/





































































































































































































































































Author: Першин С.В.
Tags: деятельность и организация общая теория связи и управления (кибернетика) общая и теоретическая биология механика судостроение гидробионика
ISBN: 5-7355-0003-1
Year: 1988




Text
С. В. Першин
основы
ГИДРОБИОНИКИ
С. В. Першин
ОСНОВЫ
ГИДРОБИОНИКИ
Ленинград
«Судостроение»
1988
ББК 22.253
П26
УДК 007:573.6
Рецензенты:
д-р бпол. наук П. П. Гамбарян, д-р техн, наук проф. Ю. И. Фаддеев
Научный редактор автор предисловия лауреат Ленинской премии, акад.
АН УССР Г. В. Логвинович
Pershin S. V. Fundamentals Hydrobionics. Leningrad.: Sudo-
stroenie, 1988. 264 p, ill. ISBN 5-7355-0003-1
Complex hydrodynamic and biological research data of many years in the
field of hydrobionics are summarized by the author.
To the end of technical modelling of adaptation mechanisms and non-
statlonary swimming of sea animals, e. g. whales-llke ones, have been analized.
Particular attention have been paid to the morphostructure of swimming bodies,
the skin damping and the decrease of dolphins' hydrodynamic resistance. The
self-regulation oT whales-llkc flipper elasticity has been considered alongside
with the optimization of their flapping flipper propulsion.
Designated for researchers In cybernetics, biology, bionics, hydrodynamics,
water transport and shipbuilding. May be of use for post-graduates and stu-
dents at institutes of higher education.
Першин С. B.
П26 Основы гидробионики. — Л.: Судостроение, 1988.—
264 с., ил.
ISBN 5-7355-0003-1
Обобщены многолетние комплексные гидродинамические и биологи-
ческие исследования в области гидробионики. С целью технического мо-
делирования приведен анализ адаптационных приспособлений и неста-
ционарного плавания водных животных (скоростных рыб и китообраз-
ных). Особое внимание уделено закономерностям морфоструктурной
организации плавающих тел, свойству демпфирования кожного покрова
и снижению гидродинамического сопротивления дельфинов. Рассмотрена
авторегуляция упругости плавников китообразных и оптимизация машу-
щего плавникового движителя.
Для специалистов, работающих в области кибернетики, биологии,
бионики, гидродинамики, водного транспорта и судостроения. Может
быть использована аспирантами и студентами вузов.
п 3605030000—023 , ао
|1 ——----------- О'” оВ
048(01)—88
ББК 22.253
ISBN 5-7355-0003-1 © Издательство «Судостроение», 1988 г.
ПРЕДИСЛОВИЕ НАУЧНОГО РЕДАКТОРА
Автором монографии собран, систематизирован, обработан
и представлен обширный фактический материал по функцио-
нальной морфологии, особенностям нестационарного плавания
и гидробионике многих видов водных животных. Очень важно,
что разрозненные и часто противоречивые сведения, публикуе-
мые различными авторами, в монографии критически пере-
смотрены, оценены и систематизированы с позиций гидроди-
намики морских животных.
Проблемы гидродинамики морских животных, особенно
крупных и быстроходных, представляют как теоретический, так
и практический интерес. Например, данные о строении кож-
ного покрова некоторых видов китов и дельфинов, приспособ-
ленном к управлению пограничным слоем, наряду с рацио-
нальной формой тела свидетельствуют о существенном сниже-
нии их гидродинамического сопротивления, что интересно и
важно в практическом отношении.
Надежные количественные характеристики по сопротивле-
нию морских животных получить весьма трудно, поскольку
режимы движения многообразны, а методы контроля недоста-
точно надежны. Поэтому приведенные в монографии количест-
венные результаты в некотором отношении еще подлежат уточ-
нению. Ввиду того, что гидробионты представляют объекты,
в которых реализуется формирование обтекания упругого эла-
стичного тела, совершающего нестационарные движения, изу-
чение движения в жидкости таких тел составляет новый и важ-
ный раздел механики, сулящий открытие и других новых за-
кономерностей.
Гидродинамика гидробионтов интересна также в отношении
акустического излучения. К сожалению, фактических данных
в этой области очень мало.
Наконец, гидробионты, как и все живые существа, являют
собой примеры «машин» прямого эффективного изотермиче-
ского преобразования химической энергии в механическую при
температуре, близкой к температуре окружающей среды. Это
1* 3
свойство заслуживает изучения и попыток технического повто-
рения.
К настоящему времени из всего обширного комплекса ин-
тересных проблем гидробионики оценочные теории разрабо-
таны лишь для пропульсивных характеристик.
Бытует мнение, что живой организм в результате эволюции
наилучшим образом приспосабливается к обитанию в своей
среде. Например, акула и дельфин являются «идеальными»
подводными пловцами. Однако приспособление к среде — про-
цесс асимптотический. Поэтому совершенное приспособление
возможно лишь в том случае, если физические условия среды
обитания были достаточно долго неизменными в масштабе
жизненного цикла данного организма. Быстрые изменения фи-
зических характеристик внешней среды могут привести к «не-
поспеванию» адаптации и даже к гибели целых видов. Этому
положению противоречит, пожалуй, только быстрая адаптация
многих видов болезнетворных микробов к антибиотикам, что,
по-видимому, объясняется высокой частотой их размножения.
Из высказанных соображений напрашивается и обратный вы-
вод: если наблюдаются гидробионты, близкие к «идеальным»
в отношении, например, подводного плавания, то с некоторым
основанием допустимо предположить, что условия их обитания
очень длительное время оставались неизменными. Некоторой
иллюстрацией этого тезиса служат акулы и кистеперые ры-
бы — свидетели очень древних эпох.
Мне представляется, что развитие в количественном отно-
шении изложенных соображений может привести к дальней-
шему развитию теории эволюции.
Монография С. В. Першина содержит огромный фактиче-
ский материал, а также ряд обобщений и систематизаций, от-
ражающих взгляды ее автора. При научном редактировании
мною были высказаны автору некоторые пожелания и реко-
мендации, направленные в основном на достижение большей
строгости изложения. Все это автором монографии было твор-
чески учтено. В целом монографию С. В. Першина следует
оценить очень высоко, и она будет полезна многим научным
работникам, инженерам и студентам.
ОТ АВТОРА
Особый интерес к направленному изучению жизненных
функций и адаптационных приспособлений животных в значи-
тельной мере обусловлен тем, что их организмы представляют
собой сложную самоуправляемую, регулируемую систему. Не-
обходимость этой регуляции вызывается и определяется неста-
ционарным взаимодействием организмов с внешней средой и
ее переменными факторами. Регулируемые биологические си-
стемы не только нестационарны по существу и имеют колеба-
тельный характер процессов, но и обычно существенно нели-
нейны. В процессе длительной эволюции и естественного от-
бора наиболее выживающих особей регуляция у организмов
ведет к целенаправленности и оптимизации действий.
В гидробионике, одной из новых специализаций бионики,
получившей развитие в нашей стране, научный и прикладной
интерес применительно к судостроению представляет изучение
регуляции и оптимизации плавания, свойственные крупным
скоростным рыбам и китообразным, в том числе дельфинам.
В настоящей монографии в научных целях и для возможного
гидродинамического моделирования детально исследуются на
основе системного подхода своеобразна^ морфология, неста-
ционарная и нелинейная кинематика, динамика и биоэнерге-
тика поступательного движения, близкого к периодически регу-
лярному движению многих видов рыб, дельфинов и китов. Рас-
сматриваются также используемые китообразными в активном
плавании уникальные авторегуляции: самонастройка демпфи-
рования кожного покрова для снижения гидродинамического
сопротивления и регулируемый гидроупругий эффект в плав-
никах.
Настоящая монография является обобщением и развитием
многолетних исследований автора по гидробионике китообраз-
ных и рыб, результаты которых были ранее доложены на все-
союзных конференциях и региональных семинарах и опубли-
кованы в ряде статей в трудах этих конференций, в академи-
ческой печати, республиканских сборниках и других печатных
5
изданиях. Использованы совместные с другими специалистами
(гидромеханиками и биологами) научные открытия, изобрете-
ния и связанные с ними статьи, в которых автор принимал
активное творческое участие.
Гидробионика создается в нашей стране трудами многих
ученых, гидродинамиков и биологов. Многочисленные литера-
турные источники приведены в статьях автора, а также в пре-
дыдущем научном обзоре автора [50]. При подготовке предла-
гаемой монографии работы прошлых лет нами были вновь
рассмотрены, уточнены и дополнены по результатам более позд-
них исследований. Содержание книги и степень подробности
изложения рассматриваемых проблем, естественно, ограни-
чены общим объемом книги. Однако автор стремился к тому,
чтобы монография смогла представить интерес для широкого
круга специалистов судостроения, водного транспорта, био-
ники, кибернетики — для гидродинамиков и представителей
технических дисциплин, для биологов различного профиля,
а также аспирантов и студентов соответствующих специаль-
ностей.
ВВЕДЕНИЕ
Краткие сведения о бионике. Термин «бионика» происходит
от греческого слова bion — элемент, ячейка жизни. Принято
считать, что бионика оформилась как научное направление и
получила свое название на первом национальном симпозиуме
в г. Дэйтоне (США) в сентябре 1960 г. Этот симпозиум про-
ходил под девизом «Живые прототипы — ключ к новой тех-
нике». В качестве эмблемы бионики были приняты скальпель
и паяльник, соединенные знаком интеграла, как объединение
анализа и синтеза. Биолог устанавливает качественные зако-
номерности функционирования живых организмов, математик
обобщает их аналитически, инженер строит физическую мо-
дель для последующего использования в технике. Таким обра-
зом, к бионике привлекаются ученые и инженеры разных спе-
циальностей; их совместный труд обогащает исследования, что
является одним из больших достоинств бионики.
В нашей стране зарождение и становление бионики обя-
зано работам ныне покойных акад. А. И. Берга и член-корр.
АН СССР Б. С. Сотскова. Естественно, что с течением вре-
мени исследования по бионике расширялись и углублялись,
менялось и само представление о ней. В настоящее время та-
кому представлению наиболее полно соответствует следующее
определение предмета: бионика является одним из новых на-
правлений науки, занимающихся использованием принципов
организации и функционирования живых организмов и их эле-
ментов для совершенствования существующих и создания
принципиально новых технических систем.
Методологические вопросы бионики — закономерность ее
возникновения, место в системе наук, предмет, задачи, методы
и этапы исследований подробно анализируются на основе клас-
сических положений Фридриха Энгельса о формах движения
материи в брошюре, изданной Казанским государственным
университетом [7]. В ней зарождение бионики рассматривается
как закономерный результат действия объективного закона ус-
коряющегося развития науки и тенденции к интеграции знаний.
7
Попытки выявить функциональные основы изучаемых в при-
роде явлений и затем создавать на этой основе соответствую-
щие физические модели и производственные образцы предпри-
нимались учеными в разных странах уже давно, задолго до
возникновения бионики как самостоятельной науки. Например,
известный русский изобретатель А. М. Игнатьев, биолог по об-
разованию, увлекшийся решением инженерных задач, в ре-
зультате проведенных им в 20-е годы биологических и техни-
ческих опытов установил, что у белки тонкий зуб-резец имеет
слоистое строение, благодаря чему при стачивании не тупится.
На этом принципе им были предложены самозатачивающиеся
многослойные режущие инструменты, изготовляемые из мате-
риалов различной износоустойчивости, в которых слои, воспри-
нимающие большую нагрузку, делались более прочными.
В настоящее время для исследований по бионике привле-
кается комплекс научных дисциплин биологического, техниче-
ского и физико-математических циклов. Бионика тесно связана
также с синтетическими науками, прежде всего с кибернети-
кой — наукой о закономерностях управления и связи в живых
организмах, машинах и обществе.
В кибернетике установлено, что организм представляет со-
бой высокоорганизованную самоуправляемую систему. Важ-
нейшая особенность организма — наличие в нем нелинейных
причинно-следственных зависимостей, что является необходи-
мым условием осуществления принципов управления. В орга-
низме имеется биологическая информация, основной источник
которой — внешняя среда. Задача управления состоит в том,
чтобы возможно эффективнее отвечать на изменения во внут-
ренней и внешней среде организма. Это осуществляется в ор-
ганизме на основе обратных связей, таких причинных зависи-
мостей, когда результат предыдущего действия воздействует
на последующее течение процесса. Отсюда проистекает целе-
направленность процесса управления, достижение цели запро-
граммированного полезного эффекта.
В нашей стране исследования по бионике координируются
единым научным центром в Президиуме Академии наук СССР.
Имеется соглашение по исследованиям в бионике между стра-
нами — членами СЭВ. Итоги исследований по бионике неодно-
кратно подводились на всесоюзных конференциях в Москве
(декабрь 1963 г., декабрь 1965 г., май 1968 г., июнь 1973 г.)
и на Международных конференциях по бионике стран — членов
СЭВ (сентябрь 1975 г., г. Варна, НРБ; сентябрь 1978 г., Ле-
нинград). Каждая из этих конференций способствовала выяв-
лению научных сил, работающих в области бионики, развер-
тыванию и усилению научных связей, а также обмену инфор-
мацией по бионике. Конференции формировали важнейшие
направления предстоящих биологических исследований, комп-
лексные работы с творческим участием ученых разных специ-
8
альностей. Материалы конференций по бионике опубликованы
издательством «Наука» в сборниках: Бионика. 1965; Вопросы
бионики. 1967; Проблемы бионики. 1973.
Поскольку в технике занимаются получением, передачей,
преобразованием и использованием веществ, энергии, инфор-
мации, то и в бионике можно различать вещественные, энерге-
тические и информационные аспекты исследований. В настоя-
щее время наибольшее развитие получили исследования
информационных процессов, что непосредственно связано с ин-
тенсивными запросами и затратами в области радиоэлектро-
ники, измерительной и вычислительной техники, систем авто-
матического управления.
Анализ показывает, что в природе имеется большой вы-
бор образцов для бионического изучения и создания на основе
моделирования объектов новой техники. К таким природным
образцам относятся отдельные организмы, их чувствительные
и управляющие органы, системы и средства связи, принципы
и функции взаимодействия с. внешней средой живой материи
на разных уровнях. Разнообразными свойствами, достойными
заимствования, обладают организмы и их элементы всех
трех видов среды земной оболочки—воздушной, водной и
твердой, причем не только животные, но и растения. Изучение
этих свойств представляет тем больший интерес, чем большим
множеством взаимосвязанных объектов природы они приобре-
тены в течение длительной эволюции и чем больше они опти-
мизировались.
Исследованным в бионике биологическим прототипам отве-
чают разнообразные по назначению и характеру использова-
ния новейшие объекты техники — от специальных датчиков, ин-
дикаторов, приборов и устройств до (сложнейших машин и
больших систем. Все это конкретно иллюстрирует материаль-
ное единство живой и мертвой природы, которое в сущности
является методологическим основанием возникновения и ин-
тенсивного развития бионики. Исторически известно, что чело-
век всегда учился у природы.
Гидробионика — новое комплексное научное направление.
В современный период научно-технической революции, ввиду
разнообразия областей исследований и технических приложе-
ний, бионика сразу начала развиваться по нескольким направ-
лениям. Одним из таких самостоятельных поисковых направ-
лений в бионике, которое отличается своими изучаемыми
живыми объектами (бионтами) и средой их обитания, своими
целями, задачами и методами исследований, является гидро-
бионика. Этот термин применяется нами систематически с 60-х
годов.
Гидробионика есть новое комплексное научное направление
по изучению принципов устройства, структуры и функций ор-
ганов локомоции водных животных (гидробионтов) с целью
9
Рис. 1. Внешний вид гидробион-
тов: а — рыба (лосось); б — дель-
фин беломордый; в — моллюск
(кальмар).
совершенствования пропульсации и маневренности технических
средств движения в жидкой среде, а также для создания прин-
ципиально новых систем. Особое значение гидробионика при-
обретает в области судостроения и водного транспорта, а также
в связи с развивающимся освоением Мирового океана и воз-
можным широким использованием его богатейших ресурсов.
Данное понимание гидробионики связано с содержанием
предмета, которое складывается в отечественной научной ли-
тературе по бионике. В ней для последующего моделирования
технических объектов перемещения в водной среде преимуще-
ственно изучают плавание в природе относительно крупных и
быстро плавающих гидробионтов — рыб, дельфинов, кальмаров
(рис. 1).
Гидробионика оформилась как самостоятельное научное на-
правление в основном в последние 25 лет. Однако исследова-
ния подобного рода выполнялись достаточно давно. Например,
в печати известны работы акад. В. В. Шулейкина и его учени-
ков по изучению плавания рыб и дельфинов, выполненные
в 30-е и последующие годы.
Институтом гидромеханики АН УССР, начиная с 1965 г.,
ежегодно выпускаются межведомственные тематические сбор-
ники статей «Бионика» (г. Киев, «Наукова думка»), В сбор-
никах печатаются результаты разнообразных теоретических
и экспериментальных исследований плавания рыб, дельфи-
нов, кальмаров, в процессе которых изучались морфологиче-
ские качества, покровные образования, кинематика, динамика
и энергетика, гидродинамическое сопротивление и движитель-
ный комплекс этих животных.
О зарубежных исследованиях по гидробионике сведения
приведены в работах [50, 57, 74, 91]. В США стали усиленно
интересоваться исключительной приспособленностью к жизни
в воде и плаванием водных животных в последние десятиле-
тия. Из тематики гидробионического направления американ-
ские ученые уделяют внимание изучению гидродинамических
особенностей морских животных, исследованию механизма дви-
10
жения их, физиологической и биохимической оценке способно-
стей морских животных погружаться на большую глубину, раз-
работке конструкций искусственных жабр.
Необходимо подчеркнуть непосредственную связь гидробио-
•ники с гидродинамикой, которая уже следует из целевого
назначения гидробионики. Однако и в самой теоретической
гидродинамике в последнее время проявляют повышенный ин-
терес к исследованиям нестационарного плавания водных жи-
вотных, создается новое направление «биогидродинамика».
Для повышения эффективности транспортных судов, в том
числе танкеров, и повышения устойчивости их на волнении
в разных странах появились многочисленные проектные, рас-
четные и экспериментальные разработки новых быстроходных
полупогруженных судов необычной формы, напоминающих вод-
ных животных, плавающих вблизи свободной поверхности.
В различных вариантах намечают делать основной подводный
корпус хорошо обтекаемой формы для сравнительно неболь-
шой глубины погружения, а поэтому облегченной конструкции.
Такой корпус сочетают с надводной легкой надстройкой, кото-
рая позволяет основному корпусу постоянно сообщаться с ат-
мосферой. Подобные факты свидетельствуют о настойчиво про-
должающихся поисках новых подходов и инженерных решений
в области судостроения, как, впрочем, и в других областях. Это
также подтверждает актуальность и своевременность развития
гидробионики.
В гидробионике могут представлять интерес также некото-
рые исследования аэробионтов, особенно в части изучения
принципов нестационарного машущего полета в воздухе, когда
изменение плотности воздуха незначительное и им можно пре-
небречь. Ярким примером гидробионического подхода в иссле-
дованиях механики полета в природе, использованного
задолго до обособления гидробионики в самостоятельное науч-
ное направление, являются некоторые ранние работы родона-
чальника космонавтики К. Э. Циолковского. В конце прошлого
века он изучал полет насекомых и птиц. Особо следует отме-
тить его работу «Аэроплан или птицеподобная (авиационная)
летательная машина». В этой работе им предложено буквально
следующее: «Мы будем подражать птице только отчасти, на-
сколько можем... Пусть снаряд (самолет — С. П.) имеет
форму застывшей парящей птицы, но вместо головы вообра-
зим два гребных винта, вращающихся в обратные стороны и
дающие тягу в одном направлении. Мускулы животного мы
заменим... двигателями. Вместо хвоста устроим двойной
руль — в вертикальной и горизонтальной плоскостях...
Двойной руль, двойной винт и неподвижность крыльев при-
думаны нами не ради выгоды и экономии работы, а единст-
венно ради исполнимости конструкции». (См. К. Э. Циолков-
ский. Собр. соч. Т. I. М.: изд-во АН СССР, 1951, с. 43—44).
11
Как видно, К. Э. Циолковский, отправляясь от изучения
полета аэробионтов, еще в прошлом веке предвосхитил созда-
ние в авиации современного моноплана со свободнонесущим
крылом (в то время строили только громоздкие многоплановые
«летающие этажерки») и при этом показал классический об-
разец технического решения. В технике не обязательно копи-
ровать сами образцы природы. Новые объекты техники могут
выглядеть внешне и конструктивно совсем иначе, чем природ-
ные прототипы. Например, скрепленное с фюзеляжем жесткое
авиационное крыло значительно отличается по виду и струк-
туре от машущего складного перьевого крыла птицы, хотя
принцип создания подъемной силы у них одинаковый.
Следует подчеркнуть также тесную взаимосвязь между гид-
робионикой и биологией, которая определяется общими
объектами исследований — гидробионтами, изучением их пла-
вательных возможностей. Активное творческое участие специа-
листов-биологов в комплексных гидробионических исследова-
ниях совершенно необходимо, особенно для правильной орга-
низации и обоснования методов исследований с целью изуче-
ния условий жизни гидробионтов в природе. Однако одни
биологические исследования плавания водных животных заве-
домо недостаточны и не могут подменить гидробионические ис-
следования. Биологические исследования нередко имеют общий
описательный характер, в то время как для гидробионики не-
обходимы целенаправленные эксперименты и соответствующие
им данные. В биологической литературе часто отмечают «тор-
педообразную» форму тела хорошо плавающих водных жи-
вотных. Между тем рыбы и китообразные не имеют цилиндри-
ческой формы торпеды с поперечными сечениями в виде круга.
В действительности их форма значительно сложнее, чем у тор-
педы и вообще у тела вращения. Более того, в гидробионике
иногда не удается использовать даже количественные биоло-
гические измерения. Так, для многих видов китообразных опуб-
ликованы многочисленные индексы — соотношения различных
размеров тела, которые имеют систематический характер.
И все же по этим данным невозможно точно установить даже
главные измерения этих животных, которые с позиций гидро-
динамики оказываются определяющими для плавания их
в водной среде.
Отметим и обратный выход гидробионики в биологию —
развитие гидробионики несомненно способствует расширению
и дальнейшему прогрессу исследований в области биологии,
что подтверждается опытом. Участие специалистов-биологов
в комплексных гидробионических исследованиях приобщает их
в большей мере к применению количественных методов, ши-
рокому использованию в экспериментах новейшей контрольно-
измерительной аппаратуры, углубленному применению матема-
тического аппарата и теории моделирования. Все это содейст-
12
вует внедрению результатов исследований в народное хозяй-
ство.
Гидродинамические и биологические особенности плавания
водных животных. В современной гидробионике наибольший
интерес вызывает изучение плавания крупных водных живот-
ных, таких как рыбы, китообразные, моллюски. Все рыбы и
китообразные (частично и моллюски) не имеют формы тела
вращения, что связано с характером их плавательных изгибно-
колебательных движений тела и машущего хвостового плав-
ника — вертикального у рыб и горизонтального у китообраз-
ных. Средняя плотность тела водных животных близка к плот-
ности воды, поэтому они более или менее уравновешены
статической «архимедовой» силой плавучести. При наличии
небольшой положительной или отрицательной плавучести по-
следняя нейтрализуется в движении подвижными грудными или
брюшными плавниками, которые при соответствующих углах
атаки создают динамическую подъемную (или топящую) силу.
Одновременно эти плавники служат активными рулями глу-
бины и стабилизаторами движения.
Созданные природой движители существенно отличаются
от технических винтовых судовых движителей, приводимых во
вра.щение отдельным двигателем. Сила тяги у рыб и китооб-
разных создается единым гибким двигательно-движительным
колебательным комплексом. Он состоит из сильно деформируе-
мого мышцами тела, совершающего изгибно-колебательные
движения в виде поперечной волны, зарождающейся у головы
или в средней части тела и последовательно распространяю-
щейся вдоль тела до машущего хвостового плавника. Послед-
ний работает как нестационарная упругая гидродинамическая
поверхность, у которой обе стороны рабочие; причем в гидро-
динамическом следе за телом Образуется своеобразная вихревая
система. Головоногие моллюски создают силу тяги за счет
реакции периодически забираемой и выбрасываемой ими струи
воды, т. е. их движитель работает как нестационарный водо-
мет с пульсирующим режимом. Фазы забора и выбрасывания
воды у кальмара осуществляются с применением усилий, соз-
даваемых кольцевыми мышцами.
Особенно перспективными объектами исследований в гид-
робионике являются дельфины. Это животные высокого эво-
люционного уровня развития. Они от природы любознательны
и сообразительны, дружелюбно относятся к человеку, сравни-
тельно быстро привыкают к содержанию в неволе и относи-
тельно легко поддаются тренировке (дрессировке). Поэтому
дельфины служат хорошими лабораторными животными и мо-
гут быть надежными помощниками человека по связи под во-
дой, поиску различных объектов, по транспортировке предме-
тов. Высокая эффективность быстрого плавания дельфинов
обусловлена их хорошообтекаемой формой тела, мощным
13
движительным комплексом и регулируемым снижением гидро-
динамического сопротивления.
Качественная гидродинамическая характеристика основных
групп гидробионтов приведена в табл. 1.
Природные мышечные двигатели работают при низкой тем-
пературе: у млекопитающих (гомойотермных животных) при
температуре около 37 °C и у рыб (пойкилотермных животных)
при температуре воды, в которой они плавают. Это имеет боль-
шое значение в биоэнергетике.
Отметим некоторые основные положения (принципы) эво-
люционной теории в биологии, с проявлением которых прихо-
дится встречаться в исследованиях по гидробионике.
В биологическом принципе конвергенции —
сходства признаков — утверждается, что в длительно и устой-
чиво однородных условиях обитания животные разных видов
под действием естественного отбора сближаются по некото-
рым признакам, непосредственно связанным с внешней средой.
Например, известна конвергенция сельдевой акулы, ихтиозавра
и обыкновенного дельфина. Хотя эти водные животные систе-
матически весьма отдалены и относятся даже к разным клас-
сам, нетрудно заметить присущие им общие черты. Все они
имеют хорошо обтекаемую форму тела и весьма сходные прин-
цип действия и компоновку движителей, свойственные быстро
плавающим животным. В гидробионике подобную широкую об-
щность морфологических приспособлений водных животных
можно рассматривать как явно выраженное направление оп-
тимизации внешних форм для быстрого и эффективного пла-
вания. Отсюда следует также целесообразность сравнительных
исследований плавания различных видов животных, что ши-
роко используется в гидробионике.
Существует биологический принцип диверген-
ции — расхождения признаков: в пределах вида нет совер-
шенно одинаковых организмов, первоначально однородные
группы их в процессе естественного отбора в несколько отлич-
ных условиях среды специализировались в разных направле-
ниях. В результате появились разновидности и новые виды
с более выраженными различиями в строении, функциях и об-
разе жизни. Поэтому механическое подобие гидробионтов мо-
жет выполняться только приближенно, и в гидробионических
исследованиях иногда неизбежно довольно большое расхожде-
ние в экспериментальных данных, с чем необходимо считаться.
Важным является биологический принцип эмб-
риогенеза. Оказывается, в процессе эмбрионального раз-
вития зародышей общие признаки большой группы животных
обнаруживаются раньше, чем специальные. Поэтому последо-
вательность появления различных признаков у эмбриона в ос-
новном соответствует исторической последовательности появле-
ния этих признаков у предков данного животного, и их можно
14
Таблица 1. Качественная гидродинамическая характеристика крупных гидробионтов
Характеристики Рыбы К нтообр а з ные Головоногие моллюски
угревидные скомброндные дельфины КИТЫ
Угри, вьюны, сарганы, сабля-рыба Пресноводные в морские: карповые, окуневые, щуковые, осетровые, лососевые, тресковые, сельдевые, тунцовые, акуловые Низкоскоростные: морская свинья, белуха; высоко- скоростные — бело- бочка, продельфины, китовидный, афалина, гринда, косатка Низкоскоростные — гладкий, горбатый, серый, кашалот, высокоскоростные — полосатики: сейвал, финвал, синий Кальмары, каракатицы, осьминоги
Форма тела Вытянутая — цилиндрическая или сжатая с бо- ков Разнообразная — не- симметричная и симмет- ричная, в виде крыла малого размаха и близ- кая к телу вращения Хорошо обтекае- мая, в том числе ла- минаризованного профиля Варьирует от большеголовых глад- ких китов до вытя- нутых полосатиков Кальмары в форме тела вра- щения, карака- тицы в форме крыла
Состояние поверхности тела Гладкая или с мелкой, погру- женной в кожу чешуей Чешуйчатая поверх- ность, покрытая сли- зью; у быстрых рыб ме- стами гладкая Гладкая, у китов шероховатая, демпфи- рующий кожный покров сложной ячеистой структуры с продольными дермальными ва- ликами Гладкая (рас- тяжимая, весь- ма упругая)
Тип движи- теля; преиму- щественный маневр У г р е в и д- н ы й Подвижно все тело, периоди- чески пробегает волна Скомброид- н ы й Вертикальный хвос- товой плавник и кормо- вая часть тела; пере- мещение преимущест- венно в некотором слое воды Плавниковый Горизонтальный машущий хвостовой плавник. Основной маневр в вертикальной плоскости (всплывают на поверхность воды и ныряют на глубину: дельфины — до 300 м, киты — до 1500 м) Реактив- ный Импульсный; исключительная маневренность в любом направ- лении
Продолжение табл. 1
Характеристики Рыбы Китообразные Головоногие МОЛЛЮСКИ
угревидные скомброидные дельфины киты
Угри, вьюны, сарганы, сабля-рыба Пресноводные и морские: карповые, окуневые, щуковые, осетровые, лососевые, тресковые, сельдевые, тунцовые, акуловые Низкоскоростные: морская свинья, белуха; высоко- скоростные — бело- бочка, продельфины, китовидный, афалина, гринда, косатка Низкоскоростные — гладкий, горбатый, серый, кашалот; высокоскоростные — полосатики: сейвал, финвал, СИНИЙ Кальмары, каракатицы, осьминоги
Внешние органы стаби- лизации и уп- равления дви- жением Изгибающийся корпус тела и сплошные, окай- мляющие тело вертикальные плавники Подвижные спинные, брюшные и анальные плавники, убирающие- ся стреловидные боко- вые плавники, хвосто- вые костистые кили Вертикальный спинной плавник-стабили- затор, изгибающийся двухлопастный хвосто- вой плавник-руль, парные веслообразные боковые плавники стреловидной формы в плане, ламинаризованного профиля (у скоростных видов) Поворотное сопло под раз- личными углами к оси тела и бо- ковой плавник
Основные гидробиони- ческие аспек- ты Хорошообтекаемая форма тела. Высокие пропульсивные качества движительного комплекса. Биологическая слизь, снижаю- щая гидродинамическое сопротивление Оптимальная по сопротивлению форма. Демпфирующая способность кожного по- крова дельфинов и возможная ламинариза- ция пограничного слоя. Регулируемый гид- роупругий эффект плавников у всех кито- образных Импульсная схема движения, система водо- проводящих от- верстий^и сопел
проследить в эмбриогенезе. Этот принцип ниже неоднократно
используется.
Отметим еще принцип мультифункционально-
сти— наложения функций. В процессе эволюционного разви-
тия органов животных происходит расширение и усложнение
их функций. У органов могут появиться дополнительные, пер-
воначально не свойственные им функции. Примеры подобного
рода неоднократно будут встречаться в дальнейшем изло-
жении.
Для сопоставления между собой некоторых функциональ-
ных систем гидробионтов по их относительной массе в орга-
низмах приведем табл. 2. Обращает на себя внимание отно-
сительно большая масса мышц-двигателей. У различных гид-
робионтов относительная масса мышц различается почти в два
раза, что существенно сказывается на их биоэнергетике. Это
необходимо учитывать в гидробионических расчетах.
Таблица 2. Относительная масса некоторых функциональных
систем гидробионтов (в процентах от общей массы)
Гидробионты (общая масса, кг)
Системы и органы Рыбы (МО-* — 3-102) Дельфины (3-101 — Ы0») Киты (3-10‘— 1-10»)
Системы управления: мозг сердце Опорный аппарат — скелет Двигатель — мышцы Движитель — машущий хвостовой плавник 0,15—0,70 0,07—0,50 33,00—67,00 0,70—2,00 0,25—1,60 0,45—0,90 11,00—14,00 30,00—40,00 0,90—1,50 0,02—0,05 0,30—0,75 15,00—25,00 25,00—45,00 0,70—1,60
Основные критерии моделирования плавания гидробионтов.
В природе существует большое число видов животных для вы-
бора в качестве прототипов гидробионтов, различающихся раз-
мерами, скоростью и гидродинамической эффективностью пла-
вания. Так, даже среди крупных гидробионтов насчитывается
около 102 видов китообразных и около 2-Ю4 видов рыб. Есте-
ственно, что за длительный период эволюции (порядка ,пХ
Х107 лет) гидробионты адаптировались к водной среде и мно-
гие их функции, в том числе гидродинамические, оптимизиро-
вались по определенным параметрам. Однако эта оптимизация
достигнута в интересах существования и прогрессивного раз-
вития самих живых организмов и не всегда может представ-
лять интерес для моделирования и применения в технике.
Отметим общую принципиальную трудность изучения меха-
низма плавания гидробионтов, например, таких как рыбы и
китообразные. Поскольку у них двигатель и движитель рабо-
17
тают нераздельно и составляют единый изгибно-колебательный
двигательно-движительный комплекс, то вследствие изгибных
деформаций на поверхности тела гидробионта одновременно
развиваются движущая сила и сила сопротивления. Поэтому
в отличие от технических объектов у гидробионтов отсутствует
четкое разделение гидродинамических функций на пассивный
корпус, который испытывает только сопротивление движению,
и активный движитель, создающий силу тяги. К тому же гид-
робионт является самодвижущейся саморегулируемой систе-
мой и попеременно настраивается на режимы движения в со-
ответствии с внешними условиями окружающей среды.
Комплексное изучение механизма плавания живых организ-
мов в настоящее время затруднено как в отношении матема-
тического описания с учетом присущих им гидродинамических
эффектов, так и в отношении экспериментальных исследований,
поскольку необходимо иметь новую специальную измеритель-
ную аппаратуру. В целях технического моделирования целе-
сообразно изучать влияние отдельных факторов механизма
плавания гидробионтов с применением системного подхода ис-
следований. Учитывая, что взаимодействие гидробионтов свод-
ной средой имеет механическую природу, обобщенный теоре-
тический анализ в сочетании с натурными опытами и модель-
ными гидродинамическими испытаниями плавания гидробион-
тов следует проводить с общих позиций теории механического
подобия и размерностей нестационарных движений. При этом
важнейшее значение имеют безразмерные числа (критерии) и
те характерные параметры, через которые эти критерии выра-
жаются.
Нестационарность плавания гидробиоцтов характеризуется
прежде всего неравномерным поступательным движением
центра тяжести тела (ЦТ) с переменной по времени t мгно-
венной скоростью U, так что U=U (/).
Безразмерная мгновенная скорость U движения гидроби-
онта, как и обычно, выражается с помощью длины тела L
U=U/^L.
Здесь g — ускорение силы тяжести (g = 9,81 м/с2). В качестве
длины тела для рыб и китообразных наиболее часто прини-
мают так называемую зоологическую длину, считая от кончика
носа до развилки хвостового плавника, или длину собственно
тела до плавника.
Важнейшей характеристикой гидробионта является средняя
скорость плавания Uo в данном режиме на участке пути дли-
ной $
U0 = -Uu(t)dt = ^. (1)
t о t
18
Соответственно безразмерная средняя скорость плавания оп-
ределяется выражением
£7оо=
В биологических работах и вообще в литературе часто встре-
чается термин «относительная скорость» плавания животного,
которая выражает среднюю скорость плавания соответственно
длине тела в секунду, т. е. Uq = UqIL. Эта величина размерная
1с-1], однако ее применение оправдано, так как для водных жи-
вотных она является характерной. У многих видов животных,
не слишком малых (микроскопических) и не слишком больших
(длиной в несколько метров), максимальная относительная ско-
рость плавания нередко составляет с-1, так что абсо-
лютная скорость Um= 10L. Это удобно для сравнения и выяв-
ления скоростных и нескоростных гидробионтов, а также для
различения переменных режимов плавания одного и того же
гидробионта.
Исследуя различные быстрые режимы плавания гидробион-
тов, особо выделяют среднюю максимальную скорость плава-
ния в течение нескольких минут или нескольких секунд (им-
пульсную скорость) и среднюю крейсерскую скорость плавания
в течение десятков минут и даже часов. Это обстоятельство
каждый раз следует отмечать.
Степень неравномерности плавания гидробионта может быть
оценена абсолютным ускорением движения W=dU (О/d/ и его
безразмерной величиной
W = —------dU(^ (2)
U2(t) dt
Однако для использования этого выражения требуется большая
информация по кинематике нестационарного плавания, что
в экспериментах не всегда достижимо.
В соответствии с теорией механического подобия явлений
при изучении нестационарного плавания гидробионтов исполь-
зуются два определяющих гидродинамических критерия (без-
размерных числа)—критерии Рейнольдса и Струхаля. Они те-
перь выводятся из дифференциальных уравнений движения вяз-
кой несжимаемой жидкости, физически представляют собой от-
ношения соответствующих сил, рассчитанных на единицу массы
жидкости, и определяются через характерные параметры дви-
жения.
Критерий Рейнольдса Re, как известно, представляет собой
отношение конвективных (переносных) сил инерции и сил вяз-
кости (внутреннего трения) жидкости и определяется выраже-
нием
Re = l/0£/v. (3)
Здесь Uo, L обозначены, как ранее; v — кинематический коэф-
19
фициент вязкости воды, зависящий от ее температуры. Обычно
в расчетах принимают v^l,30- 10-6 м2/с, что примерно соответ-
ствует средней годовой температуре воды Мирового океана
около 10 °C. Критерий Рейнольдса часто обозначают с индек-
сом, например Rei..
Критерий Струхаля Sh характеризует нестационарность дви-
жения, отношение локальных (местных) и конвективных сил
инерции жидкости. Он определяется выражением
ShA = An/U0. (4>
Здесь А— характерный линейный размер; п — характерная ча-
стота колебаний гидробионта. Применительно к плаванию рыб
и китообразных обычно принимают: А — максимальный размах,
т. е. удвоенная амплитуда колебаний точки у развилки хвосто-
вого плавника (иногда длина тела); п — частота полных коле-
баний хвостового плавника, считая сверху вниз и в обратном
направлении.
Численные значения критерия Струхаля по формуле (4)
обычно близки к известным значениям этого критерия в слу-
чаях срыва вихрей за неподвижными и колеблющимися плохо-
обтекаемыми твердыми цилиндрическими телами, когда в рас-
чет их поперечного обтекания принят поперечный размер тела
(диаметр цилиндра), что удобно для единообразия. Кроме этого
очевидно: в критерии Струхаля числитель представляет собой
половину средней поперечной скорости Ко движения хвостового
плавника гидробионта. Поэтому
ShA = O,5Vo/£7o, или V0/(/0 = 2ShA.
Отсюда усматривается простой физический смысл критерия
Струхаля. Этот критерий может выражаться также по формуле
(2), но численное значение его будет другим.
В теоретических исследованиях нестационарного движения
критерий Струхаля часто определяют через круговую частоту
колебаний (о = 2лм и вместо параметра А вводят в рассмотре-
ние длину тела L (или хорду крыла Ь), так что
Sha = (oL/i70- (5)
Связь между значениями ShA по формуле (4) и ShM по фор-
муле (5) устанавливается непосредственно по этим выраже-
ниям.
Критерии Рейнольдса Re и Струхаля Sh, взятые вместе, оп-
ределяют динамическое подобие нестационарного плавания гид-
робионтов. Однако следует подчеркнуть, что строго динамиче-
ское подобие возможно лишь при условии соблюдения геомет-
рического и кинематического подобия «натуры» и «модели» и
сохранения равенства (idem) чисел Рейнольдса и Струхаля для
натуры и модели Re = idem, Sh = idem. Между тем оба эти усло-
вия одновременно несовместимы. По Рейнольдсу скорость модели
20
и натуры изменяется не так, как по Струхалю (при прочих рав-
ных условиях). Поэтому динамическое моделирование гидроби-
онтов возможно лишь приближенно, в каждом случае отдельно-
по главным действующим силам.
Чтобы показать, насколько рыбы и китообразные соответ-
ствуют техническим объектам по основным параметрам плава-
ния, приведем содержащий большую информацию четырехпара-
метрический график L—Um, ReL—Um на логарифмической сетке
(рис. 2), который был разработан нами еще в работе [47]; здесь
он несколько дополнен. На графике каждая точка для китооб-
разных отвечает видам животных средней величины и макси-
мальной скорости плавания по многочисленным промысловым
данным, а для рыб точки нанесены по параметрам единичных
особей, по данным экспериментов разных авторов или по на-
турным наблюдениям различными методами. Для технических
объектов размер в длину и скорость движения показаны по дан-
ным первоисточников.
/7,2'^,
0,1
0,2
0,5. 1,0
Рис. 2. Гидродинамические параметры ви-
дов гидробионтов и технических объектов.
Четырехпараметрический график.
/ — рыбы: а — форель, б — лосось, в — ваху,
г —барракуда, д — тунец, е— меч-рыба, лс—
акула-мако, з — голубая акула; 2 — китооб-
разны е — дельфины: и — белобочка, к — афа-
лина, л — гринда, м — косатка, н —белуха; киты: о — полосатики, сейвал, фин-
вал, синий, п — серый, горбатый, кашалот; 3 — транспортировщики легководола-
зов; 4 — самоходные обитаемые аппараты; 5 — сверхмалые подводные лодки;
6 и 7 — зарубежные подводные лодки и торпеды [47]. КГ и ОБ — области пара-
метров гидродинамических моделей, испытываемых в кавитационных трубах
и опытовых бассейнах.
21
Рыбы и китообразные характеризуются в основном своими
областями значений характерных параметров и критериев. Ры-
бам свойствен большой диапазон абсолютных максимальных
скоростей плавания, обычно при сравнительно небольших раз-
мерах, за исключением нескольких видов, в том числе крупней-
ших акуловых рыб. Напротив, китообразные значительно
больше рыб по размерам, но диапазон максимальных скорос-
тей плавания у них ограничен. При каждой данной длине жи-
вотного абсолютная максимальная скорость плавания изменя-
ется у разных рыб в семь и более раз, а у китообразных только
примерно в два раза. С увеличением размеров на один порядок
максимальная скорость плавания повышается у разных видов
рыб примерно в восемь раз, а у китообразных всего на 20—
25%, так что абсолютная максимальная скорость плавания
дельфинов и китов оказывается почти одинаковой. При этом
имеют значение отмеченное ранее различие в относительной ос-
нащенности тела мышцами у рыб и китообразных, а также
различные гидродинамические режимы в пограничном слое
у дельфинов и китов. На графике ясно выделяются скоростные
и тихоходные виды рыб и китообразных.
Сопоставляя основные параметры плавания рыб и китооб-
разных с гидродинамическими моделями твердых тел, испыты-
ваемых в кавитационных трубах и опытовых бассейнах (на рис. 2
они показаны по нашим данным в виде областей КТ и ОБ),
а также с параметрами технических объектов различного назна-
чения, видим их взаимное соответствие в широком диапазоне
чисел Рейнольдса (Re=l-106—2-Ю8). Отсюда также следует,
что рыбы и китообразные являются перспективными объектами
исследований в гидробионике и могут служить некоторыми ана-
логами для технических объектов.
ГЛАВА I
ФУНКЦИОНАЛЬНО-МОРФОЛОГИЧЕСКИЕ СВОЙСТВА
КРУПНЫХ ГИДРОБИОНТОВ
И ИХ ГИДРОДИНАМИЧЕСКОЕ ЗНАЧЕНИЕ
§ 1. Общая гидробионическая характеристика
эффективно плавающих китообразных
Китообразные — самые крупные и скоростные, наиболее спе-
циализированные вторичноводные млекопитающие высокого
уровня эволюционного развития. Они встречаются во всех мо-
рях и океанах, всю жизнь активно проводят в воде, но дышат
.22
атмосферным воздухом, поэтому периодически всплывают на
поверхность воды. Многие из них ежегодно совершают для на-
гула жира и размножения продолжительные сезонные мигра-
ции, протяженность которых составляет по несколько тысяч ки-
лометров.
Подавляющее большинство видов китообразных исследовано
недостаточно. Более основательно изучены промысловые виды
дельфинов (белобочка, афалина, морская свинья, белуха) и
киты (кашалот, финвал, синий). В последние десятилетия ин-
тенсивно исследуют и другие виды китообразных, в том числе
дельфинов (продельфинов, гринду, косатку) и китов (малого по-
лосатика и серого). Особи их содержат и дрессируют в неволе,,
в океанариумах. Во многих океанариумах США, Японии и дру-
гих стран наиболее распространены афалины.
Внешний вид китообразных (рис. 3) показывает, что они
хорошо приспособлены к активному плаванию в водной среде.
Тело их обтекаемой формы, вытянутое, стройное, эллиптиче-
ского сечения, суживается от груди к хвосту. Шея очень корот-
кая, не разграничивает голову и туловище; лишь у некоторых
видов китообразных заметно едва намечающееся в этом месте
сужение. Задняя оконечность тела китообразных (хвостовой
стебель) сжата с боков и часто имеет сверху и снизу ясно выра-
женные кили, что гидродинамически целесообразно при машу-
щих движениях стебля в вертикальной плоскости. Расположен-
ный в конце стебля горизонтальный хвостовой плавник (глав-
ный машущий движитель) обладает в плане дельтовидной
формой со срединной выемкой на задней кромке, которая раз-
деляет его на две лопасти — левую и правую. Грудные подвиж-
ные парные плавники китообразных имеют веслообразный вид
и являются рулями глубины. Все плавники этих животных ха-
рактеризуются хорошо обтекаемой формой в плане и в про-
филе. Кожа тела и плавников китообразных снаружи гладкая,
на ней нет волос и слизистых выделений; лишь у тихоходных,
видов кожа обрастает паразитами и тогда становится шерохо-
ватой.
Рис. 3. Обыкновенный дельфин-белобочка в подводном плавании в море:
а — продольный вид; б — вид сзади.
23-
Рис. 4. Крупнейший скоростной дель-
фин-косатка выпрыгивает из воды.
шены выступающего рострума,
Отметим различное строе-
ние головной части у дельфи-
нов. Обыкновенный дельфин-
белобочка и продельфин, ко-
торых относят к наиболее бы-
строходным животным, имеет
длинный узкий рострум с вы-
тянутыми челюстями, на про-
филе головы сильно выделя-
ется клюв (см. рис. 3). У афа-
лины, более крупного и отно-
сительно быстроходного дель-
фина прибрежных вод, клюв
умеренный. Косатка — самый
крупный и скоростной вид
~дельфинов — имеет широкий
рострум (рис. 4). В отличие
от всех перечисленных видов
мелкие дельфины, морские
свиньи и крупные белухи ли-
клюв у них отсутствует. Нали-
чие более или менее выдающихся спереди рострума и клюва
у дельфинов («хватальщиков») объясняется в основном биоло-
гически — приспособлением к штучному захвату корма. Од-
нако, несомненно, отмеченные выступающие части на носовом
участке тела дельфинов не причиняют ущерба их гидродинами-
ческим качествам. Это следует как из общего биологического
принципа мультифункциональности органов животных, так и
непосредственно из того факта, что рострум и клюв встреча-
ются у быстроходных дельфинов.
Спинной плавник — стабилизатор плавания китообразных
в вертикальной плоскости — имеется у подавляющего большин-
ства видов. Однако он отсутствует у весьма резвого скоростного
северного китовидного дельфина и у менее быстроходных дель-
финов— большой мор'ской свиньи и крупной белухи, а также
у нескоростного усатого серого кита. Можно полагать, что от-
сутствие спинного плавника у скоростного дельфина компенси-
руется хорошо выраженными верхним и нижними килями на
хвостовом стебле большого удлинения.
Своеобразной особенностью отличаются скоростные усатые
киты-полосатики: сейвал, финвал, синий. Они имеют на наруж-
ной поверхности горла и брюха многочисленные (порядка не-
скольких десятков и до 120) продольные, параллельные, хорошо
выделяющиеся кожные складки в виде выступающих валиков
и расположенных между ними впадин глубиной до 0,06 м.
Обычно наличие этих складок биологи объясняют облегчением
расширения и сужения горла и брюха у полосатиков, поскольку
они очень прожорливы и заглатывают большую массу планк-
24
Рис. 5. Внешние контуры тела и плавников медленно плавающих млекопи-
тающих сиреновых: а — карибский ламантин; б — дюгонь.
тона. Неясно гидродинамическое значение кожных складок
у полосатиков.
Зубатый кит-кашалот с очень крупной головой — один из
самых тихоходных, однако ныряет глубже всех китообразных
за крупными кальмарами, которые составляют его основную
пищу. Туши кашалотов, запутавшихся в телефонно-телеграфных
трансатлантических кабелях, находили на глубине до 1,5 км.
Дельфины почти всех скоростных видов — хорошие прыгуны.
Они выпрыгивают из воды на несколько метров над ее поверх-
ностью. Обитатель тропических вод — длинномордый продель-
фин, выпрыгивая из воды, вращается в воздухе и делает до
двух с половиной оборотов вокруг своей продольной оси, за
что и получил название «вертящегося» дельфина. Очевидно,
это свидетельствует о его высокой скорости плавания и манев-
ренности. Однако и тихоходный усатый горбатый кит с отви-
сающим брюхом выпрыгивает из воды. Дельфины же морская
свинья и белуха не делают прыжков из воды.
Чтобы выделить высокие внешние гидродинамические каче-
ства китообразных, приведем для сравнения контуры тела и
плавников мирных тропических животных — ламантина и дю-
гоня (рис. 5). Это крупные вторичноводные млекопитающие из
рода сиреновых, длиной до 3—4 м, массой до 500 кг. Как и
дельфины, они приспособлены к постоянной жизни в воде,
имеют единый изгибно-колёбательный двигательно-движитель-
ный комплекс с одним мощным хвостовым плавником — глав-
ным движителем. Ламантины и дюгони — травоядные живот-
ные. Они питаются прибрежными морскими водорослями и по-
тому не нуждаются в большой скорости плавания. Однако им
приходится много плавать, так как эти животные поедают
в сутки своей малокалорийной пищи до 20 % массы тела. По
рисунку видно, что морда у них тупорылая, плохообтекаемая,
тело утолщенное, к тому же поверхность тела грубошерохова-
тая для усиления теплоотдачи. Животные имеют хвостовой
25
плавник с малым относительным удлинением (около 1,5), ме-
нее эффективный у ламантина и более эффективный у дюгоня.
“Соответственно скорость плавания ламантина и дюгоня в 2—
3 раза меньше, чем у дельфинов.
Количественная информация о более изученных 18 видах
китообразных приведена в табл. 3. Она разработана нами по
многократно сопоставленным между собой разрозненным био-
логическим данным. В таблице приведена так называемая «зоо-
логическая длина» китообразных, которую измеряют в биологи-
ческих исследованиях от кончика носа до развилки хвостового
плавника. Максимальная Lm и средняя L зоологическая длина
каждого вида определены по многочисленным промысловым
измерениям, иногда для нескольких десятков тысяч животных.
Средние абсолютные размеры частей тела и плавников дельфи-
нов и китов измерены на сравнительно небольшом числе особей
каждого вида. Соответственно приближенными являются и сред-
ние относительные размеры особей: относительное удлинение
тела Л = Л//Д_относительное отстояние наибольшего сечения от
досика тела x = x/L, относительный размах хвостового плавника
l = l!L и относительная высота спинного плавника h = h/L. Мак-
симальная скорость плавания Um по видам дельфинов и китов
указана кратковременная, импульсная, сохраняемая в течение
нескольких секунд и достигаемая ими, по наблюдениям на про-
мыслах, при мгновенном удалении уплывающих животных в со-
стоянии наивысшего напряжения, когда они слегка ранены или
крайне испуганы. Максимальное число Рейнольдса определено
по значениям L и Um. Несмотря на приближенные данные по
скоростям плавания, достоинство их состоит в том, что они
получены путем наблюдений в море на диких животных.
Большое значение имеют относительные габаритные раз-
меры плавников у китообразных. Можно заметить корреляцию
между размером хвостового плавника и поперечными размерами
тела. Относительный размах хвостового плавника немного
уменьшается в связи с ростом числа Рейнольдса животного во
всем диапазоне / = 0,24—0,22. Исключение составляют быстро-
ходная косатка и тихоходные киты — серый и кашалот, у ко-
торых / = 0,26—0,29, т. е. заметно выше. Это можно объяснить
большой маневренностью глубоко ныряющей косатки и особо
глубоким нырянием кашалота. Важно отметить, что у всех
дельфинов и китов абсолютный размах хвостового плавника —
главного движителя — превышает наибольшую ширину тела.
Иными словами, хвостовой плавник выступает за габариты
тела.
Средняя относительная высота спинного плавника резко от-
личается у дельфинов и китов. Она очень сильно варьируется
также у различных видов дельфинов. Наибольшая средняя
26
Таблица 3. Сравнительная гидродинамическая оценка
некоторых видов китообразных
Виды дельфинов и китов Зоологи- ческая длина, м Относительные размеры Максималь- ные скорости Число Рейнольдса 10"' Re
тела плавников
максимальная средняя L удлинение ЫН отстояние x/L размах хвосто- вого 1/L высота спин- ного h/L абсолютная м/с относительная UJL, с-1 т *
Дельфины Обыкновенный — 2,6 1,6 5,3 0,42 0,24 0,10 13,2 8,2 16
белобочка (Delphi- nus delphis) Полосатый про- дельфин (Stenella coerulealbus) —
2,4 1,8 4,6 0,42 0,23 0,08 — —
Продельфин (Ste- iiella attenuate) 2,1 1,9 5,7 0,40 0,22 0,10 — — —
Северный кито- видный (Lissodel- phis borealis) 2,5 — 6,5 0,45 0,19 0 — — —
Афалина (Tur- siops truncatus) 3,6 2,3 5,1 0,40 0,23 0,09 12,5 5,4 22
Тихоокеанский 2,5 2,2 6,0 0,38 0,25 0,10 — — ——
короткоголовый (Lagenorhynchus ob- л
liquidens)
Гринда (Globice- 6,0 4,0 5,2 0,38 0,23 0,07 11,3 2,8 35
phalus melas) Косатка (Orcinus 10,0 6,0 — — 0,28 0,16 13,4 2,2 62
Бесперая морская 1,6 1,2 4,5 0,40 0,26 0 6,8 5,2 6,3
свинья (Neomeris phocaenoides)
Морская свинья (Phocaena phocaena) 1,8 1,4 4,8 0,38 0,24 0,06 8,6 6,1 9,2
Белокрылая мор- 2,1 1,7 5,0 0,40 0,24 0,08 10,4 6,1 13
ская свинья (Phocae- noides dalli) t i
Белуха (Delphi- nap terus leucas) 6,0 4,5 4,3 0,40 0,24 0 6,1 1,35 21
Киты
Сейвал (сайдя- ной) (Balaenoptera borealis) 18 15 5,8 0,38 0,25 0,035 15,3 1,02 180
Финвал (сельдя- 24 20 6,2 0,41 0,21 0,025 13,0 0,65 200
ной) (Balaenoptera physalus)
27
Продолжение табл. 3
Виды дельфинов и китов Зоологи- ческая длина, м Относительные размеры Максималь- ные скорости Число Рейнольдса 10-в Re
тела плавников
максимальная Lm средняя L удлинение L/H отстояние x/L размах хвосто- вого 1/L высота спин- ного h/L абсолютная м/с т ' относительная "Л- с-
Синий (голубой) (Balaenoptera mys- culus) 30 24 6,3 0,43 0,22 0,013 15,0 0,63 280
Горбатый (Megap- tera nodosa) 18 12,5 4,5 — — 0,022 7,5 0,60 72
Серый (Eschrich- tius gibbosus) 15 12 4,9 0,32 0,26 0 7,0 0,58 65
Кашалот (Physe- ter catodon) 20 15 5,2 0,31 0,28 0,025 7,2 0,48 83
относительная высота спинного плавника наблюдается у круп-
ной скоростной косатки. Для косатки характерно еще половое
различие: /г = 0,10 у самок и Л = 0,16 у самцов; максимальное
значение для самцов /i = 0,22. Напротив, у китов средняя относи'
тельная высота спинного плавника весьма небольшая. Это сви-
детельствует о большой устойчивости движения крупных китов.
Один из первых вопросов, который приходится решать ис-
следователям и конструкторам при моделировании, обычно свя-
зан с выбором соответствующих форм тела объектов определен-
ного назначения. Из теоретических и экспериментальных иссле-
дований в гидродинамике известно, что для стационарно движу-
щихся в реальной вязкой жидкости твердых тел не существует
универсальной оптимальной внешней формы, обладающей ми-
нимальным гидродинамическим сопротивлением при заданном
полезном объеме. Каждому диапазону чисел Рейнольдса и ха-
рактеру движения тела отвечает своя, более или менее отли-
чающаяся, оптимальная форма. Тем больший интерес пред-
ставляет установить, в чем состоят выработанные в процессе
длительной эволюции особенности внешней формы тела живых
дельфинов, нестационарно плавающих при числах Рейнольдса
порядка Rer,= 107.
Поскольку тело дельфина по форме не является телом вра-
щения, а его наружная поверхность сложная, изменяется по
длине, высоте и ширине, то ясное представление о ней можно
получить только графически в виде распространенного в судо-
строении теоретического чертежа. Теоретические чертежи дель-
це
финов в трех проекциях необходимы для подробной характери-
стики формы тела и изготовления по ним жестких или гибких
моделей, которые продувают в аэродинамических трубах или
буксируют в опытовых гидродинамических бассейнах с целью
получения основных экспериментальных гидродинамических ха-
рактеристик этих моделей.
Опыт показывает, что подробный обмер живого дельфина
в естественных условиях и последующее построение на основе
такого обмера теоретического чертежа представляет нелегкую
задачу не только ввиду сложной криволинейной формы тела.
Вследствие периодических изменений формы тела в нестацио-
нарном цикле плавания заранее не очевидно, какую форму сле-
дует выбрать и зафиксировать. Поэтому обычно составляют как
бы обобщенный теоретический чертеж выровненного плаваю-
щего животного, с прямой продольной осью тела и нулевым уг-
лом атаки хвостового плавника, например, по обмерам на плаву
в небольшой ванне. Иногда принимают дополнительные допу-
щения о форме сечений тела дельфина или даже ограничива-
ются изображением только внешних обводов тела, его профилей
в вертикальной и горизонтальной плоскостях.
Возможно также построение профиля тела вращения, экви-
валентного телу дельфина по его натурным фотографиям сбоку
и сверху [76]. Вначале по несимметричному контуру дельфина
с изогнутой средней линией в вертикальной плоскости строят
симметричный контур с прямолинейной средней линией. Затем,
пользуясь тем, что в горизонтальной плоскости дельфин имеет
симметричный контур с прямой средней линией, наносят соот-
ветственно вертикальный симметричный контур и по ним обоим
проводят осредненный профиль тела вращения. Как показывает
•опыт, такие многократные построения различаются незначи-
тельно и вполне оправданы для технических целей.
Геометрические характеристики формы тела всех трех черно-
морских видов дельфинов — белобочки, афалины и морской
свиньи — приведены в работе [10]. Для обмеров были отобраны
здоровые самцы средней упитанности. Обмеры периметров их
тела произведены во многих поперечных сечениях и в положе-
нии на плаву. Эти данные приняты в расчет в качестве исход-
ных параметров. Теоретические чертежи строили для «обоб-
щенных» дельфинов на основе допущения об эллиптических се-
чениях тела, что значительно ближе к действительности, чем
допущение о круговых сечениях тела дельфина.
В результате обработки проведенных измерений с примене-
нием статистических методов и изучения геометрических харак-
теристик в строении тела черноморских дельфинов были обна-
ружены следующие особенности. По длине тела дельфина (от
начала лобной подушки до дыхала и от начала основания спин-
ного плавника до конца анального отверстия) имеются два
аномальных, в сравнении с хорошо обтекаемой гидродинамиче-
29
ской моделью, участка небольшой протяженности. На этих
участках средняя линия профиля дельфина отклоняется от об-
щей прямой, однако не более чем на 1 % длины тела, что объ-
ясняется асимметрией расположения внутренних органов дель-
фина. Кроме того, выявлена возможность существования пере-
менного по длине тела положения наибольшего сечения на
участке длиной около 7 %, в зависимости от режима плавания.
По нашему мнению, эта особенность весьма интересная и имеет
существенное значение для гидродинамической характеристики
исследованных видов дельфинов. Вместе с этим выполненное
сравнение фотографий поперечных срезов тела дельфина и со-
Рис. 6. Теоретические чертежи черноморских дельфинов [11, 62]: а — бе-
лобочка (обыкновенный); б—афалина; в — азовка (морская свинья).
30
ответствующих эллиптических сечений обобщенного дельфина
показало небольшое расхождение в площадях сечений, менее
1 % (только на коротких аномальных участках до 5%). Таким
образом, геометрические характеристики обобщенного «эллип-
тического* дельфина в пределах обычной точности расчетов сов-
падают с действительными их характеристиками.
Теоретические чертежи черноморских дельфинов достаточно
полно и точно представлены в работах акад. В. Е. Соколова
с соавторами [11, 62]. Для построения теоретических чертежей
гипотетически было выбрано положение дельфина при свобод*
Таблица 4. Безразмерные геометрические характеристики
черноморских дельфинов [11, 62]
Характеристики Относительные значения по теорети- ческим чертежам
белобочка афалина морская свинья
Длина выступающего рострума /р/LK 0,092 0,050 0
Длина хвостового плавника (кор- невой хорды) Zxn/LK 0,110 0,104 0,130
Наибольшая высота корпуса (без спинного плавника) H/La 0,215 0,209 0,176
^Наибольшая ширина корпуса 0,191 0,175 0,160
Отстояние наибольшей ширины от кончика носа xglL* 0,426 0,410 0,404
Степень эллиптичности корпуса Bl Н 0,89 0,84 0,91
Относительное удлинение корпуса LKlB Площадь плавника: 5,23 5,00 5,04
хвостового Sxn/Lx 0,0115 0,0237 0,0210
спинного Scn/Lx 0,0085 0,0113 0,0068
грудного Srn/£2 0,0076 0,0075 0,0096
Общая площадь смоченной поверх- ности корпуса с рострумом О/£х 0,432 0,520 0,445
Площадь миделевого сечения 0,0338 0,0378 0,0429
Водоизмещение №7£х 0,0170 0,0218 0,0160
Коэффициент общей полноты б = = WILBH 0,346 0,540 0,568
Коэффициент формы т) .= 0,154 0,150 0,142
Отношение коэффициентов полез- ного объема и гидродинамического сопротивления (Сх = 0,03), k = т)/Сх 5,14 5,00 4,20
Примечание. Дк — длина корпуса без рострума и хвостового плавника; Дд —общая длина тела животного.
31
ном скольжении по инерции в горизонтальной плоскости, когда
тело животного вытянуто в длину (рис. 6). Однако фактически
обмеры тела дельфинов проводили хотя и на плаву, но в не-
большой ванне, в расслабленном состоянии мышц животного.
Геометрические характеристики формы тела черноморских дель-
финов, рассчитанные по теоретическим чертежам, приведены
в табл. 4.
Резюмируя вышеизложенное, отметим следующие особенно-
сти формы тела дельфинов по сравнению с внешней формой тех-
нических объектов (в том числе дирижаблей, фюзеляжей само-
летов, подводных судов) при числах Рейнольдса порядка Re=
= 1074-108. У всех дельфинов и китов диаметральные в верти-
кальной плоскости обводы тела имеют несимметричную форму
с несколько искривленной средней линией, а в горизонтальной
плоскости — симметричную форму с прямой средней линией. По-
перечные сечения тела непрерывно изменяются от носа к хво-
сту; цилиндрическая вставка в средней части тела отсутствует.
Скоростные дельфины имеют сечения тела, близкие к эллипсу;
степень эллиптичности корпуса В/Я=0,844-0,91. Наибольшие
отклонения формы сечений тела от эллипса встречаются на
носовом, заостренном и хвостовом, килеватом участках. Отно-
сительное отстояние наибольшего сечения от носа тела обычно
х^0,40. Относительное удлинение корпуса, как правило, Х =
= £/Вл*5,0. Наблюдаются ламинаризованные формы тела у ско-
ростных дельфинов. Обобщенный коэффициент формы черно-
морских дельфинов т) = ^2/3/Й = 0,15, что несколько выше, чем
у так называемого «торпедообразного» цилиндрического тела
с полусферическим оголовком и конусообразной хвостовой ча-
стью, и гидродинамически целесообразно для достижения мак-
симально возможного полезного объема при относительно ма-
лом гидродинамическом сопротивлении. Сказанное подтверж-
дается экспериментами по распределению давления на телах
вращения одинаковой относительной толщины, но различных
очертаний, в том числе очертаний носовой части [76].
§ 2. Оптимизация формы в плане машущего
плавникового движителя китообразных
Все плавники китообразных — машущий хвостовой двухло-
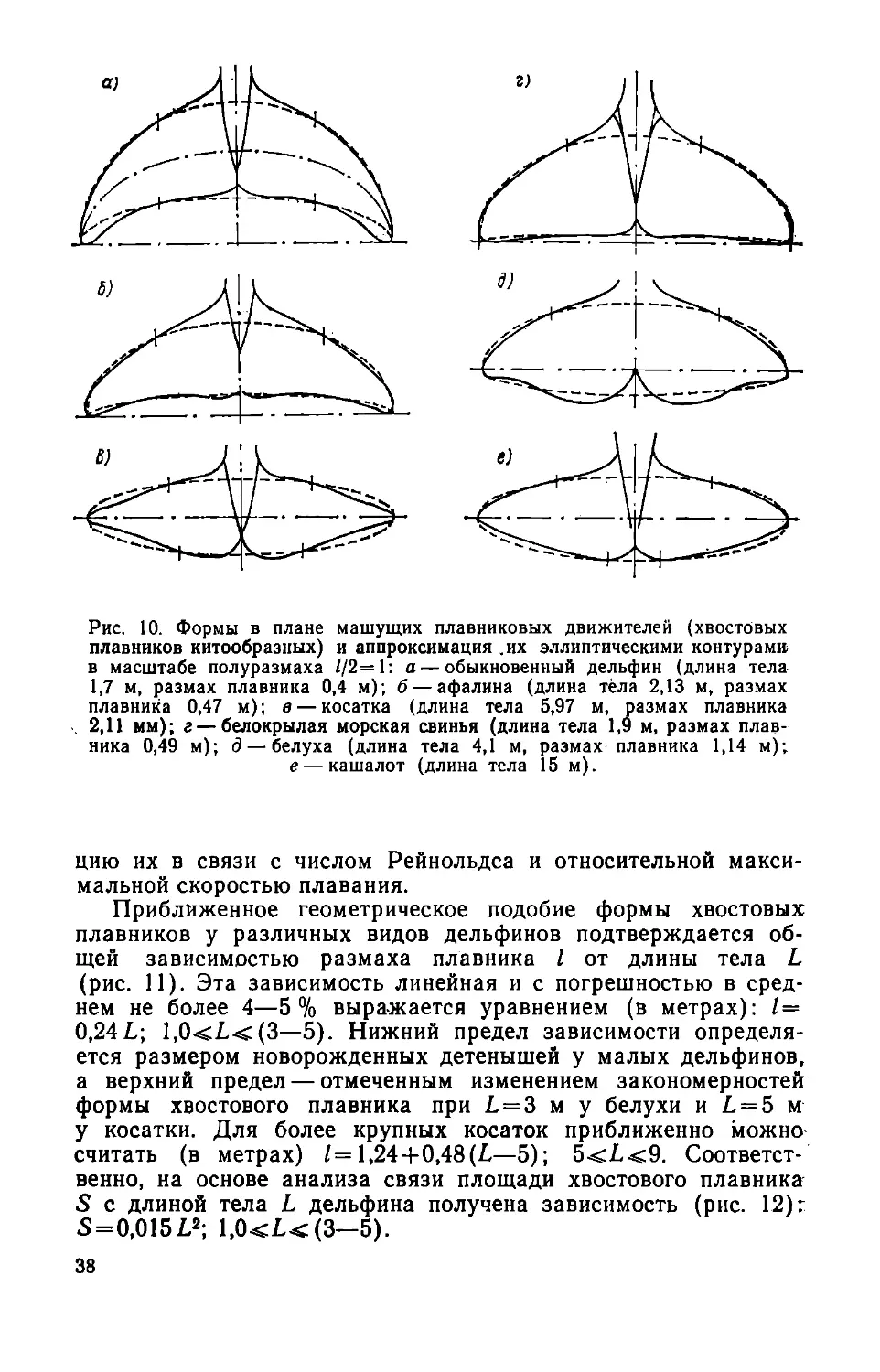
пастный, неподвижный спинной и подвижные парные грудные
однолопастные — представляют собой сплошные упругие под-
водные крылья сложной формы в плане, с криволинейными
очертаниями передней и задней кромок. Для сравнения на
рис. 7 приведены масштабные очертания плавников двух ско-
ростных черноморских обыкновенных дельфинов средних раз-
меров. Контуры плавников получены нами путем обмеров об-
водов тушек, замороженных на плаву, но не фиксированных
в формалине. Характерно, что несмотря на известное разное
32
О.........100 ....... ZOO......"» мм
Рис. 7. Масштабные контуры
в плане плавников черноморских
дельфинов-белобочек. Длина тела
1,67 м (/) и 1,88 м (2): а — хво-
стовые; б — спинные; в — грудные
плавники.
Рис. 8. Плавники крупнейших ско-
ростных дельфинов-косаток (масш-
табные контуры их в плане): а —
спинной; б — грудной; в — хвостовой
плавники.
э — эмбрион длиной 0,88 м; взрослые
дельфины длиной: / — 2,53 м; 2 — 3,56 м;
<3 — 5,18 м; 4— 5,97 м; 4'— 6,45 м.
функциональное назначение этих трех видов плавников и раз-
личное расположение их вдоль тела (спинной и грудные при-
мыкают к телу в средней части, а хвостовой плавник располо-
жен за телом), важнейшие передние кромки лопастей всех
плавников в основной своей части имеют практически близкие
очертания. Исключение составляют корневые плавные «за-
лизы» в местах сопряжения плавников с телом дельфина, кото-
рые больше различаются у разных плавников. Кроме того, не-
одинаковы углы установки плавников к потоку.
Приведенные сопоставления подтверждаются и другими экс-
периментальными данными о плавниках малых дельфинов, что
позволяет утверждать о приближенном геометрическом подобии
очертаний плавников у взрослых особей одного и того же вида.
Тем самым определяющими параметрами для формы плавников
являются длина тела и скорость плавания дельфинов, т. е. в ко-
нечном счете значение числа Рейнольдса и относительная ско-
рость плавания — главные гидродинамические характеристики
вида.
Особый интерес представляет анализ формы в плане плав-
ников самого крупного скоростного дельфина-косатки, по-
скольку с возрастом длина ее тела увеличивается в несколько
раз. Пользуясь данными обмеров и графическими материалами
о плавниках косаток различных размеров, помещенными в ра-
боте [75], мы привели их к одному масштабу и представили
в единообразном виде (рис. 8). Анализируя эти формы, заме-
чаем, что по мере роста крупного дельфина очертания его плав-
2 С. В. Першин 33
ников существенно изменяются. При длине тела косатки до 3 м
очертания хвостового и спинного плавников напоминают тако-
вые у обыкновенного дельфина (см. рис. 7). Однако с ростом
тела передняя кромка у хвостового плавника косатки стано-
вится положе, изменяется форма выемки задней кромки, появ-
ляются дополнительные приросты площади лопастей. При длине
тела косатки свыше 5—6 м передняя кромка хвостового плав-
ника делается почти прямолинейной, с небольшой постоянной
положительной стреловидностью, а задняя кромка становится
криволинейной, с переменной отрицательной стреловидностью;
обе лопасти вытягиваются и приобретают почти симметричную
форму относительно поперечной оси по размаху плавника. Это
ведет к увеличению относительного удлинения хвостового плав-
ника, что гидродинамически целесообразно. Одновременно ста-
новится симметричным, в форме равнобедренного треугольника,
высокий спинной плавник косатки.
Указанные тенденции в изменении формы хвостового плав-
ника по мере роста косатки характерны, хотя и в меньшей
степени, для хвостового плавника крупного нескоростного дель-
фина-белухи.
