/



















































































































































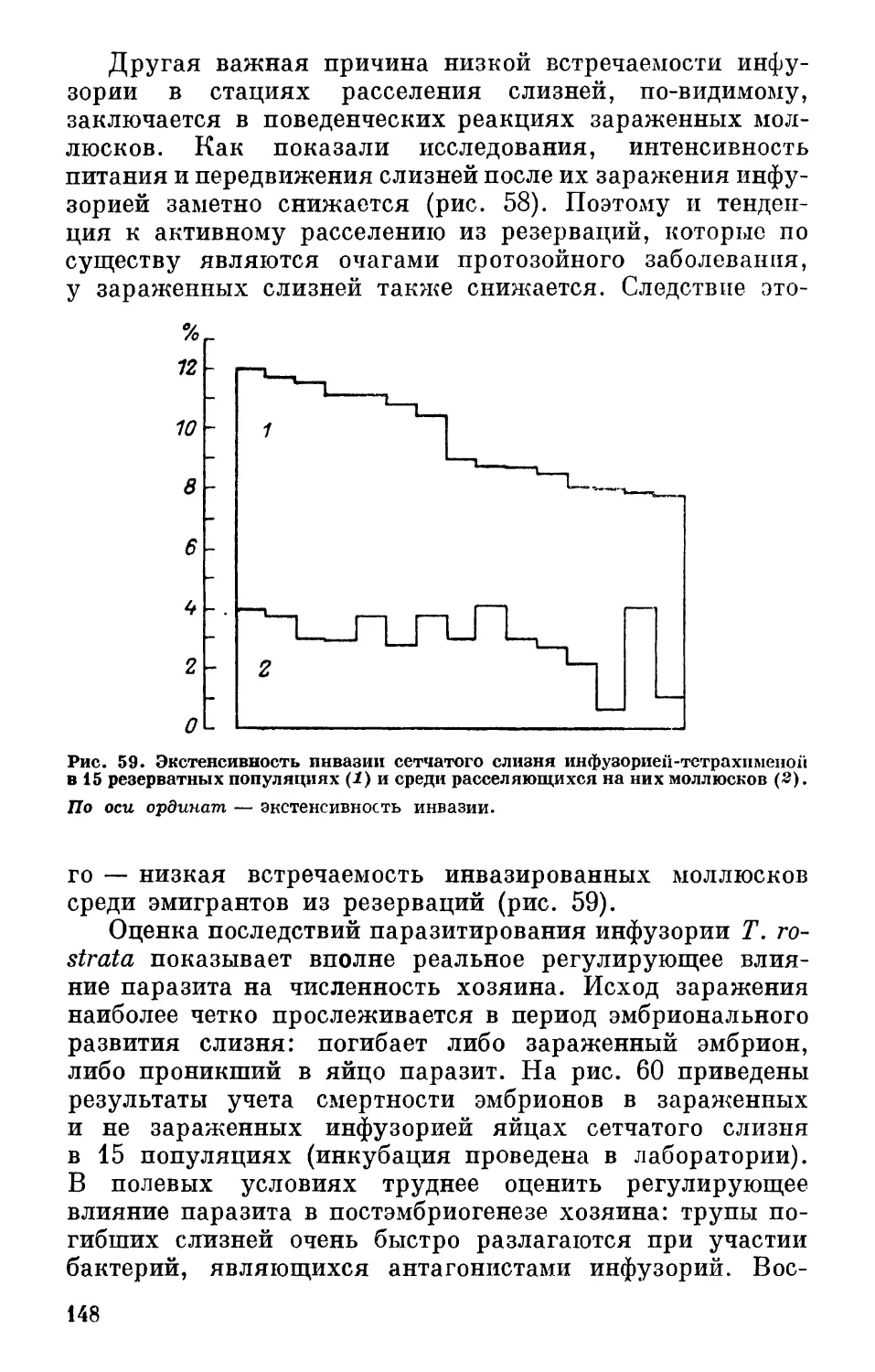











































Author: Лихарев И.М. Шапиро Я.С.
Tags: повреждения растений вредители растений болезни растений защита растений сельское хозяйство вредители
Year: 1987




Text
И.М. ЛИХАРЕВ
Я. С. ШАПИРО
СЛИЗНИ-ВРЕДИТЕЛИ
СЕЛЬСКОГО ХОЗЯЙСТВА
ИЗДАТЕЛЬСТВО-НАУКА-
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
АКАДЕМИЯ НАУК СССР
Серия
«Наука — сельскому хозяйству»
И. М. ЛИХАРЕВ
Я. С. ШАПИРО
СЛИЗНИ—ВРЕДИТЕЛИ
СЕЛЬСКОГО ХОЗЯЙСТВА
НЕЧЕРНОЗЕМНОЙ ЗОНЫ
в
Ленинград
ИЗДАТЕЛЬСТВО «НАУКА»
Ленинградское отделение
1987
УДК 632.64 (470.31)
Лихарев И. М., Шапиро Я.С. Слизни — вредители сельского
хозяйства Нечерноземной зоны.—Л.: Наука, 1987. — 190 с.
Книга посвящена безраковинным моллюскам — слизням, среди
которых многие виды являются серьезными вредителями культур-
ных растений и промежуточными хозяевами ряда гельминтов, опас-
ных для домашних животных. Приведено описание морфологии
и биологии слизней, изложены современные методы их учета,
прогнозирования численности и вредоносности. Даются описание и
определительные таблицы. Рассмотрены методы ограничения чис-
леннрсти — агротехнический, механический, биологический и хи-
мический. Описаны современные препараты для борьбы со слиз-
нями — моллюскоциды, методы учета их эффективности и способы
безопасного применения. Библиогр. 109 назв. Табл. 15. Ил. 60.
Ответственный редактор
Я. И. СТАР'ОБОГАТОВ
Рецензенты
И. Я. Поляков, А. Н. Голиков
На обложке: Слизень сетчатый (см. с. 118).
Л
3803040000-504
054 (02)-87
85-87НП
(£) Издательство «Наука», 1987 г.
ВВЕДЕНИЕ
Обычно первое знакомство со слизнями происходит
в теплую и сырую погоду на поле или огороде, где произ-
растает какая-либо излюбленная ими сельскохозяйствен-
ная культура (например, капуста или земляника) и когда
их вредная деятельность особенно бросается в глаза.
У новичков сразу же возникает ряд вопросов: что это за
животные, откуда они появляются и как защитить от них
урожай. Надо признать, что на эти вопросы не всегда удо-
влетворительно отвечают наши справочники, посвящен-
ные вредителям сельского хозяйства.
Недостаток сведений о слизнях и слабая разработан-
ность мер борьбы с ними объясняются, во-первых, тем,
что видовой состав фауны вредных слизней и основные
особенности экологии и физиологии этих животных в на-
шей стране стали основательно изучаться лишь в послед-
ние 20 лет и эти данные не нашли еще отражения в обще-
принятых руководствах по вредителям сельского хозяй-
ства. Между тем биология этих моллюсков1 резко отли-
чает их от основных групп прочих вредителей растений —
членистоногих, фитогельминтов, грызунов. И поэтому
научно обоснованное регулирование их численности
с целью снижения потерь урожая требует разработки
весьма специфических методов, в полной мере учитыва-
ющих эти отличия. Не случайно потерпели неудачу по-
пытки бороться со слизнями с помощью неорганических,
хлорорганических и фосфорорганических пестицидов,
которые достаточно эффективны против вредных насеко-
мых.
Во-вторых, в организациях службы защиты растений
существует известная недооценка роли слизней как вреди-
телей культурных растений. Отчасти это связано с тем,
что значительную часть убытков от этих моллюсков при-
1 Моллюски являются предметом изучения особого раздела зооло-
гии — малакологии.
3
иисыйают деятельности других вредителей (напрймёр,
листогрызущим гусеницам насекомых).
Слизни принадлежат к обширному классу брюхоно-
гих моллюсков (в мировой фауне в морях, пресных водах
и на суше обитает около 100 000 видов). Одной из глав-
ных особенностей большинства видов этого класса явля-
ется присутствие наружной раковины, в которой распо-
лагается большая часть внутренних органов и в которую,
в случае какой-либо опасности, моллюск полностью втя-
гивает свое мягкое тело. Но как раз именно этой особен-
ности слизни и лишены. В процессе эволюции этих жи-
вотных все внутренние органы погрузились в толщу ноги,
а раковина либо полностью исчезла, либо сильно реду-
цировалась и полностью обросла покровами спины.
Свое название — «слизни» 2 — эти животные получили
из-за сильного развития слизистых желез в покровах их
тела, экскрет которых — слизь — обильно покрывает по-
верхность тела и играет важную роль в жизни этих мол-
люсков.
Большинство наземных слизней — растительноядные
животные (полифаги и микофаги), хотя имеются и хищ-
ные группы. В нашей стране установлено 70 видов слиз-
ней, принадлежащих к 23 родам и 8 семействам (Лиха-
рев, Виктор, 1980). Среди них 23 вида в большей или мень-
шей степени наносят вред широкому кругу сельскохозяй-
ственных культур.
В СССР в условиях открытого грунта вредная деятель-
ность слизней наносит заметный ущерб народному хозяй-
ству в северо-западных, западных и центральных обла-
стях европейской части Нечерноземья, а также в сухих
и влажных субтропиках Южного берега Крыма, Кавказа
и Средней Азии. В сооружениях защищенного грунта
(в теплицах и оранжереях) слизни могут вредить повсе-
местно.
В отечественной литературе первая обстоятельная пуб-
ликация о вредной деятельности слизней датируется
1910 годом и принадлежит бывшему студенту Петровско-
Разумовской академии, а впоследствии — выдающемуся
2 В русской литературе их иногда называют «голые слизни». Это
название неправомочно, так как возникло от неправильного пере-
вода с немецкого слова «Nacktschnecken» — «голые улитки». Ниже
понятие «улптка» применяется к брюхоногим моллюскам с наруж-
ной раковиной, а «слизни» — к лишенным ее животным.
4
советскому ученому Н. И. Вавилову. Позднее весьма об-1
стоятельные исследования по биологии и вредоносности
слизней, их распространению на территории европейской
части СССР провели Гриванов (1935, 1936) и Соколов
(1933, 1935). Они же предложили ряд мер по борьбе
с этими вредителями. В 1954 г. И. М. Лихаревым была опу-
бликована брошюра, посвященная слизням — вредите-
лям сельского хозяйства, которая в доступной для мас-
сового читателя форме обобщала известные в то время све-
дения об этих вредителях и защите от них урожая.
За 30 лет, прошедших с тех пор, как в отечественной,
так и в иностранной литературе появилось много новых
данных о биологии и хозяйственном значении слизней,
об эффективных мерах борьбы с этими моллюсками. Эти
сведения разбросаны в многочисленных и подчас трудно
доступных изданиях. Обобщение и доведение их до широ-
кого круга читателей — цель данной книги.
Как известно, успешная борьба с вредителями должна
быть основана на правильном определении и глубоком
знании биологии и экологии вредящих видов. Именно
поэтому большое место в настоящей работе отведено опи-
санию морфофизиологических особенностей слизней, их
экологии, а также диагностике вредящих и сопутству-
ющих видов этих моллюсков. Эти материалы необходимы
для организации научно обоснованных мероприятий по
борьбе со слизнями на сельскохозяйственных землях.
Разработка и внедрение эффективных мероприятий по
регулированию численности вредных организмов претер-
певает постоянное развитие и совершенствование. Учиты-
вая это, авторы не ограничились констатацией устояв-
шихся и хорошо проверенных методов и приемов, а по-
пытались осветить также и вопросы проблемного харак-
тера, нуждающиеся в настоящее время в решении или
апробировании на практике. Прежде всего это касается
прогнозирования численности и вредоносности слизней
в сельскохозяйственном ландшафте, поиска новых, более
совершенных химических средств защиты от них растений,
разработки наиболее прогрессивного биологического ме-
тода, основанного на использовании в борьбе с вредите-
лями их естественных врагов.
По этой же причине значительное место в книге уде-
лено описанию методов исследования биологии, экологии
и вредоносности слизней, оценки эффективности защитных
мероприятий.
5
Видовой состав, биология и характер вредной деятель-
ности слизней европейского Нечерноземья отличается от
таковых от слизней южных областей, которые в настоя-
щее время изучены хуже. Предлагаемая работа посвящена
слизням европейского Нечерноземья. Если же читателя
заинтересуют более подробные сведения о слизнях, жи-
вущих на территории нашей страны, то он сможет почерп-
нуть их из недавно вышедшей в свет монографии И. М. Ли-
харева и А. Й. Виктора «Слизни фауны СССР и сопредель-
ных стран» (1980).
Незадолго до выхода в свет настоящего издания
Госагропром СССР утвердил новый перечень и регламент
применения пестицидов на период 1990 года. Читателям
книги следует учесть некоторые изменения в регламенте
использования описанных в книге химических препаратов.
ЗНАЧЕНИЕ СЛИЗНЕЙ КАК ВРЕДИТЕЛЕЙ
КУЛЬТУРНЫХ РАСТЕНИЙ
Слизни — это многоядные вредители, повреждаю-
щие около 150 видов и разновидностей культурных расте-
ний — полевых, кормовых, овощных, технических, пло-
дово-ягодных, цветочно-декоративных, выращиваемых
в условиях открытого и защищенного грунта. Основные
культуры, повреждаемые слизнями в центральных, запад-
ных и северо-западных областях европейской части СССР,
приведены в табл. 1.
Обширный диапазон повреждаемых культур и широкое
географическое распространение слизней определяют их
значительную вредоносность. Крупные потери от слизней
песет сельское хозяйство многих стран северной и северо-
западной Европы. Так, в Англии и Уэльсе в 1967 г.
только по картофелю и озимой пшенице убытки исчисля-
лись соответственно в 645 700 и 191 700 фунтов стерлин-
гов (Shotton, 1971). Считается, что ежегодно в этих обла-
стях слизни уничтожают 36 тыс. т картофеля (Runham,
Hunter, 1970).
Нередко большие потери от слизней несет сельское
хозяйство ряда областей СССР. Так, в дождливые 1922—
1923 гг. на территории нынешней Ленинградской области
ими было уничтожено до 20 тыс. га посевов озимой пше-
ницы, а в Новгородской области — до 32 тыс. га (Соко-
лов, 1933). В 1957 г. в Орловской области слизни нанесли
большой ущерб урожаю томатов и капусты (Бодренков,
1963), а в 1962 г. в Ленинградской области —- капусте,
картофелю, землянике и другим культурам (Дмитриева,
1963). В Новгородской области в 1980 г. в ряде хозяйств
потери капусты от слизней достигали 4 т с 1 га, а на юге
Архангельской области поврежденность той же культуры
составляла 35—40 %.
Ущерб, причиняемый слизнями растениям, культиви-
руемым в научных целях, весьма значителен, хотя его
дорой трудно выразить экономически» Примером может
7
Таблица i
Культурные растения, повреждаемые слизнями (оригинальные данные)
X озяйственно-биол о- гические группы и подгруппы купьтур Культуры Органы растений, повреждаемые сапенями
I. Зерновые: хлебные и Пшеница, рожь, Прорастающие се-
крупяные ячмень, овес, гре- мена, молодые ли-
бобовые чиха Горох, бобы, фа- стья и стебли Прорастающие се-
II. Технические: жиромаслич- соль, чина, че- чевица Лен, рапс, рыжик мена, листья и молодые стебли, цветки, незрелые плоды Проростки, моло-
ные эфиромаслич- Кориандр, мята дые листья и стебли Все надземные ор-
ные крахмалонос- Картофель, топи- ганы в молодом возрасте Проросткп, моло-
ные намбур (земля- дые листья и стеб-
наркотические ная груша) Табак, махорка ли, клубни Проросткп, моло-
лекарственные Валериана, подо- дые листья и стеб- ли Все надземные ор-
III. Кормовые: корнеплоды рожник, мак, пижма, ромашка, шалфей Свекла, морковь, ганы в молодом возрасте Проросткп, листья,
однолетние бо- брюква, репа Впка, сераделла корнеплоды Прорастающие се-
бовые травы многолетние Клевер (все виды) мена, листья, мо- лодые стеб л iii цветки, незрелые плоды Все надземные орга-
злаковые травы Люцерна, донник, эспарцет Тимофеевка, костер, ны То же в молодом воз- расте Проростки, молодые листья и стебли
райграсе, овся- ница, полевица, мятлик, лисохвост
Таблица 1 (продолжение)
Хозяйственно-биоло- гические группы и подгруппы культур Культуры Органы растений, повреждаемые слизнями
IV. Овощные: капустные корнеплодные клубнеплодные плодовые зеленные пряновкусовые многолетние V. Ягодные VI. Цветочно-де- коративные: однолетники многолетники Капуста белоко- чанная Капуста цветная, савойская, брюс- сельская, коль- раби Свекла, морковь, петрушка, редис, репа, брюква, сельдерей Картофель Томат, огурец, ка- бачок, патиссон, тыква Салат листовой и кочанный, шпи- нат Мята,укроп, ко- риандр Щавель Спаржа Земляника, клуб- ника Малина, смороди- на, крыжовник Календула, астра, петуния, циния, настурция, лев- кой, табак, го- рошек душистый Аконит, колоколь- чик, флокс, ге- оргин, хризанте- ма, гладиолус, ирис, васили- стник, ландыш, крокус, примула, незабудка Проростки, листья, стебли То же и цветоносные побеги Проростки, листья, корнеплоды Проростки, моло- дые листья и стебли, клубни Прорастающие се- мена, плоды, ре- же — молодые листья, стебли, цветки Все надземные ор- ганы То же в молодом возрасте Все надземные ор- ганы То же и корневище Ягоды, реже — мо- лодые г листья, стебли, цвето- носы и цветки Прикорневая по- росль, реже — ягоды Все надземные ор- ганы То же
9
Таблица 1 (продолжение)
X овяй ственно-биол о- гические группы и подгруппы культур Культуры Органы растений, повреждаемые слизнями
оранжерейные растения VII. Древесно-ку- старниковые VIII. Грибы Калла, гербера, ор- хидея, гвоздика ремонтантная, папоротник Клен, вяз, ясень, осина, тополь, ольха, сирень, жасмин Шампиньоны Все надземные ор- ганы Прорастающие семе- на, молодые ли- стья и побеги Мицелий и плодовые тела
служить уничтожение слизнем крапчатым (Ытах таси-
latus) ценных растений из семейства Орхидных в оранже-
рее Ботанического института им. В. Л. Комарова АН СССР
в 1984 г.
Несмотря на многоядность, у слизней есть ряд излюб-
ленных растений, которые повреждаются ими особенно
часто и сильно. Так, в областях Нечерноземной зоны наи-
более часто от этих вредителей страдают картофель, бело-
кочанная и цветная капуста, салат, различные корне-
плоды, горох, фасоль, томаты, огурцы, земляника, многие
цветочно-декоративные культуры. Особенно ощутимый
вред они причиняют озимой пшенице и ржи.
Повреждения, причиняемые слизнями, очень харак-
терны и легко отличимы от следов деятельности других
сельскохозяйственных вредителей. В листьях они обычно
выгрызают неправильной формы отверстия, оставляя
нетронутыми лишь черешок и крупные жилки. На корне-
плодах, клубнях картофеля, плодах земляники, томатов
и огурцов они выгрызают различной формы и размеров
каверны, обычно расширяющиеся внутрь. У капусты они
повреждают не только поверхностные и приземные листья,
но и выгрызают глубокие углубления в кочане. В зернах
злаков они выгрызают как зародыш, так и эндосперм
(рис. 1).
Особенно часто слизни повреждают всходы многих
культурных растений. Опасность таких повреждений за-
ключается в том, что поврежденные растения, как правило,
погибают. Проростки могут быть погрызены слизнями
еще до их появления на поверхности почвы, что наблюда-
10
Рис. 1. Повреждение различных растений слизнями.
(7 — морковь, б — клевер, в — капуста, г — земляника, д — проросток р;кп.
ется обычно при неглубокой заделке семян или посеве
их в рыхлую почву без последующего прикатывания
грунта. При этом слизни проникают к набухающим семе-
нам и могут их повредить еще до прорастания. Именно
поэтому от слизней серьезно страдают молодые всходы ози-
мых, которые нередко после нападения моллюсков уже
не могут оправиться и гибнут. Даже относительно круп-
ные всходы не всегда восстанавливаются после таких
повреждений. Уязвимые фазы развития озимых (прораста-
ние семян, всходы) приходятся на конец лета—начало
осени, когда слизни особенно многочисленны и прожор-
ливы. Нередко поэтому поврежденные моллюсками участ-
ки озимей приходится пересевать или занимать под
другие культуры.
Столь же опасны повреждения, причиняемые слизнями
прорастающим семенам и всходам овощных культур, осо-
бенно при безрассадном способе их выращивания. При
высокой численности вредителей их деятельность может
привести к полной гибели растений также и в условиях
рассадной культуры. Это наблюдается при раннем появ-
лении слизней на полях и благоприятных для них погод-
ных условиях в весенне-летний период.
Опасны также и повреждения слизнями сочных частей
растений — ягод, плодов, клубней, корнеплодов, коча-
нов, что наблюдается как в период вегетации растений,
так и во время их уборки и хранения. Как правило, это
приводит к загниванию сочных органов растений. Отчасти
это связано с разрушением покровных тканей, предохра-
няющих от проникновения вглубь возбудителей бактери-
альных и грибных гнилей. Кроме того, экспериментально
доказано участие слизней в переносе фитопатогенных ми-
кроорганизмов — грибов, бактерий, вирусов, вызываю-
щих серьезные болезни культурных растений. Слизни —
переносчики таких патогенных организмов, как грибки
Botrytis cinerea Pers., возбудители ложной мучнистой
росы капусты (Peronospora brassicae Сайт.) и бобов (Р. vi-
ciae Gum.), Х-, S-, М-вирусы картофеля (Runham, Hun-
ter, 1970; Шапиро, 1978).
Участие слизней в распространении инфекционных
болезней растений связано также и с тем, что эти моллю-
ски весьма охотно питаются растительной тканью, пора-
женной патогенными грибами — возбудителями гнилей
и плесеней (Botrytis, Mucor, Rhizopus, Aspergillus, Pent-
cillium и др.), а также бактериями. Кроме того, отмечено
12
питание слизней непосредственно мицелием, плодовыми
телами и спорами паразитических грибков. Перенос же
инфекции от растения к растению происходит обычно
чисто механически — в результате попадания инфекцион-
ного начала на слизистые покровы моллюска.
Нередко растения, поврежденные слизнями даже в сла-
бой степени, впоследствии теряют товарные качества и ста-
новятся непригодными для реализации или закладки
на хранение. Этому способствует также и загрязнение
растений слизью и экскрементами моллюсков.
Слизни как вредители культурных растений имеют
важное значение не только в условиях крупного сельско-
хозяйственного производства (т. е. в совхозах и колхо-
зах'), но также на небольших по площади опытных полях,
в теплицах сельскохозяйственных станций и других уч-
реждений, на приусадебных участках и в коллективных
садах.
В последние годы в связи с расширением производства
сельскохозяйственных продуктов в хозяйствах частного
сектора заметно возросло отрицательное значение слиз-
ней. С одной стороны, это связано с тем, что здесь выращи-
ваются в основном овощные и ягодные культуры, боль-
шинство которых сильно повреждается слизнями. Так,
например, около 80 % ягод земляники, потребляемых
жителями Ленинграда и Ленинградской области, выра-
щивается в хозяйствах частного сектора, а между тем
эта культура повреждается моллюсками особенно ин-
тенсивно. С другой стороны, на индивидуальных участ-
ках создаются более благоприятные для вредителей
микроклиматические и кормовые условия в связи с неболь-
шим размером таких участков, часто слабой их мелиори-
рованностыо, большой пестротой растительного покрова
и высокой засоренностью сорняками. Наконец, в условиях
высокой раздробленности земельного массива трудно бы-
вает организовать эффективную борьбу со слизнями.
Поэтому нередко поселения слизней на приусадебных
участках и в коллективных садах отличаются высокой
численностью и стабильностью. Почти ежегодно моллю-
ски причиняют здесь серьезный ущерб выращиваемым
растениям.
Разные виды слизней приурочены к вполне определен-
ным биотопам, и поэтому их вредная деятельность имеет
свои особенности. В Нечерноземной зоне и в Прибалтике
первостепенное значение имеют слизни из семейства агрио-
13
лимацид (Agriolimacidae). Слизни-арионы (Arionidae) оби-
тают обычно в садах и огородах и редко повреждают поле-
вые культуры. Вредная деятельность слизней из семей-
ства лимацид (Limacidae) проявляется чаще в сооружениях
закрытого грунта и в хранилищах. Хозяйственное значе-
ние отдельных видов рассматривается далее при их опи-
сании.
ОСНОВНЫЕ ОСОБЕННОСТИ МОРФОЛОГИИ
И ФИЗИОЛОГИИ СЛИЗНЕЙ
Внешний вид
Тело слизней сильно вытянуто в длину, так что при
рассматривании сверху боковые контуры его кажутся
почти параллельными. Впереди, к голове, линии тела
плавно и закругленно смыкаются, сзади — сходятся под
более или менее острым углом (у большинства) или тоже
плавно и закругленно смыкаются (все виды рода арион).
В целом тело слизней, особенно во время ползания, имеет
цилиндрическую или слабо веретеновидную форму, упло-
щенную снизу, т. е. со стороны подошвы. При сокращении
они приобретают более вздутый и массивный веретеновид-
ный или булавовидный облик, за счет сжатия в длину
и втягивания головы и шеи внутрь тела.
Благодаря отсутствию спирально закрученной наруж-
ной раковины тело слизней, в отличие от улиток, было бы
внешне строго симметричным, если бы не положение на
правой стороне дыхательного отверстия легкого (пневмо-
стом) и слабо заметных выходных отверстий: кишечника
(анус), мочеточника и полового. Тело состоит из следую-
щих основных отделов (рис. 2): голова, шея, мантия,
спина и нога с подошвой. Участки верхней поверхности
ниже мантии называются боками.
Среди слизней встречаются как «гиганты», длина кото-
рых при движении превышает 20 см (слизень черный —
Limax cinereoniger), так и «карлики» — не более 2.5 см
(слизень гладкий — Deroceras laeve).
Голова у слизней построена по тому же плану, что
и у родственных им наземных легочных улиток (рис. 3).
Впереди, в центре «лицевой» поверхности, располагается
14
рот с двумя ротовыми лопастями по сторонам и складча-
тыми верхней и нижней губами. Снизу, между головой
Рис. 2. Основные отделы тела слизня.
1 — голова, 2 — шея, 3 — мантия, 4 — спина, 5 — нога, 6 — пневмостом,
7 — бока. Прерывистой линией обозначено положение внутренней раковины
под мантией.
и передним концом подошвы, имеется широкая щель,
в которую открывается наружу ножная железа. Несколько
1 — глазное щупальце, 2 — рот, 3 — губное щупальце, 4 — ротовая лопасть,
5 — выходное отверстие ножной железы, 6 — половое отверстие, 7 — боковые
доли подошвы, 8 — срединная доля подошвы, 9 — нога, 10 — первая коль-
цевая борозда, 11 — оторочка, 12 — вторая кольцевая борозда, 13 — боковая
борозда, 14 — капюшон мантии, 15 — затылочные борозды.
отступя от ротовых лопастей, на голове располагается
пара небольших губных щупалец. Выше и немного по-
15
зади губных находится пара более крупных глазных щу-
палец.
Сзади голова плавно переходит в шею — участок тела
между головой и мантией. На правой стороне шеи, чуть
выше ноги находится половое отверстие. У большинства
видов слизней оно располагается немного позади правых
щупалец, но у видов рода арион оно сильно смещено назад
к дыхательному отверстию.
Мантия имеет вид плоского утолщения на верхней сто-
роне тела. В старой литературе ее нередко называют щит-
ком. Чаще всего она удлиненно-овальная, реже более
или менее круглая. Вблизи от правого края мантия про-
низана дыхательным отверстием (пневмостом), которое
с краем мантии связано глубокой вырезкой или мантий-
ной щелью. Сюда открываются наружу задняя кишка —
анусом, и мочеточник — выделительной порой.
Задняя половина мантии плотно прирастает к спине,
передняя же образует широкую свободную (т. е. не при-
росшую к телу) лопасть — капюшон. Он играет защитную
роль, так как при сокращении животного закрывает шею
и голову слизня. Кроме того, периодически втягивая шею
под капюшон, слизень очищает с помощью слизи эту часть
тела от пыли и других раздражающих частиц, в частности
от пестицидов.
Участок верхней стороны тела, лежащий позади ман-
тии, называется спиной. К заднему концу она снижа-
ется и сужается. У многих видов вдоль средней линии
спины тянется киль. У одних он имеет вид четкой муску-
листой складки, простирающейся от мантии к заднему
концу тела, а у других — это легкая угловатость, порой
заметная только у заднего конца тела. У видов рода арион
всякие следы киля отсутствуют, и спина в поперечном се-
чении закруглена. Другой характерной особенностью
арионов является присутствие на заднем конце тела желе-
зистого органа — хвостовой ямки, лежащей над ногой
и прикрытой треугольной лопастью (рис. 4). Перед копу-
ляцией хвостовая ямка набухает, выделяет специфиче-
скую слизь, сигнализирующую партнеру о готовности
к спариванию.
Нога у слизней располагается под внутренностями
и является мускулистым утолщением нижней стенки тела.
Внешне от остального тела она отграничена кольцевой
бороздой. Нижнюю поверхность ноги называют подош-
вой. В целом нога повторяет общие очертания тела.
16
У большинства видов при рассматривании сверху нога
почти не выступает за боковые контуры спины и шеи,
но у арионов она заметно шире верхней стороны тела.
Как сверху, так и снизу покровы слизней обладают
своеобразным рельефом, слагающимся из различной формы
возвышений — морщин и углублений — борозд. Рельеф
верхней стороны (кроме мантии) образован сетью бороз-
док, между которыми возвышаются ряды морщин. Основу
згой сети составляют бороздки, довольно регулярно ради-
Рис. 5. Рельеф по-
дошвы слизней.
1 — продольные бо-
розды, 2 — попереч-
ные бороздки.
ально расходящиеся от мантии к ноге. Число их более
или менее постоянно для каждого рода, а иногда и для
определенного вида. На шее наиболее характерны две
пары бороздок. Одна пара — затылочные бороздки, кото-
рые простираются параллельно посередине шеи. Другая
пара — боковые бороздки, идущие наклонно от мантии
до губных щупалец. Как уже упоминалось, над ногой
по всему периметру тела, с перерывом только у головы,
располагается четкая кольцевая борозда. У многих видов
над ней бывает вторая кольцевая борозда, но менее чет-
кая, чем первая.
Между радиальными и продольными бороздами распо-
лагаются короткие и неглубокие бороздки; эти борозды
и бороздки создают общую сеть. В ячеях сети располага-
ются возвышения — морщины — гексагональной или по-
лигональной формы. При жизни этот рельеф обычно
2 И, М, Лихарев и Я. С. Шапиро 17
менее четок, чем после фиксации, когда в связи с общим
сокращением тела он выражен сильнее.
