/





























































































































































































































































































































































































































































































































































































































Author: Харрисон Дж. Уайнер Дж. Тэннер Дж. Барников Н. Рейнолдс В.
Tags: антропология биология
Year: 1979




Text
HUMAN BIOLOGY
An Introduction to Human Evolution, Variation, Growth and Ecology
Second edition by
G. A. HARRISON
Department of Biological Anthropology, University of Oxford
J. S. WEINER
M. R. C. Environmental Physiology Unit, London School of Hygiene and Tropical Medicine, University of London
J. M. TANNER
Department of Growth and Development, Institute of Child Health, University of London
N. A. BARNICOT
Late of the Department of Anthropology, University College, University of London
With a chapter on “ The evolution of Human society”
by
U. REYNOLDS
Department of Biological Anthropology, University of Oxford
Oxford University Press 1977
ДЖ. ХАРРИСОН
ДЖ. УАЙНЕР
ДЖ.ТЭННЕР
Н. БАРНИНОТ
В. РЕЙНОЛДС
Перевод с английского Е. 3. ГОДИНОЙ, канд. физ.-мат. наук А. И. К03ЛЕНК0ВА и канд. мед. наук В. И. САМОЙЛОВА под редакцией проф. В. В. БУНАКА
ИЗДАТЕЛЬСТВО «МИР»
МОСКВА 1979
УДК 572.0
Настоящее—второе — издание значительно дополнено и переработано по сравнению с первым, выпущенным в свет в переводе на русский язык в 1968 г. (Харрисон Дж., Уайнер Дж., Таннер Дж., Барникот Н. Биология человека — М.: Мир, 1968). В монографии рассмотрены общие вопросы теории эволюции, история приматов, происхождение человека, генетика человека и изменчивость (изменчивость размеров и формы тела, пигментации, состава крови и т. д.), рост, физическое и умственное развитие. Заключительная часть посвящена вопросам экологии.
Предназначена для биологов всех специальностей, врачей, антропологов, этнографов, археологов, экономистов и социологов. Может служить пособием для преподавателей средних школ и техникумов, а также при чтении лекций о происхождении человека и па антирелигиозные темы.
Редакция литературы по биологии
2605070000
21001-127
Б 041 (01)-79 127’79
© 1964, 1977, Oxford University Press
© Перевод па русский язык, «Мир», 197!
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
Оксфордский университет выпустил в свет новое издание книги «Биология человека», которая уже приобрела достаточно широкую известность. В новое издание внесены существенные добавления и изменения, особенно в первую, четвертую и пятую части. В первой части описаны новые археологические находки и даны сравнительные характеристики всего материала в целом. Авторы придерживаются концепции, близкой к теории Ле Гро Кларка. В настоящем издании авторы учли возросший в последнее время интерес к биологическим условиям жизни ранних сообществ. Для освещения этой проблемы был привлечен оксфордский ученый В. Рейнолдс. Написанная им небольшая глава носит характер предварительного сообщения, однако весьма существенно, что эта развивающаяся ветвь биологии человека отражена в настоящем комплексном издании.
Н. Барникот, автор третьей части, в которой рассмотрена биологическая изменчивость популяций, расширил материал предыдущего издания; это особенно касается учения о группах крови и о белках плазмы. Излагая данные об изменчивости признаков в конкретных популяциях, автор подчеркивает, что человеческие расы не являются генетически однородными группами и разграничение их ненадежно. Эта точка зрения становится как будто преобладающей в зарубежной литературе, но все же далеко не всеобщей.
По сходству генов многих локусов Барникот строит две филе-тические схемы группы популяций. Хотя методика филетического анализа по факторам, контролирующим группы крови, разработана недостаточно полно, схемы Барникота, как и описание редких компонентов крони, существенно пополняют имеющиеся сведения по этому вопросу.
Дж. Харрисон, как и в предыдущем издании, написал две части, весьма различные по содержанию: его перу принадлежит часть I «Эволюция человека», основанная на данных палеоантро
5 Предисловие редактора перевода
пологий (то, что у нас принято называть «антропогенезом»), и часть II «Генетика человека». Достоинство этих двух частей — точность изложения, строгая обоснованность характеристик, полная корректность и вместе с тем простота математических выкладок.
Четвертая часть, написанная виднейшим специалистом в области учения о росте и конституции Дж. Тэннером, содержит свежий материал и хорошо отражает современное состояние проблем, касающихся закономерностей роста, формирования органов и систем, а также анализа конституциональных типов. Кратко, но очень содержательно охарактеризованы типы темпераментов и их связь с телосложением.
Пятая часть «Экология человека» написана Дж. Уайнером, который внес значительный вклад в построение законченной системы экологии человека, суммируя накопленные, порой разнородные, сведения. Собранный им материал охватывает, пожалуй, все основные проблемы и создает прочную основу для понимания этой новой области биологии человека, хотя рассмотрено лишь одно из существующих направлений экологического изучения человека.
Авторы монографии использовали материалы, найденные и разработанные ими самими или взятые из первоисточников. Некоторые вопросы, рассмотренные в данной книге, получили в специальной литературе также и иное освещение. Об этом подробно сказано в комментариях редактора, которыми был снабжен перевод первого издания.
Не будучи учебником по какому-то определенному разделу биологии, данная книга вполне может служить учебным пособием, так как объединяет несколько самостоятельных областей исследования, соподчинив их одной задаче. Книга заслуживает особого внимания генетиков, антропологов, врачей и всех, кто интересуется современным состоянием биологии человека.
В. Бунак
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
Что такое «биология человека»? Скрывается ли за этим термином лишь попытка дать новое наименование старым и хорошо знакомым вещам или же он действительно выражает новую концепцию в понимании естественной истории человечества?
Мне кажется, слово «концепция» здесь наиболее уместно. Биология человека — не столько научная дисциплина, сколько определенная точка зрения на самое интересное и важное из животных. Биология человека рисует портрет человечества на том же холсте, который служит для изображения других представителей животного царства. Это наука о людях, их происхождении, эволюции и географическом распространении, об увеличении численности человеческих популяций и их структуре в пространстве и времени, о человеческом организме, его развитии, включая изменение размеров и формы тела. Биология человека изучает наследственность человека, его генетическую систему, сущность и значение врожденных различий между индивидуумами, экологию и физиологию человека, а также те средства, которые человек использует в борьбе с враждебным окружением. Биология человека рассматривает также особенности его поведения — не случайные различия в поведении отдельных индивидуумов, а общие аспекты поведения, например эволюцию и роль семейных отношений, проявлений любви, истинной или скрытой агрессивности и т. п.
Наконец, одна из важнейших задач биологии в применении к человеку состоит в том, чтобы описать или разъяснить природу, происхождение и развитие системы связи между человеческими существами и основанную на ней негенетическую преемственность.
Человек — наиболее удобный объект изучения для биолога, в особенности для начинающего. Многие законы биологии могут быть столь же хорошо усвоены на примере человека, как и на примере других животных, а некоторые из этих законов даже легче разъяснить, изучая человека.
8
Предисловие к первому изданию
Человек — величайший дилетант среди животных. Там, где другие животные в силу узкой специализации ограничили свой образ жизни частными узкими рамками, человек сумел сохранить свой статус дилетанта. Человеческие существа независимы в биологическом смысле настолько, насколько могут быть независимы животные. Как млекопитающее, человек — существо «простое»: у него нет каких-либо существенных отличий — анатомических или генетических, а также особенностей роста и развития, которые не были бы предвосхищены в эволюции низших приматов. Бинокулярное зрение и перекрест зрительных трактов, увеличение коры головного мозга, освобождение передних конечностей для манипуляций, увеличение периода детства — все это отнюдь не новые качества, свойственные исключительно человеку, но лишь высшее проявление тех тенденций развития, которые мы находим у человекообразных обезьян.
Человек как биологическое существо добился решающего успеха лишь недавно, лишь в самое последнее время. Люди обычно забывают или недостаточно ясно представляют себе, насколько непрочным было их положение на Земле в первые тысячелетия их существования. Биолог многое почерпнет, изучая биологию человека: прежде всего он должен усвоить тот основной урок, что различие, которое он имеет обыкновение делать между Природой — чуждой и дикой — и Человеком и его деятельностью, лишь препятствует пониманию сущности той и другого.
Биология человека — весьма полезный предмет для студентов-медиков и врачей. Так называемые «биологические принципы» — предмет многочисленных толков и дискуссий в среде старшего и младшего поколений медиков — с научной точки зрения не выдерживают никакой критики. Наиболее абсурдна укоренившаяся у медиков вера в то, что всякого рода естественные предрасположения и приспособления организма почти совершенны и что болезни и другие недостатки — лишь часть чрезмерно затянувшегося «искупления» за совершенный человеком «грех» — разрыв с природой и переход к неестественному для него образу жизни. В связи с этим стоит серьезно задуматься над совершенствованием нашей системы медицинского образования и перестройкой ее в соответствии с принципами биологии человека. Несомненно, в медицинском образовании имеются пробелы, которые нельзя устранить лишь перетасовкой учебных программ. Многое ли из того материала, который читатель найдет в данной книге, можно отыскать в медицинских руководствах или конспектах лекций? Если такой материал и входит в учебники, то, как правило, его помещают не там, где ему надлежит быть, т. е. не в начале медицинского курса (где для него всегда можно было бы найти достаточно места путем сокращения традиционного описания акул и дождевых червей, которое уже много лет мало что дает
Предисловие к первому изданию 9
будущему медику). В то же время генетика человека если и входит в программу, то в лучшем случае как факультативный курс, который вдобавок читают в те годы, когда студент-медик занят в основном клинической практикой; сведения по демографии даются вперемежку с санитарией; на экологию человека смотрят как на предмет, который, хотя и может помочь врачу понять трудности, испытываемые переселенцами в чужой стране, едва ли заслуживает места в привычном для нас учебнике, справочнике или курсе лекций.
Я уверен в том, что эта отличная книга, написанная авторами — пионерами в области биологии человека, сделавшими крупный вклад в те отрасли науки, которые они представляют в соответствующих частях книги,— окажет большое влияние на развитие всех разделов науки о человечестве.
П. Медавар
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
Со времени выхода в свет первого издания книги в 1964 г. был сделан серьезный шаг вперед в нашем понимании биологической организации человеческих популяций как прежних времен, так и современных. И хотя в книге, носящей характер вводного курса, рассматриваются скорее принципы, чем фактические детали, многие разделы в ней заметно устарели. Во втором, исправленном издании мы постарались в целом привести книгу в соответствие с современным уровнем знаний; основательной переделке подверглись, в частности, главы, относящиеся к эволюции приматов и человека, генетической изменчивости на молекулярном и биохимическом уровне, а также природе процессов роста и адаптации. В книгу включена новая глава, написанная д-ром В. Рейнолдсом, которая посвящена поведению приматов и происхождению человеческого общества. Мы постарались тем не менее сохранить первоначальную структуру и общий объем книги.
Нам хотелось бы вновь поблагодарить сотрудников издательства Oxford University Press за постоянную помощь и проявленное терпение при подготовке рукописи к печати.
Мы выражаем также свою глубокую признательность коллегам покойного проф. Барникота, которые помогли внести необходимые изменения в написанные им ранее главы книги, а именно: д-ру Дж. Берду, проф. Уолтеру Бодмеру и его супруге, Давиду Коулмену, д-ру Ирвину Джонсону, д-ру А. Е. Муранту, д-ру М. Ф. Пе-рутцу, д-ру Прайсу Эвансу, д-ру Дж. X. Ренвику, проф. А. Дж. Стейнбергу, А. Стивенсу, Д. Тиллсу и госпоже X. Уэймс. Тони Ли подготовил все схемы в разделе книги, написанном проф. Бар-никотом. Госпожа Мэри Тайнгейт заслуживает особой благодарности не только за то, что напечатала весь текст книги, но и за помощь, которую она оказывала нам на различных этапах работы. Мы признательны многим авторам и издательствам (упомянутым в соответствующих местах книги) за разрешение воспроизвести необходимый иллюстративный материал.
Предисловие ко второму изданию
11
Проф. Н. А. Барникот закончил переделку своей части книги лишь за несколько недель до своей смерти весной 1975 г. Мы отдаем дань уважения нашему покойному коллеге, чья эрудиция и смелость всегда служили для нас источником вдохновения. Постигшая нас утрата невосполнима не только с точки зрения дальнейшей работы над книгой, но и для всей биологической антропологии Великобритании.
Оксфорд 1976 г,
Дж. А. Харрисон Дж. С. Уайнер Дж. М. Тэннер
ЧАСТЬ I
ЭВОЛЮЦИЯ ЧЕЛОВЕКА ДЖ. ХАРРИСОН, ДЖ. УАЙНЕР, ДЖ. ТЭННЕР И В. РЕЙНОЛДС
Глава 1
ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
Принципы эволюции
Эволюционная теория, правильность которой подтверждена многочисленными фактами, исходит из того положения, что все организмы родственны друг другу и имели общих предков на той или иной стадии развития жизни. Органический мир в прошлом и настоящем со всей сложностью и многообразием представляет в конечном счете продукт единого эволюционного процесса. Природу этого процесса впервые осознали со всей ясностью Чарлз Дарвин и Альфред Уоллес.
Сущность дарвинского учения состоит в следующем. Всем популяциям присуща способность к избыточному воспроизведению. Однако численность популяций, по крайней мере в течение достаточно длительного срока, остается более или менее постоянной. Из числа родившихся выживает только часть, и лишь наиболее приспособленные участвуют в продолжении рода, что и составляет сущность понятия «борьба за существование». В природе нет абсолютно одинаковых индивидуумов, и в пределах каждой популяции существует значительная изменчивость. В силу этого можно ожидать, что некоторые признаки индивидуумов окажутся более полезными для создания жизнеспособного потомства. Если эти признаки передаются по наследству, то это означает, что они с большей частотой будут встречаться в следующем поколении и что дифференциальные плодовитость и выживаемость привели в конце концов к генотипическому изменению популяции. В этом заключается роль естественного отбора в процессе эволюции. Хотя такие изменения в каждом последующем поколении относительно невелики, тем не менее эволюцию всего живого можно объяснить именно действием естественного отбора в условиях постоянно возникающих изменений, особенно если учесть длительность этого процесса. Современная наука считает, что жизнь зародилась по меньшей мере около 1000 миллионов лет назад.
Динамику отбора удалось проследить, изучая естественные популяции растений и животных и опираясь при этом на научные
14
Часть I. Эволюция человека
гипотезы, выдвинутые эволюционной теорией. Природа наследственной изменчивости, общая для всех популяций, в настоящее время в достаточной степени выяснена. Гораздо менее понятен механизм возникновения изменчивости. Как будет видно из дальнейшего, возникновение изменчивости частично можно объяснить рекомбинацией наследственных факторов, уже существующих в данной популяции. Однако главной причиной появления всех новых изменений в организмах, безусловно, являются генные и хромосомные мутации. Природа же их пока не ясна. Известно лишь, что воздействие среды само по себе не ведет к появлению таких мутаций, которые дают преимущество организму в данной среде. Напротив, большая часть мутантных генов вредна с точки зрения борьбы за существование и, следовательно, элиминируется естественным отбором; лишь случайно могут возникнуть мутации, обеспечивающие селективное преимущество и потому закрепляющиеся в популяции. Изменения в развитии индивидуума, вызываемые ими, по-видимому, незначительны. Именно этим можно объяснить длительность и постепенность эволюционных изменений. Внезапное появление новых растительных форм при полиплоидии можно объяснить тем, что в этих формах необычный состав генов передается сразу большому числу особей в результате вегетативного размножения. Однако в эволюции животного мира резкие изменения подобного рода, по-видимому, не играли существенной роли. Редкость мутаций обеспечивает устойчивость генетической структуры популяций и ограничивает скорость эволюционного процесса.
Эволюция и экология
Если за тот или иной промежуток времени одна биологическая форма эволюционировала медленнее другой, то из этого не следует делать вывод, что причина кроется в недостаточно быстром появлении новых изменений, т. е. в генетической «инерции» данной формы. Скорее всего фактором, определяющим скорость эволюционных изменений в популяциях на протяжении достаточно длительного периода времени, является характер действовавших сил отбора. Причина того, что современные простейшие не эволюционировали, скажем в человека, кроется не в том, что их генетические потенции более ограниченны, чем у тех простейших, которые были предками человека, а в том, что в данной группе естественный отбор не действовал в указанном направлении. Для целого ряда экологических ниш степень организации, свойственная простейшим, является наилучшей, и в этих нишах действие отбора состояло в том, чтобы сохранить и усовершенствовать данный тип организации. Отсюда ясно, что в то время как происхождение изменений в организмах не обусловлено непосредственно
Глава, 1. Закономерности эволюции 1S
влиянием окружающей среды, весь ход эволюционного процесса способствует тому, чтобы сделать организмы более приспособлевр-ными для использования существующих условий окружающей среды.
Требования, которые предъявляют различные условия среды к организмам, зачастую взаимно исключают одно другое. Так, животное, в совершенстве приспособившееся к обитанию на деревьях, не в состоянии в то же время быстро бегать, хорошо рыть норы или плавать, хотя, конечно, может случиться, что в некоторых конкретных условиях животному полезно владеть всеми этими навыками одновременно. Специфичность адаптации имеет далеко идущие эволюционные последствия. Если в силу естественного отбора организм приспособился к определенной нише, он может перейти в другую только при сочетании определенных условий, так как необходимые приспособительные признаки могут появиться и эволюционировать только в данной нише. Форма, никогда не занимавшая данную нишу, не имеет свойственной этой нише адаптации и потому вряд ли сможет успешно конкурировать с формами, ее занимавшими. Сказанное означает также, что биологические формы, занявшие определенную экологическую нишу, приспосабливаются к условиям жизни в ней. Если такое приспособление достигнуто, то исчезновение определенных природных условий ведет к вымиранию данной формы. Бывает, однако, что адаптация, необходимая для определенной экологической ниши, в то же время облегчает выживание формы во многих других нишах, и, следовательно, делает возможным их заселение. Это происходит в том случае, если преимущества, связанные с общей приспособленностью данной биологической формы, перевешивают тот ущерб, который обусловлен отсутствием специальных адаптаций. Высшие млекопитающие вытеснили предшествующие формы, по-видимому, благодаря своей большей способности к эффективному воспроизведению потомства. Сумчатые млекопитающие выжили только в Австралии и в меньшей степени в Южной Америке. Австралия отделилась от остального материка Евразии до того, как появились плацентарные млекопитающие; высшие млекопитающие были завезены в Австралию европейцами. Как мы увидим далее, начало эволюции человеческого мозга связано с некоторой специальной адаптацией — приспособлением к древесному образу жизни. Однако именно эта адаптация открыла в дальнейшем много различных эволюционных возможностей и в значительной мере обусловила замечательную способность человека изменять и подчинять себе окружающую природу.
На данном этапе важно подчеркнуть различия между приспособительными признаками, специфичными для частных условий существования, и приспособлениями, которые представляют собой разные степени эволюционной «специализации». Шея жира
16 Часть I. Эволюция человека
фа, копыто лошади, структура конечностей кистеперых рыб, мозг человека — все это специфичные черты в том смысле, что они являются отличительными признаками рассматриваемых биологических форм. Но в то время как первые два признака, по-видимому, предоставили их обладателям лишь ограниченные эволюционные возможности и потому могут быть названы специализациями, два последних открыли огромные возможности для эволюционного развития. Трудность в распознавании специализаций
Рис. 1.1. Схемы, иллюстрирующие различные положения барабанной косточки (кольца) по отношению к слуховой капсуле (Le Gros Clark, 1959).
а. Примитивные млекопитающие: кольцо находится снаружи, дно барабанной полости не окостеневшее.
б. Лемуроподобные: кольцо окружено костной капсулой, в. Лориевые и «платирриновые» приматы: кольцо расположено на поверхности и участвует в образовании наружной стенки капсулы, г. Катарриновые, а также современный долгопят (Таrsius): кольцо выступает наружу, образуя слуховой проход в форме трубки.
состоит в том, что мы может делать выводы только на осно-вании законченного этапа эволюции. Например, в девонском периоде кистеперых рыб можно было рассматривать как в высшей степени «специализированные» формы, но фактически эта группа дала начало всем четвероногим.
Характер специализации определяется не столько генетическими потенциями организма, сколько экологической обстановкой, в которой он существует. В начале третичного периода, когда на суше не существовало крупных хищников, способных быстро передвигаться, формы, жившие на деревьях, могли легко заселить всю сушу. В середине третичного периода эти формы не могли выжить без некоторых дополнительных приспособлений, которые компенсировали бы недостаток скорости, поскольку к этому времени постоянная борьба за существование в сухопутной фауне привела к появлению быстро передвигающихся по земле плотоядных животных. Одна из главных причин, почему направления эволюционного развития, как правило, необратимы, состоит в том, что эволюция сама по себе связана с изменениями в окружающей организм среде. Другая причина заключается в
Глава 1. Закономерности эволюции
17
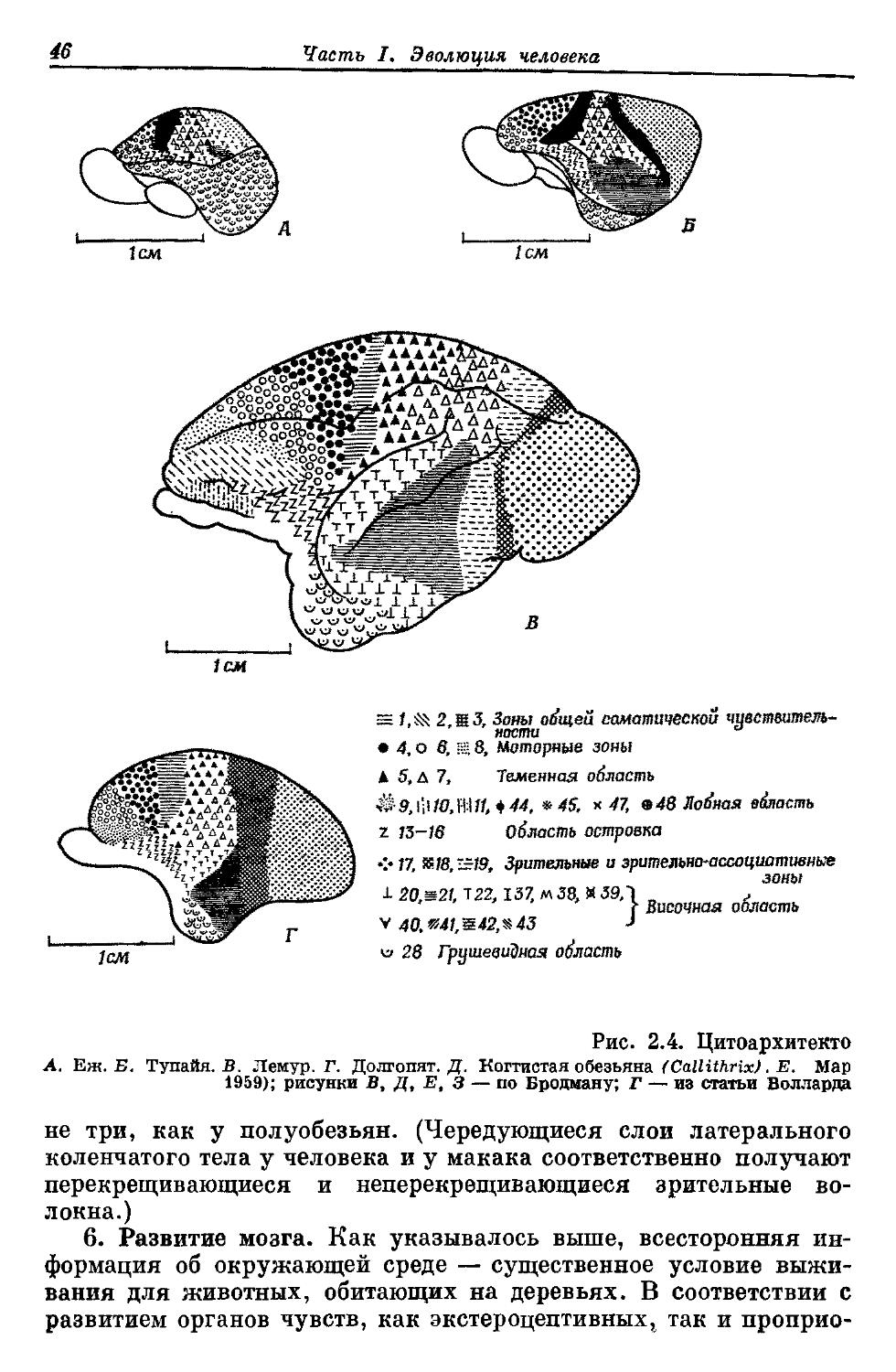
том, что каждая стадия эволюционного развития обеспечивает большую ценность приобретенных организмом признаков для выживания, чем предшествующая. В этом можно видеть одну из важнейших особенностей эволюции. Если у организма имеется несколько путей или способов удовлетворить требованиям окружающей среды и организм «выбирает» один из них в силу того, что подходящие изменения возникают первыми, то эволюционное развитие формы пойдет по пути совершенствования и усложнения признаков, соответствующих выбранному направлению, даже если этот путь сулит и в близком и в более далеком плане более ограниченные эволюционные возможности, чем другие пути развития. Организмы не могут «выжидать», пока возникнет наиболее подходящая вариация; механизм отбора начинает действовать на данную вариацию сразу же, как только она появилась. У лемуров барабанная косточка подвешена в слуховой капсуле в экто-тимпаническом кольце, а у высших антропоидов она находится в конце наружного слухового прохода, вытянутого в форме длинной трубки, и участвует в образовании наружной стенки полости. Если объяснять эти различия только как выражение разных способов защиты барабанной косточки, то трудно понять, как одно состояние могло трансформироваться в другое, поскольку на промежуточных этапах, например у лориобразных лемуров и обезьян Нового Света, эта косточка защищена в меньшей степени (рис. 1.1).
Эволюция и развитие индивидуума
Те особенности индивидуума, которые дают ему эволюционные преимущества по сравнению с другими, отчетливо проявляются во время его роста и развития. Можно сказать, следовательно, что эволюционные изменения происходят путем преобразования индивидуального развития. Сходство между развитием индивидуума (онтогенез) и его эволюционной историей (филогенез) было признано еще древними анатомами, а позже легло в основу известного закона, сформулированного Геккелем («онтогенез повторяет филогенез»). Если бы этот закон был строго верен, то изменение индивидуального развития в процессе эволюции осуществлялось бы только путем гиперморфоза, т. е. добавлением дополнительных стадий в конце онтогенеза предков. Такой гипермор-фоз действительно имеет место, но он никоим образом не является единственным способом изменения развития индивидуума в эволюционном процессе. Любая стадия в развитии может изменяться, если в результате такого изменения индивидуум станет более приспособленным к внешним условиям. Следует, однако, учитывать, что на ранних фазах индивидуального развития различные животные более сходны между собой, чем на поздних (закон
18
Часть I. Эволюция человека
Бара). Причина этого ясна. Изменение в развитии организма, происшедшее и установившееся на некоторой стадии, несомненно, влияет на все последующие стадии. Поэтому взрослое животное как бы накапливает эффекты, связанные с предшествующими изменениями индивидуального развития его предков, тогда как на более ранней стадии развития, которой предшествует меньше стадий, этих модифицирующих влияний будет меньше. Отсюда следует, что взрослые формы различных животных должны сильнее отличаться друг от друга; яйцеклетки, бластулы и гаструлы всех животных весьма сходны, тогда как взрослые формы отличаются бесконечным разнообразием. Вместе с тем существуют и такие модификации, которые затрагивают лишь ранние стадии; у взрослых же форм они утрачиваются. Их роль состоит только в том, что они увеличивают возможности выживания организма в период его развития. Эволюционные изменения, приводящие к таким адаптациям на личиночной или эмбриональной стадии развития, называются ценогенеэом. Ценогенез сыграл особо важную роль в эволюции насекомых, у которых раннее развитие часто протекает без связи с дифференцировкой личиночных форм; однако ценогенез наблюдался также и у других групп животных. Примером ценогенетической адаптации, имевшей весьма серьезные последствия, может служить эволюция оболочек плода с постепенным превращением в плаценту млекопитающих. Форма головы у большей части взрослых млекопитающих длинная и узкая, не резко отличается от головы пресмыкающихся. Однако голова плода млекопитающих имеет шаровидную форму, что облегчает прохождение его по родовым путям.
В настоящее время общепризнано, что эволюционное изменение онтогенеза может происходить в направлении, обратном тому, которое предсказывал Геккель; вместо добавления новых стадий к процессу индивидуального развития предков имеет место сокращение сроков развития. Для того чтобы такое сокращение могло произойти, необходимо, чтобы организмы достигали половой зрелости на все более ранних морфологических стадиях. При этом естественный отбор должен благоприятствовать изменениям, приводящим к более раннему половому созреванию, если в результате этого организм сможет избежать такой специализации взрослых форм, которая, будучи вызвана изменением окружающих условий, начала бы ограничивать его способность к выживанию. Эволюционные изменения, состоящие в сохранении инфантильных признаков, известны под названием неотении. Само происхождение хордовых, вероятно, обязано сочетанию ценогенеза и неотении, так как можно предполагать, что хордовые произошли от личиночных форм иглокожих. Происхождение формы головы у человека, по-видимому, объясняется сохранением головы, характерной для плода. В строении человеческого тела можно обнару-
Глава 1. Закономерности эволюции
19
Рис. 1.2. Схемы, иллюстрирующие положение головы по отношению к туловищу (De Beer, 1958).
а. Эмбрион собаки, б. Взрослая собака, в. Эмбрион человека, г. Взрослый человек. Изгиб, характерный для эмбриона, сохраняется у взрослого человека.
жить много типично «инфантильных» черт, возникновение которых обусловлено неотенией; черты эти следующие: изгиб оси черепа, характерное расположение большого затылочного отверстия, уплощение и небольшой размер лицевого скелета по сравнению с мозговым, отсутствие волосяного покрова (рис. 1.2).
Типы эволюционных изменений
Можно считать, что эволюционные изменения складываются из двух разных процессов, протекающих одновременно. Во-первых, это изменения, протекающие в одной филогенетической линии со сменой поколений; такие изменения можно назвать фи-летическими (близким по смыслу понятием является анагенез Ренша, который, кроме того, предполагает эволюционный прогресс в одной линии форм). Во-вторых, могут иметь место «разрывы» линии происхождения от предков и тогда происходит эволюция большого числа одновременно развивающихся форм. Такой тип изменений называют кладогенезом.
20 Часть I. Эволюция человека
Очевидно, что дифференциация популяций может происходить лишь тогда, когда они изолированы друг от друга; в противном случае мутантные гены, появляющиеся в одной популяции, распространяются и на другие. При филетическом изменении само время отделяет последующие популяции от предыдущих; для кладогенеза же необходимы другие изолирующие факторы, и в первую очередь пространственное разобщение популяций. Последнее может осуществляться самыми различными способами. Но если единая популяция разделится на две (или более) пространственно разобщенные популяции, то, какова бы ни была причина разобщения, эти популяции будут в дальнейшем эволюционировать независимо, так как мутантные гены, возникшие в одной из них, остаются в ее пределах. При сохранении пространственного разобщения генетическая дифференциация популяций может зайти настолько далеко, что в будущем, даже если контакт между ними восстановится, гибридизация не будет происходить. Иными словами, если в действие вступают эндогенные факторы, то для поддержания кладогенеза более не нужны какие-либо внешние изолирующие факторы. При достижении такой стадии развития каждая популяция превращается в новый вид. Майр определяет вид как «группу фактически или потенциально скрещивающихся естественных популяций, которые физиологически изолированы от других таких групп». Две популяции, пространственно перекрывающиеся, т. е. симпатрические, но не скрещивающиеся между собой, могут рассматриваться как «хорошие» виды. Если же две популяции остаются пространственно изолированными (аллопатрическими), то, очевидно, критерий скрещивания, по которому можно было бы распознать вид, уже неприменим. В этом случае для оценки степени изменений, связанных с репродуктивной изоляцией, обычно используют степень различия двух близких сим-патрических видов. Этот же критерий применим и для распознавания палеонтологических видов.
Имея в виду рассмотренные выше общие понятия, мы сможем теперь перейти к описанию некоторых основных черт эволюции, установленных при изучении всего многообразия как существующих, так и вымерших животных форм.
Адаптивная радиация
В окружающей организмы живой и неживой природе происходят непрерывные изменения, которые открывают перед животными все новые и новые эволюционные возможности. Эволюция наземных животных стала возможной в результате заселения суши растениями. Упадок рептилий в конце мезозойской эры, по-видимому вызванный климатическими изменениями, открыл для ранних млекопитающих бесчисленные экологические пиши. Мле
Глава 1. Закономерности эволюции
21
копитающие смогли противостоять климатическим изменениям: благодаря постоянству температуры тела. В конце третичного периода материки Северной и Южной Америки, до того времени изолированные, вновь соединились, и сухопутный мост между ними дал возможность северным формам использовать большое число южных экологических ниш. (Конечно, геологические изменения открыли пути для переселения животных и в обратном направлении, на север, но лишь немногие формы, подобно опоссуму, оказались биологически приспособленными для использования условий новой среды.) Далее, как уже указывалось выше, некоторые частные адаптации могут случайно приобрести более общую ценность для организма. В таком случае они дополнительно открывают организму, обладавшему такой адаптацией, много новых экологических ниш. Если перед группой животных открывается большое число эволюционных возможностей, то развитие группы идет в сторону образования множества одновременно эволюционирующих различных форм. Заселяются все наличные ниши, и формы, занимающие их, приобретают посредством естественного отбора необходимые специальные адаптации. В этом заключается процесс адаптивной радиации. Первыми позвоночными, которые приобрели независимость от водных условий жизни, были пресмыкающиеся, и в течение мезозойской эры происходила их адаптивная радиация и приспособление к многочисленным нишам, существовавшим тогда на суше. Эта радиация породила не только большое разнообразие травоядных и плотоядных форм динозавров, от небольших текодонтов до гигантских бронтозавров и тиранозавров, но также ящериц и змей, крокодилов, птеродактилей и даже такие группы, как черепахи, ихтиозавры и плезиозавры, которые вновь переселились в море. Само происхождение птиц и млекопитающих следует рассматривать как часть этой огромной радиации рептилий.
В третичном периоде радиацию рептилий сменила радиация млекопитающих. В небольшом масштабе мы можем проследить ее на примере сумчатых млекопитающих Австралии, лемуров Мадагаскара и небольшой группы птиц — так называемых дарвиновых вьюрков на Галапагосских островах. В Австралии основная группа сумчатых явилась родоначальником не только самых разнообразных наземных травоядных и плотоядных форм, но и форм роющих, планирующих, обитающих на деревьях, в воде и т. д. Радиация на Мадагаскаре привела к появлению таких необычных форм лемуров, как Megaladapis и Daubentonia. На Галапагосских островах естественный отбор, действуя на одну и ту же анцестральную группу, способствовал возникновению множества различных видов вьюрков, каждый из которых приспособился к определенному, имеющемуся на островах источнику пищи. Приводя эти частные примеры, мы вовсе не хотим сказать, что
22
Часть I. Эволюция человека
адаптивная радиация имела ограниченное значение в эволюции. Весь кладогенез — это адаптивная радиация в широком смысле слова. Однако последний термин обычно применяют в тех случаях, когда единственный род животных приводит более или менее одновременно к появлению большого разнообразия отличных друг от друга форм, каждая из которых приспособилась к определенной экологической нише.
Конвергенция и параллелизм
Строение тела животного определяется его образом жизни. Животное, которое постоянно роет землю, не может быть крупт ным; оно должно обладать обтекаемой формой тела и другими приспособительными признаками. Если такое животное не в состоянии прокапывать себе путь телом или пропускать частички почвы через кишечник, то оно должно обладать достаточно сильными конечностями для того, чтобы рыть землю. Глаза не только утрачивают свое функциональное значение, но даже становятся недостатком: зрение у таких животных либо вовсе утрачено, либо сильно ослаблено. Это лишь некоторые из обязательных признаков или следствий подземного образа жизни. Если бы мы захотели охарактеризовать другие аспекты данной экологической ниши, например климатические условия или источники пищи, то обнаружили бы и другие ограничения. Те, кто изучает эволюцию, давно уже поняли, что достаточно полная характеристика условий окружающей среды позволяет также описать в деталях строение животного, приспособившегося к этой среде. Исключением будут лишь те случаи, когда у животного имеется не один, а несколько способов использовать данную нишу и если инерция, селекционная или генетическая, задержала приобретение животным наиболее эффективных форм адаптации. И в том и в другом случае очевидно, что если одна и та же экологическая ниша существует в двух или более местах, животные, которые ее занимают, оказываются сходными независимо от того, связаны ли они близким родством. Когда две формы более схожи, чем их предки, эволюцию этих форм называют конвергентной. Если они приобретают одни и те же характерные приспособительные признаки независимо, но эти признаки делают их не более похожими друг на друга, чем были похожи их предки, то их эволюцию можно считать параллельной. Различие — только один из признаков конвергенции, тогда как параллелизм — всего лишь предельный случай конвергенции. Одним из примеров конвергенции является сходство формы тела у китов и рыб. Такая форма связана с необходимостью быстро передвигаться в воде. Установлено, что такой же формой тела обладали и ихтиозавры. Следствием одинакового образа жизни является также общность многих признаков сумчатых и плацен
Глава 1. Закономерности эволюции 23
тарных хищных. Млекопитающие «кроты» в своей эволюции дважды претерпели изменение как семейство отряда насекомоядных и один раз в составе отряда сумчатых. Эволюция обезьян Нового Света происходила почти, если не совершенно, изолированно от обезьян Старого Света. Их происхождение являет пример параллелизма, поскольку их «дообезьяньи» предки также были весьма сходны.
Число подобных примеров можно было бы увеличить ввиду чрезвычайно бюлыпой распространенности конвергенции в эволюционном развитии. Вместе с тем эта распространенность является причиной многих трудностей при попытке определить эволюционную историю группы, о которой имеются весьма скудные палеонтологические сведения: ведь, поскольку существует конвергенция, нельзя автоматически считать сходство показателем близкого родства в эволюции. Правда, в некоторых случаях, например в эволюции китов и рыб, наличие конвергенции легко установить; так, несмотря на конвергенцию, киты сохранили многие признаки млекопитающих. Но если бы мы не знали о существовании амфибий и пресмыкающихся, то могли бы легко заключить, что киты представляют собой наиболее примитивных млекопитающих. Кроме того, киты и рыбы сохранили различие не только потому, что для установления полного сходства необходима была бы очень резкая степень изменений (киты, подобно всем другим млекопитающим, не имели предков среди рыб), но и в связи с тем, что в морях и океанах существуют экологические ниши, которые могут быть лучше использованы морскими млекопитающими, нежели рыбами. Конвергенцию особенно трудно обнаружить в случае слишком незначительных различий между предками конвергентных форм. Пример этому можно найти, изучая внутривидовую изменчивость человека в зависимости от географических условий. Можно предположить, например, что темнокожие народы Меланезии и Африки приобрели многие черты сходства от своих общих предков. Но не исключено также, что они эволюционировали конвергентно, в результате большого сходства сил отбора в двух географических зонах.
Направления эволюционного развития
В целом можно считать, что межвидовая борьба все более обособляет экологические ниши, а внутривидовая совершенствует их использование. Для филетического изменения характерно образование адаптаций, все более ограничивающих условия существования, и появление линий форм с различными тенденциями развития. Это легко проследить на примере эволюции лошади, развитие которой характеризовалось постепенным увеличением
24
Часть I. Эволюция человека
размеров тела, уменьшения числа пальцев, «моляризацией» премоляров, усовершенствованием строения коренных зубов, увеличением их длины. Сходные тенденции можно обнаружить и в любой другой группе животных при наличии достаточного количества палеонтологических данных. Само существование таких тенденций помогает объяснить ход эволюции. Если, например, для нас неясно происхождение какой-либо линии форм, но из возможных групп предков у одной обнаруживаются в начальной стадии признаки, которые затем усиливаются в рассматриваемой линии, то, вероятнее всего, именно эта группа является предковой формой.
Многие склонны считать существование направленной эволюции свидетельством того, что организмы вступают на тот или иной путь эволюции под влиянием некоего эндогенного стимула. Вымирание таких форм, как саблезубый тигр или ирландский лось, приписывалось действию именно таких эндогенных стимулов, выводивших тенденцию развития за пределы приспособительной изменчивости. Было высказано предположение, что клыки саблезубых тигров стали в конце концов настолько длинными, что мешали животным хватать добычу. В то же время указывалось, что клыки саблезубого тигра Eusmilus, жившего в олигоцене, были не менее длинными, чем у тигра Smilodon, жившего в плейстоцене. Симпсон (Simpson) замечает по зтому поводу: «Совершенно нелогично считать в конечном счете неэффективным механизм, который просуществовал без заметных изменений в обширной группе и был в высшей степени эффективным в течение 40 млн. лет».
При обсуждении возможных направлений эволюции стоит указать на существование определенных «ростовых» соотношений между отдельными частями организма (аллометрические соотношения). Если одна какая-либо структура претерпевает изменение, другие также автоматически изменяются на величину, определяемую этими соотношениями. Конечно, если в определенной среде необходимо определенное изменение соотношений роста, такое изменение возникнет. Большую часть эволюционных изменений в зубном аппарате лошади можно непосредственно связать с изменением размеров тела, однако у тех лошадей, которые перешли от ощипывания листвы и побегов к питанию травой, установилось новое соотношение между высотой коронки зубов и размерами тела: стирание зубов, вызванное пережевыванием травы, привело к развитию гипсодонтии (высокой коронки зубов).
До сих пор нет неоспоримых данных в пользу ортогенеза, т. е. действия эндогенных эволюционных стимулов, но зато можно считать твердо установленным, что эволюционные тенденции часто направлены в сторону специализации признаков. В развитии ирландского лося все стадии в целом были приспособительными. Однако вымирание этого вида, без сомнения, связано с наличием у животного больших рогов, по-видимому развившихся в резуль
_______________Глава 1. Закономерности эволюции 25
тате соревнования самцов внутри вида. Даже незначительное изменение внешних условий превратило этот признак в невыгодный.
Преадаптация
Эволюция группы ограничена не только возможностями, которые ей предоставляет среда, но также и способностью группы воспользоваться этими возможностями раньше других. Не удивительно, что родоначальниками первых наземных животных стали прибрежные формы, так как они обитали в среде, наиболее благоприятной для заселения суши. Это не значит, однако, что все прибрежные формы способны были воспользоваться такой возможностью. От пластинчатожаберных моллюсков, например, не произошло ни одно наземное животное, хотя моллюски получили не меньше таких же экологических возможностей, чем другие группы. Отсюда еще не следует, что в организации животных имеются некоторые присущие им ограничения, при всех обстоятельствах препятствующие их переходу в другие экологические условия. Причина того, что пластинчатожаберные не дали ни одной наземной формы, вероятно, заключалась в том, что естественный отбор гораздо легче мог видоизменить другие формы таким образом, чтобы они смогли занять наземные экологические ниши и воспрепятствовать в дальнейшем их колонизации извне. Тем не менее можно все же сказать, что пластинчатожаберные были менее преадаптированы к существованию на суше по сравнению, например, с брюхоногими моллюсками или кистеперыми рыбами, которые нуждались в относительно меньшем изменении их организации для развития подходящих адаптаций. В самой преадаптации нет ничего мистического. Следует ожидать, что некоторые условия делают животных более приспособленными для использования предоставляющихся им эволюционных возможностей. От степени преадаптации во многом зависит то, какая именно группа животных изберет тот или иной способ существования. Как мы увидим далее, эволюция человека как наземного животного во многом связана с приспособлением к жизни на деревьях, которым обладали его предки. Это приспособление оказалось преадаптацией для определенного типа наземного существования.
Явление преадаптации помогает установить эволюционные отношения между животными формами. Если две формы избрали себе почти одинаковый образ жизни, как, например, лемуры и лори, то они смогли сделать это, вероятно, потому, что обладали сходными преадаптациями. Последние они могли унаследовать от общих предков или приобрести конвергентно. Даже те формы, условия жизни которых существенно различались, могли обла
26
Часть I. Эволюция человека
дать одинаковой преадаптацией в том случае, если выживанию этих форм в разных условиях способствовала эволюция сходных признаков. Мы увидим впоследствии, что много сходных признаков у обезьян и человека развивалось параллельно и для совершенно различных целей. Уже одно то, что происходили сходные изменения, предполагает наличие общих преадаптаций.
Одной из наиболее общих преадаптаций является, по-видимому, небольшой размер тела животного. Большинство крупных радиаций возникло от сравнительно небольших животных. У таких животных время жизни поколения обычно невелико, и в силу этого отпадают ограничения, связанные с генетической инерцией, и облегчается более быстрый эволюционный процесс.
Темп эволюции
На основании изучения существующих биологических форм и ископаемых остатков можно сделать определенный вывод, что скорость появления филетических изменений и скорость кладогенеза непостоянны. Некоторые формы оставались более или менее неизменными на протяжении целых геологических периодов, тогда как другие эволюционировали чрезвычайно быстро. Далее, отдельные линии в течение одного периода времени почти не изменяются, а затем внезапно делают эволюционный скачок. Современный бра-хиопод Lingula, живущий на деревьях, практически неотличим от своего кембрийского предка Linqulella. Скелет опоссума почти не изменился за 80 млн. лет, тогда как Eohippus за 60 млн. лет превратился в Equus. Парнокопытные эволюционировали гораздо медленнее в период от эоцена до миоцена, чем в последующие эпохи.
Скорость эволюционных изменений в группе часто выражают числом новых биологических родов, появляющихся в единицу времени, например за миллион лет. Такая характеристика является, однако, довольно грубой, так как в отличие от вида систематическая категория «род» во многом произвольна, и различия, которые в одной группе можно рассматривать как родовые, могут не быть таковыми в другой. Однако даже такая приближенная характеристика позволяет оценивать и сопоставлять скорости эволюционных процессов, причем эти оценки согласуются с полученными более объективными методами.
Новый биологический род возникает как путем филетических изменений, так и путем кладогенеза. Если учитывать только первый путь, то можно установить, что скорость эволюции у копытных животных в течение третичного периода составляет один род за 8 млн. лет. Например, в родословной лошади появлению рода Equus предшествовало 8 родов, т. е. один род за 7,5 млн. лет. Скорость эволюции многих других групп млекопитающих, включая приматов, по-видимому, аналогична. А вот формы, подобные ежу
Глава 1. Закономерности эволюции
27
и тупайе, все еще очень похожи на тех древних насекомоядных, от которых произошли все высшие млекопитающие.
Как мы уже указывали, различие в скорости эволюционных процессов не означает, что медленно развивающиеся формы обладают большей генетической инерцией. Если животное в совершенстве приспособилось к некоторой экологической нише, естественный отбор будет сохранять его неизменным, пока остается неизменной ниша. Можно, следовательно, предположить, что ниши, занятые опоссумом и ежом, не изменились радикальным образом со времени возникновения зтих форм.
До сих пор речь шла только о скорости эволюции различных животных, однако и разные части одного и того же животного могут развиваться с разной скоростью (соматическая эволюция). Например, нога лошади Hipparion очень похожа на ногу ее миоценового предка, а зубы сильно отличаются. (Интересно отметить, что другие лошади раннего плиоцена достигли различных стадий развития; у Hypohippus в отличие от Pliohippus ни ноги, ни зубы почти не изменились.) Другой пример различия в скорости соматической эволюции мы находим у Necrolemur, у которого все основные черты совпадают с признаками примитивного долгопята, однако зубы отличаются от анцестрального типа значительно больше, чем у любого другого долгопятоподобного. Как мы увидим, эволюцию человека можно подразделить на три большие фазы: 1) приобретение навыков хождения на двух ногах, 2) уменьшение размеров челюстей, 3) увеличение мозга. Есть все основания допустить, что эти три фазы не очень четко разграничены, однако в настоящее время найдены ископаемые формы, вполне приспособленные к прямохождению, но с такими особенностями черепа, которые мало изменились по сравнению с анцестральным антропоидным типом. Даже у современного человека высокий уровень развития мозга сочетается с рядом примитивных черт, например с примитивной общей морфологией передних конечностей. Вместе с тем следует учитывать, что, хотя различные системы органов могут эволюционировать с разной скоростью, животное на любом этапе развития должно быть наилучшим образом приспособлено к условиями внешней среды. Это сразу ограничивает возможное число независимых изменений в организме. Отдельные части единой функциональной системы должны развиваться совместно; например, эволюция верхней челюсти млекопитающего не может происходить со скоростью, совершенно отличной от скорости развития нижней челюсти. Кстати, функциональная несовместимость челюстей явилась одним из фактов, благодаря которому была выяснена фальсификация ископаемых остатков в Пильтдауне (Англия).
28
Часть I. Эволюция человека
Таксономия
Задача таксономии состоит в том, чтобы идентифицировать, описать, дать наименование и классифицировать организмы. Каждое животное после его открытия относят к определенному виду, вид — к роду, семейству, отряду, классу и типу. Все эти категории являются обязательными; другими (подотряд, подсемейство, надсемейство и т. д.) можно пользоваться, если информация о группе, выраженная основными категориями, оказывается недостаточной (см., например, классификацию приматов, гл. 2).
Различают три основных типа классификаций: 1) описательную, 2) фенетическую, или естественную, и 3) филетическую (филогенетическую). Единственная цель описательной классификации — идентифицировать различные группы; согласно этой классификации, даже сильно различающиеся формы могут быть сгруппированы вместе только на основании общности одного важного признака. Примером могут служить зоны с различной фауной и флорой, введенные для идентификации животных и растений, относящихся к определенным регионам или странам. В противоположность этому в фенетической классификации организмы располагаются в соответствии со степенью общего сходства; эту классификацию до Дарвина называли естественной. Согласно фенетической классификации, в одну группу попадают наиболее сходные формы. При определении фенетического родства возникают главным образом методические трудности. Для классификации необходимы методы, с помощью которых частные различия в большом числе признаков комбинируются в единое выражение общего различия, а такая процедура стала объективно возможной лишь в последние 10—20 лет благодаря развитию современных методов статистики, например методов многомерного анализа. Так как не все признаки можно выражать в одинаковых единицах, возникает необходимость приведения их к некоторой абстрактной шкале. Но, пользуясь такими сложными понятиями, как структура, физиология, поведение и генетика организма, часто бывает трудно установить, в чем именно сопоставим^ различные формы. Предполагают, например, что независимую информацию о фенетических связях могут дать признаки, которые лишь математически коррелируют друг с другом. Тем не менее можно в конце концов подойти к составлению фенетической классификации животных так же, как к сравнению ряда неодушевленных предметов, без учета природы того или иного сходства или различия. Филетическая классификация в отличие от фенетической пытается сгруппировать организмы согласно их эволюционным связям. В тех случаях, когда в ходе эволюционного процесса все линии форм диверги-руют с одинаковой скоростью, мера общего сходства будет также мерой эволюционной связи и фенетическая классификация будет
Глава 1. Закономерности эволюции
29
совпадать с филетической. Однако, хотя степень сходства между формами часто указывает на близость их филетического родства, мы уже убедились в том, что эволюция приводит как к конвергенции линий, так и к их дивергенции и что скорость изменений в разных линиях и в одном и том же ряду поколений в разное время редко бывает одинаковой. Это означает, что формы могут быть весьма сходны, но не потому, что они имеют общего предка, а потому, что эволюционируют конвергентно; большие различия могут указывать либо на медленную дивергентную эволюцию от отдаленного общего предка, либо на быструю дивергенцию от недавнего предка.
Рис. 1.3. Схема двух типов эволюционной связи.
Форма Б связана с формой А «отцовским родством», а с формой В — кладогенетическим.
Так как скорость дивергентной эволюции не постоянна, существуют два различных типа эволюционных соотношений; непонимание этого в прошлом часто приводило к недоразумениям. Все те черты сходства между организмами, которые нельзя связать с конвергенцией, возникают путем наследования их от общих предков, и в качестве одного из аспектов филетического родства можно рассматривать, следовательно, степень, в какой различные формы в результате развития от общих предков обладают одинаковыми признаками. Этот тип связи называется прямым, или «отцовским родством». Однако формы могут обладать малым отцовским родством и все же быть достаточно близкими в том смысле, что в недавнем прошлом они имели общего предка. Такая ситуация возникает при быстрой эволюции по типу кладогенеза. Этот вид генеалогической связи между формами называют «кладо-генетическим родством». Приводимая на рис. 1.3 схема иллюстрирует различие между этими двумя типами эволюционных взаимоотношений.
Очевидно, что в том случае, когда данные палеонтологии о какой-либо группе животных недостаточно полныг исследователе
so
Часть I. Эволюция человека
желающий выявить филетические связи в группе, должен установить, какова вероятность конвергенции и каков темп эволюционных изменений и вообще имели ли место те или иные изменения. При этом он может опираться на ряд общих принципов, из которых мы приводим наиболее важные.
1. Если по данным палеонтологии одна форма предшествует другой, то мало вероятно, чтобы вторая могла быть предком первой. Не исключена, однако, возможность, что установленная палеонтологами временная последовательность форм только кажущаяся, и в действительности более поздняя форма долгое времй существовала в районе, где условия для фоссилизации были мало благоприятными или же где не производилось раскопок. (Принципиальное значение данных о времени появления различных форм состоит в том, что эти данные показывают, в каком направлении происходит эволюция.)
2. Если две сходные формы обитают в областях, которые в течение продолжительного времени были географически разобщены, вероятность того, что они развивались конвергентно, выше, чем если бы эти формы проживали в смежных географических областях.
3. Если две сходные формы населяют экологические ниши одинакового типа, то вероятнее всего, что эти формы эволюционируют конвергентно.
4. Если, напротив, различие двух форм можно приписать различию в их экологии, то следует предположить, что такие различия развивались очень быстро.
5. Если какой-либо признак представляет ценность для выживания в различных жизненных условиях, т. е. имеет общую ценность, то его утрата становится маловероятной и, следовательно, форма, обладающая этим признаком, обычно не бывает анцест-ральной по отношению к форме, лишенной такого признака.
6. Если в ряде форм обнаруживается прогрессирующая функциональная тенденция, то такие формы скорее всего являются стадиями одной родословной или же стадиями близких родословных.
7. Если у двух форм развивается одна и та же функция, но разными путями, то мало вероятно, что одна форма произошла от другой, в особенности в тех случаях, когда промежуточные стадии обладали меньшей приспособленностью.
8. Если некоторая частная функция определяет собой природу большого числа признаков, то формы, которые имеют один из этих признаков, приобретают другие либо вследствие конвергенции,; либо как наследие от общих предков. Такие признаки всегда являются функциональными коррелятами рассматриваемых форм и не дают никакой информации о филетической связи между этими формами.
Глава 1. Закономерности эволюции 31
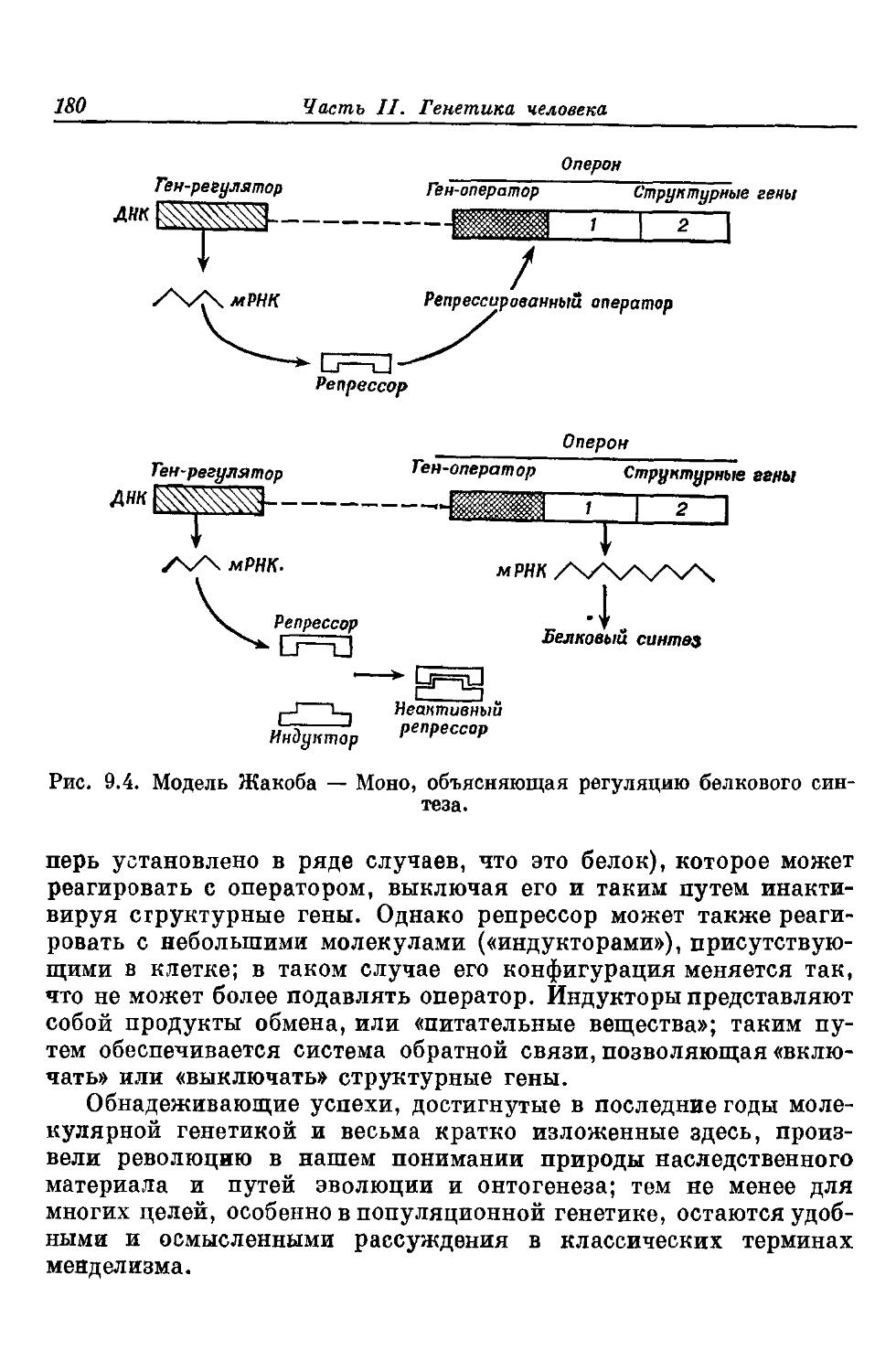
Приведенные здесь соображения показывают, насколько необходимо знание всех признаков организма, в особенности функциональных и экологических, для того чтобы можно было выявить с достоверностью эволюционные отношения в группе, о которой имеются только скудные палеонтологические данные. Если знанием таких признаков мы не располагаем, то следует воспользоваться фенетической классификацией. Вместе с тем мы имеем достаточно сведений о ближайших родственных человеку формах, как существующих, так и вымерших, чтобы оправдать попытки вскрыть ход и закономерности эволюции человека.
Глава 2
ПРИМАТЫ
Характеристика приматов
Внезапный упадок рептилий в конце мезозойской эры открыл перед ранними млекопитающими, которые в то время составляли немногочисленную группу, большие эволюционные возможности. В течение всего третичного периода происходила адаптивная радиация млекопитающих, освоение свободных экологических ниш (табл. 2.1) и совершенствование условий дальнейшего развития. Исходной группой для высших, или плацентарных, млекопитающих были насекомоядные — отряд, который в настоящее время представлен такими формами, как еж, землеройка и крот. К приматам относится большая часть тех млекопитающих, которые перешли к обитанию на деревьях и приобрели соответствующие адаптации. Сходство этих адаптаций у различных приматов, вне всякого сомнения, обязано их древесному образу жизни. Однако те животные, которые воспользовались открывшимися перед ними путями эволюции, должны были обладать для этого соответствующими возможностями, унаследованными от общих предков. Именно поэтому отряд приматов можно считать единой как филетической, так и фенетической группой.
К этому отряду относятся лемуры Мадагаскара, лори, современным местом обитания которых является тропическая Африка и Азия, долгопяты, единственный сохранившийся род которых населяет Индокитай, обезьяны Нового Света, обезьяны Старого Света, человекообразные обезьяны (в том числе гиббоны и три большие обезьяны — орангутан, шимпанзе и горилла) и человек (рис. 2.1). До недавнего времени в отряд включали также тупайю, живущую в Индии и Юго-Восточной Азии. Она близка к лемуру по многим морфологическим признакам, таким, как строение медиальной стенки глазницы или общее строение области среднего уха. Однако, целый ряд особенностей указывает также на раннее разделение филогенетических линий тупайи и приматов, например тот факт, что среднее ухо, при всем внешнем сходстве с ухом лемура, сформировано из других косточек. Можно, по-видимому, считать, что приматы в своей эволюции прошли уровень организации, свойственный тупайе.
Число различных классификаций приматов весьма велико. В основу принятой нами системы, включающей большинство известных родов, как существующих, так и вымерших, положена
Таблица 2.1
Эволюция позвоночных
Г ео хронологическая шкала
Время, млн. лет Эры Периоды Эпохи Эволюция позвоночных
0.01 2 5 23 36 54 65 135 165 205 230 280 325 360 425 505 2000 Кайнозойская Четвертичный Голоцен (современная) Плейстоцен Бесчелюстные рыбы (Agnotha) Панцирные рыбы ( Placoderml) ^Хрящевые рыбы (Chondrichthyes) Sa o' с £ S' Е Ш д. Кистеперые рыбы (Crossopt е гуд и) ] Амфибии 1 1 igaantnoiPMiqwjadn Птицы 1 Сй 1 % (3 Б о I Приматы ]
Третичный Плиоцен Миоцен Олигоцен Эоцен Палеоцен
Мезозойская Меловой Юрский Триасовый
Палеозойская Пермский Каменноугольный Девонский Силурийский Ордовикский Кембрийский
Архейская
Протерозойская
Таблица 2.2 Jj
Классификация приматов (по Е. Симонсу, 1972)
Отряд: приматы (Primates)
Подотряд: полуобезьяны (Prosimi)
Секция: плезиадапидоподобные * (Plesiadapiformes)
Семейство: плезиадапиды * (Plesiadapidae) — палеоцен — эоцен
Подсемейство: плезиадапины * (Plesiadapinae) Подсемейство: саксонеллины * (Saxonellinae) Семейство: карполестиды * (Carpolestidae) Plesiadapis * Saxonella * — палеоцен — эоцен Carpolestes*
Семейство: паромомиды * (Paromomyidae) Подсемейство: паромомины * (ParOmomyinae) Подсемейство: фенаколеумурины * (Phenacoleumu-linae) Семейство: пикродонтиды * (Picrodontidae?) — меловой—эоцен Paromomys *, Purgatorius * Phenacolemur * — палеоцен Pierodus *
Секция: лемуроподобные (Lemuriformes) Надсемейство: адапоиды (Adapoidea) Семейство: адипиды * (Adipidae) Подсемейство: адапины * (Adapinae) Подсемейство: нотарктины* (Notharctinae) Надсемейство: лемурообразные (Lemuroidea) Семейство: лемуриды (Lemuridae) Подсемейство: лемурины (Lemurinae) Подсемейство: хирогалпны (Cheirogaleinae) Семейство: индриды (Indriidae) Подсемейство: ипдрины: (Indriinae) Подсемейство: гадропитеципы * (Hadropitbecinae) — эоцен Adapts*, Pronycticebus * Notharctus *, Pelycodus * — голоцен Lemur, Hapalemur Cheirogaleus, Microcebus — плейстоцен, голоцен Propithecus, Indri Hadropithecus *
Подсемейство: археолемурины* (Archaeolemurinae)
Семейство: добентониды (Daubentoniidae)
Семейство: мегаладапиды * (Megaladapidae)
Секция; лориподобные (Lorisi formes)
Семейство: лоризиды (Lorisidae)
Подсемейство: лоризины (Lorisinae)
Подсемейство: галагины (Galaginae)
Секция: тарзиеподобные (Tarsiiformes)
Семействе: тарзииды (Tarsiidae)
Подсемейство: тарзиины (Tarsiinae)
Подсемейство: микрохорины* (Microchoerinae)
Семейство: анаптоморфиды * (Anaptomorphidae)
Подсемейство: анаптоморфины * (Anaptomorphinae)
Подсемейство: омомины* (Omomyinae)
Подотряд: высшие приматы (Anthropoidea)
Надсемейство: широконосные «платирриновые»
(Ceboidea)
Семейство: цебусовые (Cebidae)
Подсемейство: ночные обезьяны (Aotinae)
Подсемейство: чертовы обезьяны (Pithecinae)
Подсемейство: ревуновые (Alouattinae)
Подсемейство: капуциновые (Cebinae)
Подсемейство: цебупитекины * (Cebupithecinae)
Подсемейство: паукообразные (Atelinae)
Табл. 2.2' (продолжение)
Archaeolemur *
— голоцен — плейстоцен Daubentonia Megaladapis •
— мноцен—голоцен Loris, Nycticebus, Arctocebus, Perodicticus Galago
— ср. эоцен—голоцен Tarsius Necrolemur *, Pseudoloris *
— эоцен — олигоцен Tetonius*, Anaptomorphus * Omomys *, Hemiacodon *, Ro-oneyia *, Teilhardina *
— олигоцен—голоцен ffomunculus *, Aotus, Callicebus Cacajao, Pithecia Alouatta Cebus, Neosaimiri * Cebupithecia * Ateles, Lagothrix .. 8
Табл. 2.2 (продолжение)
Подсемейство: каллимиковые (Callimiconinae) Семейство: игрунковые (Callithricidae) Семейство: ксенотризиды * (Xenothricidae) Надсемейство: узконосые «катарриновые» (Cercopit-hecoidea) Семейство: мартышкообразные (Cercopithecidae) Подсемейство: мартышковые (Cercopithecinae) Подсемейство: парапитековые * (Parapithecinae) Подсемейство: тонкотелые (Colobinae) Надсемейство: ореопитековые * (Oreopithecoidea) Семейство: ореопитекиды * (Oreopithecidae) Надсемейство: гоминоиды (Hominoidea) Семейство: хилобатиды (Hylobatidae) Подсемейство: плиошиековые * (Pliopithecinae) Подсемейство: гиббоновые (Hylobatinae) Семейство: понгиды (Pongidae) Подсемейство: дриопитековые * (Dryopithecinae) Подсемейство: не установлено Подсемейство: понгинные (Ponginae) Подсемейство: гигантопитековые * (Gigantopit-hecinae) Семейство: гоминиды (Hominidae) Звездочкой отмечены ископаемые формы. Callimico — голоцеп Callithrix — плейстоцен Xenothrix * — олигоцен, голоцен Масаса, Papio. Cercopithecus, Libypithecus * Parapithecus *, Apidium* Presbytis, Colobus, Mesopit-hecus * Oreopithecus *, Mdbokopithecus * — миоцен — ниж. плиоцен — олигоцен—голоцен Pliopithecus *, Limnopithecus *, Aelopithecus * Hylobates, S ymphalangus — олигоцен—голоцен Dryopithecus *, Aegyptopithe-cui, Propliopithecus * Oligopithecus * Pongo, Pan, Gorilla Gigantopithecus * — миоцен — голоцен Ramapithecus *, Australopithe- cus *, Homo
Глава 2. Приматы
37
система Симонса — одна из последних и наиболее признанных классификационных схем (табл. 2.2).
Классическое определение приматов принадлежит Миварту (Mivart, 1836), который дал такую характеристику отряда: плацентарные млекопитающие, обладающие ключицей, глазницами, окруженными костным кольцом, зубами, по крайней мере трех типов; мозг всегда имеет затылочную долю и шпорную борозду; большой палец, по крайней мере на одной паре конечностей, противопоставляется всем остальным; первый палец стопы имеет плоский ноготь или лишен ногтя; отчетливо выражена слепая кишка; свисающий половой член, семенники спущены в мошонку, у самок — одна пара грудных молочных желез.
Ни один из перечисленных признаков в отдельности нельзя считать определяющим. Большая часть их имеется в основном стволе млекопитающих, и в таком смысле эти признаки примитивны. Все черты, отмеченные Мивартом, в какой-то степени характерны для многих других форм млекопитающих, в особенности для некоторых древесных сумчатых или насекомоядных. Более того, не у всех форм, относимых к отряду приматов, имеются все указанные признаки. Особенно отличаются Daubentonia. Распространено мнение, что они возникли лишь как сравнительно недавняя ветвь лемуров на Мадагаскаре. Далее, если включить в отряд приматов семейство Tupaiidae, критерии Миварта станут еще менее определяющими. Очевидно, их следует рассматривать лишь в совокупности, так как у любой частной формы некоторые из признаков могут отсутствовать.
С общебиологической точки зрения гораздо большую ценность представляет изучение эволюционных путей развития приматов. Направления их эволюции, которые характеризуют группу в целом, определяются почти исключительно их древесным образом жизни. Даже формы, которые в настоящее время живут на земле, сохранили черты, свидетельствующие о древесном образе жизни. Изучение этих черт, унаследованных от предков, очень важно для правильного понимания всех биологических особенностей человека.
Проследим основные тенденции, которые в какой-то степени проявились у всех приматов и которые связаны с наследованием ими признаков, характерных для древесного образа жизни.
1. Развитие хватательной функции конечностей. Животные, проводящие большую часть времени на деревьях, должны обладать конечностями, приспособленными для передвижения по ветвям. Животные, подобные белке, используют для этой цели острые когти; у приматов, однако, развитие конечностей пошло по другому пути. Их руки и ноги приобрели способность крепко схватывать предметы, главным образом за счет увеличения независимой подвижности пальцев и в первую очередь большого пальца стопы и первого пальца кисти. У разных форм в разной степени эти
38
Часть I. Эволюция человека
пальцы противопоставляются остальным. Противопоставляющимся считается палец, который может поворачиваться вокруг своей оси таким образом, чтобы его ладонная поверхность была обращена к ладонной поверхности остальных пальцев. У обезьян Нового Света первый палец кисти лишь частично противопоставляется остальным. У человека большой палец стопы вовсе не противопоставляется другим; необходимость в этом отпала с развитием прямохождения.
Несмотря на указанное различие, у всех приматов по сравнению с другими плацентарными млекопитающими хватательная способность конечностей увеличена. Эта способность наиболее
Глава 2. Приматы
99
Б
Игрунка Церкоце^
Горилла
40
Часть I. Эволюция человека
развита у существующих форм лемуров и лори, у которых четвертый палец нередко длиннее третьего, благодаря чему кисть крепче удерживает предметы. У представителей подсемейства Lorisinae второй и третий пальцы заметно укорочены, чтобы не препятствовать хватательной функции.
С отмеченной особенностью в строении конечностей приматов тесно связана и другая: замена когтей плоскими ногтями. И здесь, конечно, мы находим заметные различия у разных форм. У тех представителей семейства Tupaiidae, которые обитают на деревьях, и у ископаемой формы Plesiadapis все пальцы снабжены когтями; у Daubentonia ноготь имеется только на большом пальце стопы; у других лемуров единственный коготь на втором пальце ноги используется в целях туалета. Ногти обезьян Нового Света, в особенности игрунок, сильно сжаты. Несмотря на эти различия, у приматов имеется ярко выраженная тенденция к образованию ногтей, опять-таки обусловленная особенностями их жизни (рис. 2.1).
Подошвенные подушечки на лапах приматов расширены, кожа на них более грубая. У первобытных млекопитающих подушечки были отделены друг от друга и четко очерчены. Такая картина сохранилась у большинства стопоходящих форм. У приматов наблюдается слияние подушечек с образованием обширной трущейся поверхности, в которую входят и плоские ногти.
У некоторых обезьян Нового Света получил развитие хвост с сильной мускулатурой, дающий животному дополнительную возможность висеть на ветках деревьев.
Способы передвижения у приматов весьма разнообразны. К ним относятся: а) хождение на четырех ногах, характерное для низших обезьян; б) двуногая походка у человека; в) медленное лазание по деревьям (как у лори); г) брахиация и раскачивание на руках (гиббоны пользуются этим способом локомоции в 80% случаев; он характерен также для орангутана, паукообразных обезьян и в меньшей степени для шимпанзе); д) вертикальное передвижение посредством прыжков с дерева на дерево с цеплянием за ветки — способ, типичный для большинства полуобезьян (Намье и Уолкер определяют его как «метод передвижения по деревьям посредством прыжков, при которых обе задние конечности создают толчковое усилие, необходимое для локомоции; и до, и после прыжка туловище остается в вертикальном положении, причем животное предпочитает использовать вертикальную опору»). Многие из современных видов приматов, включая полуобезьян, значительное время проводят на земле; другие, подобно человеку, гориллам и бабуинам (и родственным им видам), вновь вернулись к преимущественно наземному образу жизни. Такой переход обычно ассоциируется с передвижением на четырех конечностях, за HCKnioneHHeMj конечно^ человека с его прямохождением. Харак
Глава 2. Приматы
41
терно, что, совершая прыжки на деревьях, приматы на земле стремятся выпрямить свое тело и начинают передвигаться посредством прыжков на двух ногах. Следует заметить также, что большие африканские обезьяны, перейдя к наземной жизни, обычно передвигаются на четырех конечностях, но опираются больше на костяшки пальцев, чем на поверхность ладоней,— этот метод иногда называют «суставной ходьбой».
2. Развитие передних конечностей как органов исследования предметов. Жизнь на деревьях сложна и полна неожиданностей. Приматы вынуждены вести очень подвижный образ жизни, и уже в силу этого их конечности должны быть более развиты и приспособлены к разнообразным движениям, чем у большинства других млекопитающих. Исходя из этого, можно понять, почему у приматов сохраняется ключица и отчетливо выражена способность к пронации и супинации. Понятно также, что в сложной обстановке, характерной для древесного образа жизни, готовность к разнообразным реакциям стала условием выживания; у приматов передние конечности служат не только средством передвижения и опоры, но выполняют и другие функции, входя в состав сложной экстероцептивной системы, служащей для исследования незнакомых предметов; передние конечности используются также для собирания плодов и поднесения пищи ко рту. Приматам становится ненужной удлиненная челюсть, свойственная другим животным. Очевидно, далее, что происхождение человеческой руки с ее сноровкой и тонким осязанием непосредственно связано с приспособлением к древесному образу жизни у предков человека.
3. Развитие пищеварительной системы травоядного типа. Среди факторов, направляющих ход эволюции животных, важная роль принадлежит питанию. Происхождение приматов можно связать в конечном счете с употреблением пищи, находимой на деревьях. Почти все приматы являются либо всеядными, либо растительноядными животными. Некоторые, однако, подобно Daubentonia, питаются исключительно насекомыми. По строению пищеварительного тракта приматы мало отличаются от насекомоядных, за исключением некоторых обезьян Старого Света, у которых имеются либо защечные мешки, либо дольчатые желудки. Хорошо развитая слепая кишка, по-видимому, сохранилась вследствие употребления вегетарианской пищи. Вместе с тем такая пища определяет форму зубов приматов, которые хотя и не столь высокоспециализированы, как у парнопалых и непарнопалых копытных, но по сравнению с зубами насекомоядных уменьшены в числе и обладают сложной структурой. Зубная формула древних млекопитающих имеет вид: 1 у : С у : РМ : М . Соответствующее ей число зубов сохраняется только у ископаемого Ana-gale, жившего в олигоцене и относимого иногда к приматам. У всех
42
Часть I. Эволюция человека
остальных число зубов уменьшено; в основном это касается резцов и премоляров. У сохранившихся форм тупайи — только два верхних резца, тогда как у Notharctus и Adapts потерян также нижний резец. Наиболее общая формула для ныне живущих полуобезьян у: Т: 4: 4 ’ ^на хаРактеРна также для обезьян Нового Света, за исключением игрунок, у которых обычно не прорезывается последний коренной зуб. Дальнейшее уменьшение числа премоляров мы наблюдаем у обезьян Старого Света, человекообразных обезьян и человека. Зубная формула приобретает вид 2 . _2_. 3
2 : 1 : 2 ; 3 •
Соответствующим образом возрастает и сложность строения зубов, в первую очередь премоляров и моляров. У древнейших
Протокоиулюс
Лротоконус
Наружный .циигулпм
Параконус
Метаконус
Метаконулюс
‘Гипоконус
Метаконид
Параконид I Эняюконид
ГЬпоконулид
Протоконид ГЬпокониЭ
А Б
Рис. 2.2. Жевательные поверхности коронок коренных зубов примата (обобщенный тип).
А. Левый верхний моляр. Б. Левый нижний моляр.
млекопитающих моляры трехбугорчатые, а премоляры имеют конусовидную форму. При окклюзии нижние зубы заходят в промежутки между верхними, образуя режущую поверхность вдоль передней и задней границ зубов. Иными словами, моляры и премоляры относятся к секториальному типу, приспособленному для раскусывания насекомых. Для пережевывания растительной пищи необходимо по возможности лучшее соприкосновение жевательных поверхностей зубов. Этому способствует развитие заднего внутреннего бугорка гипоконуса на верхних молярах и «пятки», или талонида, на нижних. Талонид переходит в конце концов в тригонид, из которого развиваются энтоконид, гипоко-нид, а часто гипоконулид. Таким образом, талонид противостоит задней части верхнего моляра. Параконид, напротив, постепенно исчезает, так как перестает участвовать в окклюзии. Верхние зубы становятся четырехбугорчатыми, нижние же имеют либо четыре, либо пять главных бугорков (рис. 2.2). У приматов наблюдаются все указанные стадии в развитии зубов, однако в це
Глава 2. Приматы.
43
лом можно сказать, что для низших приматов характерны трехбугорчатые, а для антропоидов — четырехбугорчатые зубы.
Аналогичным функциональным изменениям подвергаются и премоляры. В основной группе насекомоядных они имеют один бугорок конической формы. Подобная картина в особенности характерна для передних зубов многих лемуров и Tarsius. У всех Anthropoidea премоляры обычно имеют два бугорка, за исключением переднего нижнего премоляра, который вместе с верхним клыком рассекает куски пищи и обычно является однобугорковым зубом секториального типа. Тенденция к превращению премоляров в моляры еще более заметна у лориобразных, особенно у Galaginae, у которых задние премоляры обладают тремя или даже четырьмя бугорками.
Следует также отметить, что у большинства приматов, особенно у Anthropoidea, имеются длинные режущие клыки, используемые для защиты и нападения. Современный человек в этом отношении составляет исключение. Однако данные палеонтологии указывают на то, что у предков человека клыки были сильнее развиты. У существующих видов лемуров и лори в отличие от ранних ископаемых форм нижние клыки малозаметны: они вместе с длинными нижними резцами наклонены вперед и образуют своеобразную «зубную гребенку». Верхние резцы не участвуют в раскусывании пищи, и вследствие этого у некоторых форм они сильно уменьшены в размерах или вовсе отсутствуют. Характерная особенность многих древнейших форм полуобезьян, например принадлежащих к семейству Presiadapidae, состоит в заметном увеличении центральных резцов как верхней, так и нижней челюсти с тенденцией к удлинению последней. При этом обычно наблюдается и параллельное уменьшение размеров боковых резцов, клыков и задних премоляров, которые иногда полностью исчезают.
4. Редукция органов обоняния. Среду, в которой обитают приматы, в отличие от наземных условий обитания, не назовешь «миром запахов». В отличие от других наземных млекопитающих у приматов можно отметить прогрессирующую редукцию органов обоняния. Наиболее ярким свидетельством этого является потеря ринария (rhinarium) — голой и богатой железами кожи на верхней губе, которая соединяет ноздри со складкой слизистой, прикрепляющей губу к деснам. Ринарий имеется у лемуров и лори, которых поэтому называют стрепсириновыми приматами. Верхняя губа у них абсолютно неподвижна. У долгопята и у антропоидов губная часть среднего носового отростка, образующая ринарий, перекрывается боковыми отростками верхней челюсти. Отходящие от них мышцы подходят к верхней губе и обеспечивают ее подвижность, необходимую для выразительных движений. Такие формы получили название гаплориновых. Различие стрепсириновых и гаплориновых форм дополняется изменением в
44
Часть I. Эволюция человека
строении носовой полости. У лемуров и лори, имеющих удлиненную морду, носовые полости сравнительно велики и имеют сложное строение вследствие развития системы носовых раковин. У долгопятов и антропоидов носовые полости и раковины, а следовательно, и выстилающий их эпителий не столь сильно развиты.
5. Увеличение остроты зрения. У животных, обитающих на деревьях, естественный отбор благоприятствовал развитию зрения. У всех приматов орган зрения стал доминирующим экстеро-рецептором, что нашло свое отражение как в размерах глаз и их расположении, так и в дифференцировке сетчатки.
Сетчатка позвоночных характеризуется наличием рецепторов двух типов — палочек и колбочек. Палочки чувствительны при слабой освещенности и играют важную роль в сумеречном, или скотопическом, зрении. С их помощью, по-видимому, фиксируется движение объекта. На колбочки, напротив, действует яркий свет; их функция состоит в оценке тонких деталей формы предметов. На это указывает тот факт, что каждая колбочка имеет свой собственный зрительный нерв в противоположность палочкам, многие из которых иннервируются одним волокном. Колбочки, кроме того, имеют отношение к цветовому зрению. Палочки преобладают в сетчатке всех приматов, располагаясь главным образом по ее периферии. У тех животных, у которых имеются колбочки, они занимают центр сетчатки. Область, где сконцентрированы колбочки, соответствует точке наибольшей остроты зрения и называется желтым пятном (macula lutea). Здесь обычно находится небольшое углубление (центральная ямка), вызванное отсутствием поверхностных элементов эпителия сетчатки, которыми в других местах повсюду покрыты фоторецепторы.
У разных приматов сетчатка неодинакова как по составу фоторецепторов, так и по их строению. У лемуров, лори, Tarsius и у обезьяны Нового Света Aotes имеются только палочки, так что сетчатка у этих форм не имеет ни желтого пятна, ни центральной ямки. У Tarsius, однако, в извилистых складках центральной области имеется слой палочек. У всех представителей Anthropoidea (за исключением ночных обезьян) имеются также колбочки, а желтое пятно и центральная ямка в различной степени дифференцированы (рис. 2.3). Один из этапов такой дифференцировки ясно виден у Callithrix, у которого желтое пятно нечетко выражено, а ямка находится в зачаточном состоянии. Однако зто различие между полуобезьянами и высшими приматами не означает, что у древнейших приматов были только палочки, а колбочки развились в отряде вторично. Следует учитывать, что все существующие формы полуобезьян и Aotes ведут в основном ночной образ жизни и именно с этим связано отсутствие колбочек. Высшие приматы, по всей видимости, развились из низших еще до того, как последние перешли к ночному образу жизни, и потому у них сох
Глава 2. Приматы
45
ранился «дневной» тип сетчатки. Последний мы находимг в частности, у тупайи.
Глаза у приматов сравнительно большие, особенно у Tarsius. У последнего с наибольшей силой проявилась тенденция, характерная вообще для всех долгопятоподобных и, по-видимому, связанная с их способом передвижения. Долгопяты совершают большие прыжки с дерева на дерево, а для прыжков, очевидно, необходима большая острота зрения. Огромные глазницы мы находим
I_________|_________1, -1____________I_________I_________I
О too 200 300 400 500 600
мкм
Рис. 2.3. Центральная ямка сетчатки глаза человека (Polyak, «The vertebrate visual system», Chicago Univ. Press, 1957).
На дне ямки видны только колбочки, лежащие очень близко к поверхности. По периферии от наружного слоя среди массы колбочек (кружки) появляются палочки (зачернены).
даже у некоторых эоценовых форм, таких, как Hemiacodon. Тенденция к увеличению размеров глаз проявляется у всех приматов. Пожалуй, еще более выражена тенденция к перемещению глазниц с боковой поверхности черепа на переднюю. У первобытных млекопитающих глаза расположены сбоку и поля зрения не перекрываются. Сходную картину мы находим у лемуров, хотя их глаза несколько сдвинуты вперед. У высших приматов зрительные оси более или менее параллельны. Благодаря перекрыванию полей зрения возможен стереоскопический эффект, столь необходимый животным, обитающим на деревьях. Свидетельством развития стереоскопии у Anthropoidea может служить тот факт, что у всех представителей этого подотряда примерно половина волокон зрительного тракта перекрещивается. У лемуров перекреста вообще нет, а у Tarsius, глаза которого совершенно неподвижны, перекрест выражен в слабой степени. Вследствие такого расположения волокон наружное коленчатое тело среднего мозга имеет 6 слоев, а
46
Часть I. Эволюция человека
= t.m 2, S3, Зоны о/нцец соматической чувствительности
• 4,0 в, g 8, Моторные зоны
А 5, д 7, Теменная область
&9,\\\1О№11,444, * 45. К 47, 9 48 Лобная область
z 13-16 Область островка
.;• 17. №18,3=19, Зрительные и зрительно-ассоциативные зоны
J- 20,=2/, 122, 137, М 38, *39,1 ,
f Височная область
У 40,«41,S42,^43 J
w 28 Грушевидная область
Рис. 2.4. Цитоархитекто
А. Еж. Б. Тупайя. В. Лемур. Г. Долгопят. Д. Когтистая обезьяна (Callithrix). Е. Мар 1959); рисунки В, Д, Е, 3 — по Бродману; Г — из статьи Болларда
не три, как у полуобезьян. (Чередующиеся слои латерального коленчатого тела у человека и у макака соответственно получают перекрещивающиеся и неперекрещивающиеся зрительные волокна.)
6. Развитие мозга. Как указывалось выше, всесторонняя информация об окружающей среде — существенное условие выживания для животных, обитающих на деревьях. В соответствии с развитием органов чувств, как экстероцептивных, так и проприо-
Глава 2. Приматы
47
нические поля мозга.
пышка Ж. Шимпанзе. 3. Человек. А и Б — из книги Ле Гро Кларка (Le Gros Clark-(Woolard, J. Anat., 60; 1952); Ж — из статьи Уокера (Walker, J. Anat, 13, 1938).
цептивных, у приматов происходит усовершенствование областей головного мозга, связанных с сенсорным восприятием. У приматов должно быть также развито весьма совершенное управление движениями и чувство равновесия. В связи с этим происходит усложнение двигательных центров коры головного мозга и мозжечка. В общем отмечается тенденция к увеличению мозга, заметная уже у наиболее примитивных приматов, таких, как Adapts, если сравнивать их с насекомоядными. Эта тенденция прогрессирует у
48
Часть I. Эволюция человека
всех представителей отряда и достигает высшей степени у человека. Укажем вкратце основные морфологические изменения,-связанные с увеличением мозга (рис. 2.4).
I. Увеличение переднего мозга и особенно неопаллиума.
II. Увеличение числа борозд в коре неопаллиума; возникновение, в частности, истинной сильвиевой борозды.
III. Увеличение затылочной доли мозга, которая все более выступает назад и обособляется от расположенной по соседству теменной области вследствие образования задней шпорной борозды (ср. определяющие признаки приматов по Миварту); эта доля связана с интерпретацией зрительных образов и ассоциациями.
IV. Усложнение прецентральной коры: лобная доля управляет мышечной деятельностью и голосовым аппаратом, префронтальная выполняет ассоциативную функцию.
V. Усложнение височной доли, особенно у высших приматов; роль ее состоит в более совершенном различении звуков, необходимом для голосовой сигнализации.
VI. Усложнение мозжечка и тех областей мозга, которые связывают мозжечок с двигательными центрами коры.
VII. Редукция обонятельного мозга (риненцефалона).
В целом усложнение коры головного мозга у приматов связано с ее доминированием над деятельностью низших мозговых центров. Лучше всего это видно на примере регуляции движений. Если у собаки или кошки полностью удалить кору головного мозга, то животное будет в состоянии ходить или стоять почти так же, как нормальное. У приматов же прецентральная зона коры почти полностью регулирует функцию скелетных мышц; удаление больших полушарий у обезьян сопровождается полным или частичным параличом. Благодаря доминирующей роли коры приматы достигли в высшей степени совершенной мышечной координации, хотя, возможно, за счет несколько большей уязвимости.
7. Изменения в строении черепа. Все те изменения мозга и органов чувств, которые характерны для приматов, а также привычка приматов принимать сидячее положение и исследовать предметы с помощью конечностей — все это нашло свое отражение в строении черепа (рис. 2.5). Освоение сидячей позы привело к тому, что череп стал располагаться под некоторым углом к позвоночному столбу. Такому положению черепа способствовало перемещение затылочных мыщелков (а следовательно, и затылочного отверстия) кзади и книзу. Это наиболее сильно выражено у человекообразных обезьян и особенно, в связи с прямохождением, у человека, но заметно у всех приматов. Результатом приспособления к сидячему положению явилось также смещение лицевого черепа по отношению к мозговому, причем эта тенденция прогрессирует далее у высших антропоидов. У четвероногого животного лицевой череп составляет продолжение мозгового, а у всех приматов лицо
Глава 2. Приматы
49
Рис. 2.5. Черепа приматов (Le Gros Clark, 1959).
А. Птилоцеркус. Б. Галаго. В. Долгопят. Г. Капуцин. Д. Хульман. Е. Гиббон. Ж. Шимпанзе. 3. Проконсул.
50
Часть I. Эволюция человека
располагается ниже мозгового черепа, образуя четко выраженный базикраниальный угол (см. рис. 1.2). Образование этого угла явилось следствием изменения функции кисти, укорочения морды и уменьшения роли обоняния. Этим же можно объяснить относительно небольшой размер лицевого скелета приматов. Интересно, что, когда некоторые приматы перешли к передвижению по земле на четырех конечностях, как это вторично случилось с бабуинами и близкими к ним родами обезьян, у них снова произошло удлинение морды, но базикраниальный изгиб сохранился.
Увеличение размера глаз и перемещение их вперед на лицевую сторону черепа, очевидно, отразилось на положении и размерах глазниц. У всех приматов, за исключением некоторых самых ранних форм, получила развитие задняя (боковая) стенка глазницы: в результате кость полностью окружает глазную впадину. Хотя эта стенка имеется не только у приматов, ее развитие является отличительной особенностью таких групп, как долгопяты и высшие приматы, у которых глазница более или менее полностью отделена от височной ямы вследствие разрастания скуловой кости большого крыла основной кости (т. е. алисфеноида первобытных млекопитающих).
Благодаря увеличению глаз и смещению их кпереди у приматов в образовании медиальной стенки глазницы позади слезной кости кроме лобной и верхнечелюстной участвуют и другие кости. У лемуроподобных, как правило, две названные кости отделены отростком небной, который вытянут в направлении слезной кости; у всех остальных приматов в медиальную стенку глазницы входит бумажная пластинка решетчатой кости.
Увеличение мозга вызывает увеличение размеров лобной, теменной и затылочной костей, образующих верхние и боковые части мозгового черепа.
8. Сокращение числа одновременно рождаемых детенышей. Забота о новорожденных представляет особые трудности для животных, постоянно обитающих на деревьях. Поэтому можно думать, что чем меньше численность потомства, тем больше шансов успешно его вырастить. У всех приматов существует явная тенденция производить на свет одновременно не более двух или трех потомков, а у многих рождается только один. Этот факт имел далеко идущие эволюционные последствия. Если в утробе матери развивается сразу несколько зародышей, то между ними возникают конкурентные взаимоотношения в том смысле, что более быстро развивающиеся получают преимущество перед развивающимися медленнее. Если самка вынашивает одного или максимум трех детенышей, то отбор, благоприятствующий быстрому развитию, будет ослаблен. Аналогичная ситуация сохраняется и после рождения потомства, до тех пор пока выживание детенышей зависит от материнской заботы. Можно считать^ таким образомг что
Глава 2. Приматы
51
уменьшение численности потомства способствует увеличению времени развития. Для приматов характерно увеличение периода до наступления половой зрелости и продолжительности необходимого материнского ухода. Указанные факторы составляют предпосылку для пластичности поведения, основанного на обучении. Тенденция к увеличению периода созревания достигает наивысшего развития у человека как следствие его социальной организации. Тем не менее особенности поведения человека во многом становятся понятными, если иметь в виду образ жизни его далеких предков; жизнь на деревьях — один из немногих способов существования, при котором сокращение числа одновременно рождаемых детенышей повышает приспособленность к воспроизведению вида.
Стадии эволюции приматов
Из сказанного следует, что отличительные признаки приматов можно понять, исходя из требований, которые предъявлял или предъявляет древесный образ жизни, и что эволюционное превосходство человека коренится в унаследованных им эволюционных преимуществах. Рассмотрим теперь кратко ход эволюции приматов.
Происхождение отряда во многом остается неясным. Обычно считают, что приматы произошли от древнейших насекомоядных. Однако у ранних полуобезьян слишком мало специфических черт сходства с современными насекомоядными, если не считать некоторой общности в строении скелета. Весьма вероятно, что в меловом периоде произошло ранее разделение общего ствола млекопитающих на две ветви: одна привела в конце концов к приматам, а также к копытным, Condylartha, и ежам, другая — к «истинным» насекомоядным и тупайям, а наряду с ними — к креодонтам и некоторым другим основным группам млекопитающих. Purgato-rius из мелового периода, по-видимому, самая древняя из форм, которую несколько произвольно относят к приматам. Она остается пока единственным представителем приматов в мезозойской эре. Зато к началу палеоцена идентифицировано довольно много различных форм как в Европе, так и в Северной Америке; в настоящее время их относят к отряду приматов главным образом из-за особенностей строения коренных зубов. Тем не менее передние зубы у многих из этих форм весьма специфичны. Близких форм пока не обнаружено только в Азии и Африке, вероятно, потому, что на этих континентах не производились достаточно интенсивные расколки, а условия фоссилизации были здесь хуже» чем в Европе. Ранние приматы и те их потомки, которые не претерпели в дальнейшем заметных изменений^ составляют отряд полуобезьян (рис. 2.6).
2
3
Геологические периоды (млн. лет)
18
13
18
Голоцен
(совр. эпоха)
Плейстоцен
Плиоцен
Миоцен
Олигоцен
Рис. 2.6. Генеалогическая схема полуобезьян.
Глава. 2. Приматы
53
Рис. 2.7. Черепа некоторых наиболее известных древних полуобезьян.
А. Плезиадапис (обратите внимание на отсутствие посторбитальной кости). Б. Нотарктус. В. Адапис. Г. Некролемур. Д. Тетониус. Е. Рунейя.
Из палеоценовых отложений известны три семейства: Plesia-dipidae, Paromomyidae и Carpolestidae. Их идентифицировали в основном по остаткам зубного аппарата и фрагментам челюстей. Представителей первых двух семейств находят как в Европе, так и в Северной Америке; они существовали на протяжении почти всего эоценового периода. Формы, относящиеся к Carpolestidae, обнаружены только в Северной Америке; они вымерли в начале эоцена. К эпохе раннего эоцена относится также расцвет двух важных семейств — Adapidae и Anaptomorphidae. Первое из них включает одно подсемейство в Старом Свете и одно — в Новом. Весьма вероятно, что адапиды были предками современных лемуров. Анаптоморфиды были первыми представителями тарзие-
54 Часть I. Эволюция человека
вых. Их находят преимущественно в отложениях Северной Америки, однако известны также две формы в Старом Свете: Вегги-vius из подсемейства Anaptomorphidae и Tielhardina, которую относят к подсемейству Omomyinae. Семейство Anaptomorphidae существовало всю эпоху эоцена, а отчасти и в миоцене. В середине эоцена оно объединяется с Microchoerinae (в Старом Свете). Это последнее подсемейство Симонс включает в семейство тарзиевых, представители которого, вероятно, были предками современных долгопятов. На рис. 2.7 представлены для сравнения черепа некоторых из этих древнейших полуобезьян наряду с более поздними формами.
Хотя эоценовая эпоха была временем наибольшего обмена элементами фауны между Евразией и Северной Америкой,, имеется мало данных о том, что в этом обмене участвовали какие-либо роды подотряда низших приматов. Тем не менее сходство между формами Старого и Нового Света очень велико.
В конце эоценовой эпохи число ископаемых остатков полуобезьян резко уменьшается. В более поздних отложениях Северной Америки и Европы они полностью исчезают. Отчасти такое уменьшение — только кажущееся, поскольку ископаемые остатки третичного периода из тропической Африки и Азии немногочисленны и плохо изучены. Возможно, однако, что эоцен был эпохой расцвета подотряда полуобезьян и что с тех пор он начал приходить в упадок. Упадку полуобезьян, вероятно, способствовала конкуренция с быстро эволюционировавшими грызунами, но, без сомнения, также и прогресс их собственных потомков — высших антропоидов (рис. 2.8), стоящих на более высокой ступени организации. Показательно, что только на Мадагаскаре, отрезанном от Африканского материка еще до того, как его заселили «обезьяны», виды полуобезьян все еще многочисленны. Именно здесь произошла их вторичная радиация и возникли такие своеобразные формы, как Daubentonia (руконожка) и Megaladapis.
Степень организации, свойственная обезьянам, эволюционировала как в Новом, так и в Старом Свете. На определенном этапе эволюции к ней довольно близко подошли также формы, населявшие Мадагаскар. Действительно, лемуроид Archaeolemur из плейстоценовых отложений Мадагаскара имеет много признаков, характерных для обезьян. Также Branisella и Dolichopithecus — представители обезьян олигоценовой эпохи, найденные в Новом Свете. Остатки их, однако, слишком фрагментарны, чтобы можно было с уверенностью определить их эволюционный статус. До сих пор существовало мнение, что обезьяны Старого и Нового Света имели независимых предков на уровне полуобезьян. Такой вывод основывался отчасти на предположении, что древнейшие формы обезьян Старого Света обладали такими прогрессивными признаками, как наличие наружного слухового прохода и умень-
Нового
Геологические периоды (млн. лет)
Обезьяны
Hytobates
Gorilla Chimpanzee
sapiens
Homo erectus
Aegyptopttnecus q Propliopithecus
_ „ Dolichopithecus
Branisella
Homunculus
Apidium Pargptfhecus V • Oligopithecus
Рис. 2.8. Генеалогическая схема антропоидов.
Голоцен (совр. зпоха)
Плейстоцен
Плиоцен
Миоцен
Олигоцен
Эоцен
56
Часть I. Эволюция человека
шение числа премоляров до двух. Сейчас установлено, однако, что это не так. Более того, опираясь на теорию дрейфа материков, можно высказать предположение о существовании некоего общего антропоидного предка для форм Старого и Нового Света. Независимо от того, был ли этот предок антропоидом или полуобезьяной, можно почти не сомневаться в том, что эволюция в двух фаунистических зонах происходила во многом параллельными путями.
Есть все основания полагать, что антропоиды Старого Света произошли от какой-то формы, подобной древнему предшественнику долгопята Teilhardina. Их ранние остатки обнаружены в олигоценовых отложениях в Египте, близ Эль-Файюма, сохранившихся с той эпохи, когда область была покрыта влажными лесами. В настоящее время в этих отложениях обнаружено большое число остатков приматов, представляющих различные роды, которые включают и ранних «обезьян» (Apidium и Parapithecus), и первобытных «человекообразных приматов» (Qligopithecus, Aeolopithecus, Propliopithecus, Aegyptopithecus). Долгие годы ископаемый материал из Эль-Файюма был крайне беден и состоял из разрозненных фрагментов, что служило причиной различных спекулятивных предположений об их филогенетическом статусе. Лишь недавние раскопки дали нам более обширный материал, касающийся некоторых из упомянутых выше родов. Среди антропоидов Старого Света особый интерес представляют Apidium и Parapithecus, отличающиеся наличием трех премоляров как на верхней, так и на нижней челюсти. Тем не менее по остаткам черепа у Apidium и морфологии коренных зубов у обеих указанных форм можно заключить, что оба рода наверняка относились к антропоидам. Более того, типичный для церкопитекоидов признак, а именно билофодонтная (в начальной стадии) форма коренных зубов, т. е. соединение передних, равно как и задних, бугорков в поперечном направлении с образованием общей пластинки, указывает на принадлежность их к этой группе. Правда, следует заметить, что некоторые особенности зубного аппарата свидетельствуют о родстве с семейством Oreopithecidae, а не с церкопитекои-дами.
Особенно неясен эволюционный статус Oligopithecus. Найдена лишь часть нижней челюсти с большим числом примитивных признаков, во многом свойственных семейству Omomyinae. Вместе с тем олигопитек имел лишь два премоляра, а строение зубов у него — явно антропоидного типа, несмотря на небольшие размеры нижнего клыка. Коренные зубы несколько билофодонтны, но общее мнение сводится к тому, что олигопитека следует рассматривать как небольшую человекообразную обезьяну; она относилась к весьма широко генерализованному типу и обладала^ по-видимому, большими эволюционными возможностями.
Глава 2. Приматы,
57
У элопитека также найдена одна нижняя челюсть с зубной формулой 2 : 1 : 2 : 3. Он был, подобно олигопитеку, животным небольшого размера, но имел слишком большой для своего размера клык, а также премоляр отчетливо секториального типа. В отличие от антропоидов Старого Света третий коренной зуб у него заметно меньше — признак, который часто находят у гиббона, а нередко и у человека. Симонс, впервые обративший внимание на эту особенность, считает элопитека анцестральной формой гиббона. Фактически известен ряд форм соответствующего геологического возраста, представляющих морфологический переход от элопитека к современным гиббонам. У представителей этого ряда клыки становятся более длинными, передние нижние премоляры ближе к секториальному типу, V-образная челюсть превращается в U-образную. Как указал Ле Гро Кларк, существование такого ряда еще не означает, что нам известен истинный филогенез гиббонов, но можно быть уверенным в том, что ряд предков гиббонов, каков бы он ни был, должен был пройти через стадии, сходные с теми, которые представлены цепью форм Aelopithecus — Limno-pithecus — Pliopithecus (еще обладавший хвостом) — Hylobates — Symphalangus. Представляет интерес раннее отделение линии гиббонов от лгнии остальных представителей Hominoidea, тем более что гиббоны обладают многими особенностями в строении конечностей, характерными для больших обезьян. Поскольку таковые не встречаются у ранних форм какой бы то ни было группы, очевидно, что они должны были возникнуть в позднейших формах параллельно, подобно тому как это случилось в эволюции паукообразных обезьян в Южной Америке.
Предметом многих споров явился Propliopithecus, известный по найденной уже давно нижней челюсти и, возможно, по обнаруженному недавно коренному зубу. У проплиопитека много примитивных черт в строении зубов, и до открытия элопитека его обычно считали анцестральной формой гиббонов. Он мог быть также предком современных гоминид. Исходя из одних лишь фрагментарных остатков вряд ли стоит отводить проплиопитеку строго определенное место в филогенетическом ряду и делать серьезные эволюционные выводы. Ведь в конце концов многие ископаемые роды приматов вообще не дали никаких новых форм. Несмотря на огромные эволюционные возможности, которыми обладали антропоиды олигоценовой эпохи, проплиопитек мог изчезнуть, не оставив потомков. Лучше всего считать его одной из обобщенных форм больших обезьян. Ситуация с египтопитеком проще: обнаружены не только челюсти и отдельные зубы, но и хорошо сохранившийся череп, а возможно, и несколько костей скелета, относящихся к тому же индивидууму. У египтопитека мы находим именно такое сочетание примитивных и более поздних признаков, которое можно ожидать у переходных форм между низшими
58 Часть I. Эволюция человека
обезьянами и антропоидами. Он несколько превосходил по размерам проплиопитека, но был меньше человекообразных обезьян более поздней эпохи. В отличие от последних египтопитек имел заметно удлиненный лицевой скелет с хорошо развитыми верхними челюстями и относительно небольшой черепной коробкой. Глазницы отделены от височной ямы, как у обезьян Нового Света, и по аналогии с ними слабо развит наружный слуховой проход. Наряду с этим египтопитек имел большие верхние, но малые нижние клыки; строение коренных зубов такое же, как у больших обезьян. Насколько можно судить по фрагментам скелета, египтопитек, вероятно, хорошо бегал и, возможно, передвигался, цепляясь за ветки деревьев в лесах, покрывавших район Эль-Файюма в олигоцене.
Адаптивная радиация антропоидов достигла большого развития к началу миоцена. По остаткам челюстей и зубов идентифицированы многочисленные роды цебоидов: Homunculus из Аргентины, Cebupithecia, Neosaimiri и Strirtonia из Колумбии. В миоценовых пластах Африки найдены также цереопитекоиды, например Prohylobates и Victortapithecus. В Восточной Африке в слоях нижнего миоцена обнаружены остатки Limnopithecus, в различных местах Старого Света, преимущественно в Европе,— остатки Pliopithecus, существовавшего также в плиоценовом периоде. В миоценовых отложениях Европы, Индии и Восточной Африки также найдено немало остатков человекообразных обезьян; несмотря на значительные различия в размерах, их относят сейчас к одному роду Dryopithecus, правда выделяя в последнем большое число различных видов. Специфические признаки этого рода будут рассмотрены ниже, здесь стоит заметить только, что дриопитеки, вероятно, некоторая обобщенная форма антропоидов, не имеющая той специализированной структуры конечностей, которую мы находим у современных горилл, шимпанзе н орангутана. По многим признакам они действительно напоминают обезьян Старого Света, эти признаки следует рассматривать как примитивные. Без сомнения, дриопитеки были анцестральной группой по отношению к ныне существующим группам больших обезьян. Но еще более важно, что некоторые из дриопитеков и в особенности ряд форм из нижнего миоцена, найденных в Восточной Африке и ранее относимых к одному роду Proconsul, могли быть очень близкими к предкам первых гоминид.
В начале плиоцена церкопитекоиды и гоминоиды были, пожалуй, более широко распространены, чем в настоящее время. В плиоценовых слоях Старого Света найдено много ископаемых остатков обезьян; дриопитеки продолжали существовать и в эту эпоху. В месторождениях бурого угля в северной Италии, относящихся к плиоцену, вновь был обнаружен Oreopithecus. Этот последний род, известный по хорошо сохранившемуся скелету^
Глава 2. Приматы, 59
отличается многими необычными признаками в строении коренных зубов, которые позволяют с уверенностью отделить его от остальной филогенетической линии гоминоидов. Тем не менее ряд особенностей в строении таза и конечностей указывает, по-видимому, на то, что ореопитек обладал зачатками прямохождения. Из плиоцена и позднего миоцена известны также первые австралопитеки и Ramapithecus, играющие важную роль в оценке разделения филогенетических ветвей гоминид и понгид.
Глава 3
ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ И ЧЕЛОВЕК
Современные высшие обезьяны передвигаются по ветвям деревьев при помощи рук; человек — наземное двуногое. Оба способа передвижения связаны с вертикальным положением тела, и те черты сходства обезьяны и человека, в основе которых лежит прямохождение, могли произойти как от общих предков, так и вследствие параллельной эволюции.
У животных, сохранивших при движении горизонтальное положение тела, внутренности поддерживаются главным образом за счет фиксации к задней стенке брюшной полости. При прямохождении такая система фиксации, очевидно, недостаточна. Поэтому как у обезьян, так и у человека имеется еще и особое прикрепление внутренностей к диафрагме. Дополнительную опору снизу создает диафрагма таза, образованная мышцами — дериватами отводящих мышц хвоста; последний утратил свою балансирующую функцию и постепенно редуцировался.
В поперечном сечении грудная клетка четвероногого обычно напоминает сердце: она вытянута в дорсовентральном направлении, но сжата и уплощена с боков. Это не только придает животному необходимую устойчивость благодаря низкому положению центра тяжести, но и обеспечивает ему лучшую механическую опору, так как передние конечности могут подводиться под туловище и двигаться продольно в одной плоскости с задними. Вертикальное положение туловища, очевидно, не налагает подобных ограничений на форму грудной клетки. У человекообразных обезьян и у человека оно связано с расширением диапазона движений передних конечностей. Хотя более широкий диапазон движений обусловлен, вообще говоря, различными причинами, с ним связан ряд новых особенностей формы грудной клетки: последняя сжата в переднезаднем направлении и увеличена в поперечнике.
Когда передние конечности опираются на землю или цепляются за ветви деревьев, мышцы плечевого пояса осуществляют наряду с локомоторными движениями и дыхательные движения грудной клетки. Дыхательная функция этих мышц отходит на задний план, когда передние конечности освобождаются от опоры; у человека и обезьян более важное значение для дыхания приобретают межреберные мышцы и особенно диафрагма.
Глава 3. Человекообразные обезьяны и человек
61
С освобождением передних конечностей связано также более интенсивное развитие грудных мышц. Отдельные костные сегменты грудины сливаются, образуя сплошную кость. Последнее более характерно для больших обезьян, лазающих по деревьям, чем для человека. Однако эта тенденция наблюдается у обеих групп и обусловлена одними и теми же причинами. Развитие грудных мышц служит хорошим примером гиперморфоза, так как сегменты грудины появляются в ходе эволюции как отдельные кости и у всех остальных обезьян обычно не сливаются.
У проноградных животных позвоночный столб образует дугу со среднегрудным антиклинальным позвонком и длинным изогнутым поясничным отделом. Переход к прямохождению связан с рядом существенных изменений в строении позвоночника. Прежде всего возникает необходимость в специальных механизмах, обеспечивающих поддержание равновесия головы на позвоночнике. Этому способствует образование лордоза — изгиба кпереди — в нижнем шейном отделе. Лордоз сопровождается смещением вперед большого затылочного отверстия. Обеспеченное таким образом равновесие головы на позвоночнике позволяет удерживать ее с помощью относительно более слабой мускулатуры; менее развитыми становятся затылочные гребни, и меньше выступают остистые отростки шейных позвонков. По мере того как возрастает стабилизирующая роль грудино-сосцевидной мышцы, получают развитие сосцевидные отростки. Все указанные изменения происходят в значительно большей степени у гоминид, чем у любых известных видов обезьян: однако общность тенденций развития у тех и других по сравнению с четвероногими животными очевидна.
Поясничная область — одно из «слабых мест» при прямохождении. Не удивительно, что относительная длина ее уменьшена как у обезьян, так и у человека. Это уменьшение более заметно у обезьян, так как при передвижении по деревьям при помощи рук у них реже возникает необходимость вытягивать ноги в одну линию с осью позвоночника, а следовательно, для них не столь важен изгиб поясничной области. Последняя укорачивается за счет включения поясничных позвонков в крестец.
У обезьян, передвигающихся на четырех конечностях, крестец состоит в среднем из трех позвонков, у гиббона их обычно четыре, тогда как у человека и больших человекообразных обезьян насчитывается пять или шесть крестцовых позвонков. Слияние поясничных позвонков с крестцом отчасти можно объяснить увеличением размеров тела, которое произошло в ходе эволюции понгид и гоминид, однако нет никакого сомнения в том, что оно связано также с переходом к прямохождению, при котором вся тяжесть тела переносится на таз. Опорной функции таза способствует его вертикальное сжатие, боковое уширение и увеличение переднезаднего размера крыльев подвздошных костей. У обезьяны эти особенности
62
Часть I. Эволюция человека
строения таза выражены лишь в слабой степени, свидетельствуя о том, что многие отличительные особенности строения человеческого таза определяются в первую очередь прямохождением (рис. 3.1).
У понгид, как и у низших приматов, вертлужная впадина находится на уровне нижнего края крестца, у человека она расположена выше — на уровне крестцово-подвздошного сочленения, что обеспечивает устойчивость в тазобедренном суставе. Седалищный бугор у человека приближен к вертлужной впадине, что способствует функции мышц, прикрепляющихся на верхнем эпифизе большой
Рис. 3.1. Безыменные кости некоторых приматов.
1 — узконосая обезьяна; 2 — шимпанзе; 3 —'австралопитек из Штеркфонтейна; 4 — современный человек (крылья подвздошных костей параллельны плоскости рисунка).
берцовой кости. Обе отмеченные особенности человека связаны с передвижением на двух ногах.
Если многие черты сходства обезьян и человека, как мы видим, связаны с вертикальным положением тела, то многие различия прямо или косвенно обусловлены неодинаковым способом локомоции (см. рис. 2.1). У обезьян сильнее всего отклонились от исходной антропоидной формы передние конечности, а у гоминид изменения строения проявились главным образом в нижних конечностях. У обезьян-брахиаторов передние конечности сильно удлинены, особенно предплечье; кисть имеет форму крюка с укороченным большим пальцем. Это укорочение сказывается и в строении мускулатуры: у трех больших обезьян не всегда имеется длинный сгибатель первого пальца, а его короткие глубокие мышцы, за исключением аддуктора большого пальца, слабо развиты или вовсе отсутствуют. Интересно, что такие изменения в мускулатуре произошли также у других брахиаторных форм — у A teles и у не-
Глава 3. Человекообразное обезьяна и человек
63
Рис. 3.2. Скелет стопы некоторых приматов (Gregory, Mem. Am. Mus. nat. Hist., 1920).
1 — человек; 2 — горилла; 3 — церкопитек; 4 — капуцин; 5 — нотарктус.
которых представителей подсемейства тонкотелов, но не у гиббонов. Своеобразие гиббонов в этом отношении Штраус объясняет тем, что способ брахиации у них несколько иной.
Многие отличительные признаки человека касаются строения нижних конечностей. Об особенностях в строении таза было ска-
64
Часть I. Эволюция человека
зано выше. Следствием прямохождения является относительное удлинение бедра и голени и многочисленные изменения в скелете стопы, связанные с переносом на стопу всей тяжести тела (рис. 3.2): развитие «пятки», продольного и поперечного сводов предплюсноплюсневой области и большого пальца ноги, который располагается в одну линию с остальными пальцами и имеет с ними общую подошвенную поверхность. С таким строением связана утрата большим пальцем независимости движений, а также плоская поверхность сочленения плюсны с клиновидной костью предплюсны. Большой палец сохраняет лишь ограниченную подвижность в плюсно-предплюсневом суставе и небольшое отведение в плюсне-фаланговом суставе. У человека первый палец стопы, который первоначально был самым длинным, становится лишь частью опорной поверхности стопы, полностью утрачивая хватательную функцию. Несмотря на указанные особенности, нога человека в некоторых отношениях даже более примитивна, чем нога больших обезьян. Так, средний предплюсневой сегмент не подвергается продольному укорочению; подошвенная мышца, даже если она теряет связь с подошвенным апоневрозом, обычно сохраняется; сухожилия сгибателя первого и второго пальцев всегда тесно соединены; почти всегда имеется квадратная подошвенная мышца.
Различия в способе передвижения современных обезьян и человека в какой-то мере определяют и другую группу различий в их строении. У обезьян, живущих на деревьях, передние конечности служат для опоры и движения; обезьяны, живущие на земле (такие формы, как взрослые гориллы, значительную часть времени проводят на земле), обычно ходят на четырех конечностях. Передние конечности, следовательно, не свободны от выполнения локомоторных функций. Не удивительно, что хотя обезьяны используют для приема пищи передние конечности, для защиты и нападения у них служат большие челюсти. У человека эту роль выполняет рука, освобожденная от локомоторной функции. Поэтому не удивительно, что у человека в отличие от обезьяны челюсти и клыки малы, резцовые кости сращены с верхнечелюстными. Можно думать также, что отсутствие надглазничного валика у современного человека связано с относительно малыми размерами челюстей, так как функция валика, по крайней мере частично, заключается в том, чтобы выдерживать давление, возникающее при раскусывании твердых предметов. Исчезновение надглазничного валика связано также с увеличением лобных долей мозга и соответственно с более высокой лобной костью. Характерные сагиттальные гребни у взрослых шимпанзе, горилл и орангутанов появляются по той причине, что на боковой поверхности крыши черепа оказывается недостаточно места для прикрепления мощных височных мышц, развитие которых зависит, конечно, от размеров челюстей. Мы видим, таким образом, что изменение механизма локомоции оказывает
Глава 3. Человекообразные обезьяны и человек
65
влияние на развитие многих органов и систем и что, очевидно, детальный анализ функций этих систем весьма важен для понимания эволюционных связей.
Правда, многие различия в строении челюстей обезьяны и человека (наличие «обезьяньей полки» у обезьяны и подбородка у человека) нельзя непосредственно связать с изменениями в строении и функциях передних конечностей. Нужно учитывать и другие факторы: характер пищи (лишь отдаленно связанной со способом локомоции), роль языка и т. д. Тем не менее можно с уверенностью сказать, что морфологические признаки всегда необходимо рассматривать в тесной связи с различными и взаимодействующими друг с другом функциональными факторами.
Главные различия между обезьянами и человеком касаются строения конечностей, челюстей и мозга. Ниже мы более подробно рассмотрим эволюцию мозга у приматов. Существуют, однако, и другие различия, которым многие, особенно Вуд-Джонс (Wood Jones), придают важное филетическое значение. Вуд-Джонс отмечает, в частности, что у человека слезная и решетчатая кости сходятся на медиальной стенке глазницы, а у обезьян эти две кости, как правило, разделены лобно-максиллярным соединением. У человека в передней черепной яме решетчатая кость соединяется с клиновидной, а у обезьян между ними находятся лобные кости. У человека в области птериона сходятся теменная кость и большое крыло клиновидной, у обезьян — лобная кость с чешуей височной кости. Вуд-Джонс обращает внимание также на следующие факты: у человека отсутствуют седалищные мозоли, почка состоит из многочисленных пирамидок, от дуги аорты отходят безыменная, левая общая сонная и левая подключичная артерии, тогда как у обезьян левая общая сонная артерия отходит от безыменной. По всем этим признакам человек более примитивен, чем человекообразные и мартышковые обезьяны, и это дало основание Вуд-Джонсу утверждать, что человек непосредственно произошел от долгопята и не имел общих предков с обезьянами Старого Света и человекообразными обезьянами.
Шульц (Schultz), однако, указал, что многие из отмеченных Вуд-Джонсом различий не абсолютны и что последний недостаточно учитывал индивидуальную изменчивость, которая наблюдается у различных видов. Область птериона у многих обезьян, особенно орангутана, сохраняет примитивное расположение, а некоторые человеческие черепа имеют так называемый «церкопи-текоидный» птерион. У гиббона очень малы седалищные мозоли. По всем этим признакам шимпанзе, орангутан и горилла фактически занимают промежуточное положение между остальными обезьянами Старого Света и человеком. Наличие такой изменчивости у разных видов означает, что либо указанные выше раэли-
66
Часть I. Эволюция человека
чия не имеют большого филетического значения, либо отклонения возникли сравнительно недавно.
Необходимо указать далее на то, что сами признаки, которые, по мнению Вуд-Джонса, исключают родство больших обезьян и человека, отсутствуют у некоторых ископаемых родов больших обезьян. Так, установлено, что дриопитековые (см. рис. 2.5) обладали примитивным расположением черепных костей, таким же, как у долгопятов и человека. По-видимому, ни один из признаков, найденных у этого антропоида, жившего в раннем миоцене, не исключает возможности того, чтобы считать его древним предком гоминид. В пропорциях конечностей мы не находим брахиаторной специализации, характерной для современных обезьян. Можно думать, что для дриопитека характерна брахиация, как и для некоторых из ныне живущих обезьян, но он сохранял достаточно генерализованное строение, оставлявшее возможность дальнейшего развития в направлении понгид и гоминид. На черепе Dryopit-hecus africcmus не было надстроенных образований (надглазничного валика). Носовое отверстие было примитивным, как у церкопите-ковых обезьян. Обезьянья полка не была развита, резцы малы, а область сращения челюстей узкая. У современных человекообразных обезьян, наоборот, резцы великииимеют лопатоообразную форму, зубная дуга U-образна в отличие от человека, у которого резцы малы, а зубная дуга имеет параболическую фо'рму. Морфология зубов у дриопитека, если не учитывать некоторых незначительных деталей, в целом примитивна. В отличие от обезьян Старого Света моляры у них не билофодонтны, клыки, по всей видимости, велики, а передние нижние Премоляры режущие. Но именно такая картина характерна для ранних форм Anthropoidea, и, как мы уже говорили, все данные указывают на то, что предки человека обладали большими клыками. У современного человека клык, особенно молочный, часто выступает над остальными зубами; постоянный клык, как правило, имеет длинный корень и прорезывается позже. Твердо установлено, что некоторые ископаемые формы гоминид имели большие клыки, а иногда и диастему.
Укажем основные характерные признаки, отличающие понгид и гоминид, согласно данному Ле Гро Кларком таксономическому определению этих двух семейств.
Семейство Pongidae — особая радиация Hominoidea, отличающаяся от Hominidae следующими основными направлениями в эволюции: изменения в строении скелета, связанные с приспособлением к брахиации и проявляющиеся особенно ярко в удлинении верхней конечности в целом и ее отдельных частей; развитие большого пальца стопы, хорошо противопоставляющегося остальным пальцам; модификация деталей строения костей верхних и нижних конечностей, связанная с их большей подвижностью и развитием мускулатуры, необходимой для брахиации; тенденция к отно-
Тлава 3. Человекообразные обезьяны, и человек
67
Рис. 3.3. Вид черепа сбоку.
а. Горилла, б. Австралопитек, в. Homo erectus, г. Неандерталец из Ля-Шапелль-о-Сен. д. Штейнгеймский человек, е. Современный человек.
сительному уменьшению первого пальца передней конечности; сохранение основных пропорций таза, характерных для четвероногих млекопитающих. Особенности черепа (рис. 3.3): заметный прогнатизм с сохранением (на более поздней стадии) лицевой части резцовой кости и с убегающим подбородком; развитие (у более крупных видов) массивных челюстей и сильных мышечных гребней на черепе; усиленное развитие выйной части затылочной кости и относительно высокое расположение иниона; затылочные мыщелки располагаются сзади по отношению к слуховым отверстиям; весьма незначительный изгиб базикраниальной оси и небольшая высота крыши черепа; объем мозговой коробки без заметной тенденции к увеличению; прогрессирующая гипертрофия резцов с расширением области симфиза нижней челюсти и с образованием в конечном счете обезьяньей полки; увеличение размеров сильных конических клыков, смыкающихся в диастеме и обнаруживающих заметный половой диморфизм; отчетливо выраженный сектори-альный тип первого нижнего премоляра с развитием сильного переднего корня; позадиклыковые зубы образуют параллельные или слегка дивергирующие ряды; первый молочный моляр с одним бугорком; постоянный клык без тенденции к ускоренному прорезыванию.
Семейство Hominidae (рис. 3.3) — особая радиация Homi-noidea, отличающаяся от Pongidae следующими тенденциями
68
Часть I. Эволюция человека
развития: прогрессивные изменения скелета, связанные с приспособлением к прямохождению, в частности значительное увеличение относительной длины нижней конечности, изменение пропорций и морфологических деталей строения таза, бедра и стопы в соответствии с новыми механическими требованиями, связанными с вертикальным положением тела и прямохождением и соответствующим развитием мускулатуры; сохранение хорошо развитого первого пальца верхней конечности; окончательная утрата большим пальцем стопы способности противопоставляться прочим; усиление изгиба базикраниальной оси с увеличением объема мозговой коробки; смещение вперед затылочных мыщелков; уменьшение площади выйной области на чешуе затылочной кости и низкое расположение иниона; раннее развитие в онтогенезе непирамидального по форме сосцевидного отростка; уменьшение прогнатизма и раннее исчезновение лицевой части резцовой кости; уменьшение клыков, приобретающих лопатообразную форму, слабо смыкающихся или вовсе несмыкающихся и не обнаруживающих заметного полового диморфизма; исчезновение диастемы; замена режущих первых нижних премоляров двухбугорчатыми (с более поздним вторичным уменьшением язычного бугорка); изменение характера окклюзии, приводящее к стиранию коронок зубов до относительно ровной плоской поверхности уже на ранней стадии снашивания; образование равномерно закругленной зубной дуги; заметная (на более поздних стадиях эволюции) тенденция к уменьшению размеров коренных зубов; постепенное ускорение замены молочных зубов и прорезывания постоянных моляров; «моляриза-ция» первого молочного моляра; заметное и быстрое (на поздних стадиях эволюции) увеличение объема мозговой коробки, связанное с уменьшением челюстей и площади прикрепления жевательных мышц; развитие подбородочного выступа.
Найденные в Индии, в горах Сивалик, фрагменты верхней и нижней челюстей обезьяны, которую долгое время относили к дриопитековым, имеют исключительно большое значение для понимания ранней стадии дифференциации гоминид. В настоящее время установлено, что эти фрагменты принадлежат особой форме, получившей название Ramapithecus brevirostis. В строении челюстей и морфологии зубов рамапитека имеется много черт сходства с более поздними формами, несомненно относящимися к гоминидам. Ископаемые остатки этой обезьяны, найденные в Восточной Африке и очень сходные с остатками, обнаруженными в Индии, первоначально приписывали «кенияпитеку».
С гоминидами их сближают следующие признаки: короткий лицевой скелет, небольшие резцы и клыки, а также форма, относительные размеры, общая морфология и расположение коренных зубов и толщина эмали. Зубная дуга имеет параболическую форму, передние нижние премоляры — лишь частично секториального типа.
Глава 3. Человекообразные обезьяны и человек 69
Можно предположить, что челюсти совершали вращательное движение, способствующее истиранию пищи, а не обычное движение вверх и вниз, как у современных человекообразных обезьян. Геологическое датирование этих форм указывает на то, что они существовали в эпоху верхнего миоцена и в плиоцене. Согласно гипотезе Джолли, отличительные особенности строения зубов гоминид обусловлены изменением характера питания: вместо фруктов и плодов рамапитек стал употреблять в пищу более твердые и грубые семена растений или трав. Отсюда можно предположить, что он освоил экологическую нишу, подобную современной саванне. Остатки фауны, связанные с рамапитеком (в Индии и Восточной Африке), указывают на то, что он обитал в основном в густых лесах; не исключено, однако, что окружающая среда представляла собой смешанный тип обитания — леса и саванны.
Определение родства на молекулярном уровне
В последние годы разработано много методов определения родства приматов на молекулярном уровне. Первый из этих методов, значительно усовершенствованный к настоящему времени,— серологический; в организме путем иммунизации вызывают образование антител к тому или иному белку, например к альбумину одного из видов, а затем проверяют способность этих антител перекрестно реагировать с аналогичным белком, выделенным у других предположительно родственных видов. Более надежную информацию можно получить, однако, другим методом, основанным на определении последовательности аминокислот в белках различных биологических видов, поскольку этот метод выявляет «химическое родство» целиком, а не только в отношении компонентов, ответственных за антигенную специфичность. Комбинируя такие методы, как гидролиз и хроматография, можно не только идентифицировать аминокислоты, входящие в состав белка, но и установить их последовательность в полипептидной цепи, что позволяет оценить сходство белков, выделенных у различных видов. Принцип состоит в сравнении аминокислотных последовательностей в аналогичных белках, принадлежащих двум видам, и подсчете числа различающихся аминокислот. Усовершенствованная методика состоит в определении минимального расстояния между участками цепи, где произошли мутации, т. е. минимального числа нуклеотидов, подвергшихся изменению в процессе преобразования кодона одной аминокислоты в кодоны других аминокислот. Такого рода оценки были сделаны у приматов для ряда белков, в том числе гемоглобина, цитохрома с и фибринопептидов. Третий метод состоит в определении комплементарности ДНК, выделенной от разных видов. В основе лежит следующий принцип: отдельные цепи молекул ДНК, принадлежащих разным биологи
70
Часть I. Эволюция человека
ческим видам, при наличии гомологичных участков способны реассоциировать в двухценочечную молекулу.
Указанные серологические и биохимические данные можно дополнить результатами цитогенетического анализа, который выявляет черты сходства в наборе хромосом и их структуре у различных видов.
Применение описанного молекулярного подхода в таксономии приматов дает нам картину фонетического родства тех или иных видов. Детальное сравнение с морфологическими признаками позволяет уточнить картину связей и подтверждает, в частности, особое место, которое занимает среди приматов тупайя. Кроме того, молекулярный подход указывает на более близкое родство лемуров и лори, чем это предполагает формальная таксономия, а также на родство долгопята с человекообразными обезьянами. Новейшие молекулярно-биологические данные свидетельствуют также о более выраженных различиях между орангутаном, с одной стороны, и гориллой и шимпанзе, с другой, по сравнению с различиями между двумя последними формами и человеком.
Предпринимались попытки реконструировать филогенез приматов на основании данных о структуре белков. Однако из-за отсутствия ископаемого материала возникают значительные трудности, связанные в первую очередь с вынужденным предположением о постоянной скорости молекулярной эволюции любой выбранной группы белков. Это предположение более или менее оправдывается для периодов времени, сравнимых с геологическими эпохами, о чем свидетельствует хронология известных филетических изменений в дивергентной эволюции. Тем не менее вряд ли разумно пользоваться указанным предположением в широких масштабах и прежде всего потому, что оно не учитывает глубокой связи темпов молекулярной эволюции с механизмом естественного отбора.
Глава 4
РАЗВИТИЕ ГОМИНИД
Плейстоценовая эпоха
Большая часть известных нам остатков гоминид обнаружена в слоях, которые относят к плейстоцену. Формы, соответствующие началу этой эпохи, еще сохраняли много родственных черт с ископаемыми человекообразными обезьянами. Человек, анатомически сходный с современным, появился к концу этой геологической эпохи. Период времени, в течение которого происходила эволюция человека, охватывает примерно 2—3 миллиона лет. За это сравнительно короткое время на Земле произошли существенные изменения климата, почвы, флоры и фауны, оказавшие заметное влияние на популяции гоминид и их образ жизни (табл. 4.1).
Основное горообразование третичного периода закончилось примерно к концу плейстоцена, характеризовавшегося климатическими колебаниями, связанными, по-видимому, с изменением солнечной радиации. Но какова бы ни была их причина, можно считать установленным, что климат на Земле в плейстоценовую эпоху был то теплым, то холодным. Это отражалось прежде всего на размерах арктической ледяной шапки и оледенении гор в северном полушарии. Активность ледников в южном полушарии, например в Новой Зеландии, также была подвержена колебаниям, однако ледяной покров Антарктиды оставался гораздо более постоянным в течение всего плейстоцена и происходившие с ним изменения никогда не оказывали такого сильного влияния на климат южного полушария, какое оказывала арктическая ледяная шапка на климат северного.
В северном полушарии сменились четыре больших периода оледенения, разделенные тремя межледниковыми эпохами, в течение которых климат становился значительно мягче. Из межледниковых периодов самым продолжительным был средний, который получил название большого межледниковья. Последовательные эпохи наступления ледников в Альпах получили названия гюнц, миндель, рисе и вюрм. Каждый период, соответствовавший такому наступлению, в свою очередь прерывался по крайней мере одним интерстадиальным периодом с более сухим климатом; вюрм-ское оледенение прерывалось двумя интерстадиалами. Одновременно с наступлением альпийских ледников разрастался ледяной покров, центр которого находился в Скандинавии, так что между двумя областями оледенения оставался лишь узкий коридор.
Шкала времени (тыс лет)
Плиоцен
Сухо
Африка
Сухо ,////$ ложно", '
>< рлажро,
5
Аббевиль
Ашель
Клектон
ОлЪувей (Африка)
Техника леваллуа
с
Ното sapiens
Питекантроп
и т
е о
П а
Часть I. Эволюция человека
Гб
* £
Австралопитек
Классический
неандерталец
Прогрессивный неандерталец
Современный человек
Р9 0\ Й S Р
Р9
Глава 4. Развитие гоминид
73
Периоды наибольшей активности ледников в Северной Америке совпадают с периодами похолодания в Европе.
В популярной литературе плейстоцен обычно называют «ледниковой эпохой». Однако сейчас принято относить границу между плиоценовой и плейстоценовой эпохами ко времени значительно раннему, чем начало оледенения. Эта граница определяется как время появления ископаемых форм таких родов, как Elephas, Bos, Equus и Camelus. В начале указанной эпохи, в так называемом нижнем плейстоцене, эти млекопитающие вместе с большим числом вымерших теперь третичных видов составляли фаунистический комплекс, известный под названием виллафранкской фауны. Виллафранкским ярусом называли доледниковый период плейстоцена, однако сейчас полагают, что виллафранкский тип фауны существовал в Европе также и во время первого наступления ледников. Можно, следовательно, считать, что нижнеплейстоценовая эпоха длилась до конца гюнцского оледенения.
В течение ледниковых периодов огромные массы воды, превратившиеся в ледники, были изъяты из круговорота, вследствие чего понизился уровень океана. В результате произошло углубление рек, которые в своем нижнем течении должны были прокладывать путь по отложенным ранее слоям аллювия. Остатки этих древних отложений ныне сохранились в виде террас по берегам рек. Повторные наступления и отступления ледников привели к образованию серий таких террас, являющихся характернейшей особенностью геологии плейстоцена. Аналогично образовались береговые линии морей и озер, ныне высоко поднятые над их современным уровнем.
Известны и другие геологические данные, свидетельствующие о климатических условиях в тех или иных районах в различное время. Ископаемые мерзлые почвы свидетельствуют об условиях, типичных для современной тундры, тогда как погребенные почвы, образовавшиеся в результате химического выветривания, указывают на существование более мягких климатических условий. Одним из наиболее характерных отложений плейстоценовой эпохи на Евразийском материке является лёсс. Он представляет собой принесенную ветром пыль, состоящую из частичек окололед-никовой породы, которая накапливалась в определенных местах в больших количествах за относительно короткое время.
О климатических условиях можно судить также по ископаемой фауне и флоре. В этом отношении особенно полезным оказался пыльцевой анализ, так как каждый вид растений имеет характерную для него форму пыльцевых зерен, а ботаникам хорошо известны климатические условия, которым отдает предпочтение тот или иной вид растений. Той же цели во многом служит изучение раковин моллюсков. Типичными для определенных частных экологических условий являются многие виды млекопитающих. В
74 Часть I. Эволюция человека
начале плейстоцена по Европе бродил Elephas meridionalis (южный слон). Эволюция этой формы в течение плейстоценовой эпохи, по-видимому, привела к появлению двух других форм: Elephas antiquus (древний слон) и Elephas primigenius (мамонт). Оба эти вида легко определить, в особенности по характерному для них строению коренных зубов. Мамонт обитал в степях и в тундре, Elephas antiquus — в лесных и лесостепных районах. Последний найден только в отложениях межледникового периода. Шерстистого носорога (Ticho rhinus antiquitatis) следует отнести к тому же биому, что и мамонта, тогда как другие виды носорогов (Di-cerorhinus etruscus и D. merchii) жили в более теплом климате. Как и сегодня, дикая лошадь населяла открытые степные районы, а северный олень — тундру.
В тропических областях в плейстоценовую эпоху происходила смена влажных (плювиальных) периодов засушливыми (межплювиальными), но соответствие этих периодов периодам оледенения на севере в точности не установлено. У нас нет также достаточных оснований предполагать синхронность наступления плювиальных периодов в тропиках и в странах Средиземноморья. Об отсутствии такого соответствия можно только пожалеть, так как все палеонтологические, археологические и даже физиологические данные указывают на то, что эволюция гоминид началась в тропических областях, расположенных за пределами зоны, непосредственно испытавшей влияние оледенения. У нас нет сведений о том, что гоминиды обитали в Европе или в основной части азиатского материка в нижнем плейстоцене; они впервые заселили эти области лишь во время межледниковых или межстадийных фаз. По-видимому, человек смог приспособиться к холодному климату лишь в конце плейстоцена. В дальнейшем, когда речь пойдет об определении возраста ископаемых остатков гоминид или остатков каь менных орудий, мы будем, как это принято, основываться на последовательности ледниковых и межледниковых фаз и плювиальных и межплювиальных периодов.
Геохронология
Очевидно, что для установления наиболее вероятных эволюционных соотношений необходимо уметь определять возраст ископаемых остатков. В том случае, когда мы не располагаем абсолютной шкалой времени, часто оказывается возможным установить, что данная форма существовала раньше другой, но позже некоторой третьей формы. Если же надо получить информацию о темпах эволюционных изменений, то совершенно необходимо определение абсолютного возраста, т. е. установление геохронологии.
Известно большое число методов определения как абсолютного, так и относительного возраста пород, минералов, ископаемых
Глава 4. Развитие гоминид 7S
остатков. Мы дадим лишь краткий обзор некоторых из них, отсылая читателя за более полными сведениями к специальной литературе по геохронологии.
Наиболее важной п роблемой в хронологии является установление последовательности событий и соотношения возрастов различных пород, слоев и залегающих в них остатков. Часто случается, что в одних в тех же ярусах (в одном месте) находят остатки, относящиеся по крайней мере к двум различным возрастам. Одни имеют возраст, одинаковый с возрастом самих отложений, а другие являются лишь более поздними включениями. Если речь идет о деятельности человека, такие остатки могут быть и преднамеренными захоронениями. Задача геохронологии в таком случае состоит в том, чтобы правильно учесть все эти разнородные условия осадконакопления.
Если климатические условия, характерные для данных отложений, определены (по фауне, флоре или характеру почвы), это сразу сужает границы вероятного возраста ископаемых остатков, относящихся к плейстоцену. Иногда удается получить даже более точные данные, особенно в том случае, когда отложения содержат несомненные постплейстоценовые остатки. Наиболее яркий пример представляет чередование пластов в пещерных отложениях Санта-Фе и Албукерке в Америке. В этих пещерах самый верхний слой состоит из рыхлой пыли, в которой найдены глиняные изделия современных индейцев племени пуэбло. Ниже залегает более плотный отвердевший слой углекислого кальция, который мог образоваться только в плювиальном периоде. Третий слой представляет костеносную брекчию, в которой найдены остатки костей вымерших форм верблюдов, лошадей и наземных ленивцев, а также орудия типа Фолсом. Этот пласт отделен от нижележащих слоев, содержащих ископаемые остатки животных и орудия типа Сандиа, прослойкой бледного желто-коричневого цвета, не содержащей каких-либо остатков животных или орудий. Наличие этого желто-коричневого слоя представляет особый интерес, так как именно такие слои всегда осаждаются из просачивающихся в породу вод, содержащих особые гуминовые кислоты — продукты медленного разложения хвои. Единственное время, когда можно было бы предполагать распространение хвойных лесов в этой юго-западной части США,— это период наступания ледников на севере. Можно, следовательно, полагать, что желто-коричневый слой образовался во время последней фазы наступания последнего ледника и что культура Сандиа относится к заключительному «интерстадиалу».
Возраст ископаемых остатков, найденных в отложениях речных террас, можно определить примерно таким же способом. Из сказанного выше очевидно, что террасы представляют собой образования, соответствовавшие периодам отступания ледников, и что
76
Часть I. Эволюция человека
самые высокие террасы — самые древние, а самые низкие — наиболее молодые. Зависимостью между климатическими условиями и образованием речных террас неоднократно пользовались при определении возраста различных остатков.
Ценным источником сведений о возрасте ископаемых остатков гоминид являются другие окаменелости, залегающие в тех же слоях. Отложения, содержащие сходные ископаемые остатки, обычно имеют одинаковый возраст, и по одним из них можно определить возраст других. Правда, при этом необходимо соблюдать сугубую осторожность, так как формы, которые исчезли (и считаются вымершими) на одной территории в какой-то определенный период, могут длительное время существовать в других районах. И наоборот, могут пройти тысячелетия, прежде чем та или иная форма, появившаяся в некоторой зоогеографической области, расселится в другие (если она вообще расселится).
С этими оговорками можно все же утверждать, что различные животные формы, в том числе и некоторые млекопитающие, могут служить своего рода «индикаторами возраста», по крайней мере в пределах определенных географических зон. Так, костные остатки, принадлежащие саблезубому тигру (Machairodus), южному слону (Elephas meridionalis) и этрусскому носорогу (Dicerorhinus etruscus), найдены в Европе только в нижнем плейстоцене; мамонт (Elephas primigenius) является примером вида, жившего только в верхнем плейстоцене.
Существует несколько способов определения гомогенности образцов, найденных в одних и тех же отложениях. Вейнер (Weiner), Окли (Oakley) и Ле Гро Кларк (Le Gros Clark) воспользовались ими для того, чтобы доказать факт фальсификации пильтда-унских остатков. Один из наиболее распространенных приемов — это так называемая «фторовая проба». В костях, длительное время находившихся в земле, основная минеральная соль — гидроксиапатит Са5(РО4)3ОН — постепенно превращается во фторапатит Ca5(PO4)3F вследствие поглощения фтора из просачивающихся вглубь вод. Скорость превращения зависит, конечно, от содержания в воде фтора, но в любом данном отложении количество фтора в костях является функцией геологического возраста. Поэтому если установлено, что кости содержат различные количества фтора, то те из них, в которых фтора больше, либо пролежали в земле дольше, либо попали в данный пласт откуда-то извне.
Определять относительный возраст можно также, сравнивая содержание органического азота или органического углерода в различных костях, найденных в одном отложении. Более древние кости содержат меньше азота и углерода, так как соответствующие органические вещества постепенно разлагаются.
Многие ранние попытки определения минимального абсолютного возраста основывались на оценках скорости накопления речных
Глава 4. Развитие гоминид
77
наносов или образования последовательных слоев почвы. Поскольку, однако, скорость осаждения зависит от многих факторов, такие оценки неизбежно оказывались весьма приближенными. В последнее время удалось более точно определить скорость образования некоторых особых видов отложений, таких, как сталагмиты и сталактиты; в случае флювиогляциальных отложений слои осадков представляют годичные образования. Зимой таяние ледников происходит медленно, количество осадков, образующихся за это время года, невелико, и состоят они из мелких частиц. Напротив, наносы летнего периода более мощные и представлены гравием и моренным материалом. Каждое годичное отложение можно, следовательно, отличить от отложений предшествовавших и последовавших лет; особенно удобны в этом отношении ленточные глины — тонкие прослойки, откладывающиеся ежегодно. Подсчет количества прослоек вблизи современного края ледника и на всей площади, которую ранее покрывал ледник, может помочь определить абсолютный возраст некоторой формации. Этот метод, впервые примененный в Швеции де Геером, широко используется в Европе и Америке для определения возраста остатков.
Несколько похож на него по общему принципу метод дендрохронологии. В географических зонах со сменой времен года концентрические кольца на поперечном срезе ствола позволяют определить возраст дерева. Толщина кольца зависит от того, насколько благоприятен был данный год для роста дерева: тесное расположение нескольких соседних колец указывает на то, что условия роста в течение ряда лет были неблагоприятными. Отметка на срезе живущего дерева может оказаться где-нибудь в середине ствола, а на ископаемом стволе — на периферии. Сравнивая два образца древесины, можно определить полное число колец, которое оказывается больше, чем на любом из взятых в отдельности образцов. Серия образцов разного возраста дает возможность установить возраст археологической находки, обнаруженной вместе с ископаемыми остатками древесины. Методы, основанные на подсчете слоев ленточных глин и определении возраста древесины, позволили установить возраст ископаемых остатков, соответствующих недавним археологическим стадиям, т. е. последним 5 тысячам лет.
Наконец, укажем на целый ряд изотопных методов определения абсолютного геологического возраста. Наибольшую ценность представляет радиоуглеродный метод, впервые разработанный Либби. С помощью этого метода удается датировать ископаемые остатки, возраст которых составляет примерно 50—60 тысяч лет. Изотоп углерода С14 непрерывно образуется в атмосфере вследствие действия космического излучения на атмосферный азот. Химически он ведет себя как обычный углерод С12 и, следовательно, включается во все живые ткани. После смерти организма дальней
78
Чаешь I. Эволюция человека
ший обмен углерода прекращается и С14 медленно распадается с периодом полураспада 5568+30 лет. Содержание С14 в органическом веществе является, таким образом, функцией его возраста: чем меньше содержание С14, тем больше возраст. Ошибка в определении возраста составляет от ±100 до ±1200 лет (в зависимости от возраста), и надежно определяемый возраст не превышает 20—30 тысяч лет. Тем не менее радиоуглеродный метод широко и с успехом применялся главным образом для датировки остатков из верхнего плейстоцена и более поздних отложений.
Если возраст отложений превосходит полмиллиона лет, то используются другие изотопные методы, основанные на накоплении изотопов свинца и радиоактивных минералов, содержащих уран и торий, или измерении скорости распада радиоактивного калия, превращающегося в аргон. Последний метод основан на том, что в природном калии присутствуют микроколичества изотопа К40, который распадается, превращаясь в Са40 и Аг40 с периодом полураспада 1,3X10® лет. Этим методом была подтверждена установленная ранее продолжительность различных эпох третичного периода (табл. 2.1), а также показано, что возраст, соответствующий рубежу плиоцена и виллафранка, превосходит миллион лет, а возможно, даже достигает двух или трех миллионов лет.
Радиоизотопные методы сыграли исключительную роль в определении возраста находок в Олдувайском ущелье (Танзания). В этом районе в результате размытия берега в речном каньоне обнажился ряд слоев, относящихся к эпохе позднего плейстоцена. Основанием служит базальтовый слой вулканического происхождения; его возраст, оцененный калиево-аргоновым методом, равен приблизительно 2 млн. лет. Выше расположено пять пластов (I—V). Древнейший из них — пласт I — также вулканического происхождения, содержит ценный ископаемый материал — большое число остатков гоминид. Возраст одной из прослоек вблизи середины этого пласта (маркирующего слоя А), определенный калиево-аргоновым методом,1,75 млн. лет.Пласт II также содержит остатки гоминид, однако он образовался из материала, принесенного подземными потоками, и озерных отложений, поэтому его возраст нельзя оценить изотопными методами. Толщина его местами близка к толщине пласта I, однако процесс формирования занял несравненно больше времени. Район Олдувайского ущелья представляет собой часть Большой Рифтовой долины (Восточная Африка), простирающейся на север в Кению и Эфиопию. В южной части Эфиопии в нее входит долина реки Омо, впадающей в озеро Рудольфа в Кении. В этой долине также сделано немало важных находок, возраст которых оценен радиоизотопными методами.
Весьма перспективен новый метод определения возраста костей, сохранивших белковый материал,— так называемый метод
Глава 4. Развитие гоминид
79
рацемизации аминокислот. В живом организме 20 аминокислот имеют определенную стереоконфигурацию — все они являются левовращающими (L). После смерти происходит постепенное изменение конфигурации аминокислот, т. е. увеличение относительной! доли правовращающих молекул. Соотношение между обеими формами зависит от времени, прошедшего с момента смерти организма. Для определения возраста пока что использовали в основном одну аминокислоту — изолейцин. Было показано, что оцененный этим методом возраст остатков из Олдувая согласуется с определенными изотопными методами. Следует, однако, учитывать один существенный фактор, а именно температурную историю образца, так как с ростом температуры увеличивается скорость рацемизации.
Австралопитек
Обширные раскопки, проведенные Дартом, Брумом и Робинсоном (Dart, Broom a. Robinson), в травертиновых пещерных отложениях Ботсваны и Трансвааля, привели к открытию огромного количества ископаемых остатков, которые приписывают одной из ранних форм гоминид. Этим остаткам, извлеченным из различных пещер, были даны разные родовые или видовые наименования: Australopithecus africanus из Таунгса, A. prometheus из Макапан-сгата, Plesianthropus transvaalensis из Штеркфонтейна, Рагап-thropus robustus из Кромдраай, Paranthropus crassidens и, на более высоком уровне развития, Telanthropus capensis из Сварткранса.
Между этими формами действительно существуют определенные различия. Однако у форм, найденных в пещерах Таунгса, Ма-капансгата и Штеркфонтейна, они столь малы, что отнесение их даже к различным видам никак не оправдано. Большинство авторитетов квалифицируют их теперь как один род Australopithecus. Вместе с тем различия между указанной группой и той, которую называют Paranthropus, весьма значительны. В частности, Paranthropus имеет более массивный и прочный череп, обычно с отчетливо выраженным сагиттальным гребнем, совершенно плоское или даже «блюдцевидное» лицо и низкий лоб. Относительные размеры зубов не такие, как у всех других гоминид: моляры и премоляры велики, а резцы и клыки слишком малы. Эти различия в зубном аппарате Робинсон объясняет тем, что Paranthropus был травоядным, тогда как все остальные известные нам гоминиды были всеядными. Однако различия между всеми найденными в Южной Африке остатками кажутся нам не более значительными, чем различия, которые имеются в пределах любого другого рода млекопитающих. Поэтому в дальнейшем мы будем говорить только об одном роде Australopithecus, включающем два вида: A. africanus и A. robustus (Paranthropus).
80
Часть I. Эволюция человека
В последнее время внимание ученых привлекли раскопки в Восточной Африке — в Кении, Танзании и Эфиопии, где различными исследователями, и в первую очередь Лики и членами его семьи, было открыто большое количество ископаемых форм. Каков бы ни был точный таксономический статус этих форм, их вернее всего следует отнести к австралопитекам в широком понимании этого термина. Одна из первых находок была сделана в Олду-вайском ущелье в Танзании: вблизи основания пласта I был обнаружен почти полностью сохранившийся череп. Его идентифицировали сначала как Zinjianthropus boisei, однако в настоящее время большинство специалистов пришли к заключению, что остатки принадлежат скорее местной форме Australopithecus robustus. В отложениях вблизи Пенини, в долине реки Омо, а также к востоку от озера Рудольфа было найдено еще несколько представителей этого вида. Вместе с остатками Australopithecus robustus в Олдувае в пластах I и 1Г нашли еще одну форму гоминид, которую первоначально отнесли к роду Ното, присвоив наименование Homo habilis. Ископаемый материал включал несколько образцов, в том числе фрагменты черепа, которому дали условное наименование «Twiggy», или образец ОН24. У последнего много общих признаков с одним из наиболее хорошо сохранившихся черепов Australopithecus africanus (Штеркфонтейн 5) из Южной Африки, однако есть и отличия: челюсть и коренные зубы короче, а субназальный прогнатизм не так сильно выражен, затылочная область более круглая, а затылочное отверстие в основании черепа смещено еще дальше вперед. Из других остатков восточноафриканских австралопитеков отметим челюсть из Лотагема и фрагмент плечевой кости, найденный в Канопой (Кения). В самое последнее время большое внимание специалистов привлекли две важные находки: 1) к востоку от озера Рудольфа обнаружена значительная часть мозговой коробки и лицевого скелета индивидуума, которому присвоили кодовое название ER14,0 (его эволюционный статус будет рассмотрен ниже); 2) в Афаре (область Хаггар в Эфиопии) Иохансон (Johanson) нашел почти полный скелет; описания его пока не дано, но предполагается, что он принадлежит австралопитеку (он фигурирует под именем «Lucy»).
Ле Гро Кларк формально определяет Australopithecus как род, относящийся к Hominidae и отличающийся следующими признаками: относительно малый объем мозговой коробки — от 450 до 700 см3; сильно развитые надглазничные валики; тенденция к образованию у крупных индивидуумов низко расположенного сагиттального гребня в лобно-теменной области (не связанная с образованием высоколежащего затылочного гребня); затылочные мыщелки расположены значительно позади середины длины мозговой коробки, а в поперечном сечении — на уровне слуховых отверстий; малый размер выйной области на затылочной кости, как
Глава 4. Развитие гоминид
81
у Ното; появление как у взрослых, так и у не достигших зрелости индивидуумов пирамидальных сосцевидных отростков; типичное для гоминид строение нижнечелюстной ямки с выраженным развитием позадисуставного отростка у некоторых индивидуумов; массивные челюсти, обнаруживающие значительную индивидуальную изменчивость по абсолютным размерам; отсутствие подбородочного выступа; область симфиза относительно прямая и почти вертикальная; параболическая форма зубной дуги без диас-темы; лопатообразные клыки, стертые и уплощенные на вершине; сравнительно большие премоляры и моляры; передний нижний премоляр с двумя почти одинаковыми бугорками; выраженная «моляризация» первого молочного моляра; прогрессирующее увеличение размеров постоянных нижних моляров — от первого до третьего; типичный для гоминид скелет конечностей, отличающийся, однако, от Ното по многим деталям (рис. 3.1), а именно: удлинение кпереди верхней передней подвздошной ости, относительно малая поверхность крестцово-подвздошной области, низкое положение (у некоторых индивидуумов) седалищного бугра, заметное увеличение кпереди межмыщелковой вырезки бедра, расширение в медиальном направлении головки таранной кости.