Хвостовой плавник у китообразных совершает с помощью
хвостового стебля при поступательном движении животного
большие вертикальные машущие колебания, амплитуда которых
примерно равна высоте тела. Поэтому большое гидродинамиче-
ское значение имеет еще форма поперечных сечений хвостового
стебля у китообразных. Анализ данных показывает, что у боль-
шинства видов дельфинов и китов поперечные сечения хвосто-
вого стебля симметричные чечевицеобразные, однако они встре-
чаются и в форме толстого профиля Н. Е. Жуковского. Оче-
видно, это зависит от эффективности взмахов хвостового плав-
ника вверх и вниз, от того, одинаковые они или различные, и
обусловлено несимметричным строением тела китообразных
в вертикальной плоскости.
Чтобы выявить направление оптимизации формы в плане
машущего плавникового движителя китообразных, воспользу-
емся указанным во введении биологическим принципом эмбрио-
генеза. Имеется в виду, что в эмбриональный период зародыши
животных как бы повторяют внешние формы многовекового
хода развития и совершенствуются по опыту жизни многочис-
ленных предшествующих поколений. Для анализа применим
данные трех независимых биологических исследований 200 эм-
брионов черноморского обыкновенного дельфина-белобочки1 и
243 эмбрионов кита-финвала2, а также 227 эмбрионов кита-ка-
шалота 3. В каждом из этих исследований приведены различ-
1 См. статью М. М. Слепцова (Бюлл. МОИП. Отд. биол. 1940. № 49.
С. 5—6).
2 S. Ohsumi (SRWRT, 1960. № 15).
3 См. книгу А. А. Берзина «Кашалот» (М., 1970, 367 с.).
34
ные формы развития хвостовых плавников эмбрионов рассмат-
риваемых видов, отвечающие известной длине их тела. Сопо-
ставляя их, можно выявить 8 однотипных для всех китообраз-
ных форм развития плавниковых движителей, которые пока-
заны на рис. 9.
Как видно, хвостовые плавники эмбрионов различных ви-
дов дельфинов и китов по мере роста последовательно при-
обретают формы в плане аналогичные таковым у наземных
длиннохвостовых млекопитающих, полуводных (бобры, выдры,
каланы) и водных (ламантины, дюгони) животных, которые
генетически не родственны между собой. Отмеченный общий
путь развития обусловлен биологическим принципом конвер-
генции с универсальным приспособлением животных к все бо-
лее возрастающим скоростям плавания, наивысшими из кото-
рых среди млекопитающих обладают наиболее специализиро-
ванные в плавании вторичноводные китообразные.
В эволюции форм хвостового плавника китообразных как
машущего движителя большое значение имели следующие ва-
риации форм: 1) локализация плавника в самом конце лате-
рально уплощенного хорошо обтекаемого хвостоврго стебля
что с удалением от тела животного привело к увеличению ам-
плитуды колебаний и углов атаки плавника, а потому и к ро-
сту тяги движителя; 2) существенное изменение формы в плане
плавника — постепенное уширение его поперек тела и повыше-
ние относительного удлинения, а также последовательное из-
менение стреловидности и плавные криволинейные очертания
передней и задних кромок; это повысило гидродинамическое
качество плавника как крыла сравнительно малого размаха;
3) дальнейшая оптимизация формы хвостового плавника в на-
правлении тонких изменений выпуклости — вогнутости очерта-
ний обеих кромок, увеличение выемчатости задней кромки и
обособление левой и правой лопастей, заострение их концевых
частей, что повысило пропульсивные свойства всего движи-
тельного комплекса китообразных, применительно к каждому
виду и его особенностям.
Обращают на себя внимание следующие данные исследо-
ваний. Размеры взрослых дельфинов и китов различаются
между собой на порядок величины, а однотипные формы хво-
стового плавника, наблюдаются у эмбрионов при длине тела
одного порядка, в том числе к концу современной 8-й формы
плавника длина эмбрионов дельфина-белобочки, кита-сейвала
и кита-кашалота почти одинаковая — около 1 м. Между тем
рождаются детеныши и сразу начинают плавать при сущест-
венно различной длине тела — у малых дельфинов 1,0—1,2 м,
и у китов 5,0—6,5 м. Нами были изучены формы в плане хво-
стовых плавников крупных эмбрионов китов (сейвала и фин-
вала), близких к предродовым формам, которые удалось по-
лучпт|| в 1970—1971 гг. с известной советской китобойной базы
2*
35
36
Рис. 9. Схема последовательной смены восьми форм в плане хвостового
плавника у эмбрионов китообразных, отражающая конвергентную законо-
мерность эволюции формы: а — черноморский обыкновенный дельфин; б —
скоростной кит—антарктический финвал; в — нескоростной кит-кашалот.
«Юрий Долгорукий». Оказалось, что форма в плане хвосто-
вых плавников у крупных эмбрионов сейвала и финвала от
8-й формы до предродовой практически почти не изменяется.
В этом более позднем периоде эмбриогенеза плавники лишь
пропорционально увеличиваются в размерах; соблюдается гео-
метрическое подобие.
Таким образом, эволюция и оптимизация формы в плане
хвостовых плавников китообразных в эмбриональный период
развития разделяются на две существенно различные фазы:
1-я фаза, назовем ее «исторической», состоит из указанных
выше восьми ранних форм, когда у хвостовых плавников эм-
брионов форма в плане изменяется качественно от наземных
к водным животным и формируются собственно китообразное
формы плавников; 2-я фаза, «современная», состоит из одной
формы, при которой хвостовые плавники эмбрионов лишь по-
добно увеличиваются в размерах вплоть до рождения дете-
ныша. Пользуясь имеющимися данными по росту эмбрионов
финвалов, мы получили, что 1-я фаза эмбрионального разви-
тия плавников продолжается около 4,5 мес., т. е. менее поло-
вины периода эмбриогенеза, который у китообразных состав-
ляет 10,5—11 мес.
Теоретические соображения и экспериментальные данные
дают основание полагать [44]: различные сложные формы
в плане хвостовых плавников разных видов взрослых китооб-
разных приближенно можно аппроксимировать с помощью эл-
липтических крыльев, получаемых при скольжении границ ис-
ходного кругового крыла. Эллиптические в плане крылья
имеют различные очертания передней и задней кромок, пере-
менную по размаху прямую и обратную стреловидность. Об-
щими свойствами этих крыльев являются эллиптическое рас-
пределение по размаху длины местных хорд и закономерное
распределение в широких пределах относительного удлинения.
Нами выполнен анализ форм в плане хвостовых плавни-
ков— машущих движителей — и приближенная аппроксимация
их эллиптическими крыльями для шести видов китообразных,
достаточно разнообразных и типичных форм (рис. 10). Для
удобства сравнения полуразмах каждого плавника (большая
полуось эллипса) приняты за единицу длины //2=1 и все очер-
тания крыльев представлены в одном и том же относительном
масштабе. Основные очертания передней и задней кромок
плавников действительно эллиптические, однако от них значи-
тельно отличаются большие «зализы» сложной формы при со-
пряжении плавника с окончанием несущего машущего хвосто-
вого стебля. Важно отметить, что за счет общего осреднения
границ эллиптическими крыльями удовлетворительно аппрок-
симируются площадь и относительное удлинение реальных хво-
стовых плавников китообразных. При этом характер измене-
ний формы в плане хвостовых плавников выражает оптимиза-
37
Рис. 10. Формы в плане машущих плавниковых движителей (хвостовых
плавников китообразных) и аппроксимация .их эллиптическими контурами
в масштабе полуразмаха //2=1: а — обыкновенный дельфин (длина тела
1,7 м, размах плавника 0,4 м); б — афалина (длина тела 2,13 м, размах
плавника 0,47 м); в — косатка (длина тела 5,97 м, размах плавника
2,11 мм); г—белокрылая морская свинья (длина тела 1,9 м, размах плав-
ника 0,49 м); д — белуха (длина тела 4,1 м, размах плавника 1,14 м);
е — кашалот (длина тела 15 м).
цию их в связи с числом Рейнольдса и относительной макси-
мальной скоростью плавания.
Приближенное геометрическое подобие формы хвостовых
плавников у различных видов дельфинов подтверждается об-
щей зависимостью размаха плавника I от длины тела L
(рис. 11). Эта зависимость линейная и с погрешностью в сред-
нем не более 4—5 % выражается уравнением (в метрах): / =
0,24L; 1,0<£<(3—5). Нижний предел зависимости определя-
ется размером новорожденных детенышей у малых дельфинов,
а верхний предел — отмеченным изменением закономерностей
формы хвостового плавника при L = 3 м у белухи и L = 5 м
у косатки. Для более крупных косаток приближенно можно
считать (в метрах) 1= 1,24+0,48(L—5); 5<L<9. Соответст-
венно, на основе анализа связи площади хвостового плавника
S с длиной тела L дельфина получена зависимость (рис. 12) г
S = 0,015L2; l,0<L<(3—5).
38
Рис. 11. Зависимость размаха хво-
стового плавника от длины тела
у дельфинов различных видов и
размеров, Z/L.
J— обыкновенный дельфин; 2 — афа-
лина; 3 — морская свинья; 4 — атлан-
тическая белуха; 5 — крупные эмбри-
оны кита-сейвала.
Рис. 12. Зависимость площади
хвостового плавника от длины те-
ла у различных видов дельфинов,
включая крупные эмбрионы, S(L).
1 — морская свинья; 2 —афалина;
3 — обыкновенный дельфин; 4 — ат-
лантическая белуха; 5 — косатка.
Сопоставляя графики на рис. 11 и 12 и определяющие их
зависимости, найдем для относительного размаха X хвостовых
плавников дельфинов среднее значение
Х = —= (0,24L)2 =з,85; 1,0 sg 3,0. (6)
S 0,015La
Таким образом, X~4 = const есть оптимальное значение от-
носительного размаха хвостовых плавников малых дельфинов
при всем разнообразии их форм в плане.
У крупной косатки при длине тела L = 6,0 м относительное
удлинение хвостового плавника Х=5,75. У крупных китообраз-
ных в онтогенезе при увеличении с возрастом длины тела
в несколько раз повышаются не только абсолютные, но и от-
носительные размеры хвостового плавника, что всегда следует
иметь в виду.
39
§ 3. Гидродинамический анализ профилей плавников
дельфинов и китов
Для анализа использован следующий оригинальный мате-
риал лаборатории функциональной морфологии Зоологического
института АН СССР. Детально исследовали целый хвостовой
плавник дельфина (белокрылой морской свиньи), добытый в на-
учной экспедиции у Командорских островов в июне 1967 г.
Дельфин имел общую длину 1,91 м, массу 100 кг, максимальную
высоту тела 0,385 м и наибольшую ширину тела 0,30 м. Кроме
этого, исследовали профили продольных вырезок хвостовых
плавников двух видов китов — сейвала и кашалота. Животные
были добыты китобойной базой «Юрий Долгорукий» на про-
мысле в Антарктиде в начале 1972 г. Сейвал имел общую
длину 12,5 м и общую массу 13,5 т, а кашалот — длину 13,2 м,
массу 27,8 т. Длина хорды профилей у плавников сейвала со-
ставляла 0,790 м, у кашалота 0,928 м.
Плавник дельфина и продольные вырезки плавников китов
были на месте добычи заморожены в естественном состоянии
и поступили в лабораторию практически недеформированными,
так как они в отличие от тела животного имеют нерастяжимую
внешнюю сухожильную оплетку. Профили сечений определяли
после оттаивания материала, до фиксации формалином. Про-
фили плавника дельфина измеряли по внешним контурам уве-
личенных фотографий срезов, а вырезки плавников китов —
непосредственным скицированием с натуры. Толщину профилей
инструментально измеряли по ортогональным хорде линиям,
проведенным через каждые 5 % длины хорды; дополнительные
точки проставлялись в носовой части профилей. Профили се-
чений плавников у китообразных симметричные. В тех слу-
чаях, когда плавники тушек были несколько изогнуты в тонкой
хвостовой части, профили выпрямляли до симметричных. По-
грешность определения относительных ординат у{Ь профилей
не превышала ± Ду = 0,14-0,2 % [46].
В результате произведенных измерений построили для
удобства анализа эпюрные профили сечений исследованных
плавников, у которых масштаб чертежа по оси ординат у
в 5 раз крупнее, чем по оси абсцисс х. Симметричные профили
хвостовых и других плавников дельфинов и китов сравнивали
с классическими аэродинамическими симметричными профи-
лями Н. Е. Жуковского НЕЖ-1 и ламинаризованным профи-
лем NACA-663 соответствующей толщины. Для этих профилей
опубликованы данные аэродинамических продувок.
Дельфин-белокрылая морская свинья достаточно крупный,
плавает со сравнительно высокой скоростью. С позиции гидро-
динамики его можно считать «типичным» (но не следует сме-
шивать с малым дельфином — с морской свиньей, см. табл. 3).
На рис. 13 показан в масштабе чертеж правой лопасти хво-
40
Рис. 13. Масштабный чертеж хвостового плавника белокрылой
морской свиньи (размах /=0,49 м). Форма в плане и профили
1—5 продольных сечений.
тт — линия, проходящая через точки хорды наибольшей толщины про-
филей; к к. — линия, проходящая через точки хорды профилей, начиная
с которых кормовые части профилей представляются утолщенными для
прочности плавника по сравнению с таковыми у классических аэроди-
намических профилей.
стового плавника и 5 профилей (1—5), измеренных по срезам
на одинаковом расстоянии от продольной оси плавника.
В верхней части рисунка приведен график двух зависимостей:
Ь/1 — кривая изменения относительной длины хорды профилей
с увеличением отстояния срезов от продольной оси плавника
и 2i/max/& — практически прямая, показывающая изменение
максимальной относительной толщины профилей с увеличе-
нием того же отстояния срезов 1—5.
Анализ профилей продольных сечений хвостового плавника
белокрылой морской свиньи показывает следующее. С удале-
нием от продольной оси плавника максимальная относитель-
ная толщина профилей уменьшается с 19,6 до 15,7 % (и ме-
нее) длины хорды данного плавника, а отстояние наибольшего
сечения от носика профилей увеличивается от 32 до 35 % длины
хорды, т. е. профили сечений в концевой части лопасти за-
метно тоньше и несколько более вытянуты. Профили 1 и 2
и кормовой части имеют точки перегиба, а у последних профи-
41
± y/b, %
Рис. 14. Эпюрные профили 1, 3, 5 продольных сечений хвосто-
вого плавника белокрылой морской свиньи и классические про-
фили, пересчитанные па максимальную толщину профиля 3:
а3— обобщенный симметричный НЕЖ-1; б3— ламинариэованный
симметричный профиль NACA-663.
лей 4 и 5 их нет, вследствие относительно большего утолще-
ния концевой части менее толстых профилей. Таким образом,
форма несущей поверхности хвостового плавника дельфина
весьма сложная, с тонкими взаимопереходами, оптимально вы-
работанными в процессе длительной эволюции природного ма-
шущего движителя.
Эпюрные профили 1, 3, 5 продольных сечений хвостового
плавника белокрылой морской свиньи показаны вместе с клас-
сическими аэродинамическими профилями а3 и Ь3 на рис. 14.
Средний профиль 3 практически совпадает с обобщенным про-
филем Н. Е. Жуковского а3 той же максимальной относитель-
ной толщины 18,1 % на первых от носика 40 % и последних
30 % длины хорды, т. е. на большей части длины. В пределах
порядковых 40—70 % длины хорды средний профиль 3 лопа-
сти плавника оказывается несколько более полным, чем аэро-
динамический профиль НЕЖ-1. Сравнительно с ламинаризо-
ванным профилем NACA-663 той же относительной толщины
(на рисунке Ь3) исследуемый профиль 3 значительно полнее
в носовой части и особенно в кормовой.
Профиль продольного сечения хвостового плавника кита-
сейвала имеет максимальную относительную толщину 18,2%,
отстояние от носика сечения с наибольшей относительной тол-
щиной затянуто в корму до 35 % длины хорды. Этот профиль
менее полный, чем у дельфина и классический НЕЖ-1. В носо-
вой части он приближается по форме к ламинаризованному
профилю NACA-663, а в кормовой части почти совпадает с ним.
42
Лишь начиная с 75 % длины хорды профиль сечения хвосто-
вого плавника сейвала существенно утолщен, очевидно, для
повышения механической прочности плавника. Внешне, визу-
ально этот профиль весьма изящный. Напомним, что сейвал —
наиболее скоростной вид китов;, максимальное число Рей-
нольдса по длине его тела ReL~ 1,8 • 108, а хвостового плавника
по длине хорды Reb«9* 106.
Исследованный профиль продольного сечения плавника ка-
шалота имел максимальную относительную толщину 19,0%,
которая отвечает отстоянию от носика профиля всего на 27 %
длины хорды.’ Вид профиля своеобразный. По сравнению со
всеми другими профилями он заметно полнее в носовой части,
намного менее полный на второй трети длины, а в концевой
части снова утолщенный. Эту необычную форму профиля
можно объяснить тем, что хвостовой плавник кашалота имеет
в плане, к задней кромке, увеличивающие его площадь «при-
ливы», и потому профиль получается удлиненным, не харак-
терным для других китообразных. Если представить себе этот
профиль без приливов площади плавника, несколько укоро-
ченным, при тех же абсолютных толщинах, то его относитель-
ные ординаты увеличатся, относительная абсцисса максималь-
ного сечения сдвинется от носика в корму и очертания рас-
сматриваемого профиля приблизятся к обобщенному профилю
НЕЖ-1. Кашалот — относительно медленно плавающий вид
китов, но наилучший ныряльщик на большие глубины.
На рис. 15 показаны для сравнения эпюрные профили про-
дольных сечений хвостовых и спинных плавников разных ви-
дов дельфинов, построенные по нашим данным для профиля 3
хвостового плавника белокрылой морской свиньи и по дан-
ным работы [82] для профилей А, В, С, которые частично нами
Рис. 15. Сравнение эпюрных профилей хвостовых и спинных
плавников различных видов дельфинов.
43
пересчитаны на другие относительные толщины и обозначены
с соответствующими подстрочными индексами. Исходный про-
филь А среднего сечения хвостового плавника скоростного
обыкновенного дельфина, имеющего максимальную относи-
тельную толщину 21,1 % и ёе отстояние от носа на 32 % длины
хорды, на рисунке обозначен Ав, так как пересчитан по мак-
симальной толщине профиля В сечения спинного плавника
скоростного тихоокеанского короткоголового дельфина. Про-
филь В имеет максимальную относительную толщину 19,3 %,
которая отстоит от носа на 33 % длины хорды. Для сечения
спинного плавника белокрылой морской свиньи построен про-
филь С с максимальной относительной толщиной 15,2%, на
расстоянии от носа 36 % длины хорды. На рисунке приведены
также профили: Зс и ас — обобщенные профили НЕЖ-1, пе-
ресчитанные на толщину 15,2% профиля С; ав — профиль
НЕЖ-1, пересчитанный на толщину 19,3 % профиля В. Анализ
графика показывает, что профили средних сечений машущих
хвостовых и немашущих спинных плавников одного и того же
или близких по скоростным характеристикам видов дельфинов;
очень сходны между собой и с классическим аэродинамиче-
ским симметричным профилем НЕЖ-1, за исключением участ-
ков профилей на второй трети длины хорды.
В работе [82] утверждается, что названные выше профили
А, В, С хвостового и спинного плавников скоростных видоа
дельфинов близки по форме к малоизвестным специальным
авиационным профилям Бортмана FX-05-191 и Эпплера ЕА-6,
спроектированным для диапазона чисел Рейнольдса Reb=
= (0,5—3,0) X106. Эти профили отличаются плавным возра-
станием давления к задней кромке, что препятствует отрыву
пограничного слоя. Такие профили допускают большие углы
атаки, необходимые дельфинам при маневрировании хвосто-
вым плавником, без угрозы возникновения чрезмерного пика
давления вблизи передней кромки, отрыва потока и кавитации.
Они не вызывают увеличения сопротивления с изменением от-
носительных толщин до 0,5 % длины хорды; сопротивление не-
много возрастает лишь при увеличении толщины профиля на
0,8—1,0%.
Для выявления особенностей профилей сечений плавников;
дельфинов были выполнены также теоретические расчеты рас-
пределения гидродинамического давления на изолированных
профилях [82]. Установлено, что на рассмотренных выше сим-
метричных профилях А, В, С минимум давления при нулевом
угле атаки расположен между 14—16 % длины хорды от но-
сика профиля. Безразмерные коэффициенты гидродинамиче-
ского давления этих профилей соответственно равны: р = —0,74;
—0,66; —0,46, т. е. достаточно высокие. Анализ расчетных дан-
ных показал, что плавники А, В, С не кавитируют вблизи сво-
бодной поверхности при угле атаки а = 0° до скоростей обтека-
44
пня потоком соответственно: 16,3; 17,3; 20,7 м/с. На плавнике
А обыкновенного дельфина кавитация на различных углах
атаки а = 2, 4, 6° начинается соответственно при скорости 13,9;
11,7, 9,9 м/с. Реальные углы атаки для плавников дельфина
на высоких скоростях не превышают 3—4° и лишь на малых
скоростях плавания достигают 20°. Естественно кавитация не
возникает. В противном случае кавитация вызывала бы не
только повышение гидродинамического сопротивления, но
также и болевые ощущения у животного. Увеличение же до-
полнительного сопротивления на больших углах атаки при ма-
лых скоростях плавания несущественно для дельфина.
В работе [62] приведены профили продольных сечений хво-
стового, спинного и грудного плавников черноморского дель-
фина-афалины. Утверждается, что они практически совпадают
с известными симметричными профилями NACA с большой
максимальной относительной толщиной 18—26%, отстоящей
па 25—35 % от передней кромки профиля. Как известно, про-
фили NACA имеют оптимальные обводы. Они обладают ма-
лым гидродинамическим сопротивлением и в то же время со-
здают значительную ^поперечную силу — в данном случае силу
тяги машущего плавника.
Профили сечений плавников китообразных имеют сходство
с формой симметричных обводов их тела в горизонтальной
плоскости как по типу профиля, так и по сравнительно боль-
шой толщине, однако в соответствии с поправкой на число
Рейнольдса. Так, форма тела скоростного короткоголового
дельфина в горизонтальном разрезе ламинаризованная; мак-
симальная относительная толщина 15%, отстоящая на 45 %
длины хорды от носа тела, близка к ламинаризованному про-
филю NACA-66. Лопасть же спинного плавника этого вида
дельфина близка по форме сечения к 19 %-ному обобщенному
симметричному профилю НЕЖ-1. Это соответствует макси-
мальным числам Рейнольдса для тела ReL~l,5-107 и для про-
филя плавника Ret,~l,l • 106 данного вида дельфина. Показа-
тельным является указанная выше ламинаризованная форма
профиля продольного сечения хвостового плавника скоростного
кита-сейвала, число Рейнольдса плавника которого приближа-
ется к Re& = 107.
§ 4. Оптимизация формы тела и машущего
плавникового движителя у скоростных рыб
Формы тела и машущего плавникового движителя вырабо-
таны в природе в процессе эволюционных приспособлений рыб
к условиям плавания в водной среде в течение многих миллио-
нов лет. У многих видов рыб нестационарным движителем яв-
ляется не только вертикальный машущий хвостовой плавник,
по и упругое тело с цикличным изменением формы. При вол-
45
нообразных движениях тела и колебаниях хвостового плав-
ника в горизонтальной плоскости они обтекаются с перемен-
ными углами атаки. Однако для сопоставления с погружен-
ными твердыми телами, которые на прямолинейном ходу
обычно имеют установившийся режим движения с нулевым уг-
лом атаки, можно изучать закономерности развития основной
формы выровненного тела скоростных рыб, отвлекаясь от не-
стационарности движения и изменений конфигурации тела.
Для сравнительного гидродинамического анализа автором
книги были использованы известные расчетные и эксперимен-
тальные данные различных авторов по стационарному обтека-
нию с нулевым углом атаки моделей твердых тел, форма ко-
торых приближается к соответствующим у скоростных рыб.
В качестве исходных материалов по рыбам послужили квали-
фицированные рисунки и описания их в биологических спра-
вочниках, монографиях и статьях. В общей сложности были
просмотрены и сопоставлены между собой более тысячи точ-
ных рисунков и описаний рыб из различных морей и океанов.
Кроме того, неоднократно были осмотрены также долговре-
менные коллекции рыб Зоологического института АН СССР.
Чтобы наглядно показать влияние формы тела и формы
Рис. 16. Влияние внешнего строения нескоростных видов рыб на зна-
чение безразмерной максимальной скорости плавания =
а — рыбы, имеющие различное относительное удлинение L///=var при
схожих хвостовом и других плавниках; б — рыбы с разными формами
хвостового и других плавников при L//7 = idem.
/—форель радужная (L/H«4.6; Отах =2.0); 2 — плотва (Л/Я=3,3; ^max =
—0,8); 3 — карась (L/H = 2,2; l/max =0.6); 4 — сельдь атлантическая (L//f=4,6;
Цтах = 0,9); 5 — треска (Т-/Я=4,5; ^max =0,7); б — бычок (L/W=4,5; t/max =0,5).
46
плавникового движителя рыб на достигаемую ими максималь-
ную скорость движения, вначале приведем два характерных
примера, в каждом из которых представлены по три хорошо
изученных небольших пресноводных или морских рыбы с по-
степенно увеличивающимися невысокими скоростями (рис. 16).
Рост относительного удлинения тела рыбы при почти неизмен-
ной форме хвостового плавника или, напротив, изменение
формы хвостового и других плавников рыб при сохранении
одного и того же относительного удлинения телд приводит
в каждом из случаев к увеличению безразмерной максималь-
ной скорости в два-три раза. С учетом крупных скоростных
рыб общее различие рыб по абсолютному значению макси-
мальной скорости плавания превышает порядок величины.
Форма, размеры тела и плавников и максимальная ско-
рость движения рыб, очевидно, взаимосвязаны. Показанная
на рис. 16 смена формы хвостовых плавников является не
только характерной, но и отвечает следующей общей законо-
мерности: по мере увеличения скорости движения различных
видов рыб форма хвостовых плавников их последовательно из-
меняется в определенном порядке — сперва однолопастная,
вначале овальной формы, затем треугольной; потом двухло-
пастная, с выемкой, все меньшей стреловидности и большего
относительного удлинения, с острыми концами. Эта закономер-
ность подтверждается и в онтогенезе известных видов рыб.
Гидродинамически подобное изменение формы хвостового
плавника можно объяснить последовательным уменьшением
профильного и индуктивного сопротивления его как машущего
крыла конечного размаха с четкой фиксацией центров мощных
пропульсивных вихрей, срывающихся с острых концов плав-
ника у скоростных рыб.
«Скоростными» в биологической литературе обычно назы-
вают такие рыбы, которые плавают с максимальной скоростью
более десяти значений длины тела в секунду. Большинство рыб
плавает со скоростью в несколько раз меньшей. Поскольку
максимальная скорость движения рыб не пропорциональна
длине в первой степени, длина рыб изменяется в широком диа-
пазоне, а скорость плавания существенно зависит от продол-
жительности плавания, приведенное сравнение страдает"неко-
торой неопределенностью. Виды скоростных рыб достаточно из-
вестны, но экспериментальных данных по ним очень мало.
Внешнее строение рыбы определяется прежде всего приспо-
соблениями к движению в воде, и это является наиболее об-
щей основой исторического развития рыб [3]. Нами сделана
попытка выяснить основные признаки скоростных рыб малого
сопротивления. В результате изучения и обработки имеющихся
данных можно отметить большой ряд таких признаков.
Основными признаками скоростных рыб являются сле-
дующие:
47
1. Хорошообтекаемое тело, симметричное относительно вер-
тикальной плоскости. Обычно это не тело вращения, так как
форма тела рыбы постепенно трансформируется по всей длине
и поперечное сечение изменяется от вертикального овала за
головой до горизонтального в хвосте. Характерными являются:
а) наибольшее сечение отнесено назад, относительное от-
стояние его от передней точки x = x/L>0,35;
б) плавное очертание тела в вертикальной и горизонталь-
ной проекциях нередко напоминает ламинаризованный про-
филь, даже при малом относительном удлинении L/H<5;
в) тонкий, уплощенный и с острыми кромками хвостовой
стебель, хорошо приспособленный для чаСтых колебаний в го-
ризонтальной плоскости с наименьшим сопротивлением.
Главные измерения тела могут быть существенно различны:
относительное удлинение (по высоте Н) L/H = 3,54-7,0; относи-
тельная высота (по ширине В) ЩВ= 1,34-1,8.
Классическими примерами скоростных костистых рыб яв-
ляются тунцовые рыбы—скумбрия, пеламида, тунец, ваху. Ту-
нец, обладающий коротким толстым телом и негибким позво-
ночным столбом, приспособлен к машущим колебаниям хво-
стового плавника малой амплитуды, но большой частоты; ваху,
имеющая длинное тонкое тело, гибкий позвоночный столб и
короткие позвонки, приспособлена к изгибным колебаниям
тела большой амплитуды. Как видно, параметры изгибных ко-
лебаний скоростных костистых рыб бывают различными, что
обусловлено формой их тела. Аналогичные примеры можно
привести для хрящевых акуловых рыб (рис. 17).
2. У скоростных рыб хорошо обтекаемы все местные детали
формы. У этих рыб нет резко выделяющихся выступов и ис-
точников локальных возмущений, которые могут вызвать боль-
Рис. 17. Типичные виды скоростных рыб: а — костистые; б—аку-
ловые.
/ — тунец обыкновенный (утолщенный); 2 — ваху (удлиненная); 3 — белая;
4 — мако.
48
шие местные сопротивления и нарушение устойчивости погра-
ничного слоя при быстром плавании: а) глаза их относительно
малых размеров и имеют обтекатели; это важно, так как в го-
ловной части тела пограничный слой тонкий; б) различные
плавники (спинные, боковые, грудные) сравнительно неболь-
шие; перья их наклонены по течению; при быстром плавании
спинные плавники убираются (заваливаются) в специальные
продольные углубления в спинной части тела, а у тунца
даже боковые плавники прижимаются заподлицо с поверх-
ностью тела в имеющиеся на теле сбоку неглубокие карманы;
в) поверхность жабр с преджаберными крышками снаружи
гладкая, и выброс воды через жабры при дыхательных актах
синхронизирован с колебаниями хвостового плавника и изги-
бами тела, что встречается только у скоростных рыб (у мед-
ленно плавающих рыб такая синхронизация дыхания с изги-
бами тела не обнаружена).
3. В диффузорной части тела скоростных рыб имеются та-
кие органы, которые могут служить элементами механического
управления пограничным слоем: а) гидродинамическая ре-
шетка из пяти-десяти пар малых, специальной формы поворот-
ных дополнительных плавничков на спинной и брюшной пока-
тых сторонах тела — от наибольшего сечения до хвостового
плавника. В работе [33] их назначение объясняется снятием от-
рицательной подсасывающей силы; б) продольные парные,
ножеобразные костистые кили с обеих сторон на конце хвосто-
вого стебля, а иногда и посредине хвостового плавника.
Отметим еще «корсет» из более грубой кожи с чешуей на
в общем гладкой поверхности тела тунцовых рыб (перед наи-
большим сечением). Можно представить, что назначение «кор-
сета» подобно действию известного в гидродинамике кольца
Прандтля при смене критических режимов обтекания шара.
4. Вертикальные хвостовые плавники двухлопастные, пере-
менной стреловидности, имеют большую выемчатость и ост-
рые концы с относительным удлинением плавника до А = 8 и
более (у нескоростных рыб 1—3).
Существенными являются еще следующие характеристики
хвостовых плавников: а) чем выше скорость рыбы, тем больше
острые концы лопастей хвостового плавника выступают за пре-
делы корпуса тела в наибольшем сечении; б) высокочастот-
ный хвостовой плавник скоростных рыб жесткий, негнущийся,
без внутренней мускулатуры для управления каждой лопастью;
хвостовые же плавники удлиненных рыб, тем более нескорост-
ных (с малым числом колебаний в секунду), гибкие, расслаб-
ленные.
Из вышеизложенного следует, что вполне определенные
внешние признаки скоростных рыб малого сопротивления пред-
ставляют несомненный интерес для анализа ходкости погру-
женных твердых тел. Однако совокупность таких признаков
49
подчинена функциям плавания водного животного и основной,
особенности нестационарного движения — волнообразным из-
гибным колебаниям тела и машущим колебаниям плавнико-
вого движителя.
Кроме отмеченных выше двух групп обычного вида хорошо-
обтекаемых скоростных рыб — удлиненных и утолщенных,
тунцовых и акуловых, особо выделяется группа крупных мор-
ских хищных рыб надсемейства ксифиоидных, которые отлича-
ются своими выступающими впереди головы костными ростру-
мами. Это скоростные меч-рыба, копьеносцы, марлины и па-
русники. Наибольшей максимальной скоростью плавания,
кратковременно до 30—35 м/с, обладает меч-рыба. Она имеет
общую длину до 3 м, из них до */з занимает клиновидный уп-
лощенный меч (рис. 18). Неоднократно предпринимались по-
пытки объяснить необычайно высокую скорость плавания меч-
рыбы, в том числе и наличием выступающего рострума. Од-
нако экспериментальная проверка этих гипотез затруднена
из-за недостаточно точных исходных натурных данных.
Гидродинамической функции выступающего рострума меч-
рыбы посвящена работа Л. Ф. Козлова, И. В. Леоненко (Био-
ника. Киев. 1973. Вып. 7. С. 8—14). Однако ее выводы не объ-
ясняют высокой скорости плавания меч-рыбы.
В работе [23] также выполнены оригинальные квазистацио-
нарные расчеты пограничного слоя и эксперименты, которые
позволили утверждать, что оптимальная область чисел Рей-
нольдса твердых тел, где может проявляться «положительный
эффект меч-рыбы», составляет Re~4• 1064-8• 107. Этот вывод.
Рис. 18. Предельно скоростной вид меч-рыбы (профили сбоку, сверху; сече-
ния).
50
согласуется с действительностью, но все же не раскрывает за-
гадку меч-рыбы.
В работе М. А. Басина и др. (Тр. Ленингр. общ-ва естество-
испытат. 1976. Т. 73. Вып. 5. С. 70—78) пластмассовая модель
меч-рыбы длиной 1 м (масштаба 1 :3) подробно испытана
в аэродинамической трубе, а также в опытовом и кавитацион-
ном бассейнах, при различных скоростях обтекания потоком,
разных углах атаки и различной глубине погружения под сво-
бодной поверхностью воды. Применительно к ходкости твердых
тел можно сделать интересный вывод, что тела с обводами
меч-рыбы имеют преимущества по сравнению с телами враще-
ния традиционных обводов как благодаря уменьшению общего
сопротивления на большой скорости движения в погружен-
ном состоянии (заметно ниже сопротивления трения экви-
валентной пластины при турбулентном режиме), так и благо-
даря уменьшению волнового сопротивления при движении
вблизи свободной поверхности.
Тело меч-рыбы не имеет формы тела вращения, как это
обычно принимают в расчетах и экспериментах. В отличие от
других работ нами в гидробионическом плане впервые после-
довательно анализирован весь комплекс известных в настоя-
щее время достоверных факторов о меч-рыбе [42]. Меч-рыба
рассмотрена в качестве предельно скоростного гидробионта.
При сопоставлении ее с таковыми приняты во внимание как
гидродинамические закономерности, так и закономерности био-
логической природы. Отметим некоторые из них.
Необходимой предпосылкой высокой скорости плавания
меч-рыбы является необычайно мощный двигательно-движи-
тельный комплекс, создающий большую силу тяги. Это опре-
деляется прежде всего высокой энерговооруженностью меч-
рыбы. Мышцы составляют 67 % всей массы тела, что уже ли-
митируется биологически. Даже в сравнении со скоростными
гидробионтами — дельфином-белобочкой и акулой мако — энер-
говооруженность меч-рыбы выше почти в два раза. А у подав-
ляющего большинства рыб мышцы составляют 30—45 %
массы тела.
Хвостовой плавник меч-рыбы — высоко эффективный ма-
шущий' движитель по ряду показателей. Относительная пло-
щадь несущей поверхности хвостового плавника у меч-рыбы
наибольшая (в сравнении с другими ксифиоидными рыбами)
и значительно превышает таковую у скоростного обыкновен-
ного тунца соответствующей длины. Относительное удлинение
хвостового плавника меч-рыбы около 7, 5, т. е. весьма большое
для водного животного. Хвостовой плавник меч-рыбы имеет
размах, значительно превышающий наибольшую высоту тела.
Однако он не выступает за концы спинного и грудного плав-
ников, поэтому хвостовой плавник у меч-рыбы все же на-
ходится в пределах вихревых слоев других плавников, что
51
повышает эффективность машущего движителя [84]. Вместе
с тем относительно длинная хвостовая часть тела меч-рыбы и
жесткий практически несгибаемый скелет позволяют делать хво-
стовому плавнику большие поперечные размахи относительно
ЦТ тела. Это создает большую ометаемую площадь движителя.
Частота колебаний хвостового плавника также относительно
высокая, поскольку сам плавник жесткий и несгибаемый.
На рис. 19 приведено сравнение формы в плане машущих
плавниковых движителей — хвостовых плавников скоростных
дельфинов (белобочки и афалины) и скоростных рыб (пела-
миды и меч-рыбы). Машущий движитель мечнрыбы «вписыва-
ется» в таковой у дельфина при одинаковом приведенном раз-
махе. Из гидродинамической теории волнового движителя
в идеальной жидкости известно, что для его эффективности
особое значение имеет форма передней кромки и большая вы-
емчатость задней кромки. Выемка плавника заполняется вих-
ревыми слоями, и он работает для создания силы тяги, как
безвыемчатый, а гидродинамическое сопротивление умень-
шается в реальной жидкости. Это наблюдается и у меч-
рыбы.
Резко выделяющийся рострум у меч-рыбы несомненно
имеет различное биологическое и гидродинамическое функцио-
нальное назначение. Возможно стабилизирующее влияние
длинного мечевидного выступа у меч-рыбы на колебательное
движение ее при мощных взмахах хвостового плавника. Это
подтверждается увеличением относительной длины меча в он-
тогенезе, по мере роста молодой меч-рыбы и с повышением
скорости плавания.
Форма тела меч-рыбы хорошо обтекаемая и фактически
подобная встречается в технике. Например, схожую с ней (но
без приставки) форму имеют крупные планеры, в том числе
известный в свое время лучший рекордный паритель Генераль-
Рис. 19. Сопоставление в плане форм хвостовых плавников скорост-
ных видов дельфинов и рыб.
/ — обыкновенный дельфин; 2 — афалина; 3— пеламида; 4— меч-рыба (плавники
приведены к одному размаху).
52
Таблица 5. Сравнительная гидродинамическая оценка некоторых видов скоростных рыб
(обработка по различным источникам)
Виды рыб Максимальные значения Относительные размеры тела Хвостовой плавник Максимальные скорости Число Рейнольдса IO'1 Re
Длина L, м Масса т, кг Удлинение L/H । Отношение Н/В ф X X сс о Размах 1/L Удлинение F/S абсолютная ит- м/с относитель- ная UmIL, С-1
Форель радужная (Salmo 0,4 0,6 4,5 1,8 0,35 0,24 2,0 4,0 10,0 1,2
irideus)
Луфарь (Pomatomus sal- 0,6 2 4,5 1,8 0,43 0,24 2,6 5,5 9,0 2,5
tatrix)
Барракуда (Sphyraena barracuda) Ваху (Acantocybium 1,5 20 7,0 — 0,50 0,22 2,3 12,0 10,0 11,0
2,0 50 6,7 — 0,45 0,23 6,0 12,0 12,0 9,0
solandri)
Скумбрия (Scomber scombrus) Пеламида (Sarda sarda) 0,6 2 4,7 1,7 0,37 0,23 4,3 5,5 9,0 2,5
0,8 6 4,4 1,6 0,45 0,30 5,0 9,0 11,0 5,5
Тунец обыкновенный Thunnus thunnus) 3,0 500 3,5 1,3 0,45 0,31 7,5 25,0 12,5 38,0
Меч-рыба (Xiphios gla- 3,0 400 5,3 1,6 0,55 0,41 7,5 30,0 10,0 70,0
di us)
Марлин белый (Makaira 2,8 350 5,6 — 0,35 0,37 8,3 — — —
albida)
Акула мако (Isurus oxyrin- chus) Акула голубая (Carchar- hinus glaucus) 3,5 350 5,9 — 0,44 0,27 4,0 15,0 7,5 23,0
3,2 200 6,5 — 0,45 0,33 4,2 10,0 5,0 15,0
ного авиационного конструктора О. К. Антонова — металличе-
ский планер А-15 с ламинаризованным профилем-крыла.
Таким образом, феномен меч-рыбы состоит не только
в обычно отмечаемых выделяющемся роструме и необычайно
высокой для животных максимальной скорости плавания. Да-
лее будут показаны еще особенности кожного и слизистого
покровов меч-рыбы, которые могут способствовать существен-
ному снижению гидродинамического сопротивления.
В заключение краткого обзора гидродинамически эффек-
тивных морфологических качеств скоростных рыб приведем
•сравнительную оценку некоторых видов по данным исследова-
ний (табл. 5). В частности, для оценки максимальной скоро-
сти плавания использованы материалы экспериментов при
-спортивном лове крупных скоростных рыб с помощью специ-
ально оснащенного спиннинга. Табл. 5 составлена примерно
по тому же плану, что и табл. 3 для китообразных. Это удобно
для сопоставления между собой различных крупных скорост-
ных гидробионтов и выбора прототипов для моделирования
среди рыб и китообразных.
Морфологические данные по машущим плавниковым дви-
•жителям скоростных дельфинов и рыб достаточно хорошо со-
гласуются. Приведем график зависимости относительного уд-
линения хвостовых плавников этих гидробионтов от числа
Рейнольдса по длине тела (рис. 20). С повышением числа Рей-
нольдса в два порядка величины относительное удлинение
плавниковых движителей у гидробионтов увеличивается почти
в 3 раза. Наличие двух параллельных прямых на полулога-
рифмическом графике обусловлено различием относительной
площади хвостовых плавников, их жесткости и частоты машу-
щих колебаний у разных гидробионтов. В частности, хвосто-
вые плавники дельфинов характеризуются относительно боль-
Рис. 20. Относительное удлинение машу-
щих плавниковых движителей скоростных
видов рыб и дельфинов в функции числа
Рейнольдса по длине тела, X(Rei.).
Рыбы (см. табл. 5): / — форель радужная;
2 — луфарь; 3 — барракуда; 4 — акула-мако; 5 —
•скумбрия; 6 — пеламида; 7 — ваку; 8 — тунец
обыкновенный; 9 — меч-рыба. Черноморские
дельфины: Аз — азовка; Бб — белобочка;
Аф — афалина; Кос — атлантическая косатка.
шой площадью и малой
частотой колебаний.
Машущие плавнико-
вые хвостовые движи-
тели, как показывает ана-
лиз, обладают следую-
щими достоинствами:
1) быстро и плавно вклю-
чаются и выключаются,
сообщая телу большие ус-
корения и замедления
в короткое время; 2) хо-
рошо работают при пере-
менной нагрузке; 3) одно-
временно могут исполь-
зоваться как органы уп-
равления движением.
54
Однако плавниковые движители рыб и китообразных»
в сравнении с применяемыми в технике, имеют следующие-
существенные особенности и недостатки; а) сложность ре-
верса — непосредственно они не пригодны, для заднего хода
(дельфины назад не плавают); б) для более эффективной ра-
боты колебательные движения этих движителей сочетаются
с волнообразными движениями перемещающегося тела; в) при
моделировании, с увеличением размеров и массы твердых тел
могут возникать затруднения в управлении кинематикой дви-
жения при необходимой частоте колебаний, в соответствии
с числом Струхаля. Тем не менее отмеченные в гидробиониче-
ском плане недостатки плавниковых движителей технически,
преодолимы.
§ 5. Гидробионические закономерности строения покровов
скоростных рыб с регулируемым
сопротивлением плаванию
Для проведения комплекса исследований [22] авторами ис-
пользовались разнообразные речные, морские и океанические-
рыбы (20 видов, около 100 экземпляров), в том числе: скорост-
ные костистые — форель, щука, ставрида, пеламида, ваху, пят-
нистый и желтоперый тунцы, парусник, меч-рыба; скоростные-
хрящевые — акулы голубая, серая, мако (макрелевая), а для
сравнения и нескоростные—налим, ерш, окунь, носарь, гу-
стера, плотва, смарида, акула черноморский катран. Длина
этих рыб варьировала от 0,3 до 2,6 м, числа Рейнольдса со-
ставили широкий диапазон 2-Ю5—5-Ю7. У большинства эк-
земпляров исследовали все три вида покровов — кожный, че-
шуйный и слизистый, изучали их гистологическую структуру,
свойства, взаимосвязь, для чего изготовили более 4 тыс. пре-
паратов. Рыбы были отловлены, зафиксированы и заморожены
во время экспедиционных выездов на р. Днепр, Черное море
и в Атлантический океан.
Кожный покров рыб и его производные — чешую и слизь,
до последнего времени изучали, за редкими исключениями,
порознь и на малых нескоростных костистых рыбах, у которых
число Рейнольдса ReLclO5. По данным литературы, а также-
собственных исследований, гистологическая структура кожи’
этих рыб достаточно однообразная, несмотря на различие
в форме тела и скоростях плавания. Кожа нескоростных ко-
стистых рыб, как и у всех позвоночных животных, состоит из-
трех основных слоёв: 1) эпидермис—наружный тонкий слой;
2) кориум — собственно кожа, толстый средний слой, состоя-
щий из коллагеновых и эластиновых пучков и волокон; 3) суб-
кутис — нижний слой рыхлой соединительной ткани, часто со-
держащий многочисленные жировые клетки. Основное функ-
циональное назначение кожи нескоростных рыб — защита тела
55
от механических и других повреждений, образование чешуи и
слизи.
Гистологическое строение кожного покрова скоростных рыб
авторы изучали детально, по единой топографической схеме,
на 15—30 участках тела, расположенных в различных попереч-
ных сечениях, и в каждом сечении на спине, сбоку и брюшной
стороне тела. Материал для гистологического изучения фик-
сировали в 10 %-ном формалине, гистологические срезы кожи
рыб делали толщиной 5—10 мк (для акул 20—25 мк). Кожа
скоростных рыб состоит из тех же трех основных слоев, что
и у нескоростных, однако гистологическое строение ее значи-
тельно сложнее, выделяются различные подслои, в том числе
и новой структуры. Таким образом, характер развития кожи
у скоростных рыб дивергентный; у различных видов костистых
скоростных рыб наряду с общей структурой кожи имеются
свои особенности. Кроме того, гистологическое строение кожи
скоростных рыб различается еще вдоль тела и в разных ме-
стах сечений. Далее анализируется структура кожи в основ-
ном на участке тела, расположенном в средней части туло-
вища рыбы.
Обобщенный гистологический разрез кориума исследован-
ных экземпляров скоростных костистых рыб представляется
в следующем виде (от эпидермиса вглубь кожи): пограничная
/базальная мембрана (толщиной в тысячные доли миллимет-
ров), специфический волокнистый пигментный слой (сотые
доли миллиметров); соединительнотканный слой (десятые
доли миллиметров); ряд пластов коллагеновых пучков.и воло-
кон (общей толщиной 1 мм и более); у некоторых видов рыб
слой с жировыми клетками.
У скоростных акул многочисленные пласты коллагеновых
пучков и волокон расположены параллельно поверхности тела
в виде мелкой сетки, причем направление волокон в смежных
пластах ортогональное и составляет с продольной осью тела
угол около 45° (в хвостовой части тела угол уменьшается до
35—40°). Диагональное расположение по правой и левой спи-
ралям под углом к продольной оси тела коллагеновых воло-
кон, армирующих кожу акул, является оптимальным в усло-
виях изгибно-колебательных движений тела плавающей рыбы.
Это следует из теории сопротивления материалов, армирован-
ных кордом полимерных изделий.
Анализ изменения гистологического строения кожи кости-
стых и хрящевых рыб в связи с ростом числа Рейнольдса от
2- 105 до 5- 107 позволяет выделить важную общую тенденцию,
а именно: эпидермис кожи становится все более многослой-
ным, развивается многослойный секретирующий эпителий, ко-
торый в состоянии выделять в пограничный слой большое
количество слизи. По числу слоев клеток эпидермиса скорост-
ные рыбы располагаются примерно в следующий ряд: пела-
56
мида 2—3 слоя, желтоперый тунец 4—5 слоев, парусник 7—9
слоев, меч-рыба 8—10 слоев, акулы серая и мако 10—12 слоев,
что соответствует последовательности расположения этих рыб-
по величине [22].
Важной механической характеристикой кожного покрова
скоростных рыб, в том числе и для суждений о возможных ме-
ханизмах снижения гидродинамического сопротивления, явля-
ется локальная масса кожи, определяемая ее толщиной. С ро-
стом числа Рейнольдса относительная толщина кожи у многих
крупных скоростных рыб, костистых и хрящевых, уменьшается
от t!L~\• 10_3 до 7-Ю-4, т. е. в полтора раза (рис. 21). Од-
нако имеются и взаимно исключающие отклонения: у пятни-
стого тунца и голубой акулы кожа почти втрое толще нор-
мальной, присущей большинству крупных скоростных рыб,
а у тонкотелой, удлиненной ваху — более чем вдвое тоньше
нормальной, при одном и том же числе Рейнольдса. Таким об-
разом, среди крупных скоростных рыб можно выделить по от-
носительной толщине кожного покрова три типа рыб: с тол-
стой, средней и тонкой кожей. Это подтверждает дивергентное
развитие кожи скоростных рыб. Точка разветвления на рис. 21
приходится на пеламиду, имеющую промежуточную структуру
кожного покрова.
К наиболее многочисленной группе скоростных рыб с от-
носительной нормальной толщиной кожного покрова принад-
лежат следующие исследованные виды: форель, пеламида,
желтоперый тунец, парусник, меч-рыба, акулы серая и мако.
По полученным качественным данным о коже рыб к этой
группе относятся, кроме того, виды чешуйных тунцов при
длине тела Л>1,0 м (обык-
новенный, длинноперый,
большеглазный), а также
различные виды марлинов
и копьеносцев, т. е. в дан-
ную группу входят наибо-
лее крупные скоростные
рыбы. Общим признаком
для покровов разнообраз-
ных видов скоростных рыб
является наличие чешуи и
обильной слизи.
К скоростным рыбам
со сравнительно толстой
кожей относятся, помимо
пятнистого, полосатый и
макрелевидный тунцы и,
кроме голубой акулы, грен-
ландская акула (по каче-
ственным данным). Общими
Рис. 21. Зависимость относительной тол-
щины кожного покрова рыб от числа
Рейнольдса, (t/L) =f(Rez,).
Костистые рыбы: / — налим: 2 — фо-
рель; 3— пеламида; 4 — пятнистый тунец;
5 — желтоперый тунец; 6 — парусник; 7 — меч-
рыба. Акулы: 3 — катран черноморский;
9 — серая; 10 — голубая; //—мако. Для срав-
нения: 12 — обыкновенный дельфин; 13 —
рыба-ваху.
57
признаками покровов названных видов тунцов являются:
гладкая поверхность кожи, отсутствие чешуи и сплошного сли-
зистого покрова. Характерно, что все виды бесчешуйных тун-
цов, не имеющих слизистого покрова, малых размеров (£<
<1,0 м). По сравнению с более крупными чешуйными тунцами
покрытыми обильной слизью, они менее скоростные. Естест-
венным продолжением по числу Рейнольдса и относительной
толщине кожного покрова этих видов тунцов и акул является
обыкновенный дельфин: Reb = 2-107 и tJL^9 • \0~3 (см. точку/2
на рис. 21), а у дельфина обнаружено свойство самонастройки
демпфирования гладкой кожи при отсутствии слизи.
Чешуйный покров имеет подавляющее большинство рыб.
Чешуя вместе с кожей образует наружный скелет рыб, выпол-
няет защитную и другие функции организма. В соответствии
о образом жизни, размерами тела и скоростью плавания рыб
она различается у разных видов рыб по расположению на по-
верхности тела, залеганию в коже, строению, форме и разме-
рам чешуй. Чешуйный покров изучался вместе с кожным и
слизистым покровами на фиксированных экземплярах указан-
ных выше видов рыб. С помощью бинокулярной лупы и мик-
рофотографий исследовали топографию чешуйного покрова,
взаимное расположение и относительные размеры чешуй. В ре-
зультате выявлены гидродинамическая функция и закономер-
ности изменения чешуйного покрова рыб, включая скоростные
виды.
Отметим особенности чешуйного покрова некоторых круп-
ных скоростных костистых рыб. У пеламиды чешуя мелкая, ре-
дуцированная; поверхность рыбы, покрытая слизью, на ощупь
совершенно гладкая. У желтоперого и других чешуйных тун-
цов чешуя более крупная, однако сами эти рыбы больших
размеров, и их чешуя утоплена в соединительной ткани. У па-
русника, у марлинов и коньеносцев чешуйный покров не
сплошной, а с промежутками и видоизменяется по телу рыбы.
У взрослой меч-рыбы обнаружена рельефная поверхность
кожи с продольными складками и впадинами и как бы вторич-
ная мелкая шероховатость, образованная небольшими чешуй-
ками. Продольные складки кожи следуют от головы рыбы до
стебля хвостового плавника, сначала увеличиваясь в ширину
и в высоту до середины длины тела, а затем уменьшаясь. Они
также изменяются в размерах от спины, к боку и затем к жи-
воту. Рельеф кожи меч-рыбы отчасти выравнивается покры-
вающим слоем слизи.
Тело акул покрыто плакоидной чешуей, расположенной
диагональными рядами, по направлениям спиралей соедини-
тельнотканных волокон кориума. На пластинке плакоидной
чешуи обычно расположены продольные ребра жесткости —
кили. Число килей варьирует у разных видов от одного до
трех-четырех. Все это придает плакоидной чешуе резко выра-
58
женную шероховатость, подобно наждачной бумаге. Однако
форма элементов шероховатости такова, что они хорошо об-
текаются по потоку.
Чешуйный покров медленно плавающих и скоростных акул
существенно различается по плотности расположения в плане
и размерам плакоидных чешуй и килей. Так, у черноморского
катрана чешуя редкая и крупная (на миделе метровой акулы
10 шт/мм2). У скоростной акулы мако чешуя расположена ча-
сто, соприкасается между собой и образует сплошной панцирь
со строго диагональными рядами, мелкая (на миделе полу-
тораметровой акулы 50 шт/мм2). Высота килей чешуй соот-
ветственно уменьшается от 100 до 25 мк. Кроме того, у ско-
ростных акул кили последующих чешуй имеют направление
предыдущих и на всей поверхности тела как бы образуются
почти сплошные продольные микроребра, напоминающие
продольные складки кожи меч-рыбы.
Несмотря на большое разнообразие чешуйного покрова по
многим признакам, характерной общей тенденцией является
относительное уменьшение размеров чешуи с ростом числа
Рейнольдса рыб.
Пользуясь гистологическими исследованиями чешуйного по-
крова акул, приведем ре-
зультаты гидродинамиче-
ской оценки чешуи по тео-
рии пограничного слоя
(рис. 22). Анализ графика
показывает, что относи-
тельные размеры выступов
чешуи kIL находятся меж-
ду значениями допустимой
высоты шероховатости при
турбулентном и (условно)
ламинарном пограничном
слое, а именно (й/А)ь^
^.klL^{klL)a- С ростом
числа Рейнольдса высота
выступов чешуйного по-
крова все более приближа-
ется к наименьшим допу-
стимым значениям. Инте-
ресно, что последним для
акул отвечает относитель-
ная высота собственно
чешуй, а общая высота
(чешуя + киль) находится
на уровне соответствую-
щих значений для чешуи
костистых рыб.
Рис. 22. Гидродинамическая оценка че-
шуи рыб по относительной высоте до-
пустимой шероховатости (•) при лами-
нарном (а) и турбулентном (б) режи-
мах пограничного слоя [22].
Костистые рыбы: / — лещ; 2— карась;
3 — сазан; 4 — плотва; 5 — скумбрия; 6 —
ставрида; 7 — лосось; 8 — пеламида; 9 — угорь..
Акулы: 10 — катран; 11 — серая; 12 — голу-
бая; 13— мако (ч — чешуя, к —киль).
59
Другой характерной общей тенденцией в изменении чешуй-
ного покрова с ростом числа Рейнольдса рыб является усиле-
ние направляющего действия чешуи. Выше отмечена общая
лродольная ребристость чешуйного покрова крупных скорост-
ных костистых рыб и акул; у последних она особенно четко
выражена килями [21]. На поверхности акул ясно выделяются
и местные зоны изменения ориентации чешуй и килей по от-
ношению к продольному направлению выровненного тела рыбы
на 10—15° и до 40—45° у жаберных щелей и грудных плав-
ников, у спинного и брюшных плавников, в хвостовой части
тела. В чешуйном покрове костистых скоростных рыб, срав-
нительно с малыми нескоростными рыбами, появляется извест-
ное новообразование в виде мелкочешуйного грудного корсета.
Он расположен сразу за жаберными крышками и имеет свое-
образное сложное очертание, с размытой границей у пеламиды
и четко выраженной у чешуйных и бесчешуйных тунцов. У не-
которых видов тунцов весьма развитый корсет покрывает до
7з поверхности тела.
Из изложенного следует, что гидродинамическая функция
чешуи рыб не определяется только ее шероховатостью, как это
часто принимают. От обычной шероховатой поверхности твер-
дых тел чешуйный покров скоростных рыб существенно отли-
чается следующими свойствами: правильным шахматным рас-
положением единообразных и одноразмерных чешуй в плане,
упорядочивающим вихреобразование в пограничном слое;
пл&вными очертаниями чешуй в профиле, чем обеспечивается
безотрывное обтекание, например, хорошо известной формы
циклоидной чешуи; направляющим действием микроэлементов
чешуи акул, в том числе килей, имеющих ясно выраженное
общее продольное направление и гидродинамически оправдан-
ные местные области изменения ориентации; упругостью че-
шуй, которая проявляется в условиях изгибно-колебательных
движений тела; размеры выступов чешуй находятся в интер-
вале допустимых значений по теории пограничного слоя; че-
шуя скоростных рыб всегда покрыта эпидермисом и слизью,
выделяемой секреторными клетками. Отмеченные гидродина-
мические свойства чешуи позволяют считать ее органом регу-
лирующего влияния на пограничный слой скоростных рыб.
Экспериментальные данные по микроразмерениям чешуй
акул показывают, что площадь $ чешуй и расстояние X между
килями чешуй у скоростных акул намного меньше, чем у мед-
ленно плавающих; абсолютные значения этих величин непре-
рывно снижаются с ростом числа Рейнольдса рыб. Однако от-
носительные значения размеров чешуй стремятся к некоторым
предельным значениям: отношение длины I к ширине b чешуи
а отношение расстояния X между килями к их высоте d,
X/d<2,6. Эти соотношения показательны в связи с некоторым
условием устойчивости пограничного слоя при образовании
60
пространственных возмущений в виде продольных ячейковых
конвективных вихрей, у которых длина волны возмущений по-
перек основного потока равна Л, а глубина ячеек d. Теоретиче-
ски найдено и подтверждено экспериментально, что в случае
одной свободной границы и другой твердой (как на ячейке
чешуи) критическое значение, при котором ячейковые вихри
еще не распространяются на всю глубину пограничного слоя,
равно %/d = 2,66. Это значение поразительно точно соблюдается
в предельном чешуйном покрове скоростной акулы мако.
Слизистый покров рыб ранее привлекал внимание исследо-
вателей, однако его изучали в основном на малых нескорост-
ных костистых рыбах, вне тесной связи с кожным и чешуйным
покровами. Некоторые авторы отрицали наличие слизи у акул.
В литературе установилось мнение, что слизистый покров рыб
имеет мультифункциональное назначение защитного характера
и предохраняет кожу от загрязнения.
На исследованных костистых рыбах и акулах, включая
крупные скоростные виды, изучали: типы секреторных, слизе-
образующих клеток эпидермиса кожи; химический состав
рыбьей слизи; количественное распределение секреторных кле-
ток на поверхности тела; изменение факторов слизистого по-
крова с ростом числа Рейнольдса рыб. Химический состав сек-
реторных клеток определяли с применением гистохимических
реакций, используя окраски: сулемабромфеноловый синий,
тиомин, альциановый синий. Для изучения топографии секретор-
ных клеток была разработана специальная методика приготов-
ления препаратов эпидермиса. Толщину слизистого покрова
определяли на живой рыбе с помощью специального устрой-
ства. Слой слизи рыб оказался толщиной в среднем 40—45 мк,
с колебаниями от 25 до 60 мк (отчасти вследствие погрешно-
сти метода).
В коже различных видов рыб выделяют в основном три
типа секреторных, слизеобразующих клеток: бокаловидные,
колбовидные и зернистые. Они различаются по расположению
в различных слоях эпидермиса, форме клеток и размерам, внут-
реннему строению и химическому составу.
Обнаружено, что производство слизи и способ подачи ее на
поверхность тела зависит от размеров и скорости плавания
рыб. На пресноводных небольших и медленно плавающих ры-
бах встречаются два-три типа секреторных клеток, у рыб сред-
них скоростей плавания один-два, а у скоростных, костистых
и акул — всего один тип слизеобразующих клеток — зерни-
стый. Таким образом, с повышением скоростей плавания, а зна-
чит, и чисел Рейнольдса рыб происходит как бы узкая спе-
циализация секрета. В соответствии с характером секреторных
клеток синтезированная в них слизь поступает на поверхность
тела двумя путями: 1) продвижение клетки с секретом к по-
верхности эпидермального слоя, где и происходит полное
61
разрушение ее; 2) подача слизи из более глубоких слоев эпи-
дермиса по межклеточным ходам.
Показательно закономерное распределение по длине тела
количества на единицу площади и размеров секреторных кле-
ток в эпидермисе кожи некоторых небольших костистых рыб
(рис. 23). Как видно, это распределение связано с формой
тела и скоростью плавания рыб, особенностями формирования
их пограничного слоя. За жаберными щелями, где выбрасыва-
ются струи воды при дыхании рыбы, а также в диффузорной,
резко суживающейся хвостовой части тела, с большой ампли-
тудой изгибно-колебательных движений, возможны сильные
возмущения и даже отрыв пограничного слоя. Поэтому в этих
местах у рыб могут образоваться поля с повышенной секретор-
ной активностью, в которых сосредоточится большое количе-
ство секреторных клеток наибольших размеров (пики на гра-
фиках рис. 23).
Весьма развит слизеобразующий аппарат в хвостовой ча-
сти тунцов с циклоидной чешуей. Здесь почти все эпителиаль-
ные клетки, кроме растущего базального слоя эпидермиса
кожи, превращаются в секреторные; образуется сплошное мно-
горядное поле слизистых зернистых клеток.
Особенно мощное развитие секреторного аппарата уста-
новлено для скоростной меч-рыбы. На всей поверхности тела
и головы экземпляра длиной 2,6 м даже невооруженным гла-
зом видны поры в количестве 0,8 шт/мм2 и диаметром в свету
I
20
10
0
0 0,2 0,4 0,6 0,8, 1,0
х/Ь
Рис. 23. Закономерность распре-
деления по длине тела слизеобра-
зующих клеток в эпидермисе
кожи различных костистых рыб
[22].
1 — пеламида; 2 — щука; 3 — плотва;
---О-------поперечный размер кле-
ток d, мкм; -----•------количество
клеток л. шт/мм3.
62
0,07—0,2 мм, в среднем 0,1 мм (сбоку тела). При небольшом
механическом надавливании из пор выделяется слизистое ве-
щество. Поры ведут в обнаруживаемые в процессе препариро-
вания верхних слоев кожи внутренние каналы, которые распо-
ложены на глубине 1 мм параллельно поверхности кожи,
имеют диаметр в среднем 0,5 и не более 1,0 мм (рис. 24).
Между внутрикожными слизистыми полостями встречается
много анастомозов, особенно на голове и жаберных крышках.
На спине и животе меч-рыбы полости переплетаются и обра-
зуют своеобразную сеть, а на боковой поверхности тела они
почти параллельны и направлены ортогонально наружным
продольным складкам и впадинам кожи [27].
У меч-рыбы обнаружены не только общие лево- и право-
сторонняя внутрикожные слизеобразующие системы, которые
имеются у всех ксифиоиДных видов» но также и местные си-
стемы регуляции слизи за жаберными щелями и вдоль спины,
встречающиеся только у меч-рыбы. За жаберными щелями на
поверхности кожи обнаружены области своеобразных струк-
тур в виде системы крипт. Их выстилающий эпителий содер-
жит большое количество слизистых клеток. За толстым спин-
ным плавником, который у меч-рыбы не убирается, вдоль
средней линии спины в шахматном порядке по обе стороны
имеющегося здесь кожного валика расположены последова-
тельно одна за другой наполненные слизистым веществом
Рис. 24. Система слизистых каналов в верхнем покрове кожи меч-
рыбы: а — выводные поры слизи на поверхности тела; б — препари-
рованный продольно-складчатый слой кожи с поперечными слизистыми
каналами.
63
внутрикожные ампулы. Они обнаружены препарированием
кожи спины на глубине 10—12 мм от поверхности в количестве
2x20 шт. Длина каждой ампулы до 30 мм, диаметр до 13 мм
(у экземпляра рыбы длиной 2,60 м). Ампулы между собой не
сообщаются, но открываются выводящими продольными ще-
лями на поверхность кожи спины. Стенки ампул и выводящих
щелей выстланы многослойным секретирующим эпителием,
а внутри ампул содержится резерв слизи. Для усиления сек-
реторной активности ампул выстилающий эпителий имеет
криптообразные выросты.
Гидродинамическую функцию слизи, направленную на сни-
жение сопротивления крупных скоростных рыб, можно проде-
монстрировать также путем сравнения слизистого покрова
медленно плавающей и скоростной акул. Для черноморского
катрана характерны крупные бокаловидные клетки, которые
расположены в один ряд, плотно и по всей поверхности тела
рыбы. Напротив, у скоростной атлантической акулы мако сли-
зистые клетки мелкие и зернистого типа; они расположены
в диффузорной части тела, где заполняют собой почти всю
толщу эпидермиса, как у чешуйных тунцов [21]. Детальными
исследованиями слизистого покрова четырех видов акул опро-
вергнуто встречающееся в литературе мнение об отсутствии
у них секреторных клеток.
Эффективность рыбьей слизи для снижения гидродинами-
ческого сопротивления доказывают экспериментально также
другие разносторонние, выполненные разными средствами ис-
следования. Выявлены некоторые дополнительные особенно-
сти свежеубитых рыб, в частности, активность только их
слизи.
Определенный интерес представляют сравнительные экспе-
рименты по изучению снижения сопротивления трения при тур-
булентном режиме течения водного раствора слизи пресновод-
ных и морских видов рыб, проведенные с помощью портатив-
ного прибора — реометра [77]. Трубка из нержавеющей стали
последнего имела внутренний диаметр 0,58 мм, а скорость те-
чения раствора слизи в ней достигала 13,7 м/с. Наибольший
эффект снижения сопротивления (до 66 %) показали растворы
слизи 11 скоростных морских рыб (барракуд) длиной 0,66—
0,79 м (рис. 25). Авторы исследования не получили заметного
снижения сопротивления раствора слизи тунца «калифор-
нийский бонито» и выразили свое удивление по этому поводу.
Однако названный вид относится к бесчешуйным тунцам и, по
нашим данным, полученный результат не является неожидан-
ным. Одновременно подтверждается вывод о необходимости
разделения тунцов на две существенно различные группы по
характеру покровов.
В ряде статей приведены экспериментальные данные о био-
химизме гидродинамически активных биокомплексов слизи-
64
стых покрытий кожи, ха-
рактерных для некоторых
морских рыб, о структуре
биокомплексов и аминокис-
лотном составе их, о зна-
чении отношения содержа-
ния основных белковых
к кислотным компонентам
[39]. Интересны электрон-
ные микрофотографии слизи
сравнительно быстро пла-
вающих рыб — кефали и
ставриды (рис. 26), кото-
Рис. 25. Снижение сопротивления тре-
ния морской воды растворенной в ней
слизью морских рыб.
По горизонтальной оси — концентрация слизи
в растворе, %; по вертикальной оси — сниже-
ние сопротивления сравнительно с чистой во-
дой, %. Рыбы: 1 — тихоокеанская барра-
куда; 2 — палтус; 3 — тихоокеанская макрель;
4 — калифорнийский тунец; 5 — моллюски;
6 — морская водоросль.
рые дают представление
о размерах, форме и строе-
нии частиц биокомплексов
слизи этих рыб в виде длин-
ных цепочек.
Наблюдается следующая
закономерность в покровах
скоростных рыб, которые
отчасти выполняют функ-
цию снижения гидродинамического сопротивления. С ростом
числа Рейнольдса до ReL~ 1 • 108 у костистых и у хрящевых рыб
основное значение для возможного воздействия на погранич-
ный слой приобретает мультифункциональный слизистый
Рис. 26. Электронные микрофотографии слизистого вещества кожного
покрова кефали (а) и ставриды (б).
3 С. В. Першин
65
покров. Это обусловлено дивергентным развитием кожного по-
крова скоростных рыб с преимущественно мощным многослой-
ным секретирующим эпителием и увеличением содержания
диффузионно активной биополимерной слизи в коже хвостовой
части акуловых, чешуйных тунцовых, и на всей поверхности
ксифиоидных видов рыб. Особенно выделяется предполагае-
мой гидродинамической функцией слизистый покров меч-рыбы,
у которой также впервые обнаружены покрытая слизью рель-
ефная поверхность кожи с продольными складками и впади-
нами, слизеобразующие лево- и правосторонняя системы вну-
трикожных полостей с выводными порами, отдельные места
регуляции выделения слизи, с разветвленными криптами за
жаберными щелями и резервными ампулами вдоль спины.
ГЛАВА II
КИНЕМАТИКА НЕСТАЦИОНАРНОГО ПЛАВАНИЯ
ГИДРОБИОНТОВ И РЕГУЛИРУЮЩЕЕ ЗНАЧЕНИЕ
КРИТЕРИЯ СТРУХАЛЯ
§ 6. Экспериментальные методы и средства исследований
кинематики плавания гидробионтов
Кинематика плавания гидробионтов сложная ввиду неста-
ционарности различных режимов плавания многочисленных
видов водных животных, адаптированных к разным экологиче-
ским условиям. В настоящее время кинематика плавания вод-
ных животных теоретически не определяется. Эксперименталь-
ные исследования кинематики необходимы как для понимания
физики процесса и механизма плавания, так и в целях получе-
ния исходных количественных данных для расчета гидродина-
мических характеристик гидробионтов и моделирования. Ско-
рость плавания водных животных представляет интерес для
промысловой практики.
Методы и средства исследований кинематики гидробионтов
зависят от способов плавания различных видов, размеров их и
скоростей плавания, т. е. от чисел Рейнольдса. Имеются в виду
крупные гидробионты, такие как рыбы и китообразные. По-
этому для исследования их кинематики частично можно при-
менять и обычно принятые в экспериментальной гидродинамике
твердых тел средства исследований и типы установок. Однако
известные жесткие требования к корректному проведению гид-
родинамических экспериментов в гидробионике необходимо до-
полнить существенными физиологическими условиями содер-
66
жания живых организмов, в том числе по обеспечению их
нормального дыхания и вообще их жизненного ритма.
Исследуемый гидробионт в процессе эксперимента должен
находиться в условиях, максимально близких к естественным
для него в природе, и адаптироваться в экспериментальной ус-
тановке. Необходимо обеспечить адекватные природным фак-
торы среды — пресную или морскую воду, содержание в ней
солей и растворенйоро воздуха, соответствующую температуру
воды, а также достаточный рабочий объем экспериментальной
установки, начальную турбулентность водной среды, надлежа-
жащее освещение и т. п.
В экспериментальных исследованиях кинематики плавания
гидробионтов применяют как прямые, так и обратные методы.
К прямым методам относятся натурные исследования
в условиях моря или бухты (с использованием сетевых заграж-
дений), полунатурные эксперименты в специально оборудован-
ных океанариумах и бассейнах, а также эксперименты с ис-
пользованием малых установок — гидролотков и аквариумов.
В этих условиях подопытные гидробионты могут плавать
в свойственных им естественных режимах. Исследователи про-
водят визуальные наблюдения, надводную и подводную кино-
съемку плавания гидробионта, регистрируют проплывание его
на оборудованной дистанции с применением телеметрической
аппаратуры и т. п. Изучают регулярное нестационарное пла-
вание гидробионта и кратковременные переходные процессы.
Достоинством исследований гидробионтов в спокойной воде
является более естественное плавание, без особых искусствен-
ных побуждений животного человеком. Существенный недоста-
ток их — не вполне направленный характер эксперимента, по-
скольку заранее трудно предусмотреть, какие явления удастся
зафиксировать. Однако этот недостаток частично можно устра-
нить предварительной тренировкой животных, особенно дельфи-
нов. Известны большие достижения в дрессировке дельфинов
для выполнения ими обусловленных тренерами действий,
с применением поощрительной подкормки животных. К тому же
дрессировка осуществляется в морских искусственных океана-
риумах за относительно короткий срок (за несколько недель
или месяцев). Для дрессировки животных служит и советский
Батумский океанариум. Он открыт для посещения многочислен-
ных любителей природы и животных.
Классическим примером исследований плавания дельфинов
непосредственно в море являются подводные киносъемки
группы исследователей под руководством известного француз-
ского океанографа Жака Ива Кусто. Он создал несколько ки-
нофильмов и издал ряд книг по изучению Мирового океана.