Поверхность мантии имеет свой рельеф. У одних слиз-
ней (большинство наших видов) при жизни по пей про-
бегают волны концентрических сокращений в виде мор-
щин. После фиксации они исчезают и мантия становится
гладкой. У других (виды рода арион) — при жизни на по-
верхности мантии возникают и исчезают (пробегают) мно-
гочисленные сосочки, которые после фиксации обычно
исчезают совсем, и на их месте остается зернистый рельеф.
В отличие от верхней стороны тела подошва имеет бо-
лее простой рельеф (рис. 5) или она гладкая (виды рода
арион). У остальных слизней рельеф подошвы слагается
из двух глубоких продольных борозд, разделяющих по-
верхность подошвы на три доли: срединную и две боко-
вые. После фиксации в результате сокращения тела на
подошве появляются многочисленные поперечные бороздки.
В жизни слизней рельеф верхней стороны тела играет
важную роль. Благодаря морщинам увеличивается по-
верхность тела, что особенно важно для этих животных,
у которых значительную роль играет дыхание через кожу,
поэтому чем крупнее слизни, тем крупнее и резче рельеф
их покровов. Кроме того, бороздки служат путями, по
которым стекает слизь, смачивающая поверхность тела.
В отличие от наземных улиток, поверхность тела ко-
торых обычно окрашена в черный, темно- или сЬетло-серые
тона без всяких следов рисунка, окраска слизней более
разнообразна, как по цветовой гамме, так и нередко за
счет темного рисунка на светлом фоне. Обычно молодые
слизни окрашены светлее взрослых.
Окраска создается различными пигментами, которые
либо концентрируются в особых клетках кожи — хро-
матофорах, или свободно лежат в виде гранул между клет-
ками покровов. Наиболее распространены черный, крас-
ный и желтый пигменты. Различные сочетания их и глу-
бина залегания в толще покровов могут создавать много-
численные вариации фоновой окраски и рисунка.
Наиболее обычные типы окраски — черная, темно-се-
рая, коричневая и серовато-желтая. Рисунок может сла-
гаться из пятен: крупные или мелкие пятна, пятна-
брызги, пятна-точки. Мелкие пятна могут сливаться
в более крупные, нередко с неправильными контурами.
Слияние пятен может привести к образованию парных
(1 или 2 пары) цветных полос вдоль спины (особенно вдоль
18
киля), а на мантии — вдоль ее боковых краев и посере-
дине.
Подошва обычно окрашена значительно светлее верх-
ней стороны тела, или она лишена пигмента, белая. Сле-
дует учесть, что окраска слизней сильно изменчива и за-
висит от возраста, образа жизни и местообитания.
Внутреннее строение
Нередко близкородственные виды слизней (в частно-
сти, родов арион и дероцерас) внешне выглядят очень одно-
образно. Поэтому для точного определения видовой при-
надлежности таких слизней требуется учитывать некото-
рые особенности их внутреннего строения, в первую оче-
редь план организации кишечника и половой системы.
В этих случаях необходимо вскрывать этих животных,
чтобы разобраться в основных особенностях их анатомии.
Вскрытие желательно проводить на материале, свеже-
фиксированном 70 %-ным спиртом и предварительно про-
шедшим анестезию (Лихарев, Виктор, 1980). После фикса-
ции слизней тщательно промывают водой до полного уда-
ления слизи с поверхности тела.
Наиболее удобно начинать вскрытие, взяв слизня
в руки и нанося разрезы препаровальными ножницами
или ланцетом. При работе с мелкими экземплярами лучше
проводить вскрытие под бинокуляром. Обычно слизней
вскрывают с левой стороны (рис. 6), начиная разрез стенки
тола от головы выше кольцевой борозды и немного не до-
водя его до заднего конца тела. Затем делают два попереч-
ных надреза: один позади глазных щупалец, другой —
через задний конец на правую сторону. Все надрезы нужно
делать, оттягивая кожу кверху, чтобы не задеть внутрен-
ние органы.
Дальнейшее вскрытие проводят в неглубокой препаро-
вальной ванночке, дно которой залито смесью парафина
с воском и черным сапожным кремом. Слизня прикреп-
ляют ко дну ванночки с помощью булавок, вкалывая
их в ногу. Пинцетом отгибают направо верхнюю стенку
тела животного, попутно подрезая тонкие пленки и тяжи,
связывающие внутренности со стенкой тела, а также мел-
кие нервы и сосуды. Отогнутый участок стенки тела за-
крепляют с помощью булавок. Следует принять меры,
чтобы не оборвать заднюю кишку у места ее входа в ле-
гочную полость и аорту. При невозможности сохранить
2*
19
эти части в неповрежденном виде их надо аккуратно пере-
резать, чтобы были видны оба конца этих органов. Затем
закрепленного слизня заливают 70 %-ным спиртом так,
чтобы он был полностью погружен в жидкость.
Открывшаяся картина естественного положения внут-
ренних органов уже достаточно информативна (рис. 7).
В первую очередь бросается в глаза крупная буроватая
мелкозернистая печень, большая часть которой распола-
гается в задней половине тела, а меньшая — впереди.
Рис. 6. Основные разрезы для вскрытия слизней.
Печень в значительной степени закрывает многие другие
органы.
Видимые участки пищеварительного тракта, кроме бе-
лой и мускулистой глотки, лежащей в голове, отличаются
тонкими стенками, сквозь которые просвечивает жидкое
содержимое желтоватого, зеленоватого или красноватого
цвета (в зависимости от характера пищи). В передней
половине тела располагается большинство половых орга-
нов, отличающихся от печени и кишки окраской и тол-
стыми железистыми (белковая железа, семяяйцевод и др.)
или мускулистыми (пенис) стенками. Гонада не всегда
видна снаружи, так как может быть закрыта печенью
и кишкой. Обычно хорошо видны, особенно снизу, участки
белковой железы.
На внутренней стороне отвернутого участка кожи
сквозь нижнюю стенку легкого просвечивают органы ман-
тийного комплекса. Видны сердце (особенно его желудо-
чек), легочная сеть сосудов и желтоватая почка. Позади
мантийного комплекса крепится основание мощного му-
20
скула — переднего ретрактора тела, который впереди
дает ветви к глотке (не видны) и к щупальцам (частично
п — аорта, г — гонада, гл — глотка, же — желудочек сердца, з — зоб, зк —
задняя кишка, п — пенис, печ_— печень, рщ — ретрактор левого глазного
щупальца, ск — слепая кишка,” сод — семяяйцсвод, стт — верхняя стенка
тела, цне — мозг.
видны). У видов, кишечник которых построен по двухпет-
левому типу, основание ретрактора располагается справа
от задней кцшки, а у видов с трехцетдевцм кишечником
21
кишка огибает мускул спереди, затем тянется назад
и вновь поворачивает вперед к мантийному комплексу,
углубляясь внутрь легочной полости. Эта третья петля
всегда хорошо видна, так как она полностью лежит на
верхней стороне внутренностного мешка, непосредственно
под верхней стенкой тела. Иногда при отворачивании этой
стенки кверху она отделяется от остальных органов и оста-
ется при отвернутом участке покровов.
Дальнейшее исследование отдельных органов следует
вести путем освобождения их от различных связок и пле-
нок, мелких нервов и кровеносных сосудов. Для знаком-
ства со строением гениталий надо надрезать участок кожи
вокруг полового отверстия, перерезать ретрактор правого
глазного щупальца и оттянуть пенис и переднюю часть
женских путей (вместе с семяприемником) несколько
в сторону.
Строение мантийного комплекса органов имеет немало-
важное значение. Чтобы лучше его разглядеть, следует
подрезать нижнюю стенку легкого и отвернуть ее на-
право. Если подрезать мантию по ее заднему краю, то
откроется доступ в полость, в которой лежит рудиментар-
ная раковина.
После изучения внутреннего строения слизня можно
поместить все отпрепарированные органы на свое место
и возвратить верхнюю стенку в исходное положение.
В таком виде вскрытый слизень сохраняется в спирту.
Покровы. Кожа слизней снаружи покрыта одно-
слойным цилиндрическим эпителием, местами несущим
микроскопичёские респички. Изнутри к эпителию приле-
гают слои подкожной мускулатуры и соединительной
ткани. В отличие от улиток покровы слизней особенно
богаты слизистыми железами, которые вместе с ножной
железой обильно выделяют слизь. Практически вся по-
верхность тела слизней продуцирует слизь, но некото-
рые участки — мантия, район дыхательного отверстия,
голова, кольцевые борозды — в этом отношении играют
особенно заметную роль.
Слизь представляет собой смесь различного типа секре-
ций. Основную долю в ней занимает вода — более 90 %.
В сухом остатке слизи 77 % составляют белки, 15 —
углеводы и 8 % — неорганические вещества.
Слизь играет очень важную роль в жизни слизней.
Основное ее назначение — увлажнять покровы, что осо-
бенно важно для поддержания кожи в таком состоянии,
22
чтобы животное^ -могло дышать через нее й одновременно
впитывапЛкапельно-жидкую воду. 'Кроме того, в условиях
перегрева и низкой влажности воздуха выделение слизи
и связанное с этим испарение воды способствуют времен-
ному понижению температуры тела. Так, например, пока-
зано, что некоторые виды арионов способны при темпера-
туре 33.7 °C и влажности воздуха 24 % поддерживать
некоторое время температуру тела на уровне 21 °C, т. е.
значительно ниже верхней летальной границы.
Известно, что в жаркое время дня и в засушливое лето
многие слизни прячутся в различные укрытия (под камни,
Рис. 8. Схема системы ретракторов тела слизня (правые ретракторы глотки и
щупалец не изображены).
гл — левый глоточный, пр — половой ретрактор, щ — левый щупальцевый.
в трещины почвы и т. п.), где окружают себя чехлом из
вязкой слизи и пережидают неблагоприятное время.
Очень важную роль слизь играет при движении. Вяз-
кий секрет ножной железы, выделяясь у переднего края
подошвы, предохраняет ее от трения, создавая «дорожку»,
по которой движется животное. В свою очередь, жидкая
слизь, выделяемая покровами подошвы, способствует
скольжению животного по «дорожке».
Помимо указанных главных функций, слизь имеет
и ряд дополнительных. Стекая по поверхности тела, она
смывает посторонние твердые частицы, нейтрализует раз-
личные раздражающие вещества, смывает споры болезне-
творных грибков и микробов. У многих слизь служит
успешной защитой от хищников, как в силу своих ядови-
тых свойств (у некоторых видов), так и из-за вязкости
ее, затрудняющей действие нападающего.
Мускулатура. Слагается из кожно-мускульного
мешка и системы ретракторов, втягивающих голову и шею
внутрь тела. Различают ретракторы глотки, щупалец
и особый половой ретрактор (рис. 8).
Раковина. Одно из главных отличий слизпей от
улиток — это отсутствие на поверхности тела раковины.
Рудимент ее лежит в особой полости в толще мантии.
У слизней из рода ариоп от раковины сохранилось лишь
несколько известковых зерен.
Чаще всего раковина имеет вид округло-прямоуголь-
ной или неправильно-овальной пластинки (см. рис. 37),
состоящей из двух частей: небольшой гладкой эмбриональ-
ной и более крупной дефинитивной. Последняя сверху
имеет более или менее заметную концентрическую исчер-
ченность — линии нарастания.
Легкое и мантийный комплекс ор-
ганов. Под приросшей к спине задней частью мантии
располагается легкое — относительно небольшая полость,
служащая для дыхания и открывающаяся наружу дыха-
тельным отверстием — пневмостомом. Гладкая и муску-
листая нижняя стопка легкого называется диафрагмой.
Поочередное сокращение и расслабление диафрагмы, из-
меняя объем легочной полости, приводит к засасыванию
или выталкиванию воздуха через ппевмостом, который,
в свою очередь, попеременно то раскрывается, то замы-
кается.
Большая часть верхней стенки (потолка) легкого по-
крыта сложной системой крупных и мелких, переплета-
ющихся между собой складок. В толще их залегает сеть
кровеносных сосудов, в которых и происходит газообмен,
т. е. насыщение крови кислородом. Установлено (Run-
ham, Hunter, 1970), что у легочных улиток легкое обеспе-
чивает около одной трети общей потребности в кислороде.
Остальное покрывается за счет кожного дыхания. Так
как дыхательная поверхность легкого слизней относи-
тельно меньше, чем у улиток, кожное дыхание у них
играет еще более значительную роль.
Кровеносная система. Как и у всех брюхо-
ногих моллюсков, кровеносная система незамкнутого
типа. Она слагается из трех отделов: сердце и артериаль-
ная сеть сосудов; система полостей между органами, на-
зываемая гемоцелем, состоящая из крупных полостей —
синусов и мелких — лакун; венозная сеть сосудов.
Артериальная сеть начинается короткой аортой, отхо-
дящей от желудочка (рис. 9). Аорта распадается сначала
на крупные, а затем на более мелкие артерии, заканчива-
ющиеся очень тонкими сосудами, которые иногда назы-
вают капиллярами. Стенки артериальных сосудов снаб-
24
жены кольцевой мускулатурой, которая помогает сердцу
проталкивать кровь в органы.
В отличие от артериальной сети полости гемоцеля ли-
шены собственных стенок. То же касается большинства
венозных сосудов, за исключением сосудов легкого и ман-
тии, где стенки очень тонкие и лишены мускулатуры.
У слизней сердце пульсирует более интенсивно, чем
у улиток. У крупных видов частота биения сердца ниже,
чем у мелких. С возрастом сердце тоже пульсирует мед-
леннее. Кроме сердца и мускульной выстилки артерий,
в кровообращении слизней большую роль играют движе-
ния диафрагмы, глотки и местные сокращения мускула-
туры стенок тела (Duval А., 1982).
Дыхательную функцию крови выполняет растворен-
ный в ней белок гемоцианин. Благодаря ему насыщенная
кислородом кровь сердца и артерий окрашена в синий
цвет, а отработанная венозная кровь — бесцветная.
Кровь слизней (гемолимфа) занимает значительную
часть массы тела — около 40 % (Martin et al., 1958).
Это объясняется не только теми важными и разносторон-
ними функциями, которые играет кровь в обмене веществ,
но и еще двумя особенностями, специфическими только
для слизней. Во-первых, в гемолимфе сохраняется резерв
свободной воды, необходимой для поддержания водного
баланса для активной жизнедеятельности. Во-вторых,
гемолимфа и гемоцель обеспечивают нормальную форму
животного и расположение органов внутри тела, т. е. вы-
полняют опорную функцию (гидроскелет). В частности,
приток и отток крови способствуют прониканию слизней
в узкие убежища, а также втягиванию и выворачиванию
половых органов при копуляции и всего переднего конца
тела.
Выделительная система. Слагается из
почки и мочеточника. Последний нередко открывается
наружу через мочевой пузырь. Почка лежит на верхней
стенке (на «потолке») легочной полости вблизи от ее зад-
него края. Мочеточник и мочевой пузырь располагаются
рядом с прямой кишкой и открываются наружу вблизи
пневмостома.
Физиологические и биохимические аспекты процессов
выделения у слизней изучены слабее, чем у других мол-
люсков. Наиболее точные исследования проведены на сет-
чатом слизне (Runham, Hunter, 1970). В моче этого живот-
ного азот представлен главным образом пуринами,
?5
гла
Рис. 9. Артериальная система кровообращения слизня сетчатого.
а — аорта (главный ствол), аз — задняя аорта, ап — передняя аорта, асж —
артерия слюнных желез, асп — артерия семяприемника, бж—белковая
железа, г — гонада, газ — радияя артерия гениталий, гап — передняя арте-
рия гениталий, гл — глотка, гла — артерия глотки, з — зоб, зап — задняя
артерия печени, к — кишка, мп — мочевой пузырь, на — артерия ноги, п —
печень, па — артерия пениса, пап\— передняя артерия печени, пч — почка,
с — сердце, сж — слюнные железы, сод — семяяйцевод, цнс — мозг.
& именно мочевой кислотой и ксантином (около 92 %).
Как и у других наземных моллюсков (подобно насекомым
и птицам), у слизней в мочеточнике происходит извлечение
воды из жидкой мочи и продукты выделения выходят
наружу в виде «сухой мочи». Тем самым достигается (еще
одним путем) экономия водных ресурсов — важнейшая
проблема у всех наземных моллюсков.
Пищеварительная система. Подобно
большинству брюхоногих моллюсков у слизней пищевари-
тельный тракт имеет вид более или менее спирально за-
Рис. 10. Основные типы челюсти растительноядных слизней.
I — у арионов, II — у остальных видов.
крученной петли. Впереди он начинается ртом, ведущим
в ротовую или предглоточную полость; за нею следует
глотка, в которую открывается пара слюнных желез;
от глотки тянется назад узкий пищевод, переходящий
в зоб и желудок; в последний открываются протоки двух
долей печени; за желудком начинается кишка, которая,
образуя несколько петель и извивов, открывается наружу
заднепроходным отверстием или анусом, лежащим рядом
с пневмостомом.
На границе ротовой полости с глоткой располагается
хитиноидная челюсть, имеющая форму дуги с острым внеш-
ним краем. У видов рода арион снаружи она покрыта
поперечными ребрышками, а у остальных растительнояд-
ных слизней она гладкая с тупым выступом на середине
внешнего края (рис. 10).
Глотка представляет собой крупный орган с мускули-
стыми стенками. Системой коротких мускулов она связана
с передней и нижней стенками тела. Кроме того, два длин-
ных ретрактора соединяют ее с основной системой ретрак-
торов тела.
В полость глотки снизу выступает крупный язык, по-
крытый сверху роговой мембраной, на которой распола-
гаются многочисленные ряды мелких хитиноидных зубов.
27
Эгп мпмГ||нин1 и uyoi.i и совокупности образуют терку или
радулу (рос. II). У растительноядных слизней радула
(•лужиг для размельчения пищевого объекта путем скоб-
ления но его поверхности и подачи частиц пищи внутрь
глотки.
Короткий и узкий пищевод соединяет глотку с длин-
ным и вздутым зобом. Здесь пищевые частицы подверга-
ются дальнейшему размельчению путем перетирания друг
о друга и обрабатываются многочисленными энзимами.
Сзади к зобу прилегает сравнительно' небольшой желудок,
в который двумя притоками впадает крут»пая печень.
Рис. 11. Фрагмент радулы приона бурого ?(х 500, фот. Юнгблута).
У наземных моллюсков печень выполняет несколько
функций. Как и в зобе, здесь продолжается переварива-
ние частиц пищи путем обработки их энзимами и всасыва-
ние стенками специальных клеток. Кроме того, в печени
происходит накопление запасов гликогена, кальция и ре-
зервной воды. Некоторую роль печень играет в выделе-
нии продуктов обмена веществ.
От заднего конца желудка начинается кишка. Почти
не меняясь в диаметре, она делает несколько петель и из-
вивов среди других органов; под самый конец, перед вхо-
дом в легочную полость, она несколько сужается, и этот
участок называют прямой кишкой; пройдя через легкое,
прямая кишка открывается наружу рядом с пневмостомом.
Форма пищеварительного тракта (кишечника) расти-
тельноядных слизней имеет важное значение в систематике
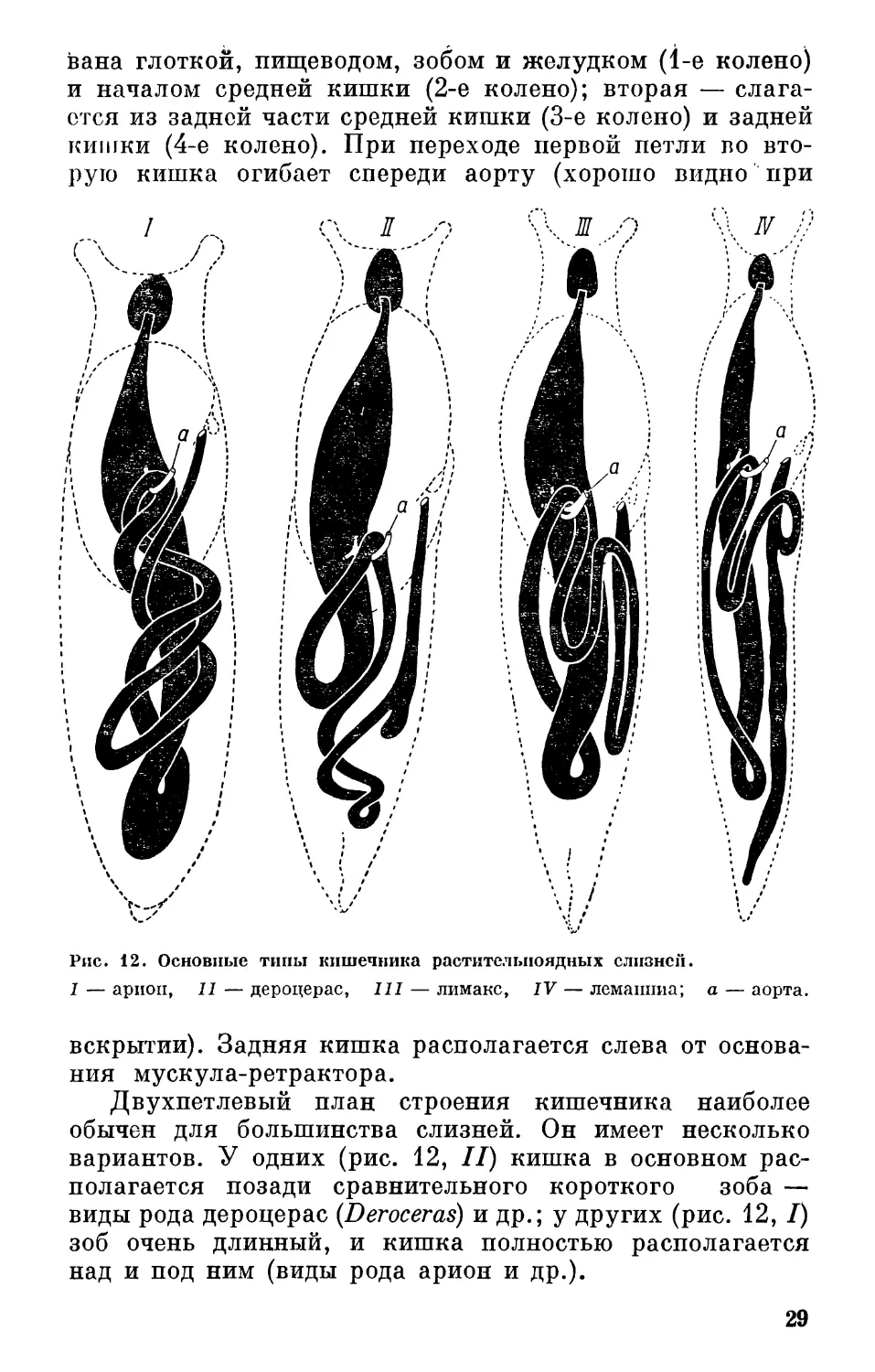
и определении этих животных. Здесь можно выделить два
основных типа строения кишечника (рис. 12).
1. Двухпетлевый кишечник. Слагается из двух петель.
Каждая петля состоит из двух колен: первая — образо-
28
вана глоткой, пищеводом, зобом и желудком (1-е колено)
и началом средней кишки (2-е колено); вторая — слага-
ется из задней части средней кишки (3-е колено) и задней
кишки (4-е колено). При переходе первой петли во вто-
рую кишка огибает спереди аорту (хорошо видно при
Рис. 12. Основные типы кишечника растительноядных слизней.
I — ариоп, II — дероцерас, III — лимакс, IV — лемаппиа; а — аорта.
вскрытии). Задняя кишка располагается слева от основа-
ния мускула-ретрактора.
Двухпетлевый план строения кишечника наиболее
обычен для большинства слизней. Он имеет несколько
вариантов. У одних (рис. 12, II) кишка в основном рас-
полагается позади сравнительного короткого зоба —
виды рода дероцерас (Deroceras) и др.; у других (рис. 12, I)
зоб очень длинный, и кишка полностью располагается
над и под ним (виды рода арион и др.).
29
2. Трехпетлопый кишечник (рис. 12, III). Если в пер-
вом типе задняя кишка располагается слева от основания
мускула ретрактора тела, то в семействе лимацид (Lima-
cidae) она огибает его спереди и образует третью петлю,
лежащую справа от упомянутого мускула на поверхности
внутренностного мешка.
Функция кишки сложная. С одной стороны, здесь
продолжается усвоение переваренных элементов пищи,
с другой — обезвоживание непереваренных остатков и фор-
мирование фекального шнура.
У некоторых видов с двух- и с трехпетлевым типами
кишечника задняя кишка имеет ориентированный назад
слепозамкнутый выступ, называемый слепой кишкой
(рис. 12, IV). Функция этого органа не изучена. При
вскрытии слепая кишка всегда хорошо заметна, так
как располагается на поверхности внутренностей, непо-
средственно под верхней стенкой тела.
Нервная система. Как у всех легочных мол-
люсков, нервная система слизней слагается из двух основ-
ных отделов: мозга и периферической системы нервов;
в некоторых органах (нога, кишечник, гениталии) нервы
образуют сложные сети — нервные плексусы. Мозг распо-
лагается позади глотки, вокруг пищевода и слагается
из пяти парных ганглиев — церебральные, буккальные,
педальные, плевральные, париетальные и одного непар-
ного абдоминального ганглия. Ганглии связаны друг
с другом нервными перемычками.
В связи с более высокой активностью по сравнению
с улитками мозг слизней отличается большей концентра-
цией ганглиев и увеличением размеров церебральных
ганглиев (рис. 13).
Органы чувств. Так как у слизней вся по-
верхность тела открыта прямому воздействию внешних
факторов, покровы этих животных обладают многочислен-
ными чувствительными клетками. Особенно много таких
клеток на голове, шее и подошве. Однако ведущее значе-
ние имеют специальные органы чувств, расположенные
на глазных и губных щупальцах.
Каждое щупальце представляет собой полый цилинд-
рический выступ головы с несколько вздутой вершиной
(особенно у глазных щупалец), в который заходит участок
головного синуса гемоцеля. Внутри щупальца залегает
ветвь мускула-ретрактора тела, которая передним кон-
цом срастается с вершиной щупальца. При сокращении
30
этого мускула вершина втягивается, а затем и все щу-
пальце погружается внутрь тела. Другие мускулы, кре-
пящиеся у основания щупальца, управляют движением
всего органа. Обратное выворачивание наружу обеспе-
гб
Рис. 13. Мозг слизня сетчатого.
га — абдоминальный ганглий, гб — буккальные ганглии, гп — педальные
ганглии, гпл — плевральные ганглии, гпр — париетальные ганглии, гц —
церебральные ганглии, ноб — обонятельный нерв глазного щупальца, ст —
статоцист.
чивается расслаблением мускулатуры и притоком гемо-
лимфы внутрь щупальца. Внутри каждого щупальца за-
легает нерв, а у глазного щупальца два нерва — обоня-
тельный и зрительный.
В верхнем конце каждого глазного щупальца распола-
гаются глаз и обонятельный ганглий. У слизней обоняние
развито лучше, чем у улиток. Так, некоторые видьГрода
31
арион способны различать запах съедобных грибов па рас-
стоянии 120 см и более. Губные щупальца также способны
различать запахи, но на значительно меньшем расстоянии.
Глаза слизней представляют собой полые овальные
пузырьки. Передняя стенка глаза прозрачная, а задняя
(ретина) слагается из многочисленных пигментных кле-
ток, среди которых регулярно расположены светочув-
ствительные клетки. Последние заканчиваются нейро-
фибриллами, которые, сливаясь друг с другом, образуют
зрительный нерв. В отличие от улиток, слизни кроме
основной обладают еще дополнительной ретиной, распо-
лагающейся в передней части глаза и служащей для вос-
приятия инфракрасного участка спектра (Newell, Ne-
well, 1968). В центре глаза находится хрусталик.
Как строение глаз, так и опыты по влиянию света на
поведение слизней показывают, что эти животные не спо-
собны различать детали предметов, и их глаза рассчитаны
па определение интенсивности освещения.
Губные щупальца’ также обладают органами обоня-
ния. Кроме того, па их поверхности располагаются сен-
сорные клетки, выполняющие роль органов вкуса. Ту же
роль выполняют и ротовые лопасти, однако главное их
назначение — осязание, особенно поверхности пищевого
объекта.
Важным органом чувств являются статоцисты. Это
парные сферические образования, лежащие на верхней
стороне педальных ганглиев. Они служат органами чув-
ства равновесия и обеспечивают ориентацию животного,
особенно при движении по наклонной поверхности и при
закручивании тела спиралью.
Половая система. Как и все легочные мол-
люски, слизни являются гермафродитными животными:
половая железа (гонада) продуцирует как спермин, так
и яйцеклетки. Обычно спермин формируются раньше.
Соответственно половой аппарат слагается из мужских
и женских половых путей и органов. Их строение отли-
чается значительным разнообразием и нередко имеет
большое значение при определении этих моллюсков
(рис. 14).
Гонада слизней имеет вид компактного округлого или
удлиненного органа, лежащего в задней половине тела
среди петель кишечника и лопастей печени или же запол-
няющего задний конец внутренностного мешка. От го-
нады отходит вперед гермафродитный проток/ нередко
32
Рис. 14. Гениталии слизня сетчатого.
ап — атриум половой, бж — белковая железа,
г — гонада, гп — гермафродитный проток, п —
пенис, пж — пениальная железа, рп — половой
ретрактор, с — семяпровод, сод — семяяйцевод,
сп — семяприемник, я — яйцевод; справа — ва-
рианты пенальной железы.
3 И. М. Лихарев и Я. С. Шапиро
дающий многочисленные извивы, особенно при подходе
к белковой железе. Здесь он впадает в сложно устроенный
многокамерный орган — квадривий. В нем происходит
оплодотворение яйцеклеток и формирование яиц, а также
через него транзитом проходит вперед своя сперма.
В квадривий открывается проток крупной языковидной
белковой железы, секрет которой формирует белковую
питательную оболочку яйца.
От квадривия простирается вперед длинный семяяйце-
вод. Он слагается из толстого фестончатого женского ка-
нала и тонкого прямого мужского канала. Внешнюю
стенку мужского канала образует лентовидная предста-
тельная железа (простата).
План строения гениталий до разделения семяяйце-
вода на семяпровод и яйцевод у всех слизней сравнительно
однообразный. С момента разделения половых путей об-
наруживаются определенные различия, особенно заметные
на органах, непосредственно участвующих в копуляции.
Как мужские, так и женские половые пути впадают
в атриум, который открывается наружу половым отвер-
стием. Недалеко от атриума находится семяприемник,
служащий для принятия и более или менее длительного
хранения спермы партнера, а также для резорбции
избыточной спермы. Этот орган состоит из резервуара
и протока. Последний у одних видов впадает во влага-
лище, у других — в основание пениса, а у третьих —
непосредственно в атриум.
Семяпровод — тонкий, нередко более или менее изви-
той проток, который, обособившись от семяяйцевода,
вначале обычно тянется вдоль яйцевода; вблизи от ат-
риума он изгибается назад, простирается вдоль пениса
и впадает в его задний конец.
У видов рода арион пениса нет, но передняя часть семя-
провода заметно утолщается за счет развития мощной
кольцевой мускулатуры и железистого эпителия, пре-
вращаясь в эпифаллус. В этом органе формируется спер-
матофор — своеобразный удлиненный пакет с хитиноид-
ными стенками, в котором при копуляции передается
сперма.
