/






























































Author: Каппуччинелли П.
Tags: биология клетки и субклеточных частиц цитология биохимия биофизика молекулярная биология
Year: 1982




Text
Motility
of Living Cells
P. Cappuccinelli
Professor of Microbiology,
Institute of Microbiology,
School of Medicine.
University of Sassari,
Sardinia. Italy
Chapman and Hall
London and New York
П. КАППУЧЧИНЕЛЛИ
ПОДВИЖНОСТЬ
живых
КЛЕТОК
Перевод с английского
канд. биол. наук Н. А. Гавеловой
под редакцией
д-ра биол. наук Б. Ф. Поглазова
«Мир»
Москва 1982
УДК 576.30
Каппуччинелли П.
К20 Подвижность живых клеток: Пер. с англ. — М.:
Мир, 1982.— 124 с., ил.
В небольшой по объему, но очень содержательной монографии
итальянского исследователя излагаются основные данные о природе
и механизмах движения клеток. Автор рассматривает как подвижность
бактерий, так и механизмы движения эукариотических клеток: микро-
трубочки и физиологическую роль тубулинов, мнкрофиламеиты и акто-
миозиновую систему, перемещение с помощью ресничек и жгутиков,
амебоидное движение.
Для цитологов, биохимиков, биофизиков, а также медиков, пре-
подавателей и студентов биологических специальностей.
Редакция литературы по биологии
2001040000
21003—113
041(01)—82
К
113—82, ч. 1
© 1980 Р. Cappuccinelli
© Перевод иа русский язык, «Мир»,
1982
ПРЕДИСЛОВИЕ
РЕДАКТОРА ПЕРЕВОДА
Исследование механизмов двигательных реакций у живых существ
началось с изучения поперечнополосатых мышц, обладающих ярко
выраженной способностью к сокращению. Именно из мышц были
впервые выделены основные сократительные белки актин и миозин,
регуляторные белки и т. д. Особенно интенсивно эти исследования
стали развиваться после того, как В. А. Энгельгардт и М. Н. Люби-
мова открыли АТРазную активность миозина, в связи с чем прояс-
нились энергетические механизмы движения.
Впоследствии оказалось, что актин и миозин присутствуют во
всех живых клетках без исключения. Сейчас считается установлен-
ным, что в основе механизма биологической подвижности в самом
широком плане лежит одна и та же универсальная реакция взаимо-
действия между миозином, актином и АТР.
Наряду с этим были обнаружены и другие двигательные систе-
мы: во-первых, система тубулин—динеин, свойственная немышечным
клеткам, и, во-вторых, вращательный жгутиковый аппарат бактерий,
который, как предполагают, не требует для своей работы АТР.
Попытки как-то суммировать и обобщить накопленный в этой
области материал делались и раньше. Но в настоящее время, когда
исследования биологической подвижности развиваются особенно
быстро, такая обобщающая работа представляется очень важной и
актуальной. В связи с этим предлагаемая книга П. Каппуччинелли,
несомненно, очень ценна для уяснения общих закономерностей по-
движности живых клеток.
Большой интерес представляют данные, касающиеся движения
бактериальных жгутиков, особенно тот факт, что вращение жгутика
осуществляется за счет траисмембраниого потенциала. Не менее
важны структурные, биохимические и генетические исследования ту-
булиновой двигательной системы и данные о движении с участием
актиновых микрофиламентов.
Следует отметить, что книга написана просто, в легкой и до-
ступной манере; привлечены наиболее показательные примеры, очень
наглядны иллюстрации. Монография воспринимается как единое
целое, читается легко и с интересом. Такое изложение материала
возможно только при очень глубоком знании проблемы. Важно, что
автор непосредственно участвовал в ее экспериментальной разра-
ботке; это придает книге особую ценность.
Книга П. Каппуччинелли будет, без сомнения, интересной и по-
лезной как для специалистов, работающих в данной области, так и
для более широкого круга биологов.
Б. Ф. Поглазов
БЛАГОДАРНОСТЬ
Я должен особенно поблагодарить мою жену
Марию за замечательные рисунки, друзей —
Теда Вайнера и Сальвадора Рубино — за по-
мощь в подготовке книги к печати и мою со-
баку Келли за то, что она не съела рукопись
за то долгое время, пока мы над ней работа-
ли (смею надеяться — не из-за неудобовари-
мое™ ее содержания).
1. ВВЕДЕНИЕ
С давних времен философы проницательно отмечали,
что жизнь в ее разнообразных проявлениях представля-
ет собой непрерывное движение. Все во Вселенной — от
космических объектов до атомов — находится в состоя-
нии движения в той или иной его форме. Способность
двигаться — одно из неотъемлемых свойств всего живого
на Земле..
В XVII в. во времена Антони ван Левенгука, кото-
рый создал первый простой микроскоп, понятие подвиж-
ности было, в сущности, синонимом жизни. С помощью
своего примитивного инструмента Левенгук наблюдал,
как микроорганизмы (которые он называл «анимальку-
ли») активно плавают в капле воды, и на основании
этого утверждал, что. «они, по-видимому, живые».
Теперь мы знаем, что движение в такой форме нель-
зя считать необходимым критерием жизни, хотя для
многих живых организмов это свойство первостепенной
важности.
Принимая во внимание общее значение подвижности
в мире живого, основное внимание в данной книге мы
уделили рассмотрению подвижности на клеточном уров-
не. Здесь она проявляется в различных формах: это и
движение клетки как целого, т. е. ее перемещение в
пространстве, и движение внутри клетки ее органелл и
других компонентов. У прокариот подвижность проявля-
ется главным образом в движении клетки как целого,
так как внутриклеточный транспорт осуществляется у
них гораздо проще, чем у эукариотических организмов.
В клетках эукариот более сложная система внутрикле-
точных органелл, а также необходимость управлять
мембранными процессами потребовали развития более
8
1. Введение
сложной, со многими внутренними связями системы
внутриклеточной подвижности. В клетках высших орга-
низмов выработались также эффективные системы для
передвижения клетки как целого. Способность к переме-
щению в пространстве необходима как отдельной клет-
ке, для того чтобы реагировать на окружающую обста-
новку, так и клеткам многоклеточного организма для
межклеточных взаимодействий. Несмотря на различия
между этими двумя типами подвижности, молекулярные
механизмы, лежащие в их основе, в принципе одина-
ковы.
Одна из важнейших черт передвижения клеток — это
направленность движения. Действительно, клетки спо-
собны ощущать изменения в окружающей среде и реа-
гировать на них, меняя направление своего движения.
Такая способность активно реагировать на стимулы, по-
ступающие из внешней среды, помогает клетке питаться,
облегчает взаимодействие с другими клетками и дает
ряд других преимуществ.
Хотя изучение клеточной подвижности относится к
одной из старейших областей науки, понимание молеку-
лярных механизмов движения пришло только в самое
последнее время благодаря применению современных ге-
нетических и биохимических методов. Вместе с тем мно-
гое все еще остается спорным; в особенности это отно-
сится к механизмам регуляции подвижности. Однако
есть основания надеяться, что идущие сейчас интенсив-
ные исследования позволят разрешить многие из этих
проблем в течение ближайших лет.
В следующей главе мы рассмотрим подвижность
прокариот, в частности жгутиковых бактерий. Другие
интересные аспекты подвижности у простейших, например
скользящие движения миксобактерий или движения ти-
па ввинчивания, свойственные лептоспирам, обсуждаться
не будут. Остальные главы посвящены системам под-
вижности у эукариот. Основное внимание мы уделим
новейшим исследованиям на молекулярном уровне.
Поскольку ни одну из тем нельзя здесь рассмотреть
исчерпывающе, в конце каждой главы дается список
литературы, который поможет читателю получить более
полные сведения по интересующим его вопросам.
1. Введение
9
Рекомендуемая литература
Dobel С. (1932). Antony van Leewenhoek and his “Little Animals”,
Constable, London. Reprinted in paperback by Dover, New York
(I960).
Книга представляет собой собрание писем А. ваи Лененгука
в Лондонское Королевское Общество, в которых он присущим ему
очаровательным слогом описывает наблюдения, сделанные им с
помощью своего микроскопа.
Интересные обзоры, касающиеся различных типов движения
клеток и возможных способов его осуществления, можно найти в
следующих сборниках:
Aspects of Cell Motility (1968). Symp. Soc. Exp. Biol., XXII, Cambrid-
ge University Press, Cambridge.
Molecules and cell movement (1975) (ed. S. Inoue and R. F. Ste-
phens), Raven Press, New York.
2. подвижность
ПРОКАРИОТИЧЕСКИХ
ОРГАНИЗМОВ
Ученых всегда восхищала подвижность простых организ-
мов, таких, как бактерии. Действительно, бактерии
весьма привлекательный объект для изучения подвиж-
ности и связанных с ней проблем: их клеточная органи-
зация относительно проста (по сравнению с клетками
эукариот), биохимические процессы хорошо изучены;
кроме того, они быстро и в больших количествах раз-
множаются, так что представляют собой идеальную сис-
тему для изучения биологических проблем с помощью
генетических методов.
Объединение биохимических и генетических подхо-
дов оказалось весьма плодотворным для изучения под-
вижности на молекулярном уровне, а также структур,
ее осуществляющих, и уже получены многообещающие
результаты.
У бактерий нет того разнообразия различных типов
подвижности, которое характерно для клеток эукариот.
Бактерии движутся всегда с помощью особой структу-
ры — так называемого жгутика, который может вра-
щаться подобно пропеллеру, тем самым обеспечивая
продвижение клетки.
У большинства бактерий жгутик прикреплен к клетке
только одним концом, а вся остальная часть его свобод-
на. Однако у некоторых бактерий (например, у спиро-
хет) жгутик прикреплен к телу клетки по всей его дли-
не с помощью наружной мембраны. В обоих случаях
подвижность клетки обусловлена вращательными дви-
жениями жгутика.
Бактерии способны управлять движениями жгутиков.
Они могут ощущать и как-то учитывать концентрации
веществ, с которыми они вступают в контакт, и соот-
2. Подвижность прокариотических организмов
11
ветственно на них реагировать. Эту способность учиты-
вать информацию можно рассматривать как примитив-
ный вариант системы памяти.
Хотя движение клетки не относится к насущно необ-
ходимым условиям ее существования (не двигающиеся
штаммы способны к росту и размножению), подвиж-
ность широко распространена у бактерий, особенно у
свободно живущих форм. В некоторых условиях свой-
ство подвижности может давать определенные преиму-
щества бактериям, позволяя им, например, уходить из
среды, содержащей токсические вещества, или, наобо-
рот, двигаться по направлению к источнику пищи.
Ниже мы рассмотрим свойства жгутиков бактерий с
морфологической и биохимической точек зр,е.ния. Мы об-
судим также вопрос о генетической регуляции сборки
жгутика, а также о том, как функционирует система
хемотаксиса и как она определяет движение бактерии.
2.1. Структуры,
обеспечивающие подвижность
Жгутики бактерий — это длинные гибкие нитевидные,
выросты, исходящие из тела бактерии. Они могут быть
одиночными, или же пучки их находятся на одном или
на обоих полюсах клетки. Жгутики могут покрывать
всю поверхность клетки, так что бактерия выглядит во-
лосатой1 (рис. 2.1). Характер расположения жгутиков
относится к генетически стабильным свойствам, по нему
жгутиковых бактерий делят на Eubacteriales (с беспоря-
дочным распределением жгутиков) и Pseudomonadales
(жгутики располагаются на полюсах клетки).
Из-за малого диаметра (120—250 нм) жгутики очень
трудно разглядеть с помощью обычных методов свето-
вой микроскопии. Однако можно применить некоторые
специальные приемы. Так, например, жгутики можно
окрасить раствором, содержащим осаждающийся реа-
гент, например танин; танин откладывается на поверх-
1 В этом случае их называют ресничками. — Прим. ред. пере-
вода.
12
2. Подвижность прокариотических организмов
Рис. 2.1. Распределение жгутиков у бактерий: а — бактерия с одним
жгутиком (вверху) и с пучком жгутиков, отходящих от одного из
полюсов клетки (внизу); б — бактерия со многими жгутиками, рас-
пределенными по всей поверхности клетки.
ности каждого жгутика, увеличивая его диаметр, что
позволяет наблюдать жгутики в обычном световом мик-
роскопе. Особенно хорош метод темного поля; при этом
жгутики четко видны как светлые линии на черном фо-
не. В этих условиях жгутики выглядят тонкими нитями,
длина которых обычно превышает длину кл,етки, из ко-
торой они исходят. В ранних работах подвижность час-
то изучали на родобактериях Chromomatium okenii. Бы-
ло показано, что на, одном из полюсов этой большой
(9—15 мкм) клетки находится группа из приблизитель-
но 40 жгутиков длиной около 25 мкм каждый. Хорошо
известная кишечная бактерия Е. coli имеет, как прави-
ло, 6 жгутиков, каждый из которых в 2—3 раза длиннее
самой клетки.
В микроскоп видна еще одна важная морфологичес-
кая черта жгутиков, а именно их форма. Жгутики всег-
да волнообразно или спиралевидно изогнуты, что харак-
терно для всех штаммов бактерий, причем жгутики от-
ходят от тела клетки под самыми разными углами,
по-видимому, без всякой закономерности.
Более точные данные о морфологии жгутиков могут
быть получены методом электронной микроскопии.
2. Подвижность прокариотических организмов
13
В ставших классическими исследованиях Де Памфилиса и
Адлера [35, 36], а также Диммита и Симона [38] на
Е. coli и В. subtilis не только выявлены детали ультра-
структуры жгутиков, но и заложена основа для понима-
ния механизма подвижности бактерий. Эти авторы раз-
работали методику получения интактных жгутиков в
довольно больших количествах. Для этого применяли,
ферментативное переваривание клеточной стенки, затем
лизировали клеточную мембрану детергентом и отделяли
жгутики от остатков клеток с помощью дифференциаль-
ного и градиентного центрифугирования в CsCl. Этот
метод позволяет удалить бактериальные оболочки и по-
лучить неповрежденные жгутики вместе со структурами,
которые прикрепляют их к клетке. Электронно-микроско-
пические исследования такого материала прояснили
многие структурные особенности жгутиков. Прежде все-
го удалось различить три самостоятельные субструкту-
ры: филамент (тело жгутика), крюк и базальное тело.
Филамент и крюк находятся снаружи от клеточной обо-
лочки, а базальное тело соединяет их с клеткой. В нор-
ме у Е. coli филамент имеет диаметр 20 нм и волнооб-
разную форму (длина волны 2,3 мкм), однако получены
мутанты с совершенно другими свойствами (рис. 2.2).
Существуют, например, прямые мутанты, жгутики ко-
торых практически лишены извивов и извилистые мутан-
ты с устойчивым признаком сильно выраженной волно-
образности жгутика (длина волны 1,1 мкм) [72]. Было
обнаружено, что переход от жгутиков с нормальным ша-
гом суперспирали (2,3 мкм) к извилистой форме'
(1,1 мкм) происходит также и в обычных условиях — в
Рис. 2.2. Типы волнообразности жгутиков: а — прямой мутант;
б — извилистый мутант; в — нормальный жгутик.
14
2. Подвижность прокариотических организмов
крюк -*•
кольцо Р
кольцо L -►
ось
кольцо S-+-
КОЛЬЦО М-к
Рис. 2.3. Схематическая модель базального тела жгутика у грам-
отрицательной бактерии (Е. coll)', элементы базального тела соот-
несены со структурами клеточной оболочки [35, 36].
процессе движения бактерии; такой переход, как будет
показано ниже, играет особую роль в плавательных дви-
жениях бактерии.
Крюк — это структура, находящаяся в основании фи-
ламента, обычно он изогнут углом или плавно. Наруж-
ный диаметр крюка несколько превышает диаметр фила-
мента; общая длина его — около 900 нм. Функция крю-
ка еще не вполне понятна, хотя можно предполагать,
что он играет роль гибкого сочленения, обеспечивая
эффективную передачу движения от базального тела к
филаменту и одновременно придавая структуре фила-
мента устойчивость по отношению к изменениям в окру-
жающей среде. Стержень, выходящий из проксималь-
ного конца крюка, проходит сквозь клеточные оболочки
и соединяется с базальными структурами.
Базальное тело — это сложная структура, ответ-
ственная не только за прикрепление жгутика к бактери-
альной клетке, но также и за создание движения. Иссле-
дование его ультраструктуры (рис. 2.3) показало, что у
грамотрицательных бактерий (т. е. у бактерий, которые
имеют наружную липополисахаридную мембрану и слой
пептидогликанов внутри клеточной стенки) в базальном
2. Подвижность прокариотических организмов
15
Рис. 2.4. Схематическая модель базального тела жгутика у грам-
положительной бактерии (В. subtilis)-, элементы базального тела
соотнесены с клеточной мембраной и клеточной оболочкой [35, 36].
теле есть четыре кольца, почти одинаковых по диамет-
ру. Самое нижнее, или кольцо М (membrane), прикреп-
лено к цитоплазматической мембране и из него выходит
стержень жгутика. Кольцо S (supermembrane) располо^
жено как раз над цитоплазматической мембраной и, по-
видимому, не соединено ни с какими другими клеточными
структурами. Остальные два кольца, Р (от peptidogly-
can, т. е. на уровне пептидогликанного слоя) и L (от
lipopolysaccaride, т. е. на уровне липополисахаридной
мембраны), погружены в соответствующие слои клеточ-
ной стенки; они фиксируют стержень, проходящий сквозь
бактериальную стенку. У грамположительных бактерий
(например, В. subtilis) базальное тело имеет сходную
структуру, но несколько упрощенную по сравнению с
грамотрицательными бактериями. В нем имеется только
одна пара колец, расположенных у основания стержня
и, по-видимому, аналогичных кольцам S и М у Е. соН;их
обозначают соответственно теми же буквами (рис. 2.4).
У В. subtilis кольцо М находится в цитоплазматической
мембране, а кольцо S прикреплено к внутренней части
пептидогликанного слоя, где оно, вероятно, контактиру-
ет с молекулами тейхоевых кислот (тейхоевые кислоты
16
2. Подвижность прокариотических организмов
входят в состав однослойной клеточной стенки грампо-
ложитёльных бактерий). Колец, фиксирующих положе-
ние стержня в клеточной стенке, не обнаружено.
Поскольку исследования ультраструктуры показали,
что структура базального тела у грамположит,ельных и
грамотрицательных бактерий в принципе одна и та же
и что у тех и других есть кольца S и М, можно предпо-
лагать, что одно из этих колец или они оба участвуют в
генерации движения. Полученные в последнее время
данны,е позволили лучше понять связь между структу-
рой жгутика и движением и, похоже, указывают на то,
что именно кольцо М ответственно за генерацию дви-
жения.
2.2. Молекулярная
структура жгутика у бактерий
Упомянутый выше метод выделения интактных жгу-
тиков обеспечил возможность не только изучить их мор-
фологически, но также иссдедовать химическую струк-
туру, что необходимо для лучшего понимания всей слож-
ности механизма функционирования этих органелл и
регуляторных процессов в них.
С химической точки зрения филамент жгутика по-
строен из идентичных субъединиц, состоящих из един-
ственного белка — флаг,е,длина; этот белок легко отде-
лить от структуры базальное тело —крюк, снизив pH
или повысив температуру среды с интактными жгутика-
ми до 60°С [3]. Флагеллиновые субъединицы способны
ассоциировать in vitro с образованием филаментов,
весьма сходных с теми, которые наблюдаются in vivo.
Химическая структура флагеллина специфична для каж-
дого штамма бактерий, и антитела, выработанные про-
тив' одного бактериального штамма, не дают реакции
преципитации с флагеллином из других штаммов; это
позволяет использовать антигенные свойства жгутиков
для классификации бактерий. Чтобы понять, как связа-
ны молекулярное строение, форма и антигенные свой-
ства жгутика, определили аминокислотную последова-
тельность флагеллинов из некоторых бактерий [31, 76];
2. Подвижность прокариотических организмов
17
однако эти исследования пока не решили эту проблему.
Как и предполагали, молекулярная масса флагеллино-
вых субъединиц у разных бактерий различна; так у
Е. coll она равна 54 000 и не превышает 40000 у некото-
рых других видов. Методом оптической дифракции на
электронно-микроскопических негативах показано, что
жгутик представляет собой полый цилиндр; стенка ци-
линдра образована одиннадцатью продольными рядами
флагеллиновых субъединиц [78]. Каждая субъединица
смещена в своем ряду на некоторый угол относительно
продольной оси, благодаря чему возникает та или иная
спиральная структура (волнообразная форма) жгутика;
величины этих углов могут быть различны в спиральных
структурах различных типов.
Биохимические исследования области крюка и ба-
зального тела вначале осложнялись тем, что эти струк-
туры составляют только около 2% массы интактного
жгутика, из-за чего было трудно набрать достаточное
для работы количество материала. Затем, однако, у
Е. coli, а также у Salmonella были обнаружены два
типа мутантных штаммов, из которых у одного интакт-
ные базальные структуры не имели филаментов [178], а
у другого ненормально увеличенные поликрюковые
структуры содержали значительное количество белка.
В первом случае базальные тела можно получить из
изолированных м,е,мбран, просто лизируя их детерген-
том, а во втором случае поликрюковые структуры легко
отделить от клеток и очистить. Методом электрофореза
в полиакриламидном геле в присутствии додецилсульфа-
та натрия этот материал можно разделить на полипеп-
тиды и исследовать их далее по отдельности. Как пока-
зали такие исследования, крюк построен, по-видимому,
из одинаковых субъединиц, содержащих только один
полипептид; у Е. coli и Salmonella субъединицы имеют
одинаковую мол. массу — 42 000, а у В. subtilis — 30 000.
Базальное тело, как можно предполагать, судя по его
ультраструктуре, и в химическом отношении устроено
значительно более сложно. У Е. coll в базальном теле
обнаружены по меньшей мере 9 различных полипепти-
дов с мол. массами от 9000 до 60000 (табл. 2.1); кроме
того, еще 3 более слабо связанных компонента, по-види-
18
2. Подвижность прокариотических организмов
мому, теряются в процессе очистки [65]. Базальные
структуры грамположительных бактерий изучены гораз-
до хуже., однако можно предполагать, что они устроены
не так сложно.
Таблица 2.1
Белки и полипептиды жгутика Е. coll (из [135])
Компонент Локализация Геи
54 000 (флагеллин)
42 000
60 000
39 000
31 000
27 000
20 000
18 000
13 000
11 000
9 000
Филамент
Крюк
Базальное тело
То же
»
»
»
»
»
»
hag
fla, область I
То же
»
»
»
Не определен
То же
»
»
2.3. Регуляция синтеза и сборки
бактериального жгутика
Структура и сборка бактериального жгутика регули-
руются с помощью набора генов, которые определены и
изучены, особенно у Е. coli [65, 135] и Salmonella [71].
Кроме того, имеются также гены, которые участвуют в
контролировании активности жгутика и переориентации
движения клетки в ответ на физико-химические стимулы.
В следующем разделе мы рассмотрим пути генетиче-
ской регуляции структуры жгутика, а также механизмы
управления синтезом и сборкой субструктур жгутика.
Исследование всей цепи генетических регуляторных про-
цессов еще не завершено, однако отдельные ее звенья
уже удалось выяснить, и это будет способствовать реше-
нию проблемы в целом.
2. Подвижность прокариотических организмов
19
2.3.1. Жгутиковые гены у Е. coli
Гены, связанные с жгутиковой подвижностью Е. coli
(и других микроорганизмов), можно разделить на три
группы в зависимости от их функций. Первая группа
включает гены fla и hag, все конечные продукты кото-
рых имеют отношение только к структуре жгутика:
клетки с одним (или более) дефектным геном fla (fla~-
мутанты) не способны построить нормальный жгутик.
Вторая группа — это гены mot, которые отвечают за
вращательные движения жгутика. Мо/_-клетки имеют
филаменты с нормальной структурой, которые не могут
двигаться. Третья группа — гены che, назначение кото-
рых— обеспечить соответствующую направленность
движения клетки в ответ на различные стимулы (так-
сис). С71е~-клетки не способны к хемотаксису и не могут
двигаться направленно. Для всех этих генов (их около
30) выяснена их локализация на генетической карте
хромосомы Е. coli и большинство генных продуктов
идентифицировано (см. рис. 2.5).
В данном разделе нас в основном интересует функ-
циональная организация генов fla и hag, однако мы
рассмотрим и более общие вопросы — о построении ге-
нетической карты и идентификации генных продуктов —
для всех тр,ех групп генов. Роль продуктов генов mot
и che будет обсуждаться в разделах, посвященных
функции бактериальных жгутиков и регуляции клеточ-
ного движения.
Если локализацию генов в бактериальной хромосоме
установили с помощью стандартных генетических мето-
дов, то для идентификации генных продуктов потребо-
вался более изощренный метод — метод клонирования
генов. Например, фрагменты генома Е. coli, несущие
жгутиковые гены, включали либо в фаг X, либо в коли-
циногенный (Col) фактор и в таком виде вводили в
клетки, способные воспроизводить только приобретенный
геном. Под действием жгутиковых генов, введенных в
пермиссивные клетки, синтезировались соответствующие
полипептиды, которые изолировали и разделяли с по-
мощью метода двумерного гель-электрофореза; получен-
ную электрофореграмму сопоставляли с электрофоре-
20
2. Подвижность прокариотических организмов
Tp*L
<•«>*)
ch«D- 64000,65000
Рис. 2.5. Локализация на генетической карте Е. coli генов, контро-
лирующих жгутиковую активность. Область I включает гены, рас-
полагающиеся между генами gal и trp (от fla VK до fla Т). Гены
между fla И и uvr С относятся к области II; остальныек обла-
сти III. Против каждого гена указана молекулярная масса (если
она известна) соответствующего генного продукта. Стрелками
отмечены сотранскрибируемые гены и направление транскрип-
ции [135].
граммой полипептидов из очищенных жгутиковых струк-
тур, и это позволило выяснить, какие гены кодируют
каждую из структур. Было показано, что ген hag коди-
рует флагеллин, ген fla К — субъединицы крюка, а все
остальные гены группы fla кодируют белки базального
тела. Кроме того, продукт гена flat необходим для экс-
прессии других жгутиковых генов. Шесть жгутиковых
генов fla локализованы в области I хромосомы Е. coll и
находятся на карте между генами gal и trp. Остальные
гены fla находятся в областях II и III между генами
zwf и sup D вместе с генами che и mot (рис. 2.5).
О том, как регулируется экспрессия жгутиковых ге-
нов, известно очень немногое. Некоторые Наблюдения
поддерживают представление о том, что репликация од-
ного из генов fla служит триггером для экспрессии ос-
тальных генов. Так, например, продукт гена fla I необ-.
ходим для репликации ряда оперонов групп fla — hag,
2. Подвижность прокариотических организмов
21
mot и che. Однако другие подобные активаторы пока
еще не идентифицированы [135].