Киносъемки выполнены группой Жака Ива Кусто из подвод-
ной наблюдательной камеры, предусмотренной в носовом от-
секе корабля и снабженной иллюминаторами, и с помощью
3‘ 67
Рис. 27. Выборочные кадры подводной киносъемки плавания в море группы
дельфинов с синхронной перекладкой хвостовых плавников в крайних ниж-
нем и верхнем положениях (в том числе в противофазе).
акванавтов, плавающих в воде с кислородными приборами.
Такие киносъемки дают большую и достоверную информацию.
Отдельные кадры киносъемки показаны на рис. 27.
Систематические исследования плавания дельфинов в ис-
кусственном прибереговом, частично остекленном, бассейне
провели сотрудники Института гидромеханики АН УССР [55,
56]. Большим преимуществом подобного рода экспериментов
являются контролируемые условия, удобство применения раз-
нообразной аппаратуры. Возможно и проведение исследований
пограничного слоя на теле дельфина. Плавательные движения
различных видов небольших рыб в неглубоком лотке с водой
уже давно зафиксированы в серии исследований известного
английского биолога Грея [74].
Наибольшее развитие исследования кинематики плавания
мелких рыб получили обратными методами, т. е. в об-
ращенном движении в замкнутых гидродинамических трубах,
в которых циркулирует вода, а рыба плавает навстречу по-
току со скоростью потока и потому находится на одном и том
же месте остекленного рабочего участка трубы [68]. При этом
используют так называемую оптомоторную реакцию рыбы, при
которой она следит своими глазами за видимым неподвижным
ориентиром, например, линиями координатной сетки, нанесен-
ной черной краской на днище трубы. Обычно прозрачный ра-
бочий участок трубы снабжают двумя плоскими зеркалами,
поставленными под углом. Плавательные движения рыбы фик-
сируют киносъемкой одновременно в вертикальной и горизон-
тальной плоскости (рис. 28). Исследование кинематики пла-
вания рыб в циркуляционных гидродинамических трубах имеет
то преимущество, что можно свободно управлять режимами
плавания рыбы путем регулирования скорости набегающего
68
Рис. 28. Рыба-елец, плавающая в био-
гидродинамической трубе с отражаю-
щими зеркалами (вертикальный н го-
ризонтальный виды).
Обращают внимание изгибы тела и хвосто-
вого плавника, приоткрытые жаберные
крышки, примкнутые к телу спинной и
брюшной плавники.
потока воды. Модели рыб и
дельфинов испытывают в
опытовых бассейнах и для
продувки в аэродинамиче-
ских трубах.
В гидродинамических
трубах не выполняются
полностью условия обраще-
ния движения при данных
числах Рейнольдса, когда
скорость плавания рыбы
приравнивают средней ско-
рости потока воды на рабо-
чем участке трубы. Дело
в том, что начальная турбу-
лентность потока в гидро-
динамической трубе обычно
в несколько раз выше, чем в естественных условиях плавания
рыбы в море, и сам поток в трубе всегда ' турбулентный.
Между тем пограничный слой на рыбе, плывущей в море,
может быть и ламинарным, хотя бы на части длины тела. Воз-
можно, несоответствие режимов пограничных слоев в прямом
и обращенном движении не имеет большого значения для не-
которых гидробионических исследований, однако это экспери-
ментально не доказано на одних и тех же живых объектах.
Можно полагать, что рыба преодолевает повышенную турбу-
лентность потока воды в трубе за счет увеличения своих энер-
гозатрат при каждой данной скорости, но тогда она быстрее
устает, чем при плавании в естественных условиях с той же
скоростью, и не достигает полных скоростных характеристик.
Автором предложена многоцелевая экспериментальная ус-
тановка для биогидродинамических исследований крупных
гидробионтов, включающая последовательно соединенные ме-
жду собой остекленные прибереговой двухкамерный морской
бассейн с продольной перегородкой и П-образную в плане
гидродинамическую трубу с двумя рабочими участками.
§ 7. Нормирование максимальных скоростей плавания
китообразных в природе и в неволе
Максимальные скорости плавания рыб и китообразных, как
и других водных животных, необходимо знать для промысло-
вого улова и разработки рациональных методов, средств и
орудий промысла, а также для расчетной оценки гидродина-
мических и энергетических возможностей плавания гидробион-
тов и моделирования. К настоящему времени проведенные ис-
следования неполны и недостаточно точны, не всегда согласу-
ются между собой, а иногда противоречивы.
69
Существуют различные методы определения максимальной;'
скорости плавания водных животных. На рис. 29 показаны
результаты определения максимальной скорости плавания ваху
и небольшого тунца распространенным способом: фиксируется
длина разматываемой лески с попавшей на крючок рыбой.
В данном случае леска имела метровые метки, которые нано-
сились с помощью металлического порошка, отмечались маг-
нитным датчиком; сигналы записывались на осциллографе [94].
Метод регистрации простой и удобный, но не вполне надежный.
Животное может вести себя необычно из-за болевых ощуще-
ний: удаляться от места поимки не по прямой линии, а меняя
направление, и т. п.
Максимальная скорость движения самой скоростной меч-
рыбы определена акад. А. Н. Крыловым расчетом по силе ее
удара в борт судна. Известен факт, когда меч-рыба пробила
медное покрытие толщиной 0,05 м, дубовую обшивку и шпан-
гоут толщиной 0,30 м, в котором застрял сломанный ее меч.
Столь мощный удар сравним с действием снаряда малой кора-
бельной пушки, и расчет показал, что скорость меч-рыбы была
более 25 м/с.
Максимальная скорость плавания рыб и китообразных бо-
лее точно регистрируется в контролируемых в эксперименталь-
ных условиях — в океанариумах и в гидродинамических лотках
и трубах. Однако не всегда есть уверенность в том, что под-
опытные гидробионты плавали в эксперименте с наивысшей
энергоотдачей и действительно показали максимальную ско-
Рис. 29. Осциллограммы импульсных рывков, попавших на крю-
чок рыб, записанные по разматыванию размеченной рыболовной
лески, U(t): а — ваху длиной 1,13 м; б — тунец длиной 0,98 м.
70
рость, даже в случаях испытания дрессированных дельфинов
[83].
Максимальные скорости плавания китообразных в природе
многократно определяли с быстроходных промысловых судов,
идущих в море параллельным курсом с животными и в спе-
циальных морских исследовательских экспедициях [79]. В ра-
боте [45] автором предложено нормировать максимальную ско-
рость плавания дельфинов и китов с учетом основных опреде-
ляющих факторов. Важнейшие из этих факторов следующие:
вид животного, который гидродинамически характеризуем из-
вестной формой Ф тела; длина его тела L, м; продолжитель-
ность плавания с данной скоростью Т, с; биологическое состоя-
ние животного — обозначим его условно как факторы Б. К по-
следним относятся: воздействие врагов и стада, условия внеш-
ней среды (температура, соленость, прозрачность воды и др.).
Биологические факторы частично определяются качественно;
все другие можно оценить количественно. В дальнейшем нами
применяется системный подход с раздельным учетом различ-
ных действующих факторов.
Осредненную за данную продолжительность плавания Т
максимальную скорость t7max животного с формой тела Ф, дли-
ной тела L в известных условиях обитания Б назовем норми-
рованной, так что
£/тах = £/(Ф, L, Т, Б).
Если один из факторов не будет учтен, то определение мак-
симальной скорости плавания гидробионтов окажется непол-
ным, что нередко встречается в литературе. Именно этим об-
стоятельством, кроме погрешностей методов измерения скоро-
стей, объясняются неувязки и противоречия в опубликованных
данных о скорости плавания гидробионтов.
В табл. 3 указана кратковременная импульсная максималь-
ная скорость плавания в море некоторых видов китообразных,
которая осреднена в течение нескольких секунд, например, при
стремительном рывке уплывающего легко раненого животного.
Поскольку для развития скорости в броске имеет значение
масса животного, то приближенно можно считать в данных
случаях для малых дельфинов Т=3с, для больших дельфинов
(косатка, белуха) Т=5 с и для китов 7=10 с. Практически
это осредненные абсолютные значения максимальной скорости
плавания дельфинов и китов.
Чтобы оценить влияние на скорость плавания китообразных
длины тела, на рис. 30 построена зависимость для относитель-
ной максимальной скорости UmIL=^(L) 18 видов. Различием
T-^idem пренебрегаем, что дает погрешность не более ±5%.
Разной является и (форма тела, фактор Ф у разных видов,
поэтому геометрическое подобие тоже не соблюдается. Таким
образом, на рассматриваемом графике сопоставляются между
71
Рис. 30. Зависимость относитель-
ной максимальной скорости пла-
вания в море диких китообразных
от длины их тела
Скоростные (/) и нескоростные (//)
виды дельфинов н китов (табл. 3):
1 — дельфин-белобочка; 2 — морская
свинья; 3 — белокрылая морская сви-
нья; 4 — афалина; 5 — гринда; 6 — ко-
сатка; 7 — кит-сейвал; 8 — кит-финвал;
9 — синий (голубой); 10 — бесперая
морская свинья; Л — белуха; 12 — се-
рый кит; 13 — кашалот; 14 — кит гор-
батый.
собой не динамически подобные
системы, а лишь относительная
быстроходность дельфинов и ки-
тов скоростных/ и нескоростных
// видов. При отмеченных усло-
виях, если скорость Um измерять
в метрах на секунду, а длину L
в метрах,
Um/L^ksL~^,
Um&ksL°-™ (7>
Здесь размерный коэффици-
ент
k =
s L
= 5,5—12,0.
L=1
Этот коэффициент представляет
собой как бы «потенциальную
быстроходность» эмбрионов дли-
ной А=1 м (постоянной вели-
чины) для различных видов ки-
тообразных, выраженную в раз-
мерностях длины и времени,
т. е. в метрах и секундах (см.
рис. 30).
Чтобы определить величину
ks~const для каждого данного вида китообразных воспользу-
емся следующими данными. Ранее было показано, что в одной
из предродовых форм развития эмбрионов малых дельфинов и
крупных китов, как раз при длине эмбриона около 1 м, сфор-
мировывается форма тела и машущего плавникового движи-
теля. Они запрограммированы предшествующей эволюцией для
различных видов эмбрионов китообразных, которые после
дальнейшего роста рождаются в воде и вслед за первыми ды-
хательными актами в воздухе начинают активно плавать.
На основании графика рис. 30 приближенно получим срав-
нительную потенциальную быстроходность различных видов ки-
тообразных:
Дельфины, киты
Белобочка, сейвал ....................12
Афалина, косатка, синий кит ...............11,4
Гринда, финвал ............................10,0
Морская свинья .............................8,2
Белуха, серый кит, кашалот .................5,5
Согласно формуле (7) длина тела мало влияет на абсолют-
ную максимальную скорость плавания китообразных. Это имеет
большое биологическое значение для совместного плавания
72
семей взрослых китообразных с малыми детенышами, в отли-
чие от рыб, у которых молодь группируется отдельными стаями.
Важнейшим фактором, влияющим на максимальную ско-
рость плавания гидробионтов, является продолжительность пла-
вания Т на данном режиме, что обусловлено интенсивностью
энергоотдачи животных. Так, импульсная и суточная активная
мощность у китообразных отличается более чем на порядок
величины. Поэтому часто встречающиеся термины — «макси-
мальная», «спринтерская», «крейсерская» скорость плавания
водных животных — без дополнительного указания продолжи-
тельности плавания не являются вполне определенными и нуж-
даются в уточнении.
В работе автора [45] после тщательного сопоставления и
графо-аналитического анализа данных различных источников
были систематизированы осредненные значения максимальной
скорости плавания различных видов дельфинов и китов в море
в течение продолжительного времени. Выделены следующие
градации продолжительности плавания Т: 10—29 мин — на-
пуганные животные, 2—4 ч — преследуемые судном, одни
сутки — мигрирующие. Эти материалы существенно подтверж-
даются для синего кита при 7=10 и 120 мин данными антарк-
тической научно-исследовательской экспедиции и для тихооке-
анского дельфина-белобочки при Т = 2—8—10—20—25 мин дан-
ными морской межведомственной экспедиции [79]. Значения
максимальной скорости вместе с рассмотренными выше им-
пульсными максимальными скоростями плавания при Т = 3—
—5—10 с приведены на логарифмическом графике (рис. 31) Для
Рис. 31. Зависимость абсолютной максимальной скорости регулярного
плавания гидробионтов от продолжительности плавания.
Китообразные: дикие дельфины ~ белобочка 1, косатка 2, белуха 3;
киты: синий 4, серый и кашалот 5, 6\ дрессированные в неволе — продельфины
7, тихоокеанская афалина 8, тихоокеанский короткоголовый 9.
Эксперименты с рыбами в гидродинамических трубах; а —
импульсный, б — продолжительный, в — мигрирующий режим. Спортивное
плавание (СП) чемпионов: г —с ластами; д — способом «кроль» и
вольным стилем.
Л — лососевая нерка длиной 0,18 м; Ф — форель радужная длиной 0,24 м.
73
китов трех видов — синего, серого, кашалота и для дельфинов
трех видов — белобочки, косатки, белухи. Среди названных ви-
дов китов и дельфинов имеются мелкие и крупные, скоростные
и нескоростные. Для них получены достаточно хорошо согла-
сующиеся зависимости Um=Um(T).
Кроме того, на графике нанесены экспериментальные
точки — результаты обстоятельных физиологических исследова-
ний Бреттом плавания лососевой рыбы Л нерки длиной L =
= 0,18 м на максимальных скоростях продолжительностью
в широком интервале времени Т= 14-3,6 • 104 с [69], а также
аналогичные данные для форели радужной Ф длиной 7 = 0,24 м
[68]. Эксперименты выполнены в специально построенных не-
больших циркуляционных биогидродинамических трубах с ос-
текленным рабочим участком. На графике приведены и данные
по спортивному плаванию способом «кроль» и вольным стилем
рекордсменов мира и олимпийских чемпионов на дистанциях
от 25 до 1500 м, а также по плаванию с ластами на аналогич-
ных дистанциях, в пределах Т= 10ч-103 с.
Примечателен тот факт, что для названных видов китооб-
разных, кроме синего кита при продолжительности плавания
7= (3—10) 4-104 с и для указанных малых скоростных рыб при
7=104-104 с приближенно справедлива аппроксимирующая за-
висимость одного и того же вида:
Um = kTT-°-w, 10 < Т < 10*. (8)
Здесь постоянный показатель степени — 0,073 характеризует
одинаковую интенсивность снижения осредненной максималь-
ной скорости (Um, м/с) с увеличением продолжительности пла-
вания (7, с) гидробионтов, а размерный коэффициент kT оп-
ределяет скорость каждого данного гидробионта при условно
фиксируемой начальной продолжительности 7=1 с. Единооб-
разная интенсивность снижения значений осредненной макси-
мальной скорости с увеличением продолжительности периода
плавания гидробионтов как бы запрограммирована для значе-
ний 7=104-104 с. Данные при 7>104 с более разнообразны и
менее надежны; они не представляют большого интереса для
гидробионики. Указанные выше закономерности по осреднен-
ным максимальным скоростям плавания дельфинов и китов
относятся к естественным природным условиям моря. Они полу-
чены по различным источникам и согласуются между собой.
Это дает основание пользоваться расчетной зависимостью
Um(T) по формуле (8) для других видов дельфинов, например,
афалины и морской свиньи, в соответствии с их абсолютными
максимальными значениями Um= 12,5 и 8,6 м/с при 7 = 3 с
(см. табл. 3).
На рис. 31 нанесены экспериментальные'точки, характери-
зующие дрессированных в неволе дельфинов при следующих
условиях: продельфин (стенэлла) длиной 1,86 м, массой 53 кг
74
при 7 = 2,0 с и Um= 11,5 м/с испытан в лагуне на дистанции
25 м; тихоокеанская афалина «Кейки» длиной 1,91 м, массой
89 кг, Т = 6,6—7,5—10—50 с, Um= 10,3—8,3—7,0—6,1 м/с испы-
тана в лагуне и открытом море с оборудованными дистанциями
61 и 320 м; тихоокеанский короткоголовый дельфин по кличке
«Нотти» длиной 2,09 м, массой 91 кг при 7 = 2,0 с и Um=
= 7,76 м/с испытан в опытовом бассейне 96X3,6X2,0 м при глу-
бине воды 1,5 м с нагоном поверхностной волны.
Как видно, значения максимальной скорости плавания дрес-
сированных в неволе дельфинов не вполне взаимно соответ-
ствуют и значительно ниже рассмотренных для диких живот-
ных в естественных условиях. Это и понятно, если иметь в виду
ограниченные условия искусственного кратковременного экспе-
римента, сам факт содержания в неволе и снижение тонуса
жизни дельфинов, лишение их главного стимула для высоких
скоростей плавания в природе — добычи пищи и спасения от
врагов. К тому же дрессировка может и не соответствовать
привычным естественным режимам плавания дельфинов —
столь высоко организованных и эволюционно адаптированных
к определенным условиям.
§ 8. Анализ кинематики цикла нестационарного
регулярного плавания крупных гидробионтов
Рассмотрим основные элементы кинематики типичных ре-
жимов нестационарного регулярного плавания гидробионтов,
имея в виду, что морфология тела и плавников у них связаны
между собой и с образом жизни, а потому и р плаванием.
В соответствии с тремя распространенными формами плавания
крупных гидробионтов — угревидной, скомброидной, плавнико-
вой— последовательно обратимся к отвечающим им экспери-
ментальным данным.
Поскольку угревидная и скомброидная формы плавания яв-
ляются ближайшими разновидностями главного, волнового,
способа плавания гидробионтов, на рис. 32 для сравнения пока-
заны результаты киносъемки сверху плавания в неглубоком го-
ризонтальном лотке со стоячей водой двух типичных рыб: мед-
ленно плавающего угря и сравнительно быстроходной скумб-
рии [74]. На дне лотка нанесена масштабная координатная
сетка; длина стороны квадрата равна 25,4 и 76,2 мм, отсчет
времени можно вести по интервалам между кадрами кино-
грамм— соответственно 0,09 и 0,05 с.
В процессе плавания обеих рыб тело поперечно изгибается,
вдоль тела в направлении, обратном его поступательному про-
движению со скоростью U, распространяется локомоторная
волна с фазовой скоростью c>U. Волна тела выделяется на
рисунках своими экстремумами. Она создается внутренними
мускульными усилиями рыб; зарождается в головной части
76
а)
Рис. 32. Кинограммы плавания в спо-
койной воде угревидной и скомброидной
рыб. Плавание угря (а) и скумбрии
(б) различается по амплитуде локомо-
торной волны тела и эффективности
действия хвостового плавника.
с небольшой амплитудой аг
и продвигается к хвосту
с большой амплитудой
ах>аг. На теле рыбы од-
новременно может нахо-
диться и более одной волны
числом m = LI'k, если длина
волны X<L. Каждый эле-
мент тела и хвостового
плавника рыбы в попереч-
ном волновом движении на-
ходится под переменным со
временем углом атаки и вы-
зывает в водной среде эле- _
ментарную реакцию. Сумма
реакций по всей длине
рыбы создает силу тяги.
Одновременно видим су-
щественное различие в пла-
вании угревидной и скомб-
роидной рыб. Поскольку
тело угря относительно
длинное (L///>15), близ-
кое к цилиндру, Н^В (Н —
наибольшая высота, В —
наибольшая ширина тела), эта рыба не имеет в конце выра-
женного хвостового плавника. Сила тяги угря создается ис-
ключительно локомоторной волной всего тела, а также про-
дольных плавников сверху и снизу тела, с относительно боль-
шой переменной по длине тела амплитудой a/L. Напротив,
скумбрия относительно толще и короче (L/H — 5), тело хорошо
обтекаемое и суживается к хвосту, а хвостовой плавник ярко
выражен. Поэтому у скумбрии изгибается только задняя часть
тела; хвостовой плавник’ создает значительную силу тяги,
уменьшается амплитуда локомоторной волны и рыскания рыбы
из стороны в сторону. У скумбрии alL меньше, чем у угря.
Скорость плавания рыбы и фазовая скорость распростране-
ния локомоторной волны определяются по кинограмме, пока-
занной на рис. 32:
£7 = Sj/г, с = (52—53)//, (9)
поэтому
с Sa — S3
U sx
(10>
Здесь Si, 5г, s3 — показанные на рисунке отрезки, a t — рассмот-
ренный промежуток времени, отсчитываемый по интервалам
кинокадров. В частности, для обоих рассмотренных выше слу-
76
чаев плавания рыб получено: угорь речной L = 0,07 м, U =
= 0,04 м/с, Re = 2,45-103, с/(7=1,55; скумбрия-макрель L =
= 0,33 м, t/ = 0,42 м/с, Re=l,2-105, с/£/=1,81.
В современной гидробионике наибольший интерес представ-
ляет плавниковая форма плавания самых скоростных рыб и
всех видов китообразных. Они также совершают волнообразные
движения телом, однако имеют весьма развитый машущий
хвостовой плавник, который создает в основном силу тяги.
Плавник имеет характерную форму: «серповидную» у скорост-
ных тунцовых и ксифиоидных видов рыб (меч-рыбы, парус-
ника, марлинов) и «полулунную» у китообразных (дельфинов
и китов). Машущий серповидный плавник скоростных рыб
жесткий и узкий, большого относительного удлинения (свыше
пяти-шести) и сравнительно высокочастотный, а полулунный
плавник дельфинов и китов упругий и широкий, дельтавидной
формы в плане с относительным удлинением около Х~4 и
низкочастотный (п^.4).
К настоящему времени изучена кинематика нестационар-
ного регулярного плавания дельфина, хотя еще несколько де-
сятилетий назад в литературе встречались самые разнообраз-
ные толкования по данному вопросу издавна интересовавшему
многих исследователей. Утверждали даже, что дельфины якобы
совершают винтообразные движения своим двухлопастным
хвостовым плавником, но это не подтвердилось новейшими
данными. Большую ясность внесли комплексное изучение ана-
томии хвостового отдела китообразных и натурные наблюдения,
главным образом, киносъемка плавания дельфинов в море и
в океанариумах. Рассмотрим обработанные кинограммы.
Рис. 33. Результаты обработки киносъемки медленного плавания черно-
морских дельфинов в большом кольцевом остекленном бассейне (Ret.«
«2- 10е): а — белобочки длиной 1,32 м; б — азовки длиной 1,33 м.
1-4— траектории движения характерных точек (/ — развилка хвостового плавника;
2 — вершина спинного плавника; 3 — оконечность грудного плавника; 4 — нос тела).
77
На рис. 33 показаны в масштабе траектории движения ха-
рактерных точек тела двух видов черноморских дельфинов:
обыкновенного (белобочки) и азовки (морской свиньи). Траек-
тории получены с применением киносъемки их медленного пла-
вания в большом кольцевом, с остекленным участком, бас-
сейне Морского гидрофизического института АН УССР [43]. Оба
дельфина одинаковых размеров, но скоростная белобочка со-
вершает колебания с меньшей амплитудой, чем нескоростная
азовка. В обоих случаях волна колебаний длиной % имеет наи-
больший размах А у развилки хвостового плавника, наимень-
ший размах колебаний у верхней точки спинного плавника и
у нижней точки грудного плавника, промежуточное значение раз-
маха— у точки в носовой части тела. По малым колебаниям
крайних точек спинного и грудного плавников можно судить
о том, что ЦТ тела дельфина перемещается почти прямоли-
нейно. В данных случаях относительные размахи A/L указан-
ных точек хвостового плавника, ЦТ тела и носа дельфинов
соотносятся примерно так: п: Ацт : Ав= 1,0 : 0,05 : 0,4. Как
видно, волна деформаций различных точек всего тела в плава-
нии дельфина имеет сложный характер. Однако траекторию
цикла колебания центра давления (ЦД) хвостового плавника
приближенно можно описать простой гармоникой типа у =
=А sin л(%А).
Подводная киносъемка плавания обыкновенного дельфина
в море (рис. 34) показывает, что при поступательном переме-
щении его хвостовой стебель делает плавные взмахи вверх-вниз,
поворачиваясь вокруг некоторого центра вращения, а обе ло-
пасти хвостового плавника вращаются за стеблем, опаздывая
по фазе примерно на угол л/2. В крайних верхнем и нижнем
положениях конца хвостового стебля плавник перекладывается
всего за 0,02—0,04 с, чем обеспечивается в любой момент
времени необходимый угол атаки хвостового плавника для соз-
дания положительной силы тяги, особенно при прохождении
хвостовым стеблем среднего положения при выровненном теле
дельфина. Продолжительность времени опускания хвостового
плавника от крайнего верхнего до крайнего нижнего положения
не обязательно равна таковой при возвратном движении. Ампли-
туда опускания хвостового плавника может быть больше ампли-
туды подъема его над средним положением, что, вероятно, за-
висит от несимметричного профиля дельфина в вертикальной
плоскости. Колебания носовой части дельфина по сравнению
с колебаниями хвостового плавника происходят со смещением
по фазе.
Чтобы более детально характеризовать кинематику плавни-
ковой формы движения гидробионтов, когда главным движите-
лем является полулунной формы хвостовой плавник, приведем
некоторые результаты исследований [24] плавания в био-
гидродинамическом канале тренированного черноморского дель-
78
Рис. 34. Кинограмма одного цикла подводного плавания в море обыкно-
венного дельфина (сверху вниз). Частота движительного комплекса
п=2 Гц; число Струхаля 5Ьд=0,37.
79
Рис. 35. Кинематические характеристики плавникового движителя
афалины длиной ~2,5 м по результатам обработки киносъемки
в биогидродинамическом канале [24]: а — траектория условного ЦД
плавника и положения корневой хорды плавника в кадрах; б —
изменения мгновенных значений продольной U(t) и поперечной У(/)
составляющих скорости плавника; в — мгновенные изменения угла
перекладки плавника Р(/), угла скоса потока у(<) и угла атаки
плавника a(t').
фина-афалины (рис. 35). Для анализа отбирали только ре-
жимы прямолинейного движения дельфина, когда хвостовой
плавник не совершал вращательных движений вокруг корневой
хорды. Последняя проектируется на вертикальную продольную
плоскость движения в виде прямого отрезка в каждом кадре,
положение которого определяется продольным расстоянием s,
пройденным хвостовым плавником, и амплитудой а его коле-
баний, тем самым — пространственными координатами х, у (см.
рис. 35, а). На рисунке показаны также номера кадров кино-
съемки и текущее время t, с.
Кривые изменения составляющих скорости движения ЦД
плавника U(t) и V (/) определены (см. рис. 35, б и а) согласно
кинограммам по временным производным:
U = ds/dt, V = da!dt. (11)
80
Мгновенная скорость движения ЦД хвостового плавника
= + (12)
На рис. 35, а и в показаны углы перекладки плавника р,
т. е. мгновенные углы наклона корневой хорды к направлению
поступательного движения ЦТ тела дельфина; углы скоса по-
тока у — углы наклона мгновенного вектора скорости к направ-
лению поступательного движения ЦТ тела дельфина; углы
атаки а между мгновенным вектором скорости и корневой хор-
дой плавника. При этом углы р определены графически, непо-
средственно по рис. 35, а, углы у получены из соотношения со-
ставляющих скоростей
tgy=— V/U, (13)
углы атаки найдены как разность двух углов
а = у—р. (14)
Анализируя приведенные характеристики видим, что посту-
пательная скорость U (/) движения дельфина неравномерная,
причем изменение скорости за цикл плавания обусловлено ко-
лебаниями двигательно-движительного комплекса. Лопасти
хвостового плавника в течение одного периода дважды пере-
секают с углом атаки почти прямолинейную траекторию посту-
пательного движения ЦТ дельфина и дважды находятся в край-
них верхнем и нижнем положениях с перекладкой через нулевой
угол атаки. Поэтому скорость поступательного движения ЦТ
дельфина за один период соответственно два раза изменяется:
от максимальной к минимальной и, наоборот, от минимальной
к максимальной. Она больше, когда хвостовой плавник дви-
жется вниз, и меньше при подъеме плавника. Перекладкой
плавника в крайних верхнем и нижнем положениях обеспечи-
вается всегда положительная сила тяги. Наибольшие ускоре-
ния наблюдаются при прохождении хвостовым плавником с уг-
лом атаки срединного, выровненного положения тела дельфина.
В данном случае, при плавании дельфина с небольшой средней
скоростью Ц0 = 2,3 м/с, мгновенные значения скорости отлича-
лись всего до ±3%, однако на скоростных режимах пульса-
ции скорости могут достигать ДСЦ/) = ± 15 %. Кривая измене-
ния поперечной составляющей скорости V(/) (см. рис. 35, в)
естественно имеет такой же характер, что и кривая амплитуд
a(t), но сдвинута по фазе на угол л/2. Экстремумы кривой
V (t) приходятся на моменты прохождения хвостовым плавни-
ком срединного, выровненного положения дельфина. Отноше-
ние max V/Uo составляет около 0,87.
Угол атаки a(t) в течение каждого полупериода колебаний
хвостового плавника дельфина имеет явно выраженный макси-
мум в виде площадки при значениях max V. Обычно а<10°,
в среднем же а = 54-6°'. Из сопоставления рис. 35, а и в
81
следует, что фазовый угол сдвига вертикальных и угловых коле-
баний хвостового плавника составляет 6 = л/2. Это соответ-
ствует максимальному коэффициенту полезного действия (кпд)
движителя типа «машущее крыло».
Как показано в работе [57], фазовая скорость распростране-
ния локомоторной волны тела дельфина при активном плава-
нии в действительности не постоянная. В области хвоста она
в полтора раза выше, чем в области головы животного (при
средней скорости плавания дельфина {70=4,3 м/с).
§ 9. Система пространственно-временных характеристик
нестационарного плавания гидробионтов
Кинематику плавания гидробионтов для единообразного
представления, использования в теоретических расчетах и мо-
делирования целесообразно выразить в виде системы простран-
ственно-временных характеристик. Такая система характеристик
впервые была разработана автором в работе [43]. Теперь эта
система уточнена и дополнена с учетом более поздних исследо-
ваний. Имеется в виду плавание крупных гидробионтов с из-
гибно-колебательным движительным комплексом, а точнее,
плавание угревидных и скомброидных рыб, и дальнейшая мо-
дификация плавникового машущего движителя с серповидным
хвостовым плавником у скоростных рыб и хвостовым плавни-
ком полулунной формы у дельфинов.
В нестационарном плавании гидробионтов необходимо раз-
личать регулярное'плавание и переходные процессы. Под ре-
гулярным плаванием понимается достаточно продолжи-
тельное, сравнительно с длительностью одного цикла, посту-
пательное перемещение гидробионтов, обычно с относительно
умеренными параметрами нестационарности. Переходные
процессы, например, трогание с места и быстрая остановка,
а также различные кратковременные процессы (импульсные
броски, повороты, заныривание, выпрыгивание из воды и т. п.)
нередко резко нестационарные, с относительно большими ли-
нейными и угловыми ускорениями, подлежат отдельному изу-
чению. Далее рассматривается только регулярное нестационар-
ное плавание гидробионтов. Оно обычно периодическое и прак-
тически гармоническое (в механике такое движение называют
установившимся).
Для характеристики кинематики регулярного нестационар-
ного плавания гидробионтов важнейшими служат следующие их
параметры и критерии: длина тела L; высота (ширина) тела
Н (В); размах колебаний (удвоенная амплитуда) А машущего
плавникового движителя (обычно точки развилки хвостового
плавника), а также относительный размах A/L; текущее время
t; продолжительность цикла т и общая продолжительность пла-
вания Т в данном регулярном режиме; частота колебаний
82
п=\]х и круговая частота co = 2nn движителя; средняя скорость
плавания Uo и относительная скорость UolL\ фазовая скорость
распространения с пропульсивной волны тела и отношение
cJUo, длина локомоторной волны \ = Lln и число m = LD. этих
волн, укладывающихся на длине тела; безразмерные критерии
Рейнольдса ReL и Струхаля ShA-
Взаимосвязь указанных параметров и критериев составляет
по результатам экспериментальных исследований систему одно-
параметрических и двухпараметрических, размерных (частных)
и безразмерных (обобщенных, линейных и нелинейных) про-
странственно-временных~зависимостей. Они необходимы, в част-
ности, для применения важнейших формул гидродинамической
теории движения удлиненных тел [33, 34, 84, 98], согласно ко-
торой, например, средняя сила тяги и гидробионта Т =
= /гр(о2Л2/2[1—t/o/c], а кпд движителя г] = 0,5[ 14- ий]с\. Отсюда
видно значение достаточно точного определения таких размер-
ных параметров гидробионтов и плавникового движителя, как
А, I, со, которые входят в формулу тяги во второй степени,
а также £70, с отношения U0/c.
Ввиду сложности экспериментов с животными гидробионика
располагает ограниченным числом детальных систематических
исследований кинематики плавания различных видов гидроби-
онтов с применением инструментальных измерений (табл. 6).
Эти исследования использованы далее для анализа. До сих пор
систематически изучали плавание только рыб небольших раз-
меров и не самых скоростных. Ограниченными были: частота
колебаний, скорость плавания и числа Рейнольдса, далеко не
достигающие максимальных значений для большинства видо'в
исследованных рыб. Плавание дельфинов-афалин эксперимен-
тально изучается пока также только на малых скоростях.
В экспериментальных исследованиях плавания гидробион-
тов (см. табл. 6) широко применяли киносъемку. Однако усло-
вия проведения экспериментов были различными и даже
несхожимы. Плавание сарганов, озерной форели, кефали, пела-
миды и луфаря изучали в циркулирующем потоке биогидроди-
намической трубы высокой турбулентности с остекленной приз-
матической рабочей частью размером 1,7X0,3X0,3 м3 [28]. На-
против, плавание форели радужной, ельца и карася изучали
в невозмущенной воде вращающегося кольцевого аквариума
среднего диаметра 0,67 м, площадью поперечного сечения
0,06x0,08 м2 [68]. Макрель японскую испытывали в циркуля-
ционной биогидродинамической трубе с рабочей частью длиной
0,80 м и диаметром 0,41 м, причем были приняты меры к сни-
жению турбулентности потока [78]. Плавание тихоокеанского
тунца исследовали в океанариуме размером 33,0X16,3X11,0 м3,
с остекленными окнами. Условия плавания тунцов-скипджек и
тунцов-альбакор были естественными, непосредственно в море,
однако рыбы, плавали в стаях [100]. Тунцы названных трех
83
Таблица 6. Систематические экспериментальные исследования кинематики плавания гидробионтов
Виды рыб и дельфинов Число особей N Длина тела Lt CM Частота колеба* ний л, Гц Относительный размах A/L Наибольшие скорости Число Рейнольдса 10~5 Re Формула скорости У0 = *Ь(п-п0) Источник
абсолют- ная м/с относи- тельная UJL. с-1 k Ло
Сарган (Belone belone euxini) 5 40—48 2,4—6,4 0,14 1,10 2,9 5,5 0,50 1,00 [28]
Форель озерная (Salmo trutta) 4 20—22 2,6—8,2 0,23 1,10 5,4 2,7 0,75 1,00 [28]
Кефаль черноморская (Mugil cephalus) 5 24-30 2,5—8,8 0,22 1,75 5,5 4,4 0,75 1,00 [28]
Пеламида (Sarda sarda) 2 16 4,0—11,6 0,24 1,30 7,0 1,7 0,79 1,50 [28]
Луфарь (Pomatomus saltatrix) 1 42 1,5—5,3 0,26 1,45 3,5 6,6 0,71 0,50 [28]
Форель радужная (Sal- mo irideus) 4 4—29 2,0—23,5 0,18 3,18 16,0 8,1 0,75 1,33 168]
Елец (Leuciscus leucis- cus) 4 5—24 1,0—25,0 0,18 2,20 20,0 4,6 0,75 1,33 [68]
Карась серебряный (Carassius auratus) 5 5—23 1,0—15,5 0,20 0,88 9,7 4,3 0,75 1,33 [68]
Макрель японская (Trachurus symmetricus) 6 5—27 2,2—20,5 0,22 2,12 17,0 5,0 0,78 1,49 [78]
Тунец тихоокеанский (Sarda chiliensis) n 45—57 1,8—6,4 — 3,70 6,5 22,0 0,64 —0,77 [86]
Тунец скипджек (Eut- hunnus pelamis) n 57 3,0—21,0 — 5,80 10,2 37,0 0,48 —4,28 [100]
Тунец альбакор (Thun- nus albacores) n 52 2,0—13,0 — 5,40 10,5 31,0 0,57 —4,32 [100]
' Дельфин-афалина (Tur- siops truncatus) 6 235—265 0,8—3,2 0,20 6,50 2,8 140,0 0,95 0,25 [26]
видов среднескоростные. Плавание дельфинов-афалин испыты-
вали в береговом морском металлическом бассейне с остеклен-
ной частью стенки, общей длиной 65 м и площадью поперечного
сечения 4,0 м2 [26]. По-видимому, условия плавания были для
них стеснительными, тем не менее дельфины выделяются сво-
ими параметрами даже при относительно малой скорости пла-
вания.
Анализируя данные табл. 6, можно заметить, что исследо-
ванные виды гидробионтов реагировали на внешние условия
эксперимента. Показательны для сравнения примерно схожие
по способу плавания скомброидные виды рыб, которых испыты-
вали как в биогидродинамической трубе высокой турбулентно-
сти, так и во вращающемся кольцевом аквариуме со спокой-
ной водой. Создается впечатление, что у этих видов рыб едино-
образно запрограммирована по длине тела и скорости плавания
частота колебаний движителя, но более вариабельна амплитуда
колебаний при различных внешних условиях, в данных случаях,
по степени турбулентности воды.
Основные типы пространственно-временных характеристик
регулярного нестационарного плавания гидробионтов. При ука-
зании сравнимых условий обозначим водных животных одного
вида формой их тела Ф, а одинаковые значения параметров —
idem.
Р а з м е р н о - с к о р о ст н а я характеристика Uo(L)
(рис. 36, а) показывает, что поступательная средняя скорость
плавания гидробионтов данного вида прямо пропорциональна
длине тела, однако при одинаковом числе колебаний движи-
теля
= Ф, n = idem.
В данной линейной характеристике коэффициент пропорцио-
нальности fei варьирует по видам гидробионтов и частоте коле-
баний, Й1(Ф, п). Таким образом, выражение скоростных ка-
честв гидробионтов разных видов и размеров без одновре-
менного соблюдения условия равенства значений частоты
движителя некорректно.
Ч а стот н о - с ко р о ст н а я характеристика Uo(n)
(рис. 36, б) показывает, что средняя скорость плавания гидро-
бионтов данного вида прямо пропорциональна частоте колеба-
ний движителя при одной и той же длине тела
[/0 = ^2(«'—«о). Ф, L = idem.
Характеристика линейная, коэффициент пропорционально-
сти k2 варьирует с видом гидробионтов и длиной тела й2(Ф, L).
Однако прямые не проходят через начало координат; по экстра-
поляции значению О0 = 0 отвечает некоторое начальное значе-
ние п0~ 1-5-4. Вблизи начала координат характеристика нели-
нейная. Это можно объяснить неустойчивостью движения на
85-
Рис. 36. Пространственно-временные ха-
рактеристики плавания рыб и дельфинов
в различных условиях (см. табл. 6): а —
размерно-скоростная характеристика
Uo(i-) форели; б — частотно-скоростная
U0(n) характеристика макрели; в — от-
носительная частотно-скоростная — раз-
мерная характеристика Uo/L=f(n)
1— сарган; 2— форель, кефаль, пеламида, лу-
.-фарь; 3 — елец; 4 — макрель японская; 5 —
тунец тихоокеанский; 6 — тунец-скипджек;
7 — тунец-альбакор; 8 — дельфин-афалина
(6 особей).
малых скоростях; теорети-
чески в окрестности нуле-
вой точки имеется особен-
ность. Начальное значение
по также варьирует у раз-
ных видов.
Относительная ча-
стотно - скоростная
характеристика
U0IL=f(n) (рис. 36, в) яв-
ляется обобщенной по ли-
нейным первым двум, по-
этому также линейная
Uo/L = k (tv—п0), Ф = idem.
На графике опущены
экспериментальные точки,
число которых около 550.
Выделяются угревидные
рыбы — сарганы и дель-
фины, схожие характери-
стики имеют скомброидные
рыбы, выше отмечались
особые условия плавания
в стае тунцовых рыб. Зна-
чения й(Ф, L) и п0(Ф,
приведены в табл. 6.
Для форели радужной
можно привести размерную
зависимость, определяющую
максимальное число коле-
баний хвостового плавника
в функции длины рыбы:
Птах = 13,5/j/T, 0,04 < L 0,29.
Размерности здесь следующие: nmax — в герцах, L — в метрах.
По-видимому, указанную зависимость приближенно можно
применять и для других видов скоростных рыб при длине их
до 2—3 м.
Выше отмечалось большое значение амплитуды колебаний
машущего плавникового движителя. Приведем соответствующие
характеристики.
Размерно-амплитудная характеристика A(L)
(рис. 37, а)
A=k3L, Ф = 1дегп, и > и0.
Эта характеристика линейная, прямая по экстраполяции, про-
ходит через начало координат, однако у различных видов гид-
36
Рис. 37. Амплитудные характеристики плавания рыб в биогидродина-
мических трубах (см. табл. 6): а — размерно-амплитудная А(£), раз-
ные виды; б — относительная амплитудно-частотная A/L=f(n),
А — размах колебаний; /—3 — длина форели соответственно 0.29; 0,23 и 0,135
робионтов наклон прямой может несколько различаться,.
&з(Ф). На графике для исследованных гидробионтов прове-
дена средняя прямая (fe3~0,20) при условии, что не приняты
во внимание нелинейные участки характеристик вблизи нулевой
точки; для рыб п0~4—5 Гц. По данным экспериментам выде-
ляются: луфарь большей амплитудой и форель радужная мень-
шей амплитудой (см. табл. 6).
Относительная амплитудно-частотная ха-
рактеристика Я/£=/(я) (рис. 37, б)
Л/Г « const; Ф, Г = idem, п^п0.
У каждой рыбы относительная амплитуда колебаний прак-
тически постоянная, однако она может быть разной у рыб од-
ного и того же вида, но различной длины, и разброс экспери-
ментальных точек большой, т. е. характеристика не очень
устойчивая. До значений п^4—5 Гц характеристика нели-
нейная.
Для крупных гидробионтов с малым числом колебаний дви-
жителя, в том числе дельфинов, у которых вообще п<4—5 Гц,
амплитудно-частотная характеристика может быть другого, бо-
лее сложного вида.
В плавании гидробионтов с изгибно-колебательным комп-
лексом по поверхности их тела пробегает пропульсивная волна,
различная у угревидных, скомброидных рыб и у дельфинов
с полулунным плавниковым движителем, поэтому необходимы
обобщенные безразмерные волновые характеристики плавания
гидробионтов,, справедливые, для определяющих критериев по-
добия в достаточно Широком интервале значений.
87
3 S
88
Число пропульсивных воли
m(ReL) (рис. 38, а), укладывающихся на
длине тела гидробионтов, в зависимости от
их числа Рейнольдса
т = Rep, Ф = idem,
где /г4(Ф), Р~—0,074. Характеристика нели-
нейная, типа показательной функции. В рас-
смотренных случаях т = 1,104-2,50; наиболь-
шее число волн у угревидной рыбы (сар-
гана), наименьшее у дельфина с плавниковым
движителем, промежуточные значения —
у рыбы скомброидного типа (форели и ке-
фали).
Относительная скорость про-
пульсивной волны c/£70 = fr(Re)
(рис. 38, б) существенно различается в зави-
симости от числа Рейнольдса и вида гидро-
бионта, принадлежности его к угревидному
или скомброидному типу плавания. Для уг-
ревидных рыб приближенно
c/U0 « const = 1,55, 1 • 10s < Re < 2- 10е.
т. е. характерно автомодельное по Рейноль-
дсу значение.
Для скомброидных рыб и дельфинов
ориентировочно справедлива нелинейная за-
висимость типа показательной функции
c/t/0 = ^5Re’, const, —0,075.
Универсальная критериаль-
ная зависимость ShA(Reb) (рис. 38, в),
по имеющимся данным, занимает на лога-
рифмической сетке клиновидную область,,
в которой в общем случае
Sh4=f(ReL, Ф, L), 1-109< ReL ЫО7.
По верхней границе области ShA^O^l.
Здесь расположены экспериментальные точки
для рыб угревидного типа. Вблизи границы
находятся точки для разных видов скомбро-
идных рыб, которые плавалй в условиях про-
мыслового эксперимента в воде, в поле элек-
трического тока. Эти рыбы, как и рыбы при
угревидной форме плавания, имели большую
амплитуду колебаний.
Основную часть графика занимают экс-
периментальные точки для скомброидных
8»
рыб 1—9, в которой число Струхаля тем больше, чем больше
длина рыбы данного вида. Каждому виду Ф скомброидной
рыбы определенной длины L отвечает своя показательная кри-
вая
Sh?1=ARe'’, k = const, г = const, Ф, L = idem.
Своеобразно расположены экспериментальные точки для
дельфинов при Rei>106. Это, видимо, связано с отмеченной
выше амплитудно-частотной характеристикой их плавания.
Существенно, что нижняя граница клиновидной области
ShAfRer) прослеживается по ряду экспериментальных точек
для различных гидробионтов единообразно при всех числах
Рейнольдса, поэтому на логарифмической сетке можно про-
вести одну прямую
8Нл = ^Ре£, k = const, s«—0,079.
Сопоставляя между собой все три волновые характеристики
для соответствующих по Рейнольдсу Reb значений т, c/Uo,
StiA, отметим их общее взаимное соответствие. В частности,
показатели степени примерно одинаковые —0,08),
т. е. указанные функции изменяются приблизительно обратно
12/-——
пропорционально у ReL — зависимости слабые.
§ 10. Гидробионические аспекты спортивного плавания
способом «брасс»
Спортивное плавание имеет свои особенности. В спортивном
плавании достигают наибольшей средней скорости на каждой
данной установленной дистанции. Спортсменов обучают пла-
вать различными способами по правилам, унифицирующим
плавательные движения тела и конечностей. Нестационарное
плавание квалифицированных спортсменов, тем более олимпий-
ских чемпионов, характеризуется четко выраженными циклами,
периодичность которых, возможно, не уступает таковой у на-
стоящих гидробионтов.
В плавании брассом на груди, т. е. с чередованием пла-
вательных движений рук и ног, попарно и симметрично с обеих
сторон тела, пловцы не достигают скоростей, доступных в слу-
чае применения других спортивных способов плавания («кроль»,
«дельфин»). Однако брасс имеет важное значение как естест-
венный, древнейший и самый массовый, способ плавания, ко-
торым плавают все: мужчины, женщины и дети всех возрастов.
Многовековой опыт показал большое прикладное значение
брасса. В природе аналогичное плавание встречается у ля-
гушки; неслучайно в народе издавна брасс называют плаванием
«по-лягушачьи». Однако лягушка активно пользуется только
одной парой кратковременно действующих движителей — зад-
ними лапами, а передние ее лапы не делают собственно греб-
ковых движений.
90
Ниже рассмотрим экспериментальные исследования спор-
тивного плавания брассом с привлечением около 80 спортсме-
нов различной квалификации — от начинающих плавать и
третьеразрядников до мастера спорта международного класса
и олимпийских чемпионов-брассистов [36]. Применяли разно-
образную методику и технику исследований плавания брассом
в спортивных бассейнах, в том числе визуализацию вызванных
плаванием спортсмена течений воды, надводную и подводную
фотографию, синхронную киносъемку в двух проекциях. Ма-
териалы исследований обработали с применением теории подо-
бия и методов математической статистики.
Отметим высокую нестационарность плавания брассом. На
рис. 39 показаны в безразмерном виде графики скорости пла-
вания спортсменов-брассистов высшей квалификации. Графики
составлены по результатам обработки киносъемки. Спортсмены
проплывали перед киноаппаратом в свойственном каждому из
них режиме с максимальной абсолютной средней скоростью
около (?о~1>4О м/с. Максимальная относительная мгновенная
скорость внутри цикла после гребка ногами и руками не пре-
вышала t/m/(70=l,4-l,6. Минимальное значение относитель-
ной мгновенной скорости в начале цикла, в подготовительной
фазе перед гребком ногами (когда руки вытянуты, почти пря-
мые, ноги согнуты в тазобедренном и коленном суставах) всего
771 = О,4О. У неквалифицированных спортсменов мгновенная ско-
рость внутри цикла снижается до нуля.
Рис. 39. Изменение мгновен-
ной относительной скорости
спортивного плавания брас-
сом в период цикла U(t).
I—11I — общие точки (/((); /—
4 — олимпийские чемпионы, ре-
кордсмены мира — брассисты;
S — аналогичный график плава-
ния гидробионта лягушки (про-
тотип).
Рис. 40. Внутрицикловые изменения абсо-
лютной скорости спортивного плавания
брассом по фазам: толкание ногами (/),
свободное скольжение (2), гребок руками-
(<3), подготовительная фаза (4).
А—тренированный брассист, чемпион мира; В —
нетренированный по брассу, но квалифицирован-
ный пловец; С — ранее тренированный, но уже
неквалифицированный пловец; D — нетренирован-
ный и неквалифицированный пловец.
91
Специалисты считают, что брасс — многовариантный спо-
соб плавания. Это зависит от влияния разных факторов. По-
этому, несмотря на общие правила, стиль плавания одного
рекордсмена отличается от другого. Для исследованных нами
рекордсменов выделяется несколько графиков изменения ско-
рости в цикле плавания брассом (см. рис. 39), а именно:
с быстрым разгоном и продолжительным замедлением (кривая
1 — женщина), с затянутым разгоном, но быстрым замедлением
(кривые 2, 3, 4 — мужчины). Пловцы различаются относитель-
ной мощностью гребковых движений рук-ног и более или ме-
нее своевременным гребком руками после гребковых движений
ногами. Однако опыт показывает, что квалифицированных
спортсменов нецелесообразно заставлять плавать одинаково,
копируя друг друга. Это обстоятельство наводит сомнения на
правильность дрессировок дельфинов для скоростного плава-
ния, приемы плавания которых детально не изучены.
Несмотря на отмеченные индивидуальные различия рекорд-
сменов, для всех четырех безразмерных кривых изменения ско-
рости внутри цикла плавания брассом характерно то, что прак-
тически все эти кривые проходят через три общие точки U(t).
Это обусловлено общностью нормативных требований для од-
ного и того же способа плавания.
На рис. 40 кривыми А, В, С, D представлены размерные
графики изменения мгновенной скорости в течение одного цикла
плавания брассом спортсменов различной квалификации и раз-
личной подготовленности, т. е. натренированности непосред-
ственно перед проведением испытаний. Основное значение для
более высоких результатов плавания имеет высокая квалифи-
кация спортсменов (кривые А и В), которые плавают брассом
со значительно большими скоростями при меньшей продолжи-
тельности циклов (большей частоте), чем неквалифицирован-
ные спортсмены (кривые С и D). Меньшее значение имеет
подготовленность, натренированность спортсменов перед испы-
таниями (сравните соответственно кривые А и В, С и D).
У неквалифицированного по этому способу и нетренированного
спортсмена (кривая D) мгновенная скорость в цикле падает
до нуля. Таким образом, в сравниваемых случаях достижения
в плавании спортсменов определяются в основном совершен-
ством способа плавания и эффективностью использования ими
движителей.
Поскольку несущими поверхностями движителей в плавании
брассом являются ладони и стопы спортсмена, на основании
анализа кинограмм были составлены схемы следов их в двух
плоскостях (рис. 41). Спортсмен массой 78 кг, ростом 1,89 м,
плавал брассом со средней скоростью 1,32 м/с. Продолжитель-
ность анализируемого цикла 1,12 с. «Шаг пловца», т. е. рас-
стояние, проплываемое спортсменом за один цикл s=l,47 м.
Изображения на схемах представлены относительно неподвиж-
92
Рис. 41. Гидробионическая мо-
дель плавания брассом: схема
траекторий несущих поверхно-
стей двух пар движителей —
конечностей спортсмена: а —
вид сбоку; б — вид снизу.
Сплошные линии — след ладоней,
пунктирные линии — след стопы
(составлено по киносъемке).
Рис. 42. Безразмерная харак-
теристика техники плавания
брассом l/s = f (l/L).
/ — олимпийские чемпионы и ре-
кордсмены мира; 2 — мастера
спорта международного класса и
мастера спорта СССР; 3 — канди-
даты в мастера спорта и спорт-
смены-брассисты I разряда; 4 —
спортсмены-брассисты II и III раз-
рядов.
ной системы отсчета, в линейном масштабе шага пловца, ко-
торый принят за безразмерную единицу длины s=l,00. Следы
ладоней и стоп показаны в проекциях сбоку и снизу. Сплошные
линии обозначают границы конечностей, причем толстыми
сплошными линиями выделены гребковые фазы движения. На-
правления движения ладоней и стоп указаны стрелками. На
схемах, кроме участков поступательного движения ладоней и
стоп спортсмена, резко выделяются петлеобразные следы их,
когда выполняются гребки. Совокупность этих схем можно
рассматривать как некоторую гидробионическую модель пла-
вания брассом. Такая модель показывает, что в плавании брас-
сом обе пары разновременно работающих движителей ладоней
и стоп пловца, симметричных относительно продольной оси
движения, создают интенсивные парные вихревые образова-
ния. Вращение воды в них с каждой стороны тела происходит
в одном направлении, противоположном вращению воды с дру-
гой стороны тела. Эти вихри неравномерно перемещаются
с поступательным потоком воды и нестационарно взаимодей-
ствуют с телом пловца и друг с другом. Таким образом, мерой
совершенства техники плавания брассом может служить про-
дольное расстояние между осями вихревых образований, соз-
данных гребковыми движениями ног и рук спортсменов.
93
Однако экспериментальное определение указанного расстоя-
ния для многих пловцов различной квалификации затрудни-
тельно. Поэтому практически воспользовались измерением по
кинокадрам в неподвижной системе отсчета (борта плаватель-
ного бассейна) взаимного отстояния в плавании двух харак-
терных фиксированных точек конечностей. В качестве послед-
них выбрали: конец среднего пальца кисти и точку на колецном
суставе, траектории которых за один цикл также имеют петли
и расстояние между которыми определяется достаточно точно.
Это расстояние обозначили через I и условно назвали «харак-
терной длиной», поскольку, можно полагать, оно пропорцио-
нально расстоянию между осями вихревых образований. Назва-
ние характерной длины оправдано еще тем, что существует
тесная связь между средней величиной 7, полученной экспери-
ментально для каждой группы брассистов различной квалифи-
кации, и нормативами времени Т проплывания дистанции спорт-
сменами соответствующих разрядов.
Определяющей оказалась безразмерная характеристика
l/s = f(l/L) (рис. 42). Здесь / — характерная длина; $ — шаг за
цикл плавания; L — рост спортсмена. На графике явно выде-
ляются области, соответствующие группам спортсменов опре-
деленной квалификации, причем эти области тем компактнее
и выражают тем более тесные зависимости, чем выше мастер-
ство брассистов. Численные значения безразмерных параметров
l/s и 1/L стремятся к своему нижнему значению, которое до-
стигается спортсменами высшей квалификации (рекордсменами
мира). Тем самым степень совершенства техники плавания
брассом получила количественное выражение, указывающее,
в каком направлении следует работать спортсмену для дости-
жения высокого результата. Эта зависимость получена впер-
вые и качественно может иметь более общее гидробионическое
значение, например, для исследований плавания гидробион-
тов— ластоногих, некоторые виды которых также имеют по
две пары движителей (конечностей).
Показательно, что безразмерное число l/s формально имеет
смысл критерия Струхаля. Действительно, I/s = l[U0Т=nl/Uо,
где п — частота циклов. Поэтому справедлива обобщенная кри-
териальная характеристика между числом Струхаля Shi и чис-
лом Рейнольдса ReL = U0L/v, где значение кинематического
коэффициента вязкости воды при средней температуре воды
в спортивных бассейнах / = 25°С равно v = 0,90’10~6 м2/с, так
что ReL= (1,84-3,0) • 10е в исследованных случаях. График
Sh/(Rei.) имеет вид, аналогичный таковому для волнового пла-
вания гидробионтов, что обусловлено общей гидродинамической
природой нестационарных движений в сплошной среде. Од-
нако численные значения критерия Струхаля для плавания
брассом непосредственно не сравнимы с другими. Они выра-
жены через иной характерный параметр длины. Тем не менее,
94
если для менее квалифицированных спортсменов Sh/~0,6, то
для спортсменов высшей квалификации Sh;~0,21.
В заключение подчеркнем, что в экспериментальных иссле-
дованиях нестационарного плавания спортсменов брассом не
только освещены такие влияющие факторы, как квалификация
спортсменов, степень их натренированности, индивидуальные
различия, но и получена количественная зависимость для
оценки степени мастерства пловцов, определены направления
совершенствования техники плавания брассом. Одновременно
проведенные исследования характеризуют весьма ограничен-
ные естественные плавательные возможности человека как гид-
робионта.
ГЛАВА III
БИОГИДРОДИНАМИКА
НЕСТАЦИОНАРНОГО ПЛАВАНИЯ ГИДРОБИОНТОВ
ПРИ БОЛЬШИХ ЧИСЛАХ РЕЙНОЛЬДСА
§ 11. Гидробионические области
и общие принципы гидробионики
Изучение функционально-морфологических свойств и кине-
матических характеристик плавания различных видов водных
животных показывает, что следует различать между собой не-
сколько гидробионических областей, гидробионты каждой из
которых обладают своими биологическими и гидродинамиче-
скими особенностями и вместе с тем выражают некоторые об-
щие принципы движения организмов в водной среде. Эти об-
ласти, с одной стороны, отражают известные в биологии эко-
логические ниши организмов, а с другой стороны, позволяют
различать гидробионты по гидродинамическим признакам —
типам движителей, способам плавания, режимам течения в по-
граничном слое.
В соответствии с гидродинамической теорией подобия для
выделения гидробионических областей воспользуемся основ-
ными параметрами и критериями подобия [41], На логарифми-
ческой сетке четырехпараметрического графика (рис. 43) каж-
дая точка отвечает измеренным и полученным из наблюдений
значениям длины тела L и максимальной скорости плавания
Um, одного животного или группы водных животных опреде-
ленного вида и одновременно — значениям числа Рейнольдса
Re и относительной скорости плавания Um. Как видно, ис-
пользованными данными охватывается весьма широкий диапа-
95
Рис. 43. Гидробионические об-
ласти водных животных, раз-
личных по типам движителей,
способам плавания, режимам
течений в пограничном слое,
в зависимости от числа Рей-
нольдса и относительной мак-
симальной скорости (L—Um,
Re-Um).
/ — мельчайшие организмы: а —
жгутиковые бактерий; сперматозо-
иды и др.; б — ресничные (про-
стейшие); 2 — планктонные, рако-
образные организмы: а — медленно
плавающие, б — быстро плавающие;
3 — рыбы костистые и акуловые:
а — угревидные, б — скомброидные,
в— с серповидным хвостовым плав-
ником; 4 — китообразные, низкоско-
ростные и высокоскоростные: а —
дельфины, б — киты; 5 — моллюски
(кальмары).
зон изменения всех четырех параметров, например, для чисел
Re он составляет 14 порядков величины. И поскольку число
Струхаля Sh функционально связано с числом Re при неста-
ционарном движении организмов, то фактически, хотя и не-
явно, учитывается влияние числа Sh.
Область 1 отвечает весьма малым числам Рейнольдса
Re=10-6-T-l, когда определяющее значение в плавании имеют
силы вязкости среды, а силы инерции пренебрежимо малы;
образуется так называемое ползущее течение. Явно выделя-
ются две подобласти: а) весьма малых чисел Re= 10~64-10~3,
при которых плавают мельчайшие жгутиковые организмы, от
бактерий до сперматозоидов, каждый из которых имеет по 1—
96
5 движительных органелл-жгутиков, волнообразно извиваю-
щихся за телом (клеткой) организма; б) подобласть малых
чисел Re=10-2-T-l характерна для реснитчатых организмов —
простейших, на поверхности которых расположены 102—103 и
более волосоподобных органелл-ресничек, совершающих коор-
динированные гребные движения весельного типа [49]. В под-
области 1 а существенно различаются числа Струхаля удли-
ненных спирохет и относительно быстро плавающих сперма-
тозоидов (см. приложение).
Область 2 условно средних чисел Рейнольдса, Re = 1 ч-
4-102 — промежуточная область течений, где одновременно ска-
зываются существенно силы вязкости среды и силы инерции.
К данной области относятся планктонные, ракообразные орга-
низмы, которые нередко двигаются рывками, скачкообразно,
с относительной средней скоростью: a) Um< 10 с-1; б) Um>
>10 с-1. В отношении гидродинамики эта область наиболее
сложная и мало исследованная. В работе Н. Н. Хмелевой
(Бионика. Киев. 1973. Вып. 7. С. 37—44) описана скорость
движения равноногих черноморских раков-идотей длиной от 2
до 22 мм (на рис. 43 подобласть 2, точки обозначены ромби-
ками). Характерно, что с увеличением размеров этих организ-
мов повышается не только абсолютная, но и относительная
скорость плавания, вплоть до t7m=10 с-1.
Область 3 сравнительно высоких ' чисел Re= 103-ь 107
с преобладающим значением сил инерции принадлежит волно-
образно плавающим костистым и акуловым рыбам и имеет
подобласти: а) угревидные рыбы; б) скомброидные и в) с сер-
повидным хвостовым плавником. Подробные данные о рыбах
приведены выше. Хотя угревидные рыбы и часть скомброидных
рыб плавают при одних и тех же числах Re= 103ч-105„ они
различаются абсолютными и относительными значениями ско-
рости плавания, а в соответствии с разными формами волно-
вого плавания — и числами Струхаля (см. рис. 38,в). Кинема-
тика их существенно различна.
Область 4 высоких чисел Re= 107-=- 108 определяет плава-
ние китообразных: а — дельфинов, б — китов с полулунным
хвостовым плавником. Как отмечалось, дельфины й скоростные
рыбы имеют ламинаризованную форму тела; они используют
свойственные им активные способы снижения гидродинамиче-
ского сопротивления, что представляет собой особый интерес
для исследований в гидробионике.
Область 5 относится к моллюскам, в данном случае
к кальмарам, отличающимся своим реактивным движением по
типу водомета. В сущности кальмары применяют два различ-
ных способа плавания: для быстрого броска они пользуются
реактивным движением с периодическим набором и выбросом
воды, а для медленного плавания — волнообразным движением
4 С. В. Першин 97
бокового ромбовидного плавника. Однако кальмары еще мало
исследованы.
Согласно данным рис. 43, вся совокупность эксперименталь-
ных точек закономерно группируется ортогонально изолиниям
равных значений Re = const, т. е. в направлении наибольших
градиентов чисел Рейнольдса. Одновременно с ростом разме-
ров и абсолютных скоростей плавания организмов в 7 поряд-
ков величины, а чисел Re в 14 порядков величины относи-
тельные значения скорости плавания организмов изменяются
в интервале двух порядков (1 —100) величины. Тем самым ве-
личина Um=UmIL является некоторым более устойчивым опре-
деляющим и регулирующим критерием, что отчасти можно
объяснить особенностями биологий организмов, их анатомии и
биоэнергетики.
Обращает на себя внимание широкое, почти безраздельное
распространение волнообразного плавания организмов от мель-
чайших простейших с числами Re=10-6 до гигантов китов
Re=108, хотя формы плавания постепенно модифицируются —
это и отражают различные гидробионические области. Отме-
ченные выше признаки областей позволяют заключить, что
границы таковых определяются гидродинамическими условиями,
когда ползущее течение жидкости сменяется промежуточной
областью, за > которой наступает практически гармоническое
движение гидробионтов, вначале с ламинарным, а затем с тур-
булентным пограничным слоем.
Обобщая вышеизложенное и известные данные, можно на-
звать следующие достаточно общие принципы гидробионики.
Принцип прогрессивного плавания. В биологическом про-
грессе водных животных с увеличением их размеров и повыше-
нием уровня организации существенно растет максимальная
абсолютная скорость плавания. Это обусловлено тем, что эф-
фективная мощность водных животных пропорциональна массе
мышц, т. е. кубу линейных размеров, а гидродинамическое
сопротивление движению пропорционально площади смоченной
поверхности — квадрату линейных размеров. Для скоростных
водных животных характерна высокая относительная доля
мышц и оптимальная по гидродинамическому сопротивлению
форма тела. Отсюда усматривается взаимосвязь биоэнергетики
с биогидродинамикой водных животных и необходимость изуче-
ния ее в гидробионике.
Принцип цикличности движения. Плавание водных живот-
ных всегда нестационарное, обычно периодическое, близкое
к гармоническому, однако с многими модификациями. Принцип
можно объяснить как жизненно важными биологическими свой-
ствами регулирования параметров живых тканей, обусловлен-
ными циклами энергообмена, актами дыхания и кровообраще-
ния, так и механической эффективностью нестационарного пла-
вания, что представляет особый интерес для гидробионики.
98
Принцип унификации движителя. Среди водных животных
наиболее распространены волнообразнодействующие движи-
тели с упругим изгибно-колебательным комплексом различной
структуры, которые при увеличении размеров и скорости пла-
вания все более локализуются в хвостовой части тела. Это
может быть обусловлено существенной особенностью привод-
ных двигателей — мышц животных, действующих антагонисти-
чески с поперечными укорочением-удлинением, а также отсут-
ствием вращающихся элементов у движителей животных, недо-
пустимых по конструкции разделенных между собой узлов и
деталей.
Принцип относительного гидродинамического соответствия.
С увеличением числа Рейнольдса последовательно выделяются
гидробионические области водных животных с закономерно из-
меняющимися характеристиками движителей и способов плава-
ния в соответствии с гидродинамическими режимами течений
в пограничных слоях и вихревых следах. Однако критические
значения чисел Рейнольдса — границ этих областей, обуслов-
ленные условиями гидродинамической устойчивости режимов
течений, в плавании живых организмов могут существенно от-
личаться от таковых для твердых тел. Здесь может сказываться
еще и масштабный эффект. Характерные примеры — повыше-
ние критических чисел Рейнольдса при плавании водных жи-
вотных с ламинаризованной формой упругих тел и активным
снижением гидродинамического сопротивления, что обуслов-
лено наличием биологической слизи у скоростных рыб и демп-
фирующего кожного покрова у скоростных дельфинов.
Высокие параметры плавания гидробионтов можно рассмат-
ривать как результат дивергенции и конвергенции в природе,
последовательной оптимизации движения деформируемых уп-
ругих живых тел с активными внешними поверхностями.
§ 12. Гидробионт как адаптационная колебательная система
Своеобразие плавания гидробионтов по сравнению с движе-
нием в воде твердых тел побуждает к разностороннему выяв-
лению и объяснению механических особенностей явления.
В большом цикле теоретических работ проф. Я. И. Грдины
по аналитической механике живых организмов, опубликован-
ных еще в 1910—1916 гг. и не посвященных непосредственно
водным животным, в общем виде было показано, что следует
различать активное и пассивное движение живых организмов
и что активное движение их всегда нестационарно. Нестацио-
нарность и нелинейность движения организмов, по нашему мне-
нию, объясняют широкие возможности регуляции плавания гид-
робионтов.
Естественно допустить, что организмы представляют собой
адаптационные колебательные системы, которые оптимально
4* 99
регулируются применительно к условиям внешней среды.
В частности, рассмотрим плавание гидробионтов с позиций тео-
рии механических колебаний [54].
Для простоты представим скоростного гидробионта в виде
колебательной механической системы с одной степенью сво-
боды — упругой балки, которая совершает вынужденные мы-
шечными усилиями изгибные колебания в демпфирующей вод-
ной среде. Пусть приведенная масса системы будет A4np; она
включает кроме собственной массы М гидробионта еще и при-
соединенную массу т, т. е. Мпр = М + т. Приведенный коэффи-
циент жесткости упругой системы, обозначим knp. Демпфирова-
ние колебаний обусловлено вязкостью жидкости и образую-
щимися вихревыми системами. Демпфирование будем считать
пропорциональным скорости деформации с осредненным за
цикл колебаний коэффициентом пропорциональности /?.
В рассматриваемой модели положим, что колебание системы
происходит под воздействием переменной силы F(t), изменяю-
щейся по гармоническому закону
F(t) = Fmeiat. (15)
Здесь Fm — амплитуда возмущающей силы, приведенная
к точке приведения; со — угловая частота вынужденных коле-
баний системы;
gi<o< _ cos sin со/, i = '—1.
Обобщенная схема колебательной системы показана на
рис. 44, а. Дифференциальное уравнение движения системы
имеет вид
MnP^ + R^- + kppy=Fme^*. (16)
dr d t
Это линейное полное дифференциальное уравнение второго
порядка с постоянными коэффициентами, решение которого из-
вестно. Нас интересует поведение системы при установившихся
колебаниях, т. е. частное решение уравнения (16), которое
может быть представлено в форме y = Cei<-at~С помощью урав-
нения (16) найдем
у = itaCe1 l'at~ф>, у—'—со2Се‘<р).
В результате получим
(—MnPco2 + i<oR + fenp)y =Fmeia>t.
Следовательно, амплитуда колебаний
г/ = -^еПш'"ф), (17)
соХ
где
ф = arctg —----------------------
—25----Л4прш
О)
Х=у Я2+(мПрС0_^ру . (18)
юо
Здесь X выражает пол-
ное сопротивление системы,
которое включает: R — ак-
тивное сопротивление си-
стемы, вызванное трением,
т. е. демпфирующей силой;
Л4Пр(о — инерционное, а
ЛПр/(о — упругое сопротивле-
ние системы. Инерционное
и упругое сопротивления
(названия условные) всегда
Рис. 44. Колебательная система с од-
ной степенью свободы как простейшая
механическая модель гидробионта: а —
теоретическая обобщенная схема; б —
составляющие приведенного реактивного
сопротивления.
44,6): Z = A4np<o—Лпр/ш. Активное
имеют противоположные
знаки.
Алгебраическая сумма
инерционного и упругого
сопротивлений называется
реактивным сопротивле-
нием Z системы (см. рис.
сопротивление R и реактивное Z сдвинуты по фазе на угол л/2.
При малой величине ш, когда АПр/со>Л1прСО, преобладающим
членом является fcnp/(i> и поведение колебательной системы оп-
ределяется ее упругостью. Напротив, при большой со, когда
Мпрсо^йпр/®, поведение системы обусловлено массой. В случае
Mnp = knp/to реактивное сопротивление Z = 0, полное сопротивле-
ние колебательной системы определяется только одним демпфи-
рованием R и по формуле (18) оказывается наименьшим. Это
случай резонанса системы.
Резонансные колебания наблюдаются при
(о2 = ЛПр/Л1пр =
где
X = VKnp/Afnp. (19)
Здесь X есть угловая частота собственных свободных колебаний
системы; Х = 2лпКр (7гкр— критическая частота колебаний, при
которой амплитуда колебаний системы имеет явно выраженный
экстремум).
Средняя за период т=2л/ш мощность, развиваемая силой F,
определяется формулой
iV = J_f yFd/ = J_ZL^=_!_{/2m(D2R. (20)
то 2 № 2
Таким образом, средняя мощность, отдаваемая при колеба-
ниях установившегося режима, пропорциональна квадрату ам-
плитуды колебаний и квадрату частоты возмущающей силы.
Для упругой системы при заданной силе Fm максимальная
мощность получается при резонансе, когда X=R. С ростом
101
реактивного сопротивления Z средняя затрачиваемая мощность
уменьшается, но уменьшается, и амплитуда скорости ут, а при
заданной угловой скорости а> уменьшается и амплитуда коле-
баний ут.
Для механической системы сумма кинетической и потенци-
альной энергии остается постоянной, поэтому внешняя энергия
расходуется только на преодоление активного сопротивления R.
В живом организме мускулатура совершает работу и потреб-
ляет энергию как при «разгоне» массы /Ипр, так и при ее «тор-
можении». Этим она отличается от простой упругости. Резо-
нанс упругой системы не вполне эквивалентен свойствам жи-
вого организма. Гидробионт или его органы должны настраи-
ваться на близкий к наивыгоднейшему режиму плавания, при
котором частота колебаний примерно равна частоте свободных
колебаний. Последняя у гидробионтов может варьироваться
с изменением жесткости тела.
Скоростной гидробионт — обыкновенный двухметровый ту-
нец— совершает миграции большой протяженности (порядка
нескольких тысяч километров) с невысокими скоростями, но
способен кратковременно плавать с большой скоростью — до
25 м/с. На рис. 45 видны белые и красные мышцы этой рыбы.
В отличие от нескоростных холоднокровных рыб с температу-
рой тела, равной температуре воды, в которой они плавают,
температура мышц у тунца в середине тела превышает темпе-
ратуру воды до 12 °C; наблюдаются большие градиенты темпе-
ратуры от центра сечения тела к периферии. При отмеченных
выше свойствах мышц плавающий тунец с помощью своей мощ-
ной кровеносной системы в состоянии варьировать изгибной
жесткостью тела на различных скоростях плавания.
