекреационного
использования. Гниль распространяется в стволе в среднем на
5-6 метров, но может достигать 14 метров, занимая до 40 %
объема ствола (Журавлев, Соколов, 1969). Пищевая
специализация — факультативный сапротроф.
Phellinus tremulae (Bond.) Bond, et Boris.— ложный
осиновый трутовик поражает осину в возрасте 25-30 лет и старше.
Вызывает трухляво-волокнистую гниль, отделенную от здоровой
древесины темной зоной шириной 2-3 мм. Гниль занимает всю
центральную часть ствола по диаметру вплоть до заболони,
выходит через отмершие сучья, где и образует плодовые тела. При
сильном развитии может занимать всю длину ствола. Осина
относится к числу клоновых пород, клоны генетически
однородны, а в природных условиях древостой могут состоять из набора
разных по устойчивости, а в данном случае по иммунитету к
поражению дереворазрушающими грибами, клонов (Стороженко
и др., 1987). В любых достаточно больших по площади массивах
лесов с присутствием значительных по площади осинников,
встречаются не поражаемые ложным осиновым трутовиком
древостой, составляющие один клон. Неустойчивые клоны
поражаются ложным осиновым трутовиком до 60-80 %. Пищевая
специализация — факультативный сапротроф.
Inonotus obliquus (Pers.: Fr.) f. sterilis (Vanin) Nicol.— чага,
стерильная форма скошенного трутовика (J. cuticularis). Наряду
с настоящим и ложным трутовиком один из наиболее
распространенных на березе трутовиков. Кроме березы, может поражать
ольху. Вызывает белую трухляво-волокнистую гниль смешанно-
-150-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
го типа. В стволе располагается локально, распространяясь на
1-2 метра от плодового тела. От желтовато-белой до ржаво-бурой
окраски гниль пронизана темными линиями. Пораженность
березовых старовозрастных древостоев и примеси этой породы в
составе коренных лесов может составлять 15-20 %. Гриб
проникает в ствол дерева через механические повреждения, отмершие
или обломанные сучья. Пищевая специализация —
факультативный сапротроф.
На хвойных породах наиболее распространены следующие
виды дереворазрушающих грибов.
Комплекс рода Heterobasidion sp.— корневая губка включает
несколько видов, которые поражают практически все хвойные
и многие лиственные породы (Журавлев, Соколов, 1969).
К. Korhonen и Т. Niemela разделили род Heterobasidion на три
интерстерильные группы: S, Р и F (не скрещивающиеся между
собой) — и на четыре вида — Heterobasidion annosum (Fr.) Bref. s.
str.; H. parviporum Niemela & Korhonen; H. abietinum Niemela &
Korhonen и Н. insulare (Murr.) Ryv. По данным К. Корхонена
(2004) в России встречаются два вида. На текущий период
Я. annosum (P интерстерильная группа) отмечен на сосне и
березе; Н. parviporum (S интерстерильная группа) отмечен на ели.
Н.И. Федоровым и М.В. Житниковой (2002) подтверждены эти
сведения в Белоруссии. Остальные два вида пока не встречены в
России. Н. abietinum обнаружен в южных районах Украины и
странах Средиземноморья, а Н. insulare — в странах Восточной
Азии (Японии, на Филиппинах, в Индии и Непале). (Korhonen,
2004)
В России грибы этого рода причиняют огромный вред
искусственным насаждениям, вызывая очаговое поражение сосновых
и еловых культур. Гниль от корневой губки на сосне поражает в
основном корни, поднимаясь в ствол только до уровня корневой
шейки. На ели гниль, проникая в ствол через корни, может
подниматься по стволу на высоту 8-10 метров. Гниль коррозионного
типа, ситовая, по диаметру занимает всю центральную часть
ствола. Поражая корни, гниль способствует вывалу деревьев и
образованию большого количества валежа. Плодовые тела
образуются на выворотнях в затененных местах, редко на стволах у
корневой шейки. Наиболее опасный и вредоносный дереворазру-
шающий гриб искусственных неустойчивых по структурным па-
-151-
В. Г. Стороженко
раметрам древостоев. В коренных древостоях естественного
происхождения вредоносность гриба значительно ниже, и, несмотря
на то, что фон его в отдельных случаях присутствует в
древостоях постоянно, его агрессивность сдерживается присутствием в
составе микокомплекса подстилки и верхних слоев почвы
большой плотности и числа антагонистов, препятствующих его
распространению и воспитанию вирулентности штаммов.
Ризосфера искусственных регулярных древостоев, как правило, не
содержит такого числа и состава антагонистов (Василяускас, Ка-
жемекене, 1977; 1986; Тиунова, 1990). В коренных ельниках и
сосняках величины присутствия этого возбудителя в общем
поражении разновозрастных древостоев могут достигать 20 %, но
очагового распространения он никогда не образует.
По пищевой специализации гриб относится к
факультативным сапротрофам, так как инфекция в виде продуцирующих
плодовых тел и вегетативного плодоношения (конидиальное спо-
роношение) может присутствовать в пораженных if
вывалившихся стволах (буреломе и ветровале) в течение 15-20 лет после
перехода пораженных деревьев в вал еж. В древостоях хвойных
формаций с продвижением от зоны смешанных лесов к северной
подзоне тайги закономерно преобладание корневой губки в
составе возбудителей, вызывающих коррозионные гнили,
сменяется преобладанием в коррозионном поражении другими
трутовиками, также вызывающими коррозионные гнили: на ели —
Phellinus chrysoloma (Fr.) Donk. (syn. Porodaedalia chrysoloma
(Fr.) Fiasson & Niemela) и на сосне — Phellinus pini (Thore.: Fr.)
Ames. (Стороженко, 1992)
Phaeolus schweinitzii (Fr.) Pat.— трутовик швейница, один из
наиболее распространенных возбудителей гнили сосны, в
меньшей степени ели, пихты, лиственницы и еще реже зафиксирован
на дубе, лещине, черемухе. В древостоях появляется в возрасте
спелости и перестоя. В коренных условно-одновозрастных
перестойных естественных сосняках Серебряного бора (Московская
обл.) наряду с корневой губкой в общем поражении древостоев
трутовик занимает около 10 %. Но в отдельных случаях пора-
женность им перестойных древостоев может достигать 30 % и
более. По данным СИ. Ванина пораженность сосны в Хреновском
бору достигала 27 %, Л.В. Любарский приводит сведения о
пораженное™ пихты на Дальнем Востоке этим возбудителем до
-152-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
35 %, а лиственницы даурской — до 33 %. Гриб поражает корни
и комлевую центральную часть ствола, не поднимаясь выше
2-3 метров по высоте. Вызывает бурую деструктивную гниль.
Плодовые тела располагаются или на земле, выходя из гнилых
корней, в пределах 1-2 метров от ствола или на корневых лапах.
Карпофоры однолетние, появляются в июне и к началу осени
достигают больших размеров — до 20-30 см в диаметре. В ствол
гриб проникает через поранения корней и комлевой части
стволов и опасен в зонах рекреационного воздействия. Пищевая
специализация — от факультативного сапрофитизма до
факультативного паразитизма.
Phellinus pini (Thore.: Fr.) Ames.— сосновая губка относится
к наиболее распространенным возбудителям стволовых гнилей
сосны, реже кедра и лиственницы. Вызывает светло-коричневую
или охряно-песочного цвета коррозионную центральную гниль
стволов. Расположение по высоте ствола — от комля до уровня
живой кроны. По данным О.Н. Ежова (1998) гниль от сосновой
губки может занимать более 30 % объема ствола, а от объема
кряжа с гнилью — 80 %. В сосняках поражение от сосновой
губки можно фиксировать с возраста спелости, а в перестойном
возрасте поражение может достигать 20-25 %. В Архангельской
области перестойные сосняки поражены до 10,4 % (Ежов, Мин-
кевич, 1998), в Беловежской Пуще (Белоруссия) пораженность
достигает 49,8 %, в Германии — до 55 % (Федоров, 1984), в При-
ангарье — до 28,4 % (Татаринцев, 1996). В тех же сосняках
Серебряного бора к возрасту перестоя поражение сосновой губкой
составляет около 10 %. В общем поражении эта величина
приближается к 30 %. Как и вышеперечисленные виды грибов, в
ствол проникает через механические повреждения естественного
и антропогенного происхождения. Пищевая специализация —
факультативный сапротроф.
Onnia triqueter (Lantz.: Fr.) Imaz.— еловый трутовик
вызывает типичную коррозионную ситовидную светло-охряную, чаще
эксцентрично расположенную по диаметру, но иногда
центральную гниль стволов ели, пихты, реже сосны и лиственницы.
Поражает комлевую часть ствола, опускаясь в корни и поднимаясь
до 3-4 метров по высоте (Гречкин, 1964; Муравьева, 1964;
Журавлев, Соколов, 1969; Черемисиной и др., 1970; Стороженко,
2002). Эксцентрическое расположение по диаметру при сильном
-153-
В. Г. Стороженко
развитии гнили является причиной образования конусовидных
некротических зон со смолотечениями, сужающимися от комля
к высоте выклинивания III стадии гнили. На этих зонах, как
правило, располагаются однолетние плодовые тела гриба. В
коренных ельниках тайги гриб является одним из основных
возбудителей, вызывающих коррозионные гнили, наряду с еловой
губкой. В спелых и перестойных условно-коренных условно-од-
новозрастных ельниках гриб может образовывать небольшие по
площади очаги поражения. В естественных ельниках 90-100 лет
в Подушкинском лесничестве Москворецкого леспаркхоза, где
нами зафиксированы такие очаги, пораженность этим грибом
составляет до 30 % от общего 30-процентного поражения
ельников. Пищевая специализация — факультативный сапротроф.
Phellinus chrysoloma (Fr.) Donk. (syn. Porodaedalia
chrysoloma (Fr.) Fiasson & Niemela) — еловая губка вызывает
типичную коррозионную охряно-светло-коричневую ситовидную
центральную гниль ели, лиственницы, пихты, ре1ке сосны.
Поражает стволовую часть деревьев, но может опускаться до
уровня корневой шейки и подниматься в ствол и в крону, поражая
нижние ветви. Плодовые тела многолетние, сидячие, если
располагаются на вертикальной поверхности ствола, или
распростертые, если располагаются на ветвях или лежачем стволе. Как и
еловый трутовик, этот гриб составляет заметную часть от общего
поражения ельников, особенно в возрасте перестоя. В редких
случаях может образовывать небольшие по площади очаги
поражения или повышенные уровни поражения, достигающие 20 %
от числа деревьев. Проникает в ствол через обломанные или
отмершие сучья, поранения стволов и комля. По зонам
растительности участие гриба в общем поражении коренных
разновозрастных ельников увеличивается от южной тайги к северной, где
гриб занимает преимущественное по сравнению с корневой
губкой положение (Стороженко, 1990; 2002; Мухин, Ушакова,
2002). Опасен в зонах рекреации в старовозрастных древостоях,
где деревья подвержены бурелому в местах расположения гнили.