Синтез жгутиковых органелл контролируется факто-
рами окружающей среды, такими, как температура, и
управляется катаболитами: он угнетается глюкозой и
рядом других катаболитов и стимулируется сАМР [7,
39]. Этот факт не относится к числу неожиданных, так
как в других бактериальных системах метаболизма
сАМР действует как неспецифический генный актива-
тор, снимающий так называемую катаболитную репрес-
сию [ИО]. Все полученные к настоящему времени данные
указывают на то, что действие сАМР обусловлено обра-
зованием комплекса между сАМР и соответствующим
белком-рецептором. Этот комплекс, по-видимому, связы-
вается с жгутиковыми генами и активирует их реплика-
цию. Центр связывания известен под названием центра,
определяющего жгутиковый синтез (на рис. 2.5 обозна-
чен cfs — constitutive flagellar synthesis). На карте он
расположен вблизи гена fla I, а возможно, входит в его
состав.
Процессы сборки и роста филамента бактериального
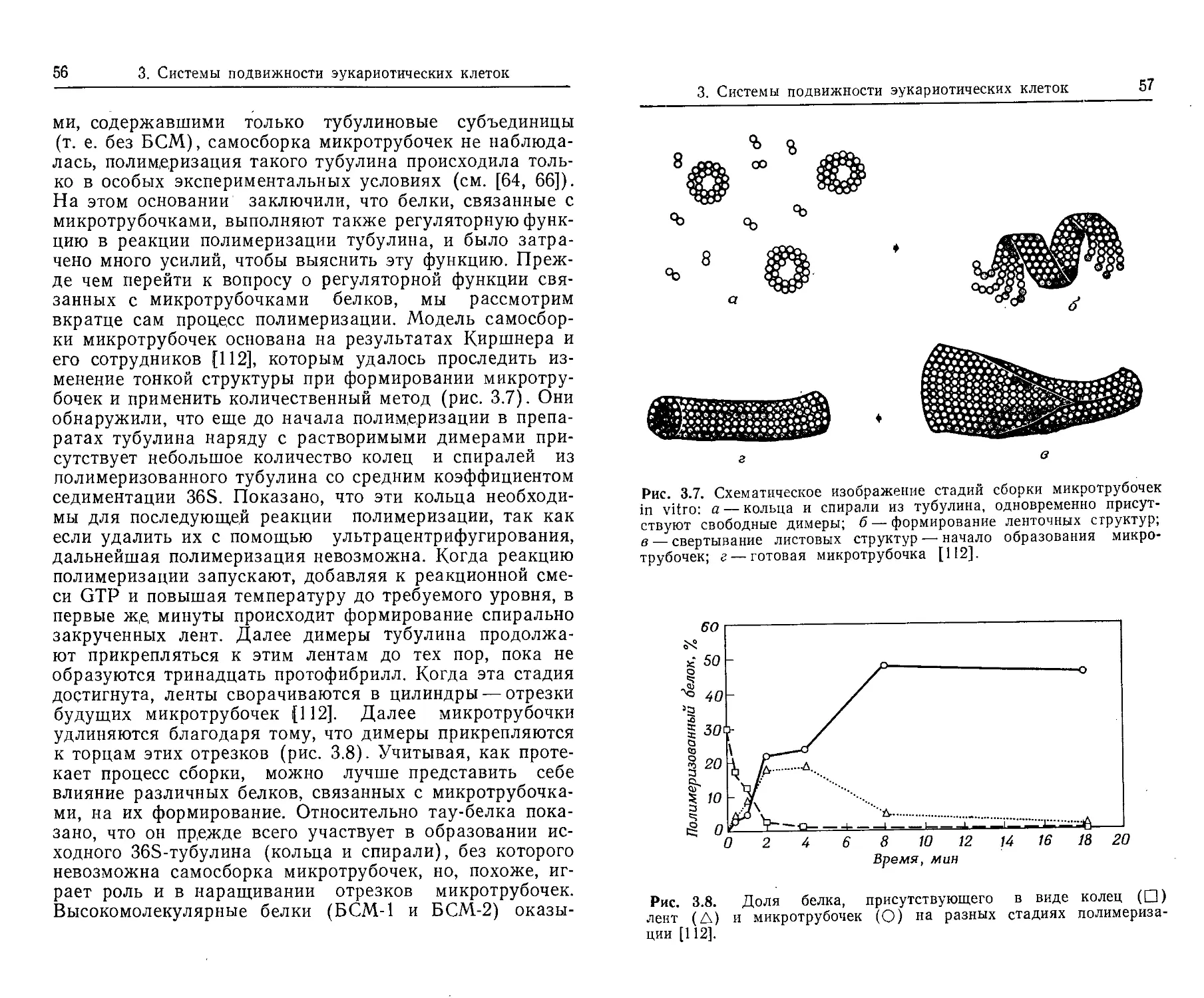
жгутика выяснены достаточно детально. Рост начинает-
ся с дистального конца крюка, к нему прикрепляются
специфической мРНК, которая управляет синтезом флагеллина.
Флагеллииовые субъединицы по центральной полости направляются
к фронту роста жгутика, где встраиваются в филамент.
22
2. Подвижность прокариотических организмов
первые флагеллиновые субъединицы, которые затем слу-
жат основой для сборки волнообразно изогнутого полого
цилиндра. Методом электронно-микроскопической радио-
автографии с использованием радиоактивного лейцина
было показано, что у В. subtilis филаменты удлиняются
путем присоединения субъединиц к их дистальным кон-
цам [44]. Лучше всего с экспериментальными данными
согласуется следующая гипотеза: флагеллиновые субъ-
единицы синтезируются в теле клетки и затем транспор-
тируются к точке роста через центральную полость (ка-
нал) филамента (рис. 2.6).
Никаких запасов резервного флагеллина в цитоплаз-
ме у бактерий не обнаружено; это означает, что каждая
вновь синтезированная субъединица немедленно транс-
портируется к точке роста.
2.4. Функция
бактериальных жгутиков
В этом разделе мы рассмотрим, как движется бак-
териальный жгутик, как генерируется движущая сила и
как она передается к филаменту.
При наблюдении за перемещением жгутиковой бак-
терии в микроскоп ясно видно, что тело бактерии вра-
щается в направлении, противоположном направлению
вращения жгутика. К сожалению, такие прямые наблю-
дения не могут дать убедительных доказательств в поль-
зу того или иного типа движения. До недавнего времени
общепринятым было представление о том, что бактерия
перемещается благодаря волнообразным движениям
жгутика, возникающим у его основания и распростра-
няющимся вдоль него. Правильную интерпретацию дан-
ных относительно характера бактериального движения
предложили Берг и Андерсон в 1973 г. [20]; они посту-
лировали, что движение бактериального жгутика явля-
ется вращательным. Правильность этого предположения
подтвердили остроумные эксперименты Силвермана и
Симона [134]. Поликрюковых мутантов Е. coll (такие
мутанты не способны синтезировать филамент и облада-
ют ненормально длинными крюками) культивировали в
2. Подвижность прокариотических организмов
23
специальной среде, способствовавшей тому, что у каж-
дой бактерии вместо обычных шести образовывалась
только одна поликрюковая структура. Ясно, что такие
клетки не могут передвигаться в среде из-за отсутствия
филаментов. Однако, когда с помощью антител, выра-
ботанных на белки крюка, удавалось приклеить крюки
к предметному стеклу (при этом тело бактерии остава-
лось не связанным со стеклом), было видно, что клетки
вращаются со скоростью 10—20 об/с. Эти опыты пока-
зали, что именно крюк (к которому в норме прикреплен
филамент) совершает вращательные движения вокруг
оси, перпендикулярной к клеточной стенке. Этот «ротор»
обладает, очевидно, достаточно большой мощностью;
чтобы вращать всю клетку бактерии, когда сам он за-
креплен. Подобные результаты были получены также на
мутантах с прямыми жгутиками (см. рис. 2.2.) и послу-
жили непосредственным доказательством того, что дви-
жение жгутиков — вращательного типа. Прикрепленные
бактерии могут вращаться как по часовой стрелке, так
и против; это позволяет предполагать, что изменение
направления вращения жгутика может лежать в основе
ориентации движения клетки.
Легко представить, как вращение единственного жгу-
тика (действующего как винт) может приводить в дви-
жение клетку; значительно труднее понять, что проис-
ходит с многожгутиковой бактерией, у которой множест-
во жгутиков исходит из разных точек бактериальной
стенки. Наблюдения с помощью микроскопа показали,
что у плавающих многожгутиковых бактерий жгутики
собраны сзади в один пучок, который вытянут вдоль
продольной оси тела бактерии. Каждый жгутик в пучке
вращается синхронно с остальными жгутиками, хотя и
независимо от них. В нормальных условиях между жгу-
тиками нет никаких соединительных структур или мости-
ков. Более того, если у нормально подвижной Е. coli
соединить соседние жгутики с помощью бивалентных
антител против флагеллина, это полностью прекращает
движение клетки.
Сопоставляя изложенные выше данные об ультра-
структуре и особенностях движения бактериального
жгутика, можно представить себе следующий механизм.
24
2. Подвижность прокариотических организмов
Кольцо М, находящееся в липидном слое клеточной
мембраны, способно вращаться. Поскольку оно соедине-
но со стержнем, вращательное движение сообщится
стержню и через него передастся за пределы клетки на
полужесткий спиральный филамент. Вращающийся фи-
ламент действует подобно винту самолета и заставляет
клетку плыть. Остальные структуры, в частности кольца
Р и L, не имеют прямой связи с вращающим мотором,
а обеспечивают, по-видимому, прохождение стержня
сквозь бактериальные оболочки.
И наконец, нужно выяснить вопрос об источнике
энергии, которая питает этот «ротор». Ларсен и сотр.
[83] в свое время показали, что АТР не является пря-
мым источником энергии для движения бактериальной
клетки; к этому выводу авторы пришли, изучая под-
вижность с помощью различных специфических ингиби-
торов, а также у мутантов Е. coli и Salmonella typhimu-
rium с нарушениями на различных этапах окислительного
фосфорилирования. Похоже, что источником энергии
служат какие-то интермедиаты окислительного фос-
форилирования, один или несколько. Недавние исследо-
вания, проведенные на стрептококках [93] и на Rhodos-
pirilium rubrum [60], подтвердили эти результаты и по-
казали, что вращение кольца М и, следовательно, самого
жгутика зависит от перемещения протонов из наружной
среды внутрь клетки через клеточную мембрану, т. е.
обусловлено работой протонного насоса. Однако, чтобы
выяснить, как именно за счет переноса протонов сквозь
клеточную мембрану приводится в действие «мотор»
жгутика, необходимы дальнейшие исследования.
2.5. Траектории и изменения
направления движения у бактерий
Жгутик следует считать очень эффективным винтом,
поскольку в нормальных условиях он может обеспечить
невероятно быстрое направленное движение клетки. Так,
например, Е. coli, длина которой не превышает 2 мкм,
может за 1 с покрыть расстояние по крайней мере
20 мкм, что сравнимо с бегом лошади со скоростью
2. Подвижность прокариотических организмов
25
180 км/ч. В микроскоп можно наблюдать, что в отсут-
ствие привлекающих или отталкивающих стимулов клет-
ка Е. coli движется прямолинейно, временами резко из-
меняет направление движения, беспорядочно кидается в
разные стороны, затем снова движется прямолинейно, и
так далее; в результате получается хаотическое переме-
щение в пространстве. Необходимо подчеркнуть, что без
дрожания (тамблинга), т. е. хаотических метаний в раз-
ные стороны, бактерия не способна изменить направле-
ние своего движения.
Наблюдения над одножгутиковыми и многожгутико-
выми бактериями показали, что механизм изменения
направления движения всегда требует смены направле-
ния вращения жгутиков на обратное; это может осу-
ществляться различными способами. Тейлор и Кош-
ланд [155] исследовали, как плавают одножгутиковые
бактерии Pseudomonas citronellotis, и показали, что эти
бактерии движутся по прямой то «головой» вперед под
действием толкающего усилия, развиваемого жгутиком,
то «пятятся», когда направление вращения жгутика
меняется на обратное и жгутик развивает тянущее уси-
Рис. 2.7. Схематическое изображение движения бактерии с • одним
жгутиком: а — бактерия плывет вперед благодаря вращению жгу-
тика; б — жгутик перестает вращаться и клетка останавливается;
в — вращение жгутика в обратном направлении вызывает движение
«задом наперед». Ломаная линия в нижией части рисунка это
траектория хаотического движения одножгутиковой бактерии, кото-
рая движется вперед и назад попеременно.
26
2. Подвижность прокариотических организмов
лие, которое заставляет бактерию двигаться назад. Это
повторяется многократно, и, так как каждый раз ори-
ентация клетки меняется на угол, который может дости-
гать 180° (но чаще бывает меньше), в результате полу-
чается ломаная траектория хаотического движения
(рис. 2.7). Во время прямолинейного движения бактерии
вперед жгутик вращается против часовой стрелки, а ког-
да клетка плывет назад, жгутик вращается по часовой
стрелке.
У свободно плавающих многожгутиковых бактерий
не обнаруживается столь четко выраженного «обратного
хода». Когда клетка 5. typhimurium или Е. coll плывет
в каком-то направлении, все жгутики, как мы уже ви-
дели, держатся вместе, образуя компактный пучок. Пу-
чок разваливается, когда направление вращения жгути-
ков меняется на обратное, и они разлетаются в разные
стороны на короткий промежуток времени, в течение ко-
торого происходит случайная переориентация оси клет-
ки. Этот период переориентации соответствует тамблин-
гу — неупорядоченному метанию бактерии из стороны в
сторону. Вращение жгутиков в том же направлении про-
должается, они снова собираются в пучок и клетка вновь
приобретает способность плыть прямолинейно, но уже в
новом направлении (рис. 2.8). Когда происходит тамб-
линг, это проявляется в случайно направленной подвиж-
ности в отсутствие каких-либо стимулов к хемотак-
сису [91].
Когда вращение жгутика меняется на обратное, ме-
няется не только направление движения бактерии, но
также и спиральная конфигурация филамента — проис-
ходит переход от спирали с большим шагом к более за-
крученной форме. Это предполагает, что меняется упа-
ковка субъединиц флагеллина, что сопровождается
структурным переходом от левозакрученной спирали к
правозакрученной. Такой переход может быть обуслов-
лен напряжением кручения, возникающим при изменении
направления вращения жгутика; при этом процесс рас-
пространяется от основания жгутика, где напряжение
кручения наибольшее, к его концу [92]. Как именно эти
изменения влияют на подвижность, можно пока что толь-
ко гадать.
2. Подвижность прокариотических организмов
27
Рис. 2.8. Схематическое изображеиие движения миогожгутиковой
бактерии: а — бактерия плывет вперед, жгутики собраны в плот-
ный пучок; при изменении направления вращения жгутики разле-
таются во все стороны (б), после чего снова образуют плотный
пучок (в). Ломаная линия — это траектория хаотического движения
многожгутиковой бактерии.
2.6. Управление движением
у бактерий
В среде (жидкой или плотной, типа агарового геля)
с градиентом концентрации аттрактанта или репеллента
жгутиковые бактерии движутся направленно — они пе-
ремещаются к тем областям, где концентрация аттрак-
танта повышена или репеллента понижена. Это явление
называется таксис. Оно наблюдается у диких штаммов
жгутиковых бактерий в ответ на химические стимулы
(хемотаксис), изменение температуры (термотаксис) или
освещенности (фототаксис) 1 [5, 162]. Эволюционные
преимущества, которые дает система, обеспечивающая
направленное движение к источнику пищи или от ток-
сических веществ, совершенно очевидны: жгутиковые
бактерии более конкурентоспособны по сравнению с
микроорганизмами, не имеющими жгутиков.
1 Известны также другие виды таксиса. — Прим. ред. перевода.
28
2. Подвижность прокариотических организмов
Движение бактерии в присутствии химического сти-
мула (хемотаксис) можно исследовать с помощью ми-
кроскопа, снабженного специальной приставкой, которую
Говард Берг [19] приспособил для наблюдения и регист-
рации движений одиночной клетки в пространстве. Ока-
залось, что траектория движения бактерии по направле-
нию к аттрактанту состоит из участков направленного
движения, между которыми находятся области тамблин-
га, т. е. неупорядоченных метаний из стороны в сторо-
ну, подобно тому, что наблюдается при ненаправленной
подвижности. Однако частота метаний меньше, а от-
дельные пробеги дольше по сравнению с ненаправлен-
ным движением. В результате происходит хемотаксис
по ломаной траектории. Когда бактерия удаляется от
репеллента, все наоборот: частота метаний повышена, а
прямолинейные пробеги укорочены. Сходный тип под-
вижности наблюдается у микроорганизмов с полярно
расположенным жгутиком, с тем только отличием, что
вместо беспорядочных метаний они делают «обратный
ход». Из этих экспериментов ясно, что направленная
подвижность осуществляется посредством усиления или
ослабления тамблинга и изменения длины прямолиней-
ных пробегов в направлении к действующему стимулу
или от него.
Бактерии способны ощущать не только пространст-
венный градиент активных веществ, но и изменение их
концентрации во времени; в ответ на быстрое введение
в среду обитания различных доз репеллентов или ат-
трактантов они изменяют характер своего движения
[90]. При этом возникает преходящее состояние двига-
тельного возбуждения, которое проявляется в учащении
беспорядочных метаний; по истечении какого-то времени
бактерии возвращаются в «спокойное» состояние. Это
означает, что они реагируют именно на изменение кон-
центрации химического вещества во времени, но не на
абсолютное ее значение. Более того, бактерии обнару-
живают своего рода кратковременную память. Если бак-
терий стимудировать, добавив в среду аттрактант в оп-
ределенной концентрации, а затем восстановить преж-
нюю нестимулирующую концентрацию, то повторное —
через короткий промежуток времени после первой сти-
2. Подвижность прокариотических организмов
29
муляции—добавление аттрактанта до ранее эффектив-
ных концентраций уже не вызовет соответствующей ре-
акции. Это указывает на то, что бактерии способны не
только ощущать, но и «запоминать» (на короткое вре-
мя) силу стимулирующего сигнала (см. [5, 47]).
Определив главные особенности хемотаксического уп-
равления бактериальным движением, мы перейдем к
анализу элементов системы управления и попытаемся
выяснить, как они связаны с направлением движения
жгутиков. Многие из результатов, которые мы обсудим,
получены в опытах с мутантами, дефектными в том или
ином аспекте проявления хемотаксиса; таких мутантов
обычно называют с/1е_-мутантами. У Е. coli система уп-
равления включает комплекс генов che, располагающих-
ся на хромосомной карте в трех разных областях, от-
ветственных за различные звенья реакции хемотаксиса.
С помощью метода, описанного выше для системы генов
fla, были найдены генные продукты большинства из
них — это полипептида с мол. массами от 8000 до
76 000; их функции также выяснены.
На рис. 2.9 изображена блок-схема системы хемотак-
сиса бактерий: ряд рецепторов, воспринимающих вход-
ные сигналы, передает их на систему переработки сиг-
налов, после чего они поступают в систему управления,
которая посылает сигнал, воздействующий на движение
жгутиков.
Наиболее полные сведения имеются о входных эле-
ментах системы хемотаксиса: известны количества раз-
личных хеморецепторов, их химическая природа, а так-
же функции. Хеморецепторы узнают аттрактанты сами
по себе — нет необходимости в предварительной модифи-
кации молекулы аттрактанта. Хеморецепторы отличают-
ся от других рецепторов бактериальной клетки (напри-
мер, от рецепторов, «узнающих» бактериофаг, бактерио-
цин и другие раздражители) уникальной способностью
информировать жгутики об обнаружении соответствую-
щего химического раздражителя. У Е. coli идентифици-
ровано более 20 различных типов хеморецепторов; одни
из них обеспечивают положительный хемотаксис (движе-
ние по направлению к аттрактанту), другие — отрица-
тельный хемотаксис (уход от репеллента). К первой
30
2. Подвижность прокариотических организмов
рецепторы
Рис. 2.9. Гипотетическая блок-схема хемотаксиса у бактерий. Рецеп-
торы, связав стимулирующую молекулу, передают сигнал на пере-
рабатывающие системы, от которых он поступает в систему управ-
ления, где трансформируется в командный сигнал, вызывающий
необходимые изменения вращения жгутиков.
группе относятся рецепторы к сахарам и аминокислотам,
ко второй — реагирующие на жирные кислоты, спирты,
индольные соединения, на изменение pH и концентрации
ионов металлов. Примечательно, что большинство ат-
трактантов используется в метаболизме, тогда как ре-
пелленты, как правило, представляют опасность для
клетки. По крайней мере в одном случае, а именно для
глюкозы, специфичности рецептора по отношению к ат-
трактанту нет, поскольку этот же сахар может воспри-
ниматься также рецепторами, «узнающими» галактозу
и маннозу.
2.6.1. Природа хеморецепторов
Хеморецепторы — это белки, которые по крайней ме-
ре периодически должны находиться вне клеточной
мембраны, чтобы взаимодействовать с внешними раздра-
2. Подвижность прокариотических организмов
31
жителями. Наряду со своей специфической функцией —
хеморецепцией — некоторые из этих белков играют так-
же роль переносчиков, необходимых для транспортиров-
ки определенных молекул через клеточную мембрану.
У Е. coli, например, белок-переносчик галактозы а-ме-
тилгалактозидной транспортной системы одновременно
выполняет функцию хеморецептора по отношению к D-
галактозе, D-глюкозе и D-фукозе [83]. Другой пример —
белок-переносчик рибозы у 5. typhimurium, который яв-
ляется также и хеморецептором того же сахара [15].
Белки-переносчики легко выделить из бактерий с по-
мощью осмотического шока, поскольку они находятся в
периплазматическом пространстве (между цитоплазма-
тической мембраной и клеточной стенкой) и связаны с
клеточной мембраной очень слабо. Взаимодействие этих
белков с сахарами, которые они связывают, можно, сле-
довательно, изучать in vitro.
Чтобы функционировать как хеморецепторы, белки-
переносчики должны уметь взаимодействовать с элемен-
тами системы, перерабатывающей сигналы от рецепто-
ров и передающей их жгутикам (рис. 2.10). Было выска-
зано предположение, что система переработки сигналов
активируется в результате конформационных изме-
нений рецепторного белка при контакте его со стимули-
рующим веществом. Эта гипотеза получила эксперимен-
передача к жгутику
Рис. 2.10. Функция рецепторов галактозы и рибозы у Е. coli. Свя-
зав соответствующий сахар, рецептор стимулирует систему перера-
ботки сигналов и одиовремеино участвует в переносе молекулы
сахара внутрь клетки.
32
2. Подвижность прокариотических организмов
тальное подтверждение пока только в случае белка, свя-
зывающего галактозу. Показано, что образование комп-
лекса белка с субстратом сопровождается конформаци-
онными изменениями белка [25].
Еще один рецептор к сахарам у Е. coll оказался фер-
ментом II- глюкозофосфотрансфер азной системы; этот
фермент прочно связан с клеточной мембраной и ката-
лизирует фосфорилирование сахаров фосфорилирован-
ным белком [8]. Сравнительно недавно было показано,
что у Е. coli и 5. typhimurium функцию рецептора по
отношению к двухвалентным катионам осуществляет
связанная с мембраной Mg2+, Са2+-зависимая аденозин-
трифосфатаза (Mg2+, Са2+-АТРаза), которая участвует
в синтезе АТР, в конечных этапах окислительного фос-
форилирования, а также в некоторых других метаболи-
ческих процессах [179].
2.6.2. Система
переработки сигналов
Природа системы, перерабатывающей сигналы от хе-
морецепторов и передающей их жгутикам, почти совер-
шенно неизвестна. Первые шаги в этом направлении уда-
лось сделать благодаря открытию Адлера и Даля [6]:
клеткам Е. coll для осуществления хемотаксиса требует-
ся L-метионин. Гетеротрофные по метионину мутанты
Е. coll (не способные синтезировать метионин) нужда-
ются в постоянном наличии этой аминокислоты в среде
их обитания; в противном случае они теряют способ-
ность к тамблингу и перестают реагировать на аттрак-
танты и репелленты. Метионин расходуется на образо-
вание S-аденозилметионина, роль которого как донора
метильных групп при метилировании белков хорошо из-
вестна [16]. В одной из своих последних работ Спрингер,
Гой и Адлер [143] приводят данные о том, что метили-
рование некоторых белков цитоплазмы — так называе-
мых метил-акцепторных хемотаксических белков
(МХБ) — необходимо для переработки информации, по-
ступающей от хемосенсорных элементов. МХБ образуют
по крайней мере две функциональные единицы, каждая
из которых необходима для переработки сигналов от
2. Подвижность прокариотических организмов
33
двух различных групп хемосенсоров (идентифицируемых
по их чувствительности к разным молекулам). Кроме
того, было показано, что продукты генов che D и che М
метилируются в ответ на хемотаксические стимулы, в
связи с чем их, следовательно, нужно отнести к МХБ.
Для метилирования продуктов гена che D и гена cheM
требуется экспрессия двух других генов группы che —
гена che X и гена che W [136].
Переработанный входной сигнал, вероятно, непосред-
ственно передается жгутикам как команда изменить ско-
рость и направление вращения «ротора». Этот сигнал
может воздействовать на жгутик также и опосредован-
но — через белки, находящиеся в клеточной мембране,
которые управляют вращательным механизмом клетки.
Оказалось, что эти белки — конечные продукты генов
mot А и mot В; они входят в состав цитоплазматической
мембраны и локализованы, по-видимому, вокруг базаль-
ного тела жгутика [137].
Рекомендуемая литература
Приведенные ниже работы относятся главным образом к общим
аспектам жгутиковой подвижности у бактерий:
Berg И. С. (1975). How Bacteria Swim, Sci. Amer., 233, 36—44.
Silverman M., Simon M. I. (1977). Bacterial flagella, Ann. Rev.
Microbiol., 31, 397—419.
Simon M., Silverman M., Matsumura P., Ridgway H., Komeda У.,
Hitmen M. (1978). Structure and function of bacterial flagella.
In: Relations between Structure and Function in the Prokaryotic
Cell (ed. R. Y. Stanier, H. J. Rogers, J. B. Ward), Cambridge
University Press, Cambridge.
MacNab R. M. (1978). Bacterial motility and chemotaxis. Molecular
biology of a behavioural system, CRC Crit. Rev. Biochem., 5,
291—342.
Генетика подвижности рассмотрена в следующих работах:
lino Т. (1977). Genetics of structure and function of bacterial flagel-
la, Ann. Rev. Genet., 11, 161—182.
Parkinson J. S. (1977). Behavioral genetics in bacteria, Ann. Rev.
Genet., 11, 397—414.
Хемотаксису посвящены следующие обзоры:
Adler J. (1975). Chemotaxis in bacteria, Ann. Rev. Biochem., 44,
341—356.
34
2. Подвижность прокариотических организмов
Chet I., Mitchell R. (1976). Ecological aspects of microbial chemo-
tactic behaviour, Ann. Rev. Microbiol., 30, 221—239.
Frere J. M. (\ЪП). La chimiotaxie ches les bactbries, Bull. Inst.
Pasteur, 75, 187—202.
