















































































































































































































































































































































Author: Афанасьев Ю.И. Юрина Н.А.
Tags: анатомия анатомия человека сравнительная анатомия вирусология происхождение и эволюция, генетика гистология цитология учебное пособие эмбриология
ISBN: 5-225-00002-9
Year: 1989




Ю. И. АФАНАСЬЕВ, Н. А. ЮРИНА.
ГИСТОЛОГИЯ.
УЧЕБНИК ААЛ МЕДИЦИНСКИХ
ИНСТИТУТОВ.
МОСКВА, ИЗДАТЕЛЬСТВО "МЕДИЦИНА"
1Р8? Г.
ОЗНАКОЛИТЕАЬНАЯ ВЕРСИЯ. КО/ИЖЕРЧЕСКОЕ использование аанного хчевного пособия
ЗАПРЕЩЕНО СОГААСНО ЗАКОНОААТЕАЬСТВХ ОС АВТОРСКОЕ ПРАВЕ
^официальный сайт <Lpfj .
они ал
ой
УЧЕБНАЯ ЛИТЕРАТУРА
Для студентов медицинских институтов
Гистология
Под редакцией
Ю. И. АФАНАСЬЕВА, Н. А. ЮРИНОЙ
Издание четвертое,
переработанное и дополненное
Допущено Главным управлением учебных
заведений Министерства здравоохранения СССР
в качестве учебника для студентов
медицинских институтов
U
Ж
Москва
« Медицина »1989
Отсканировано
в Рязани 2006
ЬБК 2S 3
Г 51
УДК 611-018 + ЮА-0Ч1 Н/С075.8)
VAs\Q>
Ю. И. Афанасьев, Н А Юрина, Б В. Алешин, Е. Ф. Котовский, Ю <•
Я А Винников, В Г. Елисеев, Г С Катинас, М Я Субботин, Т. Г.
Т Н. Радостина
Рецензент: А. А. Колосова, проф., зав. кафедрой гистологии и эмбр,
Ростовского ордена Дружбы народов медицинского ин т.
Гистология: Учебник/Ю. И. Афанасьев, Н. А. Юр
Г 51 Б, В. Алешин и др.; Под ред. Ю. И. Афанасьева, Н. А. К
ной — 4-е изд. перераб. и доп. — М.: Медицина, 198*
672 с.: ил. (Учеб. лит. Для студ. мед. ин-тов)
ISBN 5-225-00002-9
В четвертом издании (третье вышло в 1983 г.) на совреме! ювж
жены основы гистологии, цитологии и эмбриологии. Предста юг*
дения о строении клетки и ее производных, строении, функции i и
в эволюционном и возрастном аспекте. Все термины приведе> потв
с международными гистологической и эмбриологической номенкла.
4107000000 -201
Г ---------------62—88 ББк
039(01)—89
С Издательство «Медицина, Москва, 1983
С Издательство «Медицина», Москва, 1989, с изменен
ISBN 5-225-00002-9
ПОСВЯЩАЕТСЯ
Учителю и соратнику, организатору
авторского коллектива учебника
заслуженному деятелю науки РСФСР
профессору
ВЛАДИМИРУ ГРИГОРЬЕВИЧУ
ЕЛИСЕЕВУ
ПРЕДИСЛОВИЕ К ЧЕТВЕРТОМУ ИЗДАНИЮ
Гистология, цитология и эмбриология, как и другие фундамен-
;льные науки, формируя научное материалистическое мировоззре-
ие о закономерностях строения и жизнедеятельности организма
ловека, являются важнейшими учебными дисциплинами. Буду-
им врачам необходимо хорошее знание строения клеток и тканей
>ганов, являющихся структурной основой всех видов жизнедея-
лыюсти организма. Значимость гистологии, цитологии и эмбри-
логии для врачей возрастает еще и потому, что для современной
гедицины характерно широкое применение цитологических и
истологических методов при проведении анализов крови, костно-
о мозга биопсии органов и пр.
Четвертое издание учебника исправлено и обновлено. Структу-
<а кни! и осталась в основном прежней, но в каждую главу внесены
вменения в соответствии с новыми данными науки, рецензиями
пожеланиями коллег и студентов.
В отличие от предыдущего издания упразднена глава «Введение
учение о тканях», но введена новая — глава XII «Общие
инципы организации тканей», которая заключает раздел «Общая
стология». Такое расположение этой главы, по нашему мнению,
легчает студентам понимание обобщений сложных теоретиче-
IX вопросов.
Главы написаны по единому плану в основном прежним автор-
им коллективом, за исключением главы X «Мышечные ткани»
1рэф. Г. С. Катинас). Глава XII «Общие принципы организации
аней» написана проф. Ю. И. Афанасьевым в соавторстве с
°ф. Г С. Катинасом. В переработке глав «Органы чувств»,
I система», кроме авторов предыдущего издания, приняли
*ас1 ооф. Ю. И. Афанасьев и проф. Н. А. Юрина. В главе
•и <тельная система» подглаву «Зубы» написал проф.
Il знасьев, «Печень» — проф. Е. Ф. Котовский, «Поджелу-
'"1 слеза» — проф. Ю. И. Афанасьев и проф. Е. Ф. Котов-
ий.
В данном издании учебника сокращены материалы, которые
1гаются в смежных дисциплинах, носят описательный харак-
• В главе V «Основы эмбриологии» сокращено изложение
териала по вопросам органогенеза, который кратко дан в разде-
«Чистная гистология». В главе XIII «Нервная система» также
<енынем объеме изложены вопросы о микроскопическом строе-
нии органов. В книге использованы русские эквиваленты терм»
Международной гистологической номенклатуры, утвержден
Всесоюзным съездом анатомов, гистологов и эмбриологов.
В учебнике нашли отражение особенности преподавания
логии, цитологии и эмбриологии на различных факульт ета\
медицинского института (лечебном, педиатрическом, стомато логи-
ческом и санитарно-гигиеническом).
В отличие от предыдущих изданий учебник иллюстр иров
не только черно-белыми и цветными рисунками, но цвет ным
микрофотографиями препаратов и электронными микрофото^р
фиями, среди которых многие сделаны авторами и их сотрудникагЛ
Авторы выражают искреннюю благодарность всем коллега'м
гистологам, цитологам и эмбриологам, предоставившим для bkj но
чения в учебник оригинальные микрофотографии и схемы, а такж
коллегам-морфологам за рецензии и сделанные замечания —
профессорам В. И. Архипенко, К. А. Зуфарову, А. И. Брусилов-
скому, ст. науч. сотр. С. Л. Кузнецову.
Авторы и редакторы глубоко признательны сотрудникам
кафедр гистологии и эмбриологии I ММИ им. И. М. Сеченова
доц. А. Н. Яцковскому, старшим преподавателям Л. П. Бобовой
В. Л. Горячкиной, Е. А. Хачатурян, ассистенту Т. В. Боронихи
ной, канд. биол. наук. Н. Н. Босовой, ст. лаб. Л. В. Кушко, Н. Б. Ле
бедевой и Университета дружбы народов им. П. Лумумбы — до
центам А. И. Радостиной, Л. С. Румянцевой, В. А. Ремизовой
ст. лаб. Н. Н. Тупиковой за большую помощь при подготовь
учебника к изданию.
Проф. Ю. И. Афанасъ
Проф. Н. А. Юри>
Глава I
ГИСТОЛОГИЯ, ЦИТОЛОГИЯ И ЭМБРИОЛОГИЯ
ИХ СОДЕРЖАНИЕ, ЗАДАЧИ И СВЯЗЬ С ДРУГИМИ
МЕДИКО-БИОЛОГИЧЕСКИМИ НАУКАМИ.
ЗНАЧЕНИЕ ДЛЯ МЕДИЦИНЫ
Организм человека и животных представляет собой целостную
истему, в которой можно выделить ряд иерархических уровней
рганизации живой материи: клетки — ткани — морфофункцио-
нальные единицы органов — органы — системы органов. Каждый
ровень структурной организации имеет морфофункциональные
особенности, отличающие его от других уровней.
Гистология (от греч. hist os — ткань, logos — учение) — наука
о строении, развитии и жизнедеятельности тканей животных
организмов.
Таким образом, гистология вместе с другими фундаменталь-
ными медико-биологическими науками изучает закономерности
структурной организации живой материи. В то же время в отли-
чие от других биологических наук основным предметом гистоло-
гии являются именно ткани, представляющие собой систему сле-
дующей за клеточным уровнем организации живой материи в
целостном организме. Поэтому тканям присущи общебиологиче-
ские закономерности, свойственные живой материи, и вместе с тем
собственные особенности строения, развития, жизнедеятельности,
внутритканевые (внутриуровневые) и межтканевые (межуровне-
вые) связи. Сами ткани служат элементами развития, строения и
жизнедеятельности органов и их морфофункциональных единиц.
Ткани представляют собой систему клеток и неклеточных
структур, объединившихся и специализировавшихся в процессе
эволюции для выполнения важнейших функций в организме.
Для каждой из 5 основных тканевых систем (нервная ткань,
мышечная ткань, эпителиальная ткань, соединительная ткань,
ровь) характерны присущие именно им особенности строения,
развития и жизнедеятельности. Предметом общей гистоло-
гии, или собственно учения о тканях, являются общие законо-
мерности, присущие тканевому уровню организации, и отличитель-
ные особенности конкретных тканей; предметом частной
истологии — закономерности жизнедеятельности и взаимо-
действия различных тканей в органах на более высоких уровнях
рганизации. Частная гистология служит основой для изучения
икроскопического строения морфофункциональных единиц
рганов и органов в целом.
Курс гистологии включает в себя также цитологию —
чение о клетке и эмбриологию — учение о зародыше. Эти
самостоятельные курсы предшествуют общей и частной гисто-
логии.
4
5
Цитология (от греч. kytos — клетка, logos — учение) — наук»
о развитии, строении и жизнедеятельности клеток.
Цитология составляет необходимую часть гистологии, так как
клетки являются основой развития и строения тканей. В разделе
общей цитологии рассматриваются общие принципы строе-
ния и физиологии клеточных структур. Частная цитоло-
гия изучает особенности строения специализированных клеток
в различных тканях и органах. Цитология в последние годы обо-
гатилась многими научными открытиями, внесшими существенный
вклад в развитие биологических и медицинских наук и практику
здравоохранения. Новые данные о структуре ядра, его хромосом-
ного аппарата легли в основу цитодиагностики наследственных
заболеваний, опухолей, болезней крови и многих других болезней.
Раскрытие особенностей ультра структуры и химического состава
клеточных мембран является основой для понимания закономер-
ностей взаимодействия клеток в тканевых системах, защитных
реакциях и др. В медицинской практике широко используется
цитодиагностика. Клетки здорового и больного организма изуча
ются в мазках крови и костного мозга, спинномозговой жидкости
слюны, мочи, в образцах различных органов, взятых при биопсии.
Эмбриология (от греч. embryon — зародыш, logos — учение)
учение о зародыше, о закономерностях его развития.
В курсе эмбриологии, преподаваемом в медицинском вузе
основное внимание обращается на закономерности эмбриональной
развития человека. Знакомство будущего врача с особенностям»
эмбриогенеза человека имеет большое значение для формирования
его научного материалистического мировоззрения и для практиче
ской деятельности. Сравнительная эмбриология дает богаты»
фактический материал, подтверждающий, что развитие человек;
есть результат эволюции живой природы. Особое внимание в курс
эмбриологии обращается на источники развития и механизмь
образования тканей (гистогенез) на определенном этапе эмбриоге-
неза. Закономерности гистогенеза определяют морфофункциональ
ные особенности тканевых структур в постнатальном онтогенезе
в частности их способность к регенерации. Поэтому изучени
основных этапов эмбрионального развития предшествует изучении
тканей. Таким образом, объединение гистологии, цитологии »
эмбриологии в один предмет не формально, а отражает внутрен
ние естественные связи между ними.
Гистология с цитологией и эмбриологией, как и другие биоло
гические науки, решает главную задачу — выяснение структурно»
организации процессов жизнедеятельности и в связи с этим -
возможности целенаправленного воздействия на них.
Познание сущности жизни должно вестись на основе научно
методологии диалектического материализма. Необходимое!
применения законов философии в естественных науках обоснован
в трудах классиков марксизма-ленинизма. В. И. Ленин указыва.
что «... без солидного философского обоснования никакие есте
ственные науки, никакой материализм не может выдержать
борьбы против натиска буржуазных идей и восстановления бур-
жуазного миросозерцания. Чтобы выдержать эту борьбу и про-
вести ее до конца с полным успехом, естественник должен быть
современным материалистом, ... должен быть диалектическим
материалистом»1.
Предмет гистологии с цитологией и эмбриологией предостав-
ляет студентам богатые возможности для раскрытия на конкрет-
ном фактическом материале законов материалистической диалек-
тики: взаимосвязи строения и функции, общего и частного, един-
ства и борьбы противоположностей. Овладеть этими категориями
и уметь применять их в своей практической деятельности при
анализе сложных процессов в организме человека — одна из
важных задач в подготовке врача.
Познание закономерностей строения клеток, тканей и органов
должно вестись в связи с их функциями. Взаимоотношения
между структурой и функцией рассматриваются с позиций закона
диалектического материализма о единстве материи и ее движения.
Поэтому структура включает в себя понятия и морфологического
строения и функции.
Изучение каждой структуры должно проводиться с историче-
ских позиций, основывающихся на эволюционном учении Ч. Дар-
вина, согласно которому все составные части человеческого орга-
низма рассматриваются как результат филогенетического разви-
тия. Теории развития тканей (параллельных рядов А. А. Заварзина
и дивергентного развития Н. Г. Хлопина) устанавливают основные
закономерности формирования тканей в филогенезе.
Исследование различных уровней организации живой материи
в целостном организме должно базироваться на системном анали-
зе, так как всякая структура является сложной системой, взаимо-
действующей с другими структурными элементами одинакового
или различного уровня организации. Системный анализ позволяет
выявить корреляции, характерные для внутриклеточных, тканевых
и органных систем, установить закономерности взаимодействия
части и целого и др. Вот почему задачей гистологии является не
только описание строения и функционального назначения струк-
тур, но и установление связей между ними, раскрытие закономер-
ностей их развития.
Для познания закономерностей развития, строения и функции
клеток, тканей и органов в современной гистологии широко
применяются экспериментальные методы исследования, позволяю-
ие вести наблюдения на живых объектах, моделировать различ-
ью процессы. Изучение микроструктур ведется на молекулярном,
Уоклеточном, клеточном и тканевом уровнях с помощью микро-
копирования в различных системах светооптических и электрон-
ных микроскопов, методов цито- и гистохимии, авторадиографии,
иометрии. Количественный анализ структур включает применение
Ленин В. И. Поли. собр. соч., т. 45, с. 29—30.
6
7
математического моделирования, использование ЭВМ, специализи
рованных автоматических устройств.
Современные гистология, цитология и эмбриология внося?
существенный вклад в разработку теоретических и прикладных
аспектов современной медицины и биологии.
К фундаментальным теоретическим проблемам
относятся:
— изучение закономерностей цито- и гистогенеза, строения и
функции клеток и тканей;
— изучение закономерностей дифференцировки и регенерации
тканей;
— выяснение роли нервной, эндокринной, иммунной систем
организма в регуляции процессов морфогенеза клеток, тканей и
органов;
— исследование возрастных изменений клеток, тканей, ор-
ганов;
— исследование адаптации клеток, тканей и органов к дей-
ствию различных биологических, физических, химических и
других факторов;
— изучение процессов морфогенеза в системе мать — плод;
— исследование особенностей эмбриогенеза человека.
Актуальными прикладными проблемами являются
исследование клеточной и тканевой совместимости при перели-
вании крови, трансплантации тканей, при действии стрессовых
факторов, изучение регенерационных возможностей тканей в
различных условиях, разработка морфологических тестов для
оценки возрастных изменений и др.
Прогресс современной гистологии в большей степени определя-
ется тем, что она основывается на достижениях физики, химии,
математики, кибернетики. Внедрение новейших методов исследо-
вания обусловило бурное развитие гистологии с цитологией и
эмбриологией. Курс гистологии с цитологией и эмбриологией
тесно связан с преподаванием других медико-биологических
наук — биологии, анатомии, физиологии, биохимии, патологиче-
ской анатомии, а также клинических дисциплин. Так, раскрытие
основных закономерностей структурной организации клеток явля
ется основой для изложения вопросов генетики в курсе биологии.
С другой стороны, изложение вопросов, касающихся эволюции
живой материи, в курсе биологии является необходимой предпо-
сылкой для изучения различных уровней организации живой
материи в организме человека. Изучение закономерностей разви-
тия и строения органов в курсе анатомии базируется на данных
гистологического анализа. В настоящее время, когда исследова-
ния клеточных и тканевых структур ведутся на субклеточном и
молекулярном уровнях с применением биохимических методов,
отмечается особенно тесная связь гистологии, цитологии и эмбри-
ологии с бирхимией. В преподавании, научных исследованиях и
клинической диагностике широкое применение нашли цито- и
гистохимические данные. Знание нормальной структуры клеток.
ней и органов является необходимым условием для понимания
ТКханизмов изменений в них в патологических условиях. Поэтому
Отология с цитологией и эмбриологией тесно связана с патологи-
ческой анатомией и многими клиническими дисциплинами (внут-
енние болезни, акушерство и гинекология и др.).
Р Таким образом, гистология с цитологией и эмбриологией
занимает важное место в системе медицинского образования,
закладывая основы научного структурно-функционального подхода
в анализе жизнедеятельности организма человека в норме и пато-
логии.
Глава II
МЕТОДЫ ИССЛЕДОВАНИЯ в гистологии, цитологии
и ЭМБРИОЛОГИИ
В современной гистологии, цитологии и эмбриологии применя-
ются разнообразные методы исследования, позволяющие всесто-
ронне изучать процессы развития, строения и функции клеток,
тканей и органов.
Главными этапами цитологического и гистологического анали-
за являются выбор объекта исследования, подготовка его для
изучения в микроскопе, применение методов микроскопирования,
качественный и количественный анализ изображений.
Объектами исследования служат живые и мертвые
(фиксированные) клетки и ткани, их изображения, полученные в
световых и электронных микроскопах или на телевизионном
экране дисплея. Существует ряд методов, позволяющих проводить
анализ указанных выше объектов.
МЕТОДЫ ИССЛЕДОВАНИЯ ЖИВЫХ КЛЕТОК И ТКАНЕЙ
Изучение живых клеток и тканей позволяет получить наиболее
полную информацию об их жизнедеятельности — проследить дви-
жение, процессы деления, разрушения, роста, дифференцировки
и взаимодействия клеток, продолжительность их жизненного
цикла, реактивные изменения в ответ на действие различных
факторов.
Прижизненные исследования клеток в организме (in vivo).
Одним из прижизненных методов исследования является наблю-
дение структур в живом организме. С помощью специальных
просвечивающих микроскопов-иллюминаторов, например, можно
изучать в динамике циркуляцию крови в микрососудах. После
проведения анестезии у животного объект исследования (напри-
мер, брыжейка кишечника) выводят наружу и рассматривают в
микроскопе, при этом ткани должны постоянно увлажняться
физиологическим раствором. Однако длительность такого наблю-
дения ограничена. Лучшие результаты дает метод вживле-
ния прозрачных камере организм животного. Наиболее
9
8
удобным органом для вживления таких камер и последующего
наблюдения является ухо какого-либо животного (например, кро-
лика). Участок уха с прозрачной камерой помещают на предмет-
ный столик микроскопа, и в этих условиях изучают динамику
изменения клеток и тканей в течение продолжительного времени.
Таким образом могут изучаться процессы выселения лейкоцитов
из кровеносных сосудов, различные стадии образования соедини-
тельной ткани, капилляров, нервов и другие процессы. В качестве
естественной прозрачной камеры можно использовать глаз экспе-
риментальных животных. Клетки, ткани или образцы органов
помещают в жидкость передней камеры глаза в угол, образован-
ный роговицей и радужкой, и могут наблюдаться через прозрач-
ную роговицу. Таким образом была произведена трансплантация
оплодотворенной яйцеклетки и прослежены ранние стадии разви-
тия зародыша. Обезьянам были пересажены небольшие кусочки
матки и изучены изменения слизистой оболочки матки в различ-
ные фазы менструального цикла.
Широкое применение нашел метод трансплантации
клеток крови и костного мозга от здоровых животных-доноров
животным-реципиентам, подвергнутым смертельному облучению.
Животные-реципиенты после трансплантации оставались живыми
вследствие приживления донорских клеток, образующих в селезен-
ке колонии кроветворных клеток. Исследование числа колоний
и их клеточного состава позволяет выявлять количество родона-
чальных кроветворных клеток и различные стадии их дифференци-
ровки. С помощью метода колониеобразования установлены
источники развития для всех клеток крови.
Исследования живых структур в культуре клеток и тканей
(in vitro). Этот метод является одним из самых распространенных.
Выделенные из организма человека или животных клетки, малень-
кие образцы тканей или органов помещают в стеклянные или
пластмассовые сосуды, содержащие специальную питательную
среду — плазму крови, эмбриональный экстрат и стимуляторы
роста клеток. Различают суспензионные культуры
(клетки взвешены в среде) и мо послойные культуры
(эксплантированные клетки образуют на стекле сплошной слой).
Обеспечивается стерильность среды и температура, соответствую-
щая температуре тела. В этих условиях клетки в течение длитель-
ного времени сохраняют основные показатели жизнедеятель-
ности — способность к росту, размножению, дифференцировке,
движению. Такие культуры могут существовать многие дни, меся-
цы и даже годы, если обновлять среду культивирования и переса-
живать жизнеспособные клетки в другие сосуды. Некоторые виды
клеток благодаря изменениям в их геноме могут сохраняться и
размножаться в культуре практически бесконечно, образуя непре-
рывные клеточные линии. В разработку методов культивирования
клеток и тканей большой вклад внесли А. А Максимов, А. В. Ру-
мянцев, Н. Г. Хлопин, А. Д. Тимофеевский, Ф М. Лазаренко.
В настоящее время получены клеточные линии фибробластов.
10
миоцитов, эпителиоцитов, макрофагов и др., которые существуют
многие годы.
Использование метода культивирования позволило выявить ряд
закономерностей дифференцировки, злокачественного перерож-
дения клеток, клеточных взаимодействий, взаимодействий клеток
с вирусами и микробами. Показана возможность хрящевых клеток
формировать в культуре межклеточное вещество, клеток надпочеч-
ников — продуцировать гормоны. Культивирование эмбриональных
тканей и органов дало возможность проследить развитие кости,
кожи и других органов. Разработана методика культивирования
нервных клеток.
Особую значимость метод культуры тканей имеет для прове-
дения экспериментальных наблюдений на клетках и тканях чело-
века. Взятые из организма человека клетки при пункции или
биопсии могут в культуре тканей использоваться для определения
пола, наследственных заболеваний, злокачественного перерожде-
ния, выявления действия ряда токсических веществ. В последние
годы клеточные культуры широко применяются для гибриди-
зации клеток. Сущность метода гибридизации заключается
в том, что из клеток любых типов и стадий развития можно
получить клеточные гибриды при воздействии ряда факторов —
инактивированного вируса парагриппа, полиэтиленгликоля и др.
В образованных гибридах формируются гетерокарионы, содержа-
щие два вида генотипа. Изучение белковых продуктов активности
генов позволяет устанавливать локализацию генов в составе хро-
мосомы. На основе метода гибридизации создан метод получе-
ния моноклональных антител, позволяющий выявлять различные
стадии развития иммунокомпетентных и других клеток.
Необходимо помнить, однако, и о недостатках метода, которые
нужно учитывать при оценке полученных результатов. Изоляция
клеток из целостного организма ведет к ряду изменений условий
их существования: утрачиваются взаимосвязи с другими клетками
и тканями, действие комплекса нейрогуморальных факторов регу-
ляции и др. Для устранения этого недостатка применяют культи-
вирование in vivo, когда образцы тканей и органов помещают в
камеры из пористого материала, которые подсаживают в участок
тела животного (в брюшную полость, под кожу и др.).
Витальное и суправитальное окрашивание. При витальном
(прижизненн ом) окрашивании клеток и тканей краситель
вводят в организм животного, при этом он избирательно окраши-
вает определенные клетки, их органеллы или межклеточное
вещество. Например, трипановый синий или литиевый кармин
выявляет фагоциты, ализарин — новообразованный матрикс кости.
Суправитальным окрашиванием называют окра-
шивание живых клеток, выделенных из организма. Таким спосо-
бом выявляют молодые формы эритроцитов — ретикулоциты
крови (краситель бриллиантовый крезиловый голубой), митохон-
дрии в клетках (краситель зеленый янус), лизосомы (краситель
нейтральный красный).
и
ИССЛЕДОВАНИЕ МЕРТВЫХ (ФИКСИРОВАННЫХ) КЛЕТОК И ТКАНЕЙ
Основным объектом исследования здесь являются гистологи-
ческие препараты, приготовленные из фиксированных структур.
Препарат может представлять собой мазок (например, мазок
крови, костного мозга, слюны, спинномозговой жидкости и др.),
отпечаток (например, селезенки, тимуса, печени), пленку
из ткани (например, соединительной или брюшины, плевры, мяг-
кой мозговой оболочки), тонкий срез. Наиболее часто для изу-
чения используется срез ткани или органа. Гистологические
препараты могут изучаться без специальной обработки. Например,
приготовленный мазок крови, отпечаток, пленка или срез органа
могут сразу рассматриваться под микроскопом. Но вследствие
того, что структуры имеют слабый контраст, они плохо выявля-
ются в обычном световом микроскопе и требуется использование
специальных микроскопов (фазово-контрастные и др.). Поэтому
чаще применяют специально обработанные препараты- фиксиро-
ванные, заключенные в твердую среду и окрашенные.
Процесс изготовления гистологического препарата для световой
и электронной микроскопии включает следующие основные этапы:
I) взятие материала и его фиксация, 2) уплотнение материала,
3) приготовление срезов, 4) окрашивание или контрастирование
срезов. Для световой микроскопии необходим еще один этан —
заключение срезов в бальзам или другие прозрачные среды (5).
Фиксация обеспечивает предотвращение процессов разложения,
что способствует сохранению целостности структур. Это достига-
ется тем, что взятый из органа маленький образец либо погружают
в фиксатор (спирт, формалин, растворы солей тяжелых металлов,
осмиевая кислота, специальные фиксирующие смеси), либо под-
вергают термической обработке. Под действием фиксатора в
тканях и органах происходят сложные физико-химические из-
менения. Наиболее существенным из них является процесс необра-
тимой коагуляции белков, вследствие которого жизнедеятельность
прекращается, а структуры становятся мертвыми, фиксированными.
Фиксация приводит к уплотнению и уменьшению объема кусочков,
а также к улучшению последующей окраски клеток и тканей.
Уплотнение кусочков, необходимое для приготовления срезов,
производится путем пропитывания предварительно обезвоженного
материала парафином, целлоидином, органическими смолами.
Более быстрое уплотнение достигается применением метода замо-
раживания кусочков, например, в жидкой углекислоте.
Приготовление срезов происходит на специальных приборах —
микротомах (для световой микроскопии) и ультрамикротомах
(для электронной микроскопии).
Окрашивание срезов (в световой микроскопии) или напыление
их солями металлов (в электронной микроскопии) применяют для
увеличения контрастности изображения отдельных структур при
рассматривании их в микроскопе. Методы окраски гистологических
структур очень разнообразны и выбираются в зависимости от
12
задач исследования. Гистологические красители подразделяют на
кислые, основные и нейтральные. В качестве примера
можно привести наиболее употребительный краситель азур II,
который окрашивает ядра клеток в фиолетовый цвет, и кислый
краситель — эозин, окрашивающий цитоплазму в розово-желтый
цвет. Избирательное сродство структур к определенным красите-
лям обусловлено их химическим составом и физическими свой-
ствами Структуры, хорошо окрашивающиеся кислыми красителя-
ми, называются оксифильными, а окрашивающиеся основными —
базофильными. Структуры, воспринимающие как кислые, так и
основные красители, являются нейтрофильными (гетерофильны-
ми). Окрашенные препараты обычно обезвоживают в спиртах
возрастающей крепости и просветляют в ксилоле, бензоле, толуоле
или некоторых маслах. Для длительного сохранения обезвожен-
ный гистологический срез заключают между предметным и по-
кровным стеклами в канадский бальзам или другие вещества.
Готовый гистологический препарат может быть использован для
изучения под микроскопом в течение многих лет. Для электронной
микроскопии срезы, полученные на ультрамикротоме, помещают
на специальные сетки, контрастируют солями марганца, кобальта
и др., после чего просматривают в микроскопе и фотографируют.
Полученные микрофотографии служат объектом изучения наряду
с гистологическими препаратами.
ИССЛЕДОВАНИЕ ХИМИЧЕСКОГО СОСТАВА И МЕТАБОЛИЗМА КЛЕТОК
И ТКАНЕЙ
Цито- и гистохимические методы. Эти методы позволяют
выявлять локализацию различных химических веществ в структу-
рах клеток, тканей и органов — ДНК, РНК, белков, углеводов,
липидов, аминокислот, минеральных веществ, витаминов, актив-
ность ферментов. Эти методы основаны на специфичности реакции
между химическим реактивом и субстратом, входящим в состав
клеточных и тканевых структур, и окрашивании продуктов хими-
ческих реакций. Для повышения специфичности реакции часто
применяют ферментативный контроль. Например, для выявления
в клетках рибонуклеиновой кислоты (РНК) часто используют
галлоцианин — краситель с основными свойствами, а наличие
РНК подтверждают контрольной обработкой рибонуклеазой,
расщепляющей РНК. Галлоцианин окрашивает РНК в сине-фиоле-
товый цвет. Если срез предварительно обработать рибонуклеа-
зой, а затем окрасить галлоцианином, то отсутствие окрашивания
подтверждает наличие в структуре рибонуклеиновой кислоты.
Описание многочисленных цито- и гистохимических методов
дается в специальных руководствах.
Метод радиоавтографии. Этот метод дает возможность наибо-
лее полно изучить обмен веществ в разных структурах. В основе
метода лежит использование радиоактивных элементов (например,
фосфора — 32Р, углерода — |4С, серы — 35S, водорода — 3Н) или
13
меченных им соединений. Радиоактивные вещества в гистологи-
ческих срезах выявляют с помощью фотоэмульсии, которую нано-
сят на препарат и затем проявляют. В участках препарата, где
фотоэмульсия соприкасается с радиоактивным веществом, про-
исходит фотореакция, в результате которой образуются засвечен-
ные участки (треки). Этим методом можно определять, например,
скорость включения меченых аминокислот в белки, образование
нуклеиновых кислот, обмен йода в клетках щитовидной железы
и др.
Метод дифференциального центрифугирования. Этот метод
основан на применении центрифуг, дающих от 20 000 до 150 000
об/мин. Такие центрифуги позволяют отделять и осаждать различ-
ные компоненты клеток — ядра, митохондрии и др. и определять
их химический состав.
Интерферометрия. Этот метод позволяет оценить сухую массу
и концентрацию плотных веществ в живой и фиксированной
клетках. С помощью этого метода, например, можно устано-
вить суммарное содержание белков в живых и фиксированных
клетках.
В настоящее время наряду с качественными методами разра-
ботаны и применяются количественные гистохими-
ческие методы определения содержания различных веществ
в клетках и тканях. Особенность количественно-гистохимических
(в отличие от биохимических) методов исследования заключается
в возможности изучения локализации химических компонентов
в конкретных структурах клеток и тканей.
Цитоспектрофотометрия — метод количественного изучения
внутриклеточных веществ по их абсорбционным спектрам.
Цитоспектрофлюориметрия — метод количественного изучения
внутриклеточных веществ по спектрам их флюоресценции или по
интенсивности флюоресценции на одной заранее выбранной волне
(цитофлюориметрия).
Современные микроскопы — цитофлюориметры позволяют об-
наружить в различных структурах малые количества вещества
(до 10 ,4—10 16 г) и оценить локализацию исследуемых веществ
в микроструктурах.
В последние годы сочетание гистохимических методов с
методом электронной микроскопии привело к развитию нового
перспективного направления — электронной гистохи-
мии. Этот метод позволяет изучать локализацию различных
химических веществ не только на клеточном, но и на субклеточном
и молекулярном уровнях
Методы иммунофлюоресцентного анализа. Они эффективно
используются в современной гистологии. Эти методы применяются
для изучения процессов дифференцировки клеток, выявления в
них специфических химических соединений и структур. Они осно-
ваны на реакциях антиген — антитело. Каждая клетка организма
имеет специфический антигенный состав, который главным образом
определяется белками. Продукты реакции можно окрашивать и
выявлять в люминесцентном микроскопе. На рис. 13,Б демонстри-
руется выявление актина и тубулина в клетке с помощью метода
иммунофлюоресцентного анализа.
МЕТОДЫ МИКРОСКОПИРОВАНИЯ ГИСТОЛОГИЧЕСКИХ ПРЕПАРАТОВ
Световая микроскопия. Микроскопирование — основной метод
изучения препаратов — используется в биологии уже более
300 лет. С момента использования первых микроскопов в иссле-
довательских целях они постоянно совершенствовались. Современ-
ные микроскопы представляют собой разнообразные сложные
оптические системы, обладающие высокой разрешающей способ-
ностью и позволяющие изучать очень тонкие детали строения
клеток и тканей. Размер самой маленькой структуры, которую
можно видеть в микроскопе, определяется наименьшим разре-
шаемым расстоянием (d0). В основном оно зависит от
длины световой волны X, и эта зависимость приближенно выража-
ется формулой d 0— 1/2^- Таким образом, чем меньше длина
световой волны, тем меньше разрешаемое расстояние и тем мень-
шие по размерам структуры можно видеть в препарате.
Для изучения гистологических препаратов чаще применяют
обычные световые микроскопы различных марок, когда в качестве
источника освещения используют естественный или искусственный
свет (рис. 1,А,Б). Минимальная длина волны видимой части спек-
тра света соответствует примерно 0,4 мкм. Следовательно, для
обычного светового микроскопа разрешаемое расстояние равно
приблизительно 0,2 мкм (do = '/г- мкм = 0,2 мкм), а общее
увеличение (произведение увеличения объектива на увеличение
окуляра) достигает 2500 раз.
Ультрафиолетовая микроскопия. Это разновидность световой
микроскопии. В ультрафиолетовом микроскопе используют более
короткие ультрафиолетовые лучи с длиной волны около 0,2 мкм.
Разрешаемое расстояние здесь составляет приблизительно 0,1 мкм
(d0= */2 • 0,2 мкм — 0,1 мкм). Полученное в ультрафиолетовых
лучах невидимое глазом изображение преобразуется в видимое с
помощью регистрации на фотопластинке или путем применения
специальных устройств (люминесцентный экран, электронно-опти-
ческий преобразователь).
Флюоресцентная (люминесцентная) микроскопия. Явления
флюоресценции заключаются в том, что атомы и молекулы ряда
веществ, поглощая коротковолновые лучи, переходят в воз-
бужденное состояние. Обратный переход из возбужденного состо-
яния в нормальное происходит с испусканием света, но с другой,
большей длиной волны. В флюоресцентном микроскопе в качестве
источников света для возбуждения флюоресценции применяют
ртутные или ксеноновые лампы сверхвысокого давления, облада-
ющие высокой яркостью в области спектра 0,25—0,4 мкм (ближ-
ние ультрафиолетовые лучи) и 0,4—0,5 мкм (сине-фиолетовые
лучи). Длина световой волны флюоресценции всегда больше длины
15
Рис. 1. Микроскопы для биологиче-
ских исследовании.
А — световой биологический микроскоп
♦Биолам-С»: / основание; 2 — тубусо-
держатель, 3 наклонный тубус; 4
окуляр; 5 — револьвер, 6 объективы;
7 — столик; 8 — конденсор с ирисовой
диафрагмой; 9 — винт конденсора; 10 —
зеркало; // — микрометрический винт;
12 — макрометрический винт; Б элек-
видеосигнала.
тронный микроскоп ЭМВ-100АК с ав-
томатизированном системой обработки
изображений: 1 колонка микроскопа
(с электронно-оптической системой и
камерой для образцов); 2 — пульт уп-
равления; 3 — камера с люминесцент
ным экраном; 4 — блок анализа изобра
жений; 5 — датчик
k ® ® ®
HIIIIIIIIIIIBMIIII )
Б
волны возбуждающего света, поэтому их разделяют с помощью
светофильтров и изучают изображение объекта только в свете
флюоресценции. Различают собственную, или первич-
ную^ наведенную, или вторичную, флюоресцен-
цию. Любая клетка живого организма обладает собственной
флюоресценцией, однако она часто бывает чрезвычайно слабой.
Вторичная флюоресценция возникает при обработке препаратов
специальными красителями — флюорохромами. Например,
при обработке препаратов чаще всего употребляется флюорохром
акридиновый оранжевый. В этом случае ДНК и ее соединения
в клетках имеют ярко-зеленое, а РНК и ее производные — ярко-
красное свечение. Таким образом, спектральный состав излучения
несет информацию о внутреннем строении объекта, его химиче-
ском составе. Вариант метола флюоресцентной микроскопии, при
котором и возбуждение, и излучение флюоресценции происходят
в ультрафиолетовой области спектра, получил название метода
ультрафиолетовой флюоресцентной микроскопии.
Фазово-контрастная микроскопия. Этот метод служит для
получения контрастных изображений прозрачных и бесцветных
объектов, невидимых при обычных методах микроскопирования.
Для изучения препаратов в обычном световом микроскопе необхо-
димая контрастность структур достигается с помощью окрашива-
ния. Метод фазового контраста обеспечивает необходимую кон-
трастность изучаемых неокрашенных структур за счет специальной
кольцевой диафрагмы, помещаемой в конденсоре, и так называ-
емой фазовой пластинки, находящейся в объективе. Такая конс-
трукция оптики микроскопа дает возможность преобразовать не
воспринимаемые глазом фазовые изменения прошедшего через
неокрашенный препарат света в изменение его амплитуды, т. е.
яркости получаемого изображения. Повышение контраста позволя-
ет видеть все структуры, различающиеся по показателю прелом-
ления. Разновидностью метода фазового контраста является метод
фазово-темнопольного контраста, дающий негативное по сравне-
нию с позитивным фазовым контрастом изображение.
Кроме перечисленных методов, для специальных целей при-
меняются микроскопия в темном поле при изучении живых
объектов, в падающем свете для рассмотрения толстых объектов,
поляризационная микроскопия для изучения архитектоники гисто-
логических структур. Описание этих методов и соответствующих
приборов приводится в специальных руководствах.
Электронная микроскопия. Большим шагом вперед в развитии
техники микроскопии было создание и применение электронного
микроскопа (см. оис. 1, Б). В электронном микроскопе использу-
ется поток электронов с более короткими, чем в световом микро-
скопе, длинами волн. При напряжении 50 000 В длина волны
электромагнитных колебаний, возникающих при движении потока
электронов в вакууме, равна 0,0056 нм. Теоретически рассчитано,
что разрешаемое расстояние в этих условиях может быть около
0,002 нм, или 0,000002 мкм, т. е в 100 000 раз меньше, чем в
световом микроскопе. Практически в современных электронных
микроскопах лучших марок разрешаемое расстояние составляет
0,1—0,7 нм.
С помощью просвечивающего электронного
микроскопа можно получить лишь плоскостное изображение
изучаемого объекта. Для получения пространственного представ-
ления о структурах используют растровые электронные
микроскопы, способные создавать трехмерные изображения.
Растровый электронный микроскоп работает по принципу скани-
рования электронным микрозондом исследуемого объекта, т. е.
последовательно «ощупывает» остро сфокусированным электрон-
ным пучком отдельные точки поверхности. Для исследования
выбранного участка микрозонд двигается по его поверхности под
действием отклоняющих катушек (принцип телевизионной раз-
вертки). Такое исследование объекта называется сканирова-
нием (считыванием), а рисунок, по которому движется микро-
зонд, — растром. Растровый электронный микроскоп не ре-
гистрирует электроны, проходящие сквозь объект, а собирает все
электроны (вторичные), выбитые из атомов образца потоком
электронов микрозонда (первичные электроны). Полученное изо-
бражение выводится на телевизионный экран, электронный луч
которого движется синхронно с микрозондом.
Главными достоинствами растровой электронной микроскопии
являются большая глубина резкости (в 100—1000 раз больше,
чем у световых микроскопов), широкий диапазон непрерывного
изменения увеличения (от десятков до десятков тысяч раз) и
высокая разрешающая способность.
МЕТОДЫ АНАЛИЗА ИЗОБРАЖЕНИЙ КЛЕТОЧНЫХ И ТКАНЕВЫХ
СТРУКТУР
Полученные изображения микрообъектов в микроскопе, на
телевизионном экране дисплея, на электронных микрофотографи-
ях могут подвергаться специальному анализу — выявлению мор-
фометрических, денситометрических параметров и их статистиче-
ской обработке.
Морфометрические методы позволяют определять с помощью
специальных сеток (Е. Вейбеля, А. А. Глаголева, С. Б. Стефанова)
число любых структур, их площади, диаметры и др. В частности,
в клетках могут быть измерены площади ядер, цитоплазмы, их
диаметры, ядерно-цитоплазматические отношения и др. Существу-
ет ручная морфометрия и автоматизирован-
ная морфометрия, при которой все параметры измеряются
и регистрируются в приборе автоматически.
В последние годы все большее распространение получают
автоматизированные системы обработки изображений (АСОИз),
позволяющие наиболее эффективно реализовать перечисленные
выше количественные методы для изучения клеток и тканей. При
этом аналитические возможности количественной микроскопии
18
I
дополняются методами анализа и распознавания образов, основан-
ными на обработке с помощью электронных вычислительных
машин (ЭВМ) информации, извлекаемой из изображений клеток
и тканей. По существу можно говорить об устройствах, не только
усиливающих оптические возможности зрительного анализатора
человека, но и многократно расширяющих его аналитические
возможности. Высказывается мнение, что АСОИз совершает
такой же переворот в морфологии, какой около 300 лет назад
произошел благодаря изобретению светового, а около 35 лет
назад — электронного микроскопа, поскольку они не только неиз-
меримо повышают производительность труда исследователя и не
только объективизируют наблюдения, но и позволяют получать
новую информацию о не выявляемых ранее процессах, численно
моделировать и прогнозировать их развитие в клетках и тканях.
Вместе с тем участие в эксперименте ЭВМ требует от исследо-
вателя нового подхода к его проведению, владения навыками
составления алгоритмов процесса исследования, точности рассуж-
дений и в конечном итоге повышения научно-методического
уровня исследования.
Одним из методов, существенно расширивших число решаемых
морфологических задач, является оптико-структурный машинный
анализ (ОСМА), предложенный в 1965 г. К. М. Богдановым.
В 1978 г. автор метода был удостоен Государственной премии
СССР. С появлением ОСМА сделан качественно новый шаг в раз-
работке единой методологии количественного анализа микрострук-
тур на основе статистических характеристик. В последнее время
ОСМА нашел эффективное применение в исследовательской прак-
тике и народном хозяйстве.
На рис. 2 представлена выпускаемая в СССР фирмой «ЛОМО»
автоматизированная система обработки изображений «Протва-
М11». Система предназначена для проведения комплексных иссле-
дований клеток и тканей с использованием методов абсорбцион-
ной, флюоресцентной микроскопии и радиоавтографии.
Входящий в состав системы специальный сканирующий опти-
ческий или электронный микроскоп осуществляет последователь-
ный просмотр изображения препарата по двум координатам, прео-
бразуя его в цифровую форму, и вводит в ЭВМ, которая в свою
очередь производит цифровую обработку изображения и выдает
информацию о геометрических и других характеристиках анали-
зируемого объекта. С помощью цветного дисплея исследователь
может «препарировать» изображение, выделяя лишь те структур-
ные составляющие, которые его интересуют. Входящие в состав
ЭВМ емкие накопители информации на магнитных дисках или
лентах позволяют запоминать как сами изображения, так и ре-
зультаты их обработки для последующего хранения и документи-
рования.
Использование методов автоматизированного анализа микро-
объектов рассмотрим на примере обработки изображения лейке
цита крови (рис. 3). Сканирующий микроскоп-фотометр позволяет
19
Рис. 2. Автоматизированная система обработки изображений «Протва-М 11».
/ — сканирующий микроскоп фотометр; 2 — дисплей; J — ЭВМ.
построчно «просматривать» значения оптической плотности с
шагом, заданным исследователем. В результате оптический сигнал,
соответствующий оптической плотности объекта, преобразуется в
цифровую форму. Полученная цифровая матрица подлежит препа-
ровке с помощью специального математического аппарата. Вначале
убирается фон и вычленяется «чистый» объект — изображение
клетки (1а), затем из изображения клетки выделяется любая
интересующая исследователя деталь, например цитоплазма (1б) и
ядро (1в). ЭВМ рассчитывает и строит гистограммы опти-
ческой плотности для всей клетки (Па), для ее цитоплаз-
мы (Пб) и ядра (Ив). В полной гистограмме уже можно выделить
фазы цитоплазмы (б) и ядра (в), а также получить их отдельно
для каждой структуры (Пб, Пв).
По гистограммам рассчитываются параметры I поряд-
к а: среднее и интегральное значение оптической плотности, дис-
персия, асимметрия, эксцесс и др. По изображению объекта по-
лучают морфометрические параметры: площадь,
периметр, диаметр, ядерно-цитоплазматическое отношение, коэф-
фициент формы и др.
Следующим этапом обработки изображения является построе-
ние двухмерных диаграмм взаимозависимости оптичес-
кой плотности для всей клетки (см. рис. 3, III), ее цитоплазмы (IПб)
и ядра (Шв).Так же как и в первом случае, на диаграмме всей
клетки (П1а) можно выделить фазу цитоплазмы (1Пб) и ядра
(Шв). Данные диаграммы позволяют рассчитать гистограммные
20
Рис. 3. Схема автоматизированной обработки изображения клетки.
Изображение лейкоцита (а), его цитоплазмы (б) и ядра (в). / — цифровое изображение;
// — гистограммы оптической плотности; /// двухмерные гистограммы зависимости зна-
чений оптической плотности.
параметры II порядка: гомогенность, локальный контраст,
энтропию и др. Полученные таким образом параметры представ-
ляют многомерный «портрет» клетки и имеют конкретное числовое
выражение. Они могут быть подвергнуты различным методам
статистической обработки, позволяют предельно точно классифи-
цировать микрообъекты, выявлять особенности их структуры, не
обнаруживаемые визуально.
Г л а в a III
КРАТКИЙ ОЧЕРК РАЗВИТИЯ гистологии, цитологии
И ЭМБРИОЛОГИИ
СТАНОВЛЕНИЕ ГИСТОЛОГИИ, ЦИТОЛОГИИ
И ЭМБРИОЛОГИИ КАК НАУК
Развитие гистологии. «Развитие науки движется толчками в
зависимости от успехов, делаемых методикой». Это определение
И. П. Павлова полностью относится к гистологии, цитологии и
эмбриологии. Успехи гистологии с момента зарождения и по
настоящее время прежде всего связаны с развитием техники, опти-
ки и методов микроскопирования. Успехи же в микроскопических
исследованиях давали возможность накопить новые факты и сде-
лать теоретические обобщения. В связи с этим в истории учения
о тканях и микроскопическом строении органов можно различить
три периода: 1-й домикроскопический (продолжительностью около
2000 лет), 2-й микроскопический (около 300 лет), 3-й электронно-
микроскопический (около 40 лет).
Первый период (домикроскопический), наиболее продолжи-
тельный по времени (с IV века до н. э. и до середины XVII века),
является собственно предысторией гистологической науки, осно-
ванной на макроскопической технике. В этот период фактически
создавались лишь общие представления о тканях организма как
об «однородных» частях организма, отличающихся друг от друга
физическими свойствами («твердые», «мягкие»), удельным весом
(«тонущие в воде», «нетонущие») и пр. Но так как представления
о тканях в то время складывались лишь на основании анатомиче-
ского расчленения трупов, то все классификации тканей строились
на их внешнем сходстве и отличиях. Вследствие этого в одну
группу попадали иногда такие различные ткани, как нервная и
соединительная (нерв и сухожилие). Вот почему, когда в конце
первого периода, а именно в середине XVII века, английским
физиком Р. Гуком был усовершенствован микроскоп1 (1665), поз-
воливший изучить строение тканей, начинается второй период в
учении о тканях, породивший разработку технических методов
исследования невидимых невооруженным взглядом структурных
единиц тканей и накопление фактического материала об их строе-
нии. В этот период «зуд познания», по выражению М. Мальпиги,
и «желание постичь дела творца» (Н. Грю) побуждали многих
исследователей к микроскопическим исследованиям.
Первые микроскописты второй половины XVII века — физик
Р. Гук, анатом М. Мальпиги, ботаник Н. Грю, оптик-любитель
А. Левенгук и др. с помощью микроскопа описали строение кожи,
селезенки, крови, мышц, семенной жидкости и др. Каждое иссле-
дование по существу являлось открытием, которое плохо ужива-
Создание первой модели микроскопа относится ко второму десятилетию
XVII века
лось с метафизическим взглядом на природу, складывавшимся
веками. Случайный характер открытий, несовершенство микроско-
пов, метафизическое мировоззрение не позволили в течение 100 лет
(с середины XVII века до середины XVIII века) сделать существен-
ные шаги вперед в познании закономерностей строения животных
и растений, хотя и делались попытки обобщений (теории «волок-
нистого» и «зернистого» строения организмов и др.).
В конце XVIII — начале XIX века трудами многих отечествен-
ных (петербургских), а также голландских ученых и мастеров
были созданы ахроматические микроскопы, которые сделали более
достоверными микроскопические наблюдения и позволили перейти
к систематическому изучению структурных элементов самых раз-
нообразных животных и растительных организмов.
Применение ахроматического микроскопа в научных исследова-
ниях послужило новым импульсом к развитию гистологии. В на-
чале XIX века сделано первое изображение ядер растительных
клеток. Я. Пуркине (в 1825—1827 гг.) описал ядро в яйцеклетке
курицы, а затем ядра в клетках различных тканей животных.
Позднее им было введено понятие «протоплазма» (цитоплазма)
клеток, охарактеризована форма нервных клеток, строение желез
и др. Р. Броун сделал заключение о том, что ядро является обя-
зательной частью растительной клетки. Таким образом, постепен-
но стал накапливаться материал о микроскопической организа-
ции животных и растений и строение «клеток» (cellula), названных
так еще Р. Гуком.
Завершением этого периода являются исследования А. Дютро-
ше, П. Ф. Горянинова, Г. Валентина (ученика Я. Пуркине),
Я. Генле (ученика И. Мюллера), М. Шлейдена и особенно Т. Шван-
на, который обобщил все предыдущие исследования и сформули-
ровал клеточную теорию (1838—1839). Т. Шванн рас-
сматривал клетку как универсальный структурный компонент
животного и растительного мира. Это поставило на материалис-
тическую основу биологию и патологию.
Создание клеточной теории оказало огромное прогрессивное
влияние не только на развитие биологии и медицины, но также
и на развитие философии диалектического материализма. Ф. Эн-
гельс назвал клеточную теорию в числе трех великих открытий
наряду с законом постоянства энергии и эволюционным учением
Ч. Дарвина.
В середине XIX века начался период бурного развития описа-
тельной гистологии. На основе клеточной теории был
изучен состав различных органов и тканей, их гистогенез, что
позволило уже тогда создать в основных чертах микроскопи-
ческую анатомию и уточнить классификацию тканей с
учетом их микроскопического строения (А. Келликер и др.). Но
научная мысль во второй половине XIX века не могла плодотворно
развиваться без дальнейших успехов в гистологической технике
и методах микроскопического исследования. В этот период были
введены в практику и усовершенствованы водные и масляные
23
иммерсионные объективы, изобретен микротом, применены новые
фиксаторы (формалин, осмиевая кислота, хромовая кислота).
Весьма плодотворным оказался метод импрегнации солями сереб-
ра, разработанный итальянским ученым К. Гольджи, описавшим
пластинчатый комплекс (внутриклеточный сетчатый аппарат).
Этот метод и его модификации позволили провести фундамен-
тальные исследования нервной системы (Р. Кахаль) и создать
основы нейрогистологии. Признанием научных заслуг
К. Гольджи и Р. Кахаля явилось присуждение им в 1906 г. Нобе-
левской премии. В последней четверти XIX века были открыты
органеллы клетки.
Благодаря успехам, достигнутым в области изучения строения
клетки, в конце XIX века были заложены основы цитоло-
гии. Но микроскопирование фиксированных клеток не позволяло
говорить о процессах жизнедеятельности в них. Поэтому внимание
ученых привлекли методы культивирования клеток и тканей
(И. П. Скворцов, Р. Гаррисон, А. Каррель и др.). Однако это не
означало, что исследование тканей на гистологических препаратах
потеряло смысл и значение.
Методы прижизненного введения красителей, примененные
многими исследователями в то время, введение инородных тел в
организм и другие методы сделали возможным изучение физиоло-
гии гистологических структур. В 1900 г. Н. М. Гайдуковым был
предложен метод микроскопирования живых объектов в темном
поле зрения. В это же время был изобретен микроманипулятор,
с помощью которого можно было производить операции на
отдельных клетках (удаление ядер, разрезы клеток и др.) в целях
выяснения роли и значения их в жизнедеятельности организма.
Развитие эмбриологии. Эмбриология, изучающая закономер-
ности пренатального развития организмов, имеет еще более
продолжительную историю своего формирования как науки.
Тайна зарождения, развития и становления различных живых
существ, возможности создания условия для проявления этих
процессов (по крайней мере у птиц) возникали еще в древности.
Так, упоминания о выведении цыплят в искусственных условиях
(инкубаторы) в древнем Египте, а затем в Индии, Китае имеются
в трудах греческих философов. Задолго до нашей эры появились
упоминание о плаценте в связи с рождением ребенка и некоторые
другие сведения.
Однако первые медицинские эмбриологические наблюдения и
формирование важных эмбриологических представлений, по-види-
мому, принадлежат Гиппократу (IV век до н. э.) и его последова-
телям («О природе женщины», «О семимесячном плоде», «О сверх-
оплодотворении», «О семени», «О природе ребенка» и др.). Многие
высказывания врачей того этапа развития медицины скорее
представляли умозрительные заключения, которые тем не менее
были близки к истине. Например, утверждение «о высыхании»
зародыша по мере его развития, т. е. об уменьшении содержания
воды в нем, или о необходимости смешения мужского и женского
24
семени (мужские и женские половые клетки были обнаружены с
помощью микроскопа соответственно лишь в XVII и XIX столе-
тиях).
Современник Гиппократа, Аристотель в своих сочинениях
«О возникновении животных» и др. по существу положил начало
общей и сравнительной биологии. Предложенная им классифика-
ция животных по эмбриологическим признакам явилась итогом
научного анализа рассматриваемых им в 5 книгах вопросов
(«О происхождении семени», «О формах матки у различных жи-
вотных», «О живорождении и ящеророждении» и др.). Следует
заметить, что уже Аристотелем был поднят вопрос о механике
развития и сформировано положение об эпигенезе (от греч.
epi — над и genesis — происхождение). Отстаивая идею развития,
Аристотель основывался на неверных умозрительных заключениях
о том, что зародыш развивается из женской крови («материи») и
внесенного мужчиной семени («души»), одухотворившего эту
кровь. Подобные идеалистические рассуждения о нематериальном
факторе (энтелехии) существовали долго и после Аристотеля в
связи с сильным влиянием теологии на мировоззрение ученых,
пытавшихся разобраться в причинности развития и конечной
цели.
До середины XVII века история эмбриологии не была ознаме-
нована существенными достижениями. Известно, что некоторые
конкретные описания зародышей, их временных и постоянных
органов были сделаны в разных странах.
В эпоху Возрождения определенный вклад в эмбриологию
внес В. Гарвей — автор открытия кровообращения, который,
проанализировав развитие зародышей, описал их в книге «Зарож-
дение животных» (1651). Он высказал ряд принципиально важных
утверждений. В частности, Гарвей отрицал возможность самоза-
рождения и утверждал тезис о развитии животных только из
яйца («Живое — из яйца»). Он первый высказал предположение,
которое позже было подтверждено, что «пятно» на желтке яйца
птиц «есть начало цыпленка», а прыгающая «кровяная точка»
является зачатком сердца. Гарвей правильно в принципе тракто-
вал значение раннего развития крови как элемента, обеспечиваю-
щего трофику зародыша. «Жизнь заключается в крови, ... кровь
возникает прежде, чем начинает существовать какая-либо часть
тела, и она является перед всеми прочими частями плода перво-
рожденной», — утверждал Гарвей. Несмотря на то что Гарвей
тяготел к витализму, он стремился проникнуть в причинно-след-
ственные отношения. Он писал: «В порождении животных всякое
исследование надо вести от причин, в особенности от материаль-
ной и действующей».
Острая борьба мировоззрения разыгралась во второй половине
XVII века, когда с диссертацией «Теория зарождения» (1759)
выступил молодой немецкий ученый К. Ф. Вольф (1733—1794).
Он подверг резкой критике взгляды преформистов и обосновал
теорию эпигенеза. Согласно теории префоризма, развитие по
25
существу представляло развертывание в пространстве заложен-
ных при сотворении жизни готовых частей организма. Теория же
эпигенеза, напротив, отстаивала новообразование органов, пол-
ностью отрицая предопределенность, или преформацию.
К. Ф. Вольф впервые наблюдал у зародышей животных образова-
ние органов из листовидных пластинок (зародышевых листков),
описал развитие сердца у цыпленка, развитие почки (ряд структур
назван его именем) и др. Несмотря на то что первая работа
К. Ф. Вольфа была враждебно встречена в академических кругах,
прогрессивные идеи ее нашли позднее отражение в трудах отече-
ственных эмбриологов— X. И. Пандера (1794—1858), К. Э. Бера
(1792—1876) и в эволюционном учении Дарвина, появившемся
100 лет спустя (1859) после опубликования диссертации К. Ф. Воль-
фа. Роль К. Ф. Вольфа в развитии науки высоко оценил Ф. Эн-
гельс. Он писал: «...К. Ф. Вольф произвел в 1759 г. первое напа-
дение на теорию постоянства видов, провозгласив учение об эво-
люции»1. В 1768 г. К. Ф. Вольф по приглашению Петербургской
академии переехал из Германии в Россию, где и протекала вся
его дальнейшая деятельность.
Однако эти теории представляли две противоположные край-
ности и объективно отображали лишь определенные стадии
эмбриогенеза, хотя в ходе развития зародыша имеют место как
периоды полипотентности (от лат. poly — много, potentio — воз-
можность), так и жесткой предопределенности (преформации)
развития клеток и тканей.
Соотечественник К. Ф. Вольфа, А. Галлер, занимавшийся
широким кругом научных проблем в области эмбриологии и физи-
ологии, выпустил ряд диссертаций, утверждавших преформизм в
процессе эмбрионального развития (1750—1767). Вместе с тем
А. Галлер и его сотрудники провели тщательные морфометри-
ческие исследования растущего зародыша. Впоследствии использо-
вание морфометрических показателей стало одним из распростра-
ненных объективных подходов для изучения тканей и органов.
В развитии эмбриологии, как и гистологии, начиная с XVII
века, значительную роль сыграли успехи в технике исследования,
в новых методических приемах, позволивших подняться над
схоластикой. В частности, использование увеличительных стекол,
микроскопов во второй половине XVII в. существенно обогатило
науку. Так, Р. де-Грааф и Я. Сваммердам описали в 1670 г. шаро-
видные полости в яичнике (граафовы пузырьки), которые ими
были неправильно отождествлены с яйцеклетками, а вскоре (1677)
любознательный человек и искусный шлифовальщик увеличитель-
ных стекол А. Левенгук и студент-медик Гам описали мужские
половые клетки, назвав их «семенными животными» — спермато-
зоидами.
С помощью микроскопа вновь были изучены, описаны и зари-
сованы стадии развития цыпленка. Однако небольшие увеличения
Маркс К.. Энгельс Ф. Сеч., т. 20, с. 354.
26
микроскопа, а главное — метафизический характер мышления и
предвзятость были характерны для ряда исследователей (М. Маль-
пиги, Н. Мальбранш, Я. Сваммердам и др.).
ГИСТОЛОГИЯ И ЭМБРИОЛОГИЯ В РОССИИ КАК ПРЕДМЕТ ПРЕПОДАВАНИЯ.
ОТЕЧЕСТВЕННЫЕ ГИСТОЛОГИЧЕСКИЕ ШКОЛЫ ВТОРОЙ ПОЛОВИНЫ XIX
И НАЧАЛА XX ВЕКА
Отечественная гистология и эмбриология формировались в тес-
ной связи с развитием мировой науки, с прогрессом техники
микроскопических исследований.
Если не считать отдельных гистологических исследований,
проведенных соотечественниками на заре развития микроскопии,
то началом русской гистологии надо считать 30—40-е годы XIX
века. Сначала гистология преподавалась в виде курса в програм-
ме смежных дисциплин — анатомии, физиологии, а в 60-х годах
XIX века были учреждены кафедры гистологии и эмбриологии
одновременно в Московском (1864) и Петербургском (1864)
университетах, а несколько позднее в Харьковском (1867), Казан-
ском (1868) и Киевском (1868) университетах.
Очень скоро все эти кафедры стали центрами крупных гистоло-
гических исследований и школами подготовки кадров. Первыми
руководителями кафедр и основоположниками отечественной гисто-
логии как самостоятельной науки были А. И. Бабухин, Ф. В. Ов-
сянников, Н. М. Якубович, М. Д. Лавдовский, Ф. Н. Заварыкин,
К. А. Арнштейн, П. И. Перемежко, Н. А. Хржоншевский.
Московская школа гистологов была создана одним из крупных
представителей материалистического направления в естествозна-
нии второй половины XIX века А. И. Бабулиным (1827—1891).
Большое внимание в ней уделялось вопросам гистогенеза и гисто-
физиологии различных тканей, особенно мышечной и нервной,
вопросам теории микроскопа. А. И. Бабухину принадлежит
открытие происхождения и выяснение гистофизиологии электри-
ческих органов рыб; им проводились исследования развития и
строения сетчатки глаза, развития осевых цилиндров нервных
волокон и др. Позднее под руководством И. Ф. Огнева (1855—
1927), ученика и преемника А. И. Бабухина, в круг изучаемых
кафедрой вопросов были включены исследования по влиянию
различных внешних и внутренних факторов (лучистая энергия,
темнота, голодание) на гистоструктуру и физиологию клеток,
тканей и органов. Это гистофизиологическое нап-
равление, положенное в основу исследований московской
школы гистологов, дало много ценного для понимания развития
и функций тканей и органов. Тесная связь гистологии и физиоло-
гии выгодно характеризует развитие научной медицинской мысли
в России во второй половине XIX века. Оно особенно проявилось
в связи с критикой чисто морфологического «целлюлярного» нап-
равления в зарубежной науке и развитием идей нервизма в России.
27
А. И Бабухин
(1827 -1841)
Ф В. Овсянников
(1827—1906)
В Петербургском университете
курс гистологии читал акад.
Ф. В. Овсянников (1827—1906)
сначала на кафедре анатомии и
физиологии, а с 1894 г. на само-
стоятельной кафедре гистологии.
Ф В. Овсянников — один из осно-
воположников гистофизиологиче-
ского направления в морфологии,
автор интересных исследований
нервной системы и органов чувств
различных животных. Большой
вклад в развитие нейрогистологи-
ческих исследований этой кафед-
ры внес А. С. Догель (1852—1922),
ранее работавший в Казани и
Томске. Ему принадлежат класси-
ческие работы по строению вегета-
тивной нервной системы и класси-
фикации ее нейронов, иннервации
органов чувств. А. С. Догель осно-
вал в 1915 г. журнал «Архив ана-
томии, гистологии и эмбриологии»
в России.
Курс гистологии и эмбриоло-
гии в Медико-хирургической ака-
демии (ныне Военно-медицинская
академия им. С. М. Кирова) в Пе-
тербурге впервые начал читать в
40-х годах XIX века зав. кафедрой
сравнительной анатомии и физио-
логии эмбриолог К. Э. Бэр.
С 1852 г. гистология и эмбриоло-
гия были выделены в специальный
курс, который читал Н. М. Якубо-
вич (1817—1879), прославивший-
ся изучением тонкого строения
центральной нервной системы.
В 1868 г. при академии была уч-
реждена самостоятельная кафедра
гистологии и эмбриологии. Весо-
мый вклад в развитие этой кафед-
ры и отечественной гистологии внес М. Д. Лавдовский (1846—
1902), известный своими исследованиями ганглиозных клеток
мочевого пузыря, регенерации и дегенерации нервных волокон
после травмы. Под редакцией М. Д. Лавдовского и Ф. В. Овсянни-
кова было создано в 1887 г. первое в России фундаментальное
руководство по гистологии. Велики заслуги в развитии отечест-
венной и мировой науки А. А. Максимова (1874—1928),
28
возглавлявшего кафедру гистоло-
гии в академии после М. Д. Л де-
довского. Его исследования соеди-
нительной ткани и крови, а также
процессов кроветворения не поте-
ряли значения и поныне. Эмигри-
ровав за границу, А. А. Максимов
оказал огромное влияние на раз-
витие американской гистологиче-
ской школы. Учебник гистологии,
созданный А. А. Максимовым,
был одним из лучших, поэтому не-
однократно переиздавался не толь-
ко в нашей стране.
Основателем Казанской школы
К. А. Арнштейном (1840—1919) и
его учениками собран богатейший
материал по морфологии концевых
нервных волокон и нервных узлов в
различных тканях и органах (в мо-
чевом пузыре, мочеточнике, поло-
вых органах, роговице, легком, пи-
щеводе, коже и др.). Разработан-
ный А. С. Догелем метод окраски
нервной ткани позволил успешно
исследовать различные отделы нер-
вной системы и создать капиталь-
ные труды по нейрогистологии.
Работы по исследованию нервной
системы быстро выдвинули Казан-
скую лабораторию в ряды перво-
классных лабораторий Европы.
В 1888 г. А. С. Догель основал
кафедру гистологии в Томском
университете, которой с 1895 г.
руководил другой ученик К. А. Арн-
штейна А. Е. Смирнов (1859—
1910). Под его руководством ка-
федра гистологии при Томском
университете оформилась в само-
стоятельную научную нейрогисто-
к. Э. Бэр
(1792—1876)
М. Д. Лавдовскйй
(1846—1902)
логическую школу.
Кафедру гистологии в Киевском университете возглавил в
1868 г. П. И. Перемежко (1833—1893). Исследования гистологов
киевской школы были направлены на изучение развития зароды-
шевых листков эмбриона, глаза, надпочечников, селезенки, попе-
речнополосатой и гладкой мускулатуры, а также строения различ-
ных органов — печень, щитовидная железа, поджелудочная желе-
за, костный мозг, кровеносные сосуды и др. П. И. Перемежко
29
К. А. Арнштейн
(1840 -1919)
П. И. Перемежке
(1833—1893)
описаны фигуры митотического
деления клеток.
Кафедру гистологии и эмбрио-
логии в Харьковском университете
возглавил Н. А. Хржонщевский
(1836—1917). Ему принадлежат
оригинальные работы о строении
надпочечных желез, легких, пече-
ни, о кровоснабжении почки и др.
Исследования, проводимые
Н. А. Хржонщевским и сотруд-
никами, были основаны на гисто-
физиологическом подходе.
Одновременно с развитием гис-
тологии бурного расцвета достигла
в середине XIX века эмбриология.
Продолжая исследования, начатые
К. Ф. Вольфом, русские академи-
ки X. И. Пандер и К. Э. Бэр рас-
крыли очень важную биологиче-
скую закономерность в развитии
зародышей — образование заро-
дышевых листков. X. И. Пандер
заметил, что еще до появления за-
кладок нервных органов в зароды-
ше образуется два листка, а позд-
нее к ним присоединяется третий.
К. Э. Бэр проследил развитие за-
родышевых листков и образование
из них различных органов у мле-
копитающих. Он установил, что у
различных животных есть много
общего в ранних стадиях развития
их зародышей, и в своих обобще-
ниях приблизился к эволюционно-
му пониманию развития животно-
го мира. С помощью микроскопа
К. Э. Бэр обнаружил в описанных
ранее граафовых пузырьках яйце-
клетку млекопитающих (1827).
Трудами К. Ф. Вольфа, X. И. Пан-
дера и К. Э. Бэра были заложены
эмбриологии.
основы современной
Эволюционное учение Ч. Дарвина явилось предпосылкой для
дальнейшего развития материалистической теории в вопросах эмб-
риологии. Крупнейшие русские ученые И. И. Мечников (1845—
1916) и А. О. Ковалевский (1840—1901), вооруженные эволюци-
онным учением Дарвина, внесли много нового в эмбриологию.
Классическими исследованиями И. И. Мечникова и А. О. Ковалев-
10
ского по сравнительному изуче-
нию беспозвоночных и низших
позвоночных установлено, что у
разных классов и типов животных
есть много общего, что все они в
своем развитии проходят сходные
этапы, в частности стадию образо-
вания зародышевых листков. Этим
было окончательно доказано един-
ство животного мира. Исследова-
ния И. И. Мечниковым и А. О. Ко-
валевским микроскопического
строения многих животных в даль-
нейшем легли в основу эволю-
ционной гистологии и
эмбриологии.
В области эмбриологии конец
XIX — начало XX века ознамено-
вались также развитием экспери-
ментальных методов (В. Ру,
X. Шпеман и др.), позволивших
заложить основы нового направле-
ния — механики разви-
тия. В этот период произошло
сближение цитологии и эмбриоло-
гии на основе исследования роли
клетки в развитии и наследствен-
ности (А. Вейсман, Т. Морган
и др.).
РАЗВИТИЕ ГИСТОЛОГИИ,
ЦИТОЛОГИИ
И ЭМБРИОЛОГИИ В СССР
В советский период на основе
учения диалектического материа-
лизма нашли свое дальнейшее раз-
витие цитология, эмбриология, об-
щая и частная гистология. В этот
Н. А. Хржонщевский
(1836—1917)
период широкое развитие получи- Б и Лав нтьев
ли гистохимические, радиоавто- (1892 1944)
графические, люминесцентные и
другие специальные гистологические методы. В последние десяти-
летия с успехом применяется электронная микроскопия.
Советская гистология за годы своего существования развивалась
по нескольким направлениям. Большое внимание было уделено во-
просам нейрогистологии, особенно в связи с разработкой учения
И. П. Павлова. Казанской нейрогистологической школой был со-
бран богатейший материал по морфологии концевых нерв-
31
А. А. Заварзин
(1886—1945)
Н I Хлопин
(1897—1961)
ных волокон и нервных узлов в раз-
личных органах и тканях (в пище-
варительном тракте, мускулатуре,
эпителии, железах и др.). А. Н. Ми-
славский подготовил плеяду та-
лантливых нейрогистологов. Из
них особое значение имела дея-
тельность Б. И. Лаврентьева.
Б. И. Лаврентьев и его сотруд-
ники (Е. К. Плечкова и др.) разра-
батывали вопросы гистофизиоло-
гии вегетативной нервной системы,
интернейрональных синапсов, раз-
личных рецепторов, антагонистиче-
ской иннервации. Под руководст-
вом Б. И. Лаврентьева было создано
экспериментальное гистофизиоло-
гическое направление в отечест-
венной нейрогистологии. Исследуя
живые нервные клетки, Б. И. Лав-
рентьев наблюдал изменения си-
напсов при раздражении нервов.
Примененный им метод перерезки
нервов нашел широкое распростра-
нение при изучении источников ин-
нервации органов и тканей. Поль-
зуясь этим методом, Б. И. Лаврен-
тьев доказал несостоятельность
теории фибриллярной непрерыв-
ности и утвердил нейронную
теорию. Применение современ-
ных методов исследования (люми-
несцентной, электронной микро-
скопии, гистохимии и др.) позволи-
ло раскрыть тонкие механизмы
функции и реактивные изменения
тканевых элементов нервной систе-
мы в условиях экспериментальных
патологических воздействий на ор-
ганизм. Перспективные исследова-
ния в области регенерации ткане-
вых элементов нервной системы и пересадки головного мозга
проводятся в ряде научно-исследовательских институтов.
Советские гистологи уделяют особое внимание вопросам связи
нервной системы с органами, а также вопросами корреляции
нервной и эндокринной систем в жизнедеятельности организма.
Большой вклад сделан советскими гистологами в разработку функ-
циональной гистологии эндокринной системы (А. В. Немилое,
А. В. Румянцев и др.).
P
1
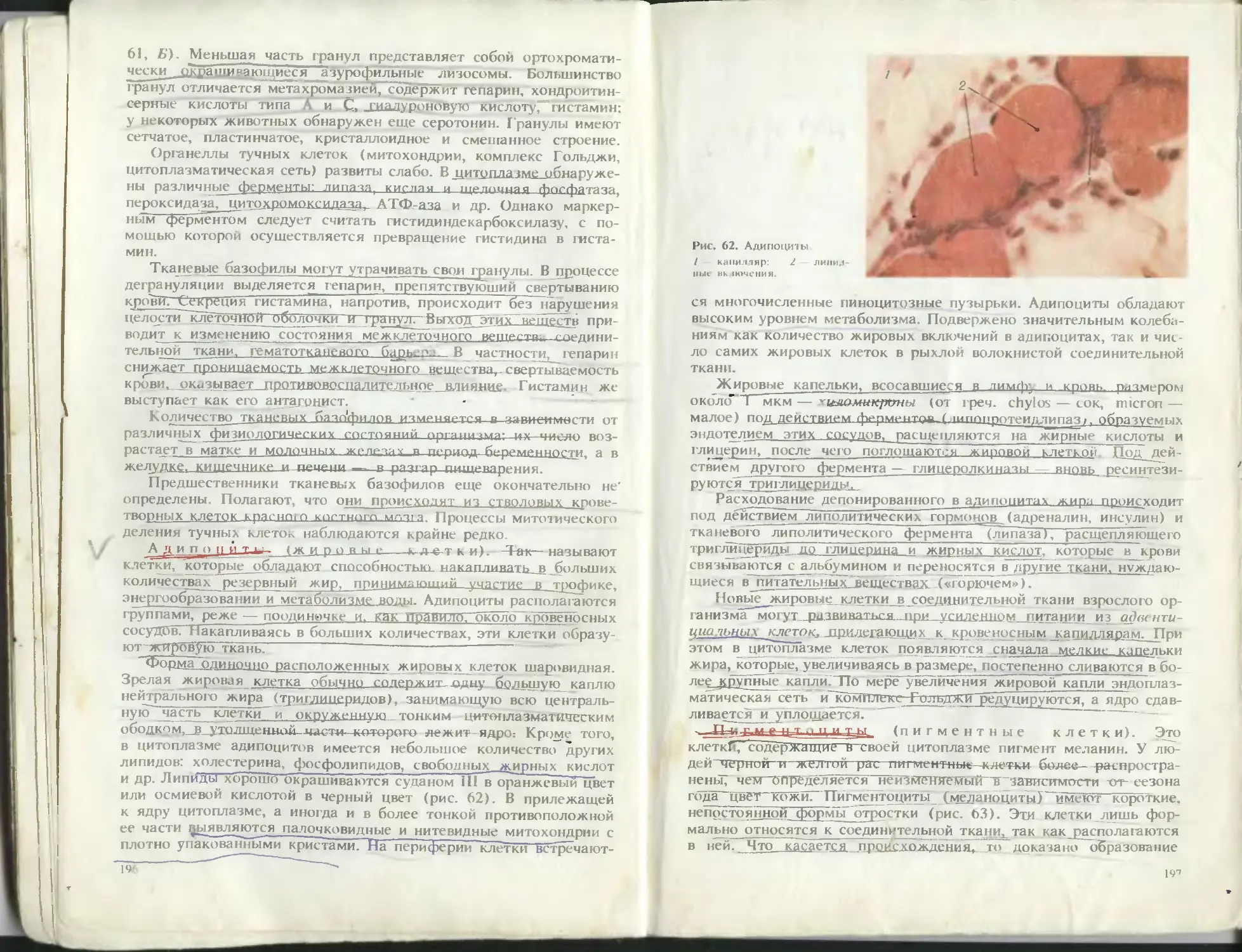
S И 'li ’а."в
32
Начатое еще А. А. Максимовым
изучение соединительной ткани
приобрело широкий размах в совет-
ское время. Оно ведется в основ-
ном по двум направлениям. Первое
направление в изучении соедини-
тельной ткани выражается в широ-
ких сравнительно-гистологических
исследованиях соединительной
ткани и крови (С. В. Мясоедов,
А. А. Заварзин, Ф. М. Лазаренко,
Е. С. Данини, Г. В. Ясвоин и др.).
«Очерки по эволюционной гистоло-
гии крови и соединительной ткани»,
а также «Очерки по эволюционной
гистологии нервной системы»
А. А. Заварзина представляют со-
бой развернутое теоретическое обо-
снование эволюционного направле-
ния в гистологии. Основной задачей
эволюционной гистологии А. А. За
варзин считал выяснение общих за-
кономерностей филогенетической
дифференцировки разновидностей
специализированных клеток в пре-
делах каждой ткани («эволюцион-
ное расщепление») при сохранении
ограниченного числа морфофунк-
циональных типов тканей («теория
параллельных рядов»). Крупное
теоретическое обобщение в области
изучения развития тканей («теория
дивергентного развития») сделал
Н. Г. Хлопин.
Второе направление — изучение
гистофизиологии соединительной
ткани различных органов, системы
крови и других органов, а также из-
менений их под влиянием нервных
и эндокринных факторов (В. Г. Ели-
сеев, Т. А. Григорьева и др.). С эти-
ми направлениями логически свя-
В. Г. Елисеев
(1899—1966)
П. Г. Светлов
(1892—1974)
зано изучение гистогенеза соединительной ткани. В Советском Сою-
зе стали впервые систематически изучаться вопросы коррелятивной
связи соединительной ткани и эпителия. Большую роль в этом
сыграли методы культивирования ткани (А. В. Румянцев, Ф М. Ла-
заренко, Н. Г. Хлопин и др.). В частности, А. В. Румянцев и другие
гистологи проследили индуктивное влияние переходного эпителия
мочевого пузыря на соединительную ткань.
2 ГИСТОЛОГИЯ
33
Большие успехи достигнуты в разработке гистофизиологии
мышечной ткани, в изучении гистогенеза и регенеративных воз-
можностей органов. Советские гистологи опровергли теории о
неспособности тканей высокоорганизованных животных к регене-
рации и на примере восстановительных процессов в поперечно-
полосатой мышечной ткани убедительно показали пути и способы
ее осуществления.
Наряду с этим вскрыты основные закономерности физиоло-
гической и репаративной регенерации ряда внутренних органов.
В 40-е годы XX века были внесены существенные коррективы
и в научные направления в области цитологии. Основным в совет-
ской школе цитологов стало изучение функционального значения
органелл, включений, их цитотопографии при различных физио-
логических состояниях клетки, а также вопросы цитохимии и
механизма деления клеток, вопросы клеточной адаптации (Д. Н. На-
сонов, В. Я. Александров, Н. К. Кольцов, П. В. Макаров, А. Г. Гур-
вич, Б. В. Кедровский, Г. И. Роскин, В. Я. Рубашкин, Л. Б. Ле-
винсон и др.). Ценными для развития цитофизиологии явились
работы Д. Н. Насонова, В. Я. Александрова по прижизненному
изучению органов и тканей. На основании этих опытов Д. Н. На-
соновым и В. Я. Александровым была создана теория паранекроза.
Современная цитология, а точнее биология клетки, разрабаты-
вает проблемы ритмической деятельности клеток (суточные,
сезонные), тканевоспецифической регуляции размножения клеток,
дифференцировки, реактивных изменений под влиянием факторов
внешней среды и др. Результаты экспериментальных цитологи-
ческих исследований находят воплощение в медицинской практике
(цитодиагностика болезней, цитоиммунологические пробы и др.).
В области эмбриологии нашли отражение эксперимен-
тальные методы, позволяющие уточнить представления об органи-
заторах зародышевого развития, нейрогуморальной регуляции и
влиянии факторов внешней среды на процессы эмбриогенеза.
В 30—40-е годы успешно разрабатывались вопросы эволюци-
онной эмбриологии большим отрядом советских эмбрио-
логов во главе с академиком А. Н. Северцовым. В настоящее
время это направление продолжает разрабатываться эмбриоло-
гами в различных лабораториях нашей страны.
А. Н. Северцов (1866—1936) и его сотрудники получили
большой сравнительно-эмбриологический материал о развитии орга-
нов, который позволил решить вопрос об эволюции всех низших
позвоночных от древнейших предков хордовых и до появления
первых наземных четвероногих. Установлено два типа закладки
изменяющихся во время эволюции органов. Один тип характери-
зуется поздними прибавлениями к зародышевым признакам пред-
ков. При другом новые изменения органа возникают на ранних
стадиях развития, перестраивая дальнейший ход последнего.
Это послужило основанием уточнить биогенетический закон Мюл-
лера — Геккеля о том, что онтогенез повторяет филогенез. Ока-
залось, что этот закон справедлив лишь по отношению к живот-
34
ным первого типа развития. Такое представление о взаимоотно-
шениях между онтогенией и филогенией вошло в науку под назва
нием филэмбриогенеза.
Отклонения в темпах последовательного индивидуального
развития органов по сравнению с очередностью эволюционного
развития, по мнению академика П. К. Анохина, обусловлено раз-
витием функциональных систем организма первоочередной важ-
ности, обеспечивающих кровообращение, акт приема пищи и др.
Большое значение в развитии эмбриологии сыграли работы
Д. Ф. Филатова (1876—1943) и П. П. Иванова (1872—1942).
Д. Ф. Филатов изучал значение обмена веществ в ускорении
органогенезов, гетерогенную индукцию конечностей и ее значение
для теории органогенезов. П. П. Иванов внес вклад в разработку
ряда важнейших эмбриологических проблем, таких как взаимо-
действие эмбрионального развития и регенерации, влияние факто-
ров среды на дифференцировку тканевых зачатков. Он показал
наличие двух организаторов, стимулирующих органогенез заро-
дыша — головного и туловищного, создал теорию развития сег-
ментированных животных. Эти и другие положения нашли отра-
жения в его фундаментальном учебнике по общей и сравнительной
эмбриологии (1937, 1945).
П. Г. Светлов (1892—1974) —ученик и последователь П. П. Ива-
нова, уделил внимание изучению роли ряда экологических факто-
ров (температура, голодание, ионизирующая радиация и др.) в
ходе эмбриогенеза. Им установлены критические периоды разви-
тия у всех животных, включая млекопитающих, во время которых
зародыши оказываются легкоранимыми (см. гл. V). Теория крити-
ческих периодов, разработанная П. Г. Светловым, имеет большое
значение для биологии и медицины, так как позволяет прогно-
зировать возможность возникновения патологии развития и уродств.
Современный, третий период развития гистологии, цитологии
и эмбриологии характеризуется новым методическим уровнем
исследований, широким использованием электронной микроскопии,
метода замораживания-скалывания, электронно-микроскопической
цитохимии и других методов.
Научно-технический прогресс, успехи развития методов иссле-
дования позволили дойти до макромолекулярного уровня органи-
зации клеток и неклеточных структур, уточнить представления о
процессах дифференцировки, регенерации, передаче наследствен-
ности признаков и др. Благодаря этому созданы основы ультра-
микроскопической цитологии и гистологии и разрабатываются
проблемы молекулярной биологии.
2*
цитология
Глава IV
УЧЕНИЕ О КЛЕТКЕ (ОСНОВЫ
ОБЩЕЙ ЦИТОЛОГИИ)
Основой строения эукариотических* организмов является наи-
меньшая единица живого — клетка (cellula).
Клетка — это ограниченная активной мембраной, упорядочен-
ная, структурированная система биополимеров, образующих ядро
и цитоплазму, участвующих в единой совокупности метаболиче-
ских и энергетических процессов, осуществляющих поддержание
и воспроизведение всей системы в целом.
Кроме клеток в организме находятся их производные, которые
не имеют клеточного строения.
Содержимое клетки отделено от внешней среды или от сосед-
них клеток плазматической мембраной (плазмолеммой). Все эука-
риотические клетки состоят из двух основных компонентов: ядра
и цитоплазмы. Цитоплазма неоднородна по своему составу и стро-
ению и включает в себя гиалоплазму (основную плазму), в которой
находятся включения и органеллы. Все они, дополняя друг друга,
выполняют внутриклеточные функции, необходимые для существо-
вания клетки как целого, как элементарной живой единицы. Изу-
чением общих черт строения и функционирования клеток и их
производных занимается наука цитология. Она исследует
отдельные клеточные структуры, их участие в общеклеточных
физиологических процессах, пути регуляции этих процессов, вос-
произведение клеток и их компонентов, приспособление клеток к
условиям среды, реакции на действия различных факторов, пато-
логические изменения клеток. Изучение цитологии имеет большое
прикладное значение, так как практически при всех заболеваниях
происходят нарушения функций клеток.
КЛЕТОЧНАЯ ТЕОРИЯ
История вопроса. Клеточная теория — это обобщенное пред-
ставление о строении клеток как единиц живого, об их воспроиз-
ведении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточ-
ной теории предшествовал довольно длительный (более 300 лет)
период накопления знаний о строении различных одноклеточных
1 Эукариотические, собственно ядерные организмы — основная масса животных
и растений, за исключением бактерий и сине-зеленых водорослей, не имеющих
оформленного ядра, — прокариотических организмов.
36
и многоклеточных организмов, растений и животных. Этот период
связан с применением и усовершенствованием различных опти-
ческих методов исследований.
Первым, кто наблюдал наименьшие единицы в составе много-
клеточных, был Роберт Гук (1665). С помощью увеличительных
линз в срезе пробки он обнаружил «ячейки», или «клетки».
Его описания послужили толчком для появления систематических
исследований строения растений и животных.
М. Мальпиги (1671), Н. Грю (1671), Ф. Фонтана (1671) под-
твердили его наблюдения и показали, что разнообразные части
растений состоят из тесно расположенных «пузырьков», или «ме-
шочков». Позднее, в 70-е годы XVII века, оптик-любитель А. Ле-
венгук открыл с помощью микроскопа мир одноклеточных орга-
низмов. Но эти и другие многочисленные исследования не привели
еще в то время к пониманию универсальности клеточного строения
животных и растений и к правильным представлениям об органи-
зации клетки. Прогресс в изучении морфологии клетки связан с
успехами микроскопирования в XIX веке. К тому времени изме-
нились взгляды на строение клеток. Главной составной частью
клеток стала считаться не клеточная стенка, а содержимое самой
клетки — ее протоплазма (Я. Пуркине, 1830). В протоплазме было
открыто ядро как постоянный компонент клетки (Р. Броун,
1833). Многочисленные данные, касающиеся строения животных
и растений, позволили подойти к обобщениям, которые впервые
были сделаны Т. Шванном (1838) и легли в основу сформулиро-
ванной им клеточной теории. Его главным достижением является
утверждение, что клетки, из которых состоят как растения, так
и животные, принципиально сходны между собой (гомологичны)
и возникают единообразным путем. Заслуга Т. Шванна заключа-
лась не в том, что он открыл клетки как таковые, а в том, что он
оценил их значение как основного структурного компонента орга-
низма. Дальнейшее развитие и обобщение эти представления
получили в работах Р. Вирхова (1858).
Создание клеточной теории стало важнейшим событием в
биологии, одним из решающих доказательств единства происхож-
дения всей живой природы. Клеточная теория оказала значитель-
ное влияние на развитие биологии и медицины, послужила глав-
ным фундаментом для становления таких дисциплин, как эмбрио-
логия, гистология и физиология. Она дала основы для материа-
листического понимания жизни, для объяснения эволюционной
взаимосвязи организмов, для понимания индивидуального разви-
тия. Как считал Ф. Энгельс, открытие клетки стало главным фак-
том, революционизировавшим физиологию и сделавшим возмож-
ным сравнительную физиологию.
Основные положения клеточной теории сохранили свое значе-
ние и в настоящее время, хотя почти за 150-летний период были
получены новые сведения о структуре, жизнедеятельности и разви-
тии клеток. В настоящее время клеточная теория гласит:
1) клетка является наименьшей единицей живого, 2) клетки раз-
37
ных организмов сходны по своему строению, 3) размножение
клЬток происходит путем деления исходной клетки, 4) многокле-
точные организмы представляют собой сложные ансамбли клеток
и их производных, объединенные в целостные интегрированные
системы тканей и органов, подчиненные и связанные между собой
межклеточными, гуморальными и нервными формами регуляции.
1. Клетка — наименьшая единица живого.
Представление о клетке как о наименьшей самостоятельной живой
единице было известно из работ Т. Шванна, Р. Вирхова и др.
Полностью сохранила свое значение в определении живою
формулировка Ф. Энгельса: «Жизнь есть способ существования
белковых тел, и этот способ существования состоит по своему
существу в постоянном самообновлении химических составных
частей этих тел»1. Действительно, все проявления жизни связаны
с участием белков. Белки — функционирующие молекулы, обла-
дающие сложной организацией и строгой функциональной специ-
фичностью. Теперь уже хорошо известно, что эта функциональная
специфичность белков определяется нуклеиновыми кислотами, ко-
торые несут в себе информацию о строении тех или других белков.
Живому свойствен ряд совокупных признаков: способность к
воспроизведению (репродукции), использование и
трансформация энергии, метаболизм, чувстви-
тельность, адаптация, изменчивость. И такую со-
вокупность этих признаков впервые можно обнаружить только
на клеточном уровне. Именно клетка как таковая является наимень-
шей единицей, обладающей всеми вместе взятыми свойствами, отве-
чающими определению «живое».
У животных, кроме отдельных клеток, встречаются так назы-
ваемые симпласты, синцитии и межклеточное вещество. Симплас-
ты — это крупные образования, состоящие из цитоплазмы со
множеством ядер. Примерами симпластов могут быть мышечные
волокна позвоночных, наружный слой трофобласта плаценты и др.
Если проследить за развитием таких «неклеточных» форм, то лег-
ко убедиться в том, что они возникают вторично за счет слияния
отдельных клеток или же в результате деления одних ядер без
разделения цитоплазмы, без цитотомии.
Синцитии (соклетия) характеризуются тем, что после деле-
ния исходной клетки дочерние остаются связанными друг с другом
с помощью тонких цитоплазматических перемычек. Такие синци-
тии можно наблюдать при развитии сперматогониев. Часто они
встречаются в тканях высших растений, где клетки могут быть
связаны с помощью цитоплазматических мостиков (плазмодесмы).
Есть примеры безъядерных клеток — эритроциты млекопитаю-
щих. Это — элементы, имеющие в своем составе клеточную мемб-
рану и цитоплазму. Они обладают ограниченными функциональ-
ными потенциями, лишившись способности к самообновлению
и саморепродукции в связи с утратой ядра.
1 Энгельс Ф. Анти-Дюринг. — В кн. Маркс К., Энгельс Ф. Соч., т. 20, с. 82.
38
Межклеточное вещество представляет собой продукт жизнедея-
тельности определенных групп клеток (например, основное веще-
ство и волокна соединительной ткани).
2. Сходство клеток разных организмов по
строению. Клетки могут иметь самую разнообразную внеш-
нюю форму: шаровидную (лейкоциты), многогранную (клетки
железистого эпителия), звездчатую и разветвленно-отростчатую
(нервные и костные клетки), веретеновидную (гладкая мускулату-
ра, фибробласты), цилиндрическую (кишечный эпителиоцит),
уплощенную (эндотелиоцит, мезотелиоцит) и др. Однако при изу-
чении клеток органов различных растений или животных обращает
на себя внимание существование общего плана организации их,
несмотря на то, что по внешнему виду они отличаются друг от
друга (рис. 4). Одновременно это сходство указывает на общность
происхождения всех эукариотических организмов.
Рис. 4. Ультрамикроскопическое строение клетки животных организмов (схема).
3 — ядро; 2 — плазмолемма; 3 — микроворсинки; 4 — агранулярная эндоплазматическая
сеть; 5 — гранулярная эндоплазматическая сеть; б — комплекс Гольджи; 7 — центриоль и
микротрубочки клеточного центра; 8 — митохондрии; 9 цитоплазматические пузырьки-
Ю — лизосомы; 11 — микрофиламенты; 12 — рибосомы; 13 — выделение гранул секрета.
39
Клеточные функции можно подразделить на две основные
группы: обязательные и необязательные (фа-
культативные). Обязательные функции, направленные на
поддержание жизнеспособности самих клеток, осуществляются
постоянными внутриклеточными структурами — органеллами.
Различие клеток в многоклеточных организмах, обусловленное
специализацией их функций, связано с развитием особых функ-
ционзльных клеточных структур — органелл специального значе-
ния. Так, в мышечной клетке, кроме структур, являющихся при-
надлежностью всех клеток (мембранные органеллы, рибосомы
и др.), встречаются в большом количестве фибриллярные компо-
ненты — миофибриллы, имеющие специфическую пространст-
венную организацию и обеспечивающие характерную для этой
клетки функцию — движение. В нервных клетках, обладающих
специальными концевыми структурами {синапсами), общекле-
точные компоненты, такие как эндоплазматическая сеть, фибрил-
лярные элементы, имеют своеобразную композицию в цитоплазме,
что также связано с особенностью ее функционирования. Они
обеспечивают генерацию и проведение нервных импульсов. Таким
образом, структурная неоднозначность клеток сопряжена с разно-
образием их специальных функций, осуществляемых данной клет-
кой как бы на фоне общих, обязательных проявлений клеточной
жизнедеятельности
Каким образом возникает это разнообразие, сегодня еще до
конца не ясно. Индивидуальное развитие, от одной клетки до
многоклеточного зрелого организма — результат последовательно-
го, избирательного включения работы разных генов в различных
клетках, называемого дифференцировкой.
Сходство в строении — гомология клеток как данного организ-
ма, так и разных организмов — определяется одинаковостью
общеклеточных функций, направленных на поддержание жизни
самих клеток и на их размножение. Разнообразие же в строении
клеток — это результат их функциональной специализации.
3. Размножение клеток путем деления ис-
ходной клетки. Т. Шванн в своих обобщениях подчеркивал
одинаковость принципа развития клеток как у животных, так и у
растений. Однако следует заметить, что первоначальная разработ-
ка этого принципа основывалась на ложном тезисе о развитии
клеток из неклеточной «бластемы». Сформулированное позднее
Р. Вирховым положение «всякая клетка от клетки» можно считать
биологическим законом. Размножение клеток, прокариотических
и эукариотических, происходит только путем деления исходной
клетки, которому предшествует воспроизведение ее генетического
материала (репродукция ДНК). У эукариотических клеток единст-
венно полноценным способом деления является митоз или мейоз
(при образовании половых клеток). При этом образуется специ-
альный аппарат клеточного деления, клеточное веретено, с по-
мощью которого равномерно и точно по двум дочерним клеткам рас-
пределяют хромосомы, до этого удвоившиеся в числе. Митоз на-
40
блюдается у всех эукариотических, как растительных, так и жи-
вотных, клеток. Современная наука отвергает иные пути образова-
ния клеток и увеличения их числа.
4. Клетки как части целостного организма.
Каждое проявление деятельности целого организма, будь то реак-
ция на раздражение или движение, иммунные реакции и многое
другое, осуществляется специализированными клетками. Однако,
хотя клетка и является единицей функционирования в многокле-
точном организме, деятельность ее не обособлена.
Многоклеточные организмы представляют собой сложные ан-
самбли специализированных клеток, объединенных в целостные,
интегрированные системы тканей и органов, подчиненные и свя-
занные межклеточными, гуморальными и нервными формами регу-
ляции. Вот почему мы говорим об организме как о целом, а о
клетках — как об элементарных единицах его, специализирован-
ных на выполнении строго определенных функций, осуществляю-
щих их в комплексе со всеми элементами, входящими в состав
сложно организованной живой системы многоклеточного единого
организма. Расчлененность функций организма дает ему большие
возможности для приспособления с целью сохранения вида, раз-
множения отдельных индивидуумов.
СТРУКТУРНЫЕ КОМПОНЕНТЫ КЛЕТКИ
Цитоплазма
Цитоплазма (cytoplasma), отделенная от окружающей среды
плазмолеммой, включает в себя гиалоплазму, находящиеся в ней
обязательные клеточные компоненты — органеллы, а также различ-
ные непостоянные структуры — включения.
Г иалоплазма
Г иалоплазма (от греч. hyalinos — прозрачный) — основная
плазма, или матрикс цитоплазмы, представляет собой очень важ-
ную часть клетки, ее истинную внутреннюю среду.
В электронном микроскопе матрикс цитоплазмы имеет вид
гомогенного или тонкозернистого вещества с низкой электронной
плотностью. Гиалоплазма является сложной коллоидной системой,
включающей в себя различные биополимеры: белки, нуклеиновые
кислоты, полисахариды и др. Эта система способна переходить
из золеобразного (жидкого) состояния в гелеобразное и обратно.
В организованной, упорядоченной многокомпонентной системе
гиалоплазмы отдельные зоны могут менять свое агрегатное состоя-
ние в зависимости от условий или от функциональной задачи; в
бесструктурной на взгляд гиалоплазме могут возникать и распадать-
ся различные фибриллярные, нитчатые комплексы белковых моле-
кул. В состав гиалоплазмы входят главным образом различные
глобулярные белки. Они составляют 20—25% общего содержания
белков в эукариотической клетке. К важнейшим ферментам гиало-
41
плазмы относятся ферменты метаболизма сахаров, азотистых ос-
нований, аминокислот, липидов и других важных соединений.
В гиалоплазме располагаются ферменты активации аминокислот
при синтезе белков, транспортные (трансферные) РНК (тРНК).
В гиалоплазме при участии рибосом и полирибосом (полисом)
происходит синтез белков, необходимых для собственно клеточных
нужд, для поддержания и обеспечения жизни данной клетки.
Клеточные мембраны. Структурно-химическая
характеристика мембран клеток
Общей чертой всех мембран клетки является то, что они пред-
ставляют собой тонкие (6—10 нм) пласты липопротеидной приро-
ды (липиды в комплексе с белками) (рис. 5).
Основными химическими компонентами клеточных мембран
являются липиды (~40%) и белки (~60%); кроме того, во мно-
гих мембранах обнаружены углеводы (5—10%).
К липидам относится большая группа органических веществ,
обладающих плохой растворимостью в воде (гидрофобность) и
растворимостью в органических растворителях и жирах (липо-
фильность). Состав липидов очень разнообразен. Характерными
представителями липидов, встречающихся в клеточных мембранах,
являются фосфолипиды (глицерофосфатиды), сфингомиелины и
из стероидных липидов — холестерин.
Особенностью липидов мембран является разделение их моле-
кул на две функционально различные части: гидрофобные непо-
лярные, не несущие зарядов «хвосты», состоящие из жирных
кислот, и гидрофильные, заряженные полярные «головки». Это
определяет способность липидов самопроизвольно образовывать
двухслойные (билипидные) мембранные структуры толщиной
5—7 нм. Различные клеточные мембраны могут •значительно
отличаться друг от друга по липидному составу. Они различаются
и набором белковых молекул.
Многие мембранные белки состоят из двух частей, из участков,
богатых полярными (несущими заряд) аминокислотами, и участ-
ков, обогащенных неполярными аминокислотами: глицином, ала-
Рис. 5. Строение клеточной
мембраны (схема).
I — липиды; 2 — гидрофобная
зона бислоя липидных молекул;
3 — интегральные белки мембра-
ны; 4 — полисахариды гликока-
ликса.
42
ничом, валином, лейцином. Такие белки в липидных слоях мемб-
ран располагаются так, что их неполярные участки как бы погру-
жены в «жирную» часть мембраны, где находятся гидрофобные
участки липидов. Полярная (гидрофильная) же часть этих белков
взаимодействует с головками липидов и обращена в сторону вод-
ной фазы.
Кроме таких интегральных белков, существуют белки, частично
встроенные в мембрану — полуинтегральные и примембранные, не
встроенные в билипидный слой. По биологической роли белки
мембран можно разделить на белки-ферменты, белки-переносчики,
рецепторные и структурные белки.
Углеводы мембран входят в состав не в свободном состоянии,
они связаны с молекулами липидов или белков. Такие вещества
называются соответственно гликолипидами и гликопротеинами.
Количество их в мембранах обычно невелико.
Как бы ни было велико различие между мембранами по коли-
честву и составу их липидов, белков и углеводов, мембраны обла-
дают рядом общих свойств, определяемых их основной структу-
рой. Все мембраны являются барьерными структурами, резко
ограничивающими свободную диффузию веществ между цитоплаз-
мой и средой, с одной стороны, и между гиалоплазмой и содер-
жимым мембранных органелл — с другой. Особенность же спе-
цифических функциональных нагрузок каждой мембраны опреде-
ляется свойствами и особенностями белковых компонентов, боль-
шая часть из которых представляет собой ферменты или фермент-
ные системы. Большую роль в функционировании мембран играют
гликолипиды и гликопротеиды.
Плазмолемма. Барьерн о-p ецепторная
и транспортная система клетки
Плазмолемма (plasmalemma), или внешняя клеточная мембрана,
среди различных клеточных мембран занимает особое место. Это
поверхностная периферическая структура, не только ограничиваю-
щая клетку снаружи, но и обеспечивающая ее непосредственную
связь с внеклеточной средой, а следовательно, и со всеми вещест-
вами и стимулами, воздействующими на клетку.
Химический состав плазмолеммы. Основу плазмолеммы состав-
ляет липопротеиновый комплекс. Она имеет толщину около 10 нм
и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный
слой — гликокаликс (Glycocalyx). Толщина этого слоя около 3—
4 нм, он обнаружен практически у всех животных клеток, но
степень его выраженности различна. Гликокаликс представляет
собой ассоциированный с плазмолеммой гликопротеиновый комп-
лекс, в состав которого входят различные углеводы. Углеводы
образуют длинные, ветвящиеся цепочки полисахаридов, связанные
с белками и липидами, входящими в состав плазмолеммы
(см. рис. 5). При использовании специальных методов выявления
полисахаридов (краситель рутениевый красный) видно, что они
образуют как бы чехол поверх плазматической мамбраны.
В гликокаликсе могут располагаться белки, не связанные непо-
средственно с билипидным слоем. Как правило, это белки-фермен
ты, участвующие во внеклеточном расщеплении различных ве-
ществ, таких как углеводы, белки, жиры и др.
Функции плазмолеммы. Эта мембрана выполняет ряд важней-
ших клеточных функций, ведущими из которых являются функция
разграничения цитоплазмы с внешней средой, функции рецепции
и транспорта различных веществ как внутрь клетки, так и из нее.
Рецепторные функции связаны с локализацией на плазмолемме
специальных структур, участвующих в специфическом «узнавании»
химических и физических факторов. Клеточная поверхность обла-
дает большим набором компонентов — рецепторов, определяющих
возможность специфических реакций с различными агентами.
Рецепторами на поверхности клетки могут служить гликопротеиды
и гликолипиды мембран (см. рис. 5). Считается, что такие чувст-
вительные к отдельным веществам участки могут быть разбро-
саны по всей поверхности клетки или собраны в небольшие зоны.
Существуют рецепторы к биологически активным веществам —
гормонам, медиаторам, к специфическим антигенам разных клеток
или к определенным белкам.
С плазмолеммой связана локализация специфических рецепто-
ров, отвечающих за такие важные процессы, как взаимное рас-
познавание клеток, развитие иммунитета, рецепторов, реагирующих
на физические факторы. Так, в плазмолемме светочувствительных
клеток животных расположена специальная система фоторецеп-
торных белков (родопсин), с помощью которых световой сигнал
превращается в химический, что в свою очередь приводит к генера-
ции электрического импульса.
Выполняя транспортную функцию, плазмолемма обеспе-
чивает пассивный перенос ряда веществ, например воды, ионов,
некоторых низкомолекулярных соединений. Другие вещества про-
никают через мембрану путем активного перено са против
градиента концентрации с затратой энергии за счет расщепления
АТФ. Так транспортируются многие органические молекулы (са-
хара, аминокислоты и др.). Эти процессы могут быть сопряжены
с транспортом ионов, в них принимают участие специальные бел-
ки-переносчики.
Крупные молекулы биополимеров практически не проникают
сквозь плазмолемму. В ряде случаев макромолекулы и даже их
агрегаты, а часто и крупные частицы попадают внутрь клетки в
результате процессов эндоцитоза (рис. 6, А, Б). Эндоцитоз фор-
мально разделяют на фагоцитоз (захват и поглощение клеткой
крупных частиц, например бактерий или даже фрагментов дру-
гих клеток), и пиноцитоз (захват макромолекулярных соеди-
нений).
Эндоцитоз начинается с сорбции на поверхности плазмолем-
мы поглощаемых веществ. Связывание их с плазмолеммой опреде-
44
Рис. 6. Эндоцитоз. Разные типы (Л, /й образования пиноцитозных пузырьков.
/ — сорбция частиц на поверхности плазматической мембраны; 2 — погружение частиц в
цитоплазму; 3 — первичные лизосомы.
ляется наличием на ее поверхности рецепторных молекул. После
сорбции веществ на поверхности плазмолемма начинает образо-
вывать сначала небольшие впячивания внутрь клетки. Эти впя-
чивания могут иметь вид еще незамкнутых округлых пузырьков
или представлять собой глубокие инвагинации, впячивания внутрь
клетки. Затем такие локальные впячивания отшнуровываются от
плазмолеммы и в виде пузырьков свободно располагаются под ней.
В дальнейшем эндоцитозные пузырьки могут сливаться друг
с другом, расти и в их внутренней полости, кроме поглощенных
веществ, начинают обнаруживаться гидролитические ферменты
(гидролазы), поступающие сюда из лизосом (см. ниже). Эти фер-
менты расщепляют биополимеры до мономеров, которые в результа-
те активного транспорта через мембрану пузырька переходят в гиа-
лоплазму. Таким образом, поглощенные молекулы внутри мемб-
ранных вакуолей, образовавшихся из элементов плазмолеммы,
подвергаются в н у т р и к л е т о ч н о м у пищеварению.
Плазмолемма принимает участие в выведении веществ из клет-
ки (экзоцитоз). В этом случае внутриклеточные продукты (бел-
ки, мукополисахариды, жировые капли и др.), заключенные в ва-
куоли или пузырьки и отграниченные от гиалоплазмы мембраной,
подходят к плазмолемме. В местах контактов плазмолемма и
мембрана вакуоли сливаются и содержимое вакуоли поступает
в окружающую среду.
Процесс эндоцитоза и экзоцитоза осуществляется при участии
связанной с плазмолеммой системы фибриллярных компонентов
цитоплазмы — таких, как микротрубочки и сократимые микрофи-
ламенты. Последние, соединяясь с определенными участками
плазмолеммы, могут, изменяя свою длину, втягивать мембрану
внутрь клетки, что приводит к отделению от плазмолеммы эндоци-
тозных вакуолей. Часто, непосредственно примыкая к ней, микро-
филаменты образуют сплошной, так называемый кортикальный
слой.
Плазмолемма многих клеток животных может образовывать
выросты различной структуры. У ряда клеток такие выросты вклю-
чают в свой состав специальные компоненты цитоплазмы (микро-
45
трубочки, фибриллы), что приводит к развитию специальных
структур — ресничек, жгутиков и др.
Наиболее часто встречаются на поверхности многих животных
клеток микроворсинки. Эти выросты цитоплазмы, ограниченные
пла змолеммой, имеющие форму цилиндра с закругленной верши-
ной. Микроворсинки характерны для клеток эпителиев, но обнару-
живаются и у клеток других тканей. Диаметр микроворсинок
около 100 нм. Число и длина их различны у разных типов клеток.
Возрастание числа микроворсинок приводит к резкому увеличению
площади клеточной поверхности. Это особенно важно для клеток,
участвующих во всасывании. Так, в кишечном эпителии на 1 мм2
поверхности насчитывается до 2 • 108 микроворсинок.
Межклеточные соединения (контакты)
Плазмолемма многоклеточных животных ор1анизмов принима-
ет активное участие в образовании специальных структур — меж-
клеточных соединений (junctiones intercellulares), обеспечивающих
межклеточные взаимодействия. Различают несколько типов таких
структур (рис. 7). Простое межклеточное соединение (junctio
intercellutaris simplex) — сближение плазмолемм соседних клеток
на расстояние 15-—20 нм. При этом происходит взаимодействие
слоев гликокаликса соседних клеток. Плотное соединение (запи-
рающая зона) (zonula occludens) — зона, где слои двух плазмо-
лемм максимально сближены, здесь происходит как бы слияние
участков плазмолемм двух соседних клеток. Роль плотного замы-
кающего соединения заключается в механическом соединении
клеток друг с другом. Эта область непроницаема для макромоле-
кул и ионов и, следовательно, она запирает, отграничивает меж-
клеточные щели (и вместе с ними собственно внутреннюю среду
организма) от внешней среды.
Часто встречается, особенно в эпителии, особый тип соедине-
ния — пятно сцепления, или десмосома (desmosoma). Эга струк-
тура представляет собой небольшую площадку, иногда имеющую
слоистый вид, диаметром до 0,5 мкм, где между мембранами
располагается зона с высокой электронной плотностью. К плазмо-
лемме в зоне десмосомы со стороны цитоплазмы прилегает участок
электронно-плотного вещества, так что внутренний слой мембраны
кажется утолщенным. Под этим утолщением находится область
тонких фибрилл, которые могут быть погружены в относительно
плотный матрикс. Функциональная роль десмдсом заключается
главным образом в механической связи между клетками.
Щелевидное соединение, или нексус (nexus), представляет
собой область протяженностью 0,5—3 мкм, где плазмолеммы раз-
делены промежутком в 2—3 нм. Со стороны цитоплазмы никаких
специальных примембранных структур в данной области не обна-
руживается, но в структуре плазмолемм соседних клеток друг
против друга располагаются специальные белковые комплексы
(коннексоны), которые образуют как бы каналы из одной клетки
46
Рис. 7. Межклеточные соединения (схема).
/ простое соединение; 2- пальцевидное соединение; 3 — дес-
мосома; 4 — плотное соединение; 5 — щелевидное соединение.
в другую. Этот тип соединения встречается во
всех группах тканей. Функциональная роль ще-
левидного соединения заключается, по-видимо-
му, в переносе ионов и мелких молекул (моле-
кулярная масса 2 • 1О'!) от клетки к клетке.
Так, в сердечной мышце возбуждение, в основе
которого лежит процесс изменения ионной про-
ницаемости, передается от клетки к клетке
через нексус.
Синаптические соединения, или синапсы
(synapsis). Этот тип соединений характерен для
нервной ткани и встречается в специализиро-
ванных участках контакта как между двумя
нейронами, так и между нейроном и каким-либо
иным элементом, входящим в состав рецептора
или эффектора (например, нервно-мышечные,
нервно-эпителиальные синапсы). Синапсы —
участки контактов двух клеток, специализиро-
ванных для односторонней передачи возбужде-
ния или торможения от одного элемента к
другому (см. гл. XI).
Органеллы цитоплазмы
Органе плы — постоянно присутствующие и обязательные
для всех клеток микроструктуры, выполняющие жизненно важные
функции.
Классификация органелл. Различают мембранные орга-
неллы — митохондрии, эндоплазматическую сеть, аппарат
Гольджи, лизосомы, гладкую эндоплазматическую сеть (к кате-
гории мембранных органелл относится и плазмолемма); н е м е м-
бранные органеллы: свободные рибосомы и полисомы,
микротрубочки, центриоли и филаменты (микрофиламенты, про-
межуточные филаменты). Во многих клетках органеллы могут
принимать участие в образовании особых структур, характерных
для специализированных клеток. Так, реснички и жгутики обра-
зуются за счет центриолей и плазматической мембраны, микро-
ворсинки — это выросты плазматической мембраны с гиалоплаз-
мой и микрофиламентами, акросома спермиев — это производное
элементов аппарата Гольджи, «эллипсоид» зрительных клеток —
скопления митохондрий и пр.
Мембранные органеллы
Мембранные органеллы представляют собой одиночные или
связанные друг с другом отсеки цитоплазмы, отграниченные мемб-
раной от окружающей их гиалоплазмы, имеющие свое собственное
47
содержимое, отличное по составу, свойствам и функциям от других
частей клетки, т. е. это замкнутые, закрытые объемные зоны —
компартменты (англ, compartment—купе). В гиалоплазме мемб-
ранные органеллы распределены закономерно. Эндоплазматиче-
ская сеть, различные вакуоли, возникающие из нее, составляют
вакуолярную систему цитоплазмы, систему синтеза и внутрикле-
точного транспорта веществ. Кроме того, в ее состав входят
комплекс Гольджи, лизосомы, аутолизосомы и пероксисомы. Для
всех элементов вакуолярной системы характерно наличие одной
ограничивающей мембраны. Митохондрии отделены от гиалоплаз-
мы двумя мембранами (двухмембранные органеллы).
Эндоплазматическая сеть
Эндоплазматическая сеть была открыта К. Р. Портером в
1945 г. Этот компонент цитоплазмы представляет собой совокуп-
ность вакуолей, плоских мембранных мешков или трубчатых
образований, создающих как бы мембранную сеть внутри цито-
плазмы. Различают два типа — гранулярную и гладкую эндоплаз-
матическую сеть.
Гранулярная эндоплазматическая сеть (reticulum endoplas-
maticum granulosum) на ультратонких срезах представлена замкну-
тыми мембранами, которые образуют на сечениях уплощенные
мешки, цистерны, трубочки. Ширина полостей цистерн значитель-
но варьирует в зависимости от функциональной активности клетки.
Наименьшая ширина их — около 20 нм, но они могут достигать
диаметра в несколько микрометров. Отличительной чертой этих
мембран является то, что они со стороны гиалоплазмы покрыты
рибосомами (рис. 8).
Гранулярная эндоплазматическая сеть бывает представлена
редкими разрозненными цистернами или их локальными скопле-
ниями. Первый тип гранулярной эндоплазматической сети харак-
терен для малоспециализированных клеток или для клеток с
низкой метаболической активностью. Скопления эндоплазматиче-
ской сети являются принадлежностью клеток, активно синтези-
рующих секреторные белки. Так, в клетках печени и некоторых
нервных клетках гранулярная эндоплазматическая сеть собрана
в отдельные зоны. В клетках поджелудочной железы гранулярная
эндоплазматическая сеть в виде плотно упакованных друг около
друга мембранных цистерн занимает базальную и околоядерную
зоны клетки. Рибосомы, связанные с мембранами эндоплазма-
тической сети, участвуют в синтезе белков, выводимых из данной
клетки («экспортируемые» белки). Кроме того, гранулярная эндо-
плазматическая сеть принимает участие в синтезе белков — фер-
ментов, необходимых для организации внутриклеточного метабо-
лизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети,
могут, минуя гиалоплазму, транспортироваться в вакуоли комп-
лекса Гольджи, где они часто модифицируются и входят в состав
48
Рис. 8. Строение гранулярной эн-
доплазматической сети.
А — схема; Б — электронная микро-
фотография участка среза печеноч-
ной клетки. I — рибосомы; 2 —
пластинки; 3 — внутренние полости
цистерн; 4 — отщепляющиеся мем-
бранные пузырьки, лишенные рибо-
сом.
либо лизосом, либо секре-
торных гранул, содержи-
мое которых остается
изолированным от гиало-
плазмы мембраной. В ряде
случаев внутри самих ка-
нальцев или вакуолей гра-
нулярной эндоплазмати-
ческой сети может проис-
ходить модификация бел-
ков, например связывание
их с сахарами (глюкози-
лирование), или конденса-
ция синтезированных бел-
ков с образованием круп-
ных агрегатов — секретор-
ных гранул.
В гранулярной эндо-
плазматической сети про-
исходит синтез мембран-
ных интегральных белков,
которые встраиваются в
толщу мембраны.
Итак, роль грануляр-
ной эндоплазматической
сети заключается в синте-
зе на ее полисомах эк-
спортируемых белков, в их
изоляции от содержимого
гиалоплазмы внутри мем-
бранных полостей, в тран-
спорте этих белков в другие участки клетки, в химической моди-
фикации таких белков и в их локальной конденсации, а также
в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum
endoplasmaticum nongranulosum) также представлена мембранами,
образующими мелкие вакуоли и трубки, канальцы, которые могут
ветвиться, сливаться друг с другом. В отличие от гранулярной
эндоплазматической сети на мембранах гладкой эндоплазмати-
ческой сети нет рибосом. Диаметр вакуолей и канальцев гладкой
эндоплазматической сети обычно около 50—100 нм.
44
Гладкая эндоплазматическая сеть возникает и развивается за
счет гранулярной эндоплазматической сети.
Деятельность гладкой эндоплазматической сети связана с мета-
болизмом липидов и некоторых внутриклеточных полисахаридов.
Гладкая эндоплазматическая сеть участвует в заключительных
этапах синтеза липидов. Она сильно развита в клетках, секретиру-
ющих такие категории липидов, как стероиды, например, в клетках
коркового вещества надпочечников, в сустентоцитах семенников.
Тесная топографическая связь гладкой эндоплазматической
сети с отложениями гликогена (запасной внутриклеточный поли-
сахарид животных) в гиалоплазме различных клеток (клетки
печени, мышечные волокна) указывает на ее возможное участие в
метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплаз-
матическая сеть способна депонировать ионы кальция, необходи-
мые для функции мышечной ткани (см. гл. X).
Очень важна роль гладкой эндоплазматической сети в дез-
активации различных вредных для организма веществ за счет их
окисления с помощью ряда специальных ферментов. Особенно
четко она проявляется в клетках печени. Так, при ряде отравлений
в клетках печени появляются ацидофильные зоны (не содержащие
РНК), сплошь занятые гладким эндоплазматическим ретикулумом.
Комплекс Гольджи (внутренний сетчатый аппарат)
В 1898 г. К. Гольджи, используя свойства связывания тяжелых
металлов (осмия или серебра) с клеточными структурами, выявил
в нервных клетках сетчатые образования, которые он назвал
внутренним сетчатым аппаратом (apparatus reticularis internus),
который позднее стали называть комплексом Гольджи (complexus
Golgiensis). Подобные структуры затем описаны во всех клетках
эукариот.
При рассмотрении в электронном микроскопе комплекс Гольд-
жи представлен мембранными структурами, собранными вместе в
небольшой зоне (рис. 9). Отдельная зона скопления этих мембран
называется диктиосомой. Таких зон в клетке может быть несколько.
В диктиосоме плотно друг к другу (на расстоянии 20—25 нм) рас-
положены 5—10 плоских цистерн, между которыми располагаются
тонкие прослойки гиалоплазмы. Каждая цистерна имеет перемен-
ную толщину; в центре ее мембраны могут быть сближены (до
25 нм), а на периферии иметь расширения, ампулы, ширина которых
непостоянна. Кроме плотно расположенных плоских цистерн, в зоне
комплекса Гольджи наблюдается множество мелких пузырьков
(везикул), которые встречаются главным образом в его перифе-
рических участках. Иногда видно, как они отшнуровываются от
ампулярных расширений на краях плоских цистерн. Принято раз-
личать в зоне диктиосомы проксимальный и дистальный участки.
В секретирующих клетках обычно комплекс Гольджи поляризован:
50
3
Рис. 9. Комплекс Гольджи.
А — нервная клетка спинного моз>а, импрегнация серебром по методу Гольджи: / — ядро;
2- ядрышко; 3 комплекс Гольджи. Б — схема ультрамикроскопического строения
(трехмерная реконструкция). В — комплекс Гольджи на ультратонком срезе (печеночная
клетка). /— пузырьки; 2 — трубочки: 3—уплощенные мешочки (цистерны); 4- пла-
стинки гранулярной эндоплазматической сети.
его проксимальная часть обращена к ядру, в то время как дисталь-
ная — к поверхности клетки.
В клетках отдельные диктиосомы могут быть связаны друг с
другом системой везикул и цистерн, примыкающих к проксималь-
ному концу скопления плоских мешков так, что образуется рыхлая
трехмерная сеть, выявляемая в световом микроскопе.
Комплекс Гольджи участвует в сегрегации и накоплении про-
дуктов, синтезированных в цитоплазматической сети, в их хими-
51
Гольджи заключается в
Рис. 10. Участие клеточных структур в белковой
секреции (схема).
I — поступление аминокислот из гемокапилляра к ри
босомам гранулярной эндоплазматической сети; 2 —
синтез и сегрегация белков; 3 — переход белков в
вакуоли комплекса Гольджи; 4 — отщепление от ком-
плекса Гольджи пузырьков с секреторными продук-
тами; 5 - экструзия, выход секрета из клетки.
ческих перестройках, созревании; в
цистернах комплекса Гольджи происхо-
дит синтез полисахаридов, их комплек-
сирование с белками, что приводит к
образованию мукопротеидов, и, главное,
с помощью элементов аппарата Гольд-
жи происходит процесс выведения гото-
вых секретов за пределы клетки. Кроме
того, комплекс Гольджи обеспечивает
формирование клеточных лизосом.
Секреторная функция комплекса
том, что синтезированный на рибосомах
экспортируемый белок отделяется и накапливается внутри цистерн
эндоплазматической сети, по которым он транспортируется к зоне
мембран пластинчатого комплекса (рис. 10). Затем накопленный
белок может конденсироваться, образуя секреторные белковые
гранулы (как это наблюдается в поджелудочной, молочной и
других железах), или оставаться в растворенном виде (как им-
муноглобулины в плазматических клетках).
В дальнейшем от ампулярных расширений цистерн комплекса
Гольджи отщепляются пузырьки, содержащие эти белки. Такие
везикулы также могут сливаться друг с другом и увеличиваться в
размерах, образуя секреторные гранулы. После этого секреторные
гранулы начинают двигаться к поверхности клетки, соприкасаются
с плазмолеммой, с которой сливаются их собственные мембраны,
и таким образом содержимое гранул оказывается за пределами
клетки. Морфологически этот процесс называется экструзией (вы-
брасывание, экзоцитоз), напоминает пиноцитоз только с обратной
последовательностью стадий.
Нужно отметить, что с самого момента образования до выве-
дения из клеток секретируемые продукты отделены мембраной от
гиалоплазмы. Следовательно, мембраны комплекса Гольджи выпол-
няют сегрегирующую роль при образовании клеточных секретов.
В зоне комплекса Гольджи могут происходить многие метаболи-
ческие процессы. Здесь большинство белков подвергается моди-
фикации, некоторые их аминокислоты фосфорилируются, ацетили-
руются или глюкозилируются. Во многие секреторные продукты
входят сложные белки — гликопротеиды и мукопротеиды (муци-
ны) — белки, связанные в единую цепь с сахарами и полисахарида-
ми разной природы. Синтез этих полисахаридов идет в комплексе
Гольджи.
В пузырьках комплекса Гольджи иногда происходит накопле-
52
ние ресинтезированных молекул липидов и образование сложных
белков липопротеидов, которые могут транспортироваться пузырь-
ками за пределы клетки.
Мембраны комплекса Гольджи образуются при участии грану-
лярной эндоплазматической сети.
Лизосомы
Лизосомы (lysosomae) — это разнообразный класс шаровидных
структур размером 0,2—0,4 мкм, ограниченных одиночной мембра-
ной. Характерным признаком лизосом является наличие в них гид-
ролитических- ферментов — гидролаз (протеиназы, нуклеазы, глю-
козидазы, фосфатазы, липазы), расщепляющих различные биопо-
лимеры. Лизосомы были открыты в 1949 г. де Дювом.
Среди лизосом можно выделить по крайней мере 3 типа: пер-
вичные лизосомы, вторичные лизосомы (фаголизосомы и аутофаго-
сомы) и остаточные тельца (рис. 11). Разнообразие морфологии
лизосом объясняется тем, что эти частицы участвуют в процессах
внутриклеточного переваривания, образуя сложные пищеваритель-
ные вакуоли как экзогенного (внеклеточного), так и эндогенного
(внутриклеточного) происхождения.
Первичные лизосомы представляют собой мелкие мембранные
пузырьки размером около 0,2—0,5 мкм, заполненные бесструктур-
ным веществом, содержащим гидролазы, в том числе активную кис-
лую фосфатазу, которая является маркерным для лизосом фер-
ментом. Эти мелкие пузырьки практически очень трудно отличить
от мелких везикул на периферии зоны комплекса Гольджи, которые
также содержат кислую фосфатазу. Местом ее синтеза является
гранулярная эндоплазматическая сеть, затем этот фермент появля-
ется в проксимальных участках диктиосом, а затем в мелких везику-
лах по периферии диктиосом и, наконец, в первичных лизосомах
Таким образом, весь путь образования первичных лизосом очень
сходен с образованием секреторных (зимогенных) гранул в клетках
поджелудочной железы, за исключением последнего этапа — вы-
брасывания из клетки.
Вторичные лизосомы, или внутриклеточные пищеварительные
вакуоли, формируются при слиянии первичных лизосом с фагоци-
тарными вакуолями (фагосомами) или пиноцитозными вакуолями,
образуя фаголизосомы, или гетероф>агосомы, а также с изменен-
ными органеллами самой клетки, подвергающимися перевариванию
(аутофагосомы). При этом ферменты первичной лизосомы получа-
ют доступ к субстратам, которые они и начинают расщеплять.
Вещества, попавшие в состав вторичной лизосомы, расщепляются
гидролазами до мономеров, которые транспортируются через мем-
брану лизосомы в гиалоплазму, где они реутилизируются, т. е. вклю-
чаются в различные обменные процессы.
Однако расщепление, переваривание биогенных макромолекул
внутри лизосом может идти в ряде клеток не до конца. В этом случае
в полостях лизосом накапливаются непереваренные продукты. Та-
53
Рис. 11. Строение ли юсом.
4 - схема участия структур клетки в образовании лизосом и во внутрик неточном пище-
варении: 1 образование из гранулярной эндоплазматической сети мелких пузырьков,
содержащих гидролитические ферменты; 2 — перенос ферментов в комплекс Гольджи:
3 — образование первичных лизосом; 4 выделение и использование (5) гидролаз при
внеклеточном расщеплении 6 эндоцитозные пузырьки; 7 - слияние первичных лизосом
и эндоцитозных пузырьков; 8 образование вторичных лизосом (фаголизосом); 9—
телолизосомы; 10 - экскреция остаточных телец; II — слияние первичных лизосом с
разрушающимися структурами клетки; 12 — аутофагосома. Б электронная микрофото-
(рафия среза вторичных лизосом (обозначены стрелками).
кая лизосома носит название «телолизосома», или остаточное те ль-
це (corpusculum residuale). Остаточные тельца содержат меньше
гидролитических ферментов, в них происходит уплотнение содер-
жимого, его перестройка. Часто в остаточных тельцах наблюдается
вторичная структуризация неперевариваемых липидов, которые об-
разуют слоистые структуры. Там же происходит отложение пиг-
ментных веществ. Так, у человека при старении организма в клет-
ках мозга, печени и в мышечных волокнах в телолизосомах про-
исходит отложение «пигмента старения» — липофусцина.
При участии лизосом в переваривании внутриклеточных эле-
ментов (аутолизосомы) они могут обеспечивать модификацию про-
дуктов, приготавливаемых самой клеткой, например, с помощью
54
гидролаз лизосом. В клетках щитовидной железы гидролизуется
тироглобулин, что приводит к образованию гормона тироксина,
который затем выводится в кровеносное русло.
В аутофагосомах обнаруживаются фрагменты или даже целые
цитоплазматические структуры, например митохондрии, элементы
цитоплазматической сети, рибосомы, гранулы гликогена и др., что
является доказательством их определяющей роли в процессах де-
градации.
Функциональное значение аутофагоцитоза еще не ясно. Есть
предположение, что этот процесс связан с отбором и уничтожением
измененных, поврежденных клеточных компонентов. В этом случае
лизосомы выполняют роль внутриклеточных «чистильщиков», уби-
рающих дефектные структуры. Интересно, что в нормальных усло-
виях число аутофагосом увеличивается при метаболических стрес-
сах, например при гормональной индукции активности клеток пе-
чени. Значительно возрастает число аутофагосом при различных
повреждениях клеток; в этом случае аутофагоцитозу могут под-
вергаться целые зоны внутри клеток. Увеличение числа аутолизо-
сом в клетках при патологических процессах — обычное явление.
Пероксисомы
Пероксисомы (peroxysoma) — небольшие (размером 0,3—
1,5 мкм) овальной формы тельца, ограниченные мембраной, содер-
жащие гранулярный матрикс, в центре которого часто видны крис-
таллоподобные структуры, состоящие из фибрилл и трубок (серд-
цевина). Пероксисомы, вероятно, образуются на расширенных сто-
ронах цистерн эндоплазматической сети. Они особенно характерны
для клеток печени, почек. Во фракции пероксисом обнаруживаются
ферменты окисления аминокислот, при работе которых образуется
перекись водорода, а также выявляется фермент каталаза, разру-
шающая ее. Каталаза пероксисом играет важную защитную роль,
так как Н2О2 является токсическим веществом для клетки.
Таким образом, мембранные органеллы клетки, составляющие
вакуолярную систему, обеспечивают синтез и транспорт внутрикле-
точных биополимеров, продуктов секреции, выводимых из клетки,
что сопровождается биосинтезом всех мембран этой вакуолярной
системы. Производные вакуолярной системы — лизосомы и пе-
роксиомы — участвуют в деградации экзогенных и эндогенных суб-
стратов клетки.
Митохондрии
Митохондрии (mitochondriae) — органеллы синтеза АТФ. Их
основная функция связана с окислением органических соединений
и использованием освобождающейся при распаде этих соединений
энергии для синтеза молекул АТФ. Исходя из этого, митохондрии
часто называют энергетическими станциями клетки, или органелла-
ми клеточного дыхания.
55
Термин «митохондрия» был введен Бенда в 1897 г. для обозначе-
ния зернистых и нитчатых структур в цитоплазме разных клеток.
Митохондрии можно наблюдать в живых клетках, так как они об-
ладают достаточно высокой плотностью. В живых клетках мито-
хондрии могут перемещаться, сливаться друг с другом, делиться.
Форма и размеры митохондрий животных клеток разнообразны,
но в среднем толщина их около 0,5 мкм, а длина — от 1 до 10 мкм.
Подсчеты показывают, что количество их в клетках сильно варьиру-
ет — от единичных элементов до сотен. Так, в клетке печени они
составляют более 20% общего объема цитоплазмы и содержат
около 30—35% общего количества белка в клетке. Площадь поверх-
ности всех митохондрий печеночной клетки в 4—5 раз больше по-
верхности ее плазматической мембраны.
Обычно митохондрии скапливаются вблизи тех участков цито-
плазмы, где возникает потребность в АТФ. Так, в сердечной мышце
митохондрии находятся вблизи миофибрилл. В сперматозоидах
митохондрии образуют спиральный футляр вокруг оси жгутика
и т. д. Увеличение числа митохондрий в клетках происходит путем
деления, или почкования, исходных митохондрий.
Митохондрии ограничены двумя мембранами толщиной около
7 нм (рис. 12. А). Наружная митохондриальная мембрана (mem-
brana mitochondrialis externa) отделяет их от гиалоплазмы. Обычно
она имеет ровные контуры и замкнута, так что представляет собой
мембранный мешок. Внешнюю мембрану от внутренней отделяет
межмембранное пространство шириной около 10—20 нм. Внутрен-
няя митохондриальная мембрана (membrana mitochondrialis inter-
na) ограничивает собственно внутреннее содержимое митохондрии,
ее матрикс (matrix mitochondrialis). Характерной чертой внутренних
мембран митохондрий является их способность образовывать мно-
гочисленные впячивания внутрь митохондрий. Такие впячивания
чаще всего имеют вид плоских гребней, или крист (crista).
Матрикс митохондрий имеет тонкозернистое строение
(рис. 12, Б) в нем иногда выявляются тонкие нити (толщиной
около 2—3 нм) и гранулы размером около 15—20 нм. Нити мат-
рикса митохондрий представляют собой молекулы ДНК, а мелкие
гранулы — митохондриальные рибосомы.
Основной функцией митохондрий является синтез АТФ, про-
исходящий в результате процессов окисления органических суб-
стратов и фосфорилирования АДФ. Начальные этапы этих слож-
ных процессов совершаются в гиалоплазме. Здесь происходит
первичное окисление субстратов (например, сахаров) до пирови-
ноградной кислоты (пирувата) с одновременным синтезом неболь-
шого количества АТФ. Эти процессы совершаются в отсутствие
кислорода (анаэробное окисление, гликолиз). Все последующие
этапы выработки энергии (дыхания) — аэробное окисление и син-
тез основной массы АТФ — осуществляются с потреблением кисло-
рода и локализуются внутри митохондрий. При этом происходит
дальнейшее окисление пирувата и других субстратов энергетиче-
ского обмена с выделением СО2 и переносом протонов на их акцеп-
56
3
1
1
Рис. 12. Ультрамикроскопическое строение митохондрии.
д — схема. Б — электронная микрофотография среза митохондрии печеночной клетки;
I наружная митохондриальная мембрана; 2 — внутренняя митохондриальная мембра-
на; 3 — кристы; 4 — митохондриальный матрикс.
торы. Эти реакции осуществляются с помощью ряда ферментов так
называемого цикла трикарбоновых кислот, которые локализованы в
матриксе митохондрии.
В мембранах крист митохондрии располагаются системы даль-
нейшего переноса электронов и сопряженного с ним фосфорили-
рования АДФ (окислительное фосфорилирование). При этом про-
исходит перенос электронов от одного белка-акцептора электронов
к другому и, наконец, связывание их с кислородом, вследствие
чего образуется вода. Одновременно с этим часть энергии, вы-
деляемой при таком окислении в цепи переноса электронов, за-
пасается в виде макроэргической связи при фосфорилировании
АДФ, что приводит к образованию большого числа молекул
АТФ — основного внутриклеточного энергетического эквивалента.
Именно на мембранах крист митохондрии происходит процесс
окислительного фосфорилирования с помощью здесь расположен-
ных белков цепи окисления и ферментов фосфорилирования АДФ,
АТФ -синтетазы.
Выявлено, что в матриксе митохондрии локализуется автоном-
ная система митохондриального белкового синтеза. Она представ-
лена молекулами ДНК, свободными от гистонов, что сближает их
57
-
с ДНК бактериальных клеток. На этих ДНК происходит синтез
молекул РНК разных типов: информационных, трансферных
(транспортных) и рибосомных. В матриксе митохондрий наблю-
дается образование рибосом, отличных от рибосом цитоплазмы.
Эти рибосомы участвуют в синтезе ряда митохондриальных бел-
ков, не кодируемых ядром. Однако такая система белкового синте-
за не обеспечивает всех функций митохондрии, поэтому автоно-
мию митохондрий можно считать ограниченной, относительной.
Малые размеры молекул митохондриальных ДНК не могут оп-
ределить синтез всех белков митохондрий. Показано, что боль-
шинство белков митохондрий находится под генетическим контро-
лем со стороны клеточного ядра и синтезируется в цитоплазме.
Наиболее вероятно, что митохондриальная ДНК кодирует лишь
немногие митохондриальные белки, которые локализованы в мем-
бранах и представляют собой структурные белки, ответственные за
правильную интеграцию в митохондриальных мембранах отдель-
ных функциональных белковых комплексов.
Митохондрии в клетках могут увеличиваться в размерах и
числе. В последнем случае происходит деление перетяжкой или
фрагментация исходных крупных митохондрий на более мелкие,
которые в свою очередь могут расти и снова делиться.
Немембранные органеллы
Рибосомы
Рибосомы fribosomae) —элементарные аппараты синтеза бел-
ковых, полипептидных молекул — обнаруживаются во всех клет-
ках. Рибосомы — это сложные рибонуклеопротеиды, в состав ко-
торых входят белки и молекулы РНК примерно в равных весовых
отношениях. Размер функционирующей рибосомы эукариотиче-
ских клеток 25 X 20 X 20 нм. Такая рибосома состоит из большой
и малой субъединиц. Каждая из субъединиц построена из рибо-
нуклеопротеидного тяжа, где рРНК взаимодействует с разными
белками и образует тело рибосомы.
Различают единичные рибосомы и комплексные рибосомы
(полисомы). Рибосомы могут располагаться свободно в гиалоплазме
или быть связанными с мембранами эндоплазматической сети.
В малоспециализированных и быстрорастущих клетках в основном
обнаруживаются свободные рибосомы. В специализированных клет-
ках рибосомы располагаются в составе гранулярной эндоплазмати-
ческой сети. Степень интенсивности синтетической деятельности
свободных рибосом меньше, а образуемые белки используются
в основном на собственные нужды клетки. Связанные рибосомы
обеспечивают синтез белков «на экспорт», т. е. на обеспечение нужд
организма. Содержание РНК и соответственно степень белковых
синтезов коррелируют с интенсивностью базофилии цитоплазмы.
Опорно-двигательные структуры клетки. Цитоскелет
Микротрубочки
В цитоплазме клеток, кроме мембранных структур и органелл,
встречается большое количество различных фибриллярных об-
разований, выполняющих разнообразные функции.
К таким фибриллярным компонентам относятся микротрубоч-
ки (microtubuli) белковой природы. В цитоплазме они могут об-
разовывать временные сложные образования, например веретено
клеточного деления. Микротрубочки входят в состав сложноор-
ганизованных специальных ор<анелл, таких как центриоли и ба-
зальные тельца, а также являются основными структурными эле-
ментами ресничек и жгутиков.
Микротрубочки представляют собой прямые, неветвящиеся
длинные полые цилиндры. Их внешний диаметр составляет около
24 нм, внутренний просвет имеет ширину 15 нм, а толщина стен-
ки — 5 нм. Стенка микротрубочек построена за счет плотно уло-
женных округлых субъединиц величиной около 5 нм. В электрон-
ном микроскопе на поперечных сечениях микротрубочек видны
большей частью 13 субъединиц, выстроенных в виде однослойно-
го кольца. Микротрубочки, выделенные из разных источников
(реснички простейших, клетки нервной ткани, веретено деления),
имеют сходный состав и содержат белки — тубулины.
Очищенные тубулины способны при определенных условиях
собираться в микротрубочки с такими же параметрами, какие
характерны для микротрубочек внутри клеток. Добавление алка-
лоида колхицина предотвращает самосборку микротрубочек или
приводит к разборке уже существующих. Деполимеризация ту-
булинов или торможение их полимеризации также вызывается по-
нижением температуры, но после повышения температуры до 37"С
снова происходит самосборка микротрубочек. Деполимеризация
тубулинов и исчезновение микротрубочек происходит и при дейст-
вии на живую клетку колхицина или охлаждения.
Полагают, что в клетке тубулины существуют в двух формах —
свободной и связанной. Сдвиг равновесия между этими формами
может привести или к диссоциации микротрубочек, или к их рос-
ту. Ни тубулины в чистом виде, ни построенные из них микро-
трубочки не способны к сокращению, они не обладают АТФ-азной
активностью. Скорее всего они выполняют роль каркасных струк-
тур. В клетках микротрубочки принимают участие в создании ря-
да временных (цитоскелет интерфазных клеток, веретено деле-
ния) или постоянных структур (центриоли, реснички, жгутики).
Микротрубочки интерфазных клеток
Практически во всех эукариотических клетках в гиалоплазме
можно видеть длинные неветвящиеся микротрубочки. В больших
количествах они обнаруживаются в цитоплазматических отростках
нервных клеток, фибробластов и других изменяющих свою форму
59
A
Б
Рис. 13. Микрофиламенты.
А — схема; Б — микрофотографии (иммунофлюоресцентный анализ); / — микротрубочки в
культуре клеток фибробластов мыши (тубулин); // — актиновые микрофиламенты в
культуре клеток; Ш - промежуточные микрофиламенты а культуре клеток эмбриональ-
ной почки свиньи (аиментин).
клеток. Они могут быть выделены сами или можно выделить об-
разующие их белки: это те же тубулины со всеми их свойствами
(рис. 13, А, Б).
Главное функциональное значение таких микротрубочек цито-
плазмы заключается в создании эластичного, но одновременно
устойчивого внутриклеточного каркаса (цитоскелета), необходимо-
го для поддержания формы клетки.
Действие колхицина, вызывающего деполимеризацию тубули-
нов, сильно меняет форму клеток. Так, если отростчатую и плос-
кую клетку в культуре фибробластов обработать колхицином, то
она теряет полярность и сжимается. Точно так же ведут себя
другие клетки: колхицин прекращает рост клеток хрусталика, от-
ростков нервных клеток, образование мышечных трубок и др.
Так как при этом не исчезают элементарные формы движения,
присущего клеткам, в частности пиноцитоз, ундулирующие дви-
жения мембран, образование мелких псевдоподий, вероятнее все-
го, роль микротрубочек заключается в образовании каркаса для
поддержания формы клеточного тела, для стабилизации и укрепле-
ния клеточных выростов.
Создавая внутриклеточный скелет, микротрубочки могут быть
факторами ориентированного движения клетки в целом и ее внут-
риклеточных компонентов, задавать своим расположением векторы
для направленных потоков разных веществ и для перемещения
крупных структур. Разрушение микротрубочек колхицином нару-
шает транспорт веществ в аксонах нервных клеток, приводит к
блокаде секреции и т. д. С цитоплазматическими микротрубочка-
ми связаны специальные белки, участвующие в механическом
переносе отдельных внутриклеточных компонентов: микровакуо-
лей, рибосом, митохондрий и др.
В неделящейся (интерфазной) клетке система микротрубочек
развивается в связи с особой клеточной органеллой — центри-
олью, которая является местом, где происходит начальная поли-
меризация тубулинов и рост микротрубочек цитоскелета.
Центриоли
Этот термин был предложен Т. Бовери в 1895 г. для обозна-
чения очень мелких телец, размер которых находится на границе
разрешающей способности светового микроскопа. В некоторых
объектах удавалось видеть, что мелкие плотные тельца — цент-
риоли (centriolum), обычно расположенные в паре — диплосома
(diplosoma), окружены зоной более светлой цитоплазмы, от ко-
торой отходят радиально тонкие фибриллы (центросфера). Со-
вокупность центриолей и центросферы называют клеточным цент-
ром (cytocentrum). Эти органеллы в делящихся клетках прини-
мают участие в формировании веретена деления и располагаются
на его полюсах. В неделящихся клетках центриоли часто опре-
деляют полярность клеток эпителия и располагаются вблизи комп-
лекса Гольджи.
61
I
Рис. 14. Строение клеточного
центра в полюсе митотического
веретена клетки.
А — схема; Б - электронная микро
фотография; / — активная материн-
ская центриоль, окруженная тонко-
фибриллярным матриксом, от кото-
рого отходят микротрубочки по-
лярной лучистости (2); 3 — неактив-
ная дочерняя центриоль.
Тонкое строение центриолей удалось изучить только с по-
мощью электронного микроскопа. Основой строения центриолей
являются расположенные по окружности 9 триплетов микротрубо-
чек (triplomicrotubuli), образующих таким образом полый цилиндр.
Его ширина около 0,2 мкм, а длина — 0,3- 0,5 мкм (хотя встре-
чаются центриоли, достигающие в длину нескольких микрометров)
(рис. 14).
Кроме микротрубочек в состав центриоли входят дополнитель-
ные структуры — «ручки», соединяющие триплеты. Системы мик-
ротрубочек центриоли можно описать формулой: (9x3) 4-0, под-
черкивая отсутствие микротрубочек в ее центральной части.
Обычно в интерфазных клетках всегда присутствуют две цент-
риоли, располагающиеся рядом друг с другом, образуя диплосо-
му. В диплосоме центриоли располагаются под прямым углом по
отношению друг к другу. Из двух центриолей различают мате-
ринскую и дочернюю. Обе центриоли сближены и расположены
так, что конец дочерней центриоли направлен к поверхности ма-
теринской центриоли.
Вокруг каждой центриоли расположен бесструктурный, или
тонковолокнистый, матрикс. Часто можно обнаружить несколько
дополнительных структур, связанных с центриолями: спутники
(сателлиты), фокусы схождения микротрубочек, дополнительные
микротрубочки, образующие особую зону, центросферу вокруг
центриоли.
При подготовке клеток к митотическому делению происходит
удвоение центриолей. Этот процесс у различных объектов про-
исходит в разное время — в течение синтеза ядерной ДНК или
после него. Он заключается в том, что две центриоли в диплосоме
62
расходятся и около каждой из них возникает заново по одной но-
вой дочерней, так что в клетке перед делением обнаруживаются
две диплосомы, т. е. четыре попарно связанные центриоли. Этот
способ увеличения числа центриолей был назван дупликацией.
Важно отметить, что увеличение числа центриолей не связано с
их делением, почкованием или фрагментацией, а происходит пу-
тем образования зачатка, процентриоли, вблизи и перпендикулярно
к исходной центриоли.
Полагают, что центриоли участвуют в индукции полимериза-
ции тубулином при образовании микротрубочек. Так, в интерфа-
зе именно в связи с центриолью происходит рост микротрубочек
клеточного каркаса. Перед митозом центриоль является одним из
центров полимеризации микротрубочек веретена клеточного деле-
ния. Центриоль — центр роста микротрубочек аксонемы ресничек
или жгутиков. Наконец, она сама индуцирует полимеризацию ту-
булинов новой процентриоли, возникающей при ее дупликации.
Реснички и жгутики
Это специальные органеллы движения, встречающиеся в не-
которых клетках различных организмов. В световом микроскопе
эти структуры выглядят как тонкие выросты клетки. В основании
ресничек fcilium) и жгутика f flagellum) в цитоплазме видны хо-
рошо красящиеся мелкие гранулы — базальные тельца (corpusculum
basale). Длина ресничек 5—10 мкм, а длина жгутиков может до-
стигать 150 мкм (см. рис. 43, В).
Ресничка представляет собой тонкий цилиндрический вырост
цитоплазмы с постоянным диаметром 200 нм. Этот вырост от
основания до самой его верхушки покрыт плазматической мембра-
ной. Внутри выроста расположена аксонема («осевая нить») —
сложная структура, состоящая в основном из микротрубочек.
Проксимальная часть реснички (базальное тело) погружена в
цитоплазму. Диаметры аксонемы и базального тельца одинаковы
(около 150 нм).
Базальное тельце по своей структуре очень сходно с центри-
олью. Оно также состоит из 9 триплетов микротрубочек, имеет
«ручки». Часто в основании реснички лежит пара базальных те-
лец, располагающихся под прямым углом друг к другу подобно
диплосоме — центриоли.
Аксонема (filamentum axiale) в своем составе имеет в отличие
от базального тельца или центриоли 9 дублетов микротрубочек с
«ручками», образующих стенку цилиндра аксонемы. Кроме пе-
риферических дублетов микротрубочек, в центре аксонемы распо-
лагается пара центральных микротрубочек. В целом систему мик-
ротрубочек реснички описывают как (9x2)+ 2 в отличие от
(9x3) +0 системы центриолей и базальных телец. Базальное
тельце и аксонема структурно связаны друг с другом и составляют
единое целое: две микротрубочки триплетов базального тельца
являются микротрубочками дублетов аксонемы.
63
I
Свободные клетки, имеющие реснички и жгутики, обладают
способностью двигаться, а неподвижные клетки движением рес-
ничек могут перемещать жидкость и корпускулярные частицы.
При движении ресничек и жгутиков длина их не уменьшается,
поэтому неправильно называть это движение сокращением. Тра-
ектория движения ресничек очень разнообразна. В различных
клетках это движение может быть маятникообразным, крючкооб-
разным, воронкообразным или волнообразным.
Основной белок ресничек — тубулин — не способен к сокраще-
нию, укорочению. Вероятным кандидатом на роль сократимого
белка считается белок «ручек» — динеин, так как он обладает
АТФ-азной активностью. В последние годы для объяснения спо-
соба движения ресничек и жгутиков используется гипотеза «сколь-
зящих нитей». Известно, что сокращение мышечных волокон про-
исходит за счет встречного скольжения фибрилл двух мышечных
белков: миозина и актина; при этом также не происходит собст-
венно укорачивания или сокращения отдельных мышечных бел-
ковых фибрилл. Предполагается, что незначительные смещения
дублетов микротрубочек друг относительно друга могут вызвать
изгиб всей реснички, а если такое локальное смещение будет про-
исходить вдоль жгутика, то может возникнуть волнообразное его
движение.
Другие фибриллярные структуры цитоплазмы
Кроме микротрубочек, к фибриллярным компонентам цито-
плазмы эукариотических клеток относятся микрофиламенты (mic-
rofilamenti) толщиной 5—7 нм и так называемые промежуточные
филаменты, или микрофибриллы (microfibrillae), толщиной около
10 нм (см. рис. 13, III).
Микрофиламенты встречаются практически во всех типах кле-
ток. По строению и функциям они бывают разные, однако от-
личить их морфологически друг от друга трудно. Располагаются
микрофиламенты в кортикальном слое цитоплазмы, непосредствен-
но под плазмолеммой, пучками или слоями. Их можно видеть в
псевдоподиях амеб или в движущихся отростках фибробластов,
в микро ворсинках кишечного эпителия. Микрофиламенты часто
образуют пучки, направляющиеся в клеточные отростки.
Сеть микрофиламентов выявлена в большинстве клеток. Они
отличаются по химическому составу. В зависимости от их хими-
ческого состава они могут выполнять функции цитоскелета и
участвовать в обеспечении движения. Эта сеть — часть цитоске-
лета. С помощью иммунофлюоресцентных методов четко пока-
зано, что в состав микрофиламентов кортикального слоя и пучков
входят сократительные белки: актин, миозин, тропомиозин, а-ак-
тинин. Следовательно, микрофиламенты не что иное, как внутри-
клеточный сократительный аппарат, обеспечивающий не только
подвижность клеток при активном амебоидном их перемещении,
но, вероятно, и большинство внутриклеточных движений, таких
64
как токи цитоплазмы, движение вакуолей, митохондрий, деление
клетки.
Промежуточные филаменты, или микрофибриллы, тоже бел-
ковые структуры. Это тонкие (10 нм) неветвящиеся, часто распо-
лагающиеся пучками нити. Характерно, что их белковый состав
различен в разных тканях. В эпителии в состав промежуточных
филаментов входит кератин. Пучки кератиновых промежуточ-
ных филаментов в эпителиальных клетках образуют так называе-
мые тонофибриллы, которые подходят к десмосомам. В состав
промежуточных филаментов клеток мезенхимальных тканей (на-
пример, фибробластов) входит другой белок — виментин,
в мышечные клетки — десмин, в нервных клетках в состав их
нейрофиламентов также входит особый белок.
Роль промежуточных микрофиламентов скорее всего опорно-
каркасная, однако эти фибриллярные структуры не так лабильны,
как микротрубочки.
В последнее время с помощью иммуноморфологических ме-
тодов стало возможным определить тканевое происхождение тех
или иных опухолей именно по белкам их промежуточных фила-
ментов, что очень важно для правильного выбора типа химиоте-
рапевтических противоопухолевых препаратов.
Включения
Включения цитоплазмы (inclusion's c>toplasmicae)—необя-
зательные компоненты клетки, возникающие и исчезающие в
зависимости от метаболического состояния клеток.
Различают включения трофические, секреторные,
экскреторные и пигментные. К трофическим включе-
ниям относятся капельки нейтральных жиров, которые могут на-
капливаться в гиалоплазме. В случае недостатка субстратов для
жизнедеятельности клетки эти капельки могут резорбироваться.
Другим видом включений резервного характера является глико-
ген — полисахарид, откладывающийся также в гиалоплазме
(рис. 15). Отложение запасных белковых гранул обычно происхо-
дит в связи с активностью эндоплазматической сети. Так, запасы
белка вителлина в яйцеклетках амфибии накапливаются в вакуолях
эндоплазматической сети.
Секреторные включения — обычно округлые образования раз-
личных размеров, содержащие биологически активные вещества,
образующиеся в клетках в процессе жизнедеятельности (см.
гл. VI, XVIII).
Экскреторные включения не содержат каких-либо ферментов
или других активных веществ. Обычно это продукты метаболизма,
подлежащие удалению из клетки.
Пигментные включения могут быть экзогенные (каротин, пы-
левые частицы, красители и др.) и эндогенные (гемоглобин, гемо-
сидерин, билирубин, меланин, липофусцин). Наличие их в цито-
65
3 гистология
/
Рис. 15. Включения глико-
гена в клетках печени.
Окраска по Бесту.
/ — ядро; 2 — цитоплазма;
3 — гликоген.
плазме может изменять цвет ткани, органа временно или постоян-
но. Нередко пигментация ткани служит диагностическим призна-
ком.
ЯДРО
Ядро (nucleus) клетки — система генетической детерминации
и регуляции белкового синтеза.
Роль ядерных структур в жизнедеятельности клеток
Ядро обеспечивает две группы общих функций: одну, связанную
собственно с хранением и передачей генетической информации,
другую — с ее реализацией, с обеспечением синтеза белка.
Хранение и поддержание наследственной
информации в виде неизменной структуры ДНК связаны с
наличием так называемых репарационных ферментов, ликвидирую-
щих спонтанные повреждения молекул ДНК. В ядре происходит
воспроизведение или редупликация молекул ДНК, что дает возмож-
ность при митозе двум дочерним клеткам получить совершенно оди-
наковые в качественном и количественном отношении объемы гене-
тической информации.
Другой группой клеточных процессов, обеспечиваемых актив-
ностью ядра, является создание собственно аппарата белкового син-
теза (рис. 16). Это не только синтез, транскрипция на
молекулах ДНК разных информационных РНК, но и транскрип-
ция всех видов транспортных и рибосомных РНК. В ядре проис-
ходит также образование субъединиц рибосом путем комплексиро-
вания синтезированных в ядрышке рибосомных РНК с рибосомны-
ми белками, которые синтезируются в цитоплазме и переносятся t
ядро.
Таким образом, ядро является не только вместилищем генетиче-
ского материала, но и местом, где этот материал функционирует и
воспроизводится. Вот почему выпадание или нарушение любой из
66
о
Рис. 16. Белковый синтез в клетке (схема).
перечисленных выше функций гибельно для клетки в целом. Все это
указывает на ведущее значение ядерных структур в процессах син-
теза нуклеиновых кислот и белков.
Структура и химический состав клеточного ядра
Ядро неделящейся, интерфазной клетки обычно одно на клетку
(хотя встречаются и многоядерные клетки). Ядро состоит из хрома-
тина, ядрышка, кариоплазмы (нуклеоплазмы) и ядерной оболочки,
отделяющей его от цитоплазмы (рис. 17).
Хроматин
При наблюдении живых или фиксированных клеток внутри ядра
выявляются зоны плотного вещества, которые хорошо воспринима-
ют разные красители, особенно основные. Благодаря такой способ-
ности хорошо окрашиваться этот компонент ядра и получил назва-
ние «хроматин» (от греч. chroma — цвет, краска). В состав хрома-
тина входит ДНК в комплексе с белком. Такими же свойствами
обладают и хромосомы, которые отчетливо видны во время митоти-
ческого деления клеток. В неделящихся (интерфазных) клетках
хроматин, выявляемый в световом микроскопе, может более или
«7
3*
Рис. 17. Ультра микроскоп и чес кое строение ядра интерфазной клетки.
А - схема; Б — электронная микрофотография участка ядра; / — ядерная оболочка (две
мембраны, перинуклеарное пространство); 2- комплекс поры; 3 конденсированный
хроматин; 4 диффузный хроматин; 5 — ядрышко (гранулярная и фибриллярная части);
6 — межхроматиновые гранулы РНК; 7 — перихроматиновые гранулы; 8 — кариоплазма.
менее равномерно заполнять объем ядра или же располагаться
отдельными глыбками.
Хроматин интерфазных ядер представляет собой хромосомы,
которые, однако, теряют в это время свою компактную форму,
разрыхляются, деконденсируются. Степень такой деконденсации
хромосом может быть различной. Зоны полной деконденсации и
их участков морфологи называют эухроматином (euchromatinum).
При неполном разрыхлении хромосом в интерфазном ядре видны
участки конденсированного хроматина, иногда называемого гетеро-
хроматином (heterochromatinum). Степень деконденсации хромо-
сомного материала — хроматина в интерфазе может отражать
функциональную нагрузку этой структуры. Чем «диффузнее» рас-
пределен хроматин в интерфазном ядре, тем интенсивнее в нем син-
тетические процессы.
Максимально конденсирован хроматин во время митотического
деления клеток, когда он обнаруживается в виде плотных хромосом.
В этот период хромосомы не выполняют никаких синтетических
функций, в них не происходит включения предшественников ДНК
и РНК
68
Таким образом, хромосомы клеток могут находиться в двух струк-
турно-функциональных состояниях: в активном, рабочем, частично
или полностью деконденсированном, когда с их участием в интер-
фазном ядре происходят процессы транскрипции и редупликации, и
в неактивном, в состоянии метаболического покоя при максималь-
ной их конденсированности, когда они выполняют функцию распре-
деления и переноса генетического материала в дочерние клетки.
Наблюдения за структурой хроматина с помощью электронного
микроскопа показали, что как в препаратах выделенного интерфаз-
ного хроматина или выделенных митотических хромосом, так и в
составе ядра на ультратонких срезах всегда видны элементарные
хромосомные фибриллы толщиной 20—25 нм.
В химическом отношении фибриллы хроматина представляют
собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в
состав которых входят ДНК и специальные хромосомные белки —
гистоновые и негистоновые. В составе хроматина обнаруживается
также РНК. Количественные отношения ДНК, белка и РНК состав-
ляют 1:1,3:0,2. Обнаружено, что длина индивидуальных линейных
молекул ДНК может достигнуть сотен микрометров и даже санти-
метров. Среди хромосом человека самая большая первая хромосома
содержит ДНК с общей длиной до 7 см. Суммарная длина молекул
ДНК во всех хромосомах одной клетки человека составляет около
170 см, что соответствует 6 • 10—12г.
В хромосомах существует множество мест независимой реплика-
ции ДНК — репликонов. ДНК эукариотических хромосом представ-
ляют собой линейные молекулы, состоящие из тандемно (друг за
другом) расположенных репликонов разного размера. Средний раз-
мер репликона около 30 мкм. В составе генома человека должно
встречаться более 50 000 репликонов, участков ДНК, которые син-
тезируются как независимые единицы. Синтез ДНК как на участках
отдельной хромосомы, так и среди разных хромосом идет неодно-
временно, асинхронно. Так, например, в некоторых хромосомах че-
ловека (1, 3, 16) репликация наиболее интенсивно начинается на
концах хромосом и заканчивается (при высокой интенсивности
включения метки) в центромерном районе (см. ниже). Наиболее
поздно репликация заканчивается в хромосомах или в их участках,
находящихся в компактном, конденсированном состоянии. Таким
примером может являться поздняя репликация генетически инакти-
вированной Х-хромосомы у женщин, формирующей в клеточном
ядре компактное тельце полового хроматина.
Белки хроматина составляют 60—70% от его сухой массы. К ним
относятся так называемые гистоны и негистоновые белки. Негисто-
новые белки составляют 20% от количества гистонов. Гистоны —
щелочные белки, обогащенные основными аминокислотами (глав-
ным образом лизином и аргинином). Очевидна структурная роль
гистонов, которые не только обеспечивают специфическую укладку
хромосомной ДНК, но и имеют значение в регуляции транскрипции.
Гистоны расположены по длине молекулы ДНК не равномерно, а в
виде блоков. В один такой блок входят 8 молекул гистонов, обра-
69
зуя так называемую нуклеосому. Размер нуклеосомы около 10 нм.
При образовании нуклеосом происходит компактизация, сверхспи-
рализация ДНК, что приводит к укорачиванию длины хромосомной
фибриллы примерно в 5 раз. Сама же хромосомная фибрилла имеет
вид нитки бус или четок, где каждая бусина — нуклеосома (см.
рис. 17). Такие фибриллы толщиной 10 нм дополнительно продоль-
но конденсируются и образуют основную элементарную фибриллу
хроматина толщиной 25 нм.
Негистоновые белки интерфазных ядер образуют внутри ядра
структурную сеть, которая носит название ядерный белковый мат-
рикс, представляющий собой основу, определяющую морфологию и
метаболизм ядра.
В ядрах, кроме хроматиновых участков и матрикса, встречаются
перихроматиновые фибриллы, перихроматиновые и интерхромати-
новые гранулы. Они содержат РНК и встречаются практически во
всех активных ядрах, представляют собой информационные РНК,
связанные с белками, — рибонуклеопротеиды (информосомы).
Матрицами для синтеза этих РНК являются разные гены, разбро-
санные по деконденсированным участкам хромосомных (хромати-
новых) фибрилл.
Особый тип матричной ДНК, а именно ДНК для синтеза рибо-
сомной РНК, собран обычно в нескольких компактных участках,
входящих в состав ядрышек интерфазных ядер.
Ядрышко
Практически во всех живых клетках эукариотических организ-
мов в ядре видно одно или несколько обычно округлой формы телец
величиной 1—5 мкм, сильно преломляющих свет — это ядрышко,
или нуклеола (nucleolus). К общим свойствам ядрышка относится
способность хорошо окрашиваться различными красителями, осо-
бенно основными. Такая базофилия определяется тем, что ядрышки
богаты РНК. Ядрышко — самая плотная структура ядра — являет-
ся производным хромосомы, одним из ее локусов с наиболее высо-
кой концентрацией и активностью синтеза РНК в интерфазе. Оно не
является самостоятельной структурой или органеллой.
В настоящее время известно, что ядрышко — это место образо-
вания рибосомных РНК (рРНК) и рибосом, на которых происходит
синтез полипептидных цепей в цитоплазме.
Образование ядрышек и их число связаны с активностью и чис-
лом определенных участков хромосом — ядрышковых организато-
ров, которые расположены большей частью в зонах ' вторичных
перетяжек; количество ядрышек в клетках данного типа может из-
меняться за счет слияния ядрышек или за счет изменения числа хро-
мосом с ядрышковыми организаторами. При исследовании фиксиро-
ванных клеток вокруг ядрышка всегда выявляется зона конденсиро-
ванного хроматина, часто отождествляемая с хроматином ядрышко-
вого организатора. Этот околоядрышковый хроматин, по данным
электронной микроскопии, представляет собой интегральную часть
70
сложной структуры ядрышка. ДНК ядрышкового организатора
представлена множественными (несколько сотен) копиями генов
рРНК: на каждом из этих генов синтезируется высокомолекулярный
предшественник РНК, который превращается в более короткие мо-
лекулы РНК, входящие в состав субъединиц рибосомы.
Схему участия ядрышек в синтезе цитоплазматических белков
можно представить следующим образом: на ДНК ядрышкового ор-
ганизатора образуется предшественник рРНК, который в зоне яд-
рышка одевается белком, здесь происходит сборка рибонуклеопро-
теидных частиц — субъединиц рибосом; субъединицы, выходя из
ядрышка в цитоплазму, участвуют в процессе синтеза белка.
Ядрышко неоднородно по своему строению: в световом микро-
скопе можно видеть его тонковолокнистую организацию. В элект-
ронном микроскопе выявляются два основных компонента: грану-
лярный и фибриллярный. Диаметр гранул около 15—20 нм, толщина
фибрилл — 6—8 нм.
Фибриллярный компонент может быть сосредоточен в виде цен-
тральной части ядрышка, а гранулярный — по периферии. Часто
гранулярный компонент образует нитчатые структуры — нуклеоло-
немы толщиной около 0,2 мкм. Фибриллярный компонент ядрышек
представляет собой рибонуклеопротеидные тяжи предшественни-
ков рибосом, а гранулы — созревающие субъединицы рибосом. В зо-
не фибрилл можно выявить участки ДНК ядрышковых организато-
ров.
Ультраструктура ядрышек зависит от активности синтеза РНК:
при высоком уровне синтеза рРНК в ядрышке выявляется большое
число гранул, при прекращении синтеза количество гранул снижает-
ся, ядрышки превращаются в плотные фибриллярные тельца базо-
фильной природы.
Действие многих веществ (актиномицин, митомицин, ряд канце-
рогенных углеводородов, циклогексимид, гидрооксимочевина и др.)
вызывает в клетках падение интенсивности ряда синтезов и в первую
очередь активности ядрышек. При этом возникают изменения в
структуре ядрышек: их сжатие, обособление фибриллярных и грану-
лярных зон, потеря гранулярного компонента, распад всей структу-
ры. Эти изменения отражают степень повреждения ядрышковых
структур, связанных главным образом с подавлением синтеза рРНК.
Ядерная оболочка
Ядерная оболочка (nucleolemma) состоит из внешней ядерной
мембраны (m. nuclearis externa) и внутренней мембраны оболочки
(т. nuclearis interna), разделенных перинуклеарным пространством,
или цистерной ядерной оболочки (cisterna nucleolemmae). Ядерная
оболочка содержит ядерные поры (pori nucleares).
Мембраны ядерной оболочки в морфологическом отношении не
отличаются от остальных внутриклеточных мембран. В общем виде
ядерная оболочка может быть представлена как полый двухслойный
мешок, отделяющий содержимое ядра от цитоплазмы.
71
Рис. 18. Строение ком-
плекса поры (схема).
/ — перинуклеарное про-
странство; 2 — внутренняя
я дерна я мембрана; 3 — на-
ружная ядерная мембрана;
4 — периферические грану-
лы; 5 — центральная грану-
ла; 6 — фибриллы, отходя
щие от гранул; 7 диафрагма
поры; 8 — фибриллы хрома
тина.
Внешняяя мембрана ядерной оболочки, непосредственно контак-
тирующая с цитоплазмой клетки, имеет ряд структурных особеннос-
тей, позволяющих отнести ее к собственно мембранной системе
эндоплазматической сети: на ней со стороны гиалоплазмы располо-
жены многочисленные полирибосомы, а сама внешняя ядерная мем-
брана может прямо переходить в мембраны эндоплазматической се-
ти. Внутренняя мембрана связана с хромосомным материалом ядра.
Наиболее характерными структурами ядерной оболочки являются
ядерные поры. Они образуются за счет слияния двух ядерных мем-
бран. Формирующиеся при этом округлые сквозные отверстия -поры
(annulus pori) имеют диаметр около 80—90 нм. Эти отверстия в
ядерной оболочке заполнены сложноорганизованными глобулярны-
ми и фибриллярными структурами. Совокупность мембранных пер-
фораций и этих структур называют комплексом поры (complexus
pori) (рис. 18). Такой сложный комплекс поры имеет октагональ-
ную симметрию. По границе округлого отверстия в ядерной оболоч-
ке располагается три ряда гранул по 8 в каждом: один ряд лежит со
стороны ядра, другой — со стороны цитоплазмы, третий расположен
в центральной части поры. Размер гранул около 25 нм. От этих гра-
нул отходят фибриллярные отростки. Фибриллы, отходящие от пе-
риферических гранул, могут сходиться в центре и создавать как бы
перегородку, диафрагму поперек поры (diaphragma pori).
Размеры пор у данной клетки обычно стабильны, так же как от-
носительно стабилен размер ядерных пор клеток разных организ-
мов. Число ядерных пор зависит от метаболической активности
клеток: чем интенсивнее синтетические процессы в клетках, тем
больше пор на единицу поверхности клеточного ядра. Так, у эрит-
робластов (клеток-предшественников ядерных эритроцитов) низ-
ших позвоночных животных во время интенсивного синтеза и на-
копления гемоглобина обнаруживается в ядре около 30 ядерных
пор на 1 мкм2. После того как эти процессы заканчиваются, в яд-
рах зрелых клеток — эритроцитов прекращается синтез ДНК и
72
РНК и количество пор снижается до 5 на 1 мкм2. В ядерных обо-
лочках полностью зрелых сперматозоидов поры не обнаружива-
ются
Из многочисленных свойств и функциональных нагрузок ядер-
ной оболочки следует подчеркнуть ее роль как барьера, отделя-
ющего содержимое ядра от цитоплазмы, ограничивающего свобод-
ный доступ в ядро крупных агрегатов биополимеров, регулирующе-
го транспорт макромолекул между ядром и цитоплазмой. Одной из
важных функций ядерной оболочки следует считать ее участие в
создании внутриядерного порядка — в фиксации хромосомного
материала в трехмерном пространстве ядра. В интерфазе часть
хроматина структурно связана с внутренней ядерной мембраной.
Описаны случаи примембранной локализации центромерных и те-
ломерных участков интерфазных хромосом.
ВОСПРОИЗВЕДЕНИЕ КЛЕТОК
Клеточный цикл
Один из постулатов клеточной теории гласит, что увеличение
числа клеток, их размножение происходят путем деления исходной
клетки. Обычно делению клеток предшествует редупликация их
хромосомного аппарата, синтез ДНК. Это правило является об-
щим для прокариотических и эукариотических клеток. Время су-
ществования клетки как таковой, от деления до деления или От
деления до смерти, обычно называют клеточным циклом
(cyclus cellularis).
Во взрослом организме высших позвоночных клетки различных
тканей и органов имеют неодинаковую способность к делению.
Встречаются популяции клеток, полностью потерявшие свойство
делиться. Это большей частью специализированные, дифференци-
рованные клетки (например, зернистые лейкоциты крови). В орга-
низме есть постоянно обновляющиеся ткани — различные эпите-
лии, кроветворные ткани. В таких тканях существует часть клеток,
которые постоянно делятся, заменяя отработавшие или погибаю-
щие клеточные типы (например, клетки базального слоя покровно-
го эпителия, клетки крипт кишечника, кроветворные клетки кост-
ного мозга). Многие клетки, не размножающиеся в обычных усло-
виях, приобретают вновь это свойство при процессах репаративной
регенерации органов и тканей. Размножающиеся клетки обладают
разным количеством ДНК в зависимости от стадии клеточного
цикла. Это наблюдается при размножении как соматических, так
и половых клеток.
Как известно, половые мужские и женские клетки несут еди-
ничный (гаплоидный) набор хромосом и, следовательно, содержат
в 2 раза меньше ДНК, чем все остальные клетки организма. Такие
половые клетки (сперматозоиды и ооциты) с единичным набором
хромосом называют гаплоидными. Плоидность обозначают буквой
и. Так, клетки с 1 и гаплоидны, с 2 и диплоидны, с 3 п триплоидны
73
и т. д. Соответственно количество ДНК на клетку (с) зависит от
ее плоидности: клетки с 2 п числом хромосом содержат 2 с коли-
чества ДНК. При оплодотворении происходит слияние двух кле-
ток, каждая из которых несет 1 п набор хромосом, поэтому обра-
зуется исходная диплоидная (2 п, 2 с) клетка-зигота. В дальней-
шем в результате деления диплоидной зиготы и последующего де-
ления диплоидных клеток разовьется организм, клетки которого
(кроме зрелых половых) будут диплоидными.
При изучении клеточного цикла диплоидных клеток в их попу-
ляции встречаются как диплоидные (2 с), так и тетраплоидные
(4 с) и интерфазные клетки с промежуточным количеством ДНК.
Такая гетерогенность определяется тем, что удвоение ДНК проис-
ходит в строго определенный период интерфазы (periodus interrru-
toticus), а собственно к делению клетки приступают только после
этого процесса.
Весь клеточный цикл состоит из 4 отрезков времени: собствен-
но митоза (М), пресинтетического (G,), синтетического (S) и
постсинтетического (G2) периодов интерфазы (рис. 19). В G,-ne-
риоде, наступающем сразу после деления, клетки имеют диплоид-
ное содержание ДНК на одно ядро (2 с). После деления в период
G, в дочерних клетках общее содержание белков и РНК вдвое
меньше, чем в исходной родительской клетке. В период G, начи-
нается рост клеток главным образом за счет накопления клеточ-
ных белков, что определяется увеличением количества РНК на
клетку. В этот период начинается подготовка клетки к синтезу
ДНК (S-период).
Обнаружено, что подавление синтеза белка или иРНК в G,-ne-
риоде предотвращает наступление S-периода, так как в течение
G,-периода происходят синтезы ферментов, необходимых для
образования предшественников ДНК (например, нуклеотид-
фосфокиназ), ферментов метаболизма РНК и белка. Это совпа-
дает с увеличением синтеза РНК и белка. При этом резко повы
шается активность ферментов, участвующих в энергетическом
обмене.
В следующем, S-периоде происходит удвоение количества ДНК
на ядро и соответственно удваивается число хромосом. В разных
клетках, находящихся в S-периоде, можно обнаружить разные ко-
личества ДНК — от 2 до 4 с. Это связано с тем, что исследованию
подвергаются клетки на разных этапах синтеза ДНК (только при-
ступившие к синтезу и уже завершившие его). S-период является
узловым в клеточном цикле. Без прохождения синтеза ДНК неиз-
вестно ни одного случая вступления клеток в митотическое де-
ление.
Единственным исключением является второе деление созрева-
ния половых клеток в мейозе, когда между двумя делениями нет
синтеза ДНК.
В S-периоде уровень синтеза РНК возрастает соответственно
увеличению количества ДНК, достигая своего максимума в G2-ne-
риоде
74
Рис. 19. Клеточный цикл
(схема) Объяснение в те-
ксте.
Постсинтетическая (G2) фаза <^ще называется премитотической.
Последним термином подчеркивается ее большое значение для
прохождения следующей стадии — стадии митотического деления.
В данной фазе происходит синтез иРНК, необходимый для про-
хождения митоза. Несколько ранее этого синтезируется рРНК
рибосом, определяющих деление клетки. Среди синтезирующихся
в это время белков особое место занимают тубулины — белки
митотического веретена.
В конце С2-периода или в митозе по мере конденсации мито-
тических хромосом синтез РНК резко падает и полностью прекра-
щается во время митоза. Синтез белка во время митоза понижает-
ся до 25% от исходного уровня и затем в последующих периодах
достигает своего максимума в С2-периоде, в общем повторяя харак-
тер синтеза РНК.
В растущих тканях растений и животных всегда есть клетки,
которые находятся как бы вне цикла. Такие клетки принято назы-
вать клетками Сс-периода. Именно эти клетки представляют собой
так называемые покоящиеся, временно или окончательно перестав-
шие размножаться клетки. В некоторых тканях такие клетки мо-
гут находиться длительное время, не изменяя особенно своих мор-
фологических свойств: они сохраняют в принципе способность к
делению, превращаясь в камбиальные, стволовые клетки (напри-
мер, в кроветворной ткани). Чаще потеря (хотя бы и временная)
способности делиться сопровождается появлением способности к
специализации, к дифференцировке. Такие дифференцирующиеся
клетки выходят из цикла, но в особых условиях могут снова вхо-
дить цикл. Например, большинство клеток печени находится в Go-
периоде; они не участвуют в синтезе ДНК и не делятся. Однако
при удалении части печени у экспериментальных животных, мно-
гие клетки начинают подготовку к митозу (Gf-период), переходят
к синтезу ДНК и могут митотически делиться. В других случаях,
например в эпидермисе кожи, после выхода из цикла размно-
жения и дифференцировки клетки некоторое время функцио-
нируют, а затем погибают (ороговевшие клетки покровного эпи-
телия).
75
Деление клеток
Митоз
Митиз (mitosis), кариокинез, или непрямое деление, — универ-
сальный, широко распространенный способ деления клеток. При
этом конденсированные и уже редуплицированные хромосомы
переходят в компактную форму митотических хромосом, образует-
ся веретено деления, участвующее в сегрегации и переносе хромо-
сом (ахроматиновый митотический аппарат), происходит расхож-
дение хромосом к противоположным полюсам клетки и деление
тела клетки (цитокинез, цитотомия).
Процесс непрямого деления клеток принято подразделять на
несколько основных фаз профаза, метафаза, анафаза, телофаза
(рис. 20)
Профаза. После окончания S-периода количество ДНК в
интерфазном ядре равно 4 с, так как произошло удвоение хромо-
сомного материала. Однако морфологически регистрировать удвое-
ние числа хромосом в этой стадии не всегда удается. Собственно
хромосомы как нитевидные плотные тела начинают обнаруживать-
ся микроскопически в начале процесса деления клетки, а именно
в профазе митотического деления клетки. Если попытаться под-
считать число хромосом в профазе, то их количество будет равно
2 п. Но это ложное впечатление, потому что в профазе каждая из
хромосом двойная, что является результатом их редупликации в
интерфазе. В профазе эти сестринские хромосомы тесно соприка-
саются друг с другом, взаимно спирализуясь одна относительно
другой, поэтому трудно увидеть двойственность всей структуры в
целом. Позднее хромосомы в каждой такой паре начинают обособ-
ляться, раскручиваться. Двойственность хромосом в митозе наблю-
дается у живых клеток в конце профазы, когда видно, что общее
их число в начинающей делиться клетке равно 4 п. Следовательно,
уже в начале профазы хромосомы состояли из двух сестринских
хромосом, или, как их еще называют, хроматид. Число их (4 п) в
профазе точно соответствует количеству ДНК (4с).
Параллельно конденсации хромосом в профазе происходят ис-
чезновение, дезинтеграция ядрышек в результате инактивации ри-
босомных генов в зоне ядрышковых организаторов.
Одновременно с этим в середине профазы начинается разру-
шение ядерной оболочки: исчезают ядерные поры, оболочка распа-
дается сначала на фрагменты, а затем на мелкие мембранные
пузырьки. Меняются в это время и структуры, связанные с синте-
зом белка. Происходит уменьшение количества гранулярного эндо-
плазматического ретикулума, он распадается на короткие цистер-
ны и вакуоли, количество рибосом на его мембранах резко падает.
Значительно (до 25%) редуцируется число полисом как на мем-
бранах, так и в гиалоплазме, что является признаком общего паде-
ния уровня синтеза белка в делящихся клетках.
Второе важнейшее событие при митозе тоже происходит во
время профазы — это образование веретена деления. В профазе
76
Рис. 20. Митоз клетки (схема).
/ интерфа ia; 2 — профаза: 3 — метафаза: 4 — анафаза: 5 — телофаза: 6 — ранняя
интерфаза.
уже репродуцировавшиеся в S-периоде центриоли начинают рас-
ходиться к противоположным концам клетки, где будут позднее
формироваться полюса веретена. К каждому полюсу отходит по
двойной центриоли, диплосоме. По мере расхождения диплосом
начинают формироваться микротрубочки, отходящие от перифери-
ческих участков одной из центриолей каждой диплосомы.
Сформированный аппарат деления в животных клетках имеет
веретеновидную форму и состоит из нескольких зон: двух зон цен-
тросфер с центриолями внутри них и промежуточной между ними
зоны волокон веретена. Во всех этих зонах имеется большое число
микротрубочек (рис. 21).
Микротрубочки в центральной части этого аппарата, в собст-
венном веретене деления, так же как микротрубочки центросфер,
возникают в результате полимеризации тубулинов в зоне центри-
олей и около специальных структур — кинетохоров, расположен-
ных в области центромерных перетяжек хромосом. В веретене
деления принято различать два типа волокон: идущие от полюса к
центру веретена и хромосомные, соединяющие хромосомы с одним
из полюсов.
В индукции роста микротрубочек веретена в зоне полюса деле-
ния принимает участие одна из центриолей диплосомы, а именно
материнская. Такое новообразование и рост нитей (пучков микро-
трубочек) веретена происходят в профазе митоза.
В то же время видны появляющиеся на хромосомах в местах
первичных перетяжек (см. ниже) пластинчатые кинетохоры, около
которых позднее также появляются микротрубочки, идущие в на-
правлении полюсов деления. Таким образом, у животных клеток
центриоли и хромосомные кинетохоры являются центрами органи-
зации микротрубочек веретена деления.
Метафаза занимает около трети времени всего митоза.
Во время метафазы заканчивается образование веретена деления,
а хромосомы выстраиваются в экваториальной плоскости веретена,
образуя так называемую метафазную пластинку хромосом, или ма-
теринскую звезду. К концу метафазы завершается процесс обособ-
Рис. 21. Строение митотического вере-
тена (схема).
/ — хромосомы; 2 — клеточный центр; 3 —
центриолярные микротрубочки; 4 — к ине-
тохорные микротрубочки.
ления друг от друга сестринских
хроматид. Их плечи лежат па-
раллельно друг другу, между
ними хорошо видна разделяю-
щая их щель. Последним мес-
том, где контакт между хрома-
тидами сохраняется, является
центромера.
Анафаза. Хромосомы
все одновременно теряют связь
друг с другом в области центро-
мер и синхронно начинают уда-
ляться друг от друга по направ-
лению к противоположным
полюсам клетки. Скорость дви-
жения хромосом равномерная,
она может достигать 0,2—
0,5 мкм/мин. Анафаза — самая
короткая стадия митоза (не-
сколько процентов от всего
времени), но за это время происходит ряд событий. Главным из
них является обособление двух идентичных наборов хромосом и
перемещение их в противоположные концы клетки.
Движение хромосом складывается из двух процессов, расхож-
дения их по направлению к полюсам и дополнительного расхожде-
ния самих полюсов.
Предположения о сокращении микротрубочек как о механизме
расхождения хромосом в митозе не подтвердились, поэтому мно-
гие исследователи поддерживают гипотезу «скользящих нитей»,
согласно которой соседние микротрубочки, взаимодействуя друг с
другом (например, хромосомные и полюсные) и с сократительны-
ми белками, тянут хромосомы к полюсам.
Телофаза начинается с остановки разошедшихся диплоид-
ных (2 п) наборов хромосом (ранняя телофаза) и кончается нача-
лом реконструкции нового интерфазного ядра (поздняя телофаза,
ранний G(-период) и разделением исходной клетки на две дочерние
(цитокинез, цитотомия). В ранней телофазе хромосомы, не меняя
своей ориентации (центромерные участки — к полюсу, теломер-
ные — к центру веретена), начинают деконденсироваться и увели-
чиваться в объеме. В местах их контактов с мембранными пузырь-
ками цитоплазмы образуется новая ядерная оболочка. После за-
мыкания ядерной оболочки начинается формирование новых яд-
рышек. Клетка переходит в новый С,-период.
Важное событие телофазы — разделение клеточного тела, ци-
78
тотомия, или цитокинез, который происходит у клеток животных
путем образования перетяжки в результате впячивания плазмати-
ческой мембраны внутрь клетки. При этом в кортикальном, под-
мембранном слое цитоплазмы располагаются сократимые элемен-
ты типа актиновых фибрилл, ориентированные циркулярно в зоне
экватора клетки. Сокращение такого кольца приведет к впячива-
нию плазматической мембраны в области этого кольца, что завер-
шается разделением клетки перетяжкой на две.
При повреждении митотического аппарата (действие холода
или агентов, вызывающих деполимеризацию тубулинов) может
произойти или задержка митоза в метафазе, или рассеивание хро-
мосом. При нарушениях репродукции центриолей могут возникать
многополюсные и асимметричные митозы и т. д. Нарушения цито-
томии приводят к появлению гигантских ядер или многоядерных
клеток.
Морфология митотических хромосом
Как интерфазные, так митотические хромосомы состоят из
элементарных хромосомных фибрилл — молекул ДНП. В послед-
нее время принято считать, что на каждую хромосому приходится
одна гигантская фибрилла ДНП, сложно уложенная в относитель-
но короткое тельце — собственно митотическую хромосому. Уста-
новлено, что в митотической хромосоме существуют боковые петли
этой гигантской молекулы дезоксирибонуклеопротеида. Боковые
петли хромосом в вытянутом состоянии могут достигать 30 мкм.
При их компактизации (спирализации) образуются структуры
промежуточного характера — так называемые хромонемные фиб-
риллы. Взаимодействие этих компонентов хромосом друг с другом
и их взаимная агрегация приводят к конечной компактизации хро-
матина в виде митотической хромосомы.
Морфологию митотических хромосом лучше всего изучать в
момент их наибольшей конденсации, в метафазе и в начале анафа-
зы. Хромосомы в этом состоянии представляют собой палочковид-
ные структуры разной длины с довольно постоянной толщиной.
У большинства хромосом удается легко найти зону первичной
перетяжки (центромеры), которая делит хромосому на два пле-
ча (рис. 22). Хромосомы с равными или почти равными плечами
называют метацентрическими, с плечами неодинаковой длины —
субметацентрическими. Палочковидные хромосомы с очень корот-
ким, почти незаметным вторым плечом называют акроцентриче-
скими. В области первичной перетяжки расположен кинетохор. От
этой зоны во время митоза отходят микротрубочки клеточного ве-
ретена, связанные с перемещением хромосом при делении клетки.
Некоторые хромосомы имеют, кроме того, вторичные перетяжки,
располагающиеся вблизи одного из концов хромосомы и отделяю-
щие маленький участок — спутник хромосомы. Вторичные пере-
тяжки называют, кроме того, ядрышковыми организаторами, так
как именно на этих участках хромосом в интерфазе происходит
79
Рис. 22. Строение хромосомы»
Хромосома в световом микроскопе (Л) и ее схематическое изображение (£); хромосома
при дифференциальной окраске (В) и ее схематическое изображение (Г); Д— хромосома
в сканирующем электронном микроскопе; Е — хромосома в трансмиссионном мегавольт
ном электронном микроскопе; 1 — теломеры; 2 — центромеры, J — плечи хромосомы.
образование ядрышка. В этих местах локализована ДНК, ответ-
ственная за синтез рибосомных РНК.
Плечи хромосом оканчиваются теломерами -— конечными уча-
стками. Размеры хромосом, как и их число, у разных организмов
варьируют в широких пределах.
Совокупность числа, размеров и особенностей строения хромо-
сом называется кариотипом данного вида.
При специальных методах окраски хромосомы неравномерно
воспринимают красители: вдоль их длины наблюдается чередова-
ние окрашенных и неокрашенных участков — дифференциальная
неоднородность хромосомы. Важно то, что каждая хромосома
имеет свой, неповторимый рисунок такой дифференциальной
окраски. Применение методов дифференциальной окраски позво-
лило детально изучить строение хромосом. Хромосомы человека
80
принято подразделять по их размерам на 7 групп (А, В, С, D, Е, F,
G). Если при этом легко отличить крупные (1, 2) хромосомы от
мелких (19, 20), метацентрические от акроцентрических (13), то
внутри групп трудно различить одну хромосому от другой. Так в
группе С6 и С7 хромосомы схожи между собой, так же как и с
Х-хромосомой. Дифференциальное окрашивание позволяет четко
отличить эти хромосомы друг от друга.
Эндорепродукция
Эндорепродукция — образование клеток с увеличенным содер-
жанием ДНК. Появление таких клеток происходит в результате
полного отсутствия или незавершенности отдельных этапов митоза.
Существует несколько моментов в процессе митоза, блокада кото-
рых приводит к его остановке и появлению полиплоидных клеток,
т. е. клеток с увеличенным числом хромосомных наборов. Блокада
может наступить при переходе от С2-периода к собственно мито-
зу, остановка может произойти в профазе и метафазе, в последнем
случае часто нарушается функция и целость веретена деления.
Наконец, следствием нарушения цитотомии также может явиться
появление полиплоидных клеток — одноядерных и двуядерных.
При блокаде митоза в самом его начале, при переходе его от
G2 к профазе, клетки приступают к следующему циклу репликации,
приводящему к прогрессивному увеличению количества ДНК в яд-
ре. При этом не наблюдается никаких морфологических особен-
ностей таких ядер, кроме увеличения их объема.
Появление полиплоидных соматических клеток может происхо-
дить в результате блокады деления клеточного тела. В печени
взрослых млекопитающих встречаются, кроме диплоидных, тетра-
и октаплоидные (8 и) клетки, а также двуядерные клетки разной
степени плоидности. Процесс полиплоидизации этих клеток проис-
ходит следующим образом. После S-периода клетки, обладающие
4 с количеством ДНК, вступают в митотическое деление, проходят
все его стадии, включая телофазу, но не приступают к цитотомии.
Таким образом, образуется двуядерная клетка (2 X 2 п). Если она
снова проходит S-период, то оба ядра в такой клетке будут содер-
жать по 4 с ДНК и 4 п хромосом. Такая двуядерная клетка входит
в митоз, на стадии метафазы происходит объединение хромосом-
ных наборов (общее число хромосом равно 8 и), а затем — нор-
мальное деление, в результате которого образуются две тетрапло-
идные клетки. Этот процесс попеременного появления двуядерных
и одноядерных клеток приводит к появлению ядер с 8 и, 16 п и да-
же 32 п количеством хромосом. Подобным способом образуются
полиплоидные клетки в печени, в эпителии мочевого пузыря, в
пигментном эпителии сетчатки, в ацинарных отделах слюнных и
поджелудочной желез, мегакариоциты красного костного мозга.
Необходимо отметить, что полиплоидизация соматических клеток
встречается на терминальных периодах развития клеток, тканей и
органов. Она большей частью характерна для специализированных,
81
дифференцированных клеток и не встречается при генеративных
процессах, таких как эмбриогенез (исключая провизорные орга-
ны) и образование половых клеток; нет полиплоидии среди стволо-
вых клеток.