Когда Дарт (Dart) в 1925 г. описал первого найденного им в Таунгсе Australopithecus, а именно часть черепа и слепок полости черепа индивидуума, не достигшего зрелости, общее мнение свелось к тому, что найденные остатки — не более как остатки еще не известного ископаемого понгида. Однако после дальнейших раскопок и открытий большинство специалистов пришли к выводу, что A ustralopithecus относится к гоминидам. Мнение изменилось потому, что череп, несмотря на его небольшой объем и общее внешнее сходство с черепом обезьяны, имел многие признаки, характерные для гоминид. Дальнейшее изучение скелета конечностей показало, что Australopithecus был наземным существом и передвигался на двух ногах. В приведенном выше описании рода Ле Гро Кларк явно отметил признаки, которые прямо указывают на выпрямленное положение тела и передвижение на двух ногах: характерное строение таза, положение затылочных мыщелков и выйных линий, развитие сосцевидного отростка. Строение челюстей и зубов дополняет общую морфологическую картину, характерную для гоминид. Особенно типична форма постоянного и молочного клыков и нижних передних премоляров, а также форма всей зубной дуги. Строение сочленения нижней челюсти с черепом и характер стирания коренных зубов указывают на то, что у австралопитека, так же как и у других гоминид, челюсти в процессе жевания совершали вращательные движения; у обезьян же из-за смыкающихся клыков возможно только вертикальное движение челюстей. У современного человека в противоположность австралопитеку суставной отросток превратился в небольшой
82
Часть I. Эволюция человека
бугорок, а размер моляров постепенно уменьшается от первого к третьему; промежуточная картина обнаруживается у других гоминид. Отсутствие подбородка, как мы видели, является примитивным признаком гоминид, и ему нельзя придавать особого значения. Однако у австралопитека отсутствует обезьянья полка, а симфиз более вертикальный, чем у понгид. Эти два признака очень важны в филетическом отношении. Развитие надглазничного валика найдено у разных гоминид. Наличие этого признака следует ожидать у форм, обладающих большими челюстями и низко посаженными лобными костями. Австралопитек представляет исключение среди гоминид в том отношении, что у некоторых индивидуумов имеется сагиттальный гребень. Хотя этот гребень и отличается от сагиттального гребня обезьян тем, что не примыкает к затылочному гребню, некоторые палеонтологи считают, что наличие этого гребня исключает австралопитека из последующей эволюции в направлении гоминид. Однако наличие такого гребня, по-видимому, связано с небольшим размером мозговой коробки и большими челюстями, и можно, следовательно, ожидать, что он исчез бы в ходе дальнейшей эволюции, если бы она привела к увеличению мозговой коробки и к уменьшению челюстей. Подобные тенденции действительно характерны для филогенеза гоминид.
Не приходится удивляться тому, что у австралопитека мозговая коробка мала, ибо малый размер коробки и мозга — анцест-ральный (предковый) признак; на некоторой стадии эволюции гоминиды должны были иметь небольшой мозг. Этот признак придает австралопитеку внешнее сходство с обезьянами, однако и этот признак свидетельствует об эволюции в направлении гоминид. Ле Гро Кларк показал, что та часть мозговой коробки, которая расположена выше уровня глазниц, у австралопитека больше, чем у гориллы, шимпанзе и орангутана. Это служит дополнительным указанием на принадлежность австралопитека к гоминидам.
В этой связи особый интерес представляют данные о том, что австралопитеки (по крайней мере некоторые из них) уже использовали примитивные каменные орудия. Их обнаружили вместе с остатками скелетов в Олдувайском ущелье и Сварткрансе. Имеются указания на то, что южноафриканские австралопитеки в качестве орудий применяли кости, рога и зубы животных.
Приведенное выше определение рода Australopithecus (принадлежащее Ле Гро Кларку) было сформулировано до того, как была получена большая часть ископаемого материала из Восточной Африки. В связи с этим уместно задать вопрос, подпадают ли вновь найденные остатки под это определение, нуждается ли оно в изменении или же для указанных остатков следует ввести дополнительную таксономическую категорию. Все специалисты, по-видимому, согласны друг с другом в том, что остатки, приписывав-
Глава 4. Развитие гоминид
83
мые «Ното habilis» из Олдувайского ущелья, и в особенности те из них, которые извлечены из пласта II, в морфологическом отношении представляют шаг вперед по сравнению с материалом, найденным в Южной Африке. Об этом говорят не только форма и величина зубов, но и имеющиеся указания на большой объем мозговой коробки. Основываясь на этих фактах, Лики (Leaky) и его сотрудники квалифицировали новый вид «habilis», отнеся его к роду Ното. В настоящее время мы располагаем таким великолепным материалом, как череп ER1470 (иэ района к востоку от озера Рудольфа) с объемом мозговой коробки не менее 800 см3, превосходящим объем коробки у некоторых Homo erectus. Этот череп характеризуется целым рядом других прогрессивных признаков, например круглой формой (рис. 4.1). Здесь стоит заметить, что более тщательный анализ остатков «Тelanthropus capensis» из Сварт-кранса, включающих часть черепа и нижнюю челюсть, также свидетельствует о наличии более развитых гоминидных признаков по сравнению с другими австралопитеками Южной Африки. Эти признаки сближают «телантропа» с более поздними формами Ното erectus. По-видимому, именно телантроп был «автором» простых орудий, найденных в Сварткрансе.
Ситуация в целом достаточно хорошо иллюстрирует трудности, которые возникают при попытке описать непрерывный процесс эволюции с помощью дискретных таксономических категорий. Если известные ископаемые остатки разделены морфологическими «разрывами», то эти разрывы можно использовать для разграничения таксонов. Однако по мере накопления данных таксономические категории становятся все менее отчетливыми. Ведь в конечном счете введение тех или иных таксонов в палеоантропологию основывается в значительной степени на субъективной оценке величины морфологических вариаций во временной последовательности форм, которую можно считать совместимой с идентификацией отдельных видов. По мнению специалистов, морфологические различия между формой Australopithecus africanus, представленной первоначально найденными остатками в Южной Африке, и наиболее ранними представителями Homo erectus на Яве недостаточны, чтобы можно было признать эти две формы отдельными видами. С этой точки зрения новые находки следует относить либо к одному, либо к другому таксону, и те, кто сталкивается с подобной необходимостью, обычно относят ископаемый материал к Australopithecus africanus. С другой стороны, специалисты, признающие видовую категорию «Ното habilis», более склонны теперь относить этот вид к австралопитеку (Australopithecus habilis),) а не к Ното. При этом учитывается большее сходство A. habilis с ископаемыми остатками австралопитеков, чем с Homo erectus. В обоих указанных случаях идентификация форм, подобных ER1470, вызывает значительные трудности. Правда^ следует заме-
Рис. 4.1. Генеалогическая схема гоминид, показывающая связь между важнейшими ископаемыми формами.
Нижний плейстоцен [ Средний плейстоцен | Верхний плейстоцен
Глава 4. Развитие гоминид
85
тить, что при сравнительном анализе отдельных образцов недостаточно учитывается вариабельность в пределах популяции. Череп ERj470 мог принадлежать, например, индивидууму мужского пола необычно больших размеров. Ясно, однако, что если отнести его к австралопитекам, то определение этого рода придется несколько изменить, чтобы включить в него случайные формы со столь большим объемом черепа. Существует и другое мнение, а именно, что ER1470, «телантропа» и ряд других, еще не описанных детально находок из Восточной Африки следует отнести к Ното (а возможно, и к Homo erectus, что привело бы к значительному увеличению древности этого вида).
Дело, конечно, заключается не столько в таксономии, сколько в правильном понимании филогенетической истории гоминид. Находки в Восточной Африке во многом способствовали прояснению этого вопроса, и прежде всего потому, что оказалось возможным определить абсолютный возраст отложений, в которых были найдены ископаемые формы. Что касается остатков гоминид из Южной Африки, то их датирование всегда вызывало серьезные трудности вследствие того, что они были извлечены из отложений невулканической природы. Возраст пласта I и наиболее глубоких слоев в пласте П в Олдувайском ущелье составляет от 1,9 до 1,6 миллионов лет. Отложения к востоку от озера Рудольфа, в которых был обнаружен ER1470, а также Australopithecus robustus, образовались 2—2,6 миллиона лет назад. Возраст фрагментов плечевой кости из Канопой, вероятно принадлежавших австралопитеку^ оценивается в 4 миллиона лет, а возраст челюсти, найденной в Лотагеме,— не менее чем в 5 миллионов лет. Датирование материала из Южной Африки, основанное главным образом на анализе связанной с ним фауны, показывает, что его возраст соответствует указанному весьма широкому временному интервалу. Возраст остатков Australopithecus africanus оценивают в 3—2,5 миллиона лет; представители A ustralopithecus robustus из отложений в Кром-драае и Сварткрансе несколько «моложе», но и их возраст датируется по крайней мере временем образования пласта I в Олдувае. Обе формы долгое время считали значительно более древними.
Из всей совокупности известных нам данных можно сделать следующий вывод: мы наверняка имеем дело с длительно существовавшим родом, который на протяжении большей части своей истории был представлен двумя видами — А. africanus и А. robustus,, нередко обитавшими в пределах одного ареала. Последний вид в конце концов исчез, тогда как первый постепенно эволюционировал в направлении Ното, пройдя через ряд форм, подобных Homo hdbilis. И если верно то, что именно А. africanus впервые стал изготовлять простейшие орудия, то можно сделать вывод, что он в конце концов перешел к охотничьему образу жизни. Возможно, однако, что Ното происходит от одной из известных форм австрало
86
Часть I. Эволюция человека
питеков и что в нижнем плейстоцене одновременно существовали не две, а три предковые линии. Даже если это так, можно утверждать, что предки человека прошли через «австралопитекоидную» фазу развития, во многом сходную с A. africanus. Менее ясен вопрос о филетической связи А. africanus и А. robustus. Разделение линий понгид и гоминид должно было произойти не 5 миллионов лет назад, а гораздо раньше. Можно думать, что предком австралопитека была одна из форм, подобных рамапитеку.
Ното
Все прочие ископаемые формы гоминид в настоящее время, как правило, относят к одному-единственному роду Ното, подразделяя его на два вида: Homo erectus (ранее его включали в род Pithecanthropus'), теперь вымерший вид, и Homo sapiens, который включает все современные человеческие популяции и ряд предшествующих ископаемых форм. Ниже мы приводим формальное определение рода, основанное на определениях Ле Гро Кларка, а также Лики, Тобиаша и Напье (Leaky, Tobias a. Napier).
Homo — род семейства Hominidae; его характерные признаки: относительно большой объем мозговой коробки, обычно от 800 см3 до 1600 см3; костные гребни на черепе развиты в различной степени, височные гребни по высоте расположения варьируют, но никогда не достигают средней линии; посторбитальное сужение иногда выражено, но слабее, чем у австралопитека; надглазничная область лобной кости изменяется в широких пределах от массивного выступающего надглазничного валика до полностью отсутствующего выступа и очень гладкой надбровной области; лицевой скелет не вогнутый (и не «блюдцевидный»); для него характерны слабый прогнатизм и ортогнатизм; контур переднего симфиза может заметно отступать назад, но иногда наклонен вперед, подбородочная кость иногда полностью отсутствует, но может образовывать более или менее развитый подбородочный треугольник; зубная дуга округлая, у большинства представителей рода без диастемы; первый нижний премоляр двухбугорчатый; коренные зубы различной величины, но меньше, чем у австралопитека; клыки и резцы малы; тазовый пояс и скелет конечностей приспособлены к прямохождению; рука способна крепко схватывать предметы.
Homo erectus
В табл. 4.2 перечислены ископаемые остатки, которые обычно относят к этому виду, с указанием географического местоположения находок.
Глава 4. Развитие гоминид
87
Таблица 4.2
Ископаемые остатки гоминид
Страна Место находки Что найдено Общепринятое наименование Возраст находки или геологический горизонт
Ява Ява Ява Ява Ява Ява Ява Ява Ява Китай Китай Алжир Марокко Танзания Триниль Сангиран » » » Мо дж оперто Сангиран » » Чжоу-коу- дянь Лантянь Тернифин Сиди-Абдер-Рахман Олдувай Крыша черепа и бедренная кость Крыша черепа Теменная кость Фрагменты четырех черепов Задняя часть крыши черепа и верхняя челюсть Череп ребенка Фрагмент нижней челюсти То же » Примерно 14 индивидуумов (в основном черепа и зубы) Череп и челюсть 3 нижние челюсти и небольшой обломок теменной кости Нижняя челюсть Фрагменты крыши черепа и несколько костей скелета Питекантроп I (Р. erectus) Питекантроп II (Р. erectus) Питекантроп III (Р. erectus) Питекантроп V — VIII Питекантроп IV ' (Р. robustus) Н. modjokertensis Питекантроп IV Р. dubius Мегантроп (Meganthropus palaeo]avanicus) t Синантроп (Sinanthropus pekinensis ) Лантяньский человек Атлантроп ( Atlanthropus mauritanicus ) A. mauritanicus ► Триниль-ский, 500-700 тыс. лет Джетисский нижний уровень, 1,9.10е 300 тыс. лет Начало среднего плейстоцена Верхний пласт II, 700 тыс. лет (пласт IV)
88
Часть I. Эволюция человека
Табл, 4.2 (продолжение)
Страна Место находки Что найдено Общепринятое наименование Возраст находки или геологический горизонт
Германия Гейдельберг Нижняя челюсть Homo heidelbergen-sis Гюнцмин-дельское межледниковье
Греция Петралоиа Череп Ср. плейстоцен
Венгрия Фертесзол- лош Затылочная кость Мипдель-ский ин-терстади-ал
Таблица 4.3
Сопоставление яванского человека с пекинским
Признак Яванский человек Пекинский человек
Объем мозговой коробки Наибольшая длина мозговой коробки Высота мозговой коробки Лоб Лобные синусы Надглазничный валик Нёбо Тело нижней челюсти Подбородочный выступ Коренные зубы Примерно 900 см3 190 мм 105 мм Очень покатый Умеренно развиты Не отделен от чешуи лобной кости Гладкое Массивное Отсутствует Большие 1075 см3 194 мм 115 мм Менее покатый Слабо развиты Отделен от чешуи лобной кости углублением Негладкое Не слишком массивное Слабо выражен Не очень большие
Первые остатки Homo erectus были найдены в 1891 г. на Яве Дюбуа (Dubois), который ввел для их обозначения родовое название Pithecantropus. Более поздние находки были сделаны на Яве Кенигсвальдом, Джекобом и Сартоне (Koenigswald, Jacob a. Sartone) в основном в районе Сангирана. Ископаемые остатки обнаружены в двух геологических горизонтах: 1) тринильском, относящемся к среднему плейстоцену, абсолютный возраст которого
Глава 4. Развитие гоминид
89
оценен в 700 тыс. лет у основания и в 500 тыс. лет — у вершины, и 2) джетисском, соответствующем нижнему плейстоцену; возраст последнего у основания, вблизи места находки черепа ребенка, определен в 1,9 млн. лет и явно завышен. Найденные в Триниле формы весьма однородны, и средний объем мозговой коробки (для 5 черепов) составляет 860 см3. В их число включен, однако, и недавно найденный, хорошо сохранившийся череп (образец VIII) с объемом мозговой коробки 1029 см3. Джетисские формы во многих отношениях более примитивны, чем тринильские, например у них имеется небольшая диастема в верхней челюсти. Размер мозговой коробки для одного из черепов равен 750 см3. К остаткам, найденным в Джетисе, относят и несколько обломков челюстей, ранее известных под названием Meganthropus pala-eojavanicus. Они сходны с соответствующими фрагментами австралопитеков, особенно A. robustus, и многие исследователи включили их в этот род. Однако из-за фрагментарного характера остатков невозможно с уверенностью определить таксономический статус «мегантропа», хотя,.конечно, более убедительное доказательство существования в Азии форм, близких к австралопитекам, представляло бы исключительный интерес. Что касается остальных находок в Джетисе, то они близки к тринильским формам, хотя и имеются некоторые отличия, связанные с разным возрастом указанных форм. И те и другие, несомненно, относятся к одному виду.
Ископаемые остатки из нижней пещеры Чжоу коу-дянь близ Пекина (их раньше относили к роду Sinanthropus) характеризуются более прогрессивной морфологией, чем яванские остатки, и относятся они к более позднему времени, чем формы, найденные на Яве. Объем мозговой коробки варьирует от 915 до 1225 см3. Вейденрейх провел детальное сравнение всех ископаемых, обнаруженных на Яве и вблизи Пекина. Из 120 морфологических признаков синантропа 74 можно было использовать для прямого сравнения. При этом оказалось, что по 57 признакам (из 74) обе формы весьма сходны; выраженные различия отмечены лишь по 4 признакам. Перечень основных различий приведен в табл. 4.3; они указывают на то, что таксономические различия двух форм соответствуют вариациям в пределах одного вида.
Ископаемые остатки Homo erectus в Европе и в Африке крайне скудны. Они относятся примерно к тому же периоду, что и пекинская или тринильская формы, и сходны с последними по многим признакам. Найденная в Фертесзоллоше затылочная кость отличается рядом прогрессивных признаков, и оцененный по ней объем мозговой коробки (1350—1400 см3) превышает минимальный объем, характерный для современного человека. Полагают, что человек из Фертесзоллоша был переходной формой от Н. erectus к Н. sapiens. Тем не менее для лучше сохранившегося черепа из Петралоны объем мозговой коробки составляет лишь 1120 см3.
90
Часть I. Эволюция человека
Хотя мы знаем очень мало об этих западных формах, имеющиеся данные говорят о том, что они в целом соответствуют яванским и пекинским формам и относятся к одному с ними виду. При этом следует учитывать возможность значительной географической изменчивости в пределах вида.
Приведем определение вида, основанное на ранее сформулированном Ле Гро Кларком определении питекантропа (когда последнего еще относили к отдельному роду).
Homo erectus — вид из рода Ното; его характерные признаки: объем мозговой коробки в среднем 1000 см3; заметное уплощение черепного свода, небольшая выпуклость лба; массивные надглазничные валики; заметное посторбитальное сужение; совпадение опистокраниона с инионом; наличие сагиттального гребня и выраженный вертекс; сосцевидный отросток различен у разных форм, но обычно мал; толстая стенка черепа; тимпанальная пластинка утолщена и имеет тенденцию к горизонтальному расположению; широкие, плоские носовые кости; тяжелая нижняя челюсть без подбородочного выступа; тяжелые зубы с хорошо развитым базальным цингулюмом; клыки, иногда выступающие и смыкающиеся; небольшая диастема в верхней челюсти; первый нижний премоляр с двумя почти равными бугорками; моляры с отчетливо дифференцированными бугорками и выраженной вторичной морщинистостью эмали; второй верхний моляр больше первого, а третий нижний может превосходить по длине второй; кости конечностей ничем не отличаются от костей Homo sapiens.
У Homo erectus мы ясно обнаруживаем как «примитивные», так и более «прогрессивные» признаки и можем рассматривать его морфологический статус как промежуточный между Austra~ lopithecus и Homo sapiens.
Основные примитивные признаки Н. erectus таковы:
1. Кости черепа очень толстые.
2. Сильно развитый надглазничный валик, не прерывающийся на всем протяжении.
3. Очень покатый лоб.
4. Мощный затылочный гребень, простирающийся в сосцевидную область и создающий угловатый контур затылочной кости.
5. Небольшие сосцевидные отростки.
6. Наибольшая ширина черепа на уровне слуховых отверстий.
7. Характерное расположение тимпанальной пластинки.
8. Широкие носовые кости.
9. Выраженный субназальный прогнатизм.
10. Массивная нижняя челюсть.
1.1. Отсутствие выдающегося вперед подбородка.
12. Образование нескольких подбородочных отверстий.
13. Крупные верхние резцы.
Прогрессивные признаки Н. erectus следующие:
Глава 4. Развитие гоминид
91
1. Объем мозговой коробки превосходит минимальные величины для Н. sapiens; выпуклый черепной свод.
2. Увеличенный по сравнению с австралопитеком изгиб лицевой части черепа по отношению к мозговой коробке; передняя черепная яма простирается над глазницами.
3. Относительные размеры лицевого черепа меньше, чем у австралопитека.
4. Большое затылочное отверстие расположено ближе к центру, чем у австралопитека.
5. Нижнечелюстное сочленение такое же, как у Homo sapiens.
6. Зубная дуга параболической формы.
7. Морфология зубов ближе к Homo sapiens, чем к австралопитеку.
8. Начальная стадия развития подбородка (у пекинского человека).
9. Кости конечностей по размерам и пропорциям аналогичны костям Homo sapiens.
Особенность, которая бросается в глаза,— это резкое отличие формы черепа у Homo erectus от таковой у австралопитека, не исключая и наиболее развитых его форм, подобных «Н. habilis» (из Олдувая). Мозговая коробка у Н. erectus уплощена и расширена, а челюсти меньше, чем у австралопитека. Исходя из указанных различий, Лики предложил исключить вид Н. erectus из прямой предковой линии Н. sapiens. Мы считаем, однако, что для этого нет достаточных оснований. Все имеющиеся данные указывают на то, что общая морфология скелета у Н. erectus и современного человека одинакова. Н. erectus обладал всеми необходимыми приспособлениями для прямохождения.
Н. erectus является промежуточной формой между австралопитеком и Н. sapiens не только в отношении общей морфологии, но и хронологически. Если допустить, что абсолютный возраст основания джетисского горизонта определен правильно, то отсюда следует, что формы в Олдувае и на Яве существовали примерно в одно и то же время. Однако, как мы указывали выше, возраст джетисских остатков явно переоценен. Древнейшие остатки Ното erectus, по-видимому, не «старше» 1 миллиона лет.
Мы не можем сказать, где именно произошел переход от австралопитека к Н. erectus, тем более если согласиться с тем, что «ме-гантроп» представлял собой азиатскую разновидность австралопитека. Было бы наивно полагать, что такой переход мог совершиться лишь у нескольких популяций в пределах ограниченного и изолированного географического региона. Принято считать, однако, что предки гоминид обитали в Африке и что именно на этом континенте происходила начальная стадия их эволюции. В пользу этого говорят находки ряда последовательных форм австралопитеков на Африканском материке и близкое родство чело
92
Часть I. Эволюция человека
века и современных человекообразных обезьян, обитающих в Африке. Тем не менее стоит вспомнить о рамапитеке, существовавшем в Азии. Не исключено, что на Азиатском континенте происходила параллельная эволюция гоминид в направлении Ното. Наиболее вероятно, что более поздние формы австралопитеков или ранние формы Н. erectus распространились из Африки в Азию. Каков бы ни был истинный ход эволюции гоминид, можно быть уверенным в том, что с появлением Н. erectus гоминиды значительно расширили область своего обитания в Старом Свете, постепенно заселив более удаленные от экватора области и страны умеренного пояса. Имеются указания на то, что пекинский человек умел добывать огонь.
Homo sapiens
Кроме ископаемых остатков гоминид, которых мы относим к роду Australopithecus и Homo erectus, было найдено много других (рис. 4.1), и почти каждой вновь открытой форме давалось свое особое видовое или даже родовое название. Специалисты, занимающиеся вопросами классификации, под впечатлением различий, которые в зоологическом материале обычно наблюдаются между близкими формами, сконструировали множество запутанных филогенетических интерпретаций поздних фаз эволюции человека. Так, некоторые французские антропологи, анализируя найденные в Европе ископаемые костные остатки человека, относимые к периоду от последней стадии первого наступания вюрмских ледников до конца плейстоценовой эпохи, выделяли четыре или пять различных расовых типов. Морант, однако, показал, что общая изменчивость в пределах указанных форм была не более выражена, чем в пределах одной популяции эскимосов, и оказалась заведомо меньшей, чем у жителей Лондона в XVII столетии. Мы хотим подчеркнуть, что при любых таксономических и, следовательно, эволюционных определениях необходимо всегда учитывать изменчивость, которая является неотъемлемой чертой всех естественных популяций.
Различия между разными ископаемыми остатками гоминид, найденными в плейстоцене, но не относящимися к Homo erectus, более выражены, чем между представителями этого вида. Однако у всех этих форм имеется много общих признаков, и поэтому нет особых оснований выделять несколько видов. Этот единственный вид получил название Homo sapiens. Мы приводим в несколько сокращенном виде определение этого рода, данное Ле Гро Кларком.
Homo sapiens — вид Ното; его характерные признаки: большой объем мозговой коробки — в среднем больше 1100 см3; надглазничные гребни развиты в различной степени; для лицевого скелета характерен ортогнатизм или слабый прогнатизм; затылоч
Глава 4. Развитие гоминид
93
ные мыщелки расположены приблизительно на середине длины черепа; височные гребни по высоте расположения варьируют, но никогда не достигают средней линии; подбородочный выступ развит в различной степени; зубная дуга округлая, без диастемы; первый нижний премоляр имеет два бугорка, причем язычный редуцирован; размер коренных зубов варьирует в широких пределах, но последний моляр обычно несколько редуцирован; клыки относительно малы и не перекрываются уже после начальной стадии стирания; скелет конечностей приспособлен к прямохождению.
При рассмотрении ископаемых остатков Homo sapiens удобно выделить, с одной стороны, формы, близкие к современному человеку, и формы, которые отличаются от современного человека в большей степени, чем различаются представители различных рас живущих ныне людей. В табл. 4.4 представлен перечень основных находок ископаемых остатков Н. sapiens, относящихся к плейстоцену, с указанием места находки. Список не претендует на полноту, но включает все наиболее известные остатки, геологический возраст которых определен достоверно. Самые ранние из этих остатков датируются ранним верхним плейстоценом. Что касается Западной Европы, где, как мы видели, геохронология остатков наиболее надежно установлена, то представители современного человека впервые появились здесь в конце первой стадии вюрмско-го оледенения. Для остатков, найденных в Комб-Капелль, радиоуглеродным методом установлен возраст порядка 30 тыс. лет. Эти остатки связывают с поздней ориньякской эпохой верхнего палеолита. В других областях найдены первые еще более древние остатки, несомненно принадлежащие Н. sapiens. Возраст обнаруженного в древесном угле черепа Нья с острова Борнео (Калимантан) составляет примерно 40 тыс. лет, возраст торфяных отложений Южной Африки, где был найден флорисбадский череп, определен в 35—47 тыс. лет. Для черепов в районе реки Омо установлен возраст порядка 37 тыс. лет, другие ископаемые остатки Н. sapiens в Южной Африке, по-видимому, значительно древнее — около 60 тыс. лет. Следует заметить, что датирование последовательности археологических находок в Африке во многом еще не завершено, и возможно, что эти находки значительно старше, чем ранее предполагалось.
Предшественником современного человека в Западной Европе был неандерталец, который один населял эту область во время первого вюрмского оледенения (70—40 тыс. лет назад).
Остатки неандертальца, относящиеся к указанному времени, весьма однородны и характерны (табл. 4.5). От современного человека неандерталец отличается прежде всего тем, что имеет низкий свод черепа, покатый лоб и тяжелый сплошной надглазничный валик. Для него характерен пяткообразно нависающий затылок,
Таблица 4.4
Остатки скелета современного человека (относящиеся, по-вндимому, к верхнему плейстоцену)
Страна Место находки Содержание находки Эпоха Примечания
Европа Англия Павиленд, близ Суонси Скелет мужчины (без че- Ориньякская (крес-
Чехословакия Франция Брно Пршедмост (Моравия) Шанселад, Дордонь репа) Брюнн 1, часть черепа молодого мужчины + фрагменты скелета Части черепов 20 индивидуумов Череп мужчины фраг- свельян) Ориньякская Мадленская Ранее предполагалась
Германия Комб-Капелль, округ Монферран, Дордонь Кро-Маньон, округ Танак, Дордонь Ложери-Бас, округ Тайак, Дордонь Солютре (Сона и Луара) Оберкассель близ Бонна менты скелета индивидуума 35—40 лет Череп мужчины+фрагменты скелета индивидуума 40—50 лет Пять скелетов -|- остатки скелетов детей Фрагменты черепа четырех индивидуумов Остатки примерно 60 индивидуумов Фрагменты скелета двух индивидуумов Ориньякская (Ча-телль-перрон) Ориньякская Мадленская Ориньякская и со-лютрейская Мадленская близость к эскимосскому типу —* Старик, типичный представитель «кроманьонской расы»
Италия
Грот Детей, Гримальди, Ривьера
Африка Северная Африка
Западная
Африка Восточная
Африка
Афалу-бу-Рюмель, близ
Бужи, Алжир
Мехта-эль-Арби, близ
Константины, Алжир Асселар, сев. Тимбукту
Стоянка Бромхед, Эль-ментейта, Кения
Пещера Гэмбл, Эльмен-тейта, Кения
Канам, бухта Кавирон-до, Кения
Каньера, бухта Кави-рондо, Кения
Долина р. Омо, югозап. часть Эфиопии
Остатки скелета, относящиеся к верхнему плейстоцену и к современному периоду. Древнейшие из них принадлежат взрослой женщине и молодому мужчине
Остатки 50 индивидуумов
Остатки 30 индивидуумов
Череп и фрагменты скелета
Остатки 30 индивидуумов
Остатки 5 индивидуумов
Передняя часть нижней челюсти
Остатки 4 черепов
Фрагменты 2 черепов и костей конечности
Ориньякская Ранее предполагалась близость к негроидному типу
Иберо-Маврская (оранская) Иберо-Маврская (капспйская) —
— Рассматривается как негрский тип
Считаются не негрским по типу
Кения То же
Кафуанская (?)
Позднеашельская » Один из черепов вполне современного типа, у других примитивные признаки. Возраст 37 тыс. лет
Страна Место находки Содержание находки
Южная Африка Олдувай, долина Серенгети, Танзания Боскоп, округ Потче-фстрём, Трансвааль Полный скелет Череп и скелет
Азия Кэп-Флетс, близ Кейптауна Фиш-Хук, близ Кейптауна Флорисбад, близ Блумфонтейна Река Матвее, западнее Порт-Элизабет Спрингбок Флэте, сев. Претории Фрагменты скелета 3 индивидуумов Череп и скелет Крыша черепа Остатки скелетов 26 индивидуумов Череп и скелет
Китай Ява Чжоу-коу-динь, близ Пекина Вадиак близ Триниля Остатки скелетов 7 индивидуумов в верхней пещере Два черепа
Табл. 4.4 (продолжение)
Эпоха
Примечания
Мезолит (?) Стиллбей (?) Так называемый боско-поидный тип, близкий к бушменам и готтентотам Боскопоиды
Стиллбей »
Африканский ранний средний каменный век Средний и поздний африканский каменный век Африканский средний каменный век Предполагается родство с австралоидным типом Боскопоиды
Верхний палеолит —
— Очень похожи на череп из Кейлора
Калимантан
Пещера Нья
Череп
Австралазия Австралия Кейлор, близ Мельбурна Кау-Свэмп, сев. Виктория Крыша черепа и обломки бедренных костей Разнообразные остатки
Нов. Гвинея Озеро Южн. Аитапе Мунго, Уэльс Нов. Скелет Часть крыши черепа
Америка США Мексика Нэтчез, Миссисипи Тепекспан, близ Мехико Таз Череп и фрагменты скелета
Возраст по С14 40 тыс. мет. Родство с меланезийско-тасманий-
ским типом
Кварцитовые сколы
Возраст ио С14 6546 ± ±250
Возраст 8 тыс. лет до н. э., сходство с человеком Соло
Возраст 23,5 тыс. лет до н. э.
Возраст по С14 11 003± ±500
98
Часть II. Генетика человека
Таблица 4.5
Наиболее известные ископаемые остатки классических неандертальцев
Страна Место находки Содержание находки Эпоха
Бельгия Энгис I, провинция Льеж Ла-Нолетт, провинция Намюр Спи, провинция Намюр Фрагменты черепа ребенка Фрагменты пижней челюсти + фрагменты скелета Фрагменты двух черепов мужчин1 и фрагмент большой берцовой кости Мустьер-ская Мустьер-ская
Англия Залив Сент-Брилейд, Джерси Зубы, фрагменты большой берцовой кости + фрагмент детского черепа Поздняя ле-валлуа-мустьер-ская
Франция Ла-Шапелль-о-Сен, Корреза Ла-Ферраси, Дордонь Ла-Мустье, Дордонь Ла-Кина, долина Шарент Череп мужчины и почти полный скелет Остатки черепов шести индивидуумов, один почти полный скелет мужчины Неполный череп мужчины Фрагменты черепа женщины 4- фрагменты скелета Мустьер-ская То же » ъ
Германия Неандерталь, близ Дюссельдорфа Крыша черепа мужчины и несколько костей скелета —•
Гибралтар Форбс-Кверри, Башня Дьявола Крыша черепа женщины+ +кости черепа ребенка Верхне-му- стьерская
Италия Гора Чирчео, южнее Рима Крыша черепа мужчины + + части нижней челюсти другого индивидуума Мустьер-ская
Ирак Шанидар Остатки скелета трех взрослых и одного ребенка Му с тверская
Израиль Гора Кармел, пещера Мугарет Табун Пещера Амуд Остатки черепа и скелета взрослой женщины + 4- нижняя челюсть и много других фрагментов скелета Почти полный череп и скелет Леваллуа-мустьер-ская
СССР Тешик-Таш, Узбекская ССР Фрагменты черепа ребенка 8—10 лет —ч
1 Определение пола как в данной таблице, так и в табл. 4.6 весьма условно.
Глава 4. Развитие гоминид
99
к которому прикреплялись сильно развитые мышцы шеи. Большое затылочное отверстие смещено назад, а сосцевидные отростки меньше, чем у современного человека. Длинный и узкий лицевой скелет неандертальца составляет меньший угол с основанием черепа; прогнатизм заметно выражен: скуловые дуги несильно выдаются. Клыковая ямка отсутствует. Челюстной симфиз наклонен назад вследствие того, что подбородочный выступ не развит, но, как и у современного человека, подбородочно-язычные мышцы отходят от подбородочных бугорков. Скелет туловища в основных чертах похож на скелет современного человека, а объем мозговой коробки так называемых «классических неандертальцев» также довольно велик (1300—1600 см3), хотя по форме их череп существенно отличается от черепа современного человека.
Ископаемые остатки, во многом сходные с классическими неандертальцами и приблизительно одного с ними возраста, обнаружены также на Ближнем и Среднем Востоке: в Шанидаре (Ирак), в пещере Мугарет Табун на горе Кармел и в пещере Амуд (Израиль) и в Тешик-Таше (Узбекская ССР). Можно отметить систематические различия форм из разных географических районов: так, например, у ближневосточных форм отсутствует типичный для неандертальцев признак — «пяткообразпо» нависающий затылок, а у человека из пещеры Амуд обнаружены большие сосцевидные отростки. Все эти формы, без сомнения, родственны западным неандертальцам.
Отмеченные здесь признаки, характерные для неандертальца, в той или иной степени имеются у большей части ископаемых форм Homo sapiens, относящихся к последнему межледниковью и найденных в Европе и соседних с ней областях. Но если рассматривать признаки, которые отличают классического неандертальца от современного человека, то ранние формы оказываются более «нео-антропными», чем формы, которые позже заменили их в Западной Европе. Ранние формы более разнородны, однако у всех таких форм в отличие от классического неандертальца мозговая коробка уже, короче, имеет более выпуклые боковые стенки и более высокий изгиб лобной кости. Имеется тенденция к разделению надглазничного валика на центральные и боковые элементы, к разрастанию затылочных костей и образованию наружного выступа затылочного бугра с плохо развитым затылочным гребнем. Изгиб основания черепа больше, а лицевой скелет меньше, чему классических форм. Скуловые кости имеют более выраженную переднебоковую ориентацию и более четко отделены от верхних челюстей, на которых намечается клыковая ямка. По всем этим признакам так называемые «прогрессивные неандертальцы» в большей степени сходны с современным человеком, чем классические формы. Места соответствующих находок указаны в табл. 4.6.
Исключение из этого общего правила составляют ископаемые,
100
Часть II. Генетика человека
Таблица 4.6
Остатки Homo sapiens из Европы, Северной Африки и Ближнего Востока, относящиеся к верхнему плейстоцену
Страна Место находки Содержание на-ходки Геологический ярус Эпоха
Марокко Джеб ель Игхуд 2 черепа Вюрм, 45 тыс. лет до н. э. (?) Леваллуа-мусть ер-скан
Палестина Джебель Кафза Полный череп+ части других индивидуумов Вюрм То же
Израиль Пещера Муга-рет-эс-Схул Фрагменты по меньшей мере 10 индивидуумов Вюрм, 36 тыс. лет до н. э. » »
Югославия Крапина, близ Загреба Фрагментарные остатки 13 индивидуумов Ранний вюрм? Мустьерская
Германия Эринге дорф, Части черепа жен- Рисс-вюрм- Домустьер-
близ Веймара щппы (Эрингс-дорф I) и 2 нижние челюсти + остатки скелета ребенка (Эрингс-дорф II) ское межледниковье ская
Италпн Саккопасторе, близ Рима Череп женщины (Саккопасторе I) и части черепа мужчины (Саккопасторе II) То же Мустьерская
Чехослова- Гановцы Слепок полости )>
кия черепа
Франция Фонтешевад, округ Шареп Лобная кость одного индивидуума (I), верхняя часть черепного свода другого индивидуума (П) » Тайякская
Глава 4. Развитие гоминид
101
Продолжение
Страна Место находки Содержание находки Геологический ярус Эпоха
Франция Обри Суар ля Шез, округ Шарен Затылочная, теменная и височная кости Поздний рисе
Франция Лазарет, близ Ниццы Теменная кость Рисе
Франция Пещера Араго, Тотанель Передняя часть крыши черепа, лицевой скелет и 2 нижние челюсти Ранний рисе
Монморан, Нижняя челюсть Миндель- Домустьер-
Верхняя Гаронна п зубы рисе? ская
Германия Штейнгейм, близ Штутгарта Череп женщины Миндель-рисское межледниковье
Англия Сванскомб, Кейт Затылочные и теменные кости То ясе Средне-ашельская (ручные рубила)
найденные Ламли (Lumleys) на юге Франции в пещере Араго и близ Обри Суар. Их датируют рисской фазой оледенения. Они сохраняют многие палеоантропные черты, и, пожалуй, по ряду признаков даже более примитивны, чем остатки человека из Фер-тесзоллоша. Особого внимания, возможно, заслуживает тот факт, что формы, найденные в южной Франции, подобно классическим неандертальцам, относятся к периоду оледенения, а не к межледниковой фазе. На существование уже в то время форм с более выраженными неоантропными признаками указывают остатки из Штейп-гейма и Сванскомба, датируемые вторым, или великим, межледниковьем. В отложениях 100-футовой террасы на Темзе близ Сванскомба были найдены затылочная и теменные кости и вместе с ними орудия среднеашельской эпохи (ручные рубила). От соответствующих костей современного человека кости из Сванскомба отличаются только толщиной. Череп, найденный в Штейнгейме, несмотря на наличие тяжелого надглазничного валика, также обнаруживает много прогрессивных черт. Очевидное родство с неандертальцами можно найти у некоторых форм, существовавших в пос
102
Часть II. Генетика человека
ледний межледниковый период (например, у черепов из Эрингс-дорфа и Саккопасторе). Вместе с тем у фонтешевадского человека (Франция) не было массивного надглазничного валика. Он сохранился, однако, у ископаемых форм, найденных в пещере Схул на горе Кармел (Израиль) ив Джебель Кафза (Палестина), хотя эти формы по большинству признаков неотличимы от современных. В настоящее время принято считать, что более поздние формы представляют конечную стадию дифференцировки скелета человека, если сравнивать его со строением скелета у современного человека.
Прежде чем рассматривать отношение описанных выше европейских и ближневосточных форм к конечной фазе эволюции человека, перечислим плейстоценовые ископаемые остатки, найденные в других частях света. Африканский материк доставил наиболее богатый материал. В Африке были найдены не только различные виды австралопитеков, но также скелетные остатки сильно отличающегося от них родезийского человека, единственный череп которого был найден в пещере холма Брокен-Хилл в Северной Родезии.
Возраст этих остатков определен в 30 тыс. лет, но, возможно, эта оценка занижена. В строении лицевого скелета, несмотря на то, что он очень велик и не имеет клыковой ямки, нет существенных отличий от скелета современного человека. Мозговая коробка, однако, низка, лоб очень покатый. Надбровные дуги массивны и образуют сплошной выступ над глазницами. Все кости настолько толстые, что внутренний длинный диаметр мозговой коробки составляет лишь 81 % его наружного диаметра. Несмотря на указанные палеоантропные черты, большое затылочное отверстие расположено почти так же, как у современного человека.
Задние мышцы шеи прикрепляются к затылочной кости выше, чем у современных форм, но затылок не так уплощен, как у классических неандертальцев. Поэтому следует признать, что найденные в той же пещере кости конечностей вполне современного типа принадлежат тому же индивидууму. До тех пор пока был известен только один череп родезийского человека, можно было бы подозревать, что он представляет какую-то патологическую форму, но обнаружение другого черепа близ бухты Салданьи, почти неотличимого от черепа из Брокен-Хилла, позволило исключить зто предположение.
Упомянем еще так называемого Africanthropus. Эта форма, вероятно, не относится к Homo sapiens, но остатки, обнаруженные в Танзании, слишком плохо сохранились, чтобы им можно было дать обоснованное таксономическое определение.
В Африке было найдено много других ископаемых остатков гоминид, но все они очень похожи на современного человека. Большое эволюционное значение приписывалось, в частности, остаткам,
Глава 4. Развитие гоминид
103
открытым в Канаме и Канжере, поскольку первоначально полагали, что они очень древние. Однако все данные, полученные к настоящему времени, указывают на то, что этот, а также весь другой ископаемый материал, несомненно относящийся к Homo sapiens, не древнее верхнего плейстоцена. Многие формы относятся, по-видимому, к более позднему времени.
Многочисленные ископаемые остатки, вероятно относящиеся к плейстоцену, обнаружены в Восточной и Юго-Восточной Азии и Австралии. Но, за исключением тех, которые были открыты в Нган-донге на острове Ява, в террасах реки Соло, все остальные следует отнести к Homo sapiens. Найденные в Нгандонге одиннадцать черепов, все без лицевого скелета, и две большие берцовые кости, согласно Кенигсвальду, относятся к верхнему плейстоцену. Все черепа — плоской формы, с необычайно толстыми стенками, каждый имеет сильно развитый надглазничный валик. В расположении большого затылочного отверстия мы видим ряд особенностей, но если не считать небольшого объема мозговой коробки (1150— 1300 см3) и того факта, что берцовые кости тонкие, с прямым диафизом, общее строение скелета соответствует неандертальскому типу.
В последние десятилетия большое количество ископаемых остатков человека обнаружено в Северной и Южной Америке; первоначально их считали очень древними. Все они, по-видимому, принадлежат Homo sapiens, причем многие являются сравнительно недавними формами. Тем не менее на основании археологических данных можно считать, что на Американском континенте человек появился в середине последнего ледникового периода, а скелеты, обнаруженные вТепекспане (Мексика), определенно имеют возраст около 10 тыс. лет.
Эволюционные связи между различными формами Ното служили предметом живой полемики. В настоящее время большинство специалистов согласны с тем, что все формы Homo sapiens произошли от Homo erectus. Основные изменения касались размеров мозга, а значит, и размеров черепа. Кроме того, у гоминид отмечается общая тенденция к уменьшению челюстей и более совершенному равновесию головы на позвоночнике.
При рассмотрении последних фаз эволюции гоминид важно иметь в виду, что между популяциями, по-видимому, существовал интенсивный поток генов, который в противовес тенденциям к дивергентному развитию приводил к эволюции по сетевидному типу. Наиболее вероятно, однако, что в Африке в условиях сравнительной изоляции Homo erectus эволюционировал в родезийского человека, а человек, остатки которого найдены в районе реки Соло, был прямым потомком восточных питекантропоидов. Несколько сложнее ситуация в Евразии. На рис. 4.2 схематически представлены три наиболее приемлемые интерпретации имеющихся данных. Рис. 4.2 указывает на раннее происхождение современного человека и со-
Современный человек
Классический неандерталец
Современный человек
Классический неандерталец
Человек из Фонтешевада
Прогрессивный неандерталец
Человек из Сванскомба Штейнгеймский человек
Родезийский
человек
Человек „Соло”
Питекантроп
Современный человек
. 4.2. Три различные филогенетические схемы эволюции и дифференц
Глава 4. Развитие гоминид
105
вершенно независимую эволюцию неандертальцев. В основе этой точки зрения лежит несомненное сходство сванскомбского и фонтешевадского человека с современным. Ее разделяли многие в то время, когда еще не был доказан факт фальсификации остатков в Пильтдауне и когда саму стадию Homo erectus в дифференциации Homo sapiens считали маловероятной.
Указанная схема игнорирует, однако, большое сходство сван-скомбских остатков и соответствующих костей штейнгеймского человека. Многие уверены сейчас в том, что сванскомбский человек принадлежал к прогрессивным неандертальцам, хотя и представлял собой наиболее неоантропный вариант группы. Если это соответствует действительности, то не остается никаких данных, которые свидетельствовали бы о том, что современный человек появился раньше последнего межледниковья: следует предположить, что его предками были прогрессивные неандертальцы, крайние стадии дифференциации которых найдены в Схуле, Джебель Кафза и Фонтешеваде. Эта наиболее широко распространенная сейчас точка зрения схематически представлена на рис. 4.2,6. Согласно третьей схеме, в противоположность двум предыдущим прогрессивный и классический неандертальцы происходят от совершенно различных предков. Основанием для того, чтобы считать одну из этих групп предком другой, послужила их непрерывная временная последовательность в одной и той же географической области. С этой точки зрения трудно, однако, объяснить возвратный характер главной линии эволюции гоминид, который допускает такая связь. Можно предположить, что палеоантропные черты классических неандертальцев представляют собой адаптации к климату холодных стран, поскольку практически все их остатки были открыты в той части Европы, которая была изолирована от Азии обширным ледяным покровом, распространившимся с Альп и Скандинавского полуострова. Но вместе с тем палеоантропные признаки, по всей видимости, являются примитивными. В таком случае существует как временной, так и морфологический разрыв между классическими неандертальцами и Homo erectus. Если классический неандерталец был северной формой, то не удивительно, что его ранние остатки не были обнаружены: северные области материка Евразии пока мало исследованы. В этой связи следует обратить внимание на палеоантропные признаки у форм, найденных в Араго и Обри Суар.
У нас нет, конечно, достаточно полных сведений о том, что помогло современному человеку вытеснить классических неандертальцев, когда в конце первого вюрмского ледникового периода он проник в Западную Европу. Важно, однако, иметь в виду, что ножи, резцы и наконечники стрел и копий верхнего палеолита, изготовленные пришельцами, были гораздо более совершенными, чем орудия, которые характеризуют мустьерскую технику класси
106
Часть I. Эволюция человека
ческих неандертальцев. Вероятно также, что их более высокая материальная культура отражала и более высокую степень социальной организации.
Невозможно с определенностью сказать, какая из существующих рас более всего похожа на первые полностью дифференцированные популяции современного человека; однако широкая распространенность австралоидного типа как у ископаемых, так и у ныне живущих популяций, его большая морфологическая близость к анцестральным стадиям дают основание полагать, что этот тип обладает наибольшим сходством с первобытной формой современного человека. Как бы то ни было, этот тип не был обнаружен в Европе, хотя в популяциях верхнего палеолита были выделены (и почти наверняка ошибочно) различные другие предки современных нам рас. Стоит заметить также, что древнейшие из ископаемых форм, обнаруженных в Австралии, например в районе озера Мунго, по всей видимости, имеют более грацильный скелет, чем менее древние формы, представленные, например, группами, населявшими болотистый район Коу.
Подразделение ветвей родословного древа (рис. 4.2) на дискретные таксоны неизбежно оказывается условным. В прошлом классическому неандертальцу, человеку «Соло», родезийскому человеку и даже некоторым формам прогрессивных неандертальцев приписывали самостоятельный эволюционный статус. Подобная практика приводит, однако, к многочисленным трудностям. Поэтому в настоящее время принято включать все эти формы в один-единст-венный вид Homo sapiens, подразделяя его на большое число подвидов, таких, как Н. neanderthalensis, Н. rhodesiensis, Н. soloensis; возможно, что к этим подвидам следует отнести и основные географические расы — как вымершие, так и ныне живущие.
Глава 5
ПРОИСХОЖДЕНИЕ ЧЕЛОВЕЧЕСКОГО ОБЩЕСТВА
Введение
При обсуждении проблемы происхождения человеческого общества мы вынуждены в значительной степени опираться на умозрительные гипотезы и предположения. Эти гипотезы, однако, отнюдь не всегда беспочвенны, порой они основаны на вполне реальных данных. Возникает вопрос: какого рода информация наиболее важна для понимания существа проблемы? По-видимому, наибольшую ценность представляют фактические данные, касающиеся поведения приматов (отличных от человека) в сообществе, а также австралопитеков и древнейших людей (полученные главным образом при изучении ископаемых остатков). Первая группа данных позволяет более четко представить характер организации сообщества у приматов; вторая дает нам возможность сказать нечто разумное об экологических и физических условиях, в которых существовали наши гоминидные предки. Изучая поведение различных видов приматов, мы получаем представление о различных формах организации сообщества и можем сопоставить их с учетом особенностей самих приматов и экологических условий, в которых они существуют, с тем, что известно об условиях жизни наших далеких предков. В этом состоит преимущество изучения приматов по сравнению с изучением организации современного человеческого общества. Развивая эту мысль, можно сказать, что анализ структуры сообществ приматов, вероятно, имеет большую ценность для.создания моделей организации сообществ догоминидных форм приматов, нежели изучение ныне живущих примитивных сообществ охотников или собирателей. Действительно, организация сообществ претерпела коренные изменения в ходе эволюции, в основном в результате усложнения структуры мозга и совершенствования речевых навыков. В то же время на примере приматов мы видим внутригрупповые отношения, основанные на системе сигналов и процессах обучения, не связанных с использованием речи. Что касается сведений об ископаемых остатках предковых форм и экологических условий, в которых они существовали, то зти сведения слишком скудны, чтобы можно было на их основе создать картину общественного уклада догоминидных форм или ранних гоминид. Немногочисленные твердо установленные факты представляют с этой точки зрения особую ценность.
108
Часть I. Эволюция человека
Изложение в данной главе построено по следующему принципу. Вначале рассмотрены основные данные, касающиеся организации сообществ у некоторых видов приматов. Далее сформулирован ряд общих идей, вытекающих из изучения организации сообществ приматов; кратко анализируются условия обитания и данные об ископаемых остатках ранних гоминид. В заключение сделана попытка объединить результаты изучения современных форм приматов и догоминйдных предковых популяций для того, чтобы выработать некоторые общие концепции, касающиеся структуры древних человеческих сообществ.
Сообщества приматов
Простые павианы (Papio anubis) и макаки-резусы (Масса mulatto)
Внутригрупповое поведение этих двух видов обезьян Старого Света изучали достаточно подробно в течение длительного времени. Павианы — весьма преуспевающий вид приматов, широко распространенный в настоящее время на обширной территории африканских саванн. Макаки водятся в Индии, в лесах и не заселенных человеком районах.
Павианы ведут наземный образ жизни и питаются растительной пищей. Это не означает, конечно, что они никогда не взбираются на деревья или скалы и вообще не употребляют мясную пищу или насекомых. Им приходится время от времени делать то и другое. Тем не менее их пища в основном растительного происхождения; они расстались с типичным для приматов древесным образом жизни.
Павианы живут большими группами — от 8 до 185 особей в группе; средная численность группы зависит от места обитания и составляет 30—50 животных. На большинстве обследованных территорий такие группы были несколько обособлены от остальных; однако в лесном районе, где вел наблюдения Rowell (1966), группы оказались менее изолированными от соседних, и животные, в особенности самцы, время от времени переходили из одной группы в другую. Более типичной является, однако, другая ситуация, когда стадо павианов держится в основном обособленно, избегая контактов и смешивания с другими группами, и лишь случайно встречается с ними у источников воды, но затем группы расходятся без обмена особями. Свой обычный день павианы проводят, странствуя немногочисленной группой в пределах занятой ею территории и возвращаясь к определенному месту, называемому «центром обитания» и выбранному для постоянного ночлега. Участки, занимаемые различными группами, могут, вообще говоря, перекрываться.
Глава 5. Происхождение человеческого общества J09
Тем не менее группы обычно держатся поодаль, стараясь избегать друг друга. Стадо проходит за сутки в среднем расстояние около 3 км. Группа состоит из равного числа самцов и самок. Однако самки развиваются быстрее, достигая зрелости к 31/2—5 годам, тогда как у самцов половая зрелость наступает лишь к 7—10 годам. В связи с этим отношение полов у взрослых особей таково: на одного зрелого самца приходятся 2—3 зрелые самки.
У павианов резко выражен половой диморфизм. Самцы вдвое или даже втрое крупнее самок. Это объясняется, по всей видимости, влиянием хищников. Крупный, сильный самец помогает защитить стадо от нападения хищников. Однако чем крупнее самцы, тем больше общая потребность в пище для группы с данным числом особей; выгодно поэтому, чтобы вся сила и агрессивность, необходимые для защиты от врагов, сосредоточились у представителей одного пола, именно у самцов; самки могут быть более миниатюрными и получить защиту от самцов. В результате снижается общая биомасса группы, а также потребность в пище и увеличивается способность группы к выживанию. Отбор на агрессивность среди самцов может оказать существенное влияние на характер организации сообщества у данного вида.
Мы подходим в итоге к необходимости рассмотрения иерархии в группе. Агрессивный характер взрослых самцов-павианов, а также замкнутость общественных групп приводят к установлению доминантной иерархии у данного вида обезьян. Эта доминантность может проявляться в различных формах. Смысл термина можно уяснить, проследив, в чем именно выражается «успех» самца, занимающего главенствующее положение в группе. Мы можем наблюдать, в какой степени ему отдают предпочтение самки при спаривании; следить за жестами «покорности» со стороны других обезьян; изучать агрессивные действия вожака по отношению к другим членам группы; наблюдать, в какой степени он проявляет себя как вожак, угрожая другим приматам или другим животным, приближающимся к группе; наконец, мы можем оценить его роль в качестве защитника, вокруг которого в момент опасности собираются самки с детьми.
Простой линейный закон соподчинения необязательно соблюдается в стаде, включающем несколько взрослых самцов. В этом случае некоторые из них действуют совместно как единая ячейка. Chance (1967) изучал группу самцов, образовавших доминантное ядро в стаде макаков. Все члены такого ядра — обязательно взрослые самцы, каждый из которых тщательно следит за действиями других членов своей группы. Самцы совместно останавливаются, согласуют свои движения в группе и управляют поведением других членов. Доминантное положение этих животных обусловлено в значительной степени их связями друг с другом. De Vore а. Hall (1965) показали, что при уходе одного из членов доминирую
ПО
Часть I. Эволюция человека
щей пары павианов оставшийся член пары может попасть в подчинение самцу, стоявшему ранее на более низкой ступени иерархии. Главенствующее положение самца в группе требует постоянного подтверждения силой и может иногда изменяться при возникновении ссор в группе. Всевозможные угрозы и контругрозы, переадресация угроз другим индивидуумам, особые жесты, с помощью которых животное одновременно выражает свое послушание вожаку,— все это элементы весьма сложной системы внутригрупповых взаимоотношений, посредством которых устанавливается и поддерживается общественный статус индивидуумов в группе.
У самок также существует определенная иерархия. Подобно самцам, они участвуют в ссорах, но ранг самок, вероятнее всего, связан с астральным циклом и определяется также их связями с определенными самцами, прежде всего с вожаком группы. Вдобавок к этому у макаков положение самок (а косвенно и самцов) в сообществе зависит от их относительного положения в иерархии матерей. Изучение поведения японских макаков и резусов показывает, следовательно, что наряду с тем местом в группе, которое определяется конкуренцией особей, положение в сообществе определяется принадлежностью особи к определенной линии, доминирующие члены которой уже занимают высокое положение.
Доминантность в группах макаков и павианов выражается также в пространственных взаимоотношениях животных: доминирующие особи обычно занимают место в центре группы, а младшие по рангу — на периферии. В результате не достигшие зрелости самцы оказываются более удаленными от центра, чем другие члены группы, а у некоторых видов макаков они даже покидают группу и возвращаются (иритом далеко не всегда) лишь через несколько лет. Такого правила как будто не существует у павианов, но молодые самцы держатся на периферии и исполняют обязанности «часовых», предупреждая группу о приближении хищников или о других непредвиденных событиях.
Важными элементами организации сообщества наряду с системой подчинения являются половое поведение и половые связи членов группы. Самкам павианов свойствен 35-дневный астральный половой цикл. Ко времени овуляции у них происходит набухание кожи вокруг наружных половых органов. В типичной группе Papio anubis главенствующий самец спаривается с самкой лишь в течение 3—4 дней, соответствующих пику ее эстрального цикла. После спаривания пара распадается, и нет никаких указаний на то, что члены пары сохраняют близость в течение длительного времени, что отличает их от гамадрилов, у которых, как правило, самцы и самки образуют устойчивые пары, и от макаков, у которых такие постоянные пары также встречаются довольно часто. В некоторых областях (Кения) спаривание носит заметно выраженный сезонный характер, в других (Южная Африка) этого не наблюдается.
Глава 5. Происхождение человеческого общества
111
Другой формой общественных взаимоотношений является связь матери с детенышем. Новорожденный имеет темную окраску кожи; рано развивающиеся рефлексы обеспечивают подвешивание его к телу матери со стороны ее живота. После рождения детеныша мать переходит в центр группы и остается там под защитой сильных самцов вместе с другими самками, имеющими или не имеющими детенышей. В первые несколько дней после рождения малыша осматривают и другие самки, однако он крепко цепляется только за свою мать. В первые месяцы жизни детеныш начинает постепенно перебираться с живота на спину матери, садится на нее верхом, пробует пищу, которую она ест, совершает кратковременные вылазки для обследования интересующих его предметов. Позднее, примерно к 6 месяцам, его кожа светлеет; он начинает есть более твердую пищу и играть со своими сверстниками. Детеныш находится под защитой матери и взрослых самцов весь период кормления (до 11—15 месяцев). После отнятия от груди он занят играми, которые принимают все более грубый характер, что может вызвать неудовольствие взрослых. Взрослые самцы становятся все менее терпимыми к шалостям подрастающих молодых самцов — в конечном счете действия «подростков» начинают вызывать у них постоянное неудовольствие. После окончания периода кормления матери возвращаются к своим обычным занятиям и перестают защищать детенышей в их играх и ссорах.
Часто наблюдаются дружеские отношения между животными, выражающиеся во взаимном обыскивании, очистке шерсти — гру-минге (grooming). Партнер побуждает к обыскиванию, приняв особую позу. Взаимное обыскивание более характерно для самок; они проводят за этим занятием больше времени, причем чаще обыскивают друг друга, чем самцов; нередко объектом груминга становятся взрослые самцы, особенно те из них, которые играют главенствующую роль в группе. Самки обыскивают часто других самок, у которых наступил период течки, или молодых матерей. Животные, находящиеся на нижних ступенях иерархии, по-видимому, чаще «обслуживают» животных более высокого ранга. Груминг создает и укрепляет взаимные связи в сообществе. Павианы используют для общения самые разнообразные звуки. Их можно разделить на категории в соответствии с характером ситуации и предполагаемыми мотивами общения. Можно выделить, например, звуки, предшествующие агрессивным актам (лай, рев, рычание), сигналы тревоги (пронзительный крик), призывание полового партнера (урчание), звуки, выражающие дружеское расположение (пощелкивание и похрюкивание). Нелегко дать всем этим звукам простые наименования и распределить по классам: во-первых, в большинстве случаев каждому звуку соответствуют определенные градации и близкие по звучанию сигналы могут использоваться для разных целей и наоборот; во-вторых, возникают трудности с описанием
112
Часть I. Эволюция человека
звуков. Правда, здесь во многом помогает использование современных средств звукозаписи.
Наряду со звуковой сигнализацией в стаде приматов существует сложная система связи посредством мимики и жестов. Мимика запугивания выражается в том, что животное широко открывает рот, подергивает головой, взлохмачивает шерсть. Испытывая страх, обезьяны оскаливаются и припадают к земле; дружеские чувства они выражают причмокиванием и другими знаками расположения. Наконец, существует и связь посредством прямого соприкосновения: через укусы, поглаживания и т. д.
Гамадрилы
Куммер (Kummer, 1968) и его сотрудники изучали в течение ряда лет организацию сообщества гамадрилов в Эфиопии. Предпочтительным местом обитания этих животных являются открытые и полузасушливые пространства, гораздо более бесплодные, чем те, в которых обитают простые павианы. Обезьяны этого вида живут большими группами — от 12 до 750 особей. Среднее число членов группы 136. Такие большие стада можно встретить, однако, только в ночное время в местах, где гамадрилы располагаются на ночлег. Днем стада разбиваются на множество мелких групп, самостоятельно занятых поисками пищи. Такие небольшие группы состоят из одного взрослого самца, одной или нескольких самок, их потомства и нескольких более старых животных, сопровождающих группу. Небольшие группы, возглавляемые единственным самцом,— характерная основная ячейка в сообществе гамадрилов, отличающая его организацию от наблюдаемой у простых павианов. У последних отсутствует тенденция к подразделению па малочисленные группы. Все члены основной ячейки согласованно перемещаются по территории и остаются вместе на протяжении всей жизни. Самец подгоняет отстающих самок угрожающими криками. Оп угрожает им также в тех случаях, когда они вступают в общение с другими самцами. Угрожая самкам, он поднимает брови, пристально смотрит на них, делает выпады в их сторону, иногда преследует и кусает их в шею. Самки в ответ собираются вместе и издают характерный кашляющий звук — «стаккато», если угрозы выражаются в мягкой форме, и резкие вопли, если самец их кусает. До кровопролития дело доходит редко. Столкновения, заканчивающиеся укусами, также довольно редки: в среднем самец кусает один раз в день. Самка принадлежит одному самцу, который постоянно занимает позицию впереди самки, в противоположность другим самцам, которые держатся поодаль или сзади. Если к группе с одним главенствующим самцом приближаются другие самцы, все самки группы устремляются к своему господину. Если самец теряет из виду одну из самок, он упорно ищет ее; Куммер наблюдал поиск,
Глава. 5. Происхождение человеческого общества
113
длившийся 12 мин. Группа никогда не покидает больных или отставших животных в противовес простым павианам, которые часто бросают на произвол судьбы больных и отставших от группы.
Копуляция ограничена членами основной единицы. В сообществе гамадрилов имеются также самцы, у которых нет самок. Они образуют группы «холостяков», никогда пе спаривающихся с самками. Если в ячейке, возглавляемой одним самцом, имеется лишь одна самка, спаривание происходит лишь в период эструса. В период беременности и кормления спаривания не происходит. Ограничения на спаривание касаются лишь взрослых самцов. Молодые самцы и самцы, только что достигшие половой зрелости, спариваются с любыми самками. Это иногда вызывает неудовольствие вожака, который в подобных случаях нападает па провинившуюся самку, не атакуя, однако, самца, спарившегося с ней. Как следствие подобные спаривания происходят обычно втайне от вожака группы. Ранние копуляции практикуют как молодые самки, так и молодые самцы.
У гамадрилов, как и у простых павианов, дружеские чувства часто выражаются взаимным обыскиванием (груминг). Этот вид общения проявляется у обоих полов, но в большей степени у самок. Располагаясь на почлег на скалистых уступах, самки отдают этому занятию до 28% своего времени. Обыскивание связано с определенными позами партнеров, животное как бы предлагает другому принять участие в совместном туалете. Объектом обыскивания бывает чаще всего вожак группы. Он нередко оставляет без внимания молодых самок, приглашающих его к взаимному обыскиванию. Предпочтение отдается тем членам группы, с которыми он проводит большую часть времени.
Самки нередко ссорятся друг с другом, оспаривая, например, право обыскивать самца. Если ссора принимает затяжной характер, самец может вмешаться и положить ей конец, укусив одну из самок. Самцы стараются избегать друг друга; вожаки держатся на расстоянии не менее 1,5 м и никогда не обыскивают друг друга. Изредка они вступают в драки, принимающие форму «ритуальных» стычек с укусами и пинками и стоянием друг против друга. Такие драки длятся не более 10 с и заканчиваются бегством одного из участников. Как и у собак, явным признаком «покорности» у гамадрилов служит подставленная для укуса шея. По наблюдениям Куммера, из 13 стычек 10 возникли из-за самок, а 3 — «по ошибке». Подобные «ошибки» связаны с неправильным истолкованием угрозы, адресованной не данному самцу, а другому животному. Драки между самцами не всегда связаны с «незаконными» совокуплениями. Нередко они возникают в результате нападения самца на чужую самку: атакующий в этих случаях получает отпор со стороны хозяина самки. Такие стычки бывают из-за самок, у
lit
Часть l. Эволюция человека
которых не наступил период эструса, так что обладание самкой — это не только половые отношения.
Большой интерес представляет механизм зарождения групп, возглавляемых одним самцом. Куммер наблюдал у молодых самцов стремление схватить и удержать при себе в течение получаса темнокожих детенышей. Такой тип поведения он назвал «похищением детей» («киднеппинг»). Похищение превращается в «удочерение»: если молодой самец получает в свое распоряжение самку детского или совсем юного возраста (от года до полутора), он носит ее с собой и остается при ней постоянно. Такая пара удивительно прочна, самка активно «ухаживает» за самцом. Из-за незрелости молодой самки половая связь между ними на первых порах отсутствует. Наказывая самку, самец никогда не кусает ее в шею. Временами самка подвешивается к самцу, как если бы он был ее матерью. Мотивы такого поведения самца не половые, а скорее псевдо-материнские. Формирующаяся таким образом постоянная пара становится нередко ядром группы с одним главенствующим взрослым самцом. На ранних стадиях связи самец в случае ухода юной самки не кусает ее в шею, но возвращает к себе, притягивая руками. Что помогает самцу увести из семьи самку в незрелом возрасте? Ответ пока не ясен. Обратим, однако, внимание на два обстоятельства. Во-первых, мать перестает защищать детеныша, начиная примерно с 6-месячного возраста, и, во-вторых, взрослые вожаки-самцы никогда не носят на себе юных самок. Так что молодых самок легко похитить (речь идет об обезьянах только данного вида). Подобная система обладает очевидным селективным преимуществом: она оставляет свободными молодых животных для связи с другими группами, позволяя избежать нежелательных последствий внутригруппового спаривания. У «похищенных» молодых самок примерно до 4-летнего возраста не появляется потомства, хотя половую жизнь они начинают вести значительно раньше.
Группы во главе с одним самцом формируются и другим путем. Самец, находящийся в расцвете лет, иногда уводит самку от более старого вожака. Последний позволяет самке уйти к молодому самцу без каких-либо мер возмездия. Что касается самок, то их переход — дело сравнительно простое, поскольку у них в большей степени выработана привычка следовать за самцом вообще, нежели привычка соблюдать верность одному избранному самцу.
Более крупная общественная единица у гамадрилов — это большое стадо животных, которое совместно располагается на ночлег, но в дневные часы разделяется, как было сказано выше, на семьи с одним вожаком. Стадо разделено также на группы средних размеров; члены каждой из них стремятся держаться вместе. Эти группы можно разграничить искусственным путем, положив, например, пищу в месте стоянки стада. Между группами вспыхивают стычки, и победители забирают себе весь корм, заставляя побеж
Глава 5. Происхождение человеческого общества
115
денных ретироваться. Одно такое столкновение вспыхнуло естественным образом на скале, выбранной для ночлега. Куммер полагает, что дружеские взаимоотношения между самцами одной группы основаны на связях между самцами-сверстниками, установившихся еще в ранние годы их жизни, до того как каждый из них возглавил семейные ячейки.
Во время поиска корма в дневное время нередко образуются также группы с двумя самцами — более старшим по возрасту и молодым. Они совершают все переходы совместно. Куммер наблюдал 8 таких групп. Один из самцов шел впереди, другой замыкал группу, самки держались посредине. Путь, который выбирала группа, представлял определенный компромисс между направлениями движения обоих вожаков, но решающая роль все же принадлежала старшему по возрасту. Куммер назвал такую систему согласованных перемещений ID-системой, обозначив символом I инициативную роль более молодого самца, а символом D определяющую роль старшего по возрасту самца в принятии окончательного решения.
Шимпанзе (Pan troglodytes)
Жизнь шимпанзе в естественных условиях изучали во многих лесных районах. Длительные наблюдения над группами, занятыми поисками корма, проводились также в национальном парке Гомбе и в других районах западной Танзании. В данном разделе книги мне хотелось бы обратить внимание читателя на особенности поведения лесных шимпанзе, в первую очередь тех, которые обитают в тропическом лесу Будонго в Уганде.
Характер взаимоотношений в сообществе шимпанзе отличается от описанного в стадах павианов и макаков. У шимпанзе не существует замкнутых групп с большим числом особей, как у простых павианов и резусов, а также небольших семейных ячеек под главенством одного вожака, как у гамадрилов. Отличительная особенность их сообщества — весьма гибкий характер групп, члены которых собираются вместе, пребывают на одной территории в течение нескольких часов или дней, а иногда и больше, а затем расстаются, чтобы встретиться вновь через некоторое время. Такой тип организации сообщества свойствен по крайней мере шимпанзе, населяющим влажные тропические леса. В других районах Африки, например в лесах Танзании, шимпанзе живут, вероятно, более обособленными группами. Тем не менее ряд японских авторов обращают внимание на постоянные переходы самок из одних групп в другие в сообществах лесных шимпанзе (Kawanaka a. Nishida, 1974). Для того чтобы определить, подчиняется ли такое поведение каким-либо принципам организации сообщества или же, напротив, формирование и перестройка групп происходят совершенно слу
116
Часть I. Эволюция человека
чайно, было предпринято исследование процесса дробления и слияния групп шимпанзе в лесу Будонго. Анализ показал, что чаще всего встречаются группы четырех типов. Это, во-первых, группы взрослых самцов, не включающие самок и молодых животных; во-вторых, группы матерей, состоящие из самок с детенышами, но без «подростков» и бездетных матерей (иногда с примкнувшим к ним пожилым самцом); в-третьих, группы, объединяющие взрослых животных обоего пола, а часто и самок в период эструса, но не подростков; и, наконец, смешанные группы из животных обоего пола и любых возрастных классов.
Функционально эти группы можно дифференцировать следующим образом. Группы взрослых самцов обычно очень подвижны, создают много шума и стремятся первыми обнаружить в лесу новые источники пищи. Группы матерей, напротив, относительно инертны, стараются не создавать шума и обычно остаются на одном месте; переход на новую стоянку, которую они выбирают, ориентируясь по крикам других животных, совершается очень быстро и почти бесшумно. У матери обычно два, а то и три детеныша. Самый маленький прицепляется к животу, следующий по возрасту сидит на спине, а старший шествует позади матери или рядом с ней. Известно, что детеныши шимпанзе остаются с матерью до 5 лет. Группы взрослых обоего пола включают самцов и самок, проявляющих наибольшую половую активность. Она отчасти связана с происходящими у самок в период течки изменениями наружных половых органов, способствующими привлечению самцов. Совокупляется с самкой только один из них. Смешанные группы из особей обоего пола и разного возраста обычно кормятся на деревьях, богатых пищей, например на фиговых деревьях. Такие группы, как правило, представляют собой большие скопления животных, объединившихся для совместного пропитания.
Перечисленные выше группы, конечно, не следует абсолютизировать. Правильнее считать их наиболее вероятными формами организации сообщества у шимпанзе, обитающих во влажных лесах, подобных лесному массиву Будонго в Уганде. Возможно, что в других районах тип организации будет иным; исследования, проводимые в Танзании и в ряде других областей, помогут пролить дополнительный свет на эту проблему. Рейнолдс и Рейнолдс (Reynolds U. a. Reynolds F.) оценили общую численность популяции шимпанзе, обитающих в условиях влажного тропического леса, в 70—80 особей. Такая популяция соответствует «сообществу», постоянно живущему в данном лесном районе. Полагают, что и в других изученных местах популяции насчитывают по 70—80 животных. Из-за интенсивных контактов между сообществами трудно установить точные границы территории, занимаемой данным сообществом, или строго оценить его численность. «Плотность населения» в лесу Будонго составляет примерно 10 шимпанзе на 1 кв. ми
Глава 5. Происхождение человеческого общества
117
лю, что близко к максимальной плотности, когда-либо зарегистрированной для этого вида животных.
Между описанным выше типом организации сообщества и формой отношений у шимпанзе, населяющих смешанную зону лесов и саванн (Kawanaka a. Nishida), существуют, по-видимому, вполне реальные различия. В последнем случае шимпанзе образуют группы, четко отделенные друг от друга, хотя и не «замкнутые» (см. ниже). Места обитания различных групп заметно перекрываются. Однако между группами существует определенная иерархия, и группа, занимающая подчиненное положение, вынуждена покидать общую территорию при появлении главенствующей группы. Создается впечатление, что одних лишь громких звуков, совместно издаваемых животными доминирующей группы, достаточно, чтобы вызвать беспокойство, а затем и бегство членов подчиненной группы. Ситуация, очевидно, полностью отлична от той, которую мы наблюдаем у лесных шимпанзе: там подобные звуки, по-видимому, лишь привлекают животных.
Особенно интересно, что антагонизм между различными группами шимпанзе, живущими на границе лесов и саванн, по всей видимости, перерастает в антагонизм взрослых самцов, составляющих ядро таких групп. Самцы двух изученных групп не покидали их в течение 8 лет, тогда как самки довольно часто переходили из группы в группу обычно в период, когда они могли принимать самцов. Эти особенности поведения имеют большое значение для понимания организации сообществ у ранних гоминид (см. ниже).
Гудэлл (Goodall) и другие подробно описали внутригрупповое поведение шимпанзе. Животные используют для общения богатый и сложный арсенал мимики и жестов, звуковых и тактильных сигналов. Среди них можно выделить, как обычно, сигналы, выражающие агрессивность, материнскую заботу и половое влечение. Еще одну важную группу сигналов составляют различные знаки приветствия: к ним относятся особая мимика и телодвижения при встрече партнеров, взаимные прикосновения, чаще всего руками. Ряд движений, выполняемых преимущественно взрослыми самцами — вожаками групп, имеет целью привлечь внимание животных к наиболее крупным самцам, занимающим центральное место в группе. Интересное явление, описанное уже довольно давно в литературе (Garner, 1896), получило название «карнавала». Последний представляет собой длительную вспышку радостного возбуждения, которая охватывает большую группу шимпанзе, собравшихся для встречи других групп. Карнавал сопровождается шумными возгласами, «барабанной дробью», пританцовыванием и сотрясением ветвей деревьев. Термин «барабанная дробь» требует некоторого разъяснения. Звук, похожий на стук барабана, производится ударами ладоней по стволам деревьев, например по стволу железного дерева. Иногда шимпанзе бьют по стволам подош
118
Часть 1. Эволюция человека
вами ног. Барабанная дробь разносится на большие расстояния и служит для связи с другими группами животных в условиях ограниченной видимости. Как указывают Рейнолдс и Рейнолдс (1965), громкие крики и шум, а также барабанная дробь, производимые большим числом обезьян, — зто своеобразная система дальней связи, посредством которой шимпанзе оповещают своих сородичей о наличии обильных источников пищи. В основе этого предположения лежит тот факт, что шимпанзе всегда спокойны, если фруктов мало, однако, если деревья богаты плодами, шимпанзе издают большой шум, привлекающий других обезьян из окружающего леса.
Экологические и филогенетические детерминанты организации сообществ
Многие авторы пытались обобщить наблюдения, касающиеся организации сообществ у приматов. Одни делали упор на экологические факторы, другие — на филогенетические. De Vore (1963) построил демографическую таблицу изученных к тому времени видов приматов — от древесных форм до тех, кто в основном перешел к наземному образу жизни. Он пришел к выводу, что 1) территория, занимаемая наземным видом, значительно превышает территорию, на которой обитает древесный вид, и что 2) приматы, ведущие наземный образ жизни, живут более многочисленными и лучше организованными группами, чем приматы, проводящие большую часть времени на деревьях. Крук и Гартлан (Grook a. Gartlan, 1966) составили таблицу «степеней приспособленности» приматов к различным условиям обитания, включая тропический лес, саванну, степи и районы с засушливым климатом. Лесные виды, как правило, живут небольшими группами. Вид, обитающий в открытой местности, состоит из более многочисленных групп с несколькими самцами в каждой и стремится сохранить за собой определенную территорию; однако в степных районах с засушливым климатом наблюдается тенденция к формированию групп под главенством одного самца. Половой диморфизм и функциональная дифференциация выражены тем сильнее, чем выше степень территориальной обособленности вида и чем более рассеяны источники пищи.
Многие считают, что внутригрупповое поведение приматов и структура их сообществ в гораздо большей степени определяются филогенетическими факторами, чем экологическими. Штрузакер (Strnhsaker, 1969) изучал жизнь обезьян, населяющих леса Западной Африки, главным образом церкопитековых, а также некоторых других видов (дрилл, мангабей, колобус). Оказалось, что большинство лесных видов обезьян, ведущих древесный образ жизни, а также все церкопитековые живут небольшими группами, в каждой из
Глава 5. Происхождение человеческого общества
119
которых имеется лишь один взрослый самец. Штрузакер пришел к заключению, что «филогенетический вклад каждого вида в данную экологическую среду неодинаков, и это различие необходимо учитывать при рассмотрении связи экологических факторов со структурой сообщества. Всякая попытка понять эволюцию общественных отношений у приматов должна основываться на анализе не только экологических, но и филогенетических факторов».
Как мы увидим ниже, вопрос об относительной роли экологических и филогенетических детерминантов структуры сообщества играет важную роль при выборе конкретного вида приматов в качестве модели, изучение которой может привести к более глубокому пониманию структуры общества древних людей. Учитывать и те и другие факторы, безусловно, необходимо. Наибольшую ценность с экологической точки зрения представляют данные, указывающие на способность определенного вида приматов изменять организацию сообщества при изменении условий окружающей среды. Примером могут служить лангуры (Presbytis entellus), а также простые павианы (Papio anubis), если сравнивать между собой группы, обитающие в лесах, в саванне и в лесостепной зоне. У других видов организация сообщества отличается, однако, большим постоянством. Так, у церкопитековых в группе почти всегда находится лишь один взрослый самец независимо от того, обитает ли группа в лесу или в саванне. Структура сообщества у гамадрилов также не отличается гибкостью: хотя они и не живут в лесах, но сохраняют разделение на группы с одним взрослым самцом даже в неволе. Клаттон-Брок (Clatton-Brock) призывал с большой осторожностью относиться к попыткам объяснить организацию сообществ приматов одними лишь экологическими факторами, характеризующими давление среды на популяцию. Сравнив между собой два вида колобусов, он обратил внимание на то, что эти виды по-разному используют одно и то же местообитание в зависимости от своих привычек и наклонностей (в частности, от характера питания), определяемых не средой, а филогенезом. По мнению Клат-тон-Брока, такие категории, как «лес», «саванна», «лесостепь» (определяющие тип местообитания), а также термины, характеризующие тип питания («насекомоядные», «всеядные», «питающиеся плодами», «питающиеся листвой»), фактически не дают однозначной характеристики экологических условий, в которых существует данный вид. Эти категории обычно маскируют важные различия в пищевом рационе, относящиеся как к отдельным видам пищи, так и к их распределению, а также различия в привычках, связанных с приемом пищи у разных видов.
Отметим в заключение, что все сделанные до сих пор попытки установить прямую связь между структурой сообществ приматов, с одной стороны, и филогенетическими или экологическими факторами, с другой, неизменно наталкивались на значительные трудное-
120
Часть I. Эволюция человека
ти. Мы вновь приходим к необходимости более тщательного изучения специфических адаптаций, которыми обладают определенные группы приматов в характерных для них условиях обитания.
Экология, и организация сообществ у ранних гоминид
Выше была подробно описана морфология австралопитека. Изучение скелета показывает, что австралопитек передвигался на двух ногах, а его руки были свободны от локомоторных функций и, вероятно, использовались для ношения а, возможно, также для бросания различных предметов и манипулирования. Объем мозга был примерно такой же, как у современных больших обезьян, и, следовательно, значительно уступал объему мозга у последовавших за австралопитеком форм Ното. Тем не менее Холловей (Holloway, 1974) на основании изучения слепков черепа пришел к выводу, что мозг австралопитека имел типичные для человеческого мозга пропорции; мы, вероятно, вправе считать, что по своим умственным способностям австралопитек превосходил современных человекообразных обезьян. Имеются указания на то, что австралопитеки обладали некоторой степенью полового диморфизма, что, возможно, свидетельствует о начальной стадии разделения труда в сообществе.
Ряд исследователей, опираясь на данные об ископаемой фауне и флоре и в значительной степени на собственное художественное воображение, пытались воссоздать картину естественной среды, в которой обитал австралопитек (Howell). Эту среду называли «саванной», «местностью открытого типа», «лесостепью», «парковой территорией». Ясно, по-видимому, лишь то, что австралопитек не был видом, живущим в лесах, и потому все известные ископаемые остатки его обнаружены на территории, свободной от леса. (Заметим в этой связи, что простые павианы, которых нередко характеризуют как вид, обитающий в саванне, населяют также тропические леса.) Потеря волосяного покрова и развитие системы потоотделения в качестве механизма теплоотдачи, вероятно, увеличили потребность в воде у ранних гоминид, заставив их поселиться ближе к рекам и озерам. Именно вблизи воды, в Олдувей-ском ущелье и в районе озера Рудольфа, были сделаны основные находки ископаемых остатков древних гоминид в Восточной Африке. Места, богатые водой, обычно привлекают много видов животных, что облегчает поимку дичи и сбор падали.
Глинн Исаак (Cl. Isaak, 1970) проанализировал археологический и палеонтологический материал, собранный в местах стоянок австралопитеков в Южной и Восточной Африке. Ископаемые остатки из Макапансгата (Южная Африка) включают кости австралопитеков, а также и многих видов животных. Однако из-за
Глава 5. Происхождение человеческого общества
121
трудностей, связанных со стратиграфией, нельзя с определенностью сказать, доставил ли австралопитек убитых животных к местам стоянки или же кости животных постепенно скопились там под действием каких-либо иных факторов. Первое предположение, однако, остается весьма спорным. Что касается находок в Восточной Африке, то здесь связь ископаемых остатков австралопитека с остатками различных животных прослеживается более четко. В Олдувайском ущелье найдены кости гоминид, кости некоторых видов животных и специально заготовленные камни. Исаак различает три типа стоянок: стоянки, где убивали животных и готовили пищу, места постоянного пребывания австралопитека и «мастерские». Первые содержат кости многих видов животных, но мало посторонних предметов, вторые — и кости, и посторонние предметы в достаточных количествах: в мастерских найдены главным образом посторонние предметы. Последних особенно много на некоторых стоянках, таких, как FLKi, где был открыт Ztnjanthropus (2659 предметов), и стоянка ВКП, где насчитали 7223 различных предмета. Среди остатков животных, найденных вместе с австралопитеком, чаще всего встречаются кости Dinotherium, антилопы и дикой свиньи, а также остатки более мелких существ: пресмыкающихся, лягушек, птиц и грызунов. По этим остаткам нельзя судить, какое время австралопитек отдавал собственно охоте. Скорее всего он вообще не охотился в прямом смысле этого слова. Тем не менее можно думать, что австралопитеки, подобно современным шимпанзе, могли в определенных обстоятельствах действовать достаточно слаженно, чтобы успешно поймать, разделать и съесть добычу: обезьяну, речного кабана или бушбака (Teleki, 1973).
Мы подошли к последнему пункту наших рассуждений, а именно к вопросу о том, как наилучшим образом объединить фактические данные и теории, вытекающие из изучения ныне живущих видов приматов (помимо человека), со сведениями, полученными при изучении ископаемых остатков австралопитека, с тем чтобы воссоздать картину общественной жизни древних людей.
На начальной стадии мы, очевидно, имеем дело с каким-то видом, произошедшим от понгид (дриопитековых), т. е. филогенетически этот исходный вид ближе к понгидам, чем к церкопитеко-вым. Существенной чертой дивергентного развития в направлении гоминид является переход к прямохождению на двух ногах, что следует рассматривать как адаптацию к наземному образу жизни на территориях открытого типа и вблизи источников воды. Если бы предками австралопитека были необычные виды простых обезьян, то мы имели бы все основания использовать для модельных исследований современные виды нечеловекообразных обезьян, приспособленных к жизни в саванне. Мы могли бы строить свои выводы на результатах наблюдений за обезьянами вида патас, простыми павианами, гамадрилами или геладами. Однако эти выводы
122
Часть 1. Эволюция человека
неизбежно отличались бы значительной неопределенностью в силу того, что типы организации сообществ у разных видов и внутри указанных видов весьма разнообразны. Ради простоты мы могли бы, конечно, рассматривать некоторые «идеальные» типы внутригрупповых отношений.
Модель, основанная на изучении павианов и макаков, приводит к заключению, что в открытой местности, подобной современной саванне, древние гоминиды жили большими группами, тесно связанными между собой. У них был явно выражен половой диморфизм и обусловленное им разделение труда; самцы обеспечивали защиту самок и молодых животных от хищников. Объектами конкуренции взрослых самцов были привлекательные в половом отношении самки, а также скудные ресурсы (например, немногочисленные примитивные инструменты — камни с острыми краями) и предпочитаемые места стоянок. Конкуренция не была, однако, единственным фактором, определявшим положение индивидуума в сообществе; оно зависело в какой-то степени и от родственных связей по материнской линии. Странствуя по территории своего местообитания, группа время от времени собиралась у источников пищи; молодые самцы выполняли функции разведчиков и часовых, физически сильные самцы поддерживали внутренний порядок в группе и защищали ее от хищников.
К несколько иной картине жизни сообществ древних гоминид мы приходим в результате изучения гамадрилов (а также гелад или обезьян вида патас): основное отличие от рассмотренной выше ситуации состоит в существовании в группе небольших подгрупп, состоящих из одного взрослого самца, одной или нескольких зрелых самок и их потомства. Мы вплотную подошли к введению такого элемента взаимоотношений, как формирование постоянных супружеских пар. Речь идет о тенденции разного пола к устойчивым взаимоотношениям, более длительным, чем период половой активности или даже время совместного ухода за потомством. Такая система неизбежно ведет к ограничениям половой деятельности членов группы, а также к формированию «холостяцких» групп, состоящих из самцов без партнерш.
Можно построить еще немало подобных «обезьяньих» моделей, и они действительно были построены. Придавая важное значение филогенетическим факторам, мы должны, однако, особенно внимательно отнестись к анализу данных, полученных при изучении понгид в качестве модельного сообщества. На примере шимпанзе мы видим еще одну, третью, возможность приспособления к условиям жизни в саванне. Она основана на принципе так называемого «открытого» сообщества, т. е. такой его структуры, которая допускает значительно большую свободу перехода индивидуумов из одной группы в другую, чем это подразумевалось в рассмотренных выше моделях. Как следствие, состав групп в таком сообществе
Глава 5. Происхождение человеческого общества
123
остается постоянным лишь в течение сравнительно короткого времени. Самцы объединяют свои усилия в поисках пищи, держатся вместе некоторое время, но затем расходятся, подбирая себе пари среди сексуально активных самок также на ограниченный период времени. Детеныши находятся под присмотром матерей, однако по мере того как дети взрослеют, матери становятся все более мобильными и, наконец, оставляют своих детей. Родственные связи — один из принципов организации сообщества; потомство даже в зрелом возрасте поддерживает контакты с родней по материнской линии. Объединение (и распад) группы диктуется в основном пищевыми потребностями ее членов. Следует учитывать и другой фактор — так называемое «чувство сообщества». На территории, бедной пищевыми ресурсами, плотность населения очень низка (скажем, 1—2 особи на 1 кв. милю), и сообщества фактически представляют собой отдельные группы, или «трибы». В такой системе ни доминантная иерархия, ни половая разобщенность не играют существенной роли в структуре связей сообщества. Можно думать, что возникновение «семейной ячейки» в такой системе было связано либо с постепенно усиливающейся тенденцией к формированию постоянных пар в противовес беспорядочным половым отношениям, либо с экономическими причинами. Семейная ячейка никогда не была ничем иным, кроме как санкционированным обществом экономическим образованием, и ее формирование и сохранение скорее противоречило, чем соответствовало естественным наклонностям взрослых гоминид.
В этой связи заслуживают особого упоминания данные японских авторов, на которые мы ссылались выше. Если в качестве модели отношений между членами сообщества у ранних гоминид использовать сообщество шимпанзе, обитающих в саванне, а не в тропических лесах (такой выбор кажется наиболее оправданным как с филогенетической, так и с экономической точки зрения), то можно выделить наиболее важную особенность организации сообщества, а именно тенденцию к образованию квазизамкнутых групп, основанных на взаимном антагонизме взрослых самцов и в то же время полной свободе передвижения из группы в группу взрослых самок, способных в данный момент принимать самцов. По указанным выше причинам такая модель предпочтительнее других. Вдобавок она устанавливает столь желательное соответствие между данными, полученными при изучении современных видов приматов, и характерным признаком древних человеческих сообществ, отмеченным рядом антропологов (например, Levi-Strauss) — обменом женщинами между группами. Возможно, что древний человек не изобрел такой обмен, а лишь продолжил уже существующую практику, превратив обмен в систему и подчинив ее определенным правилам, подобным различным запретам кровосмешения и узаконенным системам родства.
124
Часть I. Эволюция человека
Мы рассмотрели четыре типа сообществ приматов, каждый из которых послужил рабочей моделью для создания картины общественной жизни древнего человека. Основные черты этих четырех модельных систем представлены на рис. 5.1. Можно было воспользоваться данными, относящимися к другим видам приматов, например гиббонам и сиамангам, основной структурной единицей которых служат малочисленные семейные ячейки и которых отличает высокая степень территориальной обособленности. Модельные системы, выбранные нами, заметно отличаются друг от друга. Все они (за исключением последней,, описанной лишь недавно) использовались различными авторами в качестве основных моделей для
Г
Рис. 5.1. Модели организации сообществ древнего человека.
А. Модель, основанная на изучении сообществ простых павианов и (или) макак. Б. Модель, основанная на изучении сообщества гамадрилов. В. Модель, основанная на изучении сообщества лесных шимпанзе. Г. Модель, основанная на изучении шимпанзе, живущих в лесостепной саванне. Сплошной линией показана граница замкнутой группы, прерывистой — граница открытой группы.
анализа организации древних человеческих сообществ. Удастся ли когда-нибудь исключить из рассмотрения некоторые из этих моделей, признав их абсолютно «нерабочими», или найти богатый ископаемый материал и места стоянок древних людей, изучение которых позволило бы сделать однозначные выводы об организации их сообществ, сказать трудно. Такими возможностями и такими данными мы пока не располагаем.
И наконец, несколько слов о языке. Развитие системы свя-зи, основанной на использовании символических категорий, коди-
Глава 5. Происхождеиие человеческого общества
125
рованных словами, привело в конечном счете к образованию сообщества того типа, которое отличает современного человека. Одно из предположений, не вытекающее непосредственно из изучения модельных сообществ современных приматов, состоит в том, что в этих сообществах для связи не используются понятия, так что речь может идти лишь о моделировании догоминидной стадии организации сообщества. Возникает вопрос: когда появился язык, а вслед за ним и соответствующие возможности для перестройки сообщества? Мы не можем категорически утверждать, что у австралопитека отсутствовал язык понятий. Знание объема мозговой коробки (или, точнее, относительного объема мозга) не дает нам достаточных сведений на этот счет, поскольку остаются неопределенными характер организации мозга, его строение и степень сложности. Насколько можно судить об этих признаках по ископаемым остаткам, включая изучение слепков черепа, мозг австралопитека был подобен мозгу человека. Эксперименты последних лет показали, что после интенсивного обучения шимпанзе способны делать символические знаки руками, а также манипулировать пластмассовыми кружками или пуговицами разной формы и разного цвета, причем характер манипулирования позволяет предположить, что у них определенно имеется некоторая, хотя и слабо выраженная связь при помощи символов (Gardner a. Gardner, 1969; Premak a. Premak, 1972). Возможно, что по своим способностям австралопитек превзошел в этом отношении шимпанзе.
Гораздо более вероятно, однако, что речевая система связи впервые появилась у Homo erectus — первого из гоминид с объемом мозга, значительно превосходящим объем мозга австралопитека. Действительно, можно доказать, что именно развитие речевых навыков и обусловленное ими давление сил отбора качественно нового типа привело к увеличению размеров мозга у Homo erectus и к еще большему увеличению мозга у Homo sapiens. Объяснялся ли человек первоначально посредством жестов и лишь потом освоил речь, как утверждают Хьюес (Hewes, 1973) и ряд других исследователей, или же в своем развитии он миновал фазу жестикуляции, сразу перейдя к совершенствованию навыков речи при одновременном развитии «речевых» областей коры (поле Брока и поле Вернике),— эти вопросы до сих пор служат предметом полемики. Неясно также, были ли у всех гоминид, начиная с Homo erectus, речевые способности столь же выражены, как у современного человека. Согласно Либерману и др. (Lieberman et al., 1972), голосовой тракт неандертальца был неприспособлен к произнесению всей гаммы речевых звуков и в лучшем случае мог обеспечить ему «возможность лишь очень медленной связи». Фальк (Falk, 1975), однако, оспаривает этот вывод, полагая, что упомянутые авторы поместили реконструированную гиоидную кость слишком высоко» и вопрос требует дальнейшего изучения.
126
Часть I. Эволюция человека
Список литературы
Bartholomew G. A. and Birdswell J. В. (1953). Ecology and the proto-hominids. Am. Anthrop. 55, 481—498.
Beach F. A. (1947). Evolutionary changes in the physiological control of mating behaviour in mammals. Psychol. Rev. 54, 297—315.
Birdsell J. B. (1972). Human evolution. Rand McNally and Co., Chicago.
Cain A. J. (1954). Animal species and their evolution. Hutchinson’s University Library, London.
Cain A. J. and Harrison G. A. (1960). Phyletic weighting. Proc. zool. Soc. Lond. 135, 1-31.
Campbell B. G. (1966). Human evolution. Heinemann Educational Books, London.
Campbell B. G. (1967). Human evolution: an introduction to man’s adaptation. Heinemann, London.
Chance M. R. A. (1967). Attention structure as the basis of primate rank orders. Man 2, 503-518.
Clutton-Brock T. (1974). Primate social organisation and ecology. Nature, Lond. 250, 539—542.
Cold Spring Harb. Symp. quant. Biol. (1950). 11, Origin and evolution of man.
Coon C. S. (1963). The origin of races. Jonathan Cape, London.
Crook J. H. and Gartland J. S. (1966). Evolution of primate societies. Nature, Lond. 210, 1200—1203.
Day M. H. (1965). Guide to fossil man. World Publication Co., Cleveland.
De Beer G. R. (1951). Embryos and ancestors (revised edn). Oxford University Press, London.
De Vore I. (1963). A comparison of the ecology and behaviour of monkeys and apes. In Classification and human evolution (ed. S. L. Washburn). Viking Fund Publ. Anth. Volume 37, pp. 301—319.
De Vore I. and Hall K. R. L. (1965). Baboon ecology. In Primate behavior (ed.
I. De Vore), pp. 20—25. Holt, Rinehart and Winston, New York.
Dolhinow P. and Sarich V. M. (eds.) (1971). Background for man: readings in physical anthropology. Little, Brown and Co., Boston.
Falk D. (1975). Comparative anatomy of the larynx in man and the chimpanzee: implications for language in Neanderthal. Am. J. Phys. Anth. 43, 123—132.
Fox R. (1967). In the beginning: aspects of hominid behavioural evolution. Man, 2, 415-433.
Garner R. L. (1896). Gorillas and chimpanzees. Osgood, London.
Gardner R. A. and Gardner В. T. (1969). Teaching sing language to a chimpanzee. Science, N. Y. 165, 664—672.
Goodall J. (1965). Chimpanzees of the Gombe Stream Reserve. In Primate behavior (ed. I. De Vore), pp. 425—473. Holt, Rinehart and Winston, New York. Goodall J. van Lawick (1968). The behaviour of free-living chimpanzees in the Gombe Stream Reserve. Anim. Behav. Monogr. 1.
Hewes G. (1973). Primate communication and the gestural origin of language.
Current Anthrop. 14, 5—12.
Holloway R. L. (1974). The casts of fossil hominid brains. Sci. Am., July.
Howell F. C. (1966). Early man. Time Life, New York.
Howells W. W. (1967). Mankind in the making. Pelical, London.
Howells W. W. (1973). Evolution of the genus Homo. Addison-Wesley. Reading, Mass.
Hulse F. S. (1971). The human species (2nd edn.). Random House, New York.
Huxley J. S. (1942). Evolution, the modern synthesis. Allen and Unwin, London.
Isaac G. (1970). The diet of early man: aspects of archaeological evidence from lower and middle Pleistocene sites in Africa. Wld. Arch. 2, 278—299.
Глава 5. Происхождение человеческого общества 127
Jolly С. J. (1970). The seed eaters: a new model of hominid differentiation based on a baboon analogy. Man 5, 5—26.
Jolly A. (1972). The evolution of primate behavior. Macmillan, New York.
Kawanaka K. and Nishida T. (1974). Recent advances in the study of inter-unit-group relationships and social structure of wild chimpanzees of the Mahali Mountains. Symp. 5th Cong. Int. Primat. Soc., 173—186.
Kummer H. (1968). Social organisation of hamadryas, baboons, p. 189. University of Chicago Press.
Le Gros Clark W. E. (1948). Observations on certain rates of somatic evolution in the Primates, pp. 171—180. Robert Broom Commemorative Volume of the Royal Society of South Africa.
Le Groc Clark W. E. (1955). The fossil for human evolution. Chicago University Press.
Le Gros Clark W. E. (1967). Man-apes or ape-men? Holt, Rinehart and Winston, New York.
Le Gros Clark W. E. (1971). The antecedents of man (3rd edn). Edinburgh University Press.
Le Gros Clark W. E. (1973). History of the Primates (10th edn). British Museum (Nat. Hist.), London.
Lieberman P., Crelin E. S. and Klatt D. H. (1972). Phonetic ability and related anatomy of the newborn and adult human. Neanderthal man and the chimpanzee. Am. Anthrop. 74, 287—307.
Mayr E. (1965). Animal species and evolution. Harvard University Press.
(Э. Майр, Зоологический вид и эволюция. М.: «Мир», 1968.)
Napier J. (1971). The roots of mankind. Allen and Unwin, London.
Oakley К. P. (1969). Frameworks for dating fossil man (3rd edn). Weidenfeld and Nicolson, New York.
Oakley К. P. (1973). Man the tool-maker (5th edn). British Museum (Nat. Hist.), London.
Pfeiffer J. E. (1969). The emergence of man. Harper, New York.
Pilbeam D. (1972). The ascent of man: an introduction to human evolution. Macmillan, New York.
Premack A. J. and Premack D. (1972). Teaching language to an ape. Scient. Am., October, 92—99.
Reynolds V. (1966). Open groups in hominid evolution. Man 1, 441—452.
Reynolds V. and Reynolds F. (1965). Chimpanzees of the Budongo Forest. In Primate behavior (ed. I. De Vore), pp. 368—424. Holt, Rinehart and Winston, New York.
Rowell T. E. (1966). Forest living baboons in Uganda. J. Zool. 147, 344—364.
Sanderson I. T. (1950). The physical distinctions of man. Proc. Am. phil. Soc. 94, 428—449.
Sanderson I. T. (1957). The monkey kingdom. Hamish Hamilton, London.
Simons E. L. Primate evolution: an introduction to man’s place in nature. Macmillan, New York.
Simpson G. G. (1950). The meaning of evolution. Oxford University Press, London.
Simpson G. G. (1953). The major features of evolution. Columbia University Press, New York.
Simpson G. G. (1961). Principles of animal taxonomy. Columbia University Press, New York, and Oxford University Press, London.
Sneath P. H. A. and Sokal R. R. (1973). Numerical taxonomy. The principles and practice of numerical classification. Freeman, San Francisco.
Spuhler J. N. (ed.) (1959). Evolution of man’s capacity for culture. Human Biol. 31, 1—54. (Papers by Hackett, Harlow, Sahlins).
Struhsaker R. T. (1969). Correlates of ecology and social organisation among African Cercopithecines. Folia Primat. 11, 80—118.
128 Часть I. Эволюция человека
Teleki G. (1973). The predatory behavior of wild chimpanzees. Bucknell University Press, Lewisburg.
Washburn S. L. and De Vore I. (1961). Social behaviour of baboons and early man. In Social life of early man. Viking Fund Publications, Anthropology Volume 31, pp. 91—104.
Washburn S. I. (1963). Classification and human evolution. Aldine, Chicago.
Weiner J. S. (1956). The evolutionary taxonomy of the Hominidae in the light of the Piltdown investigation. Selected papers of the Fifth International Congress of Anthropological and Ethnological Sciences, pp. 741—752.
Weiner J. S. (1971). Man’s natural history. Weidenfeld and Nicolson, London.
Weiner J. S. (1958). The pattern of evolutionary development of the genus Homo. S. Afr. J. med. Sci. 23, 111—120.
ЧАСТЬ II
ГЕНЕТИКА ЧЕЛОВЕКА
ДЖ. ХАРРИСОН
Глава в
ЗАКОНЫ МЕНДЕЛЯ И ЧЕЛОВЕК
Введение
За те 70 лет, которые истекли после вторичного открытия законов Менделя, генетика не только стала важнейшей самостоятельной отраслью науки, но и проникла во все биологические дисциплины. Генетика стала краеугольным камнем и в биологии человека, так как в конечном счете только взаимодействие наследственных задатков и среды определяет все признаки индивидуума и, следовательно, изменчивость в пределах популяции, а именно изменчивость в первую очередь и интересует исследователя биологии человека. Своими успехами на ранних этапах развития генетика была обязана исследованиям, проводившимся на растениях и «низших» животных, для которых характерна небольшая продолжительность жизни одного поколения и при изучении которых можно применять экспериментальные скрещивания. Исходя из исследований популяции человека, вряд ли удалось бы открыть законы наследственности, но когда было показано, что законы Менделя приложимы и к генетике человека, исследователи биологии человека получили возможность внести свой весьма существенный вклад в развитие общей генетики. Ведь и биология индивидуума и, что более важно, структура и история популяций человека изучены значительно лучше, чем для любых других организмов. Со времени выхода в свет первого издания этой книги генетика человека достигла замечательных успехов; это особенно касается молекулярной, биохимической, клеточной и медицинской генетики. Тем не менее большие области здесь остались неизученными, в первую очередь генетические аспекты механизмов развития и популяционная генетика. Вместе с тем основные законы, как и прежде, выдержали испытание временем, и в том, что касается исходного элементарного уровня, преобладающая часть написанного в первом издании остается справедливой и но сей день.
130
Часть II. Генетика человека
Корпускулярная наследственность
Первый закон Менделя — закон расщепления признаков — гласит, что наследуемые признаки регулируются парами дискретных факторов. Члены пары отделяются друг от друга (сегрегируют) при образовании первичных половых клеток и расходятся по разным гаметам. Эти факторы воссоединяются в пары при оплодотворении, допускающем их объединение в определенных пропорциях. Вследствие этого определяемые «ими признаки также способны сегрегировать, так как они будут появляться в последующих поколениях с определенной частотой.
Примером действия этого закона может служить открытие природы и характера наследования одной из систем групп крови человека — системы MN. Группы крови определяются сложными веществами, входящими в состав мембран эритроцитов. Систему MN открыли Ландштейнер и Левин в 1927 г. Эти авторы вводили кроликам кровь человека и затем исследовали способность кроличьей сыворотки агглютинировать эритроциты людей. Оставляя в стороне некоторые технические детали, систему MN можно выявить следующим образом. Если кроличья антисыворотка (против крови человека) прореагировала с пробой крови одного человека («истощение» сыворотки), то иногда в такой сыворотке еще остаются антитела (агглютинины), способные агглютинировать эритроциты другого человека. Если для истощения сыворотки взять эритроциты второго индивидуума, то в сыворотке останутся антитела, способные агглютинировать эритроциты первого.
Результаты этих экспериментов можно объяснить следующим образом: эритроциты человека несут на своей поверхности либо один из двух антигенов (гемагглютиногенов), либо оба эти антигена (М и N). Соответствующих антител в сыворотке человека нет, однако они образуются в организме у кролика, если ввести ему определенный тип крови человека. Так, если для инъекции взять кровь, содержащую антигены М и N, то образуются антитела обоих типов. Если полученную сыворотку смешать с избытком крови, содержащей антиген N, то на агглютинацию эритроцитов будут израсходованы соответствующие антитела (анти-N). Анти-М не примут участия в реакции. Они останутся в сыворотке и сохранят способность реагировать с эритроцитами, содержащими антиген М. Напротив, если первоначально для истощения антисыворотки взять эритроциты группы М, то в сыворотке останутся анти-N.
Таким образом, в соответствии с системой MN люди делятся на три группы: у одних эритроциты несут антиген М, у других — антиген N, а у третьих — оба антигена, М и N. Следовательно, в популяции возможно 6 различных типов брака. Эти типы брака представлены в табл. 6.1 с указанием групп крови рожденных в
Глава 6. Законы Менделя и человек
131
этих браках детей и соотношения индивидуумов с различными группами крови.
Таблица 6.1
Тип брака Группы крови детей
МхМ м
NxN N
MxN MN
MNxM 1 1 уМ yMN
MNxN 1 1 yMN — N Ы di
MNxMN 11 1 -5-м 4-mn 4n 4 2 4
В отдельных небольших семьях эти соотношения часто неразличимы, но при достаточно большой численности семьи было бы найдено близкое соответствие указанным в таблице соотношениям. Такой семье генетически эквивалентна группа детей, рожденных в браках одного из шести типов. Суммарные данные о подобных браках выявляют ожидаемые соотношения.
Тип наследования антигенов MN можно объяснить, исходя из того, что их наличие у данного индивидуума определяется единичными факторами, сохраняющими свою идентичность из поколения в поколение. Любой индивидуум обладает двумя такими факторами, получая их по одному от каждого из родителей. Эти факторы могут быть или одинаковыми или различными. Обозначив эти факторы символами и .4gN, генетические основы результатов, приведенных в табл. 6.1, можно представить следующим образом (табл. 6.2).
Поведение этих факторов в процессе их наследственной передачи согласуется с их локализацией в хромосомах клеточного ядра. Об их положении в ядре свидетельствует независимость передачи от пола индивидуума, несмотря на то что в яйцеклетке значительно больше цитоплазмы, чем в сперматозоиде. О связи рассматриваемых факторов с хромосомами свидетельствует полное соответствие между наблюдаемым в микроскоп поведением хромосом во время формирования половых клеток (гамет) и поведением этих факторов, проявляющимся в типе наследования. За некоторыми исключениями, ядра всех соматических клеток содержат одинаковое число хромосом, и это так называемое диплоидное число оказывается величиной, постоянной для данного вида. Трудности в изготовлении хороших цитологических препаратов хромосом
132
Часть II. Генетика человека
Таблица 6.2
Генотип родителей Генотип детей
д§-м д^м х л gM^ gM 4gM4gM
4gN^gNxXgN4gN 4gN4gN
AgMAgMX Ag^AgN 4gM4gN
Ag^Ag^xAg^Ag^ 4- 4gM4gM 4 4gM4gN Z Z
4gM4gNX4gN4gN -= 4gM4gN 4- 4gN4gN z z
4gM4gNx4gM4gN 4-4gM4gM 1 4gM4gN 1 AgN4gN 4 Z 4
человека повлекли за собой в прошлом споры об истинном числе хромосом в ядрах клеток у человека. Теперь с достоверностью установлено, что их 46 (рис. 6.1).
Хромосомы, которые во время гаметогенеза образуют парыг называются гомологичными. Как правило, члены пары морфологически идентичны. Если не считать возможных небольших различий в расположении отдельных участков хромосом, то это утверждение безоговорочно правильно лишь в отношении хромосом женского организма, каждая из которых представлена дважды в ядрах всех соматических клеток. В клетках мужского организма, кроме 22 пар гомологов, имеется пара хромосом, у которых гомологичен в лучшем случае небольшой участок. Одна из этих двух хромосом идентична 23-й паре хромосом женского организма; это так называемая Х-хромосома. Другая хромосома значительно меньше, встречается она исключительно у мужчин; ее называют Y-хромосомой. X- и Y-хромосомы — это половые хромосомы; все прочие хромосомы называются аутосомами.
В результате мейоза в гаметах оказывается по одному члену из каждой пары гомологичных хромосом; следовательно, гамета содержит половинное число хромосом соматической клетки — гаплоидный набор. При оплодотворении восстанавливается диплоидный набор хромосом.
Тип наследования группы крови системы MN легко объяснить, допустив, что соответствующие наследуемые факторы находятся в одной паре хромосом. При генотипе одна из пар-
ных хромосом несет фактор а ее гомолог — фактор 24gN. Члены каждой пары могут с равной вероятностью попасть в ту или иную гамету, так что половина гамет организма, принадлежащего к типу ,4gM.4gN, содержит только Ag*\ тогда как другая половина — только ^4gN. В случае брака индивидуума такого
133
134
Часть II. Генетика человека
типа с другим, который несет только один из факторов —
или т. е. гаметы которого все одинаковы, равновероятно
слияние как двух одинаковых, так и двух разных гамет. Ребенок от такого брака с равной вероятностью может быть сходным с одним или с другим из родителей. Иными словами, в этом браке половина детей будет сходна с одним из родителей, а другая половина — с другим.
Родитель 4gM4gN
Родитель 4gM4gM Соотношение гамет 4gM v^gM 4-^N
Соотношение зигот 4-4gM4gM 4- 4gM4gN
Черточками под символами представлены хромосомы-носители этих факторов.
В случае брака между индивидуумами с генотипами AgMAgN у обоих родителей с одинаковой частотой образуются гаметы обоих типов, так что возможно рождение детей трех генотипов — AgM и AgN.
Родитель 4gM4gN
Соотношение гамет l^gM ±4gN
Соотношение зигот
|4gM 1 1 “4 -j-
Родитель 4gM4gN yAgN у 4gM4gN 4gN4gN
Определения
Здесь уместно привести определение ряда генетических терминов. Факторы AgK и AgN называются генами. Гены — зто наследуемые частицы, определяющие характерные признаки организ-
Глава 6. Законы Менделя и человек
135
ма. То место, которое ген занимает в хромосоме, называется локусом. Два гена, занимающие гомологические локусы, могут заменять друг друга; такие гены называются аллеломорфами, или аллелями. Если в гомологичных локусах находятся одинаковые гены (безразлично или j4gNj4gN), тдкой индивидуум именуется
гомозиготным. Если же эти гены различны (^gMHgN), то индивидуум называют гетерозиготным. Брак между гомо- и гетерозиготными индивидуумами можно назвать возвратным скрещиванием, а между двумя гетерозиготными — интеркроссом. Характерные признаки индивидуума, такие, например, как агглютиногены его эритроцитов (М, MN или N), образуют его фенотип. Генетическая конструкция индивидуума (Ag^Ag^, Ag^Ag® или же _4gM4gN) — это его генотип.
В системе MN все генотипы различаются и фенотипически. Однако во многих других случаях различные генотипы могут иметь одинаковое фенотипическое проявление. Некоторые люди способны ощущать вкус сильно разведенных растворов фенилтиомочевины и близких ей соединений, тогда как другие лишены этой способности (см. гл. 18). Людей первого типа называют «ощущающими вкус» (англ, «testers»), второго типа — «не ощущающими вкуса» («поп-testers»). Способность ощущать вкус фенилтиомочевины наследуется подобно группоспецифическим антигенам системы MN при участии двух аллелей Т и t (от taste — вкус). Наличие только двух (а не трех) типов людей объясняется тем, что гетерозиготы Tt способны ощущать вкус фенилтиомочевины подобно гомозиготам ТТ (табл. 6.3). Иначе говоря, эти два генотипа, Tt и ТТ, фенотипически неразличимы. Способность ощущать вкус фенилтиомочевины называется поэтому доминантным признаком по отношению к неспособности его ощущать, и обратно, неспособность ощущать вкус этого вещества — рецессивный признак по отноше-
Таблица 6.3
Фенотипы родителей Генотипы родителей Фенотипы детей Генотипы детей
Ощущающийх Ощу- ТТхТТ Ощущающие ТТ
щающий TTxTt » 1/2 ТТ, 1/2 Tt
TtxTt 3/4 ощущающие, 1/4 неощущающие 1/4 ТТ, 1/2 Tt, 1/4 tt
Ощущающийх Не- TTxtt Ощущающие Tt
ощущающий Ttxtt 1/2 ощущающие, 1/2 неощущающие 1/2 Tt, 1/2 tt
Неощущающий х Неощущающий ttxtt Неощущающие tt
136
Часть II. Генетика человека
нию к способности ощущать его вкус. Другим примером доминантности и рецессивности у человека может служить выделение (секреция) или невыделение группоспецифических веществ АВО в жидкости тела, в том числе в слюну. Одни люди выделяют большие количества групповых веществ АВО, другие же выделяют их мало или вовсе не выделяют. Способность к секреции — доминантный признак, отсутствие такой способности — рецессивный (гл. 12). Термин доминантный и рецессивный желательно применять, говоря о признаках, а не о генах, так как у одного и того же гена некоторые эффекты могут быть доминантными, а другие — рецессивными. Нередко, однако, эти термины используют в применении к самим генам; при этом следует всегда помнить^ что это делается лишь для краткости.
Аллелизм
Очевидно, изучению доступна генетическая детерминированность лишь тех признаков, которые проявляются не менее чем в двух взаимоисключающих вариантах. Если бы у всех людей кровь принадлежала к группе М, то нельзя было бы выяснить тип наследования этого признака; это оказалось возможным лишь благодаря наличию группы N. Многие признаки обнаруживаются у людей лишь в одном варианте: все люди оказываются гомозиготными по генам, контролирующим эти признаки. Вместе с тем возможно существование более двух аллелей, определяющих возникновение того или иного варианта какого-то определенного признака, хотя, как правило, каждый данный индивидуум обладает, разумеется, только двумя соответствующими аллелями. Так, группы крови системы АВО определяются четырьмя часто встречающимися и несколькими редкими аллелями (гл. 18).
Естественно, что при наличии только одного гена возможен только один генотип, при наличии двух аллелей одного гена возможны уже 3 генотипа — два гомозиготных и один гетерозиготный, при наличии трех аллелей число возможных генотипов увеличивается до 6 — три гомозиготных и три гетерозиготных:
Д1Д1 дм2 4343
4l42 4U3
4243
При наличии п аллелей число возможных генотипов равно п + (п —1) + (п—2) + (п—3). .. + {п (n—1)}.
Число возможных фенотипов, очевидно, зависит от наличия доминантности. Если эффект А1 доминирует над эффектом А2, а эффект А2 —над А3, то будут возможны только 3 фенотипа — ведь все гетерозиготы будут проявляться так же, как AM1 или как А2А2.
Глава 6. Законы Менделя и человек
137
Хромосомный набор
Вышеупомянутые 23 пары хромосом у человека удалось классифицировать, отнеся их в различные группы в зависимости от размеров и положения центромер. На рис. 6.1 представлен кариотип нормального мужчины; гомологичные хромосомы здесь подобраны парами, и аутосомы расположены по порядку соответственно 7 группам (от А до G). На обычных цитологических препаратах идентифицировать некоторые пары хромосом нередко удается лишь с трудом; обычно для того, чтобы с уверенностью найти именно парные хромосомы, приходится их располагать в виде кариотипа. Группа А образована тремя парами довольно крупных хромосом с центромерой, расположенной примерно посередине хромосомы и делящей хромосому на два плеча примерно равной длины. В группу В входят две пары столь же крупных хромосом, однако в них центромера расположена гораздо ближе к одному концу хромосомы; таким образом, плечи каждой хромосомы этой группы оказываются неравными. Для 7 пар, образующих группу С, характерна средняя длина и небольшое более или менее выраженное отклонение положения центромеры от срединного (субмедианное положение центромеры). К этой группе относят и Х-хромосому. Группа D состоит из двух пар хромосом средней длины, как и хромосомы группы С; однако их центромеры расположены очень близко к одному концу хромосомы, тем не менее короткое плечо четко различимо. Такие хромосомы называют акроцентрическими. Три пары небольших хромосом отнесены в группу Е, хотя в одной паре центромеры расположены строго посередине хромосом, тогда как в двух других парах они локализованы субмедианно. Хромосомы двух пар, входящих в группу F, еще меньше и их плечи еще короче; их центромеры занимают срединное положение на хромосоме. Наконец, группа G состоит из двух пар очень небольших аутосом, центромеры которых расположены акроцентрично, оставляя тем не менее короткое плечо. К этой группе относят также Y-xpo-мосому. Иногда все хромосомы со срединным, субмедианным и дистальным расположением центромер называют «метацентрическими» в отличие от «акроцентрических».
Идентификация каждой отдельной хромосомы облегчается наличием у некоторых из них характерных сужений или миниатюрных терминальных образований, сателлитов. Недавно было показано, что при окраске в определенных условиях некоторые флуоресцентные красители типа акрихина неодинаково окрашивают хромосому по ее длине, вследствие чего на ней образуются флуоресцирующие поперечные полосы, позволяющие различать пары хромосом. Этот метод весьма облегчил точную идентификацию отдельных хромосом.
138
Часть II. Генетика человека
Анализ родословных
Если не считать систем групп крови, то наши знания по генетике человека касаются главным образом редко встречающихся признаков, которые наследуются по простому менделевскому типу. Многие из этих признаков имеют важное медицинское значение; их редкость в значительной мере связана с тем, что индивидуум, обладающий таким признаком, оказывается не так хорошо приспособленным, как индивидуум с альтернативным вариантом этого признака. Поэтому часто носителей такого патологического признака называют пораженными, а их здоровых родственников — непораженными. Ген, определяющий тот или иной патологический признак, обозначают каким-нибудь особым, соответствующим диагнозу символом, а его нормальный аллель может быть обозначен знаком «+» независимо от специфики изучаемого признака.
Характер наследования того или иного признака выясняется путем анализа семей, в которых проявляются два или более вариантов этого признака. Удобная форма сводки данных о проявлении того или иного признака в семьях — это так называемые родословные (рис. 6.2). Существует несколько способов представления родословных; пример, приведенный на рис. 6.1, позволяет понять, по какому принципу они строятся.
Доминантное наследование
Признак наличия так называемых, «шерстистых (woolly) волос», наследование которого представлено в родословной на рис. 6.2, не играет существенной роли в биологии индивидуума; несколько факторов, по-видимому, обусловили его крайне редкую встречаемость в любой популяции. Однако в семье, родословная которой представлена на рис. 6.2, этот признак встречается очень часто. Такая, ситуация типична для многих наследуемых состояний, имеющих значительно более важное значение с точки зрения медицины, но в этих случаях далеко не всегда удается получить столь же полное ретроспективное описание семей.
Анализ родословной показывает, что во всех случаях у одного из родителей пробанда обнаруживался данный признак, который поэтому никогда не «перескакивает» через поколение. У двух родителей, лишенных данного признака, никогда не рождаются дети с шерстистыми волосами независимо от того, сколь близкими родственниками приходятся им «пораженные» индивидуумы. В 20 браках между пораженными и непораженными родилось 38 пораженных детей и 43 непораженных. Отношение этих двух чисел достоверно не отличается от 1. Наконец, можно заметить, что шерстистые волосы обнаруживаются у мужчин и женщин с примерно одинаковой частотой и что данный признак с равной ве-
плизкородапвенный у\ Идентичные ’—г-' Неидентичные
брак Q-Q близнецы | близнецы
Глава 6. Законы. Менделя и человек
Рис. G.2. Родословная, иллюстрирующая доминантное наследование у человека (Mohr, J. Hered., 1932).
Признак «шерстистые волосы» в одной норвежской семье.
140
Часть II. Генетика человека
роятностыо передается потомству матерью и отцом; эти факты свидетельствуют о том, что рассматриваемое свойство волос не принадлежит пп к «сцепленным с полом», ни к «ограниченным полом» признакам (см. гл. 7).
Отношение 1 : 1 в потомстве типично для возвратного скрещивания. Если бы шерстистые волосы были рецессивным признаком, то отношение 1 :1 в потомстве было бы возможно лишь при браке типа -\-wXww (где w — ген шерстистых волос, а «+» его «нормальный» аллель). Но если бы этот признак был рецессивным, то в данной родословной можно было бы себе представить браки типа + +Хшш и типа-(-шХ +ш. От браков первого типа не должны были бы рождаться дети с этим признаком, несмотря на то что у одного из родителей он имелся. В приведенной родословной мы находим только один случай отсутствия шерстистых волос у родителя ребенка, имеющего данный признак, причем отношение 1 : 1 при этом не нарушалось. При браке второго типа (Ц-шХ 25% детей должны были бы иметь данный признак, несмотря на отсутствие его у родителей. В родословной же у всех детей с шерстистыми волосами этот признак обнаружился по крайней мере у одного из родителей.
Теперь представим себе, что ген шерстистых волос проявляется у гетерозигот, т. е. в единичной дозе. Возвратное скрещивание в этом случае можно было бы представить так: И7-(-х++. При этом 50% детей окажутся носителями признака. У 50% признака не будет; в небольших семьях тот или иной тип может случайно отсутствовать. В типичном случае данный признак должен обязательно обнаруживаться хотя бы у одного из родителей пораженного ребенка, так как ген W, проявившийся у ребенка и полученный им от одного из родителей, должен был обязательно проявиться и у родителя. Или, иначе говоря, у двух родителей, лишенных данного признака, не может родиться ребенок с шерстистыми волосами, так как ни у одного из таких родителей не может быть гена W. Все сказанное полностью соответствует тому, что мы наблюдаем в приведенной родословной. Отсюда можно сделать вывод, что ген шерстистых волос проявляется у гетерозигот.
В медицинской генетике принято говорить, что шерстистые волосы — признак, доминантный по отношению к нешерстистым волосам, или, наоборот, нешерстистые волосы рецессивны по отношению к шерстистым. Строго говоря, это не точно. Прежде чем прийти к заключению о доминантности шерстистых волос, следует убедиться в том, что гетерозиготный индивидуум ТУ-}-неотличим от гомозиготного WW. Поскольку шерстистые волосы — это весьма редко встречающийся признак, брак между двумя соответствующими гетерозиготными индивидуумами представляется маловероятным, а ведь только при таком браке возможно
Глава 6. Законы Менделя и человек
141
возникновение гомозиготного индивидуума WW. В рассмотренной родословной браки между людьми с шерстистыми волосами отсутствуют; до сих пор такие браки вообще не описаны. Поэтому нет и сведений о том, как выглядят индивидуумы, гомозиготные по этому признаку. Возможно, что они неотличимы от гетерозиготных, но равновероятно и некоторое различие между ними. Тип наследования в данном случае может оказаться доминантным, подобным наследованию способности ощущать вкус фенилтиомочевины, но может быть промежуточным или подобным наследованию групп крови системы MN. Последнее не является строго промежуточным. У гетерозиготного индивидуума образуются .антигены обоих типов, так что эти признаки обычно называют кодоминантными,.
Анонихия, ахондроплазия, врожденная устойчивая куриная слепота, хорея Гентингтона и эллиптоцитоз — все эти признаки наследуются аналогично шерстистым волосам. Анонихия — это недоразвитие ногтей. Часто анонихия сопровождается деформацией кистей и стоп, например отсутствием пальцев (одного или более). Ахондроплазия — это форма карликовости, при которой голова и туловище достигают обычных размеров, а конечности сильно укорочены вследствие нарушения роста эпифизарных хрящей длинных трубчатых костей. Деформируется также и основание черепа, которое тоже формируется из хряща. При врожденной устойчивой куриной слепоте резко нарушено сумеречное (скотопическое) зрение. Этот дефект оказывается врожденным и с возрастом не прогрессирует. Иначе обстоит дело с хореей Гентингтона, которая представляет собой прогрессирующую дегенерацию элементов нервной системы, приводящую в конце концов к гибели пораженного. Обычно (хотя и не всегда) это заболевание впервые проявляется в возрасте 30—40 лет. При эллипто-цитозе (овалоцитозе) не менее 20, а иногда до 50% эритроцитов имеют форму эллипса; при этом отмечается нерезко выраженная анемия.
Из рассмотрения этих примеров видно, что заболевания, которые наследуются по простому менделевскому типу, встречаются во всех системах органов и неодинаковы по степени тяжести. Другие два патологических состояния — брахидактилия и множественная телеангиэктазия — интересны тем, что были описаны у лиц, по-видимому гомозиготных по аномальному гену. Брахидактилия — это укорочение пальцев на руках и ногах; описан случай, когда от брака пораженного мужчины с его двоюродной сестрой (вероятно, также пораженной тем же дефектом) родился ребенок с аномалией всего скелета, умерший вскоре после рождения. У супругов, страдавших множественной телеангиэктазией, родился ребенок, умерший в возрасте 2 1/2 месяцев при явлениях множественных наружных и внутренних кровотечений. Это позво
142
Часть II. Генетика человека
ляет предположить, что последние-два состояния не строго доминантны, причем у гомозигот болезнь протекает тяжелее, чем у гетерозигот. Но дети от брака между индивидуумами, страдавшими ахондроплазией, оказывались пораженными не более, чем их родители. Так же обстоит дело с таким состоянием, как врожденная катаракта. Поэтомуможнопредположить, что эти состояния наследуются-по доминантному типу, однако, поскольку количество пораженных мало, моглю-случиться-, что гомозиготное состояние никогда не реализовалось. Вовможно также, что гомозиготы по этим признакам настолько мало жизнеспособны, что соответствующая беременность прерывается в самом начале. Только статистический анализ достаточно большого числа случаев позволит решить, какое из этих предположений правильно.
У человека известно-более ста редких признаков, которые наследуются по простому доминантному типу или по меньшей мере проявляются в гетерозиготном состоянии. Вполне возможно, что проявление многих из- этих признаков генетически осложнено; возможно также, что существует много других признаков, наследуемых по такому же типу, которые по тем или иным причинам не были или не могли быть изучены генетически. К последней категории относятся, например, заболевания, летальное действие которых проявляется- до достижения детородного возраста и которые не могут, естественно, передаваться по наследству.
Рецессивное наследование
Рассматривая родословную семьи с признаком шерстистых волос, мы сосредоточили на нем внимание как на примере особенностей доминантного наследования. Как указывалось выше, признак нешерстистых волос, напротив, наследуется по рецессивному типу. Но здесь речь идет о нормальном типе. Для того же чтобы показать в сравнимых условиях особенности рецессивного наследования, мы рассмотрим другой редкий признак — альбинизм. Цвет кожи, волос и глаз- у людей весьма вариабелен не только в различных популяциях, но и в пределах одной и той же популяции. Эти варианты наследуются сложным образом. У отдельных лиц меланин не образуется или образуется в очень малых количествах, так что кожа и волосы оказываются совсем светлыми, а глаза — красными (пигмент не маскирует кровеносных сосудов сетчатки). Пример соответствующей родословной представлен на рис. 6.3. Эта родословная резко отличается от родословной семьи с шерстистыми волосами в первую очередь по частоте появления пораженных. Отметим также, что у детей пораженного индивидуума может вовсе не быть изучаемого признака, который, несмотря на это, появляется в последующем поколении. Иными словами, у родителей с нормальным развитием пигментации могут быть дети-
Глава 6. Законы Менделя и человек
143
альбиносы. Но в таких случаях в семье почти наверняка окажутся и непораженные дети: ведь подобные браки относятся к типу +а X 4*а, а в среднем лишь 25% детей должны быть альбиносами. Если семьи невелики, то вполне возможно, что от брака двух гетерозиготных индивидуумов не родятся дети-альбиносы и поэтому такая семья не будет включена в статистические данные. Вследствие этого нарушится ожидаемое соотношение (3 : 1). Подобного нарушения можно избежать; чаще всего с этой целью из анализа ис-
Рис. 6.3. Родословная, иллюстрирующая рецессивное наследование у человека.
Альбинизм (Pearson, Nettleship a. Hspcr, 1311 —1913).
ключается пробанд, т. е. пораженный индивидуум, впервые привлекший внимание к семье, -в которой имеется изучаемая аномалия.
Дети-альбиносы могут родиться и от браков типа ааХаа и ааХ-\-а. При первом из них все дети, очевидно, окажутся альбиносами; при втором типе (возвратное скрещивание) в среднем 50% детей будут альбиносами, а другие 50% — непораженными гетерозиготами по данному гену, т. е. «носителями». Самый частый тип брака с участием альбиноса — это (++Хаа); все дети от такого брака — носители. Почти наверняка это и имело место в поколении I (1 и 2), в поколении II все дети (1—12) гетерозиготны. Случилось так, что один из них женился на носительнице из другой семьи, и от этого брака родился ребенок-альбинос (III, 1). Некоторые из сестер и братьев этой девочки (или сокращенно ее «сиблинги», или «сибсы»), вероятно, были носителями, тогда как другие нет. Вероятность для одного из сибсов оказаться носителем характеризуется отношением 2:1, так как из 3/4 непораженных детей, родившихся от брака между индивидуумами, не состоящими в родстве (+ах+«), в среднем 2/3 будут гетерозиготными, а 1/3 — гомозиготной (++)• Представляется вероятным, что, подобно своей пораженной альбинизмом бабке, женщина-аль
144
Часть II. Генетика человека
бинос из поколения III вышла замуж за непораженного — ведь ни один из ее детей (поколение IV) не оказался альбиносом. Если бы их отец был носителем, то в среднем 1/2 детей оказалась бы альбиносами, причем все они были бы гетерозиготными по данному гену.
Примерами других состояний, наследуемых по рецессивному типу, могут служить алкаптонурия, фенилкетонурия, врожденный ихтиоз и глухонемота. Алкаптонурия и фенилкетонурия относятся к так называемым «врожденным ошибкам обмена», привлекшим к себе внимание биохимиков и генетиков. Для пораженных алкаптонурией характерно выделение с мочой алкаптона (гомо-гентизиновой кислоты). Вследствие окисления этого вещества моча таких детей на воздухе быстро темнеет. Это состояние часто распознают уже у новорожденного по характерным пятнам на пеленках. Гомогентизиновая кислота оказалась промежуточным продуктом распада двух аминокислот — фенилаланина и тирозина; в настоящее время считают, что у пораженных алкаптонурией отсутствует определенный фермент, катализирующий превращение гомогентизиновой кислоты в малеилацетоацетат (оксидаза гомо-гентизиповой кислоты). Подобная же ситуация отмечается у больных фенилкетонурией: отсутствие фенилаланингидроксилазы исключает возможность превращения фенилаланина в тирозин; поэтому фенилаланин накапливается в крови и превращается в ряд продуктов, в частности в фенилпировиноградную и фенилмолочную кислоты и в фенилацетилглутамин, выводимые с мочой. Однако в отличие от гомогентизиновой кислоты, которая не оказывает явного неблагоприятного влияния на организм, одно или несколько из этих веществ, присутствуя в необычно высокой концентрации, оказываются токсичными для ткани мозга; поэтому дети с фенилкетонурией отстают в умственном развитии. В последнее время это состояние стало объектом особого внимания; было показано, что если с самого раннего возраста свести до минимума количество фенилаланина, поступающего в организм ребенка, то можно избежать нарушения функций мозга. Поэтому для медицины весьма важно научиться распознавать носителей гена фенилкетонурии, с тем чтобы подвергнуть их детей соответствующему обследованию и в случае необходимости рекомендовать им подходящую диету. Эта цель в какой-то мере достигнута; показано,; что после приема с пищей стандартной дозы фенилаланина концентрация его в плазме у гетерозигот оказывается примерно вдвое выше, чем у лиц, не являющихся носителями соответствующего гена. Правда, пока удается выявить не все случаи гетероэигот-ности по данному гену.
Врожденный ихтиоз — заболевание чрезвычайно редкое; оно протекает столь тяжело, что чаще всего пораженный ребенок рождается мертвым или умирает сразу после рождения. При наиболее
Глава 6. Законы Менделя и человек
145
тяжелой форме ихтиоза вся поверхность кожи покрыта обширными ороговевшими пластинами с глубокими трещинами между ними; кожное дыхание оказывается невозможным. В отличие от ихтиоза генетически обусловленная глухонемота встречается довольно часто и не влечет за собой серьезной инвалидизации, по крайней мере в современном обществе. Это редкая ситуация для рецессивного признака, носитель которого лишь в исключительных случаях имеет плодовитость, достигающую */8 плодовитости непораженного. Глухота развивается вследствие поражения внутреннего уха, а отсутствие слуха с самого рождения влечет за собой немоту. С генетической точки зрения весьма интересен факт высокой частоты браков между глухонемыми.
Здесь уместно подчеркнуть, что даже состояние, представляющее собой определенную нозологическую единицу, не всегда определяется одним и тем же геном. Так, альбинизм, обычно наследуемый у человека как рецессивный признак, в некоторых семьях оказывается доминантным. Аналогичным образом обстоит дело и с такими состояниями, как алкаптонурия и глухонемота. Более того, частые браки между глухонемыми позволили установить, что даже рецессивная форма этого состояния определяется различными генами: в таких браках иногда рождаются здоровые дети.
Глава 7
ГЕНЕТИКА ПОЛА
Генетическое определение пола
Прежде чем приступить к рассмотрению механизмов наследования признаков, которые контролируются генами, локализованными в половых хромосомах, уместно рассмотреть способ генетического определения пола у человека. Генетическая основа пола зависит от хромосомного состава зиготы: если зигота содержит две Х-хромосомы, то формируется организм женского пола, если же в зиготе имеется одна Х-хромосома и одна Y-хромосома, то развивается мужской организм. Разумеется, как и другие генетически определяемые признаки, оба зти фенотипа окончательно реализуются лишь по истечении определенного периода развития и дифференцировки; в случае определения пола этот период оказывается весьма длительным, а соответствующий процесс — сложным. Вначале эмбрион анатомически нейтрален: гонады состоят из коркового и мозгового вещества, имеются закладки мюллерова и вольфова протоков, наружные половые органы неразличимы. Позднее, если эмбриону свойственна генетическая конституция XX, начинает пролиферировать корковое вещество гонад, мюллеровы протоки дифференцируются с образованием влагалища, матки и яйцеводов, а вольфовы протоки дегенерируют; формируются характерные для женского пола наружные половые органы. При генетической конституции XY сохраняется и дифференцируется только мозговое вещество гонад, из вольфовых протоков развивается сложная система протоков, по которым семя выводится наружу; мюллеровы протоки остаются в зачаточном состоянии; формируется половой член и мошонка, в которую спускаются яички. Наконец, в юношеском возрасте завершается созревание и избирательный рост половых желез и вторичных половых органов, специфических для каждого пола; различия касаются, в частности, формирования молочных желез, таза, развития волосяного покрова.
Результаты опытов по введению половых гормонов, кастрации и пересадке половых желез четко свидетельствуют о роли эндокринной системы в определении вторичных половых признаков у млекопитающих. Дифференцировка этих соматических тканей не детерминирована «безоговорочно» генетической конституцией особи. Однако один из моментов развития генотипа оказывается решающим; в этот момент^ по-видимому х определяется, какая часть
Глава 7. Генетика пола
147
развивающейся половой железы будет пролиферировать — корковое или мозговое вещество.
Для определения пола существенно, что в женском организме возникают яйцеклетки только одного типа — содержащие только Х-хромосому; в мужском же организме формируются сперматозоиды двух типов — содержащие Х-хромосому и содержащие Y-хромосому. Если яйцеклетку оплодотворяет сперматозоид, содержащий Х-хромосому, то родится девочка, если же оплодотворение осуществляет сперматозоид, содержащий Y-хромосому, то родится мальчик. Таким образом, у человека, как и у других млекопитающих, гетерогаметен мужской пол и в определенном смысле можно считать, что пол ребенка «зависит» от отца. Заметим, что у животных некоторых видов (птицы, чешуекрылые) ге-терогаметны женские особи, а мужские — гомогаметны.
Некогда считали, что у человека, как и у дрозофилы и у некоторых других видов, Y-хромоеома как таковая не имеет значения в детерминировании пола; решающим считали баланс между числом Х-хромосом и аутосом. У дрозофилы преобладающая часть генов (если не все гены), определяющих мужской пол, расположена в аутосомах, а гены, детерминирующие женский пол# находятся главным образом в Х-хромосоме. Если зигота содержит только одну Х-хромосому, преобладающим оказывается эффект генов, определяющих мужской пол; при наличии же двух Х-хромосом число генов, определяющих женский пол, оказывается вдвое больше и равновесие сдвигается в сторону женского типа. У дрозофилы в отсутствие Y-хромосомы и при наличии лишь одной Х-хромосомы формируется морфологически совершенно нормальная мужская особь. У человека же недавно удалось доказать роль Y-хромосомы в детерминировании пола. Описано состояние интерсексуальности, известное под названием синдрома Тернера: наружные половые органы и вторичные половые признаки потомства сформированы по женскому типу, но остаются недоразвитыми; половые железы недифференцированы. Число хромосом при синдроме Тернера равно 45. Соответствующая генетическая конституция — ХО; Y-хромосома отсутствует. Формирование организма при синдроме Тернера по женскому типу ясно свидетельствует о том, что Y-хромосома играет важную роль в детерминировании мужского пола. Правильность подобного взгляда подтверждается фактом существования у человека интерсексуальности иного типа — так называемого синдрома Клейнфельтера. Такие больные выглядят почти как нормальные мужчины, но у них отмечается азооспермия и малые размеры половых желез. У большей части этих больных обнаруживается 47 хромосом (вместо 46) и соответствующая генетическая конституция — XXY. Известны также случаи генетической конституции XXXYe XXXXY и даже XXXXXY. Следовательно^ Y-хромосома столь
148
Часть II. Генетика человека
эффективно детерминирует мужской пол, что даже при наличии двух и более Х-хромосом формируется почти нормальный мужской организм.
Интерсексуальность такого рода возникает вследствие нерас-хождения половых хромосом в мейозе. Так, если в женском организме не расходятся Х-хромосомы, то образуются яйцеклетки двоякого типа: не содержащие ни одной Х-хромосомы или содержащие их две. В мужском организме нерасхождение половых хромосом приводит к образованию сперматозоидов типа XY и 00. Здесь следует отметить, что известны случаи возникновения других аномальных по половым хромосомам генотипов, например XXX, ХХХХ, XXYY и XYY. Индивидуумы с аномалией XXX — морфологически нормальные женщины, способные к деторождению; правда, некоторые из них оказываются умственно отсталыми. Умственное развитие индивидуумов с более высоким числом Х-хромосом серьезно нарушается. При аномалии XXYY обнаруживаются признаки, характерные для синдрома Клейнфель-тера. Имеются некоторые основания для предположения (правильность которого далеко не доказана), что индивидуумы с XYY при их тенденции к необычно высокому росту обнаруживают предрасположенность к антиобщественному поведению.
Численное соотношение полов
Соотношение полов принято выражать числом мужчин, приходящихся на каждые 100 женщин в данной популяции. Соотношение полов в момент зачатия, при рождении и по достижении зрелости называют соответственно первичным, вторичным и третичным. При нормально протекающем мейозе образуется равное число сперматозоидов с X- и Y-хромосомой; можно считать поэтому, что зачатие девочки или мальчика равновероятно. Однако вторичное соотношение полов неизменно оказывается высоким: мальчиков рождается больше. Для белого населения Америки этот показатель равен 106, а для американских негров — 103, в Греции он достигает 113; даже на Кубе (одной из стран с минимальным вторичным соотношением полов) он все же составляет 101. В популяциях, обитающих на больших высотах над уровнем моря, соотношение полов оказывается особенно высоким. Можно предположить, что это преобладание мальчиков связано с более высокой смертностью эмбрионов женского пола. Однако в действительности наблюдается обратная картина: чаще гибнут плоды мужского, а не женского пола, по крайней мере на поздних сроках беременности. Нет оснований полагать, что на более ранних сроках развития дело обстоит иначе, так как на протяжении любого другого периода жизни мужские индивидуумы менее жизнеспособны, чем женские. Следует поэтому допустить, что первичное соотно
Глава 7. Генетика пола
149
шение полов еще более сдвинуто в сторону преобладания мужского пола. Объясняется это, по-видимому, естественным отбором, который направлен к тому, чтобы обеспечить создание равного количества особей обоего пола к моменту наступления половой зрелости.
Фишер подчеркивает, что воспроизведение потомства требует со стороны родителей затраты материала, энергии и времени. Эти затраты оказываются напрасными, если потомство не доживает до зрелости и в свою очередь не производит потомства; биологическую ценность этих затрат можно охарактеризовать числом потомков, которые сами стали родителями. За период, когда родители расходуют энергию на рождение и воспитание детей, умирает больше мальчиков, чем девочек, так что затраты на каждого уже выращенного мальчика в среднем больше, а на каждого новорожденного мальчика — меньше, чем затраты на каждую девочку соответствующего возраста. Рассмотрим теперь моногамную популяцию, в которой первичное соотношение полов равно 100. Расходы, затраченные на мальчиков, умерших за тот период, когда дети требуют заботы со стороны родителей, оказываются непроизводительными; еще большими оказываются непроизводительные расходы на девочек, которые, достигнув зрелости, не смогут вступить в брак вследствие гибели части мальчиков. В таких условиях в преимущественном положении оказываются те родители, у которых рождается больше мальчиков, поскольку затраты родителей на сына, который станет отцом, в среднем меньше затрат на дочь, которая станет матерью. Иначе говоря, при равных расходах можно воспроизвести больше отцов, чем матерей. Таким образом, соотношение полов, при котором на 100 девочек приходится более 100. мальчиков, представляет собой селективное преимущество, и любой обеспечивающий это соотношение генетический механизм будет распространяться в популяции. Соотношение полов будет, таким образом, возрастать до тех пор, пока затраты родителей на воспроизведение будущих матерей не станут эквивалентными по биологической ценности затратам на воспроизведение будущих отцов. Это равновесие для белого населения Америки соответствует 106, т. е. уже упоминавшемуся вторичному соотношению полов. Это соотношение ниже для американских негров и для кубинцев, по-видимому, в связи с тем, что общая детская смертность здесь выше или по меньшей мере была выше, что вело к сглаживанию различий в смертности мальчиков и девочек.
Как именно обеспечивается отклонение первичного соотношения полов от 100, неизвестно. Вполне возможно, что сперматозоид, содержащий Y-хромосому, более подвижен, более жизнеспособен или обладает большей способностью оплодотворить яйцеклетку, чем сперматозоид, содержащий Х-хромосому. Известно,j что у некоторых млекопитающих существует определенная связь
150
Часть II. Генетика человека
между pH крови отца и соотношением полов: низкое значение pH благоприятствует рождению дочери. Все признаки такого рода могут, разумеется, находиться под генетическим контролем; действительно, известны гены, которые в природных условиях контролируют соотношение сперматозоидов обоих типов в популяциях. Так, у Drosophila pseudo-obscura имеется ген, элиминирующий почти все сперматозоиды с Y-хромосомами; такая ситуация довольно широко распространена в природе.
Наследование, сцепленное с полом
Кроме генов, определяющих пол, в половых хромосомах расположены и другие гены; признаки, контролируемые этими генами, связаны по характеру наследования с полом. Такие гены могут быть расположены в Х-хромосоме (в таком случае говорят об X-сцепленном наследовании) или в Y-хромосоме (Y-сцеиленное наследование). Первая форма сцепления встречается гораздо чаще второй, несомненно, потому, что Х-хромосома достаточно велика. Особенности Х-сцепленного наследования легко понять, если вспомнить, во-первых, что каждый мужчина получает Х-хромосому от матери и передает ее всем своим дочерям, ню ни одному из сыновей и, во-вторых, что каждая женщина получает по одной X-хромосоме от каждого из родителей и передает либо одну, либо другую из них своим сыновьям и дочерям.
Примером признака, сцепленного с иолом, может служить слепота на красный и зеленый цвет; этот признак контролируется геном, локализованным в Х-хромосоме (гл. 18). У женщиной рецессивен по отношению к нормальному цветовому ярению. Нарушение восприятия цвета развивается у женщин, только если они гомозиготны по гену цветовой слепоты. Термины «доминантный» и «рецессивный» в применении к этому признаку у мужчин в таком случае теряют смысл: ведь у мужчин только одна Х-хрюмосома и, следовательно, только один ген цветового зрения (гемизиготность). В табл. 7.1 приведены 6 возможных типов брака между женщинами трех возможных генотипов и мужчинами двух возможных генотипов. Ген нормального цветового (colour) зрения обозначен буквой С, соответствующий аллель цветовой слепоты — буквой с.
Следует обратить внимание на две особенности рецессивного Х-сцепленного наследования, которые обнаруживаются при рассмотрении таблицы. Во-первых, среди пораженных цветовой слепотой мужчины встречаются гораздо чаще, чем женщины; это не удивительно, так как соответствующий ген у мужчин находит свое выражение всегда, а у женщин данный патологический признак наблюдается лишь при наличии двух генов цветовой слепоты. Следовательно, женщинаг пораженная цветовой слепотой, может
Глава 7. Генетика пола
151
Таблица 7.1
Генотипы родителем Генотипы детей
матери ОТЦЫ сыновья дочери
сс сс Сс Сс сс сс Горизонтальной ч с с С с С с ертой отмечены сл с с ЛЛ £ — 1С-с 2 2- С с учаи проявления цветово сс Сс 1 1 -^СС-^Сс Z А 1 г 1 УСс -2 - Сс сс t слепоты.
родиться только от отца с тем же дефектом зрения, так как, чтобы быть гомозиготной по гену цветовой слепоты, женщина должна унаследовать от отца Х-хромосому с этим геном. От генотипа ее матери зависит, разовьется ли у нее в действительности цветовая слепота. Во-вторых, цветовое зрение мужчины зависит только от генотипа его матери, от которой он получает свою единственную Х-хромосому. Так, половина сыновей женщины-носительницы будет поражена цветовой слепотой независимо от генетической конституции ее супруга. Эта особенность весьма существенна с точки зрения медицины. Типичная особенность наследования рецессивного сцепленного с полом и сравнительно редкого признака такого типа, как цветовая слепота,— это чередование поколений пораженных цветовой слепотой мужчин и женщин-носительниц соответствующего гена; ясно, что если пораженный цветовой слепотой мужчина вступит в брак со здоровой гомозиготной женщиной, то все его дочери окажутся носителями гена цветовой слепоты, но ни один из его сыновей не будет поражен; половина его внуков будет поражена и унаследует цветовую слепоту от своих матерей.
Известен еще один дефект, который обычно наследуется как рецессивный признак, сцепленный с полом. Речь идет о гемофилии — одной из форм нарушений свертывания крови. Существуют 2 формы сцепленной с полом гемофилии: классический вариант — гемофилия А, возникающая вследствие недостаточности «антиге-мофильного глобулина», и встречающаяся реже гемофилия В — болезнь Кристмаса„при которой отсутствует в крови «плазменный
Королева Виктория
Лрини Альберт
С?----
ФридрихШ Германский
Ж И, Принцесса тании Алиса
&—
Леопольд
Принцесса Беатриса
Королевские Королевские дома дома
Германии Великобритании
Королевские дома Греции и Югославии
Королевские дома Швеции и Дании
Генрих Прусский
Принцесса Ирена
Принц Фредерик
Николай 1Г.
Россия
Принцесса Алиса
АльфонсоЛШ Испанский
$ <
Королева Леопольд Морис Эна Маунтбзттен Маунтбзттен
Принц Принц
Вальдемар Генрих
ф $ ф ф г
Царевич Алексей
Виконт
Трематон
Герцог Астурийский
Принц Гонзоло
Больные гемофилией © Женщины* мужчины т носители
© Генотип точно неизвестен
Рис. 7.1. Родословная потомков королевы Виктории. Гемофилия.
Глава 7. Генетика пила
153
компонент тромбопластина». По сравнению с цветовой слепотой гемофилия — более серьезный патологический признак; поэтому он встречается в определенной популяции значительно реже — пораженным оказывается примерно 1 мужчина из 10 000. Случаев гемофилии у женщин не описано; одна из причин этого кроется в том, что возникновения гемофилии у женщины следует ожидать с частотой (1/10 000)2, т. е. 1/100 000 000. Причины такого различия между частотами поражения мужчин и женщин станут ясными, если рассмотреть поведение соответствующих генов в популяции.
Гемофилия получила широкую известность в связи с тем, что это заболевание наблюдалось у нескольких членов двух царствующих домов Европы: ген гемофилии был унаследован ими от королевы Виктории — носительницы данного гена (рис. 7.1).
Если какой-либо признак, сцепленный с Х-хромосомой, проявляется в гетерозиготном состоянии, то в отличие от рецессивного признака .он будет чаще встречаться у женщин, чем у мужчин, поскольку 2/3 Х-хромосом в каждой данной популяции принадлежат женщинам. Таких патологических признаков известно немного, и все они встречаются редко. К ним относится коричневая эмаль зубов и одна из форм фосфатемии, проявляющаяся симптомами рахита, не поддающимися терапии витамином D. Поскольку речь идет о редко встречающемся признаке, то практически возможны два типа брака: между гетерозиготной женщиной и непораженным мужчиной и между пораженным мужчиной и непораженной женщиной. Гетерозиготная женщина передает свою Х-хромосому с соответствующим патологическим геном поровну своим сыновьям и дочерям; поэтому половина дочерей и половина сыновей от такого брака оказываются пораженными, а половина — здоровыми. Иначе говоря, данная аномалия наследуется при браках такого типа подобно аутосомным доминантным признакам. Однако при браках второго типа пораженный мужчина передает свою Х-хромосому только дочерям и все дочери оказываются пораженными, а сыновья нет.
Полное сцепление с Y-хромосомой выявляется довольно легко. Y-хромосома имеется только у мужчин, и поэтому те признаки, которые контролируются расположенными в ней генами, обнаруживаются только у мужчин. Кроме того, патологический признак, имеющийся у отца, должен передаваться всем его сыновьям, поскольку все они получают от него Y-хромосому. Несмотря на легкость распознавания такого сцепления, пока еще однозначно не доказано, что и в норме в Y-хромосоме человека имеются гены, контролирующие какие-либо иные признаки, кроме пола. Одним из наиболее наглядных примеров признака, сцепленного с Y-хромосомой, считается наличие пучка длинных жестких волос на мочке уха. Особенно часто этот признак встречается в семьях индейцев, но описан и в других популяциях; он наблюдается толь
154
Часть II. Генетика человека
ко у мужчин, по крайней мере в некоторых родословных этот признак отмечался у всех сыновей имеющих его отцов Правда, в других семьях наследование этого признака происходит менее однозначно, по-видимому, вследствие того, что соответствующий ген не всегда получает выражение в фенотипе. Однако результаты анализа единичных семей следует интерпретировать с осторожностью, ибо всегда есть опасность ошибочно истолковать такого рода наблюдения. Ведь не исключено, например, что в упомянутой выше семье сцепление данного признака с Y-хромосомой представляет собой аномалию, возникшую вследствие того, что фрагмент какой-то другой хромосомы оказался транслоцированным на Y-хромосому.
В настоящее время накопилось много фактов, свидетельствующих в пользу гипотезы, впервые выдвинутой Мэри Лайон. Согласно этой гипотезе, в женском организме из двух Х-хромосом, находящихся в каждой клетке тела, метаболическую функцию несет лишь одна. Инактивация другой Х-хромосомы происходит на ранних стадиях эмбриогенеза; в соматической клетке Х-хромосома остается инактивированная не только в течение жизни этой клетки, но и в дочерних клетках, возникающих в результате ее деления. Тем не менее в момент инактивации чисто случайным образом определяется, какая из Х-хромосом инактивируется; благодаря этому приблизительно в половине клеток регуляция осуществляется одной Х-хромосомой, а в другой половине клеток — другой Х-хромосомой. Таким образом, все ткани женщины, гетерозиготной по сцепленному с полом признаку (за исключением яичников), образованы мозаикой из клеток двух типов: клетки одного типа представляют собой продукт одного из генов, а клетки другого типа — продукт аллеломорфа. Такая гетерогенность клеточных продуктов была обнаружена, например, для вариантов глюкозо-6-фосфатдегидрогеназы (Г6ФД), детерминация которых сцеплена с полом.
В неделящихся клетках инактивированную Х-хромосому часто удается наблюдать в виде темноокрашенного хроматинового тельца вблизи ядерной мембраны. Это тельце называют тельцем Барра, по имени открывшего его исследователя; про содержащую это тельце клетку говорят как о «хроматин-положительной». Наличие или отсутствие тельца Барра характеризует набор Х-хромосом, а следовательно, и пол индивидуума; при этом отпадает необходимость в анализе кариотипа делящихся клеток. В случаях многих аномалий, затрагивающих Х-хромосому, число телец Барра в клетке, которое всегда на единицу меньше числа Х-хромосом, позволяет определить природу этих аномалий. Так, типичный индивидуум с синдромом Тернера (набор ХО) оказывается аналогичным хроматин-отрицательному мужчине. При синдроме Клейн-фельтера XXYY имеется одно тельце Барра, тогда как при XXX их
Глава 7. Генетика пола
155
два, а при ХХХХ — 3. Среди населения Великобритании около 0,2% мужчин и 0,08% женщин оказываются носителями аномального числа телец Барра.
Наследование, ограниченное полом
Из того, что некий признак встречается чаще у представителей того или иного иола, еще не следует, что наследование этого признака сцеплено с полом. Наклонность к преждевременному облысению — это доминантный признак, встречающийся только у мужчин; синдром Лоуренса — Муна — Бидла и одна из форм олигофрении — это рецессивные признаки, которые обнаруживаются чаще у мужчин, чем у женщин. Однако соответствующие этим признакам гены локализованы в аутосомах, что следует из характера их передачи. Неодинаковое же распределение этого признака между представителями разных полов обусловлено тем, что женский генотип создает среду, которая подавляет или ослабляет выражение этих генов. Обычно говорят, что наследование таких признаков ограничивается или регулируется полом.