Кроме того, работы по различным аспектам подвижности бакте-
рий можно найти в книге, посвященной примитивным системам по-
движности:
Cell Motility (ed. R. Goldman, T. Pollard, J. Rosenbaum), Cold
Spring Harbor Laboratory, Cold Spring Harbor (1976), Section 1,
pp. 29—113.
3. СИСТЕМЫ
подвижности
ЭУКАРИОТИЧЕСКИХ
КЛЕТОК
Системы, предназначенные для обеспечения подвижнос-
ти у эукариотических организмов, имеют сложную струк-
туру, однако у всех изученных клеток они построены из
удивительно похожих элементов. Компоненты систем под-
вижности могут, очевидно, приспосабливаться к любым
ситуациям, будь то свободно живущая амеба, клетка
млекопитающего или клетка цветка. Сходство в строе-
нии основных компонентов подвижности указывает на
то, что они кодируются генами, которые сохранились не-
изменными на протяжении всей клеточной эволюции.
По морфологическим свойствам и по белковому со-
ставу все структуры, создающие движение у эукариоти-
ческих клеток, можно отнести к двум основным систе-
мам. Первая система строится из микротрубочек, основ-
ной компонент которых тубулин, а вторая — из микро-
филаментов, содержащих главным образом актин. Эти
две системы ответственны за различные типы движе-
ния. Микротрубочки связаны с движением ресничек и
жгутиков, миграцией пигментных гранул в хроматофо-
рах, изгибанием аксостилей и т. д., микрофиламенты —•
с амебоидным движением, токами цитоплазмы, цитоки-
незом и т. п. Некоторые другие типы движения, напри-
мер расхождение хромосом при делении клетки, пока
еще не удалось с определенностью связать с микротру-
бочками либо с микрофиламентами.
В системах подвижности, кроме основных белковых
компонентов, идентифицированы также и многие другие
белки. Некоторые из них непосредственно взаимодей-
ствуют с тубулином (динеин) и с актином (миозин) при
генерации движения. Многие другие выполняют регуля-
торные функции либо образуют соединения между мик-
36
3. Системы подвижности эукариотических клеток
ротрубочками, микрофиламентами и другими структура-
ми клетки. Хотя в морфологическом и биохимическом
аспектах система микротрубочек и система микрофила-
ментов существенно различаются, некоторые данные ука-
зывают на то, что функционально они могут перекры-
ваться и находиться в кооперативных отношениях.
Так, например, разветвленная сеть микротрубочек внут-
ри клетки может служить каркасом для прикрепления
микрофиламентов либо, хотя это менее вероятно, обе
системы могут действовать кооперативно, развивая при
этом сокращающую силу.
Определив в самых общих чертах компоненты систем
подвижности, мы перейдем в следующем разделе к бо-
лее детальному рассмотрению их свойств, взаимосвязей
и механизма действия.
3.1. Микротрубочки
Микротрубочки предстали как особые внутриклеточ-
ные структуры (особенно внутри жгутика) благодаря
применению методов электронной микроскопии. В 1946 г.
Джакус и Холл [75] продемонстрировали наличие одина-
ковых по диаметру трубчатых структур в ресничках
Paramecium. Впоследствии трубчатые структуры обна-
ружили почти во всех клетках. Стали считать, что они
характерны для эукариотического уровня организации.
Однако микротубулярные структуры были обнаружены
и у прокариот — в цитоплазме спирохет [69]. Морфологи-
чески и биохимически эти микротрубочки подобны тем,
которые наблюдаются в эукариотических клетках [94].
Это открытие поддержало «экзогенную гипотезу» проис-
хождения микротрубочек эукариот, согласно которой
реснички и жгутики клеток высших организмов считают-
ся приобретенными извне — путем симбиоза клеток, ра-
нее не имевших жгутиков или ресничек, со спирохетами,
содержавшими микротрубочки.
Микротрубочки представляют собой длинные полые
цилиндры, наружный диаметр которых около 24 нм, а
внутренний— 15 нм. В большинстве клеток длина тру-
бочек обычно не превышает нескольких микрон, хотя в
некоторых специализированных клетках, например в мо-
3. Системы подвижности эукариотических клеток
37
Рис. 3.1. Электронные микрофотографии микротрубочек в краевых
областях эритроцитов тритона: а — продольный срез, на котором
видна линейная упаковка субъединиц в микротрубочках; б — по-
перечный срез; видно, что стенка каждой микротрубочки образова-
на тринадцатью субъединицами. Негативное окрашивание. X 200 000.
(Снимки любезно предоставлены д-ром G. Monaco.)
торных нейронах центральной нервной системы, трубоч-
ки могут быть длиной в несколько сантиметров. Стенка
микротрубочки (около 5 нм толщиной) построена из
продольно ориентированных протофибрилл. Протофиб-
риллы состоят из глобулярных субъединиц (их очень
хорошо видно в электронный микроскоп при негативном
окрашивании), содержащих только один белок—тубу-
лин (рис. 3.1). Помимо того что микротрубочки имеют
прямое отношение к подвижности клетки, они участву-
ют и в других процессах, более или менее связанных
с подвижностью, например в поддержании формы клет-
ки, во внутриклеточном транспорте веществ, в секреции
клеточных продуктов, в движении хромосом при делении
клетки и, возможно, в осуществлении сенсорных связей,
а также в перемещении компонентов клеточной мембра-
ны (107, 149]. Микротрубочки могут быть рассеяны по
всей цитоплазме, а могут быть собраны в организован-
ные структуры.
38 3. Системы подвижности эукариотических клеток
Чтобы понять, как клетка движется с помощью мик-
ротрубочек, мы рассмотрим их организацию в ресничках
и жгутиках эукариотических клеток, в сократимом аксо-
стиле некоторых жгутиконосцев, а также в микротубу-
лярной сети цитоплазмы животных клеток.
3.1.1. Микротрубочки
в ресничках и жгутиках
Эукариотические реснички и жгутики представляют
собой специализированные структуры, выступающие за
пределы клеточной поверхности и способные двигаться.
Эти два типа органелл идентичны по своей структуре; в
функциональном отношении между ними обнаруживают-
ся лишь незначительные различия. Жгутики и реснички
удается легко и в довольно больших количествах отде-
лить от клеток, иногда просто энергичным встряхивани-
ем клеточной суспензии. Это облегчило исследование их
структуры и химического состава.
Структурная организация ресничек и жгутиков прак-
тически во всех исследовавшихся клетках отличается
удивительным единообразием: центральную пару микро-
трубочек окружает кольцо из микротрубочек (обычно
парных) [139]. Микротрубочки связаны между собой по-
перечными мостиками; вся конструкция называется ак-
сонемой жгутика. Расположение микротрубочек и мости-
ков в аксонеме изучено в настоящее время довольно по-
дробно; оно схематически представлено на рис. 3.2.
В центре находится пара микротрубочек, окруженных
общей оболочкой из белковых колец, плоскость которых
наклонена к оси аксонемы. Стенка каждой из централь-
ных микротрубочек образована тринадцатью продольны-
ми рядами (протофибриллами) белковых субъединиц.
Эту центральную структуру окружают девять располо-
женных по кольцу пар микротрубочек. В каждом
дублете одна микротрубочка (основная или А) имеет
полностью замкнутую стенку, состоящую из 13 прото-
фибрилл (рис. 3.3); другая микротрубочка (микротру-
бочка В) имеет серповидное сечение и примыкает к мик-
ротрубочке А, так что ее стенка состоит из меньшего
3. Системы подвижности эукариотических клеток
39
Рис. 3.2. Схематическое изображение аксонемы жгутика. Показана
взаимосвязь между элементами структуры на разных уровнях: а —
на конце жгутика, б — в средней части и в — на уровне базаль-
ного тела.
40
3. Системы подвижности эукариотических клеток
Рис. 3.3. Схематическое изображение расположения тубулиновых
субъединиц в микротрубочке: а — поперечный срез; б — вид сбоку.
числа протофибрилл, обычно из 10. Каждая микротру-
бочка А соединена с микротрубочкой В соседнего дуб-
лета с помощью мостика из белка, называемого некси-
ном. От обращенной внутрь поверхности микротрубочки
А отходит по направлению к центру белковый выступ —
так называемая радиальная спица. Вдобавок каждая
микротрубочка А имеет двойной ряд боковых ручек
(их длина около 14 нм), построенных из белка (динеи-
на) и направленных к соседнему дублету. Все эти вы-
ступы расположены с определенной периодичностью от-
носительно оси микротрубочки А и, как будет видно из
дальнейшего, играют очень важную роль в генерации
движения.
Описанное выше строение аксонемы соответствует
всей средней части жгутика, т. е. за исключением его
концевой части и основания, где аксонема устроена не-
сколько иначе. На самом конце центральная пара мик-
ротрубочек обычно длиннее, чем окружающие ее пери-
ферические дублеты, так что жгутик на конце заострен.
Нередко микротрубочки В бывают короче микротрубо-
чек А, и периферические дублеты вблизи конца жгути-
ка переходят в одиночные трубочки (рис. 3.2). На оди-
ночных микротрубочках А в этой области жгутика вы-
ступов нет [129].
3. Системы подвижности эукариотических клеток
41
Специализированная структура — базальное тело, —
из которой исходит жгутик, находится в цитоплазме
клетки у основания жгутика. Она может быть различна
в клетках разных типов, однако при этом основные, об-
щие черты сохраняются. В базальном теле ресничек
Tetrahymena. piriformis наряду с микротрубочками А и
В имеются еще микротрубочки С (они образованы де-
сятью протофибриллами), так что по периферии рес-
нички располагаются не дублеты, а триплеты микротру-
бочек. Эти триплеты связаны' между собой боковыми
выступами (поперечными мостиками) и прикреплены к
центральному кольцу радиальными спицами (рис. 3.2).
Базальное тело реснички или жгутика по своей структу-
ре похоже на центриоль животных клеток. И базальное
тело, и центриоль действуют как организаторы ассоциа-
ции микротрубочек; в первом случае образуется реснич-
ка или жгутик, во втором — митотическое веретено.
Еще одна интересная часть жгутика — это так назы-
ваемый воротничок, кольцевой валик, окружающий ос-
нование жгутика там, где он выходит из клетки. В элект-
ронный микроскоп на поперечных срезах воротничка в
этой области видны волокнистые соединения между пе-
риферическими дублетами жгутика и внутренними слоя-
ми клеточной мембраны. Они жестко закрепляют микро-
трубочки в клеточной мембране и участвуют в движении
жгутика.
Подвижность ресничек и жгутиков обусловлена свой-
ствами именно аксонемы, но не других компонентов этих
органелл (т. е. клеточной мембраны, цитоплазмы и др.).
Это наглядно продемонстрировали Джиббонс и Джиб-
бонс [55], которым удалось получить интактные аксоне-
мы из хвостов сперматозоидов морского ежа (мембраны
лизировали неионным детергентом тритон Х-100) и вы-
звать движение аксонем, добавив АТР в качестве источ-
ника энергии1.
1 Этот эффект впервые продемонстрировали на глицеринизиро-
ванных моделях ресничек мерцательного эпителия В. Я- Александров
и Н. И. Арронет (ДАН СССР, 1956, НО, 457—460). — Прим. ред. пе-
ревода.
42
3. Системы подвижности эукариотических клеток •
3.1.2. Микротрубочки
сократимого аксостиля
Сократимый- аксостиль некоторых жгутиковых — еще
один пример подвижной структуры, в которой способ-
ность к движению обусловлена микротрубочками. Аксо-
стиль— это палочковидная или же лентообразная орга-
нелла, которая проходит через всю цитоплазму от одно-
го конца клетки до другого (рис. 3.4). Эту структуру
впервые описал Грасс, который изучал симбиотических
жгутиковых, живущих в кишечнике термитов и древояд-
ных тараканов [61]. Аксостиль способен совершать вол-
нообразные движения, причем волны, возникающие на
его переднем конце, распространяются по всему аксости-
лю к его противоположному концу. В результате форма
клетки непрерывно меняется и именно благодаря движе-
нию своей поцерхности клетка перемещается [62]. Аксо-
стили можно извлечь, лизировав клетки с помощью три-
тона Х-100, и затем очистить от других клеточных ком-
Рис. 3.4. Сократимый аксостиль Metamonadida: а — расположение
аксостиля в теле клетки; б — схематическое изображение параллель-
ной упаковки микротрубочек.
3. Системы подвижности эукариотических клеток
43
понентов дифференциальным центрифугированием. Изо-
лированные аксостили, как и аксонемы, способны дви-
гаться при добавлении АТР [99].
Что касается ультраструктуры, аксостиль состоит из
параллельных листов, каждый из которых построен из
множества микротрубочек. Число листов, образующих
аксостиль, и число микротрубочек в каждом листе
варьируют в зависимости от вида микроорганизма. От-
носительно небольшие аксостили состоят всего из двух-
трех листов по 20—30 миктротрубочек в каждом, тогда
как более крупные аксостили построены из 20—30 лис-
тов, в каждом из которых около 150 микротрубочек. Это
означает, что суммарное число микротрубочек может
варьировать от менее чем 100 до более чем 5000. Листы
в аксостиле находятся на расстоянии приблизительно
30 нм друг от друга, а соседние микротрубочки одного
и того же листа — около 40 нм (от центра до центра)
[99]. Белковые мостики, расположенные в определенном
порядке вдоль микротрубочек, соединяют их между со-
бой в пределах каждого листа, а также, возможно, со-
единяют и трубочки соседних листов. Микротрубочки,
находящиеся в соседних листах, соединяются, кроме то-
го, динеиновыми ручками (см. рис. 3.4,6). Такая струк-
тура подвижна, так как построенные из микротрубочек
листы могут скользить друг относительно друга.
Аксонемы в аксоподиях солнечника [161], ротовом же-
лобке ресничной инфузории Nassula [163], щупальцах со-
сущих инфузорий [67, 164], а также микротрубочки в
хвостах сперматозоидов червеца Pianococcus [125] тоже
построены из связанных между собой микротрубочек,
образующих упорядоченные структуры. (Для более де-
тального ознакомления с перечисленными выше приме-
рами читателю следует обратиться к оригинальным ра-
ботам, приведенным в списке цитируемой литературы.)
3.1.3. Цитоплазматические
микротрубочки животных клеток
За последние десять лет существенно возрос интерес
к изучению микротрубочек и микрофиламентов в меди-
кобиологическом аспекте, что способствовало быстрому
44
3. Системы подвижности эукариотических клеток
развитию исследований тонкой структуры и биохимии
этих объектов в клетках млекопитающих. Благодаря
применению двух методических приемов возможности
выявления таких легко повреждаемых структур, как ци-
топлазматические микротрубочки, чрезвычайно расши-
рились. Один из этих приемов — альдегидная фиксация
(в частности, глутаральдегидом), которая позволяет хо-
рошо сохранять цитоплазматические органеллы [127].
В большинстве исследованных клеток нашли обра-
зования, весьма сходные с микротрубочками жгутиков
по своей тонкой структуре. Оказалось, что митотическое
веретено состоит в основном из микротрубочек, они. рас-
сеяны также и в цитоплазме, где их распределение за-
висит от типа клетки. Обычные методы электронной
микроскопии не позволяют получить общую картину
распределения микротрубочек в цитоплазме интерфаз-
ной клетки; приходится применять трудоемкий метод
трехмерной реконструкции по серии последовательных
срезов. Такую реконструкцию удалось осуществить с по-
мощью флуоресцирующих антител на тубулин: клетки,
приклеенные к предметному стеклу, обрабатывают флуо-
ресцирующими антителами, которые специфически
окрашивают только тубулин, это позволяет увидеть ци-
топлазматические микротрубочки в микроскоп в ультра-
фиолетовом свете [51, 172]. Вначале для выработки анти-
тел использовали тубулин из хвостов сперматозоидов
морского ежа Strongylocentrotus purpuratus (эти анти-
тела дают перекрестную реакцию преципитации с тубу-
лином млекопитающих, что указывает на высокую кон-
сервативность аминокислотной последовательности тубу-
лина в процессе эволюции). В последнее время для этого
стали применять высокоочищенный препарат тубулина
из мозга млекопитающих; с помощью выработанных
против него антител удалось подтвердить и дополнить
ранее полученные данные о распределении микротрубо-
чек в цитоплазме различных клеток.
В качестве примера in vivo мы рассмотрим микроту-
булярную структуру в культуре животных клеток. В ци-
топлазме фибробластов в состоянии интерфазы обнару-
живается огромное количество тончайших волоконец
одинакового диаметра. Часто эти волоконца радиусами
3. Системы подвижности эукариотических клеток
45
Рис. 3.5. Распределение микротрубочек в цитоплазме клетки на раз-
ных фазах жизненного цикла (данные получены с помощью флу-
оресцирующих антител): а — интерфаза: микротрубочки образуют в
цитоплазме тонкую сеть; б — профаза: флуоресценция сосредото-
чена в области центриолей; в — метафаза иг — анафаза: флуорес-
цируют микротрубочки митотического веретена; д — дочерние клетки
расходятся: флуоресценция видна только в соединительном мостике
между ними.
расходятся от ядра, почти достигая поверхности клетки;
иногда они образуют неупорядоченные пучки, которые
располагаются параллельно цитоплазматической мемб-
ране, что особенно заметно в местах выпячивания псев-
доподий (рис. 3.5). Если разрушить микротрубочки кол-
хицином, снижением температуры или повышением дав-
ления, волоконца исчезают и клетка теряет свою форму.
Когда действие этих ингибиторов прекращается, струк-
тура микротрубочек восстанавливается и клетка вновь
обретает нормальную форму (171, 172]. В клетке, при-
ближающейся к моменту деления, флуоресценция цито-
плазмы (обусловленная флуоресцирующими антителами
на тубулин микротрубочек) сгущается возле ядерной
мембраны — в двух противолежащих областях, соот-
46
3. Системы подвижности эукариотических клеток
ветствующих центриолям; у клеток в прометафазе вид-
ны две флуоресцирующие звезды. По мере развития ми-
тоза происходит дальнейшее сгущение флуоресценции,
и g метафазе интенсивно светится лишь митотическое ве-
ретено клетки, а в экваториальной плоскости — там, где
расположены сконденсировавшиеся хромосомы, флуорес-
ценция отсутствует. Флуоресценция веретена начинает
уменьшаться с наступлением анафазы (так как разру-
шается веретено), и к моменту разделения двух дочер-
них клеток светится только мостик между клетками, в ко-
тором видно много микротрубочек. Когда дочерние клет-
ки окончательно отделяются друг от друга, восстанавли-
вается исходная картина распределения тонких цито-
плазматических микротрубочек [51, 171, 172]. Эти ре-
зультаты указывают на то, что в клетках млекопитаю-
щих существует цикл микротрубочек, параллельный
циклу развития клетки; в соответствии с нуждами клет-
ки микротрубочки образуют те или иные структуры, не-
обходимые на данном этапе развития клетки.
Цитоплазматические микротрубочки, конечно, не
могут генерировать движение клетки, как это делают
микротрубочки жгутиков или аксостиля. Однако на том
основании, что в некоторых типах культур клеток под
действием ингибиторов, вызывающих деструкцию микро-
трубочек, нарушается способность клеток двигаться на-
правленно [22, 58], было сделано предположение, что ци-
топлазматические микротрубочки все же имеют значе-
ние для подвижности клетки [59]. Что касается различ-
ных типов амебоидного движения, то здесь совершенно
ясно, что если цитоплазматические микротрубочки и
принимают в них какое-то участие, то только вспомога-
тельное.
3.2. Состав микротрубочек
Основной белок, из которого построены микротрубоч-
ки,— это тубулин; другие белковые компоненты присут-
ствуют в меньших количествах и их называют белками,
связанными с микротрубочками.
3. Системы подвижности эукариотических клеток
47
3.2.1. Тубулин
Тубулин имеет изоэлектрическую точку в области
pH 5,2—5,4. Он обычно выделяется в виде димеров с
мол. массой 115000; это так называемый 68-тубулин.
При обработке димеров тубулина такими денатурирую-
щими агентами, как мочевина, гуанидин или додецил-
сульфат натрия, образуются 35-субъединицы, называе-
мые а- и р-тубулин. Они имеют одинаковую мол. массу
55000, но различаются по аминокислотному составу [27]
и по последовательности аминокислот [89]. Как недавно
показали Брайан и сотр. [28], основное различие .между
субъединицами обусловлено не посттрансляционной мо-
дификацией (хотя она и может происходить), поскольку
каждая из субъединиц кодируется своей мРНК (эти
мРНК удалось выделить из мозга куриных эмбрионов)..
Посттрансляционная модификация тубулина тем н,е
менее имеет место. Тубулин может связываться с гуани-
новым нуклеотидом (например, с GTP), для которого в
каждом димере есть два центра связывания с разным
сродством [21, 150]. Возможно также фосфорилирование
сериновых остатков тубулина протеинкиназой, активи-
руемой сАМР. Показано, что это происходит в р-субъ-
единицах цитоплазматического тубулина [43] и в «-субъ-
единицах тубулина жгутиков Chlamydomonas [114].
Третий вид модификации — это специфическое присоеди-
нение тирозина (или фенилаланина) к С-концу «-субъ-
единицы [18]. Эту реакцию катализирует специальный
фермент — тубулин :тирозин лигаза, который широко
распространен в тканях птиц и млекопитающих [122].
Значение двух последних превращений пока неясно. Что
касается связывания GTP, то это обычное свойство ту-
булина, имеющее отношение к самосборке микротру-
бочек.
Помимо нуклеотидов, фосфата и, аминокислот, тубу?
лин специфически связывает двухвалентные катионы,
например Mg2+ (1 моль димеров тубулина Прочно свя-
зывает по крайней мере 1,0 моль Mg2+ [108]); кроме
того, в определенных стехиометрических соотношениях
он связывает такие алкалоиды, как колхицин [175] и
винбластин [176]. Последнее имеет существенное значе-
48
3. Системы подвижности эукариотических клеток
ние, так как эти алкалоиды избирательно угнетают раз-
личные проявления активности микротрубочек, что ши-
роко используется для исследования реакций, протекаю-
щих с участием этих структур, в опытах как in vitro,
так и in vivo.
Исследование тубулина из клеток различных типов
показало, что большинство его физико-химических
свойств высококонсервативно, т. е. мало изменялось в
процессе эволюции. Все изученные тубулины из различ-
ных источников имеют одинаковую молекулярную массу
и одну и ту же электрофоретическую подвижность. Бо-
лее того, по предварительным данным об аминокислот-
ной последовательности а- и р-субъединиц тубулина из
аксонемы хвостов сперматозоидов морского ежа и цито-
плазматического тубулина из мозга куриных эмбрионов
в первых 25 остатках N-конца а-субъединиц нет ника-
ких различий, а в р-субъединицах обнаружена замена
только одной аминокислоты (остаток 7) [89]. Иммуноло-
гическими методами также подтверждается отсутствие
видовой специфичности тубулинов различного происхож-
дения; показано, что есть перекрестная реакция между
тубулином млекопитающих и тубулинами морского ежа
[33, 51], одноклеточной двухжгутиковой водоросли
Chlamydomonas reinhartii [115] и миксамеб слизистого
гриба Dictyostelium discoideum [30]. Тубулин из мозга
млекопитающих может также сополимеризоваться (т. е.
образовывать гибридные микротрубочки) с тубулином
дрожжей [170], плесневых грибов [132] и миксомицетов
[29]. Эти результаты демонстрируют, что тубулины раз-
личного происхождения химически очень похожи, хотя
и не идентичны.
Другой интересный вопрос: возможно ли, что в пре-
делах одной и той же клетки есть различные классы ту-
булина, которые могут быть нужны для построения раз-
личных структур? Такая возможность предполагает, что
либо есть различные наборы тубулиновых генов, либо
происходят различные посттрансляционные модификации
исходно одинаковых молекул. Действительно, на разно-
образных объектах удалось продемонстрировать присут-
ствие в одной клетке не вполне одинаковых тубулинов.
У амебоидных простейших Naegleria gruberi, которые
3. Системы подвижности эукариотических клеток
49
в условиях эксперимента могут синхронно дифференци-
роваться, переходя в жгутиковую форму, Фалтон и его
сотрудники [50, 81, 82] обнаружили антигенную неиден-
тичность жгутикового и цитоплазматического тубулинов
и показали, что жгутиковый тубулин синтезируется
de novo, когда начинают формироваться жгутики. Сте-
фенсу [148] удалось разделить микротрубочки А и В из
внешних дублетов хвостов сперматозоидов морского ежа
и показать, что а- и р-субъединицы в микротрубочках А
и В имеют разный аминокислотный состав и по-разному
расщепляются трипсином. Эти результаты согласуются
с гипотезой о том, что в клетке есть различные тубули-
ны, которые имеют разную структуру и разные свойства
при агрегации; это, вероятно, необходимо для выполне-
ния микротрубочками различных функций. Если это
так, то тубулин обнаруживает удивительное сходство с
другим белком, участвующим в осуществлении подвиж-
ности, а именно с актином. Для актина небольшие хи-
мические вариации обнаружены не только в связи с
его видовыми особенностями, но также и в пределах
одной и той же клетки.
3.2.2. Белки,
связанные с тубулином
Связанные с тубулином белки — это необходимые
компоненты микротубулярной системы; они участвуют
в построении элементов ее структуры и в осуществлении
ее функций. Они могут принимать участие в генерации
движения, в процессе сборки (либо дезагрегации) мик-
ротрубочек или же соединяют микротрубочки с другими
структурами клетки (см. табл. 3.1). Таким образом
взаимодействующие с тубулином белки образуют с ним
единую систему с точки зрения как структуры, так и
функции. Большая часть данных о белках, связанных с
тубулином, получена при исследовании аксонем жгути-
ков и ресничек и цитоплазматических микротрубочек
из мозговой ткани.
Аксонема ресничек и жгутиков представляет собой
весьма сложную структуру, построенную из многих бел-
50
3. Системы подвижности эукариотических клеток
Таблица 3.1
Некоторые из белков, взаимодействующих или микротрубочками с тубулином .
Белок Вид активности Источник белка
В аксонеме жгутиков: Динеин 1 Г Динеин 2 J Нексин АТРазная активность; Хвосты сперматозои- генерация движения дов морского ежа Образование мостиков между соседними
В цитоплазматических микротрубочках: дублетами
Высокомолекуляр- ные белки(БСМ1 (БСМ2 Инициирование сбор- Мозг телят ки и регулирование длины микротрубо- чек (?)
Тау-белок Содействие самосбор- То же ке тубулина в коль- ца или микротру- бочки (?)
Протеинкиназа Фосфорилирование » субъединиц тубулина
ков. Поэтому при электрофорезе аксонем из хвостов
сперматозоидов морского ежа в полиакриламидном геле
в присутствии такого денатурирующего агента, как до-
децилсульфат натрия, удается идентифицировать по
крайней м,ере 20—30 полипептидных полос. Две из них
наиболее интенсивны: одна соответствует тубулину (ко-
торый в этих условиях мигрирует в виде субъединиц с
мол. массой около 55 000) и другая — высокомолекуляр-
ному белку (мол. масса >300 000) динеину. Жгутиковый
динеин, впервые очищенный Джиббонсом [56], образует
ручки на микротрубочках А периферических дублетов
аксонем (см. рис. 3.2). Он обладает аденозинтрифосфа-
тазной активностью. В хвостах сперматозоидов морско-
го ежа (так же, как и в других системах) присутствуют,
по-видимому, два изофермента динеина, которые можно
разделить методом дифференциальной солюбилизации в
солевых растворах. Динеин 1 существенно преобладает,
3. Системы подвижности эукариотических клеток
51
и именно из него состоят ручки микротрубочек. Второй
изофермент — динеин 2—содержится в гораздо мень-
ших количествах; он, вероятно, непосредственно не
участвует в генерации движения, а играет в этом про-
цессе регуляторную роль. Этот вопрос нуждается, одна-
ко, в дальнейшей проверке [57]. Ряд данных указывает
на то, что динеин благодаря своей АТРазной активности
играет основную роль в активном скольжении соседних
дублетов микротрубочек друг относительно друга, кото-
рое делает возможны^ движение жгутика.