Сравнительно медленно плавающая рыба — мерланг —
обычно плавает с волнообразными изгибами тела и траектория
движения, ее хвостового плавника близка к синусоиде
(рис. 46, а). В остром опыте, после ампутации хвостового
плавника, рыба делает машущие движения; траектория кон-
чика ее хвоста резко отлична от синусоиды (рис. 46, б). Ско-
рость плавания рыбы при этом уменьшилась всего на 10 %, но
гармонические колебания сменились на другие, напоминающие
колебания релаксационного типа [74]. Число Струхаля ShA
резко повысилось, а потому должен был существенно изме-
ниться коэффициент полезного действия гидробионта.
У быстро плавающей рыбы скомброидного типа (форели)
сила тяги создается преимущественно хвостовым плавником,
амплитуда колебаний которого значительно больше таковой
у головы (рис. 47, а). Напротив, в электрическом поле воды, ис-
кусственно созданном в опытах по электролову рыбы, та же
форель плавала с большими изгибами всего тела [37], как угре-
видная рыба (рис. 47, б). Скорость плавания форели при этом
практически не изменилась, но число Струхаля соответственно
102
Рис. 45. Поперечное сечение
тела высокоскоростной рыбы —
обыкновенного тунца: а—ло-
кализация красных (темных)
мышц; б — распределение тем-
пературы.
Рис. 46. Плавание пелагической
рыбы (мерланга) по данным
киносъемки: а — с хвостовым
плавником; б — после его ам-
путации; вверху — совмещен-
ные виды колебаний, внизу —
траектории движения задней
кромки.
Sh^l.OO
повысилось с 0,18 до 0,42, следовательно, изменился кпд си-
стемы. В природе подобным образом, с сильными изгибами
всего тела, скомброидные лососевые рыбы преодолевают поро-
жистые участки рек.
Как видно, можно привести различные случаи плавания раз-
ных видов гидробионтов, подтверждающие, что гидробионт
представляет собой адаптационную колебательную систему
с переменной жесткостью тела и возможным резким измене-
нием режима и даже способа плавания.
В работе [58] на основе квазистационарного подхода рас-
смотрены кинематические условия для получения оптимальных
пропульсивных характеристик машущего движителя, так ши-
роко распространенного у водных животных. Кинематические
103
Рис. 47. Различные колебания тела и траектории движения крайних то-
чек головы и хвоста одной и той же рыбы — форели радужной: а — при
обычном скомброидном плавании; б — при угревидном плавании в элект-
рическом поле (покадровая обработка киносъемки).
условия максимума силы тяги Т и кпд т] машущего движителя
различны.
В оценочном расчете плавникового движителя дельфина [31]
рассматриваются дорезонансный, резонансный и зарезонансный
режимы плавания. Резонансной частоте колебаний плавнико-
вого движителя соответствует наибольший гидродинамический
кпд его, а возможно и наибольшее значение общего кпд си-
стемы гидробионта.
Интересный пример экспериментальной гидробионической
резонансной установки с принудительно колеблющейся упругой
машущей лопастью, работающей в закрытом призматическом
канале как воздушный насос (колебательный вентилятор), рас-
смотрен в работе [48]. Упругая лопасть в виде деревянной пла-
стины из сошлифованного шпона одним своим концом крепи-
лась на опоре с помощью стальных пружин, что превращало
машущую лопасть с пружинами в колебательный контур с оп-
ределенной постоянной собственной частотой колебаний. Она
104
имела типичную резонансную
амплитудно-частотную характе-
ристику, резонанс наступал при
частоте колебаний п = 4,5 Гц
(рис. 48). Особенностью этой ус-
тановки, кроме упругого крепле-
ния машущей лопасти, был еще
уравновешивающий инерцион-
ные силы дополнительный меха-
низм, который включал ряд де-
талей, связанных с ведущим тол-
кающим пластину штоком, в том
числе, закрепленную на коро-
мысле сменную регулировочную
металлическую массу. Путем за-
крепления на коромысле регу-
лировочной массы в различных
положениях, изменения значе-
ния этой массы и смены упругих
Рис. 48. Резонансная амплитудно-
частотная характеристика машу-
щей лопасти.
Размерные характеристики А(п): I,
2 — амплитуды (полуразмахи) влево и
вправо; 3, 4 — полный размах по сумме
(1) + (2). Безразмерная характеристика
А(п)—5‘ параметры А, п отнесены
к их резонансным значениям (Дтах=1
пкр=1’-
элементов-пружин можно регу-
лировать приведенную к штоку
массу и коэффициент жесткости,
а потому и собственную частоту
всей колебательной системы и
настраивать ее в резонанс при
различной частоте колебаний
машущей лопасти в интервале
п=1-4-6 Гц. Это снижало подво-
димую мощность при каждой данной частоте колебаний ма-
шущей лопасти на некоторых режимах в несколько раз. Такова
высокая эффективность регулирования данной колебательной
системы.
§ 13. Теоретическое моделирование волнового плавания
гидробионтов и проблема волнового движителя
Далее рассмотрим основы биогидродинамики волнового пла-
вания угревидных и скомброидных рыб, а также дельфинов.
В отличие от наиболее распространенных в технике водного
транспорта движителей гребных винтов и применения обладаю-
щих хорошими маневренными качествами крыльчатых движите-
лей в природе преобладает волновое плавание крупных водных
животных. Поэтому в гидробионике одновременно с изучением
возникает и проблема моделирования нового типа волнового
движителя.
Волновым движителем, в соответствии с исследованиями
плавания гидробионтов, назовем хорошо обтекаемое поступа-
тельно перемещающееся удлиненное гибкое тело, которое при-
105
нудительно деформируется по закону поперечной бегущей
волны, распространяющейся от передней кромки тела к задней.
Длина волны равна или менее длины тела, амплитуда ее соиз-
мерима с поперечными размерами тела и увеличивается к зад-
ней кромке, фазовая скорость распространения волны превы-
шает скорость поступательного движения тела в обратном на-
правлении. Принцип действия волнового движителя гидродина-
мически основан на законе сохранения импульса, по которому
отброшенная бегущей волной тела жидкость уносит с собой им-
пульс, а само деформирующееся тело приобретает противопо-
ложный импульс. Создается сила тяги, работой которой пре-
одолевается сопротивление жидкости и восполняется часть
энергии, затраченной на создание бегущей волны тела.
В общем виде принцип создания силы тяги при волновом
движении гибкого тела в сплошной среде теоретически изучали
в Институте гидродинамики СО АН СССР академики М. А. Лав-
рентьев и М. М. Лаврентьев. (Журнал ПМТФ. 1962. Вып. 4.
С. 3—9). Исследована задача движения в жидкости несжимае-
мой деформируемой материальной кривой — цилиндрической
пластины конечной длины с постоянной плотностью на единицу
длины, которая принята в качестве математической модели
движущегося тела. Движение определяется характером внут-
ренних сил, которые выражаются функционалом от координат
точек пластины в функции длины дуги кривой, ее кривизны и
времени. Эта функция при плавании живых организмов опреде-
ляется напряжением мышц, создающим известного вида изги-
бающие мышечные усилия, которые распространяются вдоль
тела в виде бегущей волны. Принцип создания силы тяги при
волновом движении деформируемого тела в сплошной среде де-
монстрируется следующим примером: передняя часть изгибаю-
щегося тела как бы создает криволинейный канал в. жидкости,
а сила тяги получается за счет реакции жидкости при деформа-
ции в основном в задней части тела, с концевой кромки кото-
рого непрерывно сбегают вихри.
Модель — тонкая гибкая пластина или профиль системати-
чески и детально изучалась применительно к плаванию рыб на
основе аэрогидродинамической теории плоского нестационар-
ного движения крыла во многих работах, в том числе и в ряде
работ исследовательского центра ВМС США [89, 92, 98, 99
и др.].
Данную модель можно рассматривать как первое прибли-
жение весьма сложной пространственной задачи теоретического
моделирования волнового плавания гидробионтов, поскольку
исследуется обтекание пластины (профиля) плоским потоком
жидкости.
Постановка задачи следующая. Для удобства решения рас-
сматривается обращенное движение. Гибкая тонкая пластина
бесконечной ширины, которой заменяется плавающая рыба,
106
имеет конечную безразмерную длину L = 2b = 2. Пластина рас-
положена в плоскости хОу вдоль оси х от х= — 1 до х— +1. Де-
формация пластины у = у(х, t) малая и волнообразно изменя-
ется со временем t, поперечная волна перемещается вдоль
хорды в положительном направлении оси х с постоянной фазо-
вой скоростью с. Плоский потенциальный поток набегает на де-
формированную пластину в том же направлении с постоянной
скоростью Uo. Пластина и след за ней заменяются вихревыми
слоями переменной по абсциссе и времени интенсивности. Гра-
ничные условия непроницаемости поверхности пластины лине-
аризуются. В процессе решения удовлетворяется условие Жу-
ковского — Чаплыгина о конечной скорости на задней кромке
пластины.
Волнообразные колебания пластины, моделирующие форму
деформаций оси тела рыбы, задают в виде бегущей волны
у = у (х, t) = а (х) cos (ах—и/); —1^х< + 1. (21)
Здесь а(х) — переменная по длине хорды амплитуда колебаний
нейтральной оси пластины; а=2л/Х— волновое число (X —
длина волны); (о = 2л/т=2лп— круговая частота (т — период
волны: п—число полных колебаний снизу вверх и обратно
в единицу времени). Фазовая скорость распространения волны
вдоль пластины с=пХ=ш/а = Х/т.
Прогрессивное увеличение амплитуды бегущей волны от го-
ловы к хвосту рыбы определяется многочленом второй степени
а(х) =а0 + а1х + аах2, (22)
или в экспоненциальной форме
а (х) = аеЬх. (23)
Коэффициенты а0, Oi, az, a, b — постоянные, находятся при об-
работке материалов исследования плавания рыб.
Результаты теоретических исследований показывают, что оп-
ределяющими параметрами волновых движителей являются:
закон изменения амплитуды а(х) вдоль пластины (линейный,
квадратичный и др.), волновое число а и так называемая «ре-
дуцированная частота» <j=®/.U0. Как обозначено выше, полу-
хорда b = L/2=l, поэтому о представляет собой видоизмененное
число Струхаля Shr,. Действительно,
<т =---=------= л5Пл, Shr=------
uo 2Ua L л
Наибольший интерес для гидробионики представляют сред-
ние за_период колебаний значения коэффициентов: Ст — силы
тяги Т, СР — мощности Р и кпд т). Учитывая, что в условиях
плоской задачи — в данном случае характерная площадь пла-
стины в расчете на единицу ширины h=\ — будет S = Lh = 2,
выражения для названных коэффицентов запишем в следую-
Ю7
щем феномологическом виде (опуская формулы в явном виде
через специальные функции, которые приведены в работах):
Сг=—^-г=ф1[а(х), а, ст],
Ср=—^з- = ф2[а(х), а, о],
Т) = = Фз [а (х), а, а].
Р
(24)
В выражении для т] в числителе указана полезная буксировоч-
ная мощность, а в знаменателе подведенная мощность, которая
в идеальной жидкости включает еще потерянную среднюю ки-
нетическую энергию вихревого следа деформируемой пластины.
Значения расчетного коэффициента силы тяги Ст для волно-
образно деформируемой твердой пластины достаточно высокие:
по формулам (24) порядка нескольких сотых единицы. Вели-
чина Ст существенно зависит как от вида функции изменения
амплитуды а(х) и ее максимальной величины, так и от значе-
ний волнового числа а и безразмерного ст. При данном законе
а(х) величина CT = f(a, о) тем выше, чем больше ст и чем
меньше а. Для постоянных значений а зависимость CT = f(a)
графически имеет следующий вид (по оси абсцисс ст, по оси ор-
динат Ст): вначале, при о<а имеется «провал» кривой с не-
большими отрицательными значениями Ст<0, при а = а всегда
Ст = 0 («критическая» точка), затем для всех ст>а встречаются
только положительные значения Ст>0. Область малых отрица-
тельных значений Ст, т. е. силы сопротивления, а не тяги, опреде-
ляется тем, что в этих случаях фазовая скорость с распростране-
ния бегущей волны меньше скорости Uo набегающего потока
жидкости; когда с/[/0<1, пластина тормозится. Критиче-
ской точке ст = а отвечает значение c—U0. При с/С0> 1 значения
Ст>0, тяга положительная. Значительная доля энергии (де-
сятки процентов) теряется в вихревом следе пластины,- Коэф-
фициент полезного действия т] волнообразно деформируемой
пластины по формулам (24) для всех значений а>0 имеет ве-
личину т|>0,50 и достигает т) = 0,804-0,90.
Подробно результаты теоретических исследований плоского
волнообразного движения тонкой гибкой пластины в идеальной
жидкости изложены И. М. Петровой. Здесь ограничимся рас-
смотрением аналогичных результатов для профилирован-
ных тел.
Теоретическое исследование движителя в виде волнообразно
деформируемых профилей Н. Е. Жуковского конечной толщины
е = 0,1; 0,2; 0,3, выполненное с применением функций комплекс-
ного переменного при тех же предпосылках, что и для пла-
108
стины е = 0, показало аналогичные результаты [92]. Однако чис-
ленно коэффициент силы тяги Ст и кпд т] профиля несколько
меньше, чем для пластины, и тем меньше, чем толще профиль.
При квадратичном законе изменения амплитуды и волновом
числе а = л значение CT = f(a) профиля с относительной толщи-
ной е=0,20 ниже, чем у пластины, примерно на 30 %. В этом
случае кпд т] — 0,75 (рис. 49).
Экспериментальные исследования модели — тонкой гибкой
пластины [80] — были поставлены специально с целью проверки
теоретических решений плоской задачи для пластины в реаль-
ных условиях вязкой жидкости.
Сравнение теоретических значений коэффициента силы тяги
с экспериментальными показывает качественное соответствие
почти во всех исследованных вариантах, и тем более хорошее,
чем меньше амплитуда колебаний а(х) и чем больше значения
волнового числа а, особенно при а = л. Этот случай отвечает ра-
венству длины бегущей волны — длине пластины (X = L), что
часто встречается у гидробионтов, в том числе у рыб.
В работе [90] на основании аналогичных предыдущим теоре-
тических исследований, экспериментов по визуализации потока
и натурных наблюдений сделана попытка качественно предста-
вить картину плавания рыбы с учетом влияния в реальных ус-
ловиях пограничного слоя (рис. 50). Показаны направления
Рис. 49. Теоретические значения средних за цикл коэффициентов силы тяги
Ст, мощности Ср и кпд т] волнообразно деформируемых в идеальной жидко-
сти гибких профилей Н. Е. Жуковского относительной толщины е в функ-
ции безразмерной частоты колебаний о и волнового числа а=л, для формы
хорды с переменной по квадратичному закону амплитудой у(х) =0,023 +
+ 0,042х + 0,034х2, (еаксп = 0 — эксперимент с резиновой пластиной).
109
Рис. 50. Теоретическая модель периодических деформаций тела и обтекания
рыбы через каждые л/4 цикла волнового гармонического плавания с бегу-
щей волной, амплитуда которой увеличивается к хвосту [90] (а), и сводная
схема (б).
движения выделенных- сечений тела рыбы и течения жидкости
относительно этих сечений, а также области вихреобразований,
которые образуются с обеих сторон рыбы и последовательно
перемещаются к ее хвосту. В результате за плывущей рыбой
образуется пространственная вихревая цепочка с мощными дис-
кретными пропульсивными вихрями, являющаяся обобщением
известной плоской пропульсивной вихревой дорожки В. В. Го-
лубева.
ПО
Другой подход в теоретическом исследовании и оптимизации
волновогЬ движителя пластины для достижения наивысшего
кпд при заданной тяге с учетом упругих свойств движителя
предложен В. В. Беляевым и Р. А. Грунфестом. Рассмотрено
плоское обтекание идеальной жидкостью колеблющейся упру-
гой тонкой пластины. Передней кромке пластины задано попе-
речное перемещение и вращение, которые изменяются по гар-
моническому закону, со сдвигом фаз угловых колебаний отно-
сительно поперечных, а задняя кромка свободна и на ней
выполняется условие Жуковского — Чаплыгина. Предполагая
колебания пластины малыми, а движение жидкости потенци-
альным, с линией разрыва касательных скоростей за пластиной,
реализующей вихревой след, формулируется смешанная крае-
вая задача для потенциала скорости, в Которой функция дви-
жения пластины связана с интегрированием динамического
уравнения изгибающейся пластины как балки. Пластина на-
гружена определяемой в задаче распределенной нагрузкой
q(x)=pn—рв, выражающей разность значений давления, дей-
ствующего со стороны жидкости на нижней и верхней поверх-
ностях пластины. Краевая задача сводится к сингулярному ин-
тегральному уравнению Фредгольма 1-го рода, решение кото-
рого представлено в виде ряда. Расчет выполнен на ЭВМ.
Показано, что оптимальные варианты идеализированного
волнового движителя упругой пластины зависят от трех без-
размерных комплексных параметров: числа Струхаля Shb, от-
носительной плотности y = pi/i/pa, где pi — плотность материала
пластины, р — плотность воды, h — толщина и a = Lf2 — полу-
длина пластины, а также от параметра изгибной жесткости
пластины A4 = pico2a4/i/£)'(D — цилиндрическая жесткость). На
эффективных режимах кпд пластины т)>0,80, скорость переме-
щения упругих волн вдоль пластины с =1,2 Оо. Существует не-
большой участок пластины (в рассмотренном примере у попе-
речного сечения на 0,7 ее длины), который в течение всего пе-
риода колебаний не нагружен и не создает тяги, а потому его
можно делать менее широким для уменьшения сопротивления
трения. Этот -результат может объяснить существование зау-
женного переходного участка от тела к хвостовому плавнику
у многих скоростных рыб и других водных животных. Для каж-
дого фиксированного параметра жидкости А* существует свое
значение числа Sh*, при котором достигается максимальный
кпд т]* и его регулирование. Увеличение жесткости пластины
и соответствующее повышение скорости ведет к уменьшению
тахт). Отсюда усматривается значение упругости волнового
движителя. Для повышения равномерности подвода энергии
к двигателю рекомендуется использовать две одинаковые пла-
стины, колеблющиеся со сдвигом фаз 6 = л/2.
Из вышеизложенного следует, что достоинство простейшей
теоретической модели волнового движителя в виде тонкой гиб-
111
кой пластины состоит в детальном изучении теоретическим пу-
тем влияющих факторов и определяющих параметров и в полу-
чении числовых результатов при известных упрощающих пред-
посылках. Однако экспериментальная проверка таковых в ре-
альных условиях пока выполнена только при малых числах
Рейнольдса ReL= 1044-105, что в природе отвечает плаванию не-
больших и не самых скоростных рыб..
Модель — удлиненное гибкое тело успешно применена в про-
странственной теории волнового плавания водных животных
акад. АН УССР Г. В. Логвиновичем [33, 34].
Постановка задачи следующая [33]. Волнообразное движе-
ние тонкого деформируемого тела, например, рыбы рассматри-
вается в инерциальной системе координат Oxyz, движущейся
вместе с телом в направлении оси Ох — основного поступатель-
ного движения с постоянной скоростью t/0 = const (рис. 51).
Продольная криволинейная ось $ тела отклоняется от оси Ох
в плоскости хОу на малую деформацию т)=т)(х, t). Абсцисса
носа тела хг, хвоста хь длина тела Ь = Хг—Xj. Поперечные сече-
ния тела.образованы эллипсами с большой полуосью R = R(x),
параллельной оси z. Нос тела острый, /?2(хг)=0, хвост, как
у рыбы, вырождается в плоскость, на задней кромке R\(x\) =
= Ri. В виду тонкости тела, по всей его длине dR/dx — dR/ds—
малая величина. Оригинально применяется метод плоских сече-
ний с использованием концепции «пронизываемого слоя», не-
подвижного относительно покоящейся жидкости. Тело, проходя
через некоторый плоский, ортогональный оси х, пронизываемый
слой, порождает в нем' поперечное, почти плоское течение, близ-
кое к течению идеальной жидкости. На задней кромке хвоста
тела, при x=xi выполняется условие Жуковского — Чаплыгина
о конечной скорости жидкости. За телом остается след, экви-
валентный пелене сбегающих вихрей. Это приближает задачу
к реальным условиям.
Для решения вводятся в рассмотрение известные обобщен-
ные характеристики вызванного телом нестационарного движе-
ния идеальной жидкости. В плоскости пронизываемого слоя
Рис. 51. Модель удлиненного гибкого
тела в пространственной теории
Г. В. Логвиновича — волнового плава-
ния водных животных (ПС — «попереч-
ный пронизываемый слой» жидкости).
с абсциссой х, в расчете на
единицу толщины слоя име-
ются: удельная присоеди-
ненная масса /п*(х); удель-
ный импульс (количество
движения) m*(x)vn, на-
правленный вдоль нормаль-
ной скорости vn в точке на
криволинейной оси тела;
удельная кинетическая
энергия m*(x)On2/2. Удель-
ная присоединенная масса
для эллиптического сечения
112
такая же, как и для кругового, т* (х) =рл/?2(х), где р—плот-
ность жидкости, и в данном случае есть функция только х
(или s). Нормальная к оси s тела скорость vn = vn(x, t) при
неподвижном пронизываемом слое относительно покоящейся
жидкости, определяется линеаризованным выражением
ип =
dt дх
(25)
Действующими силами на элемент длины dS тела со стороны
жидкости в пронизываемом слое будут: элементарная нормаль-
ная сила dFn и элементарная продольная сила dP.
Элементарная нормальная сила dFn просто выражается по
теореме импульсов
d f" - —77|m’ М”-’1 ds—77 (I> ("77“ ('«-57)]ds-
(26)
Элементарная продольная сила dP в данном случае ориги-
нально определяется как направленная вперед подсасывающая
сила, вызванная нестационарным избыточным «круговым дав-
лением» dQ эллиптического сечения, действующим на скошен-
ной боковой поверхности заостренного спереди тела вследствие
бокового обтекания тела.
Доказывается, что элементарная подсасывающая сила для
эллиптического сечения рассчитывается, как и для кругового,
и равна
dQ dR v2 dR v2„ dm* (s)
dP =---------ds = — 2nPp —------ds=------2------ids.
d s ds 2 ds 2 ds
(27)
Проекции на оси координат Ох и Оу значений dFn и dP по
(26) и (27) дадут элементарную силу тяги dFx и ортогональ-
ную к ней dFy,
dFx=—dFn—---dP,
дх
dFy = dFn + dP^--
дх
(28)
Интегрируя эти выражения по всей длине тела Ь = х2—Хь
можно получить мгновенные результирующие составляю-
щие сил.
Мгновенная результирующая, составляющая силы тяги,
обусловленная импульсами, по формулам (26) и (28) будет
<29>
из
Мгновенная результирующая составляющая силы тяги, обус-
ловленная подсасывающей силой, по формулам (27) и (28)
ха
Р =----L С и0 dx (30)
2 J dx V dt dx )
Xl'
С задней кромки хвоста тела, при х = х\, в след на единицу
пути тела будет переходить кинетическая энергия
Е=±т*(х)(^--и0^-'\2 , х = Х1. (31)
2 \ dt dx )
Чтобы по полученным выражениям для /, Р, Е фактически
выполнить интегрирование, необходимо знать законы деформа-
ции оси тела т] = т] (х, t) и изменения вдоль тела присоединен-
ной массы т* (х).
Волнообразные колебания тела в общем случае определя-
ются зависимостью
П(*> 0 = TloWsin^-j---(32)
Здесь т)о(х) — амплитуда колебаний точек оси тела в предпо-
ложении, что длина тела Хч—х1 = Ь = 2л1т, где I — приведенная
длина волны деформации, 1=Ё/2лт = К/2л, (X — длина волны;
т— число волн, укладывающихся на длине тела); с.— фазовая
скорость волны, бегущей по телу назад. При этом период коле-
баний тела т = 2л//с.
Амплитуда колебаний оси тела в общем случае может быть
представлена рядом
По = а0 + (хг1— х) + а2(х2—х)2 + . . .
Далее рассматривается простейший случай, когда ампли-
туда (полуразмах) волнообразных колебаний тела одинакова
во всех его точках
По = flto = const. (33)
В этом случае решение существенно упрощается. Нормальная
скорость по выражениям (25), (32) и (33) будет
un = ^(C^.t/0)cos(^---^^y (34)
Тогда среднее за период колебаний значение квадрата нор-
мальной скорости оказывается постоянной величиной в интер-
вале Хч—х
v2 = у-1 v2 d t = A- (с— (/0)2 = const. (35)
При таком периодическом движении среднюю за период ре-
зультирующую составляющую силы тяги, обусловленную им-
пульсами, можно вычислить не интегрируя по длине тела вы-
114
ражение (29) и вместо формулы (30) использовать характери-
стику гидродинамического следа. Жидкость на единицу длины
следа содержит на задней кромке хвоста тела (хвостового
плавника рыбы), т. е. при x=xlt секундный импульс m*(xl)vni,
который направлен вдоль нормальной составляющей скорости
хвоста. Проекция на ось Ох потока импульсов, стекающих
с хвоста за единицу времени, взятая с обратным знаком, опре-
деляет мгновенную результирующую, составляющую силу
тяги I, обусловленную потоком импульсов, стекающим в след
/ = m*(Xi)t/of-5~ Х = Х" <36)
k dt дх / дх
Среднее за период значение результирующей, составляющей
силы тяги, обусловленной импульсами, будет
7 = -L f iAt = т\ (хх) Uo , (37)
то c — Uo
Среднее за период значение результирующей подсасываю-
щей силы в данном случае, при ц2п=const по формуле (35),
в соответствии с выражением (30), будет постоянным, и закон
распределения масс по длине тела оказывается несущест-
венным
т
_ 1 Г . С 1 -9 dm* (х) j 1 • , < — 2 г.
Р = \ dM — v2n —77^ dx = — пц (xx) vn = E. (38)
V 1 I U A At
о X,
Одновременно получено по формуле (31), что Р=Е.
В общем случае закон изменения присоединенных масс
может иметь существенное значение для результирующих сил.
Подставляя в выражения (37) и (38) значение удельной
присоединенной массы срыва mi*(xi) = рл/?2ь амплитуду коле-
баний плавника а0 и величину v2n по формуле (35), оконча-
тельно получим расчетные формулы.
Средняя импульсная составляющая сила тяги
/ = V т‘ (77 ~ 0 ^° = 4 Р^2 • -V <39)
2 k I J \Ua ) 2 sh£
Средняя подсасывдющая составляющая сила тяги
р = т т*1 (-гУ (4- - О2 =4- pR2a^2 -4 (Sh*~1)а-
4 к I ) к Uo J 4 sh?
(40)
Средняя общая сила тяги
Т = Т+ Р=^-т\(- 1^о = ~рЯ?аоо>2 f 1------------------’
4 к U ) 4 I Shx )
(41)
115
Средний идеальный гидродинамический кпд, как отно-
шение полезной работы A = TU0 к полной затраченной работе,
включая энергию, оставленную в следе, будет
TU$ -|- EUо
(42)
Выше нами введено специальное выражение числа Стру-
халя через длину волны \
о с л А,
Shi =--------=------
А и0 и0
(43)
и, по-прежнему, и = 2л/г, п — частота колебаний. Поскольку
рассмотрено поступательное равномерное передвижение со
скоростью Uo = const волнообразно колеблющегося тела гидро-
бионта, средней силой ,тяги Т в реальной жидкости преодоле-
вается среднее сопротивление W жидкости движению тела
- — и2
T = W = CxSp _Г°_, (44)
2
где S — площадь смоченной поверхности тела. Безразмерный
коэффициент Сх силы сопротивления
С — 2Т — п
х~ SpU2 ~ 25
1
с2
(45)
Этот коэффициент можно представить в функции числа Рей-
нольдса гидробионта ReL = (J0L/n.
Как видно по расчетным формулам (39) — (45), сила тяги и
кпд волнового движителя гидробионта определяются его гео-
метрическими размерами и кинематическими данными; сущест-
венное значение имеет число Струхаля. Коэффициент полезного
действия этого движителя теоретически всегда больше */г и до-
стигает высоких значений, близких к единице. В режиме плава-
ния с высоким кпд, когда iq>0,9, подсасывающая сила не
превышает 20 % импульсной составляющей силы тяги.
Анализируя линейную теорию и результаты в целом, заме-
тим, что при указанных предпосылках и упрощениях, ее авто-
ром получено весьма простое решение пространственного дви-
жения гидробионта с волновым движителем при интегральной
оценке по следу за телом, известной своей общностью. Изло-
женная теория позволяет развить ее с учетом более сложных
конфигураций и законов деформаций тела гидробионтов [28].
Решение задачи значительно упрощено за счет принятого
выше допущения о постоянстве амплитуды колебаний тела гид-
робионта ?]o = ao = const вдоль тела, что, строго говоря, в при-
роде не наблюдается. Несколько более подходит это допущение
для угревидного способа плавания рыб. Однако, учитывая сде-
116
ланную выше обобщенную оценку по гидродинамическому
следу, принимая во внимание размер хвостового плавника,,
можно полагать изложенное решение и расчетные формулы
справедливыми также для скомброидных рыб, у которых хвос-
товой плавник сравнительно небольшого относительного удли-
нения, например Х<2.
Для скоростных гидробионтов, таких как крупные тунцовые
рыбы "и дельфины, имеющих серповидные и полулунные хвосто-
вые плавники относительно большого удлинения Х>2, вывод и
расчетные формулы гидродинамики также предложены акад..
Г. В. Логвиновичем .[33]. Анализ показывает: если длина локо-
моторной волны существенно превышает размах плавника
(2л/^>2/?1), то активные силы, действующие на хвостовой плав-
ник, в первом приближении можно вычислить пользуясь гипо-
тезой квазистационарности. Хвостовой плавник можно рас-
сматривать как крыло большого удлинения, для которого из
аэродинамики известны теоретические значения коэффициента
подъемной силы Сь = [2л1/(2 + Х)]а и коэффициента индуктив-
ного сопротивления С, = С2ь/лХ,. Окончательно получены сле-
дующие выражения.
Средняя ''за период сила тяги плавника скоростного гидро-
бионта
* / \ 2 ( / \ z х 2-»
— ( ап \ с ( с \ 1 (с » л
Т =-----!— -5- ---------------1-------!----------1 1/2
1 + X/2 k / ) ( ий к U0 ) 1 + Х/2 к Uo J ) °'
(46)
Средний идеальный гидродинамический кпд
ПГйО—7-)' <47>
Сравнивая между собой формулы (41) и (46), а также (42)
и (47), замечаем, что при относительном удлинении хвостового
плавника Х = 2 эти формулы соответственно совпадают, а для
Х>2 кпд в случае серповидного и полулунного плавников
скоростных рыб и дельфинов получается более высоким, напри-
мер, при Х = 4, когда c/U0=l,5, приблизительно на 8%, а когда
с/[7о=2,О примерно на 12 %, хотя при выводе формулы (47) не
учтены подсасывающие силы.
Чтобы сопоставить между собой известные в литературе
значения коэффициентов гидродинамического сопротивления
плавания различных гидробионтов, хотя и полученные разными
методами исследований, нами составлен сводный график Сх =
= f(Re) (рис. 52). Показаны следующие значения коэффициен-
тов: расчетные по теории тонкого тела и экспериментальным
данным кинематики плавания живых рыб и дельфинов (кривые
/—13), экспериментальные по испытаниям живых рыб в био-
гидродинамической трубе в комплексе с физиологическими
117
Re
Рис. 52. Теоретические и экспериментальные значения коэффициента сопро-
тивления гидробионических объектов в функции числа Рейнольдса, Cx(Re).
Расчеты по внешней гидродинамике тонкого тела рыбы: 1—4—
морская форель длиной соответственно 0.1; 0,2; 0,4 и 0,6 м [95]; разные виды рыб:
5 — пеламида 0,16 м; 6 — озерная форель, £=0,21 м; 7 и 8 — сарган длиной соответ-
ственно 0,42 и 0,48 м; 9 — луфарь, £«=0,42 м [28, 56]; 10—12 — соответственно почти
равномерное движение, ускоренное, активное плавание и замедленное, пассивное инер-
ционное плавание афалины длиной 2,5 м. Расчет по внутренней динамике
рыбы: 13 — сайда, £=0,22 м [93]. Испытания живых рыб в биогидродинамической
трубе: 14 и 15 — байкальские омуль. £=0.3 м, и хариус, £=0,29 м [35]. Испытания
мертвых (снулых) рыб: 16, 17— омуль, хариус (буксировка в бассейне); 18— щука,
£ = 0,73 м; 19 — скумбрия, £ = 0,42 м; 20 — сайда, £ = 0,85 м; 2/—лосось, £=1,05 м [91].
Буксировка тушки дельфина в бассейне: 22 — азовка, £ = 1 м [28]. Испытания жестких
моделей: в гидролотке (23— японская скумбрия, £=0,4 м) и в опытовом бассейне
(24 — афалина, £=2,15 м) [88]. Тела вращения: 25 — тело, £=0.3 м, £/D=5,9; 26 — тело,
£=0,5 м, £/D = 6,0 [35]. Продольное обтекание плоской гладкой пластины: Л —лами-
нарный, П — переходный, Т — турбулентный режимы.
исследованиями их энергообмена (14—15): по испытаниям мерт-
вых (снулых) рыб и дельфина (16—22), по испытаниям жест-
ких моделей рыбы и дельфина (23—24) и хорошо обтекаемых
тел вращения (25—26). В больших интервалах изменялись как
числа Рейнольдса Re = 3-1044-1 • 107, так и коэффициенты соп-
ротивления Сх= 1 • 10-3ч-1 • 10-1, что позволяет получить доста-
точно надежные интересные выводы.
Широкое изменение (в функции указанных чисел Рей-
нольдса) коэффициентов гидродинамического сопротивления
рыб и дельфинов, которые получены расчетным путем по тео-
118
рии тонкого гибкого тела, свидетельствует о том, что изложен-
ный выше приближенный метод оценки Г. В. Логвиновича су-
щественно отражает влияние главных параметров кинематики
гидробионтов на их гидродинамику плавания. Это подтвержда-
ется также плавными взаимопереходами всех кривых графика
на рис. 52 — от меньших к большим значениям чисел Рей-
нольдса, а также соответствием в одинаковых интервалах чи-
сел Re расчетных и экспериментальных значений Сх для рыб
(кривые 5—9 и 14—17) и дельфинов (кривые 10—12 и 22 и 24).
Сказанное подкрепляется и соответствующими значениями
Cx(Re) испытаний жестких моделей (кривые 23—26), хорошо
согласующихся.
Показательны расчетные кривые (1—4) на рис. 52 для че-
тырех морских форелей длиной соответственно 0,1: 0,2; 0,4 и
0,6 м. В работе [95] аналогичные расчеты произведены по ре-
зультатам теории тонкого гибкого тела и экспериментальным
кинематическим данным. При одних и тех же числах Рей-
нольдса форель разной длины имеет существенно различные
(в 2—5 раз) значения Сх, причем тем большие, чем крупнее
рыба. Это несомненно свидетельствует о существовании в гид-
родинамике плавания гидробионтов заметного масштабного эф-
фекта. Косвенно данный вывод подтверждается и расчетами:
для различных видов рыб — пеламиды, озерной форели, сарга-
нов и луфаря (кривые 5—9).
Расположение на графике кривых 7—8 для угревидной
рыбы (саргана) среди кривых для скомброидных рыб (фо-
рели 6 и луфаря 9) подтверждает сказанное выше о возмож-
ности расчета по методу Г. В. Логвиновича гидродинамики пла-
вания не только угревидного, но и скомбриодного типа гидро-
бионтов с плавниками относительно небольшого удлинения.
Кривые 5—9 единообразно рассчитаны по формуле (45) и од-
нообразным экспериментальным кинематическим данным, ко-
торые рассмотрены в § 9 (см. также рис. 38).'
Судя по графику на рис. 52, расчетные значения коэффици-
ентов сопротивления Сх живых рыб при активном плавании
с относительно большими колебаниями хвостовой части тела
получены значительно более высокие, чем обычно для жестких
твердых тел. Кроме того, кривые Cx(Re) для рыб более резко
снижаются. Однако в рассмотренных случаях числа Рейнольдса
были порядка 104—105, т. е. невысокие, а режим потока в био-
гидродинамических трубах при экспериментальном изучении
кинематики плавания небольших рыб был заведомо турбулент-
ным, причем с высокой начальной турбулентностью. Разуме-
ется, у скоростных крупных рыб, использующих свою биополи-
мерную связь для снижения гидродинамического сопротивле-
ния в более благоприятных природных условиях, соотношения
могут быть существенно иные. Отчасти это подтверждается и
при изучении небольших скоростных рыб (кривые 14 и 15).
119
Особого внимания заслуживают расчетные кривые 10—12
для плавания крупных живых дельфинов-афалин длиной 2,5 м,
имеющих Re= (44-9) • 106, тем более в сопоставлении с букси-
ровкой в бассейнах тушки дельфина-азовки длиной 1 м (кри-
вая 22) и жесткой модели афалины длиной 2,15 м (кривая 24).
Во-первых, ясно, что в данном случае дельфины плавали в пе-
реходном режиме; коэффициент гидродинамического сопротив-
ления дельфина значительно ниже, чем у модели, тем самым
расчетно-экспериментальным путем подтверждается существо-
вание известного «парадокса» Грея (1936 г.) о плавании дель-
финов с малым гидродинамическим сопротивлением. Во-вто-
рых, коэффициент сопротивления дельфина при активном пла-
вании с ускорением (кривая 11) ниже, а при замедленном инер-
ционном плавании 12 выше, чем при равномерном движении 10.
Влияние затяжного по времени положительного и отрицатель-
ного ускорения плавания дельфинов, когда U=U(t) определя-
лось из уравнения RX = T—Ri, где Rx— среднее за период
колебаний гидродинамическое сопротивление дельфина; Т—по-
прежнему среднее значение силы тяги; Ri — значение инерци-
онного сопротивления. Последнее рассчитывали по второму за-
кону Ньютона Ri = ma(l+kn)dU/At. Здесь пга — масса дель-
фина, — безразмерный коэффициент присоединенной массы,
значение которого принято как для равновеликого эллипсоида
вращения в поступательном неравномерном движении вдоль
длинной оси (Ац~0,05). В экспериментах по кинематике пла-
вания дельфинов ускорения и замедления движения в данных
случаях были: dt7/d£ = 4- (0,104-0,70; 0,754-1,50) м/с2 и AU/dt =
= — (0,104-0,20) м/с2. Средняя за цикл колебаний полная сила
тяги дельфина определялась по формуле (46) и идеальный гид-
родинамический кпд — по формуле (47). Расчетный идеаль-
ный кпд имел значения т]~0,97, что, надо полагать, не-
сколько завышено. В расчетах кпд гидробионтов по теории
тонкого гибкого тела часто встречается величина т] >0,804-0,85.
К настоящему времени появилось много зарубежных печат-
ных работ по изучению гидродинамики волновых колебаний
тонкого гибкого тела, в том числе применительно к плаванию
рыб с высоко эффективным полулунным хвостовым плавником,
с учетом влияния боковых плавников у рыб, по оптимизации
движения и т. п. Различные аспекты и литература приведены
в обобщающих аэрогидродинамических работах Лайтхилла
1975—1977 гг. [84, 85], Ньюмена и др. [87, 98, 99].
Таким образом, по проблеме волнового движителя выпол-
нено уже большое число теоретических и отчасти эксперимен-
тальных исследований, что позволяет делать приближенную
оценку работы волновых движителей, упрощает и сокращает
необходимые экспериментальные исследования.
120
§ 14. Теоретические и экспериментальные исследования
эффективного машущего плавникового движителя
В отличие от удлиненного комплекса — пропульсирующее
гибкое тело и плавник волнового движителя водных живот-
ных — машущим плавниковым движителем скоростных гидро-
бионтов назовем гидродинамическое крыло, выделяющееся от-
носительно большим размахом и обычно с переменной стрело-
видностью в плане, которое совершает гармонические колеба-
ния и создает большую силу тяги с высоким кпд.
Практическое применение машущего крыла в технике более
перспективно в водной среде, нежели в воздушной. В авиации
не применяется машущее крыло в качестве одновременно несу-
щей поверхности и движителя не только из-за специфических
конструктивных сложностей, но также и ввиду трудностей обес-
печить его прочность вследствие больших инерционных сил,,
обусловленных частыми колебаниями массы крыла. Напротив,
в более плотной среде воды для создания силы тяги, а при не-
обходимости и подъемной силы требуется машущее крыло зна-
чительно меньших размеров,, с небольшой амплитудой и низкой
частотой колебаний. При этом инерционные силы, обусловлен-
ные массой самого крыла, оказываются существенно меньшими,
сравнительно с гидродинамическими силами, действующими на
крыло в воде.
Теоретические исследования машущего крыла уже давно
проводят в нестационарной аэродинамике идеальной несжимае-
мой жидкости, однако обычно в условиях плоской задачи и не-
больших (теоретически бесконечно малых) амплитуд колеба-
ний. Важным этапом изучения в ЦАГИ им. Н. Е. Жуковского
аэродинамических эффектов вибрирующего крыла была работа,
выполненная М. В. Келдышем и М. А. Лаврентьевым примени-
тельно к проблеме флаттера крыла с малыми флюктуациями
тонкой пластины, при ее поступательном равномерном движе-
нии с поперечными вращательными колебаниями и изменением
угла атаки по гармоническому закону. Впервые было показано,
что среднее за период колебаний значение подъемной силы ма-
шущей пластины совпадает с подъемной силой, определяемой
по формуле Н. Е. Жуковского для среднего угла атаки, и что
существуют режимы колебаний, при которых средняя горизон-
тальная сила тянет вперед, т. е. является подсасывающей
силой тяги.
Известна монография акад. А. И. Некрасова по плоской тео-
рии крыла в нестационарном потоке. Установлено, что выра-
женные через специальные функции сложные формулы для сил
и моментов при различном движении тонкого профиля пред-
ставляются через соответствующие суммы трех составляющих:
квазистационарной, зависящей от присоединенных масс и вих-
ревого следа. Максимум силы тяги крыла достигается при раз-
121
ных колебаниях в функции числа Струхаля. В случае малых
чисел Струхаля (большие скорости движения) этот максимум
может быть при совместных машущих и крутильных колеба-
ниях со сдвигом фаз на 90°.
Коэффициент полезного действия вибрирующего профиля
как движителя зависит от числа Струхаля и для поступатель-
ных колебаний крыла больше, чем для вращательных. В даль-
нейшем теория малых плоских колебаний тонкого крыла в иде-
альной жидкости была развита, в работах акад. Л. И. Седова.
Вихревая теория толстого машущего крыла в плоскопарал-
лельном потоке несжимаемой жидкости предложена в ряде
оригинальных работ известного аэродинамика В. В. Голубева.
В основе его теории лежит гипотеза о том, что при колебаниях
крыла в реальной жидкости на его поверхности происходит на-
копление пограничного слоя и в крайних положениях крыла от
него отходят дискретные вихри конечной интенсивности, кото-
рые образуют позади крыла пропульсивную двойную вихревую
дорожку с шахматным расположением вихрей, обратных по на-
правлению вращения вихрям в двойной вихревой дорожке Кар-
мана, возникающей в результате преодоления сопротивления
жидкости. Расчетные формулы названной теории удовлетвори-
тельно согласуются с поставленными экспериментами [50]. Од-
нако эта теория разработана применительно к возвратно-посту-
пательным колебаниям крыла вверх и вниз с постоянным углом
установки, когда скорость подъема и опускания крыла постоян-
ная. Большое внимание изучению машущего с большими ам-
плитудами крыла уделил М., К. Тихонравов. Он детально про-
анализировал качественные дискретные схемы вариантов меха-
ники взмаха и составляющих аэродинамических сил птичьего
крыла.
Рассмотрим более подробно современные теоретические ра-
боты по исследованию оптимизации действия машущего крыла
как движителя. В работе [13] устанавливается оптимальный за-
кон совместных поступательно-крутильных колебаний машу-
щего крыла для получения силы тяги с достаточно высоким
кпд. Рассматривается тонкое слабо изогнутое крыло с дли-
ной полухорды Ь, которое совершает движение в плоской си-
стеме координат хОу, связанной со средним положением про-
филя и началом в середине хорды (рис. 53), по закону
у = w (I) + a (t) (х0<—х) = 4>а0 Re (-у - -у + • (48)
Здесь Хо — абсцисса оси вращения крыла; w(t) — поступатель-
ное перемещение оси вращения параллельно оси Оу, t — время;
а (0 —Угол поворота крыла (а>0, если передняя кромка выше
задней); о» — круговая частота; р, — сдвиг фаз между поступа-
тельными и крутильными колебаниями; х = о»о/(bcto)—отноше-
ние амплитуд: ао и соо — амплитуды колебаний; Re — значение
122
реальной части выражения
(48). В общем случае хо = О
и р,5^=0.
Крыло обтекается без-
граничным потоком идеаль-
ной несжимаемой жидко-
сти, который в бесконечно-
сти имеет скорость Uo,
направленную вдоль оси
Ох. Далее приближенно
Рис. 53. Теоретическая схема движения
машущего крыла с совместными посту-
пательно-крутильными колебаниями [13].
рассматривается случай ао<С1, ®о/^<С1 и используются изве-
стные зависимости линейной теории крыла в плоском нестаци-
онарном потоке применительно к рассматриваемому закону
колебаний. Эти зависимости выражаются через специальные
функции Теодорсена (мы их опускаем). Колеблющееся крыло
создает силу тяги T=Q—а У, где Т, Q, Y — соответственно
средние за период т = 2л/со сила тяги, подсасывающая и подъ-
емная силы, действующие _на единицу длины крыла. Как
обычно, средняя сила тяги Т и затрачиваемая на поддержание
колебаний крыла средняя мощность N, равная работе всех
сил на соответствующем перемещении за единицу времени для
элемента крыла единичной длины, выражаются через безразмер-
ные коэффициенты
Т = jipbCrUo, 1
N = npbCNUo. j
(49)
У движителя кпд т] есть отношение средней полезной
к средней затрачиваемой мощности
ТЦЛ _ Ст
N Cn
(50)
Эффективность машущего крыла как движителя определя-
ется коэффициентом тяги Ст и кпд т). Рассматривается опти-
мизация работы машущего крыла движителя по закону коле-
баний крыла, при котором значение Ст достигает максимума.
Вводится условие ограниченности амплитуды колебаний
max [to 4- а (х0-—x)] = Ab, А= const, |х0| (51)
и принимается число Струхаля в виде
Sha =(ob/U0. (52)
Дальнейшее исследование ограничивается двумя важными
случаями: 1) Shw > 1; 2) 5ЬШ 1.
Случай Sha 1 соответствует малым скоростям Uo движе-
ния. Тогда т] = 0,5 независимо от конкретного закона колебаний
крыла и, если выполняются условия (51), значение max Ст
123
достигается при чисто поступательных колебаниях крыла
(х —оо, aax-+wa/b = A)
max Сг = -^- A2Sh^. (53)
В случае чисто крутильных колебаний (х = 0, а0=А/(1 + |х0|/Ь),
если ось вращения расположена на крыле, | х01 < b
Cr< —A2Sh2. (54)
т 64 “ '
Случай Sho <1 (большие скорости движения) при рассмот-
рении асимптотических приближений позволяет сделать вывод,
что когда Sho->0 для чисто поступательных колебаний, т)->-1.
Величина Ст в общем случае при заданных Sho и максималь-
ной безразмерной амплитуде колебаний А зависит от трех па-
раметров: х, р,, Хо/b, определяющих закон колебаний крыла; за-
висимость Ст и Хо/b проявляется через условие (51). Оказыва-
ется, что по параметру р, значения max Ст и г] достигаются при
р. = л/2, а минимум при ц = —л/2.
Затем анализ проведен для оптимального случая, когда
сдвиг по фазе между поступательными и крутильными колеба-
ниями крыла ц = л/2. Анализ показывает, что единственным су-
щественным параметром, определяющим закон колебаний
крыла при р. = л/2 и Shw <С 1, является отношение амплитуд ко-
лебаний n = wo/(aob). Максимальный кпд при фиксирован-
ном числе Струхаля ShM достигается для чисто поступатель-
ных, а минимальный — для чисто крутильных колебаний. Мак-
симальной сила тяги будет для Shm 1 при приближенном от-
ношении амплитуд
х = х* = л/2 + ф25Ьш + У(л/2 + ф2 ShJ2 + ф2, (55)
где ф = 1 4- | х01 lb. При совместных поступательно-крутильных
колебаниях с отношением амплитуд х>х* имеется существен-
ный выигрыш в силе тяги по сравнению с режимом чисто посту-
пательных колебаний, который тем больше, чем меньше Shffl
и чем ближе х и х*.
Для эффективности машущего крыла как движителя, когда
можно достигнуть высокого значения как силы тяги, так и
кпд, необходимо выполнить три условия: Sha <g 1, р = л/2,
х>х*. При заданной частоте колебаний число Струхаля опре-
деляется из условия равенства силы тяги, развиваемой движи-
телем, силе сопротивления всего тела (аппарата).
Систематические теоретические исследования гидродина-
мики тонкого колеблющегося крыла (пластины) различной
формы в плане и относительного размаха выполнены примени-
тельно к анализу нестационарного плавания китообразных и
скоростных рыб в отделе прикладной математики и теоретиче-
124
ской механики Кэмбриджского университета (Англия). Здесь
обобщены известные работы Лайтхилла 1960—1975 гг. и By
1961—1971 гг. В работе М. Ю. Чопра [72] рассмотрено тонкое
прямоугольное крыло с хордой 2с конечного размаха 2s с ок-
ругленной передней и острой задней кромками. Крыло совер-
шает в идеальной жидкости поступательные движения со ско-
ростью Uo и поперечные возвратно-поступательные и враща-
тельные со сдвигом фаз на 90°, волновые колебания малой ам-
плитуды, при среднем нулевом значении подъемной силы. Уч-
тены гидродинамические эффекты не только в направлении по-
тока, но также и по размаху крыла. В среднем положении
крыло расположено в плоскости хОу: ось х — по хорде, ось у—
по размаху, ось z—перпендикулярно координатной плоскости;
начало координат в центре крыла. Поперечные смещения крыла
от среднего положения 2 = 0 определяются зависимостью
z = f(y) [h—ia (х—&)] eia>t,
где h — амплитуда поступательных колебаний; а — амплитуда
крутильных колебаний; х = Ь — абсцисса оси вращения; <о —
круговая частота колебаний,
00
Цу) = Z “n cos р = -^- ’ —4s<t/<4s.
n=0 4s
Значения ап определяются по сформулированным в работе ус-
ловиям.
В результате трудоемких математических выкладок опреде-
лены: сила тяги, потребная мощность, энергия, теряемая
в следе, и кпд крыла в функции относительного размаха, по-
ложения оси вращения, числа Струхаля Sho и обобщенного
параметра 6, выражающего отношение угла наклона хорды
плавника к углу наклона траектории оси вращения
Shffl = (ос/£/0, Q = aU0/ha.
Численные результаты решения, полученные с применением
ЭВМ, представлены в виде графиков для безразмерного коэф-
фициента Ст средней силы тяги, обусловленной поперечными
силами и кпд в интервале относительных размахов крыла-
пластины Х = 44-8
С Т
Т. 2рс sh2<£>2
Т] = ТС0/Е.
Некоторые графики воспроизведены на рис. 54. Значения Ст
и изменяются в зависимости от значений 0: когда Ст увели-
чивается, т] уменьшается и, наоборот, с уменьшением Ст воз-
растает Т).
125
Рис. 54. Теоретические значения коэффициентов силы тяги Ст
и кпд Г] для тонкого крыла — пластины относительного размаха
(Х.=4) с возвратно-поступательными и крутильными колебаниями
и сдвигом фаз на 90°, при различных положениях оси вращения
(х=6) в функции числа Струхаля Sh© и обобщенного пара-
метра 0 = 0; 0,2; 0,4; 0,6; 0,8 [73]: а—Ь = 0 (половина хорды);
б—Ь = Чг (три четверти хорды).
В обобщающей теоретической работе [73] исследованы ана-
логичные гидродинамические характеристики крыльев фигур-
ных в плане форм (рис. 55), приближающихся к лунообразным
формам хвостовых плавников-движителей быстро плавающих
животных. Сопоставлены полученные численные результаты
для крыльев различных форм и с прямоугольным крылом.
В частности, установлено, что искривление передней кромки
крыльев в связи с наличием
локальной стреловидности при-
водит к относительному умень-
шению части силы тяги, обус-
ловленной разряжением в окре-
стности передней .кромки. Это
препятствует отрыву потока у пе-
редней кромки со значитель-
ным ухудшением характеристик
крыла. Однако слишком боль-
шая стреловидность, превышаю-
щая 30°, также приводит к ухуд-
шению эффективности крыла.
Наиболее близким к оптималь-
ному по эффективности крылу из
Рис. 55. Теоретически исследован-
ные машущие пластины с перемен-
ной стреловидностью кромок [73].
Наиболее эффективная форма В2 напо-
минает плавниковый движитель скоро-
стных гидробионтов.
126
исследованных форм оказывается В2, которое имеет умеренную
локальную стреловидность передней и задней кромок и по
форме в плане напоминает высокоэффективные плавниковые
движители скоростных гидробионтов. Тем самым теоретически
подтверждается оптимальная форма последних.
Экспериментальные исследования машущего крыла, в том
числе применительно к изучению эффективных плавниковых
движителей скоростных гидробионтов, проводились в разное
время [14, 24, 25, 58, 76].
Исследования немецких ученых кратко представлены в из-
вестной монографии Хертеля [76]. Показано, что тело анестизи-
рованных и мертвых рыб (щуки, форели) при буксировке в опы-
товом бассейне имеет почти в 5 раз большее сопротивление,
чем жесткая модель при тех же числах Рейнольдса. Это обус-
ловлено режимом автоколебаний с большой амплитудой упру-
гого, но мертвого длинного тела рыбы, при отсутствии стаби-
лизирующего движения активного действия плавников у живой
рыбы. Исследованы закономерности вихреобразований за хвос-
товым плавником нестационарно плавающих морских живот-
ных и для жестких моделей. Путем киносъемки выявлено, что
в зависимости от режимов движения морским животным при-
сущи следующие три системы вихреобразований
в следе. При ускоренном движении рыбы, так называемом ак-
тивном ударе хвостового плавника, за телом образуется про-
пульсивная вихревая дорожка с двумя рядами шахматно рас-
положенных вихрей; между вихрями возникает волнообразной
формы кильватерная струя, в которой жидкость отбрасывается
назад, в сторону, противоположную движению рыбы. При рав-
номерном движении рыбы, «нейтральном» ударе хвостового
плавника, вихри расположены за телом рыбы не в шахматном
порядке, а чередуются с взаимно обратным вращением в одном
продольном ряду, нет ярко выраженной кильватерной струи.
При замедленном движении рыбы, так называемом пассивном
ударе хвостового плавника, образуется двухрядная вихревая
дорожка и обратная кильватерная струя, в которой жидкость
течет в направлении движения рыбы. Эти исследования прове-
дены при малых числах Рейнольдса; тем не менее они наглядно
показывают тесную взаимосвязь сил тяги и сопротивления
с различными системами вихреобразований.
Указанные три режима вихреобразований в следе, характер-
ные для плавания морских животных, были искусственно вос-
произведены на экспериментальной установке с небольшой
жесткой моделью машущего плавника длиной профиля 0,22 м
и толщиной 0,08 м при числах Рейнольдса порядка Re~104.
Приведены также результаты испытаний о влиянии на силы,
действующие на плавник, угла и скорости перекладки плавника
при крутильных колебаниях. Если, угол перекладки увеличи-
вать медленно, то коэффициент подъемной силы Су плавника
127
увеличивается практически линейно до своего критического
максимального значения, предшествующего отрыву погранич-
ного слоя (аКр~12°), и обтекание можно рассматривать как
стационарное. Если же угол перекладки изменять быстро, то
безотрывное обтекание плавника и линейное изменение Cv
может затягиваться до значительно большего критического угла
(акр~40°), при этом нестационарное значение Су повышается,
например, в 3—4 раза. Это объясняется тем, что вследствие
быстрой перекладки плавника не создаются условия для от-
рыва пограничного слоя.
Наиболее полные экспериментальные исследования гидро-
динамических характеристик машущих крыльев проведены
в ЦАГИ им. Н. Е. Жуковского [14]. Показано, что машущие
крылья могут выполнять одновременно функции как движи-
теля, так и несущей поверхности с достаточно высоким кпд —
порядка ц = 0,6-:-0,7 и выше, которое близко к кпд обычного
гребного винта.
В эксперименте прямоугольное в плане машущее крыло
с хордой b двигалось с постоянной горизонтальной скоростью
Uo, одновременно совершая гармонические поступательные вер-
тикальные колебания с амплитудой а и крутильные угловые ко-
лебания вокруг горизонтальной оси с амплитудой 0, при угло-
вой частоте колебаний о) = 2лп. Отметим некоторые особенности
обработки результатов испытаний.
Для определения безразмерных коэффициентов средних за
период гидродинамических характеристик машущего крыла
введено амплитудное значение U скорости движения крыла, ко-
торое пригодно и для случая U0=Q (на стопе)
U = ^U20 + (a<i>)2. (56)
Полезной мощностью Nn, отдаваемой машущим крылом,
считали ту, которая определяется уменьшением сопротивления
и появлением средней силы тяги Т машущего крыла по сравне-
нию с сопротивлением Хо этого крыла при стационарном обте-
кании его со скоростью Ue под средним углом атаки ао. Сред-
няя за период колебаний полезная мощность машущего крыла
#„ = (7—Хо) Uo. (57)
Безразмерные коэффициенты средней силы тяги и средней
затрачиваемой мощности N = Na + N& крыла площадью S = bl
будут
Ст=—, CN =---------------- (58)
pU2S pU2Saa>
Для машущего крыла кпд
Т] = -^- = (Т—Х0)и0 _ (59)
N Na + Np
128
В работе приведены графики значений пропульсивных ко-
эффициентов Ст, CN, т] в функции характерных кинематических
параметров — относительной скорости колеблющегося крыла
Лр и числа Струхаля Sh^,
Лр = Ц0/Д®. 5Ьш = <о6/{/0. (60)
Относительная скорость Лр характеризует максимальный нак-
лон траектории оси вращения крыла к направлению поступа-
тельного движения. Этот параметр аналогичен относительной
поступи гребного винта и удобен в случае машущего крыла
с двумя степенями свободы (с поступательными и угловыми ко-
лебаниями) тем, что экспериментальные точки пропульсивных
коэффициентов в зависимости от ?.р ложатся на одну кривую
при различных значениях Uq, относительной амплитуды а = а/Ь
и круговой частоты а> колебаний.
Испытания машущих крыльев проводили в опытовом бас-
сейне ЦАГИ на специально изготовленной колебательной уста-
новке, размещенной на буксировочной тележке. С помощью ус-
тановки сообщали крылу вертикальные колебания с амплиту-
дой а=15, 30, 50, 70 мм при среднем угле атаки ао = О-?5° и уг-
ловые колебания с амплитудой р<20° с любым заданным углом
сдвига б по фазе и частотой и<3,5 Гц. Испытаны две модели
существенно различных типов крыльев с профилями NACA-0015
и ЦАГИ КВ-1-7, при одном и том же значении относительного
размаха К = 11Ь = А, прямоугольной формы в плане, со скруглен-
ными боковыми кромками. Стационарные гидродинамические
характеристики обоих крыльев приведены на рис. 56 при оди-
наковом относительном погружении крыльев под поверхностью
воды Я=/7/6 =1,65. Профиль NACA-0015 с хордой 6 = 0,175 м,
относительной толщиной с=15%, оптимальный по аэродинами-
ческому качеству K, = CVICX и как сравнительно толстый, пред-
почтителен для колеблющегося крыла, которое работает в ши-
роком диапазоне углов атаки. Профиль ЦАГИ КВ-1-7 с хордой
6 = 0,15 м относительной толщиной с = 7 % выбран из соображе-
ний сохранения устойчивого положения при изменении углов
атаки и соответствует малой толщине крыльев, обычно приме-
няемых для судов на подводных крыльях. Ось вращения крыла
расположена на четверти хорды от носка (приблизительно
в ЦД крыла).
Основные выводы работы сводятся к следующему. Наилуч-
шие пропульсивные качества по силе тяги и кпд достига-
ются в случае машущих колебаний крыла с двумя степенями
свободы, когда угловые колебания опережают поступательные
по фазе б = л/2. Тогда горизонтальная проекция мгновенной
подъемной силы У/ в течение всего периода колебаний направ-
лена в сторону движения крыла, т. е. является силой тяги, ко-
торая большую часть времени превышает горизонтальную про-
5 С. В. Першин
129
ские характеристики моделей двух пря-
моугольных в плане крыльев относи-
тельного размаха Х=4 с профилями
NACA-0015 толщиной 15%, с хордой
6=0,175 м и ЦАГИ-КВ-1-7 толщиной
7 %, с хордой 6 = 0,15 м, испытанных
как машущие крылья в опытовом бас-
сейне при относительном погружении
Я=Я/6=1,65 [14].
екцию мгновенной силы
Xi — силы сопротивления.
Другая составляющая силы
тяги обусловлена дополни-
тельной подсасывающей си-
лой, возникающей на но-
сике машущего крыла, как
это показано эксперимен-
тами с горизонтально дви-
жущимся с постоянной
скоростью машущим кры-
лом, совершающим только
одни угловые колебания
вокруг некоторой оси.
Средний угол атаки ао су-
щественно влияет на про-
пульсивные характеристики
машущего крыла; с увели-
чением ао сила тяги и кпд
уменьшаются в связи с рос-
том сопротивления крыла.
Увеличение несущих
свойств данного крыла,
т. е. средней за период
вертикальной силы, приво-
дит к снижению его про-
пульсивных характерис-
тик— силы тяги и кпд. Из
двух испытанных машущих крыльев более толстое крыло
с профилем NACA-0015 имеет значительно более высо-
кие пропульсивные качества сравнительно с тонким крылом
ЦАГИ КВ-1-7. Напомним, что у китообразных машущие плав-
никовые движители относительно толстые (см. § 3).
Основными безразмерными кинематическими параметрами,
определяющими пропульсивные характеристики крыла, колеб-
лющегося с двумя степенями свободы являются: относительная
скорость движения kp=U0/aa, амплитуда угловых колебаний
Р, сдвиг фаз между угловыми и вертикальными колебаниями б.
При изменении относительной амплитуды вертикальных посту-
пательных колебаний в диапазоне а = а/Ь = 0,14-0,47, в интер-
вале относительной скорости Хр = 24-7, кривые действия машу-
щего крыла Ст(А,р), Cn(%p), т] (%р) практически не зависят от
относительной амплитуды а. Некоторые результаты испытаний
обоих моделей приведены на рис. 57.
С целью обоснования методики приближенной оценки про-
пульсивных характеристик машущего крыла по гипотезе стацио-
нарности, проведены эксперименты по измерению мгновенных
130
Рис. 57. Пропульсивные характеристики
двух машущих крыльев с совместными воз-
вратно-поступательными вертикальными и
крутильными колебаниями. Видно сущест-
венное различие характеристик обоих испы-
танных крыльев.
/-NACA-0015, <3 = 0,285 ; 2 - ЦАГИ-КВ-1-7, ~а-
=0,467; в обоих случаях одинаковые значения:
00=3,7°, 0=3°, б=Л/2, Я-1,66 [14].
Рис. 58. Результаты экспери-
ментов по измерению мгновен-
ных нестационарных значений
(/) коэффициентов сопротивле-
ния Сх и подъемной силы Cv
машущего крыла с профилем
ЦАГИ-КВ-1-7 при заданных:
</ = 0,467; а0=3,7°; Р=3°; 6=
=л/2; Я =1,65, в сравнении со
стационарными испытаниями
(2) [14].
значений подъемной силы и сопротивления колеблющегося
крыла. Линейный характер зависимости CVi(a,) сохраняется
в значительно более широком диапазоне изменения мгновенного
угла атаки а< = 0ч-20°, чем при стационарном обтекании
(рис. 58). Мгновенные значения коэффициента сопротивления
Cxt, соответствующие максимальным мгновенным углам атаки
at, оказываются близкими к стационарным значениям только
при малых мгновенных углах атаки а><8°. С увеличением
а/>8° существенно увеличивается нестационарное сопротивле-
ние крыла в сравнении со стационарным, что и понятно при
конечных параметрах колебаний машущего крыла и его широ-
ком следе.
Для оценки гидродинамического взаимодействия двух кры-
льев в опытовом бассейне ЦАГИ проведены эксперименты, в ко-
торых за машущим крылом КВ-1-7 на различных расстояниях
устанавливалось другое, стационарное крыло аналогичных раз-
меров в плане, с углом атаки, соответствующим нулевому зна-
чению его подъемной силы. Как показали эксперименты, вто-
5* 131
рое крыло в этом случае не оказывает заметного влияния на
гидродинамические характеристики первого, машущего крыла,
даже при малых расстояниях между ними.
Ряд экспериментальных исследований машущих крыльев
провели в Институте гидромеханики АН УССР [24,25, 58 и др.].
В работе [58] обращено внимание на гидродинамические осо-
бенности машущего крыла как движителя в сравнении с греб-
ным винтом. У гидробионтов ометаемая площадь Fo плавнико-
вого движителя в несколько раз может превышать площадь
миделевого сечения FM тела (F0/FM = 5-i-8), а у подводных су-
дов соотношение обратное: Fo/FM = 0,2-i-0,5. Визуализация вих-
ревого следа за машущим крылом в эксперименте показала,
что с концов его сходят два вихревых шнура, которые затем
замыкаются при смене циркуляции. Замкнувшиеся вихревые
шнуры образуют в следе за крылом вихревую систему взаимно
наклоненных замкнутых, приближенно прямоугольных вихре-
вых шнуров, которые расположены на плоскостях ломаной про-
дольной поверхности, имеющей те же период и амплитуду, что
и колеблющееся крыло. Как известно, за винтом след состоит
из спиральных вихревых шнуров, сходящих с кромок лопасти,
и прямоугольного осевого вихревого шнура, сходящего со сту-
пицы винта. Приближенная оценка вызванных скоростей от
вихревых следов машущего крыла и винта показывает, что
вследствие указанных различий по форме и интенсивности вих-
рей в следе за движителем потери на создание вызванных ско-
ростей у машущего крыла меньше, чем у винта, особенно
вследствие наличия у последнего осевого вихревого шнура. На-
против, машущим крылом при особенностях его вихревой си-
стемы используется часть кинетической энергии следа.
Экспериментальное исследование тяги машущего крыла
с симметричным профилем NACA-0015, хордой £> = 0,14 м и раз-
махом / = 0,42 м, заглубленного под поверхность воды в ней-
тральном положении на глубину Н = Н[Ь=\,4, проведено в гид-
ролотке с поперечным сечением рабочей части ВхЯ = 0,62х
Х0.42 м при скорости набегающего потока Uo = 0,314-0,67 м/с.
С помощью специально изготовленной колебательной экспери-
ментальной установки крыло было подвержено совместным
возвратно-поступательным поперечным колебаниям с амплиту-
дами ао = 0,06 и 0,10 м, а также вращательным колебаниям
с угловыми амплитудами 0 = 04-18°, с шагом Др = 3,6° и 6°, при
частоте колебаний п= 1=2,5 Гц. Показано, что увеличение ам-
плитуды угловых колебаний 0о при постоянной амплитуде Яо
поперечных колебаний крыла и частоте п повышает силу тяги
только до определенного максимума, отвечающего оптималь-
ному значению 0о для каждого режима работы колеблющегося
крыла. Приведем типичные цикловые графики, построенные по
анализу осциллограмм при следующем режиме колебаний
крыла: а = 0,06 м, 0=14°, ф = 36°, п=2,0 Гц, С/о = О,31 м/с
132
(рис. 59). Обращает на себя вни-
мание фазовый сдвиг изменения
мгновенной суммарной верти-
кальной силы по отношению
к мгновенной силе тяги кото-
рый обусловлен наличием верти-
кальных инерционных сил Р/ ма-
шущего. крыла.
Квазистационарный расчет
изолированного машущего дви-
жителя по известным для него
стационарным значениям аэро-
гидродинамических коэффициен-
тов подъемной силы Су(а) и
силы сопротивления Сх(а) явля-
ется простым инженерным мето-
дом приближенной оценки про-
пульсивных и прочностных ха-
рактеристик нестационарных ре-
жимов работы этого движителя
[14, 24, 31, 58]. В квазистацио-
нарном приближении не учиты-
вается предыстория движения и
предполагается, что гидродина-
мические силы, действующие на
машущий движитель в данный
Рис. 59. Мгновенные изменения
сил, действующих на машущее
крыло с профилем NACA-0015, за
период поступательно-крутильных
колебаний [25]: а — y(t)—траек-
тория средней точки хорды про-
филя; б — T(t)—сила тяги; в —
вертикальные силы.
Pj, — суммарные значения в воде; Ру—
инерционные силы в воздухе; —
вертикальная составляющая гидродина-
мической СИЛЫ, Ру = “ Ру»
момент времени, определяются
значением мгновенного угла атаки ос* и вектора суммарной
мгновенной скорости движения крыла W=U0+V, где V —
мгновенная вертикальная составляющая скорости движения
машущего крыла. При этом не принимаются во внимание инер-
ционные силы и сдвиг гидродинамической силы по фазе при
нестационарном движении крыла (см. рис. 59), что допустимо
при некоторых режимах работы. Равнодействующая гидродина-
мических сил может быть приложена в центре статического
давления крыла, положение ЦД мало зависит от угла атаки.
Заданными для расчета машущего движителя (по конст-
руктивным, технологическим и предварительным теоретическим
соображениям) являются: геометрические параметры — форма
в плане, профиль и толщина крыла, длина хорды (или средней
хорды) b и размах I, тем самым удлинение к = 1[Ь и площадь
S = bl крыла; кинематические параметры — поступательная ско-
рость движения Uo, амплитуда возвратно-поступательных вер-
тикальных колебаний оси вращения а, амплитуда угловых ко-
лебаний р0 при вращении крыла, угол сдвига фаз между вра-
щательными и поступательными колебаниями <р, частота ко-
лебаний п, Гц и угловая частота щ=2лп (а также средний угол
атаки ао, если машущий движитель одновременно выполняет
133
ные и крутильные
Рис. 60. Теоретическая схема машу-
щего крыла с поступательными и уг-
ловыми колебаниями в системе коор-
динат (угол сдвига фаз ф=л/2).
а, Р — амплитуды колебаний; Uo—const,
V(0, UP(/) — скорости равномерного дви-
жения по горизонтали, мгновенная по вер-
тикали и суммарная; ар Рр Vp — мгно-
венные значения угла атаки, угла пере-
кладки крыла и угла скоса потока; Т,
Р — мгновенные значения силы тяги и по-
перечной вертикальной силы.
и функцию несущей поверхности); динамические характери-
стики крыла — стационарные зависимости Сх(а), Су(а) и ка-
чество К=Су(а)/Сх(а). Последние получают в результате ста-
ционарных продувок модели данного крыла-движителя в аэро-
динамической трубе, или при буксировке в опытовом бассейне
(см., например, рис. 56), а при отсутствии таковых выбирают
по справочным данным. В необходимых случаях выполняют
пересчет справочных данных на крыло другого удлинения, вво-
дят поправку на форму крыла и т. п.
Требуется определить средние за период колебаний т=1/л
основные гидродинамические характеристики машущего дви-
жителя: среднюю силу тяги Т и среднее значение кпдтр
Расчетная схема приведена на рис. 60. Начало системы коор-
динат £От] помещено в ЦД крыла, на оси его вращения в сред-
нем положении. Поскольку используется гипотеза стационар-
ности, на расчетной схеме показаны скорости обращенного
движения.
Пусть машущий движитель, в том числе гидробионта, со-
вершает, как обычно, совместные гармонические поступатель-
: оптимальным углом сдвига
фаз <р = л/2. Вертикальные ко-
лебания определяются за-
висимостью g = acosco/. Угол
р(7) перекладки хорды крыла
0i= Pocos (a7 + q>). Поперечная»
вертикальная скорость V(t)
колебаний крыла будет У =
= i(t)=—acosincot Истинная
мгновенная скорость W набе-
гающего на крыло потока (см.
рис. 60)
W = ^U20 + V2.
Вертикальные колебания
крыла l(t), движущегося го-
ризонтально с постоянной
скоростью Uq=const, приво-
дят к появлению переменного
по времени мгновенного угла
скоса потока
*=arct« (--£-)=
. . / аш sin at \
.. =arcte(—— )•
который характеризует угол
134
наклона касательной к траектории оси вращения крыла. При
этом мгновенное значение угла атаки определяется выражением
а, = у,—р,-.
Каждому значению угла атаки а, соответствуют мгновен-
ные значения подъемной силы У,- и силы сопротивления Xi
крыла, которые определяются в скоростной системе координат
Xt = Cx£p-^-S, (61)
^ = C^P-y-S’ (62)
где Cxi и CVi — коэффициенты силы сопротивления и подъем-
ной силы крыла, взятые при стационарном обтекании с углом
атаки а», (здесь р — плотность жидкости). Проектируя эти силы
на горизонтальную ось От) и вертикальную ось О|, получим
мгновенные значения горизонтальной оси тяги Tt (или силы
сопротивления) и вертикальной силы Pi крыла
Тi = (Су{ sin ye— Cxi cos y() p S, (63)
w?