Глава 8
НЕЗАВИСИМОЕ РАСПРЕДЕЛЕНИЕ И СЦЕПЛЕНИЕ
Независимое распределение
Нами был рассмотрен принцип наследования единичных признаков, теперь можно обсудить наследование двух признаков, передающихся совместно. Согласно второму закону Менделя, признаки наследуются независимо друг от друга. На уровне генов это означает, что при одновременном расщеплении двух или более пар генов аллели каждой пары распределяются по гаметам независимо от других пар.
Это можно проиллюстрировать на примере образования гамет у индивидуумов, гетерозиготных по гену ощущения вкуса фенилтиомочевины (tester) и гену секреции (secretor) группоспецифических антигенов (генотип TtSese'). При независимом распределении у таких индивидуумов должны с равной частотой возникать 4 типа гамет: TSe, tSe, Tse и tse. Другими словами, ген Т так же часто оказывается связанным с геном Se, как и ген t, и оба они с той же частотой могут быть связаны с геном se. В случае брака двойной гетерозиготы с двойной рецессивной гомозиготой возможны зиготы четырех типов, причем, как видно из схемы, частоты появления каждого типа одинаковы (если семья достаточно велика, для того чтобы ошибка выборочного метода не играла существенной роли).
TtSese (ощущающий вкус, секретор)
Соотношение гамет */tTSe «/4<Se »/4Tse */4««е
ttsese (не tse 1/iTtSese Соотношение зигот 1/ittSese (не ^iTtsese 1/ittsese (не
ощущающий вкуса, несек-ретор) (ощущающий вкус, секретор) ощущающий вкуса, секретор) (ощущающий вкус, несекре-тор) ощущающий вкуса, несек-ретор)
При сочетании двойных гетерозигот, каждая из которых с равной частотой образует 4 типа гамет, в потомстве возможны те же 4 типа детей, причем частота их будет соответствовать хорошо известному менделевскому соотношению: 9 (ощущающие вкус
Глава 8. Независимое распределение и сцепление
157
фенилтиомочевины, секреторы): 3 (не ощущающие вкуса, секретеры): 3 (ощущающие вкус, несекреторы): 1 (не ощущающие вкуса, несекреторы). Это соотношение определяется доминантностью обоих признаков, благодаря чему четыре разных генотипа, TTSeSe, TtSeSe, TTSese и TtSese, определяют один фенотип (ощущение вкуса, секреция); генотипы TTsese и Ttsese — другой фенотип (ощущение вкуса, отсутствие секреции); генотипы ttSese и ttSeSe — третий фенотип (отсутствие ощущения вкуса, секреция) и лишь один генотип ttsese— один фенотип (отсутствие того и другого признака). Если же гетерозиготы по обоим признакам отличаются от соответствующих гомозигот, то все девять получающихся генотипов должны быть фенотипически различимыми.
Соотношение 1 : 1 : 1 : 1, наблюдаемое при возвратном скрещивании с двойным рецессивом,— результат попарного перемножения членов двух отношений 1:1, получаемых для каждого единичного признака, а соотношение 9 : 3 : 3 : 1 есть результат попарного перемножения членов отношения 3:1. Если имеются три доминантных признака, то после скрещивания они дадут фенотипы с частотой (9 : 3 : 3 : 1)Х(3 : 1)=27 : 9 : 9 : 9 : 3 : 3 : 3 : 1. Таким образом, можно предсказать соотношение для любого числа признаков, рассматриваемых в совокупности.
Независимое распределение признаков объясняется тем, что контролирующие их гены локализуются в различных хромосомах. Распределение членов одной пары хромосом по гаметам в большинстве случаев происходит независимо от распределения членов другой пары. Так, ген, контролирующий способность ощущать вкус, и ген, определяющий способность к секреции, расположены в различных парах хромосом. Следовательно, вероятность встречи хромосомы, несущей ген Se, с хромосомой, несущей ген Т или t, определяется исключительно законом случая. То же справедливо и для гена se.
Сцепление
Если первый закон Менделя универсален, то во втором законе постоянно встречаются исключения. Причина этого станет сразу ясной, если вспомнить, что каждая хромосома несет множество генов и что гены, локализованные в одной хромосоме, не могут распределяться независимо; будучи связаны в единую генетическую структуру, такие гены имеют тенденцию наследоваться совместно. Рассмотрим для иллюстрации гипотетический пример скрещивания ААВВ Xaabb, в котором два рассматриваемых локуса расположены в одной хромосоме. Если между гомологичными хромосомами не происходит обмена участками, то гибриды первого поколения, получаемые при таком скрещивании (АаВЬ), способны производить только два типа гамет (АВ и ab) вместо четырех, ожи
158
Часть II. Генетика человека
даемых при независимом распределении, поскольку попадающая в гамету хромосома с геном А обязательно несет ген В, а хромосома, несущая ген а„ всегда имеет и ген Ъ.
Родители
Pi Р2
АВ а Ъ
X ~ —
АВ а Ь
Гаметы
А В а Ъ
Fj-гибрид
А В
а Ь
Гаметы
1 А В.
2 +
1
2 а Ь
При скрещивании родительских типов ААЬЪУааВВ гибриды первого поколения будут генотипически АаВЬ, как и в первом
А Ъ
примере, но поскольку гены в них расположены по схеме ==, то а В
они будут образовывать гаметы АЪ и аВ. Гены, локализованные в одной хромосоме, называются сцепленными. Если, как в первом случае, оба доминантных гена расположены в одной хромосоме, то говорят, что они находятся в «фазе притяжения», или в цис-фазе; если же, как во втором случае, доминантные гены находятся в разных хромосомах данной пары, то говорят, что они находятся в «фазе отталкивания», или в транс-фазе. Ситуация осложняется тем, что сцепление редко бывает полным. В большинстве организмов происходит обмен материалам между гомологичными хромосомами; этот обмен происходит во время профазы первого деления мейоза, что можно наблюдать под микроскопом по образованию хиазм между парами хроматид. Следовательно, каждая конкретная хромосома при передаче из поколения в поколение не остается идентичной. Вследствие перекреста (кроссинговера) двойная гетерозигота может образовывать все четыре возможных типа гамет вместо двух родительских типов, даже когда гены оказываются в одной хромосоме. В упрощенном виде это представляет схема на стр. 159. Каждый из четырех получившихся продуктов расщепления входит в гаметы, но сочетается неодинаково: гаметы АЬ и аВ называются рекомбинантными, так как они образуются вслед-
Глава. 8. Независимое распределение и сцепление
159
ствие рекомбинации хроматид, а нантными: А В гаметы А АВ В и ab — А нерекомби- в
А в- А в—* - А В А ъ
а ь а ь а X ь и в
а ъ а ь а ь
Хотя кроссинговер привадит к образованию тех четырех типов гамет, сцепление между веками можно нее же выявить, так как рекомбинантные гаметы образуются обычно в меньшем числе, чем нерекомбинантные. Во-внрвых, для того чтобы возникли рекомбинанты, кроссинговер должен .обязательно иривгаойти между двумя сцепленными генами. В приведенной выше схеме локусы А и В расположены в противоположных концах хромосомы, однако если бы локус В оказался ближе к локусу ^4, чем точка мере-креста, то образование соотнеижвующих реком^вныйнтных гамет было бы невозможно:
А В В В
А В ____________, Л В
a bx а Ь
Во-вторых, между хроматидами мажет ироиаайти больше одного кроссинговера. Если локусы .отстоят досшатзадво далеко друг от друга, то второй кроссинговер может воостамолит* иервомачаль-ное сцепление генов, разделенных в результате л^рвого кроссинговера:
А В А В
А Д Л -В
а X X Ъ а Ь
а Ь а а
Нечетное число кроссинговеров между двумя генами, несомненно разъединяет их, а четное число кроссинговеров сохраняет их первоначальное сцепление. Сцепление двух генов, расположенных столь далеко друг от друга, что их распредел>ение происходит практически независимо, может быть обнаружено только с по
160
Часть II. Генетика человека
мощью промежуточных генов, сцепленных с обоими исследуемыми генами.
Появляется все больше данных, согласно которым у женщин кроссинговер происходит гораздо чаще, чем у мужчин, однако при стандартных условиях частота кроссинговера между двумя сцепленными генами оказывается постоянной. Оставив без обсуждения некоторые оговорки, можно сказать, что кроссинговер возможен с равной частотой в любой точке вдоль хроматиды; следовательно, частота, с которой два сцепленных гена расходятся в результате кроссинговера, является, как правило, функцией расстояния между этими генами вдоль хромосомы. Чем ближе друг к другу расположены гены, тем реже между ними происходит кроссинговер и, следовательно, тем реже возникают рекомбинантные типы гамет. На этом основано построение хромосомных карт, которые уже составлены для некоторых видов животных и растений.
Гены, расположенные в хромосоме близко друг к другу и поэтому редко разделяемые кроссинговером, называются тесно сцепленными. Гены, расположенные далеко друг от друга, так что их распределение происходит почти независимо, называются слабо сцепленными.
В генетике растений и животных простейшим способом выявления сцепления и определения частоты кроссинговера между двумя генами является возвратное скрещивание двойной гетерозиготы с двойной рецессивной гомозиготой. При независимом распределении, как уже упоминалось, все четыре возможных фенотипически различных генотипа возникают о равной частотой. При наличии сцепления соответствующих генов два фенотипа, аналогичные фенотипам дважды гетерозиготных родителей, встречаются в потомстве чаще, чем два фенотипа, отличные от родительских (сами родительские фенотипы встречаются в потомстве с равной частотой). Частота фенотипов, отличных от родительских, отражает частоту рекомбинаций и, следовательно, частоту кроссинговера.
У человека выявить сцепление генов труднее пз-за невозможности произвольных скрещиваний. Нельзя получить информацию о сцеплении и путем исследования корреляции между признаками в той или иной популяции, так как из-за кроссинговера гены одинаково часто будут находиться в фазе сцепления и в фазе отталкивания (исключение составляют лишь тесно сцепленные гены, не разделяемые кроссинговером). Так, в той или иной популяции отношение числа ощущающих вкус фенилтиомочевины секретеров к числу не ощущающих ее вкуса секретеров будет одинаковым независимо от того, сцеплены или не сцеплены соответствующие гены. Вместе с тем часто бывает известна генетическая конституция родителей в семьях, анализируемых на сцепление, так что
Глава, 8. Независимое распределение и сцепление
161
анализ потомства может быть использован для определения сцепления у человека. Трудность заключается в том, что у людей семьи, как правило, немногочисленны, и приходится комбинировать данные о многих семьях. Даже в случае подходящих браков необходимы данные о нескольких поколениях, а их часто не хватает. Последнюю трудность можно до некоторой степени обойти, применив непрямые методы, в частности наиболее простой из них — метод пар сибсов, предложенный Пенроузом; этот метод позволяет ограничиться данными об одном поколении.
Рассмотрим еще раз брак типа AaBbXaabb, в котором имеет место сцепление двух доминантных генов. Генотипы двойных гетерозигот могут быть ABlab или АЫаВ. В первом случае в отсутствие кроссинговера в потомстве встретятся либо ABlab, либо ablab. Назовем признак, определяемый аллелями А и а, первым, а признак, контролируемый аллелями В и Ь, вторым. Дети с генотипом АВ/ab будут сходны между собой по первому и по второму признакам, так же как и дети с генотипом aWab. В то же время дети с генотипом АВ/ab будут отличаться по обоим признакам от детей с генотипом ablab. Аналогичным образом обстоит дело и во втором случае — у родителей АЫаВ. Дети с генотипом АЫаЬ или аВ/ /аЪ будут либо одинаковы по обоим признакам, если будут иметь один и тот же генотип, либо при разных генотипах будут различаться по обоим признакам. Стало быть, в обоих случаях сходство сибсов по одному признаку при различии их по второму может возникнуть только в результате кроссинговера. Рекомбинантные потомки при браке типа ABlabXablab будут иметь генотип либо АЫаЬ, либо aB/ab. Эти два рекомбинанта различаются по обоим признакам, но от нерекомбинантного потомка ABlab рекомбинант АВ/аЬ отличается лишь по второму признаку, оставаясь сходным с ним по первому, а от аЫаЪ отличается по первому признаку, сохраняя сходство по второму.
Таким образом, ясно, что, сравнивая по отдельным признакам сибсов в потомстве от брака с двойным гомозиготным рецессивом, можно установить, что при наличии сцепления соответствующих генов частота случаев сходства или несходства сибсов сразу по обоим признакам будет выше, чем частота случаев сходства по одному признаку и различия по другому, поскольку последние случаи возникают лишь в результате кроссинговера. Это справедливо как для браков типа ABlabXablab, так и для браков тина ABIaBXablab, так что можно комбинировать результаты обследования различных семейств.
Другие типы браков приводят к тем же результатам, что и при браке двойной гетерозиготы с двойным рецессивом. Для выявления сцепления, очевидно, необходимо, чтобы хотя бы один из родителей был гетерозиготен по обоим признакам. Это единственный генотип, при котором кроссинговер приводит к образованию
162
Часть II. Генетика человека
гамет, отличающихся от нерекомбинантных. В том случае, когда один аллель в каждом из рассматриваемых локусов относится к категории редких (как это часто бывает в действительности), двойные рецессивы также будут редки. Если, например, гены а и В часто встречаются в популяции, а их аллели А иЬ — редко, то сибсы с фенотипически выявляемым расхождением обеих пар аллелей могут возникать, как правило, в результате брака типа Аа/ВЬ Хаа/ВЬ. В этом случае опять получается, что при наличии сцепления отношение числа пар сибсов, различающихся или сходных сразу по обоим признакам, к числу пар сибсов, различающихся только по одному признаку, будет больше, чем можно ожидать, исходя из независимого распределения, хотя в этом случае возникает некоторое число нерекомбинантных потомков, отличающихся от других нерекомбинантных потомков по одному признаку, но сходных по другому. Если мы построим схему всех возможных сравнений сибсов, то при наличии сцепления в клетки 1 и 4 попадет большее число случаев, чем в клетки 2 и 3.
Первый признак
СХОДСТВО различие
Второй Сходство 1 2
признак Различие 3 4
Степень отклонения чисел в клетках этой схемы от ожидаемых в случае независимого распределения можно использовать для вычисления частоты кроссинговера. При независимом распределении (исключая случайные различия) можно ожидать, что сибсы, сходные по первому признаку, будут различны или сходны по второму с той же частотоГй, с какой сходство и различие по этому признаку встречается в общей популяции. Аналогично у сибсов, различающихся по первому признаку, сходство и различие по второму признаку будет встречаться с той же частотой. Возвращаясь к числам в клетках схемы, можно сказать, что при независимом распределении отношение чисел в клетках 1 : 2 будет равно (1+3) : (2+4), а отношение 3 : 4 также будет равно (1+3): (2+4). Ожидаемые значения могут быть также получены из 1 : 3=(1 + +2) : (3+4) и 2 : 4=(1+2) : (3+4).
Этот способ выявления сцепления не очень эффективен, он был усовершенствован Пенроузом. Вместе с тем метод пар сибсов указывает на характер проблемы и пути ее решения.
Применение другого метода, известного под названием «метода коэффициента сходства», можно в общем виде проиллюстрировать
Глава 8. Независимое распределение и сцепление 163
приведенным ниже примером. Рассмотрим простую родословную:
Ла ВЬ аа ЪЪ
Ла ВЬ аа ЪЪ
У отца с двойной гетерозиготностью оба локуса, если они сцеплены, могут находиться в фазе притяжения (АВ lab) или в фазе отталкивания (Ab/аВ). Ясно, что первый сын получил от отца аллели АВ, а второй ab. Иными словами, если гены находились в фазе притяжения, ни один из сыновей не получил соответствующего рекомбинантного набора от своего отца; если же они находились в фазе отталкивания, то оба сына произошли из рекомбинантных гамет. Пусть х — показатель рекомбинации. Вероятность рекомбинантного генотипа у первого сына равна г/2х и у второго сына также 1/2х. Таким образом, общая вероятность получения такого потомства в этой родословной, в которой гены отца находятся в фазе отталкивания, равна (1/2я;)2. Вместе с тем, если гены находятся в фазе притяжения и не произошло рекомбинации, вероятность получения данного генотипа у первого сына равна 1/г(1 — х) и второго V2(l—х), так что общая вероятность равна [1/2(1 — 2)]2. Поэтому общая вероятность получения результатов, представленных в этой родословной, в фазе отталкивания или притяжения, равна
Если принять а:=0,5, что имеет место при независимом распределении, вероятность равна 1/1в; если 2=0 при полном сцеплении, вероятность равна 1/1в. Иными словами, вдвое вероятнее, что описанный результат соответствует полному сцеплению, а не независимому распределению. Такой подход применим и для анализа более обширных родословных с выводом соответствующих формул; подставляя различные значения х от 0,5 до 0, можно определить, какое из них наиболее соответствует анализируемой родословной и, следовательно, характеризует степень сцепления.
Несмотря на существование в настоящее время этого и других изящных методов, число известных примеров сцепления аутосомных генов у человека невелико, хотя и растет. Весьма тесно сцепление в пределах ряда серологических систем — например систем групп крови MNS и Rh, вариантов иммуноглобулинов Gm, системы тканевых антигенов HL-A, генов, определяющих р- и S-цепи полипептидов в молекуле гемоглобина; рекомбинации внутри этих групп редки. В некоторых семьях отмечено сцепле
164
Часть II. Генетика человека
ние эллиптоцитоза с системой Rh (частота кроссинговера около 3%), тогда как в других семьях эти два признака наследуются независимо. Следовательно, гены, по крайней мере в двух локусах, могут вызывать эллиптоцитоз. Синдром дефекта ногтей и надколенника — патологическое состояние, нри котором наряду с прочими отклонениями имеет место дистрофия ногтей, недоразвитие надколенника и появление «рожек» на подвздошных костях — сцеплен с системой групп крови АВО (частота кроссинговера около 10%). Локус, определяющий систему групп крови Lutheran, сцеплен с локусом, контролирующим секрецию группоспецифических веществ системы АВО, с частотой кроссинговера около 15%.
Система трансферрина сцеплена с вариантами сывороточной холинэстеразы с частотой кроссинговера 16%. Большинство случаев аутосомного сцепления, обнаруженных к настоящему времени, затрагивает лишь 2 локуса; однако есть основания считать, что локус «деспирализации хромосом» сцеплен с системой групп крови Duffy, а последняя весьма тесно сцеплена с одним из типов врожденной катаракты.
Известно несколько других случаев аутосомного сцепления благодаря открытию четких флуоресцирующих полос на окрашенных соответствующим образом хромосомах; вероятно, скоро будут обнаружены многие подобные случаи. При этом появится возможность установить, в какой именно хромосоме расположена группа сцепления. Главным образом благодаря анализу вариантов аномальных хромосом уже удалось отнести некоторые гены к соответствующим хромосомам. Локус «деспирализации», с которым связан дефект спирализации части плеча хромосомы в цитогенетических препаратах, локализован в хромосоме 1; из формального анализа сцепления следует, что гены системы групп крови Duffy и врожденной катаракты также локализованы в этой хромосоме. Изучение ряда транслокаций показало, что локус, контролирующий а-полипептид молекулы гаптоглобина, расположен в хромосоме 16; опыты с культивируемыми гибридными клетками человека и мыши показали, что у человека ген, контролирующий синтез тимидинкиназы, находится в хромосоме 17.
Установление сцепления между генами Х-хромосомы теоретически представляется более простым, чем выявление сцепления в аутосомах, поскольку сцепление с полом, несомненно, свидетельствует о локализации генов в данной хромосоме. Кроме того, у мужчин невозможно участие этих генов в кроссинговере. К сожалению, кроме сравнительно недавно обнаруженного локуса недостаточности глюкозо-6-фосфатдегидрогеназы, часто встречающейся у жителей Африки и Средиземноморья, и локусов системы групп крови Xg, в этой хромосоме отсутствуют локусы, несущие аллели обычных генов (гены-маркеры), и, следовательно, отсутствуют те исходные точки, от которых ведется отсчет при определении
Глава 8. Независимое распределение и сцепление
165
частоты кроссинговера. Тем не менее теперь можно попытаться составить предварительную карту взаимного расположения генов в некоторых частях Х-хромосомы.
Xg 1с Оа
11
_.J_______I___L_
18
G6PD
Xm Cd Ср На
7 12
Обозначения генов: Cd—дейтеранопия, Ср—протанопия, G6PD —глюкозоб-фосфатдегидрогеназа, На—гемофилия А, 1с—ихтиоз, Оа — альбинизм (глазной), Xm —один из белковых антигенов сыворотки.
Следует отметить, что эта карта не является окончательной; с суммой современных знаний совместимы и другие варианты взаимного расположения генов. Подчеркнем также, что анализа родословных было бы недостаточно для того, чтобы установить, что локусы Xg и G6PD находятся в одной хромосоме, не окажись они сцепленными с полом.
Глава 9
ГЕНЫ И ИХ ДЕЙСТВИЕ
Действие генов
То, что мы рассматривали до сих пор, можно назвать цитологическими механизмами наследственности; о том, каким именно образом гены определяют возникновение того или иного признака, говорилось мало. Лишь при обсуждении развития пола было отмечено, что установление определенного генотипа и его фенотипическое проявление разделены длительным процессом дифференцировки. Действительно, очень редко удается доказать прямую связь между наличием какого-то определенного гена и развитием соответствующего конкретного признака. По большей части признак возникает в результате взаимодействия нескольких генов, а один ген проявляет свое действие в нескольких признаках. Примеры такого плейотропизма уже приводились. Так, было показано, что при фенилкетонурии симптом экскреции фенилпировиноградной кислоты сочетается с умственной отсталостью. Выражение того или иного гена может быть еще более разнообразным. Например, при синдроме Лоуренса — Муна — Бидла наличие одного гена в двойной дозе проявляется в тучности, умственной отсталости, полидактилии и недоразвитии половых органов (гипогенитализм). При арахнодактилии — синдроме удлинения пальцев рук и ног — наряду с основными имеются и другие аномалии скелета, а также аномалии глаз и сердца. Морфогенетическая связь этих множественных аномалий с изменением одного гена не во всех случаях прослежена полностью. Однако все признаки гипогенитализма при синдроме Лоуренса — Муна — Бидла определяются одним и тем же фактором — недостаточностью гонадотропинов, а вместе с тем известно, что и другие факторы, в том числе поражение гипоталамуса, приводящее к тучности, уменьшают секрецию этих гормонов. У крыс встречается наследуемый по Менделю синдром поражения почти всей костной системы и внутренних органов; было показано, что вся совокупность патологических признаков связана с нарушением образования хрящевой ткани. Подобные ситуации показывают, что чем ближе мы подходим к непосредственному действию гена, тем сильнее сужается класс определяемых этим геном продуктов, так что есть основания полагать, что каждый ген контролирует синтез только одного белка, чаще фермента. Это гипотеза «один ген — один белок»; теперь ее предпочитают формулировать как гипотезу «один ген —
Глава 9. Гены и их действие
167
один полипептид», но и в таком виде она иногда нуждается в дополнительных пояснениях, например в случае синтеза молекул иммуноглобулинов у млекопитающих; тем не менее известно много фактов в ее пользу, полученных главным образом при изучении обмена у бактерий и грибов. У человека единственное непосредственное действие гена фенилкетонурии — это недостаточность или отсутствие фенилаланин — гидроксилазы. Все другие симптомы фенилкетонурии можно объяснить, исходя из этого изменения. Это не означает, что ген фенилкетонурии в отличие от своего аллеля дикого типа не образует никакого продукта. Просто продукт действия этого гена не обладает способностью превращать фенилаланин в тирозин. Такой эффект может быть вызван совершенно незначительным изменением в химической структуре фермента. Теперь легко понять, почему взаимозамещение аллелей, контролирующих группы крови, оказывает такое локальное действие. По-видимому, группоснецифические вещества по своему происхождению очень близко стоят к непосредственным продуктам действия генов, и, кроме того, эаи вещества не оказывают существенного влияния на формирование других признаков.
Поскольку первичным действием каждого гена является синтез определенного полипептида, то легко понять, что большинство признаков зависит от совместного действия многих генов. Развитие любой морфологической системы определяется огромным количеством биохимических реакций, и изменение одной из этих реакций способно в той или иной степени изменить весь курс морфогенеза. Так, например, функция зрения определяется не единичным геном, а целой группой генов, продукты действия которых сложным образом взаимодействуют между собой в течение всей жизни индивидуума и обеспечивают развитие и поддержание функций глаз и мозга. Нарушение целостности этой системы под действием генетических факторов или факторов внешней среды может привести к слепоте. Вот почему так много генов могут обусловить слепоту. Как с генетической, так и с клинической точки зрения слепота представляет собой гетерогенный признак. Развитие хрусталика определяется одними генами, а развитие сетчатки или роговицы — другими. Слепота же может возникнуть при дефектах любой из этих, а также многих других структур.
В ряде случаев генетика может улавливать более тонкие различия, чем клиническая медицина. Рассмотрим случай пигментного ретинита, при кото-ром пигмент мигрирует из тапетума в сегчатку, где его можно увидеть при помощи офтальмоскопа. Перемещение пигмента приводит к ночной слепоте и постепенному сокращению поля зрения. В тяжелых случаях наступает полная слепота. Степень поражения зрения различна, но выделить какие-либо четкие типы не удается. Есть основания предполагать, что развитие пигментного ретинита определяют по крайней
168
Часть II. Генетика, человека
мере три различных гена: два аутосомных (один рецессивный и один доминантный) и сцепленный с полом рецессивный ген. Клинически удается выделить только случаи пигментного ретинита, определяемые аутосомным рецессивным геном; в этих случаях ретинит сопровождается глухотой. Не известно, является ли действие этих генов кумулятивным; поскольку гены пигментного ретинита относятся к редко встречающимся, до сих пор не было описано случаев одновременного присутствия нескольких генов пигментного ретинита у одного индивидуума, хотя теоретически такие случаи вполне возможны.
Полигенная наследственность
Представление о кумуляции действия генов чрезвычайно важно для понимания наследования таких признаков, как рост, вес, телосложение и умственные способности. Распределение этих признаков в семьях и популяциях принципиально отличается от распределения таких признаков, как рассмотренные выше — группы крови или патологические состояния, которые распадаются на дискретные классы. Мы знаем, что кроме очень высоких и очень низкорослых людей встречаются люди любого промежуточного роста; различия в росте, стало быть, не качественные, а количественные, и любое разделение по росту на классы в этом смысле произвольно. При изучении таких признаков применяется непрерывная шкала измерений.
Одной из наиболее важных особенностей количественно варьирующих признаков оказывается то, что распределение их частоты в популяции близко к нормальному, или гауссовому (рис. 9.1). Это распределение описывается симметричной одномодальной кривой и часто встречается при статистическом анализе явлений. Ось симметрии этой кривой проходит через вершину и делит площадь под кривой пополам; точка на оси абсцисс, через которую проходит ось симметрии, определяет среднее значение данного распределения. Степень отклонения от среднего значения измеряется такими показателями, как дисперсия, квадратный корень из дисперсии (квадратичное, или стандартное, отклонение) и коэффициент изменчивости (отношение квадратичного отклонения к средней). Если распределение несимметричное и, стало быть, не строго нормальное, то его характеризуют другими усредненными величинами — модой и медианой. Мода — это абсцисса вершины кривой, т. е. наиболее часто встречающееся значение; медиана — это точка на оси абсцисс, через которую проходит перпендикуляр, делящий площадь под кривой пополам. Если в той или иной популяции произвести измерения некоторого признака у большого числа индивидуумов (как это обычно делается в биометрии), то распределение измерений в выборке (если она не
Глава 9. Гены и их действие
169
очень мала) будет близко к нормальному. Это распределение будет характеризоваться своим собственным средним значением и стандартным отклонЬнием. Однако маловероятно, что среднее значение для выборки будет совпадать со средним значением для всей популяции; это отличие связано с ошибкой выборочного метода, которая тем больше, чем меньше выборка. Величина ошибки определяется стандартным отклонением от среднего. Более подробное изложение этих приемов можно найти в учебниках по статистике.
Рис. 9.1. Гистограмма, иллюстрирующая распределение по длине тела 117 мужчин и соответствующая гауссова кривая (Bailey, Statistical methods im biology, 1959).
Сразу после вторичного открытия законов Менделя считали, что наследование признаков, характеризующихся непрерывной изменчивостью, нельзя объяснить участием только дискретных элементов, необходимо учитывать какие-то формы слитной наследственности. Но в случае неразделяемой смеси наследственного материала изменчивость в популяции должна была бы в каждом последующем поколении уменьшаться вдвое. Этого, как мы знаем, не происходит; в настоящее время известно, что возникновение новых вариантов, которбе будет рассмотрено позднее, не согласуется с представлением о слитной наследственности. Вместе с тем можно показать, что если тот или иной признак определяется совместным действием многих генов, то легко себе представить возможность его непрерывной изменчивости подобно тому, что происходит с таким признаком, как рост человека.
Сказанное легко объяснить на примере двух несцепленных локусов, гены которых контролируют не самостоятельные признаки, а компоненты какого-то одного признака. Пусть, например, действие гена А проявляется в образовании одной единицы какого-либо измерения, а действие его аллеля А' — в образовании
170
Часть II. Генетика человека
двух единиц. Пусть, далее, гены В и В' оказывают такое же действие на это измерение, причем вся система аддитивна. Тогда у двойной гомозиготы ААВВ при измерении этого признака получим 4 единицы, а у другой двойной гомозиготы А'А'В'В' — 8 единиц. В отсутствие доминантности двойная гетерозигота АА'ВВ' будет фенотипически строго промежуточна между двумя гомозиготами и даст 6 единиц. Рассмотрим теперь скрещивание двух гетерозигот. На схеме цифрами обозначено количество единиц признака, производимых каждым генотипом. Из схемы видно, что при скрещивании возникает 5 фенотипических классов, встречающихся в отношении 1 (4 единицы): 4 (5 единиц): 6 (6 единиц): 4 (7 единиц): 1 (8 единиц).
АА'ВВ'X АА'ВВ'
6 6
Гаметы
АВ А'В АВ’ А'В'
Гаметы Зиготы
АВ ААВВ АА'ВВ ААВВ' АА'ВВ'
4 5 5 6
А'В АА'ВВ А'А'ВВ АА'ВВ' А'А'ВВ'
5 6 6 7
АВ' ААВВ' АА'ВВ' ААВ'В' АА'В'В'
5 6 6 7
А'В' АА'ВВ' А'А'ВВ' АА'В'В' А'А'В'В'
6 7 7 8
Другими словами, распределение изменчивости данного измерения является симметричным, причем промежуточный фенотип встречается чаще крайних. Мы получили нечто напоминающее нормальное распределение. Если различия между двумя крайними фенотипами (в нашем случае между фенотипами с 4 и 8 единицами) определяются не двумя парами генов, а многими, причем каждый отдельный ген определяет только часть единицы признака, то число классов промежуточных фенотипов сильно возрастает. Это означает, что различие между фенотипически смежными классами соответственно уменьшается.
Ясно, что чем больше число генов, влияющих на изменение того или иного признака, тем труднее" провести грань между со
Глава, 9. Гены и их действие
171
седними классами; при очень большом числе генов распределение частот становится практически непрерывным, но и та дискретность изменения признака, которая теоретически должна бы проявиться, полностью маскируется взаимодействием генотипа с внешней средой (см. гл. 10). Такие факторы, как неравноценность действия отдельных генов и доминантность, наменяют форму кривой распределения и тем самым усложняют картину, но ни в коей мере не опровергают основного принципа, а именно того, что наследование количественно изменяющихся признаков можно объяснить, исходя из представления о дискретных элементах — генах. Гены, каждый из которых дает слабый эффект, но действие которых кумулятивно при определении какого-либо признака, носят название полигенов, а наследование таких признаков называется поливенным. Полигены обнаруживаются не только в эухромати-новых участках хромосомы, где локализованы так называемые «главные» гены, но и в гетерохроматиновых участках. Это было показано на разных примерах; наиболее яркий из них — влияние отсутствия Y-хромосомы на некоторые количественные признаки. У дрозофилы Y-хромосома практически полностью гетерохрома-тична. Генетический анализ признаков, определяемых полигенами, значительно сложнее, чем выяснение условий наследования признаков, определяемых одним или несколькими главными генами. Вместо того чтобы измерять частоту появления в потомстве различных классов признака при изучении полигенной наследственности, исследователи используют параметры кривой распределения и данные о природе и степени сходства между родственниками. Большую трудность представляет установление шкалы, по которой данные гены были бы аддитивными; нет оснований полагать, что замещение того или иного гена его аллелем проявится одинаково на шкале, используемой для измерения признака. Часто приходится трансформировать эту шкалу в логарифмическую или в шкалу обратных величин. Тем не менее несмотря на трудности такого рода, для некоторых количественно изменяющихся признаков у растений и животных удалось выделить генетические компоненты и компоненты, определяемые влиянием внешней среды, а также определить такие параметры, как общая доминантность, сцепление и минимальное число генов (эффективные факторы). Это оказалось возможным благодаря широкому применению контрольных скрещиваний с особями, общая генетическая конституция которых хорошо известна. В генетике человека такие тесты, конечно, невозможны, что особенно осложняет изучение наследования количественных признаков. Цвет кожи в гибридных популяциях негров и европейцев относится к количественным признакам. Штерн (Stern) предпринял попытку установить число генов, определяющих цвет кожи, сравнивая частоту распределения вариантов цвета кожи американских пег-
172
Часть II. Генетика Человека
ров с модельным распределением, вычисленным для различного числа пар генов. Частоты аллелей, использованные в модели, вычислялись по степени метизации западноафриканских негров и европейцев в популяции американских негров. Основываясь на ряде допущений, таких, как равенство и аддитивность эффекта отдельных генов, Штерн пришел к выводу, что в определении различий цвета кожи между «черными» африканцами и «белыми» европейцами принимают участие 3—4 пары генов. Весьма вероятно, что в определении различий в росте и аналогичных антропометрических признаках даже в пределах одной популяции принимает участие гораздо большее число генов.
Многое о наследовании количественных признаков можно почерпнуть при изучении сходства индивидуумов в пределах одной семьи. Такое изучение часто помогает выявить, обусловлены ли данные различия генетически или они связаны с действием факторов внешней среды. Каждый из родителей передает ребенку половину своих хромосом, так что у родителей и детей половина генов оказывается общей. В поколениях внуков, с одной стороны, и дедов и бабок, с другой — общей оказывается только одна четвертая часть генов, так как у каждого ребенка — две бабки и два деда. Точно так же сибсы в среднем схожи по половине генов, по которым их родители гетерозиготны, поскольку каждый ребенок с равной вероятностью может наследовать как ген, уже переданный сибсу, так и его аллель. По той же причине каждый ребенок будет иметь четвертую часть общих генов с дядьями и тетками и лишь восьмую часть общих генов с двоюродными братьями и сестрами. В общем случае с уменьшением родства на одну степень число общих генов уменьшается вдвое. Это означает, что если гены, ответственные за какой-либо количественный признак, обладают аддитивным действием и не обнаруживают доминантности, то в среднем половина генов каждого индивидуума делает его похожим на родителя или сибса, тогда как по второй половине генов он похож на любого из этих родственников не более, чем на случайного постороннего человека. Данный индивидуум будет иметь не только половину генов, общих с каждым из родителей и с сибсами, но будет также иметь одну четверть генов, общих с дедами, бабками, дядьями и тетками, и одну восьмую часть генов, общих с двоюродными братьями и сестрами. Приведем теоретический пример: пусть рост человека определяется исключительно генетическими факторами и соответствующие гены оказывают аддитивное действие без доминантности. Если средний рост в популяции 170 см и браки заключаются независимо от роста супругов, то у мужчин ростом 178 см средний рост сыновей будет составлять 174 см, так как от отца, рост которого на 8 см выше среднего для данной популяции, сын получит половину генов, тогда как вторая половина генов будет такой же, как у популяции в
Глава 9. Гены и их действие
173
среднем. Точно так же внуки и племянники будут иметь средний рост 172 см, а правнуки и двоюродные братья — 171,5 см. Полезно заметить, что кроме регрессии от отцов к детям можно вычислить регрессию от сыновей к отцам. Так, у упомянутых выше индивидуумов с ростом 178 см средний рост отцов составлял 174 см, так как в этой группе половина генов, обеспечивших ее членам превышение над средним ростом на 10 см, была передана от отцов.
Влияние доминантности зависит от частоты данного гена. В общем случае это легко показать, рассматривая единичный локус с двумя аллелями, один из которых доминантен. Если доминантный ген встречается редко и гомозиготы очень редки, то два других генотипа фенотипически различимы и сходство между родственниками оказывается таким же, как и в отсутствие доминантности. Вместе с тем если доминантный ген встречается часто, то сходство между родителями и ребенком может вовсе отсутствовать: ведь если оба родителя гетерозиготны, они могут отличаться от гомозиготного рецессивного ребенка в такой же степени, как и любой случайный индивидуум. Так же обстоит дело со сходством между данным индивидуумом и его дедами, бабками, дядьями, тетками и т. д. Однако сходство между сибсами сохраняется, так как если ребенок гомозиготен по рецессивным признакам, то вероятность того, что и другой ребенок, родившийся от гетерозиготных родителей, будет иметь тот же генотип, составит 25%. Таким образом, в зависимости от частоты гена фенотипическое сходство между родственниками может меняться от значений, вычисленных для случая отсутствия доминантности, до 1/4 для сибсов и нуля для прочих родственников. В общем виде степень родства между родителями и потомством выражается формулой q: (1—q), а между сибсами 14-3^.74 (1+д), где q — частота данного рецессивного аллеля в популяции.
Тот факт, что количественный признак определяется многими локусами, усложняет картину наследования. Действительно, в одних локусах могут существовать доминантные аллели, в других доминантность может быть неполной, а в третьих вовсе отсутствовать, так что можно говорить лишь об общем влиянии доминантности. Может существовать сцепление между полигенами и неаддитивность их действия, связанная с. взаимодействием локусов. По этим причинам изучение генетического определения количественных признаков у человека все еще находится в зачаточном состоянии.
Генный комплекс
Явление взаимодействия локусов даже в случае относительно простого наследования признака часто встречается в мире растений и животных. Пурпурная окраска цветков душистого горош
т
Часть II- Генетика человека
ка зависит от наличия определенных генов в двух локусах. Если оба гена отсутствуют, цветки остаются белыми. Такие факторы называются «комплементарными», и наиболее вероятным биохимическим объяснением данного явления может быть предположение о необходимости продукта одного гена для действия другого. Бывают и такие случаи (как при определении оперения у кур породы леггорн белые), когда действие гена в одном локусе препятствует выражению гена в другом. Эта особого рода «доминантность» известна под названием эпистаза.
В настоящее время накапливаются данные, согласно которым взаимодействия локусов аналогичного типа встречаются и у человека. В качестве примера упомянем взаимодействие локусов групп крови 0 (секретеры) и Льюис, которые ответственны за пути биосинтеза групповых веществ крови систем АВО и Льюис и в особенности влияют на специфичность этих веществ в продуктах секреции (см. гл. 17). Особенностью этой сложной ситуации является отсутствие образования группоспецифических веществ крови А или В (в системе АВО) у гомозигот hh, даже если у них есть гены, контролирующие синтез А или В.
Другой важный тип взаимодействия локусов известен как эффект положения. В этом случае на выражение гена влияют его положение в хромосоме или природа соседних генов. Поразительным примером может служить система групп крови Rh (гл. 17): лица с комплексом CDEIcDe имеют те же гены, что и с комплексом CDelcDE, однако у первых образуется большое количество антигена Е и мало С, а у вторых — много С и мало Е. Таким образом, если С и Е находятся в одной и той же хромосоме, подавлен синтез вещества С, а если С и Е расположены в разных хромосомах, подавлен синтез Е.
Известно немало примеров взаимодействия локусов у человека. Генное взаимодействие приводит к тому, что тот или иной проявляющийся определенным образом ген у некоторых индивидуумов не получает фенотипического выражения. Подобным примером может служить полидактилия, которая наследуется, как правило, по доминантному типу. Тем не менее она иногда встречается у детей совершенно здоровых родителей. В этом случае остается предположить, что действие гена подавляется остальной частью генотипа. Перед нами явление, именуемое неполной пенетрантностью, когда один и тот же ген в одних случаях получает выражение в фенотипе, а в других не получает. К этому явлению близка так называемая изменчивая экспрессивность действия гена. Так, при брахи-дактилии пальцы рук и ног, как правило, укорочены и снабжены перепонками, однако один или несколько симптомов данного синдрома могут отсутствовать. Эти явления показывают, что возникновение того или иного признака связано с действием многих генов. Генотипы в популяции формируются под действием естест
Глава 9. Гены и их действие 17S
венного отбора, так что замещение того или иного определенного гена, как правило, оказывает постоянное влияние на формирование какого-то нормального признака. Это не означает, однако, что рассматриваемый ген целиком ответствен за данный признак. Не удивительно, следовательно, что вредные гены, для которых данный генотип не подготовлен, зачастую характеризуются изменчивым и многоступенчатым проявлением.
В настоящее время имеется достаточно указаний на то, что основой для общего развития индивидуума и, следовательно, для его эволюционной приспособленности-является всесторонне приспособленный генный комплекс. Нетрудно представить себе, как благодаря естественному отбору происходит образование такого комплекса. При возникновении нового гена, даже если его общее действие является благоприятным для организма, некоторые его проявления оказываются вредными. Индивидуумы, генотип которых не обеспечивает подавления нежелательных проявлений нового гена, оставят меньше потомков, чем те, у которых нежелательные эффекты подавляются. Этот механизм подавления затем совершенствуется благодаря естественному отбору, и вредное действие нового гена, ведущее к снижению жизнеспособности или фертильности (или той и другой), будет полностью элиминировано. Гены, обеспечивающие подавление нежелательного действия других генов, образуют группу так называемых генов-модификаторов, которые «встроены» в генотип благодаря действию отбора. Некоторые из них, по-видимому, лишены каких бы то ни было других функций. Кроме генов, подавляющих вредное действие, несомненно, существуют гены, усиливающие благоприятные проявления других генов. На этой основе могло возникнуть и само явление доминантности. Вновь возникающий ген, будучи, естественно, редким, существует только в гетерозиготах. Если этот ген оказывает вредное действие, то естественный отбор приведет к подавлению его действия посредством генов-модификаторов. Другими словами, ген превращается в рецессивный, так что аллель дикого типа автоматически становится доминантным. Если же новый ген обеспечивает организму преимущество, то благодаря естественному отбору это преимущество закрепится и у тех индивидуумов, которые имеют новый ген в одной дозе, т. е. данный ген превратится в доминантный. Тот факт, что вредный ген, возникающий в результате мутации, оказывается обычно рецессивным с самого начала, не противоречит высказанной гипотезе, поскольку любая мутация, как будет показано далее, явление повторяющееся, и ген, возникший в данный момент, мог неоднократно возникать и в прошлом, что сопровождалось образованием модифицирующей системы, подавляющей его действие в гетерозиготном состоянии. Данные, подтверждающие эту точку зрения, были получены при гибридизации различных популяций живот-
176
Часть II. Генетика человека
ных, обладающих одним и тем же доминантным геном. В тех случаях, когда доминантность в родительских популяциях была обусловлена различными модифицирующими системами, в гибридной популяции она полностью исчезала.
Природа гена
Генетическая информация закодирована в дезоксирибонуклеиновой кислоте (ДНК) хромосом. ДНК образована последовательностью нуклеотидов; каждый нуклеотид состоит из сахара дезоксирибозы, фосфорной кислоты и азотистого основания. Азотистые основания нуклеотида неодинаковы: это может быть один из двух пуринов, аденин (А} или гуанин (Г), и один из двух пиримидинов, тимин (Т) или цитозин (Ц). ДНК всех высших организмов состоит из двух цепей нуклеотидных последовательностей, связанных друг с другом основаниями, причем аденин всегда связан с тимином, а гуанин—с цитозином. Спаренные основания образуют как бы перекладины веревочной лестницы, а дезоксирибоза и фосфорная кислота друг за другом — боковые канаты. В дейст-
ф ф ф ф
. ф
^д-ц-г-д^ С Z-ф
^Д-А-Т-Д^ С У?
^Д-Г-Ц-Д^ С ~?Ф
^д-ц-г-дС,
ф.
ф-С
Т?д-А
фС
ФС
2?д-д
ф
Г-ДСА ф
Т-Д^Л ф «~ДСА ф
Г-ДС
ф_ ф
д-д-г-д^ ф
Л^Л-А-Т-Д ФГ ф
^д-г-ц-д"" ф-^
ф_ ф
>-ц-г-дс фС ^ф
^д-д-т-дС ф-^ ф
д-г-ц-д^.^ ф Г-Д^
ф ^.ф
ф ф
^Д-a-t-aZ ф д-т-ц-д^ ф^
д—ц—г—дС
ф д-ц-г-дс^ ФС ф
Д-А-Т-ДЦ ФС. ф
^Д-г-и-дС ф-^. ф
^д-ц-г-д^.
Рис. 9.2. Репликация ДНК,
Глава 9. Гены и их действие
177
Б
Двойная спираль ДНК
А
УУГ
УУА
МРНК
ЦЦГ
АЦА
АУГ
Б
Рис. 9.3. Процесс синтеза белка.
А, Код ДНК. В. мРНК, образованная соответственно той цепи ДНК, которая представлена непрерывной линией. В. Образование полипептида на рибосоме.
вительности, как впервые было показано в замечательной работе Уотсона и Крика, эти цепи изогнуты, образуя «двойную спираль», и структуру ДНК правильнее сравнивать не с веревочной, а с винтовой лестницей. Схематически эта структура представлена на рис. 9.2, где также показана способность ДНК к репликации, необходимой в ходе клеточного деления. После разобщения цепей образуются две новые цепи, каждая комплементарная по отношению к одной из исходных, так как одно основание точно предопределяет, с каким основанием оно образует пару.
Генетическая информация, заключенная в ДНК, передается из клеточного ядра в цитоплазму с помощью матричной рибонуклеиновой кислоты — мРНК (рис. 9.3). РНК также состоит из последовательности нуклеотидов, но в их состав входит сахар рибоза, а место тимидина (содержащегося в ДНК) занимает урацил. В ходе синтеза мРНК информация, содержащаяся в ДНК, транскрибируется: считывается лишь одна цепь ДНК и РНК получает последовательность оснований, комплементарную по отношению к этой цепи; лишь место аденина ДНК при транскрипции занимает урацил. В цитоплазме мРНК присоединяется к рибосомам, которые в свою очередь содержат рибосомную РНК (рРНК), и, находясь здесь, направляет течение процесса синтеза белка. Главной
178
Часть II- Генетика человека
единицей, или «словом», в этой информации служит триплет-3 нуклеотида, 3 основания которого определяют одну из 20 аминокислот, из которых состоят белки. Эти триплеты, которые можно получить в числе 64 вариантов, называют кодонами; главным образом благодаря работам Ниренберга установлено соответствие между кодонами и кодируемыми ими аминокислотами (табл. 9.1.). Следует отметить, что если некоторые аминокислоты кодируются только одним кодоном (например, метионину соответствует ТАЦ в ДНК и АУГ в РНК), то другие кодируются двумя и более кодонами (например, фенилаланину соответствует в ДНК ААА и ААГ); такой код называют вырожденным или избыточным. Отметим, что 3 кодона — АТТ, АТЦ и особенно АЦТ — в ДНК (УАА, УАГ и УГА в РНК) не кодируют аминокислот. Они, по-видимому, должны означать точку в сообщении, отделяя синтез одной полипеп-тидной цепи от синтеза другой.
Генетический код (ДНК-код)
Таблица 9.1
Первая буква Вторая буква Третья буква
А Г Т Ц
А ААА 1 АГА ч АТА ч АЦА ч ААГ / Фен АГГ 1 АТГ / Тир АЦГ )^ИС ААТ j лей АГТ ( Сер АТТ \ «Стоп» АЦТ «Стоп» ААЦ 1 АГЦ) АТЦ / АЦЦ Три А Г T ц
Г ГАА ч ГГА ч ГТА ч ГЦА ч ГАГ 1 ГГГ 1 ГТГ ( Гие ГЦГ 1 ГАТ [ Леи ГГТ | Пр0 ГТТ 1 Глн ГЦТ [ АрГ ГАЦ J ГГЦ) ГТЦ J ГЦЦ) А Г T Ц
Т ТАА ч „ ТГА ТТА ч ТЦА ч „ грдр | Иле 'ррр ТТГ | Асн ТЦГ j ' TAT 1 Мет ТГТ Тре ТТТ 1 див ТЦТ ) Арг ТАЦ J ТГЦ ТТЦ 1 ТЦЦ f А Г Т ц
Ц ЦАА ч ЦГА ч ЦТА ч ЦЦА ЦАГ 1 ЦГГ 1 ЦТГ ) Асп ЦЦГ ЦАТ (Вал ЦГТ [ Ала ЦТТ ( Глу ЦЦТ Гли цац; цгц; цтц / ццц J А Г Т Ц
Глава 9. Гены и их действие
179
В цитоплазме имеется и РНК третьего типа — транспортная РНК (тРНК), или растворимая РНК. Каждая молекула этой РНК специфична для определенной аминокислоты и содержит антикодон для кодона, определяющего ту же аминокислоту в мРНК. Так, например, некоторые молекулы имеют антикодон ААА и переносят аминокислоту фенилаланин к тем участкам мРНК на рибосоме, которые имеют кодон УУУ; другие тРНК имеют антикодон УГУ и переносят аминокислоту треонин на кодон АЦА в составе мРНК и т. д. Таким образом, аминокислоты собираются и связываются, образуя полипептиды в последовательности, которую кодирует мРНК, а следовательно, и ДНК. Если не считать того, что урацил занимает место тимина, триплеты в РНК соответствуют триплетам ДНК.
Первой аминокислотой, включающейся в полипептид, всегда оказывается метионин. В ходе дальнейшего синтеза этот концевой метионин может быть отщеплен, и у многих белков не метионин занимает первое положение в полипептидной цепи. Тем не менее триплет ТАЦ в ДНК*(АУГ в мРНК) можно считать инициирующим кодоном (старт) в отличие от терминирующих кодонов (стоп). Следует, однако, отметить, что метионин может содержаться и в середине полипептида; имеются 2 вида тРНК — для терминального и для «внутреннего» метионинов.
Всякая снабженная ядром клетка взрослого организма содержит полный набор хромосом и генетическую информацию, полученную зиготой; тем не менее клетки дифференцируются, различаясь по форме и активности и обеспечивая функции различных тканей. В ткани одного типа одни гены активны, а другие — нет, тогда как в других тканях другие наборы генов диктуют, какие полипептиды будут синтезированы. Есть основания считать, что в большинстве тканей активно лишь около 5% генов; правда, в ткани мозга этот процент возрастает до 12. Жакоб и Моно предложили модель, объясняющую регуляцию активности генов и дифференцировку тканей; модель эта получила убедительное экспериментальное подтверждение, по крайней мере в опытах на микроорганизмах (рис. 9.4). Модель постулирует участие генов двоякого типа: структурные гены, которые кодируют полипептиды, синтезируемые на рибосомах, и гены, определяющие активность структурных генов; последние делятся на операторы и регуляторы. Считается, что структурные гены, участвующие в кодировании полипептидов, относящихся к единому метаболическому процессу, часто расположены по соседству (по мнению Шеппарда, это, по всей видимости, обусловлено естественным отбором); ген-оператор, контролирующий их активность, расположен в непосредственной близости от этих структурных генов. Такой комплекс называют опероном. Ген-регулятор, расположенный на расстоянии от оперона, образует вещество, репрессор (те-
180
Часть Л. Генетика человека
Ген-рееулятор
Оперон
Ген-оператор Структурные гены
Репрессор
________Оперон
Ген-оператор Структурные гены
—> | 2 I
I
мРНК
Белковый синтез
-J L_ Неактивный Hri^p РепР^сор
Рис. 9.4. Модель Жакоба — Моно, объясняющая регуляцию белкового синтеза.
перь установлено в ряде случаев, что это белок), которое может реагировать с оператором, выключая его и таким путем инактивируя структурные гены. Однако репрессор может также реагировать с небольшими молекулами («индукторами»), присутствующими в клетке; в таком случае его конфигурация меняется так, что не может более подавлять оператор. Индукторы представляют собой продукты обмена, или «питательные вещества»; таким путем обеспечивается система обратной связи, позволяющая «включать» или «выключать» структурные гены.
Обнадеживающие успехи, достигнутые в последние годы молекулярной генетикой и весьма кратко изложенные здесь, произвели революцию в нашем понимании природы наследственного материала и путей эволюции и онтогенеза; тем не менее для многих целей, особенно в популяционной генетике, остаются удобными и осмысленными рассуждения в классических терминах менделизма.
Глава 10
ВЛИЯНИЕ ВНЕШНЕЙ СРЕДЫ
Фенотип индивидуума в целом зависит не только от генотипа, но и от факторов внешйей среды, в которой существует индивидуум. В течение всей жизни происходит взаимодействие между этими двумя группами факторов, которые в конечном счете и детерминируют все биологические признаки индивидуума, причем обе эти группы имеют одинаково важное значение. Иногда, например в случае групп крови, различия между индивидуумами имеют исключительно генетическую природу, поскольку при всех условиях среды, совместимых с жизнью, данный генотип характеризуется одним и тем же проявлением. Хотя подобные случаи служат предметом особого внимания, так как они убедительно иллюстрируют законы Менделя, тем не менее они представляют исключения, и даже в этих случаях чисто генетическими являются лишь различия между признаками, тогда как для выявления самих признаков необходимы определенные условия внешней среды. И наоборот, различия, обусловленные исключительно факторами среды, касаются признаков, определяемых генетически. Так, различия в числе эритроцитов у индивидуумов часто связаны с высотой местности над уровнем моря. Однако сама способность к изменению числа этих клеток в зависимости от парциального давления кислорода в атмосферном воздухе обусловлена генетически. Здесь приведены два крайних случая; чаще же даже различия между индивидуумами определяются обеими группами факторов — генетическими и факторами внешней среды. Различия в росте обусловлены генетически, но они подвержены также влиянию внешней среды, поскольку рост существенно зависит от характера питания и, по-видимому, также от климатических условий. В подобных случаях приходится прибегать к анализу того, что Добжанский назвал «нормой реакции» организма. Он определил эту концепцию как «полный спектр, весь репертуар различных путей развития, которые могут выявиться у носителей данного генотипа в любой среде».
Далее, характер изменений, вызываемых обеими группами факторов, часто бывает сходным. Фактор внешней среды и аномальные гены порой вызывают весьма сходные эффекты. Например, у женщин, перенесших краснуху на ранних сроках беременности, часто родятся глухонемые дети или дети с врожденной катарактой, причем эти аномалии неотличимы от соответствующих аномалий, обу
182
Часть II. Генетика человека
словленных генетически. Такие обусловленные факторами внешней среды изменения, сходные с изменениями генетической природы, получили название фенокопий. Более важное значение имеет сходство изменений, определяемых полигенами, с нормальной изменчивостью, обусловленной факторами внешней среды.
Как уже говорилось, полигонная наследственность обусловливает более или менее симметричное одномодальное распределение частот вариантов признака. Изменчивость, обусловленная факторами внешней среды, дает аналогичную картину, что можно видеть при анализе зависимого от влияния среды признака в генетически гомогенной экспериментальной популяции растений или животных. Диапазон изменений под действием факторов внешней среды зависит не только от степени изменений внешних условий, но и от уровня, относительно которого происходят эти изменения, а также от генотипа, с которым взаимодействует среда. Так, колебания температуры около 21° не вызывают такой фенотипической изменчивости в росте мышей, как такие же колебания около 32°С. Если две генетически гомогенные популяции, одна из которых гетерозиготна по большинству локусов, а другая гомозиготна по этим локусам, поместить в одинаковые условия, то вызываемые средой различия окажутся в гетерозиготной популяции менее выраженными, чем в гомозиготной.
Мерой, широко употребляемой при изучении роли факторов наследственности и среды в определении фенотипической вариабельности, является наследуемость. Наследуемость — это доля наблюдаемой вариансы признака в популяции, которую можно отнести на счет генетических причин. Есть два типа показателей наследуемости: широкая наследуемость, общий вклад генетических факторов, и узкая наследуемость, которая складывается лишь из аддитивных компонентов генетической вариансы, т. е. является следствием усредненного действия отдельных генов на признак независимо от наличия других генов. Дополнительные компоненты общей генетической вариансы возникают вследствие доминантности и эпистаза (взаимодействия локусов друг с другом) и, лишь когда ими можно пренебречь, величины узкой наследуемости соответствуют величинам широкой наследуемости.
Интерпретация величин наследуемости затрудняется возможным вмешательством главным образом двух феноменов: ковариацией факторов среды и наследственности и их взаимодействием. Ковариация возникает, если определенные фенотипы обнаруживают тенденцию выбирать особые участки разнообразной среды, действию которых подвержена популяция. Вариацию, возникающую по этой причине, при применении многих методов определения относят к генетическому компоненту, что, несомненно, неправильно. Вместе с тем взаимодействие генетических факторов и факторов среды, характеризующееся различиями в ответах раз-
Глава 10. Влияние внешней среди
183
них генотипов на одни и те же условия среды, часто относят к средовому компоненту. Величина этих компонентов, несомненно, может быть значительной, особенно для таких признаков, как особенности поведения человека; однако определение этой величины часто оказывается трудной задачей.
Рассматривая показатели наследуемости, важно понимать, что они относятся лишь к определенной популяции и к определенной среде, для которых зти показатели получены. Качественные и количественные изменения генетического состава популяции или характера среды также могут повлиять на наследуемость.
Таким образом, показатель наследуемости не имеет общего значения и не позволяет экстраполировать от изученной ситуации для одной популяции к другой. Тем не менее если признак имеет тенденцию к проявлению постоянно высокой (или низкой) наследуемости в ряде различных популяций и сред обитания, разумно допустить, что он стабилен (или лабилен) по отношению к факторам среды в пределах колебаний интенсивности этих факторов для определенной популяции.
Иногда полученные данные приложимы лишь к наименьшей популяционной единице — семье; для данных другого типа удается охарактеризовать влияние генетический гетерогенности и гетерогенности среды на семьи, составляющие одну популяцию, в пределах которой возможны браки (между семьями и внутри семей); однако в общем случае невозможно переносить ситуацию, обнаруженную внутри одной популяции, на объяснение различий между популяциями. Иными словами, признак может быть высокопасле-дуемым в обеих популяциях, и тем не менее всякое различие между этими популяциями все же может быть полностью обусловлено факторами среды. Это положение вполне совместимо со сделанным выше заявлением, согласно которому признаки с высокой наследуемостью в одной популяции часто обнаруживают высокую наследуемость и в других, поскольку диапазон факторов среды в пределах популяций часто отличается от диапазона факторов среды между популяциями если не качественно, то хотя бы количественно.
У экспериментальных растений и животных, па которых можно проводить массовые опыты по скрещиванию, удалось определять наследуемость многих количественных признаков, т. е. оценивать ту долю изменчивости, которая обусловлена действием генов. Для такой оценки используются анализ сходства близкородственных организмов и измерение ответа признака на искусственный отбор. Остальная часть общей фенотипической изменчивости связана, естественно, с влиянием среды. По вполне понятным причинам в применении к человеку анализ взаимодействия генетических факторов и факторов внешней среды не может быть проведен с такой полнотой. Здесь приходит на помощь так называемый «близнецовый метод»*
184
Часть tj. Генетика человека
дополненный данными о сходстве родственников другой степени родства и приемных детей с их биологическими родственниками или родственниками по воспитанию.
Близнецы бывают двух типов: близнецы одного типа возникают как результат независимого оплодотворения двух различных яйцеклеток; такие близнецы называются неидентичными, двуяйцевыми (дизиготними)', генетически они сходны между собой не более, чем обыкновенные сибсы. Близнецы другого типа возникают при дроблении одной оплодотворенной яйцеклетки. Такие близнецы называются идентичными, однояйцовыми (монозиготными); они генетически идентичны, если только в одном из зародышей после разделения не произойдет соматической мутации. Различия между однояйцовыми близнецами лкогут быть связаны, следовательно, только с влиянием среды, тогда как различия между двуяйцовыми близнецами могут быть вызваны обеими группами факторов. Вследствие полной генетической идентичности реакция однояйцовых близнецов на воздействия среды более однородна, чем у дву-яйцовых, но все же различия в вариабельности двух типов близнецов имеют в основном генетическую природу при одинаковых условиях внешней среды. Различия, обусловленные влиянием внешней среды (а не только генетические различия), между родственниками нередко менее выражены, чем между индивидуумами, не находящимися в родстве; это объясняется тем, что условия существования у членов одной семьи обычно более сходны, чем во всей популяции в целом. Бывают, однако, случаи, когда члены одной семьи с самого детства оказываются в различных условиях, и если это происходит с идентичными близнецами, то такая ситуация предоставляет исследователям идеальную возможность определить долю участия факторов внешней среды в возникновении фенотипических различий в данной популяции.
Наиболее распространенный метод оценки относительной роли генетических факторов и факторов в возникновении различий между индивидуумами — это сопоставление так называемой кон-кордантности идентичных и неидентичных близнецов. Близнецы называются конкордантными, если они одинаковы по исследуемому признаку, и дискордантными, если они различаются по этому признаку. Так, при наличии у обоих членов пары одного и того же заболевания или в случае отсутствия у обоих этого заболевания (в определенных условиях) говорят о конкордантности; если же один близнец заболевает, а другой нет, то пару называют дискор-дантной. Конкордантность однояйцовых близнецов по такому инфекционному заболеванию, как корь, весьма высока, но такой же высокой конкордантностью обладают двуяйцовые близнецы. Высокая конкордантность в этом случае означает только, что при заболевании одного индивидуума инфекция легко передается близнецу. По таким заболеваниям, как туберкулез и рахит, конкордант-
Глава 10. Влияние внешней среды
185
ность однояйцовых близнецов значительно выше, чем двуяйцовых, так что, хотя одно из заболеваний является в основном инфекционным, а другое связано с условиями питания, в обоих случаях важную роль играет генетически обусловленная восприимчивость к этим заболеваниям.
Для количественных признаков их распределение по парам однояйцовых (монозиготных, MZ) близнецов, которое характеризуется либо коэффициентом корреляции, либо средней дисперсией внутри пар, можно сопоставить с распределением по парам двуяйцовых (дизиготных, DZ) близнецов. Основная формула для оценки наследуемости имеет следующий вид:
DZ—, DZ ’
где DZ — дисперсия (варианса) для дизиготных близнецов, а MZ — для монозиготных.
Если это отношение равно 1, то обсуждаемое различие имеет целиком генетическую природу, а если оно равно 0, то определяется исключительно факторами внешней среды. Другой, более надежный способ определения роли факторов среды сводится к сравнению различий между однояйцовыми близнецами, которые выросли вместе (В), и такими же близнецами, которые были рано разлучены и выросли отдельно (О). Если результат сравнения выразить в форме
О-В О ’
то приближение этого отношения к 1 будет означать, что рассматриваемый признак определяется в основном факторами внешней среды, которые сильнее варьируют в масштабе всей популяции, чем в масштабе одной семьи.
Применяя эти методы анализа к таким признакам, как вес тела, удалось показать, что различиям в условиях среды принадлежит важная роль в фенотипической изменчивости данной популяции. Рост оказался гораздо менее лабильным признаком. Эти же методы были применены для оценки различий в поведении; в табл. 10.1 приведены некоторые средние данные о степени сходства между близнецами и сибсами в отношении коэффициента интеллекта (КИ) в различных популяциях Европы и Америки. Однако интерпретация подобных результатов связана с рядом противоречивых суждений частично из-за трудностей измерения таких сложных признаков, как умственные способности и свойства личности, частично вследствие ограничений, налагаемых близнецовыми методами; некоторые из этих ограничений становятся особенно серьезными, когда речь идет об особенностях поведения.
18S
Часть II. Генетика человека
Таблица 10.1
Обобщенная картина сходства по умственному развитию по данным различных исследований1)
Степень родства Генетическая корреляция Обнаруженные коэффициенты корреляции
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0
Индивидуумы, не состоя- 0,00
щие в родстве, но воспитанные вместе Приемный родитель и 0,00
ребенок Родитель и дитя 0,50
Сибсы, воспитанные раз- 0,50
дельно Сибсы, воспитанные вме- 0,50
сте Однополые дизиготные 0,50
близнецы Монозиготные близнецы, 1,00
воспитанные раздельно Монозиготные близнецы, 1,00
воспитанные вместе
>) Горизонтальные линии соответствуют диапазону сходства в различных иссле* дованиях. Вертикальными линиями обозначены средние значения для всех обобщенных результатов. По сводным данным (Erlenmeyer-Kirnling a. Jarvik, 1963).
Во-первых, не во всех случаях удается точно установить тип зиготности. Определение типа зиготности основано на изучении сходства нескольких различных, генетически обусловленных признаков. Идентичность или очень большое сходство всех рассматриваемых признаков указывает на монозиготность, тогда как четкое различие хотя бы по одному из них может служить доказательством дизиготности.
Наиболее подходящие для такого сравнения признаки должны удовлетворять следующим требованиям: тип их наследования должен быть точно известен; определяющие их гены должны обладать полной пенетрантностью и постоянной экспрессивностью при любых условиях внешней среды; альтернативные аллели должны достаточно часто встречаться в популяции. Значение типа наследова
Глава 10. Влияние внешней среды
187
ния позволяет определить вероятность идентичности двуяйщовых близнецов по рассматриваемому признаку. Группы крови и подобные им признаки идеально подходят для такого сравнения. Сопоставление по группам крови в сочетании с анализом таких признаков, как дерматоглифические узоры, которые наследуются по сложному типу, но весьма устойчивы по отношению к действию факторов среды, практически исключает возможность ошибочного диагноза типа зиготности. В прошлом за основу сравнения обычно брали антропометрические данные и такие признаки, как цвет глаз и форма волос, что часто приводило к ошибкам. Если в том или ином конкретном случае диагноз особенно важен, то его можно установить методом пересадки кожи. Успешная пересадка кожи возможна только между индивидуумами с одинаковым генотипом. Здесь следует упомянуть о том, что близнецы могут возникать также при оплодотворении различными сперматозоидами двух половин предварительно разделившейся яйцеклетки, а также при оплодотворении второго полярного тельца. Важно помнить, что в некоторых случаях однояйцовые близнецы оказываются зеркальным изображением друг друга. Нельзя упускать из виду возможность соматической мутации, которая произошла на достаточно раннем этапе развития, но после разделения близнецов. Быть может, еще более важным, по меньшей мере для особенностей поведения, является тот факт, что реакции генетических идентичных однояйцовых близнецов на воздействие внешней среды более сходны, чем реакции генетически различных двуяйцовых близнецов; по той же причине однояйцовые близнецы имеют тенденцию выбирать сходные условия внешней среды. Иначе говоря, существуют взаимодействия и корреляции «ген — среда», размах которых почти невозможно оценить. Следует также помнить, что все типы близнецов отличаются от других детей тем, что в материнском организме они делили друг с другом общую среду и в той или иной мере конкурировали друг с другом во время внутриутробного развития и в раннем детстве. Близнецы служат объектом особого внимания родителей. Эффект того факта, что ребенок является одним из близнецов, может быть учтен при сравнении сходства двуяйцовых близнецов между собой и сходства сибсов. Однако для этого нужны сведения о сибсах в одном и том же возрасте, которые нередко бывает трудно получить. Другим опасным источником ошибок является тенденциозность, особенно сказывающаяся, если выборка близнецовых пар производится несистематизированно; в таких условиях часть двуяйцовых близнецов остается неучтенной, особенно если они дискордант-ны и если один член пары умер. И наконец, близнецы встречаются нечасто (один раз на 90 рождений), и при рассмотрении редких генетических аномалий трудно набрать число пар близнецов, достаточное для исследования.
Как впервые предложил Вейнберг, число однояйцовых близне
188
Часть II. Генетика человека
цов в популяции можно определить путем вычитания из общего числа пар близнецов удвоенного числа пар разнополых близнецов, поскольку разнополые двуяйцовые близнецы должны встречаться с той же частотой, что и однополые. Оказалось, что около четверти всех близнецов монозиготные.
Интересно отметить, что само рождение близнецов, и особенно двуяйцовых, подвержено влиянию как генетических факторов, так и факторов среды. Вероятность рождения близнецов того и другого типа зависит от возраста матерей. Частота рождения однояйцовых близнецов зависит от возраста матерей линейно, тогда как частота рождения двуяйцовых близнецов характеризуется максимумом, который приходится на возраст около 37 лет, Вероятность рождения двуяйцовых близнецов зависит также от числа предыдущих родов, причем для каждых последующих родов вероятность рождения близнецов возрастает. Естественно, что для оценки влияния числа предыдущих родов следует предварительно исключить влияние возраста матери. Во время последней войны частота рождения двуяйцовых близнецов во многих странах Европы уменьшилась; это, по-видимому, связано с ухудшением условий питания. Для матери двуяйцовых близнецов вероятность рождения еще одной двойни выше; для сестер матерей близнецов вероятность рождения близнецов выше, чем для других женщин. Имеются также расовые различия в частоте рождения близнецов: у негров она выше, а у японцев ниже, чем у европейцев (гл. 18).
Совершенно иной метод получения сведений о механизме действия среды на онтогенетическое развитие — это изучение мигрантов из одной экологической ситуации в другую. Этот подход нашел широкое применение в антропометрических исследованиях Боаса по отношению к мигрантам из Европы в Северную Америку; недавно его модифицировали для анализа вариаций физиологических параметров. Он оказался особенно полезным при изучении физиологических последствий проживания на различных высотах над уровнем моря, поскольку весьма резкие изменения условий среды происходят в горных районах в пределах географически малых расстояний, а часто и в пределах популяций, между которыми существует столь интенсивный миграционный обмен, что генетические различия не могут возникнуть. Сравнивая коренных горцев с недавними мигрантами из низин в горы и коренных жителей низин с недавними мигрантами с гор в низины, удается идентифицировать не только немедленное влияние изменений высоты на физиологические и онтогенетические параметры, но и любые генетические компоненты различий между коренными жителями гор и низин, если они были изолированы друг от друга. Главная трудность здесь заключается в том, что мигранты часто не образуют репрезентативной выборки, по крайней мере в отношении некоторых признаков, что не позволяет характеризовать популяцию, из которой они произошли.
Глава 10. Влияние внешней среды,
189
Реакции индивидуума на изменение внешней среды могут быть либо благоприятными (дающими какие-то преимущества), либо неблагоприятными. В качестве примера неблагоприятной реакции можно привести реакцию организма на большие дозы ионизирующего излучения. Те индивидуумы, которые реагируют на облучение слабее, окажутся в более выгодном положении в случае воздействия массивных доз излучения. Хотя подобные реакции индивидуумов можно считать генетически обусловленными (в широком смысле), тем не менее они не составили особой эволюционной категории. Действительно, если усиленная радиация станет постоянно действующим фактором, то благодаря естественному отбору произойдет повышение резистентности индивидуумов к облучению (если, конечно, они раньше не погибнут!). Способность индивидуума к защите от действия неблагоприятных факторов среды носит название гомеостаза. Поскольку среда, окружающая индивидуум, постоянно изменяется, состояние гомеостатического механизма также изменяется, обеспечивая тем самым желаемый «буферный» эффект. Если различия между индивидуумами связаны с различиями в их определяемых средой гомеостатических состояниях, то такие различия являются адаптивными. Способность организма адаптивно реагировать на изменения среды следует отличать от адаптации, происходящей на генном уровне; назовем эту способность приспособляемостью. Примером приспособляемости у человека могут служить: изменение числа эритроцитов в зависимости от высоты местности над уровнем моря, интенсивности основного обмена в зависимости от температуры среды и изменение цвета кожи в зависимости от интенсивности солнечной радиации. В большинстве случаях приспособляемость выражается в обратимых физиологических процессах и в особенностях поведения, но возможно, что и такие необратимые процессы, как рост и развитие, также являются приспособительными.
Глава 11
ГЕНЕТИЧЕСКАЯ СТРУКТУРА
ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ L СИСТЕМЫ БРАКОВ
Популяции
Изучение генетической структуры популяций у человека только начинается, тем не менее эта область генетики столь важна для понимания многих проблем биологии человека, что даже то малое, что уже выяснено, заслуживает весьма внимательного рассмотрения. Мы коснемся здесь лишь нескольких самых простых вопросов общей генетики популяций.
Обычно термином «популяция» пользуются в том или ином географическом или экономическом контексте, говоря о популяции как о населении некоего города, местности, среды обитания. Такая популяция может вполне соответствовать популяции в генетическом смысле; однако генетика придает первоочередную важность родственным связям, а не географической близости. Поэтому удобно различать популяции в зависимости от распространения тех или иных генов, как это сделал Добржанский, определив менделе-вскую популяцию как «репродуктивное сообщество разнополых и перекрестно оплодотворяющихся особей, обладающих неким общим генетическим фондом».Подобно тому как географическая популяция страны слагается из популяций областей, так любая менделевская популяция состоит из других популяций меньшей численности. Самая крупная менделевская популяция — это вид; ведь каждая особь потенциально (если не фактически) обладает всеми теми генами, которыми обладает вид; различные виды имеют различные генофонды, так как представители разных видов не скрещиваются между собой. Нередко в пределах вида имеются различные подвиды или расы; это менделевские популяции более низкого ранга, которые в свою очередь могут состоять из еще более мелких групп. Различные расы удается распознать благодаря генетическим различиям между особями, входящими в их состав; однако для опреде-: ления границ популяции в первую очередь существенно, насколько с точки зрения размножения они изолированы друг от друга. Изолирующие факторы могут быть чисто географическими (например, большое расстояние или другие препятствия к передвижению); однако, поскольку речь идет о человеке, это могут быть и факторы социального или религиозного порядка, так что даже жители одного небольшого района могут часто образовывать ряд совершенно изолированных групп — изолятов. Изолятами обычно именуют небольшие группы, входящие в состав популяции, однако
Глава 11. Генетическая структура популяций. I.
191
размер их может быть и довольно большим при условии, что в отношении размножения соответствующие группы остаются некой отдельностью. Именно сохранение из поколения в поколение постоянства круга вступающих между собой в брак и обусловливает генетические различия между популяциями: ведь по причинам, которые будут рассмотрены ниже, гены, закрепившиеся в одной группе, имеют тенденцию ограничиваться именно этой группой. По крайней мере в прошлом расстояния и другие географические особенности служили наиболее устойчивыми барьерами на пути к смешению популяций; в этом кроется одна из причин широкой географической вариабельности человека как вида.
Неизбирателъные браки
Одна из простейших систем скрещивания, возможных в популяции разнополых особей,— это та, для которой характерна равная вероятность скрещивания любого индивидуума одного пола с любым индивидуумом противоположного пола в этой же популяции. Такое случайное скрещивание получило название панмиксии. Строго панмиктических групп в популяции человека не существует; даже если в группе, гомогенной по происхождению и изолированной от прочих групп, каждый из индивидуумов имеет равную возможность избрать для вступления в брак любого индивидуума противоположного пола, все равно предпочтения в выборе и социальные факторы воспрепятствуют чисто случайному заключению браков. Тем не менее существующие гены, аналогичные тем, которые определяют группы крови, обычно редко принимаются во внимание при подборе брачных пар и распределяются среди жителей некоторой весьма ограниченной местности так, как если бы подбор пар происходил более или менее случайно.
Влияние случайного подбора пар на распределение генов в той или иной популяции играет основополагающую роль в генетике популяций. Для рассмотрения только этого влияния следует допустить, что генотипы не различаются по жизнеспособности и плодовитости, что мутаций не происходит и что популяция достаточно велика, чтобы обеспечить случайность в подборе гамет. Для понимания последствий случайного подбора пар удобно рассмотреть процесс формирования некой панмиктической группы в результате гибридизации двух популяций — одной, которая содержит только один из пары генов, например А, и другой, которая содержит только соответствующий аллель, например А'. Все индивидуумы в этих исходных популяциях, очевидно, гомозиготны по рассматриваемому локусу соответственно для одного из двух рассматриваемых генов. В гибридной популяции окажутся не только гомозиготы обоих типов, но и гетерозиготы; возникает вопрос: какой будет частота этих трех генотипов при условии случайного подбора пар?
192
Часть II. Генетика человека
Ответ определяется, по-видимому, величиной пропорционального представительства обеих исходных популяций, образующих вследствие смешения новую популяцию. Предположим, что гомозиготная популяция АА дает долю р всех родителей гибридной популяции,, а оставшуюся долю q дает гомозиготная популяция А'А'. Тогда при условии одинакового соотношения мужчин и женщин в каждой популяции частоты браков различного типа будут p2(AAxAA)t 2pq(AA XA'A') и q2(A'A'xA'A'), как легко можно видеть из схемы:
р (АА) р2 (ААхАА) pq (А'А'ХАА) д(А'А') рд(ААхА'А') q2 (А' А' х А’А')
При браках между гомозиготными родителями одного типа могут, разумеется, родиться дети только того же типа, тогда как при браках между гомозиготами различного типа дети будут только гетерозиготными, так что соответствующие частоты генотипов детей будут р2АА, 2pqAA', q2A'A':
Частоты браков различного типа Частоты различных генотипов детей
р2 (ААхАА) Р2 (А А)
2pq (АА ХА'А') 2рд (АА')
q2 (А' А' ХА' А') q2 (А’А’)
Следовательно, именно таким будет состав родителей в следующем поколении. Таким образом, вместо двух генотипов родителей в смешанной популяции окажутся три генотипа. Количественный результат этого изменения вычисляется аналогичным образом: сначала вычисляют частоты различных типов брака, а на основе этих частот — частоты различных генотипов детей (схема 11.3).
Глава 11. Генетическая структура популяций. I.
193
Генотипы мужчин Генотипы женщин \ Частоты браков различного типа
р2 (АА) 2pq (АД') 92 (А'А')
р^(ААхАА) 2р39 (ААхАА’) р2д2 (А А хА'А')
2р? (АА’) 2р3д (АА' ХАА) 4р2д2 (АА’хАА’) 2pq3 (АА'хА'А')
q2(A’A') p2q2 (А' А' Х-АА) 2pq3 (А'А ХАА') д* (А'А' ХА'А')
На этой схеме три типа браков появляются дважды; рассматривая частоту генотипов детей в таких браках, соответствующие перекрестные браки суммируют:
Частота браков различного типа Частоты различных генотипов детей
АА АА' А'А'
(ААхАА) р* (ААхАА') ^р’г1 (АА ХА'А') 2p2q2 (АА’хАА’) 4р2д2 (АА' хА' A') 4pq3 (А'А' хА'А') д* Р1 2p3q 2p3q 2p2q2 p2q2 2p2q2 p2q2 2pq3 2pq3 9*
Суммарная частота различных генотипов детей p2 (p2 + 2pg + q2) + 2pq (p2 + 2pq + q2) + + q2 (p2 + 2pq-i q2)
Суммарная частота каждого из трех генотипов детей определяется суммированием результатов различных браков по каждому из столбцов таблицы. Множитель (p2-\-2pq-\-q2) имеется в каждой колонке; поэтому три различных генотипа встречаются с частотами р2(АА), 2pq(AA') и q2(A'A'). С такой же частотой встречаются различные генотипы родителей. Таким образом, если рассматривать одни лишь следствия чисто случайного скрещивания, то частота, с которой каждый из трех генотипов встречается в некоторой популяции, остается постоянной из поколения в поколение, а именно р2для АА, 2pq для АА' и q2 для А 'А'; после смешения популяций равновесие устанавливается за время жизни одного поколения. К этому в сущности и сводится закон Харди — Вейнберга.
194
Часть II. Генетика человека
Факт сохранения постоянства частот генотипов при смене поколений одновременно означает также, что частота обоих типов генов осталась прежней. Вспомним, что первоначально мы обозначили буквамир и (/частоты гомозигот АА и А'А' (соответственно) среди родителей вновь образованной популяции. Каждый из этих индивидуумов обладает двумя генами одного и того же типа; поэтому буквы р и q характеризуют также частоту обоих генов А и А ' в родительской популяции. В первом поколении частоты генотипов изменяются вследствие образования нового генотипа — гетерозиготы АА', однако частоты генов не меняются; поэтому их можно по-прежнему обозначать в каждом поколении через р и q.
Частоты генов чаще выражают в десятичных дробях, а не в обычных дробях и не в процентах. Так, если четверть всех генов определенного локуса в данной популяции принадлежит к типу А, то говорят, что частота этого гена равна 0,25. Если А' — единственный аллель гена А, то его частота должна быть 0,75, так как р+?=1.
Нередко возникает необходимость установить частоту определенных генов в данной популяции. В тех случаях, когда гетерозигота фенотипически отличима от обеих гомозигот, возможен прямой подсчет частоты обоих аллелей. Примером может служить наследование групп крови системы MN. Каждый индивидуум, кровь которого принадлежит к группе М, обладает двумя генами Ag^, а индивидуум, кровь которого относится к группе MN,— только одним геном Поэтому частота гена Ag1^ в данной популяции равна
2M-J-MN
2Т ’
где М и MN означают соответственно число индивидуумов с группами крови М и MN в этой популяции, а Т — общее число индивидуумов в популяции. Частоту гена 4gN можно подсчитать или по аналогичной формуле, или же путем вычитания частоты гена из единицы. Если же гетерозигота неотличима от гомозиготы, только что описанный способ явно неприменим и частоту генов приходится определять косвенным путем. Рассмотрим описанный выше признак — способность ощущать вкус фенилтиомочевины; при неизбирательных браках три генотипа будут встречаться с частотой р2(ТТ) : 2pq(Tt) : q2(tt). Частота гена Т, т. е. р, равна квадратному корню из частоты индивидуумов ТТ, а частота гена t, т. е. q, равна квадратному корню из частоты индивидуумов tt. Вспомним, что индивидуумы tt отличаются от гетерозигот, а ТТ не отличаются. Квадратный корень из частоты индивидуумов, не ощущающих вкуса фенилтиомочевины, в данной популяции равен поэтому частоте гена, определяющего отсутствие ощущения вкуса фенилтиомочевины (д); р получают, вычитая q из единицы.
Глава 11. Генетическая структура популяций. I.195
Тот же принцип можно применить и в тех случаях, когда приходится иметь дело не с парой аллелей, а с тремя и более аллелями. В системе групп крови АВО А и В доминантны по отношению к О, а между А и В нет отношений доминантности, т. е. гетерозигота АВ может быть обнаружена, а гетерозиготы АО и ВО нет. Поэтому в отсутствие других аллелей (например, А2) возможны только четыре различных генотипа. Пусть р — частота гена A, q — частота В и г — частота О, а p+q+r=1- Тогда при условии неизбирательных браков частота г равна квадратному корню из частоты индивидуумов, кровь которых относится к группе О. Величину q можно вычислить следующим образом:
д2 + 2д + г2 = /а + /о,
где /д и /о — частоты индивидуумов с группой крови А и 0 соответственно. Отсюда
(р + г)2 = /а+/о
Р+г = К/а+/о •
Но
р + ?+г = 1; поэтому
1—q — //а +/о и ______
q = 1—Г/а +/о •
Аналогично можно показать, что р=1—V/b+Zo. В реальной ситуации эти формулы приходится несколько модифицировать, но основной принцип метода соответствует изложенному.
Для описания различий между популяциями чаще приводят именно частоты генов (а не фено- и даже генотипов), знание которых существенно для решения многих проблем генетики. В особенности их необходимо знать для определения частоты гетерозиготных носителей некоторых вредных генов. Как мы видели в случае системы из двух аллелей, частота гетерозигот при панмиксии равна 2pq. Следовательно, максимальная частота гетерозигот достигается в популяции, где частоты обоих генов равны 0,5. В этом случае 50% индивидуумов будут гетерозиготными. Со снижением частоты одного из генов снижается частота не только гомозигот по этому гену, но и гетерозигот.
Рецессивный ген, определяющий ту или иную аномалию, может быть весьма редким в популяции, и тем не менее частота его носителей может быть сравнительно высокой. Например, альбинизм встречается с частотой 1 : 20 000. Следовательно, частота гена альбинизма q=^l : 20 000«0,00709, а частота аллеля дикого типа р =0,99291. Последнюю величину можно принять равной 1, тогда 2pq=2 х 0,00709 =0J014181 т. е. примерно 1 : 70.
196
Часть II. Генетика человека
Инбридинг и аутбридинг
Отклонения от панмиксии могут происходить в двух основных направлениях. Люди, состоящие между собой в родстве, могут вступать в брак или чаще или реже, чем при чисто случайном подборе пар. В первом случае говорят об инбридинге, во втором — об аутбридинге.
Аналогична инбридингу по последствиям система положительного ассортативного скрещивания; это система, при которой индивидуумы, сходные фенотипически, а следовательно, имеющие тенденцию к генетическому сходству, вступают между собой в брак чаще, чем по закону случая. В применении к человеку это справедливо главным образом в отношении умственного развития и роста: браки между индивидуумами, сходными по этим признакам, отмечаются чаще, чем можно было бы ожидать при случайном подборе пар. Отрицательное ассортативное скрещивание имеет место при предпочтительном вступлении в брак несходных индивидуумов. Имеются указания, что рыжеволосые вступают в брак друг с другом реже, чем следовало ожидать при случайном подборе пар. Следствия такого отрицательного ассортативного скрещивания в общем случае такие же, как при аутбридинге, однако ассортативное скрещивание (как отрицательное, так и положительное) затрагивает лишь генетические системы, контролирующие те признаки, по которым направляется ассортативное скрещивание, тогда как ин- и аутбридинг, если их рассматривать как ассортативное скрещивание по признаку получения потомства, затрагивают весь геном.
Самый тесный инбридинг — самоопыление — наблюдается у многих растений; па этом примере легче всего продемонстрировать последствия инбридинга. При самоопылении гетерозиготы АА' одна половина потомства оказывается гетерозиготной, а другая — гомозиготной (АЛ или А'А'). После самоопыления гомозиготы, разумеется, образуются тоже гомозиготы, но и половина потомства гетерозигот оказывается гомозиготной. Таким образом, при самоопылении частота гетерозигот в популяции непрерывно снижается — вдвое за одно поколение. В конце концов гетерозиготы практически исчезают из популяции, которая оказывается состоящей из гомозигот двух типов. Сказанное о единичном локусе, несомненно, распространяется на все локусы, так что тенденция к гомозиготности будет возрастать с той же скоростью; при исключении прочих факторов создается ситуация, при которой каждый индивидуум дает потомство, идентичное с ним по всем генам. Другое следствие инбридинга — это возникновение ряда совершенно различных линий. Так, исходя из двойной гетерозиготы АаВ -Ъ, можно получить следующие 4 линии: ААВВ, ААЬЬ, ааВВ и aabb. Темп снижения гетерозиготности как таковой не зависит от числа рас
Глава 11. Генетическая структура популяций. I.
197
сматриваемых генных пар, однако вероятное число гомозиготных индивидуумов и число индивидуумов, гетерозиготных по 1, 2, 3 и т. д. локусам, в любом поколении зависит от исходного числа пар и может быть вычислено путем развертки двучлена 1+ (2Г—1)”, где г — порядковый номер рассматриваемого поколения, а п — число генных пар.
Заметим, что при данном рассуждении исходили из допущения, что каждый член некоторого поколения участвует в размножении и все потомство достигает зрелости. При избирательном подборе пар темп приближения к полной гомозиготности, очевидно, зависит от генотипа исходного индивидуума. Если в качестве примера взять тот крайний случай, когда лишь один индивидуум — и притом гомозиготный — был родителем следующего поколения, то, очевидно, все последующие поколения с этого момента будут гомозиготами того же типа. Вместе с тем, если этот единственный родитель гетерозиготен, частота гетерозигот в дальнейшем будет снижаться. В случае самооплодотворения можно утверждать, что ни один индивидуум не может быть более гетерозиготным, чем его родитель; гетерозиготность индивидуума может быть равной гетерозиготности родителей или меньше ее, гомозиготность же всегда сохраняется.
Сказанное не относится к следующей форме тесного инбридинга — по типу брат — сестра или родитель — потомок; при подобных типах браков элиминирование гетерозигот требует большего числа поколений; так, 6 поколений самооплодотворения в этом отношении эффективнее 17 поколений браков по типу брат — сестра. Столь тесный инбридинг (брак брата и сестры) в человеческом обществе допускался лишь в очень редких случаях (в династии Птолемеев и в государстве инков); в племени эрнадан с Малабар-ского берега старшая дочь в семье обычно становилась второй женой отца. Однако обычно самый тесный инбридинг, допускаемый в человеческом обществе,— это браки между племянницей и дядей (племянником и теткой) или между двоюродными братом и сестрой. Брак между племянницей и дядей запрещен во многих государствах, а более чем в трети штатов США запрещены и браки между двоюродными сибсами. Длительное поддержание браков последнего типа создает тенденцию к общей гомозиготности, однако темп снижения частоты гетерозигот существенно ниже, чем при браках типа брат — сестра или родитель — потомок.
Браки между родственниками (близкородственные браки) имеют немаловажное значение с медицинской точки зрения, так как вероятность ситуации, при которой оба супруга обладают одинаковыми генами, будет выше, если они состоят в близком родстве. Разумеется, родство — это понятие весьма неопределенное. В эволюционном смысле все люди состоят в родстве; на некотором этапе существования человечества все они имели общих предков.
198
Часть II. Генетика человека
В этом легко убедиться, как только мы обратимся к какой-либо конкретной популяции. Если бы предки нынешних ее членов не состояли в родстве, то численность этой популяции в прошлом должна была бы быть нереально большой. Ведь у каждого из ныне живущих индивидуумов было 2 родителя, 4 прародителя, 8 пра-прародителей и т. д., или в общем виде 2" предков п поколений назад. Если принять, что за столетие сменяется в среднем 4 поколения, то это будет означать, что 1000 лет назад у индивидуума, предки которого не состояли в родстве, имелось 240 (миллион миллионов!) предков. Но в X в. н. э. все население земного шара не превышало 200 млн. человек, а в более ранние периоды истории человечество было еще малочисленнее! Поэтому при рассмотрении степени родства приходится исходить из признания существования некой гипотетической исходной популяции, в которой индивидуумы не состоят в родстве.
Следует различать гены, «идентичные по свойствам» и «идентичные по происхождению». Так, два гена AgM могут быть идентичными по свойствам, т. е. они способны взаимно замещать друг друга с идентичным генетическим эффектом, но в то же время они оба могли либо возникнуть при репликации одного гена в каком-то предшествующем поколении («идентичны по происхождению»), либо не иметь общего источника в столь близком прошлом; такие гены в смысле происхождения неидентичны («независимы»). Неидентичные по свойствам гены, разумеется, независимы (если исключены мутации).
Генетическое родство двух индивидуумов можно выразить как вероятность наличия у каждого из них в определенном локусе генов, идентичных по происхождению. Выше было показано, что этот коэффициент родства между родителями и детьми или между братьями и сестрами равен (гл. 9). Коэффициент родства между дедом (или бабкой) и внуками, а также дядей (теткой) и племянниками равен 4/4, а между двоюродными сибсами — Vs- Это значит, что если один из двоюродных сибсов является носителем ка-кого-то рецессивного гена, то вероятность наличия этого гена у другого равна */8- Мы принимаем величину вероятности максимальной потому, что данный двоюродный сибс может оказаться носителем рассматриваемого гена и в силу простой случайности. Если этот ген встречается редко, то это мало вероятно. Тем не менее в любом случае вероятность рождения пораженного ребенка резко возрастает, если носитель гена вступает в брак с двоюродным братом (или сестрой), а не с индивидуумом, не состоящим с ним в родстве, так как вероятность вступления в брак с гетерозиготным индивидуумом в общей популяции равна 2pq. Можно добавить, что для двоюродного брата (сестры) пораженного индивидуума вероятность оказаться носителем соответствующего гена равна */4 (если не выше), поскольку наверняка известно, что оба родителя пора
Глава 11. Генетическая структура популяций. I.
199
женного индивидуума гетерозиготны и один из них приходится ему дядей (теткой).
Рассматривая эти примеры, мы не только не принимали в расчет независимые гены, но и допускали, что генотип одного из вступающих в брак двоюродных сибсов нам известен; но в случае, когда один из супругов является носителем гена, это условие в действительности не выполняется. Поэтому мы вынуждены поставить вопрос в более общем виде: какова вероятность рождения ребенка с рецессивным патологическим признаком у вступивших между собой в брак двоюродных сибсов?
Рис. 11.1. Линии родства при браке между двоюродными сибсами.
Штрихами обозначены линии происхождения от деда, точками — от бабьи.
Чтобы ответить на этот вопрос, необходимо определить, какова вероятность того, что у определенного индивидуума в данном локусе окажутся два гена, идентичные по происхождению. Это так называемый коэффициент инбридинга (F); для потомков двух полностью неродственных родителей он равен 0. В общем / 1 \ п + п'+ 1 ,
случае F=( у) (1-J-FJ, где п и п —число поколении, счи-
тая от общего предка до родителей индивидуума, a Fz — коэффициент инбридинга для общего предка. Если этого предка можно считать неинбредным, то FZ=Q и тогда F— + + . Величина (1—F) и представляет собой вероятность того, что этот индивидуум имеет в данном локусе два независимых гена.
Двоюродные сибсы имеют двух общих предков среди прародителей (дедов и бабок). Возможные их родословные представлены на схеме (рис. 11.1).
Рассмотрим некий ген, носителем которого является дед; вероятность его передачи его сыну равна %, его внуку — У4 и (далее
200
Часть II. Генетика человека
по этому пути) потомству этих двоюродных сибсов — Vs- Однако этот же ген от деда может быть передан его дочери, внучке и потомству двоюродных сибсов с той же вероятностью 1/8. Тогда вероятность «встречи» этих генов равна J/8 Х1/8=1/64- Тем не менее существуют 4 гена, которые могут передаваться таким способом: тот, что был рассмотрен в этом примере, его аллель у деда и соответствующие 2 — у бабки. Поэтому существует вероятность 4х 1/в4==1/1в> что ребенок двоюродных сибсов будет гомозиготным по 2 генам, идентичным по происхождению. Соответственно существует вероятность 15/1в того, что ни один ген не «встретится» с таким же геном. Тот же результат получим, применив вышеприведенную общую формулу.
Два поколения отделяют двоюродных сибсов от их дедов по обеим возможным линиям родства, так что если сами прародители неипбредны, то для потомства двоюродных сибсов
/1Х2+2+1
I = uJ +
(общий предок мужчина)
( -
\ 2
(общий предок женщина)
2 4- 2 4- X
1
16'
Следовательно, в такой ситуации для ребенка, рожденного в браке между двоюродными сибсами, вероятность идентичности происхождения обоих генов данного локуса равна 1/18, а вероятность независимости этих генов — 15/1в. Для рецессивного гена, частота которого в данной популяции равна q, вероятность идентичности происхождения двух генов ребенка двоюродных сибсов равна (1/16) q. Вероятность того, что гены не «встретятся», равна (15/ie) (ft однако один ген может совпасть и с другим таким же геном независимо от происхождения с вероятностью (15/16) q X q. Вероятность гомозиготности ребенка равна сумме этих двух 115 1
вероятностей, т. е. q -f- yyy q2 — yg q (1 4- 15 q).
Описанные только что закономерности изложены в применении к бракам между двоюродными сибсами и к передаче рецессивных генов, так как именно такие близкородственные браки чаще всего наблюдаются в популяциях человека, а линии родства имеют исключительно большое значение в медицинской генетике; однако те же закономерности сохраняют силу в отношении всех близкородственных браков и всех генов. Речь идет при этом о законах распределения генов в потомстве, а не о природе или проявлении генов.
Коэффициент инбридинга популяции можно представить как среднюю величину коэффициентов для всех индивидуумов, образующих популяцию. Коэффициенты инбридинга были получены для различных групп, в особенности небольших и изолированных, таких, как островные популяции и некоторые религиозные секты.
Глава 11. Генетическая структура популяций. I.
201
К самым высоким показателям относятся величины, найденные для членов секты самаритян,— средний коэффициент инбридинга у них равен 0,0434, т. е. почти эквивалентен целой популяции, инбредной подобно потомству двоюродных сибсов. Далее можно заметить, что коэффициент инбридинга ближе к минимальным значениям, особенно если генеалогия известна лишь для нескольких поколений, поскольку невозможно установить, до какой степени сами общие предки были инбредны.
Индивидуумы, рожденные в браках между двоюродными сибсами, в какой-то мере инбредны; браки между кузенами, разумеется, возможны в популяции, для которой характерны неизбирательные браки. Частота таких браков в большинстве западноевропейских стран не превышает 1 %. Однако в определенных популяциях соответствующая цифра значительно выше, например в сельских районах на севере Швеции, в высокогорных альпийских деревнях Швейцарии и в еврейских общинах многих городов Германии. Здесь нередко частота браков между кузенами достигает 6, а порой и 12%. Отсюда не следует, что в этих популяциях такие браки считаются предпочтительными; просто эти популяции слишком малы. Если группа, из которой выбирается супруг, мала, то составляющие эту группу индивидуумы неизбежно окажутся в более близком родстве, чем члены более многочисленной группы. В частности, в малой группе чаще встречаются двоюродные родственники и неизбирательные браки чаще заключаются между родственниками. Если же в небольших швейцарских общинах частота родственных браков относительно низка, надо думать, что жители умышленно избегают заключения такого рода браков; следовательно, подбор пар не случаен. Наличие связи между частотой браков между кузенами и размером изолята позволило Дальбергу создать метод определения величины изолята.
Пусть в некоторой популяции в каждом браке рождается в среднем х детей. Тогда у каждого индивидуума имеется (х—1) дядьев или теток со стороны каждого из родителей, всего 2(х—1). Поскольку в среднем каждый из дядьев и теток имеет х детей, число двоюродных сибсов у каждого индивидуума равно 2(х— 1) х. Половина из них, т. е. х(х—i), принадлежит к полу, противоположному полу данного индивидуума, т. е. составляет то число, из которого последний может выбрать себе пару. Пусть N — число индивидуумов в изоляте; тогда N/2 — число индивидуумов противоположного пола, т. е. общее число возможных супругов. Тогда вероятность с брака между двоюродными сибсами составит
. л(ж—1)
А72 •
Отсюда
_2ж(а-—1)'
202
Часть II. Генетика человека
Биологические последствия различных систем браков
В небольших популяциях, несмотря на случайный выбор супруга, отмечается тенденция к возрастанию инбредности. Значение этого факта легко понять, если вспомнить, что у многих видов растений и животных продолжительный инбридинг часто (хотя и не всегда) ведет к снижению жизнеспособности и плодовитости. Одно из возможных объяснений состоит в том, что вредные рецессивные гены (которые при аутбридинге, или случайном скрещивании, содержатся в популяции в скрытом виде) переходят в гомозиготное состояние. Теоретически возможно, что ни один из этих генов не закрепится ни в одной линии; однако если неблагоприятные последствия связаны во многими локусами (как это обычно и бывает), то вероятность этого весьма мала, особенно для популяции, самый размер которой ограничивает число устанавливающихся линий. Предполагают также, что «инбредная депрессия» витальных свойств популяции представляет собой прямое следствие утраты гетерозиготности как таковой; гетерозиготам, обладающим двумя различными генами в некотором локусе, свойственна большая «биохимическая пластичность», чем гомозиготам. В пользу этого свидетельствует немало фактов; тем не менее ясно, что организмы способны приспосабливаться к гомозиготности, ведь многие из них размножаются в условиях самооплодотворения без каких бы то ни было вредных последствий. Поэтому основой приспособленности организмов можно считать специфическую природу генов и способность их к интеграции в сбалансированный генотип.
На протяжении небольшого периода времени самооплодотворение обеспечивает немалые преимущества, особенно для неподвижных форм, таких, как растения. Комбинированное влияние инбридинга и естественного отбора ведет в конце концов к формированию популяции, все индивидуумы которой гомозиготны в отношении одинаковых генов. Отсюда вытекает высокая вероятность того, что каждый индивидуум будет иметь фенотип, наиболее соответствующий преобладающим условиям внешней среды, причем способность такой популяции реагировать на изменения среды низка. Для популяции же с высокой генетической вариабельностью более высока вероятность выживания в ходе эволюции; хотя из-за неизбежной фенотипической вариабельности число индивидуумов с оптимальным фенотипом невелико, популяция в целом оказывается в некотором смысле преадаптированной к изменениям среды. Поэтому можно ожидать, что при постоянстве условий внешней среды самооплодотворяющиеся виды будут успешно конкурировать с видами, у которых более выражен аутбридинг; однако в природе условия внешней среды непрерывно изменяются, и поэтому у боль
Глава 11. Генетическая структура популяций. I. 203
шинства организмов не имеет места самооплодотворение. Огромные эволюционные преимущества двуполости и перекрестного скрещивания заключаются в том, что при этом обеспечивается возможность возникновения большого числа комбинаций различных генов. Это весьма обобщенное изложение проблемы призвано показать, почему популяции генетически гетерогенны; более частные ситуации, иллюстрирующие это положение, будут рассмотрены ниже. В частности, можно ожидать, что в некоторой популяции имеется более одного гена для большинства полигонных локусов, поскольку именно степень гетерозиготности по этим локусам существенна для распределения вариантов количественных признаков (гл. 9). Иногда такое распределение оказывается тем механизмом, который обеспечивает идеальный компромисс между формированием определенного фенотипа, с одной стороны, и фенотипической вариабельностью, с другой; вот почему изменения условий внешней среды вызывают изменение тех признаков, которые способны варьировать количественно. Для популяций, для которых характерен аутбридинг, это означает, что большинство индивидуумов в популяции будет гетерозиготно по многим поли-генным локусам, а именно эти индивидуумы (а не редкие в таких популяциях гомозиготы), вероятно, окажутся ближе всех к оптимальному генотипу. Гомозиготы можно рассматривать как неизбежный побочный продукт в процессе формирования количественных признаков. Далее, широкая распространенность гетерозигот создает условия для возникновения в них путем естественного отбора эффективных механизмов гомеостаза. Не удивительно поэтому, что инбридинг генетически сформированных таким образом организмов будет давать прежде всего формы с низкой жизнеспособностью — ведь это формы, лишь изредка возникающие в естественных условиях и неспособные создать фенотип, адекватный среде.
Общая структура человеческих популяций в ходе большей части эволюции — это структура, типичная для небольших долго существовавших изолятов, которая время от времени нарушалась в связи с миграциями, вторжениями или смешением. Именно в этом контексте следует воспринимать особенности эволюции человека с характерными для нее периодами дифференцировки в пределах малых по-пуляций, прерываемой периодами притока генов. Однако в настоящее время в связи с прогрессом техники круг возможных браков значительно расширился, замкнутость изолятов нарушается во всем мире и, по-видимому, навсегда. Почти повсеместно этот процесс зашел столь далеко, что ничего подобного обособленным популяциям не осталось. Однако в условиях, препятствующих повышению плотности населения, а также в достаточно прочных социальных и религиозных группах (особенно при сохранении примитивной техники) этот процесс только еще начинается. Дарлингтон указы
204
Часть II. Генетика человека
вал, что население подобных длительно существующих изолятов, по-видимому, не испытывает неблагоприятных последствий инбридинга потому, что оно в какой-то мере приспособилось к гомозиготности. Тем не менее есть основания считать, что широко распространенное в Европе увеличение роста, которое началось в XX в., частично связано с разрушением изолятов. Нет оснований утверждать, что данный признак обязательно связан с общим повышением приспособленности, хотя вполне возможно, что это так и есть. У растений и животных при инбредной депрессии обычно понижается скорость роста и уменьшаются размеры взрослой особи; гибридизация таких инбредных форм не только восстанавливает их жизнеспособность, но и ускоряет рост. Повышение жизнеспособности и плодовитости вследствие гибридизации известно под названием гибридной мощности, или гетерозиса. Гетерозис наблюдается не только при гибридизации форм, ранее размножавшихся строго инбредно, но и при скрещивании различающихся подвидов растений и животных. Если рассматривать лишь жизнеспособность как таковую, то последняя может возрасти даже при гибридизации хороших видов. Это явление, казалось бы, подтверждает воззрение, согласно которому всякая гетерозиготность способствует выживанию. Однако в подобных случаях жизнеспособность особей второго поколения оказывается меньше не только жизнеспособности особей первого поколения, но и жизнеспособности родительских подвидов. Гибриды второго поколения между инбредными линиями менее жизнеспособны, чем гибриды первого поколения, поскольку в целом они менее гетерозиготны, но все же их приспособляемость выше, чем у их инбредных родителей. В настоящее время невозможно решить, настолько все сказанное применимо к человеку. Во многих областях земного шара браки между представителями различных больших рас стали обычным делом. Несмотря на всякого рода ограничения, большие смешанные группы имеются в Южной Африке и на юге США; межрасовые браки широко распространены также в Южной Америке и на островах Полинезии (особенно на Гавайях). Большинство непредубежденных исследователей не нашли при этом реального снижения приспособленности, а на острове Питкерн, где мятежники с «Баунти» взяли себе жеп-полинезиек, отмечаются некоторые признаки гетерозиса.
Глава 12
ГЕНЕТИЧЕСКАЯ СТРУКТУРА
ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ II. ИЗМЕНЕНИЯ ЧАСТОТ ГЕНОВ
Хотя система браков, господствующая в пределах той или иной популяции, предопределяет частоту различных генотипов (причем инбридинг создает тенденцию к гомозиготности, а аутбри-динг — к гетерозиготности), тем не менее на частоты генов система браков как таковая не влияет. Изменение системы браков не влечет за собой ни утраты, ни приобретения генов. Этот факт не означает, что система скрещивания не имеет значения для эволюции, но поскольку эволюция подразумевает некоторую замену одного набора генов другим, к основным механизмам эволюции относятся те факторы, которые вызывают изменение частот генов. Известны четыре таких фактора: гибридизация, мутация, дрейф генов и естественный отбор. Благодаря первым двум явлениям в популяцию поступают новые гены, а два последних определяют их дальнейшую судьбу.
Гибридизация
Изменение частоты того или иного гена в популяции вслед за появлением в ней индивидуумов из другой популяции зависит от числа иммигрантов и от частоты этого гена среди них. Если частота двух аллелей А и А ' равна соответственно рх и 7, в исходной популяции и р2 и q2 среди иммигрантов, а вновь образованная популяция состоит из доли х исходной популяции и доли у иммигрантов, то частота аллеля А становится равной хр^уРъ, а аллеля А '—хц^ург- Если в исходной популяции один из аллелей отсутствует (например, q'1=0), то частота А ' после смешения популяций, очевидно, равна yq2. Влияние смешения на частоту генов легко подсчитать; однако чаще приходится решать, сколь велика примесь в уже существующей гибридной популяции. Сделать это нетрудно, если известны частоты некоторого варьирующего гена как в родительской, так и в гибридной популяции. Вклад исходной популяции в гибридную равен {qk—q2) : (д2—?2), где qk — частота гена в гибридной популяции, qt — его частота в исходной популяции, a q2 — в группе иммигрантов. Вклад иммигрантов равен (?* — ?i) : (?2 — ?i), или ! — Qi)- (?i — Qi)- Разумеется, величины q± и q2 следует определить, изучая популяции, генетически подобные тем, которые положили начало гибридной популяции
206
Часть II. Генетика человека
и не претерпели с тех пор существенных изменений. Таким способом было установлено, что примесь от белых у американских негров составляет 25%, а у бразильских — 40%. Когда две человеческие популяции оказываются в контакте, они далеко не сразу превращаются в единую панмиктическую популяцию, особенно если они сильно различаются по физическим признакам или образу жизни, что неизбежно ведет к ограничению случайного подбора брачных пар. В течение некоторого времени различные группы живут рядом, в одной и той же местности, и тем не менее остаются более или менее изолированными друг от друга в генетическом отношении. Однако в конце концов смешанные браки неизбежны, что ведет к потоку генов из одной популяции в другую. Скорость этого потока можно вычислить, применив уравнение
(1— ' ' 31 —«2
правая часть которого равна доле примеси на данный момент (см. выше), а к — число поколений, при образовании которых имел место поток генов. Модальная скорость потока т — это доля генов в смешанной популяции, которые поступают за одно поколение из популяции с частотой гена q2. Этот метод позволил установить, что скорость потока генов из популяции белых в популяцию негров в США колеблется от 0,02 до 0,025 (т. е. 2—2,5%), а в Бразилии — от 0,045 до 0,055.
Две генетически различающиеся популяции могут быть географически удалены друг от друга, но связаны рядом промежуточных популяций, через которые осуществляется поток генов. Создается, так сказать, географический градиент частоты генов между крайними популяциями. Этот градиент исчезает лишь после того, как обе крайние и все промежуточные популяции окажутся идентичными. Подобные градиенты частот генов называются гено-клинами, тогда как градиенты вариантов тех признаков, которые обусловливаются соответствующими генами, называют фенокли-нами. Клины, или клинальная изменчивость, часто встречаются в популяциях человека. К наиболее изученным примерам клина относятся распределение в Европе гена группы крови В (ген В), частота которого постепенно снижается в направлении с востока на запад. Это явление обычно считают следствием притока генов из Азии, чему способствовали неоднократные вторжения в Европу с Востока.
Мутация
Гибридизация — это лишь способ передачи уже существующих генов из одной популяции в другую; в результате гибридизации могут возникнуть совершенно новые комбнации генов; возникно
Глава 12. Генетическая структура популяций. II. 207
вение же действительно новых генов возможно только путем мутации. Мутацию определяли как возникновение наследуемого изменения', в широком смысле это понятие приложимо как к отдельным генам, так и к целым хромосомам. В результате мутации возникает мутантный ген или мутантная хромосома, дающая начало мутантному признаку. Генные мутации могут возникнуть в любой момент, по их появление более вероятно в делящейся, а не в «покоящейся» клетке; при гаметогенезе появление мутаций наиболее вероятно во время мейоза. В генетическом отношении важны именно те мутации, которые возникают при гаметогенезе, но и соматические мутации имеют определенное значение (в частности, медицинское). Мутантный ген воспроизводится с высокой точностью до тех пор, пока он не изменится в результате новой мутации.
Причины возникновения генной, или «точковой», мутации еще далеко не выяснены; тем не менее раскрытие генетического кода позволило выяснить ее природу. В типичном случае она возникает при замене одного азотистого основания другим в одном из триплетов ДНК. Поскольку код избыточен, некоторые такие замены не влияют на синтез белка; однако нередко замена одного основания влечет за собой включение в полипептидную цепь другой аминокислоты. Так, если в триплете ЦТТ цитозин замещен тимином с образованием ТТТ, то на соответствующем месте в синтезируемом полипептиде вместо глутаминовой кислоты окажется лизин. Оказалось, что именно такого рода изменение ведет к образованию 0-цепи гемоглобина С (НЬ-С) в отличие от 0-цепи нормального гемоглобина взрослых (Hb-А) (см. гл. 16). Иногда замена основания влечет за собой сигнал стоп при считывании кода, например если аденин заменяет цитозин в ЦТТ с образованием АТТ. В таком случае будет протекать неполный синтез исходного полипептида — образуется лишь та его часть, которая предшествовала сигналу стоп. Возможна также вставка совершенно нового основания или делеция ранее существовавшего основания. Подобные изменения приводят к так называемому сдвигу рамки и могут вызвать изменение всех последующих аминокислот в цепи. Например, вставка дополнительного цитозина перед ЦТТ (глутаминовая кислота) — ААА (фенилаланин) приведет к тому, что код будет прочтен так: ЦЦТ (глицин) — ТАА (изолейцин) — А--------.
Генная мутация представляет собой обратимый процесс. Так, если ген А мутирует в аллель А', то существует равновероятная возможность того, что А' мутирует обратно в А. Часто мутацию называют случайным процессом, но это не означает, что число путей, по которым может мутировать определенный ген, неограниченно велико; это лишь означает, что соответствующее изменение не связано с изменением окружающей среды. Естественно, когда речь идет о замене оснований в ДНК, возможности хоть и многочисленны, но число их конечно; последствия изменений сильно зависят от
208
Часть II. Генетика человека
кодона, в котором произошла мутация. Обычно мутантный ген влияет на развитие той же системы признаков, что и ген, из которого он возник. Однако он влияет на эту систему либо иным способом, либо в иной степени. Так, ген, определяющий цвет глаз, может путем мутации превратиться из гена карих глаз в ген голубых глаз; вероятность превращения данного гена, скажем, в ген, контролирующий группу крови, чрезвычайно мала.
Хромосомные мутации известны в двух основных формах: изменение числа целых хромосом и изменение числа и расположения генов в пределах одной хромосомы. При увеличении числа наборов хромосом возникает так называемая полиплоидия. Последняя представляет собой следствие аномального деления клетки в мейозе, так что возникают гаметы, имеющие, например, диплоидное число хромосом. При оплодотворении такой гаметы другой гаметой с нормальным, гаплоидным, числом хромосом образуется триплоид-ная зигота; если же и вторая гамета, участвующая в оплодотворении, имеет диплоидный набор хромосом, возникает тетраплоидная зигота. Если меняется лишь число хромосом одного или нескольких типов, а не число наборов, то говорят о гетероплоидии, или полисомии.
В настоящее время хорошо изучен пример патологии, возникающий вследствие полисомии у человека; это синдром Дауна. У индивидуумов с этим синдромом, как правило, отмечается замедление роста и развития, аномальный дерматоглифический рисунок, наличие характерной кожной складки в углу глаза (эпи-кант), различные патологические состояния, в том числе предрасположение к лейкозу и выраженная умственная отсталость. Вероятность рождения ребенка с таким синдромом резко повышается с возрастом матери от 0,04% у женщин моложе 30 лет до 3,15% у женщин старше 45 лет. Этот синдром обычно развивается вследствие того, что одна из хромосом группы G представлена трижды, а не дважды во всех клетках тела в результате нерасхождения хромосом в мейозе или какого-то иного нарушения клеточного деления. В течение ряда лет считали, что речь идет о хромосоме 21, и до сих пор этот синдром называют «трисомией по хромосоме 2Ь, хотя теперь точно установлено, что аномалия касается хромосомы 22.
Изменение числа или расположения генов возможно одним из следующих четырех путей. Первый — утрата части хромосомы (делеция). Подобным примером может служить синдром, известный как «cri du chat» (кошачий крик); свое название синдром получил в связи с более или менее постоянным симптомом: новорожденный с такой патологией нередко издает жалобный крик; для синдрома характерны серьезные нарушения физического и умственного развития. У пораженных утрачено короткое плечо хромосомы 5; они гетерозиготны по данной аномалии.
Глава 12. Генетическая структура популяций. II,
209
Второй путь изменений — в каком-то смысле обратная ситуация — это увеличение длины хромосомы вследствие повторения некоторого сегмента (дупликация). Известны механизмы, благодаря которым может осуществиться полная или почти полная дупликация структурного гена. Убедительно показано, что гены, кодирующие |3- и 6-цепи гемоглобина А и гемоглобина Аа и тесно сцепленные друг с другом, возникли вследствие дупликации одной исходной последовательности ДНК; дупликация сыграла существенную роль в эволюции различных иммуноглобулинов и некоторых их участков. Дупликация — это, несомненно, тот процесс, с помощью которого может увеличиваться общее количество ДНК; она же создает новые гены, которые могут подвергнуться мутационным изменениям и будут «предложены» для отбора, тогда как старые гены продолжают выполнять свои исходные необходимые функции. Дупликация, вероятно, играет крайне важную роль в ходе эволюции.
Третий путь изменений — перенос части хромосомы на другую хромосому негомологичной пары (транслокация); примеры транслокации известны и в генетике человека. Так, часть хромосомы 21 может транслоцироваться на другую аутосому, например 14. У такого индивидуума никаких отклонений от нормы нет, так как в его клетках содержится полный комплект хромосом (одна хромосома 14, одна хромосома 21 и одна слитная хромосома 14-21); однако если при мейозе хромосома 14-21 попадет в ту же гамету, что и хромосома 21, а затем произойдет оплодотворение нормальной гаметой, содержащей одну хромосому 21, то хромосома 21 окажется в зиготе, представленной трижды, и разовьется синдром Дауна. Таким образом синдром Дауна может наследоваться по Менделю, и не у всех индивидуумов с синдромом Дауна имеется отдельная добавочная хромосома.
Наконец, некий сегмент хромосомы может оказаться перевернутым (инверсия). Так, вместо прежнего порядка локусов, например а, Ь, с, d, е, f, g, возможен иной порядок — a, b, с, f, е, d, g. Такого рода изменения хромосом часто встречаются у дрозофилы, где их легко обнаружить и где они имеют огромное значение для формирования эволюционной приспособленности организма. Почти с полной уверенностью можно говорить о существовании подобных мутаций и у человека, хотя их обнаружению и препятствуют малые размеры хромосом.
Генные мутации появляются с определенной, характерной для них частотой. Для единичного локуса эта величина составляет в среднем 1 : 100 000 половых клеток, хотя и существуют значительные колебания этой величины от локуса к локусу. Для каждого локуса в отдельности мутация — чрезвычайно редкое событие; однако у человека насчитывается от 5000 до 120 000 генов
210
Часть II. Генетика человека
(если не более), и поэтому для генотипа в целом мутация — явление совсем не редкое.
На частоту мутаций влияют многие факторы. Известно, что в сперматозоидах молодых самцов дрозофилы больше мутантных генов, чем в сперматозоидах старых самцов; у человека влияние возраста на частоту мутаций замечено, например, в отношении ахондропластической карликовости: вероятность рождения больного ребенка у более пожилых супругов выше, чем у молодых. Некоторые гены Х-хромосомы мутируют в мужском организме чаще, чем в женском. Подобное влияние пола в отношении сцепленных с полом генов нетрудно распознать, сопоставляя частоту рождения гемизиготных сыновей и гетерозиготных дочерей у родителей, у которых нет мутантных генов. Если сцепленный с полом ген мутирует с одинаковой частотой в мужском и женском организме, то следует ожидать, что отношение числа гемизиготных сыновей к числу гетерозиготных дочерей равно 1:2 — ведь принадлежащая мужчинам треть Х-хромосом популяции передается ими дочерям, а принадлежащие женщинам 2/я Х-хромосом передаются поровну их сыновьям и дочерям. Если мутация возникает только у женщин, то сцепленный с полом мутантный ген с одинаковой вероятностью будет передан дочерям и сыновьям и отношение числа гемизиготных сыновей к числу гетерозиготных дочерей окажется равным 1:1. Если же сцепленный с полом мутантный ген появляется только в мужских гаметах, то вначале он будет обнаружен только у гетерозиготных дочерей. Анализ такого рода позволил установить, что некоторые сцепленные с полом мутации, по-видимому, возникают в мужском организме в 10 раз чаще, чем в женском.
При наличии достаточных демографических данных (например, в Дании) нетрудно определить частоту мутаций доминантного признака с полной пенетрантностью, поскольку у индивидуумов, обладающих мутантным геном, последний проявляется сразу же. Частоту мутаций вычисляют, исходя из частоты рождения детей с мутантным признаком у родителей, не имеющих данного признака. В Копенгагене на 94 075 родов зарегистрировано 10 случаев ахондроплазии. Поскольку в двух случаях подобная аномалия имелась и у родителей, то частота вновь возникших случаев составляет 8 : 94 075, т. е. примерно один на 12 000. Так как каждый индивидуум обладает двумя аллелями, частота мутаций для гена дикого типа к аллелю ахондроплазии составляет один на 24000. Это довольно высокий показатель; некоторые факты позволяют полагать, что ахондроплазия может возникать вследствие мутаций более чем одного локуса. Возможно также, что ахондроплазия иногда оказывается рецессивным признаком или фенокопией. Тем не менее подобный «прямой» метод остается наиболее правильным путем определения частоты мутаций для доминантного признака. Этот метод явно неприменим, когда речь идет о рецессивном при
Глава 12. Генетическая структура популяций. II. 211
знаке; ведь если вследствие мутации возникает рецессивный ген, то он получит выражение только при сочетании с другим геном такого же типа. Если же частота этого гена низка (а она будет оставаться низкой в популяции до тех пор, пока поддерживается лишь за счет возникновения новых подобных мутаций), то появлению гомозиготного рецессивного индивидуума должна предшествовать смена многих поколений.
Имеется и другой, «непрямой» метод определения частоты той или иной мутации, основанный на возникновении равновесия между появлением новых генов в популяции вследствие мутации и их элиминацией под действием естественного отбора. Большинство мутантных генов оказываются вредными для организма. Это и не удивительно, если вспомнить, что характер мутации не зависит от внешней среды. Генотип — это высокоинтегрированная система взаимодействующих элементов, и случайные изменения в ее составе скорее всего повлияют на нее отрицательно. Поэтому отбор действует в направлении элиминации большинства мутантных генов из популяции, и присутствуют они в ней только, потому, что постоянно возникают вследствие новых мутаций. Частота мутантных генов в популяции будет поэтому постоянной, если скорость их появления будет равна скорости их элиминирования. Равновесие, разумеется, будет достигнуто лишь в случае, если частоты мутации и элиминации были равны в течение некоторого времени, п есть основания считать, что если в настоящее время величина этих показателей изменилась, то, возможно, в прошлом они были более или менее постоянными в течение достаточно длительного периода времени.
Рассмотрим редко встречающийся доминантный аномальный признак, сохраняющийся в популяции только вследствие появления новых мутаций. Если этот признак встречается в популяции из N родителей с частотой х, то число пораженных родителей будет равно xN. Все они гетерозиготны по мутантному гену, так что xN их генов будут мутантными и xN — дикого типа. Число непораженных родителей равно N — xN с общим числом генов дикого типа 2 (N — xN). Поэтому общее число аллелей дикого типа, которые могут мутировать, равно xN -[-2N—2xN=2N—xN=N (2—x). Если частота данной мутации равна и, то число новых мутантных генов среди потомства этих родителей равно uN (2— х). Поскольку частота данного признака весьма низка, то влияние х на величину этого выражения настолько мало, что им можно пренебречь, и тогда частота вновь возникающих мутантных генов равна 2Nu. Способность мутантного гена к самовоспроизведению / определяется как частота, с которой этот ген передается следующему поколению по сравнению с аллелями дикого типа. Поэтому частота его элиминации равна (1—/). Поскольку частота мутантных генов в родительской популяции равна Nx, то число му-
212
Часть II. Генетика человека
таитных генов, утрачиваемых популяцией за время одного поколения, равно (1—f)Nx.
В случае установившегося равновесия число вновь возникающих мутантных генов равно числу элиминированных, т. е. 2Nu= (l—f)Nx, где и=-^(1—f)x. В случае рецессивного признака в состоянии равновесия н = (1—f)x, так как утрачиваются два мутантных гена при элиминировании каждого индивидуума. Для полностью сцепленных с Х-хромосомой рецессивных признаков м=-д-(1—f)x', где х'—частота пораженных данной аномалией мужчин, так как мутантный ген подвержен действию отбора главным образом в организме мужчин, а они являются носителями трети всех Х-хромосом в данной популяции.
В качестве примера применения непрямого метода вернемся к рассмотрению ахондроплазии. Как говорилось выше, в Дании на 94 075 родов было зарегистрировано 10 случаев ахондроплазии, так что частоту этой аномалии в популяции (х) можно считать равной 10/94075. Показатель способности мутантного гена к самовоспроизведению можно определить, сравнив число детей, родившихся у карликов, с числом детей, родившихся у их здоровых сибсов. В то время в Дании было зарегистрировано 108 карликов, у которых было 27 детей, причем только 10 из них унаследовали карликовость, а 17 оказались нормальными; следовательно, их соотношение существенно не отличалось от 1,0. У 457 здоровых сибсов было 582 ребенка. Поэтому способность мутантного гена к самовоспроизведению следует считать равной (27/108) • (457/582) = 0,196. Способ нахождения последней величины станет более ясным, если проследить за следующим рассуждением. В среднем 108 генов ахондроплазии в одном поколении представлены в следующем поколении 13,5 гена (х/2- 27); 582 нормальных аллеля из 914 (2-457) перенесены из того же поколения в следующее. Отсюда относительная способность мутантного гена к самовоспроизведению равна (13,5/108) • (914/582)=0,196. Таким образом, по сравнению с нормальным аллелем лишь 19,6% мутантных генов передаются из одного поколения последующему, а элиминируется 80,4%; (1—/) =0,804. Поэтому показатель частоты мутаций и равен
i/2• 0,804- (10/94075) да 0,000043,
т. е. 4,3 гаметы из каждых 10° гамет оказываются носителями нового мутантного гена ахондроплазии, приблизительно 1 на 23 000. Получено, таким образом, довольно близкое соответствие между результатами применения прямого и непрямого методов.
Можно привести примеры частоты мутаций и для некоторых других локусов: 4 на 105 для гена ретинобластомы, 3 на 105 для ге
Глава 12. Генетическая структура популяций. II.
213
на гемофилии и от 0,8 до 1,2 на 105 для гена эпилойи; возможно, эти показатели представляют собой максимальные величины; относительная частота этих аномалий сделала их наиболее удобными для исследования. Частоты мутаций такого же порядка обнаружены у растений и животных. Если частоту мутаций отнести к числу поколений, то полученные показатели оказываются близкими для всех организмов. Большие различия в продолжительности жизни одного поколения влекут за собой столь же большие различия между видами в частоте мутаций за единицу времени.
Как уже говорилось, характер мутаций не зависит от внешней среды, однако такие факторы внешней среды, как ионизирующие излучения и некоторые химические вещества, увеличивают частоту мутаций. К ионизирующим излучениям относятся рентгеновские и ультрафиолетовые лучи, а-и |3-частицы, а также нейтроны. На пойкилотермных животных мутагенное действие оказывают также инфракрасные лучи; в эксперименте в качестве химических мутагенов часто используют производные иприта. Мутагенный эффект различных типов излучений зависит от их проникающей способности, но все типы ионизирующего излучения способны вызывать мутации любого типа — как генные, так и хромосомные. Химические же мутагены отличаются некоторой избирательностью действия как в смысле типа вызываемых мутаций, так и в отношении поражаемого мутацией локуса.
Мутагенное действие излучений кумулятивно, так что не обязательно вся необходимая доза должна быть получена одномоментно. Однако недавно было показано, по крайней мере на гонадах мыши, что при хроническом облучении частота мутаций вчетверо меньше, чем при однократном применении той же дозы. Тем не менее для многих организмов и в довольно широком диапазоне доз число мутаций пропорционально полученной дозе.
Несмотря на то что действие излучений кумулятивно, естественный радиоактивный фон — около 90—150 мрад в год — недостаточно высок, чтобы им можно было объяснить частоту спонтанных мутаций; в связи с этим удалось обнаружить гены, которые сами по себе повышают частоту мутаций. Предполагают, что эти гены создают для остального генотипа такую химическую среду, в условиях которой его стабильность снижается. Важно отметить при этом, что поскольку частота спонтанных мутаций хотя бы частично подвержена генному контролю, эта частота подвержена и влиянию естественного отбора.
Воздействие на человека высоких доз коротковолновых излучений вызывает развитие лучевой болезни. Если человек получает одномоментно дозу около 600Р, то примерно через 2 ч у него появляется тошнота; затем следует бессимптомный период, после чего развивается понос, рвота и ангина, повышается температура, наступает истощение; примерно через 2 недели облученный поги
214
Часть II. Генетика человека .
бает от анемии, сепсиса и множественных кровоизлияний. Для лиц, подвергающихся опасности облучения в производственных условиях, рассчитана максимально допустимая доза — ОДР в неделю. Однако при этом расчете, по-видимому, не учитывалась опасность мутаций; ведь считается, что доза 80 Р на одно поколение удваивает частоту мутаций, а, как уже говорилось, генетический эффект излучения кумулятивен.
Мутантные гены чаще всего бывают рецессивными. Объясняется это вредным действием большинства мутаций; поскольку мутации повторяются, есть все основания считать, что они происходили в данной популяции и в прошлом. В большинстве генотипов действие мутаций смягчалось благодаря присутствию дикого аллеля в геноме (см. гл. 9). Как полагает Мёллер, для того чтобы под влиянием повышенного радиоактивного фона даже в наиболее тесно инбредных популяциях возник индивидуум, гомозиготный по тому или иному рецессивному мутантному гену, должно пройти не менее 600 лет; возможно, пройдет несколько тысяч лет, прежде чем появится такой гомозиготный организм. Генетический эффект облучения редко проявляется сразу, однако не следует недооценивать грозящей будущим поколениям опасности накопления вредных генов в популяции.
Постоянное присутствие в популяции вредных мутантных генов, которых на весь геном приходится немало, рассматривается как своеобразный груз, который приходится нести данной популяции. Однако кроме этого мутационного груза следует иметь в виду и другой, возникший вследствие сегрегации в полиморфных системах гомозигот, имеющих меньшую приспособленность, чем гетерозиготы, например в случае локуса, направляющего синтез нормального и серповидноклеточного гемоглобина. Вместе с мутационным грузом этот сегрегационный груз образует «генетический груз». Насколько сегрегационный груз действительно является грузом, еще спорно; нелегко на деле провести различие между сегрегационным и мутационным компонентами, особенно для человека. Те, кто считает полезной концепцию груза, оценивают его величину в различных человеческих популяциях, сравнивая смертность в потомстве от близкородственных браков (которая по упомянутым выше доводам, вероятно, выражает груз) со смертностью в потомстве не стоящих в родстве родителей. По этим оценкам будто бы следует, что общая величина груза в обеих группах одинакова, хотя вклады различных локусов различаются.
Вследствие повторения мутации соответствующий ген постоянно вводится в популяцию, но даже если мутантный ген, как это изредка случается, окажется не лишающим организм каких-либо преимуществ, давление мутации приведет лишь к весьма медленному увеличению частоты этого гена, поскольку мутации — это не только весьма редкое, но еще и обратимое явление. Для того
Глава 12. Генетическая структура популяций. II.
215
чтобы некий ген распространился в популяции в течение какого-то обозримого периода времени, этому должны содействовать другие факторы.
Дрейф генов
Один из путей относительно быстрого изменения частот генов — это случайное распределение генов, или так называемый дрейф генов. Возможности влияния случайности в качестве эволюционного фактора математически изучил Райт; дрейф генов часто называют эффектом Райта.
Как мы упоминали выше, вид обычно распадается по условиям размножения на множество небольших более или менее изолированных единиц. Воспроизведение одного поколения потомства одним поколением родителей неизбежно влечет за собой некий подбор родительских гамет; чем меньше популяция, тем меньше возможности выбора. Рассматривая закон Харди — Вейнберга, мы исходили из допущения, что гаметы, несущие тот или иной из двух аллелей (а или Л), воспроизводятся с частотой соответственно р и <7 и комбинируются в зиготах в том же соотношении. Однако если набор гамет, образующих поколение, мал, то частоты обоих типов могут случайным образом сильно отклониться от ожидаемых. Это положение можно проиллюстрировать следующим примером: наполним мешок черными и белыми шарами (взяв тех и других поровну) и затем возьмем оттуда горсть. Будем считать, что шары аналогичны двум аллелям в популяции; р=0,5 и <7=0,5. Весьма мало вероятно, чтобы в каждой горсти оказалось поровну черных и белых шаров. При этом чем меньше шаров мы берем из мешка, тем сильнее будет отклонение от соотношения 1:1. Этот же прием можно повторить, наполнив мешок па этот раз шарами в таком же соотношении черных и белых, какое было обнаружено в первой горсти. Возьмем теперь горсть из такой смеси. Отклонение от соотношения 1 : 1 окажется еще более выраженным, причем в том же направлении, что и в первой горсти. Правда, направление отклонения может быть и противоположным, но при повторении данного приема достаточное число раз и при исследовании малых выборок в конце концов в выборке начнут появляться только шары одного типа. На языке генетики это означает, что один аллель популяция утратила, а другой в ней фиксирован. Теперь только мутация может привести к восстановлению утраченного аллеля. Таким образом, в небольших популяциях могут возникнуть значительные колебания в частоте генов, причем гены могут фиксироваться или утрачиваться случайным образом.
Число индивидуумов, способных оставить потомство,— не единственный фактор, определяющий стабильность популяции по отношению к случайным колебаниям. Второй фактор — это вариабель
216
Часть II. Генетика человека
ность в размере семьи. Если небольшое число случайных сочетаний приводит к рождению многочисленного потомства, а прочие скрещивания дают небольшое число потомков, то генетическая гетерогенность потомства,вероятно, будет меньше, чем она была бы, если бы то же самое общее число потомков появилось вследствие одинакового участия всех различных сочетаний.
Оба эти фактора объединяются в выражении «эффективная численность популяции» N. Математически величина N выражается формулой
А = (4А'-2):(2 + о|),
где N' — число индивидуумов в поколении родителей, а о| — дисперсия числа гамет, вносимых различными родителями и порождающих следующее поколение.
В прошлом человеческие популяции были малочисленными и более изолированными, чем большинство современных популяций. Численность человеческих сообществ в палеолите точно не известна, но, по всей вероятности, она не превышала нескольких сотен индивидуумов; именно такова численность современных сообществ охотников и собирателей (коренное население Австралии и бушмены). Подобные популяции состоят из представителей двух или трех поколений; в них входят также не производящие потомства индивидуумы; можно полагать, что размеры семей в таких популяциях подвержены случайным колебаниям. Вероятно, эффективная численность популяций человека на протяжении значительной части периода эволюции была немногим более 100. Значительное возрастание численности популяций мало влияет на ее эффективную численность, если за этим возрастанием следует соответствующее снижение.
Райт (Wright) показал, что темп фиксации того или иного из аллелей, происходящей исключительно за счет случайных колебаний, можно выразить следующим уравнением:
Lr = Lae~T^,
где La и LT — это число нефиксированных генов в исходном поколении и в поколении Т соответственно, а е — основание натуральных логарифмов. Поэтому если в некой популяции, эффективная численность которой равна 100, имеется ряд локусов, занятых парами аллелей, то в порядке случайных колебаний половина локусов перейдет в фиксированное состояние через 139 поколений, а половина оставшейся части — еще через 139 поколений. Эту величину находят простым расчетом, если принять LTILa равным 0,5. Тогда величина Т (число поколений) находится из уравнения
Лг:Ао = О,5 = е-т/2оо.
Глава 12. Генетическая структура популяций. II.
217
Структура популяций человека в прошлом создавала идеальные условия для дрейфа генов; однако остается спорным вопрос о том, оказал ли дрейф генов существенное влияние на дифференциацию человечества. В приведенных выше рассуждениях исходили из допущения, что два или более аллеля данного локуса имеют равную приспособленность. Однако вопрос о том, насколько это всегда соответствует действительности, остается спорным. Более сложный математический анализ и новые данные о полиморфизме белков в пределах одного вида и у разных видов позволили ряду авторов настаивать на существовании значительной доли нейтральных генов. Однако, как мы неоднократно подчеркивали, маловероятно, чтобы замена одного аллеля другим в комплексе адаптированных друг к другу генов оказалась нейтральной и не повлияла бы на выживание. Малое селективное преимущество одного гена перед другим выявить очень трудно, особенно у человека, для которого характерна большая продолжительность жизни одного поколения; а вместе с тем (за исключением самых малочисленных популяций) это преимущество предотвращает случайный дрейф генов; весьма вероятно, что селективное значение замены одного аллеля другим с эволюционной точки зрения весьма велико. По-видимому, следует признать правильным предположение, согласно которому различия в частоте некоторых групп крови между близкими селениями в отдаленных уголках земного шара возникли вследствие дрейфа, тем более что факторы, регулирующие эти гены, по-видимому, действуют периодически. Возможно, что именно дрейф генов послужил причиной того, что частоты генов в существующих в Америке небольших религиозных изолятах немецкого происхождения отличаются от соответствующих частот в исходной популяции (в Германии) и в соседней популяции (в Америке). С предположением стохастической или вероятностной природы многих популяционных феноменов увеличивалась изощренность нашего эволюционного мышления, особенно при создании математических моделей генетической динамики человека в малых группах; тем не менее большинство специалистов придерживаются в настоящее время того мнения, что дрейф не оказал серьезного влияния на длительную эволюцию человека или какого-либо другого организма. Есть, однако, обстоятельства, при которых случайность оказывается существенным фактором в генетике популяций. Во-первых, создающий эволюционные преимущества ген может неоднократно возникать путем мутаций, прежде чем он окажется фиксированным; будучи первоначально очень редким, он легко случайно утрачивается. Весьма вероятно, что такой мутант даже не попадет в зиготу. Во-вторых, если небольшая группа иммигрантов колонизирует новую территорию, то возможно, что именно в этой группе какой-то ген случайно отсутствует или встречается столь редко, что легко может быть утрачен. И если мутация не создаст его вновь,
218
Часть II. Генетика человека
все потомки этой группы будут лишены данного гена, а генный комплекс приспособится к его отсутствию. Это явление известно под названием «явления родоначальника». Возможно (хотя и не вполне вероятно), что отсутствие гена В в большинстве популяций коренного населения Америки причинно связано именно с этим явлением.
Отбор
В предыдущих разделах книги постоянно упоминалось о процессе естественного отбора; остановимся на нем более подробно. Естественный отбор формирует генотипы организмов таким образом, что они дают фенотипы, приспособленные к среде обитания. Однако естественный отбор не действует на генотипы непосредственно; он оказывает свое влияние через фенотипы. Если некий индивидуум обладает тем или иным признаком, который обеспечивает ему большую жизнеспособность или плодовитость по сравнению с другими членами данной популяции, то можно ожидать, что такой индивидуум оставит более многочисленное потомство. Иными словами, он будет более приспособлен к преобладающему комплексу условий среды. Соответствующий признак можно назвать адаптивным. Если различие в признаках имеет генетическое происхождение, то гены, определяющие этот признак, будут преимущественно представлены в следующем поколении и передадут это преимущество тем потомкам, у которых они окажутся. Таким образом, с эволюционной точки зрения между фенотипом и генотипом существует реципрокное взаимодействие; в результате гены, определяющие некоторые преимущества, распространяются, замещая менее приспособленные аллели, и соотношение различных фенотипов в популяции изменяется. Напротив, само возникновение вредного гена создает тенденцию к его элиминации путем естественного отбора (см. гл. 10). Обе эти ситуации взаимозависимы: распространяется ген, обусловливающий некоторое преимущество, и утрачивается его дефектный аллель. Эффект, который некогда создавал преимущество, может стать неблагоприятным либо потому, что новый ген создает еще большие преимущества для обитания в той же среде, либо потому, что сама среда изменилась и предъявляет новые требования. Рассматривая относительные приспособительные свойства генов, необходимо учитывать селективную роль среды (см. гл. 10) именно в данный момент, поскольку ген, превосходящий некий аллель в условиях одной среды, может оказаться уступающим ему в другой. Существенно, что отбор в только что изложенном смысле оказывает свое действие лишь через посредство репродуктивной приспособленности отдельных индивидуумов или их гамет. Следует, однако, отметить, что некий ген может получить преимущества, даже если он снижает
Глава 12. Генетическая структура популяций. II.
219
вероятность выживания некоторого определенного носителя этого гена. Это произойдет в том случае, если снижение приспособленности индивидуума с избытком компенсируется выживанием его родственников, вероятно имеющих тот же ген. Так, генетически определяемый признак, который обусловливает альтруистическое поведение родителей по отношению к ребенку, может иметь приспособительное значение, и соответствующие гены распространятся в популяции.'Именно это произойдет, когда выигрыш ребенка вдвое превзойдет проигрыш одного из родителей, поскольку половина генов родителя и ребенка одинаковы. Такого рода отбор, представляющий собой в расширенном виде дарвиновский отбор, называют «родственным отбором». Он же входит в одну категорию с вызвавшим много споров «групповым отбором».
Рассматривая групповой отбор в общем виде, можно представить себе, что популяции как таковые обладают наследственно закрепленными свойствами, например гауссовым распределением количественно варьирующих признаков, которое способствует генетической вариабельности, а последняя в свою очередь способствует их закреплению в эволюции. Труднее объяснить случаи, когда, казалось бы, потребности популяции в целом противоречат тому давлению отбора на отдельные индивидуумы, которое заставляет их стремиться к максимальной интенсивности репродукции; подобный конфликт возникает, например, при таком регулировании численности популяции, которое препятствовало бы чрезмерной эксплуатации среды. Если групповой отбор по признаку регулирования численности популяции вообще имеет место (а многие отрицают его существование), то неизвестен механизм, по которому он осуществляется у всех видов, кроме человека. В случае человека его уникальная способность к небиологическому наследованию, при котором все члены группы, а не только родственники получают преимущества благодаря альтруистическому поведению, интересы индивидуума могут быть подчинены интересам группы. Спорным остается лишь вопрос: как часто это происходит в действительности?
В популяции будут распространяться как доминантные, так и рецессивные гены, если они обеспечивают некие преимущества, однако при условии одинаковой приспособительной ценности доминантный ген должен распространяться значительно быстрее. Это, по-видимому, связано с тем, что для доминантного гена действие отбора распространяется и на гетерозиготы, а для рецессивного — только на гомозиготы. В момент своего первого появления в популяции ген встречается редко, и большинство носителей оказываются гетерозиготными по данному гену. Вместе с тем по той же упомянутой выше причине вредный доминантный ген элиминируется из популяции быстрее, чем рецессивный. Разумеется, если доминантный ген, распространяясь в популяции, приближается
220
Часть II. Генетика человека
к фиксированному состоянию, то вытесняемый им рецессивный ген становится очень редким, а темп его утраты замедляется, достигая той же скорости, с какой возрастает частота редкого рецессивного гена, создающего эволюционные преимущества. Таким образом, общее время, необходимое для полной фиксации доминантного гена, равно времени фиксации рецессивного гена равной адаптивной ценности.
Эти несколько неопределенные общие замечания приобретут большую ценность, если мы рассмотрим конкретный случай изменения генотипа и частот гена в случае, когда пара аллелей, находящихся в равновесии согласно закону Харди — Вейнберга, подвергается действию отбора вследствие различной приспособительной ценности их генотипов. Простейшей ситуацией будет та, при которой одна гомозигота всегда погибает, не достигнув репродуктивного возраста, а другая гомозигота и гетерозигота имеют одинаковую приспособительную ценность, поскольку один из аллелей доминантен во всех своих проявлениях. Если при этом аллели А и а появляются в определенном положении с частотами р и q соответственно, то имеющиеся три генотипа возникают с частотой р2(АА), 2pq(Aa) и q2(aa). После того как отбор устранит из популяции все гомозиготы аа, частота гомозигот АА оказывается равной p2/(p2A-2pq), т. е. pl{pA-2q), а частота гетерозигот Аа — равной 2pq/(p2-\A2pq), или 2q/(p+2q), т. е. их исходной частоте, деленной на величину p2+2pq новой популяции.
Подставляя 1 — q вместо р в выражение р/(р-|-2д), соответствующую частоту можно выразить как (1—q)/(i.+q). Сходным образом частота А а может быть представлена как 2t?/(l+1?). Все гены а, остающиеся в популяции, присутствуют в составе гетерозигот, у которых половина генов принадлежит к этому типу. Поэтому вновь получаемая частота гена а равна <?/(1+?). Частота соответствующего аллеля А равна 1— [9/(1+?)], т. е. 1/(1 +?)• При условии случайного скрещивания частота этих же трех генотипов в следующем поколении будет равна (1/1-(-д')2ЛЛ, 2(1/1 A-qY,qH+q)Aa и (q/l+q)2aa.
Частоты гена а в ряду следующих друг за другом [т. е. от q к qHl + q)} поколений образуют гармонический ряд, общий член которого выражается формулой
qn = qd^+nqQ'), где qa — исходная частота гена, п — число поколений, подвергшихся влиянию отбора, a qn — частота того же гена после п поколений отбора.
Применение этой формулы позволяет убедительно показать, как мала та скорость, с которой некий редкий рецессивный ген элиминируется из популяции, даже если его отрицательное влияние весьма велико. Одновременно оно доказывает неэффективность
Глава 12. Генетическая структура популяций. II.221
большинства самых строгих евгенических мер, направленных на избавление популяции от того или иного нежелательного рецессивного признака. Рассмотрим в качестве примера, какое влияние могла бы оказать стерилизация всех носителей редкого рецессивного аномального признака, производимая до наступления фертильного возраста на протяжении 25 поколений. Пусть исходная частота данного признака равна 1/10 000; следовательно, частота гена равна 0,01. Подставив эту величину в приведенную выше формулу,^ получим
1/100 1
?25—1 + 25(1/100) —125 ’
Иными словами, после 25 поколений такого отбора частота данного гена уменьшилась всего на 1/5!
Генотип редко бывает вовсе лишен приспособленности, и распространение или утрата генов в естественных условиях происходит при частичном воздействии отбора. Относительную приспособленность генотипов можно выразить долей потомства, получаемого от каждого из генотипов, по сравнению с потомством генотипа, обладающего максимальной приспособленностью. Мера отклонения менее приспособленных генотипов от приспособленности, принятой за единицу, называется коэффициентом отбора и обозначается буквой з. В только что рассмотренном случае полного отсутствия приспособленности рецессивной гомозиготы з=1. Если же изменение признака не влияет на выживаемость, то з=0 и частота генов остается постоянной из поколения в поколение. Таким образом, коэффициент отбора з и коэффициент приспособленности / в сумме дают единицу, т. е. з=1—/; выше, при рассмотрении непрямого метода определения частоты мутаций, мы сосредоточили свое внимание на приспособленности организма, обладающего мутантным геном, а не на мутантном генотипе в целом.
Действие частичного отбора можно описать также, исходя из превосходства доминантного гена над соответствующим рецессивным аллелем при условии равенства приспособленности гетерозиготы и гомозиготы АА. Такая ситуация представлена в табл. 12.1.
После отбора ген а существует в выживших гомозиготах и в гетерозиготах, так что его частота равна pqA~q2(l—s)/(l—sq2). Если и в этом выражении вместо р подставить 1 — q, то получим (1—q)qA~q2(l—з)/(1— sq2), или после упрощения q(i—sg)/(l—sq2). Поэтому изменение частоты гена а вследствие отбора, т. е. Aq, равно q—q(l—sq)/(l—sq2). Это выражение равно ^(1 — sq2)—g(l — sq)/(l—sq2), что после упрощения дает sq2(l—q)/(l—sq2). Снижение частоты гена а, разумеется, означает соответствующее увеличение частоты его аллеля А, и, подставляя 1—р вместо q в приведенном выше выражении, получаем величину изменения pt
222
Часть II. Генетика человека
Таблица 12.1
Генотипы Общее число
АА Аа аа
Частота до воздействия отбора Р3 2РЧ ч3 1
Относительная приспособленность 1 1 1-S
Частота после воздействия отбора п Р3 2pq ч3 (1-5) 1 —sq2
") Приведенную частоту рецессивных гомозигот после воздействия отбора можно, разумеется, представить в виде g2 —sg2; этот прием позволит показать, почему общее число после отбора равно 1 — sg2, ведь р2 + 2рд + д2 = 1.
т. е. Ap=s(l—р)2р/[1—s(l—р)2]. Если в популяции ^=0,5, а s=0,l, то в соответствии с приведенным выше уравнением снижение частоты гена а за одно поколение составит 0,0128, что представляет собой весьма существенное изменение, но если исходное q очень мало, то снижение оказывается ничтожным.
У человека не известно ни одного гена, который вследствие благоприятного влияния естественного отбора распространялся бы теперь в популяции, вытесняя свой некогда господствовавший аллель. Это и пе удивительно, если учесть большую продолжительность жизни одного поколения у человека и трудность обнаружения даже существенных благоприятствующих воздействий. Люди не обращаются к врачу, если их приспособленность выше средней! Однако подобное распространение гена, несомненно, наблюдается в некоторых популяциях растений и животных, и эффективность отбора в изменении генотипа отлично видна на примере одомашнивания или в специально поставленных опытах. Имеются серьезные основания полагать, что частота некоторых генов в популяциях человека меняется под влиянием отбора, особенно в популяциях, обитающих в среде, которая в недалеком прошлом претерпела серьезные изменения; однако до сих пор не описано ни одного такого бесспорного случая, если не считать тех, в которых лечебные мероприятия улучшили приспособляемость некогда весьма дефектных генотипов. Правда, во многих популяциях наблюдаются изменения таких признаков, как рост, форма головы и нёба, но эти изменения не обязательно происходят вследствие отбора, и многие из них вообще не имеют генетической природы. Анализ изменения таких признаков осложнен тем, что большинство их у человека относится к категории полигенных, и среда обитания непосредственно участвует в регуляции развития фенотипа. Все же можно полагать, что отбор продолжает действовать даже в обществах с высокоразвитой техникой. Пенроуз показал, что в Англии не менее 15% беременностей прерываются в самом
Глава 12. Генетическая структура популяций. II.
223
начале; 3% оставшегося числа беременностей оканчиваются рождением мертвого ребенка, а в период новорожденности гибнет еще 2%; 3% не доживают до половой зрелости, а из выживших до достижения этого возраста 20% не вступают в брак; из вступивших в брак 10% остается бездетными. Разумеется, генетические последствия этих «репродуктивных смертей» остаются нераспознанными; вне всякого сомнения, в этом процессе играют роль и различия в приспособленности индивидуумов.
Полиморфизм
В процессе распространения того или иного главного гена в популяции за счет его аллеля признак, определяемый соответствующим локусом, будет существовать в виде по меньшей мере двух, а возможно, и трех вариантов. Такая ситуация может служить примером полиморфизма. Форд определил полиморфизм как «сосуществование в одной сфере обитания двух или более обособленных форм какого-либо вида в таких соотношениях, что частота самой редкой из них все же слишком велика, чтобы ее можно было отнести за счет многократного мутирования». Такое определение исключает, во-первых, различие между популяциями, известное под названием политипизма', во-вторых, непрерывную изменчивость, которая, часто имея в основном те же причины, представляет собой ситуацию, прямо противоположную фенотипической изменчивости; в-третьих, варианты, возникающие исключительно вследствие мутации (например, ахондроплазия или альбинизм).
Полиморфизм, возникающий в ходе вытеснения одного гена другим, преходящ, поскольку, как только обладающий преимуществами аллель фиксируется в данной популяции, последняя становится мономорфной в отношении соответствующего признака. Однако многие признаки в популяции остаются полиморфными более или менее постоянно. Одним из наиболее ярких примеров подобного «сбалансированного полиморфизма» является пол. Другие примеры, относящиеся ко многим популяциям человека, кгсаются систем групп крови, ощущения вкуса фенилтиомочевины, секреции группоспецифических антигенов, состава гемоглобинов, эритроцитарных ферментов сывороточных белков (см. гл. 16—18). Если исключена гибридизация, то частоты генов, определяющих вариабельность соответствующих признаков, остаются более или менее постоянными на протяжении длительного времени; о древности полиморфизма некоторых систем групп крови свидетельствует обнаружение их у человекообразных обезьян. Одно время такой полиморфизм считали следствием дрейфа генов; однако, как уже упоминалось, два гена никогда не бывают настолько равноценными в смысле приспособленности, чтобы дрейф мог вызвать наблюдающиеся вариации. Теперь общепризнано^
224:
Часть II. Генетика человека
что сбалансированный полиморфизм является результатом равновесия между противоположными силами отбора. Впервые это отметили Фишер, Форд и Холдейн. Действие противоположно направленных сил отбора может проявляться различными путями. Наиболее важной представляется такая ситуация, при которой один из пары аллелей имеет сравнительные преимущества в своем проявлении при условии его малой частоты, однако начинает обусловливать относительную неблагоприятность, если достигает высокой частоты. Подобная ситуация автоматически возникает в том случае, когда гетерозигота оказывается более приспособленной, чем любая из гомозигот. Если отбор благоприятствует гетерозиготам, то он будет способствовать сохранению в популяции обоих аллелей. Рассмотрим этот процесс подробнее (табл. 12.2). Пусть sr и з2 — коэффициенты отбора для данных двух гомозигот.
Таблица 12.2
Генотипы Общее число
АА Аа аа
Частота до воздействия отбора Относительная приспособленность Частота после воздействия отбора Р2 (1-^) Р2(1-Я) 2pq 1 2pg 92 (l-s2) g2 (l-s2) 1 1 —SiP2 —s2g2
Частота гена а после действия отбора равна
1—SjP2 —s2g2 ’ ' 1— SiP2 — s2q2
Поэтому изменение частоты вследствие отбора равно
q — [q (l—s2q)/(l — 8гр2 — ад2)].
Целесообразно произвести дальнейшее упрощение:
д Р9(«29 — Sip) д_1-«1Р2-зд2 ’
q(l — s2g) g(l —stp2 —s2g2 —1 |-s2g) _g[ — stp2 + s2g (1 — 9)1 .
—1—srp2 —s2g2 1— SiP2 — s2g2 1—s^2 —s2g2 ’
подставляя p вместо 1—q, получим
pg(s2g —stp) 1— s^2 — s2q2
Глава 12. Генетическая структура популяций. II.
225
Отметим, что если s^q^s^p, то выражение будет положительной величиной и частота гена а возрастет по сравнению с его аллелем, тогда как в случае, если частота этого гена будет умень-
шаться. В каком бы направлении ни происходило изменение частоты этого гена, оно будет продолжаться до тех пор, пока не возникнет ситуация, когда s.tq=stf). После этого изменение частоты прекратится, система окажется сбалансированной. Частоту интересующего нас гена при условии наступившего равновесия можно вычислить по формуле
(М = siP) = [М = «1(1 — 2)1 = iq = «i/(«i !' «2)1, а частоту аллеля А при равновесии — аналогично по формуле
P = s2/(«i + «2).
Отсюда можно видеть, что когда гетерозиготы обладают определенными преимуществами, частота генов в двухаллельной системе в момент равновесия определяется относительной приспособленностью соответствующих гомозигот.