Пищевая специализация — от факультативного сапрофитизма
до факультативного паразитизма.
Climacocystis borealis (Fr.) Kotl.et Pouz.— северный трутовик
вызывает светло-бурую по внешним признакам призматическую
гниль ели, сосны, пихты, лиственницы, что позволяет относить
-154-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
его к грибам, вызывающим деструктивную гниль древесины.
В.А. Соловьев, однако, относит его к разрушителям лигнина
клеточных стенок, то есть к грибам, вызывающим коррозионные
гнили. Имеет наибольшее распространение на ели. Поражает
комлевую часть ствола, заходя в корни и поднимаясь вверх по
стволу до 2-3 метров. На месте выхода гнили к заболони и лубу
образуется, как и в случае с еловым трутовиком, конусовидный
потек смолы, на котором появляются в середине лета однолетние
плодовые тела. В естественных коренных разновозрастных
ельниках поражение грибом составляет 5-7 % от общего. Не
образует очагов поражения, но в отдельных случаях может составлять
значительное поражение условно-коренных условно-одновозрас-
тных еловых спелых и перестойных особенно рекреационно
нагруженных древостоев. В ельниках Правдинского лесхозтехни-
кума, подверженных III стадии рекреационной дигрессии,
поражение северным трутовиком в отдельных местах достигает 60 %.
В ствол проникает через поранения корней и комлевой части
стволов, что делает его опасным в зонах рекреационного
воздействия. Пищевая специализация — от факультативного
паразитизма до факультативного сапрофитизма.
Перечисленные возбудители гнилевых болезней составляют
основу биотрофного поражения лесов Русской равнины.
Ксилотрофный комплекс дереворазрушающих грибов
представлен в лесных микоценозах значительно более широко,
нежели биотрофный. В большинстве своем микологи, занимающиеся
изучением дереворазрушающих грибов, относят к составу кси-
лотрофов и биотрофов, что на наш взгляд, неверно по
обозначенным выше причинам. Тем не менее и большинство микологов
определяют в формировании комплексов ксилотрофов ряд эколо-
го-биологических закономерностей (Бондарцева, 1992, 2000;
Мухин, 1990, 1993 и др.). Наши исследования состава и
закономерностей функционирования этой группы грибов микоценозов
недостаточно обширны и за более подробными сведениями мы
отсылаем заинтересованного читателя к указанным авторам.
Вместе с тем наши совместно с М.А. Бондарцевой
наблюдения и учеты ксилотрофов в коренных древостоях позволили
определить наиболее часто встречающиеся виды. Эти скромные
данные представлены в таблице 36.
-155-
В. Г. Стороженко
Таблица 36
Встречаемость некоторых видов ксилотрофов на валеже в
еловых древостоях Центрально-лесного биосферного заповедника
(% встреченных от всего количества наблюдений)
."№
1
2
8
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24,
Вид гриба
Fomitopsis pinicola (Sw.: Fr.) Karst.
Trlchaptum abietinum (Dicks.: Fr.)
Fomitopsis rosea (Alb.et Schw.) Karst.
Antrodia seriates (Fr.) Donk.
Armillaria sp.
Sceletocutis amorpha (Fr.) Kotl. et Pouz.
Onnia triqueter (Lentz.: Fr.) Imaz.
Phellinus nigrolimitatus (Rom.) Bourd. et Galz.
Gloeophyllum odoratum (Wulf .:Fr.) Imaz.
Otigoporus stipticus (Pers.rFr.) Gilb.et Ryv.
Antrodia sinuosa (Fr.) Karst
Phellinus chrysoloma (Fr.) Donk.
Pycnoporellus fulgens (Fr.) Donk.
Inonotus weirii (Murr.)Kotl. et Pouz.
Onnia leporina (Fr.) Jahn.
Ph. microporus (Pil.) Parm.
Oligoporus tephroleucus (Fr.) Gilb.et Ryv.
Climacocystis borealis (Fr.) Kotl. et Pouz.
Gloeophyllum abietinum (Fr.) Karst.
Phellinus isabelinus (Fr.) Bourd. et Galz.
Sceletocutis stellae (Pil.) Kell.
J. separabelina (Pouz.) Ryv.
Junghuhnia collabens (Fr.) Ryv.
J. luteoalba (Karst.) Ryv.
Встречаемость
77
43
25
16
7
6
6
6
5
5
5
5
4 1
5
4
4
4
4
3
2
2
2
1
1
Учеты ксилотрофов на валеже в древостоях других зон
растительности показали, что их основной состав варьирует
незначительно и представленные сведения можно принять за основу с
вариациями по лесным формациям.
Существуют гнилевые фауты, природа которых еще не
выяснена. Например, не определены возбудители мокрой темно-
бурой гнили комлевого расположения у ели, которая в общем
поражении ельников может составлять до 5-7 %, или
возбудители комлевой серо-черной гнили, переходящей в дупло, у
липы, поднимающейся в отдельных случаях до 1,5 по высоте
ствола и т.д.
-156-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
В то же время, безусловно, существуют консортивные связи
между грибами биотрофной и сапротрофной групп, которые во
многом определяют их поведение во взаимоотношениях с авто-
трофами.
Почти все дереворазрушающие биотрофные виды обладают
факультативными свойствами и при отмирании деревьев
продолжают какое-то время функционировать на валеже, на мертвом
субстрате. Некоторые виды, обладающие высокой патоген-
ностью по отношению к автотрофам, переходя вместе с ними в
валеж, могут формировать плодовые тела на разлагающемся
субстрате в течение длительного времени. Карпофоры корневой
губки, одного из наиболее активных дереворазрушителей биотро-
фов, мы находили на валеже третьей стадии разложения, то есть
на стволах не менее 20 лет после вывала. Приблизительно те же
периоды сапротрофного питания можно проследить и у
типичных биотрофов — елового трутовика, еловой губки, северного
трутовика и некоторых других биотрофов. Широкая пищевая
пластичность этих видов определяется адаптационной
способностью их ферментативной системы при изменении
физиологической активности субстрата (Бондарцева, 2000)
Безусловно, существуют и консортивные взаимоотношения
между видамютрибов в структуре самого микоценоза. Эти
взаимоотношения не менее важны для функционирования биогеоценоза,
чем любые другие. Одной из важных особенностей консортивных
взаимоотношений в структуре лесного микоценоза являются
антагонистические связи между отдельными видами грибов био-
трофного и сапротрофного комплексов. Именно эти
взаимоотношения определяют активность и воспитание патогенности у био-
трофных видов дереворазрушителей. Многие виды ксилотрофов
проявляют в разной степени антагонистические свойства к
наиболее активным биотрофам, ограничивая их патогенность.
Наиболее наглядно антагонистические свойства
прослеживаются при совместном росте колоний испытуемых видов грибов в
чистых культурах. Нами проведены опыты по совместному
выращиванию в чистых культурах наиболее активных биотрофов и
некоторых ксилотрофов. В качестве тест-объекта принят один из
видов рода Heterobasidion, скорее всего Н. parviporum (N. et
Korh.) — корневая губка, выделенная из образца гнили ели.
Степень совместного влияния учитывалась по методике В. Рипачека
(1967): активный антагонизм — полное подавление роста коло-
-157-
В. Г. Стороженко
нии тест-объекта, нарастание на неё, лизис мицелия; пассивный
антагонизм — сдерживание роста колонии тест-объекта, зона
ингибирования; отсутствие антагонизма — взаимное
прорастание мицелиев колоний без изменения их свойств. В итоге
получены следующие данные. Активный антагонизм к корневой
губке проявили Fomitopsis pinicola, Trichaptum abietinum,
Penlophora gigantea, Inonotus weirii, Trichaptum fusco-violaceum.
Эти виды являются активными антагонистами корневой губки,
препятствуя её распространению по площади биогеоценоза.
Причем /. weirii оказался наиболее активным по отношению к
корневой губке. Этот сапротроф вызывает типичную коррозионную
гниль светлой окраски и может быть использован в качестве ино-
кулюма при изготовлении биопрепарата для ограничения
распространения корневой губки в древостоях ели.
Пассивный антагонизм проявляли Fomitopsis cajanderi,
Gloeophyllum sepiarium, G. odoratum, Stereum sanguinolentum,
Fomitopsis rosea, Coreolus zonatus. Эти виды сапротрофов
являются конкурентами корневой губки в обладании субстратом и
также способны ограничить её распространение в биогеоценозе.
Не проявляли антагонизма к корневой губке, взаимно
проникая в колонии друг друга, Antrodia serialis, Phellinus microporus,
Oxyporus populinus, Phellinus nigrolimitatus.
Обратный антагонизм осуществляла корневая губка к
Skeletocutis amorpha.
Биотрофы Climacocystis borealis, Onnia triqueter, Phellinus
chrysoloma проявляли пассивный антагонизм к корневой губке.
Естественно, в природных условиях состав антагонистов к
активным видам биотрофов значительно шире, особенно в
прикорневой сфере деревьев (Тиунова, 1990).
Несмотря на присутствие антагонистических свойств
некоторых биотрофов к корневой губке и видам опенка, обладающих
факультативными свойствами, в природных условиях
некоторые из них способны к совместному поселению на деревьях.
Известны одновременные поселения на дубе серно-желтого и
ложного дубового трутовиков, на сосне корневой и сосновой губок,
на ели корневой губки и опенка и др.
Подводя итог разделу о видовом составе и особенностям
распространения основных видов биотрофов в коренных лесах,
можно сформулировать некоторые закономерности участия этой
группы дереворазрушающих грибов в формировании лесных со-
-158-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
обществ и определить возможность отнесения этих
закономерностей к одним из критериев устойчивости лесов.
1. В устойчивых коренных лесах видовой состав грибов дере-
воразрушающего биотрофного комплекса сбалансирован по
соотношению видов различной пищевой специализации.
2. Виды биотрофов, обладающих способностью к увеличению
патогенности и агрессивности и как результат к очаговому
распространению, не могут использовать эту способность и
присутствуют в биогеоценозе как рядовые биотрофы, находясь в составе
других дереворазрушителей биотрофного поля биогеоценоза.
3. В составе ксилотрофного комплекса дереворазрушающих
грибов присутствует большая группа видов, проявляющих
антагонистические свойства разной степени активности к
патогенным видам биотрофов, в определённой степени осуществляющих
контроль за их распространением по площади лесных сообществ
в микогоризонтах подстилочного, корневого, комлевого и
стволового слоев микоценоза.