РЕАКЦИЯ КЛЕТОК НА ВНЕШНИЕ ВОЗДЕЙСТВИЯ
Организм и его клетки постоянно подвергаются воздействию
самых разнообразных химических, физических или биогенных
факторов. Эти фактоРы могут вызывать первичное нарушение од-
ной или нескольких клеточных структур, что в свою очередь при-
водит к функциональным нарушениям. В зависимости от интенсив-
ности поражения, его длительности и характера судьба клетки
может быть различна. Измененные в результате повреждения
клетки могут адаптироваться, приспособиться к повреждающему
фактору, репарировать повреждения, реактивироваться после сня-
тия повреждающего воздействия или измениться необратимо и по-
гибнуть. Исходя из этого функциональные и морфологические
картины клеток в этих состояниях очень разнообразны. На раз-
личные факторы при обратимом повреждении клетки отвечают ря-
дом изменений. Проявлением общеклеточной реакции на повреж-
дение является изменение способности клетки связывать различ-
ные красители. Так, нормальные клетки, поглощая из внеклеточ-
ной среды растворенные в ней красители, откладывают их в виде
гранул. Такое гранулообразование происходит в цитоплазме, ядро
при этом остается бесцветным. При повреждении клеток многими
физическими (нагревание, давление) или химическими факторами
(изменение pH среды, добавление спирта или какого-либо иного
денатурирующего агента) гранулообразование прекращается, цито-
плазма и ядро диффузно окрашиваются проникшим в клетку кра-
сителем. Если действие фактора обратимо и при устранении его
клетка возвращается к норме, то снова восстанавливается ее спо-
собность к гранулообразованию. При различных повреждениях
клеток значительно падает окислительное фосфорилирование: пре-
кращается синтез АТФ и растет потребление кислорода. Для по-
врежденных клеток характерно усиление гликолитических процес-
сов, падение количества АТФ, активация протеолиза. Совокуп-
ность неспецифических обратимых изменений цитоплазмы, возни-
кающих под воздействием различных агентов, была обозначена
термином «паранекроз» 1Насонов Д. Н., Александров В. Я., 1940].
При различных воздействиях на клетку наиболее частым измене-
нием структуры ядра является конденсация хроматина, что может
отражать падение ядерных синтетических процессов. При гибели
клетки происходят коагуляция хроматина, собирание его в грубые
агрегаты внутри ядра (пикноз), что часто завершается распадом на
части (кариорексис) и растворением ядра (кариолизис). Ядрышки
при подавлении синтеза рРНК уменьшаются в размерах, теряют
гранулы, фрагментируются.
К наиболее часто встречающимся изменениям ядерной оболочки
82
относятся расширение (отечность) перинуклеарного пространства,
извитость контура ядерной оболочки, что нередко сочетается с пик-
нозом ядра. На ранних этапах повреждения клетки часто приобрета-
ют шаровидную форму и теряют многочисленные клеточные вырос-
ты и микроворсинки. В дальнейшем, наоборот, изменения плазмо-
леммы сводятся к появлению на поверхности клеток различных вы-
ростов или мелких пузырей. На начальных стадиях нарушения окис-
лительного фосфорилирования происходит сжатие митохондриаль-
ного матрикса и некоторое расширение межмембранного простран-
ства. В дальнейшем этот тип рекции митохондрий может смениться
их набуханием, что особенно часто встречается при самых различ-
ных патологических изменениях клеток. Митохондрии при этом
принимают сферическую форму и увеличиваются в размерах, проис-
ходит обводнение матрикса, он становится светлым. Набухание
митохондрий, как правило, сопровождается редукцией числа и раз-
мера крист. При необратимом повреждении митохондрий происхо-
дит разрыв их мембран, матрикс смешивается с гиалоплазмой.
Система эндоплазматического ретикулума чаще всего подверга-
ется вакуолизации и распаду на мелкие пузырьки. При этом на
мембранах гранулярного ретикулума уменьшается число рибосом,
что однозначно указывает на падение белкового синтеза. Цистерны
комплекса Гольджи также могут увеличиваться в объеме или рас-
падаться на мелкие вакуоли. В поврежденных клетках происходит
активация их лизосом, увеличивается число аутофагосом. При тяже-
лых клеточных повреждениях мембраны лизосом разрываются и ли-
зосомные гидролазы начинают разрушать сами клетки — происхо-
дит лизис клеток.
Поврежденные клетки резко снижают митотическую активность,
часто задерживаются на разных стадиях митоза главным образом
из-за нарушения митотического аппарата, очень чувствительного к
изменениям внутриклеточной среды.
Развитие процесса повреждения клеток останавливается после
неблагоприятного воздействия. Если изменения в клетке не зашли
слишком далеко, происходит репарация клеточных повреждений,
возврат клетки к нормальному функциональному уровню. Так,
в ряде случаев повреждения клеток, связанные с набуханием мито-
хондрий и с фрагментацией эндоплазматического ретикулума,
оказываются обратимыми. Процессы восстановления внутриклеточ-
ных структур называют внутриклеточной регенерацией.
Репарация клеток бывает полной, когда восстанавливаются все
свойства данных клеток, или неполной. В последнем случае после
снятия действия повреждающего фактора нормализуется ряд
функций клеток, но через некоторое время они уже без всякого
воздействия погибают. Особенно часто это наблюдается при пора-
жениях клеточного ядра.
Повреждение клеток внешними и внутриорганизменными факто-
рами может привести к нарушениям регуляции их метаболизма.
При этом происходит интенсивное отложение или же, наоборот,
резорбция ряда клеточных включений. Кроме того, наблюдается
83
нарушение регуляции проницаемости клеточных мембран, что при-
водит к вакуолизации мембранных органелл. В патологической
анатомии такие изменения в структуре клеток называют дистро-
фиями. При жировой дистрофии в клетках накапливаются жиро-
вые включения. Жировая инфильтрация, когда клетка, поглощая
жиры, неспособна к их утилизации, приводит к накоплению жиро-
вых капель в цитоплазме. Часто в цитоплазме измененных клеток
обнаруживаются скопления липопротеидных комплексов, имеющих
вид многослойных мембранных пластов. Нарушение регуляторных
процессов метаболизма сахаров приводит к патологическому
отложению и накоплению гликогена, что, вероятно, связано с
недостаточностью фермента, расщепляющего гликоген (глюкозо-
6-фосфатазы). Часто в измененных клетках животных происходит
отложение различных пигментов, белковых гранул и др.
Особой формой патологического нарушения регуляторных
процессов могут быть нарушения специализации, одним из которых
является злокачественный опухолевый рост. Опухолевые клетки
характеризуются безудержностью, неограниченностью размноже-
ния, нарушением уровня специализации, изменениями строения
клеток, относительной автономностью от регуляторных влияний
со стороны организма, способностью к метастазированию. Все эти
свойства опухолевые клетки сохраняют от поколения к поколе-
нию, т. е. свойства злокачественности являются наследственной
особенностью таких клеток. Вот почему считается, что раковые
клетки являются мутантами, обладающими измененной генетиче-
ской структурой; именно изменением генотипа клетки можно
объяснить непрерывную передачу дочерним клетками дефектной
(в отношении регуляции) информации.
При необратимом повреждении клетки гибнут. Дать определе-
ние момента клеточной смерти очень трудно (так же, как и при
смерти целого организма), так как умирание — это не одномо-
ментное явление, а процесс. При необратимом повреждении разво-
рачивается ряд последовательных событий, приводящих к разруше-
нию клеток. В самом начале изменения клеток имеют характер
обратимых, паранекротических. Отличие состоит в том, что после
снятия воздействия они не исчезают, а прогрессируют. Явным
признаком гибели клетки является активация внутриклеточных
гидролитических ферментов. Они активируются в гиалоплазме и
начинают расщепление белков, липидов и др., при этом разрушают-
ся внутриклеточные мембраны, в том числе и мембраны лизосом.
Все это приводит к лизису, разрушению клеток, но это уже отно-
сится к посмертным изменениям.
ЭМБРИОЛОГИЯ
Глава V
ОСНОВЫ ЭМБРИОЛОГИИ
СВЯЗЬ ИНДИВИДУАЛЬНОГО И ИСТОРИЧЕСКОГО РАЗВИТИЯ ОРГАНИЗМА
Эмбриология (от греч. embryon — зародыш, logos — учение) —
наука о зародыше, о закономерностях его развития.
Медицинская эмбриология изучает закономерности развития
зародыша человека, структурные, метаболические и функциональ-
ные особенности плацентарного барьера (система мать — плацен-
та — плод), причины возникновения уродств и других отклонений
от нормы, а также механизмы регуляции эмбриогенеза. Пути и
методы влияния на эмбриогенез исследуются главным образом в
условиях эксперимента на животных, а также в клинических усло-
виях при патологии беременности. Одним из актуальных аспектов
современной эмбриологии является изучение источников и меха-
низмов развития тканей (гистогенез).
В понятие эмбриогенеза включают период от момента опло-
дотворения до рождения (для живородящих животных), вылупле-
ния из яиц (для яйцекладущих), окончания метаморфоза (для
животных с личиночной стадией развития).
Эмбриогенез — часть индивидуального развития, т. е.
онтогенеза. Он тесно связан с прогенезом (развитие и
созревание половых клеток) и ранним постэмбриональ-
ным периодом.
Актуальными задачами эмбриологии являются изучение влия-
ния различных эндогенных и экзогенных факторов, роли микро-
окружения на развитие и строение половых клеток, развитие и
взаимоотношение тканей, органов и систем, исследование меха-
низмов, контролирующих репродуктивную функцию и обеспечи-
вающих гомеостаз зародышей человека и млекопитающих, и дру-
гих факторов, изучение критических периодов развития. Частным,
но очень важным вопросом современной эмбриологии является
культивирование яйцеклеток, зародышей и имплантация их в мат-
ку. Знание условий и факторов оплодотворения и эмбриональ-
ного развития позволяет врачам решать такие практически важные
проблемы, как искусственное оплодотворение женщин при
бесплодных браках, цитодиагностика патологии беременности
и др.
Изучению эмбриогенеза человека предшествует краткое изло-
жение основ сравнительной эмбриологии, так как в ходе истори-
ческого развития млекопитающих сложились основные этапы,
последовательность и закономерности эмбриогенеза.
85
Процесс эмбрионального развития человека является результа-
том длительной эволюции и в определенной степени отражает
черты развития других форм животного мира. Некоторые ранние
стадии развития человека очень сходны с аналогичными стадиями
эмбриогенеза более низко организованных хордовых животных.
Идея связи индивидуального и исторического развития была
обоснована в начале XIX столетия. В частности, К. Бэр, изучая
в сравнительном аспекте развитие некоторых позвоночных, пришел
к заключению, что у большой группы животных на ранних ста-
диях развития проявляется больше сходства, чем частных, инди-
видуальных различий. По мере увеличения сроков эмбрионального
развития это частное, индивидуальное вырисовывается все более
отчетливо (закон зародышевого сходства). Ф. Мюллер, изучая
развитие личиночных стадий ракообразных, также обнаружил
сходство некоторых личиночных форм с вымершими ракообраз-
ными. Ч. Дарвин, придавая большое значение явлениям зароды-
шевого сходства, считал это одним из доказательств общности
происхождения животного мира.
В конце 60-х годов XIX столетия Э. Геккель сформулировал
биогенетический закон, согласно которому индивидуальное раз-
витие зародыша есть сжатое, сокращенное повторение истори-
ческого развития, иначе онтогенез повторяет в краткой форме
филогенез. Идея биогенетического закона сыграла большую роль
в развитии не только эмбриологии, но и эволюционного учения.
Вместе с тем формулировка биогенетического закона не отра-
жает влияние факторов окружающей среды, экологических усло-
вий, которые имеются в действительности и оказывают влияние на
эмбриогенез. А. Н. Северцов, продолживший в 20—30-е годы
XX столетия разработку биогенетического закона, пришел к
заключению, что эволюционный процесс совершается не путем
накопления изменений признаков взрослых животных, как счита-
ли Ч. Дарвин и Э. Геккель, а путем суммирования изменений,
появляющихся у зародышей (теория филэмбриогенеза). Биоло-
гически важная перемена в условиях существования (среда)
данного вида животных согласно воззрениям А. Н. Северцова,
является стимулом к изменению его организации; характер же
изменения среды, количественное и качественное соотношение
между изменением среды и морфофункциональными изменениями
организма определяет направление, в котором будет происходить
эволюция изменяющегося вида в данную эпоху.
ОСНОВЫ СРАВНИТЕЛЬНОЙ ЭМБРИОЛОГИИ
Прогенез
Половые клетки (гаметы)
Зрелые половые клетки в отличие от соматических содержат
одиночный (гаплоидный) набор хромосом. Все хромосомы гаметы,
за исключением одной половой, называются аутосомами. В муж-
К6
ских половых клетках у млекопитающих содержатся половые
хромосомы либо X, либо Y, в женских половых клетках — только
хромосома X. Дифференцированные гаметы обладают невысоким
уровнем метаболизма и неспособны к размножению.
Мужские половые клетки
Мужские половые клетки — сперматозоиды, или спермии,
развиваются в очень большом количестве: выделяющаяся при
эякуляции семенная жидкость содержит несколько миллионов
сперматозоидов. Они невелики по размерам. У человека их размер
достигает 70 мкм. Сперматозоиды обладают способностью к
активному движению. Скорость их движения у человека 30—
50 мкм/с. Мужские половые клетки имеют жгутиковую форму.
Строение. В сперматозоиде различают головку и хвост (рис. 23).
Головка сперматозоида (caput spermatozoidi) включает небольшое
плотное ядро, окруженное тонким слоем цитоплазмы. Ядра спер-
матозоидов характеризуются высоким содержанием нуклеопрота-
минов и нуклеогистонов. Передняя половина ядра покрыта плоским
мешочком, составляющим «чехлик» сперматозоида. В нем у
переднего полюса располагается акросома (от греч. асгоп —
верхушка, soma —тело). Чехлик и акросома являются производ-
ными комплекса Гольджи. Акросома содержит набор ферментов,
среди которых важное место принадлежит .гиалуронидазе и про-
теазам, способным растворять оболочки, покрывающие яйцеклетку.
Важно отметить, что у высших позвоночных способность к опло-
дотворению (явление капацитации сперматозоидов) приоб-
ретается постепенно, по мере их продвижения по репродуктивному
женскому тракту.
За головкой имеется кольцевидное сужение. Головка так же,
как и хвостовой отдел, покрыта клеточной мембраной.
Хвостовой отдел (flagellum) сперматозоида состоит из связую-
щих, промежуточных, главной и терминальной частей.
В связующей части (pars conjungens) или шейке (cervix)
располагаются центриоли — проксимальная и дистальная, от
которой начинается осевая нить (ахопета). Промежуточная часть
(pars intermedia) содержит 2 центральных и 9 пар перифери-
ческих микротрубочек1, окруженных расположенными по спирали
митохондриями (митохондриальное влагалище — vagina mitochond-
rialis). Именно митохондрии обеспечивают энергией двигательную
активность сперматозоидов, нарушение которой нередко связано с
поражением процесса энергообразования в митохондриях. Движе-
ния хвостового жгутика бичеобразны. Они обусловлены последо-
вательным изменением белков микротрубочек (динеин и др.). Эти
белки обладают АТФ-азной активностью и расщепляют АТФ,
1 У многих животных между центральными и периферическими микротру-
бочками имеется еще •) одиночных фибрилл.
87
Рис. 23. Строение мужских половых
клеток.
А — спермин в двух плоскостях; Б — уль-
трамикроскопическое строение спермиев;
В — фрагмент главного отдела хвоста; Г —
фрагмент дистального отдела хвоста. I —
головка; II — хвост; а — связующий отдел
(шейка); б—промежуточный отдел; в —
главный отдел; г — дистальный отдел; / —
цитолемма; 2 — акросома; 2а — акросо-
мальный пузырек; 3 — ядро; 4 — прокси-
мальная центриоль; 5 — дистальная цент
риоль; 6 — митохондрии; 7 — осевая нить;
8 — циркулярные фибриллы; 9 — перифе-
рические микротрубочки; 10 — централь-
ные микротрубочки.
вырабатываемую митохондриями. Освобождаемая при этом
энергия используется для сокращения белков и обеспечения
подвижности спермиев в жидкой среде. Среди факторов, влияющих
на скорость движения, большое значение имеют степень зрелости
спермиев, температура и pH среды.
Главная часть (pars principalis) по строению напоминает
ресничку. Окружена тонкофибриллярным влагалищем (vagina
fibrosa). Терминальная, или конечная, часть (pars terminahs) содер-
жит единичные сократительные филаменты.
Сперматозоиды животных отличаются друг от друга соотно-
шением указанных отделов и главным образом формой головки.
Продолжительность жизни и оплодотворяющая способность
сперматозоидов после эякуляции в определенных оптимальных
условиях неодинаковы у различных животных У млекопитающих
они варьируют от нескольких часов до нескольких суток. В кислой
88
среде сперматозоиды быстро утрачивают способность к движению,
оплодотворению и склеиваются. Способность к оплодотворению
зависит также от концентрации сперматозоидов в семенной жид-
кости, продолжительности их пребывания в эякуляте и др.
Женские половые клетки. Классификация
Яйцеклетки, или овоциты (от лат. ovum — яйцо), созревают
в неизмеримо меньшем количестве, чем сперматозоиды. Для неко-
торых млекопитающих количество созревающих в течение всей
жизни яйцеклеток исчисляется сотнями. У других позвоночных их
может быть гораздо больше (например, у рыб и амфибий). Как
правило, яйцеклетки имеют шаровидную форму, больший объем
цитоплазмы, чем у спермиев, они не обладают способностью
самостоятельно передвигаться.
Характерным для яйцеклеток является наличие желтка
(lecithos) (белково-липидных включений) в цитоплазме. В зави-
симости от количества желтка размеры яйцеклеток колеблются от
нескольких микрометров до нескольких сантиметров (например,
яйцеклетки птиц, акуловых рыб). Яйцеклетки классифицируют
на безжелтковые (алецитальные), маложелтковые (олиголеци-
тальные) и многожелтковые (полилецитальные). Маложелтковые
яйцеклетки подразделяются на первичные (у примитивных
хордовых, например ланцетника) ивторичные (у млекопитаю-
щих и человека). Количество желтка в цитоплазме находится в пря-
мой зависимости от условий развития животного (во внешней
или внутренней среде) и продолжительности развития во внешней
среде (рис. 24).
Как правило, в маложелтковых яйцеклетках желточные вклю-
чения (гранулы, пластинки) распределены равномерно, поэтому они
называются еще изолецитальными (греч. isos — равный). У боль-
шинства полилецитальных яйцеклеток желток в большей или мень-
шей степени сосредоточен у одного полюса (вегетативного), а орга-
неллы — у противоположного (анимального). Такие яйцеклетки
называются телолециталъными (греч. thelos—конец), а если
жедток находится в центре клетки — центролециталъными. Среди
телолецитальных различают умеренно телолецитальные — мезо-
лециталъные (например, у амфибий) и резко телолецитальные
(например, у птиц).
У животных, ведущих наземное существование, организация
яйцеклеток сложная. В частности, у пресмыкающихся и птиц
яйцеклетки резко телолецитальны. Размеры яйцеклетки большие.
Наземное развитие привело к возникновению вторичных и третич-
ных оболочек, предохраняющих яйцеклетку от повреждающего
действия механических, температурных и других факторов окру-
жающей среды (пресмыкающиеся, птицы).
У плацентарных млекопитающих в связи с внутриутробным
развитием и питанием за счет материнского организма отпала необ-
ходимость создания сколько-нибудь значительных запасов желтка
89
Рис. 24. Строение женских половых клеток.
А — различные типы яйцеклеток: а — первично изолецитальная у ланцетника; б — умеренно
телолецитальная у лягушки; в—резко телолецитальная у птицы; г—вторично изолеци-
тальная у человека; I — ядро; 2 — цитоплазма, 3 — желточные зерна; 4 — желточные пла-
стинки (масштаб не соблюден). Б— схема микроскопического (а) и ультрамикроскопи-
ческого (б) строения яйцеклетки. / — ядро; 2 — цитоплазма с желточными включениями;
3 — кортикальные гранулы: 4 — цитолемма; 5 — микроворсинки цитолеммы; 6 — блестя-
щая оболочка; 7 — фолликулярные клетки с отростками, формирующими лучистый венец.
в яйцеклетке. Поэтому вторично в эволюции появились маложелтко-
вые яйцеклетки. Исключением являются яйцеклетки представите-
лей примитивных млекопитающих (клоачные, отчасти сумчатые).
Эти животные сохраняют многие черты своих предков — пресмы-
кающихся, в том числе и резко телолецитальные яйцеклетки.
Яйцеклетка плацентарных млекопитающих относительно неболь-
шая, диаметром 50—150 мкм, окружена прозрачной зоной (zona
pellucida) и слоем фолликулярных клеток, принимающих участие
в ее питании (см. рис. 24).
Строение. Яйцеклетка содержит ядро, цитоплазму (ооплазму),
включающую в том или ином количестве питательный материал —
90
желток и оболочки. Все яйцеклетки имеют цитолемму (оволем-
му), или первичную оболочку, а многие еще окружены вторичной
(углеводно-белковой) и некоторые — третичной (скорлуповой,
подскорлуповой) оболочками. Строение яйцеклеток характеризу-
ется полярностью, которая выражена тем сильнее, чем больше
желтка в клетке, например, у птиц. Та часть яйцеклетки, в которой
накапливается желток, составляет вегетативный полюс, а проти-
воположная, куда смещается ядро, — анималъный. Поверхность
яйцеклетки покрыта микроворсинками.
Ядро женской половой клетки имеет гаплоидный набор хромо-
сом. В период роста ооцита в ядре происходят интенсивные синтети-
ческие процессы амплификации генов синтеза РНК — образо-
вание многочисленных копий с тех участков хромосомной ДНК,
которые кодируют рибосомную РНК. Копии ДНК замыкаются в
кольца и смещаются к периферии ядра. На них возникают новые
копии ДНК, которые в виде ядрышек выходят в цитоплазму, где
становятся центрами усиленного синтеза рРНК и иРНК. Большая
часть копий ДНК оказывается заблокированной белковыми молеку-
лами (информосомы) до наступления оплодотворения.
Особенностью ооцитов является накопление огромных запасов
химических компонентов аппарата трансляции: ^рибосом, иРНК,
тРНК, количество которых в сотни и тысячи раз может превы-
сить содержание их в соматических клетках. В цитоплазме яйце-
клеток накапливаются также запасы разнообразных белков:
гистонов, структурных белков рибосом, тубулина, липофосфопро-
теидов желтка.
Среди органелл в яйцеклетках разных животных хорошо развита
эндоплазматическая сеть. Количество митохондрий умеренно.
Комплекс Гольджи на ранних стадиях развития яйцеклетки распо-
лагается около ядра, а в ходе созревания яйцеклетки смещается на
периферию цитоплазмы. Здесь располагаются небольшие корти-
кальные гранулы (granula corticalia), содержащие гликозаминогли-
каны. В цитоплазме яйцеклеток млекопитающих постоянно выяв-
ляются мультивезикулярные тельца. Из включений ооплазмы особо-
го внимания заслуживает желток — питательный материал, во
"многом определяющий характер эмбриогенеза. Желток выявляется
в виде гранул или более крупных шаров и пластинок, образованных
фосфолипидами, протеинами и углеводами. Структурной единицей
желтка является комплекс липовителлина (липопротеида) и фос-
фовитина (фосфопротеина). Каждая пластинка состоит из более
плотной центральной и более рыхлой периферической зон, снаружи
ограничена осмиофильной мембраной. Плотная зона образова-
на молекулами фосфовитина, имеет вид кристаллической решетки.
Образуется желток при непосредственном участии эндоплазматиче-
ской сети и комплекса Гольджи.
В процессе роста и созревания яйцеклеток в яичнике они окру-
жаются слоем плоских или кубических клеток, называемых фолли-
кулярными. За счет деятельности ооцита и фолликулярных клеток
вокруг яйцеклетки образуется зона, богатая гликозаминогликанами.
91
У млекопитающих она называется прозрачной зоной (zona pellu-
cida).
Фолликулярные клетки посылают через прозрачную зону длин-
ные отростки, направленные к ооциту. В свою очередь цитолемма
ооцита имеет микроворсинки, располагающиеся между отростками
фолликулярных клеток (см. рис. 24, Б). Фолликулярные клетки
выделяют вещества, которые поглощаются яйцеклеткой и способ-
ствуют ее росту. Фолликулярный эпителий выполняет также за-
щитную функцию.
Эмбриогенез
Развитие зародыша происходит стадийно с постепенными ка-
чественными и количественными изменениями. Различают следую-
щие стадии: оплодотворение, дробление и образование
бластулы, гаструляция и дифференцировка заро-
дышевых листков с образованием зачатков тканей (гис-
тогенез), органов (органогенез) и систем органов
(системогенез) плода.
Оплодотворение
Оплодотворение (fertilisatio) — слияние мужской и женской
половых клеток, в результате чего восстанавливается диплоид-
ный набор хромосом, характерный для данного вида животных,
и возн1 кает качественно новая клетка — зигота (опчодотворен-
ная яйцеклетка или одноклеточный зародыш).
В зиготе масса ядра увеличивается вдвое, а объем цитоплаз-
мы практически остается тот же, особенно при оплодотворении
телоцитальных яйцеклеток. Оплодотворению предшествует осе-
менение — излияние семенной жидкости в половые пути при
внутреннем оплодотворении или в среду, где находятся яйцеклет-
ки, при наружном оплодотворении.
В процессе оплодотворения различают три фазы: 1) дистант-
ное взаимодействие и сближение гамет; 2) контактное взаимодей-
ствие и активизация яйцеклетки; 3) вхождение сперматозоида в
яйцо и последующее слияние — сингамия.
Первая фаза — дистантное взаимодействие — обеспечи-
вается совокупностью ряда неспецифических факторов, повышаю-
щих вероятность столкновения половых клеток. Важную роль в
этом играют химические вещества, вырабатываемые половыми
клетками, — гамоны: гиногамоны (I, II), вырабатываемые яйцеклет-
ками иандрогамоны (I, II), продуцируемые спермиями. Гиногамоны
I (низкомолекулярные вещества небелковой природы, выделяемые
яйцеклетками, активизируют движение спермиев. Гиногамо-
ны II (фертилизины) — видоспецифические белки, вызываю-
щие склеивание спермиев при их реакции с комплементарным
андрогамоном II, встроенным в цитолему спермия. Склеивание
92
спермиев предохраняет яйцеклетку от проникновения многих спер-
миев.
Андрогамоны I — антагонисты гиногамонов 1 — вещества не-
белковой природы, подавляют подвижность спермиев.
Вторая фаза — контактное взаимодействие гамет и про-
никновение спермия в яйцеклетку — осуществляется с помощью
акросомы и ее ферментов спермолизинов. Плазматические мем-
браны в месте контакта половых клеток сливаются и происходит
плазмогамия — объединение цитоплазм обеих гамет.
У млекопитающих при оплодотворении в яйцеклетку проника-
ет лишь один сперматозоид. Такое явление называют моноспер-
мией. У беспозвоночных животных, рыб, хвостатых амфибий,
рептилий и птиц возможна полиспермия, когда в яйцеклетку про-
никает несколько сперматозоидов, однако сливается с ядром яйце-
клетки ядро только одного спермия. Оплодотворению способст-
вуют тысячи других принимающих участие в осеменении сперма-
тозоидов. Ферменты, выделяемые из акросом, — спермолизины
(трипсин, гиалуронидаза), разрушают лучистый венец, расщепляют
гликозаминогликаны вторичной (блестящей) оболочки яйцеклет-
ки. Отделяющиеся фолликулярные клетки склеиваются в конгло-
мерат, который вслед за яйцеклеткой перемещается по трубе бла-
годаря мерцанию ресничек эпителиальных клеток слизистой обо-
лочки.
Третья фаза. В ооплазму проникает головка и проме-
жуточная часть хвостового отдела. После вхождения спермато-
зоида на периферии ооплазмы происходит уплотнение ее (кор-
тикальная реакция) и образуется оболочка оплодотворения
(рис. 25).
Как показано на беспозвоночных, механизм кортикальной ре-
акции включает: приток ионов натрия через участок мембраны спер-
матозоида, встроенный в поверхность яйцеклетки после заверше-
ния акросомальной реакции. В результате отрицательный мем-
бранный потенциал клетки становится слабоположительным. При-
ток ионов натрия обусловливает высвобождение ионов кальция
из внутриклеточных депо и увеличение его содержания в цито-
плазме яйцеклетки. Вслед за этим начинается экзоцитоз корти-
кальных гранул. Освобождающиеся из них протеолитические фер-
менты разрывают связи между блестящей оболочкой (или жел-
точной оболочки у амфибий и птиц) и плазмолеммой яйцеклет-
ки, а также между спермиями и прозрачной оболочкой. Кроме то-
го, выделяется гликопротеид, связывающий воду и привлекающий
ее в пространство между плазмолеммой и блестящей оболочкой.
Вследствие этого формируется перивителлиновое пространство.
Наконец, выделяется фактор, способствующий затвердению про-
зрачной оболочки и образованию из нее оболочки оплодотворе-
ния (membrana fertilisationis).
Кортикальная реакция — один из механизмов, препятствую-
щий проникновению в яйцеклетку других сперматозоидов.
Проникновение сперматозоида через несколько минут значи-
93
Рис. 25. Стадии оплодотворения и начала дробления (схема).
I — ооплазма; 1а — кортикальные гранулы; 2 — ядро; 3 — блестящая оболочка; 4 — фол-
ликулярный эпителий; 5 — спермин; 6 — направительные тельца; 7 — деление созревания
ооцита; 8 бугорок оплодотворения; 9 — оболочка оплодотворения; 10 — женский про-
нуклеус; II— мужской пронуклеус; 12—синкарион; 13— первое митотическое деление
зиготы; 14 — бластомеры.
тельно усиливает процессы внутриклеточного обмена, что связано
с активизацией ферментативны систем яйцеклетки, в частности
окислительно-восстановительных, а позднее — белковых синтезов.
Зигота. Ооплазматическая сегрегация. Образование
мужского и женского пронуклеусов
Вслед за проникновением спермия в яйцеклетку и усилени-
ем окислительно-восстановительных реакций начинается интенсив-
ное перемещение составных частей цитоплазмы (ооплазмы) с
образованием зон повышенной концентрации желточных и пиг-
ментных гранул, органелл, что носит название ооплазмати-
ческой сегрегации. Методом маркировки установлено,
что в ходе дальнейшего развития каждый участок оплодотворен-
ной яйцеклетки даст начало определенной структуре зародыша.
94
Такие участки называются презумптивными (от лат. praesumptio —
предположение, основанное на вероятности).
Попавшая в яйцеклетку головка спермия поворачивается на
180г, ядро постепенно набухает, округляется, хроматин разрых-
ляется и оно превращается в мужской пронуклеус. Центриоли,
внесенные мужской половой клеткой, становятся при этом цент-
ром движения внутри оплодотворенной яйцеклетки (зиготы).
Ядро женской половой клетки, имеющее также гаплоидный на-
бор хромосом, набухает, превращается в женский пронуклеус.
Пронуклеусы сближаются. При этом в них происходит реплика-
ция ДНК. В конце сближения происходит спирализация хромо-
сом, образование метафазной пластинки из двух гаплоидных про-
нуклеусов. Объединение двух пронуклеусов — синкарион
(от греч. sin — связь, karyon — ядро) — приводит к восстановле-
нию характерного для данной особи животного или человека
диплоидного набора хромосом. Таким образом, зигота приобре-
тает гены, унаследованные от обоих родителей. В реализации
наследственной информации, кроме ядер, половых клеток, важная
роль принадлежит цитоплазме клетки. Об этом свидетельствуют
эксперименты с пересадкой ядер соматических клеток в яйце-
клетку. При этом пол развивающегося организма зависит от по-
ловых хромосом. При слиянии яйцеклетки со сперматозоидом,
несущим хромосому X, образуется женская особь, а при слиянии
со сперматозоидом, имеющим хромосому Y, — мужская особь.
Дробление
Дробление (fissio) — последовательное митотическое деление
зиготы на клетки (бластомеры) без последующего роста их до
размеров материнской.
Вследствие фактического отсутствия интерфазы G|-периода,
во время которого происходит рост клеток, образовавшиеся в
результате деления клетки гораздо меньше материнской, поэтому
и величина зародыша в целом в этот период независимо от со-
ставляющих его клеток не превышает величину исходной клет-
ки — зиготы. Все это позволило назвать описываемый процесс
дроблением, измельчением, а клетки, образующиеся в результате
дробления, — бластомерами (от греч. blastos — зачаток, nwros —
часть).
На ранних стадиях все бластомеры сохраняют способность
к развитию при определенных условиях в самостоятельный ор-
ганизм, или, как принято говорить, они тотипотентны. Дробление
(уменьшение размеров бластомеров) продолжается до тех пор,
пока не восстановится характерное для соматических клеток дан-
ного вида животного соотношение ядра и цитоплазмы. После
этого наступает дерепрессия синтеза белка, и каждая дочерняя
клетка увеличивается до размеров материнской.
Дробление зародыша происходит неодинаково у различных
Q5
позвоночных, что определяется прежде всего количеством и ха-
рактером распределения желтка в яйцеклетке.
Существует определенный строгий порядок появления борозд
дробления. Борозды и плоскости попеременно проходят через
анимальный и вегетативный полюса клетки (меридианное на-
правление), поперечно (широтные) или параллельно поверхности
(тангенциальные). Чем больше желтка в яйцеклетке у различных
видов животных, тем менее полно и менее равномерно происходит
дробление (рис. 26).
Первично олиголецитальные изолецитальные яйцеклетки дро-
бятся полно и равномерно. В мезолецитальных яйцеклетках дроб-
ление полное, но неравномерное, так как в вегетативной части,
где сосредоточен желток, дробление происходит медленнее, чем
на анимальном полюсе, и неполно. В резко телолецитальных яйце-
клетках дробление частичное — меробластическое. Например, у
птиц дробится лишь часть яйцеклетки у анимального полюса1. Для
вторично олиголецитальных, изолецитальных яйцеклеток пла-
центарных млекопитающих и человека характерно полное,
или голобластическое, асинхронное, неравно-
мерное дробление. Дробление происходит во время дви-
жения зиготы по яйцеводу, причем количество бластомеров на-
растает в неправильном и притом у различных животных не-
одинаковом порядке (2, 3, 5, 10, 13, 17 и т. д.). В результате
дробления образуется многоклеточный зародыш, сначала в форме
плотного скопления клеток (морула), а затем в виде пузырька
с небольшой полостью — бластоциста (бластула) (см. рис. 26).
Дробление зародышей требует соблюдения оптимальных усло-
вий среды (химический состав, осмотическое давление, темпера-
тура, содержание кислорода и др.) Зародыши обладают высокой
чувствительностью к химическим, физическим и другим повреж-
дающим факторам, которые могут привести к мутациям.
Бластула (blastula) имеет стенку — бластодерму и полость —
бластоцель, заполненную жидкостью — продуктом секреции блас-
томеров. В бластодерме различают крышу, образовавшуюся за
счет раздробившегося материала анимального полюса, дно —
из материала вегетативного полюса и краевую зону, расположен-
ную между ними.
При полном равномерном дроблении (например, ланцетника)
бластула имеет однослойную бластодерму, а бластоцель находит-
ся в центре. Такая бластула называется целобластулой. В резуль-
тате полного неравномерного дробления (минога, лягушка) об-
разуется бластула с многослойной бластодермой и эксцентрично
расположенным бластоцелем — амфибластула. Крыша такой блас-
тулы, состоящая из мелких бластомеров, сравнительно тонкая,
1 Дробление центролецитальных яйцеклеток (у беспозвоночных) мероблас-
тическое, поверхностное. Оно заканчивается образованием перибластулы, т. е.
бластулой с бластомерами, расположенными по всей поверхности, окружающими
неразделенный желток.
96
г
Рис. 26. Схема типов дробления, бластул, гаструл (рис. Ю. И. Афанасьева).
IA—1Г — полное равномерное дробление у животных с первично олиголецитальными
яйцеклетками (ланцетник); 1Д— бластула; 1Е— начало гаструляции путем инвагина-
ции; ПА—ПГ—полное неравномерное дробление у животных с умеренно телолециталь-
ными яйцеклетками (амфибии); ПД — бластула; НЕ — начальный этап гаструляции путем
эпиболии; П1А—111 Г — неполное, частичное дробление у птиц и животных с резко тело-
лецитальными яйцеклетками; 1ПД — бластула; I11E — гаструляция путем деламинации,
IVА 1УГ — полное неравномерное дробление у человека и млекопитающих животных;
IVД — бластула; IVE— гаструляция путем деламинации (масштаб не соблюден)
4 ГИСТОЛОГИЯ
а дно, образованное крупными перегруженными желтком бласто-
мерами, массивное.
Дискобластула пресмыкающихся и птиц представляет собой
зародышевый диск, распластанный на желтке. Зародышевый диск,
образованный бластомерами, соответствует крыше и краевой зоне
целобластулы, желток — дну бластулы, а узкая щель между ни-
ми — бластоцелю.
У млекопитающих в результате полного и асинхронного дроб-
ления образуется зародышевый пузырек, называемый бластоцис-
той (от греч. blastos— зачаток, kystis— пузырь). В нем различают
стенку — трофобласт (от греч. troplw — пища) и небольшое скоп-
ление бластомеров в виде узелка на внутренней поверхности тро-
фобласта — эмбриобласт. На этой стадии развития зародыш мле-
копитающих соответствует стадии бластулы других животных, но
не гомологичен ей, так как стенка бластоцисты в построении тела
зародыша участия не принимает.
Гаструляция. Образование осевых зачатков органов
Гаструляция (от лат. g aster— желудок)—сложный процесс
химических и морфогенетических изменении, сопровождающийся
размножением, ростом, направленным перемещением и дифферен-
цировкой клеток, в результате чего образуются зародышевые
листки, наружный (эктодерма), средний (мезодерма) и внутрен-
ний (энтодерма) —источники зачатков тканей и органов.
В начале гаструляции образуются наружный и внутренний за-
родышевые листки (ранняя гаструляция), а затем (поздняя гаст-
руляция) хордомезодермальный зачаток. Зародышевые листки
располагаются послойно: снаружи — эктодерма, изнутри — энто-
дерма, между ними — хорда и мезодерма. В ходе эмбриогенеза,
начиная с этой стадии, помимо размножения и роста
клеток, постепенно происходят их перемещение и диф-
ференцировка, представляющие разные стороны единого
поступательного процесса, в результате которого формируется ор-
ганизм. На стадиях бластулы и ранней гаструлы можно марки-
ровать клетки и проследить их последующее развитие.
Способы гаструляции. Гаструляция совершается 4 основными
способами: путем иммиграции, инвагинации, эпи-
бол и и и деламинации. В результате образуется гаструла
(см. рис. 26). При иммиграции часть бластомеров стенки
бластулы перемещается внутрь зародыша, образуя внутренний
слой — энтодерму. Инвагинация, или впячивание (in —
в, лат. vagina— влагалище), например у ланцетника, заключается
в том, что часть стенки (дно) вдавливается внутрь бластулы.
Эпиболия — обрастание (от греч. epibole — накладывание,
набрасывание) быстро делящимися клетками одного участка
стенки бластулы других участков, где темп дробления замед-
98
лен, например у амфибии. Это происходит в тех случаях, когда
бластомеры вегетативной области перегружены желтком, медленно
делятся и не могут инвагинироваться. При деламинации,
или расщеплении (от лат. de — отделение, lamina — пластинка),
например у птиц, бластомеры стенки бластулы (зародышевого
щитка) делятся тангенциально, что приводит к образованию двух
слоев клеток — первичной эктодермы — наружного и первичной
энтодермы — внутреннего. У позвоночных имеется комбинация
двух или трех из указанных путей, но с преобладанием какого-
либо одного.
В гаструле ланцетника можно выделить первичный наружный
зародышевый листок (эктодерма), который образовался из крыши
бластулы. Он включает в себя закладки эктодермы и нервной
пластинки; первичный внутренний зародышевый листок (энтодер-
ма), образовавшийся из дна бластулы и включающий в себя за-
кладки энтодермы, мезодермы и хорды; щелевидный бластоцель
и новую полость — гастроцель, открывающуюся во внешнюю сре-
ду первичным ртом, или бластопором (от греч. blastos — зачаток,
poros — отверстие). Первичный рот ограничен 4 губами: дорсаль-
ной, соответствующей спинной стороне зародыша, вентральной,
или брюшной, и лежащими между ними боковыми губами. В со-
став губ бластопора входит клеточный материал краевой зоны
бластулы. Далее зародыш растет в длину, причем губы бластопо-
ра постоянно сближаются.
Гаструляция у птиц с меробластическим типом дробления и
дискоидальной бластулой включает два основных механизма —
деламинацию и иммиграцию. В первой стадии гастру-
ляции в результате деламинации образуются первичный наруж-
ный листок (эпибласт) и первичный внутренний листок (гипо-
бласт) (рис. 27, I, II). Эпибласт включает в себя зачатки нервной
пластинки, мезодермы, и хорды.
Клетки гипобласта центральной части диска мелкие, содер-
жат мало желточных включений. В будущем из них образуется
зародышевая энтодерма (кишечная). Периферическая зона ги-
побласта представлена крупными перегруженными желтком клет-
ками, формирующими внезародышевую энтодерму (желточную).
Центральная часть диска (просвечивающая область) выглядит
прозрачной: здесь выделяется еще овальное поле, занимающее ее
центральную часть. Это зародышевый щиток, из которого возни-
кает тело зародыша.
Во второй стадии гаструляции (иммиграция) материал с пер-
вичного наружного листка по краям зародышевого диска пере-
мещается по направлению к заднему концу зародыша (см. рис. 27,
//). В месте встречи двух клеточных потоков возникает вытянутое
скопление клеток — первичная по тоска, которая на переднем кон-
це заканчивается плотным первичным (головным) узелком. Кпе-
реди от первичного узелка, располагаясь между внутренним и
наружным зародышевыми листками, врастает клеточный тяж —
хорда, а из области первичной полоски по сторонам ее — два ме-
99
Рис. 27. Гаструла у птиц (по А. Г. Кнорре).
/ — ранняя стадия (деламинация и миграция): 1 — эпибласт; 2 — гипобласт; II схема
перемещения клеточного материала в бластодиске куриного зародыща в стадии закладки
осевых органов. А — образование первичной полоски и первичного узелка; Б — образова-
ние хордомезодермального зачатка; 1 — эктодерма; 2 — материал будущей нервной пла-
стинки; 3 — материал будущей хорды; 4— первичный (головной) узелок; 5—первичная
ямка; 6 — первичная полоска; 7—первичная бороздка; 8 — хорда; 9 — мезодерма; крас-
ные стрелки обозначают направление перемещения материала в составе наружного листка,
а синие — в составе среднего листка.
зодермальных пласта. Таким образом, первичный узелок соот-
ветствует дорсальной губе, а первичная полоска — боковым гу-
бам бластопора. Образование нервной трубки и дифференцировка
мезодермы совершаются так же, как и у других позвоночных.
Таким образом, у птиц хордомезодермальный зачаток развивается
вне связи с энтодермой (см. рис. 27, И; рис. 28). Сходные про-
цессы имеют место у рыб и рептилий.
Описанные особенности гаструляции сохраняются и у млеко-
питающих. В бластоцисте, стенка которой образована трофоблас-
том, прилегающая к нему изнутри группа клеток — эмбриобласт —
уплощается и соответствует зародышевому щитку описанных жи-
вотных. Путем деламинации из эмбриобласта образуются два
100
Рис. 28. Образование осевых зачатков у зародыша курицы (по А. Г. Кнорре).
А, Б, В — три последовательные стадии: / — нервный желобок; 2 — нервная трубка;
3 — эктодерма; 4 — хорда; 5 — мезодерма; 6 — сомиты; 7 — висцеральный листок сплан-
хнотома; 8 — париетальный листок спланхнотома; 9 — кишечная энтодерма.
листка — наружный (эпибласт), обращенный к трофобласту, и
внутренний (гипобласт), под которым находится жидкость по-
лости бластоцисты. В наружном листке зародышевого щитка,
как и у птиц, происходят перемещения материала, приводящие
к образованию первичной полоски и первичного узелка, из кото-
рых в дальнейшем формируются соответственно мезодерма и
хорда.
Факторы, вызывающие гаструляцию. Способы гаструляции за-
висят от предшествующих стадий развития и в конечном счете
от степени накопления желтка в яйцеклетке. На примере живот-
ных с умеренно телолецитальными яйцеклетками видно, что тем-
пы деления клеток (бластомеров) неодинаковы: на ани-
мальном полюсе они выше, чем на вегетативном. На стадии блас-
тулы краевая зона оказывается областью наиболее быстро раз-
множающихся клеток, что приводит к созданию определенного
101
повышенного давления, а затем к наползанию клеток в области
краевой зарубки на материал дна бластулы. При этом клетки кры-
ши и краевых зон бластулы отличаются примерно в 4 раза более
высоким уровнем метаболизма по сравнению с клетками дна.
Этот дорсовентральный метаболический градиент
определяет в дальнейшем дорсовентральный и краниокаудальный
градиенты дифференцировки. Ускоренное размножение клеток бо-
ковых губ бластопора способствует образованию мезодермы и
прорастанию ее в щель между экто- и энтодермой амфибий.
Определенную роль в процессе гаструляции играет разность
поверхностного натяжения клеток, при которой сме-
щение одних клеток увлекает за собой другие. Состояние неоди-
накового поверхностного натяжения может поддерживаться а к-
тивными амебоидными движениями клеток.
Несомненным является также индуктивный фактор.
Так, если, маркировав каким-либо красителем клетки зародыше-
вых листков, диссоциировать зародыш, а затем перенести в ис-
ходные условия, то клетки эктодермы выстраиваются в один
пласт, а клетки энтодермы — в другой; между ними залегают
клетки мезодермы. Согласно теории организационных
центров, предложенной Г. Шпеманом, в определенных пунктах
зародыша возникают организующие факторы (индукторы), кото-
рые оказывают индуцирующее влияние на другие участки зародыша,
обусловливая их развитие в определенном направлении. Так, ор-
ганизатор, находящийся в дорсальной губе бластопора, индуци-
рует участок эктодермы, обусловливая его дифференцировку в
нервную пластинку; это организатор 1-го порядка. В свою очередь
в нервной пластинке возникает организатор 2-го порядка, который
вызывает превращение участка самой нервной пластинки в глазной
бокал и т. д. Опыты Г. Шпемана и X. Мангольд с пересадкой
материала дорсальной губы от одного зародыша другому на вен-
тральную сторону тела привели к появлению животного, у которо-
го были две закладки хордомезодермы и две закладки нервной
системы. В настоящее время выяснена химическая природа
многих индуцирующих факторов (белки, нуклеопротеиды, стеро-
иды и др.). Установлена роль межклеточных взаимо-
действий, осуществляемых с помощью щелевых контактов.
Дифференцировка зародышевых листков. Мезенхима
Под дифференцировкой понимают изменения в
структуре клеток, связанные с их функциональной специализа-
цией, обусловленные активностью определенных генов. В резуль-
тате репрессии и депрессии различных генов возникают морфо-
логические и химические отличия между имеющими одинаковый
геном клетками организма. В развивающемся организме диффе-
ренцировка сопровождается определенной организацией или раз-
мещением специализирующихся клеток, что выражается в уста-
102
новлении определенного плана строения в ходе онтогенеза (мор-
фогенеза).
Различают 4 основных этапа дифференцировки. Первый этап —
оотипическая дифференцировка, когда материал
будущих зачатков представлен презумптивными участками цито-
плазмы яйцеклетки или зиготы. Например, у амфибий материал
будущей хордомезодермы находится в участке цитоплазмы, на-
зываемом серым серпом. Второй этап — бластомерная
дифференцировка. У некоторых животных различие в
клеточном материале устанавливается очень рано: уже первые
бластомеры отличаются друг от друга. В стадии бластулы блас-
томеры крыши, дна и краевой зоны зародыша еще более разли-
чаются. Третий этап — зачатковая дифференциров-
к а, которая выражается в появлении обособленных участков —
зародышевых листков (стадия ранней гаструлы) и четвертый —
гистогенетическая дифференцировка зачатков
тканей (стадия поздней гаструлы), когда в пределах одного
зародышевого листка появляются зачатки различных тканей,
например в сомитах мезодермы. В основе гистогенетической
дифференцировки лежит процесс дифференцировки и специализа-
ции клеток зародышевых листков. При этом формируются
диффероны — последовательные ряды клеток, каждый из которых
развивается из общего предшественника. Стволовой дифферон —
совокупность клеток, развивающихся по общей схеме, включает
в себя исходные полипотентные клетки (стволовую и полуство-
ловую клетки), из которых формируются унипотентные клетки и
бласты, а из последних образуются через ряд стадий созревания
зрелые клетки.
В процессе гаструляции и дифференцировки зародышевых
листков выделяется осевой комплекс эмбриональ-
ных зачатков органов.
Зародышевые листки являются эмбриональными источниками
развития тканей (гистогенез), из совокупности и взаимо-
действия которых развиваются органы (органогенез). В процессе
эмбрионального гистогенеза происходят клеточное размножение,
рост, миграция, дифференцировка клеток, установление межкле-
точных и межтканевых взаимодействий и гибель клеток.
Каждый зародышевый листок дифференцируется в определен-
ных направлениях, и у большинства позвоночных это происходит
сходным образом (схема 1).
При дифференцировке первичной эктодермы образуются нерв-
ная трубочка, ганглиозные пластинки, плакоды, кожная экто-
дерма, прехордальная пластинка, а также внезародышевая экто-
дерма. Дифференцировка первичной энтодермы приводит к
образованию зародышевой (кишечной) и внезародышевой (жел-
точной) энтодермы.
При дифференцировке мезодермы дорсальный отдел ее сначала
подразделяется на сомиты, начиная с головного конца. В каждом
сомите из наружной части дифференцируется дерматом — ис-
юз
Схема I. Дифференцировка зародышевых листков (по В. В. Яглову).
Эмбриональные Части Тканевые производные
зачатки
Продолжение
Эмбриональные
зачатки
Тканевые производные
А Дифференцировка первичной эктодермы
Эмбриональные
зачатки
Тканевые
производные
точник соединительнотканной части кожи — дермы, из внутрен-
ней (медиальной) —склеритом — источник хрящевой и костной
тканей, из центральной — миотом — источник скелетной мышечной
ткани. Из сегментных ножек (нефрогонотом) закладывается
эпителий почек и гонад.
Вентральная мезодерма — спланхнотом — расщепляется на два
листка, из которых образуются наружные оболочки многих внут-
ренних органов (серозные оболочки).
В процессе развития зародыша очень рано образуется мезен-
хима, представляющая собой скопления отростчатых клеток. Она
появляется на ранних стадиях, тотчас после сформирования за-
родышевых листков, заполняя промежутки между ними (рис. 29).
Мезенхима представляет собой эмбриональный зачаток многих
тканей и органов (см. схему 1, В). У позвоночных главным источ-
ником мезенхимы служит мезодерма (из области спинных
сегментов, из стенок спланхнотома). В образовании мезенхимы
принимают участие и другие закладки. Так, из нервной пластинки
формируется мезенхима, которую называют эктомезенхимой,
или нейромезенхимой. Она принимает участие в образовании
мозговых оболочек и др.
Провизорные органы (внезародышевые органы)
Провизорные, или временные, органы, развивающиеся в про-
цессе эмбриогенеза вне тела зародыша, выполняют многообраз-
ные функции, обеспечивающие рост и развитие самого зароды-
ша. В связи с тем что некоторые из этих органов окружают
зародыш, распространено и другое название — зародышевые обо-
лочки. К ним относят: желточную, амниотическую, серозную обо-
лочки, аллантоис, хорион, плаценту. В эволюции они появились не-
одновременно.
Желточный мешок
В ряду хордовых животных внезародышевые органы впервые
появляются у рыб в виде желточного мешка, депонирующего
желток, используемый зародышем в процессе развития. Его фор-
мирование начинается на стадии ранней гаструлы, когда во внут-
реннем листке можно выделить зародышевую (кишечную) энто-
дерму и расположенную по периферии диска внезародышевую жел-
точную энтодерму. Своим свободным краем желточная энтодерма
образует край обрастания, который начинает надвигаться на жел-
ток. После возникновения хордомезодермального зачатка между
экто- и энтодермой прорастают париетальный и висцеральный
листки мезодермы. Желток обрастает всеми четырьмя листками.
Зародыш приподнимается над диском и отделяется от желтка
туловищной складкой. При образовании туловищной складки
зародышевая энтодерма, до того распластанная на желтке, сво-
рачивается в кишечную трубку. Зародыш связан с желточным
106
Рис. 29. Мезенхима у зародыша морской свинки в стадии 12 сомитов
(по А. А. Максимову).
1 — нервная трубка; 2 — эктодерма; 3 — сомит; 4 — мезенхима, образовавшаяся из меди-
альной части сомита; 5 — аорта; 6 — кровяные клетки; 7 — стейка кишки; 8 — хорда;
9, 11 — висцеральный и париетальный листки мезодермы; 10— целомическая полость.
мешком полым канатиком — желточным стебельком. Желточный
мешок у рыб выполняет трофическую функцию. Другая
функция мешка — кроветворная — заключается в образова-
нии в мезодерме его стенки клеток крови.
С выходом животных на сушу (у пресмыкающихся и птиц)
в связи с развитием зародыша под скорлупой появляются новые
зародышевые органы: амнион, серозная оболочка и аллантоис.
Как и у рыб, у пресмыкающихся и птиц возникают туловищные
складки, отделяющие зародыш от желточного мешка. Желточный
мешок у них также выполняет трофическую и кроветворную
функции.
Амнион
Позднее в эмбриогенезе пресмыкающихся и птиц за счет экто-
дермы и париетального листка мезодермы формируются амнио-
тические складки, растущие в направлении дорсальной поверх-
ности зародыша. По мере роста головного конца зародыша
амниотические складки как бы надвигаются спереди на зародыш,
причем он одновременно вдавливается в желток. Нарастающие
на зародыш складки смыкаются и оба листка — эктодерма и
107
4
1
8
I
Рис. 30. Схема развития внезародышевых органов у птиц (I) и млекопитаю-
щих (II).
А — сближение амниотических складок и начало образования аллантоиса; Б, В — сфор-
мированные внезародышевые органы; / —тело зародыша; 2 — туловищная складка; 3 —
амниотические складки; 4 — полость амниона; 5 — серозная оболочка; 6 — аллантоис;
7 — желточный мешок; 8 — амниотическая оболочка; 9 — кишка; 10 — хорион. Красная
линия —эктодерма, желтая — энтодерма, синяя — мезодерма.
прилегающий к нему париетальный листок мезодермы — сраста-
ются с одноименными листками противоположной стороны. Из
двух листков складок при этом образуются две оболочки —
амниотическая, или водная, обращенная к зародышу, и серозная',
наружная (рис. 30, I).
Амниотическая оболочка на ранних стадиях отделена от тела
зародыша узкой щелью, которая позднее превращается в запол-
ненную жидкостью амниотическую полость. Эта жидкость, выра-
батываемая клетками эктодермы амниотической оболочки, обра-
щенной в полость амниона, содержит белки, углеводы. Жид-
кая среда амниона обеспечивает условия для свободного раз-
вития зародыша, а также амортизации возможных сотрясений
и ударов.
Не следует путать с серозной оболочкой, покрывающей снаружи большинство
внутренних органов.
108
Серозная оболочка
Серозная оболочка образуется одновременно с амниотической
оболочкой у пресмыкающихся и птиц. Серозная оболочка участ-
вует в снабжении эмбриона кислородом, что позволяет рассмат-
ривать ее как провизорный орган дыхания у яйце-
кладущих позвоночных.
Аллантоис
В отличие от амниона развитие аллантоиса начинается в
каудальном отделе самого зародыша в виде выроста вентральной
стенки задней кишки, образованного энтодермой и висцеральным
листком мезодермы. Проксимальная часть аллантоиса располага-
ется вдоль желточного стебелька, а дистальная, разрастаясь,
врастает в щель, между амнионом и серозной оболочкой. Это
орган газообмена и выделения: по сосудам, обра-
зующимся в мезодерме аллантоиса, доставляется кислород; в
аллантоис выделяются продукты обмена веществ зародыша.
Сращенную серозную оболочку и аллантоис у птиц и пресмы-
кающихся часто называют хорионом, или хориоаллантоисом.
У млекопитающих внезародышевые органы пред-
ставлены хорионом, амнионом, желточным мешком и аллантоисом
(рис. 30, II).
Хорион
Хорион, или ворсинчатая оболочка, развивается из трофо-
бласта и внезародышевой мезодермы. Первоначально трофобласт
представлен оболочкой с первичными ворсинками, за счет которых
после имплантации зародыша устанавливается связь с материнс-
ким организмом. Со времени появления в эмбриобласте внеза-
родышевой мезодермы (у человека — на 2—3-й неделе развития)
она подрастает к трофобласту и образует вместе с ним вторичные
эпителиомезенхимальные ворсинки. С этого времени трофобласт
превращается в хорион, или ворсинчатую оболочку.
Внедряясь в слизистую оболочку матки, хорион образует,
вместе с ней плаценту.
Плацента
Функции плаценты многообразны: трофическая, депо-
нирующая, дыхательная, выделительная, эндо-
кринная, защитная. По строению различают 4 типа пла-
цент: эпителиохориальную, десмохориалъную, эндотелиохориаль-
ную и гемохориальную (рис. 31, I, А, Б, В, Г), по характеру тро-
фики — два типа. В плаценте 1-го типа (М. Я. Субботин) хорион
поглощает из материнских тканей преимущественно белки и рас-
щепляет их до полипептидов и аминокислот; синтез эмбриоспе-
109
Рис. 31. Типы плацент и плацентарный барьер (Рис. Ю. И. Афанасьева)
/ — типы плацент: А — эпителиохориальная плацента (у свиньиI; Б— десмохориальная
плацента (у жвачных); В — эндотелиохориальная плацента (у хищных);Г — гемохориаль-
ная плацента (у человека и обезьяны); / — трофобласт; 2 — эмбриональная соединитель-
ная ткань хориона с сосудами зародыша; 3 — эпителий матки; 4 — соединительная ткань
слизистой оболочки матки с материнскими сосудами: 5 — материнские сосуды, контак-
тирующие с трофобластом; 6 — сосуды матери, открывающиеся в лакуны; 7 — кровенос-
ные лакуны (по Флугфельдеру); 8— фибриноид; 9 — мышечная оболочка; 11—плацен-
тарный барьер.
цифических белков происходит главным образом в печени эмбрио-
на. К этому типу относятся диффузные эпителиохориальные
плаценты, у которых ворсины хориона, врастая в отверстия
маточных желез, контактируют с эпителием этих желез (напри-
мер, у верблюда, лошади, свиньи и китообразных — дельфина,
по
кита); множественные десмохориалъные плаценты, в которых
хорион частично разрушает эпителий маточных желез и ворсины
врастают в подлежащую соединительную ткань, например у жвач-
ных парнокопытных млекопитающих (коровы, овцы). В плаценте
1-го типа обеспечивается донашивание зародыша до такого состо-
яния, что к моменту родов он уже способен к самостоятельному
питанию и передвижению.
В плацентах 2-го типа хорион усваивает из материнских
тканей преимущественно аминокислоты и синтезирует эмбрио-
специфические белки; эмбрион получает таким образом готовые
белки, которые использует для строительства собственных тканей.
К таким плацентам относятся эндотелиохориальная, образованная
ворсинами, расположенными в виде пояса в средней части хорио-
на, которые разрушают эпителий и соединительную ткань и кон-
тактируют с эндотелием сосудов. Плацента подобного рода харак-
терна для хищных (кошачьи, псовые, куницеобразные) и ластоно-
гих (тюлени, моржи) млекопитающих. Гемохориальная плацента,
свойственная насекомоядным (крот, еж, выхухоль), рукокрылым
(летучая мышь), грызунам (крыса, бобр), зайцеобразным (кро-
лик), приматам и человеку, при своем образовании разрушает
стенку сосудов матки и ворсинки хориона входят в контакт не-
посредственно с материнской кровью. Этим и обусловлено назва-
ние данной плаценты. Синтез эмбриоспецифических белков у
животных и человека, имеющих плаценту 2-го типа, происходит
преимущественно в хорионе и поэтому с рождением уровень
синтетических процессов резко уменьшается. Естественно, что
такие зародыши после рождения сравнительно долгое время
метаболизируют лишь материнское молоко и неспособны само-
стоятельно питаться.
Через плаценту проходят из крови матери аминокислоты,
глюкоза, липиды, электролиты, витамины, гормоны, иммуноглобу-
лины, вода, кислород, а также лекарственные средства и вирусы.
Из эмбриона в кровь матери выделяются продукты метаболизма
и углекислый газ.
Кровь матери и плода никогда в норме не смешивается, благо-
даря наличию гематоплацентарного барьера. Он состоит из эндо-
телия сосудов хориона, его базальной мембраны, окружающей
этот сосуд рыхлой волокнистой соединительной ткани, базальной
мембраны трофобластического эпителия, цитотрофобласта, синци-
тиотрофобласта. Одной из важных функций этого барьера является
обеспечение иммунологического гомеостаза в системе мать —
плод.
Критические периоды развития
В ходе онтогенеза, особенно эмбриогенеза, отмечаются перио-
ды более высокой чувствительности развивающихся половых
клеток (в период прогенеза) и зародыша (в период эмбриогенеза).
Впервые на это обратил внимание австралийский врач Норман
ill
Грегг (1944). Советский эмбриолог П. Г. Светлов (1960) сформу-
лировал теорию критических периодов развития и проверил ее
экспериментально. Сущность этой теории заключается в утвержде-
нии общего положения, что каждый этап развития зародыша в
целом и его отдельных органов начинается относительно коротким
периодом качественно новой перестройки, сопровождающейся
детерминацией, пролиферацией и дифференцировкой клеток.
В это время эмбрион наиболее восприимчив к повреждающим
воздействиям различной природы (рентгеновское облучение, ле-
карственные средства и др.).
Такими периодами в прогенезе является спермио- и овогенез
(мейоз), а в эмбриогенезе — оплодотворение, имплантация, во
время которой происходят гаструляция, дифференцировка зароды-
шевых листков и закладка органов, период окончательного созре-
вания и формирования плаценты, становление многих функцио-
нальных систем, рождение.
Среди развивающихся органов и систем человека особое место
принадлежит головному мозгу, который на ранних стадиях высту-
пает в роли первичного организатора дифференцировки окружаю-
щих тканевых и органных зачатков (в частности, органов чувств),
а позднее отличается интенсивным размножением клеток (при-
мерно 20 000 в минуту), что требует оптимальных условий трофики.
Повреждающими экзогенными факторами в критические пе-
риоды могут быть химические вещества, в том числе многие ле-
карственные, ионизирующее облучение (например, рентгеновское
в диагностических дозах), гипоксия, голодание, наркотики, ни-
котин, вирусы и др.
Химические вещества и лекарства, проникающие через пла-
центарный барьер, особенно опасны для зародыша в первые
месяцы беременности, так как они не метаболизируются и накап-
ливаются в повышенных концентрациях в тканях и органах за-
родыша. Наркотики нарушают развитие головного мозга. Голода-
ние, вирусы вызывают пороки развития и даже внутриутробную
гибель.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ЧЕЛОВЕКА
В процессе эмбрионального развития человека сохраняются
общие закономерности развития и стадии, характерные для поз-
воночных животных. Вместе с тем появляются особенности, от-
личающие развитие человека от развития других представителей
позвоночных; знание этих особенностей необходимо врачу. Про-
цесс внутриутробного радвитиядзародыша щ^овека продолжается в
среднем 280 суток (10 лунных месяцев). Эмбриональное разви-
тие_дюловека можно разделить на три периода: начальный
(1-я неделя развития). з а рдэ д ы щ с взд й (2 8-я неделя раз-
вития), п л о д н ы й (с 9-й недели развития до рождения ребен-
ка). К_конш1 зародышевого периода заканчивается закдадка ос-
новных эмбриональных зачатков тканей и органов и заро ыш
112
приобретает основные черты, характерные для человека. К 9-й не-
деле развития (начало 3-го_ме£яца) длина зародыша составляет
40 мм, а масса около_5 г. В курсе эмбриологии человека, изучае-
мом на кафедре гистологии и эмбриологии, основное внимание
уделяется особенностям половых клеток человека, оплодотворе-
ния и развития человека на ранних стадиях (начальный и заро-
дышевый периоды), когда происходят образование зиготы, дроб-
ление, гаструляция, формирование зачатков осевых органов и за-
родышевых оболочек, гистогенез и органогенез, а также взаимо-
действия в системе мать — плод. Процессы формирования систем
органов у плода подробно рассматриваются в курсе анатомии.
Прогенез
Половые клетки
Мужские половые клетки. Спермии человека образуются в
течение всего активного полового-периода^в больших количествах.
Продолжительность развития зрелых сперматозоидов из родо-
начальных клетбк^^сперматоРбнТГй — составляет около 72 дней.
Подробное описание—процессов спсрматогенеза дается в главе
XXII. С фор м ирова н ный сперматозоид имеет размер около 70 м к м
и состоит из головки и хвоста (см. рис. 23). В ядре сперматозои-
да человека содержится 23 хромосомы, одна из которых является
половой (X илй~У~) остальные — аутосомами. Среди спермиев
50% содержат Х-хромосому—и 50% — Y-хромосому. Показано,
что масса "Х-хромбСоМЫ больше массы Y:хромосомы, поэтому
спермии, содержащие Х-хромосому, менее по вижны, чем содер-
жащие_У^хромосому.
У человека объем эйякулята в норме составляет около 3 мл;
в нем содержится в среднем 350 млн. сперматозоидов. Для обеспе-
чения оплодотворения общеб^количество сперматозоидов в сперме
должно быть не менее-150 млн., а концентрация их в 1 мл —
не менее 60 млнГВ половых путях женщины после копуляции их
число уменьшается по направлению от влагалища к дистальному
концу маточной трубы. Благодаря высокой подвижности спермато-
зоиды при оптимальных условиях могут через 30 мин — 1 ч дости-
гать полости маткина через I1 '2—2 ч находиться в дистальной
(ампулярной) части маточной трубы, где происходят встреча с
яйцеклеткой и оплодотворение. Спермии сохраняют оплодотво-
ряющую способность до 2 сут.
Женские половые клетки. Образование женских половых кле-
ток (овогенез) совершается в яичниках циклически, при этом в
течение овариального цикла каждые 24—28 дней образуется, как
правило, один овоцит 1-го порядка (см. гл. XXII). ^Вышедший
из яичника при овуляции овоцит 1-го порядка имеет диаметр око-
ло 130 мкм и окружен плотной блестящей танои, или мембраной,
и вен ом фолликулярных клеток, число которых достигает 3—
4 тыс. Он подхватываетсяГбйХримками маточной трубы (яйцевода)
ИЗ
Рис 32. Яйцеклетка и зигота человека (по Б. П. Хватову).
А яйцеклетка человека после овуляции: / цитоплазма; 2 — ядро, 3 блестящая обо-
лочка; 4 фолликулярные клетки, образующие лучистый венец. Б — зигота человека в
стадии сближения мужского и женского ядер (пронуклеусов): / — женское ядро, 2
мужское ядро.
и продвигается по ней. Здесь и заканчивается созревание по-
ловгПГ клетки. При этом в результате второго деления созрева-
ния образуется овоцит 2-го порядка (яйцеклетка), который утра-
чивает центриоли и тем самым способность к делению. В ядре
яйцеклетки человека содержится 23~хромосомы; одна из них явля-
ется^половой Х-хромосомой.
Яйцеклетка женщины (как и млекопитающих животных) вто-
рично изолецитального. jHna, содержит небольшое количество
желточных зерен, более-или менее равномерно расположенных в
орплазме (рис. 32, А, Б). Свой резерв .питательных веществ яйце-
клетка человека .обычно расходует в течение 12—24 ч после ову-
ляции, а затем погибает, если не будет оплодотворена.
Эмбриогенез
Оплодотворение
Оплодотворение происходит в ампулярной части яйцевода.
Оптимальные условия для взаимодействия сперматозоидов с яйце-
клеткой обычно создаются в пределах 12 ч после овуляции. При
осеменении многочисленные спермин приближаются к яйцеклетке
и вступают в контакт с ее оболочкой. Яйцеклетка начинает со-
вершать вращательные движения вокруг своей оси со скоростью
4 вращения, в минуту. Эти движения обусловлены влиянием биения
‘жгутиков сперматозоидов и продолжаются около 12 ч. В процессе
взаимодействия мужской и женской половых клеток в них проис-
ходит ряд изменений. Для спермиев характерны явления к а п а-
цщтаци.и и акросомальная реакция. Капаци-
114
т а ц и я представпяр'и сабаи^прпцесс активации спермиер, кото-
рыи происходит—В .яйцеводе под влиянием слизистого декрета его
железистых клеток. В механизмах ткапаЦитации ольшое Значение
принадлежит1 ГИртональным факторам1 прежде всего прогестерону
(гормон желтого тела), активизирующему_секрецию железистых
клеток яйцеводов. После капацитации следует акросомаль-
jta я. Р-еа kllh я, при которой происходит выделение из сперма-
тозоидов ферментов — гиалуронидазы и трипсина, играющих
важную роль в процессе оплодотворения. Гиалуронидаза расщеп-
ляет гиалуроновую кислоту, содержащуюся в блестящей зоне.
Трипсин расщепляет белки цитолеммы яйцеклетки и клеток лу-
ДДСТОГО, рунца. JB резупьтате происходят дйссЛЦИаЦТГя ТГ-уДаЛСПТте
клеток лучистого венца, окружающих яйцеклетку, и растворение
блестящей зоны. В яйцеклетке цитолемма в области прикрепле-
ния спермия образует приподнимающий бугорок, куда входит один
сперматозоид, при этом за счет кортикальной реакции (см. выше)
образуется плотная оболочка — оболочка оплодотворения, препят-
ствующая вхождению других спермиев и явлению полиспермии.
Ядра женской и мужской половых клеток^превращаются в про-
нуклсусы, сближаются, наступает стяя^^инкариона (см. рис. 32, Б).
возникает зигота и к концу 1-х суток после оплодотворения начи-
нается д р о б~л е н и е.
Пол будущего ребенка определяется комбинацией половых
хромосом в зиготе. Если яйцеклетка оплодотворена спермато-
зоидом с половой хромосомой X, то в образующемся диплоидном
наборе хромосом (у человека их 46) содержатся две Х-хромосомы,
характерные для женского организма. При оплодотворении спер-
матозоидом с половой хромосомой Y в зиготе образуется комби-
нация половых хромосом XY, характерная для мужского организ-
ма Таким образом, пол ребенка зависит от половых хромосом
отца. Так как число образующихся сперматозоидов с X- и Y-
хромосомами одинаково, число новорожденных девочек и мальчи-
ков должно быть равным. Однако в связи с большей чувствитель-
ностью эмбрионов мужского пола к повреждающему действию
различных факторов число новорожденных мальчиков немного
меньшед-Чем девочек: на 100 мальчиков рождаются 103 девочки.
В медицинской практике выявлены различные виды патологии
развития, обусловленные аномальным кариотипом. Причиной по-
добных аномалий является чаще всего нерасхождение в анафазе
половинок половых хромосом в процессе мейоза"женских половых
клеток. В результате этого в одну клетку попадают две хромосомы
и формируется набор половых хромосом XX, а в другую не попа-
дает ни одна. При одлодотворении таких яйцеклеток спермиями
с X или Y-половыми хромосомами могут образоваться следующие
кариотипы: 1) с 47 хромосомами, из них—хромосомы- X (тип
XXX) — сверхженский jun, 2) кариотип OY (45 хромосом) —
нежизнеспособный- 3) кариотип XXY (42_хромосом) - мужской
организм с рядом нарушений — уменьшены мужские половые же-
лезы, отсутствует сперматогенез, увеличены молочные железы
115
(синдром Клайнфельтера); 4) тип ХО (45 хромосом) —женский
органи 1.м"с~рядсм_и1м£нений — невысокий рост, недоразвитие по-
ловых—органов____(яичника, матки,„ яйцеводов), отсутствие мен-
струаций и вторичных половых признаков (синдром Тернера).
Дробление
Дробление зародыша человека начинается к концу J-х суток
и продолжается в течение 4~сут после"оплодотворения, по мере
продвижения зародыша по яйцеводу к~матке. Движение зародыша
обеспечи вается перистальтически ми сокращениями мускулатуры
яйцевода, мерцанием ресничек его эпителия, а также перемеще-
нием секрета желез маточной трубьТГПитание зародыша осуще-
ствляется за счет небольших запасов желтка в яйцеклетке и,
возможно, содержимого маточной трубы.
Дробление зиготы человека полное неравномерное
асинхронное. В течение ТОрнь(Я“ суток оно происходит
Медленно." Первое деление завершается_через^30...ч: при этом бо-
розда дробления проходит по меридиану и образуется два бласто-
мера” (рис. 33, А). За стадией двух бластомеров следует стадия
трехбластомерод. Через 40 ч образуются 4 клетки.
С первых же делений, формируются два мида. бластомеров:
«темные» и «светлые». «Светлые» бластомеры дробятся быстрее и
располагаются одним слоем вокруг «темных», которые оказывают-
। ся^в середине зародыша. Из поверхностных «светлых» бластомеров
в дальнейшем возникает”грофобласг^ связывающий зародыш с
материнским организмом и обеспечивающий его питание. Внутрен-
ние^Темные» бластомеры формируют ^.мбриобласт — из него об-
разуются тело, зародышам все остальные внеза] одышевые ор-
ганы, кроме трофобласта. Начиная с трех суток дробление идет
быстрее и на 4-е сутки „зародыш состоит из 7—12 бластомеров.
Уже через 50—60 ч образуется морула, а на 3—4-е сутки начи-
нается формирование бластоцисты — полого пузырька, заполнен-
ного жидкостью (рис. 33, Б).
Бластоциста в течение 3 сут находится в яйцеводе, через
4—4*/? сут "она состоит иТ58 клеток, имеет хорошо развитый
трофобласт и расположенную внутри клеточную массу эмбриоб-
ласта. Через 5—5‘/2 сут бластоциста попадает в матку. К этому
времени она увеличивается "в размерах благодаря росту числа
бластомеров до 107 клеток и объема жидкости вследствие усилен-
ного всасывания трофобластом секрета маточных желез, а также
активной выработке жидкости самим трофобластом. Эмбриобласт
располагается в виде узелка зародышевых клеток, который при-
креплен изнутри к трофобласту на одном из полюсов бласто-
цисты.
Всгечение около 2 еут (с 5-х по 7-е сутки) зародыш проходит
стадию свободной бластоцисты. В этот период в трофобласте и
эмбриобласте происходят изменения, связанные £ подготовкой к
внедрению зародыша в стенку матки — имплантации.
116
Рис. 33. Зародыш человека на ранних стадиях развития (по Гертигу и Рокку).
А — стадия двух бластомеров; Б — бластоциста: / — эмбриобласт; 2 — трофобласт; 3 —
полость бластоцисты.
Бластоциста покрытаоболочкой оплодотворения. В трофобласте
увеличивается количество лизосом, в которых накапливаются
ферменты, обеспечивающие разрушение (лизис)_тканей матки и
тем “Самым' способствующие внедрению зародыша в толщу слизи-
стой оболочки матки. Появляющиеся в трофобласте выросты раз-
рушают оболочку оплодотворения. Зародышевый узелок упло-
щается и пр вращается в зародышевый щиток, в котором начи-
нается подготовка к первой фазе гаструляции. Гаструдяция осу-
ществляется путем деламинации с образованием двух
листков: наружного — ЭПйЬласта~ и внутреннего — гипобласта
(рис. 34).
Имплантац и я (н и д а ц и я) — внедрение зародыша в
стенку матки — начинается с 7-х суток после оплодотворения и
продолжается около ~4О~ч7~П0И~ймпЛаптащии зародыш полностью
погружается в ткани слизистой оболочки маткиГРазлйчаются две
ста дии имплантации:- адгезия (прилипание) и иЗГв а з и я
(проникновение). В п е р в о й стадии трофобласт прикрепля-
ется к слизистой обоу1<)чке'матки“й в нём начинают-дифферен-
цироваться два слоя — цитотрофобласт и симпластотрофобласт,
или плазмодиотрофобласт. Во время втор о и с т а д и и'сймплас-
тотрофоблалт, продуцируя протеолитические ферменты, разрушает
слизистую оболочку матки. При этом формирующиеся ворсинки
трофобласта, в недр я яр ь в матку, последовательно разрушают ее
эпителий, затем подлежащую соединительную ткань и стенки со-
судов,_и трофобласт вступает в непосредственный контакт с
кровью материнских сосудов. Образуется имплантационная ямка,
в которой вок г зародыша появляются участки кровоизлияний.
Трофобласт вначале (первые 2 нед) потребляет продукты распада
материнских тканей (ги стиотрофны й_т и п питания),
затем питание-зародыша осуществляется непосредственно из ма-
нт
Рис.34. Схема дробления, гаструляции и имплантации зародыша человека.
1 — дробление; 2 — морула; 3 бластоциста; 4 полость бластоцисты; 5 — эмбриобласт;
6 — трофобласт; 7 — зародышевый узелок; а — эпибласт; 6 — гипобласт; 8 — оболочка
оплодотворения; 9—амниотический (эктодермальный) пузырек; 10— внезародышевая
мезодерма; // —эктодерма; 12— энтодерма; 13— цитотрофобласт; 14 — синцитиотрофо-
бласт; 15 — зародышевый диск; 16 — лакуны с материнской кровью; 17 — хорион; 18 — ам-
ниотическая ножка; 19 — желточный пузырек; 20 — слизистая оболочка матки.
теринской крови (гематотрофный тип питания).
Из крови матери зародыш получает не только все питательные
вещества, но и кислород, необходимый для дыхания. Одновре-
менно в слизистой оболочке матки усиливается образование из
клеток соединительной ткани богатых гликогеном децидуальных
клеток. После того как зародыш полностью погружается в имплан-
тационную ямку, отверстие, образовавшееся в слизистой оболочке
матки, 1аполняется кровью и продуктами разрушения ткани сли-
зистой оболочки матки. В последующем дефект слизистой обо-
лочки покрывается регенерирующим эпителием.
Период имплантации является первым критическим-
периодом развития зародыша. Гематотрофный тип пита-
ния, сменяющий гистиотрофный, сопровождается переходом к
качественно новому этапу эмбриогенеза — ко второй фазе гастру-
ляции и закладке внезародышевых органов.
Гаструляция
Гаструляция у человека осуществляется в две фазы. Первая
фаза предшествует имплантации или идет в процессе ее, т. е.
118
совершается на 7-е сутки, а вторая фаза начинается только на
14" -15-е сутки. В период между этими фазами активно форми-
руются внезародышевые органы, обеспечивающие необходимые
условия для развития зародыша (см. рис. 34).
Первая фаза гаструляции происходит путем
деламинации, при этом клетки эмбриобласта расщепляются
на два листка — наружный — эпибласт (включает материал экто-
дермы, нервной пластинки, мезодермы и хорды), обращенный к
трофобластуТТГ внутренний — гипобласт (включает материал заро-
дышевой и внезародышевой энтодермы), обращенный в полость
бластоцисты. На 7-е сутки развития обнаруживаются выселив-
шиеся из зародышевого щитка клетки, которые располагаются
в полости бластоцисты и формируют внезародышевую мезо-
дерму (мезенхиму) К 11-м суткам она заполняет полость бласто-
цисты. Мезенхима~подрастает к трофобласту и внедряется в него,
при этом формируется хорион — ворсинчатая оболочка зародыша
с первичными хориальными ворсинками (рис. 35, А. Б).
Внузародышевая мезодерма участвует в формировании закла-
док амниотического (вместе с эктодермой) и желточного (вместе
с’энтодермой) пузырьков. Края эпибласта растут по мезодермаль-
ной закладке и формируют амниотический пузырек, дно которого
обращено к энтодерме. Размножающиеся клетки энтодермы обра-
зуют к 13—14-м суткам желточный пузырек. У человека желточ-
ный мешок практически не содержит желтка, но заполнен сероз-
ной ЖИДКОСТЬЮ.
К 13—14 суткам зародыш имеет следующее строение. Трофо-
бласт вместе с подстилающей его внезародышевой мезодермой
образует хорион (рис. 36, А, Б). В части зародыша, которая об-
ращена в глубь стенки матки, располагаются прилегающие друг
к другу амниотический пузырек и желточный пузырек. Эта часть
прикреплена к_хориону при помощи амнГС&тической. или зароды-
шевой, ножки, образованной внезародышевой мезодермой. При-
легающие друг- к-другу дно,амниотического и крыша желточного
пузырьков образуют зародышевый щиток. Утолщенное дно амнио-
тического_пузырька представляет собой_эпибласт, а остальная
д часть егсустенки — внезародышевую .эктодерму. Крышу желточно-
\ го пузырька образует гидобласт, а стенку его вне щитка — внс-
у зародышевая..энтодерма.
U Та_ким образом, у человека вранние периоды эмбрио-
х) генеза хорошо развиты внезародышевые час-
ти — хорион, амнион и желточный мешок.
Вторая фаза гаструляции начинается на 14—15-е
сутки и продолжается до 17-х суток развития. Она становится
возможной лишь после описанных процессов формирования вне-
зародышевых органов и установления гематотрофного типа пита-
ния. В эпибласте клетки интенсивно делятся и смещаются по на-
правлению к це'йтру~и вглубь, располагаясь между наружным и
внутренним зародышевыми листками. В результате процесса
иммиграции клеточного материала образуется первичная по-
\ . 119
Рис. 35. Зародыши человека 7’/2 и 11 сут развития в процессе имплантации в
слизистую оболочку матки (по Гертигу и Рокку).
А -7 / сут развития; Б — 11 сут развития; / — эктодерма зародыша; 2- энтодерма
зародыша: 3 амниотический пузырек; 4 — вне зародышевая мезодерма 5 — цитотрофо-
бласт; 6 симпластотрофобласт; 7 маточная железа; 8 лакуны с материнской кро-
вью; 9 — эпителий слизистой оболочки матки; 10 — собственная пластинка слизистой обо-
лочки матки; // — первичные ворсинки.
4
O®O
о©
go
о e
6^-^е «rzs’ ,
*z**4* 7?
ет® »Ms W<
Рис. 36. Схема строения двух-
недельного зародыша челове
ка. Вторая стадия гаструля
ции.
А — поперечный срез зародыша;
Б — зародышевый щиток. Вид
со стороны амниотического пу-
зырька. / — хориальный эпите-
лий; 2 — мезенхима хориона;
3 — лакуны, заполненные мате-
ринской кровью; 4 — основание
вторичной ворсины; 5 — амнио-
тическая ножка; 6 — амниоти-
ческий пузырек; 7 — желточный
пузырек; 8 — зародышевый щи-
ток в процессе гаструляции; 9 —
первичная полоска; 10 — зача-
ток кишечной энтодермы; 11 —
желточный эпителий; 12 — эпи-
телий амниотической оболоч
ки; 13 — первичный узелок; 14—
прехордальный отросток; 15 —
внезародышевая мезодерма;
16 — внезародышевая эктодер-
ма; 17 — внезародышевая энто-
дерма; 18 — зародышевая экто-
дерма; 19 — зародышевая энто-
дерма.
лоска, соответствующая по своим потенциям боковым губам
бластопора, и первичный узелок — аналог дорсальной губы. Ямка,
находящаяся на вершине узелка постепенно углубляется и,
прорываясь через эктодерму, превращается в гомолог нейрокишеч-
121
ного канала ланцетника. Клеточный материал эпибласта, располо-
женныикпереди от первичного узелка, через дорсальную губу
смещается в пространство между дном амниотического пузырька
и крышей желточного, давая хордальный отросток. Одновременно
с этим клеточный материал первичной полоски ложится в виде
мезодермальных крыльев в околохордальное положение. Заро-
дыш приобретает трехслойное строение и почти не отличается
от Зародыша птиц на сходной стадии эмбриогенеза.
К этому же времени относится и появление зачатка аллан
тоиса. Начиная с 15-х суток в амниотическую ножку из заднего
отдела кишечной грубки врастает небольшой пальцевидный вы-
рост — аллантоис. Таким образом, к концу второй фазы гастру-
ляции завершается закладка всех зародышевых листков и всех
внезародышевых органов.
На 17-е сутки продолжается за к л а д ка _з а ча т ко в осе-
вых органов.. В этой стадии видны все три зародышевых лист-
ка. В составе эктодермы клеточные элементы располагаются в не-
сколько слоев. Из_зоны головного узелка наблюдается массовое
выселение клеток^ которые, располагаясь между экто- и энтодер-
мой, и~~йбразуют зачаток хорды. Стенки амниотического пузырька
и желточного мешка на большем протяжении двухслойны. В стен-
ке желточного мешка происходит образование кровяных остров-
ков и первичных кровеносных сосудов.
Связь тела эмбриона с хорионом осуществляется за счет сосу-
дов, ТТрорастающих в стенку аллантоиса и ворсинки хориона.
Наружный зародышевый листок в головном конце образован од-
ним слоем клеток, наиболее высоких по медиальной оси зароды-
ша. При переходе в эктодерму амниотического пузырька клетки
его уплощаются. В переднем краниальном отделе можно видеть
первичную полоску и первичный узелок. Полость плодного пузыря
выстлана хорошо развитым наружным листком мезодермы (сома-
топлевры), которая образует также основу хориальных ворсин.
Стенки желточного мешка и амниотического пузырька выстланы
однослойным эпителием (соответственно эктодермального и экто-
дермального происхождения) и висцеральной экзоцеломической
мезодермой. •
Питание и дыхание эмбриона происходит посредством ал-
лантбхориона__Первичные ворсийкй омываются материнской
кровью.
“Начиная_с 20—21-х суток происходит обособление тела заро-
дыша от внезародышевых органов и окончательное формирование
осевых зачатков. Йзменения_в самом зародыше раньше всего
выражаются в дифференцировке мезодермы и расчленении части
ее на сомиты. Поэтому данный период называют сомитным
в отличие от предыдущего, пресомитно г о Периода закладки
осевых зачатков эмбриона.
Обособление тела зародыша .от внезародышевых (провизор-
ных) органов рройсходит путем образования чудовищной складки,
которая на 2б^е сутки достаточно отчетливо выражена. Зародыш
122
все более отделяется от желточного мешка, пока не оказывается
связанным с ним стебельком, при этом формируется кишечная
трубка, '
Дифференцировка зародышевых зачатков
Дифференцировка эктодермы. Л е й р ул я-
ц и я — процесс образования нервной трубки — протекает во вре-
мени неодинаково в различных частях зародыша. Замыкание
нервной трубки начинается в шейном отделе, затем распростра-
няется, кзади и несколько замедленнее — в краниальном направ-
лении, где формируются мозговые пузырьки. Примерно на 25-е
Сутки__нервная трубка полностью_замыкается^ с внешнегГ"срёдой
сообщаются только два незамкнувшихся отверстия на перед-
нем и заднем концах — передний и задний невропрры. Задний
невропор соответствует нейрокишечному каналу. Через—5 -6. сут
оба невропора зарастают. При смыкании боковых стенок нерв-
ных валиков и .образовании нервной трубки появляется группа
эктодермальных__кпето~ образующихся ' в__области среди нения
нейральной и остальной (кожной) эктодермы. Эти клетки, сначала
располагающиеся в виде продольных~рядов по обе стороны между
нервной трубкой и поверхностной эктодермой, образуют нервный
гребень. Клетки нервного гребня способны к миграциям. В туло-
вище мигрирующие^ клетки образуют два главных потока: одни
мигрируют в поверхностном слое, дерме, другие — в брюшном
направлении, образуя парасимпатические и симпатические ганг-
лии и мозговое вещество надпочечников. Часть клеток.летается
в области нервного гребнд^-формируя гангпиозные пластинкгц ко-
торые сегментируются и дают начало. спинномозговым. узлам
Хордальный отросток — провизорный орган — рассасывается.
Д и ф ф ер ё н ц и р ов’к а мезодермы начинается с 20-х
суток "эмбриогенеза* Дорсальные участки мезодермальных листков
разделяются на- плотные сегменты, лежащие по сторонам от
хорды - самитьц Процесс сегментации дорсальной мезодермы и
образования сомитов начинается в головной части зародыша и
быстро распространяется в каудальном направлении* На-22-е.лутки
развития у эмбриона имеется 7 пар сегментов, на 25-е — 14. на
30-е -— 30 и на 35-е сутки — 43—44 пары. В отличие от сомитов
вентральные отделы мезодермы (епданхнотом) не сегментируются,
а расщепляются на два листка — висцеральный и париетальный.
Неболышай участок мезодермы, связывающий сомиты со спланх-
нотомом, разделяется на сегменты — сегментные'ножки_(^нефро-
гонотом). На заднем конце зародыша сегментации этих отделов
не происходит.--Здесь взамен сегментных ножек располагается
несегментированный нефрогенный зачаток (нефрогенный тяж).
В процессе-деффе^нцировки'мёзодермы из дерматома и скле-
ротома возникает“;эмбриоиальньц1 -зачаток -соединительной тка-
ни — мезенхима. В образовании мезенхимы принимают участие
и другис^зародышевые листки, хотя преимущественно она возни-
123
кает из мезодермы. Часть мезенхимы развивается за счет клеток,
имеющих эктодермальное происхождение. Участие в образовании
мезенхимы принимает и зачаток энтодермы головного отдела
кишечной трубки.
Дифференцировка энтодермы, выделение кишеч-
ной энтодермы начинается с момента появления туловищной
складки. Последняя, углубляясь, отделяет зародышевую энтодерму
будущей кишки от внезародышевой энтодермы желточно-
го мешка. В задней_части зародыша_в_со_став образующейся киш-
ки входит и тот участок энтодермы, из которого возникает экто-
дермальный вырост аллатгоиса. В начале 4-й недели на переднем
конце зародыша образуется эктодермальное впячивание — рото-
вая ямка. Углубляясь^ямка _доходит до._переднег..а. .конца кишки
и после прорыва разделяющей их мембраны превращается в рото-
вое отверстие будущего ребенка.
Кишел-ная-Т-Рубка образуется первоначально как часть
энтодермы желточного мешка, затем в состав ее переднего отдела
включается материал прехордальной пластинки. Из материала пре-
хордальной пластинки развивается в дальнейшем многослойный
эпителий переднего отдела пищеварительной трубки и ее произ-
водных. Мезенхима кишечной трубки преобразуется в соедини-
тельную ткань и гладкую мускулатуру.
Анатомическое формирование органов (ор-
ганогенез) совершается параллельно процессам гисто-
генеза (образование тканей).
Внезародышевые органы человека
Хорион
/3- /cf-iZC
Ворсинчатые разрастания трофобласта, именуемые позднее
хорионом, состоят из лвух~труктурных компонентов — эпителия
и внезародышевой мезенхимы. Слизистая оболочка в той части,
которая после импланта щи войдет в состав плаценты — decidua
basalis (основная отпадающая оболочка), разрастается сильнее,
чем в других участках — decidua parietalis (пристеночная отпа-
дающая оболочка) и decidua capsularis (сумочная отпадающая
оболочка, отделяющая зародыш от полости матки) (рис. 37, А, Б).
В дальнейшем это различие выступает все более отчетливо, причем
ворсины в области пристеночной и сумочной оболочек—вообще
исчезают, а в области основной отпадающей оболочки заменяются
сильно разветвленными вторичными ворсинами, строму которых
образует соединительная ткань с кровеносными сосудами. С этого
’момент*| тгтрЙЧП ptrywnrnn rr д— «wraana — летлистый (chorion
frondosum) и гладкий (chorion laeve). В области расположения
ветвистого хориона формируется плацента. За счет основной
отпадающей_ оболочки ^образуется - м a-те р иссека я- часть
1 См. учебник анатомии.
124
Рис. 37. Зародыш человека 9'/2 нед
А: / — амнион; 2 — хорион; 3 — желточный
мешок; 4 — пуповина' 5 — формирующаяся
плацента.
/> схема взаимоотношения зародыша, внезародышевых органов и оболочек матки.
/ — брюшина, 2 — decidua basalts, 3 — полость амниона; 4 — полость желточного мешка;
5 — экстраэмбриональный целом (полость хориона); 6 decidua capsularis; 7 — decidua
parietalis; 8 — полость матки; 9 — шейка матки; 10 - эмбрион; // — межворсинковое про-
странство; 12 — ворсинки хориона; 13 — гладкий хорион; 14 — аллантоис; /5 мезенхима
пупочного канатика (по Гамильтону, Бойд и Моссман)
плаценты (partes maternae), а ^а счет ветвистого хориона —
ее плодная часть (partes fetaIesT7 ВЕёвйс-фыи хорион к
3 мес приобретает вместе с основной отпадающей оболочкой
типичную для сформированной плаценты дискоидальную форму.
Плацентация у человека совершается в течение 3—6-й недели
внутриутробного развития и совпадает с периодом формирования
зачатков органов. Этот период является—вторым—к~рл т и ч е-
с к и-м в эмбриогенезе челове к а, так как различные
патогенные воздействия в это время наиболее часто могут вызвать
нарушения.
Детское место, или плацента
П лднентя — внезародышевый орган, за счет которого ус та на в-
ливается связь зародыша с организмом матери—Плацента чело-
века относится к типу дискоидальных гемохориальных ворсинча-
тых плацент
Это важный временный орган с многообразными функциями,
обеспечивающий связь плода с материнским организмом. Плапен-
та выполняет трофическ у ю,...а кскретор ну_ю(ддя пло-
да ), эндокринную (вырабатывает хориальный гонадотропин,
прогестерон, плацентарный лактоген, эстрогены и др.) защит-
н ю (включая иммунологическую защиту) Однако через плацен-
ту (через гематоплаце тарный баръер)_л£Г¥м проникают алкоголь,
наркотические и лекарственные_вещества, никотин, а также многие
гормоны из крови матери в кровь плода.
125
lUttKI.Vl
В плаценте различают зародышевую, или плодную, часть (pars
fetalis) ухматерНйСк^кГ' или маточную (pars materna) Плодная
часть представлена ветвистым хорионом и приросшей к нему ам-
ниотической оболочкой, а материнская — видоизмененной базалfa-
ной ч~гсТСкгэндометрия (decidua basalis).
Р~а з в и т и е л л ац е н т ы_начинается на 3-й неделе, когда
во вторичные (эпителисмезенхимные ворсины ~начйнают врастать
сосуды и образовываться третичные ворсцны. На 6~8-и неделе
вокруг сосудов дифференцируются макрофаги, фибробласты,~кол-
лагеновые волокна7~В дифференцировке фибробластов и синтезе
коллагена важную роль играют витамины С и А- без ллгтаточ-
ного поступления которых в организм беременной женщины нару-
шается прочность связи зародыша с материнским организмом
и создадутся-угроза самопроизвольного аборта.
Параллельно увеличивается активность гиалуронидазы, за счет
которой и происходит расщепление молекул гиалуроновой кис-
лоты.
Уменьшение вязкости основного вещества создает наиболее
благоприятные условия для обмена веществ между тканями ма-
тери и плода. В основном веществе соединительной ткани хориона
содершит| ч. шащ'тр цц»» чиигстно гиалуцщщнпй и хондроитин-
серной кислот, с которыми связана регуляция проницаемости, пла-
центы.
Формирование коллагеновых волокон в ворсинах совпадает
по времени с усилением протеолитической активности трофоблас-
тического эпителия (цитотрофобласта) и его производного (син-
цитиотрофобласта).
С развитием плаценты происходят разрушение слизистой обо-
лочки матки и смена гистиотрофного питания на
гем адг.о т р о ф н о е. Это означает, что ворсины'хбриона- омы-
ваются__кровью матери, излившейся из разрушенных сосудов
'Зародышевая, или плодна я, часть планенты к концу
3 - го месяца представлена ветвящейся хориальной пласти нкой,
состiianigM из an.uu uujnnii <го шаненовой) соединительной гкани,
покрытой—цито- и ечнпитиотрофобластом-^ Ветвящиеся ворсины
хориона (стволовые, или якорные, ворсины) хорошо развиты лишь
со стороны, обращенной к миометрию. Здесь они проходят~через
всю толщу плаценты и своими вершинами погружаются в базальную
часть разрушенного~эндометрйя. ———
Хориальный эпителий, или цитотрофобласт, на ранних стадиях
развития, представлен однослойным ^эпителием с овальными ядра-
ми. Эти, клетки размножаются митотическим путем. Иа-них-разви-
вается синцитиотрофобласт — многоядерная структура, покрываю-
щая.редуцирующийся цитотрофобласт. В синцитцццюфобласте
со де р жи тся большое количество различных- протепдитиче с к и х и
окислительных ферментощДАТФ азы, щелочная и кислая фосфа-
тазы, 5-нуклеотидазы, ДПН-диафоразы, глюкозо-6-фосфатдегид-
рогеназы (Г-6-ФДГ), а-ГФДГ, сукцинатдегидрогеназа—СДГ,
126
цитохромоксидаза — ЦО, моноаминоксидаза — МАО, неспецифи-
ческие эстеразы, ЛДГ, НАД и НАДФ-диафоразы и др. — всего
около 601, что связано с его ролью в обменных процессах между
организмом матери и плода. В цитотрофобласте и в синцитии
выявляются пиноцитозные пузырьки, лизосомы и другие органел-
лы. Начиная со 2-го месяца хориальный эпителий истончается
и постепенно заменяется-синцитиотрофиблас Юм. В этот период
синцитиотрофобласт по толщине превосходит—цитотрофобласт,
на 9—10-й неделе синцитий истончается, а количество ядер в нем
увеличивается^ На поверхности синцития, обращенной в ла куны,
ПОЯВЛЯЮТСЯ ми/чацир-ненгнте-мжфнворсинки в виде щеточной ка-
емки_(рис. 38).
Между синцитием и клеточным трофобластом имеются щеле-
видные субмикроскопические пространства, доходящие местами
до базальной мембраны трофобласта, что создает условия для
двустороннего проникновения трофических веществ, гормонов
и др. между синцитием и стромой ворсин.
Во_лвторой_половине беременности, и особенно в конце ее,
трофобласт мутями сильно истончается и ворсины покрылаются
фибринополобной оксифильной массой, являющейся, по-видимому,
продуктом свертывания плазмы и распада трофобласта («фибри-
ноид Лангханса»).
С увеличением срока беременности ^уменьшается количество
макрофагов___и коллагенпродуиируюших _ дифференцированных
фибробластов, появляются фиброциты. Количество коллагеновых
волокон, хотя и нарастает, но до конца беременности в большин-
стве ворсин остается небольшим.
Структурно-функциина вьмпй—^дишщей сформированной пла-
центы является котиледон, образованный стволовой ворсиной и
ее вторичными и третичными (конечными4-яаздетвяениями. Общее
количество котиледонов в плаценте достигает200.
М ат еринская часть плаце н т ы представлена ба-
зальной пластинкой и соединительнотканными септами, отделяю-
щими_ котиледоны друглтгщруга, а~ такжё лдкундлш, заполненными
материнской кровью. Н мёстах контакта стволовых ворсин с от-
падающей оболочкой встречаются также трофобластические клет-
ки (периферическии трофобласт).
Уже на ранних стадиях беременности ворсины хориона раз-
рушают наружные, т. е. ближайшие к плоду, слои основной отпа-
дающей оболочки, и на их месте образуются заполненные мате-
ринской кровью лакуны, в которые свободно свисают ворсины
хориона Глубокие неразрушенные^ части отпадающей оболочки
вместе с трофобластом образуют базальную пластинку.
Базальный_слой_ эндометрия (stratum basale endometrii) —
соединительная-.ткань слизистой оболочки матки, содержащая де-
цидуальные _ клетки. Эти крупные, богатые гликогеном клетки
соединительной _ткани расположены—в—глубоких-слоях слизистой
оболочки матки. Они имеют чёткие ' границы, округлые ядра и
оксифильную цитоплазму. В базальной пластинке, чаще в месте
127
6
Рис. 38. Строение плаценты (I) и хориальных ворсин (II).
/ — строение плаценты (по Ю. И. Афанасьеву, Е. Ф Котовскому). /, 3 — плодная часть
плаценты; 1а — слизистая соединительная ткань; 2 — гемохориальное пространство, за-
полненное материнской кровью; 3 — ворсинки хориона; За — фибриноид Рора; 4 — маточ-
ная часть плаценты (базальный слой слизистой оболочки матки); 4а — перегородка между
лакунами; 5 — мышечная оболочка матки; 6 — пупочный канатик; 7 — эпителий амниона;
А — собственный слой амниотической оболочки; 9 — амниотическая оболочка; 10 — мезен
хима (соединительная ткаиь) хориона; // — цитотрофобласт; 12—синцитиотрофобласт;
12а фибриноид Ниттабуха; 13 — хориальная пластинка; 14 — кровеносный сосуд; 15 —
децидуальные клетки; 16 — кровеносный сосуд в мышечной оболочке; А — пупочная ар-
терия; В — пупочные вены
Рис. 38 (продолжение).
// — схема строения хориальных ворсин в I и 111 триместрах беременности: А — 8—9 нед;
Б 40 нед развития; 1 — меж ворсинковое пространство; 2 — микроворсинки; 3 — сим-
пластотрофобласт; 4 — ядра симпластотрофобласта; 5 цитотрофобласт; 6 — ядро цито-
трофобласта; 7 — базальная мембрана; 8 — межклеточное пространство; 9 — фибробласт;
10— макрофаги (клетки Кащенко—Гофбауэра); 11 —эндцтелиоцит; 12— просвет крове
носного сосуда; 13 — эритроцит; 14 базальная мембрана гемокапилляра (по Э. М. Швир-
сту).
прикрепления ворсин к материнской части плаценты, встречаются
скопления клеток периферического цитотрофобласта. Они напоми-
нают децидуальные клетки, но отличаются более интенсивной
базофилией цитоплазмы. Аморфная субстанция (фибриноид Рора)
находится на поверхности базальной пластинки, обращенной к
хориальным ворсинам. Трофобластические клетки базальной плас-
тинки вместе с фибриноидом играют существенную роль в обеспе-
чении иммунологического гомеостаза в системе мать — плод.
Часть основной отпадающей оболочки, расположенной на гра-
। < и не ветвистого и гладкого хориона, т. е. по краю пла це нта р но го
дискщ ппи развитии плаценты не разрушается. Плотно прирастая
к хориону- она образует замыкающую пластинку, препятствующую
истечению крови из лакун плаценты.
Кровь в лакунах-'нёпрёрьтвно обновляется. Она поступает из
маточных артерий, входящих сюда из мышечной оболочки матки.
Эти артерии идут по плацентарным перегородкам и открываются
в лакуны. Материнская кровь оттекает от ппаненты по венам,
берущим -начало от лакун-крупными отверстиями.
Кровь матери и кровь плода ци к лирует по самостоятельным
сосудистым системам и не смешивается муж. собой,""Гелгохо-
5 ГИСТОЛОГИЯ
129
риальныи барьер, разделяющий оба кровотока, состоит из эндоте-
лия~сосудбв плода, окружающей^ сосуды соединительной ткани,
эпителия хориальных ворсин Iцитотрофобласт, синцитиотрофо-
бласт , а, кроме того, из фибриноида, который местами покры-
вает ворсины снаружи.
Формирование плаценты заканчивается в конце 3-го месяца
беременности.
Сформированная к этому времени плацента обеспечивает окон-
чательную дифференцировку и бурный рост образовавшихся в
предыдущем периоде зачатков органов плода.
Желточный мешок /Л-CL -/3 - 't'K'-C-
Жедточный мешок образован внезародышевой энтодермой и
внезародышевой мезодермой, принимает активное участие в пита-
нии и дыхании эмбриона человека очень недолго. После образо-
вания3;улшвищнои^кладкйижелто.иный мешок оказывается связан-
ным с кишкой желточным стебельком. Сам желточный мешок
смещается в пространство между мезенхимой хориона и амниоти-
ческой оболочкой. Его основная роль — к р о в е т в 6 р н а я.
В качестве кроветворного органа он функционирует до 7—8-й не-
дели. щЗзатем подвергается обратному развитию. В составе пупоч-
ного канатика остаток желточного мешка позднее обнаруживается
в виде узкой трубочки. В стенке^ДкелтрчногсГмешка формируются
первичные половые клетки — гонобласты, мигрирующие из него с
кровью в зачатки 'половых желез. ~
Амнион
Амнион очень быстро увеличивается в размерах и к концу
7-й недели его соединительная .ткань входит в контакт с соедини-
тельной тканью хориона При этом эпителий амниона переходит
на амниотическую ножку, пр>-м|>ащамш|укц-а imi/iupp а ^уточный
канатик, и_в области пупочного кольца смыкается с эктодермаль-
ным покровом кожи эмбриона.
Амниотическая оболочка образует стенку резервуара, в кото-
ром находитеяллод. „Основная его функция — выработка около-
плодных вод, обеспечивающих среду для развивающегося организ-
ма и предохраняющих его от механического повреждения. Эпите-
лий амниона, обращенный в его полость, выделяет околоплодные
воды, а также принимает участие и-обратмдм ыасина.чии их. Ам-
ниотическая жидкость создает необходимую для развития заро-
дыша водную среду, поддерживая до конца беременности необхо-
димый состав и концентрацию. солей в околоплодной жидкости
(см. рис. 37, А). Амнион выполняет также защитную функцию,
предупреждая попадание в плод вредоносных агентов.
Э п ителйй'враннихстапиях на всем протя жен и и однослойный
плоский, образован крупными полигональными, тесно прилегаю-
щими друг к другу клетками^в которых постоянно происходит ми-
во
тоз. На 3-м месяце эмбриогенеза эпителий преобразуется в
призматический. Эпителий плацентарного- диска призматический,
местами многорядный. На ^оаеруно<ти эпителия имеются микро
ворсинки. В цигоплазме всегда содержатся небольшие капли ли-
пидов, зерна гликогена и гликозаминогликаны. В апикальных
частях клеток имеются различной величины вакуоли, содержимое
которых выделяется в полость амниона. Эпителий внеплацентарного
амниона кубический- В эпителии амниона, покрывающем плацен-
тарный диск вероятно, имеет место преимущественно сеуэеция,
а в эпителии—внеплацентарного амниона — преим щественно ре-
зорбция околоплодных вод.
В__строме амниотической оболочки различают базальную
мембрану, слой плотной соединительной ткани и губчатый слой
рыхлди~соединитёльной~ткани1 связывающей амнион с хорионом.
В сдое плотной соединительной ткани можно выделить лежащую
под базальной мембраной бесклеточную часть и клеточную часть.
Последняя состоит из нескольких слоев фибробластов, между
которыми находится густая сеть плотно прилежащих друг к другу
то н к их пучков коллагеновых—и ретикулярны х~волокон,~образу ю -
щих неправильную решетку, ориентированную параллельно по-
верхности оболочки.
Губчатый с 1ой образован пнем», 4«г пмзистлй» >_£оедини-
тельной тканью. Редкие пучки коллагеновых волокон, являющиеся
продолжением тех, которые залегают в слое плотной соединитель-
ной ткани, связывают амнион с хорионом. Связь эта очень не-
прочная, и поэтому обе оболочки легко отделить друг от друга.
В основном веществе соединительной ткани много гликозамино-
гликанов.
Аллантоис
Аллантоис представляет собой небольшой пальцевидный отро-
сток энтодермы, врастающий в амниотическукгттожкуг~У "человека
аллантоисупе достигает "большого развития^ но его значение в обе-
спечении питания и дыхания~зародыша все же ведико, _так как
по нему к хориону растут трупы, конечные разветвления которых
залегают в.лрдме ворсин. На 2-м месяце эмбриогенеза аллантоис
редуцируется. /г xz
СЫ • ьъс 7
Пупочный канатик
Пупочный канатик образуется в основном из мезенхимы, на-
ходящейся ЕГамПиотичесиий "ножку и желточном стебельке. В его
формировании принимают участие также-алламтоис-и растущие по
нему сосуды. С поверхности все эти образования окружены ам-
ниотизтеской оболочкой. Желточный стебе пек и аппантоис быстро
редупир)клея. л а пупочном канатике нлнщюжденного обнаружи-
ваются лишь их остатки.
Сформированный пупочный канатик — упругое соединительно-
5*
131
тканное образование, в котором проходят две пупочные артерии
и пупочная вена. Он образован типичной студенистой (слизи-
стой) тканьк>, в которой содержится огромное количество гиалу-
роновой кислоты. Именно эта ткань получившая название варто-
нова студня, обеспечивает тургор и упругость канатика (см. рис. 71).
Покрывающая поверхность канатика амниотическая оболочка
срастается с его студенистой тканью.
Значение этой ткани чрезвычайно велико. Она предохраняет
пупочные сосуды от сжатия, обеспечивая тем самым непрерывное
снабжение эмбриона питательными веществами, кислородом. На-
ряду ТГэтим студенистая ткань тгрепятствует'ттроникновенйю вре-
доносных агента ич плаценты к эмбриону внееосудистым путем
и выполняет, таким образом, защитную функцию.
На. основании изложенного можно отметить о с н о в н ы е
о с о б е н н о с т и ранних_стад и-й—p-а з в и т и .я_з_а р о д ы ш а
ч е ло в е к а: 1) асинхронный тип полного дробления и обра-
зование «светлых» и «темных» бластомеров; 2) раннее обособление
и формирование внезародышевых органов; 3) раннее образование
амниотического пузырька~ПЙ~отсутствие ~~ амниотических складок:
4) наличие двух фаз гаструляции — деламинации и иммиграции,
в течение" которых происходит также развитие провизорных
органов; 5) интерстициальный тип имплантации; 6) сильное
развитие'амниона, хориона и слабое развитие желточного мешка
и аллантоиса^ ' ' -
Система мать — плод
Система мать — плод возникает в процессе беременности и
включает в себя две подсистемы — организм матери и организм
плода, а также плаценту, являющуюся связующим звеном между
ними.
Взаимодействие между организмом матери и организмом плода
обеспечивается прежде всего нейрогуморальными механизмами.
При этом в обеих подсистемах различают следующие механизмы:
рецепторные, воспринимающие информацию, регуляторные, осуще-
ствляющие ее переработку, и исполнительные.
Рецепторные механизмы организма матери
расположены в матке в виде чувствительных нервных окончаний,
которые первыми воспринимают информацию о состоянии раз-
вивающегося плода. В эндометрии находятся хемо, механо- и
терморецепторы, а в кровеносных сосудах — барорецепторы. Ре-
цепторные нервные окончания свободного типа особенно много-
численны в стенках маточной вены и в децидуальной оболочке в
области прикрепления плаценты. Раздражение рецепторов матки
вызывает изменения интенсивности дыхания, уровня кровяного
давления в организме матери, направленные на обеспечение
нормальных условий для развивающегося плода.
Регуляторные механизмы организма мате-
р и включают отделы центральной нервной системы (височная
132
доля мозга, гипоталамус, мезэнцефальный отдел ретикулярной
формации), а также гипоталамоэндокринную систему. Важную
регуляторную функцию выполняют гормоны: половые, тироксин,
кортикостероиды, инсулин и др. Так, во время беременности про-
исходит усиление активности коры надпочечников матери и по-
вышение выработки кортикостероидов, которые участвуют в ре-
гуляции метаболизма плода. В плаценте вырабатывается хорио-
нический гонадотропин, стимулирующий образование адрено-
кортикотропного гормона гипофиза, который активизирует дея-
тельность коры надпочечников и усиливает секрецию кортикосте-
роидов.
Регуляторные нейроэндокринные аппараты матери обеспечи-
вают сохранение беременности, необходимый уровень функциони-
рования сердца, сосудов, кроветворных органов, печени и оптималь-
ный уровень обмена веществ, газов в зависимости от потребно-
стей плода.
Рецепторные механизмы организма плода
воспринимают сигналы об изменениях организма матери или соб-
ственного гомеостаза. Они обнаружены в стенках пупочных
артерий и вены, в устьях печеночных вен, в коже и кишечнике
плода.
Раздражение этих рецепторов приводит к изменению частоты
сердцебиения плода, скорости кровотока в его сосудах, влияет на
содержание сахара в крови и т. д.
Регуляторные нейрогуморальные механиз-
мы организма плода формируются в процессе развития.
Первые двигательные реакции у плода появляются на 2—3-м ме-
сяце развития, что свидетельствует о созревании нервных центров.
Механизмы, регулирующие газовый гомеостаз, формируются в
конце II триместра эмбриогенеза. Начало функционирования
центральной эндокринной железы — гипофиза — отмечается на
3-м месяце развития. Синтез кортикостероидов в надпочечниках
плода начинается со второй половины беременности и увеличи-
вается с его ростом. У плода усилен синтез инсулина, который
необходим для обеспечения его роста, связанного с углеводным и
энергетическим обменом. Следует отметить, что у новорожденных,
родившихся от матерей, страдающих сахарным диабетом, когда
снижена выработка инсулина, наблюдается увеличение массы тела
и повышение продукции инсулина в островках поджелудочной
железы.
Действие нейрогуморальных регуляторных систем плода на-
правлено на исполнительные механизмы — органы
плода, обеспечивающие изменение интенсивности дыхания, сердеч-
но-сосудистой деятельности, мышечной активности и т. д. и опре-
деляющие изменение уровня газообмена, обмена веществ, термо-
регуляции и других функций.
Как уже указывалось, в обеспечении связей в системе мать —
плод особо важную роль играет плацента, которая способна не
только аккумулировать, но и синтезировать вещества, необходимые
для развития плода. Плацента выполняет эндокринные функции,
вырабатывая ряд гормонов: прогестерон, эстроген, хорионический
гонадотропин, плацентарный лактоген и др. Через плаценту между
матерью и плодом осуществляются гуморальные и нервные связи.
Существуют также экстраплацентарные гуморальные связи через
плодные оболочки и амниотическую жидкость.
Гуморальный канал связи — самый обширный и информатив-
ный. Через него происходит поступление кислорода и углекислого
газа, белков, углеводов, витаминов, электролитов, гормонов, анти-
тел и др. В норме чужеродные вещества не проникают из организ-
ма матери через плаценту. Они могут начать проникать лишь в
условиях патологии, когда нарушена барьерная функция плаценты.
Важным компонентом гуморальных связей являются иммунологи-
ческие связи, обеспечивающие поддержание иммунного гомеостаза
в системе мать — плод.
Несмотря на то что организм матери и плода генетически
чужеродны по составу белков, иммунологического конфликта
обычно не происходит. Это обеспечивается рядом механизмов,
среди которых существенное значение имеют: 1 — синтезируемые
синцитиотрофобластом белки, тормозящие иммунный ответ мате-
ринского организма; 2 — хориональный гонадотропин и плацен-
тарный лактоген, находящиеся в высокой концентрации на по-
верхности синцитиотрофобласта, принимают участие в угнетении
материнских лимфоцитов; 3 — иммуномаскирующее действие гли-
копротеинов перицеллюлярного фибриноида плаценты, заряженно-
го так же, как и лимфоциты омывающей крови, отрицательно;
4 — протеолитические свойства трофобласта также способствуют
инактивации чужеродных белков. В иммунной защите принимают
участие и амниотические воды, содержащие антитела, блоки-
рующие антигены А и В, свойственные крови беременной, и
не допускают их в кровь плода в случае несовместимой бере-
менности.
Показана определенная взаимосвязь гомологичных органов ма-
тери и плода: поражение какого-либо органа матери ведет к нару-
шению развития одноименного органа плода. В эксперименте на
животных установлено, что сыворотка крови животного, у которо-
го удалили часть какого-либо органа, стимулирует пролиферацию
в одноименном органе. Однако механизмы этого явления изучены
недостаточно.
Нервные связи включают плацентарный и экстраплацентарный
каналы: плацентарный (у плода — интерорецептивный) — раздра-
жение беро- и хеморецепторов в сосудах плаценты и пуповины,
а экстраплацентарный (у плода — экстерорецептивный) — поступ-
ление в центральную нервную систему матери раздражений, свя-
занных с ростом плода и др. Наличие нервных связей в системе
мать — плод подтверждается данными об иннервации плаценты,
высоком содержании в ней ацетилхолина, отставании развития
плода в денервированном роге матки экспериментальных живот-
ных и др.
134
В процессе формирования системы мать — плод существует
ряд критических периодов, наиболее важных для установления
взаимодействия между двумя системами, направленных на создание
оптимальных условий для развития плода.
В онтогенезе человека можно выделить несколько критических,
периодов развития: в прогенезе, эмбриогенезе и постнатальной
'^з^и. К ним относятся: 1) развитие половых клеток — овоге-
.¥з и сперматогенез; 2) оплодотворение; 3) имплантация (7—
8-е сутки эмбриогенеза); 4) развитие осевых зачатков органов и
формирование плаценты (3—8-я неделя развития); 5) стадия уси-
ленного роста головного мозга (15—20-я неделя); 6) формирова-
ние основных функциональных систем организма и дифференци-
ровка полового аппарата (20—24-я неделя); 7) рождение; 8) пе-
риод новорожденное™ (до 1 года); 9) половое созревание (11 —
16 лет).
ОБЩАЯ ГИСТОЛОГИЯ
Глава VI
ЭПИТЕЛИАЛЬНЫЕ ТКАНИ
Эпителиальные ткани, или эпителии (epithelia), покрывают по-
верх ногти тела, слизистых и серозных оболочек внутренних ор-
ганов_(желудка, кишечника, мочевого пузыря и др.), а также
образуют большинство желез. В связи с этим различают покров-
ный и железистый эпителий^
U ркровный' эпителий является пограничной тканью. Он отделя-
ет организм (внутрёнгпою~среду) иг внешней среды, но одновре-
менно с этим участвует- в обмен*»—веществ^ организма"г окружаю-
щей .средой, осуществляя функции поглощения веществ (всасыва-
ние) и выделения продуктов обмена (экскрецию). Напдумер,
через кишечный эпителий всасываются в кровь и лимфу продукты
переваривания нищи, которые Служат источником энергии и строи-
тельным материалом для организма, а через почечный эпцтелий
выделяется ряд продуктов азотистого обмена, являющихся шла-
ками для организма. Кроме этих функций, покровный эпителий
выполнят ттажную ЦДД^тную функцию, 7Трёдохраняя~подлежащие
ткани организма от различных внешних воздействий — химиче-
с к и х, механических, инфекционных и др. Например, ко жныйэпи-
телий является мощным барьером для микроорганизмов и многих
ядов к нец, эпителий, покрывающий внутренние ррганы, нахо-
дящиеся в полостях тела, адздает уедовия для их подвижности,
напри мер для сокращения сердца, экскурсии легкий и т.
^Келеэистый_ эпителий осуществляет секреторную функцию,
т. е. образует и выделяет специфические продукты — секреты,
которые ибТЮльзуютея—в—процессах,—про [екающих в организме.
Напри мер, секреТ~ поджелудочной железы участвует в перевари ва-
нии белков, жиров й углеводов в тонкой кишке.
ИСТОЧНИКИ РАЗВИТИЯ ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ
Эпителии развиваются из всех трех зародышевых листков
начиная! с~ 3 4 и нед*аи=^эмбриональниги—развития—человека.
В зависимости от эмбрионального источника различают эпителии
эктодермального, мезодермального и эктодермального происхож-
дения.
Строение. Эпителии участвуют в построении многих органов, в
связи с чем обнаруживают большое разнообразие морфофизио-
логических свойств. Некоторые из них являются общими, позво
ляющими отличать эпителии от других тканей организма.
Эпителии предет вляю собой дд^сты клеток — дрителлоцитов
(рис. 39), которые имеют неодинаковую форму и строение в раз-
136
Go Ба 6$
Рис. 39.4^троение эпителиальной клетки (схема).
/ ядро; 2 — митохондрии; 2а комплекс Гольджи; 3 — тонофибриллы, 4 — структуры
апикальном поверхности клеток: 4а микроворсинки, 46 - щеточная каемка 4в рес-
нички; 5 - структуры межклеточной поверхности: 5а - плотные контакты, 56 десмо-
сомыг 6 - структуры базальной поверхности клеток: ба — инвагинация цитолеммы. 66 —
полудесмосомы; 7 — базальная мембрана; 8 — соединительная ткань; 9 — кровеносные
капилляры.
личных видах эпителия. Между клетками, составляющими эпите-
лиальный пласт7~Ттет~-меж клеточного—вещества и клетки тесно
связаны дру_____ругом омощ ю различных контактов —
со-н. и др.. Эпителии пяспплягаю^гя на
базальных мембранах (пластинкахХ- Базальные мембраны имеют
толщину около 1 мкм и состояти^амррфногр^веТцества и^фибрил-
лярных структур. В базальной мембране содержатся углевОДно-
белково-липйдные комплексы, от которых зависит ее избиратель-
ная проницаемость для веществ. С базальной мембраной клетки
эпителия могут быть связаны полудесмосомами, аналогичными по
строёйИТо половинкамтгеимосом.
Эпителии не содержат кровеносных сосудов. Питание эплте-
лиоцитов"осщубствЛяется диффузно ;рехбазальную мембрану со
стороны подлежащей Соединительной ткани, с которой эпителий
находится в тесном взаимодействии. ЭпителйгГ обладают поляр-
ност^ю7~т. е. базальные и апикальные отделы всего^эпит'ёлиальнЬго
пларта и составляющих -его клеток имеют разнбе строение. Эпите-
137
лиям присуща внрркая способность к регенерации. Восстановление
эпителия происходит за счет митотического деления и дифферен- I
ци[овки стволовых клеток. *
КЛАССИФИКАЦИЯ
Существует несколько классификаций эпителиев, в основу ко-
торых положены различные признаки: происхождение, строение,
функция. Из них наибольшее распространение получила морфоло-
гическая классификация, учитывающая отношение клеток к ба-
за л ьной мембране и их форму чп <'н<>боптЙ7-янн>тлП|ОЙ~Чггг-Лг<т.
apex — вершина) части эпителиального пласта (схема 2).
Схема 2. Морфологическая классификация эпителиев.
В морфологической классификации отражено строение эпите-
лиев, зависящее от их функции.
Согласно этой классификации, прежде всего различают дд-
н о с л'Синые и многослойные эпителии. В "первых все
клетки питслии сьязамы з. базальной мёмбранон~~но ВТ piix^c ба-
за льной мембраной непосредственно связан лишь один ни ж ни й
слой к пеллк..д_остальные слои такой связи лишены и соединены
друг с другом. В со тветствии с формой клеток, составляющих
эпителий, они подразделяются на плоские, кубические и при ма-
138
тическце. (цилиндрические). При этом в многослойном эпителии
учитывается лишь форма наружных слоев ю^ТОЮ-Надримегь-эпи-
телий роговицы — многослойный плоский, хотя нижние сдои его
состоят из клеток призматической и крылатой формы.
Однослойный эпителии может быть однорядным и многоряд-
ным. Уо норя кого эпителия все клетки имеют одинаковую фор-
му^— плоскую, кубическую или призматическую и, следовательно,
их ядра лежат на одном уровне, т о -в один ряд. Такой эпителий
называют еще изоморфным (от греч. isos — равный). Однослойный
эпи гудий. имевший клетки рч»‘{-умы и ьц. пнд. м ц.а ко-
торых лежат на разных уровнях, т. е. в несколько рядод.__црсит
название многорядного, или псевдомнагос дойного.
Многослойный эпителий бывает ороговевающим, неороговеваю-
щим и перехолнь1м7~Э|]^гте7гий, в которомпротекают процессы
ороговения^ связанные с превращением клеток верхних слоев в ро-
говые чешуйки, называют многослойным плоским ороговевающим.
При отсутствии ороговения эпителий- является многослойным
плоским неороговевающим.
Переходный эпителий выстилает органы, подверженные си л ь-
ному растяжению — мочевой пузырь, мочеточники и др. При изме-
нении объёма"оогана толщина и строение эпителия такжТ изме-
няются? -------~ '
Наряду с морфологической классификацией используется о н-
т о ф~иУГб генетическая к ~л а с стт ф> и к а ц и-я, —розданная
советским гистологом Н. Г. Хдопиным. В основе ее лежат особен-
ности развития эпителиев Пиз~тканетГых зачатков. Она включает
эпидермальный (кожный), энтеродермальный (кишечный), цело-
нефродермальный, эпендимоглиальный и ангиодермальный типы
эпителия.
' Эпидермальный тип эпителия образуется из эктодермы, имеет
многослойное или многорядное строение, приспособлен к выполне-
нию прежде всего защитной фунмТии Чнапример многослойный
плоский ороговевающим эпителий кожи).
Энтеродепмальный тип эпителия развивается из энтодермы,
я вл яется по строению однослойным призматическим, осуществля-
ет процессы всасывания веществ (например, однослойный каем-
чат.ый эпителий тонкой кишки), выполняет железистую функцию.
। Цело!<ефродер.мальцый тип эпителия име£т__мезодермальное
происхождение, по строению он однослойный, плоский, кубиче-
скйй~или призматический, выполняет-главным образом барьерную
или экскреторную функГ1ИЮ(ттаггрИмёр7ПГлоскйй эпителий сероз-
ных оболочек — мезотёлий7 кубический и призматический эпите-
лии в мочевых канальцах~пбчек).
Q Эпендимоглиальный тип—предетавлетгспециальным эпителием,
выстилающим, например, цр-огти »«а?Гз Истпипигом ого образона-
ния является нервная трубка.
j К ангиодермальному типу относят эндотелиальную выстилку
кровенбсных~сисудов, имеющую мезенхимное происхождение. По
строению эндотелии является однослойным ским эпителием.
139
СТРОЕНИЕ РАЗЛИЧНЫХ ТИПОВ ПОКРОВНОГО ЭПИТЕЛИЯ
Однослош । bi и п nix цьц 4ни-гс.ий (epithelium simplex squamosum).
Э|у_т вид эпителия 11[»-11 r.-in i^M в организме и и мезо-
телием.
Эндотелий (entothelium) выстилает кровеносные и лимфатиче-
ские Сосуды," а~также камеры серлпа—Он представляет собой пласт
ш ских-клеток—оидогвдиоцитпа, лежащих в один слои на ба зль-
HOji мембране,- Энло и ц < др <ают< я отцесчтеанион бедно-
стью органелл и ирису к-нн+ем w~диТоТТЛазме пиноциточных ве-
зикул.
Эндотелий участвует в обмене веществ и газов (О?, СО2) М£жду_
кров"ыо~и другими тканями организма. При его 'повреждении воз-
можно~изменение кровотока в сосудами образо1вание~в~йх просвете
сгустков крови—тромбов^
Мезотелий. (mesothelium) покрывает серозные оболочки
(листки плевры, висцеральную и париетальную брюшину, около-
сердечную сумку й др.). Клетки мезотелия — мезотелиоциты
плоские, имеют полигональную форму и неровные края (рис. 40, А).
На месте залегания ядер клетки несколько утолщены. Некоторы;
и' । и д ржат не о/mo, а два или даже три ядра,________ ой
п о вс р хности клетки имеются- щиничные—микроворсицкиГ Чере з
мезотелий происходил выделение и всасывание серозной ~жПд-
|уостй7~благодаря его ладкои поверхности легк осугцес вляется
скольжение внутренних органов. Мезотел и; р_____твует образо-
ванию соединительнотканнькх спаек между органами брюшной
и .трудной полостей, развитие которых возможно при нарушении
его целостности.
Однослойный кубический эпителий (epithelium simplex cuboi-
deum). Oi выстилает часть- почечных уяю.'Ч.црп (прокеимлл пые
и дистальные). Клетки проксимальных канальцев имеют щеточную
каемку и з< у__________р норТЬ-________рченность )бусловлена
концентрацией митохондрий в базальных отделах клеток и нали-
чием здесь глубоких складок плазмолеммы. Эпителий почечных
канальцев выполняет функцию обратного всасывания (реабсорб-
ция) рядя_н£дцестн из первичной мочи в кровь.
Опнослоииыи—призматическим—эпите и ift (epithelium simplex
columnarе). Этол вид эпителия характерен для среднего отдела
пи ще ва р ительной системы. Он выстилает внутрен нюю поверх но с ть
желудка, тонкой и толстой кишки. желчн£1ТД_пузыпя ряда про-
токов печени и поджелудочной железы.
В желудке в однослойном призматиче к м эпителии все клетки
являются железистыми, продуцирующими слизь, которая защища-
ет стенку желудка от грубого влияния комков пиши и перевари-
вающего действия желудочного сока_ Кроме того, через эпителий
желудка всасываются в_кровь- вода-и-некоторые-еоли.
В тонкой кишке—однослойный призматический эп и те л и й
(«к а е м ч а т ы й» эпи т е л и й) активно выполняет функцию
всасывания. Эпителий образован призматическими эпителиоцита-
140
mi|, среди которых располагаются бокаловидные клетки (рис. 40, Б).
Эпителиоциты имеют хорошо выраженную исчерченную (щеточ-
ную) всасывающую каемку, состоящую из множества микровор-
синок. Они участвуют в ферментативном—ресщеплении—пищи
(присдо ц р ) и всасывании пбряэлняв1пихся про-
дуктов _в кровь и лим$ ot в д е клетки выдел. ох- слизь.
Пок| ыв I ащищает—его и подле жащ е ткани от
механических и химических воздействий.
На ряд, и ддц. мчат д ми и,—ви1ь^н>виян1ами—члстж ими имеются
базалы1о-.1срписгые щИчьрннпые клетки нсскопь^ц.»—иидов (ЕС,
D, S, J и др.) и апикально-зернистые жел зистые слетки. Выде-
ляемые в кровь гормоны эндокринных клеток принимают участие
в регуляции функции органов пищеварительного аппарата.
MiioropHAiibitj (псевдомногослойный) эпителий (epithelium
pseudostratificatum). В и ае_ воздухоносные пути — носовую по-
лость, трахею, бронхи, ? также р°д других органо L В воздухонос-
ных путях эгорялный—эпителий—является ~реснитчатым, или
мерцательным. В нем различают—4 ^ида - клеток;—реснщчатые
(мерцательные) клетки, короткие и длинные вставочные клетки,
с илистые (бокаловидные)—клетки (рис. 41; см. рис. 42, В), а
также база тьно-зе пнистые д коминые) клетки. Вставочные
клетки, вероятно, являются стволовыми, способными делиться
и превраща______реснитчатые и слизистые клетки.
К базальной мембране вставочные клетки прикрепляются ши-
рокой проксимальной частью. У мерцательных клеток эта часть
узкая, а их широкая дистальная часть обращена в просвет органа.
Благодаря этому в эпителии можно выделить три ряда ядер:
ни2КНШ4—и—средний—ряды-^_ядра_ вставочных клеток, верхний
ряд — ядра мерцательных клетокГДо поверхности эпителия вер-
шины вставочных клеток не—доходят, поэтому она образована
только дистальными частями мерцательных клеток,__докрытыми
141
Рис. 41. Строение многорядного
реснитчатого эпителия (схема).
/ — мерцательные реснички; 2
бокаловидные клетки; 3 — мерца-
тельные клетки; 4, 5 — вставочные
клетки; 6 базальная мембрана;
7 — соединительная ткань.
многочисленными ресничкамш_£лдзпстые клетки имеют бокадо-
визн Подули овоитную форму и выделяют муцины на поверхность
пласта.
Попавшие вместе с воздухом в дыхательные пути частицы
пыли оседают на слизистой поверхности эпителия и движением
его мерцательных рес ни чд;к_Ш1стщчщ+но-выталкиваются^ в носовую
полость и далее во внешнюю среду. Кроме мерцательных, вставоч-
ных и слизистых эпителиоцитов в лдат^чии воздухоносных путей
обнаружено несколько видов ^ц^окринных, базально-зернистых_
клеток (ЕС-, Р-, D-клетки). Эти клетки выделяют в кровеносные
сосуды биологически активные вещества — гормоны, с помощью
которых осуществляется местная регуляция дыхательной системы.
Многослойный плоский неороговевающий эпителий (epithelium
stratificatum squamosum noncornificatum). Покрывае_т снаружи
гови . глаза выстилает полости рта и пищевода. В нем различают
три слоя: базальный, шиповатый (промежуточный^—и. плоский
(поверхностный) (рис. 42, А), разад^ц^й.^слой^ состоит ид. рпите-
лиОцитов прнзматической формьГГ васполасающихся на базальной
мембра~не7 именная стволовые клетки^ способные к ми-
тотичес1^муптёл^Ги;а.^а_счет вновь образованных клеток^Дступа-
юп1и влиф(Беренцнр'>ику| происходит смена элите лиоцитов^ыше-
лежаших слоев эпителия- ^устоиф из клеток
неправильной многоугольной—формы- В оазальном а шиповатом
слоях в эпителиоцитах хорЗшо развиты тонофибриллы (пучки
тонофиламент), а между эпителиоцитами--деемосомы и другие
виды контактов—слои—эпителия образована царскими
КЛИКАМИ-, Заканчивая гвпи*жвшеиный цикл, они-отмирают и от-
падают с поверхности эпителия.
Многослойный плоский ороговевающий эпителий (epithelium
stratificatum squamosum comificatum). Покрывает поверхность
КО4ЧУ обвали, есы э п ц д р о м и с -. и мпзшлм шмшгхплит.процесс
превращения (трансформация) эпителиальных клеток в роговые
uemyitKH - ороговение. При этом в клетках синтезируются и все
больше накапливаются специфические белки—(-кератиныЦ—а сами
' -----------------
142
Рис. 42. Строение эпителиев.
А — микрофотография многослойною, плоскою неороговевающего эпителия роговицы
глаза (окраска гематоксилин-зозином) 1 слой плоских клеток; 2 шиповатый слой;
J — базальный слой: 4 базальная мембрана; 5 — соединительная ткань. Б схема
строения многрслдиного плоского ррогове мающего эци^с^и?.: / — роговой слой; 2 — бле-
стящий слой; 3 — 1ернистыи слой; 4 шиповатый слой; 5 — базальный слой; 6 - базаль-
ная мембрана; 7 — соединительная ткань; 8 пигментоцит. В — псевдомногослойный
(однослойный мноюрядный) реснитчатый эпителий трахеи (окраска гематоксилин-эози-
ном): / — реснички; 2— ядро реснитчатых и вставочных эпителиоцитов; 3 базальные
эпителиоциты; 4 соединительная ткань. Г—однослойный микроворсинчатый (каемча-
тый) эпителий тонкой кишки (окраска гематоксилин-эозином): / — каемка; 2 — ядро ка-
емчатого эпителиоцита; 3 — бокаловидная клетка; 4 — соединительная ткань. Д — желе
зистый эпителий. Сложная смешанная слюнная железа. ШИК-реакция.
клетки из нижнего слоя постепенно перемещаются в вышележа-
щие7}тои эТ1Ителияг“В эпидермисекожи пальцев, ладоней и подошв
различают J осТГОвных^'слоев: базальный; щщюву гыи, зернистый,
блестящий _и~(~рис.~ 42, Ё). Кожа остальных участков тела
имеет эпидермис, в котором..одсутствуе7 блестящий слой. *
Базальный. слои состоит из цилиндрических по форме эпите-
лиоцитов. В ИХ~ЦИтопла ме синтезируются специфические бёлкн,
143
формирующие тонофиламенты. Здесь же находятся стволовые
клетки. Стволовые клетки'деЛятся, после чего часть новообразован-
ных клеток "ди ффе ренци ру е ге я -и—перемещается в вышележащие
слои. Поэтому базальный слой называют ростковым, или зачат-
ковым (stratum"germinativurn). Шиповатый слой образован клет-
ками Многоугольной формы, которые прочно связаны между собой
многочисленными десмосбмами. В месте десмосом на поверхности
клеток имеются мельчайшиё~~6ь1 посты — «шипики», направленные
навстречу друг к другу. Они хорошо заметны при расширении
межклеточных пространств или при сморщивании клеток. .Д^цито-
плазме шиповатыг клеток тонофиламенты образуют пучки—jo-
нофзибриллы. Кроме эпителиоцитов в базальном и шиповатом слоях
присутствуют отростчатые по форме пигментные клетки мела
ноциты, содержащие гранулы чёрного- пигмента" —^меланина,
а также—эпидермальные макрофаги — дендроциты и лимфоциты,
образующие в эпидермисё~~мГг’гную гигтему иммунного надзора.
Зернистый слой состоит из уплощенных клеток, в'цитоплазме ко-
то рых,содержатся тонофибриллы и зерна кератогиалина. Керато-
гиалин ~— фибриллярный белок, способный в дальнейшем в клет-
ках вышележащих_слоев превращаться в элеидин, а затем в кера-
тин — роговое вещество. Блестящий слой образован плоскими
клеткамд- Их цитоплазма содержит сильно преломляющий свет
элеидин,—который представляет собой комплекс кератогиалина
с тонофибриллами. Роговой слой очень мшиньш в коже пальцев,
ладоней, подошв и относительно тонкий в -остальных участках
кожи. Ло мере того как клетки из блестящего слоя перемешаются
в роговой, в них при участии, лизосом постепенно исчезают ядра
и органеллы, а комплекс кератогиалина. с тонофибриллами пре-
вращаетстгв кератиновыефибриллы и клетки становятся роговыми
чешуйками, по форме напоминающи]Щ1_д1лоские многогранники.
Они заполнены кератином—(рОГбвым веществом), состоящим из
плотно упакованных кератиновых фибрилл, и пузырьками воздуха.
Самые наружные роговые чешуйки под влиянием ферментов лизо-
сом утрачивают связь друг с другом и постоянно отпадают с по-
верхности эпителия. На смену им возникают новые за счет раз-
множения, дифференцирования и перемещения клеток из ниже-
лежащих слоев. Роговой слой эпителия отличается значительной
упругостью и плохой теплопроводностью, что имеет значение для
защиты кожи от механических воздействий и для процессов тер-
морегуляции организма.
Переходный эпителий (epithelium transitionaleЪ Этот вид эпите-
лия типичен для мочеотводящих органов — лоханок почекГ^моче-
точников,—мочевого- пузыря, стенки которых подвержены значи-
тельному растяжению при заполнении м~очбй7 В~~нем "различают не-
сколько слоев клеток — базальный, промежуточный, поверхностный
(рис. 43,“"Х V).
Базальный слой образован мепкими—округлы ми (темными)
клетками, ^промежуточном слое располагаются клетки различной
полигональной фор'мы. Поверхностный слой состоит из очень круп-
144
Рис. 43. Строение переходною эпи-
телия (схема).
ных, нередко дву- и трехъядерных клеток,, имеющих куполообраз-
ную И ПИ уц-ал111.енную ф1рму ц от состояния СИНКИ
органа.-’-ИрИдДастяжении стенки вследствие заполнения органа
мочой и- его- поверхностные
клетки уплошайУгся. По нремя сокращений стенки органа толщина
эпителиального пласта резко—возрастает. При этом_ некоторые
клеткй~в промежуточном' слое «выдавливаются» кверху и принима-
ют гру1.ие"идную форму, а расположенные над ними поверхност-
ные клетки — куполообразную форму. Между поверхностными
клетками обнаружены плотные контакты, имеющие значение для
предотвращения проникновения жидкости через стенку органа
(например, мочевого пузыря).
Регенерация. Покровный эпителий, занимая пограничное поло-
жение, постоянно испытывает влияние внешней среды, поэтому
эпителиальные клетки сравнительно быстро изнашиваются и по-
гибают.
Исходником их восстановления являются стволовые клетки
эпителия. Они сохраняют способность к делению в течение всей
жизни организма. Размножаясв^'часть <щовь образованных клеток
вступает в дифференцировку й~~п ре вращаете я в^эпнтелио! мты
подобные утраченным. Стволовые клетки в многослойных эпите-
лиях находятся в базальном (зачатковом) слое, в МНашрядных
эпителиях к ним относятся вставочные (короткие) клетки, в одно-
слойньГХ~~ЭПителиях они располаТаются в определенных участках,
например в тонкой кишке в эпителии крипт, в желудке в эпителии
шеек собственных жёлез и т. д. Высокая способность эпителия
к 'фиЗИило! и четкий рё генерации служит основой- для~~быстрого
восстановлении—его В патологических условиях (репаративная
регенерациях.
Васкуляризация. Покровные эпителии не имеют кровеносных
сосудов, за исключением сосудистой полоски (stria vascularis)
145
внутреннего уха. Питание к эпителиям поступает от сосудов, рас-
положенных в подлежащей соединительной ткани.
Иннервация. Эпителий хорошо иннервирован. В нем имеются
многочисленные чувствительные нервные окончания — рецепторы
Возрастные изменения. С возрастом в покровном эпителии
наблюдается ослабление процессов обновления.
СТРОЕНИЕ ЖЕЛЕЗИСТОГО ЭПИТЕЛИЯ
Железистый эпителий (epithelium glandulare) состоит из желе-
зистых, или секреторных, клеток — гландулоцитов^ Они осущест-
вляют'синтез а также- выделение специфических продуктов —
секретов-на поверхность кожи, слизистых оболочек и в полости
ряда внутренних органов [внешняя (экзбкринкая) секрецйяГдын
в кровь и лимфу ^внутренняя (эндокринная) секреция!.
Путем секреции в организме выполняются многие важные фун-
к ци и; образование молока слюны, жедудочТТОгО и ки щечного со к а,
желчи, эндокринная (гуморальная) регуляция и др.
Бол ьшинствб-жёлезистых ~клеток~с ~внеш ней секрецией (экзо-
кринные) отличаются наличием секреторных включений в цито-
плазме, развитой эндоплазматической сетью, полярным распо-
ложением органелл и секреторных гранул.
Секреция. (от лат.__secreiio — отделение) является сложным
процессом, включающим -4 фазы:—длт.д.п шение исходных
продуктов гландулопитами, синтез—и—на к о п. н и е в них
секрета, -выделение секрета из гландулоиитов — экструзия
и восстановл с_ни е их- структуры^ Эти фазы могут проис-
ходить в гландулоцитах циклично, т. е. одна за другой, в виде так
называемого секректорного цикла. В других слуяаях__они совер-
шаются одновременности. характерно..для диффузной или спон-
танной секреции.
П е р в а я фаза секреции заключается в том, что из крови
и лимфы в железистые- клетки со стороны базальной поверхности
поступают^различные неорганические соединения, вода и низкомо-
лекулярные органические вещества:_аминокисдптыг моносахариды,
жирные кислоты и УД. Иногда путем пиноцитоза в клетку прони-
кают брлее крупные молекулы органически х веществ, на пример
белки. Во второй фазе из этих продуктов в_эндоплазма-
тической сети синтезируются секреты, причем Белковые с участием
гранулярнои^~эндоплазматической сети, а небелковые с участием
агранулярн нд плазматической се.т(г_ интезируемый секрет по
эндоплазматической сети перемещается в зону комплекса Гольджи,
где постепенно накапливается; подвергается химической пере-
стройке и оформляется в виде гранул. В третьей ф а,з е об-
разовавшиеся секректорные гранутГы выделяются из клетки. Выде-
ление секрета~происходйт- неодинаково, в_. связи—с. чеКс различают
три типа секреции: 'м е ро кринов ый (эккриновый), п о-
К Ринов ый и голокриновый (рис. 44, А, Б, В). При
мерокриновом типе секреции железистые клетки полностью со-
146
...-it nW* Л
’’ •7.’
Рис. 44. Различные типы секреции (схема).
Л — мерокриновый; Б — апокриновый; В — голокриновый типы; / — мало дифференци-
рованные клетки; 2 — перерождающиеся клетки; 3 — разрушающиеся клетки.
храня ют свою структуру (например, клетки слюнных желез). При
апокриновом типе секреции происходит частичное разрушение
железистых клеток (например; клеток молочных желез), т. е.
вместе с секреторными, продуктами отделяются либо апикальная
ч асть ци то плазмы железистых клеток (макроа покури нова я секре-
ция) и ли верхушки микроворсинок (микроапокриновая^екреция).
Голокриновый тип секреции сопровождается накоплением жира
в цитоплазме и полным разрушением желе5УГСтых клеток (напри-
мер, клеток сальных желез кожи).
Четвертая Фаз а~секреции заключается в восстановле-
нии исходного состояния железистых клеток. Чаще всего, однако,
восстановление клеток происходит "по" мере их разрушения.
Гландулоциты лежат на базальной мембране. Форма их весьма
разнообразна и~меия>ттзг-п~^ятпггимости от фазы секреции. Ядра
бывают обычнокрупными, с изрезанной поверхностью, что при-
дает им неправильную форму. В цитоплазме глаидулоцитов, кото-
рые вырабатывают секреты белкового характера (например, пище-
варительные ферменты), хорошо развита гранулярная эндоплаз-
матическая еть. В клетках, синтезирующих небелковые секреты
(липиды, стероиды), выражена агранулярная цитоплазматическая
сеть. Комплекс Гольджи обширный. Его форма и расположение
в клетке меняются в зависимости_от^фазы секреторного процесса.
^Митохондрии, как~~правило, многочисленны.. Они накапливаются
I в местах наибольшей активности клеток, т. е. там, где образуется
• “Секрет.~В~~цитог1дазмо клеток—обычнп^ттри^утствуют секреторные
грану ы, размер и строение которых зависят от химического сос-
тава секрета. Число их колеблется в связи i _ф1азами_секреторного
процесса. В цитоплазме некоторых глаидулоцитов (например,
участвующих в образовании соляной кислоты в желудке) обнару-
живаются внутриклеточные секреторные канальцы — глубокие впя-
чивания цитолеМмЬ, СТенки которых покрыты микроворсинками.
Цитолемма имеет, различное строение на боковых, базальных
и апикальных поверхностях клеток. На боковых поверхностях она
образует десмосомы и плотные 'замыкательные контакты (терми-
147
нальные перемычки). Последние окружают верхушечные (апикаль-
ные) части клеток, отделяя таким образом межклеточные щели от
просвета железы. На базальньи лщипхностях клеток цитолемма
образует небольшое число узких складок, проникающих в цито-
плазму. 1'акие складки особенно хорошо развиты в клетках желез,
выделяющих секрет, богатый солями, например в протоковых
клетках слюнных желез. Апикальная поверхность клеток покрыта
микроворсинками.
В железистых .клетках- хорошо заметна дифферен-
цировка. Она обусловлена направленностью секреторных про-
цессов, например, при внешней секреции от базальной к апикаль-
ной части клеток.
ЖЕЛЕЗЫ
Железы (gianduiae) выполняют в организме секреторную функ-
цию. Большинство из—них—являются- производными жёле з и сто го
эпителия. Вырабатываемые в железах секреты имеют важное зна-
чение для процессов- пищеварения, роста, развития, взаимодей-
ствия с внешней средой л др. Многие железсамостоятельные,
анатомиич г - .. д т,це органы (наппимер, поджелудочная
железа? крупные слюнные железы, щитовидная железа). Другие
железы являются лини частью органов (например, железы же-
лудка).
~~Ж£Д£ЗЫ подразделяют на две группы: железы внутренней сек-
['ецци, или эндокринныё, И ЖСЛС7Ы ЬИСТннсЙ секреции, или экзо-
кринные (рис. 45, А, Б, В).
~Эпдокриннь|£ железы выражать!вают высокоактивные вещест-
ва — цдсСУДающиё непосредственно в кровь. Вот почему
эти *елезь; тс^лько^ич железистых клеток и не имеют вы-
вод ных протоков. К ним Относят гипофиз, эпифиз,^.щитовидную
и околощитовидную железы, надпочечники, островки поджелудоч-
ной железы и“др. Все они входят в состав эндокринной системы
организма, которая вместе с нервной системой выполняет регу-
дирующую функцию^
ЭкзоКринпЫе железы вырабатывают секреты, выделяющиеся во
внен1нкур, грелу, т.Т!'ГсГ*повег>хность кожи или в полости органов,
выстланные эпителием. В связи с этим они состоят из дву.х.дастец:
. l крет.рных, или концевых. отдему (portiones terminalae) и <<ы-
протоков (ductus excretorii). Концевые отделы образованы
гл а н/|улопитамИ- /Драящими на базальной мембране. Выводи ые
Протоки выстланы раздичнымц видами рпитёдиев в зависимостй'От
происхождения ^гелёзГ"В железах, образующихся из энтеродер-
мальнбго эпителия (например в поджёлУдочнойЭкеяезе), они вы-
стла мы длмгм-яайшлып irjrfiM—nmi шт lyiimiMliwiniH зш!Телием,
а в железах, развивающихся из эктодермального эпителия (напри-
мер, в Сальных железах кожи), — многослойным неороговевающим
Строение эндокринных желез описано в гл. XVII.
14К
Рис. 45. Схема строения эк-
зокринных и эндокринных
желез (рис. Е. Ф Котов- 1 2 5 4
ского):
А — экзокринная железа; Б
эндокринная железа: / — кон-
цевой отдел; 2 - секреторные
гранулы; 3 — выводной проток
экзокринной железы; 4 — по-
кровный эпителий; 5 — соедини-
тельная ткань; 6 — кровеносный
сосуд; В разновидности экзо-
кринных желез: / — простые
трубчатые железы с неразвет-
вленными концевыми отделами:
2 — простая альвеолярная же-
леза с неразветвленным кон-
цевым отделом; 3 — простые
трубчатые железы с разветвлен-
ными концевыми отделами; 4 —
простые альвеолярные железы с
разветвленными концевыми от-
делами; 5 — сложная альвео-
лярно-трубчатая железа с раз-
ветвленными концевыми отде-
лами; 6 - сложная альвеоляр-
ная железа с разветвленными
концевыми отделами.
эпителием. Экзокринные железы чрезвычайно разнообразны, от-
личаются друг от друга строением, типом секреции, т. е. способом
выделения секрета и его составом. Перечисленные признаки поло-
жены в основу классификации желез. По-сдроению экзокринные
железы^ццдразделяцц на. следу^ьур^ виды ~ (схема .^).
Пдостые ^железы имеют неветвящийся выводной проток, слож-
Uhi? железы *— ветвящийся (см. рис. 45, В). В него~~открываются
в неразветвленных железах по одному, а в разветвленных оКелезах
по нескольку концевых отделов, форма которых может быть в виде
трубочкй~либо мешочка (альвеола) или промежуточного между
ними типа?- “ ' ------------------
14*0
Схема 3. Морфологическая классификация экзокринных желез.
В некоторых железах* производных эктодермального (много-
слойного) эпителияг наг!ример в слюнных, помимо секреторные
клеток, встречаются эпителиальные клетки, обладающие способ-
ностью сокращаться,^ лтгтпэтгг/тглраЖныё клетки. Эти клетки,
имеющие щростчатую форму, охватывают концевое отделы. J3 их
нитопдадйге^присутсхвуют микроФиламенты. солержашие сократи-
тельные белки. Миоэпителиальные клетки при сокращении сдавли-
вают концевые отделы и, следовательно, облегчают выделение из
них секрета: '
Химический состав декрета» может быть различным, в связи
с этим экзокринные железы подра3деля1отся_на_оелко<>ы<; (сероз-
ные), слизистые, белковое,дизцстые ,1см. рис. 42, Д) и сальные,
В .мешанных железах кюхут присутствовать два вида секретор-
ных клеток^—"бёлковы^ исдиздетыь.Они образуют З1ибо по отдель-
ности концевые отделы (чисто белковые и чисто слизистые), либо
вместе смешанные концевые отделы (белково-слизистые). Чаще
всего в состав секрёторного продукта входят белковые и слизистые
компоненты лишь с преобладанием одного из них.
Регенерация. В железах в связи с их секреторной деятель-
ностью постоянно происходят.процессы физиологической регене-
рации В мерокриновых и апокриновых железах, в которых нахо-
дятся долгоживущие клетки, восстановление исходного состояния
iландулонитов после выделения из них секрета происходит путем
внутриклеточной регенерации*) а иногда и путем размножгния.
железаХ-ВОсстановление Осуществляется за счет
размножения специальных, стволовых клеток. Вновь образовавши-
в железистые клетки, .(клеточная регенерация).
Васкуляризация. Жрдезы обйАыi. 1 снабжены кровеносными со-
с^дами Среди них имеются артериоло-венулярные анастомозы
и вены, снабженные сфинктерами (замыкающие вены). Закрытие
150
анастомозов и сфинктеров замыкающих вен приводит к увеличе-
нию давления в капиллярах и обеспечивает выход из них веществ,
используемых гландулоцитами для образования секрета.
Иннервация. Осуществляется симпатической и парасимпа-
тической нервной системой. Нервные волокна следуют в соеди-
нительной ткани по ходу кровеносных сосудов и выводных про-
токов желез, образуя нервные окончания на клетках концевых
отделов и выводных протоков, а также в стенках сосудов.
Кроме нервной системы, секреция экзокринных желез регули-
руе сц гуипмлЬНЫМЦ ^факторами, т. е. гормонами эндокринных
желез.
Возрастные изменения. Впожилом возрасте .изменения в же-
лезах могут проявляться а снижении секреторной активности же-
лезистых клеток и изменении состава вырабатываемых секретов,
а также в ослаблении процессов регенерации и в разрастании со-
единительной ткани (стромы желез).
Глава VII
КРОВЬ И ЛИМФА. КРОВЕТВОРЕНИЕ
ПОНЯТИЕ О СИСТЕМЕ КРОВИ
Среди производных мезенхимы особое место занимают ткани
и органы системыПкрови, включающей в себя к ровьи лимфу,
о р га ны к~р~о ветворения и иммунопо э з_а. (красный
к ост ный мозг.тймус, селезенка^-Димфати ческие узлы, скопления
лимфоидной ткани). В системе крови все элементы взаимосвязаны
гистогенетически и функционально и подчиняются общим законам
нейрогуморальной регуляции. Исходя из этого понимание вопросов
развития, строения и функции отдельных элементов системы крови
возможно лишь с позиций изучения закономерностей, характери-
зующих систему в целом.
Весу клетки крови (hemocyti) развиваются из общей тишпо-
тентнои стволовой клетки, дифференцировка_которей-в-различные
виды клеток крови определяется как- м пиро о-кр ужен и е м
(ретикулярная ткань кроветворных органон!, так и дейсттц^м-спепи-
фических гемопоэтинов. Процессы разрушения и новообра-
зования клеток крови в оптимальных условиях сбалансированы и,
следовательно, поддерживается постоянство количества и состава
крови. Тесное взаимодействие между органами гемопоэза и иммуно-
поэза осуществляется путем миграции, циркуляции и рециркуля-
ции клеток крови, нейрогуморальной регуляции кроветворения
и распределения крови.
Кровь и лимфа являются особыми разновидностями тканей
мезенхимного происхождения, образующими внутреннюю
среду о рга’ни з м а _Свместе о рыхлой волокнистой сое-
динительной тканью). Филогенетически кровеносные сосуды и цир-
кулирующая в них кровь развиваются раньше лимфатических сосу-
151
дов и лимфы. Кровь_и лимфа имеют жидкую консистенцию и сос-
тоят из Двух основных компонентов: межклеточного вещества
(плазмы) и взвешенных в лей форменных элементов. Обе ткани
функционально тесно взаимосвязаны;—в них. Происходит постоян-
ный обмен форменными элемент тми, а также ынцветвамщ находя-
щимися в жидкой межклеточной среде — плазме. В последние
годы установлен факт рециркуляции лимфоцитов из крови в лимфу
и из лимфы в кровь.
Кровь и лимфа генетически и функционально тесно связаны
с соединительной^гканью. Начальные стадии развития всех клеток
крови Происходят в специализированных видах соединительной
ткани — лимфоидной и миелоидной, а дифференцированные формы
поступают в кровь и циркулируют там, выполняя основнькГфунк-
ции длительное время (эритроциты) nMgn цфдмий срок (пейкп-
। ситы) Последние далее мигрируют ич-жррнч н окружающую с ое ди -
нительную-ткань, где—осуществляют- свои основные специальные
функции.
Популяция клеток крови — обновляющаяся, с коротким циклом
развития, где зрелые формы являются конечными (погибающими)
клетками.
КРОВЬ
Кровь (sanguis) состоит из межклеточн о г о веще-
ств JT— пл а з м ы к р о в К и взвешенных в ней форменных
э л е м е н то в: эритроцитов».лейкоцитов и тромбоцитов (кровяные
пластинки). Объем плазмы равен 55—-60%, а форменных элемен-
тов — 40—45% крови. Кровь ворганизмечеловека составляет от
5 до 9% массы тела. В среднем в теле человека с массой тела 70 кг
содержится около 5 — 5,5 л крови.
Функция крови. Основными функциями крови являются:
1) транспортная, 2) защитная, 3) гомеостатическая, 4) дыхатель-
наЯт-5) трофическая^
Транспортная и трофическая функции заклю-
чаются в переносе веществ, получаемых организмом с пищей, про-
дуктов обмена? гормонов ^других биологически активных веществ,
д ы х а тельная ф у н к п и я —хв доставке кислорода из легких
в др^-ие органы и удалении углекислоты, 3 а щитная функ-
ц и я~состоит в~ обеспечении гуморального и клеточного иммуни-
rejjfl и др. Вместе с нервной и эндокринной системами кровь прини-
мает участие в поддержании г о м е о ст а з а — постоянства внут-
ренней срёдьГорганизмами том числе имунного гомеостаза.
Плазма крови
Плазма крови (plasma sanguis) представляет собой вещество
жидкой консистенции. Она содержитбО—93%7водыи 7—1()%~ту-
хого . вещества, в котором около 6,6—8,5% составляют белки
и 1,5—3,5% — другие органические и минеральные соединения.
152
К основным белкам плазмы крови относятся альбумины, глобулины
и фибриноген. Из фракции глобулинов выделены антитела. Плазма
кроцц_имеет pH около_7Д6. Подробное описание химического сос-
тава плазмы крови дается в учебниках биохимии и физиологии.
Форменные элементы крови
Эритроциты
Эритроциты (erythrocytus), или красные кровяные тельца, чело-
века и млекопитающих представляют собой безъядерные клетки,
утратившие в процессе фило- и онтогенеза ядро и большинство
органелл. Основная функция эритроцитов — транс пор тиров-
к а к ислорода и утл е к -и-с л о т-ы—и^-таким- образом, учас-
тие в обеспечении дыхательной функции. Эта функция обеспечи-
вается^ специальным пигментом — гемоглобином сложным-бел-
ком, имеющим_в своем составе железо. Кроме того, эритроциты
участвуют в транспорте аминокислот, антител, токсинов и ряда
лекарственных веществ, адсорбируя их на поверхности цйто-
леммы.
Количество эритпоцитпн у трлгплгп мужчины копебпется от
3,9 • 1012 до 5,5 • 101 в 1_д, а_у женщины — от 3,7 • 1012 до 4,9 • 1012
в „ 1л. Число эритропитон у здоровых людей ~может варьировать
в зависимости от возраста, гормонального фона, эмоциональной
и мышечной нагрузки, действия экологических факторов и др. б*час-
тности, женские половые гормоны тормозят развитие эритроцитов,
что приведет; Фолее'тптзкому-еодержатптю'эритроцитбвГу" женщин.
Форма и строение. ^Эритроциты у человека и млекопитающих
в токе крови (т. е. во взвешенном состояиии) обычно имеют форму
двояковогнутых дисков и называются дискоцитами (рис. 46, А, Б,
В). Дискоциты составляют около 80% от общего числа эритроцитов
крови—Исследования в сканирующем электронном микроскопе вы-
явили и другие формы эритроцитов — планоциты (с плоской по-
верх ностью), стоматоциты (куполообразные), седловидные, двухъ-
ямочные, сферопиты (шаровидные)^ эхиноциты (шиловидные).
Последние_два.вида относятся-К стареющим формам эритроцитов
(рис. 46, Г). Деформация дискоцита в эхиноцит наблюдается в
клетках при уменьшений содержания АТФ. Значительное измене-
ние формы эритроцитов отмечаетсяпрй-движении их через крове-
носный капилляр. Диаметр эритроцитов у человека колеблется от
7,1 до 7,9 мкм, толщина эритроцитов в краевой зоне — Г,9—2,5 мкм,
вщлГТре'= 1 мкм. В нормальной крови указанные размеры -имеют
75% всех эритроцитов — нормоциты (normocytus); большие разме-
ры (свыше 8,0 мкм) — 12,5% —макроциты (macroctttus).'y осталь-
ных-эритроцитов диаметр может быть 6 мкм и менее — микроци-
ты (microcytus). Поверхность отдельного эритроцита у человека
приблизительно равна 125 мкм2, а объем — 90 мкм3. Если считать,
что среднее количество крови у человека равно 5,5 л, то общая повер-
//
т л
ь I
Рис.
46. Форменные элементы крови человека.
А — мазок крови: / — эритроцит; 2 — сег-
ментоядерный нейтрофильный гранулоцит;
3 — палочкоядерный нейтрофильный грану-
лоцит; 4 — юный нейтрофильный грану-
лоцит^ — эозинофильный (ацидофильный)
гранулоцит; 6 — базофильный гранулоцит;
7 — большой лимфоцит; 8 — средний лим-
фоцит; 9 — малый лимфоцит; 10 — моноцит;
II — тромбоцит (кровяные пластинки). Б —
ретикулоциты (по Г. А. Алексееву и
И. А. Кассирскому). Зернисто-сетчатая суб-
станция имеет вид клубка (/), отдельных
нитей в виде розетки (II), (ill), зерны-
шек (/И). В — свежая кровь: / — эри-
троциты (дискоциты); а) — эритроциты с
выростами цитоплазмы (эхиноциты);
6) эритроцитарные группы — «монетные»
столбики эритроцитов; 2 — эозинофильный
лейкоцит; 3 — нейтрофильный лейкоцит;
4 — лимфоцит; 5 — моноцит; 6 — тромбо-
циты (кровяные пластинки); 7 — нити
фибрина.
г
Рис. 46 (продолжение).
Г—эритроциты в сканирующем электронном микроскопе (препарат А. В. Корнева):
/ — диско ниты-нормоциты; 2 — дискоциты-микроциты; 3 — дискоциты-макроциты; 4 — ре-
тикулоциты; 5 — сфероцит; 6 — эхиноцит.
155
хность эритроцитов, циркулирующих в крови, составит около
3500—3700 м2.
Плазмолемма (plasmolemma) эритроцитов под электронным
микроскопом имеет толщину около 20 нм. На наружной поверх-
ности ее расположены фосфолипиды, сиаловая кислота, антиген-
ные олигосахариды, адсорбированные протеины, на внутренней
поверхности — гликолитические ферменты, натрий- и калий-АТФ-
азы, гликопротеины, гемоглобин. Являясь полупроницаемой мемб-
ранойТ'бболочка-эритроцита обеспечивает активный перенос через
мембрану ионов натрия и калия, О-, и СО, и других веществ. Внут-
реннее содержимое эритроцита (гиалоплазма) под электронным
микроскопом выглядит электронно-плотным, содержит многочис-
ленные гранулы гемоглобина размером 4—5 нм? В эритроцитах
содержится около 60% воды и 40% сухого остатка. 95% сухого
остатка составляет гемоглобин и 5% — другие вещества. У челове-
ка содержится два типа гемоглобинов — НПА (характерен для
взрослых), НЬР“(характерен для эмбрионов). Их белковые части
отличаются по составу аминокислот. К моменту рождения ре-
бенка HbF составляет 80%, а НЬА — 20%. У взрослых людей
основную часть составляет НЬА (98%) и меньшую—HbF (не
более 2%). При ряде заболеваний (гемоглобинозы, или гемогло-
бинопатии) в эритроцитах появляются другие виды гемоглоби-
нов, белковая часть которых по сравнению с НЬА и HbF характе-
ризуется другим составом аминокислот. Основной путь обмена
энергии в эритроцитах.— гликолиз, в процессе которого происхо-
дит образование АТФ и НАД-Н2. Энергия гликолиза обеспечивает
активный транспорт катионов через плазмолемму, поддержание
оптимальногб^ботношения концентрации калия, р натрия в плазме
крови и эритроцитах^ а также поддержание формы и сохранение
целостности мембраны эритроцита. НАД-Н2 поддерживает актив-
ное__функциональное состояние гемоглобина, предотвращая его
окисление в метгемоглобин. В эритроцитах имеются также фер-
менты пентозно-фосфатного цикла. Наличие железа в гемоглобине
обеспечивает желтую окраску эритроцитов свежей кровц, а в боль-
шом количестве эритроциты придают крови красный цвет. При
окрашивании препаратов крови зрелые эритроциты проявляют
оксифилию (см. рис. 46, А). Наряду со зрелыми эритроцитами
в нормальной крови содержится 1—5% молодых форм/бедных
г£мщуюбинолц_Они обладают спосооностью'окрашиваться и кислы-
ми, и основными красками, т. е. являются полихроматофильнцмиг
При суправитальном окра ш ива н и и брилл иа нтовьгмгкрезйло вым си-
ним в их цитоплазме выявляются зернисто-сетчатые структуры
(substantia reticularis). Исходя из этого молодые формы эритро-
цитов называют рехшсулгнштамл (erylhrocytus reticulatus) (см. рис.
46, В). Электронно-микроскопически показано, что ^зернистогсет-
чатые структуры представляют собой остатки органелл, содержа-
щих рибосомальную РНК^. эндоплазматической сети, рибосом, а
также митохондрий. В ретикулоцитах в незначительной степени
осуществляется^ синтез белка, (глобина), гема, пуринов, пиридин-
156 г I о V
И (ч^
^7 М1С1ЦИ
нуклеотидов, фосфатидов, дипидов, однако РНК в них не синте-
зируетсял Продолжительност ь жизни э р и т р оТГи-
т о в составляет около 120 дней- В ~организме ежедневно разруша-
ется около 200 млн. эритроцитов. При этом гемоглобин распада-
ется на глобин и железосодержащую геминовую группу. Освобо-
дившееся железо используется для образования новых эритроци-
тов, При старении эритроцитов отмечается уменьшение активности
различных ферментов, в частности гексокиназы, глюкозо-6-фос-
фатдегидрогеназы, что приводит к снижению интенсивности глико-
лиза и реакций пентозного цикла. Имеет место также уменьшение
содержания липидов и АТФ и вследствие этого увеличение
чувствительности к осмотическому лизису и механическим воз-
действиям. В результате нарушения проницаемости -мембраны
наблюдается выход ионов калия в плазму и увеличение в эритро-
цитах содержания натрия. В Фтярвющи'х ~эритропитах отмечается
нарушение газообменнои функции из-за уменьшения способности
к восстановлению метгемоглобина в гемоглобин.
Общая характеристика и классификация. Лейкоциты (leucocy-
tus), или белые кровяные клетки .периферической крови позвоноч-
ных и человека характеризуются активной подвижностью и весьма
разнородны по морфологическим признакам-и биологической роли.
Все лейкоциты подразделяют иа две большие- группы: ££1р_н_ид>
т ы е л е й к о и и т~ы-~или г и а н у угОТГи т ы (granulocytus), и
н е з е р нистые лейкоциты, или а гранулоциты
(agranulocytus).Группа зернистых^ лейкоцитов характеризуется
наличием в цитоплазме специфической зернистости и сегменти-
рованными ядрамц.~Т1ри окраске крови смесью кислого (эозин)
и основного (азурТ красителей по методу Романовского — Гимзы
зернистость в одних лейкоцитах обнаруживает сродство к кислым
красителям, и такие лейкоциты называются эрчинпфи гц.ныыи., или
аиидофилъншиЦвцругцх.— к основным красителям — базофиль-
_ ные лейкоциты: зернистость третьих обнаруживает сродство к кис-
лым и основным красителям, .такие лейкоциты называются нейт-
рофильными, или гетерофильными.
Группа н езернис ты х л е й к о ц и ддхв отличается отсут-
ствием специфической зернистости в цитоплазме и несегментиро-
ванными ядрами. Они полрязлеаяются-на лимфошцы-и. монепиты "
(см. рис. 46, А), имеющие разные морфологические и функцио- <
нальные показатели.
Jjrx- лейкоциты имеюддшаревидную-форму. У взрослого чепове-
ка их насчитывается - IO!I Q (1. 1fl2 ц 1 -в кропи Количество
лейкоцитовмо же т значительно, ^сняться в зависимости от приема
пщци. физического л умственною надрь крниц и др. Лейкоциты \
способны _к активному перемещению, их движение осуществляется
путем образования псевдоподии, при этом у них может резко из-
меняться форма телаТгзгдод. Лейкоциты способны проходить меж-
157
ду_клетками эндотелия капилляров и перемещаться по основному
веществу соединительной ткани, проникать через базальные
мембраны и между клетками эпителия. Скорость движения лей-
коцитов зависит_от различных условий: консистенции среды, ее
pH “температуры _шлр. Направление движений лейкоцитов опреде-
ляется, по-видимому-различнымп-факторамщ—иэ которых опреде-
ленную роль играет хгмртдлсис- (от араб, kimia — химия и греч.
taxis — расположение )7х. е .“Движение под влиянием химического
раздражителя. В этом отношении большуюфоль игразот_цродукты
распада тканей, которые и являются таким -Химическим раздражи-
телем. Важное значениёвнаправленном движении и взаимодей-
ствии клеток имеют специальные рецепторы плазмолеммы лейко-
цитов. Током крови лейкоциты разносятся по всему организму,
выселяются в ткани и органы, где и проявляют наибольшую актив-
ность. Определенные типы лейкоцитов участвуют в защитной фун-
кции, обеспечивая фагоцитоз микробов, инородных веществ и про-
дуктов распада клеток, а также участвуют в формировании гумо-
рального и клеточного иммунитета.
Гранулоциты (зернистые лейкоциты)
eur tit f лет /ifnucyw
Нейтрофильные гранулоциты (granulocytus neutrophilicus) —
нейтрофильные лейкоциты, или нейтрофилы, имеют округлую фор-
му, их диаметр в капле свежей крови около 7—9 мкм. На стекле при
изготовлении мазка они несколько распластываются и их диаметр
составляет 10—12 мкм. В крови взрослого человека нейтрофилов
содержится больше, чем других лейкоцитов; их относительное
количество дФслйТ^£Т~ф5^г75%~"от~общего числа лейкоцитов,
_ЦитоцлаДМа-нейтро«Ьилов слабооксиФильна—в пей содержится
мелкая зернистость, плохо заметная не только На свежих, но и на
фиксированных, окрашенных препаратах. Количество гранул в
каждой клетке может быть от 50 до 200. При окраске по методу
Романовского—Гимзы зернистость принимает розово-фиолетовый
цвет. Зернистостью занята не вся цитоплазма — поверхностный
слои ее в ви.у: чзкой ц гпмлпрнным .содержит тон-
кие филаменты. Этот слой играет главную роль при амебоидном
движении клетки, участвуя в образовании псендоро дий. _В зависи-
мости от строения “и химического состава различают два “о с-
н о вных типа гранул — азурофилвные-луаыуды (granulum
azurophilicum) и~ нейтрофильные гранулы^ (granulum neutrophi-
iicum) Азурофильиыв гранулы, появляются в процессе разви-
тия нейтрофила раньше, поэто‘му их называют первичными. Их
больше в малоспециализированных клеткахфв "процессе-специа-
лизации их число уменьшается, В зрелВХ~Р~^'
азурофильных гранул составляет лишь 10—20% от общего числа
гранул. Их размеры варьируют от 0,4 до 0,8 мкм. Эти гранулы
представляют гмбой |ызтн>идл1ост1. иеовичных лизосом- О-ЧСМ сви-
детельствует содержание в них типичных для лизосом гидролити-
ческих Ферментов ^ кислой фосфатазы, fl-глюкуронидазы, кислой
m л, ,1
[5-глицерофосфат дегидрогеназы, кислой пррдха <4, арилсульфатазы
и "др. у^арактернгг татгже наличие миелопероксидазы и муро-1
мйдазы (лизоцима)_, имеюшеи выра'женное бактерицидное дей-
ствие. Гра'нулы имеют круглую или овальную Форму, и для них .ха-'
рактёрна большая электронная плотность, чем у специфических
нейтрофильных гранул. Специфические нейтрофильные гранулы
появляются в процессе развития нейтрофила позже азурофильных,
поэтому их называют вторичными. Их количество возрастает в
процессё~специализации клетки. В зрелом_нейтрофиле оно состав-
ляет 80—90% от всего числагранулГЗрелые нейтрофильные гра-
нулы имеют диаметр 0,1—0,3 мкм, округлую, овальную или ганте-
левидную форму, ртличаются разнообразием ультраструктуры. Не-
зрелые гранулы имеют большой размер (0,2—0,4 мкм), как прави-
ло, окруедую-ФормугШодержат более электронно-плотную центра-
льную часть. Дпя чх- химического состава хдрактерно отсутствие
л и зосо мальных ферментов и пероксидазы _и наличие шелоч нои
Фосфатазы- ф фагоцитинов^обладаю- .
щих антибактериальной^активностью, лактоферрина, лизоцима;
аминопептидаз и др. Таким образом, маркерами спетщфических
гранулмогут служить П1рпочная фосфа^^аЗт-катишжые-бёдкй^ а
а зубрильных грянул —кислая фосфатаза и пероксидаза. Ij нито-
плазЯ(Гнеитрофило;Гслабо представлены органеллы — .имеется не- /
много митохондрий, небольшой комплекс Гольджи, инеддаедстре-
чЗКУгся~дтеДуЦированные элементы эндоплазматической сети. Ха-
рактерно наличие включений — гликогена, липидов и др.
Ялрд нейтрофильных лейкоцитовСодержат плотный хроматин,
особенно цо прпифррии. в котором трудно различить, ядрышки.
форм ^Гя лес, непл и на KnHgi_ поэтому их называют также полиморфно-
ядерными лейкоцитами. Зрелые нейтрофилы имеют сегментиро-
ванные ядра, состоящие ИЗ 2—по.шеедолск.Связанных очень
тонкими, иногда почти незаметными перемычками. Это так назы-
ваем ые непрофильные ^оануцоциты (graпи 1осу-
tus neutrophilicus segmentonuclearis) (см. рис. 46, А, рис. 47)._Они
составляюцодавляющую часть нейтрофильных лейкоцитов
65%J. Меньше содержится палочкоядерных нейтрофильных гра-
"иулоццгоб (3—5% 17 Ядра эТй х кл етшТиме ют - вил изогнутпи~па-
лочки, или подковы, или буквы S. Еще_реже—встречаются юные
нейтрофильные, гранулоциты (granulocytus neutrophilicus juveiru-
lis) (0—0,5%) c^ бобовидными ядрами. Соотношение этих трех
видов лейкоцитов имеет определенное диагностическое значение
в клинике. Например,увеличение юных и палочкбядерных форм
свидетельствует об усилении кроветворения вследствие потери кро-
ви _цди наличия очага воспаления.^ строении, ядершеитрофилов
у Человека существуют пазличЙЯ в'Тавйси мости от пола. В ядрах
большинства нейтрофилов женщин -имеются_дколоядерные при-
датки, или сателлиты ядра. Полагают, что в околоядерном придат-
ке заключена _рдна из X-хромосом, поэтому-его называют также
половылкхроматином. Околоядерный придаток как правило, имеет
форму ба абанной~'тталочки—{см. рис. 47, Л). Наличие или отсут-
159
Рис. 47. Схема ультрамикро
скопического строения грану-
лоцитов (по Н. А. Юриной и
Л. С. Румянцевой).
А сегментеядерный нейтро-
фильный гранулоцит; Б — эози-
нофильный (ацидофильный)
гранулоцит; И — базофильный
гранулоцит: / сегменты ядра;
2 — тельце полового хроматина;
3 — первичные (азурофильные)
гранулы; 4 вторичные (спе-
цифические) гранулы; 5 — зре-
лые специфические гранулы, со-
держащие кристаллоиды; 6
гранулы базофила различной
величины и плотности: 7 — пе-
риферическая зона, не содер-
жащая органелл; S. микро вор-
синки и псевдоподии.
ствие вышеописанных ядерных придатков позволяет определять
в мазках вероятную долевую лринаддежность крови, что имеет
большое практическое значение.
Нейтрофильные еранулопиты---шутнижные клетки—Они могут—
мигрировать идЗхрове-нЬсных" сосудоб и передвигаться,. .к истоя=—
нику раздражения. Если в очаге воспаления имеются микробы—
то нейтроФиль! их фагоцитируют, И. И. Мечников назвал их мик
ptrfw-wnu. Нейтрофилы обладают высокой способностью к а -
г о ц и т о з у, После nornop'^piin бактерий и други х_д.асхиц в_них
формируются фдгосолщ,-сливающиеся вначале сосвецифияескими
гранулами, ферменты которых (щелочная фосфатаза, лизоцим,
лактбфёрин) действуют в нейтральной или .тедоыной среде около
3 мин. После этого происходит слияние фагосом с пелвичными
гранулами~~Т7Пгзосома1Уги2~ и формирование—фдеолизосом, в кото-
рыхсоздаются-мшмальн_ые усплпчп /снижение pH .др4,0—5,0)
для"действия гидролитических ферментов. разрушающих-фагоци-
тЧфокянтттЛе~ микроорганизмы. Фа_г идит а_р ная актив-
но с т’ь- нейтрофилов количественно выражается процентом
160
ф а г cm-и. тирующих клеток и фагоцитарн ы м и н -
Дек сом (количество частиц, поглощенных одной клеткой).
Процент "фагоцитирующих нейтрофилов у здоровых людей
18—45 лет составляет от 68,5 до 99,3%, а фагоцитарный индекс —
от. 12 до 23,
В. нейтрофилах обнаружены кейлоны - специфические-веще -
,ства. подавляющие синтез ДНК в клетках гранулоцитарного ряда и
оказывающие регулирующее действие на процессы пролиферации
и дифференцировки лейкоцитов.
Продолжительность жизни нейтрофмиль ок» <n сут- При этом
в кровяном русле они находятся 8—12 ч, а далее выходят в
соединительную ткань, где проявляется их максимальная функцио-
нальная активность.
Эозинофильные (ацидофильные) гранулоциты (granulocy-
tus eosinophilicus), или эозинофилы. Это более крупные клетки,
чем нейтрофилы; диаметр их -в капле свежей крови около 9—
10 мкм, а на мазках — около J2—14_ мкм. Количество эозино-
фильных лейкоцитов в периферической крови колеблется_в. лреде-
лах 1—5% от общего числа лейкоцитов. В цитоплазме имеется
два типа гранул.. Характерным признаком является^ наличие, ^спе-
цифических окуифидьдых гранул овальной или полигональной
формы, размером около 0,5—1,5 мкм (см. рис. 46, А). ОВгриФиль-
ность гоацул обусловлена содержанием в них основного белка,
богатого аргинином. Электронно-микроскопически в экваториаль-
ной плоскости эозинофильных гранул выявляются единичные или
множественные кристаллоидные структуры, которые имеют вид
пластинчатого тела (см. рис. 47, Б). Кристаллоидные структуры
погружены в аморфный тонкозернистый матрикс. В гранулах
содержатся большинство гидролитических ферментов, пероксида-
за, характерные для азурофильных гранул, (лизосом)_нейтрофи-
лов, поэтому их рассматривают как разновидность лизосом. Сог-
ласно другому мнению, их относят к пероксидазосомам-JlepQK-
с и да за в незрелой грануле располагается по всей ее плошали.
в зрелых — лищь_до_лериферии гранулы (рис. 48). Активность
кислой фосфатазы выявляет также w цу nonfpvпостно1' части.
В гранулах отсутствуют лизоцим и фагоцитин. Кроме того, эозино-
фильные гранулы обладают активностью эстеразы, гистаминазы.
Второй тип гранул имеет меньшие размеры —0,1—0,5 мкм, округ-
лую *форму, гомогенную или зернистую ультр^стпуктуру- Они со-
держат. кислую фосфатазу и аридсульфатазу. Число этих гранул
в процессе специализации эозинофила уменьшается. Их рассмат-
ривают как стадию развития специфических оксифильных гра-
нул . Органеллы эозинофила—в—цитоплазме слабо—пазвиты.
Различают три [я^овидности-фстедии-ратвитни) аизинофилов/
сегментоядерные, палочкоядерщ^ ц юцще. Ядро сегментоядерных
эозинофилов, как правило, с^Ётоит из двух сегментов (реже из
трех), соединенных между собой тонкими перемычками. И-зредка
встречакц£я_палочкоядерные йюные формы, ядра которых имеют
форму, сходную с ядрами-нейтрофилов гоотлетствующих стадий
6 гистология
161
Рис. 48. Гранулы эозинофильных гранулоцитов. Реакция на пероксидазу. Элек
тронная микрофотография V 12 000 (по Д. Байнтону и М. Фарквару).
1 ядро; 2 пероксидаза в зрелых гранулоцитах; 3 — кристаллический центр зрелых
гранул < отрицательной реакцией на пероксидазу
развития. Ядра эозинофилов имеют в своем составе в основном
гетерю хроматин. ядрышки не видны. Эозинофильные лейкоциты
тдёнее подвижны, чем нейтрофилы. Они могут м и три ро вать из
кровяного русла в ткани, передвигаясь к источнику раздражения.
(Положительным хемотаксическим влиянием на эозинофилы обла-
дает ряд вещестн:гистамин, лимфокины (выделяются стимули-
рованными лимфоцитами), иммунные комплексы, состоящиеиз ан-
тигенов и антител и др. Эозинофилы имеют положительный хемо-
таксис к пептиду, выделяемому тканевыми базофилами при разру-
шении их гранул (эозинофильный хемотаксический фактор ана-
филаксии). Выявлено, что в областях развития реакции антиген —
антитело в очагах воспаления образуются вещества, оказывающие
стимулирующее влияние на выход эозинофилов из костного мозга
в циркулирующую кровь и оттуда в ткани.
Эозинофильные гранулоциты способны к фагоцитозу,
однако^их фагоцитарная активность ниже, чем у нейтрофилов. Они
принимают_уяас!ие. в а щит нъГдс р е. а кц иях организма
н а чужеродны й_£_е л о к, в а л л е р г и ч е с к2и х и а на-
ф и-л-а-к-тщцгес к и х_реакциях. Введение чужеродного белка
приводит к j/велцчению числа-эозинофилов - эозинофилии вслёд-
ствие мобилизации резервов этих клеток из костного мозга. Эозино-
филы участвуют в метаболизме гистамина: они инактивируюГ^гп с
помощью фермента гистаминазы, обнаруживающего высокую ак-
тивность в эозинофилах; не обладая способностью синтезировать
гистамин, они могут накапливать его, фагоцитируя гйстаминсо-
держащиё~гранулы, выделяемые базофилами и тучными клетками,
а также адсорбировать его на цитолемме, содержащей специ-
фические рецепторы к гистамину. Кроме того, эозинофилы выра-
батывают специальный фактор, тормозящий освобождение "гиста-
мина" из "базофилов и тучных клеток. Ферменты эозинофилов
разрушают*вещества, высвобождаемые тканевыми базофилами при
иммунных реакциях (аллергия, анафилаксия): арилсульфатаза
разрушает анафилаксии; гистаминаза — гистамин. Возрастание в
крови уровня адреналина, адренокортикотропного гормона гипо-
физа и глюкокортикоидных гормонов надпочечников вызывает
падение числа эозинофилов в крови — эозинопению. Содержание
эозинофилов в крови—в-течение суток— -колеблется (максимум*—
ночные часы, минимум — утренние)- Определяется это уровнем
секреции глюкокортикоида — гидрокортизона в организме. Время
нахождения эозинофилов в кровяном русле —3—8 ч, в последую-
щем онщмигрируют в соединительную*ткань"органовх где функцио-
нируют^.
Базофильные гранулоциты (granulocytus basophilicus). Ба
зофильные гранулоциты, или базофилы, имеют , иаметр около
9 мкм в капле свежей крови и около 11 —12 мкм в мязке^ В крови
человека ггни~составляют~~0^5^1 % от общего числа лейкоцитов.
Цитоплазма базофилов заполнена крупными, округлыми или поли-
гональными базофильными гранулами (granulum basophilicum),
размер КОторых варьирует от 0,5 до 1,2 мкм. Гранулы обладают
метах рдлш рце р'Метахромазия зерен связана с_наличием в них
гликозаминогликана — гепарина. Гепарин легко раство-
ряется в воде, и поэтому в препаратах, "фиксированных-плохо
обезвоженным метиловым спиртом, на месте зерен в цитоплазме
образуются просветления. При фиксации клеток абсолютным ме-
тилов ым спиртом зерна~хороши сохраняются. Вгранудах содер-
жатся также гистамин, серотонищ пероксидаза, киспая фосфатаза,
гистидиндекарбоксилаза (фермент синтеза гистамина) и др.
Изучение базофилов с помощью электронного микроскопа по-
казало неоднородность гранул по плотности, что, вероятно, отра-
жает различную степень их зрелости или функционального состоя-
ния (см. рис. 47, В). Помимо специфических базофильных__гранул,
в базофилах содержател и азурдфильные грану„ы ( титтмы)
Ядра* базофилов чаще слабодольчатые, реже — сферические,
окрашиваются гораздо менее интенсивно, чем ядра нейтрофилов
или эозинофилов^ В цитоплазме выявляются все виды основных ор-
ганелдЗ
Функция базофилов заключаетеявуце участии я метаболизме
гис та м ййД~И"ГС~ГТа р и~н~аП>азофильные грану попиты, так же
как "и тучные клетки соединительной ткани, выделяя гепарин и
г ис.тамйн, принимают участиеврегуляции дроне ссо в сверты ва н и я
крови и проницаемости сосудов,. Базофил^^^частвутот в иммуно-
логических реакциях организма, в частности в реакнтуяхаллерги-
ческого характера. БазофИЛБгреагируКйГ-Да^комплекс антиген —
1 Метахромазия — свойство клеток и тканей окрашиваться в тон, отличающийся
от цвета красителя. '
6*
163
антитело. Они легко присоединяют особые антитела — иммуно-
глобулины (IgE), циркулирующие в крови. Чужеродные белки
(антигены), вызывающие образование IgE-антител, действуя на
базофилы, приводят к их дегрануляции и высвобождению гистами-
на, обусловливающего резкое расширение сосудов, появление
отеков и др. Фагоцитарная активность базофилов выражена слабо.
Агранулоциты (незернистые лейкоциты)
Лимфоциты (lymphocytus). В крови взрослых людей лимфоциты
составляют 20—35%. Величина лимфоцитов значительно варьирует
от 4,5 до 11) мкм. В зависимости от размера всветовом мик р о-
с о п е различают лимфоциты: малые — диаметром 4,5—6 мкм,
средние — диаметром 7—10 мкм и большие — диаметром 10 мкм и
более (см. рис. 46, Л). Большие лимфоциты встречаются в крови
новорож leuuiix д-Асдсй. > шхутутвуют. Для лймфо-
цитов характерно наличие интенсивно окрашенного ядра округлой
или бобовидной формы и О1нш телБПо~нёбольшогб ободка базо-
фильной цитоплазмы, В цитоплазме некоторых________лимфоцитов
содержится небольшое количество азурофильных гранул (ли-
зосом)^
Э л е к т р о н н о-м икроскопичес к и у взрослых людей в
популяции лимфоцитов выделено 4 Типа клеток: 1) малые светлые
лим фоциты, 2) малые темные лимфо ц и т ы, 3) средние лимфоц и -
ты, 4) плазмоциты (лимфоплазмоциты).
Малые хае 1лые лимфоциты ееетавлятпт бод^иую часть (около
70—75%)'всех лимфоцитов крови человека. Их диаметр около
7 мкм, ядернсьЦИТОп'лазматичёское отношение сдвинуто в пользу
ядер. Ядро округлой формы может иметь небольшие впячивания;
хроматин конденсирован по периферии ядра (рис. 49, А) В светлой
цитоплазме"расположены 'небольшое количество—свободных рибо-
сом и полисом, слабо выраженные элементы гранулярной эндо-
плазматической сети, центросомы, комплекс Гольджи, митохондрии
В цитоплазме много везикул и мультивезикулярных телец,
встречаются лизосомы. Органеллы обычно расположены около
выемки ядра.
Малые темные лимфоциты составляют около 12—13% лимфо-
цитов крови, и>Г диаметр 6—7"~мкм. Ядерно-цитоПлазматическое
отношение еще более сдвинутоТГГГОЛЬзу ялра7~Хроматцн выглядит
плотным, ядрышко крупное. Цитоплазмарюкружающая ядро узким
ободком, имеет высокую электронную плотность (темная),. в ней
расположено большое количество рибосом. Митохондрий немного,
их светлый матрикс выделяется на тёмном фоне цитоплазмы. Дру-
гие органеллы встречаются редко.
Средние лимфоциты составляют около 10—12% лимфоцитов
крови человека. Они имеют диаметр окол<у НХлпсм—Ядра этих клеток
обычно округлые, иногда бобовидные^часто видны пальцевидные
впячЗвания ядернои оболочки. Хроматин более рыхлый, участки
конденсированного хроматина видны около ядернои оболочки,
164
Рис. 49. Ультрамикроскопическое
строение а!ранулоцитов (схема)
(рис. Н. А. Юриной, Л. С. Румянце-
вой): А — лимфоцит; Б- моноцит;
/ ядро; 2 - рибосомы; 3 мик-
роворсинки; 4 — лизосомы, 5 — ком-
плекс Гольджи; 6 митохондрии;
7 — пиноцитозный пузырек; В —
дифференцировка моноцита в мак-
рофаг (по АН. Радостиной): /—
моноцит; // — дифференцирующий-
ся макрофаг; 7//. IV — зрелые мак
рофа! и.
ядрышко хорошо выражено. В цитоплазме расположены удлинен-
ные “канаЛЪЦы гранулярной—эндоняазматич^ской =сети-. .имеются
элементы агранулярной сети, свободные рибосомы и полисбмы.
ЦёнтрТГСТУШ~й комплекс Гольджи “обычно расположены рядом
с областью инвагинации ядерной мембраны. Митохондрии более
мелкие. Лизосомы встречаются в небольшом количестве.
I. П шямациты (лимфо плазмоциты) составляют в крови человека
I около 1—2%. Характерным признаком этих- клетСГК- является
концентрическое расположеаиесвикруг ядра канальцев гранулярной
эндоплазма i ическби~сети
Среди лимфоцитов по путям дифференцировки и роли установ-
лении -^защитных реакций выделены два основных, вида ,— Т- и
В-лимфоциты.
Л Тимусзависимые лимфоциты (Т-лимфоциты), образующиеся из
ствоЯовых клеток костного мозга в тимусе, о~беспечивают
0 р е акции к л е т о ч нтггс и~м м /1ГЙ т е т а и., р е г у л~я~Т[ и ю
гуморального им м у н ит е т а,В популяции Т-лимфоцитов
икЛй^нолигичсСкрми методами выявлены следующие две основные
группы лимфоци.тов.
Перцую__груш «у—составляют литотомичгские Т-лимфоциты
(киллеры)), являющиеся эффекторными клетками клеточного
иммунитета.