Можно привести только один пример определяющей роли гетерозиготы в поддержании полиморфизма в человеческой популяции — это аномальные гемоглобины. Известно, что отбор влияет и на группы крови, и на ощущение вкуса фенилтиомочевины, и на секрецию, однако те его факторы, которые выявлены до сих пор, действуют либо не в должном направлении, либо недостаточно интенсивно, чтобы они могли поддержать полиморфизм. Многие из них действуют лишь в зрелом возрасте, а различия в смертности оказывают свое действие на ход эволюции, разумеется, только в том случае, если они влияют на фактическую плодовитость. Сложнее обстоит дело с трехаллельными системами или с другими особыми генетическими ситуациями. Тем не менее можно предсказать, что если факторы отбора не устранятся вследствие прогресса техники и медицины, то рано или поздно будут обнаружены факторы, действовавшие в направлении сохранения полиморфизма популяций человека путем сохранения преимуществ, присущих гетерозиготам; это предсказание можно признать обоснованным, поскольку, по-видимому, отбор как таковой связан с повышением приспособленности гетерозигот. Как было показано выше, эффекты генов множественны, и даже ген, суммарный эффект которого создает преимущества для индивидуума, может оказывать некоторое неблагоприятное действие. Отбор ведет к превращению благоприятных эффектов в доминантные, а неблагоприятных — в рецессивные (см. гл. 9). Поэтому гетерозигота, образованная Двумя генами,- неблагоприятные эффекты которых рецессивны, будет обладать только благоприятными свойствами. Некоторые данные, главным образом касающиеся степени полиморфизма в ферментных системах и группах крови, свидетельствуют о том,
226
Часть II. Генетика человека
что для 16% всех локусов структурных генов у человека характерен полиморфизм. Здесь уместно упомянуть о том, что некоторые явно «вредные» рецессивные гены присутствуют в определенной популяции не в результате многократного мутирования, а вследствие преимущества гетерозигот по этим генам. Если такая возможность реальна, то описанные выше непрямые методы определения частоты мутаций, по всей вероятности, весьма неточны.
Кроме преимущества гетерозигот можно представить себе и другие ситуации, при которых силы отбора направлены противоположно друг другу. Например, некий генотип может иметь относительные преимущества, если он встречается в популяции сравнительно редко, при высокой же частоте оказывается неблагоприятным. Иными словами, природа отбора по какому-либо гену может зависеть от частоты этого гена в популяции. Такой отбор называют зависимым от частоты. Довольно близка ситуация, когда среда обитания популяции сама по себе гетерогенна и ее различные части благоприятствуют различным генотипам. Таким образом, благодаря генетической вариабельности становится возможной более эффективная эксплуатация различных экологических ниш, зависящих от распределения популяции по территории. Такую форму отбора называют диверсифицирующим отбором, или дисруптив-ным отбором-, вероятно, он весьма важен для создания полиморфного разнообразия по меньшей мере некоторых видов. О том, насколько он функционирует в отношении такого вида, как человек с его высокой индивидуальной подвижностью и большой пластичностью поведения, можно лишь догадываться.
Обобщая, скажем, что естественный отбор в виде направленного отбора представляется главной, если не единственной, силой в эволюционном процессе; тем не менее он оказывает также консервативное влияние, поддерживая генетический status quo и уровень равновесия путем нормализующего отбора против нежелательных мутантных генов, путем балансирующего отбора на гетерозиготность и путем диверсифицирующего отбора в гетерогенных средах обитания.
Политипизм
В приведенном выше изложении эволюционных механизмов наше внимание было сосредоточено главным образом на том, как меняется частота генов в пределах популяции; однако те же факторы (мутация, отбор и в меньшей степени дрейф генов) могут быть причиной возрастания многообразия в популяциях. Популяции, обитающие в различных местностях, подвергаются действию различных факторов отбора. Не существует двух мест обитания, которые были бы идентичны на протяжении сколько-нибудь дли
Глава 12. Генетическая структура популяций. II.
227
тельного времени; но даже если бы они и оказались таковыми, совершенно невероятно, чтобы в результате их действия возникли полностью идентичные генные комплексы, поскольку привнесение новых вариантов, даже не отличающихся по своей генетической природе, неизбежно будет происходить в различном порядке, а отбор сразу же начинает действовать на все то, что оказывается в сфере его действия. Если дифференциация началась, то ее последующее течение зависит от тех изменений, которые уже произошли. Избежать адаптации в той или иной форме невозможно, если единственный путь избежать ее лежит через этап, на котором индивидуумы оказались бы менее приспособленными, чем в настоящее время, несмотря на то что преимущества, создаваемые альтернативным признаком, весьма значительны, а исходное состояние предков обеспечивает значительно меньшую приспособленность. Стало быть, если популяции изолированы друг от друга, то они будут иметь различные гены, а если изоляция продлится достаточно долго, эти популяции превратятся в различные подвиды и в конце концов в различные виды, конечно, при условии, что в течение всего этого времени обе популяции остаются репродуктивно изолированными независимо от географического положения их места обитания. Однако возникновение некоторого разнообразия возможно и при неполной изоляции. Существенную роль в этом играет сравнительная интенсивность давления отбора и потока генов. Если дифференцирующий эффект отбора сильнее унифицирующего действия потока генов, то произойдет дивергенция популяций. Высокий географический градиент давления отбора может привести к возникновению «клина» частоты генов или различных вариантов признаков в такой же мере, как и центробежные потоки генов. Конечно, дивергенция будет происходить быстрее, если популяции полностью изолированы друг от друга. Однако более тонкая дифференциация вида как целого будет идти быстрее в тех случаях, когда периоды изоляции и дивергенции признаков сменяются периодами гибридизации. Такого рода сетчатой эволюцией характеризуется и происхождение человека; благодаря ей поддерживается единство рода человеческого как вида.
В изолированных популяциях с неизбежностью устанавливаются генетические различия, причем степень различий оказывается функцией продолжительности времени изоляции; отсюда, однако, не обязательно следует, что две популяции, сходные генетически или фенотипически, произошли в недалеком прошлом из одного и того же источника или что две различные популяции произошли из него в весьма отдаленном прошлом. Дивергенция популяций происходит очень быстро, если на них действуют эффективные и противоположно направленные силы отбора. Если же популяции подвергаются действию одинаково направленных сил отбора, то они окажутся, по крайней мере фенотипически, подобными
228
Часть II. Генетика человека
независимо от того, насколько различными они были в прошлом. Отсюда, разумеется, не следует, что они и генетически однородны.
Мы уже говорили о том, что отбор оказывает свое влияние именно на фенотип, и обычно имеется несколько возможных генетических путей, ведущих к возникновению одного и того же фенотипа. Следовательно, если в единичной популяции не происходит фенотипических изменений, то на этом основании еще нельзя сделать вывод, что в ней не происходит и генетических изменений. Фенотипическая изменчивость может иногда затухать, генотипическая, по-видимому, происходит всегда. Однако фенотипические различия должны воспроизводить генетические различия при условии, что они не возникли под влиянием среды. Биологическое значение различий между популяциями обычно выступает более отчетливо, чем вариабельность в пределах единичной популяции, поскольку удается получить некоторые сведения о действующих силах отбора, исследуя корреляцию между географической изменчивостью признака и географическим распределением некоторых факторов внешней среды.
Список литературы
Baiema С. J. (ed.) (1971). Natural selection in human populations. Wiley, New York.
Boyce A. J. (1976). Chromosome variations in human evolution. Symposium of the Society for the Study of Human Biology. Taylor and Francis, London.
Cavalli-Sforza L. L. and Bodmer W. F. (1971). The genetics of human populations. Freeman, San Francisco.
Crawford M. H. and Workman P. L. (1973). Methods and theories of anthropological genetics. University of New Mexico Press.
Crow J. F. and Kimura M. (1970). An introduction to population genetics theory. Harper and Row, New York.
Dobzhansky Th. (1962). Mankind evolving. Yale University Press, New Haven, Connecticut.
Dobzhansky, Th. (1970). Genetics of the evolutionary process. Columbia University Press.
Harrison G. A. and Boyce A. J. (1972). The structure of human populations. Oxford University Press, London.
Lerner I. M. (1968). Heredity, evolution and society. Freeman, San Francisco. Li С. C. (1955). Population genetics. University of Chicago Press.
Mather K., Jinks J. L. (1971). Biometrical genetics (2nd edn). Chapman and Hall, London.
McKusick V. A. (1969). Human genetics (2nd edn). Prentice-Hall, New Jersey. (Имеется nep. 1-ro изд. В. Маккьюспк, Генетика человека. М.: «Мир», 1967.)
Medawar Р. В. (1960). The future of man. Methuen, London.
Roberts J. A. F. (1973). An introduction to medical genetics (6th edn). Oxford University Press, London.
Schull W. J. and Neel J. V. (1965). The effects of inbreeding on Japanese children. Harper and Row, New York.
Spuhler J. N. (1967). Genetic diversity and human behavior. Aldine, Chicago. Stern C. (1973). Principles of human genetics (3rd edn). Freeman, San Francisco. Strickberger M. W. (1968). Genetics. Macmillan, New York.
Часть III
БИОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ В ПОПУЛЯЦИЯХ СОВРЕМЕННОГО ЧЕЛОВЕКА
Н. БАРНИКОТ
Глаеа 13
ВВЕДЕНИЕ
Предыдущие части книги были посвящены эволюции и основам генетики человека. Здесь мы рассмотрим изменчивость современного человека как вида. Различия в размерах и телосложении, в цвете кожи и других признаках между обитателями главных континентов земного шара достаточно хорошо известны и привлекали внимание антропологов еще в конце XVIII в.; в течение последних 50 лет иммунологические и биохимические исследования выявили в дополнение к этим явным различиям многочисленные невидимые различия, что весьма способствовало изучению человеческих популяций. Географическая дифференциация — хотя и главная, но не единственная тема данной части книги. Можно ожидать, что в сложных сообществах существуют биологические различия между экономическими и другими составляющими их элементами. Изучение подобных различий может иметь важное значение, например, для медицины.
Люди отличаются друг от друга в самых разных аспектах и образуют весьма разнообразные группы, рассеянные по всему земному шару. Описание этих различий имеет смысл лишь в том случае, если оно приведет к пониманию того, как осуществляется эта региональная дифференциация и каково ее биологическое значение в прошлом и настоящем. Палеонтология и археология дают наиболее прямые и адекватные сведения о событиях прошлого, и можно надеяться на создание с их помощью цельной картины позднейших фаз эволюции человека; однако пока сведения эти носят фрагментарный характер и оказываются далеко не полными. Археологи находят, как правило, только кости и зубы и лишь редко какие-либо другие ткани. Поэтому развитие знаний в данной области происходит крайне медленно и получаемый материал весьма ограничен. Археология дает нам также некоторые сведения о таких имеющих возможное биологическое значение переменных величинах, как численность популяций, их возрастная и
230
Часть III. Биологическая изменчивость популяций
половая структура, климатические условия и способы, которыми люди добывали себе пропитание.
Генетика находится в самом центре эволюционных проблем, поскольку гены — это материальный субстрат той связи, которая существует между поколениями, и филогенетические изменения зависят от изменений свойств и частот генов. Точность, с которой мы можем описать генотип, во многом зависит от характера признаков, выбранных нами для изучения. Мы еще не достигли наивысшей точности, которая позволила бы нам сравнивать строение генов человека на основе химического анализа; тем не менее анализ белков как непосредственных продуктов действия генов заметно приблизил нас к этому идеалу. За последние два десятилетия с помощью довольно простых биохимических методов были открыты многие наследственные варианты белков.
Понятно, что такие биохимические признаки обладают большой притягательной силой для тех, чья цель — сравнение популяций точными методами на уровне генов, к тому же с привлечением математического аппарата эволюционной генетики для интерпретации результатов. Вот почему в этой части книги много места уделяется биохимической генетике.
У человека наследование большинства признаков (в том числе умственных способностей, оцениваемых с помощью стандартных тестов, восприимчивости к ряду болезней и многих других) еще нельзя проанализировать в столь же точных биохимических терминах. Обычно в определении этих признаков участвует много генов, а на вариабельности признаков сказываются условия внешней среды. К таким признакам относятся размеры тела и цвет кожи, изучаемые классической антропологией. Сказанное, разумеется, не означает, что подобные признаки" и их изменчивость не интересуют исследователей биологии человека, однако их значимость для эволюционной генетики ограниченна, так как мы не можем идентифицировать индивидуальные гены, о которых идет речь.
В данной части книги речь пойдет о биологической изменчивости человека; необходимо поэтому определить границы биологии человека. Люди отличаются друг от друга по строению тела и по многочисленным биохимическим и физиологическим признакам. Такую изменчивость мы без колебаний отнесем к области биологических исследований, проводимых теми же методами, какие применяются при изучении животных. Но, кроме того, люди говорят на разных языках, подчиняются разным законам, имеют разные обычаи и верования и весьма сильно различаются по характеру и масштабам своей деятельности. Несомненно, что различия социального характера могут оказаться столь же важными для выживания, как и поддержание нормальных физиологических функ
Глава 13. Введение
231
ций организма; это нельзя упускать из виду при углубленном изучении биологии человека. Необычайное развитие средств связи, техники и форм общественной жизни присущи только человеку. Сама сложность проблемы требует проведения исследований по многим направлениям, с которыми не приходится иметь дело биологу, изучающему животных, и которые обычно не относят к области биологии.
Особенности культуры передаются из поколения в поколение благодаря обучению и условиям жизни в определенной социальной среде, а не по законам биологической наследственности; они могут изменяться гораздо быстрее признаков, закодированных в геноме и регулируемых естественным отбором. Тем не менее способность овладеть языком или культурным наследием общества, несомненно, зависит и ог особенностей мозга, хотя мы пока очень далеки от понимания неврологических основ обучения и памяти. Гены определяют развитие и функциональную деятельность структур мозга; об этом наглядно свидетельствуют случаи умственной отсталости, связанной с некоторыми мутациями. Тем не менее ткани, которые развиваются согласно «инструкциям» генома, не статичны, а обладают способностью реагировать в определенных пределах, приспосабливаясь к изменениям среды; по-видимому, зто положение распространяется и на высшую нервную деятельность организма. Созревание умственных способностей, несомненно, зависит от влияния социальной среды; то, чему человек научается, зависит от того, чему его учат, накоплению каких знаний способствуют. Анализируя развитие разнообразных культур, авторитетные исследователи изучают географические и исторические обстоятельства, а не генетически обусловленную изменчивость умственных способностей, играющих роль в развитии культуры. Если такой взгляд верен, то биология в общепринятом смысле этого слова не способна обеспечить большого вклада в изучение культурно-социологического аспекта антропологии.
В последнее время этологические исследования животных, особенно приматов, стимулировали поиски общих элементов поведения, которые человек мог унаследовать от своих отдаленных предков. Но даже если поведение современного человека в каких-то отношениях и пронизано подобными скрытыми тенденциями, перспективность такого подхода к объяснению богатства и разнообразия человеческой культуры представляется маловероятной.
Из сказанного, разумеется, не следует, что, изучая биологию человека, можно пренебрегать социальными факторами. Начнем с практического примера. Человеческие популяции часто неоднородны; они разделены на более или менее четкие социальные подгруппы, различающиеся по материальному обеспечению, роду деятельности, религии, политическим взглядам, языку,
232
Часть III. Биологическая изменчивость популяций
этническому происхождению. Такие подгруппы могут различаться, кроме того, по генетическим характеристикам и экологическим особенностям вроде характера питания и опасности заболеть той или иной болезнью. Если биолог не осведомлен о такой неоднородности, его выборка из популяции окажется непредставительной и он получит при сравнении ошибочную информацию.
Система браков определяет распределение генов в следующем поколении. Как мы отмечали, у человека подбор супружеских пар ограничен социальными и географическими преградами. В некоторых обществах браки или половые сношения между представителями различных этнических групп, например между «черными» и «белыми», запрещены законом, тогда как в других существуют более или менее серьезные препятствия к бракам между приверженцами различных религий. Даже там, где законодательство или обычаи не накладывают таких ограничений, люди часто предпочитают вступать в брак с представителями своей социальной среды и имеют для этого больше возможностей. Такое поведение затрудняет поток генов между группами. Кроме того, разные общества допускают различную степень родства между вступившими в брак. Браки между близкими родственниками повышают вероятность того, что копии генов от общего предка попадут в одну зиготу. Это влияет на гомозиготность популяции (правда, в очень малой степени) и повышает частоту редких рецессивных аномалий. Социологи уделяют немало внимания родственным отношениям, ставя, однако, 'акцент на предписываемые обществом правила, а не на реальные генетические последствия. Эта ситуация может служит показательным примером расхождения интересов и отсутствия контактов между биологами и социологами.
Проблемы естественного отбора привлекают наше внимание к причинам различий в смертности и фертильности. Во всех густонаселенных областях земного шара до недавнего времени (а в некоторых развивающихся странах и до сих пор) первое место среди причин смерти занимали инфекционные болезни, вызываемые вирусами, бактериями и простейшими. Эксперименты на животных показали, что гены оказывают влияние на восприимчивость к этим болезням; есть основания считать, что это относится и к человеку. Частота того или иного заболевания в определенной местности зависит и от особенностей климата и от таких факторов, как размер и структура данного общества, способ размещения и сооружения поселений, характер питания, гигиенические навыки и многие другие стороны жизни. Выбор сферы обитания и преобразование среды под влиянием сельского хозяйства также могут повлиять на вероятность заболевания и создать благоприятные условия для размножения и распространения возбудителей заболевания. История знает много примеров распространения эпидемий армиями и паломниками; массовая гибель американских индейцев и жителей островов
Глава 13. Введение
233
Тихого океана от инфекционных болезней, занесенных европейцами-колонистами, может служить печальным примером теневых сторон контактов между культурами. Общеизвестно, что недоедание, особенно в раннем детстве, резко повышает смертность от инфекционных болезней. Большую роль здесь играют особенности ведения сельского хозяйства, практика вскармливания младенцев, а также различного рода «табу» в отношении некоторых видов пищи.
В развитых странах, где главными причинами смерти служат болезни пожилого возраста, различия в фертильности, вероятно, представляют больше возможностей для действия отбора, чем различия в смертности. Хотя в некоторых странах основное влияние на фертильность оказывают инфекционыые болезни, в других более важное значение имеют экономические и религиозные факторы, определяющие регуляцию рождаемости. Биолог, изучающий воспроизведение потомства у человека, вряд ли может игнорировать сложные проблемы, связанные с культурными и социальными различиями.
Расы и популяции
Общепризнано, что все люди принадлежат к одному виду, Ното sapiens. Этот вид, как, впрочем, и большинство видов животных, географически неоднороден. Нет убедительных оснований считать, что такая географическая дифференциация связана с каким-либо снижением плодовитости вследствие смешения между группами в связи с генетически обусловленными причинами. Нет также разумных причин считать, что межпопуляциоиные гибриды менее приспособлены, если исключить обстоятельства их взаимного социального положения.
В XVIII в. Блуменбах (Blumenbach) ввел классификацию людей на 4 главные группы; при этом он был достаточно осторожен, чтобы указать на существование промежуточных подгрупп. С тех пор было опубликовано много более изощренных классификаций, которые основывались на немногих легко наблюдаемых внешних признаках, таких, как цвет кожи, форма головы и форма волоса. Некоторые народности Азии, например, объединялись в монголоидную расу, отличающуюся слабо пигментированной кожей, прямыми волосами, округлым черепом, плоским лицом с выступающими скулами и характерным разрезом глаз (складка эпиканта). Однако у некоторых народов Юго-Восточной Азии и у американских индейцев одна часть этих признаков встречается редко или отсутствует полностью, а другая часть встречается чаще, но в слабо выраженной форме. Эти группы не укладываются в простые схемы классификации.
234
Часть III. Биологическая изменчивость популяций
Разумеется, можно создать и для них дополнительные подгруппы, однако поскольку все популяции окажутся различными, если их достаточно подробно изучить, то неясно, где же остановится это формирование все новых подгрупп. Следует также отметить, что создатели классификаций в зоологии стремятся построить их на генетических различиях или по крайней мере не на таких различиях, которые явно находятся под влиянием местных условий среды обитания. А известно, что у человека рост и другие размеры тела зависят от подобных различий окружающей среды.
Подразделение людей на монголоидов, европеоидов, негроидов и т. д., несомненно, выражает некоторые реально существующие особенности изменчивости человека, но выражает их весьма несовершенно и таит в себе источник ошибок. Само перечисление расоводиагностических признаков маскирует тот факт, что все популяции очень изменчивы. Границы изменчивости таких количественных признаков,как рост или цвет кожи, для которых характерно непрерывное распределение в разных популяциях, часто перекрываются. В подобных случаях различия между популяциями по рассматриваемому признаку имеют статистический смысл, как различия между средними значениями, хотя при этом неучтенной остается немало информации. Нанеся на карту ряд популяционных средних величин, мы можем лучше оценить картину географического распределения характерных линейных величин и часто обнаруживаем градиенты изменения (клины), а не четкие границы. При этом следует помнить, что физические различия могут существовать между экономическими, религиозными или иными социально определенными группами, живущими в одном и том же месте; понятно, что эти различия трудно или невозможно представить на географической карте.
Различия между популяциями по дискретным признакам (например, по отсутствию или наличию определенного гена) следует оценивать соответствующими подсчетами, а не измерениями. Может оказаться, что ген встречается повсеместно, но частота его меняется или же ген может практически отсутствовать в одних областях и встречаться более или менее часто в других. И в этом случае отнесение индивидуума к той или иной группе определяется с той или иной вероятностью, и если мы нанесем частоты генов на географическую карту, то обнаружим градиенты этих частот.
Классификация по дискретным категориям может оказаться удовлетворительной, если соответствующие природные группы четко различимы. Для человеческих популяций такой ситуации не существует. Всякое точное определение таксономических групп с неизбежностью оказывается произвольным, поскольку различия между популяциями по природе своей количественные, статистические, а географические границы недостаточно определены.
Для простоты мы пользовались единичными признаками, ил
Глава 13. Введение
235
люстрируя общую природу популяционных различий; если же нас интересует родственность популяций, то правильнее изучать много признаков, чтобы избежать возможных ошибок. У таксоно-мистов принято создавать так называемые естественные классификации. Если мы измерили большое число определенных признаков или определили их частоту в ряде различных популяций, то нам нужен такой способ оценки различий между популяциями, который позволил бы учитывать всю эту информацию. Существуют статистические методы, с помощью которых эти сведения могут быть сведены к измерению «таксономических расстояний» между двумя популяциями, а индивидуумы отнесены к той или другой популяции с наименьшей ошибкой. Такие методы многих переменных обсуждаются в гл. 14. Использование большого числа признаков осложняет задачу обеспечения географической репрезентативности; нередко распределения для разных признаков не совпадают, и границы становятся еще более туманными.
В зоологии слово «раса» иногда употребляют для обозначения подвидовых групп; хотя это слово не имеет официального статуса, его не избегают. В применении к человеку этот термин так часто используют неправильно, что некоторые авторитетные исследователи предпочитают вовсе не применять его. Например, когда говорят о германской или еврейской расе, смешивают биологические и культурные различия. Эти группы ограничены рамками культуры; называя их расами, мы создаем априорное впечатление, что они различаются также и генетически, не говоря о том, в чем и в какой степени. Такие неверные концепции легко приводят к расистским доктринам, настаивающим на чистоте и превосходстве определенных групп. Ни одна природная популяция не является чистой, т. е. генетически однородной, хотя некоторые популяции могут быть менее других затронуты недавним притоком генов. Вопрос о превосходстве неизбежно влечет за собой социально-этические оценки, а па практике связан со стремлением одной группы доминировать над другой и эксплуатировать ее.
В этой части книги мы часто, хотя и риекуем услышать упрек в однообразии, будем пользоваться термином популяция. Это слово достаточно нейтрально и не уточняет само по себе, чем именно различаются группы. Мы вольны определить популяцию географическими, политическими, лингвистическими, генетическими и любыми другими терминами, соответствующими проводимой работе. Генетиков, например, нередко интересуют группы, члены которых чаще вступают в брак между собой, чем с членами других групп. Образование и сохранение таких групп может быть обусловлено географическими и социальными факторами. Люди, живущие в одной деревне, в одном городе, в одной общине, вероятнее всего, вступят в брак с другими членами той же популяции, однако некоторые будут вступать в брак п с «чужаками», живущи
236 Часть III. Биологическая изменчивость популяций
ми на том или ином расстоянии от этой популяции. Поэтому границы общины, в пределах которой заключаются браки, диффузны. На практике биологи обычно формируют выборки из таких признанных и имеющих свое название общественных групп, как нация, племя, каста; главное, что необходимо,— это определить критерии выборки по всем аспектам.
Глава 14
РАЗМЕРЫ И ФОРМА ТЕЛА
А нтропометрия
Необходимость проведения точных измерений в дополнение к словесному описанию форм и размеров тела была осознана уже в начале девятнадцатого столетия, когда «физическая» антропология выделилась в самостоятельную науку, В недавнее время главным ее стержнем стала антропометрия. Пропорции человеческого тела издавна привлекали внимание художников, но их задачей было не систематическое описание реально существующих вариантов, а определение идеальных пропорций. Не удивительно, что антропологи, которых особенно интересуют проблемы эволюционного развития, придают большое значение измерениям скелета, так как именно по фрагментам костей мы можем судить о размерах и форме тела представителей древнейших популяций. Эта особенность нашла свое отражение в тех стандартных измерениях, которые производятся для сравнения живых людей; большинство этих измерений определяют размеры скелета. Однако измерения мягких тканей, по которым можно оценить количество жира и мышц, не менее важны, чем измерения скелета. Кроме того, в прикладной антропометрии, имеющей отношение к вопросам изготовления одежды, мебели, транспортных средств, могут потребоваться дополнительные нестандартные данные.
Формы тела человека и его скелета весьма сложны. При большой затрате сил и времени некоторые их детали могут быть выражены метрически, однако для многих целей можно ограничиться небольшим числом измерений, выбранных так, чтобы охарактеризовать основные особенности формы тела.
В принципе нет ничего проще, чем произвести линейные или круговые измерения при помощи линейки, калипера или сантиметровой ленты, но для получения достоверных результатов необходимо детально разработать методику измерений. При измерении расстояния между крайними точками тела или той или иной костью следует точно определить положение этих точек с учетом позы обследуемого в момент измерения, так как многие размеры зависят от позы. Точность измерительных инструментов должна периодически проверяться. Несмотря на многочисленные попытки установить международные стандарты по технике антропометрии, зачастую невозможно сопоставить данные разных исследователей вследствие отсутствия информации об используемых ими методах.
238
Часть III. Биологическая изменчивость популяций
Статистические методы столь же важны для антропометрических исследований, как и «внешняя» анатомия или остеология, поскольку без них трудоемкий сбор результатов даст очень мало полезной информации. Человеческие популяции слишком изменчивы; поэтому для описания наших данных в компактной форме мы нуждаемся в статистических методах, и, поскольку обычно приходится полагаться на ограниченные выборки, характеризуя целую группу, эти методы позволяют нам учитывать элемент случайности при формировании выборки. Здесь мы лишь укажем некоторые наиболее важные концепции и методы, отослав читателя к соответствующим работам.
Длина тела
Мы начинаем с измерения роста, поскольку этот параметр определяется наиболее часто и позволяет проиллюстрировать различные приемы исследования метрических признаков. Отчеты о призванных на военную службу содержат многочисленные данные о росте молодых мужчин в разных странах и его изменении в течение значительного периода времени.
Метод измерения. Обычный прибор для измерения роста — вертикальная шкала с перемещающейся по ней поперечной рейкой, которая может быть наложена на голову. Измеряемый снимает обувь и, выпрямившись, становится под рейку спиной к шкале, так чтобы пятки были сдвинуты вместе, а зрительная ось была горизонтальна. Чтобы сгладить влияние сутулости на результат измерения, исследователю следует голову измеряемого взять позади ушей и приложить к ней давление, направленное вверх.
Измерить рост очень просто, однако анатомически это сложный признак, включающий размеры ног, таза, позвоночника и черепа; вклад каждого из них неодинаков у разных индивидуумов, а в среднем — ив разных популяциях. Рост человека дает нам некоторое общее представление о величине его тела, однако в последнем случае вес тела был бы лучшим показателем, поскольку он зависит и от поперечных размеров. Ясно, что одни люди высоки и худы, тогда как другие — высоки и толсты. Связь между ростом и весом или между ростом и одним из поперечных размеров может служить для оценки изменчивости телосложения (см. часть IV).
Тот эмпирически установленный факт, что вес тела, превышающий средний для популяции вес тела при данном росте, коррелирует с меньшей продолжительностью предстоящей жизни, выдвинул эту зависимость в число важных факторов при страховании жизни.
Связь между ростом и другими антропометрическими показателями можно представить в виде таблицы корреляции (табл. 14.1). Произведя различные измерения у большой группы людей, мы мо
Глава 14. Размеры и форма тела
239
жем подвергнуть результаты статистическому анализу, чтобы установить, связана ли большая величина одного из показателей, скажем высокий рост, с большой или малой величиной какого-либо другого показателя; в первом случае говорят о положительной корреляции, во втором — об отрицательной. Удобной мерой такой взаимосвязи может служить коэффициент корреляции г, который принимает значения от +1,0 до —1,0. Не удивительно, что длина ног и туловища характеризуется высокой корреляцией с ростом, тогда как корреляция роста и поперечных размеров тела значительно ниже. Вес тела, как и следовало ожидать, характеризуется высокой положительной корреляцией со всеми другими размерами.
Таблица 14.1
Усредненные корреляции между размерами тела 2400 мужчин в возрасте 17—38 лет
(английские военно-воздушные силы)
Рост стоя Рост сидя Длина руки Длина ноги Длина бедра Окружность живота Окружность бедра Окружность 1 плеча О о
Рост стоя 0,732 0,677 0,864 0,608 0,321 0,490 0,386 0,627
Рост сидя 0,732 •— 0,421 0,498 0,201 0,173 0,417 0,384 0,548
Длина руки (от кисти до плеча) 0,677 0,421 — 0,683 0,447 0,266 0,483 0,363 0,466
Длина ноги 0,864 0,498 0,683 — 0,817 0,312 0,424 0,304 0,556
Длина бедра 0,608 0,201 0,447 0,817 — 0,515 0,440 0,234 0,525
Окружность живота 0,321 0,173 0,266 0,312 0,515 — 0,667 0,526 0,709
Окружность бедра 0,490 0,417 0,483 0,424 0,440 0,667 — 0,562 0,785
Окружность плеча 0,386 0,384 0,363 0,304 0,234 0,526 0,562 — 0,681
Вес 0,627 0,548 0,466 0,556 0,525 0,769 0,785 0,681 —
Статистическое распределение значений длины тела
Метрические признаки популяции, например рост, можно представить в виде гистограммы. Изменчивость роста называют непрерывной, поскольку рост человека в определенных пределах может принимать любое значение, причем соответствующие величины
240
Часть III. Биологическая изменчивость популяций
не распадаются на определенное число четко различимых классов, не диктуемых ограниченной точностью измерений.
На рис. 14.1 распределение роста в большой группе молодых англичан, призванных на военную службу в 1939 г., представлено в виде гистограммы. Индивидуумы разного роста объединены в группы с интервалом в 2,5 см; число людей в каждой группе представлено площадью соответствующего прямоугольника. Распределение имеет колоколообразную форму с наиболее многочисленной,
Рис. 14.1. Распределение роста молодых англичан, призванных на военную службу в 1939 г. (Martin, 1949).
Сплошная прямая соответствует значению х, прерывистые вертикальные линии — стандартному отклонению о Представлена также гауссова кривая, вычисленная исходя из среднего и стандартного отклонения (х = 169 см, ст» 6,24).
или модальной, группой около середины и более или менее симметричным убыванием численности по обе стороны от максимума. Эта кривая напоминает гауссову, или нормальную, кривую. Такое сходство чрезвычайно удобно, поскольку свойства гауссова распределения хорошо изучены и могут быть использованы для описания распределения вариантов. В некоторых случаях, например при измерении веса, распределение бывает асимметричным. В других случаях вершина кривой может быть уплощенной или кривая имеет слишком длинные «хвосты». При асимметричном распределении кривую можно приблизить к гауссовой, откладывая на оси абсцисс логарифмы измеренных величин. Надобность в таком выражении возникает, например, при измерении толщины кожной складки (при помощи специального инструмента — калипера).
Для многих целей наиболее удобной характеристикой может служить среднее арифметическое значение. В случае гауссовой кривой эта величина совпадает с модой, а также с медианой. Часто возникает необходимость охарактеризовать изменчивость признака в какой-либо группе. На первый взгляд кажется, что для этого можно воспользоваться разностью между самым большим и самым малым значениями. Но при такой оценке используются толь
Глава 14. Размеры и форма тела
241
ко два крайних значения, которые могут очень сильно и притом случайно варьировать. Наиболее употребительный статистический критерий — это среднеквадратичное, или стандартное, отклонение. На гауссовой кривой вертикальные прямые, проведенные на расстоянии среднеквадратичного отклонения по обе стороны от среднего, делят площадь под кривой таким образом, что около 2/3 (68%) располагаются между этими прямыми. Это означает, что вероятность каждого варианта оказаться за пределами этого интервала равна примерно */3 (или */t) для каждого конца распределения. И далее, вероятность того, что данный вариант будет отстоять от среднего больше чем на два среднеквадратичных отклонения, равна приблизительно 5%.
Мы предпринимаем измерения с целью охарактеризовать всю популяцию в целом, однако по вполне понятным причинам обычно можем исследовать только небольшую выборку; следовательно, информация о распространении признака во всей популяции будет ограниченной. Если выборка мала (скажем, меньше 100), то гистограмма примет, вероятно, неправильную форму, и мы не сможем сказать, является ли распределение гауссовым. Вообще чем меньше выборка, тем больше вероятность того, что среднее значение и среднеквадратичное отклонение, вычисленные для этой выборки, будут отличаться от соответствующих величин, характеризующих всю популяцию. Мы можем, однако, количественно оценить степень этого отличия, вычислив так называемую среднеквадратичную ошибку среднего.
Сравнение популяций
Рис. 14.2 иллюстрирует распределение роста в двух популяциях. Одна из них — пигмеи группы бамбути из лесного района Итури в Заире; это, по-видимому, самые малорослые люди в мире. Вторая группа — скотоводческое племя тутси из находящейся по соседству Руандийской Республики — самые высокие люди на земном шаре. Различие между средним ростом бамбути (144 см) и тутси (176,5 см) составляет 32,5 см. В обеих популяциях наблюдаются вариации, причем крайние отклонения до некоторой степени перекрываются. Приведенный пример представляет собой крайний случай. Обычно разница между средними значениями двух распределений бывает значительно ниже, а степень перекрывания распределений значительно больше. Индивидуумы, попавшие в зоны перекрывания, не могут быть отнесены к той или иной популяции на основании только роста. Если выборки малы или изменчивость признака в популяциях слишком велика, наши оценки средних будут соответственно неточны и не исключена возможность того, что наблюдаемые различия между средними связаны не с реальными различиями между популяциями, а со случайным характером вы
242
Часть III. Биологическая изменчивость популяций
борок. Мы должны, следовательно, оценить вероятность того, что две выборки, средние значения для которых различаются на данную величину, представляют собой случайные выборки из одной популяции (так называемая нуль-гипотеза). Соответствующие методы, описание которых можно найти в специальных учебниках, состоят в сопоставлении различия средних со стандартной ошибкой этого различия. Если выборки малы, то следует применять критерий Стьюдента как более чувствительный.
Рис. 14.2. Распределение по росту мужчин и женщин пигмеев из района Итури (Л) (данные Gusinde, 1948) и мужчин из племени тутси (Б) (данные J. Hiernaux).
Средние показаны прерывистыми вертикальными линиями.
На рис. 14.2 показано распределение результатов измерения роста женщин и мужчин из племени бамбути. Средний рост женщин (137 см) на 4,9% ниже соответствующей величины для мужчин. Во всех популяциях рост женщин в среднем меньше роста мужчин на 5—8%.
Географические различия в росте
Мы располагаем многочисленными данными о росте жителей на всем земном шаре. Зачастую, однако, эти данные основаны либо на слишком малых выборках, либо на выборках, не представительных для популяции в целом. Во многих странах отмечаются различия в росте не только между жителями различных местностей, но и между представителями разных социально-экономических групп, а также между жителями городов и сельских местностей. Анализ призывных контингентов, произведенныйМартиномв Анг
Глава 14. Размеры, и форма тела
243
лии в 1939 г. (Martin), показал, что в сельских местностях средний рост мужчин составляет 172 см, тогда как в районах с преобладающим городским населением средний рост был равен 171 см. Различия в росте между основными областями Англии невелики: от 173 см в графствах близ Лондона до 171 см в Ланкашире. Тремольер, изучая призывные контингенты во Франции, обнаружил, что территориальные различия в росте были незначительны, тогда как у мужчин различных профессий средний рост различался более чем на 2 см. Несомненное увеличение скорости роста, наблюдаемое в экономически развитых странах в последнее столетие, приводит к увеличению среднего роста к моменту достижения зрелости. Эти изменения следует учитывать при сравнении роста населения различных районов, измеренного в разное время. В целом изменчивость тела человека на земном шаре не подчиняется каким-либо характерным закономерностям; в большинстве районов встречаются люди как высокого, так и низкого роста. Относительно высокий средний рост (170—175 см) характерен для жителей северо-западных районов Европы (Скандинавия) и для населения Албании. В некоторых областях Испании и южной Италии средний рост ниже 160 см. Лопари, а также ряд сибирских народностей еще ниже ростом. В Африке мы обнаруживаем резкие контрасты роста — от очень высоких тутси и представителей нильских племен, таких, как нуэры и динка, до миниатюрных пигмеев и бушменов; среди последних отмечаются значительные колебания от группы к группе. Бушмены с юга пустыни Калахари, ныне весьма многочисленные, относятся к числу самых низкорослых (145— 150 см); у населения более северных районов средний рост достигает 155 см. К очень низкорослым принадлежат местные племена Андаманских островов, Малайи и Филиппин, так называемые негритосы. В Новом Свете, особенно в Северной Америке, сравнительно недавно происходило перемещение племен с их исконных земель; многие племена при этом вымерли или сильно смешались с другими народностями. Тем не менее самые высокорослые группы (170 см и выше) сосредоточены в областях Великих равнин на восточном побережье Северной Америки и в Патагонии, а самые низкорослые — в тропиках Центральной Америки и в северных областях Южной Америки. Такое распределение приводится как пример градиента, адаптивного по отношению к климату (правило Бергмана). Существенная отрицательная корреляция между ростом и средней годовой температурой установлена для различных районов земного шара.
Определение роста по величине длинных костей
При изучении скелетов, добываемых при археологических раскопках, а также в судебной медицине рост индивидуумов определяют по длине костей конечностей. Для этого прежде всего необ
244
Часть III. Биологическая изменчивость популяций
ходимо установить связь между ростом и, скажем, длиной бедренной кости для достаточно большой случайной выборки. Построив график зависимости этих двух переменных, мы обнаружим, что точки не ложатся точно на прямую линию (или на какую-либо правильную кривую) вследствие индивидуальной вариабельности переменных. Однако существует статистический метод анализа регрессии, который позволяет вывести уравнение прямой линии или в случае необходимости любой другой кривой, на которую паилучшим образом ложились бы экспериментальные точки. Критерий соответствия сводится к тому, что отклонения значений от прямой или кривой должны быть минимальными. С помощью уравнения регрессии можно вычислить наиболее вероятный рост индивидуума, имеющего данную длину той или иной кости. Троттер и Глезер (Trotter a. Gleser), основываясь на большом материале, собранном в Америке, получили следующие линейные выражения для роста мужчин 18—30 лет по максимальной длине высушенной бедренной кости:
рост (см) = 2,38 X длина бедренной кости (см) 4- 61,41 ± 3,27.
Эта величина имеет определенные пределы точности, соответствующие среднеквадратичной ошибке ±3,27 см. Следовательно, рост может быть определен с точностью 3 см лишь в 68% случаев. Точность определения можно слегка повысить, если учитывать вместе с длиной бедра и длину большой берцовой кости (учет длины костей рук не влияет на точность определения роста).
Следует отметить, что оценки роста и других метрических показателей, основанные на методах анализа регрессии, оказываются наиболее точными в применении к той популяции, для которой была получена линия регрессии. Для каждого пола необходимо особое уравнение; уравнения, полученные для европейцев, мало применимы для других популяций.
Например, у американских негров отношение длины пог к росту выше, чем у европейцев, и относительно длиннее дистальные сегменты конечностей. Применение уравнений регрессии, полученных для определения роста в популяциях современного человека, вряд ли применимы при изучении ископаемых гоминид.
Длина туловища и конечностей
Наиболее широко применяемый метод измерения длины туловища — это определение роста сидя. Измеряемый садится на горизонтальную поверхность (например, на стол), и измеряется расстояние от макушки до плоскости сиденья. Измерение производится при помощи вертикальной градуированной стойки с передвижной поперечной планкой (антропометр). Следует добиться того, чтобы спина была максимально выпрямлена; для этого надо при
Глава 14. Размеры и форма тела
245
поднять голову; бедра должны быть горизонтальны, чтобы избежать отклонений назад или вперед.
Разница между общим ростом и ростом сидя часто применяется как условная мера длины нижних конечностей, которую трудно измерить непосредственно, поскольку положение большого вертела бедренной кости трудно определить с достаточной точностью.
Рис. 14.3. Схема, иллюстрирующая средние пропорции тела в двух популяциях.
Осевая пиния соответствует росту сидя; биакромиальное расстояние отмечено на произвольной высоте; относительная длина сегментов верхних конечностей отмечена одинаково на всех схемах. За длину нижних конечностей принята разность между ростом стоя и сидя. 1 — австралийцы (данные Albie, 1957); II — индейцы майя (данные Steggerda, 1932):
Длина ноги, определенная таким образом, несколько меньше ее анатомической длины, так как вертлужная впадина оказывается несколько выше уровня сиденья.
Имеются некоторые различия между популяциями по отношению длины туловища к росту. У коренного населения Австралии и у многих африканских негров туловище относительно корот-
кое, кормическии индекс —г--------X100) у них равен 45 и
г 'общий рост ' J 1
50% соответственно, тогда как у китайцев, эскимосов и американских индейцев эта величина достигает 53—54% (рис. 14.3).
Измерения длины различных сегментов ноги, в частности длины бедра на живом человеке, не отличаются точностью и не очень удобны, зато длину высушенных костей измерить легко: обычно для этого применяется остеометрическад доска с фиксированной планкой на одном конце (в котором укрепляется головка кости) и скользящей рейкой (устанавливаемой против другого конца кости). Длина большой берцовой кости по отношению к длине бедра (берцово-бедренный индекс) имеет тенденцию к увеличению (85% и выше) в популяциях, для которых характерна относительно большая длина ног.
246
Часть III. Биологическая изменчивость популяций
Общую длину руки измеряют от самой латеральной точки у края акромиального отростка лопатки до кончика самого длинного (третьего) пальца. Рука с ладонью, повернутой внутрь, вытягивается вдоль тела так, чтобы локоть, кисть и пальцы были максимально разогнуты в суставах. Длина руки, измеренная таким образом, больше анатомической длины, так как акромион лежит выше головки плечевой кости. Длина плеча измеряется от акромиальной точки до проксимального края головки лучевой кости, которую легко прощупать в области локтевого сгиба, а длина предплечья — от головки до вершины шиловидного отростка локтевой кости. Для популяций, для которых характерна большая относительная длина берцового сегмента, как правило, характерна и большая длина предплечья по отношению к плечу.
Поперечные размеры
Назовем три поперечных размера тела человека, наиболее доступных для измерения: ширина плеч, расстояние между акромиальными отросткамиДбиакромиальный диаметр); этот размер искажается, если плечи отведены назад, поэтому следует установить позу так, чтобы этот размер был наибольшим: плечи следует расслабить, опустив их вниз и немного вперед. Два измерения производятся в области тазового пояса: тазогребневой диаметр — расстояние между наиболее латеральными точками на гребнях подвздошных костей — и биспинальный диаметр — расстояние между передними верхними подвздошными остями, которые можно прощупать. Оба измерения надо производить при значительном надавливании, продавливая жировую ткань насколько возможно; даже в этом случае измерения менее точны, чем измерения, сделанные на скелете.
Круговые измерения иногда производятся при помощи сантиметровой ленты на определенном уровне грудной клетки, живота пли бедер.
Показатели отношения поперечных размеров к росту существенно различаются в разных популяциях. Схема на рис. 14.3 показывает средние пропорции тела мужчин и женщин одного из племен Центральной Австралии и индейцев майя с полуострова Юкатан. Для австралийцев с их относительно более узкими плечами и бедрами характерен линейный тип строения тела. Половые различия в строении бедер у них менее .выражены, чем у европейцев.
Антропометрия головы
Мозговая коробка. Наибольшее значение долгое время придавали измерениям длины и ширины головы. По этим показателям вычисляли головной индекс х 100^, предложенный
Глава 14. Размеры и форма тела
247
шведским анатомом Ретциусом более 100 лет назад как показатель формы головы и быстро получивший признание как способ классификации популяций.
Максимальная длина, определяемая обычно калипером, измеряется от глабеллы (костного вздутия между надбровными дугами) до опистокраниума (наиболее удаленной точки затылка). Максимальная ширина, измеряемая под прямым углом к сагиттальной плоскости, обычно определяется в области теменных бугров.
Статистическое распределение величин головного индекса непрерывно и приблизительно соответствует гауссову. У живого этот индекс в среднем на 2 единицы выше, чем при измерении непосредственно на черепе. Весь диапазон изменений головного индекса делят обычно на три группы: долихоцефалы (ниже 75), мезоцефалы
Таблица 14.2
Средний головной индекс в различных популяциях
Популяции Число обследованных Средний индекс Популяции Число обследованных ' Средний ин-; деке
Черногорцы 100 88,6 Марокканцы 5 210 74,3
Албанцы 112 86,4 Корейцы 522 83,4
Норвежские лопари 254 85,0 Чукчи (Сибирь) 148 82,0
Армяне 234 83,5 Китайцы (Восточный Ки- 359 81,7
Немцы 925 82,5 тай)
Голландцы 4 600 80,3 Японцы 6 000 80,8
Норвежцы (Тромс) 548 81,0 Айны (Япония) 95 77,3
Норвежцы (Хедмарк) 988 77,7 Майя (Юкатан) 133 85,8
Шотландцы (северо- 320 78,1 Эскимосы (юго-западная 61 80,7
восток Шотландии) Аляска)
Португальцы 11 658 76,4 Сиу (центральные районы 537 79,6
Бедуины руала 270 75,0 США)
Негритосы (Филип- 147 82,7 Отомн (южная Мексика) 178 77,9
пины) Эскимосы (восточная Грен- 225 76,7
Семанг (Малайя) 103 79,0 ландия)
Брамины (Бенгалия) 100 78,7 Гавайцы 203 84,0
Наири (Малабарский 175 73,2 Население Фиджи 133 81,5
берег) Маори 421 77,7
Балуба (Конго) 367 81,6 Население Новой Каледо- 185 76,5
Пигмеи (Итури) 386 76,5 НИИ
Ибо (Нигерия) 2 603 76,4 Австралийцы (область Арн- 236 71,8
Кикуйю (Кения) 384 76,0 хема)
Сомалийцы 244 74,3
248
Часть III. Биологическая изменчивость популяций
(75—80) и брахицефалы (выше 80) — и сравнивают частоты этих групп в различных популяциях. Однако искусственное «разрезание» такого непрерывного распределения па произвольно выделенные категории может лишь ввести в заблуждение, и нет оснований рекомендовать такой прием. Этот индекс может служить лишь грубой мерой формы черепа в горизонтальной плоскости: черепа с одинаковым индексом могут существенно различаться по своей форме в целом. Не удивительно, что изучение наследуемости такого произвольно выделенного признака не дало четких результатов.
В Европе среднее значение индекса (75—80) обнаружено у жителей Британских островов, части Скандинавии и в некоторых районах Средиземноморья. У жителей Западной и Центральной Европы среднее значение головного индекса обычно выше 80 или даже 85 (табл. 14.2). У большинства жителей Африки индекс ниже 75, однако для некоторых племен Центральной Африки характерен индекс 80 и выше. Индекс около 80 обнаружен у населения многих районов Китая, Японии и на островах Индонезии; у жителей Новой Гвинеи и некоторых других частей Меланезии индекс несколько ниже, тогда как у аборигенов Австралии найдены особенно низкие значения. У гренландских и канадских эскимосов средний индекс около 75, тогда как эскимосы Аляски относятся к брахицефалам. Археологические находки из многих различных районов показали, что для современных популяций характерна тенденция к более выраженной брахицефалии. Подчеркнем, что, как и при определении других размеров тела, все популяции весьма вариабельны, включая по три квадратичных отклонения с обеих сторон от среднего.
Емкость мозговой коробки. Этот параметр, представляющий особый интерес для палеоантропологов в связи с его низким значением у ранних гоминид плейстоцена, вряд ли имеет какую-либо ценность при исследовании современных популяций. Существует немало разногласий по поводу методики измерения емкости мозговой коробки. Согласно общепринятому методу, в полость черепа засыпают горчичное семя и затем измеряют его объем. Емкость мозговой коробки у животного можно определить по формуле, основанной на измерении длины, ширины и высоты черепа.
Среднее значение для мужчины-европейца — 1450 см3; у женщины емкость мозговой коробки на 10% меньше. Несколько большие значения известны для неевропейских народов, например для эскимосов. К самым низким значениям относится объем черепа у коренных жителей Австралии (менее 1300 см3).
Увеличение размера мозга считается характерной особенностью ранних этапов эволюции человека; в современных же популяциях соотношение объема мозга и интеллекта остается неясным. В пределах европейской популяции диапазон изменений объема
Глава 14. Размеры и форма тела
249
мозга очень широк: отклонения от среднего на 400 см3 совместимы с нормальными и даже выдающимися умственными способностями. Индивидуальные различия объема мозга в какой-то степени зависят от размера тела. Сомнительно, чтобы средние различия по этому показателю между современными популяциями имели какую-либо связь с различиями в умственных способностях.
Высоту мозговой коробки, знать которую необходимо для определения емкости, можно измерить: это расстояние между базисном (самой передней точкой большого затылочного отверстия на средней линии) и брегмой (точкой смыкания сагиттального и венечного швов). Согласно другому методу, череп ориентируют в так называемой франкфуртской плоскости (проходящей через самые нижние точки глазницы и самые верхние точки барабанного кольца, окружающего слуховой проход). Измерение аурикулярной высоты можно производить между теменем и порионом. С помощью специальных инструментов высоту черепа можно приближенно измерить и на живом.
Лицо. Длина лица обычно измеряется от назиона (у края носа) до самой нижней точки подбородка на средней линии (общая высота лица). Назион соответствует точке соединения верхнего края носовых костей с лобными на средней линии; правда, у живого этот шов трудно прощупать. Поэтому вместо него можно использовать горизонтальную линию, проходящую через нижние края орбитальной части верхних век. Верхняя высота лица измеряется от назиона до альвеолярной точки — самой нижней точки костной стенки между центральными верхними резцами или самой нижней точки десны на живом.
Высота носа измеряется от назиона до точки соединения носовой перегородки с верхней губой или (на черепе) до средней точки линии, проходящей через одну из двух самых нижних точек грушевидного отверстия по любую сторону носовой ости.
Чаще всего измеряют следующие поперечные размеры лица: межскуловой (расстояние между самыми удаленными точками скуловых дуг в плоскости,расположенной перпендикулярно сагиттальной) и бигониалъный (расстояние между углами нижней челюсти). Ширина носа определяется как расстояние между наиболее удаленными точками крыльев носа, а на черепе — как максимальная ширина грушевидного отверстия. Средний индекс ширины носа
/ширина носа jgg\ д0Стигает Ю4 у пигмеев Итури, превосхо-\ высота носа J J J г
дит 90 у многих африканских негров, у жителей Меланезии и у аборигенов Австралии. У эскимосов, напротив, нос, особенно измеренный на черепе, необычайно узкий. Было показано, что если американских негров сгруппировать в соответствии с долей евро
250 Часть III. Биологическая измениивость популяций
пейцев среди их предков, то обнаруживаются соответствующие различия в индексе ширины носа. Следует, однако, помнить, что обмеры носа таят в себе источники значительных ошибок.
Мышцы, жировая ткань и кости
Различия в количестве жира и мышц зависят главным образом от характера питания и физической нагрузки. Эти различия имеют важное значение, особенно при диетологическом и других направлениях физиологических исследований.
Методы измерения содержания жира путем определения удельного веса, применяемые в лабораторных условиях, неудобны для измерений на больших контингентах. При сопоставлении данных, полученных при лабораторных исследованиях, с результатами, полученными другими методами, было показано, что содержание жира можно определить путем измерения толщины кожной складки. Измерения, проводимые специальным инструментом, обеспечивающим постоянное давление, можно производить в разных областях тела, но наиболее правильное представление о содержании жира дает измерение толщины складки на уровне середины плеча над трехглавой мышцей и чуть ниже угла лопатки. Приближенную оценку развития мышц можно получить путем измерения окружности предплечья и голени после внесения поправки на толщину жирового слоя. Развитие жировой, мышечной и костной тканей можно определить на основании измерения ширины тени от этих компонентов на рентгенограмме голени. По диаметру стенок бедренной кости на середине ее длины, измеренному рентгенографически, можно оценить вес скелета; следует иметь в виду различия в компактности костных структур.
До сих пор мы располагаем довольно скудными данными о соотношении компонентов человеческого тела, хотя в этом направлении и проделана немалая работа как в Европе, так и в Америке. По данным Ньюмена, содержание жира у американских военнослужащих (средний возраст 20,7 лет) составляет в среднем 7,4% веса тела. Кривые, характеризующие распределение показателей процентного содержания жира, сильно асимметричны и имеют длинный «хвост» в области высоких значений; содержание жира может достигать 25% и более. В хорошо питающихся группах населения Европы наблюдается повышение содержания жира с возрастом.
Значение антропометрии для проблеял «наследственность и среда»
Часто молчаливо принимается, что различия между популяциями в размерах скелета мало зависят от окружающей среды и обусловлены только генетически. Трудно себе представить, чтобы при
Глава 14. Размеры и форма тела
251
изменении среды пигмеи Заира могли превратиться в негров тутси или динко. Но это крайний случай; некоторые менее выраженные различия между популяциями можно связать с влиянием окружающей среды.
Наследование антропометрических признаков, насколько оно изучено, зависит от многих генов; о некоторых особенностях по-лигенной наследственности мы упоминали (см. часть II). Корреляционный анализ сходства близнецов и родственников прямо указывает на то, что индивидуальные различия в росте действительно обусловлены генетически. Существуют, однако, значительные трудности в интерпретации этих корреляций для человеческих популяций. Конкретные данные, подтверждающие «наследуемость», в данном случае достоверны только для той популяции, для которой они получены. Относительно ограниченный диапазон изменчивости под влиянием среды для западноевропейских популяций может привести к недооценке значения среды при попытках экстраполяции выводов на человечество в целом.
Многочисленные данные указывают на то, что средний рост взрослого человека за последнее столетие в экономически развитых странах значительно увеличился. Заметные различия в среднем росте представителей разных социальных слоев наблюдались во многих европейских странах. Эти различия в большей мере связаны с условиями среды, и в частности с условиями питания, чем с этническим расслоением или дифференциальной миграцией. Эти вопросы будут более полно рассмотрены в части IV.
Другой подход к проблеме «наследственность и среда» состоит в сопоставлении потомства групп, мигрировавших в новые, более благоприятные условия, с потомством родительской популяции, оставшейся на прежнем месте. Имеется несколько исследований такого рода. Первой появилась работа Боаса (Boas, 1911). Боас показал, что выросшие в США потомки сицилийцев и евреев — выходцев из России отличались антропометрически от родительских популяций в Европе. Наиболее полное исследование, касающееся японских иммигрантов на Гавайях, провели Шапиро и Хьюлз (Shapiro a. Hulse). Иммигранты нанимались рабочими на плантации, а затем выписывали к себе невест из японских деревень. Оказалось, что их дети, выросшие на Гавайях, в среднем (с поправкой на возраст) сильно отличались от родителей по ряду антропометрических признаков (табл. 14.3).
Мужчины, например, были на 4,1 см выше, однако некоторые размеры, например длина головы, оказались уменьшенными. Были обнаружены также различия между первым поколением иммигрантов и их сверстниками, родившимися в тех же японских деревнях, откуда прибыли их родители. Как ни интерпретировать это явление, ясно, что сопоставление групп лиц, родившихся и выросших в новой стране, С оставшимися на родине может ввести иссле-
Сопоставление антропометрических данных о мужчинах-японцах, живущих в Японии (А), иммигрировавших на Гавайи (Б) и родившихся на Гавайях (В)
Таблица 14.3
А Б В
Метрические признаки Число обследованных среднее значение стандартное отклонение число обследованных среднее значение стандартное отклонение д о S 1 ай число обследованных среднее значение стандартное отклонение разность В-Б
Возраст, годы 172 35,55 0,64 178 40,60 0,55 +5,05 188 26,15 0,26 -14,45
Вес, кг 143 54,39 0,35 174 56,30 0,38 +1,91 185 57,84 0,42 +1,54
Рост, см 171 158,39 0,28 178 158,72 0,26 +0,33 188 162,83 0,26 +4,11
Рост сидя, см 171 84,50 0,20 178 83,10 0,19 -1,40 187 85,48 0,18 +2,38
Длина бедра, см 170 35,78 0,17 177 37,52 0,15 +1,74 186 37,72 0,16 +0,20
Длина голени, см 171 31,71 0,10 178 32,65 0,10 +0,94 188 33,93 0,10 +1,28
Биакромиальный размер, см 171 39,53 0,10 178 40,28 0,09 +0,75 187 41,35 0,09 +1,07
Длина головы, мм 172 189,70 0,35 178 189,38 0,32 —0,32 188 186,54 0,32 -2,84
Ширина головы, мм 172 151,90 0,26 178 152,72 0,27 +0,82 188 155,08 0,28 +2,36
Верхнелицевая длина, мм 172 72,66 0,23 178 81,40 0,25 -1-8,74 187 81,62 0,25 +0,22
Длина носа, мм 172 47,88 0,19 176 50,24 0,20 +2,36 188 50,22 0,18 -0,02
Ширина носа, мм 172 36,50 0,14 176 35,80 0,15 -0,70 188 34,56 0,14 -1,24
Глава 14. Размеры и форма тела
253
дователя в заблуждение, если учитывать только различия условий в период роста и формирования организма. Исследования Ласкера (Lasker) на мексиканских иммигрантах в США показали, что (как и следовало ожидать) наиболее выраженные изменения антропометрических показателей отмечались у тех, кто иммигрировал в детском возрасте. Следовательно, факторы, влияющие на процессы роста, несомненно, участвуют в изменении антропометрических признаков. Ясно, что миграция в другую страну связана с изменением характера действия многих факторов внешней среды. Еще не выяснено, какие из этих факторов оказывают наиболее сильное влияние на размеры тела; однако улучшение питания и снижение частоты инфекционных и паразитарных заболеваний, по-видимому, относятся к числу наиболее важных.
Многомерный анализ
Мы сравнили рост пигмеев итури и тутси. Различие между средними значениями дает нам показатель их несходства в том отношении, который зрительно можно представить как расстояние между пиками распределений. Поскольку распределения основаны на выборках, этот показатель представляет собой лишь примерную оценку истинного различия популяций; тем не менее его адекватность может быть оценена по величине стандартной ошибки.
Определенный метрический признак может представлять интерес сам по себе; если же мы хотим сравнить в целом две популяции, то нам следует исходить не из одного, а из многих признаков. Тогда для обобщения результатов нам понадобится мера расстояния, учитывающая много переменных (признаков). Нелегко установить, как эти признаки должны быть выбраны, однако их число должно быть достаточно большим, чтобы не был придан необоснованно большой вес одному из признаков. Кроме того, они не должны сильно коррелировать друг с другом — в противном случае большая доля информации будет повторяться. (Слово «информация» применено здесь в разговорном, а не в строго математическом смысле.) Если нас инетересуют общие различия, то размеры тела — не самый удачный из признаков, поскольку они подвержены влиянию среды. Лучше сравнить частоты аллелей для ряда хорошо известных генных локусов, которые, например, будут рассмотрены при обсуждении биохимической изменчивости (гл. 16) и групп крови (гл. 17). Нам следует допустить, что эти локусы, которые оказались хорошо изученными благодаря удобству работы с кровью, представительны в отношении генома в целом.
Две или более популяции, для которых были измерены по две переменных, можно представить геометрически (рис. 14.4). Они изображаются точками на плоскости, положение которых определяется значениями координат (х и у). Расстояние d между любой
254 Часть III. Биологическая изменчивость популяций
парой точек можно найти по теореме Пифагора. Такой способ представления данных можно распространить на трехмерный случай (но не на большое число измерений), хотя само понятие о расстоянии и математический аппарат, использующийся для его определения, остаются приложимыми и к очень большому числу измерений (признаков).
Рис. 14.4. Два индивидуума (А и Б), у которых количественно оценены два признака (х и у).
d — «расстояние».
К числу первых и простейших показателей «многомерного расстояния» относится коэффициент сходства рас (С#) Карла Пирсона (К. Pearson). По существу, он представляет собой среднее квадратов разностей между рядом средних величин для двух популяций. Различные измерения весьма разнятся по величине (например, рост и высота носа), поэтому удобно пронормировать их, выразив каждый в единицах стандартного отклонения. В такой стандартизованной форме С// для популяций можно записать как
£(ж2//—я»)/»»
где x-tj — среднее значение для признака i в популяции /, п — число признаков, а У, означает суммирование по всем признакам. Как показал Пенроуз, ценность Сн заключается в том, что его можно подразделить на две нескоррелированные компоненты — размер Cq и форму C2Z, измеряющие соответственно различия в общем размере и различия, остающиеся после устранения влияния фактора размера. Эти показатели легко вычислить; обычно вычисленные результаты хорошо согласуются с результатами более строгих, но и более трудоемких методов. Нанеся на график показатели размера и формы для ряда популяций, как это сделано на рис. 14.5, получаем наглядную картину сходства и различий.
Глава 14. Размеры и форма тела
255
Недостатком коэффициента 0# является то, что он не учитывает корреляций между показателями и, следовательно, включает лишнюю информацию. Этого недостатка нет у обобщенного расстояния D2 по Махаланобису, так как благодаря преобразованию переменных влияние таких корреляций устраняется. Вычисление этого показателя оказалось делом весьма трудоемким, по здесь помогло внедрение в практику ЭВМ.
1,ООг-Это
S. -
О)
5
S 0,8 -
Са
Си
• Са
0.6 -
Эм
0,4 -
0,2
- По о
Ри
БрЖ л Ба
Ма
Ег о оГрМ
Др
ЭТ а ГрЖ 0,2
Ки о
Де
। ° । । I I I I I
0,4 0,6 0,8 1.00
Разность коэффициентов формы
J______1
/,20
J 7,40
L
Рис. 14.5. Отличие некоторых народов Средиземноморья от мужчии-этрусков (Barnicot a. Brothwell, Medical biology a. Etruscian origins, 1959).
Статистические данные о размерах и форме основываются на определении трех размеров черепа — длине, ширине и высоте. ЭТ — этруски, мужчины; По — помпейцы; Эм — эми-лпйцы; Са — сардинцы; Кр — жители Крита; Ри — римляне; Си — сиенцы; Ма — мальтийцы, Гр — греки; Си — сицилийцы; Ки — киприоты (бронзовый век); ГрМ — греки микенской культуры; ГрЖ — греки (век железа); Ке — кефалонцы; Ту — турки; Ег — египтяне; Ба — баски; БрЖ — британцы (век железа); Эт — этруски, женщины;
Аш — ашанти.
Располагая данными о большом числе популяций и рассчитав величины D2 для каждой пары популяций, мы получим таблицу (матрицу) значений; при этом наглядно представить общую картину межпопуляционных различий довольно трудно. Точки, соответствующие популяциям, располагаются в многомерном пространстве, и мы не можем нанести их непосредственно на двумерный график, не исказив расстояний и не утеряв часть информации. Однако созданный Крускалом (Kruskal) метод многомерного шкалирования — одно из средств преодолеть эту трудность. Разброс точек в многомерном пространстве концентрируется в основном по двум-трем осям, для выявления которых существует ряд сходных мето
256
Часть III. Биологическая изменч ивость по пуля ций
дов. Это позволяет представить положения популяций на одном или большем числе двумерных графиков. Один из таких методов — «анализ главных компонентов». В простейшем случае можно представить себе набор рассеянных на плоскости точек, которые соответствуют средним значениям для ряда различных популяций; точки эти находятся более или менее в пределах эллипса. Существует некий вектор, проекции точек на который дают максимальный
Рис. 14.6. Точки, соответствующие ряду популяций, охарактеризованных средними значениями переменных х и у.
Прямая линия — главный компонент. Точки, соответствующие данным, имеют максимальную дисперсию в направлении этой линии; сумма квадратов расстояний от точек до этой же линии минимальна.
разброс (дисперсию) (рис. 14.6). Таким вектором является большая ось эллипса. Если точки разбросаны в более чем двумерном пространстве, мы можем поступить аналогичным образом, найдя вначале вектор, дающий наибольшую дисперсию, а затем — ортогональный вектор, который не коррелирует с первым, и так далее. Нередко оказывается, что первые 2—3 компонента дают большую часть дисперсии и достаточны для разумного представления данных.
Несколько иной подход к многомерным сравнениям — применение «дискриминантных функций». Эту статистическую процедуру разработал Фишер, чтобы решить задачу отнесения какого-либо индивидуума к одной из двух популяций на основании множественных измерений (исходя из того предположения, что он действительно принадлежит к одной из них). Обычная форма дискриминантной функции — это линейная комбинация (у=с1ж1+с2а;2... спхп) измерений с соответствующими весами с, выбранными таким образом, чтобы вероятность неправильной классификации свести к минимуму. Таким способом множество измерений заменяется одним
Глава 14. Размеры и форма тела
257
суммарным значением. Легко видеть, что если у нас будет результат лишь одного измерения, то правильная классификация индивидуума будет зависеть от степени перекрывания распределений для двух популяций (рис. 14.7), а степень перекрывания в свою очередь — от величины отношения различия d их средних к стандартным отклонениям ст. Дискриминантная функция максимизирует это отношение и таким образом делает межпопуляционное расстояние как можно большим при учете разброса величин вокруг
А. Два гауссовых распределения, перекрывающиеся в заштрихованной области; d — расстояние между средними. Б. Расстояние d уменьшено, область перекрывания возросла и различить распределения стало труднее, о — стандартное отклонение.
средних. Этот метод может быть обобщен в приложении к большему числу популяций, что позволит решить, к какой из ряда различных популяций следует отнести, например, тот или иной череп; однако в этом случае однозначного решения не существует. Тем не менее мы можем рассчитать последовательность функций, вклад которых в эффективное разделение популяций в многомерном пространстве становится все меньше и меньше. Расстояния между популяциями, полученные методом множественной дискриминации, численно не равны Z)2, однако могут быть простым способом связаны с Z)2 с помощью некоторого масштабного множителя.
Определение расстояний в многомерном пространстве на основе данных о частотах (например, о частотах генов), а не на основе непрерывного ряда размеров порождает ряд теоретических проб
258
Часть III. Биологическая изменчивость популяций
лем; в этой связи было предложено несколько методов. На практике все они дают сходные результаты.
Когда мы используем D2 или дискриминантные функции, подразумевается, как само собой очевидный факт, существование раздельных групп. Однако иногда может возникнуть ситуация, когда мы охарактеризовали выборку индивидуумов, получив большое число измерений (или зарегистрировав наличие или отсутствие множества признаков) у каждого; теперь мы хотим расположить их систематически на основании сходства признаков. Для этого существует несколько методик. Результаты такого определения «кластеров» часто представляют в форме дерева, «дендрограммы», в которой два индивидуума (или две популяции) присоединяются к общей точке ветвления (узлу), если они похожи друг •на друга больше, чем на какого-либо другого индивидуума в этой выборке (см. гл. 17, рис. 17.8). Решить, распадается ли выборка на две или более группы, нелегко, даже если мы рассматриваем только один размер: если распределение содержит две (или больше) четкие моды, возникает уверенность в наличии более чем одной группы; однако при значительном перекрывании такое решение принять гораздо труднее.
Детали методов многомерного анализа сложны; мы попытались показать лишь общие очертания некоторых главных концепций. Следует подчеркнуть, что хотя статистика, несомненно, помогает нам извлечь максимум информации из наших наблюдений, интерпретация полученных данных остается задачей биологов. В некотором смысле эти методы упрощают наши задачи, заменяя бесконечную массу сведений немногими величинами. Однако эти производные величины получены сложным путем, и бывает нелегко понять их биологический смысл. Сравнивая популяции, рекомендуется рассматривать отдельные признаки, индексы и другие простые значения одновременно с применением более тонких методов. Подобно большинству других статистических методов, многомерные методы требуют определения упрощающих допущений относительно свойств полученных данных; например при применении метода Z)2 допускается, что совместное распределение измерений нормально (многомерная нормальность), а дисперсии и ковариации одинаковы для всех популяций. Эти допущения оправданны не для всех биологических данных; если эти методы применять механически без правильного понимания их возможностей, то полученные результаты лишь введут в заблуждение.
Глава IS
ПИГМЕНТАЦИЯ И НЕКОТОРЫЕ ДРУГИЕ МОРФОЛОГИЧЕСКИЕ ПРИЗНАКИ
Никакие естественные различия не бросаются в глаза так, как различия в пигментации. Хотя географическое распределение таких признаков, как цвет волос, глаз и кожи, известно давно, накопление систематических точных данных об этом распределении наталкивается на серьезные технические трудности. Химический анализ самих пигментов, естественно, очень трудное дело, а это ограничивает возможности биохимического подхода к решению проблемы.
Цвет кожи
Цвет кожи обусловлен двумя основными факторами: наличием крови в мельчайших сосудах дермы и темного пигмента меланина в эпидермисе. При низком содержании меланина цвет кожи целиком зависит от присутствия крови в сосудах дермы; он меняется в зависимости от количества кровеносных сосудов и степени окислен-вости гемоглобина. При темной коже зависимость цвета от кровоснабжения в большей или меньшей степени маскируется меланином.
Меланин образуется в особых, снабженных отростками клетках — меланоцитах, расположенных в базальном слое эпидермиса. Эти клетки вырабатывают гранулы пигмента (меланина) диаметром около 0,5 мк и при помощи своих отростков транспортируют эти гранулы в клетки эпидермиса. Число меланоцитов на единицу поверхности кожи в разных участках тела различно, но в каждом данном участке у негров и у белых оно одинаково; следовательно, различия в цвете кожи зависят не от числа меланоцитов, а от содержания в них меланина. У негров гранул больше и они крупнее. У белых в базальном слое эпидермиса встречаются единичные коричневатые гранулы, у'негров же этот слой окрашен в интенсивный черный цвет; гранулы обнаруживаются также в мальпигиевом и роговом слоях, эпидермиса.
Меланины — широко распространенная группа пигментов в растительных и животных тканях. Пигменты этой группы представляют собой нерастворимые темноокрашенные комплексы, образующиеся путем полимеризации окисленных фенольных соединений. У человека и многих других организмов меланин образуется
260
Часть III. Биологическая изменчивость популяций
из аминокислоты тирозина через ряд последовательных стадий, причем некоторые из этих стадий катализируются ферментом тирозиназой. Образовавшийся в результате индол-5,6-хинон путем полимеризации превращается в меланин. Устойчивость меланинов к химическим воздействиям затрудняет их анализ. Имеются,
Длина волны, нм
Рис. 15.1. Рефлектометрические кривые для кожи светлых европейцев (7) и негров (II) в видимой и ближней инфракрасной областях спектра (Barnicot,
Man, 1957).
Обратите внимание на впадину между 500 и 600 нм, обусловленную гемоглобином; на кривой I она более выражена; соответственно в инфракрасной области спектра кривые сильно сближаются.
однако, данные о том, что меланины, синтезируемые в лабораторных условиях, различаются по структуре в зависимости от условий их синтеза. Гранулы меланина в тканях содержат в своем составе белки, что еще более затрудняет изучение химии этих природных пигментов.
Методы измерения. Для сравнительного изучения популяций, а также для генетических исследований, в которых сопоставляются члены одной семьи, желательно иметь метод количественной оценки содержания меланина. Прямое химическое определение пигмента в коже, подобное определению концентрации тех или иных веществ в крови и моЧе, оказывается невозможным, однако существует способ количественного определения влияния меланина на цвет кожи — так называемая рефлектометрия (спектрофотометрия в отраженном свете). Принцип этого метода состоит в следующем. Пучок световых лучей направляется на кожу, и количество диффузно отраженного света сравнивается с количеством света, отраженного от чистой белой поверхности (стандарт) при тех же условиях. Измерения проводятся во всем диапазоне види
Глава 15. Пигментация и некоторые другие признаки 261
мой области спектра (400—750 ммк), а если нужно, то в инфракрасной и ультрафиолетовой областях. Результаты измерений представляют обычно в виде кривой, выражающей долю отраженного света для каждой длины волны (по сравнению со стандартом). Примеры таких кривых для темной и светлой кожи приведены на рис. 15.1. Первая кривая с увеличением длины волны (от голубого конца спектра к красному) круто идет вверх, за исключением зеленой области, где образуется впадина, что связано с поглощением света гемоглобином. На второй кривой эта впадина менее выражена, и кривая идет более полого, слегка поднимаясь к красному концу. Большое количество меланина в этом случае в основном и определяет характер отражения света, и кривая напоминает кривую отражения суспензии пигмента. Изменения кровоснабжения относительно слабо влияют на отражение у красного конца спектра, и поэтому измерения в этой области могут служить удовлетворительной мерой влияния меланина на цвет кожи.
В настоящее время благодаря наличию дешевых портативных рефлектометров данный метод вытесняет более грубые старые методы. Простые, но менее точные приборы позволяют отнести каждого индивидуума к одной из нескольких произвольно выделенных степеней, как-то: светлые, смуглые или темные. Поскольку специальных стандартов для обозначения границ выделенных групп не существует, то разные исследователи могут отнести одного и того же человека к различным группам. Это делает невозможным надежное сравнение данных. Для устранения подобных недостатков были предприняты попытки использовать стандарты из различных материалов, раскрашенных под цвет кожи; этот путь оказался не очень удачным, так как достичь полного сходства стандартных образцов с кожей не удается. Точность измерений в значительной мере зависит от освещения, которое не всегда воспроизводимо в полевых условиях. Даже при самых благоприятных условиях измерений остается опасность ошибок, зависящая от различной способности обследующих сопоставлять цвета.
Действие солнечного света увеличивает количество меланина в коже. Чтобы уменьшить различия, вызываемые окружающей средой, измерения обычно проводятся на участках кожи, защищенных от солнечных лучей и легкодоступных наблюдению, например на внутренней поверхности руки. Конечно, перед измерением руку следует вымыть.
Наследование цвета кожи. По генетике цвета кожи опубликовано сравнительно мало работ. Рефлектометрические обследования детей от смешанных браков между неграми и белыми показали, что четкого расщепления, по-видимому, не происходит. Общепризнано, что наследование цвета кожи определяется не одним геном, но мнения относительно их действительного числа противоречивы. Статистическое распределение вариантов цвета кожи,
262
Часть III. Биологическая изменчивость популяций
измеряемого рефлектометрически в однородной популяции, оказывается непрерывным, т. е. аналогичным распределению роста и других метрических признаков. Средняя интенсивность пигментации у гибридов первого поколения (мулатов) близка к среднеарифметической величине содержания меланина в коже родителей. Такая же зависимость имеет место при возвратном скрещивании (т. е. при браках мулатов с членами родительских популяций). Было высказано предположение, что различия в цвете кожи в потомстве от брака между коренными жителями Австралии и европейцами можно объяснить, исходя из существования всего лишь нескольких генов, контролирующих цвет кожи; это предположение требует, однако, подтверждения с помощью более точных методов. Суеверие, согласно которому у светлокожих родителей, имеющих негритянских предков, может родиться «черный» ребенок, преувеличено, если не основано на недоразумении. Не существует рецессивных генов, оказывающих решающее влияние на усиление пигментации, и если цвет кожи родителей обусловлен аддитивным влиянием нескольких генов, то в их потомстве мы можем наблюдать отдельные случаи, когда ребенок будет несколько темнее родителей. Здесь многое зависит от того, что считать черным.
В некоторых странах не приняты браки между людьми с разным цветом кожи. Так, в Южной Африке и в некоторых штатах США браки между неграми и белыми запрещены законом. В Индии высшие слои в сложной системе эндогамных каст отличаются в целом более светлой кожей; эти различия поддерживаются специальными ограничениями, касающимися браков. Возможно, что в некоторых странах, где правящая верхушка состоит из белых, менее пигментированная кожа приобретает особую ценность и местные жители предпочитают выбирать в жены женщин с более светлой кожей.
Функция пигмента кожи; естественный отбор. Ультрафиолетовые лучи, оказывающие повреждающее действие на ткани, сильно поглощаются меланином. Темнокожие люди встречаются в основном в тропических районах; можно думать, что такое распределение является результатом естественного отбора, поскольку пигментированная кожа лучше защищает от солнечного света. Из этого географического правила есть, конечно, исключения. Американские индейцы расселены по всему континенту, однако они не сильно различаются по цвету кожи; темнокожие представители коренного населения Тасмании жили в умеренном климате. Такие исключения обычно объясняются тем, что эти народности переместились на новое место сравнительно недавно и естественный отбор не внес еще заметных изменений в характер пигментации. Кроме того, не для всех тропических районов характерна высокая интенсивность солнечной радиации (взять хотя бы тропические леса в период дождей). Вместе с тем она может быть иногда достаточно высокой и в Арктике благодаря отражению лучей от
Глава 15. Пигментация и некоторые другие признаки
263
поверхности снега. Мы не располагаем достаточными данными для сравнения интенсивности ультрафиолетового излучения в различных географических районах.
В областях с интенсивной солнечной радиацией, таких, как Южная Африка, Австралия и часть южных районов США, случаи рака кожи (эпителиомы) среди людей, проводящих значительную часть времени под лучами солнца, встречаются гораздо чаще у светлокожих. Правда, у негров рак кожи нижних конечностей встречается довольно часто, но он, по-видимому, связан с травмой, вызывающей изъязвления. Опухоли под действием солнечной радиации возникают главным образом в преклонном возрасте и не отличаются злокачественностью; тем не менее без соответствующего лечения они могут оказать значительное влияние на смертность в данной популяции.
Под действием ультрафиолетовых лучей из некоторых стероидов, находящихся вблизи поверхности кожи, образуется витамин D. В связи с этим предполагали, что депигментация у жителей северных областей земного шара благоприятствует этому полезному действию солнечного света. Тот факт, что рахит — заболевание, связанное с недостаточностью витамина D,— особенно часто встречается в популяции негров, иммигрировавших на север, нельзя, однако, объяснить на основании только этого предположения; дело в том, что условия питания в этих популяциях также неудовлетворительны. Мы пока точно не знаем, образуется ли витамин D в тех слоях кожи, которые надежно защищены пигментацией от ультрафиолетовых лучей.
Черные поверхности обычно поглощают видимый свет, энергия которого выделяется при этом в виде тепла; следовательно, тело темнокожего человека должно поглощать избыточное количество тепла. Перегрузка теплорегулирующих механизмов даже при интенсивном солнечном освещении в общем невелика, но все-таки она может, по-видимому, стать существенным отрицательным фактором, если индивидуум попадет в условия, близкие к предельно допустимым для организма.
Согласно общепринятому мнению, при обычной температуре поверхности тела излучение тепла практически не зависит от цвета кожи. Если зто так (а это мнение оспаривается рядом исследователей), то темная кожа не обладает преимуществом перед светлой как фактор охлаждения за счет излучения.
Цвет волос
Цвет волос зависит от гранул пигмента, содержащихся в ороговевших кортикальных клетках волоса. Эти гранулы образуются в меланоцитах, располагающихся в эпителии волосяной луковицы, прилежащем в фолликулу.
264
Часть III. Биологическая изменчивость популяций
Рис. 15.2. Рефлектометрические кривые для человеческих волос различного цвета (Barnicot, Man, 1957).
А. Европеец-альбинос. Б. Европеец-блондин. В. Европеец с ярко-рыжими волосами; виден выраженный подъем кривой, начиная с 520 нм. Г. Европеец с каштановыми волосами. Д. Африканский негр.
Объективно измерить цвет волос можно с помощью той же реф-лектометрии. Как показывает кривая на рис. 15.2, даже у альбиносов волосы не бывают чисто белыми. Все цвета волос, за исключением рыжих, составляют непрерывный ряд от темного до светлого, что соответствует уменьшению концентрации одного и того же пигмента—меланина. Хотя мы и не можем вовсе исключить возможность различий в химической структуре пигмента, есть все основания утверждать, что различия в цвете волос обусловлены чисто количественными изменениями в содержании пигмента. В светлых волосах гранул меланина меньше, они мельче и не так интенсивно заполнены пигментом, как в темных. Кривая отражения ярко-рыжих волос отличается по форме и резко изгибается
Глава 15. Пигментация и некоторые другие признаки
265
вверх в зеленой области спектра. Пигмент рыжих волос также образует гранулы, но, по-видимому, он отличается по составу от меланина.
Распределение пигмента. У большинства жителей земного шара волосы темные, и лишь на северо-западе Европы преобладают люди со светлыми и рыжими волосами. Следует отметить, однако, что даже в темноволосой популяции на самом деле отмечаются значительные различия в количестве пигмента, содержащегося в волосах. Эти различия трудно оценить на глаз или даже с помощью рефлектометра, когда цвет приближается к черному. Описаны интересные примеры изменчивости цвета волос в разных районах одной страны. Широко известно, что численность блондинов на севере Италии значительно выше, чем в южных областях. Параллельно отмечается соответствующее изменение других признаков. Эти данные были получены много лет назад при помощи субъективных методов, но, по-видимому, правильно передают характер региональных различий. В Великобритании по данным рефлектометрии рыжеволосые наиболее часто встречаются на севере и на западе; такое распределение совпадает примерно с распределением группы крови О (восточная Англия представляет исключение) (рис. 15.3).
Цвет волос часто меняется с возрастом. В некоторых странах южной Европы, где у взрослых преобладают черные волосы, дети часто бывают светловолосыми. В Китае и Африке волосы сильно пигментированы даже у детей. Светлые волосы нередко встречаются и у детей коренных жителей Австралии.
Наследование. За исключением наследования рыжего цвета, генетика цвета волос изучена недостаточно. Изменение цвета волос с возрастом усложняет соответствующий анализ. Различия в цвете волос, связанные с содержанием меланина, зависят, по-видимому, от многих генов, которые определяют степень потемнения волос и его скорость.
В наиболее ярко выраженных случаях рыжие волосы легко отличить от любых других, но детальное исследование больших популяций показало, что этот цвет не составляет четко ограниченной категории, а переходит в темно-рыжий и светло-рыжий оттенки Тем не менее рыжий цвет волос обычно рассматривают как отдельный класс, хотя его не всегда удается однозначно распознать (визуально или при помощи рефлектометра). Характерная форма кривой отражения может быть использована для количественной оценки «рыжести». Затем при помощи статистических методов можно произвести разделение на рыжие и нерыжие оттенки, что значительно точнее визуальной классификации, которая всегда субъективна. Возможно, что химические критерии обеспечили бы четкую классификацию соответствующих признаков. Неопределенность в отнесении того или иного индивидуума в группу рыжих затрудняет, естественно, и генетический анализ. Рыжий цвет
266
Часть III. Биологическая измени ив ость популяций
Рис. 15.3.
А. Частота встречаемости рыжих волос в Великобритании, вычисленная на основании рефлектометрических кривых (Sunderland, Ann Human Genet , 1956) Отмечаются высокие показатели для Уэльса и прилежащих районов.
волос наследуется примерно так, как рецессивный признак, контролируемый одним геном, но иногда он проявляется и у гетерозигот. Не исключено, что одинаковые фенотипы могут определяться несколькими различными генами, которые в настоящее время мы не можем различить, и что какие-то модификаторы также влияют на проявление этого признака.
Глава 15. Пигментация и некоторые другие признаки
267
Рис. 15.3.
Б. Частота встречаемости гена О системы АВО в Великобритании (Kopec, The distribution ot the blood-groups in the UK, 1970).
Цвет глаз
Цвет радужной оболочки глаза зависит, с одной стороны, от присутствия гранул пигмента (меланина), а с другой — от характера отражения света. Черный и коричневый цвета гла'з обусловлены наличием многочисленных пигментных клеток в переднем
268 Часть TII. Биологическая изменчивость популяций
слое радужной оболочки (рис. 15.4). В радужной оболочке более светлого глаза содержится значительно меньше пигмента. Часть падающего света отражается от зерен пигмента и от бесцветной, но мутной ткани наружного слоя, а часть света проходит через этот слой и поглощается темным пигментом заднего слоя эпителия. Если свет проходит через мутную среду, содержащую частицы, размеры которых сравнимы с длиной волны падающего света, то более короткие волны отражаются от частиц сильнее (рэлеевское
Рис. 15.4. Поперечный срез половины радужки глаза человека.
] — передний слой, содержащий пигментные клетки, особенно ближе к поверхности, и рыхлую соединительную ткань, 2 — круговая мышца (сужающая зрачок), 3 — радиальная мышца (расширяющая зрачок), 4 — задний слой пигментированного эпителия, 5 — цилиарное тело.
рассеяние). Этим оптическим эффектом и объясняется преобладание голубого цвета в свете, отраженном от депигментированного переднего слоя радужной оболочки. Различное содержание пигмента и определяет весь диапазон цвета глаз — от карих до зеленых и, наконец, голубых и серых. Радужная оболочка, однако, не представляет собой равномерно окрашенной поверхности. В светлых глазах ткань переднего слоя в одних местах толще, а в других тоньше, что придает глазу характерную радиальную исчер-ченюсть. Кроме того, меланин распределен неравномерно, образуя иногда хорошо заметные пятна. Ввиду этих сложностей неудивительно, что цвет глаз трудно точно определить, а это в свою очередь затрудняет генетический анализ.
Для соспоставления цвета глаз обследуемого используют специальный набор стандартных стеклянных глаз. Тот факт, что радужная оболочка представляет собой весьма малую поверхность, меняющуюся при изменении диаметра зрачка и испытывающую влияние многих других переменных, создает ряд таких затруднений при определении цвета глаз, с какими не приходится сталкиваться при определении цвета волос и кожи.
Нередко утверждают, что темный цвет глаз доминантен по отношению к светлому. Это утверждение правильно лишь в самой общей форме; четкое расщепление можно установить, если пренебречь мелкими различиями и сгруппировать цвета глаз в опре
Глава 15. Пигментация и некоторые другие признаки
269
деленные категории: коричневый, смешанный и голубой. Простота оказывается несколько искусственной, зависящей от первоначальной классификации фенотипа. По-видимому, в определении цвета глаз участвует несколько генов, одни из которых оказывают большее, а другие меньшее влияние на количество пигмента или на ход атрофии ткани. В некоторых западноевропейских популяциях темные глаза у женщин встречаются чаще, чем у мужчин, что было истолковано как указание на роль сцепленных с полом генов. Следует отметить, что в некоторых районах светлые глаза часто встречаются у темноволосых индивидуумов. Следовательно, некоторые гены влияют только на цвет глаз, тогда как другие оказывают влияние на пигментацию вообще.
Благодаря пигменту радужной оболочки свет попадает в глаз только через зрачок, что предохраняет сетчатку от повреждающего действия ультрафиолетовых лучей. Трудно сказать, оказывает ли депигментация, не выходящая за пределы нормальной для данной популяции, какое-либо заметное влияние на функцию зрения. Известно, однако, что крайний случай отсутствия пигмента — альбинизм — сопровождается светобоязнью (фотофобия) и различными дефектами зрения.
Дерматоглифические узоры на пальцах и ладони
На ладонной поверхности кисти и на подошвенной поверхности стопы имеются многочисленные тонкие эпидермальные гребни, которые образуют правильные, хотя и сложные узоры. Сходные системы дерматоглифических узоров обнаружены и у других приматов, а также у некоторых древесных сумчатых. Между гребнями на одинаковых расстояниях открываются выводные протоки потовых желез; при соприкосновении кожи с какой-либо гладкой поверхностью продукты секреции оставляют отпечаток узора; однако для четкого воспроизведения узоров, обеспечивающего детальное исследование, кожу следует окрасить специальным веществом и приложить пальцы или ладонь к бумаге. В 1892 г. Галь-тон впервые попытался классифицировать отпечатки пальцев и исследовать характер наследования дерматоглифического узора. Хотя большую часть узоров на пальцах руки можно отнести к одному из трех типов — дуги, петли и «завитки»,— имеется настолько выраженное разнообразие в деталях и в их сочетаниях, что нельзя найти двух человек (включая однояйцевых близнецов) с одинаковым дерматоглифическим узором.
Три основных типа дерматоглифических узоров приведены на рис. 15.5. Между ними нет абсолютного различия — обнаружены переходные формы, а также такие сложные системы, которые не укладываются в принятую классификацию. Простей
270 Часть III. Биологическая изменчивость популяций
ший тип узора — дуги, образованные рядом параллельно идущих изогнутых гребней, пересекающих «подушечку» пальца поперек. Петли — это системы линий, образующих при встрече Y-образную фигуру, известную как трирадиус, или дельта; различают трирадиусы лучевого и локтевого типа. Завиток состоит из двух трирадиусов и главной линии, окружающей центральный участок.
2 J2u7 2 23 и 18
Рис. 15.5. Основные типы дсрматоглифичсских узоров.
Жирными сплошными даны главные линии, образующие трирадиус. Прямые более тонкие линии проведены от трирадиуса к центру узора; учитывается число гребней, пересекаемых этой линией.
В Великобритании (согласно отчетам Скотланд-Ярда) наиболее распространенным узором являются петли (70%), затем следуют завитки (25%) и, наконец, дуги (5%). Классификация по отпечаткам пальцев усложняется тем, что они на разных пальцах различны. У европейцев завиток часто встречается на 1-м и на 4-м пальцах (35%), реже на 5-м. Дуги особенно часты на 2-м (11%) и редки на 5-м (1%). Петли лучевого типа (открытые в сторону лучевой кости) чаще встречаются на правой руке, чем на левой.
Подсчет числа трирадиусов на обеих руках дает представление об интенсивности узора. Дуги, как правило, не образуют трирадиусов и характеризуются баллом 0 (хотя на некоторых пальцах дуги бывают сжаты с боков, и в центре Образуется трирадиус).
Глава 15. Пигментация и некоторые другие признаки
271
Петли, образующие один трирадиус, считаются за 1, а завитки — за 2. Диапазон интенсивности узора может, следовательно, изменяться от 0, когда на всех пальцах имеются дуги, до 20, когда везде имеются только завитки.
Количественную характеристику узора можно получить следующим образом: провести прямую от каждого трирадиуса к центру узора и подсчитать число пересекаемых ею гребней (рис. 15.5). В случае дуг мы опять получим 0. Завитки с двумя трирадиусами дадут в среднем больше баллов, чем петли.
Изменчивость общей формы узора можно оценить стандартными методами, измеряя соотношение длины и ширины.
Дерматоглифические узоры находятся под генным контролем, но до сих пор не предложено какой-либо простой схемы их наследования. Исследования Холта по наследованию числа гребней, распределение которых для произвольной популяции оказывается непрерывным, но не гауссовым, позволили выявить корреляцию, близкую к 0,5, между сибсами, а также между родителями и детьми без каких-либо признаков доминантности. Корреляция между однояйцовыми близнецами оказалась равной 0,95±0,02, тогда как для двуяйцовых близнецов эта величина равна 0,46±0,10. Отклонение распределения числа борозд от гауссова может, по-видимому, означать, что различия следует отнести за счет небольшого числа главных генов. Если это действительно так, то количественные различия между генотипами перекрываются и отдельные модальные значения становятся неразличимыми в распределении.
При сравнении дерматоглифических узоров в популяциях мы смело можем допустить, что фенотипические различия отражают различия в распределении генов, но, как и в случае любого метрического признака, мы не можем проанализировать полученные результаты с учетом частот отдельных генов. Сложная природа дерматоглифических узоров и громадный диапазон различий, которым они подвержены, затрудняют обобщение фактов в сжатой количественной форме. Обычный метод при изучении популяции состоит в простом сопоставлении трех главных типов узоров и вычислении индекса интенсивности узора описанным выше методом. Некоторые крайние различия представлены в виде схемы (рис. 15.6). У бушменов, например, особенно часто встречаются дуги и интенсивность узора соответственно низка, тогда как у китайцев и американских индейцев наиболее распространены завитки и интенсивность узора высока.
Узор гребней на ладони также характеризуется индивидуальной и популяционной изменчивостью. Обычно различают по крайней мере пять трирадиусов, причем четыре из них находятся у основания 2—5-го пальцев, а пятый — на границе запястья. Обычнре расположение этих трирадиусов и начинающихся от них
272
Часть III. Биологическая
изменчивость популяций
Дуги, % Завитки, %
25 15 5 5 15 25
I—J—I—I—I I- । । । ।
—_ Бушмены
—i- — Пигмеи Etpe
Голландцы
Англичане
, Либерийцы Французы Венгры
1 Айны
. Итальянцы
- Жители^восточной Африки
~ Индейцы Центральной Америки
Эскимосы
Японцы
— Индейцы Северной Америки
— • Китайцы
Буряты
Рис. 15.6. Процент дуг и завитков (по горизонтали) и индекс интенсивности рисунка (по вертикали) для ряда популяций (Cummins a. Midlo, 1943).
линий показано на схеме (рис. 15.7). Дуги, петли, завитки или более сложные узоры встречаются с различной частотой на поверхности межпальцевых подушечек, на возвышениях большого пальца и мизинца. У китайцев и ряда других монголоидных народностей главные линии идут более продольно, чем у европейцев, а узоры на первой и третьей межпальцевых подушечках встречаются сравнительно редко. У бушменов мы наблюдаем противоположную картину: узор на возвышении мизинца встречается редко, а на второй межпальцевой подушечке чаще, чем в других популяциях.
Мы не знаем, какие факторы определяют эти региональные различия в частоте разных дерматоглифических узоров. Различные фенотипы, которые встречаются в нормальных популяциях, до сих пор не удалось связать с какими-либо существенными преимуществами или недостатками, что могло бы дать нам ключ к пониманию природы действия отбора. Вопрос о связи этих разли-
Глава 15. Пигментация и некоторые другие признаки
273
Рис. 15.7. Главные линии и трирадиусы ладони. Ладонные складки даны прерывистыми линиями.
чий с заболеваниями не подвергался специальному изучению. Отмечено только, что при синдроме Дауна дерматоглифические узоры явно аномальны.
Анатомические отклонения
Описаны многочисленные варианты в деталях строения скелета мышц и кровеносных сосудов; некоторые из них, как установлено, встречаются с неодинаковой частотой в различных популяциях. Большей частью эти варианты не имеют функционального значения, однако иногда они лежат в основе тех или иных нарушений. Посемейный анализ показывает, что некоторые из этих вариантов наследуются как простые менделевские признаки, однако полученные результаты, по-видимому, следует считать лишь приблизительными вследствие широкой изменчивости и трудности точного прижизненного учета подобных признаков.
Работы по изучению различий в строении зубов и скелета на чистых линиях мышей показали, что фенотипически распознаваемые различия (частью напоминающие те, которые встречаются и у человека) могут быть тем не менее генетически сложными. Их проявление в различных штаммах иногда зависит не только от генетических различий, но и от условий внутриутробного развития, характера питания и ряда других факторов. Врожденное отсут
274 Часть III. Биологическая изменчивость популяций
ствие 3-го коренного зуба, нередко встречающееся у европейцев, эскимосов и китайцев, лишь изредка можно наблюдать у африканцев. Отсутствие соответствующего зуба часто можно наблюдать и у мышей некоторых линий, но здесь это можно рассматривать просто как крайнюю степень уменьшения размера, зависящего от многих генов.
Вариации в строении позвоночника — несращение дуг в некоторых отделах (например, spina bifida), наличие добавочных ребер, измененная структура остистых отростков шейных позвонков — были изучены Олбруком (Allbrook) на скелетах африканцев. Олбрук показал, что частота этих аномалий в разных популяциях различна.
Незаращение шва между лобными костями (метопизм) встречается у жителей Западной Европы с частотой 5—10%, а у коренного населения Австралии, эскимосов и африканцев — значительно реже. Наличие добавочных косточек в области лямбдовидного шва может служить еще одним примером аномалии черепа, встречающейся с различной частотой.
Третья малоберцовая мышца, сухожилие которой можно прощупать на стопе (оно направляется от наружной лодыжки к основанию 5-й плюсневой кости), иногда редуцирована или вообще отсутствует. Посемейный анализ показал, что отсутствие этой мышцы наследуется как рецессивный признак; однако анализ осложняется существованием половых различий в степени редукции и в частоте отсутствия этой мышцы; отмечаются также различия между левой и правой сторонами тела. Данные о частоте случаев отсутствия этой мышцы следует принимать с осторожностью; все же, по-видимому, эта аномалия чаще встречается у некоторых африканских племен и у американских индейцев (15%), чем у европейцев (6—8%) или японцев (3,5%). Отмечаются также различия в размере длинной ладонной мышцы, сухожилие которой находится в центре ладонной поверхности кисти. Сухожилие этой мышцы отсутствует у 15—25% европейцев; у негров и японцев редукция этой мышцы встречается реже.
Глава 16
БИОХИМИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Введение
Эта глава посвящена главным образом изменчивости белков, так как данная область биохимии имеет непосредственное отношение к генетике. Причины такой связи уже обсуждались в разных главах этой книги, в том числе в введении к данной части, и здесь нет нужды повторяться. Еще более существенна связь генетики с нуклеиновыми кислотами (ДНК и РНК), однако они пока менее доступны для исследования. Методы гибридизации, о которых упоминалось выше, весьма полезны для оценки дивергенции целых геномов, но пока еще недостаточно чувствительны для изучения подвидовой дифференциации у человека.
Изречение, согласно которому генетическая информация становится тем яснее, чем ближе мы подходим к первичному эффекту гена, особенно приложимо к сфере биохимии. Так, нормальная изменчивость уровня сахара в крови зависит, несомненно, от закодированных в генах различий тканей, участвующих в его регуляции, но даже если бы факторы среды удалось стандартизировать, эти различия трудно проанализировать, так как регуляторный механизм состоит из многих компонентов. Вместе с тем известно много примеров серьезных нарушений обмена, наследуемых по простому менделевскому типу,— это фенилкетонурия, галактоземия и «болезни накопления», при которых липиды или углеводы накапливаются в ряде тканей в необычно больших количествах. Биохимические исследования довели диагностику этих метаболических расстройств до высокой степени точности и в ряде случаев позволили установить тот этап метаболического пути, который оказался нарушенным в результате мутации, вызвавшей нарушение синтеза определенного фермента.
Прежде чем приступить к отдельным примерам генетической изменчивости белков, полезно вспомнить некоторые их общие свойства. Белки — зто необычайно обширный класс макромолекул, молекулярный вес которых варьирует от нескольких тысяч до миллиона и выше; их свойства и функции чрезвычайно разнообразны. Существуют тысячи различных ферментов, каждый из которых служит катализатором определенной химической реакции в составе сложных метаболических путей. Существуют такие белки, как гемоглобин и трансферрин, обеспечивающие перенос определенных веществ в организме (кислорода, железа). Существу
276
Часть III. Биологическая изменчивость популяций
ют гормоны, выделяемые в кровоток эндокринными железами и вызывающие определенные ответы в тканях — мишенях. Эти белки растворимы в воде и относятся к глобулярным. Известны, кроме того, фибриллярные белки, молекулы которых имеют форму длинных нитей; некоторые из них (например, кератин волос и кожи) образуют прочные нерастворимые структуры, другие (коллаген соединительной ткани и актин мышечных волокон) значительно легче растворимы.
Все белки состоят из цепей, в которых аминокислотные остатки прочно соединены пептидными связями. Именно последовательность аминокислотных остатков в этих цепях (первичная структура белка) закодирована в генах. Участки полипептидной цепи по ее длине закручиваются в спирали (вторичная структура), а затем складываются определенным образом, образуя клубок. Такая (третичная) структура молекулы имеет решающее значение для функции. В некоторых случаях функционирующие молекулы состоят из двух или более идентичных или разных субъединиц, связанных друг с другом в более крупную (четвертичную) структуру. Конформация молекулы часто поддерживается относительно слабыми (водородными и гидрофобными) связями; в ряде случаев более прочные ковалентные связи за счет сульфгидрильных (SH) групп вносят свой вклад в стабильность молекулы. Эти разнообразные связи образуются между боковыми группами аминокислотных остатков, которые отходят от осевой цепи на одинаковом расстоянии друг от друга. Размер и химические свойства боковых групп разнообразны; одни из них гидрофобны и поэтому в случае водорастворимых белков ориентированы внутрь молекулы; другие гидрофильны и вступают в контакт с водной средой у поверхности белковой молекулы. Некоторые гидрофильные группы способны ионизироваться, получая положительный или отрицательный заряд. Изменения заряда на поверхности белковой молекулы при замене аминокислот в результате мутации позволяют обнаруживать генетические варианты белков довольно простым методом (электрофорез).
Варианты, гемоглобина (НЬ)
Гемоглобин (НЬ) займет почетное место среди тех белков, о которых пойдет здесь речь, так как его строение и функция изучены очень подробно и описано множество его вариантов. Первым из этих вариантов был открыт гемоглобин серповидных эритроцитов (Hb-S), который представляет особый интерес для популяционной генетики как пример мутантного признака, высокая частота которого в отдельных географических районах поддерживается давлением отбора.
Глава 16. Биохимическая изменчивость
277
Гемоглобин серповидных эритроцитов (Hb-S)
Начать изложение следует с упоминания о том, что в 1910 г. в Вест-Индии была впервые описана тяжелая форма врожденной анемии. Ее назвали серповидноклеточной анемией (СКА), так как в мазках крови находили эритроциты необычной удлиненной формы. Исследование семей американских негров и африканцев показало, что СКА наследуется по простому менделевскому типу: больные гомозиготны по аллелю Hbs. Большинство гомозигот умирало в раннем возрасте, до тех пор пока не стала доступной квалифицированная медицинская помощь. Дети с СКА рождаются у двух гетерозиготных родителей, каждый из которых является носителем нормального гена и гена серповидноклеточности (генотип HbAHbs). Эритроциты гетерозиготных носителей имеют нормальную форму. В условиях достаточной оксигенации эритроциты гетерозиготных носителей имеют нормальную форму; в отсутствие кислорода они становятся серповидными и возвращаются к норме при новом поступлении кислорода. В обычных условиях признак серповидноклеточности (СК) не приносит гетерозиготному носителю почти никакого вреда — он выявляется лишь при недостатке кислорода, например на больших высотах. Кроме того, в подобных случаях несколько снижена концентрационная способность почек.
Большой успех в изучении данной проблемы был достигнут в 1949 г., когда Полинг (Pauling), Итано (Itano), Сэнджер (Singer) и Уэллс (Wells) показали, что в случаях серповидноклеточности в эритроцитах содержатся гемоглобины двух типов, разделимые при помощи электрофореза. Мы уже говорили о поверхностном заряде белков; величина и знак этого заряда зависит от кислотности среды (pH). Если щелочной раствор нормального гемоглобина (pH 8,6) поместить в электрическое поле, то белок мигрирует к положительному полюсу (аноду). Если материал для этой процедуры взят у носителя признака серповидноклеточности, то от нормального гемоглобина Hb-А постепенно отделится движущийся медленнее аномальный белок Hb-S. В материале, взятом от больных СКА, обнаруживается только Hb-S и различные количества гемоглобина, характерного для плода (фетального) Hb-F, но нет Hb-А. У здорового новорожденного большая часть гемоглобина представлена Hb-F, который, однако, постепенно на протяжении первых 6 мес замещается гемоглобином взрослого типа Hb-А. При СКА и некоторых других тяжелых врожденных анемиях этот переход оказывается неполным. Интересно, что известен и другой мутантный ген, вызывающий сохранение Hb-F взрослого. Гетерозиготы по этому гену встречаются в Западной Африке с частотой около 1 %; установлено, что у взрослых гомозигот имеется только Hb-F, однако здоровье их при этом не страдает.
278
Часть III. Биологическая, изменчивость популяций
Признак серповидноклеточности возникает вследствие мутации, в результате которой синтезируется аномальный вариант НЬ; в дезоксигенированном состоянии этот НЬ менее растворим, чем нормальный; поэтому он выпадает в осадок, что влечет за собой изменение формы эритроцита. Ингрэм провел подробное исследование Hb-S; при помощи фермента трипсина молекула Hb-S была
Рис. 16.1. Пептидная карта Hb-A (по Baglioni, 1961).
На соответствующей карте Hb-S пятно, обозначенное точками, перемещается в положение, обозначенное стрелкой. Направление электрофореза — горизонтальное, хроматографии — вертикальное.
расщеплена на фрагменты (пептиды), смесь которых затем разделили, комбинируя методы электрофореза и хроматографии. Хроматография вообще получила широкое применение в биологии для разделения небольших количеств исследуемых веществ. В одном варианте этого метода полоска фильтровальной бумаги с нанесенной на нее каплей исследуемого материала погружается в подходящую смесь органических растворителей. По мере того как растворитель поднимается по бумаге, гидрофильные компоненты остаются в тонком слое воды на целлюлозе и таким образом отделяются от более гидрофобных компонентов. Ингрэму удалось таким способом разделить смесь пептидов Hb-S и после окраски, выявившей их положение, сравнить картину пятен (пептидную карту) с такой же картой для НЬ-А(рис. 16.1). Различие состояло в положении одного пептида; дальнейший анализ показал, что в этом пептиде остаток глутаминовой кислоты (Глу) заменен остатком валина (Вал). Это единственное различие между Hb-А и Hb-S; его можно объяснить заменой одного основания в соответствующем триплете гена (ЦТТ или ЦТЦ на ЦАТ или ЦАЦ). Заменой амино-
Глава 16. Биохимическая изменчивость
гп
Рис. 16.2. Сильно упрощенная модель молекулы гемоглобина.
Две а-субъединицы — светлые, две 3-субъединицы — темные. Дискообразная группа гема расположена в «кармане», образованном каждой из субъединиц; все спиральные области (А — Н) обозначены только на одной субъединице. N и С — N-конец и С-конец. Модель ориентирована так, чтобы показать близость друг к другу областей F — G, и С, важных для контакта ах- и 0,-цепей.
кислотного остатка и объясняется изменение поведения при электрофорезе: Глу заряжен отрицательно, а Вал нейтрален; правда, до сих пор неясно, каким образом такое небольшое изменение приводит к тому, что Hb-S оказывается нерастворимым в дезоксигенированном состоянии.
Гемоглобин — это не очень крупный белок; его молекулярный вес 64 000; молекула гемоглобина состоит из 4 субъединиц двух типов — а- и p-цепей, которые различаются по 57 аминокислотным остаткам, но уложены весьма одинаковым способом. Состав НЬ-А можно записать так: а2р2; в Hb-А выявлена цепь третьего тина — ^(“гТг); цепь четвертого типа — б — обнаружена в минорном компоненте НЬ-А — в НЬ-А2(а262), почти постоянно присутствующем в эритроцитах взрослых. Эти типы цепей возникли вследствие дупликации гена, за которой последовала мутационная дивергенция; таким образом, эти цепи кодируются различными генами, хотя локусы р, 6 и у тесно сцеплены. Принято нумеровать аминокислотные остатки в цепях, начиная с N-конца. Спиральные участки обозначают А — Н, также начиная с N-конца. Тогда
280
Часть III. Биологическая изменчивость популяций
можно сказать, что Hb-S отличается от Hb-А заменой Глу-> Вал в положении 6 ₽-цепи. Складывание цепей происходит так, что образуется карман, в котором плоская группа гема фиксирована многочисленными связями (рис. 16.2). Атом железа в центре группы гема служит местом связывания кислорода. Перутц (Perutz) провел большую работу, чтобы установить, какие изменения происходят с молекулой гемоглобина при связывании кислорода и его высвобождении. Этот процесс весьма сложен; связывание О2 слегка сдвигает группу гема и вызывает взаимное перемещение субъединиц, что повышает сродство других групп гема к кислороду. Эти перемещения сказываются также на заряде определенных участков молекулы и на ее способности связывать двуокись углерода (СО2). Недавно было установлено, что молекула 2,3-дифосфоглице-рата (2,3-ДФГ) способна связываться с молекулой гемоглобина вблизи от входа в центральную полость между субъединицами и таким путем способствовать стабилизации дезокси-формы. Изменение содержания 2,3-ДФГ в эритроците служит важным механизмом адаптации к недостатку О2.
Многое о том, как «работает» гемоглобин, стало известно благодаря изучению строения и свойств различных мутантов. Велик вклад сравнительных исследований и в установление того факта, что участки молекулы, связанные с выполнением важнейших функций (например, фиксация группы гема или движение в местах контакта субъединиц), редко изменяются в ходе эволюции.
Другие варианты НЬ
После того как был открыт Hb-S, началось обследование выборок из различных популяций в ходе клинических исследований или для составления карт географического распределения этого гена. В результате этой работы было обнаружено много других вариантов; некоторые из них довольно часто (5—30%) встречаются в ряде районов земного шара. Проанализировано около 80 вариантов p-цепей и 50 вариантов a-цепей; известно полдюжины вариантов Hb-F и НЬ-А2. В большинстве этих вариантов отмечается одиночная замена аминокислоты, изменяющая поверхностный заряд молекулы. Как правило, такие замены безвредны, по крайней мере в гетерозиготном состоянии; поскольку же в большинстве своем они встречаются редко, то гомозиготы обычно не обнаруживаются. Преобладание вариантов с измененным зарядом объясняется тем, что в исследованиях применяют главным образом такие методы, как электрофорез на бумаге или крахмальном геле. Лишь от четверти возможных мутаций можно ожидать изменения заряда; остальные могут быть обнаружены лишь в случае, когда они влекут за собой клинические симптомы или несомненные изменения стабильности молекулы in vitro.
Глава 16. Биохимическая изменчивость
281
НЬ-С — это вариант, мигрирующий при pH 8,6 еще медленнее, чем Hb-S (рис. 16.3), так как в нем место Глу занимает положительно заряженный остаток лизина (Лиз); замена происходит в том же участке ₽-цепи, что и в Hb-S. Этот
гемоглобин достигает довольно высоких "Г"
частот в одном небольшом районе в Запад- Н, Г 1
ной Африке. У гомозигот НЬСНЬС обнаруживаются при исследовании мазка крови J
небольшие аномалии, но это не сказыва-
ется на состоянии организма.
Другой медленно мигрирующий вариант, Hb-Е, широко распространен в Таиланде и прилежащих районах. Замена в нем соответствует ₽26Глу -> Лиз. И в зтом случае здоровье гомозигот страдает лишь незначительно, несмотря на то что замена поражает область контакта субъединиц и сродство к О2 в результате несколько снижается.
Hb-D был обнаружен в различных районах земного шара; наиболее высока его частота (1—5%) среди синдхов и гуд-жерати. Ему свойственна та же электрофоретическая подвижность, что и Hb-S,
однако он не вызывает серповиднокле-точности. Позднее изучение Hb-D в раз- Старт личных районах показало, что он гетероге-
нен; последнее хорошо иллюстрирует ограниченность метода электрофореза при «скрининге» на предмет идентификации мутантных вариантов Hb. Hb-Dpunjab (₽121Глу-> Гли) оказался идентичным
Рис. 16.3. Исследование гемоглобина методом электрофореза на бумаге (вероналовый буфер pH 8,6).
многим вариантам, которым вначале давали различные обозначе-
ния. Вместе с тем 3 другие Hb-D имеют иной состав: Hb-DIba<jan — это ₽87Тре-> Лиз, Hb-DBushman — Р16Гли->Арг, a HbDBaitimor— а26Асн-> Лиз. Гемоглобины другой группы, Hb-G, также ока
зались гетерогенными.
Среди более вредных мутантов упомянем Hb-М: у больного обнаруживается аномально высокая концентрация метгемоглобина, коричневого производного, в котором железо гема находится в окисленном состоянии [Fe(III)]. В метгемоглобине железо соединено с кислородом необратимо, и поэтому часть НЬ выключается из переноса О2; у таких новорожденных отмечается синюшность кожных покровов (цианоз). Однако этот дефект в значительной мере компенсируется и вполне совместим с жизнью. В нормальном гемоглобине остатки гистидина в положении 8 F-спирали и поло
282
Часть III. Биологическая изменчивость популяций
жении 7 Е-спирали взаимодействуют с железом гема и способствуют его удержанию в восстановленном состоянии. В четырех из^ вестных в настоящее время вариантах Hb-М эти остатки замещены тирозином (Тир) по аЕ8, аЕ7, РЕ8 и рЕ7 соответственно.
Существует, кроме того, группа мутантных гемоглобинов, у которых а- или р-субъединица нестабильна из-за того, что незаряженный остаток внутри молекулы замещен другим, с неподходящими химическими свойствами или размерами. У некоторых вариантов гемоглобина (например, HbTorino, HbBibba. HbSydney) ослаблено связывание группы гема, и она может выпасть из своего «кармана». Тогда структура молекулы гемоглобина нарушается, она «спадается», и в эритроцитах образуется осадок, так называемые тельца Гейнца. В HbGenova пролин, который в норме находится вблизи концов спиралей или в межспиральных участках, включается в состав р-спирали и разрывает ее. В Hbwien локализованный внутри молекулы Тир замещается заряженным остатком аспарагиновой кислоты (Асп), что влечет за собой заметную нестабильность молекулы и соответственно анемию даже у гетерозигот. Некоторые варианты (HbGbesapeake> HbGapetown> Hbjakkjm и HbKempsey) обладают повышенным сродством к О2 вследствие замены на месте контакта между а- и р-субъединицами (контакт ociPa); при этом наблюдается компенсаторное увеличение числа эритроцитов в периферической крови.
Кроме мутантов с вредными заменами в одном положении, существует несколько мутантов с делецией по одному или нескольким положениям, возникшей, вероятно, вследствие межгенного кроссинговера. В HbFreibUrg и в HbLeiden отсутствует один остаток, а в Hbaun нш — пять остатков в области p-цепи, важной для связывания гема.
Другие интересные варианты — это HbHariem с его двумя заменами: рбВал->Глу (как в Hb-S) и р73Асп->Асн; последняя существует независимо в одном из африканских вариантов, HbKorie-bu- На этом основании можно предположить, что НЬНаг1ет мог возникнуть вследствие простого кроссинговера между этими двумя аллелями.
Группа вредных вариантов (Lepor), по-видимому, образовалась в результате повторного кроссинговера между различными генами, занимающими соседние положения в хромосоме (межцистрон-ные кроссинговеры). HbLepOr имеет цепь, состоящую из 87 первых остатков 6-цепи и из 31 остатка p-цепи. Известны и другие примеры такого слияния (6-р или р-6), затрагивающего различные участки обеих цепей.
Некоторые из перечисленных мутантных гемоглобинов приведены в табл. 10.1. Из сказанного понятно, что эффект мутации зависит от точной локализации происшедшего изменения трехмерной
Глава 16. Биохимическая изменчивость
283
структуры молекулы и от свойств аминокислотных остатков, включенных и утраченных.
Таблица 16.1
Вариант гемоглобина Положение Замена Эффект мутации
М Boston a 58 (E 7) Гис — Тир Атом железа гема аномальной
М Iwate a87(F8) Гис —> Тир цепи стабилизирован в окис-
М Saskatoon P 63(E 7) Гис — -> Тир ленной форме, что препятст-
М Hyde Park p 92 (F 8) Гис — -> Тир вует его связыванию с О2
Torino a 43 (CD 1) Фен—»- Вал Нарушает контакт с группой гема. Нестабильность при 50°С. Анемия с тельцами включения
Hammersmith p 42 (CD 1) Фен —► Сер Карман гема доступен для воды. Нестабильность при 50°С. Анемия с тельцами включения
Sydney Вал- -* Ала Нарушает контакт с гемом. Нестабильность. Гомолитическая анемия
E P 26 (B 8) Глу —► Лиз Ослабляет контакт ajpi; снижает сродство к О2
Chesapeake a 92 (FG4) Apr — -> Лей Поражает контакт aiP2; новы-
Capetown a 92 (FG4) Apr — -*Глн шает сродство к О2
Yakima p 99 (Gl) Асп — -►Гис
Kempsey p 99 (Gl) Асп — -* Асн
Wien P 130 (H8) Тир — -> Асп Внутренние неполярные группы заменяются полярными. Нестабильность. Гемолитическая анемия
Genova Gun Hill p 28 (BIO) p 92—96 (делеция) Лей — -* Про Пролин ведет к разрыву В-спирали. Гемолитическая анемия Поражает контакты с гемом и контакты между субъединицами. Потеря гема. Гемолитическая анемия
284
Часть III. Биологическая изменчивость популяций
Талассемия
Все упомянутые до сих пор варианты НЬ отличались структурными изменениями в молекуле. Существует другая группа наследуемых вариантов, которая характеризуется частичным или полным нарушением синтеза той или иной нормальной цепи. Название thalassemia major (греческое слово таласса означает море) было дано летальной врожденной анемии, так как она часто встречалась у детей, родившихся в районе Средиземноморья; эта болезнь известна также как анемия Кули. В мазке крови таких больных обнаруживаются эритроциты необычных размеров и формы с низким средним содержанием НЬ. Компенсаторная пролиферация костного мозга приводит к утолщению костей черепа; наблюдается отставание в росте. При электрофорезе обнаруживаются НЬ-А и непостоянные количества Hb-F (10—90%).
Анемия Кули развивается у гомозигот по мутантному гену pthal, который либо тесно сцеплен с геном рА, либо является его аллелем. У гетерозиготных родителей обычно обнаруживается мягкий вариант заболевания (thalassemia minor) с весьма разнообразной картиной крови — от состояния, сходного с картиной у гомозигот, до нормального. Это непостоянство позволило заподозрить существование по меньшей мере двух типов р-талассемии, различающихся по степени выраженности эффекта мутации. Выявление гетерозигот при обследовании населения — нелегкая задача; приходится проводить целый ряд тестов: определение числа эритроцитов, их осмотической резистентности и содержания НЬ в них. Относительное увеличение содержания НЬ-А2 в среднем на 5% и во многих случаях небольшое повышение содержания Hb-F также могут быть использованы как диагностические признаки. Диагноз особенно сложен, если речь идет о популяциях с высокой частотой железодефицитных анемий, за исключением тех случаев, когда фенотипы биохимических вариантов легко различимы независимо от условий окружающей среды.
Поскольку р-талассемия довольно часто встречается в районах, где нередки и структурные варианты Hb-S, Hb-C и Hb-Е, иногда выявляются гетерозиготы с мутациями обоих типов. Клинические и биохимические проявления серповидноклеточной талассемии (генотипы pthal ps) весьма разнообразны — от слабо выраженной до тяжелой анемии, как при СКА. В зависимости от степени подавления синтеза p-цепи геном pthal клетки могут содержать в основном Hb-S или до 30% Hb-А. Симптомы НЬ-С-талассемии также разнообразны: у негров выражены слабо, а у итальянцев — гораздо сильнее. Это связывают с наличием двух различных генов — pthai+ и pthal о Hb-E-талассемия довольно часто встречается в Таиланде; нередко она приводит к состояниям, сходным с анемией Кули. Несомненно,, эти взаимодействия р-талассемии со
Глава 16. Биохимическая изменчивость
285
структурными вариантами, имеющие вредные последствия, необходимо учитывать при обсуждении влияния естественного отбора на частоты этих генов (см. ниже).
Следует отметить, что упомянутый выше HbLepOr обусловливает р-талассемическое состояние, так как синтез продукта слияния р-б протекает чрезвычайно медленно.
Как можно было ожидать, существуют и другие типы талассемии, при которых страдает синтез а-, а не p-цепи. Первой из а-талассемий была обнаружена Hb-H-болезнь. Это анемия, для которой характерно наличие больших количеств быстромигрирующего НЬ-Н. НЬ-Н представляет собой тетрамер, образованный четырьмя нормальными p-цепями (Р4). Его стабильность несколько снижена; при окраске крезиловым ярко-синим во многих эритроцитах обнаруживаются включения — преципитаты НЬ-Н. Этот гемоглобин образуется вследствие частичного подавления синтеза a-цепи; образующиеся в избытке p-цепи затем объединяются в тетрамеры. В крови, взятой из пупочного канатика новорожденных, у которых впоследствии развилась Hb-H-болезнь, был обнаружен другой быстромигрирующий гемоглобин, HbBarts> это тетрамер нормальных у-цепей (у4), т. е. эмбриональный вариант НЬ-Н.
У одного из родителей ребенка с Hb-H-болезнью обычно выявляются в мягкой форме симптомы талассемии. Трудность выявления взрослых гетерозигот по этому гену служила препятствием на пути изучения генетики и распространения а-талассемий; систематически проводимое определение количества HbBarts в крови пупочного канатика позволяет получить полезную информацию. В некоторых районах Таиланда частота этого варианта НЬ достигает 30%; его доля в составе всего гемоглобина колеблется от 1 до 90%, и по этому признаку больных можно отнести к четко различимым группам. Очень высокое содержание HbBarts отмечается в случаях гибели плода при явлениях водянки (hydrops foetalis), а содержание его около 30% характерно для случаев позднего развития Hb-H-болезни. Сопоставив различные факты, предположили, что существуют два разных гена а-талассемии: a‘hal и c4hal. Гомозиготность по ajha' приводит к hydrops foetalis, тогда как у гетерозигот a{hal a|hai развивается НЬ-Н-бо-лезнь. У гетерозигот с одним нормальным геном и одним геном a)ha' возникает во взрослом состоянии нерезко выраженная талассемия, а гетерозиготы a^hal/aA могут быть выявлены лишь по наличию в крови пупочного канатика 1—2% HbBarts.
Следует также упомянуть о весьма интересном варианте а-цепи, который приводит к развитию Hb-H-болезни, если соответствующий ген присутствует вместе с генома-талассемии. Hbconstant spring содержит 31 добавочный аминокислотный остаток, присоединенный к С-концу a-цепи, возможно, вследствие мутации
286 Часть III. Биологическая изменчивость популяций
терминирующего кодона. Такие аномальные a-цепи либо медленнее синтезируются, либо быстрее разрушаются, так что в крови обнаруживаются лишь очень небольшие количества этого варианта НЬ(<1%), поэтому их легко не заметить. В некоторых районах Дальнего Востока этот вариант, по-видимому, встречается довольно часто.
Природа генетического дефекта при классической талассемии еще не выяснена. Теоретически он может затрагивать транскрипцию, приводя к уменьшению образования мРНК, или один из компонентов, участвующих в трансляции. Накопленные в настоящее время данные свидетельствуют в пользу первого предположения; не ясно, однако, дефектна ли структура мРНК или регуляторного локуса, который регулирует активность структурного гена.
Проблема дупликации генов. Рассматривая механизм а-талас-семий, следует иметь в виду существование, по крайней мере в некоторых популяциях, двух а-локусов. Есть основания считать, что этот локус подвергся дупликации у некоторых млекопитающих, в том числе у шимпанзе; полагают, что существуют два, а быть может, и больше локусов у-цепи у человека. Шредер (Schroeder) показал, что Hb-F представляет собой смесь двух типов у-цепей, одна с глицином, другая — с аланином в положении 136.
Гетерозиготы по структурным вариантам a-цепи обычно содержат не более 25% аномального НЬ, тогда как гетерозиготы по вариантам (5-цепи достаточно часто обладают примерно равными количествами Hb-А и аномального НЬ. Объяснением этого явления может служить дупликация локуса а, но не р, хотя данные в пользу такого объяснения нельзя считать точно доказанными. Довольно убедительны наблюдения, касающиеся одной семьи в Венгрии, где произошло расщепление по двум вариантам а-це-пей. У некоторых индивидуумов имелись оба эти варианта вместе с Hb-А; следовательно, присутствовали три различных а-локуса. Вместе с тем гомозиготы по a-цепи варианта Hb-JTongariki, который впервые был найден с умеренной частотой на одном из островов Новых Гибридов, были лишены Hb-А. Этот факт несовместим с гипотезой о двух локусах, если не допустить, что вариантный ген тесно сцеплен с локусом а-талассемии, который полностью подавляет образование нормальных а-цепей.
Популяционная генетика и географическое распределение вариантов НЬ
Известно более 130 структурных вариантов НЬ, однако только три из них (Hb-S, НЬ-С и Hb-Е) обнаруживаются в достаточно обширных районах земного шара с частотой гетерозигот не менее 10%. Во многих популяциях жаркого влажного пояса Африки
Глава 16. Биохимическая изменчивость
287
Рис. 16.4. Частота гена серповидноклеточности Hbs в различных частях Старого Света (Allison, Genetical variation in human population, 1961).
серповидноклеточность встречается с частотой 20—30%; сравнительно высокие показатели отмечены в районе Средиземноморья и у различных народностей Индии (рис. 16.4). В северной Европе и на восток от Индии частота гена серповидноклеточности низка; в Новом Свете он почти наверняка отсутствовал до появления там африканских рабов. Hb-С часто встречается в довольно ограниченном районе Западной Африки с центром в Верхней Вольте и северной Гане, а НЬ-Е — исключительно часто в Таиланде и соседних странах, но по направлению к Бангладеш и границам Юго-Восточной Азии его частота снижается.
Частоты гетерозигот около 5% встречаются локально и для нескольких других вариантов, например Hb-D на северо-западе субконтинента Индии, НЬ-К — в некоторых районах Западной и Северной Африки, НЬ-0 — в Индонезии на Целебесе. Преобладающее большинство вариантов НЬ встречается редко и не достигает частоты гена, равной 1%, что иногда произвольно принимают за нижний предел полиморфизма.
Частоту генетических вариантов иногда выражают через частоту генов, а иногда через частоту гетерозигот (признаков); важно не смешивать эти два способа выражения; показатель частоты генов используется главным образом в теоретических исследованиях, однако он выводится из данных о фенотипе и может оказаться неточным, если не установлены соотношения фенотипа и генотипа. Отметим, что в простых случаях, когда одна пара аллелей имеет
288 Часть III. Биологическая изменчивость популяций
частоту не более 10%, частота гена составляет примерно половину частоты гетерозигот, поскольку большинство вариантных генов находится в гетерозиготном состоянии; однако с увеличением частоты гена величина ошибки, связанной с пренебрежением данными о гомозиготах, возрастает.
В связи с высокой частотой гена Hbs в определенных районах перед исследователем сразу же возникает такой вопрос. Поскольку носители гена серповидноклеточности умирают, не достигнув половой зрелости, особенно в условиях отсутствия медицинской помощи, со смертью каждого такого больного из популяции элиминируются два гена Hbs. Можно было бы ожидать, что за несколько столетий в результате такого отрицательного отбора частота этого гена снизится до очень низких значений. Такая утрата генов Hbs может, однако, компенсироваться, если приспособленность гетерозигот выше, чем нормальных гомозигот. В таком случае частоты генов достигают равновесия, которое имеет тенденцию восстанавливаться после отклонений, вызванных, например, притоком генов Hbs или НЬА извне. Равновесие такого типа называют устойчивым, а полиморфизм — сбалансированным. Когда мы говорим о большей «приспособленности» гетерозигот по сравнению с гомозиготами, мы имеем в виду большую вероятность для них передать гены следующему поколению. Это связано с более высокой вероятностью дожить до репродуктивного периода, или с повышенной фертильностью, или с сочетанием обоих факторов.
Ген Hbs, несомненно, часто встречается в определенных тропических районах и связан с поражением эритроцитов. Возникло, естественно, предположение, что гетерозиготы по этому гену менее восприимчивы к малярии, которая широко распространена в этих районах и занимает далеко не последнее место в перечне причин смерти. Возбудителем заболевания является паразит, который проникает в эритроцит, где проходит часть его жизненного цикла. Во многих тропических областях наиболее распространенным возбудителем малярии, нередко летальной, является Plaslodium falciparum', он вызывает трехдневную так называемую «тропическую» форму болезни. В более влажных районах перенос паразитов комарами рода Anopheles осуществляется более или менее равномерно на протяжении всего года (голоэндемичное состояние); в районах, для которых характерен сезон дождей, перенос паразитов происходит главным образом во влажные периоды. В голоэндемических районах почти все дети, достигшие двухлетнего возраста, инфицированы; с возрастом, по мере созревания иммунологических механизмов, число паразитов в крови уменьшается.
В 1954 г. Аллисон, обследуя население Восточной Африки, обнаружил, что у детей с серповидноклеточностью Р. falciparum встречается реже, а число паразитов на единицу объема крови в
Глава 16. Биохимическая изменчивость
289
среднем меньше, чем у детей с нормальными эритроцитами. Эти данные, подтвержденные более поздними исследованиями, сви-детельствуют в пользу идеи о том, что серповидноклеточность связана с определенной степенью резистентности к малярии; однако они не означают ничего определенного с точки зрения отбора, поскольку в них ничего не сообщается относительно различий в смертности. В экономически слаборазвитых странах с высокой заболеваемостью часто трудно определить точную причину смерти в раннем детстве, хотя резкое снижение детской смертности, которое обычно наблюдается после ликвидации в стране малярии, почти не оставляет сомнений в том, что малярия — одна из основных среди этих причин. Существует, однако, особенно тяжелая форма болезни — церебральная, при которой паразиты проникают в мозг; причину смерти в таких случаях можно установить с большей точностью. Интересно, что частота серповидноклеточнос-ти среди умерших от этой формы малярии исключительно низка.
Если носители гена серповидноклеточности реже умирают от малярии, то доля таких носителей должна быть в старших возрастных группах выше, чем в первый год жизни. Величину ожидаемого изменения можно вычислить из отношения pe/qe = t/S, где ре и qe — равновесные частоты НЬА и Hbs, a t и S — коэффициенты отбора против гомозигот — носителей серповидноклеточности и нормальных соответственно. Если, например, Hbs поддерживается в равновесии при частоте гена 15%, а приспособленность больного СКА принять за нулевую (£=1,0), то приспособленность у нормальных будет примерно на 17% ниже, чем у индивидуумов с признаком серповидноклеточности. Показатели относительной приспособленности, полученные в полевых исследованиях, разумно согласуются с ожидаемыми.
Имеются некоторые данные о более высокой фертильности женщин с серповидноклеточностью, так что различия в приспособ-бленности, возможно, не целиком обусловлены детской смертностью. Полагают, что инвазия плаценты малярийными паразитами ведет иногда к гибели плода. Если признак серповидноклеточности связан с некоторой резистентностью к малярии, то можно полагать, что у женщин — носительниц этого гена подобные случаи гибели плода должны встречаться реже.
Географическое распределение Hbs в основном согласуется с «малярийной гипотезой»: этот признак особенно часто встречается в районах, эндемичных по малярии, или там, где малярия была ликвидирована лишь в самое последнее время. Не следует ожидать точного соответствия,, поскольку недавние миграции населения могут изменить локальную картину генных частот, причем равновесные уровни еще не успели восстановиться. В Восточной Африке частота этого гена нижег а среди пастушеского хамитского насе
290 Часть III. Биологическая изменчивость популяций
ления долины Нила он и вовсе отсутствует. Возможно, это объясняется их происхождением из районов, менее всего пораженных малярией. Полагают, кроме того, что скот отвлекает комаров от людей. Возможно, малярия стала серьезной проблемой во влажных тропиках только после того, как дождевые леса стали доступны для земледелия и плотность населения здесь возросла. Быть может, в некоторых либерийских племенах именно этим следует объяснить низкую частоту серповидноклеточности; следует, правда, заметить, что для пигмеев эфе из лесов Итури характерна высокая частота данного признака. В Индии наивысшие частоты Hbs найдены у некоторых народностей, которые занимаются охотой и подсечно-огневым земледелием, а также в некоторых низших кастах хинди, которые частично происходят из этих народностей. Изолированные районы с высокой частотой серповидноклеточности, например некоторые районы Греции и определенные оазисы Южной Аравии, возможно, коррелируют с очагами малярии.
В целом частота Hbs у американских негров около 5%, что почти вдвое меньше, чем во многих районах Западной Африки; имеются, однако, более или менее изолированные группы, в которых частота этого гена достигает 10%. Когда рабов вывезли из Западной Африки в Новый Свет, где малярия встречается реже, гетерозиготы, по-видимому, утратили селективное преимущество и частота гена снизилась вследствие отбора; однако неопределенность ряда положений усложняет интерпретацию данных. Исходные частоты генов точно не известны, так как они неодинаковы в различных районах Западной Африки, а рабов вывозили из разных районов. Кроме того, несомненно, произошло смешение с европейцами и индейцами, что также должно было снизить частоту Hbs. Это смешение, несомненно, было неодинаковым в разных районах. В принципе его можно оценить, изучая число генов с весьма разнящимися частотами в родительских популяциях, не подвергшихся столь сильному влиянию отбора, как Hbs. Так, частоты генов групп крови 7?0 и Fyk у африканцев и европейцев весьма различны; однако соответствующие данные весьма скудны и недостаточно точны. Было бы также неоправданным упрощением полагать, что селективное преимущество гетерозигот по Hbs в Новом Свете полностью утратилось. Вопрос о существовании какой-либо формы малярии в доколумбовой Америке не решен, тем не менее наличие Р. falciparum в Центральной и Южной Америке — факт твердо установленный; встречался он до самого последнего времени и на юге США.
Ген НЪС встречается с высокой частотой (10—15%) в одном ограниченном районе Западной Африки. Не ясно, связан ли он с резистентностью к малярии или с каким-либо другим фактором отбора. Гомозиготность по этому гену не наносит серьезного вреда организму, поэтому небольшого преимущества гетерозигот вполне
Глава 16. Биохимическая изменчивость
261
достаточно для поддержания довольно высокого равновесного уровня; однако обнаружить его было бы нелегко. Ситуация еще более усложняется тем, что НЬС встречается в тех же районад, для которых характерна высокая частота его аллеля НЬ3. Гетерозиготы Hb3 Hbc явно несколько менее приспособлены, однако не в такой степени, как гомозиготы Hbs НЪ3. Смогут ли эти три аллеля (ЯЬА, НЪ3 и НЬС) сосуществовать в равновесном состоянии или же НЪ3 или НЪС будет элиминирован, зависит от относительной приспособленности различных генотипов; ее величину трудно определить с необходимой точностью. В тех районах Западной Африки, где встречаются и НЪ3 и НЪС, отмечена отрицательная корреляция между их частотами: частота НЪ3 низка там,: где высока частота НЬС. Возможно, распространяясь, вытесняет Hb\ однако точно это не установлено. Мы уже упоминали болезни,, обусловленные взаимодействием ^-талассемии со структурными вариантами p-цепи. Существование Р-талассемии в Западной Африке еще более осложняет адекватную интерпретацию.
Варианты глюкозо-6-фосфатдегидрогеназы (Г6ФД)
Зрелый эритроцит получает энергию для удовлетворения своих небольших метаболических потребностей в результате анаэробного расщепления глюкозы до лактата (гликолитический путь Эмбдена — Мейергофа). Г6ФД действует на боковой петле (гек-созомонофосфатный шуит, ГМШ) этого основного пути. Этот фермент катализирует превращение глюкозо-6-фосфата в 6-фосфо-глюконат в присутствии кофермента НАДФ, который в ходе реакции восстанавливается до НАДФ-Н. Данная реакция сопровождается восстановлением трипептида глутатиона, что в свою очередь существенно для стабилизации мембраны эритроцита. Недостаточность активности Г6ФД часто сочетается со склонностью к гемолизу (разрыву мембраны эритроцитов под действием ряда препаратов и инфекционных агентов). В своей активной форме фермент представляет собой димер с молекулярным весом 100 000; считается, что его субъединицы идентичны и синтез их контролируется одним геном.
Во время войны в Корее было замечено, что у многих американских солдат-негров после приема противомалярийного препарата примахина наступал гемолиз. Позднее установили, что повышенная чувствительность к примахину связана с врожденной недостаточностью Г6ФД — признаком, сцепленным с Х-хромосомой. Массовое электрофоретическое обследование гемолизатов с применением избирательной окраски на ГбФД показало, что у жителей многих тропических районов Африки очень часто встречаются два варианта фермента (примерно по 20% каждый). При pH 8,6 оба они мигрируют к аноду быстрее нормальной формы
292
Часть III. Биологическая изменчивость популяций
[Gid(Bt)]. По данным лабораторных исследований активность одного из них, Gd(A+), снижена незначительно, а другого, Gid (А—), весьма резко (8—20% нормальной), что сочетается с чувствительностью к примахину и ряду других веществ. По-видимому8 мутантный фермент разрушается особенно быстро в зрелых эритроцитах; его дефицит обнаруживается главным образом в старых клетках; активность фермента в лейкоцитах и других тканях нормальна.
Более тяжелая форма недостаточности Г6ФД, называемая GidMediterranean или Gd(B—), часто встречается в ряде районов Греции, на о. Сардиния, на Ближнем и Среднем Востоке и к востоку от Индии. Наиболее высокая из известных частот (50—60%) обнаружена в еврейских изолятах, прежде живших в Курдистане; у европейских евреев она гораздо ниже. При исследовании гемолизатов активность фермента оказывается менее 7% нормы; она снижена также в лейкоцитах.. Активность фермента у гетерозиготных женщин занимает промежуточное положение между его активностью у гемизиготных мужчин и в норме, тогда как у гомозиготных женщин отмечается приблизительно столь же тяжелое поражение, как и у гемизиготных мужчин. По крайней мере в части популяций недостаточность Г6ФД такого типа сочетается с повышенной частотой желтухи у новорожденных и случаев гемолиза при различных инфекциях или при потреблении в пищу конских бобов Uicia faba. Заболевание фавизм было известно в ряде районов Средиземноморья со времен древности; без лечения оно нередко приводит к смерти. Восприимчивость к фавизму, по-видимому^ зависит и от других факторов, которые, однако, еще не изучены.
Если вспомнить о гемоглобинах, то нет ничего удивительного в том, что описано более 70 вариантов Г6ФД. Для их идентификации необходим арсенал современных лабораторных методов,; в том числе электрофоретическое исследование белков, определение активности в отношении различных субстратов при различных значениях pH, определение констант сродства и тесты на термостабильность. Важно помнить, что активность in vitro, не всегда надежно коррелирует с активностью фермента в интактных клетках. Концентрация Г6ФД в крови невелика (2 мг/л), и поэтому трудно получить достаточно материала для определения аминокислотной последовательности вариантов; тем не менее есть основания считать, что Gid(A+) отличается от нормального фермента Gd(Bt) лишь по одной аминокислоте.
Большинство вариантов Г6ФД встречается редко, и активность большой их части снижена. Тщательное изучение населения одной из областей Греции, где частота резко выраженной недостаточности превышала 20%, показало, что 70% случаев приходилось на вариант GdMe(jiterranean и лишь с небольшой частотой встречались 3 других варианта, связанные с тяжелой формой недоста
Глава 16. Биохимическая изменчивость
293
точности, 3 варианта со слабо выраженной недостаточностью и 3 — с нормальной активностью фермента. Недостаточность Г6ФД обнаружена примерно у 5% жителей южных районов Китая; здесь обнаружены по меньшей мере 3 различных варианта — чаще всего это Gdcanton- Другой вариант, GdMarkham, по-видимому, широко распространен в равнинной части Новой Гвинеи. Хотя! как правило! гемолиз, связанный с недостаточностью
Рис. 16.5. Частоты талассемии и случаев недостаточности Г6ФД у мужчин в различных районах Сардинии (Bernini и др., Accad. Naz. Lincei, 1960).
Видна корреляция между возникновением этих состояний и высотой над уровнем моря; в низинных районах наиболее распространена малярия; встречаемость талассемии соответствует частоте фенотипа.
Г6ФД, бывает преходящим и возникает лишь под влиянием определенных факторов среды, существуют редкие варианты, при которых гемолиз имеет хронический характер (не-сфероцитарные гемолитические анемии).
В результате изучения этого фермента можно сформулировать два положения: состояния, связанные с недостаточностью Г6ФД, весьма неоднородны; определенные варианты фермента характерны для некоторых районов земного шара.
Локус Г6ФД находится в Х-хромосоме; поэтому соответствующие дефекты полностью выражены у гемизиготных мужчин, и их частота позволяет прямо оценить частоту гена. Как мы видели, некоторые из часто встречающихся типов недостаточности оказываются при определенных обстоятельствах вредными для орга
294 Часть III. Биологическая изменчивость популяций
низма. Возникает естественный вопрос, каким образом в некоторых популяциях оказались столь высокими частоты соответствующих генов и установилось ли равновесие частот. Изучение динамики частот генов сцепленных с полом рецессивных признаков усложнено необходимостью учитывать относительную приспособленность трех женских и двух мужских генотипов. От значений этой приспособленности зависит, может ли быть достигнуто устойчивое равновесие при промежуточных частотах (т. е. между О и 1). В настоящее время мы можем определить эти значения для фенотипов Г6ФД лишь весьма приблизительно. Сбалансированный полиморфизм теоретически возможен при наличии преимуществ у гетерозиготных женщин или в ситуации, когда наличие гена создает преимущество индивидуумам одного пола, но оказывается вредным для индивидуумов другого пола; даже при выполнении этих условий различия в приспособленности должны находиться в определенных пределах.
Высокая частота недостаточности Г6ФД обычно обнаруживается в районах, которые в настоящее время высоко эндемичны по тропической малярии или были такими в недалеком прошлом. В Западной Африке и на о. Сардиния, например, такая географическая корреляция четко выражена, что позволяет признать возможную защитную роль данного признака в отношении этой болезни (рис. 16.5). В крови гетерозиготных женщин часть эритроцитов дефектна по Г6ФД, а часть нормальна, поскольку инактивация Х-хромосомы происходит случайным образом. Можно изучить восприимчивость к Р. fasciparum клеток этих двух типов у одного и того же индивидуума; установлено, что в дефектные клетки Р. fasciparum проникает реже. Таким путем оказываются, по-видимому, защищенными как женщины, так и мужчины; однако зто преимущество в какой-то мере утрачивается мужчинами и гомозиготными женщинами вследствие других^ повреждающих эффектов данного мутантного аллеля.
Варианты других эритроцитарных ферментов
Известен целый ряд ферментов, которые могут быть разделены методом электрофореза на крахмальном геле и избирательно окрашены. Упомянем лишь о некоторых из них, изученных в достаточно широком диапазоне популяций. Обычно обнаруживаются 1—2 частых вариантных аллеля и отдельные редкие аллели, выявляемые при массовых обследованиях. Несомненно, усовершенствование методов позволит обнаружить еще большую гетерогенность. Гомозиготные фенотипы обычно характеризуются единичными полосами или рядом полос соответственно числу изоферментов.
Глава 16. Биохимическая изменчивость
295
Гетерозиготность иногда проявляется в простом сложении полос, образуемых двумя составляющими аллелями; однако в некоторых случаях картина усложняется образованием гибридных молекул.
Кислая фосфатаза эритроцитов
Этот фермент отщепляет фосфат от различных органических фосфатов, в том числе от фенолфталеинфосфата. Фенолфталеин является индикатором, который краснеет в щелочной среде, что
_________Начала
А БА В СА СВ С
Рис. 16.6. Кислая фосфатаза эритроцитов (Giblett, Genetic markers in human blood, 1969).
Окраска крахмального геля, при электрофорезе применен сукцинатный буфер.
позволяет простым способом локализовать активность кислой фосфатазы в геле. На рис. 16.6 показаны некоторые из наиболее часто встречающихся фенотипов, которые можно объяснить как результат расщепления по трем аллелям, которые принято было обозначать РА, Рв и Рс. Картина фенотипов несколько меняется в зависимости от примененной буферной системы; тем не менее в последнее время показано, что в синтезе кислой фосфатазы могут участвовать 3 разных локуса; их теперь обозначают АСР^, АСР^ и АСР?. Прежде считали, что кислая фосфатаза встречается только в эритроцитах, однако при использовании подходящего субстрата этот фермент удается обнаружить и в других тканях; физиологическая роль кислой фосфатазы остается неясной. Ферментативная активность, которую можно связать с продуктами этих трех аллелей, возрастает в порядке АСР^, АСР?, АСР%. Тем не менее распределение активности в случайной выборке из популяции унимодальног так как диапазоны активности для разных
296
Часть III. Биологическая изменчивость популяций
фенотипов широки и сильно перекрываются, так что реально существующая гетерогенность маскируется.
Как правило, аллель АСР® оказывается самым частым (60— 80%), а АСР^ — самым редким. Аллель АСРрчаще всего встречается в Европе (4—9%) и реже во всех прочих популяциях. Среди европейцев частота АСР^ колеблется от 35 до 38%, причем более высокие величины отмечены в некоторых популяциях лопарей, тогда как в Африке, насколько можно судить по имеющимся данным, частоты этою аллеля значительно ниже. Среди аборигенов Нового Света частоты АСР^ сильно варьируют (25—67%). Описан также ряд редких аллелей.
6-Фосфоглюконатдегидрогеназа (6ФГД)
Этот фермент следует за Г6ФД в реакциях гексозомонофасфаг-ного шунта; он катализирует окисление 6-фосфоглюконата до ри-булозо-5-фосфата и при этом генерирует НАДФ-Н. По-видимому,
Рис. 16 7. Часто встречающиеся фенотипы Г6ФД (Giblett, 1969).
А Лейкоциты В Эритроциты В случае лейкоцитов гомозиготы I и III дают единичную полосу, у гетерозигот II имеются слабее выраженные полосы, соответствующие по положению полосам у гомозигот, и третья промежуточная полоса, образованная рекомбинацией субъединиц Картина полос в случае эритроцитов сложнее.
повсеместно часто встречается только один вариантный аллель PGDC. Картина фенотипов лейкоцитарных экстрактов довольно проста (рис. 16.7, А); для гомозигот по нормальным аллелям PG©А характерна одна полоса; для гомозигот PGDC — также одна, но расположенная ближе к катоду; в случае гетерозигот имеется дополнительная резко выраженная полоса с промежуточной подвижностью, которую сНязывают с рекомбинацией между субъединицами молекул. При анализе эритроцитов отмечается несколько
Глава 16 Биохимическая изменчивость
397
Начало
АК1 АК2-1 АК2
Рис. 16.8. Три самых распространенных фенотипа аденилаткиназы (АК) у человека (Giblett, 1969).
Гомозигота АК2 встречается редко, для гетерозигот характерно сложение полос АК1 и АК2
более сложная картина для гомозигот PGDQ и гетерозигот PGDk/PGDc (рис. 16.7, Б). Активность продукта аллеля PGDC несколько понижена и менее стабильна по сравнению с нормальным ферментом. Известны другие редкие аллели; некоторые из них связаны с более или менее выраженной недостаточностью активности фермента.
Частота PGDC в Европе колеблется около 3%; в некоторых популяциях Африки и Дальнего Востока она заметно выше. Высокие частоты описаны для Бутана (21%) и для южноафриканских банту (15%).
Аденилаткиназа (АН)
Этот фермент присутствует, кроме эритроцитов, в различных тканях; он катализирует превращение аденозинтрифосфата в аденозиндифосфат и монофосфат. При стандартном электрофорезе на крахмальном геле самый частый фенотип (АК1) представляется в виде ряда полос, выраженность которых по направлению к аноду снижается (рис. 16.8). Вариантный аллель АК2 дает фермент более катодного типа; обычно этот аллель обнаруживается в гетерозиготной форме (АК2—1), так как этот аллель вообще редко встречается. В большинстве европейских популяций частоты АК2 варь-
298
Часть III. Биологическая изменчивость популяций
2 2-1 1
Рис. 16.9. Часто встречающиеся фенотипы фосфоглнжомутазы у человека (Giblett, 1969).
Картина складывается из набора полос, расположенных ближе к катоду (соответственно гену PGM,), и из набора полос, расположенных ближе к аноду (ген PGM»). На схеме представлены только варианты по локусу PGM!. Видно, что соответствующая гетерозиготе картина складывается из компонентов гомозигот 1 и 2.
ируют от 2 до 5%; в Африке и во многих азиатских популяциях этот аллель встречается реже; однако в некоторых популяциях Индии описаны частоты около 10%. Естественно, эти сравнительно низкие величины для АК2 имеют весьма высокую стандартную ошибку, так как размер выборки не превышает нескольких сотен.
Фосфоглюкомутаза (ФГМ)
Этот фермент, катализирующий превращение глюкозо-1-фос-фата в глюкозо-6-фосфат, также содержится в ряде тканей. Его синтез регулируется тремя локусами, которые не относятся к тесно сцепленным. Продукты этих локусов, ФГМ t и ФГМ2, проявляются как два раздельных набора изоферментных полос при ис
Глава 16. Биохимическая изменчивость
299
следовании гемолизатов стандартными методами. Продукт третьего локуса, ФГМ3, как правило, неопределим в эритроцитах, поможет обнаруживаться в виде набора более анодных полос в экстрактах тканей, например плаценты. Для каждого из этих трех локусов
известны вариантные аллели, однако часто встречаются лишь некоторые из них. На рис. 16.9 показаны фенотипы, сочетающиеся с часто встречающимися аллелями PGM\ и PGM{. Частота PGM{ в Англии составляет 76%; если исключить несколько аберрантных частот для небольших изолятов, то большинство величин для других популяций лежит между 65 и 85% с тенденцией к большим значениям у аборигенов Африки и Америки. Частота этого аллеля среди лопарей иногда низка (45—79%), но зависит от изучаемой группы. Редкие варианты PGM$ и PGM[
с частотой 1% каждый обнаружены в Китае. Ни один из четырех или более вариант- ц. пых аллелей PGM3 не встречается со значительной ча- h стотой; PGM* обнаруживается с частотой около 1%в ряде выборок африканцев; он вна-
чале был зарегистрирован как вариант Atkinson.
Лактатдегидрогеназа (ЛДГ)
_ .5(A)
Рис. 16.10. Лактатдегидрогеназа эритроцитов человека (Giblett 1969).
Видны 5 полос; полоса 1 соответствует 4 Б-субъединицам, полоса 5 — 4 Л-субъединицам. Промежуточные (2—4) полосы соответствуют комбинациям субъединиц.
Этот фермент представляет собой тетрамер, образованный двумя субъединицами А и В, которые представляют собой продукты различных локусов. Обычная картина ЛДГ для препаратов, полученных из эритроцитов, состоит из 5 полос (рис. 16.10). Полоса, самая близкая к аноду, состоит из 4 субъединиц А, а самая близ
кая к катоду — из 4 субъединиц В; прочие 3 полосы содержат в разных соотношениях обе субъединицы (АААВ, ААВВ иАВВВ). Относительная выраженность полос весьма заметно варьирует от ткани к ткани. Описаны электрофоретические варианты обеих субъединиц — А и В; по-видимому, они нигде не встречаются с достаточно высокой частотой: в выборках американских негров и индусов было найдено по 1—2% вариантных фенотипов, распределение которых детально ещё не изучено.
SCO
Часть III. Биологическая изменчивость популяций
УаЧаПО
АВА 2-1 2
Рис. 16.11. Три наиболее часто встречающихся фенотипа аденозиндезами-назы человека (Spencer et al., Ann. Hum. Gen;, 32, 1968).
Аденозиндезаминаза (АДА)
Три чаще всего встречающихся фенотипа этого фермента представлены на рис. 16.11. Частота аллеля ADA? по всей территории Европы близка к 5% , заметно выше у итальянцев и греков и еще выше в различных еврейских популяциях, а также у индейцев изученных групп (11—14%). И в этом отношении опять лопари с их частотой ADA? (10—16%) отличаются от большинства других народностей Скандинавии; правда, этот аллель довольно часто встречается у финнов (10%). Найдено несколько популяций в Африке с низкой частотой ADA? (0—3%); невысок этот показатель также у эскимосов Гренландии и у японцев (3%).
Другие ферменты
Кратко упомянем несколько других ферментов. Малатдегидрогеназа (МДГ) присутствует в растворимой форме в митохондриях. Интересно, что несколько ее вариантов изредка встречаются повсюду, что может разочаровать тех, кто ожидал увидеть большую изменчивость. Описан ряд вариантных аллелей фосфогексоизомеразы
Глава 16. Биохимическая изменчивость
301
(ФГИ), но и они все довольно редко встречаются: тип 3—1 найден с частотой 1% у индусов, а тип 4—1 — с частотой 0,8% у японцев. Локус фосфоглицераткиназы (ФГК) сцеплен с X-хромосомой; вариантные аллели PGKP и PGK1 встречаются с небольшой частотой в Микронезии и на Новой Гвинее (7 и 5% соответственно); по-видимому, варианты этого фермента повсюду редки. Вариантный аллель глутаматпируваттрансферазы ОРТ*- обнаружен у африканцев. В тканях плода образуется плацентарная щелочная фосфатаза; обнаружены три часто встречающихся ее аллеля РР, PP, РР (прежние Pls, PlF, Pl1). Частота РР колеблется между 26 и 29% в Европе и значительно меньше у африканцев, китайцев и японцев (2—4%); вместе с тем Р/8чаще встречается у китайцев и японцев (21—24%), чем у европейцев (3—9%) и африканцев (4%). Это весьма изменчивый фермент; дополнительные редкие аллели встречаются в Европе с суммарной частотой 2,5%. В различных популяциях описан ряд редких вариантов эритроцитарной ангидразы угольной кислоты, катализирующей разложение бикарбоната с выделением СО2. У японцев обнаружена мутация по гену каталазы — фермента, катализирующего разложение перекиси водорода; фермент в этом случае практически отсутствует.