Пенис слизней очень разнообразен по внешнему и внут-
реннему строению. У одних он имеет вид длинной и строй-
ной трубки, у других он мешковидный, а у третьих вздут
сзади или у середины. Он может быть прямым или более
или менее изогнутым или даже спирально закрученным,
34
Внутренние стенки пениса (при копуляции они ста-
новятся наружными) обычно имеют различные складки.
Одна из них может быть особенно мускулистой и приобре-
тать облик конического или листовидного выступа, ко-
торый сильно набухает и служит для возбуждения парт-
нера перед копуляцией. Такое образование называют сти-
мулятором. Другие внутренние складки пениса служат
для направления спермы внутрь партнера.
Снаружи пенис нередко имеет различные придатки.
У слизней из рода дероцерас придатки на пенисе мягкие,
с тонкими железистыми стенками; их обозначают как
пениальные железы. Функция этих желез неясна, но так
как при копуляции они тоже выворачиваются, не исклю-
чено, что и они играют роль в стимуляции партнера.
Кроме уже упомянутых придаточных органов, на пе-
нисе встречаются и другие образования.
Подавляющее большинство слизней имеют более или
менее хорошо развитый пенис. Только у видов из рода
арион он полностью отсутствует, и эпифаллус непосред-
ственно открывается в атриум.
Участок женских половых путей после разделения
семяяйцевода обозначают как яйцевод. Если семяприем-
ник открывается в женские пути, то участок между его
протоком и атриумом называют влагалищем (см. рис. 45, в).
Передние участки половых путей снабжены особыми
мускулами, служащими для втягивания половых орга-
нов внутрь тела после копуляции. Наиболее важным из
них является половой ретрактор. Задний конец этого
мускула крепится к стенке тела вблизи от мантийного
комплекса или же непосредственно к диафрагме легкого.
Передний конец чаще всего прикреплен к пенису, если же
его нет, то к другим совокупительным органам — эпи-
фаллусу, яйцеводу, семяприемнику. Кроме полового рет-
рактора, нередко имеются еще 1, 2 или более коротких
мускула, связывающих концевые участки гениталий с пе-
редней стенкой тела.
3*
35
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
Копуляция. У большинства слизней копуляция
обычно происходит ночью или в сумерки, реже в сырые
пасмурные дни. Ей предшествует серия поведенческих
реакций, называемых брачными играми или танцами.
Продолжительность как брачных игр, так и копуляции
различна у разных видов. В общей сложности они могут
занимать от трех минут до суток.
Брачным играм предшествует поиск готового к копуля-
ции партнера. При этом важную роль играет обоняние
и слизь, которую оставляет после себя животное, так как
замечено, что при попадании на след партнера слизень
неоднократно задерживается, чтобы съесть оставшуюся
после него слизь. У арионов в этом отношении особенно
важную роль играет хвостовая железа, которая к началу
размножения сильно вспухает и начинает выделять круп-
ные капли густой слизи. По-видимому, слизь содержит
какие-то вещества, сигнализирующие о готовности к
копуляции и вызывающие определенные реакции у
партнера.
После встречи начинаются брачные танцы. В наиболее
полном виде они состоят из трех фаз: преследование,
кружение и соединение. По-видимому, одной из главных
задач брачных танцев является синхронизация определен-
ных физиологических процессов у партнеров. В это время
сперма обильно поступает в семяяйцевод и семяпровод,
простата обогащает ее своим секретом, а у видов, раз-
множающихся с помощью сперматофоров, в эпифаллусе
заканчивается их формирование. Выворачивание копуля-
тивных органов также начинается уже в конце брачных
танцев и обычно идет синхронно у обоих партнеров. В те-
чение всех фаз брачных танцев происходит взаимное ощу-
пывание партнеров с помощью щупалец и поедание слизи
как с поверхности тела, так и с субстрата. После оконча-
ния преследования слизни серповидно изгибаются, так
что их половые отверстия прижимаются друг к другу,
и в таком положении они начинают кружиться (рис. 15).
В это время они выделяют большое количество слизи,
густо покрывающей площадку, на которой происходит
копуляция. К концу кружения, а у некоторых видов и
раньше, начинается выворачивание атриумов и пенисов
и набухание стимуляторов (если они имеются). При этом
партнеры все время ощупывают друг друга вывернутыми
36
участками гениталий; особенно активны стимуляторы и
другие придатки пениса и атриума.
Затем прекращается кружение, слизни плотно прижи-
маются друг к другу популяционными органами и сильно
сжимают тела. При этом происходит выталкивание и
передача спермы либо в виде желеобразных капель, либо
внутри сперматофоров. В обоих случаях сперма подается
Рис. 15. Брачные игры и копуляция у слизня Штуранпя (по Косипьской).
А — фаза распознавания (преследования), Б — фаза быстрого кружения,
В — фаза замедленного кружения, Г — предкопуляционная фаза, Д — копу-
ляция (фаза неподвижности), Е — разделение партнеров (завершающая
фаза).
внутрь семяприемника, щелевидный вход в который не-
редко заметен снаружи. После завершения копуляции и
разъединения партнеров некоторые слизни съедают слизь,
оставшуюся на месте копуляции.
Описанный ход брачных игр и копуляции с некоторыми
вариациями обычен для многих видов слизней. В частно-
сти, так идет копуляция у арионов и дероцерасов.
Большинство видов совершают брачные игры и копуля-
цию на горизонтальных поверхностях: на почве, верхней
37
стороне камней и т. п. и значительно реже на наклон-
ных поверхностях, например на стволах деревьев. Совсем
иначе ведут себя слизень большой {Limax maxima х), сли-
зень черный {Limax cinereoniger) и некоторые другие виды
того же рода. Они копулируют в подвешенном состоянии
над землей, прицепившись к веткам деревьев или к ниж-
ней стороне нависающих над землей скал, к потолкам
гротов и т. п.
При встрече эти слизни также ощупывают и облизывают
друг друга. Затем они сплетаются телами и, одновременно
приклеиваясь с помощью слизи к ветке или к камню
задними концами, повисают в воздухе вниз головами.
У слизня большого партнеры повисают на слизистом шнуре
длиной 15—45 см. Сплетаясь, они постепенно выворачи-
вают пенисы, которые, в свою очередь, переплетаются,
но в обратном направлении, чем туловища.
Нередко в поведении партнеров наблюдаются некото-
рые различия в активности. Так, у слизня большого один
копулянт более активен и в этом отношении напоминает
самца: он начинает преследование, а после завершения
копуляции съедает слизистую нить; другой менее активен:
он следует за первым на дерево или стену и сразу уходит
после обмена спермой (Langlois, 1963).
Известно, что иногда у некоторых слизней обмен
спермой не всегда обоюдный: только один из партнеров
передает другому свою сперму. Их роли могут меняться
при последующих встречах (Williamson, 1959).
Оплодотворение и формирование
яиц. Когда после копуляции сперма попадает в семя-
приемник партнера, она освобождается от слизи или
оболочки сперматофора.
По-видимому, значительная часть спермы партнера
активно проникает из семяприемника по семяяйцеводу
в квадривий, где и происходит перекрестное оплодотворе-
ние. Экспериментально показано, что чужая сперма, еще
находясь в семяприемнике, стимулирует овогенез и овуля'
цию (поступление зрелых яйцеклеток в квадривий). После
оплодотворения неиспользованная «чужая» сперма рас-
творяется секретом стенок семяприемника, а также,
вероятно, и квадривия.
Время, проходящее между копуляцией, оплодотворе-
нием яйцеклетки и откладкой яиц, различно у разных
групп слизней: так, у сетчатого слизня на это требуется
8—10 дней, а у некоторых арионов несколько недель.
38
После оплодотворения, еще в квадривии, каждую зи-
готу окружает слой питательного белка — продукта вы-
деления белковой железы, образующего основную массу
будущего яйца. В дальнейшем, продвигаясь вперед по жен-
скому каналу семяяйцевода, окруженная белком зигота
последовательно одевается внутренней студневидной и
внешней оболочками. На границе между белком и внутрен-
ней оболочкой возникает перивителлиновая мембрана.
Внутренняя оболочка состоит из мукополисахаридов и
небольшого количества кальция, а внешняя — из поли-
сахаридов, протеина и кальция (Garrick, 1938; Bayne,
1968; Kosinska, 1980).
У большинства слизней яйца прозрачные, но у не-
которых арионов внешний слой заполнен кристаллами
углекислой извести, что делает яйца непрозрачными.
Откладка яиц. Обычно размножение и откладка
яиц более или менее приурочены к определенным сезонам
года, но нередко аномальные погодные условия могут
изменить сроки. Так, во Франции арион бурый (Arion
subfuscus) обычно размножается осенью, но если созрева-
ние запаздывает из-за очень сухого лета, то большая часть
популяции не откладывает яйца до будущей весны.
Слизни откладывают яйца в щели почвы, у подножия
травянистых растений, под камни и другие предметы, ле-
жащие на земле, где имеется достаточная влажность.
Нередко перед откладкой яиц в почву слизни расширяют
избранную ямку или щель. Чаще всего каждый слизень
откладывает яйца порциями (кладками), и не в одно место,
а в несколько расположенных по соседству. Число яиц
в каждой кладке, по-видимому, сильно варьирует в за-
висимости от возраста, питания, физиологического состоя-
ния слизня и погодных условий. Однако общее число яиц
более или менее постоянно. Так, слизни сетчатый и Штура-
ния в среднем откладывают за сезон до 500 яиц (Carrick,
1938; Kosinska, 1980), слизень желтоватый и арион бу-
рый — до 340 (Kunkel, 1916).
Постэмбриогенез и рост. Даже из яиц
одной кладки слизни вылупляются не одновременно.
Уже на этой стадии они заметно различаются по размерам
и дальнейший рост также идет неравномерно.
В довольно обширной литературе анализируется не
только влияние питания и абиотических факторов на рост
слизней, но и связь между общим ростом этих животных
и формированием гениталий, созреванием половых про-
39
дуктов, наступлением зрелости и размножением. В пост-
эмбриогенезе рост слизней слагается из следующих фаз:
инфантильная, ювенильная, взрослая и старческая (Run-
ham, Hunter, 1970; Kosinska, 1980). Эти фазы характери-
зуются темпом роста тела (вес, размеры) и состоянием
гениталий (табл. 2).
Инфантильная фаза отличается высоким темпом роста
животного, зачаточными гонадой и половым трактом;
только семяяйцевод начинает дифференцировку. С наступ-
лением ювенильной фазы темп роста падает, а гонада
начинает интенсивно увеличиваться в размерах и диффе-
ренцироваться на отдельные доли: к концу фазы масса
ее достигает максимума. Внутри гонады идет интенсивный
сперматогенез (сперматоциты превращаются в сперматиды).
Кроме того, происходит дифференциация белковой же-
лезы, а к середине фазы заканчивается формирование
семяяйцевода.
Взрослая фаза знаменуется остановкой роста тела.
При этом завершается формирование всех органов поло-
вого тракта. В начале этой фазы оканчивается спермато-
генез и гонада вместе с ее протоками заполняется зрелыми
спермиями. Вслед за этим начинается овогенез, а масса
гонады и ее размеры уменьшаются. Завершается созрева-
ние белковой железы, которая к началу этой фазы начинает
функционировать. Простата, начавшая выделять секрет
еще в ювенильной фазе, заканчивает этот процесс к се-
редине взрослой фазы, когда начинается развитие желез
женского канала семяяйцевода. К концу взрослой фазы
приурочены копуляция и откладка яиц.
У однолетних слизней (виды родов арион и дероцерас)
вслед за взрослой фазой наступает фаза старения. Нередко
она очень коротка и заканчивается смертью животного.
Лишь при благоприятных условиях, особенно в лабора-
ториях, эта фаза может растянуться и даже сопровож-
даться ростом тела животного (Kosinska, 1980).
У многолетних слизней после периода покоя гонада
вновь повторяет свой цикл (Abeloos, 1943, 1945).
Описанная связь между ростом и репродуктивным
циклом может несколько варьировать в разных группах
слизней, причем расхождения связаны с особенностями
физиологии. Это заметно, если сравнить арионов с деро-
церасами. Если у первых мужская и женская функции
гениталий четко разграничены во времени, то у вторых
они частично совпадают или перекрывают друг друга
40
Таблица 2
Изменения в половой системе в течение п остэмбриогевеза Deroceras reticulatum (по: Runbam, Laryea, 1968)
Органы Фаза роста (стадия зрелости)
инфантильная (предзр ювенильная | взрослая елость) (зрелость) 1 старения
Гонада: стадия вес проток Белковая железа Семяяйцевод Семяприемник Пенис Не <- дифференцп > рована <— Нарастает —> Макс *- if “* *- СПврМа‘ -> Спермин -> Овоциты тоциты тиды 1 Не <- дифферен- цирована
„ 1 Ч- . ЧОТТАТТпЛТУ ЛПДПИ/И! Без ** спермы * ия > ия
Не <- дифференци > рована <- Дифференциация * Пуст Дифферен —> циация Сек | > < про ой i < Созревание — Секрец :реция Секрец
статы > * женского i >< Со спермой—> сан а ла Без спермы *
— Оощи [й рост '* с Рост стимул - pf П ТТЛ Г Т СТ ТТ ТТ СТ - - штора > дка > 1
< Яйцекла
bo взрослой фазе. Именно в силу того, что арпонам трё-
буется больше времени на завершение репродуктивного
цикла, они дают только одно поколение в вегетационный
сезон, а при неблагоприятных условиях размножение
переносится на весну следующего года (Ghevallier, 1969,
1971). Слизень сетчатый и слизень Штурания, напротив,
при благоприятных условиях способны давать в год до
2—3 генераций, которые повторяют жизненный цикл,
каждый раз завершая его размножением, откладкой яиц
и смертью родительских особей.
Известно много примеров, указывающих на связь
роста и репродуктивного цикла слизней с питанием и
такими важными абиотическими факторами, как влаж-
ность, температура и световой режим. Кроме того, на рост
и репродуктивный цикл влияют и внутренние механизмы.
В этом отношении очень показательны эксперименты, про-
веденные Лавиолетом (Laviolette, 1950, 1954), установив-
шим, что в крови многих слизней имеются гормоны, вы-
деляемые гонадой и контролирующие развитие белковой
железы и полового тракта. Природа этих гормонов пока
неясна, но влияние гонады видно из описанной выше связи
главных фаз роста слизней с состоянием гениталий
(табл. 2).
В свою очередь, созревание гонады контролируется
другими физиологическими механизмами. Эксперимен-
тально доказано, что нейросекреторная деятельность мозга
способствует усилению овогенеза, а какие-то клетки глаз-
ных щупалец секретируют фермент, тормозящий овогенез
и способствующий сперматогенезу (Pelluet, Lane, 1961;
Wattez, 1978). По-видимому, именно этот эндокринный
механизм вызывает более раннее созревание мужских
гамет (протерандрию) слизней и определяет упомянутые
выше различия в сроках наступления овогенеза у арионов
и дероцерасов.
Жизненный цикл. Широкий круг вопросов,
связанных с фенологией роста и размножения, для многих
слизней пока еще недостаточно изучен или полностью
неизвестен.
Из слизней, обитающих в европейской части СССР,
наиболее полно изучены жизненные циклы у слизня
сетчатого (Дмитриева, 1969) и слизня Штурания (Ko-
sinska, 1980).
По основным особенностям жизненного цикла и дли-
тельности жизни слизней можно разделить на однолетние
42
и многолетние виды. Однолетние живут 3—5 либо 12—
18 мес и имеют только один период размножения. Отложив
яйца, взрослые слизни гибнут. Многолетние живут 2.5—
3 года и имеют два периода размножения; после второго,
отложив яйца, животные тоже вскоре умирают.
Среди однолетних слизней наблюдается несколько
типов жизненного цикла.
1. Циклы дероцерасов (рис. 16, I). Эти слизни живут
около 5 мес. Благодаря этому генерации, начавшие свою
жизнь весной, успевают закончить цикл за один вегета-
ционный сезон. При благоприятных климатических и
погодных условиях и хорошем питании эти "виды могут
дать вторую и третью генерации, жизненные циклы кото-
рых заканчиваются нередко уже в следующем вегетацион-
ном сезоне.
Наглядным примером такого типа жизненного цикла
служит слизень сетчатый. В центральных областях евро-
пейской части СССР он дает за вегетационный сезон одну
генерацию, на зимовку остаются яйца. В северо-западной
зоне, в частности в Ленинградской области, при благо-
приятных погодных условиях — дождливое лето, поздние
осенние заморозки — может появиться и вторая генера-
ция, если не всюду, то в отдельных биотопах. В этом
случае на зимовку остаются не только яйца, но и молодые
слизни (Дмитриева, 1969). Наконец, в Англии с ее влаж-
ным и теплым климатом и мягкими короткими зимами этот
слизень размножается почти круглый год.
По-видимому, еще более коротким жизненным циклом
обладает слизень гладкий. По нашим данным, он присту-
пает к размножению в середине второго месяца жизни.
Не случайно, что этот вид — единственный слизень, жи-
вущий в тундре Евразии и Северной Америки в условиях
очень короткого и прохладного лета.
Короткий жизненный цикл дероцерасов тесно связан
с некоторыми особенностями физиологии этих животных.
Они отличаются высоким темпом роста и более ранним,
чем у других слизней, наступлением зрелости, в част-
ности, более ранним началом овогенеза (см. выше). Благо-
даря этому между копуляцией, оплодотворением и отклад-
кой яиц проходит значительно меньше времени, чем у дру-
гих слизней (например, у арионов). Кроме того, у деро-
церасов эмбриональное развитие идет значительно быстрее,
чем у других видов. Так, у слизня сетчатого при влаж-
ности около 100 % и температуре воздуха 15 °C молодь
43
I
Рис. 16. Схемы жизненных циклов у слизней.
1 — слизень сетчатый; II — арион бурый. Пружками обозначены периоды
яйцекладки.
вылупляется пз яиц па И—13-й день после яйцекладки
(Дмитриева, 1969).
2. Циклы слизней из рода арион. Эти виды живут
12—18 мес (рис. 16, II). Чаще всего, а в европейской
части СССР всегда, жизненный цикл, начавшись с вылуп-
ления из яиц осенью, через полтора года заканчивается
размножением и откладкой яиц. Поэтому обычно зимуют
как молодые, так и почти взрослые слизни. Только в таких
странах, как Франция и Англия, при благоприятных по-
годных условиях (в частности, при безморозной зиме) и
хорошем питании в отдельных популяциях эти слизни
завершают жизненный цикл за 12 мес, начав его осенью
и закончив откладкой яиц осенью следующего года. Но и
в этих странах 1.5-летний жизненный цикл достаточно
обычен.
Столь растянутый во времени жизненный цикл этих
животных, в первую очередь, связан с темпом роста и раз-
вития половой системы. У арионов темп роста значи-
тельно ниже, чем у дероцерасов и зрелость наступает
позднее. Кроме того, овогенез заметно отстает от спермато-
генеза, а поэтому откладка яиц начинается позднее и
сильно растянута во времени. Следует учесть, что и
эмбриональное развитие у арионов занимает около ме-
сяца, что значительно длиннее, чем у дероцерасов.
В заключение остановимся на жизненных циклах
многолетних слизней. Наши представления о них исходят
из того, что мы знаем о слизнях черном, большом и желто-
ватом. Жизненные циклы слизня большого и слизня
черного очень сходны. Эмбриональное развитие длится
минимум 18—21 день. Вылупление из яиц происходит
осенью. Первый год жизни эти животные только растут,
но не размножаются. Первая копуляция обычно про-
исходит в июне—июле второго года жизни, на 21—22-й мес
после вылупления. Вторая копуляция происходит на
третьем году жизни, обычно в мае—июне. Откладка яиц
тянется до двух месяцев. Продолжительность жизни обоих
видов около 3 лет.
Жизненный цикл слизня желтоватого отличается более
раннпм наступлением половозрелости — первая копуля-
ция и откладка яиц отмечены в конце первого года жизни.
В отношении дальнейшего размножения этого слизня
мнения расходятся. Кюнкель (Kunkel, 1916) считает, что
размножение происходит еще два раза — на втором и
третьем году жизни. Фрёмминг (Fromming, 1954) с ним
45
но согласен и утверждает, что слизень с небольшими
перерывами откладывает яйца почти до самой гибели.
Он объясняет это тем, что данный вид живет в подпольях
и подвалах различных строений и на его половом цикле
не сказываются ни изменения погоды, ни сезоны года.
Оба автора единодушны, что слизень живет 2.5—3 года.
Связь гаметогенеза и формирования полового пути
с общим ростом у многолетних слизней не прослежена.
ПИТАНИЕ
Подавляющее большинство слизней — растительнояд-
ные полифаги с большей или меньшей склонностью к сапро-
и капрофагии или к микофагии. Основой питания этих
животных служат листья, стебли, плоды, луковицы, кор-
неплоды и подземные клубни, зерна многих диких и куль-
турных растений (табл. 1, 3).
Наблюдения, сделанные непосредственно в полевой
обстановке, и лабораторные опыты показали, что слизни
используют в пищу очень широкий ассортимент растений,
как в свежем виде, так и на различных фазах гниения.
Обычно отдается предпочтение сравнительно сочным и
мягким частям растений — листовая паренхима, мякоть
сочных плодов и клубней; слизни избегают участков
с жесткими покровами или с крупными сосудисто-волокни-
стыми пучками.
/ Выбор пищи в значительной степени зависит от того,
что имеется в непосредственной близости от убежищ,
в которых скрываются слизни на дневное время, а также
от их возраста — взрослые слизни охотнее поедают более
грубую пищу, чем молодые, что, вероятно, связано с тем,
что глотка их имеет более мощную мускулатуру и радулу.
Только что вылупившиеся слизни вначале питаются
остатками своих же яиц и невылупившимися яйцами
из той же кладки, а затем переходят к питанию гумусом
и разлагающимися остатками растений. Лишь с возрастом
свежий растительный корм^ занимает все большее место
в их рационе.
Даже у такого в основном растительноядного живот-
ного, как слизень сетчатый, в питании заметное место
занимает животный белок, в первую очередь различные
тли и мелкие двукрылые (Pallant, 1969). Еще бблыпую
46
Таблица 3
CopHbie растения, повреждаемые слизнями (оригинальные данные)
Биологические группы
и нодгругшы сорняков
Виды сорняков
I. Малолетние:
эфемеры
яровые ранние
яровые поздние
озимые
зимующие
II. Многолетние:
размножающиеся преиму-
щественно семенами
размножающиеся семе-
нами и вегетативно:
корневищные
корнеотпрысковые
Звездчатка (мокрица)
Овсюг обыкновенный, марь бе-
лая, горец вьюнковый, гор-
чица полевая, репа полевая
Пикульнпк красивый, щирица
пониклая
Костер ржаной, костер полевой
Ярутка полевая, василек си-
ний, трехреберппк непаху-
чий, пастушья сумка
Одуванчик обыкновенный, по-
дорожник большой, репейник
Хвощ полевой, мать-и-мачеха
Сурепка обыкновенная, вьюпок
полевой, льнянка обыкновен-
ная
долю продукты животного происхождения занимают у не-
которых арионов. С другой стороны, такие лесные виды,
как арион бурый, малаколимакс нежный и слизень чер-
ный, не являясь чистыми микофагами, охотно поедают
шляпочные грибы и лишайники.
В связи с тем что активность слизней чаще всего при-
ходится на ночи и сумерки, они в основном и питаются
в это время. Наибольшая прожорливость слизней совпа-
дает с периодом интенсивного роста перед размножением
и резко снижается к началу копуляции и в период яйце-
кладки. В это время слизни почти не питаются.
Количество потребляемой пищи тесно связано также
с температурой и влажностью. При влажности воздуха
90—100 % у слизня сетчатого наибольшая прожорли-
вость наблюдается при температуре 17—19 °C и суточная
прибавка массы тела в среднем составляет 22.7 мг (Дми-
триева, 1969). При 24—25 °C потребление пищи сокра-
щается в три раза. Лабораторные опыты показали (Pal-
lant, 1970; Пахорукова, 1976), что у этого слизня, как
и у близкого вида — слизня полевого, при питании наи-
более излюбленной пищей (салатом) процент усвоения
47
ее очень высокий в среднем составляет 70—85 % от всего
съеденного корма. Характерно, что при столь высоком
уровне усвоения пищи на рост идет только 27 % ассими-
лированной энергии. Последнее понятно, так как слизни
затрачивают много энергии на движение и регулирование
водного баланса.
СЛИЗНИ И СРЕДА ОБИТАНИЯ
Влияние окружающей среды на слизней довольно
сложное. Оно проявляется в том, что все основные физио-
логические функции и поведенческие реакции, а также
численность и распределение на местности прямо или
косвенно зависят от сочетания определенных внешних
факторов, которые для удобства анализа их влияния
на слизней могут быть подразделены на три основные
группы: абиотические, биотические и антропогенные.
К анализу этих факторов и их роли в жизни слизней мы
и переходим,
1. Абиотические факторы
В жизни всех наземных моллюсков и особенно слизней
из абиотических факторов наибольшую роль играют тем-
пература и влажность, выступающие нередко в тесной
взаимосвязи. Они в значительной степени определяют ход
физиологических процессов, поведение и активность этих
животных. Остальные факторы — осадки, свет, почва и др.
в первую очередь сказываются на слизнях через изменения
режима температуры и влажности.
Если у улиток крупная, нередко толстостенная наруж-
ная раковина способствует защите от перегрева, отражая,
подобно экрану, избыток солнечной энергии, а при на-
ступлении засухи в известной степени защищает мягкие
части тела от высыхания, то слизни лишены подобной
защиты. В то же время важная роль открытых покровов
в кожном дыхании этих животных повышает их уязви-
мость к потере влаги. Поэтому главной защитой от вы-
сыхания и перегрева тела является выделение слизи,
которая поддерживает кожу в увлажненном состоянии.
Кроме того, слизь играет важную роль в движении (с. 22)
и в защите от врагов. Однако расходование слизи сопря-
48
Жено с сокращением водного баланса и с необходимостью
все время ого восстанавливать.
Поэтому в отличие от многих улиток (особенно ксеро-
бионтных), которые при угрозе перегрева и высыхания
ищут защиту внутри раковины, закрывая вход в нее
эпифрагмой, слизни всегда стремятся найти убежище
с подходящим микроклиматом, где, сжавшись и тем самым
уменьшив поверхность тела, окружают себя чехлом
из слизи и пережидают неблагоприятное время. Более
высокая подвижность слизней, чем улиток, позволяет
им быстрее найти такое укрытие.
Температура. Влияние этого фактора сказы-
вается как на поведении, так и на ходе отдельных жизнен-
ных процессов. Исключительно важно влияние темпера-
туры на интенсивность дыхания слизней, а следовательно,
на общий метаболизм этих животных. Специальные экспе-
рименты показали, что адаптация слизней к повышению
температуры состоит из двух этапов (Runham, Hunter,
1970). Первый, сравнительно короткий, сопровождается
сильным нарастанием скорости дыхания, которая через
несколько часов резко снижается до определенного уровня,
более высокого, чем до начала опытов. Второй этап более
растянут во времени и приводит к дальнейшему снижению
скорости дыхания, нередко до исходного уровня.
Отчетливо прослеживается влияние температуры на
интенсивность питания и рост слизней. Так, у слизня
сетчатого наиболее оптимальный режим питания при-
ходится на температуру 17—19 °C (Дмитриева, 1969).
При 25 °C интенсивность питания и рост животного
снижаются, и длительное пребывание в таких условиях
приводит его к гибели.
Очень велико влияние температуры на размножение
слизней. Отмечено, что низкие температуры (ниже 10 °C)
задерживают развитие гонады, в то время как высокие
(25 °C н выше) ведут к уменьшению числа ооцитов или
к их полному исчезновению (Runliam, Hunter, 1970).
У слизня сетчатого температура 17—19 °C наиболее благо-
приятна для созревания половых продуктов, а 10—
15 °C — для откладки яиц; при 5 °C кладка практически
останавливается (Дмитриева, 1969).
Слизни сравнительно слабо устойчивы к отрицатель-
ным температурам и гибнут по прошествии определенного
срока пребывания в таких условиях (Pinder, 1969). Так,
среди сетчатых слизней, находившихся при температуре
4 И. М. Лихарев а Я. С. Шапиро
49
—3.5 °C в течение 45 ч, после переноса в комнатную тем-
пературу ожило около 50 % особей. При —4.5 °C слизни
погибали через 12 ч. А температуру —2.5 °C в течение
3 недель перенесли без потерь.
Е. В. Шиков (устное сообщение) проводил весной
размораживание вмерзших в лед
наземных моллюсков.
При этом многие улит-
ки оживали, а слизни
нет.
Если взрослые
слизни не обладают
достаточной устойчи-
востью к отрицатель-
ным температурам,
то их яйца в этом
отношении гораздо
выносливее. Так,
у слизня сетчатого
яйца выдерживают
охлаждение до —11 °C
в течение двух суток
и в течение суток —
до —1G °C. Развитие
Рис. 17. Зависимость про-
должительности развития
яиц от температуры у слизня
сетчатого.
яиц начинается уже при температуре 5 °C, а 15 °C является
оптимальной температурой, так как при наиболее корот-
ком сроке развития (11 дней) отрождается почти 100 %
молоди (Дмитриева, 1969). Ниже оптимальной темпера-
туры не только увеличивается срок развития яиц, но и
уменьшается процент вылупления молоди, при более вы-
сокой температуре (17—25 °C) происходит небольшое
удлинение инкубационного периода и заметное увеличение
процента неразвившихся и погибших яиц (рис. 17).
Влажность. Как у всех наземных моллюсков,
роль влажности в жизни слизней очень велика. Это
в первую очередь связано с тем, что в теле этих животных
вода занимает очень большую долю. Так, у арионов она
составляет около 86 % массы тела (Martin et al., 1958).
Есть основания считать, что у других слизней тело со-
держит еще больше воды.
50
В течение жизни слизня постоянно происходит расхо-
дование и пополнение водного баланса, особенно когда
животное находится в активном состоянии. Вода тратится
не только на выделение слизи для смачивания поверх-
ности тела, на движение и защиту от механических раз-
дражителей и врагов, но и на все основные жизненные
функции. Даже при содержании слизней в условиях
100 %-ной влажности воздуха и без пищи у них происхо-
дят периодические колебания массы тела в пределах
15—30 %. Они связаны с изменениями водного баланса,
происходящими из-за расхода воды, главным образом
на движение (Barnes, Weil, 1944, 1945; Dainton, 1954).
Восстановление водного баланса происходит в первую
очередь за счет поглощения воды через кожу; другим источ-
ником является пища.
Многие слизни способны короткое время переносить
большие потери воды. Так, слизень сетчатый и слизень
большой выдерживают потери воды до 50 % от массы тела
и при возвращении к условиям 100 %-ной влажности
восстанавливают свой водный баланс за несколько часов.
Подобно взрослым слизням, их яйца и эмбрионы спо-
собны кратковременно выдерживать значительное высыха-
ние — потерю до 80 % воды (Bayne, 1968). С другой
стороны, полное погружение в воду на 4 сут слабо сказы-
вается на ходе развития яиц слизня сетчатого, но более
длительное пребывание значительно снижает процент
вылупившейся молоди (Arias, Crowell, 1963). Косиньска
(Kosinska, 1980) установила, что яйца влаголюбивого
слизня Штурания могут нормально и без запоздания
развиваться в воде, но только при температуре до 20 °C.