Ко второй группе относятся Т-лимфоциты, оказывающие ре-
гул и рующее влияние на В-лимфоциты. Среди них разли ча ют
Т-хелперы и Т-супрессоры. Т-хелпёры (помощники) обладают спо-
собностьюспецифически распознавать антиген и усиливатьобразо-
ваниё антител. Т-супрессоры" (угнетающие) подавляют-'способ-
ность В-лимфоцитов участвовать в выработке антител. Действие
Т-лимфоцитов на В-клетки опосредуется с “помощью особых раст-
вори мых-веществ — лимфд'Кинов, вырабатываемых ими при 'дейст-
вщцант и ге но в.
В лимфдциты образуются из стволовых клеток костного мозга
в фабрициевой сумке (bursa Fabricios) у птиц, а у чёловёка, возмож-
но, в эмбриональной печени, а у взрослого — в костном мозге.
Их главндя_ функция — обе с-н-о-ч-е-н-и-е—i-y-м-е-р-а-л-ьн ого
и м му ни те тц. Образующиеся_из В-лимфоцитов эффекторные
клетки — плазмоциты вырабатывают особые защитил? белки —
иммунагуюбулинъ^фшттитела ), которые- -инету па ют-в кровь.
Четкие морфологические различия между Т- и В-лимфоци-
тами пока не установлены. Исследования, проведенные с помо-
щью электронного микроскопа, показали, что js В-лимфоцчтах
более развита гранулярная эндоплазматическая сеть, а в Т-лим-
фо I щтах — более многочисленные лизосомы- Морфо метр и чёс к и й
анализ выявляет различия в размерах клеток, их ядер, ядерно-
цитоплазматических отношениях, содержании и распределении
гетерохроматина и эухроматипа ядер. Т-лимфоциты и их ядра
имеют меньшие размеры, большее содержание гетерохроматина.
Имеется ряд различий в ферментном профиле Т- и В-лимфоцитов.
Выявление Т- и В-лимфоцитов и их популяций проводится с по-
166
мощью иммунологических и иммуноморфологических методов ис-
следования, многие из которых основываются на характеристи-
ке специфических особенностей строения их мембран. Мембрана
лимфоцитов характеризуется__большой динамичностью, так как
химические компоненты ее (белки и др,) могут перемещаться
внутри мембраны при действии ряда факторов, например анти-
генов.— Перемещающиеся молекулы могут концентрироваться
на поверхности клеток, формируя «рецептор ы», обеспечи-
вающие- взаимодействис с антигС11ами~ тгрн Иммунных "реакциях.
Существование такого «рецептора»_ временное, иногда всего не-
сколько часов. Роль рецепторов для антигена на В-лимфоцитах
выполняют поверхностные иммуноглобулины (Sig).
Мембраны В-лимфоцитов имеют разнообразные поверхностные
рецепторы на антиген, которые определяют гетерогенность по-
пуляции В-клеток. Каждый В-лимфоцит отличается спецификой
и классом своего поверхностного иммуноглобулина. У взрослого
человека » периферической-крови содержится W—20% лимфо-
цитов, имеющих-поверхностные иммуноглобулиньщ (Sig). Около
2/я из них имеют IgM, около ’/з — IfiG, а содержание клеток cJgA,
IgD и IgE обычно ire превышает t Т%. Поверхностные иммуно-
глобулины обычно выявляют иммунофлюоресцентным методом.
Кроме того, в мембране В-лимфоцитов обнаружен рецептор
для комплемента (С3) и Fc-рецептор’, оба они имеют белковую
природу. Выявлены также антиген, специфический для В-клеток
(MBLA), и специфические аллоантигены (HL В). Указанный
специфический антиген MBLA выявлен у 17 25% лимфоцитов
крови.
Химическая природа рецепторов для антигена в Т-лимфоци-
тах пока не установлена. В мембранах Т-лимфоцитов, помимо ре-
цепторадля антигенаТ"имеется рецептор для некоторых клеток,
например для эритроцитов барана (Е-рецептор), обеспечиваю-
щий реакцию розеткообразования, Fc-рецептор, который способен
связывать иммунные комплексы и обеспечивать кооперативные
взаимодействия Т- и В-клеток. Выявлены также антигены гистосо-
вместимости (HLA-антигены) — мембранные компоненты Т кле-
ток донора, ответственные за индукцию иммунного ответа (оттор-
жение клеток) в организме реципиента, а также специфиче-
ский 0-антиген, который способен перемещаться на один из
полюсов поверхности лимфоцита и располагаться там в виде
шапочки. 50—90% лимфоцитов периферической крови содержат
0-антиген. Продолжительность жизни лимФопитов варьирует от
нескольких^недель до нескоЛътагЧ'"лёт> Большую часть в перифе-
рической крови"доставляют долгоживущие [месяцы и годы)
рециркулирующие Т-лимфоциты, меньшую часть — ко-
ротко ж ивущйё (недели"и месяцы) tt-лймфОЦитьг.~ С)>еди—клеток,
имеющих морфологию малых лимфоцитов, имеются циркулирую-
щие стволовые клетки крови и соединительные ткани, поступаю-
щие сюда из костного мозга. Впервые эти клетки были описаны
А. А. Максимовым и обозначены им как «подвижный мезен-
Л
167
химный резерв». Из этих стволовых клеток, поступающих в ге-
мопоэтические органы, дифференцируются различные клетки кро-
ви, а из поступающих в соединительную ткань — фибробласты,
тучные клетки, макрофаги и др.
Моноциты (monocytus). В капле свежец—кровц^ эти _клетки
лишь немного крупнее других лейкоцитов (9—12 мк^г), в мазке
крови дщцильно распластываются по стеклу и размер их дости-
гает 4"#-—20 мк^Т/В крови человека количество моноцитов колеб-
лется в пределах. Я.% от общего числа лейкоцитов.
Ядра- моноцитов разнообразной тПГзмёнчивой Псонфигурашш:—»
встречаются бобовидные, подковообразные, реже — дольчатые яд-
ра с многочисленными выступами и углублениями. Хроматин
рас се я н мелкими зернами по всему ядру, но_обычна- ши х
количествах он располагается под ядерной мембраной (см. рис. 49,
Б}. В ядре моноцита содержится одноТппг~Несколько маленьких
ядрышек.
Цитоплазма моноцитов менее базофильна, чем цитоплазма
лимфоцитовПТри окраскепо методу Романовского — Гимзы она
имеет бледно-голубой цвет, но по периферии окрашивается не-
сколько темнее, чем около ядра; в ней содержится различное
количество очень мелких азурофильных зерен (лизосом).
Характерно наличие пальцеобразных выростов_цитоппазмы и
образование фагоцитарных вакуолей. В цитоплазме расположено
множество пиноцитозных везикул. Имеются короткие канальцы
гранулярной эндоплазматической сети, а также небольшие по
размеру митохондрии. Моноциты относятся к макрофаги-
ческой системе организма, или к так называемой
мононуклеарной'фагоцитарной системе (МФС).
Клетки этой системы характеризуются происхождением из про-
моноцитов костного мозга, способностью прикрепляться к поверх-
ности стекла, активностью пиноцитоза и иммунного фагоцитоза,
наличием на мембране рецепторов для иммуноглобулинов и комп-
лемента. Моноциты циркулирующей крови представляют собой
подвижный пул относительно незрелых клеток, находящихся на
пути из костного мозга в ткани. Время пребывания моноцитов в
крови варьирует от 36 до 104 ч.
-—"Моноциты, выселяющиеся в тканщ-нревращаютоя в лшкрс^фаги.
при этом у них появляются большое количество лизосом, фаго-
сомы, фаголизосомы (рис. 49, В).
Тромбоциты (кровяные пластинки)
Тромбоциты (thrombocytus) в свежей крови человека имеют
вид мельчайших бесцветных телец округлой, овальной, веретено-
видной или неправильной формы. Они представляют собой отде-
лйвшиеся от гигантских клеток костного мозга — так называемых
мег<тк<777ш?щегтд»г4?езъядсрные фр<н менты ИХ цитоплазмы размером
2—3 mjcm.
Благодаря способности к агглютинации, склеиванию в мазках
168
Рис. 50. Ультрамикроскопическое строение тромбоцита (кровяной пластинки)
(схема).
/ — гранулы; 2 (лыбки гликогена; 3 - эндоплазматическая,
5 — вакуоли; 6 — отростки кровяной пластинки.
:лазмзтическгя сеть; 4 митохондрии-
Ice-rtf - io&fc®
крови они обычно встречаются группами, поэтому подсчет кровя-
ных пластинок затруднен и приводимые количества их в 1 л ко-
ле бл!отся от200 » 1()9 по-300 « И.)'1-
Кажлау нпаотимкА > -т.чп и < । и(hyalomerus), являю-
щегося основой пластинки, и грдд.уд£шдш. (granulomerus) — зер-
нышек образующих скопление в центре пластинки или разбросан-
ных _по гиаломеру. !'ранулОМер, хотя и~ окрашивается азуром;._н£
Содержит'я дерного вещества — хроматина; гистохимическая реак-
ция на ДНК в нем отрицательная. Под электронным микроскопом
в .Грану номере видны окруженные мембраной округлые плотные
гранулы диаметром около 0,2 мкм, химическая природа которых
недостаточно выяснена (рис. 50). Они называются . а -гранулами
в отличие от второй группы____гранул-—содержащих—еерето ни н
(5-гидрокситриптамин). Кроме того, в грануломере обнаружива-
ются митохондрии и различные количества гранул гликогена.
В гиаломере нет i ранул, при больших увеличениях электрон-
ного микроскопа видны тонкие филаменты. Кроме того, имеются
циркулярно расположенные пучки, состоящие из 10—15 микро-
трубочекТ которые помогают поддерживать форму_ пластинок.
Кровяные пластинки могут образовывать большое количество от-
ростков различного размера и толщины. При окрашивании по
методу Романовского— Гимзы различают 5 видов кровяных плас-
тинок:^/ юные, с базофильным гиаломером и единичными азуро-
фильными Транулами;(<2У)зрелые, со слабооксифильным гиаломе-
ром и выраженной аэурб фильнОй зернистостью; ($)/< стары е,
более темные, сине-фиолетового оттенка с темночриолётбЯой
зернистостью; 4) дегенеративные с серовато-синеватым
гиаломером и с серовато-фиолетовой зернистостью; 5) гигант-
с к и е (формы раздражения), размер которых в 2—3 раза пре-
169
вышает нормальные, с розовато-сиреневым гиаломером и фиоле-
товой зернистостью.
Кровяные пластинки принимают участие в процессе с в е р-
т ы в а н и я кр &ви. Эта функция спределяется их способность^
быстрр-рагпадаться, склеиваться в конгломераты, вокруг которых
возникают нити фибрина В процессе свертывания крови кро-
вяные пластинки выделяют ряд веществ (фосфолипиды, липопро-
теиды и др.) и различные ферменты (тромбокиназа, пептидаза,
нуклеотидаза, кислая фосфатаза, каталаза и др.).
П ро дол ж ительность жизни кровяных пластинок—около 5—
8 днёш
.. ——'
Гемограмма. Лейкоцитарная формула
В медицинской практике анализ крови играет большую роль.
При клинических анализах исследуют химический состав крови,
определяют количество эритроцитов, лейкоцитов, гемоглобина, ре-
зистентность эритроцитов, быстроту их оседания — скорость осе-
дания эритроцитов (СОЭ) и др. У здорового человека формен-
ные элементы крови находятся в определенных количественных
соотношениях, которые принято называть гемограммой, или фор-
мулой крови. Важное значение для характеристики состояния
организма имеет так называемый дифференциальный подсчет лей-
коцитов. Определенные процентные соотношения лейкоцитов на-
зывают лейкоцитарной формулой.
Возрастные изменения крови
Число эритроцитов в момент рождения и в первые часы жиз-
ни выше, чем у взрослого человека, и достигает 6,0—7,0 • 10|2/л.
К 10—14 сут оно равно тем же цифрам, что и во взрослом орга-
низме. В последующие сроки происходит снижение числа эритро-
цитов с минимальными показателями на 3—6-м месяце жизни
(физиологическая анемия). Число эритроцитов становится таким
же, как и во взрослом организме, в период полового созревания.
Для новорожденных характерно наличие анизоцитоза (разнооб-
разие размеров) с преобладанием макроцитов, увеличенное со-
держание ретикулоцитов, а также присутствие незначительного
числа ядросодержащих предшественников эритроцитов.
Число лейкоцитов у новорожденных увеличено и достигает
10—30 • 109/л. В течение 2 нед после рождения число их падает
до 9—15 • 109/л. Количество лейкоцитов достигает к 14—15 годам
уровня, который сохраняется у взрослого. Соотношение числа
нейтрофилов и лимфоцитов у новорожденных такое же, как и у
взрослых. В последующие сроки содержание лимфоцитов воз-
растает, а нейтрофилов падает, и, таким образом, к 4-м суткам
количество этих видов лейкоцитов уравнивается (первый фи-
зиологический перекрест лейкоцитов). Дальнейший рост
числа лимфоцитов и падение нейтрофилов приводят к тому, что
170
на 1—2-м году жизни процент лимфоцитов составляет 65, а ней-
трофилов — 25. Новое снижение числа лимфоцитов и пов! шение
нейтрофилов приводят к выравниванию обоих показателей у 4-лет-
них детей (второй физиологический перекрест).
Постепенное снижение содержания лимфоцитов и повышение ней-
трофилов продолжаются до полового созревания, когда количество
этих видов лейкоцитов достигает нормы взрослого.
ЛИМФА
Лимфа (лат. lympha — влага) представляет собой слегка жел-
тбватую-- жидкость белково й природы, проте ка ющую в лимфа -
тических капиллярах и сосудах. Она состоит из лимфоплазмы
(plasma lymphae) и форменных элементов. По химическому со-
ставу лимфоплазмаблизка к плазме крови, но содержит меньше
белка. Среди фракций белка альбумины преобладает над гло-
булинами. Часть белка составляют ферменты: диастаза, липаза и
гликолитические ферменты. Лимфоплазма содержит также ней-
тральные жиры, простые сахара, NaCl, Na2CO3 и др., а также раз-
личные соединения, в состав которых входят кальций, магний,
железо.
Ф о р менные эдеме и м ф_ы представлены гла fl-
ным образом лимфоцитами а также моноцитами и дру-
гими видами лейкоцитов, иногда в ее составе обнаруживаются
эритроциты. Лимфа образуется, в лимфатических капиллярах тка-
ней и органоЬ, кудапод влиянием различных факторов^ в част-
цости-лд^отичёсКйго^и гидростатического ' давления, из чуканей
Ростоянно поступают различные компоненты лимфоплазмы. Из
капилляров лимфа перемещается и периферические лимфатические
сосуды, по ним — в лимфатические узлы, затем в крупные лимфа-
тические срсу/тбГТГ~вливается в кровь. Состав лимфы постоянно
меняется. Различают периферическую лимфу (до лим-
фатических узлов), промежуточную (после прохождения
через лимфатические узлы) и центральную (лимфу груд-
ного и правого лимфатического протоков). Процесс лимфообра-
зования тесно связан с поступлением воды и других веществ из
крови в межклеточные пространства и образованием тканевой
жидкости.
КРОВЕТВОРЕНИЕ (ГЕМОЦИТОПОЭЗ)
Г е мо цито п о э_зо м (haemocytopoesis) называют развитие
крови. Разпичают а. м б'р иональный гТмоц й~ТТПт -о э-а,-
который происходит в эмбриональный период и приводит к раз-
витию крови как ткани, и постэмбриональны и гемо-
ц и т о п о э з, который представляет собой процесс физиологи-
ческой регенерации—крови. Развитие эритроцитов называют
эритроцитопоэзом, развитйё'гранулоцитов - грануло-
171
ц и поэз о м, тромбоцитов — тромбоцитопоэзом, разви-
тие моноцитов — моноцитопоэзом, развитие лимфоцитов
и иммуноцитов — лимфоцит о- и иммуноцитопоэзом.
Эмбриональный гемоцитопоэз (развитие крови как ткани)
Развитие крови как ткани происходит у эмбриона сначала в
стенке желточного мешка, затем в печени, костном мозге и лим-
фоидных органах (тимус, селезенка, лимфатические узлы).
Кроветворение в стенке желточного мешка. У человека оно
начинается в конце 2-_й — в начале 3-й недели эмбрионального
развития, В мезенхиме стенки желточного мешка обособляются
зачатки сосудистой крови, или кровяные островки. В них мезен-
химные клетки округляются, теряют отростки и преобразуются
в стволовые клетки, крови (СК). Клетки, ограничивающие кровя-
ные островки, уплощаются, соединяются между собой и образуют
эТЩбтедиальную выстилку будущего сосуда. Часть стволовых кле-
ток дифференцируется в первичные клетки крови (бласты), круп-
ные клетки с базофильной цитоплазмой и ядром, в котором
хорошо заметны крупные ядрышки (рис. 51, А, Б, В, Г). Боль-
шинство первичных кровяных клеток митотически делится и пре-
вращается в первичные эритробласты, характеризующиеся круп-
ным размером (мегалобласты). Это превращение совершается в
связи с накоплением гемоглобина в цитоплазме бластов, при этом
сначала образуются полихроматофильные эритробласты, а затем
оксифильные эритробласты с большим содержанием гемоглобина.
В некоторых первичных эритробластах ядро подвергается кари-
орексису и удаляется из клеток, в других ядро сохраняется.
В результате образуются безъядерные и ядросодержащие пер-
вичные эритроциты, отличающиеся большим размером по сравне-
нию с нормоцитами и поэтому получившие название мегалоци-
тов. Такой тип кроветворения называется мегалобластическим.
Он характерен для эмбрионального периода, но может появлять-
ся в постнатальном периоде при некоторых заболеваниях (зло-
качественное малокровие). Наряду с мегалобластическим в
стенке желточного мешка начинается нормоб л астич е-
t к 6 ё к р б в"ет в о р е н и е, при котором из бластов обра-
зу ГОТСя- вторичные эритробласты; сначала они превращаются в по-
лихроматофильные эритробласты, далее в нормобласты, из ко-
торых образуются вторичные эритроциты (нормоциты); размеры
последних соответствуют эритроцитам (нормоцитам) взрослого
человека (см. рис. 51, В). Развитие эритроцитов в стенке жел-
точного мешка происходит “внутри первичных кровеносных со-
судов, т. е. и н т р а в а~?Т у л я р н о. Одновременно экстраваску-
лярно из бластов, расположенных вокруг сосудистых стенок,
дифференцируется небольшое количество гранулоцитов — нейтро-
филов и эозинофилов. Часть стволовых клеток остается в недиф-
ференцированном состоянии и разносится током крови по раз-
личным органам зародыша, где происходит их дальнейшая диф-
172
Рис. 51. Эмбриональный гемоцитопоэз.
А поперечный срез кровяного островка в'/^-дневного зародыша кролика: / полос-ть
сосуда; 2 — эндотелий: 3 — интраваскулярные кровяные клетки; 4 — делящаяся кровяная
клетка; 5 — формирование первичной кровяной клетки; 6 — энтодерма; 7 — висцеральный
листок мезодермы. Б — кроветворение в стенке желточного мешка зародыша морской
свинки: / - мезенхимные клетки; 2 — эндотелий стенки сосудов; 3 — первичные кровяны.
клетки-бласты; 4 — митотическое деление бластов. В - развитие вторичных эритробластов
в сосуде 13 /?-дневного зародыша кролика: / — эндотелий; 2 — проэритробласты;
3 — базофильные эритробласты; 4 полихроматофильные эритробласты; 5 — оксифильные
эритробласты (нормобласты); 6 - оксифильный эритробласт с пикнотическим ядром;
7— обособление ядра от оксифильного эритробласта (нормобласта); 8 вытолкнутое
ядро нормобласта; 9 — вторичный эритроцит. Г — кроветворение в костном мозге зародыша
человека величиной 77 мм. Экстраваскулярное развитие клеток крови: / — эндотелий сосуда;
2__бласты; 3 — нейтрофильные гранулоциты; 4 — эозинофильный миелоцит (по А А Мак
симову).
ференцировка в клетки крови или соединительной ткани. После
редукции желточного мешка основным кроветворным органом вре-
менно становится печень.
Кроветворение в печени. Печень закладывается примерно на
3—4-й неделе эмбриональной жизни, а на 5—й неделе она ста-
новится центром кроветворения. Кроветворение в печени проис-
173
ходит экстраваскулярно, по ходу капилляров, врастаю-
щих вместе с мезенхимой внутрь печеночных долек. Источником
кроветворения в печени являются стволовые клетки, мигриро-
вавшие из желточного мешка. Из стволовых клеток образуются
бласты, дифференцирующие во вторичные эритроциты. Процесс их
образования повторяет описанные выше этапы образования вто-
ричных эритроцитов. Одновременно с развитием эритроцитов в
печени образуются зернистые лейкоциты, главным образом ней-
трофильные и эозинофильные. В цитоплазме бласта, становящей-
ся более светлой и менее базофильной, появляется специфиче-
ская зернистость, после чего ядро приобретает неправильную
форму. Кроме гранулоцитов, в печени формируются гигантские
клетки — мегакариоциты. К концу внутриутробного периода кро-
ветворение в печени прекращается.
Кроветворение в тимусе. Тимус закладывается в конце 1-го
месяца внутриутробного развития, и на 7—8-й неделе его эпите-
лий начинает заселяться стволовыми клетками крови, которые
дифференцируются в лимфоциты тимуса. Увеличивающиеся в чис-
ле лимфоциты тимуса дают начало Т-лимфоцитам, заселяющим
Т-зоны периферических органов иммунопоэза.
Кроветворение в селезенке. Закладка селезенки происходит в
конце 1-го месяца эмбриогенеза. Из вселяющихся сюда стволовых
клеток происходит экстраваскулярное образование всех
видов форменных элементов крови, т. е. селезенка в эмбриональ-
ном периоде представляет собой универсальный кро-
ветворный орган. Образование эритроцитов и гранулоци-
тов в селезенке достигает максимума на 5-м месяце эмбриоге-
неза. После этого в ней начинает преобладать лимфоцитопоэз.
Кроветворение в лимфатических узлах. Первые закладки лим-
фатических узлов человека появляются на 7—8-й неделе эмбрио-
нального развития. Большинство лимфатических узлов развивает-
ся на 9—10-й неделе. В этот же период начинается проникнове-
ние в лимфатические узлы стволовых клеток крови, из которых
на ранних стадиях развития дифференцируются эритроциты, гра-
нулоциты и мегакариоциты. Однако формирование этих элемен-
тов быстро подавляется образованием лимфоцитов, составляющих
основную часть лимфатических узлов. Появление единичных лим-
фоцитов происходит уже в течение 8—15-й недели развития,
однако массовое «заселение» лимфатических узлов предшественни-
ками Т- и В-лимфоцитов начинается с 16-й недели, когда фор-
мируются посткапиллярные венулы, через стенку которых осу-
ществляется процесс миграции клеток. Из клеток-предшествен-
ников дифференцируются лимфобласты (большие лимфоциты), а
далее средние и малые лимфоциты. Дифференцировка Т- и В-лим-
фоцитов происходит в Т- и В-зависимых зонах лимфатических
узлов.
Кроветворение в костном мозге. Закладка костного мозга
осуществляется на 2-м месяце эмбрионального развития. Первые
гемопоэтические элементы появляются на 12-й неделе развития;
174
в это время основную массу их справляют эритробласты и пред-
шественники-гранулшдитов. Из стволовых клеток в костном мозге
формируются все форменные элементы крови, развитие которых
происходит экстраваскулярно (см. рис. 51, Г). Часть
стволовых клеток сохраняется в костном мозге в недифференци-
рованном состоянии, они могут расселяться по другим органам и
тканям и являться источником развития клеток крови и соеди-
нительной ткани. Таким образом, костный мозг становится цент-
ральным органом, осуществляющим универсальный ге-
мопоэз, и остается им в течение постнаталыюй жизни. Он
обеспечивает стволовыми клетками тимус и другие гемопоэти-
ческие органы.
I
Постэмбриональный гемоцитопоэз
Постэмбриональный гемоцитопоэз совершается в специали-
зированных гемопоэтических тканях — миелоидной (textus myeloi-
deus), где происходит образование эритроцитов, гранулоцитов,
тромбоцитов, моноцитов, и в ""лимфоидной, где происходит диф-
ференцировка и размножение Т- и В-лимфоцитов и плазмоцитов.
Постэмбриональный гемопоэз представляет собой процесс физио-
логической регенерации крови (клеточное обновле-
ние), который компенсирует физиологическое разрушение диф-
ференцированных клеток. Миелопоэз происходит в миелоидной
ткани, расположенной в эпифизах “трубчатых и полостях многих
губчатых костей (см. гл. XVI). Здесь развиваются форменные
элементы крови: эритроциты, гранулоциты, моноциты, кровяные
пластинки, предшественники лимфоцитов. В миелоидной ткани на-
ходятся стволовые клетки крови и соединительной ткани. Пред-
шественники лимфоцитов постепенно мигрируют и заселяют та-
кие органы, как тимус, селезенка, лимфатические узлы и др.
Лимфопоэз происходит в лимфоидной ткани (textus lymphoi-
deus), которая имеет несколько разновидностей, представленных
в тимусе, селезенке, лимфатических узлах. Она выполняет три
основные функции: образование лимфоцитов, обра-
зование плазмоцитов и удаление клеток и про-
дуктов их распада.
Миелоидная и лимфоидная ткани являются разновидностями
соединительной ткани, т. е. относятся к тканям внутренней сре-
ды. В них представлены две основные клеточные линии — клетки
ретикулярной ткани и гемопоэтические. В ретикулярной
ткани различают следующие типы клеток: фибробластоподоб-
ные (выполняющие опорную функцию) и макрофагические. По-
следние обеспечивают процессы фагоцитоза, в частности разру-
шение погибающих клеток крови и использование продуктов раз-
рушения их для образования новых гемопоэтических элементов
Г е м о п о э тические элементы, заселяющие ретику-
лярную ткань, развиваются нт пшипптентных стволовых клеток
крови, которые дифференцируются в унипотентные полусаволо-
175
вые _клетки, дающие начало определенномувиду форменных эле-
ментов крови.ТТроцесс~дифференцировки стволовых клеток обу-
словлен действием многих факторов, среди которых важное место
занимает микроокружение: оба структурных компонен-
та — ретикулярные и гемопоэтические элементы — функциони-
руют в неразрывной связи.
ДляГвсёх'разновидностей лимфоидной ткани характерно также
наличие стромальных ретикулярных элементов и гемопоэтических,
образующих единое функциональное целое. В тимусе имеется
сложная строма, представленная как соединительнотканными,
так и ретикулоэпителиальными клетками. Эпителиальные клетки
секретируют особое вещество — тимозин, оказывающее влияние
на дифференцировку из стволовых клеток Т-лимфоцитов.
В лимфатических узлах и селезенке специализированные ретику-
лярные клетки создают микроокружение, необходимое для про-
лиферации и дифференцировки в специальных Т- и В-зонах Т- и
В-лимфоцитов и плазмоцитов.
Стволовые клетки крови являются полипотентными предшест-
венниками всех клеток крови и относятся к самбподдерживающей-
ся популяции клеток. Они редко делятся. Впервые представление
о рбдоначальных клетках*крови сформулировал в начале XX века
А. А. Максимов, который считал, что по своей морфологии они
сходны с лимфоцитами. В настоящее время это представление
нашло подтверждение и дальнейшее развитие в новейших экспери-
ментальных исследованиях, проводимых главным образом на мы-
шах. Выявление стволовых клеток крови стало--возможным при
применении метода к о л о н и е о б р а з о в а н и я.
В селезенке смертельно облученных животных можно на-
блюдать~образовгтие~колоний_кроветворных клеток из введенных
в их кровь к теток чро°“ ,Л кроветворных органов животных-до-
иорон- Каждая стволовая клетка образует одну колонию и назы-
вается колониеобратуюшей единицей (КОЕ). Подсчет колоний по-
зволяет судить о количестве стволовых клеток, находящихся во
введенной взвеси клеток. Таким образом, было установлено, что
у мышей на 105 клеток костного мозга приходится около 50 ство-
ловых клеток, в селезенке — 3,5 клетки, среди лейкоцитов кро-
ви — 1,4 клетки. Исследование чистой фракции стволовых клеток
с помощью электронного микроскопа показало, что по ультра-
структуре они очень близки к малым темным лимфоцитам.
Исследование клеточного состава колоний позволило выявить
две линии их дифференцировки. Одна линия дает начало полипо-
тентной полустволовой клетке JT1CK)—предшественнице грану-
лоцитарного, эритропитарылт моноцитарного и мегакариоцитарно-
го рядов гемопоэза (КОЕ — ГЭММ). Вторая линия дает_начало
полипотентной полустволовой клетке — предшественнице лимфо-
поэза-(рис. 52).
В ряду полустволовых клеток первой линии в последние годы
выделены КОЕ — ГМ, дающие начало моноцитам и гранулоцитам,
КОЕ — ГнЭ, являющиеся общим источником для дифференциров-
\ 176
ки нейтрофильных гранулоцитов и эритроцитов, и КОЕ — МГЦЭ,
из которых образуются мегакариоциты и эритроциты.
Из ПСК дифференцируются унипотентные_предщественники
для всех видов клеток крови. Методом- колониеобразования оп-
ределены унипотентные предшественники моноцитов (КОЕ — М),
нейтрофильных ГраПулДцйтов (КОЕ—ГнУ7~эозинофилов (КОЕ—
Эо), базофилов (КОЕ—Б), эритроцитов (КбЕ—Э), мегакариоци-
тов (КОЕ—МГЦ). В лимфопоэтическоуГряду выделяют унипотент-
ные клетки — предшественницы для В-лимфоцитов и соответст-
венно-для Т-лимфоцитов. ПотгипотептнБге стволовыеТ’пблустволо-
вые и унипотентные клетки морфологически не различаются.
Дифференцировка подипо.тентных полустволоных клеток в уни-
потентны£_определяется—действием—ряда специфических факто-
ров — эритропоэтинов (для эритробластов), гранулопоэтинов (для
миелооластов),~лимфопоэтинов (для лимфобластов), тромбопоэ-
тинов (для мегакариобластовХ и др.
Из каждой унипотентной клетки происходит образование блас-
та для конкретного вида клеток. Бласты дают начало морфологи-
чески идентифицируемым стадиям дифференцировки гемопоэти-
ческих клеток.
Процессы гемоцитопоэза представлены на схеме (см. рис. 52).
При развитии конкретных видов клеток в процессе миелопо-
эза можно выявить ряд морфологических особенностей.
Эритроцитопоэз
Развитие эритроцитов протекает по схеме: СК — ПСК (КОЕ-
ГЭММ — КОЕ-ГнЭ или КОЕ-МГЦЭ) — унипотентные прел-
звественники (КОЕ — Э) — проэритробласт — эритробласт (ба-
зофильный,пол их рома тофил ьный, оксифильный) — ретикулоцит ^
эритроцит. В клетке происходят изменения: уменьшаются ее
размеры умгныпае.тся, уплотняется и исчезает ядро, ослабляется
базофилия цитоплазмы в связи с уменьшением содержания PFTK,
накапливается гемоглобин и появляется связанная с ним__окс и -
филия цитоплазмы- (рис. 53).
Проэритроб часты- (proerythroblastus) представляют собой клет-
ки,“обладающие крупными округлыми или овальными ядрами, с
мелко распыленным хроматином и -хорошо выраженными ядрыш-
ками. Слегка базофильная цитоплазма узким ободком окружает
ядро. Величина этих клеток около 15 мкм. Они располагаются
поодиночке или группами" (по 2—4 клетки). Характерным при-
знаком этих клеток является способность к интенсивной проли-
ферации — деление происходит каждые 8—Т2 ч. В результате ми-
тотического деления проэритробластов появляются более мелкие
(10—12 мкм) округлые клетки. Они отличаются тем, что их пол-
ностью гомогенная цитоплазма имеет резко выраженную базо-
филию, связанную с тгакоплением в—ней РНК. Ядрсг“округлое,
несколько более интенсивно окрашенное. Эти клеточные формы по-
лучили название базофильных эритробластов (erythroblastus ba-
моиобласт I миелобласты
про моноцит . про миелоциты
эритробласт мег а карио -
бласт
полихрома-
тофильный
эритробласт
оксифильный
эритро бласт
*
I
мегакариоцит
1
тромбоциты.
эритроцит кровяные
пластцнни
I
Рис. 52. Постэмбриональный гемоцитопоэз (схема Н. Юриной).
I VI — стадии дифференцировки клеток крови: I IV морфологически неидентифици-
л
>
>4
Рис. 53. Схема ультрамикроскопическо-
го строения последовательных стадий
дифференцировки проэритробласта в
эритроцит (по В. Блюму и Д. Фоссету
с изменениями)
А — базофильный эритробласт; Б — поли-
хроматофильный эритробласт; В — окси-
фильный эритробласт; Г — ретикулоцит;
Д — эритроцит: / — ядро; 2 — рибосомы и
полирибосомы; 3 — митохондрии; 4 — гра-
нулы гемоглобина
sophilicus). При исследовании под электронным микроскопом в
цитоплазме базофильных эритробластов выявляются небольшое
количество митохондрий различных размеров, диффузно распо-
ложенные рибосомы и несколько пузырьков эндоплазматической
сети. Базофильные эритробласты энергично размножаются и дают
ряд еще более мелких клеточных форм размером 8—10 мкм,
цитоплазма которых постепенно теряет базофилию и начинает
окрашиваться не только основными, но и кислыми красителями
благодаря накоплению гемоглобина. По признаку окрашиваемо-
сти цитоплазмы эти клеточные формы называются полихромато-
фильными эритробластами (erythroblastus polichromatophilicus)
(см. рис. ^2). 1Готличие от базофильных эритробластов в ядре у
полихроматофильных эритробластов ядрышки, как правило, исчеза-
ют, а хроматин в виде довольно крупных глыбок неправильной фор-
мы равномерно заполняет все ядро. После размножения и диффе-
ренцировки нескольких генераций полихроматофильных эритро-
бластов, постепенно накапливающих гемоглобин, образуется клетка
с отчетливо выраженной оксифильной цитоплазмой — ацидофиль-
ный эритробласт (erythroblastus acidophilicus). В начальных ста-
диях образования ацидофильные эритробласты еще могут делить-
ся митотически, но очень скоро они теряют эту способность, так
180
Рис. 54. Развитие эритроцитов в печени плода человека Электронная микро
фотография (по Замбони).
Л, В — 15-недельного, z 6000; В — 20-недельного. X 15 000= / —эксцентрично расположен
ное ядро эритробласта; 2 — обособление пикиотического ядра нормобласта; 3 — отделение
пикнотического ядра с узким ободком цитоплазмы от нормобласта; 4 — ретикулоцит с еди-
ничными органеллами (указано стрелками).
как в их ядрах возникают процессы, ведущие к постепенной их
деструкции и исчезновению. Ядра в таких клетках утрачивают
правильную округлую форму, пикнотизируются и удаляются из
клетки, при этом клетки превращаются сначала в ретикулоцит,
а затем в эритроцит (см. рис. 52, 53; рис. 54).
У взрослого организма потребность в эритроцитах обычно
обеспечивается за счет усиленного размножения полихромато-
фильных эритробластов. Но всякий раз, когда потребность орга-
низма в эритроцитах возрастает (например, при потере крови),
эритробласты начинают развиваться из предшественников, а по-
следние — из стволовых клеток.
В норме из костного мозга в кровь поступают только эритро-
циты и ретикулоциты.
ранулоцитопоэз
| Источником для гранулоцитопоэза являются также СК и ПСК
^см. рис. 52), одновременно начинающие дифференцироваться
через ряд промежуточных стадий в трех различных направлениях
и продуцирующие клетки трех видов: нейтрофилы, эозинлфипы и
базофилы. Основные ряды для каждой из групп гранулоцитов
(слагаются из следующих клеточных форм: _СК — ПСК (КОЕ —
ГЭММ — КОЕ — ГМ; КОЕ — ГнЭ) — унипотентные ~предшест-
.ренники (КОЕ — Б, КОЕ — Эо, КОЕ — Гн) — миедпйпаст —
про миелоцит — миелоцит ^метамиелоцит палочко>ии^рый гра-
нулоцит и сТгментоярерный главупгшидч
По мере созревания гранулоцитов клетки уменьшаются в раз-
мерах, изменяется форма ядер от округлой до сегментирован-
1Ной, в цитоплазме накапливается специфическая зернистость
Цсм. рис. 52; рис. 55).
Миелобласты (myeloblastus), дифференцируясь в направлении
того или иного гранулоцита, дают начало промиелоцитам fpro-
myelocytus) (см. рис. 52). Это крупные клетки, содержащие оваль-
ное или округлое светлое ядро, в котором имеется несколько
ядрышек. Около ядра располагается ясно выраженная центросома,
хорошо развиты комплекс Гольджи, лизосомы. Цитоплазма слегка
базофильна. В цитоплазме накапливаются первичные (азурофиль-
ные) гранулы, которые характеризуются высокой активностью
кислой фосфатазы, т. е. относятся к лизосомам. Промиелоциты
делятся митотически. Специфическая зернистость отсутствует.
Нейтрофильные, или гетерофильные, миелоциты ('myelocytus
neutrophilicus) имеют размер от 12 до 18 мкм. Эти клетки энергич-
но размножаются митозом. Цитоплазма их становится диффузно
ацидофильной, в ней появляются наряду с первичными вторичные
(специфические) гранулы, характеризующиеся меньшей электрон-
ной плотностью. В миелоцитах обнаруживаются все органеллы.
Количество митохондрий невелико. Эндоплазматическая сеть со-
стоит из пузырьков. Рибосомы располагаются на поверхности
мембранных пузырьков, а также диффузно в цитоплазме. По мере
182
Рис. 55. Схема процесса дифференцировки нейтрофтьного гранулоциту в костном
мозге (по Д Байнтону, М. Фарквару, Дж. Элиоту с изменениями).
А — миелобласт; Б — промиелоцит; В — миелоцит; Г — метамиелоцит; Д палочкоядер-
ныи нейтрофильный гранулоцит (нейтрофил); Е — сегментоядерный нейтрофильный гра-
нулоцит; /—ядро; 2 — первичные (азурофильные) гранулы; 3 — комплекс Гольджи;
4 вторичные специфические гранулы.
размножения нейтрофильных миелоцитов округлое или овальное
ядро становится бобовидным, начинает окрашиваться темнее, хро-
матиновые глыбки становятся грубыми, ядрышки исчезают.
Такие клетки уже не делятся. Это — метамиелоциты (meta-
myelocytus) (см. рис. 55). В цитоплазме увеличивается число спе-
цифических гранул. Если метамиелоциты встречаются в перифе-
рической крови, то их называют юными формами. При дальней-
шем созревании их ядро приобретает вид изогнутой палочки. По-
добные формы получили название палочкоядерных лейкоцитов.
Затем ядро сегментируется и клетка превращается в сегментоядер-
ный, нейтрофильный лейкоцит. Полный период развития нейтро-
фильного гранулоцита составляет около 14 сут, при этом период
пролиферации составляет около 7,5 сут, а постмитотический пе-
риод дифференцировки — около 6,5 сут.
183
Эозинофильные миелоциты (см. рис. 52) представляют собой
клетки округлой формы размером около 14—16 мкм По характеру
строения ядра они мало отличаются от нейтрофильных миелоцитов.
Цитоплазма их заполнена характерной эозинофильной зернисто-
стью. В процессе созревания миелоциты митотически делятся, а
ядро приобретает подковообразную форму. Такие клетки называ-
ются эозинофильными метамиелоцитами. Постепенно в средней
части ядро истончается и становится двудольчатым, в цито-
плазме увеличивается количество специфических гранул. Клетка
утрачивает способность к делению. Среди зрелых форм различают
палочкоядерные и сегментоядерные лейкоциты с двудольчатым
ядром.
Базофильные миелоциты (см. рис. 52) встречаются в мень-
шем количестве, чем нейтрофильные или эозинофильные миелоци-
ты. Размеры их примерно такие же, как и эозинофильных миело-
цитов; ядро округлой формы, без ядрышек, с рыхлым расположе-
нием хроматина. Цитоплазма базофильных миелоцитов содержит
в широко варьирующих количествах специфические базофильные
зерна неодинаковых размеров, которые проявляют метахромазию
при окрашивании азуром и легко растворяются в воде. По мере
созревания базофильный миелоцит превращается в базофильный
метамиелоцит, а затем в зрелый базофильный лейкоцит.
Все миелоциты, особенно нейтрофильные, обладают способ-
ностью фагоцитировать, а начиная с метамиелоцита приобретают
подвижность.
У взрослого организма потребность в лейкоцитах обеспечи-
вается за счет размножения миелоцитов. При особых состояниях
организма миелоциты начинают развиваться из миелобластов, а
последние из унипотентных и полипотентных стволовых клеток.
Мегакариоцитопоэз. Тромбоцитопоэз
''-ч Кровяные пластинки также образуются в костном мозге. Этот
процесс слагается из следующих стадий: СК — ПСК (КОЕ —
МГЦЭ) унипотентный предшественник (КОЕ — МГЦ) — мега-
кариобласт — промегакариоцит — мегакариоцит — тромбоциты
(см. рис. 52).
Мегакариобласт (megacaryoblastus) представляет собой круп-
ную клетку (диаметром около 16 мкм) с лапчатым ядром и харак-
терным для него диффузным расположением хроматина. Цито-
плазма его базофильна При дальнейшем развитии, на стадии про-
мегакариоцита и мегакариоцита (megacaryocytus) происходит зна-
чительное увеличение (полиплоидизация) и сегментация ядра и
обогащение клетки цитоплазмой. Размеры клетки могут достигать
нескольких десятков микрометров (см. рис. 52), поэтому в маз-
ках костного мозга их легко определить даже при малых увеличе-
ниях. Цитоплазма содержит много центриолей, разбросанных в
углублениях ядерной поверхности и других органелл, а также
очень мелкую базофильную, полихроматофильную или оксифиль-
1К4
Рис. 56. Схема ультрамикроскопического строения мегакариоцита
(рис. НА. Юриной, Л. С. Румянцевой).
/ — ядро; 2 — канальцы эндоплазматической сети; 3 — гранулы; 4 — формирующиеся
кровяные пластинки.
ную зернистость. Характерным для них является наличие каналов
агранулярной эндоплазматической сети. По ним происходит от-
щепление кровяных пластинок от поверхности мегакариоцита, бо-
гатой псевдоподиями. В области псевдоподий и происходит отделе-
ние участков цитоплазмы, т е. кровяных пластинок (рис. 56).
Моноцитопоэз
Образование моноцитов происходит из стволовых клеток кост-
ного мозга по схеме: СК— ПСК (КОЕ — ГЭММ; КОЕ —
ГМ) —унипотентный предшественник моноцита (КОЕ—М) —
монобласт (monoblastus) — промоноцит — моноцит (monocytus)
Г Лимфоцитопоэз и иммуноцитопоэз
Лимфоцитопоэз проходит следующие стадии СК — ПСК —
унипотентный предшественник лимфоцита—лимфобласт (lym-
phoblastus) — пролимфоцит — лимфоцит. Особенностью лимфоци-
топоэза является способность дифференцированных клеток (лим-
фоцитов) дедифференцироваться в бластные формы
185
Процесс дифференцировки Т-лимфоцитов в периферических
органах приводит к образованию из унипотентных предшествен-
ников Т-бластов, из которых формируются сначала большие (lym-
phocytus magnus), средние (lymphocytus medius), а затем малые
эффекторные лимфоциты (lymphocytus parvus) — киллеры, хелпе-
ры, супрессоры, а также Т-клетки памяти (см. гл. X).
Процесс дифференцировки унипотентных предшественников
В-лимфоцитов приводит к образованию плазмоб ластов (plasino-
blastus), затем проплазмоцитов и, наконец, плазмоцитов fplasmo-
cytus) и В-клеток памяти. Более подробное изложение процессов
образования иммунокомпетентных клеток дано в гл. IX.
С - ХЗС
Глава VIII
СОЕДИНИТЕЛЬНЫЕ ТКАНИ'
Эту группу составляют: собственно соединительные ткани,
соединительные ткани со специальными свойствами и скелетные
соединительные ткани—(.хрящевая—и—костная). Соединительные
ткани характеризуются разнообразием клеток и хорошо развитым
межклеточным нгпих-тмом, не—волокон и основного
аморфного вещества. Физико-химические особенности межклеточ-
ного вещества и строение его в ' значительной—мере__одределя-
ют функциональное значение разновидностей Средийительных
тканей.
Функции. Соединительные ткани выполняют м е х а н и ч е с-
к у ю, о п о р н ую и формообразующую функции —
входят в состав капсулы и стромы многих органов и, др.; з а-
щ и т ну ю , осуществляемую путем механической защиты (фас-
ции, хрящи, кости); фагоцитоза и” выработки иммунных тел;
п л а с т и ч е с к у ю,~ выражающуюся в активном участии в про-
цессах адаптации ~к меняющимся условиям существования, ре-
генерации, заживлении ран; т ро~ф~и ч е с~к~у-ю,—связанную с ре-
гуляцией। питания структур данного региона, их участием в обмене
веществ и поддержании гомеостаза внутренней среды организма.
СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Классификация. В основу классификации положен принцип со-
отношения клеток й~"межклеточных „структур, а также степень
упорядоченности расположения соединительнотканных волокон.
Собственно соединительную ткань подразделяют на волокнистые
соединительные ткани и соединительные ткани со специальными
свойствами.
Волокнистые соединительные ткани подразделяются на рыхлую
и плотные, а последние — на неоформленную и оформленную.
Рис. 57. Рыхд^я волокни-
стая соединительная ткан^
Микрофотография.
А — фибробласт; Б мак-
рофаг:, I коллагеновые
волокна; 2 — эластические
волокна; 3 — основное амор-
фное вещество; 4 — отростки
фибробласта; 5 — вакуоли
макрофага.
Волокнистые соединительные ткани
Рыхлая волокнистая соединительная ткань
Рыхлая волокнистая соединительная ткань (textus connectivus
collagenosus laxus) обнаруживается во все^х-органах^-так-как она
сопровождает кровеносные и лимфатические сосуды и образует
строму многих, .органов? Несмотря на наличие органных особен-
ностей, строение рыхлой волокнистой соединительной ткани в
различных органах имеет сходство (рис. 57, А, Б). Она состоит
из клеток и межклеточного вещества.
Клетки
Различают: фи Араб пасты, макрофаги, плазмоциты,—тканевые
базофилы (тучные клетки), адипоциты, пигментоциты, адвентици-
алы1Ы£~клетки, перициты .сосудов, а также лейкоциты, мигриро-
вавшие сюда из крови. Клетки соединительной ткани гетерогенны
по происхождению.
187
Ф и б ор бла^с т ы (Фибпобластоииты) (от лат. fibra — волок-
но, греч. biastos^- росток, зачаток) — наиболее многочисленная
группа клеток, различных по степени дифференцировки,, характе-
ризующаяся прежде всего Iспособностью синтезировать фибрил-
лярные белки (коллаген, эластин) и гликозаминпгликаны__(куль-
фатировддшые-и-несулвфатированиьк^ё^йоследующим вьщелением
их в межклеточное вещество. Развиваются—н эмбриогенезе из
мезенхимных клеток, а после рождения из .стволовых .клеток.
В процессе дифференциравки_образуется—рад клеток (диф-
ферон); стволовые- клетки. по пустволовые клетки-предшественни-
ки, малоепецивяызырованньи^--диффе-ренцырованные-фибробласты
(зрелые, активно функционирующие), ^цброцыгьсДдефиадгивные
формы клеток), а также миофибробластыи фиброкласты. С дея-
тепьностьюфибробластов связано образование основного вещест-
ва и~волокон, заживление ран, развитие рубцовой ткани, образо-
вание соединительнотканной капсулы вокруг инородного тела и др.
Малоспёциализированные фибробласты —-_малоотростчатые
клетки с округлым или овальным~ядром и небольшим ядрышком,
базофильной цитоплазмой, богатой РНК. В эмбриогенезе’ разви-
ваются из мезенхимы. Размер клеток не превышает 20—25 мкм.
В цитоплазме обнаруживается большое количество свободных
рцбосом_(рис. 58, ЛГТъ'^Л'Эндоплазматическая сеть и митохон-
дрии развиты слабо. Комплекс Гольджи представлен скоплениями
коротких трубочек и пузырьков. Радиоавтографически показано,
что на этой стадии цитогенеза фибробласты обладают очень низ-
ким уровнем синтеза и секреции белка. 1^алоспециализирован-
ные фибробласты способны к размножению митотическим путем.
Они могут возникать также из клеток-предшественников. Пола-
гают, -что существует две популяции фибробластов^^крроткожи-
нущие (лргкпплп u ДОЛП131 ивущие (шч колько -месяцев)
Специализированные (зрелые) фибробласты круиыее по разме-
ру и в распластанном виде на пленочных препаратах могут
достигать 40—50 мкм и более (см. рис. 57). Это активно функци-
онирующие клетки. Ядра у таких клеток дягтлые. овальные содер-
жат 1—2 крупных ядрышка. Цитоплазма слабо _базофильна,
соде’ржит хорошо развитую гранулярную эндоплазматическую
сеть, которая местами контактирует с цитолеммой (см. рис. 58;
рис. 59, А, Б, В). Комплекс Гольджи в виде цистерн и пузырьков
распределен по всей клетке. Митохондрии и лизосомы развиты уме-
ренно.
Биосинтез РНК, коллагеновых, эластиновых белков и гликоз-
амиТюглйка’нов'й протеогликанов", необходимых для формирова-
ния основного—вещества и-волокон, в зрелых фибробластах осу-
ществляется довольно интенсивно,’'особенно в условиях понижен-
ной концентрации кислорода. Один из гидролитических фермен-
тов—"коллагеназа — расщепляет внутри клетки незрелый колла-
ген, чем, по-видимому, регулируется на клеточном уровне интен-
сивность секреции коллагена.
В цитоплазме фибробластов, особенно в периферическом слое,
188
A
В
Рис. 58. Схема ультрамикроскопиче
ского строения фибробласта на раз
ных стадиях дифференцировки (рис
Ю. И Афанасьева).
Л - малоелециалнитрованный; Б
молодой; В — зрелый; I — ядро; 2 —
комплекс Гольджи; 3 — митохондрия;
4 — рибосомы и полирибосомы; 5 —
гранулярная эндоплазматическая сеть;
6 — сократительные филаменты; 7 -
цитолемма; 8 — коллагеновые фиб-
риллы.
располагаются микрофиламенты толщиной 5—6 нм, содержащие
бел~кгг тигта- а~кТИн;Г и^миочинаГ что обусловливает способность
этих клеток к движению.
В обычных условиях фибробласты обладают незначительной
подвижностью и слабой фагоцитарной актив1ЮСТЬКк_Движ£ние
фибробластов становится возможным только ihujh их t нмзывания
с опорными фибриллярными структурами (фибрин, соединитель-
нотк~Зтжыв-чюлоктГа) с помощью фибронектина — гликопротеина,
си итерированного фибробластами и другими клетками, обеспечива-
ющего адгезию клеток и неклеточных структур. Во время движе-
ния фибробласт уплощается, а поверхность его может 10-кратно
увеличиться.
Фиброциты — дефинитивные формы развития фибробластов.
Они имеют веретеновидную форму И крылониднЫеотростки. Содер-
жат небольшое число органелл, вакуолей,’ липидов и гликогена.
Синтез, коддагена и других веществ у этих клеток резко снижен.
Установлено, что фибробласты могут превращаться в миофиб-
робласщ, функционально сходные с гладкими мышечными клет-
ками. но в отличие от последних имеющие хорошо развитую
эндоплазматическую ~сёть (<ж "рис. 59~, 7>);—Такие • Клетки
наблюдаются в грануляционной ткани в условиях раневого про-
цесса и в матке при развитии беременности.
В период инволюции органов (например, матки после оконча-
ния беременности) в соединительной ткани обнаруживаются
189
Рис. 59. Фибробласт, миофибробласт и фиброкласт. Микрофотографии.Х 24 000.
А — фибробласт: / ядро; 2 — гранулярная эндоплазматическая сеть; 3 — пиноцитозные
пузырьки: 4 комплекс Гольджи; 5 митохондрии; 6 цитолемма; 7 — коллагеновые
фибриллы (препарат А. И Радостииой); Б - -миофибробласт: / — ядро; 2 — гранулярная
эндоплазматическая сеть; 3 рибосомы; 4 —» комплекс Гольджи; 5 — сократительные
филаменты; 6 - цитолемма.
Рис. 59 (продолжение)
В- фиброкласт: /— ядро; 2 — гранулярная эндоплазматическая сеть; 3 рибосомы;
4— лизосомы; 5— фа го л изо со мы с фрагментами коллагеновых фибрилл {препараты
А, Б. Шехтера).
i jsiulPOKJi/acTbi — клетки с более высокой фагоцитарной и гидроли-
тической активностью,—принимакЯцйе —участие в «рассасывании»
межклеточного вещества.-^- ни-х-содержатсяв большом количестве
лизосомы (см. рис. 5!Х,“В).
а крофа г и (макрофагоциты) (от греч. makros — боль-
шой, длиннйи, fagos—пракирдющий) —блуждающие, активно
фагоцитирующие клетки, богатые органеллами для внутриклеточ-
ного переваривания поглощенного материала и синтеза антибак-
териальных и других Ьйблогически_активнь1х~ вёшёств~Тпироген‘,
интерферон, лизоцим и др.). Поглощенные чужеродные белки —
антигены, т. е. любые вещества, против которых организм выра-
батывает антитела после проникновения в~него антигена, макро-
фаги^ переводят^ИЗ корпускулярной фТГрмуфрмелекулярную.
Вступая в контщсг_с_иммунокомпетентлыми клетками Тлимфопи-
тами), макрофаги передают им антигенную информацию, необхо-
димую Для~Т5бразования антител^ - --- -
Макрофаги образуются из стволовой гемопоэтической клетки.
Переходные формы развития от промоноцита до зрелого макро-
фага показаны на рис. 49, В. Полное обновление макрофагов
в рыхлой волокнистой соединительной ткани экспериментальных
Вещество, повышающее температуру тела при воспалении.
191
A
Рис. 60. Макрофаги.
Д — макрофаги с частицами
туши из очага асептического
воспаления подкожной сое
дииительиой ткани крысы
(препарат Ю. И. Афанасье-
ва); Б — электронная микро-
фотография макрофага
X 18 000 (препарат А. И. Ра
достииой): 1 ядро; 2 —
первичные лизосомы; 3
вторичные лизосомы; 4 —
профили канальцев эндопла-
зматической сети; 5 — ми-
кровыросты периферического
слоя цитоплазмы.
животных осуществляется примерно в 10 раз быстрее, чем фибро
бластов.
Макрофагов много в участках, богато снабженных кровенос-
ными сосудами? количество их значительней возрастает при вос-
палении.
Форма макрофагов различна^встречаются клетки уплощенные,
округлые, вытянутые й~неправильной формы. Их границы всегда
четко^очёрчёны? а края неровные (см. рис. 57; рис. 60, X).JJh-
192
толемм макрофагов образует-глубокис складки и /i/iiwhw-ми кро-
выросты, с помощью которых эти клетки захватывают инородные
частицы. На поверхности плазмолеммы макрофага имеются реце-
пторы длзСллухолевых клеток и эритроцитов, Т и В лимфоцитов,
антигенов,—ИМД1Уноглобулинов. Наличие рецепторов к иммуноглсы
булинам обусловливает их участие в иммунных реакциях
Обычно макрофаги имеют одно ядро, за исключением некото-
рых видов макрофагов — гигантских клеток инородных тел и ос-
теокластов, образующихся путем слияния нескольких одноядер-
ны х.макротЬагов. Ядра макрофагов небольшого размера, округлой,
овальной. _шт . бобовидной формыГПЗ "них" содержатся крупные
л ыбки хроматина. Цитоплазма бдзофиль на, богата лизосом а м и,
фаюсомами и пиноцитознымц пузырьками содержит умеренное
количество митохондрий, гранулярной эндоплазматической сети,
комплекса Гольджи, включения гликогена, липидов и др. (рис.
60, Б).
Макрофаги секретируют в межклетолноо-воинч^гво-биологичес-
ки активные факторы и ферменты (интерферон^лизоцим, пироге-
ны, цротеазы, кислые гидролазы и др.), чем обеспечиваются их
разнообразные защитные функции.
В функциях, выполняемых макрофагами, важная роль при-
надлежит —вырабатываемым ими медиаторам-монокинам: ццтер-
л е йкцну 1, активирующему синтез ДНК в лимфоцитах; фа кто ру,
активирующему выработку иммуноглобулинов 6-лимфоцитами;
факторам, стимулирующим дифференцировку Т- и R-лимфбцитов-
факторам, вызывающим хемотаксис Т-лимфоцитов и активность
Т-хелперов, цитолитическим факторам, избирательно разрушающим
опухолевые клетки и др. Контакт макрофагов с антигенами резко
усиливает поглощение ими кислорода, расход глюкозы, липидный
обмен и фагоцитарную активность.
уПонятие о макрофагической системе. К этой системе относят
совокупность всех .клеток, обладающих способней тью захват;,1вать
из ткайевой жидкости" ганизма инородные частицы, погибающие
клетки, неклеточные структуры,бактерии и~др. Фагоцитирован-
ный "Материал подвергается ннутр’й"клетки ферментативному рас-
щепденйю~Г«завёршенный фагоцитоз»), благодаря чему тиквиди-
руютсл врёдные~д?тя иранизма аТёнтЫ, возникающие местно или
проникающие извне. К таким клеткам относятся макрофаги (гис-
тиоциты ) рыхлой неофф7млснтютг~вп.ТОкнйстби~ соединительной
тканщ звездчатые клетки синусоидных сосудов печени, свободные
и фиксироватптае~макрофаги кроветворных органов (костного моз-
га, селезенки, лимфатических узлов), макрофаги легкого, вбспали-
тельных экссудатов (перитонеальные макрбфагйТ7осте<и<ласхы, ги-
гантские клетки инородных тел и глиальные макрофа!И нервной
ткани (микроглия)-фсе уши способны к активному фагоцитозу,
имеют на своей поверхности "рецепторы к иммупоглебулинам
и происходят из прэмоноцдсшв костного мозга и Моноцитов
крови. В отличие от таких «профессиональных» фагоцитов спо-
собность к факультативному поглощению может быть выражена
7 ГИСТОЛОГИЯ
>91
независимо от указанных циторецепторов у других клеток
(фибробласты, ретикулоциты, эндотелиоциты, нейтрофильные лей-
коциты). Но эти клетки не входят в состав макрофагической
системы.
И. И. Мечников первымцрищел к мысли о том, что фагоцитоз,
появившийся в эволюции как форма внутриклеточного пищеваре-
ния и закрепившийся за_многими клетками, одновременно явля-
ется важным защитным механизмом. Он обосновал целесообраз-
ность Ъбъёдинения их в одну систему и предложил назвать ее
макрофагической. Г)1акрофагическая система представляет собой
мошныйзащитный аппарат? поинимаюшии участие как—в общих,
так и в местных защитных реакциях организма. В целостном
организме макрофагическая система регулируется как"местными
механизмами, так нервной й эндокринной“системами;
В ЗО^^б-гГТбдаходту'защитную систему называли ретикулоэн-
дотелиальной системой. В последнее время ее называют систе-
мой мононуклеарных фагоцитов, что, однако, неточно характери-
зует эту систему клеток в связи с тем, что среди клеток, вхо-
дящих в эту систему, есть и многоядерные (остеокласты).
Пл а з м а т и чес кие клетки (плазмоциты). Эти
клутки обеспечивают гуморальный иммунитет. Они синтезируют
антитела — гамма-глобулины (белки) вырабатывающиеся при по-
явлении в организме антигена и обезвреживающие его (см. рис.
93). Плазматические клетки обычно встречаются в рыхлой волок-
нистой соединительной ткани собственного слоя слизистой оболоч-
ки кишки, в'“са.льнике~в'интерстициальной соединительной ткани
различных желез (молочных, слюнных и др.), в лимфатических
узлах, селезенкЕт-Костном мозге и др. Они образуются из В-лим
фоцитов w
“Величина плазмоцитов колеблется от 7 до 10 мкм. Форма
клеток округлая или -овальная—Ядра—относительно небольшие,
круглой или овальной формы» „расположены эксцентрично^ Цито-
плазма резко базосЬидьна,содержит хорошо развитую концентри-
чески расположенную—трануля.рцу.КЭ„Эндоплазматическую сеть, в
которой синтезируются белки (антитела). Выявление различных
антител возможно при применении фтюоресцентных красителей.
I Базофилии лишена только небольшая светлая зона около ядра,
обрщгуюЩггя -гат—пазьгваемую сферу? йлй'двбрик. Здесь обнаружи-
*'ваются центрИОли~и~комплёкс Гольджи.
Т к а н е в ы е б а з о rh и л ы (тучные клетки). Этими
терминами называют клетки, в цитоплазме которых находится
специфическая зернистость?напоминающая гранулы базофильных
лейкоцйтовГТканевыё^ба’зофилы являются регуЗТяторамй'мёстного
гркгёостаЧа спади ннтезпщойодтаттил Они Принимают участие в по-
нижении свертывания крови, повышений~проницаемости гемато-
тканёвого~барьера, в процессе воспаления, иммуногенеза и др.
У человека тканевые базофилы обнаруживаются всюду, где
имеются прослойки рыхлой вотокнистой соединительной ткани.
Особенно много тканевых базофилов в стенке органов желудочно-
194
Рис. 61. Тканевый базо-
фил.
А в подкожной соедини-
тельной ткани: / ядро; 2 —
метахроматические гранулы
в цитопл зме; Б схема
ультра микроскопического
строения (рис Ю И. А фа
насьева) 1 ядро; 2 —
комплекс Гольджи; J — ли-
зосома; 4 - митохондрии:
эндоплазматическая
<etb. 6 микроворсинки;
'етерогенные гранулы;
. екреторные гранулы в
межклеточном веще^гве.
кишечного тракта, матке, молочной железе, тимусе, вило псовой
железа миндалинах. Они часто располагаются группами по ходу
кровеносных сосудов микрОДиркуля г<'пно£О__русла —капилляров,
артериол, венутг’и мелких лимфатических сосудов (рис. 61, Л).
Форма тканевых базофилов разнообразна. Она можёт быть не-
правильнаЯт~евальная;ттнпгда~с" корП’гкйми широкими отростками,
что обусловлено-енособностью их к амебоидным движениям. У-че-
ловека размер-такихлслеток-кгптеблётся~6т*1 до 14~мкм в ширину
и до 22 мкм в длину. Ядра лаброцитов сравнительно невелики,
обычно округлой или овальной формы с плотно расположен-
ным хроматином. В цитоплазме расположены многочисленные i ра-
нульг^Величина и количествсГтранул варьируют; Еранульг различ-
ны по плотности и составу; их диаметр около 0,3— 1 мкм (рис.
14
61, Б). Меньшая часть гранул представляет собой ортохромати-
чески _од<ратирак)щиеся_азурофильные лизосомы. Большинство
гранул отличается метахромазией, содержит гепарин, хондроитин-
серные кислоты типа А и С, гиалу'ро новую кис лоту “гистамин;
у некоторых животных обнаружен еще серотонин. Гранулы имеют
сетчатое, пластинчатое, кристаллоидное и сметанное строение.
Органеллы тучных клеток (митохондрии, комплекс Гольджи,
цитоплазматическая сеть) развиты слабо. В цитопла зме обнаруже-
ны различи ые ферменты: липаза, кислая и щепочная фосфата за,
пероксидаза, цитохромоксидаза, АТФ-аза и др. Однако маркер-
ным ферментом следует считать гистидиндекарбоксилазу, с по-
мощью которой осуществляется превращение гистидина в гиста-
мин.
Тканевые базо ^илы могут утр чивать свои гранулы. В процессе
дегрануляции выделяется гепарин, препятствующий свертыванию
кровиГ~Секреция гистамина, напротив, происходит безоару шени я
I Гелости~клеточной оболочки И гранулу В ьгхдд~этих~ веществ п р и -
водит к изме нению состояния межклеточного веществ- < пепин и -
тельной ткани, гематотканевого бадьегы. В частности,_гепарин
снижает проницаемость межклеточного вещества,-свертываемость
крови,_оказываех_лротивовррцадИ1ельное_ивлиянц£ Гистамин же
выступает как его антагонист.
Количество тканевых базофилов изменяется в аавиеимнсти от
различных физиологических состояний пргяииша- число во з-
растает в матке и молочных, же летах в. период, беременности, а в
желудке, кишечнике, н neueim _—в разгар пищеваре н и я.
Предшественники тканевых базофилов еще окончательно не'
определены. Полагают, что они происходят из стволовых крове-
творных клеток-красного костного-мг.31 а Процессы митотического
деления тучных клеток наблюдаются крайне редко
А д и п о пит г4-_ (жировые-----кл-ст к и). 7ак-- называют
клетки, которые обладают способностью. накапливать_в больших
количествах резервный жир, принимающий участие в трофике,
энергообразовании и метаболизме воды. Адипоциты располагаются
групнами, реже — поодиночке и, как ГГравило~,~околб~~кровеносных
сосудов. Накапливаясь в больших количествах, эти клетки образу-
ют~жтгровую тканы '
~ Форма~~одиночно расположенных жировых. кл< ток шаровидная.
Зрелая жировая клетка обычно солержит-ддну большую каплю
нейтрального жира (триглицеридов), занимающую всю централь-
ную часть клетки и окруженную тонким цитоплазматическим
ободком,_-В_утолщеннай-Амсти—которого лежит-ядро: Кроме того,
в цитоплазме адипоцитов имеется небольшое количество других
липидов: холестерина, фосфолипидов, свободных жирных киелот
и др. Липиды хорошо Опрашиваются Суданом III в оранжевый цвет
или осмиевой кислотой в черный цвет (рис. 62). В прилежащей
к ядру цитоплазме, а иногда и в более тонкой противоположной
ее части щяявляются палочковидные и нитевидные митохондрии с
плотно упакованными кристами. Йа периферии клетки встрёчают-
75б ~ "
Рис. 62. Адипоциты
/ капилляр: 2 липид-
ные вк мочения.
ся многочисленные пиноцитозные пузырьки. Адипоциты обладают
высоким уровнем метаболизма. Подвержено значительным колеба-
ниям как количество жировых включений в адипоцитах, так и чис-
ло самих жировых клеток в рыхлой волокнистой соединительной
ткани.
Жировые капельки, всосавшиеся в лимфу и кровь, ра.змером
около""! мкм — хиломикроны (от греч. chylos — сок, micron —
малое) под действием ферментов (зипппротеидлипаз,, образуемых
эндотели м сих сосудов, расщепляются на жирные кислоты и
глицерин, после чего поглощаются жировой клеткой Под дей-
ствием другого фермента — глицуролкиназы — вновь, ресинтези-
руются триглицериды.
Расходование депонированного в адипоцитах жира происходит
под действием липолитических гормонов (адреналин, инсулин) и
тканевого липолитивеского фермента (липаза), расщепляющего
триглиериды до 1лицерина и жирных кислот, которые в крови
связываются с альбумином и переносятся в другие ткани, нуждаю-
щиеся в~питательных веществах («горючем»).
Новые жировые клетки в соединительной ткани взрослого ор-
ганизма могут развиваться—при усиленном питании из адвенти-
циальцых клеток, _цридегаю1цих_к. кровеносным капиллярам. При
этом в цитоплазме клеток появляются сначала мелкие капельки
жира, которые, увеличиваясь в размере, постепенно сливаются в бо-
лее крупные капли. По мере~увеличения жировой капли эндоплаз-
матическая сеть и^кдмТТпетсеЧ^лтджп редуцируются, а ядро сдав-
ливается и уплощается.
—» н т , и и Т N (пигментные клетки). Это
клеткй,'соде’рЖащие в~своей цитоплазме пигмент меланин. У лю-
дей~черной тт желтой рас пигмент-ньнклетки более—рас простра-
не ньф чем-определяется неизменяемый в зависимости ют- еезона
года “цвет кожй. Пигм< нтоциты__fмеланоциты)“имеют короткие,
непостоянной формы отростки (рис, ЬЗ). Эти клетки лишь фор-
мально относятся к соединительной ткани^ так как располагаются
в ней. Что касается происхождения, то доказано образование
19-7
Рис. 63. Пигментоцигы в коже человека
(африканца).
/ эпителии: 2 волокна рыхлом волокни-
i гои «единительнои ткани: 3 - пигментные
клетки
их H i неРвных гребней, а не из
мезенхим.ьд_
А д и циальные
к л е т к и. Это малоспециали-
зйро ванные клетки, сопровож-
дающие кровеносные сосуды.
Они имеют уплощенную или ве-
ретенообразную форму со сла-
бобазофильной _ цитоплазмой,
овальным ядром и слаборазви-
тыми-органеллами. В процессе
дифференцировки эти клетки
могут превращаться, по-види-
мому, в фибробласту, миофиб-
робдасты. и адипоциты. Многие
авторы отрицают существование адвентициальных клеток как
самостоятельного клеточного типа, считая их малодифференци-
рованпыми клетками фибробластическою ряда.
Пер иц и г ы - клетки,-екружающиекровеносные капилляры
(см. гл. XV).
Межклеточное вещество
М,еж.клеточное_ведцество (substantia inlercellularis) соединитель-
ной ткани состоит из коллагеновых—ретикулярных, эластических
волокон, а также из основного вещества. Межклеточное вещество
как у^зародышей, так и у взрослых образуется, с одной стороны,
путем сёкреций7Ъсуществляемой~<юе®Гй'йтель11ЬГкаИнЫМи клетка-
ми, а с другой — замечет плазмы крови, поступающей в межкле-
точные пространства.
У зародышей человека образование межклеточного вещества
происходит начиная с 1—2-го месяца внутриутробного развития.
В течение жизни межклеточное вещество постоянно обновляет-
ся — резорбируется и восстанавливается.
Коллагеновые волокна (fibrae collagenosae). В составе разных
видов соединительной ткани они определяют их прочность. В рыхлой
неоформленной волокнистой соединительной ткани коллагеновые
волокна располагаются вфазличных направлениях в виде волнооб-
разно изогнутых спйралевилно скрученных округлых иди уплощен-
ных тяжей толщиной I—3 мкм и более. Длина их неопределенна.
Внутренняя структура, коллагеновою волокна определяется фиб-
риллярным белком — КОЛ-ШЮНЮИ, который синтезируется в-рибосо-
мах гранулярной эндоплазматической сети фибробластов.
;оч
Различают 12 Типов коллагена, отличающихся молекулярной
организацией, органной и тканевой принадлежностью. Коллаген I
типа встречается главньна_с>брязом в соелиниТВ^ЕЙГТъани кожи,
когти? роговице, глаза, склере, стенке артерий и др., II типа — в
гиалиновом и фиброзном хрящах, в стекловидном теле, III типа —
в дерме кожи плода, в стенке крупных кровеносных сосудов, в
ретикулярных волокнах, IV типа — в базальных мембранах, в кап-
суле хрусталика. В отличие от других типов коллагена, он содер-
жит гораздо больше боковых углеводных цепей,ф также гидроок-
силизина и гидрооксипролина, В последние годы выделен еще
V ти п. кол ла гена, располагающ идея вокруг.сдГнтезируЖпих eirr кле-
ток фибробластов, эндотелиальных л гладких мышечных клеток в
виде экзоцитоскелета. Коллаген 6—12 типов изучен неполно.
Молекулы коллагена, экстрагированные из волокон, имеют дли-
ну около 280 нм и ширину 1,4 нм. Они построены из трипле-
тов — трех полипептидных цецочек (g-цепочки) предшественника
коллагена — проколлагена, свивающихся в единую спираль, еще
находясь в клетке. Это — первый, молекулярный уровень органи-
зации коллагенового волокна. Каждая цепочка-Проколлагена со-
де ржиттгаборы из трех^разлйчных аминокислот, много кратно и за -
кономернр повторяющихся на протяжении ее длины. Первая ами-
нок и слота в таком наборе может быть любой, вторая прол и н
или лизин, третья — глицин. В течение нескольких минут пролин
и лизин окисляются в гидрооксинролин и гидрооксилиз.щ — мар-
керные аминокислоты зрелого коллагена. Пэоколлаген секретиру-
ется в межклеточное вещество.
Второй, надмолекулярный, внеклеточный организации
представляет агреттфОванные в длину и поперечно связанные с по-
моа(ью~в ^дородных связей молекулы коллагешщСначала образу-
ются протОфибриллЫ7~а~5^6 протофибрилл, скрепленных между
собой боковыми связями, составляют микрофибриллы (рис. 64,
А, Б) толщиной около 10 нм. Они различимы в электронном мик-
роскопе в виде слабоизвилистых нитей.
При участии гликозаминогликанов и гликопротеинов, также
секретируемых'фиброблЗс'гбм, образуются пучки фибрилл — тре-
тий, фибриллярный, уровень организации. Они представляют собой
поперечно 'исчерченные структуры ~т0лЩИ1юй в среднем 50—109 нм
Период повторяемости темных и светлых участков 64 нм (рис.
65, 66). Темные полосы образованы более густым асимметричным
расположением вторичных тонких поперечных линий в периоде,
обусловленных расположением полярных аминокислот в молеку-
лах коллагена.
В состав коллагенового волокна (толщиной 1 —10 мкм) в зави-
симости о7~т0потрафии~вХ0дят от нескольких фибрилл до несколь-
ких десятков - четвёртый, волоконный~уровень организации. Во-
локна "складываются в"пучки толщиной до 150 мкм.
Коллагеновые волокна отличаются малой растяжимостью и
большой проТптостБкптгг разрьгвг'&'вщге'до.'гйщна- сухожилий в ре-
зультате-набухания увеличивается на 50%, а в разбавленных кис-
199
Рис. 64. Уровни структурной органича-
ции коллаieнового волокна (схемы),
А: / — полипелтидная цепочка; Н — моле-
кулы коллагена (тропоколлаген);/// — про-
тофибриллы; IV фибрилла минимальной
толщины, у которой становится видимой
поперечная исчерченность; V - коллагено-
вое волокно: /> спиральная структура
макромолекулы колла!ена (по Ричу); мел-
кие светлые кружки - тицин, крупные
светлые кружки пролин, заштрихованные
кружки гидроксипролин.
1~3мнм
fl
Рис. 65. Колла! еновая фибрилла. Элек-
тронная микрофото! рафия. 130 000
(препарат Н.П. Омельяненко).
П период.
1
ta
Рис. 66. Образование
Р. Кристичу с изменением).
/ фибробласт; 2 — почипептилные цепочки; 3 - молекулы
тропоколлагена; 4 - гликозаминогликаны, 5 — полимериза-
ция молекул тропоколлагена; 6 — протофибрилла; 7 пучок
протофибрилл (коллагеновая фибрилла); 8 молекула элас-
тина; 9 — эластическая фибрилла.
межклеточного вещества (по
лотах и щелочах — в 10 раз, но при этом она
укорачивается на 30 % Способность к ~набу-
ханию больше выражена у молодых волокон.
При варке коллагеновые волокна образуют
клейкое вещество (греч. kolla — клеи), отку-
да и появилось название этих волокон.
Ретикулярные волокна (fibrae reliculares).
Они "относятся к типу коллагеновых волокон,
таьГ~как~Е~~их состав входят бслок -коллаген
(IH типа) и повышенное клпииягтвп vr,свопов, которыехинтези-
руются ретикулярными клетками органов кроветворения и обра-
зу юТп-рехшфПу (ретикулум), откуда и берут свое название.
Эластические волокна (fibra elasticae). Наличие их в_соеди-
нительной ткани определяет ее эластичность и растяжимость. По
прочности эластические вопокна-уступают коллагеновым. Форма
волокон округлая или уплощенная. В рыхлой волокнистой сое-
динительной ткани они широко анастомозируют друг с другом.
Толщина эластических волокон обычно меньше коллагеновых, но
201
А схема:/ центральная гомогенна» часть, 2 — микрофибри.шы (рис. Ю. И. Афанагье-
ва). Б - электронная микрофотш рафия. X45000 (препарат В. П Слюсарчука). I — цент-
ральная гомогенная часть; 2 — микрофибриллы на периферии волокна; 3 — комплекс
Гольджи фибробласта; 4 — эндоплазматическая сеть; 5 центриоль.
может достигать нескольких микрометров (например, в выйной
связке).
Основным химическим компонентом эластических волокон яв-
ляетсзГ 1лобулщ>нь1ибелок.элдсгп/4синтсзируемый -фибробласта-
ми. (первый, молекулярный, уровень организации). По сравнению с
коллагеном в этом белке содержится значительно меньше поляр-
ных аминокислот, гидрооксилизина, гидрооксипролина при боль-
шом содержании—яролин»—и—слили на- -Кроме того^—для -эластина
характерно_отсу1£твие нистина и наличие пну у производных ами-
нокислот —и изпбесиалтп -ттторыо участвуют в ста-
билизации^ молекулярной структуры эластина и придании ему спо-
собцо£ти_к растяжению, эластичности. Молекулы эластина, имею-
щие форму глобул, диаметром 2,8 нм вне клетки соединяются в це-
почки, называемые эластиновыми прото фибриллами толщиной 3—
3,5 нм (второй, надмолекулярный, уровень организации). Эласти-
новые филаменты окружены гликопротеином и образуют микро
фибриллы (рис. 67, А, Б) толщиной 8—10 нм (третий, фибрилляр-
ный, уровень организации). Чем больше аморфного вещества, тем
более «зрелым» считается волокно (четвертый, волоконный, уро-
вень организации). В отличме-от-коялагеццвых волокон, в эласти-
ческих нет_структур.. с. поперечной иечерченноетыо на -их протя-
202
жении, Наиболее зрелые эластические волокна содержат около
90% эластических белков (эластина) аморфного компонента
Кроме зрелых эластических волокон, различают элауниновые
и окситалановые волокна. В элау ни новых волокнах соотношение
микрофибрилл и аморфного компонента примерно равное, а окси-
талановые волокна состоят только из микрофибрилл.
Изучение распределения различных типов эластических воло-
кон показывает, что микрофибриллярный компонент сильнее раз-
вит там, где требования к механической прочности выше, чем к
эластичности.
Электронная микроскопия позволила установить, что эласти-
ческие волокна в центре содержат аморфный компонент, а по пе-
риферии — микрофибриллярный.
^Аморфный компонент межклеточного вещества
Клетки и волокна соединительной ткани заключены в аморф-
ный компонент, или основное вещество (substantia fundamentalist
Эт<1 — <ТУ1И.-.Ю| И1Ч.ЧЬ rnnprvj м цыыя I в образовании кото-
рой важную роль играют фибробласты. Основное вещество участ-
вуех-в-три нспорте метябл Титов. межлу Ючётк~а1МЙ~И"КривЬ'Ку^В меха-
нической, опорной.—защитной функциях.- В—его-госта в^ входят
сульфатированные гликозаминогликаны, такие как хсшдрцитинсер-
ная кислота (хондроитинсульфагы), дерматансульфат, кератиты
судьфдт, гепарицсульфат и др., которые обычно более или менее
прочно связаны с белками, образуя протеогликаны, а также не-
су л ьфатированные—(гиалуроновая кислота). Они обусловливают
консистенцию и функциональные особенности основного вещества.
Кроме указанных компонентов, в состав основного вещества
входят липиды, альбумины и глобулины кррвщ мИнёраЛЬНЙбДвеще-
ства (соли натрия, калия, кальция и др.).
Количество основного вещества в различных участках соедини-
тельной ткани неодинаково. Около капилляров и мелких сосудов,
в участках, содержащих жировые прослойки, основного вещества
мало, а на границах с тканями другого происхождения, например
с эпителием, его много. В детском возрасте основного вещества
больше, чем в зрелом и пожилом.
Аморфный компонент межклеточного вещества участвует в ме-
таболизме воды, регуляции ионного состава, в связывании кле-
ток и волокон, адгезии клеток и др.
Физико-химическое состояние межклеточного вещества в зна-
чительной~мере определяет функциональные особен11ости^соедини-
тельной ткани. Чем плотнее межклеточное вещество, тем сильнее
вы раженЭ Механическая, опорная функция? Трофич ёск а яфу н к ци я,
напротив, лучше' обеспечивается полужидким по консистенции
межклеточным веществом_____.(интерстициальная соединительная
ткань, окружающая кровеносные сосуды) благодаря протеоглика-
нам аморфного компонента межклеточного вещества, обладающим
гидрофильностью. Под влиянием эндогенного и экзогенного гис-
203
тамина, i иалуронидазы происходит повышение проницаемости
аморфного компонента межклеточного вещества. Повышение кон-
центрации гликозаминогликанов, снижение активности гиалурони-
дазы, напротив, понижают его проницаемость.
Плотная волокнистая соединительная ткань
Плотная волокнистая соединительная ткань (textus connectivus
collagenosus compactus) характеризуется относительно большим
количеством плотно расположенных роликеш и незначительным
количеством клеточных элементов и о новного аморфного вещест-
ва между ними.
В зависимости от характера расположения волокнистых струк-
тур она подразделяется на плотную неоформленную и плотную
оформленную соединительную ткань. Плотная неоформленная сое-
динительная ткань характеризуется неупорядоченным расположе-
нием волокон. В плотной оформленной волокнистой соедини-
тельной ткани расположение волокон строго упорядочено и в
каждом случае соответствует тем условиям, в каких функциони-
рует данный орган. Оформленная волокнистая соединительная
ткань встречается в сухожилиях и связках1, в фиброзных мембра-
нах.
Сухожилие (tendo). Оно состоит из толстых, плотно
лежащих параллельных пучков коллагеновых волокон. Между эти-
ми пучками располагаются фиброциты и небольшие количества
фибробластов и основного аморфного вещества. Тонкие пластин-
чатые отростки фиброцитов входят в промежутки между пучками
волокон и тесно соприкасаются с ними. Фиброциты сухожильных
пучков часто называются сухожильными клетками (tendinocyti).
Каждый пучок коллагеновых волокон, отделенный от сосед-
него слоем фиброцитов, называется пучком первого порядка.
Несколько пучков первого порядка, окруженных тонкими прос-
лойками рыхлой волокнистой соединительной ткани, составляют
пучки второго порядка. Прослойки рыхлой волокнистой соедини-
тельной ткани, разделяющие пучки второго порядка, называются
эндотенонием. Из пучкоЬ второго порядка слагаются пучки третье-
го порядка, разделенные более толстыми прослойками рыхлой
соединительной ткани (перитеноний). Иногда пучком третьего
порядка является само сухожилие, В крупных сухожилиях могут
быть и пучки четвертого порядка. В перитенонии и эндотенонии
проходят кровеносные сосуды, питающие сухожилие, нервы и
проприоцептивные нервные окончания, посылающие в централь-
ную нервную систему сигналы о состоянии натяжения ткани
сухожилий.
К плотной оформленной волокнистой соединительной ткани
относится и выйная связка, только пучки ее образованы
эластическими волокнами и нечетко подразделены.
' По некоторым классификациям сухожилия и связки относятся к скелетным
тканям в связи с их выраженной основной механической функцией.
'04
Некоторые сухожилия в местах прикрепления к костям заклю-
чены во влагалища, построенные из двух волокнистых соедини-
тельнотканных оболочек (vagina), между которыми находится жид-
кость (смазка), богатая гиалуроновой кислотой.
Фиброзные мембраны. К этой разновидности плотной
волокнистой соединительной ткани относят фасции, апоневрозы,
сухожильные центры диафрагмы, капсулы некоторых органов,
твердую мозговую оболочку, склеру, надхрящницу, надкостницу, а
также белочную оболочку яичника и яичка и др. Фиброзные
мембраны трудно растяжимы вследствие того, что пучки колла-
геновых волокон и лежащие между ними фибробласты и фибро-
циты располагаются в определенном порядке в несколько слоев
друг над другом. В каждом слое волнообразно изогнутые пучки
коллагеновых волокон идут параллельно друг Другу в одном
направлении, не совпадающем с направлением в соседних слоях.
Отдельные пучки волокон переходят из одного слоя в другой,
связывая их между собой. Кроме пучков коллагеновых волокон,
в фиброзных мембранах есть эластические волокна. Такие фиброз-
ные структуры, как надкостница, склера, белочная оболочка
яичка, капсулы суставов и др., характеризуются менее правиль-
ным расположением пучков коллагеновых волокон и большим
количеством эластических волокон по сравнению с апоневрозами.
Соединительные ткани со специальными свойствами
К таким тканям относят ретикулярную, жировую, слизистую и
пигментную. Эти ткани характеризуются преобладанием одно-
родных клеток, с которыми обычно связано само название этих
разновидностей соединительной ткани.
Ретикулярная ткань
Ретикулярная соединительная ткань (textus reticularis) - - это
разновидность соединительной ткани, имеющая сетевидное строе-
ние и состоящая из ретикулярных клеток и ретикулярных (арги-
рофильных) волокон (рис. 68, А, Б). Большинство ретикулярных
клеток связано с ретикулярными волокнами и стыкуется друг
с другом отростками. Волокна и отростчатые клетки образуют
рыхлую сеть (reticulum), в связи с чем эта ткань получила
свое название.
Ретикулярная ткань образует строму кроветворных органов и
микроокружение для развивающихся в них клеток крови. Разли-
чают ретикулярные фибробластоподобные клетки, связанные с
волокнами, фагоцитирующие клетки моноцитарного происхожде-
ния и малоспециализированные клетки
Ретикулярные волокна являются производными ретикулярных
клеток (см. рис. 68, Б). Они обнаруживаются при импрегнации
солями серебра, поэтому называются еще аргирофильными (от
греч. argentum — серебро). Ретикулярные волокна по химическому
20'
Рис. 68. Ретикулярная
ткань.
А — схема строения ретику-
лярной клетки и ретикуляр-
ных волокон: / ядро рети-
кулярной клетки; 2 от-
ростки цитоплазмы; 3 ре-
тикулярные волокна. <900.
В микрофотография рети-
кулярных волокон лимфати-
ческого узла. Импрегнация
нитратом серебра. / — рети-
кулярные волокна. ' 120.
составу близки к коллагеновым, но отличаются от них меньшей
толщиной, ветвистостью и анастомозами
В группе аргирофильных волокон различают собственно ретику-
лярные волокна и преколлагеновые волокна. Собственно ретику-
лярные волокна — дефинитивные, окончательные образования,
содержащие коллаген III типа. Преколлагеновые волокна представ-
ляют собой начальную форму образования коллагеновых волокон
в эмбриогенезе и при регенерации. Ретикулярные волокна отли-
чаются высокой концентрацией серы, липидов и углеводов по
сравнению с коллагеновыми волокнами. Под электронным микро-
скопом фибриллы ретикулярных волокон имеют не всегда четко
выраженную исчерченность с периодом 64—67 нм.
Ретикулярные волокна устойчивы к действию слабых кислот и
щелочей и не перевариваются трипсином. По растяжимости эти
волокна занимают промежуточное положение между коллагено-
выми и эластическими.
Жировая ткань
Жировая ткань (textus adiposus) — это скопления жировых
клеток, встречающихся во многих органах. Различают две разно-
видности жировой ткани белую и бурую. Эти термины условны и
№
5
Рис. 69. Схема строения
белой жировой ткани (рис.
Ю. И. Афанасьева).
А - адипоциты с удаленным
жиром в световом оптиче-
ском микроскопе; Б - уль-
трймчкросконическое строе-
ние липоцитов. / — ядро жи-
ровой клетки; 2 - крупные
капли липидоз; 3 - нервные
волокна; 4 — гемокапилля-
ры; 5 митохондрии.
отражают особенности окраски клеток. Белая жировая ткань
широко распространена в организме человека, а бурая встречается
главным образом у новорожденных детей. У некоторых животных
она хорошо выражена (у грызунов и зимоспящих) в течение всей
жизни.
Белая жировая ткань у человека располагается под кожей,
особенно в нижней части брюшной стенки, на ягодицах и бедрах, где
она образует подкожный жировой слой, в сальнике, брыжейке и
ретроперитонеальной области. Жировая ткань более или менее
отчетливо делится прослойками рыхлой волокнистой соединитель-
ной ткани на дольки различных размеров и формы. Жировые
клетки внутри долек довольно близко прилегают друг к другу
(рис. 69, А, Б). В узких пространствах между ними располагаются
фибробласты, лимфоидные элементы, тканевые базофилы. Между
жировыми клетками во всех направлениях ориентированы тонкие
коллагеновые волокна. Кровеносные и лимфатические капилляры,
располагаясь в прослойках рыхлой волокнистой соединительной
ткани между жировыми клетками, тесно охватывают своими
петлями группы жировых клеток или дольки жировой ткани.
В жировой ткани происходят активные процессы обмена жирных
кислот, углеводов и образование жира из углеводов. При распаде
207
Рис. 70. Схема строения
бурой жировой ткани
<рис. Ю. И. Афанасьева).
А адипоциты с удален-
ным жиром в световом оп-
тическом микроскопе; —
ультра микроскопическое
строение липоцитов. / яд-
ро адипоцита: 2 мелко раз-
дробленные липиды; 3 -
многочисленные митохонд-
рии; 4 - гемокапилляры; 5
нервное волокно.
жиров высвобождается большое количество воды и выделяется
энергия. Исходя из этого жировая ткань играет не только роль
депо субстратов для синтеза макроэргических соединений, но и
косвенно — роль депо воды.
Во время голодания подкожная и околопочечная жировая ткань,
жировая ткань сальника и брыжейки быстро теряют запасы жира.
Капельки липидов внутри клеток измельчаются и жировые клетки
приобретают звездчатую или веретено видную форму. У животных,
откармливаемых после голодания, в цитоплазме жировых клеток
наблюдается вначале накопление гликогена. Впоследствии он ис-
пользуется жировыми клетками при синтезе липидов. Однако не
всякая жировая ткань легко отдает липиды при голодании. В области
орбиты глаз, в коже ладоней и подошв она теряет лишь небольшое
количество липидов даже во время продолжительного голодания.
В этих случаях жировая ткань играет преимущественно
механическую роль, а не обменную. В этих местах она разделена
на мелкие дольки, окруженные соединительнотканными волокнами.
Бурая жировая ткань обнаружена у новорожденных детей и
у некоторых животных на шее, около лопаток, за грудиной,
вдоль позвоночника, под кожей и между мышцами. Она состоит из
жировых клеток, густо оплетенных гемокапиллярами. Эти клетки
принимают участие в процессах теплопродукции.
Адипоциты бурой жировой ткани имеют множество мелких
жировых включений в цитоплазме (рис. 70, А). По сравнению с
клетками белой жировой ткани здесь обнаруживается значительно
больше митохондрий (рис. 70, Б). Бурый цвет жировым клеткам
придают железосодержащие пигменты — цитохромы митохондрий.
Окислительная способность бурых жировых клеток примерно в
20К
20 раз выше белых и почти в 2 раза превышает окислительную
способность мышцы сердца. При понижении температуры окру-
жающей среды активность окислительных процессов в бурой
жировой ткани повышается. При этом выделяется тепловая энер-
гия, обогревающая кровь в кровеносных капиллярах. В регуляции
теплообмена определенную роль играют симпатическая нервная
система и гормоны мозгового вещества надпочечников — адрена-
лин и норадреналин, который через циклический аденозинмоно-
фосфат стимулирует активность тканевой липазы, расщепляющей
триглицериды на глицерин и жирные кислоты. Последние, накап-
ливаясь в клетке, разобщают процессы окислительного фосфори-
лирования, что приводит к высвобождению тепловой энергии,
обогревающей кровь, протекающую в многочисленных капиллярах
между липоцитами. При голодании бурая жировая ткань изме-
няется меньше, чем белая.
Слизистая ткань
Слизистая, или студенистая, соединительная ткань (textus
mucosus) встречается у зародыша- Классическим объектом
для ее изучения является пупочный канатик человеческого плода
(рис. 71). Клеточные элементы здесь представлены в основном
мукоцитами — клетками типа фибробластов. Между клетками этой
ткани в первой половине беременности в большом количестве
обнаруживается гиалуроновая кислота. Это обусловливает желе-
образную консистенцию основного вещества. Фибробласты студе-
нистой соединительной ткани слабо синтезируют фибриллярные
белки. На поздних стадиях развития зародыша в студенистом
веществе появляются рыхло расположенные коллагеновые фиб-
риллы, количество которых увеличивается по мере развития заро-
дыша
Рис. 71. Студенистая
ткань из пупочного кана-
тика человека.
/ мукоциты; 2 — студне
образное межклеточное ве
щес.тво
Пигментная ткань
Пигментная соединительная ткань (textus connectives pigmen-
tosus) — это ткань, в которой содержится много пи! ментных
клеток — меланоцитов (cellulae pigmentosae). К ней относятся
соединительнотканные участки кожи в области сосков, в мошонке,
около анального отверстия, а также в сосудистой оболочке и ра-
дужке глаза, родимых пятнах
СКЕЛЕТНЫЕ ТКАНИ
К скелетным тканям (textus sceletales) относят хрящевые и
костные ткани, выполняющие прежде всего опорную, защитную,
механическую функции, а также принимающие участие в водно-
солевом обмене веществ.
Хрящевые ткани
Хрящевые ткани (textus cartilaginei) входят в состав ряда
органов дыхательной системы, суставов^ межпозвонковых дисков
и др. Они состоят из клеток (хондроцитов и^хондробластов) и
большого количества межклеточного вещества, отличающегося
упругостью Именно с этим связана опорная функция^лрящевых
тканей.
В свежей хрящевой ткани содержится около 70—80% воды,
10—15% органических веТпеств и 4—7% солей^В сухом веществе
хрящевой ткани от .50 до ТО % приходится на содержание к ллаге-
на. Собственно хрящевая ткань не имеет кровеносных сосудов, а
питательные вешества-ли{}зфунди-руют из окружающей ее надхрящ-
ницы.
Классификация. Различают гри вида хрящевой ткани:
гиалиновую, япшйййгтую. Такое по1раз~леление
хрящевых тканей основано на структурно-функциональных осо-
бенностях строения их межклеточного вещества;--
Хондрогистогенез
Развитие хрящевой ткани осуществляется как у эмбриона, так
и в постэмбриональном периоде при регенерации.
Эмбриональный хондрогистогене з.1 Источни-
ком развития хрящевых тканей является мезенхима. В тех местах
тела зародыша, где образуется хрящ, мезенхима сначала уплот-
няется, клетки теряют свои отростки, усиленно размножаются и
плотно прилегают друг к другу, создавая определенное напряже-
ние — тургор. Такие участки называют хондрогенным
зачат ко м, или хондрогенным островком (рис. 72,
А, Б, В)1Мезенхимные клетки, входящие в их состав, дифферен-
цируются в хондробласты (хондробластоциты) — клетки, обра-
зующие хрящевую ткань. В цитоплазме таких клеток увеличи-
вается количество свободных рибосом, появляются участки грану-
210
Рис. 72. Схема развития 1иачипо-
вою хряща (рис. Ю. И. Афа-
насьева).
А — хондрогенный островок; Л'
первичная хрящевая ткань; В — ста-
лия дифференцировки хондроцитов.
/ мезенхимные клетки: 2 ми-
готически делящиеся клетки; J —
оксифильное межклеточное веще-
ство; 4 — изогенная группа хряще
вых клеток; 5 — базофильное меж-
клет »чное вещество; 6 молодые
хондроциты; 7 — кровеносный ка-
пилляр.
лярной эндоплазматиче-
ск ои сети.ТПВ следу юще и
стадии, называемой ста-
дией образования пер-
вичной хрящевой
ткани, клетки централь-
ного участка (первичные
хондроциты) округляются,
увеличиваются в размере,
в их цитоплазме развива-
ется гранулярная эндо-
плазматическая- сеть, с
участием которой проис-
ходят синтез и секреция
фибриллярных белков
(колдагена). Образующее-
ся таким образом межкле-
точное вещество отличает-
ся оксифилией. В дальней-
шем процессе /^стадии
дифференцировки
хрящевой ткани —
хцядродигы приобретают
способность синтезиро-
вать, кроме упомянутых
ранее фибриллярных бел-
ков, гликозаминогликаны,
главным образом сульфа-
тированные (хондроитин-
сульфаты), связанные с
неколлагеновыми белками
(протеогликаны).
По периферии хрящевой закладки, на границе с мезенхимой
формируется надхрящница — оболочка, покрывающая снаружи
развивающийся хрящ, состоящая из наружною волокнистого и
11
внутреннего хондрогенного (камбиального) слоев. В хондрогеннои
зоне клетки интенсивно делятся, дифференцируются хондро-
блаыы, которые сохраняют способность к синтезу ДНК, размно-
жению, а также к синтезу компонентов межклеточного вещества
(см. ниже). В процессе секреции продуктов синтеза и наслаивания
на уже имеющимся хрящ по его периферии клетки сами «заму-
ровываются» в продукты своей деятельности. Так происходит
рост хряща способом наложения, или аппозиционный
рост.
“Хрящевые клетки, лежащие в центре молодого развивающегося
хряща, сохраняют способность в течение некоторого времени
делиться митотически, оставаясь в одной лакуне (изогенные груп-
пы клеток; от греч. isos—равный, genesis — развитие). За счет
увеличения количества этих клеток происходит увеличение массы
хряща изнутри, что называется интерстициальным рос-
том. Интерстициальный рост наблюдается в эмбриогенезе, а
также при регенерации хрящевой ткани.
По мере роста и развития хряща центральные участки его
все более отделяются от питающих сосудов и начинают испыты-
вать затруднения в питании, осуществляемом диффузно со сторо-
ны сосудов надхрящницы. Вследствие этого хондроциты теряют
способность размножаться, некоторые из них подвергаются
разрушению, а протеогликаны превращаются в более простои
оксифильный белок’— альбумоид.
Клетки хрящевой ткани
R холе развития хрящевой ткани образуется диф ф е р о н;
ствотшые к-.четки, по’‘устно чавые (прехондробласты), хондроблас-
ты (хондробластоциты), хондроциты.
Хондроциты (от греч. chondros — зерно, хрящ, cytos — клет-
ка) — основной вид клеток хрящевой ткани. Они имеют оваль-
ную, округлую или полигональную форму в занисимсши от сте-
пени- дифференцировки- Расположены в.особых долостях (лаку-
налЦ^зйежклеточном веществе в одиночку или группами. Группы
клеток, лежащие в общей полости, называются и з о г е иными.
Они происходят путем деленияГоднЪй клетки Грис. 73 У. Различают
три типа хондроцитов в изогенных группах.
Первый тип хондроцитов характеризует! я высоким ядерно-ци-
топЛазматичАким лтищигнигле развитием вакуолярных элементов
пластинчатого комплекса, наличием митохондрий и свободных
рибосом в цитоплазме. В этих клетках нередко наблюдаются
карт ины деления, что"позволяет рассматривать их как источ ник
репр_одукции изогенных групп клеток (рис. 74, А, Б, В). Количест-
во хондроцитов первого типа преобладает в молодом, развиваю-
щемся хрящеТ^торОй ТИП' хбндрбцйтов отличается снижением
ядерно-цитоплазматического отношения, ослаблением с и нте за
ДНК, сохранением высокого уровня синтеза РНК, .интенсивным
Рис. 73. Изогенные группы хондроцитов суставного хряща. Сканирующая элек-
тронная микрофотография, z 6000 (препарат М. Н. Павловой).
/ хондроциты; 2 — коллагеновые волокна межклеточного вещества
тов комплекса Гольджи, которые обеспечивают образование и
секрецию гликозаминогликанов и протеогликанов в межклеточное
вещество. Цитолемма и кариолемма обычно извилист>Д!Д\р е т и й
тип хондроцитов отличается самым низким >с1ерно-цитоплаз-
матическим отношением'—сильным развитием й упорядоченным
расположением гр?н} ^прний-ондопня мил 1.тчггкои сети Эги_клетки
сохраняют способность к образов? “дю и секреции бепка но снижа-
ется- синтез гликозаминогликанов.
'Xotulpofi ласты (от греч. ctiondros — хрящ, blastos — зачаток) —
это молодые уплощенные___клетки,- способные—к, пролиферации и
синтезу межклеточного вещества хряща Оми яд л я ются. потомками
стводовых клеток.
Цитоплазма хондробластов имеет хорошо развитую грануляр-
ную и агрЗмулярную Эндоплазматическую сеть, комплекс Гольджи.
При окрашивании цитоплазма хондробластов оказывается базо-
фильной в связи с богатым содержанием IfeHK. При участий
хондробластоцитов происходит периферический (аппозиционный)
рост хряща. Хондробластоциты в процессе развития хряща
превращаются в хондроциты.__
6
Межклеточное вещество хряща
Рис. 74. Схема трех (Л. Б, В) типов
хондроцитов (рис. Ю. И. Афанасье-
ва).
/ митотически делящаяся клетка;
2 - эндоплазматическая сеть; 3 — ми-
тохондрии; 4 гликоген; 5 — межкле-
точное вещество.
В м.уиг к-неточном веществе хряща (substantia fundamentals
cartilaginea) органические компоненты предстдддзшы—главным об-
паум (улками липинами гпикмаминиглттми м ппотеоглика-
нами. Концентрация последних в этой разновидности соедини-
тельной ткани является наибольшей. .В большом количестве со-
держатся фибриллярные белки (главным образом коллаген И типа)
Ориентация волокон определяется направлением силовых линий,
возникающих' при деформациях хряща в процессе функциониро-
вания органа. Слой межклеточного вещества, прилегающий к кле-
точной полости и обра уюший ее стенку, отличается ( льшим
светопреломлением, содержит войлокообра зную сеть ' фи бри лл.
Его иногда называют капсулой хрящевой клетки.
Межклеточное вещество обладает..в'ыТюкой. гидрофильностью,
что обусловливает ее плотносты-лупгор и..способствует диффузии
питательных веществ, воды н го юй.. Газы и многие метаболиты
также свободно диффундируют через него. Однако крупные, белко-
вые молекулы, обладающие антигенными свойствами, не проходят.
Этим объясняется успешная Трансплантация—в -клинике (пере-
садка от одного человека к другому) участков хряща.
Гиалиновая хрящевая ткань
Бопьщинство даслей-чл^люта разнивающется_а.мбдир1>а состоит
из гиалиновой-..хрлщевой- ткани (textus cartilaginous hyalinus).
Этухряшевую ткань называют еще стекловидной (греч. hyalos —
стекло) в связи с ее прозрачностью и голубовато-белым цветом.
Во взрослом органИТме гиал~йновая~Тк^нГ.~встречается в местах
соединения ребер с грудиной, в гортани, в-.воздухоносыых путях,
на суставных поверхностях костей.
Строение гиалиновой хрящевой ткани различных органов имеет
много общего, но в то же время содержит признаки органоспе-
цифичности, проявляющиеся в расположении клеток, строении
межклеточного вещества. Гк тьшая часть—встречающейся в орга-
низме человека гиалиновой хрящевой ТКДИИ покрыта надхрящни-
цей (perichondrium) и представляет собой—вместе с пластин-
кой хрящевой ткани анатомические образования — хрящи.
В надхрящнице выделяют два слоя: наружный состоит из
волокнистой соединительной ткани с к овеносны ми сосудами;
внутренний, преимущественно клеточный, содержит хондробласты
и их предшественники^^дрехожЗдойласты. Под надхрящницей в
поверхностном _слое хряща располагаются молодые хондрТУЦиты
веретенообразной формы, длинная ось которых'направлена-вдоль
поверхности хряща (рис. 75, А, Б). В более глубоких слоях
хрящевые клетки приобретают овальную или округлую форму.
В связи с тем что синтетические и секреторные процессы у этих
клеток ослабляются, они после деления далеко не расходятся, а
лежат компактно, образуя так называемые изогенные группы из
2—4 хондроцитов^.
Клетки окружены волокнистым каркасом (капсулой!—из_кШ1ла-
геновых-волоконх_црош1танмы-м--аморфным. веществом (см рис. 73),
причем вокруг молодых хрящевых клеток межклеточное вещество
оксифильно. Более дифференцированные хрящевые клетки и изо-
генные группы, кроме оксифильного перицеллюлярного слоя,
имеют базофильную зону межклеточного вещества. Эти свойства
объясняются неравномерным распределением химических компо-
нентов межклеточного вещества — белков и гликозаминогликанов.
Однако не все хрящи одинаково Так, структурной
особенностью гиалинового хряща суставной поверхности является
отсутствие надхрящницы на поверхности, обращенной в полость
сустава. Суставной хрящ состоит из трех нечетко очерченных зон.
В цар жнби~~зоне "располагают я мелкие -уплощенные -малоспе-
циализированные хондроциты. В средней зоне клетки более
крупные, округлые, расположёны в виде.колонок пёрпёвликуляр-
но "поверхности, Глубокая^ зона суставного—хряща состоит из
кальцинированнрго хряща. Тодыш—в__этой-зоне-обнаруживаются
кровеносные сосуды.
Питание суставного хряща лишь частично осущесглл_яется из
сосудов^ глубокой зоны, а"в основном из синовиальной-жидкости
полостщсу става
Эластическая хрящевая ткань
В ряде органов, где хрящевая основа подвергается изгибам
(в ушной раковине, рожковидных и клиновидных хрящах гортани
и др.)ГрасТ1благается эластическая~хрящёвая ТК~атть"Пе?апх~сгтЛае1-
215
Рис. 75. Гиалиновым хрящ.
А микрофотография гиалинового
хряща трахеи: / надхрящница;
2 молодые хондроциты; 3 — основное
вещество с расположенными внутри
него изогенными группами хондроцитов
(4) /;— схема .троения надхрящницы
(рис. Ю. И. Афанасьева): / наруж^
ный волокнистый слой: 2 внутренний
клеточный слой; 3 хрящевая ткань.
neus elasticity). В свежем, нефиксированном состоянии эласти-
ческая хрящевая ткань желтоватого-цве!а и не такая прозрачна я,
как гиалиновая.
По общему плану строения эластический хрящ сходен с гиали-
новым. Снаружи он покрыт надхрящницей,., Хрящевые клетки
(мополые и специализированные хондроциты) располагаются в
капсулах поодиночке или образуют изогенные группы. Одним из
главных отличительных_нри знаков эластического хряща является
наличие л—е-го—меж к ле-точ ном—веществе наряду—т кпттЛсГНПТо выми
216
Рис. 76. Эластический хрящ ушной
раковины. Микрофотография.
/ июгенные группы хондроцитов; 2
пластические волокна
водок нами—эластических—воло-
кон, I ipo н и эыва ющих
точ ное вещество во все х на -
правлениях (рис. 76). Н.хдоях,
Прилежащих к надхрящнице,
эластические волокна без пере-
рыва~ переходят в эластические
волокна надхрящницы. Лилия
дств; гтпТкТГгена Yr хиидроитин-
сульфатов в эластическом хря
ще меньше, чем в гиалиновом.
В эластическом хряще ни-
кбгда не происхо дит лбы шест-
вления----
Волокнистая хрящевая ткань
Волокнистая -хрящевая ткань
(textus cartilagineus fibrosa)
встречается между позвонками дисков, полуподвижных сочленени-
ях, в местах, где совершается переход волокнистой соединительной
ткани (сухожилия, связок) в гиалиновый хряну^де ограничен
ные движения сопровождаются сильными натяжениямйТ'-.>//т£
Межклеточное вещество содержит параллельно наиравлеиные
коллагёййвые пучки;Щ<6торыс постепенно разрыхляются и пепе-
хоДят в гиалиновый хрящ. В нем содержатся полости, в которых
заключены хрящевые клетки. Последние располагаются поодиноч-
ке или образуют изогенные группы. Цитоплазма клеток часто
ва ку о л и зи рова на. П о направлению от гиалиновое. хряща к сухо-
жилию волокнистый хрящ становится все более_похожим на
су хОЖТТлие. На > ранице хряща и сухожилия—вместо столби ков
сухожильных клеток между коллагеновыми пупками, впаянными
в ос новное вещество, лежат столбики сдавленных хрящевых
клеток, которые без какой-либо границы переходят в настоящие
с у х о ж ильные клетки, расположенные- в плотной -соедините л ь н о й
ткани (рис. 77).
—Возрастные изменения. Цо мере старения организма в хпяше-
вой ткани уменьшается концентрация протеогликанов и свя зан-
ная с ними гидрофильность. Ослабляются процессы размножения
хондробластов и молодых хондроцитов. В цитоплазме этих клеток
уменьшается объем комплекса Гольджи, гранулярной эндоплазма-
тической сети, митохондрий и снижается активность ферментов.
Одновременно увеличивается количество гликогена лизосом необ-
ходимых для резорбции дистрофически измененных клеток и
неклеточного вещества. Часть лакун после гибели хондроцитов
217
заполняется аморфным веществом и коллагеновыми фибриллами.
Мес тами в межклеточном веществе об наруживаюг с я сггЛ о же н и я
солей кальция («омелёниё-хряща»!, вследствие чего хрящ стано-
вится!мутным" непрозрачным, приобретает твердость и ломкость.
В результате появляющееся нарушение трофики центральных
участков хряща лиожет-привести' R врастанию в них кровеносных
сосудов с последующим костеобразованием.
Регенерация. Физиологическая регенерация хояшевой ткани
осуществляется за~счет малоспециализированных клеток надхрящ-
ницы и хряща путем размножения и дифференцировки прехондро-
блас^1.н2^бндро5да£1Ов^.идиака -этот- процесс.. иде_т очень мед-
ленно.
Костные ткани
Костные ткани (textus ossei) -^специализированный тип
соединительной ткани с высокой минерализацией межклеточного
вещества, содержащего- около 70% неорганических соединений,
главным образом фосфатов кальция. Из этих тканей построены
кости скелета, которые из всёх~разнбвидноетей соединительных
тканей обладают наиболее выраженными опорной;-метанивеской,
защитной “функциями ДЛЯ внутренних органов, а также я вл я юте я
депег ссщей~кальтпгя--фо£фора -и-др-Органические вещеслво (мат-
рикс костной ткани) представлено в основном белками.—и- липи-
дами. Органические и неорганические компоненты в сочетании
друг с другом дают очень прочную опорную ткань, отличающуюся
способностью сопротивляться растяжению, сжатию и др.
Несмотря на высокую степень минерализации, в костных
тканях црсшсходят-постоянное-обновление входящих в-их_.состав
веществ^ постоянное разрушение и созидание, адаптивные пере-
стройки к изменяющимся условиям функционирования. Морфо-
функнионяльные свойства костной ткани меняются в зависимости от
возраста, мышечной деятельности, условий питания, а также под
влиянием деятельности .желез, внутренней секреции, иннервации и др.
218
Классификация. Существует пва.-л< ыовныу_ища кост-
ной ткани — ретикулофиброзная и пластинчатая. _Эти разновид-
ности костной ткани различаются по структурныкГй~фйзическим
свойствам,- которые обусловлены главным образом строением
межклеточного вещества.
Остеогистогенез
Различают эмбриональн ый и постэмбриональ-
ный остеогистогенез. У эмбриона костная—ткаттв—развивается
из мезенхимы "Двумя спбсо'бдмИ^ДХ1тепосредственно из мезенхимы
(п рямой о сте о г и с т о ген е з) и 2) из мезенхимы на
месте ранее развившейся хрящевой модели кости (непря мой
о с те п г и сто I е н е з). Постэмбриональное развитие кости
осуществляется при регенерации и эктопическом остеогистогенезе.
Развитие кости у эмбриона
Прямой остеогистогенез. Такой способ остеогенеза характерен
для развития грубоволокнистой костной ткани при образовании
плоских костей, например покровных костей черепа. Этот процесс
наблюдается в основном в течение первого месяца внутриутробно-
го развития и характеризуется образованием сначала первичной
«перепончатой», остеоидной костной ткани с последующей импрег-
нацией (отложением) солей кальция, фосфора и др. в межклеточ-
ном веществе. В первой стадии — образование скелетоген-
ного островка — в местах развития будущей костй~происходят оча-
говое размножение мезенхимных клетрк_и васкуляризация скелетр^^^*^
геццого островка Трис. 78, А, Б, В, Г). В о второ й_с ра д и и,
заключающейся, д_диффгренцировке—клеток островков, образуется
окси фильное межклеточное вещество с коллагеновыми фибрилла -
ми — органическая матрица костной ткани (остеоидная стадия).
Разрастающиеся волоюза_раздвигают_клетки,_ которые, не теряя
своих отростков, остаются связанными друг с.другбмГБ "основном
веществе появляются мукопрртеиды (оссеомукоид), цементирую-
щие волокна в одну прочную масху. Некоторые клетки, дифферен-
цирующиеся в пстеоцихы,—уже в этой стадии могут оказаться
включенными в толщу волокнистой массы. Другие, располагаю-
щиеся по поверхности, дифференцируются в остеобласты. В тече-
ние некоторого времени остеобласты, располагаются по одну
сторону волокнистой_ массы, но вскоре коллагеновые волокна
появляются и с других сторон, отделяя остеобласты друг от друга.
Постепенно эти клетки оказываются - шмурованными» в межкле-
точном веществе,~теряют способность размножаться и превра-
щаютсяв остеоциты. В то жё""врёмя“из окружающей мезенхимы
образуются новые генерации остеобластов, которые наращивают
кость снаружи (аппозиционный рост).
Третья стадия — кальцификация (импрег нация солями)
межклеточного вещества. Остеобласты выделяют фермент щелоч-
ную фосфатазу,_расщеплягощую содержащиеся в периферической
"ЧЧ
/7
Рис. 78. Прямой (пере
пончатыи) остеогене!
(схема).
Л — остеогенный островок;
/, остеоидная l гания;
В оссификация межкле-
точного вещества-
в
I
крови глицерофосфаты на углеводные соединения (сахара) и
4юсфо^^КИ£ДаЯ'СПос^еДня^^тУпает к Реакцию с солями
кальция, который осаждается в основном веществе- й~ волокнах
сначала в виде соединений кальция, формирующих аморфные от-
ложения |Са3(РО4)2], в дальнейшем из него образуются кристаллы
гидрооксиапатита [СЭ|0(РО4)Ь(ОН)2].
Кальцификацию оссеоида связывают с так называемыми мат-
риксными пузырьками типа лизосом диаметром до 1 мкм (подобные
пузырьки обнаружены и при минерализации органической матри-
220
Рис. 78 ( продол дени ( )
/ «прямой» ocreoi енез в
плоской кости (микрофото-
Iрафия). / мезенхимные
клетки; 2 кровеносные
капилляры; 3 остеобла
сты; 4 остеоид; 5 ми-
нерализованное межклеточ
нее вешество: 6 — остео-
класт.
цы развивающегося зуба). Эти пузырьки обладают высоком актив-
ностью щелочной фосфатазы и пирофосфатазы, содержат липиды
накапливают на внутренней поверхности, покрывающей их мемб-
раны, кадацдй
Одним из посредников кальцификации является остеонек-
тин — гликопротеин, избирательно связывающий соли кальция и
ЙЬрсФора с коллагеном. *
R рртупктатг кальцификации <'£рз?у1РТЯП ‘ i .- ru.nQ и 'ины. '
или балки. Затем от этих перекладин ответвляются выросты,
соепиняюшиеся~ меж1у собой-и образующие широкую сеть. Прост-
ра нстна между ре ре юта ди на мм—оказываются занятыми соеди н и -
теньной нопокнигтл;. тканью_с_прохолян|ими в ней кровеносными
сосудами.
^моменту завершанин-гнетогенеэа-но-периферии зачатка кос-
ти, в эмбриональной соединительной ткани появляется большое
количество волокон и остеогенных клеток. Часть этой волокнистой
ткани, прилегающей непосредственно к костным перекладинам,
превращается~в периост (periosteum), который обёсцечиваст_тр£>-
фикул регенерацию кости (см. ниже). Такая кость, появляющая-
ся на ранних стадиях эмбрионального развития и состоящая из
перекладин ретикулофиброзной костной ткани, называется первич-
ной губчатой костью. В более поздних стадиях развития она заме-
няется1мооичмпй. гу&1ааа«&«ДЙ1ЬЮ ВзИСЛЫх? -которая- от-ли чается
от первой тем, что построена из пластинчатой_к.осхысш-ткани (ч е т-
в е р та я стадия остеогенеза).
Вазвитие ттластинчатой-костно# ткани. тесно свазаыо_£..процес-
сом разрушения отдельных участков костили, врастанием кровенос-
ных сосудов в тол1Цу_Т^тикудофиброзной кости! В этом процессе
как ё^пёрйод эмбрионального остеогенеза, так и после рождения
принимают участие остеокласты (см. рис. 78, В).
Костные”пластинки обычно образуются вокруг кровеносных со-
судов путем дифференцировки прилегающей к ним мезенхимы.
На такую пластинку накладывается слой новых остеобластов, и
А Б В Г
Рис. 79. Схема «непрямого» (хрящевого) остеогенеза. Образование хрящевой
модели кости и перихондральной костной манжетки (рис. Ю. И. Афанасьева).
/__первичная хрящевая модель трубчатой кости; 2 — надхрящница; 3 хрящевая ткань,
4__ перихондральная костная манжетка; 5 — надкостница; 6 — колонки хрящевых кле
ток; 7 зона пузырчатых клеток; 8 — врастающая в хрящ мезенхима < дифференци-
рующимися остеокластами (9) и кровеносными капиллярами (10); // остеобласты,
12 — эндохондрально образованная костная ткань; 13 — точка окостенения в эпифизе.
возникает новая пластинка. Коллагеновые волокна в каждой плас-
тинке ориентированы под углом к волокнам в предыдущей плас-
тинке. Таким образом, вокруг сосуда возникает подобие костных
цилиндров, вставленных один в другой (первичные остеоны).
С момента появления остеонов ретикулофиброзная костная ткань
перестает развиваться и заменяется пластинчатой костной ткапью,
Со стороны, надкостницы формируются общие, или генеральные,
пластинки, охватывающие всю кость снаружи. Так развиваются
ц л_о с к и е_ к_РХТ и- в дальнейшем образовавшаяся в эмбрио-
нальном периоде кость подвергается перестройке: _ра утушаются
первичные остеоны и развиваютсд_щовые-генёраЦВи_остеонов. Та-
кая перестроикТГкости практически продолжается всю жизнь.
Таким образом, в отличие от хрящевой ткани кость всегда рас-
тет способом наложения новой ткани на уже имеющуюся, т. е.
аппозиции, а оптимальное кровоснабжение необходимо для
дифференцировки клеток скелетогенного островка.
Непрямой остеогистогенез. На 2-м месяпе эмбрионнльнаго раз-
вития в местах будущих трубиатых костей закладывается из ме-
зенхимы хрящевой зачэ гцк, который очень быстро при ним ает фо р-
му-будущей кости (хрящевая модель) (рис. 79, А, Б, В, Г). Зача-
ток состоит из эмбрионального гиалинового хряща, покрытого
надхрящницей. Некоторое время он растет как"^за счет клеток,
образующихся со стороны надхрящницы, так и за счет размноже-
нил клеток -во~~Бнутренних участках?"
Окостенение диафиза. Развитие—кости на месте хряща начина-
ется с диафиза. Этот процесс обусловлен разрастанием- кровенпг-
ных сосудов и_доявлением в надхрящнице,прилежащей к средней
’части^диафиза, остеобластов, образующих в виде манжетки снача-
ла ретикулофиброЗНузСХостную ткань- (ггеф ггх ондральное
окостенение), а затем пластинчатую.
.2 Образование костной манжетки нарушает питание хряща.
Вследствие этого.в центре диафизарной части хрящевого зачатка
возникают дистрофические изменения. Хондроциты вакуолизиру-
ются, их ядра пикнотизируются, образуются так называемые пу-
пырчатые хондроциты. Хрящ в этом месте перестает расти. Тзх
как соседние неизмененные дистальные отделы диафиза пролоп-
жают свой рост, хдццроциты на границе эпифиза и диафиза соби-
раются в колонки, направление которых совпадает с длинной осью
будущей кости. Таким образом, в колонке хондроцитов имеются
два_2противополржн<Г направленных процесса — размножение и
рост в дисткльных~отделах диафиза И~дис i рофические чгроцессы- в
его прокеимальн£ум_гитц?де Одновременно мезкду набухшими клет-
ками происходит отложение минеральных солей, обусловливающее
появление резкой базофилии и хрупкости хряща.
С момента разрастания сосудистой сети и появления остео-
бластТггзгатгардщницд Нф^Рррайваедся, превращаясь в надкостни-
цу. В дальнейшем кровеносные сосуды с окружающей их мезен-
химой, остеогенными клетками и остеокластами врастают через
отверстия костной манжетки и входят в соприкосновение с обыз-
вествленным хрящом (см. рис. 79, Г). П.н! влиянием ферментов,
выделяемых осгеокл.дтями^ происходит растворение (хондролиз)
обызвествленного межклеточного вещества. Диафизарный хрящ
разрушается, в нем возникают удлиненные пространства, в кото-
рых поселяются рстеоциты, образующие на поверхности остав-
шихся участков обызвествленного хряща костную ткань (см.
рис. 79).
Первичный, или диафизарный, центр окостенения. Процесс от-
ложения кости внутри хрящевого. заилтка-4Юлудил название" эн-
fl о хондральн о г о, или энхондраль ног о, окосте-
н е ни я (греч. endon — внутри).
Одновременно с процессом развития энхондральной кости no-
fl вл я юте я и признаки ее разрушения остеокластами. Вследствие
разрушения энхондральной костной ткани образуются еще боль-
шие полости и пространства (полости резорбци rtjn, на конец, воз-
ни кает костнол<озгошя полость. Из проникшей сюда мезенхимы
образуется строма костного мозга,~в которой поселяютсзг~стволо-
вые клетки крови и^соединительнойиткашы В это же время по пе-
риферии диафиза со стороны надкостницы нарастают все новые и
новые перекладины костной ткани, образующейся из надкостницы
(см. ниже). Разрастаясь в длину по направлению к эпифизам и
увеличиваясь в толщину, они образуют плотный слой кости. Даль-
нейшая организация периостальной кости протекает иначе, чем
энхондральной костной ткани. Вокруг сосудов ._юхк>рью- -идут по
длинной оси зачатка кости из прилегающей к ним мезенхимы, на
месте"разрушающейся ретикулбфйброзной кости начинают образо-
вываться концентрические пластинки, состоящие из параллельно
ориентированных тонких коллагеновых волокон и цементирующего
межклеточного вещества. Так возникают первичные остеоны. Про-
свет их широк, границы пластинок нерезко контурированье Вслед
за появлением первой генерации остеонов со стороны периоста
начинается развитие общих пластинок, окружающих кость в об-
ласти диафиза. Приблизительно в этот же период периостальная
кость смыкается с энхондральной и образуется компактное вещест-
во диафиза кости.
Окостенение эпифизов. Вслед за диафизом центры окостенения
появляются в эпифизах. Этому предшествует сначала дифферен-
цировка хондроцитов, их гипертрофии, сменяемая ухудшением пи-
та нйяГ~Дис грсгфПеи^й кальцинацией. В дальнейшем" отмечаете я
процесс окостенения, подобный описанному выше. Оссификация
сопровождается врастанием в эпифиз сосудов.
В промежуточной области между диафизом и эпифизами, а
также на поверхности эпифизов сохраняется хрящевая ткань.
Клетки костной ткани
Костные ткани содержат три вида клеток: остеоциты, остео-
бласты^ и Остеокласты.
В _ход£~развитйя костной ткани образуются два дифферона:
первый - стволовые.остеогенныс-клелки, пилуство.човые е.ромаль-
ные клетки. остеобласты- ocTeoipiri.i Второй днффероп гематоген-
ного происхождения: стволовая кроветворная клетка, полустволо-
вая кроветворная клетка предшественница миелоидных клеток и
макрофагов, унипотентная колониеобразующая моноцитарная
клетка — монобласт — промонОцит — моноцит — остеокласты (мак-
рофаги) . ' ' '
Qr тр о ци ты (от греч. osteon — кость, cytus — клетка) —
костные гпртги. Это преобладающие по количеству дефинитивные
клетки костной ткани, утратившие, способность к делению. Они
имеют отростчатую форму, ипмплктипр, относительно крупное ядро
и стабобазофильиуюцигшыгазму (рис. 80, А, Б). Органеллы развиты
слабо. Наличие центриолей в остеоцитах не установлено.
Костные клетки лежат влшегдыт полостях, или лакунах, которые
повторяют контуры остеоцита Длина полос гей Колеблется от 22 до
55 мкм, а ширина — от 6 до 14 мкм. Канальцы костных полостей за-_
полнены тканевой жшнч<югьиханастом<)зирую1- между собой и с пе-
ри васкулярными пространствами сосудов, заходящих bnyipr> пости.
Обмен веществ между остеоцитами и кровью^ осуществляёТГяДерез
ткано "УЮ HMOLlb.
Остеобласты, остеобластов идг ы (от греч. oste-
on — кость, blastos — зачаток). Это молодые клетки, создающие
костную ткань. В сформировавшейся кости они встречаются толь-
ко"в глубоких слоях надкостницы и в местах регенерации костной
’24
I
Рис. 80. Схема строения остеонита (рис.
К). И. Афанасьева).
А на светооптическом уровне; Б — на субми-
кроскопическом уровне: / — отростки остеоцитов;
2 ядро; 3 — эндоплазматическая сеть; 4 — ком-
плекс Гольджи; 5 — митохондрии; 6 — оссеоидное
(иеобызвествленное) вещество кости по краям
лакуны, в которой расположены остеоциты.
ткани после ее травмы. В образую-
щейся кости они—покрывают почти
непрерывным слоем всю поверхность
развивающейся костной ба пки
(рис? 81, Л). Остеобласты бывают
различной формы: кубической, пира-
мидальной иди .утловатай. Размер их
тела около 15—20 мкм. Ядро округ-
лой или овальной формы, часто рас-
полагается эксцентрично, содержит одно или несколько ядрьипек.
В цитоплазме остеобластов хорошо развиты гранулярная эндоплаз-
матическая сеть, митохондрии и комплекс Гольджи (рис. 81, Б).
В ней выявляются в значительных количествах РНК и высокая
активность щелочной фосфатазы. Остеобласты так же. как и остео-
циты, в которые они превращаются, — неделящиеся клетки.
О стер кл а с т ы (о с те о к л а сто ци ты) (от греч. oste-
on — кость и ciao — раздрг|бля'ть{ разбивать). Это клетки гемато-
генноицриролы способные рЯзрушйть Лйм-хмаЛ-п мх..... ц"т хрящ и
крсть.. Диаметр их достигает 90 мкм и более, и дни содержат от 3 до
нескольких д^сятков_ядеп (рис. 82, А, Б). Цитоплазма слабобазо-
фильна, иногда оксифильна, Нд стороне прилегания остеокласга
к .разруицщдщкйовёцхиостн раз, нчают w зпиьГ Первая зона, на-
иболее обширная, богата*. цитинлазматичгскими наростами (гоф-
"ированная каемка), яцдяется областью абсорбции и секреции гид-
ролитических ферментов. Вторая зона -^з5ттз~дтлотнпггг--прилета-
ния остеокласта _к_кО£ТНОЙ поверхности, окружая~~первую7~как бы
герметизирует область действия ферментов. Эта зона цитоплазмы
светлая, содержит мало органелл, за исключением микрофиламент,
состоящих из актина.
Периферический слой цитоплазмы над гофрированным краем
содержит многочисленные мелкие пузырька и более крупные —
вакуоли. Полагают, что остеокласты выделяют СО2 в окружающую
в гистология
225
Рис. 81. Схема строе-
ния остеобласта (рис.
Ю. И. Афанасьева).
А — на светооптическом
уровне; Б — на субмикро-
скопическом уровне; / — яд-
ро; 2 — цитоплазма; 3 — раз-
витая гранулярная эндоплаз-
матическая сеть; 4 — осте-
оид; 5 — минерализованное
вещество костной ткани.
Рис. 82. Схема строе-
ния остеокласта (рис.
Ю. И. Афанасьева).
А — на светооптическом уро-
вне; Б — на субмикроскопи-
ческом уровне; / — ядро; 2—
гофрированный край остео-
класта; 3 — светлая зона;
4 — лизосомы; 5 — зона ре-
чорбнии межклеточного ве-
щества; 6 — минерализован-
ное межклеточное вещество.
фермент карбоангидраз», обнаруживаемый здесь, способ-
ствует образованию кислоты (Н,,СО^), разрушении) органической
матрицьГко'сти и~растворению _кальци£дв1х._£РДей Остесдддст богат
митохондриями шллзоеомамн. Гранулярная эндоплазматическая
сетк-развита ОТНосйтелТно слабо. В том месте, где остеокласт со-
п ри касаетс я скостным веществом, в последнем образуется ла куна.
Дифференцировка остеокластоцитов зависит от воздействия лим-
фокинов, которые вырабатываются Т-лимфоцитами.
Межклеточное вещество
Межклеточное вещество (substantia intercellularis) состоит из
ос новного~веществаТУмп|‘>егймрлнаннпго неорганическими сол ям и,
в котбром~располагаются коллагеновые волокна, образующие не-
большие~пучки77?ни 111,114.41 tn таггп 1 Tiifff ВОлОкна мо-
гут иметь беспорялсшнде (д ретику лофиб розной костнои ткани}
или строголриел1тмравам»&&—{-»-пласт-инчашй..костной ткани) на-
правление.-
В основном веществе костной ткани по сравнению с хрящевой
содержится относительно небольшое количество хондроитинсер-
226
ной кислоты, но много лимонной и других кислот, образующих
комплексы с кальцием, импрегнирующим органическую матрицу
кости. Основное вещество кости содержит кристаллы гидроксиапа-
тита, упорядоченно расположенные по отношению к фибриллам
органической матрицы кости, а также аморфный фосфат кальция.
В костной ткани обнаружено более 30 микроэлементов (медь,
стронций, цинк, барий, магний и др.), играющих важнейшую роль
в метаболических процессах в организме.
Ретикулофиброзная костная ткань
Строение. Ретикууюфцбдщная (грубоволокнистая) костная
ткань (textus osseus reticulofibrosus) встречается главным образом
у зародышей. У взрослых ее можно обнаружить на месте зарос-
ших черепных швов, в местах прикрепления сухожилий к костям.
Беспорядочно расположенные коллагеновые волокна!образуют
в ней толстые--пучки-;--отчетливо заметные даже при сравнительно
небольших увеличениях микроскопа (рис. 83).
В основном вещесьве ретикулофиброзной-КОСТВОЙ^ткашр нахо-
дятся удлиненно^овалызой формы костные полости, или лакуны, с
длинными анастомозйрующими _ канальцами, в которых лежат
костные клетки — остеоциты с_их_отростками. С поверхности гру-
боволокнйстая кость покрыта надкостницей.
Пластинчатая костная ткань
Строение. Пластинчатая костная ткань (textus ossues lamella-
ris) — наиболее распространенная разновидность костной ткани во
взрослом организме. Она состоит из костных пластинок (lamellae
osseae), образованных~~костными клетками и минерализованным
аморфным веществоМ~С~КОЛЛа1|ен1>вым1Г~ВОлокна~ми, ориентирован-
ными в определенном направление В соседних пластинках волок-
на обычно имеют разное направление, благодаря чему достигается
большая прочность пластинчатой костной ткани (рис. 84). Цз_этой
ткани построены компактное и губчатое. вещества в^ольшинстве
плоских и трубчатых костей скелета.
Гистологическое строение трубчатой кости
Трубчатая кость в основном построена из пластинчатой кост-
ной ткани, за исключением" бугорков.
Строение диафиза. Компактное вещество, образующее диафиз
кости, состоит из костных лдастинок, толщина которых колеблет-
ся от 4 до 12—15 мкм. Костные пластищшдрасполагаются в опре-
деленном порядке, образуя-слажные, системы—В_диафизе различа-
ют три слоя: наружный слой общих пластинок, средний, образо-
ванный — концентрически напластованными вокруг сосудов кост-
ными пластинками —рстепиаии и называемый остеонным слоем,
и внутренний слой общих пластинок (рис. 85).
8'
227
г
Рис. 83. Схема строения ретикулофиброзной костной ткани (рис. Ю. И. Афанась-
ева, Ю. А. Рунихина).
/ — пучки переплетающихся коллагеновых волокон, 2 — остеоцитьг
Рис. 84. Схема строения пластинчатой костной ткани (рис. Ю. И. Афанасьева).
/ — костные пластинки; 2 остеоциты; 3 - контакты отростков остеоцитов; 4 коллаге-
новые волокна, ориентированные в пределах каждой костной пластинки параллельно.
228
Рис. 85. Схема строения трубчатой кости (по В. Г. Елисееву, Ю И Афанасьеву,
Е. Ф Котовскому).
А — надкостница; Б компактное вещество кости; В — эндост: Г — костномозговая по-
лость. / — слой наружных общих пластинок; 2 — остеон; 3 канал остеона; 4 — вставоч-
ные пластинки: 5 — слой внутренних общих пластинок; 6 — костная трабекула губчатой
ткани; 7— волокнистый слой надкостницы; 8— кровеносные сосуды надкостницы; 9—
прободающий канал; 10 — остеоциты.
Наружные общие пластинки не образуют полных колец вокруг
диафта -кости, перскрываюня на поверхности следующими слоя-
ми пластинок. Ннутреннйё~о^щие пластинки хорошо развиты толь-
ко~там, Тде~компактное вещество кости непосредственнсГграничит
с костномозговой полостью. В тех же местах, где компактное ве-
щество переходит в гуТУчатое,~етб~~внутрснние общие пластинки
продолжаются в пластинки перекладин губчатого вещества.
В наружных общих Пластинках залегают прободающие каналы,
по которБгм из надкос ГНйцьГвнутрь~костивходят~сосуды. Со сторо-
ны надкостницы^ в кость под разными углами проникают коллаге-
новые_волокна._Эти—волокна получили название прободающих во-
локон. Чаще всего они разветвляются^голько в наружном слое об-
щих пластинок, но могут проникать—и_в_средний остеонный слой,
однако они никогда не входят в пластинки остеонов. В растущих
229
5
Рис. 86. Остеон. Окраска
по методу Ш моря я.
/ канал остеона; 2 ос-
теоциты; 3 — костные плас-
тинки.
костях пробрдаюшие_волокна выражен.ьсхорощощ_могут.быть лег-
коббнаружены на препаратах—В-костях старых людей они кальци-
нируются частично или полностью и делаются почти невидимыми.
В среднем слое кпгтныр пластинки располагаются главным об-
разом в остеонах, формируя остеонные пластинки, а также вста-
вочные пластинки, лежащие между остеонами. Остеоны яв-
ляются_____с тр jк ту р н ой ед и н и ц е й к_о мцик т н о г о
вещества трубчатой кости (рис. 86). Каждый остеон
отграничен от соседних.лстеснов-так называе.мой_стйпой линией,
образованной основным веществом, цементирующим их Впент-
ральном канале остеона проходят кровеносные сосуды с сопровож-
дающей их соедините ч,ной тканыа И остеогенными клетками.
В диафизе длинной кости остеоны расположены преимущест-
венно параллёЛБНО длинной оси. Каналы ОСтеонов анастомозируют
друг с другом; в местах анастомозов прилежащие к ним пластинки
изменяют свое направление (см. рис. 84). Такие каналы называют
прободающими, или питательными. Сосуды, расположенные—в_ка-
налах остеонов, сообщаются друг с другом и с сосудами костного
мозга и надкостницы.
Надкостница, или периост (periosteum). В надкостнмщ£_разли-
чают два слоя: наружный (волокнистым!—и лиутреиинм—4жпеточ-
ный). Наружный СЛОи образован в основном волокнистой соедини-
тельнри-тканыо (рис7"87). Клеточный слой содержит остеоген-
ные ютегки ДостёоблггггыУ азличной степени дифференцировки и
остеокласты. ЧёреТ'наДкпстницу^про'ходят^питающиех.ость сосуды
и нервы которые разветвляйте я во внутреннем слое. Надкостница
связывает кость-с ТГкружающими ..тканями и принимает участие в
ее трофике, развитии, росте и регенерации.
Эндост (endosteum) — очень тонкая и нежная, оболочка, вы-
стилающая кость со стороны- костпо гоммоз га. Как и периост, она
образована волокнистой - соединительной- тканью. Содержит остео-
генные к летк йТлаДа к же"бстуокла сты.
230
Рис. 87. Надкостница (рис. Ю. И. Афанасьева).
/ — наружный волокнистый слой: 2 — внутренний слой; 3 — остеогенные клетки; 4 —
костная ткань.
Васкуляризация. Кровеносные сосуды образуют во внетреннем
слое надкостницы густую сеть. Отсюда берут начало тонкие арте-
риальные веточки, которые проникают в костный мозг через пита-
тельные отверстия и принимают участие в образовании питающей
его сети капилляров. Лимфатические сосуды располагаются глав-
ным образом в наружном слое надкостницы.
Иннервация. В надкостнице миелиновые и безмиелиновые нерв-
ные волокна образуют рыхлое сплетение. Часть волокон сопровож-
дает кровеносные сосуды и проникает с ними через питатель-
ные отверстия в одноименные каналы, а затем в каналы остеонов
и далее достигает костного мозга. Другая часть волокон заканчи-
вается в надкостнице свободными нервными разветвлениями, а
также участвует в образовании инкапсулированных телец.
Рост трубчатых когтей. Рост костей — процесс очень медлен-
ный. Он начинается у человека с ранних эмбриональных стадий и
кончается в среднем к 20-летнему возрасту. В течение всего пери-
ода роста кость увеличивается как в длину, так и в ширину.
Рост трубчатой кости в длину обеспечивается наличием мета-
эпифизарной хрящевой пластинки роста, в которой проявляются
два противоположных гистогенетических процесса. Один — это
разрушение эпифизарной пластинки и другой, противоположный
ему, — непрестанное пополнение хрящевой ткани путем новообра-
зования клеток Однако с течением времени процессы разрушения
начинают преобладать над процессами новообразования клеток,
вследствие чего хрящевая пластинка истончается и исчезает.
В метаэпифизарном хряще различают пограничную зону, зону
столбчатых клеток и зону пузырчатых клеток.
Пограничная зона, ближе всего расположенная к эпи-
физу, состоит из округлых и овальных клеток и единичных изоген-
ных групп, которые обеспечивают связь хрящевой пластинки с
231
костью эпифиза, В полостях между костью и хрящом находятся
кровеносные капилляры, обеспечивающие питанием клетки глубже-
лежащих зон хрящевой пластинки.
Зона столбчатых клеток содержит активно размно-
жающиеся клетки, которые формируют колонки, расположенные
по оси кости, и обеспечивают ее рост в длину. Проксимальные кон-
цы колонок состоят из созревающих, дифференцирующихся хря-
щевых клеток. Они богаты гликогеном и щелочной фосфатазой.
Обе эти зоны наиболее реактивны при действии гормонов и
других факторов, оказывающих влияние на процессы окостенения
и роста костей.
Зона пузырчатых клеток характеризуется гидрата-
цией и разрушением хондроцитов с последующим эндохондраль-
ным окостенением. Дистальный отдел этой зоны граничит с диа-
физом, откуда в нее проникают остеогенные клетки и кровеносные
капилляры.
Продольно ориентированные колонки энхондральной кости яв-
ляются по существу костными трубочками, на месте которых бу-
дут формироваться остеоны (см. ниже).
Впоследствии центры окостенения в диафизе и эпифизе слива-
ются и рост кости в длину заканчивается.
Рост трубчатой кости в ширину осуществляется за счет периос-
та. Со стороны периоста очень рано начинает образовываться кон-
центрическими слоями тонковолокнистая кость. Этот аппозицион-
ный рост продолжается до окончания формирования кости
(рис. 88, А, Б, В). Количество остеонов непосредственно после
рождения невелико, но уже к 5 годам в длинных костях конечнос-
тей количество их значительно увеличивается.
Перестройка кости и факторы, влияющие на структуру
костей
В костной ткани в течение всей жизни человека происходят
взаимосвязанные процессы разрушения и созидания. Перестройка
остеонов всегда связана с разрушением первичных остеонов и одно-
временным образованием новых остеонов как на месте разрушен-
ных, так и со стороны периоста. Разрушение первичных остеонов
начинается только после образования остеокластов. Под влиянием
остеокластов, активированных различными факторами, костные
пластинки остеона разрушаются и на его месте образуется полость.
Этот процесс называется резорбцией (от лат. resorptia —
рассасывание) костной ткани. В образовавшейся полости
вокруг оставшегося сосуда появляются остеобласты и начинается
построение новых пластинок, концентрически наслаивающихся
друг на друга. Так возникают вторичные генерации остеонов,
которые отличаются от первичных остеонов хорошо выраженными
границами костных пластинок Примыкая друг к другу, остеоны
образуют компактное вещество кости.
232
Рис. 88. Диафиз бедренной кости 2-летнего (Л), 13-летнего (Б) и 25-летнего (В)
человека. Микрорентгенограммы (препарат А. Н. Полякова).
/ — наружные общие костные пластинки; 2 остеоны различной степени оссификации;
3 — внутренние общие костные пластинки.
Между остеонами располагаются так называемые вставочные
пластинки. Они представляют собой остатки разрушенных остео-
нов ранних генераций. Процесс перестройки остеонов не приоста-
навливается и после окончания роста кости. Одной из причин,
вызывающих последующую перестройку кости, является измене-
ние физической нагрузки на кость в течение жизни.
Среди факторов, влияющих на перестройку костной ткани, су-
щественную роль играет при различных деформациях ее так назы-
ваемый пьезоэлектрический эффект. Оказалось, что в костной
пластинке (живой или выпиленной из кости) при изгибах появля-
ется определенная разность потенциалов между вогнутой и вы-
пуклой стороной. Первая заряжается отрицательно, а вторая —
положительно. В живой кости на отрицательно заряженной по-
верхности всегда отмечается процесс аппозиционного новообразо-
вания костной ткани, а на положительно заряженной, напротив,
часто наблюдается ее резорбция с помощью остеокластов. Уста-
новлено также влияние искусственно созданной разницы электро-
потенциалов на регенерацию костной ткани и трофику, что нахо-
дит применение в хирургической клинике при лечении переломов.
Отсутствие физической нагрузки на костную ткань (продолжи-
тельная иммобилизация, пребывание в состоянии невесомости
и др.) приводит к повышению функций остеокластов и выведению
солей.
233
На структуру костной ткани и костей оказывают влияние вита-
мины (С, D, А), гормоны щитовидной, околощитовидной и других
эндокринных желез. В частности, при недостаточном количестве
витамина С в организме (например, при цинге) подавляется обра-
зование Коллагеновых волокон, ослабляется деятельность остео-
бластов, уменьшается их фосфатазная активность, что практиче-
ски приводит к остановке роста кости вследствие невозможности
образования костных пластинок вокруг пролиферирующих остео-
бластов. В этих условиях в отдельных участках костей значитель-
но уменьшается количество солей кальция, что при веди т к умень-
шению прочности кости. При дефиците витамина D'(рахит) не
происходит полная кальцификация органической матрицы кости,
что приводит к размягчению костей (остеомаляция). В условиях
гипервитаминоза А усиливается функция остеокластов и связанная
с этим деструкция костей.
Существенное значение для развития и роста костей имеют и
эндокринные факторы. Гормон околощитовидной железы парати-
рин оказывает сильное влияние на рост и состояние костной ткани.
При избытке этого гормона наблюдается резорбция кости и обра-
зование фиброзной ткани, содержащей большое количество остео-
кластов, что приводит к патологическому состоянию, известному
под названием фиброзного остита. Тирокальцитонин щитовидной
железы действует диаметрально противоположно паратирину. При
гипофункции щитовидной железы и снижении концентрации йоди-
рованных гормонов (тироксин и др.) замедляется рост длинных
трубчатых костей в результате подавления активности остеоблас-
тов и торможения процесса -оссификации. Регенерация кости в
этих случаях протекает слабо и неполноценно. Определенную роль
играет и соматотропный гормон гипофиза, под влиянием которого
осуществляется синтез белков в костных клетках. В случае тести-
кулярной недоразвитости или препубертатной кастрации задержи-
вается окостенение метаэпифизарной пластинки, вследствие чего
руки и ноги у такого индивидуума становятся непропорционально
длинными. При раннем половом созревании отмечается остановка
роста из-за преждевременного диафизо-эпифизарного сращения
костей. При недостатке эстрогенов после наступления климактери-
ческого периода у женщин иногда развивается остеопороз, кото-
рый излечивается женскими половыми гормонами.
Соединения костей
Две кости могут иметь соединения непрерывные (синдес-
мозы, синхондрозы и синостозы) и прерывные (суставы).
Синдесмозы — соединения с помощью плотной волокнистой
соединительной ткани, пучки которой в виде прободающих воло-
кон внедряются в костную ткань. Примером таких соединений яв-
ляются швы теменных костей черепа, соединительнотканная мем-
брана между лучевой и локтевой костями.
Синхондрозы (симфизы) — соединения при помощи хряща,
234
например межпозвоночный диск. Они состоят из наружного фиб-
розного кольца и внутренней части, называемой пульпозным яд-
ром. Обе эти части нерезко отделены и незаметно переходят друг
в друга. Пульпозное ядро располагается во внутренней зоне меж-
позвоночного диска. В разные возрастные периоды оно имеет раз-
личное строение. В возрасте до 2 лет оно представляет полость с
гомогенным содержимым, в котором находятся только отдельные
клетки. В последующие годы жизни эта полость разделяется на
отдельные камеры. С 6—8-летнего возраста в пульпозном ядре от-
мечается появление, а затем и нарастание количества коллагено-
вых волокон и хрящевых клеток. С 15 лет нарастание волокон и
хрящевых клеток еще более усиливается, и в возрасте 20—23 лет
пульпозное ядро приобретает характерный вид волокнистого хря-
ща. Примером другого, более плотного соединения может быть
лобковый симфиз. К синхондрозам относятся также соединения
эпифиза и диафиза с помощью метаэпифи за рного хряща.
Синостозы — плотные соединения костей без волокнистой сое-
динительной ткани, например тазовые кости.
Прерывные соединения, или суставы (диартрозы), состоят из
сочлененных поверхностей, покрытых хрящом, а в некоторых слу-
чаях из хрящевого промежуточного мениска и суставной сумки.
Суставная капсула состоит из наружного фиброзного и внутренне-
го синовиального слоев. Под последним понимают пласт специфиче-
ски дифференцированной соединительной ткани, содержащей кро-
веносные и лимфатические сосуды, нервные волокна и окончания.
Пограничное положение этой соединительной ткани, несвойствен-
ное другим производным мезенхимы, постоянное растяжение, сме-
щение и давление в связи с участием в локомоторной функции
сочленения определяют рост и ее структурные особенности.
В синовиальной оболочке млекопитающих и человека различа-
ют два волокнистых коллагеново-эластических слоя — поверх-
ностный и глубокий и выстилающий полость покров-
ный слой (рис. 89, А, Б, В). Резкой границы между слоями не су-
ществует. В крупных сочленениях отмечается богатый жировой
тканью подсиновиальный слой, граничащий с фиброзной капсулой.
Коллагеновые и эластические волокна поверхностного слоя ориен-
тированы в направлении длинной оси сочленения. В глубоком слое
они расположены под углом к волокнам поверхностного слоя.
Покровный слой синовиальной оболочки состоит из клеток —
синовиоцитов (см. рис. 89). Различают три вида синовиоцитов:
А-клетки, или макрофагальные синовиоциты, В-клетки, или сино-
виальные фибробласты, которые обладают способностью к выра-
ботке и секреции гиалуроновой кислоты — специфического компо-
нента синовиальной жидкости, и С-клетки — промежуточные фор-
мы, совмещающие в себе признаки А- и В-клеток.
Васкуляризация. Кровеносные сосуды проникают в синовиаль-
ную оболочку со стороны подлежащих тканей и распределяются в
ее толще, включая и покровный слой, где они располагаются непо-
средственно под синовиоцитами. Таким образом, синовиальная по-
235
Рис. 89. Синовиальная оболочка (рис. В. Н. Павловой).
А покровный слой; Б — поверхностный коллагеново эластический слой; В — глубокий
коллагеново-эластический слой. / — синовиоциты; 2 — основное вещество; 3 кровенос-
ные капилляры; 4 — нервные волокна и окончания; 5 лимфатические капилляры; 6
коллагеновые волокна; 7 — эластические волокна; 8 артериолы и венулы.
лость отделена от кровеносного русла только клетками, основным
веществом соединительной ткани и эндотелием самих капилляров.
Для эндотелия гемокапилляров синовиальных оболочек характерны
фенестры и способность к фагоцитозу. Лимфатические капилляры
располагаются всегда глубже кровеносных в пределах поверхност-
ного волокнистого слоя.
Иннервация. Синовиальная оболочка богато иннервирована во-
локнами афферентной и эфферентной (симпатической) природы.
Глава IX
ИММУННАЯ СИСТЕМА И КЛЕТОЧНЫЕ ВЗАИМОДЕЙСТВИЯ
В ИММУННЫХ РЕАКЦИЯХ
ОЫЦАЯ ХАРАКТЕРИСТИКА. ОПРЕДЕЛЕНИЕ ПОНЯТИИ
( И м м у н и т е т это защита организм а—о_т._в сего
г е не тически чужеродного — микробов^_вщ1усов, от
чужих "клеток или генетически измененных собственных клеток.
Иммунная система объединяет органы и ткани я которых про-
исходит образование и взаимодействие клеток — иммуноци-
236
то в, выполняющих функцию р.п познана» »я генетически чужерод-
ных субстанции (антигенов) и осуществляющих специф» че-
скую реакцию.
Иммунная система обеспечивает поддержание генетической це-
лостности и постоянства внутренней среды организма, выполняя
фу н к цию распознавания «своего» и «чужого». В организме взро с -
лого _человека она представлена: красным костным мозгом —
источником стволовых клеток для иммуноцитов, центральными ор-
ганами лимфоцитопоэза (тимус), периферическими органами лим-
фоцитопоэза (селезенка лимфатические узлы, скопления лимфоид-
HoiFткани в органах , УГфо_цитацр кровили лимфы, а также по-
пуляциями лимфоцитов и плазмоцитов, проникающими во все со-
единительные и эпителиальные ткани. Все органы иммунной систе-
мы функционируют как единое целое благодаря нейрогумораль-
ным механизмам регуляции, а также постоянно совершающимся
процессам миграции и рециркуляции клеток по кровеносной и
лимфатической" системам. Главными клетками, осуществляющими
контроль и иммунологическую защиту в организме, являются лим-
фоциты а также плазматическисллетки и макрофаги.
Постоянно перемещающиеся лимфоциты осуществляют «им-
мунный надзор». Они способны «узнавать» чужие макромолекулы
бактерий и клеток различных тканей многоклеточных организмов
и осуществлять специфическую защитную реакцию.
Для понимания роли отдельных клеток в иммунологических
реакциях необходимо прежде всего дать определение некоторым
понятиям иммунитета.
Антигены — это сложные органические вещества, способные
при поступлении в организм человека и животных вызыватьТпе ци -
фическйи иммунный ответ. Свойствами антигенов обладают бак-
терии, вирусы паразиты.~чужербдные клетки и ткани/ мутационно
изменившиеся собственные клетки тела (напримео, паковые), про-
дукты_ жизнедеятельности чужеродных клеток — белки, полисаха-
риды, полипептиды, а также искусственные высокополимерные со-
единения.
Антитела — это сложные белки, находящиеся, в иммуненяобу-
пиновой фракции плазмы крови человека, и теплокровных -живот-
ных, синтезируемые плазматическими клетками под воздействием
различных антигедав. Антитела способны специфически соеди-
няться с соответствующими антигенами. Такая реакция наблюда-
ется, например, при попадании в организм большинства бактери-
альных антигенов. Обнаружение антител в- глобулиненюй -фракции
белков крови обусловило их название — иммуноглобулины (1g).
Изучено несколько классов иммуноглобулинов (IgG, IgM IgA,
JgE, IgD). Иммуно? mfiy пин О—(IgG) является главным в плазме
крови чедодека. I;_G -.ттитела очень ->,)у[<-кгцнны «щи нммтрализа-
ции бактериальных токсинов и защите организма от вирусов, но
малоэффективна\П|Тй~актИВЯции комплемента. (системъгГбёлков
плазмы, необходимых для лизиса чужеродных тел). IgG-антите-
237
ла вырабатываются при первичном иммунном ответе позднее дру-
гих иммуноглобулинов. Илциунаглабуиин М (IgM) не так специфи-
чен, как IgG, к антигенным детерминантам и очень эффективен в
нейтрализации токсических антигенов. Однако IgM эффективен в
активации комплемента, в лизисе чужеродных клеток, более акти-
венв агглютинации антигена и опсонизации (облегчение захвата и
переваривания бактерий фагоцитами). Иммуноглобулин A (IgA)
содержится в различных секретах (слезы, слюнаф. муцинах дыха-
тельных путей и просвете кишечника. IgA характеризуется высо-
ким содержанием углеводов и устойчив к действию протеолити-
ческих ферментов. Он обеспечивает защиту эпителиальных поверх-
ностей слизистых оболочек. Иммуноглобулин Е (IgE) вызывает
выделение гистамина тучными клетками и ответствен за некоторые
аллергические реакции. Функция иммуноглобулина D (IgD) недо-
статочно изучена. Его относят к эмбриональному виду глобулина.
Он присутствует на поверхности лимфоцитов новорожденных; у
взрослых IgD обнаружен совместно с IgM на поверхности 20%
всех лимфоцитов.
Комплементом является группа белков, содержащихся в све-
жей сыворотке крови человека "и животных л активизирующихся,
когда антитело связывается с антигеном. Этот процесс приводит к
лизису определенных типов клеток (лизис, опосредованный комп-
лементом) или к образованию биологически активных веществ из
белков комплемента, которые, прикрепляясь к бактериям, облегча-
ют их фагоцитоз нейтрофилами. Последние вещества называют
опсонинами.
Согласно клонально-селекционной теории иммунитета, в орга-
низме'существуктг-мнОгбчйсленныё группы (клоны) лимфоцитов,
генетически запрограммированные реагировать на один_ или не-
сколько антигенов.ТТоэтому каждый конкретный антиген оказыва-
ет избирательное действие~стимулируя только те лимфоциты, ко-
торые имеют сродство к его Тюверхностным детерминантам.
Г----При первой встрече т~ антигеном (п е~р тп, чный ответ)
лимфоциты"- стимулируются—и- -подвергаются г fra деформа-
ции в бластные ф оф м ы, которые способны к пролифе-
рации и дифференцировке в иммуноциты. В результате пролифе-
рации увеличивается число лимфоцитов соответствующего клона,
«узнавших» антиген. Дифференцировка приводит к появлению
двух типов клеток — эффекторных и клеток памяти.
Эффекторные клетки непосредственно участвуют в ликвидации
или обезвреживании чужеродного материала. К эффекторным
клеткам относятся активированные лимфоциты и плазматические
клетки. Клетки памяти — это лимфоциты, возвращающиеся в не-
активное -состая1шщ.ко_десущие информацию (память) о встрече
с конкретным антигеном. При повторном введении данного ан-
тигена они "способны обеспечивать быстрый иммунный ответ
большей интенсивности (вторичный ответ) вследствие
усиленной пролиферации лимфоцитов и образования иммуно-
цитов.
238
В зависимости от механизма уничтожения антигена различают
клеточный иммунитет и гуморальный иммунитет.
При точном иммунитете эффекторными клетка-
ми являются цитотоксические Т-лимфоциты, или лимфоциты-
киллеры (убийцы), которые непосредственно участвуют в уничто-
жении чужеродных клеток других органов или патологических
собственных (например, опухолевых) клеток и выделяют литиче-
ские вещества. Такая реакция лежит в основе отторжения чуже-
родных тканей в условиях трансплантации или при действии на
кожу химических (сенсибилизирующих) веществ, вызывающих
повышенную чувствительность (гиперчувствительность замедлен-
ного типа).
При гуморальном иммунитете эффекторными
клеткамй 'являются плазматические клетки, которые синтезируют
и выделяют в кровь антитела.
ХАРАКТЕРИСТИКА ИММУНОКОМПЕТЕНТНЫХ КЛЕТОК
Лимфоциты. Разпичают две основные разновидности лимфоци-
тов: В-лимфоциты и Т-лим оциты.
Стволовые кпетки и клетки-предшественники В-лимфоцитов
образуются в костном мозге. У млекопитающих и человека здесь
же происходит дифференцировка В-лимфоцитов, характеризую-
щаяся появлением у клеток иммуноглобулиновых рецепторов.
Далее такие дифференцированные В-лимфоциты поступают в пе-
риферические лимфоидные органы: селезенку, лимфатические
узлы, лимфатические^ узелки-пищеварительного тракта. В этих
органах при действии_антигенов происходит пролиферация и
дальнейшая специализация В-лимфоцитов с образованием эффек-
торных клеток — плазмоцитов и В-клеток памяти.
Т-лимфоциты также развиваются из стволовых клеток костно-
мозгового происхождения. Последние переносятся с током крови
в тимус, превращаются в бласты, которые делятся и дифферен-
цируются в двух направлениях—Одни-блаеты-образуют-популяцию
лимфоцитов- обладающих специальными рецепторами, восприни-
мающими 2зужероднь1е_ антигены. Дифференцировка этих клеток
происходит под влиянием индуктора дифференци-
ровки, вырабатываемого и выделяемого эпителиальными элемен-
тами тимуса. Образующиеся в результате Т-лимфоциты (анти-
генреактивные лимфоциты} 'заселяют специальные Т-зоны (тимус-
зависимые) в периферических лимфоидных органах. ~Там под
влиянием антигенов они могут подвергаться трансформации в
Т-бласты, Пролиферировать и дифференцироваться в эффекторные
клетки, участвующие в трансплантационном (Т-киллеры) и гумо-
ральном иммунитете (Т-хелперы и Т-супрессорыУ7 а также в
Т-клетки памяти. Другая" часть потомков Т-бластов дифферен-
цируется=гс'"образованием клеток, несущих'рецепторы к антигенам
собственного организма. Эти клетки подвергаются разрушению.
Таким образом, необходимо различать антигеннеза-
239
Рис. 90. Развитие иммунокомпетентных клеток (по М. Куперу, А. Лоутону с из-
менением).
А — схема развития и дифференцировки лимфоцитов млекопитающих; Б — антигеннеза-
висимая и аитигеизависимая дифференцировка В-лимфоцитов и плазмоцитов
ви си м у ю и антигензависимую пролиферацию,
дифференцировку и специализацию В- и Т-лим-
фоцитов (рис. 90).
▲нтигеннезависимая пролиферация и дифференцировка
Эти процессы генетически запрограммированы на образование
клеток, способных давать специфический тип иммунного ответа
при встрече с конкретным антигеном благодаря появлению на
плазмолемме лимфоцитов особых «рецепторов». Она совершается
в центральных органах иммунитета (тимус, костный мозг или
240
фабрициева сумка у птиц) под влиянием специфических факто-
ров, вырабатываемых клетками, формирующими микроокружение
(ретикулярная строма или ретикулоэпителиальные клетки в ти-
мусе).
.х'^Антигензависимая пролиферация и дифференцировка Т-
и В- лимфоцитов
Эти процессы происходят при встрече с антигенами в пери-
ферических лимфоидных органах, при этом образуются эффек-
торные клетки и клетки памяти (сохраняющие информацию о
действовавшем антигене).
Образующиеся Т-лимфоциты составляют пул долгоживущих,
рециркулирующих лимфоцитов, а В-лимфоциты — короткоживу-
щих клеток.
Т-лимфоциты играют важную роль как в клеточном, так и в
гуморалъйом иммунитете. Действующий антиген связывается с
«рецептором» на цитолемме Т-лимфоцита и оказывает на него сти-
мулирующее влияние, приводящее к превращению малого лимфо-
цита "в большой Т-лимфоцит (Т-лимфобласт).~ способный к
пролиферации. Превращение малого лимфоцита в Т-лимфобласт
сопровождается увеличением размера клетки и ядра, для которого
характерно диффузное расположение хроматина (эухроматино-
вое ядро), размер ядрышка также увеличивается. В цитоплазме
накапливается большое число полирибосом, увеличивается комплекс
Г ольджи.
В случае форм ирования клеточного иммуни-
тета (например, при "трансплантации клеток и тканей) при
действии тканевых антигенов дифференцировка Т-лимфобластов
приводит к появлению т^тотоксических^^лимфоцитов- (Т-киллеры)
и Т-клеток памяти (рис. 91). Цитотоксические лимфоциты спо-
собны разрушать чужеродные клетки (клетки-мишени) или непо-
средственно или посредством выделяемых ими особых веществ —
медиаторов (лимфокинов). При. прямом действии Т-киллера на
клетки-мишени _у последних_происходит повреждение -цито леммы
и клетки разрушаются. Лимфокины, выделяемые активированными
Т-лимфоцитами, представляют собой растворимые вещества
(молекулярная масса 8000—80 000), лишенные‘антигенной Специ-
фичности. Имеется несколько видов лимфокинов, обладающих
способностью влиять на интенсивность миграции макрофагов,
вызывать разрушение (лизис) клеток, стимулировать трансформа-
цию и пролиферацию лимфоцитов, переносить информацию о
неспецифическом антигене циркулирующим лимфоцитам и др.
Описанные изменения Т-лимфоцитов могут наблюдаться при
реакциях гиперчувствительности замедленного
типа (местная воспалительная реакция, возникающая при
повторном введении антигена), при развитии аутоиммун-
ных заболеваний, сопровождающихся разрушением собст-
венных клеток организма, а также при развитии опухолевых
новообразований.
241
Тимус
Костный мозг
Кровь
Мон
тх
тс
плазмоцит
л
Неактивный
нокрофаг
нтигено
Периферические
лимфоидные
органы
лв
ПТ
активация ,
активныи
макро-
фаг
В-влас
Б
Рис. 91. Иммуноцитопоэз и кооперация клеток.
А схема иммуноцитопоэза (рис. Н. Ю. Юриной): СК — стволовые клетки; ПСК — по-
лустволовые клетки: Л В лимфоциты костного мозга; ЛТ лимфоциты тимуса; В
В-лимфоииты; ВП — В-лимфоциты памяти; Т - Т-лимфоциты; ТЭ — Т-эффекторы; ТХ —
Т-хелперы; ТС — Т-супрессоры; ТП — Т-лимфоциты памяти; ТК — Т-киллеры; Мои —
моноциты. Б схема взаимодействия Т- и В-лимфоцитов и макрофагов (по Р В. Петрову);
В — В-лимфоциты; Т Т-лимфоциты; М — макрофаги. / антиген; 2 — рецепторы Т-лим-
фоцитов; 3 — рецепторы В-лимфоцитов; 4 — рецепторы макрофага, способные удерживать
Fc-фрагменты тяжелой цепи иммуноглобулинов (IgT); ИИ — индуктор иммунопоэза;
АТ — продуценты антител.
Таким образом, эффекторные Т-клетки (киллеры) обеспечи-
вают генетическое постоянство внутренней среды организма,
разрушая чужеродные клетки.
При. формировании----г у м.о рального____иммунитета
большинство растворимых и других антигенов также оказывает
242
Рис. 91 (продолжение)
В. Г электронные микрофотографии макрофага {МУ в кооперации с лимфоцитом (Л>
и тканевым базофилом (Т£) (препарат А. И. Радостиной).
I стимулирующее влияние на Т-лимфоциты; при этом формируются
/ Т-лимфоциты-хелперы, которые выдел я ют^Тме диаторы (лимфо-
/ кины), взаимодействующиес^В^лимфоцитамн и вызывающие их
/ трансформацию в В-бласты, специализирующиеся в секретирую-
щие антитела плазматической клетки (см. рис. 91). Пролифе-
рация стимулированных антигеном Т-лимфоцитов приводит также
к увеличению числа ~клётОк~которые превращаются в~неактивные
малые лимфоциты, сохраняющие в течение нескольких- лет инфор-
мацию о данном антигене и поэтому называющиеся Т-клетками
памяти. При повторной встрече с этим же антигеном Т-клетки
памяти первыми распознают его и отвечают описанными выше
пролиферацией и дифференцировкой, но эти процессы более
интенсивно выражены (вторичный ответ).
Т-хелпер обусловливает специализацию В-лимфоцитов в на-
правлении^формирования антителообразующих плазмоцитов, кото-
рые обеспечивают «гуморальный иммунитет», вырабатывая и
выделяя Bj<poBb иммуноглобулины. Вот почему медиатор, выде-
ляемый Т-хелпером, называют индуктором иммунопоэ-
з а. Одновременно В-лимфоцит получает антигенную информацию
от макрофага,_который захватывает антиген, перерабатывает его
и передает В-лимфоциту. На поверхности В-лимфоцита располо-
\ жено большое количество иммуноглобулиновых рецепторов
\\ (50 000—150 000) (рис. 92).
'^__>Таким. образом, для обеспечения иммунологических реакций
необходима кооперация деятельности трех основных типов клеток:
В-лимфоцитов, макрофагов и Т-лимфоците. Механизмы этой
кооперации можно представить следующим образом (см. рис. 91).
Молекулы антигена соединяются с иммуноглобулиновыми ре-
цепторами Т-лимфоцитов и в комплексе с ними отделяются от
клетки (происходит как бы снятие рецепторов с поверхности
Т-лимфоцитов). В образованном комплексе рецептор — антиген на
свободном конце находятся Fc-фрагменты тяжелых цепей имму-
ноглобулинов. На цитолемме макрофага имеются специальные
рецепторы, способные присоединять Fc-фрагменты. Макрофаги
после переработки захваченных антигенов передают их В-лимфо-
циту. Передача большого количества переработанных макрофагом
антигенов обеспечивает пролиферацию и дифференцировку
В-лимфоцитов в направлении образования плазмоцитов, выраба-
тывающих специфические антитела на конкретный вид антигена.
Плазматические клетки могут образовываться и при отсутствии
антигенов, но в этом случае необходимо наличие индукторов
иммунопоэза, вырабатываемых Т-лимфоцитами. Тогда сформиро-
вавшиеся плазмоциты синтезируют неспецифические гамма-гло-
булины. Действие антигенов в отсутствие индуктора иммунопоэза
вызывает гибель части лимфоцитов и появление невосприимчи-
вости к данному антигену (толерантность). Без Т-хелперов
В-клетки утрачивают способность формировать антителообразую-
нще клетки.
\ Выявлены также Т-супрессоры, которые подавляют способ-
Рис. 92. В-лимфоциты.
/ — В-лимфоциты мыши с
меченными 1/,1 антителами
(—►) против иммуноглобули-
нов кролика (по Унанье);
// — В-лимфоциты человека
с иммуноглобулинами на по-
верхности цитолеммы (-«—),
выявленными с помощью ан-
тисыворотки человека, ме-
ченной пероксидазой. < 3000
(препарат В. П. Слюсарчу-
ка).
ность лимфоцитов участвовать в выработке антител и таким
образом -обеспечивают им му но л о г и че с к у к> i о те р а н т-
ность, т. е. нечувствительность _к_£>пр£Д£ленным антигенам. Они
регулируют количество образующихся плазматических клеток и
количество антител, синтезируемых этими клетками. Оказалось,
245
что тормозить выработку, антител-можеч и особая субпопуляция
В-лимфоцитов, которые получили название В-супрессоров. Показа-
но, что Т- и В-супрессоры могут действовать подавляюще также
на реакции клеточного иммунитета.
Дифференцировка клеток плазматического ряда. Этпт процесс
проходит в несколько этапов и продолжается _в_тече_ние суток.
Из стимули рванных, Лт лимфоцитов образуются В-лимфобласты,
которые размножаются, часть из них приобретает способность
к синтезу антител и становится пдазмобяастами, превращающими-
ся в последующем в проплазмоциты и_пдазмоциты (рис. 93).
Плазмобласт (plasmoolastus) — крупная клетка, характеризует-
ся наличием большого количества рибосом и небольшим числом
уплощенных цистерн гранулярной эндоплазматической сети.
Ядро содержит деконденсированный хроматин (эухроматин) и
1—2 больших ядрышка.
П роплазмоцит характеризуется меньшим размером тела кле-
ток, увеличением количества концентрически расположенных
узких канальцев гранулярной эндоплазматической сети. Ядро
лежит эксцентрично, хроматин более компактный, расположен
группами около ядерной мембраны (имеет вид спиц колеса).
Около ядра видна зона более светлой цитоплазмы, в которой
расположен увеличенный комплекс Гольджи.
Плазмоцит (plasmocytus) характеризуется появлением боль-
шого количества расширенных цистерн гранулярной эндоплазма-
тической сети, заполненных продуцируемыми клеткой иммуно-
глобулинами. Ядро компактное, расположенное эксцентрично.
Процесс.плазмоцитогенеза сопровождается потерей способности
клеток к ^лению и движению и уменьшением количества поверх-
ностных иммуноглобулинов (Гцйтолемме. Про/юлж.ительностьТжиз-
ни плазмоцитов составляет несколько недель. Лимфобласты и не-
зрелые плазматические клетки из лимфатических узлов, где они
образуются, способны проникать в выносящие лимфатические
сосуды и заселять соседние лимфатические узлы. Часть образован-
ных из них мелких клеток, напоминающих по виду лимфоциты,
проникает в кровеносные сосуды. Они имеют центрально располо-
женное ядро, окруженное узким ободком цитоплазмы, в которой
видна развитая гранулярная эндоплазматическая сеть. Эти клетки
получили название лимфоплазмоцитов.
Синтез иммуноглобулинов (антител) происходит при участии
информационной РНК на рибосомах гранулярной эндоплазмати-
ческой сети В-лимфоцитов и образуемых из них плазмоцитов.
Синтезированные молекулы поступают в просвет цистерн.
В В-лимфоцитах первые антитела выявляются в перинуклеарных
цистернах В процессе дифференцировки плазмоцитов антитела
находятся во всех цистернах гранулярной эндоплазматической
сети.
В зрелых плазмоцитах антитела в перинуклеарных простран-
ствах отсутствуют и исчезают из некоторых цистерн гранулярной
сети. К полипептидным частям тяжелых цепей антител присоеди-
246
Рис. 93. Дифференцировка плазмоцитов (по Вайсу).
/ — предшественник плазмоцита (ПСК); 2 — плазмобласт; 3 — юный плазмоцит; 4
плазмоцит с расширенными цистернами эндоплазматической сети, содержащими иммуно-
глобулины (тельце Русселя); 5 зрелый плазмоцит.
няются углеводы (N-ацетилглюкозамин), и этот комплекс транс-
портируется в комплекс Гольджи, где и происходит присоедине-
ние дополнительных углеводов (галактоза). Из пластинчатого
комплекса антитела переносятся на поверхность клетки и выде-
ляются. Наиболее раннее выделение антител на месте действия
антигена осуществляется лимфоцитами. Плазмоциты начинают
секретировать антитела несколько позднее, но в гораздо большем
количестве. Одна плазматическая клетка может продуцировать
3000 молекул антител в сутки. Часть, иммуноглобулинов может
депонироваться'в растянутых цисдерыах-Ера чу пирной энп.оппазма-
TMMgCKOH сети.
Макрофаги
Макрофаги играют.важную, роль как в естественном, так и в
приобретенном иммунитет£_<2рг.анизма. Участие макрофагов в ес-
тественном иммунитете проявляется в их-способности к фагоци-
247
тозу и в синтезе ряда активных веществ — пищеварительных
ферментов, компонентов системы комплемента, фагоцитина, ли-_
зоцйма, интерферона, эндогенного пирогена и др., являющихся
основными факторами естественного иммунитета. Их роль~в при-
обретенном иммунитете заключается в пассивной передаче анти-
гена иммунокомпетентным клеткам (Т- и В-лимфоцитам), в
индукции специфического ответа на антигены. Макрофаги также
участвуют в обеспечении иммунного гомеостаза путем контроля
над размножением клеток, характеризующихся рядом отклоне-
ний от нормы (опухолевые клетки).
Для оптимального развития иммунных реакций при действии
большинства антигенов необходимо участие макрофагов как в
первой индуктивной фазе иммунитета, когда они стимулируют
лимфоциты, так и в его конечной фазе (продуктивной), когда
они участвуют в выработке антител и разрушении антигена. Анти-
гены. фагоцитированные макрофагами, вызывают более сильный
иммунный ответ по сравнению с теми, которые не фагоцитированы
ими. Блокада макрофагов введением в организм животных взвеси
инертных частиц (например, туши) значительно ослабляет
иммунный ответ. Макрофаги способны фагоцитировать как раст-
воримые антигены (например, белки), так и корпускулярные.
КорпусKy^rpntJe_aHTWTVHU ~тллывакн более сильнпй~тгммунный
ответ Некоторые виды антигенов, например пневмококки, _со-
держащие на поверхности углеводный компоненту____могут быть
фагоцитированы после_Тфедварительндй опсонизации. Фагоцитоз
значительно облегчается», если антигенные детерминанты чужерод-
ных клеток опсонизированы, т. е. соединены с антителом или
комплексом “антитела-и комплемента. Процёсс~опсонйзадии-юбес-
печиваётся присутствием__на_мембране макрофага рецепторов,
которые связывают часть молекулы антитела (Fc-фрагмент) или
часть комплемента (F3). С мембраной макрофага у человека не-
посредственно могут связываться только антитела класса IgG,
когда они находятся в комбинации с соответствующим антигеном.
Иммуноглобулины класса М могут связываться с мембраной
макрофага в присутствии комплемента. Макрофаги способны
«распознавать» растворимые антигены, например гемоглобин.
В механизме распознавания антигена выделяют два этапа, тесно
связанных друг с другом. Первый этап заключается в фагоцитозе
и переваривании антигена. Во втором этапе в фаголизосомах
макрофага накапливаются полипептиды, растворимые антигены
(сывороточные альбумины) и корпускулярные бактериальные
антигены. В одних и тех же фаголизосомах может быть обнару-
жено несколько введенных антигенов. Изучение иммуногенности
различных субклеточных фракций выявило, что наиболее активное
антителообразование вызывает введение в организм лизосом.
Антиген обнаруживается также в мембранах клеток. Выделяемая
макрофагами большая часть переработанного материала антиге-
нов оказывает стимулирующее влияние на пролиферацию и диф-
ференцировку клонов Т- и В-лимфоцитов. Небольшое количество
248
антигенного материала может длительное время сохраняться в
макрофагах в виде химических соединений, состоящих не менее
чем из 5 пептидов (возможно, в связи с РНК).
В В-зонах лимфатических узлов и селезенки имеются спе-
циализированные макрофаги (дендритные клетки), на поведхнос-
ти многочисленных отростков которых сохраняются многие анти-
гены, попадающие в организм и передающиеся соответствующим
клонам В-лимфоцитов" В '('-зонах-лимфатических фолликулов
расположёны интердигитирующие клетки, влияющие на дифферен-
цировку клонов Т-лимфоцитов.
Таким образом, макрофаги принимают непосредственное ак-
тивное участие в кооперативном взаимодействии клеток (Т- и
В-лимфоцитов) в иммунных реакциях организма (см. рис. 91, Б).
Участие тканевых базофилов и эозинофилов в иммунных
реакциях
При первичном и особенно при повторном введении антигенов
наблюдаются увеличение числа и массовая дегрануляция тканевых
базофилов. Высказывается предположение, что дёгрануляция
обусловлена соединением антигена с антителами, фиксированными
на цитолемме. При этом выделяются содержащиеся^ гранулах
биологически активные вещества (гистамин, серотонин, гепарин),
которые могут оказывать неспецифическое стимулирующее влия-
ние на^ процёссй пролиферации и дифференцировки иммуноком-
петентных клеток 1- и В-лимфоцитов. Появление в тканях избыт-
ка гистамина приводит-к-увеличетптю числа эозинофилов, которые
участвуют в его разрушении. Введение в организм большинства
антигенов сопровождается увеличением числа эозинофилов в тка-
нях и регионарных лимфатических узлах. В ранней (индуктивной)
фазе иммунной реакции, когда происходит «распознавание анти-
гена», эозинофилы, как и тканевые базофилы, принимают участие
в активизации макрофагов (см. рис. 91, В). В продуктивной фазе
иммунитета (выработка антител) эозинофилы выполняют дезин-
токсикационную функцию, участвуя в фагоцитозе и разрушении
комплекса антиген — антитело.
МЕХАНИЗМЫ ИНТЕГРАЦИИ ЭЛЕМЕНТОВ ИММУННОЙ СИСТЕМЫ
Иммунная система функционирует как единое целое благодаря
наличию центральных нейрогуморальных и местных факторов,
регулирующих процессы пролиферации и дифференцировки
клеток, упорядоченную миграцию стволовых клеток и лимфоцитов,
осуществляемую через кровь и лимфу (рис. 94). В эмбриональный
период переключение кроветворения из желчного мешка в печень
и далее в костный мозг осуществляется посредством переселения
стволовых клеток через кровоток. В конце эмбрионального и в
первые дни постэмбрионального развития содержание стволовых
клеток в крови в десятки раз больше, челГ у взрослых. У взрослых
249
Рис. 94. Миграция и рециркуляция клеток в иммунной системе (схема).
СК — стврловая клетка; ЛТ — лимфоцит тимуса; Т — Т-лимфоцит; ЛБ — лимфоцит
бурсы; В — В-лимфоцит; ВБ — В-бласт; ТБ — Т-бласт; ВП — В-лимфоциты памяти;
ТП — Т-лимфоциты памяти; ТЭ, ВЭ эффекторные Т- и В-лимфоциты, ПК — плазмо-
цит; IgA; IgG; IgM, IgE; IgD - иммуноглобулины.
основным поставщиком стволовых клеток становится костный
мозг, при этом в кровь мигрирует за сутки около 2% всех стволо-
вых клеток костного мозга. При действии антигенов их число
увеличивается в Десятки раз. Предполагают, что процесс миграции
стволовых клеток из костного мозга и их рециркуляция находятся
под контролем гормонов гипофиза и надпочечников. Глюкокорти-
250
коидные гормоны коры надпочечников препятствуют избыточной
миграции стволовых клеток и тем самым предохраняют их от «пере-
расходования». Стволовые клетки, мигрирующие в тимус и неиз-
вестный у человека аналог фабрициевой сумки, подвергаются
антигеннезависимой пролиферации и дифференцировке в иммуно-
компетентные Т- и В-лимфоциты. Далее Т- и В-лимфоциты по-
ступают в кровь и заселяют Т- и В-зоны в лимфатических узлах,
селезенке. Отсюда образующиеся Т- и В-эффекторные клетки
(цитотоксические лимфоциты и плазмоциты), а также Т- и
В-клетки памяти снова поступают в кровь и лимфу, и, свободно
передвигаясь, как бы «патрулируют» организм. При повторной
встрече со «своим» антигеном они обеспечивают более сильный
и быстрый иммунный ответ организма.
Различают два типа миграции клеток иммунной системы:
медленный и быстрый. Продолжительность медленной миграции
измеряется неделями. Медленный тип миграции харак-
терен для стволовых клеток и Т- и В-лимфоцитов, заселяющих
периферические лимфоидные органы. Быстрый тип мигра-
ции (несколько часов) характерен для постоянно рециркулирую-
щих долгоживущих лимфоцитов памяти. Они мигрируют из крови
в периферические лимфоидные органы и из них обратно в кровь.
Популяция быстро рециркулирующих лимфоцитов содержит
главным образом Т-лимфоциты. Большинство медленно мигрирую-
щих клеток относится к В-клеткам. При остром иммунном ответе
происходит миграция предшественников эффекторных цитотокси-
ческих лимфоцитов и плазмоцитов через кровь и лимфу во все
отделы иммунной системы и соединительную ткань, где обеспечи-
ваются условия для иммунного, ответа. Число рециркулирующих
лимфоцитов в крови человека составляет около 1О10.
Рециркулирующие лимфоциты представляют со-
бой основную часть популяции малых лимфоцитов, большинство
из которых являются долгоживущими Т-лимфоцитами; меньшее
число составляют В-лимфоциты.
Рециркулирующие лимфоциты имеют оптимальную возмож-
ность к встрече с любой чужеродной мишенью, которую они сами
непосредственно уничтожают, вырабатывая антитела.
Лимфоциты при помощи рецепторов осуществляют «проверку»
всех тканей организма, выявляя чужеродный материал. При попа-
дании антигена в периферические лимфоидные органы в зонах
его расположения концентрируются Т-лимфоциты. В этих же
зонах несколько позднее появляются В-лимфоциты. Происходит
взаимодействие макрофагов, Т- и В-лимфоцитов и кооперативное
распознавание антигена, приводящее к дифференцировке из В-
клеток памяти и плазмоцитов, продуцирующих антитела.
Лимфоциты памяти живут в течение 20 лет и более. Приобре-
тенное свойство памяти лимфоцитов наследуется.
Таким образом, процессы миграции и рециркуляции клеток
иммунной системы обеспечивают поддержание иммунного гомео-
стаза.
251
Глава X
МЫШЕЧНЫЕ ТКАНИ
Ведущая функция мышечных тканей (textus muscularis) —
обеспечение перемещения в пространстве организма в целом и его
частей. Это достигается .тем, что элементы этих тканей способны
изменять свою форму под влиянием пусковых импульсов — сокра-
щатьсяГСвойством изменения формы обладают клетки и других
тканей. Но в мышечных тканях эта способность становится глав-
ной их функцией. Мышечные сокращения всегда сопровождаются
изменениями мембранных потенциалов, регистрация которых слу-
жит важным средством современной диагностики.
ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА
И КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ
Строение элементов мышечных тканей: удлиненная форма,
продольное расположение миофибрилл, обеспечивающих сократи-
мость — соответствует их ведущей функции. Изменение формы
сократимых элементов мышечных тканей является следствием
взаимодействия молекул сократительных белков — актина и мио-
зина, котороё~оСущесгвляется с участием других белков и ионов
кальция. Гак как для сокращения требуется много энергии, струк-
турные элементы мышечных тканей богаты-митохондриями. В их
цитоплазме содержится также много гликогена и миоглобина.
Морфофункциональная классификация. Все мышечные'ткани
составляют морфофункциональную группу, а в зависимости от
структуры органелл сокращения ее. делят на две подгруппы.
Первая подгруппа — гладкие (неисчерченные) мьццечные^тка-
ни (textus muscularis nonstriatus) — характеризуется^гем, что нити
актина (актиновые миофиламенты) и _нити миозина (миозиновые
миофйламенты), присутствующие в цитоплазме гладких мышеч-
ных клеток —«миоцитов. формируют сократительн ыем иофибри л -
лы^не_имеющие поперечной исчерченности. При специальных окрас-
ках они видны й под световым микроскопом в виде продольных
равномерно окрашенных по всей длине (гладких) нитей.
Вторая подгруппа — поперечнополосатые (исчерченные) мы-
шечные ткани (textus muscularis striatus) —состоят из мышечных
элементов, постоянно имеющих в составе специальны)?'органелл
и актиновые, и миозиновые протофибриллы, формирующие мио-
фибриллы.__Взаимное—расположение актиновых и миозиновых
протофибридд—создает—поперечную исчерченность. Исчерченные
мышечные ткани сокращаются быстрее, чем гладкие.
ИСТОЧНИКИ РАЗВИТИЯ МЫШЕЧНЫХ ТКАНЕЙ
Известны 5 источников развития мышечных тканей, и соответ-
ственно последние тгодразделяются на Зттютосенетииеских типов,
м е з е н х имные, эпидермальн ы е, н е йра л ь н ы е,
252
цело мл ч е с к ие_и со м-а т.н ч е с к и е (миотомные). Первые
три типа <.>цюсл1ся_с1ЮД1руш1с гладких мышечных тканей, чет-
вертый и гтятый — к подгруппе поперечнополосатых тканей.
ГЛАДКИЕ МЫШЕЧНЫЕ ТКАНИ
Мышечная ткань мезенхимного происхождения
Гистогенез. Стволовые клетки этой ткани и клетки-предшест-
венники на этапах эмбрионального развития пока точно не отож-
дествлены. По-видимому, они родственны предшественникам
фибробластов соединительной ткани и располагаются, будучи уже
детерминированными, в составе мезенхимы. Подобно фиброблас-
там они синтезируют гликозаминогликаны и молекулы коллагена,
из которых уже вне клетки осуществляется сборка матрикса
базальной мембраны и волокон. У дефинитивных клеток (миоци-
тов) синтетическая способность снижена, но не исчезает пол-
остью.
Строение и функциональные особенности. Гладкий,,
представляет собой._вг.ре.теповинную клетку длиногГ20— 50(Г—мкм,
толщиной 5—8 мкм (рис. 95). Ядро палочковидной формы нахо-
дится_в_ее центральной части. Когда миоцит сокращается*. оно
изгибается и~даже~^акррШ~наетч.я. Органеллы общего значения,
среди которых много митохо*ндрий, сосредоточены чжоло-тюяюсов
ядра Тв эндоплазме). Комплекс Гольджи и эндоплазматическая
сеть, особенно гранулярная, развиты слабо, что свидетельствует
о малой интенсивности синтетических функций. Рибосомы в боль-
шинстве случаев расположены свободно. Цитолемма образует
многочисленные впячивания — пиноцитозные 'пузырьки и кавеолы.
Посредством их в цитоплазму^дшстаВдщш-ся. в частнрсти.~йоны
кальция^
Актиновые миофиламенты идут в цитоплазме преимущественно
продольно или под углом к длинной оси клетки, образуя трехмер-
ную сеть. Места прикрепления их. к. цито лемме. либа_друг к другу
выглядят- на электронных микрофотографиях как электронно-
плотные тельца. Они состоят из белка а-актинина. В тех плотных
тельцах, которые прикрепляются к плазмолемме, обнаружен вин-
кулин. Иногда они заметны как темные пятнышки и при светооп-
тическом изучении клеток. Миозиновые миофиламенты распола-
гаются в цитоплазме миоцита-продольно. При сокращении наблю-
дается перераспределение актиновых и миозиновых-нитей относи-
тельно друг друга. Благодаря межмолёкулярным воздействиям с
гиозином параллельные актиновые нити смещаются навстречу
друг другу7~энёргия тяги передается- на цйтолемму. и конфигура-
ция клетки изменяется.
Каждый миоцит окружен базальной мембраной. В ней есть от-
верстия^ области~Тоторых между-соседним и мйдиитамгГЪбразу-
ются щелевидные соединения (нексусы), обеспечивающие функ-
циональные взаимодействия миоцитов в ткани.. В базальную мем-
брану вплетены многочисленные ретикулярные фибриллы. На кон-
251
Рис. 95. Строение неисчерче иного
гладкого миоцита /схема/.
Л, В — при расслаблении; Б, Д — при
наибольшем сокращении; Г — при не-
полном сокращении. / - цитолемма;
2 плотные тельца; J — ядро; 4 — эн-
доплазма; 5 — сократительные комп-
лексы; 6 митохондрии; 7 — базаль-
ная мембрана; 8 — актиновые (тонкие)
миофиламенты; 9 - миозиновые (тол-
стые) миофиламенты. В, Г, Д — увели-
ченное изображение участка, обведен-
ного рамкой на фрагментах А и Ь.
цах клеток цитолемма обра-
зует узкие тонкие впячива-
ния, в которые заходят эти
фибриллы, так что на них
передается усилие, разви-
ваемое при сокращении.
Вокруг мышечных клеток
ретикулярные, эластические и
тонкие коллагеновые волокна
образуют трехмерную_сеть —
эндомизий, которая объеди-
няет__ соседние миоциты
(рис. 96).
Регенерация. Физиологи-
ческая регенерация гладкой
мышечной ткани проявляется
обычно в условиях повышен-
ных функциональных нагру-
зок преимущественно в фор-
ме компенсаторной гипертро-
фии. Наиболее отчетливо это
наблюдается в мышечной
оболочке матки во зремя бе-
ременности. В миоцитах ин-
тенсифицируются синтетиче-
ские процессы, активизиру-
ются ядра, увеличивается ко-
личество элементов эндо-
плазматической сети, коли-
чество рибосом и комплекс
Гольджи. Некоторые миоци-
ты начинают делиться. Не
исключена также возмож-
ность превращения миофибробластов в миоциты. При репаратив-
ной регенерации восстановление ткани возможно скорее всего
за счет этих же двух источников.
254
Рис. 96. Схема строения неначер-
ченной гладкой мышечной ткани на
светооптическом (А) и ультра микро
скопическом (Б) уровнях.
А: 1 — гладкий миоцит; 2 — ядро; 3 —
пучки миофиламентов; 4 — сарколемма;
5 — зндомизий; 6 — нерв; 7 — кровенос-
ный капилляр (по Крёлиигу и Грау).
Б: I — верете невидные гладкие миоци-
ты; 2 — цитоплазма миоцита; 3 — ядра
миоцитов; 4 — цитолемма; 5 - базаль-
ная мембрана; 6 — поверхностные пино-
цитозные пузырьки: 7 межклеточные
соединения; 8 — нервное окончание, 9
коллагеновые фибриллы; К) — микрофи
ламенты (по Р. Кристичу с измене
ниями).
6
Рис. 97. Миоэпителиальные клетки в концевом отделе слюнной железы
(рис. Г. С. Катинаса).
А — поперечный срез; Б — реконструкция. 1 — ядра миоэпителиоцитов, 2 отростки
миоэпителиоцитов; 3 — ядра секреторных эпителиоцитов, 4 — базальная мембрана.
Мышечная ткань эпидермального происхождения
Элементами этой ткани являются миоэпителиальные клетки,
развивающиеся из кожной эктодермы. Они располагаются в црто-
вых, молочных, слюнных и слезных железах, дифференцируясь
пДТОвременно с их секреторными эпителиальными клетками из
общих предшественников. Миоэпителиальные клетки непосредст-
венно прилежат к эпителиальным, так что от соединительной
ткани их отделяет общая базальная мембрана.
Миоэпителиальные клетки имеют звездчатую форму
и охватывают отростками концевые отделы и мелкие выводные
протоки желез; их называют также корзинчатыми (рис. 97).
В центральной части клетки располагаются ядро и органеллы
общего значения, а в отростках — сократительный аппарат, орга-
низованный так же, как в неисчерченных миоцитах мезенхимного
происхождения. Сокращаясь, отростки способствуют выведению
секрета железы. В ходе регенерации и миоэпителиальные, и секре-
торные эпителиальные клетки развиваются из общих малодиффе-
ренцированных клеток эпидермальной природы.