4. В целом, сбалансированный по видовому составу и
пищевой специализации микоценоз устойчивых коренных лесов, в
том числе комплексы дереворазрушающих грибов биотрофной и
ксилотрофной групп, соответствует динамическим
характеристикам биогеоценозов, имеет вполне определенную структуру и
иерархическую подчиненность видов, осуществляя контроль за
текущим отпадом нужного количества деревьев из состава
древостоя. В то же время при изменении структур фитоценозов и
потере ими устойчивости состав микоценоза оперативно
меняется в область увеличения массы активных видов
дереворазрушающих грибов, увеличения их агрессивности и патогенности и
распространения по площади лесных сообществ. В определенном
смысле в биогеоценозе всегда существует структура, призванная
контролировать это качество (устойчивость). Добавим однако,
что это не единственная из состава гетеротрофных организмов
структура, к числу таковых можно отнести зооценоз и в
частности энтомокомплексы биогеоценозов.
Таким образом, после того как мы в каждой из
характеристик структурного строения фитоценозов и микоценозов
обозначили основные закономерности их функционирования, по
которым можно характеризовать устойчивые лесные сообщества и
конкретные параметры этих позиций, вполне возможно
составить свод основных критериев устойчивости лесов.
- 159-
10. КРИТЕРИИ УСТОЙЧИВОСТИ ЛЕСНЫХ
СООБЩЕСТВ
Оценивая структуру работы, можно представить себе весь
набор искомых критериев по составляющим эту работу разделам,
но для завершенности изложения мы все же выделим эту
позицию в отдельный раздел. Итак, ниже приводится весь набор
критериев, выделенных нами (но, естественно, далеко не всех
вообще), по которым мы можем отнести лесные сообщества к
устойчивым. Следует добавить, что обозначены только сами критерии
без каких-либо градаций. Все отмеченные показатели присущи
лишь максимальным значениям этих позиций, характерным
для наиболее «выработанных», климаксовых сообществ.
1. Соответствие состава лесного биогеоценоза условиям
произрастания, коренному экотопу, то есть климату,
геоморфологии местности, почвенным условиям (в понимании Braun-
Blanquet, 1964).
Автотрофная составляющая биогеоценоза
2. Оптимальный породный состав фитоценоза по всем
биогоризонтам, включая подрост, подлесок, напочвенный покров.
3. Сложность структурного строения фитоценоза:
3.1. Непрерывный возрастной ряд в возрастной структуре
древостоя от последнего поколения (подрост) до первого
поколения (деревья предельного возраста для конкретных условий)
3.2. Соизмеримые величины объемов деревьев в возрастных
поколениях.
3.3. Мозаичное или групповое размещение деревьев,
относящихся к одному возрастному поколению, по площади
биогеоценоза (мозаики возрастных поколений). Оптимальный размер
мозаик в устойчивых климаксовых лесах тайги составляет 600-
1000 м2 каждая:
1) размещение деревьев по площади устойчивых коренных
разновозрастных древостоев неравномерно, но эта неравномер-
-160-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ность носит системный, постоянный для всего сообщества
характер. Такое размещение можно рассматривать как
равномерно-агрегированное;
2) равномерно-агрегированное размещение деревьев в таких
сообществах обеспечивает полную занятость площади
биогеоценозов коренных девственных сообществ, непрерывность
горизонтальной структуры и тем самым непрерывность существования
сообществ, то есть является одним из важнейших качеств
постоянно функционирующих устойчивых лесов.
4. Непременным условием существования устойчивого
лесного сообщества является присутствие и достаточное количество
естественного возобновления коренных и сопутствующих пород
разного возраста. Процентное соотношение подроста разного
возраста имеет закономерность стремительного убывания
количества благонадежного подроста от возраста всходов до возраста
конца последнего поколения (период 40-50 лет), когда
количество подроста этого возраста составляет от 0,1 до 0,5 от
количества учтенных деревьев на 1 гектар в зависимости от подзоны и
типа леса.
5. В устойчивых сообществах показатели состояния древосто-
ев сопоставимы по зонам растительности с некоторым
ухудшением средних показателей к северным лесам и сообществам
дигрессивных фаз динамики.
6. В коренных ельниках соотношения запасов древостоев с
объемами древесного отпада в биогеоценозах сравнимы по зонам
растительности. В сосняках эти значения имеют, во-первых,
меньшие абсолютные величины древесного отпада, во-вторых,
гораздо более значительные расхождения в величинах
соотношений. Это связано с двумя причинами: 1) почти обязательным
воздействием пожаров, в результате которых выгорает валеж, 2)
неравномерностью процесса усыхания деревьев и появления в этой
связи древесного отпада.
7. Относительные величины объемов древесного отпада,
представляющие собой отношения объемов древесного отпада к
запасам древостоев и соответственно выделяемого при его
разложении углерода, соизмеримы во всех зонах растительности.
Колебания этих величин зависят только от фазы динамики
биогеоценозов.
- 161-
В. Г. Стороженко
Гетеротрофная (грибная) составляющая
биогеоценоза
8. Устойчивость растительного сообщества непосредственно
связана со структурным строением биогеоценоза и микоценоза.
Чем сложнее структурное строение биогеоценоза, чем
многообразнее его функциональное строение, тем сложнее и
разнообразнее (в пределах оптимальности) по видовому и функциональному
составу микоценоз и тем устойчивее это растительное
сообщество.
9. Грибная биота — ценотическая структура лесного
сообщества, обладающая морфологическим, экологическим и
функциональным строением, как и фитоценоз, и формирующаяся
вместе с ним по определённым законам совместной динамики
развития.
10. В лиственных сообществах зоны лесостепи и смешанных
лесов наибольшие уровни поражения дереворазру*нающими
грибами биотрофного комплекса коренных биогеоценозов
естественного происхождения отмечаются в дубовых и сосновых древосто-
ях глубоко перестойного возраста (дубовых, сосновых,
смешанных), но не более 50 % по числу деревьев.
В сообществах хвойных пород коренных экотопов, начиная
от зоны смешанных лесов и кончая подзоной северной тайги,
средние значения пораженности как еловых, так и сосновых
лесов, во-первых, имеют в пределах формаций сопоставимые
величины, во-вторых, лежат в пределах 30 %. Более значительные
величины отмечены в древостоях дигрессивных фаз динамики,
где в первых возрастных поколениях сосредоточено большое
количество перестойных деревьев. В этих же сообществах
прослеживается вполне объяснимая и оговоренная выше
закономерность нарастания пораженности по мере увеличения возраста
деревьев.
11. В устойчивых коренных лесах видовой состав грибов де-
реворазрушающего биотрофного комплекса сбалансирован по
соотношению видов различной пищевой специализации.
Общей для всех лесов закономерностью качественных
характеристик комплексов биотрофных дереворазрушающих грибов
явилось стремление к балансу возбудителей, вызывающих
деструктивные и коррозионные гнили в древостоях климаксовых
-162-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
фаз динамики, то есть к 1. С продвижением в область демутации
или дигрессии этот баланс нарушается в сторону преобладания
количества деревьев, пораженных видами грибов, вызывающих
коррозионные или деструктивные гнили. В фазах демутации
видовой состав возбудителей, как правило, состоит из меньшего
числа видов дереворазрушителей — биотрофов с преобладанием
наиболее активных, вызывающих коррозионные гнили. С
углублением в фазу дигрессии возрастает не только количество
деревьев с присутствием деструктивных гнилей, но и общий видовой
состав различных биотрофных дереворазрушающих грибов, во
многих случаях с преобладанием возбудителей, вызывающих
деструктивные гнили.
12. В коренных устойчивых разновозрастных лесах деревья,
пораженные дереворазрушающими биотрофными грибами,
независимо от величины общего поражения древостоя, относительно
равномерно распределяются по площади сообщества. Очагового
поражения деревьев и активизации распространения какого-то
одного вида биотрофа в таких лесах нет.
Виды биотрофов, обладающие способностью к наращиванию
патогенности и агрессивности и, как результат, к очаговому
распространению, не могут использовать эту способность и
присутствуют в биогеоценозе как рядовые биотрофы, находясь в общем
составе вместе с другими дереворазрушителями биотрофного
поля биогеоценоза.
13. В составе ксилотрофного комплекса дереворазрушающих
грибов присутствует большая группа видов, проявляющих
антагонистические свойства разной степени активности к
патогенным видам биотрофов, в определённой степени осуществляющих
контроль за их распространением по площади лесных сообществ
3 микогоризонтах подстилочного, корневого, комлевого и
стволового слоев микоценоза.
14. В целом сбалансированный по видовому составу и
пищевой специализации микоценоз устойчивых коренных лесов, в
том числе комплексы дереворазрушающих грибов биотрофной и
ксилотрофной групп, соответствует динамическим
характеристикам биогеоценозов, имеет вполне определенную структуру и
иерархическую подчиненность видов, осуществляя контроль за
текущим отпадом нужного количества деревьев из состава
древостоя.
-163-
В. Г. Стороженко
Таким образом, если суммировать перечисленные выше
(далеко не все существующие в лесных биогеоценозах) критерии,
определяющие устойчивое лесное сообщество, можно
сформулировать итоговый вывод о том, что устойчивое лесное
сообщество — это сообщество, в котором потоки вещества и энергии
сбалансированы в консортивных цепях, объединяющих все его це-
нотические структуры, как автотрофов, так и гетеротрофов.
Проведенные исследования имеют существенное прикладное
значение для разделения лесов по устойчивости, при котором
возможно прогнозировать их будущее развитие и определять ле-
сохозяйственные мероприятия, направленные на воспитание у
лесных сообществ этого качества.
11. ГРАДАЦИИ УСТОЙЧИВОСТИ ЛЕСНЫХ
СООБЩЕСТВ
Для цельности восприятия мы вначале повторим в несколько
сжатой форме основные положения, на которых базируется
наша парадигма устойчивых лесов.
Как было упомянуто выше понятие «устойчивости лесов»
известно еще с конца XIX века. Наши великие предшественники
А.Ф. Рудзкий (1881), Г.Ф. Морозов (1929), В.Н. Сукачев (1972) и
другие определили, что устойчивость как качество леса — это
производное от сложности его структурного строения. И в общем
эта концепция принята в лесоведении, подтверждена многими
исследователями и исследованиями, в том числе и нами в
настоящих исследованиях и ранее (Стороженко, 1992). Суть этой
концепции изложена в теории «климакса», разработанной Ф. Кле-
ментсом (1936), или теории динамического развития и «выра-
ботанности лесов», разработанной В.Н.Сукачевым (1973),
в которых коренные разновозрастные леса, развивающиеся
эволюционным путем, в последних стадиях сукцессии,
рассматриваются как сообщества в наибольшей степени обладающие этим
качеством — устойчивостью. Все остальные леса, какого бы
происхождения и каких бы структурных характеристик они ни
были, относятся лишь к градациям устойчивости. При этом надо
отметить, что для лесов всех зон растительности и формаций эти
градации должны быть идентичны, однако наполнение их
разное. Если принять как позиционное положение о том, что
устойчивость, как качество эндогенного свойства лесных сообществ,
основана на структурных характеристиках биогеоценозов и её
наивысшие показатели соответствуют показателям наиболее
сбалансированных, коренных сообществ, то градации устойчивости
лесов напрямую соотносятся с положением биогеоценозов в сук-
цессионном ряду, могут быть сравнимы с градациями
структурного строения лесов (Дыренков, 1984) и иметь следующие
определения по степени убывания качества устойчивости: абсолютно-
устойчивые, устойчивые, относительно-устойчивые,
неустойчивые и абсолютно-неустойчивые.