Как правило, слизни не могут длительное время пере-
носить влажность ниже 90 %. Вначале всякое снижение
влажности вызывает кратковременное повышение подвиж-
ности, которое слизень использует для поиска укрытия.
Если таковое не будет найдено, то слизень впадает в оце-
пенение и вскоре погибает. Снижение подвижности свя-
зано с тем, что потеря воды вызывает повышение осмоти-
ческого давления крови, что, в свою очередь, влечет
подавление нервной активности педальных ганглиев (Кег-
kut, 1959). Разбавление крови водой способствует нараста-
нию активности этих ганглиев и тем самым увеличивает
подвижность животных.
|На рост слизней влажность оказывает влияние, анало-
гичное температуре. Питание прекращается, если влаж-
4*
51
ность воздуха ниже 100 %. В сухое время они прячутся
в укрытия — различные щели в почве, под камни и дру-
гие предметы на почве, под густой полог растений, и их
активность на поверхности почвы прекращается или
ограничена особо влажными местами, например берегами
водоемов. По-видимому, для поддержания нормального
водного баланса слизням не столь важна влажность воз-
духа, как наличие на почве водной пленки, из которой
они и черпают необходимую влагу, впитывая ее через
кожу. Другим источником воды служит пища.
В итоге можно сделать вывод, что, кроме высокой водо-
проницаемости покровов тела, слизни лишены специаль-
ных механизмов, контролирующих их водный баланс.
Отсюда их высокие требования к влажности окружающей
среды.
Осадки. Высокие требования слизней к влажности,
особенно к влажности почвы, делает понятным значение
осадков в жизни этих животных. Не случайно, что именно
в западном Закавказье, в той области, где выпадает наи-
большее для территории СССР количество осадков (до
3000 мм за год), обитает самая своеобразная и богатая
видами фауна слизней.
Общее количество осадков и характер их распределе-
ния в течение года наряду с другими факторами в значи-
тельной степени определяют жизненный цикл, характер
распределения на местности, численность и географиче-
ское распространение слизней (Соколов, 1935; Гриванов,
1936; Дмитриева, 1969). Сравнительная редкость и не-
продолжительность летних засух на северо-западе, западе
и в центре европейской части СССР способствуют тому,
что обитающие здесь слизни достаточно активны.
Реакция слизней на выпадение дождя в значитель-
ной степени определяется температурой воздуха. Если
дождю сопутствует оптимальная температура, то слизни
становятся активными и выползают на поверхность даже
в дневное время. Если же температура неблагоприятна,
то слизни остаются в укрытиях. Однако в тех случаях,
когда дождь начинает затапливать их убежища, слизни
выходят наружу и спасаются от воды на возвышенных
местах. В такое время в лесу многие слизни подымаются
вверх по стволам деревьев и кустов, а также на траву,
а на открытых местах заползают на верхние стороны
камней и отвесные склоны скал.
Не меньшую роль, чем дождь, имеет снежный покров:
52
во-первых, как резерв влаги для будущей весны и лета;
во-вторых, благодаря низкой теплопроводности снег явля-
ется своеобразным теплоизолятором, предохраняющим
почву от сильного охлаждения и резких колебаний тем-
пературы. Поэтому имеется прямая связь между толщиной
снежного покрова и успешной зимовкой слизней и,
в особенности, их яиц (Дмитриева, 1969).
Ветер. Всякое движение воздуха приводит к усиле-
нию испарения с поверхности тела слизней. Поэтому они
заметно реагируют на направление и силу ветра (Kalmus,
1942; Dainton, 1954). Очень слабый ветер обычно вызывает
лишь вытягивание щупалец в его сторону; при более силь-
ном ветре слизни начинают двигаться, причем всегда
по ветру. Последнее связано со стремлением в первую
очередь защитить от высыхания головной конец и щу-
пальца — наиболее чувствительные части тела. Во всех
случаях слизни стараются укрыться от ветра в каком-
нибудь убежище.
Свет. Отношение слизней к этому фактору достаточно
сложно. В первую очередь оно выражается в наличии
определенного суточного ритма активности этих живот-
ных, который связан с влажностью и температурой.
Повышение активности совпадает с периодами, когда оба
фактора наиболее близки к оптимуму. Поэтому максимум
активности слизней чаще всего приходится на утренние
и вечерние часы.
Слизни особенно чувствительны к инфракрасному
участку спектра, влияние которого они стараются осла-
бить, прячась в укрытия. Как уже указывалось (с. 32),
рецептором инфракрасных лучей является дополнитель-
ная ретина глаза.
Общий характер изменчивости окраски многих видов
в значительной степени зависит от освещенности биотопов.
Подмечено, что в популяциях, населяющих открытые
места (поля, огороды, луга), преобладают особи с более
сильным развитием меланина, придающего темный цвет
покровам тела. Их относительно меньше в популяциях,
живущих в затененных биотопах (леса, кустарники и
т. п.), где преобладают светлоокрашенные особп.
Отношение к свету не ограничивается его влиянием
на активность и окраску слизней. Показано, что опре-
деленное количество света необходимо для успешного
созревания половых продуктов (Pelluet, Henderson, 1954;
Sokolove, McCrone, 1978). С другой стороны, длительное
53
освещение высокой интенсивности вызывает торможение
гаметогенеза, а слабый свет ускоряет созревание спермиев.
Полный мрак приводит к резорбции гамет, и гонада при-
обретает облик, характерный для стареющего животного.
Почва. Химические и физические особенности почв
заметно влияют на численность слизней и их распределе-
ние на местности. Наиболее благоприятными для этих
животных являются щелочные и нейтральные почвы.
Кислые почвы они явно избегают, хотя в этом отношении
более устойчивы к ним, чем улитки. Особенно большое
значение имеет количество гумуса в почве. Его роль много-
гранна: с одной стороны, он является источником пищи,
особенно для молоди; с другой — присутствие гумуса
увеличивает влагоемкость почвы; наконец, обилие орга-
ники в почве способствует образованию густого травяного
покрова, который предоставляет слизням и хорошее
убежище и источник пищи.
Пожалуй, большее значение, чем химические, имеют
для слизней физические особенности почв — влагоемкость,
структура и микрорельеф. Они явно предпочитают влаго-
емкие и тяжелые суглинистые и глинистые почвы и из-
бегают водопроницаемых, легких каменистых, песчаных
и супесчаных почв, в первую очередь из-за их непостоян-
ной и низкой влажности. Последняя в особенности опре-
деляет отношение слизней к почвам. Как известно, для
этих животных совершенно необходимо присутствие на по-
верхности почв водной пленки — главного источника
поддержания водного баланса слизней. Только наличие
такой пленки позволяет им вести активный образ жизни,
когда влажность воздуха меньше 100 %, Однако на торфя-
ных болотах слизни обычно не живут, за исключением
ариона бурого, но и он встречается здесь в небольшом
количестве.
Очень большое значение в жизни слизней, особенно
на открытых биотопах, имеет микрорельеф почвы. Участки
со сглаженным рельефом, где мало убежищ в виде ямок
и щелей в почве или под камнями, неблагоприятны для
этих животных, особенно если растительный покров в та-
ких местах редкий. С другой сторопы, обилие таких
убежищ в сочетании с достаточно густым растительным
покровом (например, на полях и огородах) способствует
высокой численности небыстрому расселению слизней,
в частности такого серьезного сельскохозяйственного
вредителя, как слизень сетчатый.
54
2. Биотические факторы
Растительность, основные ланд-
шафты и биотопы. Окружающий слизней расти-
тельный мир — это в первую очередь важнейший элемент
среды обитания, а затем уже источник пищи.
Растительный покров создает под своим пологом осо-
бый микроклимат, порой сильно отличный от макро-
климата и значительно более благоприятный для многих
моллюсков. Растения не только защищают живущих под
ними животных от иссушающего воздействия солнца и
ветра, но и, благодаря испаряющей работе листьев,
создают псд своим пологом более высокую влажность
воздуха и почвы, чем на открытых местах.
В этом отношении очень показательно сравнить,
с одной стороны, широколиственный лес с хорошо разви-
тым подлеском и густым травянистым покровом и, с другой
стороны, расположенный по соседству сухой луг с редким
травянистым покровом. Первая ассоциация в силу благо-
приятного микроклимата, создающегося под ее пологом,
обилия и разнообразия экологических ниш, как правило,
отличается высоким разнообразием фауны слизней, от-
дельные виды которой могут достигать большой числен-
ности.
Иная ситуация на сухом лугу. Здесь фауна слизней
значительно беднее и численность отдельных видов ниже.
Моллюски находят убежища только там, где травяной
покров образует сгущения — в прикорневой части расте-
ний, а также в отдельных углублениях микрорельефа и
под камнями.
Большинство видов слизней обитает в лиственных,
особенно в широколиственных, и в смешанных лесах,
в затененных местах с богатой и достаточно влажной под-
стилкой или под густым травяным покровом, по оврагам,
ущельям и вдоль лесных речек и ручьев. В чисто хвойных
насаждениях обитают лишь немногие виды (например,
арион бурый, малаколимакс нежный), и то лишь при
достаточной влажности и наличию шляпочных грибов и
лишайников, которыми они питаются. При этом числен-
ность этих слизней здесь заметно ниже, чем в смешанных
лесах или е хвойном лесу с хорошо развитым лиственным
подлеском л травяным покровом.
Отношение слизней к тому или иному типу леса в зна-
чительной мере определяется обилием и разнообразием
55
убежищ, где животные могут укрыться от жары п мо-
розов. Наиболее универсальным убежищем служит лист-
венная подстилка, которая для многих видов одновременно
является источником пищи или субстратом, на котором
развиваются грибы, употребляемые данными слизнями
в пищу. Хвойная подстилка неблагоприятна для назем-
ных моллюсков, как в силу ее большей сухости, так и
непригодности в пищу (жесткость и высокое содержание
смолистых веществ).
Некоторые виды лесных слизней часто встречаются
на стволах деревьев — слизень черный и леманниа каемча-
тая. Здесь они питаются древесными лишайниками и гри-
бами, а также гниющей древесиной. Нередко на этих же
деревьях они находят укрытие в дуплах, щелях коры и
в моховом покрове. Однако их нельзя считать чисто дре-
весными обитателями, так как они обитают и под де-
ревьями — в подстилке, в гнилом валежнпке, под корой
пней и гниющих на земле стволов, под камнями, и в ще-
лях скал.
Наиболее богаты слизнями участки лиственного и сме-
шанного леса, находящиеся в тенистых сырых оврагах и
ущельях, вблизи от водоемов или со скалами и камени-
стыми осыпями. В этих осыпях, среди влажных камней,
чередующихся с гниющей подстилкой и слоями перегноя,
особенно много влаголюбивых улиток и слизней.
Пойменные леса и кустарники, периодически подвер-
гающиеся паводкам, обычно слабо заселены слизнями.
Причина этого в прямом воздействии паводка, когда этих
животных сносит водой и они гибнут от длительного пре-
бывания в воде. Кроме того, паводок разрушает почвен-
ный слой и подстилку, лишая слизней как убежища, так
и источника пищи.
Сравнительно бедны слизнями дубовые леса, как чи-
стые дубняки, так и дубовые леса с примесью других
древесных пород (липа, граб). По-видимому, это связано
с их большой сухостью, особенно с сухостью подстилки.
Кроме того, вероятно, лиственный опад дуба менее при-
годен в пищу моллюскам из-за высокого содержания
в нем дубящих веществ. Поэтому не только слизни, но и
большинство улиток избегают селиться в чисто дубовой
подстилке.
Как для лесных видов слизней, так и для видов, на-
селяющих открытые пространства, наиболее благоприятны
скальные участки и каменистые осыпи, в особенности
56
сложенные из известняковых и других щелочных горных
пород. Не только химизм этих субстратов благоприятен
для слизней, но и обилие в них всевозможных щелей,
углублений и других карстовых образований, в которых
накапливается гумус и поддерживается свой более ста-
бильный микроклимат, порой сильно отличающийся от на-
ружного.
В отличие от лесной, фауна слизней открытых про-
странств — лугов, степей — значительно беднее видами.
В задерненных сухих и травянистых биотопах, в особен-
ности по южным склонам, лишенным осыпей и имеющим
слабо расчлененный рельеф, без отдельных камней и скал,
популяции слизней отличаются низкой численностью.
Обычно тут обитают только некоторые виды родов деро-
церас и арион. Более влажные открытые биотопы — сырые
луга — обладают аналогичной фауной, но численность
популяций здесь заметно выше.
В биотопах с постоянной высокой влажностью, где
свободная вода имеется на почве в течение всего вегета-
ционного периода, — в заболоченных низинах, на зарос-
ших густой травой берегах водоемов, — фауна слизней
отличается большим однообразием. Наиболее характер-
ным видом таких мест является широко распространенный
слизень гладкий, который в жаркое время года становится
настоящим амфибионтом, нередко обитая совместно с амфи-
бионтными водными моллюсками, например с прудовиком
малым.
Будь то на равнине или в горах, наличие достаточного
числа подходящих убежищ, в которых слизни могут
успешно зимовать, пережидать засуху или скрываться
на дневное время, является важнейшим моментом, опре-
деляющим их численность, способность к расселению и
даже географическое распространение. Такие убежища,
как камни, щели скал и каменистые осыпи, позволяют
многим слизням жить даже в условиях жаркого климата
полупустынь Кавказа и Средней Азии.
Обширные открытые пространства, занятые планта-
циями, полями и огородами, где почва периодически под-
вергается глубокому разрыхлению и покрыта густым
покровом культурных растений, заселяются некоторыми
синантропными слизнями (с. 60). Обилие зеленой пищи
и убежищ во взрыхленной почве и под пологом растений,
гниющих растительных остатков, высокая влажность
почвы и воздуха — все это способствует высокой числеп-
57
ности этих видов. Отсюда проистекает в вред, который
они наносят многим культурным растениям.
< Если избыток тепла и низкую влажность слизням
удается нейтрализовать, подгоняя свою активность и
жизненный цикл к благоприятным сезонам и к ночам,
а остальное время скрываясь в хорошо защищенных
убежищах, то общий недостаток тепла является для них
непреодолимой преградой, ограничивающей их возмож-
ности распространения в области с холодным климатом.
В тех случаях, когда нехватка тепла достигает крайних
пределов, единственный путь преодоления этой пре-
грады — обитание в жилище человека или в теплицах.
Однако этот путь открыт только некоторым синантропным
и антропохорным видам (см. ниже).
Хищники, паразиты и болезни. Слизни
имеют довольно много врагов, в том числе и хищников.
Ими питаются многие позвоночные животные, однако
специфических «слизнеедов» среди них нет. Слизней охотно
поедают ежи, кроты, землеройки и некоторые мышевид-
ные грызуны; среди птиц — грачи, галки, скворцы и не-
которые чайки, а из домашних птиц — куры и утки.
Слизни также входят в пищевой рацион многих лягушек,
жаб, ящериц и змей.
Среди беспозвоночных животных слизнями питаются
многие насекомые, особенно часто жуки-жужелицы (Са-
rabidae). Почти исключительно питаются наземными мол-
люсками, в том числе и слизнями, хищные личинки жуков-
светляков (Lampyridae), а также некоторых жуков-
мягкотелок (Cantharidae). Однако как взрослые насекомые,
так и их личинки заметно предпочитают улиток слизням.
По-видимому, это объясняется тем, что улитки не спо-
собны так успешно защищаться от нападения с помощью
слизи, как это делают взрослые и здоровые слизни, связы-
вая ею ноги и ротовые придатки хищных насекомых.
К тому же личинки жуков, поедая улитку, нередко
используют ее раковину как убежище. Если жуки и их
личинки нападают на слизней, то чаще всего на молодых
или ослабленных взрослых особей.
На стадии личинки питаются слизнями некоторые
серые мясные мухи (Sarcophagidae), и в особенности
мухи-сциомизиды (Sciomyzidae).
Хищники, нападающие на слизней, известны и среди
других групп беспозвоночных: некоторые губоногие многое
ножки (Chilopoda), отдельные сенокосцы (Opiliones),
58
Слизни служат хозяевами (факультативными, про-
межуточными или основными) для многих паразитов. Так,
в пищеварительном тракте, печени или в почке некоторых
слизней обнаружено несколько видов инфузорий и кокци-
дий.
Многие слизни являются промежуточными хозяевами
ряда дигенетических сосальщиков (Trematoda) из семейств
Dicrocoeliidae, Brachilaimidae, Brachycoeliidae. Взрослые
черви этих групп паразитируют у домашних и диких
млекопитающих и птиц.
Слизни являются промежуточными хозяевами для не-
большого числа видов ленточных червей (Cestoda). Из них
в хозяйственном отношении особенно важна Davainea
proglottina — возбудитель опасного кишечного заболева-
ния домашних и диких птиц — давайнеоза.
Паразитами слизней являются многие виды круглых
червей (Nematoda). Роль слизней в жизненном цикле этих
гельминтов различна. Для тех видов нематод, взрослая
фаза которых протекает в почве, в слизнях происходит
только развитие личинки (Mermithidae). После выхода
наружу червь достигает половой зрелости и размножается.
У других круглых червей наземные моллюски, и в част-
ности слизни, являются промежуточными хозяевами,
в которых происходит развитие личинки до инвазионной
стадии, пригодной для заражения окончательного хозяина
(млекопитающие, птицы).
В легком и на поверхности тела многих слизней можно
встретить микроскопического клеща Ricardoella limacum.
Он питается некоторыми клеточными элементами соедини-
тельной ткани и крови слизней.
Не только сами слизни служат объектами нападения
хищников и заражения паразитами, но и их яйца. Однако
наибольший урон яйцекладкам наземных моллюсков,
особенно в теплые и влажные дни, наносят паразитические
грибки. Они могут покрывать поверхность яиц наподобие
густого войлока, проникать внутрь и в конечном счете
сильно снижать процент вылупившейся молоди.
Имеются отрывочные данные о болезнях слизней,
вызываемых грибами, бактериями и, возможно, вирусами.
Однако природа их пока не изучена.
Несмотря на то что хищники, паразиты и болезни
должны играть важную роль в регуляции численности
слизней, особенно в природных ассоциациях, достаточно
обоснованных данных об этом пока еще нет. Тому при-
59
чиной — скрытный образ жизни большинства слизней, быст-
рая их перевариваемость в желудках хищников, наши
слабые знания о биологии паразитов и о болезнях слиз-
ней и, наконец, чисто методические трудности накопления
соответствующей информации.
Какого-либо заметного предпочтения у хищников и
паразитов к определенным видам слизней не прослежи-
вается. Больше того, все упомянутые враги слизней с оди-
наковым успехом нападают или паразитируют в теле мно-
гих наземных улиток. По-видимому, главную роль играют
экологические факторы, обеспечивающие контакт между
слизнями и хищниками или с паразитами, находящимися
в инвазионной стадии развития.
3. Антропогенные факторы
Помимо природных (абиотических и биотических)
экологических факторов, важное влияние на слизней
оказывает хозяйственная деятельность человека. Это
влияние проявляется как прямо, так и опосредованно —
через изменение природных факторов среды. Под воздей-
ствием антропогенных факторов формируется животное
население, которое резко отличается от обитателей при-
родных ландшафтов. Эти отличия, относящиеся к видо-
вому составу и к экологии отдельных видов и их популя-
ций, определяют вредное значение некоторых видов слиз-
ней и динамику их численности. Вначале мы остановимся
на особенностях формирования фауны слизней антропо-
генных ландшафтов, в том числе и сельскохозяйственных,
а затем проследим динамику популяции вредящих видов
на сельскохозяйственных землях.
Синантропные и антропохорные
виды. В процессе хозяйственной деятельности человек
сознательно или невольно разрушает или преобразует
в выгодном для себя направлении природные ландшафты
или биотопы. На их месте создаются культурные (поля,
огороды, плантации) или другие антропогенные ланд-
шафты и биотопы (вторичные леса и луга, пустоши). При
этом значительная часть видов животных и растений,
исходно населявших природные биотопы, уничтожается
или вытесняется с освоенной территории, а другая часть
сохраняется и нередко даже занимает господствующее
положение. Оставшиеся виды обычно отличаются довольно
широкой экологической валентностью и, в особенности
60
в условиях монокультурного хозяйства, Их популяций
могут достигать высокой численности, в то время как
в природных сообществах они занимали довольно скром-
ное место. Такие виды принято называть синантропными.
Крайнее проявление синантропии можно видеть у жи-
вотных, которые обитают в полном отрыве от природы —
в жилье человека и в таких вспомогательных сооруже-
ниях, как погреба, подвалы и склады, или в столь искус-
ственных условиях, как оранжереи и теплицы. Упомянутые
проявления синантропии встречаются и у слизней.
Как первый шаг к синантропии можно рассматривать
слизней, которые обитают в первичных биотопах, не
затронутых или почти не измененных человеком, и во
вторичных биотопах, которые возникли в результате
сукцессии на месте первичных биотопов, уничтоженных
человеком, — во вторичных лесах, лугах и т. п. В куль-
турные биотопы эти виды проникают сравнительно редко
и в небольшом количестве, поэтому как вредители сель-
скохозяйственных культур они имеют третьестепенное
значение. В Нечерноземной зоне Европы к этой группе
относятся: арион бурый, слизень гладкий и слизень по-
левой.
Другие синантропы — это слизни, которые явно пред-
почитают антропогенные, особенно культурные, биотопы,
где в благоприятные годы они дают вспышки численности
и могут наносить существенный вред многим сельскохо-
зяйственным культурам. Они обитают и в природных био-
топах, как первичных, так и вторичных, но там их по-
пуляции не отличаются столь высокой численностью, как
на полях и огородах. Как правило, эти слизни и являются
серьезными вредителями сельского хозяйства. К ним от-
носятся арион полосатый, слизень Штурания, слизень
сетчатый.
К третьей группе принадлежат слизни, которые на
территории СССР обитают чаще всего непосредственно
внутри домов (например, в подвалах), во вспомогатель-
ных помещениях (погреба, продуктовые склады), в пар-
ках, садах (особенно в ботанических садах), в оранжереях
и теплицах; в ближайших природных биотопах, в том
числе и во вторичных, они отсутствуют. Чаще всего их
появление в таких местах связано с непроизвольным за-
возом, и поэтому их можно рассматривать как антропо-
хорные виды. Сюда относятся бётгерилла светлая, сли-
зень большой, слизень желтоватый, слизень крапчатый,
61
леманниа валенсийская. Многие из них (кроме бётгериллы)
могут наносить существенный вред растениям и запасам
пищевых продуктов.
Некоторые антропохорные виды могут исходно оби-
тать на территории СССР и в природных биотопах. Однако
непосредственных контактов между «дикими» и «культур-
ными» популяциями нет, почему можно считать, что по-
следние возникли в результате завоза человеком. Так,
бётгерилла светлая в природных условиях обитает в за-
падной Грузии и Абхазии. В антропогенных условиях
этот вид встречен в оранжереях Москвы и Челябинска,
в парках Ленинграда, Выборга и Львова. Слизень боль-
шой, встреченный в парках и садах многих крупных го-
родов европейской части СССР, в природе живет на Вос-
точных Карпатах. Слизень крапчатый широко распро-
странен в лесах Крыма и Кавказа, но, кроме того, обна-
ружен в погребах и оранжереях Ленинграда, Пушкина
(Ленинградская область), Севастополя, Орджоникидзе и
Тбилиси.
На этих примерах достаточно четко видно, что упомя-
нутые антропохорные виды могут быть завезены человеком
очень далеко от их естественного ареала. У остальных же
слизней этой группы их «родина» находится за пределами
СССР. Так, у слизня желтоватого, который обнаружен
в некоторых городах европейской части Союза, природный
ареал находится в западном Средиземноморье; у леман-
нии валенсийской, встреченной в садоводствах Ленин-
града, Ростова-на-Дону и Ташкента, природный ареал
находится на Пиренейском полуострове и в близких
к нему областях Франции.
Границы между упомянутыми тремя группами синан-
тропных слизней в известной степени условны. Сильное
антропогенное воздействие на природу в некоторых
областях и районах, при котором даже от вторичных
природны?< ассоциаций остается очень мало, приводит
к тому, что некоторые виды первой группы сохраняются
только в парках, садах и на лугах-пастбищах (арион
бурый, слизень полевой).
Некоторые виды второй группы, явно тяготеющие
к полям и огородам, завезены далеко от их естественного
ареала. Именно с этим связано, в частности, сравнительно
недавнее появление слизня сетчатого на Кавказе и в юго-
восточном Казахстане, а слизня Штурания — в Средней
62
Азии. По-видимому, эти виды завезены в указанные
области в последние 30—40 лет.
Показательно, что европейские по происхождению
слизни (например, арион бурый и полосатый, слизень
сетчатый, большой и желтоватый, леманниа валенсийская
и др.) к настоящему времени завезены и укоренились
во многих странах умеренных поясов северного и южного
полушарий, где активно выступают как вредители сель-
ского хозяйства. А слизень гладкий завезен даже во мно-
гие страны тропического пояса. По-видимому, завоз
европейских слизней на другие материки и на океаниче-
ские острова начался еще во времена парусного флота.
Быстрому расселению слизней путем непроизвольного
завоза в первую очередь благоприятствует значительная
устойчивость яиц этих животных к колебаниям темпера-
туры и влажности. С другой стороны, полифагия и не-
прихотливость в выборе пищи также явно способствуют
завозам. И наконец, на новые места слизни попадают
с привычными для них полевыми и огородными куль-
турами.
Попытаемся определить условия, способствующие ста-
новлению синантропии среди слизней (Лихарев, 1965).
Совершенно очевидно, что основным фактором, стимули-
ровавшим появление синантропных животных и растений,
была активная хозяйственная деятельность человека,
приведшая к сокращению первичных природных ланд-
шафтов и биотопов, а местами и к их полному исчезнове-
нию или существенной перестройке. При этом основная
масса видов была оттеснена в уцелевшие от антропо-
генного воздействия резерваты. Но некоторые, и таких
было немало, не только сохранились в новых условиях,
но и стали процветать. Последнему способствовало созда-
ние человеком больших площадей, засеянных определен-
ными культурными растениями (монокультуры), предо-
ставившими этим видам обильную и высококалорийную
пищу.
Но не только непосредственное воздействие хозяй-
ственной деятельности людей способствовало отбору из
дикой фауны видов, которые мы называем синантропными.
Определенные биологические особенности этих животных,
в свою очередь, сыграли в этом важную роль.
Для слизней, живущих на полях и огородах, где пе-
риодически происходит вспашка и уборка урожая, при
которых резко меняется весь комплекс условий жизни,
63
большое значение имеет длительность жизненного цикла.
Только достаточно короткий жизненный цикл, в условиях
умеренного климата занимающий не более 5—7 месяцев,
может позволить этим животным уцелеть и дать потом-
ство за период от вспашки почвы и посадки (или посева)
до уборки урожая. Не случайно на полях и огородах
обитают только виды с таким циклом — слизень сетча-
тый и другие виды рода дероцерас.
Причины, вызвавшие появление антропохорных ви-
дов, несколько иные. Как правило, эти теплолюбивые
южные виды, поселяясь на севере, далеко от своих при-
родных ареалов, в непосредственной близости от чело-
века — в домах, погребах, парках, оранжереях, — на-
ходят здесь не только обильный корм, но и главное, бла-
гоприятный микроклимат (в первую очередь температуру),
которого они не имели бы, поселившись где-нибудь по-
близости в природе.
ДИНАМИКА ПОПУЛЯЦИЙ СЛИЗНЕЙ
В АГРОЛАНДШАФТЕ И ЕЕ ПРОГНОЗИРОВАНИЕ
Сильный антропогенный пресс, который испытывает
фауна сельскохозяйственного ландшафта, определяет ее
четкую поляризацию. При этом выявляются синантроп-
ные и так называемые мизантропные виды, а появление
исторически молодых типов биоценозов (агробиоценозов)
приводит к формированию весьма своеобразных популя-
ций тех видов, которые уцелели в процессе сельскохозяй-
ственного освоения территории или проникли в агро-
ландшафт позднее (в том числе завозные виды). Если
в агробиоценоз внедряется вид-фитофаг, его популяции
часто наносят вред выращиваемым растениям. Для успеш-
ного контролирования численности вредящих видов не-
обходимо ясно представлять происхождение, функцио-
нальное значение, структуру и динамику как отдельных
популяций этих видов, так и всей их совокупности в сель-
скохозяйственном ландшафте (Поляков, 1968, 1973).
Все сказанное в полной мере относится к слизням,
и поэтому для успешной борьбы с ними необходимо учи-
тывать общие закономерности стадиального распределе-
ния и динамики популяций этих вредителей на сельско-
64
хозяйственных землях. Важность такого подхода опре-
деляется тем, что слизни заселяют практически все основ-
ные типы сельскохозяйственных угодий: поля с однолет-
ними пропашными культурами, посевы зерновых злаков
и многолетних кормовых трав, плодово-ягодные насажде-
ния, пастбища и сенокосные луга. Кроме того, эти моллю-
ски’__постоянные члены биоценозов с дикой травянистой
и древесно-кустарниковой растительностью, граничащих
с сельскохозяйственными угодьями.
Современный агроландшафт Нечерноземья характери-
зуется высокой экологической неоднородностью, связан-
ной с особенностью рельефа региона и с многоотраслевым
характером сельскохозяйственного производства. Этому
экологическому разнообразию биотопов соответствуют
популяции слизней, весьма различающиеся по своей
структуре, фенологии и динамике. Различно и их функ-
циональное значение в многолетней динамике числен-
ности вредителей на сельскохозяйственных землях: одни
популяции являются резерватными, выполняют функцию
сохранения вида в ландшафте во время ухудшения эко-
логической обстановки; другие популяции появляются
только в сезоны с благоприятными для слизней погод-
ными условиями.
К числу резерваций слизней относятся практически
все антропогенные несельскохозяйственные в узком смысле
стации: луга-пустыри в селах и деревнях, межи, перелоги,
залежные земли, граничащие с сельскохозяйственными
угодьями и испытывающие антропогенное влияние (леса,
рощи, лесопарки, кустарниковые заросли, заболоченные
участки и т. п.). Здесь слизни обычно живут из года в год,
а метеорологические особенности сезона хотя и влияют
на численность моллюсков, но почти никогда не приводят
их популяции к гибели благодаря «буферным» свойствам
этих стаций. Среди сельскохозяйственных угодий устой-
чивыми поселениями слизней отличаются многолетние
пастбища и сенокосы, сады с междурядьями, засеянными
многолетними травами, посевы многолетних травосмесей
g HQjigBbix и кормовых севооборотах, земляничные план-
тации и некоторые другие.
Стации, заселяемые слизнями лишь в благоприятные
для них сезоны, — это в первую очередь участки с разно-
образными однолетними пропашными культурами, а также
сады с систематически культивируемыми междурядьями,
5 И. М. Лихарев и Я. С. Шапиро 65
поля с легкими (песчаными и супесчаными) и хорошо
дренированными почвами и другие.
Причины успешного освоения слизнями экологически
различных стаций агроландпгафта объясняются рассмот-
ренными выше морфофизиологическими адаптациями этих
моллюсков, проявляющимися на видовом уровне. При-
чины же принципиальных различий между популяциями,
населяющими разные агробиотопы, становятся понят-
ными в результате анализа влияния на них антропогенных
факторов. Последнее важно для прогнозирования числен-
ности и распространения слизней на сельскохозяйствен-
ных землях и, следовательно, для организации эффектив-
ной борьбы с вредящими видами.