Мышечн^ ткань ы^ир н >оисхождения
Миоциты £т&/тка^л развеваются из клеток нейрального зачат-
ка в составе стенки глазного бокалаТПо строению это — неисчер-
ченные миоциты с соответственно организованным сократитель-
ным аппаратом. Они входят в состав двух мышц радужки глаза —
расширяющей и суживающей зрачок.
Гладкие мышечные ткани в составе органов
В составе органов гладкие (неисчерченные) миоциты объединя-
ются в пучки, между которыми располагаются тонкие прослойки
рыхлой волокнистой соединительной ткани (перимизий), а сово-
256
купность'пучков образует мышцу, которая окру.кена более толсты
ми прослойками волокнистой соединительной ткани (эпимизий).
В них проходят кровеносные сосуды, которые доставляют питание
гладкой мышце, и нервные волокна, оканчивающиеся не непосред-
ственно на миоцитах, а между ними. Поэтому после поступления
от нервных центров управляющих импульсов выделенные порции
медиатора диффузно распространяются между клетками и воз-
буждают сразу многие из них. Гладкие мышцы присутствуют в
стенках многих внутренних органов, образуя в них слои и оболоч-
ки (например, в органах дыхания, желудочно-кишечного тракта,
в кровеносных сосудах и др.).
ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЕЧНЫЕ ТКАНИ
Сердечная поперечнополосатая мышечная ткань
Гистогенез. Источники развития сердечной поперечнополоса-
той мышечное—скани (textus muse u laris striatus cardiacus) —
симметричные участки висцерального листка спланхнотома (цело-
мической выстилки) ншейной части тела зародыша, называемые
миоэпикардиальной пластинкой. Большинство ее клеток диффе
ренцируется в сердечные миоциты (кардиомиоциты), остальные —
в клетки мезотелия эпикарда (см. гл. XV). Возможно, что и те,
и другие имеют общие клетки-предшественники. Выходе гистоге-
неза дифференцируются несколько видов карлиомионитов; сокра-
тительные, проводящие,—переходные (промежуточные), .д-.дакже
секреторные- (см. гл. XV).
Строение сократительных кардиомиоцитов. Клетки имеют
удлиненную (100—150 мкм), близкую к цилиндрической форму.
Их концы_соединяются друг с'другом, так что цепочки кардио-
миоцитов составляют так называемые функциональные волскна
тол щи но и ~ ГО—207л км, а области контакта ~образуют',нГ'тг7?7?7Ч ные <
диски (см. ниже). Кардиомиоциты могут ветвиться и образуют
пространственную сеть (см. гл. XV). Их боковые поверхности по-
крытьц базальной мембраной, в которую снаружи вплетаются
тонкие р тикулярныеи коллагеновые, волокна.
Ядро (или ядра, если их два) овальной формы, располагается
в Антральной части клетки (рис. 98)7 Большинство ядер поли-
плоидно. У полюсов ядра в цитоплазме сосредоточены органеллы
общего значения, включая клеточный центр, комплекс Гольджи,
слаборазвитую гранулярную эндоплазматическую сеть, отдельные
лизосомы. Агранулярная эндоплазматическая сеть хорошо разви-
та. Она формирует субсарколемма льные цистерны, прилежащие
к Т7системам. а также анастомозирующие между собой трубочки
L-систем Здесь же имеются включения гликогена и липидов
Включения миоглобина распределены по цитоплазме равномерно.
Митохондрии образуют цепочки вцкруг специальных органелл
миофибрилл.
Миофибриллы построены из постоянно существующих упоря-
доченно^ расположенных нитей, актинии миозина — сократителг
9 гистология
Рис. °S. Сердечные миоциты (кардиомиоциты) (рис. Г. С. Катинаса).
! хема ультрамикрпскопического строения трех видов кардиомиоцитов: проводящих
( промежуточных (/>), рабочих (В) I бз зальная мембрана; 2 ядра клеток: 3
миофибриллы- 4 — соединение рабочих кардиомиоцитов (вставочный диск) 5 - соеди-
нения промежуточного кардиомиоцита с рабочим и проводящим кардиомиоци >ами
О соединения проводящих кардиомиоцитов; 7 — поперечная трубочка (органеллы
оощегт значения не показаны). II — схема сокращения саркомера: А расслаб-
ленно. ...стояние; Б состояние сокращения: 1 - юлофрагма (Z-линия); 2 — тонкие
(актиновые) миофитаменты: 3 - толстые (миозиновые) миофиламенты
пых—бедков. Для их закрепления служат особые структуры —
телофрагмы и_л«уЗ/2Ф.0&з\КЫ, построенные из других'бёлков. Тело- z
фратмы-представляют собой сети ич белковых молекул, натянутые
поперек—клетки—и—прикрепленные—к читпцрммр. На протольном
срезе —кардиомиоцита они выглядят линиями толщиной около
100 нм, получившими название__Z чиций. Участок миофибпиплы
между двумя_телафрагмами называется ^аркомчрйм 7см рис. 98).
По середине его располагается прополfa-
ном cneael- От мезофрагмы гу сторону тепофрп^мы отхотят ыити
миозина, а от телофрагмы навстречу им — ни,ти-_актипа- Они
встречаются и на некотором расстоянии идут параллельно, -причем
кажди.4. толстыи~Дмиотииовый4-филамент-еопров<Уждает^и б тон-
кими (актиновыми) миофиламентами.
Ha ju?£3£_ саркомера участок, занятый М-линией и прилежащи-
ми зонами, в которых расползаются только миозиновые нити,
носит название Н полосы (светлой зоны), а участок, в котором
располагаются нити миозина и частййн<Г~актйна, — А-полосы
(А-дискаК~ Участки ~ двух соседних саркомеров, разделенные
Z-линией, которые содержат только нити- актина, составляют вмес-
те / полосу (I-<)uckJ. Названия__поло£_возникли -потому, что из-за
различной молекулярной организации в области 1-полосы прелом-
ление лучей .поляризованного света идатропное, а в области
же А-полосы: — а н изот р о ли О е.___Между миофибриллами
располагаются митохондрии и агранулярная эндоплазматическая
сеть. Митохондрии, разделяющие миофибриллы, очень крупные и
образуют трехмерную сеть (см. рис. 99, ///) с более плотным
расположением в области 1-диска.
Ццдолемма- на уровне телофрагм образует глубокие каналооб-
разные впячивания, именуемые поперечны ми.^грубоика ми (tubulus
transversus) или Т-трубочками', в них заходит и базальная мембра-
на. Такая система имеет чрезвычайно важное функциональное
значение, обеспёчиваябыстрое проведение потенциала”действия
к каждой миофиЬрцдле. Канальцы агранулярной-эндоплазмати-
ческой сети в цитоплазме.между миофибриллами располагаются
преимущественно продольно,.анастомозируя друг с другом, дости-
гают Т-трубочеки_щд некотором расстоянии лдут параллельно
последним.
В области вставочных дисков разные участки концов кардио-
миоцитов оканчиваются не на одном уровне. Вы тупающая часть
однлго_кардиомиоцита вдвинута в углубленную часть последующе-
го. Контактирующие поверхности клеток образуют пальцевидные
соединения (интердигйтации), а также многочисленные песмосо-
мы. В них вплетаются актиновые нити ближайшего саркомера
миофибриллы. Боковые поверхности выступов (параллельные
длиннику клетки) тоже~~сбпрйкасаются ц__ объединяются много-
численными нексусами. В области интердигитаций и десмосом
кардиомиоциты прочно связываются один с другим, что обеспечи-
вает развитие единого усилия при сокращении многих соседних
клеток. В области нексусов осуществляются ионные и химические
взаимодействия, что, в частности, способствует синхронизации
сокращения кардиомиоцитов.
Гистофизиология сокращения. Когда кардиомиоцит расслаб
лен, в канальцах агранулярной эндоплазматической “сети аккуму-
лируются- И кальцию Иод влиянием потенциала~дёйствия,
который распространяется по щи гоЛёмМё й_ТГ-труббчкам7 ионы
калБциятзсвобождатОтся,““поступают к миофибриллам и иниции-
руют сократительныи~акт:~они взаимодействуют с регуляторными
белками =дроттониЯом и тропомиозином, подле пего актиновые
и миозиновые миофиламенты получают возможность взаимодей-
ствовать друг с другом специализированными боковыми цепочка-
ми и перемещаться друг другу' навстречу. Так как при этом концы
актиновых нитей приближаются к М-линии, Н-полоса становится
9’
I
-
3
т
I—
t
Jt
>-
Я.
У
э-
ит
ИЯ
го
ак
но
261
уже_,_в равной мере будет уже и 1-полоса в связи с тем, что концы
миозиновых нитей сближаются с телофрагмами. В, то же время
ширина A-полосы не меняется- В результате взаимного встречного
перемещения нитей актина и миозина телофрагмы сближаются,
а поскольку они прикреплены к цитолемме, укорачивается и весь
кардиомиоцит.
Особенности строения проводящих кардиомионитов. Клетки_
значительно крупнее рабочих кардиомиоцитов (длина около
J 00 мкм, а толщина около~50 мкм)', чти позволяет легко отличить
их на препаратах.
Цитоплазма содержит все органеллы общего значения; мито-
хондрии мелкие, распределены равномерно. Мирфибриллы немно-
гочисленны и лежат по периферии клетки. Цитолемма не образует
Т-систем. Проводящие кардиомиоциты соединяются в волокна
друг с другом не только концами, но и боковыми поверхностями.
Между— кардиомиоцитами формируются вставочные диски,
более просто устроенные, чем между сократительными кардиомио-
цитами. Интердигитации, десмосомы и нексусы в них обнаружива-
ются реже.
Основная функция этих кардиомиоцитов состоит в том, что
они воспринимают управляющие сигналы от пейсмекерных элемен-
тов и передают информацию к сократительным кардиомиоцитам.
Волокна, гси-тавленмые из проводящих кардиомиоцитов, ветвятся
между сократительными рабочими кардиомиоцитами и доставляют
им возбуждающий импульс.
Возможности регенерации сердечной мышечной ткани^ В дефи-
нитивном состоянии сердечная мышечная ткань не сохраняет ни
стволовых клеток, ни клеток-прелшёственников. поэтому^ если
кардиомиоциты гибнут (инфаркт) вследствие травмы или прекра-
щения поступления по кровеносным сосудам питательных веществ
и кислорода, то они не восстанавливаются.
Скелетная поперечнополосатая мышечная ткань
Гистогенез. Источником развития элементов скелетной _попе-
речнополосатой мышечной ткани (textus muscularis striatus sceleta-
lis) являются клетки миотомии Одни из них дифференцируются
на месте, другие же мигрируК5г~иТ~мйотомов в мезенхиму?~Они
уже детерминированы в направлении развития элементов мышеч-
ной ткани, хотя внешне не отличаются от других клеток мезенхи-
мы. Их дифференцировка продолжается в местах закладки буду-
щих мышц. Дифференциация клеток-предшественников сходна
как в области самих миотомов, так и после миграции. При этом
возникают две линии дифференцировки. Клетки одной из них
сливаются, образуя симпластические структуры — мышечные тру
бочки (миотубы). В них происходит дифференцировка специаль-
ных органелл — миофибрилл, которые сначала располагаются под
плазмолеммой, а затем заполняют большую часть миотубы. Ядра,
напротив, из центральных отделов смещаются к периферии, что
260
свидетельствует о возникновении дефинитивного состояния —
формировании миосимпласта. Клетки другой линии остаются
самостоятельными, дифференцируясь в миосателлитоциты.
Структурные элементы скелетной мышечной ткани (рис. 99,
/, II, Ill, IV; 100). Основным элементом является ,мь>-
шечцое волокно, образованное ми оси мп частом и миосатвлчитоци
тому Rnnnvun окружено сарколеммой—Опа образована—боддльирд
мембраной, в которую нипетепы рет-и-кулярные и тонкие коллаi ено-
вые волокна, продолжающиеся в окружающую соединительную
ткань,.и плазмолеммой симпласта.
Миосимпласты покрыты плазмолеммой. По ней распространяет-
ся мембранный потенциал действия, переходящий на-мембрану
Т-трубочек.~Под плазмолеммой ।
[giWlfOlBI
ядер может достигать в миосимпласте нескольких десятков тысяч.
Их форма вытянутая. 11оскольку~симпласт не "клетка, термин
«цитоплазма» не применяют, а говорят «саркоплазма» (от греч.
sarcos — мясо).
У полюсов ядер располагаются органеллы общего значения:
комплекс Гольджи, участки агранулярной эндоплазматической
сети, митохондрии. Гранулярная эндоплазматическая сеть развита
слабо, так как синтез белка в дефинитивном миосимпласте обеспе-
чивает лишь пластические потребности. Специальные органеллы
представлены миофибричламц. Вдоль миофиирилл и по ходу
телофрагм выявлены—бедки____(титин_и_небулин), отличающиеся
эластичностью.—С ними связывают остаточное напряжение при
расслаблении.
Механизм сокращения мыщечных волокон такой же, как и в
кардиомиоцитах (см. выше).
'Мирфибриллы лежат обособленно друг от друга, каждая из
них окружена вытянутыми продольно и анастомозируюши ми
между собой петлями специализированной агранулярной эндо-
плаз ма ти ческой сети (саркоплазматической еогц!_Между ми о-
фибриллами располагаются также многочисленные вытянутые
крупные митохондрии (см. рис. 99, III). На уровне телофрагм по
обе^стороны каждой из них в толщу миосимпласта проникают
Т.-трубочки. Они тоньше, чем у кардиомионитов, базальная мемб-
рана в них не проникает Рястом с ними участки саркопдазмати-
ческой сети_сд.и ва юте я пруг_.е другом и образуют конечные и и стер-
нь/ которые располагаются параллельно Т-трубочкам и сопро-
вождают каждую из них с двух сторон, так что образуют. т/?ддс)ы.
Функциональная роль саркоплазматической сети та же. что и у
с оотнетст ву ющихс тру к ту р.дкардиоми оци то в.
Большую роль в деятельности мышечных волокон играют вклто-
чения, в первую очередь миоглобина и гликогена. Гликоген служит
лсновным_источникоя энергии, необходимой как для совершения
мышечной работы, ТЭК И ДЛЯ поддержания т.еплояоее-6'апанса всего
оргашцма.
Миосателлитоциты__прилежат к поверхности сими лас та, та к
что их плазмолеммы соприкасаются. С одним симпластом связано
261
2
I
7 8 ю
is течй итог
Рис. 99. Скелетное мышеч-
ное волокно (миосим-
пласт).
/ — строение мышечного во-
локна на светооптическом
уровне. А — схема, Б — ми-
крофотография, / — плазмо-
лемма; 2 — саркоплазма;
J — ядро; 4 — миофибрил-
лы; 5 — анизотропный диск
(полоска А): 6 изотроп-
ный диск (полоска I); 7 -
телофрагма (линия Z); 8
светлая зона (полоса Н), в
середине которой проходит
меюфрагма (линия М); 9 —
миомер (саркомер); 10—
миосателлитоци г; / / — су-
хожильные волокна (по
А. Н. Студите кому).
значительное количество сателлитрцитов, Каждый миосателлито-
н.ит^-.од ноя дерна я клетка. Ядро мельче, чем ядро,.миосим11ласта,
и более округлое. Митохондрии и эндоплазматическая сеть распре-
делены в цитоплазме равномерно, комплекс Гольджи и клеточный
центр расположены рядом с ядром. Специальных органелл нет.
Миосателлитоциты — камбии nhHhie зплшшты скелетной мышечной
ткани.
Типы мышечных волокон. Разные мышечные волокна в силу
специфических фуйКтдеотгальтпях^СЙиггятаан^ условий их
деятельности обладают неодинаковой силой, скоростью и длитель-
ностью сокращения, а также утомляемостью;-й~в связи с этим
различаются по строению. На светооптическом уровне различают
красные мышечные волокна (I тип), белые мышечные волокна
(И тип) и Ъереходные .формы Каждый из этих типов волокон
характеризуется особенностями ультраструктуры и метаболизма.
Особенности ультраструктуры проявляются степенью развитости
саркоплазматической сети, протяженности контакта Т-трубочек с
этой сетью, структуры телофрагм, группировкой актиновых и
миозиновых нитей в миофибриллы, количеством митохондрий и
др. Различно в мышечных волокнах и содержание включений
262
Рис. 99 (продолжение)
11 — схема ультрамикроскопического
строения скелетного мышечного волок
на (по Р. Кристичу с изменением)
1 — саркомер; 2 — анизотропный
диск (полоска А); 2а изотропный
диск (полоска I); 3 — мезофра<ма (ли
ния М) в середине анизотропного
диска А; 4 телофрагма (линия Z)
в середине изотропного диска: 5 — ми-
тохондрии; 6 — саркоплазматический
ретикулум; 6а — конечная цистерна;
7 — поперечная трубочка; 8 триада;
9 cap*.лемма- III схема простран-
ственного расположения митохондрий
в мышечном волокне. Верхняя и ниж-
няя плоскости рисунка соответствуют
анизотропным дискам саркомеров (по
Л. В. Бакеевой, В. П. Скулачеву,
Ю. С. Ченцову). IV - соединения мы-
шечных волокон коллагеновыми фиб-
риллами. Сканирующая электронная
микрофотография X 2600 (препарат
Ю. А. Хорошкова). 1 - мышечные во
локна; 2 коллагеновые фибриллы
Рис. 100. Поверхностный участок мышечного волокна (миосимпласта) и миосател-
литоцита. Электронная микрофотография (препарат В. Л Горячкиной. С. Л. Куз-
нецова ).
1 базальная мембрана; 2 плазмолемма; 3 — ядро мышечного волокна; 4 — ядро
миосателлитоцита: 5 — миофибриллы; 6 — канальцы агранулярной эндоплазматической
(саркоплазматической) сети; 7 — митохондрии; 8 — гликоген.
миоглобина, гликогена, липидов. Неодинакова в них и активность
ферментов. На ее определении гистохимическими методами осно-
ваны способы выявления различных мышечных волокон (рис. 101).
Н а иболее распространено определен ие акдшшлсти аденозин-
трифосфатазы (АТФазы) миоТИТСг—и су к ци натдегидроге на з ы
-<С'Д1 ). 1'ак называемыё~вО2Юкна I типа содержат~АТФазУ медлен~
HOffrgfiHia. им ^присущи вьцююГя активность СД1 (фермент аэ-
робного окисления) и высокое солепжяцие миоглобина и гликоген
па Волокна Н типа додержат 4ТФазу. миозина быстрого типгц___
активн^т)7с'ДТг^ни^Г^ЮК^аклт0^1ИЙ гликогена ОольШе, мибглс>
бина меньше. По степени активности ДЫкательныхферкГеПтов
между крайними формами волокон встречаются переходные; по
этим свойствам они образуют как бы непрерывный спектр, наподо-
бие света, в котором есть волны с любой возможной длиной
между крайними значениями. В мышечной ткани такие волокна
располагаются мозаично (см. рис. 101).
Свойства мышечных волокон меняются при изменениях нагру-
зок (спортивных, профессиональных экстремальных воздейст-
виях). Эти изменения при восстановлении обычных условий
деятельности обратимы. При некоторых заболеваниях (мышечная
атрофия, дистрофия, последствия денервации и т. д.) мышечные
волокна с различными исходными свойствами изменяются неоди-
264
Рис. IОI. Активность сукцинатде-
гидро1еназы в мышечных волокнах
разного типа в скелетной мышечной
ткани (препарат И П Рехачевой).
/ высокая; 2 — средняя; .? - низкая,
наково, что позволяет уточ-
нять диагноз, для чего иссле-
дуют биоптаты скелетных
мышц.
Регенерация скелетной
мышечной ткани. В миосим-
пласте нет цитоцентров, его
ядра неспособны делиться.
Камбиальными _ элементами
служат миосателлитоциты.
П ри перерезке мышечного
волокна на некотором протя-
жении от места травмы возни-
кает полное разрушение' с а р-
крлеммы, саркоплазмы и м и о -
сателлитоцитов, за пределами
ж е этой зоны волокно сох р а -
н я.ет жизнеспособность. М а к -
рофаги фагоцитируют не к ро-
тизированные фрагменты. Восстановление структуры мышечных во-
локон осуществляется посредством двух механизмов: реактивных
изменений сохранившейся части миосимплаета и -размножения
миосателлитоцитов.
1Г~ц|Ипгимппасте благодаря активизации комплексов Гольджи
у си л и ваетс я сборка мембранных структур, и на поврежденны х
концах восстанавливается целость плазмолеммы, активизируется
синтез эндоплазматической сетью веществ, необходимых для
построения саркоплазмы. Концы миосимпластов утолщаются и
растут навстречу друг Другу; образуются так называемые мышеч
ные почки. Миосателлитоциты, сохранившиеся рядом_с^поврежде-
нием, делятся. Одни из них мигрируют к концам поврежденных
волокон и включаются в мышечные почки. Другие сливаются (так
же, "как миобласты в ходе гистогенеза) и образуют мыше.члые
трубочки, которые затем дифференцируются в миосимпласты.
Таким _рбразом^-ири—регенерации—восстанавливается не только
целость поврежденных мышечных волокон, но и возникают новые.
Регенерация тем интенсивнее, чем больше освобождается из-под
базалртои~мембранБГ~Мйосателлитоццтов. А. Н. Студитскии Пбка-
зал, что, если мышпу измельчить до кашеобразной массы, а эту
массу—поместить в фасциальное ложе, огупич тьлмгт, и neivHepa-
ция как.налгканевлм так и на органном уровне. При таком_пов-
ре ж де н и и ка ждое волокно распадается на множество отдел ьн ы х
фрагментов й~ соответственно в зоне повреждения содержится
очень много миосателлитоцитов — основных источников регене-
раций; '
Регенерация на тканевом уровне, вплоть до формирования
мышечных волокон, осуществляется благодаря внутритканевым
локальным и внешним (гормональным) регуляторным механизмам.
Если затем не произойдет, реиннервации волокон, восстановле-
ния на органном уровне не будет, если же возникает связь мышеч-
но'го волокна с терминальными ветвлениями отростков нервных
кл етокр~на ч и наетия согласованна я сократительная ~де я тел ь ность
многих мышечных волдкон'Тнесогласованные сокращения воз-
можны в симпластё сразу же после регенерации миофибрилл),
затем восстанавливается структура мышцы как органа. Следует
учитывать, что регенерация мышечной и соединительной тканей
осуществляется параллельно, но последняя протекает быстрее.
Территория между концами поврежденных мышечных волокон
заполняется соединительнотканным регенератом раньше, чем
сблизятся концы мышечных волокон, возникает рубец.
Мышца как орган
Между мышечными волокнами находятся тонкие прослойки
рыхлой волокнистой соединительной ткани — эндомизий. Его
ретикулярные и коллагеновые волокна переплетаются с волокнами
сарколеммы, что способствует объединению усилий при сокраще-
нии (см. рис. 99, IV). На конце каждого мышечного волокна
плазмолемма образует узкие глубокие впячивания, куда проникают
коллагеновые и ретикулярные волокна. Последние пронизывают
азальиую мембрану и образуют петлю, которая межмолекулярны-
ми связями крепится к плазмолемме как раз в том месте, где
изнутри с ней контактируют актиновые нити саркомеров. Поки-
нув базальную мембрану, ретикулярные волокна переплетают-
ся с коллагеновыми, последние в свою очередь переходят в су-
хожилия.
Мышечные волокна разных типов в определенных сочетаниях
группируются в пучки, между которыми располагаются более
толстые прослойки рыхлой волокнистой соединительной ткани —
перимизии. В нем содержатся также и эластические волокна.
Соединительная ткань, окружающая мышцу в целом, называется
эпимизием.
Васкуляризация. Артерии, вступающие в мышцу, ветвятся в
перимизии. Рядом с ними много тканевых базофилов, регулирую-
щих проницаемость сосудистой стенки. Капилляры располагаются
в эндомизии. Они идут большей частью вдоль мышечных волокон,
анастомозируя друг с другом. Венулы и вены лежат в перимизии
рядом с артериолами и артериями. Здесь же проходят и лимфа-
тические сосуды.
Иннервация. Нервы, вступающие в мышцу, содержат как
эфферентные < двигательные^—iuk—и афферентные (чувствитель-
ные) волокну. Отросток- :ервной клетки, -Врннееящнн эфферент-
Lgz
Рис. 102. Схема построения мышечных волокон
с ядерной цепочкой (Л) и ядерной сумкой (/5)
(рис. Г. С. Катинаса).
I — ядра; 2 — миофибриллы (органеллы общего
значения не показаны).
ный нервный импульс, проникает через
ба зальную мембрану и ветвится между
ней^ иплазмолеммой симпласта, участвуя
в образовании так_называемой двига-
тел!м6й,_лти.лш:арной. Пышки (более
детально ее строение рассмотрено в
гл. XIII). Нервный импульс освобождает
здесь химические вещества — медиато-
ры, которые вызывают возбуждение
(потенциал действия), распространяю-
щееся по плазмолеммесимпласта.
Итак, каждое мышечное волокно
иннервируется самостоятельно и окру-
жено сетью гемокапилляррв.^Этот ком п -
леке образует морфофункциональную
единицу скелетной, мышцы—=— мион;
иногда мионом называют само мышечное волокдо, jrro не соответ
ствует Международной гистологическом-номенклатуре.
Чувствительные нервные окончания располагаются не на рабо-
чих мышечных волокнах, а связаны со специализированными
мышечными волокнами в так называемых лишенных веретенах,
которые расположены~в перимизии (см. гл. XI).
Специализированные мышечные волокна веретен значительно
тоньше рабочих. Они подразделяются на два типа: волокна с
ядерной сумкой и волокна с ядерной цепочкой (рис. 102). В пер-
вых ядра симпласта образуют скопления в утолщенной средней
части волокна. В волокнах с ядерной цепочкой ядра симпласта
расположены также преимущественно в его средней части, но одно
за другим, при этом они лежат в центре волокна. Рядом со скопле-
нием ядер локализованы органеллы общего значения. Миофиб-
риллы находятся в концах волокна. Сарколемма волокна соеди-
няется с соединительнотканной капсулой нервно-мышечного вере-
тена.
На мышечных волокнах веретен, как и на рабочих, образуются
двигательные нервные окончания (моторные бляшки), так что
они сокращаются под влиянием управляющих нервных им-
пульсов.
Их сокращение не развивает большой силы и практически не
суммируется с усилием, развиваемым рабочими мышечными волок-
нами, а лишь натягивает капсулу изнутри. Подробнее строение
мышечных веретен и их роль в pel ул я ци и мышечной деятельности
описаны в следующей главе.
267
Глава XI
НЕРВНАЯ ТКАНЬ
Значение нервной ткани. -ворганизмещггределяеття’Основными
свойствами нервных клеток (нейронов, неиронитгнф воспринимать
раздражение, приходить в состояние возбуждения, вырабатывать
импульс и передавать его. Нервная ткань (textus nervosus) осуще-
ствляет регуляцию деятельности" тканей и органов, их взаимо-
связь и •связь "0 окружающей ~ средой.
Нервная ткань состоит из нейронов (neuronum),.выполняющих
специфическукт~функцию, и нейроглии (neuroglia), обеспечиваю-
щей существование и специфичсскую. функдию -1 leppi нях-хлеток и
осуществляющей опорную, трофическую, разграничительную, сек-
реторну юТГзащитну ю "функций.
•РАЗВИТИЕ НЕРВНОЙ ТКАНИ
Нервная ткань развивается из дорсального утолщения эктодер-
мы— нервной пластинки (рис. 103). Края пластинки утолщаются
и приподнимаются как нервные валики. Между ними образуется
нервный желобок. Нервные валики сближаются и сливаются. Тогда
нервная пластинка замыкается в нервную трубку и отделяется
от лежащей над ней эпидермальной эктодермы. Часть клеток
нервной пластинки не входит ни в состав эпидермальной экто-
дермы, ни в состав нервной трубки и располагается между ними
в виде рыхлого скопления клеток — нервного гребня. Клетки
гребня начинают мигрировать в латеральном и вентральном нап-
равлениях. Кдетки гребня головного отдела участвуют л формиро-
вании ядер черепных не; вов, вторым источником развития кото-
рых являются и нейральные плакоды (см. ниже). В туловищном
отделе клетки гребня распадаются на два потока клеток. Один
из них, поверхностный, распространяется между эктодермой и
мезодермой и дает начало пигментным клеткам кожи Другой
направляется~~вглубь и вентрально, проходя между сомитом и
нервной трубкой, а также между мезенхимными^ клетками, коТО-
рые выселяются из сомита. Из этих клеток формируются~Пейроны
спиналь ых ганглиев и ганглиев автономной нервной системы,
а также нейроглия — леммоциты.
Нейральными плакодами называются утолщения эктодермы по
бокам головы. У высших позвоночных они не очень четко отгра-
ничены, однако миграция клеток из этих утолщений и их участие
в формировании ганглиев V, VII, IX и X пар черепных нервов
доказаны экспериментально.
Нервная трубка на ранних стадиях эмбриогенеза представляет
собой многорядный нейроэпителий, представленный вентрикуляр-
ными или нейроэпителиальными клетками Вентрикулярные клетки
имеют цилиндрическую форму. Их апика.1 ьные отростки, грани-
чащие с полостью нервной трубки, соединены щелевыми контак-
тами. Базальные концы соприкасаются с субпиальной погранич-
268
Рис. 103. Схема формирования нервной трубки зародыша цыпленка (по А. Г. Кнор-
ре).
Л — стадия нервной пластинки; Б — замыкание нервной трубки; Б — обособление нерв-
ной трубки и ганглиозной пластинки от эктодермы; / — нервный желобок; 2 нервные
валики; 3 — кожная эктодерма; 4 — хорда; 5 — мезодерма: 6 ганглиозная пластинка:
7 — нервная трубка; 8 — мезенхима.
ной мембраной. Для вентрикулярных клеток характерно цикличес-
кое перемещение ядер: ядра премитотических клеток лежат глу-
д боко во время профазы приближаются к поверхности, кариокинез
\ совершается вблизи желудочковой поверхности, а ядра дочерних
клеток опять уходят вглубь. Пролиферативная активность вентри-
кулярных клеток снижается в процессе эмбрионального развития
и после рождения не наблюдается.
Морфологически сходные, вентрикулярные клетки неоднород-
ны по способности к дифференцировке в различные типы клеток зре-
лой нервной ткани. Часть из них дает начало нейроцитам, другая —
глиальным клеткам: эпендимоцитам астроцитам и олигодендрогли-
оцитам. В некоторых областях мозга, где гистогенез совершается
особенно интенсивно, вентрикулярные клетки утрачивают цилин-
269
дрическую форму и способность к перемещению ядер, но сохраня-
ют высокую пролиферативную активность. Эти клетки называются
субвентрикулярными и экстравентрикулярными нейрогерминатив-
ными (камбиальными) клетками. В дальнейшем они дают начало
некоторым типам нейроцитов и глиальным клеткам. Суб- и эк-
стравентрикулярные клетки существуют еще некоторое время
после рождения. Так, экстравентрикулярная камбиальная зона
мозжечка исчезает у человека к 20 мес постнатального онтогенеза.
В отличие от юных нейронов (нейройласащ), которые утрачи-
вают способность к делению сразу после начала mhiрации из
вентрикулярной, суб- и экстравентрикулярной герминативных зон
мозга, незрелые глиальные клетки (глиобласты) сохраняют высо-
кую пролиферативную активность после завершения процессов
миграции. Не утрачивают ее полностью и зрелые астроциты и
оли годендроглио ци ты.
По мере дифференцировки нейробласта изменяется субми-
кроскопическое строение его ядра и цитоплазмы. В ядре возни-
кают участки различной электронной плотности в виде зерен и
нитей. В цитоплазме выявляются в большом количестве канальцы
и цистерны эндоплазматической сети, уменьшается количество
свободных рибосом и полисом, хорошего развития достигает ком-
плекс Гольджи. Специфическим признаком нам-зыш-йон <iепиа-
лизации нервных клеток спечует считать появление « ы* и.итлплаз-
ме тонких фибрилл — пучков нейрофиламентов и микротрубочек.
Количество неирофиламентов в процессе специализации увеличи-
вается. Тело нейробласта постепенно приобретает грушевидную
форму, а от его заостренного конца начинает развиваться отро-
сток — нейрит. Позднее дифференцируются другие отростки —
дендриты. Нейробласты превращаются в зрелые нервные клетки —
нейроны. Между нейронами устанавливаются синаптические кон-
такты.
В отличие от клеток макроглии — эпендимоцитов, астроцитов
и олигодендроцитов, развивающихся из нейро эктодермы, микроглия
имеет иной источник развития (см. ниже).
НЕЙРОНЫ
Нейроны^ или нейрациты (neuronum, neurocytus), различных
отделов нервной системы значительно отличаются друг от~друга
по функциональному значению и морфологическим особенностям.
В зависимости от функции нейроны делят на_р-е ц&нторн ы е
(чувствительные, или афферентные), а с с о ц и а т-и-в-н-ы-е- и
э ф^фе к т о р н ы е_(эфферентные). Первые генерируютлервный
импульс под влиянием различных воздействий внешней-или внут-
ренней средыорганизма. Ассоциативные_(вставочные) нервные
клетки осуществляют различные связи между нейронами. Эффек-
торные нейр^ны^ередаюхлозбуждение на гкани рабочих орга-
нов, побуждая их к действию. Размер нейроцитов широко варьиру-
ет. Так, например, диаметр тела клеток зернистого слоя мозжечка
770
Рис. 104. Схема нейрона (по И. Ф Иванову).
/ - тело нейрона: 2 осевой цилиндр: 3 мие-
линовая оболочка в разрезе; 4 ядра нейролем-
моцнтов; 5 — миелиновый слои, 6 — насечка
миелина; 7 — узловой перехват нервного волокна;
8 иервное волокно, лишенное миелина; 9 —
нервно-мышечное (двигательное) окончание; 10 —
миелиновые нервные волокна, обработанные ос-
миевой кислотой.
Рис. 105. Схематическое изображение типов
нервных клеток (рис. Т. Н. Радости ной.
Л. С. Румянцевой).
А униполярный нейрон; Б — псе вдоу ни поляр-
ный нейрон; В — биполярный нейрон; Г — муль-
типолярный нейрон.
колеблется от 4 до 6 мкм, размер тела гигантских пирамидных
клеток коры больших полушарий головного мозга (клетки Беца)
достигает 130 мкм. Столь же разнообразна и специфична для раз-
личных отделов нервной системы форма нейронов (рис. 104).
Xарактерной чертой ддя-всех зрелых- ней врио в-являедса. наличие
у дргх отростков. Эти отростки обеспечивают проведение нервного
импульса по телу человека из одной его части в другук^_подчас
весьма удаленную, и поэтому длина их—колеблетея в больших
пределах — от нескольких микрометров до 1 —1,5 м.
271
По функциональному значению отросткинейроно дделятся на
два вида. Одни выполняют функцию отведения нервного импульса
обычно от тел нейронов (рис. 105) и шиываются аксонами
(от греч. axon — ось), или_/щЙ£дгалщ^_Нейрпт заканчивается
концевым аппаратом или на другом нейроне, или на тканях рабо-
чего органа — на мышцах, железах. Второй вид отростков нервных
клеток на^1ва^тся~~Э?н<)ригЗЛ/п~Тот греч. dendron—дерево)._В
большинстве случаев они сильно ветвятся, чем и определяется
и х на зван ие. Количество и длина- дендритов, характер их ветвле-
ния специфичны для различных типов нейроцитов. Так, дендриты
двигательных нейронов спинного мозга короткие и относительно
мало ветвящиеся. Обильные ветвления придают дендритам груше-
видных клеток^корьГТюзжечка характерный вид дерева е пышной
кроной. Дендриты пирамидных нейронов коры большого мозга
отходят от верхушки пирамиды и ее боковых поверхностей и также
имеют характерный вид. Дендриты клеток-зерен коры мозжечка
очень короткие и распадаются на концах на несколько коротких
терминалей. Дендриты проводят импульс к телу нейрона. По коли-
честву отростков нейроны делятся натри групцы^трптиолярные —
клетки с одним отростком, биполярные — клетки с двумя отро-
стками^ мультйполярные — клетки.—имеющие три и больше от-
ростков (см. рис. 105). Последняя группа наиболее_распростране-
на у. млекопитающих животных и человека._Из многих отростков
такого нейрона один представлен нейритом, тогда как все осталь
ные являются дендритами. Биполярные клетки имеют два отро-
стка^- нейрит и дендрит. Истинные биполярные клетки в теле
человека встречаются редко. К ним относятся часть клеток сет-
чатки глаза, спирального ганглия внутреннего уха и некоторые
другие. Однако по существу своего строения к биполярным клет-
кам должна быть отнесена большая группа афферентных, так на-
зываемых псевдоуниполярных нсдронон краниальных и спиналь-
ных нервных узлов. Псевдоуниполярными они называются потому,
что нейрит и дендрит этих клеток начинается ~с~общего выроста
тела, создающего впечатление одного отростка, с последующим
Т-образным делением его., Истинных униполярных клеток, т, е.
клеток с одним отростком — нейритом, в теле человека нет. Уни-
полярную форму Имеют лишь нейробласты.
Ядро нейрона. Нейроны человека в пода вл я юще м бол ьши нс тве
содержат одно ядро, расположенное в центре, реже — эксцентрич-
но. Двуядерные нейроны и тем более многоядерные встреча юте я
к райне редко. Исключение сос тавляют нейрон ы некоторых га н гл и -
ев вегетативной нервной системы, например в предстательной же-
лезе и шейке матки, где можно иногда наблюдать нейроны, содер-
жащие до 15 ядер.
Форма ядра нейронов округлая. В соответствии с высокой
активностью метаболизма нейроцитов хроматин в их ядрах дис-
пергирован. В ядре имеется 1, а иногда 2—3 крупных ядрышка.
Усиление функциональной активностй~нейронов обычно уопровож-
даетсд. увеличением объема и количества ядрышек.
Рис. 106. Схема ультраструктурной органтации нервной клетки коры головного
мозга позвоночных (по И Г. Павловой).
/ — плазмолемма; 2 — ядро: 3 — гранулярная эндоплазматическая сеть (хроматофиль-
ная субстанция); 4 — комплекс Гольджи; 5 — лизосомы; 6 — митохондрии; 7 — нейро-
филаменты; 8 — микротрубочки; 9 — дендрит; 10 аксодендритические синапсы;
// — аксосоматические синапсы.
Цитоплазма нейрона. В соответствии с высокой специфичностью
функциональной активности нейронов они имеют специализиро-
ванную плазмолемму, их цитоплазма богата органеллами. В цито-
плазме представпены эндоплазматическая Сеть. рибосомы, мито-
хондрии, комплекс Гольджи, клеточный центр, лизосомы, нейроту-
булы и неирофиламенты (рис. 106).
Плазмилемма нейронов, ьуоме функции, типичной для цито-
лемм ы любой клетки, характеризуется способностью проводить
возбуждение. Сущность этого процесса сводится j< быстрому, пере-
мещению локальной деполяризации плазмолеммы по ее дедридам
к перикариону и аксону. Деполяризация определяется проникнове-
273
нием через ппазмопямму и клетку иплов натрия (Na ), что ме
няет знак заряда внутренней поверхности мембран на положитель-
ный. _Это в свою очередь повышает проходимость ионов натрия
на смежном участке и выход ионов~калия TIC') на внешнюю пОверх-
ность мембраны в поляризованном участке, в котором при этом
восстанавливается исходный уровень рдзности^гютенциалов. Ско-
рость движения волны деполяризации поверхностей мембраны
определяет быстроту передачи нервного импульса.
При окрашивании нервной ткани анилиновыми краситеЛями
(тионин, толуидиновый синий, крезиловый фиолетовый и др.)
в цитоплазме ..нейронов выявляется в вине бязофипьных глыбок
и зерен различных размеров и формы хроматофильная субстанция
(substantia cpromatophilica). Базофильные, глыбки локализуются
в перикарионах и дендритах нейронов, но никогда не обнаружива-
ются~~в~нейритах и их конусовидных основаниях (segmentum ini-
tiale) (рис i07, А, Б). Базофильные глыбки цитоплазмы нейронов
характеризуются высоким содержанием. 'рйбо11укттелггрттгеипов.
Электронно-микроскопическими исследованиями показано, что_им
соответствуют участки цитоплазмы, содержащие скопления упло-
щенных цистерн гранулярной эндоплазматической сети, располо-
женных~~параллелвно друг~~другу7 Степень ориентации цистерн в
нейронах различных типов неодинакова. Максимально упорядочен-
но они располагаются в нейронах спинного мозга.,В целом, эндо-
плазматическая сеть в нервных клетках — структура подвижная,
меняющаяся в соответствии с фуНкциональным~сбстоянием клетки.
Так, в моторных клетках спинною мозга глыбки' хроматофиль-
ной с£б танцйтфупные—неправильной у зватой формы, распо-
ложены они наиболее плотно~~вокруг ядраУ Ближе х__периферии
тела клетки_И_&-Д£НДРИ1ах_онгуо6ычно~мельче^ несколько вытянуты
в длин ' и лежат реже. В чувствительных нейронах спинальных
ганглиев глыбки имеют вид мелкой пылевидной зернистости.' Хро-
матофйльное вещество в глыбках большинства узлов вегетативной
нервной системы представлено мелкими зернами, расположенными
в цитоплазме неравномерно, и образует сеточку (узлы пограничного
симпатического ствола, верхний шейный узел). В других ганг-
лиях хроматофильная субстанция состоит из грубых глыбок,
заполняющих все тело клетки (узлы солнечного сплетения, звезд-
чатый узел) и ее дендриты. Обилие |ранулярной эндоплазмати-
ческой сети в нейронах соответствует высокому уровню синтети-
ческих процессов в цитоплазме и, в частности, синтеза белков,
необходимых для поддержания массы их перикарионов и отрост-
ков. Для аксонов, не имеющих органелл, синтезирующих белок,
характерен постоянный ток цитоплазмы от перйокариона к тер-
миналям со скоростью 1—3 мм в сутки. Это медленный ток,
несущий белки, в частности ферменты', нёобходимые~ДТЯ~-еинтеза
медиаторов в окончаниях аксонов. Кроме того, существует б ы -
с т р ы й ток (5—10 мм/ч), транспортирующий главным образом
компоненты, необходимые для синаптической______функции. Суще-
ствует также и ток белков, например. ацетТТпхппирдгтгрязы —
274
Рис. 107. Хроматофильная субстанция и нейрофибриллярный аппарат в нейронах.
А хроматофильная субстанция (субстанция Ниссля); окраска толуидиновым синим по
методу Ниссля; Б хроматофильная субстанция; окраска метиловым геленым-пиронипом
(по методу Браше). 1 — глыбки хроматофильной субстанции; 2 — аксональный холмик;
3 — аксон; 4 — дендриты; В нейрофибриллы, импрегнация нитратом серебра (препараты
Ю. И. Афанасьева, Е. Ф Котовского. Е. А. Хачатурян, Г. А. Косолапова!.
фермента, разрушающего нейромедиатор ацетилхолин, в направ-
леншГТёрминалеидендрйТОВ. Этот дендритн ы й т.р а_и спорт
совершается соткоросгью 3”мм/ч. Помимо тока веществ от перика-
риона 1Г терминалям аксонов и дендритов наблюдается-и обрат-
ный — ретроградный ток, посредством которого ряд
г.омпоненТов^цитоптгазмьивозвращается из окончании в-тела клет-
ки. ТГтра нс порте веществ по_отро£ткам нейроцитов участвуют
эндоплазматическая сеть, ограниченные мембраной пузырьки и
грануды, микротрубочки и актиномиозиновая система цитоскелета.
Комплекс Гольджи в нервных клетках при световой микро-
скопии виден как скопление различных по форме колечек, извитых
нитей, зернышек. Ультраструктура комплекса Гольджи обычна.
Клеточный центр чаще всего располагается между ядром и ден-
дритами. В нейробластах он находится со стороны растущего
отростка (аксона). Митохондрии рас положе ны_как_в теле нейрона,
так и во всех отростках. Особенно богата митохондриями цито-
плазма нейроцитов 5~концёвых аппаратах цтррстков, в частности
в области синапсов. Кристы в митохондриях нейроцитов обычно
слабо развйтьГгГ могут быть направлены продольно.
Нейрофибриллы (neurofibrilla). При импрегнации нервной тка-
ни серебром в цитоплазме нейронов выявляются нейрофибриллы,
образующие плотную сеть в перикарионе клетки и ориентирован-
ные параллельно в составе дендритов и нейритов, включая' их
тончайшие концевые ветвления (см. рис. 107, В). Методом элек-
тронной микроскопии установлено, что нейрофибриллам соответ-
ствуют пучки нейрофиламентов диаметром 6—10 нм й цейротубул
(нейротрубочек) диаметром 20—30 нм, расположенных в пери-
карионе и дендритах между хроматофильнымн глыбками и ориен-
тированных параллельно аксону.
?75
СЕКРЕТОРНЫЕ НЕЙРОНЫ
Способность синтезировать и секретировать биологически ак-
тивные вещества, в частности медиаторы, свойственна всем нейро-
цитам. Однако существуют нейроциты, специализированные
преимущественно для выполнения этой функции — секреторные ней
роны (neuronum secretorium), например клетки нейросекретор-
ных ядер гипоталамической области головного мозга. Секреторные
нейроны имеют ряд специфических морфологических признаков.
Этц^--крупные нейроны. Хроматофильная субстанция преимуще-
ственно располагается~по~~периферии тела кпеток К"цитоплазме
нейронов и в аксонах находятся различной величины гранулы
секрета"121 неиросекрСлЛ (subsiantia neurosecretorial, содержащие
белок, а В некоторых случаях липиды и „полисахариды. Транулы
нейросекрета выводятся в кровь или мозговую жидкость. Многие
секреторные нейроны имеют ядра неправильной формы, что сви-
детельствует об их высокой функциональной активности. Нейро-
секреты выполняют роль нейрорегуляторов, участвуя во~~~взаимо-
действии нервной и гуморальной систем интеграции.
НЕЙРОГЛИЯ
Неиршлия—(neuroglia)__выполняет в нервной ткани опорную,
ра згра н и чительную- трофическую, секреторную и защитную функ-
ции. Рее клетки нейроглии п.ч, wT.a. на различ-
ных вида: гаиоииты (н чк.,МН >1ИН > н иихрчгшп. Среди ГМИОПИТОВ
ра з л и ча ют эпендимоциты, астроциты и олигодендроци т ы.
Эпендимоциты (ependymocyti). Они образуют плотный слой
клеточных элементов., высти лающих ^спинном-оуовой, канал) и все
желудочки мозга. В npniie.ee гистогенеза нервной naw эпенди-
моциты дифференцируются первыми нзглиобчастав нервной труб-
ки (см. рис. 103) и выподняюх-в-этой стадии - развитиа_разгра-
ничительную и опорную функции., Вытянутые тела глиобластов
на внутренней поверхности нервной трубки образуют слой| эпите-
лиоподобны^клеток1 (рис. 108). На поверхности клетбк,"обращен-
Ю) в полобхьТюа'наДа нервной трубки, дифференцируютсТГреснич
ки£ которые, очевидно, своим мерцапиемтпособствуют движению
цереброспинальной жидкости. Базальные"—концы эпендимоцитов
снабжены длинными отростками, которые, разветвляясь, перёсека
ют всю нервную трубку^ образуя есттаддерживакиций аппарат, Эти
отросТкй, достигая внешней поверхности нерЪой~труб1Ш;~принимают
участие образований поверхностной глиальной пограничной
мембраны (fnembrana tirriitans gTiae superficial^), „отделяющей
вещество трубки от других тканей. В постэмбриональном перио-
де развития реснички в эпендимоцитах постепенно утрачиваются
и сохраняются лишь в некоторых местах ~ценграттБной— нервной
системы, например в водопроводе среднего мозга.
Не которые эпендимоциты выполняют секреторную фу н к ци ю,
выделяя различные активные вещества прямо в полость мозговых
желудочков или кровь Например, в области задней комиссуры
276
головного мозга эпендимоциты образуют особый «субкомиссураль-
ный орган», выделяющий секрет, возможно, участвующий в регуля-
(ции водного обмена.
Эпендимоциты, покрывающие сосудистые сплетения желудоч-
ков мозга, кубической формы. У новорожденных они имеют на
своей поверхности реснички, которые позднее редуцируются. На
базальном полюсе цитоплазма образует многочисленные и глубо-
ие складки. Цитоплазма содержит крупные митохондрии и раз-
личные включения, жир, пигмент и др. При введении в кровяное
русло животного трипанового синего она накапливается в цито-
плазме этих клеток, что подтверждает мнение об активном участии
эпендимоцитов в процессах образования цереброспинальной жид-
кости и регуляции ее состава.
Астроциты (astrocyti). Они образуют опорный аппарат цент-
ральной^ нервной системы. Э^о мепкие кпетхи с многочисленными
расходящимися во все стороны отростками. Различают два вида
астроцитов: протоп —и долокнцс^ые. Между ними
имеются и переходные формы. Протоплазматические-астроциты
(astrocyti protoplasmatici) нежат преимушественно/н сером веще-
s-rG 277
стце центральной нервной системы—Олт-характеризуются нали-
чием крупного ^крухзшго^светлот -ядра- и—множеством сильно
разветвленных коротких отростков. Цитоплазма содержи! неболь-
шое количество цистерн эндоплазматической сети, свободных
рибосом и микротрубочек, богата митохондриями. Протоплазмати-
ческие астроциты несут разграничительную и трофическую функ-
ции. ' "
~*~ Волокнистые астроциты (astrocyti fibrosi) располагаются глав-
ным образом в белом веществе мозга. Эги клетки -имеют-2б— 40
гладкоконтуриро ванных, длинных, слабовегвяшихс.я отростков,
которые формирую! i лиальнБГё~волокна7 образующие а_совоку п-
ностибтлотную сеть — поддерживающий аппарат мозга. Отростки
астроцитов на кровеносных сосудах и на поверхности мозга свои-
ми концевыми расширениями фбрмИрую’Г~и?р>п^гГ-у глиа-
льные пограничные мембраны (membrana limitans gliae perivascu-
laris). При электронно-микроскопическом исследовании фиброз-
ные астроциты характеризуются светлой цитоплазмой. Их ядерная
оболочка образует иногда глубокие складки, а кариоплазма харак-
теризуется равномерной электронной плотностью. В цитоплазме
имеются многочисленные фибриллы диаметром 8—9 нм, выходя-
щие пучками в отростки.
Основная функция астроцитов— опорная и и <оляция нейронов
от внешних влияцищ-что необходимо-для осуществления специфи-
ческой деятельности нейронов.
Олигодендроциты (oligodendrocyti). Это самая многочисленная
группа jcneTOK нейроглии (см. рис. 108). ОлигодендроцитыГокружа
ют тела_нейронов-в-неит-рапьной и перферической нервной системе,
находятся,в составе оболочек нервных волокон и в нервных окон-
чаниях. В разных отделах нервной системы олигодендроциты
имеют различную форму. В сером веществе мозга они небольшого
размера; От тед клеток, имеющиурнальную или угловатую форму,
отходиз несколько коротких и слаборазвёт вленньГХ отростков.
Особенности Строения олигодендроцитов в составе перифериче-
ских нервных узлов, волокон и окончаний будут описаны в со-
ответствующих разделах.
Изучение методом электронной микроскопии показало, что по
плотности цитоплазмы клетки олигодендроглии приближаются к
нервным и отличаются от них тем, что не содержат нейрофил''-
ментов. Функциональное значение этих клеток очень разнообраз-
но. Они_ выполняют трофическую функцию, принимая участие
в обмене веществ нервных клетбк';~ОлИ1'0Дендр01ГИТЬ1йграют зна-
чительную роль щобразевйпии~ттбодочек вокруг отрбст’ков клсток,
при этом они называютсгГнейролеммоцитами (леммоцитьГ^~шван-
новские клетки). В .процессе пегенерапий~и регенерации нервных
волокон этим клеткам также принадлежит важная роль.
Микроглия (microglia). Вопрос o’ происхождении микроглии,
так же как и о ее макрофагальной природе, в настоящее время
дискутируется. Согласно одной из гипотез, клетки микроглии
являются глиальными макрофагами и происходят от_ промоноци-
тов костного мозуа, Клетки микроглии небольших размеров,
преимущественно отростчатой формы, способны, к амебоидным
движениям. Два^три более крупных отростка имеют на своей
поверхности короткие вторичные и третичные ветвления. Ядра
клеток вытянутой или треугольной формы, богаты хроматином
(см. рис. 108). При раздражении клеток микроглии их форма
меняется, отростки втягиваются, клетки -Приобретают рпецифи-
ческий характер^ округляются. В таком виде они называются
зернйстым1Гиларами.
НЕРВНЫЕ ВОЛОКНА
Отростки нервных клеток, обычно покрытые оболочками, назы-
ваются нервными волокнами (neurofibra). В различных отделах
нервной системы оболочки нервных волокон значитёльно~0тлича-
ются друг от~друга пб С ное му строению, поэтому в соответствии
с особенностямтг-их—етроения—все нервные волокна делятся на
две основные""группы — миелиновые и безмиелиновые (рис. 109,
А, Б). Те и другие состоят из отростка нервной клетки, который
лежит в центре~волокна ~и~поэтОму~Знадываётся осевым цйливдром
(cylindraxis), и оболочкщ оЕразованной клетками олигодендроглии,
которые здесь называются нейролеммоцитами (шванновские
клетки).
Безмиелиновые нервные волокна
Безмиелиновые нервные волокна (neurofibra amyelinata) на-
ходятся^-прёим^щёСТвеиий.. В. составе- вегетативной нервной систе-
мой Клетки олигодендроглии оболочек безмиелиновых нервных
волокон, располагаясГ'ПЛбтно,"образуют"тяжщ в которых на опре-
деленном расстоянии друг от друга видны овальные ядра. В нерв-
ных волокнах внутрёшшх~брганов, как правило, в таком тяже
располагается не один, а несколько (10—20) осевых цилиндров,
принадлежащйх~различнБгм“ нейронам. Они могут, покидая одно
волокно, переходить в смежное. Такие волокна, содержащие не-
сколько осевых цилиндров, называются волокнами радельного
типа. При электронной ~мйкроскопии безмиелиновых нервных во-
локон видно, что по мере погружения осевых цилиндров в тяж
леммоцитов последние одевают мХ как муфтой. Оболочки леммопи-
тов при этом прогибаются, плотно охватывают осевые цилиндры
и, смыкаясь над ними, образуют глубокие гидаДКгГ-МЩДнепкото-
рых и располагаются отдельные осевые цилиндры,—Сближенные в
области складки участки оболочки _нейролеммоцита образуют
(Двоенную мембрану — мезаксон, на которой как бы подвешен
осевой цилиндр "Тем. рис. 109, Б). Оболочки чейролеммоцитов
очень тонкие, поэтому нц._ме.заксона, ни—границ этих клеток под
световым 'микроскопом нельзя рассмотреть, и оболочка безмие-
линрвьпГ нервных волокон в этих условиях выякпяется как одно-
родный~ТЯЖ ЦИ1Ч)П; азмьь_<юдевающий1>. осевые цилиндры С повер-
хности каждое нервное волокно покпыто базальной мембраной.
279
Рис. 109. Схема строения нервных волокон на светооптическом (Л, Б) и ультра-
микроскопическом (а, 6} уровнях (рис. Т. Н. Радости ной, Ю. И. Афанасьева,
JI. С. Румянцевой).
А, а миелиновое волокно; Б, б — безмиелиновое волокно. / осевой цилиндр; 2 — миели-
новый слой; 3 соединительная ткань; 4 — насечка миелина; 5 ядро нейролеммо-
цита; 6 — узловой перехват; 7 - микротрубочки; 8 - неирофиламенты; 9 — митохондрии:
10 - мезаксон; // базальная мембрана.
Миелиновые нервные волокна
Миелиновые нервные волокна (neurofibra myelinata) __встре-
чаются как в центральной, так и в периферической нервной систе-
ме. Они значительно толще безмиелиновых нервных .валикон.
Диаметр поперечного сечения их колеблется от 1 до 20 мкм. Они
также___СОСТОЯТ__из_осевого.—цилиндра, «одетого»___оболочкой из
нейролеммоцитов, но диаметр осевых цилиндров этого типа воло-
кон значительно толще 7ГтФггдпхгка=*^*жнее-, В-ет
гптйанном
миелиновом вбл.окне принято различать лна слоя оболочки: внут-
ренний,__£шюе—тпистыи, — миелиновый слой (stratum myelim)
(см. рис. 109, Л) и наружный, тонкий, состоящий из цитоплазмы
и ядер нейролеммоцитов — нейролемму (neurolemma
280
Рис. 110. Схема развития
миелинового волокна (со-
ставлена Т. Н. Радости ной
по схеме Робертсона).
А — поперечные срезы по-
следовательных стадий раз-
вития (по Робертсону); Б —
трехмерное изображение
сформированного волокна
/ дубликапия оболочки
нейролеммоцита (мезаксон);
2 — аксон; 3 — насечки
миелина; 4 — пальцевидные
контакты нейролеммоцита в
области перехвата; 5 — ци-
топлазма нейролеммоцита;
6 — спирально закрученный
мезаксон (миелин); 7 — яд-
ро нейролеммоцита.
Миелиновый слой содержит в своем составе липиды, а поэтому
при обработке волокна осмиевой кислотой он интенсивно окра-
шивается в темно-коричневый цвет. Все волокно в этом случае
представляется однородным цилиндром, в котором на определен-
ном расстоянии друг от друга располагаются светлые линии — на-
сечки миелина (incisura myelini). Чере < некоторые интервалы (от
нескольких микрометров до нескольких миллиметров) встречаются
участки волокна, лишенные миелинового слоя, — узловые перехва-
ты (nodus neurofibrae). Перехваты соответствуют границе смежных
ней роле м мо ци то в. Отрезок волокна, заключенный, между см е ж н ы м и
перехватами, называется межузловым сегментам, а_еса_иболочка
представлена одной глиальной клеткой.
В процессе развития миелинового волокна осевой цилиндр,
погружаясь в нейролеммоцит, прогибает его оболочку, образуя глу-
бокую складку, при этом формируется мезаксон (рис. 110, А, Б).
При дальнейшем развитии мезаксон удлиняется, концентрически
наслаивается на осевой цилиндр и образует вокруг-него плотную
слоистую~зону — миелиновый спой (см. рис. 109, ПО). Под эле-
ктронным "микроскопом каждый завиток мезаксона виден как
светлый слой, имеющий ширину около 8—12 нм и'соответствую-
щий липидным слоям двух листков плазмолеммы нейролеммоцита.
По середине и по поверхности его видны тонкие темные линии,
образованные молекулами белка.
281
У '
Наружным слоем (не Про лемма) называется периферическая
зона нервного волокна,, содержащая оттесненную сюда цитонлаз-
му нейролеммоцитов (шванновских клеток) и их ядра. Эта зона
при обработке волокна осмиевой кислотой остается светлой.
В области насечек между завитками мезаксона имеются зна-
чительные прослойки цитоплазмы, а соответствующие клетс 'ые
мембраны здесь находятся на большом расстоянии (см. рис 09,
Л). При осмировании волокна эти участки не окрашиваются.
На продольном сечении вблизи перехвата видна облает в
которой завитки мезаксона последовательно контактируют с ?се-
вым цилиндром. Места прикрепления самых глубоких завг "ков
его наиболее удалены от перехвата, а все последующие завитки
закономерно расположены ближе к нему. Это легко понять, если
представить себе, что наслоение мезаксона идет в процессе роста
осевого цилиндра и «одевающих» его нейролеммоцитов. Естест-
венно, что первые слои мезаксона оказываются короче, чем по-
следние. Края двух смежных леммоцитов в области перехв? а
образуют пальцеобразные отростки, диаметр которых равен 50 1 и.
Длина отростков различна. Снаружи миелиновое нервное волок но
покрыто базальной мембраной, связанной с плотными тяжами
коллагеновых фибрилл, ориентированных продольно и не преры-
вающихся в перехвате.
Осевой цилиндр нервных волокон состоит из неиррплазмы
цитоплазмы нервной клетки, содержащей прлцппьно ориентиров?ч-
ные_ нейрофиламенты и нейротубулы. В нейроплазме осе во э
цилиндра лежат.МИТОхондрии, которых.Дольше в- неносредственнсй
близости к перехватам—и особенно -много-в -концевых аппарат, .<
волокна.
С поверхности осевой цилиндр покрыт мемЬраной — аксолем-
мой„ обесдедивающей проведение 11ервного4змдульС^~Скорость пе-
редачи импульса миелиновыми волокнами больнку-чём-бёзкгиели-
новыыц,_ Тонкие волокна, бедны»—м-иелином.
клиновые
как толстые миелиновые — со скорость
В безмиелиновом волокне--волна' "депгтярпзлции мембраны
идет по_тееи_плйЯоД£ММз:<_лце црерываясь*__а в миелиновом
волокне возникает только в области перехвата. Таким образом,
для миелиновых--волокон—характерно—сальтаторноё Проведение
возбуждения, т. е. прыжками. Между перехватами по аксолемм^
идет электрический ток, скорость которого выше, чем прохожде-
ние волны деполяризации.
РЕГЕНЕРАЦИЯ НЕЙРОНОВ И НЕРВНЫХ ВОЛОКОН
Нейроны являются несменяемой клеточной популяцией. Им
свойственна только внутриклеточная физиологическая регенера-
ция, заключающаяся в непрерывной смене структурных белков
цитоплазмы.
Отростки нейронов и соответственно периферические нервы
282
обладают способностью к ре генерации в случае их поврежде-
ния При этом регенерации нервных волокон предшествуют явле-
ния дегенерации. Нейролеммоциты периферического отрезка волок-
на уже в первые сутки резко активизируются. В цитоплазме нейро-
лем лОцитов увеличивается количество свободных рибосом и поли-
сог г эндоплазматической сети. В цитоплазме нейролеммоцитов
об(1 зуется значительное количество шарообразных слоистых струк-
тур различных размеров. Миелиновый слой как обособленная зона
ней юлеммоцита исчезает. В течение 3—4 сут нейролеммоциты
значительно увеличиваются в объеме. Нейролеммоциты интенсивно
размножаются. К концу 2-й недели миелин и частицы осевых ци-
линдров рассасываются. В резорбции продуктов принимают
участие как глиальные элементы, так и макрофаги соединительной
ткани.
Осевые цилиндры волокон центрального отрезка образуют
на к >нцах булавовидные расширения — колбы роста и врастают в
лентовидно расположенные нейролеммоциты периферического от-
ре жа нерва и растут со скоростью 1—4 мм в сутки. Рост нервных
волокон замедляется в области терминалей. Позднее происходит
миелинизация нервных волокон и восстановление терминальных
структур.
НЕРВНЫЕ ОКОНЧАНИЯ
Все нервные волокна заканчиваются концевыми аппаратами,
которые получили название нервных окончаний (terminationes пег-
vofum). По-функциональному значению нервные окончания мож-
но разделить на три ipyrfnbiT £или._эффщ<торы).
рецепторные (аффекторные или чубстецг£^дшые)_и. концевые ап-
параты, образующие___межнейронные___синапсы, осуществляющие
связь нейронов между собой.
Эффекторные нервные окончания
Они бывают двух типов — двигательные и секреторные. Д в и-
г а т е л ь н ы е—н-е-рв-тг ьге" “о к о н ч а н и я — это концевые ап-
параты нейритов двигательны5Г~клетик~соматической или вегета-
тивной нервной системы. ПрйТТх~~участии нервный импудьс_пере-
даеГтся на ткани рабочих органов. Двигатедьные_3?кончания в
поперечнополосатых мышцах называются нервно-мышечными
окончаниями (terrhTnatio neurorrniscuraris). Они представляют со-
бой окончания нейритов клеток двигательных ядер передних рогов
спинного—мезга—или моторных—ядер головного моТТа.’ Нервно-
мышечное 'окончание состоит из концевого ветвления осевого
цилиндра нервного волокна и специализированного участка мы-
шечного волокна (рис. 111). .Миелиновое нервное волокно, подой-
дя к мышечному волокну, теряет миелиновый слой и прогружа-
ется в мышецное волокно, вовлекая—за Дщбош1его~Ш1азмолемму.
Соединительнотканные элементы при этом переходят в наружный
слой оболочки мышечного волокна. Плазмолеммы терминальных
2X3
Рис. 111. Схема ультрамикроскопического строения нервно-мышечного окончания.
/ — цитоплазма нейролеммоцита 2 — ядро нейролеммоцита; 3 — плазмолемма нейро-
леммоцита; 4 осевой цилиндр нервного волокна; 5 — аксолемма; 6 постсинапатиче-
ская мембрана (сарколемма); 7 — митохондрии в аксоплазме; 8 — синаптическая
щель; 9 митохондрии мышечного волокна; /О пресинаптические пузырьки; // пре-
синаптическая мембрана (аксолемма); /2 — сарколемма; 13 ядро мышечного волокна;
14 — миофибрилла.
ветвей аксона и мышечного волокна разделены синаптической
щелбЮ шириной около 50 нм. Кроме того, мем рана мышечного
волокна сама образует—многочисленные.__складки, формирующие
вторичные синаптические щели эффекторного окончания.
В области окончания мышечное волокно не имеет типичной по-
перечной йсчерченности и характеризуется обилием митохондрий,
скоплением круглых или слегка овальных ядер. Саркоплазма с
митохондриями и ядрами в совокупности образует постсинапти-
ческую часть синапса. ---------
Терминальные ветви нервного волокна в мионевральном си-
на псе Jxapaктеризудшл о им. шс м митохшлчрйй и унлтчисле н н ы м и
пресинаптическими пузырьками, содержащими характерный для
этого вида окончаний медиатор-----ацётйлхоли'п. 11ри—возбужде-
нии ацетилхолин поступает через пресинаптниеекую--мембрану в
синаптическую щель на холинопеиепторы постсинаптической (мы-
шечной) мембраны^ вызывая ее. возбуждение (волну деполяри-
зации).
Постсинаптическая мембрана моторного нервного оконча-
ни я содержит фермент ацетилхолинэстеразу, разруша ющий
медиатор и ограничивающий этим срок его действия. Двигатель-
ные тщрвные окончания в гладкой мышечной ткани построены
проще. Здесь тонкие пучки аксшчт и и их одиночные терминали,
следя между mhiiipiiuumb—клетками,—образуют четкообразные
расширения ^Хларикозы); содержащие -холинергические или адре-
нергические пресинаптические пузырьки.
С е к р еторные нервные окончани я имеют прос-
тое строение. Они п ре дета вл я юх.-сабой—концевые, утолщения,
284
или четковидные расширения воддкда,_С_ синаптическими пузырь
камкС содержащими главным образом ацетилхолин.
Рецепторные (чувствительные) нервные окончания
Эти_нервые окончания — рецепторы — рассеяны по всему
организму и воспринимают различные раздражения как из_внеш-
ней среды, так и от внутренних органов. Соответственно выделяют
две большие группы рецепторов: экстеро рецепторы и интероре-
цепторы. В зависимости от специфичности раздражения, воспри-
нимаемого- да иным видом рецептора, все чувствительные о конча-
ния разделяют на механорецептлры, барорецепторы, хеморецеп-
торы, терморецепторы и др.
По особенностям строения чувствительные окончания подраз-
дели ются на свободные нервные окднчцндя (tefTninatio nervi
libera), т.е. состоящие только из конечных ветвлений осевого
цилиндра, и несвободные, содержащие в своем составе все компо-
ненты нервного волокна, а именно ветвления осевого цилиндра
и клетки гпич. Несвободные-,окончания, кроме того, могут быть
покрыты соепинительнот-ка-ннди капсулой,..JI тогда ОНИ называются
uHjiancyMipoeattHbtMU (corpusculum nervosum capsulatum). Несво-
бодные—нервные окончания, не имеющие соединительнотканной
капсулы, носят название неинкапсу лированных (corpusculum
nervosum noncapsulatum).
/) Для эпителия характерны свободные окончания. В этом случае
миелиновые нервные волокна подходят к эпителиальному пласту,
теряют миелин, а осевые цилиндры проникают в эпителии и рас-
падают£я._там „между. клетками на тонкие—терминальные ветви.
В многослойном эпителии есть окончания, в состав которых вхо-
дят7~кроме терминалей отростков нервных клеток^специфически
измененные эпителиальные клетки — осязательные эпителиоциты
(epitheliocytus tactus). От других клеток эпителия они отличают-
ся светлой цитоплазмой, наличием осмиофильных гранул-диамет-
ром 65—180 |1м и уплощенным темным ядром. Концевые нервные
веточки подходят к таким клеткам и расширяются, образуя диско-
видные концевые структуры, связанные с основаниями осязатель-
ных эпителиоцитов.
М Очень разнообразны рецепторы__в^соединительной ткани. Ог-
ромное большинство их представляет собой разной етепени-слож-
ности 'ветвления осевого цилиндра. В состав таких.—концевых
аппаратов, как правило, вх~одят нейролеммоциты, которые сопро-
вождают все ве'1ВЛения_долокна1„(рис. 112). Инкапсулированные
рецепторы соединительной ткани при всем их_разнообразии всегда
состоят пт ветвления—осевого цилиндра и глиальных клеток. Сна-
ружи такие рецепторы покрыты соединительнотканной капсулой.
Примером подобных окончаний могут служить весьма распрОстра-
ненные у человека пластинчатые тельца (corpusculum lamellosum).
В центре такогшельца—располагается внутреняяя луковица (bul-
bus internus], образованная видоизмененными леммоцитами.
?ks
Рис. 112. Рецепторные нервные окончания в соединительной ткани (по А. П. Мас-
лову).
Чувствительные миелиновые волокна около пластинчатого тельца
теряют миелин и проникают во внутреннюю колбу {рис. 113,
А). Снаружи тельце покрыто слоистой капсулой, образованной
фибробластами и спиоальцо ориентированными колла ге н о вы м и
волокнами..С функцией пластинчатых телец связывают восприятие
давления. Их много в глубоких слоях кожи и во всех внутрен-
нихорганах7 “
К чувствительным инкапсулированным окончаниям относятся
и осязательные тельца fcorpusculum tactus), находящиеся~~в со-
ставесосочков кожи. Они состоят из олигодендроглиоцитов, рас-
положе иных перпендикулярно к -Дл~и~нТ|7Г(Г'осй~тельпа. “Ми ел и новое
нервное волокно, проникая в тельце, лишается миелина и распада-
ется на~ несколько конечных ветвей, контактирующих с поверх-
ностью глиальных клеток. Соединительнотканная капсула тельца
очень тонкая и состоит преимущественно из коллагеновых волокон
(рис. 113,Б). ------------------
К рецепторам скелетных мышц и сухожилий лтчос^чч-я-ттервно-
мышечные веретена (fusus neu romuscu laris), регистрирующие из-
менения длины МЫ1пгчны2С^)лсыгш.-ц.-скорост:ь_этмх. изменений,
и нервно-сухожильные веретена (fusus neurotendineus), реагирую-
щие на напряже ние, прилагаемое к сухож илию при сокраще ни и
мышц. Веретено состоит из нескольких (до 10—12) ощ^их и
коротких поперечнополосатых мьпиечных волокон, заключенных
в растяжимую соединительнотканную капсулу. — и н т пафузальных
286
Рис. 113. Схема ультрамикроскопического строения капсулированных нервных
окончаний.
А — пластинчатое тельце: / — слоистая капсула; 2 — внутренняя луковица; 3 — дендрит
чувствительной нервной клетки: 4 — спиральные коллагеновые волокна; 5 — фиброциты;
6 — вторичночувствующие клетки с ресничками; 7 синаптические контакты аксонов
вторично чувствующих клеток с дендритами чувствительной нервном клетки (по
А А. Отелину, В. Р. Машанскому, А. С. Миркину). Б — осязательное тельце: / - капсула-
2 специальные клетки; 3 — нервные терминали; 4 — миелиновое нервное волокне
5 - опорные (поддерживающие) фибриллы; 6 эпителий (по Р. Кристичу с изменением).
волокон (от лат. fusus — веретено). Остагтьныеволшаш мышцы ле-
жат~зЗ пределами капсулы и называются ж^рафу-шляными (рис.
114,Л,£). И нтрафу зальные волокна имеют актиновые и миозино-
вые миофиламенты только на концах, которые .и сокращаются.
Рецепторной частью интрафузального мышечного волокна являет-
ся центральная, несокращающаяся часть. Различаютинтр^Гфучаль-
ные волокна двух типов—волокна с ядернои сумкой (bursa nuclea-
ris) и волокна с ядернои цепочкой (vinculum nucleare). Волокон
с ядернои сумкой в веретене 1—3. В центральной расширенной
части они содержат много .ядер. Волоконс~ ядсрттпй"цепот'кой в
веретене может насчитываться J 7. Они вдвое тоньше и вдвое
кор<тче,"чем волокна с ядерной сумкой, и ядра в них расположены
цепоч к ои по всей рецепторной оБласткь К йнтрафу за л ь н ы м
мышечным волокнам подходят афферентные волокна двух-'типов.
R
Рис. 114. Схема строения нервно-мышечного веретена.
А — моторная иннервация интрафуза'льных и экстрафузальных мышечных волокон
(по А. Н. Студитскому); Б сриральные афферентные нервные окончания вокруг интра-
фузальных мышечных волокон в области ядерных сумок (по Р. Кристичу с изменением).
/ — нервно-мышечные эффекторные окончания экстрафузальных мышечных волокон;
2 - моторные бляшки интрафузальных мышечных волокон; 3 — соединительная ткань;
4 — ядерная сумка; 5 чувствительные кольцеспиральные нервные окончания вокруг
ядерных сумок; 6 — скелетные мышечные волокна; 7 — нерв.
Первичные волокна диаметром 17 мкм образуют окончания в виде
спирали- кольцеспиральные окончания (terminatio nervi annulo-
spiralis) как на волокнах с ядерной сумкой, так_И на волокнах с
ядерцрй цепочкой. Вторичные волокна диаметпом 8 мкм иннерви-
рукгГтолокна с ядерной цепочкой. По обеим сторонам от кольцеспи-
рального окончания онщдбразуют гроздьевидные окончания (ter-
minatio nervi racemosa).
При L4t t lau.iri’HH (н in (М,Ц|*| ЦИИ) МШИ»1' у тытчивается
и длицд И1П-)>афузя.1кн>4х ылплклн, регистрируется рецептора-
288
Х4И, К-ольцеспиральные окрнчавия-реагируют на изменение длины
мышечного волокна и _на скоростьэто1чт-изменения,-гроздьевидные
окончания — только на изменение длины. При внезапном растя-
жении из кольцеспиральных окончаний в спинной мозг поступает
сильный сигнал, который вызывает резкое сокращение мышцы,
с которой поступил сигнал — динамический рефлекс на растяжение.
При медленном, длительном растяжении волокна возникает статиче-
ский сигнал на растяжение, передаваемый как от кольцеспираль-
ных, так и от гроздьевидных рецепторов. Этот сигнал может под-
держивать мышцу в состоянии сокращения в течение нескольких
часов.
Интрафузальные волокна имеют также эфферентную иннерва-
цию. К ним подходят тонкие моторные волокна, оканчивающиеся
аксомышечным синапсом на концах мышечного волокна. Вызывая
сокращение концевых участков интрафузального волокна, они
усиливают растяжение центральной рецепторной его части, повы-
шая реакцию рецептора.
Нервно-сухожильные веретена обычно располагаются в месте
соединения мышцы с сухожилием. Коллагеновые пучки сухожи-
лия, связанные с 10—15 мышечными волокнами, окружены
соединительнотканной капсулой. К нервно-сухожильному веретену
подходит толстое (диаметром ~ 16 мкм) миелиновое волокно,
которое теряет миелин и образует терминали, ветвящиеся между
пучками коллагеновых волокон сухожилия. Сигнал с нервно-сухо-
жильных веретен, вызванный напряжением мышцы, возбуждает
тормозные нейроны спинного мозга. Последние тормозят соответ-
ствующие двигательные нейроны, предотвращая перерастяжение
мышцы.
Межнейронные синапсы '
Поляри заци я проведения нервного импульса по цепи нейронов
определяется их специализированными контактами — синапсами
(от греч. synapsis — соединение). Различают синапсы с химиче-
ской передачей — химические синапсы и электрической переда-
чей — электрические синапсы (беспузырьковые). Последние у
высших животных встречаются редко.
В синапсах с химической передачей веточки аксона нейрона
образуП>>Т~ето пресынатическуп) Часть (pars presynaptica), взаимо-
действующую с Плазмолеммой другого нейрона — его постсинап-
тическои частью (pars postsyiiapticay.T'HHaiicbi-весьмарачнообраз-
ны й характерны для различных отделов нервной системы (рис.
115,Л, Б, В, Г).
По локализации различают аксосоматические синапсы (терми-
нальные-ветви нейрона оканчиватотстгна'ТёлёУтругого), аксоденери-
тические синапсы Ггёрмйнальные ветви аксона_______одного нейрона
вступают в синаптическую связь с дендритом другого) и аксоаксо-
нальные синапсы (терминали аксона одного нейрона оканчиваются
на аксоне другого). Последним прмпи.-мпАстгя фуцкгия передачи
to гистология
289
возбуждения, а торможения импульса, полученного чеоез аксосома-
тические и аксодендритические синапсы от каких-либо других ней-
ронов.
Треси на пт и челка я часть характеризуется скоплени-
ями пресинаптических пузырьков и митохондрий. Пресинаптиче-
ские пузырьки содержат медиаторы, наиболее распространенными
из которых являются ацетилхолин (холинергические синапсы)
и норадреналин (адренергические_синащ;ыЬ_Ррль медиатора могут
играть и другие биологически активные вещестиаг дофамин, глицин,
гамма-аминомасляная кислота, глутаминовая кислота, вещество
Р, серотонин, гистамигГй др.ТТз'них дофамин, глицин и гамма-ами-
номасляная кислота являются дормозными медиаторами. В х о л и-
нергических синапсах пресинаптические пузырьки мел-
кие (30—50 нм) и прозрачные.—Одновременно здесь может содер-
жаться~нёскОЛБКСГТ<рупных электронно-плотных пузырьков (80—
150 нм). Значение последних до настоящего времени не вполне
ясно. Предположительно с ними связывают начилие в синапсах
биогенных аминов.
Пресинаптические пузырьки______а дренергических си-
напс <7н~имеют размер 50—*)0 нм и характеризуются наличием
в каждом тггнИХТТлОтИой гранулы. На внутренней сторонеjipec-инап-
тической мембраны (аксолеммы терминали аксона) располагаются
коцуцовидныеГ электронно-плотные образования. Каждый-конус
связан с 5 соседними плотными тяжами таким образом, что между
конусовидными уплотнениями тяжами образуются участки
треугольной формы, через которые, как полагают, и выходит содер-
ж и мое си наптических.. пузырьков. Распространение потенциала
действия на пресинаптическую мембрану вызывает слияние мембран
нескольких пресинаптических пузырьков с пресинаптическои мем-
бран ой~й~излйяние медиатора в синаптическую щель, ширина
которой около 20-нм. Сами пресинаптические пузырьки остаются
в пресинаптическои части и нескольке>~Раз~по~вторно заполняются
медиатором.
Влияние медиатора на постсинаптическую часть зависит от при-
сутствия н£ГТТ<)СГО1натггической~-ме мбра не (плазмолемма нейрона-
исполнителя) особого белка — рецептора__медиатора. В-п-о с т си-
нап т и ческой части часто наблю ; юте я субмембранные уплот-
нения цитоплазмы в области синапса — постсинаптические
уплотнения, иногда — субмембранные цистерны гладкой эндоплаз-
матической сети. Функциональное значение этих структур не выяс-
нено.
Рис. 115. Строение синапсов.
А — схема цитотопографии синапсов; Б — схема строения синапсов: а тормозного
типа; б — возбудительного типа; в -электрического (беспузырькового) типа В — схема
строения синаптических пузырьков: а- холинергических (светлых), б — адренергиче-
ских (плотных), в — пуринергических, г—пептидергических (по Л. Д. Маркиной);
Г — электронная микрофотография аксодендритического синапса (препарат И- Г. Павло-
вой): / аксосоматический синапс; 2—аксодендритические синапсы, 3 аксоаксональ-
ный синапс; 4 дендриты; 5 — дендритный шипик; 6 — аксон; 7 синаптические пузырьки;
8 пресинйпгическая мембрана; 9— постсииаптическая мембрана; 10 - синаптическая
шель; II — постсинаптические уплотнения.
290
10‘
18
Рис. 116. Простая рефлекторная дуга (рис. В. Г. Елисеева, Ю. И. Афанасьева,
Е. Ф. Котовского).
/ - чувствительная нервная клетка; 2 — рецептор в коже; 3 — дендрит чувствительной
клетки; 4 — плазмолемма нейролеммоцита; 5 — ядра нейролеммоцитов; 6 — миелиновый
слой: 7 — узловой перехват нервного волокна; 8 — осевой цилиндр; 9 — насечкв мие-
лина; /О — аксои чувствительной клетки; // — двигательная клетка; 12 — дендриты
двигательной клетки; 13 — иейрит двигательной клетки; 14 — миелиновые волокна;
15 — эффектор иа мышце; 16 — спинномозговой узел; 17 — дорсальная ветвь спинномоз-
гового нерва; 18 — задний корешок; 19 — задний рог; 20 — передний рог; 21 — передний
корешок; 22 — вентральная ветвь спинномозгового нерва.
В возбуждающем синапсе взаимодействие медиатора с рецеп-
торным белком на постсинаптической мембрЯ'нетта'Тывает повыше-
ние про ни ца емости2мембракьЬ—в_резу льТате: л его ~ быстрый приток
ионов натрия в клетку снижает отрицательный потенциал-покоя.
Потенциал покоя (—70 mV) создается в покоящемся нейроне в
результате выведения натрия из клетки под действием—Анатрий-
калиевого насоса». Когда потенциал снижается ДО —59 mV, возни-
кает возбуждение. Воздействие__на постсииаптическую мембрану
медиатора тормозящего синапса повышает отрицательный потенци-
ал, в результате чего нейрон становится, менее чувствительным к
действию возбуждающих синапсов.—Электрические синапсы могут
быть или типа щелевого-контакта е-шириной щели-между двумя
292
клетками около 2 рм или щель между _двумя нейронами может
отсутствовать^ клетки сопри каса ются наружным и_ поверхности ми
плазмолеммы. Импульс в электрических синапсах может переда-
ваться в обоих направлениях. Химические синапсы проводят воз-
буждение только водном направлении пт-концевого апггарата ак-
сона одного нейрона на другой нейрон. Это весьма важное свойство
синапса лежит в основе динамической поляризации рефлектор-
ных дуг.
ПОНЯТИЕ О РЕФЛЕКТОРНЫХ ДУГАХ
Нервная ткань входит в состав нервной системы, функционирую-
щей по рефлекторному принципу, морфилш ическим субстратом
которого является рефлекторная дуга. Рефлектбрная~дуга"представ-
ляет собой цепь нейронов, связанных друг с другом синапсами и
обеспечивающих проведение нервного импульса от рецептора
чувствительного нейрона до эфферентного окончг ни я в рабочем
органе.
Самая простая рефлекторная дуга состоит из двух нейронов —
ч у в с т BjLLe_i-b_H ого- идо ич-н тельп о г о (рис. 116). В по-
давляющем большинстве случаев между чувствительными и двига-
тельными-нейронами включены вставочные, или а с с о ц и а-
т и в н ы е, нейроны У высших животных рефлекторные дуги
состоят обычно из многиХ—нейроноал имеют—значительно более
сложное строение, чем на приведенном рисунке.
Глава XII
ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ
Ткань — это_возникшая_ь ходе эволюции ч.чиыя система
организма, состоящая из одного или нескольких лиФФероно кле-
ток и их~ производных, обладающая специфическими функциями
благодаря кооперативной ^деятельности всех ее элементов.
ТКАНЬ КАК СИСТЕМА
Любая ткань — сложная система, элементами которой служат
клеки и их производные. Сами ткани тоже являются элементами
морфофункцйбнальных единиц, а последние выступают в роли
элементов органов. Поскольку по отношению к системе высшего
ранга в нашем случае — организму) системы более низких рангов
рассматриваются как частные, то и о тканях следует говорить
как о частных системах.
В любой системе все ее элементы упорядочены в пространстве
и функционируют согласованно друг с другом; система в целом
обладает при этом свойствами, не присущими ни одному из ее
элементов, взятому в отдельности. Соответственно И В каждой
293
ткани ее строение и функции несводимы к простой сумме свойств
отдельных входящих в нее клеток и их производных.
Ведущими элементами тканевой системы являются клетки.
Кроме клеток; различаю 1 клеточные производные и межклеточное
вещество:——----------------
И р о и з к о д н ы м клет о к относят симпласты (мыше-
ч ные. ролокна, наружная часть трофобласта) и-сшщисии. ~t сое дине н-
ные друг с другом цитоплазматическими мостиками развивающиеся
мужские половые клетки, пульпа эмалевого органа)^ постклеточные
структуры (эритроциты, тромбоциты, роговые чешуйки эпи^ ермиса
и т. д.).
Меж клеточное вещество подразделяют на основное
и на япяпкни. Оно может бьи- ЛДсШЕЩЛСДк гелекгилй быть
минерализрванным. Среди волокон различают обычно три вида:
ретикулярные1, коллагеновые, эластические.
Клетки всегда находятся но взаимодействии друг с другом и с
межклеточным веществом. При этом формируются различные
структурные объединения. Клетки могут лежать в меж1Цтёточном
веществе на расстоянии друг от друга и взаимодействовать через
него без непосредственных контактов (например, в ^рыхлой волок-
нистой соединительной ткани), соприкасаясь отростками (ретику-
лярнаятканьГили образуя сплошные~клеточные массыТили пласты
(эпителий^ эндотелий).
~Ксе межклеточные взаимодействия, как непосредственные, так и
через межклеточное вещество обеспечивают функционирование
ткани как единой системы. Только на основе системного подхода
возможно изучение тканей, понимание общей гистологии.
Взаимосвязь тканей. В состав органов входят различные ткани.
Одни из них во многих случаях образуют строму (остов), представ-
ленную соединительной тканью, другие — п«ренГсцму,_Строма и
паренхима всегда "тёс но взаимосвязаны и обеспечива ют вып о л не н и е
основной функции органа^ Так, доказаны взаимоотношения эпите-
лия ТГсо^ДИНТГгельтюйттани: эпителий стимулирует синтез .коллаге-
на, а последний влияет на секреторную деятельность железистого
эпителия.
Многие клетки или их производные в одних тканях отделены
от соседних ткаяей'Оо.зальнои мембраной (базальной пластинкой) —
слбжТПДМ. д и нам и ЧИым~ акта вн ы м угле водно-белково-ли п и д ны м
комплексом, выполняющим барьерную и оргайизуюТцую функции.
Базальная мембрана состоит из матрикса и коллагена IV типа с
высоким содержанием гидрооксипролина, гипрооксилцдиндТГ осо-
бенно углеводов. Базальная мембрана обладает свойствами полу-
проницаемости.
Согласованная деятельность различных тканей и органов обе-
спечивается нервной, эндокринной и иммунной системами, которые
называются и нт еграционн ы м и.
1 В состав ретикулярных волокон входит белок коллаген, поэтому они род-
ственны коллагеновым волокнам.
294
РАЗВИТИЕ ТКАНЕЙ
Свойства любой ткани несут на себе отпечаток всей предыдущей
истории ее становления. Под развитием живой системы понимаются
ее преобразования и в филогенезе, и в онтогенезе. Ткани как
системы, состоящие из клеток и их производных, возникли исто-
рически с появлением многоклеточных организмов. Уже у низ-
ших представителей животного мира, таких как губки и кишеч-
нополостные, клетки имеют различную функциональную специали-
зацию и соответственно различное строение, так что могут быть
объединены в различные ткани. Однако признаки этих тканей еще
не стойки, возможности превращения -клеток и соответственно
одних тканей в иные достаточно широки. По мере исторического
развития животного мира совершалось закрепление свойств от-
дельных тканей, а возможности их взаимных превращений ограни-
чивались, количество же тканей одновременно постепенно увеличи-
валось в соответствии со все более возрастающей специализацией.
Онтогенез. Детерминация и коммитирование.
Развитие организма начинается с одноклеточной стадии — зиготы.
В ходе дробления возникают бластомеры, но совокупность бласто-
меров еще не ткань. Бластомеры, по крайней мере на начальных
этапах дробления, еще не имеют стойкой детерминированности, и
если отделить их один от другого, то каждый может дать начало
возникновению полноценного самостоятельного организма (меха-
низм возникновения монозиготных близнецов). Постепенно на
следующих стадиях происходит ограничение потенций. В основе
его лежат процессы, связанные с блокированием отдельных компо-
нентов генома клеток и детерминацией, т. е. определением даль-
нейшего пути развития клеток на генетической основе.
Ограничение возможностей путей развития вследствие детерми-
нации определяется термином «коммитирование». Оно совершает-
ся ступенчато. Сначала соответствующие преобразования генома
касаются крупных его участков. Затем все более детализируются,
поэтому вначале детерминируются наиболее общие свойства клеток,
а затем и более частные.
Эмбриональные зачатки. Как известно, на этапе гаструляции
возникают эмбриональные зачатки (см. гл. V). Клетки, которые
входят в их состав, еще не окончательно детерминированы, так что
из одного зачатка возникают клеточные совокупности, обладающие
разными свойствами. Следовательно, один зачаток служит источ-
ником развития нескольких тканей.
Эмбриональные зачатки имеют различную пространственную
организацию в теле зародыша. Они могут представлять собой груп-
пировки, где клетки тесно прилежат друг к другу (сосредоточенная
форма), например кожная эктодерма, кишечная энтодерма. У не-
которых зачатков их части расположены в различных местах тела
зародыша (расчлененная форма), например нейральный зачаток,
представленный нервной трубкой, нейральными гребнями и плако-
дами. Клетки некоторых зачатков рано выселяются в мезенхиму i
*95
не образуют компактных группировок (диффузная, рассредоточен-
ная ф°Рма>> например клетки ангиобласта, из которых впослед-
ствии развивается эндотелий сосудов.
ТЕОРИЯ ЭВОЛЮЦИИ ТКАНЕЙ
Последовательная ступенчатая детерминация и коммитирова-
ние потенций однородных клеточных группировок — дивергентный
процесс. В общем виде эволюционная концепция дивер-
гентного развития тканей в филогенезе и в онтогенезе
была сформулирована Н. Г. Хлопиным. Современные генетические
концепции подтверждают правоту его представлений. Именно
Н. Г. Хлопин ввел понятие о генетических тканевых. типах. Кон-
цепция В. Г. Хлопина хорошо отвечает на вопрос, как и к а к и-
м и путями происходило развитие и становление тканей, но не
оста на вливается на причинах, определяющих пути ра звития.
Причинные аспекты развития тканей раскрывает-теория п а-
р а ллелизмов А. А. Заварзина.-Он обратив внимание на_схрд-
ство строения тканей, которые выполняют одинаковые функции,
у животных, принадлежащйх2даже2к.~весъма--уй&ленным-друг от
друга эволюционным группировкам. Современные электронно-мик-
роскопические исследования показали также чрезвычайное-сход-
ство ультраструктур клеток, выполняющих сходные функции, у
всех животных, независимо от их таксономического положения.
Вместе с тем известно, что, когда эволюционные ветви только
расходились, у общих_дредков таких специализированных -тканей
филогенетического древа самостоятельно, как бы параллельно,
возникали одинаково организованные ткани, выполняющие сход-
ную функниюГТТричиной этого является естественный отбор: если
возникали какие-то организмы, у которых соответствие строения
И функпйи клеток, тиачез, .>ргаи..ы uqpyi. нее
жизнеспособны. Теория А. А. Заварзина отвечает на вопрос, поче-
му развитие тканей шло тем, а не иным путем, раскрывает казу-
альные аспекты эволюции тканей.
Концепции А. А. Заварзина и Н. Г. Хлопина, разработанные
независимо одна от другой, дополняют друг друга и былиi объеди-
нены А. А. Брауном и В. П. Михайловым: сходные ткане-
в ы е структуры “возник алй параллельно в ~то д е
д и вергентного
развития.
ПРИНЦИПЫ КЛАССИФИКАЦИИ ТКАНЕЙ
Общность дефинитивного строения тканей, которые обладают
сходными функциональными признаками, позволяет объединять
их в 4 морфофу н кч и она льн ые группы: эпителии —
в связи—с выполнением прежде всего барьерных (пограничных)
функций; ткани внутренней среды (кровь, лимфа, соединительные
ткани) — в связи с обеспечением гомеостаза, трофической^ защит-
296
ной, опорной функций, мышечные — в связи с обеспечением под-
вижности тела и ~~неирам>ные — в связи ~с осуществлением
интегративных реакций на основе генерации возбуждения и
проведения ^егбГН то Же время- ткани, развивающиеся йз~~Пдтгого
зачатка, обладают такими—общими свойствами, которые не всегда
за метны в обычных условиях, но могут проявляться при па то л о ги и
или в ходе регенерации после повреждения. Так, при регенерации
многорядного эпителия трахеи возникают многослойные плоские
структуры, а при регенерации многослойного плоского неорогове-
вающего эпителия роговицы — многослойный плоский ороговева-
ющий эпителий; гладкие миоциты мезенхимного происхождения
и фибробласты, также развивающиеся из мезенхимы, близки меж-
ду собой, а в некоторых опухолях (фибромиомы) могут встречать-
ся многообразные переходные формы между ними. Наличие общих
свойств у тканей, развившихся из одного эмбрионального зачат-
ка, позволяет объединять их в единый гк а н е в о й—тип.
В предалах одной ткани могут быть разновидности. Так, выде-
ляют ретикулярную ткань с клетками микроокружения, которую
называют миелоидной тканью (в красном костном мозге) или лим-
фоидной тканью (например, в лимфатических узлах, селезенке).
Морфофункциональная и генетическая классификация допол-
няют друг друга. После того как в результате второй фазы гастру-
ляции возникают три зародышевых листка, каждый из них~сбдер-
жит разные эмбриональные зачатки. Ткани, развивающиеся из
какого-либо зачатка, в соответствии с морфофункциональными
признаками могут относиться к разным группам. Так как сово-
куппость клеток, входящих в один эмбриональный зачаток, служит
источником развития нескольких тканей, в ходе гистогенеза-еовер-
шается дальнейшая детерминация. Она охватывает меньшие участ-
ки генома, чем это было в ходе образования зачатков, так что
отличия между тканями, принадлежащими к одному типу, менее
резки, по сравнению с тканями, принадлежащими к разным типам.
Это^цроявляется, в частности, в том, что в рамках одного типа
возможны случаи превращения ткани в другую (метаплазия),
например многорядного эпителия в многослойный к ходе репара-
тивной регенерации после травмы.
ТКАНЕВЫЙ ГОМЕОСТАЗ
Каждая ткань в зрелом организме выполняет специфические
функции, которые определяются как деятельностью клеток, так и
свойствамитпг производных. Контроль функциональной активнос-
ти осуществляют регуляторные механизмы. Одни из них~обёспечи-
вают необходимые связи частей в пределах самснГ тканевой систе-
мы~(например, кейлоны), другие — в рамках межтканевых и более
высоких межсистемных отношении (например, гормональные,
иммунпьгг, 1 >ервные_>Т~Фу~|ПппттпптБнаъ активность тканей не оста-
ется нетменной. а втеб дате ы> мок рут некоторого сТуднего уровня.
Эти колебания — проявления биологических ритмов —
297
имеют равный период (время полного цикла). ^Внутриклеточным
пррцессам свойственна ритмика 4- периодами порядка несколькох
минут — одного-двух часов, внутрисистемным тканевым — не-
скольких часов; процессам на организменном уровне присущи
колебания с периодом около суток, известны и более длительные
циклы (околонедельные, окологодовые). Поэтому при анализе
ритмики всегда выявляется сложный спектр.
Изменения в обменных реакциях и-специфических функциях
отражают либо процессы адаптации, либо процессы патологиче-
ской изменчивости ткани, дезадаптации. При этом -тканевые .клет-
ки могут терять часть своих специфических структур и переходить
в особое -.едифференцированное состояние, когда сшиприобре-
таютспособность развиваться в некоторых других направлениях
в зависимости от создавшихся условий. В результате указа иных
причин могут возникать структуры,~не свойственные исходным
тканям. Подобного рода превращения Iwrntuitiwu) небезгранич-
ны. Qhm известны только пределах пинию тканещд, д_ипа- Не
исключено, что успехи генной инженерии позволят в дальнейшем
расширить эти возможности.
Как и организм в целом, ткани реагируют на изменение внеш-
них условий активными реакциями. "При этом может изменяться
и йх пространственная организация, и функциональная актив-
ность, включая параметры ритмики (изменения спектра, уровня,
амплитуды колебаний и т. д.).
^нкщ1Ональная~акТйвность тканей контролируется благодаря
согласованной деятельтгос’пг-ретуляторньгххгеханизмов гомеостаза
на собственном тканевом (в том числе кейлонами), межтканевом
(генотропные активаторы) и организменном уровнях (эндокрин-
ная. иммунологическая, нервная регуляция). Кроме поддержания
подвижного равновесия функциональной активности, тканевый
гомеостаз ббёспечйваёт-в—дрганйзтае—1Г_сохранение общей массы
клеток, а следовательттиртт соотношение^междуч целом делящихся,
дифференцированных и гибнущих клеток? Изучение этих законо-
мерностей составляет предмет кинетики клеточных популяций.
ОСНОВЫ КИНЕТИКИ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ
Каждая ткань имеет или имела в эмбриогенезе стволовые клет-
ки — наименее дифференцированные и наименее коммитирован-
ные, которые, видимо, детерминируются в составе эмбриональных
зачатков к концу второй фазы гаструляции. Они обр зуют само-
о ю ржииаюшуюся популяцию, их потомки способны лифекф' нци-
роватьгя в нескольких направлениях под влиянием микроокруже-
нил__(факторов дифферепцировки), образуя клётки-предшествен-
ники и, далее, функционирующие дифференцированные клетки.
Таким образом, стволовые клетки пол иг патентны. Они делятся
редко, пополнение убыли зрелых клеток ткани, если это необхбдй-
мо, осуществляется в первую очередь за счет клеток следующих
генераций—Ск ггетлУ-прелщественникбвГ По сравнению~сб“ всеми
29К
другими клетками данной ткани стволовые клетки наиболее устой-
чивы к повреждающим воздействиям.
Хотя в состав ткани входят не только клетки, именно они
являются ~вё(Гущими элементами системы,—тТе. определяют ее
основные свойства—И х разрушение приводит к деструкции систе-
мы и, как правило, их гибель (особенно если затронуты стволо-
вые клетки) делает ткань нежизнеспособной.
Есди одна из стволовых клеток вступает на путь дифферен-
пиации. то в результате последовательного ряда коммитирующих
митозов возникаю! сначала полу стволовые, а затем й дифферен-
цированные клетки со специфической функцией. Выход'стволивьй
клетки из популяции служит сигналом (механизм неясен)._для
деления другой стволовой клетки по типу некоммитирующего
митоза. Общая численность стволовых клеток в итоге восстанавли-
вается; в условия^—нормальной жизнедеятельности онаТохраняет-
ся приблизительно постоянной. Совокупность ТПГ’ГЛК, ртгтпиваю
Н1ихся_ из ЦЛШ1ГП ьила гтио.ншых » четок. составляет стволовой
диЗтферон, Иногда в образовании ткани участвуют различные
диффероны. Так, в состав эпидермиса, кроме кератиноцитов,
вхГЩятклётки, развивающиеся в нейральном гребне и имеющие
другую детерминацию-(меланоциты^-,-я также кпетки. развиваю-
щиеся путем лиф^решившим х-тнлъ.иой клетки крови, т. е. при-
надлежащие уже .к третьему дифферону (внутриэпидерминальные
макрофаги, или клетки Лангерганса).
Дифференцированные клетки наряду с выполнением своих
специфических функций способны синтезировать особые веще-
ства — кейлоны, тормозящие интенсивность размнб~жгийя"клеток-
прсдшественнйкОь ~й~СТВ&Лбвых?~ЕсЛИ в СИЛу_Таких-либо~причин
количество дифференцированных функционирующих Клеток умень-
шается (например, пос пе-трянмы), .тррмотаттгее-действие зсей попов
ослабевает .и численность попуаяции восстанавливается^ Кроме
ке й ло но в — местных ре гуляторов, клеточное размножение контро-
лируется гормонами; одновременно продукты жизнедеятельности
клеток регулируютактивность желез внутренней секреции. Если
какие-либо клетки под воздействием внешних повреждающих фак-
торов претерпевают мутации, они элиминируются из тканевой
системы вследствие иммунологических реакций.
Выбор пути дифференциации клеток определяется межклеточ-
ными взаимодействиями. Влияние микроокружения изменяет ак-
тивность генома дифференцирующейся клетки, активируя одни и
блокируя другие гены. У клеток, уже дифференцированных и
утра 1 и вшил способность к дальнейшему размножению, строение
и функция тоже могут изменяться (например, у гранулоцитов
начиная со стадии метамиелоцита), ^акой процесс не приводит
к возникновению различий среди потомков клетки и для него ботьше
подходит название «спе иализа ия» .
1 Понятия «дифференциация» и «специализация» не всеми гистологами трак-
туются одинаково Приведенные формулировки отражают точку зрения авторов
главы
299
РЕГЕНЕРАЦИЯ ТКАНЕЙ
* Знание основ кинетики клеточных популяций необходимо для
«понимания теории регенерации, т. е. восстановления структуры
биологического объекта после ее разрушения. Соответственно
уровням организации живого различают к п е т о ч н у ю (внутри-
клеточную) , т к а н е в у ю, органную регенерацию.
Предметом обшей гистологии является регенерация^ на^тканевом
уродне^-Вазличают регенерацию- ф изиологическую, кото-
рая совершается постоянно в здоровом организме,^ р е п а р а-
Т и в Н у Ю — вслепствие повреждения,^ разных тканёй~возмож-
ности регенерации неодинаковы.
В ряде тканей гибель -клеток,генетически запрограммирована
и совершается постоянно (в многослойном пллском-ероговенаю-
щем эпителии кожи, в однослойном каемчатом эпителии тонкой
кишки, в крови!.. За счет непрерывного размножения, в первую
очередь полустволовых клеток-предшественников, количество
клеток в популяции пополняется и постоянно находится в состоя-
нии ра в новесия. Наряду с запрограммированной фичипплгической
гибелью клеток во всех тканях ироне конит-и-неаянрограммирона н-
ная.— от случайных причин: интоксикаций (в том числе..и алко-
гольной), воздействийпостоянного природного радиационного
фона, космических лучей. Хотя в ряде тканей запрограммйрован-
нок гибели нет, но в.де'чени_е_всей жязии_в_них сохраняются ство-
ловые и полустволовые клетки. В ответ на случайную гибель возни-
кает их размножение и популяция восстанавливается. Именно
так функционируют остеокласты и остеобласты костной ткани.
У взрослого человека в тканях, где стволовых клеток не остается,
регенерация на тканевом уровне невозможна, она происходит
лишь на клеточном уровне.
ЧАСТНАЯ ГИСТОЛОГИЯ
Глава XIII
НЕРВНАЯ СИСТЕМА
-У Нервная система обеспечивает регуляцию всех жизненных про-
цессов в организме и его взаимодействие с внешней средой. Ана-
томичКк>Гнёрвную~сисгему~дёлят на ц е н т р а ль н у ю и п ер и-
Ф е р и ч е с к у ю. К первой относят~~гсоговнбц и спинной мозг,
вторая_объединяет пе рифе рическиенервные узлы, стволы и окон-
чания^ Такое деление нервной системы условно и допускается
лишь из методических соображений. Морфологическим субстратом
рефлекторной деятельности нервной системы являются рефлектор-
ные дуги, представляющие собой цепь нейронов различного функ-
ционального" значения, тела которых расположены в разных отде-
лах нервной системы, как~в периферических узлах, так и в сером
веществе центральной нервной системы.
С физиологической точки зрения нервная система делится на
с о мат и ч е с к у ю , иннервирующую все тело, кроме внутренних
органов, сосудов и желез, и автономную, или вегетатив-
н у юТрегул ирующую пеятепьность перечисленных органов.
Развитие. Нервная система развивается из нервной трубки
и ганглиозной пластинки. Из краниальной части нервной трубки
дифференцируется головной мозг и органы чувств. Из туловищ-
ного отдела нервной трубки и ганглиозной пластинки формируются
спинной мозг, спинномозговые и вегетативные узлы и хромаффин-
ная ткань организма. Особенно быстро возрастает масса клеток в
боковых отделах нервной трубки, тогда как дорсальная и вентраль-
ная ее части не увеличиваются в объеме и сохраняют эпендим-
ный характер. Утолщенные боковые стенки нервной трубки делят-
ся продольной бороздой на дорсальную — крыльную пластинку и
вентральную — основную. В этой стадии развития в боковых стен-
ках нервной трубки можно различить три зоны: эпендиму, высти-
лающую канал, плащевой слой и краевую вуаль. Из плащевого
слоя в дальнейшем развивается серое вещество спинного мозга, а
из краевой вуали — его белое вещество. Нейробласты передних
столбов дифференцируются в двигательные нейроны ядер перед-
них рогов. Их аксоны выходят из спинного мозга и образуют
его передние корешки. В задних столбах и промежуточной
зоне развиваются различные ядра вставочных (ассоциативных)
клеток. Их аксоны, поступая в белое вещество спинного мозга,
входят в состав различных проводящих пучков. В задние рога
входят нейриты чувствительных клеток спинномозговых ганглиев.
Одновременно с развитием спинного мозга закладываются
спинномозговые и периферические вегетативные узлы. Исходным
материалом для них служат клеточные элементы ганглиозной пла-
301
Рис. 117. Строение спин
номо и оного ганглия.
Л продольный разрез гаи
глия и корешков: Б — псев-
доуниполярный нейрон;
стинки, дифференцирующиеся в нейробласты и глиобласты, из
которых образуются клеточные элементы спинномозговых ганглиев.
Часть их смещается на периферию в места локализации вегетатив-
ных нервных ганглиев и хромаффинной ткани.
ЧУВСТВИТЕЛЬНЫЕ УЗЛЫ
Чувствительные узлы лежат по ходу задних корешков спинного
мозга либо черепномозговых~нервов.————
Сццнномозговой узел окружен соединительнотканной капсулой.
От капсулы_в паренхиму узла проникают тонкие_________прослойки
соединительной тканщ которая образует его остов и проводит кро-
ве нос н ые сосудБП
Нейроны'спинномозгового узла располагаются__£руплами, пре-
имущественно по перифирии органа, тогда как его центр состоит
302
Рис. 117 (продолжение).
В — ультраструктурная ор-
ганизация псевдоуннполяр-
ного нейрона и его микро-
окружения (по Р. Кристичу
с и1менением): / — задний
корешок; 2 ганглий; 3 —
псевдоу ниполярные нейро-
ны; 4 — нервные волокна;
5 — соединительнотканная
капсула; б — передний коре-
шок: 7 — леммоцит; 8 Т-
образное разветвление от-
ростка; 9 — глиоциты ган-
глия (мантийные глиоциты);
10 — базальная мембрана;
// ге\:о*?пмлчяры; I? —
миелйновые и безмиелиновые
нервные волокна.
главным образом из отростков, этих клеток. Дендриты идут в
составе чутитвительнои части смешанных спинномозговых нервов
на периферию и заканчиваются там рецепторами. Нейриты в со-
вокупностй~образуют заднйё~корешки, несущие нервные импульсы
или в серое вещество спинного мозга, или по его заднему канати-
ку в^зродОлговаТый мил. Биполярные клетки у~нйз~шйЗСпозвоночных
сохраняются в тёчение~всей жизни. Биполярными являются и аффе-
рентные нейроны некоторых черепных нервов (gang), spirale cochle-
are). В спинномозговых узлах высших позвоночных животных и че-
ловека биполярные нейроны в процессе созревания становятся
псевдоуниполярными. ОтрОсТКи клеток постепенно сближаются, и
их основания: сливаются (рис. 117, А, Б, В,). Вначале удлинен-
ная часть тела (основание отростков) имеет небольшую длину,
но со временем, разрастаясь, она многократно обвивает клетку
и часто образует клубок. Существует и другая точка зрения на
процесс формирования псевдоуниполярных нейронов: аксон отрас-
тает от удлиненной—части тела- -неироцита__после формирования
дендрита. Дендриты нейриты-клетак. в узле и за его пределами
покрыты оболочками из нейролеммоцитов. Нервные клетки спин-
номозговых узлов окружены слоем клеток глии, которые получили
101
здесь название мантийных глиоцитов, или глиоцитов ганглия
(gliocyti ganglii) (см. рис. 117, А, Б). Их можно узнать по круг-
лым ядрам клеток, окружающих тело нейрона. Снаружи глиаль-
ная оболочка тела нейрона покрыта тонковолокнистой соедини-
тельнотканной оболочкой. Клетки этой оболочки отличаются
овальной формой ядер.
ПЕРИФЕРИЧЕСКИЕ НЕРВЫ
Периферические нервные стволы — нервы — состоят из миели-
новых и безмиелиновых волокон и соединительнотканных оболо-
чек. В некоторых нервах встречаются одиночные" нервные клетки
и мелкие ганглии. На поперечном срезе нерва видны сечения осе-
вых цилиндров нервных волокон и одевающие их глиальные оболоч-
ки. Между нервными волокнами в составе нервного ствола распо-
лагаются тонкие прослойки соединительной ткани —• эндоневрий
(en d on е или гл). Пуч ки 'нёрвных~~вдлокон~ одеты периневри ем (peri-
neurium). ПериневрнЙ^С£1СХС1ИТ_из чередующихся слоев плотно рас-
положенных клеток и тонких фибрилл. Таких слоев в периневрии
толстых яервов~нескилыог"(5- ЬугФйЙрилльГ"ориентированы вдоль
нервагНаружмая оболочка нерннОго ствола — эпиневрий (epineuri-
um) — представляет -собой волокнистую соединительную ткань,
богатую фибробластами, макрофагами и жировыми клетками. Сое-
динительнотканные оболочки нерва содержат кровеносные и лим-
фатические сосуды и нервные окончатлпг~~В ~этгинсврий~ поступает
по всейдлинё’ нерва ольшое коли чествсГа настомози рующи х меж-
ду собой кровеносных сосудов. Из эпиневрия артерии проникают
в периневрий и эндоневрий (рис. 118, 4, Б).
СПИННОЙ мозг X-
Спинной мозг состоит из двух симметричных половин, отгра-
ниченных друг от друга спереди глубокой серединной щелью, а
сзади — соединительнотканной перегородкой (рис. 119). На све-
жих препаратах спинного мозга невооруженным глазом видно, что
его вещество неоднородно. Внутренняя часть органа темнее — это
его серое вещество (substantia grisea)T На’ периферии спиннЪго
мозга располагается более светлое белое вещество (substantia alba).
Серое вещество на поперечном сечении мозга видно в виде буквы
«Н» или бабочки. Выступы серого вещества принято называть
рогами.—Различают передние, иди вентральные, задние, или дор-
сальные, Л4—боковые, ияи-яатер«лъпые,-ро£а- (cornu ventrale, cornu
dorsale, cornu laterale).
Серое вещество спинного мозга состоит из нейронов, безмие-
линовых и тонких миелиновых волокон и нейроглии. Основной
составной частью серого вещёстват~отличающей его от белого, яв-
л я ются мульти полярные ней ро ньь “
Белое веществб’спинного'мозга образуется совокупностью про-
дольяо~~ориентирбванньпг преимущественно миелиновых волокон.
304
Рис. 118. Строение нерва.
Л — микрофотография (по
В. В. Яглову): / эпинев-
рий; 2 — периневрий; 5 —
эндоневрий; 4 нервные
волокна миелиновые и без-
миелиновые; 5 — кровенос-
ные сосуды; Б — электрон
ная микрофотография (пре-
парат Т. Н. Радостиной):
/ — миелиновые нервные
волокна; 2 — безмиелиновые
нервные волокна; 3 — про-
слойки соединительной тка-
ни (эндоневрий).
Пучки нервных волокон, осуществляющие связь между различны-
ми отделами нервной системы, называются проводящими путями
спинного мозга: ~
Т' НёщхмГйтьк Кц<лч<иг сход1Г1.1е .по размерам, тонкому строению и
Функциональному значению, лежат в сером веществе группами,
котопые называются ядпами. Спели нейронов спинного мозга мож-
cytus radiculatus), нейриты-зсотор^х покидают спинной мозг в сос^
таве его передних корешков, внутренние клетки (neurocytus $
internus), отростки которых заканчиваются синапсами в пределах
серого вещества спинного мозса^ и:’лучковые клетки (neurocytus
funicularis), аксоны которых проходят в белом веществе обособ-
ленными пучками в»локон несущими нервные импульсы~от~опре-
деленных ядер спинного мозга в его' другие сегменты йлИ в тгоот-
305
22
Рис. 119. Строение спин-
ного мозга.
А — схематический разрез
спинного мозга и спиналь-
ного ганглия (рис. Т. Н Ра
достиной и Л. С. Румянце-
вой): /. 2 рефлекторные
пути сознательных проприо-
цептивных ощущений и ося
зания; 3, 4 рефлекторные
пути проприоцептивных им-
пульсов; 5 — рефлекторные
пути температурной и боле
вой "чувствительности; 6 —
задний собственный пучок;
7 боковой собственный пу-
чок, д передний собственный пучок; 9 задний спинномозжечковый путь; 10 — передний
спинчомозжечковый путь; // — спиноталамический путь; 12 тонкий пучок; /3— клино-
видный пучок; 14 — руброспинальный путь; /5 таламоспинальный путь; 16 вестибулсЖ
спинальный путь; /7 -- ретикулоспинальный путь; 18 тектоспинадьный путь; /9 корти-
ко-спиназьный (пирамидный) боковой путь; 20 - кортико-спинальный (пирамидный)
передний путь; 21 - собственное ядро заднего рога; 22 грудное ядро; ~3. 24 ядра
промежуточной зоны; 25— боковое ядро (симпатическое); 26— ядра переднего рога;
Б микрофотография двигательных нейронов переднего рога.
ветствук>щие отделы головного мозга, образуя_гпроводящие пути.
Отдельные~участкй серого вещества спинного мозга значитель-
но отличаются друг от друга по составу нейронов, нервных воло-
кон и нейроглии.
-у-В _з а_д н их . р о г а_х различают: губчатый слой, желатиноз-
ное вещество. сабстве иное ядро_ заднего рогсГ~й гру<Гное_ ядро.
Между задними и боковыми рогами серое-вещество вдается тяжами
в белое, вследствие чего образуется его сетеобразное разрыхление,
получившее название сетчатого образования.
з()б
у) Г^Лшггып спои зп.лнмх рогов характеризуется широкопетлис-
тым глиальным остовом^вкотором содержится большое количе-
ст’во мелки -< встарочных-нейро+юв.
й "В желатинозном веществе преобладают глиальные элементы.
НервныеПслетки здесь мелкие и количество их незначительно.
Задние рога богаты диффузно расположенными вставочными
клетками. Это мелкие мультиполярные ассоциативные и комиссу-
ральные клетки, аксоны которых заканчиваются в пределах серо-
го 'вещества спинного мозгй той же стороны ^ассоциативные
клетки) или противоположной стороны (комиссуральные клетки).
Нейроны губчатой зоны, желатинозного вещества и вставочные
клетки осу ществляют связь между чувствительнымгГклетками спи-
нальных ганглиев и двигательны^~клёткамй~'/пёредни)Г~ рогов,
замы кая' местные рефлекторные дугг:.$В середине заднего рога рас-
полагяется£o6creeHHQg^adj2oзаднего_рща. Онрсостоит из вставоч-
нну нейронов, аксоны которых переходйт через переднюю белую
спайку, на противоположную^сторону спинного мозга в боковой ка-
натик белого вещества, где они входят в состав 'вентрального
спинномозжечкового и спиноталамического путей щ_ца.правлдются
в мозжечок, «-зрительный бугор (см. рис. 119).
ц).\Груднос ядро состоит из крупных вегавочных нейронов с силь-
но разветвленными дендритами И» акелны выходят в боковой кана-
тик белого вещества той же стороны и в составе дорсального спин-
номозжечкового пути полнимаются__к_мозжечку.
В промежуточной зоне различают|)медиальное промежуточное
ядро, нейриты клеток которого присоединяются к вентральному
спинномоз~жечковому пути той же стороны.—И ^латеральное про-
межуточное ядро, расположенное в б о к о в и х_р О.гдл и_пред-
ста^ляюш е с ц_____группу ассоциативных клеток симпатической
рефлекторной дуги. Аксоны этих клеток покидают мозг вместе с
Lсоматическими двигательными волокнами в составе передних ко-
ре шков й бособляются от них в виде белых соединительных вет-
вей симпатического ствола.
' +В п е р ел.н и.х-ро-1-а х распшшже.цы-£амые крупные, нейроны
сптЦщгр мозга, которые имеют диаметр 100—140 мкм и образуют
значительные по объему ядра. Это, так же как и нейроны ядер боко-
вых рогов, корешковые клетки, поскольку их—нейриты составляют
основную массу волокон передних корешков. В составе смешан-
ных сГГИ)п1омозговь1х нервов они поступают на периферию и обра-
зуют моторные окончания в скелетной мускулатуре. Таким обра-
зом^эти ядра представляют собой моторные соматические центры.
Различают в передних рогах медиальную и латеральную группы
моторных клеток. Первая иннёрвирует-мышцы-туловища и развита
хорошо на всем протяжении спинного мозга. Вторая находится в
области щейного~и поясничного утолщений и иннервирует мышцы
конечностей.
В сером веществе спинного мозга рассеянных Тучковых нейро-
нов много. Аксоны этих клеток вы ход ягщ белое вещество и сразу
же делятся на более длинну.ю восходящую и более короткую нис-
307
с
ходящую ветви. В совокупности эти волокна образуют собственные,
или осповные?_пучки_белого, вещества, непосредственно прилегаю-
щие к серому веществу. По своему ходу они дают много коллате-
ралей, которые^ как и самшетвщ-заканчиваюФсяеинаттсами на дви-
гательных,клетках передних-регов-4—5 смежных сегментов спин-
ного мозга. Собственных пучков три-лары.
Глиоциты спинного мозга. Спинномозговой канал выстлан эпен.
димоцитами, участвующими в выработке~~спиннрмочТовпй _жидкл-
сти. От "периферического концд_эпендимоцита отходит длинный
отро~сток7~вхЬдящий в состав наружной погряничной мем раны
сщцшоПЕмозга.
Основную часть остова серого вещества составляют протоплаз-
матические и волокнистые астроциты. Отростки волокнистых аст-
роцитов бБГхидяг за пределы серого вещества и^вместёТГэлемента-
ми соединительной ткани принимают участие в образовании пере-
городок^ белом веществе-и глиальных-мембран вокруг "кровенос-
н ы х сосудов и на поверхности спинного мо зга. Ол и го де н дрогл и я
входит в состав оболочек нервных водбкон. Микроглия поступает
в спиннои мозг по мере врастания в него кровенОсных~сбсупрв и
распределяется в сером и белом веществе.
ГОЛОВНОЙ мозг
В головном мозге различают серое и белое вещество, но рас-
пределение этих двух составных частей здесь значительно слож-
нее^ чем вдзпинном мозге. Большая часть серого вещества_головно-
го М£зга располагае~тся~~на~пбвёрхности большого мозга и в моз-
же ч ке, образуя wwtopy. Меньшая часть обра.зуе г м н от они еле н н ые
я дра_£твс да-мозга.
Ствол мозга. Проводящие пути и детали строения ствола мозга
изложены в курсах нормальной анатомии и неврологии. В состав
ствола мозга входят продолговатый мозг, мост, мозжечок и струк-
туры среднего И промежутоиного ядра серлгл неществя
ствола мозга состоят из мультиполярных нейронов. Различают
ядра черепных нервов и переключательные яЛТюГК первым отно-
сят ядра подъязычного, добавочного, блуждающего, языкоглоточ-
ного, преддверно-улиткового нервов продолговатого..мозг-я; отво-
дящёгоГТПЩВвого. троиннпппго-нерппр моста -К_чиспу' вторых от-
носятся нижнее, медиальное добавочное и заднее добавочное
оливные ялш продолговатого мозга; верхнее оли в ное ядро, ядра
трапециевидного тела и ядро латеральной петли моста; зубчатое
ядро, пробковидное ядро, ядро шатра, шаровидное ядро мозжечка;
красное ядро среднего мозга и др.
Продолговатый мозг. Продолговатый мозг характеризуется
присутствием перечисленных выше ядер черепных нервов, которые
коНПеитиируЮтсялпеймушественнб в ё1ч> дорсальной части, обра-
зующей дно£У желудочка, Из^числа~пёрек.1юча'1,г.тннътх ядер сле-
дует—отмстить нижние оливы. Они содержат крупные мультипо-
лярные нервные клетки, нейриты которых образуюТсинаттгИческие
ДО
связи с клетками мозжечка и зрительного бугра.В;_нижние_.Ш1ивы
поступаютврлокна от длозжё'жа-. -красного ядра, ретикулярной
формации и спинного мозга, с которыми нейроны нижних олив
связащ>Г особыми волокнами. В центральной области-продолгова-
того мозга располагается важный координационный аппарат го-
ловного мозга — ретикулярная формация.
Ретикулярная "формация начинается в верхней части спинного
мо зга и тянется через пралодговатыи~ мозг, мост, сред ни и мозг,
центральные части зоительного буграГгипоталЛмус и другие облас-
ти, соседние со зрительным бугром. ^1ногочислещ1ые_нервные во-
локна идут в ретикулярной формации в различных на правде идя х и
<в~ббвб купности образу ют сеть. В этой~сети ра спол а га юте я_ ме л к ие
группы мультиполярных нейронов., Нейромы-варьируют по разме-
рам от очень маленьких.до очень больших. Мелкие нейроны_сос-
тавляющие большинство, имеют короткие аксоны, образующие
множество контактов в самой ретикулярной формации. Большие
нейроны характеризуются тем, что их аксоны часто образуют би-
фуркации с одним ответвлением,.идущим вниз вТпингкнГ мозг и
другим — вверх в зрительный бугор или другие базальные области
промежуточного мозга и в большой мозг. Сенсорные волокна рети-
кулярная формация получает из многих источников, таких как
спиноретикулярный тракт, вестибулярные ядра, мозжечок, кора
большого мозга, особенно ее двигательная область, гипоталамус
. и другие соседние области. Ретикулярная, формация представляет
I собой сложный рефлекторный центр и принимает участники конт-
роле над тонусом мышц и стереотипными движениями. 1
Белое вегцёство ~ в~ продолговатом мозге занимает прей му щест-
ве нно вентролатеральное поло же ни е. Qc новные пучки миелиновы х
нервных волокон представлены кортикогспкнальными______пучками
(пирамиды продолговатого,мозга). лежащими в его, вентральной
чаерц Блоковых областяЗГтесполагаются'веревчатые тела, образо-
ванные волокнами спинно-мозжечковых путей. Отсюда эти волокна
поступаютллшзжЕВЗк.'Отростки нейронов ядер клиновидного и тон-
кого пучкбВ’В виде внутренних дуговых волокон пересекают рети-
кулярную формацию, перекрещиваются по средней линии, образуя
шов, и направляются к зрительному бугру.
Мост делится на дорсальную (покрышковую) и вентральную
части. Дорсальная часть содержит волокна проводящих путей про-
долговатого мозга, ядра V—VIII черепных нервов, ретикулярную
формацию моста. В вентральной части располагаются собственные
ядра моста и волокна пирамидных путей, идущие продольно. Ядра
моста построены из мультиполярных нейронов, размеры и форма
которых в различных ядрах различны. К переключательным ядрам
задней части моста относятся верхнее оливное ядро, ядра трапе-
циевидного тела и ядро латеральной петли. Центральные отростки
нейронов улиткового узла заканчиваются на переднем и заднем
улитковых ядрах продолговатого мозга. Аксоны нейронов передне-
го улиткового ядра заканчиваются в верхнем оливном ядре и яд-
рах трапециевидного тела. Аксоны верхнего оливного ядра, заднего
309
улиткового ядра и ядер трапециевидного тела образуют латераль-
ную петлю. В состав последней входят также клетки ядра лате-
ральной петли и их отростки. Латеральная петля заканчивается
в первичных слуховых центрах — нижнем холмике крыши сред-
него мозга и медиальном коленчатом теле.
Средний мозг состоит из крыши среднего мозга (четверо-
холмия), покрышки среднего мозга, черного вещества'"иножек
мозга7~Четверохолмие состоит из пластинки крыши, двух ростра-
"льных- {верхних) и двух каудальных (нижних) холмиков. Ростра-
льные холмики (звено зрительного анализатора) характеризуют-
ся послойным расположением нейронов, каудальные (часть слу-
хового анализатора) построены по ядерному принципу. В по-
крышке среднего мозга находится до 30 ядер и в том числе
красное ядро. Красное ядро состоит из крупноклеточной и м<мнт
клеточной частей. Крупноклеточная часть .получает импульсы из
базальных ганглиев конечного мозга и передает сигналыпо руб-
роспинальному тракту в спинной мозц^а^др коллатералям_рубро-
ci шпального тракта —в ретикулярную формацию. Мелкие нейро-
ны краского»! дра возбуждаю.тся .импульсами из мозженюа по
церебеллорубрадьному тракту и посылают импульсы в рети-
кулярную формацию. Черное вещество получило свое название
в связи с тем, что в его мелких веретенообразных нейронах
содержится меланин. Ножки мозга образованы миелиновыми во-
локнами, идущими от коры большого мозга.
Промежуточный мозг. В промежуточном мозге преобладает по
объему зрительный бугор. Вентрально от него располагается бо-
гатая мелкими ядрами гипоталамическая (подбугорная) область.
Зрительный бугор содержит много ядер, отграниченных друг от
друга прослойками белого вещества. Ядра связаны между собой
ассоциативными волокнами. В вентральных ядрах таламической
области заканчиваются во сходя пЩеднувсТвительные пути. От них
нервныеттстпутГьсыТТере даются >ре. Нервные импульсы к зритель-
ному бугру иЗ"головного мозга идут по экстрапирамидному двига-
тельному пути.
В каудальной группе ядер (подушка зрительного бугра) закан-
чиваются волокна зрительного пути.
Гипоталамическая область — важный вегетативный центр
головного мозга, регулирующий температуру, кровяное давление,
водный, жировой обмен и др. Гипоталамическая область у челове-
ка состоит из 7 групп ядер (см. гл. XVII).
Мозжечок
Мозжечок представляет собой центральный орган ‘равновесия
и координации" движений. Он связан-со С'ГВОлбм мозга афферен-
тными и эфферентными—проводящим и "пу ч ка ми,' образу ющи ми в
совокупности три пары йОЖетгмпзжечкаг'Ш поверхности мозжеч-
ка мчого извилин и бороздок, которые значительно увеличивают
ее площадь (у взрослых людей 975—1500 см2). Борозды и изви-
310
лины создают на разрезе характерную для мозжечка картину
«древа жизни». Основная масса серого вещества в мозжечке
располагается на поверхности и образует его кору. Меньшая часть
серогоГЪещества лежит глубоко в белом веществе в виде цен-
тральных ядер. В центре каждой извилины имеется тонкая
прослойка белого вещества, покрытая слоем серого вещества —
корой.
Вскоре мозжечка различают три сюя- наружный—молекуляр-
ный ("stratum moleculare), средний — ганглионарный слой или слой
грушевидных нейронов (stratum neuronum piriformium) и_внутрен-
ний-— г^рч'кти (stratum granulosum) (рис. 120, 1,11). Jjjyiue-
видные нейроны (neuronum piriforme) имеют нейриты, KOTopjye^ по-
кидая кору мозжечка, образуют начальное звено его эфферентных
тормозных путей. В га н г л и о н а р tw м с^л о е клетки распо-
лагаются строго~в~~одйн~ряд~Тсм. рис. 120,//). От их крупного
(60x35 мкм) грущевидного_теда а -молекулярный слои отходит
2—3 дендрита, которые, обильно ветвясь, пронизывают всю толщу
^молекулярного слояТ ~ТТсе ветви—дендритов располагаются только
i ОДНОЙ—плоскости,—перпенди кулярной к. напра вле ни ю из вил и н,
поэтому при поперечном и продольном сечении извилин дендриты
грушевидных клеток, ^н^глядят различно. От основания тел этих
клеток отхотят нейриты, проходящие через зернистый слой коры
мозжечка в белое вещество и заканчивающиеся на клетках ядер
мозжёчк£>Л5 пределах зернистого, слоя! от них отходят коллате-
рдди,-которые, возвращаясь в 1англщунарный~~сл6й; вступают в си-
наптическую .связь с. соседними грушевидными нейронами.
М о л е к у л я р н ы й с ло^ содержит дваткнявинх. нита ней-
ронов: корзинчатые и звездчатые. нейроны/(neuro-
num corbiferum) находятся в нижней трети молекулярного _слоя.
Это неправильной формы мелкие клет|\Ц размером около 10—
20 мкм. Их тонкие длинные дендриты'-етвятгя преим-ущеетвен1у^^
в плоскости, распрдож.еииой-1)опере-иыа ж-извилине. Длинные*'?!^-
Рифы клеток всегда идутдюперек-иавилины и. параллельно поверх-
ности ьад_грушевидными нейронам^ Онибтдают коллатерали,'
спекающиеся-к-телам грушевидных нейронов^и совместно с дру-
гими воюкнамигттгптгбплётая'Трушевидные нейроны, формируют
на_цих характерную-структуру корзинок^ нервных волокон (corbis
neurofibrarum). Активность нейритов корзинчатых-нейронов- вызы-
ваес-торможение грущщшдцых нейронов.
^вездчатые нейроны (neuronum stellatum) лежат выше корзий-
чатых й^УйаюТ двух типов.' Мелкие звездчатые унейроны снабжены
тонкими кб'рбткимиЗдендритами и' слаборазветвленными нейрита-
ми, образующими синапсы~~нд2дёнДрттта^~'груше видных клеток.
Крупные звездчатые нейроны вЪтличие, 0ТдЛ)^£ких имеют длинные
и сильно разветвленные дендриты и нейриты. Ветви-их нейритов к
соединяются с дендритами грушевидных клеток, но некоторые из
них достигают тел грушевидных клеток й’вхсдят в-состав так на-
зываемых корзйноКи. Корзинчатые, и звездчатые । ейроны молеку-
лярного слоя представляют собой единую систему вставочных ней-
•II
Рис. 120. Строение мозжечка.
/ микрофотография' А молекуляр-
ный слои; Б — ганглионарный слой
(слой грушевидных нейронов); В — зер-
нистый слой: / — тела 1рушевидных
нейронов; 2 - разветвления дендритов
грушевидных нейронов. // — схема
синаптических связей нейронов в коре
мозжечка (по Сентаготаи): А — изви-
лина мозжечка (пунктиром на про-
дольном сре*е ограничены участки,
показанные на рис. Б и Б) ; Б — клубо-
чек зернистого слоя; В - синапсы в
молекулярном слое. / — грушевидный
нейрон; 2 — дендриты грушевидных
нейронов; 3 — нейрит грушевидного
нейрона; 4—корзинчатые нейроны; 5 —
дендриты и 6 — аксон корзинчатого
нейрона; 7 — звездчатый нейрон; 8 —
большие звездчатые нейроны; 9 — ден-
дриты большого звездчатого нейрона;
10 — аксон большого звездчатого ней-
рона: // — зерновидные нейроны
(клетки-зерна); 12 аксон клетки-
зерна; 13 — дендриты клеток-зерен;
14 — моховидные (лазящие) нервные
волокна; 15 — синаптические пузырьки
В черный цвет окрашены тормозящие
нейроны.
/л£&
понов передающую тормозные-нервные импульсы на..дсндриты и
тепа грУУ11еви,пныу,ипогиппи, поперечной извилинам.
бчень богат нейронами зернистый слой. Он характери-
зуется особыми' клеточными формами, получившими-.названи£-зер-
иовипных нейронов, или к.летакт.зере.н. (neuronum granuliformis).
У них маленький по объему (5—8 мкм в диаметре) бедный цито-
плазмой перикарион с круглым крупным ядром. Клетка имеет?!—
4 коротких дендрита, заканчивающихся в этомже_£лое концевыми
ветвлениями в виде лапки птицы. Вступая в синаптическую связь с
окончаниями приходящих в мозжечок возбуждающих афферент-
ных (моховидных) волокон, дендриты клеток-зерен образуют ха-
ракте 1ные структуры, именуемые клудочкулш^огшечка (glome-
rulus"'cerebellaris).
Нейриты клеток-зерен проходят в молекулярный слой и в нем
Т-образно пелятся~на две ветви, ориентированные параллельно по-
ве"рхнос1 и коры вдоль извилин мозжечка. Преодолевая большие
расстояния, ^эти_варалледьные подокна _пеаесекаютветю1СНИЯД.енд-
ритов многих грушевидных клеток и образуют с ними и дендрита-
тЯ^ТорзйнчатъиГи звездчатых -нейронов синапсы. Таким образом
^нейриты клеток-зерен-Передают возбуждение. гюлученное__ими от
моховидных-волоксшу на значительное расстояние многим груше-
видным клеткам.
Вторым типом клеток зернистого слоя мозжечка являются тор-
мозные звездчатые неиооцы (neuronum stellatum mag-
num). Различают два вила таких клеток: с корохкимИ-И. длинными
нейритами?//ейрощу с короткими нейритами (neuronum stellatum
breviacsonicum) лежат вблизиганглйондрного слоя. Их разветвлен-
ные таымты распростраИЯУбтТй в молекулярном слое и образуют
синдцсы с2~ПйрЙЛЛелвными—воЗЯгёййми_— а ксонами_ кде1ою.зере н.
С '^Неипитьгнйттрзтст^зер'нистыи'слой к Тслубочкам мозжечка и
закачиваются синапсами на концевых ветвлениях дендритов кле-
1Рк<зебён проксимальнее синапсов моховидных волокон. Возбуж-
дение звездчатых нейронов может блокировать импульсы, посту-
пающие по моховидны^» волокнам,'^Немногочисленные звездчатые
нейроны с длинными_11Сир-1^аМ1г (пёпГопит stellatum longiaxsoni-
гтптЬ^ймеют оби льновегвяшиеся в зернистом слое дендриты и ней-
риты, вы холящие н белое вещество. Предполагают, что эти клетки
обеспечиваютсвчзхьМ£жду-радли.чнымцобластями коры мозжечка.
Третин-вид-клеток составляют веретеновидные -горизонтальные
KsieiKOL (neuronum fusiformie horizontale). Они_встречаются пре-
имущественно .между—зернистым и. гднглионарным-сдоями, имеют
небольшое аытянутое_.тело, -от которого—в- -обе._стороны отходят
длинные горизонтальные дендриты,_заканчивяюшиеся _н ганглио-
нарном и зернистом слоях.Нейриты же этих клеток дают коллате-
раЛй 5* зернистый слой «и.у’ходЯТ д белое-вещеетво.
---Афферентные волокна, поступающие в кору мозжечка, пред-
ставле_ны двуЮя виттамиГ^— моховиднылаГи так называемыми лазя-
щими волбюшКГи._Моха5ц<)нь<е волокна идут'в^еоитаве^едиаомоз
жечкового и мостомозжечкового путей____и опосредованно через
клетки-зерна оказывают на_грушевидные клетки возбуждающее
действие/Они заканчиваются._в_ клубочках (glomerulus) зернистого
слоя_мозжёчка, где вступают в контакт с дендритами клеток-Уёрен.
Каждое волокно дает ветви к многим клубочкам мозжечка, и каж-
дый клубочек получает ветви от многих моховидных волокон. Ней-
риты клеток-зерен по параллельным волокнам молекулярного слоя
передают импульс дендритам грушевидных;-корзинчатых, звезд-
чатых нейронов, больших' звёздчатых нейронов зернистого слоя.
Лазящие валснсна поступают-'в~ к'орумозжечка, по-видимому, по
спийно-мозжечковому и вестибуломозжечковому пу ям.—Они пере-
с’ёкатот'зернисгыи слой, прйлегают_к грушевидным нейронам и сте-
лются пр их ендритам. заканчиваясьла их поверхности-синапсами.
Лазящие волокна передают возбуждение непосредственно^груше-
видным нейронам. Дегенерация грушевидных нейронов ведет к рас-
стройству .координации движений.
Таким образом, возбуждающие импульсы, поступающие в кору
мозжечка, достигают грушевидных нейронов или непосредственно
по лазящим волокнам, или по параллельным волокнам клеток-зерен
(см. рис. 120). Торможение — функция звездчатых нейронов мо-
лекулярного слоя, кбрзинчатых нейронов, а такжё~больших звезд-
чатых нейронов ~зернистого-слоя. Нейриты двух первых, следуя
поперек извилин'й-тормозя-дкгивность грушевидных клеток, огра-
ничивают их возбуждение узкими дискретными зонами коры. По-
ступление в кору мозжечка возбуждающих сигналов по моховид-
ным волокнам, через клетки-зерна и параллельные волокна может
быть прервано тормозными синапсами больших звездчатых нейро-
нов, локализованными на концевых ветвлениях дендритов клеток-
зерен проксимальнее возбуждающих синапсов.
Кора мозжечка содержит различные глиальные эде-
ме йты.“В ’зёрнистём-сЛй?-имеются’ волокнистые и плазматиче-
ские астроциты. Ножки отростков волокнистых астроцитов образу
ют периваскулярныё мембраны. Во всех слоях в мозжечке имеют-
ся элементы олигодендроглии-Особенно богаты этими клетками
зернистый слой и белое вещество мозжечка. В ганглионарном слое
между грушевидными нейронами лежат глиальные клетки схем-
ными ядрами. Отростки "Этих клеток направляются к поверхности
коры и'образуют^гЛиаль1П>генолбкна молекулярного слоя мозжечка,
поддерживающие ветвления дендритов грушевидных клеток (glio-
fibra sustentans). Глиальные макрофаги в большом количестве со-
держатся в молекулярном и ганглионарном слоях.
Кора большого мозга
Развитие коры больших полушарий (неокортекса) млекопита-
ющих и человека в эмбриогенезе происходит из вентрикулярной
герминативной зоны конечного мозга, где расположены малоспе-
циализированные пролиферирующие клетки. Из этих клеток диф-
ференцируются нейроциты неокортекса. При этом клетки утра-
чивают способность к делению и мигрируют в формирующуюся
314
корковую пластинку вдоль вертикально ориентированных волокон
эмбриональных радиальных глиоцитов, исчезающих после рожде-
ния. Вначале в корковую пластинку поступают нейроциты будущих I
и VI слоев, т.е. наиболее поверхностного и глубокого слоев коры.
Затем, как бы раздвигая эту первичную корковую закладку, в нее
встраиваются в направлении изнутри и кнаружи последовательно
нейроны V, IV, III и II слоев. Этот процесс осуществляется за счет
образования клеток в небольших участках вентрикулярной зоны в
различные периоды эмбриогенеза (гетерохронно). В каждом из
этих участков образуются группы нейронов, последовательно вы-
страивающихся вдоль одного или нескольких волокон радиальной
глии в виде колонки (рис. 121, 7). Подобные, так называемые онто-
генетические колонки в дальнейшем служат основой для формирова-
ния функциональных интегративных единиц неокортекса: мини- и
макроколонок. Для установления сроков формирования в эмбрио-
генезе различных групп нейронов применяют радиоизотопный ме-
тод (рис. 121, 11).
Строение. Кора большого мозга представлена слоем серого ве-
щества толщиной около .З -мм. Наиболее сильно развита она в пе-
редней центральной извилине, гдё~толщина корьГдостигает 5 мм.
Обилие борозд И-Извилин значительно увеличивает площадь серого
вещества головного мезга. В коре содержится около 10—14 млрд,
нервных клеток. Различные участки ее, отличающиеся друг от дру-
га некоторыми особенностями расположения и строения клеток
(цитоархитектоника), расположения волокон (миелоархитектони-
ка) и функциональным значением^ называются полями. Они пред-
ставляют собой ме.с.та-высшего анализа и-еинтеза нервных импуль-
сов. Резко очерченные границы между ними отсутствуют. Для ко-
ры характерно расположение клеток и волокон слоями (рис. 122, А,
Б, В).
Цитоархитектоника коры большого мозга. Мультиполярные
нейроны коры весьма разнообразны по_форме._ Среди них можно
выделить. ; j гвездчатые, веретенообразные, паукообраз-
ные и гори юнтадьные нейроны. Пирамидные нейроны составляют
основную и наиболее специфическую для коры большого мозга
форму (см. рис. 122, Л). Размеры их варьируют от 10 до 140 мкм.
Они имеют вытянутое треугольное тело, вершина которого обращена
к -поверхности коры. От вершины и боковых поверхностей тела
отходят дендриты, заканчивающиеся—а--различных слоях серого
вещесгва. От основания пирамидных клеток берут начало 'веири1
т« "в одних" кдетках .короткие,.образующие ветвления в пределах
данного участка коры, в других — длинные, поступающие в белое
вещество.
Пирамидные клетки разли ных. сдоев коры отличаются раз-
мерами и имеют различное функциональное значение. Мелкие
клетки представляют собой .вставочные нейроны, нейриты которых
связывают отдельные участки корь одного^ полушария (ассоциа-
тивные нейроны) или двух, полушарий (комиссуральные нейроны).
Эти клетки встречаются в разных количествах во всех слоях коры.
315
Рис. 121. Эмбриогенез коры полушарий большого мозга (неокортекса).
/ схема пространственно-временного образования нейронов в неокортексе млекопи-
тающих (представлены две соседние колонки коры): А — кора (корковая пластинка);
Б белое вещество; А — вентрикулярная герминативная зона; Г, Д две рядом лежа-
щие колонки неокортекса; / VI формирующиеся слои коры: / мало дифференциро-
ванные делящиеся клетки; 2 — радиальные эмбриональные глиоциты; 3 — молодые ней-
роны, мшрирующие в корковую пластинку; 4 — группы нейронов, последовательно обра-
зующиеся в различные сроки эмбриогенеза (по К. Ю. Резникову с изменением).
Особенно богата ими кора большого мозга человека. Нейриты
крупных пирамид принимают участие.д.образовании пирамидных
путей, проецирующих импульсы в соответствующие центры ствола
и спинного мозга.
Нейроны коры_расположены нерезко отграниченными слоями.
К а ж дыи~с пой., х а ра к те рй^ется~ттреоб71а даниекг какого-либо одного
вида клеток. В двигательной зоне коры различают 6 основных_слр-
ек^\.^^мплекулярнын (lafnTna riiolecularis), 11 — наружный зернис-
rww_(lamina granularis externa). III — пирамидных neupoHVir (lami-
na piramidalis), IV — внутренний зернистый (lamina granularis •
interna), V — ганглионарный (lamina ganglionaris), — слой
полиморфных клеток (lamma multiformis) (см. рис. 122, Л).
316
Рис. 121 (продолжение)
И — радиоавтограф неокор-
текса новорожденной мыши,
получившей Н-тимидин на
16-е сутки эмбриогенеза. По-
лутонки й срез, окрашенный
толуидиновым синим, v 2000.
/ — группы меченных изото-
пом нейронов, образовавших-
ся на 16-е сутки эмбриоге-
неза; 2 нейроны бет метки,
образовавшиеся в другие сро-
ки эмбриогенеза (препарат
К. Ю. Резникова, Г. Д. На-
заре вской).
В период эмбрионального развития первыми на_6-м месяце
дифференцируются У~и VI слои, а 11,111 и IV слои развиваются прзд-
нее — на^8_-м_месяце внутриутробного развития.
Молекулярный слой коры содержит небольшое количество мел-
ких ассоциативны хкдеток веретеновидной формы (см. рис. 122, А,
В). Их нейриты, проходят параллельно поверхности мозга в соста-
ве тангенциального сплетения нервных волокон молекулярного
слоя._Однако основная масса волокон этого сплетения представле-
на ветвлениями дендритов нижележащих слоев.
Наружный зернистый г/юй-обрптовптгмепк'ими нейронами диа-
метром около 10 мкм, имеюгцими округлую, угловатую и пирами-
да льную Форму. и—шездчатыми-шейро-цйИяи:—
Дендригы этих клеток поднимаются в молекулярный слой.
Нейриты или уходят в белое вещество, или^ образуя дуги, так-
же поступаю!- д_тангенциа пьное сплетение волокон молекуляр-
ного слоя.
М7
Рис. 122. Схема цито- и
миелоархитектоники коры
полушарий большого моз-
га человека.
А - схема расположения
клеток (цитоархитектоника);
Б — схема расположения
волокон (миелоархитектони-
ка):
Z7
Рис. 122 Г (продолжение)
Г — электронная микрофотография белого вещества полушарий большого мозга (препарат
И. Г. Павловой): 9 — миелиновые волокна; 10 — безмиелиновые возокна.
Рис. 122 В (продолжение).
В — схема межнейрональных связей в коре полушарии большого мозга. /, 11, 111, IV, V, VI —
слои (пластинки) коры. Афферентные волокна (обозначены красным): 1 — кортико-корти-
кальные; 2 — специфические; 2а — зона распространения специфических афферентных
волокон; 3 — пирамидные нейроны (обозначены зеленым); За — заторможенные пирамид-
ные нейроны (обозначены зеленым с черным пунктиром); 4 — тормозные нейроны и их
синапсы (обозначены черным цветом); 4а — клетки с аксональной кисточкой; 46 — малые
корзинчатые клетки; 4в — большие корзинчатые клетки; 4г — клетки, формирующие аксо-
аксональные синапсы; 4д — клетки с двойным букетом дендритов (тормозящие тормозные
нейроны); 5 — шпиковые звездчатые клетки (окрашены в желтый цвет), возбуждающие пи-
рамидные нейроны непосредственно и путем стимуляции клеток с двойным букетом дендри-
тов; б — наружный главный слой волокон; 7 — полоска внутренней зернистой пластинки:
8 — полоска внутренней пирамидной пластинки;
(
Самый широкий слой коры большого мозга — пирамидный. Он
особенно хорошо развит в прецентральной извилине. Величина пи-
рамидных клеток последовательно увеличивается в пределах 10—
40 мкм от наружной зоны этого слоя к внутренней.'От верхушки пи-
рамидной клетки отходит главный дендрит, который располагается
в молекулярном слое. Дендриты, берущие начало от боковых по-
верхностей пирамиды и ее основания, имеют незначительную ДДИ-,
ну ио(^зуют~синапсь1 со смежными клетками-эдрго слоя. Нейрит'
пирамиднбгГ клетки всегда отходит от ее основания. В мелких
.клетках он остается в пределах коры; аксон ^ке^принадлежащий
крупной пирамиде, обычно формирует миелцндвое-асооциативное
|^пй2ко1йиссуральнае--вояок11П.- идущее в белое вещество.
Внутренний зернистый слои в некоторых полях коры развит
<5чень сильно (например, в зрительной зоне коры). Однако он мо-
жет почти отсутствовать (в прецентральной извилине). 1Этот слой
образован мелкими звездчатыми нейронами. В его состан~Входит
большое количество горизонтальных волоко^.
Ганглионарный слой коры образован крупными пирамидами,
причем область прецентральной извилины содержит пн антские пи-
рамиды, описанные впервые киевским анатомом В. А. Бецем в
1874 г. (клетки Беца). Это очень крупные клетки, достигающие в
высоту 120 мкм и в ширину 80 мкм. В отличие от других пирамид-
ных клеток коры гигантские пирамиды характеризуются наличием
крупных глыбок хроматофильного вещества. Нейриты клеток этого
слоя образуют главную часть кортико-спинальных и_кортико-нук-
леарныл_ путей и оканчиваются синапсами на клетках моторных
ядер.
Прежде чем пирамидный путь покинет кору, от него отходит
множество коллатералей. Аксоны от гигантских пирамид Беца да-
ют коллатерали, посылающие тормозящие импульсы в саму кору.
Коллатерали волокон пирамидного пути идут в полосатое тело,
красное ядро, ретикулярную формацию, ядра моста и нижних олив.
Ядра моста и нижйих олив передают сигнал в мозжечок. Таким
образом, когда пирамидный путь передает сигнал, вызывающий
моторную активность, в спинной мозг, одновременно сигналы по-
лучают базальные ганглии, ствол мозга и мозжечок. Помимо кол-
латералей пирамидных путей, существуют волокна, которые идут
непосредственно от коры к промежуточным ядрам: хвостатому те-
лу, красному ядру, ядрам ретикулярной формации ствола мозга
и др.
i Слои полиморфных клеток образован нейронами различной,
преимущественно веретенообразной, формы. Внешняя зона этого
слоя содержит более крупные клетки. Нейроны внутренней зоны
мельче и лежат на большом расстоянии друг qt друга. Нейриты
клеток полиморфногохлоя-улодят в белое вещество в составе эф-
ферентных путей головного га. Дендриты достигают молеку-
лярного СЛОЯ~~КО рЫ.Д
Крупные пирамидные клетки являются основными нейронами, к
которым по центрифугальным волокнам приходят импульсы из
дру| их отделов центральной нервной системы и передаются через
синапсы на их дендриты и тела. От больших пирамид импульс ухо-
дит по аксонам, формирующим центрипетальные эфферентные пу
ти. Внутри коры между нейронами формируются сложные связи^
(см. рис. 122, В).
Исследуя ассоциативную кору, составляющую 90% неокортек-
са, Сентаготаи и представители его школы установили, что струк-
турно-функциональной единицей неокортекса является модуль
вертикальная колонка диаметром около 300 мкм. Модуль орга-
низован вокруг кортико-кортикального волокна (см. рис. 122,
В, 7), представляющего собой волокно, идущее либо от пирамид-
ных клеток того же полушария (ассоциативное волокно), либо от
противоположного (комиссуральное). В модуль входят два тала-
мо-кортикальных волокна (2) специфических афферентных во-
локна, оканчивающихся в IV слое коры на шипиковых звездчатых
нейронах и базальных дендритах пирамидных нейронов. Каждый
модуль, по мнению Сентанотаи, подразделяется на два микромоду-
ля диаметром менее 100 мкм. Всего в неокортексе человека при-
мерно 3 млн. модулей. Аксоны пирамидных нейронов модуля про-
ецируются на три модуля той же стороны и через мозолистое тело
на два модуля противоположного полушария. В отличие от специ-
фических афферентных волокон, оканчивающихся в IV слое коры,
кортико-кортикальные волокна образуют окончания во всех слоях
коры и, достигая 1 слоя, дают горизонтальные ветви, выходящие
далеко за пределы модуля. Помимо специфических афферентных
волокон, на выходные пирамидные нейроны (см. рис. 122, В, 3)
возбуждающее влияние оказывают шипиковые звездчатые нейроны
(см. рис. 122, В, 4). Различают два типа шипиковых звездчатых
клеток: 1) шипиковые звездчатые нейроны фокального типа, обра-
зующие множественные синапсы на апикальных дендритах пира-
мидного нейрона (правая сторона рисунка), и 2) шипиковые звезд-
чатые нейроны диффузного типа, аксоны которых широко ветвятся
в IV слое и возбуждают базальные дендриты пирамидных нейро-
нов. Коллатерали аксонов пирамидных нейронов (не показанные
на схеме) вызывают диффузное возбуждение соседних пирамид.
Тормозная система модуля представлена следующими типами
нейронов: 1) клетки с аксональной кисточкой (см. рис. 112, В, 5)
образуют в I слое множественные тормозные синапсы на горизон-
тальных ветвях кортико-кортикальных волокон; 2) корзинчатые
нейроны — тормозные нейроны, образующие тормозящие синапсы
на телах практически всех пирамид. Они подразделяются на малые
корзинчатые нейроны (см. рис. 122, В, 6), оказывающие тормозя-
щее влияние на пирамиды II, III и V слоев модуля, и большие кор-
зинчатые клетки (7), располагающиеся на периферии модуля и
имеющие тенденцию подавлять пирамидные нейроны соседних мо-
дулей; 3) аксоаксональные нейроны (см. рис. 122, В. 8), тормозя-
щие пирамидные нейроны II и III слоев. Каждая такая клетка об-
разует тормозящие синапсы на начальных участках аксонов сотен
нейронов II и III слоев. Они тормозят, таким образом, кортико-
11 гистология
321
кортикальные волокна, но не проекционные волокна нейронов V
слоя; 4) клетки с двойным букетом дендритов (см. рис. 122, В <,)
располагаются во II и III слоях и, тормозя практически все тормоз J
ные нейроны, производят вторичное возбуждающее действие на пи-
рамидные нейроны. Ветви их аксонов направлены вверх и вниз и рас
пространяются в узкой колонке (50 мкм). Таким образом, клетка с!
двойным букетом дендритов растормаживает пирамидные нейроны
в микромодуле (в колонке диаметром 50—100 мкм). Мощный воз
буждающий эффект фокальных шипиковых звездчатых клеток
объясняется тем, что они одновременно возбуждают пирамид-
ные нейроны и клетку с двойным букетом дендритов. Таким обра-
зом, первые три тормозных нейрона тормозят пирамидные клетки,
а клетки с двойным букетом дендритов возбуждают их, тормозя
тормозные нейроны.
Система тормозных нейронов играет роль фильтра, тормозящего
часть пирамидных нейронов коры.
Кора различных полей характеризуется преимущественным
развитием тех или других ее слоев. Так, в моторных центрах коры,
например в передней центральной извилине, сильно развиты III, V
и VI слои и плохо выражены II и IV. Это так называемый агра-
нулярный тип коры. Из этих областей берут начало нисхо-
дящие проводящие пути центральной нервной системы. В чувстви-
тельных корковых центрах, где заканчиваются афферентные
проводники, идущие от органов обоняния, слуха и зрения, слабо
развиты слои, содержащие крупные и средние пирамиды, тогда как
зернистые слои (II и IV) достигают своего максимального развития.
Это гранулярный тип коры.
Миелоархитектоника коры. Среди нервных волокон коры полу-
шарий большого мозга можно выделить: ассоциативные волокна,
связывающие отдельные участки коры одного полушария, комис-
суральные, соединяющие кору различных полушарий, и проекцион-
ные волокна, как афферентные, так и эфферентные, которые свя-
зывают кору с ядрами низших отделов центральной нервной систе-
мы (см. рис. 122, Б). Эти волокна в коре полушарий образуют
радиальные лучи, заканчивающиеся в пирамидном слое. Кроме уже
описанного тангенциального сплетения молекулярного слоя, на
уровне внутреннего зернистого и ганглионарного слоев расположе-
ны два тангенциальных слоя миелиновых нервных волокон —
внешняя и внутренняя полосы, которые, очевидно, образуются
концевыми ветвлениями афферентных волокон и коллатералей
нейритов клеток коры, таких как пирамидные нейроны. Вступая в
синаптические связи с нейронами коры, горизонтальные волокна
обеспечивают широкое распространение в ней нервного импульса.
Строение коры в различных отделах большого мозга сильно варьи-
рует, поэтому детально^ изучение ее клеточного состава и хода
волокон является предметом специального курса. Кора полушарий
головного мозга содержит мощный нейроглиальный аппарат, вы-
полняющий трофическую, защитную, опорную и разграничитель-
ную функции.
322
АВТОНОМНАЯ (ВЕГЕТАТИВНАЯ) НЕРВНАЯ СИСТЕМА
Часть нервной системы, контролирующая висцеральные санк-
ции организма, такие как моторика и секреция органов пищевари-
тельной системы, кровяное давление, потоотделение, температура
тела, обменные процессы и др., называется автономной, или веге-
тативной, нервной системой. По своим физиологическим осо-
бенностям и морфологическим признакам вегетативная нервная
система делится на симпатическую и парасимпатическую. В боль-
ши нстве случаев обе системы одновременно—принимают участие в
иннервации органов.
Вегетативная нервная система состоит из центральных отде-
лов2 представленных ядрами головного и сгтинпогтгмозга; тгпери-
ферических: нервных стволов, узлов (ганглиевУ и сплетений:
Ядра^центрального отдела вегетативной нервной системы нахо-
дятся в среднем~гГпрбдблговатом мозге, а также в боковьогрогах
грудных^- поясничных й крестцовых сегментов оГйнного мозга.
К симпатической нервной системе опюсятся^ вегетативные ядра
боковых рогов грудного и верхнёпбясничного опделов спинного
мозга, к парасимпатической — вегетативные ядра ТП~УП~, IX И X
пар черепных нервов и вегетативные ядра крестцового отдела
спинного мозга. Мультиполярные нейроны ядер центрального от-
дела представляют собой ассоциативные нейроны рефлекторных
дуг вегетативной нервной системы (рис. 123). Их_Ц£ЙВИты покида-
ют центральную нервную систему через передние_корещки спинно-
го мозга и Ли черепные нервы и оканчиваются синапсами на нейро-
нах одного из периферических вегетативных ганглиев. Это пре-
ганглионарные волокна вегетативной нервной системы; ^обычно
миелиновые .Пре ганглионарные волокна симпатической и парас и м-
патической вегетативной нервной системы — холинергические. Их
терминали содержат мелкие светлые синаптические пузырьки (40—
60 нм) и одиночные крупные темные везикулы (60—150 нм).
Периферические узлы вегетативной нервной-системы лежат как
вне ор7анбв~ (симпатические паравертебральные древертебралfa-
ные ганглии, парасимпатические узлы головы), так иjg.стенке ор-
ганов в составе интрамуральных нервных сплетений пищевари-
тельного тракта,__£ердид,. матки, мочевого, пузыря и др.
Паравертебральные ганглии расположены по обе стороны по-
звоночника^ со своими СоедгшцУ^КныКнГствЬлами образуют сим-
патические цепочки.
Пре ее рте бральные ганглии образуют кпереди дт брюшной аор-
ты и ее главных ветвей брюшное сплетение, в состав которого вхо-
дят чревный, верхнийТУрыжеечный и нижний брыжеечный ганглии.
Вегетативные ганглии снаружи покрыты соединительнотканной
капсулойГТГрОслОиКТГ соединительной тка'нй'проникают в паренхи-
му узла, образуя его остов. Узлы состоят из мультиполярныХ~нерв-
ных клеток, весьма разнообразных по форме и величине. Дендри-
ты нейронов многочисленны и сильно ветвятся. Аксрньг_в_составе
постганглионарных (обычно безмиелиновых) волокон поступают в
323
I
7
Рис. 123. Простая вегетативная рефлекторная цуга (рис. В. Г. Елисеева, Ю. И. Афа-
насьева, Е. Ф. Котовского).
/ — спинной мозг; 2 спинномозговой узел; 3 — передний корешок; 4 — задний рог; 5 —
боковой рог; 6 передний рог; 7 - чувствительный (афферентный) нейрон соматической
и симпатической нервной системы; 8 — центральный (эфферентный) нейрон вегетативной
нервной системы; 9 — двигательный (эфферентный) нейрон передних рогов; 10 — узел сим-
патического ствола; // — узел солнечного сплетения; 12 — интрамуральный узел (узел
нервного сплетения пищевода); 13, 14. /5 — периферические (афферентные) нейроны
вегетативной нервной системы; 16 — преганглионарные волокна эфферентного пути веге-
тативной нервной системы; 17 — постганглионариые волокна эфферентного пути вегета-
тивной нервной системы; 18 афферентный путь соматической нервной системы; 19 — стен-
ка пищевода; 20 - поперечнополосатая скелетная мышечная ткань; 21 — микроскопиче-
ское строение периферических узлов вегетативной нервной системы; 22 — мульти полярная
нервная клетка; 23 — клетка глии; 24 — нервное волокно.
соответствующие внутренние органы. Каждый нейрон и его отро-
стки- окружены—глиальной—оЙозочкрй. Наружная поверх нос ть
глиальной оболочки покрыта базальной мембраной, кнаружи от
324
которой расположена тонкая соединительнотканная-еболочка^Пре-
ганглйонарные во.юьла. лсхуная в ..>>.< »*-!<*! оу к >иппг ганглий, закан-
чиваются на дендритах или перикарионах нейроноваксодендритиче-
скими либо аксосоматическими синапсами. Синапсы микроскопи-
чески выявляются в виде варикозов волокна или терминальных
утолщений. Электронно-микроскопически пресинаптических полюс
характеризуется типичными для холинергических синапсов про-
зрачными^ мелкими синаптическими пузырьками (40—60 нм!, и
оди1ючными крупными (80—150 нм) темными везикулами.
Цитоплазма нейронов симпатического ганглия содержит кате-
холамины, о чем свидетельствуют наличие мелких гранулярных
пузырьков и различная степень флюоресценции на препаратах, об-
работанных формальдегидом по методу Фалька, их перикарионов
и отростков, в том числе аксонов, поступающих в виде постганг-
лионарных волокон в соответствующие органы (рис. 124, А, Б).
В составе симпатических ганглиев имеются небольшие группы
м е л к и~ х, гранулосодержащих, интенсивно флюоресцирующих
клеток (МИФ-клетки). Они характеризуются короткими отростка-
ми и обилием в цитоплазме гранулярных пузырьков, соответствую-
щих по флюоресценции и элктронно-микроскоггической характе-
ристике пузырькам клеток мозгового вещества надпочечника.
МИФ-клетки окружены глиальной оболочкой. На телах МИФ-кле-
ток, реже на их отростках, видны холинергические синапсы, обра-
зованные терминалями преганглионарных волокон. ^МИФ-клетки
раосматриваются как внутриганглионарная тормозная_ система.
О уч. ^.»?бу.>г.'С*и‘-'^ >ными холинергичеекими- ги ыюкна-
ми. выделяют катехоламины. Последние, распространяясь диффуз-
но или по сосудам ганглия, окязырвют тормозящее ивияние_ня си-
н а птическую передачу с 11 регадгдирцдрных волокон на_лери фе р и -
ческие нейроны ганглия.
Ганглии парасимпатического отдела вегетативной нервной сис-
темы, содержащие ее второй нейрон, лежат или вблизи иннервиру-
емого органа, или в его интрамуральных нервных сплетениях. Пре-
ганглионарные волокна заканчиваются на телах нейронов, а чаще
на их дендритах холинергическими синапсами. Аксоны этих кле-
ток (постганглионарные волокна) следуют в мышечной ткани ин-
нервируемых органов в виде тонких варикозных терминалей и
образуют мионевральные синапсы. Их варикозные расширения
содержат холинергические синаптические пузырьки. Холинергиче-
ские нейроны и их отростки выявляются реакцией на ацетилхо-
линэстеразу по методу Келле в различных модификациях (см.
рис. 124, Б).
Интрамуральные сплетения. Значительное количество нейронов
вегетативной нервной системы сосредоточено в нервных сплетени-
ях самих иннервируемых органов: в пищеварительном трактеГсер-
дце. мочевом пузыре и др. ~ ~
Ганглии интрамуральных сплетений, как и другие вегетативные
узлы,^содержэт. кроме эфф>~ре»^цчу. нейронов, рецепторные и ас-
социативные клетки местных рефлекторных—дуг—Морфологически
325
Рис. 124. Нейроны и нервные волокна
вегетативной нервной системы. А —
нейроны вегетативного ганглия: I —
длинноаксонный нейрон; 2 — аксон;
3 — равноотростчатые нейроны;
4 — ядра глиоцитов. Б — адренер-
гические нейроны; В — нервные во-
локна (препараты Т. Н. Радости-
ной). Метод Фалька.
в интрамуральных нервных сплетениях различают три типа клеток.
Длцнноакс<>нныёэД)фер'ентные неир()ньГ1кАетку\ 1-го типа) имеют
много коротких ветвящихся лецпритов и ппинный нейрит, уходя-
щий за пределы ганглия (см. рис. 124, А), Равноотростчатые (аф-
ферентные) нейроны___(клетки—2=со—хила)_содержат несколько
отростков_Цо__морфологическим признакам нельзя определить,
который 1£3_ниха кернита к как отростки, не разветвляясь, уходят
далеко от тела клетки. Экспериментально установлено, что их нейри-
ты. образуют синапсы на клетках 1-го типа. Клетки 3-го типа_1дссо-
циативные) посылают свои отростки в соседние ганглии, где они за-
ка нч и ва юте я на дендритах их нейронов'
Специфическими особенностями отличается интрамуральная
система желудочночщшЕчнбго тракта (энтеральная система).
Рис. 125. Схема интраму-
ральных нервных сплете-
ний пищеварительного
тракта (по А. Д. Ноздра-
чеву).
/ — слизистая оболочка; II —
подслизистая основа; III —
мышечная оболочка; IV —
серозная оболочка; / — под-
слизистое сплетение; 2 —
мышечно-кишечное сплете-
ние; 3 — симпатический
ганглий; 4 — постганглио-
нарные симпатические во-
локна; 5 — афферентные
волокна (дендриты нейронов
спинального ганглия); 6 —
симпатические преганглио-
нарные волокна; 7 эффе-
рентные волокна блуждаю-
щего нерва; 8 - афферент-
ные волокна местных реф-
лекторных путей.
В стенке пищеварительной трубки расположены три нервных спле-
тения: подсерозное, межмышечное и подслизистое, содержащие
скопления нервных клеток^ связанные пучками нервных волокон
(рис. 125). Наиболее массивное нервное сплетение — межмышеч-
ное — расположено, между'лродольнымл. циркулярным мышечными
слоями, Электронно-микроскопически и гистохимически в межмы-
шечном сплетении установлены холинергические нейроны, возбуж-
дающиедвигательную активность кишечника, и тормозные нейро-
ны,“представленные адренергическими и_иеадренергическими (пу-
ринергическими) нейроцихами. Морфологически пуринергиче-
ские нейроциты характеризуются содержанием в перикарионе и
отростках крупных (размером 80—120 нм) электронно-плотных
гранул. В составе интрамуральных вегетативных ганглиев содер-
жатся и пептидергические нейроны, выделяющие ряд гормонов
(вазоинтестинальный пептид, вещество Р, соматостатин и др.).
Считается, что эти нейроны осуществляют нервные и эндокринные
функции, а также модулируют функциональную деятельность эн-
докринных аппаратов различных органов.
Постганглионарные волокна нейронов интрамуральных сплете-
ний в мышечной ткани органа образуют терминальное сплетение,
тонкие стволы которого содержат несколько, .варикозно рагшйрен-
ных аксонов. Карйко^ные расширения (0,5—2 мкм в диаметре)
327
сод^жах_гшна»ти*+еекие—пузырьки и митохондрии. Межварикоз-
ные участки (шириной 0,1—0,5 мкм) заполнены нейротрубочками
и нейрофиламентами. Синаптические пузырьки холинергических
мионевральных синапсов мелкие светлые (размером 30—60 нм),
адренергических—мелкие гранулярные (размером 50—60 нм).
ОБОЛОЧКИ ГОЛОВНОГО И СПИННОГО МОЗГА
Головной и спинной мозг покрыты тремя оболочками: мягкой,
непосредственно прилегающей к тканям мозга, паутинной и твер-
дой, которая граничит с костной тканью черепа и позвоночника.
Мягкая мозговая оболочка непосредственно прилежит к ткани
мозга и отграничена от нее краевой глиальной мембраной. В рых-
лой волокнистой соединительной ткани оболочки имеются большое
количество кровеносных сосудов, питающих мозг, многочисленные
нервные волокна, концевые аппараты и одиночные нервные клетки.
Паутинная оболочка представлена тонким слоем рыхлой волок-
нистой соединительной ткани. Между ней и мягкой мозговой обо-
лочкой лежит сеть перекладин, состоящих из тонких пучков кол-
лагеновых и тонких эластических волокон. Эта сеть связывает
оболочки между собой. Между мягкой мозговой оболочкой, повто-
ряющей рельеф ткани мозга, и паутинной, проходящей по возвышен-
ным участкам, не заходя в углубления, располагается подпаутин-
ное (субарахноидальное) пространство, пронизанное тонкими кол-
лагеновыми и эластическими волокнами, связывающими оболочки
между собой. Субарахноидальное пространство сообщается с же-
лудочками мозга и содержит цереброспинальную жидкость.
Твердая мозговая оболочка образована плотной волокнистой
соединительной тканью, содержащей много эластических волокон.
В полости черепа она плотно сращена с надкостницей. В спинно-
мозговом канале твердая мозговая оболочка отграничена от пери-
оста позвонков эпидуральным пространством, заполненным слоем
рыхлой волокнистой соединительной ткани, что обеспечивает ей
некоторую подвижность. Между твердой мозговой и паутинной
оболочками располагается субдуральное пространство. В субду-
ральном пространстве содержится небольшое количество жидкости.
Оболочки со стороны субдурального и субарахноидального про-
странства покрыты слоем плоских клеток глиальной природы.
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ НЕРВНОЙ СИСТЕМЫ
Изменения в центральной нервной системе в раннем постна-
тальном онтогенезе связаны с ее созреванием. У новорожденных
для корковых нейронов очень характерно высокое ядерно-цито-
плазматическое отношение. С возрастом размеры нейронов уве-
личиваются за счет увеличения объема цитоплазмы. При этом
наиболее быстро (в первые 3 мес жизни) увеличиваются размеры
пирамидных нейронов II и IV слоев. Более медленное увеличение
характерно для клеток-зерен и малых пирамид IV слоя. Увели-
чивается число синаптических контактов.
128
У взрослых людей, по сравнению с новорожденными, умень-
шается число нейронов в коре на единицу объема. Уменьшение
зависит от гибели части нейронов, но главным образом от разра-
стания нервных волокон и нейроглии, что приводит к увеличению
толщины коры и механическому «раздвиганию» нейронов. У но-
ворожденных в нейронах средней лобной извилины отсутствует
базофильное вещество, количество хроматофилыюго вещества в
нейронах увеличивается у ребенка 3—6 мес, а в двухлетнем воз-
расте достигает уровня взрослых. Формирование миелиновых
оболочек вокруг аксонов в ряде областей коры (средняя и нижняя
лобные извилины, средняя и нижняя височные извилины и др.)
происходит после рождения ребенка.
Изменения в центральной нервной системе в старческом воз-
расте связаны прежде всего со склеротическими изменениями
сосудов мозга. В старости мягкая и паутинная оболочка мозга
утолщаются. В них могут появиться отложения извести. Наблю-
дается атрофия коры больших полушарий, прежде всего лобной
и теменной долей. Уменьшается число нейронов на единицу
объема коры, зависит это главным образом от гибели клеток.
Нейроны уменьшаются в размере, частично теряют базофильное
вещество, ядра уплотняются, их контур становится неровным.
Быстрее других изменяются пирамиды V слоя двигательной зоны
коры и грушевидные клетки коры мозжечка. В нейронах раз-
личных отделов нервной системы накапливаются гранулы липо-
фусцина.
КРОВОСНАБЖЕНИЕ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ.
ГЕМАТОНЕЙРОНАЛЬНЫЙ БАРЬЕР
Кровоснабжение спинного мозга осуществляется через перед-
ние и задние корешковые артерии, входящие с передними и
задними корешками и образующие артериальную сеть в мягкой
мозговой оболочке. Здесь формируются продольные артерии, из
которых главная — передняя спинальная артерия, проходящая в
передней срединной щели.
Капиллярная сеть в сером веществе более густая, чем в белом.
Вены спинного мозга не сопровождают артерии. Мелкие вены,
идущие с периферии спинного мозга и из передней срединной
щели, образуют сплетение в мягкой мозговой оболочке, особенно
густое на дорсальной поверхности спинного мозга, откуда кровь
оттекает в вены, сопровождающие вентральные и дорсальные
корешки.
Артериальное кровоснабжение головного мозга осуществляется
за счет внутренних сонных и позвоночных артерий, которые сли-
ваются в основании мозга в базилярную артерию. Ветви этих
артерий проходят в мягкую мозговую оболочку, и отсюда мелкие
веточки следуют в вещество мозга. Капиллярная сеть в сером
веществе головного мозга также более густая, чем в белом. Капил-
ляры мозга имеют непрерывную эндотелиальную выстилку и
329
Рис. 126. Схема строения гематоэнцефалического барьера.
/ — эндотелий гемокапилляра; 2 — базальная мембрана; 3 — тело астроцита; 4 — плас-
тинчатые окончания отростков астроцитов; 5 — нейрон; 6 — отростки нейронов; 7 —
оли годе ндрогл иоци т.
хорошо развитую базальную мембрану. Здесь происходит изби-
рательный обмен веществ между нервной тканью и кровью, в ко-
тором принимает участие так называемый гематоэнцефалический
барьер. Избирательность обмена веществ между тканью и кровью
обеспечиваете^, помимо морфологических особенностей самих
капилляров (сплошная эндотелиальная выстилка с хорошо раз-
витыми десмосомами, плотная базальная мембрана), также и тем,
что отростки глиоцитов, прежде всего астроцитов, образуют н'
поверхности капилляров слой, отграничивающий нейроны от не
посредственного соприкосновения с сосудистой стенкой (рис. 126
Г л а в а XIV
ОРГАНЫ ЧУВСТВ
р^ОБЩАЯ ХАРАКТЕРИСТИКА И КЛАССИФИКАЦИЯ
Органы уувств (organa sensuum), по определению И. П. Пав-
лова^ это периферические части анализаторов. Анализато-
Р ы ~ сложные структурно-функциональные* системы," осуществля-
ющие связь центральной нервной системы с внешней и внутрен-
ней средой. В системе каждого анализатора различают три части:
периферическую, в которой происходит рецепция, или восприятие,
330
пром? суточную — проводящие пути и подкорковые образования,
по которым передаются импульсы, и центральную — кору большо-
го мозга, где происходит окончательный анализ и синтез воспри-
нятого ощущения.
Органы чувств воспринимают специфические раздражения, пре-
образуют их в нервный импульс и передают информацию, закоди-
рованную в серии нервных импульсов, через промежуточные час-
ти анализаторов-в-центральные.
Раскрытие закономерностей цитофизиолоши рецепторных кле-
ток и структуры анализаторов является основой для материалис-
тической теории познания, согласно которой человеческие ощуще-
ния, понятия, научное познание в ц0юм являются отражением
объективно существующей реальности. Критикуя идеалистические
теории агностицизма (непознаваемости внешнего мира), в част-
ности теорию символов Г. Гельмгольца, В\И. Ленин писал: «Наши
ощущения отражают объективную реальность, т. е. то, что сущест-
вует независимо от человечества и от человеческих отношений»1.
Классификация органов чувств. Различают три Основных
типа органов чувств. К первому типу относятся_овр_гоа_ц__^^^^=и_21
и сГрт~а~н—ег б^п£адд~ЗП~'В~~йх составе находятся особые рецеп-
1Ррныё 51^1ш^'1сдетки~ГнейросёнсорныёУ, которые названы п е р-
в и_ч н о-ч у в с т р у ю щ ими. Источником их развитоя яиля юте я
нёрвныё~элементы, образующиеся из эмбриональной нервной плас-
тинки. ЭТИ Клетки имеют специализированные периферические
отростки—дендриты, воспринимающие колебания световых во л н
или молекул пахучих веществ, а также центральные отростки —
не ириты, поезеоторым .возбуждение в виде импульсов передаете я
Ко второму „типу относятся органы вкуса- равное-
с и я и слуха. Эти органы ~заклалыняютс.я в эмбриогенезе
в сосТаде„зктодермь1 из особых ее утолщений — ц ла код. В этих
органах чувств основным воспринимающим элементом являются
специализированные эпителиальные клетки (сенсоэпитедиальные)^
Эт них~преОбразованное—раздражение—передается Нервным клет-
,ам, которые поэтому названы вторично-чувствующими^ Дендриты
червных клеток воспринимают возбуждение, возникающее всёнсо-
пителиальных клетках-под вездействием вкусовых веществ либо ко-
лебаний воздушной’ или жидкой’ среды,ттгпе’рёдают его в проме-
жуточные части соответствующих анализаторов, т. е. вкусового,
слухового или вестибулярного“нервепт"—————
К .третьему типу с невыраженной четкой органной органи-
зацией относится группа рецепторных инкапсулированных и
неинк^псулиррвяпньПГнервных~окончании, например пластинчатые
нервные тельца, луковицеобразные тельца, отдельные клёткщ яв-
ляющиеся тоже периферическими частямй соответствующих ана-
лизаторов (давления, осязания и др.).
1 Для всех воспринимающих клеток характерно наличие спе-
Ленин В. И. Поли. собр. соч., т. 18, с. 246. \
131
\.1
циализированных структ р, обеспечивающих восприятие специфи-
ческих~раздражёнйи~^- ресничек (киноцилий), связанных с базаль-
ными тельцами, или микроворсинок (стереоцилий). В плазмолемму
ресничек и микроворсинок вмонтированы молекулы особых фото-,
хемст- и механоре'цепторных белков, которые кодируют энергию
стимула в специфическую информацию клетки. Образующиеся
биопотенциалы"поступают в центральную нервную систему, где
происходит декодирование стимула
ОРГАН ЗРЕНИЯ
Глаз (oculus) представляет собой_____п_е р_и.ф е_р и н е скую
часть з р и те льного а н я п и за т п.р а Он—состоит из
глазного яблока (Jmlbus___oculi), содержащего фоторецепторные
клетки, соединенного—посредством зрительного...нерва—с- мозго^л,
и вспомог а т е л ь н о го а п п ар а таг включающего веки,
слезный аппарат и-цоперещюнеяееатые глазадаигате пьные мыш-
цы.
Глазное яблоко образовано тремя оболочками, фиброзной
(склера и роговица), сосудистой и внутренней (сенсорной )—м их
производными (радужка, ресничное тело), а также хрусталиком,
жидкостью передней и задней калацгглаза, стекловидным телом.
В глазном яблоке—различают три основных функциональных
аппарата. ~ д~ии п т р и ч е е к~ и'щ йлй “свети пр е л о м л я-
ю щ и и (роговица, жидкость передней и задней камер глаза,
хруогалйк. стекловидное___тело), аккомодаци сГн"ньГй а п-
п а рат (радужка, ресничное тело с “ресничным пояском) и
р е цеп торный аппарат (сетчатка). Склера выполняет
защитную и опорную функции. Ц
Развитие. Глаз гдзных-источников (рис. 127).
Сетчатка л_ зри гель:ши нерп ф<>рмируёмз_я_льГЗ>чагАа_ззервной
систем ы —-нервной трубки ь киле пыпччи^нмм ы х
глазными пузырьками, сохраняющих связь с эмбриональным моз
гом при помощи полых глазных стебельков. Вдоль стебелька »
глазнби пузырек проникают сосуды. Передняя часть__ЕДазноп
пу^йрька 'ВБЛптивае1'СЯ:~;внутрБ:ге'|г) полости, благодаря чему я
приобретает форму двустенного глазного бокала. Часть эктодермы,
расположенная напротив отверстия глазцого2бока^за, утолщается,
инвагинирует и отшнуровывается, давая начало зачатку хрустали-
ку^ Эти изменения эктодерма претерпевает под влиянием индук-
торов дифференцировок, образующихся в глазном пузырьке.
Первоначально х русталик имеет вид полого эпителиального
пузырика Затем~клетки эпителия его задней стенки удлиняются
и превратаются~в~так называемые хрусталиковые волокну, запол-
няющие полость пузырька. В процессе развития внутренняя
сте нка~глазного бокала преобразуется в сетчатку, а нару ж на я
в пигментныи~слои сетчатки. Из~нейвобластов вну-тредней стен-
ки глазного бокала"0~бразу38тся колбочконесущие и пазючконесу-
щие фоторецепГОршзте—тяпгмёнты й другие нейроны сетчатки.
332
Рис. 127. Развитие глаза.
Д, />, В—сагиттальные разрезы эмбриональною вмазка глаза на различных стадиях
развития / — эктодерма; 2 - хрусталиковая плакода — будущий хрусталик; 3 — глазной
пузырек; 4 — сосудистая выемка; 5 — наружная стенка глазного бокала будущий
пигментный слой сетчатки; 6 — внутренняя стенка глазного бокала — будущая сетчатка;
7 — стебелек — будущий зрительный нерв; 8 — хрусталиковый пузырек.
происхождению явля-
Стебелек глазного-бокала пронизывается нейритами образующих-
ся в сетчатке ганглиозные клеток. Эти нейриты и формируют
зрительный нерв, направляющийся в зг. Из окружшощей~чаз-
ной бокал мезенхимы формируются сосудистая оболочка и склера.
В передней' части глаза склепа переходит в покрытую много-
слойным плоским эпителием прозрачную роговицу. Сосуды и ме-
зенхима, проникающие в ранних стадиях развития внутрь глазного
бокала, совместно с эмбриональной сетчаткой принимают участие
в образовании стекловидного тела и радужки. Мышца радужки,
суживающая зрачок, развивается из краевого утолщения наруж-
ного и внутреннего листков глазного бокдла. Мышечные клетки,
расширяющие зрачок, развиваются из наружного—листка. Таким
образом7~обе мышцы радужки по своему
, ются нейральными.
iv ксдфа Lcicrv
Строение глазного яблока и
Фиброзная оболочка .(tunica fibrosa bulbi). Эта'обрлочка обра-
зует наружнукГчасть глаэ^ и представлена склерой, ^закрывающей
большую-поверхность глаз^Ц и переходящей в передней его части
в роговицу. \
Склера (sclera). Это плотная соединительнотканная ободочка
толщиной 0,3 0,4 мм в задней_наг1и vC0,6 мм вблизи роговицы.
Состоит _из расположенных параллельно поверхности^ глаза со-
едини тельнотканных пластинок. всн1ержащих коллагеновые волок-
на (рис. 128), между которы\иАнах дя _уплацщщцоц_ формы
фибробласты ц. отдепы4ыа алд<тмиАгкие подокна—Пу коллаге-
новых волокон, .Hcjan’iniieb—Нропопются в собственное вещество
роговицы. Прозрачная роговина довольно резко переходит в непроз-
рачную склеру в области лимба Здес\ наружный слой склеры
частично-побывает )фай роговицы. ЭНйтелий роговицы в ее
краеной зоне—поётеФемио переточит в э^итедим—конъющцивы
глаза, В _т£дни_склеры—на—месте—соединения ее т, роговицей
имеются небольшие—цепряиипьной формы разве гвденные нопости
Рис. 128. Строение переднего отдела глазного яблока (схема).
/ роговица; 2 — передняя камера глаза; 3 — радужка; 4 — задняя камера глаза;
5 - хрусталик; б ресничный поясок (циннова связка), 7 — стекловидное тело; 8
гребенчатая связка; 9 венозный синус склеры; 10 — цилиарное тело; а — отростки
цилиарного тела; б — цилиарная мышца; 11 — склера; 12 — сосудистая оболочка; /? —
зубчатая линия; !4 — сетчатка.
копцпде, сообщаясь между собой, образуютчгчрзныц синус. скле
ры_ (шлеммов канал), Внутренняя Поверхность склеры контакти
рует с ралужкой, образует так называемое пространство радужно-
роговичного угла, в котором расположена гребенчатая связка.
Через эту область проходит отток водянистой, влаги из передней
камерьГ~глаза~к”вёнозному Минусу. С наружной поверхности склеры
располагаются конъюнктива (в области перехода в роговицу) и
глазсщвигательныё^мъптщгт- .
Сосудистая оболочка (tupica vasculbsa bill bi). Представлена
собственно сосудистой оболачКой рединным—|е.твл|, радиккой.
Собственно сосудистая оболочки (choroidea) осуществляет пита-
ние сетчатки. В ней, начиная \царужи, различают пластинки:
над сосудистую, сосудистую,''сосудцсто-капиллярную и -базальный
комплекс. / \
Н а д~с о с у д и с т а я п л /с т и\, к a (lamina suprachoroidea)
представляет собой самый наружныйХ слой сосудистой оболочки,
лежащей на границе со "склерой?-ОнДу^брзгзована рыхлой, волок-
нистой соединительной ржанью, соДерждЩбй~~ббльшое количество
эластических волокон, фибробластов и пигментных клеток (мела-
ноцитов V~
С ос у д и с т а я/ пластинка (lamina^ vasculosa) состоит из
переплетающихся” вртерйй~й""вен, между ко >рыми располагается
пыхлая волокттисч'ая'~соединительная ткань, содержащая-большое
Зл
количество пигментных клеток. Здесь также залегают отдельные
пучки Гладких миоцитов.
С ocV ди ст о-к ап >игля р н а я пл а С т и_н к a (lamina cho-
nocapillans) содержитгемокапи пляры, отличающиеся неравномер-
ным калиб^о>1^Часть их принадлежит к капиллярам синусоид-
ного типщЛЙвжду капиллярами располагаются уплощенные фиб-
робластвг X.
Ба за л ь н ыКк о м п л е к с (complexus basalis) — очень тон-
кая полоска (1—4 мкм), располагающаяся между сосудистой обо-
лочкой и пигментным сйоем сетчаткй.~Впетт~различают три слоя.
Наружный — эластический^—слой. содержит тонкие^эластические
волокна, являющиеся продолжением волокон сосудисто-капилляр-
ной пластинки. В утренним^ бхмее широким, сост и _из волокнисто-
го (фиброзного) слоя.Дретийхлой-тюставдяет-базальная мембрана.
Диоптрический (светопреломляющий) аппарат глаза
Диоптрический аппарат глаза образован системой прозрачных
структур и сред, преломляющих свет.
- Роговица (cornea). Это оболочка, толщина которой колеб-
лется от 0,8 до 0,9 мм в центре и составляет 1,1 мм на пери-
ферии. Особое строение роговицы и химический состав делают ее
прозрачной. Радиус кривизны роговицы составляет около 7,8 мм,
показатель преломления — 1,37. В роговице различают 5 слоев:
передний эпителий, переднюю пограничную пластинку, собствен-
ное е цество роговицы, заднюю пограничную пластинку, задний
эпителий (рис. 129).
Перед ний эпителий (epithelium anterius) — мнргослой-
HbiujiJiQci непрптвевающии, общей толщиной до 50 мкм, состоит
из 5—6 слоев. В эпителии роговицы располагаются многочислен-
ные свободные нервные окончания, обусловливающие высокую
тактильную*чувствительность и формирование рефлексов роговн-
иы. Поверхность роговицы увлажнена секретом слезныхЛ-ЬТЩъюн-
ктидальных желцзГЭпйтелий роговицы отличается высокой реге-
не рационнои"СПОСобностью и проницаемостью для”разнообразных
жидких и газообразных веществ. 11бслёднёё~свойств6^йспользуется
во врачебной практике при введении лекарственных препаратов.
Передний эпителий роговицы продолжается в многослойный
плоский эпителии конъюнкхцвьн- Базальная мембран _ переднего
эпителия состоит из-электронно-светлого и электронно-темного
слоев (см. рис. 129, Л).
Перед н я я ддср,а ни ч и а.я пласт и н к a (lamina limi-
tans апТег1ог7~^ежит~тго^базальной_мембранощ- Л меет- -фибрил-
лярцое строение. 'Толщина~пластинки ко геблется от до 9 мкм.
clrto с тТв-н нее. в е щ е с т в о о_о_т_н и ц ы (substantia
propria corneae) состоит из правильно чередующихся взаимно
пересекающихся под уцзом—тонких соединительнотканных плас-
тинок. Каждая пластинка состоит из параллельно расположен-
ных пучков коллагеновых волокон различном толшйны. В плас-
2
Рис. 129. Роговица глаза.
А — интактная роговица:
/ — многослойный плоский
неороговевающий эпителий;
2 - базальная мембрана пе-
реднего отдела роговицы; 3—
собственное вещество; 4 —
задняя пограничная мембра-
на; 5 — задний эпителий
роговицы; Б — роговица с
трансплантатом (Т) (препа-
рат В. В. Веретенниковой).
тадкаХ- и__между ними располагаются отростчатые плоские
клетки, являющиеся разновидностями фибробластов. Клетки и
пластинки погружены в аморфное вещество, богатое гликозами-
ногликанами (в основном кератинсульфатами), которое обеспе-
чивает^, прозрачность собственного вещества роговицы. Собствен-
ное вещество роговицы не имеет кровеносных сосудов. В области
радужно-роговичного угла оно продолжается в непрозрачную
наружную обблочкуглаза -^склеру.
3 а дня я ~п о г~р?а~н и ч-н-а-я—н-ж.а с т и н к a (lamina limitans
posterior) имеет толщину от 5 до 10 мкм. Она иредставлстииколлаге-
новыми волокнами диаметром 10 нм, погруженными в аморфное
вещество.
Задний эпителий (epithelium posterius) состоит из
плоских полигональных- клеток. Ядра клеток отличаются много-
образием формы.
Питание роговицы происходит за счет диффузии питательных
веществ из передней камеры глаза и кровеносных сосудов лимба,
в с амой роговице кровеносных сосудов нет\ Лимфати че с к ая
система роговицы формируется из узких лимфатических щелей,
сообщающихся с ресничным венозным сплетением.
В случае воспаления кровеносные капилляры вместе с клетками
46
(лейкоциты, макрофаги и др.) проникают из области лимба в
собственное вещество роговицы, вызывая ее помутнение и орого-
вение (бельмо). Для восстановления прозрачности роговицы бель-
мо удаляют и производят пересадку донорской роговицы (см.
рис. 129, Б).
Хрусталик (lens). Это прозрачное двояковыггуюпое телсд форма
которого меняется_во время аккомодации глаза к видению близ-
ких и овдале иных—объектов.-- Вместе--е--роговицей, стекловидным
телом хрусталик составляет основную светопреломляющую среду.
Радиус кривизны хрусталика варьирует от 6 до 10 мм, показатель
преломления составляет Хрусталик покрыт прозрачной кап-
сулой толщиной 11 —18 мкм. Его передняя стенка-, призе>ка1пая
к капсу. с. rocTopTj<4j^i£HnrapHiiuiо наптшп л/шо-
(epithelium lentis).
По направлению к экватору эпителиоциты становятся выше и
образуют ростковую зону г руг та п и к а, Эта зона пос-
тавляет в течение всей жизни новые клетки как на переднюю,
так Я—на заднюю поверхность хрусталика—Новые—эпителиоциты
преобразуются^ так называемые хрусталиковые .волокна (fibrae
lentis). Каждое,.волокно представляет собой прозрачную шести-
угол ь ну юл ризму. В цитоплазме хрусталиковых волокон находится
прозрачный бел&к — кристалин. Волокна дклеиваютсл друг.с дру-
гом -особым веществом, которое имеет такой же, как и они,
коэффициент преломления. Центрально расположенные волокна
теряют свои ядра, укорачиваются и, накладываясь друг ня друга,
образуют ядро хрусталика.