-165-
В. Г. Стороженко
Опираясь на выдвинутые ранее положения об
основополагающем значении для отнесения биогеоценозов к определенной
степени устойчивости критериев соответствия экотопу породного
состава фитоценоза, в состав которого обязательной структурой
входит достаточное количество возобновления, первыми самыми
общими ключевыми позициями разделения биогеоценозов на
устойчивые и неустойчивые являются наличие в биогеоценозе
именно этих структур и условий: 1) соответствие породного
состава фитоценоза коренному экотопу; 2) присутствие
достаточного количества возобновления коренной и сопутствующих
пород.
При этом следует принимать во внимание, что в состав древо-
стоев коренных формаций, лесов естественного происхождения
во всех лесорастительных зонах, неотъемлемой составляющей
входят так называемые «пионерные» породы — в основном
береза, осина. Например, в составе девственных коренных ельников
их присутствие может доходить до 2-4 единиц Большее или
меньшее присутствие этих пород в составе лесов указывает на
определенное положение биогеоценоза в сукцессионном ряду.
Несомненно то, что отнесение лесного сообщества к
определенной градации устойчивости регламентируется его
происхождением и структурным строением, то есть теми параметрами,
которые описаны выше. Но важной составляющей при этом
является и учет параметров его санитарного состояния, некоторые
из них тесно связаны с устойчивостью. Такие показатели как
усыхание в различной степени крон деревьев или их отмирание в
результате воздействия различных факторов (грибные болезни,
энтомовредители, спонтанные техногенные или антропогенные
воздействия, изменение условий роста в результате
климатических катаклизмов), могут кардинально менять положение
лесного сообщества в сукцессионном ряду или приводить к смене
коренного биогеоценоза на производный. Поэтому одновременно с
учетом всех лесоводственных и структурных характеристик
биогеоценозов при отнесении их к градациям устойчивости
необходимо принимать во внимание и их санитарное состояние — балл
категорий состояния основных лесообразующих пород,
присутствие очагов болезней или энтомовредителей, объемы древесного
отпада, степень изменения анатомических или морфологиче-
-166-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ских показателей растений фитоценоза, рекреационной
дигрессии и захламленности территорий в техногенно и антропогенно
нарушенных лесах. Ниже приводится предлагаемая нами
определительная таблица разделения лесов по градациям устойчивости.
В приведенной таблице обозначены общие характеристики
устойчивых лесов. Тем не менее, каждая характеристика имеет
конкретные числовые выражения, которые приведены в
разделах работы, изложенных выше. Это замечание имеет важное
значение как ответ оппонентам, ищущим конкретное
подтверждение позициям устойчивых лесных сообществ, обозначенным в
таблице. Именно на констатации количественных и
качественных параметров этих характеристик и получены выводы,
позволившие разделить леса по градациям устойчивости.
В практическом применении наиболее показательно и
эффективно разделение лесов по устойчивости провести в древостоях
различного происхождения, состава, рекреационных и техноген-
нных нагрузок. Леса Серебряноборского лесничества Института
лесоведения РАН, примыкающие к кольцевой дороге г. Москвы
Таблица 37
Определительная таблица разделения лесных биогеоценозов
по градациям устойчивости
Градации устойчивости
Абсолютно-устойчивые
сообщества
Описание лесных сообществ
Соответствие характеристик почвенно-гидрологи-
ческих условий составу и структуре фитоценоза.
Баланс воспроизводимой автотрофами и
разлагаемой грибами биомассы. Непрерывный возрастной
ряд древостоя от последнего поколения (подрост
разного возраста) до первого поколения (деревья
предельных для данного экотопа и природной
зоны возрастов) в близких показателях
соотношений объемов деревьев в возрастных поколениях;
мозаичная горизонтальная структура фитоценоза
по возрастному признаку; полное использование
площади биогеоценоза коренными и
сопутствующими породами разного возраста; достаточное
для непрерывной смены поколений количество
возобновления разного возраста коренных и
сопутствующих пород в оптимальных соотношени-
-167-
В. Г. Стороженко
Табл. 37 (продолжение)
Градации устойчивости Описание лесных сообществ
ях; присутствие древесного отпада разных стадий
разложения в близких соотношениях объемов ва-
лежа в стадиях разложения; оптимальный по
качественным и количественным параметрам состав
и структура грибных комплексов биотрофов и
ксилотрофов, энтомокомплекса и животных —
коренные разновозрастные леса климаксовых и
близких к ним фаз динамики.
Устойчивые сообщества Коренные и условно-коренные сообщества, имею-1
щие в составе древостоев (в том числе в
структурах возобновления), кроме коренных и
некоренные породы или их преобладание; имеющие
неполный ряд возрастных поколений или
упрощенную возрастную структуру древостоев (услов-
но-одновозрастную, относительно-разновозрастную);
имеющие неполное или неоптиИгальное по
объемным характеристикам присутствие валежных
структур; отсутствие или нарушенные мозаичные
структуры древостоев по возрастным,
горизонтальным и вертикальным параметрам,
нарушенные по известным параметрам грибные
комплексы биотрофов и ксилотрофов, но имеющие
достаточное для формирования последующих
поколений возобновление коренных пород — коренные
разновозрастные леса дигрессивных или демута-
ционных фаз динамики, в том числе находящиеся
на начальных этапах коренных сукцессии, и
условно-коренные леса тех же характеристик.
Относительно- Условно-коренные древостой естественного про-
устойчивые сообщества исхождения и искусственные насаждения,
выросшие или высаженные в условиях коренного экото-
па, не имеющие в первом ярусе коренных пород
или имеющие их в количестве, недостаточном для
формирования основного полога; древостой с
измененными лесоводственными
характеристиками, не соответствующими коренному экотопу, но
при наличии подроста коренных пород в
достаточном для формирования в перспективе коренного
древостоя; как правило, не имеющие очагового
распространения биотрофных видов грибов.
Например, лиственные леса естественного проис-
-168-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Табл. 37 (продолжение)
Градации устойчивости
Описание лесных сообществ
Неустойчивые
сообщества
Абсолютно-неустойчивые сообщества
хождения, возникающие на площадях вырубок,
сплошных ветровалов и т.д., или насаждения
искусственного происхождения, в коренных
условиях роста, простых возрастных и горизонтальных
структур, но имеющие подрост коренных пород в
достаточном для формирования в перспективе
коренного разновозрастного древостоя. Такие леса
могут быть отнесены к сообществам,
находящимся на начальных этапах коренных сукцессии.
Естественные условно-одновозрастные, одновоз-
растные или искусственные леса некоренных или
коренных пород, не имеющие возобновительных
структур коренных пород до периода распада
основного полога древостоя, с неопределенной
перспективой формирования подроста в обозримом
времени; с вероятностью или присутствием
очагового распространения отдельных видов биотроф-
ных грибов. Например, древостой с плотным, за-
дернелым напочвенным покровом, с сомкнутым
ярусом подлеска других пород (лещины, рябины
и т.д.) или культуры сосны и ели без подроста с
очаговым поражением болезнями или без
такового, обреченные на распад и переформирование
через смену пород. В перспективной динамике не
просматривается возможность отнесения их к
начальным этапам коренных сукцессии, напротив
возможна смена биогеоценоза.
Среди лесов естественного и искусственного
происхождения абсолютно-неустойчивые встретить
крайне трудно, поскольку в естественно
возникающих лесах условия экотопа диктуют
формирующемуся фитоценозу определенный набор видов,
соответствующих текущему этапу сукцессии, а
искусственные леса создаются, как правило, с
учетом хотя бы нескольких природных факторов
соответствия. Исключение могут составлять
посадки экзотов или посадки пород в нетипичных
для их развития экотопах.
-169-
В. Г. Стороженко
и частично входящие в черту города, испытывающие и
техногенное влияние самого мегаполиса, и влияние выбросов
автотранспорта, и огромные рекреационные нагрузки, вполне подходят
как полигон для практического испытания нашей разработки с
применением вышеизложенной методики.
В каждом квартале лесничества были обследованы наиболее
крупные по площади выделы, в которых определялись лесовод-
ственные параметры биогеоценозов — состав и структура
древесного полога, подлеска, подроста, тип леса, покров,
происхождение древостоев; определялись параметры санитарного состояния
насаждений — вычислялся средний балл состояния по каждой
породе в древостое (Санитарные правила..., 1992) и причины,
вызвавшие это ослабление, объемы валежа, присутствие очагов
распространения возбудителей болезней деревьев, энтомовреди-
телей, величина текущего отпада; степень рекреационной
дигрессии (Полякова, 1980) и захламленности лесов, по
разработанной нами методике.
Коренными породами, слагающими леса Серебряноборского
лесничества, являются сосна, липа, дуб, ель (в восточных
кварталах с более богатыми почвами). Ель как лесообразующая
порода исчезла из состава древостоев лесничества. В лесах
пограничного с Серебряноборским лесничеством Подушкинского
лесничества сохранились устойчивые, условно-коренные, условно-
одновозрастные ельники с примесью дуба, липы, березы. Леса
Серебряноборского лесничества также следует считать условно-
коренными и в большинстве своем условно-одновозрастными,
соответствующими коренным экотопам.
Распределение обследованных выделов по градациям
устойчивости приводится в таблице 38.
Таким образом, на территории Серебряноборского
лесничества нет абсолютно-устойчивых сообществ, устойчивые
биогеоценозы тоже редки. Понятно, что это связано с нарушением
структур лесов в результате их длительного хозяйственного
использования или не вполне целенаправленного ведения лесного
хозяйства. Наиболее значительные площади лесов относятся к
категории относительно-устойчивых, имеющих надежный
подрост коренных пород и определенные перспективы сохранения
или формирования коренного леса. При этом необязательным
-170-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Таблица 38
Распределение обследованных древостоев Серебряноборского
лесничества по градациям устойчивости
Градации
устойчивости
Абсолютно
устойчивые
Устойчивые
Относительно
устойчивые
Неустойчивые
Абсолютно-
| неустойчивые
Распределение по преобладающим породам, %
Сосна
0
4,3
37,8
16,5
0
Береза
0
0
7,9
9,8
0
Липа
0
0
9,1
0
0
Дуб
0
0
3,7
3,0
0
Осина
0
0
1,8
6,1
0
Всего
0
4,3
60,3
35,4
0
условием является сохранение в будущем той же главной
породы. Основные массивы сосновых, дубовых, березовых и
осиновых лесов при естественном ходе формирования, а вернее
переформирования, состава фитоценоза в будущем сменятся на
липовые с участием перечисленных пород. По составу
возобновительных структур, пород второго яруса и подгона биогеоценозы,
отнесенные к градации относительно-устойчивых, в ожидаемой
перспективе будут формировать основной полог с преобладанием
пород, представленных в таблице 39.