Опять-таки на материале из хвостов сперматозоидов
морского ежа (очень удобный объект, дающий возмож-
ность выделять довольно большое количество аксонем)
удалось охарактеризовать другой белок, также связан-
ный с микротрубочками А. Это растворимый в кислой
среде-полипептид с мол. массой около 165000, который
получил название нексин [146], потому что он связывает
два соседних (т. е. ближайших, по-английски next) дуб-
лета. Он, вероятно, ограничивает степень смещения двух
соседних дублетов при их скольжении друг относитель-
но друга во время движения жгутика.
Вовсе не просто разобраться в химическом составе и
функциональной роли других элементов аксонемы жгу-
тиков. Есть два подхода к решению этой задачи. Можно
очистить аксонемы с помощью физико-химических мето-
дов, а можно изучать химический состав и ультраструк-
туру у мутантных организмов с нарушенной активностью
жгутиков. Необходимо отметить, что второй подход, хо-
тя он и представляется более разумным и плодотвор-
ным, трудно применить в случае сложных организмов.
Линку [87] с помощью ряда сложных методов (диффе-
ренциальное растворение элементов аксонемы, электрон-
но-микроскопическое наблюдение происходящих струк-
турных изменений, электрофоретический анализ хими-
ческого состава) удалось получить микротрубочки А
вместе с прикрепленными к ним дополнительными
структурами. При этом он показал, в частности, что
10 минорных белков наружных дублетов локализовано
именно в микротрубочках А. Изучение мутантов Chla-
mydomonas с парализованными или аномально двигаю-
щимися жгутиками позволило соотнести соответствую-
52
3. Системы подвижности эукариотических клеток
щие структуры с отдельными белками. Так, например,
сопоставив электрофореграммы аксонем диких штаммов
и мутантов, лишенных радиальных спиц, удалось уста-
новить, что основной материал спиц — это белок с мол.
массой 118 000. Изучение мутантов без центральных
микротрубочек и без окружающей их оболочки позволи-
ло идентифицировать три полипептида с мол. массами
более 220 000; эти белки образуют оболочку [177].
Вполне вероятно, однако, что состав аксонемы слож-
нее, чем получается по данным описанных выше мето-
дов, так как разрешающая способность электрофореза
могла быть недостаточна. Действительно, применив дву-
мерную изоэлектрофокусировку в сочетании с гель-
электрофорезом в додецилсульфате натрия, в составе
аксонемы диких штаммов Chlamydomonas удалось вы-
явить по крайней мере 100 полипептидов. С помощью
того же метода исследовали мутантов с аксонемой без
радиальных спиц и обнаружили, что у таких организмов
не хватает 12 белков [116]. Эти результаты показывают,
как сложна эта система белков и как много надо сде-
лать для выяснения ее состава.
С другой стороны, оказалось приятной неожидан-
ностью, что система белков, связанных с цитоплазматиче-
скими микротрубочками, как сейчас представляется, не
так сложна, как в жгутиках. Возможно, однако, что мень-
шая сложность белков, связанных с микротрубочками,
свойственна именно клеткам нервной системы, которые
пока что были практически единственным объектом та-
кого рода исследований. Современные представления о
свойствах тубулина основываются на данных о тубулине
из мозговой ткани, который иссл.едован лучше всего, так
как был найден простой метод его получения в доволь-
но больших количествах из мозга млекопитающих
(экстракты которого содержат более 20% тубулина от
общего количества белка) с помощью многократно по-
вторяемых циклов полимеризации — деполимеризации
микротрубочек при изменении температуры [133, 174].
При получении тубулина методом полимеризации — де-
полимеризации микротрубочек другие белки также под-
вергаются очистке и выделяются вместе с тубулином в
стехиометрических соотношениях. Было показано, что
3. Системы подвижности эукариотических клеток
53
эти сопутствующие белки (сокращенно их называют
БСМ — белки, связанные с микротрубочками) связаны
не только с интактными микротрубочками в клетках
центральной нервной системы, но и с цитоплазматичес-
кими микротрубочками и микротрубочками митотичес-
кого веретена клеток других типов.
Хотя в разных лабораториях условия очистки не со-
всем одинаковы и есть существенные различия между
выделяемыми препаратами белков, связанных с
микротрубочками, удается достаточно надежно иденти-
фицировать и охарактеризовать две группы БСМ
(см. табл. 3.1). Наиболее обширную из них составляют
высокомолекулярные белки, подвижность которых при
электрофорезе в полиакриламидном геле соответствует
мол. массе около 300 000 [24]. На эти белки в некоторых
препаратах микротрубочек приходится около 20% от со-
держания тубулина. По данным электронной микроско-
пии высокомолекулярные БСМ образуют тонкие высту-
пы, равномерно распределенные вдоль микротрубочек
[101]. По высокой молекулярной массе этих белков и их
способности образовывать боковые ручки их можно бы-
ло бы считать похожими на динеин. Однако в отличие
от последнего в них не обнаружена АТРазная актив-
ность и нет указаний на то, что они участвуют в движе-
нии микротрубочек. Другая группа БСМ — белки с мол.
массой около 70000. В некоторых лабораториях получе-
ны препараты тубулина, в которых этим белкам соответ-
ствует одна полоса при электрофорезе и они названы
тау-белком [173]. Показано, что флуоресцирующие анти-
тела против тау-белка локализуются в цитоплазмати-
ческих микротрубочках; этот белок способствует, по-ви-
димому, их сборке.
Стимулируемая циклическим АМР протеинкиназа,
ответственная, по-видимому, за посттрансляционное
фосфорилирование тубулина, также может быть отнесе-
на к БСМ, поскольку показано, что она связывается с
тубулином в определенных стехиометрических соотноше-
ниях, которые не изменяются в течение нескольких цик-
лов очистки путем полимеризации — деполимеризации
микротрубочек [140].
54
3. Системы подвижности эукариотических клеток
3.3. Самосборка микротрубочек
и ее регуляция
В любой живой клетке происходит агрегация тубу-
лина с образованием микротрубочек и распад этих
структур. Вопрос о том, как регулируются процессы
сборки и деструкции микротрубочек имеет первостепен-
ное значение для понимания многих проявлений жизне-
деятельности клетки.
3.3.1. Самосборка in vitro
Изучение in vitro полимеризации тубулина, выделен-
ного из нервных клеток, оказалось ценным источником
информации для понимания механизма сборки микро-
трубочек in vivo и путей управления этим процессом.
В этом разделе мы прежде всего рассмотрим сборку
микротрубочек in vitro и затем обсудим, что может про-
исходить в живой клетке.
Если нейротубулин находится в растворе в виде ди-
меров в присутствии связанных с ним белков, то в
определенных условиях легко идет процесс самосборки.
Полимеризация происходит в условиях умеренной ион-
ной силы, при слабо кислом pH (около 6,6—6,7), в при-
сутствии нуклеотидов (GTP) и ионов магния. Направ-
ление реакции зависит также от температуры: in vitro
при физиологических температурах идет полимеризация
тубулина с образованием микротрубочек, а снижение
температуры до 0°С вызывает их быструю деполимери-
зацию (рис. 3.6). Реакцию полимеризации можно ре-
Рис. 3.6. Реакция полимеризации димеров тубулина с образованием
микротрубочек. Указано влияние двухвалентных ионов и темпе-
ратуры.
3. Системы подвижности эукариотических клеток
55
гистрировать спектрофотометрически — по изменению
светорассеяния — или же с помощью вискозиметра — по
изменению вязкости раствора. Образование микротру-
бочек можно наблюдать также в электронный микроскоп
[53, 142]. Ясно, что в такой простой системе in vitro лег-
ко можно порознь оценить эффект различных парамет-
ров, влияющих на реакцию полимеризации. В частности,
было установлено, что полимеризации способствуют вы-
сокие концентрации сахарозы или глицерина, тогда как
полианионы (РНК, алкалоиды типа колхицина) угнета-
ют полимеризацию. С помощью упомянутых методов де-
тально исследовали влияние различных ионов и GTP.
Эти данные представляют особый интерес в связи с тем,
что такие вещества можно рассматривать как возмож-
ные регуляторы полимеризации тубулина in vivo.
Высокие концентрации двухвалентных катионов угне-
тают самосборку микротрубочек. Особенно активным ин-
гибитором является Са2+: в концентрациях выше
1,0 мМ он угнетает полимеризацию на 50%. GTP необ-
ходим для полимеризации в концентрациях, по крайней
мере эквимолярных количеству димеров тубулина. По-
видимому, GTP гидролизуется при полимеризации тубу-
лина: недавно была обнаружена ОТРазная активность
растущих концов микротрубочек [34]. Однако GTP мож-
но заменить его негидролизуемыми аналогами (напри-
мер, б'-гуанозинметилендифосфонатом), при этом поли-
меризация тубулина все равно стимулируется. Это по-
зволяет предполагать, что роль GTP заключается в том,
чтобы поддерживать определенное конформационное
состояние тубулина, благоприятное для его полимериза-
ции. Расщепление концевой фосфатной связи GTP, ве-
роятно, сдвигает равновесие реакции в сторону образо-
вания микротрубочек [17]. Однако GTP не используется
при этом в качестве источника энергии, поскольку, как
показывают эксперименты с негидролизуемыми аналога-
ми GTP, сборка микротрубочек не требует затраты
энергии.
Приведенные выше данные получены на таких препа-
ратах нейротубулина, которые содержали также сопут-
ствующие белки (белки, связанные с микротрубочками).
В экспериментах со специально очищенными препарата-
56
3. Системы подвижности эукариотических клеток
ми, содержавшими только тубулиновые субъединицы
(т. е. без БСМ), самосборка микротрубочек не наблюда-
лась, полимеризация такого тубулина происходила толь-
ко в особых экспериментальных условиях (см. [64, 66]).
На этом основании заключили, что белки, связанные с
микротрубочками, выполняют также регуляторную функ-
цию в реакции полимеризации тубулина, и было затра-
чено много усилий, чтобы выяснить эту функцию. Преж-
де чем перейти к вопросу о регуляторной функции свя-
занных с микротрубочками белков, мы рассмотрим
вкратце сам процесс полимеризации. Модель самосбор-
ки микротрубочек основана на результатах Киршнера и
его сотрудников [112], которым удалось проследить из-
менение тонкой структуры при формировании микротру-
бочек и применить количественный метод (рис. 3.7) . Они
обнаружили, что еще до начала полимеризации в препа-
ратах тубулина наряду с растворимыми димерами при-
сутствует небольшое количество колец и спиралей из
полимеризованного тубулина со средним коэффициентом
седиментации 36S. Показано, что эти кольца необходи-
мы для последующей реакции полимеризации, так как
если удалить их с помощью ультрацентрифугирования,
дальнейшая полимеризация невозможна. Когда реакцию
полимеризации запускают, добавляя к реакционной сме-
си GTP и повышая температуру до требуемого уровня, в
первые ж,е минуты происходит формирование спирально
закрученных лент. Далее димеры тубулина продолжа-
ют прикрепляться к этим лентам до тех пор, пока не
образуются тринадцать протофибрилл. Когда эта стадия
достигнута, ленты сворачиваются в цилиндры — отрезки
будущих микротрубочек [112]. Далее микротрубочки
удлиняются благодаря тому, что димеры прикрепляются
к торцам этих отрезков (рис. 3.8). Учитывая, как проте-
кает процесс сборки, можно лучше представить себе
влияние различных белков, связанных с микротрубочка-
ми, на их формирование. Относительно тау-белка пока-
зано, что он прежде всего участвует в образовании ис-
ходного ЗбБ-тубулина (кольца и спирали), без которого
невозможна самосборка микротрубочек, но, похоже, иг-
рает роль и в наращивании отрезков микротрубочек.
Высокомолекулярные белки (БСМ-1 и БСМ.-2) оказы-
3. Системы подвижности эукариотических клеток
57
Рис. 3.7. Схематическое изображение стадий сборки микротрубочек
in vitro: а — кольца и спирали из тубулина, одновременно присут-
ствуют свободные димеры; б — формирование ленточных структур;
в — свертывание листовых структур—’Начало образования микро-
трубочек; г — готовая микротрубочка [112].
Рис. 3.8. Доля белка, присутствующего в виде колец (□)
лент (Л) и микротрубочек (О) на разных стадиях полимериза-
ции [112].
58
3. Системы подвижности эукариотических клеток
вают сходное действие; известно, что они увеличивают
не только начальную скорость самосборки, но и общее
число готовых микротрубочек, т. е. ускоряют наращива-
ние отрезков. Есть даже данные о том, что ускорение
роста отрезков — основной вид регуляторной активности
высокомолекулярных белков [141]. Возможно, что дру-
гая их функция заключается в формировании связую-
щих мостиков между микротрубочками и другими эле-
ментами каркаса клетки, например микрофиламентами
и промежуточными филаментами.
Многообещающий подход к проблеме самосборки
микротрубочек и ее регуляции заключается в изучении
действия на системы in vitro некоторых внутриклеточ-
ных структур — так называемых центров организации
микротрубочек, которые способствуют сборке микротру-
бочек в клетке. Этот вопрос еще недостаточно исследо-
ван, однако похоже на то, что эти структуры, будучи
выделены из клетки, сохраняют способность функциони-
ровать сходным образом и в системах in vitro. Это по-
казано для суспензий базальных тел жгутиков и кине-
тохор, полученных из метафазных хромосом.
3.3.2. Самосборка микротрубочек in vivo
Плодотворный метод изучения сборки микротрубочек
(в частности, исследование влияния условий внешней
среды, таких, как температура или давление, а также
фаз жизненного цикла клетки и связанных с ними собы-
тий) — прямое наблюдение в микроскоп формирования
митотического веретена в процессе клеточного деления.
С помощью высокочувствительной поляризационной оп-
тики некоторые исследователи смогли непосредственно
наблюдать микротубулярные структуры, которым свой-
ственно двойное лучепреломление (поляризованные лу-
чи на микротрубочках раздваиваются). Важные резуль-
таты получили Инуи и сотр. [73] на яйцеклетках морских
иглокожих, а также другие исследователи, изучавшие
другие объекты.
Было показано, что некоторые химические вещества
(D2O, а также некоторые гликоли), равно как и повы-
шение температуры, усиливают двойное лучепреломле-
3. Системы подвижности эукариотических клеток
59
ние митотического веретена, т. е. способствуют полиме-
ризации. С другой стороны, низкая температура, повы-
шенное давление и ингибиторы микротрубочек (колхи-
цин, винбластин) уменыпаШт двойное лучепреломление.
Такие наблюдения позволили даже измерить скорость
самосборки микротрубочек (обычно 1,5 мкм/мин).
Нецелесообразно обсуждать здесь все эксперимен-
тальные данные, поэтому мы перейдем к тем важным
заключениям, которые были сделаны на основании при-
веденных выше результатов и других исследований in
vivo. В клетке есть два типа регуляции самосборки
микротрубочек. Один из них — временной контроль: по-
лимеризация или деполимеризация тубулина приуроче-
на к определенным моментам жизненного цикла клет-
ки. Другой тип — пространственный контроль, который
необходим для того, чтобы содержащие микротру-
бочки структуры формировались правильно. Некоторые
данные указывают на то, что для временного контроля
количество тубулина, находящегося в клетке, не имеет
значения; иначе говоря, клетка це начинает продуциро-
вать микротрубочки просто потому, что в ней накопилось
много тубулина. Содержание тубулина довольно постоян-
но на протяжении всего жизненного цикла клетки [124], и,
когда нужно производить много микротрубочек (напри-
мер, при росте нейронов в нейробластоме), необходимость
в дополнительном синтезе тубулина, по-видимому, не воз-
никает [97]. Следовательно, полимеризацией тубулина
или разрушением микротрубочек в соответствии с по-
требностями клетки управляют другие факторы. Были
попытки идентифицировать эти факторы с определенны-
ми белками, например с высокомолекулярными белка-
ми, связанными с микротрубочками; регулирующая роль
также приписывалась изменению обменного фонда ио-
нов кальция; предполагался еще более сложный конт-
рольный механизм с участием сАМР. Однако до сих
пор никаких четких данных о механизме регуляции
сборки микротрубочек получить не удалось. В отличие
от животных клеток у некоторых низших эукариот сбор-
ка микротрубочек, похоже, связана с синтезом тубули-
на de novo. Это, по-видимому, имеет место при регенера-
ции жгутиков у Chlamydomonas и ресничек у Tetrahy-
60
3. Системы подвижности эукариотических клеток
тепа и наверняка происходит при росте жгутика у
Naegleria.
Механизм пространственного контроля сборки микро-
трубочек предполагает, что внутри клетки есть некие
центры, способные организовать полимеризацию тубули-
на в нужном месте, а также система, контролирующая
наращивание микротрубочек, их ориентацию и связь с
другими структурами клетки. Иначе говоря, эти-две сис-
темы должны предопределять все детали конструкции
строящейся микротрубочки, а также ее местоположение
и ориентацию в клетке. «Ядра конденсации микротрубо-
чек», более известные под названием «центры органи-
зации микротрубочек» [97], широко распространены во
всех клетках. Морфологически они детально описаны,
однако об их составе, к сожалению, известно очень не-
многое. Митотические центры (полюса митотического ве-
ретена), кинетохоры хромосом и вообще все те струк-
туры, из которых исходят пучки микротрубочек, следу-
ет рассматривать как центры организации микротру-
бочек.
Наряду с регуляцией процесса полимеризации тубу-
лина при сборке микротрубочек должен быть механизм,
задающий тип укладки микротрубочек в соответствии с
архитектурой той или иной структуры—аксонемы, ак-
состиля, митотического веретена и т. и.. Были предложе-
ны две гипотезы, объясняющие многообразие возникаю-
щих структур [156] (см. рис. 3.9).
Согласно одной из них, особенности формирующейся
из микротрубочек структуры обусловлены образованием
специфических мостиков между уже готовыми рядом
расположенными микротрубочками. Тип и число возни-
кающих мостиков определяют окончательное располо-
жение микротрубочек в формирующейся структуре.
Можно предполагать, что именно так строятся аксо-
подии некоторых солнечников.
Согласно другой гипотезе, конструкция предопреде-
лена самим местоположением центров конденсации каж-
дого элемента будущей структуры, так что сборка
микротрубочек осуществляется в заранее «запланиро-
ванных» местах. Этот механизм, по-видимому, действует
при сборке ресничек и жгутиков: круговое расположение
3. Системы подвижности эукариотических клеток
61
Рис. 3.9. Два пути формирования микротубулярных структур: а —
тип структуры определяется после сборки микротрубочек; б — ми-
кротрубочки растут в соответствии с типом будущей структуры,
заданным расположением базальных тел [165].
микротрубочек в аксонеме предопределяется в самом
начале морфогенеза базального тела, когда начинается
сборка девяти триплетов на равных расстояниях друг
от друга строго по окружности [165].
3.4. Как микротрубочки
генерируют движение?
Микротрубочки могут генерировать движение с по-
мощью по крайней мере двух различных механизмов: за
счет активного скольжения или же путем изменения сво-
ей длины.
Модель скольжения для движения ресничек и жгути-
ков предложил Петер Сатир [128, 130]; в дальнейшем
эта модель получила так много независимых подтверж-
62
3. Системы подвижности эукариотических клеток
дений в разных лабораториях, что теперь ее можно счи-
тать доказанной. По этой модели основные рабочие эле-
менты — микротрубочки и динеиновые ручки, а энергия
для перемещения получается при гидролизе АТР.
Данные о том, что динеин (белок с АТРазной актив-
ностью, из которого построены боковые ручки перифери-
ческих дублетов в ресничках и жгутиках) непосред-
ственно участвует в генерации движения, были впервые
получены на препаратах аксонем из хвостов спермато-
зоидов морского ежа. После того как радиальные спи-
цы и нексиновые мостики избирательно переваривались
трипсином, аксонемы с интактными боковыми ручками
вце, еще могли двигаться [152]. Дальнейшее подтвержде-
ние получено при исследовании спермы у человека. При
одной из форм бесплодия, известной как синдром Карта-
генера, сперматозоиды не способны двигаться. Оказа-
лось, что при этом периферические дублеты в аксонеме
не имеют динеиновых ручек [9, 111].
Скольжение дублетов аксонемы удается наблюдать
непосредственно под микроскопом (по методу темного
поля), если аксонемы предварительно частично перева-
рить трипсином. После добавления АТР можно видеть,
что продольные нити микротрубочек становятся длиннее
и тоньше. Эти эксперименты показывают, что, когда
нексиновые мостики и радиальные спицы переварены
трипсином, соседние дублеты имеют возможность сколь-
зить друг относительно друга и делают это до тех пор,
пока це, перестанут перекрываться, т. е. пока не разде-
лятся полностью. Какую же роль играют динеиновые
ручки в этой модели? Динеиновые ручки могут образо-
вывать жесткие мостики между микротрубочкой А, из
которой они исходят, и микротрубочкой В соседнего дуб-
лета. При гидролизе АТР мостик разрывается и в тот
же момент изменяется ориентация динеиновой молекулы
(ручки). Это вызывает скольжение. В результате повто-
рения таких циклов соседние дублеты заметно скользят
друг относительно друга и в конечном счете могут пол-
ностью разделиться (рис. 3.10). Этот процесс можно
сравнить с тем, как гусеница (дублет с динеином) ползет
по веточке (соседний дублет), попеременно перемещая
ноги (динеиновые ручки) до тех пор, пока не доползет
3. Системы подвижности эукариотических клеток
63
Рис. 3.10. Механизм скольжения микротрубочек в аксонеме. Дуб-
лет 1 скользит вдоль дублета 2 за счет движения динеиновых
ручек.
до конца веточки и не упадет с нее. Если дублеты соеди-
нены между собой нексиновыми мостиками и связаны с
радиальными спицами, то при скольжении они не разде-
ляются, а изгибаются; это, однако, уже другой вопрос,
и мы вернемся к нему, когда будем рассматривать ме-
ханизм движения ресничек и жгутиков.
Движение может также генерироваться путем управ-
ления полимеризацией и деполимеризацией микротрубо-
чек. Примеры такого типа движения — удлинение и уко-
рочение аксоподий у солнечников, а также, по-видимому,
движение хромосом. Солнечники имеют длинные высту-
пы, лучами исходящие из тела, которые придают им вид
солнышка (название Heliozoa произошло от греческого
слова helios — солнце). Внутри выступов микротрубоч-
ки образуют каркас характерной спиральной конструк-
ции. Когда солнечник Actinophrys движется по плоской
поверхности (рис. 3.11), то его аксоподии, которые вы-
ступают вперед по направлению движения, укорачива-
ются со скоростью около 20 мкм/мин, в то время как ак-
соподии, обращенные назад, удлиняются со скоростью
64
3. Системы подвижности эукариотических клеток
изображение солнечника, катящегося с
Рис. 3.11. Схематическое
помощью аксоподий.
Рис. 3.12. Сокращение аксоподий у солнечников сопоставлено со
стимуляцией. Вверху. Centrohelidae; внизу. Actinophryidae.
3. Системы подвижности эукариотических клеток
65
около 7 мкм/мин. Эти изменения длины обусловлены
частичной деполимеризацией или дополнительной поли-
меризацией микротубулярных осей аксоподий; деполи-
меризация сопровождается сокращением аксоподия, по-
видймому, за счет поверхностного натяжения клеточной
мембраны. У солнечников Actinophrys сокращения
крайне медленны из-за того, что в концевом участке
каждой аксоподий должна произойти деполимеризация
очень большого числа (сотен) микротрубочек [169].
У более мелких солнечников Centrohelidae в аксоподиях
находится только по шесть микротрубочек и сокращения
происходят очень быстро, по-видимому, в связи еще и с
тем, что при воздействии на аксоподию соответствующе-
го стимула деполимеризация микротрубочек происходит
по всей их длине, в результате чего вся структура ру-
шится мгновенно (рис. 3.12). Удлинение, однако, осу-
ществляется более медленно — приблизительно с такой
же скоростью, как и в аксоподиях Actinophrys sol.
Хотя в настоящее время молекулярный механизм
расхождения хромосом все еще остается предметом дис-
куссий, многие данные указывают на то, что в нем
может участвовать процесс полимеризации — деполиме-
ризации микротрубочек митотического веретена. Предло-
жена более или менее удовлетворительная модель рас-
хождения хромосом, однако согласие пока не достигну-
то, а недавнее открытие актомиозиновых нитей в мито-
тическом веретене еще более запутало этот и без того
сложный вопрос. Поэтому целесообразно отложить об-
суждение до получения новых данных.
3.5. Проблема
промежуточных филаментов
Филаменты диаметром около 10 нм (т. ,е. тоньше, чем
микротрубочки, и толще, чем микрофиламенты) — оди-
ночные или в виде небольших пучков — были обнаруже-
ны в цитоплазме различных клеток уже на заре приме-
нения электронной микроскопии в биологии. Относи-
тельное количество этих структур зависит от типа клет-
66
3. Системы подвижности эукариотических клеток
ки; в клетках эпидермиса они представлены широко и
известны под названием тонофиламентов; в клетках
других типов, например в клетках центральной нервной
системы, они многочисленны в аксонах и называются
нейрофиламентами. В других клетках млекопитающих
этих филаментов не так много, но они все же встреча-
ются [52].
Химический состав промежуточных филаментов еще
не вполне ясен; они, по-видимому, построены из одного
или нескольких основных б,едков. В препаратах проме-
жуточных филаментов из почечных клеток новорожден-
ных хомячков (из клеток типа 21) при электрофорезе в
полиакриламидном геле четко разрешаются две полосы,
очень похожие по соответствующей им молекулярной
массе на полосы двух тубулиновых субъединиц [145].
Есть, однако, данные и о том, что белки промежуточных
филаментов составляют гомогенный класс белков, от-
личных от других структурных белков клетки. Антитела,
выработанные против промежуточных филаментов одно-
го типа клеток, перекрестно реагируют с промежуточ-
ными филаментами клеток других типов, но не с микро-
трубочками или микрофиламентами.
В этой книге мы упоминаем о промежуточных фила-
ментах в связи с тем, что а) эти структуры могут быть
соединены с микротрубочками и б) они могут играть
какую-то роль в подвижности клеток млекопитающих.