Варианты сывороточных белков
Сыворотка крови, т. е. жидкость, остающаяся после удаления фибрина из свернувшейся плазмы, содержит весьма сложную смесь белков. С помощью электрофореза на бумаге их можно грубо разделить на несколько больших фракций: альбумин; а±-, а2-, р- и у-глобулины. Фракция у-глобулинов состоит из белков антител (иммуноглобулинов); их варианты подробно описаны в гл. 17. Электрофорез сыворотки на крахмальном или акриламидном гелях дает лучшее разрешение различных белковых компонентов, поскольку мельчайшие поры в этих средах служат ситами, разделяющими белки в зависимости от их молекулярного веса и электрического заряда.
Гаптоглобин
Гаптоглобин — зто а2-глобулин, способный связывать свободный оксигемоглобин. Комплекс НЬ—Нр можно обнаружить, окрашивая гели на НЬ реагентами типа о-толидин — перекись водорода. Три фенотипа, наследуемые по простому менделевскому типу (рис. 16.12), обнаруживаются практически во всех популяциях, хотя и с весьма различной частотой. Тип 1 — у гомозигот по аллелю Нр1; для него характерна одна полоса; тип 2 — у гомозигот по аллелю Нр2 — дает ряд полос, находящихся ближе к катоду, что представляется странным, если считать, что соответствую
302 Часть III. Биологическая изменчивость популяций
щий ген кодирует, как обычно, полипептидную цепь одного типа. У гетерозигот 2—1 для гаптоглобина характерна полоса, типичная для Нр1, и ряд расположенных ближе к катоду полос, отличающихся от полос типа 2 по положению и сравнительной выраженности. Оказалось, что аллель Нр2 действительно кодирует белок одного вида, однако его молекулы склонны «слипаться» друг с другом (полимеризоваться), давая таким образом ряд полимеров
Рис. 16.12. Результаты исследования сыворотки человека методом электрофореза на крахмальном геле (Barnicot, 1961).
Система с трис-боратиым буфером; горизонтальный метод. Для насыщения гаптоглобинов к сыворотке был добавлен избыток гемоглобина. Показаны три обычных типа гаптоглобинов — 1—1, 2—1 и 2—2, а также модифицированный 2—1. В образце с модифицированным типом 2—1 показан вариант трансферрина Т/D,, следующий за Т/С. Глобулин, основная масса которого располагается позади старта, не показан.
возрастающего молекулярного веса. Молекула гаптоглобина состоит из двух легких (а) и двух тяжелых (Р) цепей. Описанная выше изменчивость возникает вследствие изменений в a-цепях; однако описаны также немногочисленные редкие варианты Р-цепи.
Молекулярный вес a-цепи Нр2 почти вдвое выше молекуяр-ного веса a-цепи Нр1. Анализ аминокислотных последовательностей показал, что a-цепь Нр2 действительно состоит из двух а-цепей Нр1, соединенных «конец в конец», причем в месте соединения отсутствует какой-то участок последовательности. Поэтому можно предположить, что ген Нр2 образовался вследствие крое-
Глава 16. Биохимическая изменчивость
303
синговера между двумя неправильно конъюгировавшими генами Яр1. У других приматов обнаруживается вариант с одной полосой, напоминающий Нр1. Поэтому представляется вероятным, что этот кроссинговер затронул ветвь гоминид; тот факт, что ген Нр2 обнаруживается у населения всего земного шара, позволяет считать, что этот ген присутствовал у самых ранних предков человека.
Другой фенотип, сходный с типом 2—1, со слабо выраженными катодными полосами встречается довольно часто (10%) у африканцев и называется модифицированным типом 2—1 (2—1М). В некоторых тропических популяциях у многих людей вообще не удавалось обнаружить Нр. Оказалось, что это — временное состояние, возникшее вследствие гемолиза при малярии или от других причин; комплекс НЬ—Нр выводился из кровотока и содержание Нр восстанавливалось лишь постепенно за счет синтеза в печени. Известны, однако, и случаи агаптоглобулинемии без явной экзогенной причины; по-видимому, иногда она бывает врожденной.
Если подвергнуть Нр-1 электрофорезу в кислых гелях, содержащих мочевину в высоких концентрациях (для разрушения водородных связей) и меркаптоэтанол (для расщепления дисульфидных связей), a-цепи разделяются и движутся к аноду с различной подвижностью. Таким путем было показано, что популяции полиморфны по цепям Нр-1 двух типов, определяемых двумя аллелями HpalF и Нра15 и различающихся лишь по одной аминокислоте. В Западной Европе аллель Hpals обычно встречается чаще (20—25%), чем HpalF (10—15%); у китайцев, японцев, эскимосов и австралийских аборигенов Hpa1F встречается редко. У нигерийцев обнаружены более высокие частоты HpalF (47%), чем HpaiS (26%). Такая изменчивость Дра-цепи являет хороший пример скрытого разнообразия, выявляемого соответствующей модификацией лабораторного метода.
На рис. 16.13 приведена карта мира с указанием частот Нр1. Высокие частоты (^60%) обнаружены у многих народностей тропической Африки, в Центральной и Южной Америке, на Новой Гвинее и в Полинезии; зона сравнительно низких частот простирается от Ближнего Востока до Индии и Юго-Восточной Азии, а также включает Австралию. Не ясно, подвержены ли аллели Нра давлению отбора, и если да, то каким образом оно осуществляется. Гемоглобин, связанный с Нр, не может проникнуть через клубочковый фильтр, что в какой-то мере обеспечивает защиту от повреждения почечных канальцев и от потери железа; не ясно, однако, различаются ли фенотипы Нр по эффективности этой защиты и обладают ли гетерозиготы преимуществом перед гомозиготами. Гемолиз, вызванный малярией или другими болезнями, повсеместно встречается в Африке, где отмечается высокая частота Нра.1-, однако это же состояние часто обнаруживают в тех областях Азии, где частоты Нра1 несомненно низки.
Рис. 16.13. Частоты генов Яр1 и Яр3 и распространение их по земному шару (Walter a. Steegmuller, 1969).
Глава 16. Биохимическая изменчивость
305
Трансферрин (Tf)
Трансферрин -(молекулярный вес 70 000) — это fJ-глобулин, переносящий железо из мест разрушения эритроцитов и из кишечника в костный мозг, где синтезируется НЬ. Каждая молекула способна присоединить два атома железа; возможно, молекула Tf состоит из двух субъединиц.
По различиям в электрофоретической подвижности выделено около 20 врожденных вариантов Tf. Такие различия, вероятно, зависят от присутствия ряда аллелей; тем не менее семьи с расщеплением двух таких вариантов обнаруживаются редко, поскольку варианты в большинстве своем редки. Наиболее частый тип трансферрина TfG характеризуется одной полосой, тогда как у гетерозигот по одному из медленных (D) или по одному из быстрых (В) вариантов отмечается добавочная полоса. Полосы трансферрина можно избирательно идентифицировать, добавив к сыворотке радиоактивное железо (89Fe) и поместив срез геля после электрофореза на фотографическую пластинку. Когда после соответствующей экспозиции пластинка будет проявлена, участки контакта с меченым Tf будут обнаружены в виде темных полос.
Медленный (катодный) вариант TfDi довольно часто встречается в популяциях Африки (частота гена 1—5%), а также в Австралии, Новой Гвинее и на соседних островах. В некоторых группах австралийских аборигенов частота гена достигает 20%. Анализ пептидных карт показал, что африканский и австралийский TfDx имеют одинаковую аминокислотную замену. Не ясно, возникла ли одна и та же мутация независимо в этих двух частях света с последующим увеличением частоты благодаря отбору или дрейфу по каждой из них или же обе части света получили их от какого-то отдаленного общего предка. Другой вариант, TfDChi, электрофоретическая подвижность которого весьма схожа с подвижностью TfDj, впервые был описан у китайцев; он обнаружен в различных популяциях Юго-Восточной и Восточной Азии, у вед-дахов Цейлона, у некоторых племен Индии, а также в ряде популяций американских индейцев. Большинство вариантов В встречается редко, однако TfB0_1 был найден у 8% индейцев навахо на юго-западе США. До сих пор экспериментальным путем не удалось обнаружить каких-либо различий в способности транспортировать железо между часто встречающимися типами Tf.
Сывороточный альбумин
Те, кто предпочитал выраженный полиморфизм, когда-то считали альбумин «скучным» белком с редко встречающимися вариантами. Затем было установлено, что у индейцев наскапи и монтанье Лабрадора весьма часто (9—14%) встречается быстрый (анодный)
ЗОв Часть III. Биологическая изменчивость популяций
вариант этого белка. Оказавшийся идентичным ему аллель dZNaskapi, теперь переименованный в AZAlgonkln, был обнаружен с меньшей частотой у различных других племен, говорящих на алгонкинском языке (кри, оджибве, черноногие), у атапасков (сиу и тлинкиты), а также у эскимосов Аляски (но не Гренландии). Этот ген распространен главным образом на севере США и Канады; тем не менее он также встречается у навахо и апачей, родственных по языку атапаскам. Навахо, апачи и другие юго-западные племена, например зуньи и пима, также отличаются довольно высокой частотой другого варианта альбумина, обнаруженного у мексиканских индейцев и названного альбумин Mexico.
В различных частях земного шара — Малайзии, Индии, Африке, Европе и на Новой Гвинее — описано более десятка редких вариантов альбумина.
«,-Антитрипсин (система Pi)
Этот белок — ингибитор активности трипсина и ряда других протеолитических ферментов. Возможно, его физиологическая роль состоит в блокировании протеолитических ферментов, высвобождаемых лейкоцитами в очагах воспаления. В подкисленных крахмальных гелях этот белок дает ряд полос, расположенных ближе к аноду по отношению к альбумину. В большой выборке норвежцев с заметной частотой встречались четыре аллеля: Pim (95%), РР (2,3%), PP(2%)nPif (1,3%); обнаружены были также два других редких аллеля. В настоящее время мало известно о распространении этих аллелей на земном шаре; у лопарей, финнов и в некоторых азиатских выборках РР не найден. У гомозигот по РР отмечается явная недостаточность антитрипсиновой активности; в меньшей мере она ослаблена у гетерозигот РР Pim и у носителей аллеля РР. У гомозигот по РР отмечено предрасположение к развитию хронической бронхэктатической болезни легких, которая у них обычно возникает в возрасте около 40 лет; возможно, что такое предрасположение имеется также у гетерозигот. Эти вредные эффекты, по-видимому, влекут за собой более или менее выраженный отбор против данных генов, однако начинается заболевание в столь позднем возрасте (по отношению к репродуктивному периоду), что давление отбора выражено слабо.
С3-компонент комплемента
Комплементом иммунологи именуют фактор свежей сыворотки, необходимый для разрушения чужеродных антигенов под влиянием антител. Оказалось, что это сложная система, состоящая из 11 белков; ее компоненты последовательно вступают в действие после того, как первый из них присоединится к молекуле анти
Глава 16. Биохимическая изменчивость
307
тела, связавшей экзогенный антиген. Полиморфизм одного из компонентов комплемента — (С3) можно оценить при электрофорезе сыворотки на агарозном геле в условиях высокого напряжения. Известно два часто встречающихся аллеля, и С|, и около десяти редких. С3 встречается с частотой 15—25% в большинстве изученных европейских популяций. До сих пор не обнаружено генетических вариантов с нарушенной активностью комплемента.
Компонент Gc (группоспецифический)
Этот белок выявляется при электрофорезе на крахмальном, а лучше на акриламидном геле при щелочном pH в виде двух и более слабых полос, расположенных несколько ближе к катоду,
А Предметное стекло покрыто тонким слоем агара на буферном растворе. Сыворотку помещают в небольшую лунку у края пластинки. Затем пластинку соединяют с содержащими буферный раствор сосудами и создают электрическое поле. Происходит разделение сывороточных белков на пять фракций — одну альбуминовую и четыре глобулиновых. После окраски хорошо видно положение белковых фракций. Б. Сыворотка, исследуемая на наличие Gc-компонента, вносится в 2 лунки. После электрофореза (в агаре) вдоль средней линии вырезают канавку и заполняют ее анти-йс-сывороткой На месте контакта антигена и антител, диффундирующих навстречу друг другу, возникают полосы (дуги) преципитации. На стекле видны результаты исследования гомозиготы (одна дуга) и гетерозиготы (две перекрывающиеся дуги).
чем полоса альбумина. Для точной его идентификации служит реакция со специфическими антителами к Gc. Генетические варианты выявляются методом иммуноэлектрофореза; вначале образец сыворотки грубо фракционируют методом электрофореза на тонком слое агара; затем в агаре параллельно направлению миграции белка прорезают щель, которую заполняют специфической антисывороткой. Там, где встречаются диффундирующие навстречу друг другу антиген и антитела, появляются дуги преципитации (рис. 16.14). Во всех популяциях обнаруживаются три фенотипа
308 Часть III. Биологическая изменчивость популяций
соответственно двум часто встречающимся аллелям — Gc1 и Gc2. Частота Gc2 в Европе колеблется около 25%, но оказывается ниже (около 10%) у лопарей, африканцев и аборигенов Австралии. Велика его изменчивость у американских индейцев; у навахо аллель Gc2 встречается с частотой 2%, а у шавантов в Бразилии — 69%. Другой аллель, GcChippewa, был обнаружен в некоторых популяциях североамериканских индейцев, и еще один, GicAborigen. в Австралии и на Новой Гвинее; последний неотличим существующими методами от аллеля, встречающегося у африканцев. Интересно, что локус Gc тесно сцеплен с локусом сывороточного альбумина; выдвинуто предположение, согласно которому он и возник из гена альбумина путем дупликации. В самое последнее время появилось мнение, что белок Gc, возможно, участвует в транспорте витамина D; не исключено, что эти представления могут привести к пониманию действия отбора на данную систему.
Сывороточная псевдохолинэстераза
Если препараты, полученные при электрофорезе сыворотки человека на щелочном крахмальном геле, обработать смесью а-нафтилацетата с подходящим азокрасителем, то проявится ряд из четырех окрашенных полос — изоферменты псевдохолинэстеразы. Этот фермент, гидролизующий определенные органические эфиры, присутствует в различных тканях, но не в эритроцитах: он отличается от ацетилхолинэстеразы, которая расщепляет нейромедиатор ацетилхолин в нервно-мышечном соединении и присутствует также в эритроцитах. Было отмечено, что в некоторых случаях, когда вводится перед хирургическим вмешательством миорелаксант суксаметоний (сукцинилдихолин), развивается длительный паралич и нарушение дыхания. Подробное изучение вопроса с применением различных субстратов и ингибиторов ферментов показало, что в этих случаях речь идет о гомозиготах по редкому аллелю Е[ псевдохолинэстеразы, что приводит к снижению активности фермента. У гетерозигот по этому аллелю и нормальному аллелю E'i наблюдается промежуточный уровень активности. Частота E'i в различных европейских популяциях составляет около 1,5%, а у жителей Африки, Восточной Азии, у американских индейцев и аборигенов Австралии значительно ниже. Третий аллель, Е1, отличается способностью снижать чувствительность фермента к ингибирующему действию фтора; существует также редкий аллель Ef, который в гомозиготном состоянии обусловливает практически полное отсутствие активности псевдохолинэстеразы.
Глава 17
ГРУППЫ КРОВИ
Введение
Различия в групповой принадлежности крови зависят от различий в строении определенных сложных углеводных компонентов мембраны эритроцитов; это относится по крайней мере к группам АВ О и Lewis, которые наиболее изучены с химической точки зрения. Поэтому рассмотрение групп крови отдельно от других биохимических различий представляется несколько искусственным; тем не менее такое рассмотрение удобно в связи с тем, что тесты на групповую принадлежность крови главным образом иммунологические (серологические), а также потому, что химическая природа многих группоспецифических веществ еще не известна.
Групповые вещества крови представляют собой антигены, т. е. при введении индивидууму, лишенному именно этого антигена, вызывают образование специфических антител. Многие из этих антигенов представляют собой белки, однако большинство близки к углеводным антигенам, описанным у бактерий. Антитела — это белки (иммуноглобулины), которые выделяются в плазму крови особыми антителообразующими клетками. Группоспецифические антитела, используемые для выявления антигенов групп крови, обычно обнаруживаются в сыворотке беременных женщин (иммунизированных антигенами собственного плода) или в сыворотке индивидуумов, которым многократно переливали кровь. Иногда такие антитела возникают «спонтанно» у индивидуумов с особыми группами крови; некоторые же (например, анти-М и анти-N) получают путем иммунизации кроликов эритроцитами человека.
Главный метод определения различий по группам крови — агглютинация. Если отмытые эритроциты смешать с сывороткой, содержащей специфические антитела, то они покрываются белком антител и агглютинируют. Однако некоторые специфические антитела, связываясь с эритроцитами, не вызывают их агглютинации; такие антитела называют неполными. Даже в таких условиях агглютинацию можно вызвать, проводя реакцию в среде с высоким содержанием белка, после предварительной обработки эритроцитов протеолитическими ферментами или в присутствии анти-у-глобулинов. Последний метод (тест Кумбса) основан на том, что неполные антитела представляют собой у-глобулины. Поэтому анти-у-глобулин связывается с ними и вызывает агглютинацию.
310
Часть III. Биологическая изменчивость популяций
Оказалось, что некоторые реагенты на групповые вещества крови присутствуют в экстрактах семян некоторых бобовых. Эти так называемые лектины также являются глобулинами. Ценный анти-О-реагент получен из семян Ulex europaeus, анти-А — из Dolichos biflorus, а анти-N — из Vieta graminea.
Наследование различных вариантов групп крови удивительно точно следует законам Менделя. Этим можно воспользоваться при решении вопроса об отцовстве, для различения монозиготных близнецов от дизиготных, в работах по изучению сцепления генов. Многие различия по группам крови встречаются в разных популяциях с высокой и неодинаковой частотой; поэтому они представляют ценность и для антропологии. Более того, многие из них могут быть определены сразу после рождения и впоследствии изменяются лишь при нескольких редких заболеваниях. Вначале казалось, что правило «один ген — один антиген» приложимо и в этих случаях; однако с развитием молекулярной генетики и после описания ряда необъяснимых явлений стала очевидной ограниченность этого правила. Известно, что структурные гены представляют собой участки ДНК, в которых закодированы аминокислотные непоследовательности полипептидных цепей белковых молекул. Поэтому углеводные антигены, выявляемые при тестировании групп крови, не могут быть прямыми продуктами генов. Соответствующие гены, как полагают, контролируют ферменты (гликозилтрансферазы), участвующие в синтезе углеводных цепей. Далее, гены состоят из множества азотистых оснований, способных к мутациям. Возможно, что мутация, затрагивающая тот или иной фермент, приводит к изменению не одной антигенной специфичности групповых веществ крови. Ниже мы приведем несколько примеров, когда определенная группа антигенных специфичностей наследуется как целое. Такую ситуацию можно теоретически объяснить, постулируя существование ряда тесно сцепленных генов; однако если сцепление столь тесно, что кроссинговер практически никогда не наблюдается, этот постулат остается лишь удобным предположением, не проясняя истинной структуры этих генов. Серологические методы чувствительны и высокоэффективны, но для понимания природы генов они дают нам меньше сведений, чем химия. К сожалению, почти ничего не известно о физиологическом значении групповых веществ крови, а ведь знание их функций позволило бы успешно вести поиски агентов, избирательно действующих на эти вещества. Можно надеяться, что ведущееся в настоящее время углубленное изучение клеточных мембран позволит в ближайшем будущем ликвидировать наше невежество в этом вопросе.
В 1968 г. Рзйс и Сенджер в своей ставшей классической монографии перечислили 14 различных систем групп крови, т. е. группоспецифических веществ^ которые контролируются независи
Глава 17. Группы крови
311
мыми генными локусами. Кроме того, существует немалое число антигенов групп крови, которые не варьируют (общие антигены), а также антигенов, найденных лишь в одной или нескольких семьях (личные антигены).
Система
Эта система была обнаружена первой; в 1900 г. Ландштейнер показал, что сыворотка некоторых индивидуумов вызывает агглютинацию отмытых эритроцитов ряда других индивидуумов. Это открытие позволило разработать метод безопасного переливания крови, а также предоставило в распоряжение генетиков и антропологов набор просто наследуемых вариантов, довольно часто встречающихся в большинстве популяций, легко обнаруживаемых и практически не зависящих от пола, возраста и окружающей среды. Затем лишь в 1924 г. Бернштейн установил тип наследования групп крови системы АВО, существо которого представлено в табл. 17.1. Эритроциты могут быть носителями антигенов А,
Таблица 17.1
Генетическая и серологическая характеристики групп крови системы АВО
Фенотипы Генотипы Антигены эритроцитов Антитела в сыворотке
Группа А АА и АО А Анти-В
Группа В ВВ и ВО В Анти-А
Группа О 00 '— Анти-А и анти-В
Группа АВ АВ А и В Отсутствуют
или В, или А и В, или же ни одного из них; таким образом, мы получаем четыре фенотипа: группы А, В, АВ и О; наследование определяется тремя аллелями1— А, В и О, которые при расщеплении дают 6 генотипов — АА, АО, ВВ, ВО, АВ, 00. Присутствие антигена А можно определить, смешивая клетки с сывороткой, содержащей анти-А, после чего они агглютинируют; антиген В определяется сходным образом с помощью анти-В. Истинный анти-0 не найден; поэтому мы не можем серологически отличить гомозиготы А или В от гетерозигот А или В (АО и ВО). Отсюда также следует, что мы не можем определить частоты этих генов в
1 В первом издании этой книги мы пользовались символами GA и GB, предложенными Фордом, однако такие обозначения не получили общего признания, и поэтому мы вернулись к менее обоснованному, но более распространенному способу — применяются те же символы, что и для антигенов,— А, В, О, но при обозначении генов они даются курсивом.
312
Часть III. Биологическая изменчивость популяций
популяции путем простого подсчета; для этого приходится применять косвенные методы, исходя из допущения, что в данной выборке для этих аллелей соблюдается равновесие Харди — Вейнберга.
Анти-А, как правило, присутствует в сыворотке индивидуумов, лишенных этого антигена, а анти-В — у тех, кто лишен антигена В. По-видимому, эти естественно встречающиеся антитела образуются в ответ на A-подобные и В-подобные антигены бактерий и других организмов, проникающие в пищеварительный тракт человека вскоре после рождения.
Группу А можно подразделить на группы А х и А2. Эритроциты Аз слабо реагируют с обычным анти-А, однако у нас нет специфических анти-Аз, чтобы идентифицировать этот вариант. Правда, в организме некоторых А2- и А2В-индивидуумов образуются специфические анти-А х, которые можно применить для субтипирования А-клеток. Тем не менее генотипы А Мз серологически неотличимы от АгАг или АгО, а А2А2 — от А2О. Описаны различные другие количественные варианты группы А (например, A-intermediate), довольно часто встречающиеся у африканцев.
Фактор секреции и химические свойства антигенов АВН и Lewis
В Западной Европе примерно у 75% индивидуумов, принадлежащих к группам А, В или АВ, эти антигены содержатся в слюне и других секретах слизистых. Способность к секреции групповых антигенов в водорастворимой форме наследуется как доминантный признак, контролируемый геном Se, который сегрегирует независимо от локуса АВО. В некоторых популяциях (американские индейцы и аборигены Австралии) этот ген встречается еще чаще, несекреторы (sese) соответственно редки.
Секретеров группы О можно выявить по выделению антигена, обозначаемого Н, который связывается с лектином анти-Н, получаемым из семян Ulex europaeus. Антиген Н не специфичен только для группы О; он образуется также при других фенотипах АВО, хотя и в меньшем количестве. Это вещество-предшественник, которое превращается в антиген А или В в присутствии генов А или В; его образование определяется геном Н, наследуемым независимо от генов АВО. Таким образом, существует взаимодействие между локусами Н и АВО'. //-локус образует вещество, на которое могут воздействовать продукты локуса АВО. Известны редко встречающиеся семьи, у отдельных членов которых нет антигенов АВН ни на эритроцитах, ни в секретах (фенотип Bombay). Такие индивидуумы, по-видимому, гомозиготны по редкому аллелю h, и у них не образуется антиген Н.
Антигены Lewis Lea и Leb впервые были описаны как эритроцитарные антигены, контролируемые локусом Le, наследуемым
Глава 17. Группы, крови
313
независимо от локуса АВО. Тогда же было замечено, что индивидуумы с клетками Le(a—Ь+) относятся обычно к АВН-секре-торам, а с клетками Le (a-f-b—) — обычно к АВН-несекреторам. Тем не менее у индивидуумов Le(a—Ь+) в секретах содержится как Lea, так и Leb. По-видимому, антигены Lewis исходно представляют собой водорастворимые антигены, которые связываются с эритроцитами лишь при достаточно высокой концентрации в плазме. Образование Lea контролируется геном Le; при наличии этого гена происходит секреция Lea независимо от типа секреции АВН у данного индивидуума. Специфичность Leb зависит не только от гена Let но и от генов Н и Se.
Таблица 17.2
Взаимосвязь между антигенами эритроцитов и секрета слизистых и генотипами (локусы Н, Se и Le)
Генотипы Антигены клеток ABH LeaLeb Антигены секрета слизистых ABH LeaLeb
Локус Н Локус секретера Локус Lewis
НН или SeSe LeLe + - + + + +
Hh SeSe Lele
НН или sese LeLe + + - - + -
Hh Lele
hh SeSe LeLe a
SeSe LpZ» l Фенотипы - + - - + -
hh sese lele / ВотЬаУ - . .
НН или sese lele -1
Hh
НН или SeSe lele _| . -1
Hh Sese
Эти довольно сложные отношения между генотипами по локусам Н, Se и Le л антигенами в эритроцитах и секретах представлены в табл. 17.2. Их легче понять в свете химических исследований антигенов АВН и Lewis. Эта работа касается главным образом водорастворимых форм, поскольку их легче исследовать, нежели антигены эритроцитарной мембраны, связанные с липидами. Эти водорастворимые антигены представляют собой крупные молекулы, состоящие в основном (на 80—90%) из углеводов, которые в форме боковых цепей из остатков сахаров присоединены к поли-пептидной основе. Главным образом благодаря работам Моргана (Morgan) и Уоткинса (Watkins) было установлено, что специфичность антигенов Lewis и АВН определяется схемой расположения углеводных остатков на свободном конце цепи (рис. 17.1). Полагают, что соответствующие гены кодируют ферменты (трансфера-
314
Часть III. Биологическая изменчивость популяций
Рис. 17.1. Строение и взаимопревращение антигенов групп крови А, В, Н и Le (Watkins, Science, N. Y., 152, 1966).
Специфичность определяется терминальными структурами углеводных цепей; представлены только эти структуры. Стрелками указаны эффекты отдельных генов.
зы), которые катализируют присоединение определенных углеводных остатков; строение этих ферментов до сих пор не выяснено. Существуют два весьма сходных вещества-предшественника; оба они могут быть превращены в антиген Н при наличии генов Н. Тем не менее образование фукозилтрансферазы, участвующей в синтезе водорастворимого антигена Н, каким-то образом зависит от присутствия гена Se. Гены А т В кодируют синтез трансфераз, присоединяющих соответственно N-ацетилгалактозамин и галактозу в качестве концевых компонентов Н-цепи. В присутствии гена Le к предшественнику 1 в качестве предпоследнего остатка присоединяется фукоза с образованием антигена Lea; Leb образуется путем соответствующего превращения антигена Н.
Географическое распределение групп крови А±А2В0
Распределение групп системы АВО в разных странах изучено значительно подробнее, чем распределение других систем. Несомненно, это связано с простотой получения соответствующих сывороток и определения групп, а также с тем, что эта система была обнаружена первой. В странах с высоким уровнем медицинского обслуживания имеются многочисленные контингенты доноров, что
Глава 17. Группы, крови
315
позволяет проводить подробные региональные сравнения (см., например, на рис. 15.3, Б).
Картина широкого распространения аллелей О, В и А по всему свету представлена на контурных картах (рис. 17.2, 17.3 и 17.4). Частоты этих аллелей в сумме дают 1 (100%); поэтому между этими частотами существует некоторая корреляция: заметное повышение одной должно сопровождаться снижением одной или двух других. Удивительно, что ни А, ни Б не превышают 50%, тогда как О кое-где достигает 100% (в некоторых племенах южноамериканских индейцев, которым удалось избежать смешения). Можно поэтому допустить существование сил отбора, ограничивающих относительные частоты этих аллелей. Действительно, частота О превышает 80% в большей части Нового Света (хотя у эскимосов Аляски и крайнего севера Канады эти величины ниже). Другая крайность отмечается в широкой зоне низких частот О, простирающейся от Восточной Европы через большую часть Центральной Азии вплоть до части побережья Тихого океана. В Европе частоты О на Британских островах выше на северо-западе и в Ирландии. Сравнительно высокие частоты (75 %) обнаруживаются в Исландии, которая, согласно письменным источникам, была в IX веке колонизирована выходцами из Норвегии. Тем не менее ни в одной области Норвегии частота эта не превышает 66%. Такое расхождение, вероятно, возникло потому, что древние скандинавы брали с собой исландских рабов, доля которых в последующем росте населения оказалась непропорционально большой. Частота гена секреции в Исландии (36%), Северной Ирландии (45%) и Шотландии (46%) по сравнению с Норвегией (53—59%) явно снижена. Ниши с высокой частотой О обнаруживаются и у других народов Европы, например у басков, родственные связи языка которых непонятны, у сардинцев, у племен берберов в Атласских горах и у жителей некоторых районов Кавказа. Частота О довольно высока также в Аравии; однако говорящие по-арабски племена берберов отличаются от арабов другими особенностями групп крови; это пример того, что распространение языка происходит независимо от потока генов. Заканчивая рассмотрение карты гена О, отметим его относительно’высокую частоту в Северной Австралии по сравнению с другими областями этой страны и с Новой Гвинеей. У племени тиви на островах залива Карпентария частота гена О достигает 95%. Несомненно, это пример дрейфа генов в малой общине или же эффект родоначальника; на различных островах этого района обнаружены и другие примеры аберрантных частот генов.
Карта гена В (рис. 17.3) до некоторой степени представляет картину, обратную карте гена О: высокие частоты обнаруживаются в обширных районах востока Центральной Азии с максимумом 25—30% в области Гималаев. Однако в Новом Свете и в Австралии
Рис. 17.2. Частота гена О групп крови системы АВО (Mourant, Kopec a. Sobczak, The distribution of human blood groups, 1976).
Рис. 17.3. Частота гена В групп крови системы АВО (Mourant et al., 1976).
Рис. 17.4. Частота гена А групп крови системы АВО (Mourant et al., 1976).
Глава 17. Группы, крови
319
этот аллель встречается редко (5%) или вовсе отсутствует. В направлении от Юго-Восточной Азии -к Индонезии частота гена обнаруживает тенденцию к снижению; заметный спад происходит на пути между Новой Гвинеей и Австралией, где ген В обнаруживается, да и то с низкой частотой, в районе мыса Йорк. Столь же редко встречается этот ген в Полинезии. К западу от Центральной Азии частота гена снижается, следуя неправильному геноклину; она особенно низка у басков. По-видимому, народы Азии, впервые проникшие в Новый Свет, характеризовались низкой частотой гена В в отличие от современных обитателей Восточной Азии; однако по крайней мере у одного народа Восточной Сибири, у чукчей, частоты генов А и В низки. В Африке частоты гена В низки у бушменов и гораздо выше у готтентотов.
Обратившись к карте гена А (рис. 17.4), мы отмечаем его довольно высокие частоты в Европе с рядом пятен особенно высокой частоты в Лапландии и в Армении. В Новом Свете картина распределения гена А до некоторой степени обратна выявленной для гена О. Ген А встречается редко или отсутствует в Южной и Центральной Америке; гораздо чаще он обнаруживается в Северной Америке, достигая 25% и более в ряде племен североамериканских индейцев. Ген А часто встречается также среди эскимосов, живущих на пространстве от Аляски до. Гренландии. В Австралии снова отмечается сходство распространения генов А и О: у живущих южнее племен с низкой частотой гена О явно повышена частота гена А, достигающая в некоторых случаях 40% и более.
Распространение подгруппы А2 оказывается гораздо более ограниченным по сравнению с Ах. Частота соответствующего гена близка к 10% в Европе и Африке и падает до 5% в Индии и Юго-Восточной Азии. В других частях света этот ген встречается редко или отсутствует. Частота гена А2 заметно повышена (25—37%) у лопарей, хотя и не одинакова в разных их группах; она довольно высока и у финнов. Это один из нескольких примеров необычной частоты генов у лопарей, происхождение которых остается невыясненным.
Интересно также обратить внимание не на локальное распределение частот аллеля, а на места с его максимальными частотами; при этом приходится принимать во внимание величину популяций. В некоторых австралийских племенах частота гена А очень высока, однако их вклад во всемирный итог пренебрежимо мал, так как эти племена невелики. Мак-Артур и Пенроуз (McArthur a. Penrose) подсчитали средние частоты генов для различных регионов и обнаружили, что на всем земном шаре они составляют для О — 62,5%, для А — 21,5% и для В— 16,2%. Эти авторы предлагают усредненные величины в качестве удобного стандарта, в сравнении с которым можно оценивать отклонения, имеющиеся в местных популяциях.
320 Часть III. Биологическая изменчивость популяций
Группы АВО и естественный отбор
Тот факт, что огромное большинство популяций во всем мире полиморфно по двум, а обычно даже по трем аллелям АВО, позволяет полагать, что эти вариации существовали в течение тех многих тысячелетий расселения человека по земному шару, когда популяции были невелики и были склонны утрачивать аллели вследствие дрейфа; тем не менее мы не знаем, насколько этот процесс препятствовал току генов между группами людей в те давние времена.
Антигены, близко напоминающие антигены АВН человека, встречаются у человекообразных приматов, которые также полиморфны по двум или трем аллелям. На этом основании можно полагать, что полиморфизм по АВО существовал уже гораздо раньше, когда произошла дивергенция линий гоминид и понгид.
Описание случаев эритробластоза новорожденных, вызванного присутствием у плода группоспецифического антигена системы Rhesus, отсутствующего у матери, стимулировало поиски подобных эффектов крови в системе АВО. Оказалось, что сходные заболевания новорожденных, связанные с несовместимостью по антигенам системы АВО, действительно встречаются, хотя и редко. Вполне возможно, что иммунологическое повреждение происходит на гораздо более ранних этапах беременности и остается незамеченным, если не предприняты специальные исследования для его обнаружения. В целом, если пренебречь незначительными флуктуациями, можно ожидать, что частоты генов АВО в большой выборке новорожденных будут теми же, что и у родителей. Поэтому можно проверить, существует ли значимый недостаток определенных фенотипов в потомстве определенных браков в системе АВО. В ряде исследований групп крови в больших выборках матерей и их детей были выявлены значительные потери некоторых генотипов, особенно в случаях гена О у матерей; однако в других работах этот результат подтвердить не удалось. Разумеется, следует помнить, что для убедительной демонстрации отбора порядка 1—5% могут понадобиться очень большие выборки, порядка 15—20 тыс. индивидуумов. Тем не менее и такое давление отбора может оказать существенный эволюционный эффект. Отбор по несовместимости матери и плода обязательно повлечет за собой элиминацию гетерозиготного потомства, так что равновесие частот генов будет неустойчиво, если отсутствует какой-либо компенсирующий механизм типа преимущества гетерозиготного состояния в более позднем возрасте.
Следует рассмотреть и возможность того, что определенная группа системы АВО способна обеспечивать защиту от каких-то болезней. Имея в виду такую возможность, проводили сравнение больших выборок больных с каким-то определенным патологичес-
Глава 17. Группы крови
321
ним процессом и контрольных популяций, из которых происходят эти больные. Более высокая частота группы А была обнаружена среди больных раком желудка, а группы О — среди больных с язвой двенадцатиперстной кишки (табл. 17.3). Сходные, хотя и менее убедительные результаты получены для некоторых других болезней.
Таблица 17.3
Частоты групп крови О и А у больных с язвой двенадцатиперстной кишки (F. Roberts, Brit. med. Bull., 1959)
Город Величина выборки Частота группы О(%) Частота группы А(%)
больные контроль больные контроль больные контроль
Лондон 946 10000 56,6 45,8 32,9 42,2
Ливерпуль 1059 15 377 59,6 48,9 29,0 39,1
Глазго 1642 5 898 57,7 53,9 31,5 32,3
Копенгаген 680 14 304 50,3 40,6 38,4 44,0
Вена 1160 10000 41,0 36,3 41,3 44,2
Айова 1301 6 313 53,7 45,8 36,3 41,6
Подобная связь группы крови с тем или иным патологическим состоянием не может считаться свидетельством причинной связи между ними. Если популяция неоднородна, в ней может существовать подгруппа с высокой частотой группы О, особенно предрасположенная к язве двенадцатиперстной кишки по какой-то другой причине. Однако величина некоторых из этих эффектов и тот факт, что сопоставимые результаты были получены в весьма отдаленных друг от друга местах, противоречат попыткам выдвинуть в качестве объяснения социальную стратификацию. Механизм, посредством которого групповая принадлежность крови влияет на предрасположение к тому или иному заболеванию, все еще остается нераскрытым; отметим, что в случае болезней желудочно-кишечного тракта соответствующие антигены присутствуют в секретах слизистых, по крайней мере в случае секреторов. Болезни типа язвы двенадцатиперстной кишки обнаруживают тенденцию к появлению на поздних этапах репродуктивной жизни; поэтому селективный эффект на группы АВО должен быть небольшим.
Антигены, сходные с антигенами АВО человека, обнаруживаются на поверхности различных бактерий. Поэтому представляется вероятным, что у индивидуумов, принадлежащих, например, к группе А, снижена способность к образованию антител против микроорганизмов с A-подобными антигенами. Действительно, утверждали, что индивидуумы с группой А более восприимчивы к
322 Часть III, Биологическая изменчивость популяций
оспе (если судить главным образом по реакциям на вакцинацию); однако эти данные не получили подтверждения; до сих пор не выяснено, имеется ли антиген А у вируса оспы, если исключить случаи, когда его культивируют на A-положительном курином эмбрионе. Области сравнительно низких частот О-генов и высоких частот 5-генов в Азии приблизительно совпадают с резервуаром грызунов — носителей чумы. Возбудитель чумы Pasteurella pestis обладает Н-подобным антигеном; из этого делают вывод, что индивидуумы с группой О могут оказаться особенно восприимчивыми к чуме. Имеются основанные на опытах указания на то, что принадлежность к тому или иному фенотипу по АВО может влиять на восприимчивость к вирусу простудного катара.
Итак, накоплено много данных о роли системы АВО как фактора отбора, однако эти данные часто неоднозначны и четкая картина путей поддержания полиморфизма еще не сформировалась.
Система Rhesus (Rh)
В 1940 г. Ландштейнер и Винер (Landsteiner a. Wiener) обнаружили, что сыворотка кролика, иммунизированного эритроцитами макаки-резуса, агглютинирует эритроциты 85% белых американцев. Оказалось, что способность реагировать с анти-Rh наследуется как доминантный признак. Два аллельных гена Rh и rh образуют два Rh-положительных генотипа (RhRh и Rhrh) и один Rh-отрицательный (rhrh).
Позднее в крови беременных женщин и индивидуумов, подвергшихся неоднократным переливаниям крови, были обнаружены антитела, которые давали практически те же результаты. С помощью соответствующих антисывороток, полученных от людей, можно выявлять антиген D, который, хотя и отличается от антигена обезьян-резусов, тем не менее близок ему. Следовательно, Rh-положительные генотипы можно обозначить по-новому: DD и Dd, а Rh-отрицательный как dd.
Медицинский аспект системы Rhesus прояснился в 1939 г., когда в результате работы Левина и Стетсона (Levine a. Stetson) стало ясно, что эта система связана с возникновением тяжелого заболевания новорожденных. Оказалось, что оно возникает в тех случаях, когда лишь мать — носитель Rh~ (dd), а отец — Rh+ (DD или Dd). В определенной части таких браков плод гетёрози-готен (Dd) и, следовательно, является носителем антигена D, которого нет у его матери. Нарушение’целостности плаценты при родах приводит к тому, что В+-клетки плода попадают в кровоток матери и иммунизируют ее к D-антигену. При последующих беременностях соответствующие анти-В-антитела способны проникать
Глава 17. Группы, крови
323
сквозь плацентарный барьер и оказывать' повреждающее действие на эритроциты плода, у которого развивается тяжелая анемия (гемолитическая болезнь новорожденных). Если отец гетерозиготен (Dd), вероятность этого осложнения меньше, поскольку в среднем половина потомства будет иметь генотип dd, т. е. будет лишена D-антигена.
Примем частоту индивидуумов Rh~ в стране приблизительно равной 16%, тогда частота d будет равна 40%, а вероятность возникновения гемолитической болезни при случайном формировании брачных пар составит 9%. Однако частота этой болезни составляет всего 1 : 150—1 : 200; следовательно, должны существовать другие факторы, которые снижают опасность возникновения этого заболевания. Один из таких факторов — групповая принадлежность родителей по системе крови АВО. Если они несовместимы и по этой системе, то попавшие в организм матери клетки, плода будут разрушаться ее антителами и не смогут вызвать анти-D -иммуниз ации.
До последнего времени единственным средством спасения таких новорожденных было быстрое переливание крови. В 1965 г. Кларк (Clark) и его сотрудники пришли к выводу, что попавшие в организм матери В+-клетки плода могут быть разрушены путем введения соответствующих доз антисыворотки (анти-D). Этим методом удается успешно предотвращать иммунизацию матери.
Система Rh и естественный отбор
Мы уже упоминали об иммунологическом отборе, направленном против гетерозиготных плодов, когда обсуждали* систему АВО. В случае системы Rh с помощью такого же механизма элиминируются генотипы Dd. Отбор против гетерозигот обеспечивает равновесие, только если частоты обоих аллелей точно равны 50%; в противном случае аллель, встречающийся с меньшей частотой, постепенно элиминируется. Во всех популяциях, за исключением басков, частота d ниже 50%; возможно, что в этом случае полиморфизм преходящий и частота d снижается. Если это так, то остается объяснить прежде всего, как могли быть достигнуты столь высокие локальные частоты этого гена. Предполагают, что баски представляют собой остатки доисторической западноевропейской популяции с высокой частотой гена d; позднее произошло их смешение с восточными народностями, для которых характерна очень высокая частота гена D. Тем не менее возможно, что равновесие в действительности устойчиво, по-видимому, благодаря каким-то неизвестным преимуществам гетерозигот Dd в более старшем возрасте.
324
Часть III. Биологическая изменчивость популяций
Дальнейшее усложнение системы Rh
До сих пор мы говорили о системе Rh как о состоящей лишь из антигенов D и d, действительно наиболее важных с точки зрения медицины. Однако дальнейшее изучение сывороток людей выявило наличие и других антигенов. Обнаружив определенные закономерности в результатах серологических исследований, Фишер в 1943 г. предложил схему, объясняющую генетическую структуру этой системы. Он постулировал существование трех весьма тесно сцепленных генных локусов, каждый из которых мог быть занят одним из пары аллелей: D или d, С или с, Е или е. Это создает возможность возникновения 8 типов генных триплетов! или гаплотипов:
обозначения по Фишеру—Рейсу:
CDE, CDe, cDE, cDe, cde, Cde, cdE, CdE\ обозначения по Винеру: 7?z, /?[, Rz, r, r', r", ry.
Фишер полагал, что каждый ген продуцирует 'свой особый антиген, и предсказал открытие шести соответствующих антисывороток. Это предсказание сбылось, за одним исключением: сыворотка анти-d не была найдена, а возможно, и не существует. Фишер также отметил, что наиболее часто встречаются типы cDe{Ri), cDE(R2), cDe(R2) и cde, и предположил, что типы, встречающиеся реже, возникли в результате редких кроссинговеров. Действительно, без кроссинговера нет генетических способов отличить друг от друга гипотетические триплеты и единичные гены. Винер не принял концепцию Фишера, предпочитая признавать существование набора из 8 аллелей со сложными свойствами, определяющими антигенность. Его обозначения удобны своей простотой; здесь они приведены вместе с символами Фишера и Рейса. Несомненно, раскрытие химической структуры (которая до сих пор неизвестна) этих антигенов снимет указанные противоречия.
Читатель понимает, что, поскольку гены-триплеты расположены в одной хромосоме, генотип какого-либо индивидуума может быть, например, CDelcde(Rx/r) или cDEjcDe (R2/Ra), т. е. может содержать 6 генов. Тесты с применением 5 доступных сывороток необязательно позволят однозначно определить генотипы, поскольку они не дают сведений о том, как эти 6 генов расположены в виде пары гаплотипов; такое определение возможно лишь в случаях типа (с+), (С—), (D—), (e-j-), (Е—), которые должны относиться к гомозиготам cdejcde. В каждом отдельном случае генотип обычно удается выяснить по семейному анамнезу; однако на материале популяции можно сомневаться в наличии редко встречающихся гаплотипов.
Глава 17. Группы крови
325
Исследование Rh-фактора позволило обнаружить много любопытных серологических феноменов; не все они полностью объяснимы. Упомянем лишь те немногие, которые существенны для изучения популяций. Определенные D-антигены слабо реагируют с некоторыми анти-В-сыворотками, их обозначили Du; более того, выраженность реакции непостоянна, что позволяет предполагать 'существование серии различных Би-антигенов. Du часто встречается в Африке; такой индивидуум может быть ошибочно принят за D-отрицательный. Описана анти-У-сыворот-ка, которая дает положительные реакции с клетками лишь у 1% белых американцев и американских индейцев, но реагирует с клетками 40% жителей Западной Африки. Объясняется это, невидимому, тем, что анти-V реагирует с клетками, которые содержат как с, так и аллели е, е3 в одной хромосоме. Обнаружены и другие антисыворотки такого рода: анти-Се и анти-се.
Сообщалось о редко встречающихся индивидуумах, имеющих только| D-антиген (—D—), и о других, не дающих реакции ни е одним; из Rh-антигенов («Rh-нулевые»); среди случаев последнего типа, по крайней мере в одном, родители были Rh+; соответствующие антигены у их ребенка, по всей видимости^ подверглись супрессии.
Географическое распределение Rh-групп
Система Rh с ее 8 гаплотипами и набором разнообразных аллелей в каждом локусе создает простор для изменчивости; это иллюстрирует табл. 17.4, где приведены частоты вариантов локуса Rh в различных популяциях. Например, частота CDe(Rl) колеблется от 5% в некоторых популяциях Африки до 90% и более во многих племенах Новой Гвинеи. Отмечается тенденция повышенной частоты этого гаплотипа в районе Средиземноморья по сравнению с другими областями Европы; cde (г) здесь встречается соответственно редко. Эта тенденция, заметная у сардинцев, сохраняется в Северной Индии; на Дальнем Востоке cde встречается редко. Cw, аллель гена С, необычно часто встречается у лопарей, финнов и латышей. cDE (R2) заметно чаще обнаруживается у американских индейцев и весьма часто в Полинезии и Юго-Восточной Азии. Замечательная особенность cDe(R0) состоит в том, что его частота в Африке заметно выше, чем в любом другом районе. Частоты, превышающие 90%, были зарегистрированы для народностей, столь соматически различных, как, например, определенные племена из долины Нила и бушмены. По мере приближения к Среднему Востоку частоты Ro снижаются, хотя и остаются равными 50—60% в Эфиопии, 15—20% в некоторых племенах Йемена и 10—16% в районах южнее Каспийского моря'. Умеренно высокие частоты Ro были обнаружены также у некото-
326
Часть III. Биологическая изменчивость популяций
Таблица 17.4
Частоты вариантов локуса Rh в различных популяциях
Число обследованных CDE (Bz) CDe (Hl) cDe (H2) cde (Но) Cde (г') CdE (ry) cdE (И cde (r>
Англичане 1798 0,27 41,43 14,50 2,62 1,19 0,00 0,97 39,03
Итальянцы (г. Ми- 772 0,35 47,56 10,77 1,63 0,67 0,33 0,69 38,00
лан)
Баски 383 0,00 37,56 7,07 0,50 1,47 0,00 0,25 53,15
Сардинцы 107 — 66,84 8,84 2,12 0,00 — 0,00 22,19
Норвежские лопа- 183 0,00 52,46 18,37 10,34 0,00 0,00 0,00 18,83
ри Пакистан (с.-з. Па- 253 0,63 66,69 7,78 0,00 0,50 0,00 0,00 24,40
кистан) Китайцы (южный 250 0,49 75,91 19,51 4,09 0,00 0,00 0,00 0,00
Китай) Австралийцы 234 2,08 56,42 20,09 8,54 12,87 0,00 0,00 0,00
Новокаледонцы 147 0,00 80,27 12,71 7,02 0,00 0,00 0,00 0,00
Североамерикан- 241 4,06 47,81 38,84 0,00 0,00 0,00 3,42 9,87
ские индейцы Индейцы чиппева 161 2,04 33,67 53,03 0,00 0,00 0,00 3,22 8,03
Нигерийцы (ю.-з. 145 0,00 9,51 8,28 61,91 w 1,87 0,00 0,00 18,43
Нигерия) Банту Южной Аф- 644 0,00 2,81 4,27 73,95 К 7,07 0,00 0,00 11,84
рики Бушмены 232 — 9,04 1,96 j 89,00D 0,00 — 0,00 0,00
*) В том числе сРие
рых разрозненных народностей (негритосы Малайи и некоторые племена северной Австралии).
Что касается более редких гаплотипов, то довольно часто CDE (Rz) встречается в так называемых веддоидных племенах Индии и в некоторых австралийских племенах, но не среди самих веддахов. Редкий тип cdE(r') удивительно часто обнаруживают у айнов (северная Япония).
Мы упомянули выше о «сложном» антигене V (ces). Как и в случае cDe, высокие частоты этого антигена характерны для Африки, где частоты соответствующих генов колеблются от 20 до 30%, падая до 5—10% в Аравии.
Глава 17. Группы крови
327
1 Система MNS
Группы крови MN были открыты Ландштейнером и Левином в 1927 г. в результате иммунизации кроликов эритроцитами человека. Кроличьи антисыворотки до сих пор широко применяются для работы с этой системой, хотя пришлось ввести методику очистки; непростой задачей оказалось приготовление антисывороток, которые реагируют с нужным антигеном и достаточно сильно и достаточно специфично. Компоненты этой системы наследуются по простому менделевскому типу: три фенотипа — М, MN и N — удается распознать по отдельности соответственно трем генотипам — ММ, MN и NN (или Agh\ Ag1* AgN и AgN AgN).
Карта мира для гена М (рис. 17.5) показывает существование ряда выраженных региональных контрастов — от 20% в Новой Гвинее и ряде районов Австралии до 90% в некоторых частях Нового Света. Районы со сравнительно высокой частотой М имеются также в Аравии, на северо-востоке Сибири и в Юго-Восточной Азии. На большей части территории Западной Европы частоты М близки к 50%; они ниже в некоторых группах лопарей и выше в соседней Финляндии. Сардинцы с их высокой, как у басков, частотой О резко отличаются от последних высокой частотой М (75%).
В 1947 г. концепция простоты системы MN была поколеблена вследствие открытия анти-Э-антисыворотки. Она обнаруживает антиген, явно отличный от М и от N, но каким-то образом связанный с ними. Если бы наследование групп S и MN было независимым, следовало бы ожидать одинаковую (в пределах ошибки метода) долю 3+-индивидуумов в каждом из фенотипов М, MN и N; однако данные одной британской выборки показывают, что зто не так: доля S+-M гораздо выше, чем следовало ожидать при случайном их сочетании:
Анти-S М + MN N
Наблюдалось (-)-) 93 15 1
Ожидалось (+) 82 26 J lUo
Наблюдалось (—) 52 30 ) Q9
Ожидалось (—) 63 19 J
145 45 1£О
Анализ семей показал, что М и S сегрегируют попарно. Име-ется, следовательно, ситуация, подобная группам Rh. Анти-s-
Рис. 17.5. Частота гена, М групп крови системы MN (Mourant et al., 1976).
Глава 17. Группы, крови
329
сыворотка, обнаруживающая аллельный антиген s, была открыта позже. Теперь, применяя четыре антисыворотки, можно различить четыре генные пары, или гаплотипа: MS, Ms, NS, Ns. Анти-s — весьма редкая антисыворотка; ее нечасто применяют при массовых обследованиях населения; в таком случае не удается отличить гетерозигот типа MSMs, однако можно идентифицировать MsMs по отсутствию реакции с анти-5. Частота S в Европе обычно близйа к 30—35% ,чаще всего в сочетании с М. Частота S еще выше на Среднем Востоке (40%); здесь, а также в Индии MS представляет собой наиболее частое сочетание. В Восточной Азии частоты S гораздо ниже 5—20%; в этом районе М обычно сочетается с $. В Австралии 5 практически отсутствует, несмотря на то что в отдельных районах Новой Гвинеи частота этого гена достигает высоких показателей.
Дальнейшее развитие исследований этой системы привело"к открытию анти-U. Было обнаружено, что эта антисыворотка не реагирует с клетками некоторых американских негров, у которых обычно (но не всегда) отвечалась также отрицательная реакция «а S и s. Теперь принято считать, что в локусе 5 в таких случаях имеется аллель 5“, супрессирующий выражение другого аллеля, если таковой присутствует. 5“ обнаруживается весьма часто в определенных районах Африки; в одной выборке пигмеев Конго 36% индивидуумов не реагировали с анти-U. Ген Su не был обнаружен у австралийских аборигенов.
Система MNS в действительности еще сложнее, .но мы не будем рассматривать ее подробно. С одной стороны, имеются различные редкие гены, которые являются аллелдми М, N или S; с другой стороны, существуют дополнительные специфичности, связанные с этой системой, подобно тому как MN ассоциируется с Ss. Одна из этих специфичностей — Хеншоу — была обнаружена в крови одного нигерийца с помощью кроличьей антисыворотки; она, как оказалось, широко распространена в Африке, но встречается редко (в 5% и реже) и в сочетании с NS, по крайней мере в Западной Африке.
Система Р
Успехи в изучении этой системы после того, как Ландштейнер открыл ее, служат прекрасным примером серологического детектива; тем не менее о ней мало что можно сказать из-за тех методических трудностей, которые затрудняют проведение массовых обследований. Вначале одна антисыворотка, анти-Р, позволила различать два фенотипа — Р(+) и Р(—). Спустя много'лет оказалось, что сыворотка к весьма широко распространенному антигену Tja способна агглютинировать клетки Р(+) и Р(—), но не клетки очень редкого фенотипа Tj(a—). Эта ситуация аналогична
330
Часть III. Биологическая изменчивость популяций
ситуации с AiA2O в системе АВО, и поэтому Р(+)иР(—-) переименовали соответственно в Рх и Р2. Поэтому анти-Tj0 (PPJ ведет себя аналогично анти-А и агглютинирует клетки Ах и А2, а исходная анти-Р — аналогично анти-А п агглютинируя только клетки Рх. Фенотип Tj(a—) считают гомозиготным по очень редкому аллелю р системы Р.
Несомненно, существуют большие пазличия в частотах генов Р. Частота Р2 в Европе около 50% (в Финляндии несколько ниже); в Западной Африке она ниже 5%; другая крайность — частота, равная 75—80%,— отмечается в Китае и Японии.
Система Lu (Lutheran)
В системе Lutheran первыми были открыты антитела, которые давали положительную реакцию в 4% случаев при обследовании англичан. Позднее были найдены антитела другого типа, реагирующие с Биа-отрицательными клетками. Используя антисыворотки обоих типов, можно поэтому выявлять три фенотипа — Lu(a+h—), Lu(a+b+) и Lu (а—b+), соответствующие трем генотипам — LuaLua, LuaLub и LubLub. Наблюдали также очень редкий генотип Lu (а—b—), который определяется независимым локусом, способным блокировать образование веществ Lu.
Ген Lua обнаружен в Европе, а также в Африке, но встречается там редко. Еще реже он встречается в Бирме, редко обнаруживается в Индии и практически отсутствует в Австралазии. С низкой частотой этот ген наблюдается среди американских индейцев; возможно^ он был внесен к ним после открытия Америки европейцами.
Система Kell
Антиген К, проявляя сходство по распространению с антигеном Lua, нигде не встречается достаточно часто; вне Европы и Среднего Востока он наблюдается редко или отсутствует. Самая большая частота (10%) отмечена в Аравии; при переходе к Западной Евройе она надает до 5% и ниже. Этот ген не часто обнаруживают в Африке и Индии; он практически отсутствует на Дальнем Востоке, в Австралии и Новом Свете. Известна антисыворотка, позволяющая обнаружить антиген к, образуемый аллельным геном. Несовместимость по К и к иногда обусловливает гемолитическую болезнь новорожденных.
Подробное изучение системы Kell показало, что, подобно системам Rh и MNS, она представляет собой ассоциацию локусов или антиген образующих сайтов. Было показано, что примерно у 2% белых американцев имеется антиген- Кра, определяемый тем же локусом, что и к. У американских негров Кра встречается
Глава 17. Группы крови
331
очень редко; они являются носителями антигена Sutter, Js®, который, как было показано, также связан с системой Kell. Таким образом, эта система содержит аллельные пары Кк, КраКрь и JsaJsb-, однако обнаружена лишь половина из возможных 8 гаплотипов, поскольку К и Кра свойственны почти исключительно белым, а их смешение началось слишком недавно, чтобы вследствие кроссинговера могли возникнуть все возможные рекомбинанты.
Jsa встречается с частотой около 9% среди американских негров; в Африке и соседних регионах его частота гораздо выше, чем где бы то ни было еще.
Система Duffy
Первый из антигенов системы Duffy Fya был открыт в 1950 г.,, а годом позже была выделена сыворотка, с помощью которой можно выявлять аллельный продукт Fy6. Расщепление пары аллелей дает 3 фенотипа: Fy(a+b—), Fy(a+b+) и Fy(a—Ь+). Тем удивительнее было обнаружить, что около 70% американских негров не реагируют ни на одну из этих сывороток и относятся к генотипу Fy(a—b—). Возможно, ген Fy контролирует антиген, сыворотка к которому еще не найдена, возможно также, что ген Fy супресси-рует образование антигена своим партнером.
Вероятно, Fy надежнее других известных генов позволяет обнаружить различие между европеоидами и африканцами. В одной выборке западных пигмеев частота Fy составила 100%; величины порядка 90% отмечены у ряда народностей в Центральной Африке. В Бейе на южном берегу Красного моря частота Fy равна 78%, в Аравии — 50%, среди курдов северного Ирана и' Ирака — 15—50%, тогда как у жителей Западной Европы частота этого гена 3% и менее. За пределами Европы и Среднего Востока Fy, вероятно, встречается редко, однако данных по этому вопросу недостаточно.
Частота гена Fya (рис. 17.6) также весьма широко варьирует, достигая 100% среди некоторых австралийских аборигенов; она высока также среди американских индейцев и индусов. В Западной Европе частота этого гена ниже (35—45%); она повышена у лопарей (54—82%). Интересно, что в Африке Fya обнаруживается с частотой 5—10% у бушменов.
Система Diego
Антиген Diego был впервые обнаружен в одной венесуэльской семье, и долгое время его считали «частным» антигеном с очень ограниченным распространением в отдельных семьях. В настоящее время выяснено, что в некоторых племенах южноамериканских
Рис. 17.6. Распространение на земном шаре гена Еуг системы Duffy (Baraicot, 1963). На карте приведены неопубликованные данные.
Глава 17. Группы крови
333
индейцев число Diego-положительных индивидуумов достигает 30%. Некоторые данные о распределении этого гена приведены в табл. 17.5. Несколько меньшие частоты наблюдаются у североамериканских индейцев, и еще меньшие — у китайцев и японцев. Этет антиген встречается на о. Калимантан, но до сих пор не был обнаружен в Меланезии, Полинезии и в Австралии, а также в Европе или Африке. Создается впечатление, что антиген Diego распространен среди восточных популяций, классифицируемых как монголоидные, однако-распределение его частот не соответствует распределению различных морфологических признаков, на которых основана классификация типов. Ген достигает наивысшей частоты у некоторых народов, у которых монголоидные признаки отсутствуют.
Таблица 17.5
Частота фенотипа Di (а+) в различных популяциях
Популяции Число обследован НЫ1 Частота фенотипа, % Популяция Число обследованных Частота фенотипа, %
Каингапги (Бразилия) 48 45,8 Лопари, Норвегия 433 0,0
Каража (Бразилия) 36 36,1 Полинезийцы 80 0,0
Карибы (Венесуэла) 121 36,5 Население Новой Бри- 74 0,0
Индейцы майя 363 17,6 тании
Гуахибо (Венесуэла) 76 14,5 Аборигепы Австралии 162 0,0
Японцы 65 12,3 Белое население США 1000 0,0
Индейцы чиппева 148 10,8 Индийцы 75 0,0
Корейцы 277 6,1 Африканцы (Либерия, 775 0,0
Гуахиро (Венесуэла) 152 5,3 Берег Слоновой Ко-
Индейцы апачи 73 4,1 сти)
Эскимосы, Аляска 241 0,8 Бушмены 114 0,0
Эскимосы 156 0,0
Антиген Xg
В 1962 г. был открыт антиген Xga, который, несомненно, контролируется геном Xga, расположенным в Х-хромосоме. Оказалось, что частота Xg3 среди европейцев высока — она составляет 68%; эта величина вычислена по данным о числе мужчин Xg(a+). Около 90% женщин также являются носителями этого гена, из них около 45% гетерозиготны, что можно использовать для изучения сцепления.
К сожалению, пригодность Xga для изучения сцепления оказалась ограниченной: соответствующий локус расположен вблизи
334 Часть III. Биологическая изменчивость популяций
короткого плеча Х-хромосомы и поэтому весьма отдален от большинства других локусов. Это разочарование компенсируется, пожалуй, существенным вкладом Xga в«изучение различных аберраций Х-хромосомы, при которых оказывается аномальным число или строение этой хромосомы (гл. 7). Например, оказывается возможным, применяя Xga в качестве маркера, установить, от кого из родителей исходит избыточная хромосома при синдроме Клейнефельтера (XXY).
Данные о частотах Xga в разных популяциях приведены в табл. 17.6.
Таблица 17.6
Частоты Xg а в разных популяциях (Simons, Nature, Lend. 227, 1363, 1970)
Популяция Число обследованных Частота Xga. %
Новая Гвинея 263 85
Аборигены Австралии 352 79
Американские индейцы навахо 308 77
Сардинцы 322 76
Израиль 201 68
Северная Европа 5388 66
Индийцы (Бомбей) 100 65
Китайцы (материк) 171 6)
Китайцы (Тайвань) 178 53
Негры и выходцы с Ямайки (Нью-Йорк) 219 55
Аборигены Тайваня 164 38
Любопытно, что антиген, неотличимый от Xga, был обнаружен у гиббонов (Hylobates lar lar), но его нет у понгид и у некоторых изученных видов нечеловекообразных обезьян. У гиббонов этот антиген также сцеплен с полом (насколько можно судить по небольшой выборке).
Построение филогенетических схем
Главная цель работы по сбору сведений о частоте генов среди населения земного шара — проследить эволюционные связи популяций и понять, как создалась их современная мозаика. Предполагается, что популяции, разделившиеся недавно, сохраняют много общего в распределении частот генов; если же они изолированы друг от друга в репродуктивном отношении, то возникает все большая дивергенция под действием различных эволюционных факторов. Ход эволюции удобно изображать в
Глава 17. Группы крови
335
виде дерева, концы живых ветвей которого соответствуют современным популяциям. В случае внутривидовой эволюции остается возможность потока генов между популяциями, что замедляет дивергенцию. В экстремальном случае возможно полное смешение популяций и образование петли на родословном дереве. Вообще популяции, находящиеся в отдаленном родстве, могут становиться
Рис. 17.7. Гипотетическое дерево' эволюционных потомков, соединяющее 4 популяции — а, Ь, с и d, для которых известны частоты двух генов — А и В (Cavalli-Sforza et al., 1964).
в процессе эволюции все более и более сходными, по крайней мере в определенных отношениях, в результате адаптации к одним и тем же условиям.
В течение последних 10 лет применение ЭВМ сделало возможной реконструкцию наиболее вероятной модели эволюционного дерева, которое могло бы привести к возникновению наблюдаемой в настоящее время мозаики генных частот. Здесь мы можем лишь поверхностно рассмотреть эту работу; приведем результаты ее лишь потому, что создатели этого направления исследований — Кавалли-Сфорца (Cavalli-Sforza) и Эдвардс (Edwards) — выбрали в качестве объекта частоты групп крови человека.
На рис. 17.7 представлены четыре популяции, положение которых на плоскости определяется частотой встречающихся у них двух генов. Если принять во внимание возникновение данного распределения вследствие дивергенции во времени, то приведенное здесь дерево интуитивно представляется наиболее вероятным. Однако желательно было бы использовать данные о возможно большем числе локусов и аллелей, так что популяции
336
Часть III. Биологическая изменчивость популяций
следует рассматривать как точки многомерного пространства (гл. 14).
Простейшая и математически наиболее привлекательная модель дивергенции генных частот учитывает только случайный дрейф генов; однако это касается дивергенции уже разделенных популяций, а не самого разделения. Как правило, в пашем распоряжении почти нет сведений о времени и причинах такого разделения, так что в этой модели исходят из предположения о его случайном характере.
о,о
0,2 -
0,4 -
0,6 -
0,8 -
{0 -
7,2 -
1,4 -
Англичане
Эфиопы
I Банту жители Гоны
----1 1--------
I Горка I (Непал) Ледда ] (Цейлон)
Турки
Лопари
Корейцы
Эскимосы
жители Новой Гвинеи
Австралийцы
Маори
Индейцы
Сев Америки Индейцы Южн. Америки
Рис. 17.8. Наиболее вероятное дерево связи 15 популяций, основанное на генных частотах для 5 систем групп крови (Cavalli-Sforza et al., 1964).
На шкале обозначено число генов, замещенных за определенное время
Даже при применении этой относительно простой модели число возможных вариантов такого дерева увеличивается очень быстро с увеличением числа популяций. По-видимому, нахождение наиболее вероятного дерева (а иногда целой их группы) классическими методами наталкивается на определенные трудности, и авторы прибегли к допустимым приближениям; в частности, считали, что наиболее вероятное дерево должно быть близко к такому, у которого длина путей эволюции наименьшая.
На рис. 17.8 представлено эволюционное дерево для 15 популяций, построенное на основе данных о 5 локусах групп крови (в системах АВО, Rh, MNS, Fy, Di). Вертикальная шкала — не время, а число генных замещений, которое (если действует лишь дрейф генов) зависит от (эффективного) размера популяции и от времени, протекшего с момента разделения популяций.
Глава 17. Группы крови
337
Ясно, что эта модель сильно упрощает ситуацию; многие биологи скажут, что отбор играет важную, но не единственную роль в географической дифференциации человека. Весьма важная особенность дрейфа — это его влияние на все локусы; после коррекции различий, обусловленных частотами генов, дрейф вызывает постоянный рост вариабельности генных частот. Отбор же имеет тенденцию влиять неодинаково на различные локусы, быстро меняя частоты одних генов и стабилизируя частоты других. Если удается оценить ожидаемую вариабельность генных частот вследствие дрейфа, то расхождения в соответствующих значениях для каждого конкретного локуса позволяют решить, повлиял ли отбор на данный локус. Как указывают авторы, способность дрейфа вызвать дифференциацию генных частот довольно велика. Они полагают, что вариабельность, обнаруженная ими в полученной модели, могла возникнуть на протяжении полумиллиона лет, даже если размер популяции (эффективный) достигал 10 000.
Группы Gm и Inv
Иммуноглобулины (1g)
Факторы Gm и Inv представляют собой серологически выявляемые варианты белков антител (иммуноглобулинов), содержащихся в сыворотке. Иммуноглобулины — это класс физиологически очень важных и крайне сложных белков. Поэтому вначале мы сообщим некоторые самые общие сведения об их строении и функциях.
В постнатальном периоде образуются антитела к самым разнообразным микроорганизмам, чужеродным белкам и сложным углеводам, попадающим в ткани тела. Замечательное свойство антител заключается в их способности связываться более или менее избирательно с антигенами, вызвавшими их образование. Происходит это, вероятно, благодаря тому, что связывающая ан-1иген часть молекулы иммуноглобулина довольно строго соответствует определенным участкам (антигенным детерминантам) поверхности антигена. Известно, что аминокислотная последовательность этих связывающих антиген участков крайне вариабельна; это и не удивительно, если учесть существование множества самых разнообразных антигенов. Считают, что такая вариабельность кодируется генами; не ясно, однако, какая часть соответствующей генетической информации передается через гаметы, а какая созда-' ется в соматических клетках индивидуума. Согласно наиболее распространенной «клональной теории», антиген не служит матрицей, на которой формируется антитело; само присутствие антигена каким-то способом стимулирует пролиферацию именно тех
338 Часть III. Биологическая изменчивость популяций
предо уществующих «клеток, которые закодированы так, что образуют антитела, соответствующие антигену.
Схематически молекула 1g представлена на рис. 17.9. Она имеет форму буквы Y и состоит из двух идентичных половин, каждая из которых содержит тяжелую (Heavy — Н) и легкую (Light — L) цепи, соединенные в вертикальном участке Y. Обе половины, Н- и L-цепи, удерживаются рядом главным образом дисульфидными связями. Вариабельные участки L- и Н-цепей
Рис. 17.9. Схема молекулы иммуноглобулина.
Каждая половина состоит из легкой (L) и тяжелой (Н) цепей. Цепи удерживаются вместе дисульфидными связями. Черным обозначены вариабельные (V) участки L- и Н-цепей, белым — неизмененные участки (С).
(обозначенные на схеме V) образуют N-концевые половины вертикального участка Y. Поэтому каждая молекула 1g содержит два идентичных участка связывания антигена; электронно-микроскопически показано, что такая «двухвалентность» приводит к образованию агрегатов, состоящих из молекул антитела и антигена. Остальные участки L- и Н-цепей по своей аминокислотной последовательности гораздо менее вариабельны (constant, С). Тем не менее удается различить несколько структурно неодинаковых классов и субклассов этих цепей; существует дополнительный полиморфизм ограниченных участков цепей определенных типов, создающий ту или иную специфичность по Gm и Inv. У человека и высших позвоночных известны два типа L-цепей (х и л) и 5 классов Н-цепей (у, а, ц, 6, е). Последние характеризуют 5 классов молекул Ig(IgG, IgA, IgM, IgD, IgE), различающиеся по иммунологическим свойствам. IgG, составляющий основную массу иммуноглобулина нормальной сыворотки, способен проникать через плацентарный барьер; большая часть 1g новорожденного — это IgG его матери. Молекула IgM (молекулярный вес 900 000) состоит
Некоторые свойства классов иммуноглобулинов
Таблица 17.7
Класс igG IgA IgM IgD IgB
Цепи (X ИЛИ X) 3 Уз (х или л)2 осз [(х ИЛИ Х)з Из]5 (х или X) з 6а (х или Х)а еа
Молекулярный вес 150000 152 000 или 385 000 900000 175000 190000
Содержание углеводов, % 2,5 5-10 5-10 11,5
Средняя концентрация в сыворотке взрослых, мг/мл 12,0 1,8 1,0 0,03 0,0003
Функциональная характеристика Связывает комплемент; проникает через плаценту Содержится в секретах Связывает ко*м-племент; характерен для раннего ответа; обладает высокой агглютинирующей способностью Характерен для некоторых форм аллергии
340
Часть III. Биологическая изменчивость популяций
из 5 субъединиц, каждая примерно той же величины, что и единичные молекулы какого-либо другого класса; обычно антитела этого класса первыми образуются в ответ на антигенный стимул. IgA обнаруживается главным образом на слизистых и содержится в их секрета®:. Некоторые свойства этих цепей приведены в табл. 17.7. На рис. 17.9 цепи изображены прямыми, однако в действительности они имеют перегибы в определенных участках, стабилизируемые дисульфидными связями.
Специфичность Gm (теперь их насчитывается 20) определяется лишь вариантами Н-цепей IgG. Распознают четыре подкласса тяжелых цепей IgG(yGi, yG2, yG3, yG4). Некоторые специфичности Gm связываются с yGi, другие — с yG3, одна-две — с yG2; до сих пор не известны специфичности, связанные с yG4 (табл. 17.8).
Таблица 17.8
Взаимоотношения некоторых Gm-факторов с подклассами Н-цепей
Подкласс Н-цепей IgG Факторы Gm
1, 2, 3, 4, 17
yG2 23
?G3 5, 6, 11. 13, 14, 21
?g4 Неизвестны
В нескольких случаях удалось установить, какие аминокислотные замены лежат в основе специфичностей Gm. Первым %ыл открыт фактор Gm 4 (Grubb, 1956); в положительной по этому фактору молекуле в двух соседних участках Н-цепи находятся аспарагин и лейцин, в отрицательной — глутаминовая кислота и метионин. Специфичности Inv определяются L-цепями; известно, что в молекулах Inv3-положительных молекулах в положении 191 лейцин замещен валином. Кратко упомянем несколько других вариантов цепей Ig: Isf расположен на Н-цепях IgGj, Oz — на цепях 0Х, Ат — на Н-цепях IgA.
Несомненно, белки Ig нормальной сыворотки чрезвычайно неоднородны по строению; практически невозможно выделить чистые фракции в количествах, достаточных для химического анализа. Однако клетки некоторых редко встречающихся лимфоидных опухолей (миелом) синтезируют большие количества гомогенных Ig; в некоторых подобных случаях с мочой выделяются свободные L-цепи (так называемый белок Бенс-Джонса). Полагают, что эти опухоли образуются путем пролиферации из одной антителообразующей плазматической клетки. Одна плазматическая клетка способна в определенный момент вырабатывать Ig только
Глава 17. Группы крови
341
одного класса и подкласса; если же больной гетерозиготен по факторам Gm, то в Н-цепях, образующихся в его организме, обнаруживаются факторы, кодируемые только одним из двух аллелей. Такое «выключение аллеля» напоминает сходный процесс инактивации одной из двух Х-хромосом у женщин.
При рассмотрении гемоглобинов мы видели, что структурно различные цепи (а, р, у) кодируются разными' генами (цистро-намц). То же, по-видимому, имеет место и в случае классов и подклассов L- и Н-цепей иммуноглобулинов. Существует точка зрения, согласно которой имеются отдельные гены У и С, кодирующие соответственно вариабельные и константные участки цепей; в таком случае для образования одной молекулы 1g клетка должна иметь 4 гена (KL, Ен, Cl и Сн).
Изучение наследования единичных факторов Gm, например Gmj, выявило ситуацию, характерную для случая пары кодоминантных аллелей. Если же провести тесты на несколько различных факторов, то окажется, что некоторые из них передаются вместе, единой группой; с этим явлением мы уже встречались, рассматривая группы крови систем Rh и MNS. В Европе широко распространены и передаются единой группой комплексы Gm (1, 17, 21), Gm (1, 2, 17, 21) и Gm (3, 5, 13, 14). Известно, что факторы 1, 2, 3 и 17 расположены на yGt, а 5, 13, 14 и 21 — на yG3; следовательно, они должны контролироваться различными локусами. Эти локусы, по-видимому, тесно сцеплены; другие комбинации факторов, которые могли бы образоваться путем межцистрон-ных кроссинговеров, в данной популяции встречаются редко. Сообщалось о случаях таких кроссинговеров в отдельных семьях; возмоя^но, что некоторые из этих комбинаций, присутствующих в различных популяциях, возникли именно таким путем. Комбинации факторов Gm, по-видимому, передаются участками ДНК, содержащими несколько цистропов; поэтому правильнее говорить об этих комбинациях как о гаплотипах. Этот термин означает, что некий набор факторов, как правило, передается как единое целое гаплоидным хромосомным набором гаметы.
Обнаружение факторов Gm и Inv
Обычный метод выявления этих факторов — торможение гемагглютинации. Вначале эритроциты обрабатывают неполными Rh-антителами, несущими искомый фактор Gm. Затем, если добавить антитела, специфичные в отношении этого фактора, происходит агглютинация. Соответствующие антисыворотки можно получать от больных ревматоидным артритом и от некоторых здоровых индивидуумов. Если исследуемую сыворотку добавить к смеси антисыворотки и обработанных эритроцитов, то при наличии того же фактора Gm исследуемая сыворотка будет конкурировать за
342
Часть,-!!!. Биологическая изменчивость популяций
антцтела, нанесенные на эритроциты, и тогда агглютинации не произойдет. Для некоторых факторов Gm соответствующие антисыворотки были получены путем иммунизации кролика и обезьяны Н-цепями; однако классический метод до сих пор остается" более чувствительным.
Географическое распределение факторов Gm и Inv
До сих пор применяются две системы обозначений этих факторов, что отнюдь не способствует прояснению довольно запутанной ситуации; обозначения эти сопоставлены в табл. 17.9.
Таблица 17.9
Сопоставление прежних и новых обозначений для факторов Gm и Inv
Новое обозначение Прежнее обозначение Новое обозначение Прежнее обозначение
Grrij а 15 £
2 X 16 t
3 bw—b2 17 Z
4 f 18 Rouen 2
5 b = b3 19 Rouen 3
6 с = с5 20 20
7 Г 21 g
8 е 22 У
9 р 23 n
10 а b8
11 р ь° c3
12 У Invj 1
13 Ь3 2 a
14 Ь4 3 b 1
Число популяций, исследованных в отношении 9 факторов Gm, "еще не очень велико; тем не менее данные, сведенные в табл. 17.10, свидетельствуют об удивительно больших различиях между коренным населением главных географических областей, а в некоторых случаях — между популяциями, живущими в одной и той же области. Некоторые гаплотипы часто встречаются в одвдх популяциях и редки или отсутствуют в других. Так, Gm (3, 5, 13, 14) распространен среди европеоидов, но редко встречается или отсутствует в других местах, если только там не произошло смешения с европеоидами. Известны 4 гаплотипе, частота которых достигает высоких значений среди африканских негров или в популяциях, которые происходят от них. Интересно, что в 10%
Цлава 17. Группы крови
343
популяций Gm(l, 5, 13, 14, 17) (один из африканских гаплоди-пов) находят среди курдских евреев в Ираке, а среди евреев ашкенази — в 2%. Заметим, что у бушменов наблюдается аберрантная картина, в том числе гаплотип Gm (1, 5, 17), который характерен лишь для них и для народностей, с которыми они смешивались (рис. 17.10). У них есть и другой гаплотип, Gm (1, 13, 17),
Таблица 17.10
Часто встречающиеся гаплотипы Gm в различных популяциях, тестированные по 9 факторам (Steinberg, 1973)
ABCDEFGHI J К Гаплотипы
Европеоиды Монголоиды Айны Новая Гвинея Австралийцы Африканцы (негроиды) Бушмены + + + + + + + + + + + + + + + + + + + + + + + + + А (1, 1/, 21) В (1, 2, 17, 21) С (3, 5, 13,'14) D (1, 5, 13,14,17) Е (1, 5, 14, 17) F (1, 5, 6, 17) G (1, 5", 6, 14, 17) Н (1, 13, 17) I (1, 3, 5, 13, 14) J (2, 17, 21) К (1, 5, 17)
который встречается в Восточной Азии и совершенно нехарактерен для африканцев.
Из табл. 17.10 следует, что тестирование по многим факторам Gm расширяет возможности этого метода в разграничении популяций друг от друга. Так, Gm (1, 5) встречается во многих районах мира; если же провести исследование и на дополнительные факторы, то такая комбинация будет встречаться в четырех различных гаплотипах в Африке и еще в одном гаплотипе в Восточной Азии и Океании. Интересно, что у европейцев факторы Gm (1) и Gm (5) сегрегируют независимо.
Табл. 17.10 дает полное представление о глубоких различиях по гаплотипам Gm между регионами, однако различия в частотах гаплотипов описаны также и для более ограниченных областей. Несколько лет назад был обнаружен клин возрастания частоты Gm(l) в направлении к северу Европы. При обследовании аборигенов Австралии обнаружено, что в некоторых племенах на севере Квинсленда с заметной частотой встречается гаплотип Gm (1, 5, 13, 14), отсутствующий в племенах центральных и западных пустынь Австралии. Эти и некоторые другие данные о факторах
344
Часть III- Биологическая изменчивость популяций
Рис. 17.10. Карта Южной Африки с указанием процента примеси бушменов в различных племенах (Jenkins et al., Am. J. phys. Anthrop., 32, 1970).
Определено по частотам гаплотипа Gm(l,13).
Gm соответствуют концепции о притоке генов в Австралию через Торресов пролив. В самой Новой Гвинее отмечается немалая вариабельность частот гаплотипов. В долине реки Маркхейм частоты определенных гаплотипов коррелируют с делением племен по языку — говорящих на меланезийском и на не-австронезийских наречиях; зти данные свидетельствуют в пользу концепции о приходе племен, говорящих на меланезийских наречиях, из Юго-Восточной Азии.
Частота Gm (1, 13) была использована при картировании примеси бушменов-готтентотов у южноафриканских банту (рис. 17.10).
Стейнберг (Steinberg), перу которого принадлежат многочисленные антропологические работы по факторам Gm, определил при
Глава 17. Группы крови
345
месь европеоидных компонентов и койсанских компонентов в народности сидамо на юго-западе Эфиопии; соответствующие zвеличины оказались равны 40 и 12%. Он обследовал также айнов на о. Хоккайдо в Японии, которые являются носителями свойственного только им гаплотипа Gm (2, 17, 21), и пришел к заключению, что в его выборке среди айнов оказалось около 30% японской примеси.
Если в ходе дальнейших исследований подтвердится впечатление о том, что популяции различаются сильнее по частотам гаплотипов Gm, чем по прочим генетическим маркерам, то возникнет вопрос: почему эти различия сохраняются, несмотря на пот^к генов и рекомбинацию, стремящиеся разорвать связи, обусловленные сцеплением? Очевидный, хотя до сих пор и полученный умозрительным путем ответ — наличие эффективных региональных сил отбора. Привлекательным, но недоказанным предположением представляется ассоциация определенных гаплотипов с наличием антител, эффективных против ряда болезней.
Известны три фактора Inv — Inv (1), Inv (2) и Inv^3). Inv (1) и Inv (2) тесно связаны при наследовании, a Inv (3) наследуется самостоятельно, почти всегда в отсутствие первых двух. Популяционные исследования в основном проводились с применением сывороток анти-Inv (1) и анти-Inv (2). Во всех исследованных популяциях Inv (1, 2) был обнаружен с заметной частотой. Его частота довольно низка (<20%) только в Европе и на Среднем Востоке; максимума (94%) она достигает в некоторых племенах индейцев Венесуэлы.
Система тканевой совместимости (HL-A)
Хорошо известно, что кожа и другие ткани, трансплантированные от одного индивидуума другому, обычно вызывают иммунологическую реакцию, приводящую к гибели трансплантата. Подобное отторжение трансплантата, как правило, оказывается менее выраженным, если реципиент и донор состоят в близком родстве. Важным фактором такой несовместимости тканей оказались врожденные различия по тканевым антигенам HL-A; образуемая ими система усиленно изучается с целью добиться приживления трансплантатов путем соответствующего подбора донора и реципиента по антигенам. Антигены системы HL-A обнаруживаются в большинстве тканей, но не в эритроцитах; тем не менее уместно упомянуть их в этой главе.
Антисыворотки к антигенам HL-A обычно готовят из крови беременных, подвергшихся иммунизации антигенами плода; в отличие от некоторых антител к групповым веществам крови антитела, образующиеся в этом случае, не приносят вреда плоду. Эти антигены определяются с помощью теста на цитотоксичность. Лимфоциты исследуемых индивидуумов смешивают со специфичес
346
Часть III. Биологическая изменчивость популяций
кой анти-НЬ-А-сывороткой в присутствии комплемента (гл. 16). При наличии соответствующих антигенов клетки оказываются поврежденными и их можно отличить от неповрежденных клеток, применив определенные красители.
Антигены HL-A контролируются двумя различными, но тесно сцепленными локусами LA и4. У европейцев известно по меньшей мере 14 различных антигенов, контролируемых локусом LA, и 17 — локусом они определяются рядом аллелей по каждому локусу. Вследствие тесного сцепления локусов определенные парни генрв (например, 1 (LA) и 8 (4), 1 (LA) и 5 (4)) в данной популяции наследуются вместе. Все сказанное в этом отношении о гап-
Таблица 17.11
Географическая изменчивость частот некоторых аллелей
N 1 Локус LA М Локус 4
2 3 9 10 5 7 8 12 13
Англия 100 18 29 14 8 3 7 4 17 13 18 1
Германия 442 14 26 16 9 6 5 7 15 10 12 4
Франция 152 12 32 13 12 7 5 7 12 5 21 3
Исландия 116 11 34 18 12 3 6 2 23 9 13 1
Страна Басков 142 9 28 8 13 9 7 И 11 8 24 0
Сардиния 200 3 32 3 11 1 10 8 3 3 3 2
Лопари Скольт 178 1 28 37 16 7 8 9 23 2 1 1
Арабы 181 16 20 14 10 6 7 20 3 4 5 2
Пенджабцы 150 15 11 7 10 6 15 19 4 13 9 2
Индусы 153 11 17 10 18 2 14 12 10 3 13 3
Малайцы 137 3 15 2 40 1 16 6 5 0 3 8
Новая Гвинея (Восточное плоскогорье) 214 0 0 0 76 3 4 0 0 0 0 5
Австралийцы (уолбири) 177 0 13 0 31 43 10 0 0 0 0 21
Японцы 127 0 27 1 37 14 4 18 5 0 8 1
Китайцы (Кантон) 166 1 31 1 17 5 28 6 1 0 1 12
Айны 127 0 29 1 31 17 3 И 1 0 5 0
Тибетцы 138 9 29 4 33 6 16 21 0 0 2 1
Папуасы 105 0 51 1 37 1 0 5 1 1 2 0
Гватемала 131 б 51 0 22 1 0 1 0 1 1 0
Ивечуа (Перу) 111 1 59 1 22 0 1 3 4 0 1 1
Эскимосы (Гренландия) 177 1 16 1 65 1 2 17 1 1 2 0
Эскимосы (Канада) 336 0 28 0 63 0 0 15 10 0 0 0
Замбия 172 2 14 5 11 5 0 1 10 2 19 1
Ши (Заир) 144 7 26 4 13 9 2 17 12 1 16 1
Бушмены куне 84 5 17 8 14 7 0 9 19 4 6 2
Глава 17. Группы крова
347
лотипах Gm относится также к ассоциированным гаплотипам HL-A. Можно ожидать, что со временем’их ассоциация нарушится в результате кроссинговера, и с наступлением равновесия они станут неразличимыми в данной популяции. Сохранение их до настоящего времени позволяет допустить существование сил отбора, благоприятных для некоторых ассоциаций аллелей.
Частоты некоторых аллелей HL-A подвержены географической изменчивости. В табл. 17.11 приведены некоторые примеры для небольшого числа аллелей. Большинство выборок невелико, и соответственно стандартные ошибки частот аллелей велики; тем не менее определенные тенденции несомненны. Для Европы показатели довольно единообразны, хотя у лопарей, басков и жителей Сардинии обнаруживаются отклонения, как, впрочем, и в случае ряда других генов. Некоторые аллели (например, 1, 3, 7, 8 и 12) встречаются в Восточной Азии, в Австралазии и у американских индейцев гораздо реже, чем среди европейцев. Вместе с тем аллель 9 встречается явно чаще у аборигенов Нового Света, особенно у эскимосов. Как правило, величины для африканцев менее отличаются от таковых для европейцев, хотя теперь известны некоторые антигены, характерные для этого района. Следует отметить большую изменчивость у племен Австралии и Новой Гвинеи; такая локальная вариабельность всегда затрудняет представление данных в сжатой форме. Система HL-A с ее выраженным полиморфизмом создает замечательные возможности для изучения родства популяций.
В настоящее время нам известно недостаточно о природе сил отбора, действующих на систему HL-A; однако некоторые факты позволяют выдвинуть интересные гипотезы. Дивергенция частот определенных аллелей найдена в выборках больных некоторыми нечасто встречающимися заболеваниями, например лимфогранулематозом и красной волчанкой. Имеются также указания на тесное сцепление локусов HL-A с локусами, модифицирующими эффективность иммунологического ответа на некоторые вирусы.
Глава. 18
ДРУГИЕ ВИДЫ ИЗМЕНЧИВОСТИ
Ощущение вкуса фенилтиомочевины
В 1931 г. Фокс случайно подметил, что есть люди, которые не ощущают вкуса синтетического препарата — фенилтиомочевины, другие же считают его горьким, подобно хинину. Позднее было показано, что способность ощущать вкус этого соединения наследуется по законам Менделя как простой доминантный признак. Самый простой способ испытания — это дать человеку в рот кристаллик фенилтиомочевины или пропитанную раствором бумажку. Более совершенный способ — это применение ряда различных концентраций вещества в воде. Применение последнего способа показало, что лица, ощущающие вкус фенилтиомочевины, сильно различаются по чувствительности к ней. Диагноз наличия или отсутствия признака весьма субъективен. Для дальнейшего усовершенствования теста можно использовать методику, разработанную Харрисом и Кальмусом (Harris a. Kalmus): испытуемому предлагают отобрать растворы фенилтиомочевины из ряда образцов, половина которых содержит только воду. Если этот тест Повторять, увеличивая каждый раз концентрацию фенилтиомочевины, то можно выявить пороговую концентрацию, при которой впервые появляется ощущение вкуса препарата. Распределение пороговых величин в популяции можно затем представить в виде гистограммы (рис. 18.1). При этом обнаруживается бимодальное распределение, в котором одна мода характеризует ощущающих, а другая — не ощущающих вкуса фенилтиомочевины; обычно наблюдается взаимное перекрывание обеих групп, связанное с трудностью отнесения некоторых лиц к той или иной категории. Дальнейшее уточнение теста достигается путем внесения поправок на несколько ббльшую чувствительность к горькому вкусу, которая оценивается в контрольных опытах с бруцином. Отсутствие способности ощущать вкус фенилтиомочевины наследуется как рецессивный признак; имеются, однако, указания на тот факт, что порог чувствительности к этому веществу у гетерозигот (Tt) выше, чем у гомозигот (ТТ).
Распределение. Методом множественных проб было исследовано небольшое число популяций. Некоторые результаты этого исследования представлены в табл. 18.1. Доля лиц, не ощущающих вкуса фенилтиомочевины, составляет на северо-западе Европы 35—40%; в районе Средиземного моря этот показатель значитель-
Глава 18. Другие виды изменчивости
349
но ниже. Среди африканцев, китайцев, японцев, североамериканских индейцев и лопарей данный фенотип встречается значительно реже.
Влияния отбора. Бимодальное распределение касается способности ощущать вкус не только фенилтиомочевины, но и многих других, родственных ей соединений. Однако наиболее четкое
Рис. 18.1. Распределение индивидуумов с различными порогами ощущения вкуса фенилтиомочевины в трех популяциях (Barnicot, Ann. Eugen, 1950). Номер раствора соответствует номеру последовательного двукратного разведения 0,13%-. ного раствора фенилтиомочевины. Форма распределения не ощущающих вкуса (область разведений от 5 до <1) ясно видна на верхней схеме. А. Англичане (155). В. Африканцы (74). В. Китайцы (06).
разделение двух фенотипов характерно именно для фенилтиомочевины. В химической структуре подобного рода соединений важную роль, по-видимому, играет группа — С—N.
II
S
Аналогичные производные мочевины, в которых атом серьь(З) замещен атомом кислорода (О), не обладают интересующей нас биологической активностью.
Некоторые производные тпомочевины, например тиоурацил, как известно, применяются в клинике для подавления функции щитовидной железы. В связи с этим возникла мысль о том, что по-
350
Часть III. Биологическая изменчивость популяций
Таблица 18.1
Частота отсутствия способности ощущать вкус фенилтиомочевины в различных популяциях
Популяция Число обследованных Процент лиц, не ощущающих вкуса фенилтиомочевины Популяция Число обследованных Процент лиц, не ощущающих вкуса фенил-тиомоче-вины
Хинди 489 33,7 Японцы 295 7,1
Датчане 251 32,7 Лопари 140 6,4
Англичане 441 31,5 Негры Западной 74 2,7
Испанцы 203 25,6 Африки
Португальцы 454 24,0 Китайцы 50 2,0
Негритосы (Ма- 50 18,0 Индейцы Южной 163 1,2
лайя) Америки (Бра-
Малайцы) 237 16,0 зилия)
диморфизм в отношении данного гена каким-то образом связан с функцией щитовидной железы. Соответствующие исследования ощущения вкуса при ряде заболеваний этой железы позволили установить различие в частоте ощущения вкуса фенилтиомочевины при некоторых формах зоба.
Поскольку индивидуумы, неспособные ощущать вкуса фенилтиомочевины, обнаружены и среди шимпанзе, было высказано предположение, что данный тип полиморфизма поддерживается, начиная с отдаленных этапов эволюции человека, вероятно, вследствие гетерозиса.
Цветовая слепота
Известны случаи аномального восприятия цвета, в частности слепота на красный и зеленый цвета; она служит примером сцепленного с полом наследования у человека. Пораженные этим дефектом склонны путать красный и зеленый цвета, а в более тяжелых случаях вообще их не различают. Углубленные исследования показали, что существуют две физиологически различные формы слепоты на красный и зеленый цвета — «протанопическая» и «дей-теранопическая», причем каждая из них может быть различной степени тяжести. Дефекты наиболее тяжелого типа называют «протанопией» (слепота на красный цвет) и «дейтеранопией» (слепота на зеленый цвет). Соответствующие, но менее тяжелые дефекты называют «протаномалией» и «дейтераномалией».
Почти окончательно установлено, что протанопический' и дейтеранопический дефекты возникают вследствие мутаций по
Глава 18. Другие виды изменчивости
351
двум различным, но весьма тесно сцепленным локусам Х-хромосомы. Если это так, то можно ожидать обнаружения в редких случаях мужчин с дефектами обоих типов, а также семей, в которых у женщины с нормальным цветоощущением будут сыновья протанопы, дейтеранопы и здоровые. Последняя ситуация может возникнуть, если мать гетерозиготна по обоим локусам и дает нормальную хромосому путем кроссинговера. Обе эти ситуации описаны, хотя диагностировать смешанные дефекты нелегко. Полагают, что различия в тяжести этих дефектов связаны с существованием двух или большего числа аллельных вариантов по каждому локусу.
Механизм действия мутаций цветовой слепоты неизвестен, так как трудно получить для детального биохимического исследования такой материал, как сетчатка глаза человека. Результаты некоторых изящных оптических экспериментов позволяют предполагать, что у протанопов в рецепторах колбочек отсутствует пигмент, чувствительный к кра’сному цвету; действительно, их чувствительность к свету красной части спектра очень низка. У дейтеранопов, напротив, сохранена чувствительность к свету в красной и зеленой областях спектра; возможно, у них дефект затрагивает какой-то более высокий уровень организации нервной системы. Следует также упомянуть аутосомно наследуемые де-
Таблица 18.2
Частота различных аномалий цветового зрения у европейцев (Wright, 1953)
(Частота в популяции, % 1 - у мужчин------------------у женщин
1. Монохрематизм 2. Дихроматизм Протанопия Дейтеранопия Тританопия 3. Аномальный трихроматизм Протаномалия Дейтераномалия Тританомалия Встречается очень редко 2,105 1.0 1.1 0,005 5,9 1,0 4,9 Встречается очень редко Встречается очень редко 0,06 0,02 0,01 0,03 0,40 0,02 0,38 Встречается очень редко
Приблизительная суммарная частота аномалий 8,0 0,46
352
Часть III. Биологическая изменчивость популяций
фекты цветоощущения в 'Желто-голубой области («тританопичес-кие» дефекты), которые, однако, встре'чаются редко. Еще реже обнаруживается колбочковый монохроматизм, при котором полностью нарушена способность воспринимать цвета. Примерные частоты этих разнообразных нарушений в западноевропейских популяциях представлены в табл. 18.2. Заметим, что чаще всего отмечается дейтераномалия. Частота дефектов заметно выше среди мужчин, чем среди женщин, чего и следовало ожидать от сцепленного с полом рецессивного признака. Частота дефекта у мужчин прямо соответствует частоте гена q; при установившемся равновесии частота встречаемости пораженных женщин (гомозигот) должна быть равна q.
Подробное исследование аномалий цветового зрения включает измерение чувствительности к свету, определение способности сравнивать цвета и отличать друг от друга образцы различного цвета. Это весьма трудоемкие исследования, требующие сложного оборудования. Поэтому для массового обследования населения необходимы более простые и быстрые тесты. Широко применяются специальные таблицы, например хорошо известный набор Ишиха-ра. Правда, применяя их, легцо упустить случаи слабо выраженных аномалий. Исследуемого просят назвать ряд цветных цифр на цветном фоне или проследить ход извилистой линии (при обследовании неграмотных). И цифры, и фон образованы из цветных точек главным образом разных оттенков красного или зеленого; они подобраны так, что страдающий цветовой слепотой не способен различить цифру или, видя только часть ее, ошибочно принимает эту цифру за другую. Эти тесты следует проводить при рассеянном дневном свете; при ином освещении иногда получают ошибочные результаты.
Удобным прибором для обнаружения менее серьезных дефектов цветового зрения служит аномалоскоп Нагеля; он приспособлен для полевых исследований. Лица с нормальным цветовым зрением способны составить желтый цвет спектра из смеси монохроматических цветов — красного и зеленого. Кроме того, они могут составить любой цвет, смешивая не более трех цветов; обычно это правильно подобранные красный, зеленый и синий. Поэтому их называют «трихроматами». Протанопы и дейтеранопы сопоставляют все цвета лишь с двумя цветами; их называют «дихроматами». Работая с аномалоскопом, испытуемый смотрит через окуляр на круглое поле, половина которого освещена желтым светом; вращая калиброванные рукоятки, он может добавлять красный или зеленый свет к другой половине поля до совпадения обеих половин по цвету. Доля красного и зеленого, подбираемая нормальными индивидуумами, довольно постоянна, а дихроматами — вариабельна: протаномалы «добавляют» больше красного, а дейтераномалы — больше зеленого. Ощущение красного и зеле
Глава 18. Другие «иды, изменчивости
353
ного цветов у них ослаблено, но не отсутствует, и поэтому их называют «аномальными трихроматами». Интерпретируя результаты, полученные с помощью аномалоскопа, необходимо соблюдать особую осторожность при исследовании популяций, не знакомых с подобными приборами и не привыкших к точной подгонке цвета.
В табл. 18.3 представлены данные о частоте случаев слепоты' на красный и зеленый цвета в выборках мужчин из некоторых популяций, обследованных быстрыми методами. Для Европы получены величины около 7—8%; у различных народностей Восточной Азии они несколько ниже. В Индии (насколько можно судить по довольно малым выборкам) в некоторых районах отмечаются большие различия между кастовыми группами; различаются также частоты для популяций хинди и ряда других племен. Показатели для субэкваториальной Африки, Нового Света и Австралазии сравнительно ниже (1—3%). Можно полагать, что для народностей, занимавшихся охотой и собирательством, цветовая
Таблица 18.3
Частота случаев цветовой слепоты у мужчин в различных популяциях
Популяция Размер выборки Цветовая слепота Автор
Арабы (друзы) 337 10,0 Kalmus, 1961
Норвежцы 9047 8,0 Waaler, 1927
Шведы 2 000 8,0 Von Planta, 1928
Немцы 6863 7,7 Schmidt, 1936
Бельгийцы 9 540 7,4 De Laet, 1935
Англичане 16180 6,6 Grieve, 1946
Иранцы 947 4,5 Plattner, 1959
Индийцы (Андра Пра- 292 7,5 Dronamraju, 1961
деш)
Китайцы (Пекин) 1164 6,9 Chang, 1932
Китайцы (все) 36 331 5,0 Chun, 1958
Тибетцы 241 5,0 Tiwari, 1969
Японцы 259000 4,0 Sato/ 1935
Мексиканцы 571 2,3 Garth, 1933
Индейцы навахо 535 1,1 Garth, 1933
Эскимосы 279 2,5 Skeller, 1954
Сваны 409 3,0 Squires, 1942
Хуту 1000 2,9 Hiernaux, 1953
Тутси 1000 2,5 Hiernaux, 1953
Народности Заира • 929 1,7 Appelman, 1953
Аборигены Австралии 4455 1,9 Mann, 1956
Островитяне Фиджи 608 0,8 Geddes, 1946
354 Часть III. Биологическая изменчивость популяций
слепота у мужчин была неблагоприятным селективным фактором; с переходом к земледелию и развитием цивилизации давление такого отрицательного отбора ослабляется. Однако данные об охотниках и собирателях скудны; не ясно, могло ли давление мутаций создать столь большой прирост генных частот за период менее 10 000 лет.
С помощью аномалоскопа обследовано лишь небольшое число популяций, и потому имеющиеся данные об относительных частотах аномалий цветового зрения на земном шаре весьма скудны. Наиболее вариабельным типом представляется дейтераномалия — от 5% среди европейцев до 1—2% в Уганде, на Новой Гвинее и в Таиланде (Adam, 1970); можно, однако, ожидать, что снижение давления отбора более всего скажется на тяжелых аномалиях.
Частота рождения двоен
В демографической и медицинской статистике собраны обширные данные о рождении двоен, и, хотя частота их рождения низка, удалось проследить различия между популяциями. Частота рождения дизиготных близнецов колеблется в весьма широких пределах, тогда как монозиготные близнецы появляются в различных популяциях с поразительно близкой частотой. Вероятность рождения дизиготных близнецов увеличивается с возрастом матери и числом предшествующих родов, поэтому более полные сведения удается получить, если принимать во внимание оба эти фактора. Необходимо тщательно следить за тем, чтобы избежать ошибок выборочного метода; например, более высокая частота случаев многоплодия среди женщин, обращающихся за акушерской помощью, может привести к необоснованному преувеличению данных о частоте двоен, если данные акушерских клиник будут считаться представительными для той или иной области. Установление типа зиготности в каждом отдельном случае требует применения сложных методов, в том числе определения групп крови; но определить частоту двоен того и другого типа в популяции можно при помощи специального расчета по методу Вейнберга. Разнополые близнецы всегда относятся к категории дизиготных; при соотношении полов, равном 1,0, можно ожидать, что число однополых дизиготных близнецов равно числу разнополых; путем вычитания числа разнополых двоен из общего числа однополых получим примерное число монозиготных двоен.
Табл. 18.4, заимствованная из работы Балмера, свидетельствует о наличии четких различий в частоте дизиготных двоен в разных странах Европы и даже в пределах одной страны. Сообщают о весьма высокой частоте дизиготных двоен в Африке, на о. Ямайка и среди американских негров. Данные о частоте двоен в Африке и на о. Ямайка не корригированы с учетом возраста мате
Глава 18, Другие вида изменчивости,
3S5
рей и числа предшествующих родов; однако внесение соответствующих поправок, вероятно, лишь несколько увеличило бы эти показатели. Частота дизиготных двоен в Японии, напротив, значительно ниже, чем в Европе.
Рождение дизиготных двоен явно имеет тенденцию к концентрации в определенных семьях, однако никакого простого закона наследования этого признака установить не удалось. Во всяком случае, географические различия в стандартизованной частоте рождения дизиготных двоен можно считать предварительным указанием на существование соответствующих генетических различий.
Таблица 18.4
Число двоен на 1000 родов в различных популяциях (Bulmer, Ann. Hum. Geuet., 1980)
Страна Число двоен Страна Число двоен
ДИЗИГОТ- НЫХ монозиготных диэигот-ных монозиготных
Нигерия 39,9 5,0 Англия и Уэльс
Родезия 26,6 2,3 (1946—1955) 8,9 3,6
Заир 18,7 3,1 Швеция (1946—1955) 8,6 3,2
о. Ямайка 13,4 3,8 Италия (1949—1955) 8,6 3,7
США 11,8 3,9 Франция (1946—1951) 7,1 3,7
Греция (1931—1938) 10,9 2,9 Испания (1951—1953) 5,9 3,2
Япония (1926—1931) 2,7 3,8
Распределение редких наследственных заболеваний
Многие из рассмотренных нами биохимических и серологических вариантов встречаются довольно часто, во всяком случае в некоторых популяциях; этим облегчается сравнение частот этих вариантов в разных географических областях даже на основе небольших выборок. Труднее получить данные о частоте редко встречающихся заболеваний; тем не менее ясно, что некоторые из них распределены неравномерно и представляют интересный объект для генетики популяций. Тяжелые наследственные болезни, будучи даже весьма редкими, обычно служат предметом особого внимания врачей, а редкие заболевания с доброкачественным течением порой выявляются случайно при массовых анализах крови или мочи, проводимых с какой-либо целью. Многие популяции расположены слишком далеко от центров квалифицированной медицинской помощи, которые могли бы обеспечить сбор надежных
356
Часть III. Биологическая ивменчцвостъ популяций
данных; несмотря на это, о географических различиях в частоте той или иной наследственной болезни можно судить по данным об иммигрантах из интересующих нас областей в страны с более развитой сетью медицинских учреждений. При систематическом изучении частоты редких заболеваний в популяции в целом следует с особой тщательностью подходить к статистической оценке результатов, чтобы исключить вероятность ошибки выборки.
Случаи редких рецессивных заболеваний часто бывают сосредоточены в некоторых ограниченных областях, где малые размеры и изолированность сообщества неизбежно увеличивают частоту близкородственных браков. Если такой изолят представляет собой потомство небольшого числа предков, среди которых случайно окажется носитель определенного гена, то локальная частота этого гена может оказаться здесь значительно выше, чем в популяции в целом. В результате создаваемой инбридингом гомозиготности вредный рецессивный ген подвергается более жесткому отбору вплоть до полной элиминации, тогда как в обычной популяции некоторого избытка гетерозигот оказывается достаточно для поддержания равновесия на уровне низкой частоты данной аномалии. В малой популяции существенное значение приобретают также случайные колебания в передаче генов от одного поколения к следующему.
Известна детская форма амавротической идиотии — тяжелого неврологического расстройства, наследуемого по рецессивному типу и ведущего к преждевременной гибели пораженного. Частота этой аномалии в ряде европейских стран достигает 1/10*000; оказалось, что большая часть пораженных этим дефектом — евреи. Оказалось также, что весьма редкая и, по-видимому, не ведущая к серьезной патологии аномалия, известная под названием Z-ксилокетозурии (появление в моче необычного углевода), также поражает главным образом евреев, в большинстве своем выходцев из западных областей России. Другая форма наследственной умственной отсталости — фенилкетонурия, напротив, редко встречается у евреев; она распространена во многих районах Европы, причем есть основания предполагать существование географических различий в ее частоте; гораздо реже эта аномалия встречается у американских негров, у африканцев и японцев. Редко встречающаяся наследственная недостаточность фермента каталазы в эритроцитах до сих пор наблюдалась только у японцев. Характерная депигментация альбиносов, гомозиготных по некоему рецессивному гену, весьма облегчает их выявление в популяциях пигментированных индивидуумов; и действительно, они были описаны в большинстве частей света. Дефекты зрения обычно заставляют таких альбиносов обратиться к врачу в тех местностях, где для этого имеются необходимые условия. Частота случаев альбинизма в ряде европейских стран составляет 1/10 000—
Глава 18. Другие виды, изменчивости
357
1/20 000; в Западной Африке она, по-видимому, в 3—5 раз выше, в некоторых индейских племенах Панамы, обитающих на островах сравнительно небольшими деревенскими общинами, частота этой аномалии достигает 1/200.
В последнее время интерес к генетическим последствиям воздействия ионизирующих излучений повлек за собой настоятельную необходимость уточнить сведения о частоте редких наследственных аномалий, в том числе тех, которые обычно именуют врожденными уродствами. К этой группе относятся заячья губа, конская стопа, врожденный вывих бедра, разного рода полидактилии и пороки развития центральной нервной системы (анэнцефалия и spina bifida). Общая частота появления врожденных уродств в европейских популяциях и среди американских негров и японцев довольно постоянна (примерно 1—1,5%), однако по частоте отдельных перечисленных выше форм эти же популяции различаются. Различия эти, несомненно, обусловлены целым комплексом причин; несмотря на тенденцию к семейной концентрации некоторых уродств, для большинства из них четких генетических закономерностей не установлено. Некоторые аномалии могут быть сгруппированы на основе фенотипического сходства, несмотря на то что генетически они явно гетерогенны. Упомянутые различия между популяциями в смысле частоты аномалий могут быть отражением различий не только в частотах определенных генов, но и в характере влияния факторов внешней среды на проявление этих генов и на возможность возникновения фенокопий.
Пенроуз собрал данные о распределении в Европе случаев анэнцефалии (недоразвитие мозга); он установил, что частота этой аномалии колеблется в пределах от 1,0 до 0,32% в Ирландии и в ряде западных районов Великобритании; в большинстве стран континентальной Европы она в 10 раз ниже, а на юго-востоке Франции эта аномалия встречается еще реже. Сирл (Searle), изучив данные о ряде этнических групп в Малайе, обнаружил колебание соответствующих частот в различных группах от нуля (китайцы группы хакка) до 0,65% (сикхи). Причины анэнцефалии совершенно неяснь^, однако, рассматривая ее распространение по популяциям в комплексе с другими данными, можно предположить, что в возникновении этой аномалии играют роль как генетические факторы, так и факторы внешней среды.
Толерантность к лактозе
Тонкий кишечник новорожденных млекопитающих содержит фермент лактазу, расщепляющий характерный для молока сахар, лактозу, на глюкозу и галактозу. Ко времени прекращения грудного вскармливания количество этого фермента резко снижается. У многих детей аналогичный процесс происходит на 4-м году жиз
358 Часть III. Биологическая изменчивость популяций
ни. Люди в этом отношении отличаются от прочих млекопитающих тем, что научились (вероятно, 10 000 лет назад) употреблять в пищу молоко коров и некоторых других одомашненных копытных; для ряда народностей свежее молоко составляет существенный компонент рациона взрослых. Было замечено, что в некоторых районах Азии и Африки у многих индивидуумов при употреблении в пищу молока возникают боли в животе и понос. Эти симптомы развиваются вследствие брожения нерасщепленной лактозы в толстом кишечнике. В Западной Европе и в определенных пастушеских племенах (тутси и фулани в Африке), где пьют много свежего молока, отсутствие толерантности к лактозе встречается гораздо реже. Существует простой количественный тест на толерантность к лактозе: испытуемому дают лактозу в дозе 2 г на 1 кг веса тела (всего не более 50—100 г) и через определенные промежутки времени определяют содержание в крови глюкозы. Толерантность к лактозе считается пониженной, если повышение уровня глюкозы в крови не достигает 20 мг на 100 мл; не у всех таких испытуемых при проведении теста развиваются кишечные симптомы. Коровье молоко содержит 4,5—5% лактозы; поэтому тест-доза эквивалентна 3 л молока, т. е. очень высока.
Оказалось, что в составе популяций китайцев, японцев, таиландцев, эскимосов, а в Африке в племенах ибо, йоруба, ганда и у бушменов 80% индивидуумов не переносят лактозу, тогда как для тутси, фулани и жителей Западной Европы соответствующие показатели лежат между 5 и 25%.
Возникает вопрос, связана ли толерантность к лактозе с индукцией относительно высокого уровня лактазы под влиянием систематического потребления в пищу молока или же это наследственный признак. Изучение семей в Нигерии, Израиле и других странах позволяет предполагать, что толерантность к лактозе наследуется как простой доминантный признак; однако непонятно, каков механизм действия соответствующего гена; возникает мысль об аналогии с генетическим признаком сохранения у взрослого Hb-F. Возможно, что в тех популяциях, у которых свежее молоко служит одним из основных продуктов питания, подобная мутация обеспечивает определенное селективное преимущество. Однако не все так просто; по некоторым данным, у взрослых с частичной или полной недостаточностью лактазы (судя по результатам описанного выше теста или определения активности фермента в биопсийных препаратах тонкого кишечника) удается выработать толерантность к лактозе. Поскольку такая адаптация не сопровождается повышением уровня лактазы, можно допустить, что она обусловлена изменением кишечной флоры. Здесь уместно заметить, что у некоторых народностей вошло в обычай потреблять в пищу кислое молоко, в котором большая часть лактозы уже расщеплена благодаря активности бактерий.
Глава 18. Другие виды изменчивости
359
Влажная и сухая ушная сера
В западноевропейских популяциях у 90% индивидуумов (и более) ушная сера (cerumen) представляет собой желтую и вязкую массу, у остальных она сероватая и сухая, содержит меньше липидов. Этот любопытный признак контролируется парой аллелей, причем сухой тип соответствует гомозиготности по рецессивному признаку. Влажный тип редко встречается в Северном Китае (4, 2%) и в Корее (7, 6%), тогда как на юге Китая он обнаруживается чаще (21—42%). Подробные исследования были проведены у индейцев Центральной и Северной Америки; частота влажного типа у них широко варьирует — от 36,7% у навахо до 93,3% у цельтали майя (табл. 18.5). Весьма мало известно об этом признаке у африканцев; влажный тип у них, по-видимому, встречается по меньшей мере столь же часто, как и у европейцев. Функциональное значение такого диморфизма неизвестно.
Таблица 18.5
Частота признака влажной ушной серы в различных популяциях (Matsunada, Ann. Hum. Genet., 1962; Petrakis, Nature, Load., 1969)
Популяция Численность выборки Частота признака,%
Европеоиды 368 98,7
Негры 51 100,0
Цельтали майя 296 93,3
Чоктавы 1> 432 79,2
Айны 30 86,7
Меланезийцы 732 72,2
Аборшены Тайваня 1420 71,4
Чилкотин 1> 261 66,7
Ноотка 1> 244 63,6
Сиу 1> 147 63,3
Микронезийцы 458 62,9
Алеуты 140 51,4
Папаго 437 50,3
Китайцы 169 41,5
Жители о-ва Рюкю 898 37,5
Навахо 183 36,7
Китайцы (Южный Китай) 463 21,2
Корейцы 381 7,6
Китайцы (Северный Китай) 216 4,2
>) Американские индейцы
360
Часть III. Биологическая изменчивость популяций
Инактивация, изониазида
Выше упоминалось о двух врожденных, биохимических признаках, обнаруженных практической медициной при лекарственной терапии: характерная для негров недостаточность фермента Г6ФД, которая проявляется при лечении примахином, и дефектный тип сывороточной псевдохолинэстеразы при лечении гексаме-тонием. Ниже приводится еще один пример.
Препарат изониазид вошел в практику лечения туберкулеза в 1952 г. Было подмечено, что скорость его выведения с мочой подвержена резко выраженным индивидуальным колебаниям. Систематическое исследование показало, что если вводить стандартную дозу препарата и через 6 ч определять его уровень в плазме, то на гистограмме уровней для случайной выборки обнаружатся два распределения с частичным наложением друг на друга. Анализ родословных показал, что медленная инактивация наследуется как простой рецессивный признак.
Немногочисленные сравнительные данные (табл. 18.6) свидетельствуют о том, что у 50% европейцев отмечается медленная инактивация; отсюда следует, что частота этого аллеля 70—80%. Возможно, что у африканцев эта величина еще выше. Вместе с тем в различных популяциях Юго-Восточной и Восточной Азии соответствующие частоты явно ниже (30—60%), как и в выборках эскимосов и североамериканских индейцев.
Таблица 18.6
Инактивация изониазида
Популяция Численность Частота аллеля медленной инактивации, %
Американцы белые 122 73,0
негры 41 7.6,5
Суданцы 102 79,8
Индийцы (Мадрас) 321 78,1
Бирманцы 121 61,2
Таиландцы 108 54,2
Китайцы 60 52,1
Японцы 209 28,9
Айны 86 30,8
Эскимосы (Канада) 216 22,4
Североамериканские индейцы (сиу) 15 45,8
Глава 18. Другие виды, изменчивости
361
Полиморфизм хромосом, человека
Возможность систематически изучать кариотип человека появилась впервые примерно 20 лет назад; вскоре стало ясно, что некоторым хромосомам присуща значительная вариабельность, не сопровождающаяся какими бы то ни было аномальными проявлениями. Особенно разительным примером может служить длина Y-хромосомы: она может быть очень крупной, подобно членам группы G (13—15), и очень маленькой, подобно короткому акро-центрику. При обычных методах окрашивания хромосомы имеют гомогенную структуру; однако в 1970 г. при исследовании хромосом, окрашенных акрихином (quinacrine, Q), в ультрафиолете были обнаружены четкие поперечные полосы. Дистальная область длинного плеча хромосомы сильно флуоресцировала; оказалось, что вариабельность длины этой хромосомы зависит от количества Q-хроматина, или, как его назвали, гетерохроматина, который иногда практически отсутствует. Работы по сравнению разных популяций немногочисленны; тем не менее установлено, что длинная Y-хромосома довольно часто встречается у японцев, а короткая — у аборигенов Австралии. При Q-окрашивании обнаруживаются также сильно флуоресцирующие полосы в центромерной области хромосомы 3 и в коротких плечах и сателлитах хромосом групп D и G. Обнаружена также изменчивость размера этих полос. Разработаны методы преимущественной окраски гетерохроматина вблизи центромеры (С-полосы). Особенно широкие С-полосы видны в хромосомах 1, 9 и 16; для них также характерна вариабельность. Еще окончательно не выяснено, распадаются ли эти различия в количестве гетерохроматина по четким категориям или образуют ряд постепенных изменений. Применение этих недавно сделанных открытий для сравнения разных популяций — дело будущего.
Умственные способности
На протяжении столетий путешественники интересовались, восхищались или возмущались образом жизни чуждых им обществ, сопоставляя его с привычным для них образом жизни у себя на родине. Было бы опрометчиво поддаться соблазну и приписать эти различия, иногда совпадающие с поразительным внешним несходством, генетическим различиям в умственной деятельности. Огромные изменения, которые могут произойти при переносе индивидуума в раннем возрасте в иную культурную среду, предостерегают от поверхностных заключений в этом вопросе. Вместе с тем ясно, что генетические факторы могут серьезно влиять на умственное развитие; об этом свидетельствуют многочисленные примеры наследственной умственной отсталости. Закономерности наследования умственных способностей в пределах нормальных
362 Часть III. Биологическая изменчивость популяций
колебаний изучали с помощью различных методов, отвечающих требованиям анализа полученного наследования. Изучение корреляций менаду родственниками и между одно- или двуяйцовыми близнецами, а также сопоставление однояйцовых близнецов, воспитывавшихся врозь или вместе, доказало роль генетических факторов в возникновении нормальных различий между индивидуумами. Этот вывод, однако, применим только к относительно гомогенной популяции западноевропейских стран, в которых были проведены эти исследования, и мало говорит о том, какова роль среды в возникновении различий, существующих между весьма несхожими культурами. Естественно ожидать, что гены, контролирующие функцию мозга или другие функции, связанные с умственной деятельностью, могут в различных местностях варьировать по частоте; однако можем ли мы надеяться, что нам удастся выявить эти «естественные» различия доступными нам методами?
Вера во врожденную интеллектуальную ограниченность некоторых людей имеет длинную историю. Там, где чувство приверженности какой-то идее переходило известные границы, слишком часто страдала научная объективность.
С целью избежать субъективных суждений при оценке уровня интеллекта или темперамента психологи создали многочисленные стандартные тесты. Мы не можем рассмотреть их здесь детально; одни из них предназначены для оценки памяти, общего уровня знаний и знакомства с общественной жизнью, владения речью и способности к решению задач, другие служат для оценки состояний эмоциональной сферы. В применении к людям, воспитанным так, как это принято в Западной Европе, эти тесты оказались полезным средством для определения их соответствия тем или иным профессиональным требованиям или для диагностики психических заболеваний; однако применение этих тестов для обследования популяций, живущих в совершенно иной среде, наталкивается на весьма серьезные трудности.
Применимость этих тестов ограничивается, в частности, необходимостью знать язык, на котором они составлены, но и после перевода они могут оказаться бесполезными, поскольку предусматривают знакомство со сведениями и концепциями, характерными лишь для общества определенного типа. Для того чтобы преодолеть эти затруднения, были созданы не словесные тесты, а такие, при проведении которых испытуемого просят обнаружить связь между абстрактными узорами, расположить в определенном порядке цветные кубики, нарисовать человечка, проложить маршрут по лабиринту ит. п.; утверждали, что такие тесты позволят избежать влияния различий в культуре. Однако опыт работы с африканцами, американскими индейцами, аборигенами Австралии и т. п. заставляет серьезно усомниться в правильности подобного
Глава 18. Другие виды изменчивости
363
утверждения. Вполне возможно, что низкие оценки оказываются следствием не умственной неполноценности, а неопытности в обращении с кубиками и другими игрушками, в интерпретации символических изображений или в различении цветов. Вообще сомнительно, чтобы можно было создать тест, «свободный от влияния культуры»; скорее всего такая идея сама по себе абсурдна'.
Такого рода тесты могут дать более или менее полноценную информацию в случае, когда мы сравниваем различные этнические группы людей, воспитанных в идентичных условиях. В этом направлении большая работа была проделана по изучению американских негров. Результаты суммировали в виде так называемого коэффициента умственного развития (intelligence quotient, I. Q.). Нередко оказывалось, что у негров величина этого коэффициента равнялась 80, а у белых — 100. Однако в пределах обеих популяций вариабельность была весьма значительной, как это имеет место в отношении метрических признаков; отмечалось также значительное перекрывание соответствующих кривых распределения. У негров величины этого коэффициента оказываются выше в тех группах, члены которых воспитывались в более благоприятных условиях; при этом они могли оказаться выше, чем у белых, находившихся в худших условиях. Некоторые авторы убеждены в реальном характере различий в среднем уровне интеллекта у белых и негров, не обусловленных факторами среды; однако другие берут под сомнение правильность их выводов. В США все еще наблюдается расовая сегрегация; поэтому сомнительно, чтобы при интерпретации подобных результатов можно было исходить из того, что сопоставляемые группы находятся в одинаковых условиях среды.
Проделанная предварительная работа позволяет прийти к выводу, согласно которому неевропейские народности в реально существующих условиях разнообразия культур часто считаются не приспособленными для выполнения такого рода задач, которые кажутся существенными для европейцев; это и не удивительно. Вопрос, который предстоит решить главным образом экспериментально, сводится к тому, в какой степени представители этих народностей, если они того захотят, смогут сравняться с европейским стандартом или превзойти его, получив соответствующее воспитание и моральную поддержку.
Список литературы
Общие вопросы
Buettner-Janushh J. (1973). Physical anthropology—a perspective. Wiley, New , York.
Burnet A. M. (1973). Genes, dreams and reahites. Penguin, Harmondsworth. Cavalli-Sforza L. L. and Bodmer W. (1971). The genetics of human populations.
Freeman, New York.
Ebling J. (ed.). Racial variation in man. Institute of Biology, Blackwells, Oxford.
364 Часть III. Биологическая изменчивость популяций
Haldane J. В. S. (1956). The argument from animals to men: an examination of its value in anthropology. Huxley Mem. Lecture. J. R. anthrop. Inst. 86, 1.
Harrison G. A. (ed.) (1961). Genetical variation in human populations. Perga-mon, Oxford.
Harrison G. A., Peel J. (1969). Biosocial aspects of race. Blackwells, Oxford.
Hulse F. (1971). The human species. Random House, London.
Garn S. (ed.) (1968). Readings on race. Thomas, Springfield, Illinois.
Roberts D. F. and Harrison G. A. (eds.) (1959). Natural selection in human populations. Pergamon, Oxford.
Roberts D. F., Sunderland E. (eds.). Genetic variation in Britain. Taylor and Francis, London.
Young J. Z. (1971). An Introduction to the study of man. Clarendon Press, Oxford.
Антропометрия
Barnicot N. A. and Brothwell D. R. (1959). The evaluation of metrical data in the comparison of ancient and modrn bones, CIBA Symposium on medical biology and Etruscan origins, p. 131. Churchill, London.
Shapiro H. L. (1939). Migration and environment. Oxford University Press, London.
Trevor J. C. (1953). Race crossing in man: the analysis of metrical characters. Eugenics Lab. Mem. XXXVI. Galton Laboratory, London. Cambridge University Press.
Weiner J. S. and Lourie J. A. (1969). Human biology—a guide to field methods. IBP Handbook no. 9, Blackwell, Oxford.
Статистика
Cavalli-Sforza L. L., Barrai I. and Edwards A. W. F. (1964). Analysis of human evolution under random genetic drift. Cold Spring Harb. Symp. quant. Biol., 29, 9-20.
Hill A. B. (1971). Principles of medical statistics. Oxford University Press, London.
Moroney M. J. (1969). Facts from figures. Penguin, Harmondsworth.
Sneath P. H. and Sokal R. R. (1973). Numerical taxonomy, Chapter 5. Freeman, New York.
Snedecor G. W. and Cochran W. G. (1967). Statistical methods (6th edn). Iowa State University Press.
Van der Geer J. P. (1971). Introduction to multivariate analysis for the social sciences. Freeman, New York.
Weiner J. S. and Huizinga J. H. (eds.) (1972). The assessment of population affinities in man. Oxford University Press, London.
Биохимическая изменчивость
Allison A. C. (1965). Population genetics of abnormal haemoglobins and G6PD. In Abnormal haemoglobins in Africa (ed. J. H. Jonxis). Blackwells, Oxford.
Bodmer W. F. (1972). Evolutionary significance of the HL—A system, Nature, Lond. 237, 139.
Giblett E. R. (1969). Genetic markers in human blood. Blackwells, Oxford.
Grubb R. (1970). The genetic markers of human immunoglobulins. Molecular biology, Biochemistry and Biophysics, Vol. 8. Chapman and Hall, London.
Harris H. (1969). Human biochemical genetics. Cambridge University Press.
Lehman H. and Huntsman R. G. (1974). Man’s haemoglobins. North—Holland, Amsterdam.
Lehman H. and Carrell R. W. (1969). Variation in the structure of human haemoglobin. Br. med. Bull. 25, 14—23.
Livingstone F. B. (1967). Abnormal haemoglobins in human populations. Aldine, Chicago.
Глава 18. Другие виды изменчивости
365
Группы крови
Fraser Roberts J. А. (1959). Some associations between blood groups and disease, Br. med. Bull. 15, 129.
Kopec A. C. (1970). The distribution of the blood groups in the United Kingdom. Oxford University Press, London.
Mourant A. E. (1959). Blood groups, genetical variation in human populations (ed. G. A. Harrison), p. 1. Pergamon, Oxford.
Mourant A. E. (1976). The distribution of the human blood groups. Oxford University Press.
Mourant A. E., Kopec A. C. and Sobczak K. (1958). The ABO blood groups. Comprehensive tables and maps of world distribution. Blackwell Scientific Publications, Oxford.
Race R. R. and Sanger R. (1968). Blood groups in man. Blackwells, Oxford.
Robinson M. G., Tolchin T., and Halpern C. (1971). Enteric bacterial agents-and the ABO blood groups. Am. J. Human Genet. 23, 135—145.
Sheppard P. M. (1959). Blood groups and natural selection. Br. med. Bull. 15, 134.
Vogel F. (1970). ABO blood groups and disease. Am. J. Human Genet. 22, 464. Weiner A. S. (1970). Blood groups and disease. Am. J. Human Genet. 22, 4.
Особые виды чувствительности
Kalmus H. (1965). Diagnosis and Genetics of Defective Colour vision, Pergamon, Oxford.
Kretchmer N. (1972). Lactose and lactase. Sci. Am., Oct. 1972.
Wright W. D. (1953). Defective colour vision. Br. med. Bull. 9, 36.
Дерматоглифические узоры
Cummins II. and Midlo C. (1962). Fingerprints, palms and seles. Dover, New York.
Holt S. B. (1968). The Genetics of dermal ridges. Thomas, Springfield, Illinois.
Penrose L. S. (1969). Dermatoglyphics. Sci. Am., December.
Пигментация
Barnicot N. A. (1957). Human pigmentation. Man, no. 144.
Harrison G. A. (1961). Pigmentation. In Genetical variation in human population (ed. G. A. Harrison), p. 99, Pergamon, Oxford.
Врожденные аномалии
Adam A. (1973). Genetic diseases among Jews. Esrael. J. mod. Sci. 9, 1383.
Carter С. O. (1969). Genetics of common disorders. Br. med. Bull. 25, 52.
Умственное развитие
Bodmer W. F. and Cavalli-Sforza L. L. (1970). Intelligence and Race. Sci. Am., October 1970.
Butcher H. J. (1968). Human intelligence. Methuen, London.
Jensen A. R., Kagan J. S., Hunt J. MeV., Crou> J. F. et al. (1969). Environment, heredity and intelligence Harvard Educational Review Reprint Series No. 2.
Penrose L. S. (1972). The genetics of mental defect (4th edn). Sidgwick and Jack-son, London.
Vernon P. E. (1969). Intelligence and cultural environment. Methuen, London.
ЧАСТЬ IV
РОСТ И КОНСТИТУЦИЯ ЧЕЛОВЕКА ДЖ. ТЭННЕР
Глава 19
КРИВАЯ РОСТА ЧЕЛОВЕКА
Изучение роста чрезвычайно важно для выяснения механизмов эволюции, так как эволюция морфологических признаков осуществляется всегда путем изменения генетически обусловленного роста и развития. Кроме того, изучение процесса роста существенно для понимания индивидуальных различий формы и функции у человека, поскольку многие из этих различий обусловлены различиями в относительной скорости роста отдельных частей тела.
На рис. 19.1 показана кривая роста длины тела мальчика; измерения проводили каждые 6 мес с момента рождения до 18 лет. На верхнем графике нанесены абсолютные размеры длины тела в соответствующие годы, на нижнем — прирост длины тела от одного возраста к другому. Если представить себе процесс роста как некую форму движения, то верхний график даст нам представление о пройденном пути, а нижний — о скорости. Понятно, что скорость роста лучше отражает состояние ребенка в каждый данный момент, чем абсолютные значения длины тела, так как они во многом зависят от роста ребенка в предыдущие годы. Содержание в крови и тканях различных биохимических веществ изменяется с возрастом параллельно изменению скорости роста. В некоторых случаях природу физиологических процессов лучше отражает ускорение, а не скорость. Можно, например, думать, что резкое увеличение секреторной активности эндокринных желез у подростков более отчетливо проявляется в ускорении роста (см. рис. 20.3).
Данные, приведенные на рис. 19.1, относятся к одной из самых ранних публикаций по вопросу о росте. Эти измерения были проведены в 1759—1777 гг. графом Филибертом де Монбейяром, наблюдавшим за развитием своего сына, и опубликов’аны Бюффоном, другом де Монбейяра, в приложении к «Естественной истории». График показывает, что скорость роста длины тела с возрастом неуклонно уменьшается, но в самом конце ростового периода эта тенденция неожиданно прерывается. В это время (в данном случае
Рис. 19.1. Рост длины тела сына де Монбейяра с момента рождения (1755 до 18 лет (Tanner, 1962).
А. Абсолютный рост. Б. Скорость роста (годичный прирост длины тела).
в возрасте 13—15 лет) наблюдается отчетливо выраженное уск< рение роста, получившее название пубертатного скачка рост? который мы рассмотрим подробнее в следующей главе. По некой
Часть ГУ. Рост, и конституция Человека рым данном, небольшое увеличение скорости роста (появление дополнительного зубца на общей кривой) наблюдается между 6 и 8 годами. Его называют иногда «полуростовым» скачком. Хотя рис. 19.1 как будто бы подтверждает эту точку зрения, при многих других индивидуальных исследованиях детей от 3 до 13 лет в большинстве случаев этот скачок не выявляется. По-видимому, если он и имеет место, то у меньшинства детей.
Пренатальный рост
Аналогичный тип кривой скорости роста характерен и для внутриутробного развития. На рис. 19.2 приведены кривые, описывающие изменения абсолютных значений и скорости роста длины тела для внутриутробного периода и первых двух лет жизни. Максимальная скорость роста плода характерна для первых четырех месяцев внутриутробного развития. (Возраст в период внутриутробного развития обычно отсчитывается от первого дня последней менструации; это в среднем на две недели раньше действительного оплодотворения, но зато это единственный надежный ориентир.)
Таким же образом изменяется и вес плода, с той разницей, что максимальная скорость достигается несколько позже — обычно на 34-й нед. внутриутробного развития. Как свидетельствует целый ряд данных, в период между 36 и 40 нед. (момент рождения) скорость роста плода несколько замедляется, так как к этому времени полость матки оказывается целиком заполненной. Скорость роста близнецов замедляется раньше — в тот период, когда их общий вес становится равным весу одиночного 36-недельного плода. Таким образом, вес и размеры тела новорожденного зависят не столько от генотипа, сколько от внешней среды, каковой в данном случае является материнский организм. Если генетически крупный ребенок развивается в матке у небольшой женщины, замедляющие механизмы способствуют успешным родам. Сразу же после рождения скорость роста вновь увеличивается, особенно у генетически крупного ребенка. Прибавка в весе достигает максимума через два месяца после рождения.
На приведенных кривых (рис. 19.2) это замедление роста отражается более низкими показателями, приходящимися на 9-й и 10-й месяцы, и низкими скоростями роста в интервалах от 8-го до 9-го и от 9-го до 10-го месяца. Увеличение скорости роста сразу после рождения сглаживает кривую. Однако мы не можем быть абсолютно уверены в этом, так как данные по пренатальному и постнатальному росту получены на разных детях, и ошибки выборочного метода могут создать видимость снижения и последующего увеличения скорости роста.
Глава 19. Кривая роста человека
369
Рис. 19.2. Рост длины тела в пренатальном и раннем постнатальном периодах (Thompson, 1942).
А. Абсолютный рост. Б. Скорость роста.
В первые два месяца внутриутробного развития скорость роста длины тела не очень велика. Это стадия эмбриона с характерными для нее процессами «регионализации»— дифференцировки гомогенного целого на отдельные части, такие, как голова, конечности и т. д., и гистогенеза — дифференцировки клеток с образованием специализированных тканей, таких, как мышечная или нервная. В то же время благодаря дифференциальному росту клеток и кле
370
Часть IV. Рост и конституция человека
точным миграциям каждая часть приобретает совершенно определенные очертания. Этот процесс, известный под названием морфогенеза, продолжается до взрослого состояния, а в некоторых частях тела — и до старости. Но в основном он завершается на 8-й неделе внутриутробного развития, и к этому времени эмбрион приобретает характерные признаки человека.
Большая скорость роста плода объясняется главным образом тем, что клетки все еще продолжают делиться. По мере роста плода число митозов во всех тканях уменьшается, и принято считать, что после 6-го месяца внутриутробного развития не происходит (или почти не происходит) образования новых мышечных и нервных клеток (если не считать клеток нейроглии). В это же время скорость роста линейных размеров резко падает. Мышечные и нервные клетки плода существенно отличаются от аналогичных клеток ребенка или взрослого. Клетки того и другого вида содержат мало цитоплазмы вокруг ядра. Мышечная ткань плода отличается высоким содержанием межклеточного вещества и воды. Дальнейшее развитие мышечных клеток заключается в построении цитоплазмы, включении солей и образовании актомиозина. Клетки становятся больше, межклеточное вещество почти исчезает, уменьшается содержание воды. Этот процесс активно протекает до трех лет, а затем замедляется. Скорость его вновь возрастает в период полового созревания, особенно у мальчиков, когда под влиянием андрогенных гормонов происходит включение новых веществ в состав фибрилл. В это же время происходит увеличение абсолютного количества ДНК, что свидетельствует о дальнейшем появлении ядер (поскольку ДНК содержится в ядрах клеток почти в постоянной концентрации). Таким образом, в фибриллах в течение всего периода роста происходит увеличение числа ядер. В нервных клетках происходит синтез цитоплазмы, появляются нуклеопротеидные включения и усиленно растут аксоны и дендриты.
Таким образом, постнатальный рост, по крайней мере некоторых тканей, сводится к развитию и увеличению уже существующих клеток, а не к образованию новых. Исключение составляет жировая ткань. Число жировых клеток продолжает увеличиваться вплоть до начала пубертатного периода, но темпы прироста постепенно уменьшаются. Однако и для этой ткани количественно более существенным аспектом постнатального роста является увеличение размеров каждой клетки.
Подбор кривых роста (выравнивание результатов измерений)
Рост — это в общем чрезвычайно регулярный процесс. Вопреки все еще распространенному мнению он никогда не осуществляется скачками. Чем точнее будут наши измерения (например, если нам
Глава 19. Кривая роста человека
371
удастся свести к минимуму влияние суточных колебаний роста), тем более регулярным будет расположение точек. На рис. 19.3 нанесены точки, изображающие результаты измерений длины тела у одного ребенка, проведенных одним и тем же наблюдателем через каждые 6 мес от 3 1/2 до 10 лет. Ясно видно, что легко подобрать гладкую кривую, на которую хорошо укладываются эти точки.
Рис. 19.3. Соответствие кривой вида y=a+bi-|-clni индивидуальным данным (Tanner, 1960).
Длину тела девочки измеряли каждые 6 мес с 3 1/2 до 10 лет. Харпенденовское исследование роста.
При обследовании группы детей отклонения от таких подобранных кривых редко превышают 6 мм, причем точки одинаково часто располагаются выше и ниже кривой. Однако у многих детей отмечены регулярные сезонные вариации роста, при которых скорость роста увеличивается весной и уменьшается осенью. Эти шестимесячные колебания усложняют ход общей ростовой кривой. Эффект подобных сезонных колебаний настолько велик, что скорости роста, взятые за временной промежуток меньше года, не могут считаться пригодными для медицинских целей. Средний ребенок в Англии растет втрое быстрее в тот период, когда скорость роста у него максимальная, по сравнению с периодом минимальной скорости роста.
Было сделано много попыток математически описать процесс роста человека и животных. Большинство из них не принесло ничего, кроме разочарования, потому что новые данные не соот
372
Часть IV.. Рост и конституция Человека
ветствовали уже выведенным кривым, а часть этих попыток относилась к области фантазии, так как модели оказывались настолько сложными и содержали так много параметров, что их биологическая интерпретация становилась совершенно невозможной. Задача состоит в том, чтобы найти формулу или формулы, содержащие относительно небольшое число констант, каждая из которых имела бы определенный биологический смысл. При этом полученные математические выражения должны, разумеется, обеспечивать совпадение с эмпирическими данными (в пределах ошибок измерений). Затруднения, возникающие при решении этой задачи, заключаются в том, что речь идет о подборе кривых для описания таких измерительных признаков, которые в большинстве своем биологически сложны. Так, например, длина тела складывается из длины ног, длины туловища и высоты головы, причем характер изменения этих признаков с возрастом неодинаков. Даже когда речь идет о таких сравнительно однородных признаках, как длина большой берцовой кости или ширина икроножной мышцы, пока еще трудно выдвинуть четкие биологические предпосылки, которые объясняли бы форму подобранной кривой. Если исходить из предположения, что в основе роста лежит непрерывное деление клеток, мы придем к заключению, что величина прироста должна быть пропорциональна измеренному признаку (dyldt = ky, или у~ =e?+bt), где у представляет собой измерение в возрасте t, а к, а и Ъ — константы. Если же допустить, что рост происходит в результате непрерывного добавления или включения во все клетки определенного количества какого-то неделящегося вещества, то это приведет к заключению о постоянстве прироста. Наиболее сложная, но вместе с тем и наиболее вероятная гипотеза состоит в том, что с возрастом постепенно уменьшается доля делящихся клеток или клеток, в которые происходит включение нового вещества, а также изменяется скорость этих процессов. Математическая интерпретация такого предположения далеко не однозначна.
Тем не менее подбор кривой, наилучшим образом описывающей результаты измерений, проведенных на одном индивидууме,— это единственный способ получения максимальной информации о характере индивидуального роста. Это становится все очевиднее по мере разработки таких проблем, как влияние патологических процессов на скорость роста или генетические основы различных типов роста. По всей вероятности, для характеристики всего периода постнатального роста недостаточно одной кривой. В настоящее время наиболее пригодными по крайней мере для большинства признаков, в том числе для роста и веса, оказываются две кривые. Кривая
у = а±Ы Ц-clog t, по-видимому, хорошо подходит для возрастного интервала от
Глава 19. Кривая роста человека
378
нескольких месяцев с момента рождения до начала подросткового периода, т. е. приблизительно до 10 лет. Такая кривая приведена на рис. 19.3. Пубертатному скачку роста соответствует кривая — симметричная S-образная экспоненциальная кривая, описываемая уравнением
y = P + (K/l+ea~bt),
где Р — наименьшая асимптота и К — общий прирост за весь подростковый период. Другая подобная кривая — это кривая Гом-пертца, искаженная экспонента. На рис. 19.4 изображена кривая
Рис. 19.4 Соответствие кривой Гомпертца (сплошная линия) индивидуальным данным (Tanner, 1963).
Длину тела девочки измеряли каждые 3 мес на протяжении пубертатного периода.
Гомпертца, описывающая изменения длины тела девочки во время пубертатного периода.
Результаты продольных и поперечных исследований
Вышеупомянутые кривые приложимы к данным о росте отдельных индивидуумов. Усредненные годичные величины, полученные в результате массовых однократных обследований детей, в целом не дают такой кривой. Эти два вида исследований называ; ют соответственно продольным и поперечным. При поперечном исследовании каждого ребенка измеряют только один раз, так что, например, группа восьмилетних детей отличается по составу от группы семилетних. При продольном исследовании, напротив, одного и того же ребенка измеряют каждый год, и между группа
374 Часть IV. Рост и конституция человека
ми восьмилетних и семилетних детей нет разницы по составу. Продольное исследование можно проводить в течение нескольких лет. Существуют краткосрочные продольные исследования, продолжающиеся не более двух лет, а также продолжительные исследования, охватывающие весь период от рождения до зрелости, во время которых одни и те же дети измеряются один или несколько раз в течение года от рождения до 20 лет. Практически не удается измерять в течение длительного времени одну и ту же группу детей.
Рис. 19.5. Соотношение между индивидуальной и средними скоростями роста (Tanner, 1962).
А. Кривые скорости роста нанесены по хронологическому возрасту Б Кривые построены в соответствии с временем наступления максимальной скорости роста.
Состав группы неизбежно меняется. Это так называемое смешанное продольное исследование. Чтобы извлечь из этих данных максимум информации, необходимо применить ряд специальных статистических приемов. В прошлом этого вообще не понимали и нередко отбрасывали больше трех четвертей полезных данных смешанного исследования. Один из частных видов смешанного исследования заключается в том, что параллельно проводится несколько непродолжительных продольных исследований, которые перекрывают друг друга. Так, для того чтобы охватить весь возрастной интервал, можно взять следующие возрастные группы: 0—6, 5—11, 10—16 и 15—20 лет. Однако в том случае, если выборки были недостаточно хорошими, могут возникнуть сложности при соединении отдельных частей исследования.
Каждый из этих методов находит свое применение, но позволяет получить различные данные о характере роста, и обращаться с этими данными следует по-разному. Поперечные исследования
Глава 19. Кривая роста человека
375
быстрее, проще, дешевле и охватывают больше детей. Они позволя-ют нам получить представление об абсолютном росте, установить нормальные стандарты длины тела и веса для данной популяции. Но у них есть один недостаток: они не вскрывают индивидуальных различий в скорости роста и во времени наступления отдельных фаз, например, пубертатного периода. А между тем именно эти различия проливают свет не только на тонкие механизмы генетической регуляции роста, но и на соотношения между ростом, воспитанием, развитием интеллекта и поведением. Продольные исследования требуют большой затраты времени и труда со стороны тех, кто их выполняет, и тех, кто служит объектом. Не подкрепленные результатами поперечных исследований и экспериментами на животных, они теряют с годами свою ценность и превращаются в бесплодное коллекционирование цифр. Но продольные исследования необходимы.
Результаты поперечных исследований могут иногда ввести в заблуждение. На рис. 19.5 показано усреднение индивидуальных кривых, различающихся по времени наступления пубертатного скачка роста. На рис. 19.5, А приведен ряд индивидуальных кривых скорости роста, охватывающих возрастной промежуток от 6 до 18 лет, причем начало скачков роста у разных индивидуумов приурочено к разным возрастам. Усредненная кривая, показанная пунктиром, была получена путем сложения всех индивидуальных значений для каждого возраста (5, 6, 7 лет и т. д.) с последующим делением этих сумм на пять — обычный подход к материалу при поперечном исследовании. Совершенно очевидно, что суммарная кривая никоим образом не отражает «средней» скорости роста, напротив, она искажает ее. Она сглаживает скачок роста и «растягивает» его во времени. То же самое неизбежно происходит и со средними, вычисленными для каждого возраста при поперечном исследовании. Они не отражают скорости и интенсивности индивидуального скачка роста. На рис. 19.5, Б те же самые кривые нанесены таким образом, что их точки максимальной скорости совпадают. В данном случае усредненная кривая вполне удовлетворительно характеризует группу. В этом случае шкала времени изменена: по оси абсцисс нанесен уже не хронологический возраст, а приведены величины, показывающие, насколько быстро протекает процесс развития. Другими словами, учитывается истинный статус развития ребенка или его физиологический статус. Мы еще остановимся на этом в гл. 21.
Это как раз та проблема, которая успешно разрешается путем подбора кривых, наилучшим образом описывающих индивидуальные данные. Допустим, что у двух детей пубертатные скачки роста идентичны по форме и интенсивности, но различаются по возрасту наступления. Если кривая подобрана достаточно точно, то один или несколько из ее параметров будут отражать крутизну скачка,
376
Часть IV. Рост и конституция человека
другой — высоту пика и т. д. Все эти параметры будут у обоих детей одинаковы, так как они не зависят от возраста. Следующий параметр кривой роста должен выразить время начала скачка. И здесь выявляется различие между детьми по величинам, характеризующим опережение или запаздывание по отношению к их хронологическому возрасту.
Сравнивая различные группы детей по подобранным кривым, следует помнить об одном весьма важном моменте. Чтобы получить, скажем, среднее значение параметра а кривой роста для группы детей, нужно определить значение параметра а для каждого индивидуума и потом усреднить их (если, конечно, кривая характеризуется достаточно сложным уравнением). Если же выводить уравнение для средних результатов измерений, проведенных в каждом возрасте, то получится совершенно другое и в большинстве случаев ошибочное среднее значение параметра. Таким образом, правильную групповую кривую можно построить только по средним значениям параметров, рассчитанным на основании подобранных индивидуальных кривых.
Кривые роста для различных тканей и различных частей тела
Кривые роста большинства скелетных и мышечных размеров примерно повторяют ход кривой роста длины тела. То же самое можно сказать и о размерах отдельных органов — печени, селезенки, почек. Однако существует целый ряд других тканей и органов, кривые роста которых существенно отличаются по типу. К ним относятся мозг, череп, органы размножения, лимфатическая ткань и подкожная жировая клетчатка.
Эти различия изображены графически на рис. 19.6, где нанесены размеры, достигаемые различными тканями к каждому возрасту в процентах от общего прироста за период с момента рождения до наступления зрелости. Изменения длины тела и большей части других его размеров характеризуются одной «общей» кривой. Рост органов размножения, как внутренних, так и наружных, идет по кривой, в принципе сходной с общей кривой, но явно отличной от нее. В предпубертатном периоде рост этих органов замедлен, а в периоде полового созревания сильно ускорен. По сравнению с костной системой они более чувствительны к одним гормонам и менее чувствительны к другим.
, Мозг (а вместе с ним череп, глаз и ухо) развивается раньше любой другой части тела (подробнее см. в гл. 22); он имеет характерную кривую постнатального роста. У новорожденного мозг уже достигает 25% своего дефинитивного веса, у 5-летнего ребенка — 90%, у 10-летнего — 95%. Таким образом, если мозг и имеет какой-либо пубертатный скачок, то лишь очень незначительный.
Глава 19. Кривая роста человека
377
Возраст, годы
Рис. 19.6. Кривые роста различных тканей и органов тела, иллюстрирующие четыре основных типа роста (Tanner, 1962).
По оси ординат достигнутая величина признака выражена в процентах к общему приросту от рождения до зрелости (20 лет), так что величина признака в 20-летнем возрасте соответствует 100%. I — лимфоидный тип: тимус, лимфатические узлы, лимфатические массы кишечника; II — мозговой и головной тип: мозг и его части, твердая мозговая оболочка, спинной мозг, глаз, размеры головы; III — общий тип: тело в целом, внешние размеры (за исключением головы), органы дыхания и пищеварения, почки, аорта н легочная артерия, мышечная система, объем крови; IV — репродуктивный тип: яички, придаток, пред-, стательная железа, семенные пузырьки, яичники, фаллопиевы трубы.
Небольшой, но выраженный скачок наблюдается для продольного и поперечного диаметров головы, но скорее всего его следует отнести за счет утолщения костей черепа и мягких тканей головы, а также за счет развития воздухоносных пазух. Кривая роста лица занимает промежуточное положение между кривыми роста верхней части черепа и остального скелета. К моменту рождения размеры лица ближе к дефинитивным, чем длина тела. Однако пубертатный скачок выражен довольно резко, причем значительнее всего для нижней челюсти. Таким образом, голова в целом опережает в своем развитии остальные части тела, а ее верхняя часть — глаза и мозг — опережает нижнюю.
Лимфатическая ткань миндалин, червеобразного отростка, кишечника и селезенки характеризуется совершенно иной кривой роста (рис. 19.6). Эта ткань достигает максимального развития еще до наступления подросткового периода. Затем, вероятно, под
200
Возраст, годы
Глава 19. Кривая роста человека
379
непосредственным влиянием половых гормонов, она пЬдвергается некоторому обратному развитию до уровня, характерного для взрослого.
Подкожный жировой слой также имеет своеобразную и довольно сложную кривую роста. Толщину этого слоя можно определить рентгенологически или при помощи специального инструмента, предназначенного для измерения жировой складки, отделенной от подлежащих мышц. На рис. 19.7 (А и Б) представлены оба типа кривых увеличения кожных складок, измеренных на задней поверхности плеча (над трицепсом) и на спине (под нижним углом лопатки). Отложение подкожного жира начинается у плода на 34-неделе внутриутробного развития и продолжается до момента ррждения и затем в течение 9 мес первого года жизни (это относится к «среднему» ребенку; у некоторых детей максимальная толщина подкожного жирового слоя наблюдается в 6 мес, у других — в год и даже в 15 мес); начиная с 9 мес, когда скорость роста подкожного слоя равна нулю, количество подкожного жира постепенно уменьшается, т. е. скорость роста становится отрицательной. Так продолжается до 6—8-летнего возраста, когда она вновь начинает увеличиваться.
Следует отметить, что речь шла лишь о толщине жирового слоя. Уменьшение толщины отнюдь не предполагает уменьшения площади поперечного сечения, занятой жиром. Жир — это кольцо вокруг мышечно-костного стержня, размеры которого с возрастом увеличиваются. Если площадь поперечного сечения жира остается постоянной, то ширина жирового кольца будет уменьшаться просто за счет увеличения мышечно-костного стержня. Тем не менее, судя по измерениям, произведенным на рентгенограммах, в раннем детском возрасте имеет место истинное уменьшение площади поперечного сечения жировой ткани. Это уменьшение в меньшей степени выражено у девочек, чем у мальчиков, так что в возрасте старше одного года количество жира у девочек больше.
С семилетнего возраста начинается увеличенное отложение жира как у девочек, так и у мальчиков. Однако у мальчиков-подростков количество жира на конечностях уменьшается (рис. 19.7, А и Б). Так продолжается почти до двадцати с лишним лет. Отложение жира на туловище у мальчиков (на рис. «подлопаточная»
Рис 19.7.
А. Кривые абсолютного увеличения слоя подкожной жировой ткани (Tanner, 1962). Толщину складки измеряли калипером Харпендена над трицепсом (а) и под лопаткой (б). За период с момента рождения до 1 года данные получены только продольным методом на 65 девочках (1) и 74 мальчиках (II); с 2 до 7 лет — данные Лондонского центра по изучению детей, причем с 4 до 6 лет данные были получены только продольным методом на 57 девочках (I) и 59 мальчиках (II), а затем средние годичные прибавки вычитались или прибавлялись для получения средних арифметических в 2, 3 и 7 лет; с 5 до 16 лет — данные Лондонского городского совета, полученные методом поперечного исследования; численность каждой возрастной группы: от 5 До 14 лет — 1000—1600, 15 лет — 500 и 16 лет — 250. Б. Кривые скорости роста слоя подкожной жировой ткани, измеренного над трицепсом (а) и под лопаткой (6). Источники данных те же, что и для рис. 19.7, А.
380
Часть IV. Рост и конституция человека
складка) происходит без заметного уменьшения — можно говорить лишь о временной остановке постоянного прироста. У девочек отложение жира на конечностях слегка приостанавливается, но не уменьшается, а количество жира на туловище все время возрастает вплоть до 14-летнего возраста.
Поскольку вес тела складывается из веса различных тканей, его кривая роста часто менее показательна, чем кривые, характеризующие отдельные компоненты. Тем не менее индивидуальные кривые скоростей роста веса и длины тела, как правило, совпадают. Хотя по весу можно в какой-то мере судить о здоровье ребенка, следует помнить, что увеличение веса может идти как за счет роста костей и мышц, так и за счет простого накопления жира. Если ребенок старшего возраста не прибавляет в весе, это означает лишь, что нужно обратить больше внимания на его питание и тренировку, если же отсутствует рост в длину или увеличение мышечной массы, то необходимо всесторонне исследовать ребенка.
Упорядоченность и неупорядоченность процесса роста
Мы видели, что увеличение того или иного отдельного размера — зто в высшей степени регулярный процесс. Кроме того, мы убедились, что различные ткани и области человеческого тела растут с различной скоростью. Именно благодаря этому различию в скоростях роста в процессе эмбриогенеза плод приобретает типичные для человека очертания, а в процессе дальнейшего роста ребенок превращается во взрослого. Совершенно очевидно, что это высокоорганизованные процессы, так как начало одного из них зависит от наступления определенной стадии другого или, более вероятно, нескольких других. Выяснить, как осуществляется эта организация или, вернее, в каких терминах мы можем лучше всего ее описать,— одна из основных задач в процессе изучения роста.