В некоторых случаях процесс формирования может быть
скорректирован или ускорен хозяйственными методами.
По лесоводственным характеристикам в ожидаемой
перспективе основные массивы лесов сменятся по главному эдификатору
на липовые с примесью дуба, клена, березы и сосны. Восстанов-
Таблица 39
Ожидаемая смена эдификатора в древостоях
обследованных выделов
Эдификатор
существующего
биогеоценоза
Сосна
Береза
Липа
!Дуб
Осина
Эдификатор будущего биогеоценоза, % от площади,
занятой существующими выделами
Липа
61,3
69,2
100,0
83,3
66,6
Дуб
29,0
30,8
0
16,7
33,4
Сосна
9,3
0
0
0
0 '
-171-
В. Г. Стороженко
ление чистых сосняков, как и ельников, не предвидится. Только
9,3 % существующих сосняков имеют перспективы в следующем
поколении иметь в первом ярусе сосну с примесью других пород.
Значительные площади лесов в лесничестве заняты
неустойчивыми насаждениями — 35,4 % от всех обследованных.
Причем, среди них наиболее представлены сосновые леса, не
имеющие подроста коренной породы. В составе этих лесов у 22,2 %
древостоев зафиксированы очаги усыхания, вызванные
распространением такого биотрофа, как корневая губка (Heterobasidion
annosum (Fr.) Bref.). Все они относятся к искусственным древо-
стоям.
Состояние древостоев в целом оценивалось по состоянию
отдельно каждой породы, слагающей древостой. Средний балл для
смешанного по породному составу выдела не исчислялся из-за
больших расхождений в значениях ослабленности отдельных
пород. Средние показатели состояния отдельных пород по
лесничеству приведены в таблице 40.
Таблица 40
Средние показатели состояния основных лесообразующих
древесных
Породы
Показатели
! состояния
(баллы)
Сосна
1.4
Дуб
3,5
Липа
1,3
Береза
1,9
Осина
2,2
Наиболее устойчивой, наименее ослабленной, а
следовательно, в наибольшей степени перспективной основной лесообразую-
щей породой будущего является липа. Сосна, несмотря на
довольно высокие показатели состояния, имеет малые шансы на
сохранение в будущем лидирующих позиций по упомянутым
выше причинам. Искусственные регулярные посадки сосны
также мало перспективны из-за сохраняющегося постоянного фона
корневой губки, угрозы поражения древостоев этим
возбудителем уже в молодом возрасте и распада его к возрасту приспева-
ния.
Береза имеет довольно низкий средний показатель
состояния, определяемый в основном низкими показателями состоя-
-172-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
ния старовозрастной березы как в составе древостоев других
пород, так и чистых старовозрастных березняков. Причин
ослабления березы, по нашему мнению, несколько. Во-первых, не
вполне соответствующие этой породе довольно легкие песчаные и
супесчаные почвы, во-вторых, сокращенный в этой связи
жизненный цикл и преждевременное старение в составе древостоев,
в-третьих, повышенный фон загрязнения атмосферы вредными
выбросами мегаполиса, в-четвертых, воздействие поздневесен-
них заморозков нескольких лет.
Осина по показателям состояния значительно ослаблена по
тем же причинам, что и береза, к которым следует добавить
значительное поражение древостоев ложным осиновым
трутовиком — Phellinus tremulae (Bond.) Bond, et Boris. При
обследованиях древостоев лесничества в двух случаях обнаружены
устойчивые клоны осины без каких-либо признаков поражения этим
биотрофным грибом.
В катастрофическом положении по показателям состояния
находится дуб. Среднее значение усыхания крон составляет
60 %, максимальные достигают 100 %. Одной из основных
причин усыхания крон является многократное, в течение многих
лет, объедание листвы комплексом листоверток — зеленая
дубовая; палевая дубовая; боярышниковая; розанная (золотистая);
пестро-золотистая.
Некоторое участие в ослаблении дуба играет мучнистая роса
(Microsphaera alphitoides Gr. Et Maubl.), поражающая листья, и
ряд видов дереворазрушающих грибов, вызывающих гнили
стволов — ложный дубовый трутовик (Phellinus robustus (Karst.)
Bourd. et Galz.; серно-желтый трутовик (Laetiporus sulphureus
(Bull.: Fr.) Murr.; Inonotus dryadeus (Pers.: Fr.) Murr. и др. Тем
не менее, по-видимому, существуют и другие, пока не
определенные причины, влияющие на массовое усыхание крон дуба на
огромных территориях Русской равнины и Европы.
Описывая представленным выше образом древостой самых
различных характеристик, открывается возможность
объективно, с учетом параметров самого сообщества определять
целесообразность, виды и интенсивность хозяйственного вмешательства в
процессы формирования биогеоценозов.
Применительно к обследованным лесам Серебряноборского
лесничества, учитывая их огромное значение, как лесов, выпол-
- 173-
В.Г. Стороженко
няющих важные средообразущие, санитарно-гигиенические,
эстетические функции, можно определить следующие общие
положения хозяйственной деятельности.
В группе устойчивых древостоев (4,3 % от всех
обследованных) основным видом хозяйственной деятельности должны
быть, во-первых, поддержание сложных структур возрастного,
горизонтального и породного строения древостоев,
способствующих сохранению устойчивости биогеоценозов, во-вторых,
содействие появлению и свободному росту и развитию
возобновительных структур коренных пород в достаточном для формирования
последующих поколений количестве. При этом необязательно
искусственно поддерживать присутствие в подросте породы,
слагающей в текущий период основной полог и являющейся эдифи-
катором биогеоценоза, поскольку это может нарушить
естественный ход сукцессионного развития сообщества.
В группе относительно-устойчивых биогеоценозов (60,3 %)
хозяйственное вмешательство должно быть направлено на
формирование последующих поколений коренных пород,
сохранение подроста этих пород по параметрам оптимального
количества и разнообразия состава.
В группе неустойчивых древостоев (35,4%) единственным
мероприятием, с помощью которого можно избежать
неблагоприятной смены пород при распаде основного полога, является
высадка подпологовых культур коренных пород, способных
переносить относительное затенение — ель, липа, дуб. К числу
неустойчивых насаждений в Серебряноборском лесничестве почти
поровну относятся сосняки — 16,5 %, березняки и осинники —
15,9 %. Неустойчивые сосняки представляют собой, как
правило, чистые древостой с густым подлеском из кустарниковых и
подлесочных пород — лещины, рябины, бузины, черемухи и т.д.
Возобновление сосны в них практически невозможно, поэтому во
избежание нежелательной смены пород необходимо проводить
опытные посадки других коренных пород: липы, дуба, ели.
В таких древостоях неизбежна смена эдификатора. Одним из
вариантов воспроизводства на таких территориях сосновых лесов
может быть сплошная вырубка перестойного древостоя,
корчевка и посадка культур этой породы. Однако этот путь в зоне
интенсивного рекреационного использования лесов мало пригоден
-174-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
и к тому же чреват появлением в создаваемых культурах очагов
корневой губки или опенка.
Разработанная нами методика позволяет оценить
перспективы формирования биогеоценозов и определить возможности
повышения их устойчивости хозяйственными методами в
соответствии с коренными экотопами. Особенно это важно в лесах,
предназначенных для долговременного функционирования,
выполняющих важные водоохранные, санитарно-гигиенические,
различные экологические функции, подверженных
рекреационным и техногенным воздействиям с измененными параметрами
фитоценозов, пораженных грибными болезнями или энтомовре-
дителями, нуждающихся в оптимизации структур и функций.
Подводя итог проведенным исследованиям, хотелось бы
выразить обоснованную надежду, что нам удалось ответить по
крайней мере на три позиции, вынесенные в качестве основных
задач этой работы и обозначенных в её начале. Конечно,
хотелось бы и далее продлить наши исследования в область
прикладного их использования, тем более что на этом пути открывается
широчайшее поле применения в интересах оптимальной
структуризации лесного хозяйства с учетом не только хозяйственного
использования леса, но и максимального использования его эко-
системных функций.
-175-
ЗАКЛЮЧЕНИЕ
У заинтересованного читателя, одолевшего эту книгу, могут
возникнуть вопросы, касающиеся вообще целесообразности
исследований в области устойчивости лесных сообществ, особенно
в наш такой прагматичный период истории. Зачем исследовать
какие-то девственные леса, которых на Земле осталось и так
немного, да и последние скоро вырубят. Зачем говорить о какой-то
устойчивости лесов, когда у всей страны на глазах происходит
их хищническое уничтожение при полном хаосе в лесном деле?
Забота об экологическом здоровье нации удовлетворяется
поддержанием относительного порядка в парках, лесопарках и
пригородных лесах больших городов. Состояние, структура и запасы
остальных лесов пока мало кого волнуют и рассматриваются как
возобновляемый ресурс для удовлетворения потребностей
человека.
Однако в экологической политике многих стран все более
отчетливо проявляется осознание необходимости соизмерения
потребностей человечества и возможностей природных
экосистем удовлетворять эти потребности.
В осознании важности этих проблем и заключается
актуальность таких исследований. Если рассматривать нашу планету
как один биогеоценоз, то баланс между геоценозом,
фитоценозом, микоценозом и зооценозом в нем формировался в течение
долгого исторического периода и каждый организм, в том числе
и человек, адаптирован именно к этому балансу. И важным с
этих позиций становится приведение в известность параметров
этого баланса, пока еще сохранились в отдельных регионах
нашей страны не затронутые антропогенным воздействием
устойчивые леса.
Не может вызывать дискуссий тезис о том, что устойчивость
лесных сообществ относится к разряду эндогенных качеств,
имеющих определенные характеристики, параметры которых
можно измерить или описать. Лесные сообщества в рамках лесного
биогеоценоза имеют свои вполне определенные границы, в пере-
-176-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
делах которых и определяются параметры этого качества.
Именно этот биогеоценотический уровень сообществ рассматривается
в настоящем издании. Вместе с тем, по нашему глубокому
убеждению, те же или близкие качества, удовлетворяющие условиям
устойчивости, вполне применимы и к более сложным по
организации, структурным характеристикам и масштабам системам.