В пользу первого предположения свидетельствуют дан-
ные электронной микроскопии (обнаружены тонкие мос-
тики из фибриллярного материала, которые связывают
стенки микротрубочек с промежуточными филаментами),
а также тот факт, что ингибиторы митоза (например,
колхицин) вызывают перераспределение промежуточных
филаментов in vivo. Второе предположение подтвержда-
ется тем, что, по данным электронной микроскопии,
локализация промежуточных филаментов в цитоплазме
лейкемических клеток различна в разных фазах их дви-
жения [45]. Чтобы выяснить свойства и функции этих
наименее изученных элементов структурного скелета
клетки, нужны, однако, дальнейшие исследования.
.3. Системы подвижности эукариотических клеток
67
3.6. Микрофиламенты
Микрофиламенты эукариотических клеток представ-
ляют собой длинные нитевидные структуры толщиной
5—7 нм, находящиеся в цитоплазме. Они состоят глав-
ным образом из актина (хотя можно обнаружить и дру-
гие белки), в связи с чем их часто называют актиновы-
ми филаментами (или нитями). Они иногда образуют
группы или пучки, а в некоторых высокоспециализиро-
ванных клетках, например в мышечных волокнах, упо-
рядоченные и стабильные структуры. Чаще, однако, из
микрофиламентов формируются нестабильные пучки или
тонкие сетчатые структуры, их форма и локализация в
клетке изменяются в зависимости от фазы жизненного
цикла клетки, ее движения и т. п. Это означает, что,
подобно цитоплазматическим микротрубочкам, микрофи-
ламенты немышечных клеток представляют собой из-
меняемые структуры, способные к самосборке или депо-
лимеризации на составляющие их молекулы в зависи-
мости от нужд клетки.
Морфологию микрофиламентов и их поведение в про-
цессе клеточного движения и цикла развития клетки
много изучали на культуре животных кдеток с помощью
флуоресцирующих антител против актина, а также про-
тив других белков (миозин, тропомиозин и а-актинин),
которые часто обнаруживаются в микрофиламентах
[84, 86]. Когда клетки находятся в состоянии покоя и
прикреплены к стенкам сосуда, в котором живут, в них
можно наблюдать длинные пучки микрофиламентов, тя-
нущиеся сквозь всю клетку, по-видимому, непосредствен-
но под клеточной мембраной. В «возбужденные» высту-
пы клетки с нестабильными, как бы волнующимися края-
ми они обычно не заходят. Пучки микрофиламентов на-
зываются стрессовыми волокнами; они типичны для
нормальных клеток и отсутствуют в клетках, трансформи-
рованных онкогенными вирусами. Когда клетки движут-
ся, в дополнение к стрессовым волокнам появляются
тонкие пучки актиновых нитей, рассеянные в цитоплаз-
ме; они особенно заметны в направленных вперед высту-
пах клеток. В процессе деления клетки флуоресценция
сосредоточена в митотическом веретене, причем картина
и
3. Системы подвижности эукариотических клеток
69
ее распределения удивительно похожа на ту, которую
создают антитубулиновые антитела в тех же условиях.
Когда дочерние клетки расходятся, микрофиламенты со-
бираются в области тяжа, соединяющего дочерние клет-
ки; там они образуют сократимое кольцо, обеспечиваю-
щее разделение клеток (рис. 3.13). Приведенное описа-
ние дает представление о том, как в соответствии с
фазами клеточного цикла может изменяться простран-
ственная организация микрофиламентов.
Существуют, однако, и такие клетки, в которых мик-
рофиламенты образуют стабильные структуры, — их
форма и относительное расположение в них микрофила-
ментов меняются редко. Не говоря уже о мышечных
клетках (им посвящена особая книга в этой серии, по-
этому мы не будем их рассматривать), хороший пример
стабильной конструкции из микрофиламентов — длин-
ные пальцеобразные выросты клеток кишечного эпите-
лия (так называемые микроворсинки; они есть и во мно-
гих других животных клетках). В каждой микроворсин-
ке пучок микрофиламентов идет параллельно оси вор-
синки — от ее кончика, к которому он прикреплен, вдоль
оси к клеточной мембране [100]. Стабильные пучки ак-
тиновых нитей можно обнаружить также в таких хоро-
шо изученных объектах, как сперматозоиды иглокожих
и ооциты морского ежа [157]. Исследование микрофила-
ментов в сперме Mytilus и в ооцитах морского ежа ме-
тодами оптической дифракции и трехмерной реконструк-
ции показало, что микрофиламенты в каждом пучке
имеют правильную гексагональную упаковку и в опреде-
ленном порядке соединены поперечными мостиками, так
что каждый пучок представляет собой паракристалли-
ческую структуру [37]. Это удивительно похоже на орга-
низацию микротрубочек в аксостил,е жгутиковых.
В электронный микроскоп нечасто можно увидеть,
что актиновые нити в немышечных клетках связаны с
более толстыми и короткими нитями миозина (они
обычно имеют 8—9 нм в диаметре и 300 нм в длину),
белка, который взаимодействует с актином при генера-
ции движения. Электронно-микроскопических данных о
миозиновых нитях в немышечных клетках мало, видимо,
потому, что на тонких срезах такие толстые нити увидеть
70
3. Системы подвижности эукариотических клеток
трудно. Если же обработать микрофиламенты антитела-
ми против миозина, то хорошо видна диффузная флуо-
ресценция, указывающая на то, что миозин действитель-
но присутствует1. Флуоресцирующие антитела против
других белков сократительной системы также выявляют
характерную периодичность цитоплазматических микро-
филаментов; так, например, участки связывания антител
против а-актинина и тропомиозина в микрофиламентах
чередуются, демонстрируя присутствие этих двух
белков.
Считается, что микрофиламенты в немышечных клет-
ках выполняют в основном две функции. Их классичес-
кая роль — генерировать движение; в частности, их счи-
тают ответственными за амебоидное движение. С другой
стороны, сокращение микрофиламентов может служить
источником силы для различных видов клеточной ак-
тивности: изменения формы клетки, токов цитоплазмы,
движения клеточных органелл, фагоцитоза, секреторной
активности, деления клетки и регуляции распре-
деления белков в мембранах. Однако накапливаются
данные о том, что микрофиламенты играют также роль
цитоскелета: жесткие пучки этих волокон могут поддер-
живать определенную форму клетки [32, 80].
3.7. Состав микрофиламентов
Чтобы лучше понять динамику микрофиламентов и
их функцию в движении немышечной клетки, мы рас-
смотрим те биохимические свойства составляющих их
белков, которые имеют наиболее прямое отношение к
этим вопросам.
1 Аналогичная ситуация в некоторых гладкомышечных клетках
навела на мысль о самосборке миозиновых нитей в этих клетках
непосредственно перед развитием сократительной реакции.—Прим,
ред. перевода.
3. Системы подвижности эукариотических клеток
71
3.7.1. Свойства актинов
немышечных клеток
Актин широко распространен во всех эукариотичес-
ких клетках. Его можно обнаружить как в растворимой
мономерной форме, так и в полимерной — в виде фила-
ментов (рис. 3.14). В некоторых активно передвигаю-
щихся клетках (амебы, макрофаги, тромбоциты) актин
преобладает среди белков клеточного экстракта: его со-
держание может достигать 20—30% от общего белка.
В менее подвижных клетках содержание актина
заметно меньше— 1—2% от общего белка.
Однако если вспомнить, что в эукариотической клетке
одновременно присутствуют многие тысячи различных
белков, то такое небольшое количество весьма сущест-
венно.
Благодаря относительно высокому его содержанию
актин удается выделить из клеточных систем. Более или
Рис. 3.14. Электронные микрофотографии микрофиламентов, обра-
зованных in vitro актином из мозга телят: а — нормальные микро-
филаменты; б — после обработки тяжелым меромиозином, который
специфически связывается с актиновыми микрофиламентами. Нега-
тивное окрашивание, X 120 000. (Снимки любезно предоставлены
д-ром G. Monaco.)
72
3. Системы подвижности эукариотических клеток
менее полно изучено уже около 30 различных актинов
из клеток самых разных типов—от миксамеб слизистых
грибов до клеток человека.
Актин можно очистить обычными методами разделе-
ния белков— ионообменной хроматографией и гель-
фильтрацией [144]. Для многих исследований вполне до-
статочна аффинная хроматография супернатанта, полу-
ченного после высокоскоростного центрифугирования
клеточного экстракта, на агарозных гранулах с кова-
лентно связанной ДНКазой I. Актин имеет большое
сродство к ДНКазе I и сорбируется на агарозных гра-
нулах, после чего его легко элюировать соответствую-
щим буфером [85]. Сродство к ДНКазе I — хорошо из-
вестное свойство актина; он образует с этим ферментом
прочный, лишенный ферментативной активности комп-
лекс. Физиологическое значение этой реакции остается,
однако, неизвестным [68].
Все изученные актины очень сходны по мол. массе
(всегда около 42 000), электрофоретической подвижно-
сти, аминокислотному составу (у всех актинов есть не-
обычная аминокислота — N-метилгистидин; ее роль в
молекуле неизвестна), по способности связывать адени-
новые нуклеотиды в эквимолярных соотношениях и по
способности к полимеризации [80]. Что касается последо-
вательности аминокислот, у актинов из разных источни-
ков обнаруживаются неболыпце, иногда только в одном
аминокислотном остатке, но существенные различия.
Различия в химическом составе выявлены у актинов не
только из разных организмов, но и из различных тка-
ней одного и того же организма. Более того, показано,
что даже в одной и той же клетке могут существовать
одновременно актины различных типов. Разницу между
ними трудно уловить — требуется сочетание двух высо-
коэффективных методов разделения: изоэлектрофокуси-
ровки (при этом белки разделяются в соответствии с их
изоэлектрическими точками) и электрофореза в полиак-
риламидном геле в присутствии додецилсульфата натрия
(белки разделяются в зависимости от размера мо-
лекулы и суммарного заряда). Такой комплексный под-
ход позволил идентифицировать три типа актина, кото-
рые получили название а-, р- и у-актин. Кроме того, мо-
3. Системы подвижности эукариотических клеток
73
гут присутствовать еще две нестабильные формы этого
белка, а-Актин свойствен полностью дифференцирован-
ной мышечной ткани, р- и у-актин — большинству изу-
ченных немышечных клеток; в культуре миобластов об-
наруживаются все три типа актина. Поскольку были
изолированы мРНК для каждого из этих актинов, мож-
но заключить, что в клетке есть по крайней мере три
различных актиновых г,ена и их экспрессия строго конт-
ролируется в соответствии с типом клетки [70, 151].
3.7.2. Белки,
взаимодействующие с актином
Актин, как известно, взаимодействует со многими
белками. В зависимости от того, какой из белков всту-
пает во взаимодействие с актином, результаты могут
быть самыми разными: либо генерируется движение,
либо меняется процесс полимеризации актина, либо
образуются связующие мостики, не исключено также
влияние на какие-то звенья метаболизма.
Чаще всего обнаруживают взаимодействие актина с
миозином. Свойства миозина из мышечных волокон изу-
чены гораздо более полно, чем миозина из це.мышечных
клеток. Миозин можно определять по его ферментатив-
ной (аденозинтрифосфатазной) активности, обычно
усиливаемой актином, и по его способности взаимодей-
ствовать с ним [32]. Стимулирующий эффект актина за-
висит от присутствия определенных ионов, поэтому раз-
личные миозины можно классифицировать как Са2+,
Mg2+- или К+-зависимые АТРазы. Что касается струк-
туры, большинство изученных немышечных миозинов
состоит из двух тяжелых полипептидных цепей с мол.
массой около 200 000 каждая и двух пар легких цепей
(мол. масса 17 000 и 20 000); в результате мол. масса
миозина составляет около 500 000. В молекуле миозина
хорошо различаются две области. Одна из них, так на-
зываемый стержень, или «хвост», состоит из «-спираль-
ных участков двух тяжелых цепей, свитых в суперспи-
раль; стержневые части миозиновых молекул способны
связываться друг с другом с образованием биполярных
74
3. Системы подвижности эукариотических клеток
молекулы миозина
полимеризация
Рис. 3.15. Образование биполярных нитей миозина.
Рис. 3.16. Схематическое изображение молекулы обычного миозина
(вверху) и миозина из Acanthamoeba (внизу).
нитей (рис. 3.15). На одном конце стержня находятся
две «головки»1, в каждой из которых .есть центр связы-
вания с актином и центр АТРазной активности. Легкие
цепи связаны с головками.
Миозин с другой структурой, названный миозин I,
обнаружен у почвенной амебы Acanthamoeba castellani.
Особенность этого белка в том, что в его молекуле толь-
1 Каждая из глобулярных головок соединяется со стержнем с
помощью гибкой полипептидной цепи, что имеет существенное зна-
чение для генерации движения. — Прим. ред. перевода.
3. Системы подвижности эукариотических клеток
75
ко одна тяжелая цепь с мол. массой 140000 и две лег-
кие цепи (14 000 и 16 000) (рис. 3.16) *. Миозин I не об-
разует биполярных нитей и активируется актином толь-
ко в присутствии кофактора [118, 119]. Из той же амебы
недавно выделили другой миозин (миозин II). Он боль-
ше похож на обычный: может образовывать биполяр-
ные нити и имеет две головки с АТРазной актив-
ностью [120].
М§,2+-зависимая стимуляция АТРазной активности
миозина актином в случае некоторых немышечных мио-
зинов требует предварительного фосфорилирования лег-
кой цепи специальной протеинкиназой, переносящей фос-
фат от донорной молекулы к легкой цепи 20000.
Это относится также к миозину I Acanthamoeba: его
кофактор недавно идентифицирован как киназа, кото-
рая специфически катализирует фосфорилирование тя-
желой цепи [95]. С генетической точки зрения система
миозинов не столь консервативна, как система актинов;
к настоящему времени выявлено около 15 типов миози-
нов, различающихся по многим свойствам.
Другие белки, взаимодействующие с актиновыми или
актомиозиновыми филаментами, можно попытаться
классифицировать в соответствии с их активностями
(табл. 3.2). Присутствие некоторых из них, например
тропомиозина и а-актинина, в составе микрофиламентов
недавно удалось увидеть непосредственно с помощью
иммунофлуоресценции [84].
Тропомиозин из тромбоцитов, мозга, поджелудочной
железы и фибробластов млекопитающих по молекуляр-
ной массе и пептидной карте подобен мышечному тропо-
миозину; он участвует в регулировании концентрации
Са2+, необходимой для «сокращения» актомиозина.
а-Актинин (или же похожие на него белки) обнаружен
(в количествах, больших по сравнению с другими объ-
ектами) в ворсинках эпителия, в культурах животных кле-
ток, а также в сперматозоидах подковообразного краба,
1 На схеме двуглавой молекулы миозина допущена ошибка:
каждая головка должна сидеть иа отдельной «шейке», которые за-
тем соединяются в общий стержень молекулы. — Прим. ред. пере-
вода.
76
3. Системы подвижности эукариотических клеток
Таблица 3.2
Некоторые белки, взаимодействующие с актином
в иемышечных клетках
Источник белка Взаимо- действую- щие белки Взаимодействующие белки Источник белка
Селезенка Профи-"! Филамин а-Актинии Макрофаги Различные си- стемы
По джелу- ЛИН G-актинид t^F-актии Гелактины I-II-III-IV A. castellani
дочная же- леза ДНКаза Тропонин Т ропомиозин Актинсвязыва- ющий белок Различные си- стемы То же Макрофаги
когда в них формируются аксонемы. а-Актинин, по-види-
мому, каким-то образом присоединяет микрофиламенты
к клеточным мембранам и, возможно, к другим структу-
рам клетки; присутствует этот белок главным образом
именно там, где есть такие контакты.
Некоторые белки угнетают полимеризацию актина
in vitro и, следовательно, могут контролировать образо-
вание актиновых филаментов in vivo. Так, например,
удалось выделить из селезенки и очистить так называе-
мый профилин, который представляет собой полипептид с
мол. массой 16 000, способный образовывать комплексы с
растворимым актином. Подобные белки, по-видимому,
широко распространены в немышечных клетках, особенно
в тех, где концентрация растворимого актина велика.
Такие белки, как филамин и актинсвязывающий бе-
лок, выделенные из макрофагов, могут связываться с
актиновыми микрофиламентами in vitro; при этом способ-
ность актина стимулировать АТРазную активность мио-
зина подавляется, что угнетает сократимость, обуслов-
ленную актомиозином. Функция этих белков in vivo не-
ясна, ио можно предположить, что в клетке они нужны
для того, чтобы некоторые актиновые филаменты играли
роль просто каркасных структур [80]. Подобную функ-
3. Системы подвижности эукариотических клеток
77
цию, возможно, выполняют и другие белки, например ге-
лактины — группа из четырех белков, выделенных из
Acanthamoeba. Гелактины превращают суспензию микро-
филаментов в гель, образуя между отдельными волокна-
ми связующие мостики. Эти белки угнетают также Mg2+-
зависимую АТРазную активность миозина (в присут-
ствии актина); характер ингибирования, похоже, тот же,
что и в случае филамина и актинсвязывающих белков.
Наконец, к числу б.едков, взаимодействующих с акти-
ном, относятся также альдолаза и некоторые другие фер-
менты гликолиза.
Описанная выше система представляется уже доста-
точно сложной, однако то, что изучено до сих пор, —
только царапины на поверхности неизведанного. Многие
предположения и теории относительно актина и взаимо-
действующих с ним белков в немышечных клетках нуж-
даются в дальнейшей разработке и проверке, прежде
чем могут быть сделаны какие-либо конкретные выводы.
3.8.. Сборка микрофиламентов
и ее регуляция
Как мы уже упоминали выше, актин может сущест-
вовать в двух функциональных состояниях: в виде раст-
воримых мономеров и в виде полимеров, когда одиноч-
ные субъединицы собираются в двухнитевые филамен-
ты. Актин в мономерной форме называется G-актин, а в
полимерной — F-актин. Двойная спираль F-актина име-
ет толщину около 7 им и шаг 68 нм (рис. 3.17).
мономеры актина
(G-актин)
актиновый филамент
( F-актин)
Рис. 3.17. Полимеризация актиновых субъединиц—.сборка микро-
филаментов.
78
3. Системы подвижности эукариотических клеток
Реакция полимеризации in vitro идет при оптималь-
ных концентрациях определенных солей и в присутствии
связанного АТР. Для этой реакции критична концентра-
ция актина, и образующийся полимер всегда находится
в равновесии с мономерным актином в соответствии с
уравнением
(актин)п (актин)„-I + актин
Критическая концентрация актина, при которой он в
соответствующих условиях полимеризуется, своя для
каждого типа актина и может служить как отличитель-
ный признак.
Поскольку во многих типах клеток концентрация не-
полимеризованного актина значительно превышает его
критическую концентрацию, должны существовать фак-
торы, способные взаимодействовать с актином и предот-
вращать тем самым его полимеризацию. Эти факторы
можно, следовательно, считать регуляторами сборки ак-
тиновых филаментов. Белки-ингибиторы полимеризации
актина обнаружены в самых различных клетках (один
из них — профилин — уже упоминался). В качестве при-
мера системы, в которой важную роль играют ингибито-
ры полимеризации актина, мы рассмотрим семенные
клетки иглокожих, а именно морского огурца (Thyone).
Эти клетки — также замечательный пример определен-
ным образом ориентированной сборки микрофиламен-
тов, так что они вполне заслуживают детального рас-
смотрения. На апикальном конце головки семенной клет-
ки Thyone есть чашевидное углубление, в котором нахо-
дится акросомальная вакуоль. Между акросомальной
вакуолью и хроматином ядра находится аморфное ве-
щество, которое представляет собой мономерный актин.
Когда сперматозоид приближается к яйцеклетке своего
вида, в первые же несколько секунд контакта головки
сперматозоида с внеклеточным веществом, окружающим
яйцеклетку, развивается акросомальная реакция. Она
заключается в быстрой сборке актиновых микрофила-
ментов в толстый пучок, исходящий из дна чащевидного
углубления и выступающий вперед приблизительно на
90 мкм (рис. 3.18). Функция этой структуры заключает-
ся в том, чтобы проткнуть желеобразное вещество, ок-
3. Системы подвижности эукариотических клеток
79
Рис. 3.18. Фазы акросомальной реакции головки сперматозоида
Thyone: а — исходное состояние; б — начало полимеризации актина;
в — готовый акросом.
ружающее яйцеклетку, что дает возможность мембра-
нам обеих кдедок слиться. Из изложенного выше ясно,
что в маленьком чашеобразном углублении головки се-
менной клетки до активизации содержится много непо-
лимеризованного актина — достаточно для сборки пучка
микрофиламентов длиной 90 мкм. Можно с большой
вероятностью предполагать, что в этом углублении обна-
ружатся белки, предотвращающие полимеризацию ак-
тина. С целью найти эти белки аморфный актин из го-
ловок клеток Thyone подвергли электрофорезу в поли-
80
3. Системы подвижности эукариотических клеток
акриламидном геле в присутствии додецилсульфата нат-
рия. Выявились три основных полипептида, один из
которых соответствует актину. Два других белка (мол.
массы 230 000 и 250 000), по-видимому, ответственны за
поддержание актина в неполимеризованном состоянии.
Обработка протеолитическим ферментом (трипсином)
позволяет отделить мономерный актин от высокомоле-
кулярных белков [158]. Акросомальную реакцию у спер-
матозоидов Thyone можно вызвать различными актива-
торами; повышение внутреннего pH освобождает актин
от связанных с ним белков, делая тем самым возможной
полимеризацию. Процесс полимеризации инициирует
специфическая структура, которую можно увидеть на
дне акросомальной чаши и которая называется актоме-
ром. Она представляет собой пучок из примерно 25 ко-
ротких микрофиламентов, заключенный в плотное
аморфное вещество. В момент активации актомер начи-
нает действовать как ядро полимеризации, из которого
вырастают микрофиламенты [160].
На основании этих и других данных высказано пред-
положение, что полимеризацией и ростом микрофила-
ментов управляют какие-то организующие центры, по-
добно тому, как центр, управляющий жгутиковым синте-
зом, регулирует сборку микротрубочек.
Специфические точки роста актиновых филаментов
обнаружены в семенных клетках Mitylus и Limulus, а
также при формировании кишечных ворсинок [150];
вполне возможно, что они есть и во многих других объ-
ектах. Действительно, в немышечных клетках часто вид-
но, что микрофиламенты прикреплены к определенным
точкам клеточной мембраны, однако пока неясно, управ-
ляют ли эти точки сборкой микрофиламентов или же
прикрепление к ним происходит уже после.
3.9. Как микрофиламенты
генерируют движение?
Микрофиламенты могут генерировать движение дву-
мя различными способами: путем скольжения — соглас-
но этому механизму актиновые и миозиновые нити
3. Системы подвижности эукариотических клеток
81
скользят друг относительно друга — или же просто пу-
тем сборки и дезагрегации пучков микрофиламентов.
Опять мы сталкиваемся с удивительным соответствием
между системой микрофиламентов и системой микротру-
бочек: обе системы генерируют движенце одними и те-
ми же способами. Это представляет интерес с эволюци-
онной точки зрения, поскольку указывает на то, что сис-
темы, которые возникли, по-видимому, независимо друг
от друга, решили проблему генерации движения одина-
ково, хотя и с помощью различных материалов.
Модель скольжения микрофиламентов, описывающая
зависимые от актомиозина движения немышечных кле-
ток, исходит из данных о сокращении мышц, модель
которого можно применить с незначительными модифи-
кациями к микрофиламентам.
В немыщечных клетках такая система подвижности
состоит из актиновых микрофиламентов, один конец
которых прикреплен к каким-либо структурам клетки (к
клеточной мембране, микротрубочкам или другим орга-.
неллам), а другой конец свободен; между свободными
концами двух противолежащих актиновых микрофила-
ментов находятся биполярные миозиновые нити. Когда
два противолежащих актиновых филамента скользят
вдоль миозиновой нити, их свободные концы сближают-
ся, а закрепленные концы тянут за собой те структуры,
к которым они присоединены. В мышцах все это проис-
ходит в структурах, специально предназначенных для
генерации движения (саркомерах). Саркомер состоит из
двух пучков актиновых нитей (они прикреплены к
Z-мембранам, ограничивающим саркомер) с миозиновы-
ми нитями между ними. В результате скольжения нитей
саркомер укорачивается, что соответствует сокращению
мышцы. В немышечных клетках такое скольжение при-
водит к сближению структур, к которым прикреплены
противоположные концы микрофиламентов (рис. 3.19).
Рассмотрим теперь, как же при взаимодействии ак-
тиновых и миозиновых нитей возникает скольжение.
Инициатором является миозин, точнее головки его моле-
кулы, где находятся центры АТРазной активности. Мио-
зиновые головки отличаются большим сродством кАТР,
и при его избытке каждая головка связывает одну мо-
82
3. Системы подвижности эукариотических клеток
молекула миозина
Рис. 3.19. Модели взаимодействия актина и миозина. Вверху, в
мышце; посередине и внизу: в немышечных клетках.
лекулу АТР. Связав АТР, миозиновая головка сразу же
переходит в активированное состояние с высоким срод-
ством к актину и прикрепляется к одной из актиновых
субъединиц ближайшего микрофиламента. Связывание с
актином немедленно вызывает гидролиз АТР, за счет
выделившейся при этом энергии головка поворачивает-
ся на небольшой угол, что немного перемещает актино-
вый филамент, к которому головка прикреплена. При
утилизации новых порций АТР такой цикл повторяется
многократно, скольжение становится заметным. Мы при-
вели весьма упрощенное описание процессов, связанных
с актомиозинзависимой сократимостью мышечных воло-
кон. Бол,ее полное описание читатель найдет в другой
книге этой же серии (R. М. Simmons „Muscle Contrac-
tion”). Этот основной механизм взаимодействия актина
и миозина можно, по-видимому, распространить также и
на немышечные клеточные системы.
ЗЛО. Регуляция скольжения белками
микрофиламентов
В мышечных клетках актиновые нити содержат
(кром,е актина) два регуляторных белка — тропомиозин
и тропонин, благодаря которым скольжение чувствитель-
3. Системы подвижности эукариотических клеток
83
но к концентрации ионов Са2+. Связывание комплекса
миозин — АТР с актином возможно только в присут-
ствии Са2+, т. е. Са2+ служит регулятором мышечного
сокращения. Концентрация Са2+ внутри саркомеров ре-
гулируется высвобождением его из саркоплазматичес-
кого ретикулума при деполяризации мембраны [41].
Если мы хотим перенести наши представления о со-
кращении мышц на немышечные клетки, вполне можно
представить, что взаимодействие между актиновыми фи-
ламентами и биполярными миозиновыми нитями регу-
лируется' таким же способом. К сожалению, не удается
показать, что в немышечных клетках миозиновых нитей
много (за исключением некоторых особых случаев, на-
пример, в тромбоцитах). Это не исключает, однако, то-
го, что они есть, но просто не видны в электронный мик-
роскоп из-за малой длины или потому, что рассеяны по
препарату. Тот факт, что миозин действительно присут-
ствует в микрофиламентах животных клеток, был проде-
монстрирован с помощью антител к миозину, меченных
флуоресцеином [48]. Возможно также, что в немышеч-
ных клетках миозиновые молекулы существуют не в ви-
де типичных нитей с выступающими головками1, а прос-
то как двуглавые мономеры, сохраняющие способность
связывать актиновые микрофиламенты. Другая модель,
которую предложили Марута и Корн [95], предполага-
ет, что одиночные миозиновые молекулы присоединены к
актиновому филаменту стержневыми участками тяже-
лых цепей, так что свободные головки могут взаимодей-
ствовать с соседними актиновыми филаментами. В этом
случае подвижность обеспечивалась бы непосредственно
скольжением двух актиновых микрофиламентов друг от-
носительно друга, т. е. так, как скользят микротрубочки
в ресничках и жгутиках. Согласно этой модели, одно-
главый миозин, который, по-видимому, не способен об-
разовывать биполярные нити (как, например, миозин I
Acanthamoeba), тоже мог бы участвовать в генера-
ции движения (см. рис. 3.19).