Большая часть морфологических различий между фетальными, ювенильными и дефинитивными формами отдельных видов и родов возникает за счет различий в скоростях роста отдельных частей тела. Для человека (в отличие от человекообразных обезьян) характерна большая длина нижних конечностей относительно туловища или верхних конечностей. Это объясняется тем, что начиная с ранних стадий внутриутробного развития относительная скорость роста нижних конечностей выше. У человека по сравнению с другими приматами место сочленения черепа с позвоночным столбом несколько сдвинуто вперед. Однако к моменту рождения это различие не выражено. У всех антропоидов относительное положение сочленения остается примерно одинаковым, но у низших и человекообразных обезьян постнатальный рост головы ха-
Глава 19. Кривая роста человека
381
растеризуется значительно большей скоростью в той части, которая лежит кпереди от затылочных мыщелков, и, таким образом, сочленение несколько отодвигается назад. У человека скорости роста предмыщелковой и позадимыщелковой частей головы почти одинаковы и положение сочленения существенно не меняется. Различия в скоростях роста отдельных частей играют важную роль в механизмах морфогенеза.
«Канализирование»
Несмотря на важность проблемы, мы все еще слишком мало знаем о закономерностях роста. Несомненно, существуют регуляторные механизмы, которые удерживают процессы развития в определенных рамках. Ибо на протяжении сложного и длительного процесса, который отделяет первичное химическое действие генов от окончательной формы взрослого организма, есть все условия для самых разнообразных, пусть незначительных, отклонений и нарушений, которые могли бы в дальнейшем прогрессивно умножаться. Так, например, трудно себе представить, чтобы при делении яйца на две части, из которых образуются идентичные близнецы, в каждую попала точно половина всего количества цитоплазмы. Столь же мало вероятно, чтобы в двух таких одновременно развивающихся организмах концентрации продуктов гена были абсолютно идентичными; если же возникают те или иные различия, то по мере дальнейшего развития они должны были бы усугубиться. А между тем на самом деле однояйцовые близнецы настолько сходны между собой, что только точными измерениями удается выявить разницу между ними. Именно это сходство, а не различие нуждается в объяснении.
Принято считать, что процессы дифференцировки и роста относятся к категории самостабилизирующихся, или направленных. Теперь, когда мы начинаем все глубже проникать в динамику сложных систем, состоящих из многих взаимодействующих элементов, мы понимаем, что способность к достижению сходных конечных форм посредством различных путей отнюдь не является исключительным свойством живого организма. Многие сложные системы, в особенности так называемые «открытые», т. е. находящиеся в постоянном взаимодействии с другими окружающими их системами, обладают способностью к подобной внутренней регуляции, которая является логическим следствием их организации, даже если они состоят из самых простых, неживых компонентов.
Способность стабилизировать ход развития и возвращаться к предонределенной кривой роста даже в том случае, когда нарушена, если можно так выразиться, траектория развития, характерна для всего периода роста и отчетливо проявляется в реакции молодых животных на болезни и голод. Уоддингтон (Waddington)
382
Часть IV. Рост и конституция человека
Рис. 19.8. Кривые длины тела (Z) и скорости роста длины тела (77) у девочки с двумя периодами отсутствия аппетита (обозначены черным), вслед за которыми наступает «наверстывающий» рост (Prader et al., J. Pediat., 1963).
назвал это свойство канализированием, или гомеорезом. (В отличие от гомеостаза, т. е. поддержания постоянства условий в стабильных системах, гомеорезом называют поддержание постоянства в развивающихся системах.)
Необычно высокая скорость роста, наблюдаемая в некоторые периоды, получила название «наверстывающего» роста (Prader,, Tanner, von Hamack, 1963). На рис. 19.8, II приведен пример подобного «наверстывающего» роста. Аналогичный «наверстывающий» рост наблюдается, например, после успешного лечения состояния гипотиреоза или после удаления кортизолпродуцирующей опухоли, вызывавшей задержку в росте. Скорость роста в начале восстановительного периода может втрое превышать соответствующую данному возрасту. Для описания подобного явления специалисты по питанию иногда пользуются термином компен
Глава 19. Кривая роста человека
383
саторный рост. Однако этот термин впервые был применен для обозначения совершенно иного явления — заместительного роста органов или частей. Так, после удаления одной почки другая гипертрофируется, или, как принято говорить, проявляет компенсаторный рост. «Наверстывание» может быть полным или неполным. Если стресс был тяжелым и его воздействие пришлось на ранние периоды жизни животного, тогда «наверстывающий» рост, хотя и продолжается в течение некоторого периода времени, может оказаться недостаточным для полного восстановления нормальной кривой роста. Механизм подобной регуляции далеко не выяснен.
Градиенты роста
Одно из проявлений организации роста — это наличие градиентов созревания. На рис. 19.9 показан один из таких градиентов. Разберем более простой случай (рис. 19.9, Л): длину стопы, длину голени и длину бедра у мальчиков для каждого возраста выражают в процентах от дефинитивного значения. В любом возрасте стопа оказывается ближе к дефинитивному состоянию, чем голень, а голень ближе, чем бедро. Поэтому говорят, что в нижней конечности между быстрее созревающей дистальной частью и отстающей в росте проксимальной существует градиент созревания. На рис. 19.9, Б показан аналогичный градиент для верхней конечности; вместе с тем иллюстрируется тот факт, что независимо от дистальнопроксимального градиента девочки во всех возрастах ближе к зрелости, нежели мальчики (см. гл. 20).
Имеется множество других градиентов. Некоторые из них существуют лишь кратковременно и распространяются на небольшие участки, тогда как другие охватывают целые системы и наблюдаются во время всего периода роста. Так, голова во всех возрастах опережает в своем развитии туловище, а туловище опережает конечности. Однако в пределах самого туловища этот «кефало-каудальный» градиент не обнаруживается. Его заменяют более сложные градиенты, распространяющиеся на ограниченные области. Градиенты созревания мозга, имеющие огромное значение для теории и практики воспитания, описываются в гл. 22.
Множество химических реакций, протекающих в процессе дифференцировки и роста, должны самым точным образом сочетаться между собой. Так, для того чтобы обеспечить нормальную остроту зрения, рост хрусталика должен быть гармонично согласован с ростом глазного яблока в глубину. Нужно ли удивляться, что эта координация не всегда осуществляется и что у большинства людей отмечается некоторая степень близорукости или дальнозоркости. По-видимому, многие признаки лица и головы контролируются самостоятельными генами, которые не оказывают су-
384
Часть IV. Рост и конституция человека
2 4 6 8 10 12 14 16 18
Возраст, еоЪы
Рис. 19.9. Градиенты созревания верхних (Л) и нижних (5) конечностей (Tanner, 1962).
По оси ординат отложена длина сегментов конечностей, выраженная в процентах к дефинитивной длине. Кисть во всех возрастах ближе к окончательному размеру, чем предплечье, а предплечье ближе, чем плечо, независимо от половых различий в степени зрелости. Смешанные продольные данные. 1 — кисть; 2 — предплечье; з — плечо; 4 — стопа;
S — голень; 6 — бедро.
щественного влияния на другие близкие признаки. Однако в общем отдельные части лица составляют единое целое. Это происходит потому, что конечные стадии роста пластичны, и при взаимной подгонке частей, например верхней и нижней челюстей, вступает в действие взаимная регуляция, под влиянием которой происходят отклонения от первоначального генетически обусловленного характера роста отдельных частей.
Эти регуляторные силы не всегда действуют успешно. Если равновесие генетических сил нарушается с самого начала, то нормального развития не произойдет. Так, если при репликации одной из хромосом в оплодотворенном яйце окажется аномальное число генов или если нарушится их правильное распределение, то возникнут разнообразные аномалии развития, которые обычно приводят к выкидышу, но иногда — как в случае трисомии по небольшой 21-й хромосоме, обусловливающей синдром Дауна,— к нарушениям физического и умственного развития.
Однако совершенно ясно, что не только такие аномалии, но и многие индивидуальные морфологические, а возможно, и функциональные различия возникают как следствие дифференциальной
Глава 19. Кривая роста человека 385
изменчивости скоростей роста различных структур. Интересно, что совсем недавно ряд психологов выдвинули предположение, что некоторые индивидуальные особенности в структуре личности могут возникать аналогичным образом. Более того, полагают, что некоторые психологические аномалии, т. е. чрезмерные с точки зрения цивилизации отклонения от среднего (подобно крайней степени близорукости), обусловлены нарушением согласованности скоростей развития разных структур и функций. Это может произойти либо по генетическим причинам — например, ребенок случайно несет относительно дисгармоничный набор генов,— либо вследствие воздействия окружающей среды, когда развитие одной стороны личности стимулировалось внешними факторами уже в раннем детстве, а другая сравнительно отставала в своем развитии. Мы не располагаем точно доказанными примерами из области психологии человека, но у животных хорошо известен целый ряд фактов дисгармоничного развития, затрагивающего поведение. Например, можно экспериментально получить «эдипов комплекс» у гуся, скрестив дикого гусака с домашним. Домашний гусь несет гены раннего полового созревания, и у некоторых молодых самцов половая зрелость может наступить уже в то время, когда гусята еще ходят с гусыней. Вследствие этого молодая птица упорно пытается спариться с матерью. Поскольку у дикого гусака (отца) половая активность приходится на более поздний период весны, он подобным образом не реагирует.
Чувствительные периоды
Чувствительные, или критические, периоды представляют собой крайние примеры подобной связи событий дифференциального роста. Под «чувствительным периодом» понимают некоторую стадию ограниченной продолжительности, во время которой под влиянием специфического воздействия другой области развивающегося организма или окружающей среды возникает специфическая ответная реакция. Эта реакция может быть полезной п даже существенно важной для нормального развития, но может быть и патологической. В качестве примера нормального чувствительного периода рассмотрим дифференцировку гипоталамуса крысы в мужской или женский. Для дифференцировки гипоталамуса крысы в мужской необходимо стимулирующее действие тестостерона, приходящееся на первые 5 дней жизни. До этого периода, а также спустя пять дней после него тестостерон не оказывает никакого воздействия на гипоталамус. Как и в большинстве подобных примеров, степень чувствительности может быть выражена количественно, опа постепенно повышается до максимума, а затем вновь снижается.
386
Часть IV. Рост и конституция человека
Второй пример можно взять из классических работ Хыобеля и Визела, показавших, что для развития клеток центральной нервной системы, обеспечивающих восприятие света, котенок должен находиться на свету в течение первых недель после того, как у него открылись глаза. Если же котенка перенести на свет по истечении 3 нед, зрение у него уже не разовьется. Животные рождаются как бы подготовленными к «ожидаемой» среде, в которой все события, запрограммированные для каждого чувствительного периода, должны реализоваться.
Постпубертатный рост
Рост скелета не совсем прекращается к концу подросткового периода. У человека в отличие от других млекопитающих, таких, например, как крысы, эпифизарные хрящи длинных трубчатых костей окостеневают полностью и поэтому рост конечностей невозможен. Рост же позвоночного столба продолжается и в возрасте 20—30 лет за счет отложения новых слоев костного вещества на верхних и нижних поверхностях позвонков. Таким образом, длина тела за эти годы несколько увеличивается — в среднем на 3—5 мм. Между 30 и 45—50 годами она остается постоянной, а потом начинает уменьшаться. Эта периодичность позволяет предположить, что андрогенные гормоны, возможно, имеют важное значение в поддержании роста, так как именно эти гормоны стимулируют рост позвоночного столба у подростков (см. гл. 20). Для практических целей тем не менее полезно установить тот возраст, для которого можно констатировать фактическое прекращение роста длины тела, т. е. такой возраст, после которого прирост длины тела не превысит 2%. Продольные исследования показывают, что для мальчиков это в среднем 17/4 лет, а для девочек 16. Нормальные колебания для различных индивидуумов составляют до 2 лет в ту и другую сторону от среднего.
Большая часть размеров головы и лица после подросткового периода продолжает постоянно увеличиваться. Этот процесс происходит, хотя и очень медленно, по крайней мере до 60 лет. Между 20 и 60 годами прибавка составляет 2—4% значений для двадцатаяетнего возраста.
Кривая роста человека как характерная особенность приматов
Для человекообразных и низших обезьян характерна такая же форма кривой роста, что и для человека. По-видимому, это характерная особенность приматов, так как кривые роста таких животных, как грызуны и крупный рогатый скот, носят совершенно иной характер.
Глава 19. Кривая роста человека
387
Половое
Рис. 19.10. Кривая скорости увеличения веса мыши (Tanner, 1962).
1 — результаты получены методом поперечного сечения, оба пола объединены; 2, з — данные для самцов (18) и самок (18), полученные только продольным методом на крупных мышах линии Мак-Артур; 4, 5 — мелкие мыши линии Мак-Артур. Время полового созревания указано по Engle and Rosasco, которые приводят первую течку у альбиносов в возрасте 37 дней при среднем квадратическом отклонении в 5 дней.
До сих пор проведено мало продольных исследований, касающихся линейных измерений, на представителях животных разных видов. Поэтому при сравнении видов нам приходится использовать кривые увеличения веса, а не роста. На рис. 19.10 показана кривая скорости увеличения веса мыши. Малая продолжительность интервала между окончанием вскармливания и половым созреванием и отсутствие явного пубертатного скачка связаны с отсутствием периода низких скоростей роста между моментом рождения и наступлением зрелости. Если судить по уровню созревания органов, то мышь рождается на более ранней стадии своего развития, чем человек. Максимум на кривой скорости увеличения веса у мыши приходится на то время, которое соответствует по стадии созревания органов моменту рождения у человека, т. е. первому максимуму на кривой веса человека. Кривая для крыс сходна с таковой для мышей, за исключением того, что крысы рождаются еще менее зрелыми. Морские свинки рождаются более зрелыми, и максимум скорости роста у них действительно приходится на момент рождения.
У шимпанзе, напротив, кривая имеет совершенно иной вид и полностью напоминает кривуюг характерную для человека (рис.
388
Часть IV. Рост и конституция человека
Рис. D.11. Кривая скорости увеличения веса шимпанзе (Tanner, 1962). Возразч первой менструации в среднем 8,8 года при индивидуальных колебаниях от 7,0 до 10,8 года. I — самцы, II — самьи.
19.11). Первый пик на кривой скорости увеличения веса обезьяны наблюдается незадолго до рождения или в момент рождения; затем происходит постепенное уменьшение скорости, охватывающее длительный период от конца вскармливания до полового созревания. Для обезьян, особенно самцов, характерен выраженный пубертатный скачок. Для макаков-резусов характерна сходная кривая, правда с меньшей продолжительностью интервала между вскармливанием и половым созреванием. Выраженность подросткового скачка и степень наблюдаемых в это время половых различий варьируют от вида к виду.
Увеличение времени между окончанием вскармливания и половым созреванием, а также появление пубертатного скачка роста можно рассматривать как этап в эволюции приматов. Весьма существенным изменением является, по-видимому, некоторая отсрочка в наступлении полового созревания, так как другие млекопитающие продолжают расти относительно дольше после достижения половой зрелости. Поиски непосредственной причины отсрочки, вероятно, приведут нас к механизмам гипоталамуса, так как начало пубертатного скачка определяется именно функцией мозга. Это дополнительное время, необходимое для созревания мозга, у приматов создается за счет увеличения периода между окончанием вскармиливания и половым созреванием п задержки
Глава 19. Кривая роста человека 389
созревания гипоталамуса до тех пор, пока ассоциативные зоны коры не достигнут соответствующей стадии созревания.
Этот процесс заходит особенно далеко у низших и человекообразных обезьян и у человека. По крайней мере некоторые эволюционные причины этого достаточно ясны. По-видимому, обучение, и в частности обучение сотрудничеству в групповой или семейной жизни, протекает гораздо успешнее, если оно происходит в то время, когда отдельные особи относительно послушны и еще не пытаются соперничать со взрослыми самцами. Существование групп, состоящих из послушных, но умеренно активных членов, само по себе могло быть эволюционно выгодным.
Глава 20
ПУБЕРТАТНЫЙ СКАЧОК РОСТА И ОПРЕДЕЛЕНИЕ ВОЗРАСТА ПО СТЕПЕНИ РАЗВИТИЯ
Пубертатный скачок роста — это общее явление, варьирующее по интенсивности и продолжительности у разных детей. Максимальная скорость роста длины тела в среднем составляет около 10 см в год для мальчиков и немного меньше для девочек. У мальчиков пубертатный скачок роста происходит в среднем между 12% и 15% годамиг а у девочек — на два года раньше.
Рис. 20.1. Пубертатный скачок роста длины тела у девочек (/) и мальчиков (II).
Максимум скорости роста приходится в среднем на 12—13 лет у девочек и на 14—15 у мальчиков. Средние прибавки нанесены в центре полугодовых периодов.
На рис. 20.1 приведены кривые скорости роста, иллюстрирующие половые различия между группой мальчиков с максимумом скорости в 14—15 лет и группой девочек с максимумом в 12—13 лет. Эти группы были подобраны таким образомл чтобы по возможности
Глава 20. Пубертатный скачок роста
391
избежать ошибок «растягиванием» максимума во времени (см. рис. 19.5). Различия в размерах тела у взрослых мужчин и женщин в значительной степени зависят от времени наступления, продолжительности и интенсивности пубертатного скачка роста. До этого скачка различия в длине тела у мальчиков и девочек не превышают 2% , а после него они достигают в среднем 8%. Различия возникают также вследствие более позднего наступления пубертатного скачка роста у мальчиков, в связи с чем увеличивается продолжительность периода предпубертатного роста, хотя и протекающего с малой скоростью; имеет значение и интенсивность самого скачка.
Практически пубертатный скачок касается почти всех костных и мышечных размеров, хотя и не в равной мере. Скачок роста длины тела происходит главным образом за счет роста туловища, а не конечностей. Скачок в увеличении мышечной массы происходит через 3 мес, а скачок веса — через 6 мес после скачка роста длины тела.
Пубертатный скачок распространяется и на сердечную мышцу и на все остальные органы. Некоторое пубертатное ускорение роста имеет место, по-видимому, даже у такого органа, как глаз, который к моменту рождения ребенка оказывается ближе всего к дефинитивному состоянию. О наличии подобного скачка свидетельствует выраженный миопический сдвиг, приуроченный именно к данному периоду. Миопия неуклонно прогрессирует начиная с 6-летнего возраста (или даже раньше) вплоть до достижения взрослого состояния. Резкий сдвиг, характерный для пубертатного периода, можно объяснить тем, что ускорение роста глазного яблока в направлении зрительной оси выражено cnnbHeet чем в вертикальном направлении.
Точно не установлено, наблюдается ли пубертатный скачок в развитии мозга. Незначительный пубертатный скачок имеется в росте лицевого скелета. Что касается индивидуальной изменчивости в смысле возникновения пубертатного скачка, то она довольно сильно выражена, и у некоторых детей вообще не происходит ускорения роста многих измерительных признаков лица и головы, в том числе и гипофизарной ямки. Однако, как правило, в пубертатном периоде челюсть удлиняется, утолщается и выступает вперед. Профиль делается более прямым, резцы обеих челюстей— более вертикальными, а нос — более выступающим. Все эти изменения сильнее выражены у мальчиковг чем у девочек.
Половые различия
Многие половые различия в величине и форме тела, наблюдаемые во взрослом состоянии, в большинстве своем возникают благодаря дифференциальному характеру пубертатного роста. М-ы уже говорили о больших размерах тела у мужчин. Столь характерные
392
Часть IV. Рост и конституция человека
признаки, как относительно широкие плечи у мужчин и бедра у женщин, обусловлены в основном специфическим стимулирующим действием андрогенов (в первом случае) и эстрогенов (во втором) на клетки хряща. Усиленное развитие мышц у мужчин и ряд других приводимых ниже физиологических особенностей также возникает за счет стимулирующего действия андрогенов.
Далеко не все половые различия развиваются подобным образом. В частности, более длинные относительно туловища нижние конечности могут быть следствием увеличения периода предпу-бертатного роста у мужчин, так как именно в это время ноги растут несколько быстрее туловища. Другие половые различия проявляются еще раньше. У мужчин относительная длина предплечья больше, чем у женщин. Это различие наблюдается уже у новорожденного, а затем постепенно усиливается с возрастом. Причина его, возможно, кроется в том, что у мужчин на ранних стадиях внутриутробного развития на данном участке происходит отложение несколько большего количества ткани или несколько более активной ткани. Аналогичные соотношения наблюдаются и у других приматов.
Подобным образом, вероятно, возникают и половые различия в относительной длине второго и четвертого пальцев. У женщин чаще, чем у мужчин, второй палец бывает длиннее четвертого. Эти соотношения также устанавливаются еще до рождения. Однако самое удивительное из всех предпубертатных половых различий — это более раннее созревание женщин.
Развитие системы органов размножения
Пубертатный скачок в росте костных и мышечных размеров тесно связан с интенсивным развитием репродуктивной системы, которое происходит в это же время. На рис. 20.2, Б схематически представлен пубертатный период у мальчиков. Сплошные прямоугольники соответствуют периодам ускоренного роста пениса и яичек, а горизонтальные линии с оценочными баллами (2—5) отражают появление и дальнейший рост волос на лобке. В каждом случае приводятся средние значения возраста. Представление об индивидуальных отклонениях от этих средних можно получить по цифрам, расположенным под каждым показателем и обозначающим размах изменчивости возраста проявления для того или иного признака. Так, ускорение роста пениса начинается в среднем в 12 Vg лет, но иногда происходит очень рано — в 10 1/2 лет, а иногда поздно — в 14 лет. Следует отметить, что у некоторых мальчиков ускорение роста длины тела и пениса начинается после того, как у рано созревающих мальчиков этот период уже закончился. В любой группе мальчиков 13—14-летнего возраста можно отметить широкий диапазон колебаний — от полной зрелости до
Глава, 20. Пубертатный скачок роста
393
Рис. 20.2. Схема, иллюстрирующая последовательность наступления пубертатных явлений (Marshall and Tanner, Archs Dis. Childh., 45, 1970).
А. У девочек: I — скачок роста длины тела, II — наступление менструации, III — развитие молочных желез, IV — появление волос на лобке. Б. У мальчиков: I — то же, II — пенис, III — яичко, IV — общий балл, V — появление волос на лобке; стрелкой обозначен максимум увеличения мышечной силы. Представлены средние данные; приведены индивидуальные вариации возраста начала и окончания того или иного явления.
предпубертатной стадии. В связи с этим возникают сложные социальные и педагогические проблемы, выражающиеся прежде всего в психологической неприспособленности^ наблюдаемой иногда в подростковом периоде (см. гл. 22).
Последовательность процессов в пубертатном периоде варьирует гораздо меньше, чем возраст, в котором они происходят. Первый признак наступающего пубертатного периода у мальчи
394 Часть IV. Рост и конституция человека
ков — это ускоренный рост яичек и мошонки. Приблизительно в это же время начинается незначительный рост волос на лобке. Сначала они растут медленно, затем одновременно с ускорением роста длины тела и пениса увеличивается и скорость роста волос. Обычно это происходит через год после первого ускорения роста яичек. Рост яичек происходит главным образом за счет разрастания семенных канальцев. В то же время развиваются клетки Лейдига, продуцирующие андрогены.
В подкрыльцовой впадине волосы обычно появляются лишь через 2 года после начала роста волос на лобке, по как проявление индивидуальной изменчивости в отдельных случаях они появляются раньше лобковых. В перианальной области волосы вырастают несколько раньше, чем под мышкой, и отличаются от лобковых, распространяющихся на всю промежность. На лице у мальчиков волосы начинают расти приблизительно в одно время с подмышечными. Прежде всего удлиняются и темнеют волосы, растущие в углах верхней губы, а затем — расположенные медиально. В дальнейшем волосы появляются на верхних частях щек, по средней линии под нижней губой и, наконец, по обеим сторонам и вдоль нижнего края подбородка. Рост волос на теле начинается одновременно с появлением волос под мышкой и заканчивается лишь спустя некоторое время после полового созревания. Общее количество волос на теле человека определяется, по-видимому, генетически, хотя точно не известно, является ли причиной их роста природа и количество секретируемого гормона или реактивность волосяных фолликулов.
Разрастание гортани у мальчиков начинается тогда, когда рост пениса почти заканчивается. Изменение голоса происходит постепенно и часто завершается лишь после окончания подросткового периода. У мальчиков-подростков нередко встречаются некоторые изменения со стороны грудных желез: околососковый кружок увеличивается в диаметре и темнеет. У некоторых мальчиков (до 30% в большинстве изученных групп) наблюдается отчетливое возвышение соска с развитием твердой субареолярной железистой ткани. Это происходит примерно в середине пубертатного периода и длится в течение 12—18 мес, после чего у большинства мальчиков возвышение и железистая ткань спонтанно исчезают.
В медицинской, антропологической и педагогической практике нередко приходится решать вопрос о том, как далеко зашел процесс созревания. Стандарты для оценки роста волос на лобке, развития половых органов и молочных желез можно найти в книгах о развитии подростков.
Пубертатный период у девочек схематически представлен на рис. 20.2, Л. И в этом случае отмечается существенная изменчивость во времени наступления пубертатного скачка, но последовательность процессов остается довольно постоянной. Первый при-
Глава 20. Пубертатный скачок роста
395
Рис. 20.3. Кривые-скорости (4) и ускорения (Б) роста длины тела девочки от 9 до 18 лет (Israelson, в кн. Tanner, 1960).
Вычислено по данным, выравненным с помощью первой и второй производной кривой Гомпертца. I — начало развития молочных желез; II — возраст первой менструации.
знак пубертатного периода — это набухание области сосков, хотя появление волос на лобке иногда может ему предшествовать. Матка и влагалище развиваются одновременно с развитием грудных желез. Первая менструация почти всегда начинается после того, как пройдена максимальная точка скорости роста длины тела. На рис. 20.3 показаны кривые, полученные в результате дифференцирования кривой Гомпертца, построенной на основании измерения роста девочки; на рис. 20.3, А приведена кривая скорости, а на рис. 20,3, Б — кривая ускорения. Особый интерес представля
396 Часть IV. Рост и конституция человека
ет форма кривой ускорения: сначала ускорение постепенно увеличивается, затем наступает период отрицательного ускорения и, наконец, происходит постепенное уменьшение отрицательного ускорения. Точками I и II отмечено соответственно начало набухания области сосков и наступление первой менструации. Поражает точное совпадение первой точки с максимумом положительного ускорения, а второй — с максимумом отрицательного.
Начало первой менструации служит показателем относительной зрелости матки, хотя, как правило, это не означает, что становление репродуктивной функции завершилось полностью. Первые менструальные циклы часто не сопровождаются выходом яйцеклетки из яичника. В течение года после начала первой менструации, а иногда и дольше наблюдается период относительного бесплодия. Это характерно не только для человека, по и для низших и человекообразных обезьян.
IIвменения физиологических функций и развитие мышечной системы
С пубертатным скачком совпадают во времени существенные изменения физиологических функций. Они выражены гораздо резче у мальчиков, чем у девочек, и благодаря им мужчины приобретают большую физическую силу и выносливость. До наступления подросткового периода мальчики в среднем несколько сильнее девочек. Даже в это время в популяции насчитывается больше мускулистых, мезоморфных мальчиков, чем девочек. Но разница эта невелика. По окончании подросткового периода мальчики становятся значительно сильнее девочек. Это происходит за счет увеличения мышечной массы, а также, возможно, вследствие способности к проявлению большей мышечной силы на 1 г веса мышц. Сердце и легкие становятся у мальчиков относительно крупнее, кислородная емкость крови выше; повышается способность крови нейтрализовать химические продукты мышечной деятельности.
Короче говоря, за время подросткового периода мужчины становятся более приспособленными к охоте, борьбе, поднятию и переноске тяжестей и другим видам деятельности.
На рис. 20.4 приведены данные по двум силовым тестам, полученные при продольном исследовании группы мальчиков и девочек на протяжении всего подросткового периода. Силу растяжения определяли по степени растягивания динамометра перед грудью при разведении рук, а силу сжатия — при сведении рук. В качестве результата фиксировалась лучшая из трех попыток на соревнованиях школьников, проводившихся каждые полгода. Только при таком условии удавалось получить надежные максимальные значения. У мальчиков в возрасте от 13 до 16 лет наблюдался выраженный пубертатный скачок. У девочек скачок почти невыражен, хотя
Возраст, годы
Рис. 20.4. Показатели мышечной силы (Jones, 1949).
Сила сжатия правой (А) и левой (В) кисти; сила растяжения (В) и сжатия (Г) рук. I — мальчики, II — девочки. Данные смешанного продольного исследования, численность группы мальчиков 65—93, группы девочек 66—93.
398
Часть IV. Рост
и конституция человека
Рис. 20.5. Изменение содержания гемоглобина (Л) и числа циркулирующих в крови эритроцитов (Б) у детей, иллюстрирующее появление половых различий в подростковом периоде (Tanner, 1962).
1 — мальчики, 11 — девочки. Смешанные продольные данные из Mugrage and Anderson (1936, 1938), Amer. J. Dis. Child.
данные динамометрии кисти, полученные в той же самой группе, выявили некоторое ускорение в увеличении мышечной силы, которое приходилось на 12—13% лет.
Увеличение числа эритроцитов, а следовательно, и содержания гемоглобина в крови у мальчиков-подростков показано на
Глава 20. Пубертатный скачок роста
399
рис. 20.5. До наступления подросткового периода половых различий в этих показателях не существует. Поэтому в нижней части графика приведены общие данные для обоих полов. Систолическое артериальное давление постепенно повышается уже в детстве, но у мальчиков-подростков процесс этот значительно ускоряется; отмечается уменьшение частоты сердечных сокращений. У мальчиков повышается парциальное давление углекислого газа в альвеолах, а у девочек нет, что влечет за собой половые различия в парциальном давлении углекислоты в артериальной крови. Одновременно у мальчиков повышается резервная щелочность крови. Благодаря этому во время мышечной нагрузки кровь взрослого мужчины способна поглощать без изменения pH значительно больше молочной кислоты и других продуктов мышечной деятельности, чем кровь женщины, что и необходимо, поскольку мышечная масса у мужчин развита значительно сильнее. Повышение эффективности реакции на нагрузку достигается разными способами, в том числе, например, уменьшением общего количества прошедшего через легкие воздуха на каждый литр усвоенного кислорода.
В результате указанных анатомических и физиологических сдвигов в подростковом периоде мышечная сила у мальчиков резко увеличивается. Распространенное представление о том, что в этом возрасте продольный рост мальчиков опережает развитие мышечной силы, не имеет под собой научной почвы. Совершенно верно, что максимум скорости в увеличении мышечной силы достигается примерно через год после максимального ускорения роста большинства костных размеров, и в течение некоторого кратковременного периода подросток, завершив свой скелетный рост, еще не обладает силой молодого мужчины при тех же размерах и телосложении. Но это временное явление. Сила, атлетизм, физическая выносливость быстро и прогрессивно увеличиваются на протяжении пубертатного периода.
Определение возраста по степени развития и понятие физиологической зрелости
Хотя все описанные выше процессы подросткового периода связаны друг с другом и характеризуются довольно однообразной последовательностью, возраст их наступления сильно варьирует. Чтобы проиллюстрировать это, выберем три снимка из стопки фотографий нормально развивающихся мальчиков 14-летнего возраста. Один мальчик — невысокий, с детскими мышцами, неразвитыми половыми органами и отсутствием волос на теле; его легко принять за 12-летнего. Другой — практически взрослый мужчина с широкими плечами, сильными мышцами, развитыми наружными половыми органами и низким голосом. Третий занимает как бы промежуточное положение. Было бы явно нелепым считать всех
400
Часть TV. Рост и конституция человека
троих одинакового сформированными в физическом или социальном отношении, поскольку поведение в этом возрасте во многом определяется физическим статусом. Когда мы говорим, что мальчику 14 лет, мы фактически ограничиваемся очень туманной характеристикой. G точки зрения морфологии, физиологии и социологии все зависит от того, находится ли ребенок в доподростковом, подростковом или послеподростковом периоде развития.
Очевидно, необходимы какие-то другие показатели физиологической зрелости, более точные, нежели хронологический возраст, и, пожалуй, наиболее подходящим из них следует признать степень развития органов размножения. Но аналогичные различия в темпах роста, как их назвал впервые описавший их Франц Боас (F. Boas), наблюдаются в любом возрасте, хотя они менее выражены, чем в пубертатном периоде. Таким образом, мы должны найти показатель степени развития, или физиологической зрелости, применимый в течение всего периода роста. В настоящее время существуют три возможных показателя: скелетная зрелость, зубная зрелость и зрелость формы.
Скелетная зрелость
Наиболее широко распространенным показателем физиологической зрелости является степень развития скелета, определяемая рентгенологически. Оссификация каждой кости начинается с первичного центра и проходит через ряд последовательных стадий увеличения и формирования области окостенения. К этому добавляется в некоторых случаях один или несколько дополнительных центров в эпифизах, где окостенение начинается независимо от главного центра; в заключение зпифизы срастаются с телом кости, и созревание кости завершается. Все эти изменения можно легко увидеть на рентгенограмме, так как содержащие кальций зоны окостенения, непроницаемые для рентгеновских лучей, отличаются от хрящевых зон, где оссификация еще не начиналась. Последовательность стадий формирования центров окостенения в диафизе и эпифизах постоянна. Поэтому скелетная зрелость, или, как чаще говорят, костный возраст, оценивается как по числу имеющихся центров, так и по стадии развития каждого из них.
Теоретически для оценки скелетной зрелости можно использовать любую часть скелета или даже все его части. Но на практике наиболее удобными и общепринятыми областями исследования являются кисть и запястье. Получить рентгенограмму кисти очень легко; при этом не облучаются другие области тела, требуется незначительная доза рентгеновских лучей и самая простая аппаратура. Наконец, кисть — это область с большим количеством развивающихся костей и эпифизов. Принято исследовать левую руку. Ее укладывают на кассету ладонью в распрямленном состоянии и ус
Глава 20. Пубертатный скачок роста
401
танавливают трубку на расстоянии 76 см от сгиба среднего пальца.
Оценка скелетной зрелости производится путем сравнения данной рентгенограммы с рядом стандартов. Это можно сделать двумя способами. При более старом методе с использованием специальных атласов данную рентгенограмму последовательно сопоставляют со стандартными, соответствующими различным возрастам, и смотрят, с каким стандартом она приблизительно совпадает. Метод, разработанный в последнее время, устанавливает серию стандартных стадий, характерных для каждой кости, и сопоставляет ту или иную кость на данной рентгенограмме с этими стадиями. Затем каждая кость получает балл, соответствующий достигнутой стадии, а вся рентгенограмма оценивается по сумме баллов для многих центров созревания. Суммарный балл сопоставляется с распределением баллов в стандартной группе соответствующего возраста, и, наконец, определяется перцентильный статус ребенка по скелетной зрелости. (Если, например, 80% детей данного возраста имеют более низкий суммарный балл, то данного ребенка относят к восьмидесятому перцентилю.) Скелетный возраст можно определить как возраст, при котором данный балл приходится на пятидесятый перцентиль.
Зубная зрелость
Зубную зрелость можно определить путем подсчета числа прорезавшихся зубов и сопоставления его со стандартными величинами — почти так же, как это делалось в случае скелетной зрелости. Молочные зубы прорезываются с 6 мес до 2 лет и могут быть использованы как показатель физиологической зрелости в этом возрастном интервале. Прорезывание постоянных зубов может служить показателем зрелости в интервале от 6 до 13 лет. С 2 до 6 лет и после 13 лет простой подсчет зубов мало что дает в этом смысле. Однако в последнее время были предложены новые методы определения зубной зрелости с использованием стадий окостенения зубов на рентгенограммах челюстей, подобно тому как оценка скелетной зрелости производится по стадиям окостенения.
Определение возраста по форме тела
По мере того как ребенок становится старше, меняется и форма его тела благодаря различиям в скоростях роста разных частей (см. гл. 19). В принципе степень изменения формы тоже может служить мерой зрелости и практически весьма удобной. Однако такая оценка связана с некоторыми трудностями, не возникавшимп при определении склетной и зубной зрелости. Изменение формы может быть использовано для определения возраста лишь в том случае,
402
Часть IV. Рост и конституция человека
если удается найти показатель, который был бы полностью независим от дефинитивной формы (при определении скелетного и зубного возраста мы не сталкиваемся с этой проблемой, так как окончательная стадия развития этих структур одинакова у всех людей.) Именно поэтому потерпели неудачу попытки определения «возраста по длине тела» и «возраста по весу», которые предпринимались педиатрами, и «возраста по интеллекту», применяемого психологами.
Под «возрастом по длине тела» понимается тот возраст, при котором «средний» ребенок достигает данной длины тела. Допустим, что ребенок имеет высокий рост для своего возраста. Это может быть вызвано либо тем, что ребенок развивается быстрее среднего (это мы и должны выяснить), либо тем, что он в будущем станет высоким взрослым и уже сейчас обгоняет своих сверстников.
Следовательно, для того чтобы определение возраста формирования было действительно эффективным, нужно найти такую комбинацию размеров тела, которая изменялась бы с возрастом, но пе зависела от окончательных размеров и формы тела. Это математически сложная, но выполнимая задача. В настоящее время определение возраста по форме тела представляет научную проблему, а не практический метод.
Соотношения между различными оценками зрелости
Данные, приведенные на рис. 20.2, не должны заслонять того факта, что дети очень сильно отличаются друг от друга как по той скорости, с которой они проходят через различные стадии пубертатного периода, так и по степени связи между различными показателями, включая скелетную зрелость. В качестве одного крайнего варианта можно привести здоровую девочку, которая еще не начала менструировать, хотя уже достигла взрослой стадии по развитию грудных желез и волос на лобке и прошла максимум скорости роста длины тела два года назад. Однако ее костный возраст также будет запаздывать, как наступление менструации, поскольку эти два показателя очень тесно связаны. Среднее квадратическое отклонение хронологического возраста наступления менструаций составляет приблизительно 1 год; соответствующий показатель для костного возраста к моменту наступления менструаций — всего 0,7 года. Таким образом, нормальные пределы хронологического возраста наступления менструаций для Англии в настоящее время составляют 11,0—15,0 лет, а костного возраста — 11,5—14,5 лет. Костный возраст не связан с возрастом, в котором начинает развиваться грудная железа, и лишь в слабой степени связан с возрастом появления волос на лобке. Последовательность развития грудных желез и волос на лобке характеризуется значи-
Глава 20. Пубертатный скачок роста
403
Рис. 20.6. Соотношения между скелетной зрелостью и воврастом первой менструации (Tanner, 1962).
Приводится возраст скелетного развития (по стандартам Тодда) для трех групп девочек от 7 до 17 лет — с ранним (Ml), средним (М2) и поздним (М3) наступлением первой менструации. Данные смешанного продольного исследования.
тельной независимостью, так же как и рост половых органов и волос на лобке у мальчиков. Индивидуальные различия в степени связи между показателями созревания, вероятно, отражают различия в организации гипоталамо-гипофизарной системы.
Как показывает рис. 20.6, дети в течение всего периода роста или по крайней мере начиная с трехлетнего возраста имеют тенденции либо запаздывать в своем развитии, либо, наоборот, обгонять своих сверстников. На рисунке приведены данные по трем группам девочек: с ранним, средним и поздним наступлением первой менструации. У девочек, рано начавших менструировать, костная система оказывается более развитой не только во время подросткового периода, но и в предшествующие годы начиная с 7 лет. У девочек, поздно начавших менструировать, отмечается постоянное отставание по костному возрасту. Точки Ml, М2, М3 соответствуют среднему возрасту первой менструации в каждой группе.
404
Часть IV. Рост и конституция человека
У детей в возрасте от 6 до 13 лет, идущих с опережением по костному возрасту, в среднем больше прорезавшихся зубов. Аналогичным образом у детей с ранним наступлением подросткового скачка зубы прорезываются раньше (рис. 20.7). Однако, как видно из того же рисунка, здесь имеет место не столь тесная корреляция: даже при наличии по каждому полу всего лишь трех групп различной степени зрелости кривые иногда пересекаются.
Эта некоторая независимость развития зубов от общего развития тела вовсе не кажется удивительной. Зубы представляют собой часть головного конца организма, а мы уже видели (гл. 19), что голова опережает в своем росте остальную часть тела и благодаря этому кривая ее роста несколько отличается от общей кривой роста.
Очевидно, в процессе роста существует какой-то общий фактор, регулирующий созревание организма и определяющий тип развития ребенка в целом как опережающий или отстающий по окостенению скелета, росту, прорезыванию постоянных зубов, физиологическим реакциям, развитию умственных способностей и по другим психологическим реакциям. Подчиненные этой общей тенденции группы признаков, характеризующих степень зрелости на ограниченных участках, варьируют независимо друг от друга. К числу подобных признаков относятся зубы (прорезывание молочных и постоянных зубов в значительной степени не зависит друг от друга), центры окостенения и, возможно, мозг. Мы можем догадываться и о некоторых механизмах, лежащих в основе этих соотношений. Например, у детей с гипофункцией щитовидной железы запаздывают не только прорезывание зубов и развитие скелета, но и умственное развитие, тогда как в случаях преждевременного полового созревания, обусловленного мозговыми расстройствами или заболеваниями надпочечников, отмечается ускоренное созревание костной системы и половых желез, не сопровождающееся соответствующим ускорением в развитии зубов и мозга.
Длина тела в данном возрасте, выраженная в процентах от окончательной, обнаруживает довольно тесную связь со скелетной зрелостью начиная с 7 лет и дальше. Разработаны уравнения регрессии для прогнозирования окончательной длины тела, исходя из роста ребенка, хронологического и костного возраста. Для девочек следует ввести дополнительную информацию о наступлении первой менструации, а для обоих полов сделать поправку на рост родителей.
Половые различия в возрасте физиологической зрелости
По скелетной зрелости девочки в среднем опережают мальчиков в течение всего периода роста, начиная с момента рождения и до взрослого состояния. То же можно сказать и о прорезывании по-
Глава 20. Пубертатный скачок роста
405
30
L. . . ,1__I I । ! I_______!__1_____!-----1
6 7 8 9 10 11 12 13 14 15
Возраст, годы
Рис. 20.7. Общее число прорезавшихся зубов в каждой возрасте для рано (7), средне (2) и поздно (3) созревающих девочек (7) и мальчиков (77) (Tanner, 1962).
Степень зрелости группы определяется по возрасту максимальной скорости роста длины тела. Смешанные продольные данные.
406
Часть IV. Рост и конституция человека
стоянных зубов (но не молочных). По-видимому, половые различия касаются общего фактора созревания (так же как и многих частных факторов), что, надо предполагать, не может не отразиться на интеллектуальных тестах и социальных реакциях.
Половые различия в костном возрасте проявляются уже во время внутриутробного развития, причем отставание особей мужского пола, по-видимому, прямо или косвенно связано с действием генов Y-хромосомы. Доказательством этого служит то, что дети с такой аномалией хромосом, как XXY (синдром Клейнфельтера), не отличаются по скелетной зрелости от нормальных мальчиков (XY), а при аномалии ХО (синдром Тернера) скелетная зрелость соответствует таковой у нормальных девочек (XX), по крайней мере до наступления пубертатного периода. Каков механизм действия этих генов, кпд не знаем. Возможно, что замедление процесса созревания у лиц мужского пола связано с дифференцировкой яичек, начинающейся на 2-м месяце внутриутробной жизни и определяющей фундаментальное различие в сроках развития. Более вероятно, однако, что подобные различия могут возникать вскоре после этого возраста за счет секреции специфических половых гормонов гонадами и надпочечниками плода. Парадокс состоит в том, что все до сих пор известные специфические мужские гормоны оказывают ускоряющее, а не тормозящее влияние на созревание кости.
Новорожденные мальчики отстают по скелетной зрелости от девочек приблизительно на 4 нед, и в течение всего периода роста их костный возраст составляет 80% костного возраста девочек того же хронологического возраста. Именно поэтому у девочек на 2 года раньше начинается подростковый период и завершается созревание. Процентные различия в зубном возрасте не столь велики: у мальчиков он составляет 95% зубного возраста девочек того же хронологического возраста.
Половые различия в степени зрелости наблюдаются не только у человека, но и у человекообразных и низших обезьян, а также у крыс и, по-видимому, могут служить характерным признаком всех или большинства млекопитающих. Полное биологическое значение этой особенности пока не выяснено.
Физическая зрелость, умственные способности и развитие эмоциональной сферы
Ряд факторов показывает, что интеллектуальное и эмоциональное развитие до некоторой степени связано с развитием скелета. Это можно проще всего истолковать, по крайней мере в отношении интеллектуального развития, как доказательство того, что мозг, как и зубы, подвержен действию общего фактора, задающего темп развитию. Так, дети, быстрее развивающиеся физи
Глава 20. Пубертатный скачок роста 407
чески, лучше справляются с психологическими тестами, чем физически отсталые. Эти вопросы еще будут обсуждаться в гл. 22.
Почти не вызывает сомнений, что более раннее или более позднее созревание оказывает существенное влияние на эмоциональное развитие и социальное поведение ребенка, особенно во время подросткового периода. Эти проблемы также обсуждаются в гл. 22. Ясно, что существование различий в темпах развития людей имеет большое значение для теории и практики воспитания.
Глава 21
ГОРМОНАЛЬНЫЕ, ГЕНЕТИЧЕСКИЕ И СРЕДОВЫЕ ФАКТОРЫ, РЕГУЛИРУЮЩИЕ РОСТ
Эндокринная система играет огромную роль в регуляции роста и развития, будучи одним из главных факторов, при участии которых заключенная в генах информация реализуется во взрослый организм с такой скоростью и в такой форме, как это допускает данная конкретная среда.
Пренатальный период
На 9-й нед внутриутробного развития, считая с первого дня последней менструации (или на 7-й нед, считая с момента оплодотворения), под действием генов Y-хромосомы ранее недифференцированная гонада начинает превращаться в яичко. Пока не ясно, какова роль гормонов в этом процессе. На 11-й постменструальной неделе в яичке появляются клетки Лейдига, и к 12-й нед они начинают продуцировать тестостерон или родственное вещество. Вероятно, это происходит под влиянием гонадотропина хориона, содержание которого в моче матери как раз в это время достигает максимальной величины (определение гонадотропина в моче служит стандартным тестом в диагностике беременности). Под влиянием тестостерона ранее не дифференцированные наружные половые органы превращаются в пенис и мошонку.
У женщин дифференцировка яичника и наружных половых органов проходит, по-видимому, не столь бурно. В отсутствие Y-хромосомы на 9-й нед ничего не происходит, а на 10-й гонада превращается в яичник. Формирование наружных половых органов по женскому типу происходит примерно на 14-й нед, очевидно, без участия гормонов.
Нельзя не упомянуть и еще об одном аспекте половой дифференцировки, изученном пока только на животных, но имеющем большое как теоретическое, так, возможно, и практическое значение для человека. У крыс секреция клеток Лейдига воздействует не только на наружные половые органы, но и на мозг. У всех изученных млекопитающих гормональные и в значительной степени поведенческие признаки, характерные для мужского пола, зависят от структуры гипоталамуса. Если гипофиз пересадить от самки крысы взрослому гипофизэктомированному самцу, то после восстановления сосудистых связей с гипоталамусом гипо
Глава 21. Факторы., регулирующие рост
409
физ начнет секретировать гонадотропные гормоны мужского, а не женского типа. Эксперимент по пересадке гипофиза от самца к самке дает аналогичный результат.
У крыс дифференцировка гипоталамуса происходит под влиянием тестостерона, выделяемого клетками Лейдига, в первые 2— 3 дня после рождения. Это истинный чувствительный период. Введение тестостерона за несколько дней до рождения, а также спустя 5 дней после рождения гонадэктомированной крысе не вызывает дифференцировки мозга. Сигнал должен быть получен гипоталамусом точно в нужный момент. Однократная инъекция женского полового гормона па 5-й день после рождения приводит к прекращению нормальной дифференцировки по мужскому типу, а однократная инъекция тестостерона самке на 5-й день после рождения вызывает эффект «стерилизации андрогеном», и у взрослого животного женский половой цикл будет отсутствовать. Известно, что некоторые области мозга избирательно поглощают тестостерон; это области, относящиеся к половому поведению, к регуляции секреции гонадотропина. Если новорожденным самкам крыс вводили тестостерон, у них во взрослом состоянии не развивался женский тип полового поведения даже после того, как им удаляли яичники и проводили эстроген-прогестероновую заместительную терапию, так что по гормональному статусу они соответствовали нормальной самке. После удаления яичников и введения тестостерона они вели себя как самцы.
Уже сейчас ясно, что крысы в этом отношении не отличаются от других млекопитающих. До какой степени и в какие промежутки времени аналогичная ситуация имеет место у человека, мы пока не знаем. Момент рождения у крыс соответствует, вероятно, 16—18-й нед. внутриутробного развития у человека или, поскольку клетки Лейдига у человека развиваются раньше, чем у других млекопитающих, даже 14—16-й нед. Действительно, это хорошо согласуется с секрецией тестостерона у плода. Имеет ли это какое-нибудь отношение к различным типам полового поведения у человека, в настоящее время неизвестно.
Роль других эндокринных желез в пренатальном периоде не выяснена. Эстрогены материнского организма, проникая через плаценту, вызывают преходящее увеличение матки у новорожденной девочки. Щитовидная железа плода продуцирует тиреоидный гормон, необходимый для нормального развития мозга. В надпочечнике существует особая зона, хорошо развитая при рождении, а впоследствии несколько редуцирующаяся; однако вопрос о ее назначении и роли пока не ясен.
410
Часть IV. Рост и конституция человека
Постнаталъный период
Наиболее важным гормоном, воздействующим на процесс роста от момента рождения до подросткового периода, является соматотропин, или гормон роста. Он представляет собой полипептид, выделяемый гипофизом и обладающий большей степенью видовой (точнее, надвидовой) специфичности, чем остальные гормоны гипофиза. Стимуляцию роста у человека вызывают только гормоны человека или обезьяны.
Хотя соматотропин присутствует у плода, он не является необходимым для его роста. Однако начиная с момента рождения он становится жизненно важным для установления нормальной скорости роста ребенка. К 2 годам дети с изолированной недостаточностью гормона роста заметно отстают от здоровых сверстников (хотя в действительности это отставание замечают не раньше 5—6-летнего возраста, когда ребенок идет в школу или когда младшие братья или сестры обгоняют его по росту).
Гормон роста сам по себе не оказывает воздействия на хрящ. Он вызывает рост посредством стимуляции печени, которая продуцирует промежуточный гормон, называемый соматомедином. Это тоже пептид, но с меньшим молекулярным весом. Введение гормона роста при его недостаточности вызывает рост мышц с увеличением включения аминокислот, используемых для синтеза белка. Параллельно с этим происходит уменьшение количества жировой ткани, т. е. сдвиг метаболических процессов в сторону запасания белка. Дети с недостаточностью гормона роста не только низкорослые, но и толстые.
Секреция гормона роста, как и других гипофизарных гормонов, находится под контролем гипоталамуса. Ко времени написания этой книги было установлено существование ингибирующего гипоталамического гормона (соматостатин, пептид из 14 аминокислот), но гипоталамический рилизинг-гормон пока не известен.
Гормон роста выделяется на протяжении суток не непрерывно, а отдельными порциями. Физическая нагрузка, состояние тревоги, сон всегда вызывают секрецию гормона, о других факторах нам пока ничего не известно. Все еще многое неясно относительно регуляции и функции гормона роста у взрослых. Из-за прерывистого выделения трудно установить суточный ритм секреции гормона роста, и поэтому мы до сих пор не знаем, как меняется скорость секреции в детском возрасте, если такие изменения вообще имеют место. Со всей определенностью можно лишь сказать, что индивидуальные различия в длине тела в пределах нормальных вариаций не связаны с различиями в секреции гормона роста. Быть может, они обусловлены различиями в количестве сома-томедина, однако более вероятно объяснить их количественными или качественными особенностями рецепторов в клетках хряща.
Глава 21. Факторы, регулирующие рост
411
Гормон щитовидной железы играет очень важную роль на протяжении всего периода роста. Функциональная активность щитовидной железы постепенно падает начиная с момента рождения и вплоть до подросткового периода, а затем в течение года, вероятно, несколько увеличивается или по крайней мере постепенно снижается. Что касается скорости роста, то влияние щитовидной железы следует признать стимулирующим, но не контролирующим. При гипофункции щитовидной железы (гипотиреоз) рост замедляется, причем это сказывается и на склетной, и на зубной зрелости, и на развитии мозга.
Хотя нам ясно, что механизм, регулирующий скорость созревания скелета, должен иметь гормональную природу, мы пока ничего не знаем о соотношении участвующих в нем гормонов. В случае недостатка гормона щитовидной железы или гормона роста наблюдается задержка созревания, и, наоборот, половые гормоны и андрогены надпочечника вызывают его ускорение. Небольшие концентрации половых гормонов и андрогенов надпочечника содержатся в крови до наступления подросткового периода, но в какой мере различия в их количестве могут контролировать темпы роста, пока совершенно неизвестно.
Подростковый период
С наступлением подросткового периода начинается новая фаза роста, во время которой действие гормонов половых желез и надпочечников в сочетании с гормоном роста вызывает пубертатный скачок роста. По-видимому, полная реализация скачка роста зависит от присутствия гормонов того и другого типа: у мальчиков с недостаточностью гормона роста максимальная скорость роста во время пубертатного периода достигает половины нормальной величины.
Уровень двух (из трех) основных групп гормонов надпочечников в крови остается относительно неизменным начиная с момента рождения; речь идет о кортизоне и альдостероне; последний контролирует концентрацию электролитов в тканевой жидкости.
Третья группа гормонов надпочечников — андрогены — появляется, по крайней мере в заметном количестве, только в подростковом периоде. Считают, что именно андрогены вместе с гормоном роста регулируют ход всего пубертатного ускорения роста у женщин. У индивидуумов мужского пола они действуют совместно с половым гормоном — тестостероном, секретируемым яичком. Несомненно, что именно тестостерон вызывает рост мышц и увеличение числа эритроцитов у мужчин во время подросткового периода. До сих пор не выяснено, существуют ли в пубертатном
412
Часть IV. Рост и конституция человека
периоде половые различия в количестве и типах андрогенов, секретируемых надпочечником.
До сих пор нет законченной теории, которая устанавливала бы последовательность эндокринных процессов в пубертатном периоде, хотя многое в этом запутанном вопросе уже разгадано. Вся последовательность событий начинается с гипоталамуса. Хотя гипофиз содержит гонадотропины или даже обладает способностью продуцировать их и до наступления пубертатного периода, в кровоток эти гормоны не поступают из-за тормозящего действия гипоталамуса. Выброс гонадотропина осуществляется с помощью особого вещества — рилизинг-фактора лютеинизирующего гормона (РФЛГ), представляющего собой октапептид, который синтезируется некоторыми клетками гипоталамуса и достигает гипофиза через его портальную систему. Именно гипоталамус, а не гипофиз несет информацию относительно процесса созревания. В «нужный момент» при достижении определенной стадии физической зрелости и зрелости структур гипоталамуса происходит выброс РФЛГ.
То, каким образом это происходит, имеет важное значение для понимания механизмов развития. В предпубертатВом периоде у ребенка уже устанавливается и действует система обратной связи, при которой очень низкие уровни циркулирующих в крови тестостерона (у мальчиков) и эстрогена (у девочек) оказывают тормозное влияние на нейроны гипоталамуса, которые секретируют РФЛГ. Таким образом, содержание гонадотропина также поддерживается на крайне низком, хотя и заметном, уровне. Затем в пубертатном периоде происходит нечто такое, что понижает чувствительность нейронов гипоталамуса к половым стероидам. Уровень циркулирующих гормонов становится недостаточным для подавления выброса РФЛГ, и тогда возникает следующая цепь событий: РФЛГ -> гонадотропины -> эстроген (у девочек), причем концентрация эстрогена постоянно повышается до тех пор, пока не достигнет уровня, при котором выброс РФЛГ подавляется. Эта новая концентрация гормона в крови достаточна, чтобы вызвать увеличение грудных желез и другие пубертатные изменения. Однако, что именно вызывает снижение чувствительности клеток гипоталамуса, до сих пор остается загадкой. Очевидно, мы имеем дело со своеобразными внутренними часами, причем такими, которые зависят не просто от хронологического возраста или даже сроков развития в целом, а от последовательности прохождения многочисленных предшествующих событий в организме.
Механизм увеличения андрогенов надпочечника менее ясен. Так как в "подростковом периоде кортизон продолжает выделяться с прежней интенсивностью, мало вероятно, чтобы в связи с процессами полового созревания происходило увеличение секреции адре-
Глава 21. Факторы, регулирующие рост
413
нокортикотропных гормонов. По-видимому, каким-то образом повышается чувствительность клеток надпочечника, секретирующих андрогены, так что то же количество адренокортикотропина вызывает усиленное образование андрогенов. В опытах на животных было показано, что эстрогены способны повышать чувствительность клеток надпочечника. Тестостерон, по-видимому, не обладает такой способностью, так что механизм аналогичного процесса у мужчин остается неизвестным.
Многое в данном вопросе еще предстоит выяснить. Неизвестны причины усиленного накопления жира в предпубертатном возрасте, хотя по времени этот процесс как будто совпадает с усиленным выделением гонадотропина. Все еще вызывает споры вопрос об идентичности андрогенов надпочечника, стимулирующих рост волос на лобке и в подкрыльцовых впадинах у женщин. И хотя мы уже начинаем постигать тончайшие механизмы гормональных явлений в подростковом периоде, более глубокие знания буду! получены лишь после проведения продольных исследований с применением более чувствительных химических и биологических методов.
Взаимодействие наследственности и среды в процессе регуляции скорости роста
Известно множество факторов, оказывающих влияние на скорость развития. Некоторые из них носят наследственный характер и с раннего возраста способствуют ускорению или замедлению физиологического созревания. Другие (например, недостаточное питание, время года, тяжелый психологический стресс) относятся к категории факторов внешней среды и влияют на скорость роста лишь в период своего действия. И наконец, некоторые факторы, например социально-экономическое положение, представляют собой результат сложного взаимодействия наследственности и среды.
Длина тела, вес, телосложение ребенка или взрослого являются результатом не только действия, но и взаимодействия генетических факторов и факторов среды. От определенного набора генов, определяющего длину тела, очень далеко до роста данного конкретного человека. В современной генетике считается прописной истиной, что выражение любого гена зависит в первую очередь от внутренней среды, создаваемой другими генами, а затем от внешней среды. Более того, взаимодействие генов и окружающей среды может быть не аддитивным. Например, определенное увеличение пищевого рациона не обязательно вызовет увеличение роста, скажем, на 10% у всех людей независимо от их генетической конституции. Вместо этого у генетически высоких людей произойдет увеличение роста на 12%, а у генетически низкорослых — на 8%. Этот тип взаимодействия называется мулътипликаторным.
414____________Часть IV. Рост и конституция человека
Вообще говоря, одни и те же условия окружающей среды могут оказаться весьма подходящими для ребенка с одним набором генов и совершенно не подходящими для ребенка с другим набором. Таким образом, чрезвычайно трудно количественно оценить относительную роль наследственности и среды в регуляции роста и телосложения, не имея в виду определенных условий окружающей среды. В общих чертах, чем ближе условия среды к оптимальным, тем скорее проявятся потенциальные возможности генов. Однако это всего лишь общее правило, и, без сомнения, в процессах роста и дифференцировки наблюдается множество более тонких и специфических взаимодействий.
Тем не менее совершенно ясно, что наследственные факторы имеют огромное значение. Основной план роста определяется очень рано, когда развивающийся в матке зародыш находится в относительной безопасности. Зачаток кости конечности, пересажанный от плода или новорожденной мыши под кожу спины взрослого животного той же инбредной линии (что исключает образование антител), продолжает развиваться в нормальную зрелую кость. Более того, то же самое произойдет и с хрящевой манжеткой, изолированной до начала костеобразования. Итак, структура зрелой кости задолго до ее реализации в основном заложена в хрящевой модели. Последующее влияние условий окружающей среды — натяжение прикрепленных к кости мышц и связок — завершает формирование.
Генетика роста
Генетическую регуляцию скорости роста проще всего проследить на примере наследования возраста появления первой менструации. У сестер — однояйцовых близнецов этот возраст разнится в среднем на 2 мес, а у разнояйцовых — на 10 мес. Коэффициент корреляции между возрастом первой менструации у матерей и дочерей лишь немногим меньше соответствующего коэффициента для длины тела и равен приблизительно 0,4. Этот факт указывает, что индивидуальная изменчивость возраста первой менструации в европейских популяциях в значительное мере обусловлена генетически. Наследование возраста первой менструации, вероятно, идет как по отцовской, так и по материнской линии и связано не с одним, а с многими генами, каждый из которых обладает незначительным действием. Это такой же характер наследования, который типичен для таких количественных признаков, как рост, вес и другие.
Генетический контроль действует на протяжении всего периода роста. Однояйцовые близнецы в любом возрасте характеризуются одинаковой степенью скелетной зрелости. Время прорезывания молочных и постоянных зубов, а также последовательность
Глава 21, Факторы, регулирующие рост
415
Рис. 21.1. Коэффициент корреляции между ювенильными и дефинитивными значениями длины (Л) и веса (Б) тела (Tanner, 1962).
Кривые для обоих полов (III) построены на основании обследования 124 индивидуумов в Абердине. Кривые для мальчиков (I) и девочек (II) построены по результатам обследования 66 мальчиков и 70 девочек в рамках Калифорнийской программы профессиональной ориентации. Все данные получены продольным методом.
их кальцификации и прорезывания в значительной мере наследственно обусловлены. Гены, контролирующие процесс роста, могут быть самыми разнообразными: одни из них определяют, вероятно через посредство эндокринного аппарата, скорость роста всего тела, другие обусловливают в высшей степени локализованные градиенты роста, устанавливающие порядок прорезывания зубов и появления центров окостенения в костях запястья.
416 Часть IV. Рост и конституция человека
Не все гены оказываются активными к моменту рождения. Некоторые из них проявляются только в последующие годы в определенных физиологических условиях; об их действии говорят, что оно «ограничено возрастом». Вероятно, так можно объяснить характер изображенной на рис. 21.1 кривой, описывающей коэффициенты корреляции между величинами количественных признаков у ребенка в разные возрастные периоды и величиной соответствующих признаков у взрослого. Эти данные были получены в результате длительных продольных исследований. Коэффициент корреляции между длиной тела у новорожденного и взрослого очень низок, так как длина тела при рождении определяется не столько генотипом плода, сколько размерами матки. Гены, определяющие рост, начинают проявляться в первые три года жизни, и степень корреляции резко возрастает. Зато впоследствии, вплоть до подросткового периода, наблюдается лишь слабое ее увеличение. По всей видимости, интенсивность и время наступления пубертатного скачка контролируются генами, определяющими секрецию больших или малых количеств андрогенов. Действие этих генов может не проявляться вплоть до момента начала секреции андрогенов. Допубертатный и пубертатный рост в какой-то мере независимы.
Расы и экологические условия
Существование расовых различий в скорости и характере роста приводит к различиям в физическом типе между взрослыми индивидуумами. Некоторые из этих различий имеют явно генетическое происхождение, тогда как другие зависят от питания и, возможно, климата. Мы должны предположить, что в каждой из основных популяций земного шара рост ее членов был приспособлен действием отбора к условиям той среды, в которой происходила эволюция. Мы можем увидеть остатки этого процесса в современных популяциях, однако только остатки, так как сравнительно недавние миграции сильно изменили расселение народов и многие из них больше не живут в тех областях, где происходила эволюция. Действительно, существует довольно тесная положительная связь между показателем «линейности», который оценивается как относительный вес взрослого человека (вес по отношению к длине тела), и средней годовой температурой места обитания. Различия в размерах тела следует строго отделять от различий в пропорциях тела, поскольку первые из них в отличие от последних довольно легко изменяются под действием недоедания. Европеец, который голодал в детстве, в конце концов окажется невысокого роста, но у него не будет относительно коротких ног и длинного туловища, как у японца, или длинных ног и короткого туловища, как у африканца. Форма его скелета практически не изменится, по
Глава 21. Факторы, регулирующие роет
417
при достаточно продолжительном и сильном недоедании количество жира и мышц окажется резко уменьшенным.
Возрастные кривые длины тела для групп европейских, африканских и азиатских народов точно установленного происхождения, живущих в сходных и благоприятных условиях с почти оптимальным статусом питания, не позволяют обнаружить каких-либо существенных различий между неграми и европейцами. Что касается китайцев, то длина тела у них меньше и процесс роста заканчивается раньше; возможно, этим и объясняется их короткопогость. В отличие от двух групп генетически сходных популяций, живущих в различных условиях окружающей среды, общая задержка роста при недоедании очевидна. Вопреки распространенному мнению климат не оказывает прямого влияния на скорость роста. У школьниц Нигерии, получающих относительно хорошее питание, средний возраст появления первой менструации — 14,3 года, а у девочек-эскимосок — 14,4 года. У бирманских и ассамских девочек, получающих прекрасное питание и пользующихся хорошим медицинским обслуживанием, появление первой менструации в среднем падало на 13,2 года. Несмотря на очень жаркий климат (45°С), эта цифра практически не отличалась от средних данных для европейских девочек. Воздействие питания на возраст появления первой менструации (см. ниже) настолько велико, что оно полностью перекрывает возможное влияние климата, которое, если и существует, крайне незначительно.
Некоторые популяционные различия в форме тела, по-видимому, обусловлены в основном генетическими причинами. Относительный вес детей американских индейцев в возрасте от 6 до 11 лет, живущих в Аризоне, несколько выше, чем детей местных белых, несмотря на то что индейцы живут в худших экономических условиях. Это можно было бы принять за побочное действие плохого питания, когда приостанавливается рост в длину и усиливается рост костей и мышц в толщину. Но по некоторым данным, относительный вес африканских негров, и в особенности нилотов, напротив, во всех возрастах меньше, чем белых того же роста. Либо в этих двух местностях различен сам характер недоедания, либо, что гораздо более вероятно, мы имеем дело с генетическими различиями.
Генетические различия — это, без сомнения, и есть та причина, благодаря которой негры Западной и Восточной Африки и США опережают белых по скелетной зрелости начиная с момента рождения и в течение первых двух лет жизни. Это связано с более интенсивным развитием двигательной активности: негритянские дети раньше начинают сидеть и ползать. Указанное опережение в развитии исчезает примерно к третьему году жизни (по крайней ме-ре, у африканских негров) вследствие недостаточного питания. Негритянские девочки из состоятельных семей в США опережают
418
Часть IV. Рост и конституция человека
своих белых сверстниц в течение всего периода роста. Постоянные зубы у негров прорезываются в среднем на год раньше. Зубы закладываются очень рано и отличаются большей устойчивостью к голоду и болезням, чем костная система, возможно, потому, что они в меньшей степени подвержены гормональным влияниям.
Время года
При анализе многочисленных данных из развитых стран, расположенных в умеренных областях, отмечается отчетливо выраженное влияние времени года на скорость роста. В среднем самый быстрый продольный рост происходит весной, а самая интенсивная прибавка в весе — осенью. По старым западноевропейским данным, средняя скорость роста длины тела с марта по май почти вдвое превышает скорость роста за сентябрь и октябрь.
Однако время наибольшей интенсивности сезонного пика, так же как и его выраженность, сильно варьируют, и у значительного числа детей трудно уловить сезонные колебания скорости роста. Эти различия могут отражать индивидуальные колебания эндокринной активности.
Питание
Недоедание приводит к задержке роста. Об этом свидетельствует, например, влияние голода во время войн. На рис. 21.2 нанесены ежегодные величины веса и длины тела штутгартских школьников за период с 1911 по 1953 г. С 1920 по 1940 г. происходит постоянное увеличение обоих размеров во всех возрастах (обсуждение этого явления см. ниже), но в последние годы второй мировой войны наблюдается противоположная тенденция.
Дети обладают большим запасом восстановительных сил, если, конечно, неблагоприятные влияния не были очень продолжительными и не нанесли организму серьезного ущерба. В течение кратковременного периода недоедания рост замедляется. Когда же условия снова изменяются к лучшему, начинается необычайно бурный рост с быстрым восстановлением генетически обусловленной кривой роста. Во время этой восстановительной фазы вес, длина тела и развитие скелета «наверстывают упущенное», по-ви-димому, с одной и той же скоростью. У крупного рогатого скота чередование периодов хорошего питания и недоедания приводит к изменению окончательной формы и состава тканей в соответствии с продолжительностью периодов, причем быстро растущие ткани реагируют на голод сильнее. Данных о подобном влиянии недоедания на организм человека нет.
Девочки, по-видимому, лучше мальчиков «защищены» от голода и болезней, и ход кривой их роста нарушается значительно ре-
л
л
Рис. 21.2. Влияние недоедания на рост длины (Л) и веса (Б) тела (Tanner, 1962).
Данные о штутгартских школьниках (от 7—8 до 14—15 лет — народная школа, С 15—16 и старше — средняя школа) за 1911—1953 гг. Линии соединяют значения, соответствующие одному возрасту, и демонстрируют эпохальный сдвиг и влияние военных условий, I — мальчики, II — девочки.
420
Часть IV. Рост и конституция человека
же. Возможно, зто происходит потому, что две Х-хромосомы надежнее обеспечивают регуляцию, чем одна Х-хромосома и небольшая Y-хромосома.
Психологические воздействия
Мысль о том, что неблагоприятные психологические воздействия могут вызвать некоторую задержку роста, часто приходит в голову. В последние годы было достоверно установлено, что у некоторых детей под влиянием эмоционального стресса происходит задержка секреции гормона роста, что сказывается на их развитии так же, как и первичное отсутствие гормона роста. Однако после прекращения воздействия условий стресса гормон роста вновь начинает выделяться и наблюдается обычная стадия быстрого восстановления. Эти случаи сравнительно редки, хотя Нелиган (Neli-gan) показал, что среди населения Ньюкасла они встречаются в 3 раза чаще, чем случаи первичного отсутствия гормона роста.
Подобная задержка роста может наблюдаться не только в условиях крайнего стресса, и, по-видимому, именно этим можно объяснить незначительные вариации индивидуального роста, хотя убедительными доказательствами на этот счет мы, естественно, не располагаем. Однако одно подобное исследование, проведенное Виддаусоном (Widdowson), можно считать тщательно проверенным.
После войны Виддаусон изучал влияние пищевого рациона на развитие осиротевших детей, получавших скудное питание, доступное в Германии в 1948 г. Ему представился редкий случай наблюдать последствия замены воспитательницы. По плану эксперимента детям из приюта II после 6-месячпого контрольного периода давали прибавку к пищевому рациону, а затем сравнивали их рост и вес с ростом и весом детей из приюта I, не получавших такой прибавки. Однако (как видно из рис. 21.3) результат оказался прямо противоположным предполагаемому. Хотя дети из приюта II во время первого, контрольного периода лучше прибавляли в весе, чем дети из приюта I, во время второго периода, когда калорийность их рациона была увеличена на 20%, они стали хуже прибавлять в весе. Причина, по-видимому, заключалась в том, что как раз на рубеже двух периодов одна из воспитательниц была переведена из приюта I и стала во главе приюта II. Она «воспитывала» детей розгами и обычно выбирала обеденное время для публичного и часто несправедливого наказания провинившихся. Это расстраивало всех остальных детей. Исключение составляли лишь восемь ее любимцев, которых она взяла с собой из приюта I. Эти восемь всегда весили больше, чем другие, а при повышенном питании в приюте II стали прибавлять в весе еще быстрее. Тот же эффект, но менее выраженный, наблюдался и в отношении длины тела.
Глава 21. Факторы, регулирующие рост
421
4 в 12 16 20 24 28 32 36 40 4 4 48 52 Время наблюдения, недели
Рис. 21.3. Изменение веса (Л) и длины (Б) тела у детей из двух сиротских приютов (Tanner, 1962).
Увеличение пищевого рациона в приюте II отмечено вертикальной полосой, одновременно в приют II из приюта I перешла старшая воспитательница (обозначено х). Обратите внимание, что изменения роста связаны с присутствием нли отсутствием воспитательницы, а не с количеством пищи Кривые III соответствуют изменениям длины и веса тела 8 любимцев воспитательницы, которые вместе с ней были переведены из приюта II в приют I.
«Лучше скудный обед, приправленный любовью, чем жирный окорок, нашпигованный ненавистью»,— пишет Виддаусон.
Вероятно, подобным образом можно хотя бы отчасти объяснить результаты некоторых наблюдений по увеличению роста и веса школьников в учебное и каникулярное время.
422
Часть IV. Рост и конституция человека
Социально-экономические факторы; размер семьи
Дети из различных социально-экономических слоев населения во всех возрастах различаются по средним метрическим признакам; причем дети из более обеспеченных групп опережают своих сверстников на протяжении всего развития. В большей части исследований социально-экономическое положение определялось по профессии отца, хотя в последние годы стало ясно, что во многих странах этот признак больше не может служить четкой характеристикой условий жизни. Необходимым дополнением становится оценка жилищных условий, а также доля расходов на ребенка в семейном бюджете.
Разница в длине тела у детей из обеспеченных слоев населения— специалистов, квалифицированных рабочих и административного аппарата, с одной стороны, и детей неквалифицированных рабочих — с другой, составляет около 2 см в трехлетием возрасте, а в подростковом периоде достигает 5 см. Разница в весе несколько меньше, так как для детей из низших социально-экономических слоев характерен больший вес относительно длины тела. На рис. 21.4 приведены данные о росте 7-летних детей в Великобритании (родившихся на протяжении одной недели 1958 г.) в зависимости от социально-экономического положения и размера семьи.
Различия в длине тела отчасти обусловлены более ранним созреванием детей из обеспеченных классов, а отчасти более высоким ростом представителей этих классов во взрослом состоянии. Отмечается разница (на 2—3 мес) и в возрасте появления первой менструации у девочек, принадлежащих к различным социально-экономическим группам. У детей из более обеспеченных слоев несколько раньше происходит смена зубов (если исходить из усредненного варианта).
В основе этих различий лежит, по-видимому, множество причин. Наиболее важная из них — питание, а вместе с ним и все навыки регулярного приема пищи, сон, физкультура и общая организованность, которыми отличается с этой точки зрения «хороший дом» от «плохого». Описанные различия в росте в большей степени определяются укладом жизни в семье, чем ее экономическим положением; а уклад этот в значительной мере отражает интеллект и личные качества родителей. У подавляющего большинства хорошо питающихся детей такие болезни, как корь, грипп и даже излечиваемые антибиотиками воспаление среднего уха и пневмония, не вызывают заметной задержки роста, которая может наблюдаться в подобных случаях у детей, находящихся в плохих условиях в смысле питания и ухода. Возможно, что высокая частота таких болезней в менее обеспеченных и неорганизованных семьях играет определенную роль в задержке роста у таких детей, хотя это и не было
Глава- 21. Факторы, регулирующие рост
423
твердо установлено. Различия в весе и длине тела между различными слоями населения за последние 50 лет стали менее выраженными в связи с улучшением социальных условий, однако они по-прежнему зависят скорее от домашних условий и образования родителей, чем от материального положения семьи.
Не удивительно поэтому, что более «умные» дети (если судить по результатам соответствующих тестов во всех возрастах) обго-
Рис. 21.4. Различия в длине тела 7-летних детей в зависимости от положения родителей в обществе и числа детей в семье (Goldstein, Hum. Biol., 43, 1971). 2—5 — слои общества; За — отец занимается умственным трудом; зб — отец занимается физическим трудом.
пяют своих менее интеллектуальных сверстников из той же социальной прослойки. Эта связь, вероятно, представляет собой сложный комплекс воздействия генетических и средовых факторов, причем одни из них усиливают другие.
Эпохальный сдвиг
В течение последних ста лет наблюдается удивительная тенденция: дети во всех возрастах становятся все крупнее и крупнее. Эта тенденция достигает значительных размеров и стирает различия между развитием детей из разных социально-экономических слоев. На рис. 21.5 приведены данные о росте и весе шведских школьников, полученные в 1883, 1938 и 1965—1971 гг. Во всех возрастах начиная с 7 лет дети 1938 г. Kpynnee^j чем их сверстники 1883 г.j а дети 1965—1971 гг. еще крупнее.
424
Часть IV. Рост и конституция человека
Рис. 21.5. Эпохальный сдвиг в росте длины (4) и веса (Б) тела шведских детей с 1883 по 1968 г. (Ljung В., Bergsten-Brucefors A. a. Lindgren G., Ann. hum. Biol., 1, 245, 1974).
I — девочки; 11 — мальчики.
Глава 21. Факторы, регулирующие рост
425
Рис. 21.6. Эпохальный сдвиг в росте длины тела английских мальчиков с 1833 по 1958 г. (Tanner, 1962).
I — фабричные мальчики (1833); II — дети рабочих (1874); III — дети служащих (1874); IV — школьники старших классов (1878); V — дети высших слоев бирмингемского общества (1955); VI — средние данные по Англии (1958).
Британские, скандинавские, немецкие, польские и североамериканские статистики приводят сходные величины эпохального сдвига. Средняя прибавка с 1880 по 1950 г. у детей от 5 до 7 лет составила около 1 см и 0,5 кг за каждое десятилетие; у подростков она увеличилась до 2,5 см и 7 кг, а у взрослых вновь уменьшилась до 1 см. Довольно скудные данные по дошкольному возрасту свидетельствуют о том, что расхождение наблюдается уже у новорожденных, а для 2—5-летнего возраста достигает, вероятно, большей величины, чем в последующие годы. По-видимому, эта тенденция все еще продолжается в большинстве европейских стран и особенно сильно выражена в Японии. Однако в большей части обеспеченных слоев общества она, несомненно, приостановилась. Возможно, что дети из этих слоев полностью реализовали свои генетические ростовые потенции. Трудно сказать, когда началась тенденция к увеличению размеров тела. Судя по весьма любопытной серии измерений, начатых в Норвегии еще в 1741 г., с 1760 по 1830 г. произошло небольшое увеличение роста взрослых, с 1830 по 1875 г. прибавка составляла 0,3 см за 10 лет, а с 1875 г. и до наших дней — 0,6 см за десятилетие. По измерениям, начатым в Дании в 1815 г., никакой прибавки не отмечалось вплоть до 1845 г. На рис. 21.6
426
Часть IV. Рост и конституция человека
Рис. 21.7. Эпохальный сдвиг возраста первой менструации за 1840—1970 гг. (Tanner, 1962, и дополнительные данные из источников, приведенных в кн. Eveleth a. Tanner, Worldwide variation in human growth. Cambridge University Press, 1976).
I—Швеция, II—Норвегия, III — Финляндия, IV — Дания, V—Голландия, VZ — Великобритания, VII — США.
приведены данные о росте английских мальчиков начиная с 1833 по 1958 г. Эпохальный сдвиг перекрыл различия между социальными классами, и хотя они все еще существуют, средний мальчик любого возраста в наши дни выше, чем его ровесник из высших слоев общества в 1878 г.
Эта тенденция к увеличению размеров происходит, во-первых, за счет более раннего созревания, приводящего в конечном итоге к тому, что рост, характерный для взрослого, достигается несколько раньше, а во-вторых, за счет увеличения длины тела во взрослом состоянии. По большинству европейских данных, эпохальный сдвиг длины тела для взрослого составляет 1 см за десятилетие, или 2,5 см за поколение.
Ускорение роста проявилось и в эпохальном сдвиге возраста первой менструации (рис. 21.7). Во всех сериях наблюдений эта тенденция оказалась на редкость устойчивой. С 1830 по 1960 г. начало первой менструации в странах Западной Европы сдвигалось за каждое десятилетие примерно на 4 мес. В последние годы, однако, появились доказательства, что этот процесс приостановился в Осло и в Лондоне, но все еще продолжается в Швеции, Голландии и Венгрии. В большинстве европейских стран средний возраст появления первой менструации сегодня колеблется от 12,8
Глава 21. Факторы, регулирующие рост
427
до 13,2, а в Италии он несколько ниже. Соответствующая цифра для белых девочек США составила 12,8 года. Кажется вероятным, что в некоторых странах тенденция к увеличению размеров тела приостанавливается, по мере того как достигается максимальная скорость роста. Можно только догадываться, является ли это достижение максимальной скорости роста желательным с медицинской точки зрения, не говоря уже о социальном аспекте этой проблемы.
Никто не может сказать наверняка, чем вызван этот эпохальный сдвиг. Обычно, и не без веских на то оснований, говорят об улучшении питания и общих условий окружающей среды. Однако, как мы знаем, отмеченная тенденция вовсе не ограничивается менее обеспеченными слоями. И если питание действительно является причиной сдвига, то, по-видимому, речь идет не только об увеличении калорийности, но и об изменении соотношения ингредиентов в рационе и о каких-то важных пищевых факторах. Было высказано предположение, что тенденция к увеличению длины тела, возможно, имеет генетическое происхождение и вызвана неуклонно происходящим разрушением генетических изолятов, т. е. нарушением традиции к заключению браков между членами одной и той же деревенской общины. Безусловно, увеличению числа браков, выходящих за пределы замкнутой общины, способствовало, в частности, широкое распространение велосипедов в сельских местностях. Однако для проявления общей тенденции к увеличению длины тела многие контролирующие рост гены должны действовать таким образом, чтобы в среднем рост детей от брака между высокими и низкорослыми людьми хотя бы ненамного превосходил среднее арифметическое между значениями роста родителей. В настоящее время нет прямых доказательств в пользу того, что гены длины тела действуют именно таким образом, однако нет и недостатка в предположениях и догадках на этот счет. Иногда высказывают предположение, что в наши дни благодаря резкому снижению частоты инфекционных заболеваний больше людей высокого роста доживают до детородного возраста и оставляют потомство. Однако это предположение не объясняет наблюдаемую скорость эпохального сдвига, даже если бы оно было верно по существу, что весьма сомнительно.
Есть доказательства, что в связи с индустриализацией в Европе произошло значительное замедление темпов пубертатного развития, а наблюдающаяся ныне тенденция началась тогда, когда стали улучшаться условия жизни. Во всяком случае, эпохальный сдвиг в отношении как более раннего созревания, так и увеличения размеров тела — одно из самых значительных явлений в современной биологии человека, которое, несомненно, должно иметь серьезные медицинские, педагогические и социологические последствия.
Глава 22
ФИЗИЧЕСКОЕ И УМСТВЕННОЕ РАЗВИТИЕ
Совершенно ясно, что изучение роста и развития следует рассматривать как одну из ведущих наук, на которых должна основываться теория и практика воспитания. Раньше педагоги были тесно связаны с биологией человека, а некоторые из них сами были врачами или антропологами. Но в течение первой половины этого столетия бурный расцвет педагогической психологии привел к тому, что педагоги почти совсем перестали интересоваться вопросами физического развития. Теперь с этим покончено. Учителя со всевозрастающей настойчивостью требуют от специалистов по биологии человека точных и полных данных о росте мозга, о критических периодах и стадиях развития, о зависимости результатов психологических тестов от скорости созревания, о влиянии раннего или позднего созревания на эмоциональную уравновешенность, об эпохальном сдвиге и т. д.
На многие из этих вопросов пока можно дать лишь весьма схематичные ответы, так как наукой о росте и развитии долгое время пренебрегали. Тем не менее уже сейчас мы располагаем обширным арсеналом фактов и целым рядом выводов, которые имеют важное значение для педагогики. Главные среди них — это вопросы о градиентах роста мозга; об индивидуальной изменчивости в темпах роста, особенно в пубертатном периоде; о взаимосвязи между темпами умственного и физического развития.
Рост мозга
Начиная с самых первых стадий внутриутробного развития и на протяжении всего дальнейшего роста мозг по весу ближе к своему конечному значению, чем любой другой орган человеческого тела, за исключением, пожалуй, глаза. В этом смысле мозг развивается раньше, чем остальная часть тела (см. рис. 19.6). У новорожденных вес мозга составляет 25% его веса у взрослого, у ребенка 6 мес — почти 50%, в 2% года — около 75%, в 5 лет — 90% и в 10 лет — 95%. Эти величины резко отличаются от соответствующих общих показателей веса тела, составляющего у новорожденного лишь 5% общего веса тела молодого взрослого, а у 10-летнего ребенка — 50%.
Глава 22. Физическое и умственное развитие
429
Рис. 22.1. Пренатальный рост различных отделов головного и спинного мозга (Tanner, 1961).
По оси ординат отложен объем соответствующих отделов в процентах от их объема при рождении. I — средний мозг; II — большой мозг (включает полушария, полосатое тело и промежуточный мозг); III — спинной мозг; IV — варолиев мост; V — мозжечок.
Различные части мозга растут с различной скоростью и на одновременно достигают максимальной скорости своего роста. На рис. 22.1 приведены данные по пренатальному периоду роста; показано существование градиентов роста, аналогичных приведенным на рис. 19.9 для верхних и нижних конечностей, и нанесены значения веса мозга (в том числе мозолистого тела, базальных ядер, промежуточного мозга с таламусом и гипоталамусом), мозжечка, среднего мозга, моста и продолговатого мозга, спинного мозга,
430 Часть IV. Рост и конституция человека
выраженные в процентах от веса соответствующей части мозга у новорожденного. Во всех возрастах, начиная с 3-го мес внутриутробной жизни и вплоть до момента рождения, средний и спинной мозг оказываются наиболее зрелыми, за ними идут варолиев мост и продолговатый мозг. Большой мозг несколько отстает от них, но все-таки опережает мозжечок. Хотя данные по постнатальному росту этих частей мозга в большинстве случаев отсутствуют, очевидно, что подобные соотношения, как правило, сохраняются, если выразить показатели веса отдельных частей в процентах от их величины у взрослого. Иными словами, судя по известным данным, в постнатальном периоде не возникает противоположно направленных градиентов.
Спинной и средний мозг и варолиев мост первыми достигают максимальной скорости роста. Это происходит примерно на 2-м мес внутриутробного развития. К 3-му мес максимальной скорости достигает рост полушарий головного мозга, а к 6-му — рост мозжечка. Таким образом, для мозга характерно существование таких же градиентов роста и очередности в процессах созревания, как и для других частей тела.
Развитие коры головного мозга
Мы все еще слишком мало знаем о росте мозга и развитии его организации. Анатомическое изучение мозга —необычайно кропотливый процесс, и лишь у немногих исследователей достаточно мужества, упорства и возможностей, чтобы провести морфологические исследования мозга детей различного возраста. Физиологические исследования, касающиеся, например, возрастных изменений характера энцефалограмм, все еще находятся в начальной стадии.
Большую часть сведений по развитию структуры мозга мы черпаем из добросовестных работ Конела (Conel), который подверг анализу структуру коры головного мозга новорожденного, трехмесячного, шестимесячного, пятнадцатимесячного, двухлетнего, четырехлетнего и шестилетнего ребенка. Данные по пренатальному развитию структуры очень малочисленны и носят качественный характер. Что касается структурных изменений после шестилетнего возраста, то подобные данные практически отсутствуют.
Кора головного мозга может быть идентифицирована примерно на 8-й нед внутриутробного развития. Впоследствии происходит ее увеличение в ширину, и к 26-й нед она приобретает характерное строение из шести нечетко разграниченных слоев нервных клеток и одного внутреннего слоя волокон. Созревание слоев происходит неодновременно: клетки пятого слоя к моменту рождения оказываются более развитыми по сравнению с другими, за ними следуют клетки шестого, третьего, четвертого и второго слоев. Полагают, что все нервные клетки, имеющиеся у взрослого
Глава 22. Физическое и умственное развитие
431
человека, формируются во время первых 15—18 нед внутриутробного развития, за исключением, может быть, некоторых клеток мозжечка, появляющихся несколько позднее. Впоследствии происходит рост аксонов и дендритов, появление нуклеопротеидов в цитоплазме, увеличение размеров клеток и образование миелиновых оболочек аксонов. Однако новые нервные клетки уже не образуются. Клетки нейроглии, опорной соединительной ткани, продолжают появляться в течение значительно более длительного времени. Уже на ранних этапах развития их больше, чем нейронов, а впоследствии они составляют около 90% всех клеток мозга.
Анализируя эти изменения, можно выделить ряд критериев для определения зрелости различных частей коры, подобных критериям скелетной зрелости, за которые приняты изменения центров окостенения кисти и запястья. Конел выделил 9 критериев, в том числе число нейронов на единицу площади, величину нейронов, состояние вещества Ниссля и нейрофибрилл, длину аксонов и степень миелинизации.
Можно выделить два четких градиента развития: первый касается последовательности развития основных областей мозга, второй — последовательности развития центров в пределах каждой области. Ведущей частью коры является первичная моторная область в прецентральной извилине (рис. 22.2); за ней следует первичная сенсорная область в постцентральной извилине, затем первичная зрительная область в затылочной доле и первичная слуховая область в височной доле. Все ассоциативные области отстают от первичных, причем градиенты развития всегда идут в направлении от первичных зон к вторичным. Так, в лобной доле прежде всего начинают развиваться те ее части, которые находятся непосредственно впереди моторной коры, а потом уж — полюс доли. Извилины медиальной поверхности полушарий и островок развиваются в последнюю очередь.
В самой моторной зоне нейроны, контролирующие движения рук и верхней части туловища, развиваются раньше тех клеток, которые управляют функцией ног. То же самое характерно и для сенсорной области. Это согласуется, с одной стороны, с относительно большей зрелостью руки по сравнению с ногой, а с другой — с тем, что ребенок лучше владеет движениями рук.
У новорожденного кора мозга развита очень слабо; морфологическая картина соответствует малочисленности и даже полному отсутствию кортикальных функций. К концу 1-го мес жизни гистологическая картина первичной моторной области верхних конечностей и туловища свидетельствует о возможности ее функционирования, а к 3-му мес все первичные области оказываются в относительно зрелом состоянии, что связано с развитием у ребенка зрения и слуха. Однако ассоциативные области, выполняющие интегративную функцию, в это время еще не развиты. К 6 мес про-
432
Часть IV. Рост и конституция человека
Рис. 22.2. Латеральная и медиальная поверхности мозга с обозначением отделов коры и областей локализации функций (Tanner, 1961).
Глава 22. Физическое и умственное развитие
433
исходит миелинизация отдельных волокон, приходящих в кору с нижних уровней мозга, хотя внутри самой коры созревает лишь небольшое число ассоциативных волокон. В возрасте между 6 мес и 2 годами происходит дальнейшее развитие, во время которого первичная сенсорная область достигает уровня развития первичной моторной области. Однако многие области все еще находятся в незрелом состоянии, особенно область гиппокампа, поясной извилины и островка.
Начиная с момента рождения и до четырех лет, а может быть, и несколько дольше происходит постоянное увеличение числа и размеров дендритов во всех слоях коры. Кроме того, увеличиваются и усложняются как экзогенные волокна, приходящие в кору с нижних уровней мозга, так и ассоциативные волокна, расположенные в пределах отдельных зон коры и между ними. Степень взаимосвязи (т. е. возможности клетки оказывать влияние на другие клетки посредством связей с ними) увеличивается, и это, очевидно, имеет первостепенное значение в формировании более сложных функций мозга.
Исследования Яковлева и его коллег по миелинизации показали, что мозг продолжает постепенно развиваться, по крайней мере вплоть до наступления пубертатного периода, а может быть, и дольше. Миелинизация нервных волокон представляет собой только один из признаков зрелости. Волокна могут проводить импульсы и, вероятно, иногда проводят их еще до появления миелиновой оболочки. Данные о миелинизации хорошо согласуются с данными Конела о появлении нервных клеток в тех случаях, когда оба источника информации пересекаются. Как правило, волокна, несущие импульсы к определенным полям коры, миелинизируются одновременно с теми волокнами, которые проводят импульсы из этих полей к периферии. Таким образом, созревание проходит по дуге, затрагивая в первую очередь функциональные единицы, а не топографические участии.
Миелинизация целого ряда путей завершается примерно к трем или четырем годам. Волокна, связывающие мозжечок с корой мозга, которые необходимы для осуществления точного контроля над произвольными движениями, после рождения только начинают миелинизироваться; полное завершение этого процесса происходит лишь к четырем годам. Ретикулярная формация — часть мозга, особенно развитая у приматов и человека и связанная с функцией поддержания внимания и сознания, — продолжает миелинизироваться, по крайней мере вплоть до наступления пубертатного периода, а может быть, и несколько дольше. Сходным образом миелинизация продолжается и в частях переднего мозга, расположенных ближе к средней линии. Яковлев предполагает, что это может быть связано с замедленным развитием типа поведения,
434
Часть IV. Рост и конституция человека
относящегося к метаболической, висцеральной и гормональной активности во время половой жизни.
В процессе всего развития мозга, начиная с ранних стадий внутриутробной жизни, становление функций тесно связано с созреванием структуры. Волокна звуковоспринимающей системы (звукового анализатора) начинают миелинизироваться уже на 6-м мес внутриутробной жизни, но завершение этого процесса происходит постепенно, продолжаясь вплоть до 4-го года жизни ребенка. В противоположность этому, волокна световоспринимающей системы (зрительного анализатора) начинают миелинизироваться лишь после рождения ребенка, но завершение этого процесса происходит очень быстро. Яковлев считает, что во внутриутробном периоде развития звуки функционирующих материнских органов являются главными сенсорными раздражителями, если не считать антигравитационных стимулов. Очевидно, они не воспринимаются на уровне коры, но на подкорковом уровне анализатор работает. После рождения быстро начинают преобладать зрительные раздражители, так как для человека характерно развитое зрение. Вскоре кора начинает принимать эти сигналы: корковый конец зрительного анализатора миелинизируется в течение первых нескольких месяцев после рождения. Напротив, корковый конец слухового анализатора миелинизируется очень медленно, что, вероятно, связано с развитием речи.
Едва ли можно допустить, что подобная взаимосвязь между созреванием структуры и становлением функции внезапно прекращается в 6, 10 или 13 лет. Напротив, у нас есть все основания полагать, что высшие функции мозга развиваются лишь тогда, когда заканчивается созревание некоторых структур, или клеточных центров, локализованных по всей коре. Даже миллионы дендритов занимают очень небольшое пространство, и, следовательно, значительное увеличение степени взаимосвязи может проходить при общем повышении веса мозга всего на несколько процентов. Стадии умственного развития, описанные Пиаже (Piaget) и другими специалистами в области детской психологии, отличаются множеством характерных особенностей, присущих развивающимся структурам, а последовательность стадий, вероятно, зависит от постепенного созревания и организации коры (и ограничена ими).
Пока еще не ясно, в какой мере окружающая среда может влиять на созревание и организацию мозга. По мнению Кахала (Cajal) и Хебба (НеЬЬ), функциональная активность клетки стимулирует дальнейшее развитие ее связей, однако эта гипотеза не подтверждена экспериментальными данными.
Многие аспекты функционирования мозга, по-видимому, совершенно не подвержены изменениям условий окружающей среды, если последние колеблются в нормальных пределах. Так, например, дети, родившиеся до окончания нормального 40-недельного
Глава 22. Физическое и умственное развитие
435
срока внутриутробного периода, формируются в неврологическом отношении так же, как и дети того же возраста, развивающиеся в утробе матери. Недоношенные дети не начинают раньше ходить или стоять, несмотря на более продолжительное воздействие со стороны внешней окружающей среды. Это, конечно, не значит, что созревание мозга вообще не зависит от внешних условий. При некоторых состояних, таких, как недоедание или воздействие токсичных веществ, нормальный рост может нарушаться. В настоящее время все еще нельзя определенно утверждать, до цакой степени недоедание, встречающееся в некоторых слаборазвитых странах, может тормозить созревание мозга, хотя эта проблема широко дискутируется. Источником множества недоразумений послужило непонимание некоторыми экспериментаторами того факта, что голодание новорожденной мыши соответствует, логически говоря, голоданию плода человека, находящегося в середине внутриутробного развития, а не новорожденного ребенка. Поэтому большая часть экспериментальных исследований, показавших на крысах постоянное воздействие такого голодания, может относиться только к развитию детей, родившихся с низкими для их возраста весом и размерами тела вследствие заболеваний плаценты. В последние годы Суссером и Стейном было выполнено классическое исследование развития детей, которые ощутили на себе воздействие тяжелого голода в Голландии в 1944—1945 гг. В то время многие из них находились еще в утробе матери или были в младенческом состоянии (Stein, Susser, Saenger and Marolla, 1975). Обследованные в 18-летнем возрасте перед поступлением на военную службу эти юноши не отличались по размерам тела или умственному развитию от своих сверстников, которые в детстве не голодали. В настоящее время есть основания считать, что высокий потенциал «наверстывающего» роста обеспечивает полное восстановление нормальной длины тела и, вероятно, умственного развития даже после тяжелого недоедания в том случае, если условия жизни ребенка во время восстановительного периода были хорошими. Именно это последнее условие обычно редко выполняется в развивающихся странах. Суссер и Стейн великолепно подытоживают современное состояние знаний в этой области следующими словами: «По нашему мнению, следует признать, что в промышленных странах плохое питание в пренатальном периоде не может считаться существенным фактором в социальном распределении умственных способностей среди выживших взрослых. Однако его нельзя исключать в качестве возможного фактора в сочетании с плохим постнатальным питанием, особенно в экономически слаборазвитых странах» (1975).
436
Часть IV. Рост и конституция человека
Влияние темпов роста
В гл. 20 были описаны индивидуальные различия в темпе роста и отмечалось их важное значение для общественной и педагогической практики, особенно в отношении подросткового возраста. Для удобства обсуждения мы рассмотрим сначала развитие интеллекта, а потом эмоциональное развитие.
Развитие интеллекта
Накоплено много данных, свидетельствующих о том, что дети, более развитые в физическом отношении (судя по их раннему созреванию во время подросткового периода), получают в среднем более высокие баллы в психологических тестах, чем их отстающие в развитии сверстники. Эти различия невелики, но постоянны и наблюдаются у детей всех изученных возрастных групп начиная с 6)4 лет.
Таким образом, при возрастных обследованиях обнаруживаются большие возможности у детей с быстрым физическим созреванием по сравнению с медленно созревающими.
Отмечено также, что крупные, т. е. физически более развитые дети получают болеэ высокие баллы при определении так называемого коэффициента интеллекта (КИ) во всех возрастах, начиная с 6 лет, чем их низкорослые сверстники. В выборке из 6940 11-летних шотландских школьников коэффициент корреляции между длиной тела и величиной оценочного балла группового теста «Мор-рей Хаус» составил 0,25+0,01 при допущении возрастных различий от 11,0 до 11,9 лет. При переводе результатов этого теста в баллы КИ по «Терману — Мериллу» было получено среднее увеличение в 0,67 балла на каждый сантиметр длины тела. Подобная связь была обнаружена и при обследовании лондонских детей. Различия для отдельных детей могут быть весьма значительными. У 10-летних девочек обнаружена разница 9 баллов в величине КИ между теми, чья длина тела была выше значений 75-го перцентиля, и теми, у которых опа была ниже значений 15-го перцентиля. Такой размах вариаций составляет 2/3 среднего квадратического отклонения для балльной оценки этого показателя.
Обычно считалось, что у взрослых связь между длиной тела и результатами психологических тестов исчезает. Этого и следует ожидать, если коэффициенты корреляции отражают лишь взаимосвязь между параллельным ходом физического и умственного развития. Действительно, различий по длине тела между рано- и поздносозревающими мальчиками после окончания роста уже не наблюдается. Однако в настоящее время совершенно ясно, что, как это ни странно, связь между длиной тела и КИ, хотя бы частично, сохраняется и у взрослых. Мы пока не знаем, в какой мере это
Глава 22. Физическое и умственное развитие
437
обусловлено воздействием генетических, а в какой мере средовых факторов. Вполне возможно, что в качестве основного движущего фактора в данном случае выступает различная социальная мобильность.
Эмоциональное развитие
Не приходится сомневаться, что раннее или позднее созревание отражается на поведении, причем у некоторых детей в значительной степени. В мире маленького мальчика престиж и успех идут рядом с удалью, а главный инструмент в жизни — мускулистое тело. Те мальчики, которые в своем развитии опережают сверстников не только во время пубертатного периода, но и до пего, выделяются из их среды и становятся вожаками. В самом деле, мальчики с развитой мускулатурой в среднем созревают раньше других, и пубертатный скачок наступает у них раньше. И наоборот, у долговязого неразвитого мальчугана, который часто не может постоять за себя в свалках и драках, пубертатный период в среднем наступает позднее.
Видя вокруг себя успешно развивающихся сверстников, ребенок с поздним развитием в глубине души иногда начинает сомневаться, сможет ли он догнать их в физическом отношении и станет ли он настоящим мужчиной. Беспокойство и тревога, связанные с вопросами пола, проявляются большей частью лишь подсознательно и вызываются гораздо более сложными и глубокими причинами. Однако и в этом случае события подросткового периода — или их отсутствие — играют роль триггера и вызывают к жизни страхи, живущие глубоко в душе ребенка.
При раннем развитии на первый взгляд все обстоит благополучно. В большинстве работ, посвященных изучению личности детей с известными данными по физическому росту, действительно отмечается, что рано созревающие дети более общительны и уравновешенны, менее нервозны и пользуются большим успехом в обществе, по крайней мере в США (это, впрочем, может объясняться и тем, что, как правило, они относятся не к долихоморфному типу, а к мышечному). Но таких детей ждут свои трудности. Если одни горды своим новым положением, то другие испытывают чувство неловкости и замешательства. Девочка с развивающейся грудью, вызванная к доске прочесть наизусть стихотворение перед всем классом, будет сутулиться вместо того, чтобы стоять прямо. Подросток тоже может испытывать подобное смущение. Кроме того, у рано созревших детей в течение более длительного срока продолжается задержка сексуальных влечений, стремления к независимости и профессиональной ориентации. Именно с этим, по мнению большинства авторов, и связаны все те психологические трудности, которые испытывают в этот период многие юноши и девушки.
438 Часть IV. Рост и конституция человека
Таковы некоторые социальные проблемы, связанные с выраженной изменчивостью темпов роста. Практически эта изменчивость целиком биологического происхождения, и социальные мероприятия не могут существенно уменьшить ее. Это заставляет нас согласовывать педагогическую систему как в теории, так и на практике с отмеченными биологическими фактами, противопоставляя биологической изменчивости соответствующую ей социальную гибкость.
Глава 23
АНАЛИЗ И КЛАССИФИКАЦИЯ ТИПОВ ТЕЛОСЛОЖЕНИЯ
Введение
При изучении конституции человека мы пытаемся ответить на вопрос: в чем состоят стойкие различия между людьми и чем эти различия вызваны? Слово «стойкие» используется здесь в своем обычном смысле, ибо под конституциональными признаками мы подразумеваем такие аспекты структуры, функции или поведения, которые изо дня в день или даже на протяжении нескольких лет существенно не изменяются.
В любой изучаемой группе индивидуумов изменчивость признака, повторно наблюдаемого через какой-то период времени, можно разделить на межиндивидуальную и внутрииндивидуальную; к последней относятся лишь те изменения, которые происходят во времени у одного индивидуума. При правильной постановке эксперимента с применением точных статистических методов дисперсионного анализа можно разграничить и определить долю обоих источников изменчивости. К сфере интересов учения о конституции относятся только те признаки, межиндивидуальная изменчивость которых значительно выше, чем внутрииндивидуальная. Признаки, быстро изменяющиеся в ответ на воздействие окружающей среды, исключаются из рассмотрения. Признаки, связанные со стойкими индивидуальными изменениями, характерными для процессов роста, созревания и старения, так же как и неизменяющие-ся признаки, нередко конституционально обусловлены. Следует помнить также, что и характер реакции на изменение среды может быть конституциональным признаком. Количественные показатели некоторых физиологических признаков часто отличаются более выраженной внутрииндивидуальной изменчивостью в состоянии покоя, чем при максимальной нагрузке. Так, частота сердечных сокращений у индивидуума в состоянии покоя оказывается более вариабельной и менее конституционально обусловленной, чем тот же признак при максимальной нагрузке, когда конституциональные факторы приобретают большее значение. Возможно, все это приложимо и к некоторым психологическим признакам. К конституциональным признакам относятся морфологические, физиологические и психологические. Исторически сложившееся разделение на морфологию, физиологию и психологию не имеет права на существование в теоретическом учении о конституции, однако в
440
Часть IV. Рост и конституция человека
методическом отношении каждая из этих наук выступает в качестве самостоятельной основы для практических исследований.
В определении конституции нет четкого разделения на наследственность и среду, поскольку учение о конституции возникло раньше, чем генетика как наука. Несомненно, генетические факторы имеют первостепенное значение. Правда, многие конституциональные признаки имеют очень сложную генетическую основу, а некоторые из них, как можно предположить, возникают скорее как следствие ранней физической нагрузки, нежели генетического влияния. Таким образом, учение о конституции охватывает более широкую область, чем генетика человека в узком смысле слова. В наши дни, однако, генетики все больше и больше интересуются эпигенезом, т. е. развитием признаков у взрослых индивидуумов в результате многочисленных взаимодействий в процессе роста, и в этом смысле учение о конституции и генетика неразрывно переплетаются между собой.
В медицине, главной отрасли прикладной биологии человека, по мере постепенного искоренения инфекционных заболеваний все большее значение приобретают конституциональные болезни. В 1881 г. Бенеке (Beneke), одним из первых применивший действительно научные методы в исследовании конституции, сформулировал цель подобных работ в следующих памятных выражениях. «Различные конституции,— писал он,— и обусловленная ими различная степень сопротивляемости организма создают всего лишь почву для развития некоторых болезней, если индивидуум попадает в неблагоприятные условйя. Не нужно разъяснять, как важна эта точка зрения для общей гигиены и терапии. Правильно распознав различные конституциональные типы и поняв их физиологические различия, мы поможем людям благополучно пройти через все превратности жизни». Однако мы все еще слишком далеки от достижения этой цели.
Внутрииндивидуальная изменчивость морфологических признаков сравнительно невелика. Длина тела почти не изменяется за несколько недель и не очень сильно изменяется за несколько лет (исключение составляют периоды детства и старости). Некоторые физиологические признаки, например группы крови или способность ощущать вкус фенилтиомочевины, изменяются еще меньше и потому представляют собой излюбленные признаки, используемые генетикой человека. Но большая часть физиологических признаков, даже такие стабильные, как выведение с мочей кетостероидов или креатинина и температура тела, изменяются значительно сильнее. Еще менее постоянны признаки, имеющие отношение к поведению, хотя некоторые особенности реакции на обстоятельства и некоторые черты характера и темперамента сохраняются в течение всей жизни и в свете сказанного, по-видимому, являются конституциональными. В настоящее время морфология представля
Глава 23. Анализ и классификация типов телосложения
441
ет самую очевидную и простую, хотя, быть может, и не самую важную, основу конституциональных исследований. Соответственно в этом разделе основное внимание будет уделено конституциональным различиям в телосложении; связанные с ними физиологические различия будут описаны в гл. 24; там же будут рассматриваться и типы поведения, которые также связывают с телосложением.
Между людьми существуют значительные различия по форме тела, которые касаются всех уровней детализации — от размеров и формы всего тела до формы век или мизинца. Различия эти, несомненно, имеют древнее происхождение. Они представляют собой своего рода количественный сбалансированный полиморфизм, если несколько расширить значение этого термина. Подобный полиморфизм телосложения характерен и для многих животных, но мы пока не можем сказать, выражен ли он у них в большей или меньшей степени, чем у человека, так как наши знания о животных в основном базируются на искусственно поддерживаемых, а следовательно, несопоставимых породах.
Можно предположить, что те же самые силы, которые поддерживают полиморфизм единичного локуса, такого, например, как локус, контролирующий группы крови, поддерживают и количественный полиморфизм типов телосложения. В разных условиях и в разное время селективное преимущество, по-видимому, имеет либо тот, либо иной конституциональный тип. Отбор может благоприятствовать сначала одному типу телосложения, а затем другому. Так, существуют доказательства в пользу того, что высокий, худой тип негров-нилотов сформировался благодаря вполне определенным преимуществам, которыми обладает данный тип телосложения при поддержании физиологических функций в условиях жаркого климата (см. часть V). Как и в случае с группами крови, некоторые болезни «отдают предпочтение» одним типам телосложения перед другими. Так, смертность от туберкулеза среди худых и высоких людей выше, чем среди низкорослых и коренастых (см. ниже), а коронарная болезнь чаще поражает полных, мускулистых людей, нежели худых и высоких. Иногда объектом отбора служит тип телосложения в целом (как это произошло с нилотами), но чаще происходит отбор по тем или иным физиологическим признакам, характерным для данного телосложения. Очевидно, между людьми с разными типами телосложения существуют значительные физиологические различия, но мы все еще мало знаем об этом.
Половые различия в телосложении представляют собой самый удивительный пример количественно сбалансированного полиморфизма. Еще до наступления периода полового созревания появляются различия между мальчиками и девочками, правда, не одного типа телосложения (или соматотипа, см. ниже), а в частоте встречаемости различных типов телосложения (или соматотипов).
ш
Часть IV. Рост и конституция человека
Преимущественно мышечные типы встречаются среди девочек редко. (По системе Шелдона может оказаться, что до наступления периода полового созревания девочка 4-4-2 почти не отличается от мальчика 4-4-2, то таких девочек очень мало, а девочек 2-7-1 и вовсе не бывает.) Преимущество этого механизма очевидно; он как бы примиряет две противоположные тенденции: к разнообразию типов телосложения в популяции, с одной стороны, и к специализации типов для выполнения различных задач, с другой. Характер размножения приматов предполагает относительно малоподвижную самку, которая в течение значительных периодов не может принимать участия в борьбе и быстрой смене территории. Таким образом, отбор мог привести здесь к равновесию с преобладанием более подвижных и мощных типов телосложения у самцов и менее подвижных, но в остальном более выгодных типов у самок. Этот процесс не мог идти без некоторой потери физической изменчивости. Хромосомный и эмбрионный механизмы такого равновесия пока не известны; однако изучение индивидуумов с аномальным набором хромосом — XXY и ХО наводит на мысль, что гены Х-хромосомы каким-то образом тормозят развитие мышц. Индивидуумы с кариотипом XYY характеризуются тенденцией к высокорослости, однако они, по-видимому, не более мускулисты, чем обычные люди.
Классификация типов телосложения по внешней форме тела
Было предпринято множество попыток классифицировать все разнообразие типов телосложения, связав их с физиологическими функциями, повседневным поведением и восприимчивостью к заболеваниям. Конечно, не представляет никакого труда произвести ряд антропометрических измерений на том или ином человеке, а затем сопоставить каждую из полученных цифр с размахом значений, полученных по этим признакам на стандартной группе того же пола и возраста. Данный индивидуум может оказаться в 70-м перцентиле по длине тела, в 75-м — по росту сидя, в 65-м — по весу, в 80-м — по ширине головы, в 50-м — по окружности голени и т. д. (Так как распределение антропометрических признаков чаще всего является гауссовым, или нормальным, и лишь распределение толщины подкожного жира, измеренного клипером или на рентгенограммах, оказывается лог-нормальным, то о данном индивидууме можно сказать также, что длина его тела на 0,5 стандартного отклонения выше среднего арифметического и т. д.)
Один из недостатков подобной характеристики состоит в том, что она малоэффективна. Практически все размеры тела положительно коррелируют друг с другом, так что с каждым последующим измерением мы получаем все меньше дополнительной информации. Если мы измерили рост и длину ноги, то мы практически
Глава 23. Анализ и классификация типов телосложения
443
знаем длину руки, и ее измерение не скажет нам ничего нового. Следовательно, нужно провести тщательный отбор измерений и ограничить их число определением нескольких основных компонентов телосложения, не скоррелированных между собой (см. ниже), или определением количества в организме различных тканей — жировой, мышечной, костной.
Второй недостаток этой простой характеристики состоит в том, что она почти ничего не сообщает нам о форме тела, даже если эта информация содержится в цифровых результатах. Это действительно серьезный недостаток, особенно для медицинской практики. Желательно так описать телосложение человека, чтобы у слушателя возникло представление о размерах, форме и составе тела. Простое перечисление множества измерений едва ли поможет нам добиться цели, хотя практически эта задача, вероятно, осуществима.
Человеческое тело даже по своему внешнему виду характеризуется сотнями разнообразных вариаций, и каждая классификация типов телосложения неминуемо выбирает какие-то одни признаки в качестве основных, игнорируя другие. Состоятельность или несостоятельность классификации зависит от того, как она связана с другими аспектами биологии человека и как она отражает проблемы роста, эволюции, физиологии, патологии и поведения. Судя по этим критериям, заслуживают внимания четыре классификации внешней формы тела. Три из них связаны с именами их создателей: итальянского врача Виолы, немецкого психиатра Кречмера, американского психолога Шелдона. Четвертая основана на применении статистического метода, известного под названием «факторный анализ», к многочисленным антропометрическим измерениям.
Система Виолы
Эта классификация, возникшая в начале столетия, а теперь забытая и оставленная, впервые основывалась на обширной системе измерительных признаков. Для практических целей использовали 10 измерительных признаков. Из них путем простых эмпирически установленных преобразований (современная биометрия переживала в те времена период младенчества) получили четыре индекса. По каждому индексу оценивали положение индивидуума, сопоставляя его со стандартной группой того же пола и возраста, и классифицировали тип телосложения как лонгитип, брахитип, нормотип или смешанный тип. Для лонгитипа характерны длинные конечности по отношению к туловищу, относительно широкая грудная клетка и преобладание поперечных размеров над переднезадними. Для брахитипа характерны противоположные соотношения; нормотип занимал промежуточное положение. К смешанному типу относили случаи несоответствия четырех индексов, когда
444 Часть IV. Рост и конституция человека
один из индексов соответствовал одному типу, а второй — другому. (По современной терминологии смешанные типы аналогичны «дис-пл астическим».)
Эта система классификации была положена в основу огромного числа работ; опубликовано несколько сотен статей, посвященных морфологическому анализу, различиям в восприимчивости к болезням и в меньшей степени физиологическим и психологическим соотношениям. Труды Виолы были несправедливо забыты, хотя некоторая часть их все еще имеет значительную ценность.
Система Кречмера
Эта система получила более широкую известность, так как она была переведена на английский язык и вошла в руководства по психиатрии. Однако она целиком базировалась на антропоскопических наблюдениях и была менее объективна, чем система Виолы. Кречмер подробно описал три типа телосложения: пикнический, лептосомный и атлетический. Пикник — широкий с округлыми формами и большим количеством жира, сильный и коренастый. Лептосом — длинный, тонкий, вытянутый. Атлет — мускулистый, с широкими грудной клеткой и плечами и узкими бедрами.
Ныне эта система «вышла из моды», так как она несла в себе роковую ошибку (присущую также, но лишь отчасти, и системе Виолы). Кречмер полагал, что людей действительно можно классифицировать на четко разграниченные, дискретные категории и лишь небольшое число индивидуумов остается за рамками этих категорий. Эта предпосылка, широко распространенная вплоть до 1930 г., приводила в замешательство многих последователей системы, так как каждый честный исследователь должен был попросту признать, что большая часть людей не попадает в уже установленные и, очевидно, крайние варианты.
Система Шелдона
Система Шелдона, хотя и напоминала систему Кречмера, будучи тройной, а не двойной классификацией, с самого начала исходила из ставшего теперь общепринятым предположения, что существуют не дискретные «типы», а непрерывно распределенные «компоненты» телосложения.
«Концепция типов,— писал Шелдон в 1940 г.,— сыграла свою положительную роль в изучении личности, но сами типы при этом, пожалуй, больше всего напоминали шесты, поддерживающие бельевую веревку, иными словами, что-то такое, к чему можно было «подвесить» всю классификацию. По мере того как «веревка» заполнялась, представление о типах отходило на задний план и, наконец, исчезло совсем, уступив место представлению о непрерыв-
Глава 23. Анализ и классификация типов телосложения
445
ном распределении. Развитие шло от представлений о дихотомии до концепции изменчивости по различным пространственным осям».
В выборе трех компонентов Шелдон исходил из практики наблюдения. Он начал с того, что получил около 4000 стандартных фотографий студентов колледжа в обнаженном виде спереди, сбоку и сзади (фото I). Не обращая внимания на общую величину размеров тела, которую его классификация игнорирует, он пытался найти крайние варианты телосложения и выделил три таких крайних варианта, соответствующие концу кривой распределения для каждого компонента. Затем индивидуум получал оценку по каждому компоненту. Это делалось антропоскопическим способом, с использованием оценочной шкалы от 1 до 7 баллов, с равными интервалами между цифрами (это означает, что индивидуум, получивший балл 3, так же отличается от индивидуума, получившего балл 2, как последний — от индивидуума с баллом 1). Таким образом, первый из выделенных крайних вариантов оценивался как 7-1-1, второй — как 1-7-1 и третий — как 1-1-7. Компоненты получили название эндоморфного, мезоморфного и эктоморфного, отразив тем самым не слишком популярную теорию происхождения их от зародышевых листков. Вся система получила название соматотипи-рование. (Следует подчеркнуть, что этот термин надо употреблять лишь в связи с системой Шелдона; он не может служить синонимом любых конституционных классификаций.) Набор трех цифр — это соматотип человека.
Чтобы дать описание компонентов, обратимся к их крайним проявлениям. Крайний эндоморфный вариант (7-1-1) характеризуется шарообразными формами, насколько это вообще возможно для человека. У такого индивидуума круглая голова, большой живот, слабые, вялые руки и ноги, с большим количеством жира на плечах и бедрах, но тонкие запястья и лодыжки. У него относительно крупные печень, селезенка и, по-видимому, кишечник; крупные легкие и сердце несколько отличаются по форме от легких и сердца у лиц, принадлежащих к другим крайним вариантам телосложения. Подобного человека с большим количеством подкожного жира можно было бы назвать просто толстым, если бы все переднезадние размеры его тела (включая грудную клетку и таз) не превалировали над поперечными. При длительном голодании он становится, по Шелдону, просто изголодавшимся эндоморфом, но не приближается по баллам ни к эктоморфу, ни к мезоморфу. По-видимому, тучность в большей или меньшей степени сопутствует этой конституции, и накопление жира в процессе старения, как полагают, прямо связано с оценочным баллом эндоморфного компонента. Хотя соблюдение диеты и физические упражнения позволяют приостановить эту тенденцию, угроза ожирения навсегда сохраняется, тогда как у людей с высоким эктоморфным компонентом совершенно отсутствуют всякие признаки подобной тенденции. Возможно1 что
446
Часть IV. Рост и конституция человека
% ;
И
X
Фото I. Крайние варианты трех компонен-
А. Крайняя эндоморфия, соматотип 6-3-2. Б. Крайняя мезоморфия, соматотип 1 1/2-7-2,
Глава. 23. Анализ и классификация типов телосложения
447
тов соматотипов Шелдона.
В. Крайняя эктоморфия, соматотип 1 1/2-2-6 1/2. Г. Средний тип телосложения, сомато-тип 3-4-4.
448 Часть IV. Рост и конституция человека
в организме индивидуумов с высокой степенью эндоморфии содержится больше жировых клеток, чем у тех, у кого эндоморфный компонент выражен слабо, точно так же, как у людей с высокой степенью мезоморфии, по-видимому, больше мышечных клеток. Одпако это не более чем гипотеза, так как у нас нет количественных гистологических данных о числе мышечных и жировых клеток у человека.
Крайний вариант мезоморфного компонента — это классический Геркулес с преобладанием костей и мышц. У него кубическая массивная голова, широкие плечи и грудная клетка, мускулистые руки и ноги с преобладанием дистальных сегментов над проксимальными. Сердечная мышца относительно велика. Количество подкожного жира минимально, переднезадние размеры невелики.
Крайний пример эктоморфии — это долговязый человек. У него худое, вытянутое лицо, убегающий назад подбородок, высокий лоб, худая узкая грудная клетка и живот, узкое сердце, тонкие, длинные руки и ноги. Подкожный жировой слой почти отсутствует, мускулатура неразвита, но зато по отношению к общим размерам велика поверхность кожи и хорошо развита нервная система.
Вполне естественно, что большинство людей не относится к отмеченным крайним вариантам, в их телосложении в умеренной степени выражены все три компонента. Таким образом, наиболее обычными соматотипами будут 3-4-4, 4-3-3 или 3-5-2. Метод нанесения соматотипов на плоскостную схему показан на рис. 24.1.
Отдельные части тела могут различаться по степени выраженности этих признаков; несоответствие между соматотипами различных областей получило название дисплазии. Теоретически дисплазия представляет большой интерес, но практически определение ее затруднено, так как по системе Шелдона оценка дисплазии дается после определения соматотипов отдельных частей тела — головы, рук, ног, туловища и грудной клетки, а это влечет за собой больше ошибок, чем определение соматотипа всего тела.
Определение соматотипов проводится антропоскопически, по фотографиям, которые должны быть специально сделаны для этих целей. Обследуемый должен стоять в строго определенной позе на вращающейся подставке, меняющей его положение при фотографировании. Камера отодвигается на достаточно большое расстояние, чтобы избежать укрупнения на пленке ближних частей тела по сравнению с удаленными. Эта методика очень подробно разработана, и мы теперь получаем такие фотографии, на которых с помощью специальных инструментов можно точно определить размеры тела (так называемая фотограмметрическая антропометрия).
Однако при определении соматотипов подобные измерения не используются. (Правда, в книге Шелдона приводятся таблицы фотографических измерений и описывается способ определения сома-
Глава 23. Анализ и классификация типов телосложения
449
тотипа с их применением, но эта система не оправдала себя и на практике никогда не использовалась.) Соматотип определяется по фотографиям, которые при необходимости можно сравнить с фотографиями уже известных соматотипов. Кроме того, при определении соматотипа можно пользоваться таблицами индекса рост/ р/вес, приведенного для каждого соматотипа в любом возрасте в атласе Шелдона. Хотя такое определение носит несколько субъективный характер, опытные исследователи, перешедшие теперь к использованию оценочных баллов с интервалом в % и имеющие в своем распоряжении 13 баллов вместо 7 для каждого компонента, в 90% случаев дают одинаковые оценки с точностью до Уг оценочного балла. Лишь в 10% случаев результаты различаются на целый балл, и лишь в очень редких случаях приходится сталкиваться с большими различиями. Освоить технику определения соматотипов нетрудно, достаточно 1—2 мес обучения и время от времени некоторой практики.
Против системы определения соматотипов можно привести ряд возражений. Первое из них заключается в том, что компоненты не являются независимыми. Выражаясь языком многомерного анализа, они облические, а не ортогональные. Между ними существует отрицательная корреляция, так что высокий оценочный балл по одному компоненту до некоторой степени исключает высокие баллы по другим компонентам. Таким образом, существуют сомато-типы 2-3-5, 3-3-5 и 4-3-5 и не существуют 1-3-5, 5-3-5, 6-3-5 или 7-3-5. Точно так же известны соматотипы 6-4-1 и 4-4-4, но не известны 7-7-1, 5-5-5, 3-3-3 или 1-1-1. С отсутствием независимости связаны затруднения биометрического характера при трактовке корреляций соматотипа с другими переменными. Более серьезным является затруднение теоретического характера: казалось бы, естественнее думать о компонентах Шелдона как о независимых, или ортогональных. Ортогональные компоненты, конечно, более удобны. Однако если в основе соматотипических компонентов действительно лежат какие-то физиологические и генетические механизмы, то выбор между облическими или ортогональными компонентами должен основываться на биологических, а не на статистических закономерностях. Вполне возможно, что в ходе эмбрионального развития действительно формируются подобные варианты, связанные между собой отрицательной корреляцией. Однако, судя по последним приводимым ниже данным о количестве жировой, мышечной и костной тканей в разных областях тела, по крайней мере эти ткани развиваются по совершенно независимым направлениям.
Независимость таких показателей, как толщина кости и толщина мышцы (см. ниже), служит серьезным возражением в адрес концепции мезоморфности в современном ее понимании, так как последняя предусматривает как широкие кости, так и развитые мышцы. Тем не менее при оценке этого компонента используется
450
Часть IV. Рост и конституция человека
в основном размер мышц, так что толщину кости можно практически безболезненно опустить из описания.
Субъективность определения соматотипов, безусловно, служит объектом критики, хотя в каком-то отношении она является достоинством, а не недостатком системы. Шелдон писал, что исследователь может лучше всего судить о «морфогенотипе» субъекта по со-матотипу. По определению соматотип остается постоянным в течение всей жизни; меняются внешний вид и размеры тела, но не соматотип. Следовательно, измерительные критерии, используемые при определении соматотипа, должны были бы с возрастом менять-.ся. Различные болезни или гипертрофия мышц, связанная с усиленной физической нагрузкой, изменяют очертания тела, но опять-таки не соматотип. Таким образом, человек, занимающийся определением соматотипа, должен по возможности учитывать возраст, наличие или отсутствие патологических процессов и степень физической нагрузки. Это не так трудно, как кажется. Для опытного глаза не составит труда выявить гипертрофию, связанную с физической нагрузкой, а потеря в весе более 4—5 кг обязательно выявится появлением морщин и складок кожи, отчетливо заметных на фотографиях. Глаз часто говорит больше, чем измерительный инструмент.
Хотя теоретически определение соматотипа лучше всего производить после повторных наблюдений в течение нескольких лет, на практике за основу для окончательной оценки принимается тип телосложения, характерный для 20—25-летнего возраста при нормальном питании. Определение соматотипа у детей в настоящее время не отличается большой точностью, так как необходимые продольные исследования, показывающие, как меняется вид того или иного ребенка к 20 годам, почти полностью отсутствуют. Однако, когда будет накоплено достаточно много фотографий и количественных данных, мы научимся предсказывать соматотип, исследуя 5-летнего, а может, быть и 3-летнего ребенка, сославшись, правда, на некоторую степень неточности в связи с относительной независимостью изменений, характерных для подросткового периода. Определение соматотипа женщин в настоящий момент также представляет трудности из-за отсутствия соответствующего атласа. Обычно применяются те же критерии, что и при определении соматотипов мужчин, а это приводит к избытку эндоморфных типов среди женщин по сравнению с мужчинами и полному отсутствию у них баллов 6 или 7 по мезоморфному компоненту. За последнее десятилетие был предложен целый ряд модификаций методики определения соматотипов, но ни одна из них не может быть безоговорочно рекомендована. Парнелл (Parnell) предложил систему, основанную только на антропометрических измерениях, с помощью которой можно приближенно оценить соматотип. Однако этот метод оказался менее эффективным^ чем система стандартных уравнений мно
Глава 23. Анализ и классификация типов телосложения 451
жественной регрессии между соматотипом и размерами тела, разработанная Деймоном (Damon) и его коллегами. Хит и Картер (Heath a. Carter) предложили вариант, при котором допускаются оценочные баллы выше 7, для того чтобы более прямолинейно соотнести компоненты с такими признаками, как длина тела, вес, толщина жировых складок и ширина кости. Преимущества этих методов не вполне очевидны. Система Шелдона, выдвинутая в 1940 г. и проиллюстрированная вышедшим в 1954 г. атласом, очень последовательна. Независимо от ее теоретического обоснования она чрезвычайно удобна и информативна. Чтобы сразу же распознать мужчину среди сотни других, достаточно сказать о нем, что его соматотип 3-2-5.
Факторный анализ телосложения
Классификация с помощью факторного анализа вытекает из чисто количественного, измерительного подхода к определению телосложения человека. Этот подход не противопоставляется описанным выше, а дополняет их. Факторный анализ — это ветвь многомерного статистического метода, используемая для сведения множества коррелирующих между собой измерительных признаков к небольшому числу факторов, определяющих основную долю изменчивости первоначальных измерений. Факторы могут быть об-лическими (коррелирующими между собой), ортогональными или независимыми — по выбору исследователя. Исходным пунктом служит таблица коэффициентов корреляции для группы из, скажем, десятка или более измерительных признаков. Конечным результатом являются несколько, например 3, ортогональных факторов (А, Б, В), которые в совокупности определяют около 80% общей изменчивости данного десятка измерений. Факторный анализ позволяет в физической антропологии избежать широко распространенной ошибки — измерять одно и то же несколько раз.
Существует несколько разных способов факторного анализа. Применение в данной связи одного из них, наиболее употребительного, позволяет выделить общий фактор размера тела, а вслед за ним несколько групповых факторов, которые сами могут быть подразделены на более мелкие факторы. Результаты анализа различных корреляционных таблиц хорошо согласуются и определяют ряд факторов, приведенных в табл. 23.1.
В действительности дело обстоит, вероятно, не так просто, и к приведенному в таблице следует добавить факторы размера и формы головы, кисти и стопы.
Факторы представляют собой, по существу, статистические величины, такие же, как средние квадратические отклонения. A priori их нельзя непосредственно связывать с генетическими и физиологическими механизмами. Но при анализе формы тела они бу-
452
Часть IV. Рост и конституция человека
Таблица 23.1
Классификация телосложения в виде ортогональных подразделенных групповых факторов в результате применения факторного анализа11
'2. Линейность
1. Общий
размер
тела
3. Нелинейность
г 2Аа. Длина проксималь-
/2А. Длина конечно- ных сегментов
стей 2А6. '2Ба. 2Б6. Длина дистальных сегментов Длина шеи Длина грудной
2Б. Длина туловища < 2Бв. клетки Длина поясничной области
2В. Высота головы и (Различные части
лица ' ЗА. Толщина конечностей лица)
ЗБ. Толщина тулови- /ЗБа. Поперечный размер
ща 1 ЗБб. Переднезадний размер
ЗВ. Поперечные размеры головы и лица (Различные части лица)
*) С небольшими изменениями из Tanner, 1953. Growth and Constitution. В ки. «Anthropology today» (ed. A. L. Kroeber). Chicago University Press.
дут полезными лийть в том случае, если окажется, что они действительно отражают какие-то физиологические закономерности и прямо связаны с ростом. Можно надеяться, что подобные взаимоотношения удастся обнаружить, по крайней мере если при выборе измерений для факторного анализа мы будем руководствоваться четкими биологическими предпосылками. Многомерный статистический анализ может послужить основой для дальнейшего развития классификации и понимания физических различий. Одно из обнадеживающих направлений этого развития описывается в следующей главе.
Тканевые компоненты и анализ телосложения
Описанные выше классификации телосложения базировались на внешних признаках. В основу классификации могут быть положены количественные различия в структуре тканей, например относительное содержание жировой, мышечной и костной тканей у различных индивидуумов. Эти два подхода не являются чем-то
Глава 23. Анализ и классификация типов телосложении
453
независимым и взаимоисключающим. Некоторые чисто внешние признаки, например общий размер тела, могут практически не зависеть от структуры. Однако форма и структура неизбежно связаны: толстый человек, как бы он ни старался, будет всегда иметь округлые очертания, человек с развитой мускулатурой никогда не будет казаться хрупким и долговязым. Для каждой отдельной ткани соотношения между структурой и формой более независимы: у человека с определенным процентом костной ткани могут быть длинные ноги и короткое туловище и, наоборот, короткие ноги и длинное туловище.
Современные химические и физические методы позволяют определить общее содержание жира и воды в теле человека с точностью до ±15%, а количество мышечной ткани определяется в соответствующих условиях с помощью изотопа (40К) калия. Однако подобные методы позволяют лишь определить общее содержание того или иного компонента, но не указывают, где располагается, скажем, жир и как он влияет на форму тела.
Объединение обоих методов стало возможным благодаря использованию рентгеновских лучей для измерения толщины жирового, мышечного и *костного компонентов конечностей, а также благодаря изобретению специального калипера, предназначенного для измерения толщины подкожного жирового слоя. Для этого складка кожи с подкожной клетчаткой отделяется от подлежащего мышечного слоя и измеряется калипером. В некоторых областях тела это можно проделать достаточно точно.
Для рентгенограмм лучше всего выбирать плечо, голень или бедро и пользоваться при этом мягкими рентгеновскими лучами в малой дозе; свинцовый фартук должен полностью защищать от облучения половые железы. На рентгенограммах плеча толщина подкожного жирового слоя, а также мышечного и костного компонентов измеряется на уровне середины расстояния между акромионом и головкой лучевой кости. На бедре измерение производится на границе трети длины, считая вверх от основания мыщелков бедра. На голени толщина жирового слоя, мышц и большой берцовой кости (малая берцовая кость в расчет не берется) определяется на уровне максимального диаметра.
Установив корреляцию результатов измерений, произведенных по рентгенограммам, можно выявить четкие закономерности (табл. 23.2) (подобные результаты дает и анализ вычисленных площадей компонентов на поперечных сечениях конечностей). Все три измерения одной и той же ткани характеризуются положительной корреляцией друг с другом, но не связаны с другими тканями. Так, толщина мышц голени коррелирует с толщиной мышц плеча и бедра, но не связана с толщиной жирового и костного компонентов голени или другой части тела. В табл. 23.2 приведены средние коэффициенты для трех корреляций: жир — жир, мыш-
454
Часть ГУ. Рост и конституция человека
Таблица 23.2
Средние коэффициенты корреляции между толщиной подкожного жирового слоя, мышцы и костя (плечевая, бедренная и большая берцовая) по рентгенограммам плеча, бедра и голени 166 молодых женщин и 125 молодых мужчин1»
Пол Жир Мышца Костъ
Жир ж 0,66 — -
м 0,75 — —
Мышца ж 0,09 0,43 —.
м 0,08 0,49 —
Кость ж 0,09 0,13 0,37
м —0,07 0,09 0,48
») Tanner, Healy, Whitehouse (неопубликованные данные).
ца — мышца и кость — кость, а также соответствующие перекрестные коэффициенты корреляции между компонентами. Расположенные по диагонали внутритканевые коэффициенты корреляции для жира высокие, а для кости и мышц умеренные. Межтканевые корреляции, и в том числе корреляция мышцы — кость, практически отсутствуют; коэффициенты их равны нулю. Вследствие этого можно выделить три ортогональных фактора тканевых компонентов, определяющие соответственно толщину жира, мышц и костей конечностей. Как показывают дальнейшие измерения, толщина костей конечностей складывается из двух независимых элементов: ширины костномозговой полости и толщины кортикального слоя, причем последний связан скорее с развитием мышц, чем с шириной костномозговой полости.
Подобный анализ можно продолжить и дальше, включив в него ряд других измерительных признаков скелета, полученных с помощью обычной антропометрической техники, а также показатели толщины кожной складки — с помощью калипера и данные о внешней форме тела, полученные фотограмметрическим методом. Нужно вычислить коэффициенты корреляции (следует использовать логарифмы величин, характеризующих толщину складки, так как эти величины характеризуются лог-нормальным распределением) и составить матрицу для факторного анализа. По мере увеличения числа признаков картина, естественно, усложняется, но три первичных ортогональных компонента по-прежнему сохраняются. В табл. 23.3 приведен пример подобного анализа. Колонки цифр представляют собой весовые значения (нагрузки) каж-
Таблица 23.3
Факторный анализ (максимум правдоподобия) измерений 166 молодых женщин и 125 молодых мужчин. Весовые значения измерений в первых пяти ротационных ортогональных факторах 1(
I II III IV V
Ж м ж м ж м Ж м ж м
Толщина жирового слоя -0,11
плечо 0,81 0,92 0,09 0,05 0,09 0,10 0,01 0,02 -0,09
голень 0,83 0,82 -0,01 0,03 0,03 -0,7 -0,15 0,00 -0,02 0,11
бедро Толщина мышцы 0,86 0,87 0,01 -0,05 —0,10 -0,06 0,01 -0,02 0,08 0,03
плечо 0,13 0,04 0,52 0,70 0,06 -0,04 0,15 0,10 0,05 0,02
голень 0,06 0,10 0,65 0,67 -0,01 0,10 0,11 -0,22 -0,12 0,05
бедро Толщина кости —0,04 0,07 0,72 0,84 0,10 0,30 0,12 0,26 0,01 0,07
плечо -0,05 -0,08 0,12 0,09 0,28 0,28 0,51 0,56 -0,01 —0,03
голень 0,05 —0,12 0,03 -0,09 0,45 0,37 0,51 0,75 0,06 -0,03
бедро 0,10 —0,06 -0,11 -0,01 0,47 0,37 0,51 0,57 0,06 0,04
Рост сидя —0,34 —0,10 0,21 0,17 0,69 0,72 0,00 0,11 -0,23 0,02
Длина ноги (от седалищного бугра) -0,05 0,02 0,03 -0,28 0,66 0,77 0,21 -0,10 0,48 0,55
) Tanner, Healy, Whitehouse (неопубликованные данные).
Продолжение
I II III IV V
Ж м ж м Ж м Ж м Ж м
Биакромиальный диаметр -0,17 0,02 0,26 -0,16 0,63 0,75 0,07 0,07 0,22 0,11
Тазогребневый диаметр 0,02 0,04 0,18 -0,07 0,46 0,54 0,29 0,03 —0,17 0,03
Межмыщелковый диаметр плеча 0,17 0,03 0,35 0,23 0,49 0,55 0,41 0,24 —0,00 -0,12
Межмыщелковый диаметр бедра 0,40 0,31 0,36 0,30 0,53 0,53 0,28 0,22 —0,14 —0,02
Процент общей изменчивости 16,4 16,2 11,0 13,0 17,2 20,1 8,2 9,7 2,8 2,4
Средние весовые значения
жир 0,83 0,87 0,03 0,01 0,01 —0,01 -0,04 0,00 -0,01 0,01
мышца 0,05 0,07 0,63 0,74 0,05 0,09 0,13 0,05 -0,02 0,05
Все скелетные измерения 0,03 0,01 0,01 0,02 0,52 0,54 0,31 0,27 0,03 -0,5
Ширина костей конечностей 0,02 —0,09 0,16 0,00 0,40 0,34 0,51 0,63 0,04 —0,01
Длина ноги -0,05 0,02 0,03 -0,28 (0,66 0,77) 0,21 -0,10 0,48 0,55
Глава 23. Анализ и классификация типов телосложения
457
дого измерительного признака в факторах; эти весовые значения варьируют от 0 до 1 и интерпретируются подобно коэффициентам корреляции.
Жир
Все величины, характеризующие толщину слоя подкожного жира, хорошо коррелируют друг с другом, этим определяется фактор жира. Как видно из табл. 23.3, для этого фактора (I) высокие весовые значения имеют результаты измерения количества жира на плече, бедре и голени и низкие весовые значения — другие измерения. По другим данным, у женщин существует дополнительный фактор жира на туловище кроме жира на конечностях; хотя определенные данные, касающиеся выраженности этого фактора у мужчин, отсутствуют, различия в кривых роста, характеризующих увеличение жирового слоя под лопаткой и над трицепсом у мальчиков-подростков, не исключают возможности его существования. Стойкие региональные колебания, такие, например, как избыточное отложение жира на руках при его отсутствии на ногах или избыточное отложение жира на груди при его отсутствии на животе, по-видимому, не наблюдаются. Однако некоторые узколокальные колебания встречаются; так, в одних случаях происходит избыточное отложение жира на икрах, в других — в области плеч. Эти одномерные факторы известны под названием специфических. Фактор туловище — конечности и специфические локальные факторы определяют характерную форму расположения жира как у толстого, так и у худого человека. Мы можем предположить, что эти «факторы формы расположения жира» отражают количество жировых клеток в той или иной области тела, а «фактор общего количества жира» — степень заполненности этих клеток жиром. По некоторым данным, количество жира в брюшной полости у молодых людей коррелирует с количеством подкожного жира. В старших возрастных группах эта корреляция становится, по-видимому, менее тесной. Вероятно, вследствие этого фактор жира достаточно хорошо отражает содержание жира в организме. Это можно просто подсчитать, измерив две кожные складки (на задней поверхности плеча, над трицепсом и в области нижнего угла левой лщщтки), ростовые кривые которых были представлены на рис. 19.(Ги 19.7, а затем произвести логарифмическое преобразование и сложить полученные величины.
Факторный анализ (табл. 23.3) показывает, что из всех скелетных размеров лишь рост сидя и межмыщелковый диаметр бедра (ширина колена) имеют большие весовые значения в факторе жира (I). В случае ширины колена мы сталкиваемся с чисто технической причиной, поскольку какое-то количество жира входит в значение признака при его измерении. Отрицательное весовое значение та
458 Часть IV. Рост и конституция человека
кого признака, как рост сидя, отчетливо показывает, что способность к накоплению жира среди молодых взрослых сильнее выражена у индивидуумов с коротким туловищем, чем с длинным. Эмбриологические и физиологические причины этого явления пока не изучены. Приверженцы метода соматотипирования утверждают, что степень эндоморфии может быть определена по форме скелета, правда, лишь теоретически и с некоторыми трудностями. Опи считают короткое туловище и преобладание переднезадних скелетных размеров показателями эндоморфии. Возможно, что эндоморфия, как они ее себе представляют, отражает число жировых клеток, а толщина — количество жира на одну клетку.
Мышцы
Коэффициенты корреляции между толщиной мышечного слоя в различных частях тела не так высоки, как соответствующие коэффициенты для жира, но все же довольно значительны. Фактор мышц (табл. 23.3, II) можно приблизительно оцепить путем простого сложения толщины трех мышечных слоев на рентгенограммах конечностей. Как показывает фотограмметрия, этот фактор определяет развитие мускулатуры всего тела, включая туловище и спину. Мы не имеем четких данных о стойких региональных различиях, таких, например, как избыточное развитие мышц руки при слабом развитии мышц ноги, хотя узколокальные колебания встречаются.
Скелет
Со скелетом дело обстоит гораздо сложнее. Но если мы выделим ортогональные факторы (факторы III, IV и V в табл. 23.3), то результаты окажутся приемлемыми. Одна из самых больших трудностей факторного анализа заключается в том, что он не приводит к однозначному решению; даже придерживаясь ортогональности, можно выделить другие компоненты, которые с не меньшим успехом охарактеризуют первоначальные корреляции между признаками. Исследователь должен руководствоваться прежде всего биологическими соображениями. Было бы по меньшей мере неразумным искать замену таким компонентам, как жир и мышцы; другое дело — конфигурация скелета, оценка которой более субъективна и дискуссионна.
Как видно из табл. 23.3, скелет можно описать с помощью следующих факторов: общего фактора размера скелета (III), в котором учитываются все скелетные измерительные признаки; фактора ширины костей конечностей (IV), отражающего ширину диафизов длинных костей, и фактора длины конечностей (V), отражающего длину конечностей по отношению к длине туловища.
Глава 23. Анализ и классификация типов телосложения
459
Рис. 23.1. Схема последовательного определения формы тела по пяти ортогональным факторам (4, Б, В -— факторы размера и формы скелета, Г и Д — мягкие ткани) (Tanner, Healy a. Whitehouse, неопубликованные данные). А. Форма тела индивидуума определяется числом баллов по фактору общего размера скелета (остальные факторы скелета имеют средние значения). Б. Индивидуум имеет балл выше среднего по фактору ширины костей конечностей. В Индивидуум имеет балл выше среднего по фактору длины костей конечностей Г Индивидуум имеет дополнительный балл по фактору мышц. Д. Индивидуум имеет дополнительный балл по фактору жира.
Дальнейший анализ показал, что ширина костей конечностей не связана с шириной тел позвонков, так что грацильность длинных костей не зависит от грацильности позвоночника. Более того, длина туловища складывается из нескольких совершенно независимых компонентов: длины головы и шеи, длины грудной клетки (расстояние от верхней точки грудины до пупка) и длины таза (расстояние от уровня пупка до седалищного бугра). Длина грудной клетки и таза при таком измерении совершенно независимы. Тазогребневый диаметр варьирует также в значительной степени независимо от других скелетных размеров. При определении типа телосложения следует учитывать «фактор задней части тела», который используется также при оценке формы тела крупного рогатого скота. Длина шеи в большей мере коррелирует с длиной конеч
460
Часть IV. Рост и конституция человека
ностей, чем с длиной грудной клетки. Длинные руки, ноги и шея обычно встречаются в комплексе. Коррелирующие между собой размеры кисти и стопы в значительной мере независимы от перечисленных факторов. То же самое следует сказать о размере головы и отдельных размерах лица.
Различия между признаками обусловлены различиями в скоростях роста. Поскольку многие из описанных корреляций присущи не только человеку, но и животным, механизмы их возникновения можно установить, исследуя инбредные линии животных. Основываясь на табл. 23.3, можно привести в качестве примера такое объяснение механизмов корреляции. У мужчин длина ноги имеет отрицательное весовое значение в факторе мышц (—0,28). Мы знаем, что для мужчин с развитой мускулатурой, как правило, характерно раннее созревание (см. выше), а следовательно, более короткий период ускоренного роста ног по сравнению с туловищем, поэтому у таких мужчин относительно короткие ноги.
Первые, самые простые результаты анализа тканевых компонентов приведены схематически на рис. 23.1. Основываясь на ряде ортогональных, или независимых, факторов, мы можем реконструировать основные элементы телосложения (за исключением головы, кисти, стопы и различных мягких частей) в виде баллов по: а) общему размеру скелета (рис. 23.1, Л); б) ширине костей конечностей (рис. 23.1, Б, где кости конечностей несколько шире средних); в) длине костей конечностей (рис. 23.1, В, где кости конечностей несколько длиннее средних); г) толщине мышц (рис. 23.1, Г); д) толщине слоя жира (рис. 23.1, Д).
Факторы Б-\-В вместе определяют, насколько размеры конечностей выходят за пределы средних, обусловленных фактором общего размера скелета (А). Отношение факторов Б/В определяет форму конечностей, т. е. отношение их длины к ширине. Вполне естественно, что эта схема не учитывает многих признаков, в том числе формы туловища. Однако это лишь начало. Мы можем объективно измерить все факторы и выяснить эмбриологический смысл по крайней мере некоторых из них. Эти компоненты приложимы в равной мере к анализу телосложения как у мужчин, так и у женщин, хотя по некоторым из них женщины получают более высокие, а по другим — более низкие баллы, чем мужчины. Для объективного определения половых различий, о которых уже шла речь в главах о росте, необходимы новые факторы.
Глава 24
СВЯЗЬ ТЕЛОСЛОЖЕНИЯ С ФИЗИОЛОГИЧЕСКИМИ ФУНКЦИЯМИ, БОЛЕЗНЯМИ И ПОВЕДЕНИЕМ
Физиологические функции
Мы все еще поразительно мало знаем о связи между телосложением и физиологическими функциями. Однако такая связь, несоменно, должна существовать, по крайней мере между телосложением и эндокринной функцией и метаболизмом. Ведь трудно себе представить, чтобы у людей с высокоразвитым мышечным компонентом уровень эндокринной секреции был таким же, как у тех, у кого мышечный компонент развит слабо, или чтобы люди с противоположными баллами по шкале эндоморфии не отличались бы по характеру обмена веществ. Трудно предположить также, чтобы различия в структуре и функциях центральной нервной системы совершенно не были связаны с телосложением. Но вопросы эти все еще совершенно не изучены.
В старой литературе по вопросу о конституции мы находим многочисленные предположения о связи между эндокринной функцией и телосложением. На основании зтой предполагаемой связи была построена даже система классификации по «биотипам». Большинство этих предположений не имеет под собой научной основы и исходит из ненадежных клинических аналогий. Телосложение, по-видимому, действительно тесно связано с активностью эндокринной системы; оно может быть также связано с различной численностью клеточных рецепторов, которые переводят гормональные сигналы в информацию, непосредственно регулирующую жизнедеятельность клетки. Лишь в самое последнее время появились такие тесты для определения эндокринного статуса, которые позволяют выявить индивидуальные различия по отдельным показателям; методика определения рецепторных белков пока находится в стадии разработки.
Можно предположить, что с телосложением должны быть связаны только относительно стабильные физиологические функции, не изменяющиеся неделями и даже годами. Следовательно, прежде всего нужно найти поддающиеся измерению функциональные признаки с низкой внутрииндивидуальной и высокой межиндивидуальной изменчивостью. С помощью соответствующего статистического метода, предусматривающего повторные измерения в течение определенного периода времени, можно отделить стабильные компоненты от изменчивых и выяснить связь между стабильными зна
462
Часть IV. Рост и конституция человека
чениями для каждого индивидуума и различными индексами его телосложения.
Результаты подобного исследования приведены в табл. 24.1. Содержание в моче 17-кетостероидов и 17-кетогенных стероидов характеризует два различных аспекта функции надпочечников. 17-кетостероиды представляют собой конечные продукты метаболизма андрогенных гормонов надпочечников и тестостерона, выделяемого яичками. 17-кетогенные стероиды — конечные продукты метаболизма гормонов коры надпочечника, которые наряду с некоторыми другими функциями участвуют в регуляции углеводного обмена. В норме у молодых людей, живущих в обычных условиях, содержание этих веществ в моче относительно постоянно. Столь же постоянно содержание в моче креатинина—вещества, которое главным образом или даже полностью образуется в мышцах.
Таблица 24 1
Коэффициенты корреляции между стабильными (межинднвидуальными) значениями суточного содержания в моче различных химических веществ и величиной жирового, мышечного и костного компонентов конечностей, весом и длиной тела3)
Жир Мышцы Кость Длина тела Вес
17-кетостероиды 0,10 0,39 О 0,23 0,05 0,31 2>
17-кетогепные стероиды -0,12 0,10 0,49 ’> 0,29 2> 0,30 2>
Креатинин 0,16 0,77 2> 0,26 0,17 0,76 2>
5%-ный уровень значимости.
1%-ный уровень значимости
•) Tanner, Healy, Whitehouse and Edgson, 1959
Экскреция 17-кетостероидов в значительной мере коррелирует с количеством мышечной ткани в организме, причем коэффициент множественной корреляции с толщиной мышечного слоя и поверхностью тела (приблизительная мера общей мышечной массы) достигает 0,56. В процессе того же исследования была выявлена довольно неожиданная корреляция между содержанием в моче 17-кетогенных стероидов и толщиной диафизов длинных трубчатых костей, точнее шириной костномозговой полости. Экскреция этой группы гормонов в большей степени, чем экскреция 17-кетостероидов, коррелировала с общими размерами тела. Как и следовало ожидать, была выявлена тесная связь между содержанием креатинина в моче и мышечной массой.
По этим результатам еще нельзя судить о характере различий в интенсивности секреции гормонов самими надпочечниками, так
Глава 24. Физиологические функции, болезни и поведение
463
как количество стероидов, выделяемых с мочой, зависит от утилизации их в тканях, связывания их в печени и способности почек к их выведению. Можно, однако, не сомневаться, что дальнейшие исследования в этом направлении должны выявить зависимость формирования и поддержания того или иного типа телосложения от гормональных агентов и соответствующих рецепторов.
По поводу других соотношений нам почти ничего не известно. Принято считать, что у высоких индивидуумов с выраженным эктоморфным компонентом повышена секреторная активность щитовидной железы, однако это предположение пока не подтверждено убедительными исследованиями с использованием современных методов. Как показали некоторые работы, у коренастых индивидуумов артериальное давление (систолическое и диастолическое) несколько выше, чем у высоких и тонких. Однако поскольку артериальное давление определяют с помощью манжетки, то вполне возможно, что полученные данные не отражают истинного положения вещей, а являются следствием систематической ошибки измерения, обусловленной большими размерами окружности руки у коренастых людей. Было показано, что объем крови относительно веса тела несколько выше у людей с выраженным мезоморфным компонентом и ниже у индивидуумов эктоморфного и эндоморфного типов со слабо выраженным мезоморфным компонентом. Время реакции, по некоторым сведениям, уменьшается с увеличением степени эктоморфии. Однако это требует подтверждения.
Болезни
Не вызывает сомнений, что люди определенного телосложения более склонны к некоторым заболеваниям. По крайней мере в отношении одной болезни — туберкулеза легких — исследования типов телосложения до начала заболевания показали, что конституциональные различия предшествуют болезни, а не являются ее результатом.
Туберкулез легких — это болезнь, характерная для эктоморфного типа телосложения или по крайней мере для людей с небольшим весом по отношению к росту. Как показало одно из продольных исследований, первичное инфецирование туберкулезной палочкой не зависит от типа телосложения, но последующее распространение возбудителя в организме и появление через несколько лет клинических симптомов заболевания при одинаковых условиях жизни значительно чаще наблюдаются у высоких людей с относительно низким весом, чем у коренастых. По-видимому, данный тип телосложения благоприятствует заболеванию, хотя, каким именно образом, мы пока не знаем. Один из недостатков этой и подобных работ заключается в том, что при характеристике типа телосложе-
464
Часть IV. Рост и конституция человека
иия исследователи часто ограничивались измерениями роста и веса, не отделяя жировой компонент от мышечного и тяжелые кости от легких. Это исключает возможность физиологического анализа.
Инфаркт миокарда также связан с определенным типом телосложения: он чаще встречается у людей с большим весом по отношению к росту. Причины этого пока не ясны, но, по-видимому, склонность к этому заболеванию каким-то образом связана с избыточным развитием как жировой, так и мышечной ткани. В основе этих соотношений могут быть и физиологические и социальные причины. Вполне возможно, что индивидуумы мезоморфного типа чаще оказываются в таких условиях, которые способствуют развитию коронаротромбоза, например в условиях стресса. По некоторым данным, люди мезоморфного типа с нетренированными мышцами характеризуются большей восприимчивостью к болезни, чем менее мускульные, но столь же мало тренированные. Физиологический механизм подобных различий в склонности к коронаротромбозу, быть может, сводится к различной степени утилизации андрогенных гормонов при физической нагрузке или к чему-нибудь аналогичному.
Другим примером болезни, связанной с телосложением, является диабет. Известны две клинические формы диабета. Одна из них проявляется в молодом возрасте, другая характерна для средних возрастных групп. При первой форме диабета для устранения клинических симптомов требуются высокие дозы инсулина, при второй симптомы купируются малыми дозами. Физиологическая основа этих двух форм болезни, по-видимому, неодинакова; во всяком случае, наблюдаются отчетливые различия по типу телосложения. У больных поздней формой диабета более выражен эндоморфный компонент, чем у больных ранней формой.