Хрустадиклоддерживается в глазу с помощью волокон реснич-
JjOiQ ~пояс^а (zonula ciliansTT-ПбразоВЯИногб- радиально "рЗспсъ
ложенными пучками нерастяжимых волокон (связки), лрикре-
пленных с одной стороны к цилиарному телу, а с другой — к
капсуле хрусталика, благодаря чему сокращение мышц цилиарного
тела передается хрусталику. Знание закономерностей строения и
гистофйзиолоГИИ хрусталйкаТюзволило разработать методы созда-
ния искусственных хрусталиков и широко внедрить в клиническую
практику их пересадку, что сделало возможным лечение больных
с помутнением хрусталика (катаракта).
Стекловидное тело (corpus vitreum). Это прозрачная масса
желеобразного вещества, заполняющего полость междухрустали-
ком и сетчаткой.__На фиксированных препаратах стекловидное
тело имеет сетчатое строение. На периферии оно более плотное,
ч е м. в центре. Чере з стекло видное тело проходит канал —^"оста -
ток эмбриональной "сосудистой системы глаза — од сосоцкр сет-
чатки до задней поверхности хрусталика. Стекловидное тело
содержит" белок витреин___и_ -гиалуроновую—кислоту. Показатель
преломления стекловидного тела равен 1,33.
Аккомодационный, аппарат глаза
Аккомодационный аппарат глаза обеспечивает изменение фор-
мы и преломляющей силы хрусталика, фокусировку изображения
337
Рис. 130. Радужка.
/ — однослойный плоский эпителий; 2 — наружный пограничный слой; 3 — сосудистый
слой; 4 внутренний пограничный слой; 5 — заднни пигментный эпителий.
на сетчатке, а также приспособление глаза к интенсивности
освещения. l
^""Радужка (iris), [Представляет собой дисковидное образование
с отверстием изменчивой величины (зрачок) в центре. Она явля-
ется ^производным сосудистой оболочки гдазя Сзади_ радужка
покрыта пигментным эпителием..сетчатой оболочки. Расположена
между роговицей и хрусталиком на границе между передней и
задней-камерами rna^ai(рис. 130). Край радужки, соединяющий
ее с цилиарным телом, называется цилиарным-краем. Строма ра-
дужки состоит из рыхлой волокнистой соединительной ткани,
богатой£пигментныму клетками. Здесь располагаются гладкие
миоциты, образующие мыщцы~ ТУжиВаюшие и расширяющие
зрачок (m. sphinctexjmpillae, rru-ditarafor pupillae).
В радужке различают 5 слоев: передний эпителий, покрыва-
ющий переднюю поверхность радужки^ наружный пограничный
(бе ссосу дистыиУ~?лой. сосудистый слой, внутренний пограничный
слой и пигментный эпь ~влий.
Передний—а п нт е л и й (epithelium anterius iridis) пред-
ставлен плоскими полигональными клетками. -Рн—является про-
должением_эпителия покрывающего заднюю поверхность рого-
вицы.
Н а р,у_ж ный п ограни ч н ы й __с л о й (stratum externum
limitans) состоит щ основного вещества, в котором располагается
значительное количество фибробластов и много пигментных кле-
ток,_Различное__положение и количество меланинсодержащих
клеток обусловливают цвет глаз. У альбиносов пигмент отсутствует
и радужка имеет краснь1и~7щёт в связи с тем,~дтб" через ее" толщину
просвечивают сосуды. В пожилом возрасте наблюдается депигмен-
тация радужки и она делается более светлой. '
Сосудистый слой (stratum vasculosum) состоит из мно-
гочисленных сосудов, пространство между которыми заполнено
138 Т
рыхлой волокнистой соединительной тканью с пигментными
клетками. /
Внутр е н н и й п о гд? а н тгдлыш.с л о й (stratum internum
limitans) не отличается по строе.йию-ат-наружного слоя.
Задний пигментный эпителий (epithelium pos-
tering pigmentosum) является продолжением двухслойного эпи-
телия сетчатки, покрывающего цилиарное тело и отростки.
Радужка осуществляет свою функцию в качестве диафрагмы
глаза с помощью ~двух~мышц: суживающей (muse ulus sphincter
pupillae) и расширяющей (musculus dilatator pupillae) зрачок.
Мышца, расширяющая зрачок, иннервируется за счет постгангли-
онарных симпатических волокон верхнего шейного узла, а.мышца,
суживающая его, — за счет постганглионарных парасимпатических
волокон цилиарионГэтого же источника происходит ин-
нервация цилиарного тела. /
Ресничное тело (corpus cillare). Ресничное тело является
производным, сосудистой и сетчатой оболочек- Выполняет функ-
цию фиксации и изменения, кривизны хрусталика,—тем самым
участвуя в акте аккомодации. На меридиональных срезах через
глаз цилиарное тело имеет вид треугольника, который своим осно-
ванием обращен в переднюю камеру глаза. Цилиарное тело под-
разделяется—на две части: внутреннюю—цилиарную корйну
(corona ciliaris) и наружную — цилиарное кольцо (orbiculus
ciliaris). От поверхности цилиарной короны отходят по направле-
нию к хрусталику цилиарные_отростки (processus ciliares), к кото-
рым прикрепляются волокна, ресничного пояска (см. рис. 128).
Основная ча’ет|Г~цилиарнпгс> теля, за—исключением отростков,
образована ресничной, или цилиарной.—мышцей (m. ciliaris),
играющей важную роль в аккомодации глаза. Она состоит из пуч-
ков гладких мышечныГ клеток, располагающихся в_трех раз-
личных направлениях. Различают наружные меридиональные мы-
шечные пучки,~~лёжащие ««посредственно под склерой, средние
радиальные и циркулярные мышечные пучки, образующие кольце-
вой мышечный слой. Между мышечными пучками расположена
рыхлая волокнистая соединителБная~'ткатгь~~С пигментными клет-
ками. Сокращение цилиарной мышцы приводит к рассла’блению
волокон—круговой—связки ресничного пояска, вследствие чего
хрусталик.станпвится выпуклым и его преломляющая сила'увели-
чивается.
Цилиарное тело_и_-цилиарпые отростки—пекрьтл цилиарной
частью сетчатки^которая в данной области состоит из наруж-
ного листка, лежащего на цилиарной мышце. Он представлен
одним слоем кубического интенсивно пигментированного эпителия.
Внутренний листок составлен одним слоем цилиндрических кле-
ток, лишенных пигмента. Непигментированный внутренний листок
с поверхности, обращенной к центру глаза, окружен стекловидной
цилиарной мембраной. Эпителиальные клетки, покрывающие цили-
арное—тело и отростки, принимают участйе’ в образовании водя-
нистой,влаги. заполняющей обе камеры глаза.
Рецепторный аппарат глаза
/ Сетчатка (retina) — внутренняя оболочка глаза, большая часть
которой (pars optica retinae) является световоспринимающей_л
содержит фоторецепторные клетки. В зависимости от формы своих
периферических Отростков^ они называются палочковыми нейро-
сенсорными и колбочковыми нейросенсорными клетками. Мень-
шая часть, покрывающая~с~ внутренней стороны цилиарное-тело
(pars ciliarS-retinae) и-заднюю поверхно'сть радужки (pars iridica
retinae), лишена фоторецепторов.
Сетчатка состоитиз-трех радиально расположенных нейронов,
(наружного-^фоторецепторного, среднего ^ассоциативного и
внутреннёТё/^ганглионпрнр^о^ и двух нейронов, включающихся
в радиальные цепочки: на уровне контакта- первого' й второго ней-
ронов ГгГ также на—уро~внё “соединения
второго и третьего нёироИив-Ф|(алйгкрт7йныё нейроны}. Йе жду
радиально наловленными цепочками нейронов" расположены
радиальные-глиоциты. В совокупности клетки образуют несколько
сдоев сетчатки: фотосенсорный слой палочек и колБочек^на руж-
ный ядерный слои, наружный сетчатый слой, внутренний ядерный
слой, внутренний сетчатый слой, ганглиозный слой и слои нер-
вных, волокон. Ядерные и ганглиозный слои сетчатки соответствуют
телам нейронов? сетчатые ёТГОи-^их “контактам (рис. 131, А, Б,
В, Г). ------
Луч света, прежде чем попасть на светочувствительный слой
сетчатки, должен пройти через роговицу, хрусталик, стекловид-
ное тело и всю толщу сетчатки. Таким образом, сетчатка глаза
человека относится к типу так называемых инвертирован-
н ы х, т. е. таких, в которых рецепторы нейросенсорных клеток
направлены от света и являются самыми глубокими частями сет-
чатки, обращенными к пигментному эпителию сосудистой оболочки.
Снаружи к сетчатке прилежит слой пигментного эпителия.
Нейросенсорные клетки воспринимают световые лучи перифери-
ческими частями — палочками и колбочками. Ядросодержащие
части фотореце[ггорных ю1еток?3лбразуюг.л—совокупности ^наруж-
ный ядерный слой сетчатки (stratum mucleare externum). Их цен-
тральные отростки принимают участие в образовании Наружного
сетчатого слоя (stratum plexiforme externum). Периферические
отростки — дендриты палочковых нейросен с орны х
клеток, лме ют радиальную ориентацию и располагаются между
отростками пигментного эпителия-сетчатки. Каждый отросток
состоит из двух частей! наружного и- внутреннего сегментов,
соединённых ресничкой. Наружный--еегметггтгмёёт цилиндрическую
(палочковидную) форму. Он состоит из множества (до 1000)
сдвоенных мембран; которые обпазуют -накладывающиеся друг
на друга не связанные между собой замкнутые диски толщиной
140 нм и шириной до 2 мкм (см рис! ГЗТ,~S). В мембранах
наружных сегментов находится зрительный пигмент — родопсин,
состоящий из белка опсина и альдегида витамина-А —"рётйналя.
340
ccf'lcc , •fce-M’HK . ,
<1 I
Наружный и внутренний сегменты связаны ресничкой, начинаю-
щейся во внутреннем сегменте базальным тельцем.
Ядросодержащая часть палочковых нейросенсорных клеток
имеет небольшоТГоббдбк~цитоплазмы вокруг ядра. (Эт~неё~ОТходиз
тонкий центральный отросток — нейрит, заканчивающийся в на-
ружном сетчатом слое синапсом с биполярной клеткой.__Количест-
во палочковых клеток в сетчатке человека около 130 млн.
К о л б о ч ко в ы_е_нейросенсорные кл е т к и (neuro-
sensorius conifer) отличаются от палочковых клеток большим
объе_мом. строением “наружного и внутреннего сегментов. и_зри-
тельным пигментом. Наружные членики колбочковых клеток
в отличие от палочковых состоят из полудисков, образованных
в результате йнйагинации плазмолеммы (см. рис. 131, Г) Во
внутреннем-сегменте имеется участок, называемый эллипсоидом,
состоящий из липидной капли и скопления..плотно-лрилёгающих
друг к другу митохондрий. Длина колбочек в центре желтого
пятна около 75 мкм, толщина — 1 —1,5 мкм. На периферии сет-
чатки их длина несколько уменьшается, достигая 45 мкм. Ядро-
содержащие части колбочковых клеток располагаются в наруж-
ном (ядерном) слое сетчатки, ближе к наружной пограничной
мембране Они отличаются от палочковых нейросенсорных _клеток
наличием округлого, более крупного и более светло окрашенно-
го ядра. От ядросодержашей части отходит центральный отрос-
ток — аксон, формирующий синапс с дендритом биполярного
нейрона. Количество колбочковых клеток в сетчатке глаза челове-
ка составляет 6—7 млн. Они являются рецепторами дневного, т. е.
цветного, а палочковые клетки — сумеречного зре ни я.
Мембраны писков наружных сегментов колбочковых клеток
содержат, другой зрительный пигмент — йодопсин, отличающийся
по химическому составу от родопсина^ Колбочковые клетки
сетчатки человека чувствительны К трем основным цветам спектра:
синему, зепепому—и__красному Определенную роль в цветовом
во с п ри ятии могут играть также липидные каплРГэллйпсойдб в. Ц ве -
товая слепота (дальтонизм) объясняется отсутствием колбочковых
клеток одного или нескольких типов, обусловленным генетически.
При освещении зрительный пигмент распадается на свои
компоненты ^бёл'йК и ретиналв:—Распад пигмента запускает в
клетк£_цепь биохимических рёа’кцййгтгогорые приводя!1 к измене-
нию-чянмой пронипднмости. мембраны фоторецептора и во з ни к но-
вению-репепторного потенция па, Ресинтез..зрительных пигментов
происходит при темновой адаптации. При функциональном нап-
ряжении глаза распад родопсина преобладает над его ресинтезом,
что приводит к временному ослаблению зрительного восприятия.
Кратковременное затемнение создает условия для усиления фазы
ресинтеза родопсина и восстановления зрения.
Наружные сегменты образуются за счет разрастания плазмо-
леммы ресничек эмбриональных нейросенсорных клеток, обращен-
ных к пигментному слою сетчатки. При этом диски будущих
колбочковых и палочковых клеток развиваются одинаково — пу-
341
. • * —к//
— МП
Рис. 131. Сетчатка.
Л — схема нейронного
состава сетчатки. 1 -
пигментный эпителий
сетчатки; // — фотосен-
сорный слой (слой пало-
чек и колбочек); ///—
наружный ядерный слой;
/И — наружный сетчатый
слой: И внутренний
ядерный слой: VI — внут-
ренний сетчатый слой;
VII — ганглионарный
слой; VIII — слой нерв-
ных волокон; / - палоч-
ки; 2 — колбочки; 3 —
-наружный пограничный
слой; 4 — центральные отростки фоторецепторных клеток (нейриты); 5 — синапсы нейритов
рецепторных клеток с дендритами биполярных нейронов; 6 — горизонтальный нейрон;
7 — амакринный нейрон; 8 — ганглионарные клетки; 9 — радиальный глиоцит;
10 — внутренний пограничный слой; II — волокна зрительного нерва; 12 центрифу-
гальный нейрон. Б — микрофотография (препарат Ю. И. Афанасьева).
тем образования складок плазматической мембраны. Затем часть
эмбриональных колбочковых клеток претерпевает еще допол-
нительную дифференцировку, превращаясь в палочковые клетки
благодаря замыканию и отрыву их дисков от плазмолеммы. Обра-
342 I
Рис. 131 (продолжение).
Ультрамикроскопическое строение налом
новой (Я) И колбочковой нейросенсорной
(/ ) клеток (рис. Ю. И Афанасьева) I
наружный сегмент; II связующий отдел;
lit внутренний сегмент; IV перика
рион; V— аксон. / диски (в палочках) и
полудиски (в колбочках); 2 — плазмо-
лемма; 3 реснички; 4 — липидное тело;
5 __ митохондрии; б — эндоплазматиче-
ская сеть; 7 — ядро; 8 — синапс.
зование дисков индуцируется ви-
тамином А. В его отсутствие они
не развиваются, а у взрослых
при длительной недостаточности
витамина А диски разрушаются
(«куриная слепота»).
Во внутренних сегментах
нейросенсорных клеток нахо-
дятся ферментные системы,
обеспечивающие энергетический
обм^н и биосинтез основных
хщиических компонентов клет-
ки.
ядерномс пое
сетчатки (stratum nucleare in-
ternum) располагаются ассоциа-
тивные нейроны трех типов —
так называемые горизонталь-
ные, биполярные и амакринные
нервные клетки.
Г оризонтальные._____нервные
клетки (neuronum horisontalis)
располагаются в один тиди_два
дендритов, которые контактиру-
ют сдкронами фоторецепторных
1.
клеток. Их аксоны, имеющие го-
ризонтальную ориентацию, могут тянуться на довольно значитель-
ном расстоянии и вступать в контакт с аксонами как палочек, так
и колбочек. Передача возбуждения с горизонта льных... клеток _на
синапсы рецепторного и биполярного нейронов вызывает времен-
ную блокаду в передаче импульсов от Фоторецепторов (эффект ла-
терального торможения), что увеличивает контраст рассматривае-
мых объекте в.
Биполярные нервные клетки (neuronum bipolaris) соединяют
палочковые и колбочковые клетки с ганглиозными-.клетками сет-
чатки, причем иесколько палочконых клеток соединяются, с одной
би полярлоК-Д-холбочковые к-четки—контактируют^ .сортно ше ни и
1:1. Такое сочетание-обесленивает-более-высокую остроту цветового
видения пр сравнению с._черно-белым, Биполярные клетки имеют
^радиальную ориентацию. Их ядросодержащие части расдоласаются
во внутреннем ядерном слое, а дендриты — в наружном сетчатом
слое, где они образуют синапсы с аксонами нейросенсорных клеток.
Среди биполярных нейроцитов иногда встречаются клетки, ядросо-
держащисГучастки которых располагаются ближе целою ганглиоз-
ных клето^Это — центрифуга пьные Гшпоиярные Они пере-
дают импульсы в противоположном направлении — от ганглиозных
клеток к зрительным, что является морфологическим выражением
обратной_афферентации_ как_формы~ самоконтроля системы нейро-
нов. Биполярные клетки играют существенную роль в концентрации
импульсов, получаемых от нейросенсорных клеток и затем переда-
ваемых в ганглиозные клетки.
Амакринные^клетки (neuronum amacrinus) выполняют роль,
сходную с горизонтальными клетками только на уровне соединения
биполярных и мультиполяоных ганглиозных нервных клеток.
Ганглиозные, мулътиполярные клетки (neuronum multipolare) —
наиболее, крупные клетки сетчатки, В их цитоплазме хорошо выра-
жено хроматофильное ве!цество. Дендриты их-располагаются'^во
внутреннем c£z^tixiM_cjuje~7sTratum plexiforme iniernum), где они
контактируют с нейритами биполярных клеток. Тела ганглиозных
клеток образуют слой, который носит название^/ганглионарного
(stratum ganglionare). Нейриты ганглиозных клеток формируют
самый внутренний слой сетчатки слой нервных волокон (stratum
neurofibrarum), отделенных от стекловидного тела внутренним
пограничным слоем (stratum limitans interna).
Нервные волокна сетчатки, за исключением тех, которые распо-
лагаются в области центральной ямки («желтого пятна»), радиально
направлены и сходятся, _как_спип.ы в зшлесе^д. диске—зрительного
нерва («слепом пятне») сетчатки. Отсюда они, окруженные миели-
новой оболочкой, переходят в зри re -и.ныи нерв и поспе перекреста
(chiasma optica) заканчиваются в подкорковых зрительных_буграх.
Нейроглия сетчатки представлена особыми волокноподоб-
ными радиальными глиоцитами (gliocytus radialis), располагающи-
мися радиально во всей толще внутреннего листка сетчатки от на-
ружного до внутреннего пограничного слоя (см. рис. 131, А). Их
ядросодержащие части располагаются в центре внутреннего ядерно-
го слоя, а внутренние отростки образуют анугренний ппг/¥1ничнн11
слой (stratum limitans iniernum), отделяющий сетчатку отстекловил-
ногщела. Наружный пограничный слой (stratum limitans externum)
формируется на границе между слоем______палочек и колбочек и
наружным ядерным слоем благодаря плотному прилеганию
периферических концов глиоцитов друг к другу. В сетчатых слоях
клетки глии образуют при помощи своих отростков горизонтальную
пластинчатую сеть, в которой располагаются нейроны сетчатки.
На внутренней поверхности сетчатки у заднего конца оптической
оси глаза имеется -округлое и п.и_£>вальное желтое пятно диаметром
около 2 мм. Слегка углубленный центр этого образования носит
название центральной ямки (рис. 132, А). Центральная ямка — ме-
сто наилуяшего восприятия зрительных раздражений. В этой ебла-
Рис. 132. Центральная ям-
ка (А) и диск зрительного
нерва (Б).
А: I — склера, 2 — сосуди-
стая оболочка; .? - сетчатка;
4 — центральная ямка (жел-
тое пятно). Б: / — склера;
2 — сосудистая оболочка;
3 — сетчатка; 4 — диск
зрительного нерва (слепое
пятно): 5 зрительный
нерв.
сти внутренний идейный и ганглиозный СЛОИ-резка истончаются,
а несколько утолщенный—наружный—ядерный слттй "Представлен
главным образом телами колбочковых нейросенсорных клеток,
которые здесь у человека приобретают палочкообразную форму.
На месте центральной ямки все слои сетчатки, кроме наружного
ядерного слоя, раздвинуты, как бы облегчая ход световых лучей
до слоя колбочек. Аксоны нейросенсорных клеток в области цент-
ральной ямки расходятся радиально в стороны.
Кнутри от желтого пятна на сетчатке имеется вол вы шение,
образованное выходом зрительного нерва. В этой области, носящей
название диска зрительного нерва, или слепого пятна (рис. 132, Б),
все слои се тчатки отсутствуют, за исключением слоя нервных воло-
кон, которые, собираясь со всех участков сетчатки, составляют
зрительный.нерв. На месте своего перегиба волокна образуют валик,
окружающий центральное углубление. В этом месте из толщи зри-
тельного нерва на внутреннюю поверхность сетчатки выходят пита-
ющие сетчатку сосуды.
Пигмент ый слой (stratum pigmentosum) - - самый на-
ружный слой сетчатки^ состоит из призматических полигональ-
ных, главным образом^шестиугольных, клеток. Своими основаниями
клетки располагаются-на базальной мембране и, таким образом,
прилежат к сосудистой оболочке глаза. Общее количество пигмент-
ных клеток у человека варьирует от 4 до 6 млн. В центре желтого
345
пятна они более высокие^ а на периферии сетчатки уплощаются,
но становятся в несколько раз шире. Микроворсинки на апикаль-
ной поверхности пигментоцитов охватывают дистальные участки
наружных сегментов фоторецепторных клеток. Один пигментоцит
контактирует с 30—45 наружными сегментами палочковых нейро-
сенсорных клеток. Вокруг наружного сегмента палочки обнаружи-
вается 3—7 отростков пигментоцитов, содержащих меланосомы,
фагосомы и органеллы общего значения. Количество отростков
пигментоцитов вокруг каждой колбочки достигает 30—40, они
длиннее и обычно не содержат органелл^ за-исключением меланосом.
Пигментоциты участвуют в защитных реакциях, тормозящих
перекисное окисление липидов с помощью ферментов микроперок-
сисом (пероксидаза, каталаза) и функциональных групп меланосом,
адсорбирующих металлы, катализирующие перекисное окисление
липидов. Фагосомы образуются в процессе фагоцитоза участков
наружных сегментов нейросенсорных клеток. Считают, что пигмен-
тоциты являются разновидностью специализированных макрофагов
центральной нервной системы, отличающихся по происхождению
от гематогенных макрофагоцитов.
Наличие меланосом-обуславливаег_прглощепие-85-^90% света,
попадающего вхлаз. Поглощение пигментоцитами «лишнего» рассе-
янного света повышает разрешающую способность глаза и умень-
шает распад родопсина.
Морфофункциональные изменения глаза
в зависимости от интенсивности
освещения
На изменение освещенности реагируют все функциональные
аппараты глаза, особенно светочувствительная часть сетчатки (све-
товая адаптация). При этом происходит перемещение меланосом
в апикальные отростки пигментоцитов, плотно окружающих наруж-
ные сегменты нейросенсорных клеток. Наблюдаемое при этом уко-
рочение колбочковых и удлинение палочковых клеток приводит
к сильному экранированию палочковых и, наоборот, хорошему
освещению колбочковых клеток — рецепторов дневного света.
При темновой адаптации происходит перемещение меланосом в
обратную сторону — из микроворсинок в цитоплазму пигментоци-
тов. Одновременно колбочковые клетки удлиняются и экранируются,
а палочковые укорачиваются, что обусловливает осуществление
ими функции сумеречного зрения. Перемещение мелансом осуще-
ствляется с помощью микрофиламентов. На этот процесс оказывает
влияние гормон меланотропин. Кроме того, пигментоциты участвуют
в метаболизме веществ, обеспечивающих фоторецепторный процесс.
Витамин А (ретинол), являющийся необходимым компонентом
зрительного пигмента, переносится особым белком, синтезируемым
в печени (ретинелсвязывающий белок — РСБ). Комплекс РСБ —
ретинол соединяется со специфическими рецепторами пигментоци-
тов, входит в состав их плазмолеммы и поступает в цитоплазму.
346
Пигментоциты обеспечивают предотвращение утечки витами-
на А в кровь на ярком свету, снабжение нейросенсорных клеток
ретинолом для регенерации и биосинтеза родопсина.
Регенерация сетчатки. Процессы физиологической регенерации
палочковых и колбочковых клеток происходят в течение всей жизни.
Ежесуточно в каждой палочковой клетке ночью или в каждой кол-
бочковой клетке днем формируется около 80 мембранных дисков.
Процесс обновления каждой палочковой клетки длится 9—12 дней.
В одном пигментоците ежесуточно фагоцитируется около 2—4 тыс.
дисков, утилизируется 60—120 фагосом, каждая из которых содер-
жит 30—40 дисков.
Таким образом, пигментоциты обладают исключительно высокой
фагоцитарной активностью, которая повышается при усилении
функции глаза в 10—20 и более раз.
Выявлены циркадные ритмы утилизации дисков: отделение и
фагоцитоз сегментов палочковых клеток происходят обычно утром,
а колбочковых — ночью. Постоянное многодневное освещение при-
водит к торможению указанных процессов, которые вновь активи-
зируются при отключении светового раздражения.
В механизмах отделения отработанных дисков важная роль
принадлежит ретинолу (витамин А), который в больших концентра-
циях накапливается в наружных сегментах палочковых клеток на
свету и, обладая сильно выраженными мембранолитическими свой-
ствами, стимулирует указанный выше процесс. Циклические нуклео-
тиды (цАМФ) тормозят скорость деструкции дисков и их фагоци-
тоз. В темноте, когда цАМФ много, скорость фагоцитоза невелика,
а на свету, когда уровень цАМФ снижен, она возрастает.
Васкуляризация. Ветви глазничной артерии формируют две груп-
пы разветвлений: одна образует ретинальную сосудистую систему
сетчатки (центральная артерия, вена и их разветвления), васкуля-
ризирующую сетчатку и часть зрительного нерва; вторая образует
цилиарную систему, снабжающую кровью сосудистую оболочку,
цилиарное тело, радужку и склеру. Лимфатические капилляры
располагаются только в склеральной конъюнктиве, в других участ-
ках глаза они не найдены.
Л
Вспомогательный аппарат таза
К вспомогательному аппарату глаза относятся глазные мышцы,
веки и слезный аппарат.
Глазные мышцы. Они характеризуются теми же структурными
особенностями, чт'р и соматическая мускулатура (топографию
см. в учебнике анатомии).
Веки. В них различают переднюю кожную поверхность и зад-
нюю — конъюнктив^, которая продолжается в конъюнктиву глаза,
покрытую многослойным эпителием (рис. 133). Внутри века, ближе
к его задней поверхности, располагается тарзальная пластинка,
состоящая из плотной волокнистой соединительной ткани. Ближе
к передней поверхности в толще век залегает кольцевая мышца.
347
Рис. 133. Веко (сагиттальный
срез).
/ — передняя (кожная) поверх-
ность; 2 внутренняя (конъ-
юнктива) поверхность; 3 — рес-
ница; 4 - тарзальная железа;
5 — ресничная железа; б —
сальная железа.
Между пучками мышцы располагается прослойка рыхлой волокни-
стой ткани. В этой прослойке оканчивается часть сухожильных
волокон мышцы, поднимающей верхнее веко. Другая часть сухо-
жильных волокон этой мышцы прикрепляется прямо к проксималь-
ному краю тарзальной (соединительнотканной) пластинки. Наруж-
ная поверхность покрыта тонкой кожей с тонкими волосками и саль-
ными железами. По краю века располагаются в 2—3 ряда ресницы.
В воронку корня ресницы открываются выводные протоки несколь-
ких сальных желез. Одновременно туда же впадают протоки так
назваемых ресничных желез. Это видоизмененные потовые железы,
имеющие прямые концевые отделы. В толще тарзальной пластинки
заложены открывающиеся по краю века разветвленные сальные
(мейбомиевы) железы. Рудиментарное третье веко, расположенное
в медиальном углу глаза, покрыто многослойным плоским эпители-
ем, содержащим слизистые клетки.
Сосуды века образуют две сети — кожную и конъюнктиваль-
ную. Лимфатические сосуды формируют третье дополни-
тельное, тарзальное сплетение.
Слезный аппарат глаза. Он состоит из слезных желез, слезного
мешка и слезно-носового протока. Слезные железы образуются из
нескольких групп сложных альвеолярно-трубчатых желез серозного
характера. Секрет слезных желез содержит около 1,5% хлорида
натрия, незначительное количество альбумина (0,5%) и слизи.
Слезная жидкость имеет в своем составе лизоцим, оказывающие
бактерицидное действие. Стенки слезного мешка и слезно-носового,
протока выстланы двух- или многорядным эпителием, расположен'
14S
ним на рыхлой волокнистой соединительной'ткани. В слезный ме-
шок открываются мелкие разветвленные трубчатые железы.
Возрастные изменения. С возрастом ослабляется функция
всех аппаратов глаза. В связи с изменением общего метаболизма
в организме в хрусталике и роговице часто происходит уплотнение
межклеточного вещества и помутнение, которое практически необ-
ратимо. Утрачивается эластичность хрусталика и ограничивается
его аккомодационная возможность. Склеротические процессы в
сосудистой системе глаза нарушают трофику, особенно сетчатки,
что приводит к изменению структуры и функции рецепторного
аппарата. ; [ ,rij
ОРГАН ОБОНЯНИЯ Ш ‘ )
Орган-обоияния (organum olfactus) в .своем периферическом
отделе представлен ограниченным _участком слизистой оболочки
юса — обонятельной областью, покрывающей у человека верхнюю
л~отчасти среднююраковины носовой полоети и перегородки..носа.
Знешне обонятельная область отличается от респираторной части
слйзистой желтоватым цветом.
РазвИтйе. Сходно с развитием глазного бокала. Во время онто-
генеза орган обоняния, так же как и орган зрения, неразрывно свя-
!ан с центральной нервной системой, закладывая~сь в одной общей
змбрионально^нервной пластинке. При этом парная закладка орга-
га обоняния у эмбрионов занимает ее самый передний край на
границе с эктодермой. В процессе дальнейшего развития, пепифери-
ческая часть обонятельного анализатора отделяется _от._за.чатка
центра_льной~исрвиой системы и .за тем только вторично, при помощи
обо н ятельного нерва связывается с центральными частям и анали за-
тора. В момент отведения от нервной пластинки зачатки органа обо-
няния обнаруживаются на ее переднем крае в виде парных, так назы-
ваемь1х~обонят7льных ямок. Далее эти зачатки в связи (Тростом го-
ловы перемещаются в верхние и средние носовые раковины (обоня-
тельная область)-.-"У зародышей на 4-м месяце развития из
элементов, образующих стенки обонятельных ямок, формируются
поддерживающие эпителиоциты и нейросенсорные обонятельные
клетки. Аксоны обонятельных клеток, объединившись между собой,
образуют в совокупности^УО- 4Q нервных пучков {обонятельных пу-
тей - fila olTactbnay, устремляющихся через отверстия в хрящевой
закладке^'будущей-решетчатой кости к опоиятеиъны-м-^луковицам
головного^чозгд.Здесь осуществляется синаптический контакт меж-
ду терминалями^ксонов и дендритами митральных нейронов обоня-
тельных луковиц. Некоторые участки эмбриональной_эпителиопо-
добной обонятельной выстилки, погружаясь в подлежащую соедини-
тельную ткань, превращаются в обонятельные железы.
Строение. Обонятельная выстилка состоит из эпителиоподоб-
ного пласта высотой 60—90 мкм, в котором различают обонятельные
нейросенсорные, поддерживающие и базальные этштелиоциты (рис.
134, А, Б). От подлежащей соединительной ткани они отделены
349
Рис. 134. Строение обонятельного
эшфелия (схема).
А — микроскопическое строение (по
Я. А. Винникову и Л. К. Титовой):
хорошо выраженной базальной мембраной. Обращенная в носовую
полость поверхность обонятельной выстилки покрыта слоем слизи.
Рецепторные, или нейросенсорные, обонятельные клетки
(сеIIu7ae пёTlrosensoriae olfactoria)~~~располатаются ~мёжду~поддер-
живающими эпителиоцитами и имеют короткий периферический
отросток — дендрит и длинный —^центральный — аксон. Их ядро-
содержащйе части занимают, как правило, срединноеположёние
в толще обонятельной выстилки: У собак^ которые отличаются
хорошо развитым органом обоняния, насчйтывшбт~около~225~Млн.
обонятельных клеток, у человека их число значительно меньше,
но все же_до£.тигает^6 млн. (30 тыс. рецепторов на 1 мм2). Дисталь-
ные части периферических отростков обонятельных клеток "заканчи-
ваются характерны мй утолщениями — обонятельными _булавами
(clava olfactoria). Обонятельные "булавы клеток на своей округлой
верши не несут до _10—12 заостренных подвижных обонятельных
ресничек (см. рис. 134, Б). Однако обнаружены также клетки (око-
ло 10%), имеющие на своей поверхности только микроворсинки.
Цитоплазма периферических отростков содержит митохондрии и
вытянутые вдоль оси отростка микротрубочки диаметром до
20 нм. Около ядра в этих клетках отчетливо выявляется гра-
нулярная эндоплазматическая сеть. Реснички булав содержат
продольно ориентированные фибриллы: 9 пар периферических
и 2 — центральных, отходящих от базальных телец. Обонятельные
реснички подвижны и являются своеобразными антеннами для
молекул пахучих веществ. Периферические отростки обонятельных
клеток могут сокращаться под-дёйствиёмТпахучих всществ. Ядра
обонятельных клеток светлые с однииилидвумя крупными ядрыш-
ками._Базальная часть клетки продолжается в .узкий, слегка изви-
350
в
Рис. 134 (продолжение).
Б ультрамикроскопическое строение (по А. А. Бронштейну с изменением»: В — регенера-
ция обонятельных клеток (по Л. Арденсу); а, б, в дифференцирующаяся нейросен-
сорная клетка; г, д — разрушающаяся клетка, / — обонятельный эпителий: // собствен-
ная пластинка слизистой оболочки; / — обонятельные клетки; 2 периферические от-
ростки (дендриты); 3 — обонятельные булавы: 4 — центральные отростки (аксоны);
5 — обонятельные реснички; 6 — микроворсинки; 7 — поддерживающие эпителиоциты;
8 — базальные эпителиоциты; 9 малодифференцированные нейроциты; 10— базальная
мембрана; II нервные стволики-аксоны обонятельных клеток; 12 — обонятельная
железа.
351
вдццщийся аксон, который проходит между опорными клетками.
В соединительнотканном слое центральные отростки составляют
пучкбЪёзмиёлинСВпттгюбрдятелътгого нерва, которые объединяются
в 20—40 нитевидных стволиков (fila olfactoria) (см. рис. 134, А)
и через отве стия решетчатой кости направляются в обонятельные
луковицы.
Поддерживающие эпителиоциты (epitheliocytus sustentans).jJ)op-
миру1бт Многоядерный эпителиальный пласт, в котором и располага-
ются обонятельные клетки, разделенные поддёрживаюшими зпите-
лиоцитами. На апикальной поверхности поддерживающих эпители-
оцитов имеются многочисленные микроворсйнки ддйной“до 4 мкм.
Поддерживающие эпителиоциты проявляют признаки апокриновой
секреции и обладают высоким уровнем метаболизма. В цитоплазме
их имеется эндоплазматическая сеть, которая расположена главным
образом вдоль длинной оси клетки. Митохондрии большей частью
скапливаются в апикальной части, где находится также большое
число гранул и вакуолей. Внутриклеточный сетчатый аппарат распо-
лагается над ядром. В цитоплазме поддерживающих клеток
содержится коричнево желтый пигмент. В результате присутствия
Батальные^ Эпителиоциты (epilFieliocytus basales) находятся на
базальной мембране и снабжены цитоплазматическими выростами,
окружающими пучки центральных отростков обонятельных клеток.
Цитоплазма их имеет относительно ровные контуры, заполнена
рибосомами и не содержит тонофибрилл. Существует_мне_ние^что
базальные эпителиоциты служат источником регенерации рецептор
ны^клето^В-пбдл’ёжащёи”рь1хлой волокнистой ткани обонятельной
области располагаются~Котщевые етлетпл Трубчато-альвеолярных
желез (см. рис. 134), выделяющие секрет, который содержит муко-
протеиды. Концевые отделы-состоят йз_ элементов, двоякого рода:
снаружи лежаТ" более уплощенные клетки — миоэпителиальные,
внутри — клетки;техретирующие по мерркридовому_типу. Их проз-
рачный, водянистый секрет вместе с секретом поддерживающих
эпителиоцитов ^увлажняет—пиверУПОСТь'-обонятельной выстилки,
что являетсяГнеобходимым условием длзгфунк'ционирования обоня-
тельных .клеток, В_это"м~се крете, омывающём обонятельные ресни ч-
ки, растворяются пахучие вещества, прйсутствйёТсоторытолько в
этом случае и~~воспринимае-тся рецепторными, белками, вмонтиро-
ванными в мембрану ресничек обонятельных клеток.
Васкуляризация.-Слизистая оболочка полости носа обильно
снабжена кровеносными и лимфатическими сосудами. Сосуды мик-
роциркуляторного типа напоминают кавернозные тела. Кровеносные
капилляры синусоидного типа образуют сплетения, которые могут
депонировать кровь. При действии резких температурных раздра-
жителей и молекул пахучих веществ слизистая оболочка носа может
сильно набухать и покрываться значительным слоем слизи, что
затрудняет рецепцию.
Возрастные изменения. Чаще всего они обусловлены перенесен-
ными в течение жизни воспалительными процессами (риниты),
352
которые приводят к атрофии рецепторных клеток и разрастанию
респираторного эпителия.
Регенерация. У млекопитающих в постнатальном онтогенезе
обновление рецепторных обонятельных клеток происходит в течение
30 сут. В конце жизненного цикла нейроны подвергаются деструкции
(см. рис. 134, Б, В). Малодифференцированные нейроны базального
слоя способны к митотическому делению, лишены отростков. В про-
цессе их дифференцировки увеличивается объем клеток, появляется
специализированный дендрит, растущий к поверхности, и аксон,
растущий в сторону базальной мембраны. Клетки постепенно пере-
мещаются к поверхности, замещая погибшие нейроны. На дендрите
формируются специализированные структуры (микроворсинки и
реснички).
/орган ВКУСА ( И
’ Орган_вкуса (organum gustus) представлен совокупностью так
называемых вкусовых почек (caliculi gustatoriae), располагающихся
в многослоином~эпителии боковых стенок желобоватых, листовид-
ных и шляпках грибовидных_сосочков языка человека (рис. 135,
А, Б). У детей.гГиногда и у взрослых вкусовые почки могут нахо-
иитьсянагубах, наружной и внутренней поверхностях надгортанни-
ка, голосовых связках Количество вкусовых почек у человека дости-
гает 2000, из них около 50% находятся в желобоватых сосочках.
Развитие. Первые признаки развития зачатков вкусовых почек
удается обнаружить в сосочках языка человеческого эмбриона
длиной 60 мм. Источником дифференцировки клеток вкусовых луко-
в и и. является эмбриональный многослойный эпи тедий .СССР чк ов,
он подвергается счеи.мя и> ной ли4к}.еру|'иировкг пол инлупирующим
ВО3дей.ствием оуонпрчмй nep.iui.iy волокон ялуАЧНОГО, ЯЗЫКОГЛОТОч-
ного и блуждающего нервов. Таким образом, иннервация вкусовых
почек появляется одновременно с возникновением их зачатков.
Строение. Каждая вкусовая почка имеет эллипсоидную форму
и занимает всю Толщу многослойного эпиТёЛИалКнОГО пласта
сосочка. Она состоит из плотно прилежащих друг к ДРугу_40—60
клеток, среди которых различают три типа: рецепторные, поддержи-
вающие~й~базальныё клетки. От подлежащей соединительной ткани
вкусовая почка отделяется базальной мембраной. Вершина, почки
сообщается с поверхностью языка при помощи„отверстия — вкусо-
вой поры~[рогиь gustatorius) (см. рис. 135, Б). Вкусовая пора ведет
в небольшое углубление образованное апикальными поверхностями
вкусовых сенсорных клеток, — вкусовую ямку.
Вкусовые сенсорные эпителиоциты (epitheliocytus sensorius
gustatoriae) отделяются друг от друга поддерживающими эпители-
оцитами. Их ядра имеют вытянутуюовальцую форму и распола-
гаются ближе-к основанию клеток. Цитоплазма вкусовых клеток
в апикальном “отделе богата агранулярной эндоплазматической
сетью и митохондриями. На апикальном конце вкусовой, клетки
имеются микро ворсинки (см. рис. 135, Б), благодаря чему значи-
12 ГИСТОЛОГИЯ ’5’
А микроскопическое строение: / •— мно-
гослойный плоский эпителий сосочка; 2 —
пространство между сосочками; 3 — вкусо-
вая пора; 4 - клетки вкусовой почки
f> — ультрамикроскопическое строение
(рис. Р. А. Певзнер): / — рецепторный
вкусовой эпителиоцит; 2 поддерживаю-
щий эпителиоцит; 3 — базальный эпителио-
цит; 4 — эпителиоцит; 5 — микроворсин-
ки; 6 — нервные окончания; 7 — нервное
волокно; 8 — мукопротеиды.
тельно увеличивается поверхность воспринимающей мембраны.
Между микроворсинками во вкусовой ямке находится электронно-
плотное вещество с высокой активностью фосфатаз и значитель-
ным содержанием белка~и мукопротеидов, которое- “играет роль
I адсорбента~~для "вкусовых "веществу тгопадающих~“ия тюверхность
языка.
Во вкусовых точках_переднеи^2*асти языка обнаружен сладко-
чувствительный рецепторный белок, задней части — горькочув-
ствительный. Вкусовые вещества адсорбируются на примембран-
ном слое цитолеммы микроворсинок, в которую вмонтированы
специфические рецепторные белки. Одна и та же вкусовая клетка
способна воспринимать несколько вкусовых раздражений. При
адсо.рбцик-ВОЗД£Йствующихмолекул происходят конформационные
изменения-редептепшях_йЖрвь1х-Тмолёкул7тгсггпрьте'“приврдят к
локальному .изменению проницаемости мембран вкусового сенсор-
ного эпителиоцита и генерации потенциала. Этот процесс имеет
(сходство с процессом в холинергических синапсах, хотя допуска-
ется участие и других медиаторов. В каждую вкусовую почку
входит и разветвляется около 50 афферентных нервных волокон,
формирующих СИПапсы с "базальными отделами рецепторных кле-
ток.
Возбуждение из вкусовых почек передается в виде импульсов
через синапсы к нервным окончаниям,~аГот них по нервным пу-
тям —-н центральные звенья вкусового анализатора, где и форми-
руются-представления б характере-вкусовых рецепций.
Поддерживающие эпителиоциты (epilheliocytus sustentans) отли-
ваются наличием крупного ядра, хорошо выраженными элемента-
ми гранулярной и агранулярной эндоплазматической сети, комп-
:.4
юксом Гольджи, присутствием пучков тонофибрилл. Они окружа-
ют и изолируют вкусовые клетки и нервные волокна в базальном
отделе вкус'ОНОЙ почки И~прТ<11ймают участие^процессах секреции
липопротеидов. Базальные ^дигелщ?^ит.ы_расположенына базаль-
ной мембране и в отличие от сенсорных и^юддерживающих клеток
не достигают поверхности эпителиального слоя. Они представляют
собой малоспециализированные клетки, из которых, видимо, раз-
виваются поддерживающие и вкусовые сенсорные эпителиоциты.
Нервные волокна, входящие во вкусовую почку из подлежащей
соединительной ткани, идут вдоль-поддерживающих эпителиоци-
тов и заканчиваются на боковой поверхности сенсорных клеток.
Возрастные изменения. С возрастом число вкусовых
почек уменьшается и отмечается повышение вкусовых порогов
для всех вкусовых веществ, особенно для сладких.
Регенерация. Сенсорные и поддерживающие эпителиоциты
вкусовой почки непрерывно обновляются. Продолжительность
жизни их примерно 10 сут. При разрушении вкусовых сенсорных
эпителиоцитов синапсы прерываются и вновь образуются на новых
клетках.
.* П '
ОРГАН СЛУХА И РАВНОВЕСИЯ
[ПРЕДДВЕРНО-УЛИТКОВЫЙ ОРГАН)
В состав преддверно-улиткового органа (organum vestibulo-
cochleare) входят наружное., среднее и внутреннее ухо, которые в
совокупности осуществляют восприятие звуковых, гравитационных
и вибрационных стимулов, линейных и угловых' ускорений.
Наружное ухо
Наружное ухо включает ушную раковину, наружный слуховой
проход и барабанную перепонку.
Ущная"^акбвцн^^остоит_из_хонкрй пластинки эластического
хряща, покрытой кожей с немногочисленными тонкими волосами
и сальными железами. Потовых желез в ее составе мало.
Наружный слуховой проход образован хрящом, являющимся
продолжением эластического хряща ракови'ныГ Поверхность про-
хода покрыта"тонкой~ кожей, содержащей волосы и связанные
с ними сальные железы. Глубже сальных желез расположены
трубчатые церуминозные .железы (glandula ceruminosa), выделяю-
цие ушную серу. Их протоки_ открываются самостоятельно на
поверхности слухового лрохода или в выводные—протоки сальных
желез. Церуминозные железы располагаются неравномерно по
ходу слуховой трубы: во внутренних двух третях они имеются
лишь в коже верхней части трубы.
Барабанная перепонка овальной, слегка вогнутой формы. Одна
из слуховых косточек среднего уха — молоточек — сращена с
помощью своей ручки с внутренней поверхностью барабанной
перепонки. От молоточка к барабанной перепонке проходят крове-
носные 'сосуды и нервы. Барабанная перепонка в средней части
12*
355
состоит из двух слоев, образованных пучками коллагеновых воло-
кон и залегающими между ними фибробластами. Волокна наруж-
ного слоя~расположены радиально^ а внутреннего —--циркулярно.
В верхней части "барабанной перепонки количество коллагеновых
волокон уменьшается."" Эластические тонкие волокна имеются
на периферии и в центре барабанной перещщки. На наружной
ее поверхности-располагается очень тонким слоем (50—60 мкм)
эпидермис, на внутренней поверхности, обращенной в среднее
ухо — слизистая лбодонка толщиной около 20—40 мкм, покрытая
однослойным плоским эпителием.
Среднее ухо^
Среднее ухо состоит из барабанной полости, слуховых косто-
чек и слуховой трубы.
Барабанная ~полость — уплощенное пространство, покрытое
опноспойным7~ппоским, местами переходящим в кубический или
цилиндрический эпителий. На медиальной стенке барабанной
полости имеются два отверстия, или «окна». Первое — овальное
окно В нем_ располагается основание стремечка, которое удержи-
вается с помощью тонкой связки по окружности окна.jObb льное
окно отделяет барабанную полость от вестибулярной лестницы
улитки. Второе окно — круглое, находится несколько позади
овального?" Оно закрьгго~волокнистой мембраной. Круглое окно
отделяет барабанную полостъ_от_барабанной_ лестницы улитки.
Слуховые' косточки — молоточек, наковальня, стремечко как
система рычагов передают колебания барабанной перепонки на-
ружного уха к овальному окну, от которого начинается вестибу-
лярная _деггии((а-внутреннего уха.
Слуховая труба, соединяющая барабанную полость с
носовой частью"глотки; имеетгтарошо" выраженный просвет диа-
метром 1—2 мм. В области, прилежащей к барабанной полости,
слуховая труба окружен!? костной стенкой, а ближе к глотке со-
держит островки гиалинового хряща. Просвет трубы _выстлан
призматическим... многорядным реснитчатым___эпителием. В нем
имеются бокаловидные железистые клетки. На поверхности эпите-
лия открываются протоки слизистых желез. Уерез сдухсшую_трубу
регулируется-давдение воздуха—н_.^арабанноЙ_ЛСДрс ^среднего
уха.
Внутреннее ухо
Внутреннее ухо состоит из костного и расположенного.в нем
перепончатого лабиринта, в кот<5ром~наХОдятся рецепторные клет-
ки — в5л6сковые~Сежорные эпи гелиоцитьт-органа слуха неравнове-
сия. Они располбжены"~в определенных участках перепончатого
лабиринта:' слуховые рецепгорнБгегютеТки — '~в~спцральнол< органе
улитки, а рецепторные клетки органа равновесия — в_эллиптиче-
ском^и~сфёричёском мешочках и ампулярных гребешках полу-
кружных каналов.
356
Рис. 136. Схема развития слухового пузырька у эмбриона человека (по Арею с
и вменением).
А стадия 9 сомитов; В стадия 16 сомитов; В — стадия 30 сомитов; / — эктодерма;
2 — слуховая плакода; 3 — мезодерма; 4 — глотка; 5 слуховая ямка; 6 — мозговой
пузырь; 7 — слуховой пузырек.
Развитие внутреннего уха. У эмбриона человека перепонча-
тый лабиринт развивается путем впЯчиванйЯ~тподлежащую эмбри-
ональную соединительную ткань эктодермы, котораязатем замы-
каете я и образует тазе -называемый слухосоц пузырек (рис. 136).
Он_ располагается вблизи первой жаберной ш.епи пп обеим сторо-
нам закладки про.лопговатого мозга. Слуховой пузырек состоит
из многорялного эпителия, который секретируе-т.. эндолимфу, за-
полняющую просвет пузырька. Одновременно слуховой пузырек
ко нтактирует с_эмбриональным слуховым нервным ганглием,
который вскоре делится на две части — ганглий__лред^узерия и
ганглий улитки. В процессе дальнейшего развития пузырек меняет
свою форму, перетягиваясь на две части: первая — вестибуляр-
ная *~превращаегся~~в~эллинтическии тагептзчек — утрикулюс (utri-
с u lus) с полукружными каналами и их ампулами, вторая об разует
сферический мешочек — саккулюс (sacculus) и закладку улитково-
го канала. Улитковый канал постепенно растет, завитки jero увели-
чиваются и он отделяется от^эллиптического мешочка. На месте
прилегания слухового-ганглия к слуховому-пузырьку стенка по-
следнего утолщается. Волосковые сенсорные и поддерживающие
эпителиоциты органа слуха и равновесия обнаруживаются уже у
эмбрионов длиной 15— 18,5 мм. Улитковый канал вместе со спи-
ральным органом развивается в виде "трубки, которая впячивается
в завитки костной улитки. Из эпителия базальной стенки пере-
пончатого канала развивается спиральной орган, содержащий
рецепторные слуховые клетки. В это же время происходит образо-
вание синапсов между чувствительными клетками лабиринта и
периферическими отростками клеток вестибулярного и улиткового
ганглиев.
Одновременно развиваются и перилимфатические полости.
В улиткеЭМбриона длиной 43 мм ймеётся~~гюрйлимфатИческая
полость барабанной лестницы, а у эмбрионов длиной 50 мм — и
вестибулярная перилимфатическая лестница. Несколько позднее
357
Рис. 137. Строение пере-
пончатого канала улитки и
спирального органа.
А — схема; £ — спираль-
ным орган. / — перепонча-
тый канал улитки; 2 — вес-
тибулярная лестница; 3 —
барабанная лестница; 4 —
спиральная костная пластин-
ка; 5 — спиральный узел; 6—
спиральный гребень; 7 —
дендриты нервных клеток;
8 — вестибулярная мембра
на; 9 — базилярная мембра-
на; 10 — спиральная связ-
ка; / / — эпителий, высти-
лающий барабанную лест-
ницу; /2 — сосудистая по-
лоска; 13 — кровеносные
сосуды; 14 — покровная
пластинка; 15 — наружные
волосковые сенсорные эпи-
телиоциты; 16 — внутрен-
ние волосковые сенсорные
эпителиоциты, /7 — внутрен-
ние поддерживающие эпите-
лиоциты; 18 — наружные
поддерживающие эпителио-
циты; 19 — клетки-столбы;
У) — туннель.
происходят—процессы—окостенения и формирования костного
лабиринта улитки и. полукружных каналов.
Улитковый канал перепончатого лабиринта. Восприятие звуков
осуществляется в спиральном органе, расположенном по всей
лпине^-улиткового каната перепончатого лабиринта. Улитковый
канал представляет собой спиральный слепо заканчивающийся
мешок- длиной 3,5 см, заполненный эндолимфой и окруженный
снаружи_дщзилц.я</>ой. Улитковый канал илжружающие его запол-
ненные перилимфой полости барабазшоД-Шестибу лярнои лестницы
в свою очередь заключены в костную улитку, образующую у чело-
века 2 */., завитка вокруг центрального костного стержня.
Упцггковый канал плпереччом разрезе-йТййет форму тре у -
гольника (рис. 137, 4), стороны которогО-С^разоШЛаьй^СШЙуляр-
458
Рис. 138. Схема ультрамикро-
сконического строения сосу-
дистой полоски (рис.
Ю. И. Афанасьева).
/ — светлые базальные клетки;
2— промежуточные клетки; J —
темные призматические клетки;
4 - митохондрии; 5 - крове-
носные капилляры; 6 — базаль-
ная мембрана.
ной мембраной, сосудистой полоской, лежащей на наружной стен-
ке костной улитки, и базилярно11__ллх1схиикой. Вестибулярная
мембрана (membrana vestibularis) образует верх н_.е м е_дта л ь-
н у ю с т е н к у канала. Она представляет собой тонкофибрилляр-
нуга соединительнотканную, _ддаспшку, покрытую однослойным
плоским эпителием, обращенным к энпопимфе,—и эндотелием,
обращенным к перилимфе.
Нару ж,н ая стенка образована сосудистой______________полоской
(stria vascularis), .расположенной на спиральной связке (ligamen-
tum spirale). Эпителий~м1кй орядный cbiтоит_ма-ллпских .базальных
светлых клеток и высоких отрлстмад-ыу—прггчмнтчщеслшх темных
клетоКд^кйТСрые богаты митохондриями (рис. 138). Митохондрии
клеток отличаются очень высокой активностью окислительных
ферментов. Ме^жду кпетками проходят гемлкяпиппяры Пред по ла
га ют, что сосудистая полоска выполняет секреторну ю фу нк ци ю,
продуцируя эндолимфу, и играет значительную_роль в трофике
спиратгБПОгй органа:
Нижняя, базилярная, пластинка (lamina basilaris), на кото-
рой располагается спиральный орган, построена наиболее сложно.
С внутренней стороны она прйкпёпляётся2_1с'хдиральнаи-2костной
пластинке в том месте, где ее надкостница — лимб (см. рис. 1э7)
.159
делится на две части: верхнюю — вестибулярную губу и ниж-
нюю — барабанную губу. Последняя переходит в базилярную
пластинку, которая на противоположной стороне прикрепляется
к спиральной связке.
Базилярнаядндастинка представляет .собой соединительноткан-
ную пластинку, которая в виде спирали тянется вдоль всего
улиткового канала. На стороне, обращенной к спиральному орга-
ну, она покрыта базальной~мембраной эпителия этого органа.
В основе базилярной пластинки лежат тонкие коллагеновые
волок на («струны»),~которые-тянутся в виде непрерывного ра -
диального пучка от спиральной^КОСТНОЙ пластинки до спираль-
ной. связки, выступающих в полость костного канала улитки Ха-
рактерно, что длина волокон неодинакова по всей длине улиткового
канала— Более длинные (около 505 мкм) волокна находятся на
верщине_улитки, короткие (около 105 мкм) — в ее основании. Рас-
полагаются волокна в гомогенном основном веществе. Подокна
состоят из тонких фи рилл диаметром около 30 нм, анастомозирую-
щих между собой с помощью еще более тонких пучков. Со стороны
барабанной лестницы базилярная пластинка покрыта слоем плоских
клеток мезенхимной природы.
Поверхность спирального лимба покрыта плоским эпителием
Его клётки'пзбладяют-енособттогтьтсг К^ССкреции. Выстилка спи-
ральной бороздки (sulcus spiralis) представлена несколькими ряда-
ми крупных плоских полигональных клеток, которые непосредст-
венно переходят в поддерживающие эпителиоциты, примыкающие
к внутренним волосковым клеткам спирального органа.
Покровная мембрана (membrana tectoria) имеет связь с эпите-
лием вестибулярной губы. Она представляет собой лентовидную
пластинку желеобразной консистенции, которая тянется в виде
спирали по всей длине спирального органа, располагаясь над
вершинами его волосковых клеток. Эта пластинка состоит из
тонких радиально направленных коллагеновых волокон. Между
волокнами находится прозрачное склеивающее вещество, содержа-
щее гликозаминогликаны.
Строение спирального органа. Спиральный j»praH состоит из
двух групп клеток — сенсорных и поддерживающих. Каждая из
этих групп клеток подразделяется на внутренние и наружные (см.
рис. 137, Б). Границей служит туннель.
Внутренние Сенсорные во чпскдвъте эпителиоциты (epitheliocyti
sensoriae pilosae internae) кувшинообразной формы с расширен-
ным основанием лежат в один ряд. fla" поверхности их слегка
выпуклых вершин имеется от 31) до 60 коротких уплотненных
микроворсинок, способных отклоняться. — стереоци пий (рис. 139).
Они располагаются в 3—4 ряда. Стереоцилии в совокупности
о разуют пучок, в~котором длина самой высокой из них достигает
40 мкм. Ядра в этих клетках залегают в базальной части.
У человека приблизительно 3500 внутренних ~Ъолосковьгх клеток.
Апикальная часть внутренней волосковой к летки-шмсрызд кутику-
лой, через жрдопую проходят стереоцилии. В цитоплазме обнару-
1Я1
360
Рис. 139. Ультраструктурная организа-
ция внутренней (А) и наружной (Б)
слуховых волосковых клеток (схема).
/ - волоски; 2 — кутикула; 3 — митохонд-
рии; 4 — ядра; 5 — синаптические пузырьки
в рецепторных клетках; 6 светлые нерв-
ные окончания; 7 — темные нервные оконча-
ния.
живаются митохондрии, элемен-
ты гладкой и гранулярной эндо-
плазматической сети и пере-
плетающиеся актиновые и мио-
зиновые микрофиламенты. На
ружные волосковые „сенсорные
зпите шациты (epitheliocytr
sensoriae pillosae externae) име-
ют округлое основание. На сво-
ей апикальной поверхности они
несут кутикулярную пластинку
со стерео[!или_я_ми. Волосковые
клетки здесь лежат в три па-
па ллельных ряда. У человека в
верхних завитках улитки может
быть 4—5 таких рядов. Своими
основаниями волосковые клетки располагаются во вдавлениях,
образованных телами лежащих под ними поддерживающих эпи-
вых клеток. Они, как и внутренние клетки, несут на своей апикальной
поверхности кутикулярную пластинку со стереошзлиями, которые
образуют щеточку из нескольких рядов в виде буквы V (рис. 140).
Стереоцилии наружных волосковых клеток своими вершинами
прикасаются к внутренней поверхности текториальной мембраны.
С те реоцилии содержат многочисленные плотно упако ванные фиб-
риллы, имеющие в своем составе сократительный белок актомиозин,
благодаря чему после наклона они вновь принимают исходное
вертикальное положение. Киноцилия в волосковых клетках спи-
рального органа взрослых млекопитающих отсутствует.
Цитоплазма сенсорных эпителиоцитов богата окислительными
ферментами, монофосфоэстеразой, содержит РНК. Наружные сен-
сорные эпителиоциты содержат большой запас гликогена, а их
стереоцилии богаты ферментами, в том ЧИСЛУ ацетилхолин-
эстераз6иГ~Активность ферментов и других химических веществ
при непродолжительных звуковых воздействиях возрастает, а при
длительных — снижается. Наружные сенсорные эпителиоциты
значительно чувствительнее к—зву%а&1г=&1адщЕЙ12Щнтеттсивности,
чем внутренние. Высокие звуки раздражают только волосковые
клетки, расположенные на нижних завитках улитки, а низкие
звуки — волосковые клетки вершины улитки и часть клеток на
нижних завитках.
В о_ время звукового воздействия на барабанную пе ре по н ку
ее колебания передаются2ыа_мадоточек, наковальню и стремечко,
361
Рис. 140. Наружная поверхность клеток спирального органа. Сканирующая элек-
тронная микрофотография. Х2500 (препарат К. Койчева).
/ - наружные волосковые сенсорные эпителиальные клетки; 2 — внутренние волосковые
сенсорные эпителиальные клетки: 3— границы поддерживающих эпителиоцитов.
а далее через овальное окно на перилимфу, базилярную и
текЧория nt-ну ил мембря мыЭто движение строго соответствует
частоте и интенсивности звуков. При этом происходит отклоне-
ние стереопилий и возбуждение рецептор(щ1х~ю1ётокТ~~Онр сопро-
вождается взаимодействием ацетилхолина, содержащегося в эндо-
лимфе с холинорецепторным белком в мембранах стереоцилий,
где~ также локализована ацетилхолинэстераза, которая^разрушает
я идтипхлдин -Все это..„приводит возникновению рецепторного
потенциала (-микрофонный., эффект). Афферентная информация
по слуховому нерву—передается в центральные части слухового
анализатора.
Поддерживающие эпителиоциты спирального органа в отличие
от сенсорных своими основаниями непосредственно располагают-
362
ся на базальной мембране. В их цитоплазме обнаруживаются
тонофибриллы. Внутренние^ фаланговьнэпителиоциты (epithelio-
cyti phalangeae internae), лежащие под внутренними волосковы-
ми сенсорными эпителиоцитами, имеют тонкие пальцевидные
отрбстки Гфаланги)._ Этими отргосткалпг вершины рецепторных
клеток отделены друг от другй? Н спиральном органе расположены
также так называемые внутреннийи наружные ~сто.тбовьГе эпите-
лиоциты (epitheliocyti pilaris internae et externae). На месте своего
соприкосновения они сходятся под острым углом друг к другу и
образуют правильный треугольный канал — внутренний туннель
(cnniculus internus), заполненный эндолимфой Туннель тянется
по спирали вдоль всего спирального 6рГ5на~ Реновация клеток-
столбов прилежат друг к другу и располагаются на базальной
мембра"не. Через- туннель проходят безмякотныё~нёрвныё^волокна,
идущие от нейронов спирального ганглия к сенсорным клеткам.
На базилярной мембране располагаются также наружные фа-
ланеовыё^ клетки (epitheliocyti phalangeae externae). Они залегают
в 3—4 ряда в; непосредственной близости от наружных столбо-
вых клеток. Эти клетки имеют призматическую форму/_В их
базальной части располагается ядро, окруженное пучками тоно-
фибрилл. В верхней трети, на месте соприкосновения с наруж-
ными волосковыми клеткамЩ в наружных фаланговых эпите-
лиоцитах "есть чашевидноё~вдавЛёние, В кОтороё входит'основание
наружных сенсорных кдёток. Только один узкий отросток наружных
поддерживающих эпителиоцитов доходйт своей ТОНЮТЙиершиной —
фалангой — до верхней пгТверхТюсти"спирального органа. В этом
органе есть еше лва вида клеток. Наружные пограничные
эпителиоциты (cellulae epitheliocyti Inrntans externae) распола-
гаются на базальной мембране рядом с наружными фаланговыми
эпителиоТ|и^мии^б-р^ЗУКНП:Л;Т0:Ш1ЮЙ~^та~ТГ^1ИЛСТКЮ.^ПТШтелиаль-
ных клеток. На наружной поверхности клеток имеется большое
количество микроворсинок. Эти клетки отличаются- богатством
гликогена7-итаг_очевиднс1,_^бу£Л£)влецр__их_1рофи.чес^ой функцией.
Латеральнее этих клеток располагаются наружные поддерживаю-
щие (epitheliocyti sustentans externus), которые имеют кубическую
форму и, постепенно видоизменяясь, переходят в эпителий,
выстилающий сосудистую полоску.
Иннервация и васкуляризация спирального органа см. ниже.
Вестибулярная часть перепончатого лабиринта.' Это место рас-
положения рецуптороворгана равновесия. Она состоит из двух
мешочков — эллиптического (utriculus) и сферического 4t>acculus),
сообщающихся при помощи" узкого канала и связанных с тремя
полукружньгми^ каналами, локализующимися в костных КЭналах,
расположецных__в дгрех—взаимно -перпендикулярньгх-нанравде ниях.
Эти каналы на. месте соединения их с эллиптическим мешочком
(маточка) имеют расширения — ампулы. В стенке перепончатого
лабиринта в области эллиптического- и сфёричёскйш "мешочков
и ампул есть~учасТки, содержащие чувствительные (сенсорные)
клетки. B~MCfno4Kax~3TtT~участюГ~называютеж-нятндлщ. или маку
ЗЬЗ
лами± соответственно: пятно эллиптического мешочка (macula
utriculi) и пятно круглого мешочка (macula sacculi), а в ампу-
лах— гребешками, или кристами (crista ampullaris).
Стенка вестибулярной части перепончатого лабиринта состоит
из однослойного плоского ^пителия»__за исключением области
крист полукружных каналов и макул, где он превращается в
кубический и призматический.
Пятна мешочков-(макулы). Пятно выстлано эпителием, рас-
положенным на базальной мембране и состоящим из сенсор-
ных 1и~опорных клеток (рис. 141, А, Б). Поверхность эпителия
покрыта~особой студенистой отолитовой мембраной (membrana
statoconiorum), в которую включены состоящие из карбоната каль-
ция кристаллы—отолиты, или статоконии (statoconia).
Макула эллиптического мешочка — место восприятия линей-
ных ускорений, 'г. е: земного притяжения, рецептор гравитации,
связанный Tjшшепетпгем тинуса! мыпмттггВедёляющих установку
тела. Макула сферического мешочка, являясь также рецептором
гравитации, одновременно воспринимает и вибрационные колеба-
ния.
Волосковые сенсорные клетки (cellulae sensoriae pilosae) не-
посредственно обращены своими вершинами, усеянными волоска-
ми, в полость лабиринта. Основание клетки контактирует с аффе-
рентными и эфферентными нервными окончаниями. По строению
волосковые клетки подразделяются на два типа (см. рис.
141, Б). Клетки первого типа (грушевидные) отличаются округ-
лым широким основанием, к которому примыкает нервное
окончание, образующее вокруг него футляр в виде чаши. Чаша
местами образует синаптические контакты с рецепторной клеткой.
Клетки второго типа (столбчатые) имеют призматическую форму.
К основанию клетки непосредственно примыкают точечные
афферентные и эфферентные нервные окончания, образующие
характерные синапсы. На наружной поверхности этих клеток
имеется кутикула, от которой отходят 60—80 неподвижных
волосков — стереоцилии длиной около 40 мкм и одна подвижная
ресничка — киноцилия, имеющая строение сократительной рес-
нички. Круглое пятно человека содержит около 18 000 рецептор-
ных клеток, а ов&льное — около 33 000. Киноцилия всегда поляр-
но располагается по отношению к пучку стереоцилий. При смеще-
нии киноцилий в сторону стереоцилий клетка возбуждается, а
если движение направлено в противоположную сторону — проис-
ходит торможение клетки. В эпителии макул различно поляри-
зованные клетки собираются в 4 группы, благодаря чему во
время скольжения отолитовой мембраны стимулируется только
определенная группа клеток, регулирующая тонус определенных
мышц туловища. Другая группа клеток в это время тормозится.
Полученный через афферентные синапсы импульс передается
через вестибулярный нерв в соответствующие части вестибуляр-
ного анализатора.
Поддерживающие клетки (epithehocyti sustentans), распола-
364
Рис. 141. Макула.
А — строение на светоопти-
ческом уровне (схема): /—
поддерживающие эпителио-
циТы; 2 — сенсорные эпите-
лиоциты (по Кольмеру); 3 —
волоски; 4 — нервные окон-
чания; 5 — миелиновые нерв-
ные волокна; 6 — студени-
стая отолитовая мембрана;
7 — отолиты. Б — строение
на ультрамикроскопическом
уровне (схема). / — киноци-
лия; 2 — стереоцилии; 3 —
кутикула; 4 — поддерживаю-
щий эпителиоцит; 5 — чаше-
видное нервное окончание;
6 — эфферентное нервное
окончание; 7 — афферентное
нервное окончание; 8 — мие-
линовое нервное волокно
(дендрит).
гаясь между сенсорными, отличаются темными овальными ядрами.
Они имеют большое количество митохондрий. На их вершинах
обнаруживается множество тонких цитоплазматических микровор-
синок.
Ампулярные гребешки (кристы). Они_в виде поперечных скла-
док цаходятся_в каждом ампулярном расширении _полукружного
канала._Ампулярный_ гребешок выстлан сенсорными волосковыми
и поддерживающими эпителиоцита миГА1 шкальная часть этих кле-
ток окружена желатинообразным прозрачным куполом (cupula
gelatinosa), который имеет форму колокола, лишенного полости.
Его длина достигает 1 мм. Тонкое строение волосковых клеток
и их иннервация сходны с сенсорными клетками мешочков
(рис. 142). В функциональном отношении желатинозный купол —
рецептор угловых ускорений.—При тхвижепиигиловБг или ускорен-
365
лами
utric
ЛЗХ_
с
из <
Крис
куби
г
поле
ных
ПОК{
stato
ция
Ч
ных
связ
тела
грав
HHflj
ПОС]
ми,
рен1
ВОЛ1
141
лыь
око
мес
Кл<
К
аф(1
хар
име
BOjJ
рес|
НИЧ
ньг
но
НИ1>
ест
хо;
301
вре
оп^
мы
По
чер
НО1
164
Рис. 142. Схема строения ампулярного гребешка (по Кольмеру с изменением),
/ — гребешок;// — желатиновый купол; / — поддерживающие эпителиоциты; 2 — сенсорные
эпителиоциты, 3 — волоски; 4 — нервные окончания; 5 - миелиновые нервные волокна;
б — желатинозное вещество пограничного купола; 7 — эпителий, выстилающий стенку
перепончатого канала.
ном вращении всего тела купол легко меняет свое положение.
Отклонение купола под влиянием движения эндолимфы в полу-
кружных каналах стимулирует волосковые клетки. Их возбужде-
ние вызывает рефлекторным ответ той части скелетной мускула-
туры, которая корригирует положение тела и движения глазных
мышц.
Иннервация. На волосковых сенсорных эпителиоцитах спираль-
ного и вестибулярного органов расположены афферентные нерв-
ные окончания биполярных нейронов, тела которых располагают-
ся в основании спиральной костной пластинки, образуя спираль-
ный ганглии, большинство нейронов которого одето миелиновой
оболочкой. Нейриты этих клеток в составе слухового нерва несут
импульсы в центральную нервную систему. В спиральном органе
наряду с афферентной иннервацией имеется и эфферентная иннер-
вация, которая осуществляется так называемым оливокохлеарным
пучком. Те и другие волокна при переходе на базилярную
мембрану теряют свою миелиновую оболочку, окружают внутрен-
366
ние волосковые клетки, а частично проходят через туннель в
область наружных волосковых клеток. Волокна, переплетаясь,
подходят к основаниям внутренних и наружных волосковых кле-
ток и заканчиваются здесь терминалями. При этом образуется
два связанных между собой сплетения, располагающихся по спи-
рали вдоль всего улиткового канала. Одно из них лежит в области
основании внутренних волосковых клеток—внутреннее спиральное
сплетение. Другое, расположенное между наружными поддержи-
вающими клетками, называется наружным спиральным сп /ете-
нием. В области внутреннего и наружного спиральных сплетении
обнаруживается высокая активность специфической холинэстера-
зы, которая, как известно, расщепляет ацетилхолин, принимающий
участие в передаче импульсов в спиральном органе.
Васкуляризация. Артерия перепончатого лабиринта берет свое
начало от верхней мозговой артерии. Она делится на две ветви,
вестибулярную и общую кохлеарную. Вестибулярная артерия
снабжает нижние и боковые части эллиптического и сферического
чочков, а также верхние боковые части полукружных каналов,
с зуя капиллярные сплетения в области слуховых пятен. Кох-
леарная артерия снабжает кровью спиральный ганглий и через
надкостниц, вестибулярной лестницы и спиральной костной пла-
стинки прошкает до внутренних частей базальной мембраны
спирального оргзна. Венозная система лабиринта складывается
трех независимых друг от друга венозных сплетений, нахо-
дяиГ-’хся в улитке, преддверии и полукружных каналах. Лимфа-
тическ сосуды в абиринте не обнаружены. Спиральный орган
сосудов <е имеет. \
Возрас’ные измен ния. С возрастом у человека могут
возникать ни тения органа слуха. При этом изменяются от-
дельно или col *стно звукопроводящая и звуковоспринимающая
системы. Это с зано с тем, что в области овального окна
костного лабиринт появляются очаги оссификации, распростра-
няющиеся на подко. |ную пластинку стремечка. Стремечко теряет
подвижность в овальном окне, что резко снижает порог слыши-
мости. С возрастом чаще поражается звуковоспринимающий
нейросенсорный аппарат, т. е. сенсорные клетки, которые, проделав
свой жизненный цикл, гибнут и не восстанавливаются.
Глава XV
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
В составсердечно-сосудистой системы входят сердце, крове-
но<.ц.Ы° 'сосуды^ -nnii/i ггпиегуггр—гпгп’1<пт‘ Спеупиетпп система и
сердце обеспечивают распространение по организму крови, пита-
тель н ых и биологически активных веществ, газов, продуктов
метаболизма
^67
ламик
utric
лах -
с I
ИЗ (
крис*
куби
поле
ных
покр
stato
ция
НЫХ;
свял'
тела,
грав|
ния.
ПОС|
ми,
рент
волг
141,
лым
OKOI
мес
Кле
К
аф<$
xapj
име
воле
peci
нич
ны>
но |
НИИ
есл
ход
зов
вре
опр!
МЫ1
По;
чер
ног
164
КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды представляют собой систему замкнутых
трубок различного диаметра, осуществляющих транспортную
функцию, регуляцию кровоснабжения органов и обмен веществ
между-кровью и окружающими тканями.
Развитие. Первые^кровеносные сосуды появляются в мезенхиме
стенки желточного мешка на 2-й — 3-й неделе эмбриогенеза,
а также в стенке хорщща. составе—так называемых кровяных
островков. Часть мезенхимных клеток ЛО периферии островков те-
ряет связь с клетками, расположенными в центральной части, и
превращается в_ эндотелиальные клетки первичных кровеносных
сосудов, Клетки центральной части островка округляются, диффе-
ренцируются и превращаются в клетки кровщ.. Из мезенхимных
клеток, окружающих сосуд, позднее дифференцируются гладкие
мышечные клетки,—перициты и адвентициальные клетки сосуда,
а также фибробласты, синтезирующие межклеточное вещество.
В теле зародыша образуются первичные кровеносные сосуд;
имеющие вид трубочек-”и щелевидных пространств, формирующу-
ся ”из мезенхимы, клетки которой дают начало всем элеме"‘ам
сосудистой стенки. В конце 3-й недели внутриутробного-раа’ити^
сосуды зародыша начинают сообщаться с сосудами внезап’Дыше"
вых органов. Дальнейшее развитие стенки сосудов гроисходиз
после- начала циркуляции крови под влиянием те эмодинами
ческих условий (кровяное давление, скорость кров ока), которые
создаются в различных частях тела. Последнее хбетоят ,ЛЬСТво
обусловливает появление специфических особе остгУ. строения
стенки внутриорганных и внеорганных сосудов . ходе перестроек
первичных сосудов в эмбриогенезе часть из ни ре т^цируется.
Классификация и общая характеристика сосудов. В кровенос-
ной системе различают артерии, артериолы, ге {.экапилляры, вену-
лы, вены и артериоловенулярные анастомозы.,^ io середины XX ве-
ка в ангиологий гемокапилляры считались единственным связу-
ющим звеном между артериями и венами. Однако, как выясни-
лось, взаимосвязь между артериями и венами осуществляется
системой сосудов микроциркуляторного русла, включающей не
только капилляры, но и мельчайшие артерии, вены и артерйолове-
нулярные анастомозы (см. ниже)’.
По артериям кровь течет от сердца к_органам. Как правило,
эта кровь насыщена кислородом, за исключением легочной артерии,
несущей венозную кровь:- По венам кровъ-Лритекает к сердцу и
содержи мало кислофода, кроме крови в легочных венах. Гемока-
пилляры соединяют _артериаль^но~ё~звено кровеносной системы с
венозным, кроме так называемых чудесных сетей (rete mirabile),
в которых капилляры находятся между двумя олноименными
сосудами (например, в клубочках почки).
Артерии
Классификация. По особенностям строения артериибывают
трех типов: эластического, мышечного и смешан-
368 —-----------------
Рис. 143. Аорта человека; попереч-
ный срез (микрофотография).
I__внутренняя оболочка; II средняя
оболочка; /// наружная оболочка; I —
эндотелий; 2—подэндотелиальный слой;
5__эластические мембраны; 4 — глад-
кие миоциты; 5 сосуды сосудов.
•------2
3
//
н о г_о (мышечно-эластичес-
кого). Стенка всех артерий,
так же как и вен, состоит из трех
оболочек: внутренней (tunica
interna),.средней (tunica me-
dia) и___наружной (tunica
externa). Их толщина, ткане-
вый состав и функциональ-
ные особенности неодинако-
вы в сосудах разных типов.
Артерии эластического
типаг (arteriae elastotypica).
К" ним относятся сосуды
крупного калибра, такие... как
аорта и легочная артерия, в
которых кровь протекает под
высоким давлением (120—
130-ММ т. ст.) и с большой скоростью (0,5—1,3 м/с). В эти сосу-
ды кровь поступает либонепосрёдственно из сердца, либо вблизи
4
5
///
главным образом транспортную функцию. Наличие большого
количества эластических элементов (волокон,'мембран) позволяет
этим сосудам растягиваться при систоле сердца и возвращаться
в исходное положение во время диастолы. В качестве примера
сосуда эластического типа рассматривается строение аорты
(рис. 143).
Внутренняя оболочка аорты включает _ эндотелий
(endothelium), подэндотелиалЪный слой (stratum subendotheliale) и
сплетение эластических волокон (plexus fibroelasticus).
Эндотелий аорты_человека состоит из- клеток,—различных по
форме "и размерам, расположенных на__ба за пьыой мембране. По
протяженности сосуда размеры и форма клеток неодинаковы.
Иногда клетки достигают 500 мкм в длину и 150 мкм в ширину.
Чаще они бывают одноядерными, но встречаются и многоядерные.
Размеры ядер также неодинаковы. В эндотелиальных клетках
слабо” развита эндоплазматическая сеть, содержащая небольшое
количество рибосом. Митохондрии весьма многочисленны (от 200
до 700), разнообразны по форме и величине, очень много микро-
филаментов.
Подэндотелиальный слой состоит из рыхлой тонкофибрилляр-
нои соединительной ткани, богатой малоспециализированными
клетками“звездчатой формы. Толщина подэндотелиального слоя
369
в аорте значительная (15—20% от толщины стенки). В этом слое
встречаются отдельные продольно направленные гладкие мышеч-
ные клетки (гладкие миоциты). В межклеточном веществе внут-
ренней оболочки аорты и в ме н ьше й с тепе ни в дру гих оболбч ка х
при специальной обработке выявляются большое количество гли-
козаминогликанов и фосфолипиды. Основное аморфное-вещество
играет большую роль в трофике стенки сосуда. Физико-химическое
состояние этого вещества обусловливает степень проницаемости
стенки сосуда. У лиц среднего и пожилого возраста в межклеточ-
ном веществе обнаруживаются холестерин и жирные кислоты.
Глуб-ж‘* подэндогепиапьного слоя в составе внутренней оболочки
расположено густое сплетение эластических "волокон, в котором
обычно удается различить внутренний циркулярный и наружный
продол ьный'слСГи: '
Внутрённяя~оболочка аорты в месте отхождения от сердца обра-
зует три карманоподобные створки («полулунные клапаны»).
С р е д ня я оболочка »»рты состоит из большого- количе-
ства (40- ^0) эластических окончатых мембран (membranae
elasticae'Tenestratae), связанных между собой эластическими во-
локнами и образующих ёдйпый элас1ичеекий каркас- вместе с
эластическими элементами других оболочек (см. рис. 143). Между
мембранами залегают гладкие мышечные клетки, имеющие косое
по~отношению к мембранам направление, и небольшое количество
фибробластов. Такое строениесреднейоболочки делает аорту
высокоэлаегичной и._£мягчает^толчки крови, выбрасываемой в со-
суд во время сокращения левого желудочка сердца, а также обе-
спечивает поддержание тонуса сосудистой стенки во время ди-
астолы.
Н цр у ж на я оболочка аорты построена из рыхлой
волокнистой соединительной ткани с большим количеством толс-
тых э/юс/'ЦУЛг'ХИУ-и-¥ иеоттйг-лмеюших -главным
образом~Продольное направление. В средней и наружной оболоч-
КЯУ аорты, как и вообще во всех крупных сосудах, протгуфт пита-
ющие сосуды (vasa vasorum) п^цервные стволики (nervi vasorum).
Наружная оболочка предохраняет сосуд от перерастяжения и
разрывов.
Артерии смешаннег* или мышечн»-эластическ»п» типа (аа.
mixtotypicae). По строению и функциональным особенностям они
занимают промежуточное положение между сосудами мышечного
и эластического типов. К ним относятся, в частности, .сонная и
подключичная арт^рдч_Внутренняя оболочка этих сосу-
дов состоит из эндотелия, расположенного на базальной мембране,
подэндотелиалъного слоя и внутренней эластической мембраны
(membrana elastica interna). Эта мембрана располагается на грани-
це внутренней и средней оболочек и характеризуется четкой выра-
женностью и отграниченностью от других элементов сосудистой
стенки.
С редняя оболочка артерий смешанного типа состоит
из примерно равного количества гладтшх~мышечных~клеток, спи-
370
рально^ориентированных эластических водоко/1Л1-.оканчатых- элас-
тических мембран. Между гладкими мышечными клетками и элас-
тическими элементами обнаруживается^ небольшое ~ количество
фибробласте и коллагеновых волокон.
В наружной оболочке артерий можно выделить два
слоя: в нутре н н и й,^содержащий отдельные пучки~?ладких мыше ч-
ных клегок.~й~наружный, состоящий преимущественно из продоль-
но и косорасположенных пучков коллагеновых и эластических
волокон и соединительнотканных клеток. В ее составе присутствуют
сосуды сосудов и нервные волокна. Занимая промежуточное поло-
жение между сосудами мышечного и эластического типов, артерии
смешанного типа (например, подключичные) не только могут
сильно сокращаться, но и обладают выгпким» эчагшийтимим оной-
ствами, что особенно четке-проявлястся при повышении кровяного
давления.
Артерии мышечного типа (аа. myotypicae). К мим относятся
прей мущественно сосуды среднего и мелкого калибра, т. ,е. бол fa-
ши н C.TBO артерии организма (пргйрии-Тйла, конечностей и в ну тре н -
них органов).
В стенках этих артерий имеется относительно большое количе-
ство гладких мышечных клеток, что обеспечивает дополнительную
1агнетательную силу их и регулирует приток крови к органам
(рис. 144, А,Б).
В -состав внутренней об о л о ч к и.входят___эндотелий с
базальной мембраной, подэндотелиальный слой и вну_гредняя элас-
тическая мембрана.
'?5ндо1елйальнь1е клетки вытянуты вдоль продольной оси сосу-
да. Прдэндате.пиа дьный слой состоит из тонких эластических и
коллагеновых волокон, преимущественно продольно направленных,
а также малоспёциалйзированных соединительнотканных клеток.
В основном веществе подэндотелиального слоя находятсяглико-
замшпТглйканы. Подэндотелиальный слой лучше развит в артериях
среднего и крупного калибра и слабее — в мелких артериях..Кна-
ружи от подэндотелиального слоя расположена тесно связанная
с ннм~вТТу1 ренняя-элаетычеекая -мембрана. В мелких'артериях она
очень тонкая?-В более крупных артериях мышечного типа, эласти-
ческая мембрана четко выражена (на гистологических препаратах
она имеет вид извитой блестящей эластической пластинки).
С р е д н я я обо дочка артерии состоит из гладких мышеч-
ных к.четак, расположенных по пологой спирали, между которыми
находятся в небольшом числе соединительнотканные клетки типа
фибробплстпи и волокна (коллагрипп! 1й » плпотнчйские). Такое
расположение мышечных клеток обеспечивает при сокращении
уменьшение объема сосуда и проталкивание крови в дистальные
отделы. Эластические волокна сопровождают мышечные клетки,
а на граниьщс наружной и внутренней оболочками они с.'щвЯются
с их эластическими элементами. Таким обпазом. создается единый
эластический каркас, который с одной стороны, придает срсуду
эластичность при растяжении, а с другой — упругость при сдавле-
371
Рис. 144. Строение стенки артерии и вены среднего калибра (рис. Ю. И. Афа-
насьева).
А — схема: слева — артерия, справа — вена. / — внутренняя оболочка: I — эндотелий;
2 — базальная мембрана; 3 — подэндотелиальный слой; 4 — внутренняя эластическая мем-
брана; П — средняя оболочка; 5 — гладкие мноциты; 6 — эластические волокна; 7 — кол-
лагеновые волокна; /// — наружная оболочка; 8 — наружная эластическая мембрана; 9 —
волокнистая соединительная ткань; 10 — сосуды сосудов.
Рис. 144 (продолжение)
Б— эластический каркас артерии; поперечный срез (микрофотография): /— внутренняя
эластическая мембрана; 2 — наружная эластическая мембрана. В — вена; поперечный
срез (микрофотография): / —эластические волокна. Г—сосудисто-нервный пучок: / —
артерия мышечного типа; 2 — вена; 3 — нерв.
нии (см. рис. 144,Л). Эластический каркас препятствует спадению
артерий, что обусловливает их постоянное зияние и непрерывность
в них тока крови
Гладкие, мышечные .клетки средней оболочки артерий мышеч-
ного типа своими сокращениями поддерживают кровяное давле-
ние, регулируют приток крови в сосуды микроциркудяторного
русла органов. На границе между средней и наружной оболочками
располагается наружная эластическая мембрана (membrana elasti-
са externa). Она состоит из продольно идущих толстых, густо
переплетающихся эластических воликин,- ки'1'орьте иногда приобре-
тают- ВИД 'С'ПЛО'Шнои эластической пластинки. Обычно наружная
эластическая мембрана бывает тоньше внутренней и не у всех
артерий достаточно хорошо выражена.
Наружная оболочка состоит из рыхлой волокнистой
соединительной ткани, в которой соединительнотканные волокна
имеют преимущественно косое и продольное направление. В этой
оболочк£_достоянно—встречаются нервы и кровеносные сосуды,
питающие стенку
Но мере уменьшения диаметра артерии и их приближения к
артериолам все оболочки артерии истончаются. Во внутренней
оболочке резко уменьшается толщина подэндотелиального слоя и
внутренней эластической мембраны. Количество мышечных клеток
и эластических волокон в средней оболочке также постепенно
убывает. В наружной оболочке уменьшается количество эластиче-
ских волокон, исчезает наружная эластическая мембрана (см.
ниже).
МикроциркуЛяторное русло
Этим термином в ангиологии обозначается система мелких сосу-
дов, включающая~~артериолы, гемокапилляры, венулы, а также
артериоловенулярные анастомозы. Этот функциональный ком-
плекс кровеносных сосудов, окруженный~~лимфатическими капил-
лярами и лимфатическими ~С~0судамй. вместе с окружающей соеди-
нительной тканью обеспечивает регуляцию кровенаполнения орга-
нов,” транскапиллярный обмен и дренажно-депонируюшую функ-
цию” (рис. 145). Чаще всего элементы.микроциркудяторного русла
образуют густую систему я нягтомо.зон прекапиллярных, кяпиппяр-
ныХ иГпосткапиллярных сосудов, но могут быть и другие варианты
С выделением какого-либо основного, предппитмггедьноиг канала,
например анастомоза прекапиллярной артериолы и посткапилляр-
ной венулы и др. В “каждом органе адекватно его функции Суще-
ствуют специфические особенности конфигурации, диаметра и
плотности расположения сосудов микроциркудяторного русла.
Сосуды микроциркудяторного русла пластичны при изменении
кровотока. Они могут депонировать форменные элементы или быть
спазмированы и пропускать лишь плазму, изменять проницаемость
для тканевой жидкости.
Артериолы. х Это наиболее мелкие артериальные -сосуды мы-
Рис. 145. Сосуды микроциркудяторного русла (рис. Ю. И. Афанасьева).
/ артерия; 2 — вена; 3 артериолы; 4 — гемокапилляры; 5 - венулы; 6 — артериовену-
лярный анастомоз; 7 — лимфатический капилляр; 8 лимфатические сосуды.
щечного типа диаметром не более 50— 100 мкм, которые, с одной
стороны, связаны с артериями, а £ другой — постёпённсгпереходят
в капилляры (см. рис. 14бЕ В артерилпах сохраняются три обо-
лочки^ характерные для артерий вообще, однако выражены они
очець слабо7~В н у я- обол о-ч к а этих-оесудоа состоит
из эндотелиальных клеток с базальной мембраной, тонкого подэн-
дотелйальнбгГГТлия- и ТОНКОЙ"внутренней эластической мембраны.
С р ej. няя оболо чка образована 1—2 слоями _?дадких
мышечных клеток, имеющих спиралевидное направление. В прека-
пиллярных артериолах гладкие мышечные^ клетки располагаются
поодиночке. Расстояниемё'ждуштимиувёличивается в дистальных
отделах, однако они обязательно присутствуют в месте отхожде-
ния прекапилляров~от артериолы и в месте разделения прекапил-
ляра на капилляры. В артёриолаху обнаруживаются перфорации
в базальной мембранё~эндотелия и внутренней эластической мем-
бране, благодаря которым осуществляется непосредственный
тесный контакт" эндотёлйбцйтОВ и гладких мышечных клеток
(рис. 146). Такие контакты "Создают условия для передачи инфор-
мации от эндотелиЛ~к~тладким мышечным клеткам, в -частности,
при выбросе в кровь -адреналина из мозгового вещества надпо-
375
Рис. 146. Схема строения арте-
риолы (рис. Ю. И. Афанасьева).
/ — эндотелиальная клетка; 2 — ба-
зальная мембрана; 3 — эластиче-
ская мембрана; 4 — гладкая мышеч-
ная клетка; 5 - контакт гладкого
миоцита с эндотелиоцитом; б —
клетки; 7 — соединительнотканные
волокна.
чечников, который вы з ы -
вает__срк₽ащение гладких
мышечных клеток. Между
мышечнымй~клётками ар-
териол обнаруживается
небольшое количество
эластических волокон? На-
ружная эластическая
мембрана отсутствует.
Наружная о б о л о ч-
к а_представлена рыхлой
волокнистой соединитель-
ной ^тканью.
В функциональном от-
ношении артериолы яв-
ляются, по выражению И. М. Сеченова, «кранами сосудистой
системы», которые регулируют приток крови к органам благодаря
сокращению спирально направленных гладких мышечных клеток,
иннервируемых эфферентными нервными волокнами. В месте от-
хождения гемокапилляра от прекапиллярных артериол имеется
сужение, обусловленное циркулярно расположенными гладкими
мышечными клетками в устье капилляров, выполняющих роль
прекапиллярных сфинктеров.
Капилляры. Кровеносные капидпяры (vasae haemocapillariae)
наиболее многочисленные_и самые тонкие сосуды, имеющие,
однако, различный просвет. Это обусловлено как органными осо-
бенностями кЭТГИЛЛЯрОвТ'так и функциональным состоянием сосу-
дистой системы. Например, наиболее узкие капилляры (диаметром
от 4,5 до 6—7 мкм) находятся в попередшополосатых мышцах,
нервах, легких и т. д., более широкие капилляры (диаметром 7—
11 мкм) — в коже и слизистых-ободочках. В кроветворных орга-
нах, некоторых, железах внутренней секреции, печени встречаются
капилляру с широким но—денаюишмгн___н-ч протяжении сосуда
диаметром (20—30 мкм и более). Такие капилляры называются
синусоидными. Специфические вместилища—крови калил л я рн ого
ТИЦЛ-^-^даьулы —.имеются в и-...ри. тщ тепах ППЛрВОГО члена.
В большинстве случаев капилляры формируют сеть, одна ко
они могут образовывать, петли (в сосочках ножи и синовиальных
ворсинках суставов), а также клубочки (сосудистые клубочки
в .почке).
376
В капиллярах, образующих петли, выделяют артериальный и
венозный отделы. Ширина артериального отдела в среднем равна
диаметру эритроцита, а венозного — несколько больше. Количество
капилляров в разных органах неодинаково. Например, на попереч-
ном разрезе в мышце человека на 1 мм2 насчитывается от 1400 до
2000 капилляров, а в коже на той же площади — 40. В любой
ткани в обычных физиологических условиях находится до 50%
нефункиионирующих капилляров. Просвет их, как правило, сильно
уменьшен, но полного закрытия его при этом не прТЯТСТОДит. Для
форменных элементов крови эти капилля ы оказываются непрохо-
дим ыми, в то же время плазма продолжает по ним циркулировать.
Число капилляров в ..определенном органе связано с его общими
мопс^офункциональными особенностями, а_количество открытых
капилдяров зависит от интенсивности работы органа в данный
момент.
Площад^ поперечного сечения среза капиллярного русла в
любой области во много раз превышает площадь поперечного
сечения ИСХОДНОЙ артерии. В стенке капиппярон_дазличают три
тонких слоя (как аналоги трех оболочек рассмотренных выше
сосудов). Внутренний слой предотавдец эндптепиа пьными клетка-
ми, расположенными С*™ мембране средним пктлит из
перицитов', заключенных в базальную мембрану, д наружный — из
адвентициальных^ клеток и тонких коллагеновых волокон, погру-
женныхГ в аморфное вещество.
Э/Н д- о т е л и а л ь и ы-й-—е л О"Й7—Внутренняя выстилка капил-
ляра представляет собой пласт лежащих на базальной мембране
вытянутых, полигональной формы эндотелиальных хдеток с изви-
листыми границами, которые хорошо выявляются при импрегна-
ции серебром (рис. 147,4, Б, В, Г). Ядра эндотелиальных клеток
обычно уплощенные, овальной формы.
Наиболее вытянутые эндотелиоциты достигают в длину 75—
175 мкм, а наиболее короткие — 5—8 мкм. Толщина эндотелиаль-
ных клеток неодинакова. В различных капиллярах она колеблется
от 201) нм до 1—2 мкм на периферии и 3—5 мкм в околоядерных
участках. Как появино, клетки эндотелия тесно прилежат друг к
другу (см. рис. 147), часто обнаруживаются соединения, по типу
замка и зоны слипания. Поверхность эндотелиальных клеток,
обращенная к току крови, покрыта слоем гликопоотеидов (пара-
плазмолеммальный слой). Вдоль внутренней и наружной поверх-
носте й эндотелиальных клеток асполагаются пиноци то з н ые
пузырьки и кавеолы, отображающие трансэнлотепиальный транс
порт "различных веществ и метаболитов. В венозном конце их
больше, чем в артериальном Органеллы, как правило, немногочис-
ленны и расположены в околоядерной зоне. В цитоплазме эндоте-
лиальных клеток обнаруживаются микрофиламенты толщиной
около 5 нм, которые могут образовывать пучки, проходящие
1 Нередко в литературе базальную мембрану и перициты объединяют в
базальный слои
377
—г~~
Б
Рис. 147. Эндотелий капилляров (схема).
А — плоскостное изображение; Б — отвесной срез
(рис. Ю. И. Афанасьева).
/ — границы клетки; 2 — цитоплазма; 3 — ядро;
В фенестры ( <— ) в эндотел ио цн та х перитубу-
лярного гемокапилляра почки. Электронная микро-
фотография. X 20 000 (по А. А. Миронову); Г —
параплазмолеммальный слой эндотелиоцита гемо-
капилляра. Электронная микрофотография.
X 80 000 (по В. В. Куприянову, Я. Л Караганову
и В. И. Козлову). / — просвет гемокапилляра; 2 —
цитолемма; 3 — параплазмолеммальный слон; 4 —
базальная мембрана; 5 — цитоплазма перицита.
Рис. 148. Три типа капилляров (рис. Ю. И. Афанасьева).
/ — гемокапилляр с непрерывной эндотелиальной клеткой и базальной мембраной;
II — гемокапилляр с фенестрированным эндотелием и непрерывной базальной мембра-
ной; ill — синусоидный гемокапилляр с щелевидными отверстиями в эндотелии и пре-
рывистой базальной мембраной; / — эндотелиоцит; 2 — базальная мембрана; 3 — фене-
стры: 4 — щели (поры); 5 — перицит; 6 — адвентициальная клетка; 7 — контакт эндоте-
лиоцнта и перицита; 8 — нервное окончание.
в гиалоплазме. Эти структуры составляют цитоскелет, а также
обеспечивают внутриклеточные перемещения органелл.
Внутренняя поверхность эндотелия капилляра, обращенная к
току крови, может иметь субмикроскопические выступы в виде
отдельных микроы^н-м^1^, 1ИпГ>1ни»» мшпчпм концу капилляра.
В венозных отделах капилляра цитоплазма эндотелиоцитрв обра-
зует клапанообразные структуры. Эти цитоплазматические вырос-
ты увеличивают поверхность эндотелия и в зависимости от актив-
ности транспорта жидкости через эндотелий изменяют свои раз-
меры.
379
Классификация капилляров обусловлена орган-
ными особенностями их (рис. 148). Кроме описанного, наиболее
распространенного первого типа капилляров, различают капилляры
фенестрированного типа (второй тип) с локальными истончениями
(фенестрами) цитоплазмы эндотелия (капилляры клубочков поч-
ки^ ворсинок кишечника; желез внутренней секреции и др.) и
капилляры с щелевидными отверстиями в эндотелии и базальной
мем()[Х1Ш’, К5к,~ например? В селезенке. печени (третий тип).
Фенестры и в особенности щели облегчают проникновение
различных макромолекулярных—и-корпускулярных веществ через
стенку капилляров. Растяжимость эндотелия и проницаемость
для коллоидных^частиц в венозном отделе капилляра выше, чем
в артериальном Данные цитохимических исследований указывают
на"* наличие в эндотелии гликогена, фосфатаз, АТФ-азы и других
веществ, играющих большую роль в метаболизме и транспорте
веществ через клетки и выполнении других функций.
Базальная мембрана эндотелия капилляра — тонкофибрил-
лярная структура толщиной 30—35 нм, которая выполняет много-
образные функции (см. гл. VI). В базальной мембране капилля-
ров некоторых органов (мозг, надпочечники и др.) обнаружена
АТФ-аза, что указывает на активную роль мембраны в метаболиз-
ме клетки. Между эндотелиальными клетками—и—перицитами
базальная мембрана местами истончается и прерывается, а сами
клетки здесь связаны между собой путем плотных контактов
цитолеммы. Эта~область эндотелиоперицитарных контактов, воз-
можно, служит местом передачи__возбуждении-от одной клетки
другой?
П е_р и ц и т ы. Эти_соединительнотканные^ клетки имеют
отростчатую форму и в виде корзинки окружают кровеносные
капилляры; располагаясь в расщеплениях базальной мембраны
эндотелия. Плотность расположения этих клеток неодинакова в
капиллярах разных органов. .На перицитах некоторых капилляров
обнаружены эфферентные нервные окончания, функциональное
значение которых, по-видимому, связано с-регуляцией изменения
просвета капилляров (рис. 149).
А двентици альные клетки. Это малодифференциро-
ванные клетки, расположенные снаружи от перицитов. Они окру-
жены аморфным веществом соединительной ткани, в котором
находятСя тонкие коллагеновые вожухна? —'
Кровенбснйе'—капилляры осуществляют основные обме иные
процессы между кровью и тканями, а в некоторых органах (лег-
кие) участвуют в обеспечении газообмена между кровью_и воз-
духом^. Тонкость стенок капилляров, огромная площадь их сопри-
косновения с тканями (более 6000 м2), тледленньтй кровоток
(0,5 мм/с), низкое кровяное давление (20—30 мм рт. ст.) об.ееггг-
чивают наилучшие условия для обменных процессов.
С те нка каОЯТшяров тесно связана функпионапьно-и мо рфоло-
гически с окружающей соединительной тканью Изменение гостоя-
ни я ба зальной мембраны и основного вещества соединительной
380
Рис. 149. Схема строения гемо капилляра
(рис. Ю. И. Афанасьева).
/ — эндотелиоцит; 2 перицит; 3 — ба
зальная мембрана; 4 — плотный контакт ци
толеммы эндотелиальной клетки и перици-
та; 5 -- эфферентное нервное окончание на
периците.
ткани, окружающей сосуды,
быстро сказывается на прони-
цаемости капилляров, что имеет
огромное значение при фи-
зиологических и патологиче-
Ских состояниях организма.
Расщепление гиалуроновой
кислоты под влиянием гиалу-
ронидазы приводит к повыше-
нию проницаемости капилляров,
которая меняется также с изме-
нением концентрации солей
кальция в крови или при попа-
дании в кровь протеолитических
ферментов (например, при
гнойной инфекции и др.).
Изменение просвета капил-
ляров при различных физиоло-
гических и патологических уело
в значительной мере зависит
от давления крови в самих капиллярах, что связано с тонусом мы-
шечных клеток артериол и мелких вен, прекапиллярных сфинктеров,
а также артериоловенулярных анастомозов и состоянием перици-
тов.
О тродящий отдел микроциркудяторного русла начина-
ется венозной частью капилляров, для которые vap;,Штерны более
крупные микровореннки на люминальной поинрхпости .эндотелия
и складки, напоминающие створки клапанов. В эндотелиальных
клетках содержится относительно большое шзоло-митохондрий
и пинюпитозных пузырьков, лучше представлен комплекс Гольджи.
В эндотелии чаще обнаруживаются фенестры. Диаметр венозного
отдела капилляра может быть шире артериального в I1/,'— 2 раза.
Beнулы (venulae). Различают три разновидности венул: пост-
капиллярные, собирательные и мышечные. Посткапиллярные
венулы (диаметр 8—30 мкм) по своему строению напоминают
венозный отдел капилляра, но в стенке этих венул отмечается
больше перицитов, чем в капиллярах. В собирательных венулах
(диаметр 30—50 мкм) появляются отдельные гладкие мышечные
клетки и более четко выражена наружная оболочка. Мышечные
венулы (диаметр 50—100 мкм) имеют один-два слоя гладких мы-
шечных клеток в средней оболочке и сравнительно хорошо разви-
тую наружную оболочку.
381
Венозный отдел микроциркуляторного русла вместе с лимфат
ческими капиллярами выполняет дренажную функцию, регулир
гематолнмфатическое равновесие между кровью и внесосудист<
жидкостью, удаляя продукты метаболизма тканей. Через стен)
венул, так же как через капилляры, мигрируют лейкоциты. Me
ленный кровоток (не более 1—2 мм в секунду) и низкое кровян
давление (около 10 мм рт. ст.), а также растяжимость эп
сосудов создают условия для депонирования крови.
Артериоловенулярные, днастомозы (АВА) (anastomosae arteri
lovenularis). Это — соединения сосудов, несущих артериальну
кровь в вены в обход~капидлярного русла. Они обнаружены почти^
всех органах. Калибр АВА колеблется от 30 до 500 мкм, а длина м
жет достигать 4 мм. АВА способны к сокращениям 2—12 раз
минуту.
Классификация АВА. Различают две группы анаст
мозов:—1) игтмцш,ц»—АВА—(шунты), по которым сбрасывает*
чистая артериальная кровь; 2) атипичные АВА (полушунты), .
которым течет смешанная кровь.
Первая группа анастомозов (шунты) может иметь ра зли ч ну
внешнюкцформу — п ямые корот^и^. соустья, петгти, ветвящие*
соединения.
По своему строению они подразделяются на две подгрупп
а) простые АЪА; б)~АТ?АГснабженньгр специальными сцкрятитрп
ными структурами.
В. первой подгруппе истинпым ?из<-тлмпзов граница перехо
одного сосуда в другой соответствует участку, где заканчивает,
среднязшболочка артериолы. Регуляция кровотока осуществляет
гладкимц_мышечными клетками средней -оболочки са?.ши артер
олы, без специальных сократительных аппаратов (рис. 150, 1, Hi
Анастомозы второй подгруппы могут иметь специальные сокр
тительйые устройства в-виде валиков или подущек в подэндотел
ально'м~слое, образованные продольно расположенными гладки)
мышечными-клетками. Сокращение мышечных подушек, выступ
ющи*~в просвет анастомоза, приводит к прекращений) "Кровото
(см. рис. 150, II, А). К этой же подгруппе относятся АВА эпител
оидного типа (простые~и сложные) (см. рис. 150,11, Б, В). Црост)
АВА эпителиоидного типа характеризуются наличием в средней
оболочке внутренне го продольного й наружного циркуляр но го
слоев гладких мышечных клеток, которые по мере при! йжения
к венозному концу заменяются на короткие овальные светлые
клетки (Е-клетки), похожие на эпителиальные. И венозном се г-
менте АВА стенка его резко истончается. Средняя оболочка здесь
содержит лишьнезйЭЧИЗ'ельние киличесГВОТЛадхих-мылечнык кле-
ток в виде циркулярно расположенных поясков. Наружная оболочка
состоит из рыхлой соединительной ткани.
Сложные, или клубочковые, АВА эпителиоидного типа отлича-
ются от простых тем, что приносящая (афферентнаяартериола
делится на 2—4 ветви, которые переходят в венозный сегмент.
Эти ветви окружены одной обшей соединительнотканной-оболоч-
382
Рис. 150. Артериоловенулярные анастомозы (АВА) (рис. Ю. И. Афанасьева).
/ АВА без специального запирательного устройства: / артериола: 2— венула; 3—
анастомоз: 4 — гладкие мноцнты анастомоза. // — АВА со специальным запирательным
устройством: А — анастомоз типа «амыкающей артерии: Б — простой анастомоз эпите-
лиоидного типа; В — сложный анастомоз эпителиоидного типа (клубочковый): /—эндо-
телии: 2 — продольно расположенные пучки гладких миоцитов; 3 — внутренняя эластическая
мембрана; 4 — артериола; 5 венула; 6 - анастомоз; 7 эпителиоидные клетки анастомо-
за; 8 капилляры в соединительнотканной оболочке- Hi — атипичный анастомоз: / — арте-
пиола: 2 — короткий гемокапилляр: 3 — венула
<ой (ем. рис. 150, II, В). Такие анастомозы часто обнаруживаются
в дерме кожи и гиподерме, а также в параганглиях.
Broj>az ~~груиоя — 1тн.|цчные анастомозы (полушунты) —
представляет собой соединения артериол и венул, посредством
короткого сосуда капиллярного типа (см. рис. 150,111). Поэтому
сбрасываемая в венозное русло кровь является не полТЮГтью
артериальной.
АВА, особенно клубочкового типа, богато иннервированы.
Соединения артериальной и венозной систем без посредства
капилляров имеют большое значение для регуляции тока крови
через орган и для регуляции кровяного давления (общего и мест-
383
ного). Эти соединения играют определенную роль для стимуляции
венозного кровотока, артериализации венозной крови, мобилиза-
ции депонированной крови и регуляции тока тканевой жидкости
в венозное русло. Велика роль АВА в компенсаторныя-реакция''
организма при нарушении кровообращения и развитии патологи
чески х процессов.
__------------' 4 *
Вены
Вены составляют отводящее звено сосудистой системы. Отток
крови начинается по посткапиллярным венулам (см. выше). Низ-
кое кровяное давление (15—20 мм рт. ст.) и незначительная
скорость (в органных венах около 10 мм/с) кровотока определя-
ют сравнительно слабое развитие эластических элементов в венах
и большую растяжимость их. Кбличёствб~же гладких мышечных
клеток в стенке вен неодинаково и зависит от того, движется ли
в них кровь к сердцу под действием силы тяжести или против нее.
Необходимость преодоления силы тяжести крови в венах нижних
конечностей приводит к сильному развитию гладких мышечных
элементов в этих сосудах по сравнению с венами верхних конеч-
ностей, головы и шеи. Во многих венах имеются клапаны (valvulae
venosae), являющиеся производными внутренней оболочки. Вены
головного мозга и его оболочек, внутренних органов, подчревные,
подвздошные, полые и безымянные клапанов не содержат.
По .степени развития мышечных элементов в стенке вен они
могут быть разделены на две группы: вены волокнистого (безмы-
шечного) типа и вёпы мышечного типа.-Вены мышечного типа в
свою очередь подразделяются на вены со слаб ы м, с р е д н и м
и с и лдн_ы м. развитием мышечных^ элементов.
В венах, так же как и в артериях, различаюттри оболочки:
внутреннюю,' среднюю и наружную. Выраженность этих оболочек
и строение их в различных венах существенно отличаются.
Вены волокнистого типа (venae fibrotypicae). К венам этого
типа ТУтносят безмышечные вены твердой и мягкой ^мозговых
оболочек (рис.' 151), вены сетчатки глаза, костей, селезенки и
плаценты Вены мозговых оболочек и сетчатки глаза ЭтодатТГИвы
при изменении кровяного давления, могут сильно растягиваться,
но скопившаяся в них, кровь сравнительно легко под действием
собственной силы тяжести оттекает в более крупные венозные
стволы. Вены костей, селезенюГ и плаценты гак ассивны в
продвижении по ним крови. Это объясняется тем, что все они
плотно сращены с плотными элементами соответствующих орга-
нов и не спадаются, поэтому отток крови по ним совершается
зегко. -'^нпптрлиальные клетки, выстилающие эти вены, имеют
более извилистые границы, чем в артериях. Снаружи к ним щтй~
лежит базадьная-мембрдна, а затем тонкий слои рыхлой волокнис-
той соединительной ткани, срастающийся с окружающими тка-
нями^
Вены мышечного типа (venae myotypicae). Вены со сщ_а_б-ы-м
р а з в и тием мышечн ых э л е м е н т о в_различны ПО диа-
384
Рис. 151. Вены волокнис-
того типа. Тотальный пре-
парат МЯГКОЙ МО1ГОВОЙ
оболочки.
/ вена; 2 эндотелий; 3—
адвентициальные клетки; 4—
артериола.
метру. Сюда относятся вены мелкого и среднего калибра (до 1 —
2 мм), сопровождающие артерии мышечного типа в верхней части
туловища, шеи и jinpa, а также такие крупные вены, как, например,
верхняя полдя^веца. В этих сосудах к ровь в з на ч и тельной мере
продвигается пассивно вследствие своей тяжести. К этому же типу
вен мТгжно отнести и вены верхних конечностей. Стенки "таких
вен несколько тоньше соответствующих по калибру артерий, со-
держлт_м£ныпе мышечных элементов и на препаратах находятся
обычно в спавшемся состоянии.
Вены мелкого и среднего калибра со слабым развитием
мышечных элементов имеют пдохо аыраженныа ттолэп
4рТ€5ЯИл±лыи Я®, а в вредней оболочке содержится небольшое
количество МЫШЕЧНЫХ |С1етоц.ВнекдТррьтх мелких венатц например
в венах. пищеварительного тракта, гладкие мышечные клетки jj
средщей- оболочке-образуют отдельные «пояски», располагающиеся
далеко друг от друга. Благодаря такому строению вены могут сильно
расширяться -и-выполнять. депонирующую функцию. В наружной
эболоцке мелких вен встречаются-единичные продольно направлен-
ию гладкие мышечные клетки..
Среди вен крупного калибра, в которых слабо развиты мышечные
элементы, наиболее типична верхняя "ПОлая вена (рис. 152), в сред-
ней оболочке- стенки- тготорой оimcчается небольшое количество
гладких мышечных клеток. Эю обусловлено отчасти прямохож-
дением человека,-В Силу чего кровь по этой вене стекает к сердцу
благрдарзт~гобственной-тяжести;—а^такЖЕ"дыхательным движениям
грудной клетки. В начале диастолы (расслабление мускулатуры)
желудочков сердца в предсердиях появляется даже небольшое от-
рицательное кровяное давление, которое как бы подсасывает кровь
из полых вен.
Примером вены среднего калибра со с р е_д н и М._ р а з в и т и е м
м ы ш ечных элементов является плечевая вена. Эндотели-
альные клетки, выстилающие ее внутреннюю о b 0~7Г0 ч к у,
короче, чем в соответствующей артерии По<1г>ндотелиальный слои
состоит из соединительнотканных волокон и клеток, ориецтироиян-
13 гистология
385
I
Рис. 152. Вены (поперечный срез).
А вена со слабым развитием мышеч-
ных элементов (верхняя полая вена).
/ — внутренняя и средняя оболочки;
// наружная оболочка; 1 — клетки
крови в просвете вены; 2 - эндотелий,
3 - гладкие мноциты; 4 - эластические
волокна; 5 коллагеновые волокна; 6—
соединительнотканные клетки. Б— вена
с сильным развитием мышечных эле-
ментов (ннжняя полая вена человека).
I — внутренняя оболочка; // — средняя
оболочка; Hi — наружная оболочка; / —
гладкие миоциты; 2 — пучки продольно
идущих гладких миоцитов: 3 — соеди-
нительная ткань; 4 — сосуды--сосудов.
В— бедренная вена с клапаном (про
дольный срез): / — внутренняя оболоч-
ка; Н — средняя оболочка; / - створка
клапана; 2 — эндотелий; 3 — гладкие
мноциты.
ных в основном вдоль сосуда. Внутренняя оболочка этого сосуда
формирует клапанный аппарат, а также имеет в своем-Сбставе
отдельные продольно направленные гладкие мышечные клетки.
Внутронняя-эпастическая мембпана в вене ну выражена. На границе
между внутренней и средней оболочками располагается только сеть
эластических-волокон, Эла~< тичесхиегволокна внутренне й-обол о ч к и
плечевой вены, как и в артериях, связаны с эластическими волокна-
ми средней и наружнои оболоче'к и"составляют^БДиный каркас.
С~р~ё д н я я о бо дочка этой вены гораздо тоньше средней обо-
лочки соответствующей артерии. Она обБГчно состоит~йТтртрку.1Ярно
расположенных пучков ;дддких_миоцитов, рачделетшьгх~прбулойка-
mj воло^ктои .рудинитульнеи TKi}jtn£._ Наружная эластическая
мембрана в этой вене отсутствует, поэтому соедйнИТСЛыютканные
прослойки* средней оболочки переходя? непосредственно -в. рыхлую
волокнистукг соедипителъную ткань н^рру-жндхй—о&олоч к и,
В плечевой вене она очень сильно развита: ее размеры в 2—3 раза
превышают размеры средней оболочки. Коллагеновые и эластиче-
ские волокна наружной оболочки направлены преимущественно
продольно. Кроме того, в наружной обопочке-встречаются отдельные
глаПкйё^ы щечные клетки и небольшие пучки их, которые также
расположены продольно.
К венам я ьным развитием мышечных э л е-
м е н~т о~в относятся крупные вены нижней половины .туловища и
ног. Для них характерно развитие пучков гладких мышечных клеток
но нгех^ тдехЛГхЪбол очках Ш’Ииё*Гпп Tityiptiiiivn и иарт.жппйобб-
лочках они имеют продольное направление, а в средней^—нирку-
лярТгоК ' ~
Наиболее типично для этой группы вен строение бедренной
вены.~Внутренняя оболочка ее состоит из hhi~1oтелия и
па^лнПотелиального сдоя, образованного рыхлой волокнистой сое-
динительной тканью, в которой продольно залегают пучки гладких
мышечных-клеток. Внутренняя эластическая мембрана отсутствует,
однако на ее месте видны ^копления влаетичее-ких волокон.
Внутренняя оболочка бедренной вены формирует клапаны, пред-
ставляющие собой тонкие складки ее. Эндотелиальные клетки,
покрывающие клапан со стороны, обращенной в просвет сосуда,
имеют удлиненную форму и направлены вдоль створок клапана,
а на противоположной стороне^клапан покрыт эндотелиальными
клетками полигональной-формы, лежащими поперек створок. Осно-
ву клапана составляех волокнистая соедини тел ьная ткань. При этом
на стороне,обращенной к просвету сосуда, под эндотелием залегают
преимущественно эластические, волокна, а на противоположной
ст<)роне'т=г-Много коллагеновых_вшюкон. В основании створки кла-
пана может находиться некоторое количество гладких мышечных
клеток.
К.жшанп » венах способствуют току венозной- крови -к сердцу,
препятствуя ее обратному движению. Одновременно клапаны пре-
дохраняют герппе Ю из нтнней. затраты зиергмм на Ц|ГПДПЛеНие
колебательных дпн жений -крови, постоянно возникающих в венах
13'
187
под влиянием различных внешних воздействий (изменение атмос-
ферного давления, мышечное сжаТйе и т. д.) Однако наличие пучков
гладких мышечных клеток в оболочках и в створках клапанов бед-
ренной вены оказывается недостаточным для подъема крови против
сил тяжести. Существенную роль в этом играет сокращение скелет-
ной мускулатуры нижних конечностей.
Средняя оболочка бедренной вены содержит пучки
циркуляр но расположенных гладких мышечных клетшслВыте ТТёно -
бзшГя клапамасрёдняя оболочка истончается. Ниже места прикре-
пления клапана мышечные пучки перекрещиваются, создавая утол-
щение в стенке вены. В н а р у жн ой оболочке, образованной
волокнистой соединительнои^тканью, обнаруживаю!ся~тучки~про-
дольнб~расположённых гладких мышечных клеток.
Нижняя полая вена по строению резко отличается от впадаю-
щих в нее вен (см. рис. 152, Б). Внутренняя и средняя оболочки ниж-
ней полой вены развиты относительно слабо. Наружная оболочка
нижней полой вены имеет большое количество продольно располо-
женных пучков гладких мышечных клеток и по своей толщине в 6 —
7 раз превышает внутреннюю и среднюю оболочки, вместе взятые.
Между пучками гладких мышечных клеток лежат прослойки волок-
нистой соединительной ткани. Продольное расположение гладких
мышечных клеток в наружной оболочке имеет определенное физи-
ологическое значение: сокращение этих пучков не только способ-
ствует проталкиванию крови вверх (против силы тяжести), но и
приводит к образованию поперечных складок, препятствующих
обратному току крови (в нижней полой вене отсутствуют клапаны).
В устье нижней полой вены в ее наружную оболочку заходят пучки
поперечнополосатых мышечных клеток миокарда. В средней и
наружной оболочках содержатся сосуды сосудов, лимфатические
капилляры, многочисленные нервные волокна.
Особенности строения органных сосудов
Некоторые отделы сосудистой системы имеют органные особен-
ности строения артерий. Например, артерии черепа отличаются сла-
бым развитием эластических элементов в средней и наружной обо-
лочках; наружной эластической мембраны в них нет. Внутренняя
эластическая мембрана, наоборот, выражена отчетливо. Такие же
особенности существуют и у артерий головного мозга.
В пупочной артерии отсутствует внутренняя эластическая мем-
брана. В затылочной артерии сильно развиты пучки гладких мышеч-
ных клеток во внутренней оболочке. В почечной, брыжеечной, селе-
зеночной и венечной артериях пучки продольно расположенных
гладких мышечных клеток хорошо выражены в наружной оболочке.
В артериях матки, полового члена, артериях сосочковых мышц
сердца и пупочного канатика, особенно в месте его перехода в пла-
центу, пучки гладких мышечных клеток находятся и во внутренней,
и в наружных оболочках.
Некоторые вены, как и артерии, имеют ярко выраженные орган-
ные особенности строения. Так, у легочной и пупочной вен в отли-
188
чие от всех других вен очень хорошо развит циркулярный мышечный
слой в средней оболочке, вследствие чего они напоминают по своему
строению артерии. Вены сердца в средней оболочке содержат про-
дольно направленные пучки гладких мышечных клеток. В воротной
же вене средняя оболочка состоит из двух слоев: внутреннего —
кольцевого и наружного — продольного. В некоторых венах, напри-
мер сердечных, обнаруживаются эластические мембраны, которые
способствуют большей упругости и эластичности этих сосудов в
постоянно сокращающемся органе. У глубоких вен желудочков
сердца нет ни мышечных клеток, ни эластических мембран. Они
построены по типу синусоидов, имеющих на дистальном конце
вместо клапанов сфинктеры. Вены наружной оболочки сердца
содержат продольно направленные пучки гладких мышечных кле-
ток. В надпочечниках есть вены, которые имеют продольные мы-
шечные пучки во внутренней оболочке, выступающие в виде подуше-
чек в просвет вены, особенно в устье. Вены печени, подслизистой
основы кишечника, слизистой оболочки носа, вены полового члена и
др. снабжены сфинктерами, регулирующими отток крови.
ХЛ4 / С /
ЛИМФАТИЧЕСКИЕ СОСУДЫ г ! I '
Лимфатические сосуды — часть лимфатической системы, вклю-
чающей в себя еще и лимфатические узлы. В функциональном
отношении лимфатические сосуды тесно связаны с кровеносными,
особенно в области расположения сосудов микроциркуляторного
русла. "Именно здесь происходит образование тканевой жидкости
и проникновение ее в лимфатическое русло.
Через мелкие лимфоносные пути осуществляется пострян-
н а я миграция лимфоцитов из кровотока И ИХ рециркуляция из ли м -
фатическйх узлов в кровь.
Классификация. Среди лимфатических сосудов различают:
лимфатические тпиппяры интра- и экстраорганные лимфатиче-
ские сосуб)ы-31твддяшие лимфу-оторганов, и-^лавиыс-лимфатические
стралы—ттгла--грудной -проток, и правый лимфатический проток,
впадающие н крупные вены щей. По строению различают лимфати-
ческие сосуды безмышечного (волокнистого) и мышечного типов.
Лимфатические капилляры. Лимфатические капилляры — на-
чал ь ны е отдел ыл имфатической с~истемы. в котовые из тканей посту -
пает тканевая жидкость вместе с продуктами обмена веществ, а в
патологических случаях - инородные частицы и микроорганизмы.
По лимфатическому русЛу могут распространяться и клетки злока-
чественных опухолей.
Лимфатические капилляры представляют собой систему замкну-
тых сЗхднеп^Дйонцау-уплощенных Эндотелиальных трубок, анастомо-
зирую1Цйх~друг с другом и пронизывающих органы' (рис. 153, А, Б),
1 Лимфатические капилляры не обнаружены в головном мозге, селезенке,
плаценте, костном мозге, в склере глазного яблока и хрусталике, а также в эпителии
и хрящевых тканях.
389
Рис. 153. Лимфатические
капилляры (препарат
Г. В. Булановой):
А тотальный препарат
(импрегнация серебром): /
границы эндотелиальных
клеток; 2 — замкнутый
конец капилляра (по Янчо).
Б — электронная микрофо-
тография лимфатического
капилляра перикарда крысы.
X 6300. / — эндотелий; 2—
фиксирующие филаменты;
3 - просвет капилляра.
гд^хиш сопровождают гемокапипляры (см. рис. 145) . (Диаметр лим-
фа тических. капилляров а несколько раз больше, чемдёрдвсносных.
Как и в кровеносной, в лимфатической системе почти всегда имеют-
ся резервные капилляры, наполняющиеся лишь при усилении
лимфообразования. Станка лимфатических капилляров состоит из
inn w.H4.i>' кплтщг, которые в отличие от таковых в кровенос-
н ы х ка и иллярах в 3—4 раза крупнее. 1дазальна я мембранаипери ци -
уы~ в лимфатических капиллярах отсутствуют. Энд(1телш|7Й.ная
выстЯ7Пш~лцмфатиш-. j,он, капплляра тесно связана с окружающей
^соединительной тканью С помощью так называемых стропных, или
фиксирующих. Фшимснтод, которые вплетаются в коллагеновые
волокна, распоиожвнныа вдоль—адмфатических капилляров (см.
рис. 153, Б). Лимфатические капилляры и начальные отделы.отво-
дящих лимфатических"сосудов (см. ниже) обеспечивают гемато-
390
1
Рис. 154. Внутриорганный лимфатический сосуд из перикарда крысы. Электрон-
ная микрофотография. X 2700 (препарат Г. В. Булановой).
1 — эндотелий; 2 — гладкие миоциты; 3 — коллагеновые волокна адвентициальной обо-
лочки.
лимфатическое равновесие как необходимое условие микроциркуля-
ции^дГздОровом организме;--------—-----------------
Отводящие лимфатические сосуды. Основной отличительной
особенностью-строения лимфатических сосудов является ^дли.чие
в них клаианппд1_ххф1)ЩО риштой на^у.-цдой обр/ючки (рис. 154).
В местах расположения клапанов лимфатические сосуды колбовид-
но расширяются. В строении стенок лимфатические сосуды имеют
много общего с венами. Это объясняется сходством лимфо- и гемо-
динамических условий этих сосудов: наличием низкого давления
и направлением тока жидкости от органов к сердцу.
Лимфатические сосуды в зависимости от диаметра подразделя-
ются на мелкие,.средние и крупные. Как и вены, эти сосуды по свое-
му строению могут быть безмыщечными и мышечными. Ц м.е лдсп_х
с о с^дит диаметром —40 мкм, которые-являются главным
образом внутриорганными лимфатическими сосудами.'~мьПП'ечные
элементы отсутствуют и их стенка состоит из эмдотедия и сое ди-
нитедьнбтканной оболочки. ~ ~
Сред пл ей крупные лимфатические сосуды (диаметром
более 0,2 мм) имеют три хсщошо развитые оболочки: внутреннюю.
391
Рис. 155. Грудной лимфа-
тический проток. Продоль-
ный срез (препарат
В. А. Кудряшовой).
/ — эндотелий; 2 — продоль-
но ориентированные гладкие
миоциты внутренней оболоч-
ки; 3 — циркулярно ориен-
тированные гладкие миоциты
средней оболочки; 4 — ад-
вентициальная оболочка.
среднюю и наружнунр-Во внутренней оболочке, покрытой эндотели-
ем, находятся продольно и косо направленные пучкиколлагеновых
й~эпастических вл лТ>кпн. Она-формирует многочисленные клапаны.
Они, состоят из центральной соединительнотканной пластинки,
по к р ыто й с внутренней и наружной поверхностей эндотелие м. Под
эндотелием створки клапана обращенной к стенке сосуда,~расЬола-
гается эластическая мембпана. В толще центральной соединитепь-
нотканной пластинки клапана обнаруживаются пучки _сладких
мышечных клеток. На границе внутренней и средней~об7Тпочек
лежит не всегда'ч^тко ныра женная_внутренняя эластическая мем-
бра ня
Средняя ийи-юцка лимфатических сосудец слабо развита в со-
суда х голоьы. верхней части туловища и верхних конечностей.
В лимфатических СОСУПаи нижних к.щ.-иш»., т»-й ои», налГюрот..выра-
ж^на отчетливо, В стенке этих сосудов находятся пучки гладких
МышечныТ клеток, имеющие циркулярное и косое направление.
Большого~ратпИтия достигает мышечный слой в средней оболочке
коллекторов подвздошного лимфатического сплетения, околоаор-
тальных лимфатических сосудов и шейных лимфатических стволов,
сопровождающих яремные вены. Эластические волокна в средней
о бол о ч ке могут различаться по количеству, толщи не и на правд е н и ю.
Наружная обо эпика лимфатических сосудов образована рыхлой
волокнистой нгпфюрм . сосдиннтельи щ тканьк£ЗйЁшрая без
резких границ переходит в окружающую соединительную ткань.
Иногда в наружной оболочке встречаются отдельные продольно
направленные гладкие мышечные клетки.
В качестве примера строения крупного лимфатического сосуда
расемотрим О/1ИН из r ianiiux j кмфштичегких стволов — ГРУДНОЙ
лимфатический проток. Его стенка имеет неодинаковое строение
на различных уровнях. Наиболее сильного развития она достигает
на уровнещиафрагмы (рис. 155). Ца этом месте в стенке сосуда чет-
ко выделяются три оболочки, напоминающие по своему строению
оболочки нижнеи "полой вены. Внутренняя и гр а.п н я я
обо дочки выражены относительно слабо. Цитоплазма эндоте-
•“ " — Рд рдД
392 1
лиальцых клеток богата пиноцитозными пузырьками. Это указывает
на*а1сгйви™й--ч^>ансэндотелиальный транс портЭКИдкости. Базальная
часть клеток неровная. Сплошной базальной мембраны нет.
В подэндотелиальном'слое рыхло залегают пучки коллагеновых
фцбр'йлл. Несколько глубже находятся единичные гладкие мышеч-
ные клетки, имеющие во внутренней оболочке продольное, а в
средней — косое и циркулярное направление. На границе внутрен-
ней и средней оболочек иногда встречается' плотноё~~Гпявтение
тонких эластических волокон, которое ‘сравнивают е внутренней
эластической мембраной. Как и в кровеносных сосудах, эти эласти-
ческие волокна связаны с подобными элементами других оболочек
грудного протока в единый эластический каркас.
В средней обо u ci к е п и ищеуких волокон в
основгк" и ин ым наш авлснисм nvчков
гладких мышечных клеток. Наружная оболочка трудного
л и мфатического протока пУД раза топну- двух других обило чек
и содержит мощные продольно лежащие 1^уч~китлалких_мы1печных
клеток, разделенные i.noc.niiii a jH соепинит. дыюй ткани. Толщина
мышечных слоев грудного лимфатического протока, особенно в
наружной его оболочке, уменьшается в направлении тока лимфы.
При этом стенка лимфатического протока в его устье оказывается
в 2—3 раза тоньше, чем на уровне диафрагмы. .На протяжении
груд ногр л поток? встречается до 9 нрлрунчцу кпяпщрв Створки
клапанов состоят из тех же элементов, что и внутренняя оболочка
протока. У основания клапана в стенке протока наблюдается утол-
щение, образованное скоплением соединительной ткани и гладких
мышечных клеток, направленных циркулярно. В створках клапанов
имеются единичные мышечные клетки, расположенные поперечно.
Васкуляризация сосудов. Все крупные и средние кровеносные
сосуды имеют для своего питания собственную систему, носящую
название «сосуды сосудов». Они приносят артериальную кровь к
стенке сосудов из артерий, проходящих в окружающей соединитель-
ной ткани. В артериях сосуды сосудов проникают до глубоких слоев
средней оболочки. Внутренняя оболочка артерий получает питатель-
ные вещества непосредственно из крови, протекающей в данной
артерии. В диффузии питательных веществ через внутреннюю обо-
лочку артерий большую роль играют белково-гликозаминогликано-
вые комплексы, входящие в состав основного вещества стенок этих
сосудов. Кровеносные капилляры стенок артерий собираются в вены,
которые чаще всего попарно сопровождают соответствующую арте-
рию и открываются в близлежащую вену. В венах сосуды сосудов
снабжают артериальной кровью все три оболочки. Капилляры
стенок вен открываются в ту же вену. В крупных лимфатических
сосудах артерии и вены, питающие их стенки, идут раздельно.
Кроме кровеносных сосудов, в стенке артерий, вен и лимфатиче-
ских стволов находятся лимфатические сосуды.
Иннервация. Сосуды снабжаются нервами вегетативной нервной
системы. Как правило, они сопровождают сосуды и заканчиваются
в их стенке. По строению нервы сосудов являются либо миелиновы-
393
Рис. 156. Адренер! ическое нервное
сплетение артерии головного мозга
человека. Метод Фалька (препарат
Л. Д. Маркиной).
ми, либо безмиелиновыми. Чувствительные нервные окончания в
капиллярах многообразны по форме. Характерная особенность
чувствительных нервных аппаратов артериол — их большая протя-
женность и поливалентность — связь с концевыми нервными волок-
нами соседних венул и окружающей соединительной тканью. Рецеп-
торы на венулах нередко кустикообразно ветвятся. В артериях
среднего и крупного калибра разнообразные рецепторы залегают
во всех трех оболочках сосуда (рис. 156). В поверхностных слоях
наружной оболочки и в околососудистой рыхлой волокнистой
соединительной ткани есть инкапсулированные пластинчатые и
неинкапсулированные нервные окончания, образующие периваску-
лярное нервное сплетение (plexus nervorum perivascularis).
Артериоловенулярные анастомозы имеют сложные рецепторы,
расположенные одновременно на анастомозе, артериоле и венуле.
Эффекторные нервные волокна заканчиваются на гладких мы-
шечных клетках маленькими утолщениями. Эффекторы на арте-
риях и венах однотипны. По ходу сосудов, особенно крупных,
встречаются отдельные нервные клетки и небольшие ганглии
симпатической природы.
Возрастные изменения. Строение сосудов непрерывно меняется
в течение всей жизни человека. Развитие сосудов под влиянием
функциональной нагрузки заканчивается примерно к 30 годам.
В дальнейшем в стенках артерий происходит разрастание соедини-
тельной ткани, что ведет к их уплотнению. В артериях эластического
типа этот процесс выражен сильнее, чем в остальных артериях.
В основном веществе внутренней и средней оболочек накапливаются
сульфатированные гликозаминогликаны, с которыми связано, до- ;
локнробразпвание. После 60—70 лет во внутренней оболочке всех
артерий обнаружила юте я^очаговые утолщения коллагеновых воло- п
кон, в результате чего в крупных артериях внутренняя оболочка
"по размерам приближается к средней. В мелких и средних артериях
внутренняя оболочка разрастается слабее. Внутренняя эластическая
мембрана с возрастом постепенно истончается и расщепляется.
М у шеч ные к четкц-средней обо почки атрофируются, Эпа сти ческие
394
волокна подвергаются зернистому распаду и фрагментации, в то
время как коллагеновые волокна разрастаются. Одновременно с
этим во внутренней и средней оболочках у пожилых людей по явля-
ются известковые отложения, которые прогрессируют с возрастом.
В наружной оболочке у лиц старше 60—70 лет возникают продольно
лежащие пучки гладких мышечных клеток.
Возрастные изменения в венах сходны с таковыми в артериях.
Однако перестройка стенки вены человека происходит еще на пер-
вом годужидни* Так, к моменту рождения человека в средней обо-
лочке стенок бедренной и подкожных вен нижних конечностей
имеются лишьгпучки циркулярно ориентированных—мышечных
клеток. Только к моменту вставания на-ноги (к концу первого года)
и повышения дистального гидростатического давления развиваются
продольные мышечные пучки. Просвет вены по отношению к просве-
ту артерии у взрослых (2:1) больше, чем у детей (1:1). Расширение
просвета вен обусловлено меньшей эластичностью стенки вен,
возрастанием у взрослых кровяного давления.
Сосуды сосудов до возраста 50—60 лет, как правило, бывают
умеренно спазмированными, после 65—70 лет просвет их расши-
ряется.
Лимфатические сосуды многих органов у лиц старческого воз-
раста характеризуются многочисленными мелкими варикозными
вздутиям^__и выпячиваниями. Во внутренней оболочке стенок
крупных лимфатических стволов и грудниго11ротЬка’у людей старше
35 лет увеличивается количество~"коллагеновых волокон. 'ЗтОт
процесс значительно прогрессирует к 60—70 годам? Одновременно
количество мышечных клеток_и эластических волокон уменьшается.
кровеносные и особенно лимфатические сосуды значительно
изменяютсЯ~~яе~только с возрастом,—но и при функциональных
перестройках органов, например в матке и яичнике_в зависимости
от овариалыто-тиенсгруального цикла и Т. д.
Регенерация. Кровеносные и лимфатические сосуды обладают
высокой способностью к регенерации. Восстановление дефектов
сосудистой стенки после ее повреждения начинается с регенерации
и роста ее эндотелия. Уже к концу-цервых?^- началу вторых суток
тяа' месте нанесенного повреждения наблюдается многочисленное
деление эндотелиальных клеток.
Мышечные клеткй~поврёжденного сосуда, как правило, восста-
авливаются~более медленно и неполно по сравнению с другими
каневыми элементами сосуда. Восстановление их происходит
зстично путем деления миоцитов, а также ^результате дифферен-
ирлвки миофибробластов. Эластические элементы развиваются
або. В случае полного перерыва среднего и крупного сосудов
егенерации его стенки без оперативного вмешательства, как прави-
э, не наступает, хотя восстановление циркуляции крови в соответ-
вующеЧобласти можетТзаблюдаться очень рано. Это.происходит,
одной стороны, благодаря компенсатбрПОЙ~перестройке коллате-
льных сосудов, а с_другой — вследствие развития и роста новых
елких сосудов — капилляров. Новообразование капилляров начи-
395
нается с того, что цитоплазма эндотелиальных клеток артериол и
венул набухает в виде почки, затем эндотелиальные клетки подвер-
гаются делению. По мере роста эндотелиальной почки в ней появля-
ется полость. Такие слепо заканчивающиеся трубки растут навстре-
чу друг другу и смыкаются концами. Цитоплазматические перего-
родки между ними истончаются и прорываются, и во вновь образо-
ванном капилляре устанавливается циркуляция крови.
Лимфатические сосуды после их повреждения регеиерируюу
несколько медленнее, чем_кроведо£ные. Регенерация лимфати-
ческих сосудов может происходить за сдёшли почкования дистал
ных концов эндотелиальных трубок, или перестройки лимфатиче-
ских капилляров в отводящие сос ды.
СЕРДЦЕ
Сердце (сог) — основной орган, приводящий в движение кровь.
Развитие. Первая закладка сердца появляется в начале 3-й не
дели развития у эмбриона длиной 1,5 мм в виде парного скопления
мез^цхимны>.-кг^ток, которые расположены в задней части голов-
ного отдела зародышевого щитка по сторонам от средней линии
под висцеральным листком мезодермы. Позднее эти скопления
превращаются в две удлиненные трубочки, вдающиеся вместе с
прилегающими висцеральными листками мезодермы в целомиче-
скую полость тела (рис. 157, А, Б„ В). В дальнейшем мезенхимные
трубки сливаются и из их стенки образуется эндокард.
Та ^область висцеральных листков мезодермы, которая приле-
жит к этим трубкам, получила название миоэпикардца пьных плас-
тинок. Из этих пластинок дифференцируются две части: одна —
внутренняя, прилежащая к мезенхимной трубке, превращается в
зачаток миокарда, а из наружной образуется эпикард. Клетки
зачатка миокарда — кардиомиобласты — делятся, их объем увели-
чивается и на 2-м месяце развития зародыша в них появляются
идущие в разных направлениях миофибриллы с поперечной ис-
черченностью. Z-полоски появляются одновременно с саркотубу-
лярной сетью и поперечными инвагинациями клеточной мембраны
(Т-системы). В местах контакта миобластов отмечаются десмосо-
моподобные структуры, преобразующиеся позднее во вставочные
диски. В течение эмбрионального периода вставочные диски пред-
ставляют собой слабоизвитые мембраны контактирующих клеток.
В конце 2-го месяца появляются признаки формирования проводя-
щей системы, которая отличается большим количеством ядер,
замедленной дифференцировкой фибриллярного аппарата и др.
К 4-му месяцу заканчивается образование всех отделов проводящей
системы сердца. Развитие мышечной ткани левого желудочка про-
исходит быстрее, чем правого.
Клапаны сердца — предсердно-желудочковые и желудочково-
сосудистые— развиваются в основном как дупликатура эндокар-
да. Левый клапан появляется в виде эндокардиального валика, в
который позднее (у эмбриона 2 */2 мес) начинает врастать соеди-
нительная ткань из эпикарда.. На 4-м месяце внутриутробного пе-
396
Рис. 157. Развитие сердца. Попереч-
ные разрезы зародышей в трех по-
следовательных стадиях формирова
ния сердца (по Штралю, Гису и Бор-
ну).
Л — две парные закладки сердца, Б -
их сближение; В — слияние в одну не-
парную закладку; / — эктодерма; 2 —
энтодерма; 3 — париетальный листок
мезодермы; 4 — висцеральный листок
мезодермы; 5 — хорда- 6 — нервная
пластинка; 7 — сомит; <8 — вторичная
полость тела; 9 — эндотелиальная за-
кладка сердца (парная); 10—нервный
желобок; 1/ — нервные валики; 12 —
нисходящая аорта (парная); 13 об-
разующаяся головная кишка; 14 — го-
ловная кишка; 15 — спинная сердечная
брыжейка; /б — полость сердца, 17 —
эпикард; 18 — миокард; 19 — эндокард;
20 — околосердечная сумка; 2/ — пери-
кардиальная полость; 22 — редуцирую-
щаяся брюшная сердечная брыжейка.
Z/7
риода из эпикарда в створку клапана врастает пучок коллагено-
вых волокон, образующий в будущем фиброзную пластинку. Пра-
вый клапан закладывается как мышечно-эндокардиальный валик.
С 3-го месяца развития зародыша мышечная ткань правого атрио-
вентрикулярного клапана уступает место соединительной ткани,
врастающей со стороны миокарда и эпикарда. У взрослого челове-
ка мышечная ткань сохраняется в виде рудимента только с пред-
сердной стороны в основании клапана. Таким образом, предсерд-
но-желудочковые клапаны являются производными не только
эндокарда, но и соединительной ткани миокарда и эпикарда.
Аортальные клапаны имеют двойное происхождение: синусная
сторона их образуется из соединительной ткани фиброзного коль-
ца, которая покрывается эндотелием, а желудочковая — из эндо-
карда.
Развитие интрамурального нервного аппарата сердца человека
начинается па 7-й неделе, когда впервые обнаруживаются нейро-
бласты. На 9—10-й неделе отмечается их дифференцировка, а на
12-й — первые синапсы.
Строение. В стенке сердца различают три оболочки: внут-
реннюю— эндокард, с р е д и ю ю, или мышечек), — мио-
кард и наружную, или серозную, — эпикард.
Э нд~окар Д- Внутренняя оболочка сердца эндокард (endo-
сafflium»^ в^тиллет_изнутри камеры сщща, пдпи.иярные мыш-
1Щ1. сухо4Ц1Н1МЙ пичи, а такте клапаны сердца. Толщина эндо-
397
Рис. 158. Эндокард и миокард
сердца человека.
/ эндотелий; 2 подэндотелиальныи
слой; 3 — мышечно-эластическии слой:
4 — гемокапилзяры; 5 — атипичные мы-
шечные клетки (проводящие миоциты);
6 типичные кардиомиоциты миокар-
да.
карда неодинакова в различ-
ных участках. Он толще в ле-
вых камерах сердца, особен-
но на межжелудочковой пе-
регородке и у устья крупных
артериальных стволов —
аорты и легочной артерии,
а на сухожильных нитях зна-
чительно тоньше.
.Со стороны поверхности,
обращенной в полость серд-
ца, эндокард выстлан^ эндо-
телием, СОСТОЯЩИМ-ЛД-ЛОЛИ-
гональных клеток, лежащих
.на Толстой—бщадльнсш мем-
б[>ане (рис. 158). За ним следует подэТк^отелиадьный елой^обра^
зованный соедини тельной тканью, богатой малодифференцирова н -
нымй клетками. Глубже располагается .^ышечн<оэластический
слои, в которомэластическиеволокна переплетаютсяс гладкими
мышечными клетками. Эластические волокна гораздо луярте вы-
ражены в эндокарде предсердий, чем в желудочках. Гладкие
мышечные клетки сильнее всего развиты в эндокарде у места вы-
хода аорты и могут иметь многоотростчатую форму. Самый глубо-
к и й слой, эндокарда — шружн!^й—С(М’динител(>н(Нканн111и — ле ж и т
на границе с миокардом. Он состоит щз соединительной ткани,
содержащей толстые ЗЛаТТические, коллагеновые й ретикулярные
волокна. -====—- —-------= --------
Питание эндокарда осуществляется главным диффуз-
но замечет крови, находящейся в камерах сердца. КровёнДгяне
согупч имеется лишь в наружном соединительнотканном слое
эндокарда.
—К_дапаны. Между предсердиями и желудочками сердца, а так-
же желудочками и крупными сосудами располагаются клапаны.
П редсёрдно-желудочковый Татриовентрикулярный ) клапан в
левой половине сердца двустворчатый, в правой — трехстворча-
тый._ Они представляют собой покрытые эндотелием тонкие .плас-
тинки плотной волокнистой соединительной ткани с небольшим
количеством клеток (рис 159) Энпотепиапьные клетки по к ры -
вающие клапан, частично покрывают друг друга в виде черепицы
или образуют пальцевидные вдавливания цитоплазмы одной клет-
ки в другую. Кровеносных сосудов створки клапанов не имеют.
В подэндотелиальном слое выявлены тонкие коллагеновые волок-
398
Рис. 159. Предсердно-желудочковый (атриовентрикулярный) клапан сердца чело-
века (по В. В. Соколову).
/ _ предсердная сторона; II — желудочковая сторона; I — волокна миокарда в основании
створки клапана; 2 — кровеносные сосуды; 3 — эндокард левого желудочка: 4 миокард
левого желудочка.
на, которые постепенно переходят в фибрознуюпластинку створ-
ки клапана, а В Месту iipHKjirmfHHH iy.y- и трехгт1орч1нчно клапа-
нов —’ в фиброзные кольца. В основном веществе створок клапа-
нов обнаружено большое количество гликозаминогликапод
Строение гГрёДСердных ижелудочковых частей створок клапа-
нов неодинаково. <-т.ц¥>ца их имеет гладкую ло.х-рх-
HOCTi. < :rct- и 1:огпкг10Т£ли;ч1 ном ом- | -ь.. . р, 1 ое спле-
тен и е эластических волокон и пучки гладких мышечных клеток.
Количество мышечных пучков заметно увеличивается в основании
клапана. Желудочковая сторона обладает неровной поверхностью.
Она^иабхеиа^Еипостами, от которых начинаются сухожильные
нити fchordae tendineae). В этой области под эндотелием распола-
гается лишь небольшое количество эластических волокон.
На границе-между восходящей частью дуги аорты и левым же-
лудо«ПСОМ Сурдца-лпкализуютГм аариииьиьи^лцлапаны. По своему
строению они имеют много общего с предсердно-желудочковыми
клапанами и клапанами легочной артерии. На вертикальном раз-
резе в створке клапана можно различить три слоя: внутренний,
средний и наружный. Внутренний слой, обращенный к же-
лудочку сердца, представляет сибиЙгтфоДОлжение эндокарда. Эн-
лотелий этого слоя характеризуется ~на;1йЧием пучков субЯНкро-
скопических филаментов толщиной 5—8 нм и многочисленных
пиноцитозных пузырьков. В подэндотелиальном слое содержатся
399
Рис. 160. Строение кардиомиоцита.
А — схема (рис. Ю. И. Афанасьева и В. Л.
Г орячкиной);
^фибробласты с длинными тонкими отростками, которые в •виде кон-
солей поддерживают эндотелиальные клетки. К.подэндотелиально-
му слои! ipnn-жат сыютны^ iij и>.ифибрилл, идущнх
продольно и поперечно, зддсоторым следуёт~смётттатшая~ эластико-
коллагеновая прослойка. Средний слои тонкий, состоит из
рьтЯлоф нотгОкИИсюй Соединительной ткани, богатой клеточными
эле_ментами, Н7Гр~у-~Ж~ТГы й слой, обращенный к аорте, кроме
эндотелия, содержит кодддксновые волокна, которые берут начало
от фиброзного кольца вокруг аорты. "=—
Миокард. Мышечная оболочка сердца (myocardium) состо-
ит и,з тесно связанных между собой поперечнополосатых мышеч-
ные клеток, которые образуют функциональные мышечные «во-
локна», залегающие послойно (см. гл. X). Ме_жду мышечными
элементами миокарда располагаются прослойки рыхлой соедини-
тельной ткани, сосуды, нервы. Различают типичные сократите пь-
ные мышечные клетки ка[к)иомио_циты и прааадящие сердечные
миоццты, входяГцшГв состав так называемой проводящей системы
сердца.
Сердечные сократительные миоциты (myociti cardiaci) харак-
теризуются рядом структурных и цитохимических особенностей,
400
Рис. 160 (продолжение).
Б электронная микрофотография вставочного диска. / — миофибриллы; 2 — мито-
хондрии; 3 — саркотубулярная сеть; 4 — Т-трубочки; 5 — базальная мембрана; 6 — лизо-
сома: 7 — вставочный диск; 8 — десмосома; 9 — зона прикрепления миофибрилл; 10 —
щелевые контакты; // — рибосомы.
отличающих их от проводящих кардиомиоцитов и от волокон
скелетной поперечнополосатой мышечной ткани (рис. 160).
Кардиомиоциты на продольных срезах почти прямоугольной
формьцих длина колеблется от 50 до 120 мкм, ширина составля-
ет 15—20 мкм. В центральной части миоцита расположено 1^-
2 ядра .овальной йли удлиненной формы. Миофибриллы распбла-
гаютс.я продольно (см. гл. X).
В отличие от желудочковых кардиомиоцитов, форма которых
близка к цилиндрической, предсердные миоциты чаще имеют от-
рогтчзтую форму В миоцитах, предсердий меньше митохондрий,
миофибриллу саркоплазматической сети~В предсердных карлио-
401
миоцитах менее выря»ечя активность сукпинатдегидрогеназы. а
бойее высока активность ферментов, спя )анных—С—метаболизмом
цщкогена (фосфорилаза, гликогенсинтетаза и др.). Отличительной
особенностью этих кардиомиоцитов является относительно хрро-
iiici развитая гранулярная сеть и интенсивное развитие комплекса
Гольджи? Указанные выше^морфологические признаки связаны с
наличием в предсердных кардиомиоцитах специфических пред-
сердных гранул, в синтезе белковых компонентов которыхтзначале
принимают~участие гранулярная цитоплазматическая сеть, а окон-
чательное формирование гранул происходит в комплексе Гольд-
жи . С пецифические гранулы предсердий богаты гликопротеинам и.
Ряд экспериментальных данных дает (Основание предположить,
что гликопротеины предсердных гранул, поступая в кровь, связы-
вают липопротеины, проявляя тем самым ^нтитромбическое дей-
ствие. Кроме ТОГО, преасердные к-.|р>|илмщщИТЫ, подобно о'собБГМ
клеткам, '’расположенным около почечных клубочков, выделяют
вещества (натрийуретический фактор), участвующие в регуляции
.артуриальногодавления т. е. эти клетки- пбпапают трнзосенсогР
ной функцией.
Еще одной отличительной чертой пре>н ерлных кардиомиоцитов
многих млекопитающих является слабое рдзвитиеТ-системьрка-
нальцев, В тех прелсерлных миоцитах, где нет Т-системы, на пе-
риферии клеток, под сарколеммой, располагаются многочислен-
ные пиноцитозные пузырьки и канеолы. Полагают, что эти пу-
зырьки и кавеолы являются фушго.чопа льпыми- пивлогами Т-ка-
нальцев
~Каплиом,иопиты. образующие мышечные «волокна», сообщают-
ся между собой в области вставочных „дисков (disci intercalaTty.
В гистологических препаратах они имеют вид темных полосок.
Строение вставочного диска на его протяжении неодинаково (см.
рис. 160). Различают места вплетения миофибрилл
в плазмаг1вмму -ц—щелевые кшншды. Если первые два участка
диска выполняют м^аническую функцию. т“ третий осуществляет
электрическую свящ МрдИПМИРчщов. С помощью дисков кардио-
миоциты соединяются ё мышечные «пиюкиа». Между Соседними
мышечными волокнами имеются анастомозы. Продольные у боко-
вые связи~кардиомиоцитов обеспечивают функциональное единст-
во миокарда.
Мышечнь|р волокна миокарда прикрепляются к опорному ске-
летусердца, который образован фиброзными кольцами между
предсердиями и желудочками и плотной соединительной тканью
в устьях крупных сосудов. Кроме плотных пучков коллагеновых
волокон, в составе «скелета» сердца имеется много эластических
волокон, а иногда бывают даже хрящевые пластинки.
В интерстициальной соединительной ткани заключено большое
количество кровеносных и лимфатических капилляров. Каждый
миоцит ^контактирует е 2~—Э--капиллярами.
Как было отмечено выше, другой разновидностью миоцитов в
миокарде являются проводящие сердечные миоциты (myocyti соп-
402
Рис. 161. Кардиомиоциты проводящей системы сердца (по П. П Румянцеву).
/ — схема расположения элементов проводящей системы сердца; // -=- кардиомиоциты
синусного и атриовентрикулярного узлов: а — Р-клетки. б — переходные клетки; 111—
кардиомиоцит из пучка Гиса; IV — кардиомиоцит из ножек пучка (волокна Пуркинье):
/ — ядра; 2 — миофибриллы; 3 — митохондрии; 4 — саркоплазма: 5 — глыбки гликогена;
6 — промежуточные филаменты; 7 — миофиламентные комплексы.
ducens cardiacus), входящие в состав так называемой проводящей
системы сердца. ~ ~
Проводящая система сердца (systema conducens cardiacum).
Этим термином объединяют мышечные клетки, формирующие и
провсщящиё~ймиульсы К СйкРаТЙтёльным клеткам сердца. В со-
став проводящей системы входят: синусно-предсердный узел, пред-
сердно-желудочковый _ учел, предсердно-желудочковый rtyipOK
(ствол, правая и левая ножки) и его разветнЯёНИЯ,—передающие
импульсы на сократительные мышечные клетки. Различают три
403
типа мышечных клеток, которые в разных соотношениях находят-
ся в различных отделах этой системы (рисГГ61).
У зл ы проводящей с лх темы. Формировапцу^импуль-
са происходит центральную часть которого за-
нимают клетки первого типа — водители ритма, или пеисмекер-
ные клетки ~(Р~клетк_и), способный к самопроизвольным сокраще-
ниям (см. рис. 161). Они отличаются небольшими размерами,
многоугольной формой С максимальным диаметром 8 10 мкм, не-
большим количеством миофибрилл, не^имеюших уплрядочеынгГи
ориентировки. Миофиламенты в составе миофибрилл упакованы
рыхло. А- и Т-диски различаются нечетко. Митохондрии неболь-
шие, округлой или овальной формы, немногочисленные. Сарко-
плазматический ретикулум развит слабо. Т-система отсутствует,
но вдоль цитолеммы находится много пгГноцитозных пузырьков
и кавеол, которые в 2 раза увеличивают Мембранную ТТбВерхпПсть
клеток. Высокое содержание свободного кальпия-.д_цитоплазме
этих клетбк~при слабом развитии саркоплазматической сети обу-~~
слрвлива.ет способность клеток синусного уз па генерировать им -
пульсы к сокращению. Поступление необходимой энергии обеспе-
чивается преимущественно процессами гликолиза. Между клетка-
ми встречаются единичные десмосомы и нексусы. Др периферии
У зла располагаются переходное клетки —б»+лттей
части кдеток в д т fa и п ь.е н т и к у л я и о м у ч л г. р-кдеток в
атриовентрикулярном узле^ напротив, мало. Основную часть состав-
ляет второй inn — HePi’WpHl’le клетки. Это — тонкие, вытянутые
клетки, поперечное сечение которых меньше поперечного сечения
типичных сократительных кардиомиоцитов. Миофибриллы более
развиты, ориентированы параллельно друг к другу, но не всегда. От-
дельные переходные клетки ~мбгуГ~годержать короткие I-трубоч-
Переходные клетки сообщаются между собой как С Помощью
простых контактов, так и путем образования более сложных со-
единений типа’ вставочных дисков. Функциональное значение этих
клеток состоит в передаче возбуждения от Р-клеток к клеткам
пучка и рабочему миокарду.
Клетки пучка проводящей системы и его
н о ж ё~кГ Они составляют Третий тип. В функциональном отяоше-
н и и я вляются п^ре;иатчиками возбуждения or nne¥iynu*x клеток
к клеткам щцмнегп мирыпн кпиуопнклв
Мышечные клетки проводящей системы в стволе и разретвле-
н и я х ноже к стволапроводящеи системы располагаются небольши -
ми пучками, ойи~отгружены —мыиш шуюкнистой
соединительной ткани. Ножки пучка разветвляются под эндокар-
дом, а также в толще миокарда желудочков. Клетки Проводящей
системы не только разветвляются в миокарде, но и проникают в
сосочковые мышцы. Это обусловливает натяжение сосочковыми
мышцами створок клапанов (левого и правого) еще до того, как
начнется сокращение миокарда желудочков.
По стропит ч АШИ пупка пт ,ишЬ.т< я крупными раз-
мерами (15 мкм и более) в диаметре, почти полным отсутствием
404
Т-систем, тонкостью миофи^рилл, которые без определенного
порядка располагаются главным образом по перифёрйи клетки.
Ядра, как правило, располагаются эксцентрично. Эти клетки в
совокупности образуют предсердно-желудочковый ствол и ножки
пучка («волокна Пуркине»).
Клетки проводящей системы сердца богаты лабильным глико-
сен<та?легко расщепляемым амилазой. Содержание рибонуклео-
протеидО в; липидов меньше, чем в~сократительных сердечных мио-
цитах. Что же касается активности окислительно-восстановитель-
ных ферментов, то в проводящей системе сердца преобладают
энзимы, принимающие участие в анаэробном гликолизе (фосфо-
рилаза, дегидрогеназа молочной кислоты. Понижена активность
аэробных ферментов цикла трикарбоновых кислот (дегидрогена-
за янтарной, яблочной,'ct-кетоглютаровой и изолимонной кислот) и
митохондриальной цепи переноса электронов (цитохромоксидаза).
В проводящих волокнах уровень калия ниже, а кальция и натрия
выше по сравнению с сократитель1гыми кардиомиоцитамтт,-
В миокарде много афферентч<ну. -нервных во-
локон. Типичных Нёрвн<1-мы1чечн1.1ч еннлнгоя ч.лепь нет. Раздраже-
ние нервных волокон, окружающих проводящую систему, а~также
нервов, подходящих к серд-цу, П| пып<нм1>-ипмй11«»ние ритма оердеч-
ных сокращений. Это указывает на решающую рОлБ~~нервнс>й~сис-
темы в ритме сердечной деятельности, а следовательнО-Щ в переда-
че импульсов по проводящей системе.
Эпикард и перикард. Наружная оболочка сердца, или
эпикард /epicardium), представляет собой -висцеральный-.чисток
перикарда (pericardium). Эпикард образован тонкой (не более
0,3—0,4 мм) пластинкой соединительной ткани, плотно срастаю-
щейся с миокардом. Свободная поверхность ее плкрыта музоте-
ц1рм R—слепииитепьнлтканной основе эпикарда различают по-
верхностный сдой коллагеновых волокон, слой эластических
волокон, глубокий слой коллагеновых волокон и глубокий коллаге-
ново-эластический слой, который составляет до 50% всей толщи
эпикарда. На предсердиях и некоторых участках желудочков
последний слой отсутствует или сильно разрыхлен. Здесь же иног-
да отсутствует и поверхностный коллагеновый слой. Между эпи-
кардом И перикардом имеется щелевидное пространство, содержа-
щее небольшие количество 'Жидкости, ветполняющей-рель-смазки
В перикарде соединительнотканная основа развитгГ~1‘Илънее,
чем в эпикарде. В ней много эластических волокон, особенно в
глубоком еги слое; Понерхность черикардаг"ббращённая~к~~пери-
^чем. Но ходу крове-
кардиальной полости, тоже покрыта joF-
:МИ||
носных сосудов встречаются скопления жировых клеток. Эпикард
и париетальный листок перикард» имеют многочисленные нервные
окончания, преимущественно свободного типа.
Васкуляризация. Венечные артерии имеют плотный эластиче-
ский каркас, в котором четко выделяются внутренняя и наружная
эластические мембраны. Гладкие мышечные клетки в артериях
обнаруживаются в виде продольных пучков во внутренней и на-
405
Рис. 162. Адренергические
(А) и холинергические
(fi) нервные .волокна и ма
лые интенсивно флюорес-
цирующие клетки
МИФ-клетки (В, Г).
А — алюминий-формальде-
гидный метод (препарат
Р А. Стропуса); Б метод
М Карновского;
4С. 162 (предо чжение) .
— метод флюоресцентной микроскопии; Г — электронная микрофотография. X 10 000
репарат А. А. Сосунова и В. Н. Швалева). / малая интенсивно флюоресцирующая
,етка; 2 — ядро. 3 — гранулы секрета, 4 — гемокапилляр.
ружной оболочках. В основании клапанов сердца кровеносные со-
суды у места прикрепления створок разветвляются на капилляры.
Кровь из капилляров собирается в коронарные вены, впадаю-
щие в правое предсердие или венозный синус. Проводящая систе-
ма, особенно ее узлы, обильно снабжена кровеносными сосудами.
Лимфатические сосуды в эпикарде сопровождают кровеносные.
В миокарде и эндокарде они проходят самостоятельно и образуют
густые сети. Лимфатические капилляры обнаружены также в атри-
овентрикулярных и аортальных клапанах. Из капилляров лимфа,
оттекающая от сердца, направляется в парааортальные и пара-
бронхиальные лимфатические узлы. В эпикарде и перикарде на-
ходятся сплетения сосудов микроциркуляторного русла.
Иннервация. В стенке сердца обнаруживается несколько нерв-
ных сплетений (в основном из безмиелиновых волокон адренер-
гической и холинергической природы) и ганглиев. Наибольшая
. ютность расположения нервных сплетений отмечается в стенке
правого предсердия и синусно-предсердного узла проводящей сис-
темы.
Рецепторные окончания в стенке сердца (свободные и инкап-
сулированные) образованы нейронами ганглиев блуждающих нер-
вов и нейронами спинномозговых узлов (С7—Th6) и, кроме того,
407
за счет ветвления дендритов равноотростчатых нейроцитов внутри-
органных ганглиев (афферентные нейроны).
Эффекторная часть рефлекторной дуги в стенке сердца пред-
ставлена расположенными среди кардиомиоцитов и по ходу сосу-
дов органа нервными волокнами холинергической природы, об-
разованными аксонами находящихся в сердечных ганглиях длин-
ноаксонных нейроцитов (эфферентные нейроны). Последние по-
лучают импульсы по прегаш лионарным волокшем из нейро-
нов ядер продолговатого мозга, приходящих сюда в составе
блуждающих нервов. Эффекторные адренергические нервные во-
локна образованы ветвлениями аксонов нейронов ганглиев симпа-
тической нервной цепочки. На этих нейронах также синапсами
заканчиваются преганглионарные волокна — аксоны нейронов
симпатических ядер боковых рогов спинного мозга. Эффекторы
представляют собой варикозные утолщения по ходу адренергиче-
ских нервных волокон, содержащие синаптические пузырьки.
В состав нервных ганглиев сердца входят богатые катехол-
аминами так называемые малые интенсивно флюоресцирующие
клетки — МИФ-клетки (рис. 162, А, Б, В, Г), на которых обнару-
жены нервные окончания как адренергических, так и холинерги-
ческих нервных элементов сердца и которые рассматриваются как
вставочные нейроны, выделяющие свои медиаторы в кровеносное
русло.
Возрастные изменения. В течение онтогси&да млжнц-яыпепить
три периода изменения гистоструктуры сгршь'с пврно-я—д и ф-
ф е р енцировки, перил ц гтяй и л м ia и ни и пери о д
и н валипии —tn. п< п.гнчес^их элементов
сердца, начавшаяся еше в заро-тмпм-вом ш-рилче_заг..шчнп.1с»я к
16 20 годам- Существенное влияние на процессы лиФФере нци-
ровки кардиомиоцитов и морфогенез желудочков оказывает за-
ращение овального отверстия и артериального протока, которое
приводит к изменению г^кГодинаминес учу yr.mwn» — у^ены пению
давления и сопротивления в малом круге и увеличению давления
в бо7П1НЮм~ ().пноь[нмгшш отмечаются физиологическая атрофия
миокдрпа правого же.л.'ючгд и физиологическая типертрнфия ми-
окарда левого желудочка. В ходе дифференцировки сердечные
миоциты обогащаются саркоплазмой, в результате чего их ядер-
но-плазменное отношение уменьшается. Количество миофибрилл
прогрессивно увеличивается. Мышечные клетки проводящей систе-
мы при этом дифференцируются быстрее, чем сократительные.
При дифференцировке волокнистой стромы сердца наблюдаются
постепенное уменьшение количества ретикулярных волокон и за-
мена их зрелыми коллагеновыми волокнами.
В цериод между 20 и 30 годами при обычной функциональной
нагрузке сердце человека находится в стадии относительной ста-
билизации. В возрасте старше 30—40 лет в миокарде обычно на-
чинается некоторое увеличение его соединительнотканной стромы.
При Этом в~стенке сердца особенно в эпикарде- появляются, ади-
поциты.
408
Степень иннервации сердца также изменяется с возрастом. _
Максимальная плотность внутрисердечных сплетений на единицу ь
площади и высокая активность медиаторов отмечаются в период
полового созревания. После 30-летнего возраста неуклонно умень- i
шаются плотность адренергических нервных сплетений и содержа- >
ние“М5Диаторов<в них, а плотность холинергических сплетений и
количество медиаторов в ийх сохраняются тгочти ла 'исходном -
уров11е7~Нарушение равновесия в вегетативной иннервации сердца
предрасполагает к развитию патологических сисюяний.- -В—етар-
ческом возрасте уменьшается активность медиатордв ~и в холинер-
гических сплетениях сердца. >
Регенерация. У новорожденныт, а ioahawmh и в раннем дет- ।
ском возрасте, когда способные к делению кардиомиоциты еще ,
сохраняются. Dvivt|cr|4горные процесс^ 1'иепиче- 1
пием/ ,ичества___кардиомиоиитов У взрослых физиологическая
регенерация осуществляется в миокарде главным образом путем
внутрнк ||,ТО,1М ‘"I"1 ЬС i ЧЯ.1ИЧеНИЯ КОЛИЧрстьл клеток.
Клетки соединительной ткани всех оболочек пролиферируют, как
в любом другом органе.
При повышенных систематических функциональных нагрузках
общее количество клеток не возрастает, но в цитоплазме увеличи-
ваются содержание органелл общего значения и миофибрилл,
размер клеток (происходит функциональная гипертрофия); соот-
ветственно возрастает и степень плоидности ядер.
Глава XVI
ОРГАНЫ КРОВЕТВОРЕНИЯ И ИММУНОЛОГИЧЕСКОЙ
ЗАЩИТЫ
К органам кроветворения и иммунологической защиты отно-
сят: красный костный мозг, тимус (вилочковая железа), лимфа-
тические узлы, селезенку, а также лимфатические узелки пище-
варительного тракта (миндалины, лимфатические узелки ки-
шечника) и других органов. Они образуют единую с кровью
систему.
В настоящее время принято различать центральные
и периферические органы кроветворения и иммунологи-
ческой защиты.
К центральным органам кроветворения у человека относятся
красный костный мозг, тимус и пока неизвестный у млекопитаю-
щих аналог сумки Фабрициуса1. В красном костном мозге из ство-
ловых клеток образуются эритроциты, гранулоциты, тромбоциты
(кровяные пластинки), В-лимфоциты и предшественники Т-лим-
фоцитов. В тимусе предшественники Т-лимфоцитов превращаются
в Т-лимфоциты. В центральных органах происходит антигеннеза-
висимое размножение лимфоцитов.
1 Предположительно лимфатические скопления по ход} кишечника.
409
В периферических кроветворных органах (лимфатические уз-
лы, гемолимфатические узлы, селезенка) происходят размноже-
ние приносимых сюда из центральных органов Т- и В-лимфоцитов
и специализация их под влиянием антигенов в эффекторные клет-
ки, осуществляющие иммунологическую защиту. Кроме того, здесь
погибают клетки крови, завершившие свой жизненный цикл.
Органы кроветворения функционируют содружественно и обес-
печивают поддержание морфологического состава крови и иммуно-
логического гомеостаза в организме. Координация и регуляция
деятельности всех органов кроветворения осуществляются по-
средством гуморальных и нервных факторов организма, а также
внутриорганных влияний, обусловленных микроокружением.
Несмотря на различия в специализации органов гемопоэза,
все они имеют сходные структурно-функциональные признаки.
В основе их лежит ретикулярная соединительная, а иногда эпите-
лиальная ткань (в вилочковой железе), которая образует строму
органов и выполняет роль специфического микроокружения для
развивающихся клеток. В этих органах происходит размножение
кроветворных клеток, временное депонирование крови или лимфы»
Кроветворные органы благодаря наличию в них специальных фа-
гоцитирующих и иммунокомпетентных клеток осуществляют так-
же защитную функцию и способны очищать кровь или лимфу от
инородных частиц, бактерий и остатков погибших клеток. Процессы
кроветворения в эмбриогенезе проходят три этапа: мезобластиче-
ский, гепатолиенальный и юстномозговой, или медуллярный.
костный мозг
K-iCXJbiiLмозг (medula ;енгп) — центральный кпоа. t ьор.чый
1 • орган, где находился само, д. приживающаяся популяция с» во то -
вых кроветворных клеток и образуются клетки как миелоидного,
так и лимфоидногу ряда-
Развитие. Костный мозг у человека появляется впервые на
2-м месяце внутриутробного периода в ключице эмбриона, на
3-м месяце он образуется в развивающихся плоских костях —
лопатках, безымянных костях, затылочной кости, ребрах, грудине,
костях основания черепа и позвонках, а в начале 4-го месяца раз-
вивается также в трубчатых костях конечностей. До 11-й неде-
ли — это остеобластический костный мозг, который выполняет
остеогенную функцию. В данный период костный мозг накапли-
I зает стволовые клетки, а клетки стромы с остеогенными потенция-
ми создают микросреду, необходимую для дифференцировки ство-
ловых кроветворных клеток. У 12—14-недельного эмбриона че-
ловека происходит развитие кровеносных сосудов и дифференциров-
ка вокруг них гемопоэтических клеток. У 20—28-недельно-
го плода человека в связи с интенсивным разрастанием кост-
но о мозга отмечается усиленная резорбция костных перекладин
остеокластами, в результате чего образуется костномозговой канал,
а красный костный мозг получает возможность расти в направле-
но
нии эпифизов. К этому времени костный мозг начинает функцио-
нировать как основной кроветворный орган, причем большая часть
образующихся в нем клеток относится к эритроидным.
У зародыша в возрасте 36 недель в костном мозге диафиза
трубчатых кос гей обнаруживаются жировые клетки. Одновременно
появляются очаги кроветворения в эпифизах.
Строение. Во взрослом организме человека различают крас-
ный и желтый костный мозг.
Красный костный мозг (medula ossium гиЬгд) яв-
ляется~крОЬёТвормой час!ыи КЪСТНого мозга. Он заполняет губча-
тое" вещество плоских костей~и эпифизовтрубчатых__костей"и во
взрослом организме ^составляет в среднем "бкоЯб^Л—5% общей
массы тела. Красный костный мозг имеет темно-красный цвет и
полужидкую консистенцию, что позволяет легко приготовить из
неТтТ онкйе” мазки на стекле.
Через ретикулярную строму проходит множество крове-
носных сосудов микроциркуляторного русла, между которыми рас-
полагаются гемопоэтические клетки: стволовые, полустволовые
'••орфологически не идентифицируемые}, бласты, проэритроблас-
ты, ^ромиелоциты, метамиелоциты, пролимфоциты, промоноциты,
чего», триобласты, мегакариоциты, и зрелые форменные элементы
крови
Нд пее интенсивно кроветворение происходит вблизи эндос-
та, где \-,.ондентрация стволовых кроветворны* диь-таи. примерно в
в 3 pa-jgy coj ыпе, чем в центре костномозговой полости.
Гемопоэтические клетки располагаются островками. Например,
эритробласты в процессе созревания окружают макрофад. на-
копивший железо используемое эритробластами для построе 11 и я
гем и новой части гемоглобина^ Макрофаги служат своего рода
«кормильцами» для эритробластов, которые с их помощью посте-
пенно, обогащаются железом- Незрелые эритроидные клетки окру-
жены—гликопротеидамй? По мере—созревания клеток количество
гл иксш ратей до в—уменьшается. Одновременно noBbiiiiae тся . п од-
ни ЖН0£Т1, эритроидных клеток и нч F гпгуишчп ц ло,
тредщавленное гемокапиллярами, посткапиллярными сосудами си-
тусоидного типа, или синурами. Стенки синусов состоят из эндо-
телиальных уплощенныхТлеток, пронизанных щелевидными от-
ве рс тиями, или порами, в которые проникают форменные эле мен-
ты крови и плазма (рис. 163). Среди эндотелиальных клеток есть
фиксированные макрофаги.
Гоанулоцитопсэтические клетки также образуют островки, раз-
деляющиеся между собой. Незрелые клетки гранулоцитарных
рядов окружены протеогликанами. В процессе созревания грану-
иты депонируются в красном костном мозге, где их насчиты-
-сялримерно 1 3 разя больше, чем эритроцитов, и в 20 раз
ле, _Ы£м_ гранулоцитов в периферической крови.
Легакариоблас! ы й^ мегакариоциты располагаются в тесном
акте с синусами так, что периферическая часть цитоплазмы
поникает в просвет сосуда через поры. Отделение фрагмен-
411
Рис. 163. Срез красного
костного мозга.
/ адипоциты; 2 — к тетки
гемоцитопоэтических рядов;
3 ретикулярная клетка,
4 — мегакариоцит; 5 — ве-
нозный синус (капилляр си-
нусоидного типа).
гов цитоплазмы в виде тромбоцитов (кровяные пластинки) про-
исходит непосредственно в кровяное русло.
С{уди островков клеток миелоидного ряда встречаются не-
бел ьшие' скопления- -костномозговых—лимфоцитов—и—моноцитов
которые окружают кровеносный сосуд.
В обычных ~фйзиг>ппгицргуиу—утопиях через стенку син’л ов
костного мозга проникают лишь созревшие форменные элементы
крови. Миелоциты и эритробласты попадают в кровь толь о при
патологических состояниях организма. Причины такой избира-
тельной проницаемости стенки сосудов остаются недостаточно
ясными, но факт проникновения незрелых клеток в кровяное рус-
ло всегда служит верным признаком расстройства костномозгово-
го кроветворения.
Вышедшие в кровоток клетки выполняют свои функции либо
в сосудах микроцирКуля'1'upnoi'Q русла-'(эритроциты, тромбоциты),
либо при попадании в соединительную ткань (лейкоциты) и пери-
фе рйчесТСие лимфоидные органы (лимфОЦИТы). В частности, ТГлим-
фоциты мигрируют в тимуснезависимые зоны селезенки и лимфа-
тических узлов, где они клонируются на клетки иммунологической
памяти и клетки, непосредственно дифференцирующиеся в анти-
телопродуценты (плазмоциты) уже при первичном иммунном
ответе.
Желтый костный мозг (medula ossium flava) у взрос-
лых находится в диафизах трубчатых костей. В его составе на-
ходятся многочисленные жировые клетки (адипоциты). Благодаря
наличию в жировых клетках пигментов типа липохромов костный
мозг в диафизах имеет желтый цвет, что и определяет его назва-
ние. В обычных условиях желтый костный мозг не осуществляет
кроветворной функции, но в случае больших кровопотерь или рри
некоторых патологических состояниях организма в нем появляют-
ся очаги миелопоэза за счет дифференцировки приносимых сюда
с кровью стволовых и полустволовых клеток.
Резкой границы между желтым и красным костным мозгом
не существует. Небольшое количество адигоцитов постоянно
412
встречается и в красном костном мозге. Соотношение желтого и
красного костного мозга может меняться в зависимости от воз-
раста, условий питания, нервных, эндокринных и других факто-
ров.
Васкуляризация. Костный мозг снабжается кровью посредст-
вом сосудов, проникающих через надкостницу в специальные от-
верстия в компактном веществе кости. Войдя в костный мозг,
артерии разветвляются на восходящую и нисходящую ветви, от
которых радиально отходят артериолы, которые сначала переходят
в узкие капилляры (2—4 мкм), а затем в области эндоста про-
должаются в широкие тонкостенные со щелевидными порами
синусы (диаметром 10—14 мкм). Из синусов кровь собирается в
центральную венулу. Постоянное зияние синусов и наличие ще-
лей в эндотелиальном пласте обусловливаются тем, что в синусах
гидростатическое давление несколько повышено, так как диаметр
выносящий вены меньше по сравнению с диаметром артерии.
К базальной мембране с наружной стороны прилежат адвентици-
альные клетки, которые, однако, не образуют сплошного слоя, что
создает благоприятные условия для миграции клеток костного
мозга в кровь. Меньшая часть крови проходит со стороны периос-
та в каналы остеонов, а затем в эндост и синус. По мере контакта
с костной тканью кровь обогащается минеральными солями и
регуляторами кроветворения (колониестимулирующие факторы
и др.).
Возрастные изменения. Красный костный мозг в детском воз-
зсте заполняет эпифизы и диафизы трубчатых костей и нахо-
ится в губчатом веществе плоских костей. Примерно в 12—18 лет
распый костный мозг в диафизах замещается желтым. В стар-
иком возрасте костный мозг (желтый и красный) приобретает
(изистую консистенцию и тогда называется желатинозным кост-
(м мозгом. Следует отметить, что этот вид костного мозга может
гречаться и в более раннем возрасте, например при развитии
:тей черепа и лица.
Регенерация. Красный костный мозг обладает высокой физио-
ической и репаративной регенерационной способностью. Йсточ-
ом образования гемопоэтических клеток являются стволовые
тки, находящиеся в тесном взаимодействии с ретикулярной
ромальной тканью. Скорость регенерации костного мозга в зна-
тельной мере связана с микроокружением и специальными рост-
имулирующими факторами гемопоэза (см гл. VII).
ТИМУС (ВИЛОЧКОВАЯ. ИЛИ ЗОБНАЯ, ЖЕЛЕЗА)
Гимус (thymus) центральный орган лимфоиитопоэза и им-
ногенеза. Из костномозговых предшественников Т-лимфоцитов
нем происходит антигеннезависимая___дифференцировка их в
лимфоциты, разновидности которых____осуществляют'реакции
••легочного иммунитета л регулируют реакции гуморального им-
мунитета Гем. гл. VII). Удаление тимуса (тимэктомия) у ново-
Рис.
леза человек 12 вед |ш
вития (прт рвт З.С. Хль
стовой =*• Шмелеве,
И. И. К Иной).
/ соеди ельноткаь
капсула, 2 ретикутяр...
строма;. 3 —| лимфоцит
164. Ви^чковдя ж
рожденных животных вызывает резкое угнетение пр бифераии
лимфоцитов во всех лимфатических узелках кроветвбДЫХ ор '
нов, исчезновениемалых лимфоцитов_и„з_крови, резкое уменьшен*
количества гейкоцитов и другие характерные призна (атроф
органов, кровоизлияния и пр.). При этом организм азывает.
весьма чувствительным ко многим инфекционным за ’Леваниш^
не отторгает.чужеродные трансплантаты органов.
Развитие. Закладка тимуса у человека происходя1 в конц
первого месяца внутриутробного развития в эпитеди глоточно!
кишки, в области главным образом III и IV пар ж< рных карм,
нов в виде тяжей многослойного эпителия. Дистал часть за
чатков III пары, утолщаясь, образует тело тимуса, оксималь
ная вытягивается, подобно выводному протоку экзс\ шной же
дезы. В дальнейшем тимус обособляется от жабернь кармана
Правый и левый зачатки сближаются и срастаются. Р 7-й неде
ле развития в эпителиальной строме тимуса человека
первые лимфоциты. На 8—11-й неделе врастающая
ную закладку органа мезенхима с кровеносными сос
деляет закладку тимуса на дольки. На 11 — 12-й нел
эмбриона человека происходит дифференцировка Л1
на поверхности клеток появляются специфические
Т-антиген (рис. 164). На 3-м месяце происходит ди<’
ка на мозговую и корковую части, причем поел
нее инфильтрируется лимфоцитами и первоначаль
эпителиальная структура зачатка становится труд.
Эпителиальные клетки пласта раздвигаются и оста1
ними друг с другом только межклеточными мостку
тая вид рыхлой сети. В строме мозгового вещее!
своеобразные структуры — так называемые слоисч
ные тельца.
Образующиеся в результате митотического де
штелиаль
1и подраз
развитие
•оцитов, I
^епторы г
ренциров
|я обидь
типична»
злкчимои
^’связан
1риобр»
являютс
ителиа.ъ
Т-лимф.
414
Рис. 165. Схема строения и кровоснабжения дольки вилочковой железы
(рис. Ю. И. Афанасьева и Л. П. Бобовой).
/ соединительнотканная капсула; 2 - корковое вещество; 3 — мозговое вещество
дольки; 4 лимфоциты; 5 — эпителиоретикулонит тимуса; 6 — тельце тимуса; 7 меж-
дольковая волокнистая соединительная ткань; 8 — адипоцит; 9 — междольковая арте-
рия: 10— капиллярная сеть коркового вещества; 11— подкапсулярная вена; 12— капил
лярная сеть мозгового вещества; 13 — междольковая вена.
циты мигрируют затем в закладки лимфатических узлов (в ти-
мусзависимые зоны) и другие периферические лимфоидные
органы.
В функциональном отношении Т-лимфоциты подразделяются
на киллеры, хелперы, супрессоры, которые различаются по набору
антигенов и рецепторов плазмолеммы.
Строение. В основе органа лежит эпителиальная ткань, сос-
тоящая из отростчатых клеток, которая в процессе развития пре-
образовалась так, что базальный сдой эпителия с базальной мем-
браной~ обращен наружу и граничит с окружающей соединитель-
ной тканью. кцпсу той От нее
внутрь от ходяТпе ре го род к и, раздели кнцие тимус, на дольки. В
каждой л ольке различают корковое и мозговое вещества (рис. 165).
К j в о е вещество (cortex олек содержит Т-лим
эторые густо заполняют просветы сетевидного эпитет
лТп >£-aci о остова. В подкапсулярной зоне коркового-вещества нахО
дятс>‘ крупные лимфоидные клетки — лимфобласты — предшест-и
венг.щ чП^лимфоцитов, мигрировавшие сюда из красного костного
мозга” % под влиянием тимозина, выделяемого эпителиоре
тику^ядн^йы клетками стромы и макрофагами, проЛИфе-
рируки Новые генерации лймфоцйтОВ ПОЯВЛЯЮТСЯ в тимусе каж
дые 6 -9 ч. Полагают, что__Т-димфоциты коркового вещества
мигрируют вкроваток^щелхоля в мозговое вещество^тидимфа
^щты оглшаю -ч по составу рецепторов от Т-лимфоцитов моз 'о-
вого вещества. С током крови они понадают в периферические
органы лимфоцитопоэза — лимфатические узлы и селезенку, где
созревают в субклассЫ! ЗнтиГенреактивнъге киллеры, хелперы
супрессоры. Однако не все образующиеся в тимусе лимфоциты
выходят в циркуляторное русло, а лишь те, которые прошли «обу-
чение» и приобрели специфические циторецепторы к чужеродным
антигенам. Лимфоциты, имеющие циторецепторы к собственным
антигенам, как правило, погибают в тимусе, что служит проявле-
нием отбора иммунокомпетентных клеток. При попадании этих
Т-лимфоцитов ТВ кровоток развивается аутоиммунная реакция.
Клетки коркового вещества определенным образом отграниче-
ны от крови гем&тТУМмуснъш 15сттд>сраугппре7гохратгяю1ггим-7ги(^фе-
ренцируюгциеся лимфоциты коркового вещества от избытка анти-
генов, В его состав входят эндотелиальные клетки гемокапил-
ляров с базальной "мембраной, перикапиллярное пространство с
единичными лимфоцитамщ-макгюфагами и межклеточным вещест-
вом, а также эпителиоретикулоциты с их базальной мембраной.
Барьер обладает избирательной проницаемостью по отношению к
анти гену, но если последний и проходит через барьер, концентра-
ция'антигена при этом значительно снижается. При нарушении
барьера среди клеточных элементов коркового вещества обнаружи-
ваются также единичные плазматические клетки, зернистые лейко-
циты и тканевые базофилы. Иногда в корковом веществе появляют
ся очаги экстрамедуллярного миелопоэза.
М оз г о в о е _ в.£-ше с т в о (medulla) дольки на гистологи
ческих препаратах имеет более светлую окраску, так как п<
сравнению с -коркпвым~-Бещёствбм содержит меньшее количеств
лимфоцитов. Лй~м~фбцйты~этой зоны~~представляют собой"рецв
ркулируюгций пул Т-лимфоцитов и могут входить и выхолить и.
кровотПка Чёрёз~посткапиллярные венулы и лимфатические сосуды.
Количество митотически делящихся клеток в мозговом ве-
ществе примерно в 15 раз меньше, чем в корковом. Особенностью
ультрамикроскопического строения отростчатых эпителиоретику-
лоцитов является наличие в цитоплазме гроздевидных вакуолей
и внутриклеточных канальцев, поверхность которых образует
ми к ро В Ы рос+ыГ
В средней части мозгового вещества расположены слоистые
эпителиальные тельца (corpusculum thymicum). Он,, с ~азованы
концентрически наслоенными эпителиоретикулоцитами, цк Лтлаз-
ма которых содержит крупные вакуоли, гранулы кератг и
грубые пучки фибрилл (рис. 166). Количество слоистых Э1/ге-
лиальных телец и их размеры увеличиваюся с возраст®Г~К.ж в
корковом, так и в мозговом веществе часто встречаются макро-
фаги.
Васкуляризация. Внутри органа артерии ветвятся ча междоль-
ковые и внутридольковые, которые образуют дуговые вс тви. От
них почти под прямым углом отходят кровеносные капилляры,
образующие густую сеть, особенно в корковой зонС. Капилляры
коркового вещества окружены непрерывной базальной мембраной
41ь
Рис. 166. Эпителиоретикулоцит мозгового вещества тимуса, х 18 000 (препарат
Л. П. Бобовой).
I —ядро; 2— вакуоли; 3— лизосомы; 4 -митохондрии; 5 — лимфоцит, инвагинирован-
ный в эпителиоретикулоцит.
и слоем эпителиальных клеток, отграничивающим перикапиллярное
пространство. В перикапиллярном пространстве, заполненном тка-
невой жидкостью, встречаются лимфоциты и макрофаги. Большая
часть корковых капилляров переходит непосредственно в подкап-
сулярные венулы. Меныпая часть идет в мозговое вещество и на
границе с корковым веществом переходит в посткапиллярные
венулы, отличающиеся от капсулярных венул высоким призмати-
ческим эндотелием. Через этот эндотелий могут рециркулиро-
вать (уходить из вилочковой железы и вновь возвращаться)
лимфоциты. Б арьера вокруг капилляров в мозговом веществе нет.
Таким образом, отток крови из коркового и мозгового вещества
происходит самостоятельно.
Лимфатическая система представлена глубокой
(паренхиматозной) и поверхностной (капсулярной и подкапсуляр-
ной) выносящей сетью капиляров. Паренхиматозная капиллярная
сеть особенно богата в корковом веществе, а в мозговом капилля-
ры обнаружены вокруг эпителиальных слоистых телец. Лимфати-
ческие капилляры собираются в сосуды междольковых перегоро-
док, идущие вдоль кровеносных сосудов.
Возрастная и акцидентальная инволюция тимуса. Тимус достига-
ет максимального развития в паннем детском возрасте В период от
3 до 20 лет отмечается стабилизация его массы. В более позднее
14 ГИСТОЛОГИЯ
417
время происходит _обратдое_ развитие__(возрастная инв
тимуса. Это сопровождается уменьшением количества лим(
особенно в корковом веществе, появлением липидных вю
в соединительнотканных клетках и развитием жировой
Слоистые эпителиальные тельца сохраняются гораздо
В редких случаях тимус не претерпевает возрастной инь
(status thymicolymphaticus). Обычно это сопровождается дефици
том глюкокортикоидов коры надпочечников. Такие люди отли-
чаются пониженной сопротивляемостью к инфекциям и интокси-
кациям. Особенно увеличивается риск заболеваний опухолями.
Временная, быстрая, или а к и и д е н т а л ь н а я, инволю-
ция может наступить в связи с воздействием на организм рзз-
личнбгх чрезвычайно" сильных раздражителей (травма, интокси-
кация,'ТТнфекция, голодание и др.). При стресс-реакции происхо-
дят выброс Т-лимфоцитов в кровь и массовая гибёлЬ~71 и м фоцитов
в самом органе, особенно в корковом веществе. В связи с этим
становится менее заметной граница корковосо.-и. мозгового^вещест-
ва. Кромё~~лимфоцитолиза, наблюдается фагоцитоз макрофагами
внешне^ не измененных^лимфоцитов. Биологический смысл ли мфо-
цитолиза окончательно не установлен. Вероятно, гибель лимфо-
цитов является выражением селекции Т-лимфоцитов.
Одновременно с гибелью лимфоцитов происходит разрастание
эпителиальной стромы органа. Эпителиоретикулоциты набухают,
в цитоплазме появляются секретоподобные капли,"дающие "поло-
жительную реакцию на гликопротеиды. В некоторых случаях они
скапливаются между клетками, образуя подобие фолликулов.
Тимус вовлекается в стресс-реакции вместе с надпочечниками.
Увеличение в организме количества гормонов коры Надпочечника,
в первукт~очередь~~тлюкокортикоидов, вызывает очень быструю и
сильную акцидентальную_инводю1(ию тимуса.
Таким-образом, функциональное значение тимуса в процессах
кроветворения заключается в образовании тимусзависимых лим-
фоцитов, или Т-лимфоцитов, а также в селекции лимфоцитов,
регуляции пролиферации и дифференцировки в периферических
кроветворных органах благодаря выделяемому органом гормону —
тимозину. Помимо описанных функций, тимус оказывает влияние
на организм, выделяя в кровь и ряд других биологически актив-
ных факторов: инсулиноподобный фактор, понижающий содержа-
ние сахара в крови, кальцитониноподобный фактор, снижающий
концентрацию кальция в крови, и фактор роста.