к Особенности фенологии и струк-
тура популяций. Наши исследования (Шапиро,
1979, 1980, 1981) показали, что популяции слизней,
населяющие различные элементы агроландшафта, суще-
ственно различаются по своей фенологии и возрастной
структуре. Высокая экологическая лабильность агро-
биоценозов приводит к резким колебаниям плотности
популяций, а метеорологические особенности каждого
сезона вызывают определенные внутрипопуляционные из-
менения в соотношении разных возрастных групп моллю-
сков. Однако принципиальные различия в структуре
популяций, населяющих основные типы сельскохозяй-
ственных угодий и залежные земли, остаются значимыми
на протяжении длительного периода. Это объясняется
регулярно повторяющимися антропогенными воздей-
ствиями на агробиоценозы, которые зависят от технологии
возделывания культурных растений и поэтому из года
в год принципиально не меняются.
Эти воздействия проявляются в основном опосредо-
ванно — через микроклимат и пищевой режим стации.
Непосредственное влияние микроклимата стации на струк-
туру и фенологию населяющей ее популяции слизней,
а пищевого режима — на ее плотность показано в ряде
работ (Соколов, 1933; Carrick, 1938; Бодренков, 1963;
Hunter, 1968). Более всего оказывают влияние на слиз-
ней температура и влажность в местах обитания, особенно,
если они принимают экстремальное значение. Данные
рис. 18 и табл. 4 иллюстрируют эту зависимость. Если
на капустном поле зимуют лишь яйца полевого слизня,
а в саду — яйца и молодые слизни 1-го весового класса,
то на участке, заросшем кустарником, в зимующей цо-
пуляции присутствуют дополнительно моллюски 2-го ве-
сового класса. Половое созревание и откладка яиц слиз-
нями на более влажных участках сада и клеверного ноля
начинаются обычно на 1 мес раньше, чем на капустных
полях, где в результате систематической культивации
почва иссушается сильнее; во влажной подстилке участка,
заросшего кустарником, яйца слизней встречаются на
протяжении почти всего года.
Рис. 18. Встречаемость пяти весовых классов слизня полевого в различных
популяциях.
I — на капустных "полях, II — на клеверных полях, III — в садах, IV —
в кустарниковых зарослях. По оси абсцисс — весовые классы слизней (ем.
табл. 7) и месяцы, по оси ординат — частота встречаемости.
Наблюдения показывают, что сроки размножения
и эмбрионального развития слизней в общем ограничены
периодами наиболее высокой влажности почвы конкрет-
ного участка, с чем в итоге и связаны структурные осо-
бенности популяций. Значительные колебания влажности
почвы в течение сезона (например, на капустных полях)
определяют наибольшую возрастную гомогенность по-
пуляции, тогда как постоянно высокие значения этого
показателя микроклимата (в стациях с дикой раститель-
ностью и на залужаемых участках) приводят к заметному
усложнению структуры, увеличению гетерогенности по-
пуляции в результате удлинения периода размножения
моллюсков, сглаживания временного «пика» откладки
яиц.
5*
67
[ ц а 4 со 9.8 о ci тн СМ о о со
я &
ч (О о со *<г °?
к Ен сч ТГ-1 об т-i СО
я Т“< Т-< Т-1 Т-1
60 Q
Е* о тЧ СМ СО
СМ т4 Ю
V 3 «п-н СМ тЧ т-1
я я СМ Ю °.
& со ci ci ci СО
иО
&
3 я о К СО [- 00 СО
Й Е- сч СО см L"i 00
со Ф
Я Я 2 со см
Я О i к т-1 см ci см’ см
ф
и. сэ со LO
fa я со о iri см’
о я см
я
с» я si< со
TH к сч о об
ф со т-1
ю о о с Т"( о см СО СО СО т^
5 сГ4 Т-1
& н СМ
со о о о Si<
1 я о ь яиц ИЮЛ1 сч о о НО о см
Ч ь<
о тН о о о о
И я
со
Я ф
ь о я о СО о о о о
я я
к я
& 2 СЧ о о о о
3 я
см 00 °. А
на Т"( ю см’ см
к
g СО со 1О со
м ! СО см см’
о 5S о сэ со О
о СО 2 сч ю СО со
Я
О
я СМ CD О
К ю об
Ц
>4 bi ф ф
О Я- о о я
О И о к £ я е g & Ф ч к й « о и
Я 8 о СО m >> О. § 5 ф й Ф Ч з X О 45 и° X о СО
АЙ 1 к К
Аналогичное явле-
ние наблюдал Бурен-
ков (1977): на распахан-
ных участках огорода
у слизней отмечен
лишь летне-осенний пе-
риод размножения, а на
задерненных участках,
где почва иссушается
слабее, — также и ве-
сенне-летний.
На фенологию по-
пуляций слизней кроме
микроклимата сильное
влияние оказывает про-
должительность суще-
ствования конкретного
сельскохозяйственного
угодья. У всех слизней
рода дероцерас в Не-
черноземной зоне
РСФСР в течение года
развивается не менее
одной полной генера-
ции; раннее половое
созревание служит важ-
ным фактором успеш-
ного заселения ими
сельскохозяйственных
земель. Однако на раз-
ных стадиях онтогенеза
слизни реагируют на
ликвидацию (распашку)
угодья по-разному. По
нашим наблюдениям,
зяблевая вспашка ока-
зывает губительное
влияние на зимующие
постэмбриональные ста-
дии слизней. На зимую-
щие яйца отрицатель-
ное влияние этого аг-
роприема не отмечено.
Вероятно, причина яв-
68
ленпя заключатся в более сильном промерзании почвы
распаханного участка, что вызывает массовую гибель
слизней, но не опасно для их яиц. Поэтому усиление
культивации территории, повышение интенсивности зем-
леделия снижает возрастную гетерогенность популяций
слизней, тогда как в многолетних угодьях, где вспашка
ежегодно не проводится, а также на залежных участках
популяционная структура более разнообразна.
Это влияние наглядно прослеживается при многолет-
них наблюдениях на сельскохозяйственных землях, вклю-
ченных в севооборот, в процессе которого обычно проис-
ходит смена культур, отличающихся по интенсивности
культивации. На рис. 19 показана структура двух по-
пуляций сетчатого слизня, обитающих на капустных полях.
Одно из полей расположено на месте распаханного трех-
летнего клеверного поля, а второе — на месте капустного
поля. Фенология и структура популяций второго участка
типичны для однолетних угодий с пропашными культу-
турами. При замене клеверного посева капустой наблю-
дается резкое снижение возрастной гетерогенности, ха-
рактерной для популяций клеверных полей. Поэтому
относительное возрастное разнообразие «капустной» по-
пуляции первого участка — это своеобразное последействие
более благоприятного для слизней микроклимата кле-
верного поля. Когда на его месте возникают на следующий
год иные условия, определяемые агротехникой возделы-
вания более интенсивной культуры — капусты, возраст-
ная гетерогенность популяции уже не является адекват-
ной реакцией на местные экологические условия, и по-
этому гетерогенность резко снижается. Увеличение воз-
растной однородности протекает здесь на фоне необхо-
димой гибели части популяции (двух осенних генераций —
в год распашки клеверища и в первый год возделывания
на его месте капусты) и представляет вынужденную реак-
цию моллюсков на повышение интенсивности земледелия.
Сами же популяции, гомогенные по возрастному признаку,
являются единственно возможной формой существования
слизней в ежегодно распахиваемых угодьях.
Увеличение возрастной гетерогенности можно наблю-
дать на участках, засеянных многолетними травами или
оставленных под залежь. На рис. 20 показана фенология
популяций сетчатого слизня на посевах клевера разного
возраста. Структура популяции однолетнего клеверного
поля близка к структуре популяций, обитающих на еже-
69
Рис. 19. встречаемость нагл весовых классов слизня'сетчатого в популяциях
капустных полей
I — капустное поле на месте клеверного,' II — на^яесте^капустного поля’
остальные обозначения как на ряс. 18.
Рис. 20. Встречаемость пяти весовых классов слизня сетчатого в популяциях
клеверных полей.
I — однолетнего, II — двухлетнего, III — трехлетиего; остальные обозна-
чения как на рис. 18.
Рис. 21. Встречаемость пятп весовых классов слизня сетчатого в популяциях
залежных участков.
I — однолетнего, II — четырех летнего, III—семилетнего; остальные обо-
значения как на рис. 18.
годно перепахиваемых участках: вспашка, проведенная
перед посевом травосмеси, уничтожает зимующие пост-
эмбриональные стадии моллюсков. На двулетнем
поле уже появляется второе (осеннее) поколение слизней,
хотя его доля невелика — в октябре на неполовозрелых
слизней этого поколения приходится 7 %. Посевы пер-
вого и второго года различаются по густоте травостоя.
Влажность почвы и воздуха на однолетнем участке ниже,
чем на двухлетнем, и колеблется сильнее в течение сезона.
Это приводит к появлению здесь первых яйцекладок
и к вылуплению молоди слизней на 15—20 дней позднее,
чем на двухлетнем посеве. Популяция трехлетнего поля
отличается наибольшим возрастным разнообразием! на
протяжении всего сезона здесь присутствуют слизни
1-го весового класса, тогда как на двухлетнем посеве они
встречались только в течение 4 мес, а на однолетнем —
только в июне и июле. Благодаря ранним срокам размно-
жения значительная часть яиц на трехлетием поле успе-
вает завершить развитие в том же году, в результате
чего в зимующей популяции велика доля молодых слиз-
ней (86 %).
На этом увеличение гетерогенности «клеверных» по-
пуляций слизней заканчивается, поскольку трехлетнее кле-
верное поле, как правило, распахивается, в результате
чего происходит противоположный процесс — нараста-
ние однородности возрастной структуры.
Дальнейшее усложнение возрастной структуры на-
блюдается в популяции залежных участков (рис. 21).
На однолетнем участке структура и фенология популя-
ции аналогичны тем, которые характерны для полей
с однолетними культурами, т. е. на однолетней луговине
возрастное разнообразие не увеличивается. Популяция
четырехлетнего участка приближается по своей структуре
и фенологии к популяции трехлетнего клеверища: в те-
чение всего сезона здесь присутствуют слизни 1-го весо-
вого класса, появляется второе (осеннее) поколение;
в зимующей популяции имеется большая доля неполово-
зрелых слизней. Еще более разнообразной становится
популяционная структура на семилетней луговине: резко
увеличивается возрастная гетерогенность весенних и осен-
них популяций; в течение всего сезона здесь присутствуют
слизни первых трех весовых классов, причем на их долю
приходится не менее половины всех особей. Таким об-
разом, в усложнении возрастной структуры популяций
71
залежных участков прослеживаются следующие этапыз
увеличение доли самых молодых слизней (1-го весового
класса) и появление второго (осеннего) поколения; уве-
личение доли слизней 2-го и 3-го весовых классов, в ре-
зультате чего возрастает гетерогенность весенне-летних
и октябрьских популяций; сглаживание временного «пика»
и увеличение продолжительности размножения.
Итак, в процессе формирования различных по своей
структуре и фенологии поселений слизней в агроланд-
шафте выявляются две основные тенденции. Первая
проявляется в упрощении, повышении однородности воз-
растной структуры тех популяций, которые обитают на
землях, подвергнутых наиболее интенсивной обработке.
Формирование таких популяций, которые представляют
единственно возможную форму существования вида в од-
нолетних угодьях, — безусловно, вынужденная реакция
моллюсков на сильный антропогенный пресс. Вторая
тенденция — это усложнение возрастной структуры по-
пуляций, увеличение их гетерогенности. Такие поселения
характерны для экстенсивных, подверженных длитель-
ному залужению угодий и для участков с дикой расти-
тельностью. Отмеченные процессы протекают параллельно,
а их соотношение зависит в общем от способа земледелия.
Если учесть, что развитие сельскохозяйственного про-
изводства связано с его интенсификацией, то можно пред-
положить, что первое направление будет все более доми-
нировать.
Каждая из двух отмеченных тенденций имеет важное
значение для поддержания существования слизней в агро-
ландшафте. Так, способность формировать однородные
по возрасту популяции с относительно коротким перио-
дом размножения позволяет слизням успешно осваивать
однолетние, ежегодно перепахиваемые угодья интенсив-
ного типа. В то же время длительное существование этих
популяций, как будет показано ниже, тесно связано с на-
личием в ландшафте популяций экстенсивных и залежных
земель. Поэтому современное структурное и фенологи-
ческое разнообразие агроценотических популяций слиз-
ней можно рассматривать как необходимое условие и
одновременно следствие заселения этими моллюсками
сельскохозяйственных земель, а следовательно, и как
основу прогнозирования и контролирования их числен-
ности в агроландшафте.
Сезонная и многолетняя динамика
72
численлост и. Численность слизней, как и других
беспозвоночных животных, в сильной степени зависит
от абиотических факторов, которые, как было показано
выше, непосредственно влияют на плодовитость и выжи-
ваемость моллюсков. Вместе с тем важным фактором,
определяющим плотность популяции слизней в конкрет-
ном местообитании, является его пищевой режим. По-
этому соотношение в ландшафте или в отдельном агро-
массиве различных по своему значению для слизней
стаций наряду с погодными условиями определяют ди-
намику численности и размеры вредной деятельности
этих моллюсков. Вероятно, определенное регулирующее
влияние на численность слизней оказывают патогенные
для них организмы, однако их роль в этом процессе изу-
чена пока очень слабо.
Сезонное варьирование численности слизней в агро-
ландшафтах Нечерноземья может быть значительным
лишь в случае массовой гибели моллюсков (например,
в суровую зиму или засуху) или при появлении второй
(осенней) генерации. Последнее явление довольно редкое:
на северо-западе зоны, по расчетам Дмитриевой (1969),
второе поколение сетчатого слизня может появляться
в среднем 1 раз в 10 лет.
Напротив, плотность отдельных популяций слизней
часто испытывает в течение сезона сильные колебания,
причем равновероятно и уменьшение, и увеличение плот-
ности постэмбриональных стадий моллюсков. Это свя-
зано с динамикой вылупления их из яиц, с возможной
гибелью слизней до начала размножения, с естественной
элиминацией отложивших яйца особей, а также нередко
и с активным расселением моллюсков, в результате кото-
рого происходит перераспределение их между соседними
биотопами.
Динамика численности слизней в разных стациях имеет
принципиальные отличия, определяемые структурно-фе-
нологическими особенностями популяций. Эти отличия
необходимо учитывать в процессе выявления вредителей
на сельскохозяйственных землях, при составлении про-
гнозов численности и определении (сигнализации) сроков
борьбы с ними. Рассмотрим наиболее важные особенности
сезонной динамики численности слизней.
На рис. 22 показана интенсивность вылупления слиз-
ней рода дероцерас из перезимовавших яиц в конце весны—
начале лета в популяциях двух типов — населяющих
73
однолетние угодья и векультйЁй^емыё участки. В тоН
случае, когда все отложенные осенью яйца остаются
зимовать, интенсивность отрождения молоди графически
выражается кривой 1. Участок кривой а соответствует
вылуплению моллюсков из яиц, отложенных в самые ран-
ние сроки в предшествовавшем году; участок б — вы-
луплению слизней из основной массы перезимовавших
яиц; участок в — отрождению моллюсков из яиц, отло-
женных в поздние сроки. Поскольку распределение об-
щего количества яиц по срокам их откладки обычно при-
Весна Лето
ближается к нормальному, кри-
вая 1 близка к логистической.
Именно такой характер носит
динамика численности отрож-
дающейся молоди на ежегодно
распахиваемых участках или
Рио. 22. Схема динамики отрождения слиз-
ней деронерас из перезимовавших яиц.
По оси ординат — численность отродив-
шихся слизней в однолетних агробиото-
-L пах (2) и на залежных участках (Р); а—в —
различные стадии развития слизней.
на однолетней луговине, поскольку здесь, как правило,,
зимуют все отложенные в конце сезона яйца.
На залежных участках (кривая 2) отрождение слизней
начинается раньше благодаря более продолжительному
развитию яиц летом и осенью предшествовавшего года.
Последнее, как отмечалось, связано с более ранними сро-
ками размножения слизней на таких участках. Из яиц,
откладываемых в самые ранние сроки, в том же году
обычно вылупляется молодь, поэтому кривая 2 отли-
чается отсутствием пологого участка а. Вылуплению
слизней из яиц, отложенных в поздние сроки, соответ-
ствует участок б'. В целом отрождение слизней здесь
более равномерное, поскольку временной «пик» размно-
жения выражен слабее. Помимо залежных участков,
популяциями слизней с аналогичной динамикой засе-
ляются сенокосные угодья, пастбища, клеверища, засеян-
ные травами междурядья садов.
Отрождение моллюсков одного вида из перезимовав-
ших яиц во всех биотопах заканчивается приблизительно
в одни и те же сроки, что связано с весьма синхронным
74
прекращенном откладки яиц при наступлении похолода-
ния в конце осени.
В отдельные годы с засушливой весной наблюдается
массовая гибель яиц и вылупляющихся моллюсков.
Вероятность этого заметно выше на ежегодно распахивае-
мых участках или на однолетней луговине (кривая 7).
Это связано с более поздним отрождением большего коли-
чества слизней, что, в свою очередь, связано с уменьше-
нием положительного влияния на яйца и молодь талых
Рис. 24. Схема сезонной ди-
намики численности пост эмбрио-
нальных стадий слизней дсро-
церас.
Рис. 23. Схема динамики естествен-
ной гибели слизней дероцерас после
откладки яиц.
По оси ординат — численность по-
гибших моллюсков в однолетних
агробиотопах (I) и на залежных
участках (2).
1 — в однолетних агробиото-
пах, 2 — на многолетних па-
стбищах и сенокосах, 3 — на
залежных участках. По оси
ординат — численность слиз-
ней.
вод. Кроме того, в начале сезона микроклимат многолет-
них участков обычно более влажный, чем однолетних
угодий.
Естественная гибель слизней наблюдается вскоре поело
откладки ими яиц. Поскольку сроки и интенсивность
откладки в разных стациях различны, то и кривые есте-
ственной элиминации слизней выглядят по-разному
(рис. 23). Более растянут этот процесс в тех популяциях,
где слизни раньше достигают половозрелости и присту-
пают к размножению, т. е. на залежных участках и мно-
голетних угодьях (кривая 2). В однолетних же угодьях,
где временной «пик» репродукции выражен более резко,
естественная элиминация протекает более интенсивно
(кривая 2). В случае раннего осеннего похолодания ин-
тенсивность гибели моллюсков увеличивается, и это
в первую очередь отражается неблагоприятно па по-
пуляциях однолетних, ежегодно распахиваемых участков.
Как показывают многолетние наблюдения, принци-
пиальные особенности сезонной динамики различных
агроценотических популяций слизней остаются значи-
мыми из года в год. Эти различия, так же как и рассмот-
ренные структурно-фенологические особенности популя-
ций, определяются характером антропогенных воздей-
ствий. Поэтому среднемноголетние характеристики попу-
ляций, населяющих основные типы сельскохозяйственных
земель, могут служить достаточно надежным основанием
для составления оперативных прогнозов численности
слизней и планирования борьбы с ними.
На рис. 24 схематично показана сезонная динамика
численности постэмбриональных стадий слизней в угодьях
двух типов и на залежных землях. Наиболее сильно
колеблется численность моллюсков в однолетних агро-
биотопах, что связано с весьма синхронным развитием
большинства членов популяции. Здесь четко выражен
«пик» численности, приходящийся на конец лета—на-
чало осени. В многолетних угодьях в связи с более растя-
нутым периодом размножения и присутствием постэмбрио-
нальных стадий в зимующей популяции синхронность
развития особей заметно нарушена, поэтому и колебание
численности менее значительно. На залежных участках,
где обитают популяции с наиболее сложной и разнооб-
разной возрастной структурой, численность колеблется
еще слабее.
Разнообразие возрастной структуры популяций слиз-
ней удобно выразить математически, через показатель
возрастной гетерогенности (А — индекс):
/ s \-i
д= 2^1 .
\ i=l /
где Р. — доля слизней t-класса, S — число классов
слизней (с. 98). Колебание численности моллюсков можно
выразить через коэффициент вариации. Сопоставление
этих параметров весьма наглядно иллюстрирует зави-
симость динамики численности слизней от возрастной
структуры популяции (табл. 5). Эту зависимость для
76
Таблица 5
Варьирование численности и возрастная структура популяций сетчатого
слизня в некоторых антропогенных стациях (из: Шапиро, 1981)
Станин КлэФФииюн" ез<»нн<»г вар прощания чи' енноети °/о Коэффициент Н'М’о РТВ го вар п р« «ван и я «игленности °/о Показатель в'»зра тной грт роггн но. ТИ П"ПУ я- ПИИ (Д)
Капустные поля 104.78 103.31 2.04
Картофельные поля 102.24 94.77 2.06
Пастбища п сенокосы 73.42 73.97 2.35
Луга-пустыри в населенных 62.51 58.17 2.58
пунктах Злаково-разнотравные луга 53.53 56.63 2.41
Лесопарки 59.93 61.43 2.38
Залежные земли с древес- 46.68 42.11 2.67
ной растительностью Заброшенные сады 58.10 60.75 2.49
сетчатого слизня достаточно точно выражает следующее
уравнение, используемое в целях прогноза:
Угвз, = 271.46 — 84.95. Д,
где Vce3 — коэффициент варьирования плотности попу-
ляции слизня в течение одного сезона. Расчеты показы-
вают, что различия в сезонном колебании численности
постэмбриональных стадий сетчатого слизня на 88 %
зависят от структурных особенностей отдельных по-
пуляций вредителя.
Таблица 6
Динамика плотности популяций сетчатого слизня в Ленинградской
области в 1972—1979 гг. (указана среднесезонная плотность
постэмбриональных стадий, экз./м2, оригинальные данные)
Стация 1972 1973 1974 1975 1977 1978 1979
Капустные поля 0.0 0.1 3.1 5.0 15.3 13.5 28.6
Картофельные поля 0.0 0.1 2.3 3.5 9.6 7.8 16.0
Долголетние пастбища 0.2 1.0 4.6 4.8 19.7 19.9 23.7
Луга-пустыри в населен- 5.5 7.3 11.8 14.1 25.9 25.7 48.7
ных пунктах Злаково-разнотравные 4.3 5.5 6.6 8.7 14.0 12.3 23.0
сенокоспые луга Лесопарки 2.9 3.0 4.0 5.3 8.5 7.6 15.5
Залежные участки с дре- 4.5 6.1 6.9 12.3 12.4 16.2 20.1
весно-кустарниковой растительностью Заброшенные сады 10.3 12.7 15.6 28.3 46.3 55.9 63.2
77
Многолетнее варьирование численности слизней на
сельскохозяйственных землях тесно связано со структурой
агроландшафта — соотношением в нем стаций, различаю-
щихся по степени благоприятности для слизней. Значе-
ние отдельных поселений для поддержания существова-
ния вида в ландшафте прослеживается в ходе многолет-
них наблюдений за их динамикой. В табл. 6 показана
динамика плотности некоторых популяций сетчатого
слизня в течение 1972—1979 гг. в Ленинградской области.
Анализ данных показывает, что многолетняя динамика
численности моллюска, так же как и сезонная, тесно
связана с возрастной структурой отдельных популяций
(табл. 5). Эту связь выражает уравнение регрессии:
Уиного„т. = 271.31 - 82.23-Д,
где Vuноголет. — коэффициент варьирования численности
слизня по годам. Различия в многолетней динамике плот-
ности поселений моллюска на 79 % зависят от различий
в их возрастной структуре.
Биологический смысл установленных регрессий — се-
зонного и многолетнего варьирования плотности популя-
ции по индексу ее возрастной гетерогенности — состоит
в том, что возрастное разнообразие популяции в целом
увеличивает ее жизнеспособность, поскольку разные ста-
дии жизненного цикла обладают различной устойчивостью
к действию экологических факторов. Следовательно, по-
казатель возрастной структуры популяций слизней может
служить одним ив основных критериев для анализа посе-
лений моллюсков в агроландшафте и прогнозирования
их динамики.
Такой вывод подтверждается результатами многолет-
них наблюдений за пространственным распределением
и динамикой численности слизней на сельскохозяйствен-
ных землях. Наиболее резкие многолетние колебания
испытывают однородные по возрасту популяции однолет-
них агробиотопов. В этих угодьях в благоприятные се-
зоны численность моллюсков достигает экопомическп
ощутимых значений, возникает необходимость борьбы
о вредителями. Высокая плотность популяций в такие
годы определяется здесь высокой концентрацией фито-
массы кормовых для слизней растений. Такие условия
на полях Ленинградской области складывались в 1922—
1923 гг. (Соколов, 1933; ГриваНов, 1935), в 1962 и 1965 гг.
(Дмитриева, 1969) в 1977, 1979—1981 гг. (Шапиро, 1981).
В неблагоприятные сезоны популяции однолетних
угодий, особенно с пропашными культурами, часто исче-
зают вообще и их последующее возрождение происходит
благодаря иммиграции моллюсков извне (Шапиро, 1981).
Поэтому популяции слизней, обитающие в однолетних
агробиотопах с пропашными культурами, справедливо
считать временными, а заселяемые ими стации — стаци-
ями расселения.
Относительно гетерогенные популяции экстенсивных
угодий (многолетних посевов, пастбищ, сенокосов и т. п.)
и залежных земель отличаются известной стабильностью —
варьирование их плотности по годам не столь значительно.
Особенно это характерно для поселений моллюсков на
залежных участках с древесно-кустарниковой раститель-
ностью. Естественно, концентрация кормовых растений
здесь не очень высокая, однако микроклимат таких место-
обитаний наиболее благоприятен для слизней. Поэтому
плотность популяций вредителей здесь редко бывает высо-
кой, однако животные переживают в этих стациях периоды
с неблагоприятными погодными условиями. Локализа-
ция слизней в резерватных стациях наблюдалась в Ле-
нинградской области, например, в 1971—1973 гг. и
в 1983 г., когда количество осадков, выпадавших в течение
вегетационного периода, было значительно меньше нормы.
Таким образом, многолетняя динамика популяций
слизней в агроландшафте выражается, с одной стороны,
в варьировании плотности отдельных популяций, а с дру-
гой — в закономерной динамике стациального распре-
деления на заселенной моллюсками площади. В этом про-
цессе можно наметить ряд фаз многолетней динамики
популяций (рис. 25), которые важно иметь в виду при
выявлении и учете вредителей, для прогнозирования их
численности и расселения, при организации защитных
мероприятий.
В фазу депрессии (а) слизни локализованы практиче-
ски лишь в резервациях, причем их численность здесь
невелика; в эти годы на культивируемых участках мол-
люски обычно отсутствуют. При наступлении благоприят-
ных погодных условий численность слизней в резерва-
циях заметно увеличивается и одновременно начинается
их расселение на соседние поля (б). Резкого увеличения
численности слизней и заселенней ими площади в течение
79
одного вегетационного сезона не происходит — для этого
необходимо сочетание благоприятных погодных условий
в течение 2—3 лет. Это связано, во-первых, со сроками
заселения слизнями новых стаций, а во-вторых, с од-
ногодичной генерацией у главных вредящих видов. Есте-
ственно, в тех ландшафтах, где преобладают резерватные
стации, эта фаза протекает быстрее. Массовое размноже-
ние слизней (в) наблюдается при заселении ими большин-
ства сельскохозяйственных угодий и сохранении благо-
Рпс. 25. Схема многолетней динамики популяций
слизней дероцерас в агроландшафте.
По оси ординат — численность слизней (N),
усредненный показатель возрастной гетероген-
ности популяции (Д) и варьирования ее плот-
ности (V); а—г — фазы динамики численности.
приятных для моллюсков погодных условий. При резком
ухудшении метеорологической обстановки наблюдается
массовая гибель в стациях расселения и как результат —
резкий спад численности вредителей в ландшафте (г).
Как отмечалось, популяции слизней в стациях рассе-
ления и в резерватных стациях характеризуются суще-
ственными структурными различиями и разной потен-
циальной изменчивостью плотности. Поэтому в процессе
многолетней динамики численности слизней и адекват-
ного изменения их стациального распределения наблю-
даются вполне закономерные изменения обобщенных пока-
зателей возрастной структуры и плотности поселений
в агроландшафте (рис. 25). С увеличением численности
и площади заселения средняя возрастная гетерогенность
поселений слизней снижается, поскольку в ландшафте
постепенно доминируют однородные по возрасту по-
80
пуляции. А это, в’свою очередь, приводит к увеличению
показателя изменчивости плотности популяций (с. 78).
Из этого следует важный для прогнозирования вывод:
вероятность вспышки численности вредителей, равно как
и вероятность их массовой гибели, находится в прямой
зависимости от размеров заселенной территории, так как
по мере увеличения площади сельскохозяйственных уго-
дий, заселенной слизнями, усиливается влияние на их
численность экологических факторов.
Расселение слизней на сельскохо-
зяйственных землях. Как было показано
выше (с. 65), в сельскохозяйственном ландшафте наблю-
дается весьма четкая функциональная дифференциация
популяций слизней. Благодаря же расселению слизней
устанавливается контакт между их отдельными по-
пуляциями. Поэтому расселение служит не только факто-
ром освоения новых местообитаний, но и способом поддер-
жания длительного существования вида на уже заселенной
территории.
В ряде работ анализируются способы пассивного рас-
селения наземных моллюсков, в частности слизней (Ли-
харев, 1965; Rees, 1965; Хохуткин, 1967; Старобогатов,
1970; Шилейко, 1972; Шиков, 1977). Пассивный, т. е.
непроизвольный (неумышленный) занос слизней челове-
ком на сельскохозяйственные земли может происходить
при перевозке рассады растений (в основном овощных
и декоративных культур), саженцев плодово-ягодных
и декоративных деревьев и кустарников. В почвенном
комке, окружающем корневую систему пересаживаемого
растения, могут находиться как слизни (чаще молодь),
так и их яйца. Кроме того, нередки случаи заноса слизней
в сооружения защищенного грунта с тарой, полиэтиле-
новой пленкой, цветочными горшками и другими пред-
метами, на которых могут находиться моллюски. Слизней
и их яйца можно обнаружить в массе органического
удобрения (торфа, перегноя, навоза) и почвы, вместе с ко-
торыми вредители попадают на поля, в парники, теплицы,
оранжереи. Часто для укрытия зимующих растений
используют лиственный опад, старое сено, солому и дру-
гой материал, с которыми также возможен занос слизней
на культивируемый участок.
Поскольку слизни обладают относительно невысокой
мобильностью, значение таких случайных заносов нельзя
недооценивать. Нужно учитывать, что гораздо проще
6 И. М. Лихарев и Я. С. Шапиро
81
предотвратить подобные заносы, чем ликвидировать пХ
последствия.
В распространении слизней на небольшие расстояния
возрастает значение активного расселения («своим хо-
дом»). Особенно велика роль активного расселения в пе-
риод возрастания численности слизней, когда наряду
с увеличением плотности отдельных поселений моллюсков
происходит их интенсивное расползание из резерваций
на соседние участки. Вследствие активного расселения
от слизней систематически страдают культурные растения,
выращиваемые вблизи лесных и лесопарковых массивов,
заброшенных садов, пустошей с дикой травянистой рас-
тительностью и других мест скопления моллюсков.