Судя по старой итальянской литературе, случаи рака чаще встречались у брахитипов, чем у лонгитипов. Недавние исследования Деймона (Damon) и его сотрудников, по-видимому, подтверждают эту точку зрения, по крайней мере в отношении рака молочной железы.
Что касается психических заболеваний, то в этой сфере патологии отмечается явная связь с телосложением. Шизофрения чаще встречается у лиц с высоким эктоморфным компонентом; маниакально-депрессивный психоз наблюдается у людей с низкой степенью эктоморфии и высокими мезоморфным и эндоморфным компонентами; к паранойе более склонны мезоморфные типы. Все эти данные неоднократно подтверждались исследованиями с применением различных методов и могут считаться доказанными, но механизмы, лежащие в основе этих корреляций, пока не изучены. Полагают, что характер симптомов при неврозах также связан с телосложением. Истерия и депрессия более типичны для мезоморфного и эндоморфного типов, симптомы тревоги и страха — для эктоморфного.
Глава 24. Физиологические функции, болезни и поведение 465
Как мы видим, в эмпирических данных, касающихся связи типов телосложения с патологией, нет недостатка, тогда как каузальный анализ этих соотношений, а также лежащий в их основе физиологические и социологические механизмы далеко не изучены. Представляется наиболее вероятным, что в большинстве случаев телосложение является фенотипическим выражением генного комплекса, воздействующего на пенетрантность и экспрессивность отдельных генов, определяющих предрасположение к тем или иным заболеваниям. R настоящее время эта область широко открыта для изучения.
Поведение
Почти каждый, кто занимался изучением конституции человека, интересовался также и соотношением между телосложением и поведением. Старые авторы считали подобную взаимосвязь само собой разумеющейся и не видели необходимости в статистических доказательствах. Сравнительно недавно в центре внимания оказался вопрос о том, какие элементы поведения более или менее стабильны в течение жизни и связаны ли они с физическими различиями, а если такая связь вообще существует, то с какими именно различиями и насколько тесно.
Умственные способности, оцениваемые по баллам коэффициента интеллекта, остаются относительно стабильными и весьма слабо коррелируют с телосложением. В значительно большей степени выражена корреляция между телосложением и темпераментом или глубочайшими свойствами личности с присущими ей навыками поведения. Кречмер первоначально выделил два типа темперамента — циклотимический и шизотимический, более или менее соответствовавшие экстравертному и интравертному типам Юнга. Кречмер связывал их с типами телосложения: циклотимию — с пикническим телосложением, шизотимию — с лептосомным. Позже он описал и тип темперамента, характерный для атлетического телосложения.
Шелдон продолжил эту работу, использовав при описании темперамента не тип, а компонент, точно так же как и в классификации телосложения. Выделив пятьдесят признаков для определения постоянных аспектов поведения на основании 50-часового клинического обследования, он начал оценивать их у 30 человек. Все признаки разделились на три категории, и на этой основе Шелдон выделил три компонента темтерамента, каждый из которых характеризовался двадцатью признаками. Каждый признак оценивался по семибалльной шкале, а средний балл по двадцати признакам определял весь компонент. Набор признаков приведен в табл. 24.2. Компоненты темперамента получили название «висцеротонии», «соматотонии» и «церебротонии». Определив число баллов по компо-
466
Часть IV. Рост и конституция человека
Таблица 24.2
Набор признаков из шкалы темпераментов по Шелдону
№ п/п Висцеро тония Соматотония Церебротония
1 Расслабленность в осанке и движениях Уверенность в осанке и движениях Заторможенность в движениях, скованность в осанке
2 Любовь к комфорту Любовь к приключениям Необщительность, социальная заторможенность
3 Жажда похвалы и одобрения Эмоциональная черствость Скрытность, эмоциональная сдержанность
4 Легкость в общении и выражении чувств; висцерото-ническая экстра-версия Экстраверсия в поступках, но скрытность в чувствах и эмоциях; соматопическая экстраверсия Церебротоническая интраверсия
5 Общительность и мягкость в состоянии опьянения Агрессивность и настойчивость в состоянии опьянения Устойчивость к действию алкоголя и других депрессантов
6 Тяга к людям в тяжелую минуту Тяга к действию в тяжелую минуту Тяга к одиночеству в тяжелую минуту
7 Ориентация к детству и семейным взаимоотношениям Ориентация к юношеской деятельности Ориентация к поздним периодам жизни
центам темперамента у 200 испытуемых и сопоставив их с данными по компонентам соматотипов, Шелдон установил коэффициент корреляции порядка 0,8 между висцеротонией и эндоморфией, соматотонией и мезоморфией, церебротонией и эктоморфией.
Это несколько более тесная взаимосвязь, чем полагали, и работа Шелдона подверглась критике не столько из-за наличия каких-либо противоречащих ей данных, сколько из-за недоверчивости оппонентов. Здесь возникают, по-видимому, две основные проблемы. Во-первых, необходимо проследить, в какой мере оценки отдельных признаков темперамента по Шелдону согласуются друг с другом на протяжении всей жизни и как они изменяются в различные ее периоды. Необходимо изучить и генетическую основу компонентов темперамента. Классификации, построенные на признаках подобного рода, всегда представляют статичным то, что на самом деле изменчиво. Во-вторых, работа нуждается в подтверждении, при этом нужно по возможности стараться избежать «круговой поруки»,
Глава 24. Физиологические функции, болезни и поведение
467
т. е. надо сделать так, чтобы психолог и специалист по сомато-типам работали отдельно, причем психолог должен пользоваться лишь записями беседы с обследуемым и не иметь представления о его телосложении. Одно подобное исследование было осуществлено, и хотя оно касалось лишь части проблемы, а именно соотношений между доминирующей церебротонией и доминирующей экто-морфией, выводы Шелдона в этой части получили подтверждение.
Существуют две теории возможного происхождения связи между телосложением и темпераментом. Одна из них объясняет это действием плейотропных, или тесно сцепленных, генов, другая — влиянием среды на ранних этапах развития. Согласно первой теории, гены, контролирующие телосложение, контролируют, кроме того, и развитие мозга и эндокринной системы, определяющих темперамент; в другом варианте предполагается существование отдельных генов для каждого признака, которые, однако, тесно сцеплены между собой. Согласно второй точке зрения, ребенок с выраженной степенью мезоморфии, растущий среди других детей, случайно осознает, что он сильнее остальных и может их победить; впоследствии он привыкает выделяться из среды своих сверстников, потому что это ему нравится и этого ждут от него другие. Эти две теории, конечно, не исключают одна другую. Вполне возможно, что генетическая тенденция реализуется при наличии соответствующих условий.
Люди с различным телосложением обычно выбирают различные пути, и это, безусловно, свидетельствует о различных свойствах темперамента — о предпочтении того или иного образа жизни. На рис. 24.1 показано распределение соматотипов в достаточно представительных выборках — у студентов Оксфордского университета, у курсантов младших курсов Королевской военной академии в Сандхерсте и у группы будущих преподавателей физкультуры — учащихся физкультурного колледжа (Лафборо). Мезоморфный компонент оказался у курсантов более выраженным, чем у студентов Оксфордского университета, а у будущих преподавателей физкультуры более выраженным, чем у курсантов. (Использованный здесь метод представления распределения соматотипов на плоскости довольно иллюстративен, но также нуждается в применении критериев значимости к полученным величинам.)
Это простой и, по-видимому, закономерный результат. Человек вряд ли выберет карьеру преподавателя физкультуры или кадрового военного, если он недостаточно силен и ловок. Были исследованы и другие профессиональные группы. Оказалось, что люди, занятые исследовательской работой на заводе, независимо от их заработной платы, более высокие и худые, чем работающие на производстве. Водители транспорта на дальние расстояния и летчики отличаются от остальной части населения более высоким мезоморфным компонентом. Студенты Гарвардского университета1
Б Мезоморфия
Глава 24. Физиологические функции, болезни и поведение
469
Рис. 24.1. Распределения соматотипов в трех выборках.
А. Студенты Оксфордского университета. Б. Курсанты Королевской военной академии в Сандхерсте. В. Студенты физкультурного колледжа.
изучающие естественные и социальные науки, характеризуются более атлетическим телосложением по сравнению со студентами искусствоведами, филологами и философами; для будущих инженеров, медиков и зубных врачей характерна более выраженная мезоморфия и менее выраженная эктоморфия, чем для студентов физиков и химиков. Юноши-правонарушители, бесспорно, отличались развитием мезоморфного компонента выше среднего.
Все эти результаты согласуются в своих выводах. Не вызывает удивления тот факт, что некоторые очень разнородные группы, такие, как курсанты военных училищ и юноши-правонарушители, оказываются сходными, например отличаются более выраженной мезоморфией. Для обоих видов деятельности требуется сила, риск, решительность, но цель, ради которой реализуется эта энергия, различна. Вполне возможно, что энергия и напористость индивидуумов соматотонического типа — такой же врожденный признак, как и описанные этологами инстинкты. Соматотония может быть столь же инстинктивной, как и сексуальная активность, но более подверженной влиянию культуры.
470
Часть IV. Рост и конституция человека
Большинство прямых исследований, изучающих соотношения между телосложением и поведением, приводят к выводу о меньшей степени корреляции, чем это предполагал Шелдон. Однако число подобных работ невелико, и они недостаточно хорошо продуманы. По-видимому, вопрос об этой связи остается открытым, и к нему нужно относиться достаточно объективно, хотя, наблюдая за своими друзьями и близкими, мы с трудом сохраняем объективность. Повседневный опыт заставляет нас поверить в существование той общей связи, которая была описана Кречмером и Шелдоном.
Подобные представления, даже если они и ошибочны, глубоко укоренились в нашем сознании. Трудно представить себе, чтобы Лорел и Харди, сохранив свой прежний облик, поменялись характерами, так же как нельзя представить Фальстафа Мефистофелем, а Скруджа Пиквиком. Мы не можем представить себе Христа похожим на Будду. Очевидно, наше мышление несет стереотипный образ, связывающий воедино телосложение и поведение. Этот стереотип сформировался в средние века или еще раньше и с тех пор существенно пе изменился. Возможно, это лишь плод коллективной фантазии, и все-таки более вероятно искать его истоки на твердой почве биологии человека.
Список литературы
Рост
Falkner F. (ed.) Human development. Saunders, Philadelphia, 1966.
Goss R. J. (ed.) Regulation of organ and tissue growth. Academic Press, London, 1973.
Le Gros Clark W. E., Medawar P. B. (ed.) Essays on growth and form presented to D’Arcy Wentworth Thompson. Clarendon Press, Oxford, 1945.
Marshall W. A., Tanner J. M. Puberty. In «Scientific foundation of paediatrics» (ed. J. A. Davies, J. Dobbing). Heinemann, London, 1974.
Stem Z., Susser M., Saenger C., Marolla F. Famine and human development. Oxford University Press, 1975.
Tanner J. M. (ed.) Human growth. Symposium of the Society for the Study of Human Biology No. 3. Pergamon Press, Oxford, 1960.
Tanner J. M. Education and physical growth: the implications for educational practice of the facts and principles derived from the study of children’s physical growth. University of London Press, 1961.
Tanner J. M. Growth at adolescence: with a general consideration of the effects of hereditary and environmental factors upon growth and maturation from birth to maturity. 2nd ed. Blackwell Scientific Publications, Oxford, 1962.
Tanner J. M. Regulation of growth in size in mammals. Nature, Lond. 199, 845, 1963.
Tanner J. M. Human growth hormone. Nature, Lond. 237, 431—437, 1972.
Tanner J. M. Physical growth and development. In «Textbook of paediatrics» (eds. J. 0. Forfar, G. G. Arneil). 2nd ea. Edinburgh and London, Churchill and Livingstone, 1977.
Tanner J. M., Inhelder B. (ed.) Discussions on child development, vols. I—IV. A consideration of the biological, psychological and cultural approaches to the understanding of human development and behaviour. Tavistock Publications, London. 1956—1960.
Глава 24. Физиологические функции, болезни и поведение 471
Thompson D'Arcy Wentworth. Growth and form (revised edn.). Cambridge University Press, 1942.
Waddington С. H. The strategy of the genes. Allen & Unwin, London, 1957.
Телосложение
Brozek J. (ed.). Human body composition. Symposia of the Society for the Study of Human Biology, vol. 7. Pergamon, London, 1965.
Damon A., Bleibtien H. K., Elliot O., Giles E. Predicting smatotype from body measurements. Am. J. phys. Anthrop. 20, 461—473, 1962.
Heath В. H., Carter L. A modified somatotype method. Am. J. phys. Anthrop. 27, 57-74, 1967.
Petersen G. Atlas for somatotyping children. Royal Vangorcum, Assen Netherlands, 1967.
Schreider E. La biometrie. Presses Universitaires, Paris, 1960.
Sheldon W. II., Dupertuis C. W-, McDei mott E. Atlas of men. Harpers, New York, 1954.
Sheldon W. H., Hartl E. M., McDermott E. The varieties of the delinquent youth. Harpers, New York, 1949.
Sheldon W. H., Stevens S. S. The varieties of temperament. Harpers, New York, 1942.
Sheldon W. H., Tucker W. B. The varieties of human physique. Harpers, New York, 1940.
Tanner J. M. Physique, character and disease: a contemporary appraisal. Lancet, 2, 635—637, 1956.
Tanner J. M. The regulation of human growth. Child devel. 34, 817, 1963.
Tanner J. M. The physique of the Olympic athlete. George Allen and Unwin, London, 1964.
Tanner J. M., Healy M. J. R., Whitehouse R. H., Edgson A. C. The relation of body build to the excretion of 17-ketosteroids and 17-ketogenic steroids in healthy young men. J. Endocr. 19, 87—101, 1959.
Viola G. Criteres d’appreciation de la valeur physique, morphologique et foncti-onelle des individus. Biotypologie 3, 93, 1935.
Viola G. Il mio metodo di valutazionc della constituzione individuale. Endocrinol. Patol. costit. 12, 387, 1936.
ЧАСТЬ V
ЭКОЛОГИЯ ЧЕЛОВЕКА ДЖ. УАЙНЕР
Глава 25
ВВЕДЕНИЕ
Целостность, постоянство и, если оно происходит, расселение человеческих групп основаны на непрерывном взаимодействии с окружающей средой — непрерывном обмене веществами и энергией; благодаря этому обмену группа получает необходимые вещества и создает условия для своего существования. Экология в широком смысле означает динамическую взаимосвязь сообщества со средой, в которой оно находится. Приспособления, необходимые для успешного существования в определенных условиях, называются «адаптациями». Приспособляемость — свойство не только каждого из индивидуумов, образующих группу, но и всей группы в целом. Экологическое взаимодействие складывается а) из биологических процессов, влияющих на жизнедеятельность, рост и формирование членов группы, и б) из небиологических процессов, которые принимают форму культурных, технических и общественных реакций со стороны группы. Адаптация — двусторонний процесс: с одной стороны, она включает изменение физиологических и социальных функций, необходимых для того, чтобы удовлетворить требованиям окружающей среды; с другой стороны, сама среда, ее абиотические и биотические компоненты, изменяется, чтобы удовлетворить требованиям организма. Среда — это комплекс окружающих условий, в которых находится сообщество. К ней относятся абиотические и биотические компоненты, в том числе особенности рельефа, климат, средства сообщения, растительный и животный мир. Даже в самых примитивных человеческих сообществах обмен продуктами и передвижение групп приводят к столь значительному расширению контактов с другими группами и природой в целом, что понятие среда не приходится ограничивать узкими рамками их непосредственной среды обитания. Естественная среда может быть частично заменена одомашненным, искусственно созданным окружением, состоящим из жилищ, садов, сельскохозяйственных угодий, или, наконец, полностью заменена городскими условиями существования.
Глава 25. Введение
473
Человеческие популяции сильно различаются как по численности, так и по характеру организации. Мы встречаем самые разнообразные сообщества — от простейшей, сравнительно изолированной самообеспечивающейся группы, которая соответствует в животном мире дему, или естественной популяции, до более крупных комплексов демов, организованных в трибы и ассоциации триб, и на высшем уровне донациональных и наднациональных организаций.
Комплекс, состоящий из определенной группы живых существ и среды, в которой эта группа существует, часто называют экосистемой. Экосистема — одно из основных биологических понятий, применяемое для характеристики реально существующего сообщества, которое удовлетворяет свои нужды в динамической взаимосвязи со средой. Экосистема — арена «борьбы за существование», в которой успех зависит от эффективности адаптаций и в которой постоянно действует так называемый «естественный отбор». Под «экологией» человека мы понимаем экологию всей экосистемы в целом, рассматриваемой с точки зрения роли, которую играет в ней «человеческая компонента». Вид Homo sapiens представляет собой обширную сеть популяций, существующих в самых разнообразных условиях обитания. Человеческие популяции политп-пичны как по своим характерным признакам, так и в отношении экологической взаимосвязи популяций. Функциональную человеческую группу в определенной экосистеме можно рассматривать как зоологический дем. В экологическом смысле раса — более или менее искусственное понятие, основанное для многих популяций на выделении их из различных экосистем и объединении на основе общности или частоты появления некоторого числа физиологических признаков. С другой стороны, одну и ту же экосистему, например в пустынных областях, могут использовать в своих целях группы или демы, обладающие заметными генетическими различиями. Различия человеческих групп в действительности обусловлены различиями в их общих экологических взаимоотношениях, а последние лишь частично можно считать генетическими.
Процессы экологической адаптации
Взаимодействие организма со средой протекает в двух главных направлениях. Одно из них касается тех биохимических изменений в организме человека, которые обусловлены требованиями среды или возникают в процессе воздействия человека на среду. Здесь следует отличать деятельность и индивидуальные реакции мужчины, женщины и ребенка от деятельности и реакций всей группы в целом. Было предложено использовать термин аутэкология для характеристики индивидуальных реакций и термин синэкология для описания групповых реакций. Индивидуальные реакции носят
474
Часть V. Экология человека
в первую очередь физиологический характер: в силу общего соматического свойства физиологической приспособляемости организм может приобрести адаптацию, акклиматизироваться или выработать иммунитет к самым разнообразным внешним факторам — алиментарным, климатическим, патологическим.
Приспособляемость — свойство вида: все представители вида Homo sapiens способны проявить необходимую пластичность реакций в ответ на изменение внешних условий. Как мы уже указывали, эта физиологическая пластичность, характерная для человека, выработалась в ходе эволюции и представляет такой же важный продукт эволюции вида, как строение конечностей или концептуальное мышление.
Второе направление — это биологические реакции, специфичные для данного индивидуума и определяемые данным частным генотипом. Такие реакции и признаки могут быть свойственны лишь немногим индивидуумам; иногда они гарантируют особые преимущества целым популяциям (как, например, узкий нос в холодных или засушливых странах). Появление приспособительных признаков, выработка их в процессе естественного отбора сыграли важную роль в эволюции первых гоминид. Тем не менее порой бывает трудно установить ценность для выживания тех или иных «региональных» или «расовых» особенностей.
Цель и значение адаптации состоят в сохранении биологического гомеостаза, который, согласно Кеннону, представляет собой «совокупность устойчивых состояний, поддерживаемых в организме». Гомеостаз включает два взаимосвязанных процесса — достижение устойчивого равновесия и саморегуляцию. В физиологическом смысле его следует рассматривать как сохранение того постоянства внутриклеточных условий, которое необходимо для жизни организма. В растущем организме роль так называемого морфогенетического гомеостаза сводится к ограничению возможных путей роста определенным каналом. Насколько нам известно, «гомеостатически» устойчивые состояния у всех рас характеризуются определенным диапазоном изменений температуры тела, pH крови, уровня сахара, осмотического давления и других показателей, хотя тот или иной конкретный уровень в пределах данного диапазона может зависеть от конкретных условий деятельности организма или условий окружающей среды. Любая серьезная угроза нарушения гомеостатического уровня порождает компенсаторные процессы, направленные к нейтрализации такой угрозы и стабилизации внутренних параметров в пределах нормы. Таким образом, организм проявляет как инерцию, так и приспособляемость.
Действие компенсаторных процессов ограничено определенными рамками; например, существуют пределы изменения минутного объема сердца при физической нагрузке, или буферная способность крови. Превышение указанных пределов вызывает пато
Глава 25. Введение
415
логические изменения в организме. Приспособляемость организма может возрасти как по величине, так и по чувствительности в результате акклиматизации, физических упражнений, выработки определенных навыков, применения лекарственных средств и иммунизации. Индивидуальная изменчивость в приспособляемости организма, по-видимому, отражает различия в генетическом фонде индивидуумов.
Сходным образом и человеческая популяция в целом проявляет «гомеостатические» свойства, обеспечиваемые генетическим составом группы. Существует определенная тенденция к устойчивости диапазона изменений различных признаков, причем, их величины тесно группируются около некоторых средних значений. Разброс величин связан с действием многих адитивиых генов (гл. 9); среднее значение соответствует наибольшей степени гетерозиготности, крайние значения — наибольшей степени гомозиготности. Средние величины, как правило, соответствуют наиболее благоприятному выражению в фенотипе, а большие отклонения от средних — наименее благоприятным его проявлениям. Элиминация наименее приспособленных (в интересах приспособленности всей группы) не лишает группу генотипической изменчивости, так как гетерозиготы, которым особенно благоприятствует отбор, обеспечивают эту изменчивость (гл. 11). Таким образом, существует «генетический гомеостаз», поддерживающий как оптимум проявления признака, так и диапазон его изменчивости и обеспечивающий возможность генотипического сдвига в случае изменения окружающих условий. Поэтому группа способна к генетической адаптации, сохраняя при этом известную генетическую «инерцию». Генетическая изменчивость, свойственная всем скрещивающимся популяциям, имеет и другое приспособительное значение. Она дает популяции возможность выработать адаптации и использовать широкий диапазон условий окружающей среды и жизненных обстоятельств (см. также обсуждение биологического значения полигенов, гл. 9).
Помимо биологических процессов поддержанию гомеостаза способствуют различные виды общественной и технической деятельности: они дают группе возможность приспособиться к окружающей среде и подчинить ее своим потребностям. Необходимо помнить, что небиологические факторы — организация, разделение труда, нравы и обычаи общества — также служат биологическим целям. Многие общественные институты, не имеющие непосредственного отношения к удовлетворению биологических нужд членов общества, на самом деле служат биологическим целям. В этом можно убедиться, изучая производство и распределение предметов питания, размещение рабочей силы, мероприятия по созданию жилищ, заботу общества о подрастающем поколении и т. п. За примерами далеко ходить не надо. Религиозные обычаи племени
476
Часть V. Экология человека
Тсембага на Новой Гвинее играют важную роль в перераспределении земли и излишков свиного поголовья, обеспечивая группу в целом мясом и другими белковыми продуктами (Раппопорт). Ритуал гадания на лопатках у некоторых племен североамериканских индейцев служит, по мнению Мура, рандомизации мест охоты на оленей и способствует их отстрелу. Эшманн считает, что снятие половых ограничений во время некоторых сезонных церемоний у индейцев Байя в Центральной Калифорнийской пустыне объясняет колебания рождаемости, связанные с сезонными флуктуациями источников пищи. Разбросанность поселений туземцев Новой Гвинеи препятствует распространению инфекции. Мы далеки, конечно, от мысли пытаться установить простое соответствие между формами общественных институтов и биологическими потребностями общества: подобный «биологизм» ни в коей мере не может служить основой для социального анализа. Общественная организация имеет свои цели и особенности; последние нельзя сводить к чисто биологическим признакам. Для характеристики различных аспектов общественной деятельности мы должны пользоваться такими понятиями, как общественное положение, обязанности, родственные отношения, т. е. терминами, определяющими социальную структуру группы. Тем не менее среда налагает явно выраженные ограничения на материальную культуру, биологическую структуру и функции и даже на социальную организацию общества.
Разнообразие человеческих экосистем
Чтобы оценить все разнообразие человеческих экосистем, необходимо рассматривать различные комбинации природных условий, в которых обитают сообщества с разными формами хозяйства. Нам нужны такие классификации типов среды обитания и экономики, которые включали бы как экосистемы древнейших доисторических людей, как и более развитые сообщества. Простая классификация типов среды обитания должна учитывать важнейшие климатическое особенности, рельеф, характер фауны и флоры; классификация же экономических или общественных групп должна достаточно полно характеризовать образ жизни и степень сложности сообщества. Не следует стремиться к слишком подробной и сложной классификации природных условий, форм экономического уклада или экосистем, так как эти условия и формы чрезвычайно разнообразны, но в каждом конкретном случае необходим «детальный анализ любого активного сообщества» (Форд).
Среду обитания можно классифицировать в соответствии с географической широтой местности, выделив, согласно П. Джеймсу (Р. James), 10 основных типов (табл. 25.1). Исключение будет составлять лишь один, десятый тип, определяемый не широтой мест-
Глава 25. Введение
477
Таблица 25.1
Классификация типов среды обитания
I. Экваториальные леса
Густой влажный лес: вечнозеленые, высокие, образующие сомкнутый покров деревья; лес сырой, не горит и трудно поддается раскорчевке.
II. Тропические леса и кустарники
Менее густой полулиственный лес, поддающийся раскорчевке; заросли кустарника с травянистым покровом и небольшими колючими растениями; кустарник переходит с одной стороны в тропический лес, а с другой—в степи.
III. Тропические степи (саванны)
Большие степные пространства с одиноко растущими деревьями или небольшими островками леса. Богатый животный мир.
IV. Пустыни и засушливые области
Скудная растительность: а) сочные растения, запасающие воду (суккуленты) н (или) колючие растения (кактусы, алоэ); б) однолетние растения, обладающие периодом покоя; в) многолетние растения с длинными корнями. Поверхностные источники воды встречаются редко, мало тени. Животный мир беден. В оазисах и вблизи речных систем климатические условия несколько более благоприятны.
V. Леса умеренного пояса
Широколиственный лес, теряющий иа зиму листву, граничащий с хвойным лесом и вересковыми пустошами (на западном побережье Европы), теперь сильно вырублен. Богатый животный мир.
VI. Леса и кустарниики средиземноморского типа
«Маки»: чаще вечнозеленые, реже теряющие листву небольшие деревья (дубы и каштаны) и кустарники; растения с глубокими корнями и ксерофиты; островки степей. Животный мир небогат.
VII. Степи умеренного пояса
Степи, поросшие низкой травой; прерии с высокотравьем. Черноземные почвы, хорошо увлажняемые и задерживающие влагу.
VIII Северные леса (тайга)
Хвойный лес, в основном состоящий из ели, пихты, лиственницы, реже из бука, березы и тополя. Обилие озер, рек и болот. Богатый животный мир, например мыши, олени, пума в Америке, тигры в Сибири.
IX. Полярные области и тундра
Тундра начинается на окраине тайги в виде низкого леса из карликовых ивы, ясеня, ольхи, постепенно переходит в сплошной ковер, образуемый осокой, мхом и лишайниками; далее следует пустынная тундра с изолированными островками травя
478
Часть V. Экология человека
нистой тундры, весной покрывающейся цветами. Довольно богатый животный мир: рыба, морские млекопитающие, много арктических видов, например лисы, зайцы, медведи.
X. Горные районы
Растительность зависит от местоположения, высоты и ориентировки местности по отношению к солнцу. Например, вблизи экватора по мере увеличения высоты тропический лес переходит в смешанный, далее в хвойный и затем в альпийские луга.
ности, а высотой над уровнем моря. Экологические «стрессы» в каждой из этих групп неодинаковы — они зависят от самых разнообразных комбинаций алиментарных, климатических и патологических факторов. В табл. 25.1 приведены главные биотические особенности 10 различных типов обитания. Географическое распределение этих типов на разных континентах дано в табл. 25.2 (Европу и Азию следовало бы рассматривать как единый материк).
Из табл. 25.3 мы видим, что самые многочисленные современные популяции обитают в двух резко отличных географических зонах — в тропических лесах и кустарниковых зонах и в областях умеренного пояса, покрытых смешанными лесами. В последней графе таблицы показана относительная плотность населения. Весьма вероятно, что первые гоминиды появились в тропических степных и лесостепных районах. По сравнению с ними современный человек существенно расширил зону своего обитания, заселив с высокой плотностью тропические области, покрытые влажными лесами (плотность, вдвое превосходящая общую среднюю плотность), районы Средиземноморья, поросшие невысоким лесом и кустарником (плотность вчетверо выше средней), и области умеренного пояса, покрытые смешанными лесами (плотность, вшестеро превосходящая среднюю). Плотность населения довольно высока также в горных районах.
Грубой характеристикой экономического уклада может служить наиболее распространенный и типичный для данного сообщества способ производства (табл. 25.4). Он определяет как объем производимой продукции и способ организации труда, так и сложность социальной структуры сообщества.
Не следует, однако, придавать слишком большого значения каким-либо классификациям, основанным на различиях в технологии производства; способы производства редко взаимно исключают друг друга, и у групп, использующих одинаковую технику производства, социальная струкутура может заметно различаться.
Далее, как видно из табл. 25.5, 25.6 и 25.7, нельзя рассматривать «расу» как некую преграду, ограничивающую среду обитания данного сообщества или формы его хозяйственной деятельности определенными узкими рамками. Одна и та же рассовал группа участвовала нередко в исторической последовательности в форми-
Таблица 25.2
Типы среды обитания — географическое распределение
(несколько примеров)
Африка Азия Европа Сев. и Южн. Америка Австралазия
I. Экваториальные тропические леса Бассейн Конго Камерун Индия Юго-Восточная Азия Малайя Индонезия Бразилия (бассейн Амазонки) Новая Гвинея Меланезия
И. Тропические и кустарниковые леса Ангола, Уганда, Южный Судан Индия Южный Китай Юкатан Гран-Чако Полинезия Район Кейрнс
III. Тропические степи (саванны) Судан Танзания Индонезия Южная Америка Пампа, Льянос Северная Австралия
IV. Пустыни и засушливые области Сахара Калахари Намиб Аравийская Индостанская (Тар) Кызылкум Каракум Калифорнийская Атакама Парагвайская Центральная Австралийская
V. Леса умеренного пояса Китай к югу от Великой стены и к северу от лесного пояса Области между Средиземным морем, тайгой и степями Побережье, омываемое Гольфстримом Район к востоку от Миссисипи Долина реки Св. Лаврентия Северо-западное побережье Северной Америки Чили Австралия о. Тасмания Новая Зеландия
Африка Азия
VI. Леса и кустарники средиземноморского типа Южный берег Средиземного моря Пиренейский полуостров Израиль Ливан Южный берег Черного моря
VII. Степи умеренного пояса Южноафриканские степи (вельд) Трансвааль Узкая полоса к югу от гор Атлас Казахстан Маньчжурия Монголия
VIII. Тайга Сибирь
IX. Тундра Сибирь
X. Горные районы Эфиопия Басутоленд Тибет Гиндукуш
Продолжение
Европа Сев. и Южн. Америка Австралазия
Крым Северный берег Средиземного моря Калифорния до Ванкувера Средние районы Чили Область вблизи городов Перт и Аделаида
Степи Поволжья и Приуралья Черноземный пояс Прерии Пампасы Патагония
Лапландия Северная Канада Аляска Южная оконечность Южной Америки Горы Новой Зеландии
Лапландия Гренландия Прибрежные районы Антарктиды
Швейцарские и Австрийские Альпы Мексика Перу Чили Новая Зеландия (остров Южный)
Глава 25. Введение
481
Таблица 25.3
Географическое распределение популяций
Среда обитания Площадь, % от общей площади суши Население, % ОТ населения земного шара Относительная плотность населения
Засушливые земли и пустыни 18 4 0,2
Тропические леса | 15 28 1,9
Тропические кустарниковые леса
Тропические степи (саванны) 1 91
Степи умеренного пояса (континентальные)
Смешанные леса умеренного пояса 7 39 5,6
Кустарниковые районы средиземноморского 1 4 4,0
типа
Тайга 10 1 0,1
Полярные земли и тундра 16 (<1) 0,01
Горные районы 12 12 1,00
Таблица 25.4
Формы хозяйства
Собиратели. Жпвут сбором плодов, корней, меда, насекомых, мелкой дичи. Их материальная культура (одежда, жилища, домашняя утварь и т. д.) очень бедна и примитивна. Единственное домашнее животное собака.
Охотники. Живут охотой, ставят силки и капканы для дичи. Их жилища часто хорошего качества. У охотников существуют уже зачатки гончарного ремесла и ткачества. Люди палеолита и мезолита относились к этим двум категориям.
Пастухи и скотоводы. Держат крупный рогатый скот, овец и коз. Часто строят хорошие прочные жилища. Охота и возделывание земли носят подсобный характер.
Кочевники. Скотоводы, которые регулярно в течение года перемещаются с места на место в поисках пастбищ для скота (годовой цикл).
Примитивные земледельцы. Заняты садоводством и огородничеством, имеют подсобные угодья; вспомогательную роль играет собирание плодов и ягод, а также охота; иногда держат свиней и домашнюю птицу. К этой категории следует отнести сообщества эпохи неолита.
Земледельцы на более высокой ступени развития. Сельское хозяйство для них—главное средство существования. Понимают важность сохранения плодородия почв и значение севооборота. В период после неолита начинают практиковать задержание влаги и орошение почв.
Таблица 25.5
Типы среды обитания и расовые группы
Рассы
Среда обитания Негроидная Кавказоидная Монголоидная Австрало-веддоидная
I. Пустыни Бушмены Народы Средиземно- морья Монголы (Гоби) Индейцы пустыни Атакама Австралийцы
II. Тропические леса Пигмеи Индо-дравиды Южные монголоиды Малайцы Американские индейцы «Муррианцы» Ведды
III. Тропические Кустарниковые леса Банту Анголы Индо-дравиды Американские индейцы (Южная Америка) Веддоиды Индии
IV. Саванны V. Леса умеренного пояса Народы Нила Хауса Американцы испанского происхождения Народы Северной и Центральной Европы Американские индейцы (Южная Америка) Североамериканские индейцы Монголы Китая Австралийцы Полинезийцы и карпентарианцы
VI. Кустарники и низкий лес (типичный для Средиземноморья) VII. Степи умеренных широт VIII. Тайга IX. Тундра Г оттентоты Банту (сото) Средиземноморские группы Европейские группы Лапландцы Лапландцы Североамериканские индейцы Американские индейцы Монголы Маньчжурии Вогулы (остяки) Ненцы Эскимосы Чукчи Карпентарианцы Айны
X. Горы Банту Армепоидная и альпийская группы Монголоиды Тибета, Ирана и Боливии Горцы Новой Гвинеи
Примечание. Данные в таблице примеры служат иллюстрацией того, что люди любых рас способны жить в самых разнообразных условиях обитания.
Таблица 25.6
Формы хозяйства и расы
Форма хозяйства Расы
Негроидная Кавказоидная Монголоидная Австрало-веддоидная
I. Собиратели Бушмены Ведцы Сообщества мезолита Индейцы Огненной Земли Калифорнийские индейцы Австралийцы
II. Охотники и рыболовы Пигмеи Народы Западной Африки Меланезийцы (Айны) Ископаемые люди из Пржедмоста Эскимосы Американские индейцы
III. Пастухи и скотоводы Банту Масаи Готтентоты Лапландцы «Нордические» народы железного века Группы южной Манчжурии
IV. Кочевники Негроиды Сахары (народы Нила) Бедуины Туареги Монголы Народы Туркестана Чукчи
V. Примитивные земледельцы Банту Народы Новой Гвинеи Индо-дравиды Южные монголы Малайцы Меланезийцы
VI. Земледельцы на более высокой ступени развития Народы Западной Африки Народы Средиземноморья Хопи, Майя Индонезийцы Северные монголы Полинезийцы
Примечание. Приведенные в таблице примеры указывают на то, что люди всех рас имеют вполне совершенные типы хозяйства.
Таблица 25.7
Условия обитания, формы хозяйства и народы
(несколько примеров)
Собиратели ОХОТНИКИ Пастушеские племена и скотоводы Кочевники Примитивные земледельцы Земледельцы на более высокой ступени развития
I. Экваториальные леса II. Тропические Семанги Индейцы Гран- Пигмеи Меланезийцы Банту Бемба Амазонские племена Народы Новой Гвинеи Индо-драв иды Индонезийцы о. Ява Банту
и кустарниковые леса III. Тропические Чако Австралийцы Хадза (Восточ- Народы Нила Народы Южно американские группы Североамери- Хамиты
степи (саванны) IV. Засушливые земли и пустыни Бушмены Австралийцы ная Африка) Нила Бедуины канские индейцы Жители оазисов Население оазисов и речных бассей- нов
Собиратели Охотники
V. Леса умеренных широт Австралийцы Европейские народы мезолита Тасманийцы Ископаемые люди Пржед-моста
VI. Леса и кустарники средиземноморского типа Мезолитические племена Юго-Восточной Африки Индейцы Калифорнии
VII. Степи умеренного пояса Европейские народы палеолита Североамериканские индейцы
VIII. Тайга Индейцы Огненной Земли Ненцы
IX. Тундра Эскимосы
Продолжение
Пастушеские племена и скотоводы Кочевники Примитивные земледельцы Земледельцы на более высокой ступени развития
Европейцы в железном веке Китайцы Китайцы
Балканские народы Берберы Народы неолита п железного века Маори Европейцы в средние века
Монголы Буряты Монголы Индейцы племени сиу Индейские племена поуни
Лапландцы
Лапландцы
486
Часть V. Экология человека
ровании ряда отличных, сменяющих друг друга экономических систем возрастающей сложности.
Некоторые частные типы среды обитания могут, конечно, оказаться более благоприятными для развития и совершенствования определенного способа производства. И наоборот, естественная среда в отдельных случаях делает невозможным определенный тип хозяйственной деятельности. В целом, однако, на всех стадиях жизни сообщества среда обитания не налагала сколько-нибудь серьезных ограничений на экономику. Различные типы среды обитания могут использоваться по-разному. Соответствующие им экосистемы также будут заметно различаться, и в первую очередь в отношении плотности населения. Сообщества с развитой культурой земледелия могут существовать в самых разнообразных условиях, и плотность населения может быть всюду достаточно высокой.
Результатом внедрения человеческих сообществ в природные экосистемы может явиться состояние устойчивого или, наоборот, динамического равновесия. В первом случае основные биологические признаки человеческой группы остаются более или менее неизменными в течение длительного времени: сохраняются размер популяции, плотность и состав, энергетический и пищевой баланс, общий уровень жизни. Этот консерватизм отражается и на уровне общественной организации и способе производства. Та или иная близость к экологическому равновесию достигается в различных условиях среды, при различных формах хозяйственного уклада и в популяциях разной плотности. В случае динамического равновесия, характерного для большинства расселяющихся и быстро развивающихся сообществ, возрастающие потребности в продуктах питания, водных ресурсах, сырье, размещении отходов удовлетворяются за счет увеличения полезной деятельности сообщества, затраты энергии и более интенсивного использования биотических и абиотических компонентов среды. Этот тип равновесия (или та или иная степень близости к нему) особенно показателен для человеческих сообществ в эпоху индустриализации. В истории сообществ бывали, конечно, периоды, когда экологический баланс нарушался, что влекло за собой весьма серьезные социальные и биологические последствия. Цель экологического анализа состоит в том, чтобы понять биологические свойства сообщества и входящих в него семейств и индивидуумов, изучив характер процессов, поддерживающих равновесие в системе, а также процессов обмена. К таким процессам относятся передача энергии, транспортировка пищи и воды, распространение заболеваний в экосистеме.
Говоря об экологическом «успехе» того или иного сообщества, нередко используют термин «уровень жизни» (жизненный стандарт). Это понятие включает в себя совокупность биологических, медицинских или демографических факторов. Весьма общими показате
Глада 25. Введение
487
лями «уровня жизни», характеризующими «приспособленность» сообщества к условиям среды, могут служить величина популяции и ее стабильность, смертность, средняя продолжительность жизни индивидуума, состояние питания. Пожалуй, наиболее убедительным доказательством «неприспособленности» может служить уменьшение численности популяции (депопуляция). Для примера укажем на безуспешность попыток различных иммигрантских групп утвердиться на новых территориях в качестве самостоятельных и обособленных.
Для анализа «эффективности» или «емкости» экосистемы необходимо детально рассмотреть условия жизни общества, его структуру. В любом аспекте взаимоотношений между организмом и средой можно обнаружить определенные ограничения: климатические в прямом или косвенном смысле (связанные, например, с недостатком строительных материалов для постройки жилищ), ограничения, связанные с нехваткой пищевых ресурсов, со спецификой распространения болезней и т. п. Эти ограничения могут быть также следствием несовершенства социальной организации, например неправильной организации трудовой деятельности в течение годового цикла, плохого хранения продуктов питания и т. д. Несомненно, одни экологические условия значительно менее благоприятны для человека, чем другие. Так, проживание в тропической Африке всегда представляло целый комплекс существенных трудностей, связанных с климатом, питанием, болезнями. Возникновению древнейших цивилизаций на Ближнем Востоке способствовал комплекс благоприятных условий, сложившийся в речных долинах Египта и Месопотамии,— обилие воды, плодородие почвы, благоприятный климат, возможность использования рек как средств сообщения, защита от нападений извне,. которую обеспечивали окружающие пустыни.
Глава 26
ЭКОЛОГИЯ ПИТАНИЯ
Теперь перейдем к анализу экосистем как систем, в которых происходит обмен веществами и энергией, т. е. рассмотрим вопросы, связанные с экологией питания.
Энергия
Поддержание общего баланса калорий у взрослого индивидуума, исключающее резкие колебания веса тела, связано с регуляцией, осуществляемой нервной системой.
Энергия, получаемая из пищи, необходима для поддержания обмена веществ в состоянии покоя (основного обмена), для мышечной деятельности, роста и размножения, для сохранения постоянной температуры тела. Можно весьма точно указать необходимый для удовлетворения этих биологических нужд расход энергии (в калориях) для одного индивидуума, семьи или целой группы.
Приводя ниже соответствующие таблицы, мы должны сделать, однако, ряд замечаний.
1. К основному обмену относят энергию, расходуемую организмом во время сна, в покое или в сидячем положении, во время беременности и лактации, а также в процессе роста. В среднем можно принять, что организм взрослого человека в состоянии сна или «покоя» расходует примерно 300 кДж/ч. Эта величина зависит, однако, от размеров тела. Расход энергии при беременности и кормлении грудью равен 400 кДж/ч. Ребенок в зависимости от возраста расходует в состоянии покоя 150—250 кДж/ч.
2. Мы рассматриваем отдельно энергию, затрачиваемую при различных видах деятельности, связанной с добыванием пищи, чтобы показать, какая часть всех калорий расходуется на работу, цель которой — обеспечение организма необходимой энергией. Эквивалент работы в джоулях зависит от характера работы. Он может изменяться, скажем, от 450 кДж/ч, затрачиваемых при обычной пешей прогулке или деятельности, не требующей больших физических усилий (например, собирание плодов, поливка), до 1600— 2000 кДж/ч, расходуемых при тяжелой работе. Работа, сопровождаемая такой большой затратой энергии, может выполняться лишь в течение нескольких часов в день. Только для немногих видов деятельности, характерных для примитивных сообществ1 была
Глава 26. Экология питания
489
произведена подобная оценка. Вот некоторые примеры (Passmore a. Durnin, 1955):
Вид деятельности Расход энергии, кДж/ч
Переноска бревен Посадка чуфы Рыхление земли мотыгой Прополка Выкорчевывание кустарника Рубка леса 840 920 1050—1470 1340 1700 2000
Наиболее тяжелая физическая работа в промышленности Западней Европы требует затраты примерно 10 000 кДж для мужчин и 6700 кДж для женщин за восьмичасовой рабочий день (т. е. 1250 кДж/ч и 838 кДж/ч соответственно). Можно выбрать два средних «уровня» для энергетической характеристики легкой и тяжелой работы, а именно 5000 кДж и 8400 кДж за рабочий день.
3. Оценка иных видов деятельности, кроме тех, которые непосредственно связаны с добыванием пищи, с энергетической стороны весьма затруднительна. В простых сельскохозяйственных сообществах значительное время занимает домашняя работа, собирание топлива, приготовление пищи, ремонт и изготовление одежды и инвентаря, постройка жилищ, изготовление мебели и т. д. В этих видах деятельности человек затрачивает, по-видимому, примерно в 1,5 раза больше энергии, чем необходимо для поддержания обмена в состоянии покоя, т. е. 460—500 кДж. Такие работы, как размалывание риса, требуют большой затраты энергии (до 1250 кДж/ч). В некоторых странах Азии и Африки женщина нередко занята размалыванием маиса или маниока до двух часов в день (Platt, 1946). Многие виды развлечений или активного отдыха, танцы и другие церемонии требуют не меньшей затраты энергии, чем иной трудоемкий вид работы; их поэтому также следовало бы учитывать в общем балансе энергии. На холоде человек расходует избыточную энергию («дрожь» от холода) примерно на 30% выше обычной.
Можно рассчитать общий расход энергии за день для условной семьи, состоящей из родителей и троих детей, из которых двое (подростки) участвуют в некоторых видах деятельности, связанной с добыванием пищи. Максимальные и минимальные величины приведены в табл. 26.1.
Общее количество потребляемых человеком калорий зависит от ряда факторов, в частности от возраста, пола, размеров тела, а следовательно, в какой-то мере и от генетических факторов. Основной обмен для данного возраста можно выразить в зависимости от общей величины тела по известной формуле Дюбуа
^4 = ^0,425.#0,725.71^4,
490
Часть V. Экология человека
Таблица 26.1
Энергетические потребности гипотетической семьи
Основной обмен Деятельность, связанная с добыванием пищи Прочие виды деятельности Всего, кДж
Время, ч Энергия, Дж Время, ч Энергия, Дж Время, ч Энергия, Дж
мин. макс. мин. макс. МИН. макс.
Отец 8 2 350 2 560 8 5 040 9 200 8 3 790 11180 15 600
Мать 8 2 350 3 700 6 3 780 5 020 10 3 790 9 920 12 530
1-й ребенок 10 2 520 2 770 6 2 520 3150 8 3 070 8 070 .8 950
2-й' ребенок 11 2 310 2 520 4 1 680 1850 9 3030 7 020 *7 400
3-й ребенок 12 2100 2180 0 — — 12 3 530 5630 5 710
Всего — 11 630 13 730 — 12 920 19 220 — 17170 41 820 50190
Примечание. Энергия, необходимая 1 чел. в сутки: 8364 кДж—минимум (приближенно 8500), 10 038 кДж—максимум (приближенно 10 000).
где А — площадь поверхности тела в квадратных сантиметрах, W — вес в килограммах, Н — рост в сантиметрах.
Фактор размера также влияет на энергетическую «стоимость» работы, выраженную в килоджоулях. Например, пешая прогулка со скоростью 3 км/ч требует (независимо от пола и возраста) затраты Q килоджоулей в 1 мин, где Q зависит от веса тела:
Q = 0,197^ + 4,284.
В среднем можно считать, что в любом обществе на кажлого человека необходимо от 8500 до 12 750 кДж в день. Эти цифры учитывают, в частности, такие факторы, как размер тела, климат, трудность работы.
Между экосистемами существуют, однако, значительные различия в доле энергии, затрачиваемой на производство продуктов питания. Из табл. 26.1 следует, что из каждых 40 кДж, расходуемых семьей для всех ее нужд, 10 или 15 кДж (в зависимости от трудности работы) затрачиваются на деятельность, непосредственно связанную с добыванием пищи. Эта «производительная» доля общей затраты энергии, составляющая 30—40%, по-видимому, характерна для примитивных обществ, занятых в основном добыванием средств существования. Первобытный человек в этом отношении сделал значительный шаг вперед по сравнению со своими обезьяньими предками: высшие приматы (большие человекообразные обезьяны), занятые непрестанными поисками пищи,
Глава 26. Экология питания
491
затрачивают до 70% энергии на этот вид деятельности. В наиболее эффективном сельском хозяйстве довольно легкая работа, требующая 6700 кДж/день, способна обеспечить пропитание 20 или более членов сообщества. Следовательно, из каждых 40 кДж в этом случае лишь 2 кДж затрачиваются на деятельность, непосредственно
Таблица 26.2
Среднее суточное потребление жиров, углеводов и белков в разных популяциях
Жители Великобритании Кикуйю Эскимосы Жители о. Барбадос
0м о CQ О <х> р* S § [ергия, кДж эличество, г (ергия, кДж См о 8 0) tJ* Я гергия, кДж См О 03 S сг S * тергия, кДж
X Ф X Ф X ф X ф
Жиры (38 кДж/г) НО 4150 22 832 162 6120 63 2380
Углеводы (17 кДж/г) 400 6730 390 6500 59 990 416 7000
Белки (17 кДж/г) 100 1680 100 1680 377 5870 45 756
Всего — 2990 — 2158 — 3102 — 2411
связанную с производством продуктов питания. Изменение условий среды при сохранении основных форм хозяйственной деятельности может изменить «производительную» долю расходуемой энергии не более чем в 1,5 раза. В то же время изменение техники производства при неизменных условиях обитания может изменить эту долю в 10 раз и более. Для оценки усилий, затрачиваемых на добывание средств к существованию, иногда используют отношение W/С, где W — количество человеко-дней, связанных с добыванием пищи, а С — количество человеко-дней, связанных с ее потреблением. Этот показатель близок по смыслу к производительной доле расходуемой энергии и может служить лишь для грубой характеристики соответствующих затрат энергии. Для стада бабуинов отношение W/С близко к единице (что соответствует непрерывному поиску корма), тогда как для бушменов Кунга оно«0,25. В некоторых примитивных сообществах (например, на Новой Гвинее) эффективность деятельности, связанной с энергетическим обеспечением группы, гораздо выше и производительная доля усилий снижается почти до 10%.
Источники пищи и энергии чрезвычайно разнообразны. Это видно из табл. 25.7, в которой представлены 50 различных комбинаций форм хозяйства и сред обитания. Физиологическая пластич
492
Часть V. Экология человека
ность в использовании разнообразных источников питания обусловлена, во-первых, способностью человеческого организма усваивать и перерабатывать в процессе обмена три источника энергии — жиры, углеводы и белки — во всевозможных сочетаниях и, во-вторых, способностью использовать указанные вещества из огромнейшего количества пищевых продуктов.
Табл. 26.2 может служить полезной иллюстрацией физиологической (метаболической) пластичности человеческих популяций. Она показывает среднее суточное потребление на душу населения жиров, углеводов и белков в четырех популяциях, резко различающихся по типу экономики.
Мы видим, что в четырех различных экосистемах разные комбинации основных пищевых продуктов обеспечивают необходимое организму количество энергии. Различия проявляются не только в соотношении белков, жиров и углеводов, но и в том, из каких продуктов они извлекаются; ассортимент продуктов зависит от окружающей среды и способа добывания пищи. Главными источниками углеводов служат злаки, корнеплоды, плоды и сахар; источниками жира — растительные и животные жиры и масла; белки содержатся главным образом в животной пище (включая рыбу и молоко), в орехах и в бобовых растениях.
Доля энергии, поступающей в организм в виде жиров, в европейских странах варьирует от 21 % в районах Средиземноморья до 31% в странах Западной Европы и в США. Из табл. 26.2 видно, что относительное потребление энергии в виде жиров у эскимосов достигает 47%, а у кикуйю составляет лишь 10%. Доля белков в рационе различных сообществ более постоянна: относительный белковый эквивалент энергии изменяется в узких пределах — от 9 до 12%.
Предположение о том, что специфика питания у разных человеческих популяций так или иначе связана с генетическими особенностями этих групп, подтверждается пока что лишь данными для одного или двух питательных веществ. Говоря о такой связи, обычно приводят в качестве примера традиционную, богатую жирами пищу эскимосов. Такая пища мало привлекательна для европейца, в организме которого эти жиры окисляются не полностью, что ведет к накоплению кетоновых тел. Утверждают, что у эскимосов кетоз не наблюдается. Даже если они генетически более приспособлены к усвоению такой пищи (что еще нельзя считать установленным), такое превосходство весьма относительно. Было показано, что и у некоторых европейцев вырабатывается привычка к постоянному потреблению пищи, содержащей много жиров и белка. Люди, по-видимому, легко переносят (если не психологически, то по крайней мере физиологически) существенные изменения в характере рациона. В этом проявляется общее свойство Homo sapiens приспосабливаться к изменяющимся условиям среды.
Глава 26. Экология питания
493-
Баланс энергии
Группа, численность которой на данной территории остается постоянной в течение длительного времени, скажем в течение жизни нескольких поколений, очевидно, способна сохранять баланс расходуемой и потребляемой энергии. Однако поддержание такого баланса — отнюдь не простое дело, причем достигнутое в определенных условиях равновесие может оказаться весьма непрочным. Например, собиратели и примитивные земледельцы, как мы увидим ниже (гл. 29), нередко испытывают острую нехватку воды и продуктов питания.
Эффективность использования источников энергии и способность популяции сохранять энергетический баланс в системе определяются такими величинами, как производительная доля расходуемой энергии и поток энергии в сообществе. Эффективным следует считать сообщество с низкой производительной долей затрат энергии и высоким энергетическим выходом, в особенности если оно способно получать излишки энергии сверх необходимого минимума для создания запасов или торговли. При высокой производительной доле затрат энергии те же результаты могут быть достигнуты гораздо более дорогой ценой. В примитивных сообществах мы, однако, часто сталкиваемся с ситуацией, когда низкий уровень производительных затрат сочетается с низким энергетическим выходом. Этот случай требует дополнительного рассмотрения. Подобное сообщество может существовать при довольно низком уровне трудовой активности; члены его работают не больше, чем необходимо для выживания. При этом нередко достигается некий оптимум; дополнительные затраты труда в данной экологи-' ческой ситуации сопровождаются эквивалентным поступлением энергии в систему.
Пищевые вещества
Жиры, белки и углеводы помимо той роли, которую они играют в качестве источников энергии, выполняют также особые функции в процессах обмена веществ. Входящие в состав белков аминокислоты необходимы для роста тканей и их восстановления, а также для синтеза многих белков, выполняющих в организме самые разнообразные функции. Жиры необходимы не только как форма хранения энергии в организме, но и для обеспечения теплоизоляции тела. Углеводы участвуют во всех процессах превращения энергии. Содержание сахара в крови отличается, как правило, строгим постоянством.
Тот факт, что всем этим требованиям удовлетворяют самые различные соотношения углеводов, жиров и белков (табл. 26.2), вновь указывают на поразительную пластичность метаболических
494
Часть F. Экология человека
процессов. Различные вещества, участвующие в этих процессах, взаимозаменяемы: источниками углеводов могут служить и белки и жиры; их поступление в какой-то степени избавляет организм от необходимости производить соответствующий вид топлива. Если пища богата углеводами и бедна жирами, организм способен пополнять запасы жиров за счет углеводов и тем самым компенсировать недостаток жиров в пище. Правда, некоторый минимум жиров, особенно ненасыщенных, все же необходим при любых условиях. Биологическая ценность продуктов, содержащих белки животного происхождения, несравненно выше, чем продуктов, в состав которых входят только растительные белки. В животных белках содержится значительно больше так называемых «незаменимых» аминокислот. Однако, употребляя пищу, содержащую животные и растительные белки в различных соотношениях, можно обеспечить полноценное белковое питание. Это дает возможность изменять рацион в зависимости от условий обитания. Поскольку белковая недостаточность чревата серьезными последствиями для организма, определенный минимум животных белков следует считать необходимым. Потребность в белках (в граммах в сутки), согласно наиболее достоверным оценкам, такова: 30—50 г для ребенка в возрасте 4 лет, 50—80 г — в возрасте 12 лет, 50—90 г — для взрослого мужчины, 40—70 г — для взрослой женщины и 55— 90 г — для женщины во второй половине беременности или в период кормления грудью. Убыль азота за счет усиленного потоотделения в жарких странах, которую необходимо компенсировать, невелика — не более 10%. Расовых различий в интенсивности белкового обмена, по-видимому, не существует. Приводившиеся ранее оценки потребления азота у горных племен Новой Гвинеи скорее всего неточны (или же современные стандарты потребления слишком высоки), так что нет оснований считать, что мы имеем здесь дело с особым генотипическим признаком. Однако белковая недостаточность, как об этом свидетельствует, например, частота заболеваний квашиоркором после отнятия ребенка от груди,— весьма распространенное явление. Потребление белков не превышает в этом случае 30 г в сутки. Если кормящая женщина не получает с пищей нужного количества белка, то лактация заметно сокращается, однако содержание белка в молоке остается высоким. Это имеет очень большое значение для выживания, особенно если период кормления грудью чрезмерно удлинен, как это часто бывает в примитивных обществах. Длительное ограничение половых сношений в послеродовом периоде, способствующее защите ребенка, значительно чаще встречается в сообществах с недостаточным количеством белковых продуктов.
Трудности, связанные с определением относительной роли факторов среды и генетических факторов, можно проиллюстрировать на следующем примере. Для многих темнокожих народов (в том
Глава 26. Экология питания
495
числе для негров Западной Африки, аборигенов Австралии, индейцев навахо) характерно высокое содержание гамма-глобулина в сыворотке. Этот признак считают генетическим по происхождению, но, несомненно, нужно учитывать и роль других факторов и в первую очередь инфекционных заболеваний, в частности малярии. Низкий уровень альбумина, по-видимому, связан с особенностями питания.
Выраженная индивидуальная изменчивость в характере выделения аминокислоты D-фенилаланина, а также р-аминоизомасля-ной кислоты имеет, как мы видели, генетическую природу. Однако популяционную изменчивость в характере выделения р-аминоизо-масляной кислоты нельзя, по-видимому, относить за счет»особен-ностей промежуточного обмена, так как она связана главным образом с функцией почек.
Минеральные соли и вода
Среди веществ, которые мы обычно относим к этой группе, наибольшую роль в процессах жизнедеятельности организма играют железо, хлористый натрий, иод, кальций, и, конечно, вода. Эти вещества распространены в природе весьма неравномерно. В организме устанавливается устойчивое равновесие между поступлением каждого из этих веществ и его выведением. Если потребность организма в том или ином веществе резко возрастает и ее не удается удовлетворить за счет внешних источников, организм вынужден компенсировать отрицательный баланс, используя свои собственные внутренние ресурсы. Как только вещество начинает вновь поступать извне в достаточном количестве, запасы его пополняются и на некоторое время устанавливается положительный баланс. Таким образом, между поступлением и выведением минеральных солей всегда поддерживается равновесие, сколько бы солей ни поступало в организм в данных конкретных условиях.
Вода
Количество воды в организме у разных индивидуумов изменяется в довольно широких пределах и зависит в основном от содержания жиров. Если минимизировать колебания веса тела^ связанные с нарушением энергетического баланса (по возможности уравновесив поступление и расход энергии), то суточные колебания количества воды в организме будут сохраняться на уровне 1% от веса тела. Такая стабильность отражает эффективность гомеостатической регуляции, осуществляемой почками и особым механизмом, лежащим в основе чувства жажды. Мы не располагаем, однако, сколько-нибудь убедительными доказательствами
496
Часть F. Экология человека
того, что у жителей пустынь — бедуинов, ведущих кочевой образ жизни, имеется особый механизм регуляции количества воды в организме, связанный, например, со способностью почек выделять более концентрированную мочу.
Одно из важнейших экологических требований в любом сообществе — поддержание равновесия между наличными источниками воды и ее потреблением для любых нужд, включая расход воды на нужды туалета и на удаление нечистот. Зависимость численности популяции от имеющихся в ее распоряжении водных ресурсов будет рассмотрена ниже. Организм может справиться с некоторой нехваткой воды, выделяя более концентрированную мочу; однак^ способность выжить при минимальном количестве воды — около 500 мл в сутки — следует рассматривать скорее как приспособление к экстремальным условиям существования.
Кальций
Поступление кальция в организм может изменяться в широких пределах. При этом устанавливается простой баланс между потреблением и выведением. Если в пище содержится мало кальция, то интенсивность всасывания его в кишечнике повышается, что позволяет поддерживать постоянный уровень этого вещества (в организме взрослого) и даже удовлетворять потребности, связанные с ростом (в организме ребенка).
Механизм этого процесса не вполне понятен, ясно лишь, что, по-видимому, благодаря ему организм сравнительно легко приспосабливается к диете, бедной кальцием. Другой фактор, способствующий поддержанию нормального баланса кальция, связан с действием ультрафиолетового излучения солнца. Поглощение ультрафиолетовых лучей поверхностными слоями кожи содействует превращению содержащегося в подкожных тканях вещества, родственного эргостеролу, в активное начало витамина D. Искусственное ограничение доступа ультрафиолетовых лучей к более глубоким подкожным тканям у лиц с темной кожей может привести к развитию рахита или остеомаляции (как это часто бывает в странах, где женщины носят паранджу). Светлая кожа у людей в странах с недостаточным количеством солнечного света обеспечивает определенные селективные преимущества.
Во многих географических районах в организм человека поступает недостаточное количество кальция, так как пища, за исключением молока и сыра, содержит мало солей кальция. При низком содержании кальция в пище люди часто бывают низкорослыми; кости у них недостаточно длинные, развитие зубов задерживается. Можно думать, что при недостатке кальция снижается и скорость роста ребенка. Если при этом начать давать кальций в избытке, то он будет задерживаться и накапливаться в организме.
Глава 26. Экология питания
497
Имеются сведения о низком содержании кальция в богатой углеводами и бедной мясом и молоком пище сельских жителей некоторых стран, например у беднейших жителей Шри Ланки и аборигенов Ньюфаундленда.
Иногда высказывают мнение, что само существование низкорослых расовых групп объясняется чрезмерно низким поступлением кальция в организм; эта гипотеза не подкреплена, однако, надежными фактами. В связи с этим указывают обычно на то, что в местностях, где растительность бедна кальцием, водятся низкорослые породы овец (Marett). Между тем известно, что такие низкорослые народности, как эскимосы и лапландцы, получают с пищей вполне достаточное количество кальция. Пока не ясно, выработалась ли у низкорослых народов какая-либо специфическая резистентность к дефициту кальция.
Железо
Суточная потребность в железе мала и ее легко удовлетворить. Однако у ребенка, которого кормят грудью, нередко возникает дефицит железа. В организме легко восстанавливается равновесие между поступлением и выведением железа, и временный дефицит его легко восполняется за счет имеющихся запасов. Потребность в железе значительно возрастает при анемии, вызванной, например, такими паразитарными инвазиями, как малярия и анкилостомоз, которые очень широко распространены в тропических странах. Существуют формы анемий, не связанные с недостатком железа в окружающей среде; речь идет о заболеваниях генетической природы — талассемии и серповидноклеточной анемии, которые нередко детальны. В некоторых районах гены, определяющие эти формы анемий, достигают высокой частоты, так как у гетерозигот по этим генам анемия протекает в легкой форме, а вместе с тем в районах, эндемичных по малярии, присутствие этих генов в гетерозиготном состоянии обеспечивает определенное селективное преимущество (гл. 16). Необходимо следить за тем, чтобы в странах с высокой частотой анемии содержание железа в пище людей было повышено.
Железо содержится во многих пищевых продуктах, и, следовательно, географические различия в этом отношении не имеют значения. При смешанной диете с достаточным количеством овощей дефицит железа в организме развивается редко. Но все же для беременных или кормящих женщин и для детей количество железа, поступающее с пищей в определенное время года, может оказаться недостаточным.
498
Часть V. Экология человека
Хлористый натрий (соль)
Человек не может обойтись без минимальной ежедневной дозы хлористого натрия. В жарких странах значительное количество хлористого натрия выделяется с потом. Тем не менее количество соли, поступающее в организм в этих странах, ненамного выше, чем в странах умеренного пояса, поскольку в ходе акклиматизации устанавливается новое равновесие при более низком уровне поступления за счет уменьшения выведения хлористого натрия с мочой и потом. У лиц, прибывших в страну с жарким климатом, в течение некоторого времени наблюдается недостаточность хлористого натрия, но после акклиматизации у них, так же как у коренных жителей, устанавливается необходимое равновесие (на весьма умеренном уровне — около 8—10 г в сутки).
Известно, что организм человека легко приспосабливается как к низкому, так и к высокому уровню потребления соли. Эскимосы, в пище которых мало овощей, не испытывают какой-либо особой потребности в соли. Там, где вода солоновата, количество поступающей в организм соли довольно велико, как, например, у некоторых групп бедуинов. При систематическом потреблении е пищей больших количеств соли развивается привычка к «соленому». Во многих странах соль используют для консервирования пищевых продуктов.
Мы не располагаем данными, которые указывали бы на то, что потребность в соли или умение приспосабливаться к изменению наличных количеств соли различны у разных популяций. Леши (Leschi) утверждает, что у представителей некоторых племен тропической Африки уровень натрия в крови несколько ниже, чем у европейцев. Это не обязательно «расовый» признак, так как известно, что у европейцев, живущих в тропиках, уровень натрия в плазме также понижен. По-видимому, это одно из физиологических изменений, сопровождающих акклиматизацию.
Источники соли имеют довольно ограниченное распространение (главные из них — море, соляные источники и солончаки); поиски таких источников были в древности одним из факторов, который способствовал ликвидации изоляции человеческих групп. Караваны с солью еще и сейчас пересекают Сахару; широко распространены покупка и меновая торговля солью; истории известны большие соляные экспедиции, иногда даже военного характера.
Иод
Для нормального роста и жизнедеятельности организма необходим некоторый суточный минимум иода. Мы знаем, что в некоторых районах содержание иода в почве, воде и растениях оказывается недостаточным. Области земного шара, эндемичные по
Глава 26. Экология питания
499
зобу, характеризуются не какими-либо особенностями климата, а более или менее интенсивной вулканической деятельностью (в прошлом или настоящем). Способность организма приспосабливаться к недостаточному поступлению иода довольно ограниченна. Более интенсивное потребление иода из организма до некоторой степени компенсируется увеличенной активностью щитовидной железы (с характерным развитием зоба). Однако дефицит иода приводит к задержке роста и другим нарушениям в развитии скелета. У нас нет веских оснований рассматривать популяцию пигмеев как «расу» тиреоидных карликов, хотя в области Улела в Заире существует дефицит иода. У карликов, живущих в этой области, в отличие от пигмеев нарушена функциональная деятельность щитовидной железы. Тем не менее их низкий рост никак пе связан е какими-либо генетическими особенностями. До сих пор не описано этнических групп, которые не реагировали бы на недостаточное поступление иода. У народов Пенджаба, готтентотов и банту, эскимосов и европейцев — у всех развиваются клинические симптомы недостаточности иода; они могут быть обусловлены нехваткой иода в данной местности или действием различных, так называемых зобогенных факторов, нарушающих использование иода в организме.
Витамины
Организм не способен вырабатывать витамины; подобно незаменимым аминокислотам, они должны поступать извне. Поэтому диета круглый год должна быть достаточно разнообразной, чтобы обеспечить поступление хотя бы в самых минимальных количествах примерно дюжины витаминов, необходимых человеку. Организм располагает известным резервом витаминов (или их предшественников) и поэтому способен в течение некоторого времени восполнять их недостаток в пище. Последствия витаминной недостаточности обнаруживаются обычно спустя некоторое время (через недели или месяцы). Авитаминозы встречаются у представителей всех рас. Клиническая картина зависит от особенностей питания в данных конкретных условиях и от комбинации с другими формами алиментарной недостаточности. Если пища бедна продуктами животного происхождения, организм испытывает недостаток в витаминах А и D; однако эти витамины содержатся также в свежих овощах, и соответствующим изменением диеты можно восполнить их дефицит. Сказанное в полной мере относится к витаминам групп Вик витамину С. Таким образом, в самых различных условиях обитания можно обеспечить' достаточное поступление в организм этих витаминов. Лишь в тех случаях, когда пища слишком однообразна и содержит главным образом злаки, при низком содержании жиров и белков появляется ост-
Рис. 26.1. Географическое распределение болезней, связанных с алиментарной недостаточностью (по данным Американского географического общества, Атлас болезней, карта 9).
I — белковая недостаточность (квашиоркор, голодные отеки), II — недостаток солей (зоб), III — множественный авитаминоз.
Глава 26. Экология питания
501
рая витаминная недостаточность. Например, употребление в пищу больших количеств маиса связано с дефицитом никотиновой кислоты и ведет к пеллагре. При постоянном употреблении в пищу только пшеничной муки и очищенного риса развиваются симптомы недостаточности тиамина и рибофлавина. Недостаток в пище животных жиров и свежих овощей ведет к авитаминозу D, степень которого усиливается при недостаточном количестве ультрафиолетовых лучей, что может быть связано либо с атмосферными условиями, либо с ношением плотной одежды. В худшем положении оказываются темнокожие народы (гл. 27) Имеются указания на высокую частоту рахита у детей индийских иммигрантов в Англии в начале 70-х годов.
Географическое распределение болезней, связанных с алиментарной недостаточностью
Американское географическое общество составило карту, иллюстрирующую распределение на земном шаре болезней, связанных с алиментарной недостаточностью (рис. 26.1). Отмечена тесная связь таких болезней с географическими зонами, бедными продуктами скотоводства, овцеводства, рыболовства. Именно в этих областях чрезвычайно высок уровень потребления углеводов. (Изданы также карты, показывающие географическое распределение на земном шаре углеводов, белков и других пищевых веществ). В обзоре, содержащем данные по 21 стране за период 1955— 1962 гг., среди главных проблем, связанных с нехваткой продуктов питания, названы белковая недостаточность (квашиоркор), низкая калорийность пищи (маразм) и дефицит иода, анемии алиментарного происхождения, авитаминоз А, недостаточность тиамипа (бери-бери), рибофлавина, витамина С и никотиновой кислоты (пеллагра).
Зависимость характера пищи от среды обитания
Большому разнообразию человеческих экосистем соответствует и разнообразие пищевых ресурсов. Чтобы оценить все аспекты, связанные с особенностями питания, необходимо провести тщательный анализ каждого конкретного случая. Тем не менее можно сделать ряд общих замечаний о различных типах питания. С точки зрения питания полезно отметить важные различия между тремя группами: 1) охотничьими (включая рыбную ловлю) и пастушескими сообществами; 2) земледельческими группами; 3) сообществами, занятыми одновременно и земледелием и животноводством. Можно в общих чертах охарактеризовать состав пищи в этих трех группах. В первой пищу можно назвать преимущественно белковой, во второй — преимущественно углеводной с
502
Часть V. Экология человека
низким потреблением белков, а в третьей — смешанной. Отмеченные главные различия лишь приблизительно характеризуют диету. Первичные источники основных пищевых веществ в пределах каждой категории чрезвычайно разнообразны. Высокое содержание белка может обеспечить мясо диких и домашних животных; содержание жиров в такой пище может быть либо высоким, либо низким. Термин преимущественно углеводная пища отражает зависимость от одной или нескольких главных зерновых культур (маиса, пшеницы, риса), растений с клубнями, богатыми крахмалом, от тех и других одновременно или от рафинированного сахара. Потребление других продуктов питания, а именно овощей, зелени и фруктов, может изменяться совершенно независимо от того, богата пища белками или нет.
Диета примитивных сообществ обеспечивает лишь необходимый минимум пропитания — сообщество должно обеспечить себя всеми пищевыми продуктами, необходимыми в течение года, из окружающей среды. Во многих экосистемах, вероятно в большинстве из них, существует зависимость от естественной последовательности источников пищи, не всегда достаточно надежных, особенно в примитивных сообществах. Как общества, занятые сельским хозяйством (земледелием или животноводством), так и охотничьи племена и собиратели часто испытывают острую нехватку продовольствия, хотя и по разным причинам. Вареное мясо дает 850— 1250 кДж на каждые 100 г веса в зависимости от содержания жира. Для того чтобы обеспечить организм необходимым числом калорий (8370 кДж на полдня), надо около 1000 г мяса в день. Мяса убитого на охоте животного обычно хватает нескольким семьям на 2—3 дня. С точки зрения удовлетворения повседневных энергетических потребностей пища охотников и собирателей плодов гораздо менее надежна, чем при разумно организованном земледелии. Однако примитивное земледелие, хотя оно и не связано с повседневной заботой о пище, типичной для жизни охотников, почти повсеместно сопровождается сезонной нехваткой продовольствия, и земледелец, весь труд которого направлен на обеспечение себя и своей семьи продуктами питания, может в лучшем случае добиться соответствия между наличным и потребляемым количествами продуктов лишь в среднем за год. Пастушеские племена также не застрахованы от сезонной нехватки продовольствия.
Циклический и часто совершенно ненадежный с точки зрения питания характер этих двух главных типов хозяйства, связанных лишь с добыванием пищи, можно проиллюстрировать на примере эскимосов — в высшей степени искусных охотников — и крестьянской общины в Гамбии. В обоих случаях имеется явная зависимость от сезонной последовательности наличных источников пищи — появления определенных пород арктических животных
Глава 26. Экология питания
503
(для эскимосов) или созревания тех или иных сельскохозяйственных культур (для гамбийцев).
Зимой эскимосы Канады, живущие к северу от Гудзонова залива, охотятся на тюленей. Охотник ожидает добычу на плавучей льдине; часы дневного света коротки, льдину часто окутывает густой туман, за которым нередко следуют снежные бураны; температура воздуха может достигать — 35°С. Убитый тюлень почти полностью съедается; кровь и печень идут в пищу в первую очередь, а ворвань съедают лишь при самых неблагоприятных обстоятельствах. Это период острой нехватки пищи и неуверенности в завтрашнем дне. Весной начинается охота на моржей, которых бьют гарпунами из каяков. И на этот раз почти все животное, за исключением содержимого желудка, съедается. Эскимос крадется к моржам и тюленям по краю льдины и охотится с собаками за полярным медведем. Ни человек, ни собаки не употребляют в пищу печень медведя: эскимосам хорошо известно, что она токсична из-за слишком высокого содержания в ней витамина А. В давние времена еще одним источником пищи служил также мускусный бык. Именно в это время года эскимосы запасают впрок в ледяных колодцах большие количества различных пищевых продуктов и ворвани. Летом появляется растительность, и вдали от берега эскимосы устраивают засады на диких оленей в период их ежегодной миграции. Олени питаются мхом и лишайником. Эскимосы съедают кислое, богатое ферментами содержимое желудка оленей. Эскимосы охотятся и на других наземных животных — волков, зайцев, уток и гусей, а также ловят лососевых рыб. В это время года эскимос перестает быть исключительно плотоядным: ягоды (брусника, клюква), различные корни и молодые побеги карликовых ив входят в его ежедневный рацион. Состав пищи эскимосов приведен в табл. 26.3.
Отличительная особенность этой охотничьей диеты — высокое содержание белков и жиров и низкое содержание углеводов. Организм получает в достаточном количестве железо, витамины А, В и С, много витамина D, особенно если учесть употребление в пищу костей животных. Очевидно, что при таком питании ежедневно или на каждый прием пищи должно приходиться большое количество мяса. Синклер (Sinclair) приводит цифры, свидетельствующие об исключительной способности эскимосов и якутов съедать до 10 кг мяса или даже больше за один прием. Зимой нередко наступает голод, когда затруднен доступ к источникам пищи и туман препятствует охоте.
Эскимосы не страдают рахитом, цингой или другими авитаминозами; им абсолютно неизвестны болезни, связанные с белковой недостаточностю. Имеются данные о болезнях, обусловленных недостаточностью иода; описано несколько географический районов, эндемичных по зобу.
Питательная ценность диеты взрослого эскимоса (Sinclair, 1953)
Таблица 26.3
Вид пищи Вес съедаемой порции, г Калорийность, кДж Общее количество белка, г Животные белки, г Углеводы, г Жиры, г Железо, мг Кальций, мг Фосфор, мг Витамин А, ME Каротиноиды, мкг Витамин D, ME Тиамин, мг 1 Никотиновая кислота, мг ' Рибофлавин, мг Аскорбиновая кислота, мг
Мясо морских млекопитающих 860 7100 163 163 26 103 23,2 95 1 686 7 740 0 0 0,95 42,1 1,2 69
Прочая мясная пища 225 1 855 43 43 7 27 6,1 25 441 2025 0 0 0,25 11,0 0,32 18
Мойва (лососевая рыба) 620 2710 105 105 0 19 6,2 155 1500 508 0 6144 1,30 4,61 0,87 56
Прочая рыба 370 1862 61 61 0 11 3,3 67 699 56 0 0 0,15 8,5 0,18 7
Яйца 5 34 1 1 0 1 0,1 3 10 35 30 3 0,01 0 0,02 0
Ягоды 50 60 0 0 3 0 0,6 30 22 0 27 0 0,02 0,2 0,02 45
Хлеб 27 269 2 0 13 0 0,3 6 20 0 0 0 0,01 0,2 0,01 0
Ячмень н бобы 6 60 1 0 2 0 0,4 11 19 0 0 0 0,03 0,1 0,02 0
Сахар 6 100 0 0 6 0 0 0 0 0 0 0 0 0 0 0
Кофе 6,5 80 1 0 2 1 0,3 9 10 0 0 0 0,06 0,6 0 0
А. Общее количество — 14130 377 373 59 162 40,5 401 4 407 10 364 57 6147 2,78 108,8 2,64 195
Б. Оксфордская норма потребления — 12 600 72 36 432 102 10 750 1000 833 3000 200 1,2 12 1,8 30
Процент от нормы — 112 524 1036 14 159 405 54 441 1244 2 3 074 232 907 147 650
Глава 26. Экология питания
505
Пища донеолитического человека
Пища эскимосов дает представление о всех существенных особенностях питания охотничьих обществ, в том числе и человека, жившего в период палеолита и мезолита. Правда, для диеты эскимосов характерна одна особенность — очень малое потребление растительной пищи. Каннибализм неизвестен у современных или живших в недавние времена народов, занятых собиранием плодов, а также у первобытных охотников и земледельцев. Имеются сведения, что он был распространен у Homo erectus в Китае, а также у неандертальцев и некоторых других групп Homo sapiens в эпоху позднего палеолита. В общем можно сказать, что достаточный по калорийности рацион охотника, успешно использующего все доступные ему источники пищи, в целом обеспечивает организм необходимыми питательными веществами, пока сохраняется основной источник энергии. Весьма вероятно, что именно такого положения достиг человек в доисторическую эпоху, предшествовавшую появлению групп, занятых сельским хозяйством.
Pithecanthropus pekinensis, если судить по сломанным и сожженным костям, найденным при раскопках пещер, употреблял в пищу мясо ныне вымерших видов — бизона, лошади, носорога, плоскорогого оленя, бурого медведя, большерогой овцы, мамонта, верблюда, страуса, антилопы, индийского буйвола, дикого кабана, гиены и других животных. Мы достоверно знаем, что и другие охотничьи племена, как давно вымершие, так и жившие в недалеком прошлом, успешно охотились на всех этих животных. Обуглившиеся кости из пещеры Чоу-коу-тьен принадлежат также лисице, крупным диким кошкам, макакам и бабуинам. В золе костров найдены остатки растений, таких, как Celtis sinensis карликовой породы, достигающих в настоящее время 10 м в длину, и Celtis occidentalism из плодов которого длиной до 2 см получали сладкий напиток, содержащий сахарозу и гексозу и богатый витамином С. Pithecanthropus pekinensis употреблял в пищу различные бобовые растения.
Раскопки неандертальской стоянки в Крапине свидетельствуют о том, что неандертальцы пользовались огнем, а также о том, что они разбивали кости животных, чтобы добыть костный мозг. Установлено также, что они употребляли в пищу мясо лошадей, свиней, выдр, медведей и носорогов. Полагают, что как у человека из Крапины, так и у пекинского человека имел место каннибализм. В гибралтарских пещерах найдены ныне вымершие гагарки и много раковин моллюсков, в том числе двустворчатых. И птицы и моллюски употреблялись в пищу. У нас есть все основания полагать, что неандерталец мог использовать самые разнообразные источники питания, и сомнительно, чтобы исчезновение неандертальцев, если принять во внимание их широкое распространение
506
Часть V. Экология человека
и имеющиеся в их распоряжении пищевые ресурсы, можно было приписать каким-либо факторам, связанным с питанием, например с недостатком в пище витамина D.
Об охотничьей деятельности людей эпохи ориньякской культуры мы можем судить как по остаткам, так и по наскальным рисункам. Древние люди эпохи Граветт оставили огромные кучи костей мамонтов и носорогов, что свидетельствует о достаточно совершенных методах охоты. Остатки огромного количества лошадей сохранились в Сутре; в Вистонице и Пржедмосте найдены груды костей мамонтов, главным образом молодых, которых убивали, загоняя в глубокие ямы. Из костей мамонтов добывали костный мозг. В остатках культуры Капси (Северная Африка) среди убитых животных обнаружены большерогий буйвол, слон, носорог, зебра; в пищу употреблялись также яйца страуса и сухопутной черепахи. Наконечники стрел и копья мадленской эпохи свидетельствуют об интенсивной охоте на оленей и о рыбной ловле. Наглядным примером интенсивной эксплуатации экологических ниш в переходный период от собирательства к земледелию служит Центральная Америка.
Крестьянское сообщество Гамбии
Вернемся теперь к земледельцам и рассмотрим в качестве примера крестьянское сообщество Гамбии. Рис. 26.2 иллюстрирует последовательность употребления различных видов пищи — злаков, овощей, мясных и молочных продуктов — на протяжении года. Каждый столбец соответствует одной неделе, а высота столбца показывает, сколько раз в течение недели данный продукт употреблялся в пищу. Основной род пищи — злаки (финди, маис, рис, камышовое просо, сорго следуют друг за другом в течение года). Финди — ранняя кормовая культура — имеет зерна примерно такого же размера, как семена злаковых трав, и используется в пищу начиная с июля, вскоре после сбора урожая. Наиболее распространенная культура — рис; рис едят все время, пока имеются его запасы. Приблизительно в апреле, когда запасы истощаются, главными продуктами питания становятся просо и сорго. Для употребления в пищу эти злаки толкут в ступе, чтобы отделить мякину и большую часть отрубей. Рис едят в виде цельных зерен, другие злаки перемалывают в муку, причем выход муки варьирует от 70 до 85%. Молоко и яйца относятся к редким продуктам. Таким образом, в ежедневный рацион входят преимущественно углеводы и лишь небольшое количество мяса и сушеной рыбы, земляные орехи и бобы, в определенное время года — зелень и фрукты.
В табл. 26.4 показан состав пищи гамбийцев. (Для сравнения приводится состав пищи разных крестьянских сообществ, а также
Глава 26. Экология питания
507
А
Время сбора урожая
Крупы В Майе EJ Финди Сорго S Просо □ Рис
Овощи
□ Помидоры
£3 Окиа
В Сушеные листья свежие листья г-! Бобы рожно-ш вого дерева □ Земляные орехи
Мясо и другие продукты И Масло ЕЗ Молоко Сушеная рыба □ Сушеное мясо □ Свежее мясо
Рис. 26.2. Частота употребления различных видов пищи в крестьянском хозяйстве в Африке.
Каждый столбец соответствует одной неделе, а высота столбца показывает, сколько раз в неделю употреблялся в пищу данный продукт А Основные виды пищи Б. Дополнительные блюда (овощи) В. Мясные продукты.
состав пищи жителей Великобритании.) Отличие диеты гамбийцев от диеты эскимосов (табл. 26.3) очевидно. Наиболее серьезное последствие такой диеты — белковая недостаточность, поэтому в Гамбии широко распространены квашиоркор и цирроз печени. Азотистый баланс в организме поддерживается на очень низком уровне: общее выделение азота с мочой примерно вдвое меньше, чем у американских детей. Хорошо известно, что народы, главным пищевым продуктом которых является маис, страдают от недостатка витамина В. У них часто встречается пеллагра и хейлоз. У народов, питающихся рисом, весьма часты случаи бери-бери и различных стоматитов. Нередко организм испытывает недостаток в железе и кальции, но кальцификация зубов и костей в целом хорошая, вероятно из-за более эффективного усвоения кальция,
508
Часть V. Экология человека
Таблица 26.4
Содержание основных пищевых веществ в диете некоторых крестьянских сообществ
(Platt, Trans. Roy. Soc. Trop. Med. Hyg., 40, 379, 1947)
Цифры для развивающихся стран, рекомендованные на ближайшее будущее Питательные вещества, приходящиеся в день на человека
Великобритания (1944 г.) о. Барбадос (1944 Г.) Деревня в Гамбии (1945 г.) Ньясаленд (1938—1939 гг.)
1 Деревня 1 в горах Деревня у подножия I горы 1 Деревня на берегу озера Город
Эвергия (кДж) 10 500 12 250 10100 8 518 7 580 8 610 7 300 6 950
Белки, г 60 87 45 56 50 59 26 54
Жиры, г — 117 63 25 15 16 9 11
Углеводы, г — 381 416 396 357 400 387 334
Спирты, мг — — — — 2,5 8,6 1,4 0.8
Кальций, мг 800 1037 254 219 280 476 1065 702
Железо, мг 20 . 16 12 26 25 34 21 24
Витамин А, ME (как р-каро-тин) 5000 3 773 5 215 3 536 7 757 10 220 6020 6 914
Витамин Bi, мг (аневрин, тиамин) 1,6 2,0 0,8 1,8 0,8 1,2 0,7 0,8
Рибофлавин, мг 1,8 2,1 0,8 0,5 0,5 0,7 0,5 0,6
Никотиновая кислота, мг 12 19,7 7,0 14,5 9,0 11,5 9,6 7,0
Витамин С (аскорбиновая кислота), мг 30 123 69 28 86 98 120 73
имеющегося в пище. Алиментарная недостаточность особенно четко проявляется, когда ребенка отнимают от груди. До этого, несмотря на то, что пища матери бедна белками, молоко остается полноценным, а содержание белка в нем — высоким. Однако увеличение продолжительности кормления грудью до 3—4 лет приводит к прогрессирующему ухудшению качества молока. Отнятие ребенка от груди не всегда исправляет положение, так как пища ребенка теперь состоит почти целиком из углеводов — жидкой каши и пюре. На такой диете ребенок продолжает испытывать хронический недостаток белка.
Глава 26. Экология питания
509
Расход Щ Потребление
Рис. 26.3. Схема, иллюстрирующая потребление, наличные ресурсы и расход энергии в течение года (для сельских жителей) (Fox, 1953).
Как уже упоминалось, характерные трудности первобытных земледельцев, связанные с питанием, особенно резко проявляются в «голодный сезон», когда взрослые теряют в весе, рост детей замедляется и появляются клинические симптомы алиментарной недостаточности. Результат количественного анализа баланса калорий на примере хозяйства Гамбии показан схематически на рис. 26.3.
В период от марта до мая у деревенских жителей почти нет работы. Вес тела остается стационарным. Источников энергии вполне хватает, чтобы поддерживать равновесие, соответствующее небольшому расходу энергии. В мае крестьяне начинают расчищать землю, готовясь к дождливому сезону, и расход энергии возрастает. По-настоящему тяжелая работа начинается в июне, увеличивается как потребление, так и расход энергии. Из-за недостаточного питания в это время года расход энергии превышает ее поступление; начинается потеря в весе (приблизительно 20 927 кДж на каждый килограмм веса). В сентябре и октябре энергия, затрачиваемая в процессе производства сельскохозяйственных продуктов, уменьшается, но вместе с тем уменьшается и ее поступление в организм. Вес тела продолжает падать. В ноябре расход
510
Часть V. Экология человека
энергии близок к минимуму; выполняемая в это время работа наименее интенсивна. Источники питания становятся обильнее, увеличивается количество продуктов, и потребление энергии начинает превышать ее расход. В течение ноября, декабря, января и февраля вес тела возрастает: все сельскохозяйственные работы к
Рис. 26.4. Изменение веса тела в ходе сельскохозяйственного цикла (Fox, 1953).
I. Мужчины. II. Женщины.
этому времени заканчиваются, а продовольствие имеется в достатке благодаря недавно собранному урожаю.
Заметьте, что в период от начала июля до конца октября организм должен восполнять дефицит энергии за счет собственных запасов. Только используя эти запасы, организм может поддерживать равновесие между количеством поступающих и расходуемых калорий (рис. 26.4).
Пастушеские племена и народы, как правило, также испытывают сезонную нехватку продовольствия. В Судане (область Бутан) за коротким влажным сезоном следует продолжительная засуха, во время которой становится очень трудно или даже невозможно пасти скот и возделывать землю. Отсюда сезонная нехватка животных белке®!, рибофлавина, витаминов А и С* Вскоре после начал® дюдадай по
ложение меняется: скотоводы получают вдоволь молока и зелени, а несколько позже и зерна, так как многие; из них один раз в ти>д снимают урожай зерновых.
Сильные и слабые стороны питания для различию форм экосистем очевидны. Недостатки диеты охотников заключякгв®» ив в составе пищи, а в ее количестве, связанном с неиадеяпмггаи» источников питания. К особенно тяжелым последствия» ведет низкая калорийность пищи одновременно с алименгаряий недостаточностью. В противоположность скотоводам у земледельцев пища более надежна с точки зрения поступления энергии, Одатако в качественном отношении она часто неполноценна вследствие сезонных трудностей и высокого содержания утаетюдав. Лишь
Глава 26. Экология питания
511
при наличии надежной системы водоснабжения и орошения можно достичь существенного улучшения условий питания. Однако зависимость от нескольких главных зерновых культур может быть в этом случае даже более выраженной со всеми вытекающими отсюда последствиями, связанными с недостатком белков, минеральных солей и витаминов.
Преимущество смешанных типов хозяйства не подлежит сомнению. Сельскохозяйственное производство в неолите, по-видимому, приближалось к такому типу, и пища была достаточно полноценной с точки зрения состава и качества. Наиболее древняя из известных нам земледельческих культур средней Европы, так называемая «дунайская» культура, относится приблизительно к 4000 г. до н. э. Она просуществовала несколько тысячелетий. Сельскохозяйственная техника была основана на использовании орудия, похожего на деревянную мотыгу. Основными зерновыми культурами были ячмень, пшеница (однозернянка), эммер (культурная двузернянка) и другие виды пшеницы; в некоторых районах — бобы, горох, чечевица. В небольших количествах дунайцы держали скот: овец, свиней, быков. В южной части Балканского полуострова, на территории современной Греции, в период раннего неолита выращивали ячмень, пшеницу, инжир и держали крупный рогатый скот, овец и коз. Некоторую роль в хозяйстве еще играла охота (на благородного оленя). Неолитические племена фаюма (Египет) сеяли пшеницу и ячмень, охотились, удили рыбу, держали домашних животных, крупный рогатый скот и овец.
Преимущества смешанных типов хозяйства, типичных для многих земледельческих сообществ неолита, можно наглядно проиллюстрировать на примере индейцев племени яруро, населяющих плоскогорья южной Венесуэлы. Наряду с возделыванием сельскохозяйственных культур на участках, очищаемых от леса вырубкой и огнем, в их хозяйстве важную роль играет сбор плодов, раз-ведение свиней, охота и рыбная ловля, причем деятельность, связанная с добыванием пищи, носит явно выраженный сезонный характер. В сухое время года заметно возрастает сбор клубней и семян, по-видимому богатых крахмалом; фрукты собирают преимущественно в дождливый сезон. Сухой сезон — это также время интенсивной охоты, когда животные собираются по берегам рек и водоемов. Реки в это время года мелеют, освобождаются от ила и зарослей травы, что облегчает лов рыбы. Выращивание некоторых полезных культур, например ананасов и проса, в сухой сезон невозможно; уменьшается также количество сахарного тростника.
Смешанный тип хозяйства, вероятно, был наиболее распространен в районах, где не было искусственного орошения; однако появление железных орудий изменило характер хозяйства — оно стало почти полностью зависеть от интенсивного выращивания лишь нескольких основных культур.
512
Часть V. Экология человека
Физические и «расовые» различия и их связь с питанием
Антропологи часто склонны приписывать различия в физических признаках, размерах тела, форме и размерах лица и черепа действию длительных факторов — расовых или генетических. Между тем эти различия можно с успехом объяснить влиянием факторов внешней среды, в частности питания. Наблюдение над близнецами (см. гл. 10) показывает, в какой мере различия в росте или весе могут быть связаны с негенетическими причинами. Физические изменения, связанные с миграцией, вскрывают те же тенденции, какие обнаруживают группы, сходные в расовом отношении, но находящиеся в различных условиях обитания. В гл. 21 подробно рассмотрено влияние таких факторов, как экономическое положение и питание, на рост человека.
Обычно мы лишь косвенным путем можем установить, что питание — вероятная причина физических различий. Когда мы находим, что размеры тела у одной группы больше, чем у другой с близким генетическим составом, проживающей в той же зоне (но пища первой более полноценна), мы почти не сомневаемся в действии факторов, связанных с питанием. Эти факторы могут иногда играть более важную роль, чем генетические.
Хорошо аргументированный пример приводит Ёрно (Hiernaux). Представители двух генетически близких групп народности хуту в Руанде, резко различающихся по характеру питания, заметно отличаются друг от друга средними размерами тела, причем именно теми, которые у близнецов наиболее чувствительны к изменению условий внешней среды. И, наоборот, признаки с сильно выраженной наследственной компонентой оказались у обеих групп более схожими.
Более высокий рост, крепкое телосложение, большая выносливость сикхов — народности, проживающей в северной Индии,— пр сравнению с мадрасцами, населяющими южную Индию, по-видимому, непосредственно связаны с более высоким содержанием белка в рационе сикхов, состоящем из мяса, молока и молочных продуктов, тогда как пища мадрасцев — в основном вегетарианская. Мак-Каррисон (McCarrison) провел эксперименты на крысах; те из них, которые получали пищу сикхов, весили в среднем 250 г, а получавшие рацион мадрасцев — 155 г. В двух обследованных Ёрно генетически близких популяциях хуту в Руанде жители более плодородных и богатых областей весили в среднем на 7 кг больше, чем жители бедных районов, и соответственно имели большие размеры грудной клетки и бедер.
Бойд-Opp и Джилке (Boyd-Огг a. Gilks) изучали племена масаи и кикуйю в Кении. Здесь мы сталкиваемся, вероятно, с одновременным действием как генетических, так и алиментарных
Глава 26. Экология питания
513
факторов. Земледельцы кикуйю употребляют в пищу злаки, клубни некоторых растений и бобы. Масаи — скотоводы; в их рацион входит мясо, молоко, кровь буйволов. В физическом отношении эти две группы, живущие рядом в одних и тех же природных условиях, заметно отличаются друг от друга. Представители племени масаи на 7—8 см выше, а вес их на 10—11 кг больше, чем у их соседей из племени кикуйю. По мнению Бойд-Орра и Джилкса, это различие — прямое следствие существенных различий в их диете. В рационе масаи достаточно белков животного происхождения, а племя кикуйю живет в условиях почти постоянного белкового голодания.
Де Кастро (De Castro) наблюдал нечто подобное на северо-востоке Бразилии. В прибрежных районах и в засушливой области, удаленной от берега, преобладают люди высокого роста, а в зоне тропических лесов доминируют низкорослые — их средний рост ниже, чем в двух других областях. Жители прибрежных районов ловят рыбу, и потому в их пище много белка. Потребление белка довольно высоко также во внутренних областях, где основное средство существования — скотоводство, дающее много мяса, молока и сыра. Зато в зоне джунглей, где сахарный тростник утвердился в качестве монокультуры и производство его вытеснило все другие виды деятельности, диета бедна; в состав пищи входит мука из маниока с крайне низким содержанием белка. Указанными различиями в питании, особенно в потреблении белка, можно объяснить различия в размерах тела у трех групп людей, проживающих в одной и той же географической зоне.
Детально анализируя условия питания, Никольс (Nicholls) пришел к выводу, что различия в весе и росте жителей Шри Ланки зависят не от их расового происхождения, а от экономического и социального положения. Рост 15-летних мальчиков из зажиточных слоев общества на 12,5 см выше, а вес на 9 кг больше, чем у мальчиков того же возраста из бедных слоев. Различия в физических признаках у лиц, принадлежащих к двум разным общественным группам, гораздо больше, чем различия между племенами масаи и кикуйю. Мальчики особенно быстро растут в возрасте от 5 до 8 лет, но темпы роста выше для мальчиов из зажиточных слоев. Мальчики из бедных слоев особенно сильно отстают от своих товарищей в 11 —14 лет и лишь отчасти компенсируют это отставание в старшем возрасте.
Еще одним примером может служить Пуэрто-Рико. Тьем (Thieme) показал, что негры из более зажиточных слоев выше не только своих более бедных африканских сородичей, но и беднейших европейцев.
Хорошее полноценное питание — фактор, который оказывает большое влияние на рост и другие размеры тела. В то же время многие физические признаки и пропорции тела остаются неизмен
514
Часть V. Экология человека
ными при изменении диеты. Таково, например, соотношение длины рук и ног у негров. Дети японцев, живущих в Калифорнии и питающихся так же, как американцы, растут гораздо быстрее, но сохраняют отношение роста сидя к общему росту, характерное для японцев, родившихся в Японии.
Нелегко дать точное и надежное объяснение различиям в размерах тела, отмечаемым даже в пределах ограниченных территорий, с точки зрения влияния среды или генетических факторов. Частично это связано с взаимодействием факторов друг с другом, а частично объясняется сложностью генетического определения роста и другими антропометрическими признаками. Кроме того, в ряде случаев влияние климата или болезней может оказаться более существенным, чем влияние питания. (Более подробное обсуждение этого вопроса читатель найдет в части IV.)
Социальные аспекты питания
До сих мы делали упор на биологию питания, но добывание пищи — процесс общественный. Даже в самом примитивном обществе он включает в первую очередь ответственность родителей за сохранение потомства, поскольку дети в течение многих лет полностью зависят от родителей. «Питание как биологический процесс,— пишет Одри Ричардс (Audrey Richards), — определяет в большей мере, чем любая другая функция, характер общественных групп и формы, которые принимает их деятельность». Мы не собираемся здесь исследовать вопрос о том, разделяют ли антропологи точку зрения Ричардса, но нет сомнения в том, что некоторые стороны социологии питания имеют прямое биологическое значение. Из большого круга относящихся сюда вопросов мы рассмотрим следующие: 1) сведения об окружающей среде, которыми располагает данное сообщество, 2) сведения о питательной ценности пищевых продуктов, о способах их приготовления, хранения и распределения, 3) ограничения общественного и технического характера, связанные с удовлетворением пищевых потребностей.
Тщательное изучение первого вопроса показывает, что большинство первобытных или находящихся на примитивной стадии развития сообществ имеет прекрасное представление о пищевых ресурсах окружающей среды, хорошо знает, что съедобно, а что ядовито. Аборигенов Австралии мы с полным правом можем считать специалистами по экологии. У них есть имена для всех видов ботанических ассоциаций, для любого дерева или растения. Они могут подробно описать все стадии уборки урожая в любое время года. О том, насколько хорошо охотники знают дичь и ее повадки, мы можем судить, наблюдая мимику и жесты в танцах бушменов
Глава 26. Экология питания
515
или изучая наскальные рисунки древних народов. Поиски и сбор меда изображены на стенах пещер палеолита в Аранае (Испания).
Горожанину наших дней покажется необычной и даже неприятной пища многих примитивных групп. Пища, которую мы на первый взгляд считаем неполноценной, в действительности содержит, как показывает анализ, много полезных веществ, жиров, белков и витаминов. В рацион бушменов входят личинки насекомых, гусеницы, лягушки, змеи, ящерицы, саранча, муравьи и термиты.
Ряд других примеров, относящихся к разным типам экосистем, приводит Боденгеймер (Bodenheimer). Он показывает, что особо важным элементом пищи являются насекомые. В Австралии ночные бабочки, жуки и муравьи играют в питании такую же важную роль, как и в тотемических верованиях. В Африке над озером Танганьика ловят и собирают рои крошечных комаров, делают из них муку и используют для приготовления пирожных. Для многих народов Западной Африки существенно важным источником белка, жиров и калорий служат термиты.
Известно более 400 видов ядовитых растений. Разительным примером тех познаний, которыми обладает народ, находящийся на доисторическом уровне развития, о свойствах негодных в пищу и ядовитых веществ, может служить «детоксикация» клубней маниоки, проводимая индейцами Амазонки. Эти клубни содержат цианистый водород. «Детоксикация» состоит в том, что разрезанные ломтиками клубни пропитывают особым раствором; при этом цианистый водород обезвреживается; затем мякоть высушивается и растирается в порошок.
В дополнение к тому, что мы сказали выше об использовании «необычных» видов пищи, отметим, что способы приготовления пищи у многих племен и народов весьма существенны с точки зрения сохранения или даже улучшения питательных свойств пищи. Листья баобаба в африканских странах, употребляемые для приготовления супа, высушивают в тени, а не на солнце, что обеспечивает сохранение в них витаминов. Многие богатые ферментами пищевые продукты, например пиво из дурры (Мексика), имеют большую ценность как источники минеральных солей и как противоцинготные средства. Для приготовления тортиллы (особый вид хлеба) в Мексике маис пропитывают однопроцентной известковой суспензией. Концентрация кальция в таком маисе увеличивается по сравнению с обычной почти в 20 раз. Питательную ценность сои повышают, проращивая семена (которые в таком виде употребляют в пищу) или подвергая их брожению (приготовление соевого соуса). Варка пищи уничтожает различные микроорганизмы и сильно расширяет ассортимент употребляемых в пищу веществ. Большое значение имеет использование гладких, покрытых глазурью горшков и сосудов, которые регулярно моются перед едой.
516
Часть V. Экология человека
Существуют, однако, такие традиции и верования в отношении отдельных видов пищи, которые оказывают явно неблагоприятное влияние на организм. В некоторых странах, например, свежее коровье молоко и рыба считаются неподходящей пищей для молодых людей. Культ риса в ряде стран настолько велик (рис считают абсолютно необходимым для крепкого здоровья), что его вводят в пищу ребенка уже через несколько дней после рождения, вызывая расстройства пищеварения. Можно было бы привести множество таких запретов (табу) и верований. У различных племен широко распространена практика табу на определенные виды продуктов.
Здесь уместно упомянуть о каннибализме. Необходимо отличать людоедство как один из способов пропитания от людоедства как обряда, например военного обряда, когда съедается небольшая часть тела побежденного врага. Число известных нам случаев каннибализма очень мало. Гарн и Блок (Garn a. Block) утверждают, что «умеренный» каннибализм у некоторых живущих на грани голода племен Новой Гвинеи, возможно, служил источником критически необходимого минимума белков. По предположению одна из таких групп людоедов — миянмины — обитает у истоков реки Сепки и насчитывает около 100 человек. Испытывая постоянный недостаток в белковых продуктах, миянмины могли удовлетворять до 10% своей потребности в год, съедая 15 взрослых весом по 60 кг! Последнее из нападений на соседнее племя ат-бальминов в 1956 г. завершилось поимкой и съедением 16 человек, в том числе И взрослых. Из семидесяти известных примеров истинного каннибализма 20% приходится на долю охотничьих народов, находящихся на самой низкой ступени развития, преимущественно народов австралийской группы, а 50% — на долю первобытных земледельцев. Среди пастушеских племен людоедство почти неизвестно. Равным образом и приношение в жертву людей описано в основном у земледельческих народов. Нет никаких оснований полагать, что где-нибудь на земном шаре истинный каннибализм служил значительным или постоянным источником белковых продуктов.
Способ отнятия ребенка от груди оказывает влияние на здоровье, выживаемость, дифференциальный отбор внутри популяции. В западных странах постепенно переходят от грудного вскармливания к смешанной диете, состоящей из специально приготовленных продуктов, среди которых главная роль отводится коровьему молоку. «Западная» практика обеспечивает, пожалуй, самый безопасный из всех мыслимых переходов. В прошлом вскармливание и отнятие от груди сопровождались рядом ритуальных действий, и обращение с ребенком в этот критический для него период являло собой разительный контраст с современной практикой. У негров Африки ребенок после отнятия от груди иногда сразу
Глава 26. Экология питания
517
же переходит на общий для всей семьи рацион. Иногда, однако, смена питания не столь резкая: с самых ранних месяцев ребенок может получать в небольших порциях и в сильно размельченном виде «взрослую» пищу, например жидкую кашу или простоквашу (у народности талленси), кашицу из зерна дурры (у народности коса) или же предварительно пережеванную пищу и пюре из овощей (у народности алор).
Широко распространенная практика увеличения времени кормления грудью означает, что растущий ребенок не может получить необходимое ему количество белка; его потребности возрастают, а общее количество молока у матери, несмотря на то что его состав может оставаться неизменным, уменьшается. Один автор пишет, что в северо-восточной Африке после чрезмерно затянувшегося питания грудным молоком дети становятся хилыми, страдают анемией и часто умирают от поноса, связанного с употреблением малопригодных для них видов пищи после отнятия от груди. В самом переходе к диете с низким содержанием белка, состоящей в основном из углеводов (маниок или просо), кроется опасность заболевания квашиоркором. Ослабленный ребенок может оставаться на попечении старших братьев и сестер, когда мать выполняет различные сельскохозяйственные работы с другими членами племени. Приготовление каши требует тщательного перемалывания зерен в течение нескольких часов; мать же слишком устает и не может затратить необходимого для этого времени, вследствие чего дети получают недостаточно хорошо размолотую крупу; употребление ее в пищу нередко ведет к тяжелым заболеваниям, сопровождающимся поносом.
Одна из распространенных причин неполноценного питания — неэффективное или неумелое использование пищевых ресурсов. В некоторых районах Африки рогатый скот — лишь символ богатства и власти и животных используют главным образом как производителей молока, а не мяса. Из-за плохого хранения продуктов или даже полного отсутствия необходимых помещений значительная часть собранного урожая во многих крестьянских сообществах портится и пропадает. Крысы, насекомые, а также грибы способны уничтожить до 1/3 собранного урожая зерновых и бобовых культур.
В годовом сельскохозяйственном цикле рост производства может тормозиться под действием различных факторов. Экономика Гамбии служит примером устойчивости экологического равновесия и трудностей в достижении радикальных сдвигов в сторону более высокой производительности труда, которая обеспечила бы получение излишков сельскохозяйственной продукции при меньшей затрате труда. Дожди выпадают здесь между июнем и октябрем, притом довольно нерегулярно. Сев проводится поэтому вскоре после начала дождей в очень короткий срок, чтобы почва не
518
Часть V. Экология человека
успела высохнуть и затвердеть на солнце, а растения успели закончить вегетацию до окончания дождливого сезона. Дожди бывают проливными, и какие-либо работы в поле во время ливней невозможны. Вследствие этого ограничивается число рабочих дней в указанные месяцы. Религия запрещает проведение работ также и в дни религиозных праздников. В остальные дни июня и июля крестьянин вынужден работать с чрезмерной нагрузкой — общий расход энергии достигает 14 230 кДж в день. Однако в это же время года ощущается нехватка продовольствия, организм теряет в весе, и, как следствие, быстро наступает переутомление. В июле сказывается отрицательная роль и других неблагоприятных факторов; под влиянием теплового излучения, высокой температуры воздуха и большой влажности средняя эффективная температура (гл. 27) достигает 27°С. В этих условиях чрезвычайно затруднены такие виды полевых работ, как копка земли, разрыхление почвы мотыгой, посадка растений. Крестьяне стремятся обрабатывать большую по размеру площадь, и одно лишь хождение к более удаленным земельным участкам может увеличить расход энергии и одновременно сократить полезное рабочее время.
Оценка действительного количества потребляемых организмом питательных веществ в примитивных обществах весьма затруднительна. Пожалуй, наиболее разумный метод оценки — непосредственное изучение потребления продуктов одной семьей. Процедура, связанная с приемом пищи, может быть довольно сложной: завтрак, обед, ужин нередко проходят в различных, особо отведенных для этого строениях или хижинах. Это связано с обычаями гостеприимства, родственными отношениями и т. д. У зулусов первыми по традиции обедают мужчины, затем женщины и, наконец,— дети; воины получают, следовательно, более качественную пищу, что способствует сохранению их физической силы. Чтобы учитывать возможные изменения в питании, выборочные наблюдения должны охватывать полный цикл (время активной деятельности и отдыха). Такой цикл далеко не всегда занимает неделю; он может составлять и 10 дней. Сезонные колебания также весьма существенны, и их необходимо учитывать. С точки зрения питания важны также празднества и другие церемонии, когда людям предоставляется везможность потребления избыточного количества продуктов, богатых белком и витаминами.
Глава 27
КЛИМАТИЧЕСКАЯ АДАПТАЦИЯ
Для каждой среды обитания, в которой живет человек и его домашние животные и в которой развиваются выращиваемые им растения, характерен свой климатический режим. Климатические условия на земном шаре чрезвычайно разнообразны, и только благодаря наличию у человека определенных приспособлений он способен существовать в среде более разнородной, чем у всех других видов млекопитающих (за исключением, быть может, только крыс). Подобно всем живым организмам, тело человека чувствительно ко многим элементам, которые в совокупности определяют климат местности. Эта чувствительность и способность к соответствующим реакциям отвечает необходимости поддержания гомеостаза. Система терморегуляции в условиях сильной жары или холода действует таким образом, чтобы поддерживать температуру тела в относительно узких пределах (гомойотермия). В условиях жизни на больших высотах система органов дыхания поддерживает в стабильных пределах давление кислорода и углекислоты в жидкостях и тканях человеческого' тела. Повышенные требования к организму предъявляют не только тепловые факторы и низкое барометрическое давление на больших высотах, но и воздействие коротковолновой радиации (ультрафиолетового и ионизирующего излучения).
Температурная среда
«Климат» и «погода» — термины, которые мы употребляем, чтобы охарактеризовать состояние атмосферы. Климат представляет собой сезонное среднее условий погоды в данной местности. Полное описание состояния атмосферы, определяющего погоду и климат и регистрируемого через регулярные промежутки за некоторый период времени, должно включать данные о температуре и влажности воздуха, скорости и направлении ветра, величине и характере облачности, числе солнечных дней, общей дозе теплового излучения, количестве осадков в виде дождя и снега, пыли в атмосфере, а также ряд других параметров. Климат, в котором живет человек, в действительности состоит из ряда климатических «оболочек» — микроклимата его одежды, микроклимата жилых и производственных помещений и географического макроклимата.
520 Часть V. Экология человека
Среди всех географических факторов первостепенную физиологическую роль играют те, которые оказывают прямое влияние на интенсивность теплового обмена между поверхностью тела (обнаженной или закрытой одеждой) и окружающей средой. Для полной характеристики условий температурной среды непременно должны быть представлены сведения о температуре и влажности воздуха, скорости ветра и средней температуре окружающей среды. В помещении к этой среде относятся стены, печи, камины и другие поверхности, которые излучают тепло в направлении субъекта; вне помещений эту роль играют солнце, небо и почва. Все эти элементы в совокупности определяют то количество тепла, которое тело отдает во внешнюю среду. Возможна, конечно, ситуация, когда тепло, наоборот, поступает из окружающей среды в организм, добавляясь к тому «внутреннему» теплу, которое выделяется в процессе мышечной деятельности и обмена веществ.
Эффективность адаптации организма зависит от степени нарушения «гомойотермии». Адаптивные приспособления человеческого организма к температурной среде бывают трех типов (ср. со сказанным в гл. 26).
Д. Общие физиологические приспособления, связанные с функцией терморегуляторной, метаболической и циркуляторной систем, обеспечивают возможность жить и работать в самых разнообразных условиях температурной среды. Способность к таким приспособлениям — это свойство, получившее наибольшее развитие у человека как вида. Приспособления могут быть как кратковременными, так и длительными.
2. Специализированные физиологические и анатомические адаптивные реакции, в основе которых лежат особенности генотипа.
3. Культурные и социальные приспособления, связанные с обеспечением человека жильем, одеждой, теплом, системой вентиляции. Приспособления культуры к биологическим потребностям (а также к потребностям, связанным с питанием), закрепленные обычаями (в смысле Малиновского), до сих пор не были проанализированы достаточно тщательно. Об эффективности этих приспособлений можно судить на основании биологических критериев.
Эффективная адаптация человеческого организма к климату необходима для: а) обеспечения состояния комфорта; б) выполнения физической работы без повышенной утомляемости; в) выполнения различных видов квалифицированной работы, требующей внимания и сноровки, с минимумом ошибок; г) обеспечения нормальных условий для роста и развития.
Рассмотрим крайние, наиболее трудные условия среды как показатель способности человека приспосабливаться к климату. В Верхоянске —- одном из городов Восточной Сибири, находя
Глава 27. Климатическая адаптация
521
щемся вблизи Полярного круга, где зарегистрированы самые низкие температуры на земном шаре,— средняя температура самого холодного месяца, января, на 35°С ниже средней годовой (—18°С). В оазисе Ин-Салах в алжирской Сахаре температура самого жаркого месяца, июля, достигает 38°С, т. е. на 12° превышает среднюю годовую температуру. Таким образом, человеческие сообщества успешно выживают в различных областях с летней температурой от —17 до 4-38°С и зимней температурой от—16 до 4-28°С.
Несмотря на столь резкие изменения внешней температуры, внутренняя температура тела изменяется в относительно небольших пределах. Суточные колебания температуры тела не превышают 2°С. Она имеет максимальное значение вечером и минимальное около 4 ч утра. В тропических странах этот цикл сдвинут вверх примерно на 0,2°С. Подобный сдвиг имеется у всех рас: у европейцев, находящихся в Индии или Сингапуре, температура тела такая же, как у коренных жителей.
Организм не может перенести значительных отклонений от средней суточной температуры, и существование столь узкого интервала колебаний для различных условий (суточных, сезонных и географических) подразумевает наличие весьма чувствительной системы внутренней регуляции. Регуляция осуществляется в первую очередь термостатическим механизмом мозга (гипоталамусом), который чувствителен к повышению или понижению температуры тела в тех случаях, когда тело отдает или получает тепло в большом количестве. При изменении наружной температуры в определенном диапазоне, скажем 27—32°С, внутренняя (ректальная) температура тела обнаженного человека остается близкой к 37°С без заметного изменения температуры кожи (в пределах приблизительно от 30,5 до 33,3°С), так что температурный комфорт не нарушается. Однако и за пределами этого температурного диапазона при изменении температуры поверхностных частей тела обеспечивается постоянство внутренней температуры, необходимое для нормальной жизнедеятельности организма. Пределы климатических изменений, к которым может приспособиться организм, определяются взаимоотношением этих двух биологических факторов — поддержанием необходимого комфорта и сохранением теплового баланса.
Гомойотермия определяется простым уравнением:
Приток тепла — Потеря тепла =± Накопление тепла, или, точнее,
М ±C±R — Е = ±S, где М — количество тепла, производимое в процессе мышечной работы и обмена веществ, С — приток или отдача тепла за счет конвекции, R — приток или отдача тепла за счет излучения, Е —
522
Часть V. Экология человека
отдача тепла за счет испарения влажной поверхности кожи, S — суммарный теплообмен.
Хотя переменные, входящие в левую часть уравнения, могут изменяться в широком интервале, величина S большей частью остается относительно постоянной. Изменения величины S соответствуют либо увеличению, либо уменьшению теплосодержания тела, причем общее теплосодержание практически не изменяется. Именно в этом состоит пластичность реакции организма на перегрев или охлаждение. Если создается угроза равновесию, т. е. накопление или отдача тепла становятся чрезмерными, в действие вступают различные быстро действующие компенсаторные механизмы. Кроме того, в организме происходят длительные процессы, способствующие дальнейшему улучшению теплового баланса; всю совокупность их мы называем акклиматизацией. Все физиологические реакции организма на изменение климатических условий были описаны у людей различных рас.
Реакции на перегрев
Немедленная физиологическая реакция на перегрев — увеличение отдачи тепла организмом. Оно осуществляется, во-первых, через систему кровообращения и, во-вторых, путем потоотделения. Роль системы кровообращения состоит в усилении тока крови через кожу, которое становится возможным вследствие расширения кожных сосудов, а также, если необходимо, вследствие увеличения минутного объема сердца, сопровождающегося ускорением пульса. Тепло, поступающее в избытке к поверхности тела, рассеивается за счет увеличения конвекции и излучения; рассеяние тепла возрастает вследствие увеличения температуры кожи. Интенсивность конвекционной отдачи тепла на единицу поверхности пропорциональна разности температур кожи и окружающего воздуха (и квадратному корню из скорости движения воздуха). Интенсивность излучения на единицу излучающей поверхности приблизительно пропорциональна р.азности средних температур кожи и окружающей среды. Человеческая кожа, независимо от цвета, действует как абсолютно черное излучающее тело. Если указанные процессы недостаточны для сохранения теплового баланса и температура тела возрастает, начинается усиленное потоотделение. Интенсивность теплоотдачи при испарении пота зависит от разности давления паров воды на поверхности кожи и давления воздуха, от размеров увлажненной поверхности и от движения воздуха. Увеличение температуры кожи означает увеличение давления паров воды на ее поверхность. Теплоотдача за счет скрытой теплоты парообразования может сильно увеличиться вследствие как увеличения числа функционирующих потовых желез, так и прогрессирующего увеличения активности
Глава 27. Климатическая адаптация
523
каждой железы. Максимально возможная потеря воды (равная примерно 1 л в 1 я) эквивалентна отдаче 2500 кДж тепла в 1 ч. Хотя общее число потовых желез у разных людей неодинаково, мы не располагаем данными о сколько-нибудь существенных различиях между разными расовыми группами (табл. 27.1).
Таблица 27.1
Общее число потовых желез у взрослого мужчины
Хипди 6 1,51 ±0,19 Knip
Индусы 19 1,69±0,16 Weiner
Голландцы 9 1,47±0,29 Knip
Англичане 2 1,66±0,19 Weiner
Европейцы (Зап. Африка) 21 1,75 Thomson
Африканцы (Зап. Африка) 26 1,66 Thomson
Негры Суринама 8 1,69±0,14 Knip
*) Приведены средние значения в млн. ± стандартное отклонение.
Методы обследования, применявшиеся Томсоном (Thomson), не совсем идентичны методам, использованным другими исследователями. Число потовых желез на одних и тех же участках поверхности тела у представителей разных групп приблизительно одинаково. Число желез на различных участках тела располагается в порядке убывания следующим образом: на верхней конечности — тыльная сторона кисти, предплечье, плечо; на нижней конечности — ступня, нога, бедро; на туловище — жйвот и грудь. Можно со всей ясностью продемонстрировать пластичность физиологической реакции организма, если рассмотреть способ
Таблица 27.2
Число потовых желез на 1 кв. см поверхности тела у взрослого мужчины
Число обследованных Области тела
I живот 1 рука, кисть предплечье плечо тыльная сторона СТОПЫ стопа бедро
Европейцы 29 69 206 98 85 132 87 59
Голландцы 9 75 145 86 80 119 66 52
Индусы 19 89 209 97 91 152 91 62
Хинди 6 87 170 121 93 119 81 60
Африканцы (Зап. Африка) 26 94 240 109 119 175 78 85
524
Часть V. Экология человека
сохранения постоянной температуры в условиях повышенной тепловой нагрузки (табл. 27.2). Такая ситуация встречается, например, 1) в жарком и влажном климате при почти полном отсутствии ветра, 2) при жаркой сухой погоде с интенсивной солнечной радиацией и при движении воздуха, 3) в холодном сухом климате с интенсивной солнечной радиацией, 4) при выполнении тяжелой работы в условиях умеренной температуры. (Подробный анализ указанных случаев можно найти в учебниках по физиологии человека и в монографиях, список которых приведен в конце книги).
Акклиматизация к высоким температурам
При постоянном или многократном воздействии тепловой нагрузки происходит поразительное улучшение в приспособлении организма к реакции на эту нагрузку. Особенно заметно улучшается способность выполнять физическую работу. В лабораторных экспериментах испытуемые, подвергавшиеся действию тепла, в первый день прекращали работу менее чем через 1 ч после начала опыта, тогда как на 5-й день могли выполнять ту же работу в течение 4 ч. Заметно улучшалось состояние системы кровообращения — уменьшались такие показатели, как частота пульса и минутный объем. Более эффективной становилась терморегуляция. В первые дни опыта температура тела быстро и значительно повышалась, достигая довольно высоких значений, а в последующие дни она повышалась медленнее и достигала «плато», несмотря па продолжавшееся воздействие тепла. При акклиматизации уменьшалась также и температура кожи.
Было показано, что все эти изменения, наблюдавшиеся в искусственных условиях (жарких и влажных или жарких и сухих), происходят также в естественной среде в странах с жарким экваториальным или засушливым климатом. Главная причина улучшения системы регуляции состоит в том, что потовые железы становятся более чувстителышми к тепловому раздражению, их реакция ускоряется и потоотделение усиливается. Это означает, что площадь, с которой происходит испарение, увлажняется гораздо быстрее и равномернее, а теплоотдача за счет испарения возрастает, как это и должно быть в случае, если необходимо свести к минимуму увеличение температуры тела и накопление тепла.
Весь комплекс изменений, которые обеспечивают физиологическую акклиматизацию к высоким температурам, был подтвержден наблюдениями над людьми различных рас, живущими в странах с жарким климатом,— нигерийцами, китайцами, индусами, малайцами, бушменами в пустыне Калахари и южноафриканскими банту, а также над европейцами в тропиках и пустынях.
Глава 27. Климатическая адаптация
525
Тот факт, что люди различных рас обладают способностью к акклиматизации, неудивителен, если мы вспомним, что гоминиды возникли и развивались в экваториальных условиях. Человеку наших дней физиологическая акклиматизация позволяет вести активный образ жизни в самых разнообразных условиях жарких стран и дает возможность одному виду Homo sapiens обитать в самых различных тропических и экваториальных областях. Это очевидно из табл. 25.3 и 25.5, показывающих распределение популяций в различных географических районах. Указанную физиологическую пластичность дополняет естественный отбор, закрепляющий те морфологические признаки организма, которые позволяют ему приспосабливаться к жизни в разнообразных климатических условиях.
Реакции на охлаждение
Непосредственные реакции организма на охлаждение направлены к уменьшению теплоотдачи (С+7?) и к увеличению количества вырабатываемого организмом тепла (М), т. е. к сохранению гомойотермии. У человека, не защищенного одеждой, в состоянии покоя при температуре воздуха 31°С происходит повышение интенсивности обмена веществ, чтобы предотвратить падение внутренней температуры тела; это и есть критическая температура. Этот уровень критической температуры характерен для тропических животных вообще. Теплоотдача снижается за счет повышения изолирующих свойств поверхности тела. Теплопроводность кожи заметно уменьшается вследствие сужения сосудов, однако максимальный эффект достигается довольно быстро, так что ниже критической точки температура кожи непрерывно снижается с понижением температуры воздуха. Для жизни в холодном климате человек должен обладать добавочной теплоизоляцией, что позволяет снизить критическую температуру. Известно, что у арктических животных высокоэффективная теплоизоляция обеспечивается толстым слоем подкожного жира и мехом. Такой же степени изоляции человек может достичь, используя для защиты от холода шкуры животных или другой материал. Дополнительная теплоизоляция в 1 clo (изоляция, эквивалентная теплоизолирующему действию слоя неподвижного воздуха толщиной 0,6 см) понизит критическую температуру до 20°С, а в 2 clo — 10°С. При более низких значениях необходимо прогрессирующее повышение интенсивности теплообразования, чтобы человек смог находиться на холоде несколько часов подряд. Теплоизолирующая способность одежды из оленьего меха толщиной от 3,75 до 7,5 см, которую носят эскимосы, соответствует 7—12 clo. Этого достаточно для удовлетворения всех требований организма: во время сна необходимо 10—12 clo, а для активной деятельности при —40°С
526
Ча^тпь V. Экология человека
нужно 4 clo. В жилищах эскимосов днем сохраняется температура 21, а ночью 10° С. В зимнее время эскимосы проводят вне помещения 1—4 ч день, а в летнее время — от 5 до 9 ч. Только лицо, а иногда и руки остаются не защищенными от холода.
Большое значение имеет толщина подкожного жира, определяющая тот уровень, до которого падает температура кожи, подвергающейся действию холода. «Люди с большими жировыми отложениями способны к более длительному пребыванию в холодной воде.
На холоде количество вырабатываемого организмом тепла увеличивается; это может происходить непроизвольно (мышечная дрожь) или потому, что с целью согреться человек намеренно работает, движется и т. п. При дрожи вырабатывается почти втрое больше тепла, чем в состоянии покоя. Дрожь возникает вследствие уменьшения температуры кожи и последующей рефлекторной стимуляции особого центра, находящегося в гипоталамусе. Большое количество тепла вырабатывается при физической работе; это количество лимитируется только функциональной приспособленностью организма и наличием пищи. Если температура окружающего воздуха соответствует точке замерзания, то для поддержания постоянной температуры тела даже в теплой одежде, толщина которой втрое превосходит обычную, затрачивается вдвое больше энергии, чем на основной обмен. Известно, что эскимосы могут долгое время бежать за санями достаточно быстро, чтобы согреться на бегу, но вместе с тем не выбиваясь из сил; их функциональная приспособленность, измеренная обычными тестами, выше, чем у канадцев европейского происхождения (Shepherd).
Важное значение имеет реакция рук на действие холода. Сначала происходит интенсивное сужение сосудов, за ним, примерно через 5 мин, следует их расширение; в дальнейшем эти вазомоторные реакции периодически повторяются. Такое местное расширение сосудов препятствует понижению температуры ткани и отморожениям.
Акклиматизация к холоду
Имеются четкие данные, свидетельствующие о том, что акклиматизация к холоду, т. е. повышение способности переносить Холод, развивается постепенно. Тот, кто впервые попадает в Арктику, вначале надевает всю имеющуюся у него теплую одежду, однако когда приходит зима и температура воздуха падает, он уже не ищет какой-либо дополнительной защиты от холода. В экспедициях во время работы или сна люди часто надевают на себя меньше одежды в зимнее время, чем в начале похода. Было замечено, что у тех участников экспедиции, которые проводили большую часть времени в помещениях, отморожения появлялись при низких температурах, на ветру в первые 1,5 мин, а те, кто преиму
Глава 27. Климатическая адаптация
527
щественно находился на воздухе, выдерживали пребывание на ветру и на холоде до 10 мин. Привычные к холоду люди способны к более точной оценке температуры лица и ног и вовремя принимают необходимые меры, чтобы предотвратить появление отморожений (Irving). Имеются также данные, указывающие на стимуляцию процессов, связанных с поддержанием теплового равновесия. Основной обмен несколько возрастает по сравнению с его величиной в тропических условиях. У эскимосов наблюдается более заметное повышение интенсивности основного обмена (на 7—30%), чем у европейцев (8%), живущих в аналогичных условиях.
Однако пока не ясно, зависит ли увеличение основного обмена непосредственно от действия холода или от изменения функции эндокринной системы и характера питания. Общая потребляемая энергия в Арктике значительно выше, чем в тропиках, но она колеблется в широких пределах в зависимости от условий, а также от выполняемой работы. По мнению некоторых специалистов, увеличение количества тепла, вырабатываемого в процессе обмена веществ, происходит за счет более высокого потребления белка и за счет его специфического динамического действия. Создается, однако, впечатление, что жир в равной или даже в большей степени, чем белок, способствует защите от холода. Многие исследователи Арктики и лица, живущие на севере, утверждают, что в арктических условиях люди ощущают потребность в жире. Участники американской антарктической экспедиции рассказывали, что хотя дома они испытывали отвращение к жиру, в Арктике обычно ели его в больших количествах. Таким образом, и у животных и у человека два процесса играют основную роль в усилении адаптации к холоду: усиленное образование тепла в организме и увеличение толщины теплоизолирующего жирового слоя. Человек зависит, конечно, еще и от искусственной теплоизоляции.
Под влиянием акклиматизации усиленное образование тепла в организме уменьшает степень падения ректальной температуры при действии холода, а вторичное повышение температуры кожи приводит к улучшению субъективного самочувствия человека. Такие изменения температуры тела действительно наблюдались.
О наличии «локальной» акклиматизации свидетельствуют данные, приведенные на рис. 27.1. У лиц, находившихся в холодной комнате по 2 ч в день 5 дней в неделю в течение 5 нед при температуре — 15°С и тепло одетых, но без перчаток, отмечалось снижение тактильной чувствительности, которая, однако, со временем восстанавливалась. В тестах на тактильную чувствительность больше баллов набирали лица, находившиеся вне помещения, по сравнению с теми, кто большую часть времени проводил дома. Давно известно, что рыбаки Новой Шотландии могут подолгу держать руки в холодной воде.
528
Часть V. Экология человека
В экспериментах было установлено, что у группы лиц, находившихся вне помещения, температура кожи была выше, особенно после пребывания на морозе. Это может служить указанием на то, что у лиц, акклиматизированных к холоду, кровоток в коже пальцев и кистей повышен. Способность человека акклиматизироваться к холоду не вызывает сомнений. Примеры такой акклиматизации можно наблюдать в различных странах. Аборигены
Недели 1я
Время, дни
Рис. 27.1. Изменение тактильной чувствительности в процессе акклиматизации к холоду (Mackworth, Proc. Roy. Soc. В., 143, 392—407, 1955).
центральной Австралии спокойно спали без одежды при темпе-пературе воздуха около 0°С и излучательной температуре — 45°С и, по-видимому, не испытывали при этом ни малейшего неудобства.
Сами ученые, проводившие это исследование, никак не могли уснуть в подобных условиях — они все время дрожали от холода. Аборигены оказались способными выдержать гораздо большее падение температуры кожи, чем европейцы. Подобно аборигенам Австралии, бушмены Калахари спят в холодное время при слабо горящем костре под защитой одного покрывала. Большая выносливость к холоду (Дарвин наблюдал ее у индейцев Огненной Земли) в значительной степени может быть приобретена и европейцами. Норвежские студенты, прожившие 6 нед на открытом воздухе с минимальной защитой от холода, вначале не могли спать, но с течением времени стали спокойно засыпать в этих условиях.
Известно, что у эскимосов руки переносят холод гораздо лучше, чем у представителей других народов. Это подтверждается в эксперименте, в котором производили измерение температуры кожи пальцев у эскимосов, индейцев, негров и европейцев после погружения кистей рук в холодную воду. Опыт корейской войны сви-
Глава 27. Климатическая адаптация
529
Таблица 27.3
Скорость кровотока в сосудах предплечья при локальном охлаждении
Число испытуемых Число наблюдений Интенсивность кровотока, мл на 100 см’/мин Температура кожи, °C
Контрольная группа 5 122 4,7 ±0,19 32,8
Эскимосы 6 148 8,6±0,43 33,8
детельствует о том, что у негров чаще возникали отморожения. При исследовании сенсомоторных функций у молодых рабочих на северо-занаде Канады индейцы и эскимосы явно превосходили группу европейских рабочих, проживших в тех же условиях в течение 6 нед.
Способность людей, у которых выработалась акклиматизация к холоду, и в особенности эскимосов, активно работать руками в условиях низкой температуры связывают с усилением кровотока. В табл. 27.3 приводятся результаты измерения интенсивности кровотока у людей, которые погружали предплечья в воду при температуре 24°С (Brown a. Page, J. appl. Physiol., 5, 221, 1952).
Несмотря на то что щитовидная железа и надпочечники играют определенную роль в приспособлении организма к холоду, в опытах с изотопом иода 131 I не удалось выявить каких-либо различий в функции щитовидной железы между американскими индейцами, эскимосами и европейцами на Аляске (Rodahl).
Все имеющиеся данные указывают на существование приобретенной выносливости к холоду, в которой главную роль играют физиологические приспособительные механизмы. Ни одна расовая группа, за исключением, быть может, аборигенов центральной Австралии, не обнаружила качественно какой-либо выдающейся способности вырабатывать в себе выносливость к холоду. Возможно, что у негров медленнее происходят соответствующие защитные изменения.
Физические признаки, от которых зависят температурные реакции организма
Размеры и форма тела в какой-то степени влияют на интенсивность теплообмена. Теплоотдача за счет конвекции и испарения оказывается тем больше, чем больше поверхность кожи. Теплообмен за счет излучения происходит тем быстрее, чем больше площадь излучающей поверхности. Если главную роль в теплоотдаче играет потоотделение (при температуре воздуха, близкой к температуре кожи или несколько большей), то общая теплоотдача
530
Часть F. Экология человека
А
Рис. 27.2. Изменение состояния организма при физической нагрузке.
А — эксперимент 1; Б — эксперимент 2 (полный субъект устал и прекратил ходьбу спустя 63 мин после ее начала). I, II— интенсивность обмена; III— пульс; ГУ—ректальная температура; У — температура кожи. 1 — для невысокого полного субъекта;
!— для сухого худого.
Глава 27. Климатическая адаптация
531
должна коррелировать с площадью поверхности. Однако действительная величина корреляции составляет +0,8, что указывает на значительную индивидуальную изменчивость в способности к потоотделению; следовательно, телосложение— это не единственный фактор, влияющий па теплообмен.
Количество тепла, выделяемого человеком в процессе мышечной работы, тесно связано с его весом, причем это относится к самым разнообразным видам физической работы. Количество тепла, производимого на 1 кг веса тела, оказывается приблизительно одинаковым у лиц с большим и малым весом. Это количество, однако, не будет постоянным, если относить его к единице поверхности тела, так как чем меньше индивидуум, тем большая поверхность приходится на 1 кг веса тела. Последнее вытекает из того, что вес тела пропорционален кубу, а поверхность — квадрату линейных размеров тела; при данном весе у людей эндоморфного типа площадь поверхности тела меньше, чем у людей эктоморфного типа. У более крупных людей отношение веса к площади поверхности больше. Таким образом, меньшие по размерам индивидуумы обладают на единицу производимого телом количества тепла относительно большей площадью, участвующей в рассеянии этого тепла путем конвекции или испарения; отдача тепла на единицу поверхности у них меньше, что подтверждается данными прямых наблюдений, представленными на рис. 27.2.
Форма тела влияет на теплоотдачу и другим путем. И коэффициент теплоотдачи путем конвекции, и коэффициент теплоотдачи путем испарения (т. е. потеря тепла на единицу площади тела, отнесенная соответственно к единичной разности температур или давлений паров воды) становятся приблизительно постоянными, если поверхность велика. Эти коэффициенты быстро увеличиваются, если диаметр конечности меньше 10 см; так, при диаметре 7 см коэффициент, характеризующий испарение, почти вдвое выше, чем при диаметре 15 см.
Из этих анатомических соотношений вовсе не следует, что у индивидуумов, размеры тела которых меньше, потоотделение на единицу поверхности должно быть таким же, как у более крупных. Последние потребляют также больше воды, чем индивидуумы малых размеров. У человека большого веса образуется больше пота на единицу поверхности тела; потовые железы должны в этом случае быть более активными (так как количество этих желез, как мы видели, не зависит от веса). Из сказанного следует, что при высокой температуре индивидуумы меньших размеров и с более удлиненной формой тела обладают некоторыми биологическими преимуществами. В опыте Робинсона (Robinson) у индивидуума с большим весом в условиях работы в помещении с температурой 32°С и относительной влажностью 70% тепловое равновесие не наступало. Оно устанавливалось лишь после того, как влаж
532
Часть V. Экология человека
ность снижалась до 44% и обеспечивалась достаточная вентиляция. Несмотря на большую теплоотдачу на единицу поверхности, в организме такого индивидуума происходило накопление тепла (рис. 27.2).
Третий анатомический фактор — толщина подкожного жирового слоя. При высокой температуре большая часть тепла уносится на периферию усиленным током крови. Но жировой слой сравнительно беден кровеносными сосудами; толщина слоя оказывает большое влияние на общую теплопроводность.
Все рассмотренные выше факты, которые в жарких странах благоприятствуют установлению теплового баланса, оказывают прямо противоположное действие на теплообмен в районах с холодным климатом. В экспериментах установлено, что индивидуумы с эндоморфным типом телосложения и с большим слоем подкожного жира лучше переносят холод (исследование пловцов на дальние дистанции).
Примеры из антропологии
Форма и размеры тела
С точки зрения приспособления к климату имеют значение различия в характере телосложения между разными популяциями. Эти различия подчиняются экологическим правилам Бергмана (Bergmann, 1847) и Аллена (Allen, 1877), которые справедливы для всех животных популяций. Правило Бергмана утверждает, что в пределах одного политипического теплокровного вида размер тела подвида обычно увеличивается с уменьшением температуры окружающей среды. Правило Аллена гласит: у теплокровных животных, относящихся к одному виду, имеется тенденция к увеличению относительного размера сильно выступающих частей тела (таких, как уши и хвост) с увеличением температуры окружающей среды. Эти правила применимы к подавляющему большинству теплокровных видов (J. Huxley).
В ряде исследований было показано, что форма и размеры тела человека также подчиняются этим правилам. В популяциях жарких стран на всех континентах средний вес тела ниже, чем в популяциях, живущих в умеренном или холодном климате (рис. 27.3).
Робертс также показал, что отношение длины тела человека в сидячем положении к общей длине тела становится меньше при увеличении средней годовой температуры, т. е. у жителей жарких стран нижние конечности-относительно длиннее (рис. 27.4). То же можно сказать и о верхних конечностях: отношение размаха рук к длине тела у жителей жарких стран больше; меньше у них и размеры туловища. В совокупности все эти данные указывают на то, что при переходе от умеренного к жаркому климату отношение веса тела к площади его поверхности уменьшается (табл.
Глава ?7, Климатическая адаптация
533
Рис. 27.3. Регрессия веса юла относите^ ьно средней годовой температуры (Roberts, Am. J. phys. AntLiop., 11, 533—58, U5a).
1 — Для европейцев (У = 84,8—0 414 X), II — дтя американцев (У - ВО 2—0,123 X), III — для монголоидной расы (У = 60,1—0,100 X) IV—для африканцев (У = 87,8— — 0,512 X), V —средняя (У = 78,6—0,305 X).
27.4). Следует заметить, что корреляция между размерами или формой тела и средней температурой тела объясняет до 50—60% межпопуляционной изменчивости. Конечно, вариации в характере телосложения зависят и от других факторов, и в первую очередь от подвижности популяции.
Мы располагаем весьма скудными данными о толщине подкожного жирового слоя у представителей различных пародов. Установлено, что у американских негров толщина кожной складки
Табпща 27.4
Отношение веса тела (кг) к площади поверхности (м2)
Фраищ зы 38 Сома липцы 35
Албанцы 37 Мексиканцы 35
Арабы 36 Жители Андаманских островов 32
534
Часть V. Экология человека
Рис. 27.4. Регрессия роста сидя относительно средней годовой температуры (Roberts, дисс. докт.).
1 — для монголоидной расы (У = 562,8—0,578 %); II — для американцев (У = 540,8— —0,293 X); III — для африканской европеоидной расы (У = 566,4—0,772 X); IV — для негроидной расы (У = 558,7—0,750 X); V — средняя (У,= 554,9—0,639 X).
Рис. 27.5. Толщина подкожного жирового слоя у негров (Z) и у белых {II) США (Newman, Human Biol., 28, 1956).
Глава 27. Климатическая адаптация
535
Фото II. Нильский тип конституции (племя Динко).
меньше, чем у белых (рис. 27.5); у эскимосов, по-видимому, жировой слой толще, чем у негров. С этим фактом согласуется наблюдение, что у кикуйю и других африканцев эндоморфный компонент в конституции менее выражен, чем у европейцев или японцев (Danby).
Различие в характере телосложения между коренастыми эскимосами, с одной стороны, и длинными и тонкими аборигенами
536
Часть V. Экология человека
центральной Австралии или нилотами (фото II), с другой, служит разительным примером проявления правила Аллена. Правдоподобным кажется утверждение, что пигмеи и нилоты представляют два крайних примера действия правил Бергмана и Аллена.
Рост и развитие
Отмеченные различия в характере телосложения взрослых индивидуумов позволяют предположить, что и в характере роста должна проявляться некоторая зависимость от климатических условий. Тот факт, что удлиненная форма тела преобладает у людей, живущих в жарких странах, согласуется с наблюдением, что период роста в этих странах удлинен, а наступление половой зрелости несколько запаздывает. Удлиненная форма тела, т. е. относительно большая длина на единицу веса, обычно связана с задержкой развития скелета и общего физиологического созревания. К сожалению, данные, собранные в тропиках, довольно скудны, и наблюдения осложняются влиянием таких факторов, как питание, болезни, социальное положение и пр. Дети африканцев (обычно имеющие более длинные конечности, чем дети европейцев в том же возрасте) отстают в развитии скелета; у подростков отставание составляет почти год.
Монголоидный тип лица
Кун, Гарн и Бердселл (Coon, Garn a. Birdsell) выдвинули заманчивую, хотя и не вполне подтвержденную фактами гипотезу, что черты лица монголоидного типа — это специальный приспособительный признак для жизни в условиях суровых холодов. Для этого типа характерны уменьшенные надбровные дуги и лобные пазухи, более плоские и широкие глазничные и молярные об-
Таблица 27.5
Носовой индекс и климат
Коэффициент корреляции Стандартное отклонение
Показания сухого термометра 11 0,63 0,050
Относительная влажность 11 0,42 0,068
Показания сухого термометра и относительная влажность 1> 0,72 0,040
Показания влажного термометра 2> 0,77 0,034
Давление паров воды в воздухе 2) 0,82 0,027
1) Данные Томсона и Бакстона.
2) Данные Вайнера.
Глава 27. Климатическая адаптация 537
ласти (что способствует образованию жировых подушек), уменьшенный носовой выступ. Считается, что у американских индейцев и эскимосов при длительном пребывании на холоде реже возникают отморожения и другие неблагоприятные последствия охлаждения.
Форма носа
Отмечена тесная корреляция между формой носовых отверстий (характеризуемой «носовым индексом») и климатом (табл. 27.5).
Из табл. 27.5 следует, что форма поса определяется той основной функцией, которую он выполняет,— увлажнение вдыхаемого воздуха (поскольку поглощение влаги из воздуха дыхательной поверхностью зависит от давления паров); от этой функции она зависит в большей степени, чем от теплообмена. Именно этим можно объяснить, почему узкий нос чаще встречается у людей, живущих в пустынных районах, где сухость воздуха очень велика. Функция ресничек мерцательного эпителия, как известно, в большей степени страдает от сухости, чем от перегревания или охлаждения. При нормальных условиях вследствие колебательных движений ресничек слизь движется со скоростью до 10 мм/мин в направлении к глотке, вымывая из дыхательных путей осевые твердые частицы. При этом скорость движения слизи зависит от относительной влажности вдыхаемого воздуха. Низкая влажность может затруднить нормальный ход слизи. Этой чувствительностью дыхательных путей к сухости можно объяснить, почему инфекционные заболевания полости носа и придаточных пазух, а также респираторные инфекции часто встречаются у эскимосов (Bertelson). Подобные заболевания сравнительно редки у африканских негров или у аборигенов Малайи; это связывают с тем, что у них более длинные и широкие носовые ходы, затрудняющие доступ воздуха.
Роль генетических и негенетических факторов в климатической адаптации
Выше мы привели некоторые факты, свидетельствующие о том, что определенные физические особенности строения тела оказываются благоприятными для выполнения тех или иных функций, и в то же время указали географическое распределение этих признаков у разных народов. На первый взгляд это может означать, что существует географический отбор, развивающийся в направлении, указанном Дарвином, и что во многих случаях климатические различия обусловлены генетическими различиями; так, изучение близнецов показывает, что изменчивость формы и размеров тела, отложения жира, характер роста, развитие скелета и физиологическое созревание в гораздо большей степени определяются
538
Часть V. Экология человека
генетической конституцией, нежели действием факторов внешней среды. Несомненно, что некоторые различия между популяциями определяются различиями в генотипах или в каких-либо полифак-торных комбинациях. Такие полифакторные признаки, как форма носа или отношение длины конечностей к длине туловища, остаются неизменными при изменении окружающих условий. В действительности, однако, ситуация более сложна, так как влияние климата на вес тела и скорость роста у переселенцев примерно такое же, как у коренных жителей данной местпости. Вес тела у европейцев в тропиках в среднем ниже, чем у европейцев в холодных странах (Roberts, 1953). Вес тела и рост заметным образом зависят от времени года. Хорошо известно, что у животных даже в первом поколении строение тела претерпевает изменения в направлении, определяемом правилами Бергмана и Аллена. Другими словами, действие высокой температуры в период роста может привести к морфологическим и физиологическим изменениям, которые в последующем обеспечат большую сопротивляемость организма к действию высоких температур (Harrison). Благодаря способности к непосредственным и быстрым реакциям, подобно тенденции к загару, эти изменения могли, по-видимому, накапливаться путем сравнительно быстрого естественного отбора (в соответствии с принципом ассимиляции Уоддингтона), так что в некоторых популяциях соответствующие типы роста закрепляются генетически. Не удивительно в таком случае, что на всех континентах связь телосложения с климатическими условиями, устанавливаемая соотношениями Бергмана и Аллена, существует независимо от расы (Робертс). В некоторых случаях эта связь может быть в своей основе и генетической, но это вовсе не обязательно.
Что касается адаптации к действию высоких температур в самых разнообразных условиях обитания, то мы видели, что в организме происходят быстрые физиологические изменения, связанные с акклиматизацией; можно думать, что появляются также выгодные морфологические признаки, по-видимому, в результате естественного отбора. Исходя из этого, можно объяснить, почему выделение пота у коренных жителей тропиков меньше, чем у полностью акклиматизировавшихся европейцев, несмотря на то что коренные жители легче приспосабливаются к /дополнительному притоку тепла путем дальнейшей акклиматизации и повышенной активности потовых желез. Уменьшению потоотделения у коренного населения тропиков благоприятствует меньший вес и более удлиненная форма тела и конечностей. Тепло, выделяемое ими в процессе работы, сбалансировано меньшим потоотделением па единицу поверхности. Уайнер и Ли (Weiner а. Lee) иллюстрируют эту точку зрения следующими данными (табл. 27.6).
Тот факт, что каждая из больших расовых групп — «европеоидная^ «монголоидная^ «негроидная» и «австралоидная» — живет
Глава 27. Климатическая адаптация
539
Таблица 27.6
Сравнение характерных биологических параметров жителей Азии и Европы, находящихся в одинаковых условиях жаркого климата
Группа Средний вес, кг Расход калорий на 1 кг веса тела в 1 ч Расход калорий в 1 ч (на общий вес тела) Средняя площадь поверхности тела, м2 Количество выделяемого тепла, кДж/м2/ч Потеря тепла путем испарения, «Дж/м»/ч Количество пота, г/ч
Азиатская 55 4 220 1,6 137 165 440
Европейская 80 4 320 2,0 160 195 650
в самых различных климатических условиях, должен быть приписан отчасти широко распространенному явлению физиологической акклиматизации, частично — существованию различий в размерах тела (Бергман и Аллен) и других физических «расовых» различий и частично — технологическим приспособлениям к климатическим условиям.
Полезно привести некоторые примеры, иллюстрирующие степень климатической адаптации расовых групп (см. также табл. 25.5). Быть может, наиболее удивительны в этом отношении американские индейцы. Различные племена индейцев на Американском континенте живут в самых различных географических условиях: индейцы атабаски, населяющие район Большого Медвежьего озера, живут в таких же суровых условиях, как эскимосы; материальная культура жителей Огненной Земли более бедная, а климат сырой, холодный, с частыми штормами; индейцы майя на полуострове Юкатан или в Гватемале обитают в условиях жаркого влажного климата. Австралийские аборигены, подобно жителям Огненной Земли, переносят холода с минимальными защитными приспособлениями. Зимой в ночное время в центральной Австралии, в областях, где проживают племена анватьера и ильпирра, а также вблизи Алис-Спрингс бывает температура от —2 до —10°С. Днем эти же аборигены даже в зимнее время испытывают действие очень высоких температур — сухой воздух прогревается до 30—35°С. Вместе с тем в жарких влажных областях Австралии, также населенных коренными жителями, температура достигает 28—32°С при относительной влажности 70—80% . Негры населяют не только джунгли и жаркие влажные места, но и холодные высокогорные районы Басутоленда, Эфиопии и Кении. К монголоидной расе относятся не только народы, живущие вблизи Полярного круга, но и жители Центральной Азии, привыкшие к очень резким колебаниям суточной температурь^ и китайцы, живущие в Юго-Во
540
Часть V. Экология человека
сточной Азии, где климат жаркий и влажный. Очень широк в настоящее время климатический интервал обитания европейцев. В странах жаркого пояса отметим популяции иммигрантов — европейцев, обосновавшихся в бассейне Амазонки, итальянцев в северной Австралии, народность кота и других «европеоидов» в Индии, народы Северной Африки и Ирана, живущие в пустынных местностях вблизи Средиземного моря и на Ближнем Востоке. Напомним еще, что лапландцы — это европейцы, живущие в исключительно холодных условиях.
Границы биологической толерантности
Существуют пределы климатической адаптации, показателями которой служат чувство комфорта, работоспособность и состояние здоровья в данной обстановке. Эти пределы необходимо изучить в связи с различными типами среды обитания. Знание их особенно важно, когда возникает необходимость в выработке подходящих технологических приспособлений.
Границы зоны комфорта
Соотношение между субъективным ощущением тепла и температурой окружающей среды обычно выражают посредством американской шкалы эффективной температуры (ЭТ), которая позволяет оценить совместное действие температуры, влажности и скорости движения воздуха единым температурным показателем (рис. 27.6). В градусах эффективной температуры верхняя граница зоны комфорта зимой в Англии составляет 17, в Америке 22°С; благодаря акклиматизации этот предел увеличивается летом до 22 и 24°С соответственно; эти пределы устанавливаются для людей, носящих легкую одежду и выполняющих сидячую работу. Для лиц, раздетых до пояса, верхняя граница зоны комфорта и тепловой нейтральности начинается примерно с 27°С. Свыше 80% европейцев в Сингапуре чувствуют себя достаточно хорошо при эффективной температуре в пределах 23—26°С. Большая часть белых австралийцев, родившихся в Австралии, без каких бы то ни было неудобств переносят эффективную температуру 27°С. Высокие значения верхней границы зоны комфорта для тропических областей подтверждаются наблюдениями над индийцами и иранцами. На рис. 27.6 показаны зоны комфорта для различных частей света (Ellis). Приведенные данные касаются мужчин. Несколько по-иному сказывается влияние теплого климата на организм женщины.
Для многих районов характерен период в течение дня и целого сезона, когда эффективная температура выше верхней границы зоны комфорта (табл. 27.7). Эти районы тем не менее заселены с достаточной плотностью^ чему способствует совершенная система
Таблица 27.7
Тепловые характеристики различных типов среды обитания и климатических зон (даны средние месячные значения эффективной температуры, °C)
Среда обитания Зона Климатические провинции h oOhO и 1 \ ХО HKcipKO и ВПЛ I но Тсчло и CVXO 1 1егло и I влажно ОХЛЭ и | onWpuxortrf । Пр >хладно । и 1лажно OX(J и онНонох Холодно и вл ан ио Некоторые примеры
I. Пустыни и засушливые земли Жаркая Тропическая пустыня 29 — 17 — 10 — — — Сахара, Атакама, Австралия
Холодная Холодная, сухая пустыня 25 — 20 — 10 — 6 — Гоби
II. Тропические леса /Каркая Экваториальная — 28 — 21 — — — — Заир, Малайя, бассейн Амазонки
Экваториальная и муссонная — 28 — 22 — — — — Западная Индия, Танзания
» Тропическая морская и муссонная — 28 — 17 — — — — Побережье Бразилии, Мозамбик, Индокитай
» То же — 27 20 17 — — — — Побережье Квинсленда
III. Кустарник и ме шии лес в тропиках » Тропическая континентальная 28 23 20 — 17 — — — Внутренние районы Африки и Южной Америки
Тропическая континентальная п муссонная 24 22 20 17 12 — — •— Индия, Австралия (исключая тропические районы)
Среда обитания Зона К лим атические провинции Жарко и сухо ! Жарко и 1 влажно
IV. Тропические степи (саванны) Жаркая Тропическая континентальная (климат плато) 28 22
V. Леса умеренных широт Теплая Умеренно теплая 25 27
Холодная Холодная морская — 26
VI. Кустарник средиземноморского типа Теплая Умеренно теплая 28 27
VII. Степи умеренного пояса Холодная Холодная континентальная 25 20
VIII. Тайга » Холодная морская —- 22
» Холодная континентальная — —
IX. Полярныезем-ли » Арктическая тундра — —
Продолжение
Тепло и сухо Тепло и влажно I Прохладно и cvxo , Прохладно 1 и влажно , Холодно и . сухо Холодно и влажно Некоторые примеры
16 — 10 — — — Перу, Южная Африка
20 — 10 — — — Северный и Южный Уэльс, Аргентина
— 20 — 12 — — Великобритания, Германия, Новая Зеландия
16 — 10 — — — Калифорния, Средиземноморье
— — — 12 4 — Прерия, пампасы, черноземные области
— 17 •— 12 3 3 Аляска
— 17 — 12 3 — Сибирь, Канада, западная часть пров. Виннипег
25 — 12 10 2 — Гренландия
Глава 27. Климатическая адаптация
543
10
(°C)
30
25
20
15
а i 1 5
<Т) 30—-
80
Верхняя граница зоны -- комфорта (Иран)
Зона комфорта (Сингапур)—
Годовая зона комфорта ~ (Калькутта) Z
Оптимальная зона комфорта (США. лето) — Оптимальная зона ~ комфорта (США, зима Z и лето)
Зона комфорта Z "(Великобритания, зима) -
Рис. 27.6. Зоны комфорта для различных частей света (Ellis, Ann. R. Coll. Surg., 13, 1953).
Чтобы определить эффективную температуру, следует соединить прямой точки на вертикальной шкале стева и справа соответственно наблюдаемым значениям температур и найти точку пересечения этой прямой с линией сетки, определяющей скорость движения воздуха (последняя дана в фут/мин, 1 фут = 30 см).
вентиляции, устройства жилищ, форма одежды. Следует учитывать также, что акклиматизация дает возможность выполнять работу при температуре, превышающей верхнюю границу зоны комфорта. Нет сомнения в том, что особенности телосложения, выгодные для определенного климата, должны играть значительную роль в успешном приспособлении к условиям, которые обычно считаются крайне тяжелыми для жизни.