ЛИМФАТИЧЕСКИЕ УЗЕЛКИ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Лимфатические (лимфоидные) узелки (noduli lymphatici)
в стенке полых органов пищеварительного тракта и дыхательных
путей осуществляют локальную иммунную защиту и, возможно,
являются местом приобретения В-лимфоцитами костномозгового
происхождения рецепторов для распознания антигенов, попадаю-
щих в эти органы из внешней среды (аналог бурсы Фабрициуса,
418
или фабрициевой сумки). Располагающиеся здесь Т-лимфоциты
осуществляют реакции клеточного иммунитета и регулируют
деятельность В-лимфоцитов. Лимфоциты могут располагаться
между эпителиоцитами и на их поверхности, в рыхлой соедини-
тельной ткани под эпителием, где В-лимфоциты образуют лимфа-
тические узелки. Число лимфатических узелков и выраженность
в них герминативных центров непостоянны и зависят от иммунных
реакций.
Активизированные антигеном В-лимфоциты после деления в
лимфатических узелках выходят за их пределы. Часть из них
через стенку посткапиллярных венул мигрирует в кровоток.
Вернувшись после циркуляции обратно в слизистую оболочку этих
органов, они заканчивают свою дифференцировку в плазмоциты,
которые вырабатывают белковую часть молекул антител — имму-
ноглобулинов класса А. Углеводный компонент иммуноглобулинов
выделяют эпителиоциты. Сборка молекулы иммуноглобулина проис-
ходит в слизи на поверхности эпителиоцитов, где они осущест-
вляют местную антибактериальную и противовирусную защиту.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Лимфатические узлы (noduli lymphatici) располагаются по
ходу лимфатических сосудов, являются органами лимфоцито-
поэза? иммунологической защиты, и депонирования протекающей
лимфы. Это округлые или овальные весьма многочисленные
образования^ размером около 0,5—1 см. Обычно лимфатические
узлы с одной стороны имеют вдяпприир (рис. 167). В этом месте,
называемом воротами, в узел входят артерии и н£рвьц~а~Т1Бгходят
вены и выносящие-лимфатические сосуды. Сосуды, ипиносяшие
лимфу, ВХОДЯТ С ПрОТИВОПОДОЛШОЙ^—выпуклой СТОДОШЛ узла.
Благодаря такому распопошемию узла по_ходу__лимфатических
сосудов_он является—не -столько—кроветворным органом, но и
своеобразным фильтром для оттекающей от тканей жидкости
(лимфы) на пути в кровяное русло. Протекая через—димфати-
ческие узлы, лимфа очищается от инородных частиц, микроорга-
низмов и обогащается лимфоцитами и антителами.
Развитие. Лимфатические узлы впервые возникают в конце
2-го — начале 3-го месяца внутриутробного развития плода
человека. Их образование связано с размножением и накопле-
нием в определенной области вокруг кровеносных и лимфати-
ческих сосудов мезенхимных клеток. Разрастающиеся лимфати-
ческие щели в области закладки будущего узла сливаются и
образуют так называемый подкапсулярный (краевой) синус.
По периферии зачатка узла в это же время из мезенхимы
формируются поверхностная капсула и отходящие от нее внутрь
перегородки — трабекулы. От краевого синуса внутрь узла между
трабекулами отходят многочисленные анастомозирующие меж-
ду собой вокругузелковые и мозговые синусы. Эти синусы в свою
очередь разделяют мезенхимную ткань, превращающуюся в рети-
14'
414
Рис. 167. Схема строения и кровоснабжения лимфатического узла (рис. Ю. И. Афа-
насьева).
А - краевой синус; Б — посткапиллярная венула; I — соединительнотканная капсула;
2 — трабекула; 3 — приносящие лимфатические сосуды, 4 — подкапсульный (краевой)
синус; 5 — ретикулоэндотелиальные клетки (береговые клетки); 6— вокругузелковый
синус; 7 — мозговые синусы; 8 — ворота лимфатического узла; 9 — выносящий лимфа-
тический сосуд; 10 — ретикулярные клетки; 11 — лимфатические узелки; 12 — мозговые
тяжи; 13 — ретикулярные волокна; 14 — артерия лимфатического узла; 15 — вена лим-
фатического узла; 16 — трабекулярная артерия; 17 — артерии мякотных тяжей; 18а —
поверхностные и 186 — глубокие гемокапиллярные сети; /9 — вены мозговых тяжей;
20 — трабекулярная вена; 21 — макрофаги в синусах; 22 — лимфоциты и плазматические
клетки; 23 — эндотелий; 24 — щели между эндотелиальными клетками; 25 - лимфоцит,
проникающий в щель; 26 базальная мембрана; 27 — пара кортикальная зона.
кулярную, на овальные скопления и тяжи, которые заселяются
кроветворными клетками, и на 16-й неделе образуют лимфа-
тические узелки и мозговые тяжи. Одновременно появляются
ретикулярные волокна. В-лимфоциты вселяются раньше Т-лимфо-
цитов и главным образом в центральные участки лимфатического
узла (будущее мозговое вещество), а затем в самый поверхност-
ный (в дальнейшем — корковое вещество) слой. Т-лимфоциты все-
ляются в промежуточную зону между корковым и мозговым вещест-
вом (Т-зона).
Начиная с 16-й недели развития в строме закладки лимфати-
ческого узла нарастает количество макрофагов.
Входящие в узел лимфатические сосуды становятся прино-
сящими сосудами, а выходящие из ворот — выносящими.
В конце 5-го месяца эмбрионального развития лимфатические
узлы приобретают черты дефинитивного кроветворного органа.
Строение. Несмотря на многочисленность лимфатических узлов
и вариации органного строения, они имеют общие принципы
организации. Снаружи узел покрыт соединительнотканной капсу-
лой^ несколько толщенной~р~бб7Т5ГТИ ворот (см. рис. 167У. В кап-
суле много коллагеновых и мало эластических волокон. Кроме
соединиТелЬйбтканных элементов, в ней главным^образом в облас-
мтг.г«
ти ворот рас но за такт-я шшглкыки;—пучки—гладких_мышечных
клеток, особенно в узлах нижней половины туловища. Внутрь от
кап суды через относительно правильные промежутки отх одя т тон-
кие соединительнотканные перегородки, или трабекулы, аПЗСТОмо-
зирующйе между собой в глубоких частях узла. В совокупности они
составляют примерно */4 площади среза органа. На срезах узла,
проведенных через его илротд можно различить периферическое,
более плотное корковое вещество, состоящее из лимфатических
узелков, паракортикальную (диффузную) зону, а также централь-
ное светлое мозговое .вещество, образованное мозговыми тяжами
и синусами. Большая часть кортикального слоя и мозговые “Тяжи
составляют область заселения В-лимфоцитов (В-зона), а наракор-
тикальная, _Тймусзавйсймая “зона содержит преимущественно
Т-лимфоциты (Т-зона).
Корковое вещество
Лимфатические узелки. Лимфатические узелки (noduli lyinpha-
tici) представляют собой округлые образования диаметром около
0,5—1 мм (рис. 168, А, Б).
В ретикулярном остове узелков—проходят толстые, грубые,
извилистые, ретикулярные волокна, в основном циркулярно на-
правленные. В петлях ретикулярной ткани залегают лимфоциты,
лимфобласты, макрофаги ~и другие клетки. В периферической
Чс5сТИ узелков находятся малые лимфоциты в виде короны.
Лимфатические узелки покрыты ретикулоэндотелиальными
клетками, лежащими—на ретикулярных .волокнах. Среди рети-
кулоэндотелиальных клеток много фиксированных макрофагов
421 1
2
Рис. 168. Корковое (Л) и
мозговое (Б) вещество
лимфатического узла.
Л: / — капсула; 2 — прино-
сящий лимфатический сосуд;
3 — кровеносные капилляры;
4 — ретикулярные клетки в
краевом синусе; 5 — лимфа-
тический узелок; 6 — центр
размножения (реактивный
центр). Б: Z — мозговой тяж:
2 — ретикулярные клетки в
мозговом синусе; 3 — трабе-
кула; 4 — артериола; 5 — ве-
нула-
(«береговые макрофаги»). Центральная—часть узелков-обычно вы-
глядит светлой вследствие того, что она состоит из более крупных
клеток с большими светлыми ядрами: из лимфобластов, типичных
макрофагов и ретикулоподобных «дендритных—клеток», лимфоци-
„ тов..Лимфобласты обычно находятся в различных стадиях деления,
вследствие чего эту часть узелкаГ называют герминативным центром
(centrum germinale), или центром размножения. При интоксикации
организма, особенно микробного_происхожденця^ в центральной
части узелка могут появляться скопления фагоцитирующих клеток,
что указывает на высокую реактивность описываемых структур.
Поэтому данную часть узелка часто называют еще реактивным
центром.
Типичные свободные макрофаги преобразуют корпускулярный
антиген в молекулярный и концентрируют его _до количества,
способного побудить к пролиферации и дифференцировке распсло-
жен ные_рядом В-лимфопиты при участии Т-хелперов. В результа-
те этого образуются клетки памяти Т- и В-типа и иммунббласты.
Активированные антигеном В-лимфоциты по мере размножения
422
Рис. 169. Четыре стадии изменения строения лимфатических узелков.
А — начальная стадия: Б — образование центра размножения; В — формирование темной
зоны, состоящей из малых лимфоцитов вокруг светлого центра; Г — светлый (реактив
ный) центр, состоящий из ретикулярных клеток и макрофагов (по Е. Конвей).
и созревания образуют В-зону, откуда мигрируют__в мозговые
тяжи/где превращаются в плазмоциты и продуцируют антитела.
К петуи памяти с током лимфы или через посткапиллярные вены
вступают в циркуляцию и будут созревать в эффекторные клетки
после вторичной встречи с_антигеном. Макрофаги светлых центров
могут фагоцитировать также погибающие клетки, в результате чего
в их цитоплазме обнаруживаются хромофильные остаточные
тельца.
Отростчатые («дендритные») клетки реактивных центров яв-
ляются разновидностью макрофагов, способных с помощью рецеп-
торов цитолеммы к фиксации иммуноглобулинов а через них и ан-
423
тяге нов, вызвавших иммунный ответ организма. Накопленные на
их поверхности антигены активируют и вовлекают в иммунную ре-
акцию контактирующие с ними В-лимфоциты. Морфологически
«дендритные» клетки характеризуются отростчатой формой, элек-
тронно-прозрачной цитоплазмой, бедной рибосомами, лизосомами
и канальцами цитоплазматической сети. Полагают, что эти клетки
характерны для В-зон лимфатических узелков. Длительная за-
держка антигенов на поверхности дендритных клеток и наличие
клеток памяти обеспечивают более быстрый иммунный ответ при
повторной встрече с тем же антигеном.
Строение лимфатических узелков может меняться в зависи-
мости от физиологического состояния организма (рис. > 169).
Различают 4 стадии, отражающие происходящие в них процессы.
В I стадии — формирование центра размножения — в лимфатиче-
ском узелке имеется небольшой центр, состоящий преимущест-
венно из малодифференцированных клеток лимфоцитопоэтиче-
ского ряда. Некоторые из этих клеток могут быть в состоянии
митотического деления. Во II стадии у лимфатических узелков
центры крупнее и содержат большое количество митотически
делящихся клеток лимфоцитопоэтического ряда (от 10 и более
на срезе). Центральная часть узелка выглядит светлой. В III ста-
дии вокруг светлых центров появляется корона из малых лимфо-
цитов. Уменьшается число митотически делящихся клеток и коли-
чество молодых клеток лимфоцитопоэтического ряда. В IV стадии
в центре узелка фигуры митозов и макрофаги единичны. Вокруг
узелка корона из малых лимфоцитов состоит преимущественно
из клеток В-памяти. Это стадия относительного покоя.
Возникновение и исчезновение центров происходит в течение
2—3 сут.
Паракортикальная зона
На границе между корковым и мозговым веществом располага-
ется nhHnu тимусчависимая зона (paracortex). Она со-
держит главным образом Т-лимфоциты. Микроокружением для
лимфоцитов цдракпртмь a lib.ты* »1UU им дсятг и НИ Й7й1ГТЬ МЭК-
рофагов,—по.тграьши1—сиособноь'ть—и—янноч^иги-
тирукмцие -клетки», которые обладают многочисленными пальце-
видными отростками, вдавливающимися из одной клеткй'в другую.
Ядра интердигитирующих клеток неправильной формы, светлые,
с краевым расположением хроматина. В слабобазофильной цито-
плазме обнаруживаются везикулы, комплекс Гольджи, гладкая
эндоплазматическая сеть. Фагосомы встреч? отся редко. Полагают,
что эти клетки вырабатывают гликопротеиды, которые играют
роль гуморальных факторов лимфоцитогенеза. Гликопротеиды
примембранных слоев способны сорбировать и сохранять антиген
на цитоплазматических мембранах и индуцировать пролифера-
цию Т-лимфоцитов.
В паракортикальной зоне происходит пролиферация Т- леток,
424
бласттрансформация и^ифФерениирпиУй Р клетки
(клетки-киллеры и др.). Посткапиллярные венулы паракортикаль-
ной зоны являются местом проникновения в лимфатический узел
циркулирующих Т- и В-лимфоцитов. В некоторых случаях при
разрастании паракортикальной зоны лимфатические узелки слива-
ются в плато.
Мозговое вещество
От узелков и паракортикальной зоны внутрь узла, в^егс моз-
говое вещество, отходят мозговые тяжи (chordae medullana),
анастомозирующие между собой (см. рис. 168, Б). В_рснове их
лежит ретикулярная ткань, в петлях которой находятся В-лимфо-
циты, плазматичеТкиёТОгетки и макрофаги. Здесь происходят про-
лиферация и созревайиё плазматических клеток. Большая часть
иммуноглобулинов, образуемых щесь плазматическимй~клётками,
относится к классу иммуноглобулинов G (см. гл. IX). Внутри
мозговых ~ тяжей проходят ^кровеносные ~ сосуды и_____капилляры,
содержащие поры в эндотелии. Снаружи тяжи, так же как и
лимфатические узелки, покрытьг^ндотетптотТодЬбйымгГрётикуляр-
ными клетками^ лежащими на “пучках ретикулярных фибрилл
и образующими стенку синусов.
Пространства, ограниченные капсулой и трабекулами, с одной
стороны, и узелками и мозговыми тяжами — с другой, называются
синусами, являющимися как бы продолжением приносящих
лимфатических сосудов. Различают под капсульный, или краевой
сцдус— (sinus subcapsularis"^ располагающийся между капсулой
и узелками, «округузелуопые синусы (sinus corticalis perinodularis),
проходящие между узелками и трабекулами, мозговые синусы
(sinus-Hit'dullaiis), uiраничгнные грабёкулймй и мозговыми тяжами
(см. рис. 167).
Наружные клетки подкапсульного синуса, прилежащие к кап-
суле узла, расположены на базальной мембране. По строению и
функции они близки к эндотелиальным клеткам, выстилающим
приносящие лимфатические сосуды. Среди этих клеток встречают-
ся фагоцитирующие — макрофаги. Внутренние эндотелиоподобные
ретикулярные клетки, покрывающие лимфатические узелки кор-
кового вещества, не имеют базальной мембраны, а лежат на
пластинке ретикулярных фибрилл. Между клетками обнаружива-
ются щели, через которые в просвет синуса проникают лимфо-
циты. Клетки, выстилающие все остальные синусы, имеют ана
логичное строение.
М озговые тяжи вместе с окружающими их трабекула ми и
синусами образуют мозговое вещество (medulla).
По синусам коркового и мозгового вещества протекает лим-
фа. При этом она обогащается ли
ми
которые поступают
в нее в большем или меньшем количестве из узелков, паракорти-
кальнои. зоны и мозговых яжеи. Среди свободных клеточных
элементов в синусах при различных состояниях организма можно
425
обнаружить лимфоциты, плазмоциты, свободные макрофаги;
встречаются единичные зернистые лейкоциты и эритроциты. Сину-
сы выполняют роль защитных фильтров, в которых благодаря
наличию фагоцитирующих клеток задерживается большая часть
попадающих в лимфатические узлы антигенов.
Лимфатические узлы "очень чувствительны к различным внеш-
ним и внутренним факторам. Например, под действием ионизирую-
щей радиации быстро погибают лимфоциты в лимфатических
узелках, в мозговых тяжах. При недостаточной функции гормонов
коры надпочечников, наоборот, происходит разрастание лимфоид-
ной ткани во всех органах (status thymicolymphaticus).
Васкуляризация, Кровеносные сосуды проникают в лимфати-
ческие узлы через их ворота. После вхождения в узел одна часть
артерий распадается на капилляры в капсуле и трабекулах, дру-
гая заканчивается в узелках, паракортикалькой зоне и мозговых
тяжах. Некоторые артерии проходят сквозь узел не разветвляясь
(транзитные артерии).
В узелках различают две гемокапиллярные сети — поверхност-
ную и глубокую. От гемокапилляров начинается венозная система
узла, которая совершает обратный ход преимущественно отдельно
от артерий. Эндотелий посткапиллярных венул более высокий,
чем в обычных капиллярах, а между эндотелиальными клетками
имеются поры. Особенности строения эндотелия играют
определенную роль в процессах рециркуляции лимфоцитов из
кровотока в узел и обратно. В обычных физиологических усло-
виях кровь из сосудов узла не изливается в его синусы. Однако
при воспалительных процессах в синусах регионарных лимфатиче-
ских узлов часто обнаруживаются эритроциты.
Иннервация. Лимфатические узлы имеют афферентную и эффе-
рентную адренергическую и холинергическую иннервацию. В подхо-
дящих к органу нервах, а также в капсуле обнаружены интра-
муральные нервные узлы. Рецепторный аппарат хорошо выражен
во всех макромикроскопических структурах: капсуле, трабекулах,
сосудах, корковом и мозговом веществе. Имеются свободные и
несвободные нервные окончания. Внутри узелков нервные оконча-
ния не обнаружены.
Возрастные изменения. В течение первых трех лет после
рождения у ребенка происходит окончательное формирование
лимфатических узлов. Герминативные центры в узелках появляют-
ся по мере иммунизации и становления кроветворной и защитной
функции. В старческом возрасте центры снова исчезают. При
этом капсула узлов утолщается, количество трабекул возрастает,
фагоцитарная активность макрофагов постепенно ослабевает.
Некоторые узлы могут подвергаться атрофии и замещаться жиро-
вой тканью.
Регенерация. Регенерация лимфатических узлов (частичная
или полная) возможна лишь при сохранении приносящих и выно-
сящих лимфатических сосудов и прилежащей к узлу соедини-
тельной ткани. В случае частичной резекции лимфатического
426
узла репаративная регенерация его происходит через 2—3 нед
после повреждения. Восстановление начинается с пролиферации
клеток ретикулярной ткани, затем появляются очаги лимфоидного
кроветворения и образуются узелки. При полном удалении лимфа-
тического узла, но при сохранении лимфатических сосудов регене-
рация этого органа начинается с появления большого количества
очагов лимфоидного кроветворения, которые возникают из ство-
ловых кроветворных клеток. При этом приносящие и выносящие
лимфатические сосуды анастомозируют между собой в области
лимфоидного очага. В результате дальнейших преобразований анас-
томозы сосудов оказываются погруженными внутрь лимфоидного
очага и превращаются в синусы узла.
ГЕМОЛИМФАТИЧЕСКИЕ УЗЛЫ
Кроме обычных лимфатических узлов, у жвачных и млекопи-
тающих встречаются гемолимфатические узлы (nodus lymphaticus
haemalis), синусы которых содержат кровь. У человека такие узлы
бывают редко. Обычно они располагаются в околопочечной
клетчатке вдоль почечных артерий или по ходу брюшной аорты,
реже — в заднем средостении.
Развитие. Развитие гемолимфатических узлов весьма сходно
с развитием обычных лимфатических узлов (см. выше), но гемо,-
лимфатические узлы относительно долго сохраняют способность
к миелопоэзу (до рождения, а иногда и в течение нескольких лет
в постнатальном периоде).
Строение. По величине гемолимфатические узлы, как правило,
значительно меньше лимфатических. Снаружи они покрыты сое-
динительнотканной капсулой, нередко содержащей пучки гладких
мышечных клеток (рис. 170). Корковое вещество меньшего объ-
ема, лимфатических узелков немного; мозговые тяжи тоньше
и малочисленнее. Синусы гемолимфатических узлов, особенно
мозговые, бывают относительно широкими. Благодаря значитель-
ной примеси крови синусы не всегда легко отличаются от вен,
проходящих в мозговых тяжах. Критерием служат ретикулярные
клетки и ретикулярные волокна в просвете сосудов. Вопрос о
наличии соустьев между лимфатическими сосудами и венами гемо-
лимфатических узлов остается спорным. Гемолимфатические узлы
вырабатывают форменные элементы крови не только лимфоидного,
но и миелоидного ряда.
У эмбрионов и в раннем постнатальном периоде в гемолимфа-
тических узлах, кроме клеток лимфоидного ряда, составляющих
большинство клеточных элементов, обнаруживаются промиелоци-
ты, миелоциты и метамиелоциты, особенно эозинофильные, про-
эритроциты, нормоциты и даже мегакариоциты. Кровь, находящая-
ся в синусах, частично вымывается лимфой, частично подвергается
разрушению: эритроциты и их фрагменты фагоцитируются макро-
фагами, в цитоплазме которых всегда обнаруживается железо-
содержащий пигмент — гемосидерин. Истинные гемолимфатиче-
Рис. 170. Схема строения гемолимфатического узла (рис В А. Флоренсова).
/ — корковое вещество; II мозговое вещество, / — приносящий лимфатический сосуд;
2 — капсула узла; 5 — краевой синус' 4 — ретикулярные клетки в корковом веществе; 5
лимфобласт; 6— макрофаги; 7 — миелоцит; 8— ретикулярная клетка синуса; 9 — эри-
троциты в синусе; 10 — трабекула; II — артерия, 12 — вена; 13 — выносящий лимфатиче
ский сосуд, 14 — ворота узла, 15 — макрофаги, поглощающие эритроциты; 16 плазма-
тические клетки: 17 мегакариоцит; 18 межфолликулярный синус; 19 митотическое
деление лимфобластов и лимфоцитов; 20 малые лимфоциты; 21 — трабекулы. 22 — кро-
веносные капилляры.
ские узлы важно отличать от ложных, которые могут образовывать-
ся в результате всасывания крови лимфатическими сосудами из раз-
личных очагов кровоизлияний, в связи с чем она обнаруживается
в краевом синусе и приносящих лимфатических сосудах. В отли-
чие от добавочных селезенок гемолимфатические узлы имеют
приносящие лимфатические сосуды, а в просвете синусов встре-
чаются ретикулярные клетки. Кроме того, в добавочных селезен-
ках находятся специфические для селезенки структуры (централь-
ные артерии фолликулов, артериальные гильзы, венозные синусы),
чего нет в гемолимфатических узлах.
Возрастные изменения. С возрастом гемолимфатические узлы
подвергаются инволюции. Корковое и мозговое вещества замеща-
ются жировой тканью или прорастают рыхлой волокнистой соеди-
нительной тканью.
СЕЛЕЗЕНКА
Селезенка (splen, lien) — важный кроветворный и защитный
орган, принимающий участие как ~в~элйми нации ’ОТЖИватоЩих или
п о врежденных эритроцитов и тромбоцитов, так~~й в орган и за ци и
защитных реакций от экзогенных антигенов, которые не были
задержаны лимфатическими узлами"и Тфоникли в кровоток. Селе-
зенка играет активную роль в образовании гуморального и клеточ-
ного иммунитета, а также в выработке веществ,—у гнегающих эри-
тропоэз в красном костном мозге. Объем и масса этого органа
сильно варьируют в зависимости от депонирования крови и актив-
ности процессов кроветворения.
Развитие. У человека селезенка закладывается на 5-й неделе
эмбрионального периода развития в толще мезенхимы дорсальной
брыжейки. В начале развития селезенка представляет собой плот-
ное скопление мезенхимных клеток, пронизанное первичными
кровеносными сосудами. В дальнейшем часть клеток дифференци-
руется в ретикулярную ткань, которая заселяется стволовыми
клетками. На 7—8-й неделе развития в селезенке появляются
макрофаги. На 12-й неделе развития селезенки человека впервые
появляются В-лимфоциты с иммуноглобулиновыми рецепторами
Процессы миелопоэза в селезенке человека достигают максималь-
ного развития на 5-м месяце внутриутробного периода, после
чего активность их снижается и к моменту рождения прекращает-
ся совсем. Основную функцию миелопоэза к этому времени выпол-
няет красный костный мозг. Процессы лимфоцитопоэза в селе-
зенке к моменту рождения, наоборот, усиливаются.
На_ 3-м месяце эмбрионального развития в сосудистом русле
зенки появляются широкие ведозшя ину_сы, разделяющие ее
на островки. Вначале ост овки кроветворных клетоц_ располага-
Тотся равномерно вокрут артерии (Т-зона), а на 5-м месяце начи-
нается концентрация лимфоцитов и макрофагов сбоку от нее
(В-зоиаУ7 К этому времени популяция В-лимфоцитов, выявляемая
при помощи иммунологических методов, примерно в 3 раза превы-
429
Рис. 171. Схема строения селезенки (рис. Ю. И. Афанасьева).
А — опорно-сократительный аппарат селезенки—капсула и трабекулы; Б—кровообра-
щение в селезенке; В — гистоструктура селезенки; / — капсула; 2 — мезотелий; 3 — тра-
бекулы селезенки; 4 — селезеночная артерия; 5 — трабекулярная артерия; 6 — пульпарная
артерия; 7 — центральная артерия; 7а — гемокапилляры а лимфатическом узелке; 76 —
краевой синус; 8 — кисточковые артерии; 9 — гильзы; 10 — капилляр, свободно открываю-
щийся в пульпу (по теории открытого кровообращения); 1 I — капиляр, переходящий
в венозный синус (по теории закрытого кровообращения); 12 — трабекулярная вена:
13 — селезеночная вена; 14 — лимфатическое периартериальное влагалище; 15 — лимфа-
тические узелки (белая пульпа); 16 — красная пульпа; 17 — венозные синусы; 18 — рети-
кулярная ткань; 19— эритроциты и лейкоциты в красной пульпе; 20 — щели в эндотелии
синуса; 21 — ядра эндотелиальных клеток; 22 — аргирофильные волокна.
шает популяцию Т-лимфоцитов. Одновременно с развитием узелков
происходит формирование красной пульпы, которая становится
морфологически различимой на 6-м месяце внутриутробного раз-
вития.
Строение. Селезенка человека покрыта соединительнотканной
капсулой и брюшиной. Толщина капсулы неодинакова в различных
участках, селезенки. Наиболее толстая капсула в воротах селе-
зенки, через_ которые проходят кровеносные-и ““лимфатические
сосуды. Капсула состоит из плотной волокнистой соединительной
ткани, содержащей фибробласты и многочисленные коллагеновые
и эластические волокна. Между волокнами залегает небольшое
количество гладких мышечных клеток.
Внутрь от капсулы отходят перекладины — трабекулы селезен-
ки, которые вТлубоких частях органа анастомозируют между
собой, (рис. 171). Капсула и трабекулы в селезенке человека зани-
мают примерно 5—7% от общего объема органа и составляют
его опорно-сократительный аппарат. В трабекулах селезенки че-
ловека сравнительно немного гладких мышечных клеток. Эласти-
ческие волокна в трабекулах более многочисленны, чем в капсуле.
В селезенке различают белущ__н-красную пульпу (pulpa alba et
rubra lienis). B_ основе пульпы селезенки лежит ретикулярная
ткань,образую щая ее строму. Строение селезенки и соотношение
между белой и красной пульпой могут изменяться в зависимости
от функционального состояния органа.
Белая пульпа селезенки (pulpa alba). Препстапт<з?т собой сово-
купность лимфоидной те а нщ расположенной в адвентиции ее. арте-
рий. щдиде шаровидных скоплений^или узелков, и лимфатических
периартериальных влагалищ. В совокупности они составляют
примерно ’Д органа.
Лимфатические узелки селезенки (lymphonoduli splenici) 0,3—
0,5 мм в диаметре представляют собою скопления Т- и В-лимфо-
цитов, плазмоцитов ~>Т~макрофагбвТл~ петлях ретикулярной ткани
(дендритных клеток), окруженные капсулой из уплощенных рети-
кулярных клеток Через лимфатиписнкий узЩПЯ^^фж^лдИт. обычно
эксцентрично, аруерич учеика (a. lymphonoduli),-от которой QJ
ходят падиачцю капилляры (рис. 172).
В лимфатических узелках различают 4 нечетко разграниченные
зоньц,. париартериальную, центр размножения, мантийную и кра
евую, или маргинальную, зону. " '
ГТ ериартериальная зона занимает небольшой уча-
сток. узелка около артерии и образована главным образом из
Т-лимфоцисов, попадающих сюда через “гемокапилляры, отходя-
щих от артерии лимфатического узелка, и интердигитирующих
клеток. Субмикроскопические отррстки ithv к веток вытягиваются
на 1ГТ1П~П1ТГП|ИПВ JlflllTiW >»>ЛУ ЛИружмМЦИИМ МТ___лимфоци-
тами И ПЛОТНО С ними контактируют Полагают. ЧТО эти клс тки
адсорбируют антигены, поступающие сюда с кровотоком, и пере-
дают Т-лимфоцитам информацию о Состоянии микроокруже ни я,
стимулируя hy бласттрансформяпию и пролиферацию В течение
431
Рис. 172. Селезенка че-
ловека (препарат
Ю. И. Афанасьева).
/ — лимфатический узе-
лок (белая пульпа); 2—
центральная артерия; 3 —
красная пульпа; 4 — тра-
бекула; 5 — трабекуляр-
ная артерия.
1—3 сут активированные Д^-лцмФоциты остаются в этой—зоне
и размножаются. В дальнейшем они мигрируют из лериартериа ль-
ной зоны в синусы краевой зоны через гемокапилляры. Тем же
путем попадают в селезенку и В-лимфоцйты. Причина заселения
Т- и В-лимфоцитами «своих»’ зон недостаточно ясна.ТГфункцио-
нальном отношении периартериальная зона является анало-
гом паракортикальной тимусзависимой зоны лимфатических
узлов.
Центр размножен и я, или герминативный центр узел-
ка, состоит из ретикулярных пр»дифернру-юнньх-И-лим-
фобластов, дифференцирующихся антителообразующих клеток.
Кроме того, здесь нередко можно обнаружить скопления макро-
фагопитов С фагоцитированными лимфоцитами или их фрагмен-
тами-^ виде хромофильных телец и дендритные клетки. В этих
случаях центральная часть узелка выглядит светлой («реактивный
центр»).
На границе со следующей, мантийной зоной обнаруживают-
ся дифференцирующиеся плазмоциты. В функциональном отно-
шении эта область идентична герминативным центрам узелков
лимфатических узлов.
Мантийная зона окружает периартериальную зону и
центр размножения, состоит главным образом из плотно располо-
женных малых В-лимфоцитов~и небольшого количества Г-лимфо-
цитов, а также содержит плазмоциты и макррфаги. Прилегая
плотно друг к другу, клетки образуют как бы корону, расслоен-
ную циркулярно направленными толстыми ретикулярными во-
локнами.
К р а е tya я, или маргинальная, зо_ н а узелков селезен-
ки представляет собой переходную область -между-белой и красной
пульпой шириной около 100 мкм. Она состоит преимущественно
из Т- и В-лимфоцитов и единичных макррфагов, окружена краевы-
ЛШГ~иЛи маргинальными, синусоидными сосудами с щелевилными
порами в стенке.
432
Периартериальные лимфатические влагалища (vagina репаг-
terialis lymphatica) представляют собою вытянутые по ходу пуль-
парной артерии скопления В-лимфоцитов^ТГлазматических клеток,
а по периферии влагалища — малые "Т-лимфоциты.
Красная пульпа Селезенки (pilpa lienis rubra). Состоит из
ретикулярной ткани ~с~ расположенными в ней клеточными~эле-
ментами крови, придающими ей красный цвет, и многочисленными
кровеносными сосудами, главным образом синусоидною типа.
Частькрасной пульпы, расположенная между синусами, назы-
вается селезеночными, или пульпарными, тяжами (chordae
splenicae). Здесь по аналогии с мозговыми тяжами~лймфатических
узлов обнаруживаются очаги плазмоцитогенеза. В красной пульпе
задерживаются моноциты, которые дифференцируются в макро-
фаги.
Селезенка считается «кладбищем эритроцитов» в связи с тем,
что обладает спосо6ностью~понйжать осмотическую устойчивогть
старых илй Поврежденных эритроцитов. Это приводит эритроциты
к гибели. Такие эритроциты поглощаются макрофагами красной
пульпы. В результате расщепления гемоглобина поглощенных
ма_крофагами~эритроцйтов~образуЮТСЗГ и~выделяются в кровоток
билирубин И содержащий железо трансферрин. Билирубин пере-
носится в печень, где войдет в состав желчи. Трансферрин из
кровотока захватывается макрофагами костного мозга, ^которые
снабжают железом вновь^азвивающйёся'Дрйтроциты.
В селезенке депонируется кровь и скапливаются тромбоциты.
Старые тромбоциты подвергаются здесь разрушению.
Синусы красной пульпы, расположенные между селезеночными
тяжами, представляют собой часть сложной сосудистой системы
селезенки, в связи с чем их следует рассмотреть отдельно.
к Васкуляризация. J3 ворота селезенки входит селезеночная
артерия, которая разветвляется на трабекулярные артерии. На-
ружная~1оболочка артерий рыхло соединена -е—тканью трабекул
(см. рис. 17Г). Средняя оболочка четко заметна на любом срезе
трабекулярной артерии благодаря мышечным пучкам, идущим
в составе ее стенки по спирали. От трабекулярных артерий отходят
пульпарные артерии. В наружной оболочке этих артерий много
спирально расположенных эластических волокон, которые обеспе-
чивают продольное растяжение и сокращение сосудов. Недалеко
от трабекул в адвентиции пульпарных артерий появляются
периартериальные тимфатические влагалииуа и лимфатические
узелки (см. выше).
Артерияг—проходящая через -узелок (a. lymphonoduli), отдает
несколько гемокапилдяров (см. выше) и, выйдя из узелка, разветв-
ляется в виде кисточки на нескопькгсартериоп (arteriolae penicil-
laris). Дистальный конец 4зтпй..артериппы продолжается Я—Эллип-
соидную (гильзовую) артериолу (arteriola elipsoideae), снабжен-
ную_муфтой («гильзой») из ретикулярных клеток и волоконГ'Это
своеобразный сфинктер__на артериоле - У человека эТИТтгльзы
развиты очень елабо. В~эвдотелитг гильзовых—или -эллипсоидных
433
Рис. 173. Синус селезенки (по Судзуки).
I — эндотелий; 2 - щели в стенке синусоидного капилляра; 3 — эритроцит; 4 — лейкоцит;
5 - макрофаг в красной пульпе.
артериол обнаружены сократительные филаменты. Далее следуют
короткие артериальные гемокапилляры. большая часть капилляров
красной пульпы впадает в венозные синусы (за к р ы т о е к р о-
вообраще ние), однако некоторые могут непосредственно
открыва-|ьс>г*~П~ ре тикулярную ткань (открытое кровообращение).
Закрытое кровообращение — путь быстрой циркуляции и оксиге-
нации тканей. Открытое кровообращение — более медленное, обес-
печивающее контакт форменных элементов крови с макрофагами.
Синусы являются началом венозной системы селезенки. Их ди-
аметр колеблется от 12 до 40 мкм в зависимОСТй~от"Кровенапол-
нения. При расширении совокупность всех синусов занимает боль-
шую часть селезенки. Эндотелиоциты синусов расположены на
прерывистой базальной мембране(рис. 173). По поверхности
стенки синусов в виде колец залегают ретикулярные волокна.
Синусы не имеют перицитов. В стенке синусов в месте их перехода
в веньцзмеются подобия мышечных сфинктеров. При открытых
артериальных и венозных сфинктерах кровь свободно проходит
по синусам в вены. Сокращение венозного сфинктера приводит
к накоплению крови в синусе. Плазма крови пронйкаёт~сквозь
стенку синуса, что способствует концентрации в нем клеточных
элементов. В случае закрытия венозного и артериального сфинк-
теров кровь депонируется в селезенке. При растяжении синусов
между эндотелиальными клетками образуются щели, через кото-
рые кровь может проходить в ретикулярную строму (см. рис. 173).
Расслабление артериального и венозного сфинктеров, а также
сокращение гладких мышечных клеток капсулы и трабекул ведет
к опорожнению синусов и выходу крови в венозное русло.
Отток венозной крови из пульпы селезенли совершается по
системе вен. Трабекул~ярные вены ТПТШемы Собственного мышечно-
го слоя; средняя оболочка в них выражена очень слабо. Наружная
оболочка вен плотно сращена с соединительной тканью трабекул.
Такое строение вен обусловливает их зияние и облегчает выброс
крови при сокращении гладких мышечных клеток селезенки.
434
Между артериями и венами в капсуле селезенки, а также между
пульпарными артериями встречаются анастомозы.
Иннервация. В селезенке имеются чувствительные нервные во-
локна (дендриты нейронов спинномозговых узлов) и постганглио-
нарные симпатические нервные волокна из узлов солнечного спле-
тения. Миелиновые и безмиелиновые (адренергические) нервные
волокна обнаружены в капсуле, трабекулах и сплетениях вокруг
трабекулярных сосудов и артерий белой пульпы, а также в синусах
селезенки. Нервные окончания в виде свободных концевых веточек
располагаются в соединительной ткани, на гладких мышечных
клетках трабекул и сосудов, в ретикулярной строме селезенки.
Возрастные изменения. В старческом возрасте в селезенке про-
исходит атрофия белой и красной пульпы, вследствие чего ее тра-
бекулярный аппарат вырисовывается более четко. Количество
лимфатических узелков в селезенке и размеры их центров посте-
пенно уменьшаются. Ретикулярные волокна белой и красной пульпы
грубеют и становятся более извилистыми. У лиц старческого воз-
раста наблюдаются узловатые утолщения волокон. Количество мак-
рофагов и лимфоцитов в пульпе уменьшается, а число зернистых
лейкоцитов и тканевых базофилов возрастает. У детей и лиц стар-
ческого возраста в селезенке обнаруживаются гигантские много-
ядерные клетки — мегакариоциты. Количество железосодержащего
пигмента, отражающее процесс гибели эритроцитов, с возрастом в
пульпе увеличивается, но располагается он главным образом внекле-
точно.
Регенерация. Физиологическое обновление лимфоидных и стро-
мальных клеток происходит в пределах самостоятельных стволо-
вых дифферонов. Экспериментальные исследования на животных
показали возможность восстановления селезенки после удаления
80—90% ее объема (репаративная регенерация). Однако полного
восстановления формы и размеров органа при этом, как правило,
не наблюдается.
Глава XVII
ЭНДОКРИННАЯ СИСТЕМА (ЭНДОКРИННЫЕ ЖЕЛЕЗЫ)1
ОБЩАЯ ХАРАКТЕРИСТИКА
Эндокринная система совместно с нервной системой осущест-
вляет_ {^уляцик"и коорди||а11ию функций организма В состав
эндокринной системы входят специализированные эндокринные
железы, или железы внутренней секреции, инкреторные железы,
лишенные выводных протоков, но обильно снабженные сосудами
микроциркуляторного русла, в которые выделяются продукты
секреции этих желез, а также одиночные эндокринные клетки,
рассеянные по разным органам и тканям организма. Как эндо-
В скобках дано название в соответствии с Международной анатомической
номенклатурой.
435
кринные железы, так и одиночные эндокринные клетки выраба-
тывают и инкретируют (т."е. выделяют"в~кровь и_лимфу) высоко-
активные регуляторные факторы — гормоны (от греч. hormao —
возбуждаю>7-окзз1явагощйе стимулирующее или угнетающее влия-
ние преимущественно на основные ^унющи-организма: обмен
вещёствТ^соматичес кии рост, репродуктивные функции.
Природа горКкйюв. Подавляющее большинство гормонов при-
надлежит к белкам (пептиды, олигопептиды, гликолептиды)_и про-
изводным аминокислот, часть — к стероидам (половые гормоны и
гормоны коры надпочечников).
Некоторые гормоны вырабатываются только в одной эндокрин-
ной^железе (например, тироксин — только^в щитовидной железе),
другие — в нескольких органах. Например, инсулин продуцируется
главным образом 6-клетками поджелудочной железы, а также
в околоушной железе, некоторыми нейронами головного мозга.
Адренокортикотропный гормон (АКТГУ вырабатывается в перед-
ней^доле гипофиза, а также вголовном мозге_и некоторых органах
(тканевой АКТГ). Многие олигопептидные гормоны человека
и млекопитающих образуются в головном мозге и в слизистой
оболочке желудочно-кишечного тракта. Нередко одиночные эндо-
кринные клетки могут продуцировать одновременно—несколько
разных олигопептидных гормонов.
Гормонь1_рказывают свое специфическое-влияние на клетки
и органы, называемые эффекторами иди мишенями.
Клеточные рецепторы гормонов. Каждый гормон может дейст-
вовать при условии, что он распознается и связывается клеточ-
ными рецепторами, находящимися в клетках-мишенях.
При этом гормон долженестрого соответствовать рецептору (комп-
лементарно, т. е. подобно тому,_как ключ должен полностью со-
ответствовать запираемому им замку). Связывание гормона с
рецептором активирует фермент аденилатциклазу, который в свою
очередь вызывает образование циклического аденозинмонофосфа-
та (цАМФ) из"аденозинтрифосфорной~кислбты САТФ). Далее
цАМФ активирует внутриклеточные ферменты, ^что приводит
клетку-мишень в состояние функционального возбуждения.
Взаимосвязь нервной и эндокринной систем. Общим для нерв-
ных и эндокринных клеток является выработка гуморальных
регулирующих факторов. Эндокринные клетки синтезируют гор-
моны и выделяют их в кровь, а секреторные нейроны синтези-
руют нейротрансмиттеры (большинство из которых являются
нейроаминами): норадреналин, серотонин и другие, выделяющиеся
в синаптические щели. В гипоталамусе находятся секреторные
нейроны, совмещающие свойства нервных и эндокринных клеток.
Они обладают способностью образовывать как нейроамины, так и
олигопептидные гормоны (рис. 174).
Выработка гормонов эндокринными органами регулируется
нервной системой, с которой они тесно связаны. Внутри эндо-
кринной системы существуют сложные взаимодействия между
центральными и периферическими органами этой системы.
436
Рис. 174. Схема строения нервных, нейросекреторных и эндокринных клеток
(рис. Б. В. Алешина).
/— холинергический нейрон с ацетилхолиновыми пузырьками в терминалях; // — гомори-
положительные нейросекреторные клетки переднего гипоталамуса (пептидно-холинергиче-
ский нейрон), продуцирующие белковые гранулы; ///— адренергический нейрон с ipa-
нулами в терминали, содержащими белковую сердцевину, на которой аккумулируются
катехоламины; IV — нейросекреторная пептидадренергическая клетка медиобазального
гипоталамуса; V— эндокринная клетка (хромаффинная клетка мозговой части надпочеч-
ников) с секреторными гранулами, как в адренергических нейронах (III); VI — эндокрин-
ная клетка, вырабатывающая белковые гормоны (парафолликулярные клетки щитовид-
ной железы, энтериноциты слизистой оболочки желудочно-кишечного тракта и панкреа-
тические островки), содержит секреторные гранулы с белковой сердцевиной. / — перика-
рион; 2 — дендриты; 3 — аксон; 4 — терминаль аксона; 5 — зоны накопления нейросекре-
та; 6 — синаптические пузырьки; 7 — гранулы нейрогормона; 8—структура секреторных
гранул.
Клетки с выраженной эндокринной функцией могут распола-
гаться в нервной системе, образовывать самостоятельные органы
или части органов, а также находиться в виде одиночных гормоно-
продуцирующих клеток в составе неэндокринных органов. Среди
этих образований выделяют центральные и периферические от-
делы, взаимодействующие между собой и формирующие единую
систему.
437
Таким образом, эндокринная система представлена следую-
щими основными структурными компонентами.
I. Центральные_регуляторные образования эндокринной системы
1. Гипоталамус (нейросекреторные ядра)
2. .Гипофиз
3. Эпифиз
II. Периферические эндокринные железы
1. ' Щитовидная железа
2. Околощитовидные железы
3. Надпочечники: а) корковое вещество
• б) мозговое вещество надпочечников
III. Органы, объединяющие эндокринные и неэндокринные функ-
ции
1. Гонады: а) семенник
б)_яичник (см. гл. XXII)
2. Плацента (см. гл. V)
3. Поджелудочная железа (см. гл. XVIII)
IV. Одиночные гормонопродуцирующие клетки
1. Нейроэндокринные клетки группы ПОДПА (APUD)
(нервного происхождения)
2. Одиночные гормонопродуцирующие клетки (не нервного
происхождения)
ЦЕНТРАЛЬНЫЕ РЕГУЛЯТОРНЫЕ ОБРАЗОВАНИЯ ЭНДОКРИННОЙ
СИСТЕМЫ
Г ипоталамус
Гипоталамус является высшим центром эндокринных функ-
ций. Он контролирует и интегрирует все висцеральные функции
организма и объединяет эндокринные механизмы регуляции с
нервными, будучи мозговым центром симпатического и парасимпа-
тического отделов вегетативной нервной системы. Субстратом
объединения нервной и эндокринной систем являются нейро-
секреторные клетки, которые у высших позвоночных и человека
располагаются в гипоталамусе.
Медиальное возвышение (eminentia medialis) является нейро-
гемальным органом гипоталамо-гипофизарной системы. Оно обра-
зовано эпендимой, отдельные глиальные клетки которой дифферен-
цируются в танициты (tanicyti), отличающиеся разветвленными
отростками, контактирующими с клубочками первичной капил-
лярной сети портальной системы гипоталамо-гипофизарного кро-
вОобращения. В гипоталамо-аденогипофизарной системе аккумули-
руются аденогипофизотропные нейрогормоны, вырабатываемые
гипоталамусом, которые затем поступают в портальную систему
438
гипофиза. В ги11оталамо=ц£йрргипофи <арнпи си< реме аналогичным
йейрогемальным органом оказывается (задняя поля
гипофиза)^ где- аккумулируются _нонапептип'ные________нейрогормо-
ны пере,,негб~ гипоталамуса, в дальнейшем щ.имягуыс t ток
крови.
“X* е креторные нейроны (neuronum secretorium). JB се-
ром веществе гипоталамуса обособляются парные ядра (свыше
30 пар), которые группируются в его переднем, среднем (медио-
базальном и туберальном) и заднем отделах.|В переднем гипота-
ламусе- располагаются парные супраоптически^*ГГП1с^ел supr р гсТ)
и ндрааент/гш^дарные, (Т1йсТеГ~р^ау5Т1ТгтгТПаге5) ядра. Супраопти- '
ческие ядра образованы крупными холинергическими—(пептидохо-
линергическими) нейросекреторными клс , содержащими как
в перикарионах, так" и в отростках отчетливые секреторные гра-
нулы. Аксоны этих клеток проходят через медиальную эминен-
цию и „гипофизарную ножку-в задщоф долю гщтрфизаутде за-
канчиваются на"крРвеноснь1х капиллярах утолщенными-терми-
налями.
^(аравентрикулярные ядра построены более сложно. Их цент-
ральная крупноклеточная часть образована такими же крупными '
холинергическими нейросекреторными клетками, как в супраопти-
ческом ядре, и и^а^тзнамтфттдугцй'м заДПКно щнлю„„гипофиза
В обоих указанных %ддра1 крупные нейросекреторные клетки
продуцируют белковые (нонапептидные) нейрогормоны — вазо-
прессин или антидиуретический гормон \АДГ) и окситоцин.
человека выработка антидиуретичдского гормона" совершается
преимущеетвенцд д супраоптическом ядре, тогда как продукция
р^ситоиица преобладает в крупноклеточной__части паравентрику-
лярных ядер. ‘
Периферическая же часть пара^еггцжкулярнечхч-ядра состоит
из мелких адренергических нейросекреторных клеток. Аксоны
этих клеток направляются в МеД'иальнучо~5йиненцию. ~
В ядрах среднегТг— ЧтяйдйобазальПоттг-и туберального^ гипо-
таламуса их мелкие адренер! ИЧескиё (пептидоадренергические)
нейросекреторные клетки вырабатывают аденогипофизсаропиые
нейрогормоны, с помощью которых гипоталамус контролирует
гормонообразовательную деятельность аденогипофиза. Эти нейро-
гормоны по своей природе являются низкомолекулярными олиго-
пептидами и разделяются на либерины, стимулирующи^ выделение
и, вероятно, продукцию 1о|5монов передней и средней долей гипо-
физа, и статины, угнетающие функции аденогипофиза. Важнейшие
ядра ахрй чЗТГй гипоталамуса локализуются в сером 6vrn lutufrer^-
cinereum) —аркуатное или идф>ундрб\ чудное J in.arcuatus seu
infundibu laris) удУгбСбразно охватывающее гипофизарную ножку,
вентромедиальное (n. ventromedialis) и дорсомедиалыюе (n. dorso-
medialis)". Основным местом вы чаботки гипоталамических либери-
нов и статйнов оказываются аркуаттзые и вентромедиальные ядрат
а также '"мелкие “пёптидоадренергические клетки мелкоклеточной
части паравентрикулярного ядра и аналогичные клетки в сером
439
Рис. 175. Схема гипоталамо-гипофизарной системы и действия тропных гормонов
на органы-мишени (рис. Б. В. Алешина).
I — зрительная хиазма; 2 — медиальная эминенция с первичной капиллярной сетью; 3 —
полость III желудочка; проекция некоторых гипоталамических ядер на стенку III желу-
дочка; 4 — супраоптическое ядро; 5 — переднее гипоталамическое ядро (преоптическая
зона гипоталамуса); 6 — паравеитрикулярное ядро; 7 — аркуатновентромедиальный ком-
плекс медиобазального гипоталамуса; 8 — нейросекреторные пептидоадренергические
клетки медиобазального гипоталамуса, секретирующие аденогипофизарные гормоны в
первичную капиллярную сеть медиальной эминенции (2); 9 — адренергические нейроны
медиобазального гипоталамуса, дающие начало нисходящим эфферентным нервным
путям (парагипофизарная передача гипоталамических импульсов регулируемым эффек-
торам): 10— таламус; II— воронка III желудочка и гипофизарная ножка; 12— задняя
доля гипофиза: 13 — накопительное тельце Херринга (окончание аксонов нейросекре-
торных клеток переднего гипоталамуса — супраоптического и паравентрикулярного ядер
на капиллярах задней доли гипофиза); 14— средняя доля гипофиза; 15— гипофизарная
щель; 16 — передняя доля гипофиза со вторичной капиллярной сетью; 17 — портальная
(воротная) вена; 18 — туберальная часть аденогипофиза. Аденогипофизарные гормоны
и места их приложения: СТГ — стимулирует рост организма в целом и его отдельных
органов (в том числе рост скелета); АКТГ — стимулирует пучковую и сетчатую зоны
коры надпочечников; ЛГ — стимулирует овуляцию, образование желтого тела и продук-
перивентрикулярном веществе, в преоптической зоне гипоталамуса
и в супрахиазматическом ядре.
Аксоны мелких пептидоадренергических клеток, равно как и
сопровождающих их адренергических нейронов обычного типа,
направляются в медиальную эминенцию в составе тубероинфун-
дибулярного дофаминергического пучка, берущего начало от до-
фаминергических нейронов туберальных ядер (рис. 175).
Регуляция гипоталамусом периферических эндокринных
желез
Влияние гипоталамуса на периферические эндокринные^
жеЛЕзы осуществляется преимущественно {гуморально в два этапа.,
Г ипоталамическиё~либёрины активируют переднюю долю гипофи за
в аботизгср; ии соответствующих тропныххормшю.в^Аеист-
вуюйхи5Г~ПЗ~же7тезБГКПн1Гени. I‘акой__способпередачищазывается
тра н с аде ног ипофизарн ы м. Для трансаденогипофизар-
ного способа характерно каскадообразное усиление начального
гипочаламичёскОТо-импульуаГ Кроме того, гипоталамус посылает
свои эфферентныё~ймпульсы к регулируемым ффектораилрямо
по симпатическим или парасимпа_т ческим нервам последних, без
опосредовани* гйпбфизом, т.е. п а гГхКи. пофизарно. Па-
рагипофизарная регуТвщихгэндокринных эффекторов может про-
являться способностью эндокринных образований непосредствен-
но реагировать (по принципу отрицательной обратной связи) на
собственные гормоны, или иммунологические агенты, поступив-
шие в циркуляцию, или на величину эффекта, вызванного ими в
организме. Например, наблюдается ослабление секреторной ак-
тивности околощитовидных желез при повышении уровня ионов
кальция в крови или усиление секреции инсулина при повышении
концентрации глюкозы в крови Сильная и длительная гиперфунк-
ция щитовидной железы вызывается патологическими тиростиму-
лирующими иммуноглобулинами, возникающими в организме как
аутоантитела и действующими на щитовидную железу не че-
рез гипофиз, а непосредственно на клеточные рецепторы тиро-
цитов.
Нейросекреторная еятельность гипоталамуса в свою очередь
исцьц вает влияние высших отделов головного мозга, особенно
лимбической системы, миндалевидных_ядер, гиппокампа и эпифи-
за. В осуществлении этих влияний существенное значение при-
надлежит нейроаминам — катехоламинам (дофамин и норадрена-
цию последним прогестерона, стимулирует продукцию тестостерона в семеннике; ФСГ —
активирует рост фолликулов и выработку ими эстрогенов в яичнике, стимулирует спер-
матогенез в семеннике; ТТГ — активирует продукцию и секрецию тиреоидного гормона
щитовидной железы; ЛТГ — активирует выработку молока в молочных железах. Гормоны,
содержащиеся в задней доле гипофиза: ОКС — вызывает сокращение матки и отдачу
молока молочными железами; АДГ — стимулирует обратную реабсорбцию воды из пер-
вичной мочи в почках (уменьшает диурез) и одновременно повышает артериальное
давление крови.
441
лин), серотонину и ацетилхолину, содержание которых в гипотала-
мусе выше, чем в других отделах головного мозга. Кроме того, на
нейросекреторные функции гипоталамуса сильно влияют некото-
рые гормоны, особенно эндорфины и энкефалины, вырабатывае-
мые специальными нейроцитами головного мозга.
Гипофиз
Гипофиз состоит из трех долей: передней, средней
и зад ней.(1>-'1'
Развитие. Закладка гипофиза происходит у зародыша человека
на 4—5-й неделе эмбриогенеза как результат взаимодействия
двух отдельных зачатков — эпителиального И нейрального. Из эк-
тодермального эпителия, выстилающего потовую ямку зародыша,
выпячивается гипофизарныи карман!шАпрЬвляющийся к основа-
нию формирующегося головного мозга и дающий начало адено-
гипофизу. Но дифференцировка этого эпителиального кармана
начинается только после того, как он вступит в соприкосновение
с противоположно направленным выпячиванием промежуточного
пузыря зачатка головного мозга, которое в дальнейшем окажется
воронкой третьего желудочка. Базальная же часть промежуточно-
го пузыря дает начало гипоталамусу.
Дифференцировка эпителиального гипофизарного кармана начи-
нается разрастанием его передней стенки, становящейся передней
долей гипофиза. Задняя стенка того же кармана остается узкой
средней (промежуточной) долей. Между передней и средней до-
лями иногда сохраняется остаток полости гипофизарного кармана
в виде узкой гипофизарной щели. У человеческого зародыша эта
щель облитерируется и средняя доля гипофиза спаивается с пе-
редней. Нейроглия дистального конца воронки, разрастаясь, фор-
мирует заднюю долю гипофиза, или нейрогипофиз. Проксимальная
же часть воронки, суживаясь, становится гипофизарной ножкой,
связывающей гипоталамус с гипофизом.
Строение. В аденогипофизе различают переднюю долю
(lobus anteriorfT пром еж уточную час т ь (pars intermedia)
и туберал ь'н у ю часть (pars tuberalis).
Передняя доля образована разветвленными эпителиаль-
ными тяжами — трабекулами, формирующими сравнительно гус-
тую сеть. ПромёжуткиМежду трабекулами заполнены рыхлой
волокнистой соединительной тканью и синусоидными капилляра-
ми, оплетающими трабекулы. Каждая трабекула образована же-
лезистыми клетками (аденоцитами) трех родов (рис. 176).
Одни из них, располагающиеся по периферии трабекул, со-
держат в своей цитоплазме отчетливые секреторные гранулы, ко-
торые интенсивно воспринимают красители. В связи с этим такие
аденоциты именуются хромофильными эндокриноцитами fendo-
crinocytus chromophilus). Другие же клетки, занимающие середину
трабекулы, имеют нечеткие границы и их цитоплазма окраши-
вается слабо—хромофобные эндокриноциты (аденоциты) (endo-
442
crinocytus chromophobus). В свою очередь хромофильные эндо-
криноциты подразделяются на базофильные и ацидофильные по
окрашиваемости их секреторных гранул. Базофильные эндокрино-
циты f endocrinocytus basophilus) получили такое название потому,
что их гранулы окрашиваются основными красителями. Гранулы
содержат гликопротеиды, являющиеся материалом для биосинте-
за гормонов, продуцируемых этими аденоцитами. Относительное
количество этих клеток в норме составляет от_4 до 10% общего
числа аденоцитов передней доли. По размерам они сравнительно
крупные. Среди них различают две разновидности. Одни из них
характеризуются округлой или овальной формой и эксцентриче-
ским положением ядер, оттесняемых на периферию клетки сильно
развитой макулой — кольцевидной структурой, соответствующей
комплексу Гольджи, которая занимает центр клетки. Секретор-
ные гранулы имеют 200—300 нм в диаметре. Митохондрии не-
велики, но многочисленны; эндоплазматическая сеть представле-
на пузырьками с мелкозернистым содержимым. Количество таких
базофилов возрастает во время усиленной продукции гонадотроп-
ных гормонов (гонадотропинов), в связи с чем клетки данной
разновидности называются гонадотропоцитами, или гонадотроп-
ными эндокриноцитами (endocrinocytus gonadotropicus) (рис. 177,
/). Предполагается, что одни из гонадотропоцитов вырабатывают
фолликулостимулирующий гормон (фоллитро-
п и н), а другим приписывается образование лютеинизиру-
ющего (лютропина) гормона.
При недостаточности в организме половых гормонов в передней
доле гипофиза по механизму отрицательной обратной связи усили-
вается выработка гонадотропных гормонов (особенно фоллитро-
пина), в связи с чем в некоторых гипертрофирующихся гонадо-
тропоцитах развивается крупная вакуоль в области макулы, растя-
гивающая цитоплазму наподобие кольца и оттесняющая уплотняю-
щееся ядро на край клетки (рис. 177, И//). Такие трансформи-
рованные гонадотропоциты получают наименование клеток
кастрации.
Вторая разновидность базофильных клеток отличается непра-
вильной или угловатой формой (рис. 177,11). Их секреторные грану-
лы очень мелкие (диаметром 80—150 нм) и интенсивно окрашива-
ются альдегидфуксином. Они содержат меньше гликопротеинов,
чем гонадотропоциты. Эти клетки вырабатывают тиротропный гор-
мон — тиротропин, действующий на щитовидную железу, и назы-
ваются тиротропоцитами, или тиротропными эндокриноцитами
(endocrinocytus thyrotropic us).
Если в организме возникает недостаточность тироидных гормо-
нов — йодотиронинов, то происходит перестройка тиротропо-
цитов. Продукция тиротропина усиливается, вследствие чего не-
которые тиротропоциты значительно увеличиваются в объеме,
цистерны эндоплазматической сети расширяются, цитоплазма при-
обретает вид крупноячеистой сети. В этих цистернах обнаруживают-
ся альдегидофуксинофильные гранулы более крупного калибра,
443
Рис. 176. Гипофиз.
I — схема строения перед-
ней доли гипофиза (по
Б. В, Апешину).
чем в исходных тиротропоцитах. Такие вакуолизированные тиро-
тропоциты называются клетками тироидэктомии (рис. 177, VI).
Для ацидофильных эндокриноцитов (endocrinocytus acidophi-
lus) характерны крупные плотные белковые гранулы, воспринимаю-
щие кислые красители (рис. 177, 111). По размерам эти клетки не-
сколько меньше базофильных, но по количеству достигают 30—
35% всех аденоцитов передней доли гипофиза. Форма их округлая
или овальная. Ядра располагаются в центре клетки. Митохондрий
немного, но они крупные. Сильно развита гранулярная эндоплаз-
матическая сеть с параллельным расположением цитомембран, что
свидетельствует о высокой интенствности белкового синтеза. Комп-
лекс Гольджи развит умеренно и прилегает к ядру.
Ацидофильные эндокриноциты тоже представлены двумя разно-
видностями. Одни — соматотропоциты, или соматотропные эндо-
криноциты (endocrinocytus somatotropicus), вырабатывают гормон
роста, или соматотропи н, другие — маммотропоциты, или
маммотропные эндокриноциты, или пролактиноциты (endocrino-
cytus mammotropicus seu prolactinocyti), продуцируют лакто-
тропный гормон, или пролактин. Основное значение
лактотропного гормона (пролактина) заключается в активирова-
нии биосинтеза молока в молочной железе. Продукция этого
гормона усиливается у рожениц после родов, во время лактации
и вскармливания новорожденного. Кроме того, пролактин удлиня-
ет функционирование желтого тела в яичнике, в связи с чем ранее
иногда назывался лютеотропным гормоном (см. гл. XXII).
В соматотропоцитах секреторные гранулы имеют шаровидную
форму и достигают в диаметре 350—400 нм. Маммотропоциты
444
Рис. 176 (продолжение ).
// — микрофотография пе-
редней (А), средней (Б) и
задней (В) долей гипофиза.
Окраска азаном (препараты
Ю. И. Афанасьева). 1 —
хромофобные клетки; 2 -
базофильные клетки; 3 -
ацидофильные клетки; 4 —
кровеносные капилляры с
эритроцитами крови; 5 -
эпителий фолликулов проме-
жуточной доли; 6 — коллоид;
7 - питуициты.
II
отличаются еще более крупными гранулами овальной или удли-
ненной формы (500—600 нм в длину и 100—120 нм в ширину).
Еще одна группа хромофильных клеток — кортикотропные
эндокриноциты (endocrinocytus corticotropicus), или кортикотро-
поциты, локализующиеся преимущественно в центральной зоне
445
Рис. 177. Схема ультра микроскопического строения железистых клеток аденогипо-
физа (рис. Б. В. Алешина).
/ — гонадотропная фолликулостимулирующая клетка; II — тиротропная клетка; III — со-
матотропная клетка; IV — лактотропная (маммотропная) клетка; V — кортикотропная
клетка; VI — клетка тиреоидэктомии; VII — клетка кастрации; VIII — фолликулярно-
звездчатые клетки псевдофолликула. I — ядро; 2 — комплекс Гольджи; 3 — митохондрия;
4 — эндоплазматическая сеть; 5 — секреторные гранулы; 6 — вакуоль; 7 — микроворсинка;
8—полость лсевдофол яикула заполненная коллоидоподобной массой
передней доли гипофиза, продуцируют белковый адренокортико-
тропный гормон (АКТГ, или кортикотропин). Их форма непра-
вильна или угловата, клеточные ядра дольчатые, митохондрии
сильно развиты, эндоплазматическая сеть хорошо выражена.
Хотя АКТГ является белковым гормоном, кортикотропоциты не
принадлежат ни к ацидофилам, ни к базофилам^ а составляют
особую самостоятельную группу аденоцитов. Их секреторные гра-
нулы имеют строение пузырьков, одетых мембраной, и содержат
внутри плотную белковую сердцевину, причем между мембраной и
сердцевиной остается светлое пространство.
Цитоплазма хромофобных эндокриноцитов слабо воспринимает
красители и не содержит отчетливых секреторных гранул. Поэтому
хромофобные клетки, составляющие около 60%, нередко считаются
недеятельными. В действительности же группа хромофобов оказы-
вается сборной и включает аденоциты разной степени дифферен-
цировки и различного физиологического значения. Среди них
можно обнаружить клетки, уже начавшие специализироваться в
базофильные или ацидофильные аденоциты, но еще не успевшие
накопить специфические секреторные гранулы. Другие же хромо-
фобные клетки, наоборот, являются вполне специализированными,
но лишившимися своих базофильных или ацидофильных секретор-
ных гранул вследствие интенсивной или длительной секреции.
К ряду неспециализированных принадлежит лишь небольшая часть
хромофобных клеток, которые можно признать резервными. На-
конец, среди хромофобов встречаются звездчатые (фолликулозвезд-
чатые) клетки, небольшие по размерам, но обладающие длинными
ветвистыми отростками, которыми они соединяются в широкопет-
листую сеть. Некоторые из отростков проходят между соседними
аденоцитами и заканчиваются на синусоидных капиллярах. Иногда
же звездчатые клетки группируются в небольшие фолликулоподоб-
ные образования, в полостях которых накапливается гликопро-
теидный секрет. На апикальной поверхности таких звездчатых
клеток развиваются микроворсинки, вдающиеся в просвет фоллику-
ла (рис. 177, VIII).
Средняя (промежуточная) часть аденогипофиза
представлена узкой полоской эпителия. Аденоциты средней доли
способны вырабатывать белковый или слизистый секрет, который,
накапливаясь между соседними клетками, приводит к формирова-
нию в средней доле фолликулоподобных кист. От задней доли
эпителий средней доли отделяется тонкой прослойкой рыхлой со-
единительной ткани.
В средней части аденогипофиза содержится меланоцитостиму-
лирующий гормон (меланоцитотропин), а также липотропин —
гормон, усиливающий метаболизм липидов. По современным
данным, эти гормоны, а также адренокортикотропин в действитель-
ности возникают в головном мозге путем расщепления крупной
исходной молекулы церебрального пептида препроопиомеланокор-
тина на меньшие фрагменты, каждый из которых становится од-
ним из перечисленных гормонов.
447
Туберальная часть аденогипофиза — отдел, Прилежа
щий к гипофизарной ножке и соприкасающийся с нижней поверх -
ностью медиального возвышения гипоталамуса. Туберальная часть
образована эпителиальными тяжами, состоящими из кубических
клеток с умеренно базофильной цитоплазмой. От туберальных
тяжей, так же как от эпителия средней доли, отходят в переднюю
долю трабекулы; в некоторых клетках туберальных тяжей встреча-
ются базофильные гранулы, хотя в небольшом количестве.
Гипоталамо-аденогипофизарное кровоснабжение и его значение
в гипоталамической регуляции аденогипофизарного гормонопоэза
(см. рис. 175). Система гипоталамо-аденогипофизарного кровоснаб-
жения называется портальной (воротной). Приносящие
гипофизарные артерии вступают в медиальное возвышение медио-
базального гипоталамуса, где разветвляются всеть капилля-
ров (первичное капиллярное сплетение портальной системы). Эти
капилляры образуют петли и клубочки, с которыми контактируют
терминали аксонов нейросекреторных клеток аденогипофизотроп-
ной зоны гипоталамуса. Капилляры первичного сплетения соби-
раются в портальные вены, идущие вдоль гипофизарной ножки в
переднюю долю, где они распадаются на капилляры синусоидного
типа (вторичная капиллярная сеть), разветвляющие-
ся между трабекулами паренхимы железы. Наконец, синусоиды
вторичной капиллярной сети собираются в выносящие вены, по
которым кровь, обогатившаяся гормонами передней доли, посту-
пает в общую циркуляцию.
Задняя доля гипофиза, или нейрогипофиз. Задняя доля гипо-
физа обрадована в основном клеткамц^эпендимы. Они имеют от-
ростчатую или вере те но видную форму ^называются питуицитами.
Их многочисленные тонкие отростки заканчиваются “в адвентиции
кровеносных сосудов или__на базальной мембране^капилляров.
В задней доле гипофиза (в нейрогемалыюм органе гиИТЬталамо-
нейрогипофизарной системы) аккумулируются антидиуретический
гормон (вазопрессин} и окситоцин, вырабатываемые крупными
пептидохолинергическими нейросекреторными клетками переднего
гипоталамуса. Аксоны этих нейцосекреторнык_клеток^ собираются
в гипоталамо-нейрогипофизарные пучки, входят в заднюю долю
гипофиза, где заканчиваются крупными терминалями (называемы-
ми тельцами Херринга, или накопительными тельцами), контакти-
рующими Гкапиллярами (рис. 178).
Васкуляризация задней доли гипофиза обеспечивается само-
стоятельными сосудами, отходящими от нижних гипофизарных
артерий.
Иннервация. Гипофиз, а также гипоталамус и эпифиз получают
симпатические нервные волокна от шейных (преимущественно
верхних) ганглиев пограничных стволов. Однако аксоны нейронов
этих ганглиев заканчиваются не в гипоталамусе, а в ретикулярной
формации и ядрах ствола головного мозга, адренергические ней-
роны которых посылают свои аксоны в гипоталамус в составе нор-
адренергических пучков. Таким образом, влияние симпатических
448
Рис. !78. Схема контакта
терминали аксона гипота-
ламического секреторного
нейрона с гемокапилляром
задней доли гипофиза (по
Г ершенфельду)
/ нейрофибриллы; 2— ми-
тохондрии: J — гранулы ней-
росекрета; 4 синаптиче
ские пузырьки: 5 — базаль-
ная мембрана; 6 — эндоте
ооциты гемокапилляра.
импульсов достигает гипоталамус не прямо, а через возбуждение
адренергических (а также серотонинергических) мезенцефаличес-
ких и лимбических нейронов головного мозга. Эффект симпатиче-
ских импульсов проявляется в гипоталамусе ускорением кругообо-
рота нейроаминов и возрастанием содержания норадреналина и
серотонина. Кроме того, гипоталамус получает также афферент-
ную информацию по центростремительным нервам от перифери-
ческих органов.
Возрастные изменения. Дифференцировка гипофизарных адено-
цитов у зародыша начинается на 9-й неделе внутриутробного пе-
риода. Сначала появляются базофильные аденоциты, а на 4-м ме-
сяце развития плода — ацидофильные. К моменту рождения ре-
бенка дифференцировка гипофиза заканчивается Однако в пост-
натальном периоде преимущественно активируются ацидофильные
аденоциты (очевидно, в связи с обеспечением повышенной про-
дукции соматотропина, стимулирующего быстрый рост тела ребен-
ка), а среди базофилов преобладают тиротропоциты. В пубертат-
ном периоде, когда наступает половое созревание, увеличивается
количество базофильных аденоцитов.
15 ГИСТОЛОГИЯ
449
Регенерация. Аденогипофиз обладает ограниченной регенера-
торной способностью, главным образом за счет специализации
хромофобных клеток. Задняя доля гипофиза, образованная нейро-
глией, регенерирует лучше.
Эпифиз (шишковидная железа]
• Г’ ЛА’ •
Эпифиз участвует в регуляции процессов, протекающих в ор-
ганизме ритмически или циклически, например овариально-менст-
руального цикла. Ритмические колебания других периодических
функции, интенсивность которых закономерно изменяется на про-
тяжении суток, называются циркадными (от лат. circa diem — около
дня). Циркадные ритмы явно связаны со сменой дня и ночи (све-
тового и темнового периодов) и их зависимость от эпифиза сви-
детельствует, что гормонообразовательная деятельность послед-
него определяется его способностью различать смену световых
раздражений, получаемых организмом.
Развитие. У зародыша человека эпифиз закладывается на 5—6-й
неделе внутриутробного периода в виде выпячивания промежу-
точного мозга. У круглоротых, амфибий и пресмыкающихся (яще-
риц) образуется несколько таких парапинеальных вы-
ступов ив их числе непарный теменной глаз. У млекопи-
тающих и человека теменной глаз не развивается, но эпифиз со-
храняет способность реагировать на световые раздражения, которые
передаются к нему от сетчатки парных глаз по симпатическим нер-
вам.
Стенка эпифизарного дивертикула утолщается, а его просвет
облитерируется. Из того же выпячивания возникает как утолщение
эпендимы добавочный субкомиссуральный орган, в
ходе дальнейшего развития включающийся в состав эпифиза.
Строение. Снаружи эпифиз окружен тонкой соединительноткан-
ной капсулой, от которой отходят разветвляющиеся перегородки
внутрь железы, образующие ее строму и разделяющие ее паренхиму
на дольки (рис. 179). В пинеальной паренхиме различаются клетки
двух родов — секретообразующие пинеалоциты (endocrinocytus pi-
nealis) и поддерживающие глиальные (gliocytus centralis). Пинеало-
циты располагаются в центральной части долек. Они несколько
крупнее опорных нейроглиальных клеток, по форме многоуголь-
ны, имеют пузыревидные ядра с крупными ядрышками. От тела пи-
неалоцита отходят длинные отростки, ветвящиеся наподобие ден-
дритов, которые переплетаются с отростками глиальных клеток.
Отростки, булавовидно расширяясь, направляются к капиллярам и
контактируют с ними. В цитоплазме этих булавовидных расши-
рений содержатся осмиофильные гранулы, вакуоли и митохонд-
рии. Среди пинеалоцитов различают светлые пинеалоциты (endo-
cnnocytus lucidus), характеризующиеся светлой гомогенной цито-
плазмой, и темные пинеалоциты (endocrinocytus densus) меньшего
размера с ацидофильными (а иногда базофильными) включениями
в цитоплазме. По-видимому, обе названные формы являются не
450
Рис. 179. Эпифиз человека (схема).
А малое увеличение, Б большое увеличение; / — соединительнотканная капсула;
2 соединительнотканные перегородки, 3 — паренхима эпифиза; 4 - мозговой песок,
5 — кровеносные сосуды; 6 глиоциты, 7 — темные пинеалоциты; 8 — светлые пинеало-
циты.
самостоятельными разновидностями, а представляют собой клетки,
находящиеся в различных функциональных состояниях, или клетки,
- подвергающиеся возрастным изменениям. В цитоплазме пинеалоци-
тов обнаруживаются многочисленные митохондрии, хорошо раз-
витый комплекс Гольджи, лизосомы, пузырьки агранулярной эндо-
плазматической сети, рибосомы и полисомы. Глиальные клетки
преобладают на периферии долек. Их цитоплазма скудна, ядра
уплотнены. Их отростки направляются к междольковым соедини-
тельнотканным перегородкам, образуя своего рода краевую кайму
дольки.
Функции. Несмотря на малые размеры эпифиза, его функцио-
нальная'деятельность весьма сложна и многообразна. Клинические
и экспериментальные наблюдения показали, что эпифиз замедляет
ра «витие половой системы, так как разрушение этой железы, ее
недоразвитие или удаление эпифиза у инфантильных животных
в эксперименте имеют следствием наступление преждевременного
полового созревания. Ингибирующее влияние эпифиза на половые
функции обусловливается несколькими факторами. Во-первых, пи-
неалоциты вырабатывают серотонин, который в них же пре-
вращается в мелатонин. Этот нейроамин, по-видимому, ос-
лабляет или угнетает секрецию гонадолиберина гипоталамусом и
15- 451
гонадотропинов передней_доли гипофиза. В то же время пинеало-
циты продуцируют ряд белковых гормонов и в их числе а нтиго-
надотропин, ослабляющий секрецию лютропина передней доли
гипофиза. Наряду с антигонадотропином пинеалоциты образуют
другой белковый гормон, повышающий уровень ка-
лия в крови, следовательно, участвующий в регуляции мине-
рального обмена. Число регуляторных пептидов, продуцируемых
пинеалоцитами, приближается к 40. Из них наиболее важны а р г и-
н ин-вазотоцин, тиролиберин, люлиберин и даже
тиротропин. Образование олигопептидных гормонов совместно
с нейроаминами (серотонин и мелатонин) демонстрирует нейро-
эндокринную природу пинеалоцитов и их принадлежность к системе
ПОДПА (см. с. 467).
Иннервация. Симпатические нервные волокна, входящие в эпи-
физ и заканчивающиеся в паренхиме его долек, берут начало от
верхних шейных ганглиев пограничных стволов. Симпатические
импульсы усиливают в эпифизе превращение серотонина в мелато-
нин.
Возрастные изменения. У человека эпифиз достигает макси-
мального развития к 5—6 годам жизни, после чего, несмотря на
продолжающееся функционирование, начинается его возрастная
инволюция. Некоторое количество пинеалоцитов претерпевает атро-
фию, а строма разрастается и в ней увеличивается отложение
фосфатных и карбонатных солей в виде слоистых шариков, на-
зываемых мозговым песком.
ПЕРИФЕРИЧЕСКИЕ ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Щитовидная железа
Эмбриональное развитие. Зачаток щитовидной железы возника-
ет у зародыша человека на 3—4-й неделе внутриутробногошериода
как выпячивание стенки глотки между первой и второй парами жа-
берных карманов. Это выпячивание растет вдоль глоточной кишки
в виде эпителиального тяжа. На уровне III—IV пар жаберных
карманов этот тяж раздваивается, давая начало формирующимся
правой и левой долям щитовидной железы. Начальный эпителиаль-
ный тяж (ductus thyreoglossus), соответствующий выводному, про-
току, атрофируется и от него сохраняются только перешеек, свя-
зывающий у человека обе доли щитовидной железы, и прокси-
мальная часть в виде ямки (foramen coecum) в корне языка.
У большинства других млекопитающих атрофируется также дис-
тальный конец эпителиального тяжа, поэтому перешеек не разви-
вается и обе доли щитовидной железы обособляются. Зачатки
долей быстро разрастаются, образуя рыхлые сети ветвящихся
эпителиальных трабекул; из них формируются фолликулы, в про-
межутки между которыми врастает мезенхима с кровеносными со-
судами и нервами. Кроме того, у человека и млекопитающих
имеются нейроэндокринные парафолликулярные клетки, берущие
начало от нейробластов.
4S2
Рис. 180. Щитовидная железа человека (рис. Б. В. Алешина).
/ интрафолликулярный коллоид; 2 микрофолликул; 3 межфолликулярные соедини
тельнотканные прослойки; 4 капилляры; 5 - интерфолликулярный островок: 6 ре
зорбционные вакуоли.
Строение. Щитовидная железа окружена соединительнотканной
капсулой, прослойки которой направляются вглубь и разделяют
орган на дольки (рис. 180). Структурно-функдионадьными-едини-
цами (аденомерами) щитовидной железы являются фолликулы —
замкнутые шаровидные или слегка вытянутые пузыревидные обра-
зования варьирующих размеров с полостью внутри. Иногда, когда
функциональная активность щитовидной железы значительно уси-
лена (например, при тиротоксикозе), стенка фолликула образует
многочисленные ветвистые складки и контуры фолликулов приоб-
ретают звездча’тые_очертания. В просвете фолликула накапливается
коллоид — секреторный продукт эпителиальных клеток, составля-
ющих выстилку фолликула. При жизни коллоид, представляет
собой вязкую жидкость, состоящую в основном из тироглобулина.
Фолликулы разделяются прослойками рыхлой волокнистой соеди-
нительной ткани, по которым проходят многочисленные крове-
носные и лимфатические капилляры, оплетающие фолликулы, а
также нервные волокна. В этих же прослойках обнаруживаются
компактные скопления тироидных эпителиальных клеток. Кроме
того, в межфолликулярных соединительнотканных прослойках
всегда встречаются лимфоциты и плазматические клетки, а также
тканевые базофилы.
Фолликулярные эндокриноциты (endocrinocytus follicularis),
или тироциты — железистые клетки, составляющие большую часть
стенки фолликулов и экстрафолликулярного эпителия. В фоллику-
лах тироциты образуют выстилку (стенку) и располагаются в
один слой на базальной мембране, ограничивающей фолликул сна-
ружи. При умеренной функциональной активности щитовидной
железы (ее нормофункции) тироциты имеют кубическую форму и
шаровидные ядра. Коллоид, секретируемый ими. заполняет в виде
гомогенной массы просвет фолликула.
45’’
На апикальной поверхности тироцитов, обращенной к просвету
фолликула, имеются микроворсинки. Соседние клетки в выстилке
фолликулов тесно связаны между собой многочисленными десмо-
сомами и хорошо развитыми терминальными пластинками. Кроме
того, по мере возрастания тироидной активности на боковых по-
верхностях тироцитов возникаютлальцевидные выступы (интер-
дигитации!, входящие в соответствующие вдавления боковой по-
верхности. соседних клеток. —
В тироцитах хорошо развиты органеллы, особенно участвующие
в белковом синтезе. Белковые продукты, синтезируемые тироцита-
ми, выделяются в полость фолликула, где завершается образова-
ние йодированных тирозинов и тиронинов (аминокислот, входящих
в состав крупной и сложной молекулы тироглобулина). Тироидные
гормоны могут попасть в циркуляцию лишь после высвобождения
из этой молекулы (т. е. после расщепления тироглобулина). Когда
же потребности организма в тироидном гормоне возрастают и
функциональная активность щитовидной железы усиливается (ги-
перфункция щитовидной железы), тироциты фолликулов набухают,
принимают призматическую форму. Интрафолликулярный коллоид
при этом становится более жидким и пронизывается многочислен-
ными ресорбционными вакуолями. Ослабление же функциональной
активности (гипофункция щитовидной железы) проявляется, на-
оборот, уплотнением коллоида, его застоем внутри фолликулов, диа-
метр и объем которых значительно увеличивается; высота же гиро-
цитов уменьшается, они принимают уплощенную форму, а их ядра
вытягиваются параллельно поверхности фолликула. По мере уси-
ления тироидной активности количество и размеры микроворсинок
возрастают. Одновременно базальная поверхность тироцитов, почти
гладкая в периоде функционального покоя щитовидной железы,
становится складчатой, что увеличивает соприкосновение тироцитов
с перифолликулярными пространствами.
Секреторный цикл фолликулов. В секреторном цикле фоллику-
лов различают две фазы: фазу продукции и фазу выведения гор-
монов.
Фаза продукции, которой начинается секреторный цикл
тироцитов, состоит в поглощении через базальную поверхность
исходных веществ будущего секрета (аминокислот, в том числе
тирозина, йода и других минеральных ионов, некоторых углеводов,
воды), приносимых к щитовидной железе кровью. В эндоплазма-
тической сети происходит формирование молекулы тироглобулина.
Образующиеся соединения постепенно перемещаются в зону комп-
лекса Гольджи, где к полипептидной основе присоединяются угле-
водные компоненты и происходит формирование везикул, содержа-
щих тироглобулин. Затем они смещаются к апикальной мембране
тироцита, где их содержимое путем экзоцитоза поступает в по-
лость фолликула.
Йод поглощается тироцитами из крови в виде йодида (точнее,
иона йода), но так как в молекулу тирозина может включаться
только атомарный йод, то предварительно ион йода под действием
454
Рис. 181. Схема тироцита и процесса секреции (рис. Б. В. Алешина).
/ — капилляр; 2 — перикапиллярное пространство; 3 — базальные мембраны тироцита и
эндотелиоцита (стрелки показывают направление транспортировки перерабатываемых
веществ). 4— эндоплазматическая сеть и рибосомы; 5 — комплекс Гольджи; 6 — пере
мещение пузырьков секрета гликопротеида к апикальной мембране; 7 - экзоцитоз в
полость фолликула; 8 — поступление атомарного йода в полость фолликула и йодирова-
ние гликопротеида; 9 — микроворсинки; 10 — образование псевдоподий в процессе ре
зорбции коллоида; // фагоцитоз интрафолликулярного коллоида; 12— микротубулы
псевдоподии; 13 — капли фагоцитированного интрафолликулярного коллоида (интра-
целлюлярные капли коллоида); 14—лизосомы; /5 слияние коллоидной капли с лизо-
сомой; протеолиз тироглобулина и высвобождение тироидного гормона; /6 вакуоли с
продуктами протеолиза; /7 — протеолиз интрафолликулярного коллоида на апикальной
поверхности тироцита и эндоцитоз (пиноцитоз) продуктов этого протеолиза; 18—тран-
спорт пиноцитозных пузырьков; /9 — отдача тироидного гормона в капилляр; 20
митохондрии; 21 ядро тироцита; 22 — замыкающая пластинка; 23 межклеточная щель
с десмосома ми; 24 — интрафолликулярный коллоид; 25 — поступление веществ из капил
лвра в тироцит.
фермента пероксидазы окисляется в атомарный йод (I).
Этот процесс совершается на апикальной поверхности тироцита
и его микроворсинок, т. е. на границе с полостью фолликула. Здесь
же в тирозины, даже находящиеся в составе полипептидной ос-
новы молекулы тироглобулина, включается сперва по одному атому
йода (из тирозинов образуются монойодтирозины), а затем по вто-
рому, и монойодтирозины переходят в дийодтирозины, которые
объединяются попарно в тетрайодтирзнин. Наряду с тироксином
образуется также трийодтиронин. Трийодтиронин значительно ак-
тивнее тироксина.
Фаза выведения начинается ,с реабсорбции коллоида и
протекает по-разному в зависимости от степени и продолжитель-
455
Рис. 182. Схема соотношения
фолликулярных и парафоллику-
лярных клеток в щитовидной
железе (рис. Б. В. Алешина).
А — реакция восстановления окис
лов тяжелых металлов (аргирофи-
лия или осмифилия) только в па-
рафолликулярных клетках; Ь по-
глощение радиоактивного йода, ха-
рактерное только для фолликуляр-
ных клеток, / — парафол чикуляр-
ная клетка в межфолликулярнои
локализации; 2 парафолликуляр-
ная клетка в интраэпителиальной
локализации; 3 — фолликулярная
клетка; 4 — базальная клетка; 5 —•
«светлая» фолликулярная клетка;
6 — эпителиальная почка, во ши-
кающая в результате размноже-
ния базальных клеток; 7 — базаль-
ная мембрана фолликула,
ности активации щитовид-
ной желез ыЦЕсли эта акти-
вация сильна (например,
когда она вызвана избыт-
ком тиротропина), тироци-
ты приобретают все-при-
знаки, свидетельствующие
об их интенсивной фагоци-
тарной активности. Они на-
бухают, —значительно воз-
растают их объем и высота.
На апикальной поверх-
ности наряду с увеличени-
ем числа и размеров микро-
ворсинок появляются псев-
доподии. Коллоид захваты-
вается тироцитом путем
фагоцитоза. Фагоцитиро-
ванные фрагменты коллои-
да, попавшие внутрь тиро-
цита, с помощью лизосо-
мального аппарата подвер-
гаются протеолизу и из
молекул фагоцитированно-
го тироглобулина высво-
бождаются йодтирозины и
йодтиронины. Йодтирозины в цитоплазме тироцитов распадаются,
а освобождающийся йод реутилизируется в последующем гормоно-
генезе. Йодтиронины же выделяются через базальную мембрану
тироцита в ток крови или лимфы.
Фагоцитоз коллоида продолжается всего несколько часов.
456
При умеренной активности щитовидной железы или при значи-
тельной ее гиперфункции, но сохраняющейся-длительно (напри-
мер, при тиротоксикозе), образования апикальных псевдоподий и
фагоцитирования ими коллоида не наступает, а происходит протео-
лиз тироглобулина в полости фолликула и пиноцитоз (макроэндо-
цитоз) продуктов расщепления цитоплазмой тироцитов (рис. 181).
Парафолликулярные эндокриноциты (endocrinocytus para follicu-
laris), или кальцитониноциты. Во взрослом организме парафолли-
кулярные клетки локализуются в стенке фолликулов, залегая между
основаниями соседних тироцитов, но не достигают своей верхуш-
кой просвета фолликула (интраэпителиальная локализация пара-
фолликулярных клеток). Кроме того, парафолликулярные клетки
располагаются также в межфолликулярных прослойках соедини-
тельной ткани (рис. 182). По размерам парафолликулярные клетки
крупнее тироцитов, имеют округлую, иногда угловатую форму.
В отличие от тироцитов парафолликулярные клетки не поглощают
йод, но совмещают образование нейроаминов (норадренали-
на и~ серотонина) путем декарбоксилирования тирозина и
5-гидрокситриптофапа (ароматических аминокислот — предшест-
венников указанных нейроаминов) с биосинтезом белковых (олиго-
пептидных) гормонов — к а л ь п и т~сГн ина и соматоста-
тина.
ч)
5s4
Секреторные гранулы, густо заполняющие цитоплазму парафол-
ликулярных клеток, обнаруживают сильную осмиофилию и аргиро-
филию. В цитоплазме парафолликулярных клеток хорошо развиты
гранулярная эндоплазматическая сеть и комплекс Гольджи. Секре-
торные гранулы парафолликулярных клеток бывают двух родов.
В некоторых парафолликулярных клетках преобладают мелкие,
но сильно осмиофильные гранулы. Клетки данного типа вырабаты-
вают кальцитонин. В парафолликулярных клетках другого
типа содержатся более крупные, но слабо осмиофильные гранулы.
Эди клетки продуцируют соматостатин.
Васкуляризация. Щитовидная железа обильно снабжается
кровью. За единицу времени через щитовидную железу проходит
приблизительно столько же крови, как через почки, причем интен-
сивность кровоснабжения увеличивается при усилении функцио-
нальной активности щитовидной железы.
Иннервация. Щитовидная железа богата симпатическими и па-
расимпатическими нервными волокнами, но влияние прямых
нервных импульсов на деятельность фолликулов невелико и зна-
чительно перекрывается гуморальными эффектами тиротропина.
Тем не менее раздражение шейных симпатических ганглиев или
воздействие адренергическими веществами вызывает хотя и слабое,
но достоверное усиление образования и отдачи йодированных тиро-
идных гормонов, несмотря на то, что в данных условиях наступает
сужение кровеносных сосудов и уменьшение тока крови через щи-
товидную железу. Парасимпатическим импульсам принадлежат,
наоборот, угнетающие эффекты.
Парафолликулярные клетки полностью лишены зависимости от
457
Рис. 183. Дифференцировка микрофолликула. Электронная микрофотография,
(по Д. X. Хамидову и К. А. Зуфарову).
/ — ядро; 2 цитолемма; 3 — формирующаяся полость будущего фолликула; 4 цистерна
эндоплазматической сети; 5 — митохондрии.
гипофиза и гипофизэктомия не нарушает их деятельности. В то
же время они отчетливо реагируют на прямые симпатические
(активирующие) и парасимпатические (угнетающие) импульсы.
Регенерация. Паренхима щитовидной железы отличается повы-
шенной способностью к пролиферации. Источником роста тироид-
ной паренхимы оказывается эпителий фолликулов. Деление тиро-
цитов приводит к увеличению площади фолликула, вследствие
чего в нем возникают складки, выступы и сосочки, вдающиеся в
полость фолликулов (интрафолликулярная регене-
рация).
Размножение клеток может привести также к появлению эпи-
телиальных почек, оттесняющих базальную мембрану кнаружи, в
межфолликулярное пространство. С течением времени в пролифе-
рирующих тироцитах этих почек возобновляется биосинтез тиро-
глобулина, что приводит к дифференцировке островков в микро-
фолликулы (рис. 183). Микрофолликулы в результате продолжаю-
щегося синтеза и накопления коллоида в их полостях увеличивают-
ся
ся в размерах и становится такими же, как материнские (экстра-
фолликулярная регенерация). Парафолликулярные
клетки в фолликулогенезе участия не принимают.
•колощитовидные железы
Функциональное значение околощитовидных желез заключает-
ся в регуляции метаболизма кальция. Они вырабатывают белковый
гормон паратирин, который, действуя на костную ткань —
депо накопления кальция в организме, стимулирует высвобожде-
ние последнего, что приводит -к повышению уровня этого иона в
крови (гиперкальциемическое действие), а в костях наступает
частичная деминерализация. Следовательно, паратирин и кальци-
тонин парафолликулярных клеток щитовидной железы, оказываю-
щий гипокальциемизирующее действие, составляют антагонистиче-
скую пару и их взаимодействием обеспечивается постоянство
уровня кальция в крови (кальциевый гомеостаз организма).
Развитие. Околощитовидные железы закладываются у зародыша
как выступы эпителия 3-й и 4-й пар жаберных карманов глоточной
кишки. Эти выступы отшнуровываются и каждый из них развивает-
ся в отдельную околощитовидную железу, остающуюся самостоя-
тельной, даже если некоторые из них в ходе дальнейшего эмбрио-
генеза включаются в щитовидную железу.
Строение. Каждая околощитовидная железа окружена тонкой
соединительнотканной капсулой. Ее паренхима представлена тра-
бекулами — эпителиальными тяжами либо скоплениями эпители-
альных эндокринных клеток — паратироцитов (endocrinocytus рага-
thyroideus), разделенными тонкими прослойками рыхлой соедини-
тельной ткани с многочисленными капиллярами (рис. 184, А,
Б). Хотя между паратироцитами хорошо развиты межклеточные
щели, соседние клетки связаны интердигитациями и десмосомами.
Различают главные паратироциты (endocrinocytus principalis) и
оксифильные паратироциты (endocrinocytus oxyphilicus). В глав-
ных паратироцитах цитоплазма базофильна, в ее периферических
зонах рассеяны скопления свободных рибосом (полисом), что
свидетельствуете высокой интенсивности белкового синтеза. Комп-
лекс Гольджи развит хорошо и представлен уплощенными мешоч-
ками и многочисленными пузырьками. В них формируются секре-
торные гранулы диаметром 150—200 нм. Митохондрий много, они
удлиненной формы, с хорошо развитыми поперечными кристами.
При усилении секреторной активности паращитовидных желез
главные клетки увеличиваются в объеме. Среди главных паратиро-
цитов различают светлые (endocrinocytus principalis lucidus)
и темные (endocrinocytus principalis densus). В цитоплазме
светлых клеток встречаются включения гликогена. В оксифильных
(ацидофильных) паратироцитах особенно многочисленны мито-
хондрии. Перечисленные формы клеток являются не самостоя-
тельными разновидностями паратироцитов, а лишь возрастными
или функциональными состояниями последних.
459
Рис. 184. Строение около-
щитовидной железы чело-
века.
А — окраска по Маллори;
Б схема ультрамикроско-
пического строения (рис.
Ю. И. Афанасьева); / —
оксифильные паратироциты;
2 — главные паратироциты;
J — соединительнотканные
прослойки; 4 фолликул с
коллоидоподобным содержи
мым; 5 — гемокапилляры;
б адипоциты; 7 — проме-
жуточные клетки; 8 пре-
секреторные гранулы.
2 7 1
На секреторную активность околощитовидных желез не оказы-
вают влияния гипофизарные гормоны. Околощитовидная железа
по принципу обратной связи быстро реагирует на малейшие коле-
бания в уровне кальция в крови. Ее деятельность усиливается
при гипокальциемии и ослабляется при киперкальциемииДЛаратиро-
циты обладают рецепторами, способными непосредственно воспри-
нимать прямые влияния ионов кальция на них.
Васкуляризация. Сосуды околощитовидных желе» отличаются
тонкими стенками. Артерии распадаются на многочисленные ка-
пилляры, вены не анастомозируют, формируя петлистую сеть. Они
лакунообразно расширяются и собираются в субкапсулярные спле-
тения, сообщающиеся с венами щитовидной железы.
Иннервация. Околощитовидные железы получают обильную
симпатическую и парасимпатическую иннервацию. Безмиелиновые
волокна заканчиваются терминалями в виде пуговок или колечек
между паратироцитами. Вокруг оксифильных клеток нервные
терминали принимают вид корзиночек. Встречаются также инкап-
сулированные рецепторы. Влияние поступающих нервных импуль-
сов ограничивается сосудодвигательными эффектами.
Возрастные изменения. У новорожденных и детей младшего
возраста в паренхиме околощитовидных желез обнаруживаются
только главные клетки. Оксифильные клетки появляются не ранее
5—7 лет, к этому времени их количество быстро нарастает. После
20-~25 лет постепенно прогрессирует накопление адипоцитов.
В старческом возрасте венулы, извиваясь, нередко приобретают
вид кольцеобразных петель.
Надпочечники
Надпочечники — это парные органы, образованные соединением
двух отдельных самостоятельных гормонопродуцирующих желез,
составляющих к о р к о в о е и мозг о в о е вещество разного
происхождения, регуляции и физиологического значения (рис. 185).
Развитие. Закладка корковой части обнаруживается у зароды-
ша человека на 5-й неделе внутриутробного периода в виде утол-
щений целомического эпителия по обе стороны корня брыжейки.
В дйлЬнейшем эти эпителиальные утолщения, образованные круп-
ными клетками с ацидофильной цитоплазмой, собираются в ком-
пактное интерреналовое тело. Ацидофильные клетки ста-
новятся зачатком первичной (фетальной) коры будущих надпочеч-
ников. На 10-й неделе внутриутробного периода первичная кора
окружается снаружи мелкими базофильными клетками (происхо-
дящими тоже из целомического эпителия), которые дают начало
дефинитивной коре надпочечников.
Из того же целомического эпителия, из которого возникает
интерреналовое тело, закладываются таКжЗГ половые валики —
зачатки гонад, что обусловливает их функциональную взаимосвязь
и близость химической природы их стероидных гормонов.
Мозговая часть надпочечников закладывается у зародыша че-
ловека на 6—7-й неделе внутриутробного Периода. Из общего за-
чатка симпатических ганглиев, располагающегося в аортальной
области зародыша, выселяются неиробластЫ (симпатобласты),
внедряющиеся в интерреналовое тело, где размножаются и дают
начало мозговой части надпочеяников. Следовательно, железистые
(хромаффинные) клетки мозговой части надпочечников должны
рассматриваться как нейроэндокринные. У зародыша хромаффин-
ные клетки вначале содержат только норадреналин, а адреналин
появляется на более поздних стадиях эмбриогенеза.
461
Рис. 185. Схема строения надпочечника (рис. Б. В. Алешина).
А, Б — накопление липидов в адренокортикоцитах (окраска Суданом 111-гематоксили-
ном); В адренокортикоциты пучковой зоны после растворения липидов (окраска по
Маллори); /—капсула; 2 - клубочковая зона: 3 пучковая зона; 4—сетчатая зона:
5 хромаффинные клетки мозгового вещества. 6 — капли липидов в адренокортикоцитах;
7 капилляры; 8 эндотелий; 9 — соединительнотканные прослойки между тяжами
адренокортикоцитов; 10 — спонгиоциты.
Корковое вещество надпочечников
Строение. Снаружи надпочечники покрыты соединительноткан-
ной капсулой, в которой различаются два слоя — наружный (плот-
ный) и внутренний (более рыхлый). Под капсулой залегает тонкая
прослойка мелких эпителиальных клеток, размножением которых
обеспечивается регенерация коры и создается возможность воз-
никновения добавочных интерреналовых телец, иногда обнаружи-
ваемых на поверхности надпочечников и нередко оказывающихся
источниками опухолей (в том числе и злокачественных).
Корковые эндокриноциты (endocrinocytus corticalis) образуют
эпителиальные тяжи, ориентированные перпендикулярно к поверх-
ности надпочечника. В коре имеются три зоны: клубочковая,
пучковая и сетчатая. Промежутки между эпителиальны-
ми тяжами заполнены рыхлой соединительной тканью, по которой
проходят кровеносные капилляры и нервные волокна, оплетающие
тяжи.
Клубочковая зона (zona giomerulosafjofipaaoeaHa мел-
кими корковыми эндокриноцитами, которые формируют округлые
скопления («клубочки»). В этой зоне клетки содержат мало ли-
пидных включений. Их агранулярная эндоплазматическая сеть
представлена мелкими пузырьками, между которыми обнаружи-
ваются рибосомы. Митохондрии овальной или удлиненной формы
отличаются пластинчатыми кристами. Хорошо развит комплекс
Гольджи.
В клубочковой зоне вырабатывается альдостерон — ми-
нералокортикоидный гормон, контролирующий содержание натрия в
организме. Кроме того, минералокортикоиды усиливают воспали-
тельные процессы. Минералокортикоиды жизненно важны. Разру-
шение или удаление клубочковой зоны приводит к смертельному
исходу.
Между клубочковой и пучковой зонами располагается узкая
прослойка мелких малоспециализированных клеток. Она называет-
ся промежуточной или суданофобной. Предпола-
гается, что размножение клеток данной прослойки обеспечивает
пополнение и регенерацию пучковой и сетчатой зон.
Пучковая зона (zona fasciculate) занимает среднюю часть
тяжей и наиболее выражена. Корковые эндокриноциты этой зоны
(рис. 186) отличаются крупными размерами, кубической или при-
зматической формой; на поверхности, обращенной к капиллярам,
имеются микроворсинки. Цитоплазма этих клеток изобилует ка-
плями липидов. Митохондрии крупные, округлой или овальной
формы, с кристами в виде извитых и ветвящихся трубок (везику-
лярные кристы). Гладкая эндоплазматическая сеть хорошо выра-
жена. Рибосомы лежат в цитоплазме свободно.
Наряду со светлыми клетками встречаются в разном ко-
личестве темные с уплотненной цитоплазмой, содержащей мало
липидных включений, но повышенное количество рибонуклеопро-
теидов. В темных клетках, помимо развитой агранулярной эндо-
Рис. 186. Электронная микрофотография адренокортикоцита пучковой зоны.
54 000 (по В. П. Деревянко).
I — липидные включения; 2 комплекс Гольджи; 3— митохондрии; 4 (иалоплазма
5 — кариолемма; 6 — ядро.
плазматической сети, имеется гранулярная эндоплазматическая
сеть. Светлые и темные клетки представляют разные функцио-
нальные состояния одних и тех же корковых эндокриноцитов.
Полагают, что в темных клетках осуществляется синтез специфи-
ческйх-белков — ферментов, которые в дальнейшем участвуют в
образовании кортикостероидов, о чем свидетельствует обильное
содержание в цитоплазме темных клеток рибосом (рибонуклео-
протеидов). По мере выработки стероидов и их накопления цито-
плазма клеток становится светлой и они вступают в фазу вьнеления
готового секреторного продукта в циркуляцию.
464
В пучковой зоне вырабатываются глюкокортикоидные гормоны,
кортикостерон, кортизон и гидрокортизон
(кортизол). Они влияют на метаболизм углеводов, белков и
липидов и усиливают процессы фосфорилирования в организме,
чем способствуют образованию веществ, богатых энергией, высво-
бождаемой~затем- для энер!етического обеспечения всех процессов
жизнедеятельности, протекающих в каждой клетке организма.
Глюкокортикоиды форсируют глюконеогенез (образование
глюкозы за счет белков) и отложение гликогена в печени и мио-
карде, а также мобилизацию тканевых белков. Большие дозы
глюкокортикоидов вызывают деструкцию и распад лимфоцитов
и эозинофилов крови, приводя к лимфоцитопении и эозинофило-
пении, а также угнетают воспалительные процессы в организме.
С е т ч а та я_з о н a (zona reticularis). В ней эпителиальные
тяжи разветвляются, формируя рыхлую сеть. Адренокортикоциты
в сетчатой зоне уменьшаются в размерах и становятся кубиче-
скими, округлыми или угловатыми Содержание в них липидных
включений убывает, а число темных, клеток возрастает. Кристы
митохондрий в адренокортикоцитах трубчатые. Эндоплазматиче-
ская сеть в этих адренокортикоцитах преимущественно вакуоляр-
ная, в цитоплазме преобладают свободные рибосомы. Пластин-
чатый комплекс развит хорошо.
В сетчатой зоне вырабатывается андроген стероидный
г q р м о н., близкий по химической природе и физиологическим
свойствам к тестостерону семенников. Поэтому опухоли коры над-
почечников^ женщин нередка, оказщваюхся причиной вирилизма
(развитие вторичных половых признаков мужского пола, в част-
ности усов и бороды). В сетчатой зоне образуются также женские
половые гормоны (эстрогены и прогестерон), но в небольших
количествах. Иногда в сетчатой зоне на границе с мозговой частью
(мозговым веществом) сохраняются остатки фетальной коры.
Ее клетки отличаются ацидофильной цитоплазмой. Эти остаточные
образования иначе называются Х-зоной. Она постоянно обнаружи-
вается в надпочечниках самок некоторых млекопитающих, а у
самцов становится заметной после кастрации.
Мозговое вещество надпочечников
Строение. Мозговое вещество (medulla) отделено от кор
кового вещества тонкой, местами прерывающейся прослойкой сое-
динительной ткани. Эта часть надпочечников образована скопле-
нием сравнительнсгжрупных клеток округлой формы — мозговых
эндокриноцитов, или хромаффиноцитов (endocrinocytus medul-
Taris), между которыми находятся кровеносные сосуды (синусо-
иды). Различают светлые эндокриноциты, или эпинефроциты
( endocrinocytus lucidus, epinephrocytus), секретирующие адреналин,
и темные эндокриноциты, или норэпинефроциты (endocrinocytus
densus), секретирующие норадреналин. Цитоплазма клеток густо
заполнена электронно-плотными секреторными гранулами диа-
465
метром 100—500 нм, окаймленными мембраной. Сердцевина гра-
нулы заполнена белком, аккумулирующим секретируемые катехо-
ламины — норадреналин и адреналин. После обра-
ботки надпочечников раствором бихромата калия в железистых
клетках откладывается бурый осадок низших окислов хрома.
Подобным же образом эти клетки восстанавливают четырехокись
осмия и нитрат серебра, вследствие чего эти клетки получают
наименование хромаффинных, или осмиофильных, или аргиро-
фильных1. Кроме того, в мозговом веществе находятся мультипо-
лярные нейроны автономной нервной системы.
Регуляция секреторных функций коры надпочечников. Специ-
фическим возбудителем_гормонообразователыюй деятельности
пучковой 1й сетчатой зон является адено цдсфизарный адренокор-
тикотропин (АКТГ).
Регуляция клубочковой зоны более сложна. Поскольку альдо-
стерон образуется из кортикостерона, биосинтез которого стиму-
лируется АКТГ, начальные стадии генеза минералокортикоидов
подчиняются влиянию этого адецогипофизарного активатора.
Но переход кортикостерона в альдостерон определяется дополни-
тельным вмешательством ренина (гормон, вырабатываемый в
почке).
Возрастные изменения. Кора надпочечников у человека дос-
тигает полного развития в возрасте 20—25 лет, когда соотношение
ширины ее зон приближается к 1:9:3. После 59 лет ширина коры
начинает уменьшаться. В корковых эндокриноцитах постепенно
уменьшаются количество и размеры липидных включений, а сое-
динительнотканные прослойки между эпителиальными тяжами
утолщаются. Редукция поражает главным образом сетчатую и от-
части клубочковую зону. Ширина же пучковой зоны относительно
увеличивается, что обеспечивает достаточную интенсивность глю-
кокортикоидной функции надпочечников вплоть до преклонного
возраста. Мозговая часть надпочечников не претерпевает выра-
женных возрастных изменений. После 40 лет отмечается некото-
рая гипертрофия хромаффиноцитов, но лишь в преклонном воз-
расте в них наступают атрофические изменения, ослабевает син-
тез катехоламинов, а в сосудах и строме обнаруживаются призна-
ки склероза.
Васкуляризация. У мозгового и коркового вещества надпочеч-
ника имеется общее кровоснабжение. Артерии, входящие в над-
почечник, разветвляются на артериолы, образующие густую суб-
капсулярную сеть, от которой отходят капилляры, снабжающие
кровью кору. Их эндотелий фенестрирован, что облегчает поступ-
ление кортикальных стероидных гормонов из корковых эндокрино-
цитов в ток крови. Из сетчатой зоны капилляры вступают в мозго-
вую часть, где принимают вид синусоидов и сливаются в венулы,
1 Помимо мозговой части надпочечников, скопления хромаффинных клеток
обнаруживаются также в параганглиях, входящих в состав ганглиев автономной
нервной системы, в каротидных тельцах.
466
которые переходят в венознре сплетение мозгового вещества.
Наряду с ними в мозговую часть входят также артерии, берущие
начало от субкапсулярной сети. Проходя через кору и обога-
щаясь продуктами, секретируемыми адренокортикоцитами, кровь
приносит к хромаффиноцитам специальные, вырабатываемые в ко-
ре ферменты, которые активируют метилирование норадренали-
на, т. е. образование адреналина.
В мозговой части ветвление кровеносных сосудов таково, что
каждый хромаффиноцит одним концом соприкасается с артери-
альным капилляром, а другим обращен к венозному синосоиду,
в который выделяет катехоламины. Венозные синусоиды собира-
ются в центральную вену надпочечника, впадающую в нижнюю по-
лую вену. Таким образом, в циркуляцию поступают одновременно
и кортикостероиды, и катехоламины, чем обеспечивается возмож-
ность совместного действия обоих регуляторных факторов на эф-
фекторные органы или системы. По другим же венам кровь из
коры и мозгового вещества направляется в воротную вену печени,
принося в нее адреналин (увеличивающий мобилизацию глюкозы
из гликогена) и глюкокортикоиды, стимулирующие, наоборот,
глюконеогенез в печени.
Иннервация. Надпочечники получают эфферентные нервные
импульсы главным образом по чревным и блуждающим нервам.
В кору входят постганглионарные (безмиелиновые) симпатические
волокна, а в мозговую часть (как и в другие симпатические ганг-
лии) преганглионарные (миелиновые) волокна. В капсуле нередко
встречаются микроганглии и одиночные ганглиозные нервные
клетки. В корковой части, деятельность которой активизируется
аденогипофизарным АКТГ, роль прямых нервных импульсов огра-
ничивается вазомоторными эффектами. В мозговом веществе,
которое не зависит от гипофиза, регуляция его секреторной дея-
тельности осуществляется исключительно нервными импульсами,
поступающими главным образом по чревным нервам. Раздражение
последних усиливает отдачу адреналина и норадреналина хрома-
фи иными клетками в ток крови.
1
1
л
л
т
I-
и
1О
(И
.и
ы
>й
а,
>i.
э-
и
1-
'О
й
>-
И
ОДИНОЧНЫЕ ГОРМОНОПРОДУЦИРУЮЩИЕ КЛЕТКИ
Среди одиночных гормонопродуцирующих клеток различают по
меньшей мере две самостоятельные группьиВ первую группу вхо-
дят одиночные гормонопродуцирующие клетки нервного проис-
хождения — секреторные нейроциты, образующиеся из нейроблас-
тов нервного гребешка, которые сохраняют способность образо-
вывать нейроамины, сочетая ее с продукцией белковых (олиго-
пептидных) гормонов. Клетки данной группы принято объединять
в группу или серию APUD-клеток (Amine Precursors Uptake and
Decarboxylation), чему в русской транскрипции соответствует
аббревиатура ПОДПА (Поглощение и декарбоксилирование пред-
шественников аминов).
Нейроэндокринные клетки группы ПОДПА встречаются^ го-/
( '(
( 16Г
469
ловном мозге и в других органах, как эндокринных, так и неэндо-
крйнных? Примером нейроэндокринных клеток этой группы, обна-
руживаемых в -Эндокринных органах, могут служить парафолли-
кулярные клетки щитовидной железы и хромаффинные клетки
мозговой части надпочечников, а в неэндокринных — некоторые
специализированные нейроны головного мозга, желудочно-кишеч-
ные эндокриноциты в слизистой оболочке желудочно-кишечного
тракта, которые совмещают продукцию олигопептидных гормонов
с образованием серотонина, притом столь интенсивным, что ока-
зываются основным источником серотонина, содержащегося в пе-
риферической крови.
Олигопептидные гормоны, продуцируемые нейроэндокринными
клетками, оказывают местное действие на клетки органов,
в которых они локализуются, но главным образом дистант-
ное— на общие функции организма вплоть до высшей нервной
деятельности.
Нейроэндокринные клетки находятся в непосредственной и тес-
ной зависимости от прямых нервных импульсов, поступающих
к ним по симпатическими и парасимпатическим нервам, но не
реагируют на тропные гормоны гипофиза. Их состояние и актив-
ность не нарушаются гипофизэктомией.
В организме встречаются также одиночные гормонопродуци-
рующие клетки или скопления клеток, происходящие не из нейро-
бластов, а принадлежащие к другим тканям. К ним относятся
гландулоциты семенника, вырабатывающие гормон тестостерон и
клетки зернистого слоя фолликулов яичника, продуцирующие
эстрогены и прогестерон. Названные клетки продуцируют не бел-
ковые, а стероидные гормоны и активируются аденогипофизар-
ными гонадотропинами. Важнейшим отличием клеток этой группы
от нейроэндокринных клеток группы ПОДПА является то, что
они лишены способности декарбоксилировать аминокислоты и
вырабатывать нейроамины.
Глава XVIII
-L ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Пищеварительная система человека состоит из пищеваритель-
ной трубки и расположенных вне ее желез (слюнные железы, пе-
чень и поджелудочная железа), секрет которых участвует в про-
цессе пищеварения. Пищеварением называются процессы
химической и механической обработки пищи с последующим
всасыванием продуктов расщепления ее, происходящие последо-
вательно в различных отделах пищеварительной трубки. Значение
пищеварительного аппарата в организме человека как открытой
биологической системы велико — через него осуществляется по-
ступление веществ, обеспечивающих организм необходимой энер-
гией и строительными материалами для восстановления постоянно
разрушающихся и отмирающих его структур.
46 b
В пищеварительной системе условно выделяют три основных
отдела: передний, средний и задний. Передний отдел
включает ротовую полость со всеми ее структурными компонента-
ми, глотку и пищевод, В переднем отделе происходит главным
образом механическая обработка пищи. Средний отдел
состоит из желудка, тонкого и толстого кишечника, печени и
поджелудочной железы. В этом отделе осуществляются преимуще-
ственно химическая обработка пищи, всасывание продуктов рас-
щепления ее и формирование каловых масс. Задний отдел
представлен каудальной частью прямой кишки и обеспечивает
функцию эвакуации непереваренных остатков пиши из пищевари-
тельного канала.
Для понимания гистологического строения и гистофизиологии
различных отделов пищеварительной системы вначале необходимо
ознакомиться с особенностями гистогенеза и общими принципами
строения пищеварительной трубки.
Гистогенез. Эпителиальная выстилка пищеварительной трубки
и железы развиваются из энтодермы и эктодермы. Из энтодермы
формируются однослойный призматический эпителий слизистой
оболочки желудка, тонкого и большей части толстого кишечника,
а также железистая паренхима печени и поджелудочной железы.
Из эктодермы ротовой и анальной бухт эмбриона образуется мно-
гослойный плоский эпителий ротовой полости, слюнных желез и
каудального отдела прямой кишки. Мезенхима является источни-
ком развития соединителЧ н&Й^гкани и сосудов. Из висцерального
листка спланхнотома развивается однослойный плоский эпителий
(мезотелий) серозной оболочки — висцерального листка брю-
шины.
I L, ОБЩИЙ ПЛАН МИКРОСКОПИЧЕСКОГО СТРОЕНИЯ ПИЩЕВАРИТЕЛЬНОЙ
"Г ' ТРУБКИ
Пищеварительная трубка в любом ее отделе состоит из внут-
ренней слизцсхои. оболочки (tunica mucosa), подслизистой основы
(tela submucosa), мышечцой оболочки (tunica muscularis) и цзруж-
ной оболочки, которая представлена либо серозной оболочкой
(tunica serosa), либо адвентициальной оболочкой (tunica adventi-
tia) (рис. 187).
Слизистая оболочка. Свое название она получила в
связи с тем, что поверхность ее постоянно увлажняется выделяемой
железами слизью. Эта оболочка состоит, как правило, из трех плас-
тинок: эпителия, собственной пластинки слизистой оболочки (lamina
propria mucosae) и мышечной пластинки слизистой оболочки
(lamina muscularis mucosae) (см. рис. 187).
Эпите ши в переднем и заднем отделах пищеварительной труб-
ки многослойный плоский, а в среднем отделе — однослойный
призматический. Железы расположены либо эндоэпителиально
(например, бокаловидные клетки в кишечнике), либо экзоэпи-
телиально в собственной пластинке слизистой оболочки (пище-
469
I
Рис. 187. Общий план строения пищеварительной трубки.
А пищевод; Б желудок; В тонкая кишка; Г — толстая кишка. / слизистая обо-
лочка: // подели «истая основа; /// мышечная оболочка; IV — серозная адвенти-
циальная оболочка; / — эпителий слизистой оболочки; 2 — собственная пластинка слизи-
стой оболочки: 3 - мышечная пластинка слизистой оболочки; 4 — бокаловидный экюкри-
ноцит; 5 железы в собственной пластинке слизистой оболочки; 6 — железы, располо-
женные в подслизистой основе; 7 сосудистые сплетения; 8—подслизистое нервное
сплетение; 9— межмышечное иервное сплетение; 10— желудочные ямки: //—крипты;
12 — ворсинки.
вод, желудок) и в подслизистой основе (пищевод, двенадцати-
перстная кишка) или за пределами пищеварительного канала
(печень, поджелудочная железа).
Собственная пластинка слизистой оболочки лежит под эпите-
лием, отделена от него базальной мембраной и представлена рых-
лой волокнистой соединительной тканью. Здесь находятся крове-
носные и лимфатические сосуды, нервные элементы, скопления
лимфоидной ткани. В некоторых отделах (пищевод, желудок)
могут располагаться простые железы.
Мышечная пластинка слизистой оболочки расположена на
границе с подслизистой основой и состоит из 1—3 слоев, обра-
зованных гладкими мышечными клетками. В некоторых отделах
(язык, десны) гладкие мышечные клетки отсутствуют.
Рельеф слизистой оболочки на протяжении всего пищева-
рительного канала неоднороден. Поверхность ее может быть глад-
кой (губы, щеки), образовывать углубления (ямочки в желудке,
крипты в кишечнике), складки (во всех отделах), ворсинки
(в тонкой кишке).
Подслизистая основа. Состоит из рыхлой волокни-
стой соединительной ткани. Наличие подслизистой основы обеспе-
470
чивает подвижность слизистой оболочки, образование складок.
В подслизистой основе имеются сплетения кровеносных и лимфа-
тических сосудов, скопления лимфоидной ткани и подслизистое
нервное сплетение (plexus submucosus). В некоторых отделах
(пищевод, двенадцатиперстная кишка) расположены железы.
Мышечная оболочка. Состоит, как, правило^ из, _двух
слоев мышечных элементов — внутреннего циркулярного и наруж-
но го продольного/в передне мйза днем отделах пище варите льно го
канал;Г*мышечная ткань п^Фиущёственно поперечнополосатая, а в
среднем7— гладкая? Мышечные слои разделены соединительной
тканью, в которой расположены кровеносные и лимфатические
сосуды и межмышечное нервное сплетение (plexus nervorum inter-
muscularis). Сокращения мышечной оболочки способствуют пере-
мешиванию и продвижению пищи в процессе пищеварения.
Серозная оболочка. Большая часть пищеварительной
трубки покрыта серозной оболочкой — висцеральным листком
брюшины. Брюшина состоит из соединительнотканной основы, в
которой расположены сосуды и нервные элементьи-и- из—мезоте-
лия, В некоторых отделах (дищеводд-частьлрямой^ кишки) сероз-
ная оболочка отсутствует. В таких местах пищеварительнаятрубка
по к оытас наружи адвентициальной о_бдхл_о_ц jt о й, со-
стоящеи только из соединительной ткани.
Васкуляризация. Стенка пищеварительной трубки на всем
протяжении обильно снабжена кровеносными и лимфатическими
сосудами (рис. 188, А, Б). Артерии образуют наиболее мощные
сплетения в подслизистой основе, которые тесно связаны с арте-
риальными сплетениями, лежащими в собственной пластинке сли-
зистой оболочки. В тонкой кишке артериальные сплетения фор-
мируются также в мышечной оболочке. Сети кровеносных капил-
ляров располагаются под эпителием слизистой оболочки, вокруг
желез, крипт, желудочных ямочек, внутри ворсинок, сосочков язы-
ка и в мышечных слоях. Вены также формируют сплетения под-
слизистой основы и слизистой оболочки. Наличие артериолове-
нулярных анастомозов обеспечивает регуляцию притока крови в
различные участки пищеварительного тракта в зависимости от
фазы пищеварения.
Лимфатические капилляры образуют сети под эпителием, вок-
руг желез и в мышечной оболочке (см. рис. 188). Лимфатические
сосуды формируют сплетения подслизистой основы и мышечной
оболочки, а иногда и наружной оболочки (пищевод). Наиболее
крупные сплетения сосудов располагаются в подслизистой основе.
Иннервация. Эфферентную иннерв.ацию обеспечи-
вают ганглии вегетативной нервной системы, расположенные либо
вне пищеварительной трубки (экстрамуральные симпатические
'англии), либо в толще ее (интрамуральные парасимпатические
•ганглии) (рис. 189). К экстрамуральным ганглиям относятся
верхней шейный, звездчатый и другие узлы симпатической цепоч-
ки, иннервирующие пищевод, ганглии солнечного и тазового спле-
тений, иннервирующие желудок и кишечник. Интрамуральными
471
4
Рис. 188. Схема васк ia-
ризации стенки пищева-
рительной трубки (рис
Н. А. Юриной, Л. С. Ру
мянцевой).
А расположение кровенос-
ных и лимфатических сосу-
дов в стенке пищеваритель-
ной трубки (на примере
тонкой кишки): / слизис-
тая оболочка; // - подсли-
зистая основа; /// — мышеч-
ная оболочка; / — кровенос-
ные капилляры ** ще-'м»
3 — вены: 4 -
синка: 5
капилляр;
леза (крин
ческий сос} у — лимфати-
ческий узелок; 9 — сосуди-
стые сплетения в подсли-
зистой основе. Б — лимфати
ческие капилляры слизистой
оболочки желудка человека
(препарат В С. Ревазова):
1 — лимфатические капилля-
ры; 2 - железы желудка,
инъекция синей массы Геро-
та.
Рис. 189. Схема расположения нервных элементов
в стенке тонкой кишки (по Рамон-и Кахалю).
/ слизистая оболочка; // подслизистая основа;
мышечная оболочка; IV — серошая оболочка;
разветвление нервных волокон в слизистой обо-
лочке; 2 подслизистое нервное сплетение;
внутримышечное нервное сплетение.
Ill
являются ганглии межмышечного, под-
слизистого и подсерозного, или адвен-
тициального, сплетений. Описание ней-
ронного состава сплетений дано в
гл. XIII (см. рис. 125). Аксоны эффе-
рентных нейронов симпатических и па-
расимпатических сплетений иннервиру-
ют мышцы и железы.
Афферен тная иннервация
осуществляется окончаниями дендритов
чувствительных нервных клеток, находя-
щихся в составе интрамуральных гангли-
ев, и окончаниями дендритов чувстви-
тельных клеток спинальных ганглиев.
Чувствительные нервные окончания рас-
печатаются в мышцах, эпителии, волок-
нистой соединительной ткани и нервных
ганглиях. Афферентные окончания в
стенке пищеварительного канала могут
быть поливалентными, т. е. одновременно иннервировать различные
ткари — нервную, соединительную, а также кровеносные сосуды.
В эпителии слизистой оболочки и железах всех отделов пище-
варительной системы, но особенно в ее среднем отделе, имеются
одиночные эндокринные клетки. Выделяемые ими
биологически активные вещества — нейротрансмиттеры и гормоны
оказывают как местное действие, регулируя функцию желез и
гладких мышц сосудов, так и общее влияние на организм (см. рис. 211).
р ПЕРЕДНИЙ ОТДЕЛ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
Передний отдел включает в себя ротовую полость со всеми ее
уктурными образованиями, глотку и пищевод. К производным
овой полости относятся губы, щеки, десны, твердое и мягкое
>о, язык, миндалины, слюнные железы, зубы. Здесь находится
также орган вкуса.
Основная механическая функция переднего отдела оп-
ределяет особенности его строения. В частности, для переднего
отдела характерно наличие в слизистой оболочке многослойного
плоского эпителия, предохраняющего от повреждений подлежа-
щую соединительную ткань. Кроме того, здесь может происходить
начальная химическая обработка углеводов пищи амилазой и
мальтазой слюны. Ряд органов переднего отдела пищеварительной
трубки участвует в обеспечении защитной функции.
473
-j— Ротовая полость
Слизистая оболочка ротовой полости отличается следующими
особенностями: наличием многослойного плоского эпителия (тол-
щина 180—600 мкм), отсутствием или слабым развитием мышеч-
ной пластинки слизистои оболочки и отсутствием"~"в некоторых
Участках подслизистой основы. В последнем случае она твердо
сращена с-подлежащими тканями и лежит непосредственно на
мышцах (например, в языке) или на кости (в деснах и твердом
небе). Слизистая оболочка в местах расположения лимфоидной
ткани (миндалины) образует складки.
Наличие множества поверхностно лежащих мелких кровенос-
ных сосудов, просвечивающих через эпителий, придает слизистой
оболочке характерный розовый цвет. Хорошо увлажненный эпите-
лий способен пропускать многие вещества в эти сосуды, что часто
используется во врачебной практике для введения таких лекарств,
как нитроглицерин, валидол и др., через слизистую оболочку рта.
I Г убы------- 1 '
В губе различают три отдела: кожный (pars cutanea), про-
межуточный (pars intermedia) и слизистый (pars mucosa). В тол-
ще губы находятся поперечнополосатые мышцы (рис. 190).
Рис. 190. Схема микроскопического строения губы (по В. Г. Елисееву и др.).
/ — слизистая часть;// — переходная часть;/// — кожная часть; / многослойный плоский
эпи "г лий; 2 собственная пластинка слизистой оболочки с капиллярами; 3 глюниые
железы; 4 — мышцы; 5 — сальные железы; 6 волосяной мешок; 7 — эпидермис.
474
Кожный отдел губы имеет строение кожи. Он покрыт
многослойным плоским ороговевающим эпителием и снабжен
сальными, потовыми железами и волосами. Эпителий этой части
расположен на базальной мембране; под мембраной лежит рыхлая
волокнистая соединительная ткань, образующая высокие сосочки,
которые вдаются в эпителий.
II р о’м ежуточный отдел губы в свою очередь состо-
ит из двух зон: наружной (гладкой) и внутренней (ворсинчатой).
В наружной зоне роговой слой эпителия сохраняется, но стано-
вится тоньше и прозрачнее. В этой области нет волос, постепенно
исчезают потовые железы, а сохраняются только сальные железы,
открывающие свои протоки на поверхность эпителия. Сальных
желез больше в верхней губе, особенно в области угла рта. Соб-
ственная пластинка слизистой оболочки является продолжением
соединительнотканной части кожи; сосочки ее в этой зоне невысо-
кие. Внутренняя зона у новорожденных покрыта эпителиальными
сосочками, которые иногда называют ворсинками. Эти эпителиаль-
ные сосочки по мере роста организма постепенно сглаживаются
[и становятся малозаметными. Эпителий внутренней зоны переход-
сного отдела губы взрослого человека в 3—4 раза толще, чем в
наружной зоне, лишен рогового слоя. Сальные железы здесь, как
правило, отсутствуют. Лежащая под эпителием рыхлая волокнис-
тая соединительная ткань, вдаваясь в эпителий, образует очень вы-
сокие сосочки, в которых находятся многочисленные капилляры.
Циркулирующая в них кровь просвечивает через эпителий и
обусловливает красный цвет губы. Сосочки содержат огромное
количество нервных окончаний, поэтому красный край губы очень
чувствителен.
Слизистый отдел губы покрыт многослойным плос-
ким неороговевающим эпителием. Однако в клетках поверхност-
ного слоя эпителия все же можно обнаружить небольшое коли-
чество зерен кератина. Эпителиальный пласт в слизистом отделе
губы значительно толще, чем в кожном. Собственная пластинка
слизистой оболочки здесь образует сосочки, однако они менее
высокие, чем в лежащем рядом переходном отделе. Мышечная
пластинка слизистой оболочки отсутствует, и поэтому собственная
пластинка без резкой границы переходит в подслизистую ос-
нову, примыкающую непосредственно к поперечнополосатым
мышцам.
В подслизистой основе располагаются секреторные отделы слюн-
ных губных желез (gll. labiates). Железы довольно крупные, иног-
да достигают.величины горошины. .По строению это сложные аль-
веолярно-трубчатые железы. По характеру секрета они относятся
К смешанным слизисто-белковым железам. Выводные протоки их
выстланы многослойным плоским неороговевающим эпителием и
открываются на поверхности губы.
В подслизистой основе слизистого отдела губы расположены
артерии и обширное ^енозное сплетение распространяющееся и
в красную часть губы.
475
Щеки
Щеки — это мышечные образования, покрытые снаружи кожей,
а изнутри — слизистой оболочкой. В слизистой оболочке щеки
различают три зоны: верхнюю, или максиллярную (zona maxillaris),
нижнюю, или мандибулярную (zona mandibularis), и среднюю, или
промежуточную (zona intermedia). В слизистой оболочке отсутст-
вует мышечная пластинка.
Макс иллярная и мандибулярная зоны щеки
имеют строение, сходное со строением слизистой части губы.
Эпителий здесь многослойный плоский неороговевающий, сосочки
собственной пластинки слизистой оболочки небольших размеров.
В этих областях хорошо выражена подслизистая основа, в которой
находится большое количество слюнных желез щеки (gll. buccales).
Наиболее крупные из них лежат в области коренных зубов (gll.
molares). По мере удаления от ротового отверстия железы рас-
полагаются в более глубоких слоях, т. е. в толще щечных мышц
и даже снаружи от них.
Средняя, или промежуточная, зона щеки за-
нимает область шириной около 10 мм, тяну уюся от угла рта до
ветви нижней челюсти. В эмбриональном периоде и в течение пер-
вого года' жизни ребенка на этом участке имеются эпителиальные
ворсинки, такие же, как на переходной части губы. Сосочки соб-
ственной пластинки слизистой оболочки здесь, как и в переходной
части губы, больших размеров. Слюнные железы отсутствуют. Для
промежуточной части щеки характерно наличие нескольких реду-
цированных желез, таких же, как в промежуточном отделе губы.
Промежуточная зона щеки, как и промежуточный отдел губы, яв-
ляется зоной контакта кожи и слизистой оболочки ротовой по-
лости, формирующейся вследствие срастания эмбриональных за-
кладок при формировании ротового отверстия.
С подлежащими тканями слизистая оболочка щеки соединена
при помощи подслизистой основы, в которой располагается много
кровеносных сосудов и нервов. Мышечная оболочка щеки образо-
вана^ щечной мышцей, в толще которой лежат щечные слюнные
железы с белково-слизистыми и чисто слизистыми секреторными
отделами.
Десны. Твердое небо
Десны. Покрыты слизистои оболочкой, плотно сращенной с
надкостницей верхней и нижней челюстей. Слизистая оболочка
выстлана многослойным плоским эпителием, который иногда оро-
говевает. Собственная пластинка слизистой оболочки образует
длинные сосочки, глубоко вдающиеся в эпителий. Сосочки ста-
новятся ниже в той части десны, которая непосредственно при-
лежит к зубам. В соединительной ткани собственной пластинки
слизистой оболочки* Мёс+аМй имеются^оольшие скопления ткане-
вых базофилов. Основное вещество её” рёзко метахроматично,
т. е. содержит в своем составе значительное количество гликоза-
476
миногликанов. Мышечная пластинка слизистой оболочки отсутст-
вует. Десна богато иннервирована/В эпителии находятся свобод-
ные нервные окончания, а в собственной пластинке слизистой
оболочки — инкапсулированные и неинкапсулированные нервные
окончания.
I Твердое небо. Состоит из костной основы, покрытой слизистой
\ оболочкой. Подслизистая основа отсутствует, поэтому слизистая
оболочка плотно сращена с надкостницей. Слизистая оболочка
выстлана многослойным плоским неороговевающим эпителием.
В области шва твердого неба эпителий слизистой оболочки иногда
образует у'Г0ЛЩенйя,"7Гмеющие вид характерных дя/кей. У ново-
рбжденнь()Г они представлены эПЙТёЛиЛлЬными тельцами, состоя-
щими из концентрически наслоенных эпителиоцитов и достигаю-
щими величины булавочных головок. Собственная пластинка сли-
зистой оболочки образует сосочки, вдающиеся в эпителий. В ней
расположены мощные пучки коллагеновых волокон, сильно пере-
плетающиеся между со’бой и' вплетающиеся в надкостницу. Эта
особенйОСТО"в строении твердого неба лучше выражена в местах
плотных сращений слизистой оболочки с костью (например, об-
ласть шва, зона перехода в десны). В остальных местах между
собственной пластинкой слизистой оболочки и надкостницей рас-
полагается небольшая прослойка жировой ткани, соответствующая
местоположению подслизистой основы других частей ротовой по-
лости. В сре них детках тве[ дого нрба между слизистой оболоч-
кой и надкостницей располагаются группы небных слюнных же-
лез (gll.‘ palatinae). Небные слюнные железы ^дьвеолярно-труб-
чатые, разветвленные.
Мягкое небо. Язычок
Мягкое небо и язычок. Сост т из сухожильно-мышечнои ос-
новы, покрытой слизистой Oooj очкой. В мягком небе и язычке
“различают ротоглоточную (п е ре д ню_ю) и н о с о г ло-
то ч н у ю (_з а нюю поверхности. У плодов и новорож-
дённых гистологическая граница между ними проходит прибли-
зительно по линии перегиба слизистой оболочки с ротовой по-
верхности на носовую на дужках мягкого неба и на язычке.
У взрослых эта граница смещается на заднюю (носовую) поверх-
ность, так что весь язычок оказывается одетым слизистой оболоч-
кой, характерной для ротовой полости.
Слизистая оболочка ротовой поверхности
мягкого неба и язычка покрыта многослойным плоским неорого-
вевающим эпителием. Собственная пластинка слизистой оболочки
образует высокие узкие сосочки, глубоко вдающиеся в эпителий.
За ней располагается сильно развитый слой эластических, волокон.
Мышечная пластинка-е-лизнегёй оболочки отсутствует. Далее сле-
дует. хорошо выраженная по/ слизистая основа, образованная рых-
лой^ волокнистой соединительной тканью, богатой жировыми эле-
ментами, в которой расположены слизистые слюнные железы.
477
Выводные протоки этих желез открываются на ротовой поверх-
ности мягкого неба и язычка. В язычке скопления желез обнару-
живаются и внутри мышечного слоя. Поперечнополосатая мышеч-
ная ткань, составляющая основу язычка, имеет ряд особенностей.
Мышечные волокна ее ветвятся и образуют между собой анасто-
мозы.
Слизистая оболочка носовой поверхности
мягкого неба покрыта псевдомногослойным реснитчатым эпите-
лием, содержащим бокаловидные клетки, расположенные в собст-
венной пластинке слизистой оболочки. Собственная пластинка
слизистой оболочки здесь лишена сосочков и отделена от эпите-
лия хорошо выраженной базальной мембраной. На поверхности
эпителия открываются мелкие слюнные железы слизистого типа.
За собственной пластинкой следует слой эластических волокон.
Мышечная пластинка слизистой оболочки и подслизистая осно-
_ва отсутствуют. В месте перехода слизистой оболочки ротовой
поверхности неба в носовую эпителий становится сначала много-
слойным призматическим, а затем многорядным мерцательным.
Язык
Язык человека, помимо участия во вкусовом вос-
приятии, механической обработке пищи и ак-
те глотания, является органом речи. Основу языка состав-
ляет поперечнополосатая мышечная ткань соматического типа.
Язык покрыт слизистой оболочкой. Рельеф ее различен на
нижней, боковых и верхней поверхностях языка. Наиболее про-
стое строение имеет слизистая оболочка на н и ж ней
п о^£_р_х-тггге-т-и. Эпителий здесь многое пойньщ_плоский неоро-
грвевающий. Собственная пластинка слизистой оболочки вдается в
эпителий, образуя короткйе~~сосочки. За собственной пластинкой
следует подслиЗистая^основа,‘ которая прилежит непосредственно
к мьнццайСТэлагодаря наЛЙчию подсли’зйСтойПосновы слизистая
Оболочка нижней поверхности языка легко смещается.
Слизистая оболочка в е_р_х_а£ й и боковых
п о вер х н о с т е и яз ы к а неподвижно сращена сего мышеч-
ным__телом и снабжена особыми Образованиями — сосочками.
Подслизистая основа оттутствуе.т1 В языке человека имеется 4 ви-
"да сосочков: нитевидные (papillae filiformes), грибовидные (pa-
pillae fungiformes), желобоватые (papillae vallatae) и листовидные
(papillae foliatae). Все сосочки языка являются производными
слизистой оболочки и построены по общему плану (рис. 191).
Ловерхнесдъ сосочков образована многослойным плоским неорого-
вевающим или частично ороговевающимТй~иитекгз7Тных-с6сочках)
эпителием, ^жанщм^на^базальной^ёмб^де. Основу каждого со-
сочка составляет вырост (первичный сосочек) собственного со-
единигельнотканногсГслоя слизистой обшпучкигОт вершины первич-
ного сосочка отходитбт5до^и~и-белее^ТОнкйх'соединительноткан-
ных вторичных сосочков, вдающихся в эпителий. В соединительно-
Рис. 191. Схема микроскопического строения языка человека, продольный разрез
на разных уровнях (по В. Г. Елисееву и др.)
А — верхняя поверхность языка — спинка языка; Б средняя часть языка; В — нижняя
поверхность языка; /—кончик языка; // — боковая поверхность я лыка; /// — корень
языка; / — нитевидный сосочек; 2 — грибовидный сосочек; 3 — листовидный сосочек;
4 — вкусовые почки; 5 — серозные железы; 6 желобоватый сосочек; 7 — эпителий ва-
лика желобоватого сосочка; 8 — поперечнополосатая мышца; О — кровеносные сосуды;
10—смешанная слюнная железа; // — слизистая слюнная железа; — многослойный
плоский эпителий; /3 — собственная пластинка слизистой оболочки; — лимфатический
фолликул.
тканной основе сосочков языка расположены многочисленные кро-
веносные капилляры, просвечивающиеся через эпителий и при-
дающие сосочкам характерный красный цвет.
Нитевидные сосочки — самые многочисленные, равномерно
покрывают верхнюю поверхность языка, концентрируясь особенно
в углу, образованном сосочками, окруженными валом. По разме-
рам они наиболее мелкие среди сосочков языка. Длина их около
0,3 мм. Наряду с нитевидными сосочками встречаются конические
479
(papillae conicae). При ряде заболеваний процесс отторжения по-
верхностных ороговевающих эпителиоцитов может замедляться, а
эпителиальные клетки, накапливаясь в больших количествах на
вершинах сосочков, образуют мощные роговые пласты. Эти массы,
покрывая беловатой пленкой поверхность сосочков, создают кар-
тину языка обложенного белым налетом.
Грибовидные сосочки — немногочисленны и располагаются на
спинке языка среди нитевидных сосочков. Наибольшее их коли-
чество сосредоточено на кончике языка и пл-ещ_краям. Они бо-
лее крупных размеров, чем нитевидные, — Q,7—1,8 ум в длину и
около 0,4—1 мм в диаметре. Основная масса этИхТгосочков имеет
форму гриба с узким основанием и широкой вершиной Среди
них встречаются конические и линзовидные формы.
В толще эпителия находятся вкусовые почки (gemmae gusta-
toriae), располагающиеся чаще всего в области «шляпки» грибо-
видного сосочка. В срезах через эту зону в каждом грибовидном
сосочке обнаруживается до 3— 4 вкусовых почек. В некоторых
сосочках вкусовые почки отсутствуют.
Желобоватые сосочки (сосочки языка, окруженные валом)
встречаются на верхней поверхности языка в количестве от 6 до
12. Расположены они между телом и корнем языка вдоль погра-
ничной линии. Они хорошо различимы даже невооруженным гла-
зом. Их длина около 1 —1,5 мм, диаметр— 1—3 мм. В отличие от
нитевидных и грибовидных сосочков, отчетливо возвышающихся
над уровнем слизистой оболочки, верхняя поверхность этих со-
сочков лежит почти на одном уровне с нею. Они имеют узкое
основание и широкую, уплощенную свободную часть. Вокруг со-
сочка располагается узкая, глубокая щель — желобок (отсюда
название — желобоватый сосочек). Желоб отделяет сосочек от
валика — утолщения слизистой оболочки, окружающего сосочек.
Наличие этой детали в строении сосочка послужило поводом к
возникновению другого названия — «сосочек, окруженный валом».
В толще эпителия боковых поверхностей этого сосочка и окружаю-
сцего его' валика расположены многочисленные вкусовые почки.
В сое/itiнительной ткани сосочков и валиков часто встречаются
пучки гладких мышечных клеток, расположенных продольно, косо
или циркулярно. Сокращение этих пучков обеспечивает сближение
сосочка с валиком Это способствует наиболее полному соприкос-
новению пищевых веществ, попадающих в желоб, со вкусовыми
почками, заложенными в эпителии сосочка и валика. В рыхлой
волокнистой соединительной ткани основания сосочка и между
прилежащими к нему пучками поперечнополосатых волокон на-
ходятся концевые отделы слюнных белковых желез, выводные
протоки которых открываются в желоб. Секрет этих желез про-
мывает желоб сосочка и очищает его от скапливающихся в нем
пищевых частиц, слущивающегося эпителия и микробов.
Листовидные сосочки языка хорошо развиты только у детей.
Они представлены двумя группами, расположенными по правому и
левому краям языка. Каждая группа включает 4—8 параллельно
расположенных сосочков, разделенных узкими пространствами.
Длина одного сосочка около 2—5 мм. В эпителии боковых по-
верхностей сосочка заключены вкусовые почки. В пространства,
разделяющие листовидные сосочки, открываются выводные прото-
ки слюнных белковых желез. Их концевые отделы расположены
между мышцами языка. Секрет этих желез промывает узкие
пространства между сосочками. У взрослого человека листовидные
сосочки редуцируются, а на местах, где ранее располагались бел-
ковые железы, развиваются жировая и лимфоидная ткани.
Слизистая оболочка корня языка характери-
зуется отсутствием сосочков. Однако поверхность эпителия здесь
не ровная, а имеет ряд возвышений и углублений. Возвышения
образуются за счет скопления в собственной пластинке слизистой
оболочки лимфатических узелков,. достигающих иногда 0,5 см в
диаметре. Здесь же слизистая оболочка образуй! углубления —
крипты, в которые открываются протоки многочисленных слюнных
слизистых желез. Совокупность скоплений лимфоиднои ткани в
корне языка называется язычной миндалиной.
Мышцы языка образуют тело этого органа. Пучки по-
перечнополосатых мышц языка располагаются в трех взаимно
перпендикулярных направлениях: одни из них лежат вертикально,
другие продольно, третьи поперечно. Мускулатура языка разделе-
на на правую и левую половины плотной соединительнотканной
перегородкой. Рыхлая волокнистая соединительная ткань, лежа-
щая между отдельными мышечными волокнами и пучками, со-
держит много жировых долек. Здесь же расположены концевые
отделы слюнных желез языка. На границе между мышечным те-
лом и собственной пластинкой слизистой оболочки верхней по-
верхности языка имеется мощная соединительнотканная пластин-
ка, состоящая из переплетающихся наподобие решетки пучков
коллагеновых и эластических волокон. Она образует так назы-
ваемый сетчатый слой. Это своеобразный апоневроз языка, кото-
рый особенно сильно развит в области желобоватых сосочков.
На конце и у краев языка толщина его уменьшается. Поперечно-
полосатые мышечные волокна, проходя через отверстия сетчатого
слоя, прикрепляются к маленьким сухожилиям, образованным
лежащими в собственной пластинке слизистой оболочки пуч-
ками колллагеновых волокон.
Слюнные железы языка (gll. lingualis) делятся на т и
вида: белковые, слизистые и смешанные.
Белковые слюнные железы расположены поблизости от же-
лобоватых и листовидных сосочков в толще языка. Это^^простые
трубчатые разветвленные железы. Их выводные протоки открыва-
ются в ровики сосочков, окруженных валом, или между листовид-
ными сосочками и выстланы многослойным плоским эпителием,
иногда содержащим реснички. Концевые отделы представлены
разветвленными трубочками с узким просветом. Они состоят из
клеток конической формы, выделяющих белковый секрет, между
которыми проходят межклеточные секреторные капилляры.
481
16 гистология
Слизистые железы расположены главным образом в корне
языка и вдоль его боковых краев. Это одиночные простые аль-
веолярно-трубчатые разветвленные железы. Их протоки выстланы
многослойным эпителием, иногда снабжены ресничками. На корне
языка они открываются в крипты язычной миндалины. Трубчатые
концевые отделы этих желез состоят из слизистых клеток.
Смешанные железы располагаются в его переднем отделе.
Их протоки (около 6 млн.) открываются вдоль складок слизистой
оболочки под языком. Секреторные отделы смешанных желез рас-
положены в толще языка.
Кровоснабжение языка осуществляется входящими в него ар-
териями, которые обильно разветвляются и образуют широкую
сеть в мышцах языка. Они же дают ветви к поверхностным частям
языка. В сетчатом слое языка сосуды располагаются горизонталь-
но, а затем от них отходят вертикальные конечные ветви к сосочкам
слизистой оболочки. Конечные ветви формируют в соединительно-
тканных сосочках капиллярную сеть, от которой в каждый более
мелкий сосочек входит одна петля кровеносных капилляров. Кровь
от поверхностных слоев языка оттекает в венозное сплетение,
располагающееся в собственном слое слизистой оболочки. Более
крупное венозное сплетение находится в основании языка. Ма
ленькие лимфатические сосуды также образуют сеть в собствен
ной пластинке слизистой оболочки. Эта сеть связана с более кру1
ной сетью, имеющейся в подслизистой основе нижней поверхнос
ти языка.
Лимфатические сосуды в большом количестве находятся и ь
области миндалины языка.
Иннервация. Разветвления подъязычного нерва и барабанной
струны образуют многочисленные двигательные нервные оконча-
ния на поперечнополосатых мышечных волокнах. Чувствительная
иннервация передних 2/з языка осуществляется ветвями тройнич-
ного нерва, задней 1 /3 — ветвями языко-глоточного нерва. В собст-
венной пластинке слизистой оболочки языка имеется хорошо вы-
раженное нервное сплетение, от которого отходят нервные волок-
на к вкусовым почкам, эпителию, железам и сосудам. Нервные
волокна, входящие в эпителий, разветвляются среди эпителиаль-
ных клеток и заканчиваются свободными нервными окончаниями.
Иннервация вкусовых почек описана в гл. XIV.
- |/ Лимфоэпителиальное глоточное кольцо Пирогова.
Миндалины
На границе ротовой полости и глотки в слизистой оболочке
располагаются большие скопления лимфоидной ткани. В совокуп-
ности они образуют лимфоэпителиальное глоточное кольцо, окру-
жающее вход в дыхательные и пищеварительные пути. Наиболее
крупные скопления этого кольца носят название миндалин. По
месту их расположения различают небные миндалины, глоточную
миндалину, язычную миндалину. Кроме перечисленных миндалин,
4К2
в слизистой оболочке переднего отдела пищеварительной трубки
существует ряд скоплений лимфоидной ткани, из которых наибо-
лее крупными я вл-потея скопления в области слуховых труб —
трубные миндалины ч в желудочке гортани — гортанные мин-
далины.
Миндалины выполняют в организме важную защитную функ-
цию, обезвреживая микробы, постоянно попадающие из внешней
среды в организм через носовые и ротовые отверстия. Наряду с
другими органами, содержащими лимфоидную ткань, они обеспе-
чивают образование лимфоцитов, участвующих в реакциях гумо-
рального и клеточного иммунитета.
Развитие. Небные миндалины закладываются на 9-и неделе
эмбриогенеза в виде углубления псевдо много ело иного реснитча-
того эпителия латеральной стенки глотки, под которым лежат
компактно расположенные мезенхимные клетки и многочислен-
ные кровеносные сосуды. На 11—12-й неделе формируется тон-
зиллярный синус, эпителий которого перестраивается в много-
слойный плоский, а из мезенхимы дифференцируется ретикуляр-
ная ткань; появляются сосуды, в том числе посткапилдярные ве-
нулы с высокими эндотелиоцитами. Происходит заселение органа
лимфоцитами. На 14-и неделе среди лимфоцитов определяются
главным образом Т-лимфоциты (21%) и немного В-лимфоцитов
(1%). На 17—18-й неделе появляются первые лимфатические
узелки. К 19-й неделе содержание Т-лимфоцитов возрастает до
60%, а В-лимфоцитов—до 3%. Рост эпителия сопровождается
формированием в эпителиальных тяжах пробок из ороговевающих
клеток.
Глоточная миндалина развивается на 4-м месяце внутриутроб-
ного периода из эпителия и подлежащей мезенхимы дорсальной
стенки глотки. У эмбриона она покрыта многорядным мерцатель-
ным эпителием. Язычная миндалина закладывается на 5-м ме-
сяце.
Миндалины достигают максимального развития в детском воз-
расте. Начало инволюции миндалин совпадает с периодом полово-
го созревания.
Строение. Небные миндалины во взрослом организме представ-
лены двумя телами овальной формы, расположенными по рбеим
сторонам глотки между небными дужками. Каждая миндалина
состоит из нескольких складок слизистой оболочки, в собственной
пластинке которой расположены многочисленные лимфатические
узелки (noduli lymphathici) (рис. 192). От поверхности минда-
лины в глубь органа отходят 10—20 крипт (criptae tonsillares),
которые разветвляются и образуют вторичные крипты. Слизис-
тая оболочка покрыта многослойным плоским неорогове-
вающим эпителием. Во многих местах, особенно в криптах, эпите-
лий часто бывает инфильтрирован (заселен) лимфоцитами и зер-
нистыми лейкоцитами. Лейкоциты, проникающие в 1олщу эпите-
лия, обычно в большем или меньшем количестве выходят на его
поверхность и передвигаются навстречу бактериям, попадающим
483
• б*
Рис. 192. Небная миндали-
на (по Б. Б. Барышеву).
/ — многослойный плоский
неороговевающим эпителий:
2 — многослойный плоский
иеороговевающий эпителий,
инфильтрированный лимфо-
цитами; 3—собственная
пластинка слизистой оболоч-
ки; 4 — лимфатические узел-
ки; 5 — крипта.
в полость рта вместе с пищей и воздухом. Микробы в миндалине
активно фагоцитируются лейкоцитами, при этом часть лейкоцитов
погибает. Под влиянием микробов и различных ферментов, выде-
ляемых лейкоцитами, эпителий миндалины часто бывает разрушен.
Однако через некоторое время за счет размножения клеток элите
лиального пласта эти участки восстанавливаются.
Собственная пластинка слизистой о б о л о ч-
к и образует сосочки, вдающиеся в эпителий. В рыхлой волок-
нистой соединительной ткани этого слоя расположены многочис-
ленные лимфоидные узелки. В центрах некоторых узелков хорошо
выражены более светлые участки — герминативные центры. Лим-
фоидные узелки миндалин чаще всего отделены друг от друга тон-
кими прослойками соединительной ткани. Однако некоторые узел-
ки могут сливаться. Мышечная пластинка слизистой оболочки не
выражена.
Подслизистая основа, располагающаяся под скоп-
лением лимфоидных узелков, образует вокруг миндалины капсулу,
от которой в глубь миндалины отходят соединительнотканные
перегородки. В этом слое сосредоточены основные кровеносные и
лимфатические сосуды миндалины и ветви языкоглоточного нер-
ва, осуществляющие ее иннервацию. Здесь же находятся и секре-
торные отделы небольших слюнных желез. Протоки этих желез
открываются на поверхности слизистой оболочки, расположенной
вокруг миндалины. Снаружи от подслизистой основы лежат попе-
речнополосатые мышцы глотки — аналог мышечной обо-
лочки.
Глоточная миндалина. Расположена в участке дорсальной стен-
ки глотки, лежащем между отверстиями слуховых труб. Строение
ее сходно с другими миндалинами. Во взрослом организме она
выстлана многослойным плоским неороговевающим эпителием.
Однако в криптах глоточной миндалины и у взрослого иногда
встречаются участки псевдомногослойного реснитчатого эпителия,
характерного для эмбрионального периода развития.
4Н4
При некоторых патологических состояниях глоточная минда-
лина может быть очень сильно увеличена (аденоиды).
Язычная миндалина. Расположена в слизистой оболочке кор-
ня языка. Эпителий, покрывающий поверхность миндалины и
выстилающий крипты, многослойный плоский неороговевающий.
Эпителий и подлежащая собственная пластинка слизистой оболоч-
кТГТтфильтрированы лимфоцитами* проникающими сюда из лим-
|фатйческих узелков. На дне многих крипт открываются выводные
протоки слюнных желез языка. Их секрет способствует промы-
• ванию й ’Очищению крипт.
Слюнные железы
Общая морфофункциональная характеристика. В ротовую
полость открываются выводные протоки трех пар больших слюн-
ных желез:. околоушных, подчелюстных и подъязычных, лежа-
щих за пределами слизистой оболочки. Кроме того, в толще сли-
зистой оболочки ротовой полости находятся многочисленные мел-
кие слюнные железы: губные, щечные, язычные, небные.
Эпителиальные структуры всех слюнных желез развиваются
из эктодермы, как и многослойный плоский эпителий, выстилаю-
щий ротовую полость. Поэтому для строения их выводных прото-
ков и секреторных отделов характерна многослойность.
Слюнные железы представляют собой сложные альвеолярные
или альвеолярно-трубчатые железы. Они состоят из концевых
отделов и протоков, выводящих секрет (рис. 193).
Концевые отделы fportio terminals) по строению и характеру
выделяемого секрета бывают трех типов: белковые (серозные),
слизистые и смешанные (т. е. белково-слизистые).
Выводные протоки слюнных желез подразделяются на внут-
ридольковые (ductus interlobularis), включающие вставочные (duc-
tus intercalatus) и исчерченные (ductus striatus), междольковые
(ductus interlobularis) выводные протоки и протоки железы (duc-
tus excretorius seu glandulae) /см. рис. 193).
Белковые железы выделяют жидкий секрет, богатый фермен-
тами. Слизистые железы образуют более густой, вязкий секрет с
большим содержанием муцина — вещества, в состав которого вхо-
дят гликозаминогликаны. По механизму отделения секрета_______из
клеток все слюнные железы мерокриновые.
— Слюнные железы выполняют экзокринные и эндокринные
функции. Экзокринная функция заключается в ре-
гулярном отделении в ротовую полость слюны. В ее состав входят
вода (около 99%), белковые вещества, в том числе ферменты,
неорганические вещества, а также клеточные элементы (клетки
эпителия, лейкоциты).
Слюна увлажняет пищу, придает ей полужидкую консистен-
цию, что облегчает процессы жевания и глотания. Постоянное
смачивание слизистой оболочки щек и губ слюной способствует
акту артикуляции. Одной из важных функций слюны яв-
4К5
рис. 193. Схема строения слюнных желез (по В. Г. Елисееву и др.).
А — долька подчелюстной железы: Б — долька подъязычной железы; В — долька около-
ушной железы; Г — поперечное сечение различных отделов околоушной железы: / — вывод-
ной проток железы; 2 — исчерченный проток; 3 — вставочные протоки; 4 — белковый кон-
певой отдел: 5 — слизистый концевой отдел; 6 — смешанный концевой отдел (белково-
слизистый): а — мукоциты (слизистые клетки); б — сероциты (белковые клетка); 7 —
миоэпителиоциты; 8 - серозное полулуние.
ляется ферментативная обработка пищи. Ферменты слюны могут
участвовать в расщеплении полисахаридов (амилаза, мальтаза,
гиалуронидаза), нуклеиновых кислот и нуклеопротеидов (нуклеазы
и калликреин), белков (калликреинеподобные протеазы, пепсино-
ген, трипсиноподобные ферменты), клеточных оболочек (лизо-
цим). Помимо секреторной функции, слюнные железы выполняют
экскреторную функцию. Со слюной во внешнюю среду выделяют-
ся различные органические и неорганические вещества: мочевая
кислота, креатин, железо, йод и др. Защитная функция слюнных
желез состоит в выделении бактерицидного вещества — лизоцима.
Эндокринная функция слюнных желез обеспечивает-
4К6
ся наличием в слюне биологически активных веществ типа гормо-
нов— инсулина, паротина, фактора роста нервов (ФРН), фактора
роста эпителия (ФРЭ), тимоциттрансформирующего фактора
(ТТФ), фактора летальности и др. Слюнные железы активно
участвуют в регуляции водно-солевого гомеостаза.
Развитие. Закладка околоушных желез происходит
на 8-й неделе эмбриогенеза, koi да из эпителия ротовой полости
в подлежащую мезенхиму начинают расти эпителиальные тяжи в
направлении к правому и левому ушным отверстиям. От этих тя-
жей отпочковываются многочисленные выросты, формирующие
сначала выводные протоки, а затем концевые отделы. На 10—
12-й неделе имеется система разветвленных эпителиальных тяжей,
врастание нервных волокон. На 4—6-м месяце развития формиру-
ются концевые отделы альвеол, а к 8—9-му месяцу в них появля-
ются просветы. Вставочные протоки и концевые отделы у плодов
и детей до двухлетнего возраста представлены типичными слизис-
тыми клетками. Из мезенхимы к 5—5'/2 мес эмбриогенеза диф-
ференцируются соединительнотканная капсула и прослойки меж-
дольковой соединительной ткани. Сначала секрет имеет слизистый
характер. В последние месяцы развития слюна плода проявляет
амилолитическую активность.
Подчелюстные железы закладываются на 6-й неделе
эмбриогенеза. На 8-й неделе в эпителиальных тяжах образуются
просветы. Эпителий первичных выводных протоков сначала двух-
слойный, затем многослойный. Концевые отделы формируются на
16-й неделе. Слизистые клетки концевых отделов образуются в
процесе ослизнения клеток вставочных протоков. Процесс диффе-
ренцировки концевых отделов и внутридольковых протоков на
вставочные отделы и слюнные трубки продолжается в постнаталь-
ный период развития. У новорожденных в концевых отделах
сформированы элементы, состоящие из железистых клеток куби
ческой и призматической формы, образующих белковый секрет
(белковые полулуния). Секреция в концевых отделах начинаете»’
у 4-месячных плодов. Состав секрета отличается от секрета взрос-
лого человека^/I I од ъ язычные железы закладываются
^’0 неделе эмбриогенеза в виде отростков от оральны . кон-
цов подчелюстных желез. На 12-й неделе отмечается почкоьание
и ветвление эпителиального зачатка. . ♦ ’
Строение слюнных желез. Околоушная железа (gl. parotis) —
сложная альвеолярная разветвленная железа, выделяющая белко-
вый секрет в ротовуюполость, а также обладающая эндокринной
функцией. Снаружи она покрыта плотной соединительнотканной
капсулой. Железа имеет выраженное дольчатое строение. В про
слойках соединительной ткани между дольками расположены
междольковые протоки и кровеносные сосуды (рис. 194).
Концевые отделы околоушной железы белковые (се-
розные) (см. рис. 193). Они состоят из секреторных клеток ко-
нической формы — белковых клеток, или сероцитов (serocyti), и
~мйоэпителиальных клеток. Сероциты имеют узкую апикальную
487
Рис. 194. Околоушная железа (препарат Н. А. Юриной).
А — долька железы; / — междольковая рыхлая волокнистая соединительная ткань;
2 кровеносный сосуд; 3 — концевой отдел: 4 — миоэпителиальная клетка; 5 — вставоч-
ный проток; 6 — исчерченный проток; 7 междольковый проток.
часть, выступающую в просвет концевого отдела. В ней содержат-
ся ацидофильные секреторные гранулы, количество которых из-
меняется в зависимости от фазы секреции. Базальная часть клетки
более широкая, содержит ядро. В фазе накопления секрета разме-
ры клеток значительно увеличиваются, а после выделения его
уменьшаются, ядро округляется. В секрете околоушных желез
преобладает белковый компонент, но часто содержатся и муко-
полисахариды, поэтому такие железы могут быть названы серо-
мукозными. В секреторных гранулах выявляются а^а’милаза,
ДНК-аза. Цитохимически и электронно-микроскопически различа-
ют несколько типов гранул — ШИК-положительные с электронно-
плотным ободком1, ШИК-отрицательные и мелкие гомогенные
сферической формы. Между сероцитами в концевых отделах око-
лоушной железы располагаются межклеточные секреторные ка-
' ШИК — реактив Шиффа и йодная кислота, применяются для гистохимиче-
ского выявления углеводов.
488
Рис. 194 (продолжение).
Ультрамикроскопическое строение клеток различных отделов околоушной железы
(по Е. А. Шубниковой): ацинуса (F), вставочного протока (В), исчерченного протока
(Г. Д). I — секреторные гранулы; 2 — гранулярная эндоплазматическая сеть; 3 — ядро;
4 межклеточные секреторные канальцы; 5 — комплекс Гольджи; 6 — складки базальной
плазмолеммы; 7 — тонофиламенты; 8 — митохондрии; 9 — миоэпителиоцит; 10 — базаль-
ная мембрана.
пальцы, просвет которые имеет диаметр около 1 мкм. В эти ка-
нальцы выделяется из клеток секрет, который далее поступает в
просвет концевого секреторного отдела. Общая секреторная пло-
щадь концевых отделов обеих желез достигает почти 1,5 м2.
Миоэпителиальные клетки (миоэпителиоциты) составляют вто-
рой слой клеток в концевых секреторных отделах. По происхож-
дению это эпителиальные клетки, по функции — сократительные
элементы, напоминающие мышечные. Их называют также звезд-
чатыми эпителиоцитами, так как они имеют звездчатую форму и
489
своими отростками охватывают концевые секреторные отделы на-
подобие КбрзИПГТК (см. рис. 193). Миоэпителиальные клетки всегда
располагаются между базальной мембраной и основанием эпи-
телиальных клеток. Своими сокращениями они способствуют вы-
делению секрета из концевых отделов.
Внутридольковые вставочные протоки околоушной железы
начинаются непосредственно от ее концевых отделов. Они обычно
сильно разветвлены (см. рис. 193, 194). Вставочные протоки выс-
тланы кубическим или плоским эпителием. Второй слой в них
образуют миоэпителиоциты. В клетках, прилежащих к ацинусу, об-
наруживаются электронно-плотные гранулы, содержащие мукопо-
лисахариды, здесь же расположены тонофиламенты, рибосомы и
агранулярная эндоплазматическая сеть (см. рис. 194).
Исчерченные слюнные протоки являются продолжением вста-
вочных и располагаются также внутри долек. Диаметр их зна-
чительно больший, чем вставочных протоков, просвет хорошо вы-
ражен. Исчерченные протоки ветвятся и часто образуют ампуляр-
ные расширения. Они выстланы однослойным призматическим эпи-
телием^ Цитоплазма эпителиоцитов ацидофильна. В апикальной
части клеток видны микроворсинки, секреторные гранулы с содер-
жимым различной электронной плотности, комплек- Гольджи.
В базальных частях эпителиальных клеток отчетливо выявляется
базальная исчерченность, образованная митохондриями, располо-
женными в цитоплазме между складками цитолеммы перпендику-
лярно к базальной мембране (см. рис. 194). В исчерченных отделах
выявлены циклические изменения, не связанные с ритмом пище-
варительного процесса.
Междольковые выводные протоки выстланы двухслойным эпи-
телием. По мере укрупнения протоков эпителий их постепенно
становится многослойным. Выводные протоки окружены прослой-
ками рыхлой волокнистой соединительной ткани.
Проток околоушной железы, начинающийся в ее теле, прохо-
дит через жевательную мышцу, а его устье расположено на по-
верхности слизистой оболочки щеки на уровне второго верхнего
большого коренного зуба. Проток выстлан многослойным кубиче-
ским, а в устье — многослойным плоским эпителием.
Подчелюстная железа (gl. submaxilare). Это — сложная аль-
веолярная, местами альвеолярно-трубчатая разветвленная железа.
По характеру отделяемого секрета она смешанная, т. е. белково-
слизистая. С поверхности железа окружена соединительноткан-
лой капсулой.
Концевые секреторные отделы подчелюстной железы двух ти-
пов: белковые и белково-слизистые, т. е. смешанные (рис. 195).
В подчелюстной железе преобладают белковые концевые отделы.
Секреторные гранулы сероцитов дискретны и обладают невысокой ,
электронной плотностью. Часто внутри гранул содержится элек-
тронно-плотная сердцевина. Их строение сходно с концевыми от-
1 Второй слой образован миоэпителиоцитами.
490
делами околоушной железы. Концевые отделы (ацинусы) состоят
из 10 18 серомукозных клеток, из которых лишь 4—6 клеток
расположены вокруг просвета ацинуса. Секреторные гранулы содер-
жат_гликолипиды и гликопротеиды. Смешанные концевые отделы
более крупные, чем белковые, и состоят из двух видов клеток —
слизиЬтых и белковых. Слизистые клетки fmucocyti) более круп-
ные по сравнению с белковыми и занимают центральную часть
концевого отдела. Ядра слизистых клеток располагаются всегда
у их основания, они сильно уплощены и уплотнены. Цитоплазма
этих клеток имеет ячеистую структуру благодаря наличию в ней
слизистого секрета, который избирательно окрашивается муцикар-
мином и электронно-прозрачен. Небольшое количество белковых
клеток охватывает слизистые клетки в виде колпачка, или сероз-
ного полулуния (semilunium serosum). Белковые (серозные) по-
лулуния являются характерными структурами смешанных желез
Между железистыми клетками расположены межклеточные сек-
реторные канальцы. Снаружи от клеток полулуния лежат мио-
эпителиальные клетки.
Вставочные протоки подчелюстной железы менее разветвлен-
ные и более короткие, чем в околоушной железе, что объясняется
ослизнением части этих отделов в процессе развития. Клетки этих
отделов содержат мелкие секреторные гранулы, часто с мелкими
плотными сердцевинами.
Исчерченные протоки в подчелюстной железе очень хорошо
развиты, длинные и сильно ветвятся (см. рис. 193). В них часто
встречаются сужения и баллонообразные расширения. Выстилаю-
щий их призматический эпителий с хорошо выраженной базаль-
ной исчерченностью содержит желтый пигмент. Среди клеток при
электронной микроскопии различают несколько типов — широкие
темные, высокие светлые, мелкие треугольной формы (малодиф-
ференцированные) и клетки, имеющие форму бокала. В базаль-
ной части высоких клеток на боковых поверхностях расположены
многочисленные цитоплазматические выросты. У некоторых жи-
вотных (грызуны), кроме исчерченных протоков, имеются грану-
лярные отделы, клетки которых содержат хорошо развитый комп-
лекс Гольджи, расположенный в базальном отделе клеток, и гра-
нулы, содержащие трипсиноподобные протеазы, а также ряд гор-
мональных и ростстимулирующих факторов. Как установлено, эн-
докринные функции слюнных желез (выделение инсулиноподоб-
ного и других веществ) связаны с этими отделами.
Междольковые выводные протоки подчелюстной железы, рас-
полагающиеся в соединительнотканных перегородках, выстланы
сначала двухслойным, а затем многослойным эпителием. Проток
подчелюстной железы открывается рядом с протоком подъязыч-
ной железы на переднем крае уздечки языка. Его устье выстлано
многослойным плоским эпителием. Проток подчелюстной железы
сильнее разветвлен, чем проток околоушной железы.
Подъязычная железа (gl. sublinguale). Это — сложная альвео-
лярно-трубчатая разветвленная железа. По характеру отделяемого
Рис. 195. Строение подчелюстной железы.
Z — микрофотография дольки железы; I — смешанный концевой отдел: а — мукоциты; б —
сероциты, образующие серозное полулуние; 2 — миоэпителиальная клетка, 3 исчерченный
проток; 4 междольковая рыхлая волокнистая соединительная ткань (по Л. И. Фалину)
секрета это .смешанная, слизисто-белковая железа, с преоблада-
нием слизистой секреции. В ней имеются концевые секреторные
отделы трех типов: белковые, смешанные и слизистые.
Белковые концевые отделы очень немногочисленны (см.
рис. 193). Смешанные концевые отделы составляют основную
массу железы и состоят из белковых полулуний и слизистых кле-
ток. Полулуния, образованные серомукозными клетками, в них
выражены лучше, чем в подчелюстной железе. Клетки, форми-
рующие полулуния, в подъязычной железе значительно отлича-
ются от соответствующих клеток в околоушной и подчелюстной же-
лезах. Их секреторные гранулы дают реакцию на муцин. Эти клетки
выделяют одновременно белковый и слизистый секрет и потому но-
сят название серомукозных клеток. В них сильно развита грану-
лярная эндоплазматическая сеть. Они снабжены межклеточными
"секреторными канальцами. Чисто слизистые концевые отделы
этой железы состоят из характерных слизистых клеток, содержа-
492
Рис. 195 (продолжение).
П — схема ультрамикроскопического строения клеток ацинуса (Л), вставочного протока
(£>>. исчерченного протока (В) (по Е. А. Шубниковой), / — секреторные гранулы: 2 —
гранулярная эндоплазматическая сеть; 3 — ядро; 4 — межклеточный каналец; 5 — комплекс
Гольджи; 6 — складка базальной плазмолеммы; 7 — базальная мембрана; 8 — митохондрии:
9 — миоэпителиальная клетка; 10 — светлая клетка; // — темная клетка.
щих хондроитинсульфат В и гликопротеиды. Миоэпителиальные
элементы образуют наружный слой во всех типах концевых от-
делов.
В подъязычной железе общая площадь вставочных протоков
очень мала, так как они еще в процессе эмбрионального развития
почти целиком ослизняются, формируя слизистые части концевых
отделов. Исчерченные протоки в этой железе развиты слабо: они
очень короткие, а в некоторых местах отсутствуют. Эти протоки
выстланы призматическим или кубическим эпителием, в котором
также видна базальная исчерченность, как и в соответствующих
протоках других слюнных желез.
В цитоплазме эпителиальных клеток, выстилающих исчерчен-
ные протоки, содержатся мелкие пузырьки, которые рассматри-
ваются как показатель экскреции.
Внутридольковые и междольковые выводные протоки подъ-
язычной железы образованы двухслойным призматическим, а в
устье — многослойным плоским эпителием. Соединительноткан-
ные внутридольковые и междольковые перегородки в этих желе-
зах развиты лучше, чем в околоушных или подчелюстных железах.
Васкуляризация. Все слюнные железы богато снабжены сосу-
дами. Артерии, входящие в железы, сопровождают разветвления
выводных протоков. От них отходят веточки, питающие стенки
протоков. У концевых отделов мелкие артерии распадаются на
капиллярную сеть, густо оплетающую каждый из этих отделов.
Из кровеносных капилляров кровь собирается в вены, которые
следуют по ходу артерий.
Для кровеносной системы слюнных желез характерно наличие
значительного количества артериоловенулярных анастомозов. Они
располагаются в воротах железы, при входе сосудов в дольку и
перед капиллярными сетями концевых отделов. Анастомозы в
слюнных железах обусловливают возможность значительного из-
менения интенсивности кровоснабжения отдельных концевых от-
делов, долек и даже всей железы, а следовательно, изменения
секреции в слюнных железах.
Иннервация. Эфферентные, или секреторные, волокна крупных
слюнных желез происходят из двух источников: отделов парасим-
патической и симпатической нервной системы. Гистологически в
железах обнаруживаются миелиновые if безмиелиновые нервы,
следующие по ходу сосудов и протоков. Они образуют нервные
окончания в стенках сосудов, на концевых отделах и в выводных
протоках желез. Морфологические различия между секреторны-
ми и сосудистыми нервами не всегда можно определить. В экспе-
риментах на подчелюстной железе животных было показано, что
вовлечение в рефлекс симпатических эфферентных путей ведет к
образованию вязкой слюны, содержащей большое количество сли-
зи. При раздражении парасимпатических эфферентных путей об-
разуется жидкий белковый секрет. Закрытие и открытие просвета
артериоловенулярных анастомозов и конечных вен также опреде-
ляется нервными импульсами.
494
Возрастные изменения. После рождения процессы морфоге-
неза в околоушных слюнных железах продолжаются до 16—
20 лет; при этом железистая ткань преобладает над соединитель-
ной тканью. После 40 лет отмечаются инволютивные изменения,
характеризующиеся уменьшением объема железистой ткани, уве-
личением жировой ткани, сильным разрастанием соединительной
ткани. В течение первых 2 лет жизни в околоушных железах вы-
рабатывается в основном слизистый секрет, с 3-го года до глубо-
кой старости — белковый, а к 80-м годам опять преимущественно
слизистый.
В подчелюстных железах полное развитие серозных и слизис-
тых секреторных отделов наблюдается у 5-месячных детей. Мак-
симальное развитие их отмечается к 25 годам. После 50 лет на-
чинаются инволютивные изменения. Рост подъязычных желез, как
и других, наиболее интенсивно происходит в течение первых двух
лет жизни.
Регенерация. Функционирование слюнных желез неизбежно
сопровождается частичным разрушением эпителиальных железис-
тых клеток. Отмирающие клетки характеризуются крупными раз-
мерами, пикнотическими ядрами и плотной зернистой цитоплаз-
мой, сильно окрашивающейся кислыми красителями. Такие клет-
ки получили название набухающих. Восстановление клеток па-
ренхимы осуществляется главным образом путем внутриклеточной
регенерации.
I/ Зубы —
Зубы являются частью жевательного аппарата. У человека
они появляются двумя генерациями: вначале образуются выпадаю-
щие, или молочные, зубы, а затем постоянные. В лунках челюст-
ных костей зубы укрепляются плотной соединительной тканью —
периодонтом, который в области шейки зуба образует циркуляр-
ную зубную связку. Коллагеновые волокна зубной связки имеют
преимущественно радиальное направление. С одной стороны они
проникак'1 в цемент корня зуба, а с другой — в альвеолярную
кость. Периодонт выполняет не только механическую, но и тро-
фическую функцию, так как в нем проходят кровеносные сосуды,
питающие корень зуба.
Развитие. Молочные зубы у человека начинают разви-
ву гься в конце 2-го месяца внутриутробного периода, когда в
эпителии ротовой полости Возникает вестибулярная зубная плас-
тинка, отделяющая губы и щеки от десен, в результате чего об-
разуется преддверие ротовой полости.
В области закладки однокоренных зубов от дна преддверия
растет второе эпителиальное выпячивание в виде валика, превра-
щающегося в зубную пластинку (lamina dentalis). Зубная пластин-
ка в области закладки многокорневых зубов развивается самосто-
ятельно непосредственно из эпителия ротовой полости. На внут-
ренней поверхности зубной пластинки сначала появляются эпи-
495
телиальные скопления — зубные зачатки (germen dentis), из ко-
торых развиваются эмалевые органы (organum enamehum). В
дальнейшем навстречу каждой почке начинает расти мезенхима в
виде зубного сосочка (papilla dentis), вдавливающаяся в эпите-
лиальный орган (рис. 196), который становится похожим на двух-
стенныи бокал или колпачок. Впоследствии он постепенно отделя-
ется от зубной пластинки. Клетки эпителиального эмалевого ор-
гана дифференцируются на три вида: внутренние, наружные и
промежуточные. Внутренний эмалевый эпителий,
примыкающий к зубному сосочку, становится высоким и приоб-
ретает характер призматического эпителия (рис. 197, А, Б). Впо-
следствии он образует эмаль (enamelum), в связи с чем клетки
этого эпителия и получили название энамелобластов (ename-
loblasti, ameloblasti). Наружный эмалевый эпителий
в процессе дальнейшего роста органа уплощается, а клетки про-
межуточного слоя приобретают звездчатую форму вслед-
ствие накопления между ними жидкости. Так образуется пульпа
эмалево*х> органа, которая позднее принимает участие в образо-
вании кутикулы эма ш (cuticula enameli).
Мезен-.има, окружающая закладку зуба и зубной сосочек, по
мере развития зачатка зуба также подвергается значительным
изменениям. Она уплотняется и образует зубной мешочек fsaccu-
lus dentis). В конце 3-го месяца эмалевый орган полностью отде-
ляется от зубной пластинки.
К 4-му месяц) эмбрионального развития начинается гистогенез
дентина (рис. 198). Дифференцировка образователей дентина —
дентинобластов начинается раньше и активнее протекает на вер-
шине зуба, а позднее на боковых поверхностях. Дентинобласты —
клетки мезенхимной природы, высокие призматические клетки с
четко выраженной полярной дифференциацией. Их верхушечная
часть имеет отростки, через которые происходит секреция орга-
нических веществ, образующих матрицу дентина — предентин.
Преколлагеновые и коллагеновые фибриллы матрицы имеют ра-
диальное направление. Это мягкое вещество заполняет промежут-
ки между дентинобластами и внутренними клетками эмалевого
органа — энамелобластами. Количество предентина постепенно
увеличивается. Позднее, когда происходит кальцификация дентина,
эта зона входит в состав плащевого дентина. В стадии обызвест-
вления дентина соли кальция, фосфора и других минералтых
веществ откладываются в виде глыбок, которые объединяются в
глобулы. В дальнейшем развитие дентина замедляется, а около
пульпы появляются тангенциальные коллагеновые волокна около-
пульпарного дентина.
В конце 5-го месяца эмбрионального развития в предентине
зачатка зуба начинается отложение известковых солей и форми-
рование окончательного дентина. Однакр4 процесс обызвествления
предентина при этом не захватывает участки, окружающие апи-
кальные отростки дентинобластов. Это ведет к возникновению
системы радиальных каналов, идущих от внутренней поверхности
496
4
5
6
5
4
Рис. 196. Развитие и прорезывание молочных зубов и развитие постоянных зубов
(схема по А. Хэму и Д Кормаку).
Л _ £ _ последовательные стадии I закладка молочного зуба; 2 участок мезенхимы
3 _ зубной сосочек, 4 — нижняя челюсть; 5 — челюстно губное выпячивание, 6 язык
7 — закладка нижней челюсти 8 эмалеаый орган 9 эмаль. /О — дентин II пульпа.
12 — закладка постоянного эуба, 13 зона резорбции корня молочного зуба
fl
Рис. 197. Ультра микроскоп и чес кое строение эна-
мелобласюв (рис Ю. И. Афанасьева).
А — схема; / — эмаль; 2 — гранулы в дистальных от-
делах амелобластов; 3 эндоплазматическая сеть, 4
комн зеке Гольджи; 5 — ядро; 6 митохондрии.
дентина к наружной (рис. 199). Кроме
того, участки предентина на границе с
эмалью также остаются необызвествлен-
ными и носят название интерглобуляр-
ных пространств.
Параллельно развитию дентина в за-
кладке зуба идет процесс дифференци-
ровки пульпы, в которой с помощью фиб-
робластов постепенно образуется основ-
ное вещество, содержащее преколлаге-
новые и коллагеновые волокна. Гисто-
химически в периферической части пуль-
пы, в области расположения дентино-
бластов и предентина, обнаруживаются
ферменты, гидролизующие фосфатные
соединения (фосфогидролазы), благо-
даря которым фосфатные ионы достав-
ляются дентину и эмали. Образование
эмали происходит несколько позже.
I Энамелобласты — клетки эпителиальной
* природы, высокие, призматической фор-
мы, с хорошо выраженной полярной
дифференциацией. Первые зачатки эма-
ли появляются в виде кутикулярных
пластинок на поверхности энамелоблас-
тов,'обращенных к дентину в области ко-
ронки зуба (см. рис. 197). По ориентации
эта поверхность базальная. Однако с на-
чалом эмалеобразования происходит пе-
ремещение, или инверсия, ядра и орга-
нелл клетки (центросома и комплекс
ролоджи) в противоположный конец
клетки. В результате базальная часть
энамелобластов становится как бы апи-
кальной, а апикальная — базальной.
После такого изменения полюсов клеток
и питание их, очевидно, также начинает
I осуществляться со стороны промежуточ-
ного слоя эмалевого органа, а не со сто-
роны дентина В подъядерной зоне эн амелобластов обнаруживаются
большое количество рибонуклеиновой кислоты, а также гликоген и
высокая активность щелочной фосфатазы. Кутикулярные пластинки
на энамелобластах при фиксации обычно сморщиваются и видны
как штифтики или отростки.
При дальнейшем образовании эмали в прилежащих к отрост-
498
,Б
Рис. 197 (продолжение).
Б электронная микрофотография эмалевых призм. X 30 000. / — поперечно перерезан-
ные эмалевые призмы; 2 - продольно перерезанные эмалевые призмы; 3 — кристаллы
гидроксиапатитов (по Трэвист и Глимчер).
Рис. 198. Схема ультра микроскопичен
кого строения дентинобластов (рис.
Ю. И. Афанасьева).
/ - дентин; 2 дистальный отросток ден-
тинобласта; 3 предеитин; 4 — митохонд-
рии; 5 — комплекс Гольджи; 6 — эндоплаз-
матическая сеть; 7 — ядро.
кам участках цитоплазмы энаме-
лобластов появляются гранулы,
которые постепенно перемеща-
ются в отростки, после чего на-
чинаются их кальцинация и об-
разование предэмалевых призм.
При дальнейшем развитии эмали
энамелобласты уменьшаются в
размерах и отодвигаются от ден-
тина. К завершению этого про-
цесса, примерно к моменту про-
резывания зубов, энамелоблас-
ты peiKO уменьшаются и реду-
цируются, а эмаль оказывается
покрытой лишь тонкой оболоч-
кой — кутикулой, образованной
клетками промежуточного слоя
пульпы. Наружные клетки эма-
левого органа при прорезывании
зуба сливаются с эпителием дес-
ны и в дальнейшем разрушают-
ся. С появлением эмалевых
призм поверхность дентина де-
лается неровной. Частичная ре-
зорбция дентина, очевидно, спо-
собствует укреплению его связи
с эмалью и усилению кальцина-
ции эмали высвободившимися
солями кальция.
Развитие цемента происхо-
дит позднее эмали, незадолго до
прорезывания зубов, из окружающей зубной зачаток мезенхимы,
образующей зубной мешочек. В них различают два слоя: более плот-
ный — наружный и рыхлый — внутренний. В процессе развития
цемента во внутреннем слое зубного мешочка в области корня из ме-
зенхимы дифференцируются цементобласты. Цементобласты, по-
добно остеобластам и денгинобластам, синтезируют коллагеновые
белки, которые выделяют в межклеточное вещество. По мере разви-
тия межклеточного вещества цементобласты превращаются в от-
ростчатые цементоциты, которые погружаются в межклеточное ве-
щество. Цементоциты расположены в полостях и отходящих от них
канальцах. Наружный слой зубного мешочка превращается в зубную
5(10
Рис. 199. Верхушка зачатка верхнего
резца 5-месячного плода (препарат
Л. И Фалина).
/ — эиамелобласты; 2 — эмаль; 3 — дентин;
4—предентин; 5 — слой дентинобластов;
6 - зубной сосочек.
связку — периодонт. Таким об-
разом, эмалевый орган выполня-
ет прежде всего морфогенетиче-
скую роль, определяя форму
развивающегося зуба.
Закладка постоянных
зубов происходит в конце
4-го — начале 5-го месяца внут-
риутробного развития. Зачатки
постоянных зубов образуются
также из зубной пластинки и
подлежащей мезенхимы. Зача-
ток постоянного зуба находится
позади каждого зачатка молоч-
ного зуба. Прорезывание молоч-
ных зубов у ребенка начинается
на 6—7-м месяце жизни. К этому времени сформирована только
коронка зуба, а формирование корня лишь начинается. Молочные
большие коренные зубы (моляры) заменяются постоянными ма-
лыми коренными (премолярами).
Закладка постоянных больших коренных зубов происходит на
1—4-м году жизни. Сначала оба зуба (молочный и постоянный)
лежат в общей альвеоле. Затем между ними появляется костная
перегородка.
Постоянный зуб развивается очень медленно. Когда наступа-
ет время выпадения молочных зубов, т. е. в возрасте 6—7 лет,
остеокласты разрушают эту перегородку и корень выпадаю-
щего зуба, а постоянный зуб начинает усиленно развиваться. Так
же как и молочные, постоянные зубы выталкиваются (прорезыва-
ются) под давлением, которое создается в пульпе зуба в связи с
образованием основного вещества соединительной ткани. До про-
резывания зубов минеральные вещества (кальций, фосфор, фтор
и др.) и питательные вещества поступают только из крови. После
прорезывания в этих процессах возрастает роль слюны и соответ-
ственно ее химического состава.
Строение. Зуб состоит из твердых и мягких частей. В твердой
части зуба различают эмаль, дентин и цемент; мягкая часть зуба
представлена так называемой пульпой.
Эмаль (enamelum) покрывает коронку зуба. Наибольшего
развития она достигает у вершины коронки (до 3,5 мм). Эмаль
содержит незначительное количество органических веществ (около
3—4%) и неорганические соли (96—97%). Среди неорганических
SOI
веществ подавляющую часть составляют фосфаты и карбонаты
кальция и около 4% — фторид кальция. Эмаль построена из эма-
левых прим (prisma enameli) толщиной 3—5 мкм. Каждая призма
состоит из тонкой фибриллярной сети, в которой находятся крис-
таллы гидрооксиапатитов, имеющих вид удлиненных призм (см.
рис. 197)7 Призмы располагаются пучками, имеют извитой ход и
залегают почти перпендикулярно к поверхности дентина. На попе-
речном срезе~эмалевые призмы обычно имеют многогранную или
вогнутовыпуклую форму. Между призмами находится менее обыз-
вествленное склеивающее вещество. Благодаря S-образно изогну-
тому ходу призм на продольных шлифах зуба одни из них оказы-
ваются рассеченными более продольно, а другие — более попереч-
но, что обусловливает чередование светлых и темных эмалевых по-
лос. На продольных шлифах можно видеть еще более тонкие па-
раллельные линии. Их появление связывают с периодичностью
Проста и различной зональной обызвествленностью призм, а также
с отражением в структуре эмали силовых линий, возникающих в
результате действия силового фактора во время жевания.
Снаружи эмаль покрыта тонкой кутикулой (cuticula enameli),
которая на жевательной поверхности зуба быстро стирается и ос-
тается заметной лишь на его боковых поверхностях. Химический
состав эмали меняется в зависимости от обмена веществ в орга-
низме, интенсивности растворения кристаллов гидрооксиапатита и
реминерализации органической матрицы. В определенных пределах
эмаль проницаема для воды, ионов, витаминов, глюкозы, амино-
кислот и других веществ, поступающих непосредственно из полос-
ти рта. При этом большую роль играет слюна не только как источ-
ник поступления различных веществ, но и как фактор, активно
влияющий на процесс проникновения их в гкани зуба. Проницае-
мость повышается под действием кислот, кальцитонина, спирта,
дефицита в пище солей кальция, фосфора, фтора и др. Эмаль и
дентин соединяются с помощью взаимных интердигитаций.
Дентин (dentinum) образует большую часть коронки, шейки
и корня зубов. Он состоит из органических и неорганических ве-
ществ; органического вещества 28% (главным образом коллагена),
неорганических веществ 72% (главным образом фосфат кальция и
магния с примесью фторида кальция).
Дентин построен из основного вещества, которое пронизано
трубочками, или канальцами (tubuli den(inalis) (рис. 200). Основ-
ное вещество дентина содержит коллагеновые фибриллы и распо-
ложенные между ними мукопротеины. Коллагеновые фибриллы в
дентине собраны в пучки и имеют преимущественно два направле-
ния: радиальное и почти продольное, или тангенциальное. Ради-
альные волокна преобладают в наружном слое дентина — так на-
зываемом плащевом дентине, тангенциальные — во внутреннем,
околопульпарном дентине. В периферических участках дентина об-
наруживаются так называемые интерглобулярные пространства, ко-
торые представляют собой его необызвествленные участки, имею-
щие вид полостей, с неровными, шаровидными поверхностями.
502
Рис. 200. Дентин и пульпа зуба.
А — схема (рис. Ю. И. Афанасьева); Б — микрофотография дентина; поперечный срез
(препарат Б. М. Коркмазова и Р. М. Мамедовой). I — дентин; U-предентин; ///— пе-
риферический слой пульпы; IV — промежуточный слой пульпы; V — центральный слой
пульпы; / — дентинные канальцы с от ростками денгинсбластов; 2 - тела дентинобластов.
Наиболее крупные интерглобулярные пространства встречаются в
коронке зуба, а мелкие, но многочисленные находятся в корне, где
они образуют зернистый слой. Интерглобулярные пространства
принимают участие в обмене веществ дентина.
Основное вещество дентина пронизано дентинными канальцами,
в которых проходят отростки дентинобластов, расположенных в
пульпе зуба (см. ниже), и тканевая жидкость. Канальцы берут на-
чало в пульпе, около внутренней поверхности дентина, и, веерооб-
разно расходясь, заканчиваются на его наружной поверхности.
В отростках дентинобластов обнаружена ацетилхолинэстераза, иг-
рающая большую роль в передаче нервного импульса. Количество
канальцев в дентине, их форма и размеры неодинаковы в различ-
ных участках. Более плотно они расположены около пульпы. В ден-
тине корня зуба канальцы ветвятся на всем протяжении, а в ко-
ронке они почти не дают боковых ветвей и распадаются на мелкие
веточки лишь около эмали. На границе с цементом дентинные ка-
нальцы также разветвляются, образуя анастомозирующие между
собой аркады. Некоторые канальцы проникают в цемент и эмаль,
особенно в области жевательных бугорков, и заканчиваются колбо-
видными вздутиями. Система канальцев обеспечивает трофику
дентина. Дентин в области соединения с эмалью имеет обычно фес-
тончатый* край, что способствует более прочному соединению их.
Внутренний слой стенки дентинных канальцев содержит много
S03
преколлагеновых аргирофильных волокон, которые сильно минера-
лизованы по сравнению с остальным веществом дентина.
На поперечных шлифах дентина заметны концентрические па-
раллельные линии, появление которых, очевидно, связано с перио-
дичностью роста дентина.
Между дентином и дентинобластами находится полоска пре-
дентина, или необызвествленного дентина, состоящего из коллаге-
новых волокон и аморфного вещества. В опытах с применением
радиоактивного фосфора показано, что дентин растет постепенно
путем наслоения нерастворимых фосфатов в предентине. Образо-
вание дентина не прекращается у взрослого человека. Так, вторич-
ный, или заместительный, дентин, отличающийся нечеткой направ-
ленностью дентинных канальцев, наличием многочисленных интер-
глобулярных пространств, может быть как в предентине, так и
пульпе (дентикли). Дентикли образуются при нарушении обмена
веществ, при местных воспалительных процессах. Обычно они ло-
кализуются около дентинобластов, с деятельностью которых свя-
зано образование дентиклей. Источником их развития являются
дентинобласты (см. выше). Небольшое количество солей может
проникать в дентин через периодонт и цемент.
” Цемент (cementum) покрывает корень зуба и шейку, где в
виде тонкого слоя частично может заходить на эмаль. По направ-
лению к верхушке корня цемент утолщается.