Одна из причин высокой вредоносности слизней на при-
усадебных участках — весьма частое соседство последних
с резервациями этих вредителей.
Благодаря активному расселению слизни проникают
в парники, теплицы, оранжереи, подвалы с хранящейся
сельскохозяйственной продукцией.
Таким образом, важным фактором вредоносности слиз-
ней в конкретном местообитании нередко становится их
активное расселение. Поэтому регистрация, прогнозиро
вание и возможность предотвращения этого явления
имеет большое практическое значение.
Для регистрации передвижения слизней можно про-
водить их мечение посредством клеймения переднего,
свободно свисающего края мантии («капюшона») нагретой
проволочной петлей. Варьирование числа и расположе-
ния ожогов дает до 10 и более различных комбинаций,
а клеймение одного моллюска занимает не более 3—5 сек.
Метка, нанесенная таким способом, сохраняется на ман-
тии слизня пожизненно и хорошо различима. Простота
и возможность применения в полевых условиях описан-
ного метода позволяют считать его более удобным, чем
метод холодового ожога с использованием жидкого азота
(Richter, 1973, 1976). Наблюдения за мечеными слизнями
разного возраста на протяжении всей их жизни в лабора-
тории не выявили существенных отклонений в выживае-
мости, темпах роста и развития от контрольных моллю-
сков.
Активное расселение слизней зависит от комплекса
экологических факторов, среди которых погодные условия
и пищевой режим местообитания наиболее важны. Тесная
связь мобильности слизней с температурой и влажностью
82
среды обитания анализируется в ряде работ (Dainton,
1954; Runham, Hunter, 1970; Duval, 1972; Backer, 1973).
Исследования Дмитриевой и Шапиро (1975) показали,
что условия увлажнения оказывают сильное влияние на
интенсивность расселения слизней из резерватных стаций
на сельскохозяйственные земли; предложено аналитиче-
ское выражение зависимости интенсивности миграций
моллюсков от гидротермического коэффициента Селяни-
Рис. 26. Влияние влажности среды на интенсивность расселения слизня
сетчатого.
По оси абсцисс — гидротермический коэффициент (ГТК)* по оси ординат —
процентное отношение числа мигрантов из резерваций к числу слизней в
популяции-доноре, кривая — графическое изображение регрессии, точки —
экспериментальные данные.
нова (ГТК). Последующие исследования, проведенные
Я. С. Шапиро в 1977—1981 гг. в хозяйствах Ленинград-
ской области, подтвердили весьма высокую достоверность
и возможность использования названного метода в целях
прогноза изменений стациального распределения вреди-
телей в агро ландшафте. На рис. 26 приведено сравнение
данных, полученных в результате наблюдений за рассе-
лением сетчатого слизня, с теоретическими (расчетными)
значениями индекса миграции.
Наблюдения Дыовэла (Duval D., 1972) показали,
что в сухую погоду у сетчатого слизня в полевых условиях
хорошо выражен хоминг (возврат к исходной точке дви-
жения), тогда как в период дождей хоминг отсутствует
и передвижения моллюсков более свободны. Вероятно,
хоминг может быть выражен у разных особей в различной
степени и, в зависимости от условий увлажнения, охва-
83
тывать большую или меньшую часть популяции. Поэтому
наблюдается весьма плавное увеличение интенсивности
расселения слизней по мере возрастания значения ГТК
(рис. 26).
Важный фактор, определяющий направление актив-
ного расселения слизней, — это обеспеченность их кор-
мом в пределах конкретного местообитания. Благоприят-
ные для слизней кормовые условия сельскохозяйственных
угодий стимулируют расселение вредителей из биотопов
с дикой растительностью на участки с культурными рас-
тениями. Существенное влияние на направление мигра-
ции оказывает и различная привлекательность для слиз-
ней тех или иных культурных растений. Их смена в про-
цессе севооборота определяет вполне прогнозируемые
изменения в распространении и численности вредителей.
Весьма характерны миграции слизней, вызванные раз-
личиями в сроках созревания и уборки разных сельско-
хозяйственных культур, возделываемых на соседних участ-
ках. С полей, занятых рано убираемыми культурами (ран-
ними овощами, земляникой), слизни активно переползают
на участки с поздними или зимующими культурами (кор-
неплодами, поздними сортами капусты, клевером, ози-
мыми злаками). Такие миграции вредителей могут и
должны прогнозироваться.
Расселение слизней зависит, разумеется, и от физико-
химических свойств поверхности, по которой слизни
передвигаются. Известны вещества и материалы, затруд-
няющие передвижение моллюсков; на их использовании
основаны некоторые приемы борьбы со слизнями (с. 145).
Помимо условий внешней среды, на мобильность
слизней влияет состояние самих животных. Так, слизни
разного возраста обладают различной способностью к ак-
тивному расселению. Из данных, представленных на
рис. 27, следует, что с увеличением массы моллюска воз-
растает скорость его передвижения в полевых условиях,
однако достоверных различий в мобильности слизней
4-го и 5-го весовых классов не обнаружено.
Скорость расселения слизней связана с их возрастным
различием и влияет на численность. Если на залежных
участках отмечается большая доля половозрелых особей,
чем на соседних полях, то, вселившись на эти поля (обы-
чно в конце лета—начале осени) и принеся здесь потом-
ство, эмигранты оказывают влияние на численность и
84
возрастную структуру исконной для этого места популя-
ции. Это и далее приводит к последующему уменьшению
численности моллюсков на залежных землях и к ее уве-
личению на соседних полях.
Данные о заселении слизнями вновь создаваемых сель-
скохозяйственных угодий показывают, что традиционные
представления о медлительности этих животных часто
не согласуются с фактами. Так, небольшие участки с кор-
мовыми для слизне!! культурами (такими как земляника,
Рис. 27. Встречаемость пяти весовых классов сетчатого слизня на разном уда-
лении от места выпуска маркированных слизней спустя 10 сут.
По оси абсцисс — расстояние от места выпуска слизней, по оси ординат —
частота встречаемости, 1—5 — весовые классы (см. табл. 7).
салат, редис), делянки селекционных и коллекционных
посевов заселяются вредителями в течение одного сезона
на всю глубину. На больших массивах при благоприятных
погодных условиях слизни могут проникнуть за один
сезон в глубину массива на расстояние до 100 м. Эти
наблюдения согласуются с приведенными выше экспери-
ментальными данными. Они показывают, что в сельско-
хозяйственном ландшафте Нечерноземья, с характерной
для него мелкоконтурностыо полей, между отдельными
популяциями слизней устанавливаются тесные кон-
такты, определяющие целостность всей совокупности
поселений. Тесная зависимость расселения вредителей
от погодных условий позволяет прогнозировать этот про-
цесс, а знание биологии отдельных агроценотических
популяций — предвидеть влияние расселения на после-
дующую динамику численности слизней.
85
В заключение отметим, что большую помощь в прак-
тической работе по составлению прогнозов численности
и распространения вредных видов слизней может оказать
агромалакологическое картирование территории сельско-
хозяйственных угодий колхоза или совхоза или отдель-
ного агромассива. Такое картирование проводится после
ряда предварительных наблюдений.
1. Выявление резерваций слизней.
Рис. 28. Карта-схема пространственного распределения сетчатого слизня
(Ленинградская об л., 1979 г.). Максимально заселенная территория и воз-
растная гетерогенность (Д-индекс) поселений моллюска.
Эту работу особенно удобно проводить в годы с низкой
численностью моллюсков, когда практически все их насе-
ление представлено резерватными популяциями. Однако
выявить резервации слизней с известной достоверностью
можно и в годы с высокой численностью вредителей на осно-
вании названных выше геоботанических критериев,
а главное — путем определения и сопоставления индексов
возрастной гетерогенности различных популяций слизней
(рис. 28).
2. Выявление стаций расселения.
Стации расселения моллюсков можно установить с до-
статочной достоверностью лишь в годы широкого рас-
пространения вредителей на картируемой территории.
При этом важно определить наибольшую границу рас-
пространения рлизней и выявить те природно-хозяй-
86
стйёйные факторы, of которых заёйсит йх йрострайс1всп-
ное распределение (удаленность от резерваций, микро-
рельеф и физико-химические свойства почвы, степень
дренированности участка, привлекательность во вделы-
ваемых растений для вредителей и другие).
3. Выявление основных направлений расселения слиз-
ней.
Из сопоставления данных п. 1 и 2 выясняют основные
направления расселения слизней из резерваций на со-
седние поля при наступлении благоприятных погодных
условий.
Таким образом, на составленной карте отмечают ре-
зервации и стации расселения слизней с указанием их
площадей; структурно-фенологические особенности по-
пуляций вредителей в основных угодьях; направления рас-
селения из резерваций и последовательность заселения
отдельных участков картируемой территории. Особенно
важно составлять такие карты в хозяйствах, используе-
мых службой сигнализации и прогнозов в качестве ба-
зовых, где проводятся детальные учеты численности и
распространения вредителей. Агромалакологическое кар-
тирование осуществляется на основании достаточно про-
должительных и корректно выполненных наблюдений
за состоянием популяций моллюсков. Методические ре-
комендации по этим наблюдениям изложены в следующей
главе.
ВЫЯВЛЕНИЕ, УЧЕТ ЧИСЛЕННОСТИ
И ОПРЕДЕЛЕНИЕ ВРЕДОНОСНОСТИ СЛИЗНЕЙ
Успешная борьба с вредными слизнями, так же как и
с другими вредителями, в значительной степени зависит
от своевременного их выявления, установления видового
состава, учета численности и распространения на терри-
тории хозяйства, сигнализации сроков борьбы. Правиль-
ная организация защиты растений от вредителей преду-
сматривает также и разработку прогнозов их появления и
распространения. На этой основе возможна своевремен-
ная и обоснованная подготовка к проведению борьбы
с вредителями.
87
Выполнение этой работы требует постоянных наблю-
дений за территориальным распределением, динамикой
численности и вредной деятельности слизней.
Полевые исследования
Обычно используют два основных метода выявления
слизней и нанесенных ими повреждений растениям —
маршрутные обследования и детальные учеты. Последние
проводятся, как правило, на выделенных стационарных
участках (в базовых хозяйствах).
Маршрутные обследования. Осмотр тер-
ритории, заселенной вредными слизнями, нацелен на
выявление моллюсков, установление заселенных ими пло-
щадей, а также определение площади поврежденных рас-
тений. Обоснованное планирование этих обследований
обязательно должно учитывать фактическую тенденцию
в динамике численности слизней на обследуемой террито-
рии в прошедшие годы. Это необходимо для того, чтобы
рационально определить подлежащие обследованию уча-
стки, технику проведения обследования и избежать тем
самым ненужной работы.
Многолетние наблюдения позволяют выделить следу-
ющие основные фазы динамики численности и распределе-
ния слизней на сельскохозяйственных землях (с. 79).
1. Фаза депрессии — вредитель распространен только
в резервациях, и наносимый им ущерб культурным расте-
ниям хозяйственно неощутим.
2. Фаза увеличения численности и расселения — интен-
сивное размножение слизней в резервациях и расселение
на прилегающие культивируемые участки.
3. Массовое размножение — интенсивное размножение
вредителей и заселение ими обширной территории.
4. Спад численности — гибель временных поселений
слизней и резкое сокращение численности резерватных
популяций.
В годы депрессии и спада численности основное внима-
ние при проведении обследований необходимо уделять
выявлению на территории хозяйства всех резерваций
слизней. Ими обычно служат залежные и целинные уча-
стки, особенно с дикой древесно-кустарниковой расти-
тельностью, пойменные луга и другие земли, подвергну-
тые длительному задернению, а также межи, пустоши, при-
усадебные участки и коллективные сады, расположенные
88
возле агромассивов. В процессе маршрутных обследова-
ний, пользуясь земельным планом участка, следует опре-
делить приблизительную площадь резерваций вредителей.
Кроме того, необходимо оценить их кормовую «емкость»,
которая определяется биомассой кормовых для слизней
растений, в первую очередь двудольных сорняков с большой
листовой поверхностью (репейника, мать-и-мачехи, подо-
рожника, одуванчика). Необходимо также глазомерно
оценить приблизительную численность слизней и
вероятность их расселения на соседние участки с куль-
турными растениями в случае наступления благоприятных
погодных условий.
Данные, полученные при обследовании резерваций,
служат основой для принятия решения об их ликвидации
или организации борьбы со слизнями. Кроме того, их
необходимо учитывать при размещении сильно поврежда-
емых слизнями растений.
Маршрутные обследования в фазу увеличения числен-
ности и расселения слизней должны охватывать как при-
родные и антропогенные резервации моллюсков, так и
расположенные поблизости участки с культурными расте-
ниями. Следует иметь в виду, что в первую очередь слизни
заселяют участки с предпочитаемыми растениями (капу-
стой. салатом, земляникой, горохом). Поэтому появление
слизней па этих культурах свидетельствует о начале актив-
ного расселения моллюсков из резерваций на культиви-
руемые участки. Позднее вредители заселяют и другие
культуры, особенно возделываемые на тяжелых по меха-
ническому составу почвах (глинистых и суглинистых),
а также на участках со слабым дренажем.
В период массового размножения слизней (если его
не удалось предотвратить) обследуют все основные типы
сельскохозяйственных угодий (или всю территорию кол-
лективного сада), определяют площади, заселенные вре-
дителями, и масштабы их вредной деятельности. Данные
обследований наносят на земельный план и руководству-
ются ими при проведении детальных учетов численности
слизней и поврежденности растений.
Если в прошедшие годы маршрутные обследования не
проводились и, следовательно, территориальное распре-
деление и фаза динамики численности слизней не из-
вестны, обследование начинают с резерваций, а затем
переходят на прилегающие к резервациям окультуренные
участки. Если вредители будут обнаружены и там, об-
89
следуют все остальные угодья, где есть повреждаемые
слизнями растения.
Маршрутные обследования в центральных и северо-
западных областях Нечерноземной зоны РСФСР проводят
в период с мая по октябрь. Сроки и кратность обследова-
ний уточняют в зависимости от вида вредителя (или ком-
плекса видов), фазы динамики численности и простран-
ственного распределения слизней, агроклиматических
условий и преследуемых целей.
Обследования резерваций в годы депрессии популяций
слизней проводят после полного отрождения слизней рода
дероцерас из перезимовавших яиц (то есть в конце весны).
Такое обследование позволяет одновременно выявить пере-
зимовавших слизней рода арион, которые к этому времени
становятся достаточно активными. Установить начало
активного расселения слизней из резерваций, т. е. насту-
пление 2-й фазы динамики численности, позволяют обсле-
дования резерваций и прилегающих к ним полей в период
откладки яиц слизнями дероцерас (сентябрь—октябрь).
Если обследование проводится в фазу активного рас-
селения или массового размножения вредителей, стано-
вится необходимым определить наибольшую площадь,
заселенную моллюсками в текущем сезоне (на этой площа-
ди будут зимовать отложенные слизнями яйца). Для этого
обследование проводят в конце периода откладки яиц
слизнями дероцерас. В Ленинградской области у главного
вредного вида — сетчатого слизня — этот период совпа-
дает с понижением среднесуточной температуры до
4—5 °C. После этого дальнейшее расселение слизней в те-
кущем сезоне обычно не наблюдается. Во время осенних
обследований глазомерно оценить численность вредите-
лей легко, поскольку в этот период они достигают крупных
размеров и очень активны.
В годы с повышенным против нормы количеством осад-
ков и умеренно теплой погодой приблизительно со второй
половины лета начинается расселение слизней и освоение
ими новых мест обитания. Этот процесс необходимо ре-
гистрировать, а число обследований с этой целью увели-
чить до 1 —2 в месяц.
Методика маршрутных обследований природных резер-
ваций слизней и культивируемых участков, где они оби-
тают, различна. Если резервация представляет собой круп-
ный массив (лес, рощу, лесопарк, обширные кустарнико-
вые заросли или природное пастбища), то маршрут прсь
90
кладывают" на его территорий вдоль границы с сельскохо-
зяйственными землями, отступя в глубь массива на 20—
30 м. Островные резервации подлежат сплошному обсле-
дованию. Расстояние между станциями-пробами зависит
от экологической однородности участка: на больших сель-
скохозяйственных угодьях оно составляет 50—100 м, а на
территории приусадебных участков и коллективных са-
дов — 10—20 м.
В начале фазы расселения слизней из резерваций нет
необходимости в проведении сплошного обследования при-
легающих к ним полей. Обследование в этом случае на-
чинают с краевой полосы посева, прилегающей к резерва-
ции вредителей, и удаляются от нее вглубь до тех пор,
пока будут обнаруживаться слизни или следы их жизне-
деятельности. Сплошное обследование территории проводят
в фазу массового размножения вредителей и заселения
ими большой площади.
Диагностика повреждений растений слизнями в про-
цессе маршрутных обследований обычно не вызывает
трудностей. Наиболее надежные признаки — это следы
радулы моллюсков и слизь (бесцветная, молочно-белая,
желтая, оранжевая), которые остаются на растении после
питания вредителей.
Маршрутные обследования по возможности следует
проводить в те часы суток, когда активность и вредная
деятельность слизней наибольшие (рано утром по росе,
а днем — в пасмурную погоду или сразу после дождя).
При этом активно ползающих и питающихся моллюсков
легче выявить, а свежие повреждения растений диагно-
стируются с большей надежностью.
Слизни относятся к таким вредителям растений, кото-
рые даже в годы массового размножения не заселяют всю
площадь участка с кормовыми для них культурами. По-
этому в ходе обследований необходимо установить заселен-
ные и незаселенные площади (в процентах) по каждому ти-
пу угодий (овощные пропашные культуры, многолетние
травы, сенокосные луга, ягодные плантации и т. д.),
причем отдельно по каждому обследованному хозяйству.
Результаты маршрутных обследований наносят на карту.
Они служат исходным материалом для проведения более
точных учетов численности и вредоносности слизней на
конкретном участке.
Таким образом, маршрутные обследования должны
проводиться в точно установленные сроки и охватывать
91
вполне определенные места обитания слизней в зависимо-
сти от состояния их популяций. Наряду с хозяйствами,
типичными для обследуемого района, обследования сле-
дует проводить и в хозяйствах, отличающихся по микро-
климату, почвенному покрову, рельефу, общему уровню
организации производства. В этом случае можно получить
наиболее полное представление об отклонениях в терри-
ториальном распределении, фенологии и динамике числен-
ности слизней, а также о причинах этих отклонений. Осо-
бенно это важно для агроландшафтов северо-западных
областей Нечерноземья, где благоприятный для слизней
климат в сочетании с высокой экологической неоднород-
ностью сельскохозяйственных земель определяют суще-
ствование популяций слизней, принципиально различаю-
щихся по структуре и динамике численности. В любом
случае маршрутные обследования должны проводиться не
менее чем на 15—20 % площади, на которую будут затем
экстраполироваться полученные данные. Поскольку в ди-
намике численности слизней важную роль играют резер-
вации, все они по возможности должны обследоваться на
заселенность вредными моллюсками.
Помимо маршрутных обследований, проводятся деталь-
ные учеты, которые дают материал о динамике численности
слизней, наступлении определенных периодов в их жизни,
о нанесенном ими вреде. Такие учеты обычно проводят
на постоянных участках, выявленных при маршрутных
обследованиях. Рассмотрим виды детальных учетов.
Учет яиц слизней. Яйца слизней учитывают
методом почвенных проб. Пробы закладывают размером
0.1—0.25 м2. Их глубина зависит от мест обитания слиз-
ней: в природных резервациях моллюсков, а также на
других участках, подвергнутых длительному задернению,
она может быть не более 5—7 см, на пашне же увеличива-
ется до 10—15 см. Почвенные пробы помещают в пакеты
или мешочки и анализируют в лаборатории. Кроме соб-
ственно почвы, просматривают и весь находящийся на ее
поверхности субстрат (лесную подстилку, дерн, стерню,
другие растительные остатки). Количество проб зависит
от размера и экологической однородности участка: на
больших однородных сельскохозяйственных массивах за-
кладывают 10—12 проб на площади до 10 га, на приуса-
дебных участках и в коллективных садах — 5—8 проб
на каждые 100 м2 территории.
Сроки проведения учетов яиц зависят от вида слизня,
92
характера местообитания, погодных условий и задач
исследования. Осенний учет позволяет оценить запас зи-
мующих яиц слизней дероцерас, что важно для составле-
ния прогноза численности этих вредителей на будущий
год. Его проводят после завершения откладки яиц, т. е.
в конце осени. Для того чтобы установить сроки откладки
яиц, проведение учетов начинают при обнаружении в по-
пуляции первых половозрелых слизней. Особенно важно
установить эти сроки на участках, предназначенных для
посева озимых культур, поскольку в случае раннего раз-
множения моллюсков в том же году возможно появление
второго (осеннего) поколения слизней, которые останутся
здесь зимовать и сильно повредят прорастающие семена
и всходы озимых.
Весенние учеты позволяют оценить состояние перези-
мовавших яиц. Яйца слизней находятся обычно на не-
большой глубине, в течение осени и весны они могут за-
топляться водой, подвергаться нападению паразитов и
хищников, грибковым заболеваниям, а в случае суровой
и малоснежной зимы — вымерзать. Поэтому численности
жизнеспособных яиц слизней к весне может заметно сокра-
титься. Учеты яиц проводят рано весной, сразу после раз-
рушения снежного покрова, в местах осенней откладки
яиц вредителями.
Жизнеспособность яиц определяют в лаборатории.
В первой половине лета к откладке яиц приступают
слизни ариониды. Как правило, они откладывают яйца
на залежных и прилегающих к ним участках, в садах и
на огородах. Из отложенных яиц в том же году отрожда-
ется молодь, которая может нанести серьезный вред куль-
турным растениям. Для оценки этой возможности в наз-
ванные сроки следует провести учет численности яиц
арионов.
Учет слизней постэмбриональных
стадий. Он проводится так же, как и учет яиц, мето
дом почвенных раскопок. Следует в то же время иметь
в виду, что слизни, особенно молодые, могут находиться
не только в почве (на глубине до 15—20 см) и на ее поверх-
ности, но также и на растениях, в кучах ботвы и выпо-
лотых сорняков, внутри поврежденных плодов (огурцов,
томатов, яблок), корнеплодов (моркови, свеклы, репы),
клубней картофеля, а также в находящейся на почве
лиственной и травяной подстилке. Поэтому, помимо соб-
ственно Почвы, необходимо проанализировать и весь на-
ходящийся на ней мертвый и живой субстрат*
Слизней желательно доставить в лабораторию живыми.
Для этого взятые в ноле пробы помещают в мешочки из
мелкопористого материала, обеспечивающего свободный
доступ воздуха (например, из мельничного газа). Для
поддержания необходимой влажности субстрата внутрь
мешочка закладывают небольшое количество сочных ли-
стьев или сырого мха. Необходимые сведения о месте
сбора записывают на наружной этикетке. Класть такую
этикетку внутрь не следует, так как слизни могут ее съесть
или сильно загрязнить. Пробы с живыми слизнями до
доставки в лабораторию следует тщательно оберегать от
прямого солнечного света. Время от времени надо прове-
рять состояние слизней и влажность субстрата, заменять
высохшие листья свежими. Следует учесть, что в это время
главная опасность для моллюсков — это перегрев и
недостаток свежего воздуха.
При невозможности доставить пробы в лабораторию
извлечение моллюсков проводят в полевых условиях
с помощью одного из описанных ниже методов.
Сроки проведения учетов численности слизней зависят
от ряда факторов, в частности от стации обитания вреди-
телей. На участках, отведенных под озимые культуры,
учеты численности необходимо провести заблаговременно,
сразу же после уборки культуры-предшественника. В это
же время следует провести учет численности слизней на
соседних участках и оценить вероятность заселения ими
озимых. В случае высокой численности моллюсков следует
сигнализировать о проведении борьбы с ними, а после
проведения защитных мероприятий — обязательно опре-
делить их эффективность, т. е. провести повторный учет
вредителей до посева озимых культур.
Учеты численности слизней на озимых следует продол-
жать до окончания вегетации последних в текущем году,
поскольку именно осенью эти культуры наиболее сильно
повреждаются моллюсками. Весенние учеты здесь прово-
дят лишь в том случае, когда велика вероятность заселе-
ния озимого поля перезимовавшими слизнями ариони-
дами, а также если на озимых зимовали молодые слизни
дероцерас, отродившиеся из яиц осенью прошлого года.
На клеверных полях 2-го и 3-го года популяции слиз-
ней отличаются высокой возрастной неоднородностью —
наряду с яйцами здесь зимуют часто и молодые слизни
94
(с. 70); здесь же могут зимовать слизни ариониды. Вредо-
носность моллюсков на таких участках начинает прояв-
ляться уже ранней весной — в это время от них сильно
страдает отрастающий клевер. Поэтому здесь необходимо
провести ранневесенний учет вредителей и, в случае их
высокой численности, организовать борьбу с ними. Пери-
одические учеты проводятся в течение всего периода веге-
тации клевера; они позволяют судить о динамике числен-
ности слизней и своевременно принимать необходимые
защитные меры. При этом, учитывая высокое возрастное
разнообразие «клеверных» популяций слизней, следует
регистрировать их возрастную структуру, что имеет важ-
ное значение для выбора наиболее эффективного способа
применения моллюскоцида (с. 158). На трехлетием клевер-
ном поле численность слизней может быть очень высокой,
поскольку клеверные поля часто служат местами накоп-
ления вредителей. Поэтому после уборки клеверной массы
следует провести учет вредителей на этой территории,
чтобы оценить потенциальный вред, который слизни смо-
гут нанести последующей культуре этого участка и, в слу-
чае необходимости, провести уничтожение слизней.
Учеты численности моллюсков на овощных культурах
в открытом грунте начинают проводить с момента отрожде-
ния из яиц слизней дероцерас (в мае—июне). Динамику
отрождения следует прослеживать в течение одного ме-
сяца, так как в случае холодной весны оно может растя-
нуться. Если вероятность заселения овощных культур
слизнями, мигрирующими с соседних участков, велика,
то периодические учеты их численности следует проводить
вплоть до уборки урожая.
Нередко овощным культурам, выращиваемым в между-
рядьях садов, вредят обитающие здесь слизни ариониды.
Особую опасность для овощей представляют эти слизни
на приусадебных участках и в коллективных садах, где
создаются благоприятные условия для их вредоносности.
Учет численности арионид, проведенный сразу после раз-
рушения снежного покрова, позволяет оценить степень
их опасности для ранних овощей и заблаговременно при-
нять необходимые защитные меры.
Ягоды земляники, особенно выращиваемой на приуса-
дебных участках и в коллективных садах, часто повреж-
даются моллюсками, заселяющими земляничную планта-
цию в период сбора урожая. Решающая роль при защите
этой культуры от слизцей, как будет показано ниже,
принадлежит предупредительным мероприятиям. Поэтому
в конце мая—начале июня следует провести учет числен-
ности слизней как непосредственно на плантации культу-
ры, так и на соседних с ней участках в радиусе 100—150 м,
имея в виду возможность быстрого сплошного заселения
посадки мигрирующими вредителями в период созревания
ягод. Учету подлежат как перезимовавшие слизни арио-
ниды, так и отрождающиеся в этот период из яиц слизни
дероцерас. При высокой численности с ними проводят
борьбу. После уборки урожая учет повторяют и при необ-
ходимости вновь уничтожают слизней до начала их раз-
множения с целью предотвращения откладки ими яиц на
земляничной плантации и возле нее.
В коллективных садах и на приусадебных участках
учеты численности слизней имеют свои особенности. Здесь
выращивается большой ассортимент растений, сильно
различающихся по повреждаемости слизнями и срокам
прохождения уязвимых стадий развития. Кроме того,
в силу высокой экологической неоднородности индивиду-
альных участков популяции моллюсков, обитающие на
них, очень разнообразны по своей возрастной структуре
и фенологии. Площадь таких участков сравнительно не-
велика, поэтому их сплошное заселение вредителями
происходит очень быстро, а заселение отдельных гряд и
делянок — в еще более короткие сроки. Все это в сочета-
нии с мягким микроклиматом вблизи человеческого
жилья создает исключительно благоприятные условия для
массового размножения слизней, и их вредоносность на
таких участках бывает часто очень высокой. Поэтому
наблюдения за слизнями, учеты их численности здесь
следует проводить систематически, не реже одного раза
в 7—Ю дней. Особенно важны регулярные учеты вреди-
телей в сезоны с благоприятными для них погодными
условиями, поскольку во влажную погоду расселитель-
ная способность слизней резко возрастает и они могут
появиться на участке весьма внезапно и в большом коли-
честве.
Для проведения регулярных учетов слизней на неболь-
ших территориях кроме описанных приемов полезными
могут быть укрытия-ловушки, которыми могут служить
старые доски, куски фанеры, полиэтилен вой пленки,
мешковины, небольшие кучки выполотых сорняков и
ботвы, любые другие предметы, под которыми сохраняется
повышенная влажность почвы и воздуха. Укрытия-ло-
96
вушки равномерно распределяют на территории участка
на расстоянии 3—5 м, регулярно просматривают и уни-
чтожают скопившихся под ними слизней (с. 145).
Нередко слизни появляются в коллективных садах и
на приусадебных участках благодаря неумышленному
заносу с рассадой овощных, цветочных и других растений,
саженцами плодовых и кустарниковых культур, с торфом,
перегноем и другим материалом. Для предотвращения
заноса вредителей весь вновь поступающий посадочный
материал и органическое удобрение необходимо проверить
на присутствие слизней или их яиц. Следует учесть, что
значительно легче предотвратить появление на участке
моллюсков, чем уничтожить размножившихся вредителей.
Информацию о численности и вредоносности слизней
на отдельных участках нужно систематически обобщать
и оперативно решать вопрос о целесообразности истреби-
тельных мероприятий. Борьба со слизнями будет эффек-
тивной лишь тогда, когда она проводится на всей тер-
ритории садоводства, а также в расположенных возле
него природных и антропогенных резервациях этих вре-
дителей.
Учет повре ж ценности растений.
Разнообразный характер повреждений, наносимых слиз-
нями, а также широкая многоядность этих вредителей
определяют необходимость использования различных ме-
тодов учета их вредной деятельности.
При высокой численности и отсутствии эффективных
мер борьбы слизни нередко вызывают сплошную гибель
растений. В этом случае поврежденность культуры оце-
нивается обмером площади с погибшими растениями и
выражается в гектарах.
Если имеет место не сплошная гибель растений, а лишь
изреженность посева в результате уничтожения слизнями
семян, всходов или рассады, поврежденность также оце-
нивается в единицах площади, а степень изреженности
определяется по 3-балльной шкале:
балл 1 — изреженность слабая — погибло до 25 %
всходов;
балл 2 — изреженность средняя — погибло от 25 до
50 % всходов;
балл 3 — изреженность сильная — погибло свыше
50 % всходов.
Эту шкалу применяют для полевых культур сплошного
сева, а также для пропашных до проведения разреживания
7 И. М. Лихарев и Я. С. Шапиро
97
(прорывки). После прорывки изреженность пропашных
определяют по такой шкале:
балл 1 — изреженность слабая — погибло до 10 %
растений;
балл 2 — изреженность средняя — погибло от 10 до
30 % растений;
балл 3 — изреженность сильная — погибло от '30 до
60 % растений.
При изреженности пропашных растений более чем на
60 % посев считают полностью уничтоженным.