Те же критерии устойчивости, с некоторыми вариациями, могут
быть применимы к типам леса, группам типов леса,
экосистемам, ландшафтным системам, флористическим комплексам, ле-
сорастительным зонам и вообще ко всяким природным
комплексам, имеющим эволюционную историю развития, в том числе и в
целом к планете Земля, как природной системе, которая
испытывает сильнейший и все возрастающий антропогенный пресс.
Я приношу благодарность всем организациям,
предоставившим для меня возможность проводить исследования в
закрепленных за ними лесах, заповедникам, национальным паркам,
научным центрам, лесхозам и лесничествам. Особую
благодарность я приношу М.А. Бондарцевой, Л.В. Гарибовой за помощь в
определении видов некоторых трутовых грибов, В.И. Крутову и
М.Г. Романовскому за ценные замечания при редактировании и
рецензировании работы.
-177-
ЛИТЕРАТУРА
Александрова В.Д. Изучение смены растительного покрова.
Полевая геоботаника. М.-Л. Наука. 1964. Т. 3. С. 300-451.
Алексеев И.А. Научные основы лесохозяйственных мер борьбы с
корневой губкой в лесах Полесья и лесостепи УССР. Л. Авто-
реф. дисс. ... д-ра с.-х. наук. 1974. 36 с.
Алексеев И.А. Лесохозяйственные меры борьбы с корневой
губкой. М. Лесная пром-сть. 1969. 76 с.
Алексеев СВ. Возобновление ели на вырубках. М. Наука. 1978.
130 с.
Астапенко В.В. Что такое микоценоз, фитоценоз и биоценоз с
позиций системного подхода // Микология и фитопатология.
1989. Т. 23. В. 6. С. 505-509.
Атрохин В.Г., Кузнецов Г.В. Лесоводство. М. Агропромиздат.
1989. 399 с.
Баранов Н.И., Григорьев К.И. Ельники Севера. Л. ЦНИИЛХ.
1955.48 с.
Беклемишев В.Н. О классификации биоценотических симфизио-
логических связей // Бюлл. МОИП. Отд. биол. 1951. Т. 56.
В. 5. С. 3-30.
Бондарцев А.С. Трутовые грибы Европейской части СССР и
Кавказа. М.-Л. Из-во АН СССР. 1953. 1103 с.
Бондарцева М.А. Определитель грибов СССР. Афиллофоровые.
Л. Наука. 1986. В. 1. 192 с.
Бондарцева М.А. Определитель грибов России. Афиллофоровые.
СПб. Наука. 1998. В. 2. 391 с.
Бондарцева М.А. Видовой состав, распространение в лесных
биогеоценозах и экологическая функция дереворазрушающих
трутовых грибов // М. Наука. Научные основы устойчивости
грибов к дереворазрушающим грибам. 1992. С. 90-140.
Бондарцева М.А. Эколого-биологические закономерности
функционирования ксилотрофных базидиомицетов в лесных
экосистемах // М.— Петрозаводск. Грибные сообщества лесных
экосистем. 2000. С. 9-25.
-178-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Бондарцева М.А. Адаптация к субстрату как один из факторов
эволюции афиллофороидных грибов // М-— Петрозаводск.
Грибные сообщества лесных экосистем. 2004. Т. 2. С. 9-21.
Бузыкин А.И., Охонин В.А., Секретенко О.П., Хлебопрос Р.Г.
Анализ пространственной структуры одновозрастных древо-
стоев // Красноярск. Разновозрастные леса Сибири, Дальнего
Востока и Урала и ведение хозяйства в них. 1983. С. 5-20.
Бурова Л.Г. Экология грибов макромицетов // М. Наука. 1986.
222 с.
Быков Б.А. О методе переменной площадки и новом способе
оценки распределении особей // Экология. 1976. №4.
С. 29-37.
Быков Б.А. Экологический словарь. Алма-Ата. Наука. 1983.
247 с.
Бяллович Ю.П. Биогеоценотические горизонты. Бюлл. МОИП.
Отд. Биол. 1960. Вып. 3. С. 43-60.
Бяллович Ю.П. О биогеоценотической структуре центрального
слоя биосферы. Бюлл. МОИП. Отд. Биол. Т. 85. В. 3. 1980.
С. 25-39.
Вакин А.Т., Полубояринов О.И., Соловьев В.А. Альбом пороков
древесины. М. Лесная пром. 1969. 78 с.
Вакин. А.Т. Фитопатологическое состояние дубрав Теллерманов-
ского леса // Труды Ин-та леса АН СССР. 1954. Т. 16.
С.50-109.
Ванин СИ. Гниль дерева, её причины и меры борьбы. М.-Л.
Сельхозгиз. 1931. 160 с.
Василяускас А.П. Корневая губка и устойчивость экосистем
хвойных лесов. Вильнюс. Мокслас. 1989. 175 с.
Василяускас А.П., Кажемекене Б.Ю. Ризосферно-прикорневая
микрофлора сосны, ели и березы и её антагонистическая
активность к корневой губке // Вопросы защиты леса Южной
Прибалтики. Каунас. 1977. С. 121-138.
Величко И.М. Когда и как возникли растения. Киев. Наукова
думка. 1989. 159 с.
Вересова И.М. Исследования микофлоры измельченной
древесины в естественных условиях // Труды ЛТА. 1968. №110.
С. 35.
Воропанов П.В. Естественная история ельников Севера и их
внутренняя структура // Сб. трудов Поволжского лесотех-
нич. ин-та. Йошкар-Ола. 1949. № 46. С. 3-38.
-179-
В. Г. Стороженко
Воропанов П.В. Таблицы древесного отпада основных лесообра-
зующих пород СССР. М. Лесная пром-сть. 1973. 110 с.
Волков А.Д., Дыренков С.А. Возобновительная динамика
относительно разновозрастных древостоев ельников в
ненарушенных лесных массивах // Труды Петрозаводской лесной
опытной станции. Петрозаводск. 1971. В. 1. С. 159-174.
Гарибова Л.В. Обзор и анализ современных систем грибов.
Петрозаводск. 1999. 28 с.
Гарибова Л.В., Лекомцева С.Н. Основы микологии. М. 2005.
220 с.
Герасимов И.Л. Эволюция и дифференциация природы Земли //
Изб. Труды. АН СССР. М. Наука. 1990. 312 с.
Горбань А.Н., Охонин В.А. Устойчивая реализуемость
биогеоценозов в ходе сукцессии // М. Наука. Проблемы
экологического мониторинга и моделирования экосистем. Т. 9. 214-226.
Горленко М.А. Краткий курс иммунитета растений к
инфекционным болезням. М. Высшая школа. 1962. 303 с.
ГОСТ «Лесоводство. Термины и определения». 1998.
Грабарний П.Я., Комарова А.С. Статистический анализ
горизонтальной структуры древостоя. М. Наука. 1967. 120 с.
Гречкин В.П. Еловая комлевая губка // М. МЛТИ. Вопросы
защиты леса. 1974. С. 17-21.
Дворецкий М.Л. Пособие по вариационной статистике. М.
Лесная пром-сть. 1971. 104 с.
Демаков Ю.П. Диагностика устойчивости лесных экосистем.
Йошкар-Ола. Изд-во «Периодика Марий-Эл». 2000. 414 с.
Дробышев Ю.И., Короткое С.А. Устойчивость древостоев:
структурные аспекты. Реферативная информация. 2003. № 7.
С. 2-11.
Дробышев Ю.И., Кузнецов Е.В. Методика количественного
описания антропогенного влияния на устойчивость
рекреационных насаждений // М. Лесопользование и воспроизводство
лесных ресурсов. МГУЛ. 2000. В. 303. С. 122-128.
Дылис Н.В., Уткин А.И, Успенская И.М. О горизонтальной
структуре лесных биогеоценозов // Бюлл. МОИП. Отд. биол.
1984. Т. 69. В. 4. С. 65-73.
Дыренков С.А. Структура и динамика таежных ельников.
Л. Наука. 1984. 176 с.
-180-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Дутина О.П. Горизонтальная структура некоторых ассоциаций
пихтовых лесов южной тайги // Калинин. Взаимоотношение
компонентов биогеоценозов в южнотаежных ландшафтах.
1984. С. 90-102.
Ежов О.Н. Распространение сосновой губки (Phellinus pint
(Thore. ex Fr.) Pil. в средней подзоне тайги и ограничение её
вредоносности. СПб. Автореф. дисс. ... канд. биол. наук.
1998. 18 с.
Ежов О.Н., Минкевич И.И. Особенности распространения
стволовой гнили сосны // Лесной журнал. 1998. № 2-3. С. 12-17.
Еленев П.Ф. Попытка дифференциации различных
растительных остатков в связи с их микофлорой // Труды секции
микологии и фитопатологии Русского ботанического сада. 1923.
Т. 1. С. 81-100.
Журавлев И.И, Селиванова Т.Н., Черемисинов Н.А.
Определитель грибных болезней деревьев и кустарников. М. Лесная
пром. 1979. 248 с.
Замолодчиков Д.Г., Уткин А.И., Честных О.В. Коэффициенты
конверсии запасов насаждений в фитомассу основных лесооб-
разующих пород России // Лесная таксация и
лесоустройство. Красноярск. 2003. В. 1(32). С. 119-127.
Замолодчиков Д.Г., Коровин Г.Н., Уткин А.И., Честных О.В.,
Сонген Б. Углерод в лесном фонде и сельскохозяйственных
угодьях России // М. Тов. Научн. изданий КМК. 2005. 200 с.
Звягинцев В. Б. Распространение и вредоносность видов
Armillaria в лесах Беларуси // Материалы 5-й
международной конференции «Проблемы лесной фитопатологии и
микологии». 2002. С. 94-97.
Звягинцев Д.Г. Некоторые концепции строения и
функционирования комплексов микроорганизмов // Вести МГУ. Сер. 17.
Почвоведение. 1978. № 4. С. 48-56.
Ивашкевич Б.А. Девственный лес, особенности его строения и
развития // Лесное хоз-во и лесная пром-сть. 1929. № 10.
С. 35.
Ивашкевич Б.А. Основы хозяйства в защитных и водоохранных
лесах // Хабаровск. Труды ДальНИИЛХ. 1981. В. 23.
С.136-143.
Ильинский А.П. Растительность земного шара. М.-Л. Изд-во АН
СССР. 1937. 458 с.
-181-
В. Г. Стороженко
Инструкция по борьбе с корневой губкой сосны, ели и пихты в
лесах СССР. М. ЦБНТИлесхоз. 1979. 17 с.
Ипатов B.C. Некоторые вопросы теории организации
растительного покрова // Ботан. журнал. 1970. Т. 55. № 2. С. 184-195.
Ипатов B.C., Тархова Т.Н. Количественный анализ ценотиче-
ских эффектов в размещении деревьев по территории //
Ботан. журнал. 1975. Т. 60. № 9. С. 1237-1250.