Pt = (Cyi cos у. + Cxi sin y£) p —— S • (64)
Мгновенные значения кпд машущего крыла, вообще говоря,
могут быть найдены исходя из различных условий, как это
видно из предыдущего изложения. Простейшее условие опре-
деляется следующим очевидным выражением:
= TjUo = Cyj sin yi — Cxi cos у£ Uo
1 P(V Cyi cos Y( + CXi sin fl |V
Мгновенные значения Ti и гр можно использовать для по-
строения кривых действий машущего крыла в функции опре-
деляющих критериев и комплексных параметров, таких как
число Струхаля Sna =aa>/f/o, или относительная скорость Хр =
= и0[а(й, относительная поступь XP'=L/o/4an, например, СГ(А),
К (Л.) (см. рис. 57). В некоторых случаях целесообразно ввести
в рассмотрение отношение амплитуд поступательных и враща-
тельных колебаний крыла 0 = а/0&, а для сравнения результа-
тов расчетов с гребным винтом учитывать площадь гидравличе-
ского сечения машущего крыла, т. е. ометаемую площадь F =
= 2al, ввести в рассмотрение отношение площади крыла к по-
следней (n = S/F = b/2a) и т. п.
Для определения^ средних за период колебаний значений
силы тяги Т и кпд т] полупериод разбивается на k (например,
А = 6) равных промежутков времени; для каждого из них
135
определяются мгновенные значения Т{ и т], по формулам (63)
и (64), а затем они осредняются
/ ft \ _ /ft \ / ft \
T = П = (66)
С позиций гипотезы стационарности можно определить ji
оптимальные случаи работы машущего крыла, когда созда-
ются такие углы атаки, которыми обеспечиваются максималь-
ная тяга или максимальный кпд движителя.
Условие мгновенного максимума силы тяги 7\ по исследо-
ванию на экстремум выражения (63) будет
maxTt->-^ :-^-=ctgVi=-^.
dai d<Xi V
Этим определяется оптимальный по тяге угол атаки крыла
в любой момент времени, поскольку дСу^дщ и dCXi]dai есть
тангенсы углов наклона заданных стационарных кривых Су(а)
и Сх(а).
Условие мгновенного максимума кпд гц определим по выра-
жению (65) в случае малого угла у,, созу£~1 и, учитывая, что
Сх мало в сравнении с Су, а потому Cxsinyi<^Cv. Получим
CyVIUo— с
max Т]£ ~ —v -----
un = J 1 Цо
Су V k аа cos a>t
Здесь К=СУ1СХ — качество крыла. Как видно, максимальному
значению кпд отвечают максимальное значение качества крыла,
возможно большее значение амплитуды колебаний и сравни-
тельно невысокая скорость плавания.
В принципе гипотеза стационарности может применяться
в зависимости от формы профиля крыла при малых докрити-
ческих углах атаки, например, а<12—15°, и сравнительно не-
большой частоте колебаний п. Напомним, что у дельфинов
обычно а<10°, в среднем а— 6°, п^З Гц. Применение подобной
методики расчета для рассмотренного выше машущего крыла
с профилем ЦАГИ КВ-1-7 дало вполне удовлетворительное сов-
падение расчетных квазистационарных значений с эксперимен-
тальными значениями кривых действия крыла Ст(Хр), CN(XP),
К (Лр) при режимах движения с относительной скоростью ХР =
= £7о/асо^2, представляющих наибольший практический инте-
рес [14]. Применительно к машущему крылу с симметричным
профилем Бортмана ГХ-05-191 прямоугольной формы в плане,
с удлинениями Х = 2 и 4,5 и хордой Ь = 60 мм [58] получено, что
тяга его по экспериментальным данным больше тяги, рассчи-
танной с использованием гипотезы стационарности на 30—40 %,
т. е. Т3= (1,34-1,4) Тр. С увеличением числа Струхаля Sha —
= a<s)/Uo расхождение увеличивается [24]. Сделан вывод, что ги-
потезу стационарности целесообразно использовать при неболь-
ших значениях ShM для предварительной оценки гидродинами-
136
ческих характеристик машущего движителя, например, при вы-
боре профиля крыла, закона изменения угла атаки и т. п.
Интересно отметить результаты приближенного расчета про-
пульсивных характеристик хвостового плавника-движителя
дельфина по гипотезе стационарности (с применением других
допущений, в частности по форме крыла в плане и далее в про-
филе) [24]. Хвостовой плавник афалины квазистационарно рас-
считывался как жесткое машущее крыло с известными из экс-
перимента геометрическими и кинематическими параметрами, и
расчетная сила тяги при равномерном движении дельфина при-
равнивалась его общей силе сопротивления. Оказалось, что
квазистационарные коэффициенты гидродинамического сопро-
тивления дельфина, движитель которого рассчитывался как ма-
шущее крыло, во всех случаях примерно вдвое больше, чем
коэффициенты сопротивления, полученные при тех же режи-
мах плавания дельфина, движитель которого рассчитывался
как волновой, с изгибно-колебательными движениями всего
тела. Отсюда усматривается высокая эффективность плавания
удлиненного, упругого, а потому и гибкого двигательно-движи-
тельного комплекса дельфина с взаимно меньшими затратами
энергии. Как показали результаты расчета на ЭВМ гидроди-
намических характеристик жесткого машущего крыла с про-
филем NACA-0015, используя гипотезу стационарности, можно
определить среднюю тягу с погрешностью не более 10%,
в сравнении с экспериментальными данными, если угол атаки
крыла в течение всего периода колебания не превышает
15—20°.
По изучению гидродинамики машущего плавникового дви-
жителя выполнено большое число теоретических и эксперимен-
тальных исследований, которые в общем согласуются. Выяв-
лены определяющие функциональные параметры и комплексы,
оптимальные режимы работы машущих крыльев, условия их
расчета и применения. Однако непосредственное сопоставление
результатов исследований иногда затруднено различными ис-
ходными предпосылками и формой представления результатов,
что вызывает необходимость пересчетов. До сих пор в лите-
ратуре рассмотрены только изолированные машущие крылья и
только‘жесткие. Изучение упругих машущих плавниковых дви-
жителей, подобных существующим у скоростных гидробионтов,
т. е. во взаимодействии с гибким телом, составляет сложную
задачу дальнейших исследований, в том числе в области тео-
рии гидроупругости.
Таким образом, в гидробионике еще необходимо провести
немало комплексных экспериментально-теоретических исследо-
ваний реальных плавниковых движителей.
137
§ 15. Значение нестационарности движения по результатам
раздельного гидродинамического моделирования
Выше неоднократно отмечалось, что гидробионты всегда
плавают нестационарно. Естественно, возникает интерес выя-
вить значение управляемой нестационарности движения, в том
числе применительно к техническим объектам, для снижения
гидродинамического сопротивления твердых тел. С этой целью
рассмотрим некоторые теоретические и экспериментальные ис-
следования нестационарного плавания гидробионтов.
В отношении гидродинамики нестационарное плавание ги-
дробионтов вызвано их волновыми и машущими движителями
и характеризуется периодическими колебаниями скорости по-
ступательного движения и периодическими деформациями по-
верхности гибкого тела, влияющими на характеристики погра-
ничного слоя. Влияние этих разных факторов, ввиду труднос-
тей исследований, последовательно рассмотрено в ряде работ,
выполненных в Ленинградском кораблестроительном институте
(ЛКИ) [19, 20, 66].
Большой интерес вызывает следующая экспериментально-
теоретическая работа [66]. С целью выявления влияния неста-
ционарного изменения скорости поступательного движения на
сопротивление воды в опытовом бассейне ЛКИ провели спе-
циальные испытания твердого тела вращения длиной £=1,5 м,
наибольшим диаметром </ = 0,21 м (удлинение £/</=7,15), при
погружении под свободной поверхностью. Н = 0,7 м (Hjd =
= 3,33). Нестационарные испытания проведены при скоростях
буксировки тела U (/)^3,0 м/с, в диапазоне чисел Рейнольдса
Re = (1,04-5,0) 106 согласно трем законам изменения скорости:
по линейному U (t)=at с положительным ускорением а = 0,14-
4-0,5 м/с2 и с отрицательным ускорением U(t) = U0—at, а =
= 0,14-0,3 м/с2; по экспоненциальному; по синусоидальному
U (/) =А |sin со/|.
В процессе испытаний скорость движения тела и силы на
нем непрерывно записывали с помощью осциллографа, погреш-
ность в замерах сил специально спроектированными тензомет-
рическими динамометрами находилась в пределах 5%. При
обработке экспериментальных результатов испытаний пола-
гали, что в случае достаточного заглубления буксируемого тела
волновое сопротивление незначительное, и потому оно не учи-
тывалось. Полная сила сопротивления воды движению тела
вращения при нестационарном его движении R„ может быть
представлена в виде
Rh = Rhb + Rhhh, Rbohh ~ 0,
где Rbb — нестационарное вязкостное сопротивление тела;
/?нин — нестационарное инерционное сопротивление. Соответст-
138
венно для безразмерных коэффициентов сопротивления Схн =
= СНв + Снин, где Схн — коэффициент полного нестационарного
сопротивления, Снв и Снин — коэффициенты нестационарного
вязкостного и инерционного сопротивления.
Инерционное сопротивление определяли расчетным путем
RhHH — (tn + Хц)
dU(t)
dt
тле tn — масса тела; Хи — присоединенная масса. Полагали,
что коэффициент полного нестационарного сопротивления Схн
является функцией числа Рейнольдса Re, числа Фруда Fr и
безразмерного ускорения N
N = du(t) L/Lf2
В свою очередь, коэффициент нестационарного вязкостного со-
противления Снв представляли выражением
Снв = Св +ACHB(Re, Fr, N),
где Св — коэффициент вязкостного сопротивленйя, соответст-
вующий квазистационарному режиму движения тела; ДСНВ —
коэффициент дополнительного вязкостного сопротивления,
обусловленный нестационарностью движения.
Анализ полученных экспериментальных данных показал,
что линейный и экспоненциальный законы в диапазоне изме-
нения скорости 0^ U (t) ^3,0 м/с дают близкие результаты.
Положительное ускорение приводит к увеличению вязкостного
сопротивления твердого тела, и тем к большему, чем меньше
Лс отношение чисел Re/N. На-
против, отрицательное ускоре-
ние уменьшает вязкостное со-
противление твердого тела
(рис. 61).
Рис. 61. Экспериментальные значения
коэффициента дополнительного вяз-
костного сопротивления при нестаци-
онарном движении твердого тела по
линейному закону ACHB(Re/A^) [66].
J — положительное ускорение (разгон); 2 —
отрицательное ускорение (торможение).
Рис. 62. Осциллограмма нестационар-
ной буксировки твердого тела вра-
щения по синусоидальному закону
изменения скорости U(t) =А [sin <о<|
[66].
Р(0 — мгновенные изменения скорости
(масштаб времени 1 с); (?н — нестационар-
ные значения силы сопротивления.
139
В случае синусоидального закона изменения скорости, когда
тележка вместе с буксируемым телом вращения пробегала
вдоль бассейна несколько полуволн синусоиды скорости с на-
чальными моментами времени, при которых скорость умень-
шалась до нуля, максимум силы сопротивления соответствовал,
максимуму ускорения, а не скорости, как обычно полагают.
С дальнейшим ростом скорости до Umax наблюдалось умень-
шение силы сопротивления (рис. 62). Кривые осредненного за
полупериод синусоиды полного нестационарного сопротивле-
ния RH(U) на графиках расположены ниже соответствующей
стационарной кривой (до 40%). Это, возможно, объясняется,,
тем, что при отрицательных ускорениях на тело может дейст-
вовать дополнительная инерционная сила, направленная вдоль-
движения тела и обусловленная наличием пограничного слоя.
Для дальнейшего анализа механизма сопротивления воды
движению твердого тела при нестационарном движении теоре-
тически была рассмотрена задача о нестационарном турбулент-
ном пограничном слое на плоской жесткой пластине. Числен-
ное решение на ЭЦВМ «Урал-2» соответствующего дифферен-
циального уравнения в частных производных представлено'
графически на рис. 63, где кривые показывают влияние неста-
ционарности на коэффициент полного сопротивления трения
в функции безразмерного ускорения. Здесь коэффициент п =
= Cf„ICjZ есть отношение коэффициентов нестационарного пол-
ного сопротивления С/н к соответствующему стационарному С/с.
Положительное ускорение а>0 приводит к возрастанию CjHr
а отрицательное ускорение а<0 к его уменьшению по сравне-
нию с соответствующим стационарным коэффициентом С/с. Это
согласуется с приведенными выше экспериментальными резуль-
Рис. 63. Расчетные значения влияния
нестационарности на коэффициент пол-
ного сопротивления трения n=CfUlCfC
плоской гладкой пластины с турбулент-
ным пограничным слоем в функции ве-
личины N= WLIU1 [66]:
А — равноускоренное движение U(t)=at, а>0;
В — равнозамедленное движение U(t) = Uo—at',
значения а: / — 0,1; 2 — 0,5; 3—1 м/с’.
татами для твердого тела.
Нестационарные расчеты
показали также, что в не-
которых случаях наложе-
ние дополнительного гар-
монического колебания на
основе поступательное дви-
жение тела по закону
U(t) = t7o(l+osinw/) мо-
жет привести к уменьше-
нию коэффициента сопро-
тивления трения по сравне-
нию со стационарным, со-
ответствующим скорости
равномерного движения t70.
В работе [66] сделан вы-
вод, что. для уменьшения
сопротивления трения при
нестационарном поступа-
140
тельном движении твердого тела целесообразен режим, при ко-
тором короткий участок разгона тела с положительным уско-
рением в начале движения затем сменяется длинным участком
замедления тела с отрицательным ускорением.
Влияние нестационарности движения гидробионта, в том
числе периодических деформаций его гибкого тела, теорети-
чески исследовано в цикле работ Ю. И. Фаддева и др. на ос-
нове систематического применения теории движения тел изме-
няемой формы, деформация которых осуществляется по задан-
ному закону, вначале применительно к потенциальному обте-
канию, а затем и к нестационарному пограничному слою.
В работе Я. И. Войткунского, Ю. И. Фаддева (Бионика.
Вып. 10. 1976. С. 21—27) показано, что при решении чисто
потенциальной задачи произвольного движения деформируе-
мого тела в несжимаемой безграничной жидкости, для оценки
действующих на тело инерционных сил и моментов необходимо
ввести в рассмотрение так называемые обобщенные присоеди-
ненные массы
= —Р(Бф« —dS, i=l, 2, . . . , 7, £=1,2..........6.
J дп
s
Матрица | |Xife| | содержит 7X6 обобщенных присоединенных
масс. Вследствие свойства парности число независимых обоб-
щенных присоединенных масс сокращается до 21+6. В отличие
от обычных присоединенных масс, соответствующих движению
твердого тела, Х/ь для деформируемого тела зависят от вре-
мени. Присоединенные массы Xih (i, k=\, 2, .... 6) характери-
зуют движение тела как квазитвердого (в предположении его
мгновенного отвердения), а (i=7, £= 1, 2, ..., 6) отражают
непосредственный учет скоростей деформации поверхности
тела.
Для последующего расчета пограничного слоя анализирова-
лись характеристики потенциального течения при продольном
поступательном движении удлиненного эллипсоида вращения
с деформациями поверхности, симметричными относительно его
продольной оси, когда вызванное течение жидкости также осе-
симметрично. Характеристики потенциального течения опреде-
лены методом, основанным на распределении гидродинамиче-
ских особенностей по поверхности эллипсоида S (t), мгновенно
изменяющей свое положение во времени. Это приводит к реше-
нию нестационарного интегрального уравнения Фредгольма
второго рода. Деформации поверхности S (t) принимаются ма-
лыми, точки поверхности колеблются вблизи некоторых сред-
них положений, образующих условную равновесную поверх-
ность So, радиус поперечной кривизны которой изменяется на
деформационное слагаемое, являющееся периодической функ-
цией времени. Деформационное слагаемое было задано в об-
141
щем виде; варьируя входящими в его выражение параметрами,
можно получать различные режимы деформаций поверхности
эллипсоида, а именно: расширение-сжатие, перетекание, бегу-
щую волну, контрольный случай недеформируемого эллипсоида.
Численные расчеты, выполненные на ЭЦВМ «Минск-22» для
эллипсоида вращения с относительным удлинением ао/6о = 6,
показали, что во всех случаях деформации поверхности ока-
зывают значительное влияние на мгновенное перераспределе-
ние давления в течение периода деформаций.
Влияние закона периодического изменения скорости посту-
пательного движения на сопротивление трения эллипсоида вра-
щения с относительным удлинением L[d^=& выявлено расче-
тами нестационарного ламинарного пограничного слоя [19]. Из-
менение скорости поступательного движения эллипсоида было
задано по закону
Ц(/) = Ц0(1 + к Д ^sin©^, (67)
где Uo — средняя за период Т = 2л/6 скорость поступательного
движения; к — амплитуда изменения скорости; £;— коэффи-
циенты, определяющие вид графика U (/); со — круговая ча-
стота. Расчеты выполнены на ЭЦВМ «Минск-22» для девяти
вариантов нестационарного движения при одинаковой ампли-
туде к = 0,25 и варьируемых значениях коэффициентов £, и
числа Струхаля Sh = <oa/t/o. В расчетах приняты три группы
значений коэффициентов а именно: I— Bi = l, £2 = 0, £з = 0,
что обеспечивало синусоидальное изменение поступательной
скорости (СИПС) вблизи средней величины Uo', II — £i = l,
12 = 0,24, £3 = 0,055 — быстрый разгон и медленное торможение
(БРМТ); III — gi=l, £2 = —0,24, g3 = —0,055 — медленный раз-
гон и быстрое торможение (МРБТ). Графики скорости для всех
трех режимов движения эллипсоида показаны на рис. 64.
При расчете определяли мгновенные значения различных
Рис. 65. Расчетные зависимости длины
безотрывно обтекаемой части эллипсоида
и относительного коэффициента сопро-
тивления трения от времени для сину-
соидального закона изменения скорости
[19]:
1 1+0.25 sin ш/; 2-1^. З — С^.
Рис. 64. Законы изменения скоро-
сти движения эллипсоида, приня-
тые в расчетах влияния нестацио-
нарности движения на сопротивле-
ние трения [19].
/ — СИПС; 2-БРМТ; Э-МРБТ (рас-
шифровка в тексте).
142
безразмерных коэффициентов, в том числе: мгновенные отно-
сительные значения длины безотрывно обтекаемой части по-
верхности эллипсоида с полуосью а, т. е. /бо = Сбо/2а, и мгновен-
ные значения коэффициента сопротивления трения безотрывно
обтекаемой части поверхности CX6o = 2Rx/pU02S6O. Их измене-
ния за один период для одного режима движения при Sh= 1,0
представлены на рис. 65. Сделан вывод, что мгновенные инте-
гральные характеристики пограничного слоя за период изме-
нения скорости движения претерпевают значительное измене-
ние. Однако средние за период величины незначительно зави-
сят от закона движения и числа Струхаля. Таким образом,
при чисто ламинарном обтекании нестационарность поступа-
тельного движения гидробионтов слабо влияет на среднее за
период сопротивление трения воды их движению.
Оценка влияния деформаций поверхности тела на сопротив-
ление трения в настоящее время теоретически возможна только
в рамках концепции пограничного слоя и практически только
в частных случаях осевой симметрии. В работе [20] на примере
продольного движения эллипсоида вращения рассмотрено влия-
ние на распределение местного коэффициента трения по длине
тела нестационарности, вызванной в начале периодическим из-
менением скорости поступательного движения недеформируе-
мого эллипсоида, а затем и деформацией поверхности эллип-
соида при его движении с постоянной поступательной ско-
ростью. Нестационарный ламинарный пограничный слой на
деформируемом эллипсоиде вращения рассчитывался по при-
ближенному интегральному методу с помощью уравнения
импульсов, выведенного авторами для деформируемого тела
вращения. Результаты расчета представлены рядом графиков
распределения местного коэффициента трения по длине тела
в зависимости от числа Струхаля Sh = <oa/t/ и режима движе-
ния эллипсоида для различных моментов периода: 7=0, Т/4,
Т/2, ЗТ/4.
Местный безразмерный коэффициент трения Cf связан с на-
пряжением трения на стенке то известной зависимостью
Cf = , Cf--, (68)
' pU2 7Re„rH
где ReMrH — мгновенное число Рейнольдса.
Кривые распределения значений Cf(t, Sh) по длине тела при
нестационарном движении недеформируемого эллипсоида рас-
положены в различных случаях и выше и ниже известных сни-
жающихся кривых квазистационарного распределения коэффи-
циента трения, вычисленного в предположении dU/dt = O. Од-
нако существенно, что для всех рассмотренных вариантов не-
стационарного движения недеформируемого эллипсоида общим
является явно выраженный квазистационарный характер рас-
пределения Cf вблизи передней критической точки с резковы-
143
раженным наибольшим значением этого коэффициента. Напро-
тив, в случаях движения деформируемого эллипсоида с по-
стоянной поступательной скоростью в указанных выше режи-
мах БРМС (быстрое расширение —медленное сжатие) и МРБС
(медленное расширение — быстрое сжатие) перераспределение
коэффициента трения С/ происходит не только в области пред-
отрывного течения, но и вблизи передней критической точки,
причем различным образом в разные моменты периода. Так
сказывается предыстория пограничного слоя на его мгновен-
ные местные характеристики, а потому и возможности регули-
рования нестационарного сопротивления. Существенно, что по
сравнению с. нестационарностью, порождаемой колебаниями
скорости поступательного движения эллипсоида, нестационар-
ность, возникшая вследствие нормальной деформации поверх-
ности, сопровождается значительно меньшими перемещениями
положения точки отрыва пограничного слоя.
В связи с приведенными выше результатами представляет
интерес экспериментальная работа Л. Ф. Козлова, В. М. Ша-
кало, Л. Д. Бурьяновой, Н. Н. Воробьева (Бионика. Киев.
1974. Вып. 8. С. 13—16) по изучению нестационарности пла-
вания черноморской афалины. Афалина длиной 2,5 м проплы-
вала в море прямолинейный опытный участок траектории
движения длиной 75 м с малой скоростью 0,5—2,0 м/с. Иссле-
дование проводили с помощью телеметрической аппаратуры
с кабельной связью. Мгновенная скорость плавания афалины
измерялась укрепленной на ее теле с помощью присоски ми-
кровертушкой; степень турбулентности в пограничном слое из-
мерялась проволочным датчиком термоанемометра. Погреш-
ность измерений скорости не превышала 4 %. Обработка и
анализ осциллограмм показали, что плавание дельфина отли-
чается существенной нестационарностью сложного характера,
с рядом закономерностей периодического типа.
Сделан вывод, что наибольшее значение имеет колебание
скорости, связанное с колебаниями хвостового стебля и плав-
ника дельфина. Степень турбулентности в пограничном слое
афалины достигала максимума на участках ускорения и спа-
дала до минимума на участках торможения в каждой данной
осцилляции. Эти результаты согласуются с приведенными выше
данными и свидетельствуют о возможной регуляции степени
турбулентности в пограничном слое дельфина при активном
плавании.
Несколько иной подход к оценке нестационарных режимов
движения гидробионтов содержится в работе [59]. В частности,
выполнен анализ в квазистационарном приближении такого
цикла движения, когда вначале осуществляется активный раз-
гон гидробионта с положительным ускорением, а затем насту-
пает пассивное движение по инерции с замедлением, но без
затраты мускульной энергии. Из уравнения нестационарного
144
движения тела в вязкой жидкости с квадратичной зависи-
мостью сопротивления от скорости следует, что минимуму зат-
рат энергии отвечает прямолинейное движение с постоянной
средней скоростью. Однако анализ этого уравнения с разрыв-
ными коэффициентами показывает, что при той же средней
скорости возможен выигрыш энергии в случае периодического
движения с чередующимися участками активного разгона и
последующего движения по инерции.
Аналогичный результат теоретически получен Д. Вайсом
(Weihs) Для продолжительного нестационарного плавания рыб
с отрицательной плавучестью, таких как скомброидные — скум-
брия, пеламида, тунец, акулы — белая, макрелевая, мако.
Квазистационарными расчетами показано, что для рыб с отри-
цательной плавучестью энергетически выгодно цикличное двух-
фазное плавание с чередованием пассивного, без затрат энер-
гии, наклонного скольжения вниз по инерции на некоторую
глубину и последующего активного всплывания под углом
к горизонту. При этом экономия затрат энергии может дости-
гать 50 % и более по сравнению с горизонтальным равномер-
ным плаванием на то же расстояние.
Теоретически исследованы и гидродинамические аспекты
взаимодействия вихревых следов рыб в стае, плавающих в иде-
альной жидкости с одинаковой скоростью на близком расстоя-
нии. Однако в природе всегда встречаются трудности синхро-
низации плавательных движений рыб в стае вследствие их
различных размеров, частых изменений направления движения
и естественных возмущений жидкости. Известно, что крупные
рыбы редко плавают в стае.
В биогидродинамике встречаются работы, в которых иссле-
дуется эффективность использования взаимодействия нестацио-
нарного плавания гидробионтов с нестационарным движением
самой жидкой среды. Здесь отметим только работу А. И. Ко-
роткина (Бионика. Киев, 1973. Вып. 7. С. 27—31) по теорети-
ческому анализу механизма скольжения дельфинов на бегу-
щих корабельных и ветровых морских волнах. С помощью
простого уравнения профиля волны по линейной теории, прибли-
женным расчетом определены условия, при которых дельфин
может скользить, вместе с волной на поверхности моря без
затраты своей энергии, и установлена соответствующая область
параметров этих волн. Например, дельфин длиной 2 м, массой
91 кг перемещается на волне длиной Z = 9 м со скоростью U =
= 3,75 м/с, а при А, = 25 м £7 = 6,25 м/с. Полученные теоретиче-
ские соотношения в принципе можно применить в спорте для
расчета перемещения спортсмена, балансирующего на доске
на переднем склоне волны.
Таким образом, к настоящему времени получены разнооб-
разные экспериментальные и расчетные данные для объяснения
механизма сопротивления водной среды при различных режи-
145
мах нестационарного движения тел, которые проливают свет
на эту проблему и применительно к гидробионтам. Однако эти
Данные не всегда согласуются между собой и предстоит еще
многое сделать для уверенных экстраполяций от идеализиро-
ванного расчета и эксперимента к натуре с учетом масштабных
эффектов, которые недостаточно экспериментально исследованы.
§ 16. Моноласт как эффективный волновой движитель
для скоростного плавания способом «дельфин»
В связи с освоением Мирового океана актуальным является
повышение эффективности пропульсивных качеств человека
при использовании им мышечной энергии для плавания с при-
менением простейших индивидуальных технических средств
массового изготовления. Хорошо известны обычные парные
резиновые ласты промышленного производства, с помощью ко-
торых тренированный пловец в надводном или подводном пла-
вании повышает скорость плавания до 15—20 % сравнительно
с плаванием без ластов на той же дистанции. Одними из лучших
считают отечественные заводские резиновые ласты «Дельфин»
общей длиной 0,55 мм, с выступающей за галоши лопастью
шириной 0,225 м и площадью ее рабочей поверхности 0,066 м2.
Масса обоих ластов 1,9 кг, а развиваемая ими средняя сила
тяги около 21 кг.
Опыт спортивных соревнований показал, что для дальней-
шего повышения скорости плавания наиболее быстрым спосо-
бом «кроль» (вольный стиль), с чередованием плавательных
движений обеих ног пловца, рабочая поверхность ласта дол-
жна быть существенно увеличена и лопасть следует делать
более длинной, изгибающейся на волновой траектории. Так
впервые в нашей стране появились самодельные удлиненные
парные скоростные ласты с лопастью из листового стеклотек-
столита общей длиной 0,75 м, шириной лопасти 0,22 м и пло-
щадью рабочей поверхности 0,092 м2, которая почти в 1,5 раза
больше, чем у заводских резиновых ластов. В настоящее время
скоростные заплывы способом «кроль» с парными ластами вы-
полняются только с использованием подобных удлиненных ско-
ростных ластов.
В 1969 г. впервые в нашей стране спортсменами был изго-
товлен и успешно использован моноласт с большой титано-
вой тонкой лопастью для скоростного плавания способом
«дельфин», т. е. с одновременными плавательными движе-
ниями обеими ногами [51].
Идея применения моноласта состоит в том, что при спо-
собе плавания «дельфин» пловец совершает активные колеба-
тельные движения в вертикальной плоскости не только' конеч-
ностями, но и всем телом, и потому эффективно используются
самые крупные группы мышц человека — спины и живота.
146
В данном способе плавания заведомо лучше организуется и сам
поток воды вдоль спортсмена. Все это позволяет получить до-
полнительный прирост в скорости плавания. Последующий спор-
тивный опыт подтвердил целесообразность применения моно-
ласта, и теперь он широко используется на соревнованиях муж-
чинами и женщинами на всех дистанциях плавания. Моноласт
по-прежнему изготовляется спортсменами в индивидуальном по-
рядке, но с применением для рабочей лопасти листов стекло-
текстолита промышленного производства, которые специально
обрабатывают, выполняя трудоемкие технологические опера-
ции вручную.
На подводной фотографии (рис. 66) пловец зафиксирован
в почти выровненном положении тела на траектории, руки его
вытянуты. Основной движитель — моноласт большой площади,
состоящий из одной стеклотекстолитовой листовой лопасти и
пары резиновых галош, скрепленных с лопастью на клею.
Стеклотекстолит предпочтителен потому, что имеет малую
плотность — около 1,6 г/см3, является упругим материалом и
удобен в обработке. На лопасти виден ряд продольных по-
лос— это углубления, вырезанные вручную не на всю толщину
в листе стеклотекстолита при изготовлении моноласта. Спорт-
смены утверждают, что такие продольные углубления необ-
ходимы для устойчивости движения и предупреждения попе-
речного рыскания моноласта в экстремальных его положениях
на гребнях волновой траектории.
Сравнительное изучение моноластов, применяемых двад-
цатью спортсменами высокой квалификации, показал, что ло-
пасти моноластов изготовляют различной формы и размеров,
руководствуясь в основном интуицией и имеющимся опытом.
Определяющими факторами считают: пол спортсмена, его рост
и вес, нормативные спринтерские и стайерские дистанции, уп-
ругие свойства листа стеклотекстолита. Однако уверенно учи-
тывать полностью все эти факторы пока не представляется
возможным. Обычная форма лопасти моноласта трапециевид-
ная, близка к полуэллипсу, с волнистой кромкой и т. п. Раз-
меры монопласта варьируют
в пределах: общая длина
вместе с галошами 0,65—
0,90 м, размах, по ширине
0,65—0,87 м, площадь (с од-
ной стороны) 0,30—*0,45 м2,
что намного превышает та-
ковую у двух удлиненных
скоростных ластов. Относи-
тельное удлинение моно-
ласта варьирует в пределах
X = /2/F = 0,9—1,7, где / —
ширина моноласта по
Рис. 66. Спортсмен с моноластом в бас-
сейне при подводном плавании способом
«дельфин» (мгновенная съемка).
147
выходной кромке; F — несущая площадь моноласта. Общая
масса моноласта не превышает 2 кг. Замечено, что пловцы
чувствуют на дистанции каждые 0,2—0,3 кг лишней массы дви-
жителя. Спортсмен высокой квалификации в плавании на ко-
ротких дистанциях с моноластом развивает предположи-
тельно среднюю силу тяги около 300 Н. Это позволяет достигать
средней скорости плавания (в нырянии) до 2,75 м/с, что близко
к предельному значению для человека по физиологическим по-
казателям.
При продольном поступательном перемещении пловца и по-
перечных колебательных движениях туловищем и ногами
моноласт, в том числе его рабочая лопасть из листового стекло-
текстолита значительно изгибаются. Моноласт как бы вписы-
вается в волновую траекторию, являющуюся результатом на-
ложения обоих названных видов движений. Общее представ-
ление о движениях подводного пловца с моноластом в течение
нескольких циклов можно составить по циклограмме (рис. 67).
Съемка производилась при выключенном свете, а характерные
точки пловца были отмечены светящимися электролампочками,
укрепленными на его теле. Кинокамера в водонепроницаемом
боксе была снабжена обтюратором — вращающимся диском
с прорезями, что позволяло сделать процесс съемки прерывис-
тым с известными периодами времени. Один кадр плывущего
спортсмена снят при импульсной вспышке света, его антропо-
метрические размеры служат масштабом длины.
Анализ циклограммы показывает, что в то время как кисть
руки пловца следует почти по прямой линии а, точка вблизи
ЦТ тела описывает криволинейную траекторию б, приближаю-
щуюся к синусоиде сравнительно небольшой амплитуды, вы-
ходная кромка лопасти моноласта описывает криволинейную
траекторию в большой амплитуды, напоминающую синусоиду.
Как видно, при плавании с моноластом способом «дельфин»,
пловец совершает в вертикальной плоскости периодические из-
гибные колебания всем телом непрерывно, со сдвигом фаз тра-
Рис. 67. Циклограмма траекторий движения характерных точек тела спорт-
смена с моноластом в бассейне.
а — кисть руки; б — ЦТ тела; в — выходная кромка моноласта (снято неподвижной
кинокамерой с обтюратором и световой вспышкой).
148
екторий указанных точек. Таким образом, по телу пловца и
моноласту при поступательном перемещении их вперед про-
бегает назад упругая деформируемая бегущая волна с нара-
стающей амплитудой, которая является периодической и
близка к гармонической. Эта бегущая волна тела пловца за-
рождается на кисти его руки и заканчивается на выходной
кромке лопасти моноласта; ее прохождение хорошо видно ви-
зуально при наблюдении с борта бассейна. Результаты обра-
ботки подводной киносъемки приведены на рис. 68. Огибающая
колебаний различных точек вдоль тела пловца имеет явно кри-
волинейный характер, типа экспоненты.
Наибольший интерес представляют волновые траектории
двух точек в плавании спортсмена с моноластом, а именно: ЦТ
тела и середины концевой кромки лопасти моноласта. Рас-
смотрим характерные для мастера спорта международного
класса экспериментальные данные по рекордному заплыву
в спортивном бассейне на дистанции 400 м с аквалангом.
Спортсмен удерживал перед собой в вытянутых руках неболь-
шой баллон со сжатым воздухом и плыл на глубине 1,22 м
ЦТ тела под поверхностью воды. Временами продольная ось
движения была наклонена до ±3°. Поступательное продвиже-
ние пловца за один цикл, соответствующий подъему и опус-
канию моноласта, его «поступь» была s=l,51 м. Волновая
траектория ЦТ тела почти симметричная, с размахом 0,18 м
и средней крутизной ~ 1/4. Волновая траектория середины
концевой кромки лопасти моноласта явно несимметричная; ее
размах А = 0,63 м, средняя крутизна близка к единице, или
фронт волны имеет наклон под углом около 45°. В 'данном
случае оси обеих волн параллельны; они отстоят одна от дру-
гой на 0,20 м, что влияет на повышение гидродинамического
сопротивления пловца. Несовпадение осей обеих траекторий
можно объяснить неодинаковой сгибаемостью вверх и вниз ног
Рис. 68. Огибающие колебаний характерных точек спортсмена с моноластом
и отдельные кадры положений моноласта в вертикальной плоскости за один
цикл плавания (схематизировано).
Точки: а — кисть руки; б — ЦТ тела; в — выходная кромка моноласта; 1— верхняя,
и II — нижняя огибающие; 0; 16 — мгновенные положения моноласта (размеры в мм).
149-'
человека при плавании вследствие известного строения суста-
вов колена и стопы ноги.
При перемещении спортсмена в течение одного цикла пла-
вания с периодом Т = 0,67 с, т. е. с частотой п=1/Т= 1,49 Гц
затрачивалось времени на подъем моноласта (удар вверх)
7'п = 0,46 с, или 0,69 периода, а на опускание моноласта (удар
вниз) То = 0,21 с, или 0,31 периода цикла. Пловец продвинулся
за время подъема моноласта на расстояние $п=1 м, равное
около 2/з поступи, а при опускании моноласта so = 0,51 м, ’/з
поступи. Тем самым опровергается широко распространенное
среди спортсменов мнение о том, что якобы удар моноластом
вверх происходит пассивно и не имеет большого значений
в плавательном цикле.
Следует подчеркнуть неравномерность удара моноластом за
цикл. Поперечные, вертикальные составляющие скорости дви-
жения конца лопасти были в среднем при подъеме 1ГП = Д/7’П =
= 1,37 м/с, а при опускании Wo=A/To = 3,00 м/с, т. е. сущест-
венно различные. Тем не менее средние продольные, горизон-
тальные составляющие скорости поступательного перемещения
пловца за указанные части периода достаточно близкие: при
подъеме моноласта Un = sn/Tn = 2,l8 м/с и при его опускании
Uo = so/To.= 2,43 м/с.
Средняя поступательная скорость плавания в рассматри-
ваемом цикле Uq = s/T = 2,25 м/с. Число Струхаля, выраженное
через максимальный размах А колебательного движения мо-
ноласта, ShA = nA/Uo= (1,49-0,63)/2,25 = 0,41. Полученное зна-
чение числа Струхаля характерно. Оно соответствует значению
такового для угревидного плавания гидробионтов с относи-
тельно большим размахов колебательных движений хвостовым
плавником (см. рис. 38, в).
Пространственно-временные данные кинограмм позволили
определить расчетные значения длины бегущей волны %=
= 2,10 м и скорость распространения ее с = 3,0 м/с, а потому
и средние значения активной силы тяги Р и гидродинамиче-
ского кпд т] по изложенной выше теории гидродинамики де-
формируемого тела, с использованием концепции пронизывае-
мого слоя. В рассматриваемом случае Р — 380 Н и т] = 0,87.
Характер изменения мгновенной поступательной скорости
плавания спортсмена с моноластом на дистанции был выбо-
рочно определен акустическим способом с использованием эф-
фекта Допплера (рис. 69). Колебания скорости достаточно
закономерны. Абсолютное значение средней скорости заплыва
на всей дистанции 400 м точно зафиксировано, Uo= 1,94 м/с.
Для общей ориентировки и определения сопротивления
спортсмена без ластов в квазистационарном приближении нами
•обработаны имеющиеся в литературе экспериментальные дан-
ные. Первоисточниками этих экспериментов являются: под-
водная буксировка девяти-десяти спортсменов без ластов в бас-
150
U, м/с
Рис. 69. Осциллограмма мгновенной поступательной скорости спортсмена с
моноластом за 10 циклов плавания на дистанции.
сейне Львовского института физической культуры; буксировка
спортсмена без ластов в Шведском опытовом бассейне; систе-
матические испытания физиологии плавания человека швед-
скими спортивными организациями; систематические испыта-
ния энергетики плавания десяти спортсменов в Нью-Йоркском
университете (США). Графическая обработка показала удов-
летворительное соответствие экспериментальных данных по
различным источникам. Оказалось, что гидродинамическое со-
противление спортсменов при активном колебательном плава-
нии без ластов в 1,4 раза больше, чем при пассивной букси-
ровке выровненного тела с той же скоростью. Это вполне
объяснимо дополнительным сопротивлением спортсмена от попе-
речных движений тела и конечностей с большим размахом ко-
лебаний.
Полная сила гидродинамического сопротивления спорт-
смена с моноластом ростом /7=1,80 м, массой гп = 82 кг, при
средней скорости /7о=1,94 м/с, подсчитанная энергетическим
методом, оказалась равной /?~210 Н. Были определены без-
размерные коэффициенты гидродинамического сопротивления
спортсменов без ластов и с моноластами в зависимости от
числа Рейнольдса (рис. 70).
Следует обратить внимание не только на абсолютные значе-
ния коэффициентов сопротивления, но и на согласованный ха-
рактер протекания кривых, отвечающих различным представ-
ленным вариантам исследований. Это свидетельствует о досто-
верности полученных результатов.
Анализ полученных результатов показывает, что гидроди-
намический коэффициент сопротивления тела спортсмена без
ластов на порядок выше такового для технически гладкой пла-
стины с турбулентным пограничным слоем, или в 8 и более раз
выше, чем у тела вращения с турбулентным режимом при
одинаковых числах Рейнольдса. Это несомненно свидетельст-
вует об отрывном обтекании тела спортсмена. Коэффициент
гидродинамического сопротивления тела женщины заметно
ниже, чем у мужчины, и немного меньше, чем у модели-мане-
151
Сх
Рис. 70. Коэффициент гидродинамиче-
ского сопротивления тренированных
спортсменов в зависимости от числа
Рейнольдса Cx(Re)
1 — буксировка лебедкой и 2 — активное
плавание мужчин — в обоих случаях рост
1,70—1,80 м, масса около 80 кг; 3 — букси-
ровка лебедкой женщин, рост 1,64 м,
масса 58 кг; 4 — буксировка модели-ма-
некена женщины тех же размеров; 5 —
гладкая пластина и 6 — тело вращения
LfD=b с турбулентным пограничным
слоем. Расчет для спортсмена с монола-
стом: а — по энергообмену, б — по гидро-
динамике деформируемого тела.
кена. Расчетные точки а и б
для спортсмена с моноластом
согласуются с эксперимен-
тальными данными по сопро-
тивлению тела спортсменов
без ластов.
Хотя человек — плохо пла-
вающий гидробионт, моно-
ласт является перспективным
волновым движителем для
‘Скоростного плавания способом «дельфин». Нами, совместно
с другими авторами, предложена единообразная структура и
форма моноласта для промышленного изготовления [6].
Р. А. Грунферстом, Н. П. Березиной (Бионика. Киев. 1981.
Вып. 15. С. 29—39) теоретически исследованы малые колеба-
ния упругого ласта в двумерном потенциальном периодиче-
ском потоке идеальной жидкости при линейной постановке за-
дачи. Принятые допущения существенно ограничивают возмож-
ность использования полученных теоретических результатов.
В частности, авторы указывают, что применение двумерной тео-
рии допустимо для ласта большого удлинения е = Ь/2а, когда
е^Ю, что практически не встречается в плавании. В случае
переменной жесткости ласта, изменяющейся по линейному за-
кону уменьшения жесткости к задней кромке, кпд -q изменяется
значительно. К аналогичному эффекту приводит уменьшение
.плотности материала ласта.
152
ГЛАВА IV
БИОЭНЕРГЕТИЧЕСКИЕ АСПЕКТЫ
ПЛАВАНИЯ ГИДРОБИОНТОВ.
ДИАПАЗОН И ХАРАКТЕР РЕГУЛЯЦИИ
БИОЭНЕРГЕТИКИ
§ 17. Некоторые общие закономерности биоэнергетики.
Типовая зависимость «мощность—скорость гидробионтов»
Биоэнергетика плавания имеет большое значение для гидро-
бионики. Это обусловлено ее особенностями, а также тем, что
при некоторых обстоятельствах она позволяет рассчитать ак-
тивную мощность и гидродинамическое сопротивление круп-
ного гидробионта данного конкретного вида, что не всегда уда-
ется оценить с достаточной точностью при использовании дру-
гих методов расчета. В основу настоящей главы положены
некоторые из ранее опубликованных работ автора, дополнен-
ные новыми материалами.
Биоэнергетические затраты животных биологи (физиологи)
изучают преимущественно по обмену веществ — метаболизму,,
с которым связаны все жизненные процессы. При этом учиты-
вают реагирующие компоненты и калорийность их, прежде
всего потребление кислорода. В соответствии с физиологиче-
ским состоянием отдыхающего и работающего организма об-
мен может быть основным, активным и общим. Различают
также уровень обмена — энергия, расходуемая целым живот-
ным (размерность работы — Дж), и интенсивность обмена —
удельная энергия в расчете на единицу веса гидробионта
(Дж/Н). Уровень энергетического обмена гидробионта, отне-
сенный к единице времени, есть мощность (N, Вт), а интен-
сивность обмена в единицу времени — удельная мощность q,
Вт/Н). Отметим эквивалентность размерностей 1 Вт/Н=1 м/с,
что удобно для расчета удельной мощности гидробионта по
массе т без переводных множителей. Разумеется, расчетные
данные по обмену веществ всегда осреднены по времени, хотя
часто это не отмечают.
Мощность Основного обмена No выражает среднюю мини-
мальную мощность организма гидробионта в относительном по-
кое при известных внешних условиях отдыха, затрачиваемую
на «внутреннюю работу» органов и тканей (деятельность
сердца, легких, органов пищеварения и др.). Наряду с основ-
ным обменом используют более строгое понятие «стандартный
обмен» (стандартный метаболизм), который определяют по
экстраполяции, как уровень «нулевой активности» в так назы-
ваемой термонейтральной зоне, в диапазоне температуры
153
которой расход энергии не меняется. Численные значения мощ-
ности основного и стандартного обмена обычно близкие.
Мощность активного обмена Ма выражает средние допол-
нительные энергетические затраты гидробионта при мышечной
нагрузке в активном плавании, сверх мощности основного об-
мена. Величина Na вызывает наибольший интерес в гидробио-
нике. Активный обмен в несколько раз, при высоких скоростях
плавания до порядка величины и более, превышает основной
обмен.
Мощность общего обмена N представляет собой полную
среднюю мощность гидробионта в активном плавании. Прини-
мают, хотя и условно, что в плавании основной обмен гидро-
бионта такой же, как в покое. Поэтому для мощности энерге-
тического обмена имеем N=N0 + Na, (Na^N0). Соответст-
венно для удельной мощности общего q, основного q0 и актив-
ного <7а обмена гидробионта массой т справедливы соотно-
шения:
q = Nlm, qa = Nalm, qa = NJm.
Рассматривая гидробионта в плавании как живую машину,
среднюю мощность общего обмена N можно назвать физиоло-
гически располагаемой мощностью, которая химически высво-
бождается из субстрата организма посредством метаболизма.
Гидродинамически эффективная, полезная мощность Ne гидро-
бионта в горизонтальном плавании со средней поступательной
скоростью Uo = const выражается через среднюю силу тяги Т
или равную ей среднюю силу сопротивления R (при равномер-
ном движении) Ne=TUo = RUo.
Зависимость между Na и Ne определяется формулой
Ne = ПидПм^а = 'ПидМм.
где г)Ид — идеальный средний гидродинамический кпд в дан-
ном режиме плавания гидробионта, учитывающий потери мощ-
ности на кинетическую энергию в его следе (см. гл. III), т]м —
кпд мышц —двигателя гидробионта, которым учитывается эф-
фективность превращения биохимической энергии животного
в механическую работу с мощностью мышц NM. В среднем т]м~
«0,20, однако величина т]м сильно варьирует.
В настоящее время надежные, но ограниченные эксперимен-
тальные данные имеются по основному и активному обмену
у мелких рыб, птиц, в меньшей мере у млекопитающих. Су-
щественную помощь для биоэнергетического расчета плавания
крупных теплокровных гидробионтов оказывают обстоятель-
ные исследования энергетического обмена спортсменов.
Главным фактором основного обмена теплокровных (го-
мойотермных) животных является общая масса тела. Темпера-
тура тела у них, в том числе у крупных млекопитающих, под-
держивается приблизительно постоянной (/«36—37 °C), не-
154
зависимо от температуры среды. Приведем для сравнения об-
щего обмена следующие данные.
У многих видов аэробионтов-птиц, в результате современной
обработки и переоценки всех предыдущих исследований, в ко-
торых масса птиц варьировала во всем интервале значений
от мельчайшей колибри до гигантского африканского страуса,,
установлены формулы среднего стандартного обмена [50]:
для неворобьиных птиц
No 3,8т0-724, q0 = 3,8m-0’276, 3- IO-3 < т < 10в; (69)
для воробьиных птиц
Уо = 6,25m0’723, <?о = 6,25m-0’277. (70}
Здесь т — масса птицы, кг; No— мощность, Вт; q0— удель-
ная мощность основного обмена, Вт/кг. Интересно, что стан-
дартный обмен у воробьиных птиц выше, чем у неворобьиных,
в среднем в 1,65 раза, или в отношении 5/3, что пока не нахо-
дит удовлетворительного объяснения.
Для различных млекопитающих одного и того же вида,
размеры и масса которых увеличиваются с возрастом в про-
цессе роста, и для животных разных видов («от мыши до
слона») в результате экспериментальных исследований многих
авторов получены аналогичные средние зависимости основного
обмена от массы тела
7V0 = 3,4m0’74, <7o = 3,4m-0’26, 20-10-3 < m < 3,7- 10s. (71)
Важно отметить, что этим зависимостям удовлетворяют дан-
ные по основному обмену человека (мужчины и женщины мас-
сой 70—80 кг) и крупных морских гидробионтов-дельфинов
массой 50—200 кг, т. е. закономерности основного обмена ока-
зываются общими для млекопитающих.
Главные факторы основного обмена у рыб — температура
среды и масса тела. Зависимость интенсивности основного об-
мена рыб от температуры, как показано разными авторами и-
с применением различных методов, для пресноводных и мор-
ских рыб многих видов, размеров и возраста единообразно
выражается одной и той же эмпирической «нормальной кри-
вой» Крога. Эта кривая, по аппроксимации с погрешностью, не
превышающей ±10 %, нами определяется экспоненциальной
зависимостью, которая позволяет производить для рыб перерас-
чет основного обмена с изменением температуры среды:
q0 = 0,07е°’п, 5° < t < 27°, (72)
где t — температура воды, °C; q0 — удельная мощность основ-
ного обмена, Вт/кг. С повышением температуры воды потреб-
ление кислорода рыбой в единицу времени на единицу массы
резко увеличивается в основном обмене: от 5 до 25 °C —►
почти в 8 раз; от 10 до 20 °C — примерно в 3 раза.
155-
Главным фактором основного обмена у рыб является также
масса тела. В результате единообразной статистической обра-
ботки Г. Г. Винбергом многочисленных экспериментальных
данных, полученных многими авторами для пресноводных и
морских рыб (исследовано около 400 видов), масса которых
изменялась в 106 раз, можно принять одно осредненное урав-
нение основного обмена рыб в следующем виде:
No = 0,44m0’80, q0 = 0,44m~°’20, IO"6 < tn < 10. (73)
Здесь tn — масса рыбы, кг; No — мощность, Вт; qo — удельная
мощность основного обмена, Вт/кг. Данные приведены к одной
температуре £ = 20°С. Очевидно, характер зависимости основ-
ного обмена от массы тела для мощности — параболический,
а для удельной мощности — гиперболический, т. е. с увеличе-
нием массы, следовательно, размеров рыбы, q0 уменьшается.
При увеличении массы рыбы на один порядок удельная мощ-
ность общего обмена снижается в среднем на 15%, а для
уменьшения удельной мощности на один порядок масса рыбы
должна быть больше в 105 раз.
Как видно, для всех гидробионтов справедливы тождествен-
ные степенные зависимости для мощности основного обмена No
и удельной мощности основного обмена q0 от массы тела типа
N0 = amn, q0 = amn-\ где размерный коэффициент а определяет
соответствующие значения основного обмена при массе живот-
ного т=1 кг, а варьирующий для отдельных видов безразмер-
ный показатель степени п<1 характеризует интенсивность сни-
жения этих значений с увеличением массы тела т> 1 кг. В це-
лом значения данного показателя изменяются в интервале
0,67<пс1. Здесь нижнее значение п = 0,67 ограничено частным
случаем пропорциональности основного обмена площади по-
верхности тела некоторого животного (поскольку S^m2/3),
а- верхнее значение п=1 — частным случаем пропорционально-
сти основного обмена самой массе тела (ЛЛ«т), что изредка
встречается у отдельных видов. Приведенные выше устойчивые
значения показателей степени п^0,75 в выражениях (69) —
(73) — среднестатистические для основного обмена соответ-
ствующих групп животных, промежуточные между значениями
зависимости метаболизма как от поверхности, так и от массы
тела, поскольку влияют оба эти параметра.
Еще в 1915 г. Броди показал справедливость некоторого
основного принципа для общего обмена наземных млекопи-
тающих. Согласно этому принципу, средняя мощность общего
обмена N животных пропорциональна средней мощности основ-
ного обмена No, т. е.
W = kNо = kamn. (74)
Здесь безразмерный коэффициент кратности k, переменный по
величине, определяется характером выполняемой животным ра-
боты, ее активностью и продолжительностью. При этом актив-
156
ный обмен одного и того же мле-
копитающего может превышать ос-
новной обмен в fe=l,6; 4,8; 25 раз
(рис. 71). Основной обмен у афа-
лин соответствует таковому у че-
ловека (линия /). Однако при воз-
бужденном состоянии дельфинов —
морской свиньи и афалины — их
основной обмен превышает норму
более чем в два раза (линия 1').
Наиболее исследованы в по-
следнее время вариации коэффи-
циента k для горизонтального по-
лета птиц В. Р. Дольником — см.
[50]. Получено, что k = 34-5 для
облегченного полета в течение
всего дня птиц с повышенным аэ-
родинамическим качеством крыла
(стрижи, ласточки); k= 124-14 для
умеренного длительного полета
птиц в течение нескольких часов
без кислородного дебета (хорошие
летуны, перелетные птицы); k =
= 164-20 для изнуряющего кратко-
временного полета птиц с разви-
вающимся кислородным дебетом
(плохо летающие птицы).
Коэффициент кратности k по
экспериментальным данным для
наземных млекопитающих и птиц
не зависит от их массы. Следова-
ние. 71. Мощность Vo основ-
ного (линии 1, 1') и Na. актив-
ного (2—4) обмена в зависи-
мости от общей массы у теп-
локровных животных и чело-
века
С —собака; Ч —человек; Л — ло-
шадь; дельфины: МС— морская
свинья, Аф*—афалина; линия Г —
животное в возбужденном состоя-
нии.
тельно, показатель степени п в формулах искомой зависимо-
сти мощности активного обмена Wa от массы этих животных т
сохраняет ту же величину, что и для основного обмена, т. е.
п~0,75. Это справедливо и по единому значению размерно-
стей. Действительно, в соответствии с зависимостями для N,
No и Na по формуле (74) имеем
Wa = W—We = (k^-1) атп.
(75)
Здесь переменное значение множителя (k—1) есть функция
степени активного животного. Для гидробионтов этот множи-
тель выражается через среднюю скорость плавания Uo на каж-
дом данном режиме соответствующей продолжительности пла-
вания Т до утомления, и потому может иметь вид
Лк-1=ф((/(,) = ф-1(Т).
157
Таким образом, для мощности активного обмена теплокров-
ных гидробионтов расчетная формула структурно представля-
ется произведением двух функций
Na = <р (t/0) Ф (tn) = ф"1 (Т) ф (пг). (76)
При этом в ф(т) показатель степени по предыдущему известен
(п = 0,75), а функция скорости ф(£70) на основании гидродина-
мических соображений зависит от режима течения в погранич-
ном слое гидробионта и должна быть степенной функцией.
Расчетная формула устанавливается ниже с привлечением не-
обходимых дополнительных систематических эксперименталь-
ных данных по биоэнергетике.
Активный обмен рыб изучали физиологическими методами-
несколько авторов, однако только у немногих видов сравни-
тельно мелких рыб. В частности, по В. А. Матюхину [35]г
удельная мощность qa активного обмена байкальских омулей и
хариусов длиной L — 0,30 м, массой около т —0,3 кг зависит от
устойчивой скорости плавания до утомления продолжитель-
ностью примерно около часа следующим образом: 9a = 3,Ot/o1,67>
O,3<t/o<l,O, где [<7а]= Вт/кг, [(70] = м/с. В указанной работе
приведены различные сведения по методике и технике биоэнер-
гетических исследований.
Наиболее полные исследования основного и активного об-
мена у рыб выполнены Бреттом в ряде работ [69], которыми
широко пользуются. В биогидродинамической трубе-респиро-
метре систематически испытаны тихоокеанские лососи-нерки
длиной Л = 0,0774-0,54 м, массой т = 0,00344-1,43 кг, при раз-
личной температуре воды / = 0—24 °C. Они плавали до утомле-
ния с различной заданной потоком воды в респирометре сред-
ней скоростью (t/o = 0,18-ь 1,42 м/с). Установлено, что активный
обмен у испытанных рыб существенно зависит по крайней мере
от следующих четырех факторов: общей массы т, уровня ак-
тивности — скорости плавания Uo, температуры воды t, °C и
доопытного состояния рыбы. Ниже приведены соответствующие
графики зависимости по нашему пересчету.
Для молодой нерки длиной Л = 0,18 м, массой т = 0,050 кг
Бретт установил три уровня активности плавания (см. рис. 31):
а) импульсный (пиковый, рывками), длительностью Т< 10 с;
б) продолжительный (до состояния усталости), Т= 104-2• 104с;
в) мигрирующий (устойчивый — неопределенно долгий),.
Т>2-104 с. Существенно, что в импульсном режиме плавания
средняя максимальная скорость плавания Um уменьшается
с продолжительностью Т по экспоненциальной зависимости,
как это показано Бреттом. Однако в продолжительном плава-
нии нерки зависимость Um(T) может быть принята в виде сте-
пенной функции вида (10). Эта аппроксимация подтверждается
аналогичными данными по скоростям плавания форели радуж-
158
ной длиной L = 0,24 м (см. рис. 31), что упрощает возможный
расчет.
Значительное влияние на активный, а потому и общий об-
мен молодой нерки оказывает температура воды: тем больше,
чем меньше относительная скорость плавания С/0/^<3,5 с-1.
Однако при больших скоростях £/0/L>5 с-1 влияние темпера-
туры воды на обмен рыб возможно и небольшое, т. е. в скоро-
стном плавании рыбы ведут себя по активному обмену почти
как гомойотермные животные. Представляет интерес выявление
всех пяти границ, лимитирующих интенсивность общего обмена
рыбы (рис. 72).
Поскольку в экспериментах Бретта масса испытанных не-
рок изменялась в широком диапазоне, приведем соответствую-
щий график удельной мощности активного обмена (рис. 73).
На интенсивность общего обмена нерки оказывает также
влияние состояние рыбы: нормальное или возбужденное
(рис. 74). Однако и этот фактор более всего влияет на основ-
ной обмен (при нулевой скорости). По мере повышения скоро-
сти плавания его значение резко уменьшается. Значительное
увеличение основного обмена рыбы при возбужденном ее со-
стоянии хорошо согласуется с ранее отмеченным для дельфи-
нов— морской свиньи и афалины (см. рис. 71).
Рис. 72. Зависимость удельной мощ-
ности общего обмена тихоокеанского
лосося-нерки длиной 0,18 м от от-
носительной скорости плавания в те-
чение часа и температуры воды.
Лимитирующие границы: 1 — ос-
новной обмен: 2 — минимальная темпера-
ратура fmin=0°* 3 ~ максимальная *тах=
—26 °C; 4 — усталость рыбы (сносится по-
током); 5 — недостаток кислорода в воде.
Рис. 73. Удельная мощность актив-
ного обмена нерки в зависимости от
массы рыбы и скорости плавания
в закрытой биогидродинамической
трубе и открытом лотке, (m, Uo).
l — L=0,077 м; m=0,0034 кг; 2 — L-0,1 м;
m=0,0085 кг; 3 — L—0.188 м:- т=0,056 кг;
4 — 7=0.418 м; т-0,746 кг; 5 — L-0,539 м;
т=1,432 кг; Um—максимальная скорость
часовой продолжительности плавания
рыбы каждого размера.
159
Рис. 74. Влияние различного состояния
нерки длиной 0,2 м на удельную мощ-
ность общего обмена при различной ско-
рости плавания (оптимальная темпера-
тура воды 15 °C).
1 — нормальное; 2 — возбужденное состояние
рыбы; 3 — отношение 4 — интенсивность
основного обмена
Данные испытания свидетель-
ствуют о необходимости тщатель-
ной подготовки гидробионтов для
проведения биоэнергетических
исследований с учетом их физио-
Рис. 75. Типичная зависимость
мощности общего обмена гид-
робионта от средней скорости
его плавания различной про-
должительности (с разным
кпд) на примере лососевой
рыбы-нерки, N(U0).
U№ — оптимальная скорость при
Wmln;t/OK—крейсерская — по усло-
вию (Ж'«) m|n; / — «продолжитель-
ная» скорость плавания гидроби-
онта в океане 1/я а: 2 — устой-
чивая форсированная крейсерская
скорость — в реке против течения
(//«> t/oi); 3—4 — интервал кратко-
временных высоких скоростей пла-
вания гидробионта (О0 » //«>)
логического состояния.
Более общий интерес для гидробионики представляет типо-
вая зависимость мощности общего обмена гидробионтов от
скорости плавания — N(UO). Она имеет единообразный экстре-
мальный вид. Такая зависимость экспериментально получена
Бреттом [69] для проходной взрослой лососевой рыбы-нерки
(рис. 75). Оказалось, что со средней скоростью t/oi, близкой
к оптимальной средней скорости ^oo-*Vmin, нерка плавает
в море, когда мощность общего обмена ее минимальная. При
любой другой средней скорости плавания, в том числе и мень-
шей, чем оптимальная Uo<Uqo, мощность общего обмена выше.
В реке нерка плывет против течения с повышенной, крейсер-
ской средней скоростью U02>U00, при этом мощность общего
обмена значительно больше, чем при оптимальной скорости.
Аналогичные кривые N(U0) экспериментально получены для
аэробионтов — летучих мышей, птиц (волнистого попугай-
чика) [50]. Таким образом, единообразная, экстремального вида
зависимость N(U0) является типичной для аэрогидробионтов.
160
Это позволяет утверждать, что
гидробионты активно регули-
руют биоэнергетику своего пла-
вания.
Сравним удельные энергети-
ческие затраты животных на пе-
ремещение в различной среде;
представим их в виде зависимо-
сти для безразмерной относи-
тельной мощности N/WUK=f(W)
(рис. 76). Здесь W — вес тела,
Н; U к — крейсерская средняя
скорость перемещения; N — об-
щая мощность энергетических
затрат животного по Тьюкеру.
Относительная мощность на пе-
ремещение в любой среде опре-
деляется весом тела животного
и типом его перемещения (пла-
вание, полет, ходьба, бег) и
резко снижается с увеличением
веса, особенно у наземных жи-
вотных. При одном и том же ве
щих и наземных животных наименьшие удельные энергетиче-
ские затраты на перемещение наблюдаются у плавающих гид-
робионтов.
-5 -Ч -3 -2 -1 0 1 2 3 k
IgW.W
Рис. 76. Зависимость от общего
веса животных минимальной от-
носительной общей мощности на
перемещение единицы веса на
расстояние, преодолеваемое в еди-
ницу времени, \g(b'lWUw) =
= f(lg W).
/ — плавание (лососевые рыбы); // —
полет (/ — насекомые; 2 — птицы; 3 —
летучие мыши); /// — ходьба и бег
(4 — мыши; 5 — крысы; 6 — собака; 7 —
человек; 8 — лошадь).
мелких плавающих, летаю-
§ 18. Прыжки из воды скоростных гидробионтов
с максимальной отдачей мощности. Метод определения
активной мощности по киносъемке
Для дальнейших расчетов активной отдачи мощности гидро-
бионтами необходимо знать ее максимальное значение. Ниже
излагается разработанный и примененный автором простой и
удобный гидробионический метод определения осредненной
максимальной удельной отдачи мощности крупными скорост-
ными водными животными в кратковременный период им-
пульса. Метод основан на систематическом изучении и обра-
ботке последовательных кадров натурных киносъемок прыжков
и применим в принципе для исследования биоэнергетики раз-
личных выпрыгивающих из воды скоростных водных живот-
ных— дельфинов, рыб, кальмаров. Для повышения точности
расчетов следует, применять кинограммы прыжков с тем боль-
шей частотой кадров, чем меньше размеры тела водного жи-
вотного и выше скорость выпрыгивания его из воды. Метод
целесообразно использовать для качественного анализа -и ко-
личественной обработки обычных киносъемок прыжков столь
крупных водных животных, как дельфины.
6 с. В. Перши
16Г
Рис. 77. Группа одновременно выпрыгивающих дельфинов.
Прыжки дельфинов (рис. 77) представляют собой интерес-
ное явление и бывают различными. В дальнейшем будем раз-
личать одиночные прыжки дельфинов и групповые (парой,
тройкой, большой группой — в один ряд, в два ряда). Для це-
лей настоящей работы наибольший интерес представляют оди-
ночные прыжки дельфинов, которые по характеру прыжка и
значениям его основных параметров подразделим на настиль-
ные— относительно низкие, но длинные, и навесные — относи-
тельно высокие, но короткие (рис. 78). Здесь и далее мы вос-
пользовались достаточно известными работами Ж. И. Ку-
сто и др.
Полные кинограммы (кадр за кадром) прыжков показы-
вают линию горизонта, угол выхода' тела дельфина из воды и
угол, при котором он снова входит в воду, положение тела
в процессе прыжка и наибольший подъем его над водой, четкие
очаги брызг при выбрасывании дельфина из воды и при обрат-
ном погружении его воду. Все это вместе с отметками времени,
автоматически отсчитываемом кадрами, позволяет производить
достаточно детальную количественную обработку кинограмм,
в частности, для расчета импульсной мощности дельфинов.
Исходные предпосылки метода автора по расчету осреднен-
ной максимальной удельной отдачи импульсной мощности
крупных водных животных на основе киносъемки их прыжков
состоят в следующем.
При выпрыгивании в воздух Животное находится в свобод-
ном полете и на него действует в сущности только одна внеш-
няя сила — сила тяжести (силой сопротивления воздуха за-
ведомо можно пренебречь ввиду ее малой величины и непро-
должительности полета). Это соответствует движению по
параболической траектории тела, выброшенного под углом
162
и находящегося под воздействием постоянной по значению и
направлению силы тяжести. Выберем прямоугольную систему
координат xUy с началом О в точке выхода животного из воды;
ось х направим вдоль поверхности воды в плоскости прыжка
(рис. 79). Тогда, обозначив ао — угол выхода, Uo — скорость
выхода животного из воды и t —
Рис. 78. Выборочные последо-
вательные кадры киносъемки
в море одиночного навесного
прыжка дельфина.
текущее время прыжка, приняв
за начало отсчета времени мо-
мент прохождения ЦТ живот-
ного через начало координат
(g — ускорение свободного па-
дения), напишем следующие из-
вестные выражения.
Уравнения движения центра
массы в параметрической форме
х = Uot cos а0,
у = Uot sin а0'—gt2l<2.-,
траектория движения
У = х tg а0—gx2/2t/2 cos2 а0.
Максимальные размеры пры-
жка:
дальность полета хтах = / =
= (702 sin 2a0/g,
высота полета ymax=h=
= U02 sin2 a0!2g.
Рис. 79. Расчетная схема для определе-
ния основных параметров прыжков
дельфинов по натурным кинограммам.
6*
165
На увеличенной кинограмме прыжка гидробионта можно из-
мерить угол cto и относительные размеры прыжка //L, h/L, h/l,
где L — длина тела животного, считая от носа до начала хво-
стового плавника. Чтобы определить абсолютные размеры
(длину животного L) и параметры прыжка I и h, а также ско-
рость Uo, воспользуемся тем, что по кинограмме можно уста-
новить еще продолжительность выхода тела из воды при пере-
сечении им свободной поверхности /i = nit и длительность по-
лета ЦТ тела от поверхности воды до наивысшего положения
в воздухе ^2 = «2Т. Здесь т — период съемки одного кадра, на-
пример, т= 1/24 с, а П| и п2— соответствующее число кадров.
Тогда, из соотношений для хШах и t/max, считая, что во время
выхода тела из воды скорость Uo остается постоянной, получим
Цо=—!—72^=-^ = -^, (77)
Sin<Z0 /1 М]Т
L = 2g('^L-yjL. (78)
\ sina0 / L
В выражении (78) для L теперь все величины известны, что
позволяет определить по кинограмме длину тела животного и
размеры прыжка. Разумеется, расчет упрощается, если длина
тела животного заранее известна, например, при киносъемке
в океанариуме.
Как видно, определение параметров прыжка животного сво-
дится к следующему: по кинограмме находят величины aoi h/L,
l/L, h/l, t\, t2, затем расчетом по приведенным выше форму-
лам вычисляют значения L, h, I, Uo. Предложенный метод есте-
ственно, сопровождается погрешностями, обусловленными не-
точностью измерений по кинограмме, перекосом плоскости
прыжка относительно плоскости кинопленки, углом ракурса
при съемке. Целесообразно произвести контроль, который
можно осуществить по двум формулам, связывающим незави-
симо определенные величины. Теоретически должны соблюдаться
равенства: /i// = tga0/4, t2 = t/osin ao/g.
На основании вышеизложенного осредненная по времени
удельная отдача механической мощности животного в прыжке,
рассчитанная на единицу его массы, [<?м] = Вт/Н, будет
i?M= — = -J-£t/osinao. (79)
/2 2
Коэффициент k учитывает, как отмечено выше, что животное
в прыжке увлекает за собой воду. В соответствии с имею-
щимися подобными данными для твердых тел принято значе-
ние k= 1,1.
Отметим некоторые методические замечания, выработанные
при обработке киносъемки многих прыжков дельфинов. Для
164
Рис. 80. Зависимость относительной
высоты прыжка дельфина от угла
выхода из воды его тела.
а — настильные; б — навесные прыжки;
/—15 — номера исследованных прыжков.
Рис. 81. Зависимость угла выхода
из воды выпрыгивающего дельфина
от относительной длины его тела.
а — настильные; б — навесные прыжки;
1—15 — номера исследованных прыжков.
измерения углов и линейных размеров на кинокадрах послед-
ние проектировались на экран с увеличением более чем на по-
рядок величины. Обрабатывались только такие кинограммы, на
которых четко были видны: линия горизонта воды (она прини-
малась за горизонтальную линию), а также очаги брызг в на-
чале и в конце прыжка. Были отобраны и обработаны только
15 киносъемок прыжков дельфинов в некотором промежуточ-
ном масштабе. Результаты систематического изучения их кино-
грамм по всем относительным и абсолютным параметрам по-
казаны в виде графиков на рис. 80—82.
Полученные максимальные значения для дельфина, высота
прыжка которого Ь = 3,5м
и длина / = 7,9 м, конеч-
но, не являются рекорд-
ными, но, по-видимому,
близки к ним, и соответ-
ствуют встречающимся
в литературе сведениям.
Систематические исследо-
вания, с которыми можно
было бы достаточно полно
сравнить наши 'резуль-
таты, нам неизвестны.
Однако при расчете по
указанным выше форму-
лам погрешность опреде-
ления параметров рас-
Рис. 82. Абсолютные размеры (высота и
длина) прыжков дельфинов.
а — настильные; б — навесные прыжки (1—15 —
номера прыжков).
165
смотренных прыжков, как правило, не превышает 15 %, т. е.
для целей данной работы точность метода приемлема. Нами
использован обычный многотиражный кинофильм, а не специ-
альные кинограммы прыжков. При соответствующей организа-
ции киносъемок прыжков точность результатов может быть по-
вышена, тем более при работе с тренированными животными.
Все это позволяет считать, что подобные результаты надежно
можно использовать для определения удельной отдачи мощно-
сти животными в кратковременный период импульса. Поскольку
большой интерес представляют максимальные значения удель-
ной мощности, для такого расчета использованы только четыре
навесных прыжка № 12—15 (табл. 7).
Таблица 7. Импульсная удельная отдача мощности дельфина
по результатам обработки кинограмм прыжков
Номера прыжков Длина дельфина L, м Скорость выхода Uo, м/с Продолжи- тельность взлета /2, с Высота прыжка /г. м Осредненная удельная отдача мощности Вт/Н
12 2,04 8,16 0,667 2,45 4,04
13 2,45 9,05 0,876 2,87 3,60
14 1,60 7,70 0,834 2,40 3,17
15 2,06 9,00 0,960 3,50 4,01
Следовательно, при длине обыкновенного дельфина L = 2 м
и общей массе /п = 80 кг его мышечная система характеризу-
ется осредненной за время /~1 с общей отдачей максимальной
импульсной мощности Л/е=2,95-103 Вт. Это вполне соответст-
вует определенной по кислородному обмену отдаче мощности
мускульной системы тренированного спортсмена за тот же пе-
риод (см. рис. 71). Последнее и следовало ожидать, поскольку
все основные параметры активного обмена у дельфина при-
мерно те же, что и у человека.
Проходные лососевые рыбы, перемещаясь в нерестовый пе-
риод вверх по реке и преодолевая преграды в виде запруд и
плотин, могут выполнять высокие прыжки из нижнего бьефа
в верхний. В работе [35] приведены два примера прыжков
крупного лосося длиной Л = 0,91 м: на высоту fti = l,83 м со ско-
ростью (71 = 8,3 м/с и /12 = 3,05 м, (72=11,0 м/с. Последовательно
применяя указанные выше формулы, получим, что удельная от-
дача механической мощности крупного лосося в данных слу-
чаях составляла <71 = 32,5 Вт/кг и <72 = 41,8 Вт/кг. Эти резуль-
таты соответствуют рассмотренным выше для прыжков дель-
финов.
166
§ 19. Продолжительность ныряния водных млекопитающих
в связи с максимальной скоростью плавания
и относительной массой мышц
Некоторые виды морских млекопитающих характеризуются
не только высокой скоростью плавания, но и тем, что они адап-
тированы к продолжительному глубокому нырянию. Например,
из китообразных выдающимся ныряльщиком является кашалот.
Он питается глубоководными кальмарами и ныряет на глубину
до двух километров в течение одного-полутора часов.
Продолжительность ныряния водных млекопитающих об-
условлена возможностью выключения внешнего дыхания воз-
духом и обеспечивается резервами кислорода организма в мио-
глобине мышц, гемоглобине крови и в легких. Наибольшее зна-
чение имеют запасы кислорода в мышцах. Т. Н. Глазовой
показано, что у ныряющих млекопитающих концентрация гемо-
глобина крови в полтора и более раза, а концентрация миогло-
бина мышц в 5—10 раз выше, чем у неныряющих животных.