Таблица 7
Весовые классы слизней
Весовой класс Пределы весового класса по видам слизней, мг
сетчатый слизень полевой слизень
1-й Менее 50 Менее 50
2-й 50—250 50-150
3-й 251—500 151—250
4-й 501—1000 251-500
5-й Более 1000 Более 500
В тех случаях, когда слизни наносят повреждения
растениям, но не вызывают их гибели, проводится опреде-
ление частоты поврежденности (процент поврежденности
растений) и степени поврежденности (степень объедания
листвы). Для определения поврежденности листовой по-
верхности применяют следующую шкалу:
балл 1 — слабая поврежденность — объедено до 5 %
листовой поверхности;
балл 2 — заметная поврежденность — объедено от 5
до 25 % листовой поверхности;
балл 3 — средняя поврежденность — уничтожено от
25 до 50 % листвы;
балл 4 — сильная поврежденность — уничтожено
свыше 50 % листвы.
Такой шкалой пользуются при учете поврежденности
слизнями листовых овощей (капусты, салата, щавеля) и
зеленных культур (сельдерея, петрушки, укропа). При
оценке поврежденности слизнями генеративных органов
растений (плодов земляники, огурцов, томатов, цветков
декоративных растений), а также клубней картофеля,
корнеплодов свеклы, моркови, репы, редиса, луковиц и
клубнелуковиц тюльпанов, гладиолусов, гиацинтов и дру-
гих растений определяют процент поврежденных органов
от общего числа осмотренных, а степень поврежденности
оценивают по следующей шкале:
балл 1 — слабая поврежденность — объедена У4 часть
органа;
балл 2 — средняя поврежденность — объедена У4—
У2 часть органа;
балл 3 — сильная поврежденность — объедено более
У2 части органа.
Лабораторные исследования
Выделение слизней из почвенных
проб. Доставленные в лабораторию пробы следует
проанализировать возможно скорее, особенно при ис-
пользовании методов, основанных на выделении живых
моллюсков.
Слизней можно извлечь из проб методом ручного раз-
бора. Эффективность метода весьма высока, так же, впро-
чем, как и его трудоемкость. Применение этого метода
оправданно при небольшом количестве анализируемых
проб и наличии свободной рабочей силы. Почву при этом
высыпают из пакетов на поддоны, разрушают крупные
комки и тщательно просматривают под сильным источни-
ком света. Этот метод могут использовать садоводы-лю-
бители, поскольку он не требует специального оборудова-
ния; почвенные пробы в ясную солнечную погоду удобно
разбирать непосредственно на садовом участке.
При анализе большого количества проб следует приме-
нять более производительные методы. Одним из них явля-
ется промывка пробы водой через систему сит (диаметр
ячеек 5, 3 и 1 мм). После промывки каждое сито просма-
тривают и выбирают всех слизней. Для большей точности
содержимое каждого сита можно опустить в раствор суль-
фата магния или поваренной соли с плотностью 1.17.
Слизни вместе с другими органическими частицами всплы-
вают на поверхность. Недостаток метода заключается
в том, что он сопряжен с разрушением части мелких слиз-
ней, которые тем самым остаются неучтенными.
Другой способ выделения слизней — это выгонка их
на поверхность субстрата путем медленного (в течение 5—
10 ч) насыщения пробы водой, поступающей снизу. Для
7*
99
этого пробы, находящиеся в мешочке из мельничного газа,
следует поместить в поддон с водой (уровень воды в под-
доне постепенно поднимают). По мере насыщения пробы
водой слизни выползают на поверхность, где их собирают
и подсчитывают. При этом методе также возможна гибель
части мелких экземпляров, остающихся в толще субстрата.
Выделение из пробы яиц слизней также проводят ме-
тодом ручного разбора. Для этого почву рассыпают тон-
ким слоем на черной полиэтиленовой пленке или на дру-
гой темной поверхности, на фоне которой прозрачные
(на ранних стадиях развития) или светлоокрашенные
яйца слизней хорошо заметны, и просматривают под источ-
николм света. Яйца слизней удобно извлекать шпателем
или небольшой ложкой.
При большом объеме анализируемого материала вы-
борку яиц проводят путем просеивания почвы через набор
сит. Если почва очень влажная, ее следует перед просеи-
ванием подсушить. В то же время следует учитывать, что
при длительном пребывании в сухой почве яйца слизней
теряют тургор, сморщиваются и выделить их становится
невозможно. Поэтому при чрезмерном иссушении проб
их следует слегка увлажнить, в результате чего яйца
приобретут тургор и станут вновь хорошо заметны. Это
свойство яиц можно использовать в ситуации, когда не-
возможно провести быстрый анализ собранных в поле
проб. В этом случае пробы оставляют в воздушно-сухом
состоянии на срок до 1—2 мес, а затем увлажняют водой
и выделяют из них яйца описанным методом. Понятно,
что при этом яйца слизней, сохранив первоначальный
внешний вид, полностью утратят свою жизнеспособность.
Определение живой массы и воз-
раста слизней. В практике работы лабораторий
и пунктов диагностики и прогнозов станций защиты ра-
стений живую массу моллюсков можно определить сле-
дующим методом. Выделенных из проб моллюсков поме-
щают в сосуды с проложенной внутри влажной фильтро-
вальной бумагой и оставляют без пищи на 12 ч. После
этого слизней взвешивают на торзионных или аналитиче-
ских весах в зависимости от необходимой точности измере-
ния. Затем можно провести разбивку моллюсков на весо-
вые классы. Экспериментальные исследования показы-
вают, что при разбивке, указанной в табл. 7, молодые
слизни попадают в 1—3-й весовые классы, а половозре-
лые — в 4-й и 5-й.
100
Однако масса слизней одного возраста может варьиро-
вать и в более широких пределах, что связано с услови-
ями их местообитания, поэтому по массе моллюсков можно
лишь ориентировочно судить об их возрасте. Между тем
в целях сигнализации сроков полового созревания, на-
чала размножения слизней и прогнозирования их числен-
ности необходимо более точно устанавливать возраст и
возрастную структуру популяций моллюсков. Для этого
необходимо провести вскрытие и изучение продуктов,
продуцируемых половой железой моллюска. Это можно
сделать посредством взятия мазка из полости протока
гонады: мазок наносится на предметное стекло, окрашива-
ется и микроскопируется при увеличении Х140, Х280,
Х360. При этом в зависимости от стадии гаметогенеза
у взрослых слизней обнаруживают зрелые спермин или
же зрелые спермин вместе с яйцеклетками. Зрелые яйце-
клетки слизней имеют диаметр до 50 мкм (у сетчатого и
полевого слизней — 35—45 мкм), их протоплазма хорошо
окрашивается водным раствором конгорота в интенсивно-
оранжевый цвет, а ядро — в кирпичный. Спермин окра-
шиваются только в массе в бледно-оранжевый цвет. Их
длина у сетчатого и полевого слизней — до 100—140 мкм,
у арионов — до 200—250 мкм. Появление в протоке го-
нады зрелых половых продуктов (вначале спермиев, а за-
тем и яйцеклеток) свидетельствует о готовности слизней
к размножению.
Анализ большого количества живого материала поз-
воляет после приобретения некоторого опыта судить
о наступлении половой зрелости слизней по морфологии их
половых органов, не прибегая к микроскопированию. При
этом следует учесть, что морфологические особенности
гениталий слизней, указывающие на их возраст, четко
прослеживаются на свежефиксированном материале или
при вивисекции наркотизированных моллюсков.
Определение жизнеспособности
яиц. Учет численности яиц, отложенных слизнями, поз-
воляет прогнозировать наибольшую возможную числен-
ность вредителей на данном участке. Однако поскольку
часть отложенных яиц может быть неоплодотворенной,
а некоторые яйца утрачивают жизнеспособность в процессе
развития (не меняя при этом внешнего вида), то в целях
составления прогноза численности вредителей очень важно
определить фактическую жизнеспособность яиц. Для этого
яйца слизней в количестве 100 шт помещают в чашки Петри
101
с почвой, влажность которой поддерживают на уровне
85—90 % от полной влагоемкости, а температуру — 17—
19 °C. Слизней, отрождающихся из яиц, подсчитывают и
определяют процент жизнеспособных яиц. Период инку-
бации в этих условиях составит 2—4 недели в зависимо-
сти от вида моллюска.
Определение жизнеспособности яиц полезно провести
как в период их откладки моллюсками, так и непосред-
ственно перед вылуплением слизней. Сравнение получен-
ных данных позволит оценить влияние экологических
факторов на развитие эмбрионов.
В ряде случаев важно своевременно сигнализировать
о сроках завершения развития яиц и отрождения из них
моллюсков. Это необходимо, в частности, при высокой
численности вредителя, когда с ним необходимо вести
химическую борьбу (применяемые же моллюскоциды эф-
фективны только против вышедших из яиц вредителей).
Своевременная сигнализация основана на постоянных
полевых наблюдениях. Их трудоемкость можно заметно
снизить, если осуществлять дополнительный контроль за
яйцами слизней, собранными в природе и помещенными
в садки. Учитывая, что период размножения слизней на
разных участках может протекать в разные сроки, необ-
ходимо запастись достаточно большим и разнообразным
материалом, предназначенным для инкубации в садках.
В качестве садков можно использовать глиняные горшки
большого объема, которые наполняют почвой с места
сбора яиц. Яйца размещают в горшках с почвой на той же
глубине, на какой они были собраны в поле. Горшки затя-
гивают сверху мельничным газом и прикапывают в откры-
том грунте вровень с поверхностью почвы на отгороженной
площадке, например в вегетационном домике. Прикопан-
ные горшки систематически просматривают. Появление
в них первых вылупившихся слизней служит сигналом
к проведению учетов моллюсков на тех участках, где были
собраны яйца.
Содержание слизней в неволе. В про-
цессе наблюдений, проводимых за слизнями в полевых
условиях, многие явления их жизни остаются незамечен-
ными. Иногда нельзя установить вид молодых слизней,
и поэтому возникает необходимость содержать их в лабо-
ратории и доводить до хорошо диагностируемой стадии.
Содержание (воспитание) слизней в лаборатории или
в садках, выставленных в открытой природе, может быть
102
необходимым и для определения длительности развития,
числа генераций моллюсков, сроков прохождения отдель-
ных стадий жизненного цикла. Большой интерес могут
представить сведения о сравнительной повреждаемости
различных видов, разновидностей и сортов культурных
растений слизнями. Такие данные можно получить, вы-
кармливая содержащихся в неволе моллюсков интересую-
щим исследователя кормом. В садках-изоляторах, кроме
того, проводят предварительное исследование эффектив-
ности моллюскоцидов (с. 172).
Для длительного содержания слизней в неволе при-
годны различные садки-террариумы, тип и величина кото-
рых определяются поставленными задачами, размером и
числом содержащихся слизней. Следует учесть, что в сли-
шком малых садках слизни нередко нападают и калечат
друг друга. В то же время мелких моллюсков, помещен-
ных в большие садки, трудно отыскать. Удобны для со-
держания слизней стеклянные сосуды различного объема,
затянутые сверху марлей или мельничным газом (послед-
ний легче отмывать от слизи животных). На дно сосуда
помещают слой почвы, которую желательно взять с места
сбора слизней. Слой почвы должен быть достаточно тол-
стым и рыхлым, чтобы слизни могли найти или сделать
в нем углубление для откладки яиц или укрытия от днев-
ного света. Во всех случаях садки должны быть защищены
от солнца и обеспечивать свободный доступ воздуха при
высокой влажности. Садки необходимо периодически
очищать от экскрементов, слизи и остатков пищи.
Для питания растительноядных слизней можно ис-
пользовать те растения, которыми данный вид питается
в природных местообитаниях, а также специально выра-
щиваемые в лаборатории культурные растения. Хорошие
результаты получены при выкармливании слизней листь-
ями капусты, салата, клевера, гороха, плодами огурцов,
морковью, клубнями картофеля.
К содержащимся в лаборатории слизням подсаживать
собранных в природе моллюсков следует лишь после
длительного (до 2—3 недель) их пребывания в садках-
изоляторах. Это необходимо делать во избежание зараже-
ния лабораторных животных патогенными паразитиче-
скими организмами, носителем которых может быть вновь
поступивший материал.
Фиксация материала. Для умерщвления
и консервации слизней обычно применяют 70—75 %-ный
103
этиловый спирт. Этот метод вполне годится для обычных
исследований в области фаунистики, диагностики и систе-
матики слизней, так как позволяет сохранить их на
неопределенное время и проводить необходимые иссле-
дования по изучению строения этих животных. Однако
фиксация спиртом непригодна для гистологических и
цитологических исследований, которые требуют специ-
альных фиксаторов.
Перед фиксацией необходимо принять меры к тому,
чтобы слизень «заснул» в расправленном состоянии —
в таком виде он более удобен для анатомических исследо-
ваний. Для этого нужно поместить живых слизней в со-
суд с холодной, желательно кипяченой водой. Сосуд дол-
жен быть закрыт так, чтобы под крышкой не оставалось
воздуха. В воде слизни вытягиваются и через несколько
часов погибают.
Для ускорения усыпления слизня можно добавить
в воду несколько кристаллов ментола или несколько ка-
пель спиртового раствора этого вещества. Если в сосуд
с водой добавить немного нитрата кобальта [Co(NO3)2x
ХбН2О] и поместить в него слизней, то моллюски погиб-
нут через несколько минут, полностью расслабившись.
Раствор должен иметь интенсивно-розовый цвет.
После усыпления отмытых от слизи животных поме-
щают в 70—75 %-ный спирт. Перед этим крупным экзем-
плярам (длиной более 70 мм) следует ввести внутрь тела
спирт с помощью медицинского шприца. При отсутствии
шприца можно надрезать кожу слизня с левой стороны,
чтобы фиксатор свободно проникал внутрь тела.
Сосуд, в котором фиксируют и в дальнейшем сохра-
няют слизней, должен быть достаточно просторным, чтобы
слизни в нем располагались свободно в вытянутом состоя-
нии и чтобы объем спирта не менее чем в 3—4 раза превос-
ходил общий объем слизней. Во всех случаях через сутки,
а затем примерно через две недели необходимо сменить
спирт и повторять эту операцию периодически, пока спирт
в сосуде со слизнями не станет бесцветным и прозрачным.
Для изучения и демонстрации окраски и ее изменчи-
вости лучше фиксировать слизней в 4 %-ном водном рас-
творе формалина. В этом фиксаторе природная окраска
лучше сохраняется, чем в спирте. Но для препарирования
такой материал малопригоден, так как ткани становятся
неэластичными. При отсутствии спирта в крайнем случае
допустима фиксация слизней и в формалине (в указанной
104
концентрации). Однако iipii первой возможности такой
материал следует перевести в спирт, который после этого
необходимо сменить еще два раза с интервалами в 1.5—
2 недели.
Для сохранения образцов растений, поврежденных
слизнями, их засушивают между листами бумаги, поме-
щенными в гербарные сетки. Перед гербаризацией расте-
ния нужно тщательно расправить, а в необходимых слу-
чаях слегка прогладить теплым утюгом. Гербарные сетки
с растениями помещают под пресс или туго перевязывают
веревкой. Сочные части растений сохраняют залитыми
консервирующей жидкостью (например, формалином)
в стеклянной, хорошо закупоренной посуде.
Весь материал (слизней и образцы поврежденных ими
растений) снабжают этикетками. Этикетки пишут на плот-
ной бумаге тушью или простым карандашом, наклеивают
на боковую стенку банки с законсервированным материа-
лом или вкладывают в гербарий. В этикетке необходимо
указать вид моллюска или растения, время и место сбора
его, фамилию собравшего материал.
ОПРЕДЕЛЕНИЕ ВИДОВ СЛИЗНЕЙ
В Нечерноземной зоне европейской части СССР (вклю-
чая и Прибалтику) обитает 16 видов слизней. Из них
10 видов в большей или меньшей степени являются вредите-
лями многих сельскохозяйственных культур на полях,
огородах, садах, в закрытом грунте или их запасов в овоще-
и плодохранилищах. Остальные 6 видов являются безвред-
ными в силу их образа жизни (обитают в лесах) и по харак-
теру питания (питаются преимущественно шляпочными
грибами и лишайниками). Среди слизней — вредителей
культурных растений одни наносят больший ущерб, дру-
гие — меньший. Отдельные виды существенно различа-
ются по образу жизни, биологии размножения и жизнен-
ным циклам.
Поэтому для эффективной организации борьбы со
слизнями и составления научно обоснованных прогнозов
их численности очень важно знать, с каким видом имеешь
дело, т. е. необходимо точное определение. С этой целью
ниже дается таблица для определения видов слизней и
105
Гювидовые характеристики (очерки) каждого вида. Эти
очерки сопровождаются необходимыми рисунками.
Предлагаемая таблица для определения видов постро-
ена по принципу противопоставления признаков: тем при-
знакам, о которых сказано в одном пункте, в другом проти-
вопоставлены иные признаки, иногда совершенно проти-
воположного характера. Если первый пункт называется
тезой, то второй антитезой. Цифры, поставленные перед
каждым пунктом, означают их порядковую нумерацию.
Около каждого порядкового номера пункта в скобках
поставлено одно- или двухзначное число, указывающее
порядковый номер пункта, в котором приводятся противо-
положные признаки. Если признаки тезы не подходят
к определенному объекту, то надо переходить к антитезе.
Например:
1 (8). Тело слизня массивное и широкое, сзади плавно
закругленное.........................................
8 (1). Тело стройное, сзади заостренное.
В данном примере пункт 1 — это теза, а пункт 8 --
антитеза. Сравнивая определяемого слизня с тезой и ан-
титезой, надо решить, чему он больше соответствует. Если
данный слизень подходит к тезе — пункт 1, то надо пере-
ходить к следующему по порядку пункту 2 и сравнивать
слизня с признаками, приведенными в данном пункте
и в его антитезе. Если определяемый слизень более под-
ходит к антитезе 8, то далее следует переходить к следую-
щему по порядку пункту 9, опять-таки сравнивая свой
объект с признаками этой тезы и антитезы.
Определение заканчивается, когда у принятого пункта
справа стоит название вида. После этого для полной уве-
ренности в правильности определения следует прочесть
очерк по данному виду и сравнить определяемого слизня
с соответствующими рисунками,
Таблица для определения родов и видов слизней
Нечерноземной зоны европейской части СССР
1 (8). Тело массивное и широкое, сзади плавно закруг-
ленное. Задний конец ноги при движении выступает
из-под спины и имеет над подошвой хвостовую ямку,
сверху прикрытую треугольной лопастью
(см. рис. 4). Пневмостом лежит в передней половине
106
правого края маытии. Подошва не разделена двумя
продольными бороздами на три доли. (Род Арион)
2 (7). Даже у сократившихся слизней половое отвер-
стие располагается впереди мантийшй щели. Между
щелью и средней линией спины 14—16 радиальных
рядов морщин. Резервуар семяприемника яйцевид-
ный или заострен на конце.
3 (4). Мантия, кроме боковых полос, имеет мелкие тем-
ные пятна, особенно четкие после фиксации. На спине
боковые полосы имеют четкие верхние границы и
нечеткие (размытые) нижние. Эпифаллус без вздутия
у основания (вблизи атриума), расположен не-
сколько отступя от переднего конца с пояском из
темного пигмента . . . Арион окаймленный (с. 114)
4 (3). Мантия без пятен. Нижние границы боковых полос
спины всегда четкие, в то время как верхние иногда
могут быть нечеткими. Эпифаллус со вздутием
у основания и обычно без темной опояски.
5 (6). При жизни спина и мантия темно-пепельные
с коричневатым оттенком; после фиксации коричне-
вый оттенок исчезает и слизни становятся темно-
серыми или почти черными. Бока светлые, без жел-
тых или оранжевых полос. Длина при движенпи
до 25, при сокращении до 20 мм. Эпифаллус с не-
большим вздутием на границе с атриумом. Атриум
почти прямоугольный . . . Арион лесной (с. 117)
6 (5). При жизни спина и мантия светло-серые, без ко-
ричневатого оттенка. На боках ниже темно-серых
боковых полос имеется по одной желтой или оран-
жевой полоске, которые после фиксации исчезают.
Длина при движении до 50, при сокращении до
32 мм. Эпифаллус с крупным вздутием на границе
с атриумом. Атриум конический или треугольный
..........................Арион полосатый (с. 112)
7 (2). У сократившихся слизней половое отверстие
располагается позади или под мантийной щелью.
Между нею и средней линией спины 19—20 радиаль-
ных рядов морщин. Резервуар семяприемника более
или менее шаровидный, без^острой вершины . . .
..............................Арион бурый (с. 111)
8 (1). Тело стройное, сзади заостренное. Задний конец
ноги не выступает из-под спины и не имеет хвостовой
ямки. Пневмостом лежит в задней половине пра-
107
вого края мантии. Подошва двумя продольными
бороздами четко разделена на три доли.
9 (10). Тело очень стройное, узкое, червеобразно-ци-
линдрическое, в поперечном сечении круглое из-за
узкой подошвы. Спина резко сужается к острому
заднему концу, который при раздражении или после
фиксации сжимается с боков, приобретая форму
вертикального плавника. Киль простирается на
всю спину. На мантии от пневмостома отходят
вперед и пазад по одной неглубокой бороздке, кото-
рые вместе напоминают контур летящей птицы . .
......................Бётгерилла светлая (с. 128)
10 (9). Тело более массивное, цилиндрическо-веретепо-
видное, в поперечном сечении снизу заметно упло-
щенное за счет довольно широкой подошвы. Спипа
если сужена к заднему концу, то более плавно и
слабее. Киль (если имеется) не простирается на всю
спину, выражен слабее. Мантия без упомянутых
бороздок.
И (18). Морщины спины и боков короткие и плоские и
разделены неглубокими бороздками. Кишечник
двухпетлевого типа (рис. 12, /У); задняя кишка
располагается слева от основания мускула-ретрак-
тора тела. (Род Дероцерас).
12 (15). Слепая кишка хорошо развита (рис. 7, се), дли-
на ее в несколько раз больше ширины.
13 (14). Окраска слагается из светлого (кремового) фона
и темно-коричневого или почти черного рисунка
из пятен, которые в типичном случае образуют
сетчатый рисунок; у слабо пигментированных осо-
бей эти пятна разбросаны редко и не образуют сеть.
Гонада занимает задний конец внутренностного
мешка и почти не закрыта соседними органами.
Пениальная железа крупная и более или менее
разветвленная (рис. 14) ......................
.....................Слизень сетчатый (с. 118)
14 (13). Окраска от почти белой до кремовой, иногда
с легким коричневым оттенком, без темного рисунка.
Гонада лежит на значительном удалении от вершины
внутренностного мешка, сверху сильно или почти
полностью закрыта печенью п слепой кишкой.
Пениальная железа небольшая и имеет вид прямого
или изогнутого пальцевидного выступа, без всяких
108
следов ветвления (рис. 39) ...................
.........................Слизень полевой (с. 121)
15 (12). Слепой кишки нет или она представлена неболь-
шим «карманом» задней кишки, и длина этого вы-
ступа не больше его ширины.
16 (17). Пенис всегда имеется и хорошо развит. Он мешко-
видной формы, впереди несколько вздут, сзади
с 2 четкими боковыми выступами, нередко отогну-
тыми вперед (рис. 43) .......................
.....................Слизень Штурания (с. 126)
17 (16). Пенис может присутствовать (эуфаллпчпые особи),
по может и отсутствовать (афалличпые особи); встре-
чаются также экземпляры с рудиментарным пенисом
(гемифаллические). У эуфаллических особей пенис
крупный, более или менее цилиндрический, извитой
или спирально закрученный. Он слагается из 3—4
отделов (рис. 41).............Слизень гладкий (с. 123)
18 (И). Морщины спины и боков длинные, выпуклые и
разделены глубокими бороздками. Кишечник трех-
петлевого типа (рис. 12, III, IV); задняя кишка
(третья петля) располагается справа от основания
мускула-ретрактора тела.
19 (26). Имеется длинная слепая кишка (рис. 12, IV),
которая у взрослых животных простирается назад
до вершины внутренностного мешка.
20 (23). Кожа толстая. Рисунок образован многочисленны-
ми мелкими темными пятнами, которые не сливаются
в продольные полосы или ряды более крупных пятен.
Слизь желтая или оранжевая. (Род Лимакс).
21 (22). Верхняя сторона тела окрашена в серовато-зе-
леный или оливковый цвет. На этом фоне имеется
рисунок из серых пятен с нечеткими контурами
и мелкой сетки из микроскопических точек. Этот
рисунок исчезает на заметном удалении от края
подошвы. Пенис сильно извитой. Яйцевод состоит
из 2 отделов: заднего узкого и переднего вздутого,
толстостенного. В последний впадает семяприемник
.........................Слизень желтоватый (с. 133)
22 (21). Фоновая окраска желтая или серовато-желтая.
Рисунок того же типа, но темнее и более четкий.
Сетка из точек по бороздкам простирается до края
подошвы. Пенис слабо извитой, С-образпо загнут
или прямой. Яйцевод не подразделен на передний
и задний отделы, цилиндрический. Семяприемник
109
впадает в атриум между пенисом и яйцеводом . .
......................Слизень крапчатый (с. 135)
23 (20). Кожа тонкая. Рисунок образован темными про-
дольными полосами или рядами крупных пятен.
Слизь бесцветная. (Род Леманниа).
24 (25). Вид обитает только в лесах..................
....................Леманниа каемчатая (с. 137)
25 (24). В СССР вид обитает только в оранжереях . . .
.......................Леманниа валенсийская (с. 138)
26 (19). Слепой кишки нет.
27 (30). Крупные слизни — взрослые особи в движении
вытягиваются до 200 мм. Окраска верхней стороны
тела либо одноцветно-темная до черной, либо на
светлом фоне имеется рисунок из темных продоль-
ных полос или рядов пятен, а па мантии — из пятен.
Слизь бесцветная. Длина пениса не менее г/2 длины
тела. (Род Лимакс).
28 (29). Длина киля около г/2 длины спины; кожа более
прочная и толстая, с резкими и длинными морщи-
нами. Мантия одноцветная, без пятен. Боковые доли
подошвы чаще всего темные, а срединная светлая.
Щупальцы покрыты мелкими темными пятнышками.
Пенис равен или больше длины тела. Вид обитает
только в лесах................Слизень черный (с. 131)
29 (28). Длина киля около х/з Длины спины; кожа тоньше,
с менее резкими и более короткими морщинами, чем
у предыдущего вида. Мантия пятнистая. Вся подошва
светлая. Шупальца без пятнышек. Длина пениса —
около половины длины тела. На северо-западе
европейской части СССР обитает в населенных
пунктах......................Слизень большой (с. 132)
30 (27). Небольшие слизни — взрослые особи в движении
вытягиваются до 50 мм. Окраска верхней стороны
тела при жизни одноцветно-желтая, зеленовато-
или серовато-желтая, реже почти оранжевая. После
фиксации на мантии нередко заметен темный лиро-
образный рисунок, а па спине 2 продольные полосы.
Слизь желтая. Пенис короткий — длина его значи-
тельно меньше х/2 длины тела ........................
..................Малаколимакс нежный (с. 136)
110
Семейство Ариониды — Arionidae
Арион бурый — Arion subfuscus (Draparnaud, 1805)
(рис. 29, 30)
Тело удлиненное, при рассматривании сверху с почти
параллельными краями. Мантия занимает около х/3 длины
тела, овальная с широко закругленным задним концом.
Морщины спины тонкие, слабо выпуклые (объединены
в 19—20 рядов). Половое отверстие располагается непо-
средственно за мантийной щелью или под нею.
Окраска сильно варьирует как в пределах одной попу-
ляции, так и в течение жизни одной особи. Фон разных
оттенков, от коричневого до оранжевого, чаще ржавого
или серовато-коричневого цвета. Середина спины обычно
наиболее темная, даже темно-шоколадная, боковые поло-
сы неразличимы. Иногда встречаются особи с четкими поло-
сами на спине и с лирообразным рисунком на мантии. Этот
рисунок наиболее обычен для молодых особей и на ранней
фазе половой активности. Обе цветовые формы встреча-
ются совместно. Вблизи от пневмостома от ближайшей
полосы как бы отходит клин, благодаря которому почти
все отверстие окружено темным пигментом.
После фиксации оранжевый пигмент исчезает и окраска
становится более серой или черной. Подошва всегда кре-
мовая. У полосатых особей, как правило, слизь оранже-
вая или желтая, а у бесполосых — бесцветная или слабо-
окрашенная.
При движении длина тела до 80 мм.
Кишечник двухпетлевый, более или менее перекру-
ченный. Яйцевод длинный и мускулистый; задний уча-
сток его тонкий и короткий, а передний — вздутый и
толстостенный. Семяприемник с длинным протоком. Срав-
нительно длинный и тонкий семяпровод плавно переходит
в длинный и извитой эпифаллус, который на границе
с атриумом окружен заметным валиком.
В СССР очень широко распространен по лесным и ле-
состепным областям европейской части; вне СССР — почти
вся Европа.
Обитает в лиственных, смешанных и хвойных лесах.
В антропогенных биотопах живет сравнительно редко и
преимущественно в старых парках и на кладбищах.
Излюбленной пищей служат шляпочные грибы, но
питается также отмершими частями растений и трупами
111
животных. В теплые сырые годы этот слизень может
наносить некоторый ущерб полям и огородам, расположен-
ным по соседству с лесом и кустарниками.
Жизненный цикл — см. с. 44—45.
Рис. 29. Арион бурый.
Рис. 30. Передняя часть гениталий
ариона бурого.
9п — эпифаллус; остальные обозна-
чения как на рис. 14.
Арион полосатый1 — Arion fasciatus (Nilsson, 1823)
(рис. 31, 32)
Слизень заметно более сплющен и относительно более
широкий, чем другие виды арионов. У молодых экземпля-
ров по середине спины проходит слабый, но ясный киль;
у взрослых он едва заметен. Морщины тонкие и слабо
выпуклые; между мантийной щелью и средней линией
спины 14—16 рядов морщин. Половое отверстие лежит
1 До недавнего времени этот вид смешивали с арпоном окаймленным
(см. ниже).
ИЗ
впереди мантийной щели (даже при сокращении живот-
ного).
Phc.j.31. Арион полоса-
тый."
Рис. 32. Гениталии ариона полоса-
того.
эп — эпифаллус; остальные обозначе-
ния как на рис. 14.
Тело почти всегда светлое, как бы выцветшее. Фон кре-
мовый или желтовато-пепельный. Середина спины и ман-
тии несколько темнее периферии, без каких-либо пятны-
шек. Боковые полосы темно-пепельные, с четкими гра-
3 И. М. Лихарев и Я. С. Шапиро 113
йиЦамй сверху и снизу. У живых Слизней, кроме темйьП
полос, несколько ниже них заметны слабее выраженные
узкие желтые, оранжевые или красноватые полоски.
При фиксации спиртом эти полоски вместе с желтоватым
оттенком фона исчезают и тело становится светло-пепель-
ным. Подошва при жизни кремовая, после фиксации белая.
Слизь обычно бесцветная, иногда желтая.
При движении длина тела до 50 мм.
Кишечник двухпетлевого типа; сравнительно с кишеч-
ником у других видов арионов слабо перекручен.
Гениталии по сравнению с арионом лесным и окайм-
ленным заметно крупнее. Яйцевод тонкий, короткий и не
разделен на передний и задний участки, как у ариона бу-
рого. Семяпровод очень длинный, а эпифаллус необычно
больших размеров: длинный, толстый и на границе с атри-
умом имеет очень большое вздутие. Резервуар семяприем-
ника яйцевидный или клювообразно вытянутый; проток
его очень большого диаметра и перед впадением в атриум
вздут. Атриум обычно имеет вид сплющенного конуса,
узким концом обращенного наружу.