Поскольку подвижность зависит от взаимодействия
1 Не исключено, что такие нити могут самоорганизовываться
по мере надобности. — Прим. ред. перевода.
84
3. Системы подвижности эукариотических клеток
актина и миозина, факторы, регулирующие это взаимо-
действие, можно рассматривать как регуляторы клеточ-
ной подвижности. В мышечных клетках управление со-
кращением осуществляется с помощью ионов Са2+ и
системы тропомиозин — тропонин, связанной с актино-
выми нитями. В немышечных клетках регуляция еще не-
достаточно изучена. Ясно, однако, что в клетках различ-
ных типов может быть много разных регуляторных сис-
тем — одни из них основаны на действии Са2+, а другие
реализуют другие механизмы.
Ферментативная активность при взаимодействии
очищенных препаратов актина и миозина из немышеч-
ных клеток не зависит, как правило, от концентрации
Са2+, однако известны примеры Са2+-чувствительной
АТРазной активности актомиозина из тканей мозга,
из лейкоцитов, тромбоцитов и плазмодия миксомицета
Physarum polycephalum [32]. Чувствительность к Са2+
можно определить, регистрируя сокращение актомиози-
новых нитей в клеточных экстрактах (это сделано на
амебах и некоторых других клетках). Данные in vivo
о подвижности, чувствительной к Са2+, в которой участ-
вуют микрофиламенты, получены при исследовании то-
ков цитоплазмы у Amoeba proteus, Chaos carolinensis
и Physarum, а также ATP-зависимого сокращения изо-
лированных полосок щеточной каемки кишечного эпи-
телия [98, 156].
В мьцпечных клетках сокращение регулируется Са2+-
связывающим белком системы тропомиозин — тропонин,
поэтому некоторые исследователи искали подобные бел-
ки и в немышечных клетках. Белки, подобные тропоми-
озину, удалось найти в тромбоцитах, в тканях мозга, в
поджелудочной железе и в культуре фибробластов
мышц; Са2+-связывающий белок, похожий на тропонин
мышц, недавно выделили из мозга куриных эмбрионов.
Белки, придающие актомиозиновому комплексу чувстви-
тельность к Са2+, выделены из Physarum и Dictyoste-
Itum, однако эти данные нуждаются в дальнейшей про-
верке.
Наряду с кальциевой регуляцией, несомненно, суще-
ствуют и другие регуляторные системы, контролирую-
щие взаимодействие актина и миозина.
3. Системы подвижности эукариотических клеток
85
Одной из них может быть регуляция фосфорилирова-
ния миозина. Было показано, например, что в тромбоци-
тах АТРазная активность миозина, стимулированная ак-
тином, возрастает приблизительно в 5 раз, когда легкая
(17 000) цепь миозина фосфорилируется особой протеин-
киназой в присутствии АТР [4]. Это позволяет предпо-
лагать, что фосфорилирование прямо влияет на взаимо-
действие актина и миозина. Однако относительно этой
системы пока еще преждевременно делать окончатель-
ные выводы.
Из приведенных выше примеров должно быть ясно,
что в настоящий момент еще нет единой теории регуля-
ции актомиозинового взаимодействия в немышечных
клетках. Отчасти это обусловлено сравнительной скудо-
стью сведений, которыми мы располагаем по этому
вопросу, а отчасти — сложностью самой проблемы. Ве-
роятно, что в регуляции взаимодействия актина с мио-
зином в немышечных клетках участвует много систем.
Для некоторых клеток важное значение имеют ионы
Са2+, о других механизмах регуляции известно пока еще
слишком мало.
Есть еще один способ генерировать движение, кото-
рый используется не для перемещения всей клетки как
целого, а для движения отдельных ее частей (например,
мембран); речь идет о подвижности, обусловленной по-
лимеризацией и деполимеризацией пучков актиновых
филаментов. В этом случае движение обусловлено н,е.
скольжением, а ростом пучков микрофиламентов, кото-
рые при этом отталкивают ту часть клетки, которая
контактирует с зоной их роста (обратный процесс, как
можно представить себе, происходит при деструкции
микрофиламентов).
Пример движения такого типа — уже упоминавшая-
ся акросомальная реакция. В процессе этой реакции ме-
нее чем за 10 с формируется прямой пучок микрофила-
ментов, выпячивающий мембрану сперматозоида в на-
правлении к яйцу.
В цитоплазме некоторых немышечных клеток неред-
ко обнаруживают другой тип надмолекулярной органи-
зации актиновых мономеров: вместо обычных пучков
микрофиламенты образуют тонкую трехмерную сеть.
86
3. Системы подвижности эукариотических клеток
Это явление можно воспроизвести in vitro; оно известно
под названием процесса желатинизации. Такие, как их
еще называют, переходы золь—-гель имеют, по-видимо-
му, существенное значение для регуляции вязкости ци-
топлазмы и изменения формы клетки, и, хотя эти функ-
ции могут быть косвенно связаны с движением клетки,
их нельзя считать истинной подвижностью [80].
3.11. Микротрубочки,
микрофиламенты
и клеточные мембраны
Есть основания считать цитоплазматические микро-
трубочки и микрофиламенты тесно взаимосвязанными
структурами одной и той же системы. Они играют сход-
ную роль в генерации движения, встречаются в одних и
тех же клетках в роли опорных структур, ведут себя
сходным образом в зависимости от фазы клеточного
цикла и образуют морфологически похожие структуры.
К сожалению, приходится отметить, что, несмотря на
приведенные выше соображения, у нас нет прямых до-
казательств взаимосвязи между микротрубочками и
микрофиламентами. Многие данные, полученные различ-
ными методами, косвенно, но с большой вероятностью
указывают на то, что между этими двумя системами есть
связь, так что почти все признают, что такая связь на
самом деле существует.
Например, с морфологической точки зрения исследо-
вали распределение в клетке микротрубочек и микрофи-
ламентов, особенно в процессе митоза. Фудживара и
Поллард [49] с помощью антител к тубулину, меченных
флуоресцеином, и антител к миозину, меченных родами-
ном, сравнили распределение микротрубочек и микро-
филаментов в одной и той же клетке. В полностью рас-
пластанных клетках они не обнаружили морфологичес-
ких связей между этими двумя типами волокон. Однако
в митотическом веретене обусловленная миозином флуо-
ресценция так сочеталась с флуоресценцией микротру-
бочек, что это указывало на тесную взаимосвязь между
микрофиламентами и микротрубочками.
3. Системы подвижности эукариотических клеток
87
Более косвенные данные получены в опытах с инги-
биторами микротубулярных структур (колхицин или его
производное колцемид) и ингибиторами микрофиламен-
тов (цитохалазин В) [166]. В качестве примера такого
подхода мы рассмотрим влияние ингибиторов микро-
трубочек и микрофилам,ентов на явление образования
«шапки» (capping), наблюдаемое в клетках млекопи-
тающих [40; 104]. Реакция образования «шапки» возни-
кает после обработки клетки бивалентными антитела-
ми или другими лигандами, способными взаимодейство-
вать с белками на клеточной мембране. Она заключается
в том, что связываемые молекулы лиганда скапливаются
на ограниченном участке клеточной поверхности, обра-
зуя как бы шапку. Образование такой шапки угнета-
ется цитохалазином В, откуда следует, что микрофила-
менты играют активную роль в перемещении молекул в
мембране клетки [104]. Во многих системах угнетение
образования шапки удается снять колхицином или дру-
гими веществами, вызывающими деполимеризацию мик-
ротрубочек [153]. Если разрушить и микротрубочки, и
микрофилам,е.нты комбинированным действием колхи-
цина и цитохалазина В, образовавшаяся шапка, которая
обычно довольно стабильна, распадается, так что моле-
кулы, из которых она состояла, распределяются по всей
поверхности клетки. Эти результаты указывают на то,
что и микротрубочки, и микрофиламенты участвуют в
регуляции перемещения молекул в клеточной мембране:
микрофиламенты проявляют сократительную актив-
ность, а микротрубочки служат для них опорой. Можно
также предполагать, что для латерального перемещения
белков в липидном бислое мембраны необходимо пря-
мое взаимодействие между микрофиламентами и мем-
бранными белками (рис. 3.20).
Есть, однако, и более прямые данные о том, что
микрофиламенты соединены с поверхностными белками,
т. е. с клеточными мембранами. В разных лабораториях
различными методами показано, что непосредственно
под шапкой, образующейся на лимфоцитах (после обра-
ботки антителами) или на клетках гранулезы яичников
(после обработки конканавалином А), собирается много
микрофиламентов [10]. Там же накапливаются миозин и
88
3. Системы подвижности эукариотических клеток
Рис. 3.20. Предполагаемый механизм образования на поверхности
клетки «шапки» из рецепторов (а, б) и ее исчезновения под дей-
ствием веществ, разрушающих микрофиламенты (в), и веществ,
разрушающих микротрубочки (г).
тубулин. В более поздней работе применили м,етод
осаждения актина из лизатов клеток мышечным миози-
ном, после чего идентифицировали вещество, преципити-
рующе,е вместе с актином; показано, что актин осажда-
ется вместе с молекулами, обычно находящимися на
внешней поверхности клеточных мембран, такими, как
антигены Н-2 и иммуноглобулины. Количество иммуно-
глобулинов, связанных с актином, больше в клетках с
«шапками», чем в кл,е.тках с неизмененным распределе-
нием антигенов по поверхности [46]. Исходя из получен-
ных результатов, можно предположить, что агрегация
поверхностных белков в «шапку» вызывается возникно-
вением новых связей с актином. Это может служить до-
полнительным свидетельством в пользу того представ-
ления, что микрофиламенты контролируют по крайней
мере некоторые типы перемещения компонентов мемб-
раны.
Приведенные выше данные демонстрируют такж,е, что
актиновые филаменты прямо связаны с мембранами.
Результаты, полученные с помощью других методов и
на других объектах, также указывают на это. Например,
микрофиламенты часто выделяются при очистке цито-
плазматических мембран. Впервые это показано для
3. Системы подвижности эукариотических клеток
89
Acanthamoeba castellani [117], а впоследствии подтверди-
лось практически для всех типов клеток. В очищенных
препаратах мембран микрофиламенты можно увидеть в
электронный микроскоп (причем они прикрепл,ены к
внутренней поверхности мембраны); кроме того, при
электрофорезе препаратов мембран в полиакриламид-
ном геле в присутствии додецилсульфата натрия обна-
руживается актин.
Далее, есть прямые микроскопические наблюдения
примембранных областей, в которых много микрофила-
ментов. Иногда, например в случае ворсинок, очень чет-
ко видно, что микрофиламенты прикреплены к мембране
на конце ворсинки. На электронных микрофотографиях
поперечных сколов ворсинок центральный пучок микро-
филаментов соединен с мембраной ворсинки как бы
спицей.
Как микрофиламенты прикрепляются к мембране,
пока неясно. Непосредственно ли присоединены микро-
филаменты к мембранам или же их связь осуществляет-
ся через какие-то другие белки? Второе предположение
кажется более правдоподобным; наиболее вероятные
кандидаты на роль такого связующего белка — а-акти-
нин или подобные ему белки. Показано, что а-актинин
присутствует в кончике ворсинки — там, где микрофила-
менты прикрепляются к ее мембране; кроме того, он об-
наружен в цитоплазматической мембране и в мембранах
секреторных везикул вблизи микрофиламентов [80]. Та-
ким образом, предполагается, что а-актинин играет од-
ну и ту ж,е рбль и в мышечных волокнах (где он связы-
вает актиновые нити с Z-мембранами) и в немышечных
клетках. Возможно, микрофиламенты взаимодействуют
с мембранами также через посредство миозина, и есть
данные, указывающие на то, что миозин может высту-
пать как трансмембранный, белок.
Менее убедительны эксперименты, поставленные с
целью выявить прямое взаимодействие между микротру-
бочками и мембранами. По данным многих исследова-
телей, колхицин связывается с мембранами, однако из-
за того, что такому связыванию могут способствовать
самые разные факторы, это нельзя расценивать как до-
казательство присутствия тубулина в клеточной мембра-
90
3. Системы подвижности эукариотических клеток
не. Более прямые данные, указывающие на то, что ту-
булин входит в состав мембран, получены на синаптосо-
мах головного мозга [167] и на мембранах ресничек
некоторых моллюсков [147]. Во всяком случае, связь с
мембранами для тубулина, по-видимому, не столь обыч-
ное явление, как для актина.
Кратко рассмотрев данные о взаимодействиях между
микротрубочками, микрофиламентами и мембранами, и
помня о распределении элементов цитоскелета в живот-
ной клетке, мы можем обсудить роль цитоскелета и кле-
точной мембраны (рис. 3.21). Согласно предполагаемой
модели, в интерфазных кл,е,тках микротрубочки образу-
ют внутренний каркас, расходясь от центра к периферии
клетки. Этот каркас не участвует непосредственно в ге-
нерации движения, а служит опорой для расположенных
ближе к поверхности клетки структур, образованных
Рис. 3.21. Предполагаемые взаимоотношения между микротрубоч-
ками, микрофиламентами и клеточной мембраной. (По Nicholson et
al., 1977, из книги Dynamic Aspects of Cell-Surface Organization,
vol. 3, North Holland.)
3. Системы подвижности эукариотических клеток
91
микрофиламентами. Именно микрофиламенты, связан-
ные с мембранными белками и органеллами клетки, от-
ветственны за движения внутриклеточных структур и
всей клетки в целом. Эта модель отводит микротрубоч-
кам роль цитоскелета, а микрофиламентам—динамичес-
кую роль.
Рекомендуемая литература
Следующие книги по проблемам, рассмотренным в данной главе,
представляют общий интерес:
Primitive motile systems in cell biology (1964) (ed. R. D. Allen and N.
Kamiya), Academic Press, New York and London.
Поглазов Б. Ф. (1966). Структура и функции сократительных,
белков. — М.: Наука 1965 L
Cell motility (1976) (ed. R. Goldman, T. Pollard and J. Rosenbaum),
Cold Spring Harbor Laboratory, Cold Spring Harbor.
Чтобы оценить огромный прогресс, сделанный за последнее де-
сятилетие в изучении молекулярных основ подвижности клеток, по-
лезно сравнить содержание первых двух книг с тем, что изложено
в третьей.
Наряду со множеством обзоров и специальных статей читатель
может получить дополнительную информацию о микротрубочках в
следующих публикациях:
Snyder J. A., McIntosh R. (1976). Biochemistry and physiology of
microtubules, Ann. Rev. Biochem., 45, 699—720.
Gaskin F., Shelanski M. L. (1976). Microtubules and intermediate
filaments. In: Essays in Biochemistry (ed. P. N. Campbell and
W. N. Aldridge), vol. 12, Academic Press, New York and London,
pp. 115—146.
Stephens R. E., Edds К. T. (1976). Microtubules: structure, chemistry
and function, Physiol. Rev., 56, 709—777.
Dustin P. (1978). Microtubules, Springer-Verlag, Berlin, Heidelberg,
New York.
О микрофиламентах и касающихся их вопросах:
Clarke М., Spudich J. А. (1977). Nonmuscle contractile proteins: the
role of actin and myosin in cell motility and shape determination,
Ann. Rev. Biochem., 46, 797—822.
Hitchcock S. E. (1977). Regulation of motility in nonmuscle cells,
J. Cell Biol, 74, 1—15.
Korn E. D. (1978). Biochemistry of actomyosin-dependent cell moti-
lity, Proc. Natt. Acad. Sci. USA, 75, 588—599.
1 Вместо рекомендуемого автором английского перевода восста-
новлена ссылка на первоисточник. — Прим. ред.
4. ДВИЖЕНИЕ
ЭУКАРИОТИЧЕСКИХ
КЛЕТОК
Создаваемое микротрубочками и микрофиламентами
движение в эукариотических клетках может реализо-
ваться различно, приводя к движению либо всей клетки
как целого, либо только отдельных ее частей. В первом
случае происходит активное перемещение клетки, во
втором — движение ограничено какой-то частью клетки,
будь то м,ембрана, цитоплазматические органеллы или
сама цитоплазма. Перемещение клетки осуществляется
одним из двух основных способов: либо благодаря дви-
жению ресничек или жгутиков, либо амебоидным дви-
жением. Реснички и жгутики эукариотических кле-
ток — это цилиндрические органеллы, вдоль которых
при возбуждении распространяются волны, приводящие
клетку в движение, если она не закреплена. Если же
кдетка лишена возможности двигаться, биение ресничек
или жгутиков приводит в движение окружающую их
жидкость. Так действуют, например, реснички (ворсин-
ки) эпителия, который выстилает полости в теле живот-
ного. Можно было бы привести еще множество приме-
ров, однако и без них ясно, что двигаться с помощью
ресницей и жгутиков можно только в жидкой среде.
Амебоидное движение, напротив, эффективно для пере-
мещения клетки в нежидкой среде. Оно обнаружено не
только у свободно живущих микроорганизмов, но также
и у более высоко организованных живых существ, у ко-
торых амебоидное движение играет решающую роль в
развитии, особенно в процессе морфогенеза многоклеточ-
ного организма (см. книгу D. R. Garrod „Cellular Deve-
lopment”, изданную в этой же серии). Можно также
привести примеры того, что оба типа движения реали-
зуются одновременно в пределах одной клетки. Еще бо-
4. Движение эукариотических клеток
93
лее удивительны такие микроорганизмы, как почвенная
амеба Naegleria или плазмодии миксомицетов Physarum
и Didymium, которые движутся амебоидно, когда живут
в почве, и приобретают жгутики, приспосабливаясь к
водной среде. Благодаря этому свойству такие микроор-
ганизмы можно считать промежуточными формами меж-
ду а,м,е.бами и жгутиковыми.
Движения внутри клетки свойственны всем организ-
мам. Это отражает тот факт, что структурная организа-
ция живой клетки находится в состоянии динамики;
изменения в окружающей среде требуют от клетки адек-
ватной реакции, необходимым звеном которой часто бы-
вают внутриклеточные движения. Это могут быть дви-
жения клеточной мебраны (например, при фагоцитозе),
органелл внутри клетки (вакуолей, лизосом, пигментных
гранул и др.)1 или самой цитоплазмы (например, цик-
лоз в растительных клетках).
В предыдущей главе мы рассмотрели молекулярные
компоненты систем подвижности эукариотических кле-
ток; следующие разделы посвящены исполнительным и
регуляторным механизмам жгутикового и амебоидного
движения.
4.1. Движение ресничек и жгутиков
Реснички и жгутики эукариотических клеток — это
цитоплазматические выросты, способные генерировать
движение. Они состоят из оси, образованной микротру-
бочками (аксонемы) и клеточной мембраны, которая ее
окружает. Реснички'и жгутики имеют одинаковую струк-
туру и различаются главным образом по характеру дви-
жения: жгутики совершают равномерные волнообразные
движения, тогда как реснички—двухфазные биения
(быстрый рабочий гребок и медленное возвращение в
исходное положение).
1 К наиболее подвижным органеллам клетки относятся митохон-
дрии— они движутся практически непрерывно. — Прим. ред. пере-
вода.
94
4. Движение эукариотических клеток
4.1.1. Механизм движения
Согласно модели скольжения, которую предложил
Петер Сатир [129], движение ресничек и жгутиков созда-
ется деятельностью динеиновых руч,ек в дублетах микро-
трубочек в присутствии АТР. Возникающее при этом
скольжение приводит к изгибанию реснички или жгути-
ка благодаря связующим белковым элементам (радиаль-
ным спицам и нексиновым мостикам) между дублетами.
Решающее значение радиальных спиц и нексиновых мос-
тиков в трансформации скольжения в изгибание демон-
стрируется следующим фактом: если их переварить
трипсином, аксонемы таких жгутиков или ресничек те-
ряют способность к волнообразным движениям. Не по-
врежденные трипсином, аксонемы в присутствии АТР
изгибаются подобно интактным жгутикам. На основании
этих простых экспериментов можно сделать следующие
важные выводы: во-первых, именно аксонема ответ-
ственна за движение жгутика; во-вторых, движение воз-
никает в результате взаимодействия динеиновых ручек
с дублетами микротрубочек; в-третьих, белковые соеди-
нения между дублетами аксонемы обеспечивают транс-
формацию скольжения микротрубочек в изгибание жгу-
тика. Первые два пункта мы уже обсуждали в разде-
ле 3.4 главы 3, так что читатель может к этому вер-
нуться. Теперь наш интерес сосредоточится на последнем
пункте, а именно на вопросе о том, как скольжение мик-
ротрубочек в соседних дублетах трансформируется в из-
гибание жгутика. Мы рассмотрим также роль нексино-
вых мостиков и радиальных спиц в этом процессе и об-
судим, как локальный изгиб может породить волнооб-
разное движение всего жгутика.
Современные представления о роли нексиновых мос-
тиков и радиальных спиц основаны главным образом на
работе Уорнера и Сатира [168], которые с помощью
электронной микроскопии высокого разрешения изучили,
как изменяется структура латеральных ресничек в жаб-
рах моллюска Elliptic» spp., фиксируя ее в разных фазах
цикла биений. Показано, что в прямых (неизогнутых)
участках ресничек спицы не контактируют с централь-
ной оболочкой и перпендикулярны тому дублету, из ко-
4. Движение эукариотических клеток
95.
Рис. 4.1. Схематическое изображение аксонемы жгутика: а — про-
цесс изгибания; б — неизогнутая аксонема. Показаны отношения
между радиальными спицами и центральными дублетами, а также
смещения внутри дублетных пар на конце жгутика (сравните схе-
мы а и б поперечного сечения конца жгутика).
торого они исходят. Там, где аксонема изогнута, ранее
свободные концы радиальных спиц прикреплены к цент-
ральной оболочке, а сами спицы наклонены, причем угол
наклона пропорционален кривизне в данном участке
реснички. Похоже, что радиальные спицы периодически
смыкаются с центральной оболочкой и отсоединяются от
96
4. Движение эукариотических клеток
нее в соответствии со степенью изогнутости реснички.
В тех участках, где спицы прикрепляются к оболочке
вследствие скольжения микротрубочек, ресничка изгиба-
ется. В остальной ее части концы радиальных спиц не
закреплены, что допускает свободное скольжение дубле-
тов (рис. 4.1). Излишнее скольжение предотвращается
нексиновыми мостиками между соседними дублетами.
Эти мостики, по-видимому, не размыкаются, а служат
постоянными эластическими соединениями между дуб-
летами и допускают только ограниченное скольжение,.
4.1.2. Типы движений
В экспериментальных условиях изгибание реснички
или жгутика может начаться в любой точке аксонемы.
Это продемонстрировано в работе, Брокау и Джиббонса
[26] на частично лишенных мембраны хвостах спермато-
зоидов: при добавлении АТР изгибы возникали именно
в тех участках, с которых удалили мембрану. Однако в
естественных условиях изгибание чаще всего возникает
у основания жгутика или реснички, и затем волна рас-
пространяется вдоль по направлению к кончику
(рис. 4.2). Реже можно наблюдать обратное: например,
у трипаносом в норме волны распространяются по жгу-
тику от его конца к основанию. Размеры жгутиков эу-
кариотических клеток позволяют легко регистрировать
их движение с помощью светового микроскопа в сочета-
нии, например, со скоростной микрокиносъемкой (или
иным методом регистрации изображений). Движение
Рис. 4.2. Схема распространения волн вдоль жгутика при движе-
нии сперматозоида.
4. Движение эукариотических клеток
97
I »'
Рис. 4.3. Движения жгутика: а — плоские волнообразные движения;
б — винтовое движение. На рис. а' и б' показано, как движется
при этом кончик жгутика.
жгутика имеет волнообразный характер и может совер-
шаться как в одной плоскости, так и в пространстве.
В последнем случае движения жгутика ограничены ци-
линдрической поверхностью, и его кончик описывает
кривую, близкую к эллипсу (винтовое движение). Дви-
жения жгутика характеризуются симметрией и упоря-
доченностью волн. Так как волны возникают с довольно
большой частотой (20—30 раз в секунду), вдоль по жгу-
тику в норме одновременно бежит более чем одна вол-
на, так что он выглядит волнообразно изогнутым
(рис. 4.3).
Движение жгутика в жидкой среде создает силу, ко-
торая достаточна для того, чтобы толкать свободно пла-
вающий организм в направлении, противоположном на-
правлению распространения волны. Таким образом, ког-
да волна распространяется от основания жгутика к его
концу, он толкает клетку от себя, а когда волна бежит
от кончика к основанию, жгутик тянет клетку за собой.
Интересное исключение обнаруживают жгутиковые
Ochromonad, которые перемещаются в направлении рас-
пространения волны вдоль жгутика. Это связано с тем,
что жгутик этих простейших снабжен двумя рядами ни-
тевидных отростков (около 20 нм толщиной и 1 мкм
длиной), называемых мастигонемами, которые прикреп-
лены по всей длине жгутика с противоположных сторон
в одной плоскости (рис. 4.4). Согласно Яну и др. [74],
жгутик совершает биения в плоскости расположения
мастигонем. Жесткце выступы мастигонем создают по-
ток жидкости в направлении, противоположном распро-
98
4. Движение эукариотических клеток
волна
мастигонемы
направление движения клетки
Рис. 4.4. Направление перемещения клетки в результате волнооб-
разных движений жгутика. Вверху — жгутик без мастигонем, вни-
зу— с мастигонемами.
страняющейся по жгутику волне. Так как мастигонемы
развивают силу большую, чем жгутик, организм продви-
гается в направлении распространения волны вдоль
жгутика.
Движения ресничек очень разнообразны; их обычно
называют биениями. Согласно общей модели, основы-
вающейся на наблюдениях биения ресничек у разных
организмов, каждый цикл биения состоит из двух фаз.
В первой фазе (фаза рабочего гребного удара) ресничка
резко изгибается вблизи своего основания, а остальная
ее часть остается прямой или лишь слегка изогнутой.
При этом конец реснички описывает плоскую дугу по
крайней мере в 90°. Во второй фазе (восстановительное
движение) изгиб медленно перемещается (и постепенно
уменьшается) к концу реснички; в результате восстанав-
ливается исходное положение, с которого начинался ра-
бочий гребок (рис. 4.5). Рабочий гребной удар — это
более продуктивная фаза биения ресничек: гребок созда-
ет значительно больший поток жидкости, чем восстано-
вительное движение, и, следовательно, именно он опре-
деляет направление движения. Свободноживущий мик-
роорганизм перемещается в направлении, противопо-
ложном направлению гребного удара. В случае реснич-
ного эпителия или подобных структур поток жидкости,
создаваемый биением ресничек, направлен в ту же сто-
4. Движение эукариотических клеток
99
Рис. 4.5. Схематическое изображение движения реснички: а —
профиль гребного удара и восстановительного движения; б — траек-
тория движения кончика реснички (проекция на плоскость).