В процессе адаптации к водному образу жизни у водных
млекопитающих произошли изменения в структуре их органов
и тканей; имеются специальные «механизмы» экономного ды-
хания.
Существенно, что при быстром изменении глубины погруже-
ния (тем самым при резком увеличении или уменьшении гидро-
статического давления воды) у водных млекопитающих кессон-
ная болезнь не возникает. Напомним, что максимальная
продолжительность пребывания тренированного человека (до-
бытчиков жемчуга) под водой на небольшой глубине обычно
не превышает 2,5 мин.
Быстрое плавание и глубоводное погружение морских
млекопитающих представляют собой экстремальные режимы,
связанные с взаимопереходами потенциальной и кинетической
энергии. Возможные сопоставления показывают, что наиболь-
шая продолжительность Тт времени выключения внешнего ды-
хания и максимальная скорость плавания Um, а также размер
в длину .L морских млекопитающих взаимносвязаны, так что
Tm = f(L, Um) или (L, Тт). Эта зависимость показана
на рис. 83. Здесь в рассмотрение введена относительная макси-
мальная скорость плавания Um]L. Хотя имеющихся данных не-
много, однако точки графика расположены закономерно, что
позволяет сделать определенные выводы.
Продолжительность ныряния и выключения внешнего дыха-
ния у китообразных тем больше, чем меньше их двигательная
активность, выражаемая относительной максимальной ско-
ростью плавания, и чем больше размер животного. Одна из
этих двух адаптаций как бы снимает необходимость в другой
благодаря особенностям организма, обусловленным экологиче-
скими условиями и объектами питания. Кашалот может быть
167
Рис. 83. Зависимость максималь-
ной продолжительности выключе-
ния внешнего дыхания при ныря-
нии от максимальной относитель-
ной скорости плавания морских
млекопитающих, Тт (Um/L).
КТ — каспийский тюлень; дель-
фины: / — обыкновенный, 2 — бело-
крылая морская свинья, 3 — косатка,
4 — афалина; киты: 5 — финвал; 6 —
горбатый; 7 — синий, 8 — кашалот.
выдающимся ныряльщиком
вследствие его наименьшей среди
китообразных двигательной ак-
тивности при крупных размерах
тела. Напротив, дельфин-бело-
бочка как скоростной пловец ма-
лого размера не в состоянии на-
долго и глубоко погружаться
в воду. Показатели тренирован-
ного спортсмена, неадаптирован-
ного к условиям жизни в водной
среде, по продолжительности
ныряния и скорости плавания,
далеко уступают таковым у мор-
ских млекопитающих.
В табл. 2 уже обращалось
внимание на относительно боль-
шую у гидробионтов массу
мышц-двигателей и количествен-
ное различие ее у разных видов
почти в два раза. Теперь мышеч-
ная масса организма выявляется
как накопитель миоглобина для
резервов кислорода при нырянии.
Для обобщенных сопоставлений
и последующих расчетов приведем график (рис. 84), состав-
ленный нами по данным многих исследователей.
С ростом числа Рейнольдса как у рыб, так и китообразных
относительная масса мышц (с}=тм/т, где тк — масса мышц,
т — общая масса гидробионта) непрерывно увеличивается по
степенной функции, хотя и слабо: для скоростных рыб Ci~
?«Re0’05, для китообразных Ci~Re°’08. При низких числах Re по-
рядка 105 малые нескоростные рыбы (карась и окунь) имеют
относительную массу мышц Cj меньше в два раза, чем более
скоростные елец и форель. При числах Re=1064-107 у скоро-
Рпс. 84. Зависимость относительной массы мышц гидробионтов от мак-
симального числа Рейнольдса, Ci(Re).
А — рыбы: нескоростные (к — карась, о — окунь) и скоростные (е — елец, с — став-
рида) (см. табл. 5). / — форель радужная; 2 — пеламида; 3 — тунец обыкновенный;
4 — меч-рыба; 5 — акула голубая; Б — китообразные (см. табл. 3): 6 — мор-
ская свинья; 7 — дельфин-белобочка; S — афалина; 9 — белуха; /0—горбатый кит;
// — кашалот; 12 — финвал; 13 — синий кит.
168
стных костистых рыб величина С\ до двух раз больше, чем
у дельфинов и китов, и только у хрящевой голубой акулы Ci
значительно меньше, чем у скоростных костистых рыб. Среди
китообразных малым значением Ci выделяются киты — горба-
тый и кашалот, которые отличаются мощной жировой прослой-
кой у поверхности тела, особенно кашалот, глубоко ныряющий
в холодную воду и нуждающийся в повышенной теплозащите.
Приведенные данные свидетельствуют о необходимости учиты-
вать относительную массу мышц ct в расчете биоэнергетики
плавания гидробионтов.
Следует отметить различие в структуре двигательных
красных и белых . мышц у скоростных рыб и скоростных
дельфинов. У скоростных рыб красные мышцы, которые
насыщены миоглобином и длительно функционируют, вы-
деляются отдельными порциями в боковых частях тела среди
белых мышц, работающих кратковременно (см. рис. 45). Форма
порций красных мышц и расположение их (ближе к поверхно-
сти тела или глубже) различны у разных видов скоростных
рыб, но одинаковые с обеих сторон тела. Напротив, у скорост-
ных дельфинов волокна красных и белых мышц распределя-
ются довольно равномерно и диффузно в различных по функ-
циональному назначению мышцах. Двигательная мускулатура
хвостовой области дельфинов характеризуется гипертрофиче-
ским развитием двух пар мускульных комплексов: располо-
женным соответственно над и под позвоночником длиннейшей
мышцы-поднимателя и гипаксиальнои мышцы-опускателя, при-
чем масса первой в 1,5—2,0 раза больше, чем второй. Массы
мышц, осуществляющих боковые изгибы хвоста, в 15—20 раз
меньше массы мышц, двигающих хвостом в вертикальной пло-
скости (данные В. А. Родионова).
Крупные скоростные рыбы
с большим содержанием мышц,
такие как тунцовые, меч-рыба,
акула-мако, являются как бы
«теплокровными». У них темпе-
ратура глубоких мышц (изме-
рено термистером-иглой) на 5—
15 °C выше температуры окру-
жающей воды (рис. 85). Уста-
новлено, что эти виды рыб ре-
гулируют высокую температуру
своего тела, располагая мощной
противоточной кровеносной сис-
темой. По нашему -мнению, био-
энергетику указанных видов рыб
в первом приближении можно
рассчитывать как для теплокров-
ных млекопитающих.
Рис. 85. Максимальная темпера-
тура мышц tM «теплокровных»
крупных скоростных рыб, пойман-
ных при различной температуре
<в воды, /м(/в).
1 — обыкновенный тунец; 2 —акула-
мако.
169
Рыбы непрерывно дышат кислородом, отбираемым их жаб-
рами из воды. Грей [74], сравнительно исследуя морских рыб,
нашел, что относительная площадь поверхности жабр возра-
стает соответственно повышенной активности, быстроходности
рыб. У одного вида более подвижной рыбы площадь поверхно-
сти жабр оказалась больше, чем у малоподвижного вида, в рас-
чете на единицу общей массы в 10 раз и в расчете на единицу
площади поверхности тела в 15 раз.
§ 20. Метод аналогов в биоэнергетике,
саморегуляция отдачи мощности гидробионтами
и масштабный эффект
В гидробионике наибольший интерес представляет эффек-
тивное плавание крупных гидробионтов. Однако непреодолен-
ные пока трудности изучения активного обмена у дельфинов и
крупных скоростных рыб побуждают прибегнуть к возможному
использованию имеющихся аналогичных данных по активной
отдаче мощности у хорошо тренированных спортсменов. Для
использования этих данных имеются следующие основания.
Морские млекопитающие, в том числе наиболее распростра-
ненные виды дельфинов, являются, как и человек, гомойотерм-
ными организмами, у которых все три главные параметра
активного обмена одного порядка величины: постоянная темпе-
ратура тела / = 36—37 °C, масса тела около т^ЮО кг (у двух-
метрового дельфина), средняя из наибольших скоростей плава-
ния дельфинов Um^Ю м/с, т. е. такая же, как и максимальная
скорость бега человека. Крупные скоростные рыбы (см.
рис. 85), имеют высокую температуру тела, регулируют ее и по
активному обмену приближаются к теплокровным животным.
Выше было показано, что мощность основного и активного
обмена у человека изменяется в функции общей массы тела
единообразно с теплокровными животными, а основной обмен
дельфинов в нормальном состоянии соответствует этим данным
(см. рис. 71). Необходимая поправка на различие гидробион-
тов в массе тела достаточно очевидна по формуле (75). Воз-
можно дальнейшее уточнение обмена по видам с учетом от-
носительной массы мышц гидробионтов, особенно большой
у крупных теплокровных рыб (см. рис. 84).
В физиологических исследованиях спортсменов одновре-
менно и точно фиксируются и скоростные показатели, и про-
должительность упражнений, что позволяет последнюю обоб-
щенно ввести в расчет в соответствии с зависимостью (76) для
активного обмена. Это важнейшее обстоятельство подтвержда-
ется единообразным изменением скоростей плавания трениро-
ванных спортсменов и гидробионтов в диапазоне продолжи-
тельности плавания 7=104-2- 104 с (см. рис. 31).
Более того, можно показать, что в соответствии с биоэнерге-
тикой организма скоростные показатели и продолжительность
170
упражнений тренированных спортсменов и в других видах
спорта изменяются единообразно (сравнительно с плаванием
дельфинов). Для этого достаточно воспользоваться обобщен-
ными безразмерными координатами по скорости и времени.
Так, в диапазоне продолжительности упражнений 7’=10-т-
Ч-2-104 с, по официальным справочным данным спортивных
организаций, рекордсмены мира достигают следующих резуль-
татов на различных дистанциях (скорости округлены): плава-
ние со скоростями £70 = 2,3—1,6 м/с (25—1500 м); академиче-
ская гребля 5,0—4,7 м/с (500—2000 м); бег по дорожке 10,1—
4,9 м/с (100 м — 30 км); бег на коньках 13,5—11,4 м/с
(500 м—10 км); велоспорт (с места) 15,5—12,0 м/с (1—
100 км). В графическом виде на логарифмической бумаге по-
добные данные составляют серию соответствующих слегка вол-
нистых кривых Uq(T). Однако, если выбрать для единообраз-
ного представления этих различных данных общую для них
единицу продолжительности упражнений, например Т *= 100 с,
и отвечающую этой продолжительности в каждом виде спорта
среднюю максимальную скорость то в относительных
координатах Uo(T)/Uo* (Т *) =f(T/T *) получаем с погрешностью
±5% единую зависимость (рис. 86). Наклон средней линии
такой же, как и всех других на рис. 31 в рассматриваемом диа-
пазоне Т.
Разумеется, погрешность ±5% совершенно неприемлема
в спорте, где измеряют дистанции с точностью до сотых долей
метра, а результат по времени — до тысячных долей секунды,
поскольку рекордсмены выявляются из миллионов соревную-
щихся спортсменов. Для целей гидробионики показанное при-
ближенное единообразие достаточно удовлетворительное. Оно
свидетельствует о том, что затраты энергии в различных видах
упражнений почти эквивалентны при максимальных достиже-
ниях каждой данной продолжительности. Одновременно снима-
ется возможное возражение об эквивалентности, поскольку
l/om/ug
Рис. 86. Взаимосвязь средней максимально достигаемой скорости движе-
ния и соответствующей продолжительности преодоления дистанции
у спортсменов и дельфинов в безразмерных координатах, Uo(T)/Uo*(T*) =
= f(T/T*).
Спортсмены (мужчины, рекордсмены): / — плавание, вольный стиль; 2 — пла-
вание с ластами (на поверхности); 3 — академическая гребля; 4 — бег по дорожке;
5 — бег на коньках; 6 — велоспорт. Дельфины: 7 —белобочка; 8 — косатка.
171
в различных видах спорта участвуют группы мышц, различные
как по структуре, так и по их относительной массе. Очевидно,
при максимальных нагрузках имеет значение работа всего ор-
ганизма.
Важным достоинством известных надежных биоэнергетиче-
ских данных в области спорта является то, что они представ-
ляют собой отдачу мощности на внешнюю механическую ра-
боту [97], т. е. величину Л/е/т]ид = т)м^а в функции продолжи-
тельности упражнений Т. На основании вышеизложенного их
можно непосредственно использовать для определения силы
тяги или силы сопротивления в плавании гидробионтов. Эти и
другие подобные данные нами пересчитаны для удельной от-
дачи мощности в расчете на 1 Н общего веса спортсмена, по-
лагая его среднюю массу пг = 75 кг и относительную массу
мышц mM/m = 0,40. Расчетный график показан на рис. 87, где
экспериментальные данные аппроксимированы кривой 1 по
одночленному уравнению
= 1,73 [lg(l + Т)]-1'25. (80>
Здесь Т, с; qM, Вт/Н (м/с).
Для теплокровных гидробионтов, заметно отличающихся от
человека по общей массе тела и относительной массе мышц,
расчетная формула (80) в соответствии с формулой (76) и
рис. 84 приобретает с поправочным коэффициентом k оконча-
тельный вид
q'„=kq„, k = (^.^-Y025, (81>
\ m9 с0 )
где m — общая масса тела рассчитываемого гидробионта.
Рис. 87. Максимальная удельная отдача мощности на внешнюю меха-
ническую работу крупных теплокровных гидробионтов в функции продол-
жительности плавания, ^М(Т).
Обобщенные кривые: / — расчетная аппроксимация по формуле (80); 2 и
3 — физиологические оценки для спортсменов-чемпионов и хорошо тренированных
спортсменов [97]. Экспериментальные точки: гидробионты — / — прыжки
дельфинов, 2 — прыжки лососей, 3 — нерестовая миграция рыбы-кеты; спортсмены —
4 — гребля, 5 — бег по дорожке и в гору, 6 — упражнения на велоэргометре.
172
tn^lm — относительная масса его мышц. В сущности, поправоч-
ный коэффициент k приближенно выражает масштабный эф-
фект биоэнергетики теплокровных гидробионтов.
Анализируя рис. 87, замечаем, что обобщенные кривые
удельной отдачи мощности в одинаковой мере отвечают экс-
периментальным данным по различным видам спорта. Они со-
гласуются и с расчетными данными по прыжкам дельфинов и
лососей для импульсных максимальных значений удельной от-
дачи мощности кратковременной продолжительности, которая
превышает суточные значения мощности примерно в 20 раз.
При этом удельная отдача мощности хорошо тренированных
спортсменов меньше, чем у чемпионов и рекордсменов, при-
близительно на 30%, т. е. разница не столь значительная.
В соответствии с вышеизложенным график на рис. 87, а также
расчетные выражения (80) и (81) можно применять для опре-
деления отдачи мощности теплокровными гидробионтами на ме-
ханическую работу плавания.
§ 21. Квазистационарный биоэнергетический расчет
плавания дельфинов и анализ полученных результатов
Нестационарное движение гидробионтов, подробно рассмот-
ренное в предыдущих главах, характеризуется многими морфо-
логическими, кинематическими и гидродинамическими парамет-
рами, которые необходимо знать для достаточно точного инже-
нерного расчета. Однако эти параметры для многих видов
гидробионтов неизвестны, а их определение сопряжено с боль-
шими экспериментальными трудностями. Целесообразно наряду
с другими приближенными методами применить квазистацио-
нарный расчет динамики плавания гидробионтов и использо-
вать имеющееся биологические, в том числе физиологические
данные, формулы (80) и (81). В квазистационарном биоэнерге-
тическом расчете используются взаимосоответствующие средние
значения скорости плавания и удельной мощности активного
обмена, отвечающие одинаковой продолжительности пла-
вания.
В квазистационарном приближении регулярного поступатель-
ного плавания гидробионтов инерционные силы и моменты от-
сутствуют. Главный вектор осредненных внешних сил, дей-
ствующих на гидробионты, равен геометрической сумме двух
вертикальных противоположно направленных и практически
уравновешенных сил — веса G и плавучести Q — и двух гори-
зонтальных, равных и противоположно направленных сил —
тяги Р, развиваемой мышечной системой гидробионта, и гидро-
динамического сопротивления воды R. Последние определяем,
как при стационарном движении, по средней максимальной
скорости max Uo при данной продолжительности Т плавания.
В дальнейшем индекс max опущен.
173
Среднюю силу тяги Р гидробионта выразим через соответ-
ствующую по продолжительности отдачу мощности N'(T) его
мышечной системой
= (82)
67о Uo Uo
Здесь т) — пропульсивный гидродинамический коэффициент,
учитывающий потерю мощности при работе движительного ком-
плекса «тело — хвостовой плавник» в воде как вязкой жидко-
сти; q'K — удельная отдача мощности мышц в расчете на еди-
ницу веса тела при значениях qK и k по формулам (80) и (81);
О = рт£СзЬ3 — вес тела гидробионта в воздухе, где рт— плот-
ность тела; g=9,81 м/с2 — ускорение свободного падения; Сз—
коэффициент подобия масс при длине гидробионта L, считая от
кончика носа до развилки хвостового плавника.
Для доказательства подобия видов гидробионтов по общей
массе тела m = c3L3, где сз = const, приведем графики на рис. 88
и 89. Разброс точек достаточно большой, однако следует иметь
в виду, что данные получены по различным литературным ис-
точникам, а масса животного заданной длины есть переменная
величина, зависящая от пола, возраста, региона обитания, ус-
ловий питания по сезонам года и др. Тем не менее общая за-
кономерность подобия масс для разных видов гидробионтов
очевидна. Например, по рис. 88 для черноморской морской
свиньи сз= 14,6 кг/м3, для океанической косатки сз= 12,05 кг/м3,
а по рис. 89 для средиземноморских обыкновенных тунцов Сз =
= 15,75 кг/м3.
Грей [74] специально изучал характеристики формы тела
многих видов морских рыб различного систематического по-
ложения. Для 300 особей 39 видов, 25 семейств им были опре-
делены значения коэффициента k в зависимости S = km2/3, где
S — площадь поверхности, см2; Zt = const для каждого вида рыб;
m — масса тела, г. Оказалось, что значение коэффициента k
изменяется более чем в два раза: от 6=114-15 у удлиненных,
сжатых и плоских рыб (в том числе у обыкновенного угря
11,3 и для сабли-рыбы 12,7) до k = 6 4- 8 у быстроходных ском-
броидных рыб сравнительно небольшого удлинения: скумбрие-
вые 6,7—7,2, луфаревые 7,8, кефалевые 8,0 и т. д.
Силу гидродинамического сопротивления R гидробионта
считаем состоящими из трех основных составляющих: сопротив-
ления трения гладкой поверхности Fr, сопротивления формы
Рф и индуктивного сопротивления плавников РИц. Согласно за-
висимостям стационарной гидродинамики
i/2 и2
R = Fr + Рф + Рин = CxQp = С&Цр —— (83)
Здесь Сх — безразмерный коэффициент силы гидродинамиче-
174
• о 3)
А Б т,кг
а) Б)
Рис. 88. Подобие дельфинов по общей массе тела m = c3L3: а — морская сви-
нья (/); б — обыкновенный дельфин (2) и афалина (3); в — косатка (4).
А — черноморские дельфины; Б — океанические.
Рис. 89. Подобие крупных скоростных рыб по общей массе тела m = c3L3.
1 — средиземноморский обыкновенный тунец; 2 —меч-рыба; 3 — акула-мако; L — зооло-
гическая длина (от кончика носа до развилки хвостового плавника).
ского сопротивления, отнесенный к полной площади Q смочен-
ной поверхности гидробионта, включая площадь поверхности
плавников; р — плотность воды; Сг— безразмерный коэффициент
формы, так что Я = Сг£2.
Как видно, используется подобие видов гидробионтов по
смоченной поверхности тела. Оно показано для двух видов
черноморских дельфинов на рис. 90. Для морской свиньи Сг =
= 0,443, для афалины С2 = 0,412.
В дальнейшем коэффициент Сх гидробионта выражаем, как
обычно, для хорошо обтекаемых твердых тел, через безразмер-
ный коэффициент Cf трения гладкой плоской пластины, обте-
каемой в продольном направлении,
Сх = пС/. (84)
Подставляя выражение (84) в (83) и приравнивая послед-
нее выражению (82), получим С/ (или nCf = Cx)
q ^'П^?мРт?сэ^' 2r|fegMG (85)
ncflUo npQt/g
175
Рис. 90. Подобие черно-
морских дельфинов по
смоченной поверхности
тела, Q = c2L2.
1 — азовка (морская сви-
нья); 2 — афалина.
Практически можно считать, что
рт = р. Поэтому в формуле (85) эти ве-
личины сокращаются. Величина Су опре-
деляется непосредственно по известной
скорости, для чего нет необходимости
знать режим течения в пограничном слое
гидробионта. Напротив, если известен по-
следний, то пользуясь стационарными
гидродинамическими зависимостями, мо-
жно рассчитать среднюю максимальную
скорость плавания гидробионта опреде-
ленной продолжительности, отвечающей
принятому значению удельной отдачи
мощности.
В соответствии с теорией погранич-
ного слоя имеем:
при ламинарном режиме (закон
Блазиуса)
г - *-328 _ 1.328V0-5
f~ VRT “ У°Л°'5 '
при турбулентном режиме (закон
Прандтля), когда числа Рейнольдса не
очень высокие,
г 0,074 _ O,O74v0,2
5 ~ у/0,2г 0,2
у Re ио ь
Здесь v — кинематический коэффициент вязкости воды, v = f(0,
он зависит от температуры t воды, например, при t= 15°С,
v=l,14 м2/с. Области применения по числу Рейнольдса этих за-
висимостей для гидробионтов неизвестны. Однозначность реше-
ния выявляется из сравнения расчетных скоростей с наблюдае-
мыми в природе значениями. Если последние отсутствуют, рас-
чет следует вести в двух предположениях — при ламинарном и
турбулентном режимах. Подставим указанные зависимости
в формулу (85). Тогда в случае ламинарного обтекания
,,5/2 _ 1 .бт^мРт^Сэ^-1’5 _ 1,5r]kquGL°'5
° npC2v0’5 npQv0,5
В случае турбулентного обтекания
£/14/5 = 27t]fegMpTgc3L1,2 27r\kqMGL°'2
npc2v0’2 npQv0,2
(86)
(87)
Приведенные выше зависимости ясно показывают, от каких
параметров гидробионтов и физических свойств ‘воды и в какой
мере зависят коэффициенты сопротивления плаванию Сх или
С; и средняя максимальная скорость плавания Uo в данном ре-
176
жиме при продолжительности плавания Т. Можно анализиро-
вать влияние на гидродинамические характеристики гидробион-
тов размеров тела — длины L, формы тела — коэффициентов
полноты Сз и с2, скорости плавания Uo в соответствии с удель-
ной отдачей мощности kqM различной продолжительности и т. п.
Важнейшей характеристикой плавания гидробионтов явля-
ется значение коэффициентов гидродинамического сопротивле-
ния Сх, Cf = f(ReL). На рис. 91 показаны результаты наших
биоэнергетических расчетов для пяти наиболее характерных
видов дельфинов средних размеров: черноморских—мор-
ской свиньи, обыкновенного (белобочки), афалины — и океани-
ческих— белухи и косатки. Расчеты сопоставлены с экспери-
ментальными исследованиями соответствующих моделей Аэ, м и
D и натурным экспериментом и расчетом по гидродинамической
теории Г. В. Логвиновича для афалины 2. Среди указанных
видов дельфинов есть мелкие (морская свинья, белобочка) и
Рис. 91. Значения коэффициентов гидродинамического сопро-
тивления дельфинов и соответствующих моделей (С/, Ся).
Биоэнергетический расчет (Ср: 1 — обыкновенный дельфин
1=1,6 м; 2 — афалина, 1 = 2,3 м; 3 —косатка, 1 = 6 м; 4 — морская сви-
нья, 1 = 1,4 м;’ 5 —белуха, 1=4,5 м. Натурный эксперимент и
расчет по Г. В. Логвиновичу Сх (261: афалины 1=2,35—2,65 м;
ЛЭр—почти равномерное, а — ускоренное (активное), б — замедленное
(пассивное) плавание. Буксировка в опытовых бассейнах;
Дэ>м—жесткая модель афалины, 1=2,01 м; МСЭ т—тушка морской сви-
ньи, 1=1 м; D — всплывающая жесткая модель «Дельфин* ламинаризо-
ванной формы, 1=1,6 м [15(.
177
крупные (белуха, косатка), скоростные (белобочка, афалина,
косатка) и медленно плавающие (белуха). Для расчета по
формуле (85) использованы следующие данные по дельфинам:
размеры по табл. 3, скорости — см. график на рис. 31 и фор-
мулу (8), удельную отдачу мощности — см. рис. 87 и формулы
(80) и (81), массу тела и относительную массу мышц — см.
рис. 84, 88, 90. Значения п и т] приняты обычные: п= 1,20;
г, = 0,804-0,90 для разных видов.
Анализируя приведенные на рис. 91 результаты, можно сде-
лать следующие важные выводы. В целом биоэнергетический
расчет хорошо согласуется со всеми экспериментальными дан-
ными и экспериментально-теоретическими результатами по
Г. В. Логвиновичу. Последние получены для черноморских афа-
лин средней длиной £ = 2,5 м, на малых скоростях плавания
t7o = 2-=-5 м/с, т. е. заведомо при неполной удельной отдаче
мощности животными. Наш биоэнергетический расчет по афа-
лине сделан для скоростей £7о = 6-4-12 м/с определенной продол-
жительности в интервале 7' = 3=105 с, при полной отдаче мощ-
ности и является как бы продолжением расчета по Г. В. Логви-
новичу. Кривые значения С/(Re) для всех видов дельфинов
расположены соответственно их биологическим характеристи-
кам, определяющим и режим обтекания тела. Наименьшим
гидродинамическим сопротивлением обладает самый скорост-
ной малый дельфин-белобочка, наибольшим (при тех же чис-
лах Рейнольдса)—крупный медленно плавающий в холодных
водах дельфин-белуха. Кроме белобочки, эффективно плавают
афалины и косатки. Режим их пограничного слоя переходный,
а коэффициент С/ ниже, чем для гладкой пластины при турбу-
лентном режиме Т, почти в шесть раз.
В сравнении с буксируемой жесткой моделью афалины Аэ. м
и всплывающим ламинаризованным телом D гидродинамическое
сопротивление в плавании живых дельфинов указанных видов
снижается за счет присущего им способа, который надлежит
раскрыть. Снижение сопротивления скоростных дельфинов про-
исходит при относительно высоких числах Рейнольдса Re =
= 5 1064-7 • 107. Характерен экстремальный вид кривых сопро-
тивления.
С помощью справочных данных по скоростям плавания
мировых рекордсменов на различных дистанциях и графика по
коэффициентам сопротивления спортсменов (рис. 70) были
определены наивысшие гидродинамические кпд спортивного
плавания вольным стилем и способом «дельфин» с моноластом
на поверхности (рис. 92). Вторые значения выше первых в пол-
тора раза, т. е. весьма значительно. Моноласт является самым
эффективным мускульным волновым движителем человека.
В заключение приведем график (рис. 93) зависимости коэф-
фициента мощности активного обмена C№ = 2Wa/Sp£/3o от чи-
сла Рейнольдса для быстро плавающих, однако небольших
178
С,
Рис. 92. Гидродинамический кпд ре-
кордного плавания спортсменов при
средней максимальной скорости на
дистанции, T]r(t/om). Обращает на
себя внимание большое различие
в значениях кпд при спортивном пла-
вании различными стилями.
1 — плавание вольным стилем; 2 — плава-
ние способом «дельфин» с .моноластом на
поверхности.
Рис. 93. Масштабный эффект по
числу Рейнольдса безразмерного ко-
эффициента мощности активного об-
мена при различной массе рыб раз-
ных видов, Cn (т, Rcl).
Um — максимальная скорость каждой
нерки, плавающей в течение часа. 1—5 —
тихоокеанская нерка, L=0,077—0,539 м,
/п = 0,0034—1,432 кг, /=15°С при Т=60 мин
(см. рис. 73); б — черноморская кефаль,
1 = 0,102—0.134 м, /пэ=0,013—0,0225 кг. / =
= 20 °C (плавание в течение 30 мин) ГЗ];
7 — байкальские хариус и омуль, 1 =
-0.29—0.32 м. гп=0.30—0.39 кг, Г = 10°С
[351.
рыб: лосося-нерки, кефали, хариуса и омуля. График составлен
по известным данным экспериментальных определений [3, 35,
69]. Общий характер серии кривых для рыб разных размеров
таков: вначале, до l/2Um, видны участки параллельных пря-
мых, т. е. степенные зависимости для CNa, а затем до l,0t/m —
криволинейные участки-экспоненты. Тем самым определяется
сложный масштабный эффект, анализ которого усложняется
еще экспериментальными данными других авторов, получен-
ными при иной методике исследований (прямые 6 и 7 на всем
протяжении).
Поскольку CNa^Cx/r\, а общий кпд т]~0,20, график на
рис. 93 можно сравнить с верхней частью рис. 52 в интервале
Re= IO4-? 105. Теоретические значения, определенные по гидро-
динамической теории удлиненного тела, хорошо соответствуют
экспериментальным значениям Сдга(Ее) в биоэнергетике. Тем
самым все приведенные выше выводы согласуются и доста-
точно обоснованы.
179
ГЛАВА V
АВТОРЕГУЛЯЦИЯ ДЕМПФИРОВАНИЯ
КОЖНОГО ПОКРОВА У ДЕЛЬФИНОВ
И МОДЕЛИРОВАНИЕ СНИЖЕНИЯ
ГИДРОДИНАМИЧЕСКОГО СОПРОТИВЛЕНИЯ
§ 22. Свойство кожи китообразных активно регулировать
гидродинамическое сопротивление плаванию
В предыдущем изложении среди множества крупных гидро-
бионтов были выделены скоростные китообразные, которые от-
личаются хорошообтекаемой формой тела, эффективным волно-
вым плаванием на больших скоростях и относительно низким
гидродинамическим сопротивлением. Последнее их свойство
давно интересовало многих исследователей, однако обычно оно
рассматривалось в сравнительном аспекте и без достаточных
экспериментальных оснований. Кажущееся легким плавание
дельфинов обратило на себя внимание еще в конце прошлого
века, но наибольшее значение для гидробионики имеют иссле-
дования, выполненные в последние десятилетия и даже в по-
следние годы.
Повышенный интерес к проблеме отчасти возник после пу-
бликации известного «парадокса» английского зоолога Грея
[74]. Утверждалось, что гидродинамическое сопротивление пла-
ванию дельфина раз в семь меньше буксировочного сопротив-
ления твердого тела тех же размеров и формы и при той же
скорости, т. е. при одинаковом числе Рейнольдса. Убедительно
объяснить, разрешить этот «парадокс» не удавалось до послед-
него времени, хотя такие попытки предпринимались неодно-
кратно.
Немецкий аэрогидродинамик М. О. Крамер в 1960—1962 гг.
выступил в США со статьями, в том числе со статьей под гром-
ким названием «Секрет дельфина» — см. [81]. Он высказал ги-
потезу, что наружные слои кожного покрова дельфина об-
ладают свойством стабилизации ламинарного пограничного
слоя путем распределенного демпфирования возмущений на по-
верхности тела животного. Крамер не исследовал детально
строение и свойства кожи дельфинов и по существу непра-
вильно объяснил демпфирующую способность ее живых тканей.
Однако изобретенные им конструкции резиновых покрытий,
будучи наклеены на твердое удлиненное тело, снижали в экс-
периментах сопротивление трения тела до 60—70 %, при числе
Рейнольдса Re = 2-107 [81]. Насколько известно, эти демпфи-
рующие покрытия не получили распространения в инженерной
практике, а эксперименты никому не удалось повторить. Тем не
180
менее изобретения Крамера способствовали развитию интереса
к исследованию уникальных особенностей кожи дельфина.
В отличие от всех предшествовавших эпизодических и раз-
розненных исследований кожного покрова дельфинов ниже
кратко изложены гидробионические исследования, основные ре-
зультаты которых приведены в работах [8, 29, 60]; там же ука-
заны ссылки на литературные источники. Эти исследования
включали эколого-морфологическое изучение различных видов
китообразных, их биоэнергетических возможностей, детальный
анализ структурных и гистологических особенностей кожи
дельфинов, экспериментальное определение ее упругодемпфи-
рующих механических характеристик, опытное изучение погра-
ничного слоя активно плавающих дельфинов, техническое мо-
делирование гидродинамической функции кожи. Исследования
проводились в течение сорока лет, наиболее интенсивно — по-
следние 20 лет и привели к научному открытию [18].
Впервые установлено свойство кожи китообразных активно
регулировать гидродинамическое сопротивление плаванию по-
средством управления локальным взаимодействием кожного по-
крова с обтекающим потоком. Это свойство обусловлено рецеп-
торной чувствительностью кожи и вазомоторной подвижностью
кровеносной системы, т. е. способностью к быстрому изменению
циркуляции крови в кожном покрове в соответствии с режимом
плавания. Такое регулирование автоматически вызывает со-
ответствующие изменения упругодемпфирующих характеристик
различных слоев кожи, каждый из которых выполняет свою
специфическую функцию.
§ 23. Структурные и гистологические особенности
кожного покрова китообразных
Структурно-морфологические и гистологическое изучение
кожного покрова нескольких видов дельфинов и китов система-
тически было выполнено по материалам дальневосточной экс-
педиции АН СССР в Охотском море еще в 1951 г. [61]. Изу-
чали топографию кожного покрова, функциональное назначе-
ние отдельных элементов, черты сходства и различия в кожном
покрове дельфинов и китов (а также ластоногих). В 60-е годы
подобные исследования были продолжены и дополнены на Чер-
ном море, на базе Карадагской биологической станции Инсти-
тута биологии южных морей им. А. А. Ковалевского АН УССР.
Обнаружены уникальные особенности кожи китообразных как
высоко специализированных вторичноводных животных, отме-
ченные у дельфинов и крупных китов. Далее остановимся,
главным образом, на рассмотрении кожи дельфинов которую
детально изучали в институтах АН УССР.
Строение кожи дельфина (рис. 94) достаточно сложное. Его
кожа состоит из нескольких более или менее резко выражен-
181
Рис. 94. Схема строения кожи черноморского обыкновенного дельфина,
составленная по результатам детального препарирования [64]: а — про-
дольный вид, б — поперечный разрез.
/ — эпидермис; 2 —дерма; 3 — кожная мышца; 4 — подкожная жировая клетчатка;
5 —первая и вторая соединительнотканные мембраны; 6 — скелетная мускулатура.
ных слоев и тонких соединительнотканных мембран, которые
являются общим ее каркасом. В коже выделяются наружный
тонкий слой эпидермиса и мощная плотная и прочная дерма
(собственно кожа), кожная мышца и подкожная жировая клет-
чатка, которая лежит на скелетной мускулатуре. Каждый из
слоев кожи имеет в различных частях тела дельфина различ-
ные толщину, строение, плотность, упругость, вязкость и другие
механические свойства.
Специфическую структуру представляет собой кожная
мышца дельфина. Она имеет сложную конфигурацию и состоит
из двух частей: одна из них расположена на боковых сторонах
тела дельфина, другая — на брюшной стороне. Волокна тканей
кожной мышцы расположены под углом к продольной оси
тела. Это позволяет дельфину при изгибно-колебательных дви-
жениях тела изменять натяжение кожного покрова. Наличие
у дельфина наряду с кожной мышцей нескольких соедини-
тельнотканных мембран указывает на возможность регуляции
подвижки плотных слоев дермы из коллагеновых и эластино-
вых волокон на более рыхлой, податливой ткани подкожной
жировой клетчатки при плавании дельфина на различных ре-
жимах. Кожа дельфина представляет функциональную систему,
в которой воедино связаны все слои кожи.
Наибольший интерес с позиций гидробионики представляют
существенные детали верхних слоев кожи китообразных. Прин-
ципиальная схема В. Е. Соколова, показывающая строение
кожного покрова китообразных (рис. 95), давно получила ши-
рокое распространение и признание, стала классической.
Поверхность тела скоростных китообразных не имеет волос
и гидравлически гладкая. Возможно, вследствие изгибно-коле-
бательных движений тела на вогнутой стороне последовательно
возникают поперечные волнистые микроскладки, однако они
182
быстро продвигаются и раз-
глаживаются. Высота их
у живых дельфинов, веро-
ятно, не превышает 0,1 мм,
а на больших скоростях
плавания — и того меньше,
поскольку изгибаемость не-
большая. Поэтому поверх-
ностные микроскладки не
выступают за пределы ла-
минарного подслоя погра-
ничного слоя. Существенно,
что под наружным сплош-
ным тонким резиноподоб-
ным слоем эпидермиса рас-
положен ячеистый сосочко-
вый слой, в углубления
(ячеи эпидермиса) которо-
Рис. 95. Типичное строение кожного
покрова китообразных [61].
а — дермальные сосочки; б — продольные эпи-
дермальные перегородки; в — подсосочковый
слой дермы; г—подкожная жировая клет-
чатка.
го снизу входят тонкие
дермальные сосочки из
плотно переплетающихся
коллагеновых и эластино-
вых волокон. Сосочковый
слой у китообразных ги-
пертрофически развит, сравнительно с подобными слоями у дру-
гих млекопитающих и у человека, что показательно для упруго-
демпфирующих свойств кожи.
Хотя у дельфинов дермальные сосочки имеют микроскопиче-
ские размеры (порядка 1 мм в высоту и 0,1 мм в поперечнике),
число их велико: до 15—20 шт. на каждом квадратном мил-
лиметре поверхности кожи, или около 30- 10® шт. у двухметро-
вого дельфина. Важно, что каждый дермальный сосочек на-
сыщен артерио-венозными капиллярами и анастомозами (см.
рис. 95, слева) и содержит различные нервные окончания [1].
Обильная васкуляризация и иннервация дермальных сосочков
позволяет в широких пределах изменять кровенаполнение их
на различных режимах плавания и тем самым оказывает влия-
ние на упругодемпфирующие свойства кожи. Ниже расположен-
ный подсосочковый слой дермы содержит большое количество
кровеносных сосудов и нервных стволов, ответвления от кото-
рых проходят в дермальные сосочки.
Важная особенность сосочкового слоя кожи китообразных
состоит в том, что дермальные сосочки размещены не беспо-
рядочно, а упорядоченно на более плотных продольных эпидер-
мальных перегородках — валиках. Расположение последних
также имеет свою гидродинамическую функцию.
Гистологические срезы наружного покрова кожи весьма ак-
тивных дельфинов, плавающих в экваториальных водах, рода
183
Рис. 96. Микрофотографии ги-
стологических срезов наруж-
ного покрова кожи теплолюби-
вого дельфина рода стенслла
перед спинным плавником: а — поперечный срез; б — тангенциальный срез
у основания дермальных сосочков. Видна большая насыщенность всех дер-
мальных сосочков мельчайшими кровеносными сосудами, включая капилляр-
ные.
стенелла, показаны на микрофотографии (рис. 96). И в про-
дольном и в поперечном срезах хорошо видны упорядоченное
расположение дермальных сосочков, обильная васкуляризация
их и взаимосвязь между собой и с подсосочковым слоем. Ха-
рактерна овальная форма тангенциального среза дермальных
сосочков. Это может свидетельствовать о наклонном положении
цилиндрических сосочков в продольной плоскости. Обращает
на себя внимание относительно большая суммарная площадь
тангенциального среза всех дермальных сосочков по сравнению
с общей площадью среза. Отсюда ясно, что регуляция крове-
наполнения дермальных сосочков существенно влияет на уп-
ругодемпфирующие характеристики всего сосочкового слоя и
кожного покрова дельфина.
Дермальные сосочки не только расположены продольными
Рис. 97. Расположение участков кож-
ного покрова белобочки с различным
продольным наклоном дермальных со-
сочков.
J — а—10—25°; 2 — а=25—40°; 3 - а = 40—55°;
4 - а=55—80°.
рядами, но на различных
участках тела дельфина
имеют разные углы нак-
лона к поверхности эпидер-
мальных перегородок-ва-
ликов (рис. 97 и 98). У дель-
фина-белобочки эти углы
изменяются в широком ин-
тервале значений а=104-
4-80°. Наименьший наклон
дермальных сосочков (а =
= 104-28°) отмечается в пе-
редней части, сбоку и в хво-
стовой части тела, т. е. в ме-
184
Рис. 98. Продольные разрезы дермальных сосочков бе-
лобочки.
стах наибольшего гидродинамического давления и его градиен-
тов. Наибольший наклон, а = 554-80° наблюдается, в частности,
за спинным и грудными плавниками, т. е. в гидродинамической
тени. В каждом поперечном сечении тела дельфина с наимень-
шими углами наклона дермальных сосочков их будет по не-
сколько штук, причем одни сосочки будут располагаться над
другими, поэтому регулирующее их значение возрастает.
Отмеченная закономерность расположения указанных уча-
стков уже свидетельствует о ее значении для изменений уп-
ругодемпфирующих характеристик кожного покрова.
Детальное препарирование особей всех трех видов черномор-
ских дельфинов— белобочки, афалины и морской свиньи — пока-
зало сходство и различия в расположении их дермальных вали-
ков (рис. 99). Общим является то, что дермальные валики на
большей части поверхности тела дельфинов направлены в основ-
ном продольно, вдоль тела, т. е. в направлении линий тока при
среднем выровненном положении тела. Вместе с тем на отдель-
ных участках поверхности тела — в головной и концевой части,
на плавниках — направления эпидермальных валиков отлича-
ется от продольного, однако в соответствии с местными усло-
виями обтекания частей тела, при набольших изгибно-колеба-
тельных амплитудах. Различие в расположении эпидермальных
Рис. 99. Направление дермальных валиков в коже тела и плавниках дель-
финов: а — афалина; б—морская свинья.
185.
валиков у различных дельфинов состоит в разной попереч-
ной густоте их, например, у афалины и морской свиньи, что
вполне объяснимо известным различием размеров их тела и ско-
ростей плавания. Подобные факты объяснимы также с позиций
гидродинамической теории устойчивости движения. На первый
взгляд, это может показаться неоправданным, поскольку дер-
мальные валики расположены не на смоченной поверхности
тела, а в самой дерме. Однако расстояние валиков от наружной
поверхности тела всего порядка 1 мм, и потому различие в плот-
ности тканей валиков и в зазорах между ними несомненно ска-
зывается на характеристиках кожного покрова.
Анализ структурных и гистологических особенностей кож-
ного покрова китообразных приводит к выводу о его гидродина-
мической функции. В частности, у дельфинов кончики васкуля-
ризованных и иннервированных дермальных сосочков находятся
на глубине всего 0,5 мм от гладкой чувствительной внешней по-
верхности эпидермиса и в состоянии воспринимать пульсации
в пограничном слое тела, а также реагировать на них. Это сле-
дует также из применения теоретического принципа эмбриоге-
неза в биологии.
Воспользуемся результатами исследований развития кож-
ного покрова у многочисленных эмбрионов разных возрастов
скоростного кита-финвала. При общей продолжительности эм-
брионального развития зародышей китообразных около 11 мес.
дермальные валики закладываются у эмбриона на шестом ме-
сяце, а дермальные сосочки на них только на восьмом месяце
(рис. 100). В соответствии с принципом эмбриогенеза это ука-
зывает на то, что названные признаки являются специальными
(не общими). Ввиду весьма позднего эмбрионального появления
дермальных валиков и сосочков их функция связана с после-
дующими новыми условиями существования новорожденного
китообразного, т. е. с активным плаванием его в воде сразу
после рождения.
Структурные и гистологические особенности кожного по-
крова китообразных, если сравнить его, например, с кожным
покровом крупных скоростных рыб, прослеживаются и на по-
следующих, более глубоких слоях кожи (рис. 101). Общая тол-
щина кожи, выраженная в долях длины L тела животного, со-
ставляет в средней части обыкновенного дельфина около
8,5-10-3 L, а у наиболее крупных скоростных рыб (0,65ч-0,95) X
Х10-3 L, т. е. у дельфина кожа толще на порядок величины, что
отмечалось нами и выше (см. рис. 21). У дельфина резко выде-
ляются своей толщиной как по абсолютной величине (цифры на
рис. 101 слева от разрезов кожи), так и по отношению к общей
местной толщине кожи в процентах (цифры справа от разре-
зов) слои сплошного эпидермиса и расчлененного, но упорядо-
ченного сосочкового слоя кожи. Этот второй слой имеется только
в коже дельфина; у скоростных рыб его нет. Не менее мощный
186
1,65 м 51/z
3,30м 8 1,85м 9
Рис. 100. Схема развития верх-
них слоев кожи у эмбрионов
кита-финвала.
е — эпидермис, d — дермальные валики,
р — дермальные сосочки; цифры:
слева — длина тела, м; справа — воз-
раст эмбриона, мес.
1Ш ПППТПТП Г/И
а б 6
г д
Рис. 101. Послойная структура ги-
стологических срезов кожи крупных
скоростных рыб и дельфина.
1 — желтоперый тунец; 2 — меч-рыбач;
3 — акула-мако; 4 — дельфин. Слои
кожи: « — сплошной эпидермис; б —
ячеистый сосочковый; в — соединительно-
тканный; г--подкожная жировая клет-
чатка; д — слой коллагеновых пучков и
волокон.
и соединительный слой кожи дельфина, особенно мощный слой
подкожной жировой клетчатки. Указанные различия вполне по-
нятны. Снижение гидродинамического сопротивления у скорост-
ных рыб достигается благодаря использованию ими биологиче-
ской полимерной рыбьей слизи, инъектируемой в пограничный
слой обтекаемого потока, а у дельфинов — с помощью демпфи-
рующего кожного покрова, без слизистых выделений. Этому
способствует также то, что, как известно, у дельфинов давле-
ние крови- в сосудах на порядок величины выше, чем у рыб, и
потому возможно активное регулирование кожного покрова по-
средством кровеносной системы.
187
§ 24. Гидродинамическая функция
и механическая модель
демпфирующего свойства кожи дельфинов
Гидродинамическая функция кожного покрова дельфинов
установлена в результате разносторонних комплексных исследо-
ваний, из которых следует выделить также изучение обильного
кровоснабжения и иннервации различных слоев кожи. Механи-
ческие характеристики функционально-специфического кожного
покрова регулируются посредством сосудистой системы вместе
с изменением метаболизма животного на различных режимах
нестационарного плавания, при работе изгибно-колебательного
движительного комплекса. Для живых дельфинов характерен
большой диапазон вазомоторной подвижности — расширения и
сужения кровеносных сосудов, что было замечено еще в 1947 г.
Об этом пишет А. Г. Томилин (ДАН СССР. Т. 5. 1947. № 2.
С. 221—223). Специальные наблюдения провели на 28 подопыт-
ных животных на Черном море для изучения терморегуляции
поверхности тела. В зависимости от физиологического состоя-
ния дельфина часто отмечали усиленную гиперемию, времен-
ное порозовение белых участков кожи при нагреве солнцем,
тогда как в нормальных условиях кожа не имела розовой ок-
раски. .
Чтобы показать непосредственную связь высокой вазомотор-
ной подвижности кожного покрова дельфинов со скоростью
плавания, проведены наблюдения за афалиной, обученной в не-
неволе плавать по команде на различных скоростях. В 44 за-
плывах отмечали скорость плавания животного и изменения ок-
раски его брюха. Как показал анализ наблюдений, у одной и
той же афалины наступало заметное порозовение кожи при
возбужденном плавании с увеличением скорости в среднем на
20%. Характерно также, что регистрируемые средние значения
скорости плавания афалины повышались, происходило посте-
пенное «врабатывание» животного. Все это указывает на регу-
лирование скорости плавания дельфина с участием кожного
покрова, контактирующего с пограничным слоем обтекающего
потока воды.
В каждом дермальном сосочке кожного покрова просвет кро-
веносных сосудов ^изменяется в соответствии с активным мета,-
болизмом животного на данной скорости плавания. Это под-
тверждается остаточной формой дермальных сосочков в виде
расширенной капсулы, что часто наблюдается при препарирова-
нии кожного покрова трупа дельфина (рис. 102).
Много данных по строению, топографии, интраорганной вас-
куляризации и иннервации кожного покрова черноморских дель-
финов содержится в монографии [1]. Утверждается интенсивная
иннервация всех компонентов кожного покрова дельфинов, на-
личие афферентных и эфферентных связей, осуществляющих
188
Рис. 102. Капсулообраз-
ная форма цилиндриче-
ских дермальных сосоч-
ков при повышенном кро-
вотоке как проявление
вазомоторной подвижно-
сти кровеносной системы
кожного покрова дель-
фина (микрофотографии
гистологических срезов).
Обычная форма (в по-
кое) дермальных сосоч-
ков без концевых капсул.
передачу прямой и обратной информации между нервными ап-
паратами кожи и центральной нервной системой. Плотность
нервных окончаний в коже дельфина даже выше, чем у чело-
века. В эпидермисе кожи дельфина гистологически обнаружены
разнообразные рецепторы с высокой чувствительностью, в том
числе холодовые (инкапсулированные колбочки Краузе), теп-
ловые (тельца Руффини), механорецепторы тактильных ощу-
щений: прикосновения (клетки Меркеля), давления (тельца
Фатра — Пачини) и др.
Исключительный интерес представляет тот факт, что у дель-
финов и китов сосудистая система секционированная (рис. 103).
Кровоснабжение кожи, а потому регулирование сосудистой си-
стемой упругодемпфирующих характеристик кожи китообразных
осуществляется локально, посредством подводящих сосудов с ко-
нечными разветвлениями на каждом участке кожи вдоль тела
длиной, определяемой размером одного позвонка скелета. По-
скольку при активном нестационарном плавании происходят
волнообразные изгибные колебания тела дельфина в вертикаль-
ной плоскости, можно полагать, что система авторегуляции
демпфирования кожи включается последовательно, по секциям,
со смещением во времени по фазе. Тем самым в каждой секции
возможно чередование фаз наибольшей активности (работы) и
относительного отдыха (восстановления) системы авторегуля-
ции, локальное действие которой оптимизируется. Система авто-
регуляции весьма совершенная.
Значение локализации, секционирования сосудистой системы
авторегуляции демпфирования кожи китообразных подтвержда-
ется взаимосвязью между средней длиной секции L/п и относи-
тельной максимальной скоростью плавания Um]L, а также чис-
лом Рейнольдса Re для ряда дельфинов и китов (рис. 104).
У косатки — самого крупного и скоростного дельфина длиной
А = 6,0 м — средняя длина вдоль тела отдельной секции авто-
189
Рис. 103. Секционированная сосудистая система авторегуляции демп-
фирования кожи у дельфинов.
Рентгенограммы артерий эмбриона по свинцовой инъекции сбоку (вверху) и
снизу.
регуляции демпфирования кожи составляет, как правило.
Л/п = 0,12 м.
Прежде чем перейти к механическому моделированию кожи
дельфина в соответствии с теорией подобия, необходимо убе-
диться в том, что существует геометрическое подобие для важ-
нейших элементов кожи. С этой целью приведем график зависи-
мости толщины эпидермиса от длины тела китообразных — от
малых дельфинов и до крупных китов (рис. 105). Простейшая
линейная зависимость представлена в широком интервале зна-
чений p(L), и потому можно говорить о нормировании тол-
щины эпидермиса как важнейшего демпфирующего слоя кожи
для относительно быстро плавающих китообразных. Напротив,
норме не соответствуют значения толщины эпидермиса для
дельфина-белухи и кита-кашалота (толщина эпидермиса у них
больше соответственно в 4 и 2 раза). Однако это вполне объяс-
нимо образом жизни данных животных, при котором не требу-
ются высокие скорости плавания, но предъявляются повышен-
ные требования к прочности кожи. Белуха обитает в холодных
водах, часто среди льдов, а кашалот, как отмечалось выше,—
глубокий ныряльщик. Напомним, что по расчетному энергети-
190
ческому графику (рис. 91), в отличие от скоростных дельфи-
нов — белобочки, афалины, косатки, у которых пограничный
слой на значительной части поверхности тела должен быть ла-
минарным, белуха имеет целиком турбулентный пограничный
слой.
Для изучения структурных характеристик, в том числе тол-
щины отдельных слоев кожи дельфинов и возможных изменений
ее по длине тела животного, нами на основании биологических
экспериментальных данных выполнено нормирование послойной
модели демпфирующего кожного покрова дельфинов. Использо-
ваны результаты препарирования кожи черноморских дельфи-
нов: морской свиньи длиной 1,27 м и белобочки длиной 1,45 м
(рис. 106).
Как показывают исследования кожи китообразных, кожный
покров дельфинов по строению, свойствам, назначению есть
сложное живое, активное многослойное демпфирующее покры-
тие из органических полимерных материалов, свойства которых
благоприятны для регулирЪвания. Полимерам присущи: боль-
шие деформации при относительно низких напряжениях, взаимо-
связь напряжений с продолжительностью действия сил и ско-
ростью деформации, существенная зависимость механических
свойств от температуры, анизотропия свойств материала. Меха-
нические свойства полимеров определяются, в частности, удель-
ной массой, плотностью мате-
риала р, кг/м3, модулем упру-
гости Е = <з1г (ст— напряже-
ние, Н/м2, е — относительная
деформация), вязкостью ц =
Рис. 104. Зависимость продольной длины секции авторегуляции
демпфирования кожи китообразных от относительной максималь-
ной скорости плавания и числа Рейнольдса, L/n = fl,2(Um!L, Rer).
Длина секции равна L/п, где L — длина тела, п — число позвонков живот-
ного; / — скоростные, // — нескоростные дельфины и киты; /-дельфин-бе-
лобочка; 2 — полосатый продельфин; 3—морская свинья; 4 — бесперая мор-
ская свинья; 5 — афалина; 6 —белуха; 7 —гринда; 8 — косатка; 9 — серый
кит; 10 — кашалот; // — синий кит; /2 — финвал; 13 — сейвал; 14 — горбатый.
Рис. 105. Зависимость толщины эпидермиса кожи от длины тела
у дельфинов и китов, p = f(L).
МП — малый полосатик; СП — северный плавун, /-дельфин-белобочка; 2 —
полосатый продельфин; 3 — морская свинья; 4 — тихоокеанский короткого-
ловый дельфин; 5 — афалина; 6 — белуха; 7 — кашалот; 8 — финвал (см.
«табл. 3).
191
Рис. 106. Нормирование послойной модели кожи черноморских дельфинов —
морской свиньи (а) и белобочки (б) — на спине, сбоку и на груди.
п — число дермальных сосочков на площади 1 мм2 поверхности тела дельфина: т —
число дермальных валиков на ортогональном отрезке длиной 1 мм.
р — толщина слоя эпидермиса; h — высота дермальных сосочков; g — толщина подсо-
сочкового слоя дермы; г —толщина слоя подкожной жировой клетчатки; s — общая
толщина кожи.
= Е/ш, Н-см2 (ш — предельная частота колебаний), временем
(периодом) релаксации х = х\1Е. Для установления основных
свойств полимеров предложены наглядные механические мо-
дели с параллельным и последовательным соединением расчет-
ных упругих и вязких элементов; каждой модели отвечает свое
дифференциальное уравнение движения.
Гидродинамическая функция снижения сопротивления в пла-
вании дельфинов может осуществляться путем демпфирования
в пограничном слое возмущений типа волн Толмина — Шлих-
тинга с помощью верхних расчлененных слоев кожного покрова.
Для гашения трехмерных возмущений имеет значение различная
плотность отдельных элементов эпидермиса. При плавании
дельфина с высокой скоростью, когда на теле возникает значи-
тельный перепад давления, для сохранения хорошо обтекаемой
формы гибкого тела и создания оптимальных вязкоупругих
192
демпфирующих свойств податливой кожи существенное значе-
ние приобретают также более глубокие слои активно работаю-
щих тканей. На основании вышеизложенного и согласно теории
механического подобия, проведена аналогия между вязкоупру-
гими свойствами хорошо васкуляризованных тканей кожи и со-
ответствующими полимерными материалами и принята следую-
щая механическая модель кожи дельфина, находящейся под
воздействием внешнего возмущающего поля давления воды.
На рис. 107 соответственно показаны и интерпретированы
основные моделируемые слои кожи дельфина. Тонкий сплошной
слой 1 эпидермиса есть упругая мембрана массой Л4, и натя-
жением Ть Величины М\ и Т[ — переменные со временем, что
обусловлено изгибно-колебательными движениями тела при пла-
вании животного. Расчлененный сосочковый слой 2 эпидермиса
есть вязкоупругое тело с параллельным соединением полимер-
ных элементов с модулем упругости и коэффициентом вяз-
кости т]. Переменное значение модуля упругости Е\ определя-
ется различным наклоном к своему основанию и деформацией
дермальных сосочков. Переменное демпфирование слоя за счет
вязкости вызывается перетеканием крови и лимфы из капилля-
ров слоя 2 в сосуды слоя 3.
Подсосочковый волокнистый слой 3 дермы с горизонтально
расположенными коллагеновыми и эластиновыми волокнами и
кровеносными сосудами
представлен в виде после-
довательно соединенных
полимерных элементов
мембраны массой М2, натя-
жением Т2 и модулем уп-
ругости Е2. Переменная
масса М2 зависит от ампли-
туды возмущающего дви-
жения и действующего
внешнего давления. Вели-
чина Т2>Т\, а демпфирова-
нием слоя 3 пренебрегаем,
учитывая строение тканей и
кровеносной системы. Тол-
стый и мощный сетчатый
слой 4 дермы является уп-
ругим основанием («под-
ложкой») с модулем упру-
гости Ез. Показанные на
модели шарниры обуслов-
лены тем, что ввиду мало-
сти внешнего возмущаю-
щего гидродинамического
давления воды, напряже-
ние. 107. Типичный разрез (а) и меха-
ническая модель кожи дельфина как
демпфирующего покрытия (б).
С. В. Першин
193
ния изгиба слоев 3 и 4 пренебрежимо малы и не учитываются.
Не моделируются с рассматриваемой целью последующие глу-
бокие слои кожи (соединительнотканный 5, примыкающий
к кожной мускулатуре, кожная мускулатура 6, подкожная жи-
ровая клетчатка 7, соединительнотканный слой 8, примыкаю-
щий к скелетной мускулатуре).
Дифференциальное уравнение динамического равновесия
единичного элемента механической модели кожи дельфина
можно представить в виде
(Л + Т2) (Мх + М2) -
дх2 dt2 dt
—(EJ + Ez + ^e^p—визг, (88)
где t — время; х — координата вдоль потока; еу — относитель-
ная поперечная деформация кожного покрова, оиэг — напряже-
ние изгиба дермальных сосочков; индексы 1, 2, 3 указывают
различные слои кожи. В уравнении (88) некоторые коэффи-
циенты переменные. Однако при периодических изгибных коле-
баниях двигательно-движительного комплекса дельфина, когда
ЦТ его тела перемещается почти прямолинейно и равномерно,
можно воспользоваться осредненными значениями этих коэффи-
циентов. Применив характерные для пограничного слоя вели-
чины, такие как толщина пограничного слоя б, м; скорость Uo
м/с; давление pU0, Н/м2; число Рейнольдса, получим необхо-
димые для моделирования безразмерные параметры:
параметр массы
feM = (Re1/p)Mz; (89)
параметр натяжения
= TL/p,l/0; (90)
• параметр упругости
kE = E'^UQ Re2; (91)
параметр предельной частоты
\ М% / Ut> Rei \ км 7
параметр демпфирования
= d.R£6_. (93)
kMaoco
В выражениях (89)—(93) Rex = £7o-l/v, м-1 HRee = t/06/v—
числа Рейнольдса, единичное и вычисленное по толщине погра-
ничного слоя; р — плотность морской воды, кг/м3; М% —сум-
марная колеблющаяся* масса нескольких слоев, кг-с2/м3; —
натяжение кожи; ц— динамический коэффициент вязкости воды,
Н-с/м2; Uо — скорость набегающего потока (осредненная ско-
194
рость плавания дельфина); £2—жесткость элемента кожного
покрова, Н/м3; г, — вязкость или демпфирование элемента мо-
дели кожного покрова, м2/с; d — коэффициент демпфирования;
«о и Со — безразмерное волновое число и фазовая скорость воз-
мущающего движения. Указанными коэффициентами учитыва-
ются суммарные характеристики слоев кожи.
Кроме этих параметров, большое значение при моделирова-
нии имеет безразмерная скорость распространения поверхност-
ных волн в мембране
т ( Т \1'2
и0 \ м ) и.
Основное значение механической модели кожи дельфина
(см. рис. 107), дифференциального уравнения (88) и безразмер-
ных параметров [см. формулы (89) — (93)] состоит в том, что
они ясно показывают, от каких качеств кожного покрова зави-
сят его демпфирующие свойства, а также позволяют численно
выразить эти свойства. Очевидно, свойства кожных покровов,
стабилизирующие пограничный слой дельфинов, связаны со ско-
ростью плавания, и с изменением ее они должны автоматически
регулироваться. Это следует из выражений безразмерных па-
раметров, где основные механические характеристики элемен-
тов кожного покрова дельфинов взаимосвязаны. При увеличении
скорости плавания дельфина повышается натяжение кожи с по-
мощью кожной мускулатуры, изменяются кровенаполнение
капилляров дермальных сосочков и вязкоупругое свойство сосоч-
кового слоя, более жестким становится упругое основание сет-
чатого слоя дермы и др. Такая «перенастройка» стабилизирую-
щих свойств кожи дельфинов с изменением скорости плавания
в процессе эволюции животного оптимизировалась, что подтвер-
ждается соответствующими расчетами и приведенными ниже
экспериментальными данными.
§ 25. Экспериментальные упругодемпфирующие характеристики
кожи дельфинов
Приведем результаты некоторых ограниченных эксперимен-
тов, проведенных в США. В работе, выполненной по департа-
менту психологии и физиологии Калифорнийского университета,
непосредственно измеряли для сравнения микровибрации по-
верхности кожи у человека и дельфина в идентичных условиях
(рис. 108).
У покоящегося человека, в воздухе, микровибрации поверх-
ности кожи регулярные, с амплитудой 1—5 'мкм и частотой
11 Гц. У дельфина, вынутого из воды и лежащего на пенопла-
сте, т. е. тоже в воздухе, микровибрации также регулярные, но
со значительно большими амплитудами 5—20 мкм; выделяются
две частоты: 2,5 и 13 Гц. Напротив, у дельфина, плавающего
*/27*
195
5)
Рис. 108. Сравнение микровибраций поверхности кожи у человека
и дельфина: а — измерения выполнены на покоящемся человеке,
в воздухе; б — афалина в воздухе; в — афалина плавает в мелкой
воде.
в мелкой воде, микровибрации не столь регулярные. Они изме-
няются со временем (сравнить участки 1 и 2 на рис. 108,а).
Таким образом, чувствительность кожи к внешнему воздей-
ствию в воздушной среде у дельфина выше, чем у человека, на
полупорядок величины при той же частоте. В мелкой воде
у медленно плавающего дельфина экспериментальная запись
микровибраций кожи была изменчивой по амплитуде и менее
ритмичной, что объясняется переменной мышечной активностью
дельфина. Микровибрации поверхности кожи коррелируются
с состоянием мышечных тонусов, тем самым экспериментально
показана тесная связь нервной регуляции состояния эпидермиса
и более глубоких слоев тканей.
Специальные эксперименты по изучению влияния нагрузки
при местном давлении на кровоток в коже человека проведены
196
в биоинженерном центре исследований Вашингтонского универси-
тета. С увеличением внешнего давления всего на (1,33—2,00) X
X 103 Па, что соответствует венозному давлению крови, местный
кровоток резко уменьшился.
Известны также отечественные опыты в области медицины
спорта по активному управлению физиологическими процессами
в органах и системах человека с применением локального ва-
куума, осуществляемого при помощи малой барокамеры актив-
ной гиперемии. Подобные эксперименты, проводимые на чело-
веке, тем более показательны для условий работы кожи дель-
фина, которая обильнее снабжена кровеносными сосудами и
нервами и подвергается на высоких скоростях плавания резким
перепадам гидродинамического давления. Так, при средней
скорости плавания дельфина Uo= 10 м/с в носовой его части
создается избыточное против местного гидростатического давле-
ние около рС702/2 = 5,1 • 104 Н/м2, т. е. более полуатмосферного,
а в пике разряжения, примерно в наибольшем сечении тела,—
недостаточное давление, составляющее приблизительно 20 %
указанной величины.
Впервые систематически, в течение ряда лет и по широкой
программе, упругодемпфирующие характеристики кожи дельфи-
нов были экспериментально изучены В. В. Бабенко с сотруд-
никами в Институте гидромеханики АН УССР [8, 67]. Отметим
некоторые прямые результаты экспериментов с кожей дельфи-
нов, имея в виду, что важнейшими параметрами, определяю-
щими эффективность упругодемпфирующих покрытий, являются
колеблющаяся масса единицы площади, натяжение, упругость
и демпфирование.
Под колеблющейся массой М кожного покрова при данной
скорости движения животного понимается масса тех слоев, на
глубину которых распространяются возмущения пограничного
слоя, вызывающие в коже упругие волны малой амплитуды.
Глубина проникновения колебаний в кожу зависит от толщины
6 ламинарного пограничного слоя или вязкого подслоя турбу-
лентного пограничного слоя, энергии возмущающего движения
Е, значения пульсаций давления р пограничного слоя на по-
верхности кожи и спектра пульсаций в пограничном слое. Ве-
личина 6 определяет амплитуду колебаний кожного покрова и
распределение толщины кожи вдоль тела. Величина Е обуслов-
лена пульсационными скоростями в пограничном слое и рас-
пределением их по толщине 6. Параметр колеблющейся массы
для принятой механической модели зависит от единичного
числа Рейнольдса Rep в соответствии с режимом течения в по-
граничном слое дельфина, величины Л1£ колеблющейся массы
и плотности р материала кожного покрова.
Согласно гидродинамическим исследованиям турбулентных
шумов пульсации в турбулентном пограничном слое определя-
ются выражением (р2),/а/<7 = 0,008, где (р2)'/з —среднеквадра-
7 С. В. Першин 197
тичное значение пульсацией давления, q = pU02!2— скоростной
напор. Полагая скорость плавания дельфина Uo= 10 м/с, полу-
чим р = 4,0 Н/м2. В случае ламинарного пограничного слоя вели-
чина р примерно на порядок меньше. Вследствие ее малого
значения естественно считать, что при ламинарном обтекании
тела дельфина наименьшей колеблющейся массой будет масса
сплошного слоя мембраны 1 эпидермиса, т. е. Л!г =Afi. С уве-
личением скорости плавания дельфина и повышением пульса-
ций давления к этой колеблющейся массе следует добавить
массу сосочкового слоя 2 эпидермиса, а затем и массу подсо-
сочкового слоя 3, т. е. тогда =М\+М2 (см. рис. 107). Были
сделаны определения параметра колеблющейся массы в раз-
личных вариантах и для всех трех видов черноморских дель-
финов: белобочки, афалины, азовки, однако в одном попереч-
ном сечении перед спинным плавником. При подсчете пара-
метра kM в расчет вводили два варианта' скорости дельфинов;
малую скорость (суточной продолжительности) и большую (от-
вечающую продолжительности плавания 15—20 мин).
Расчеты показали, что параметры колеблющейся массы у бе-
лобочки и афалины близкие, а у азовки (морской свиньи) за-
метно отличаются, как это и должно быть по их скоростным
данным. Для кожного покрова белобочки и афалины получено:
плотность слоев 1—3 р= 10204-1030 кг/м3; осредненная толщина
/-го слоя 2-Ю-4 м и слоев /—3 (1,74-2,2) • 10-3 м; масса кож-
ного покрова единицы поверхности 1-го слоя = 0,205 кг/м2
и слоев 1—3 Мхз = 1,754-2,24 кг/м2; безразмерный расчетный
параметр колеблющейся массы для 1-го слоя kM\ = (0> 12 4-
4-0,14) • 104 и для слоев 1—3 kMZ = (1,564-1,73) 104. Путем
сопоставления с имеющимися результатами расчетов для пло-
ских демпфирующих покрытий выявлено, что колебательное
движение одного наружного слоя эпидермиса кожного покрова
дельфина не оказывает стабилизирующего влияния на погра-
ничный слой, а колебание первых трех слоев кожи является
оптимальным. В дальнейшем подобные расчеты были расши-
рены для ряда поперечных сечений тела белобочки и афалины,
с учетом и более глубоких слоев кожи.
При изучении упругости кожи живых дельфинов выяснилось,
что существующая аппаратура для определения упругости по-
лимеров и резин в данном случае непригодна. Поэтому был раз-
работан специальный прибор для измерения упругости кожи
дельфинов [28], который включал измерительный контактный
с кожей стержень и систему элементов электрической схемы
индикаторной части прибора, размещенных в небольшом гер-
метичном корпусе из оргстекла. Поскольку определить упру-
гость кожи у быстро плавающего дельфина в море в настоящее
время не представляется возможным, измерения проводили на
подопытных тренированных дельфинах различных видов. Упру-
гость измеряли во многих точках поверхности тела, располо-
198
женных на разных продольных линиях, в разных поперечных
сечениях.
В результате многочисленных экспериментов были состав-
лены схемы распределения упругости кожи по телу дельфинов
(рис. 109). Измерения показали, что модуль упругости кожи
зависит от вида дельфина, его тренированности и состояния жи-
вотного при эксперименте. Например, у только что отловленного
и, естественно, возбужденного дельфина-афалины модуль упру-
гости кожи в средней части тела Е = 4,5-105 Н/м2, а у трениро-
ванного, в спокойном состоянии £ = 2,4-105 Н/м2 в том же ме-
сте. Упругость кожи только что отловленного дельфина соот-
ветствует таковой при плавании его на высоких скоростях,
а у тренированного спокойного дельфина — близка к упругости
при медленном плавании. Согласно данным экспериментов, наи-
более упругая кожа у белобочки. Несколько менее упругая
кожа у афалины и значительно менее упругая кожа у морской
свиньи, что соответствует их скоростным возможностям. У боль-
ного дельфина упругость кожи уменьшается, а у мертвого ста-
новится одинаковой вдоль всего тела. Эксперименты показали,
что дельфины натяжением кожной мышцы могут изменять упру-
гость кожного покрова почти в два раза. Все это свидетель-
ствует об активной регуляции упругости кожи дельфинов, а сле-
довательно, и об активной регуляции демпфирования.
Сказанное подтверждается последующими исследованиями
упругости кожи живых дельфинов [8]. Учитывая влияние сум-
марной толщины расчетных слоев кожи на результаты измере-
ний ее упругости, целесообразно характеризовать упругие свой-
ства кожи жесткостью s, определяемой отношением s = E/t, где,
Рис. 109. Распределение модуля упругости кожи дельфинов по длине тела,
измеренное в различных сечениях: а — белобочка; б — афалина.
/ — дельфин только что отловлен и возбужден; 2 — тренированный дельфин в спо-
койном состоянии (одни и те же особи).
7*
199
по-прежнему, Е — осредненный модуль упругости слоев, a t —
толщина этих слоев. Для характеристики упругости кожи дель-
финов, вместо введенного ранее в рассмотрение параметра уп-
ругости kE [см. формулу (91)], можно воспользоваться другим
его выражением
k-=sT~k' (94)
где р и v — соответственно плотность и кинематический коэф-
фициент вязкости жидкости. Это позволило рассчитать распре-
деление величины kt вдоль тел белобочки и афалины и сопо-
ставить их с оптимальными значениями (рис. ПО). Расчеты
выполнены в двух вариантах: при малой средней скорости пла-
вания дельфинов иср, отвечающей суточной продолжительности
плавания, и большой крейсерской скорости УКр, соответствую-
щей активному плаванию в течение 15—20 мин. В первом слу-
чае в расчет принята толщина t трех верхних слоев кожи, во
втором — толщина всей кожи до слоя подкожной жировой клет-
чатки. На рис. ПО заштрихована область оптимальных значе-
ний ks, полученных расчетом. При медленном плавании со ско-
ростью UKp величина ks имеет неоптимальные значения по всей
длине тела рассмотренных скоростных дельфинов, а на высокой
крейсерской скорости UKP — оптимальные значения в средней
части тела. Это вполне соответствует рассчитанным биоэнерге-
тическим методом минимальным значениям коэффициента ги-
дродинамического сопротивления Сх скоростных дельфинов при
крейсерских скоростях плавания (см. рис. 91)..
Демпфирующее свойство материала или конструкции можно
определить по высоте отскока падающих грузов в простейшем
приборе [67]. Стальные шарики разных размеров и массы сбра-
сывали с некоторой фиксированной высоты на поверхность ис-
пытуемого материала посредством отключения удерживающего
электромагнита. Высоту отскока шарика достаточно точно из-
меряли оптическим способом, основанным на перемещении по
вертикальной шкале отсчета визирного кольца, через которое
выпадал шарик. Показателем
Рис. НО. Распределение параметра
упругости по телу дельфинов — бело-
бочки (/) и афалины (2) — при раз-
личных скоростях плавания.
демпфирования принимали
выраженное в процентах зна-
чение коэффициента Л = [1—
— (h/ho)]lOO, где h — высота от-
скока шарика; h0 — высота его
падения. Результаты измере-
ний показаны на рис. 111. Для
сравнения данные по кожным
покровам дельфинов, несколь^
ким эластомерам и конструю"
ционным материалам приве-
дены в виде зависимостей
200
коэффициента поглощения k от
энергии удара Vl = mgh0 (т —
масса падающего шарика).