Температурные границы при выполнении физической работы
Люди могут выполнять тяжелую физическую работу при весьма высоких эффективных температурах, если они физически сильны, молоды, тренированны и хорошо акклиматизировались. В лабораторных опытах европейцы выполняли работу с затратой 1050 кДж/ч (что соответствует умеренно тяжелой работе на производстве) по 4—5 ч подряд 5 или 6 дней в неделю в течение полугода при эффек
544
Часть V. Экология человека
тивной температуре 32—33°С. Рабочие рудников Южной и Западной Африки способны выполнять такую же работу. При этом высокая ректальная температура поддерживается на определенном уровне и выделение пота составляет 3,5—4 л в 1 ч. При более высоком уровне ректальной температуры даже у этих тренированных людей наступает нарушение кровообращения.
Таким образом, существует большой разрыв между допустимым пределом комфорта в состоянии покоя, равным примерно 26°С, и максимально возможным для тяжелой работы, скажем 32°С. Нужно признать, что эти пределы соответствуют исключительным условиям, но в то же время они указывают на поразительную способность человеческого организма приспосабливаться к действию высоких температур.
Верхний предел эффективной температуры для умеренной работы, длящейся годами в течение всего активного периода жизни человека, по-видимому, составляет от 25 до 28°С.
Температурные границы при выполнении различных видов квалифицированной работы
Большое число наблюдений (в которых испытуемые выполняли всевозможные тесты на внимание, а также такие операции, как передача телеграфных сообщений, наблюдение за приборами и т. п.) показывает, что температура, при которой эффективность работы снижается, а число ошибок начинает быстро возрастать, во всех случаях примерно одинакова. Эффективная температура порядка 27°С не оказывает заметного влияния на результаты работы, но уже при 28,5—31°С точность выполнения операций падает. Различные исследователи принимают за критическое значение температуры 28,5—29,5°С. Макуорт (Mackworth), исследовавший работу телеграфистов, установил, что более искусные работники несколько лучше переносят тепловую нагрузку, чем менее опытные.
В качестве основной критической величины для всех видов работы, как физической, так и умственной, и для большинства индивидуумов (в том числе и хорошо акклиматизировавшихся) можно принять значение эффективной температуры 28,5°С, которое несколько превышает верхний предел зоны комфорта. При этой температуре нарушается также и сон.
При низких температурах отмечается зависимость качества ручных видов работ, требующих сноровки, от температуры кожи. При акклиматизации к холоду способность к различению предметов и точность выполнения операций повышаются (27.1), что связано с улучшением местного кровообращения. Следует помнить,
Глава 27. Климатическая адаптация 545
что в этом отношении акклиматизировавшиеся европейцы схожи с эскимосами. Несчастные случаи на производстве при температурах пиже 13°С встречаются на 34% чаще, чем при 18°С. Квалифицированная работа, выполняемая голыми руками, при температуре воздуха ниже указанной качественно снижается, даже если человек носит плотную защитную одежду.
Влияние климата па состояние здоровья
Климат жарких стран может оказывать непосредственное вредное воздействие на организм (хотя гораздо более серьезны последствия заболеваний, вызываемых бактериями или паразитами, распространению которых благоприятствуют высокая температура и влажность). При эффективных температурах, превышающих 27°, человек может чувствовать некоторую слабость и утомляемость, и эти неприятные ощущения наряду с солнечным ожогом и часто появляющимся тропическим лишаем заставляют некоторых иммигрантов покидать тропики. Имеются сведения, что тропический лишай появляется и у коренных жителей жарких стран, в том числе у негритосов Малайи и арабов Адена, но неизвестно, встречается ли он у них реже, чем у переселенцев. Тепловой удар встречается весьма редко; ему подвержены и африканцы и европейцы. Возникновению теплового удара способствует сочетание определенных условий (недостаточная акклиматизация, перенесенная лихорадка, нехватка воды и тяжелая работа); тепловой удар чаще всего поражает тучных пожилых людей с нарушениями кровообращения, а также чрезмерно тепло одетых детей. Улучшение функции системы теплорегуляции, наступающее вследствие акклиматизации, а также рассмотренные ранее морфологические особенности играют положительную роль в защите организма от неблагоприятного воздействия высоких температур. Несомненно, что в жарких странах происходит интенсивный естественный отбор: в менее выгодном положении оказываются более тучные индивидуумы и лица, у которых меньше потовых желез или эти железы частично повреждены (например, вследствие солнечных ожогов). В эту последнюю категорию попадают альбиносы. Различные нарушения, связанные с недостатком воды или перегревом, по-видимому, менее опасны людям, вес и размеры тела которых невелики.
Вредное влияние холода, в частности отморожения, представляет опасность для тех, кто не имеет подходящей одежды, приспособленных к холоду жилищ, а также необходимого опыта. Возможно, что некоторые индивидуумы особенно чувствительны к холоду и плохо акклиматизируются.
546
Часть V. Экология человека
Климатические зоны
На земном шаре распределение и смена в течение года жары и холода, ясных и облачных дней, ветра и безветрия, дождей и засухи зависит от большого числа различных факторов: географической широты, удаленности места от моря, защищенности от ветров, рельефа поверхности и высоты над уровнем моря. Главный из отдельных факторов, который обусловливает существование больших климатических зон,— это широта местности. В конкретных условиях он проявляется, однако, в сложной форме вследствие взаимодействия остальных перечисленных факторов в' любом данном месте или в любое время года. Самое древнее и наиболее простое разделение земного шара на климатические пояса — жаркий, теплый, умеренный и холодный — связано с астрономическими наблюдениями за движением солнца в разных широтах. Эти пояса простираются, грубо говоря, от 0 до 30° широты (жаркий), от 30 до 45° (теплый), от 45 до 60° (умеренный) и свыше 60° (холодный).
Каждый большой пояс включает множество подпоясов, или климатических «провинций», так как влияние широты на климат может принимать различные формы в зависимости от действия таких факторов, как высота над уровнем моря, близость к морю, защищенность от ветров. Эти дальнейшие подразделения, вводимые климатологами (подробнее см. Miller, 1946), основаны на вариациях масштабов и времени наступления температурных изменений и количества осадков; они соответствуют, следовательно, особенностям рельефа провинции внутри каждого пояса, определяются различными комбинациями величин, характеризующих температуру воздуха, влажность (давление водяных паров), интенсивность солнечной радиации, скорость движения воздушных масс; эти комбинации изменяются в зависимости от времени суток и времени года, они определяют собой комплекс физиологических воздействий, характерных для данного климатического пояса. Для каждого пояса можно установить приближенную среднюю годовую эффективную температуру; например, для жаркого климата 27— 21 °C, для теплого — 21—16°С, умеренного — 15—5°С, холодного — ниже 5°С. Но эти цифры, будучи средними значениями интегральной величины (эффективной температуры), вместе с тем маскируют глубокие различия (а также перекрывание) климатических зон и различия внутри каждой зоны.
Пытаясь охарактеризовать и сравнить, скажем, 20 различных климатических провинций на земном шаре, мы убеждаемся в том, что задача значительно облегчается, если считать каждую зону состоящей из повторяющихся характерных комплексов, которые определяются в соответствии с временем года и суток.
Табл. 27.7 служит грубой иллюстрацией разнообразия климатических условий, в которых находится человек в главных провин-
Глава 27. Климатическая адаптация
547
днях (и в наиболее распространенных типах среды обитания). Из таблицы следует, что почти всюду имеются те или иные суточные или сезонные колебания и что даже в тех зонах, в которых живут большие популяции (см. табл. 25.3), колебания климатических условий могут быть достаточно сильными, а иногда и чрезвычайно резкими. Относительно небольшие климатические колебания наблюдаются лишь в зоне экваториальных муссонов, где тем не менее живут большие популяции и где высоко развито сельское хозяйство. Наивно предполагать a priori, будто существует один «идеальный» климат и что он, кроме того, непременно ограничен узкой климатической зоной, лежащей где-то вблизи изотермы 20°С.
Табл. 27.7 показывает, что для многих стран мира характерны суровые климатические условия, требующие соответствующих физиологических и технических приспособлений. В качестве таких особо трудных условий, ограничивающих физическую работу и требующих широких физиологических приспособлений (что, разумеется, увеличивает вероятность ошибок при наблюдении), можно выделить: 1) типичное для «экваториальной» зоны сочетание безветрия с высокой влажностью, температурой и интенсивной солнечной радиацией; 2) палящий зной с обжигающим, горячим ветром, температура которого выше температуры кожи (такой ветер вызывает перегрев вследствие плохой конвекции и теплопроводности); 3) сильный мороз (даже при полном безветрии), когда облачность препятствует притоку тепла (особенно плохо переносится при повышенной влажности); 4) сочетание свирепых холодов и сильных ветров, даже при сухом воздухе. Как видно из табл. 27.7, почти в каждой климатической зоне человек сталкивается с действием одного или нескольких из указанных неблагоприятных факторов. Конечно, в районах с мягким климатом, например в странах теплого пояса (Средиземноморье) или в странах с прохладным морским климатом, сильная жара или холод редки и непродолжительны. Однако в любой климатической зоне человек не гарантирован от случайностей погоды, от ухудшения климатических условий в то или иное время года. Виду Ното sapiens всегда приходилось и приходится приспосабливаться к климату и его изменениям, начиная со времени своего появления в условиях, которые были достаточно сложны и которые в конечном счете вызвали к жизни столь гибкую и совершенную систему теплорегуляции.
Культурные аспекты климатической адаптации
Если бы биологические приспособления не были дополнены на определенной стадии развития техническими средствами, человек никогда не смог бы так широко расселиться на земном шаре и
548
Часть V. Экология человека
существовать в самых разнообразных условиях. В эпоху раннего плейстоцена человек жил в тропических областях и оттуда постепенно переселялся в северные широты. Это расселение вначале происходило только во время теплых межледниковых периодов или в эпохи отступления ледников. По-видимому, только в начале четвертого (вюрмского) оледенения человек впервые прочно обосновался в областях с довольно холодным климатом, вблизи ледников. В степной зоне Европы с лессовыми почвами, где была обильная пища для травоядных животных, обнаружено большое число охотничьих стоянок и поселений. К этому времени использование огня прочно вошло в быт людей, они научились также применять орудия и древесину для создания жилищ. Костяные шила для сшивания шкур и кремневые скребки для их обработки свидетельствуют о том, что в верхнем палеолите человек располагал достаточно прочной одеждой для защиты от холода. Жилища первобытных людей соответствовали по приспособленности жилищам современных бушменов или индейцев Огненной Земли, и потому биологическая акклиматизация играла в жизни людей каменного века большую роль.
История и техника постройки жилья, отопительных приборов и вентиляции помещений, изготовления одежды — это обширный самостоятельный предмет, и мы ограничимся здесь лишь несколькими краткими замечаниями. Для защиты от суровых холодов стены домов, крыша и пол должны обладать достаточной теплоизоляционной способностью, чтобы обеспечить наилучшее использование пространства и различных обогревательных устройств; в то же время они должны обеспечивать эффективную вентиляцию (без сквозняков), которая необходима и для самих обитателей дома и для нормального сгорания топлива. Замечательным примером воплощения всех этих принципов, несмотря на различия в строительных материалах, служат традиционные зимние жилища эскимосов центральной Канады, Аляски и Гренландии. У других народов Севера, например у чукчей-оленеводов, жилища более примитивны, что связано, вероятно, с необходимостью делать их транспортабельными. Если воздух внутри помещения временами охлаждается, то происходит конденсация влаги, одежда становится сырой, промокает материал, из которого сделаны крыша и стены дома. Принципы обогрева помещений у народов Крайнего Севера служат хорошей иллюстрацией их умения приспособиться к суровым условиям окружающей среды. (Другим примером их экологической эрудиции является детальное знание повадок и морфологии диких животных). В домах эскимосов с глубокими входными «колодцами» и низкими куполообразными крышами холодный воздух поступает внутрь через отверстия в нижней части дома и достигает обитателей уже достаточно прогретым. Объем помещения, в котором теплый воздух накапливается уже за
Глава 27. Климатическая адаптация
549
пределами, доступными для обитателей дома, достаточно мал. Поток теплого воздуха наружу регулируется размерами отверстия в крыше дома.
Другой принцип — скопление большого числа людей в небольшом помещении с минимальной вентиляцией, т. е. обогревание помещения за счет тепла, выделяемого человеческим телом. Это — обычная для эскимосов практика, когда у них нет топлива. Именно благодаря такой практике им удается иногда выжить в крайне неблагоприятных условиях, которые могли бы стать для них гибельными. В какой-то степени этот же принцип используют чукчи, сохраняя тепло в обтянутых шкурой спальных «ящиках». Светильники или лампы обогревают у них помещение во время ужина, а ночью обитатели согреваются лишь собственным теплом, которое удерживают, во-первых, одеяла из оленьего меха и, во-вторых, стенки общего спального «ящика».
Приходится удивляться, почему жители Крайнего Севера совсем не используют спальные мешки, а вместо этого предпочитают тем или иным способом утеплить само помещение и место для сна и спать голыми под меховыми одеялами. Возможно, этим решается проблема, с которой всегда сталкивались полярные исследователи, а именно уменьшается накопление пота и испарений в постели.
Примером того, насколько хорошо народы Севера знают арктические условия, в которых они обитают, может служить также следующий факт: в зимнее время эскимосы не сооружают свои жилища в низинах, например в долинах рек, где может скапливаться густой холодный воздух.
По сравнению с постройками людей, живущих у Полярного круга, жилища многих других народов часто более примитивны. Индейцы Огненной Земли, где климат сырой и холодный, аборигены Австралии и бушмены в холодные ночи в пустынях укрываются одним или двумя одеялами или шкурами животных. Их простые жилища сделаны из палок, согнутых и связанных вместе и покрытых шкурами, травой или корой деревьев. Айны по традиции живут в грубо сделанных хижинах, с небольшим очагом внутри Пребывание в такой хижине требует достаточной выносливости к холоду. Сопротивляемость к холоду у айнов и аборигенов Австралии, по-видимому, намного выше, чем у эскимосов и лапландцев или даже бушменов Калахари. Последние используют топливо, заграждения от ветра, кожаные плащи, а иногда и одеяла, чтобы защититься от холода. В результате температура воздуха, соприкасающегося с кожей, близка к температуре тела (Wyndham).
Технические приспособления, которыми пользовался человек в жарких странах, совершенствовались на протяжении веков начиная с доисторической эпохи. Много веков назад возникли методы строительства, учитывавшие весь комплекс факторов в условиях жаркого сухого или жаркого влажного климата. Эти методы
550
Часть V. Экология человека
мы можем теперь оценить в свете современных представлений о физиологических принципах, стандартах и нормах, обеспечивающих комфорт и работоспособность (подробнее см. Lee, 1951). Во всех жарких странах исходят из сознания необходимости уменьшить количество тепла, выделяемого организмом человека. Этой цели служат: а) различные устройства, облегчающие физический труд, а также специальная конструкция жилищ и мест храпения запасов; б) всевозможные меры, направленные на ограничение тепла, поступающего в организм, и способствующие теплоотдаче путем излучения (навесы, тенты, карнизы на крышах, различные способы уменьшения отражательной способности почвы). В жарких сухих странах для защиты от солнца нередко строят толстостенные дома и строения с вентиляцией под крышей и регулируемой вентиляцией через двери и окна. В странах с жарким влажным климатом необходимы более открытые жилища. Особенность традиционных построек в этих странах — широкие проемы в стенах, продуваемые легким ветром и обеспечивающие проветривание помещения во всех направлениях в любое время дня и ночи. В работах Lee (1951) и Wulsin (1949) читатель найдет более полные сведения о способах постройки жилищ в странах с жарким климатом, применяемых различными народами.
Одежда и обувь, подобно жилищам, различны в странах с разным климатом и определяются требованиями теплообмена (подробнее см. Siple). Связь этих требований с характером одежды у разных народов в различных условиях обитания проанализировал Вульсин.
Коротковолновое излучение
На организм человека большое влияние оказывают ультрафиолетовые лучи солнечного спектра, а также ионизирующее излучение — космическое и испускаемое радиоактивными элементами, содержащимися в воздухе и земной коре. Хотя человек, проживший на земле целую геологическую эпоху, имел возможность выработать необходимые приспособления к ультрафиолетовому излучению и естественному радиоактивному фону, в наше время он столкнулся с новой большой экологической опасностью, обусловленной освобождением и накоплением искусственных радиоактивных веществ.
Ультрафиолетовые лучи
Ультрафиолетовые лучи (длина волны короче 0,32 мкм) вызывают загар и ожоги. С уменьшением длины волны эритематозное действие ультрафиолетовых лучей усиливается, достигая максимума при 0,28 мкм.
Глава 27. Климатическая адаптация
551
Распределение интенсивности ультрафиолетового излучения на земном шаре в точности не известно. Уровень ее в значительной мере зависит от географической широты местности. Колебания этого уровня — суточные, сезонные, а также географические — связаны в основном с присутствием в атмосфере озона, который поглощает почти все ультрафиолетовые лучи с длиной волны короче 0,285 мкм. Например, если солнце переходит из зенита в положение, в котором оно находится под углом 30° к горизонту, интенсивность ультрафиолетового излучения может уменьшиться более чем наполовину. Сезонными изменениями движения солнца можно объяснить, почему в северном умеренном поясе излучение, вызывающее эритему, может быть более интенсивным в мае, чем летом, или почему во время солнцестояния, когда солнце находится над тропиком Рака, области на экваторе и на широте 47° получают одинаковое количество солнечного света. Несмотря на то что ультрафиолетовые лучи, распространяющиеся от солнца, когда оно находится под углом к горизонту, меньшим 45°, сильно поглощаются озоном, значительная часть их все же попадает на землю, рассеиваясь и отражаясь в атмосфере, и способна вызывать у людей эритему. Молекулы газов, находящиеся в воздухе, гораздо сильнее рассеивают короткие волны, чем длинные (этим объясняется голубой цвет неба); вследствие этого в атмосферном излучении ультрафиолетовый компонент имеет большую интенсивность, чем видимая область спектра. В больших промышленных центрах ультрафиолетовые лучи очень сильно поглощаются пылью и дымом.
Загар и солнечные ожоги. В основе загара лежит ряд сложных изменений; по-видимому, главное из них — повреждение клеток эпидермиса, при котором выделяются вещества, расширяющие мелкие кровеносные сосуды; в результате возникает отечность и другие признаки воспаления. Роль адаптации состоит в повышении пороговой эритемной дозы. Острые явления уступают место загару. Даже при слабом загаре этот более высокий порог может сохраняться в течение двух месяцев. В основе защитного эффекта лежат два процесса — утолщение рогового слоя и накопление меланина. Об ограничении проникновения ультрафиолетовых лучей через эпидермис при утолщении рогового слоя свидетельствует то, что, например, у альбиносов, а также в области витилиго загар не возникает, но порог эритемной дозы выше. Для обычной кожи существенную роль должно играть также накопление пигмента меланина и миграция его из базальных клеток к поверхности. Было показано, что присутствие пигмента в роговом слое влияет на степень поглощения ультрафиолетовых лучей. Пигмент сохраняется в эпидермисе в течение многих месяцев и даже лет. На ранее загорелой и сохранившей следы загара коже под воздействием излучения с длиной волны короче 0,32 мкм происходит «вторичное» потемнение.
552
Часть V. Экология человека
Повреждение кожи при солнечном ожоге, по-видимому, затрагивает и потовые железы. В связи с этим в эритемный период часто нарушается теплорегуляция (Thomson). В пигментированной коже подобного повреждения не происходит.
Мы уже упоминали, что под действием ультрафиолетового излучения происходит активация присутствующего в коже вещества — предшественника антирахитного витамина D.
Широта, градусы
Рис. 27.7. Зависимость канцерогенного действия ультрафиолетового излучения от географической широты (Blum, Carcinogenesis by ultraviolet light.
По оси абсцисс отложены градусы северной широты, по оси ординат — частота случаев рака кожи в процентах от этого показателя на широте 32°. I — максимальное изменение в течение года интенсивности излучения; II и III — частота зарегистрированных в США случаев рака кожи у белых женщин и мужчин соответственно.
Рак кожи и ультрафиолетовое излучение. На роль солнечного света в возникновении рака кожи указывают следующие факты: а) рак кожи у белых (базальноклеточная и плоскоклеточная карцинома) встречается преимущественно на лице (95%) и на других открытых для солнечных лучей участках, а также на тыльной поверхности рук; б) он чаще встречается у лиц, работающих вне помещения; в) рак кожи наиболее распространен в областях земного шара, где интенсивность солнечной радиации выше. Например, у американцев с белым цветом кожи он встречается реже по мере удаления от экватора (рис. 27.7). Частота случаев плоскоклеточной карциномы возрастает по мере удаления от экватора гораздо быстрее, чем частота базальноклеточной карциномы. То же можно сказать и о меланоме, частота которой у белых тем выше, чем
Глава 27. Климатическая адаптация
553
ближе к тропикам. В Квинсленде (Австралия) рак кожи встречается примерно в 20 раз чаще, чем в Англии.
Антропологические соображения. Робертс и Калон (Roberts а. Kahlon) показали, основываясь на измерении отражательной способности кожи у различных популяций, что существует явно выраженная корреляция (с коэффициентом 0,9) между цветом кожи и широтой местности и значительно более слабая зависимость цвета кожи от средней годовой температуры. Это подтверждает теорию, согласно которой географические вариации цвета кожи у людей связаны с интенсивностью ультрафиолетового излучения. Наиболее сильная пигментация наблюдается в Судане, где интенсивность солнечной радиации максимальная. Очень темным цветом кожи отличаются также народы, населяющие пустыни и саванны, например аборигены Австралии, которые не носили защищающей от солнца одежды или стали носить ее лишь в последнее время. Если в той или иной стране с жарким климатом оказывается этническая группа с менее выраженной пигментацией, то обыдпо легко установить, что эта группа иммигрировала сюда сравнительно недавно. Примером могут служить жаркие пустыни Нового Света.
В умеренных широтах наименьшее количество солнечного света получают близкие к морю западные области континентов, где много облачных дней в году. Небо Арктики летом свободно от облаков и пыли, в зимнее время снег и лед отражают падающий свет, интенсивность ультрафиолетового излучения здесь высока. Как мы упоминали выше, цвет кожи у арктических народов — более смуглый, чем у людей умеренного пояса.
Пороговый уровень излучения, с которого начинается загар, у негров значительно выше, чем у белых. Эксперименты Томсона показали, что дело здесь не только в большей толщине рогового слоя (как утверждал Blum): биопсия показала, что у негров роговой слой не намного толще, чем у европейцев. Удалось установить, что у негров роговой слой содержит более темный пигмент (меланин) и обладает меньшей проницаемостью для ультрафиолетовых лучей.
У негров США и Южной Африки рак кожи встречается очень редко; что касается локализации, то частота его на открытых и защищенных одеждой участках кожи одинакова (в противоположность тому, что мы находим у европейцев). У метисов в Капской провинции эпителиома встречается гораздо реже, чем у европейцев. Почти все случаи рака кожи в Аргентине относятся к иммигрантам, и крайне редко рак этой локализации встречается у индейцев или негров. Меланома также реже встречается у негров, чем у европейцев.
Таким образом, географическая связь между интенсивностью ультрафиолетовых лучей и цветом кожи определяется, вероятнее всего, защитной ролью пигментации; темнокожие народы населяют
554
Часть V. Экология человека
в основном области с более высоким уровнем ультрафиолетовой радиации. Даже в тропиках существуют заметные различия в цвете кожи: племена джунглей имеют более светлую кожу, народы открытых пространств — более темную; таково, например, различие между племенами пигмеев и неграми банту.
Нелегко представить себе селективную роль базальноклеточной карциномы; эта опухоль развивается, как правило, в пожилом возрасте и обычно не оказывает серьезного общего влияния на организм. Иначе обстоит дело с меланомой, которая характеризуется более быстрым течением с фатальным исходом и довольно часто встречается в молодом возрасте. Несомненно, однако, что более позднее развитие базальноклеточной карциномы частично обусловлено защитным действием загара и одежды; то, что сказано об этой опухоли, приложимо только к умеренным и субтропическим зонам. При действии интенсивного ультрафиолетового излучения на незащищенную кожу у европейцев селективное преимущество, несомненно, будет на стороне индивидуумов с более темным цветом кожи благодаря их меньшей восприимчивости к раку кожи. Следует также учитывать результат непосредственного действия солнечных лучей — ожоги. В естественных условиях чрезмерный загар неблагоприятно действует на организм..
Развитие сильной пигментации можно объяснить, исходя из ее защитного действия в отношении рака и ожогов, вызываемых ультрафиолетовым излучением. Отсутствие пигментации, характерное для народов европейского происхождения, нельзя объяснить иным образом. Если речь идет не о каком-то анцестральном признаке, не имеющем в настоящее время биологического значения (что мало вероятно), то он должен давать какое-то селективное преимущество в странах с низким уровнем ультрафиолетовой радиации. Точный смысл такого преимущества неизвестен; известно, однако, что антирахитическое действие ультрафиолетовой радиации выражено тем сильнее, чем лучше она проникает в кожу. Отсутствие меланина облегчает проникновение ультрафиолетовых лучей, и в областях с низкой интенсивностью солнечной радиации этот фактор может оказаться решающим (Coon, Garn a. Birdsell, 1950).
Загар, приобретаемый людьми со светлой кожей в качестве средства защиты, можно рассматривать как фенокопию генетически определенного более темного цвета кожи у людей, живущих в тропических и экваториальных областях. Вполне возможно, что темная пигментация появилась независимо в разных районах земного шара у темнокожих «европеоидов» южной Индии и Аравии, у негров Океании и Африки, поскольку эти народы по многим генетическим признакам далеки друг от друга.
Глава 27. Климатическая адаптация
555
Ионизирующее излучение
Радиоактивный фон — это излучение природных источников. К радиоактивному фону относится космическое излучение и излучение, испускаемое природными материалами — радием и торием, находящимися в земной коре, а также радиоактивными изотопами калия (в почве и воде), углерода (в воздухе) и водорода (в водяных парах). Приходящаяся в среднем на каждого человека общая доза ионизирующего излучения, создаваемая радиоактивным фоном, составляет за 30 лет примерно 3—5 Р. На больших высотах из-за большей интенсивности космического излучения эта доза достигает 5,5 Р. В некоторых географических районах, например в штате Керала в южной Индии и в ряде районов Бразилии, в силу геологических причин естественный радиоактивный фон значительно выше. В современных условиях существенную роль играют искусственные источники излучения. Медицинское применение рентгеновских лучей увеличивает дозу облучения, приходящуюся на гонады (в США эта доза за 30 лет составляет в среднем 3 Р); другой источник искусственного излучения — светящиеся циферблаты часов. Но наиболее значительное увеличение радиоактивности на земном шаре связано с испытаниями ядерного оружия и последующим выпадением радиоактивных осадков.
Радиоактивные вещества попадают в организм человека через дыхательные пути и желудочно-кишечный тракт. Некоторые из них, например стронций-90 и цезий-137, быстро всасываются в кишечнике. Радиоактивная пыль должна учитываться наряду с другими метеорологическими факторами; в некоторых случаях дожди могут занести ее в районы, где обычно доза излучения невелика. Люди, находящиеся в домах, лучше защищены от действия излучения, чем те, которые большую часть своего времени проводят на открытом воздухе. Жители тропических стран в этом отношении оказываются в менее благоприятных условиях. Если бы испытания ядерного оружия продолжались с такой же интенсивностью, как в 1953 и 1955 гг., тридцатилетняя доза на гонады повысилась бы почти на 1 Р.
Самые серьезные биологические последствия облучения связаны с мутагенным действием ионизирующих лучей на половые клетки; вызываемые ими мутации могут быть субмикроскопическими или же представлять собой микроскопически видимые изменения в хромосомах. Каждый вид имеет свой собственный набор мутантных генов, и хотя частота спонтанных мутаций для многих видов точно не установлена, полагают, что она варьирует от 1 -10? до 1 «10* половых клеток на поколение. Лишь малую долю этих спонтанных мутаций можно отнести за счет действия радиоактивного фона.
Радиационные мутации, подобно спонтанным, бывают доми
556
Часть V. Экология человека
нантными и рецессивными; рецессивные встречаются гораздо чаще. Доминантные мутации крайне редки, но если у какого-нибудь индивидуума под влиянием облучения возникла доминантная мутация, то она проявится у всех его потомков. Рецессивные же мутации проявляются только у тех индивидуумов, которые получили одинаковые мутантные гены от обоих родителей. Поскольку в человеческом обществе близкородственные браки запрещены, эффекты, связанные с такими рецессивными мутациями, должны проявляться лишь через много поколений. Однако иногда они могут проявиться и раньше; это произойдет в том случае, если какая-либо радиационная мутация объединится (в зиготе) с такой же мутацией, возникшей в популяции спонтанно. Кроме того, рецессивные мутации, сцепленные с полом, могут проявиться у мужчины, получившего от матери Х-хромосому, несущую мутантный ген.
Среди рецессивных мутантных генов много таких, которые внешне ничем не проявляются, однако снижают плодовитость и продолжительность жизни, а также так называемых «летальных» генов, которые приводят к гибели организма в зародыше. Вот почему не наблюдается непосредственного повышения частоты явных генетических аномалий у детей, родители которых пережили взрыв атомной бомбы (в Японии). Доминантные летальные гены вызывают гибель зародыша на ранних стадиях развития, так что наступающий в связи с этим ранний выкидыш проходит незамеченным.
Реальную опасность представляют рецессивные летальные гены, которые передаются многим индивидуумам через многие поколения, прежде чем проявятся в гомозиготном состоянии. В результате естественного отбора у человека, так же как и у других видов, установился определенный уровень элиминации как рецессивных, так и доминантных вредных генов, необходимый для обеспечения выживания и приспособленности. Вредные доминантные гены элиминируются быстро; они исчезают на ранних этапах онтогенеза. Рецессивный летальный ген в конце концов также элиминируется (причем в двойной дозе), но происходит это нескоро.
Влияние искусственных источников радиоактивного излучения все более повышает вероятность появления вредных генов. По данным Американского научно-исследовательского совета в США, 2% новорожденных имеют ту или иную генетическую аномалию, проявляющуюся до' наступления половой зрелости. К этим аномалиям относятся: умственная отсталость, врожденные аномалии нервной и нейро-мышечной системы, кожи, скелета, эндокринных желез, нарушения слуха, зрения, аномалии желудочно-кишечного тракта и мочеполовой системы. Существующие генетические данные указывают на то, что частота субмикроскопических мутаций увеличивается линейно с повышением дозы из
Глава 27. Климатическая адаптация 557
лучения. Доза, которая удваивает число вредных мутантных генов, определялась разными авторами по-разному: она выше 5 Р, но не превышает 150 Ри, возможно; составляет 30—80 Р. Наиболее вероятной величиной считают примерно 10 Р. Эта величина в 3 раза превосходит естественный радиоактивный фон. Наличие такой дозы, по расчетам того же Американского научно-исследовательского совета, означало бы, что у детей ныне живущей в США популяции были бы выявлены 200 000 (10%) новых случаев наследственных аномалий. Существующая сейчас доза излучения, создаваемого искусственными источниками, оценивается примерно в 5 Р за поколение. Доза 10 Р может дать 50 000 случаев проявления наследуемых дефектов в первом поколении и в конечном счете около 500 000 на поколение. Эти цифры, конечно, не характеризуют всех возможных генетических нарушений, но они показывают масштабы медицинской и социальной опасности, связанной с этим новым экологическим фактором.
Отбор, действуя на мутации, элиминирует вредные гены и сохраняет гены, имеющие ценность для выживания, но радиация не может рассматриваться как фактор, вызывающий выгодные организму мутации. Как спонтанные, так и вызванные излучением мутации носят случайный характер, и прежде чем появятся полезные мутации, должно быть испытано и элиминировано множество неблагоприятных. Мало вероятно, чтобы в современных популяциях можно было зафиксировать «хорошие» мутации, как это достигается при скрещивании животных или в естественных условиях.
В природе выгодные организму мутации распространяются в пределах вида благодаря тому, что размер потомства у носителей мутантных генов превышает средний уровень; вредные гены, наоборот, элиминируются в результате гибели или бесплодия их носителей. Оба эти механизма, вероятно, неэффективны у тех видов, -для которых характерно немногочисленное потомство: мало вероятно, чтобы полезные мутации приводили к заметному увеличению числа потомков, а мутантные гены, которые обусловливают аномалии в физическом или умственном развитии естественный отбор, как правило, элиминирует (Auerbach).
Жизнь в высокогорных условиях
Почти 10 миллионов человек постоянно проживают в горной местности, на высоте от 3600 до 4000 м над уровнем моря. Жизнь в горах представляет целый комплекс экологических трудностей, связанных с низким барометрическим давлением (т. е. с пониженным давлением кислорода и двуокиси углерода во вдыхаемом воздухе), низкой влажностью воздуха и низкой температурой. Кроче того, в этих районах почва обычно трудна для обработки и требует от человека больших физических усилий.
558
Часть V. Экология человека
Пониженное барометрическое давление
По объему кислород составляет 21 % атмосферного воздуха; на долю кислорода приходится около 1/5 общего барометрического давления. При давлении кислорода 150 мм рт. ст. (около 20 кПа), соответствующем давлению на уровне моря, кислород диффундирует из легких в кровь со скоростью, достаточной для нормальной жизнедеятельности организма. С уменьшением давления кислорода по мере увеличения высоты скорость эта падает. Количество кислорода, необходимое для выполнения физической работы, на высоте 2400—3000 м над уровнем моря становится недостаточным, а выше 4000 м человек ощущает нехватку кислорода даже в сидячем положении. Компенсаторные реакции организма, впервые попавшего в такую обстановку, весьма ограниченны; поэтому довольно быстро наступает общая слабость. Однако с течением времени возрастает роль вторичных процессов, приводящих к физиологичес-~кой акклиматизации; становится возможной как физическая деятельность, так и постоянное пребывание на больших высотах. Известно, например, что некоторые обрабатываемые человеком земли в Тибете находятся на высоте 2700—4500 м над уровнем моря, а постоянные поселения в Андах располагаются даже на высоте 5200 м над уровнем моря, причем жители их (рудокопы) ежедневно поднимаются в рудники, находящиеся на высоте 5800 м. Согласно данным Бэйкера (Baker), на высоте более 4000 м преобладает пастбищное скотоводство, земледелие играет основную роль на меньших высотах.
Реакции организма
Непосредственная реакция на недостаток кислорода (аноксия) состоит в увеличении минутного объема воздуха. Это происходит за счет повышения частоты и глубины дыхательных движений. При этом лишь частично улучшается снабжение организма кислородом. Перенос кислорода облегчается тем, что молекулы кислорода легче диссоциируют при низком давлении кислорода. Вместе с тем при учащенном дыхании углекислый газ «вымывается» из дыхательных путей и как следствие понижается его содержание в крови. Это ведет к сдвигу кислотно-щелочного равновесия в организме в щелочную сторону — наступает алкалоз. В патогенезе горной болезни играют роль оба фактора — аноксия и алкалоз. Аноксия проявляется ощущением усталости, головной болью, потерей внимания и ощущением, похожим на опьянение; алкалоз сопровождается тошнотой, рвотой и головокружением. На начальной стадии болезни нарушается регуляция дыхания, появля
Глава 27. Климатическая адаптация
559
ется так называемое «чейн-стоксово дыхание». Самая незначительная физическая нагрузка сопровождается резким ускорением пульса. Частота сердечных сокращений в состоянии покоя и минутный объем сердца, возросшие в связи с недостатком кислорода, по мере акклиматизации постепенно возвращаются к норме.
При длительном пребывании на больших высотах организм становится значительно более выносливым. Акклиматизация сопровождается учащением дыхания (хотя у тех, кто постоянно живет в горах, частота дыхания ниже, чем у вновь акклиматизировавшихся). При этом уменьшается разница в парциальном давлении кислорода между атмосферным и альвеолярным воздухом, что ведет к повышению давления кислорода в артериальной крови. Давление углекислого газа в альвеолярном воздухе хотя и снижено, но все же выше, чем у неакклиматизиров энного индивидуума. Устранение алкалоза обеспечивается почками, которые продуцируют щелочную мочу, благодаря чему устраняется избыток щелочи и нормализуется pH крови.
Одновременно усиливается образование эритроцитов и повышается содержание в них гемоглобина, вследствие чего повышается способность крови переносить кислород. На высоте до 3500 м существует линейная зависимость между содержанием гемоглобина в крови и барометрическим давлением. На больших высотах кривая зависимости содержания гемоглобина от атмосферного давления идет вверх более круто. В силу этого, а также благодаря изменению режима дыхания количество кислорода в артериальной крови у коренных жителей гор превышает количество кислорода у тех, кто живет в низинных районах. К тому же мышцы обитателей гор, по-видимому, способны совершать большую физическую работу при пониженном давлении кислорода в крови.
У постоянно живущих в горах давление в легочной артерии выше, чем у тех, кто впервые поднялся в горы, что связано с гипертрофией правого желудочка. Ряд исследователей полагают, что указанный фактор имеет приспособительное значение, обеспечивая усиленную перфузию легких и стимуляцию газообмена. Минутный объем сердца у лиц, постоянно живущих в горах, больше, чем у вновь прибывших; концентрация лактата в крови и кислородная задолженность при больших физических нагрузках, наоборот, ниже.
Акклиматизация к жизни на высоте 4500 м требует примерно 10 дней. При длительном пребывании на высоте наступают морфологические изменения, увеличивается грудная клетка и емкость легких. Температура воздуха в горах на больших высотах очень низка: в Тибете на высоте 5000 м над уровнем моря она падает до —33°С зимой, а летом не превышает 4-13°С. Защите от холода способствуют одежда, жилище, система отопления, а отчасти и физиологическая акклиматизация организма.
560
Часть V. Экология человека
Популяции на больших высотах
Даже у людей, живущих на высоте 2000 м, можно обнаружить некоторые приспособительные изменения в организме; такие изменения имеются у людей, живущих на Кавказе, Пиренеях и даже в области Высокого Вельда в Южной Африке, причем одинаковые физиологические приспособления имеют место у разных расовых групп: у тибетцев, живущих на высоте 4500 м, у жителей высокогорных районов Кашмира (2500—3000 м) и у жителей Анд (5500 м). На всех этих высотах человек способен к акклиматизации; примером могут служить коренные жители Перу, у которых число эритроцитов в крови на 30% выше, чем у лиц, живущих на уровне моря.
Хотя все описанные реакции представляют собой вторичные процессы, можно предположить, что некоторые из них могут фиксироваться генетически в процессе естественного отбора. Для некоторых животных, например для ламы, викуньи и боливийского гуся, обитающих в высокогорной местности, характерна высокая численность эритроцитов и более высокое сродство гемоглобина к кислороду; все эти признаки сохраняются в первом поколении у лам, родившихся в местности, расположенной на уровне моря. Монде утверждает, что в результате естественного отбора большая часть испанских завоевателей, вторгшихся в Анды, вымерла, и именно те, кто в новых условиях оказался неспособным давать потомство. Чистокровные испанцы окончательно утвердились в этих районах лишь через несколько поколений. Однако мы не можем представить убедительных доказательств существования у человека особых генотипов, приспособленных к условиям жизни па больших высотах.
Работоспособность
Человек сумел выжить в значительной степени благодаря своей способности выполнять физическую работу, будь то охота, перенос тяжестей, выкорчевка деревьев, рыхление земли мотыгой и т. д. Организм человека — исключительно гибкая, гомеостатически управляемая, производящая энергию система, способная к непрерывному выполнению физической работы и к кратковременным проявлениям («вспышкам») особенно высокой активности. Эффективность системы в значительной мере обеспечивается быстрым использованием высокоэнергетических фосфатных связей и их восстановлением в процессе работы или по ее окончании. Пополнение запасов энергии с соответствующей скоростью происходит за счет источников кислорода (подробно об этом можно прочитать в любом учебнике физиологии).
Один из методов оценки способности индивидуума совершать мышечную работу заключается в измерении количества кислорода,
Глава 27. Климатическая адаптация
561
доставляемого к мышцам в процессе работы. Для сравнительной оценки удобно использовать показатель макримальногЬ потребления кислорода при максимальной интенсивности работы. Эта величина, называемая максимальной аэробной мощностью (сокращен-110 ^Огмакс), известна сейчас для многих популяций. Изучены также факторы, от которых зависит наблюдаемая вариабельность ^Огмакс-
Величина Уо2макс тесно связана с размерами тела, в частности с весом тела и массой за вычетом жира. Опа коррелирует также с
Возраст, годы
Рис. 27.8. Зависимость максимальною потребления кислорода на Г кг веса тела от возраста индивидуума и места (Shepard).
I — атлеты; II — эскимосы; III — скандинавы; IV — канадцы; V — американцы; VI — прочие.
объемом и мышечной массой нижних конечностей; для измерения Уо2макс чаще всего пользуются велосипедным эргометром. Поэтому при сравнении представителей одной или разных популяций необходимо учитывать размеры тела или объем мускулатуры. Максимальная аэробная мощность заметно снижается с возрастом (рис. 27.8). Это обусловлено до некоторой степени уменьшением повседневной активности, другими словами, снижением «тренированности» организма. Работоспособность, подобно другим характеристикам, имеющим первостепенное адаптивное значение (например, способность переносить тепловой или высотный стресс), можно улучшить ежедневно повторяющейся или все более интенсивной физической работой. Значение Уо2макс может возрастать при этом на 20%.
Те сообщества или профессиональные группы, представители которых ежедневно выполняют тяжелую физическую работу, характеризуются более высокими средними значениями Уо2макС по
562
Часть V. Экология человека
Таблица 27.8
Значения РО1!Макс для различных этнических групп ”
Этническая группа ^О2макс’ мл/кг/мин Источник данных
Народы, с более примитивной культурой
Индейцы Арктики 49,6 Andersen et al.
Лапландцы 54,0 Andersen et al.
Бушмены 47,9 Wyndham
Доробо и Туркана 46,0 di Prampero a Cerretelli
Пигмеи 47,4 Andersen
Эскимосы Канады 56,5 Shepherd a. Rode
Урбанизованные народы.
Банту 44,6 Wyndham et al.
Американцы 43,5 Taylor et al.
Англичане 41,0 Davies et al.
Канадцы 44,5 Shepherd et al.
Чехи и словаки 40,6 Scranc
Немцы 40,5 Konig et al.
Итальянцы 45,0 di Pramero a. Cerretelli
Норвежцы 44,0 Hennansen a. Anderson
Южноафриканцы 45,6 Wyndham et al.
Японцы 45,0 Ikai a. Shindo
>) Davis et al , J. Appl. Physiol , 33, 726. 1972.
сравнению с группами лиц, выполняющих относительно легкую работу или ведущих сидячий образ жизни. Например, для занятых тяжелым физическим трудом представителей племени йоруба характерно высокое значение Ро2макс (55,5 мл/кг/мин), а для тех, кто выполняет легкую работу, значение Ро2макс ниже (45,9). Из табл. 27.8 видно, что величина УО!макс У народов с примитивной культурой выше, чем у городских жителей. Общирные и хорошо поставленные обследования канадских эскимосов (см. Rode a. Shepherd) показали, что расход энергии (т. е. повседневная активность) и аэробная мощность у эскимосов (мужчин и подростков) выше, чем у канадцев европейского происхождения. Высокие показатели Ро2макс характерны также для некоторых европейских сообществ (например, для шведов Ио2макс=52,0).
Вопрос о влиянии наследственности на работоспособность индивидуума заслуживает внимания по двум причинам. Во-первых, известно, что лишь часть общей вариабельности величины Йо2мако у взрослого мужского населения можно объяснить изменением Ко,макс в результате тренировки. Во-вторых, очевидно, что атле
Глава 27. Климатическая адаптация 563
ты представляют собой весьма небольшую группу людей с особыми физическими данными. Изучение близнецов показывает, что генетическими компонентами можно объяснить свыше 80% всей изменчивости величины Уо,макс- Однако при этом роль генетических факторов явно завышена, так как условия среды у близнецов более однородны, чем у обычных популяций. Величина Vo,макс зависит от размеров тела и объема мускулатуры, поэтому диапазон изменений этой важной функциональной характеристики определяется, вероятно, совокупностью генетических и негенетических факторов, таких, как степень тренированности, питание и болезни.
Глава 28
БОЛЕЗНИ
Экологические аспекты того или иного заболевания различны в зависимости от его причин, которые распадаются на три категории.
Во-первых, непосредственной причиной нарушения нормальной жизнедеятельности организма и возникновения патологического процесса могут быть различные факторы внешней среды. Совершенно очевидна, например, связь географического распределения ряда патологических состояний — теплового удара, горной болезни, ползучей язвы роговицы — с климатическими зонами, высотой местности и интенсивностью ультрафиолетового излучения. В наши дни во многих странах к числу неблагоприятных факторов внешней среды прибавилось загрязнение воздуха дымом и вредными продуктами.
Во-вторых, биотический компонент окружающей среды может таить в себе этиологический агент в виде патогенных микроорганизмов (вирусы, бактерии, грибы, простейшие), ядовитых растений, насекомых и опасных для человека животных.
В-третьих, роль среды выражается в различных нарушениях, связанных с питанием, которые обусловлены недостатком пищевых ресурсов животного и растительного происхождения (а также минеральных солей).
Кроме болезней, возникающих непосредственно под влиянием неблагоприятных условий окружающей среды, существует большая группа заболеваний, внешнее проявление которых кажется результатом плохой приспособленности организма, его отдельных органов и систем. В действительности же болезни проявляются лишь в результате взаимодействия с внешней средой, особенно с социальным окружением. Генетический дефект может проявиться в нарушении способности индивидуума к самосохранению; разного рода психологические нарушения выражаются в неправильной оценке внешних факторов или требований окружающей среды и неправильной реакции на них.
Экология болезней — обширный и сложный предмет. Характер географического распределения частоты заболеваний можно понять, учитывая взаимодействие факторов внешней среды и биологических, в том числе и генетических факторов. При этом всегда следует помнить о принципах адаптации как биологических, так и технологических.
Глава 28. Болезни
565
Инфекционные болезни
Занимаясь охотой и ловлей диких животных, вырубая леса и осушая болота, человек одновременно вытеснял животных, которые были его серьезными соперниками в борьбе за жизненное пространство и пищу. Это нередко приводило к бессмысленному истреблению многих полезных видов животных. Вместе с тем необходимо помнить, что болезни, причина которых — контакт человека с другими живыми организмами, представляют собой фазы экологического конфликта, котоый еще до сих пор не разрешился целиком в пользу человека.
Человек все еще борется с мелкими животными, такими, как грызуны, насекомые, грибы и микроорганизмы. Некоторые из них — паразиты, живущие в пище или в жилищах, другие непосредственно паразитируют на человеческом теле или в организме и наносят ему вред.
Географический фон — самый главный фактор, который обусловливает тип и численность паразитов и патогенных организмов. В переносе микроорганизмов могут участвовать вода, воздух, насекомые и другие животные, включая человека. Связь между хозяином и паразитом может принимать сложные формы и распадаться на несколько стадий. В этой связи участвуют переносчик, промежуточный хозяин и один или несколько «резервуаров». Изучая места распространения различных паразитов, мы часто находим, что экологические взаимоотношения сильно зависят от таких физических параметров среды, как ветер, дождь, температура воздуха, влажность. Патогенные организмы могут существовать лишь в определенных условиях. Переносчику также необходимы особые условия для размножения, например клещам или мухам требуется сухой климат. В свою очередь и хозяин — носитель паразита также обитает в какой-то определенной среде, например белки в Малайе живут только во влажных лесах. Малярийному комару Anopheles для размножения нужны водоемы или лужи, а следовательно, дождь и структура почвы, задерживающая воду на поверхности. Для мухи цеце, переносчика африканской сонной болезни, необходим сравнительно плотный растительный покров.
Географическое распределение многих болезней представляет собой, по существу, распределение переносчиков этих болезней и промежуточных хозяев. Шистосомоз распространен среди популяций, обитающих по берегам рек в странах с теплым климатом, так как одна из стадий развития данного паразита проходит в организме особого вида улиток. Болезни, вызываемые риккетсиями, например пятнистая лихорадка Скалистых гор, связаны с особым переносчиком — разновидностью клеща, встречающегося главным образом в Северной Америке, Бенгалии и Северной Африке; азиат-
566
Часть V. Экология человека
екая форма, переносимая клещами, характерна для Японии, о-вов Тайвань и Океании; разновидности тифа в Европе и Центральной Азии переносятся мухами и вшами. География бруцеллеза находится в тесной связи с географией распределения молока и молочных продуктов: главный резервуар — стада крупного рогатого скота.
К инфекционным заболеваниям, тесно связанным с климатическими условиями, относится фрамбезия; 80% площади, где распространено это заболевание, соответствует области со среднегодовой изотермой 27°С; остальная площадь лежит между изотермами 21—27°С. Фрамбезия особенно широко распространена там, где годовое количество осадков колеблется от 125 до 175 мм. Не ясно, однако, почему выживанию и переносу трепонемы способствуют высокие температура воздуха и влажность. Глистные инвазии широко распространены в районах с температурой 25—30°С и годовым количеством осадков более 150 мм, а следовательно, в основном между 40° северной и 40° южной широты.
Из-за большой сложности экологических взаимоотношений лишь относительно немного видов из огромного числа паразитирующих на человеке характеризуются обитанием в определенной географической местности. По этой причине для большинства болезней, вызываемых микроорганизмами, специфику экологических связей можно установить только конкретным анализом условий в данной географической среде. Оди (Andy) приводит следующий пример. Климат Сомали сухой, большая часть страны покрыта колючим кустарником или представляет собой полупустыню. Отсюда высокая частота катаракт и глазных инфекций, возникающих при раздражении соединительной оболочки глаза песком и ярким солнечным светом а также инфекций, переносимых мухами, которые размножаются в сухом климате. Климат Сомали способствует также размножению клещей — переносчиков возвратного тифа. Реже встречается мадурская стопа — хроническое заболевание, вызываемое актиномицетами, попадающими в кожу вместе с шипами колючих растений. Сухость воздуха и частые песчаные бури способствуют развитию ангин (сомалийцы пытаются лечить их, отрезая «язычок» мягкого нёба!). Песок, сухость и яркий солнечный свет — первичные физические факторы, которые можно выделить в этом экологическом комплексе.
Примером другого комплекса может служить Арктика. Здесь среди множества различных видов комаров и других членистоногих нет ни одного переносчика инфекционных болезней. Зато в арктических районах Америки главным резервуаром возбудителей салмонеллеза, бешенства и ряда гельминтов являются собаки, которые до сих пор служат основным видом зимнего транспорта. Сезонный характер появления таких болезней связан с тем простым фактом, что пищевые отходы, обычно выбрасываемые рядом с
Глава 28. Болезни
567
жилищем, оказываются безвредными в холодное время года, но с наступлением весенней оттепели сохраняющиеся в них патогенные организмы активизируются и становятся источником инфекций.
Характер поселений и тип жилищ могут также способствовать распространению определенных видов болезней. И в холодном климате (у эскимосов), и в жарких сухих странах, где люди ведут кочевой образ жизни, в небольших жилых помещениях скапливается много людей, в первом случае — ради сохранения тепла, а во втором — из-за простой необходимости переносить с собой как можно меньше груза. Скученность в обоих случаях ведет к распространению респираторных заболеваний, в первую очередь пневмонии и туберкулеза. Меланезиец на Новых Гебридах тщательно оберегает свой дом от всякого доступа воздуха — как бы в дом не проникли злые духи! В Йемене шистосомоз получил распространение по той причине, что обряд омовения совершается в общих бассейнах.
Распространению инфекционных болезней могут способствовать расчистка местности от кустарника и зарослей и вырубка лесов, неизбежные при постройке человеческих поселений. Уничтожение лесов на холмах Шри Ланки привело к тому, что даже в сухое время года вблизи жилищ часто сохраняются водоемы, в которых размножаются комары. Некоторые виды крыс, служащие резервуаром ряда инфекций, переносимых клещами, очень редко встречаются в естественных лесах Малайи, но после вырубки лесов они появились вблизи жилищ в больших количествах. Распространению бильгарциоза в Африке, Китае и Японии способствовало создание оросительных систем.
Грандиозные эпидемии чумы и холеры всегда были большим социальным бедствием, размеры которого становились все опаснее по мере улучшения путей сообщения и увеличения контактов между популяциями. Эти эпидемии распространялись со всевозрастающей скоростью по маршрутам караванов, судов и вдоль железных дорог. Было время, когда паломничество в Мекку становилось первопричиной новых эпидемий холеры.
Биологические реакции
Сопротивляемость инфекционным болезням, подобно сопротивляемости любому другому виду стресса, определяется, во-первых, генетическими факторами, которые обеспечивают естественную невосприимчивость,, и, во-вторых, «активным» иммунитетом, приобретаемым только в результате контакта с болезнетворным агентом. (Пассивный иммунитет, приобретаемый ребенком от матери с циркулирующими в ее крови антителами, исчезает через несколько месяцев после рождения.) Способность приобретать иммунитет можно рассматривать как физиологический признак, присущий человеку как виду и приобретенный в процессе эволюции. Как го
568
Часть V. Экология человека
ворит Бернет (Burnet), в мире, кишащем микроорганизмами, каждый из которых способен инфицировать живые ткани, для выживания положительно необходимо, чтобы в большинстве случаев заражение микробом сопровождалось, во-первых, выздоровлением и, во-вторых, развитием устойчивой невосприимчивости к новой атаке той же болезни. Он указывает, далее, что формы иммунитета, характерные для человеческих существ, отражают способ обитания древесных предков человека в тропических джунглях; главными патогенными организмами в такой среде были, вероятно, простейшие и вирусы, которые переносятся насекомыми (особенно комарами), и иммунитет, вырабатываемый в таких условиях,— это иммунитет против вирусов, подобных вирусу желтой лихорадки.
При попадании патогенного агента в организм человека в последнем начинают вырабатываться специфические белковые тела, называемые антителами, которые в больших количествах, поступают в кровь. Они обладают свойством проникать в патогенный микроорганизм и действовать на него особым образом, нарушая его жизнедеятельность и размножение. Эти антитела специфичны по отношению к патогенному агенту и устойчивы. Так как они остаются в крови и тканях в значительных количествах, при повторном проникновении патогенного агента в организм борьба с этим агентом начнется немедленно и будет более эффективной. Антитела, обеспечивающие иммунитет против самых разнообразных патогенных организмов, могут вырабатываться благодаря бесчисленным модификациям молекулярной структуры основного белка (гамма-глобулина). Каждая такая модификация обеспечивает специфическое «опознавание» и защитную реакцию против определенного патогенного агента, проникающего в организм. В малочисленных сообществах инфекции, переносимые воздушным путем (скажем, грипп, простудные заболевания, корь, коклюш), затухают сами собой из-за недостаточной частоты контактов между индивидуумами. Подобные инфекции получили широкое распространение лишь после того, как человек перешел к городскому образу жизни.
В тот или иной момент времени разные популяции могут находиться в различной фазе резистентности по отношению к тому или иному патогенному организму, с которым им приходится сталкиваться. Болезнь, достигшая определенного равновесия и слабоэндемичная у одного народа, может вылиться в серьезные волны эпидемий у других популяций, которые не успели приобрести соответствующего иммунитета.
Устойчивость к болезни, обусловленная приобретенным иммунитетом, аналогична физиологической акклиматизации к высоте, холоду или высокой температуре. Все эти формы сопротивляемости — результат длительного воздействия той или иной формы стресса. Во всех этих случаях приспособляемость — это свойство
Глава 28. Болезни
569
вида, а индивидуальная способность вырабатывать защитные реакции должна иметь некоторый генетический детерминант, о котором мы, однако, очень мало знаем.
Вопрос, который должен интересовать медиков и антропологов, заключается в том, существуют ли в природе различия в восприимчивости или резистентности к инфекционным болезням у различных групп, обусловленные не приобретенным иммунитетом, а генетически контролируемыми признаками независимо от того, имел ли место недавний контакт с болезнью или нет.
Было высказано мнение (Haldane), что инфекционные болезни могли служить одним из наиболее эффективных факторов естественного отбора у человека, так как они благоприятствовали выживанию индивидуумов, гены которых обеспечивали повышенную сопротивляемость организма болезням. Селективная роль хронических или дегенеративных заболеваний (например, атеросклероза) должна быть, напротив, незначительной, так как эти болезни обычно губят организм лишь после того, как он произвел на свет потомство, разве что эти болезни каким-то образом снижают плодовитость. Генетическая сопротивляемость по отношению к инфекционным (и некоторым другим) болезням, без сомнения, существует, однако лишь в редких случаях удалось определить гены, ответственные за такую сопротивляемость, и выяснить их метаболическую защитную роль.
Малярия
Это наиболее хорошо изученный пример болезни, сопротивляемость к которой зависит от наличия определенных генов. Имеются данные, свидетельствующие о том, что наиболее часто встречающие-•ся в современных популяциях три типа полиморфизма — серпо-видноклеточность, талассемия и недостаточность глюкозо-6-фос-фатдегидрогеназы (Г6ФДГ) — обязаны своим происхождением селективному действию так называемой тропической -малярии. Природа этого сбалансированного полиморфизма уже обсуждалась в гл. 16. Географическое распределение этих признаков совпадает с распределением областей с высокой частотой малярии. На рис. 28.1 показано географическое распределение малярии (до 1930 г.), а также серповидноклеточности, недостаточности глюкозо-6-фос-фатдегидрогеназы и талассемии. В Африке наблюдается особенно тесная корреляция между частотой серповидноклеточности и недостаточностью упомянутого фермента, хотя оба признака встречаются пе у одних и тех же индивидуумов. Было установлено, что во многих районах, эндемичных по малярии, в крови детей с серповидноклеточностыо или с недостаточностью Г6ФДГ содержится меньше малярийных паразитов. Подобная устойчивость в отношении малярии способствовала выживанию детей, гетерозиготных по гену серповидноклеточности, и, по-видимому, повышала
570
Часть V. Экология человека
Рис. 28.1. Географическое распределение тропической малярии. (Л), серповидноклеточности (В),
Глава 28. Болезни,
571
талассемии (В) и недостаточности глюйозо-6-фосфатдегидрогеназы (Г).
572
Часть К Экология человека
плодовитость гетерозиготных женщин (Allison, 1960). Из этого и ряда других фактов Мотульский (Motulsky) делает вывод, что малярия служила общим селективным фактором, благоприятствующим закреплению независимых мутаций — серповидноклеточности и недостаточности Г6ФДГ. Одним из наиболее удивительных и пока не получивших объяснения фактов, указывающих на различную восприимчивость к малярийной инфекции, является слабая чувствительность негров Западной Африки к Р. vivax.
Согласно данным археологии, в странах восточного Средиземноморья в древности существовала связь между географическим распределением малярии и генов талассемии. Порезный гиперостоз был распространен среди земледельцев неолитической эпохи, расчищавших участки от леса, что способствовало размножению комаров. Гиперостоз не обнаружен, однако, в предшествущем охотничьем периоде, когда человек занимал под пашню преимущественно свободные от леса сухие земли (Angel, 1940). Хотя до сих пор не удалось идентифицировать конкретные генотипы, определяющие восприимчивость или иммунитет к таким, например, инфекционным заболеваниям, как туберкулез, корь или полиомиелит, тем не менее существуют довольно убедительные данные (Мо-тульски, 1958), свидетельствующие в пользу определенной роли генетических факторов в иммунитете к таким заболеваниям. Одна группа данных касается постепенного снижения смертности от некоторых болезней в последовательном ряду поколений, длительное время не прибег авших к каким-либо лечебным или профилактическим мерам. Известны, далее, случаи повышенной частоты того или иного заболевания в определенных семьях, несмотря на то что условия существования этих семей ничем не отличаются от условий других семей, реже поражаемых. Некоторые популяции оказываются гораздо менее восприимчивыми к определенным болезням, хотя условия возникновения и распространения этих инфекций у них одинаковы с другими популяциями. В подобных случаях нелегко отличить приобретенный иммунитет от резистентности, обусловленной генетически. Однако эксперименты на животных ясно показывают, что с помощью искусственного отбора можно вывести линии животных, обладающих повышенной резистентностью пли, наоборот, восприимчивостью ко многим болезням, в том числе и к некоторым, свойственным человеку, например к туберкулезу (у кроликов), полиомиелиту (у мышей), чуме (у крыс). Изучение близнецов дает нам еще одно подтверждение роли генетических факторов: оказывается, что монозиготные близнецы обладают гораздо более высокой конкордантпостью по наличию (или отсутствию) данной болезни, чем дизиготные.
Рассмотрим в качестве примера данные, касающиеся туберкулеза легких. Неоднократно сообщалось, что смертность от туберкулеза особенно высока в тех популяциях, которые прежде не
Глава. 28. Болезни
573
имели контакта с этой болезнью — это американские индейцы, эскимосы, полинезийцы, ирландцы, меланезийцы и бушмены Южной Африки. В третьем поколении годовая смертность в популяции американских индейцев снизилась с 10 до 0,2%. Можно думать, что среди них выживали те индивидуумы, которые обладали генетической резистентностью. Хорошо известно, что в Европе городское население менее восприимчиво к туберкулезу, чем сельское, и что смертность от туберкулеза начала падать в этих городах задолго до того, как были разработаны эффективные меры борьбы с ним.
Селекционеры сумели вывести породы кроликов с большей или меньшей восприимчивостью к туберкулезу. И еще: удалось показать, что у монозиготных близнецов конкордантность по туберкулезу значительно выше, чем у дизиготных. Фершуэр (Verschuer) установил, что средняя величина конкордантности для монозиготных близнецов составляла 53%, а для дизиготных — 22%.
Изучение близнецов указывает также на роль генетических факторов в восприимчивости к полиомиелиту: конкордантность у монозиготных близнецов равна в этом случае 35%, а у дизиготных близнецов и у остальных сибсов — 6%. Добавим также, что в США более изолированные группы гораздо чаще болеют полиомиелитом, чем популяции с большей плотностью; как и в случае туберкулеза, это объясняют генетически обусловленной восприимчивостью.
Хотя генетические факторы, без сомнения, играют важную роль в восприимчивости или иммунитете организма ко многим болезням, тем не менее роль этих факторов в географическом распределении инфекционных заболеваний, по-видимому, все же подчиненная. Аборигены Австралии, живущие на обширной территории в отдаленных областях, географически почти не связанных между собой, страдают от тех же болезней (инфекционных и других), что и европейцы (Cleland, 1928). Антитела, характерные для большинства вирусных заболеваний, поражающих городских жителей, найдены в крови популяций, сохранивших первобытную культуру, например у эскимосов и бушменов Калахари. Удалось выявить следующие причины, лежащие в основе различий в частоте и степени тяжести ряда паразитарных инфекций в различных популяциях: 1) различия в экологических условиях, способствующих возникновению и распространению инфекций; 2) различия в «уровне иммунитета» (последнее относится даже к малярии); 3) общественно-экономические факторы, например недоедание, недостаточно развитая сеть медицинских учреждений и т. п.
574
Часть V. Экология человека
Неинфекционные болезни
Географические различия в характере распространения многих неинфекционных болезней можно объяснить, только если рассматривать в комплексе факторы внешней среды и биологические реакции (врожденные и приобретенные). Тот факт, что негры в большей степени подвержены отморожениям, чем эскимосы или североамериканские индейцы, можно рассматривать и как результат недостаточной акклиматизации, и как показатель какой-то особой генетической восприимчивости. Согласно Картеру (Carter), такие аномалии центральной нервной системы, как spina bifida и анэнцефалия, этиология которых недостаточно изучена, чаще встречаются в определенных семьях или этнических группах, что указывает на важную роль генетических факторов в предрасположении к этим видам патологии. В то же время частота подобных аномалий зависит и от таких негенетических факторов, как возраст матери, а также от того, каким по счету является данный ребенок в семье.
Географии распространения рака всегда уделялось достаточно много внимания. Онкологи надеялись, что изучение местных условий, характерных для областей с высокой и низкой частотой этого заболевания, даст в руки исследователя ключ к пониманию этиологии рака. Подобный подход в целом оказался малоплодотворным: удалось решить ряд проблем, но возникло, пожалуй, не меньше новых. Те или иные формы рака зарегистрированы у всех человеческих популяций, однако существуют заметные вариации в частоте различных видов злокачественных новообразований. Точно оценить частоту случаев рака в различных сообществах довольно трудно, особенно в районах, где здравоохранение находится в самой зачаточной форме. В более развитых странах улучшение диагностики, а также средств профилактики и лечения рака и медицинской статистики приводит время от времени к необходимости пересмотра имеющихся оценок. Однако даже при учете указанных факторов мы не вправе игнорировать многочисленные различия в частоте случаев рака у разных популяций. Вероятность появления многих видов опухолей увеличивается с возрастом. Сравнительная оценка восприимчивости к раку, исходя из статистики заболеваний, у обоих полов для популяций с разной возрастной структурой оказывается поэтому весьма приближенной.
Higginson a. Oettli провели анализ зарегистрированных случаев рака у банту в Трансваале. Частота различных видов опухолей у банту была сопоставлена с наиболее вероятной оценкой частоты случаев рака у белого населения США, американских негров и популяций датчан с аналогичной возрастной структурой (табл. 28.1). Установлено, что у мужчин банту гораздо реже, чем в контрольных группах, встречается рак желудка, толстой и прямой кишки, а у женщин банту — рак молочной железы и тела мат
Глава 28. Болезни
575
ки. Рак печени (гепатома) и в меньшей степени рак пищевода, наоборот, более характерны для мужского населения банту. Частота некоторых форм рака у банту и американских негров весьма различна, однако утех и других редко встречаются рак кожи, полости рта и губ, что отчасти можно объяснить пигментацией. Среди белого населения Америки гораздо реже, чем у банту, встречается рак шейки матки. Отмечены также заметные различия в частоте рака печени, кишечника и молочной железы у белого населения Америки и у датчан.
Таблица 28.1
Число зарегистрированных случаев рака у банту в Иоганнесбурге (1953—1955 гг.) и ожидаемое число для трех контрольных популяций того же возраста11
Локализация опухоли Негры банту Американцы Датчане
белые негры
М ужчины
Полость рта 2 21,3 2,0 24,3
Язык 8 12,9 9,3 —
Пищевод 53 20,9 36,5 12,2
Желудок 41 89,0 128,4 119,4
Толстая кийка 7 75,7 54,1 43,6
Прямая кишка 5 67,8 45,7 61,0
Печень 114 13,8 25,7 1,4
Легкие (бронхи) 40 103,0 104,3 51,1
Половой член 8 3,9 11,4 3,6
Кожа 14 212,7 18,2 49,5
Женщины
Шейка матки 198 119,1 266,6 122,1
Тело матки 1 27,9 25,5 23,2
Молочная железа 50 221,9 170,7 131,8
*) Higginson a. Oettli, J. natn. Cancer Inst., 24, 589 (1960).
Можно было бы привести еще немало примеров, свидетельствующих о заметных различиях в частоте рака у разных популяций. В ряде случаев можно с большой степенью достоверности указать на фактор среды, обусловливающий относительное увеличение частоты тех или иных форм рака. Так, в некоторых районах' Индии распространен рак глотки, что, по-видимому, объясняется постоянным употреблением жевательного табака. У белых, живущих в местах с большим количеством солнечных дней в году, нередко возникает рак кожи на не защищенных от солнца местах, что, очевидно, связано с воздействием ультрафиолетовых лучей.
576
Часть V. Экология человека
Первичный рак печени, столь часто встречающийся у африканцев, по-видимому, представляет собой отдаленное последствие цирроза печени, высокая частота которого в этих популяциях в свою очередь обусловлена особенностями питания — низким содержанием в рационе животных белков и избытком углеводов. Резкие различия в распространенности коронарной болезни у разных рас на самом деле связаны с особенностями питания — с избыточным содержанием в рационе жиров. Так, у банту, употребляющих в пищу Меньше жиров, чем европейцы, коронарная болезнь встречается крайне редко.
Низкая частота сахарного диабета во многих негритянских популяциях, по-видимому, обусловлена малой калорийностью и во многих отношениях неполноценностью пищи. Тот факт, что с изменением характера питания частота диабета у негров становилась такой же, как и у белых, убедительно свидетельствует против отнесения диабета к категории «расовых» признаков. Против этого же свидетельствует и значительное снижение частоты диабета во время войн, когда пища людей малокалорийна и бедна жирами. Однако характер питания, по-видимому, способствует лишь проявлению диабета и определяет его течение. Недавно появилось сообщение, что предрасположение к диабету наследуется как простой аутосомный рецессивный ген с частотой, которая в США колеблется в пределах от 20 до 25% (Steinberg a. Wilder). О наследственной склонности к диабету свидетельствует также то, что <^реди родственников больных диабетом частота этого заболевания выше, чем в общей популяции. Показательна также конкордантность по диабету идентичных близнецов.
Известно, что многие заболевания и аномалии наследуются по простому менделевскому типу: больной индивидуум обычно гомозиготен по рецессивному гену, хотя при некоторых условиях могут играть роль и доминантные гены. Почти все «генетические» болезни очень редки, однако некоторые из них чаще встречаются у вполне определенных популяций. Талассемию и серповидноклеточную анемию связывают с наличием особых генов, определяющих синтез аномальных типов гемоглобина (см. гл. 16). Гемолитическая болезнь новорожденных — результат несовместимости по группам крови системы Rh — характерна для европейцев и не встречается у монголоидов или американских индейцев, среди которых нет Rh-индивидуумов. Язва двенадцатиперстной кишки, по-видимому, чаще встречается у людей с группой крови 0; лица с группой крови А более склонны к раку желудка.
Сравнительное изучение популяций, находящихся в резко отличных условиях окружающей среды, сыграло определенную роль в понимании природы сердечно-сосудистых болезней. Установлено, что стойкая гипертония служит фактором, предрасполагающим к инфаркту миокарда и кровоизлиянию в мозг. Для многих случаев
Глава 28. Болезни
577
так называемой «эссенциальной гипертонии» не удается найти патологические факторы, ответственные за высокое' артериальное давление. В странах с достаточно высоким уровнем жизни статистическое распределение систолического артериального давления в пределах данной возрастной группы непрерывно, но отличается от гауссова: кривая распределения асимметрична и имеет «хвост» в области высоких значений давления. Цифры, характеризующие средние в пределах популяции уровни систолического и диастолического давления, увеличиваются с возрастом, особенно у мужчин старше 50 лет. Некоторые исследователи полагают, что случаи эссенциальной гипертонии (несколько условно определяемые показателем 150/100 и выше) не следует выделять в особую группу; они представляют лишь наиболее крайние варианты общего распределения (с одним максимумом).
Непрерывный характер распределения обусловлен с этой точки зрения комбинированным действием многих генов и разноообраз-ных факторов внешней среды. Противоположное мнение состоит в том, что лица с высоким артериальным давлением, по существу, относятся к особой группе, отличие которой от других групп имеет простую генетическую основу; распределение фактически имеет два максимума, однако разделить их нельзя из-за слишком сильного взаимного перекрывания. Как утверждает Пиккеринг и Робертс (Pickering a. Roberts), эссенциальной гипертонией страдают люди, наследующие склонность к более высокому уровню артериального давления, т. е. частота гипертонии у разных популяций и варианты средних показателей давления определяются генетическими факторами. Мультифакторная природа распределения уровня артериального давления включает также и фактор среды. Например, у американских негров давление гораздо выше (и увеличивается с возрастом), чем у африканских. У представителей народности банту, проживающих в городах, артериальное давление выше, чем у живущих в сельской местности.
Средний уровень артериального давления у лиц определенной возрастной группы, а также смертность от гипертонии заметно различаются даже в пределах одного региона или одной этнической группы. В Японии оба показателя достигают максимума в северо-восточных районах (исключая остров Хоккайдо). Можно думать, что это связано в первую очередь с большой разницей температур вне и внутри помещений, а также с различиями в пищевом рационе. Сравнительно низкий уровень систолического давления характерен для аборигенов Австралии, населяющих центральную пустыню и ведущих кочевой образ жизни. Более высокое давление отмечено у племен, которые чаще вступают в контакты с европейскими переселенцами (например, среди аборигенов Квинсленда). Географические вариации в распределении артериального давления, а также само предрасположение к гипертонии, вероятно,
578
Часть V. Экология человека
---------------------------------------------------г-------- зависят от множества факторов: характера питания, физической нагрузки, температуры окружающей среды, психологического стресса и т. д.
У монозиготных близнецов в старших возрастных группах разница в уровне давления едва ли меньше, чем между сибсами и дизи-готными близнецами. Это указывает на то, что в пожилом возрасте уровень артериального давления в большей мере зависит от факторов с]5еды. Вопрос о том, каковы именно эти факторы, служит предметом живой полемики; это могут быть особенности питания (в частности, содержание в рационе жиров, солей, белков), степень физической нагрузки, а также психологический стресс. Согласно грубой оценке Пиккеринга, наследственные факторы и факторы среды играют равнозначную роль в этиологии гипертонии. То, что естественный отбор не элиминировал гипертонию как одну из причин смерти во многих популяциях, объясняется, вероятно, тем, что смерть от гипертонии обычно наступает в пострепродуктивном возрасте.
Токсичные вещества и загрязнители
Токсичные химические вещества могут попасть в организм человека с пищей и водой или -в составе лекарственных препаратов. Отсюда вовсе не следует, что человек столкнулся с опасным воздействием этих веществ лишь в наше время, которое можно назвать эрой химического синтеза. Первобытные люди были хорошо осведомлены о ядовитых свойствах некоторых видов растений, корней или ягод. Знание этих свойств составляло неотъемлемую часть их общей экологической эрудиции. Многие охотничьи племена умеют готовить яды и применять их на охоте или на войне, используя, например, отравленные наконечники стрел или копий. Современный человек сталкивается, однако, с несравненно более широким и опасным воздействием токсичных веществ на организм, в особенности тех, которые выделяются при различного рода промышленных процессах. Эти вещества заглатываются, вдыхаются или попадают в организм через кожу.
В пределах каждой популяции существует, как обычно, широкая вариабельность в индивидуальных реакциях организма на действие токсичных веществ. Как следствие толерантность или, наоборот, повышенную чувствительность к такого рода агентам можно понять, лишь учитывая взаимодействие факторов среды и биологических факторов. Последние в свою очередь бывают либо приобретенными, либо генетически обусловленными.
Значительная часть химических соединений теряет свою токсичность в организме в результате преобразования в неактивные метаболиты под действием ферментов печени либо выводится из организма с мочой (оба процесса могут происходить одновременно).
Глава 28. Болезни
579
Эти процессы следует рассматривать как первичные приспособительные реакции организма. Эффективность защитных метаболических реакций и степень выведения вредных продуктов зависят от физико-химической природы последних. Некоторые загрязнители, слабо диффундирующие или нерастворимые в липидах, сразу же выводятся из организма в неизмененном виде. Ферменты печени, осуществляющие детоксикацию, резко отличаются от метаболических ферментов. По предположению Броди, организм вырабатывает эти защитные ферменты с тем, чтобы избавиться от растворимых в липидах веществ, содержащихся в пище, таких, например, как углеводороды, алкалоиды, терпены и стероиды. Указанные ферменты неспецифичны: ведь человеку приходилось сталкиваться с великим множеством вредных веществ еще задолго до начала промышленной эры. Скорость метаболической детоксикации весьма сильно зависит от индивидуальных особенностей человека и определяется, в частности, его генофондом. Так, у мо-нозиготных близнецов скорость выведения лекарственных препаратов из организма одинакова, тогда как у дизиготных — различия ничуть не меньше, чем у любых других членов популяции.
Повышенная сопротивляемость к действию токсичных веществ ‘определяется способностью организма индуцировать определенные виды ферментов. Некоторые из химических продуктов окружающей среды сами стимулируют или ускоряют свое выведение из организма, обусловленное метаболизмом. Подобно всем явлениям, зависящим от «тренировки», наиболее активная фаза выработки ферментов наступает спустя некоторое время после начала приема внутрь химического вещества (обычно через несколько дней). Индукция фермента прекращается спустя 2—3 нед после удаления химического агента из организма. Индукция включает как повышение активности метаболических ферментов, так и их ускоренный синтез, осуществляемый главным образом печенью. Индукцию можно предотвратить, блокируя сйнтез белка, например, актиномицином D.
К числу токсичных веществ» способных индуцировать ферменты, относятся хлоросодержащие углеводороды (группы DDT), спирт, табак и многие из лекарственных препаратов. Так, «период полужизни» такого лекарственного препарата (как, скажем, фено-барбитон) спустя две недели после начала его приема снижается до 5 ч (вместо исходных 13); этот препарат индуцирует синтез в печени фермента глюкуронилтрансферазы. Существует, конечно, некоторая предельная способность реакции на индукцию ферментов, и вполне возможно, что интенсивность этой реакции определяется генетически (Lawrenke). Толерантность и различным лекарственным препаратам широко варьирует в пределах популяции, и эта вариабельность отчасти зависит от различий в реакции на индукцию ферментов.
Глава 29
СТАБИЛЬНОСТЬ ПОПУЛЯЦИЙ
Как мы видели, каждое сообщество вынуждено приспосабливаться к различным видам стресса, создаваемого самыми разнообразными факторами внешней среды. Именно экологическое взаимодействие среды и сообщества определяет численность популяции. Эта величина служит показателем того, насколько успешно сообщество подчиняет себе среду (в результате сознательной деятельности или каким-либо иным путем).
Плотность населения и его численность
Большое рзанообразие экосистем человека находит отражение в не меньшем разнообразии численности его различных популя-' ций. Данные, приведенные в табл. 29.1, могут служить некоторой иллюстрацией связи между формой, или стадией, развития экономики и плотностью населения.
Каждая экономическая группа испытывает на себе действие многих различных факторов. Плотность популяции в данных условиях среды зависит от ряда причин: 1) от характера климата, почвы и рельефа; 2) от флоры и фауны; 3) от культурного уровня, способности использовать природные ресурсы (производительность хозяйства).
В простых сообществах с относительно однородным культурным уровнем на данной территории можно установить прямую связь плотности популяции с некоторой величиной, характеризующей воздействие среды. Согласно Бёрдселу (Birdsell, 1953), в Австралии такой величиной может служить среднегодовое количество осадков, которое явно коррелирует с плотностью населения (коэффициент корреляции +0,8). Для популяций численностью около 500 человек применимо уравнение Y=7112,8 А-1'544’, где Y — размер площади, занимаемой данной популяцией, а X — среднее годовое количество осадков.
Отсюда, однако, не следует, что плотность популяции должна быть* обязательно одинакова в областях с одинаковыми условиями среды и формой хозяйства. Плотность изменяется также в зависимости от различий в биосфере и источниках пищи. Плотность населения у бушменов Калахари или у индейцев-шошонов в Неваде отличается от плотности населения у аборигенов Австралии, не
Глава. 29. Стабильность популяций
581
смотря на примерно одинаковое количество осадков, выпадающих в этих областях. Производительность хозяйства у индейцев южйой Калифорнии примерно такая же, как у аборигенов Австралии, а количество осадков в Калифорнии даже ниже, чем в Австралии. Тем не менее плотность населения у индейцев в 50 раз выше, чем у австралийцев; это можно связать с обилием особого вида растительной пищи (вблизи Калифорнийского залива произрастает около десятка различных видов агавы). Материальная культура простых сообществ — собирателей и охотников — очень бедна; в более развитых обществах возникает зависимость плотности населения от некоторых переменных, характеризующих социальную структуру популяции и технику производства, так как высокоорганизованные сообществ^ способны более эффективно использовать природные ресурсы и преодолевать налагаемые средой ограничения.
Плотность популяции
Таблица 29.1
Экономическая стадия Площадь на 1 чел. (кв. мили) Число жителем на 1 кв. милю
1. Собиратели 200—0,5 До 2
Люди верхнего палеолита (Англия) 200 —
Аборигены Австралии 25 —
Индейцы Огненной Земли 8 —
Жители Андаманских островов 0,5 —
2. Охотники и рыболовы 200—0,05 До 20
Эскимосы и индейцы Северо-Восточных территорий Канады 200 —
Эскимосы (Аляска) 30 —.
Люди мезолита (Англия) 10 —
Индейцы в пампасах 2 ——
Индейцы Британской Колумбии 0,05 —
3. Древние земледельцы 0,05—1 0,5—50
Люди неолита (Англия) — 2
4. Пастушеские и кочевые народы — 10—100
5. Земледельцы на более высокой ступени развития - —— 10—150
Люди железного века — 10
Люди средневековья — 50
Шведские крестьяне 20—150
На протяжении большей части истории человечества техника производства оставалась на крайне низком уровне: основными способами добывания средств к существованию были собирательство и охота, потребительское садоводство и огородничество.
582
Часть V. Экология человека
Вследствие этого численность населения долгое время оставалась очень низкой, а рост ее происходил медленно. Человек стал осваивать более холодные области только в конце плейстоцена, а Австралия и Америка были заселены лишь 12 000—15 000 лет назад. Ускоренный рост численности населения начался примерно 8000 лет назад с развитием земледелия, благодаря которому стала возможна жизнь в городах. В нынешнюю фазу, характеризующуюся колоссальным ростом численности населения и освоением все новых районов, человечество вступило лишь в самое последнее время, с развитием индустриализации. Можно думать, что в ту эпоху, когда человек жил охотой и собирательством (что соответствует культуре палеолита или мезолита), плотность населения была менее 1 человека на 3 кв. км. В эпоху неолита, когда человек стал обрабатывать землю, плотность возросла примерно в 10 раз; в бронзовом и железном веках — еще в 10 раз. Общее число людей в эпоху неолита определяют примерно в 5 млн., а в период появления первых крупных городов — в 20—40 млн.
Современному виду Homo sapiens потребовалось около 20 000 лет, чтобы достичь численности 200 млн. (во времена Римской империи). В последующие 1500 лет (к 1600 г. н. э.) населе-ление земного шара возросло до 500 млн., спустя еще 200 лет — более чем удвоилось (около 1 млрд, в 1800 г.). В наши дни на земле проживает около 3 млрд, человек.
На любой стадии развития географическое распределение населения на земном шаре было неравномерным. Даже сегодня половина всего населения земного шара проживает на территории, составляющей всего 5% площади обитаемой суши (табл. 25.3), а'80 % всей этой площади характеризуется средней плотностью несколько более 3 человек на 1 кв. км. Начиная с глубокой древности, со времени появления первых городов, наряду с большими популяциями, отличавшимися высокой плотностью, в различных местах земного шара существовали и небольшие сообщества.
Регуляция численности популяций
Динамика численности человеческих популяций может быть весьма разнообразной: они могут увеличиваться или уменьшаться и могут оставаться стабильными. Уже в первобытных человеческих сообществах существовала тенденция к сохранению численности популяции на определенном уровне. Здесь уместно упомянуть о системах естественной регуляции, действующих в животном мире. Основные принципы этих систем, сформулированные экологами Элтоном, Никольсоном и Соломоном (Elton, Nicholson a. Solomon), следующие:
Глава 29. Стабильность популяций
583
1. Приближение к стабильной численности популяций на самом деле означает, что колебания численности ограничены некоторыми пределами.
2. Стабильный средний уровень плотности определяется емкостью системы (примером могут служить популяции аборигенов Австралии).
3. Процессы, которые ограничивают диапазон колебаний в экосистеме постоянной емкости,— это внутренние процессы; их называют процессами, зависящими от плотности.
4. Внешние события могут резко нарушить равновесие в экосистеме, но они не участвуют в установлении стабильности.
5. Любая данная экосистема имеет «оптимальную» численность населения (Элтон, 1941), и процессы, регулирующие численность, направлены к установлению оптимума (который, однако, не всегда достигается в данных условиях).
Эти принципы применимы и к человеческим популяциям, по с известными ограничениями. Во-первых, численность человеческой популяции регулируется не только биологическими, но И культурными факторами. Действие последних может быть целенаправленным, но во многих сообществах оно настолько тесно переплетено с действием укоренившихся обычаев, что кажется столь же безликим, как и проявление действия биологических факторов. Во-вторых, со временем система становится все более способной к расширению. В самый последний период стало возможным непрерывное увеличение численности популяции без приближения к равновесию.
Регуляторные процессы, зависящие от плотности, можно разделить на две группы — «обусловленные конкуренцией» и «инверсные».
В процессах первой группы ограничивающее действие изменяется с изменением численности популяции — при возрастании численности оно усиливается, а при падении ослабевает. Так, при увеличении плотности популяции усиливается борьба за основные источники пищи, воды, топлива и сырья, за освоение территории и мест, пригодных для жилья. Увеличение плотности популяции может сопровождаться загрязнением окружающей среды или каким-либо иным ухудшением условий существования, что ведет к более частым вспышкам эпидемий и к более быстрому распространению заболеваний как путем прямых контактов между людьми, так и вследствие загрязнения воздуха, пищи, воды, домашней утвари. Ограничивающее действие этих процессов ослабевает с уменьшением численности популяции.
Ограничивающее действие регуляторных процессов второй группы («инверсных») изменяется в направлении, противоположном изменению численности популяции. Это такие процессы, интенсивность которых с увеличением плотности популяций спи-
584
Часть V. Экология человека
жается и, наоборот, с уменьшением ее повышается. По мере увеличения численности популяция становится способной более эффективно использовать природные ресурсы, более успешно добывать себе средства к существованию, в особенности пищу. Появляются лучшие возможности для совместной охоты и рыболовства, совместной расчистки земель и проведения оросительных работ, строительства жилищ и т. д. Более многочисленное сообщество может лучше организовать взаимную защиту своих членов или обеспечить наступательные действия против других групп и животных. Следует учитывать также, что с увеличением численности популяции возрастает фонд генетической изменчивости. При уменьшении плотности все эти преимущества теряют силу: малочисленное, разбросанное на большой территории сообщество имеет меньше шансов успешно конкурировать с другими популяциями и менее способно к проведению коллективных мероприятий.
В животном царстве эти две группы естественных регуляторных процессов, действующих в противоположных направлениях, могут, по-видимому, поддерживать колебания численности популяции в узких пределах, совместимых с постоянным выживанием в данной экосистеме. По аналогии можно привести данные, свидетельствующие об относительной стабильности - человеческих экосистем в неиндустриальных сообществах.
1. Археология предоставляет в наше распоряжение многочисленные примеры того, как в определенных условиях обитания техника производства может оставаться неизменной в течение длительного периода времени (Childe a. G. Clark). Анализируя медленное изменение культуры каменного века, Чайлд справедливо утверждал: «До тех пор пока достигнутая степень приспособленности к условиям среды остается удовлетворительной, сообщество стремится быть консервативным». Действительно, резкие изменения жизни народа и его материальной культуры, устанавливаемые раскопками, обычно связаны с вторжением новых популяций с иной культурой.
2. Этнография приводит данные, подтверждающие консерватизм тех современных сообществ, которые сохранили многие примитивные черты (Forde). К примеру, религиозные церемонии и традиции у австралийцев племени аранда подчиняются исключительно строгой регламентации, и нарушители всегда сурово наказываются; экономическая деятельность таких сообществ подчиняется строгим правилами и представляет результат опыта, накопленного за долгие времена путем проб и ошибок. Строгое соблюдение границ племенных территорий свидетельствует о стабильности популяции, так как ресурсы, необходимые для существования популяции, остаются неизменными в течение длительного времени.
3. Экономика примитивных сообществ почти полностью за
Глава 29. Стабильность популяций
585
висит от местных ресурсов. Выше мы подробно проанализировали равновесие, которое достигается в среднем в течение года между количеством продуктов питания, производимых крестьянской общиной, и ее нуждами (на примере Гамбии). Стабильность популяции обусловлена негибким характером экосистемы. Для собирания плодов и охоты может потребоваться почти весь наличный запас энергии, который человек способен израсходовать в процессе работы,— примерно 6270—8370 кДж в день (эти цифры представляют расход энергии при непрерывном движении). Можно рассчитать, что плотность популяции в таких экосистемах должна составлять около одного человека на 3 кв. км. Переход к сельскому хозяйству приводит вначале к увеличению плотности популяции, однако значительные излишки при этом не накапливаются. Энергия, которую необходимо затратить па обработку 1,2 га земли в один год одним человеком, примерно равна энергии, затрачиваемой при использовании для охоты 200 га. По этому расчету плотность населения на стадии первобытного земледелия при равновесии может достичь 30 человек на 1 кв. км или больше.
4. Мы располагаем весьма скудными сведениями о динамике численности населения в примитивных сообществах. Даже для относительно развитых сообществ, обладающих письменностью, часто отсутствуют соответствующие демографические данные. Наиболее надежными оказались данные о среднем числе дочерей, рождаемых одной матерью Хорошим показателем может служить общая плодовитость (С. Clark, 1958). Последняя определяется числом детей, рождаемых в среднем одной женщиной за весь детородный период. При благоприятных условиях для воспроизведения потомства (т. е. когда каждая женщина выходит замуж в молодости, а вдовы имеют возможность вновь вступить в брак), если предположить к тому же, что климакс наступает после 20 лет супружеской жизни, общая плодовитость должна составлять примерно 8. Это предполагает рождение одного ребенка за каждые 2,5 года замужества (учитывается возможность нескольких самопроизвольных абортов и периодов временной стерильности). При высокой смертности (когда лишь немногие переживают возраст 40 лет) общая плодовитость, измеряемая величиной 6—8, достаточна лишь для того, чтобы сохранить численность популяции на постоянном уровне. Известно, что в примитивных сообществах общая плодовитость часто бывает ниже 6 (табл. 29.2).
Если в таких сообществах численность остается на одном уровне в течение длительного времени, то отсюда следует, что смертность в них не столь высока, чтобы для поддержания постоянной численности плодовитость была равна 6—8, и, стало быть, происходит эффективное ограничение рождаемости. Подобная ситуация характерна для племен собирателей и первобытных земледельцев. Многочисленные данные свидетельствуют о том, что из каждых
586
Часть V. Экология человека
Таблица 29.2
Общая плодовитость (число детей у матери) 3)
Собиратели и охотники-одиночки
Австралийцы: западные области центральные области
Индейцы Огненной Земли
Охотники
Эскимосы: Гренландия
Североамериканские индейцы:
нутка
чинук омаха
Н римитивные земледельцы.
Оранг Кибу на Суматре
Айны
Земледельцы на более высокой ступени развития
Банту
Нигерийцы
Индейцы
Китайцы
Народы развивающихся стран (данные Совета по изучению популяций: Африка, Юго-Западная и Юго-Восточная Азия, 1965 г.)
Народы развитых стран (данные Совета по изучению популяций, 1965 г.)
Северная Америка
Центральная, северная и западная Европа
СССР
4—6 5 4
3—4
3 3
4—6
4 3—4
около 6 4,3 Вероятно, 6—8 Вероятно, 6—8
5,1—7,0
3,7
2,7
2,9
l) (Carr-Saunders, 1922.)
4—5 рождающихся детей лишь двое или трое достигают зрелости. Размер семьи, таким образом, довольно мал. В земледельческих сообществах с высокой плотностью смертность обычно гораздо выше и сохранение численности популяции возможно лишь при условии, что общая плодовитость составляет 6—8.
Процессы, регулирующие численность популяции
В сообществах с ограниченными ресурсами и консервативной техникой плодовитость такова, что по истечении достаточно длительного времени неизбежно наступило бы такое пложение, при котором численность популяции превысила бы уровень, соответствующий количеству ресурсов. Если же численность остается постоянной, то, по-видимому, существуют какие-то регуляторные процессы, поддерживающие стабильность. Такими ограничивающими факторами мальтузианцы считают голод, эпидемии, войны.
Глава 29. Стабильность популяций
587
Согласно Мальтусу, рост популяции подчиняется закону геометрической прогрессии, пищевые же ресурсы увеличиваются по закону арифметической прогрессии. Мальтус оправдывал «моральные» меры по ограничению рождаемости, например более поздние браки, которые должны привести к неизбежному уменьшению численности семей.
Некоторые теоретики (например, Hawley, 1950) в своем анализе динамики популяции в примитивных сообществах целиком остаются на мальтузианских позициях. По их мнению, «...численность популяции имеет тенденцию к изменениям, прямо связанным с изменением наличных источников продовольствия и материалов, необходимых для жизни сообщества... При таких условиях проблема численности популяции решается только применительно к данной местности, так как географическая изоляция ограничивает возможность получения помощи извне... Низкий уровень техники ставит популяцию в тесную зависимость от изменений в окружающей среде. Такая популяция фактически беззащитна перед неблагоприятными изменениями климата (засуха, наводнения), а также перед уничтожением источников пищи вредителями или хищниками. Таким образом, местные особенности физической и биотической среды — весьма неустойчивый фактор для групп, сохраняющих низкую и неизменную технику хозяйства.
Размер популяции автоматически приспосабливается к изменениям природных ресурсов чаще всего за счет изменения смертности. В неблагоприятные годы смертность резко возрастает, а в периоды изобилия падает. Хотя наши сведения о демографии изолированных групп фрагментарны, среднегодовая смертность для таких популяций, по-видимому, колеблется около 40 па 1000. В некоторых случаях она достигает 100 на 1000, а может также снизиться до 25. В целом, однако, изолированные популяции почти всегда живут под угрозой катастрофы. На это указывает не только высокая средняя смертность, но и малая средняя продолжительность жизни (30 лет или менее). «Запас прочности» у таких популяций иногда настолько мал, что снижения годового количества осадков всего лишь на несколько сантиметров или потери нескольких дней, необходимых для нормального развития сельскохозяйственных культур, оказывается достаточно для значительного увеличения смертности» (Хоули).
В таких условиях рождаемость и смертность велики, но и скорость обновления популяции очень высока.
«Интенсивное использование популяцией природных ресурсов поддерживается высоким постоянным уровнем воспроизведения потомства. В изолированной группе женщины почти постоянно беременны, но внутриутробная и детская смертность настолько велики, что лишь немногие доживают до зрелого возраста. Тем не менее родник жизни неиссякаем, и всегда существует некото
588
Часть V. Экология человека
рая потенциальная популяция, готовая поглотить любой избыток пищи, который может появиться в высокоурожайный год. Группа всегда может быстро возместить потери, понесенные ею в период острой нехватки пищи, если только численность ее не упала до чрезмерно низкой...» (Хоули). Концепция, к которой многие демографы пришли на основании наблюдений над жизнью простых сообществ и популяций животных, состоит в том, что популяция фактически стремится сохранить постоянную численность, и эта цель всегда более или менее достигается. Ле Гро Кларк писал (см. книгу «Наследие мальтузианства», стр. 17):
«Имеется достаточно данных, свидетельствующих о том, что за долгие годы существования первобытнообщинного строя люди приобрели смутное, но в основных чертах правильное представление о связи между занимаемой ими территорией, обычным прожиточным минимумом и численностью популяции. Они не могли, конечно, предвидеть время наступления засухи или наводнения, но всегда по возможности принимали в расчет неизбежность «голодного сезона». Они стремились ограничить рост численности либо такими грубыми, но эффективными мерами, как аборт, детоубийство или пожизненное вдовство, либо посредством различных табу, запрещавших половые сношения в определенное время года. Стихийные бедствия, вредители и болезни брали у людей свою дань, но в целом узаконенное детоубийство, аборты и табу всегда давали древнему человеку способ регулировать рост популяции».
Ограничение численности популяции за счет увеличения смертности в примитивном сообществе представляет собой регуляторный процесс, зависящий от плотности и так или иначе связанный с тенденцией к перенаселенности. Антисанитарные условия жизни и другие следствия перенаселенности особенно неблагоприятно отражаются на грудных детях и детях младшего возраста. Пока женщина работает, дети остаются без надлежащего ухода. Ребенок, которого только что отняли от груди, очевидно, более чувствителен к последствиям плохого ухода, чем более старшие дети. Примитивное сообщество постоянно нуждается в рабочей силе, и связанный с этим недостаточный уход за детьми превращается в фактор, действующий в направлении, противоположном увеличению численности популяции. Болезни, поражающие здоровых членов сообщества, наносят ущерб деятельности, связанной с добыванием пищи. Престарелые и слабые здоровьем перестают участвовать в добывании пищи, но продолжают за нее конкурировать, испытывая при этом большие лишения.
В результате рост популяции следует рассматривать как процесс, связанный с равновесием трех демографических факторов — плодовитости, смертности и миграции (более подробное обсуждение см. Nag, 1973). Хорошо документированные данные подтверждают
Глава 29. Стабильность популяций
589
факт миграции в малых масштабах, как, например, подразделение простых земледельческих сообществ па локальные группы; имеются также данные, свидетельствующие о больших переселениях популяций (и те и другие миграции следует отличать от передвижений групп, связанных с вырубкой и выжиганием растительности под посевы).
Как утверждает Карр-Саундерс (Carr-Saunders), рост популяции, если он должен соответствовать ресурсам сообщества, с необходимостью требует продуманной регуляции численности. Эта регуляция (наряду с действием естественных факторов) должна поддерживать оптимальную плотность, т. е. плотность, которая гарантирует уровень жизни, соответствующий технологии и ресурсам данного сообщества.
Теория «оптимума» подчеркивает роль процессов «естественной» регуляции в человеческих экосистемах, связанных с действием зависящих от плотности регулирующих факторов. С одной стороны, имеется тенденция к росту популяции, который обеспечил бы реальное увеличение дохода от разных видов обслуживания и количества товаров на душу населения. С другой стороны, существует стремление уменьшить прибыль, поскольку «при данном объеме естественных ресурсов и (капитального) оборудования и существующем уровне техники любое увеличение числа работников в популяции ведет к уменьшению других величин, от которых зависит уровень производства (например, земли), а следовательно, к уменьшению выработки на одного работника». Действие этих противоположных сил приводит в конечном счете к тому, что «при определенном для данного времени уровне знаний и других постоянных условиях достигается состояние, отвечающее получению максимальной прибыли: количество затрачиваемого труда таково, что и увеличение и уменьшение его ведет к снижению прибыли».
Технический прогресс делает возможным достижение нового «оптимального» уровня, однако эффективная деятельность может быть связана с выбором различных путей, а возможно, и с периодом, в течение которого не удается избежать перенаселенности. Хорошей иллюстрацией может служить производство риса. Расширение и усовершенствование действующей ирригационной системы может оказаться на данном этапе более выгодным, чем создание новых оросительных каналов. Однако, выбирая первый путь, мы не гарантируем прогресс в отдаленном будущем, и в конечном счете перед нами встанут новые проблемы. Идея о том, что в примитивных сообществах численность населения близка к оптимальной, хотя этому «оптимуму» и угрожает «давление плодовитости», предполагает наличие общественных мер по ограничению рождаемости. При этом «оптимальная» ситуация коренным образом отличается от пассивных мальтузианских условий. Пример такой об
590
Часть V. Экология человека
щественной регуляции мы находим у аборигенов Австралии и у других сообществ, ведущих собирательное хозяйство: кочевая жизнь в поисках воды и пищи связана с дальними переходами и матерям трудно переносить с собой более одного ребенка. Вследствие этого племя, переходя на новую стоянку, нередко убивает или бросает новорожденных. Численность популяции в первобытных земледельческих сообществах определяется максимальным количеством рабочей силы, которая необходима во время
Таблица 29.3
Меры по регуляции численности в различных популяциях
Детоубийство Аборт Ограничение половых сношений
Собиратели и охотники
Австралийские племена + + —
Тасманийцы + + —
Бушмены + — —
Американские индейцы + + +
Земледельческие народы,
Американские индейцы + + +
Африканцы + + +
Народы Океании + + +
*) ( + ) —зарегистрированные случаи, (—) —не зарегистрированные случаи.
сева и уборки урожая. Общество должно располагать достаточным числом рабочих для выполнения трудоемких земледельческих работ в короткие сроки. В оставшееся время года часто имеют место безработица и перенаселенность, связанные с постепенным истощением пищевых запасов. Тем не менее плотность популяции в целом соответствует средней равновесной плотности в пределах года.
Некоторые данные, собранные Карр-Саундерсом, свидетельствуют о существовании у различных народов продуманных мероприятий по регуляции численности (табл. 29.3).
Во многих других сообществах часто принимаются менее крутые меры, например отсрочка брака, узаконенная различными традициями. Интересно, что вступление в брак в возрасте до 17 лет в конечном счете ведет скорее к снижению, чем к увеличению общей плодовитости (статистические данные для Индии). Имеются
Глава 29. Стабильность популяций 591
также сведения о том, что способность к деторождению не всегда появляется одновременно с началом менструаций (Montagu). Удлинение периода кормления грудью может отсрочить зачатие до года. В некоторых сообществах практикуется воздержание от половых сношений в послеродовом периоде. Широко распространенной мерой против зачатия был и остается Coitus interruptus. В основе снижения плодовитости в развитых странах были такие меры, как воздержание и определенный ритм в половых сношениях; в настоящее время стали прибегать главным образом к механическим и химическим противозачаточным средствам.
Возможно, Карр-Саундерс преувеличивает необходимость общественных мер по ограничению рождаемости; ведь помимо высокой детской смертности следует учитывать также действие биологических факторов, которые могут приводить к бесплодию. К таким факторам относятся хронические болезни — глистные инвазии, малярия и венерические болезни. На жизнеспособность плода в утробе матери в значительной мере влияет характер ее питания и особенно периоды голодания.
Описаны также случаи резкого снижения плодовитости у некоторой части женщин в популяции, не связанной с умышленным вмешательством. Возможная, но не единственная причина этого — венерические заболевания. Гудхарт предположил, что в издавна существующих примитивных сообществах имел место отбор на низкую плодовитость и что существует равновесие между линиями с высокой и низкой плодовитостью. В группах охотников и собирателей малая численность потомства дает определенное преимущество; в сообществах, в которых имеет место высокая смертность (например, в крестьянских общинах с высокой плотностью населения, где скученность способствует распространению инфекций и где многочисленная семья имеет преимущества в сельскохозяйственном производстве), это соотношение изменяется в пользу линий с высокой плодовитостью. Гипотеза о преобладающей роли «генетических» факторов по сравнению с социальными (теория «оптимума») или факторами, обусловленными воздействием среды (мальтузианство), требует доказательств.
Часто полагают, что увеличение численности населения — нормальный признак развивающегося человеческого сообщества, в действительности же на протяжении почти всей истории человечества численность населения оставалась стабильной. Согласно теории «оптимума», численность населения может измениться с изменением характера общественной организации и ростом производительных сил; примером может служить современный период развития человечества. «Проблемы человеческой популяции в прошлом решались бессознательно или почти бессознательно. Сейчас для человечества настало время сознательно найти наилучшее решение!» — пишет Карр-Саундерс.
592
Часть V. Экология человека
Экологические показатели
Уровень жизни того или иного человеческого сообщества зависит от способа, с помощью которого данное сообщество достигает равновесия в определенных экологических условиях. Это равновесие может быть достигнуто за счет более высокой общей смертности или более высокой заболеваемости, а также за счет более напряженной работы, ухудшения здоровья или недостатка материальных благ. Для характеристики равновесия можно пользоваться различными критериями (в том числе такими, которые определяют уровень потребления, работоспособность, энергию или денежный доход, приходящийся на душу населения), а также различными демографическими показателями. Особого упоминания заслуживают два показателя — детская смертность и средняя продолжительность жизни.
Детская смертность. Наиболее важным показателем эффективности экологического контроля может служить детская смертность (смертность в первый год жизни на каждую 1000 новорожденных), поскольку она отражает отрицательные стороны общественного уклада — недоедание, перенаселенность, отсутствие необходимых санитарных условий. Это особенно чувствительный показатель, поскольку снижение детской смертности не связано непосредственно с уменьшением смертности в старших возрастных группах. В конце XIX в. в Великобритании смертность детей на первом году жизни была еще очень высокой (150 на 1000 новорожденны!), но смертность детей старше года неуклонно снижалась. Показатели детской смертности, приведенные в табл. 29.4, отражают социально-экономические условия жизни, в том числе условия питания. Во всех примитивных сообществах детская смертность неизменно высока, и, как уже упоминалось, она служит одним из факторов регуляции численности в экосистемах с ограниченной емкостью.
Средняя продолжительность жизни. Этот показатель для человеческого общества каменного века и более позднего доисторического периода определяется по скелетным остаткам. При определении возраста учитывают: 1) состояние зубного аппарата, 2) степень оссификации костей запястья, 3) степень заращения эпифизарных щелей трубчатых костей, 4) степень заращения черепных швов. Изученный скелетный материал распределяют по 4—5 возрастным группам, разграничивая их следующим образом: дети (от 0 до 12—13 лет), подростки и юноши (от 12—13 лет до 21 года), взрослые (от 21 до 40 лет), люди среднего возраста (от 40 до 59 лет), пожилые (60 лет и старше). По имеющимся данным можно примерно установить, что почти на всем протяжении истории человечества смерть наступала в значительно более раннем возрасте, чем в наше время. Не более 10% населения переживало возраст
Глава 29. Стабильность популяций
593
40 лет, и лишь 50% достигало двадцатилетнего возраста. Согласно Валлуа, из 40 известных науке неандертальцев лишь один был старше 50 лет; из 76 Homo sapiens, живших в эпоху верхнего палеолита и найденных в Европе и Азии, только 2 умерли в возрасте старше 50 лет; из 65 индивидуумов, относящихся к мезолиту, достигли 50 лет также только 2; из 94 неолитических скелетов, найденных в Силезии, лишь 4 принадлежали людям старше 50 лет. Эти цифры, по-видимому, характерны также для всех современных или живших в недалеком прошлом сообществ собирателей, охотников, первобытных земледельцев. В доземледельческой группе индейцев кноль (долина реки Пекос) смертность в возрасте до 21 года достигала 57% — примерно так же, как в Древней Греции. У индейцев, проживающих на Великих Равнинах, смертность в детском и юношеском возрасте очень высока (мохаве — 93%, дакота — 65%, семинол — 44%). Сходные цифры можно привести для африканских народов.
Известны и отдельные исключения: 20% черепов гуанчей принадлежало людям, умершим в возрасте старше 50 лет; в Меланезии (Новая Ирландия) свыше 75% людей доживало до брачного возраста.
Энжел (Angel, 1940), изучавший черепа древних греков, установил, что в древности продолжительность жизни неуклонно, хотя и очень медленно, увеличивалась. Это можно объяснить улучшением питания и условий жизни и развитием медицины, что, по данным археологов, имело место в Греции.
Пирсон (Pearson) определял по мумиям среднюю продолжительность жизни в Египте в период Римской империи, т. е. примерно 2000 лет назад. Он построил кривую, характеризующую продолжительность предстоящей жизни в зависимости от возраста; для новорожденного эта величина составила 22 года.
Продолжительность жизни в античные времена в среднем составляла 20—30 лет; для англичан, живших в XIII в.,—35 лет.
В последующие 5 столетий средняя продолжительность жизни изменилась очень незначительно. Таблица, которую составил астроном Галлей (Halley) на основании записей, сделанных в Бре-славле в 1687—1691 гг., показывает, что средняя продолжительность жизни была 33,5 года. Величину 35,5 получил Уигглесуорз (Wigglesworth), изучая статистику смертности в Массачусетсе и Нью-Гемпшире до 1789 г. Постепенное повышение уровня жизни в последние два столетия привело к увеличению средней продолжительности жизни. Из таблиц, составленных статистиком У. Фарром (W. Farr) для Англии и Уэльса и относящихся к 1838—1854 гг., следует, что средняя продолжительность жизни в то время была равна 40,9 года. В последующие годы с развитием медицины и гигиены средняя продолжительность жизни увеличилась до 49,2 года (1900—1902 гг.). В США в 1945 г. средняя продолжи
594
Часть V. Экология человека
тельность жизни достигла 65,8 года, т. е. увеличилась за 5 десятилетий примерно на 16 лет. В настоящее время средняя продолжительность жизни в странах Европы и в США достигла стабильного уровня — около 70 лет.
Несмотря на все сказанное, в нашу эпоху остаются еще большие различия в цифрах, характеризующих детскую смертность и среднюю продолжительность жизни. Из табл. 29.4 видно, что эти цифры тесно связаны с социально-экономическими условиями. Об уровне жизни можно судить по доле углеводного компонента в пищевом рационе. Страны с высоким потреблением белка и сме-
Таблица 29.4
Некоторые демографические данные за 1938 г. (приближенные)
Доля пищевого рациона, приходящаяся на углеводы (картофель, злаки), % Средняя продолжительность жизни, годы Детская смертность
мужчины женщины
Австралия 63,5 67,1 38
Новая Зеландия 65,0 67,9 50
США (белое население) 30—39 60,6 64,5 51
Канада 59,0 60,7 63
Великобритания 60,2 64,4 55
Норвегия 61,0 63,8 37
Г ермания 40—49 59,9 62,8 60
Нидерланды 65,1 66,4 37
Венгрия 48,3 51,3 131
Чехословакия 51,9 55,2 121
Бельгия 56,0 59,8 73
Франция 54,3 59,0 66
Болгария 70 79 45,9 46,6 144
Польша 48,2 51,4 140
Япония 46,9 49,6 144
Индия 26,9 26,6 167
шапной диетой характеризуются более высокими значениями экологических индексов, определяющих уровень жизни.
Экология в наши дни
Экология сегодняшнего дня — это всемирная экология. Интенсивные взаимосвязи в мире поставили перед человечеством такие проблемы, которые невозможно решить в узких территориальных рамках. К этим проблемам следует отнести в первую очередь давление в популяции, сохранение материальных ресур
Глава 29. Стабильность популяций
595
сов и источников энергии, их лучшее распределение в мире, размещение радиоактивных и иных отходов, организацию транспорта и туризма во всемирном масштабе, «расовые» проблемы, устранение войн и т. п.
Список литературы
Allison А. С. (1960). Abnormal haemoglobins and erythrocyte enzyme-deficiency traits. In Genetical variations in human populations (ed. G. A. Harrison). Pergamon, Oxford.
Angel J. L. (1940). The length of life in ancient Greece. J. Gerontol. 2, 18—24.
Baker P. and Weiner J. S. (eds.) (1966). The biology of human adaptability. Clarendon Press, Oxford.
Barmcot N. A. (1959). Climatic factors in the evolution of human populations. Cold Spring Harb. Symp. quant. Biol. 24, 115—129.
Birdsell J. B. (1953). Some environmental and cultural factors influencing the structuring of Australian aboriginal populations. Amer. Naturalist. 87, 171.
Blum H. F. (1945). The physiological effects of sunlight on man. Physiol. Rev. 25, 483.
Brass W. (ed.) (1971). Biological aspects of demography. Symposium No. 10. Society for the Study of Human Biology. Taylor and Francis, London.
Cannon W. B. (1932). The wisdom of the body. W. W. Norton, New York.
Carr-Saunders A. M. (1922). The population problem: a study in human evolution. Clarendon Press, Oxford.
Clark C. (1958). World population. Nature, Lond. 181, 1235.
Clark G. (1947). Archaeology and society. (2nd ed) Methuen, London.
Clegg E. J., Harris G. A. and Baker P. T. (1970). The impact of high altitudes on human population. Human Biol. 42, 486—578.
Cleland J. B. (1928). Disease amongst the Australian aborigines. J. trop. Med. Hyg. 31, 53-327.
Coon S. C., Garn S. M. and Birdsell J. B. (1950). Races, a study of the problems of race formation in man. Thomas, Springfield, Illinois.
Damon A. (ed.) (1975). Physiological anthropology. Oxford University Press, New York and London.
Davenport С. B. (1945). The dietaries of primitive peoples. Amer. J. phys. An-throp. 47, 60—82.
Edholrn O. G. (1967). The biology of work. Weidenfeld and Nicolson, London.
Elton C. (1949). Animal ecology. Sidgwick and Jackson, London.
Forde C. D. (1949). Habitat, economy and society. Methuen, London.
Fox R. H. (1953). A study of energy expenditure of Africans engaged in various rural activities. Ph. D. thesis, University of London.
GarlicK J. P. and Keay R. W. J. (eds.) (1975). Human ecology in the tropics. Symposium No. 16. Society for the Study of Human Biology, Taylor and Francis, London.
Garn S. M. (1961). Human races. Thomas, Springfield, Illinois.
Goodhart С. B. (1956). World population growth and its regulation by natural means. Nature, Lond. 178, 561—565.
Harrison G. A. and Walsh R. J. (eds.) (1974). A discussion on human adaptability in a tropical ecosystem. An I. В. P. investigation of two New Guinea communities. Phil. Trans. Roy. Soc. B. 268, 221—400.
Hawley A. H. (1950). Human ecology: a theory of community structure. The Ronald Press, New York.
James, Preston E. (1959). A geography of man. (2nd ed.). Ginn, London.
Katz S. H. (ed.) (1947). Biological anthropology, readings from Scientific American, W. H. Freeman.
596
Часть V. Экология человека
Lee D. Н. К. (1951). Thoughts on housing for the humid tropics. Geog. Rev. 41, 124—147.
Miller, Austin A. (1946). Climatology (4th ed). Methuen, London.
Moni N. (1973). Anthropology and population. Pop. Studies. 27, 59—68.
Montagu, Ashley M. F. (1946). Adolescent sterility. Thomas, Springfield, Illinois.
Motulsky A. G. (1958). Metabolic polymorphisms and the role of infectious diseases in human evolution. Human Biol. 30, 43—72.
Newburgh L. H. (ed.) (1949). Physiology of heat regulation and the science of clothing. Saunders, Philadelphia.
Nicholson A. J. (1933). lhe balance of animal populations. J. Anim. Ecol. 2, 132-178.
Passmore R. and Durnin J. V. G. (1955). Human energy expenditure. Physiol. Rev. 35, 801—839.
Platt B. S. (1946). The colonial nutrition problem. Proc. Nutr. Soc. 5, 1—17.
Pugh L. G. С. E. (1965). High altitudes. Chapter 9 in The physiology of human survival (eds. O. G. Edholm and A. L. Bacharach). Academic Press, London.
Richards A. I. (1932). Hunger and work in a savage tribe. George Routledge, London.
Roberts D. F. (1953). Body weight, race and climate. Amer. J. phys. Anthrop. II, 533—558.
Robinson S. (1942). The effect of body size upon energy exchanges in work. Amer. J. Physiol. 136, 363.
Schreider E. (1951). Race, constitution, thermolyse. Revue scient. Paris, 89, 110-119.
Sears P. B. (1959). The ecology of man. Smithsonian Report for 1958, pp. 375— 398.
Sinclair H. M. (1953). The diet of Canadian Indians and Eskimos. Proc. Nutr. Soc. 12, 69.
Solomon M. E. (1949). The natural control of animal populations. J. Anim. Ecol. 18, 1—35.
Weiner J. S. (1973). The natural history of man. Anchor Books, Doubleday.
Weiner J. S. (ed.) (1976). Physiological variability and its genetic basis. Symposium No. ??? Society for the Study of Human Biology. Taylor and Francis, London.
Wulsin F. R. (1949). Adaptations to climate among non-European peoples. In Physiology of heat regulation and the science of clothing, (ed. L. H. Newburgh). Saunders, Philadelphia.
Wyndham С. H. et al. (1964). Heat reactions of Caucasians and Bantu in South Africa. J. Appl. Physiology. 19, 598—606.
Young J. Z. (1971). An introduction to the study of man. Clarendon Piess, Oxford.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аборигены австралийские 245, 247, 262, 265, 274, 303, 305, 308, 312, 319, 327, 331, 343, 361, 495, 514, 528, 539, 549, 553, 574, 577
Австралопитек 79, 120, 121, 125
— безыменные кости 62
— определение Ле Гро Кларка 80, 82
— череп 67
Агаптоглобулинемия 303
Агглютинация 309
Агглютинины 130
Адапис 53
Адаптация 472
— климатическая см. Климатическая адаптация
— на эмбриональной стадии 18
— процессы 373
— специфичность 15
— ценогенетическая 18
Адаптивная радиация 20
Адаптивный признак 218
Аденилаткиназа 298
Аденозиндезаминаза 300
Аденин 176, 177
Айны 549
Акклиматизация к высоким температурам 524
— — холоду 526
Алкалоз 558
Алкаптонурия 144
Аллели 135, 163
— и естественный отбор 218, 220
Аллелизм 136
Аллеломорфы 135, 154
Аллена правило 532, 536, 538
Аллометрические соотношения 24
Аллопатрические популяции 20
Альбинизм 142,143, 145, 195, 356, 545
Альбумин 305
— Mexico 306
Альтруистическое поведение 219
Амавротическая идиотия 356
Американские индейцы 245, 262, 271, 274, 290, 308, 319, 325, 331, 333, 347, 359, 362, 417, 476, 528, 539 — негры 244, 249, 262, 290, 329, 331, 354, 357, 363, 417, 535, 553, 574
Анагенез 19
Андрогены 411
Анемия Кули 284
Анкилостомоз 497
Аноксия 558
Анонихия 141
Антигены 130, 309, 310
— Lewis 312, 313
— Xg 333, 334
ABH-антигены 312—314
Антитела 130
Антитрипсин 306
Антропоиды 43
— адаптивная радиация 58
— генеалогическая схема 55
Антропометрия 237, 238
— головы 246, 249
— многомерный анализ 253
Анэнцефалия 357, 574
Арахнодактилия 166
Ассортативное скрещивание 196
Атеросклероз 569
Атлетический тип Кречмера 444
Аутбридинг 196
Аутосомные гены 153
Аутосомы 132, 147
Аутэкология 473
Ахондроплазия 141, 210, 212
Бабуины 39
Базальноклеточная карцинома 552
Бамбути 241, 242
Банту и рак 574—575
Барабанные косточки 16, 17
Баски 323, 346
Бэра закон 18
Безыменные кости приматов 62
Белки, аминокислотная последовательность 69
— и мутации 207
Белки, модель Жакоба — Моно 179—
180
— основные свойства 276
— синтез 177, 178
— суточное потребление в разных популяциях 491, 492, 494
598
Л редметный указатель
Белковая недостаточность 494
Бенс-Джонса белок 340
Бергмана правило 243, 532, 536, 538
Бешенство 566
Биологическая изменчивость 230
Близкородственные браки 197, 214, 232
Близнецовый метод 183
Близнецы 184, 354
— болезни 572, 576
— двуяйцовые (пеидентичные) 184
— дискордантные 184
— зиготность 184, 185
— конкордантные 184
— однояйцовые 184
— частота рождения 188, 354—355
Блуменбаха классификация 233
Болезни 564— 579
— влияние факторов внешней среды 564
— инфекционные см. Инфекционные болезни
— неинфекциопные 574
— связанные с алиментарной недостаточностью 501
— экологические аспекты 564—568
Болезнь Кристмаса 151
Браки 232
— неизбирательные 191—195
— системы 202
Брахидактилия 174
Бронтозавры 21
Бруцеллез 566
Брюхоногие моллюски 25
Бушмены 243, 271, 524, 549, 574
Вегетативное размножение 14
Венерические болезни 591
Вид, определение по Майру 20
Виллафранкская фауна 73
Виллафранкский ярус 73
Висцеротония 465, 466
Витамины 499
Витамин С, недостаточность 499
— D, недостаточность 263, 499, 506
Внешняя среда 181—189
Вода, потребности организма 495
Возвратное скрещивание 135
— — соотношение полов 157
Возраст, определение по степени развития 391—407
— — — форме тела 401
Волосы, цвет 263—265
— — наследование 265
Воспроизведение потомства 149
Врожденные уродства 357
Выражение гена 174
Высокогорные условия, акклиматизация 557, 558
— — и популяции 560
— — реакция организма 558
Галаго, череп 49
Гамадрилы 112—115
Гаметы 130, 131, 156, 158, 163
— рекомбинантные, нерекомбинапт-ные 158—159
Га'плоидный набор хромосом 132
Гаплотипы 242—345
Гаптоглобин, распространение по земному шару 304
Гауссово распределение 168, 240
Геккеля закон 17
Гельминты 566
Гемагглютипоюны 130
Гемпзиготпость 150
Гемоглобин 276, 279, 280
— варианты 280—286
— географическое распределение 286
— Л 277—280, 284
— С 281, 284, 286
— Г> 281, 287
— Е 281, 284, 286
— F 277, 280, 284
— G 281
— Н 285
— HbBorts 285
— К 287
— М 281
— О 287
— S (серповидноклеточности) 276— 278, 284, 286
Гемофилия 151—153
Ген(ы) 134, 135
— взаимодействие локусов 174
— действие разных факторов 210
— доминантные 136, 175
— — фиксированное состояние 219—220
— идентичные по происхождению 198—200
— — — свойствам 198
— и зрение 167
— — цвет кожи 171, 172
— метод пар сибсов Пенроуза 161
— нейтральные 217
— ограниченные возрастом 416
— плейотропизм 467
— природа 176—180
— рецессивные 136, 175
Предметный указатель
599
— — скорость элиминации 219
— серповидноклеточности 569
— — и малярия 289—291
— способность к самовоспроизведению 211
— структурные 179
— сцепленные 158—161, 163, 164
— частота мутаций 209—212
— человека 160
Гены-модификаторы 175
Гены-операторы 179
Гены-регуляторы 179
Генетическая дифференциация 20 — инерция 14, 22, 26 — конституция 147
Генетический груз 214
— фонд 190
Генные мутации 14, 207, 209, 210, 213
Генный комплекс 173—176
Геноклины 206
Генотип 135, 136, 181, 191
— относительная приспособленность 221
— фенотип, взаимодействие 218
Геохронология 74
Гетерогаметный пол 147
Гетерозигота 135, 136, 191
Гетерозиготность 196, 197, 199
Гетерозис 204
Гетероплоидия 208
Гиббон 39, 57, 63
— череп 49
Гибридизация 205, 206
Гибридная мощность 204
Гибриды, жизнеспособность 204
Гиперморфоз 17
— и эволюция 61
Гиперостоз 572
Гипертония 577, 578
Гипогенитализм 166
Гипоталамус 409, 410, 412
Гинсодонтия 24
Гистогенез 369
Глазные инфекции 566
у-Глобулин 568
Глухонемота 144
Глюкозо-6-фосфат-дегидрогеназа и сцепленные гены 164
— недостаточность 291—294, 569, 571
Головной индекс 247
Гомеорез 382
Гомеостаз 189, 474, 475
Гоминиды, генеалогическая Схема 84
— определение Ле Гро Кларка 66 — развитие 71
Гоминиды ранние, экология 120 — экология и организация сообществ 120
Гомогаметный пол 147
Гомозиготность 196, 197, 200, 201, 203
Гомозиготный 135, 136, 191
Гомойотермия 519, 521
Гомпертца кривая 373
Гонадотропины 166
Гонады 146
Горилла 39
— скелет стопы 63
— череп 67
Гормоны роста 409—411
— щитовидной железы 411
Групповой отбор 219
Группы крови 309—347
— — Льюис 174
----АВО 136, 164, 174, 311
— — — географическое распределение 314—319
— — — и естественный отбор 320
----Duffy 164, 331
— — Lutheran 164
----MN 130—132, 194
----MNS 163
Гуанин 176, 177
Гуминовые кислоты 75
Дарвиновы вьюрки 21
Дауна синдром 209, 384
Двойная спираль 177
Двуполость, эволюционные преимущества 203
Действие генов 166, 167
«Дейтеранопическая» слепота 350
Делеция 207, 208
Дем 473
Дендрограмма 258
Дендрохронология 77
Дерматоглифические узоры 208,
269-271
Деторождение 591
Детская смертность 592
Дефекта ногтей синдром 164
Джетисские формы 89, 91
Диабет 576
— и телосложение 464
Диверсифицирующий отбор 226
Диета и обмен веществ 493
— эскимосов 491, 492, 503, 504
Дизиготные близнецы 184
Динозавры 21
Диплоидный набор хромосом 131, 132
600
Предметный указатель
Дискриминантные функции 256, 257
Дисплазия 448
Дисруптивный отбор 226
Дифференциальная плодовитость и выживаемость 13
Длина тела 238
— — методы измерения 238
— — статистическое распределение значений 239
Долгопят 38, 45
— череп 49
Доминантное наследование 138
Доминантный ген 136, 175
— — фиксированное состояние 219—220
Донеолитический человек 505
ДНК 176-179
ДНК-код 178
Древние люди эпохи Граветт 506
Дрейф генов 215—217, 226
Дунайская культура 511
Дупликация 209
Дюбуа формула 489
Естественный отбор 13, 174—175, 218—223
Жакоба — Моно модель синтеза белка 179—180
Железо, потребность организма 495, 496
Жилища и климат 548, 549
Жировая ткань 250
Жиры, методы измерения 250
— потребление в холодных условиях 527
— — и коронарная болезнь 576
— суточное потребление в разных популяциях 491, 492
Загрязнители 578
Законы Менделя 129—145
Зиготность, тесты 185, 186
Зиготы 147
Змеи 21
Зрелость зубная 401
— отношение между разными оценками 402
— оценки 402
— скелетная 400
— физиологическая, половые различия 404
— эмоциональное развитие 437
Зубная зрелость 401
— формула древних млекопитающих 41
— — полуобезьян 42
Зубы, билофодонтная форма 56
— гипсодонтия 24
Иглокожие 18
Игрунка 39
Иерархия у павианов 109, 110
Изменчивая экспрессивность действия гена 174
Изоляты 190, 203, 217, 356, 427
Изониазид, инактивация 360
Иммигранты, антропометрия 251— 253
Иммунитет 567, 568
Иммуноглобулины 337
Инбридинг 196—204
— коэффициент 199—201
Инверсии 209
Индейцы племени пуэбло 75
— — яруро 511
Индукторы 180
Инерция генетическая 22
— селекционная 22
Интеллект, наследуемость 185
— развитие 436
Интеркросс 135
Инфекционные болезни 565—569
— — биологические реакции 567— 569
— — генетические факторы 573 — — и естественный отбор 569 Иод, потребность организма 495, 498
Ионизирующее излучение 555—557 — — биологические последствия облучения 555
— — действие на частоту мутаций 213
Ирландский лось 24
Ископаемые остатки из Фертесзол-лоша 89
— — найденные на Яве, сравнение с пекинскими формами 89
— — пещеры Чжоу коу-дянь 89
— формы, открытые Лики 80
Ихтиоз врожденный 144
Ихтиозавры 21
Ишихара таблицы 352
Предметный указатель
601
Кальций, потребности организма 495, 496
Канализирование 381—383
Каннибализм 505, 516
Капуцин, скелет стопы 63
— череп 49
Катаракта 566
17-кетостероиды 462
Кикуйю 512, 513
Кислая фосфатаза 295
Кистеперые рыбы 16, 2?
Киты 23
Кладогенез 19, 22, 29
Кладогенетическое родство 29
Классификация Виолы 443
Клейнфельтера синдром 147, 148, 334, 406
Климат 519
— влияние на состояние здоровья 545
— и одежда 550
— — физическая работа 543
— постройка жилья 548, 549
— Сомали 566
Климатическая адаптация 519—
563
— — в высокогорных условиях 557
— — генетические и негенетические факторы 537
— — зависимость физических признаков организма 529
— — зоны комфорта 540
— — и работоспособность 560
— — культурные аспекты 547— 550
— — примеры из антропологии 532-545
— — характер роста 536
Климатические зоны 541, 542, 546
Клины 206, 227, 234
Кодоминантные признаки 141
Кодон 178
Кожа, цвет 259—263
— — естественный отбор 262
— — методы измерения 260
— — наследование 261
— — статистическое распределение 261
— функция пигмента 262
Колбочки 44
Конвергентная эволюция 22
Конвергенция 22—23
Копытные 26
— происхождение 51
Кормический индекс 245
Коронарная болезнь 576
Коронаротромбоз и физиология 464
Корпускулярная наследственность 130
Корь 572
Космическое излучение 555
Кошачий крик (синдром «cri du chat») 208
Коэффициент интеллекта 436
— отбора 221
— сходства, метод 162
— — рас 254
— теплоотдачи путем испарения 531
— — — конвекции 531
Красная волчанка 347
Краснуха 181
Креатинин 462
Креодонты 51
Крестец 61
Крестьянское сообщество, питание 506—511
Кривая роста 366, 371—374, 376, 390—406
Кровяное давление 577, 578,
— — и физиология 463
Крокодилы 21
Кроссинговер 158, 159
1-Ксилокетозурия 356
Культура дунайская 511
— Капси 506
— мадленская 506
— ориньякская 506
Кумбса тест 308
Куриная слепота 141
Лактатдегидрогеназа (ЛДГ) 299
Ледники Альп 71
Лейдига клетки 408, 409
Лекарственные вещества 579
Лектины 310
Лемуроиды 54
Лемуры 21, 37, 38
— и преадаптация 25
Ленточные глисты 77
Лептосомный тип Кречмера 444
Лимфогранулематоз 347
Лицо, размеры 249
Локус 135
Лонгитип Виолы 443
Лори 38
— и преадаптация 25
Лоуренса — Муна — Бидла синдром 155, 166
602
Предметный указатель
Мадрасцы 512
Мадурская стопа 566
Макаки 46
Макаки-резусы 108
Максимальная аэробная мощность 561-563
Мальпигиев слой 259
Мальтузианство 586—588
Малярия 288—291, 569
Мамонт 74, 76
Маниакально-депрессивный пси-
хоз и телосложение 464
Масаи 512
Мезоморфия 445
Мейоз, 132 148
Меланин 259—262, 265, 551
Меланома 553
Меланоциты 259
Менделевская популяция 190
Менделя второй закон 157
— первый закон 130
Менструация первая 395
— — генетические факторы 414
— — и расы 417
— — — социально-экономические факторы 422
— эпохальный сдвиг 426, 427
Метгемоглобин 281
Метод анализа главных компонентов 256
— коэффициента сходства 162
— многомерного шкалирования 255
— пар сибсов Пенроуза 161
— рацемизация аминокислот 79
Микроорганизмы 565
Минеральные соли, потребности организма 495
Многомерный анализ 253
— — и сравнение популяций 253
Мозговая коробка, высота 249
— — емкость 248
Монголоидный тип лица 536
Монголоидная раса 233
Монозиготные близнецы 184
Морфогенез 370
Морфогенотип 450
Мутантные гены 20, 556, 557
— — способность к самовоспроизведению 211
Мутации 206—214
— генные см. Генные мутации
— действие разных факторов 210
— радиационные 555, 556
— соматические 184, 187
— хромосомные 14
— частота 209—212, 556
Муха цеце 565
Мыпщы, развитие 250
Насекомые, эволюция 18
Население, плотность 580
— распределение на земном шаре 582
— Тасмании 262
— ускоренный рост численности 582
Наследование доминантное 138
— ограниченное полом 155
— рецессивное 142
• — сцепленное с полом 150
— Х-сцепленное 150—154
— Y-сцепленное 152
Наследственность и среда в антропометрии 250—253
— полигенная 168—173, 182
Наследуемость 182, 183, 251
Натрий хлористый, потребности ор-
ганизма 495, 498
Неандертальцы 67, 93, 99, 102, 105
— пища 505
— речевые навыки 125
Неврозы и телосложение 464
Негры 20, 261, 363, 417, 513, 529, 553, 577
Независимое распределение 156
Пеизбирательные браки 191—195
Пеинфекционные болезни 575
Нейтральные гены 217
Некролемур 53
Неолит 511
Неотения 18
Неполная пенетрантность 174
Пормотип Виолы 443
Нотарктус 53, 63
Нуль-гипотеза 242
Обезьяны Нового Света 23, 38, 42, 53, 58
— Старого Света 23, 41, 53, 58, 66
Оболочки плода, эволюция 18
Общая плодовитость 586
Озон 551
Окружающая среда 472
Оледенение, периоды 71
Олигофрения 155
Онтогенез 17, 180
Оперон 179
Опоссум 21, 26, 27
Организация сообществ раннпх гоминид 120
Ортогенез 24
Предметный указатель
603
Орудия типа Сандиа 75
— — Фолсом 75
Основной обмен, действие низкой температуры 527
— — энергия 488
Остатки из пещеры Амуд 98, 99
— — — Араго 101, 105
— — — Схул на горе Кармел 102, 105
— найденные в Вистонпце 506
— — — Джебель Кафза 102, 105
— — — Канаме 103
— — — Канжери 103
— — — Комб-Капелль 93
— — — Пржедмосте 506
— — — Обри Суард 101, 105
— — — Пильтдаун 27, 76, 105
— — — Сутре 506
— — — Штейпгейме 101
Остеология 238
Отбор балансирующий 226
— диверсифицирующий 226
— нормализующий 226
Отцовское родство 29
Отложения Санта-Фе 75
Павианы, иерархия 109, НО
— поведение ПО, 111, 122
— простые 108—112
Палеонтологические виды, распознавание 20
Палочки 44
Панмиксия 191, 206
Параллелизм 22—23
Паранойя и телосложение 464
Патогенные организмы 565, 567,
568
Пекинские формы 89
Пенроуза метод 161
Перегрев, реакции ораганизма 522
Пещерные отложения Альбукерк 75
— — Ботсваны 79
— — Трансвааля 79
Пикнический тип Кречмера 444
Пигментный ретинит 167
Питание, зависимость от среды обитания 501—504
— и болезни 576
— и обмен веществ 493
— — физические различия 512
— — энергия 488—493
— насекомыми 515
— социальные аспекты 514
Пищевые вещества 493—499
— — в диете крестьянских сообществ 508
Пища донеолитического человека 505
— кикуйю 512, 513
— крестьянского сообщества 506— 511
— мадрасцев 512
— масаи 512
— сикхов 512
Пластинчатожаберные моллюски 25
Плезиадапис 53
Плейотропизм 166
Плейотропные гены 467
Племя Андаманских островов 243
Плоскоклеточная карцинома 552
Пневмония 567
Погода 519
Подкожный жир и температурная среда 526
Подкожный жировой слой и теплообмен 532
— — — кривая роста 379
Подростковый период 411
Пол, генетическое определение 146
Полигонная наследственность 168— 173, 182
Полидактилия 166, 174
Полиморфизм 223—226
Полиомиелит 572
Полиплоидия 14, 208
Полисомия 208
Политипизм 226—227
Полуобезьяны 38, 44—46
— генеалогическая схема 52
— происхождение 51
— череп 53
Понгиды 62, 121
— и гоминиды по Ле Гро Кларку 66 Поперечные исследования 373—376 Популяции 190, 233—236
— биохимическая изменчивость 275
— стабильность 580—594
— филогенетическое дерево 335, 336
— численность, регуляция 582, 587, 590—591
— — теория «оптимума» 589
— — увеличение 591
— эффективная численность 216
Потовые железы 522—524, 552
Поток генов 206, 227, 232
Потоотделение 522, 524
Преадаптация 25
Предшествующие формы 15
Приматы, безыменные кости 62
— зубы 41, 42
— классификация по Симонсу 34—37
— — таблица 34
— кривые роста 386
604
Предметный указатель
Приматы, мозг, развитие 46
— органы обоняния 43
— организация сообществ 117
— передние конечности 41
— родство, определение на молекулярном уровне 69
— развитие пищеварительной системы травоядного типа 41
— — хватательной функции конечностей 37
— сетчатка и зрение 44
— скелет стопы 63
— стадии эволюции 51
— «степени приспособленности» 118
— стрепсириновые 43
— человекообразные 38
— череп 48, 49
— число одновременно рождаемых детенышей 50
Приспособляемость 189, 472
Пробанд 143
Производительная доля энергии 490, 491
Происхождение хордовых 18
Проконсул, череп 49
Проплиопитек 57
Простейшие 14
«Протанопическая» слепота 350
Псевдохолинэстераза сывороточная 308
Птеродактили 21
Птилоцеркус, череп 49
Пурины 176
Пятнистая лихорадка Скалистых
гор 565
Работоспособность 560
Радиоактивный фон 555
Радиоуглеродный метод 77
Райта эффект 215
Рак, географический аспект 574, 575
— и телосложение 464
— кожи 263, 552—554
Расы 233—236
Рахит 263
Реакция на охлаждение 525
— — перегрев 552
Рептилии, адаптивная радиация 20, 21
— упадок 20, 32
Ретинит пигментный 167
Рецессивное наследование 142
Рецессивный ген, частота элиминации 219
— признак 135, 136
Риккетсии 565
Рилизинг-фактор лютеинизирующего гормона (РФЛГ) 412
РНК 177—179
Родезийский человек 102, 103, 106
Родословные, анализ 138
Родственный отбор 219
Родство 197
— коэффициент 198
Рост, взаимодействие наследственности и среды 413—423
— влияние времени года 418
— — питания 418
— — психологического воздействия 420
— генетика 414—416
— географические различия 242
— гормоны 409—411
— градиенты 383—385
— дифференциальная изменчивость 384—385
— и социально-экономические факторы 422
— компенсаторный 382—383
— кривая для различных тканей и
частей тела 376
— кривая подкожного жирового слоя 379
— лимфатической ткани 377
— мозга 376, 428
— наверстывающий 382
— определение по величине длинных костей 243
— подростковый период 411
— популяции, сравнение 241, 242
— постнатальный 368—370, 410
— постпубертатный 386
— пренатальный 368—370, 408, 409
— продольные и поперечные исследования 373—376
— пубертатный скачок 391—407
— расы и экологические условия 416
— результаты исследований 373
— скорость и климат 417
— тканей 376
— чувствительные периоды 385
— эпохальный сдвиг 423
Рунейя 53
Сальмонеллез 566
Самаритяне, коэффициент инбридинга 201
Сванкомбский человек 101, 105
Сердечно-сосудистые болезни 576
Серповидноклеточная анемия 277,
576
Предметный указатель
605
Серповидноклеточная талассемия 284, 569
Серповидноклеточность 276, 277, 288
Сетчатка позвоночных 44
— приматов 44
Сикхи 512
Симпатрические популяции 20
Синдром Клейнфелътера 406
Синэкология 473
Система органов размножения 392— 396
— тканевой совместимости (HL-A) 345
— Diego 331
- Kell 330
— Lutheran 330
— Rhesus (Ph) 163, 322
— — генетическая структура по Винеру 323
— — — — — Фишеру — Рейсу 324
— — географическое распределение 325
— — и естественный отбор 323
- Р 329
АВО-система 320, 321
Скелетная зрелость 400
Складка эпиканта 233
Слепота 167
Смешанный тип В иолы 443, 444
Современный человек, череп 67
Созревание, градиенты 383—385 — физиологические функции и развитие мышечной системы 396—399
Соли, потребности организма 497
Солнечный ожог и загар 538, 545, 550—554
Соматическая эволюция, скорость 27
Соматические клетки 131, 132
— мутации 184, 187
Соматотипирование 445
Соматотипы 448—450
— распределение 467—469
Соматомедин 410
Соматотония 465, 466, 469
Соматостатин 410
Соматотропин 410
Сообщества приматов, примеры 108
Соотношение гамет 134, 156
— зигот 134, 156
— полов 148—150
Способность к самовоспроизведению мутантного гена 211
Среда, географическое распределение популяций 476, 479
— классификация 476, 478—487
— обитания 541, 542
— — и питание 501—504
Среда, типы и географическое распределение 479
— формы хозяйства 484
Средняя продолжительность жизни 592
Стопоходящие формы 40
Стронций-90 555
Стьюдента критерий 242
Сумчатые млекопитающие 15, 21
Х-сцепленное наследование 150— 154
Y-сцепленное наследование 152
Сывороточные белки, антитрипсин 306
— — варианты 301—308
— — гаптоглобин 301
— — С3-компонент комплемента 306
— — Gc-компонент 307
Сывороточный альбумин 305
Spina bifida 274, 574
Таксономия 28—31
Талассемия 284—286, 569, 570, 576
Текодонты 21
Телосложение и болезни 463
— и жир 457
— — кровяное давление 463
— — образ жизни 467—470
— — поведение 465—470
— — физиологические функции 461
— классификация Виолы 443
— — Кречмера 444
— — Шелтона 444
— мышцы 458
— полиморфизм 441
— половые различия 441
— признаки 439
— скелет 458
— тканевые компоненты 452—460
— факторный анализ 451
Тельца Варра 154, 155
Темперамент, классификация Кречмера 465
— набор признаков Шелдона 465, 466
Температура тела 521
Температурная среда 519—522
— — адаптация человеческого организма 520
— — и температура тела 521
Темпы роста 436
— — интеллектуальных способностей 436
--- эмоционального развития 437
Тепловой удар 545
606
Предметный указатель
Теплоотдача, влияние формы тела 531
Тернера синдром 147, 406
Тест на цитотоксичность 345
Тестостерон 408, 409, 411
Тетониус 53
Тигр саблезубый 24, 76
Тимин 176, 177
Тиранозавры 21
Токсичные вещества, сопротивляемость организма 578—579
Толерантность 'к лактозе 357
Толковые мутации 207
Транслокации 209
Трансферрин 305
«Тританопические» дефекты 352
Тропический лишай 545
Туберкулез 567, 572, 573
— легких и телосложение 463
Тупайя 27, 38, 51
Тутси 241
Углеводы, суточное потребление в разных популяциях 491, 492
Ультрафиолетовые лучи 550 — — и рак кожи 552—554
Умственные способности 361
Уоддингтона принцип ассимиляции 538
Урацил 177
Фавизм 292
Факторы Gm и Inv 337
— — — — географическое распределение 342
— — — — обнаружение 341
Фенетическая классификация 28
Фенилкетонурия 144, 166, 167, 356
Фенилтиомочевина 135, 141
— ощущение вкуса 348—350
Фенобарбитон 579
Феноклины 206
Фенокопии 182
Фенотип 135, 136, 181
— Bombay 312, 313
Ферменты 578, 579
— географическое распределение 294—301
Фертильность 232, 233
Физическая работа, температурные границы 543, 544
Филетическая (филогенетическая) классификация 28
Филетические изменения 20, 23, 26 — связи 30
Филогенез 17
Филогенетическое дерево 335, 336
Флорисбадский череп 93
Фонтешевадский человек 100, 102, 104
Форма носа и климат 537
Фосфатемия 153
Фосфоглюкомутаза 298 6-фосфоглюконатдегидрогеназа 296 Фотограмметрия 448, 458
Фрамбезия 566 «Фторовая проба» 76
Харди — Вейнберга закон 193, 215, 220
Хозяйство, формы и расы 483
Холера 567
Холод, акклиматизация 526—529, 544
Хордовые, происхождение 18
Хорея Гентингтона 141
Хроматиды 159
Хромосома(ы) 132
— акроцентрические 137
— гетерохроматиновые участки 171
— метацентрические 137
— мутации 14, 207, 208
— окрашивание по длине 137
— человека, полиморфизм 361
— эухроматиновые участки 171
Х-хромосома 132, 137, 146, 147, 149, 150, 151, 153, 164, 210
— взаиморасположение генов 165
— и Xg-антиген 333, 334
Y-хромосома 132, 137, 146, 147, 149, 150, 153, 406, 408
Хромосомный набор 137
Хульман, череп 49
Цвет глаз 267—269
Цветовая слепота 150, 151, 350—354
— — распространение 353
Цезий-137 555
Ценогенез 18
Ценогенетическая адаптация 18
Центромеры 137
Церебротония 465—467
Церкопитек, скелет стопы 63
Церкопитековые обезьяны 118, 119
Церкопитекоиды 56
Церкоцеб 39
Предметный указатель
607
Цианоз 281
Циклотимический темперамент 465
Цирроз печени 576
Цитозин 176, 177
Частота генов 194, 195, 205—228, 287
— — обусловленная отбором 225
Чейн-Стоксово дыхание 559
Человек из пещеры Схул 105
— — Сванскомба 104
• — — Фонтешевада 105
— классификация Блюмсибаха 233
— «Соло» 103, 104, 106
Человекообразная обезьяна, вертлужная впадина 62
— — грудная клетка 60
— — — мышца 61
— — и человек 60
— — — — главные различия 65
— — крестец 61
— — передние конечности, локомоторные функции 64
— — поясничная область 61
• — — челюсти и зубы 65
— приматы 38
Человеческое общество, происхождение 107
Череп, найденный у реки Омо 93
— Нья 93
Чукчи 549
Чума 567, 572
Шизотимический темперамент 465
Шизофрения и телосложение 464
Шимпанзе 115
— безыменные кости 62
— кривая роста 387
— из леса Будонго 115
— поведение 117
— череп 49
Ширина носа, средний индекс 249
Шистосомоз 565, 567
Шкала эффективной температуры 540
Штейнгеймский человек 105 — — череп 67
Щитовидная железа 350, 499, 529
Эволюционная «специализация» 15
Эмбриональная стадия, адаптация 18
Эволюционное развитие, направления 23
Эволюционные изменения, типы 19 — преимущества 17
Эволюция барабанной косточки 16,17
— и развитие индивидуума 17
— и экология 14
— конвергентная 22
— лошади 23, 24
— наземных животных 20
— насекомых 18
— оболочек плода 18
— параллельная 22
— плотоядных 16
— принципы 13
— темпы 26, 27
Экология питания 488—518
Экономический уклад и способ производства 478
Экосистемы 473
— человеческие 467, 478—487
Эктоморфия 445
Эллпптоцитоз 141, 164
Элопитск 57
Эмбдена — Мейергофа гликолитический путь 291
Эндокринная система 146
Эндоморфия 445
Энергия, баланс 493
— пищевые источники 488—493
— потребности семьи 489, 490
— производительная доля 490, 491
— расход при различных видах деятельности 489
Эпигенез 440
Эпистаз 174, 182
Эпителиома 263
Эритроциты, кислая фосфатаза 295
— число, влияние внешних условий 181
Эскимосы 274, 300, 303, 315, 347, 360, 498, 573
— головной индекс 247
— жилища 548 , 549
— работоспособность 562
— специфика питания 491, 492, 503, 504
— чувствительность к холоду 528, 537
Эссенциальная гипертония 577
Эффективная численность популяции 216
Явление родоначальника 218
Язва двенадцатиперстной кишки 576
Язык 125
Японцы, размеры тела 251, 252
Яруро 511
Ящерицы 21
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Adapidae 53 Adapinae 34 Adapis 42 Adapoidea 34 Adipidae 34 Aegyptopithecus 56 Aelopithecus 56, 57 Africanthropus 102 Alouatta 35 Alouattinae 35 Anaptomorphidae 53 Anaptomorphinae 35 Anaptomorphus 35 Anopheles 288, 565 Aotinae 35 Anthropoidea 43—45 Aotus 35 Apidium 56 Archaeolemur 54 Archaeolemurinae 35 Arctocebus 35 A teles 62 Atelinae 35 Australopithecus 90 — africanus 79, 83 — habilis 83 Cebus 35 Cercopithecidae 36 Cercopithecinae 36 Cercopithecus 36 Cheirogaleus 34 Cheirogalinae 34 Colobus 36 Daubentonia 21, 37, 40, 41, 54 Daubentoniidae 35 Dicerorhinus etruscus 74, 76 Dicerorhinus merkii 74 Dinotherium 121 Dolichopithecus 54 Dryopithecinae 36 Dryopithecus 58, 66 Elephas 73 — antiquus 74 — meridionalis 74, 76 — primigenius 74, 76 Eohippus 26 Equus 26, 73 Eusmilus 24 Galago 35
Gerruvius 54 Boss 73 Branisella 54 Galaginae 43 Gigantopithecinae 36 Gigantopithecus 36 Gorilla 36
Cacajao 35 Callicebus 35 Callimico 36 Callithricidae 36 Callithrix 44 . Camelus 73 Carpolestes 34 Carpolestidae 53 Cebidae 35 Cebinae 35 Ceboidae 35 Cebupithecia 58 Cebupithecinae 35 Hadropithecinae 34 Hadropithecus 34 Hapalemur 34 Hemiacodon 35, 45 Hipparion 27 Hominidae 36 Hominoidea 66, 67 Homo 86 — erectus 83, 86, 125 — habilis 83 — sapiens 92, 125 Homunculus 58 Hylobates 57
Указатель латинских названий
609
Hylobatidae 36 Hylobatinae 36 Hypohippus 27
Indri 34
Indriidae 34
Indriinae 34
Lagothrix 35
Limnopithecus 57, 58 Lemur 34
Lemuridae 34
Lemurinae 35
Lemuriformes 34 Lemuroidea 34 Libypithecus 36 Limnopithecus 36 Lingula 26 Lingulella 26 Loris 35 Lorisidae 35 Lorisiformes 35 Lorisinae 35
Мясяся mulatta 108 Machairodus 76 Megaladapidae 35 Megaladapis 21, 54 Mesopithecus 36 Microcebus 34 Microchoerinae 35 Mobokopithecus 36
Necrolemur 27
Neosaimiri 58
Notharctinae 34 Notharcthus 34, 42 Nycticebus 35
Oligopithecus 56
Omomyinae 35, 56
Omomys 35
Oreopithecidae 36 Oreopithecoidea 36 Oreopithecus 56, 58
Papio anubis 108, 119 Paranthropus crassidens 79 — robustus 79 Parapithecinae 36 Parapithecus 56 Paromomyidae 34, 53 Paromomyinae 34 Paromomys 34 Pelicodus 34
Perodicticus 35
Phenacolemur 34
Phenacoleumurinae 34
Picrodontidae 34
Pierodus 34
Pithecathropus pekinensis 505
Pithecia 35
Plasmodium falciparum 288
Plesiadapis 40
Plesiadapidae 53
Plesiadapinae 34
Plesiadaform.es 34
Plesianthropus transvaalensis 79
Pliohippus 27
Pliopithecinae 36
Pliopithecus 58
Pongidae 66, 67
Proconsul 58
Presbytis entellus 119
Presiadapis 43
Primates 34
Prohylobates 58
Pronyciicebus 34
Propithecus '34
Propliopithecus 56, 57
Prosimi 34
Pseudoloris 35
Purgatorius 34
Ramapithecus 59
— brevirostis 68
Rooneyia 35
Saxonella 34
Saxonellinae 34
Smilodon 24
Strirtonia 58
Symphalangus 57
Tarsiidae 35
Tarsiinae 35
Tarsiiformes 35
Tarsius 44, 45
Teilhardina 54, 56
Telanthropus capensis 79, 83
Tetonius 35
Tichorhinus antiquitatus 74
Tupaiidae 37, 40
Victoriapithecus 58
Xenothricidae 36
Xenothrix 36
Linj anthrop us 121
ОГЛАВЛЕНИЕ
Предисловие редактора перевода................................... 5
Предисловие к первому изданию.................................... 7
Предисловие ко второму изданию.................................. 10
Часть I. Эволюция человека (Дж. Харрисон, Дж. Уайнер, Дж. Тэннер и В. Рейнолдс)............................................. 13
Глава 1. Закономерности эволюции.......................... 13
Глава 2. Приматы......................... ............... 32
Глава 3. Человекообразные обезьяны и человек ............. 60
Глава 4. Развитие гоминид................................. 71
Глава 5. Происхождение человеческого общества............ 106
Список литературы................................. 126
Часть П. Генетика человека (Дж. Харрисон) ...................... 129
Глава 6. Законы Менделя и человек........................... 129
Глава 7. Генетика пола...................................... 146
Глава 8. Независимое распределение и сцепление.............. 156
Глава 9. Гены и их действие................................. 166
Глава 10. Влияние внешней среды............................. 181
Глава 11. Генетическая структура человеческих популяций. I. Системы браков........................................... 190
Глава 12. Генетическая структура человеческих популяций.
II. Изменения частот генов........................ 205
Список литературы . , , .......................... 228
Часть Ш. Биологическая изменчивость в популяциях современного человека (Н. Барникот)....................................... , 229
Глава 13. Введение......................................... 229
Глава 14. Размеры и форма тела............................ 237
Глава 15. Пигментация и некоторые другие морфологические
признаки.......................................... 259
Глава 16. Биохимическая изменчивость . , , ................ 275
Оглавление 611
Глава 17. Группы крови.................................... 309
Глава 18. Другие виды изменчивости........................ 348
Список литературы................................ 363
Часть IV. Рост и конституция человека (Дж. Тэннер) . ....... 366
Глава 19. Кривая роста человека............................ 366
Глава 20. Пубертатный скачок роста и определение возраста . . по степени развития........................................ 390
Глава 21. Гормональные, генетические и средовые факторы, регулирующие рост.......................................... 408
Глава 22. Физическое и умственное развитие................ 428
Глава 23. Анализ и классификация типов телосложения...... 439
Глава 24. Связь телосложения с физиологическими функциями, болезнями и поведением..................................... 461
Список литературы ....................................... 470
Часть V. Экология человека (Дж. Уайнер)........................ 472
Глава 25. Введение......................................... 472
Глава 26. Экология питания................................. 488
Глава 27. Климатическая адаптация.......................... 519
Глава 28. Болезни ......................................... 564
Глава 29. Стабильность популяций........................... 580
Список литературы................................. 595
Предметный указатель....................................... . 597
Указатель латинских названий ^ .... г.......................... 608
Дж. Харрисон
БИОЛОГИЯ ЧЕЛОВЕКА
Ст. научный редактор Е. А. Яновская
Мл. научный редактор 3. В. Соллертинская
Художник А. В. Шипов
Художественный редактор Л. Е. Безрученков Технический редактор Л. П. Бирюкова Корректор А. Я. Шехтер
ИБ № 1463
Сдано в набор 30.08.78.
Подписано к печати 19.12.78.
Формат 60Х901/!,.
Бумага типографская К» 1.
Гарнитура обыкновенная. Печать высокая.
Объем 19,25 бум. л. Усл. печ. л. 38,50
Уч.-изд. л. 39,49 Тираж 53000 зкз.
Изд. 4/9812
Зак. 3037. Цена 3 р. 10 к.
Издательство «Мир»
Москва, 1-й Рижский пер., 2
Ордена Октябрьской Революции
и ордена Трудового Красного Знамени Первая Образцовая типография
имени А. А. Жданова Союзполпграфпрома при Государственном комитете СССР по делам издательств, полиграфии
и книжной торговли.
Москва, М-54, Валовая, 28