восстановительное
движение
рону, что и гребные удары. В фазе восстановления рес-
ничка медленно возвращается в исходное положение в
другой плоскости, чем та, в которой совершался рабочий
гребок. Так как восстановительное движение медленно,
оно лишь незначительно смещает жидкую среду и обыч-
но никак не влияет на подвижность.
4.1.3. Координация движения
и управление им
Координация движения подвижных структур эука-
риотических клеток особенно наглядна в случае ресни-
чек. Это обусловлено тем, что реснички обычно никогда
н,е бывают одиночными, а образуют структуры, состоя-
щие нередко из тысяч плотно упакованных отдельных
органелл. Трудно представить себе, что в таких услови-
ях каждая ресничка движется более или менее незави-
симо. Действительно, биения ресничек в таких структу-
рах оказались чрезвычайно высоко координированными.
Предложены две гипотезы, которые объясняют меха-
низм согласованного движения ресничек с разных пози-
ций. Согласно «нервной» гипотезе координации, подвиж-
ность ресничек управляется своего рода нервным меха-
низмом: возбуждение передается последовательно от
одной реснички к другой по клеточным структурам. Дру-
100
4. Движение Эукариотических клеток
ресничка
базальные
структуры
„нервная”
координация
стимул
Рис. 4.6. Схема, иллюстрирующая возможные механизмы «нервной»
и механической координации биения ресничек.
гая гипотеза — теория механической координации —
предполагает, что движения соседних ресничек взаимо-
связаны благодаря тому, что механическая сила пере-
дается от одной реснички к другой через вязкую среду,
в которой они движутся (рис. 4.6). Чтобы выяснить, ка-
кая из гипотез верна, Слейг [138] поставил ряд остроум-
ных экспериментов на ресничных гребешках Pleu.ro-
branchia, применив механическое устройство для стиму-
ляции и остановки биения ресничек. Опыты показали,
что, когда между соседними ресничками вставлена тонкая
бумажная перегородка, задерживающая поток жидкости,
распространение биений немедленно прекращается.
Этот эксперимент и другие ему подобные подчерки-
вают важность потоков жидкости, вызываемых движе-
нием ресничек, для согласования их деятельности и под-
держивают гипотезу механической координации. Правда,
этот механизм реализуется, видимо, не у всех орга-
низмов. Есть данные о том, что у многих многоклеточ-
ных координация движения ресничек осуществляется по
механизму «нервной» регуляции.
При движении ресничек на поверхности ресничной
клетки наблюдаются метахронные волны (т. е. реснич-
ки движутся со сдвигом по фазе); это результат коор-
динации движения ресничек. Эти волны поэтично срав-
нивают с волнами, пробегающими по хлебному полю на
ветру. Тип координации биения ресничек (у различных
организмов описаны разные типы координации [77]) за-
висит от того, как направлены гребные удары ресничек
4. Движение эукариотических клеток
101
Рис. 4.7. Координация биений ресничек: а — симплектитовая мета-
хрония; б — антиплектитовая метахрония; в, г, д — диаплектитовая
метахроиия, г — вид в направлении распространения волны, д —
вид в направлении гребного удара. (Из Holwill М. Е. J., 1977,
Adv. Microbiol. Physiol., 16, 1—48.)
и как распространяются метахронные волны по поверх-
ности, несущей реснички. Когда метахронная волна и
гребной удар одинаково направлены, говорят о симплек-
титовой метахронии, когда они направлены в противо-
положные стороны — об антиплектитовой метахронии,
когда взаимно перпендикулярны — о диаплектитовой ме-
тахронии (рис. 4.7). Каждый из этих типов метахронии
свойствен определенной ресничной структуре.
102
4. Движение эукариотических клеток
Управление перемещением клеток, движущихся за
счет активности жгутиков или ресничек, несомненно, оп-
ределяется возможностями клетки изменять характер
движения этих органелл. Как и в случае бактерий, та-
кие изменения чаще всего вызываются различными воз-
действиями окружающей среды: химическими, электри-
ческими, механическими стимулами, изменениями осве-
щенности и проч. Стимулы окружающей среды регули-
руют как ориентацию, так и скорость передвижения
клетки.
Одна из ориентационных реакций у эукариотических
микроорганизмов изучена достаточно детально; это так
называемая реакция избегания у жгутиковых или рес-
ничных клеток. Эту реакцию вначале рассматривали
как механизм, позволяющий свободно плавающей клетке
избегать механические препятствия, которые она может
встретить на своем пути; в дальнейшем, однако, реак-
цию избегания наблюдали в ответ также и на многие
другие стимулы. У ресничных, например у инфузорий
Paramecium, временное изменение направления греб-
ного удара на обратное вызывает движение клетки
вспять. Эта реакция вызывается механическими или ка-
кими-либо физико-химице.скими стимулами. Реакция
заканчивается, когда гребные удары возобновляются в
нормальном направлении и клетка снова перемещается
вперед (рис. 4.8). Исследования Роджера Эккерта и его
сотрудников позволили прояснить некоторые вопросы,
связанные с изменением направления биения ресничек;
они показали, что направление гребного удара (прямое
или обратное) контролируется электрическим потенциа-
лом клеточной мембраны и связанной с ним проницае-
мостью мембраны для ионов кальция (см. [42] и [102]).
Согласно общепринятой в настоящее время модели, сти-
мул, действующий на хвостовой конец Paramecium, вы-
зывает деполяризацию клеточной мембраны; поток ио-
нов кальция из среды внутрь клетки увеличивается.
В результате возрастает концентрация кальция внутри
клетки и направление биения ресничек меняется на об-
ратное. Если деполяризующий стимул не поддержива-
ется (т. е. его длительность ограничена), Са2+ активно
выкачивается из клетки до тех пор, пока внутри клет-
4. Движение эукариотических клеток
103
Рис. 4.8. Фазы реакции избегания у Paramecium-, а — уход от
препятствия; б — переориентация клетки; в —• движение в новом
направлении [42].
Рис. 4.9. Последовательность фаз реакции избегания: а — деполя-
ризация мембраны в ответ на стимуляцию; б — вызванный деполя-
ризацией поток Са2+ внутрь клетки; в — аккумуляция Са2+ и из-
менение направлеияя биения ресничек на обратное; г — возобнов-
ление биения ресничек в нормальном направлении в результате
восстановления нормальной внутриклеточной концентрации Са2+
ки не восстановится его нормальная концентрация. Как
только достигается это состояние, реснички возобновля-
ют биения в нормальном направлении и клетка снова
плывет вперед (рис. 4.9).
104
4. Движение эукариотических клеток
Помимо Paramecium, Са2+-зависимая регуляция дви-
жения ресничек и жгутиков обнаружена также в спер-
матозоидах Tubularia и Elliptia и у таких жгутиковых
простейших, как Crithidia oncopelti и Chlamydomonas.
Каков механизм действия ионов кальция, в настоящее
время еще не ясно.
Способ переориентации движения клетки в ответ на
стимулы внешней среды не одинаков у различных мик-
роорганизмов. Двухжгутиковые, такие, как Chlamydo-
monas, изменяют направление плавания на обратное,
вытягивая на короткое время оба жгутика перед те-
лом клетки. Некоторые сперматозоиды на мгновение
останавливаются, движения жгутика прекращаются, го-
ловка поворачивается в нужную сторону, после чего
сперматозоид плывет в новом направлении. Трипаносо-
мы действуют иначе: у них нормальное направление рас-
пространения волны по жгутику (от конца к основанию)
меняется на обратное (от основания к концу), что изме-
няет направление их движения. Все эти способы очень
эффективны; не исключено, что в их основе лежит один
и тот же механизм.
4.2. Амебоидное движение
Термин «амебоидное движение» охватывает все фор-
мы перемещения клеток с помощью временно образую-
щихся на поверхности клетки цитоплазматических вы-
ростов, носящих общее название псевдоподии. Этот тип
движения вначале считался характерным свойством
амебоидных простейших класса Sarcodina, однако впо-
следствии стало ясно, что он присущ большинству кле-
ток, хотя и с различными модификациями. Клетка
может выпускать псевдоподии в любой среде (вода, воз-
дух, почва и др.). Однако передвижение возможно толь-
ко в том случае, когда у организма есть плотная опора.
Следовательно, амебоидно двигаться могут только те
клетки,, которые либо лежат на твердой поверхности,
либо находятся внутри многоклеточного организма, где
опору обеспечивают другие клетки и ткани.
4. Движение эукариотических клеток
105
4.2.1. Механизм
амебоидного движения
Как уже отмечалось выше, для амебоидного движе-
ния необходима система актиновых микрофиламентов,
которые могут ^взаимодействовать с миозином в присут-
ствии АТР и создавать силу, которая и вызывает дви-
жение. По общему мнению, при амебоидном движении
роль мотора играет взаимное скольжение актина и мио-
зина; гораздо менее ясно, как это скольжение связано с
формированием псевдоподий и их активностью.
Для объяснения амебоидного движения есть две ги-
потезы. Пантен [109] и Маст [96] задолго до открытия
сократимых микрофиламентов предложили идею о
сократимом эктоплазматическом мешке. Согласно этой
гипотезе, цитоплазма на периферии клетки (эктоплазма)
находится в состоянии геля и может «сокращаться». Со-
кратившийся эктоплазматический «мешок» оказывает
давление на центральную, более жидкую, часть цито-
плазмы (эндоплазму), выдавливая ее в ту область клет-
ки, где давление меньше (в сторону открытой части
«мешка»), где и образуется псевдоподия (как выдавли-
вается из тюбика зубная паста) (рис. 4.10, а). Роберт
Рис. 4.10. Токи эдоплазмы при амебоидном движении: а — по гипо-
тезе о сокращении эктоплазменного мешка; б — по гипотезе о со-
кращении фронтальной зоны.
106
4. Движение эукариотических клеток
Аллен (12] предложил гипотезу сокращения фронталь-
ной зоны. При этом он наделил свойством сократимости
уже эндоплазму, точнее ту ее часть, которая находится
во фронтальной зоне продвигающейся вперед псевдопо-
дии, считая, что при сокращении она будет тянуть за со-
бой эндоплазму из лежащих за ней областей клетки
(рис. 4.10,. б). Хотя показано, что сокращение эктоплаз-
матического м,ешка согласуется с моделью скольжения
актиновых и миозиновых нитей [123], в настоящий мо-
мент гипотеза Аллена более популярна; в ее пользу сви-
детельствуют многие экспериментальные результаты, а
также теоретические представления (см. [105] и обзор
[14]). Возможно, однако, что верны обе гипотезы, просто
они реализуются в различных системах.
Если не считать теоретические соображения, мы пока
что очень мало знаем о молекулярном механизме амебо-
идного движения. Хотя некоторые реакции этого процес-
са уже удалось расшифровать, предстоит еще большая
работа. Модель, предложенная Тейлором и его сотруд-
никами [154], представляется в этом отношении хорошо
согласующейся со всеми имеющимися морфологически-
ми и биохимическими данными. Эта модель основыва-
ется на представлении о том, что в цитоплазме все время
происходят переходы актина из мономерного состоя-
ния в полимеризованное и обратно, причем актин взаи-
модействует с миозином только в виде филаментов; при
этом развивается сокращение. После того как оно про-
изошло, актиновые филаменты деполимеризуются на
мономеры и цикл повторяется снова. Таким образом, со-
кращение предстает как многоступенчатый процесс,
включающий фазу структурирования актина (сборка
микрофиламентов; формирование из них пространствен-
ной сетки, т. е. образование геля, или желатинизация) и
фазу собственно сокращения гелеподобной структуры.
Внутри клетки область перехода золь—гель актина
совпадает с зоной превращения эндоплазмы в эктоплаз-
му. «Расслабление» актомиозиновых комплексов, сопро-
вождающееся деструкцией актиновых филаментов, похо-
же, совпадает с переходом эктоплазмы в эндоплазму.
Модель подчеркивает важность тех факторов, которые
контролируют как взаимодействие актина и миозина,
4. Движение эукариотических клеток
107
так и процесс желатинизации. Впервые это исследова-
лось на большой амебе Amoeba proteus и в дальнейшем
подтвердилось на других объектах.
4.2.2. Типы амебоидного движения
Чтобы клетка могла двигаться амебоидно, должны
выполняться два условия. Во-первых, клетка должна
иметь возможность прикрепиться к твердому субстрату
и, во-вторых, она должна уметь выпускать цитоплазма-
тические выросты — псевдоподии, в которые как бы пе-
ретекает содержимое клетки. Эти два условия в общих
чертах определяют важнейшие особенности амебоидного
движения, однако как форма и структура цитоплазмати-
ческих отростков, так и характер передвижения у раз-
личных клеток весьма различны. С морфологической и
структурной точек зр,едия выделяют по крайней мере
пять типов псевдоподий (рис. 4.11). Первый тип — это
характерные для амеб лобоподии, т. е. широкие, с
закругленным концом выпячивания клеточной мембра-
ны. На конце лобоподии можно видеть область прозрач-
ной эктоплазмы, за которой находится менее прозрачная
эндоплазма с клеточными органеллами. Обычно при
Рис. 4.11. Наиболее распространенные формы псевдоподий: а —
лобоподия; б — филоподии; в — аксоподия; г — ламеллоподия; <5 —
ризоподии (ретикулоподии).
108
4. Движение эукариотических клеток
движении клетки лобоподия прикрепляется к субстрату,
а затем содержимое клетки перетекает в нее, однако у
раковинных амеб дело обстоит иначе. Эти амебы строят
раковину из органического вещества, внутри которой на-
ходится почти вся цитоплазма. Из отверстия раковины
выступает только пальцевидная лобоподия; она прикреп-
ляется к субстрату, сокращается и, укорачиваясь, подтя-
гивает клетку к точке прикрепления.
Второй тип псевдоподий — различные формы ламел-
лоподий, встречающихся у клеток млекопитающих. Они
похожи на сильно уплощенные лобоподии (толщина слоя
цитоплазмы в них иногда менее 0,1 мкм), которые вы-
тягиваются перед клеткой в направлении ее движения.
Ламеллоподии содержат микротрубочки, микрофиламен-
ты и другие органеллы. Они вытягиваются вперед, при-
крепляются к субстрату и передвигают клетку.
Третий тип псевдоподий — так называемые филопо-
дии, они встречаются как у амеб, так и у многоклеточ-
ных. Это очень гибкие, тонкие (около 0,2 мкм в диамет-
ре) псевдоподии, внутри которых под клеточной мембра-
ной находится ось из микрофиламентов. Филоподии —
очень подвижные образования; они быстро выпускаются
клеткой и могут так же быстро втягиваться обратно.
Возможно, что они действуют как чувствительные щу-
пальца, с помощью которых клетка исследует консистен-
цию находящейся перед ней поверхности, прежде чем
наползет на нее [11]. Иногда филоподии превращаются
в псевдоподии другого типа —в лобоподии или ламелло-
подии, — более эффективные в целях передвижения.
Четвертый тип псевдоподий, ретикулоподии, называе-
мые также ризоподии или сетчатые псевдоподии, чаще
всего встречаются у простейших Foraminifera. Это тон-
кие, сильно разветвляющиеся нитевидные структуры,
образующие между собой множество анастомозов, что в
целом создает сложную трехмерную сеть вокруг клетки.
Внутри ризоподии находится ось из микрофиламентов,
в окружающей эту ось цитоплазме часто обнаружива-
ется много гранул (митохондрии и цитоплазматические
включения). Вся сеть может сокращаться и использует-
ся клеткой для передвижения.
4. Движение эукариотических клеток
109
И наконец, последний тип — аксоподий солнечников,
структура и подвижность которых уже описаны выше в
разделе о движении микротрубочек (разд. 3.4).
Самые существенные морфологические особенности
амебоидного движения мы теперь опишем более де-
тально на примере больших почвенных амеб Chaos
chaos и Amoeba proteus. Первая из них— особенно удоб-
ный объект для такого рода исследований, так как ее
большие размеры (несколько миллиметров в длину)
значительно упрощают морфологическое изучение. Маст
[96] и позднее Аллен [13] подробно описали это явление.
Движущаяся амеба представляет собой полярную струк-
туру, в которой передний полюс соответствует концу
псевдоподии, а задний полюс называют хвост, или уро-
ид. В передней части продвигающегося вперед конца
псевдоподии имеется тонкий чашеобразный слой совер-
шенно прозрачной эктоплазмы (гиалиновый колпачок)
без каких бы то ни было гранул; эта прозрачная чаша
охватывает всю переднюю часть клетки. Гранулирован-
ная эндоплазма с ядром и другими органеллами зани-
мает центральную область клетки. Гиалиновый колпа-
чок в продвигающемся вперед конце клетки периодиче-
ски выдвигается вперед вследствие того, что некоторая
часть центральной эндоплазмы псевдоподии резко
переходит в состояние геля. В эти моменты продвиже-
ние псевдоподии в направлении перемещения клетки
происходит с максимальной скоростью — около 3—
4 мкм/с. Токи эндоплазмы направлены к продвигающе-
муся вперед концу псевдоподии,вблизи гиалинового кол-
пачка общий поток распадается на боковые струи, эндо-
плазма под колпачком частично обтекает колпачок, а
частично переходит в состояние геля и в таком виде
включается в слой эктоплазмы. Разделение центрально-
го тока эндоплазмы на боковые струи имеет сходство с
фонтаном, поэтому область, в которой это происходит и
где совершается переход эндоплазмы в эктоплазму, по-
лучила название «зона фонтана» (рис. 4.12). Токи экто-
плазмы направляются по краям назад к уроиду, где про-
исходит обратный переход эктоплазмы в эндоплазму, за
что эта область получила название «зона восстановле-
по
4. Движение эукариотических клеток
Рис. 4.12. Различные области цитоплазмы у амебы [13].
ния». Отсюда эндоплазма начинает двигаться снова в
направлении к «зоне фонтана».
Наиболее важен для амебоидного движения контакт
с субстратом, на котором находится клетка. Амебы при-
крепляются к субстрату особым участком клеточной
мембраны в центральной части поверхности клетки.
Часть клетки позади точки прикрепления укорачивается,
в то время как передняя часть удлиняется в направле-
нии перемещения.
Способ перемещения больших амеб нет оснований
считать общим для всех клеток. Однако вполне вероят-
но, что молекулярный механизм один и тот же для всех
типов амебоидного движения. Наибольшее разнообра-
зие наблюдается на морфологическом уровне,. В культу-
ре клеток млекопитающих, например фибробластов,
продвигающиеся вперед псевдоподии (ламеллоподии)
имеют обычно листообразную форму с характерным
волнистым передним краем. Ведущая ламеллоподия
движется вперед, а остальная часть клетки следует за
ней. При этом, однако, не видно «фонтана» в токах ци-
топлазмы, подобного тому, какой имеется у больших
амеб. В фибробластах обычно хорошо выражена хвос-
товая область, часто типичная пальцевидная. Как пра-
вило, точек прикрепления к субстрату несколько, они
находятся на нижней стороне ламеллоподий, тела клет-
ки и хвоста [1, 2].
4. Движение эукариотических клеток
111
4.2.3. Управление
амебоидным движением
Способность двигаться направленно в ответ на по-
ступающую из окружающей среды информацию химиче-
ской или физической природы хорошо развита у эука-
риотических клеток с амебоидным типом движения.
В частности, хемотаксис (т. ,е„ направленная подвиж-
ность в ответ на химические стимулы) проявляют
самые разные клетки высших организмов, например
фибробласты, опухолевые клетки, нейроны, половые
клетки, лейкоциты и др., а также некоторые низшие ор-
ганизмы, такие, как амебоидные простейшие или миксо-
мицеты. Хотя эукариотические клетки — довольно рас-
пространенный объект при исследовании направленной
подвижности, бол,е,е или менее полные данные о хемо-
таксисе имеются только для полиморфноядерных лейко-
цитов (из клеток высших организмов) и для клеточной
формы миксомицета Dyctyostelium discoideum (из низ-
ших эукариот). D. discoideum особенно удобен для та-
кого рода исследований: с ним легко манипулировать,
его можно исследовать сразу многими методами и сис-
тема его хемотаксиса уже довольно хорошо изучена.
а. Хемотаксис у клеточной формы миксомицета
D. discoideum. D. discoideum — один из излюбленных
объектов по сравнению с клеточными формами других
слизистых грибов, хорошо изученный пример клеточной
дифференцировки. Его жизненный цикл можно считать
состоящим из трех фаз: фазы роста, фазы агрегации и
фазы морфогенетического развития (рис. 4.13). В фазе
роста одноклеточные амебы размножаются делением и
питаются бактериями. Когда запасы пищи истощаются,
начинается фаза агрегации: амебы мигрируют
по направлению к центрам, где они образу-
ют агрегаты из тысяч клеток. Далее наступает
фаза морфогенетического развития: такой агрегат раз-
вивается, дифференцируется, и образуется плодовое
(спорообразующее) тело, в котором можно различить
клетки трех типов (базальные диски, клетки стебля и
споры). Попав в подходящие условия, споры превра-
щаются в амеб, что соответствует началу нового жизнен-
112
4. Движение эукариотических клеток
импульсная
сигнализация
хемотаксис
потоки
релейная
передача
сигналов
фаза 'морфогенетического развития
Рис. 4.13. Жизненный цикл D. discoideum. Подробности в тек-
сте [10].
ного цикла миксомицета. (Дополнительную информацию
можно найти в монографии J. М. Ashworth „Cell Diffe-
rentiation”; см. также [88].) Предположение о том, что
именно хемотаксис обеспечивает агрегацию кл,е.ток
D. discoideum, было высказано еще в начале века пер-
выми исследователями, изучавшими слизистые грибы
[106, 121]. Позднее было показано, что хемотаксис вызы-
вается диффундирующим фактором, названным акразин
[23, 126], свойства которого частично описал Шаффер
[131]. И наконец], Конджен и сотр. [79] показали, что
циклический аденозинмонофосфат (сАМР) действует
как аттрактант на миксамеб D. discoideum и что в ес-
тественных условиях агрегация происходит именно как
хемотаксис на сАМР. Циклический аденозинмонофосфат
служит аттрактантом также и для других видов того же
рода. Однако некоторые родственные миксомицеты
(Polyspondylium pollidum, Р. violaceum) используют как
4. Движение эукариотических клеток
ИЗ
акразин низкомолекулярный пептид с мол. массой око-
ло 1500. Это примечательно тем, что подобный пептид
является хемоаттрактантом также и для лейкоцитов.
Процесс агрегации у D. discoideum представляет
собой сложное явление. С помощью дефектных мутантов
подсчитано, что в нем более или мецее непосредственно
участвует около 200 генов. Агрегация начинается в от-
вет на голодание; некоторые миксамебы начинают рит-
мически выделять порции сАМР вначале со скоростью
примерно 1 раз каждые 10 мин, а затем быстрее — до
1 «импульса» каждые 2—3 мин (рис. 4.14). Как регули-
руется эта скорость, пока выяснить не удалось. Она
может зависеть от периодических изменений активности
аденилатциклазы (фермента, участвующего в синтезе
сАМР), обусловленных колебаниями метаболических
процессов в клетке; эта проблема в настоящее время
еще не решена.
Выделяемый клеткой сАМР диффундирует на рас-
стояние вплоть до 100 мкм. В процессе диффузии сАМР
постепенно расщепляется присутствующими в среде фос-
фодиэстеразами, однако некоторые молекулы, избежав-
шие этого, достигают соседних миксамеб и связываются
находящимися на их поверхности хеморецепторами
(сАМР-рецепторами). сАМР-редепторы возникают на
поверхности клеток в ответ на голодание через несколь-
ко часов после исчезновения пищи.
Неясно, как эти рецепторы воспринимают хемосиг-
нал. Предложено два объяснения этого явления. Соглас-
но первой гипотезе, рецепторы способны реагировать на
градиент концентрации аттрактанта (сАМР) вблизи
поверхности клетки. Вторая гипотеза предполагает, что
миксамебы могут ощущать изменения концентрации
сАМР во времени, подобно тому, как это' наблюдается
у бактерий при действии на них химических стимулов.
Сигнал сАМР вызывает у миксамеб два типа огра-
ниченных во времени ответов: двигательную реакцию и
выделение своего собственного сАМР. Двигательная ре-
акция заключается в образовании псевдоподий в сторо-
ну сигнала, в результате вся клетка направленно дви-
жется к источнику сигнала. Перемещение продолжается
около 100 с, и за это время клетка проходит приблизи-
4. Движение эукариотических клеток
115
тельно 20 мкм (что примерно соответствует ее удвоенно-
му диаметру). После этого псевдоподии втягиваются и
движение клетки прекращается до тех пор, пока она не
получит новый хемосигнал.
Вторая ограниченная во времени ответная реакция
получила название релейного ответа. Он заключается в
продуцировании и выделении в среду собственного сАМР
через 12—15 с после получения стимула сАМ.Р. При ре-
лейном ответе сигнал усиливается, поскольку длитель-
ность испускания вторичного сАМР-импульса может до-
стигать 60 с. Периодичность испускания сигналов и их
распространение в среде, заполняемой сближающимися
миксамебами, приводят к образованию концентрических
колец из клеток, одновременно сползающихся к центру
агрегации; эту начальную фазу агрегации легко увидеть
в микроскоп.
По мере развития фазы агрегации возникают потоки
клеток, мигрирующих по направлению к центру агрега-
ции; процесс завершается, когда все миксамебы дости-
гают центра, к которому их с самого начала привлекал
аттрактант (см. [54] и [103]).
Такой объект, на котором влияние хемотаксиса на
движение клеток может быть исследовано как генетичес-
кими, так и биохимическими методами, оказывается
чрезвычайно полезным для изучения клеточной подвиж-
ности, обусловленной микрофиламентами. Можнд наде-
яться, что результаты исследования этой и подобных
систем позволят в недалеком будущем выяснить меха-
низмы хемотаксиса и подвижности эукариотических
клеток.
Рекомендуемая литература
Различные аспекты движения клеток с помощью ресничек и
жгутиков рассмотрены в:
Swimming and Flying in Nature (1975) (ed. T. Y. Wu, C. J. Brokaw
and C. Brennen), Plenum Press, New York.
Cilia and Flagella (1975). (Ed. M. Sleigh), Academic Press, London.
Eckert R. (1972). Bioelectric control of ciliary activity, Science, 176,
473—481.
Summers K. (1975). The role of flagellar structures in motility, Bio-
chem. Biophys. Acta, 416, 153—168.
116
4. Движение эукариотических клеток
Holwill М. Е. (1977). Some biophysical aspects of ciliary and fla-
gellar motility, Adv. Microb. Physiol., 16, 1—48.
Подробные обзоры по амебоидному движению можно найти в
следующем сборнике:
Molecules and Cell Movements. A SPG symposium (1975). (Ed.
S. Inoue and R. Stephens), Raven Press, New York.
Этот сборник включает ряд обзоров и многие статьи из уже
упоминавшейся книги
Cell Motility (1976). (Ed. R. Goldman, T. Pollard and J. Rosen-
baum), Cold Spring Harbor Laboratory, Cold Spring Harbor.