Как видно, диссипация энер-
гии кожным покровом у дель-
фина наибольшая и существенно
зависит от состояния животного.
Затухание возмущений происхо-
дит до 95 % в кожном покрове
здорового дельфина, у больного
дельфина не превышает 80%,
а у только что умершего — не
более 70 %. У части испытанных
эластомеров характеристика за-
тухания возмущений приближа-
ется к таковой для кожи дель-
фина, однако лишь при больших
значениях П. В исследованном
диапазоне энергия с ростом П
величина k у дельфинов моно-
тонно уменьшается, у эластоме-
ров имеется максимум в области
П= (2-10-3ч-2-10-2) Дж, а у
конструкционных материалов k
Рис. 111. Зависимость коэффици-
ента демпфирования от энергии,
сообщаемой кожным покровам
дельфинов, эластомерам и конст-
рукционным материалам &(П)
[67].
/ — здоровые; Дельфины: 2 — боль-
ной; 3 — мертвый. Эластомеры:
4 — неплотный, пористый (р = 42 кг/м3,
число пор на I см 20—26, толщина
h=l см, модуль упругости £=0,05—
0,09 МПа); 5 — плотный, малопори-
стый (р=1050 кг/м3, л=6, Л=0,8 см,
£=0,45 МПа); 6 — промежуточный ва-
риант (р=107 кг/м3, п=30—40» 6=
=0,8 см, £“=0,09—0,13 МПа). Конст-
рукционные материалы: 7 —
силикатное стекло, 8 — оргстекло.
увеличивается. Учитывая пульса-
ции скорости и давления в пограничном слое (пульсация ско-
рости при переходном режиме в 1,5—2,0 раза выше, чем при
турбулентном), можно полагать, что максимум поглощающей
способности кожного покрова дельфина приходится на область
энергии возмущений, соответствующую переходному и турбу-
лентному пограничным слоям. Отсюда усматриваются большие
возможности поглощения энергии возмущений в пограничном
слое на теле живого дельфина и достаточно широкий диапазон
активного регулирования поглощения этой энергии кожным пок-
ровом. В отличие от дельфинов испытанные образцы эластоме-
ров характеризуются максимумом поглощения энергии, кото-
рый в несколько порядков величины больше существующего
в пограничном слое. Этим также подтверждаются уникальные
упругодемпфирующие свойства кожи дельфинов.
§ 26. Опытные исследования пограничного слоя
активно плавающего дельфина
Вначале рассмотрим результаты анализа хорошообтекаемых
форм гидробионтов, который был выполнен для многих видов
скоростных рыб и дельфинов. При этом использовались изве-
стные фотографии, достаточно точные обобщенные рисунки (со-
ставленные по измерениям многих особей) и новые обмеры не-
которых видов скоростных гидробионтов.
201
Рис. 112. Ламииаризованная форма тела скоростных дельфинов: а—ки-
товидный дельфин длиной 2,08 м, массой 56 кг; б — тихоокеанский ко-
роткоголовый дельфин.
На рис. 112 показаны скоростные виды дельфинов — кито-
видный и тихоокеанский короткоголовый — и соответствующие
обводам их тела аэрогидродинамические профили. Указанные
виды дельфинов имеют ламинаризованные профили: наибольшее
сечение по крайней мере в одной главной плоскости затянуто
по расположению от носа до 45 % общей длины тела.
По изложенной в работе [44] методике, форму тел скорост-
ных гидробионтов для сравнения приближенно можно характе-
ризовать соответствующими эквивалентными телами вращения.
Максимальный оптимальный диаметр Do последних определя-
ется равенством площади круга наибольшего сечения прибли-
женного тела вращения с площадью эллипса наибольшего
сечения тела гидробионта высотой Но и шириной Во,. т. е. по усло-
вию лО02/4 = лН0В0/4. Эта единая методика позволяет прово-
дить общую оценку форм обводов скоростных гидробионтов, из-
вестных типов исследовательских подводных аппаратов [15] и
аэрогидродинамических профилей. Выполненные сопоставления
показали, что скоростные гидробионты-рыбы и китообразные —
составляют единую и стройную систему ламинаризованных тел,
последовательно сменяющихся цо характерной продольной от-
носительной координате х0 и диаметру Do в зависимости от зна-
чений критериев плавания: числа Рейнольдса Re и относитель-
ной минимальной скорости плавания Um=UmIL.
Для систематизации ламинаризованных тел скоростных ги-
дробионтов нами использована известная номенклатура лами-
нарных профилей NACA серий 63, 66, 67. Напомним, что их обо-
значение пятью цифрами дает основную характеристику лами-
нарных профилей, например, NACA-66-026 означает ламинарный
202
профиль низкого гидродинамического сопротивления движению
(первая цифра 6), у которого расчетная точка с минимальным
давлением отстоит на относительном расстоянии от передней
кромки хо = О,6 длины хорды (вторая цифра 6); профиль сим-
метричный (цифра 0 после тире), с относительной толщиной
26 % (последние две цифры). В частности, ламинарным профи-
лем NACA-66-026 определяется тело вращения, приближенно
эквивалентное ламинаризованным формам тела самого крупного
и скоростного дельфина-косатки.
Результаты рассматриваемых исследований приведены на
рис. 113, где параметры скоростных рыб и китообразных сред-
них размеров каждого вида показаны в поле координатной ло-
гарифмической сетки ReL и Um, а на соответствующих им на-
клонных прямых со значениями max, 0,5max и 0,lmax величин
Um указаны ламинарные профили эквивалентных тел вращения
(Ф — форель, 63-016; Т — тунец, 66-024: 1 — обыкновенный
дельфин, 66-021; 8 — косатка, 66-026 и т. п.). Таким образом,
Рис. 113. Закономерность изменения ламинаризованных форм
тел вращения, эквивалентных скоростным рыбам и китообраз-
ным, в зависимости от относительной максимальной скорости
плавания UmIL и числа Рейнольдса Ret.
Рыбы: Ф — форель; С — скумбрия: П — пеламида: Т — тунец обыкно-
венный. Дельфины: 1— белобочка; 2— афалина; 3— косатка. Киты:
4 — сейвал; 5 — синий. Технические аппараты: D, S3, DSRV.
Номенклатура профилей — NACA, серии 63, 66, 67.
203
скоростные рыбы и дельфины имеют ламинаризованные формы
тела. Следовательно, у них может быть ламинарный погранич-
ный слой на относительно большой части длины тела.
Анализируя график на рис. 113, сделаем следующие важ-
ные выводы. Скоростные рыбы и китообразные имеют лами-
наризованные формы тела при числах Рейнольдса 105—108, т. е.
в широком диапазоне, верхние значения которых превышают та-
ковые для используемых в технике ламинаризованных аэроги-
дродинамических жестких тел. Поскольку скоростные гидро-
бионты в нестационарном плавании совершают волнообразные
изгибные колебания с относительно большой поперечной ампли-
тудой, их ламинаризованные формы тела свидетельствуют
о том, что подобные колебания достаточно плавные и не нару-
шают устойчивой ламинаризации пограничного слоя на упруго-
вязкой поверхности живого гидробионта. Тем самым ламинари-
зованные формы тела скоростных гидробионтов являются необ-
ходимой предпосылкой авторегуляции демпфирования кожного
покрова у скоростных дельфинов и регуляции воздействия жи-
вой полимерной рыбьевой слизи у скоростных рыб как допол-
нительных средств существенного снижения гидродинамического
сопротивления.
Скоростные рыбы и скоростные китообразные характеризу-
ются своими областями значений критериев Reb и Um. Прини-
мая во внимание, что скоростные рыбы совершают изгибные
колебания в горизонтальной плоскости, а китообразные в вер-
тикальной и что плоскости этих колебаний ориентированы раз-
личным образом относительно симметричных профилей одних и
других, отметим: подобное различие не имеет видимого зна-
чения для ламинаризации пограничного слоя, по крайней мере
на поверхности живых тел гидробионтов. Приведенные выводы
могут быть использованы для моделирования эффективного пла-
вания скоростных гидробионтов.
Как известно, искусственная визуализация обтекания плыву-
щего живого дельфина вызывает большие трудности ввиду
гладкой и упругой поверхности кожи и больших колебаний хво-
стового стебля и плавника. Поэтому интересно воспользоваться
естественной визуализацией пограничного слоя дельфина пу-
зырьками воздуха, которые иногда выходят под водой из ды-
хала животного сверху в лобной части его тела. На рис. 114
показаны три подводные фотографии, сделанные тренером в из-
вестном флоридском океанариуме «Мэринленд» [70].
При внезапной остановке дельфина (нижняя фотография)
пузырьки воздуха из дыхала естественно всплывают верти-
кально вверх. В случае, когда дельфин плывет с большим уг-
лом атаки (средняя фотография), пузырьки воздуха образуют
четко выраженную сильно наклоненную короткую с изломом по-
лоску, а затем внезапно исчезают. Очевидно, у дыхала наблюда-
ется ламинарный пограничный слой, с излома полоски начина-
204
ется переходный участок,
который в месте исчезно-
вения пузырьков воздуха
заканчивается турбулент-
ным отрывом погранично-
го слоя. У двух афалин,
плывущих последователь-
но с малыми углами
атаки (верхняя фотогра-
фия), просматривается
пологая у поверхности
кожи и более длинная
с изломом полоски из пу-
зырьков вгоздуха, чем во
втором примере. Наблю-
даются удлиненные уча-
стки как с ламинарным
пограничным слоем, так
и с переходным.
Фотографии сделаны
в океанариуме, где на-
чальная турбулентность
водной среды намного
выше, чем в естествен-
ных морских условиях.
Тем не менее очевидно
наличие протяженного
Рис.- 114. Естественная визуализация по-
граничного слоя дельфина пузырьками вы-
дыхаемого воздуха. Подводные съемки
в океанариуме [70].
Сверху вниз: пара дельфинов плывет в гори-
зонтальной плоскости; дельфин плывет с боль-
шим углом атаки; один дельфин внезапно оста-
новлен другим на месте.
участка ламинарного по-
граничного слоя в голов-
ной части дельфина, ко-
торый в природе несом-
ненно развит на большей
длине. Достаточно вспом-
нить о наблюдениях за
плавающими дельфинами
в фосфоресцирующей воде, когда за обыкновенным дельфином
виден только весьма узкий вихревой след.
Наряду с оптимальной ламинаризованной формой тела ско-
ростных дельфинов отмечена целесообразность с позиций ги-
дродинамики расчленения эпидермиса их кожи продольными
рядами дермальных валиков и сосочков на периодически чере-
дующиеся в поперечном направлении микрополосы эпидермиса
различной упругости, вязкости, демпфирования. Этой своеобраз-
ной структурой кожного покрова дельфинов определяются пара-
метры системы продольных вихрей, образующихся в погранич-
ном слое (рис. 115). Согласно оценочным расчетам гидродина-
мической устойчивости такой системы продольных вихрей она
устойчива для характерных скоростей плавания дельфинов; Это
205
Рис. 115. Схемы продольных вих-
рей в пограничном слое на по-
верхности эпидермиса кожи дель-
фина.
/ — продольные ряды дермальных ва-
ликов и сосочков; 2 — продольные
ряды эпидермиса; 3 — поверхность
кожи; 4— продольные вихри в слу-
чаях одного (а} и двух (б, в) вих-
рей между смежными рядами дермаль-
ных сосочков; 5 — распределение про-
дольной составляющей скорости U(2);
6 — изменение толщины пограничного
слоя 0(2).
в пограничном слое вблизи
задерживает переход от лами-
нарного режима течения к тур-
булентному до чисел Рейнольдса
порядка Re~ 107.
В работе Е. В. Романенко,
В. Г. Янова (Бионика, 1973.
Вып. 7. С. 52—56) эксперимен-
тально установлено, что ампли-
туда пульсационных характери-
стик на поверхности кожного
покрова зависит от характера
движения дельфинов. При актив-
ном плавании дельфина уровень
некоторых пульсационных харак-
теристик может быть в несколько
раз меньше, чем при пассивном
движении по инерции.
Особый интерес представ-
ляют сопоставимые результаты
единообразной обработки экспе-
риментальных измерений. Изме-
рялась степень турбулентности
продольной составляющей пуль-
сационной скорости еи=д/(72 : U
обтекаемой поверхности жесткой
модели афалины и живых дельфинов (рис. 116). В Институте
гидромеханики АН УССР эти исследования проводились в те-
чение ряда лет [28]. В пограничном слое жесткой гладкой мо-
дели афалины длиной 1,75 м режим течения изучали при бук-
сировке ее на глубине 1,5 м в опытовом бассейне глубиной
3,0 м со скоростью от 0,5 до 4,5 м/с. Пользуясь проволочным
термоанемометром, регистрировали пульсации продольной ком-
поненты скоростей в пограничном слое в нескольких сечениях,
на расстоянии 1 мм от поверхности модели. Аналогичные ис-
следования были проведены на живых афалинах. Результаты
экспериментальных измерений представлены в виде графиков
зависимостей eu = fi,2(Uo, x/L) на рис. 116 для модели и актив-
ного плавающего дельфина.
Анализируя графики степени турбулентности в пограничном
слое жесткой модели афалины, по расположению и виду всплес-
ков значений еи замечаем, что ламинарный режим мог су-
ществовать только на головном участке модели при х/£<0,20.
В тех случаях, когда скорости были самыми малыми (L/o = O,5-5-
4-1,0 м/с), наблюдались высокие значения степени турбулент-
ности (еи=15—20%) почти вдоль всей модели. При скорости
буксировки t7o = 2-j-4 м/с достигались значения еи = 25—30 %
и уже в сечении № 5 при х/£ = 0,68, т. е. за спинным плавником,
наступал отрыв турбулентного пограничного слоя. Характерно,
206
Рис. 116. Результаты гидродинамических измерений степени турбулентности
еи продольной составляющей скорости в пограничном слое у поверхности
жесткой модели афалины (а) и живого дельфина-афалины (б) в разных
сечениях x/L, при различной.скорости Uo, т. е. eu(.U0, x/L) [29].
что в сечении № 6 при x/L = 0,80 с повышением скорости бук-
сировки жесткой модели афалины от 0,5 до 4,5 м/с степень
турбулентности в пограничном слое последовательно повыша-
лась от 13 до 34 %.
Результаты экспериментальных измерений в пограничном
слое активно плавающего дельфина-афалины качественно отли-
207
Рис. 117. Расчетные отношения
площади поверхности с ламинар-
ным пограничным слоем к общей
площади смоченной поверхности
тела дельфина при различной ско-
рости плавания, со/Й = НЦ)) [28].
1 — морская свиная длиной 1,4 м; 2 —
белобочка длиной 2,1 м.
плавающего дельфина степе
чаются от рассмотренных для мо-
дели. Во-первых, значения еи
выровнены вдоль тела живот-
ного, отсутствуют местные
всплески значений еи в сечениях
на различных относительных от-
стояниях от носа x/L = 0,125 4- 0,75
при исследованных скоростях
плавания. Это явно указывает
на локальное демпфирующее
действие кожи дельфина. Во-
вторых, значения еи в несколько
раз ниже в пограничном слое
афалины, чем на жесткой ее мо-
дели, что приводит к ламинари-
зации пограничного слоя дель-
фина. В-третьих, в отличие от
жесткой модели, у активного
> турбулентности при исследован-
ных значениях скорости уменьшается с повышением скорости,
т. е. регулирующее действие кожи все более оптимизируется.
Несомненно, это прямое экспериментальное доказательство ак-
тивного регулирования кожей скоростного дельфина режима
в пограничном слое, ведущего к локальному снижению пульса-
ций и к образованию ламинарного пограничного слоя на теле
животного.
С рассмотренными результатами экспериментов по локальной
регуляции демпфирования находятся в хорошем соответствии
приведенные выше экспериментально-теоретические оценки ин-
тегрального эффекта по снижению гидродинамического сопро-
тивления при активном плавании афалины (см. рис. 91). Суще-
ствование оптимума активного регулирования подтверждается
также дополнительными расчетами протяженности области ла-
минарного пограничного слоя на теле скоростных дельфинов,
(рис. 117).
§ 27. Техническое моделирование искусственных
демпфирующих покрытий для снижения гидродинамического
сопротивления трения
При моделировании искусственных демпфирующих полимер-
ных покрытий для снижения гидродинамического сопротивле-
ния трения их можно рассматривать как технические аналоги
детально изученных эффективных кожных покровов дельфинов.
Чем больше скорость движения, тем интенсивнее возникают
пульсации давления и скорости в пограничном слое на поверх-
ности покрытия, усиливается их взаимодействие и тем в бо-
лее глубокие слои покрытия проникают волны возмущений.
208
Отсюда следует, что эффективные искусственные демпфирующие
покрытия должны конструироваться и рассчитываться каждый
раз со строгим учетом их назначения и условий работы — ско-
рости движения, глубины погружения в жидкость, начальной
турбулентности потока, полимерных материалов, технологии
производства и т. д. Изложим некоторые общие вопросы выбора
демпфирующих покрытий [9].
Искусственное демпфирующее покрытие как технический
аналог не должно -полностью копировать сложнейшую конструк-
цию кожи дельфинов, да это в настоящее время и невозможно,
поскольку отсутствуют полимерные материалы, вполне соответ-
ствующие кожным, и имеются технологические трудности.
Важно использовать принцип активного действия прототипа и
возможность регулирования демпфирующих свойств покры-
тия.
Для приближения к цели вначале были испытаны два про-
стейших демпфирующих покрытия: пленочное, с полихлорвини-
ловой пленкой мембранного типа и вязкоэластичное из пенопо-
лиуретана, с некоторыми модификациями (рис. 118). В частно-
сти, конструкция гибкой поверхности позволяла исследовать
влияние на гидродинамическую устойчивость толщины слоя
воды под мембраной и свойства опоры ее в виде продольных
полосок из пенополиуретана. В последнем случае простая мем-
бранная поверхность превращалась в сложную мембранную, ко-
торая лучше стабилизирует ламинарный пограничный слой.
Изготовленные конструкции позволили провести сравнитель-
ные экспериментальные исследования гидродинамической устой-
чивости на мембранных и вязкоэластичных покрытиях и опреде-
лить все необходимые механические свойства их в соответствии
с ранее установленными критериями подобия. В результате про-
веденных испытаний, по ряду предусмотренных вариантов по-
крытий определены числовые значения размерных величин и
Рис. 118. Конструкции мембранной (а) и вязкоэластичной (б) демпфиру-
ющих поверхностей:
I — рама; 2 — дюралюминиевая пластина; 3 — прижимная планка; 4 — полоски пе-
нополиуретана; 5 — прижимная пластина; 6 — стержни рамы; 7—мембрана; 8 —
лист полиуретана.
209
безразмерных параметров покрытий, которые опубликованы
в работе [9].
Гидродинамические исследования указанных демпфирующих
покрытий выполнены на специальном стенде с малой начальной
турбулентностью потока е= (0,03-^0,05) % в рабочем диапазоне
небольших скоростей. На стенде можно искусственно созда-
вать гидродинамические возмущения различного вида. Сравни-
тельные экспериментальные исследования гидродинамической
устойчивости ламинарного пограничного слоя проведены при
продольном обтекании жесткой пластины и на упругодемпфи-
рующих поверхностях различных конструкций. Для всех эффек-
тивных упругодемпфирующих покрытий нарастание возмущен-
ного движения в ламинарном пограничном слое меньше и мед-
леннее по сравнению с таковым в случае обтекания жесткой
поверхности (по амплитуде более чем в два раза). Эффективные
демпфирующие покрытия повышают нижнее значение критиче-
ского числа Рейнольдса более чем в пять раз и увеличивают
протяженность переходной области.
В настоящее время теоретически и экспериментально дока-
зано, что искусственные упругодемпфирующие покрытия могут
стабилизировать как ламинарный, так и турбулентный погра-
ничный слой и снижать гидродинамическое сопротивление тре-
ния.
Приведем для примера новую конструкцию регулируемого
демпфирующего покрытия (рис. 119), которая позволяет широ-
кий выбор используемых упругих материалов (полимеров, ре-
зин), вязких наполнителей (например, силиконовых жидкостей),
переменного внутреннего статического давления, под которым
наполнители заполняют полости и каналы, с различными соче-
таниями размеров элементов и т. п. [4]. Предлагаемая конструк-
ция предназначена для стабилизации ламинарного погранич-
ного слоя и уменьшения пульсаций скорости и давления в тур-
Рис. 119. Регулируемое демпфирующее покрытие, наклеиваемое па обтекае-
мое жидкостью твердое тело.
1—3 — слои покрытия из эластичных материалов, склеенные в замок в виде гребенки;
4, 5, 7—10, 14 и 15 — выступы и впадины гребенок соответствующих слоев; 6 — наклон-
ные глухие каналы; 11, /2 —верхние и нижние продольные полости; 13 — наклонные
соединительные каналы; 16 — поперечные регулировочные секционные каналы — все ка-
налы и полости заполнены вязкой жидкостью под переменным давлением; 17 — об-
текаемое тело.
210
булентном пограничном слое, для снижения аэрогидродинами-
ческого сопротивления обтекаемых твердых тел.
Верхний, гребенчатый снизу слой покрытия (см, рис. 119)
представляет собой упругую разветвленную тонкую мембрану
с глухими наклонными каналами — вязкими аналогами вершин
дермальных сосочков кожного покрова дельфинов. Средний
слой с гребенчатой поверхностью как сверху, так и снизу и
предусмотренными зазорами позволяет образовать продольные
полости и имеет наклонные соединительные сквозные каналы,
которые заполнены вязким наполнителем, а в целом этот слой
является аналогом важнейшего расчлененного сосочкового слоя
кожи дельфина с дермальными сосочками и продольными дер-
мальными валиками. Нижний слой имеет гребенчатую поверх-
ность сверху и служит упругой подложкой. Это аналог нижеле-
жащих плотных слоев кожи дельфина. Соединение на клею
всех трех смежных слоев демпфирующего покрытия в замок
в виде гребенки с зазорами не только позволяет выделить про-
дольные валики и полости, но одновременно увеличивает по-
верхность сопряжения слоев и существенно повышает прочность
и надежность покрытия.
Диссипация энергии внешних возмущений в пограничном
слое обтекаемого твердого тела, на которое наклеивается дем-
пфирующее покрытие, может происходить за счет сообщения
колебаний и упругих деформаций массе элементов покрытия
и дросселирования при взаимодействии с вязким наполнителем
полостей и каналов слоев покрытия. Важнейшим элементом
покрытия является, также периодически расположенные попе-
речные каналы, через которые заполняются все полости и
каналы вязким наполнителем под некоторым статическим давле-
нием. Эти поперечные каналы позволяют секционировать покры-
тие по длине обтекаемого тела и регулировать внутреннее дав-
ление в каждой продольной секции в соответствии с распреде-
лением среднего внешнего гидродинамического давления вдоль
поверхности движущегося тела (или неподвижной поверхности,
обтекаемой набегающим потоком).
Изготовление подобного искусственного демпфирующего
покрытия — сложная техническая проблема. Трудности состав-
ляют как выбор соответствующих исходных материалов, так
и выполнение тонких деталей сложной композитной конструк-
ции. Однако дальнейшее продолжение экспериментально-тео-
ретических исследований в этом направлении представляется
перспективным.
211
ГЛАВА VI
АВТОРЕГУЛЯЦИЯ УПРУГОСТИ ПЛАВНИКОВ
И ОПТИМИЗАЦИЯ ЭФФЕКТИВНОСТИ
МАШУЩЕГО ДВИЖИТЕЛЯ КИТООБРАЗНЫХ
§ 28. Явление саморегулирования упругости плавников
китообразных соответственно режимам плавания
Одной из новых проблем гидробионики, интересной как в
теоретическом отношении, так и по возможным практическим
приложениям, является исследование так называемого регули-
руемого гидроупругого эффекта в плавниках дельфинов и ки-
тов. На это явление было обращено внимание в связи с изуче-
нием различных режимов плавания и поведения дельфинов,
существенно различающихся кинематикой и энергетической эф-
фективностью. Комплексные экологические, гидродинамические
и морфофункциональные (включая рентгенографические и ги-
стологические) исследования плавников многих видов дельфи-
нов и китов выполнены на базе лаборатории функциональной
морфологии Зоологического института АН СССР небольшим
коллективом ученых в 1963—1980 гг. Результаты этих исследо-
ваний опубликованы в работах [53, 63] и др. Впервые обнару-
женный авторами эффект официально зарегистрирован как на-
учное открытие под названием «Явление саморегулирования
гидроупругости плавников китообразных» [17]. Получены автор-
ские свидетельства на ряд изобретений, в частности [5]. Эти ра-
боты вызвали интерес как в нашей стране, так и за рубежом.
В адрес Зоологического института АН СССР поступили на них
запросы из США, Канады, Англии, Бельгии.
-Сущность явления состоит в следующем [17]. Саморегулиро-
вание упругости плавников китообразных возникает рефлек-
торно при изменении режима плавания. Оно обусловлено дей-
ствием впервые обнаруженных широко разветвленных комп-
лексных артерио-венозных сосудов, а в хвостовом плавнике еще
и наличием общего распределительного узла кровеносных со-
судов. Имеет значение также особое строение покровных тка-
ней плавников, которые включают покрытие из сухожильных
тканей.
Автоматическое регулирование упругости плавников с изме-
нением режима плавания происходит во всех плавниках дель-
финов и китов, хотя и с некоторыми различиями. Здесь иллю-
стрируем механизм саморегулирования упругости наиболее важ-
ного— хвостового плавника, главного движителя, на примере
обыкновенного дельфина-белобочки (рис. 120). Хвостовой плав-
ник, будучи сильно развитым выростом кожи, сохраняет ее ти-
212
личную структуру в виде
трех слоев органических
тканей: эпидермиса, дер-
мы и гиподермы. Вместе
с тем он существенно от-
личается от кожного по-
крова дельфина. Эпидер-
мис плавника значитель-
но толще, имеет вид
резиноподобного растя-
жимого слоя толщиной до
1,0—1,5 мм. Между сосоч-
ковым слоем дермы и сет-
чатым слоем гиподермы
расположены вдоль лопа-
стей плавника многочис-
ленные сухожильные
тяжи СТ диаметром до
1,0—1,5 мм, направлен-
ные от мышц туловища и
хвостовых позвонков. Су-
хожильные тяжи, плотно
прижатые друг к другу,
создают сплошное и проч-
Рис. 120. Схема регулируемого гидроуп-
ругого эффекта в хвостовом плавнике
обыкновенного дельфина.
КС — комплексные артерио-венозные сосуды в ги-
подерме; РУ — регулировочный узел кровеносной
сети; в — верхняя, н — нижняя самостоятельные
сети комплексных сосудов; СТ — сухожильные
гяжи покровной нерастяжимой оболочки плав-
ника; хп — хвостовые позвонки. / — хвостовой
стебель с магистральным комплексным крове-
носным сосудом под позвонками; 2 — двухлопа-
стный плавник с крупными комплексными
сосудами в гиподерме (по рентгенограмме); 3 —
продольный разрез хвостовой лопасти с капил-
лярными сосудами (по микропрепарату)
ное наружное покрытие плавника, практически нерастяжимое.
Сетчатая гиподерма с коллагеновыми и эластиновыми волок-
нами, которая заполняет всю область внутри сухожильной оп-
летки, в отличие от аналогичной ткани туловища, весьма бедна
жировыми клетками, но обильно насыщена кровеносными со-
судами.
В хвостовом плавнике дельфина отсутствует собственная
мускулатура и другие активные в метаболическом отношении
органы, а потому, казалось бы, этот плавник не нуждается
в обильном кровообращении. Между тем в плавнике имеется
весьма мощная и очень разветвленная кровеносная система,
которая состоит из густой сети артерий и вен всех рангов — от
крупных распределителей и собирателей диаметром 1,0—2,0 мм
до капилляров диаметром 0,01 мм. Характерной особенностью
сосудистой сети плавников дельфина, не встречающейся в та-
ком виде у других животных, является ее комплексный харак-
тер. Каждый достаточно заметный кровеносный сосуд есть ком-
плексный КС и состоит из центральной относительно крупной
толстостенной мышечной артерии с высоким артериальным
давлением крови, текущей от центра к периферии, окруженной
снаружи до 10—20 более мелкими тонкостенными венами с низ-
ким венозным давлением крови, в которых кровь возвращается
из периферии. Последние встречаются и отдельно в тканях плав-
ника, но тогда вены имеют более толстые стенки.
213
При пульсирующей работе сердца система разветвленных,
артериальных сосудов с их упругими толстыми стенками из
мышечных силовых элементов способна к ритмичным измене-
ниям объема сосудистого русла. Она в состоянии значительно-
изменять свою емкость и пропускную способность. При большой
физической нагрузке в плавании расход циркулирующей крови
может повышаться десятикратно за счет имеющихся в теле ки-
тообразных кровяных депо, с увеличением калибра сосудов и
вовлечения в действие многочисленных артерио-венозных ана-
стомозов. Особая роль в сосудистой системе отведена мельчай-
шей сети капилляров. С изменением кровесодержания в ней
может развиться осмотическое давление до нескольких атмо-
сфер. И так как кровь — практически несжимаемая жидкость,
то увеличение или уменьшение кроветока ведет к расширению-
или сокращению толстой мышечной стенки артерии. Это соот-
ветственно изменяет внешнее давление на венозную оплетку ар-
терии и режим ее работы. В действие вовлекается рассредото-
ченная в тканях гиподермы мельчайшая капиллярная сеть
с осмотическим аппаратом. В заключенной в плотную, почти не-
растяжимую сухожильную оплетку («рубашку») сетчатой гипо-
дерме создается объемное напряженное состояние, которое фун-
Рис. 121. Микрофотография попе-
речного среза тканей хвостового
плавника обыкновенного дель-
фина.
/ — эпидермис и сосочковый слой
дермы; 2 — слой сухожильных тканей;
3 — сетчатый слой дермы; 4 — одиноч-
ный кровеносный сосуд; 5 — комплекс-
ный артерио-венозный сосуд.
кционально связано с режимом
плавания дельфина; в хвостовом-
плавнике проявляется своеобраз-
ный регулируемый гидроупругий
эффект. Когда скорость плава-
ния дельфина увеличивается,
его метаболизм, а потому и цир-
куляция крови повышаются,
плавник автоматически стано-
вится жестким, упругим и, на-
оборот, с замедлением движения
упругость плавника понижается..
Это вполне отвечает наиболее
эффективной работе машущего
плавникового движителя с боль-
шими амплитудами колебаний.
Обычная частота колебаний
хвостового плавника соответст-
вует частоте пульса дельфина
(80—130 ударов в минуту).
Характерной особенностью со-
судистой сети хвостового плав-
ника дельфина (тоже не встре-
чается у других животных) яв-
ляется наличие одного общего
распределительного узла РУ кро-
веносной системы, который рас-
214
положен в конце подводящего магистрального комплексного со-
суда (см. рис. 120). Этим облегчается регулирование гидроуп-
ругого эффекта в хвостовом плавнике, тем более, что в нем
имеются две самостоятельные сети комплексных сосудов —
верхняя и нижняя.
Чтобы подчеркнуть существенное отличие тканей хвостового
плавника дельфина от таковых в коже туловища, приведем
рис. 121, который можно сравнить с рис. 95 и 107. Особо выде-
ляются в плавниках слой сухожильных тяжей и уникальные
комплексные артерио-венозные сосуды.
Ниже подробно рассмотрено саморегулирование упругости
плавников китообразных. Материалом для его изучения послу-
жили хвостовые и спинные плавники: черноморских дельфи-
нов—морской свиньи, афалины, белобочки; тихоокеанских
дельфинов — белокрылой морской свиньи и белобокого; океани-
ческих дельфинов — серого, длиннокрылого и косатки; крупных
эмбрионов китов — сейвала, финвала, кашалота и карликового
кашалота.
§ 29. Гидродинамическая целесообразность авторегуляции
упругости плавников китообразных
Для гидродинамических исследований проведем подробный
анализ результатов визуальных наблюдений, надводных и под-
водных киносъемок плавательных движений дельфинов как
в условиях содержания их в неволе, так и в естественных усло-
виях моря [52, 53]. Разнообразные режимы поведения и движе-
ния дельфинов даже визуально подтверждают, что авторегуля-
ция упругости плавников дельфинов вызывается их функцио-
нальными и физиологическими особенностями. В необычно
широком диапазоне и быстро изменяется гидродинамическая
нагрузка на плавники — от незначительной при отдыхе и сне
на плаву до резко повышенной в импульсных переходных про-
цессах и при маневрах.
Весьма показательно первое проявление регулируемого ги-
дроупругого эффекта. Оно непосредственно наблюдается в при-
роде в естественных условиях при выпрямлении плавников
с началом самостоятельного плавания новорожденного дель-
фина и с проявлением так называемой вазомоторной изменчи-
вости кровеносных сосудов плавников (рис. 122). У зародышей
дельфинов и китов до самого рождения хвостовой плавник
свернут в трубочку, а спинной прижат к спине. После рождения
животного плавники по мере заполнения всей сосудистой сети
кровью быстро выпрямляются, занимают обычное положение и
новорожденный оказывается в состоянии плавать. Характерно,
что это явление искусственно необратимое; плавники не удается
без травм возвратить у живого дельфина в первоначальное, со-
гнутое положение, настолько значительно повышается их упру-
215
Рис. 122. Первое проявление гидроупругого эффекта у новорожденных дель-
финов: а — свернутый в трубочку хвостовой плавник при рождении рас-
прямляется в воде; б — примкнутый к спине спинной плавник детеныша че-
рез полчаса выпрямляется.
гость при обильном заполнении кровью уникальной сосудистой
сети, заключенной в тканях гиподермы с нерастяжимой внешней
оболочкой из сухожильных тяжей.
Напротив, исчезновение регулируемого гидроупругого эф-
фекта и посмертное опадение стенок кровеносных сосудов
в плавниках китообразных наблюдалось многократно. Оно хо-
рошо заметно визуально и на ощупь в виде своеобразного се-
тевого рельефа на поверхности плавников. У живых китообраз-
ных поверхность всех плавников гладкая, однообразная, а у
мертвых и фиксированных — с явно выраженной системой по-
нижений в коже плавников по месту расположения крупных
комплексных сосудов. По окончании периода трупного окочене-
ния хвостовой плавник может быть согнут и даже свернут
в трубочку, как у эмбриона (это невозможно сделать у живого
дельфина).
Особый интерес представляет воздействие регулируемого
гидроупругого эффекта в спинном плавнике косатки, поскольку
это самый крупный и скоростной дельфин, а ее спинной плав-
ник выделяется большими абсолютными и относительными раз-
мерами (рис. 123). Однако максимальная высота спинного плав-
ника у косатки (см. табл. 3) не превышает высоты столба
крови, соответствующей наибольшему артериальному давлению
крови млекопитающих. В работе [53] приведен ряд фотографий
косатки, сделанных в океанариуме «Морской мир» в Сан-Диего
(США), на которых видны самые различные положения и со-
стояния спинных плавников активно действующих и пассивно
отдыхающих косаток. Подобные факты позволяют рассматри-
вать высокий спинной плавник косатки в связи с регулируемым
гидроупругим эффектом как своего рода наглядный индикатор
общего состояния дельфина и его метаболизма. Тренер косаток
216
Рис. 123. Влияние регулируемого гид-
роупругого эффекта на положение и
работоспособность высокого спинного
плавника дельфина: а — косатка
в надводном плавании активно пла-
вает в море (спинной плавник рас-
правлен); б — косатка в океанариуме
отдыхает (спинной плавник полусог-
нут, почти примкнут к телу).
в океанариуме сообщил, что
обычно большую часть вре-
мени у его питомцев спинные
плавники бывают не пол-
ностью расправлены, а более
или менее изогнуты и сло-
жены. Это свидетельствует
о некоторой гиподинамии у ко-
сатки в океанариуме, когда
океанический крупный ско-
ростной дельфин помещается
в водоем ограниченных разме-
ров. Отсюда также следует,
что в условиях океанариума
крупный скоростной дельфин
физиологически не может раз-
вивать максимальную ско-
рость плавания, такую как на
воле, в море. На это нами не-
однократно обращалось вни-
мание.
Вышеизложенное справед-
ливо не только для крупных
плавников косатки, но также
можно заметить у афалины
(рис. 124). Чтобы дельфин мог
впрыгнуть на помост, его хво-
стовой плавник должен ак-
тивно и усиленно работать — он расправлен и упругий, а спин-
ной плавник в нерабочем состоянии полусогнут. Подобные ак-
тивные действия хвостового плавника наблюдаются и в других,
случаях перемещений дельфина, в том числе при движении
«стоя на воде» и при выпрыгивании из воды в воздух. В этих
случаях хвостовой плавник испытывает в воде наибольшие на-
грузки и должен быть упругим.
Регулируемый гидроупругий эффект в плавниках китообраз-
ных срабатывает достаточно быстро, но плавно. Об этом можно
судить по анализу таких скоротечных переходных процессов
в плавании дельфина, как внезапное трогание с места и резкая:
остановка, которые происходят с большими продольными уско-
рениями и замедлениями тела и предъявляют жесткие требова-
ния в отношении прочности движителю — хвостовому плавнику.
Наблюдения показывают, что находящийся в покое дель-
фин в состоянии внезапно сделать один большой взмах хвосто-
вым плавником, который продвинет его вперед на расстояние,
равное удвоенной-утроенной длине тела. Напротив, быстро плы-
вущий дельфин, например со средней скоростью около 6—8 м/с,
пользуясь своим хвостовым плавником под большим углом
217"
Рис. 124. Различное одновременное состояние плавников у выпрыгнув-
шей из воды на помост афалины.
Хвостовой и грудной плавники расправлены и упругие, а спинной плавник полу-
согнут и ненапряженный.
атаки как гидротормозом, способен прекратить движение на
расстоянии всего лишь длины полукорпуса. При этом продоль-
ное замедление тела может превышать 10 м/с2, что в воде при
обычных условиях граничит с ударом. Очевидно, в этих усло-
виях упругость плавника должна нарастать плавно и точно ре-
гулироваться.
Авторегуляция упругости гидродинамически целесообразна
также ввиду экстремального вида амплитудно-частотной харак-
теристики колебаний хвостового плавника (рис. 125), одновер-
шинного графика зависимости между амплитудой и частотой ко-
лебаний в характерной точке развилки лопастей хвостового
плавника при регулярном поступательном плавании дельфина.
Обращают на себя внимание большие размахи колебаний и
резко выраженные абсолютные пики размахов у разных дельфи-
нов, практически при одном и том же значении частоты колеба-
ний /1=1,5 Гц. Назовем эту частоту «критической» пк. Экспери-
менты были проведены в большом кольцевом морском бассейне
Черноморского гидрофизического института АН УССР с непри-
нимавшими пищу дельфинами. По этим данным построена без-
размерная характеристика A/Am=f(n/nK), где А — размах, от-
вечающий некоторой выбранной для данного дельфина частоте
п; Ат — максимальный размах при критической частоте пк =
= 1,5 Гц того же дельфина.
Как видно, размерные и обобщенная безразмерная ампли-
тудно-частотная характеристики хвостового плавника черномор-
ских дельфинов напоминают резонансную характеристику гар-
монических вынужденных колебаний механической системы.
При наличии экстремума одному и тому же размаху колеба-
218
нии отвечают два значения частоты колеоании: одно докрити-
ческое га<пк, а другое сверхкритическое п>пк- Поскольку пер-
вому из них соответствует меньшая скорость плавания дель-
фина, при которой энергетически целесообразна малая упру-
гость лопастей хвостового плавника, а второму — большая
скорость плавания, при которой целесообразна повышенная
жесткость плавника, регулирование упругости хвостового плав-
ника для наиболее эффективной его работы становится необхо-
димым. Это хорошо согласуется с изменениями активного мета-
болизма и циркуляцией крови дельфина по мере увеличения или
уменьшения скорости плавания. Рассматриваемая регуляция
осуществляется автоматически посредством уникальных комп-
лексных артерио-венозных сосудов.
Чтобы установить, насколько часто дельфины плавают с ча-
стотой колебаний хвостового плавника, близкой к критической,
на рис. 126 показана частотно-статистическая характеристика их
плавания. Она построена по данным статистической обработки
кадров подводной киносъемки плавательных движений дель-
финов как для условий эксперимента в морском бассейне (160
случаев), так и в натурных условиях моря по известному кино-
фильму Ж. И. Кусто «В мире
Рис. 125. Экстремальная ампли-
тудно-частотная характеристика
колебаний хвостового плавника
дельфинов: а — размерная, Л(м);
б — безразмерная.
А — полный размах (удвоенная ампли-
туда), п — частота колебаний; 1 и 2 —
морская свинья длиной соответственно
1,11 и 1,35 м; 3 — белобочка длиной
1,53 м.
безмолвия» (105 случаев). На
графике отложены по гори-
зонтальной оси — абсолютные
значения частоты полных ко-
Рис. 126. Частотно-статистическая
характеристика плавания дельфинов.
1 — эксперименту в морском бассейне
(дельфины не принимали пищу и были
истощены); 2 — по данным киносъемки
в естественных условиях моря; 3 — рас-
четная кривая статистически нормального
распределения частоты.
219'
лебаний (сверху вниз и обратно) п, Гц и по вертикальной оси—
относительное число р, % случаев плавания дельфинов с интер-
валами Дп = 0,20 Гц (за 100 % принято общее число учтенных
случаев в каждом из вариантов расчета). Кроме эксперимен-
тальных кривых / и 2, на графике нанесена еще теоретическая
кривая 3 нормального распределения частоты, рассчитанная по
данным натурных морских условий. Частота колебаний хвосто-
вого плавника у дельфинов не подчиняется закону распределе-
ния случайных величин, а определяется функциональной зави-
симостью А(п). Наиболее часто дельфины плавают с частотой
колебаний хвостойого плавника, близкой к критической. Как из-
вестно, в механических системах резонансная частота энергети-
чески наиболее эффективна, она отвечает минимуму затрат
энергии колебаний.
Таким образом, авторегуляция упругости плавников китооб-
разных обусловлена многими особенностями их поведения и
плавания на различных режимах.
У рыб, артериальное давление крови которых на порядок ве-
личины меньше, чем у дельфинов, и хвостовой плавник пластин-
чатый, а не в виде толстого крыла, подобное регулирование не-
возможно. Однако они имеют раздвижной хвостовой плавник,
и обе его пластины могут сужаться или расширяться; кроме
того, частота колебаний рыб изменяется в несколько раз. Воз-
можно одновременное регулирование размаха плавника и ча-
стоты колебаний рыб на различных скоростях плавания.
§ 30. Рентгенографические исследования распределения
сосудистой сети в плавниках китообразных
Значительный интерес представляет распределение в плане
и в профиле плавников кровеносной сети, играющей столь важ-
ную роль в регулируемом гидроупругом эффекте. Исследования
выполнены рентгенографическим методом. Для этого плавники
дельфинов и эмбрионов китов освобождали от крови в крове-
носных сосудах, а затем артерии заполняли с помощью шприца
под давлением рентгеноконтрастной массой в виде свинцовых
белил, разведенных на бензине, с последующей рентгеносъемкой
плавников и отдельных срезов. Помимо ранее изученных взрос-
лых черноморских дельфинов разных видов было использовано
около 50 пдавников эмбрионов китов разного возраста. Мате-
риал был собран в арктических и антарктических экспедициях
и поступал для исследований в -лабораторию функциональной
морфологии Зоологического института АН СССР в заморожен-
ном виде.
Результаты исследований показали, что крупные кровенос-
ные сосуды, которые в плавниках китообразных обычно комп-
лексные, артерио-венозные, с многочисленными анастомозами,
220
распределяются в плане и в профиле различным образом"
в спинном и хвостовом плавниках и по-разному у дельфинов-
и китов (в зависимости от размеров и скорости плавания жи-
вотных). Здесь мы не касаемся парных грудных плавников —
рулей глубины, поскольку они, в отличие от других плавников,,
имеют твердые костные остатки, в виде фаланг пальцев, и по-
тому заведомо достаточно жесткие. Регулируемый гидроупру-
гий эффект для них не столь существенен.
Спинные плавники и обе лопасти хвостовых плавников всех
дельфинов и китов не имеют твердых костных элементов, од-
нако их гидродинамические функции существенно различаются.
Спинные плавники служат стабилизаторами курса и в прямо-
линейном движении являются симметричной конструкцией. По-
этому комплексные артерио-венозные сосуды в них расположены
только снизу вверх, в срединной плоскости тканей гиподермы
спинного плавника (рис. 127). Другой их особенностью явля-
ется то, что в спинных плавниках комплексные артерио-веноз-
ные сосуды разобщены между собой. Это одиночные крупные
сосуды, которые не составляют специальной сети разветвленной
сосудистой системы спинного плавника. Артерии комплексных
сосудов в спинном плавнике являются продолжением секцио-
нированных артериальных сосудов, расположенных в теле
Рис. 127. Артерии разобщенных крупных комплексных сосудов
в срединной плоскости спинного плавника и секционированная
артериальная сеть в спине под плавником у эмбриона длинно-
крылого дельфина.
Фото с рентгенограммы, свинцовая инъекция.
22 Г
дельфинов и китов под спинным плавником. Однако последние
в самом спинном плавнике окружаются венами и превраща-
ются в комплексные артерио-венозные сосуды.
Напротив, хвостовые плавники всех дельфинов и китов, как
активные машущие движители, имеют двухрядные — верхние
и нижние — общие сети комплексных артерио-венозных сосудов,
которые расположены в дерме вблизи сухожильной оплетки.
У скоростных дельфинов поступление и отток крови в них ре-
гулируется общим распределительным узлом (рис. 128). Можно
полагать, что двухрядное расположение сосудистой сети в хво-
стовых плавниках обусловлено тонкостями регулирования их
упругости при возвратных машущих движениях и перекладке
плавника в крайних верхнем и нижнем положениях. В кратко-
временный период перекладки плавника его нагнетательная и
подсасывающая поверхности меняются местами. В этих усло-
виях гидродинамически целесообразно, чтобы профиль лопастей
хвостового плавника был несимметричным. Не исключено, что
при перекладке хвостового плавника его выпуклость изменяется
на обратную с помощью соотвётствующей части сухожильной
оплетки и одного из двух рядов сети комплексных артерио-ве-
нозных сосудов. Однако это последнее предположение нужда-
-ется в тщательной проверке.
В лопастях хвостового плавника дельфинов имеются два
рода вен: одни — сильно анастомозированные, связанные в ком-
плекс с артериями, а другие — одиночные, толстостенные. Вто?
рые встречаются реже, но стенки их — силовые элементы. В це-
лом соотношение магистральных артериальных и венозных со-
Рис. 128. Типичная для ско-
ростных дельфинов двухрядные
(верхняя и нижняя) артериаль-
ные сети комплексных сосудов
хвостового плавника с одним
общим распределительным уз-
лом.
Фото средней части рентгено-
граммы, свинцовая инъекция.
судов у китообразных порядка
1: 5, что крайне редко встречается
у других млекопитающих (обычно
1:2).
У всех дельфинов и китов в хво-
стовых плавниках сети комплекс-
ных артерио-венозных сосудов
двухрядные (верхняя и нижняя),
однако имеются существенные раз-
личия в системе ветвления каждого
ряда сосудистой сети у мелких и
крупных, скоростных и нескорост-
ных китообразных (рис. 129). Встре-
чаются три типа ветвления: ма-
гистральный М, рассыпной Р и по-
лурассыпной ПР.
Магистральный М тип
ветвления сосудистой сети ха-
рактеризуется наличием в хвосто-
вом плавнике общего распредели-
тельного узла, от которого расхо-
222
Рис. 129. Типы строения двухрядной сети комплексных артерио-веноз-
ных сосудов в хвостовом плавнике различных видов дельфинов и ки-
тов: а — магистральный (обыкновенный дельфин); б—полурассыпной
(кит сейвал); в — полурассыпной (полярный крупный дельфин-белуха);
г— рассыпной (кашалот).
Рисунки составлены по рентгенограммам.
дятся в стороны 4 крупных распределительных сосуда: по 2
в правую и левую лопасти (из них один верхний, а другой ниж-
ний— см. рис. 129, а). От каждого из четырех распределитель-
ных сосудов ответвляются сосуды второго и третьего порядков,
так что действие любого из этих четырех распределительных со-
судов распространяется на всю площадь лопасти плавника,
снизу или сверху. Магистральный тип ветвления обнаружен по
анализу рентгенограмм хвостовых плавников у скоростных
дельфинов — морской свиньи, афалины, белобочки, белокрылой
морской свиньи, короткоголового дельфина.
Рассыпной Р тип ветвления, напротив, характери-
зуется отсутствием одного общего распределительного узла. На
магистральном комплексном сосуде имеется ряд последова-
тельно расположенных узлов, от которых отходит более десяти
самостоятельных сосудистых ветвей, достигающих удаленных
частей лопастей, каждая из которых обслуживает узкую
223
полоску тканей. Типичным для этого типа ветвления является
хвостовой плавник у кашалота (см. рис. 129,г).
Полурассыпной ПР тип ветвления — промежу-
точный между первыми двумя. Например, выделяется один
крупный распределительный сосуд, достигающий самого удален-
ного конца лопасти хвостового плавника; от него же ответ-
вляются сосуды первого и второго порядка к передней кромке
плавника, а на всей остальной части, составляющей более по-
ловины лопасти, крупные комплексные артерио-венозные со-
суды ответвляются непосредственно от магистрального. Этот
тип ветвления обнаружен в хвостовом плавнике крупного не-
скоростного дельфина-белухи и скоростного кита-сейвала (см.
рис. 129, б и в).
Существенно различаются у разных видов китообразных и
диаметры крупных комплексных сосудов в хвостовых плавни-
ках. У малых скоростных дельфинов (морская свинья, бело-
бочка) наибольший диаметр магистрального сосуда 2—3 мм,
диаметры распределительных сосудов 1,0—2,5 мм, у крупной
белухи соответственно 12 и 5^6 мм. В хвостовых плавниках
китов диаметры распределительных комплексных сосудов значи-
тельно больше: у сейвала 6—8 мм, у финвала 9—16 мм, у ка-
шалота 10—15 мм.
Сопоставляя все три типа ветвления М, Р, ПР сосудистых се-
тей в хвостовых плавниках различных видов китообразных,
можно заметить, что наиболее совершенный для регуляции уп-
ругости плавника — магистральный тип с одним общим распре-
делительным узлом. Этот тип ветвления встречается у скоро-
стных малых дельфинов, у которых лопасти хвостового плав-
ника более тонкие. Рассыпной тип ветвления сосудистой сети
с наибольшими диаметрами сосудов у кашалота обусловлен тем,
что кашалот глубоко ныряет в холодную воду и его комплекс-
ные сосуды выполняют другую функцию — теплорегуляцию ор-
ганизма. Одновременное влияние размеров и скорости плава-
ния дельфинов и китов на ветвление сосудистой сети в хвосто-
вых плавниках усматривается и при сравнении полурассыпного
типа у белухи и сейвала. Таким образом, скоростные качества
китообразных не только зависят от формы тела и плавников, но
и связаны со спецификой разветвления сосудистой системы ло-
пастей плавников, которая выражает разную степень проявле-
ния регулируемого гидроупругого эффекта в главном движителе
животного.
Тип ветвления сосудистой сети в хвостовом плавнике кито-
образных определяется уже в раннем возрасте развития эм-
бриона. Для рентгенографического исследования были исполь-
зованы плавники эмбрионов самых разных размеров. Оказа-
лось, что хорошо развитая сеть артерио-венозных комплексных
сосудов наблюдается у эмбриона кашалота с размахом хвосто-
вого плавника всего около 0,1 м. По топографии она почти
224
Рис. 130. Рентгеноспимок по свинцовой инъекции продольного среза лопасти
хвостового плавника эмбриона сейвала длиной тела 2,4 м.
Видны двухрядные (верхняя и нижняя) артериальные сети комплексных сосудов и
мощная снаружи их сухожильная (нерастяжимая) оплетка.
в таком же виде встречается и у взрослого кашалота с разма-
хом лопастей 3,0—3,5 м.
Приведем рентгеноснимок продольного среза лопасти хво-
стового плавника крупного эмбриона сейвала (рис. 130). Верх-
няя и нижняя сосудистые сети самостоятельны и могут рабо-
тать для регуляции упругости соответствующих по толщине
частей лопастей независимо. Поэтому вместе с большим или
меньшим натяжением мощной сухожильной оплетки верхней
или нижней поверхности лопастей сосудистая сеть, как отмеча-
лось выше, может способствовать изменению на обратную сто-
рону выпуклости хвостового плавника при перекладке его
в крайних верхнем и нижнем машущих положениях. Гидро-
динамически это целесообразно.
§ 31. Гистологические исследования тканей плавников
и уникальных комплексных артерио-венозных сосудов
китообразных
Из биомеханики известно, что биологическая сплошная
среда отличается от обычных инженерных материалов своей
сложной структурой, многофазностью, неоднородностью и анизо-
тропностью. Особенностью живых тканей является своеобраз-
ное изменение их упругости, а именно: малым напряжениям от-
вечают большие деформации; это нелинейные вязкоупругие
материалы экспоненциального типа. Биологические жидкости
обычно неньютоновские. Эксперименты показывают, что по
мере ветвления и уменьшения диаметра кровеносные сосуды
становятся все более жесткими. Микроскопические кровеносные
сосуды обладают значительно меньшей растяжимостью, чем
крупные. Основными мягкими биологическими тканями явля-
ются кровеносные сосуды, кожа, сухожилия, и все они встреча-
ются в плавниках китообразных в сложных структурах и со-
четаниях, например, в виде уникальных комплексных артерио-
венозных сосудов. Далее кратко рассмотрим данные структуры,
насколько это позволяют сделать выполненные в связи с об-
225
наружением регулируемого гидроупругого эффекта многочис-
ленные гистологические исследования [52].
В структуре хвостового и спинного плавника (рис. 131 и 132)
общим является следующее чередование слоев тканей от на-
ружной поверхности к середине плавника: эпидермис, сосочко-
вый слой, слой сухожильных тяжей, сетчатый слой дермы и
гиподермальные образования сердцевины лопасти плавника.
Все эти слои хорошо видны на поперечных срезах лопастей
даже невооруженным глазом, но абсолютная и относительная
мощность их у разных видов китообразных может существенно
изменяться. Различно в плавниках расположение комплекс-
ных артерио-венозных сосудов: в хвостовом плавнике в два
ряда, вблизи верхней и нижней частей сухожильной оплетки,
а в спинном плавнике в один ряд, в срединной плоскости гипо-
дермы. Это различие функционально обусловлено.
Эпидермальный резиноподобный слой плавников относи-
тельно толстый, особенно мощный на передней кромке, где он
в 2,0—2,5 раза толще, чем на других участках. Сосочковый
слой включает нижнюю часть эпидермиса и верхнюю часть-
Рис. 132. Поперечный срез лопасти
спинного плавника тихоокеанского
белобокого дельфина: а — общий
вид; б — увеличенная фотография ча-
сти среза с расположенными в один
ряд комплексными сосудами.
Рис. 131. Поперечный срез лопа-
сти хвостового плавника обыкно-
венного дельфина: а — общий вид;
б — увеличенная фотография ча-
сти среза с расположенными
в два ряда комплексными сосу-
дами (масштабная шкала в мм).
226
дермы. Дермальные сосочки ориентированы на передней
кромке плавников почти ортогонально к ней, а на несущих по-
верхностях имеют косое направление. Слой сухожильных тяжей
есть специфическое образование лопастей плавников китообраз-
ных. Тяжи расположены параллельно, они плотно прижаты
друг к другу и ориентированы вдоль лопастей плавников,
а в хвостовом плавнике — от хвостового стебля. Встречается
несколько рядов сухожильных тяжей (рис. 133). Сетчатый слой
дермы плавников относительно толще, чем в коже туловища,
с более сложным переплетением опорных элементов из кол-
лагеновых и эластиновых волокон. Волокна собраны в мощные
пучки, которые в основном имеют направление, ортогональное
вышерасположенным тяжам сухожильного слоя. Ориентирован-
ностью коллагеновых волокон сетчатый слой дермы лопастей
плавников резко отличается от того же слоя кожи тела дель-
фина, где волокна расположены под разными углами наклона
к поверхности кожи. Гиподермальный слой лопастей плавников
нечетко отделен от прилежащего слоя дермы, отчасти вслед-
ствие того, что отсутствует скопление жировых клеток.
На продольном срезе лопасти хвостового плавника иногда
ясно видно изменение расположения опорных элементов тка-
ней — коллагеновых и эластиновых волокон. В передней тол-
стой части профиля лопасти волокна направлены в разных на-
правлениях (беспорядочно), а в концевой тонкой части про-
филя— преимущественно вертикально. Это, очевидно, связано
с необходимостью иметь различную жесткость в разных частях
Рис. 133. Микрофотографии поперечных срезов тканей хвостовых
плавников тихоокеанского короткоголового дельфина (а) и карли-
кового кашалота (6).
Видны: эпидермис, сосочковый слой, сухожильный слой с одиночными кро-
веносными сосудами, сетчатый слой дермы с коллагеновыми и эластиновыми
волокнами и комплексным артерио-венозным сосудом.
227
плавника при колебательных движениях; концевая часть может
несколько изгибаться.
Сети распределительных и ответвляющихся от них комплекс-
ных артерио-венозных сосудов расположены в дермальном
слое лопастей хвостового плавника непосредственно под сплош-
ной сухожильной оболочкой. В целом они образуют густую,
сильно анастомозированную поверхностную сеть крупных со-
судов (рис. 134). При этом венозная оплетка не сплошь накры-
вает поверхность артерии, а с большими просветами. Из мел-
ких кровеносных сосудов в хвостовом плавнике распространены
сложная и разветвленная система артериальных, венозных и
артерио-венозных анастомозов и многочисленные капилляры.
Анастомозы между мельчайшими артериями и венами (арте-
риолами и венулами) выполняют транзитную роль. Анастомозы
между артериями обеспечивают более равномерную доставку
крови и выравнивают кровяное давление. Артерио-венозные
анастомозы существенно ускоряют ток крови из артериальной
системы в венозную, минуя капиллярную сеть. Таким образом,
сосудистая система плавников в состоянии весьма эффективно
выполнять функции, связанные с регулированием кроветока и
изменением давления крови, что непосредственно ведет к само-
регуляции упругости тканей плавников китообразных в соответ-
ствии с режимами плавания.
Образование и работу комплексных артерио-венозных сосу-
материале спинных плавников
дельфинов, поскольку, в отли-
чие от хвостового, в спинном
его плавнике комплексные со-
суды одиночные, не замыка-
ются в общую сеть и форми-
руются в самом плавнике.
На микрофотографиях
рис. 135 зафиксированы раз-
личные состояния комплекс-
ных сосудов спинного плав-
ника длиннорылого дельфина.
Просвет центральной толсто-
стенной мышечной артерии
может изменяться в широком
диапазоне. Резко изменяется
соотношение площадей про-
светов всех окружающих вен
и данной артерии. Тем самым
возможно широкое регулиро-
вание притока и оттока крови
в комплексном сосуде и изме-
нение давления на ткани за
пределами самого сосуда.
дов оолее удооно изучать на
Рис. 134. Продольный вид комплекс-
ного сосуда в плавнйках дельфина:
а — обобщенная схема; б — коррози-
онный препарат (шкала в мм).
/ — толстостенная артерия с ответвлени-
ями; 2 — тонкостенные вены с анастомо-
зами.
228
Рис. 135. Различные соотно-
шения просветов артерий и
вен в комплексных сосудах
спинного плавника длинно-
рылого дельфина рода сте-
нелла (микрофотографии
срезов).
Рис. 136. Различные формы
образования комплексных
артерио-венозных сосудов
в спинном плавнике серого
дельфина от основания до
вершины плавника (микро-
фотографии срезов).
Чтобы выявить, как образуются комплексные сосуды в спин-
ном плавнике дельфина на основе секционированных артерий
и вен, идущих из спинной части тела, был сделан ряд срезов
комплексного сосуда в различных его местах, считая от основа-
ния у спинной части тела и кончая вершиной в верхней оконеч-
ности плавника (рис. 136). По взаимному расположению арте-
рий и вен Комплексного сосуда можно различать несколько
форм образования такового. У основания спинного плавника
выделяется начальная форма комплексного сосуда, когда в от-
далении от артерии группируются одиночные вены, которые
разобщены тканями гиподермы с артерией и между собой.
В средней части плавника имеется промежуточная форма ком-
плексного сосуда: одиночные вены концентрически распо-
ложены вблизи артерии, но все еще с небольшими промежут-
ками, в том числе и между венами. Следующая, развитая
форма состоит в том, что образуется внешнее кольцо из вен,
непосредственно прилегающих к стенкам артерии и друг
к другу. Очевидно, начальная форма комплексного сосуда ме-
нее эффективна для саморегуляции упругости тканей, но она
расположена у основания спинного плавника, где он наиболее
толстый и мощный. А промежуточная и развитая формы обра-
зования комплексного сосуда более эффективны и располо-
жены в менее толстой и менее мощной части плавника.
229
§ 32. Теоретический анализ расположения сосудистой сети
и гидроупругого эффекта в плавниках китообразных
Сопоставляя сосудистую сеть спинных и хвостовых плавни-
ков дельфинов и китов, отмечаем, что вертикальное расположе-
ние одиночных комплексных артерио-венозных сосудов в спин-
ных плавниках полностью определяется положением артерий,
кровоснабжающих секционированные части спинной части тела.
Эти артерии тела продолжаются в спинной плавник, где после-
довательно покрываются венозной пленкой. Напротив, сосуди-
стая сеть в хвостовых плавниках — вполне самостоятельное об-
разование магистрального, полурассыпного и рассыпного типа
ветвления у различных видов китообразных, что может опреде-
ляться их гемодинамическими и гидродинамическими законо-
мерностями. Ниже остановимся на них, имея в виду возможное
моделирование регулируемого гидроупругого эффекта в плав-
никах.
В гемодинамике установлено, что кровяное давление при-
мерно одинаково у человека и у многих крупных (лошадь, со-
бака) и мелких (кролик, мышь) млекопитающих. Оно также
мало зависит от систематического их положения. Общие за-
кономерности гемодинамики о ритмичных изменениях объема
артериальных сосудов и кровяного давления в основном спра-
ведливы и для дельфинов. Упругое состояние стенок артерий
не является стабильным, но рефлекторно изменяется благодаря
активности мышечных элементов, поэтому при большой физиче-
ской нагрузке минутный расход крови повышается до 10—
12 раз за счет кровяных депо и увеличения калибра сосудистой
сети, что характеризует возможность регулирования упругости
тканей плавника. Неустановившееся пульсовое течение крови —
сложной неньютоновской жидкости — даже в крупных крове-
носных сосудах обычно происходит при ламинарном режиме.
Из общего развиваемого сердцем гидродинамического дав-
ления крови, пульсирующего от минимального 8 - 103 Па до
максимального 16- 103 Па, по расчету и экспериментальным
данным до 50 % теряется на гидродинамическое сопротивление
в концевых частях артерий — многочисленных артериолах,
длина которых до 2 мм и внутренний диаметр около 0,02 мм.
В артериолах пульсация кровяного давления сглаживается так,
что в последующих капиллярах она не превышает ~133 Па,
т. е. практически исчезает. В более многочисленных венах ско-
рость течения крови значительно меньше и потери давления на
гидродинамическое сопротивление незначительны. Именно ар-
териолы обычно служат как бы своеобразными кранами,
с помощью которых рефлекторно, вследствие упругости мышеч-
ных стенок самих сосудов, регулируется кровяное давление и
распределение кроветока. Однако в сосудистой системе плав-
ников большую роль играет еще мельчайшая сеть весьма
230
многочисленных капилляров длиной до 0,5 мм и диаметром по-
рядка 0,01 мм. В них теряется на сопротивление около 1/4 об-
щего давления. В системе капилляров преобладают явления
осмотического характера, при которых может развиваться дав-
ление, превышающее атмосферное в 7—8 раз, что несомненно
влияет на упругость тканей. Как отмечалось, в плавниках
китообразных имеется большое количество различного типа
анастомозов.
Животным, обитающим в водной среде, присуща своя ге-
модинамическая специфика. Для них характерно явление бра-
дикардии. Оно состоит в том, что у животных при погружении
в воду в связи с необходимостью задерживать внешнее дыха-
ние пульс становится реже. Например, у ластоногих, в том чи-
сле тюленей, частота пульса в воде уменьшается в 10 и более
раз. Однако в отличие от сравнительно медленно плавающих
ластоногих у быстро плавающих дельфинов частота пульса
в воде снижается не более чем в 2 раза [52]. Этим еще более
подчеркиваются исключительные свойства кровообращения
дельфинов. Наибольший размах вертикальных колебаний точки
у развилки хвостового плавника дельфина составляет около
0,4 длины тела, т. е. у двухметрового дельфина 0,8 м, что от-
вечает разнице между систолическим и диастолическим давле-
нием крови. Очевидно, это неслучайные совпадения. Между тем
тюлени, имея резко выраженную брадикардию, вынужденно
машут своими ластами, как рыбы плавниками, в горизонталь-
ной плоскости. В сущности, гемодинамическими закономерно-
стями предопределяется возможность регулируемого гидроупру-
гого эффекта в плавниках дельфинов (при отмеченных морфо-
логических особенностях их сосудистой сети и тканей в дерме
и гиподерме).
Комплексные сосуды в плавниках дельфинов впервые были
обнаружены А. Г. Томилиным более 35 лет назад. В то время
существование этих необычных кровеносных сосудов связыва-
лось только с интенсивной терморегуляцией через плавники
у китообразных. Впоследствии к подобным суждениям пришли
и американские биологи. Действительно, несомненны разная
температура поверхности плавников и туловища (в воздухе до
10°C) и более резкие, чем в туловище, колебания температуры
на одном и том же плавнике дельфина. Однако существуют
различные представления о роли комплексных сосудов в плав-
никах как органах терморегуляции у китообразных. Одни ав-
торы придают наибольшее значение отдаче избытка тепла пла-
вающим животным через плавники, поскольку температура
тела с толстым подкожным слоем жира всегда постоянная и
выше температуры окружающей воды. Напротив, другие ав-
торы относят комплексные сосуды в плавниках к органам со-
хранения тепла, уменьшающим тепловые потери через пере-
охлаждаемые тонкие плавники, почти не содержащие жира.
231
Если иметь в виду только терморегуляцию, то непонятно,
чем вызывается резкая разница в строении и расположении
сети комплексных сосудов в спинном и хвостовом плавниках
одного и того же китообразного животного, почему сильно раз-
витая сосудистая сеть в хвостовом плавнике распространяется
от внешней поверхности внутрь, какую функцию выполняет об-
щий распределительный узел комплексных сосудов у дельфи-
нов и т. п. Наши современные детальные рентгенографические
и микроскопические исследования строения тканей и сетей ком-
плексных сосудов плавников, а также специально поставленные
эксперименты ясно показали, что система комплексных сосудов
служит важнейшим органом регулирования упругости тканей
плавников. Одновременно система участвует и в терморе-
гуляции плавников. Это согласуется с обычной мультифункцио-
нальностью органов у животных.
Комплексные артерио-венозные сосуды представляют собой
сложные гемодинамические образования, для подробного ана-
лиза которых требуются самостоятельные продолжительные и
разнообразные исследования. С целью моделирования регули-
руемого гидроупругого эффекта прежде всего важно выяснить
некоторые закономерности образования сети комплексных сосу-
дов в наиболее совершенном магистральном типе ветвления —
в хвостовом плавнике, например, обыкновенного дельфина
(рис. 137). С позиций гидродинамики представляет интерес, от
чего зависит значение угла между крупными распределитель-
ными артериальными сосудами (от срединного магистрального
сосуда хвостового плавника до конца его лопасти) и многочис-
ленными боковыми ответвлениями сосудов следующего порядка
величины в обе стороны лопасти. Характерно, что угол ответ-
вления всех комплексных сосудов в точке разветвления — оди-
наковый для артерий в случае деления потока артериальной
крови и для окружающих вен, т. е. в случае соединения веноз-
ных потоков, текущих в обратном направлении. Ответы на эти
вопросы при известных упрощающих предпосылках можно по-
лучить теоретически.
В 1956 г. автором было обобщено точное решение задачи
о боковой струе. Рассмотрен простой недеформированный на-
порный трубопровод постоянного сечения с боковым отверстием
в стенке, через которое вытекает часть (или вся) протекающей
в трубопроводе жидкости. Следует учитывать две особенности
бокового истечения жидкости: 1) боковое отверстие прикрывает
противоположная ему стенка трубопровода; 2) в трубопроводе
имеются транзитные скорости, которые могут быть направлены
различным образом относительно отверстия (случаи деления
потока и соединения потоков). Поэтому в боковых струях из-
меняется как наклон струи, так и ее сужение. Эти параметры и
подлежат определению.
Постановка задачи следующая. Рассмотрено установивше-
232
Рис. 137. Рентгенограмма артери-
альной сети комплексных сосу-
дов и мелких сосудистых развет-
влений правой лопасти хвостового
плавника обыкновенного дельфина
(а) и график распределения пло-
щадей участков лопасти (б), последовательно обслуживаемых боковыми арте-
риями второго порядка, ответвляющимися справа и слева от распределитель-
ной магистрали (свинцовая инъекция, уменьшено в три раза).
На графике можно видеть резкий излом ступенчатой линии распределения
площадей в тонкой концевой части лопасти плавника, где она в состоянии
изгибаться.
№ 2, 3, 12, 15. 20 — боковые ответвления.
еся течение (слева направо) несжимаемой идеальной жидкости
из трубопровода через отверстие в боковой стенке в условиях
плоского потенциального течения с образованием свободных
границ (рис. 138). Заданными являются: относительный размер
бокового отверстия и = Ь0/В = ш0/й, где Ьо, со0 и В, Q — соответ-
ственно ширина и площадь сечения отверстия и трубопровода;
безразмерный кинематический параметр £= ±Vi/V0= ±<7i/?o,
—где Vo, Qo и ± Vi, qi — соответственно граничные зна-
Рис. 138. Зависимость угла наклона Ро и коэффициента сжатия в свободной
боковой струи от относительного размера бокового отверстия а и отношения
взаимодействующих расходов £ в трубопроводе, р0, е=Л,1(о, В).
8 С. В. Першин
233-
чения скорости и расхода жидкости до отверстия (слева) и
после отверстия (справа).
Величины +Vi и отвечают случаю деления потока
в трубопроводе и являются транзитными скоростью и расходом
последующего потока. Величины —Vt и —соответствуют со-
единению двух потоков в трубопроводе ; это скорость и расход
противотока. В свободной боковой струе скорость в «сжатом»
сечении Ус и расход <72; при этом <7о = <71Ч-<7г в случае деления
потока и ^о + <?1 = ^2 при соединении потоков. Боковая струя на-
клонена к продольной оси трубопровода под некоторым углом
₽о, который отсчитывается от оси трубопровода, 0<р0<л/2.
Степень сужения струи выражается коэффициентом сжатия
струи e = 6/feo=Wc/Q. Задача состоит в определении значения
функций Ро = Л (о, I), e = f2(o, I).
В результате применения метода годографа скоростей
с трехкратным конформным отображением известных областей
комплексных переменных на вспомогательную полуплоскость
получены следующие выражения:
_L+JL.
1-5 ’
о ест
cos 00 = —
1 -о 2 о , . В„ , 1 Г 1 , 1 Ъ 0—£) +ест
_-S1n Ь- - С05 Р. In Ctg 4- + - [- + «г
—L[_L+w, f-lin + .
Я Lea (1-Ч)2 J (1-Ч)-£ест
Численные результаты решения показаны на рис. 138. Гра-
фики для угла выхода свободной боковой струи 0О и коэффи-
циента сжатия струи е являются весьма общими и дают реше-
ния в явном виде при различных режимах работы бокового от-
верстия. Левый вертикальный разрез графиков | = — 1 отвечает
случаю симметричной схемы соединения двух противополож-
ных потоков трубопровода в боковую струю; средний разрез
графиков £ = 0 соответствует тупиковой схеме истечения, когда
весь расход трубопровода qo поступает в боковую струю. Об-
ласти графиков, расположенные слева от среднего вертикаль-
ного разреза, отвечают схеме противопотока при соединении
двух потоков у бокового отверстия в трубопроводе, а области,
расположенные справа, — транзитной схеме деления потока.
Именно последние и представляют в данном случае наиболь-
ший интерес.
Чем меньше относительный размер бокового отверстия сг и
относительный транзитный расход жидкости £, тем больше
угол ответвления 0О свободной боковой струи и коэффициент
сжатия ее е. Обращаясь к рис. 137, замечаем, что в случае ар-
териальной сети хвостового плавника дельфина этот вывод
качественно вполне оправдан. Если в начале верхнего распре-
делительного крупного артериального сосуда боковые ответвле-
ния отходят под углом до Ро~85°, то в конце углы уменьша-
234
ются до Ро~2О°. Верхний распределительный артериальный со-
суд питает кровью практически равномерно всю площадь
лопасти хвостового плавника (крестиками на рис. 137, а обо-
значены боковые ответвления нижнего распределительного'
крупного артериального сосуда, которые не принимались в рас-
чет). Тем самым закономерность ветвления магистрального
типа артериальной сетки комплексных сосудов хвостового
плавника китообразных получила достаточное разъяснение.