По химическому составу цемент приближается к кости. В нем
содержится около 30% органических веществ и 70% неорганиче-
ских веществ, среди которых преобладают соли фосфата и карбо-
ната кальция.
По гистологическому строению различают бесклеточный, или
первЦчный, цемент и клеточный, или вторичный. Бесклеточный це-
мент располагается преимущественно в верхней части корня, а
клеточный — в его нижней части (рис. 201). В многокорневых
зубах клеточный цемент залегает главным образом у разветвлений
корней. Клеточный цемент содержит клетки — цементоциты, мно-
гочисленные коллагеновые волокна, которые не имеют определен-
ной ориентации. Поэтому клеточный цемент по строению и соста-
ву сравнивают с грубоволокнистой костной тканью, но в отличие от
нее он не содержит кровеносных сосудов. Клеточный цемент мо-
жет иметь слоистое строение.
В бесклеточном цементе нет ни клеток, ни их отростков. Он со-
стоит из коллагеновых волокон и из лежащего между ними
аморфного склеивающего вещества. Коллагеновые волокна прохо-
дят в продольном и радиальном направлениях. Радиальные волок-
на непосредственно продолжаются в периодонт и далее в виде
прободающих волокон входят в состав альвеолярной кости.
С внутренней стороны они сливаются с коллагеновыми радиальны-
ми волокнами дентина.
Питание цемента осуществляется диффузно через кровеносные
сосуды периодонта. Циркуляция жидкости в твердых частях зуба
происходит за счет ряда факторов: давления крови в сосудах пуль-
SQ4
Рис 201. Клеточный
мент корня зуба.
/ — цементоциты; 2 — мине-
рализован ное ме ж кл еточ ное
вещество.
цы и периодонта, которое изменяется при перепаде температуры в
полости рта при дыхании, приеме пищи, жевании и др. Определен-
ный интерес препставляют данные о наличии анастомозов дентин-
ных канальцев с отростками клеток цемента. Такая связь каналь-
цев служит дополнительной питательной системой для дентина в
случае нарушения кровоснабжения пульпы (воспаление, удаление
пульпы, пломбирование канала корня, заращение полости и т. д.).
П ульпа (pulpa dentis), или зубная мякоть, находится в ко-
ронковой полости зуба и в корневых каналах. Она состоит из рых-
лой волокнистой соединительной ткани, в которой различают три
слоя: периферический, промежуточный и центральный (см.
рис. 200).
Периферический слой пульпы состоит из нескольких рядов
многоотростчатых клеток грушевидной формы — дентинобластов,
отличающихся выраженной базофилией цитоплазмы. Длина их не
превышает 30 мкм, ширина —- 6 мкм. Ядро дентинобласта лежит в
базальной части клетки. От апикальной поверхности дентиноблас-
та отходит длинный отросток, который проникает в дентинный ка-
налец. Полагают, что эти отростки дентинобластов участвуют в
снабжении минеральными солями дентина и эмали. Боковые от-
ростки дентинобластов короткие. По своей функции дентинобласты
сходны с остеобластами кости. В дентинобластах обнаружена ще-
лочная фосфатаза, играющая активную роль в процессах кальци-
нирования зубных тканей, а в их отростках, кроме того, выявлены
мукопротеиды. В периферическом слое пульпы находятся незре-
лые коллагеновые волокна. Они проходят между клетками и про-
должаются далее в коллагеновые волокна дентина.
В помежуточном слое располагаются незрелые коллагеновые
волокна и мелкие клетки, которые, подвергаясь дифференцировке,
заменяют отжившие дентинобласты.
Центральный слой состоит из рыхло лежащих клеток, волокон
и кровеносных сосудов. Среди клеточных форм этого слоя разли-
чают адвентициальные клетки, макрофаги и фибробласты. Между
505
лению.
Рис. 202. Нервные окончания в <убе
(по Бернику).
/ предентин; 2 нервные окончания;
3 дентинобласты.
клетками обнаруживаются
как аргирофильные, гак и
коллагеновые волокна. Элас-
тических волокон в пульпе зу-
ба не обнаружено.
Пульпа зуба имеет опре-
деляющее значение в питании
и обмене веществ зуба. Уда-
ление пульпы резко заторма-
живает обменные процессы,
нарушает развитие, рост и ре-
генерацию зуба.
Зубодесневое сое-
динение. Зуб укрепляется
не только с помощью перио-
донта в лунке челюсти, но и
благодаря сращению много-
слойного плоского эпителия
десны с кутикулой шейки зу-
ба. Эпителий зоны прикрепле-
ния более тонкий, чем на боковой (преддверной) поверхности десны.
Нарушение целостности зубодесневого соединения и образование
зубодесневого кармана могут привести к инфицированию и воспа-
Васкуляризация и иннервация. Сосуды (разветвления верхнече-
люстной артерии) вместе с нервами (разветвления тройничного
нерва) проникают в полость зуба через основной и дополнитель-
ные каналы, расположенные в корне зуба. Артерии входят в ко-
рень зуба одним или несколькими стволиками. Разветвляясь в
пульпе на множество анастомозирующих капилляров, они собира-
ются далее в вену. В пульпе обнаружено небольшое количество
лимфатических капилляров.
Нервы образуют в пульпе зуба два сплетения: более глубокое
состоит преимущественно из миелиновых волокон, более поверх-
ностное — из безмиелиновых. Терминальные разветвления рецеп-
торов пульпы нередко связаны одновременно с соединительной
тканью и сосудами пульпы (поливалентные рецепторы). Денти-
нобласты густо оплетаются тонкими окончаниями тройничного
нерва.
Вопрос о природе чувствительности дентина окончательно
не решен. Многие исследователи отрицают данные о проникно-
вении нервных окончаний в дентинные канальцы, хотя в началь-
ных отделах этих канальцев окончания иногда обнаруживаются
(рис. 202).
506
Возможно, в возникновении боли определенную роль играет
гидродинамический механизм раздражения нервных окончаний,
расположенных во внутренних отделах дентинных канальцев (пе-
редача давления столба жидкости, циркулирующей по дентинным
канальцам, на терминали чувствительных нейронов).
Возрастные изменения. На протяжении первых 12—15 лет
жизни происходит последовательная смена молочных зубов на по-
стоянные. Первым прорезывается большой коренной зуб (первый
моляр), затем центральные и боковые резцы, в 9—14 лет прорезы-
ваются премоляры и клыки и только в 20—25 лет — «зуб муд-
рости».
Одновременно с возрастом происходят постепенные измене-
ния в химическом составе и структуре зубов. Эмаль и дентин на их
жевательной поверхности стираются. Эмаль тускнеет и может да-
вать трещины, на ней откладывается минерализованный налет. Со-
держание органических соединений в эмали, дентине и цементе
уменьшается, а количество неорганических веществ возрастает. В
связи с этим ослабляется проницаемость эмали, дентина и цемента
для воды, ионов, ферментов, аминокислот и других веществ. С воз-
растом новообразование дентина почти полностью прекращается,
количество же цемента в корне зуба увеличивается. Пульпа зуба
с возрастом подвергается атрофии в результате ухудшения пита-
ния, вызванного склеротическим изменением ее сосудов. Количест-
во клеточных элементов в ней при этом уменьшается. В дентино-
бластах отмечается редукция значительной части клеточных ор-
ганелл, снижается пиноцитозная активность клетки. Дентиноблас-
ты превращаются в дентиноциты. Коллагеновые волокна грубеют.
После 40—50 лет в периодонте часто выявляются склеротические
изменения сосудов.
Регенерация. Регенерация зуба происходит очень медленно и
не полностью. При повреждении дентина или раздражениях его
кариозным процессом в зубе со стороны пульпы против очага по-
вреждения образуется небольшое количество заместительного, или I
вторичного, дентина. Этот процесс сопровождается регенерацией
периферического слоя пульпы путем дифференцировки клеточных
элементов промежуточной зоны и превращения их в дентиноблас-
ты. Показано также, что в дентинобластическом слое пульпы на
всех стадиях развития зуба содержатся клетки, обладающие спо-
собностью к пролиферации. Образование дентина происходит при
мерно через 2 нед после повреждения. Этот процесс начинается с
появления предентина. Волокна в основном веществе заместитель-^
ного дентина в отличие от первичного околопульпарного дентина,_
располагаются без определенного порядка. К концу 4-й неделг~о
предентин обызвествляется. Канальцы заместительного дентину
имеют неправильную ориентацию и очень слабо ветвятся. Цеменнг
зуба регенерирует плохо. Восстановление эмали после поврежде^
ния зуба не происходит вообще. При воздействии на эмаль патоо
генных факторов эмаль реагирует образованием зон гиперминера
лизации.
5Г
лотка
В глотке перекрещиваются дыхательный и пищеварительный
пути. В ней различают три отдела, которые имеют различное стро-
ение: носовой, ротовой и гортанный. Каждый из этих отделов от-
личается от другого строением слизистои оболочки.
Слизистая оболочка носового отдела глотки
покрыта псевдомногослойным реснитчатым эпителием, содержит
смешанные железы (респираторный тип слизистой оболочки).
Слизистая оболочка ротового и гортанного
отделов выстлана многослойным (плоским) эпителием, располага-
ющимся на собственной пластинке слизистой оболочки, в которой
имеется хорошо выраженный слой эластических волокон. В под-
слизистой основе лежат концевые отделы сложных сли-
зистых желез. Выводные протоки их открываются на поверхности
эпителия. Слизистая оболочка и подслизистая основа глотки при-
мыкают к мышечной стенке (аналог мышечной оболочки),
которая состоит из двух слоев поперечнополосатых мышц — внут-
реннего продольного и наружного кольцевого. Снаружи глотка
окружена адвентициальной оболочкой.
Пищевод
Развитие. Эпителий пищевода образуется из_прехррдальной
пддстинки, расположенной в энтодерме передней кишки, осталь-
ные слои - из окружающей ее мезенхимы. Особый интерес пред-
ставляет вопрос о развитии эпителия пищевода, который в течение
всего эмбрионального периода претерпевает многократные измене-
ния. Сначала эпителиальная выстилка пищевода представлена од-
нослойным призматическим эпителием. У 4-недельного эмбриона
он становится двухслойным. После этого наблюдается интенсивное
разрастание эпителия, которое приводит к полному закрытию про-
света пищевода. Разросшиеся эпителиальные клетки в дальнейшем
подвергаются распаду, вновь освобождая просвет пищевода. К 3-му
месяцу внутриутробного развития пищевод выстлан многоряд-
ным мерцательным эпителием. С 4-го месяца мерцательные клетки
постепенно вытесняются пузырьковидными, содержащими глико-
ген клетками, которые преобразуются в плоские клетки. Начиная
с 6-го месяца эпителий пищевода становится многослойным плос-
ким. У новорожденных в эпителии могут встречаться островки
мерцательных клеток. У взрослых эти клетки сохраняются изредка
только в протоках слизистых желез. Причины трансформации од-
ного вида эпителия в другой неясны. По-видимому, в формирова-
нии многослойного плоского эпителия в пищеводе ведущая роль
принадлежит его функции. Образование многослойного эпителия в
слизистой оболочке пищевода обеспечивает сохранность стенки
зищевода при прохождении грубых комков пищи. Железы пище-
юда появляются в конце 2-го месяца, мышечная оболочка пище-
юда — на 2-м месяце, а мышечный слой слизистой оболочки — на
1-м месяце внутриутробного развития плода.
1ок
Строение. Пищевод построен из слизисто_й оболочки, подсли-
зистой основы, мышечной и адвентициальной оболочек (рис. 203,
Л). Слизистая оболочка и подслизистая основа образуют в пище-
воде 7—10 продольно расположенных складок, вдающихся в его
просвет.
Слизистая оболочка пищевода построена из эпи-
телия, собственной и мышечной пластинок. Эпителий слизистой
оболочки — многослойный плоский неороговевающий, но в пожи-
лом возрасте его поверхностные клетки могут подвергаться орого-
вению. В составе эпителиального пласта имеется 20- 25 клеточ-
цых слоев. В пищеводе человека плоские клетки поверхностного
слоя эпителия содержат небольшое количество зерен кератогиа-
дина.
'Собственная пластинка слизистой ofолочки образует соедини- '
тельнотканные сосочки, вдающиеся в эпителий. В ней находятся
большие скопления лимфоцитов вокруг протоков слизистых желез,
образуя даже отдельные лимфатические узелки. В собственной
пластинке расположены кардиальные железы пищевода^ (glandulae
cardiacae oesophagi). Они представлены двумя группами! Одна груп-
па желез залегает на уровне перстневидного хряща гортани и 5-го
кольца трахеи,’ вторая группа находится в нижней части пищевода,
около входа в желудок. По своему строению эти железы напоми-
нают кардиальные железы желудка (отсюда их название). Это
простые разветвленные трубчатые железы (см. рис. 203, В). Кон-
цевые отделы их образованы кубическими и призматическими эпи-
телиоцитами с зернистой цитоплазмой, иногда реагирующей на му-
цин. В некоторых концевых отделах кардиальных желез содержат-
ся париетальные клетки, вырабатывающие хлориды. Самые мелкие
протоки этих желез соединяются в расширенные крупные протоки,
которые открываются на вершинах сосочков собственной пластин-
ки слизистой оболочки. Эпителий протоков призматический. Он
сходен со слизистым эпителием желудочных ямочек и дает харак-
терную реакцию на муцин. Иногда на месте расположения карди-
альных желез слизистая оболочка пищевода приобретает характер
слизистой оболочки желудка (см. ниже). Кардиальные железы по-
являются в пищеводе задолго до того, как его эпителий становится
многослойным. Кардиальные железы пищевода содержат большое
количество эндокринных клеток, располагающихся главным обра-
зом в концевых отделах, а также в выводных протоках. По цито-
химическим особенностям различают клетки трех типов. ‘ Первые
соответствуют энтерохромаффинным‘клеткам желудочно-кишечно-
го тракта, синтезирующим серотонин (ЕС^к^Вторые напоминают
энтерохромаффинеподобные клетки (ECLv, характерные для сли-
зистой оболочки тела желудка^ Природа третьих неясна. Для вра-
ча строение и функция кардиальных желез пищевода представля- (
ют интерес потому, что именно в местах их расположения часто
образуются дивертикулы, кисты, язвы и опухоли пищевода.
Мышечная пластинка слизистой оболочки пищевода состоит из
расположенных вдоль него пучков гладких мышечных клеток,
S09
Рис. 2(13. Строение пищевода.
Л — поперечный срез в верхней трети;
окруженных сетью эластических волокон. Эта пластинка начинает-
ся в виде отдельных небольших пучков на уровне перстневидного
хряща гортани; далее по ходу пищевода толщина этого слоя увели-
чивается и около желудка достигает 200—400 мкм. Мышечная
пластинка слизистой оболочки играет большую роль в проведении
по пищеводу пищи и в защите внутренней его поверхности от по-
вреждения острыми телами в случае их попадания в пищевод. Раз-
дражение стенки пищевода грубым пищевым комком вызывает
снижение тонуса мышечной пластинки, а это ведет к расширению
соответствующего участка слизистой оболочки. Таким образом,
прохождение комка пищи, содержащего грубые частицы, облегча-
ется. /
Подслизистая ос но в_а пищевода обеспечивает
большую под' ижность слизистой по отношению к мышечной обо-
лочке. Вместе со слизистой оболочкой она образует многочислен-
ные продоль зые складки, которые расправляются во время прогла-
тывания пищи. В подслизистой основе находятся собственные же-
лезы пищевода.
510
Рис. 203 (продолжение).
Б — микрофотография соб-
ственных желез пищевода;
В — микрофотография кар-
диальных желез пищевода;
продольный срез (препарат
В. А. Вдовцевой).
9
Z — многослойный плоский неороговевающий эпителий; 2 — собственная пластинка слизи
стой оболочки; J — мышечная пластинка слизистой оболочки; 4 — подслизистая основа
5 собственные железы пищевода; 6 мышечная оболочка; а — циркулярный слой
б — продольный слой; 7 — адвентициальная оболочка; 8 — выводной проток железы
9 — кардиальные железы.
Собственные железы пищевода (glandulae oesophagea propriae)
Это сложные сильно разветвленные альвеолярно-трубчатые желе
зы (см. рис. 203, Б). Их концевые отделы состоят исключительн
из слизистых клеток. Секрет изливается в мелкие выводные про
токи, которые сливаются в более крупные. Эти протоки проходя
через мышечную пластинку слизистой оболочки и образуют в соб
ственной пластинке крупные ампулообразные протоки, открывак
щиеся на поверхности эпителия. Эпителий, выстилающий мелки
протоки, низкий призматический, в более крупных протоках о:
многослойный плоский, иногда в нем встречаются реснитчатые
клетки. Собственные железы пищевода располагаются главным об-
разом на вентральной поверхности его верхней трети. Функция
собственных желез пищевода состоит в выделении слизи, постоян-
но увлажняющей поверхность слизистой оболочки и способствую-
щей прохождению пищевых комков.
Мышечная оболочка состоит из внутреннего цирку-
лярногои наружного продольного слоев, разделенных прослойкой
рыхлой волокнистой соединительной ткани. В верхней трети пище-
вода эти слои представлены поперечнополосатой мышечной
тканью, которая является продолжением мышечной ткани глотки.
В средней трети пищевода мышечная оболочка содержит как попе-
речнополосатую, так и гладкую мышечные ткани. В нижней трети
оба слоя образованы только гладкой мышечной тканью. Это об-
стоятельство может служить ориентиром для определения уровня
пищевода на гистологическом срезе. Оба мышечных слоя не всегда
лежат точно циркулярно или продольно. Во внутреннем слое встре-
чаются спиральные и косые пучки, в наружном слое отдельные
пучки также могут иметь неодинаковое расположение. -Утолщение
внутреннего циркулярного слоя мышечной оболочки на уровне
перстневидного хряща образует верхний сфинктер пищевода, а
утолшение этого слоя на уровне перехода пищевода в желудок —
нижний сфинктер пищевода. Сокращение мышечной оболочки пи-
щевода способствует проталкиванию пищи в пищеводе по направ-
лению к желудку.
Адвентициальная оболочка с одной стороны свя-
зана с прослойками соединительной ткани, расположенными в мы-
шечной оболочке, а с другой — с окружающей пищевод соедини-
тельной тканью средостения. В адвентициальной оболочке много
продольно ориентированных сосудов и нервов. Брюшной отдел пи-
щевода покрыт серозной оболочкой, образованной мезо-
телием с подлежащей соединительной тканью.
Васкуляризация. Артерии, входящие в пищевод, образуют спле-
тения в подслизистой основе (крупнопетлистые и мелкопетлистые),
из которых кровь поступает в крупнопетлистое сплетение собст-
венной пластинки слизистой оболочки. Здесь же имеется подэпи
телиальная сеть кровеносных капилляров. Венозный отток начина-
ется сетью мелких вен в собственной пластинке слизистой оболочки.
Эти вены несут кровь в венозное сплетение подслизистой основы,
а оттуда — в адвентициальную оболочку. Лимфатическая сис-
тема пищевода представлена сетями лимфатических капилляров,
расположенных в собственной пластинке слизистой оболочки, в
подслизистой основе и мышечной оболочке, а также сплетениями
лимфатических сосудов, которые лежат в подслизистой основе и
мышечной оболочке (иногда и в адвентициальной оболочке). Глав-
ным коллектором оттока лимфы из пищевода является подслизис-
тое сплетение.
Иннервация. Интрамуральный нервный аппарат образован свя-
занными между собой четырьмя сплетениями: адвентициальным —
Рис. 204. Переход пище-
вода в желудок (препарат
Н А. Юриной).
,Г -МНОГОСЛОЙНЫЙ
нео рогове ва ющий
плоский
эпителий
пищевода; 2 однослойный
призматический эпителий
желудка; 3 — желудочные
ямочки; 4 — кардиальные
железы желудка; 5 — кар-
диальные железы пищевода;
б — мышечная пластинка
слизистой оболочки.
средней и нижней третях пищевода; субад-
|аиболее развитым в
кнтициальным — лежащим на поверхности мышечной оболочки и
хорошо выраженным только в верхних частях пищевода; межмы-
иечным — находящимся между циркулярным и продольным мы-
печными слоями и состоящим из нервных волокон разного калиб-
ра, образующих широкопетлистое сплетение, в составе которого
располагаются крупные нервные узлы, и подслизистым сплетением,
, обнаруживающимся на всем протяжении пищевода и состоящим
из системы нервных стволов, в местах ветвления которых лежат
мелкие нервные узелки. Межмышечное сплетение самое мощное.
Его ганглии образованы в основном клетками I типа. Чувствитель-
ные нервные окончания выявляются в виде рыхлых или инкапсу-
лированных клубочков, кустиков, древовидных разветвлений в мы-
шечных тканях пищевода, в эпителии и соединительной ткани сли-
зистой оболочки и подслизистой основе. В интрамуральных ганг-
лиях обнаруживаются крупные инкапсулированные клубочковые
рецепторы, которые, по-видимому, являются баро- и механорецеп-
торами.
На гистологических срезах в области границы пищевода и же-
лудка виден стык многослойного плоского эпителия пищевода и
однослойного призматического эпителия желудка (рис. 204). В соб-
ственной пластинке слизистой оболочки пищевода в этой области
располагаются кардиальные железы, а в собственной пластинке
слизистой оболочки желудка наряду с желудочными железами мо-
гут находиться слизистые железы пищевода. Кольцевой слой мы-
шечной оболочки пищевода образует здесь утолщение, формируя
сфинктер.
СРЕДНИЙ И ЗАДНИЙ ОТДЕЛЫ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
В среднем отделе пищеварительного тракта происходит глав-
ным образом химическая обработка пищи под воздействием фер-
ментов, вырабатываемых железами, всасывание продуктов перева-
ривания пищи, формирование каловых масс.
17 ГИСТОЛОГИЯ ЯЗ
Желудок
мног
клеп
разо
собе
но у
щей
N
лярг
рых
водг
тка>
В cfj
реч1
оба
сто
пит
лех
чак
пу1
в на
пе|
ут<
ни
Щ‘
ле
за
Ш
ге
гц
’а
Желудок выполняет в организме ряд важнейших функг а
Главной из них является _с екреторная. Она заключается в ни
работке железами желудочного сока. В его состав входят ферм п
ты: пепсин, химозин, липаза, а также соляная к и <
лотаислизь. —•
Пепсин — основной фермент желудочного сока. Под его
влиянием происходит расщепление сложных белков пищи
на более простые — альбумозы и пептоны. Пепсин проявляет
ферментативное действие только в кислой среде, поэтому для его ак-
тивизации необходимо присутствие соляной кислоты. Липаза
выделяется в желудке в небольших количествах и участвует в рас-
щеплении жиров. Химозин присутствует в содержимом желуд-
ка только в раннем детском возрасте. Его роль заключается в ство-
раживании молока. Слизь, покрывая поверхность слизистой обо-
лочки желудка, предохраняет ее от действия соляной кислоты и от
повреждений 1Рубыми комками пищи. Полагают, что в желудке
выделяется особое вещество, которое называют антипепсином. На-
личием его в содержимом желудочного сока объясняют причину
устойчивости стенки желудка по отношению к переваривающему
действию пепсина.
Осуществляя химическую переработку пищи, желудок вместе с
тем выполняет еще некоторые важные для организма функции.
Механическая функция желудка состоит в перемешива-
нии пищи с желудочным соком и проталкивания переработанной
пищи в двенадцатиперстную кишку. В осуществлении функции
принимает участие мускулатура желудка. В стенке желудка обра-
зуется антианемический фактор, который способствует
.» поглощению витамина В|2, поступающего с пищей. При отсутствии
этого фактора у человека развивается злокачественное малокро-
вие. Через стенку желудка происходит всасывание таких
веществ, как вода, спирт, соли, сахар и др. Вместе с тем желудок
выполняет и определенную экскреторную фукцию. Осо-
бенно наглядно эта функция проявляется при заболевании почек,
когда через стенку желудка выделяется ряд конечных продуктов
обмена белков (аммиак, мочевина и др.). Эндокринная
функция желудка заключается в выработке ряда биологически
активных веществ — гастрина, гистамина, серотони-
на, мотилина, энтероглюкагона и др. Эти вещества
оказывают стимулирующее или тормозящее действие на моторику
и секреторную активность железистых клеток желудка и других
отделов пищеварительного тракта.
Развитие, Желудок появляется на 4-й неделе внутриутробного
развития, а в течение 2-го месяца формируются все основные его
отделы. Однослойный призматический" эпителий желудка развива-
ется из энтодермы кишечной трубки. Желудочные ямочки образу-
ются в течение 6—10-й недели развития плода, железы закладыва-
ются в виде почек на дне желудочных ямочек и, разрастаясь, в
44
дальнейшем располагаются в собственной пластинке слизистой
оболочки. Вначале в них появляются париетальные клетки, за-
тем — главные и слизистые клетки. В это же время (6—7-я неде-
ля) формируются из мезенхимы сначала кольцевой слой мышеч-
ной оболочки, затем — мышечная пластинка слизистой оболочки.
На 13 14-й неделе образуется наружный продольный и несколько
позднее — внутренний косой слой мышечной оболочки.
Строение. Стенка желудка состоит из слизистой оболочки, под-
слизистой основы, мышечной и серозной оболочек.
Слизистая оболочка желудка имеет неровную по-
верхность из-за наличия в ней трех видов образований: складок,
полей и ямочек. Желудочные складки (plicae gastricae) образова-
ны слизистой оболочкой и подслизистой основой. Желудочные по-
ля (areae gastricae) пре дета вляютиюбой отграниченные друг ст дру-
га бороздками участки слизистой оболочки. Они имеют многоуголь-
ную форму и размер поперечника от 1 до 16 мм. Наличие полей
объясняется тем, что железы желудка располагаются группами,
отделенными друг от друга прослойками соединительной ткани.
Поверхностно лежащие вены в этих прослойках просвечивают в
виде красноватых линий, выделяя границы между полями. Желу-
дочные ямочки (foveolae gastricae) — углубления эпителия в соб-
ственной пластинке слизистой оболочки. Они встречаются по всей
поверхности желудка Число ямочек в желудке достигает почти
3 млн. Желудочные ямочки имеют микроскопические размеры, но
величина их неодинакова в различных отделах желудка. В кар-
диальном отделе и теле желудка их глубина составляет всего 1 /4
толщины слизистой оболочки. В пилорической части желудка ямоч-
ки более глубокие. Они занимают около половины толщины всей
слизистой оболочки. На дне желудочных ямочек открываются же-
лезы, лежащие в собственной пластинке слизистой оболочки. Сли-
зистая оболочка наиболее тонкая в кардиальном отделе.
Эпителий, выстилающий поверхность слизистой оболочки же-
лудка и ямочек, однослойный призматический (рис. 205). Особен-
ность этого эпителия — его железистый характер; все поверхност-
ные эпителиоциты желудка (epitheliocyti superficiales gastrici) по-
стоянно выделяют мукоидный (слизеподобный) секрет. Каждая
железистая клетка четко подразделяется на две части — базаль-
ную и апикальную (рис. 206). В базальной части, прилежащей к
базальной мембране, лежит овальной формы ядро, над которым
располагается комплекс Гольджи. Апикальная часть клетки запол-
нена зернами или каплями мукоидного секрета. Специфичность
секрета поверхностных эпителиоцитов у человека и животных оп-
ределяется составом углеводного компонента, в то время как бел-
ковая часть характеризуется общностью гистохимических свойств.
Углеводный компонент играет определяющую роль в защитной ре-
акции слизистой оболочки желудка на повреждающее действие же-
лудочного сока. Роль поверхностных эпителиоцитов желудка за-
ключается в выработке слизи, которая служит защитой как от ме-
ханического воздействия грубых частиц пищи, так и от химическо-
fl
Рис. 205. Строение стенки дна желудка.
А — схема;
го действия желудочного сока.
Количество слизи в желудке
сильно увеличивается при попа-
дании в него раздражающих ве-
ществ (алкоголь, кислота, горчи-
ца и др.).
В собственной пластинке
слизистой оболочки расположе-
ны железы желудка, между ко-
торыми лежат тонкие прослойки
рыхлой волокнистой соедини-
тельной ткани. В ней в большем
или меньшем количестве всегда
имеются скопления лимфоид-
ных элементов либо в виде диф-
фузных инфильтратов, либо в
виде солитарных (одиночных)
лимфатических узелков, кото-
рые чаще всего располагаются в
области перехода желудка в две-
надцатиперстную кишку.
Мышечная пластинка слизи -
(стой оболочки состоит из трех
слоев, / образованных гладкой
мышечной тканью: внутреннего
' и наружного циркулярных и
среднего — продольного (см.
рис. 4 05). От мышечной плас-
тинки отдельные мышечные
клетк» отходят в соединитель-
ную ткань собственной пластин-
ки слизистой оболочки. Сокра-
щение мышечных элементов
слизистой оболочки обеспечива-
ет подвижность ее, а также спо-
собствует выведению секрета из
желез желудка.
Железы желудка (gll. gastricae) в различных его отделах имеют
неодинаковое строение. Различают три вида желудочных желез:
собственные железы желудка, пилорические и кардиальные. Коли-
чественно преобладают собственные железы желудка. Они залегают
в области тела и дна желудка (фундальные). Кардиачьные и
пилорические железы располагаются в одноименных частях же-
лудка.
Собственные железы желудка (gll. gastricae propriae) — наи-
516
4-
Рис. 205 (продолжение)
Б микрофотография.
/ однослойный призмати
четкий железистый эпителий;
2 — желудочная ямочка; 3 —
собственные железы желуд-
ка; 4 — собственная плас-
тинка слизистой оболочки;
5 — мышечная пластинка
слизистой оболочки; 6 — под-
слизистая основа: а — кро-
веносный сосуд; б жиро-
вая клетка; 7 — мышечная
оболочка; 8 — межмышеч-
ное иервное сплетение; 9
серозная оболочка.
i:
'Б-
1
более многочисленные. У человека их насчитывается около 35
млн. Площадь каждой железы составляет приблизительно 100-мм2.
Общая секреторная поверхность фундальных желез достига-
ет огромных размеров — около 3—4 м2. По строению эти железы
представляют собой простые неразветвленные, иногда очень слабо
разветвленные трубчатые железы (см. рис. 205). Длина одной
железы около 0,65 мм, диаметр ее колеблется от 30 до 50 мкм.
Железы группами открываются в желудочные ямочки. В каждой
железе различают перешеек (isthmus), шейку (cervix) и главную
часть (pars principalis), представленную телом (corpus) и дном
(fundus). Тело и дно составляют секреторный отдел железы, а
шейка и перешеек — ее выводной проток. Просвет в железах
очень узкий и почти не виден на препаратах.
Собственные железы желудка содержат.,^ основных видов же-
лезистых клеток; главные экзокриноциты, париетальные экзокри-
ноцигы, слизистые, шеечные мукоциты, эндокринные (аргиро-
фильные), недифференцированные эпителиоциты.
Главные экзокриноциты (exocrinocyti principals) располагают-
ся преимущественно в области дна и тела железы (см. рис. 207).
Ядра этих клеток имеют округлую форму, лежат в центре клетки.
В клетке выделяют базальную и апикальную части. Базальная
часть обладает выраженной базофилией. В апикальной части обна-
руживаются гранулы белкового секрета. В базальной части на-
ходится хорошо развитый синтетический аппарат клетки. На апи-
кальной поверхности имеются короткие микроворсинки. Секретор-
ные гранулы имеют диаметр 0,9—1 мкм. Главные клетки секрети-
руют пепсиноген — профермент (зимоген), который в присутствии
соляной кислоты превращается в активную форму — пепсин.
Предполагают, что химозин, расщепляющий белки молока, также
вырабатывается главными клетками. При изучении различных
фаз секреции главных клеток выявлено, что в активной фазе выра-
ботки и накопления секрета эти клетки крупные, в них хорошо
различимы гранулы зимогена. После выделения секрета величина
517
Рис. 206. Схема ультра-
микроскопического строе-
ния поверхностных эпите-
лиоцитов желудка (по
С. Ито).
I микроворсинки; 2—гра-
нулы слизистого секрета; 3—
митохондрии; 4 комплекс
Гольджи; 5 - гранулярная
эндоплазматическая сеть; 6—
базальная мембрана.
клеток и количество гранул в их цитоплазме заметно уменьшают»
ся. Экспериментально доказано, что при раздражении блуждающе-
го нерва клетки быстро освобождаются от гранул пепсиногена.
Париетальные экзокриноциты (exocrinocyti parietales) распо-
лагаются-снаружи от главных и слизистых клеток, плотно приле-
гая к их базальным концам (рис. 207). Они больше главных кле-
ток, неправильной округлой формы. Париетальные клетки лежат
поодиночке и сосредоточены главным образом в области тела и
шейки железы. Цитоплазма этих клеток резко оксифильна, имеет
зернистое строение В каждой клетке содержится одно или два
ядра округлой формы, лежащих в центральной части цитоплазмы.
Внутри клеток располагаются особые системы внутриклеточных
канальцев (canaliculis intracellulares) с многочисленными микро-
ворсинками, которые переходят в межклеточные канальцы, нахо-
дящиеся между главными и слизистыми клетками и открывающи-
518
Рис. 207. Ультра микроско-
пическое строение соб-
ственной железы желудка
(по Р. Кристичу с измене-
нием) .
I — просвет желе »ы; 2 — ше-
ечный мукоцит; 3 парие-
тальные экзокрииоциты;За
внутриклеточные ка на льны;
4 — главные экзокрииоциты;
5 — эндокриноциты, 6 ге-
мокапилляр; 7 нервное во-
локно.
еся в просвет железы (см. рис. 207). От апикальной поверхности
клеток'отходят микроворсинки. Для париетальных кдуток харак-
терно наличие многочисленных митохондрий. Роль париеталь-
ных клеток собственных желез желудка заключается в выработке
хлоридов, и.» которых образуется соляная кислота.
Слизистые клетки (mucocyti) представлены двумя видами.
Одни располагаются в теле собственных желез (см. рис. 207) и
имеют уплотненное ядро в базальной части клеток. В апикальной
части этих клеток обнаружено множество круглых или овальных
гранул, небольшое количество митохондрий и комплекс Гольджи.
Другие слизистые клетки (шеечные) располагаются только в шей-
ке собственных желез. Ядра у них уплощенные, иногда неправиль-
ной треугольной формы, лежат обычно у основания клеток. В апи-
si9
/
Рис. 208. Пилорическая
часть желудка.
1 — желудочные ямочки;
2 собственная пластинка
слизистой оболочки; 3 — мы
шечная пластинка слизистой
оболочки; 4 пилорические
железы.
калькой части этих клеток находятся секреторные гранулы. Слизь,
выделяемая шеечными клетками, слабо окрашивается основными
красителями, но хорошо выявляется муцикармином. По сравне-
нию с поверхностными клетками желудка шеечные клетки мень-
ших размеров и всегда содержат значительно меньшее количество
капель слизи. Их секрет по составу отличается от мукоидного
секрета, выделяемого железистым эпителием желудка. В шеечных
клетках в противоположность другим клеткам фундальных желез
часто обнаруживаются фигуры митоза. Полагают, что эти клетки
являются недифференцированными эпителиоцитами (epitheliocyti
nondifferentiati) — источником регенерации как секреторного эпи-
телия желез, так эпителия желудочных ямок. Эндокринные клетки
описаны ниже.
Пилорические железы (gll. pyloricae) расположены в зоне
перехода желудка в двенадцатиперстную кишку (рис. 208). Их
число составляет около 3,5 млн. Пилорические железы отличают-
_ ся от собственных желез несколькими признаками: расположены
’ более редко, гораздо сильнее разветвлены, имеют широкие про-
светы; большинство пилорических желез лишено париетальных
^-клеток.
Концевые отделы пилорических желез построены в основном
из клеток, напоминающих слизистые клетки собственных желез.
Ядра их сплющены и лежат у основания клеток. В цитоплазме
при использовании специальных методов окраски выявляется
слизь. Клетки пилорических желез богаты дипептидазами. Секрет,
вырабатываемый пилорическими железами, имеет щелочною реак-
цию. В шейке желез расположены также промежуточные — шееч-
ные —"клетки, которые были уже описаны в собственных желе-
зах желудка.
Строение слизистой оболочки в пилорической части имеет
некоторые особенности: желудочные ямочки здесь более глубокие,
чем в теле желудка, и занимают около половины всей толщины
520
слизистой оболочки. Около выхода из желудка эта оболочка имеет
хорошо выраженную кольцевую складку. Ее возникновение связа-
но с наличием мощного циркулярного слоя в мышечной оболочке,
образующей пилорический сфинктер. Последний регулирует по-
ступление пищи из жедудка в кишечник.
з Кардиальные железы (gll. cardiacae) —простые, трубчатые
железы с сильно разветвленными концевыми отделами. Выводные
протоки (шейки) этих желез короткие, выстланы призматически-
ми клетками. Ядра клеток сплющенной формы, лежат у основания
клеток. Цитоплазма их светлая. При специальной окраске муци-
кармином в ней выявляется слизь. По-видимому, секреторные
клетки этих желез идентичны клеткам, выстилающим пилориче-
ские железы желудка и кардиальные железы пищевода. В них
также обнаружены дипептидазы. Иногда в кардиальных железах
встречаются в небольшом количестве главные и париетальные
клетки.
Желудочно-кишечные эндокриноциты (endocrinocyti gastrointes-
tinales). В желудке по морфологическим, биохимическим и функ-
циональным признакам выделено несколько видов эндокринных
клеток (см. рис. 211). ЕС-клетки — самые многочисленные, распо-
лагаются в области тела и дна желез между главными клетками.
Эти клетки секретируют с-е-рот о н и н и м елатонин. Серо-
тонин стимулирует секрецию пищеварительных ферментов, выде-
ление слизи, двигательную активность. Мелатонин регулирует
фотопериодичность функциональной активности (т. е зависит
от действия светового цикла). G-клетки (гастринпродуцирующие)
также многочисленны и находятся главным образом в пилори-
ческих железах, а также в кардиальный' располагаясь в области
их тела и дна, иногда шейки. Выделяемый ими гастрин сти-
мулирует секрецию пепсиногена главными клетками, соляной
кислоты — париетальными, а также моторику желудка. При
гиперсекреции желудочного сока у человека отмечается увеличе-
ние числа G-клеток. Кроме гастрина, эти клетки выделяют эн-
кефалин, являющийся одним из эндогенных морфинов. Ему
приписывают роль медиации боли. Менее многочисленными явля-
ются Р-, ECL-, D-, Di-, А- и Х-клетки. Р-клетки секретируют
бомбезин, стимулирующий выделение соляной кислоты и пан-
креатического сока, богатого ферментами, а также усиливают
сокращение гладкой мускулатуры желчного пузыря. ECL-клетки
(энтерохромаффинноподобные) ""характеризуются^ разнообразием-
формы и располагаются главным образом в теле и дне фундаль-
ных желез. Эти клетки вырабатывают гистамин, который
регулирует секреторную активность париетальных клеток, выде-
ляющих хлориды. D- и Dj-клетки выявляются главным образом
в пилорических железах. Они являются продуцентами активных
полипептидов. D-клетки выделяют соматостат ин, ингибирую-
щий синтез белка. D,-клетки секретируют вазоинтестиналь-
ный пептид (ВИП), который расширяет кровеносные сосуды и
снижает артериальное давление, а также стимулирует выделение
521
гормонов поджелудочной железы. А-клетки синтезируют глюка-
гон, т. е. имеют сходную функцию с эндокринными А-Клетками
островков поджелудочной железы. Функция Х-клеток неизвестна.
Подслизистая основа желудка состоит из рыхлой
волокнистой неоформленной соединительной ткани, содержащей
большое количество эластических волокон (см. рис. 205). В ней
расположены артериальное и венозное сплетения, сеть лимфати-
ческих сосудов и подслизистое нервное сплетение.
Мышечная оболочка желудка относительно слабо
развита в области его дна, хорошо выражена в теле и наибольшего
развития достигает в привратнике. В мышечной оболочке различа-
ют три слоя, образованные гладкими мышечными клетками (см.
рис. 205). Наружный, продольный, слой является продолжением
продольного мышечного слоя пищевода. Средний — циркулярный,
также представляющий собой продолжение циркулярного слоя
пищевода, наибольшего развития достигает в пилорической об-
ласти, где образует пилорический сфинктер толщиной около 3—
5 см. Внутренний слой представлен пучками гладких мышечных
клеток, имеющих косое направление. Между слоями мышечной
оболочки располагаются межмышечное нервное сплетение и спле-
тения лимфатических сосудов.
Серозная оболочка желудка образует наружную часть
его стенки.
Васкуляризация. Артерии, питающие стенку желудка, проходят
через серозную и мышечную оболочки, отдавая им соответствую-
щие ветви, а далее переходят в мощное сплетение в подслизистой
основе. Веточки от этого сплетения проникают через мышечную
пластинку слизистой оболочки в ее собственную пластинку и обра-
зуют там второе сплетение. От этого сплетения отходят мелкие
артерии, продолжающиеся в кровеносные капилляры, оплетающие
железы и обеспечивающие питание эпителия желудка. Из кровенос-
ных капилляров, лежащих в слизистой оболочке, кровь собирается
в мелкие вены. Непосредственно под эпителием проходят относи-
тельно крупные посткапиллярные вены звездчатой формы (vv.
stellatae). Повреждение эпителия желудка обычно сопровождается
разрывом этих вен и значительным кровотечением. Вены слизистой
оболочки, собираясь вместе, формируют сплетение, расположенное
в собственной пластинке около артериального. Второе венозное
сплетение располагается в подслизистой основе. Все вены желудка,
начиная с вен, лежащих в слизистой оболочке, снабжены клапанами.
Лимфатическая сеть желудка берет начало от лимфатических
капилляров, слепые концы которых находятся непосредственно
под эпителием желудочных ямочек и желез в собственной пластинке
слизистой оболочки (см. рис. 188, Б). Эта сеть сообщается с широко-
петлистой сетью лимфатических сосудов, расположенной в подсли-
зистой основе. От лимфатической сети отходят отдельные сосуды,
пронизывающие мышечную оболочку. В них вливаются лимфатичес-
кие сосуды из лежащих между мышечными слоями сплетений.
Иннервация. Желудок имеет два источника эфферентной ИН-
42
нервации' парасимпатииееыш (от блуждающего нерва) и симца
тический (из пограничного симпатического стволаД,—В—стенке
желудка располагаются три нервных сплетения- межмышечное,
подели {истое и субсеро тое. Нервные ганглии малочисленны в
кардиальной области, увеличиваются в числе и размерах в направ-
лении привратника.
Ганглии .едмога митинга межмышечного сплетения построены
преймущественно из клеток I типа и незначительного количества
клеток II типа. Наибольшее количество клеток II типа наблюда-
ется в пилорической области желудка. Подслизистое сплетение
развито слабо. Возбуждение блуждающего нерва ведет к ускоре-
нию сокращения желудка и усилению выделения железами желу-
дочногб еока. Возбуждение симпатических нервов, наоборот, вызы-
вает замедление сократительной "деятельности же пудка~~й~лс пябпе-
ние желудочной секреции.
Афферентные волокна образуют чувствительное сплетение, рас-
положенное в мышечной оболочке, волокна которого осуществля-
ют рецепторную иннервацию нервных узлов, гладких мышц, сое-
динительной ткани. В желудке обнаружены поливалентные рецеп-
торы.
Тонкая кишка
В тонкой кишке подвергаются химической обработ-
к е все виды питательных веществ — белки, жиры и углеводы.
В переваривании белков участвуют ферменты энтерокиназа, кина-
зоген и трипсин, расщепляющие простые белки; эрепсин (смесь
пептидаз), расщепляющий пептиды до аминокислот, и нуклеаза,
которая переваривает сложные белки — нуклеопротеиды. Перева-
ривание углеводов обеспечивается амилазои, мальтазой, сахаразой,
лактазой и фосфатазой, а жиров — ферментов липазои. В тонкой
кишке происходит также процесс всасывания продуктов
расщепления белков, жиров и углеводов в кровеносные и лимфа-
тические сосуды. Кроме того, кишечник выполняет механи-
ческую функцию -^проталкивает химус в каудальном на-
правлении. Эта функция осуществляется благодаря перисталь-
тическим сокращениям мышечной оболочки кишечника. Эндо-
кринная функция, выполняемая специальными секретор-
ными клетками, заключается в выработке биологически активных
веществ — серотонина, гистамина, мотилина, секретина, энтероглю-
кагона, холецистокинина, панкреозимина, гастрина и ингибитора
гастрина.
Развитие. Тонкая кишка начинает развиваться на 5-й неделе
эмбриогенеза. Эпителий ворсинок, крипт и дуоденальные железы
тонкой кишки Образуются из кишечной энтодермы. На первых
этапах дифференцировки эпителий однорядный кубический, затем
он становится двухрядный призматическим и, наконец, на 7—8-й
11 еделг образуется однослойный призматический эпителии, На
8—Ю-й неделе развития возникают ворсинки и крипты. В тече-
523
ние 20—24-й недели формируются циркулярные складки. К этому
времени появляются и дуоденальные железы. Клетки кишечного
эпителия у 4-недельного эмбриона не дифференцированы и харак-
теризуются высокой пролиферативной активностью. Дифферен-
цировка эпителиоцитов начинается на 6—12-й неделе развития.
Появляются столбчатые (каемчатые) эпителиоциты, для которых
характерно интенсивное развитие микроворсинок, увеличивающих
резорбционную поверхность. Гликокаликс начинает формировать-
ся к концу эмбрионального — началу плодного периода. В это
время в эпителиоцитах отмечаются ультраструктурные признаки
резорбции — большое число везикул, лизосом, мульти везикуляр-
ных и мекониальных телец.
Бокаловидные экзокрииоциты дифференцируются на 5-й неде-
ле развития, эндокриноциты — на 6-й неделе. В это время среди
эндрокриноцитов преобладают переходные клетки с недифферен-
цированными гранулами, выявляются ЕС-клетки, G-клетки и
S-клетки. В плодном периоде преобладают ЕС-клетки, большин-
ство из которых не сообщается с просветом крипт («закрытый»
тип); в более позднем плодном периоде появляется «открытый»
тип клеток. Экзокрииоциты с ацидофильными гранулами мало
дифференцированы у эмбрионов и плодов человека. Собственная
пластинка слизистой оболочки и подслизистая основа тонкой
кишки образуются из мезенхимы на 7—8-й неделе эмбриогенеза.
Гладкая мышечная ткань в стенке тонкой кишки развивается
из мезенхимы неодновременно в различных участках ~ю1шечной
стенки; на 7—8-й неделе появляется внутренний циркулярный
слой мышечной оболочки, затем на 8—9-й неделе — наружный
продольный слой и, наконец, на 24—28-й неделе развития плода
возникает мышечная пластинка слизистой оболочки. Серозная,
оболочка тонкого кишечника закладывается на 5-й неделе эмбрио-
генеза из мезенхимы (ее соединительнотканная часть) и висце-
рального листка мезодермы (ее мезотелий).
'Строение. Стенка тонкой кишки построена из слизистой обо-
лочки, подслизистой основы, мышечной и серозной оболочек (рис.
209, А. Б, В).
Слизистая оболочка имеет характерный рельеф бла-
годаря наличию ряда образований: циркулярных складок, ворси
нок и крипт (кишечные железы). Эти структуры увеличивают
общую-поверхность тонкого кишечника, что способствует выпол-
нению его основных функций.
Циркулярные складки (plicae circulares) образованы
слизистой оболочкой и подслизистой основой.
Кишечные вор синки (villi intestinales) представляют
собой выпячивания слизистой~оболочки пальцевидной или листо-
видной формы, свободно вдающиеся в просвет тонкой кишки.
Число ворсинок в тонкой кишке очень велико. Больше всего их в
двенадцатиперстной и тощей кишках (22—40 ворсинок на I мм2),
несколько меньше — в подвздошной кишке (18—31 ворсинок на
1 мм2). В двенадцатиперстной кишке ворсинки широкие и корот-
524
кие (высота их всего 0,2—0,5 мм), в тошеи и подвздошной кишках
они несколько тоньше, но более высокие (по 0,5—1,5 мм). В обра-
зовании каждой ворсинки участвуют структурные_злементы всех
слоев слизистой оболочки. (ЩвсЫсст'
С поверхности каждая кишечная ворсинка выстлана однослой-
ным призматическим эпителием (см. рис. 209, Б). В "эпителий
различают три вида клеток: столбчатые эпителиоциты, бокаловид-
ные экзокрииоциты И эцпокринощиты. ~ Столбчатые эпителиоциты
ворсинки (epitheliocyti columnares villi) составляют основную
массу эпителиального пласта, покрывающего ворсинку. По формеА
это призматические клетки, имеющие высоту около 22—26 мкм
и ширину около 8 мкм. Они характеризуются выраженной поляр-Я
цостью строения, что отражает их функциональную спецйализа^Я|
_щпО_^^обеспечение резорбции и транспорта веществ, поступающих w
с пищей? ~
На апикальной поверхности клеток видна исчерченная каемка
(limbus""striatus)."Образованная множеством микроворсинок (рис.
210). Количество микроворсинок на 1 мкм2 поверхности клетки
составляет от 60 до 90. Высота каждой микроворсинки у человека
около 0,9—1,25 мкм, диаметр — 0,08—0,11 мкм, промежутки
между микроворсинками равны 0,01—0,002 мкм. Благодаря огром-
ному числу микроворсинок поверхность всасывания кишки увели-
чивается в 30—40 раз. В микроворсинках имеются тонкие фила-
мщггьц^ микротрубочки. В микроворсинках исчерченной каемки
обнаружено высокое содержание ферментов, участвующих в рас-
щеплении и транспорте всасывающихся веществ: фосфатазы,
нуклеозйддифосфатазы, Е-, D-гликозидазы, аминопептидазы и др.
Содержание фосфатаз в эпителии тонкой кишки превышает их
уровень в печени почти в 700 раз, причем 3/4 их количества нахо-
дится в каемке. Установлено, что расщепление пищевых веществ
и всасывание их наиболее интенсивно происходят в области исчер-ДЙ
ченной каемки. Этот__щюцесс получил название пристеноч - Иг
ного пищеварени я в отличие от полостногб?"соверша юще-
гося в просвете кишечной трубки, и внутриклеточного. Поверх-
ность микроворсинок выполняет роль пористого адсорбента фер-
ментов и пищевых веществ, а расстояние между ворсинками явля-
ется оптимальным для протекающих здесь ферментативных реак-
ций.
На поверхности микроворсинок расположен гликокаликс,, пред-
ставленный липопротеидами и гликозаминогликанами (см. рис.
210). '------------------------ ~
В апикальной части клетки имеется хорошо выраженный тер-
минальный слой, который состоит из филаментов, расположенных
параллельно поверхности клетки."для соединения апикальных
частей энтероцитов характерны замыкающие пластинки и плотные
контакте! (zoou\a adherens), которые соединены с терминальными
сетями.ЙГболее глубоких частях цитоплазмы видны хорошо разви-
тая гранулярная эндоплазматическая сеть, .лизосомы- мультиве-
зикулярные тельца, степень выраженности которых зависит от
525
A
Рис. 209. Строение тонкой кишки.
А — общий план строения (по Крёлингу и
Грау):
/ — слизистая оболочка; 11 — подслизистая
основа; 111—мышечная оболочка; IV —
серозная оболочка; 1 — кишечные ворсинки;
2 — кишечные железы (крипты); 3 — эпи-
телий; 4 — собственная пластинка слизи-
стой оболочки; 5 — мышечная пластинка
слизистой оболочки; 6 — сосудистая сеть;
7 — лимфатическая сеть; 8 — нервное во-
локно; 9 — подслизистое нервное сплетение;
10 — мышечно-кишечное нервное сплете-
ние. Б — продольный срез ворсинки (рис.
Н. А. Юриной, Л. С. Румянцевой): 1 — од-
нослойный призматический каемчатый эпи
телий; 2 — исчерченная каемка; 3 — бока-
ловидный экзокриноцит; 4 — собственная
пластинка слизистой оболочки; 5 — крове-
носный капилляр. 6 — лимфатический ка
пилляр; 7 — лимфоциты; 8 — гладкие мио-
циты.
Рис. 209 (продолжение)
В — схема строения железы
(крипты) тонкой крипты; / —
просвет кишки; 2 — столбча-
тые эпителиоциты; 3 — бока-
ловидный экзокриноцит; 4 —
аргирофильные клетки (эн-
докриноциты); 5 — недиффе-
ренцированные эпителиоци-
ты; б экзокриноциты с аци-
дофильными гранулами; 7 —
соединительная ткань соб-
ственной пластинки слизи-
стой оболочки.
в
интенсивности процесса резорбции (см. рис. 210). Ядро овальной
формы, имеет инвагинации. Митохондрии в апикальной части
менее развиты и меньших размеров, чем в базальной. Комплекс
Гольджи, как правило, находится .над ядром.
В базальной части столбчатого эпителиоцита расположено боль-
шое число рибосом, полирибосом и митохондрий- Боковые контак-
ты ^ежду базальными частями клеток осуществляются с помощью
десмосом и пальцевидных инвагинаций
— Бокаловидные экзокриноциты (exocrinocyti caliciformes) в вор-
синках расположены поодиночке среди стпдбчятыу упетенг1 (см.
рис. 209)._Число их увеличивается по направлению от двенадцати-
перстной кишки к подвздошной. По своему строению — это типич-
нвгеслизистые клетки. В них наблюдаются циклические измене-
ния, связанные с накоплением и последующим выделением слизи.
В фазе накопления секрета ядра этих клеток оказываются прижа-
тыми к их основанию, в цитоплазме же клеток над ядром видны
капли слизи. Комплекс Гольджи и митохондрии располагаются
около ядра. Формирование секрета происходит в области комплек-
са Годьджи В~Стадии накопления слизи в клетке~ойнар^тгйается
большое число” сильно-измененных митохондрий. Они крупные.
527
Рис. 210. Гликокаликс и исчерченная каемка в кишечных ворсинках кошки. Элек-
тронная микрофотография. X24 000 (no С. Ито).
1 — апикальные части двух столбчатых эпителиоцитов; 2 — микроворсинки исчерченной
каемки; 3 — гликокаликс.
светлые, с короткими кристами. После выделения секрета бокало-
видная клетка, .становится узкой, ядро~~ёе~уменьша,ется, цитоплаз-
ма освобождается от гранул секрета. £лизь. выделяемая бокало-
видными экзокриноцитами, служит для увлажнения поверхности
слизистой оболочки кишечника и этим способствует продвижению
пищевых частиц.
Под эпителием ворсинки находится базальная мембрана. да-
которой—следует рыхлая волокнистая соединительная ткань соб-
ственной пластинки слизистой оболочки. В—нойГпроходят крове-
носные и лимфатические сосуды и нервы, ориентированные вдоль
ворсинки. В строме ворсинки всегда .присутствуют отдельные глад-
кие мышечные клетки — производные мышечного слоя слизистой
оболочки. Пучки гладких миоцитов обвиты сетью ретикулярных
волокон, которые связывают их со стромой ворсинки и базальной
мембраной.
С.О кращение миоцитов с пособству ет п рота л киванию вса -
сывающихся продуктов гидролиза пищи в кровь и лимфу ворсин
кишечника. Другие пучки гладких мышечных клеток, проникаю-
щие в подслизистую основу, образуют циркулярные слои вокруг
проходящих там сосудов. Сокращение этих мышечных групп ре-
гудипует, кровоснабжение органа.
Кишечные железы (крипты) (glandulae sey cryptae intestina-
les) представляют собой трубчатые углубления эпителия, лежа-
528
»
(
I
,щиев собственной пластинке слизистой оболочки (см. рис. 209,
А, В). Их устье открывается д просвет между ворсинками. На
1 мм2 поверхности приходится, до 100 крипт, а всего в тонком
кишечнике более 150 млн. крипт. Каждая кишечная крипта имеет
длину около 0,25—0,5 мм, диаметр до 0,07 мм. Общая площадь
крипт в тонкой кишке составляет около 14 м2.
Эпителиальная- выстипка кишечных [хрипт содержит следую-
щие виды клеток: столбчатые эпителиоциты/ нёдифферёнцирован-
ные эпителиоциты, бокаловидные экзокриноциты, эндокриноциты
и jib слеридцц.иты ~с ацидофильными^ ранулами |(клетки 'Па'НётаГ.^
Столбчатые эпителиоциты составяядзт основную массу эпителиаль-
ной выстилки крипт. По сравнению с аналогичными клетками
ворсинок они более низкие, имеют более тонкую исчерченную
каемку и базофильную цитоплазму: Недиффе рёщъртяягтъп.
.эпителиоциты расположены в нижней половине крипт. В них часто
видны фигуры митоза. ^ти_элементы служГЙУ исхцчникбм регене ра -
гфикакдля^эпйтел'иальйыхклетО к ворси нок, так и для клеток крипт.
Период смены эпителия составляет около 2 сут. Бокаловидные
экзокриноциты постоянно находятся в криптах, их строение сходно
fc описанными выше. ~Эндокриноцитов в криптах значительна
Дольше, чем в ворсинках. Экзокриноциты с ацидофильными гранула-
ми (exocrinocyti cum granulis acidophilis) располагаются группами
или поодиночке на дне крипт (см. рис. 209, В). В их апикалБний—
* части видны плоТггьтё ~ йльнб преломляющие свет гранулы. Эти
гранулы .резко ацидофильны, окрашиваютсТГэбзином в "ярко крас- .
ный цвет, .растворяются в__кислотах, но устойчивых щелочам.
Цитохимически в гранулах .обнаружены белковочтодисахариДПБГй
комплекс, ферменты, лизоцим. Цитоплазма базальной частиэкЗо-
криноцитов с ацидофильной зернистостью обнаруживает значи-
тельную базофилию. Вокруг большого округлого ядра располагает-
ся немного митохондрий. Гистохимические исследования показали,
что в клетках с ацидофильной зернистостью содержится большое
количество цинка, выражена активность кислой фосфатазы, дегид-
рогеназ. Обнаружено также усиленное выделение гранул секрета из
этих клеток при введении подопытным животным пилокарпина.
После выделения секрета клетки выглядит узкими, темноокрашен-
ными. Через 1 ч после инъекции пилокарпина зернистость вновь
накапливается в цитоплазме. Полагают, что эти клетки выделяют 4
дипептидазы (эрепсин), принимающие участие в расщепле-
нии дипептидов до аминокислот. Согласно другому представлению, ।
/эти клетки выделяют секрет, который нейтрализует соляную кис-
лоту, содержащуюся в химусе.
Желудочно-кишечные эндокриноциты в кишечнике бывают не-
скольких типов. Наиболее многочисленными являются ЕС-клетки,
секретирующие с е ро то н и н, мТГТ’и л итз и ве"щ~ё с т в о
JP. А-клетки, продуцирующие энтероглюкагон, малочис-
ленны. £-клетки, вырабатывающие секретин, распределены в
разных отделах кишечника нерегулярно. Кроме того, в кишечни-
ке найдены 1-клетки, секретирующие холецистокинин
О Т О Н И н,
529
If
Типы клеток Секреторные гранулы (размеры S нм)
350
EC 200
G V®® м ®*®@ 300
0 350
% • • • ®.* • • •. • • 160
Д ®°®« 250
L ••V 400
X ® 300 • •••
$ • • ••• • •!• • 200
I • 750
К 350
N 500
PP Qo V §
P • •’?•• 120
A
и па нкреозими 11__ биологически активные вещества, оказы-
вающие стимулирующее действие на функции поджелудочной
железы и печени. Обнаружены также С> клетки, вырабатывающие
гастрин, р и П,-клетки, продуцируюцГие активные пептиды (рис.
2ТП7С^Г. --------
Для собственной пластинки слизистой оболочки (см. рис. 209,
Б) характерно содерждншц большого количества ретикудярмых-
волокон. Они образуют густую сеть по всей собственной пластин-
ке и, подходя кеэпителию участвуют в об шии базальной
мембраыыХС ретикулярными во.ГокпамТГтесно связаны отростча^
ТОТ Оетки, сходные по строению с ретикулярными клетка-
ми кроветворных органов. В собственной пластинке постоянно f
встречаются эозинофилу,,, лимфоциты, а также плазматические
кдетки.
' Мьийечная пластинка слизистой оболочки состоит из двух сло-
ев: внутреннего циркулярного и наружного (более рыхлого) —
и1род0Льн^ТГТсМГриег-20Д.'^). Толщина обоих слоев около 40 мкм.
В них имеются и косо идущие пучки мышечных клеток. От внут-
реннего циркулярного мышечного слоя отдельные мышечные
клетки отходят в собственную пластинку слизистой оболочки и в
подслизистую основу.
В слизистой оболочке много скоплений лимфоидной ткдшс—.
Одиночные (солитарные) лимфоидные узелки (nSttn+i—(yfnphatici
solitani) встречаются на всем протяжении тонкой кишки. Диаметр
их около 0,5—3 мм. Более крупные узелки, лежащие в дисталь-
ных отделах тонкой кишки, проникают в мышечнную пластинку
слизистой оболочки и располагаются частично______д подслизистой
основе. Количество одиночных лимфоидных узелков в стенке
тонкой кишки детей от 3 до 13 лет составляет около 15 000. По
мере старения организма количество их уменьшается.
Сгруппированные лимфоидные узе гиги (noduli lymphatici
aggr'egati), как правило, располагаются в подвздошной кишке,
но иногда встречаются в тощей и двенадцатиперстной кишках.
Число узелков варьирует в (ависимости от возраста: в тонкой
кишке у детей около 100, у взрослых — около 30—40, а в стар-
ческом возрасте их количество значительно уменьшается. Длина
одного сгруппированного лимфоидного узелка может быть от 2 до
12 см, а ширина — около 1 см. Наиболее крупные из них проника-
ют в подслизистую основу. Ворщшки в. слизистой оболочке в
| местах расположения сгруппированных лимфоидных узелков^как
как правило, отсутствуют.
> --------------
Рис. 211. Строение и распределение различных типов желудочно-кишечных эндо
криноцитов в пищеварительной системе человека.
Л — Международная классификация и распределение эндокриноцитов. Ультрастръктура
их гранул (по Д. Грубе и В. Форссману с изменением) Б — ультрамикроскопическое
строение желудочно-кишечных эндокриноцитов: / — ЕС-клетки с гранулами ракооб-
разной формы, 2 L-клетки с крупными электронно-плотными гранулами; 3 — D -клетки
с многочисленными мелкими гранулами; 4 — D-клетки с крупными умеренно плотными
гранулами (по С. М. Говалову, И. М. Коростышевской, М. С. Виноградовой).
531
2
3
Рис. 212. Строение двенадцатиперстной
кишки (продольный разрез).
/ слизистая оболочка,// подслизистая
основа, /// мышечная оболочка; IV
серозная оболочка, / кишечные ворсинки,
/ 2 кишечные железы (крипты); 3 мы-
шечная пластинка слизистой оболочки
4 — подслизистые (дуоденальные) железы;
5 —мышечно кишечное нервное сплетение.
Подслизистая осно-
в а нередко содержит дольки жи-
ровой ткашгГВ ней рдсгТолагают-
ся сосуды ^"подслизистое нерв-
н<5ё сплетение. В двена цати-
. пёрстной кишке находятся
I сложные трубчатые разветвлен-
{1~ыё железы подслизистой осно- м0**
• вы TglE submucosae), концевые
отделы которых занимают почти
всю толщину подслизистой осно-
вы. Дольки желез подслизистой
4 основы могут заходить и в об-
ласть пилорического сфинктера,
а иногда на несколько санти-
5
метров проникают в пилориче-
скую часть желудка и в началь-
ные отделы тощей кишки. Чаще
всего железы подслизистой ос-
новы располагаются в толще
циркулярных складок (рис.
212). По характеру выделяемого
секрета это слизистые железы.
ни имеют некоторое сходство
с пилорическими железами же-
лудка. У секреторных клеток
светлая ячеистая цитоплазма и сплющенное темное ядро, лежа-
щее у основания клетки. Выводные протоки этих желез открываются
в кишечные крипты или в пространства между ворсинками. В началь
ных отделах протоки выстланы кубическими или призматическими
клеткамиПТо"сравнению с клетками концевых отделов они более
мелкие и содержат меньшее количество слизи. В наружных отделах
выводные протоки выстланы клетками с исчерченной каемкой.
В этих железах могут встречаться также экзокриноциты с аципо-
фильными гранулами, желудочно-кишечные эндокриноциты, бпка-
ловидные и отдельные париетал’ьные клетки? Железы подслизистой
основы выполняют важную роль, участвуя в образовании кишечного
сока. В секрете этих желез найдены дипептидазы, подобные
тем, которые обнаруживаются в пилорических и кардиальных желе-
зах желудка. Секрет желез участвует в расщеплении углеводов при
помощи амилазы, а также активизирует амил75литическое действие
532
сока поджелудочной железы. Мукоиды, содержащиеся в секрете,
нейтрализуют соляную кислоту, поступающую из желудка. Считает-
ся, что в дуоденальных железах находятся эндокриноциты—< S-
клетки, вырабатывающие гормон с е кретин. Выделение секрета
железами происходит при стимуляции*1)луждающего нерва и при 2
введении соляной кислоты в двенадцатиперстую кишку. О
Мышечная оболочка тонкой кишки состоит из двух
слоев: внутреннего - циркулярного (более мощного) и наружно- .
го -^продольного (см. рис. 209, Л). Направление хода пучков
мышечных клётСПгт обоих слоях не строго циркулярное и продоль-
I ное, а спиральное. В наружном слое завитки спирали более рас-
‘тянуты по сравнению с внутренним слоем. Между обоими мышеч-
ными слоями располагается прослойка рыхлой волокнистой~срй-
динительной ткани, в ^которой находятся узлы мьппечПГРтгитттеч-
ного нервного сплетения и сосулы.
Функция мышечной оболочки заключается в перемешивании и
проталкивании химуса по ходу кишечника. В тонкой кишке раз-
j ичают сокращения двух видов. Сокращения местного характера
(бусловлены главным образом сокращениями внутреннего слоя
мышечной оболочки. Они совершаются ритмически 12—13 раз
в минуту. Другие сокращения — перистальтические — вызываются
действием мышечных элементов обоих слоев и распространяются
последовательно по всей длине кишки. Перистальтические сок-
ращения прекращаются после разрушения мышечно-кишечного
нервного сплетения. Усиление перистальтики тонкой кишки про-
исходит при возбуждении симпатических нервов, ослабление —
при возбуждении блуждающего нерва.
Серозная оболочка покрывает снаружи тонкую кишку
со всех сторон, за исключением двенадцатиперстной кишки,
которая покрыта брюшиной только спереди, а в остальных частях
имеет соединительнотканную оболочку.
Васкуляризация. Артерии, входя в стенку тонкой кишки, об-
разуют три сплетения: межмышечное — между внутренним и на-
ружным слоями мышечной оболочки; широкопетлистое — в под-
слизистой основе и узкопетлистое — в слизистой оболочке. Из
последнего выходят артериолы, образующие кровеносные капил-
ляры вокруг кишечных крипт, и по 1—2 артериолы, входящие
в каждую ворсинку ц распадающиеся там на капилярные сети.
Из кровеносных капилляров ворсинки кровь собирается в венулу,
проходящую вдрль ее оси (см. рис. 188, Л). Вены тонкой кишки
образуют два сплетения — сплетение в слизистой оболочке и
сплетение в подслизистой основе. Имеются многочисленные арте-
риоловенулярные /Анастомозы типа замыкающих артерий, регули-
рующие приток крови к кишечным ворсицкам. Во время акта
пищеварения' анастомозы между артериями и венами закрыты,
и вся масса крови устремляется в слизистую оболочку, к ее
ворсинкам. В период голодания анастомозы открыты и основная
масса крови проводит минуя слизистую обо; очку. Запирающие
вены регулируют 'объем венозного отрока от тонкой кишки. В
случае резкого переполнения эти вены могут депонировать значи-
тельные количества крови.
Лимфатические сосуды тонкой кишки представлены очень ши-
роко разветвленной сетью. В каждой кишечной ворсинке есть
центрально расположенный, слепо оканчивающийся на ее вершине
лимфатический капилляр. Просвет его шире, чем в кровеносных
капиллярах. Из лимфатических капилляров ворсинок лимфа от-
текает в лимфатическое сплетение слизистой оболочки, а из
него — в соответствующее сплетение подслизистой основы, образо-
ванное более крупными лимфатическими сосудами. В это сплете-
ние вливается также густая сеть капилляров, оплетающих оди-
ночные и групповые лимфатические узелки. Из подслизистого
сплетения отходят лимфатические сосуды, находящиеся между
слоями мышечной оболочки.
Иннервация. Афферентная иннервация осущест-
вляется мышечно-кишечным чувствительным сплетением (plexus
myentericus sensibilis), образованным чувствительными нервными
волокнами спинальных ганглиев и их рецепторными окончаниями
Ветвистые и кустиковые нервные окончания часто встречаются
в подслизистой основе и собственной пластинке слизистой оболоч-
ки. Их терминальные веточки достигают сосудов, дуоденальные
желез эпителия кишечных крипт и ворсинок. Обильные ветвле-
ния чувствительных волокон наблюдаются в подвздошной кишке
и илеоцекальной области, где преобладают кустиковидные формы
рецепторов. Отдельные рецепторы имеются в самих нервных
ганглиях.
Эфферентная иннервация осуществляется симпа-
тическими и парасимпатическими нервами. В толще стенки кишки
хорошо развиты парасимпатические мышечно-кишечное и под
слизистое нервные сплетения. Мышечно-кишечное сплетение
(plexus myentenens) наиболее развито в двенадцатиперстной
кишке, где наблюдаются многочисленные, плотно расположенные
крупные ганглии. Количество и размеры ганглиев в тонкой киш-
ке уменьшаются в каудальном направлении. В ганглиях различают
клет_ки I и II типа причем клеток I типа значительно больше.
Для тонкой кишки характерно наличие большого количества
клеток II типа по сравнению с другими отделами пищеваритель-
ной трубки. Их особенно много в двенадцатиперстной кишке,
в начальном отделе подвздошной кишки и в илеоцекальной
области.
Гистофизиология процесса всасывания в тонкой кишке. Ки-
шечные ворсинки являются главными структурами, обеспечиваю-
щими процесс пристеночного пищеварения и всасывания в тонкой
кишке. В этом процессе существенную роль играют и кишечные
крипты. Расщепленные под влиянием ферментов, пищевые вещества
проникают в столбчатые эпителиоциты ворсинок, где подвергаются
ряду дальнейших превращений. Затем они поступают через соб-
ственную пластинку слизистой оболочки в кровеносные капил-
ляры (белки, углеводы) или в лимфатические капилляры (жи-
534
ры). Гладкие мышечные клетки, расположенные в строме ворсинок,
сокращаясь, способствуют проникновению всосавшихся веществ в
расширяющиеся при этом сосуды. При расправлении кишечных
ворсинок питательные вещества снова поступают через эпителий
в их строму, а капилляры вновь наполняются кровью. Ежеминутно
каждая ворсинка совершает около 4—6 сокращений. Наиболее
активное движение кишечных ворсинок наблюдается на высоте
процесса всасывания. При голодании движение их замедляется.
Подсчитано, что одна ворсинка способна захватить в минуту
около 0,03 мм3 вещества из химуса. При наличии в тонкой
кишке более 1,4 млн. ворсинок в минуту в ней может всасываться
около 45 см пищевых веществ.
Всасывание жиров начинается на вершине кишечной
ворсинки и распространяется по направлению к ее основанию.
Вначале (через 20 мин после кормления подопытного животного)
мелкие капли жира (хиломикроны) располагаются в исчерченной
каемке столбчатого эпителиоцита, точнее в пространствах между
ее микроворсинками, а также в периферической зоне цитоплазмы
клетки под каемкой (рис. 213). Здесь они подвергаются действию
липаз. При этом образуются глицерин и свободные жирные
кислоты, которые эстеризуются с помощью холинэстеразы и
холинэстерина и превращаются в хорошо всасываемые клеткой
Эфиры холестерина. После прохождения через оболочку клетки
эфиры холестерина расщепляются, в результате чего появляются
свободные жирные кислоты, которые снова соединяются с всосав-
шимся в клетку глицерином, образуя капли жира. В синтезе
нейтрального жира из глицерина и жирных кислот, по-видимому,
важную роль играют комплекс Гольджи и митохондрии эпители-
оцита. >
Наряду с описанным процессом имеются данные о непо-
средственном всасывании эмульгированного жира столбчатыми
эпителиоцитами. Затем хиломикроны начинают перемещаться в
базальную часть клетки. Несколько позже (через 30 мин — 1ч
после начала эксперимента) крупные капли жира обнаружива-
ются и в зоне цитоплазмы над ядром, где расположен комплекс
Гольджи.
Гистофизиология процесса всасывания белков в
тонкой кишке изучена недостаточно. Белковые включения в эпи-
телии кишки удается видеть только у плодов и новорожденных.
Их появление в столбчатых эпителиоцитах, по-видимому, объяс-
няется заглатыванием плодом амниотической жидкости. Во взрос-
лом организме все белки всасываются только после их расщепле-
ния до аминокислот.
Всасывание углеводов в тонкой кишке, вероятно,
связано со способностью столбчатых эпителиоцитов синте>иро-
вать гликоген путем фосфорилирования — превращения гексоз в
эфиры фосфорной кислоты. В эпителиоцитах ворсинок тонкой
кишки зародыша имеется большое количество гликогена. У
взрослых людей гликоген в эпителиоцитах тонкой кишки появ-
S35
Рис. 213. Схема процесса всасывания жира через эпителий тонкой кишки (по Кар-
делю, Баденхаузену, Портеру с изменением).
А — столбчатый эпителиоцит после голодания животного; Б — он же после приема жирной
пиши; / — микроворсинки исчерченной каемки; 2 — гликокаликс; 3 капли липидов;
4 — эндоплазматическая сеть; 5 — митохондрии; 6 — хиломикроны; 7 — комплекс Гольджи;
8 — межклеточное пространство; 9 — капли липидов, поступающие через межклеточное
пространство в лимфатический капилляр (10).
ляется только при введении в кишечник большого количества
сахара. Он выявляется в виде гранул в исчерченной каемке
эпителиоцитов. В каемке и апикальных частях клеток содержатся
щелочная и кислая фосфатазы и фосфоамидаза, участвующие в
процессах расщепления и всасывания углеводов. Всосавшиеся
через эпителиоциты простые сахара (главным образом глюкоза)
поступают затем в кровеносные капилляры. Через эпителий
кишки происходит также всасывание воды с растворенными
в ней минеральными веществами, витаминов и некоторых других
веществ.
536
Толстая кишка
Толстая кишка выполняет важные функции — интенсив-
ное всасывание воды из химуса и формирова-
ние каловых масс. Способность к всасыванию жидкостей
используют во врачебной практике для введения больным пита-
тельных веществ с помощью клизм. В толстой кишке выделяется
значительное количество слизи, которая облегчает продвижение
содержимого по кишечнику и способствует склеиванию непере
варенных частиц пищи. Одной из функций толстой кишки явля-
ется выделительная. Через слизистую оболочку этой кишки
выделяется ряд веществ, например кальций, магний, фосфаты,
соли тяжелых металлов и т. д. В толстой кишке вырабатываются
витамин К и витамин В Этот процесс осуществляется с учас-
тием бактериальной флоры, постоянно присутствующей в ки-
шечнике. С помощью бактерий в толстой кишке происходит пере-
варивание клетчатки.
Развитие. Эпителий ободочной кишки и тазовой части прямой
кишки развивается из энтодермы. В >уожной и промежуточной
зонах анальной части прямой кишки эпителии имеет эктодер-
ма, ьное происхождение. Граница между эпителием кишечного
'и кожного типа выражена нерезко и расположена между стол-
бчатой и промежуточной зонами прямой кишки. Эпителий кишеч-
ной трубки сильно разрастается на 6—7-й неделе внутриутробной
жизни плода. Ворсинки и крипты в слизистой оболочке эмбриона
закладываются почти одновременно. Позднее сюда врастает мезен-
хима, что ведет к сильному выпячиванию ворсинок в просвет
кишки. На 4-м месяце эмбрионального развития закладка тол-
стой кишки содержит большое количество ворсинок. В дальнейшем
усиленный рост поверхности слизистой оболочки приводит к
растягиванию и сглаживанию этих ворсинок. К концу эмбриоге-
неза ворсинок в толстом кишечнике уже нет. Мышечная оболочка
толстой кишки развивается на 3-м месяце внутриутробного пе-
риода, а мышечная пластинка слизистой оболочки — на 4-м
месяце развития зародыша.
Ободочная кишка
Стенка ободочной кишки образована слизистой оболочкой,
подслизистой основой, мышечной и серозной оболочками. В
слизистой оболочке много__циркулярных складок и ки-
шечных желез (крипт), значительно увеличивающих ее повер-
хность, иГотсутствую‘т~в'ор^цнки~Д(рис. 214). Циркулярные складки
’образуются на внутренней поверхности кишки из слизистой
оболочки и подслизистой основы. Они располагаются- поперек ~и
имеют полулунную форму (отсюда название «полулунные склад-
ки»). 1Сишечные железы (крипты) в ободочной кишке развиты
брлынег чем в тонкой, расположены чаще, размеры их больше
(0,4—0,7 мм), они шцрс_содержат очень много бокаловидных
экзокриноцитов.
537
4
Рис. 214. Строение толстой кишки.
А - схема; Б микрофотография. / слизистая оболочка; II — подслизистая основа;
/// мышечная оболочка; IV — серозная оболочка: / эпителий; 2 — кишечные железы
(крипты); 3 — собственная пластинка слизистой оболочки; 4 — мышечная пластинка сли-
зистой оболочки; 5 — кровеносные сосуды; 6 одиночный лимфоидныи узелок; 7 — бокало-
видный экзокриноцит.
Эпителии слизистой оболочки однослойный призматический.
Он состоит из трех основных видов клеток: утолбчатых эпртО-—
лиоцитов, бокаловидны\ лкзикдинппитов и желудочно-кишечных
, эПДОкриноцитов. Кроме того, имеются не диффе рении ровЭТтыр
эгиЯ^тптОциты: ТУолбчать/е эпителиоцит Я располагаются на повер-
хности слизистой оболочки и в ее криптах. По своему строению
они сходны с аналогичными клетками тонкой кишки, но у них
более тонкая исчерченная каемка (см. рис. 214, Б; рис. 215)’.
Бокаловидные экзокрииоциты, выделяющие слизь, имеются в боль-
шом количестве___в. криптах,. Строение их было описано ранее.
УСооневапия кишечных кридт лежат недифференцированные .эпи-
телиоциты. В них часто видны митозы^За счет этих клетПК~ГТро-
исходит регенерация столбчатых эпителиоцитов и бокаловидных
экзокриноцитов. Кроме того, изредка встречаются эндокринные
клетки с ацидофильными гранулами.
Собственная пластинка слизистой оболочки образует тонкие
соединительнотканные прослойки между кишечными криптами. В
S38 ~
Рис. 215. Верхняя часть железы (крипты) толстой кишки собаки. Электронная микро-
фотография, Х6000 (по К. А. Зуфарову).
/ — просвет кишечной железы (крипты); 2— каемчатые столбчатые эпителиоциты; 3 -
исчерченная каемка (микроворсинки); 4 — бокаловидные экзокрииоциты; 5 лимфоцит
этой пластинке часто встречаются одиночные лимфоидные-узелки,
из которых лимфоциты мигрируют в окружающую соединительную
ткань и проникают в эпителий.
Мышечная пластинка слизистой оболочки сильнее выражена,
чем в тонкой кишке, и состоит из двух слоев. Внутренний слой
более плотный, образован преимущественно циркулярно располо-
жен ными пучками гл.адких миоцитов. Наружный слой представлен
* — 539
пучками гладких миоцитов, ориентированных^ частично продольно,
частично косо по отношению—к—оси. д<ишкш Мышечные клетки
’5"эТом-слое расположены более рыхло, чем во внутреннем.
Подслизистая основа содержит много жировых кле-
ток- Здесь располагаются сосудисты) _а также нервное подслизис-
тое сплетения. В подслизистой основе ободочной кишки всегда
очень много лимфоидных узелкод^_они распространяются сюда из
собственной пластинки слизистой оболочки.
Мышечная оболочка представлена двумя слоями
гладких мышц: внутренним - циркулярным и наружным — npoz
дольным. Наружный слой мышечной оболочки в ободочной кишке
“и^еет особое строение. Этот слой не сплошной, и пучки гладких _
миоцитов в нем собраны в три ленты, ^тянущиеся вдоль всей
ободочнойкишки. В участках кишки, лежащих между лентами,
обнаруживается лишь тонкий слой, состоящий из незначительного
количества продольно расположенных пучков гладких миоцитов.
Эти участки кишки образуют вздутия, выбухающие наружу..Между
двумя слоями мышечной оболочки есть прослойка рыхлой волок-
нистой соединительной ткани,- в которой проходят сосуды и
находится мышечно-кишечное нервное сплетение...
Се розная оболочка покрывает ободочную кишку ена-
^ужи^) Иногда она имеет пальцеобразные выросты. Эти выросты
представляют собой скопления жировой ткани, покрытые брюши-
ной.
Червеобразный отросток
Для этого органа характерно наличие больших скоплений лим-
фоидной ткани. Червеобразный отросток имеет просвет (рис. 216)
треугольной Формы у детей и круглой — у взрослых. С годами
этот просвет может облитерироваться, зарастая соединительной
тканью.
Слизистая оболочка червеобразного отростка имеет
кишечные железы (крипты), покрытые однослойным призматичес- ।
ким эпителием с небольшим содержанием бокаловидных клеток. ,
На пне кишечных крипт чяще, чем в других отделах толстой
кишки, встречаются -чкзлкринопиты с апил.офильными гранулами.
Здесь же располагают<'^ц5дифферен1шрова11ные зли-тепиапиты и
эндокринные ECL-клетки, причем их здесь сравнительно больше,
чем В криптах тонкой кишки (в среднем в каждой около 5 клеток).
Собственная пластинка слизистой оболочки без резкой грани-
цы (вследствие слабого развития мышечной пластинки слизистой)
переходит в полслизистую основу. В собственной пластинке и в
подслизистой основе располагаются многочисленные крупные
Рис. 216. Червеобразный отросток человека.
А — взрослого человека; Б - плод человека 24 нед развития (препарат Е. Л. Работнико-
вой); / — эпителий: а — инфильтрация лимфоцитами; 2— кишечные железы (крипты);
3 — собственная пластинка слизистой оболочки; 4 — лимфатические узелки; 5 — гермина-
тивные центры; б подслизистая основа; 7 мышечная оболочка; 8 — серозная оболочка.
S41
местами сливающиеся скопления лимфоидной ткани. При попада-
нии инфекции в просвет отростка всегда наступают выраженные
изменения его стенки. В лимфоидных узелках возникают круп-
ные светлые центры, лимфоциты сильно инфильтрируют соеди-
нительную ткань собственной пластинки и часть их проходит
через эпителий в просвет червеобразного отростка. В этих слу-
чаях в проедете отростка часто можно видеть отторгнутые эпи-
телиоциты и скопления погибших лимфоцитов. В подсли-
зистой основе располагаются кровеносные сосуды, и нер-*
вное подслизистое сплетение. ’И
Мышечная оболочка имеет два слоя: внутренний -и
циркулярный и наружный — продольный. Продольный мышечный
слой
отростка
отличие от соответствующего слоя
ободочной кишки. Снаружи отросток обычно покрыт серозной
оболочкой, которая образует собственную брыжейку отростк-1
Червеобразный отросток осуществляет защитную функций'
скопления лимфоидной ткани в нем входят в состав перифеф.
ческих отделов иммунной системы
Прямая кишка
I f МД!
I . »ч I
Стенка прямой кишки состоит из тех же оболочек, что и стен-
ка ободочной кишки. В тазовой части прямой кишки ее слизистая
оболочка имеет три поперечные складки, ^образовании этик
складок участвуют подслизистая основа и кольцевой слой мышёч
ной оболочкй7~Нижё этих складок располагается 8—1U продоль-
ных складок; ежду которыми видны углубления.
В~1Гна.1ьНОй части кишки различают три зоны: стол бч„
тую, промежуточную и кожную. В столбчатой
зоне дродольшяе^ск л а дк и образуют заднепроходные столбы.
В промежуточной зоне эти образования соединяются^ образуя
зону слизистой оболоч к и с_-гдадкпи поверхностью в видекольца
шириной около 1 см (zona haemorrhoidalis).
Слизистая оболочка прямой кишки состоит из
.эпителия, собственной и мьпнечной пластинок. Эпителии выверь
2£неМ—отделе прямой кишки однослойный призматиче< кий Й
Столбчатой зоне нижнего отдела - многослойный, кубически^.
в промежуточной цишгослоиный—гшосжийнеороговедающий, jb
кожной — многослойный плоский ороговевающии. Переход от
многослойного кубического эпителия~к многослойному плоскому
совершается резко в виде зигзагообразной — аноректальной линии
(linea anorectalis).
Переход в эпителий кожного типа постепенный. В эпителии
прямой кишки встречаются столбчатые эпителиоциты с исчерчен-
ной каемкой, бокаловидные экзокриноциты и эндокринные (ECL)
клетки. Последних осоосно много в столбчатой зоне. Эпителий
в верхней части прямой кишки образует кишечные крипты. Они
несколько длиннее, чем в ободочной кишке, но менее многочислен
ны. В нижних отделах кишки крипты постепенно исчезают
542
Собственная пластинка принимает участие в формировании
идялок прямой кишки. Здесь располагаются о()иночнь1Г7ПТифоид-
ные узелки и сосуды. В области столбчатой зоны в этой
пластинке залегает сеть тонкостенных кровеносных лакун, кровь
из которых оттекает в геморроидальные вены. В собственной
пластинке этой области проходят протоки желез, расположенных
в подслизистой основе.
В п р омежуточной зоне собственная пластинка содер-
ит большое количество эластических волокон, а также лимфох—».
.тов'и тканевых базофилов (тучные клетки). Здесь находятся
। Единичные сальные железы. J , t о '
"В кТТяГнои зон е, окружающей анальное отверстие, к
иьным железам присоединяются волосы Потовые железы в
'^ственной пластиИке слизистой обол. тки появляются на рассто-
йй 1 —1,5 см от ануса, представляют собой трубчатые железы^
цевые отделы которых свернуты кольцом (gll. circnmanales).
железы апокринового типа.
*1ышечная пластинки слизистой оболочки, как и в других от-
< толстой кишки, состоит из двух слоев. Пучки гладких—ш+о-
"св ее постепенно переходят в узкие продольные пучки, тяну-
. ?'.’ч до столбчатой зоны.
В подслизистой основе располагаются сосудистые
.1 нервные сплетения. Здесь же встречаются пластинчатые нер-
п-ьГе тельца. В подслизистой основе лежит сплетение гемор-
-лглальных—ве . При нарушении тонуса стенок _этих сосудов
'являю cj варикозные расширения. При патологических измене-
чизх эти образования могут быть причиной кровотечений. В
лстизистой основе столбчатой зоны прямой кишки имеется 6 8
1 (ветвленных трубчатых образований тянущихся до циркулярно-
лоя мышечной оболочки, прободающих его и слепо заканчи-
А.хщихся в межмышечной соединительной ткани. На концах их
эбоазуются ампулярные расширения, которые выстланы одним или
двумя слоями кубических клеток. Эпителий главных протоков
гих рудиментарных анальных желез (gll. anales) состоит из
.-скольких слоев полигональных клеток. Устье протока выстлано
ногослойным плоским эпителием Эти эпителиальные трубочки
зеценивают как гомологи анальных желез животных. У человека
при патологических условиях они могут служить местом образо-
вания фистул.
Мышечная оболочка состоит из двух слоев: внутрен-
него — циркулярного и наружного — продольного. .Циркулярный
слой на~~разных уровнях прямой кишки образует два утолщения,
которые выделяюся как отдельные анатомические образования
^фйнк'тёрьГугп. sphincter ani internus et externus). Последняя
1ышца в отличие от всей мышечной оболочки образована попе-
речнополосатой мышечной тканью. Наружный продольный слои
мышечной оболочки прямой кишки "в противоположность другим
отделам толстой кишки сплошной. Между обоими мышечными
^шоямй* имеется прослойка рыхлой волокнистой неоформленной
мз
риединительной-.ткани1_ в которой лежат мышечно-кишечные спле-
тения и сосуды. ~1 ~
СТ^рсГз пая оболочка покрывает прямую кишку {^верх-
ней ее части; в нижних отделах прямая кишка имеет соединительно-
тканную оболочку. '
Иннервация. В парасимпатическом мышечно-кишечном нер-
вном сплетении толстой кишки, начиная с проксимальных от-
делов, моторные нейроны I типа постепенно заменяются чувстви-
тельными нейронами II типа, которые в прямой кишке становятся
преобладающими.
Афферентная иннервация в прямой кишке выра-
жена. В толстой кишке афферентные волокна образуют в мышеч-
ной оболочке чувствительное сплетение. Чувствительные оконча-
ния имеют вид кустиков и терминалей, оканчивающихся в глад-
ких мышцах.
Печень
Печень (hepar) — самая крупная железа пищеварительного
тракта. Функции печени чрезвычайно разнообразны. В ней обез-
вреживаются многие продукты обмена веществ, инак-
тивируются гормоны, биогенные амины, а также
ряд лекарственных препаратов. Печень участвует в защитных
реакциях организма против микробов и чужеродных ве-
ществ в случае проникновения их извне. В ней образу-
ется гликоген — главный источник поддержания постоян-
ной концентрации глюкозы в крови. В печени синтезиру-
ются важнейшие белки плазмы крови: фибриноген,
альбумины, протромбин и др. Здесь метаболизируется железу
и образуется желчь — необходимая для всасывания жиров
в кишечнике. Большую роль она играет в обмене холес-
терина, который является важным компонентом клеточных
мембран. В печени накапливаются необходимые для организма
жирорастворимые витамины — A, D, Е, К и др. Кроме того, в
эмбриональном периоде печень является органом кроветво-
544
г
рения. Столь мно!счисленные и важные функции печени опре-
зеляют ее значение для организма как жизненно необходимого
1 органа.
Развитие. Зачаток печени образуется из энтодермы в конце
13-й недели эмбриогенеза и имеет вид мешковидного выпячивания
•-антральной стенки туловищной кишки (печеночная бухта),
растающего в брыжейку. В процессе роста печеночная бухта
подразделяется на верхний (краниальный) и нижний (каудаль-
ный) отделы. Краниальный отдел служит источником развития
печени и печеночного протока, каудальный — желчного пузыря
'♦i желчного протока. Устье печеночной бухты, в которое впадают
^краниальный и каудальный отделы, образует общий желчный про-
ток. Эпителиальные клетки краниальною отдела печеночной бухты
-быстро разрастаются в мезенхиме брыжейки, формируя многочи-
сленные тяжи. Между эпителиальными тяжами располагается сеть
широких кровеносных капилляров, происходящих из желточной
вены, которая в процессе развития дает начало воротной вене.
Сложившаяся таким путем железистая паренхима печени
своим строением напоминает губку. Дальнейшая дифференцировка
печени происходит во второй половине внутриутробного периода
развития и в первые годы после рождения. При этом по ходу
ветвей воротной вены внутрь печени врастает соединительная ткань,
разделяя ее на печеночные дольки.
Строение. Поверхность печени покрыта соединительнотканной
капсулой, которая плотно срастается с висцеральным листком
брюшины. Паренхима печени образована печеночными дольками
(lobuli).
Печеночные дольки — структурн о-ф ункциона л ь-
ные единицы печени (рис. 217). Существует несколько
представлений об их строении — старое, классическое, и более
новые, высказанные в середине XX столетия. Согласно клас-
сическому представлению, печеночные дольки имеют форму шес-
тигранных призм с плоским основанием и слегка выпуклой вер-
шиной. Их ширина не превышает 1,5 мм, тогда как высота, не
смотря на значительные колебания, несколько больше. Иногда
простые дольки сливаются (по две и более) своими основаниями
и формируют более крупные сложные печеночные дольки. Ко-
личество долек в печени человека достигает 500 гыс. Междолько-
вая ^соединительная ткань образует строму органа. В ней про-
ходят кровеносные сосуды и желчные протоки, структурно и
функционально связанные с печеночными дольками. У человека
междольковая соединительная ткань развита слабо, и вследствие
•того печеночные дольки плохо отграничены друг от друга. Такое
строение характерно для здоровой печени человека. Наоборот,
интенсивное развитие соединительной ткани, сопровождающееся
атрофией (уменьшением) печеночных долек, является признаком
тяжелого заболевания печени, известного под названием «цирроз».
Кровеносная система. Исходя из классического представления
° строении печеночных долек, кровеносную систему печени услов-
’8 ГИСТОЛОГИЯ
545
но можно разделить на три части: систему притока кро-
ви к долькам, систему циркуляции крови внутри
них и систему оттока крови от долек (рис. 218).
Система притока представлена воротной веной и печеночной
артерией. Воротная вена, собирая кровь от всех непарных органов
брюшной полости, богатую веществами, всосавшимися в кишечни-
ке, доставляет ее в печень. Печеночная артерия приносит кровь
от аорты, насыщенную кислородом1. В печени они многократно
разделяются на все более мелкие сосуды: долевые, сегментарные,
междольковые вены и артерии (vv. и аа. interlobulares), вокруг-
дольковые вены и артерии (vv. и аа. perilobulares). На всем протя-
жении эти сосуды сопровождаются аналогичными по названию
желчными протоками (ductuli biliferi). Вместе ветви воротной
вены, печеночной артерии и желчные протоки составляют так
называемые триады. Рядом с ними лежат лимфатические сосуды.
Междольковые вены и артерии, подразделяющиеся по размеру
на 8 порядков, идут вдоль боковых граней печеночных долек. От-
ходящие от них вокругдольковые вены и артерии опоясывают
дольки на разных уровнях.
Междольковые и вокругдольковые вены являются сосудами со
слаборазвитой_мыщечной оболочкой. Однако в местах разветвле-
ния в их стенках наблюдаются скопления мышечных элементов,
образующих сфинктеры. Соответствующие междольковые и вок-
ругдольковые артерии относятся к сосудам мышечного типа. При
этом артерии обычно в несколько раз меньше по диаметру, чем ря-
дом лежащие вены.
От вокругдольковых вен и артерий начинаются кровеносные
капилляры. Они входят в печеночные дольки и сливаются, обра-
зуя внутридольковые синусоидные сосуды, которые составляют
систему циркуляции крови в печеночных дольках. По ним течет
смешанная кровь в направлении от периферии к центру долек.
Соотношение между венозной и артериальной кровью во внут-
ридольковых синусоидных сосудах определяется состоянием
сфинктеров междольковых вен. Внутридольковые капилляры отно-
сятся к синусоидному (до 30 мкм в диаметре) типу капилляров
с прерывистой базальной мембраной. Они идут между тяжами
печеночных клеток — печеночными балками, радиально сходясь к
центральным венам (vv. centrales), которые лежат в центре
печеночных долек.
Центральными венами начинается система оттока крови от
долек. По выходе из долек эти вены впадают в собирательные,
или поддольковые вены (vv. sublobulares), проходящие в меж-
дольковых перегородках. Поддольковые вены не сопровождаются
артериями и желчными протоками, т. е. не входят в состав триад.
По этому признаку их легко отличить от сосудов системы ворот-
ной вены — междольковых и вокругдольковых вен, приносящих
кровь к долькам.
Часть кислорода поступает к печени с кровью также по воротной вене.,
546
5
Рис. 218. Схема кровенос-
ной системы печени (рис.
Е Ф. Кстовскою).
/ — воротная вена и пече-
ночная артерия; 2 — долевая
вена и артерия; 3 — сегмен-
тарная вена и артерия; 4 —
междольковая артерия и ве-
на; 5 вокругдольковая веиа и
артерия; 6 — внутридолько-
вые гемокапилляры; 7 —
центральная вена; 8 — соби-
рательная вена; 9 пече-
ночные вены; 10 печеноч-
ная долька.
Центральные и поддольковые вены — сосуды безмышечного
типа. Они сливаются и образуют ветви печеночных вен, которые в
количестве 3—4 выходят из печени и впадают в нижнюю полую
вену. Ветви печеночных вен имеют хорошо развитые мышечные
сфинктеры. С их помощью регулируется отток крови от долек и
всей печени в соответствии с ее химическим составом и массой.
Таким образом, печень снабжается кровью из двух мощных
источников — воротной вены и печеночной артерии. Благодаря
этому через печень проходит за непродолжительное время вся
кровь организма, обогащаясь белками, освобождаясь от продуктов
азотистого обмена и других вредных веществ. Паренхима печени
имеет огромное число кровеносных капилляров, и вследствие этого
кровоток в печеночных дольках осуществляется медленно, что
способствует обмену между кровью и клетками печени, выполняю-
щими защитную, обезвреживающую, синтетическую и другие важ-
ные для организма функции. При необходимости в сосудах печени
может депонироваться большая масса крови.
Печеночные дольки. Согласно классическому представлению,
печеночные дольки образованы печеночными балками и внутри-
дольковыми синусоидными кровеносными капиллярами. Печеноч-
ные балки, построенные из гепатоцитов — печеночных эпителио-
цитов, расположены в радиальном направлении. Между ними в том
же направлении от периферии к центру долек проходят кровенос-
ные капилляры.
Внутридольковые кровеносные капилляры выстланы плоскими
эндотелиоцитами. В области соединения эндотелиальных клеток
ДРУГ с другом имеются мелкие поры. Эти участки эндотелия называ-
ются ситовидными (рис. 219). Между эндотелиоцитами рассеяны
многочисленные звездчатые макрофаги (клетки Купфера), не об-
разующие сплошного пласта. В отличие от эндотелиоцитов они
имеют моноцитарное происхождение и являются макрофагами пе
i&* 547
Рис. 210. Схема ультра микроскопического строения печени (рис. Е. Ф Котов-
ского) .
/ — виутридольковый синусоидный сосуд; 2 — эндотелиальная клетка; 3 — ситовидные
участки; 4 — звездчатые макро фагоциты; 5 — перисинусоидальное пространство; 6 — ре-
тикулярные волокна; 7 — микроворсинки гепатоцитов; 8 — гепатоциты; 9 — желчный ка-
пилляр; 10 перисинусоидальные липоциты; II жировые включения; 12 — эритроциты
в гемокапилляре.
чени (macrophagocytus stellatus), с которыми связаны ее защитные
реакции. Звездчатые макрофаги имеют отростчатую форму и строе-
ние, типичное для фагоцитов. Прй"бсуществлении защитных реак-
ций они отходят от стенки кровеносных капилляров и превращаются
в свободные макрофаги. Базальная мембрана на большом протяже-
нии у внутридольковых капилляров отсутствует, за исключением их
периферических и центральных отделов. Капилляры окружены
узким (0,2 1 мкм) вокругсинусоидным пространством. Через поры
в эндотелии капилляров составные части плазмы крови могут попа-
дать в это пространство, а в условиях патологии сюда проникают и
форменные элементы. В нем, кроме жидкости, богатой белками,
находятся микроворсинки гепатоцитов, иногда отростки звездчатых
макрофагов, аргирофильные волокна, оплетающие печеночные бал-
548
'I
ки, а также отростки клеток, известных под названием перисину
соидальные липоциты. Эти клетки небольшою ра шера Г1' 10 мкм),
располагаются .между соседними гепатоцита мш_Оци постоянно со-
держат не^слйвакУЩИеся между собой мелкие капли жира, имеют
мрибосом и единичные митохондрии. Количество нерисинуtoи-
дальных липоцитов может резко возрастать при ряде хронических
заболеваний печени. Полагают, что липоциты, подобно фиброблас-
там, способны к волокнообразованию, а также к депонированию
жирорастворимых витаминов.
Печеночные балки состоят_Ц'4 гецатацитал. (см. ни же), связан—
ных~71руг~с другом десмосомами и по типу «замка». Балки анастомо-
зируют между собой, и поэтому их радиальное направление в
дольках не всегда четко заметно. В печеночных балках и а на с тома»
з5х между ними гепатоциты распблагаютсядвумя рядами, тесно
прилегающими друг к другу ."В связи с этим на поперечном срезе
каждая балка представляется состоящей из двух клеток. По анало-
гии с другими железами печеночные балки можно считать концевы-
ми отделами печени, так как образующие их гепатоциты секретиру-
ют глюкозу, белки крови и ряд других веществ.
Между рядами гепатоцитов, составляющих балку, располагают-
. ся желчные капилляры диаметром от 0,5 до 1 мкм. Этикапилляры
не имеют собствен ной стенки. Их стенка образована сопрйкасаюши-
мися~поверхностями гепатоцитов па которых имеются небольшие
углу блениядю впадающие друг с другом и вместе образующие
просвет желчного капилляра (рис. 220, А, Б). Просвет желчного
капилляра не сообщается с межклеточной щелью благодаря тому,
что мембраны соседних гепатоцитов в этом месте плотно прилегают
друг к другу, образуя замыкательные пластинки. Поверхности
гепатоцитов, ограничивающие желчные ^капилляры, имеют микро-
ворсинки, которые вдаются в их просвет? Напббьпшых^гистологйче-
’"'с^их препаратах желчные капилляры остаются незаметными и вы-
являются только при специальных методах обработки (импрегнация
серебром или инъекции капилляров окрашенной массой через
желчный проток). На таких препаратах видно, что желчные капил-
ляры слепо начинаются (на центральном концс_дтече]7^нбй балки,
идут вдоль нее, слегка изгиЬаясь и отдавая в стороны короткие сле-
пые выросты, и, наконец, на периферии печеночных долек пере-
ходят в холангиолы — короткие трубочки, узкий просвет которых
ограничен 2- 3 овальными клетками. Холангиолы впадают в меж-
дольковые желчные протоки (ductuli interlobulares).
Таким образом, желчные капилляры располагаются внутри пе-
ченочных балок, тогда как между балками проходят кровеносные
капилляры. Поэтому каждый .гепатоцит в печеночной балке имеет
две стороны. Одна сторона билиарная^-обращена к просвету
желчного капилляра- куда ик н и сеюк-цщую; желчи другая вас-
кулярная — направлена к кровеносному внутридольковому ,капил-
ляру, в который клетки выделяют глюкозу, мочевину, белки и другие
вещества. Между кровеносными и желчными капиллярами нигде
нет непосредственной связи, так как их отделяют друг от друга
549
Рис. 220. Строение долек (Л) и балок (£) печени (рис. Е. Ф. Котовского).
А — схема строения портальной дольки и ацииуса печени: / — классическая печеночная
долька; 2 портальная долька; 3 — ацинус (заштриховано); 4 — триада, 5 — центральные
вены. Б — схема строения печеночной балки: / — печеночные балки (пластинки); 2 —
гепатоцит; 3 — кровеносные капилляры; 4 — перисинусоидальное пространство; 5 пе-
рисинусоидальный липоцит; 6 желчный каналец; 7а — вокругдольковая вена, 76 —
вокругдольковая артерия; 7 в — вокругдольковый желчный проточек; 8 центральная
вена
печеночные и эндотелиальные клетки. Только при некоторых забо-
леваниях (паренхиматозная желтуха и др.), связанных с поврежде-
нием и гибелью части печеночных клеток, желчь может непосред-
ственно из желчных капилляров поступать в кровеносные. В этих
случаях желчь разносится кровью по всему организму и окрашивает
его ткани в желтый цвет (желтуха).
• Согласно другому представлению о строении Печеночных
долек, они состоят из широких пластинок (laminae hepaticae), анас-
томозир ющих^меж собой. Между пластинами располагайте*!
кровяные лакуны (vas sinusoTdem), по которым медленно циркулиру-
ет кровь. Стенка дакуи образована эндотелиоцитами и звездчащуш
макрофагоцитами. От пластин они отделены пе >илакунарным
пространством.
В последнее время в науке появилось третье представле-
ние о гистофункциональных единицах печени, отличных от класси-
ческих печеночных долек. В качестве таковых рассматриваются так
называемые портальные печеночные дольки и печеночные ацинусы.
Портальная печеночная ролька включает сегменты трех соседних
классических печеночных долек, окружающих триаду. Поэтому она
имеет треугольную форму, в ее центре лежит триада, а на периферии,
т. е. по углам, — вены (центральные). В увязи с этим в портальной
| дольке кровот л< по кровенруным капиллярам направлен от центра
' к периферии (см. рис. 220. Л). Печеночный ацинус образован сег-
* ментами двух рядом расположенных классических долек, благодаря
чему имеет форму ромба. У острых его углов проходят вены (цент-
ральные), а у тупого угла — трцада. от которой внутрь ацинуса
идут ее ветви (вокругдольковые). От этих ветвей к венам (цент-
ральным) направляются гемокапилляры (см. рис. 220, Л). Таким
образом, в ацинусе, так же как и в портальной дольке, кровоснабже-
ние осуществляется отего центральных участков к периферическим.
Печеночные клетки, или гепатоциты, составляют 60% всех кле-
точных элементов печени. Они выполняют большую часть функций,
присущих печени. Гепатоциты имеют неправильную много-
угольную форму. Диаметр их достигает 20—2J мкм. Многие из них
(до 20% в печени человека) содержат два ядра и больше. Количе-
ство таких клеток зависит от функционального состояния организ-
ма: например, беременность, лактация, голодание заметно отража-
ются на процентном содержании их в печени.
Ядра гепатоцитов круглой формы, величина их колеблется от
7 до 16 мкм. Это объясняется наличием вЬеченочных клетках наря-
ду с обычными ядрами (диплоидными) более крупных — полипло-
идных. Число этих ядер с возрастом постепенно увеличивается и к
старости достигает 80%.
Цитоплазма печеночных клеток окрашивается не только кислы-
ми, но и основными красителями, так как отличается большим
одержанием РНП. В ней присутствуют все виды общих органелл,
ранулярная эндоплазматическая сеть имеет вид узких канальцев
с прикрепленными рибосомами. В центролобулярных клетках она
расположена па аллельными рядами, а в периферических — в раз-
551
ных направлениях. Агранулярная эндоплазматическая сеть в виде
трубочек и пузырьков встречается либо в ограниченных участках цито-
плазмы, либо рассеяна по всей цитоплазме. Гранулярный вид сети
участвует в синтезе белков крови, а агранулярный в метаболизме
углеводов. Кроме того, эндоплазматическая сеть за счет образую-
щихся в ней ферментов осуществляет дезинтоксикацию вредных
веществ (а также инактивацию ряда гормонов лекарств). Около
канальцев гранулярной эндоплазматической сети располагаются
пероксисомы, "с которымгт*~СВязан обмен жирных кислот. Боль-
шинство митохондрий имеет округлую или овальную форму и раз-
мер 0,8—2 мкм. Реже наблюдаются митохондрии нитчатой формы,
длина ^оторых достигает 7 мкм и более. Митохондрии отличаются
сравнительно небольшим числом крист и умеренно плотным мат-
риксом. Они равномерно распределены в цитоплазме. Количество
их в одной клетке может меняться. Комплекс Гольджи в период
интенсивного желчеотделения перемешается в клетке к просвету
желчного капилляра. Вокруг него встречаются отдельные или не-
большими группами лизосомы, На васкулярной и билиарной поверх-
ностях клеток имеются микроворсинки.
Гепатоциты содержат различного рода включения:. глико!ен.
липйдЫ, пигменты и др., образующиеся из продуктов, приносимых
кровью. Количество их меняется в различные фазы деятельности
печени. Наиболее легко эти изменения обнаруживаются в связи с
процессами пищеварения. Уже через 3—5 ч после приема пищи
количество гликогена в гепатоцитах возрастает, достигая максиму-
ма через 10—12 ч. Через 24—48 ч после еды гликоген, постепенно
превращаясь в глюкозу, исчезает из цитоплазмы клеток. В тех
случаях, когда пища богата жирами, в цитоплазме клеток появля-
ются капли жира, причем раньше всего — в клетках, расположен-
ных на периферии печеночных долек. При некоторых заболеваниях
накопление жира в клетках может переходить в их патологическое
состояние — ожирение. Процессы ожирения гепатоцитов резко
проявляются при алкоголизме, травмах мозга, лучевой болезни и др.
В печени существует суточный ритм секреторных процессов: днем
преобладает выделение желчи, а ночью — синтез гликогена. По-ви-
димому, этот ритм ре гул и pye-тсм -при участии гипоталамуса и
гипофиза. Желчь и гликоген образуются в разных зонах печеночной
дольки: желчь обычно вырабатывается в периферическом эоне и
только затем этот процесс постепенно распространяется на цент-
ральную зону, а отложение гликогена осуществляется в обратном
направлений — оТ Щ1 ПТрПТ периферии дольки. Гепатоциты выделя-
ют в кровь глюкозу, мочевину, белки, жиры, а в"желчные к.пшдля-
ры—_желчь
Желчевыводящие пути. К ним относятся внутрипечено ч-
ные и внепеченочные желчные протоки. К внутрипечено^Г
ным принадлежат междольковые желчные протоки, а к внепеченоч-
ным — правый и левый печеночные протоки, общий печеночный,
пузырный и" общий желчный протоки. Междольковые желчные
протоки вместе с разветвлениями воротной вены и печеночной
артерии образуют в печени триады. Стенка междольковых прот< ков
состоит из однослойного кубическогб, а вболёё крупных протоках —
из цилиндрического эпителия, снабженного каемкой, и тонкого слоя
рьТУдой соединительной ткани. В апикальных от .елах"ЗТ1итеЛйёГпь-
ных клеток протоков нередко встречаются в виде зерен или_капель |
составные части желчи. На этом основании предполагают, что •
междольковые желчные протоки выполняют секреторную функцию.
Печеночные, пузырный и. обгний желчный протоки имеют примерно
одинаковое строение. Это сравнительно тонкие трубки диаметро^
около 3,5—5 мм, стенка которых образована тремя оболочками.
Слизистая о б о л о ч к а состоит из однослойного _высокощ
призматического эпителия и xopQpjo развй голо слоя соединительной
ткани (собственная пластинка). Для эпителия этих протоков ха-
рактёрно наличие в его клетках дизором и включений желчных
пигментов. Некоторые считают это доказательством резорбтивной,
т. е. всасывательной, функции эпителия протоков. В эпителии неред-
ко встречаются бокаловидные клетки. Количество их резко увели-
чивается при заболеваниях желчных путей. Собственная пластинка ,
желчных протоков отличается богатством эластических волокон, 1
расположенных продольно Ц.ЦиркУДДГЩО- В небольшом количестве
в ней имеются ^лизистые жел^ь^ Мы ш е_ч ная оболочка
тонкая, состоит из спирально расположенных. пучков гладких
миоцитов, между которыми много соединительной ткани. Мышечная
оболочка хорошо выражена лишь в определенных участках прото-
ков— в С1>нк<*~ТТУТырного протока при переходе его в пузырь и в
стенку общего желчного протока пои впадении -его в двенадцати-
перстную кишку. В этих местах.ДУЧКи гладких мцоцитов распола-
гаются главным образом циркулярно. Они образую; сфнцхтеры.
которые регулируют поступление желчи в кишечник. Адвентици-
альная оболочка состоит из рыхлой соединительной ткани.
)|<елчрый пузырь
Желчный пузырь (vesica biliaris) представляет собой тонкостен-
ный орган (толщина стенки 1,5—2 мм), вмещающий 40—70 мл
желчи. Стенка желчного пузыря состоит из трех_оболочек: слизис-
той, мьПиёчнбй и адвентициальной (рис. 721). Пузырь со стороны
брюшной полости покрыт серозной оболочкой. Слизистая
оболочка образует многочисленные складки. Она выстлана
| высокими призматическими эпителиальными клетками, имеющими
.каемку. Под эпителием располагается собственная пластинка сли-
зистой оболочки, содержащая большое количество эластических л
волокон. В области шейки пузыря в ней находятся альвеолярно-1
трубчатые железы, выделяющие слизь. Эпителий слизистой оболоч-*
ки обладает способностью всасывать воду и некоторые другие веще-
ства из желчи, заполняющей полость пузыря. Поэтому пузырная
желчь всегда более густой консистенции и более темного цвета, чем
желчь, изливающаяся непосредственно из печени. Мышечная
оболочка желчного пузыря состоит из пучков гладких миоци-
553
Рис. 221. Гепатоцит, Электронная микрофотография. Х8000 (препарат Е. Ф. Ко-
товского) .
/ — ядро; 2 — митохондрии; 3 — гранулярная эндоплазматическая сеть; 4 лизосома;
5 — гликоген; 6 — граница между гепатоцитами; 7 — желчный каналец; 8 десмосома;
О — соединение по типу «замка».
тов, расположенных в виде сети, в которой преобладает их цирку-
лярное направление.' В области шейки пузыря циркулярные пучки
мышечных клеток особенно сильно развиты. Вместе с мышечным
слоем пузырного протока они образуют сфинктер. Между пучками
554
мышечных клеток всегда имеются хорошо выраженные прослойки
рыхлой соединительной ткани. Адвентициальная обо-
лочка желчного пузыря состоит из плотной волокнистой соеди-
нительной ткани, в которой содержится много толстых эластиче-
ских волокон, образующих сети.
Иннервация. В капсуле печени находится вегетативное нервное
сплетение, ветви которого, сопровождая кровеносные сосуды, про-
должаются в междольковую соединительную ткань. Они иннервиру-
ют кровеносные сосуды, желчные протоки и гепатоциты в печеноч-
ных балках, формируя окончания в виде небольших утолщений.
По ходу нервов располагаются отдельные нервные клетки.
Возрастные изменения. С возрастом в гепатоцитах повышается
количество липофусцина, который окрашивает клетки в бурый цвет.
Число делящихся клеток резко уменьшается. Ядра гепатоцитов
увеличиваются в размерах — гипертрофируются, в них повышается
содержание ДНК и наблюдается гиперхроматоз. В старческом
возрасте отмечается разрастание соединительной ткани между пече-
ночными дольками. <
Регенерация. Печень обладает высокой способностью к физиоло-
гической и репаративной регенерации. У животных при удалении
от 50 до 70% ткани печени исходная ее масса восстанавливается
уже на 10—14-й день. Процессы регенерации происходят путем
компенсаторного увеличения размеров клеток (гипертрофии) и
размножения гепатоцитов. Стимулирует регенерацию печени пища,
богатая углеводами и белками.
Поджелудочная железа
Поджелудочная железа (pancreas) является смешанной железой,
включающей экзокринную и эндокринную части. В экзокрин-
ной части вырабатывается панкреатический сок, богатый пище-
варительными ферментами — трипсином, липазой, амилазой и др.,
поступающий по выводному протоку в двенадцатиперстную кишку,
где его ферменты участвуют в расщеплении белков, жиров и угле-
водов до конечных продуктов. В э н_д окринной части
синтезируется ряд гормонов — инсулин, глюкагон, соматостатин,
панкреатический полипептид, принимающие участие в регуляции
углеводного, белкового и жирового обмена в тканях и др.
Развитие. Поджелудочная железа развивается из энтодермы и
.мезенхимы. Ее зачаток появляется в конце 3-й недели эмбриогенеза
в виде дорсального и вентральных выпячиваний стенки туловищного
отдела эмбриональной кишки, врастающих в брыжейку. Из них
формируются головка, тело и хвост железы. На 3-м месяце плодного
периода энтодермальные зачатки начинают дифференцироваться
на экзокринные и эндокринные отделы железы. В экзокринных
отделах образуются ацинусы и выводные протоки, а эндокринные
отделы, вначале имеющие вид почек на выводных протоках, затем
отшнуровываются от них и превращаются в островки. Из мезенхимы
развиваются соединительнотканные элементы стромы, а также
сосуды.
555
Рис. 222. Строение желчного пузыря.
I — складки слизистой оболочки; 2 эпителий слизистой оболочки, 3 собственная
пластинка слизистой оболочки; 4 — мышечная оболочка; 5 — кровеносные сосуды; 6
наружная оболочка.
Строение. Пожелудочная железа с поверхности покрыта тонкой
соединительтканной капсулой, срастающейся с висцеральным
листком брюшины. Ее паренхима разделена на дольки, между
которыми проходят соединительнотканные тяжи. В них располо-
жены кровеносные сосуды, нервы, интрамуральные нервные ганг-
лии, пластинчатые тельца и выводные протоки (рис. 222). Дольки
включают экзокринные и эндокринные части железы. На долю
первой части приходится около 97%, а второй — до 3% всей
массы железы.
Экзокринная часть железы в дольках представлена
панкреатическими ацинусами, вставочными и внутридольковыми
протоками, а также междольковыми протоками и общим панкреа-
тическим протоком, открывающимся в двенадцатиперстную кишку.
Структурно-функциональной единицей экзо-
кринной части поджелудочной железы является
панкреатический ацинус (acinus pancreaticus). Он включает секре-
торный отдел и вставочный проток, которым начинается вся прото-
ковая система железы. Внешне ацинус напоминает мешочек разме-
ром 100—150 мкм. Между ацинусами располагаются ретикулярные
волокна, кровеносные капилляры, а также нервные волокна и нерв-
ные ганглии вегетативной нервной системы. Ацинусы состоят из 8—
556
2
Б
Рис. 223. Строение поджелудочной железы.
л — схема (по Ю. И Афанасьеву); Б микрофотография (по В В Яглову) 1 — долька,
экзокринные отделы (панкреатические ацинусы); 3 — панкреатический островок
внутридольковый проток; 5 кровеносные сосуды.
12 крупных экзокринных панкреатоцитов, или ациноцитов (acino-
cytus), расположенных на базальной мембране, и нескольких мелких
протоковых, или центроацинозных, эпителиоцитов (рис. 223, Б).
Экзокринные панкреатоциты (ациноциты) выполняют секретор-
ную функцию, синтезируя пищеварительные ферменты панкреати-
ческого сока. Они имеют форму конуса с суженной верхушкой и
широким основанием, лежащим на базальной мембране ацинуса.
Цитолемма на базальной поверхности клеток образует внутренние
складки, а на апикальной поверхности — микроворсинки. Между
боковыми поверхностями клеток имеются контакты типа замыка-
тельных пластинок и десмосом. Верхушечные (апикальные) части
клеток называются зимогенными зонами, а противоположные ба-
зальные части клеток — гомогенными зонами. Зимогенные зоны
клеток окрашиваются кислыми красителями, т. е. появляются
оксифильными. Зимогенная зона клеток в основном занята круп-
ными секреторными гранулами (до 80 нм в диаметре). Среди них
выделяют гранулы различной степени зрелости (плотности).
Зимогенные гранулы содержат синтезируемые в клетках ферменты
в неактивной форме, т. е. в виде зимогена. В гомогенной зоне
преобладает гранулярная эндоплазматическая сеть состоящая из
массы плоских мешочков, мембраны которых усеяны рибосомами.
В них осуществляется синтез ферментов панкреатического сока.
Обилие рибосом обусловливает базофилию этой зоны. Ядра ациноз-
ных клеток, содержащие 1—2 ядрышка, преимущественно распо-
лагаются в их базальной части.
В надъядерной части клеток расположен обширный комплекс
Гольджи. Митохондрии рассеяны по всей цитоплазме, но большин-
ство их находится под цитолеммой и рядом с комплексом Гольджи.
Они отличаются разнообразной формой.
Секреторную деятельность ациноциты осуществляю! цикличе-
ски. Их секреторный цикл, включающий фазы поглощения
исходных веществ, синтеза секрета, накопления его и затем выде-
ления по мерокриновому типу, занимает в среднем Р/г'—2 ч.
' Однако в зависимости от физиологических потребностей организма
в пищеварительных ферментах этот цикл может сократиться или,
наоборот, увеличиться.
Выделившийся из ациноцитов секрет попадает во вставочный
проток (ductus intercalatus), стенка которого состоит из мелких кле-
ток. В одних случаях они примыкают к ациноцитам сбоку, имея
общую с ними базальную мембрану, в других — проникают в центр
ацинуса, располагаясь на апикальной поверхности ациноцитов. При
такой локализации они называются центроацинозными эпителиоци-
тами (cellulae centroacinosi). Центроацинозные клетки имеют непра-
вильную, уплощенную форму, их овальное ядро окружено узким
слоем светлой цитоплазмы, бедной органеллами. На свободной,
обращенной в просвет ацинуса поверхности имеются единичные
микроворсинки.
Вставочные протоки переходят в межацинозные протоки (duc-
tus interacinosi). Стенки этих протоков выстланы однослойным
558
и
кубическим эпителием. Их цитолемма образует внутренние складки
и микроворсинки. Клетки соедниняются друг с другом с помощью
десмосом. В цитоплазме клеток много митохондрий и хорошо раз-
j вит комплекс” Гольджи. Существует мнение, что эпителиальные
Г клетки протоков продуцируют жидкий компонент панкреатического
* сока.
Межацинозные протоки впадают в более крупные внутридоль-
ковые протоки (dustus intralobulares), стенки которых покрыты
однослойным кубическим эпителием. Ядра эпителиальных клеток
крупные, в цитоплазме — немногочисленные митохондрии, слабо
выраженный комплекс Гольджи, свободные рибосомы^ гладкая *
эндоплазматическая сеть. Вокруг протоков расположена рыхлая
волокнистая соединительная ткань, в которой проходят кровеносные
капилляры и нервные волокна.
Внутридольковые протоки далее продолжаются в междольковые
протоки (dustus interlobulares), которые лежат в соединительно-
тканных перегородках (септах) между дольками. Они впадают в
общий проток поджелудочной железы, проходящий в ее толще от
хвоста к головке, где он открывается (вместе с общим желчным
протоком) в полость двенадцатиперстной кишки. Все эти протоки
। выстланы слизистой оболочкой, состоящей из высокого призмати-
’ ческого эпителия и собственной соединительнотканной пластинки.
В устье общего протока, кроме того, имеются циркулярно располо-
женные гладкие миоциты, образующие его сфинктер.
В эпителии протоков имеются бокаловидные экзокриноциты,
|Z1 а также эндокриноциты, вырабатывающие гормоны панкреозимин____
* ми ^^олицистокицин. Под их влиянием стимулируются секреторная
Активность ациноцитОв'поджелудочной железы и отделение желчи
из печени. В собственной пластинке протоков находятся мелкие
слизистые железы.
Эндокринна я часть железы в дольках представлена
островками (insulae pancreaticae), лежащими между панкреатиче-
скими ацинусами (см. рис. 223; рис. 224). Они обычно имеютоКруг-
лую или овальную форму, но наряду с этим могут встречаться
островки лентовидной и звездчатой формы. В средйРМ ил диаметр
колеблется от 100 до 300 мкм. Наибольшее количество островков
располагается в хвостовой части железы. Общее их число колеблет-
ся от 1 до 2 млн. и более, но при этом их объем не превышает 3%
объема всей железы.
Островки состоят из эндокринных клеток — инсулоцитов [in-
sulocyfi)? между которыми находятся кровеносные капилляры
фенестрированного типа, окруженные перикапиллярным простран-
ством. Именно сюда прежде всего поступают инсулярныеТормоны,
3"затем через стенку капилляров — в кровь.
Инсулоциты в отличие от ацинозных имеют меньшие размеры.
В их цитоплазме умеренно развита гранулярная эндоплазматиче-
ская сеть, но хорошо представлены комплекс Гольджи, митохондрии
(мелкие) и секреторные гранулы (рис. 225). Эти гранулы по своим
физико-химическим и морфологическим свойствам неодинаковы в
559
4
Рис. 224. Панкреатический
островок (по Блюму).
I — островковые ацидофиль-
ные клетки — А; 2 — ост-
ровковые базофильные клет-
ки — В; 3 — островковые
клетки — D, 4 — кровенос-
ные капилляры; 5 — экзо-
кринные отделы
различных клетках островков. На этом основании среди инсулярных
клеток различают 5 основных видов: В-клетки (базофильные),'^
А-клетки (ацидофильные), D-клетки Jдендритические), Огклетки г
(аргирофильные) и РР-клетки. с
В-клетки составляют основную массу клеток островков (около
70—75%). Большая часть их лежит в центре островков. Секретор-
, ные В-гранулы клеток не растворяются в воде, но полностью раство-
ряются в спирте. Они проявляют базофильные свойства, окрашива-
ясь альдегидфуксином, генциановым фиолетовым в синий цвет.
Гранулы имеют размер около 275 нм. Между их содержимым и
покрывающей мембраной имеется широкий светлый ободок. В неко-
торых гранулах содержатся плотная кристаллическая структура,
в которой обнаруживается цинк. Гранулы В-клеток состоят из гор-
Мона инсулина- синтезирующегося в этих клетках. Одним из наибо-
лее ярких эффектов инсулина является его гипогликемическое
действие, так как он способствует усвоению глюкозы крови клетками
тканей. Поэтому при недостатке инсулина, количество глюкозы в
тканях снижается, а, содержание ее в крови резко возрастает, что
приво/усг-к-еажарыцд1у мочеизнуретТию' (сахарный диабет).
А-клетки составляют примерного—25% от всей массы инсуляр-
ных клеток. В островках они занимают преимущественно перифери-
ческое положение. А-гранулы клеток устойчивы к спирту, но раство-
ряются в воде. Они обладают оксифильными свойствами, в связи с
чем окрашиваются кислым фуксином, в ярко-красный цвет. Раз-
меры гранул около 230 нм. Их плотное содержимое отделено от
окружающей мембраны узким светлым ободком. К гранулах А-кле-
ток обнаружен гормон глюкагон По своему действию он является
антагонистом инсулина. Под его влиянием в тканях происходит
S60
Рис. 225. Схема ультрам и крое конического строения клеток панкреатического
островка (рис. KJ. И. Афанасьева, Ю. А. Рунихина).
/ — А-клетки; 2 — В-клетки; 3 — D-клетки; 4 — РР-клетки; 5 — гемокапилляр; 6 —
нервные волокна; 7 рыхлая волокнистая соединительная ткань.
усиленное расщепление гликогена до глюкозы. В связи с этим в слу-
чаях его недостатка количество глюкозы в крови может сни-
жаться.
Следовательно, инсулин и глюкагон строго поддерживают посто-
янство сахара в крови и определяют содержание гликогена в тканях
(прежде всего в печени).
D-клетки, число которых в островках невелико (5—10%), распо-
лагаются в основном на их периферии, имеют грушевидную и реже
звездчатую форму. D-гранулы среднего размера (325 нм), умерен-
ной плотности и лишены светлого ободка. ^клетки секретируют
гормон соматостатин. Этот гормон задерживает выделение инсули-
на и глюкагона А- и В-клетками, а также подавляет синтез фермен-
тов ацинозными клетками поджелудочной железы. В небольшом
числе в островках находятся Огклетки, содержащие мелкие
(160 нм) аргирофильные гранулы, значительной плотности с
узким светлым ободком. Этот вид клеток выделяет вазоактивный
интестинальный полипептид (ВИП), который снижает артериаль-
ное давление, стимулирует выделение сока и гормонов поджелу-
дочной железой.
561
Рис. 226. Центроацинозный эпителиоцит. Электронная микрофотография.
X 10 000 (препарат В. В. Яглова).
/ — зимогенные гранулы; 2 — эндокринные гранулы тина А; 3 — гранулярная эндоплазма-
тическая сеть.
РР клетки (2—5%) вырабатывают панкреатический полипеп-
тид, стимулирующий выделение желудочного и панкреатического
сока. Это полигональные клетки с очень мелкими зернами в цито-
плазме (размер гранул не более 140 нм). РР-клетки обычно лока-
лизуются по периферии островков в области головки железы, а
также встречаются вне островков среди экзокринных отделов и
протоков.
Помимо экзокринных (ацинозных) и эндокринных (инсуляр-
ных) клеток, в дольках поджелудочной железы описан еще один
тип секреторных клеток — промежуточные, или ацинозно-инсу-
лярные клетки (рис. 226). Происхождение их дискуссионно. Ско-
рее всего, это самостоятельный тип клеток. Они располагаются
группами вокруг островков среди экзокринной паренхимы. Харак-
терной особенностью промежуточных клеток является наличие в
них гранул двух типов — крупных зимогенных, присущих ациноз-
ным клеткам, и мелких, типичных для инсулярных клеток (А, В,
D, РР).
Их митохондрии делятся по величине на крупные и мелкие,
а гранулярная эндоплазматическая сеть по степени развития зани-
мает промежуточное положение. Гранулы ацинозно-инсулярных
562
клеток напоминают гранулы одного из видов клеток островков —
А, В или D. В связи с этим предложено классифицировать ациноз-
но-инсулярные клетки по их гормональному профилю на три типа:
А, В и D. Большая часть ацинозно-инсулярных клеток выделяет
в кровь как эндокринные, так и зимогенные гранулы. Реже встре-
чаются клетки, из которых те и другие гранулы поступают в
выводные протоки железы.
Согласно одному из предположений, ацинозно-инсулярные
клетки выделяют в кровь трипсиноподобные ферменты, которые
освобождают из проинсулина активный инсулин.
Васкуляризация. Поджелудочная железа снабжается кровью,
приносимой по ветвям чревной и верхней брыжеечной артерий.
Разветвления этих артерий в междольковой соединительной ткани
и внутри долек образуют густые капиллярные сети, оплетающие
ацинусы и проникающие в островки. Существует мнение, что эти
капиллярные сети между собой не сообщаются. Согласно другому
предположению, в дольках железы существует портальная система
сосудов, когда приносящая артериола распадается на капилляры
островков, а затем они собираются в выносящие артериолы, от
которых начинается новая сеть капилляров, оплетающих ацинусы
экзокринных отделов железы. Оттекающая от поджелудочной
железы венозная кровь поступает в воротную вену.
Лимфатическая система начинается щелевидными
капиллярами вокруг ацинусов и островков. Лимфатические капил-
ляры вливаются в лимфатические сосуды, которые проходят
поблизости от кровеносных.
Иннервация. Эфферентная иннервация поджелудочной железы
осуществляется блуждающим и симпатическими нервами. Симпати-
ческие волокна сопровождают кровеносные сосуды, являясь по
своему значению сосудодвигательными. В поджелудочной железе
имеются интрамуральные вегетативные ганглии. Основную массу
их нервных клеток составляют холинергические нейроны. Вместе
с тем в ганглиях содержатся и пептидергические нейроны, секре-
тирующие полипептидные гормоны. Нервные волокна холинерги-
ческих и пептидергических нейронов заканчиваются на клетках
панкреатических ацинусов и вдоль капилляров проходят в остров-
ки, регулируя секреторную функцию железы.
Чувствительные нервные волокна образуют в междольковой
соединительной ткани разнообразные рецепторы, в том числе пла-
стинчатые тельца.
Возрастные изменения. В поджелудочной железе они прежде
всего проявляются в изменении соотношения между ее экзокрин-
ной и эндокринной частями. Островки наиболее сильно развиты
в железе в первые годы жизни. С возрастом их число постепенно
уменьшается.
Регенерация. Пролиферативная (митотическая) активность
клеток поджелудочной железы крайне низкая, поэтому в физиоло-
гических условиях в ней происходит обновление клеток путем
внутриклеточной регенерации.
563
Г л а в a XIX
ДЫХАТЕЛЬНАЯ СИСТЕМА
Дыхательная система обеспечивает в организме внешнее
дыхание, а также ряд важных недыхательных фун-
кций.
В ее состав входят различные органы, выполняющие возду-
хопроводящую и дыхательную (газообменную) фун-
кции: полость носа, носоглотка, гортань, трахея, внелегочные
бронхи и легкие.
Внешнее дыхание, т. е. поглощение из вдыхаемого
воздуха кислорода и снабжение им крови, а также удаление из
организма углекислого газа, является основной функцией дыха-
тельной системы. Газообмен осуществляется легкими. Среди дру-
гих функций очень важными являются: терморегуляция
и увлажнение вдыхаемого воздуха, очищение
его от пыли и микроорганизмов, депонирова-
ние кровив обильно развитой сосудистой системе, участие в
поддержании свертываемости крови благодаря
выработке тромбопластина и его антагониста — гепарина, участие
в синтезе некоторых гормонов в водно-солевом
и липидном обмен» веществ, а также в г о л о-
сообразовании, обонянии и иммунологической
защите.
Развитие. Гортань, /трахея и легкие развиваются из одного
общего зачатка, который появляется на 3—4-й неделе путем выпя-
’ чивания вентральной стенки передней кишки. Гортань и трахея
закладываются на 3-й неделе из верхней части непарного мешко-
видного выпячивания вентральной стенки передней кишки. В ниж-
ней части этот непарный зачаток делится по средней линии на
два мешка, дающих зачатки правого и левого легкого. Эти мешки
в свою очередь позднее подразделяются на множество связанных
между собой более мелких выпячиваний, между которыми врастает
мезенхима. На 8-й неделе появляются зачатки бронхов в виде
коротких ровных трубочек, а на 10—12 й неделе стенки их стано-
вятся складчатыми, выстланными цилиндрическими эпителиоцита-
ми. Из мезенхимы, окружающей растущее бронхиальное дерево,
дифференцируются гладкая мышечная ткань, хрящевая ткань, волок-
нистая соединительная ткань, бронхов, эластические, коллагеновые-
элементы альвеол, а также прослойки соединительной ткани, прора-
стающие между дольками легкого. В конце 4-го месяца эмбриональ-
ного периода дифференцируются бронхиолы, покрытые кубическим
эпителием. С 6-го месяца и до момента рождения в легких проис-
ходит процесс развития альвеолярных ходов и альвеол. В течение
всего эмбрионального периода альвеолы имеют вид спавшихся
пузырьков с незначительным просветом и толстой стенкой, состоя-
щей из цилиндрических или кубических клеток. Одновременно
из мезенхимы развивается сеть кровеносных сосудов, вместе с
которыми к легким подходят нервы. Из висцерального и париеталь-
564
I
«xtuCifXu
ною листков спланхнотома в это время образуются висцераль-
ный и париетальный листки плевры. При первом вдохе новоро-
жденного альвеолы легких расправляются, в результате чего резко
увеличиваются их полости и уменьшается толщина альвеолярных
стенок. Это способствует обмену кислорода и углекислоты между
кровью, протекающей по капиллярам, и воздухом альвеол.
ВОЗДУХОНОСНЫЕ ПУТИ
//
К ним относят носовую полость, носоглотку, гортань, трахею
и бронхи. В воздухоносных путях по мере продвижения воздуха
происходят очищение, увлажнение, приближение температуры
вдыхаемого воздуха к температуре тела, рецепция газовых темпе-
ратурных и механических раздражителей в также регуляция
объема вдыхаемого воздуха. Гортань принимает участие в звуко-
образовании. ,
Носова я полость. В ней различают преддверие и собственно
носовую полость, включающую дыхательную и обонятельную
области.
П реддверие образовано полостью, расположенной под
хрящевой частью носа. Оно выстлано многослойным плоским
эпителием, который является продолжением эпителиального по-
крова кожи. Под эпителием в соединительнотканном слое заложе-
• ньГуальные ж£ди.зы .и -корми щетинковых.. волос. Волосы носовой
‘полости задерживают пылевые частицы из вдыхаемого воздуха.
В более глубоких частях преддверия волосы становятся короче и
количество их уменьшается.
Внутренняя поверхность собственно носовой полос-
. ти в дыхательной части покрыта слизистой оболочкой, состоящей
I из многорядного призматического реснитчатого ^питепиц и соеди-
* нительнотканноц—собственной пластинки (рис. 227). В эпителии,
расположенном на базальной мембране, различают 4 вида кле-
ток: реснитчатые, микроворсинчатые, базальные и бокаловидные.
Реснитчатые клетки снабжены мерцательными ресничками высо- з
той около 3—5 мкм. Между реснитчатыми клетками располагают-
ся микроворсинчатые, с короткими ворсинками на апикальной
поверхности и базальные малоспециализированные клетки^ Бока- .
ловидные клетки являются одноклеточными слизистыми железами, *
выделяющими свой секрет на поверхность мерцательного эпи-
телия.
Собственная пластинка слизистой оболочки состоит из рыхлой
волокнистой неоформденнрй соединительной ткани, содержащей
большое количество эластических волокон. В ней залегают конце-
вые отделы слизистых желез, выводные протоки которых откры-
ваьйдя—на поверхности эпителия. Секрет этих желез, как и секрет
бокаловидных клеток, увлажняет слизистую оболочку. Благодаря
этому на ее поверхности задерживаются пылевые частицы, микро-
организмы, удаляемые затем движением ресничек мерцательного
эпителия. В собственной пластинке слизистой оболочки постоянно
56S
Рис. 227. Схема строения слизистой
оболочки носовой полости.
Хорошо развита эта оболочка и в
1 — многорядныи призматический рес-
нитчатый эпителии; 2 — соединительно-
тканная пластинка; 3 — многочислен-
ные кровеносные срсуды; 4 — железы.
встречаются лимфатические
узелки, особенно в области
отверстия слуховых труб, где
они образуют тубарные
миндалины.
Васкуляризация. Слизис-
тая оболочка носовой полости
очень богата сосудами, распо-
ложенными в поверхностных
участках ее собственной плас-
тинки непосредственно под
эпителием, что способствует
согреванию вдыхаемого воз-
духа и в холодное время года.
Артерии и артериолы носовой
полости отличаются выра-
женностью средней оболочки,
нах. В области нижней раковины
находится сплетение вен с широким просветом. При наполнении их
кровью слизистая оболочка сильно набухает, что затрудняет вды-
хание воздуха.
Лимфатические сосуды образуют густую сеть. Они
связаны с субарахноидальным пространством и периваскулярными
влагалищами различных частей мозга, а также с лимфатическими
сосудами больших слюнных желез.
Иннервация. Слизистая оболочка носовой полости обильно
иннервирована, имеет многочисленные свободные и инкапсулиро-
ванные нервные окончания (механо-, термо- и ангиорецепторы).
Чувствительные нервные волокна берут начало из тройничного
узла V пары черепных нервов. Слизистая оболочка околоносовых
пазух, в том числе лобных и верхнечелюстных, имеет ту же
структуру, что и слизистая оболочка дыхательной части носовой
полости, с той лишь разницей, что собственная соединительно-
тканная пластинка в них значительно тоньше.
Гортань. Орган воздухоносного отдела дыхательной системы,
принимающий участие не только в проведении воздуха, но и в
звукообразовании. Гортань имеет три оболочки: слизистую,
фиброзно-хрящевую и адвентициальную (рис. 228).
Слизистая оболочка (tunica mucosa), за исключе-
нием голосовых связок, выстлана многорядным реснитчатым
эпителием. Собственная пластинка слизистой оболочки, представ-
ленная рыхлой волокнистой соединительной тканью, содержит
многочисленные эластические волокна, не имеющие определенной
566
Рис. 228. Строение гортани;
фронтальный разрез (схема).
I — хрящ надгортанника; 2 —
собственная пластинка слизи-
стой оболочки; 3 — лимфатиче-
ские узелки; 4 — отдельные пуч-
ки гладких мышечных клеток
ложной голосовой связки; 5 —
ложная голосовая связка; 6 —
железы; 7 — щитовидный хрящ;
8 — желудочек гортани; 9 — ис-
тинная голосовая связка; 10—
мышцы истинной голосовой свя-
зки; 11 — многослойный пло-
ский эпителий.
ориентировки. В глубоких слоях слизистой оболочки эластические
волокна постепенно переходят в надхрящницу, а в средней части
гортани проникают между поперечнополосатыми мышцами го-
лосовых связок. «
— На передней поверхности в собственной пластинке слизистой I
оболочки гортани содержатся смешанные белково-слизистые же- •
лезы (gll. mixteae seromucosae). Особенно много их у основания
надгортанного хряща. Здесь же имеются значительные скопления
лимфатических узелков, носящие название гортанных минда-
лин.
В средней части гортани имеются складки слизистой оболочки,
образующие так называемые истинные и ложные голосовые связ-
ки. Складки покрыты многослойным плоским эпителием. В сли-
зистой оболочке выше и ниже истинных голосовых связок рас-
полагаются смешанные белково-слизистые железы. Благодаря
сокращению поперечнополосатых мышц, заложенных в толще
V*
567
Рис. 229. Строение трахеи.
/ — ели «истая оболочка; II — подсли-
зистая основа; Н1 волокнисто-мы-
шечно-хрящевая оболочка; / — псевдо-
многослойный призматический рес-
нитчатый эпителий; 2 — бокаловидные
экзокрииоциты; 3 — собственная плас-
тинка слизистой оболочки; 4 — железы
трахеи; 5 -- надхрящница; 6 — гиалино-
вый хрящ.
голосовых складок, происхо-
дит изменение величины ще-
ли между ними, что влияет на
высоту звука, производимого
воздухом, проходящим через
гортань (см. рис. 228).
Д Фиброзно-хряще-
вая оболочка состоит
из гиалцдовых и эластических
хрящей, окруженцых плот-
ной волокнистой соедини-
тельной тканью. _Она выпол-
няет роль защитно-опорного
каркаса гортани.
Адвентициальная оболочка состоит из коллагено-
вой соединительной ткани.
Гортань отделена от глотки надгортанником, основу которого
составляет эластический хрящ. В области надгортанника происхо-
дит переход слизистой оболочки глотки в слизистую оболочку
гортани. На обеих поверхностях надгортанника слизистая оболоч-
ка покрыта многослойным плоским эпителием. Собственная плас-
тинка слизистой оболочки надгортанника на его передней повер-
хности образует значительное количество вдающихся в эпителий
сосочков; на задней поверхности они короткие, а эпителий более
низкий.
Трахея. Полый трубчатый орган, состоящий из слизистои
оболочки, подслизистой основы, волокнисто-хрящевой и адвенти-
циальной оболочек (рис. 229).
I Слизистая оболочка (tunica mucosa) при помощи
тонкой подслизистой основы связана с фиброзно-хрящевой обо-
лочкой трахеи и благодаря этому не образует складок. Она выстла-
на многорядным призматическим реснитчатым эпителием, в кото-
ром различают реснитчатые, бокаловидные, эндокринные и базаль-
ные клетки.
Реснитчатые клетки призматической формы, имеют на свобод-
ной поверхности около 250 ресничек. Реснички мерцают в направ-
лении, противоположном вдыхаемому воздуху, наиболее интенсив-
но при оптимальной температуре (18—33°С) и в слабощелочной
среде. Бокаловидные клетки — одноклеточные эндощцтелиальные
56К
железы — выделяют слизистый секрет, богатый гиалуроновой и
сиаловой кислотами, на поверхность эпителиального пласта.
Их секрет вместе со слизистыми железами подслизистой основы
увлажняет эпителий и создает условия для прилипания попада-
ющих с воздухом пылевых частиц, удаляемых при откашливании.
Слизь содержит также иммуноглобулины, выделяемые плазмати-
ческими клетками, находящимися в составе слизистой оболочки,
которые обезвреживают многие микроорганизмы, попадаемые с
воздухом. /Эндокринные клетки имеют пирамидальную —форму,
округлое ядро и секреторные гранулы- Эти клетки выделяют
пептидные гормоны и биогенные амины: норадреналин, серотонин,
дофамин и регулируют сокращение мышечных клеток воздухо-
носных путей'./Базальные клетки камбиальные, имеют овальную
или треугольную форму По мере их. специализации в цитоплазме
появляются тонофибриллы и гликоген, увеличивается количество
органелл.
Под базальной мембраной эпителия располагается собствен-
ная дыастинка сдизистой оболочки, состоящая из рыхлой волок-
нистой неоформленной соединительной ткани, богатая эластичес-
кими волокнами. В отличие от гортани эластические волокна
в трахее принимают продольно^ направление. В собственной
пластинке слизистой оболочки встречаются лимфатические узелки
и отдельные циркулярно расположенные п чки гладких мышечных
клеток.
^Подслизистая ос нов a (tela submucosa) трахеи состо-
ит из рыхлой волокнистой соединительной ткани, без резкой
границы переходящей в плотную волокнистую соединительную
ткань надхрящницы незамкнутых хрящевых колец. В подслизис-
той основе располагаются смешанные белково-слизистые железы,
вы водные .протоки которых, образуя на своем пути колбообразные
расширения, открываются на поверхности слизистой оболочки.
Этих желез особенно, много в задней и боковой стенках трахеи.
Волокнист о-х р я щ е в’а* я обо л~б~ч ка (tunica fibrocar-
tilaginea) трахеи состоит из 16—20 гиалиновых хрящевых колец,
не замкнутых на задней стенке трахеи. Свободные концы этих хря-
щей соединены пучками гладких мышечных клеток, прикрепля-
ющихся к наружной поверхности хряща. Благодаря такому стро-
ению задняя поверхность трахеи оказывается мягкой, податливой,
что имеет большое значение при глотании. Пищевые комки,
проходящие по пищеводу, расположенному непосредственно по-
зади трахеи, не встречают препятствия со стороны стенки трахеи.
Адвентициальная оболочка (tunica adventitia)
трахеи состоит из рыхлой волокнистой неоформленной соедини-
тельной ткани, которая соединяет этот орган с прилежащими
частями средостения.
Кровеносные сосуды трахеи, так же как и гортани, образуют
в ее слизистой оболочке jt сколько параллельно расположенных
сплетений, а под эпителием — густую капиллярную сеть. Л и м-
фсГТ ИЧеские сосуды также формируют сплетения, из ко-
569
торых поверхностное находится непосредственно под сетью кро-
веносных капилляров.
Нервы, подходящие к трахее, содержат спинномозговые и ве-
гетативные волокна и образуют два сплетения, ветви которых
заканчиваются в ее слизистой оболочке нервными окончаниями.
Мышцы задней стенки трахеи иннервируются из ганглиев веге-
тативной нервной системы.
Функция трахеи как воздухоносного органа тесно связана со
структурно-функциональными особенностями бронхиального дере-
ва легких.
ЛЕГКИЕ
Легкие занимают большую часть грудной клетки и постоянно
изменяют свою форму в зависимости от фазы дыхания. Поверх-
ность легкого покрыта серозной оболочкой — висцеральной плев-
рой.
Строение. Легкое состоит из системы воздухоносных путей —
бронхов (бронхиальное дерево) и системы легочных пузырьков,
или альвеол, выполняющих роль собственно респираторных отделов
дыхательной системы.
Бронхиальное дерево легкого (arbor bronchialis). В его состав
входят главные бронхи (правый и левый), которые подразделяют-
ся на внелегочные долевые бронхи (крупные бронхи 1-го порядка),
разветвляющиеся затем на крупные зональные внелегочные (по
4 в каждом легком) бронхи (бронхи 2-го порядка). Внутрилегоч-
ные бронхи сегментарные (по 10 в каждом легком) подразде-
ляются на бронхи 3—5-го порядка (субсегментарные), которые
по своему диаметру относятся к средним бронхам (5—2 мм).
Средние бронхи, разветвляясь, переходят в мелкие (2—1 мм в диа-
метре) бронхи и затем в конечные бронхиолы (bronchioli ter-
minales). За ними начинаются респираторные отделы легкого,
выполняющие газообменную функцию.
Строение бронхов, хотя и неодинаково на протяжении брон-
хиального дерева, имеет общие черты. В н у т ренняя оболо-
чка бронхов — слизистая — выстлана, подобно трахее,
реснитчатым эпителием, толщина которого постепенно уменьша-
ется за счет изменения формы клеток от высоких призматических
до низких кубических. Среди эпителиальных клеток, помимо
реснитчатых, бокаловидных, эндокринных и базальных, описан-
ных выше, в дистальных отделах бронхиального дерева встреча-
ются секреторные клетки, каемчатые (щеточные), а также без-
реснитчатые клетки (рис. 230).
Секреторные клетки (cellulae secretoriae) характери-
зуются куполообразной верхушкой, лишенной ресничек и микро-
ворсинок и заполненной секреторными гранулами. Они содержат
округлое ядро, хорошо развитую эндоплазматическую сеть аграну-
лярного типа, комплекс Гольджи. Эти клетки вырабатывают
ферменты, расщепляющие сурфактант, покрывающий респиратор-
ные отделы (см. ниже). Безреснитчатые клетки (epitheliocyti
570
Рис. 230. Схема ультрамикроскопического строения эпителиальных клеток слизи
стой оболочки воздухоносных путей (рис. Ю И. Афанасьева).
/ — реснитчатые клетки; 2 — бокаловидные клетки; 3 — эндокринные клетки; 4 - базаль-
ные клетки; 5 — безреснитчатые клетки; 6 секреторные клетки; 7 — каемчатые клетки;
й — нервное волокно; 9 — базальная мембрана
aciliati) встречаются в брЛиолах. Они имеют призматическую
форму. Их апикальный конец несколько возвышается над уровнем
смежных реснитчатых клеток. В апикальной части содержатся
скопления гранул гликогена, митохондрии и секретоподобные
гранулы. Функция их неясна. Каемчатые клетки (epitheiiocyti
limbati) отличаются овоидной формой и наличием на апикаль-
ной поверхности коротких тупых микроворузнок (см. рис. 230).
Эти клетки встречаются редко. Полагают, что они выполняют
функцию хеморецепторов.
Собственная пластинка слизистой оболочки бронхов богата
продольно направленными эластическими волокнами, которые
обеспечивают растяжение бронхов при вдохе и возвращение их
в исходное положение при выдохе. Слизистая оболочка бронхов
имеет продольные складки, обусловленные сокращением косоцир-
кулярных пучков гладких мышечных клеток (мышечная пластинка
слизистой оболочки), отделяющих слизистую оболочку от подсли-
зистой соединительнотканном основы. Чем меньше диаметр бронха,
тем относительно сильнее оказывается сокращение мышечной
пластинки слизистой. В слизистой оболочке бронхов, особенно
крупных, встречаются лимфатические узелки.
В подслизистои соединительнотканной основе залегают конце-
вые отделы смешанных слизисто-белковых желез. Железы распо-
лагаются группами, особенно в местах, которые лишены хряща, а
выводные протоки проникают в слизистую оболочку и открываются
на поверхности эпителия. Их секрет увлажняет слизистую оболочку
и способствует прилипанию, обволакиванию пылевых и других
частиц, которые впоследствии выделяются наружу. Слизь облада-
ет бактериостатическими и бактерицидными свойствами. В брон-
хах малого калибра (диаметром 2—1 мм) железы отсутствуют.
571
Фиброзно-хрящевая оболочка по мере уменьшения калибра
бронха характеризуется постепенной сменой незамкнутых хряще-
вых колец (у главных бронхов) на хрящевые пластинки (долевые,
зональные, сегментарные, субсегментарные бронхи) и островки
хрящевой ткани (в бронхах среднего калибра). В бронхах сред-
него калибра вместо гиалиновой хрящевой ткани появляется
эластическая хрящевая ткань. В бронхах малого калибра фиброз-
но-хрящевая оболочка отсутствует.
Наружная адвентициальная оболочка построена из волокни-
стой соединительной ткани, переходящей в междолевую и меж-
дольковую соединительную ткань паренхимы легкого. Среди
соединительнотканных клеток обнаруживаются тканевые базо-
I филы, принимающие участие в регуляции местного гомеостаза и
.свертываемости крови.
Таким образом, бронхи крупного калибра, имеющие диаметр
соответственно от 15 до 5 мм, характеризуются складчатой сли-
зистой оболочкой, выстланной многорядным реснитчатым эпите-
лием, благодаря сокращению гладкой мышечной ткани, наличием
желез, крупных хрящевых пластин в фиброзно-хрящевой оболоч-
ке. Бронхи среднего калибра отличаются снижением высоты эпи-
телиального пласта и толщины слизистои оболочки, наличием
желез, уменьшением размеров хрящевых островков. В бронхах
малого калибра эпителий двухрядный, а затем однорядный, хряща
и желез нет, мышечная пластинка слизистой оболочки становится
более мощной по отношению к толщине всей стенки. Продолжи-
тельное сокращение мышечных пучков при патологических состоя-
ниях, например при бронхиальной астме, резко уменьшает просвет
мелких бронхов и затрудняет дыхание.
Следовательно, мелкие бронхи выполняют функцию не только
проведения, но и регуляции поступления воздуха в респираторные
отделы легких.
Конечные (терминальные) бронхиолы имеют диаметр около
0,5 мм. Сдизцстая оболочка их.выстлана однослойным кубическим
реснитчатым эпителием, в котором встречаются щеточные сек-
реторные и бес каемчатые клетки. В собственной пластинке сли-
зистой-Оболочки этих бронхнол-раслоложены продольно идущие
эластические волокна, между которыми залегают отдельные пучки
гладких мышечных клеток. Вследствие этого бронхиолы легко
растяжимы при вдохе и возвращаются в исходное положение при
выдохе.
Респираторный отдел. С т р у к ту р ц,.р^ф.у н к и и п на л ь-
н о й единицей респираторного отдела легкого
является ацинус (acinus pulmonaris). Он представляет собой
систему альвеол, расположенных в стенке распираторной брон-
хиолы, альвеолярных ходов и мешочков, которые осуществляют
газообмен между кровью и воздухом альвеол. Ацинус начинается
респираторной бронхиолой (bronchiolus respiratorius) 1-го порядка,
которая дихотомически делится на респираторные бронхиолы
2-го, а затем 3-го порядка. В просвет бронхиол открываются
572
1
I
А — схема; Б — микрофотография. I— респираторные бронхиолы 1-го порядка: 2
респираторные бронхиолы 2-го порядка; 3 - респираторные бронхиолы 3-го порядка;
4 — альвеолярные ходы; 5 — альвеолярные мешочки.
альвеолы (рис. 231). Каждая респираторная бронхиола 3-го по-
рядка—в свою очередь подразделяется на альвеолярные ходы
XSucTuli alveolares), а каждый альвеолярный ход заканчивается
двумя альвеолярными мешочками (sacculi alveolares). В устье
альвеол альвеолярных ходов имеются небольшие пучки гладких
мышечных клеток, которые на срезах видны в виде утолщений.
Ацинусы отделены друг от друга тонкими соединительнотканными
прослойками; 12—18 ацинусов образуют легочную дольку.
Респираторные бронхиолы выстланы однослойным
кубическим эпителием. Реснитчатые клетки встречаются редко
(рис. 232). Мышечная пластинка истончается и распадается на
отдельные, циркулярно направленные пучки гладких мышечных
клеток. Соединительнотканные волокна наружной адвентициаль-
ной оболочки переходят в интерстициальную соединительную
ткань.
На стенках альвеолярных ходов и альвеолярных мешочков
располагается несколько десятков альвеол. Общее количество их
у взрослых людей достигает в среднем 300—400 млн. Поверхность
всех альвеол при максимальном вдохе у взрослого человека мо-
жет достигать 100 mj, а при выдохе она уменьшается в 2—21 /2
раза.
Альвеолы разделены тонкими соединительнотканными перего-
родками, в которых проходят кровеносные капилляры. Между аль-
веолами существуют сообщения в виде отверстий диаметром около
10—15 мкм — альвеолярных пор (рис. 233, 234).
Альвеолы имеют вид открытого пузырька. Внутренняя поверх-
ность выстлана двумя основными видами клеток: респираторными
эпителиоцитами (клетки 1-го типа) и большими эпителиоцитами
573
Рис. 232. Респираторная бронхиола легкого ребенка 1‘/2 лет. Сканирующая элек-
тронная микрофотография. Х8400 (по Л. К. Романовой).
I — реснички на апикальной поверхности реснитчатых клеток; 2 — бе эре с нитчатые клет-
ки; 3 — границы между клетками.
Рис. 233. Альвеола легкого крысы. Сканирующая электронная микрофотография.
Х3500 (по Л. К. Романовой).
I — апикальная поверхность с микроворсинками альвеолоцитов 2-го типа; 2 — выделение
сурфактанта; 3 — межклеточные границы; 4 — кровеносные капилляры; 5 — пора между
альвеолами.
Рис. 234. Строение альвеол и
межальвеолярных перегоро-
док легкого (по Л. К. Рома-
новой с изменением).
А — схема: / — просвет альвео-
лы; 2 — сурфактант; 3 — гипо-
фаза сурфактанта; 4 — альвео-
лоцит 1-го типа; 5 — альвеоло-
цит 2-го типа; 6 — альвеолоцит
3-го типа; 7 — макрофаг; 8 —
просвет капилляра; 9 — клетка
эндотелия капилляра; 10 — кол-
лагеновые волокна; 11 — фибро-
бласт; 12 — пора. Б - электрон-
ная микрофотография. X 24 000.
1 —альвеолоцит 1-го типа; 2
базальная мембрана альвеоло-
цита; 3 — базальная мембрана
эндотелия гемокапилляра; 4 —
эидотелиоциты; 5 — цитоплазма
гранулоцита в просвете гемока-
пилляра; б — воздушно-кровяной
барьер.
(клетки 2-го типа). Кроме того, у животных существуют в альвео-
лах клетки 3-го типа — каемчатые.
Респираторные эпителиоциты (epitheliocyti respiratorii) име-
ют неправильную уплощенную вытянутую форму. Толщина клеток
в тех местах, где располагаются их ядра, достигает 5—6 мкм, тог-
да как в остальных участках она колеблется в пределах 0,2— ).
0,3 мк-мг На свободной поверхности цитоплазмы этих клеток
имеются очень короткие цитоплазматические выросты, обращен-
ные в полость альвеол, что увеличивает общую площадь сопри-
косновения воздуха с поверхностью эпителия. В цитоплазме их
575
Рис. 235. Сурфактантный альвеолярный комплекс легкое крысы. Электронная
микрофото!рафия. <"60 000 (по Л. К. Романовой).
! — просвет альвеолы; 2 — просвет кровеносного капилляра; 3 - воздушно кровяной
барьер; 4 — мембраны сурфактанта; 5 — гипофаза (жидкая фаза) сурфактантного
альвеолярного комплекса.
обнаруживаются мелкие- митрхондрии и пиноиитрзные п зырьки.
К безьядерным участкам эпителиоцитов 1-го типа прилежат
также безъядерные участки эндотелиальных клеток капилляров.
В этих участках базальная мембрана эндотелия кровеносного
капйГЛЛяра может вплотную приближаться к базальной мембране
эпителия. Благодаря такому взаимоотношению стенок альвеол и
капилляров барьер между кровью и воздухом (аэрогематический
барьер) оказывается чрезвычайно тонким — в среднем 0,5 мкм
(см. рис. 234, А, Б). Местами толщина его увеличивается за счет
тонких прослоек рыхлой волокнистой соединительной ткани.
Важным компонентом аэрогематического барьера является сур
фактантный альвеолярный комплекс (рис. 235). Он играет важную
роль в предотвращении спадения альвеол _рри выдохе, а также* в
предохранении их от проникновения через стенку альвеол микро-
организмов из вдыхаемого воздуха и транссудации жидкости из
капилляров межальвеолярных перегородок в альвеолы. Сурфак-
тант состоит из двух фаз /^мембранной и^^жидкой (гипофаза).
Биохимический анализ сурфактанта показал, что в его состав
входят фосфолипиды, белки и гликопротеиды.)») '
Фосфолипиды и белки образуют мембранйыи компонент,
сходный с элементарной Биологической мембраной. Гликопро-
теиды располагаются глубже и образуют гипофазу. Синтез фос-
фолипидов, идущих на построение мембран сурфактанта, -проис-
^одит в эпителиоцитах 2-го типа, которые выделяют его.
Эпителиоциты 2-го типа, или большие эпителиоциты (epithe-
jt. iiocyti magni), несколькоТкругщее uoвысоте! чем клетки 1-го типа.
В цитоплазме' выятстяютТя более крупные митохондрии, комп-
лекс Гольджи, осмиофильные тельца и эндоплазматическая сеть.
576
Кроме описанных видов клеток, в стс-нкс альвеол обнаружи-
ваются микрофаги, содержащие захваченные инородные частицы,
избыток сурфактанта. В цитоплазме макрофагов всегда находится
значительное количество липидных капель и лизосом. Окисление
I липидов в макрофагах сопровождается выделением тепла, которое
’обогревает вдыхаемый воздух. Макрофаш проникают в альвеолы
«из межальвеолярных соединительнотканных перегородок. Альвео-
лярные макрофаги, как и макрофаги других органов, имеют
костномозговое происхождение.
Снаружи к базальной мембране прилежат кровеносные ка-
пилляры, проходящие по межальвеолярным перегородкам, а также
сеть эластических волокон, оплетающих альвеолы. Кроме эласти-
ческих волокон, вокруг альвеол располагается поддерживающая
их сеть тонких коллагеновых волокон. Так как альвеолы тесно
прилежат друг к другу, то капилляры, оплетающие их, одной своей
поверхностью граничат с одной альвеолой, а другой с сосед-
ней. Это обеспечивает оптимальные условия для газового обмена
между кровью, протекающей по капиллярам, и воздухом, запол-
няющим полости альвеол.
Плевра. Легкие снаружи покрыты плеврой, называемой тегоч-
ной, или висцеральной. Висцеральная плевра плотно срастается
с легкими, эластические и коллагеновые волокна ее переходят в
интерстициальную ткань, поэтому и золировать плевру, не травми-
руя легкие, трудно. В висцеральной плевре встречаются гладкие
мышечные клетки. В париетальной плевре, выстилающей наруж-
ную стенку плевральной полости, эластических элементов меньше,
гладкие мышечные клетки встречаются редко. В процессе органо-
генеза из мезодермы формируется только однослойный плоский
эпителий мезотелий, а соединительнотканная основа плевры
развивается из мезенхимы. В зависимости от состояния легкого
мезотелиальные клетки становятся то плоскими, то высокими.
Васкуляризация. К р о воснабжение в легком осушест-
вляется по двум системам сосудов (рис. 236). С одной стороны,
легкие получают артериальную кровь из дез очных артерий, т. е.
из малого круга кровообращения^ Ветви легочной артерии, сопро-
вождая бронхиальное дерево, доходят до лсноняния альвеол., где
они образ)йот узкопетлистую капиллярную сеть альвеол. В альвео-
лярных капиллярах, диаметр которых колеблется" в "пределах
5—7 мкм, эритроциты располагаются в один ряд, что создает оп-
тимальное условие для осуществления газообмена между гемогло-
бином эритроцитов и альвеолярным воздухом. А львеоля рные
капилляры собираются в посткапиллярные венулы, которые форми-
рую т систему легочной вены. ——
Бронхиальные артерии отходят непосрегнтяаына от аорты—пи-
тает бронхи и легочную паренхиму артериальной кровью. Про-
никая в стенку бронхов, они разветвляются и образуют артериаль-
ные сплетения в их подслизистой основеЭйГслиЗИСТОй оболочке.
В слизистой оболочке брПтгхттв происходи! сообщение—сосу до в
бол ьшого~й малого"круга путем "анастомозирования ра з ветвле ни и
19 I ИСТОЛОГИЯ 577
Верхушка дольки
Рис. 236. Схема расположения сосудов в дольке легкого (по А. Хэму).
1 - конечная (терминальная) бронхиола; 2 — респираторная бронхиола; 3 — альвеолярный
ход; 4 альвеола; 5 ветви легочной артерии; 6 — ветви легочной вены; 7 междолько
пая соединительнотканная перегородка; 8 — сеть кровеносных капилляров; 9 лимфатиче-
ский сосуд; 10 — плевра.
броихиа л ьныхл-леиааных-араерийПо сткадил лярные-венулы, от-
ходящие от альвеол и бронхов, объединяются_в мелкие_вены,
которые дают начало передним и. -задним бронхиальным венам.
На уровне мелких бронхов располагаются многочисленные арте-
риоловенулярные анастомозы между бронхиальными и легочными
артериальными системами.
Лимфатическая система легкого состоит из поверх
ностной и глубокой сетей лимфатических капилляров и сосудов.
Поверхностная сеть располагается в висцеральной плевре. Глу-
бокая сеть находится внутри легочных долек, в междольковых
перегородках, залегая вокруг кровено< ных сосудов и бронхов
легкого. В самих бронхах лимфатические сосуды образуют два
анастомозирующих сплетения: одно располагается в слизистой
оболочке, другое — в подслизистой основе.
В плевре имеются кровеносные и лимфатические сосуды и
578
Рис. 237. Нервное оконча-
ние в стенке альвеолы.
Микрофотография (пре-
парат Т. Г. Оганесян).
1 альвеолы; 2 нервное
волокно; 3 — свободное нерв-
ное окончание в стенке аль-
веолы,
многочисленные нерв-
ные окончания. В па-
риетальной плевре
сосуды образуют по-
слойно три сплете-
ния.
Иннервация. Осу-
ществляется главным
образом симпатиче-
скими и парасимпа-
тическими, а также
спинномозговыми не-
рвами. Симпатиче-
ские нервы проводят
импульсы, вызываю-
щие расширение бронхов и сужение кровеносных сосудов, парасим-
патические — импульсы, обусловливающие, наоборот, сужение
бронхов и расширение кровеносных сосудов. Разветвления этих
нервов образуют в соединительнотканных прослойках легкого нерв-
ное сплетение, расположенное по ходу бронхиального дерева, аль-
веол и кровеносных сосудов (рис. 237). В нервных сплетениях
легкого встречаются крупные и мелкие ганглии, обеспечивающие, по
всей вероятности, иннервацию гладкой мышечной ткани бронхов
В легочной плевре есть два нервных сплетения: мелкопетлистое
под мезотелием и крупнопетлистое в глубоких слоях плевры.
Возрастные изменения. В постнатальном периоде дыхательная
система претерпевает большие изменения, связанные с началом
выполнения газообменной и других функций после перевязки
пуповины новорожденного. В детском и юношеском возрасте
прогрессивно увеличиваются дыхательная поверхность легких,
эластические волокна в строме органа, особенно при физической
нагрузке (спорт, физический труд). Общее количество легочных
альвеол у человека в юношеском и молодом возрасте увеличи-
вается примерно в 10 раз. Соответственно изменяется и площадь
дыхательной поверхности. Однако относительная величина респи-
раторной поверхности с возрастом уменьшается. После 50—60 лет
происходит разрастание соединительнотканной стромы легкого,
отложение солей в стенке бронхов, особенно прикорневых. Все
это приводит к ограничению экскурсии легких и уменьшению
основной газообменной функции.
Регенерация. Физиологическая регенерация органов дыхания
наиболее интенсивно протекает в пределах слизистой оболочки за
19"
579
счет малоспециализированных клеток. Восстановления же после
удаления части органа (воздухоносные пути) практически не про-
исходит. После пульмонэктомии в эксперименте в оставшемся
легком происходит компенсаторная гипертрофия с увеличением
объема альвеол в 3—4 раза с последующим размножением струк-
турных компонентов альвеолярных перегородок. Одновременно
расширяются сосуды микроциркуляторного русла, обеспечивающие
трофику и дыхание.
Главе XX
КОЖА И ЕЕ ПРОИЗВОДНЫЕ
Кожа (cutis) образует внешний покров организма, площадь
которого у взрослого человека достигает 1,5—2 м2. Из производ-
ных кожи у человека имеются потовые и сальные железы, волосы
и ногти.
КОЖА
Кожа выполняет различные функции. Она защищает подлежа
щие части организма от повреждений. Здоровая кожа непрони-
цаема для микроорганизмов, многих ядовитых и вредных_веществ'.
Кожа участвует в во д н о-с О левл м, а также в тепло-
вом обмене с внешней средой. В течение суток через кожу
человека выделяется около 500 мл воды, что составляет 1 % всего
ее количества в .организме Кроме воды, через кожу вместе с потом
выводятся различные соли, главным образом хлориды, а также
молочная кислота и продукты азотистого обмела. Окопл Я2%.всех
тепло вы х потерь организма происходит йерез кожную пове рх-
ность. В случаях нарушения этой функции (например, при дли
тельной работе в резиновом комбинезоне) могут возникнуть
перегревание организма и^тепловой удар.
В коже под действием ультрафиолетовых лучей синтези-
руется витамин________D. Отсутствие его в организме вызывает
тяжелое заболевание — рахит.
Наличие в коже обильной сосудистой сети и артериоловенуляр-
ных анастомозов определяет значение ее как депо крови.
У взрослого человека в сосудах кожи может задерживаться до
1 л крови. и %
Благодаря обильной иннервации кожный покров представляет
собой огромное рецепторное поле, в котором сосредото-
чены осязательные, температурные и болевые нервные окончания.
В некоторых участках кожи, например на голове и кистях, на
1 см2 ее поверхности насчитывается до 300 чувствительных то-
чек.
Жиры и масла способны проникать в кожу. Наиболее легко проникают в
кожу человека жир кашалота и персиковое масло, которые могут быть проводни-
ками в организм веществ, растворенных в них.
Рис. 238. Схема строения эпидермиса и дер-
мы кожи (рис. Е. Ф. Котовского)
/ эпидермис: // — дерма; /// - подкожная осно-
ва: / базальный слои эпидермиса; 2 меланоцит;
3 шиповатый слой; 4 зернистый слои; 5
блестящий слои; 6 роговой слой; 7 — базальная
мембрана; 8 сосочковый слой дермы; У сетча-
тый слои; 10 — потовая железа.
Развитие. Кожа развивается из
двух эмбриональных зачатков. Эли-
те лиальный покрбв сообразуется из
кожной эктодермы, а подлежащие
соединительнотканные слои — из
дерматомов {производных сомитов).
В первые недели развития зародыша
эпителий кожи состоит всего из од-
ного слоя плоских клеток. Постепен-
но эти клетки становятся все более
высокими. В конце 2-го месяца над
ними появляется второй слой клеток,
а на 3-м месяце эпителий становится
многослойным. Одновременно в на-
ружных его слоях (в первую очередь
на ладонях и подошвах) начинаются
процессы ороговения. На 3-м месяце
внутриутробного периода в коже за-
кладываются эпителиальные зачатки
волос, желез и ногтей. В соедини-
тельнотканной основе кожи в этот
период начинают образовываться во-
локна и густая сеть кровеносных со-
судов. В глубоких слоях этой сети
местами появляются очаги кроветво
рения. Лишь на 5-м месяце внутри-
утробного развития образование кро-
вяных элементов в них прекращается
и на их месте формируется жировая
ткань.
Строение. Кожа состоит из двух
частей — эпителиальной и соедини-
тельнотканной. Эпителий кожи назы-
вается надкожицей, или эпидерми-
сом, а соединительнотканная осно-
ва — дермой, или собственно кожей.
С подлежащими частями организма
кожа соединяется слоем жировой
ткани — подкожной основой, или
подкожной клетчаткой. Толщина ко-
жи в различных отделах тела варь-
ирует от 0,5 до 5 мм.
581
Рис. 239. Схема строения
меланоцита в эпидермисе
кожи (рис. Е. Ф Котов
ского).
/ отростки меланоцита;
2 — комплекс Гольджи; .?
премелаиосомы ; 4 — мелаио-
сомы; 5 - базальная мем-
брана эпидермиса.
Эпидермис
(epidermis) представ
лен многослойным
плоским ороговенаю-
щим эпителием. Тол-
щина его колеблется
от 0,03 до 1,5 мм и бо-
лее1. Наиболее тол-
стый эпидермис на
ладонях и подошвах.
Он состоит из многих
десятков слоев кле-
ток, которые объединены в 5 основных слоев: базальный, шипова-
тый, зернистый, блестящий и роговой (рис. 238, Л).
Непосредственно на базальной мембране, отграничивающей в
норме эпителий от дермы, лежат клетки, составляющие базальный
слой (stratum basale). Среди них различают базальные эпителио-
циты и меланоциты — пигментные клетки, количественное соотно-
шение которых примерно 10:1. Базальные эпителиоциты имеют
цилиндрическую или овальную форму, базофильную цитоплазму
и округлое ядро, богатое хроматином. В них выявляются орга-
неллы общего значения, тонофиламенты и в некоторых клетках
Iранулы темно-коричневого или черного пигмента —"меланина.
Соединяются они друг с другом и с вышележащими клеткЭМЙ~при
помощи десмосом, а с базальной мембраной — полудесмосомами.
Среди базальных эпителиоцитов присутствуют с-тволовые клет^
ки. Образующиеся при их делении дочерние клетки специализи-
руются и постепенно перемещаются в вышележащие слои эпи-
дермиса. Совокупность клеток во всех слоях эпидермиса, обра-
зовавшихся из одной стволовой клетки, т. е. ее потомки, составляет
дифферон.
Таким образом, базальный слой выполняет роль росткового
слоя. За счет его эпидермис обновляется в различных участках
кожи человека в течение 10—30 дней (физиологическая регене-
рация).
Меланоциты на препаратах, окрашенных гематоксилин-эози-
1 Некоторые авторы на основании различной толщины эпидермиса выделяют
толстую и тонкую кожу Толстая кожа покрывает небольшие участки тела
(ладони, подошвы), тогда как тонкая выстилает остальные обширные его поверх-
нос ги.
S82
ном, имеют вид светлых клеток. При импрегнации серебром у
них обнаруживаются длинные ветвящиеся отростки. Меланоциты
не имеют десмосом и лежат свободно. В их цитоплазме содер-
жатся в большом количестве зерна пигмента меланина, но слабо
развиты органеллы и отсутствуют тонофибриллы (рис. 239).
Над базальными клетками в 5— 10 слоев располагаются клетки
полигональной формы, образующие шиповатый слой (stratum
spinosum). Между клетками хорошо видны многочисленные корот-
кие цитоплазматические отростки («мостики»), в месте встречи
которых находятся десмосомы. В десмосомах заканчиваются пуч-
ки тонофиламент — тонофибриллы. Кроме эпителиоцитов, в ба-
зальном и шиповатом слое содержатся отростчатые клетки —
дендроциты. Они не образуют десмосом с окружающими клетками.
В их цитоплазме много аргентофинных гранул, имеющих вид
теннисных ракеток. Эти клетки расцениваются как внутриэпидер-
мальные макрофаги', мигрирующие в эпидермис из дермы. В эпи-
дермис проникают также Т-лимфоциты. Гранулярные дендроциты
и лимфоциты образуют в эпидермисе местную систему иммунного
надзора.
Зернистый слой (stratum granulosum) состоит из 3 4 слоев
сравнительно плоских клеток. В их цитоплазме имеются рибо-
сомы, митохондрии, лизосомы и их разновидность — кератино-
сомы (в виде слоистых телец), а также пучки фрагментированных
тоцофибрилл и лежащие рядом с ними крупные гранулы кера-
тогиалинаТТфанулы интенсивно красятся основными красителями,
состоят из полисахаридов, липидов и белков, отличающихся
высоким содержанием аминокислот — гистидина, пролина, аргини-
на, а также серосодержащей аминокислоты — цистина. Присут-
ствие в клетках зернистого слоя комплекса кератогиалина с
тонофибриллами указывает на то, что в них начинаются процессы
ороговения, так как, по мнению многих авторов, кератогиалин
является предшественником рогового вещества — кератина.
Следующий, блестящий слой (stratum lucidum), также состоит
из 3—4 слоев плоских клеток, в которых ядра подвергаются ка-
риорексису и гибнут, а цитоплазма диффузно пропитывается
белковым веществом — элеидином. Элеидин не окрашивается кра-
гтелями, но хорошо преломляет свет. Благодаря этому в блестя-
щем слое границы и структура клеток оказываются незаметными,
1 весь слой представляется в виде блестящей полосы. Полагают,
•то элеидин образуется из белков тонофибрилл и кератогиалина
путем окисления их сульфгидрильных групп. Сам элеидин рас-
матривается как один из предшественников кератина .
Самый поверхностный — роговой слой (stratum corneum)
состоит из многих слоев ороговевших клеток — роговых чешуек.
4ешуйки содержат роговое вещество — «мягкий» кератин и пу-
1 Ранее натыкались «клетки Лангерганса».
Электронно-микроскопически блестящий слой как отдельный слой не опре-
деляется.
583
Рис. 240. Схема процесса ороговения в
эпидермисе кожи (рис. Е. Ф. Котовско-
го).
/ банальный эпителиоцит; // — шипова-
тый эпителиоцит: /// 1ернистый эпите-
лиоцит: IV — роговые чешуйки; / - базаль-
ная мембрана; 2 - полудесмосомы; 3 - дес-
мосомы; 4 тоиофибриллы; 5 — лизосомы;
6 — межклеточные шели; 7 кератин.
зырьки воздуха. Кератин пред-
ставляет собой белок, богатый
серой (до 5%), очень устойчи-
вый к различным химическим
агентам — кислотам, щелочам
и др. Роговые чешуйки выглядят
как светлые плоские ячейки с
толстой (роговой) оболочкой.
Внутри них располагаются кера-
тиновые фибриллы, иногда вид-
ны остатки тонофибрилл в виде
нежной сети и светлая полость,
образовавшаяся на месте погиб-
шего ядра. Самые поверхност-
ные росовые чешуйки постоянно
отпадают — слущиваются и за-
меняются новыми, происходя-
щими из нижележащих слоев.
В процессе слущивания важная
роль нринадоежит кератиносо-
мам. Они выходят из клеток и
концентрируются в межклеточ-
ных пространствах.В результате
наблюдаются лизис (растворе-
ние) десмосом и отхождение ро-
говых чешуек друг от друга. Зна-
чение рогового слоя определяет-
ся тем, что он обладает большой
упругостью и плохой теплопро-
водностью.
Таким образом, в процессе
ороговения эпидермиса кожи
участвует ряд компонентов кле-
ток: тонофибриллы, кератогиа-
лин, керат иное омы. десмосомы
(рис. 240).
По сравнению с кожей ладоней и подошв эпидермис других
участков кожи значительно тоньше. Толщина его, например, на
волосистой части головы не превышает 170 мкм. Блестящий слои
в нем отсутствует, а роговой представлен лишь 2—3 рядами opo-i
говевших клеток — чешуек. По всей вероятности, ороговение d
84
этом случае протекает по сокращенному циклу. Следовательно,
большая часть кожи имеет эпидермис, который состоит из трех
основных слоев росткового, зернистого и рогового. При этом каж-
дый из них значительно тоньше, чем соответствующие слои эпи-
дермиса кожи ладоней и подошв.
Под влиянием некоторых внешних и внутренних факторов
характер эпидермиса может существенно изменяться. Так, на-
пример, при сильных механических воздействиях, при А-авита-
минозе, под влиянием гидрокортизона резко усиливаются процес-
сы ороговения'.
Собственно кожа (corinm), или дерма, имеет толщину
от 0,5 до 5 мм, наибольшую — на спине, плечах, бедрах. Дерма
делится на два слоя — сосочковый и сетчатый (см. рис. 238),
которые не имеют между собой четкой границы.
Сосочковый слой “(stratum papillare) располагается непосредст-
венно под эпидермисом, состоит из рыхлой волокнистой соедини-
тельной ткани, выполняющей трофическую функцию. Свое назва-
ние этот слой получил от многочисленных сосочков, вдающихся
в эпителий. Их величина и количество в коже различных частей
тела неодинаковы. Наибольшее количество сосочков высотой до
0,2 мм находится в коже ладоней и подошв. В коже лица сосочки
развиты слабо, а с возрастом могут совсем исчезнуть. Сосочковый
слой дермы определяет рисунок на поверхности кожи, имеющий
строго индивидуальный характер .
Соединительная ткань сосочкового слоя состоит из тонких
коллагеновых, эластических и ретикулярных волокон, а также из
клеток, среди которых наиболее часто встречаются фибробласты,
макрофаги, тканевые базофилы (тучные клетки) и др. Здесь также
встречаются гладкие мышечные клетки, местами собранные в не-
большие пучки, связанные с корнем волоса. Это мышца, подни-
мающая волосы. Однако имеются мышечные пучки, не связанные
с ними. Больше всего их в коже головы, щек, лба и тыльной по-
верхности конечностей. Сокращение мышечных клеток обусловли-
вает появление так называемой гусиной кожи. При этом сжима-
ются мелкие кровеносные сосуды и уменьшается приток крови
к коже, вследствие чего понижается теплоотдача организма.
Сетчатый слой (stratum reticulare), обеспечивающий прочность
кожи, образован плотной неоформленной соединительной тканью
с мощными пучками коллагеновых волокон и сетью эластических
волокон. Пучки коллагеновых волокон проходят в основном в двух
направлениях: одни из них лежат параллельно поверхности кожи,
^другие — косо..Вместе они образуют сеть, строение которой опре-
деляется функциональной нагрузкой на кожу. В участках кожи.
1 Нередко наблюдается изменение этих процессов на небольших участках
кожи, испытывающих сильное трение (мозоли), или это происходит в результате
нарушения обменных процессов в организме (юношеские бородавки).
По отпечаткам рисунка кожи пальцев, оставленных на различных предметах,
можно узнать лицо, которому принадлежат данные отпечатки. Этим широко поль-
зуются в криминалистической и судебно-медицинской практике (<)акты юскопия).
5X5
испытывающих сильное давление (кожа стопы, подушечек пальцев,
локтей и др.), хорошо развита широкопетлистая, грубая сеть
коллагеновых волокон. Наоборот, в тех участках, где кожа подвер-
гается значительному растяжению (область суставов, тыльная
сторона стопы, лицо и т. д.), в сетчатом слое обнаруживается
узкопетлистая более нежная коллагеновая сеть. Эластические
волокна в основном повторяют ход коллагеновых пучков. Их значи-
тельно больше в участках кожи, часто испытывающих растяжение
(в коже лица, суставов и т. д.). Клеточные элементы сетчатого
слоя представлены главным образом фибробластами.
В большинстве участков кожи человека в ее сетчатом слое
располагаются кожные железы — потовые и сальные, а также
корни волос.
Пучки коллагеновых волокон из сетчатого слоя дермы продол-
жаются в слой подкожной клетчатки, богатой жировой тканью.
Подкожная клетчатка (tela subcutanea) смягчает действие на
кожу различных механических факторов, поэтому она особенно
хорошо развита в тех участках кожи, которые подвергаются силь-
ным механическим воздействиям (подушечки пальцев, ступни
и т. д.). Здесь подкожная клетчатка полностью сохраняется даже
при крайней степени истощения организма. Подкожный слой обес-
печивает некоторую подвижность кожи по отношению к нижеле-
жащим частям, что в значительной мере предохраняет ее от раз-
рывов и других механических повреждений. Скопление жировой
ткани в гиподерме ограничивает теплоотдачу.
Кожный пигмент, за очень небольшим исключением,
имеется у всех людей. Он находится как в эпидермисе, так и в
дерме. Люди или животные, организм которых лишен пигмента,
называются альбиносами (от лат. aibus — белый). Кожный пиг-
мент относится к группе меланинов (от лат. melas — черный).
Образуется меланин при окислении аминокислоты тирозина под
влиянием фермента тирозиназы и ДОФА-оксидазы. Меланин обла-
дает высокой ультрафиолетовой абсорбцией, поэтому он защищает
организм от повреждающего действия ультрафиолетовой радиации.
Распределение пигмента в коже неравномерно: сильнее пигмен-
тирована кожа лица, шеи, спины, слабее — живота, ладоней и
подошв. Количество кожного пигмента в эпидермисе может
меняться в зависимости как от внешних, так и от внутренних
факторов. Например, резко возрастает оно при длительном облуче-
нии кожи солнечными лучами, в результате чего у лиц со светлой
кожей образуется загар и могут появиться веснушки. Пигментные
пятна на коже часто появляются во время беременности вслед-
ствие гормональных изменений, наступающих в организме в этот
период.
В эпидермисе пигмент находится в цитоплазме меланоцитов
и эпителиоцитов. Меланоциты представляют собой отростчатые
клетки. В их цитоплазме много рибосом и меланосом. Меланосо-
мы — структуры овальной формы, состоящие из 3—15 плотных
гранул, окруженных общей мембраной (см. рис. 239). Они отделя-
586
ются от комплекса Гольджи, где к ним присоединяются ферменты
тирозиназа и ДОФА-оксидаза. Положительная реакция на ДОФА-
оксидазу является специфической для меланоцитов. Наиболее
вероятно неврогенное происхождение меланоцитов. Они образу-
ются из меланобластов, которые выделяются в эмбриогенезе из
нервного гребня. Подсчитано, что в среднем на 10 базальных кле-
ток эпидермиса кожи человека приходится 1 меланоцит.
Эпителиоциты неспособны к синтезу меланина и поэтому не
дают положительной ДОФА-реакции. Они захватывают уже го-
товый меланин при выделении его из меланоцитов.
Внутриэпидермальные макрофаги также могут захватывать
пигмент из отростков меланоцитов, но при этом сами они никогда
не дают положительной ДОФА-реакции.
В дерме кожи пигмент располагается в цитоплазме дермаль-
ных меланоцитов — клеток отростчатой формы, однако в отличие
от меланоцитов эпидермиса они не дают положительной ДОФА-
реакции. В связи с этим пигментные клетки дермы содержат,
но не синтезируют пигмент. Каким путем попадает пигмент в эти
клетки — точно неизвестно, но предполагают, что он поступает из
эпидермиса.
Дермальные меланоциты встречаются лишь в определенных
местах кожи — в области анального отверстия и в околососковых
кружках.
Пигментный обмен в коже тесно связан с содержанием в ней
витаминов (А, С, РР), а также зависит от эндокринных факторов
гормонов гипофиза, надпочечников и др.
Васкуляризация. Кровеносные сосуды образуют в коже не-
сколько сплетений, от которых отходят веточки, питающие раз-
личные ее части. Сосудистые сплетения залегают в коже на раз-
ных уровнях.
Различают глубокое и поверхностное артериальное сплетения,
а также одно глубокое и два поверхностных венозных сплетения.
Артерии кожи берут начало из широкопетлистой сосудистой
сети, расположенной между мышечными фасциями и подкожной
клетчаткой (фасциальная артериальная сеть). От этой сети от-
ходят сосуды, которые, пройдя слой подкожной жировой ткани, на
границе ее с дермой разветвляются и образуют глубокую кожную
артериальную сеть. От нее идут веточки, снабжающие кровью
жировые дольки, потовые железы и волосы. Из глубокой кожной
артериальной сети начинаются артерии, которые проходят сетча-
тый слой дермы и в основании сосочкового слоя распадаются на
артериолы, образующие подсосочковую (поверхностную) арте-
риальную сеть. От этой сети в свою очередь отходят тонкие
короткие веточки, распадающиеся в сосочках на капилляры, име-
ющие форму шпилек, длина которых не превышает 0,4 мм. Корот-
кие артериальные веточки, отходящие от подсосочковой сети,
снабжают кровью группы сосочков. Характерно, что они не ана-
стомозируют друг с другом. Этим можно объяснить, почему иногда
покраснение или побледнение кожи происходит «пятнами». От
587
подсосочковой сети отходят также артериальные сосуды к саль-
ным железам и корням волос.
Капилляры сосочкового слоя, сальных желез и корней волос
собираются в вены, впадающие в подсосочковые венозные сплете-
ния Различают два подсосочковых сплетения, лежащих одно за
другим. Из них кровь поступает в кожное (мубокое) венозное
сплетение, лежащее между дермой и подкожной клетчаткой. В это
же сплетение отводится кровь от жировых долек и потовых желез.
Кожное сплетение соединяется с фасциальным венозным сплете-
нием, от которого отходят более крупные венозные стволы.
В коже многочисленны артериоловенулярные анастомозы,
особенно на кончиках пальцев рук и ног, в области ногтевого ло-
жа. Они имеют прямое отношение к процессу терморегуляции.
Лимфатические сосуды кожи образуют два сплете-
ния: поверхностное, лежащее ниже подсосочковых венозных спле-
тений, и глубокое, расположенное в подкожной клетчатке.
Иннервация. Кожа иннервируется как ветвями цереброспи-
нальных нервов, так и нервами вегетативной системы. К церебро-
спинальной нервной системе принадлежат многочисленные чувст-
вительные нервы, образующие в коже огромное количество чувст-
вительных нервных сплетений. Нервы вегетативной нервной си-
стемы иннервируют в коже сосуды, гладкие миоциты и потовые
железы1.
Нервы в подкожной клетчатке образуют основное нервное
сплетение кожи, от которого отходят многочисленные стволики,
дающие начало новым сплетениям, расположенным вокруг корней
волос, потовых желез, жировых долек и в сосочковом слое дермы.
Густое нервное сплетение сосочкового слоя посылает миелиновые
и безмиелиновые нервные волокна в соединительную ткань и в
эпидермис, где они образуют большое число чувствительных нерв-
ных окончаний. Нервные окончания распределены в коже неравно-
мерно. Они особенно многочисленны в участках кожи с повышен-
ной чувствительностью, например на ладонях и подошвах, на лице,
в области половых органов. К ним относятся свободные
и несвободные нервные окончания: пластинчатые нервные
тельца, концевые колбы, осязательные тельца и осязательные
диски. Считается, что чувство боли—передается_____расположен-
ными в эпидермисе свободными нервными окончагшямли _где
они доходяг дб зернистого.^слоя, а также нервными оконча-
ниями^ лежащими в^Т^ёбчковом слое дермы. Вероятно, что
свободные окончания одновременно являются терморецептора-
ми. Чувство прикосновения!—(осязание) воспринимается ося-
зательными тельцами и пипками, а также нервн ымц— сияе-
те ниями вокруг (корней) волос. Осязательные тельца находят-
ся в сосочковом сдое дермы, лиски — в ростковом слое_эпидер-
миса. Кроме того, в эпидермисе встречаются тактильные (осяза-
тельные) клетки, контактирующие с осязательными дисками.
' Функция сальных желез, очевидно, регулируется действием гормонов.
5KK
Чувство давления связано с наличием пластинчатых нервных
телец, лежащих глубоко в коже, К механорецепторам относятся
и 'концевые колбы, расположенные, в частности, в коже наружных
половых органов.
Железы кожи
В коже человека находится три вида желез: молочные, потовые
и сальные1. Поверхность железистого эпителия потовых и сальных
желез примерно в 600 раз превышает поверхность эпидермиса.
Эти кожные железы обеспечиваютхдерморегуляцию (около 20%
тепла отдается организмом путем испарения пота)А/защиту кожи
от повреждений (жировая смазка предохраняет кожу от высыха-
ния, а также от мацерации водой и влажным воздухом)^ выделе-
ние из организма некоторых продуктов обмена веществ (мочевина,
мочевая кислота, аммиак и др.).
Потовые железы (gll. sudoriferae) встречаются почти во
всех участках кожного покрова. Их количество достигает 2,5 млн.
Наиболее богата потовыми железами кожа подушечек пальцев
рук и ног, ладоней и подошв, подмышечных и паховых складок.
В этих местах на 1 см2 поверхности кожи открывается свыше
300 желез, тогда как в других участках кожи — 120—200 желез.