Кроме того интересные работы имеются в:
Allen R. D. (1972). Biophysical aspects of pseudopodium formation.
In: The Biology of Amoeba (ed. K. Jeon), Academic Press,
New York.
Allen D. A., Stromger Allen N. (1978). Cytoplasmic streaming in
amoeboid movement, Ann. Rev. Biophys. Bioeng., 7, 469—495.
Читатель, интересующийся хемотаксисом слизистых грибов, найдет
полезную информацию в следующих статьях:
Konijn Т. М. (1974). The chemotactic effect of cyclic AMP and its
analogues in the acrasiae, Antibiot, and Chemother. (Basel), 19,
369—381.
Gerisch G., Malchow D. (1976). Cyclic AMP receptors and the cont-
rol of cell aggregation in Dictyostelium. In: Advances in Cyclic
Nycleotide Research, vol. 7 (ed. P. Greengard and A. Robison),
Raven Press, New York.
Bonner J. T. (1977). Some aspects of chemotaxis using the cellular
slime molds as an example, Mycologia, 69, 443—459.
Newell P. C. (1978). Cellular communication during aggregation of
Dictyostelium, J. Gen. Microbiol., 104, 1—13.
ЛИТЕРАТУРА
1. Abercrombie М., Heaysman J., Pegrum S. (1970). Exp. Cell. Res.,
59, 393—398.
2. Abercrombie M., Heaysman J., Pegrum S. (1970). Exp. Cell. Res.,
60, 437—444.
3. Abram D„ Koffler H. (1964). J. Mol. Biol., 9, 168—185.
4. Adelstein R. S„ Conti M. A. (1975). Nature, 256, 597—598.
5. Adler J. (1975). Ann. Rev. Biochem., 44, 341—356.
6. Adler J., Dahl H. H. (1967). J. Gen. Microbiol., 46, 161—173.
7. Adler J., Templeton B. (1976). J. Gen. Microbiol., 46, 175—184.
8. Adler J., Epstein W. (1974). Proc. Nat. Acad. Sci. USA, 71,
2895—2899.
9. Afzelius B. A. (1976). Science, 193, 317—319.
10. Albertini D. E„ Clark J. I. (1975). Proc. Nat. Acad. Sci. USA. 72,
4976—4980.
11. Albrecht-Buehler A. (1976). In: Cell Motility (ed. R. Goldman,
T. Pollard and J. Rosenbaum), Cold Spring Harbor* Lab.,
pp. 247—264.
12. Allen R. D. (1961). Exp. Cell Res., 8 (suppl.), 17—31.
13. Allen R. D. (1961). In: The Cell (ed. J. Brachet and A. E. Mir-
sky), vol. 2, Academic Press, New York, pp. 135—216.
14. Allen R. D., Stromgren Allen N. (1978). Ann. Rev. Biophys.
Bioeng., 7, 469—495.
15. Aksamit R. R., Koshland D. E. (1974). Biochemistry, 13, 4473—
4478.
16. Amstrong J. B. (1972). Can. J. Microbiol., 18, 1695—1703.
17. Aral T., Kaziro Y. (1977). J. Biochem. (Tokyo), 82, 1063—1071.
18. Barra H. S., Arce C. A., Rodriguez J. A., Caputto R. (1974).
Biochem. Biophys. Res. Comm., 60, 1384—1390.
19. Berg H. C. (1971). Rev. Sci. Instrum.,, 42, 868—871.
20. Berg H. C„ Anderson R. A. (1973). Nature, 245, 380—382.
21. Berry R. W„ Shelansky M. L. (1972). J. Mol. Biol., 71, 71—80.
22. Bhisey A., Freed J. (1971). Exp. Cell Res., 64, 419—423.
23. Bonner J. T, (1947). J. Exp. Zool., 106, 1—26.
24. Borisy G. G., Olmsted J. B., Marcum J. M., Allen C. (1974).
Fed. Proc., 33, 167—174.
25. Boss IF., Gordon A. S., Hall L. E., Price D. (1972). J. Biol.
Chem., 247, 917—924.
26. Brokaw C. J., Gibbons I. K. (1973). J. Cell Sci., 13, 1—18.
27. Bryan J., Wilson C. (1971). Proc. Nat. Acad. Sci. USA, 68,
1762—1766.
118
Литература
28. Bryan R. N., Cutter G. A., Hayashi M. (1978). Nature, 272,
81—83.
29. Cappuccinelli P., Cuccureddu R., Hames B. D. (1977). In: De-
velopment and Differentiation in the Cellular Slime Moulds (ed.
P. Cappuccinelli and J. M. Ashworth), Elsevier/North Holland,
Amsterdam, pp. 231—241.
30. Cappuccinelli P., Martinotti G., Hames. B. D. (1978). FEBS
Letters, 91, 153—157.
31. Chang J. Y„ De Lange R. J., Shaper J. M., Glazer A. N. (1976).
J. Biol. Chem., 251, 695—700.
32. Clarke M„ Spudich J. A. (1977). Ann. Rev. Biochem., 46, 797—
822
33. Dales S. (1972). J. Cell Biol., 52, 748—752.
34. David-Pfeuty T., Laparte J., Pantaloni D. (1978). Nature, 272,
282—284.
35. De Pamphilis M. L., Adler J. (1971). J. Bacterio!., 105, 384—395.
36. De Pamphilis M. L., Adler J. (1971). J. Bacteriol., 105, 396—407.
37. De Rosier D., Mandelkow E., Silliman A., Tilney L., Kane R.
(1977). J. Mol. Biol., 113, 679—695.
38. Dimmit K., Simon M. (1971). J. Bacteriol., 105, 369—375.
39. Dimmit K-, Simon M. (1971). J. Bacteriol., 108, 285—286.
40. De Petris S. (1975). J. Cell BioL, 65, 123—146.
41. Ebashi S., Endo M. (I960). Prog. Biophys. Mol. Biol., 18, 123—
183.
42. Eckert R. (1972). Science, 176, 476—481.
43. Eipper B. A. (1972). Proc. Nat. Acad. Sci. USA, 69, 2283—2287.
44. Emerson S. U., Tokynasu K-, Simon M. I. (1970). Science, 169,
190______192.
45. Felix H.,Strauli P. (1976). Nature, 261, 603—605.
46. Flanagan J., Koch G. L. E. (\2il8j. Nature, 273, 278—281.
47. Frere J. M. (1977). Bull. Inst. Pasteur, 75, 187—203.
48. Fujiwara K., Pollard T. D. (1978). J. Cell Biol., 67, 125a.
49. Fujiwara K-, Pollard T. D. (1978). J. Cell Biol., 77, 188—195.
50. Fulton- C., Simpson P. M. (1976). In: Cell Motility (ed. R. Gold-
mann, T. Pollard and J. Rosenbaum), Cold Spring Harbor Lab.,
pp. 987—1006.
51. Fuller G. M., Brinkley B. R„ Boughter M. J. (1975). Science,
187, 948—950.
52. Gaskin F., Shelanski M. L. (1976). In: Essays in Biochemistry
(ed. P. N. Campbell and W. N. Aldridge), vol. 12, Academic
Press, New York,- pp. 115—146.
53. Gaskin F., Cantor C. R., Shelansky M. L. (1974). J. Mol. Biol.,
89, 737—758.
54. Gerish. G., Malchow D., Hess J. (1974). In: Biochemsitry of
Sensory Functions, Mosbacher Colloquium (ed. L. Jaenicke),
Springer-Verlag, Berlin, pp. 279—298.
55. Gibbons В. H., Gibbons I. R. (1972). J. Cell Biol., 54, 75—97.
56. Gibbons I. R. (1965). Arch. BioL, 76, 317—324.
57. Gibbons I. R., Fronk E., Gibbons B., Ogawa K. (1976). In: Cell
Motility (ed. R. Goldman, T. Pollard and J. Rosenbaum), Cold
Spring Harbor Lab., pp. 905—932.
58. Goldman R. (1971). J. Cell Biol., 51, 752—762.
Литература
119
59. Goldman R., Follet E. (1970). Science, 169, 268—288.
60. Глаголев A. H., Скулачев В. П. (1978). Nature, 272, 380—382.
61. Grasse P. P. (1956). Arch. Biol., 67, 595—609.
62. Grimstone A. V., Cleveland L. R. (1965). J. Cell BioL, 24, 387—•
400.
63. Hazelbauer G. L., Adler J. (1971). Nature New BioL, 230, 101—
104.
64. Herzhog W., Weber K. (1977). Proc. Nat. Acad. Sci. USA, 74,
1860—1864.
65. Hilmer M., Simon M. (1976). In: Cell Motility (ed. R. Goldman,
T. Pollard and J. Rosenbaum), Cold Spring Harbor Lab.,
pp. 35—45.
66. Himes R. H., Burton P. R., Gaito J. M. (1977). J. BioL Chem.,
252 6222______6228.
67. Hitchen E. T„ Butler R. D. (1973). Z. Zellforsch. milrosk, anat.,
144, 59—73.
68. Hitchcock S., Carlsson L., Lindberg U. (1976). In: Cell Motility
(ed. R. Goldman, P. Pollard and J. Rosenbaum), Cold Spring
Harbor Lab., pp. 545—559.
69. Hollande A., Gharagzoglu I. (1967). C. R. Acad. Sci., 265,
1309—1315.
70. Hunter T„ Garrels J. I. (1972). Cell, 12, 767—781.
71. lino T. (1977). Ann. Rev. Genet., 11, 161—182.
72. lino T., Mitani M. (1967). J. Gen. Microbiol., 49, 81—88.
73. Inoue S., Ritter N. (1975). In: Molecules and Cell Movement
(ed. S. Inoue and R. E. Stephens), Raven Press, New York,
pp. 3—30.
74. Jahn H. L., Landman M. D., Fonseca J. R. (1964). J. ProtozooL,
11, 291—303.
75. Jakus M. A., Hall С. E. (1946). BioL Bull., 91, 141—144.
76. Joys T. M„ Rankis V. (1972). J. BioL Chem., 247, 5180—5193.
77. Knight-Jones E. W. (1954). Q. J. Microsc. Sci., 95, 503—521.
78. Kondok H., Yanagida M. (1975) J. Mol. BioL, 96, 641—652.
79. Konijn T. M., Barckley D. S., Chang Y. Y., Bonner J. T. (1968).
Amer. Naturalist, 102, 225, 234.
80. Korn E. D. (1978). Proc. Nat. Acad. Sci. USA, 75, 588—599.
81. Kowitt J. D„ Fulton C. (1974). J. BioL Cheem., 249, 3638—3646.
82. Kowitt J. D., Fulton C. (1974). Proc. Nat. Acad. Sci. USA, 71,
2877—2881.
83. Larsen S. H., Adler J., Gargus J. J., Hokk R. W. (1974). Proc.
Nat. Acad. Sci. USA, 72, 1239—1243.
84. Lazarides E. (1976). In: Cell Motility (ed. R. Goldman, T. Pol-
lard and J. Rosenbaum), Cold Spring Harbor Lab., pp. 347—360.
85. Lazarides E., Lindberg U. (1974). Proc. Nat. Acod. Sci. USA,
71, 4742—4746.
86. Lazarides E„ Weber K. (1974). Proc. Nat. Acad. Sci. USA, 71,
2268—2272.
87. Linck R. W. (1976). J. Cell. Sci., 20, 405—439.
88. Loomis W. F. (1975). Dictyostelium Discoideum: a Developmen-
tal System, Academic Press, New York.
89. Luduena R. F„ Woodward D. O. (1973). Proc. Nat. Acad. Sci.
USA, 70, 3594—3598.
120
Литература
90. MacNab R., Koshland D. E., Jr. (1972). Proc. Nat. Acad. Sci.
USA 69 2509________2512
91. MacNab R., Koshland D. E., Jr. (1974). J. Mol. Biol., 84, 399—
406.
92. MacNab R., Orston H. K. (1977). J. Mol. Biol, 112, 1—30.
93. Manson M. D.. Tedesco P., Berg H. C., Harold F. M. Van der
Drift C. (1977). Proc. Nat. Acad. Sci. USA, 74, 3060—3064.
94. Margulis L., To L., Chase D. (1978). Science, 200, 1118—1124.
95. Maruta H., Korn E. D. (1977). J. Biol. Chem., 252, 8329—8332.
96. Mast S. D. (1925). J. Morphol. Physiol., 41, 347—425.
97. Morgan J. L„ Seeds N. IF. (1975). J. Cell Biol., 67, 136—145.
98. Mooseker M. S. (1976). J. Cell Biol., 71, 417—433.
99. Mooseker M. S„ Tilney L. G. (1973). L. Cell Biol., 56, 13—26.
100. Mooseker M. S„ Tilney L. G (1976). J. Cell. Biol., 67, 724—743.
IQl. Murphy D. B., Borisy G. C. (1975). Proc. Nat. Acad. Sci. USA,
72, 2696—2700.
102. Naitoh У, Eckert R. (1975). In: Cilia and Flagella (ed.
M. A. Sleigh), Academic Press, New York, pp. 305—355.
103. Newell P. C. (1978). J. Gen. Microbiol., 104, 1—13.
104. Nicolson G. L. (1976). Biochem. Biophys. Acta, 457, 57—108.
105. Odell G. M„ Frisch H. L. (1975). J. Theor. Biol, 50, 59—86.
106. Olive E. IP. (1902). Proc. Boston Soc. Nat. Hist, 30, 451—513.
107. Olmsted J. B., Borisy G. G. (1973). Ann. Rev. Biochem, 42,
507—540.
108. Olmsted J. B., Borisy G. G. (1975). Biochemistry, 14, 2996—
3005.
109. Pantin C. F. A. (1923). J. Marine Biol. Ass, 13, 24—69.
110. Pastan I., Perlman R. (1970). Science, 169, 339—342.
111. Pedersen H., Rebbe H. (1975). Biol. Reprod, 12, 541—544.
112. Penningroth S. M., Cleveland D. M., Kirschner M. W. (1976).
In: Cell Motility (ed. R. Goldman, T. Pollard and J. Rosenba-
um), Cold Spring Harbor Lab, pp. 1233—1255.
113. Pickett-Heaps J. D. (1969). Cytobios, 1, 257—280.
114. Piperno G„ Luck D. (1974). J. Cell Biol, 63, 271a.
115. Piperno G., Luck D. J. (1977). Biol. Chem, 252, 383—391.
116. Piperno G., Huang B., Luck D. L. (1977). Proc. Nat. Acad. Sci.
USA, 74, 1600—1604.
117. Pollard T. D„ Korn E. D. (1973). J. Biol. Chem, 248, 448—454.
118. Pollard T. D„ Korn E. D. (1973). J. Biol. Chem, 248, 4682—
4690.
119. Pollard T. D., Korn E. D. (1973). J. Biol. Chem, 248,4691—
4697.
12Q. Pollard T. D„ Porter N. E„ Stafford D. W. (1977). Cell Biol,
75 262a
121. Potts G. (1902). Flora, 91, 281—347.
122. Raybin D„ Flavin M. (1975). J. Cell Biol. 67, 356a.
123. Rinaldi R., Opas M., Hrebenda B. (1975). J. Protozool, 22,
286—292.
124. Robbins E., Shelanski M. L. (1969). J. Cell. Biol, 43, 371—373.
125. Robinson W. G. (1972). J. Cell Biol, 52, 66—83.
126. Runyon E. H. (1942). Collecting Nat, 17, 99—129.
Литература
121
127. Sabatini D., Bensch K-, Barnett R. (1963). J. Cell Biol., 17,
19—58.
128. Satir P. (1965). J. Cell Biol., 26, 805—834.
129. Satir P. (1968). J. Cell Biol., 39, 77—94.
130. Satir P. (1974). Sci. Amer., 231, 44—54.
131. Shaffer В. M. (1956). J. Exp. Biol., 33, 645—657.
132. Sheir-Neiss G„ Nardi R. V., Gealt M. A., Morris N. R. (1976).
Biochem. Biophys. Res. Coinm., 69, 285—290.
133. Shelansky M. L., Gaskin F., Cantor C. R. (1973). Proc. Nat.
Acad. Sci. USA, 70. 765—768.
134. Silverman M., Simon M. (1974). Nature, 249, 73—74.
135. Silverman M., Simon M. (1977). Ann. Rev. Microbiol., 31, 397—
419.
136. Silverman M„ Simon M. (1977). Proc. Nat. Acad. Sci. USA, 74,
3317—3321.
137. Simon M., Silverman M„ Latsumura P., Ridgway H., Kome-
da Y., Hitman M. (1978). In: Relations between Structure and
Function in the Prokaryotic Cell (ed. R. Y. Stayner, H. J. Ro-
gers and B. J. Ward), Cambridge University Press, pp. 272—287.
138. Sleigh M. A. (1972). In: Essay in Hydrobyology (ed.
R. B. Clark and R. Wootons), University of Exeter, pp. 119—136.
139. Sleigh M. A. (ed.) (1974). Celia and Flagella, Academic Press,
London.
140. Sloboda R. D„ Rudolph S. A., Rosenbaum J. L„ Greengard P.
(1975). Proc. Nat. Acad. Sci. USA, 72, 177—181.
141. Sloboda R. D., Dentler IF. L., Bloodgood R. A., Telzer B. R.,
Granett S., Rosenbaum J. L. (1976). In: Cell Motility (ed.
R. Goldman, T. Pollard and J. Rosenbaum), Cold Spring Harbor
Lab., pp. 1171—1212.
142. Snyder J. A., McIntosh J. R. (1976). Ann. Rev. Biochem., 45
699—720.
143. Springer M. J., Goy M. F„ Adler J. (1977). Proc. Nat. Acad.
Sci. USA, 74, 3312—3316.
144. Spudich J. A., Watt S. (1971). J. Biol. Chem., 246, 4866—4871
145. Starger G. M., Goldman R. D. (1977). Proc. Nat. Acad. Sci.
USA 74 2422________2426.
146. Stephens R. E. (1970). Biol. Bull., 139. 438a.
147. Stephens R. E. (1975). J. Cell Biol., 67, 418a.
148. Stephens R. E. (1975). In: Molecules and Cell Movements (ed.
S. Inoud and R. E. Stephens), Raven Press, New York, pp. 181—
206.
148. Stephens R. E„ Edds К. T. (1976). Physiol. Rev., 56, 709—777.
150. Stephens R. E., -Renaud F. L., Gibbons I. L. (1967). Science,
156, 1606—1608.
15LSfor« R. V., Coen D. M„ Rich D. (1976). Cell, 8, 521—527.
152. Summers К. E., Gibbons I. R. (1977). Proc. Nat. Acad. Sci. USA,
68, 3092—3096.
153. Sundquist K. G.. Ehrust A. (1976). Nature, 264, 4866—4871.
154. Taylor D. L. (1976). In: Cell Motility (ed. R. Goldman, T. Po-
lard and J. Rosenbaum), Cold Spring Harbor Lab., pp. 797—821.
155. Taylor B., Koshland D. E„ Jr. (1974). J. Bacteriol., 119, 640—
642.
122
Литература
156. Taylor D. L., Rhodes J. A., Hammond S. A. (1976). J. Cell
Biol., 70, 123—143.
157. Tilney L. G. (1975). In: Molecules and Cell Movements (ed.
S. Inoue and R. E. Stephens), Raven Press, New York, pp. 339—
388.
158. Tilney L. G. (1976). J. Cell Biol., 69, 73—89.
159. Tilney L. G. (1977). In: International Cell Biology (ed.
В. B. Brinkley and K. R. Porter), The Rockefeller University
Press, New York, pp. 388—402.
160. Tilney L. G. (1978). J. Cell Biol., 77, 5051—5064.
161. Tilney L. G., Byers B. (1969). J. Cell Biol., 43, 148—165.
162. Tsang N., MacNab R„ Roshland D. E., Jr. (1973). Science, 181,
60—63.
163. Tucker J. B. (1968). J. Cell Sci., 3, 493—514.
164. Tucker J. B. (1974). J. Cell Biol., 62, 424—437.
165. Tucker J. B. (1977). Nature, 266, 22—26.
166. Васильев И. M„ Гельфанд И. М. (1976). In: Cell Motility (ed.
R. Goldman, T. Pollard and J. Rosenbaum), Cold Spring Harbor
Lab., pp. 279—304.
167. Walters В. B„ Mathus A. I. (1975). Nature, 257, 496—498.
168. Warner F. D„ Satir P. (1974). J. Cell Biol., 63, 35—63.
169. Watters C. (1968). J. Cell Sci., 3, 231—244.
170. Water R. D„ Rleinsmith L. J. (1976). Biochem. Biophys. Res.
Commun., 70, 704—708.
171. Weber R. (1976). In: Cell Motility (ed. R. Goldman, T. Pollard
and J. Rosenbaum), Cold Spring Harbor Lab., pp. 403—417.
172. Weber R., Bibring T., Osborn M. (1975). Proc. Nat. Acad. Sci.
USA, 72, 459—463.
173. Weingarten M. B., Lockweed A. H., Hwo S. Y., Rirschnar M. W.
(1975). Proc. Nat. Acad. Sci. USA, 72, 1858—1862.
174. Weisenberg R. C. (1972). Science, 177, 1104—1105.
175. Wilson L., Bryan J. (1974). Adv. Cell Mol. Biol., 3, 21—72.
176. Wilson L., Creswell M., Chin D. (1976). Biochemistry, 14, 5586—
5592.
177. Witman G. B., Plummer J., Sander G. (1978). J. Cell Biol., 76,
729—747.
178. Yamaguchi S., lino T., Rurdiwa T. (1972). J. Gen. Microbiol.,
70, 299—303.
179. Zukin R. S., Roshland D. E., Jr. (1976). Science, 193, 405—408.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аденозиимонофосфат циклический
(сАМР) 21, 47, 59, 112—114
Адеиозинтрифосфат (АТР) 24, 32,
41, 43, 62, 81—82, 94, 96, 105
Аденозиитрифосфатаза (АТРаза)
К+-зависимая 73
— Mg2+, Са2+-зависимая 32, 73
Аденозиитрифосфатазиая (АТРаз-
ная) активность и динеин 51, 62
-----миозина 73—77, 81, 85
Акросомальная реакция 78—79,
85
Аксонема 38—40, 41, 49—50, 52,
61, 62, 93
Аксоподии 63—65
Аксостиль 42, 60
Актин, актиновые нити (фила-
менты) 67, 69, 71—85, 105,
106
а-Актинин 67, 70, 75, 76, 89
Актиисвязывающий белок 76
Амебоидное движение 46, 92, 104.
См. также Псевдоподии
----механизм 105
----- типы 107
— — управление 111
Базальное тело у бактерий 14—
16, 20, 33
-------эукариот 41, 58, 61
Белки, связанные с микротрубоч-
ками (БСМ) 53, 55—56
-------тубулином 49
Воротничок жгутика 41
Ворсинки (микроворсинки) 69, 83,
92
Гелактины 77
Гуанозинтрифосфат (GTP) 47, 54,
55
Динеин (динеиновые ручки) 50—
51, 62, 94
Жгутики бактерий 10
----движение 22—23
— — жгутиковые гены 19,- 33
----крюк 13, 14
----расположение 11
— — синтез 21
----структура 13, 16
— — филамент 13
---- форма 12
— — химический состав 16, 18
— у эукариот см. Реснички и
124
Предметный указатель
жгутики эукариотических кле
ток
Избегания реакция 102—103
Кальций (Са2+) 75, 83, 84, 102,
104
Крюк 14, 20, 21, 23
Мастигонемы 97—98
Мембрана 10, 14, 15, 24, 31, 33,
41, 80, 81, 86, 93
— деполяризация 102
Микротрубочки 36, 81
— белки, связанные с микротру-
бочками (БСМ) 55—56. См.
также Белки, связанные с тубу-
лином
— в ресничках и жгутиках 38
— животных клеток 43
— и движение 61
------ жизненный цикл клетки 45
— самосборка 59
---in vitro 54
— сократимого аксостиля 42
Микрофиламенты 35, 67
— и движение 80
— • — жизненный цикл клетки
67—69
— сборка 77
— состав 70
— функции 70
Миксомицеты 111
Миозин, миозиновые нити 67, 69,
73—75, 80—89, 105—106
Митотическое веретено 44, 46, 60,
65, 86
Нексин, нексиновые мостики 39,
40, 51, 94, 96
Промежуточные филаменты 58,
65—66
Протонный насос 24
Протофибриллы 37, 38, 41, 56
Профилин 76, 78
Псевдоподии 104, 107—109, ПО
Радиальные спицы 40, 52, 94—95
Реснички и жгутики эукариотичес-
ких клеток 38, 92, 93
---------- координация 99
----------— механизм движе-
ния 94
--------структура 38
----— — — типы движения 96
Саркомер 81
Скольжения модель для микрофи-
ламентов 81
-------ресничек и жгутиков 61
Сперматозоиды 78, 96, 104
Таксис и движение ресничек см.
Реакция избегания
— у бактерий 27
Тамблинг 25;—26, 28
Тау-белок 50, 53, 56
Тропомиозин 67, 75, 82, 84
Тропонин 82, 84
Тубулин 46, 47, 90
— белки, связанные с тубулином
49
— и жизненный цикл клетки 59
— полимеризация 52
125
Филамии 76
Флагеллин 16, 20, 22, 26
филамент в жгутиках бактерий
13, 23, 24
Хеморецепторы 29, 30, 113
Хемотаксис и амебоидное движе-
ние 111
— у бактерий 27—29, 32
Хемотаксические белки 32
Центриоль 41
Центры организации микротрубо-
чек 60
«Ш апка», реакция образования
87—88
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА 5
БЛАГОДАРНОСТЬ 6
1. ВВЕДЕНИЕ 7
2. ПОДВИЖНОСТЬ ПРОКАРИОТИЧЕСКИХ ОРГАНИЗ-
МОВ 10
2.1. Структуры, участвующие в подвижности 11
2.2. Молекулярная структура жгутика у бактерий 16
2.3. Регуляция синтеза и сборки бактериального жгутика 18
2.4 Функция бактериальных жгутиков 22
2.5. Траектории и изменения направления движения у бак-
терий 24
2.6. Управление движением у бактерий 27
3. СИСТЕМЫ ПОДВИЖНОСТИ
ЭУКАРИОТИЧЕСКИХ КЛЕТОК 35
3.1. Микротрубочки 36
3.2. Состав микротрубочек 46
3.3. Самосборка микротрубочек и ее регуляция 54
3.4. Как микротрубочки генерируют движение? 61
3.5. Проблема промежуточных филаментов 65
3.6. Микрофиламенты 67
3.7. Состав микрофиламентов 70
3.8. Сборка микрофиламентов и ее регуляция 77
3.9. Как микрофиламенты генерируют движение? 80
3.10. Регуляция скольжения белками микрофиламентов 82
3.11. Микротрубочки, микрофиламенты и клеточные мем-
браны 86
4. ДВИЖЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК 92
4.1. Движение ресничек и жгутиков 93
4.2. Амебоидное движение 104
ЛИТЕРАТУРА 117
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ 123
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги,
ее оформлении, качестве перевода и
другие просим присылать по адресу:
129820, Москва, И-110, ГСП
1-й Рижский пер,, д. 2,
издательство «Мир»