Казимиров Н.И. Ельники Карелии. Л. Наука. 1971. 139
Каламеэс К. А. Агариковые грибы Эстонии (Polyporales,
Boletales, Russulares, Agaricales). Автореф. дис... д-ра. биол.
наук. Таллин. 1975. 110 с.
Карелин Д.В., Уткин А.И. Скорость и параметры разложения
древесного дебриса в лесных экосистемах: результаты
анализа базы данных // М.— Петрозаводск. Проблемы лесной
фитопатологии и микологии. 2005. С. 173-178.
Картавенко Н.Т., Колесников Б.П. К вопросу о скорости
разложения порубочных остатков на сплошных вырубках //
Труды Ин-та биологии Урал, филиала АН СССР. 1962. В. 28.
С. 23.
Комин Г.Е., Семечкин И.В. Возрастная структура древостоев и
принципы её типизации // Лесоведение. 1970. № 2. С. 24-33.
Крутов В.И. Фитопатогенные микро- и макромицеты —
консорты древесных пород в Республике Карелия //
М.-Петрозаводск. Грибные сообщества лесных экосистем. 2004. Т. 2.
С. 114-130.
Корчагин А.А. Строение растительных сообществ. М.-Л. Наука.
1976. Т. 5. 283 с.
Кузнецов Л.М. Дыхательный газообмен древесного детрита в
таёжном лесу. СПб. Автореф. дисс... канд. биол. наук. 1998.
19 с.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. СПб.
1994. Ч. 1. 281 с.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. СПб.
1998. 358 с.
Курнаев С.Ф. Лесорастительное районирование СССР. М. Наука.
1973. 203 с.
Курнаев С.Ф. Основные типы леса средней части Русской
равнины. М. Наука. 1968. 355 с.
-182-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Лавренко Е.М. Основные закономерности растительных
сообществ и пути их изучения. М.-Л. Изд-во АН СССР. 1959. Т. 1.
70 с.
Любарский Л.В., Васильева Л.Н. Дереворазрушающие грибы
Дальнего Востока. Новосибирск. Наука. 1975. 164 с.
Мазинг В.В. Консорции как элемент функциональной
структуры биогеоценоза // Бюлл. МОИП. Отд. биол. 1966. Т. 27.
B. 2. С. 117-127.
Марков М.В. К вопросу о пространственных взаимоотношениях
в популяциях древесных растений // Казань. Структура и
организация популяций. 1985. С. 102-110.
Маслов А.Д. Усыхание лесов от засухи на Европейской
территории СССР // Лесоведение. 1972. N° 6. С. 77-87.
Маслов А.А. О современном применении метода блоков и метода
главных компонентов для анализа мозаичности лесных
сообществ // Бюлл. МОИП. Отд. биол. 1985. Т. 90. В. 4.
C. 107-117.
Маслов А.А. Природная динамика заповедных лесных
сообществ в центре Русской равнины. Автореф. дисс... д-ра биол.
наук. 2004. 43 с.
Миркин Б.М>, Розенберг Г.С., Наумова Л.Г. Словарь понятий и
терминов современной фитоценологии. М. Наука. 1989.
223 с.
Миркин Б.М., Розенберг Г.С. Фитоценология. Принципы и
методы. М. Наука. 1978. 210 с.
Мирчинк Т.Г. Почвенные грибы как компонент биогеоценоза //
Почвенные организмы как компонент биогеоценоза // М.
Изд-во МГУ. 1984. С. 220.
Мозолевская Е.Г., Катаев О.А., Соколова Э.С. Методы лесопато-
логического обследования очагов стволовых вредителей и
болезней. М. Лесная пром-сть. 1984. 152 с.
Мозолевская Е.Г. Усовершенствование методики обследования
санитарного состояния насаждений // М. Вопросы защиты
леса. 1973. В.41. С. 47-56.
Мозолевская Е.Г. Оценка роли стволовых вредителей в лесных
биогеоценозах // Брянск. Проблемы охраны природы и
защиты леса. 1979. С. 34-40.
Морозов Г.Ф. Избранные труды. М. Лесная пром-сть. 1970. Т. 1.
559 с.
-183-
В. Г. Стороженко
Морозов Г.Ф. Учение о лесе. М.-Л. Госиздат. 1928. 440 с.
Морозов Е.Е. Экологические условия и скорость микогенного
ксилолиза в биогеоценозах южной подзоны тайги. Автореф.
... дисс. к.б.н. СПб. 1994. 22 с.
Муравьева Н.Б. Фауты ели в насаждениях Лисинского лесхоза //
Лесной журнал. 1965. № 2. С. 17-20.
Муравьева Н.Б. Повреждения стволов ели и раневые гнили //
Лесной журнал. 1964. № 2. С. 6-9.
Мухин В.А. Экологические закономерности формирования и
структуры биоты ксилотрофных базидиомицетов Западно-
Сибирской равнины. М. Автореф. дисс... доктора биол. наук.
1990. 32 с.
Мухин В.А. Биота ксилотрофных базидиомицетов
Западно-Сибирской равнины. Екатеринбург. Наука. 1993. 231 с.
Мухин В.А., Ушакова Н.В. Базовые таксономические спектры
ксилотрофных базидиомицетов // М.—Петрозаводск.
Грибные сообщества лесных экосистем. Т. 2. 2004. С. 52-61.
Одум Ю.Р. Экология. М. Мир. 1986. Т. 1. 325 с.
Орлов А.Я. Значение елового подроста в мелколиственных лесах
южной тайги для восстановления ельников // Лесное хоз-во.
1983. №4. С. 11-14.
Плотников В.В. О горизонтальной структуре древесного яруса
лесных сообществ // Лесоведение. 1968. № 5. С. 3-11.
Побединский А.В. Сравнительная оценка одновозрастных и
разновозрастных древостоев // Лесное хоз-во. 1988. № 2.
С. 40-43.
Побединский А.В., Желдак В.И. Временное наставление по
проведению рубок в лесах, где допускаются только рубки ухода
и санитарные рубки, для равнинных лесов европейской части
РСФСР // М. Госкомлес. СССР. 1989. 23 с.
Полякова Г.А. Деградация сосняков Подмосковья под влиянием
рекреации // Лесоведение. 1980. № 5. С. 62-69.
Проскуряков М.А. Закономерности формирования
пространственной структуры древостоев темнохвойных горных лесов
Тянь-Шаня и Рудного Алтая. Красноярск. Автореф. дисс...
д-ра биол. наук. 1982. 44 с.
Работнов Т.А. О структурных элементах фитоценозов и фитоце-
нотических популяциях // Бюлл. МОИП. Отд. биол. 1965.
Т. 90. В. 1.С. 103-107.
-184-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Работнов Т.А. О некоторых терминах, употребляемых в
микологии // Микология и фитопатология. 1985. Т. 19. В. 6.
С.519-20.
Рипачек В. Биология дереворазрушающих грибов. М. Лесная
пром. 1967. 275 с.
Рожков А.А., Козак В.Т. Устойчивость лесов. М. Агропромиз-
дат. 1989. 238 с.
Рожков А.А. Оценка устойчивости и состояния лесов //
Лесоведение. 2003. № 1. С. 66-72.
Розенберг Г.С. Устойчивость экосистем и ее математическое
описание // Экологические аспекты гомеостаза в биогеоценозе.
Уфа. 1986. С. 120-130.
Романовский М.Г. Продуктивность, устойчивость и
биоразнообразие равнинных лесов Европейской России. М. Изд-во
МГУЛ. 2002. 91 с.
Рудзкий А. Лесные беседы: Для русских лесовладельцев и
лесничих // СПб. Дервиен. 1881. 207 с.
Рысин Л.П., Комиссаров Е.С., Маслов А.А., Петерсон Ю.В.,
Савельева Л.И. Методические предложения по созданию
системы постоянных пробных площадей на особо охраняемых
лесных территориях. М. Наука. 1988. 27 с.
Санитарные правила в лесах Российской Федерации. М. 1992.
18 с.
Санников Н.С., Локосова Е.И. Микроэкосистемный анализ
структурно-функциональных связей в лесных биогеоценозах //
Екатеринбург. Генетические и экологические исследования в
лесных биогеоценозах. 2001. С. 73-95.
Свирежев Ю.М., Логофет Д.О. Устойчивость биологических
сообществ. М. Наука. 1978. 352 с.
Синская Е.Н. Основные черты эволюции лесной растительности
Кавказа в связи с историей видов // Вотан, журнал СССР.
1933. В. 18. №5,6.
Скворцова Е.Б., Уланова Н.Г., Басевич В.Ф. Экологическая роль
ветровала. М. Лесн. пром-сть. 1983. 187 с.
Соколов Д.В. Корневая гниль от опенка и борьба с ней. М.
Лесная пром. 1964. 182 с.
Соловьев В.А. Микогенный ксилолиз, его экологическое и
технологическое значение // М. Наука. Научные основы
устойчивости лесов к дереворазрушающим грибам. 1992. С. 140-171.
-185-
В.Г. Стороженко
Сочава В.Б. Вопросы классификации растительности, типологии
физико-географических фаций и биогеоценозов // Труды Ин-
та биол. УФ АН СССР. 1961. Т. 27. С. 99.
Стороженко В.Г. Грибные болезни в рекреационных лесах //
Лесное хоз-во. 1987. № 3. С. 63-65.
Стороженко В.Г. Датировка разложения валежа ели //
Экология. № 6. 1990. С. 66-69.
Стороженко В.Г., Бондарцева М.А., Соловьев В.А., Кругов В.И.
Научные основы устойчивости лесов к дереворазрушающим
грибам. М. Наука. 1992. 221 с.
Стороженко В.Г. Грибные дереворазрушающие комплексы в
генезисе еловых биогеоценозов. М. Автореф. дисс. д-ра биол.
наук. 1994. 42 с.
Стороженко В.Г. Стратегии и функции грибных сообществ
лесных экосистем // Москва-Петрозаводск. Грибные сообщества
лесных экосистем. Изд-во Карельского НЦ РАН. 2000.
С. 37-42.
Стороженко В.Г. Датировка разложения крупных древесных
остатков в лесах различных природных зон // Лесоведение.
2001. № 1. с. 49-53.
Стороженко В.Г. Гнилевые фауты коренных лесов Русской
равнины. М. Изд-во ВНИИЛМ. 2002. 156 с.
Стороженко В.Г. Устойчивость лесов и грибные сообщества //
М. Материалы 5-й межд, конф. «Проблемы лесной фитопат.
и микол.». 2002. С. 235-238.
Стороженко В.Г. Показатели древесного отпада в коренных
ельниках таежной зоны Русской Равнины // М.- Петрозаводск.
Грибные сообщества лесных экосистем. 2004. С. 221-239.
Стороженко В.Г. Понятия «больного» и «здорового» лесного
сообщества // М.— Петрозаводск. Материалы 6-й межд. конф.
«Проблемы лесной фитопат. и микол.», 2005. С. 311-316.