Венозная сеть комплексных сосудов хвостовых плавников
отличается тем, что, во-первых суммарная площадь сечения
периферических вен намного больше, чем каждой одной цен-
тральной артерии, которую они окружают, а гидродинамиче-
ское давление в них, напротив, значительно меньше, и, во-вто-
рых, обратное течение крови в венах представляет более благо-
приятное по сопротивлению соединение потоков. Поэтому углы
ответвления вторичных сосудов на распределительном компле-
ксном сосуде определяются потоками артериальной системы.
Это подтверждается и малыми изменениями ро и е в условиях
даже еще менее благоприятной схемы противотока |<0 (см.
рис. 138). Достаточно серьезные допущения в использованном
выше решении — обычно принятые.
Теоретически покажем также, что при моделировании ре-
гулируемого гидроупругого эффекта в плавниках китообразных
можно в искусственных полимерных материалах изменять упру-
гость системы путем изменения внутреннего давления в жидко-
сти, заполняющей сеть каналов, образованных в материале си-
стемы. Для этого рассмотрим простейший случай однородной
прямоугольной резиновой пластины длиной L, шириной В и
высотой 2Н, в которой на расстоянии h по обе стороны от ней-
тральной оси имеется по п продольных каналов, каждый пло-
щадью со. В каналы с одного конца подается неагрессивная
жидкость, например вода, а в другом конце эти каналы глу-
хие, так что вода не течет, а находится под статическим давле-
нием. Пусть статическое давление воды в каналах верхней
зоны будет pi, а «в каналах нижней зоны р2. Для определен-
ности положим, что pi>p2 (рис. 139). Силу тяжести пластины
Рис. 139. Расчетная схема определения упругой линии прямоугольной ре-
зиновой пластины, армированной глухими каналами, заполненными под
изменяемым статическим давлением.
8*
235
S)
(pt+pjwn
M=(pi-pz)<i>nh.
Мц+tlMx
Рис. 140. Схема равновесия элемента канализированной пла-
стины (а) и эквивалентная ей расчетная схема (б).
не учитываем (например, пластина может быть подвешена
вертикально).
Найдем уравнение упругой линии резиновой пластины, ар-
мированной заполненными водой каналами. Рассмотрим равно-
весие вырезанного элемента пластины длиной dx (рис. 140, а).
Эту схему можно заменить расчетной (рис. 140, б), в которой
по оси пластины действует сила (pi+pziun и в любом сечении
постоянный момент Af=(pi—p2)(pnh = const. Точное дифферен-
циальное уравнение упругой линии пластины, как известно,
будет
1 = ± fy/dx* =
рх Vl 1 + (dyldx)*)* EI
где рх — радиус кривизны пластины в сечении х от момента
Afx = const. Таким образом,
d (dy/dx)_________(Pi — Pi) unit
V[1 + (dy/dx)*]» ~ El
(95)
После двухкратного интегрирования и определения произ-
вольных постоянных, имея в виду, что при х = 0, у = 0 dy/dx = 0,
окончательно получим
у=——! 1 _л/1-Г . (9б)
(Pl — Pi) tonh ( v L EI J J
Упругая линия пластины, в соответствии с направлением
действия момента Мх, будет изогнута в сторону, противополож-
ную большему внутреннему давлению pi>p2- При этом жест-
кость пластины EJ функционально связана с давлением р\ и
р2. Изменяя его, можно регулировать упругость системы. Дей-
ствительно, по формуле (96)
EI = [(pi—р2) ®n/i (х2 + у2)]/2у.
(97)
Эксперименты подтверждают теоретические выводы.
236
§ 33. Экспериментальное моделирование
и техническое использование гидроупругого эффекта
в плавниках китообразных
Машущий движитель — двухлопастный хвостовой плавник
крупного кита представляет собой внушительное зрелище
(рис. 141). Размах плавника у 20-метрового кита достигает
4 м и более. Плавник может быть полностью расправлен,
вплоть до заостренных оконечностей, или же последние могут
несколько обвисать, что зависит от вида китообразного и его
состояния, системы сухожильной оплетки и типа сосудистой
сети плавника. С помощью сухожильной оплетки, являющейся
плоским разветвлением в плавнике сухожильных округлых тя-
жей волокон — окончаний мощных продольных спинных и
брюшных мышц, хвостовой плавник поднимается и отпускается
в машущих движениях как единый локомоторный орган. Сосу-
дистой сетью комплексных артерио-венозных сосудов регули-
руется большая или меньшая упругость внутренних тканей бес-
костных лопастей хвостового плавника. Только в средней части
плавника проходят хвостовые позвонки.
Эффективность регулируемого гидроупругого эффекта в при-
роде можно показать сравнительными экспериментами на хво-
стовом плавнике живого и мертвого взрослого дельфина
(рис. 142). В опыте с извлеченным из воды живым дельфином
выровненный горизонтально хвостовой плавник был подвержен
нагрузке. Снизу плавника накладывали широкий ремень, кото-
рый с силой примерно 50 Н был натянут вверх динамометром.
Плавник деформировался по схеме ! так, что лишь тонкие
концы лопастей были резко загнуты вверх, а средняя часть ло-
пастей оставалась ровной, плоской и весьма упругой. Близкие
к поверхности плавника крупные кровеносные сосуды рель-
ефно выделялись. Напротив, в аналогичном опыте с хвостовым
плавником мертвого дельфина лопасти равномерно изгибались
по схеме II, т. е. весь плавник оказался податливым. Форма
Рис. 141. Фотографии с натуры хвостовых плавников пыряющих китов, под-
верженных регулируемому гидроупругому эффекту (виды сбоку и сзади)
237
I
Рис. 142. Схема форм деформации хвостового плавника морской свиньи
в воздухе от искусственной нагрузки.
/ — у живого дельфина средняя часть плавника не изгибается (проявляется гидро-
упругий эффект); II— у мертвого дельфина (через 12 ч после смерти) весь плавник
плавно изгибается; а — утолщенная часть плавника, мало деформируется; б — утон-
ченная оконечность плавника, легко деформируется.
деформированного хвостового плавника в этом случае напо-
минает таковую у эмбриона с не вполне заполненными кровью
кровеносными сосудами конечностей. В расчетном отношении
обе лопасти хвостового плавника являются консольными пла-
стинами, заделанными в продольном, наиболее мощном осевом
сечении плавника. Поскольку жесткость балки или пластины
при прочих равных условиях тем больше, чем меньше ее дефор-
мация (прогиб под нагрузкой), сопоставление схем деформа-
ции I и II на рис. 142 ясно показывает действие регулируемого
гидроупругого эффекта в хвостовом плавнике живого дельфина.
Гидроупругий эффект в спинном плавнике живого дельфина
(афалины) легко наблюдать при помещении его в ванну с во-
дой. Вначале, когда активно плававшего дельфина посадили
в ванну, поверхность его спинного плавника была ровной,
гладкой. А к исходу третьего часа пребывания дельфина
в ванне в малоподвижном состоянии на поверхности спинного
плавника его было замечено появление четко выраженных за-
павших желобков, соответствовавших залеганию наиболее круп-
ных комплексных артерио-венозных сосудов в направлении от
основания плавника к его вершине. При этом спинной плавник
был менее упругим, как бы несколько вялым. Желоба на плав-
нике оставались хорошо заметными до конца пребывания дель-
фина в ванне. Когда же афалину выпустили в бассейн и она
начала активно плавать, желоба на спинном плавнике полно-
стью расправились, исчезли.
Интересные наблюдения проведены с хвостовыми плавни-
ками тушек крупных эмбрионов (длиной 2,4 и 3,4 м), принад-
лежащих китам-сейвалам при подготовке их к рентгеносъемке.
238
После размораживания плавника и освобождения его сосудов
от свободно истекавшей крови, лопасти становятся мягкими,
наблюдается своеобразная «сетчатость» эпидермиса в плане на
спинной поверхности плавника, расправленного на столе. В на-
правлении от середины плавника к концам его лопастей виден
рельеф желобков над опавшими крупными комплексными ар-
терио-венозными сосудами. Если лопасть слегка согнуть, то
примерно в ортогональном направлении к продольной ее оси
образуются частые, через каждые I—3 мм, «морщины» эпидер-
миса. Направление последних в действительности изменяется
ют продольного-по оси плавника в средней его части до почти
поперечного в концевой части каждой лопасти, т. е. везде по
хорде профилей лопасти плавника. Если теперь удерживать на
столе только конец хвостового стебля, а сам хвостовой плавник
сдвинуть со стола (он окажется без опоры), то его лопасти под
действием собственного веса обвисают, однако различным об-
разом на разные стороны. На брюшную сторону хвостовые ло-
пасти отвисают совсем и могут быть искусственно подгибаемы
в трубочку. На спинную сторону лопасти хвостового плавника
не обвисают полностью и менее изгибаются (рис. 143). Оче-
видно, жесткость хвостового плавника в обе эти стороны суще-
ственно различная, что объясняется несимметричным распо-
ложением позвонков по толщине самого хвостового плав-
ника.
Затем инъекцировали под давлением с помощью медицин-
ского шприца рентгеноконтрастную жидкую массу (тонкотер-
тую свинцовую краску на бензине) в магистральный сосуд
хвостового плавника. Было замечено, что обвисшие лопасти
плйвника постепенно слегка расправляются по 'мере наполне-
ния сосудов краской. Начинал действовать искусственно со-
зданный регулируемый внутренним давлением гидроупругий
эффект. Если контрастную массу наливать через артерии, то
одновременно заполняются и оплетки вен комплексных сосу-
Но если наливать только вены, то контрастная масса
сосуды
ДОВ.
в артерии не поступает,
имеют такие анастомозы,
в которых кровь течет
только из артерий в вены.
В обратном направлении
течению крови препятст-
вуют имеющиеся в венах
клапаны. Судя по усилию
пальцев руки эксперимен-
татора примерно 20—
30 Н на шприц с краской
диаметром 2 см, статиче-
ское внутреннее давление
рентгеноконтрастной мас-
Следовательно, комплексные
собствен-
плавника
Рис. 143. Схемы обвисания под
ным весом лопастей хвостового
у тушки крупного эмбриона сейвала: а —
на брюшную сторону, б — на спинную сто-
рону.
23Q
сы в сосудистой сети хвостового плавника было около 0,07—
0,1 МПа.
Чтобы выявить различное воздействие на лопасти хвосто-
вого плавника его сухожильной оплетки под эпидермисом и
сети комплексных артерио-венозных сосудов в тканях плав-
ника, был проделан следующий опыт с оконечностью хвосто-
вого стебля тушки крупного эмбриона сейвала. Хвостовой сте-
бель, поперечный срез которого показан на рис. 144, закреп-
ляли на конце стола так, что лопасти хвостового плавника
свободно свешивались за краем стола. Когда в поперечном срезе
хвостового стебля пинцетами тянули за две пары округлых
тяжей сухожилий, расположенных в специальных оболочках
сверху и снизу стебля, то весь хвостовой плавник поднимался
или опускался, а его лопасти все же оставались обвисшими.
Однако когда дополнительно шприцем нагнетали под давле-
нием жидкость через магистральный сосуд хвостового стебля,
Рис. 144. Поперечный срез
хвостового стебля тушки
крупного эмбриона сейвала.
Видны: округлый позвонок
(в середине), парные сухожиль-
ные тяжи сверху и снизу по-
звонка. сосудистая сеть (маги-
стральный сосуд внизу позвонка,
мощные обводные и вертикаль-
ные артерии).
лопасти хвостового плавника рас-
правлялись — действовал гидро-
упругий эффект. Это воздействие на-
блюдалось и без натяжения сухо-
жильных тяжей хвостового стебля.
Тем не менее несомненно, что прак-
тически нерастяжимая внешняя сухо-
жильная оплетка лопастей хвостового
плавника повышает эффективность
регулирования упругости внутренних
тканей лопастей с помощью сети ком-
плексных артерио-венозных сосудов.
Систематические эксперимен-
тальные исследования, результаты ко-
торых приведены в статье С. В. Пер-
шина, Е. П. Носова, В. П. Исакова
(Бионика-1973. T. 6. АН СССР. 1973.
С. 93—99), были проведены на спе-
циально изготовленном испытатель-
ном стенде (рис. 145).
Природа гидроупругого эффекта
в деталях весьма сложная. С помо-
щью предварительного моделирова-
ния стремились экспериментально
изучить регулирование деформаций и
жесткости упрощенной модели в ста-
тических условиях нагружения. Мо-
дель изготовляли из резины. По
характеру деформаций и упругим
свойствам резина приближается к ор-
ганическим тканям и одновременно
является вероятным синтетическим
240
Рис. 145. Схема экспериментального
стенда для испытания гидроупругого
эффекта в заделанной канализирован-
ной резиновой пластине.
1 — консольная модель-пластина; 2 —
блок крепления модели с двухсторонней
подачи жидкости; 3 — гидравлический
поршневой цилиндр; 4 — контрольный ма-
нометр; 5 и 6 — перепускные трубки с за-
порными кранами; 7 и 9 — двухстороннее
весовое устройство с блоками; 8 — гори-
зонтальная шкала.
Б0мы40 20 0 20 40 60ми
hji8^— —*-Лпр
Рис. 146. Резиновая модель
пластины с канализирован-
ными двумя рядами внутри
глухих отверстий, заполняе-
мых водой под переменным
статическим давлением, и
с внешним двухсторонним
весовым устройством (см.
рис. 145).
материалом для технических изделий с использованием гидро-
упругого эффекта. Модель формовали в виде заделанной од-
ним концом пластины, в соответствии с расположением спин-
ного и обеих лопастей хвостового плавника на корпусе кито-
образных. Для заделки пластины один монолитный конец ее
был утолщен (рис. 146). Он же служил для крепления модели
на экспериментальном стенде и подачи жидкости под перемен-
ным статическим давлением.
Основная схематизация модели заключалась в следующем.
Очень сложные по строению и функциям кровеносные компле-
ксные пучки сосудов плавников имитировали в исследуемых
резиновых пластинах одиночными внутренними продольными
цилиндрическими глухими полостями, которые были располо-
241
жены в один или два ряда по толщине пластины, а концы их
соединены между собой по рядам. Полости заполняли водой
под избыточным, регулируемым гидростатическим давлением
с предварительным удалением из них воздуха. На модели не
была воспроизведена капиллярная сеть кровеносных сосудов
плавников, поскольку ее невозможно осуществить при совре-
менной технологии изготовления резины. Не воспроизводили и
покровную сухожильную оболочку плавников, хотя имитация
таковой возможна с применением кордовой ткани. Принятая
нами схематизация модели оправдана простым техническим
выполнением, причем заведомо с запасом по эффективности
против возможной после модернизации.
Схематизированные модели были в рабочей части прямо-
угольной формы как в плане, так и в поперечном сечении, сле-
дующих размеров: длина / = 200, ширина (высота) Ь = 70 мм,
толщина в двух вариантах — тонкой модели / = 5 и толстой
/=12 мм. Тонкая модель имела один ряд внутренних поло-
стей, оси которых расположены в срединной продольной пло-
скости, проходящей через нейтральную ось поперечного сече-
ния, подобно однорядному расположению крупных комплекс-
ных сосудов в спинном плавнике китообразных. Толстая
модель имела два независимые, не связанные между собой
(левый и правый) ряда полостей, расположенные вблизи обеих
боковых плоскостей пластины, на одинаковых расстояниях от
нейтральной оси (см. рис. 146), подобно двухрядному распо-
ложению комплексных сосудов в хвостовом плавнике. Диа-
метр полостей был d=l—3 мм, а число их выбирали из рас-
чета, чтобы общая площадь просветов полостей составляла от
3 до 12 % всей площади поперечного сечения пластины (учиты-
валась относительная емкость кровеносной сети в плавниках).
В качестве исходного материала использовали сырую ли-
стовую резину с относительным удлинением 560—600 и оста-
точным удлинением 8—12 %. Для формирования резиновых
пластин и вулканизации их под давлением при температуре
150 °C, специально изготовили массивные стальные разборные
пресс-формы. Сменные детали пресс-формы, в виде съемных
кондукторов и закладных стальных спиц, обеспечивали в про-
цессе изготовления пластины правильное расположение и необ-
ходимые размеры внутренних свободных полостей. В утолщен-
ном конце пластины были размещены металлические полые
штуцеры для подачи воды в полости.
Опыты проводили на испытательном стенде (см. рис. 145),
в воздушной среде. Канализированную резиновую пластину
укрепляли на стенде горизонтально, в положении на ребро,
для исключения влияния силы тяжести на учитываемую дефор-
мацию. Пластину нагружали последовательно на свободном
конце, слева и справа, переменной по знаку и величине внеш-
ней сосредоточенной горизонтальной силой Слв, Gnp. В запол-
242
ненных водой внутренних полостях пластины создавали кон-
тролируемое по манометру избыточное гидростатическое дав-
ление, причем в случае толстой пластины отдельно в левом и
правом рядах рлв, рПр- При различном сочетании нагрузок на-
блюдали за деформацией модели и непосредственно измеряли
по горизонтальной шкале стенда стрелку прогиба hлв» Л-пр конца
пластины (см. рис. 146). Основные результаты опытов следую-
щие.
Деформации и жесткость канализированных резиновых
пластин под нагрузкой существенно различались по характеру
и значениям в зависимости от расположения и числа рядов
действующих внутренних полостей и вида нагрузки пластины.
В случае тонкой пластины с одним симметрично расположен-
ным в сечении рядом внутренних полостей избыточное над ат-
мосферным гидростатическое давление р приводило всегда
к уменьшению деформации h, а потому соответствующему по-
вышению жесткости EJ пластины при сравнимой, одной и той
же внешней нагрузке от сосредоточенной силы G, справа или
слева на конце пластины. Для пластины с десятью внутрен-
ними полостями, общая площадь просвета которых (после
усадки резины) составляла 6 % or всей площади попереч-
ного сечения, отмеченные изменения деформации и жесткости
достигали 40%. Поскольку оси этих лопастей расположены
в срединной продольной плоскости пластины, по свойствам ра-
боты резины на растяжение и сжатие, полезный эффект был
обусловлен растягивающими напряжениями в пластине только
с одной стороны полостей, да и то вблизи нейтральной оси ра-
бочего сечения. Проверено, что деформации пластины под
внешней нагрузкой, но с незаполненными водой полостями, и
при заполнении полостей, но без избыточного гидростатиче-
ского давления (р = 0) одинаковые.
Наибольший интерес представляют эксперименты с толстой
пластиной, имеющей двухрядное расположение внутренних по-
лостей, вдали от нейтральной оси поперечного сечения пла-
стины. Стрелка прогиба свободного конца пластины от одной
внешней сосредоточенной силы h(G, 0) следует линейной, а от
одного избыточного гидростатического давления во внутренних
полостях h(0, р) — практически экспоненциальной зависимости
(рис. 147):
h (G, 0) = kG, h. [0, р) = ± (exp) р.
Прогиб пластины в отмеченных случаях различается по на-
правлению. От действия только внешней горизонтальной силы
Оле или Опр пластина изгибалась, конечно, в направлении этой
силы, с соответствующей стрелкой прогиба ЛЛв или Лпр. От дей-
ствия только гидростатического давления в одном ряду внут-
ренних полостей слева или справа (рЛв или рпр) пластина
243
Рис. 147. Деформации испы-
танной канализированной
резиновой модели-пластины.
h — отклонения конца пластины;
р — внутреннее статическое дав-
ление жидкости; G — внешняя
нагрузка; обозначения в индек-
сах: лв и пр —левая и правая
стороны.
изгибалась в противо-
положных направле-
ниях (Йпр или Ллв).
Такая деформация
объясняется нали-
чием поперечного мо-
мента сил, обуслов-
ленного действием
гидростатического да-
вления полостей
в направлении вдоль
пластины.
В тех случаях, ко-
гда приложены оба
вида нагрузок, спра-
ведливо наложение
деформаций, что весьма облегчает и упрощает регулирование
по заранее заданной программе деформаций, а следовательно, и
жесткости канализированных резиновых изделий. При этом бу-
дем иметь
h(G, p) = h(G, 0)±/i(0, p) — kG ±(exp)p. (98>
Здесь суммирование алгебраическое, так как в зависимости от
расположения ряда внутренних полостей относительно ней-
тральной оси сечения пластины, повышение избыточного
гидростатического давления р воды в них вызывает увеличение
или уменьшение стрелки прогиба h пластины при одной и той
же внешней нагрузке G.
В соответствии с известной из теории сопротивления мате-
риалов зависимостью для стрелки прогиба однородной пла-
стины (балки), заделанной в одном конце, относительное из-
менение жесткости EJ модели, где Е — модуль упругости ре-
зины на изгиб, / — момент инерции площади поперечного
сечения пластины, приближенно будет равно обратному отно-
шению прогибов в двух сравниваемых случаях, а именно:
(EI)n ~ GnP . G„P hQ ,
(Е1)о mhn ' mh0 hn
при условии, ЧТО Gn=G0, Рп^Ро-
244
В условиях проведенных экспериментов с резиновой пласти-
ной, которая имела 12 внутренних полостей в каждом из двух
их рядов, с общей площадью просвета сечения 4,5%, было-по-
лучено уменьшение деформации в 3,6 раза и повышение ее
в 2,0 раза, а потому и соответствующие изменения жесткости
пластины, т. е. весьма значительные, при сравнительно неболь-
шом гидростатическом давлении во внутренних полостях
0,4 МПа. Эффективность регулирования можно существенно
повысить за счет более рационального расположения и выбора
числа внутренних полостей, а также применения кордовых тка-
ней в качестве наружных слоев резиновых пластин.
Таким образом, проведенные систематические эксперименты
на упрощенных моделях — резиновых пластинах — полностью
подтвердили концепцию авторов регулируемого гидроупругого
эффекта в плавниках китообразных [17] и ясно показали суще-
ственное различие в регулировании жесткости пассивного
(спинного) и активного (машущего хвостового) плавников
этих животных. Одновременно эксперименты позволили сде-
лать ряд предложений по техническому использованию схема-
тической модели гидроупругого эффекта, которые защищены
авторскими свидетельствами на изобретения.
Следует подчеркнуть, что эксперименты проведены в стати-
ческих условиях нагружения модели. В динамических условиях
гидроупругости эффект может проявиться иначе.
ЗАКЛЮЧЕНИЕ
Гидробионика представляет собой новое интересное и важ-
ное научное направление. В нашей стране она получила раз-
витие начиная с 60-х годов. Развитию гидробионики способ-
ствовала координация совместных усилий ученых-биологов,
гидродинамиков, математиков, в частности, путем проведения
ряда всесоюзных конференций и издания ежегодных выпусков
межведомственных сборников по бионике. Советские ученые
внесли большой вклад в развитие гидробионики своими теоре-
тическими исследованиями и экспериментальными работами,
преимущественно с такими крупными гидробионтами, как ско-
ростные рыбы и дельфины. За истекшие 20—25 лет в нашей
стране определился состав специалистов-гидробиоников, энту-
зиастов своего дела, зарегистрированы научные открытия и
десятки изобретений, появились исследования, представляющие
непосредственный интерес для судостроения и водного транс-
порта.
Главными особенностями плавания водных животных-
гидробионтов, заслуживающими внимания применительно к за-
дачам техники, являются следующие: нестационарность движе-
ния (обычно гармонического типа); исключительная маневрен-
ность, практически в любых направлениях; высокая
эффективность плавания с использованием средств снижения
гидродинамического сопротивления, не применяемых в технике;
широкое регулирование обычно нелинейных функций движе-
ния. Нестационарность, маневренность и эффективность плава-
ния рыб и китообразных обусловлены волновыми движениями
гибкого упругого тела и машущими изгибными колебаниями
плавникового движителя, при этом тело и движитель совме-
щены в едином двигательно-движительном комплексе. Широкое
регулирование режимов движения приобретено живым орга-
низмом в эволюции при обитании в плотной водной среде
с многими переменными факторами внешнего воздействия.
Важнейшими проблемами исследований прикладного значе-
ния в гидробионике до настоящего времени были: изучение
246
главного — волнового —плавания и его разновидностей у из-
вестных видов крупных гидробионтов, исследование способов
снижения гидродинамического сопротивления в природе, воз-
можность применения в технике новых типов волновых и ма-
шущих движителей. В данной монографии эти и другие про-
блемы разработаны и обобщены на основе детального изучения
морфологии, кинематики, динамики и биоэнергетики плавания
крупных гидробионтов с использованием многочисленных лите-
ратурных источников. Для различных видов рыб и китообраз-
ных выявлены как общие гидродинамические закономерности,
так и существенные различия, свойственные им в связи со зна-
чениями определяющих критериев подобия Рейнольдса и
Струхаля.
В результате функционально-морфологического анализа
крупных гидробионтов получена общая качественная и количе-
ственная характеристика закономерного ряда видов рыб и
китообразных по форме тела, очертаниям машущих плавнико-
вых движителей в плане и в профиле, выраженная в табличной
и графической форме. Выявлены морфологические признаки
скоростных гидробионтов в направлении оптимизации плава-
ния в природе по гидродинамическим критериям. С повыше-
нием числа Рейнольдса хорошо обтекаемые каплевидные тела
и профили плавников скоростных гидробионтов приобретают
ламинаризованные формы до чисел Re«107 и выше. У рыб
мягкие и податливые двухлопастные лучевые пластинчатые
плавники сменяются более жесткими, а затем и крыловыми.
Впервые установлены гидробионические закономерности строе-
ния кожного, чешуйного и слизистого покровов рыб, в том
числе скоростных, с пониженным гидродинамическим сопротив-
лением при использовании ими инъекции в пограничный слой
тела биополимеров рыбьей слизи.
Кинематику нестационарного плавания и одновременно ре-
гулирующее значение числа Струхаля изучали в зависимости
от размеров тела и скоростей плавания гидробионтов, в гидро-
динамических циркуляционных трубах и путем буксировки
в опытовых бассейнах, проводили опыты в морских бассейнах и
открытых лагунах, с учетом биологических особенностей адап-
тации гидробионтов к условиям эксперимента. Широко при-
меняли подводную фотографию и киносъемку, что позволяло
определять скорость плавания, различные формы плаватель-
ных движений, частоту и амплитуду колебаний движителей
гидробионтов во взаимосвязи с существенными для биоэнерге-
тики организма его размерами и продолжительностью плава-
ния при каждом данном режиме. Выделены и получили коли-
чественную оценку угревидная, скомброидная и плавниковая
формы волнового плавания рыб. Нормированы максимальные
скорости плавания китообразных в природе и в неволе. Для
скоростного дельфина-афалины получен типичный режимный
Г" V
247
график прямолинейного нестационарного волнового плавания
с различными мгновенными характеристиками движения.
Предложена система пространственно-временных характери-
стик нестационарного плавания различных видов гидробион-
тов, в том числе критериальный график Sh(Re). Большинство
характеристик нелинейные. Все их можно использовать для
динамического расчета плавания гидробионтов. В качестве од-
ного из практических приложений рассмотрены гидробиониче-
ские аспекты спортивного плавания способом «брасс» и пред-
ложены рекомендации по улучшению техники плавания этим
наиболее распространенным способом.
Много исследований выполнено по биогидродинамике. Вы-
явлены гидродинамические области типичных групп гидробион-
тов в широком интервале чисел Рейнольдса Re = 10~®4-108, от
микроскопических простейших до гигантских китообразных.
Гидробионты каждой области обладают своими характерными
особенностями по способам плавания, типам движителей, ха-
рактеру течений в пограничном слое тела. Выделены некоторые
общие принципы гидробионики. Анализ и сопоставление харак-
терных случаев плавания гидробионтов в природе позволяет
отнести их к адаптационным колебательным системам, кото-
рым иногда свойственны энергетически эффективные режимы,
известные в технике, как резонансные.
Разработаны различные модификации математического мо-
делирования динамики волнового плавания гидробионтов. Наи-
больший теоретический интерес и возможные практические
приложения имеет примененная акад. АН УССР Г. В'. Логви-
новичем гидродинамическая модель удлиненного гибкого тела,
развитая им в пространственной теории волнового плавания
водных животных. Это позволяет по заданной кинематике дви-
жения гидробионтов рассчитывать средние значения силы тяги
и кпд и проводить сопоставления гидродинамических характе-
ристик различных видов гидробионтов. Проведены разнообраз-
ные теоретические и экспериментальные исследования гидро-
динамики изолированных машущих плавниковых движителей.
Рассмотрены случаи оптимизации работы изолированных ма-
шущих движителей и выявлены возможности проведения про-
стых расчетов в квазистационарном приближении. В результате
раздельного гидродинамического моделирования показано зна-
чение нестационарности движения, вызванной различными
причинами: неравномерным поступательным движением твер-
дого тела и деформациями разного типа гибкого тела. Иссле-
дован монопласт как эффективный волновой движитель для
скоростного плавания человека способом «дельфин».
Биоэнергетика плавания крупных гидробионтов ввиду био-
логических особенностей живых организмов представляет со-
бой сложную и трудно разрешимую комплексную проблему.
Однако некоторые положительные результаты исследования
248
этой проблемы нашли отражение в книге. В частности, выяв-
лены диапазон и характер регулирования биоэнергетики не-
которых гидробионтов. Рассмотрены прыжки дельфинов с ма-
ксимальной кратковременной отдачей мощности и продолжи-
тельное ныряние водных млекопитающих. Применен метод
аналогов в биоэнергетике и получена расчетная зависимость
саморегуляции отдачи мощности крупного гидробионта в функ-
ции продолжительности плавания; учитывается относительная
масса мышц гидробионта. Предложен квазистационарный био-
энергетический расчет плавания дельфинов и показано соответ-
ствие такового нестационарному гидродинамическому расчету.
Выявлено приближенное механическое подобие плавания видов
гидробионтов и существование масштабного динамического эф-
фекта.
В результате многолетних и разнообразных биологических
и гидродинамических исследований впервые установлено свой-
ство кожи китообразных активно регулировать гидродинамиче-
ское сопротивление плаванию. Тем самым получил более пол-
ное разъяснение известный «парадокс» Грея. Локальное
взаимодействие кожного покрова дельфинов с обтекающим
водным потоком происходит на основе рецепторной чувстви-
тельности кожи и вазомоторной изменчивости кровеносной си-
стемы, автоматически вызывающей изменения упругодемпфи-
рующих слоев кожи соответственно режиму плавания. Под-
робно изучены структурные и гистологические особенности
кожного покрова китообразных, тело которых имеет ламинари-
зованные формы. Выявлена гидродинамическая функция их
кожного покрова, разработана механическая модель демпфи-
рующего свойства кожи и установлены соответствующие крите-
рии механического подобия. Экспериментально исследованы
упругодемпфирующие характеристики кожи черноморских
дельфинов. Проведены опытные исследования пограничного
слоя у активно плавающего в морском бассейне дельфина.
Разработаны основы технического моделирования искусствен-
ных полимерных демпфирующих покрытий для снижения
гидродинамического сопротивления трения.
Впервые установлено также явление саморегулирования
упругости плавников китообразных соответственно режимам
плавания, обусловленное действием широко разветвленных уни-
кальных комплексных артерио-венозных сосудов в плавниках
и наличием практически нерастяжимой внешней сухожильной
оболочки плавников. Проведены многочисленные рентгеногра-
фические исследования распределения и строения сосудистой
сети в плавниках многих видов китообразных и показано раз-
нообразие этой сети магистрального и рассыпного типов. Вы-
полнены детальные гистологические исследования тканей плав-
ников и комплексных кровеносных сосудов, показано различие
их сетей в хвостовом и спинном плавниках дельфинов и раз-
249
личные формы образования комплексных сосудов. Сделан тео-
ретический анализ гидроупругого эффекта в плавниках и вы-
полнено экспериментальное моделирование такового. Предло-
жено технически целесообразное использование этого эффекта
в различных гибких изделиях с регулируемой жесткостью и
деформацией, выполненных из полимерных материалов, напри-
мер, резины.
Как видно, трудами многих ученых созданы прочные ос-
новы гидробионики. При моделировании изученных полезных
адаптационных приспособлений эффективно плавающих гидро-
бионтов следует иметь в виду, что эти приспособления воз-
никли в эволюционном развитии живых организмов и не всегда
могут быть применены в технике. Ламинаризованные формы
тела гидробионтов являются необходимой предпосылкой для
использования в природе любых дополнительных средств сни-
жения гидродинамического сопротивления (демпфирование
кожного покрова у китообразных или биополимерной слизи
у рыб). Однако выбор плоскости колебаний машущего плавни-
кового движителя в технических изделиях не ограничен, так
как вертикальные или горизонтальные колебания у гидробион-
тов обусловлены не гидродинамической причиной, а различ-
ными физиологическими актами дыхания водных животных.
Очевидно, существенная зависимость мощности двигательно-
движительного комплекса гидробионтов от продолжительности
плавания не является лимитирующим фактором для техниче-
ского моделирования. Исключительная маневренность многих
видов гидробионтов отчасти может быть обусловлена отсут-
ствием у них реверса движителей, который, однако, восполним
при моделировании в технических изделиях.
Касаясь возможных последующих исследований и дальней-
шего развития гидробионики, на основании вышеизложенного
отметим некоторые задачи гидродинамики. Следует продол-
жить теоретические исследования нестационарного движения
объектов с волновым и. машущим плавниковым движителем
с целью разработки практически удобных методов расчета,
в том числе с применением новейшей вычислительной техники.
Необходимо уделить большее внимание непосредственно вопро-
сам моделирования, в том числе изучению масштабного эф-
фекта для правильного внедрения в практику достигнутых
результатов. Целесообразно развивать теорию по изучению эф-
фектов гидроупругости изделий, используя при этом математи-
ческий аппарат как гидродинамики, так и теории упругости.
В экспериментальных гидродинамических исследованиях
назрела необходимость дальнейшего прогресса в нескольких
направлениях. Следует расширять выбор видов подопытных
гидробионтов с волновым и машущим движителями (с учетом
их больших размеров и возможных испытаний при высоких
скоростях плавания). Желательно также присовокупить к под-
250
робным и тщательным исследованиям крупных гидробионтов
исследования микроскопических простейших, которые играют
большую роль во многих технологических процессах. Не сле-
дует далее ограничиваться изучением только ходкости типич-
ных гидробионтов; настало время серьезно заняться и манев-
ренностью плавания их. Необходимо дальнейшее развитие
экспериментального оборудования и аппаратуры, в особенности
для изучения пограничного слоя на телах нестационарно пла-
вающих гидробионтов.
По нашему мнению, весьма желательно как в общенаучных
целях, так, возможно, и в практическом отношении экспери-
ментальное исследование плавания характерных в гидродина-
мическом отношении видов рыб в водных среде и потоке с раз-
личной начальной турбулентностью («от спокойного водоема
до бурного или порожистого водотока»). Несомненно, что по-
добные тщательные исследования плавания одних и тех же
особей в различных, даже искусственно созданных условиях
позволят получить новые сведения для изучения развития тур-
булентности.
Биологические исследования применительно к гидробио-
нике в дальнейшем могут дать полезные результаты по био-
энергетике плавания живых организмов. Необходимо также
провести систематические исследования и сравнения двигате-
лей— мышц различных гидробионтов — и конструкций при-
вода их машущих плавниковых движителей (типы сухожиль-
ных тяг, различное крепление их и т. п.).
ПРИЛОЖЕНИЕ
НЕКОТОРЫЕ ХАРАКТЕРИСТИКИ ПЛАВАНИЯ
МИКРООРГАНИЗМОВ
Микроорганизмы очень широко распространены в природе и в биохимиче-
ских технологических процессах и представляют большой интерес для изуче-
ния во многих современных отраслях знания: микробиологии, биомедицине,
биофизике, бионике, реологии, механике суспензий, физике аэрозолей, гидро-
механике. Плавание микроорганизмов происходит при весьма малых числах
Рейнольдса Re=10_,4-l и в биогидродинамике вызывает интерес природой
нестационарных медленных ползущих течений, которые сравнительно мало
изучали в классической теории. В ползущих течениях преобладающее значение
имеют силы вязкости среды, а силы инерции пренебрежимо малы, что пред-
определяет особенности плавания микроорганизмов, которые в последние годы
интенсивно изучаются. Так, в Калифорнийском технологическом институте США
была создана специальная исследовательская группа в составе крупных уче-
ных биологов и механиков, которая работает по комплексной программе. Про-
водятся биофизические наблюдения и физиологический анализ поведения про-
стейших организмов, создается и развивается математический анализ теорети-
ческих моделей их плавания, выполняется механическое моделирование вари-
антов различных механизмов плавания и др.
Литература по биогидродинамике плавания при малых числах Рейнольдса
ранее была рассмотрена и проанализирована в нашем научном обзоре [50]
(обобщающие и обзорные работы, различные исследования по многим вопро-
сам плавания жгутиковых и ресничных организмов, новые задачи в изучении
пропульсации одноклеточных организмов). В настоящей монографии плаваю-
щие микроорганизмы отнесены к гидробионической области 1 (см. с. 96—97,
рис. 43). Ниже частично приведена краткая характеристика их плавания по
работе [49] (опущены все иллюстрации). Это позволяет судить о том, на-
сколько существенно отличается плавание микроорганизмов от плавания
крупных гидробионтов — объектов исследования в данной книге.
Различные виды многочисленных микроорганизмов внешне весьма разно-
образны, однако выделяются более или менее короткое утолщенное тело
клетки и относительно длинные и тонкие органеллы-движители. Последние
представлены в основном жгутиками, ресничками, в связи с чем микроорга-
низмы соответственно называются жгутиковыми и ресничными. Поступатель-
ное движение микроорганизмов осуществляется при волновом изгибании жгу-
тиков и ресничек, в зависимости от вязкоэластических свойств самого орга-
низма и среды, в которой он плавает. С применением электронной микроскопии
показана единообразная ультрамикроструктура органелл-жгутиков и ресни-
чек, однако формы движения микроорганизмов весьма различные.
Жгутиковые микроорганизмы плавают посредством волновых движений
одного или нескольких жгутиков обычно длиной 5—100 и диаметром 0,1—
0,2 мкм, при частоте волн 10—100 Гц и амплитуде 0,1—10 мкм. Волны бывают
нескольких типов: а) плоская бегущая волна,' иногда близкая к синусоидаль-
ной, которая распространяется по жгутику от головы организма к концу жгу-
252
тика (или наоборот от конца жгутика к голове); б) винтовая бегущая волна;
в) трехразмерная волна с вариациями более простых предыдущих — нерегу-
лярная, спиральная и др.
Ресничные микроорганизмы, в отличие от жгутиковых плавают с помощью
множества, иногда порядка 10‘, более мелких волосоподобных органелл-рес-
ничек длиной 2—15 и диаметром 0,02—0,2 мкм, выступающих на поверхно-
сти микроорганизма. Каждая ресничка, последовательно изгибаясь, совершает
гребущие движения — периодические циклы колебаний, с частотой 10—20 Гц,
которые состоят из быстрой фазы активного маятникового удара вытянутой
реснички относительно точки закрепления ее на поверхности тела и медленной
фазы пассивного возвратного стелющегося движения различной формы у раз-
ных микроорганизмов. При совокупном биении множества ресничек на по-
верхности тела микроорганизма образуются координированные метахрональ-
ные волны, в жидкости создаются тангенциальные усилия. Фронт и направле-
ние распространения метахрональных волн могут быть различно ориентиро-
ваны относительно направления поступательного движения тела и направле-
ния удара ресничек. По отношению к последнему различают метахрональные
волны совпадающего, противоположного, смешанного и поперечных направ-
лений.
Следует подчеркнуть, что движение микроорганизмов весьма сложное.
Например, простейшая ресничная — инфузория парамециум — при свободном
плавании в воде имеет траекторию движения в форме вытянутой спирали,
диаметр которой может сильно варьировать под влиянием различных химиче-
ских реагентов и одновременно ее тело вращается вокруг продольной оси.
Вращение тела возможно влево и вправо, что в экспериментах наблюдается
под влиянием изменения таких факторов, как вязкость, температура, состав
солей среды.
Общее представление об основных параметрах жгутиковых и ресничных
дает четырехпараметрический график (см. рис. 43). Видно, что минимальная
относительная скорость плавания микроорганизмов Um= Um/L=0,3-10 с-1,
однако, как максимум, бывает равна и 20—100 с-1. Числа Рейнольдса, рассчи-
танные по длине тела (т. е. для жгутиковых без учета длины движителя I,
с целью единообразного представления с ресничными микроорганизмами),
Reb=10-B-*-l. Кинематический коэффициент вязкости среды принят, для срав-
нимости данных, одинаковым v=l-10-’ мг/с (для воды <=20°С).
Характерно, что различные группы жгутиковых и ресничных занимают до-
статочно компактные подобласти а и б области 1 на рис. 43. Далее кратко
остановимся только на параметрах плавания жгутиковых микроорганизмов,
которые имеют нечто общее с волновым плаванием животных при значительно
больших числах Рейнольдса, например, мелких рыб. Эффективность плавания
жгутиковых зависит от многих факторов: формы и размеров тела; числа и
размеров жгутиков, их амплитуд и частоты колебаний; формы, скорости рас-
пространения и числа одновременно бегущих волн жгутика; вязкости, темпе-
ратуры и солевого состава жидкой среды и др. Тело жгутиковых обычно при-
ближается к сфере или эллипсоиду, не всегда симметричное относительное
удлинение тела L/B= 1-5-5, т. е. небольшое, но у спирохет цилиндрической
формы удлинение достигает 40.
Частота колебаний жгутиков в вязкой среде варьирует у различных рас-
смотренных видов жгутиковых п довольно широком диапазоне «=74-70 Гц.
При этом она находится в тесной зависимости с относительной амплитудой ко-
лебаний, которая с учетом как длины тела L, так и длины жгутика I выражается
величиной X/L=0,02-5-0,22. В качестве примера приведены данные по частотно-
скоростной характеристике плавания жгутиковой бактерии-салмонеллы. Эта
бактерия имеет тело длиной £=4 и шириной В=1 мкм, жгутик длиной 1=7 и
диаметром d=0,12 мкм. Длина спиральной волны жгутика Х=2,2, радиус
волны В=0,22 мкм. В процессе поступательного плавания тело бактерии вра-
щается, при этом частота полных циклов вращения <р=1ч-5 с-1. Характерно,
что ее частотно-скоростная характеристика имеет линейный вид, причем в дан-
ном случае n/<p=4 = const.
Отметим также влияние вязкости среды ц на частоту п, амплитуду А и
253
длину волны X жгутиков двух видов сперматозоидов. Вязкость среды повы-
шали в эксперименте в широком интервале значений до 40 раз. Между тем
значения указанных параметров снижались при этом не столь сильно, менее
интенсивно п и X (в 1,5—3 раза), более интенсивно Я (в 5 раз).
Два обстоятельства указывают на эффективность плавания микроорганиз-
мов, приспособленность их к движению в условиях преобладающего влияния
сил вязкости среды. У жгутиковых микроорганизмов сравнительно высокая
относительная скорость плавания. Максимальное, резко выделяющееся значе-
ние (7m=200 с-1 (см. рис. 43). Вместе с этим у них небольшая относительная
масса движителя — жгутика т по отношению к массе тела М, например, для
микроорганизма длиной тела £=2-10-в м с длиной жгутика (=5-10-в м при-
близительно т/М= 1,5-10-3=0,15 %. Однако встречаются и другие, сущест-
венно отличающиеся соотношения.
В результате экспериментальных исследований с применением электрон-
ной микроскопии возникла проблема, почему двигательно-движительная си-
стема весьма распространенных жгутиков и ресничек самых различных клеток,
простейших организмов, сперматозоидов наиболее часто имеет девять перифе-
рических ультраструктур, которые в поперечном сечении органеллы равно-
мерно распределены по окружности через каждые 40° дуги. В нашей работе
впервые показано, что именно девять периферических ультраструктур создают
наиболее устойчивое движение этих органелл, в отличие от иного числа (5—
12) ультраструктур (см. С. В. Першин. О преимуществе в устойчивости дви-
жения органелл с девятью периферическими ультраструктурами//Доклады
АН СССР. 1986. Т. 291. Xs 2. С. 458—462).
СПИСОК ЛИТЕРАТУРЫ
1. Агарков Г. Б., Хоменко Б. Г., Хаджинский В. Г. Морфология дельфи-
нов. Киев. Наукова Думка, 1974, 167 с.
2. Алеев Ю. Г. Функциональные основы внешнего строения рыбы. М.:
АН СССР, 1963, 247 с.
3. Алексеева К. Д. Затраты энергии на движение у кефалей//Бионика.
Киев. Наукова Думка. 1972. Вып. 6. С. 7—12.
4. А. с. 413286 СССР, МКИ F 15d 1/12, Демпфирующее покрытие/В. В. Ба-
бенко, Л. Ф. Козлов и С. В. Першин (СССР). Заявка 1807892/25-8 от 11.07.72.
опубл. 30.01.74, Бюлл. № 4, 3 с.
5. А. с. 441188 СССР, МКИ ВбЗв 1/28, Крыло/С. В. Першин, Е. П. Но-
сов, В. П. Исаков и К. М. Петров (СССР). Заявка 1649717/27-11 от 26.04.71,
опубл. 30.08.74, Бюлл. № 32, 3 с.
6. А. с. 738626 СССР, МКИ А 63 В 31/08, Ласт/Е. П. Носов, Г. Н. Ор-
лов, В. И. Кебкало, С. В. Першин, М. Я- Туренко, С. А. Доос и> М. С. Ры-
бакова (СССР). Заявка 2429669/28-12 от 13.12.76., опубл. 05.06.80, Бюлл.
№ 21, 3 с.
7. Ахмеров У. Ш. Методологические вопросы бионики. Казань: Изд-во
Казан, ун-та, 1977, 69 с.
8. Бабенко В. В. Исследование упругости кожи живых дельфинов//Био-
ника. Киев: Наукова Думка. 1979. Вып. 13. С. 43—52.
9. Он же. Методика определения механических свойств и обоснование
выбора конструкции гибких покрытий//Там же. 1973. Вып. 7. С. 71—79.
10. Беляев В. В. Геометрические характеристики формы тела черноморских
дельфинов//Вопр. морск. биологии. Киев: Наукова Думка. 1969. С. 12—14.
11. Бурлаков В. Д., Грушанская Ж. Я., Соколов В. Е. Геометрические
особенности тел и теоретические чертежи двух видов черноморских дельфи-
нов в связи с их гидродинамическими характеристиками//Бюлл. Моск, об-ва
испытателей природы. Отд. биологии? 1977. Т. 82. № 2. С. 62—66.
12. Гид роб ионические исследования локомоции водных животных в связи
с задачами функциональной морфологии/Г. Б. Агарков, А. А. Вронский,
А. П. Носарь, А. П. Коваль//Бионика-78. М.—Л. 1978. Т. 1. С. 71—74,
13. Горелов Д. Н. Об эффективности машущего крыла как движителя//
//Бионика. Киев: Наукова Думка. 1976. Вып. 10. С. 49—53,
14. Гребешов Э.П., Сагоян О. А. Гидродинамические характеристики коле-
блющегося крыла, выполняющего функции несущего элемента и движителя//
//Тр. ЦАГИ (Центральн. аэрогидродинам. ин-т). 1966. № 1725. С. 3—30.
15. Грейнер Л. Гидродинамика и энергетика подводных аппаратов. Пер.
с англ. Л.: Судостроение, 1978, 384 с.
16. Грушанская Ж. Я., Короткий А. И., Тюшкевич В. А. Исследование
обтекания и гидродинамических характеристик твердых моделей плавников
дельфина//Тр. Ленингр. о-ва естествоиспытателей. 1976. Т. 73. Вып. 5.
С. 42—52.
255
17. Диплом Ns 95 на открытие СССР. Явление саморегулирования
гидроупругости плавников китообразных/С. В. Першин, А. С. Соколов,
А. Г. Томилин. Заявка № ОТ-7087 от 23.12.68, опубл. 26.07.71, Бюлл. Ns 23,
1 с.
18. Диплом № 265 на открытие СССР, Свойства кожи китообразных
активно регулировать гидродинамическое сопротивление плаванию... /
/В. Е. Соколов, В. В. Бабенко, Л. Ф. Козлов, С. В. Першин, А. Г. Томилин,
О. Б. Чернышев, Заявка № ОТ-8405 от 21.02.73, опубл. 07.05.83, Бюлл. Ns 17,
1 с.
19. Журова В. М., Фаддеев Ю. И. К вопросу о влиянии нестационарного
поступательного движения гидробионтов на сопротивление трения//Бионика.
Киев.: Наукова Думка. 1973. Вып. 7. С. 56—59.
20. Они же. Результаты расчета нестационарного ламинарного погра-
ничного слоя на деформируемом эллипсоиде вращения//Там же. 1977.
Вып. 11. С. 44—50.
21. Заец В. А. Особенности строения кожных покровов быстроплаваю-
щих акул//Бионика-1973. М. 1973. Т. 6. С. 45—50.
22. Закономерность в покровах скоростных рыб/С. В. Першин, О. Б. Чер-
нышев, Л. Ф. Козлов и др.//Бионика. Киев.: Наукова Думка. 1976. Вып. 10.
С. 3—21.
23. Золотов С. С., Ходорковский Я. С. Особенности сопротивления тре-
ния тел формы меч-рыбы//Там же. 1973. Вып. 7. С. 14—18.
24. Каян В. П. О гидродинамических характеристиках плавникЬвого
движителя дельфина//Там же. 1979. Вып. 13. С. 9—15.
25. Он же. Экспериментальное исследование гидродинамического упора,
создаваемого колеблющимся крылом//Там же. 1983. Вып. 17. С. 45—49.
26. Каян В. П., Пятецкий В. Е. Гидродинамические характеристики дель-
фина-афалины при различных режимах ускорения//Там же. 1978. Вып. 12.
С. 48—55.
27. Коваль А. П. Шероховатость и некоторые особенности строения кожи
меч-рыбы//Там же. 1972. Вып. 6. С. 73—77.
28. Козлов Л. Ф. Теоретическая биогидродинамика. Киев.: Вища шк.,
1983, 235 с.
29. Козлов Л. Ф., Першин С. В. Комплексные исследования активного
регулирования кожей дельфинов снижения гидродинамического сопротивле-
ния//Бионика. Киев.: Наукова Думка. 1983. Вып. 17. С. 3—12.
30. Кокшайский Н. В. Очерк биологической аэро- и гидродинамики. М.:
Наука, 1974, 255 с.
31. Коренная Л. И. Оценочный расчет плавникового движителя//Био-
ника. Киев.: Наукова Думка. 1974. Вып. 8. С. 49—52.
32. Она же. Установка для исследования гидромеханики гибкой пла-
стины//Там же. 1976. Вып. 10. С. 85—88.
33. Логвинович Г. В. Гидродинамика плавания рыб//Там же. 1973.
Вып. 7. С. 3—8.
34. Он же. Гидродинамика тонкого гибкого тела (оценка гидродина-
мики рыб)//Там же. 1970. Вып. 4. С. 5—11.
35. Матюхин В. А. Биоэнергетика и физиология плавания рыб. Ново-
сибирск: Наука, Сибир. отд-ние, 1973, 154 с.
36. Мосунов Д. Ф., Першин С. В. Гидробионические аспекты спортив-
ного плавания способом брасс//Бионика. Киев.: Наукова Думка. 1980.
Вып. 14. С. 34—41.
37. Нуссенбаум Л. М., Фалеева Т. И. Исследование поведения рыбы
в электрическом поле//Изв. Гос. НИИ озеровед, и рыб. хоз-ва. 1961. Т. 52.
Вып. 1. С. 1—92.
38. О подходе к оценке эффективности биомеханики плавания биообъек-
тов при оптимизации энергозатрат/В. М. Ахутин, С. П. Балицкий. А. С. Кат-
рин и др.//Бионика-78. М.— Л. 1978. Т. 1. С. 235—237.
39. О структуре биокомплексов слизистых веществ кожи быстроплаваю-
щих рыб/Е. Т. Ускова, А. В. Чайковская, Т. А. Костина. И. А. Усков//Био-
ника. Киев.: Наукова Думка. 1982. Вып. 16. С. 16—21.
256
40. Овчинников В. В. Меч-рыба и парусинковые. Калининград, 1970,
106 с.
41. Першин С. В. Биогидродинамические закономерности плавания вод-
ных животных как принципы оптимизации в природе погруженных тел//Вопр.
бионики/AH СССР. М.: Наука, 1967. С. 555—560.
'42 . Он же. Биогидродинамический феномен меч-рыбы как предельный
случай скоростных гидробионтов//Бионика. Киев: Наукова Думка. 1979.
Вып. 12. С. 40—48.
43. Он же. Гидробионические зависимости нестационарного гармониче-
ского движения рыб и дельфинов как погруженных тел переменной формы//
//Проблемы бионики/AH СССР. М.: Наука. 1973. С. 464—474.
44. Он же. Гидробионические закономерности оптимизации внешних форм
движительного комплекса китообразных//Бионика. Киев: Наукова Думка.
1983. Вып. 17. С. 13—24.
45. Он же. Гидродинамическая характеристика китообразных и норми-
рованная скорость плавания дельфинов в естественных условиях и в не-
воле//Там же. 1969. Вып. 3. С. 5—12.
46. Он же. Гидродинамический анализ профилей плавников дельфинов
и китов//Там же. 1975. Вып. 9. С. 26—36.
47. Он же. Гидродинамические аспекты изучения движения водных жи-
вотных//Бионика. М.: Наука. 1965. С. 207—215.
48. Он же. Машущее крыло как лопастная машина для принудитель-
ной циркуляции среды//Бионика. Киев.: Наукова Думка. 1981. Вып. 15.
С. 64—71.
49. Он же. К характеристике плавания микроорганиэмов//Гидродинам.
вопр. бионики. Киев: Наукова Думка. 1983. С. 28—37.
50. Он же. Плавание и полет в природе//Итоги науки и техники. Сер.
Бионика. Биокибернетика. Биоинженерия. М.: ВИНИТИ АН СССР, 1979,
155 с.
51. Першин С. В., Носов Е. П, Исаков В. П. Схематическая модель гидро-
упругого эффекта в плавниках китообразных и техническое использование
ее//Бионика-1973. М.: АН СССР. 1973. Т. 6. С. 93—99.
52. Першин С. В., Соколов А. С., Томилин А. Г. Биомеханические основы-
регулируемого гидроупругого эффекта в плавниках — крыловых несущих
поверхностях китообразных (дельфинов и китов)//Тр. ин-та/Зоологич. ин-т АН
СССР. Л.: Наука. 1974. Т. 54. С. 266—305.
53. Они же. Регулируемый гидроупругий эффект в плавниках крупней-
шего скоростного дельфина косатки Orcinus огса//Бионика. Киев: Наукова
Думка. 1979. Вып. 13. С. 35—43.
54. Першин С. В., Шаров Я. Ф., Носов Е. П. Гидробионт как адаптаци-
онная колебательная система //Там же. 1973. Вып. 7. С. 18—25.
55. Пятецкий В. Е., Каян В. П. Гидродинамические характеристики пла-
вания афалины//Там же. 1976. Вып. 10. С. 80—85.
56. Пятецкий В. Е., Каян В. П., Кравченко А. М. Экспериментальные
установки, аппаратура и методика изучения гидродинамики плавания вод-
ных животных//Там же. 1973. Вып. 7. С. 91 — 101.
57. Романенко Е. В. Теория плавания рыб и дельфинов. М.: Наука,
1986, 148 с.
58. Савченко Ю. Н. Некоторые особенности гидродинамики движителя
типа «машущее крыло»//Бионика. Киев. Наукова Думка. 1971. Вып. 5.
С. 11 — 19.
59. Савченко Ю. Н., Савченко В. Т. Бионические аспекты плавания
животных//Гидродйнам. вопр. бионики. Киев: Наукова Думка. 1983.
С 46—57.
60. Самонастройка демпфирования кожи китообразных при активном
плавании/В. В. Бабенко, Л. Ф. Козлов, С. В. Першин и др.//Бионика. Киев:
Наукова Думка 1982. Вып. 16. С. 3—10.
61. Соколов В. Е. Структура кожного покрова некоторых китообраэ-
ных//Бюлл. Моск, о-ва испытателей природы. Отд. биологии. 1955. Т. 60.
Вып. 6. С. 45—60.
257
62. Соколов В. Е., Бурлаков В. Д., Грушанская Ж. Я. Теоретический
чертеж черноморского дельфина-афалины//Там же. 1972. Т. 77. № 6.
С. 45—53.
63. Соколов А. С., Юрчинская Т. Л., Першин С. В. Особенности распре-
деления и строения комплексных сосудов, регулирующих упругость спин-
ного плавника дельфина//Тр. ин-та/Зоолог. ин-т. АН СССР. Л.: 1982. Т. 115.
С. 85—91.
64. Суркина Р. М. Строение соединительнотканного остова кожи дель-
фина//Механизмы передвижения животных. Киев: Наукова Думка. 1968.
С. 78—86.
65. Томилин А. Г. Дельфины служат человеку. М.: Наука, 1969, 246 с.
66. Шебалов А. Н. Некоторые вопросы влияния нестационарности на
«механизм» образования сопротивления//Бионика. Киев.: Наукова Думка.
1969. Вып. 3. С. 61—66.
67. Юрченко Н. Ф., Бабенко В. В. О моделировании гидродинамических
функций наружных покровов водных животных//Гидродинам. вопр. био-
ники. Киев: Наукова Думка. 1983. С. 37—46.
68. Bainbridge R. The speed of swimming of fish as related to size and
to the frequency and amplitude of tail beat//J. Exp. Biol. 1958. V. 5. N 1.
P. 109—133.
69. Brett T. R. The swimming energetics of Salmon//Scient. Amer. 1965.
V. 213. N 2. P. 80—85.
70. Caldwell D. K-, Caldwell M. C. The world of the Bottlenosed dolphin.
Philadelphia. New York. 1972. 157 p.
71. Carey F. G., Teal J. M. Regulation of body temperature by the bluefin
tunay/Comparative Biochemistry and Physiology, 1969. V. 28. N 1. P. 205—213.
72. Chopra M. G. Hydromechanics of lunate-tail swiming propulsion//,!.
Fluid Meeh. 1974. V. 64. Part 2. P. 375—391.
73. Chopra M. G., Kambe T. Hydromechanics of lunate-Tail swimming pro-
pulsion. Part 2//J. Fluid Meeh. 1977. V. 79. Part 1. P. 49—69.
74. Gray J. Animal Locomotion L. Weidenfeld and Nicholson. 1961. 479 p.
75. Grieg J, A. Nogle notiser fra et spekhuggersteng ved Bildostrommen
i januor 1904//Bergens Museums Aarbog. 1906. N 2. P. 1—28.
76. Hertel H. Structure, Form and Movement (Biology and Technology)//
Reinhold Publishing Corporation. New-York. 1966, 251 p.
77. Hoyt T. W. Hydrodynamic drag reduction due to fish slimes.//Swimm-
ing and Flying in Nature. N. Y.: Plenum Press. 1975. V. 1. P. 653—672.
78. Hunter J. R., Zweifel T. R. Swimming speed, tail beat frequency, tail
beat amplitude and sise in mackerel Trachurus symmetricus and other fishes//
Fish. Bull. V, S. 1971. V. 69. N 2. P. 253—266.
79. Johannessen C. L., Harder J. A. Sustained swimming spedds of Dol-
phins//Science. 1960. V. 132. N 3439. P. 1550—1551.
80. Kelly H. R., Rentz A. IF., Siekmann J. Experimental studies on the
motion of a flexible hydrofoil//J Fluid Mechanics, 1964. V. 19. Part 1. P. 30—
48.
81. Kramer Max O. Boundary layer stabilization by distributed damping//
Naval Engineers Journal. 1962. V. 74. N 2. P. 341—348.
82. Lang T. G. Hydrodynamic analysis of dolphin fin profiles//Nature.
1966. V. 209. N 5028. P. 1110—1111.
83. Lang T. G. Speed, power and drag measurements of dolphin and
porpoises//Swimm. and Fly. Nature. V. 2. N.-Y., L. 1975. P. 553—572.
84. Lighthill James. Mathematical biofluid dynamecs//SIAM, Philadelphia
Pa. 1975. 281 p.
85. Lighthill James. Aquatic animal locomotion. Trans. N. E. Coast Inst.
Eng. and Shipbuild. 1977. V. 93. N 6. P. 127—135.
86. Magnusson J. J., Prescott J. H. Coartship, locomotion, feeding and
miscellaneous behavior of pacific bonito//Animal Behaviour. 1966. V. 14. N 6.
P. 54—67.
87. Newman J. N., IFu T. Y. A generalized slender body theory forish-like
forms//J. Fluid Meeh. 1973. V. 57. N 4. P. 673—693.
258
88. Purves P. E., Dubek van Heel W. H„ Jonk A. Locomotion in dolphins..
Part 1//Aquat. Mammals. 1975. V. 3. N 1. P. 5—31.
89. Siekmann J. Investigation into the movement of swimming animals//
ZWDI, 1962. V. 104. N 10. P. 433—439.
90. Svanson N. M. Some observations on fish propulsion: qualitative
boundory layer considerations//Developm. Meeh. 1965. V. 2. Part I. P. 550—
564.
91. Swimming and Flying in Nature. Vol. 1. New-York, London: Plenum
Press. 1975. 421 p.
92. Uldrick J. P. On the propulsion effeciency of swimming flexible hy-
drofoils of finite thickmess. J. Fluid Meeh. 1968. V. 32. Part 1. P. 29—53.
93. Videler J. J., Hess F. Fast continuous swimming of two pelagic fre-
dators, saithe (Pollachius virens) and mackerel (Scomber scombrus): a kine-
matic analysis//.!. Exp. Biol. 1984. V. 109. P. 209—288.
94. Walters V., Fierstine H. L. Measurements of swimming speeds of
yellowfin tuna and wahoo//Nature. 1964. V. 202. N. 4928. P. 208—209.
95. Webb P. W., Kostecki P. T. The effect of size and swimming sped on
locomotor kinematics of rainbow trout//J. Exp. Biol. 1984. V. 109. P. 77—95.
96. - Whales, Dolphins and Porpoises/ Ed. K. S. Norris/ Berely and Los
Angeles: University of Californis Press. 1966. 789 p.
97. Wilkie D. R. Man as an Aero Engine//.!. Royal Aeronaut. Society,
L. 1960, V. 64. P. 477—481.
98. Wu T. Y. Hydromechanics of swimming of fishes and cetaceans//Adv.
Appl. Meeh. N.-Y., L. 1971. Vol. 11. P. 1—63.
99. Wu T. Y. Introduction to the Scaling of aquatic animal locomotion.//
Scale effects in animal locomotion/Ed. T. Pedley. London: Academic Press.
1977. P. 203—232.
100. Yuen H. S. H. Swimming speeds of yellowfin and Skipjack tuna//Trans.
of the Amer. Fish. Soc. 1966. V. 95. N 2. P. 203—209.
ПРЕДМЕТНО-ТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
Биологический принцип
----дивергенции 14
----конвергенции 14
----мультифункциональности 17
----эмбриогенеза 14
Бионика (определение) 7
— аспекты исследований 9
Вазомоторная подвижность 181, 188
Гидробионика (определение)
— методы 66, 100, 106, 112, 121, 127,
133, 153
— области 95
— объекты 13, 15
— принципы 98
Гидробионты 10, 15, 27, 53
Гидроупругий эффект 212
----моделирование 241
Грея «парадокс» 180
Давление кровяное
----величина 187, 197, 220
---- пульсация 214
Движитель гидробионтов
----волновой 105
----машущий плавниковый 121
----реактивный 13
----скомброидный 15, 76, 102
----угревидный 15, 76
Дельфины 13
— черноморские 29, 31
Исследования машущего крыла
----теоретические 121
----экспериментальные 127
Исследование гидробионтов 157
----биоэнергетические 156, 161, 170,
172
----теоретические 106, 112, 121
----экспериментальные 84, 127
Кваэистационарный расчет
----биоэнергетический 173
260
----гидродинамический 133 "
Кинематическая характеристика 79,
82, 104
----относительная амплитудно-ча-
стотная 87
------волновая 89
----размерно-амплитудная 86
----размерно-скоростная 85
----резонансная амплитудно-ча-
стотная 104
----частотно-скоростная 85
------ относительная 86
---- числа пропульсивных волн 89
---- универсальная критериальная
88, 89
Кожный покров китообразных 181,
187, 193
------демпфирование 197
------дермальные валики 185
---------сосочки 183
------механическая модель 193
------ микровибраций 195
------толщина 186, 191
Комплексные сосуды 213, 228
Коэффициент гидродинамический
----мощности 108
----сопротивления 116, 118, 177
----тяги 108
----полезного действия 116
Крамера демпфирующие покрытия
180
Логвиновича теория 112
Масштабный эффект 179
Метод аналогов биоэнергетики 170
Моделирование гидробионическое
-------- гидроупругого эффекта 240
------------демпфирующего кожного по-
крова 192
------формы тела гидробионтов 202
Моделирование гидробионтов
------гибкой пластиной 106
----профилем 106, 126
----удлиненным телом 112
Модель демпфирующего покрытия
---- вязкоэластичная 209, 210
---- мембранная 209
Моноласт (волновой движитель) 146
Мышц относительная масса 17, 168
Нестационарные режимы 138, 144
----деформации формы тела 141
-----изменения скорости 138, 142
Нормирование
— послойной кожи дельфинов 191,
192
— скорости плавания 71
Ныряние китообразных 167
Обмен гидробионтов (метаболизм)
153
----активный 156
----общий 156
----основной 154
----интенсивность 153
---- уровень 153
Оптимизация формы
----плавников в плане 33, 35, 36
----в профиле 40
----- тела 47, 202
Пограничного слоя дельфинов
---- визуализация 204
----ламинариэация 202
----нестационарность 138, 144
----определяющие параметры
194
---- пульсации 206
----степень турбулентности 207
Плавники гидробионтов
----дельфинов в плане 33, 37, 38
----в профиле 40, 44
----рыб нескоростных 47
----скоростных 49, 52
----смена формы 36, 46, 48
Подобие гидробионтов
----по массе мышц 168
----общей массе 175
----смоченной поверхности 176
Покровы скоростных рыб
----закономерность 65
----кожный 53
----слизистый 61
----чешуйный 58
Потенциальная быстроходность ки-
тообразных 72
Пропульсивные характеристики 130,
131
Присоединенные массы 141
Профили гидродинамические
----плавников 41
----тела' 30, 203
----эпюрные 42
Прыжков дельфинов
— — общая характеристика 162
----основные параметры 164
Резонансные колебания 101, 105
Рейнольдса критерий (число) 19
Саморегуляция упругости ялавни-
ков 212
Секционирование сосудистой сети 189
Скорость плавания
----безразмерная 19
---- крейсерская 73
----максимальная 70, 73, 171
----относительная 19
----средняя 18
Скоростные рыбы
----гидродинамическая оценка 47
----особенности меч-рыбы 51
Снижение сопротивления
----демпфированием 178
----рыбьей слизью 62, 64
----формой тела 202, 204
Сосудистая сеть плавников дельфи-
нов
----спинного 221
--------- хвостового 222
Спортивное плавание
----------брассом 90
----------«дельфином» с моноластом 146
Сопротивление
— инерционное 139
— трения 174
— формы 174
Стационарности гипотеза 173
Струхаля критерий (число) 20
Теоретический чертеж дельфинов 31
Тип сосудистой сети
---- — магистральный 222
------полурассыпной 224
-------рассыпной 223
Упругодемпфирующие параметры 194
--колеблющаяся масса 197
--коэффициент демпфирования
200
----— модуль упругости 199
Упругости плавников саморегуляция
-------активная 217, 218
------- исчезновение 216
------- первое проявление 215
Характеристика китообразных
----геометрическая 31
----гидробионическая 23
----формы тела 32, 113
---- плавников 33, 40
ОГЛАВЛЕНИЕ
Предисловие научного редактора................................ .... 3
От'автора ............. ...................... .... 5
Введение ......................................................... 7
Глава I. Функционально-морфологические свойства крупных гидробионтов
и их гидродинамическое значение.....................................22
§ 1 . Общая гидробионическая характеристика эффективно пла-
вающих китообразных........................................ 22
§ 2. Оптимизация формы в плане машущего плавникового дви-
жителя китообразных........................................32
§ 3. Гидродинамический анализ профилей плавников дельфи-
нов и китов................................................40
§ 4. Оптимизация формы тела и машущего плавникового движи-
теля у скоростных рыб.................................... 45
§ 5. Гидробионические закономерности строения покровов ско-
ростных рыб с регулируемым сопротивлением плаванию 55
Глава II. Кинематика нестационарного плавания гидробионтов и регули-
рующее значение критерия Струхаля ..... 66
§ 6. Экспериментальные методы и средства исследований кине-
матики плавания гидробионтов ............................. 66
§ 7. Нормирование максимальных скоростей плавания китооб-
разных в природе и в неволе................................69
§ 8. Анализ кинематики цикла нестационарного регулярного
плавания крупных гидробионтов ............................ 75
§ 9. Система пространственно-временных характеристик неста-
ционарного плавания гидробионтов ......................... 82
§ 10. Гидробионические аспекты спортивного плавания спосо-
бом «брасс»................................................90
Глава III. Биогидродинамика нестационарного плавания гидробионтов
при больших числах Рейнольдса........................................95
§ 11. Гидробионические области и общие принципы гидробио-
ники .......................................................95
§ 12. Гидробионт как адаптационная колебательная система 99
§ 13. Теоретическое моделирование волнового плавания гидро-
‘ бионтов и проблема волнового движителя..............105
§ 14. Теоретические и экспериментальные исследования эффек-
тивного машущего плавникового движителя....................121
§ 15. Значение нестационарности движения по результатам раз-
дельного гидродинамического моделирования..................138
§ 16. Моноласт как эффективный волновой движитель для ско-
ростного плавания способом «дельфин» ..................... 146
262
Глава IV. Биоэнергетические аспекты плавания гидробионтов. Диапазон
и характер'регуляции биоэнергетики..................................153
§ 17. Некоторые общие закономерности биоэнергетики. Типовая
зависимость «мощность—скорость гидробионтов» . . . .153
§ 18. Прыжки из воды скоростных гидробионтов с максималь-
ной отдачей мощности. Метод определения активной мощ-
ности по киносъемке........................................161
§ 19. Продолжительность ныряния водных млекопитающих
в связи с максимальной скоростью плавания и относитель-
ной массой мышц............................................167
§ 20. Метод аналогов в биоэнергетике, саморегуляция отдачи
мощности гидробионтами и масштабный эффект . . . .170
§ 21. Кваэистационарный биоэнергетический расчет плавания
дельфинов и анализ полученных результатов . . . .173
Глава V. Авторегуляция демпфирования кожного покрова у дельфинов и
моделирование снижения гидродинамического сопротивления 180
§ 22. Свойство кожи китообразных активно регулировать гид-
родинамическое сопротивление плаванию .................... 180
§ 23. Структурные и гистологические особенности кожного по-
крова китообразных.........................................181
§ 24. Гидродинамическая функция и механическая модель
демпфирующего свойства кожи дельфинов......................188
§ 25. Экспериментальные упругодемпфирующие характеристики
кожи дельфинов.............................................195
§ 26. Опытные исследования пограничного слоя активно пла-
вающего дельфина...........................................201
§ 27. Техническое моделирование искусственных демпфирую-
щих покрытий для снижения гидродинамического сопро-
тивления трения...........................................208
Глава VI. Авторегуляция упругости плавников и оптимизация эффектив-
ности машущего движителя китообразных..........................212
§ 28. Явление саморегулирования упругости плавников китооб-
разных соответственно режимам плавания....................212
§ 29. Гидродинамическая целесообразность авторегуляции уп-
। ругости плавников китообразных.......................215
§ 30. Рентгенографические исследования распределения сосуди-
стой сети в плавниках китообразных........................220
§ 31. Гистологические исследования тканей плавников и уни-
кальных комплексных артерио-венозных сосудов китооб-
разных ...................................................225
§ 32. Теоретический анализ расположения сосудистой сети и
гидроупругого эффекта в плавниках китообразных . . . 230
§ 33. Экспериментальное моделирование и техническое исполь-
зование гидроупругого эффекта в плавниках китообразных 237
Заключение...........................<•......................... . 246
Приложение. Некоторые характеристики плавания микроорганизмов . 252
Список литературы...................................................255
Предметно-тематический указатель....................................260
Научное издание
Серафим Васильевич Першин
ОСНОВЫ ГИДРОБИОНИКИ
Монография
Зав. редакцией Д. В. Павлов
Редактор А. И. Кускова
Переплет художника Б. Н. Осенчакова
Художественный редактор Э. А. Бубович
Технический редактор Р. К Чистякова
Корректор С. Н. Маковская
ИБ № 1189
Сдано в набор 08.09.87. Подписано в печать 11.02.88. М-27521. Формат бСГХЭО’Лв. Бу-
мага типографская Хе 2. Гарнитура шрифта литературная. Печать высокая. Усл.
печ. л. 16,5. Усл. кр.-отт. 17,0. Уч.-изд. л. 18,0. Тираж 1380 экз. Изд. № 4105—85.
Заказ Хе 2806. Цена 3 р. 60 к.
Издательство «Судостроение», 191065, Ленинград, ул. Гоголя, 8.
Ленинградская типография Хе 4 ордена Трудового Красного Знамени Ленинградского
объединения «Техническая книга» им. Евгении Соколовой Союэполиграфпрома при
Государственном комитете СССР по делам издательств, полиграфии и книжной тор-
говли. 191126, Ленинград, Социалистическая ул., 14.
Зак. 152 М.