Секрет потовых желез — пот—содержит 98% воды и 2% плот-
ного остатка, который состоит из органических и неорганических
веществ. Кроме воды, вместе с потом организм выделяет продукты
белкового обмена и некоторые соли, например хлорид натрия.
За сутки выделяется, около 500—600 мл пота2.
Потовые железы подразделяются на эккриновые (мерокрино-
вые) и апокриновые. Апокриновые железы находятся лишь в опре-
деленных местах кожного покрова, например в подмышечных
впадинах, области заднего прохода, коже лба, больших половых
губах. Развиваются они в период полового созревания организма
и отличаются несколько большими размерами. Секрет их богаче
белковыми веществами, которые при разложении на поверхности
кожи придают ему особенный, резкий запах. Разновидностью
апокриновых потовых желез являются железы век и железы, вы-
деляющие ушную серу.
Потовые железы по своему строению являются простыми труб-
чатыми. Они состоят из длинного выводного протока, идущего
прямолинейно или слегка извиваясь, и не менее длинного конце-
вого отдела, закрученного в виде клубочка (рис. 241, А, Б). Диа-
метр клубочка около 0,3—0,4 мм. Концевые отделы располага-
ются в глубоких частях сетчатого слоя на границе его с подкож-
ной клетчаткой, а выводные протоки, пройдя через оба слоя дермы
и эпидермис, открываются на поверхности кожи так называемой
1 Молочные железы будут описаны в главе XXII «Половая система».
При тяжелой физической работе, высокой внешней температуре и лихорадоч-
ных состояниях организма количество выделяемого пота может резко возрастать
до 2—4 и даже Юл в сутки.
5X9
Рис. 241. Строение желез кожи (схема).
А потовая железа; / концевой отдел; 2 — экзокрииоциты; 3 — веретеновидные эпите-
лиоциты; 4 выводиой проток; 5 — базальная мембрана. Б — сальная железа: / ба
зальная мембрана; 2 базальные себоциты; 3 сальные экзокрииоциты (себоциты) на
разных стадиях жирового перерождения: 4 выводной проток; 5 — >ладкие миоциты
мышцы, поднимающей волос
потовой__порой. Выводные протоки многих апокриновых желез
не образуют потовых пор, а впадают вместе с выводными прото-
ками сальных желез в волосяные воронки.
Концевые отделы эккриновых потовых желез имеют диаметр
около 30—35 мкм. Они выстланы однослойным эпителием, клетки
которого в зависимости от фазы секреции могут иметь кубическую
или цилиндрическую форму. В слабобазофильной цитоплазме
секреторных клеток постоянно встречаются капли жира, гранулы
гликогена и пигмента. Обычно в них содержится высокоактивная
щелочная фосфатаза. Среди секреторных клеток различают свет-
лые и темные. Светлые клетки выделяют воду и ионы металлов,
темные клетки — органические макромолекулы. Кроме секретор-
ных клеток, на базальной мембране концевых отделов располага-
ются миоэпителиоциты. Своим сокращением они способствуют
выделению секрета. Концевые отделы апокриновых желез более
крупные: их диаметр достигает 150—200 мкм. Секреторные клетки
имеют оксифильную цитоплазму и не отличаются высокой актив-
ностью щелочной фосфатазы. В процессе секреции апикальные
концы клеток разрушаются и входят в состав секрета. Функция
апокриновых потовых желез связана с функцией половых желез:
S90
н предменструальный и менструальный периоды и во время бере-
менности секреция апокриновых желез возрастает.
Переход концевого отдела в выводной проток совершается
резко. Стенка выводного протока состоит из двухслойного куби-
ческого эпителия, клетки которого окрашиваются более интен-
сивно. Проходя через эпидермис, выводной проток приобретает
штопорообразныи ход . Здесь его стенка образована плоскими
клетками. Клетки выводных протоков потовых желез обладают
слабо выраженной секреторной способностью.
Сальные железы (gll. sebaceae) наибольшего развития
достигают в период полового созревания. В отличие от потовых
желез сальные железы почти всегда связаны с волосами. Только
там, где нет волос (губы, соски и др.), они лежат самостоятельно.
Больше всего сальных желез на голове, лице и верхней части
спины. На ладонях и подошвах они отсутствуют. Секрет саль-
ных желез (кожное сало) служит жировой смазкой для волос
и эпидермиса кожи. За сутки сальные железы человека выделяют
около 20 г кожного сала. Оно смягчает кожу, придает ей элас-
тичность и облегчает трение соприкасающихся поверхностей кожи,
а также препятствует развитию в ней микроорганизмов. В отличие
от потовых сальные железы располагаются более поверхностно —
в пограничных отделах сосочкового и сетчатого слоев дермы. Око-
ло одного корня волоса можно встретить 1 — 3 железы. Сальные
железы являются простыми альвеолярными _с_ разветвленными
концевыми отделами. Секретируют они по голокриновому типу.
— Концевые отделы, диаметр которых колеблется от 0,2 до 2 мм,
состоят из двух видов клеток: малоспециализированных, способ-
ных к митотическому делению, и клеток, находящихся в разных
стадиях жирового перерождения (см. рис. 241, fi). Первый вид
клеток образует наружный, ростковый, слои концевого отдела.
Кнутри от него располагаются более крупные клетки, в цитоплаз-
ме которых появляются капли жира. Постепенно процесс синтеза
жира в них усиливается и одновременно клетки смещаются в сто-
рону выводного протока. Наконец, в результате удаления от источ-
ника питания происходит гибель клеток, которые распадаются
и образуют секрет железы. Выводной проток короткий, открыва-
ется в волосяную воронку. Стенка его состоит из многослойного
плоского эпителия. Ближе к концевому отделу количество слоев
в стенке протока уменьшается, эпителий становится кубическим
и переходит в наружный ростковый слой концевого отдела.
Волосы
Волосы (pili) покрывают почти всю поверхность кожи1 2.
Наибольшая плотность их расположения на голове, где их общее
1 Выводные протоки апокриновых желез в тех случаях, когда они открываются
на поверхности кожи, имеют в эпидермисе более прямой ход и отчетливо выраженную
стенку.
2 Лишена волос только кожа ладоней, подошв, тыльной поверхности ногтевых
фаланг пальцев рук и ног, красной каймы губ, сосков, малых половых губ, головки
полового члена и внутреннего листка крайней плоти.
591
число может достигать 100 000. Длина волос колеблется от не-
скольких миллиметров до 1,5 м. толщина — от 0,005 до 0,6 мм.
Различают три вида волос: длинные, к которым относятся
волосы головы, бороды, усов, а также располагающиеся в под-
мышечных впадинах и на лобке; щетинистые — волосы бровей,
ресниц, а также растущие в наружном слуховом проходе и в
преддверии носовой полости; пушковые, покрывающие остальные
участки кожного покрова.
Развитие. Волосы развиваются на 3-м месяце эмбриогенеза.
Эпидермис в виде тяжей врастает в дерму, образуя волосяные
фолликулы, из которых и происходит рост волос. Прежде всего
волосы появляются в области бровей, подбородка и верхней
губы. Несколько позднее они образуются в коже других участков
тела.
Перед рождением или вскоре после него эти первые волосы
(lanuga) выпадают (за исключением области бровей, век и голо-
вы) и заменяются новыми, пушковыми волосами (vallns). В об-
ласти бровей, век и головы позднее также происходит смена волос
на более грубые — длинные и щетинистые.
В период полового созревания грубые волосы появляются в
подмышечных впадинах, на лобке, а у мужчин, кроме того, на
лице, иногда на груди, спине, бедрах и т. д. Волосы, образовав-
шиеся в период полового созревания, по характеру строения
являются окончательными. В дальнейшем они подвергаются пе-
риодической смене.
Строение. Волосы являются эпителиальными придатками кожи.
В волосе различают две части: стержень и корень. Стер-
жень волоса находится над поверхностью кожи. Корень волоса
скрыт в толще кожи и доходит до подкожной клетчатки. Стержень
волоса образован корковым веществом и кутикулой. Корень длин-
ных и щетинистых волос состоит из коркового вещества, мозгового
вещества и кутикулы, в пушковых волосах — только из коркового
вещества и кутикулы.
Корень волоса располагается в волосяном мешке, или фолли-
куле, стенка которого состоит из внутреннего и наружного эпи-
телиальных (корневых) вчагалищ. Фолликул окружен соедини-
тельнотканным дермальным влагалищем (волосяной сумкой)
(рис. 242, А, Б).
Корень волоса заканчивается расширением — волосяной луко-
вицей. С ней сливаются оба эпителиальных корневых влагалища
фолликула. Снизу в волосяную луковицу вдается соединительная
ткань с капиллярами в виде волосяного сосочка. В месте перехода
корня волоса в стержень эпидермис кожи образует небольшое
углубление — волосяную воронку. Здесь волос, выйдя из воронки,
появляется над поверхностью кожи. Ростковый слой эпидермиса
воронки переходит в наружное эпителиальное корневое влагалище.
Внутреннее эпителиальное корневое влагалище на этом урбвне за-
канчивается. В волосяную воронку открывается проток одной или
нескольких сальных желез. Ниже сальных желез в косом направ-
9
О сь
Рис. 242. Строение волоса.
А схема (по А Хэму с
изменением): / — дермаль-
ное корневое влагалище (во-
лосяная сумка); 2 — наруж-
ное эпителиальное корневое
влагалище: 3 — два слоя
внутреннего эпителиального
корневого влагалища; 4 -
кутикула волоса; 5 — корко-
вое вещество волоса; 6 —
мозговое вешество волоса;
7 - волосяная воронка: 8 —
ростковый слой эпидермиса;
9 — роговой слой эпидерми-
10 — сальная железа;
- мышца, поднимающая
волос; 12 — волосяная луко-
вица; 13 — волосяной сосо-
чек. Б — поперечный срез
корня волоса (окраска азо-
кармином): / — дермальное
корневое влагалище (воло-
сяная сумка); 2— наружное
эпителиальное корневое вла-
галище; 3 — два слоя внут-
реннего эпителиального кор-
невого вла1алища: 4 — кор-
ковое и мозговое вещество
волоса
лении проходит мышца, поднимающая волос (m. arrector pili).
Волосяная луковица (bulbus pili) является матрицей, т. е. той
частью волоса, из которой происходит его рост. Она состоит из
эпителиальных клеток, способных к размножению. Размножаясь,
клетки волосяной луковицы передвигаются в мозговое и корковое
вещество корня волоса, его кутикулу и во внутреннее эпителиаль-
ное влагалище. Таким образом, за счет клеток волосяной луко-
вицы происходит рост самого волоса и его внутреннего эпители-
ального (корневого) влагалища. Питание волосяной луковицы
осуществляется сосудами, расположенными в волосяном сосочке
(papilla pili). По мере того как клетки волосяной луковицы пере-
ходят в мозговое и корковое вещество, в кутикулу волоса и внут-
реннее эпителиальное корневое влагалище, они все больше уда-
ляются от источника своего питания — сосудов волосяного со-
сочка. В связи с этим в них нарастают необратимые изменения
и связанные с ними процессы ороговения. В более удаленных от
волосяной луковицы участках клетки погибают и превращаются в
роговые чешуйки. Поэтому строение корня волоса, его кутикулы и
внутреннего эпителиального влагалища на разных уровнях неоди-
наковое. Наиболее интенсивно процесс ороговения клеток проис-
ходит в корковом веществе и кутикуле волоса. В результате в них
образуется «твердый» кератин, который отличается по физическим
и химическим свойствам от кератина эпидермиса кожи. Твердый
кератин более прочный. У человека из него, кроме того, построены
ногти, а у животных — копыта, клювы, перья. Твердый кератин
плохо растворяется в воде, кислотах и щелочах; в его составе
особенно много серосодержащей аминокислоты цистина. При
образовании твердого кератина отсутствуют промежуточные ста-
дии — накопление в клетках зерен кератогиалина и элеидина.
Во внутреннем эпителиальном влагалище и в мозговом веществе
волоса процессы ороговения протекают так же, как в эпидермисе
кожи, т. е. в клетках появляются сначала зерна кератогиалищ*
(трихогиалина)', которые затем превращаются в кератин.
Мозговое вещество волоса (medulla pili) хорошо выражено
только в длинных и щетинистых волосах. В пушковых волосах
оно отсутствует. Мозговое вещество состоит из клеток полигональ-
ной формы, лежащих друг на друге в виде монетных столбиков.
Они содержат ацидофильные, блестящие гранулы трихогиалина,
мелкие пузырьки газа и небольшое количество зерен пигмента.
Пигмент образуется в волосяной луковице меланоцитами, которые
располагаются непосредственно вокруг волосяного сосочка. Про-
цессы ороговения в мозговом веществе протекают медленно, по-
этому примерно до уровня протоков сальных желез мозговое
вещество состоит из не полностью ороговевших клеток, в которых
обнаруживаются пикнотические ядра или их остатки. Только
выше указанного уровня клетки подвергаются полному орогове-
1 Трихогиалин отличается от кератогиалина тем, что окрашивается не основ-
ными, а кислыми красками.
594
нию. С возрастом процессы ороговения в мозговом веществе
волоса усиливаются, в клетках снижается количество пигмента
и увеличивается число пузырьков воздуха — волосы седеют.
Корковое вещество волоса (сопех pili) составляет основную
его массу. Процессы ороговения в корковом веществе протекают
интенсивно и без промежуточных стадий. На протяжении большей
части корня и всего стержня волоса корковое вещество состоит
из плоских роговых чешуек. Только в области шейки волосяной
луковицы в этом веществе встречаются не полностью ороговев-
шие клетки с овальными ядрами В роговых чешуйках содержатся
твердый кератин, остатки ядер в виде тоненьких пластинок, зерна
пигмента и пузырьки газа. Чем лучше в волосе развито корковое
вещество, тем оно прочнее, эластичнее.
Кутикула волоса (cuticula pili) непосредственно прилежит к
корковому веществу. Ближе к волосяной луковице она представле-
на цилиндрическими клетками, лежащими перпендикулярно к
поверхности коркового вещества. В более отдаленных от луковицы
участках эти клетки приобретают наклонное положение и пре-
вращаются в роговые чешуйки, накладывающиеся друг на друга в
виде черепицы. Эти чешуйки содержат твердый кератин, но пол-
ностью лишены пигмента.
Внутреннее эпителиальное корневое влагалище (vagina epithe-
lialis radicularis interna) является производным волосяной луко-
вицы. В нижних отделах корня волоса оно переходит в вещество
волосяной луковицы, а в верхних отделах на уровне протоков
сальных желез исчезает. В нижних отделах во внутреннем эпи-
телиальном влагалище различают три слоя: кутикулу, внутренний
(гранулосодержащий) эпителиальный слой и наружный (бледный)
эпителиальный слой. В средних и верхних отделах корня волоса
все эти три слоя сливаются, и здесь внутреннее корневое влага-
лище состоит только из полностью ороговевших клеток, содер-
жащих мягкий кератин.
Наружное эпителиальное корневое влагалище (vagina epithe-
li/lis radicularis externa) образуется из росткового слоя эпидермиса
кожи, который продолжается вплоть до волосяной луковицы.
Клетки богаты гликогеном. Этот слой постепенно истончается и в
месте перехода в волосяную луковицу состоит всего из 1—2 слоев
клеток.
Корневое дермальное влагалище (vagina derma'is radicularis) —
соединительнотканная оболочка волоса. В ней различают наруж-
ный — продольный слой волокон и внутренний — циркулярный
слой волокон.
Мышца, поднимающая волос (m. arrector pili), состоит из
гладких мышечных клеток. У щетинковых, пушковых волос, волос
бороды и подмышечных впадин она отсутствует или развита слабо.
Мышца залегает в косом направлении и одним концом вплетается
в волосяную сумку волоса, а другим — в сосочковый слой дермы.
При ее сокращении корень принимает перпендикулярное на-
правление к поверхности кожи и в результате этого стержень
595
волоса несколько приподнимается над кожей (волосы «встают
дыбом»).
Смена волос. Продолжительность жизни волоса — от несколь-
ких месяцев до 2 4 лет, поэтому в течение жизни происходит
периодическая смена волос. Процесс этот начинается с атрофии
волосяного сосочка, в результате в волосяной луковице клетки
утрачивают способность размножаться и подвергаются орогове-
нию, что приводит к обра юванию волосяной колбы. Рост волоса
прекращается. Волосяная колба по футляру, образованному на-
ружным корневым влагалищем, смещается вверх к месту прикреп-
ления мышцы, поднимающей волос. Запустевшая часть эпите-
лиального футляра при этом спадается и превращается в клеточ-
ный тяж. На конце этого тяжа впоследствии вновь формируются
волосяной сосочек и новая волосяная луковица. Из нее начи-
нается рост нового волоса. Новый волос растет по эпителиальному
тяжу, который при этом превращается в его наружное эпителиаль-
ное влагалище. По мере дальнейшего роста новый волос вытес-
няет старый из его волосяного ложа. Процесс заканчивается
выпадением старого и появлением на поверхности кожи нового
волоса.
Ногти
Ноготь (ungus) — производное эпидермиса. Развитие ногтей
начинается на 3-м месяце внутриутробного развития. Прежде чем
появиться ногтю, на месте его будущей закладки образуется так
называемое ногтевое ложе. При этом эпителий, покрывающий
дорсальные поверхности терминальных фаланг пальцев рук и ног,
утолщается и несколько погружается в подлежащую соединитель-
ную ткань. В более поздней стадии из эпителия проксимальной
части ногтевого ложа начинает вырастать сам ноготь. Вследствие
медленного роста (около 0,25—1 мм в неделю) только к последне-
му месяцу беременности ноготь достигает кончика пальца. Но-
готь— роговая пластинка, лежащая на ногтевом ложе (рис. 243).
Ногтевое ложе состоит из эпителия и соединительной ткани. Эпи-
телий ногтевого ложа — подногтевая пластинка (hyponychium)
представлен ростковым слоем эпидермиса. Лежащая непосредст-
венно на нем ногтевая пластинка является его роговым слоем.
Ногтевое ложе с боков и у основания ограничено кожными склад-
ками— ногтевыми валиками (задним и боковыми). Ростковый
слой их эпидермиса переходит в эпителий ногтевого ложа, а
роговой слой надвигается на ноготь сверху (особенно на его ос-
нование), образуя так называемую надногтевую пластинку или
кожицу (eponychium). Между ногтевым ложем и ногтевыми вали-
ками имеются ногтевые щели (задняя и боковые). Ногтевая
(роговая) пластинка своими краями вдается в эти щели. Она обра-
зована плотно прилегающими друг к другу роговыми чешуйками,
в которых содержится твердый кератин. Ногтевая (роговая) плас-
тинка подразделяется на корень, тело и край. Корнем ногтя назы-
вается задняя часть ногтевой пластинки, лежащей в задней ног-
S96
Рис. 243. С троение hoith
(окраска гематоксилин-эо-
зином).
/ ногтевая пластинка: 2
ногтевое ложе: 3 — подиог-
тевая пластинка (ростковый
слой эпителия); 4—сосоч-
ковый слои дермы, 5 мат-
рикс; 6 - ногтевой валик;
7 надиогтевая пластинка
(надкожица).
тевой щели. Лишь небольшая часть корня выступает из-за задней
ногтевой щели (из-под заднего ногтевого валика) в виде беловато-
го участка полулунной формы — луночки ногтя. Остальная часть
ногтевой пластинки, расположенная на ногтевом ложе, составляет
тело ногтя. Свободный конец ногтевой пластинки — край —
выступает за пределы ногтевого ложа.
Соединительная ткань ложа содержит большое количество
волокон, часть которых располагается параллельно ногтевой
пластинке, а часть — перпендикулярно к ней. Последние достига-
ют кости конечной фаланги пальца и соединяются с ее надкост-
ницей. Соединительная ткань ногтевого ложа образует продоль-
ные складки, в которых проходят кровеносные сосуды. Участок
эпителия ногтевого ложа, на котором лежит корень ногтя, явля-
ется местом его роста и носит название ногтевой матрицы. В ног-
тевой матрице постоянно происходит процесс размножения и
ороговения клеток. Образующиеся роговые чешуйки смещаются в
ногтевую (роговую) пластинку, которая в результате этого уве-
личивается в длину, т. е. происходит рост ногтя. Соединительная
ткань ногтевой матрицы образует сосочки, в которых лежат
многочисленные кровеносные сосуды.
Г л IB а XXI
МОЧЕВЫЕ ОРГАНЫ
К мочевым органам относятся почки, мочеточники, мочевой
пузырь и мочеиспускательный канал. Среди них почки являются
мочеобразующими органами, а остальные составляют
мочевыводящие пути. Вместе с мочой из организма вы-
деляются свыше 80% конечных продуктов обмена веществ.
Развитие. В течение эмбрионального периода закладываются
последовательно три парных выделительных органа: передняя
почка, или предпочка (pronephros), первичная почка (mesonep-
hros) и постоянная, или окончательная, почка (metanephros).
Предпочка образуется из передних 8 10 сегментных ножек
среднего зародышевого листка. У зародыша человека предпочка
не функционирует в качестве мочеотделительного органа и вскоре
после закладки подвергается обратному развитию.
Первичная почка является главным выделительным органом в
S97
течение значительного периода развития зародыша. Она форми-
руется из большого числа сегментных ножек (до 25), расположен-
ных в области туловища зародыша. Сегментные ножки отшнуро-
вываются от сомитов и спланхнотома и превращаются в канальцы
первичной почки — метанефридии. Канальцы растут по направле-
нию к мезонефралъному протоку, образующемуся при развитии
предпочки, и вступают с ним в сообщение. Навстречу им от аорты
отходят сосуды, распадающиеся на капиллярные клубочки. Ка-
нальцы своим слепым концом обрастают эти клубочки, образуя
их капсулы. Капиллярные клубочки и капсулы вместе формируют
почечные тельца. Возникший при развитии предпочки мезонеф-
ральный проток открывается в заднюю кишку.
Окончательная почка закладывается у зародыша на 2-м ме-
сяце, но развитие ее заканчивается лишь после рождения ребенка.
Функционировать она начинает во второй половине эмбрионально-
го периода. Эта почка образуется из двух источников — мезо-
нефрального протока и нефрогенной ткани, представляющей не
разделенные на сегментные ножки участки мезодермы в каудаль-
ной части зародыша. Мезонефральный проток дает начало мо-
четочнику, почечной лоханке, почечным чашечкам, сосочковым
каналам и собирательным трубкам.
Из нефрогенной ткани дифференцируются почечные канальцы.
На одном их конце образуются капсулы, охватывающие сосудис
тые клубочки. Другим концом они соединяются с собирательными
трубками. Образовавшись, окончательная почка начинает быстро
расти и, начиная с 3-го месяца, оказывается лежащей выше пер-
вичной почки, которая во второй половине беременности под-
вергается обратному развитию.
почки
Почка (геп) — парный орган, в котором непрерывно образует-
ся моча. Образуя мочу, почки выводят шлаки, регулируют водно-
солевой обмен между кровью и тканями (поэтому при болезнях
почек, связанных с нарушением их функции, возникают отеки),
поддерживают кислотно-щелочное состояние крови, выполняют
эндокринную функцию.
Строение. Почка покрыта соединительнотканной капсулой и,
кроме того, спереди серозной оболочкой. Вещество почки подраз-
деляется на корковое и мозговое (рис. 244). Корковое вещество
(cortex renis) темно-красного цвета, располагается общим слоем
под капсулой1 (рис. 245).
Мозговое вещество (medulla renis) более светлой окраски, раз-
делено на 8—12 пирамид. Вершины пирамид, или сосочки
свободно выступают в почечные чашки. В процессе развития
почки ее корковое вещество, увеличиваясь в массе, проникает
между основаниями пирамид в виде почечных колонок. В свою
---------- —‘—______________________________________
1 В раннем эмбриогенезе почки имеют дольчатое строение. При дальнейшем
развитии основания долек сливаются, образуя корковое вещество. Только в мозговом
веществе дольчатость в виде пирамид сохраняется.
S98
Рис. 244 Строение ночки
на фронтальном разрезе
(схема).
/ — капсула ночки; 2 кор-
ки во** вещество почки; i -
мозговое вещее 1 но почки;
4 короткий (корковый)
нефрон; 5 длинны и (юк
ста медуллярный) нефрон;
6 — собирательная почечная
трубочка; 7 — почечное тель-
це; Я проксимальный ка-
налец; 9 — тонкий каналец:
10 — дистальный канадец;
II — почечная чашка
очередь мозговое вещество тонкими лучами врастает в корковое,
образуя мозговые лучи.
Строму почки составляет рыхлая волокнистая соединительная
ткань, богатая ретикулярными клетками и ретикулиновыми во-
локнами. Паренхима почки представлена эпителиальными почеч-
ными канальцами ftubuli renaies), которые при участии кровенос-
ных капилляров образуют нефроны (см. рис. 244). В каждой почке
их насчитывают около 1 млн.
Нефрон (nephronum) — структурная и функцио-
нальная ди ни ца почки. Длина его канальцев до 50 мм,
а всех нефронов в среднем и хло 100 км. Нефрон переходит в
599
Рис. 245. Корковое веще-
ство почки.
А — строение коркового ве-
щества (ШИК-реакция). Б—
щелочная фосфатаза в ще
точной каемке проксималь-
ных канальцев (окраска по
Гомори) / — почечное тель-
це; 2 — проксимальный от-
дел нефрона, J — щеточная
каемка; 4 — дистальный от-
дел нефрона.
собирательную трубку, которая продолжается в сосочковый канал,
открывающийся на вершине пирамиды в полость почечной чашеч-
ки. В состав нефрона входят: капсула клубочка (capsula glomeruli),
проксимальный извитой каналец (tubulus contortus proximalis),
проксимальный прямой каналец (tubulus rectus proximalis), тонкий
каналец (tubulus attenuatus), в котором различают нисходящую
часть (pars descendens) и восходящую часть (par ascendens),
дистальный прямой каналец (tubulus rectus distalis) 1 и дистальный
извитой каналец (tubulus contortus distalis). Тонкий каналец и
дистальный прямой каналец образуют петлю нефрона. Почечное
тельце (corpusculum renale) включает сосудистый клубочек (glo-
merulus) и охватывающую его капсулу клубочка.
Около 1 % нефронов целиком располагаются в корковом ве-
ществе, а у 80 % нефронов петли спускаются в наружную зону
мозгового вещества. Это соответственно короткие и промежуточ-
ные корковые нефроны (см. рис. 244). Остальные 20 % нефронов
располагаются в почке так, что их почечные тельца, проксималь-
1 Дистальный прямой каналец часто называют толсты» отделом восходящей
части петли.
600
ные и дистальные отделы лежат в корковом веществе на границе
с моновым веществом, тогда как петли глубоко уходят в мозговое
вещество. Это длинные, или околомозговые (юкстамедуллярные),
нефроны.
Собирате чьные почечные трубочки, в которые открываются
нефроны, начинаются в корковом веществе, где они входят в со-
став мозговых лучей Затем они переходят в мозговое вещество и
у вершины пирамид вливаются в сосочковый канал.
Таким образом, корковое и мозговое вещества почки образо-
ваны р« этичными отделами нефронов. Корковое вещество состав-
чяют почечные тельца, проксимальные и дистальные отделы неф-
ронов, имеющие вид извитых канальцев. Мозговое вещество со-
стоит из прямых нисходящих и восходящих частей петель нефро-
нов, а также собирательных трубочек и сосочковых каналов.
Васкуляризация. Кровь поступает к почкам по точечным арте-
риям, которые^ войдя в "почки, распадаются на междолевые арте
"—£ии_ faa. interlobares), идущие ТЛЕжду мозговыми пирамидами.
На границе между корковым и мозговым веществом они разветв-
ляются на дуговые артерии (aa. arcuatae). От них в корковое ве-
щество отходят междольковые артерии (aa. interlobulares). От
междо Нэковых артерий в стороны расходятся внутридольковые
артерии (aa iniralobulares), от которых начинаются~~прзгносяи<де
'"артериолы (arteriolae afferentes). От верхних внутридольковых ар-
терий приносящие артериолы направляются к корковым нефронам,
от нижних - к юкстамедуллярным нефронам. -В связи с этим в
почках условно ра зличают кортикальное кровообращение, обслу-
живающее корковые нефроны, и юкстамедуллярное, связанное с
околомозговг ми нефронами (рис. 245).
В кортикальной системе кровообращения
приносящие артериолы (arteriola afferentes) распадаются на ка-
пилляры, образующие сосудистые клубочки (glomeruli) почечных
телец корковых нефронов. Капилляры клубочков собираются в
выносящие артериолы (arteriolae efferentes), которые несколько
меньше по диаметру, чем приносящие артериолы. В капиллярах
клубочков корковых нефронов кровяное давление необычайно вы-
сокое — свыше .50 мм рт. ст. Это является важным условием для
первой фазы мо : - >бразования процесса фильтрации жидкости и
веществ из плазмы крови в нефрон'?
Выносящие артериолы, пройдя короткий путь, вновь распада-
ются на капилляры, оплетающие канальцы нефрона и образующие
перитубулярную капиллярную сеть. В этих «вторичных» капилля-
рах давление крови, наоборот, относительно низкое — около 10—
12 мм рт. ст., что способствует второй фазе мочеобразования —
процессу обратного всасывания части жидкости и веществ из неф-
рона в кровь. —-----------
Из капилляров кровь перитубулярной сети собирается в верх-
них отделах коркового вещества сначала в звездчатые вены, а
При снижении кровяного давления в органц зме может наступить олигурия
уменьшение мочеобразования
601
iClhl :^Г ‘ /
Рис. 246. Кровоснабжение нефронов.
А—схема: / — короткий (корковый) нефрон II— длинный (околомоз оной) нефро!
/, 2— междолевые артерии и вена; 4 — дуговая артерия и вена. 5 6 междольковая
артерия и вена, 7 — приносящая клубочковая артериола; 8 выносящая клубочковая
артериола; 9— клубочковая капиллярная сеть (сосудистый клубочек); 10 - неритубуляр
ная капиллярная сеть; II — прямая артериола; 12 — прямая венула.
затем в междольковые^ в средних отделах коркового вещества —
прямо в междольковые вены. Последние впадают в дуговые вены,
переходящие в междолевые, которые образуют почечные вены,
выходящие из ворот почек.
Таким образом, корковые нефроны в результате особенностей
кортикального кровообращения (высокое кровяное давление в ка-
пиллярах сосудистых клубочков и наличие перитубулярной сети
602
Рис. 246 (продолжение).
Б — инъекция сосудов почки кармином и
берлинской лазурью I — междольковая ар-
терия; 2 — приносящая клубочковая арте-
риола; 3 — клубочковая капиллярная сеть
(сосудистый клубочек) 4 — перитубуляр-
ная капиллярная сеть; 5— междольковая
вена.
капилляров с низким давлением
крови) активно участвуют в мо-
чеобразовании.
В юкстамедулляр-
ной системе кровооб-
ращения приносящие и вы-
носящие артериолы сосудистых
клубочков почечных телец око-
ломозговых нефронов примерно
одинакового диаметра или вы-
носящие артериолы даже не-
сколько шире. Поэтому кровя-
ное давление в капиллярах
этих клубочков ниже, чем в
клубочках Корковых нефронов.
' Выносящие клубочковые ар-
териолы юкстамедуллярных нефронов идут в мозговое вещество,
распадаясь на пучки тонкостенных сосудов, несколько более
крупных, чем обычные капилляры — прямые сосуды (vasa recta).
В мозговом веществе как от выносящих артериол, так и от прямых
сосудов отходят ветви для формирования мозговой перитубуляр-
ной капиллярной сети (rete capillare peritubulare medullaris). Пря-
мые сосуды образуют петли на различных уровнях мозгового
вещества, поворачивая обратно. Нисходящие и восходящие части
этих петель образуют противоточную систему сосудов, называемую
сосудистым пучком (fasciculis vascularis). Капилляры мозгового
вещества собираются в прямые вены, впадающие в дуговые вены.,.
Вследствие этих особенностей околомозговые нефроны не так
активно участвуют в мочеобразовании, как корковые нефроны.
В то же время юкстамедуллярное кровообращение играет роль
шунта, т. е. более короткого и _легкого пути, по-которому проходит
часть крови почки в условиях сильного кровенаполнения,
например при выполнении человеком тяжелой физической работы.
Гистофизиология нефрона. Нефрон начинается
почечным тельцем (диаметр около 200 мкм), представленным
сосудистым клубочком и его капсулой. Сосудистый клубочек fglo-
merulus) состоит более чем из 30 кровеносных капилляров. Их эн
доте, иальные л тетки имеют многочисленные фенестры (возможно,
кроме того и поры) диаметром до 0,1 мкм. Эндотелиальные клетки
капилляров располагаются на внутренней поверхности толстой,
трехслойной базальной мембгкцщ- С паружной^стороны на ней ле-
жит эпителий внутреннего листка капсулы клубочка (рис. 247).
603
Рис. 247. Строение почечного тельца с юкстагломерулярным аппаратом (схема/.
1—приносящая клубочковая артериола; 2 — выносящая клубочковая артериол?; 3—
капилляры сосудистого клубочка; 4 — эидотелиоциты; 5 — подоциты внутреннего листка
капсулы клубочка; 6 — базальная мембрана; 7 — мезангиальные клетки; 8 — полость кап-
сулы клубочка; 9 — наружный лиСток капсулы клубочка: 10 — дистальный каналец
нефрона; //— плотное пятно; 12— эндокриноциты (юкстагломерулярные клетки); 13—
юкставаскулярные клетки; !4 — строма почки.
Капсула клубочка fcapsula glomeruli) по форме напоминает
двухстенную чашу, образованную внутренним и наружным лист-
ками, между которыми расположена щелевидная полость — по-
лость капсулы, переходящая в просвет проксимального канальца
нефрона.
Внутренний листок капсулы проникает между капиллярами
сосудистого клубочка и охватывает их почти со всех сторон. Он
образован крупными (до 30 мкм) неправильной формы лиители-
604
Рис. 248. Схема ультрамикросконического строения фильтрационного барьера
почек (рис. Е, Ф Котовского).
/ — эндотелиоцит кровеносного капилтяра сосудистого клубочка; 2 — трехслойная мем-
брана; 3 — подоцит внутреннего листка капсулы клубочка; 4 — трабекула подоцита,
5 педикулы подоцита; 6 — полость капсулы клубочка; 7 — часть эритроцита.
альными клетками подоцитами t (podocyti). От тел подоцитов
отходят несколько больших широких отростков —цитотрабекулы,
от которых в свою очередь начинаются многочисленные мелкие
отростки —^цитоподии, прикрепляющиеся к трехслойной базаль-
ной мембране. Между цитрподиями располагаются зкие щели,
сообщающиеся через промежутки между телами подоцитов С по-
лос тью капсулы. Трехслойная базальная мембрана, являющаяся
общей для эндотелия кровеносных капилляров и подоцитов внут-
реннего листка капсулы, включает наружный и внутренний слои —
менее плотные (светлые) и средний слой — более плотный (тем-
ный). В среднем слое мембраны (lamina densa) имеются микро-
фибриллы, образующие сеточку с диаметром ячеек до 7 нм.
Все три названных компонента — эндотелий, -капилляров к iy-
бочка, додоциты внутреннего листка капсулы и общая для них
трехслоиная. базальная_мембрана — составляют ~ фвл ьт р а ц и~
о н н ы й барьер, через который из крови в полость капсулы
фильтруются составные части плазмы крови, образующие первич-
ную мочу (рис. 248).
Таким образом, в составе почечных телец находится почечный
фильтр. Он участвует в первой фазе мочеобразования — фильт-
рации. Почечный-.фильтр обладает избирательной проницаемостью,
задерживая, все.та, что больше размеров_лчеек в среднем~сЛОе
базальной мембраны. В норме через него не проходят формен-
ные элементы крови и некоторые белки плазмы крови с наиболее
крупными молекулами — иммунные тела, фибриноген и др. При
повреждении фильтра в случаях заболевания почек (например,
при нефритах) они могут обнаруживаться в моче больных.
605
Рис. 249. Схема ультрамикроскоьического строения проксимального (Л)
стальною (/») канальцев нефрона ночки (рис. Е. Ф. Котовского).
/ — эпителиоциты; 2 — базальная М1мбрана; 3— щеточная каемка; 4 — пиноцито if
пузырьки; 5 — лизосомы; 6— базальная исчер* емкость; 7 - кровеносный капилляр. 1
В сосудистых клубочках почечных телец в тех местах, куда
между капиллярами не могут проникнуть подоциты внутреннего
листка капсулы, лежит еще один вид клеток — мезангиальные.
Они вырабатывают основное межклеточное вещество — матрикс,
'вместе с которым образуют мезангии сосудистых клубочков.
Часть мезангиоцитов является Макрофагами, несущими 1а-анти-
ген. Благодаря этому создается возможность для локальной
реализации в клубочках иммуновоспалительной реакции (см.
(рис. 247). Наружный листок капсулы клубочка представлен од-
ним слоем плоских и кубических эпителиальных клеток, располо-
женных на~баУальной мембране. Эпителии наружного листка кап-
сулы переходит в эпителий проксимального отдела нефрона.
Проксимальный отдел имеет вид извитого и короткого прямо-
го канальца диаметром до 60 мкм с узким неправильной формы
/ просветом. Стенка канальца образована однослойным r*iy6riieeitww>
Эсаетичатым эпителием. Цн~~бсуществляет реабсорбцию, ”г. е. об-
ратное всасывание в кровь (в капилляры перитубулярной сети)
из первичной мочи ряда содержащихтя в ней веществ — белков,
глюкозы, электролитов, воды. Механизм этого процесса связан с
гистофизиологией эпителиоцитов проксимального отдела. Поверх-
ность этих клеток покрыта щеточной каемкой с высокой актив-
ностью щелочной фосфатазы участвующей в полном обрахшм
всасывании глюкозы (см. рис. 245). В цитоплазме клеток образу-
ются пйнбцитозные пузырьки и находятся лизосомы, богатые
протеолитическими ферментами. Путем пиноцитоза клетки погло-
щают из первичной мочи белки, которые расщепляются в цито-
плазме под влиянием лизосомальных ферментов до аминокислот,
транспортируемых в кровь перитубулярных капилляров. В своей
базальной части клетки имеют исчерненность, образованную внут-
г —
606
ренними складками цитолеммы и расположенными между ними
митохондриями. Митохондрии, содёржащйё~сукцинатдёгидрогена-
зу и другие ферменты, играют важную роль в обратном активном
всасывании некоторых электролитов, а складки цитолеммы имеют
большое значение для пассивного обратного всасывания части
воды (рис. 249, А, .
В результате реабсорбции в проксимальных отделах первич-
ная моча претерпевает значительные качественные изменения — из
нее полностью исчезает сахар и белок. При заболевании почек эти
вещества могут обнаруживаться в окончательной моче больного
вследствие поражения клеток проксимальных отделов нефронов.
Петля нефрона состоит из тонкого канальца и прямого дис-
тального канальца. В корковых^нефронах тонкий каналец имеет
Только нисходящую часть. В ю кета меду ллярных нефронах имеет-
ся также длинная восходящая часть тонкого канальца, которая
переходит затем в прямой дистальный каналец. Тонкий каналец
имеет диаметр^!3—15 мкмЗ Стенка его образована плоскими эпи-
е иальными клетками. Цитоплазма клеток светлая, бедная О| га-
прямой каналец имеет больший диаметр —
до 30 мккр Его эпителиальная выстилка такая же, как в дисталь-
ном^йзвитом канальце. Последний имеет диаметр (^)—50 мкм/)
Его стенка образована низким призматическим эпителием, участ-
вующим в реабсорбции — всасывании в кровь части электролитов.
Эпителиальные кле гки лишены щеточной к и, но в связи с ак-
тивным переносом электролитов (натрия, хлоридов и др.) имеют
выраженную базальную исчерченность. обусловленную скоплением
большого числа митохондрий в базальных участках цитоплазмы.
Прямой дистальный каналец, прилежащая половина извитого
дистального канальца почти непроницаемы для воды. В результате
абсорбции электролитов жидкость в этих канальцах становится
разведенной, а в интерстиции повышается осмотическое давле-
ние, что вызывает пассивное обратное всасывание воды из про-
светов конечной части извитых дистальных канальцев и собира-
тельных трубочек.
Собирательные почечные трубочки в верхней
корковой части выстланы однослойным кубическим эпителием, а
в нижней мозговой части — однослойным низким цилиндрическим
эпителиемПГЪпителии различают светлые й темные клетки. Свет-
лые клетки бедны органеллами, их цитоплазма образует внутрен-
ние складки. Темные клетки по своей ультраструктуре напоми-
нают париетальные клетки желез желудка, секретирующие соля-
ную кислоту (рис. 250). В собирательных трубках с помощью свет-
лых клеток завершается пассивное обратное всасывание из мочи в
кровь части-воды. Кроме тогбГ происходит подкисление мочи, что,
вероятно, связано с секреторной деятельностью темных эпителио-
цитов. ' ——________
Реабсорбция воды в собирательных трубочках (у некоторых
видов животных и в конечных частях дистальных извитых каналь-
цев) зависит от концентрации в крови антидиуретического гормо-
607
Рис. 250. Схема ультрамикроскопического строения тонкого канальца петли
нефрона (Л) и собирательной трубочки (Д) почки (рис. Е. Ф. Котовского).
I - эпителиоциты; 2 — базальная мембрана: 3 светлые эпителиоциты; 4 — темные эпи-
телиоциты; 5 микроворсинки; 6 инвагинации цитолеммы; 7 — кровеносный капилляр
на гипофиза. В его отсутствие стенка собирательных трубочек и
конечных частей извитых дистальных канальцев непроницаема
для воды. Поэтому концентрации мочи не происходит. В присутст-
вии гормона стенки указанных канальцев очень проницаемы для
воды, которая выходит пассивно путем осмоса в гипертоническую
среду интерстиция мозгового вещества, и затем в сосуды. В этом
процессе важную роль играют прямые сосуды (сосудистые пуч-
ки), которые, унося воду, поступающую из собирательных тру-
бочек, поддерживают градиент концентрации между содержимым
собирательных трубочек и окружающей их гипертонической сре-
дой.
Таким образом, мочеобразование — сложный процесс, который
осуществляется в нефронах. В почечных тельцах нефронов про-
исходит первая фаза этого процесса — фильтрация, в результате
чего образуется первичная моча вхутки). В каналь-
цах нефронов протекает вторая фаза р^Жования, т. е. реаб-
сорбция, следствием чего является качественное и количественное
изменение мочи Из нее полностью исчезают сахар и _белок, а
также вследствие обратного.всасываниябодьшей части водьГсни-
жается количество мочи (ло 15^2. п в сутки), что приводит к рез-
кому возрастанию в окончательной моче концентрации выделяе-
мых шлаков: /реатиновых тел — в 75 раз, аммиака — в 40 раз и
т. д. Заключительная (третья) секреторная фаза мочеобразования
осуществляется в собирательных трубках, где .реакция мочи ста-
новится слабокислой (рис. 251). Все фазы образования мочи—
результат активной деятельности клеток нефронов. I
608
Рис. 251. Схема строения противоточно-множительного аппарата почки.
J — почечное тельце; 2-—проксимальный прямой каналец нефрона; 3— тонкий каналец
(восходящая часть петли нефрона): 4 дистальный прямой каналец нефрона; 5 — соби-
рательная трубочка; 6 — кровеносные капилляры; 7 — интерстициальные клетки.
Эндокринная система почек. Она представлена рениновым и
простагландиновым аппаратами.
Рениновый аппарат, или юкстагломеруляр-
ный аппарат (ЮГА), т. е. околоклубочковый, секретирует
в кровь активное вещество — ренин. Он катализируад^-образование
в организме ангиотензинов, оказывающих сильное сосудосужи-
20 гистология
600
(вающее влияние, а также стимулирует продукцию гормона альдо-
эстерона в надпочечниках. Кроме того, возможно, что ЮГА при-
Гнадл^ЖИТ важная роль в выработке эритропоэтинов.
В состав ЮГА входят юкстагломерулярные клетки, плотное
Спятно и юкставаскулярные клетки (клетки Гурмагтига) (см.
урис. 247).
^^Окстагломерулярные клетки лежат в стенке приносящих
'бвыносяших артериол под эндотелием. Они имеют joea льну ю или
полигональную форму, а в цитоплазме — крупные секреторные
(рениновые? гранулы, которые не окрашиваются обычными
гистологическими методами, но дают положительную ШИК-реак-
цию.
Плотное пятно (“macula densa) —участок стенки дистального.
[отдела нефрона в том месте, где он проходит рядом с почечным
/тельцем меЖЛУ’приносящей.и. выносящей артериолами. 6 плотном
пятне эпителиальные клетки более высокие, почти лишены база л ь-
ной складчатости, а их базальная мембрана” чрезвычайно тонкая
(по некоторым данным, полностью отсутствует). Допускают, что
плотное пятно, подобно «натриевому рецептору», улавливает изме-
нения содержания натрия в моче и воздействует на околоклубоч-
ковые клетки, секретирующие ренин.
^—уЮкставаскулярные клетки лежат в треугольном пространстве
“между приносящей и выносящей артериолами и плотным пятном.
Они имеют реальную или неправильную форму, образуют далеко
простирающиеся отростки, контактирующие с клетками мезан-
гия клубочка. В их цитоплазме выявляются фибриллярные струк-
туры.
^\/Некоторые авторы причисляют к ЮГА также мезангиальные
Ллетки сосудистых клубочков. Предполагают, что клетки Гурмаг-
f тига и мезангия включаются в продукцию ренина при истощении
'.юкстагломерулярных клеток.
ростагландиновый аппарат включает интерсти-
^Цельные клетки и нефроциты собирательных трубочек.
Интерстициальные клетки, имеющие мезенхимное происхожде-
ние^ располагаются % строме мозговых пирамид. От их вытянуто-
го тела отходят отростки; некоторые из них оплетают канальцы
петли нефронов, а другие — кровеносные капилЗгяры." В Цйтоплаз-
ме интерстициальных клеток хорошо развиты органеллы и нахо-
дятся липидные (осмиофильные) гранулы. По поводу роли этих
клеток высказывают мнение, что они продуцируют один из видов
рростагландинощ который оказывает антигипертензивное действие,
(т. е. снижают^кровяное давление" Кроме этих клеток, источником
эбразования простагландинов, вероятно, являются светлые клетки
:обирательных,_трубочек.
УГаким образом, в почках существует эндокринный комплекс,
участвующий в регуляции общего и почечного кровообращения, а
через него оказывающий влияние на мочеобразование.
] Непосредственно на функцию нефронов воздействует альдосте-
/рон) (надпочечники) и вазопрессин, или антидиуретический гормон
610
(гипоталамус). Под влиянием первого гормона усиливается реаб-
сорбция натрия в дистальных отделах нефронов, а под влиянием
второго — реабсорбция воды в остальных канальцах нефронов
и в собирательных трубочках.
Лимфатическая система почки представлена
сетью капилляров, окружающих канальцы коркового вещества и
почечные тельца. В сосудистых клубочках лимфатических капил-
ляров нет. Лимфа из коркового вещества оттекает через футляро-
образную сеть лимфатических капилляров, окружающих междоль-
ковые артерии и вены, в отводящие лимфатические сосуды 1 -го по-
рядка, которые в свою очередь окружают дуговые артерии и вены.
В эти сплетения лимфатических сосудов впадают лимфатические
капилляры мозгового вещества, окружающие прямые артерии и
вены.
Лимфатические сосуды 1-го порядка образуют более круп-
ные лимфатические коллекторы 2-го, 3-го и 4-го порядка, кото-
рые вливаются в междолевые синусы почки. Из^этих сосудов лим-
фа поступает в регионарные лимфатические узлы.
Иннервация. Иннервацию почки осуществляют эфферентные
симпатические и парасимпатические нервы и афферентные заднеко-
решковые нервные волокна. Распределение нервов в почке различ-
ное. Одни из них имеют отношение к сосудам почки, другие —
к почечным канальцам. Почечные канальцы снабжаются нервами
симпатической и парасимпатической систем. Их окончания лока-
лизуются под мембраной эпителия. Однако, по некоторым данным,
нервы могут проходить через базальную мембрану и оканчиваться
на эпителиальных клетках почечных канальцев. Описаны также
поливалентные окончания, когда одна веточка нерва заканчивается
на почечном канальце, а другая — на капилляре.
Возрастные изменения. Возрастные особенности строения по-
чек указывают на то, что выделительная система человека в пост-
эмбриональном периоде продолжает свое развитие длительное вре-
мя. Так по толщине корковый слой у новорожденного составляет
всего 1 /4—1 /5, а у взрослого — 1 /2—’/з мозгового вещества Одна-
ко при этом увеличение массы почечной ткани связано не с обра-
зованием новых, а с ростом и дифференцировкой уже существую-
щих нефронов, которые в детском возрасте еще не полностью
развиты. В почке ребенка обнаруживается большое число нефро-
нов с мелкими нефункционирующими и слабодифференцирован-
ными клубочками. Толщина извитых канальцев нефронов у детей
в среднем 18—36 мкм, тогда как у взрослого она достигает 40—
60 мкм. Особенно резким изменениям с возрастом подвергается
длина нефронов. Их рост продолжается вплоть до половой зре-
лости. Поэтому с возрастом, по мере того как увеличивается масса
канальцев, количество клубочков на единицу поверхности почки
уменьшается.
Подсчитано, что у новорожденных на один и тот же объем
почечной ткани приходится до 50 клубочков, у 8—10-месячных
детей— 18—20 клубочков, а у взрослых — 4—6 клубочков.
20
611
I
МОЧЕВЫВОДЯЩИЕ ПУТИ
К мочевыводящим путям относятся почечные чашечки и ло
ханки, мочеточники, мочевой пузырь и мочеиспускательный ка-
нал, который у мужчин одновременно выполняет функцию выве-
дения из организма семенной жидкости и поэтому будет описан
в главе, посвященной половой системе.
Строение стенок почечных чашечек и лоханок, мочеточников
и мочевого пузыря в общих чертах сходно. В них различают сли-
зистую оболочку, состоящую из переходного эпителия и собствен-
ной пластинки, подслизистую основу, мышечную и наружную обо-
лочки.
В стенке почечных, чашечек и почечных лоханок вслед за пе-
реходным эпителием располагается собственная пластинка слизис-
той оболочки, незаметно переходящая в соединительную ткань
подслизистой основы. Мышечная оболочка состоит из двух тонких
слоев гладких мышечных клеток: внутреннего (продольного) и
наружного (циркулярного). Однако вокруг сосочков почечных пи-
рамид сохраняется только один циркулярный слой гладких мы-
шечных клеток. Наружная оболочка без резких границ переходит
в соединительную ткань, окружающую крупные почечные сосуды.
Мочеточники обладают выраженной способностью к растяже-
нию благодаря наличию в них глубоких продольных складок сли-
зистой оболочки. В подслизистой основе нижней части моче-
точников располагаются мелкие ал1ве >лярно-трубчатые железы,
по строению сходные с предстательной железой. Мышечная обо-
лочка мочеточников в верхней половине состоит их двух слоев:
внутреннего (продольного) и наружного (циркулярного). В ниж-
ней части мочеточников мышечная оболочка имеет три слоя: внут-
ренний и наружный слои продольного направления и средний
слой — циркулярного. В мышечной оболочке мочеточников в мес-
тах их прохождения через стенку мочевого пузыря пучки гладких
мышечных клеток идут только в продольном направлении. Сокра-
щаясь, они раскрывают отверстие мочеточника независимо от со-
стояния гладких мышц мочевого пузыря. Снаружи мочеточники
покрыты соединительнотканной адвентициальной оболочкой.
Слизистая оболочка мочев9гр пузыря состоит из переходного
эпителия и собственной пластинки. В ней мелкие кровеносные
сосуды особенно близко подходят к эпителию. В спавшемся или
умеренно растянутом состояли слизистая оболочка мочевого
цудыря имеет множество скл;с4рк рис. 252). Они отсутствуют в
переднем отделе дна пузыря, Рие в него впадают мочеточники и
выходит мочеиспускательный я1нал. Этот участок стенки мочевого
пузыря, имеющий форму треу|'Жльника, лишен подслизистой“осно-
вы и его слизистая оболочка плотно сращена с мышечной оболоч-
кой. Здесь в собственной пластинке слизистой оболочки заложены
железы, подобные железам нижней части мочеточников.
Мышечная оболочка мочевого пузыря построена из трех не-
резко отграниченных-слоев: внутреннего и наружного с продоль-
ным расположением гладких мышечных клеток и^ред+гетсг-— цир-
612
Рис. 252. Строение мочевого пузыря.
J — слизистая оболочка; 2 - переход-
ный эпителий; 3 — собственная плас-
тинка, 4 подслизистая основа; 5 -
мышечная оболочка; 6 — серозная обо-
лочка.
кулярного. Гладкие мышеч-
ные клетки часто напомина-
ют по форме расщепленные
на концах веретена. «Про-
слойки соединительной ткани
разделяют мышечную ткань
в этой оболочке_на отдельные
крупные пучки. В шейке мо-
чевого пузыря циркулярный
слои~^образует мышеЧНЫй-
сфинктер. Наружная оболоч-
ка на верхнезадней и частич-
но на боковых поверхностях
мочевого пузыря представле-
на листком брюшины (сероз-
ная оболочка), в остальной
его части она является ад-
вентициальной.
Стенка мочевого пузыря
богато снабжена кровеносны-
ми и лимфатическими сосу-
дами. Мочевой пузырь иннер-
вируется как симпатически-
ми и парасимпатическими,
так и спинальными (чувст-
вительными) нервами. Кроме того, в мочевом пузыре обнаружено
значительное число нервных ганглиев и рассеянных нейронов
вегетативной нервной системы. Особенно много нейронов у места
впадения в мочевой пузырь мочеточников. В серозной, мышечной
и слизистой оболочках мочевого пузыря имеется также большое
число рецепторных нервных окончаний.
Глава XXII <£
ПОЛОВАЯ СИСТЕМА | V
Половая система как в мужск’М', так и в женском организме
состоит из половых желе» / (гонад) и добавочных
органов полового тракта, к которым в мужском организме
относятся се мявыводящие пути, семенные пузырьки, предстатель-
ная железа и половой член, а в женском — яйцеводы, матка и
влагалище, а также наружные пололые органы. К этой же группе
у женщины могут быть причислены молочные железы, тесно свя-
занные с половой системой.
613
Рис. 253. Развитие гонад в эбриогенезе.
А схема первичном локализации гоно-
блвстов в желточном мешке зародыша и их
последующей мшраиии в зачаток гонад 1 по
Пэттену с и1меиением А Г. Кнорре): 1 —
эпителии желточного мешка; 2 мезенхи-
ма; 3 сосуды; 4 первичная почка <ме
юнефрос): 5 — зачаток гонады; 6 пер-
вичные половые клетки (тонопласты); 7 —
поверхностный эпителии.
Значение половой системы состоит в обеспечении размноже-
ния организма (т. е. его репродуктивной функции), что осущест-
вляется путем образования половых клеток — яйцеклеток и спер-
матозоидов, слиянием которых (оплодотворением) начинается
развитие зародыша. Одновременно половые железы вырабатывают
половые гормоны — андрогены и эстрогены, благодаря
влиянию которых в организме создаются условия, необходимые
для обеспечения возможности размножения.
Различия между полами предопределяется генетически через
половые хромосомы (XY у мужчин и XX у женщин). Существен-
ной особенностью функционирования женской половой системы
является цикличность ее деятельности, ее периодичность. При
этом выделение половой клетки и изменения интенсивности секре-
ции женских половых гормонов повторяются регулярно, тогда как
мужская половая система функционирует непрерывно с момента
достижения половой зрелости до старческого увядания.
Развитие. Закладка половой системы в начальных стадиях
эмбриогенеза протекает у обоих полов одинаково (индиффе-
рентная стадия), притоу в тесном контакте с развитием
выделительной системы (рис. ^З, 254). Закладка гонад становит-
ся заметна у 4-недельного зарЬдыша в виде половых валиков —
утолщений целомического эпителия на поверхностях обеих пер-
вичных почек ("mesonephros). Однако первичные половые клетки
у зародышей обоего пола — гзметобласты в пресомитных стадиях
эмбриогенеза появляются раньше (на 3-й неделе) и характери-
зуются крупными клеточными ядрами, повышенным содержанием
гликогена и высокой активностью щелочной фосфатазы в цито-
плазме. Вначале они обнаруживаются в стенке желточного мешка,
614
Рис. 253 (продолжение).
Б семенник плода челове-
ка 20 нед развития (препа-
рат О„ И. Бриндака)* /
половой тяж; 2 гоноциты;
3 - эпителий половых тя-
жей; 4 — дифференцирую-
щиеся гландулоциты, В се-
менник плота кролика 29 дней
развития (препарат
О. И. Бриндака): / кана-
дец сети семенника; 2 - се-
менной каналец; 3 — гоно-
циты; 4 эпителии каналь-
ца сети семенника; 5 — диф-
ференцирующиеся гландуло-
ци гы.
где быстро размножаются, затем в стенке задней кишки и с
кровью, протекающей по его сосудам, выселяются в толщу поло-
вых валиков. Из эпителия половых валиков образуются фоллику-
лярные клетки в яичниках или поддерживающие эпителиоциты
(сустентоциты) в семенниках, которые обеспечивают питание
созревающих гоноцитов. Эпителиоциты при участии интерстици-
альных (мезенхимных) клеток, или эндокриноцитов, осуществля-
ют выработку половых гормонов. Интерстициальные (мезенхим-
ные) клетки интенсивно размножаются на 9—10-й неделе. После
22-й недели количество их уменьшается.
От половых валиков в строму первичной почки, основа которой
образована мезенхимой, врастают половые шнуры — тяжи эпите-
лия, в которых располагаются гоноциты. Одновременно из мезо-
нефрального протока первичной почки, тянущегося от ее тела
к клоаке, отщепляется параллельно идущий парамезонефральный
проток. Дифференциация индифферентной половой железы по полу
у зародыша человека начинается на 6-й неделе эмбриогенеза.
МУЖСКАЯ ПОЛОВАЯ СИСТЕМА
Яички (семенники)
Яички, или семенники (testes), — мужские гонады, в которых
происходит образование мужских половых клеток и мужского по-
лового гормона.
Развитие. При развитии семенника по верхнему краю первич-
ной почки формируется будущая соединительнотканная капсула
семенника — белочная оболочка (tunica albuginea), которая отде-
ляет половые-шнуры от полового валика, давшего им начало.
В дальнейшем половые шнуры развиваются в семенные канальцы
(tubuli seminiferi convolut7J7*H6~часть этих шнуров превращается
в сеть семенника (rete testis). Вначале семенные канальцы и
канальцы сети семенника разобщены_и вступают в связь позже.
В постнатальном периоде в семенных канальцах гоноциты
размножаются, а эпителий половых шнуров сохраняется в качест-
ве поддерживающих клет^с._Ь тех^же половых шнурах, из которых
развивается сеть семенника, гоноциты постепенно редуцируются.
Канальцы сети семенника, приближаясь к белочной оболочке
средостения, сли ва ютея й выносящие канальцы (ductuli efferentes),
формирующиеся____путем перестройки эпителиальной выс-
• тилки канальцев первичной почки. Выносящие канальцы яичка,
собираясь, переходят далее в канал придатка семенника (ductus
epididymis), проксимальный отдел которого, многократно изви-
ваясь, образует, придаток семенника (epididymis), тогда как его
дистальная часть становится семявыносящим протоком (ductus
defferens). Парамезонефральный проток в мужском организме
атрофируется и сохраняется только верхний конец и нижний,
п ревра шаюшийся в предстательную (му ж с ку ю) маточку (utriculus
prostaticus), располагающуюся у взрослого мужчины в толще пред-
стательной железы у места впадения семявыносящего протока
616
Рис. 254. Стадии развития гонад и становления их гормональной регуляции в онто-
генезе (схема Б. В. Алешина, Ю. И. Афанасьева, О. И Бриндака, Н А. Юриной).
ТФ телоферрон; НУ—антиген; ГРПП - гормон регрессии парамезонефрального про
тока; ТС — тестостерон; Э — эстрадиол; П прогестерон; ФСГ фолликулостимулирую-
щий гормон; ФИСГ — фактор ингибирующий сперматогонии; ЛГ — лютеинизирующим
гормон; ИН — ингибин; ГЛ — гонадолиберин; АЯ — аркуатное ядро; ВМЯ — вентромеди-
альное ядро. I — парамезонефральный проток; 2 — мезонефральный проток; 3 — половые
тяжи; 4 — гоноциты; 5 — эпителий; 6 — гландулоциты; 7 — сеть семенника; 8 — выносящие
каиальцы семенника; 9 — корковое вещество яичника; 10 — мозговое вещество яичника;
II — примордиальные фолликулы; 12 — сустентоциты; 13 — сперматогонии; 14 — первич-
ные фолликулы; /5 — маточная труба; 16 — интерстициальные клетки.
в мочеиспускательный канал. Предстательная железа и -семенные
пузырьки развиваются как выросты мочепо понагл синуса
После 22-й недели гоноциты превращаются в сперматогонии,
при этом они утрачивают гликоген и высокую- активность щелоч-
ной фосфатазы. “---~
В онтогенезе эндокринная функция семенников устанавлива-
, ется раньше, чем генеративная. Мужской половой гормон — те-
стостерон начинает выра >атываться у зародыша человека
617
ГД
КГ
вь
ля
(<
С(
aj
к>
н,
2.
о
л
н
к
п
У
приблизительно с 8—10 нед внутриутробного периода. В заро-
дышевом семеннике ещё'ЦО ИЗчала^бипсидтетаг тестостерона об-
разуются поочередно несколько белковых (пептидных) андроге-
нов, принадлежащих кингибинам. Из них первым уже на
стадии индифферентной гонады появляется гормон, под влиянием
которого происходит редукция парамезонефрального протока, и
с этого момента индифферентный зачаток половой системы на-
1чинает дифференцироваться по мужскому типу. Источник выра-
ботки этого гормона еще не установлен.
Приблизительно к середине эмбриогенеза, когда в семенных
I
1
канальцах развивающегося семенника уже дифференцируется эпи-
телиаг.нерматогенный _слой, а в просвете я большом количестве
скапливают ся гоноциты, начинается выработка второго ингибина,
который, с одной стороны, угнетает размножение гоноцитов и
вызывает их разрушение, а с другой — одновременно ингибирует
секрецию ФСГ аденогипофизом. Этот пептид вырабатывается эпи-
тели^мсети яичка. -----——
Наконец, по мере приближения к пренатальному периоду эм-
бриогенеза у плода мужского пола появляется ингибин, который,
п
л
н
с
л
I
(
в
к
г
й
теряя депрессивное влияние на гоноциты, сохраняет способность
избирательно угнетать^секрецию ФСГ, действуя на соответствую-
щие центры гипоталамуса.
Строение. Снаружи большая часть семенника покрыта сероз-
ной оболочкой — брюшиной, за которой располагается плотная
соединительнотканная оболочка, получившая название белочной
(tunica albuginea) (рис. 255). На заднем крае семенника белочная
оболочка утолщается, формируя средостение (mediastinum testis),
от которого в глубь железы отходят прослойки соединительной
ткани (septula testis), разделяющие железу на дольки (около
250 долек), в каждой из которых находится 1—4 извитых семен-
ных канальца (tubuli seminiferi convoluti). Каждый семенной
каналец имеет диаметр от 150 до 250 мкм и длину от 30 до 70 см
Приближаясь к средостению, канальцы (300—450 в каждом се-
1
<i
1
1
1
<
I
<
меннике), сливаются и становятся прямыми, а в толще средосте-
ния соединяются с канальцами сети семенника (rete testis) '"у этой
сети выходит 10—12 выносящих канальцев (ductuli efferens), впа-
дающих в проток придатка (ductus epididymis).
Стенку семенного канальца образует собственная оболочка,
состоящая из базального слоя (stratum basale), миоидного слоя
(stratum myoideum) и волокнистого слоя (stratum fibrosum), выст-
ланная изнутри так называемым сперматогенным эпителием1, распо-
ложенным на базальной мембране. Базальная мембрана спермато-
генного эпителия у человека имеет толщину около 80 нм, образует
выросты, направленные как внутрь сем чных канальцев, так и в на-
ружные слои. Базальный слой (внутренний неклеточный слой), рас-
положенный между двумя базальными мембранами (сперматоген-
ного эпителия и миоидных клеток), состоит из сети коллагеновых
Термин устаревший, так как эпителий не является источником образования
сперматогенных клеток.
618
Рис. 255. Схема строения яичка (рис. Е. Ф Котовского)
А — сперматогенный эпителий в период размножения и в начале периода роста; Б — он же
в конце периода роста и в период созревания; В — период формирования; Г — строение
семявыносящего канальца яичка; Д — строение канала придатка; Е — строение семя-
выносящего канала; / оболочки яичка; // — перегородки яичка; III — дольки яичка;
IV— извитой семенной каналец: V — интерстициальная железистая ткань; VI—прямые
семенные канальцы; VII сеть семенника; VIII — выносящие канальцы яичка; IX —
канал придатка; X — семявыносящий канал: I — мезотелий; 2 — кровеносный сосуд;
3 — соединительнотканные клетки; 4 — поддерживающие клетки; 5 - сперматогонии;
6 — сперматоциты; 7 — сперматиды; 8 — сперматозоиды в просвете извитого семенною
канальца: 9 — мышечно-волокнистая оболочка выносящего канальца; 10 — реснитчатые
клетки; 11 — кубические клетки; 12 — сперматозоиды в выносящем канальце яичка;
13 — мышечно-волокнистая оболочка канала придатка яичка; 14 — двухрядный реснит-
чатый эпителий семявыносящего канала; 15 — двухрядный реснитчатый эпителий; 16
собственная пластинка слизистой оболочки; 17 — внутренний продольный слой мышечной
оболочки; 18 — средний циркулярный слой мышечной оболочки; 19 — наружный продоль-
ный слой мышечной оболочки; 20 адвентициальная оболочка.
волокон. Миоидный слои (внутренний клеточный слой) образован
особыми миоидными клетками, содержащими актиновые филамен-
ты, но отличающимися по структуре от типичных гладких мы-
шечных клеток. Миоидные клетки обеспечивают ритмические со-
кращения стенки канальцев. Волокнистый слой состоит из двух
частей Непосредственно к миоидному слою примыкает внутрен-
ний неклеточный слой, образованный базальной мембраной мио-
619
гд
•<Г
вь
л»
(<
сс
aj
ю
HI
2.
ol
Л1
Hi
к
У
9
П1
л<
р
н
1
с
4
• 1
<;
<
i
I
идных клеток и коллагеновыми волокнами. За ними расположен
наружный слой, состоящий из фибробластоподобных клеток.
В соединительной ткани между семенными канальцами рас-
положены гемокапилляры и лимфокапилляры, обеспечивающие
обмен веществ между кровью и сперматогенным эпителием. Из-
бирательность поступления веществ из крови в сперматогенный
эпителий и существенные различия в химическом составе плазмы
крови и жидкости из семенных канальцев позволили сформули-
ровать представление о гематотестикулярном барьере. Гематотес-
тикулярным барьером называется совокупность структур, распо-~
латающихся между просветами капилляров и семенных канальцев.
Эпителио-е^ерматоеенньт—слой Cepithehurrr spermatogenicum)
имеет две основных популяции клеток — поддерживающие клетки,
или сустентоциты (epitheliocytus sustentans) и сперматогенные
клетки (cellulae spermatogenicae), находящиеся на различных ста-
диях дифференцировки. Сперматогенные клетки представлены
стволовыми клетками, сперматогониями, сперматоцитами, сперма-
тидами и сперматозоидами. Обе популяции находятся в тесной
морфофункциональной связи.
Поддерживающие клетки лежат на базальной мембране, имеют
пирамидальную форму и достигают своей вершиной просвета из-
витого семенного канальца. Ядра их имеют неправильную форму.
Для них характерно наличие инвагинаций, трехчленного ядрышка
(ядрышко и две группы околоядрышкового хроматина). В цито-
плазме представлены все органеллы, особенно хорошо развита аг-
ранулярная эндоплазматическая сеть, комплекс Гольджи. Встре-
чаются также микротрубочки, микрофиламенты, лизосомы и
особые кристаллоидные включения. Обнаруживаются включения
Липидов, углеводов, липофусцина. На боковых поверхностях сус-
тентоцитов образуются бухтообразные углубления, в которых рас-
полагаются дифференцирующиеся сперматоциты и сперматиды.
Между соседними поддерживающими клетками формируются зо-
ны платных контактов, которые подразделяют сперматогенный
эпителий на два отдела — наружный базальный и внутренний
ауьлюминальньий. В базальном отделе расположены сперматогонии,
имеющие максимальный доступ к питательным веществам, по-
ступающим из кровеносных капилляров. В адлюминальном от-
деле находятся делящиеся сперматоциты, а также сперматиды и
сперматозоиды, которые не имеют доступа к тканевой жидкости и
получают питательные вещества непосредственно от поддержи-
вающих эпителиоцитов. Поддерживающие эпителиоциты создают
микросреду, необходимую для дифференцирующихся половых кле-
ток, изолируют формирующиеся половые клетки от токсических
веществ и различных антигенов, препятствуют развитию иммунных
реакций. Кроме того, сустентоциты способны к фагоцитозу де-
генерирующих половых клеток и последующему лизису их с по-
мощью своего лизосомального аппарата. Показана их способность
синтезировать андроген-связывающий белок (АСБ), который тран-
спортирует мужской половой гормон к сперматидам. Секре ия
620
АСБ усиливается под влиянием ФСГ. Поддерживающие эпителио-
циты имеют поверхностные рецепторы для ФСГ, а также рецепто-
ры для тестостерона и его метаболитов. Различают два вида под-
держивающих клеток — светлые клетки. продуцирующие фактор
(и н г и б и н), тормозящий^ секрецию ФСГ аденогипофизом, и
Темные клёткщ^продуцирующие фактор, стимулирующий деление
половых клеток. ~
Генеративная функция. Сперматогенез
Образование мужских половых клеток (сперматоге.н£ з)
протекает в извитыд семенных_канальцах и включает 4 после-
довательные стадии или фазы: размножение, рост, созревание и
формирование (рис. 256).
Начальной фазой сперматогенеза является р а з м н о ж е н и е
сперматогоний, занимающих наиболее периферическое (базаль-
ное) положение в спсрматогенном эпителии. Согласно современ-
ным представлениям, среди сперматогоний можно выделить два
типа клеток: 1) ^стволовые спе рматогонии типа А, которые под-
разделяются на две субпопуляции: долгоживущие резервные
стволовые клетки и быстрд_обновляюшиеся полуствОловые клетки,
~киТОрые делятся один раз в течение цикла сперматогенного эпите-
лия, 2) дифференцирующиеся сперматогонии типа А и типа В.
Стволовые клетки расположены в базальной части канальца
изолированно от других сперматогоний. Морфологически в попу-
ляции стволовых А-сперматогоний различают светлые и темные
клетки (см. рис. 256). Для обоих клеток характерно преобладание
в ядрах деконденсироваиного хроматина и расположение ядры-
шек около ядерной оболочки. Однако в темных клетках типа А
степень конденсации хроматина большая, чем в светлых клетках.
Темные клетки относят к «резервным» медленно обновляющим-
ся стволовым клеткам, а светлые — к полустволовым быстро об-
новляющимся клеткам. Для стволовых клеток характерно нали-
чие овальных ядер с диффузно распределенным хроматином, од-
ного или двух ядрышек, большое содержание в цитоплазме ри-
босом и полисом малое количество других органелл.
Часть стволовых клеток типа А при делении не завершают
цитокинез и остаются соединенными цитоплазматическими мости-
ками. Появление таких спаренных сперматогоний свидетельствует
о начале процессов дифференцировки мужских половых клеток.
Дальнейшее деление таких клеток приводит к образованию це-
почек или групп сперматогоний, соединенных цитоплазматически-
ми мостиками (см. рис. 256). Клетки типа В имеют более крупные
ядра, хроматин в них не дисперсный,~а собран в глыбки
В~~следующйи период сперматогоний' перестают делиться и
дифференцируются в сперматоциты 1-го порядка (период рос-
та). Синтициальные группы сперматогоний перемещаются в ад-
люминальную зону сперматогенного эпителия. В период^ роста
сперматогонии увеличиваются в объеме и вступают в первое
621
Рис. 256. Схема сперматогенеза (по И. Клермону с изменениями).
/ VI — стадии цикла развития мужских половых клеток в семенных канальцах человека
/ — соединительнотканная капсула канальца° 2 — базальная мембрана; 3 — поддержи
вающие клетки; 4 — сперматогонии: тип — светлый; тнп Ат — темный; В — тнп В;
5 — сперматоциты 1-го порядка: 5а — в пахнтене; 56 — в прелептотене; 5в — в лепто-
тене; 5г — в днплотене; 5д — в знготене; 5е — делящиеся сперматоциты I го порядка;
6 — сперматоциты 2-го порядка с интерфазными ядрами; 7 — сперматиды на различных
стадиях развития (а, б, в. г).
деление м е й о з а (редукционное деление). Профаза
первого деления длинная и состоит из 5 стадий: лептотены,
зиготены, пахитены, диплотены, диакинеза.
Перед профазой в S-периоде сперматоцита 1-го порядка проис-
ходит удвоение количества ДНК. Сперматоцит находится на ста-
дии прелептотены. В—стадии лептотены хромосомы становятся
„видимыми в виде тонких нитей. В стадии зиготены гомологичные
.хромосомы расположены парами (конъюгируют), образуя бива-
ленты. Е^: значение заключается в том, что между конъюгирую-
щими хромосомами происходит обмен генами. В результате
прогрессирующей спирализации пары конъюгирующих хромосом
продолжают укорачиваться и одновременно утолщаться — пахи-
тенная стадия (от лат. pachys — толстый). Гомологичные
хромосомы находятся в тесном контакте по всей своей длине.।
На этой стадии с помощьтсг Электронного микроскопа в сперма-
тоцитах 1-го порядка обнаружены синаптонемные комп-
622
лексы в местах соприкосновения гомологичных хромосом —
парные параллельно расположенные ленты шириной около 60 нм,
разделенные светлым промежутком шириной около 100 нм. В свет-
лом промежутке видны срединная электронно-плотная линия и
пересекающие его тонкие филаменты. Оба конца комплекса
присоединены к ядерной оболочке. У человека на этой стадии
формируется_23 синаптогенных комплекса. В диплотене хромо-
сомы, образующие каждый бивалент, отходят друг от друга, так
что становится видна каждая в отдельности, но сохраняют связь
в перекрестах хромосом. Одновременно можно видеть, что каждая
хромосома состоит из двух хроматид. Дальнейшая спирализация
приводит к тому, что пары конъюгирующих хромосом приобре-
тают вид коротких телец разнообразной формы — гак называе-
мых тетрад. Поскольку каждая тетрада образована двумя
конъюгировавшими хромосомами, число тетрад оказывается вдвое
меньше, чем исходное число хромосом, т. е. гаплоидным, — у
человека 23 тетрады. На стадии диакинеза хромосомы еще
более утолщаются, после чего клетка вступает в м е т а Ф.З з у
первого деления мейоза (или первое деление созревания)
и хромосомы располагаются в экваториальной плоскости. Вана-
фа зе обе хромосомы каждого бивалента расходятся к полюсам
клетки — по одной к каждому полюсу. Таким образом, в каждой
из двух дочерних клеток — сперматоцитов 2-го порядка содержит-
ся гаплоидное число хромосом (23 у человека)J
Второе деление созревания начинается сразу
вслед за первым, и происходит как обычный митоз без редупли-
кации хромосом. В анафазе второго деления созревания диады
сперматоцитов второго порядка разъединяются на монады, или
одиночные хроматиды, расходящиеся к полюсам. В результате
сперматиды получают столько же монад, сколько было диад в
ядрах сперматоцитов второго порядка, т. е. гаплоидное число.
Сперматоциты 2-го порядка имеют меньшие размеры, чем сперма-
тоциты 1-го порядка, и располагаются в средних и более поверх-
ностных слоях эпителио-сперматогенного слоя.
Таким образом, каждая исходная сперматогонии дает начало
4 сперматидам с гаплоидным набором хромосом. Сперматиды
больше не делятся, но путем сложной перестройки превращаются
в зрелые сперматозоиды. Эта трансформация составляет четвертую
фазу сперматогенеза —-период формирования, или
спермиогенеза (рис. 257).
Сперматиды представляют собой небольшие округлые клетки
со_сравнительно крупными ядрами. Скапливаясь около верхушек
поддерживающих клеток, сперматиды частично погружаются в их
цитоплазму, что создает условия для формирования спермато-
зоидов из сперматид. Ядро сперматиды постепенно уплотняется
и уплощается.
В сперматидах около ядра располагаются комплекс Гольджи,
центросома и скапливаются мелкие митохондрии. Процесс форми-
рования начинается с образования в зоне комплекса Гольджи
623
ГД
Ч
Bh
л>
((
се
а:
ю
Нк
2:
о<
л
н
к
"I
У
Рис. 257. Схема сперматогенеза (дифференцировка сперматиды в сперматозоид
(рис. Б. В. Алешина).
/ сперматиде, внедрившаяся в верхушку поддерживающей клетки; // VII — последо-
вательные стадии формирования сперматозоида; / — комплекс Гольджи; 2 — акробласт:
3 зачаток акросомы; 4 митохондрия; 5 ядро, 6 - центриоль; 7 — проксимальная
центриоль; 8 - дистальная центриоль; 9 — трубки акронемы; 10 — кольцо: II — микро-
трубочки; 12 — шейка; 13 митохондриальное влагалище; 14 — хвост; /5 — поддержи-
вающий эпителиоцит
п
Л1
н
а
л
е
(
ч
к
уплотненной, гранулы —^акробласта, прилежащего к поверхности
ядраТТСдальнейшем акробласт-д величиваясь в размерах, в виде
ШЗпочки охватывает ядро2а в середине акробласта дифференци-
руется уплотненное тельце. Такую структуру называют акросомдй-
Она лежит в том конце трансформирующейся сперматиды, кото-У/\
}рый обращен к поддерживающей клетке. Центросома, состоящая «
из двух центриолей, перемещается в противоположный конец
• сперматиды. "Проксимальная центриоль прилегает к поверхности
ядра, а дистальная разделяется на две части. От передней части
дистальной центриоли начинает формироваться жгутик (flagellum),
который затем становится осевой нитью развивающегося сперма-
тозоида. Задняя же половина дистальной центриоли принимает
вид колечка. Смещаясь вдоль жгутика, это колечко определяет
заднюю границу средней или связывающей части сперматозоида.
Цитоплазма по мере роста хвоста сползает с ядра и сосредо-
тачивается в связующей части. Митохондрии располагаются
спиралеобразно между проксимальной центриолью и колечком.
Цитоплазма сперматиды во время ее превращения в спермий
сильно редуцируется. В области головки она сохраняется только в
виде тонкого слоя, покрывающего акросому; небольшое количество
цитоплазмы остается в области связующего отдела и, наконец,
она очень тонким слоем покрывает жгутик. Часть цитоплазмы
сбрасывается и распадается в просвете семенного канальца либо
поглощается поддерживающими клетками. Кроме того, эти клетки
вырабатывают жидкость, накапливающуюся в просвете извитого
семенного канальца. В эту жидкость попадают, высвобождаясь из
624
верхушек поддерживающих клеток, сформировавшиеся спермато-
зоиды и вместе с ней уходят в дистальные части канальца.
Процесс сперматогенеза в целом длится у человека около
75 сут, но протекает на протяжении извитого семенного каналь-
ца волнообразно. Поэтому на каждом отрезке канальца имеется
определенный набор клеток сперматогенного эпителия.
Эпителиосперматогенный слой чрезвычайно чувствителен к
повреждающим действиям. При различных интоксикациях, авита-
минозах, недостаточности питания и других условиях (особенно
при воздействии ионизирующим излучением) сперматогенез ос-
лабляется или даже прекращается, а сперматогенный эпителий
атрофируется. Аналогичные деструктивные процессы развиваются
при крипторхизме (когда семенники не опускаются в мошонку,
оставаясь в брюшной полости), длительном пребывании организма
в среде с высокой температурой, лихорадочных состояний и осо-
бенно после перевязки или перерезки семявыводящих каналов.
Деструктивный процесс при этом поражает в первую очередь форми-
рующиеся сперматозоиды и сперматиды. Последние набухают,
нередко сливаются в характерные округлые массы — так называе-
мые семенные шары, плавающие в просвете канальца. Так как
нижние слои сперматогенного эпителия (сперматогонии и сперма-
тоциты 1-го порядка) при этом сохраняются более длительно, то
восстановление сперматогенеза после прекращения действия
повреждающего агента иногда оказывается возможным.
Деструкция, описанная выше, ограничивается только спермато-
генным слоем. Поддерживающие клетки в указанных обстоятельст-
вах сохраняются и даже гипертрофируются, а гландулоциты
часто увеличиваются в количестве и образуют большие скопления
между запустевающими семенными канальцами.
Эндокринная функция
В рыхлой соединительной ткани между петлями извитых каналь-
цев располагаются интерстициальные клетки — гландулоциты,
скапливающиеся здесь вокруг кровеносных капилляров. Эти клетки
сравнительно крупные, округлой или многоугольной формы, с аци-
дофильной цитоплазмой, вакуолизированной по периферии, содер-
жащей гликопротеидные_кклк>-чения, а также глыбки гликогена и
белковые кристаллоиды _в_виде палонек или лент. С возрастом в
цитоплаз'ме интерстициальных клеток начинает откладываться пиг-
мент. Хорошо развитая гладкая эндоплазматическая сеть, много-
численные ''митохондрии, с трубчатыми и везикулезными кристами
указывают на способность интерстициальных клеток к выработке
стероидных веществ, в данном случае мужского полового гормона.
Семявыносящие пути
Семявыносящие пути составляют систему канальцев (см.
рис. 255) семенника и его придатков, по которым сперма (сперма-
тозоиды и жидкость) продвигается в мочеиспускательный канал.
625
ГД
кр
вь
ля
(с
cd
of
л!
ш
"i
У
Отводящие пути начинаются прямыми канальцами семенника
(tubuii seminiferi recti), впадающими в сеть семенника (rete
testis), располагающуюся в Средостении (mediastinum). От этой
сети отходят 12—15 извитых выносящих канальцев ductuli
efferentes testis), которые соединяются с протоком придатка
(ductus epididymidis) в области головки придатка. Этот проток,
многократно извиваясь, формирует тело придатка и в нижней
хвостовой части его переходит в прямой семявыносящий проток
(ductus deferens), поднимающийся к выходу из мошонки, а затем
достигающий предстательной железы, где впадает в мочеиспуска-
тельный канал. ~~
Все семявыводящие пути построены по общему плану и состоят
из слизистой, мышечной и адвентициальной оболочек. Эпителий,
выстилающий эти канддьцы, обнаруживает признаки железистой
деятельности, особенно выраженной В головке придатка.
В прямых канальцах семенника эпителий образован клетками
призматической формы. В канальцах сети с£менника в эпителии
преобладают кубические и плоские клетки. В эпителии Семявыво-
дящих канальцев чередуются группы реснитчатых клеток с желё-
* зистыми клетками, секретирующими по апокриновому типу.
п
Л!
В придатке семенника эпителий протока становится двухряд-
ным. В его составе находятся высокие призматические клетки,
несущие на своих апикальных верхушках стереоцилии, а между
базальными частями этих клеток "располагаются вставочные клетки
Эпителий протока придатка принимает участие в выработке жид-
кости, разбавляющей сперму во время прохождения сперматозои-
дов, а также в образовании гликока/щкса — тонкого слоя, которым
покрываются сперматозоиды. Удаление гликокаликса при эякуля-
ции приводит к активизации сперматозоидов (капацитация).
Одновременно придаток семенника оказывается резервуаром для
накапливающейся спермы.
Продвижение спермы по семявыы т.-пцим путям обеспечивает-
ся сокращением мышечной оболочки, образованной циркулярным
слоем гладких мышечных клегок.
Проток придаткй "Дйлее" переходит в семявыносящий проток
(ductus deferens), в котором значительно развивается мышечная
оболочка, состоящая из трех слоев — внутреннего продольного,
среднего циркулярного и наружного продольного. В толще мышеч-
ной оболочки располагается нервное сплетение, образованное скоп-
лёнием’гащдиозных клеток, Иннервирующих пучки гладких мышеч-
ных клегок. Сокращениями их обеспечивается эякуляция спермы.
В связи со значительным развитием мышечной оболочки слизистая
оболочка семявыносящего протока собирается в продольные склад-
ки (см. рис. 254). Дистальный конец этого протока ампулообразно
расширен. Снаружи семяотводящие пути на всем протяжении
покрыты соединительноткандуй адвентициальной оболочкой.
Ниже-мёста соединения семявыносящего протока и семенных
пузырьков начинается семявыбрасывающий проток (ductus ejacu-
latorius). Он проникает через предстательную-железу (см. ниже)
626
и открывается в мочеиспускательный канал. В отличие от семявы-
носящего протока семявыбрасывающий проток не имеет столь
выраженной мышечной оболочки. Наружная оболочка его срастает-
ся с соединительнотканной стромой простаты.
Васкуляризация. Трофика семенника обеспечивается кровью,
которая поступает в него через ветвь внутренней семенной артерии,
входящей в состав семенного канатика в средостение, где она
разветвляется на сеть капилляров, проникающих по соединительно-
тканным перегородкам внутрь долек и оплетающих петли извитых
семенных канальцев. Вокруг этих капилляров скапливаются интер-
стициальные клетки.
Лимфатические капилляры также образуют сеть между каналь-
цами семенника, а далее образуют выносящие лимфатические со-
суды.
Иннервация. Нервные волокна, как симпатические, так и пара-
симпатические, проникают в семенник вместе с кровеносными сосу-
дами. В паренхиме семенника рассеяны многочисленные чувстви-
тельные нервные окончания. Нервные импульсы, поступающие в
семенник, способны оказывать некоторое влияние на его генера-
тивную и эндокринную функции, но основная регуляция его дея-
тельности осуществляется гуморальными влияниями гонадотроп-
ных гормонов аденогипофиза.
Возрастные изменения. Генеративная функция семенника
начинается еще в препубертатном возрасте, но в этот период
сперматогенез останавливается в начальных стадиях. Полное
завершение сперматогенеза (образование сперматозоидов) насту-
пает лишь после достижения половой зрелости — пубертатного
периода. У новорожденного семенные канальцы еще имеют вид
сплошных клеточных тяжей, состоящих из поддерживающих клеток
и сперматогоний. Такое строение семенные канальцы сохраняют
в течение первых 4 лет постнатального периода развития мальчи-
ка. Просвет в семенных канальцах появляется лишь к 7—8 годам
жизни. В это время количество сперматогоний значительно увели-
чивается, а около 9 лет среди них появляются одиночные спермато-
циты 1-го порядка, что указывает на начало второй стадии сперма-
тогенеза— стадии роста. Между 10 и 15 годами семенные
канальцы становятся извитыми: в их просветах обнаруживаются
сперматоциты 1-го и 2-го порядка и даже сперматиды, а поддержи-
вающие клетки достигают полной зрелости. К 12-—14 годам заметно
усиливается рост и развитие выводящих протоков и придатка
семенника, что свидетельствует о поступлении в циркуляцию муж-
ского полового гормона в достаточно высокой концентрации. В со-
ответствии с этим в семенниках отмечается большое количество
крупных интерстициальных клеток. Возрастная инволюция семен-
ника у мужчин происходит между 50 и 80 годами. Она проявляется
в нарастающем ослаблении сперматогенеза, разрастании соедини-
тельной ткани. Однако даже в пожилом возрасте в некоторых семен-
ных канальцах сохраняется сперматогенез и их строение остается
нормальным.
627
Параллельно прогрессирующей атрофии эпителиосперматоген-
ного слоя увеличивается деструкция гландулоцитов, вследствие чего
ослабевает продукция мужского полового гормона, а это в свою
очередь оказывается причиной возрастной атрофии предстательной
железы и частично наружных половых органов (см. ниже). С воз-
растом в цитоплазме интерстициальных клеток начинает отклады-
ваться пигмент.
Добавочные железы мужской поповой системы
К ним относят: семенные пузырьки, предстательную железу,
бульбоуретральные железы.
Семенные пузырьки
Семенные пузырьки развиваются как выпячивания стенки
семявыносящего протока в его дистальной (верхней) части. Это
| парные железистые органы, вырабатывающие жидкий слизистый
секрет, слабощелочной 'реакции богатый фруктозой, который
примешивается к сперме и разжижает ее. В стенке пузырьков
имеются оболочки, границы между которыми выражены нечетко:
слизистая, мышечная, адвентициальная (рис. 258)'. Слизистая
оболочка собрана в многочисленные разветвленные складки,
местами срастающиеся между собой, вследствие чего она приоб-
1ретает ячеистьТи вид. Слизистая оболочка покрыта однослойным
столбчатым эпителием, лежащим на тонкой базальной мембране.
* В собственной пластинке слизистой оболочки много эластических
волокон. В слизистой оболочке расположены терминальные отде-
лы желез альвеолярного типа, состоящих из слизистых экзокри-
ноцитов (exocrinocytus mucosus).
Мышечная оболочка хорошо выражена и состоит из
двух слоев гладких мышечных клеток — внутреннего циркулярного
и наружного продольного. Адвентициальная оболочка
состоит из плотной волокнистой соединительной ткани с большим
содержанием эластических волокон.
Рис. 258. Семенной пу-
зырек.
/ — слизистая оболочка;
// — мышечная оболоч
ка; /// — наружная сое-
динительнотканная обо-
лочка; / — складки сли-
зистой оболочки; 2 — се-
крет в просвете железы.
628
Предстательная железа
Предстательная железа, или простата, — мышечно-железистый
орган, охватывающий верхнюю часть мочеиспускательного канала
(уретры), в которую открываются протоки многочисленных проста-
тических желез.
Развитие. У зародыша человека оно начинается на 11 — 12-й не-
деле эмбриогенеза, при этом из эпителия уретры в окружающую
мезенхиму врастают 5—6 тяжей. В первой половине пренатального
эмбриогенеза человека из разрастающихся эпителиальных тяжей
развиваются преимущественно альвеолярно-трубчатые простати-
ческие железы, а со второй половины преобладает рост гладкой
мышечной ткани и соединительнотканных прослоек предстательной
железы. Просветы в эпителиальных тяжах появляются в конце
предплодного периода развития зародыша. Обособленно от указан-
ных желез из эпителия уретры возникают небольшие железы,
располагающиеся между предстательной маточкой и семявыбрасы-
вающим протоком.
Строение. Предстательная железа (простата) — дольчатая
железа, покрытая тонкой соединительнотканной капсулой. Ее па-
ренхима состоит из многочисленных отдельных слизистых желез,
выводные протоки которых открываются в мочеиспускательный
канал. Железы располагаются вокруг мочеиспускательного канала
тремя группами.
Самые мелкие железы лежат в составе слизистой оболочки
непосредственно вокруг мочеиспускательного канала. Вторая груп-
па в виде кольца залегает в соединительной ткани, непосредст-
венно окружающей мочеиспускательный канал, а третья группа
главных собственно предстательных желез занимает всю осталь-
ную, большую часть органа. Концевые отделы альвеолярно-труб-
чатых~"предстательных желез обраюнжны—выеоКимм__£лизистыми
экзокриноцшами (exocrinocytus mucosus), между основаниями
которых располагаются_д1едюде вставочные клетки (рис. 259).
Выводные протоки перед впадением в уретру расширяются в виде
аШтуТГНеправильной формы, выстланных многорядным призмати-
ческим эпителием. Мыщечно-эластическую строму железы (stroma
myoelasticum) образуют рыхлая волокнистая соединительная ткань
и мощные пучки "гладких мышечных.кл.еток, радиально расходя-
щиеся от центра предстательной железы и разделяющие ее на
дольки. Каждая долька и каждая железа окружены продольными и
циркулярными слоями гладких мьцдеяных клеток, которые, сокра-
щаясь, выбрасЬгватоТ'-секрет из предстательных желез в момент
эякуляции.
В месте впадения семявыносящих каналов в мочеиспускатель-
ный канал в предстательной железе расположен семенной бугорок
(colliculus seminalis). С поверхности он выстлан переходным эпите-
лием, а его основу составляют соединительная ткань, богатая элас-
тическими волокнами, и гладкие мышечные клетки. Благодаря нали-
чию многочисленных нервных окончаний семенной бугорок отли-
чается наибольшей чувствительностью. Возбуждение семенного
629
Рис. 259. Предстательная
железа.
/ — микрофотография (по
В. В. Яглову, М. Н. Кашири-
ной): 1 — концевые отделы
желез; 2 — гладкие миоци-
ты и соединительнотканная
строма.
бугорка вызывает его эрекцию, благодаря чему предотвращается
попадание эякулята в мочевой пузырь.
Позади семенного бугорка располагается предстательная маточ-
ка (utriculus prostaticus), открывающаяся на поверхность семенного
бугорка.
Функции предстательной железы многообразны. Вырабаты-
ваемый простатой секрет, выбрасываемый во время эякуляции,
разбавляет сперму, которая имеет очень густую консистенцию при
выходе из семявыносящего канала. По-видимому, предстательная
железа обладает не только внешней, но и внутренней секрецией.
Эта железа находится в зависимости от тестостерона семенников
и атрофируется после кастрации. Удаление предстательной железы
в свою очередь ослабляет в семенниках сперматогенез и продукцию
тестостерона. Кроме того, эта железа оказывает влияние на половую
дифференцировку гипоталамуса (участвует в предопределении его
дифференцировки по мужскому типу), а также вырабатывает
фактор, стимулирующий рост нервных волокон.
Возрастные изменения. Предстательная железа в течение всей
жизни человека претерпевает возрастную перестройку, связанную
со снижением образования половых гормонов и проявляющуюся
сдвигами в соотношении между железистым эпителием, соедини-
тельнотканными и гладкомышечными клетками этого органа.
Секреторные отделы предстательной железы ребенка имеют
эпителий, состоящий из двух видов клеток — высоких и низких
эпителиоцитов (см. рис. 259). Соединительная ткань образует
массивные пучки по ходу выводных протоков и значительно уплот-
няется вокруг секреторных отделов. В ней преобладают фибро-
бласты, макрофаги и коллагеновые волокна. Сравнительно мало в
строме гладких мышечных клеток.
В период полового созревания в цитоплазме железистых клеток
концевых отделов усиливаются секреторные процессы. Эпителий
становится высоким. В период наибольшей функциональной ак-
тивности (в возрасте 20—35 лет) в предстательной железе сек-
реторные элементы преобладают над соединительнотканными,
возрастает синтез гликогена, гликозаминогликанов и гли-
630
Рис. 259 (продолжение).
// схема возрастных изменений
предстательной железы (по Б. В. Тро-
ценко). А срез предстательной же-
лезы ребенка; Б срез предстательной
железы в зрелом возрасте; В срез
предстательной железы в пожилом воз-
расте. 1 концевые отделы желез
2 гладкие миоциты; 3 — фибробла-
сты: 4 — волокна соединительной тка-
ни; 5 — кубические клетки концевых
отделов; 6 базальные эпителиоциты;
7 эпителиальные клетки призмати
ческой формы; 8 — гемокапнлляры: 9 —
конкреции в секреторных отделах пред-
стательной железы (по Б. В. Троценко).
копротеидов. В дальнейшем
(в 35—60 лет) некоторые
железистые дольки начинают
атрофироваться, а соедини-
тельная ткань разрастается и
уплотняется. Железистый
эпителий постепенно стано-
вится низким (см. рис. 259).
В полости секреторных от-
делов образуются и скапли-
ваются предстательные кон-
креции, которые особенно
часто встречаются в старчес
ком возрасте.
Бульбоуретральные
железы
Бульбоуретральные желе-
зы по своему строени' вля-
ются альвеолярно-Г1”3 вчлты-
ми, открывающимися «.воими
протоками в верхней части
мочеиспускательного" канала.
Их концевые отделы и вы-
водные протоки имеют не-
правильную форму. Конце-
вые трубчато-альвеолярные
отделы местами соеди "ются
друг с другом и состоят из
слизистых клеток (exocrino-
cytus mucosus). В расширен-
ных альвеолах этих желез
эпителий бывает чаще всего
уплощенным, в остальных отделах железы — кубическим или
призШТПческим. Клетки эпителия заполнены капёльками~~^утоттда
и своеобразными палочковидными включениями. Между концевы-
631
ми отделами располагаются прослойки рыхлой волокнистой не-
оформленной соединительной ткани, содержащей пучки гладких
мышечных клеток.
Половой член
Половой член — копулятивный орган. Его основная масса обра-
зована тремя пещеристыми (кавернозными) телами, которые,
переполняясь кровью, становятся ригидными и обеспечивают
эрекцию. Снаружи пещеристые тела окружены белочной оболочкой
(tunica albuginea), образованной плотной волокнистой соединитель-
ной тканью. Эта ткань изобилует эластическими волокнами и содер-
жит значительное количество гладких мышечных клеток. В середине
нижнего кавернозного тела проходит мочеиспускательный канал,
по которому осуществляется выделение спермы. Он разделяется на
предстательную часть (pars prostatica), перепончатую часть (pars
membranacea) и губчатую часть (pars spongiosa).
Мочеиспускательный канал имеет хорошо выраженную слизис-
тую оболочку. Ее эпителий в предстательной железе переходный,
в мембранозной части — многорядный призматический, а начиная
с области ладьГЙТтдной -ЯТЯки в губчатой части эпителий уретры
становится многослойным плоским и обнаруживает признаки орого-
вения. В многорядном эпителии встречаются многочисленные бока-
ловидные и немногочисленные эндокринные клетки. Под эпителием
располагается собственная пластинка слизистой оболочки, богатая
эластическими волокнами. В рыхлой волокнистой ткани этого слоя
проходит сеть венозных сосудов, имеющая связь с полостями
кавернозного тела уретры. В слизистой оболочке мочеиспускатель-
ного канала располагаются мелкие слизистые железы. В подсли-
зистой основе имеется сеть ширТП<их-венозных сосудов.
Мышечная оболочка мочеиспускательного канала хорошо разви-
та в его простатической части, где она состоит из внутреннего
продольного и наружного циркулярного слоев гладких миоцитов.
При переходе перепончатой части мочеиспускательного канала в
его пещеристую часть мышечные слои постепенно истончаются и
сохраняются только одиночные пучки мышечных клеток.
Основа головки полового члена состоит из плотной волокнис-
той 'сйедттнйТетьЙби ткани, в которой заложена сеть анастомо-
зирующих вен, переполняющихся ~кровью во время эрекции.
В толстой стенке их располагаются продольные и циркулярно
расположенные пучки гладких мышечных клеток. Кожа, покры-
вающая головку полового члена тонкая. В ней расположены саль-
ные (препуциальные) железы (gll. sebacea preputiales).
Васкуляризация. Артерии, приносящие кровь к кавернозным
телам, имеют толстую мышечную оболочку и широкий просвет. Ар-
терия полового члена, снабжающая его кровью распадается на не-
сколько крупных ветвей, которые проходят по перегородкам пеще-
ристой ткани. При спокойном состоянии полового члена они
спиралевидно закручены и поэтому называются завитыми, или
улитковыми (аа. helicinae). Во внутренней оболочке этих артерий
632
имеются утолщения, состоящие из пучков гладких мышечных
клеток, а также коллагеновых волокон. При сокращении стенок
артерий эти утолщения оказываются своего рода клапанами,
закрывающими просвет сосуда. Вены тоже отличаются толстой
стенкой и хорошо выраженным мышечным слоем во всех трех
оболочках: продольным — во внутренней оболочке, циркуляр-
ным — в средней и продольным — в наружной адвентициальной
оболочке. Сосудистые полости кавернозных тел, сеть которых нахо-
дится между артериями и венами, имеют очень тонкие стенки,
выстланные эндотелием. Кровь из полостей уходит по небольшим
тонкостенным сосудам, которые впадают в глубокие вены. Эти
сосуды играют роль клапанов или шлюзов, так как во время
эрекции стенка вены сокращается и зажимает их просвет, что
препятствует оттоку крови из сосудистых полостей. В сосудистой
системе полового члена обнаружены также типичные артериоло-
венулярные анастомозы.
Иннервация. Симпатические безмиелиновые волокна в половом
члене образуют сплетение, иннервирующее пучки гладких мышеч-
ных клеток в стенках сосудов и в перегородках между сосу-
дистыми полостями пещеристых тел. В коже полового члена и
слизистой оболочке мочеиспускательного канала рассеяны много-
численные рецепторы. Среди них имеются свободные ветвящиеся
окончания, залегающие в эпителии головки полового члена и край-
ней плоти, а также в субэпителиальной ткани.
Особенно многочисленны и разнообразны в тканях полового
члена несвободные инкапсулированные окончания. К ним относят-
ся осязательные тельца в сосочковом слое крайней плоти и голов-
ке полового члена, генитальные тельца, пластинчатые в глубоких
слоях соединительнотканной части полового члена и в белочной
оболочке пещеристых тел.
Гормональная регуляция деятельности мужской пбловои
системы
Обе функции гонад (генеративная и гормонообразовательная)
активируются аденогипофизарными гонадотропинами — ф о л л и -
тропином (фолликулостимулирующий гормон) и л ю т р о -
пином (лютеинизирующий гормон). Фоллитропин влияет пре-
имущественно на эпителиосперматогенный слой, герминативную
функцию семенника, а функции гландулоцитов регулируются
лютропином. Однако в действительности взаимодействия гонадо-
тропинов более сложны. Показано, что регуляция герминатив-
ной функции семенника осуществляется совместным влиянием
фоллитропина и лютропина. Пептидные ингибины угнетают фол-
ликулостимулирующую функцию гипофиза (по механизму отрица-
тельной обратной связи), что приводит к ослаблению влияния,
оказываемого на семенники фоллитропином, но не препятствует
действию на него лютропина. Тем самым ингибин регулирует вза-
имодействие обоих аденогипофизарных гонадотропинов, проявля-
ющееся в регуляции ими деятельности семенника (рис. 260).
633
Рис. 260. Гормональная регуляция сперматогенеза (схема Б. В. Алешина,
К). И. Афанасьева, О. И Бриндака, Н. А. Юриной).
ВМЯ вентромедиальное ядро; АЯ аркуатное ядро; ГЛ— гонадолибернн; ЛГГ -ЛГ-
гонадотропонит; ФСГГ ФСГ-гонадотропоциты; ЛГ лютеинизирующий гормон; ТС
тестостерон; ФСГ — фолликулостимулирующий гормон; ИН ннгибин; АС Б андро-
генсвязывающий белок; / — интерстициальный эндокриноцит; 2 поддерживающий эпи-
телиоцит; 3 — сперматогонии; 4 - сперматоциты; 5 — сперматиды; 6 — сперматозоиды.
ЖЕНСКАЯ ПОЛОВАЯ СИСТЕМА
Женская половая система включает половые железы — яич-
ники и вспомогательные половые органы (яйцево-
ды, матка,, влагалище, наружные половые органы)
Яичники
Яичники (парный орган) выполняют репродуктивну
(образование женских половых клеток) и эндокринну
(выработка половых гормонов) функции.
3 5
634
Развитие. Дифференцировка яичника наступает несколько
позднее, чем семенников, и отчетливо заметна лишь к концу 7—8-й
недели эмбриогенеза.
В эмбриогенезе яичников усиленное развитие мезенхимы про-
исходит в основании тел первичных почек, при этом редуцируются
свободные концы половых шнуров и почечные канальцы, а мезо-
нефральные протоки атрофируются, тогда как парамезонефраль-
ные протоки становятся маточными трубами, концы которых рас-
ширяются в воронки, охватывающие яичники. Нижние части пара-
мезонефральных протоков, сливаясь, дают начало матке и влага-
лищу.
К концу 6-й — началу 7-й недели начинается отделение яични-
ка от мезонефрона и формирование сосудистой ножки яичника —
мезовария fmesbvarium). У 6—8-недельных эмбрионов яичник
представлен корковым веществом, а мозговое вещество развивает-
ся позже. Корковое вещество формируется при врастании половых
шнуров от поверхности эпителия полового валика. Между половы-
ми шнурами~постепенно прорастает мезенхима, расчленяющая
их на отдельные островки — первичные фолликулы. В результате
активного размножения овогоний в эмбриогенезе, особенно на
—4-м месяце развития, количество половых клеток прогрессивно
увеличивается. Около половины овогоний с 3-го месяца развития
начинает дифференцироваться в овоцит первого порядка (период
малого роста), находящийся в профазе мейоза (лептотен-
ная стадия). На этой стадии клетка сохраняется до периода
полового созревания, когда происходит завершение всех фаз
мёиоза (период большого роста). К моменту рождения
число овогоний прогрессивно уменьшается и составляет около
4—5%, большая часть клеток разрушается, основные клетки
представляют собой вступившие в период роста овоциты 1-го
порядка. Общее число половых клеток к моменту рождения со-
ставляет около 300 000—400 000. У новорожденной девочки еще
продолжается процесс врастания половых шнуров от поверхно-
стного эпителия, который прекращается к концу первого года
жизни (после формирования соединительнотканной оболочки
яичников). Мозговое вещество развивается из первичной почки
(разрастающейся мезенхимы и кровеносных сосудов мезовария).
Эндокринная функция яичников начинает проявляться при дости-
жении женским организмом половой зрелости. Первичный малый
рост фолликулов не зависит от гормонов гипофиза, большой рост
стимулируется ФСГ аденогипофиза.
Строение яичника взрослой женщины. С поверхности орган
окружен белочной оболочкой (tunica albuginea), образованной
плотной волокнистой соединительной тканью, покрытой мезотели-
ем (рис. 261). Свободная поверхность мезотелия снабжена микро-
ворсинками. В цитоплазме определяются умеренно развитая гра-
нулярная эндоплазматическая сеть, митохондрии и другие орга-
неллы. Под оболочкой располагается корковое вещество, а глуб-
же — мозговое вещество.
Рис. 261. Схема строения яичника (рис. Ю. И. Афанасьева).
/ — примордиальные фолликулы в корковом веществе: 2 — растущий фолликул; 3 — сое-
динительнотканная оболочка фолликула; 4 — фолликулярная жидкость; 5 — зрелый
фолликул; 6 — яйценосный бугорок; 7 — желтое тело; 8 — интерстициальная ткань;
9— белое тело; 10— атретический фолликул; II— поверхностный эпителий: 12 — бе-
лочная оболочка; 13 — кровеносные сосуды в мозговом веществе яичника.
Корковое вещество (cortex ovarii) образовано так
называемыми фолликулами различной степени зрелости, распо-
ложенными в соединительнотканной строме. Примордиальные
фолликулы состоят из овоцита в диплотене профазы мейоза,
окруженного одним слоем плоских клеток фолликулярного элите -
636
1 , <- г
лия и базальной мембраной (рис. 262). Ядра клеток удлиненные,
с инвагинациями. По мере роста фолликулов увеличивается раз-
мер самой половой клетки. Вокруг цитолеммы появляется вторич-
ная, прозрачная оболочка, снаружи от которой располагаются
в 1—2 слоя кубические фолликулярные-кле-тки на базальной мем-
бране. В цитоплазме этих клеток на стороне, обращенной к яйце-
клетке, хорошо развиты комплекс Гольджи с секреторными вклю-
чениями, рибосомы и полирибосомы. На поверхности клеток
видны два вида микроворсинок: одни проникают в прозрачную
оболочку, а другие обеспечивают контакт между фолликулоцита-
ми. Подобные микроворсинки имеются и на цитолемме овоцита.
В период деления созревания микроворсинки укорачиваются и
даже исчезают. Такие фолликулы, состоящие из растущего овоци-
та, формирующейся прозрачной оболе 1ки и нескольких слоев
фолликулярного эпителия, называются пе^рт^ыми^фоллику.шми.
Характерной особенностью этих фолликулой является образо-
вание прозрачной т>болочки, которая состоит из мукопротеинов и
гликозаминогликанов, секретируемых как овоцитом, так и фолли-
кулярным эпителием. "В неокрашенном виде она выглядит проз-
рачной, блестящей, поэтому и получила свое название zona pellu-
cida. По мере увеличения растущего фолликула окружающая его
соединительная ткань уплотняется, давая начало внешней оболоч-
ке фолликула (theca folliculi).
Дальнейший рост фодликула обусловлен разрастанием одно-
слойного фолликулярного эпителия и превращением его в много-
слойный эпителий, секретирующий фолликулярную жидкость
(liqor folliculi), содержащую стероидные гормоны (эстрогены).
При этом овоцит с окружающими его вторичной оболочкой и
фолликулярными клетками в виде яйценосного бугорка (cumulus
oophorus) смещается к одному полюсу фолликула. В дальнейшем
в наружную оболочку врастают многсТчйсленИЫе кровеносные
капилляры и она дифференцируется на два слоя — внутренний
и наружный. Во внутренней теке (theca interna) вокруг развет-
вляющихся капилляров располагаются многочисленные интерсти-
циальные клетки, соответствующие интерстициальным клеткам
семенника (гландулоцитам). Наружная теки (theca folliculi exter-
na) образована плотной соединительной тканью.
Клетки фолликулярного эпителия, усиленно размножаясь,
окружают овоцит многослойным эпителием, который вырабатыва-
ет фолликулярную жидкость, накапливающуюся между фоллику-
лярными клетками. Такие фолликулы называются вторичными
(folliculus secundorius). Овоцит в этом,фолликуле Уже не увеличи-
вается в объеме, хотя сами фолликулы за счет накопления в их
полостях фолликулярной жидкости резко увеличиваются. При
этом овоцит с окружающим его слоем фолликулярных клеток,
который называется луцретым венцом (corona radiata), оттесняет-
ся к верхнему, полюсу растущего фолликула. Зрелый фолликул,
достигший своего максимального развития, заполненный фоллику-
лярной жидкостью, называется третичным, или пузырчатым фол-
617
Рис. 262. Строение фолли-
кулов, овоцитов и желтого
тела яичника.
А — примордиальные фол-
ликулы (окраска гематокси-
лин-эозином): / овоцнты
1-го порядка (по В В Яг-
лову, Л. К. Линденберг,
М. Н. Каширннои). Б —
гликопротеины блестящей
оболочки. ШИ К-реакция, ок-
раска гематоксилином (по
Ю. И. Афанасьеву, П.А. Мо-
та вкину. М. Г. Шубнчу,
А. Н. Я цк о вс ком у, К. К. Ро-
га минской): / ядро с яд-
рышком; 2 — цитоплазма с
равномерно распределении
ми желтковыми включения-
ми; J блестящая оболоч-
ка; 4 фолликулярные
клетки.
лцкулом (folliculus ovaricus tertiarius). Клетки лучистого венца,
непосредственно окружающие растущий овоцит, имеют длинные
ветвистые отростки, проникающие’Через блестящую зону и дости-
гающие поверхности овоцита По этим отросткам к овоциту от
фолликулярных клеток поступают питательные_вещества, из'йттто-
’’ рых в цитоплазме синтезируются липопротеиды желтка, а также
другие вещества.
Пузырчатый фолликул достигает такого размера, что выпячи-
вает поверхность яичника, причем яйценосный бугорок с овоцитом
оказывается в выступающей части пузырька. Дальнейшее увели-
чение объема пузырька, переполненного фолликулярной жид-
костью, приводит к растягиванию и истончению как его наруж-
ной оболочки, так и белочной оболочки яичника в месте прилега-
ния этого пузырька с последующим раз ывом и овуляцией (см.
ниже).
Между фолликулами встречаются атретические тела (corpus
atreticum). Они формируются из прекративших свое развитие
на разных стадиях фолликулов (см. с. 637).
Мозговое вещество (medulla ovarii) состоит из сое-
638
в
Рис. 262 (продолжение).
В — преовуляторный фолли-
кул (окраска гематоксилин-
эозином); / — яйцеклетка,
свободно плавающая в поло-
сти фолликула; 2 — истон-
ченная стенка фолликула
(по О. В. Волковой). Г —
желтое тело (окраска гема-
токсилин-эозином) I — лю-
теиновые клетки на разных
стадиях дифференцировки;
2 — гемокапнлляры (по
Т. Н. Боровой).
динительной ткани, в которой проходят магистральные кровенос-
ные-сосуды и нервы, эпителиальные тяжи — остатки канальцев
первичной почки.
Генеративная функция. Овогенез
Овогенез протекает аналогично сперматогенезу, но с некото-
рыми особенностями. Так, первая стадия — период размно-
жения оогониев — осуществляется в период_ внутриутробного
развития, а у некоторых видов млекопитающих и в первые месяцы
постнатальной жизни, когда в яичнике зародыша происходит
деление _ оогониев - и формирование первичных фолликулов
(рис. 263). Вторая стадия — период роста — протекает в
функционирующем яичнике и состоит в превращении овоцита 1-го
порядка первичного фолликула в овоцит 1-го порядка.,в зрелом
фолликуле В ядре растущего овоцита происходят конъюгация
хромосом и образование тетрад, а в их цитоплазме накапливаются
желточные включения. Третья стадия — период созревания —
заканчивается образованием овоцита 2-го порядка и завершается
639
1-
2-
3-
4-
А
Рис. 263. Овогенез в антенатальном периоде развития (по Л. Ф. Курило).
Л1
hi
вс
Pl
Д1
А — схема стадий овогенеза: I — 6-7 нед; II — 9—10 нед; III — 12—13 нед; IV — 16 —
17 нед; V — 27—28 нед: VI — 38—40 нед.
/ — оогонии в интерфазе; 2 — оогонии в митозе; 3 — ооцит на стадии прелептотенной
конденсации хромосом; 4 — ооцит на стадии прелептотенной декомпенсации хромосом;
5 — ооцит в лептотене; 6 — ооцит в зиготене; 7 — ооцит в пахите не; 8 — ооцит в дипло-
тене; 9 ооцит в диктиотене; 10------островки половых клеток на границе коркового и
мозгового вещества; II примордиальиый фолликул; 12 первичиый фолликул;
!3 — покровный эпителий, 14 — белочная оболочка яичника; 15 — тяжи соединительной
ткани.
В<|
ОК
че
КС
НС
НУ
ш
at|
на
выходов зрелой яйцеклетки из яичника в результате овуляции. Пе-
риод созревания, как и во время сперматогенеза, включает два деле-
ния, причем второе следует за первым без интеркинеза, что при-
водит к уменьшению (редукции) числа хромосом вдвое, и набор
их становится гаплоидным. При первом делении созревания ово-
цит 1-го порядка делится, в результате чего образуются овоцит
2-го порядка и небольшое редукционное тельце. Овоцит 2-го по-
рядка получает почти всю массу накопленного желтка и поэтому
остается столь же крупным по объему, как и овоцит 1-го порядка.
Редукционное же тельце представляет собой мелкую клетку с не-
мо
631
Рис. 263 (продолжение) .
Б — схема ультраструктуры женских половых кчеток дофолликулярных стадий оогенеза
у плодов человека; / — гоноцит; II — оогонии в интерфазе; III — ооцит в прелептотенной
декомпенсации хромосом; IV — ооцит в лептотене; V — ооцит в зиготене; VI — ооцит в
па хите не: I — ядрышко; 2а хроматин; 26 — хромосомы; 3 — пери хроматиновые 1 ра-
нулы; 4 — сферы 90—120 им; 5 — скопления интерхроматиновых гранул; 6 синапто-
немный комплекс; 7 — элементарные хромосомные нити; 8 — рибосомы; 9 — митохондрии;
10 — эндоплазматическая сеть; II — комплекс Гольджи; 12 — ядерная оболочка.
21 ГИСТОЛОГИЯ
большим количеством цитоплазмы, получающую по одной диаде
от ”к"аждой тетрады ядра овоцита 1 -го порядка. При втором деле-
нии созревания в результате деления овоцита 2-го порядка обра-
зуются одна яйцеклетка и второе редукционное тельце. Первое
редукционное тельце иногда тоже делится на две одинаковые
мелкие клетки. В результате этих преобразований овоцита 1-го
порядка образуются одна яйцеклетка и три редукционных тельца.
Четвертая стадия — формирование — в овогенезе от-
сутствует.
Гоноциты, мигрирующие из первичной эктодермы через энто-
дерму желточного мешка в половые валики, трансформируются
при половой дифференцировке гонад в оогонии в яичниках. По
мере прохождения Периода размножения, после многократного
деления митозом оогонии (см. рис. 263) вступают в следующий
этап дифференцировки половой клетки — ооцит 1-го порядка,
в котором происходят важные биологические явления, специфи-
ческие для половых клеток — конъюгация гомологических ро
дительских хромосом и кроссинговер — обмен участками между
хромосомами. Эти процессы происходят в ооцитах 1-го порядка,
находящих'ся в профазе I деления мейоза. В отличие от спермато-
генеза прохождение ооцитами 1 -го порядка стадий профазы 1 мей-
оза у большинства видов млекопитающих и человека имеет место
в антенатальном Периоде. Подобно соматическим клеткам, гоноци-
ты, оогонии и ооциты 1-го порядка на стадиях профазы 1 деления
мейоза-содержат диплоидный набор хромосом. Оогония превра-
щается в ооцит 1-го порядка с момента, когда она заканчива-
ет период размножения и входит в период малого роста.
Морфологические перестройки хромосом и ядер в ооцитах при
их переходе от одной стадии профазы 1 деления мейоза в другую
аналогичны приведенным выше для сперматоцитов. В отличие от
сперматоцитов на стадии диплотены ооциты в диплотене вовле-
каются в формирование фолликуда, Именно после этой стадии
профазы 1 деления мейоза ооциты участвуют в последовательных
стадиях фолликулообразования. Ооциты в диплотене, заключенные
в первичные фолликулы, составляют пул половых клеток, из ко-
торого непрерывно часть их вступает в период большого роста.
В ооцитах^ покинувших пул первичных фолликулов и вступивших
в период б о л ь ш о го р о_с т а, происходит активный синтез р-
и и-РНК и белка, использующихся не только для роста ооцита,
но "главным образом на первых этапах развития дробящегося
эмориона. Лишь некоторые из вступивших в’ рост ооцитов и фол-
ликулов достигают преовуляторного размера, созревают и всту-
пают в метафазу второго деления созревания и могут быть оплодо-
творены. Большинство ооцитов в растущих и созревающих фол-
ликулах в разные периоды своего роста претерпевают атрезию.
Завершающие этапы периода большого роста ооцита и фолликула,
созревания и овуляции происходят циклически и зависят от цик-
лической деятельности системы гипофиз — гипоталамус —
яичники.
642
В начале большого роста фолликулярные клетки, ранее
распТхлагавшйе'СЯПВ'виде одного слоя уплощенных клеток, приобре- i
тают*"призматическую форму, делятся митозами и фолликулярный
эпителий становится многослойным, получая название зернистой
зоны (zona granulosa). Среди фолликулярных клеток преовулятор-
ного фолликула различают «темные» и «светлые» клетки. Однако
происхождение и значение их остается неясным.
Женские половые клегки, так же как и мvжcкиe, в определен-
ной степени отделены от микроокружения гематофолликулярным
барьером, который создает оптимальные условия для метаболизма
овоцитов. Он состоит из соединительной ткани (теки), сосудов
микроциркуляторного русла, базальной мембраны, фолликуляр-
ного эпителия и прозрачной оболочки
Овуляция. Наступление_овуляции — разрыв фолликула и вы-
брос овоцита 2-го- порядка в- брюшную полость—вызывается |
действием лютеинизирующего гормона (лют юпин), когда вы де- w
ленйё его гипофизом резко увеличивается^ Б предовуляторной
стадии происходит выраженная гиперемия- яичника, повышение
проницаемости гемато-фолликулярного барьера с последующим
развитием интерстициального отека, инфильтрацией стенки фол-
ликула сегментоядерными лейкоцитами. Объем фолликула и дав-
ление в нем быстро возрастают, стенка его резко истончается.
В нервных волокнах и терминалях обнаруживается в этот период
наивысшая концентрация катехоламинов. Известную роль в овуля-
ции может играть окситоцин. Перед наступлением овуляции секре-
ция окситоцина увеличивается в ответ на раздражение нервных
окончаний (располагающихся во внутренней оболочке), обуслов-
ленное повышением внутрифолликулярного давления. Кроме того,
истончению и разрыхлению фолликула способствуют протеоли-
тические ферменты, а также взаимодействие гиалуроновой кисло-
ты и гиалуронидазы, находящихся в его оболочке.
Овоцит 2-го порядка, окруженный фолликулярным эпителием,
из брюшной полости попадает в воронку и далее в просвет маточ-
ной трубы. Здесь быстро происходит второе деление созревания
и образуется зрелая яйцеклетка, готовая к оплодотворению.
Желтое тело (corpus luteum). Под влиянием избытка лютей- \/
низцрующего гормона, вызвавшего овуляцию, элементы стенки
лопнувшего зрелого пузырька претерпевают изменения, приводя-
щие к формированию желтого тела — временной добавочной эндо-
кринной Железы в составе яичника. При этом в полость запустев-
шего пузырька изливается кровь из сосудов внутренней оболочки,
целость которых нарушается в момент овуляции. Сгусток крови ,
быстро замещается соединительной тканью в центре развивающе- (
гося желтого тела. В развитии желтого тела рЗ^лйЧаются 4 стадии.
В церв'ои стадий — пролиферации и васкуляриза-
ции — происходит размножение эпителиальных клеток бывшего
зернистого слоя и между ними интенсивно врастают капилляры
из внутренней оболочки. Затем наступает вторая стадия — же-
лезистого метаморфоза, когда клетки фолликулярного
21к
643
эпителия сильно гипертрофируются и в. ПИЛнакапливается жел-
тый _ пигмент (лютеин), принадлежащий к группе липохромов.
Такие клетки называются лютеиновыми (luteocyti). Объем ново-
образующегося желтого тела быстро увеличивается, и оно при-
обретает желтый цвет, "Хорошо различимый—при-ЖИЭПП7~С этого
момента желтое тело начинает продуцировать свой гормон^—
прогестерон. переходя таким образом в третью стадикГ— рас-
цвета. Продолжительность этой стадии_различна. Если оплодо-
творения не произошло, период расцвету желтогЦ-Тела ограни-
чивается 12— 14 днями—В этом случае оно называется менструаль-
ным желтым телом (corpusluteum menstruationis). Более длитель-
но желтое тело сохраняется, если наступила берем нность —
желтое тёлсГЪеременности (corpusTuteum graviditaTiooTs).
Разница между желтым телом беременности и менструальным
ограничивается только^щительностью периода расцвета и размера-
ми 0,5—2 см в диамегреу менструального и более 5 см в диаметре
у желтого тела беременности). После прекращения функциони-
рования как желтое тело беременности, так и менструальное пре-
терпевают- инволюцию (стадию о б р атного развития).
Железистые клетки атрофируются, а соединительная ткань цен-
трального руб ,а_рггфастаётся. В ^результате на месте бывшего
желтого тела ф°РмиРУется белое тело (corpus albicans) — соеди-
* нительнотканный рубец. Оно сохраняется в яичнике на протяже-
нии нескольких лет,-"но затем рассасывается.
^‘Атрезия. Значительное число фолликулов не достигает стадии
^зрелости, а претерпевает а т р е з и ю — своеобразную перестрой-
ку деструктивного характера. Она начинается гибелью овоцита,
у"от которого сохраняется блестящая оболочка, сморщиваюпмяся.
но несколько утолщающаяся и гиалинизирующаяся. Одновременно
атрофируются—и___клетки -зернистого—слоя,—а и те рсти циал ьные
клетки оболочки при этом не только.-не. логиба ют но, наоборот,
’’Ч» усиленно- размножаются и, гипертрофируясь, начинают напоми-
л нать по форме и виду лютеиновые клетки желтого тепа, нахоля-
4 щиеся в расцвете. Так возникает атретическое тело (corpus^atre-
' ticum), внешне несколько напоминающее желтое тело, но отлива-
ющееся от послед не го наличием в центре блестящей обол очки
Тс.
овоцита.
ВЗГоде дальнейшей инволюции атретических тел на их месте
остаются скопления интерстициальных клеток.
Обильная иннервация атретических тел, а также увеличение
содержания рибонуклеопротеидов и липидов в гипертрофирую-
щихся интерстициальных клетках и возрастание активности ряда
ферментов в них, свидетельствующие о повышении метаболизма
этих клеток, указывают на высокую функциональную активность
атрезирующихся фолликулов. Так как интерстициальные клетки
участвуют в выработке овариальных гормонов, следует допустить,
что атрезия, результатом которой является увеличение количества
этих клеток, необходима для гормонообразования в яичнике
женщины.
644
Mr к с
WjA1
Эндокринные функции
В то время как семенник на протяжении своей активной дея-
тельности непрерывно вырабатывает половой гормон, для яичника
характерна циклическая_(поочередная) припугни" эстрогенов
и гормона желтого тела прогестерона.
Эс троге н ы (эстрадиол, эстрон и эстриол) обиаружива-
ются~в~жйД1<0СТИ, накапливающейся в полостях растущих и зрелых
фолликулов. Поэтому эти гормоны ранее .именовались фоллику-
лярным и, или фол ли к у л и_н а м и. Яичник начинает интен-
сивно продуцировать эстрогены при достижении женскиморганиз-
лом половой'зрёдасти,когда устанаю1ЙМ1Отсяполовые_циклы~ кото-
рые у низших млекопитающих проявляются регулярным наступле-
нием течки (oestrus) — выделением пахучей слизи из влагалища.
Поэтому гормоны, под влиянием которых наступает течка, получили
название эстрогенов.
Возрастное затухание деятельности яичников приводит к прек-
ращению половых циклов.
Васкуляризация. Для яичника характерны спиралевидный ход
артерий и вен и их обильное ветвление. Распределение сосудов
в яичнике претерпевает изменения в связи с циклом фолликулов.
В период роста первичных фолликулов в развивающейся внутренней
оболочке формируется сосудистое сплетение, сложность которого
возрастает ко^ времени овуляции и формирования желтого тела.
В последующем по мере обратного развития желтого тела сосуди-
стое сплетение редуцируется. Вены во всех частях яичника связа-
ны многочисленными анастомозами, и емкость венозной сети
значительно превышает емкость артериальной системы.
Иннервация. Нервные волокна, входящие в яичник, как сим-
патические, так и парасимпатические, образуют сети вокруг фол-
ликулов и желтых тел, а также в мозговой части Кроме того, в
яичниках обнаруживаются многочисленные рецепторы, через кото-
рые афферентные сигналы поступают в центральную нервную
систему и достигают гипоталамуса. г
Другие органы женской половой системы
Маточные трубы
Маточные трубы или яйцеводы —.парные органы, связываю-
щие брюшную нолость^ц_матку По нимполовьгёГ ХЛёТки -прохо-
дят в матку.
Развитие. Яйцеводы развиваются из верхней части парамезо-
нефральных протоков.
Строение. Стенка яйцевода имеет три оболочки: слизистую,
мышечную и серозную (рис. 264). Слизистая оболочка яйцеводов
собрана в крупные_разветщ1енные продольные складки. Она покры-
та однослойным призматическим эпителием, который состоит
из двух видов клеток — реснитчатых и железистых, секретирую-
щих слизь. Собственная пластинка слизистой оболочки представ-
лена рыхлой волокнистой соединительной тканью. Мышечная
645
Рис. 264. Маточная труба.
А _строение; поперечный срез (но Савараги, Тонака); 1 складки слизистой
оболочки, покрытой реснитчатым эпителием; 2 — собственная пластинка слизистои
оболочки: 3 — мышечная оболочка: 4 — кровеносный сосуд; 5 серошая оболочка.
£ __ сканирующая электронная микрофотография слизистой оболочки яйцевода,
мерцательные реснички; 2 апикальные поверхности секреторных клеток; 3 — капли
секрета.
оболочка, следующая за слизистой, состоит из внутреннего цирку-
лярного или спирального слоя и наружного продольного. Снаружи
яйцеводы окружены серозной оболочкой.
Проксимальный конец яйцевода расширяется в воронку и за-
канчивается бахромкой (фимбриями). В момент овуляции сосуды
фимбрий яйцеводов увеличиваются в объеме и воронка при этом
плотно охватывает яичник. Передвижение половой клетки по яй-
цеводу обеспечивается не только движением ресничек эпителиаль-
ных клеток, выстилающих полость маточной трубы, но и пери-
стальтическими сокращениями ее мышечной оболочки.
Матка
Матка — мышечный орган, предназначенный для осуществле-
ния внутриутробного развития плода.
Развитие. Матка и влагалище развиваются у зародыша из дис-
тального отдела левого и правого парамезонефральных протоков
в месте их слияния. В связи с этим вначале тело матки обнару-
живает некоторую двурогость, но к 4-му месяцу внутриутробного
развития слияние заканчивается и матка приобретает грушевид-
ную форму.
Строение. Стенка матки состоит из трех оболочек: слизистой
(эндометрий), мышечной (миометрий) и серозной (периметрий)
(рис. 265). В _р н.д о м е т р и и различают два слоя — базальный —-.
и функциональный. Строение функционального (поверхностного)
слоя зависит от овариальных гормонов и претерпевает глубокую
перестройку на протяжении менструального цикла. Слизистая
оболочка матки выстлана однослойным призматическим эпители-
ем. Реснитчатые клетки располагаются преимущественно вокруг
устьев маточных желез. Собственная пластинка слизистой оболо-
чки матки образована рыхлой волокнистой соединительной тканью.
Некоторые клетки соединительной ткани развиваются в дециду-
альные клетки крупного размера и округлой формы, содержащие
в своей цитоплазме глыбки гликогена и липопротеиновые включе- /
ния. Количество децидуальных клеток возрастает (со времени
менструации), особенно при формировании плаценты в период
беременности.
В слизистой оболочке находятся многочисленные маточные
железы,-простирающиеся-ч^реЗ” ВСТО толщу эндометрия и даже
проникающие в поверхностные слои миометрия. По форме маточ- •
ные железы относятся к простым трубчатым.
Миометрий — состоит из трех слоев гладких мышечных
клеток — внутреннего подслизистого (stratum submucosum), сред-
него__сосудистого с косопродольным расположением миоцитов
(stratum vasculosum), богатого сосудами, и наружного надсосуди-
стого (stratum supravasculosum) с косопродольным расположением
мышечных клеток, но перекрестным по отношению к подслизи-
стому слою. Такое расположение мышечных пучков имеет опреде-
ленное значение в регуляции интенсивности циркуляции крови в
течение менструального цикла.
647
Рис. 265. Стенка матки (рис. Ю И
Афанасьева).
подслизистый
6 сосудистый слой
надсосудистый слой
мезотелий; 9 ма
1 эндометрий; // — миометрии; /// —
периметрий: / — простей однослойный
призматический эпителий; 2 — соб-
ственный слой слизистой оболочки; 3 —
маточные железы (крипты); 4 — кро-
веносные сосуды; 5
слой миометрия;
миометрия; 7 —
миометрия; 8 —
точная труба
Между пучками мышечных клеток имеются прослойки соеди-
нительной ткани, Изобилующей эластическими волокнами и гис-
тиоцитами. Гладкие мышечные клетки миометрия длиной около
50 мкм во время беременности сильно гипертрофируются, достигая
иногда в длину 500 мкм. Они слегка ветвятся и соединяются отро-
стками в сеть.
Периметрий покрывает большую часть поверхности
матки. Не покрыты брюшиной лишь передняя и боковые поверх-
ности надвлагалищной части шейки матки. В формировании пери-
метрия принимают участие мезотелий, лежащий на поверхности
органа, и рыхлая соединительная волокнистая ткань, составляю-
щие прослойку, примыкающую к мышечной оболочке матки. Одна-
ко не во всех местах эта прослойка одинакова. Вокруг шейки
матки, особенно с боков и спереди, находится большое скопление
жировой ткани, которое получило название параметрия. В других
участках матки эта часть периметрия образована относительно
тонким слоем рыхлой волокнистой соединительной ткани.
Слизистая оболочка шейки матки покрыта, как и вла-
галище, много£лойным_плоским эпителием. Канал шейки выстлан
призматическим эпителием, который секретирует слизь. Однако
наибольшее количество секрета вырабатывается многочисленными
сравнительно крупными разветвленными железами, располагающи-
648
мися в строме складок слизистой оболочки. Мышечная оболочка
шейки представлена мощным циркулярным слоем гладких мышеч-
ных клеток~составляющим так называемый сфинктер матки, при
сокращении которого выжимается слизь из шеечных желез. При
расслаблении этого мышечного кольца возникает лишь своего
рода аспирация (всасывание), способствующая втягиванию в мат-
ку спермы, попавшей во влагалище.
Васкуляризация. Система кровоснабжения матки хорошо раз-
вита. Артерии, несущие кровь к миометрию и эндомстрик», в цир-
кулярном слое миометрия спиралевидно закручиваются, что спо-
собствует их автоматическому сжатию при сокращении матки.
Особенно важное значение эта особенность приобретает во время
родов, так как предотвращается возможность сильного маточного
кровотечения в связи с отделением плаценты.
Входя в эндометрий, приносящие артерии дают начало мелким
артериям двух видов, одни чз uuv, прямые не выходят за пределы
базального слоя эндометрия^ДРУгие же, спиральные, снабжают
функциональный слой.
Лимфатические сосуды в эндометрии образуют
глубокую сеть, которая через лимфатические сосуды миометрия
соединяется с наружной сетью, располагающейся в периметрии.
Иннервация. Матка получает нервные волокна, преимуществен-
но симпатические, от подчревного сплетения. На поверхности мат-
ки в периметрии эти симпатические волокна образуют хорошо
развитое маточное сплетение. От этого поверхностного сплетения
отходят веточки, снабжающие миометрий и проникающие в
эндометрий. Около шейки матки в окружающей клетчатке распо-
лагается группа крупных ганглиев, в которых, кроме симпатических
нервных клеток, имеются хромаффинные клетки. В толще миомет-
рия ганглионарные клетки отсутствуют. В последнее время получе-
ны данные, свидетельствующие о том, что матка иннервируется
как симпатическими, так и некоторым количеством парасимпати-
ческих волокон. В то же время в эндометрии обнаружено большое
количество рецепторных нервных окончаний различной структуры,
раздражение которых не только вызывает сдвиги в функциональ-
ном состоянии самой матки, но и отражается на многих общих
функциях организма; кровяном давлении, дыхании, общем обмене
веществ, гормонообразовательной деятельности гипофиза и других
эндокринных желез, наконец, на деятельности центральной нер-
вной системы, в частности гипоталамуса.
Влагалище
Стенка влагалища состоит из слизистой, мышечной и адвенти-
циальной оболочек. В составе слизистой оболочки имеется много-
слойный плоский эпителий, в ~ котором различают три слоя:
базальный, промежуточный и поверхностный, или функциональ-
ный (рис. 266); '
Эпителий слизистой оболочки влагалища претерпева-
ет значительные ритмические (циклические) изменения в по-
649
Рис. 266. Влагалище.
/ — многослойный плоский эпителий:
2 — собственный слой слизистой обо-
лочки; 3 подслизистая основа; 4 —
пучки гладкой мышечной ткани.
следовательных фазах мен-
струального цикла. В клетках
поверхностных слоев эпите-
лия (в его функциональном
слое) откладываются зерна
кератогиалина, но полного
ороговения клеток в норме
не происходит. Клетки этого
слоя эпителия богаты глико-
геном. Распад гликогена под
влиянием микробов, всегда
обитающих во влагалище,
приводит к образованию мо-
лочной кислоты, поэтому вла-
галищная слизь имеет кислую
реакцию и обладает бактери-
цидными свойствами, что
предохраняет влагалище от
развития в нем патогенных микроорганизмов. Железы в .стенке
влага пища отсутствуют. Базальная граница эпителия неровная,
так как собственная пластинка слизистой оболочки формирует
сосочки неправильной формы, вдающиеся в эпителиальный пласт.
Основу собственной пластинки слизистой оболочки составляет
рыхлая-волокнистая соединительная ткань, эластические волокна
которой'образуют поверхностную и глубокую сети. Собственная
пластинкгг^час1С_.инфи.тьхрирхется лимфоцитами, иногда в ней J
встречаются единичные лимфатические узелки. Подслизистая ос- ,
нова во влагалище не "Выражена и собственная пластинка сли-
зистой оболочки непосредственно переходит в прослойки соедини-
тельной ткани в мышечной оболочке, которая в осцащ_______
ном состоится? продольно идущих пучков гладких мышечных
клетО'к^ между пучками которых в средней части мышечной обо-
лочки имеется небольшое количество циркулярно расположенных
мышечных элементов.
Адвентициальная оболочка влагалища состоит из
рыхлой волокнистой неоформленной соединительной ткани, свя-
зывающей влагалище с соседними органами. В этой оболочке
располагается венозное сплетение.
Половой цикл
Циклическая деятельность женской половой системы (яичники,
маточные трубы, матка, влагалище), т. е. последовательные изме-
нения ее функции и структуры — о в а р и а ль н о-м е н с т р у-
650
а л ьн ый цикл регулярно повторяется в одном и том же порядке.
У женщин и самок человекообразных обезьян половые циклы
характеризуются регулярными маточными кровотечениями (мен-
струациями). У большинства женщин, достигших половой зрелости,
менструации повторяются регулярно через 28 дней. В овариально-
менструальном цикле различают три периода или фазы: мен-
струальный (фаза десквамации эндометрия), которым закан-
Ьчивается предыдущий менструальный цикл, постменстру-
альный период (фаза пролиферации эндометрия) и,
наконец, предменструальный период (функциональ-
ная фаза, или фаза секреции), во время которого происходит под-
готовка эндометрия к возможной имплантации оплодотворенной
яйцеклетки, если наступило оплодотворение.
Начало менструальной фазы определяется резким изменением
кровоснабжения эндометрия. На протяжении предыдущей пред-
менструальной (функциональной) фазы под влиянием прогестеро-
на, интенсивно секретируемого желтым телом, вступившим в этот
период в стадию расцвета, кровеносные сосуды эндометрия
достигают максимального развития. Прямые артерии дают начало
капиллярам, питающим базальный с^юй Ьндрметрия, а спирале-
видные ~артёрий7'~~разрастающиеся в этой фазе, вкручиваются
I в клубочки и образуют ГУСТУЮ сеть ютпиллярсгь ветвящихся в
' функциональном слое эндометрия. !о мере того как к концу
предменструального периода желтое теле в яичнике начинает
атрофироваться (вступает в стадию обратного развития), .прек-
ращается вступление прогестерона в циркуляцию. Вследствие это-
го начинаются спазмы спиралевидных артерий, результатом чего
оказывается значительное уменьшение притока крови к эндометрию
| (ишемическая фаза) и в нем развивается гипоксия, а вздыудах
возникают тромбы. Стенки сосудов теряют эластичность и стано-
вятся ломкими. На прямые артерии указанные изменения не
распространяются, й базальный слой эндометрия продолжает
снабжаться кровью.
В функциональном слое эндометрия вследствие ишемии начи-
наются некротические изменения. После длительного, £ШЗма
спиралевидные артерии щщдь расширяются и цриток крови к
эндометрию увеличивается. Но так как стенки этих сосудов
стали хрупкими, в них возникают многочисленные разрывы,
и в строме эндометрия начинаются кровоизлияния, образуются
гематомы. Некротизирующийся функциональный слой отторга-
ется, расширенные кровеносные сосуды эндометрия вскрываются
и наступает маточное кровотечение.
!В день менструации в организме женщины практически отсут-
ствует^ овариальные гормоны, так как секреция прогестерона
прекращается, Й гекрепйЯ Эстрогенов (которой ~препя тствовало
t желтое тело, пока оно было в расцвете) еще не возобновилась.
Но, поскольку начавшаяся регресрия_желтого_тела ттастормажи-
вает рост очередного фолликула, продукция эстрогенов стано-
вится возможной. Под цх влиянием в матке активизируется
Рис. 267. Овариально-менструальный цикл (схема).
I — менструальная; II — постменструальиая; 111 предменструальная фаза: 1 — извитая
артерия эндометрия; 2 — прямая артерия эндометрия; 3 спазм и регрессия терминаль-
ных ветвей извитых артерий (ишемическая фаза); 4 — кровоизлияние в эндометрии;
5 — примордиальный фолликул в яичнике; 6 — растущие фолликулы; 7 — зрелый (граа-
фов) фолликул; 8 — овуляция; 9 - желтое тело в стадии расцвета; 10 — обратное разви-
тие желтого тела; 11 — передняя доля гипофиза; 12 — воронка промежуточного мозга;
13 — задняя доля гипофиза; ФСГ — действие фоллитропина на растущие фолликулы;
Л Г — действие лютропина на овуляцию и образование желтых тел; ЛТГ — действие
лактотропина (пролактина) на сформированное желтое тело; Э — действие эстрогена на
матку, стимулирующее рост эндометрия (постменструальная или пролиферативная фаза);
Пг — действие прогестерона на эндометрий (предменструальная фаза).
регенерация эндометрия и усиливаемся пролиферация эпителия
за счет ДОньцдек. маточных желез, которые сохранились в ба-
зальном слое после десквамации функционального слоя. Через
2—3 дня пролиферации менструальное кровотечение останавли-
вается и начинается очередной постменструальный период. Таким
образом, постменструальная фаза определяется влиянием-эстроге-
нов, а предменструальная — влиянием -Прогестерона. Овуляция
наступает в" яичнике на 12—17-й день менструального цикла,
т. е. приблизительно посередине между двумя очередными мен-
струациями. В связи с участием гормонов яичника в регуляции
перестройки, претерпеваемой маткой, изложенный процесс обычно
называют не менструальным, а овариально-менструальным цик-
лом (рис. 267).
Постменструальный период. Этот период начинается вслед за
окончанием менструации (рис. 268). В^ этот момент эндометрий
представлен только базальным слоем, в котором остались диет ль-
ные отделы маточных желез~ Уже начавшаяся регенерация
652
Рис. 268. Строение эндометрия мат-
ки женщины в различные фазы
цикла. Микрофотографии (препара-
ты Ю. И. Ухова).
А — менструальная фаза; Б — пост-
менструальная фаза; В — предмен-
струальная фаза (20-й день менстру-
ального цикла); 1 — маточные же-
лезы (крипты); 2 — собственная пла-
стинка слизистой оболочки; 3 — под-
слизистый слой миометрия
Рис. 269. Эндометрий в фазе пролиферации. Электронная микрофотография.
X 17 000 (препарат Г. В. Космачевой).
/ — апикальная поверхность эпителия эндометрия; 2 микроворсинки; 3 — реснички
эпителиальных клеток.
функционального слоя позволяет назвать данный период п р о-
ли фТГр а т и в н о й ф а з о й (рис. 269). Она продолжается с
5-го по 14—15-й день цикла. Пролиферация регенерирующего
эндометрия наиболее интенсивна в начале данной фазы (5—11-й
день цикла), затем темп регенерации замедляется и наступает
период относительного покоя (11 — 14-й день).
Маточные железы в постменструальном периоде растут быстро,
но остаются узкими, прямыми и не секретируют.
Как уж.е упоминалось, рост эндометрия стимулируется эстро-
генами, которые продуцируются растущими фолликулами. Следо-
вательно^—во —Время лостменструальццго периода В яичнике про-
исходит рост очередного—фолликула, который достигает стадии
зрелого (третичного, или пузырчатого) к 14-му дню цикла.
Предменструальный период. В конце постменстпуального-.пери-
ода в_ яичнике наступает овуляция, а на месте лопнувшего пу-
зырчатого фолликула образуется желтое тело, вырабатывающее
прогестерон, который активирует маточные железы, начинающие
рекретир'бвЗ п.. 'Они —увеличиваются в размерах, -становятся
извитыми и часто разветвляются. Их клетки_набухают, а просветы
желез заполняются выделяемым декретом. В цитоплазме появ-
ляются вакуоли, содержащие гликоген и гликопротеиды — сна-
чала в базальной части, а затем смещающиеся к апикальному
краю Слизь, обильно выделяемая железами, становится густой
В участках эпителия, выстилающего полость матки между усть-
ями маточных желез, клетки приобретают призматическую фор-
му, а на верхушках многих из них азвивают<яi реснички. Толщи-
нТГэндометрия увеличивается по сравнению с предыдущим пост-
менструальным периодом, что обусловливается гиперемией и
накоплением отечной жидкости в tunica propria. В клетках сое-
динительнотканной стромы тоже откладываются глыбки гликогена
и капельки липидов. Некоторые из этих клеток дифференцируются
в децидуальные (см. «Плацента» в главе V).
Есди произошло оплодотворение, то эндометрий участвует в
формировании плаценты. Вели же оплодотворение не состоялось,
то функциональный слой эндометрия разрушается и отторгается
при очередной менструации.
Циклические изменения влагалища. С началом пролиферации
эндометрия (на 4—5-й день после окончания менструации),
т. е. в постменструальном периоде, во влагалище заметно набу-
хают эпителиальные клетки. На 7—8-й день в этом эпителии
дифференцируется промежуточный слой уплотненных клеток, а к
12—14-му дню цикла (к концу постменструального периода) в
базальном слое.эпителия клетки сильно набухают и увеличиваются
в .объеме. В верхнем (функциональном) слое влагалищного эпите-
лия клетки'"разрыхляются и в них накапливаются глыбки
кератогиалина. Однако процесс кератинизации не доходит до
полного ороговения.
В предменструальном периоде деформированные уплотненные
клетки функционального слоя влагалищного эпителия продолжают
отторгаться, а клетки базального слоя уплотняются.
Состояние эпителия влагалища зависит от уровня овариальных
гормонов в крови, поэтому по картине мазка, полученного с
поверхности влагалища, можно судить о фазе менструального
цикла и о его нарушениях. Во время менструации в мазке преобла-
дают эритроциты и нейтрофилы, эпителиальные клетки встреча-
ются в небольшом количестве. В начаде._достменструального
периода (в пролиферативной фазе'цикла) влагалищный эпителий
сравнительно тонок, а в мазке содержание лейкоцитов быстро
уменьшается и появляются эпителиоциты с пикнотическими ядра-
ми. К моменту овуляции (в середине овариально-менструального
цикла) такие клетки в мазке становятся преобладающими, а
толщина влагалищного эпителия возрастает. Наконец, в предмен-
струальной фазе цикла количество 'клеток с пикнотическим
ядром уменьшается, но усиливается десквамация нижележащих
слоев, клетки которых обнаруживаются в мазке. Перед наступле-
нием менструации содержание эритроцитов в мазке начинает воз-
растать.
Возрастные изменения органов женской половой системы
Морфофункциональное состояние органов женской половой
системы зависит от возраста и активности нейроэндокринной
системы.
65 е
Матка. У новорожденной девочки длина матки не превы-
шает 3 см и, постепенно увеличиваясь в течение препубертатного
периода, достигает конечных размеров по достижении половой
зрелости.
К концу детородного периода и в связи с приближением
климакса, когда гормонообразовательная деятельность яичников
ослабевает, в матке начинаются инволютивные изменения, прежде
всего в эндометрии. Дефицит лютей низирующиего гормона в
переходном (преклимактерическом) периоде проявляется тем, что
маточные железы, сохраняя еще способность к росту, уже пере-
стают функционировать. После установления менопаузы атрофия
эндометрия быстро прогрессирует, особенно в функциональном слое.
Параллельно в миометрии развивается атрофия мышечных клеток,
сопровождающаяся гиперплазией соединительной ткани. В связи с
этим размеры и масса матки, претерпевающей возрастную инво-
люцию, значительно уменьшаются. Наступление климактеричес-
кого периода характеризуется уменьшением размеров органа и
количества миоцитов в нем, а в кровеносных сосудах возникают
склеротические изменения. Это является следствием снижения гор-
монообразования в яичниках.
Яичники. В первые годы жизни размеры яичников у
девочки увеличиваются преимущественно за счет роста мозговой
части. Атрезия фолликулов, прогрессирующая в детском возрасте,
сопровождается разрастанием соединительной ткани, а после
30 лет разрастание соединительной ткани захватывает и корковое
вещество яичника.
Затухание менструального цикла в климактерическом периоде
характеризуется уменьшением размеров яичников и количества
фолликулов в них, склеротическими изменениями их кровеносных
сосудов. Вследствие недостаточной продукции лютропина овуляции
и образования желтых тел не происходит и поэтому овариально-
менструальные циклы сначала становятся ан овуляторным и,
а затем прекращаются и наступает менопауза.
Влагалище. Морфогенетические и гистогенетические про-
цессы, приводящие к формированию основных структурных эле-
ментов органа, завершаются к периоду полового созревания.
После наступления климактерического периода влагалище пре-
терпевает атрофические изменения, его просвет суживается,
складки слизистой оболочки сглаживаются, количество влагалищ-
ной слизи уменьшается. Слизистая оболочка редуцируется до 4—5
слоев клеток, не содержащих гликогена. Эти изменения создают
условия для развития инфекции (сенильный вагинит).
Гормональная регуляция женской половой системы
Как упоминалось, фолликулы начинают расти еще в яичниках
зародыша. Первичный рост фолликулов («малый рост») в яични-
ках зародыша не зависит от гормонов гипофиза и приводит
к возникновению фолликулов с небольшой полостью. Для даль-
нейшего роста («большого роста») фолликулов необходимо
656
стимулирующее влияние аденогипофизарного фрдлитгюпи.на на
выработку клетками фолликулярного эпителия (zona granulosa)
эстрогенов и добавочное влияние небольших количеств лютропина,
который активирует интерстициальные клетки (theca interna).
К окончанию роста фолликула возрастающее содержание лютро-
пина в крови вызывает овуляцию и образование желтого тела.
Фаза “же расцвета желтого тела, в течение которой оно продуциру-
ет и секретируеТ" прогестерон, усиливается и удлиняется благо-
даря добавочному влиянию аденогипофизарного пролактина.
Местом приложения прогестерона является слизистая оболочка
матки, которая под его влиянием подготавливается к восприятию
оплодотворенной яйцевой клетки (зиготы). В то же время про-
гестерон ингибирует рост новых фолликулов. Наряду с выработкой
прогестерона в желтом теле сохраняется в слабой степени выра-
ботка эстрогенов. Поэтому в конце фазы расцвета желтого тела
вновь отмечается поступление небольших количеств эстрогенов
в циркуляцию.
Наконец, в фолликулярной^жидкости растущих фолликулов и
зредых (пузырчатцх) фолликулов наряду с эстрогенами обнару-
живается еще и белковый гормон гонадокривин (по-
видимому, идентичный ингибину семенников), который угнетает
рост овоцитов и их созревание. Гонадокринин, как и эстрогены,
вырабатывается клетками зернистого слоя. Предполагается, что
гонадокринин, действуя непосредственно на другие фолликулы,
вызывает в них гибель овоцита и дальнейшую атрезию этого
фолликула. Атрезию следует рассматривать как предотвращение
образования избыточного количества яйцеклеток (т. е. суперовуля-
ции). Если же овуляция зрелого фолликула по каким-либо при-
чинам не наступит, то выработанный в нем гонадокринин обес-
печит его атрезию и ликвидацию.
Половая дифференцировка гипоталамуса. Непрерывность муж-
ской половой функции и цикличность женской связаны с особен-
ностями секреции лютропина гипофизом. В мужском организме
как фоллитропин, так и лютропин секретируются одновременно
и равномерно. Цикличность же женской половой функции обус-
ловливается тем, что выделение лютропина из гипофиза в цирку-
ляцию происходит не равномерно, а периодически, когда гипофиз
выбрасывает в кровь повышенные количества этого гормона, дос-
таточные для вызова овуляции и развития желтого тела в яичнике
(так называемая овуляционная квота лютропина). Гормонопоэти-
ческие функции аденогипофиза регулируются аденогипофизотроп-
ными нейрогормонами медиобазального гипоталамуса.
Гипоталамическая регуляция лютеинизирующей функции пе-
редней доли гипофиза осуществляется двумя центрами. Один из
них («низший» центр), находящийся в туберальных ядрах (арку-
атном и вентромедиальном) медиобазального гипоталамуса, акти-
вирует переднюю долю гипофиза к непрерывной тонической
секреции обоих гонадотропинов. При этом количество выделяемо-
го лютропина обеспечивает лишь секрецию эстрогенов яичниками
стостерона семенниками, но слишком мало для вызова овуля-
и и образования желтого тела в яичнике. Другой центр («выс-
лий», или «овуляторный») локализуется в преоптической области
медиобазального гипоталамуса и модулирует деятельность низше-
го центра, в результате чего последний активирует гипофиз к
массивному выбросу «овуляторной квоты» лютропина.
При отсутствии влияния андрогена преоптический овуляторный
центр сохраняет способность периодически возбуждать деятель-
ность «низшего центра», как это свойственно женскому полу.
Но у зародыша мужского пола благодаря наличию в его организме
мужского полового гормона данный овуляторный центр гипотала-
муса маскулинизируется. Критический период, после которого
овуляторный центр теряет способность модифицироваться по
мужскому типу и окончательно закрепляется в качестве женского,
ограничивается у человеческого плода концом внутриутробного
периода.
Наружные половые органы
Преддверие влагалища выстлано многослойным
плоским эпителием.- В' преддверие влагалища открываются две
железы преддверия (бартолиновы железы). По форме эти железы
альвеолярно-трубчатые, образованы призматическим эпителием,
секретируют слизь. В малых губах покрывающий их многослой-
ный эпителий слегка ороговевает, а базальный слой его пигменти-
рован. Основу малых губ составляет рыхлая волокнистая соедини-
тельная гканы-богатая эластическими волокнами и кровеносными
сосудами. Вней заложены многочисленные сальные железы.
Большие губы влагалища представляют собой
складки кожи с обильными прослойками жировой ткани. В боль-
ших губах много сальных и потовых желез.
Клитор по эмбриональному развитию и строению соответ-
ствует дорсальной части мужского полового члена. Он состоит
из двух эректильных пещеристых тел, заканчивающихся голов-
кой, которая покрыта многослойным плоским эпителием, слегка
ороговевающим.
Иннервация. Наружные половые органы, особенно клитор,
обильно снабжены различными рецепторами. В эпителии этих
органов разветвляются свободные нервные окончания. В соеди-
нительнотканных сосочках собственной пластинки их слизистой
оболочки находятся осязательные нервные тельца, а в дерме на-
ходятся инкапсулированные генитальные тельца. В больших губах
и клиторе встречаются также пластинчатые тельца.
Молочные железы
Молочные железы по своему происхождению представляют
видоизмененные лджные-потовые железы Но по своему функци-
ональному значению они относятся к женской репродуктивной
системе, так как__рбеспечивают первоначальное. питание ребенка
658
немедленно после рождения, когда прекращается получение им
питательных веше< тв чере< п чацентарцую-кровь (см. гл. V).
Развитие. Молочные железы закладываются у зародыша дли-
ной 8—10 см в виде Двух продольных уплотнении эпг миса
(«молочные линии»), тянущихся вдоль туловища. От этих утолще-
ний в подлежащую мезенхиму врастают эпителиальные _1яжи,
сначала плотные а затем разветвляющиеся на своих дистальных
концах — зачатки молочных желез.
В мужском организме молочные железы остаются инфантиль-
ными в течение всей жизни.
Строение. У половозрелой женщины каждая молочная железа
состоит из 15—20 отдельных железок, разделенных прослойками
рыхлой соединительной и жировой ткани. Эти железы по своему
с трое н и ю являются сложными альвеолярными, и их выводи ые
протоки- открываютс.яыы вершине соска Выводные протоки"пере-
ходят в расширенные моючные синусы (sinus lactifen), служащие
ре зер ву ара ми, в которых накапливается молоко, продуцируе мое в
альвеолах. В молочные синусы впадают многочисленные ветвящиеся
и анастомозирующие молочные протоки fdnctns lactifen), закан-
чиваюшиеся до наступления периода лактации тонкими_слепыми
трубочками — альвеолярными молочными ходами (ductuli alveo-
lares lactifen), которые во время беременности и лактации дают
начало многочисленным~альвеолам (рис. 2TU).
Молочные синусы открываются на верхушке соска, представ-
ляющето собой утолщение кожи.~Его эпидермис сильнб~пигмёнти-
рован, в базальную часть эпителиального слоя вдаются длинные
й~~частб~ разветвляющиеся сосочки дермы, в которых паспола-
гаЮТся' многочисленные инкапсулированные нервные окончания. .
Обилие ~этйх~чувствительных рецепторных окончаний связано с
тем, что выделение молока обусловлено рефлексом на раздраже-
ние этих окончаний актом сосания. Полного развития молочная
железа достигает во время беременности. С момента имплантации
зародыша в слизистую оболочку матки в дольках молочной
железы разрастаются альвеолярные ходы, на концах кот&рых
фо ми ются альв; оды—JBo второй половине игре ныост —желе-
зистые клетки начинают вырабатывать секрет и незадолго до
родов насту.цд£1_£.екрец,ия молозива (colostrum).
Интенсивная секреция полноценного молока устанавливается '
в течение первых_дней после рождения ребенка.
Выработка _мшюка—промсдлдит в альвеолах,______имеющих вид
округлых' или слегка вытянутых пузырьков. Железистые клетки
альвеол (лактоциты) соединяются с помощью замыкающих плас-
тинок и десмосом, располагаются в один слой на базальной
мембране. На апикальной поверхности лактоцитов выступают
небольшие микроворсинки. Местами у основания лактоцитов
(как и в других эктодермальных железах, например потовых или
слюнных) обнаруживаются корзинчатые (миоэпителиальные)
клетки, охватывающие своими отростками альвеолу снаружи.
Молоко — сложная водная эмульсия, в состав которой вхо-
659
Рис. 27(J. Строение аци-
нусов молочной желе-
зы (по Р. Кристичу с
изменением).
/ — ацинусы; 2 — мо-
лочный ход; 3 — внутри-
дольковый проток; 4 —
апокриновая секреция
5 — миозпителиоцит; 6-
нервное волокно; 7 — ге-
мокапилляр; 8 — лакто-
ЦИТ.
дятжл р-ов-ые—надъел ьк и (тпиглицепиды молока, а также
жирные кислоты, являющиеся предшественниками триглицеридов),
б елки (из них специфическими для молока являются казеин,
а также лактоглобулины и лактоальбумины), углеводы (в
том числе специфический для молока дисахарид'— лактоза, или
молочный сахар), соли и вода. Возможность продукции
столь многокомпонентного секрета предполагает соответствую-
щее усложнение ультраструктуры железистых клеток. В лактоци-
тах хорошо развита гранулярная и агранулярная эндоплазма-
тическая сеть, образованная канальцами и цистернами. В комплек-
се Гольджи, хорошо развитом в лактоцитах, завершаются обра-
зование и конденсация казеина, а также синтез лактозы,
чему способствует наличие специального фермента — лактосинте-
тазы. Кроме того, в лактоцитах обнаруживаются микротубульг
и микрофиламенты, особенно в апикальных частях цитоплазмы.
Предполагается, что микротубулы способствуют транспортировке
секреторных продуктов к апикальному краю лактоцита.
При выделении синтезированных компонентов м.олока жировые
капли,'достигающие больших азмеров, перемещаются к апикаль-
660
1
ной мембране и, облекаясь ею, вытягиваются над краем
п’яктопйта-'' Пр_л*ег»е_экстпузии жировая капля вместе с окружа-
ющей ее частью апикальной мембраны открывается и попадает
в полость альнеолы. В полости альвеолы капельки жираГраздроб-
ляясь, превращаются в тонкую эмульсию, к которой примешива-
ются казеин, лактоза и соль, т. е. формируется~молоко, которым
запВлнлс 1ся_цол0СГЬ аяьнееяы.
Опорожнению альвеолы и переходу молока в млечные ходы
способствует ^сокращение звездиаты.х—миоэпителиальных клеток.
По окончании периода лактации молочная железа претерпевает
инволюцию, однако часть альвеол, образовавшаяся во время
предыдущей беременности, сохраняется.
Возрастные изменения. У девочек в период полового созре-
вания начинается интенсивное развитие молочных желез. Из раз-
ветвленных железистых трубок дифференцируются секреторные
отделы — альвеолы, или ацинусы. В течение полового цикла
секреторная активность повышается в период овуляции и снижа-
ется во время менструации.
После прекращения образования овариальных гормонов с
наступлением климактерического периода или после кастрации
молочная железа претерпевает инволюцию.
Регуляция функции молочных желез. В онтогенезе зачатки
молочных желез начинают интенсивно развиваться после наступ-
ления полового созревания, когда в результате значительного
увеличения образования эстрогенов устанавливаются менструаль-
ные циклы и форсируется развитие вторичных признаков женского
пола. Но полного развития и окончательной дифференцировки
молочные железы достигают только во время беременности.
С момента имплантации зародыша в эндометрий матки в дольках
молочной железы разрастаются альвеолярные ходы, на концах
которых формируются альвеолы. Во второй половине беременнос-
ти в альвеолах начинается секреция молозива. Интенсивная сек-
реция полноценного молока устанавливается в первые дни после
рождения ребенка.
Регуляция деятельности функционирующей молочной железы
осуществляется двумя основными гормонами — аденогипофизар-
ным пролактином 1дактотропный гормон), который стимулирует
накопляющегося сначала в млечных ходах и выбрасываемого
оттуда во время лактации под влиянием окситоцина — гипота-
ламического нонапептидного нейрогормона. В свою очередь сек-
реция пролактина возбуждается тем же гипоталамическим тиро-
либерйном, который активирует тиротропную функцию гипофиза,
а_ угнетается дофамином — нейроамином, образующимся в тубе-
ральных ядрах медиобазального гипоталамуса и переносимым с
током портальной крови в переднюю дДлк) гипофиза, где этот
нейроамин действует п ямо на лактотропоциты, блокируя се к -
рецию пролактина. ~ ~ ~ ---
У-' * 1 1 1 •
предметный указатель
Агранулоциты 164
Адипоциты 196
А к со нема 63
Аллантоис 109
Амнион 107
Анализ иммунофлюоресцентный, методы 14
Антигены 237
Антигонадотропин, образование 452
Антитела 237
Артерии 368
— типа мышечного 371
— — мышечно-эластического 370
— — эластического 369
Астроциты 277
— волокнистые 278
— протоплазматические 277
Базофилы 163
— тканевые 194
Барьер гематоэнцефалический 329
Вазопрессин 439
— образование 448
Вещество межклеточное 39
— — ткани костной 226
— — хряща 214
— мозговое 638
— — надпочечников 465
Волокна коллагеновые 198
— мышечные, типы 262
— — с ядерной сумкой 267
— — — — цепочкой 267
— нервные 279
— — регенерация 282
— пре коллагеновые 206
— ретикулярные 201, 205
— эластические 201
Волосы 591
— строение 592
Гаметы см. Клетки половые
Ганглий спинальный, строение 302
Гаструляция 98
— способы 98
— — деламинация 99
— — иммиграция 99
— — инвагинация 98
— — эпиболия 98
Гемограмма 170
Гемоцитопоэз 171
— постэмбриональный 175
— эмбриональный 172
Гетерохроматин 68
Гиалоплазма 41
Гипоталамус 438
— влияние на железы эндокринные пери-
ферические 441
Гипофиз 442
— развитие 442
— строение 442
Гистология, методы исследования 9
Глаз, аппарат аккомодационный 337
— — вспомогательный 347
— — диоптрический 335
— — рецепторный 340
— — слезный 348
— развитие 332
— строение 333
— — оболочка сосудистая 334
— — — фиброзная 333
— — радужка 338
— — роговица 335
— — ‘сетчатка 340
— — склера 333
— — тело ресничное 339
— — — стекловидное 337
— — хрусталик 337
Глотка, развитие 508
Гомеостаз тканевый 297
Гормон анти диуретический 439, 448
— лактотропный 444
— лютеинизирующий, образование 443
— мелаиоцитостимулирующий 447
— половой 617
— роста, образование 444
— фолликулостимулирующий, образование
443
Гортань 566
Гранулоцитопоэз 182
Гранулоциты 158
— ацидофильные см. Гранулоциты эозино-
фильные
— базофильные 163
— нейтрофильные 158
— — палочкоядерные 159
— — сегменте ядерные 159
— — юные 159
— эозинофильные 161
Губы, развитие 474
Десны, развитие 476
Диафиз, строение 227
Дуга рефлекторная 293
Железа (ы) бульбоуретральные 631
— вилочковая см. Тимус 413
— зобная см. Тимус
— околоушные 487
— околощитовидные 459
— поджелудочная 555
— подчелюстная 490
— подъязычная 491
— предстательная 617
— — развитие 629
— — строение 629
— слюнные 481, 485
— шишковидная 450
— щитовидная 452
— — иннервация 457
— — развитие эмбриональные 452
— — регенерация 458
— — строение 453
— экзокринные 148
— эндокринные 148, 436
— — периферические 452
Желтое тело, формирование 643
Желудок 514
— железы кардиальные 521
662
1 — — пилорические 520
‘ — — собственные 516
— развитие 514
строение 515
Зародыш человека, дифференцировка мёзо-
дермы 123
— — — эктодермы 123
— — — энтодермы 124
— — дробление 116
— — имплантация 116
— — нейруляция 123
Зигота 94
— дробление 95
/бы 495
— молочные 495
— постоянные 501
— строение 501
— — дентин 502
— — пульпа 505
— — цемент 504
— — эмаль 501
Иммунитет 236
— роль макрофагов 247
F -му но глобулины 237
Иммуноцитопоэз 185
Иммуноциты 236
Кальцитонин 457
Кальцитониноциты 457
Капилляры, типы 379
Кислота соляная, образование 519
Кишка ободочная 537
— прямая 542
— толстая 537
— тонкая 523
— васкуляризация 533
— иннервация 534
— развитие 523
строение 524
Клетка (и) адвентициальные 198
— воспроизведение 73
— гормонопродуцирующие одиночные 467
— деление 76
— — митоз 76
— — — анафаза 78
— — — метафаза 77
— — — профаза 76
— — — телофаза 78
। — жировые 196
. — иммунокомпетентные 239
— интерфазные, микротрубочки 59
, — исследование, методы 9
— окрашивание витальное 11
— — суправитальное 11
— оплодотворение 92
— пигментные 197
— половые 86
— — женские 89, 113
— — мужские 87, 113
— — развитие, периоды 111
— реакция на внешние воздействия 82
— состав химический, исследование, ме-
тод(ы) 13
— — — — — гистохимические 13.
— — — — — дифференциального цент-
1 рифугирования 14
— — — — — интерферометрия 14
— — — — — радиоавтография 13
I
— — — — — цитоспектрофлюориметрия
14
— — — — — цитоспектрофотометрия 14
— — — — — цитохимические 13
— строение 36, 63
— структура 41
— тучные 194
— фиксированные, исследование 12
— эндорепродукция 81
— ядро 66
Клеточные популяции, кинетика 298
Клеточный цикл 73
Кожа, железы молочные 658
— — потовые 589
— — — апокриновые 589
— — — эккриновые 589
— — сальные 591
— развитие 580
— строение 581
— — пигмент кожный 586
— — собственно кожа 585
— — эпидермис 582
Комплекс Гольджи 50
Кортикотропоциты 445
Кости, соединение 234
— — синдесмозы 234
— — синостозы 235
— — синхондрозы 234
— — суставы 235
— трубчатые, рост 231
— — строение гистологическое 227
Кроветворение 171
К ровос на бжение ги по та л а мо-а де ногипофи-
зарное 448
Кровь 152
— изменения возрастные 170
— плазма 152
— регенерация физиологическая 175
— функция 152
— элементы, форменные 153
— — — базофилы 163
— — — гранулоциты 154
— — — лейкоциты 157
— — — лимфоциты 164
— — — моноциты 168
— — — плазмоциты 166
— — — тромбоциты 168
— — — эозинофилы 161
— — — эритроциты 153
Легкие, кровоснабжение 577
— строение 570
— — бронхиальное дерево 570
— — отдел респираторный 572
— — плевра 577
Лейкоцит(ы) 157
— базофильный 184
— зернистые 157
— незернистые 157
— нейтрофильные 183
— палочкоядерные 183
— сегментоядерные 183
Либерииы 439
Лизосомы 53
— аутофагосомы 53
— вторичные 53
— гетерофагосомы 53
— первичные 53
— фаголизосомы 53
Лимфа 171
663
— периферическая 171
— промежуточная 171
— центральная 171
Лимфоцитопоэз 185
Лимфоциты 164
— киллеры 239
— тимусзависимые 166
В-Лимфоциты 166
Т-Лимфоциты 166
Липотропин 447
Листки зародышевые 98
— — дифференцировка 102
— — — бластомерная 4 03
— — — гистогенетическая 103
— — — зачатковая 103
— — — оотипическая 103
Лютропин 443
Макрофаги 191
Маммотропоциты 444
Матка, развитие 647
— строение 647
Мегакариобласт 184
Мегакариоцитопоэз 184
Межклеточные соединения, типы 46
— — — десмосома 46
— — — плотное 46
— — — простое 46
— — — синаптические 47
— — — щелевидиое 46
Мелатонин, образование 451
Меланоцитотропин 447
Метамиелоцит (ы) базофильный 184
— эозинофильные 184
— юные 183
Миелобласты 182
— базофильные 184
— нейтрофильные 182
— эозинофильные 184
Микроглия 278
Микроскопия световая 15
— ультрафиолетовая 15
— фазово-контрастная 17
— флюоресцентная 15
— электронная 17
Микрофибриллы 199
Миндалина (ы) глоточная 484
— небные 483
— язычная 485
Миосателлитоциты 261
Миосимпласты 261
Миофибриллы 261
Миофибробласты 189
Митохондрии 55
Мозг головной 308
— — оболочка мягкая 328
— — — паутинная 328
— — — твердая 328
----- кора 314
— костный 410
— — желтый 412
— — развитие 410
— — строение 411
— спинной 304
— — нейроны 305
— — оболочки 328
Мозжечок 310
Моноцитопоэз 185
Моноциты 168
Надкостница 230
Надпочечники 461 д
— вещество мозговое 465 е
— кора 463 r-t
— развитие 461 »
— строение 462
Небо мягкое 477
— твердое 477
Нейрогипофиз 439, 448
Нейроглия 276
Нейрогормоны аденогипофизотропные 439
Нейроны 270
— мозга спинного 305
— регенерация 282
— секреторные 276
Нейрофибриллы 275
Нейроциты 270
Нерв(ы) зрительный, диск 345
— — центральная ямка 344
— периферические 304
Нервные окончания 283
— — рецепторные 285
— — эффекторные 283
Ногти, строение 596
Оболочка(и) амниотическая 108
— зародышевые 106
— серозная 109
Овогенез 639
Ойоциты см. Яйцеклетка 89
Овуляция 643
Окостенение диафиза 223
— эндохондральное 223
— эпифизов 224
Окситоцин 439
Олигодендроциты 278
Опсонины 238
Орган (ы) вкуса 353
— внезародышевые 106, 109
— — человека 124
— зрения 332
— иммунологической защиты 409
— кроветворения 409
— мочевые, пути мочевыводящие 612
— обоняния 349
— преддверно-улитковый см. Орган ри
весия
— равновесия 355
— слуха 355
— чувств 330
— — классификация 331
Органеллы мембранные 47
— не мембранные 58
Остеобластоциты 224
Остеобласты 224
Остеогистогенез 219
— непрямой 222
— прямой 219
Остеокластоциты 225
Остеокласты 225
Остеоциты 224
Паратирин 459
Паратироциты 459
Перициты 198
Пероксисомы 55
Печень 544
— пути желчевыводящие 552
— развитие 544
— система кровеносная 545
664
> v— строение 545 Ъргментоциты 197 пищеварение 468 ^Пищевод, развитие 508 * — строение 509 ''зма крови 152 лмолемма 43 \ ^змоциты 166, 194 Участника кровяная см. Тромбоцит 1ла цента 109 — строение 109 — функции 109 Полость носовая 565 — ротовая 474 Почки, система эндокринная 609 — строение 598 Препараты гистологические, микроскопиро- вание методы 15 Прогенез 86 Пролактин 444 Пролактиноциты 444 Промиелоциты 182 Протофибриллы 199 Проэритробласты 177 Пути воздухоносные 565 — — гортань 566 — — легкие 570 — — носовая полость 565 — — трахея 568 — — — сосуды кровеносные 569 — — центральная, кровоснабжение 329 — пищеварительная 468 — — отдел задний 513 — — — — железа поджелудочная 555 — — — — желудок 514 — — — — кишка ободочная 537 — — — — — прямая 542 — — — — — толстая 537 — — — — — тонкая 523 — — — — печень 544 — — — — пузырь желчный 553 — — — — — червеобразный отросток 540 — — — передний 473 — — — — глотка 508 — — — — губы 474 — — — — десны 476 — — — — железы слюнные 481, 485 — — — — зубы 495 — — — — миндалина (ы) глоточная 484 — — — — — небные 483 — — — — — язычная 485 — — — небо мягкое 477 — — — — твердое 477 - — — — пищевод 508 — — — — ротовая полость 474 — — — — щеки 476 — — — — язык 478 — — — — язычок 477 — — — средний 513 — — трубка пищеварительная, строение
Реакции иммунные, взаимодействия кле- точные 236 — — участие тканевых базофилов 249 — — — — эозинофилов 249 Рибосомы 58 Русло микроциркуляторное, сосуды 374 — — — анастомозы артериоловенулярные 382 — — — артериолы 374 — — — венулы 381 — — — вены 384 — — — капилляры 376 469 — половая 613 — — мужская, деятельность гормональ- ная, регуляция 633 — — — железа (ы) бульбоуретральные 631 — — — — предстательная 629 — — — пузырьки семенные — — — пути семявыносящие 625 — — — член половой 632 — — — яички 616 — — строение 619 — — функция 621
1 егрегация ооплазматическая 94 О дезенка 429 развитие 429 строение 431 ^>^дце, клапан (ы) аортальные 399 — атриовентрикулярный см. Сердце, клапан предсердно-желудочковый — предсердно-желудочковый 398 миокард 400 развитие 396 — строение 397 .еротонин, образование 451 ' эндоплазматическая агранулярная 49 - — гранулярная 48 > ьапсы межнейронные 289 лстема дыхательная, развитие 564 кровеносная 368 крови 151 иммунная 236 — элементы, интеграция 249 макрофагическая 193 - мать — плод 132 — нервная 301 — — вегетативная 323 — — изменения возрастные 328 — — развитие 301 — — женская 634 — — — влагалище 649 — — — — изменения циклические 654 — — — железы молочные 658 — — — матка 647 — — — период постменструальный 652 — — — — предменструальный 654 — — — половые органы наружные 658 — — — регуляция гормональная 656 — — — цикл половой 650 — — — яичники 634 — — — — функция регенеративная 639 — — — яйцеводы 645 — сердечно-сосудистая 367 — эндокринная 436 — — образования регулярные централь- ные 438 — — связь с нервной системой 437 Соматостатин 457 Соматотропин 444 Соматотропоциты 444 Сосуды лимфатические 389 — — капилляры 389 — — отводящие 391 — — проток лимфатический грудной 392 Сперматогенез 629
665
Сперматозоид, строение 87
Структуры клеточные, изображение, ана-
лиз, методы морфометрические 18
— — — — оптико-структурный машин-
ный 19
— тканевые, изображение, анализ, методы
18
Тестостерон 617
Тимус 413
Тирозин 454
Тиротропоциты 443
Тироциты 453
Ткань (и), взаимосвязь 294
— жировая 206
— — белая 207
— бурая 208
— исследование, методы 10
— классификация, принципы 296
— костная (ые) 218
— — вещество межклеточное 226
— — клетки 224
— — — остеобласты 224
— — — остеокласты 225
— — — остеоциты 224
— — пластинчатая 227
— — ретикулофиброзная 227
— метаболизм, исследование 13
— мышечная(ые) 252
— — гладкие 253
— — классификация морфофункциональ-
ная 252
— — поперечнополосатые 257
— — — сердечная 257
— — — скелетная 260
— — происхождения мезенхимного 253
— — — нейрального 256
— — — эпидермального 254
— нервная 268
— — развитие 268
— окрашивание витальное 11
— — суправитальное 11
— организация, принципы 293
— развитие 295
— скелетные 210
— соединительная (ые), вещество межкле-
точное 198
— — — — компонент аморфный 203
— — волокнистые 187
— — — плотная 204
— — — — мембраны фиброзные 205
— — — — связка выйная 204
— — — — сухожилие 204
— — — пигментная 210
— — ретикулярная 205
— — слизистая 209
— — функции 186
— состав химический, исследование, ме-
тод(ы) 13
— — — — — гистохимические 13
— — — — — дифференциального цент-
рифугирования 14
— — — — — интерферометрия 14
— — — — — радиоавтография 13
— — — — — цитоспектрофлюориметрия
14
— — — — — цитоспектрофотометрия 14
— — — — — цитохимические 13
— фиксированные, исследование 12
— хрящевая волокнистая 217
— — гиалиновая 214
666
— — клетки 212
— — — хондробласты 213
— — — хондроциты 212 '
— — эластическая 215
— эволюция, теории 296
— эпителиальные 136
— — строение 136
Тракт пищеварительный, узелки лимфати-
ческие 418 (
Тромбоцитопоэз 184
Тромбоциты 168
Узлы гемолимфатические 427
— — развитие 427
— — строение 427
— лимфатические 419
— — вещество корковое 421
— — — мозговое 425
— — зона паракортикальная 424
Ухо внутреннее 356
— наружное 355
— среднее 356
Фибрилла коллагеновая 200
Фибробласты 188
— малоспециализированные 188
— специализированные 188
Фиброкласты 191
Фиброциты 189
Фоллитропин 443
Формула лейкоцитарная 170
Хондрогистогенез 210
Хорион 109
Хорионаллантоис 109
Хроматин половой 159
Хромаффиноциты 465
Хромосомы митотические, морфология 79
Центриоли 61
Цитология, методы исследования 9
Цитоплазма 41
— включения 65
— — пигментные 65
— — секреторные 65
— — экскреторные 65
— микрофиламенты 64
— филаменты промежуточные 65
Человек, органы внезародышевые 124
— — — аллантоис 131
— — — амнион 130
— — — детское место 125
— — — желточный мешок 130
— — — плацента 125
— — — пупочный канатик 131
— — — хорион 124
— развитие эмбриональное 112
— — — периоды 112
Червеобразный отросток 540
Щеки, развитие 476
— строение 477
Эмбриогенез 85, 92
— человек 114
Эмбриология, методы исследования 9
— сравнительная 86
Эндокриноциты ацидофильные 444
— базофильные 443
— гонадотропные 443
— желудочно-кишечнб!е/521J
— кортикотропные 445 '—
— маммотропные 444
— мозговые 465
— парафолликулярные 457
— соматотропные 444
— тиротропные 443
— фолликулярные 453
— хромофильные 442
— хромофобные 442
Эндост 230
Эозинофилы 161
— палочкоядерные 161
— сегментоядерные 161
— юные 161
Эпендимоциты 276
Эпителий железистый 136
— — строение 146
— классификация 138
— многослойный 139
— — плоский неороговевающий 139
— — — ороговевающий 139
— однослойный 139
— — изоморфный 139
— — кубический 140
— — многорядный 139
— — однорядный 139
— — плоский, строение 140
— — — — мезотелий 140
— — — — эндотелий 140
— — призматический 140
— переходный 144
— — слой базальный 144
— — — поверхностный 144
— — — промежуточный 144
— покровный 136
— — васкуляризация 146
— — возрастные изменения 146
— — иннервация 146
— — регенерация 145
— тип ангио дермальный 139
— — цело нефро дермальный 139
— — энтеро дермальный 139
— — эпендимоглиальный 139
— — эпидермальный 139
Эпифиз 450
— развитие 450
— строение 450
— функции 451
Эритробласты ацидофильные 180
— базофильные 177
— полихроматофильные 180
Эритроцитопоэз 177
Эритроциты 153
— строение 153
— функция 153
Эухроматин 68
Ядро клеточное, состав химический 67
— — структура 67
— — — кариоплазма 67
— — — оболочка ядерная 71
— — — хроматин 67
— — — ядрышко 70
Язык, строение 478
Яйцеклетка (и) женщины 114
— классификация 89
— строение 90
ОГЛАВЛЕНИЕ
Предисловие к четвертому изданию — Ю. И. Афанасьев, Н. А. Юрина . . .
Глава 1. Гистология, цитология и эмбриология. Их содержание, задачи
и связь с другими медико-биологическими науками. Значение для медици-
ны — Н. А. Юрина............................................
Глава 11. Методы исследования в гистологии, цитологии и эмбриоло-
гии — Н. А. Юрина .....................
Методы исследования живых клеток и тканей...................
Исследование мертвых (фиксированных) клеток и тканей........
Исследование химического состава и метаболизма клеток и тканей . . . .
Методы микроскопирования гистологических препаратов.........
Методы анализа, изображений клеточных и тканевых структур...
Глава III. Краткий очерк развития гистологии, цитологии и эмбриоло-
гии — Ю. И. Афанасьев.......................................
Становление гистологии, цитологии и эмбриологии как наук ...
Гистология и эмбриология в России как предмет преподавания. Отечествен-
ные гистологические школы второй половины XIX и начала XX века . .
Развитие гистологии, цитологии и Эмбриологии в СССР.........
ЦИТОЛОГИЯ
Глава IV. Учение о клетке (основы общей цитологии) — Ю. С. Ченцов .
Клеточная теория ...........................................
Структурные компоненты клетки.............................
Цитоплазма ..............................................
Ядро ....................................................
Воспроизведение клеток .....................................
Клеточный цикл............;..............................
Реакция клеток на внешние воздействия.......................
МБР
ЛОГ
Глава V. Основы эмбриологии. — Ю. И. Афанасьев, Н. А. Юрина,
М. Я. Субботин ......................................................
Связь индивидуального и исторического развития организма...........
Основы сравнительной эмбриологии.....................................
Прогеиез..........................................................
Эмбриогенез.......................................................
Эмбриональное развитие человека......................................
Прогенез..........................................................
Эмбриогенез.......................................................
ОБЩАЯ ГИСТОЛОГИЯ
Глава VI. Эпителиальные ткани — Е. Ф. Котовский....................
Источники развития эпителиальных тканей............................
Классификация......................................................
I
<
3 I
5
9
9
12
13
15
18
22
22
27
31
1
36
36
41
41
66
73
73
82
85
85
86 '
86
92
112
113
114
136
136
138
Строение различных типов покровного эпителия ........................... 140
Строение железистого эпителия........................................... 146
Железы.................................................................. 148
лава VII. Кровь и лимфа. Кроветворение. — Н. А. Юрина, В. Г. Елисеев . 151
Понятие о системе крови................................................. 151
Кровь................................................................... 152
Плазма крови ........................................................ 152
Форменные элементы крови ............................................ 153
Гемограмма. Лейкоцитарная формула.................................... 170
Возрастные изменения крови........................................... 170
Лимфа................................................................... 171
Кроветворение (гемоцитопоэз)............................................ 171
Эмбриональный гемоцитопоэз (развитие крови как ткани).................172
Постэмбриональный гемоцитопоэз....................................... 175
Глава VIII. Соединительные ткани. — Ю. И. Афанасьев, В. Г. Елисеев . . '““'t'Sd .
Собственно соединительная ткань.......................................186 !.
Волокнистые соединительные ткани................................
Соединительные ткани со специальными свойствами .................265'' \
Скелетные ткани ... .......................................210. /
Хрящевые ткани...................................................21б~
Костные ткани...................................................218
Глава IX. Иммунная система и клеточные взаимодействия в иммунных
реакциях. — Н. А. Юрина.............................................236
Общая характеристика. Определение понятий.........................236
Характеристика иммунокомпетентных клеток..........................239
Антигеннезависимая пролиферация и дифференцировка..............240
Антигензависимая пролиферация и дифференцировка Т- и В-лимфо-
цитов .........................................................241
Макрофаги......................................................247
Участие тканевых базофилов и эозинофилов в иммунных реакциях . . . 249
Механизмы интеграции элементов иммунной системы...................249
. Глава X. Мышечные ткани. — Г. С. Катинас .......... 252
Общая морфофункциональная характеристика и классификация мышечных
тканей.......................-. . ...............................252
Источники развития мышечных тканей................................252
Гладкие мышечные ткани............................................253
Поперечнополосатые мышечные ткани.................................257
Глава XI. Нервная ткань. — Т. Н. Радостина ...... '...............268
Развитие нервной ткани .......................................... 268
Нейроны ..........................................................270
Секреторные нейроны...............................................276
Нейроглия........................................................ 276
Нервные волокна...................................................279
Регенерация нейронов и нервных волокон............................282
Нервные окончания.................................................283
Понятие о рефлекторных дугах......................................293
Глава XII. Общие принципы организации тканей. — Ю. И. Афанасьев,
Г. С. Катинас.....................................................293
Ткань как система.................................................... 293 у
Развитие тканей...................................................... 295 уЙЙ'
Теории эволюции тканей............................................... 296А '
Принципы классификации тканей........................................296
Тканевый гомеостаз...................................................297
Основы кинетики клеточных популяций..................................298
Регенерация тканей .................................................. 500
669
ЧАСТНАЯ ГИСТОЛОГИЯ
Глава XIII. Нервная система. — Т. Н. Радостина.......................301
Чувствительной- узлы.................................................302
Периферические нервы.................................................304
Спинной мозг.......................................................<?э0ф
Головной мозг........................................................308
Мозжечок .................................................. . . . 310
Кора большого мозга............................................ . 314
Автономная (вегетативная) нервная система........................... 323
Оболочки головного и спинного мозга..................................328
Возрастные изменения нервной системы.................................328
Кровоснабжение центральной нервной системы. Гематонейрональный
барьер ..............................................................329
/ Глава XIV. Органы чувств.— Я. А. Винников, Ю. И. Афанасьев,
Н. А. Юрина...............................................................330
Общая характеристика и классификация.................................330
Орган зренйя.........................................................332
Строение глазного яблока ........................................ 333
Орган обоняния.......................................................349
Орган вкуса..........................................................353
Орган слуха и равновесия (преддверно-улитковый орган)................355
Глава XV. Сердечно-сосудистая система. — Ю. И. Афанасьев.............367
Кровеносные сосуды.................................................. 368
Артерии ..........................................................368
Микроциркуляторное русло..........................................374
Вены .............................................................384
Особенности строения органных сосудов ........................... 388
Лимфатические сосуды.................................................389
Сердце...............................................................396
Глава XVI. Органы кроветворения и иммунологической защиты.—
Ю. И. Афанасьев......................................................409
Костный мозг........................................................ 410
Тимус (вилочковая, или зобная, железа).......................... ". 413
Лимфатические узелки пищеварительного тракта....................... 418
Лимфатические узлы...................................................419
Гемолимфатические узлы...............................................427
Селезенка............................................................429
/] Глава XVII. Эндокринная система (эндокринные железы). — Б. В. Але- .
шин...................................................................с .435-
Общая характеристика . ..............................................435
Центральные регуляторные образования эндокринной системы.............438
Гипоталамус ......................................................438
Гипофиз...........................................................442
Эпифиз (шишковидная железа).......................................450
Периферические эндокринные железы.................................. 452
Щитовидная железа.................................................452
Околощитовидные железы............................................459
Надпочечники .................................................... 461
Одиночные гормонопродуцирующие клетки ...............................467
Глава XVIII. Пищеварительная система. — Н. А. Юрина...............468
Общий план микроскопического строения пищеварительной трубки .... 469
Передний отдел пищеварительной системы..............................473
Ротовая полость..................................................474
Глотка'..........................................................508
Пищевод..........................................................508
670
|>едний и задний отделы пищеварительной системы...................513
Желудок.........................................................514
| Тонкая кишка....................................................523
। Толстая кишка...................................................537
Печень. — Е. Ф. Котовский.......................................544
। Желчный пузырь....................................................553
Поджелудочная железа. — К). И. Афанасьев, Е. Ф. Котовский .... 555
лава XIX. Дыхательная система — Ю. И. Афанасьев, Т. Г. Оганесян . 564
оздухоносные пути.................................................565
еткие............................................................ 570
'1 а в а XX. Кожа и ее производные. — Е. Ф. Котовский............. 580 И
>жа ..............................................................580
j Железы кожи.....................................................589
Волосы .................................591
Ногти............................................................596
Глава XXI. Мочевые органы — Е. Ф. Котовский.............. ... . 597
Почки.................................598
Мочевыводящие пути.‘..................612
। Глава XXII. Половая система. — Б. В. Алешин, Ю. И. Афанасьев,
Н. А. Юрина ....................... 6ГЗ
Мужская половая система............................................616
Яички (семенники)..............................................616,
Семявыносящие пути .............................................625
Добавочные железы мужской половой системы.......................628
Половой член.................................................. 632<
Гормональная регуляция деятельности мужской половой системы .
Женская половая система гС.....................................
Яичники . . -Лг............................................
Другие органы женской половой системы 3-...................
, Половой цикл -V..............................................
j Возрастные изменения органов женской половой системы.........
Гормональная регуляция женской половой системы.............
Наружные половые органы ...................................
Молочные железы ...........................................
633
634
634
655
656
658
658
Предметный указатель..........................................
662
А'чдб ник
ЮЛИЙ ИВАНОВИЧ АФАНАСЬЕВ,
НИНА АЛЕКСЕЕВНА ЮРИНА<
БОРИС ВЛАДИМИРОВИЧ'АЛ
гистология
I
!
ЗаЬ. реакцией О. В. Карева
Редактор И. Л. Червова
Редактор издательства И. Н. Кононова
Художественный редактор Т. К. Винокурова
Технический редактор В. И. Табенская /
Корректор М. П. Молокова
ИБ № J916
Сдано в набор 24.08.87. Подвижно к печати 1.12.8fc'
Формат бумагиФОХ 90'/i6./fc ‘ 'офсетн. № I. Гарнитура
тайме. Печать офсетная.уУ ^Д1еч. л. 42,0. Усл. кр.-отт.
159,25. Уч.-изд. л. 48,12. Ти
Цена 2 р. 40 к.
Ордена Трудового Кра(Хюго Знамени издательство «Меди-
цина» 101000 Москва,' Петроверигский пер., 6/8.
Ярославский полиграфкомбинат Союзполиграфпрома при
Государственном комитете СССР по делам йздательсщ,
полиграфии и книжной торговли. 150014, Ярослав^!,,
ул. С вободы, 97.
ж 75 000 экз. Заказ № 908.
/
f
i