Стороженко В.Г, Разделение лесов по градациям устойчивости.
Методика и эксперимент // М.— Петрозаводск. Материалы
6-й межд. конф. «Проблемы лесной фитопат. и микол.».
2005. С. 317-329.
Страхов В.В., Писаренко А.И., Борисов В.А. Глобализация
лесного хозяйства. М. 2001. 397 с.
Сукачев В.Н. Избранные труды. Л. Наука. 1972. Т. 1. 343 с.
Сукачев В.Н. Избранные труды. Л. Наука. 1973. Т. 2. 418 с.
-186-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Сукачев В.Н. Избранные труды. Л. Наука. 1975. Т. 3. 543 с.
Тахтаджан А.Л. Четыре царства органического мира //
Природа. 1973. № 2. С. 22-32.
Татаринцев А.И. Центральная стволовая гниль в сосняках,
пройденных низовыми пожарами // Новосибирск. Изд-во
СО РАН. Сибирский экологический журнал. 1996. № 1.
С. 81-87.
Тарасов М.Е. Оценка скорости разложения детрита в лесах
Ленинградской области // Труды СПбНИИЛХ. 2000. В. 1.
С. 31-45.
Тиунова М.И. Влияние почвенной и ризосферной микофлоры на
активность корневой губки на примере сосновых культур Бу-
зулукского Бора. Автореф. дисс... канд. биол. наук.
Свердловск. 1990. 23 с.
Ткаченко М.Е. Общее лесоводство. М.-Л. Гослесбумиздат. 1955.
599 с.
Третьяков Н.В., Горский П.В., Самойлович Г.Г. Справочник
таксатора. М.-Л. Гослесбумиздат. 1952. 853 с.
Толмачев А.И. К истории возникновения и развития темнохвой-
ной тайги. М-Л. Изд-во АН СССР. 1954. 156 с.
Федоров Н.И, Корневые гнили хвойных пород. М. Лесная пром-
сть. 1984. 159 с.
Федоров Н.И, Ярмолович В.А. Распространение и вредоносность
смоляного рака в сосновых фитоценозах Беларуси // Леса
Беларуси и их рациональное использование. Минск. 2000.
С. 235-238.
Федоров Н.И., Ярмолович В.А. Особенности эцидиального спо-
роношения грибов Cronartium flaccidum Wint. и Peridermium
pini Lev. в условиях Беларуси // М. Материалы 5-й Между-
нар. конф. «Проблемы лесной фитопат. и ми ко л. ж 2002.
С. 251-253.
Федоров Н.И., Ярмолович В.А. Смоляной рак сосны
обыкновенной в лесах Беларуси // М.— Петрозаводск. «Грибные
сообщества лесных экосистем.». 2004. Т. 2. С. 239-255.
Федоров Н.И, Житникова М.В. Корневая губка в хвойных лесах
Беларуси // М. Материалы 5-й Междунар. конф. «Проблемы
лесной фитопатологии и микологии». 2002. С. 248-250.
Федорчук В.Н., Кузнецова М.Л., Андреева А.А., Моисеев Д.В.
Резерват Вепсский лес // СПб. СПБНИИЛХ. 1998. 208 с.
-187-
В. Г. Стороженко
Цвел ев Н.Н. Проблемы теоретической морфологии и эволюции
высших растений // М.-СПб. РАН. БИН. 2005. 407 с.
Частухин В.Я. Распад растительных остатков в еловых лесах //
М. Биология почв. 1948 В. 1. С. 13-91.
Черемисинов Н.А., Негруцкий С.Ф., Лешковцева И.И. Грибы и
грибные болезни деревьев и кустарников. М. Лесная пром-
сть. 1970. 392.
Черемисинов Н.А. Микоценоз — компонент лесного
биогеоценоза // Микология и фитопатология. 1973. Т. 7. В. 1. С. 34-38.
Чмыр А.Ф. Биологические основы восстановления еловых лесов
южной тайги. Л. Изд-во Ленингр. универ. 1977. 160 с.
Чураков Б.П., Зырянова У.П., Пантелеев СВ. Аккумуляция
тяжелых металлов представителями микобиоты в лесных
экосистемах // М.— Петрозаводск. Грибные сообщества лесных
экосистем. 2004. Т. 2. С. 287-306.
Чураков Б.П., Нестеренко С.Г., Михалкин И.П.
Распространение очагов корневой губки в сосновых древостоях
Ульяновской области // М. Материалы 6-й междунар. конф.
«Проблемы лесной фитопатологии и микологии». 2005. С. 350-356.
Чураков Б.П. Фитопатогенные грибы дубовых лесов //М.
Грибные сообщества лесных экосистем. 2000. С. 292-317.
Швиденко А.З., Веневский B.C., Нильссон С. Обобщенная
оценка прироста и отпада в лесах России //М. Труды VII
ежегодной конф. МАИБЛ. 1997. С. 191-197.
Шенников А.П. Введение в геоботанику. Л. Изд-во ЛГУ. 1964.
447 с.
Шорохова Е.В. Потеря массы в результате ксилолиза после
сплошных ветровалов в ельниках Центрально-лесного
биосферного государственного заповедника // М.— Петрозаводск.
Материалы 6-й междунар. конф. «Проблемы лесной
фитопатологии и микологии». 2005. С. 361-367.
Шорохова Е.В., Соловьев В.А. Динамика углерода древостоев
коренных ельников средней тайги (резерват «Вепсский лес»)
// Лесоведение. 2002. № 1. С. 10-17.
Шорохова Е.В., Шорохов А.А. Характеристика классов
разложения древесного детрита ели, березы и осины в ельниках
подзоны средней тайги // СПб. Труды СПбНИИЛХ. 1999. В. 1.
С.17-23.
-188-
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА (Теория и эксперимент)
Шорохова Е.В., Гирфанов М.И. Ксилолиз крупных древесных
остатков в коренных среднетаежных ельниках // М.—
Петрозаводск. Грибные сообщества лесных экосистем. 2004. Т. 2.
С. 255-272.
Ainsworth J. and H. Bisby. Dictionary of the Fungi. 8th ed. By D.L.
Hawks worth, p.m. Kirk, B.C. Sutton and D.M. Pegler. CAB
International. Wallington. U.K. 1995. 616 p.
Braun-Blanquet J. Pflanzensoziologie. Wien-N.Y. 3. Springer.
1964. 865 p.
Cavalier-Smith T. Zooflagellata phylogeny and classification //
Cytology. 1995. V.37. P. 1010-1029.
Clements F.E. Plant succession. An analysis of the development of
vegetation. Wash. (D.C.) Carnegie Inst. 1916. 242 p.
Clements F.E. Nature and structure of the climax // Ecol. 1936.
V.21. N.l. 462p.
Clements F.E. Dynamics of Vegetation. New York. 1949. 296 c.
Heterobasidion annosun. Biology, Ecology, Impact and Control.
New-York. CAB International. 1998. 589 p.
Karjalainen L., Kuuluvainen T. Amount, quality and variability of
course w<aody debris within pine-dominated forest landscape //
Abstract of Nordic Symposium «Ecology Course Wood Debris in
Boreal Forest». Umea. Sweden. 1999. P. 12.
Kent B.V., Dress P.E. On the convergence of forest stand spetial
pattern over time the cases of regular and aggregated initial
special pattern // Forest Science. 1980.V. 26. N1. P. 10-22.
Korhonen K. Fungi belonging to the genera HETEROBASIDION
and ARMILLARIA in EURASIA // Материалы 6-й междунар.
конф. «Проблемы лесной фитопатологии и микологии».
2004. С. 89-114.
Korhonen К. Intersterility groups of Heterobasidin annosum.
Helsinli. 1978. 25 p.
Korhonen K. Intersterility and clonal size in the Armillariella
mellea complex // Karstenia. 18. 1978. P. 31-42.
Krankina O.N., Harmon M.E. Dynamics of the dead wood carbon
pool in northen-western Russian boreal forests // Water, Air
and Soil Pollution. 1995. V. 82. P. 227-238.
Margulis L., Schwartz K. Five Kingdoms. San Francisco. 1982.
400 p.
-189-
8. Г. Стороженко
Niemela Т., Korhonen К. Taxonomy of the genus Heterobasidion //
Heterobasidion annosum. Biology, ecology, impact and control.
CAB International. Oxon. New York. P. 27-33.
Nobles M.K. Identification of cultures of wood- rotting fungi //
Stad. In Forest. In Forest. Patol. 1948. V. 26. N. 3. P. 281-431.
Nobles M.K. Identification of cultures of wood-inhabiting
Hymenomycete // Can.J.Bot. 1965. V. 43. N. 9. P. 1097-1139.
Nordic Macromycetes. Polyporales, Boletales, Agaricales,
Russulales. Copengagen. 1992. V. 2. 474 p.
Nordic Macromycetes. Heterobasidioid, Aphyllophoroid and
Gasteromycetoid Basidiomycetes // Copengagen. 1997. V. 3.
444 p.
Rischbeth J. Stump protection against Fomes annosus. Inoculation
with Pheniophora gigantea // Ann. Appl. Biol. 1963. V. 52.
P. 63-77.
Selleck G.H. The climax consept // Botan. Rev. 1960. V. 26. N. 4.
P. 535-546.
Storozhenko V.G., Bondartseva M.A. The occurrence of wood
destroying fungi on the fallen logs in the spruce wildlife forests
of the Central-Forest reserve. //M. IV International Conf.
«Probl. Of Forest Patol. And MycoL». 1997. P. 88-90.
Stalpers J.A. Identification of wood-inhabiting Aphyllophorales in
pure culture. Stadies in Micology. 1978. N. 16. P. 248.
Tansley A.G. The classification of vegetation and the concept of
development // Jorn. Ecol. 1920. V. 8. N. 2.
Tansley A.G. The use and abuse of vegitational concepts and terms //
Jorn Ecol. V. 16. N. 3 1935. 485-930 p.
Yatskov M., Harmon M., Krankina O. A chronosequenceof wood
decomposition in the boreal forest of Russia // Can. Journ.
For.Res. 2003. V. 33. P. 1211-1226.
Vincent P.J., Hsworth J., Griffiths J.D., Collins R. The detection
of rendomness in plant pattern // J. Biogeogr. 1976. V. 3. N. 4
P.373-390.
Whittaker R.H. Communities and ecosystem. 2-nd ed. N. Y. 1975.
531 p.
-190-
В.Г. Стороженко
УСТОЙЧИВЫЕ ЛЕСНЫЕ СООБЩЕСТВА
(ТЕОРИЯ И ЭКСПЕРИМЕНТ)
Формат 60x84/16. Печ. л. 12,125.
Печать офсетная. Бумага офсетная.
Тираж 100 экз. Заказ № 385
ЗАО «Гриф и К», г. Тула, Октябрьская, 81-а