В СССР обнаружен в ряде областей Северо-Запада
и Центра европейской части. В силу отчетливого тяготе-
ния к антропогенным биотопам трудно определить, какие
местонахождения принадлежат к природному ареалу,
какие обязаны завозу. Вероятно, широко распространен
почти по всей Европе.
Явно предпочитает жизнь в культурных биотопах —
огородах, садах, полях, парках и др. Наносит существен-
ный вред многим сельскохозяйственным культурам; в этом
отношении данный слизень стоит на втором месте после
слизня сетчатого.
Арион окаймленный — Arion circumscriptus Johnston, 1828
(рис. 33, 34)
Тело удлиненное, узкое и слегка приплюснутое. Рельеф
поверхности и положение полового отверстия, как у ари-
она полосатого. Окраска очень характерная. Спина голу-
бовато-стального или пепельного цвета, иногда темно-
серая. Середина спины и мантии темная; на этом фоне за-
метны небольшие и нерегулярно лежащие темные пятны-
шки, которые более отчетливо видны после фиксации,
особенно на мантии. Боковые полосы темно-серые, иногда
почти черные; верхние границы их четкие, тогда как ниж-
114
ние размытые, так как пигмент плавно переходит на бока,
которые к подошве постепенно светлеют. Тем не менее
Рис. 33. ЛрпонХ'окайпленный*
пигмент хотя и в малом ко-
личестве, но доходит до края
ноги. Подошва при жизни
голубовато-белая, после фик-
сации кремовая.
При движении длина тела
до 40, чаще до 25 мм.
Кишечник, как у ариона
полосатого, но сильнее пе-
рекручен.
Рис. 34. Гениталии ариона
окаймленного.
ап — эпифаллус; остальные обо-
значения как на рис. 14.
Яйцевод почти такой же толщины, как и эпифаллус.
Семяпровод длинный и тонкий. Эпифаллус короткий
и тонкий, у основания без кольцевидного валика, но
с четким кольцом из темного пигмента. Семяприемник
8*
115
обычно с сильно вытянутым резервуаром и узким прото-
ком, без вздутия у основания. Атриум сильно сплющен
Рис. 35. Арион лесной.
и очень длинный, обычно
длина его в 2 раза больше
ширины.
В пределах Нечерно-
земной зоны европейской
части СССР найден пока
только в окрестностях Ле-
нинграда. Возможно, оби-
тает в природных биото-
пах Прибалтики и запад-
ных областей Белоруссии
и Украины. Вне СССР —
Рис. 36. Гениталии ариона
лесного.
ап — эпифаллус; остальные
обозначения как на рис. 14.
северная и средняя Европа и сопредельные страны.
Обитает во влажных смешанных и лиственных лесах,
особенно в ольховых. Безвреден для культурных растений.
116
Арион лесной — Arion silvaticus Lohmander, 1937
(рис. 35, 36)
Тело сильнее сплющено, чем у ариона окаймленного,
но слабее, чем у ариона полосатого. Самой характерной
является окраска. Спина темная, а бока ниже полос очень
светлые. При жизни спина и мантия темно-пепельные,
со слегка коричневым оттенком. Характерные для ариона
окаймленного черные пятнышки на спине и мантии у дан-
ного слизня отсутствуют, и спина несколько более темная.
Боковые полосы почти черные. Благодаря темной спине
верхние границы полос нечеткие, зато нижние хорошо
выделяются на фоне светло-кремовых боков. Подошва
обычно совершенно белая, но с более кремовым оттенком,
чем у ариона окаймленного. Слизь бесцветная.
После фиксации коричневый оттенок окраски спины
исчезает и она становится пепельной или черноватой, а
подошва кремовой. Контраст между темной спиной и свет-
лыми боками становится еще более резким.
При движении длина тела до 25 мм.
Гениталии данного вида по форме занимают промежу-
точное положение между арионом окаймленным и арионом
полосатым. Прежде всего это касается размеров и формы
эпифаллуса — он средней величины, без кольца из тем-
ного пигмента, с небольшим вздутием на границе с атри-
умом. Образование, сходное со вздутием эпифаллуса, но
меньших размеров, чем у ариона полосатого, имеет про-
ток семяприемника. Яйцевод короткий и толстый, как
у ариона окаймленного. Наиболее характерным является
атриум, который почти прямоугольной формы, сплю-
щен, и длина его лишь немного больше ширины.
В СССР данный вид обнаружен в Литве, в окрестностях
Ленинграда и в Карпатах. Вне СССР — значительная
часть Европы.
Обитает в смешанных и лиственных лесах, особенно
в долинах рек и вблизи водоемов.
В силу своего образа жизни культурным растениям не
опасен.
117
Семейство Агриолимациды — Agriolimacidae
Слизень сетчатый — Deroceras reticulatum (Muller, 1774)
(см. обложку и рис. 7, 14, 37)
Животное с сильно выпуклой спиной; задний конец
клиновидно сужен. Мантия занимает около 2/5 длины тела.
У взрослых слизней фон грязно-кремовый, светло-кофей-
ный или оливково-кремовый. Обычно имеется четкий
рисунок, образованный коричневыми, черноватыми или
темно-бурыми пятнами. Чаще всего эти пятна образуют
рисунок типа неправильной сетки (отсюда и название
вида). Темный пигмент в первую очередь концентриру-
ется по бороздкам. Взрослые особи обычно с пятнами,
молодые — долгое время лишены их. Интенсивность ок-
раски и плотность расположения пятен очень индиви-
дуальны, даже внутри одной популяции. Общее потемне-
ние окраски обычно происходит к концу осени. Чаще
всего пятна располагаются наиболее густо на спине и ман-
тии, а по бокам — реже. Голова и шея покрыты более
мелкими пятнами; щупальца черноватые. Подошва кре-
мовая, а у меланистических (темно-окрашенных) особей
коричневая, всегда одноцветная. Слизь бесцветная, при
раздражении особи молочно-белая.
При растяжении длина тела до 35, реже до 45 мм; при
сокращении до 25 мм.
Задняя кишка с хорошо развитой слепой кишкой,
длина которой в 2 раза и более превосходит ширину
(рис. 7).
Крупная бурая гонада в мужской фазе достигает
заднего конца внутренностного мешка и лишь по краям
слегка перекрыта печенью. У половозрелых особей
(рис. 14) пенис крупный и с четким перехватом посере-
дине, но у молодых слизней это сужение может и отсут-
ствовать. Внутри переднего отдела пениса находится
крупный конический стимулятор. Снаружи основание
стимулятора отмечено легкой вогнутостью стенки пениса.
На заднем конце пениса сидит крупная пениальная
железа, форма которой отличается исключительно боль-
шой изменчивостью. Она может быть одностебельчатой —
с лопастями или короткими веточками по краю или двух-
и трехстебельчатой, причем каждый стебель дает вторич-
ные ответвления или лопасти.
И8
В задний конец пенйса справа й немного бнизу впадает
семяпровод, а несколько впереди крепится мощный
Рис. 37. Слизень сетчатый. Внешний вид (сбоку и сверху) и его внутренняя
раковина.
половой ретрактор. Кроме последнего, имеется еще один
довольно крупный мускул, соединяющий передний отдел
пениса со стенкой тела, и несколько мускульных тяжей
между обоими отделами пениса. Семяприемник овальный
119
с четким протоком. Яйцевод и атриум цилиндрические,
причем последний довольно короткий.
Хотя данный вид несомненно европейского происхож-
дения, трудно четко охарактеризовать его природный
ареал. Это связано с его тяготением к агроценозам, лег-
костью завоза на новые места и явственной синантропией.
В европейской части СССР он обнаружен почти во всех
республиках и областях. Появление популяций этого
вида на Кавказе (Ереван, Орджоникидзе) и в Казахстане
(Алма-Ата) обязано завозу. Завезен также на другие кон-
тиненты и на многие острова.
Обитает на открытых местах, явно избегая лесов и
даже кустарников, преимущественно на глинистых поч-
вах. Обычен на лугах, на полях, огородах, пустошах,
свалках, в компостах, а в городах — в парках, садах и на
кладбищах. Днем укрывается под комками почвы, кам-
нями и другими предметами, лежащими на поверхности
земли, и в трещинах почвы.
Сроки размножения и число генераций зависят от
климата данной местности (рис. 16). На северо-западе
европейской части СССР копуляция и откладка яиц
начинаются чаще всего в конце лета и в начале осени.
Обычно здесь зимуют только яйца. При благоприятных
погодных условиях (теплое и влажное лето) размножение
начинается раньше и появляется обычно немногочислен-
ная летняя генерация слизней, часть которой после благо-
получной зимовки приступает к размножению раньше,
чем генерация, вылупившаяся из перезимовавших яиц.
Откладка яиц начинается через 10 дней после копуля-
ции. Один слизень в несколько приемов откладывает до
700 яиц. При оптимальной температуре (15—17 °C) и вы-
сокой влажности вылупление молоди происходит на 11-й
день. Однако в природе такие сроки относятся только
к летним генерациям. Обычно же яйца, отложенные
осенью, остаются в покое всю зиму и первую половину
весны, пока не установится теплая погода без ночных
заморозков.
Основной пищей слизня служат зеленые части расте-
ний, а также плоды и корнеплоды. Особенно предпочи-
тает молодые побеги и всходы различных полевых и ого-
родных культур. Большая прожорливость в сочетании
с высокой численностью в благоприятные («слизневые»)
годы, явное тяготение к культурным биотопам — вот
причины, почему этот слизень в определенных климати-
120
ческих и хозяйственных областях стал бичом полей и ого-
родов. В СССР он чаще всего вредит в западных и северо-
западных областях европейской части, особенно на тяже-
лых глинистых почвах. Известны случаи, когда этот сли-
зень местами полностью истреблял урожай капусты и
всходы озимых.
Слизень полевой, или пашенный — Deroceras agreste
(Linnaeus, 1758) (рис. 38, 39)
Тело более стройное, чем у слизня сетчатого, но мас-
сивнее, чем у слизня гладкого. Спина выпуклая, в попе-
речном сечении полукруглая. Мантия занимает около
1/3 длины тела.
Окраска от почти белой до кремовой с легким корич-
невым оттенком, без темного рисунка. Мантия и спина
немного темнее боков. Подошва тоже кремовая; ее края
немного темнее середины. Слизь бесцветная; при раздра-
жении слизень может выделять молочно-белую или мут-
ную слизь.
При растяжении длина тела до 40, при сокращении
до 35 мм.
Кишечник, как у слизня сетчатого.
Гонада у молодых более или менее треугольная,
светло-кофейного цвета, у взрослых она небольшая
овальная или лопастевидная, темно-шоколадного цвета.
В отличие от слизня сетчатого она меньших размеров
и никогда не располагается на вершине внутренностного
мешка, а лежит приблизительно на границе между задней
и средней третью тела, вблизи от слепой кишки или под
нею. У взрослых особей (с небольшой гонадой и крупной
белковой железой) пенис с перетяжкой посередине. У мо-
лодых экземпляров эта перетяжка выражена слабо или
совсем отсутствует и пенис имеет мешковидную или почти
цилиндрическую форму. Передняя часть пениса заметно
вздутая, так как внутри находится крупный конический
стимулятор. Пениальная железа имеет вид небольшого
пальцевидного или крючковидного, не ветвящегося при-
датка, лежащего у заднего конца пениса. Половой рет-
рактор крепится к пенису приблизительно у границы
его задней и средней трети. Здесь же впадает семяпровод.
Это место нередко заметно окрашено темным пигментом.
Овальный резервуар семяприемника цлавно переходит
121
в короткий проток. Атриум довольно длинный и цилин-
дрический.
В СССР
достоверно
распространен почти повсеместно. Вне СССР
известен из средней и северной Европы.
Рис. 38. Слизень полевой.
Обитает на откры-
тых местах, таких как
луга, болота, придорож-
ные канавы. Явно избе-
гает местообитаний с на-
рушенным дерновым
покровом, почему ма-
лочислен на полях,ого-
родах и в садах. Встре-
чается на лесных опуш-
ках и в зарослях ольхи,
но не углубляется
внутрь леса.
На северо-западе
европейской части СССР
откладка яиц обычно
происходит осенью и
на зимовку остаются
только яйца. В более
южных областях, кроме
Рис. 39. Передняя часть
гениталий слизня по-
левого.
Обозначения как на
рис. 14.
яиц, зимуют полувзрослые слизни, которые приступают
к размножению в конце весны—начале лета. В связи
с этим осенью можно наблюдать 2—3 генерации. Только
что вылупившиеся слизни имеют длину 3.5 мм; тело их
белое и просвечивающее.
В связи с тем что данный вид длительное время путали
со слизнем сетчатым, щироко расцространилось мнение,
122
что он является серьезным сельскохозяйственным вреди-
телем. В действительности его роль как вредителя не-
сравненно меньшая, чем слизня сетчатого, так как он
менее тяготеет к полям и огородам, да и к тому же и чис-
ленность его обычно значительно ниже. Однако в благо-
приятные годы он может существенно вредить на при-
усадебных участках и в коллективных садах.
Оба вида несомненно близкородственны и во многом
сходны, однако при внимательном изучении их всегда
можно отличить друг от друга по окраске, гонаде и пе-
ниальной железе (см. выше).
Слизень гладкий, или проворный — Deroceras laeve
(Muller, 1774) (рис. 40, 41)
Животное очень подвижное и беспокойное. Тело ци-
линдрическое и стройное, блестящее, мягкое, просвечи-
вающее, при сокращении веретеновидное с притуплен-
ными концами. Мантия относительно очень большая,
при жизни сзади закругленная, после фиксации нередко
угловатая; длина ее, как правило, не уступает длине
спины или лишь немного меньше. При ползании голова
и шея выступают вперед сильнее, чем у других видов
того же рода. Киль очень короткий и тупой и заметен
лишь при ползании. При сокращении морщины довольно
четкие и крупные.
При жизни окраска почти одноцветная, чаще всего от
красновато-коричневой до почти черной, очень редко бо-
лее светлая до темно-кремовой. При фиксации проступают
темные пятнышки, которых особенно много на мантии;
на спине их меньше и они располагаются по бороздкам.
Щупальца толстые и черные с крупными глазными взду-
тиями. Подошва относительно узкая, светлее верхней
стороны тела; срединная доля ее темнее, чем боковые,
за счет просвечивания внутренностей. Слизь жидкая
бесцветная.
При растяжении длина тела до 25, мантии до 10 мм;
при сокращении тело 13—16, реже до 22, мантия 5.5 —
7.0 мм.
^Кишечник без слепой кишки.
Очень характерной чертой этого вида является при-
сутствие даже в пределах одной популяции как особей
с нормально развитыми мужскими половыми путями —
эуфалличные особи, так и экземпляров, у которых пенис
123
и семяпровод полностью или частично редуцированы —
а фал личные и гемифал личные особи.
Небольшая темная шаровидная гонада обычно почти
полностью закрыта кишечником и печенью.
У эуфаллических особей пенис крупный, более или
менее цилиндрический, извитой или даже спирально
JSX А А) закрученный. Он слагается
из 3—4 отделов: 1) узкий
передний отдел, открываю-
щийся в атриум; 2) очень
вздутый отдел, нередко
образующий крупный выс-
туп, в котором лежит ко-
нический или полусфери-
ческий стимулятор; 3) зак-
рученный отдел, в задний
конец которого впадает
семяпровод, а немного
впереди крепится половой
ретрактор; 4) узкий сле-
пой выступ нередко на
конце с 2—3 бугорками.
Редукция пениса про-
является в различной сте-
пени. У гемифал личных
он становится коротким
изогнутым цилиндром, сое-
диняющим семяпровод
Рис. 40. Слизень гладкий.
с атриумом. У афалличных — это небольшое вздутие на
атриуме, которое не связано с семяпроводом (последний
может отсутствовать, или сохраняется лишь его началь-
ная часть, слепо замкнутая на переднем конце); имеется
слабый половой ретрактор. Афалличные особи встре-
чаются чаще, чем гемифал личные.
Яйцевод цилиндрический, разной длины. У афаллич-
пых особей он обычно короче, но несколько вздут. Атриум
цилиндрический, длинный и тонкостенный.
В СССР распространен почти повсеместно. Общее рас-
пространение — холодные и умеренные области север-
ного полушария.
124
Среды всех слизней Евразии — это один из наиболее
влаголюбивых и холодостойких видов. Обычными местами
обитания являются болота, берега небольших пресных
водоемов, как природных, так и искусственных, где мо-
жет находиться на почве, на растениях, в том числе и па
Рис. 41. Гениталии слизня гладкого.
Слева — афаллические, справа — эу фаллические:
остальные обозначения как на рис. 14.
подводных частях прибрежных водных растений. Обитает
также на влажных лугах и в сырых лесах. Довольно
долго выдерживает пребывание под водой. Встречается
в природных и в антропогенных биотопах. В обоих слу-
чаях популяции отличаются сравнительно постоянной, но
невысокой численностью.
Половое созревание наступает раньше, чем у других
видов того же рода. Откладка яиц идет в течение большей
125
части теплого времени года. Вылупление молоди проис-
ходит через 4—5 недель. Молодые слизни имеют длину
около 4 мм, бесцветные, просвечивающие. Голова и щу-
пальца светло-розовато-коричневые. Через несколько дней
пигментация усиливается, распространяясь с головы на
все тело.
Зимуют как яйца, так и слизни разного возраста.
В силу своих высоких требований к постоянной и вы-
сокой влажности роль этого вида как вредителя культур-
ных растений ограничена в основном тепличными хозяй-
ствами, в особенности работающими на гидропонике.
В подобных условиях при разведении ремонтантной гвоз-
дики в одном из цветоводческих хозяйств Ленинграда
были отмечены существенные убытки от слизня гладкого.
Кроме того, благодаря высокой подвижности этого
вида, по нашим наблюдениям, во влажных садах он не-
редко заползает на ягодные кустарники и вредит созре-
вающим ягодам.
Слизень Штуранпя — Deroceras sturanyi (Simroth, 1894)
(рис. 42, 43)
Тело не такое стройное, как у слизня гладкого, с вы-
пуклой спиной и резко суженным задним концом. Мантия
довольно крупная, занимает около х/2 длины тела; при
движении задний конец ее располагается в задней поло-
вине тела. Кожа очень тонкая и просвечивающая, со сла-
бым рельефом; после фиксации она теряет прозрачность,
рельеф сглаживается, а на боках появляются нитевид-
ные белые продольные полоски.
Окраска одноцветная без рисунка из темных пятен и
полос, довольно изменчивая в отношении оттенков. Чаще
всего грязновато-кремовая или серовато-коричневая.
Иногда встречаются целые популяции коричневого или
даже шоколадного цвета или наряду со светлыми отдель-
ные меланпстические особи с почти черной спиной. Ман-
тия и середина спины всегда окрашены несколько темнее,
чем бока и подошва. Слизь водянистая и бесцветная.
При растяжении длина тела 60—70, при сокращении
до 35 мм.
Слепая кишка либо отсутствует, либо представлена
небольшим «кармашком».
Небольшая темная гонада лежит вблизи задней кишки
или под нею. Пенис очень характерной формы, напоми-
126
нающей молоток. Впереди он несколько вздут, а задний
конец с 2 четкими боковыми выступами, нередко отогну-
тыми вперед. Эти выступы различны по форме и разме-
рам, так как один из них обычно к концу несколько су-
Рис. 42. Слизень Штурания.
Рис. 43. Гениталии слизня
Штурания.
Обозначения как на рис. 14.
жен и длиннее другого. Иногда более длинный выступ
по внутреннему краю расчленен на короткие лопасти.
Семяпровод и половой ретрактор крепятся к пенису
с правой (внешней) стороны между упомянутыми высту-
пами. Внутри переднего отдела пениса находится крупный
конический стимулятор с тупой вершиной. Семяприем-
ник с удлиненным резервуаром и тонким протоком и по
длине немного больше пениса.
Внешне особи данного вида (особенно темные) обнару-
додвают некоторое сходство со слизнем гладким: у обоих
127
слизней мантия относительно длинная; при ползании
задний конец ее располагается в задней половине тела.
Однако после фиксации оба вида обнаруживают четкие
различия: у слизня гладкого проступают темные пятнышки,
а у слизня Штурания — белые нитевидные полоски.
Светлые особи слизня Штурания на первый взгляд
можно принять за слизня полевого. Однако если у пер-
вого кожа очень тонкая, просвечивающая и со слабым
рельефом, то у второго она более толстая, не просвечиваю-
щая и с четким рельефом. Кроме того, у последнего вида
мантия относительно короче, чем у первого.
Наиболее четкие различия между упомянутыми тремя
видами слизней обнаруживаются в строении кишечника
и пениса.
В СССР слизень Штурания обнаружен сравнительно
недавно (Лихарев, Виктор, 1980). В частности, он встре-
чен в нескольких местах Московской, Калининской и Во-
ронежской областей, а также в ботанических садах и при-
городных полях и огородах Ленинграда, Челябинска,
Ростова-на-Дону и других городов. В природных биото-
пах он найден в буковых лесах близ Ужгорода и на пой-
менных лугах Верхней Волги.
Общее распространение этого вида изучено пока не-
достаточно.
По своему хозяйственному значению слизень Штура-
ния близок к слизню полевому. Он отмечен как вредитель
многолетних кормовых трав, а на приусадебных участках
и в коллективных садах вредит овощным, ягодным и де-
коративным культурам.
Семейство Бётгериллиды — BoettgeriHidae
Бётгерилла светлая — Boettgerilla pallens (Simroth, 1912)
(рис. 44, 45)
Небольшие слизни с очень стройным и узким телом;
в поперечном сечении почти круглые. Морщины длин-
ные, плоские, бороздки между ними неглубокие. Голова
и шея ^же остального тела; щупальца относительно тол-
стые и цилиндрические.
Спина резко сужается к заднему концу, который при
раздражении сильно сжимается с боков, образуя как бы
вертикальный плавник. Киль хорошо развит и простира-
ется на всю спину. Мантия удлиненно-овальная, сзади
128
Рис. 44. Бётгерилла светлая.
Рис. 45.'Гениталии бётгериллы
светлой.
в — влагалище, вт — веретено-
видное тело; остальные обозна-
чения как на рис. 14.
9 II. М. Лихарев и Я. С. Шапиро
заканчивается очень острым углом, располагающимся
на середине тела. Пневмостом лежит у задней половины
правого края мантии и окружен небольшим вздутием
или площадкой. Подошва трехдольчатая, очень узкая.
Окраска светло- или голубовато-серая или пепельно-
стальная, на спине темнее, по бокам книзу более светлая.
Молодые слизни белые или беловато-кремовые, реже со
слабым розовым оттенком. У особей, еще не достигших
половой зрелости, спина серая, а бока белые. Подошва
всегда белая или кремовая.
L
Рис. 46. Слизень черный.
При растяжении длина тела до 60, при сокращении
до 30 мм.
Для гениталиев наиболее характерно следующее: сред-
ний участок семяпровода вздут и превращен в толстостен-
ное очень мускулистое веретеновидное тело; семяприем-
ник впадает в женские пути, сильно отступая от атриума,
благодаря чему яйцевод короткий, а влагалище длинное.
В природных условиях этот вид обитает в широколист-
венных лесах Абхазии и Западной Грузии. В настоящее
время интенсивно завозится за пределы естественного
ареала. Так, в СССР он обнаружен в парках Выборга,
Ленинграда, Львова, Орджоникидзе и Еревана, в оран-
жереях Москвы и Челябинска. Кроме того, в антропоген-
ных биотопах он встречен во многих странах средней и за-
падной Европы.
Однолетние животные. Большую часть жизни прово-
дят в почве, в которой прокладывают ходы. Предпочи-
тают тяжелые глинистые почвы. Укрываются под круп-
ными камнями, глубоко погруженными в землю. Популя-
ции отличаются невысокой численностью.
Питаются мицелием грибов. Безвредны для культур-
ных растений.
130
Семейство Лимациды — Limacidae
Слизень черный — Li max cinereoniger Wolf, 1803
(рис. 46, 47)
Тело стройное, к хвосту заостренное, с довольно длин-
ным килем, равным половине длины спины. Мантия спереди
закругленная, сзади угловатая, закрывает около 2/4 жи-
вотного. По внешнему об-
лику походит на слизня
большого (см. ниже), но
кожа более толстая и проч-
ная, а морщины более длин-
ные и резкие.
Окраска сильно варьи-
рует. На востоке Европы
у этого вида большая
часть тела окрашена в чер-
ный или темно-серый цвет;
реже ниже киля на свет-
лом фоне располагаются
1, 2 или 3 пары продоль-
ных темных полос или ря-
дов пятен. Мантия темная,
одноцветная, по краям
иногда с мелкими светлыми
точками. Киль всегда свет-
лее остальных участков
спины, чаще всего белый.
Боковые доли подошвы
темнее средней, но у мо-
лодых слизней иногда вся
подошва светлая. Темные
щупальца испещрены чер-
ными или бурыми точ-
ками. Свежевыл упившаяся
молодь (длина 8—9 мм)
вначале бесцветная и про-
свечивающая, но вскоре
делается непрозрачной,
Рис. 47. Гениталии слизня чер-
ного.
Обозначения как на рис. 14.
9*
131
а под конец черной. Слизь бесцветная, довольно жид-
кая.
При растяжении длина тела до 150—200 мм.
Пенис очень длинный, равен длине тела или более ее,
образует внутри тела сложные извивы. Семяпровод и по-
ловой ретрактор крепятся к заднему концу пениса.
В пределах европейского Нечерноземья этот слизень
распространен в Прибалтике, Южной Ка-
релии, в Белоруссии, западных и цент-
ральных областях РСФСР. Вне СССР —
почти вся Европа. Обитает в смешанных
и широколиственных лесах и в хвойных
лесах с хорошим травяным покровом.
О размножении и жизненном цикле
см. с. 45.
Питается преимущественно грибами и
лишайниками. Безвреден для культурных
растений.
Слизень большой — Li max maximus
Linnaeus, 1758 (рис. 48)
Тело стройное, к хвосту заостренное,
с коротким килем. Мантия спереди зак-
ругленная, сзади угловатая; она закры-
вает около 7з животного.
Окраска слагается из желтовато- или
пепельно-серого, или грязно-белого фона
и черного рисунка из 2 или 3 пар по-
лос или продольных рядов пятен. Ман-
тия испещрена черными пятнами, нередко
образующими меандрический рисунок.
Щупальца розовато-коричневые без тем-
ных точек. Подошва белая. Слизь бесцвет-
ная, густая.
При растяжении длина тела до 130 мм.
В отличие от близкородственного
слизня черного, у данного вида пенис бо-
лее короткий; семяпровод и половой рет-
рактор крепятся немного отступя от его
заднего конца.
Рис. 48. Слизень большой.
132
В виде изолированных друг от друга популяций оби-
тает в парках и садах, а также в теплицах, погребах,
овоще- и плодохранилищах многих городов северо-за-
падных и центральных областей и республик европейской
части СССР; в природных биотопах живет только в лесах
Карпат. Как синантропный вид обитает почти но всей
Европе. Завезен па другие континенты и острова.
Главным образом вредит в овоще- и плодохранилищах.
Слизень желтоватый — Li max fiavus Linnaeus, 1758 (рис. 49)
Внешне походит на слизня крапчатого (рис. 50).
Тело веретеновидное, в поперечном сечении почти
круглое, с очень коротким и нечетким килем. Мантия
закрывает не более г/3 тела. Кожа с тонким рельефом.
Рис. 49. Передняя часть гениталий слизня желтоватого.
Обозначения как на рис. 14.
Основная (фоновая) окраска взрослых слизней оливковая
или серовато-зеленая. Рисунок слагается из очень мелких
темных точек, которые концентрируются в основном по
бороздкам, но на спине местами переходят и на морщины,
образуя серые пятна с нечеткими контурами. Еще боль-
шее развитие эти пятна имеют на мантии, где они, частично
сливаясь друг с другом, слагают меандрический рисунок.
Ниже спины пятен меньше, темные точки сохраняются
только по бороздкам, образуя тонкую сеть, и плавно ис-
чезают на некотором удалении от подошвы. Свободные от
темного пигмента участки спины имеют облик светлых
жемчужин.
133
Слпзь верхней стороны тела желтая, а на мантии почти
оранжевая, прозрачная и^жидкая. Желтоватые тона ок-
Рис. 50. Слизень
крапчатый.
раски тела в значительной степени
обязаны слизи; поэтому после фик-
сации окраска становится более туск-
лой, почти серой.
При растяжении длина тела до
120 мм.
Третья петля кишечника очень
короткая, с длинной слепой кишкой,
которая плотно прилегает к верх-
ней стенке тела и нередко прости-
рается до заднего конца полости
тела (рис. 12, IV).
Пенис цилиндрический, сильно из-
витый; семяприемник впадает в пе-
реднюю часть яйцевода.
В СССР данный вид обнаружен
в погребах, подвалах и оранжереях
Москвы, Калининграда, Тарту, Са-
ратова, Одессы и Ростова-на-Дону.
Рис. 51. Передняя часть гениталий
слизня крапчатого.
Обозначения как на рис. 14.
В сходных условиях он обитает в большинстве стран
Европы. Природный ареал — многие прибрежные страны
и острова Средиземного моря.
Антропохорный вид. Обитая в овощехранилищах и по-
гребах, может наносить вред пищевым продуктам.
134
Слизень крапчатый — Liinax maculatus (Kaleniczenko, 1851)
(рис. 50, 51)
Внешне походит на слизня желтоватого. Главные от-
личия: основная окраска более светлая — желтая или
серовато-желтая; рисунок более четкий, темно-серый или
почти черный, также сла-
гается из различных по раз-
мерам и форме темных пятен
на спине и мантии и сетки
из микроскопических точек,
но эта сеть простирается до
подошвы, чего не бывает
у предыдущего вида; щу-
пальца темно-серые или почти
черные. Как и у слизня жел-
товатого, молодь всегда ок-
рашена темнее, чем взрос-
лые особи, нередко одноцвет-
но черная или с единичными
светлыми пятнами.
При движении длина тела
до 120 мм.
Кишечник, как у слизня
желтоватого.
Яйцевод на всем протя-
жении одинаков, без вздутия
впереди; семяприемник впа-
дает в атриум между пени-
сом и яйцеводом; пенис зна-
чительно короче, С-образно
Рис. 52ЛМалаколимакс нежный.
изогнут, если же извитый, то слабее, чем у слизня жел-
товатого.
В СССР в природных биотопах (в лесах) обитает в Крыму
и на Кавказе. Кроме того, как антропохорный вид обна-
ружен в погребах и оранжереях Пушкина (Ленинград-
ская область), Ленинграда, Севастополя, Орджоникидзе
и Тбилиси. Вне СССР установлен на Британских островах,
в Румынии, Болгарии и Турции,
135
Может наносить некоторый вред пищевым запасам.
В теплицах и оранжереях отмечен как вредитель овощ-
ных и декоративных растений.
Малаколимакс нежный — Malacolimax tenellus (Miiller, 1774)
(рис. 52, 53)
Тело сравнительно уплощенное, мягкое, с тонкой про-
свечивающей кожей. Мантия закрывает менее уз тела,
сзади со слабо выраженным углом. Морщины плоские,
бороздки неглубокие.
Рис. 53. Передняя часть гениталий малаколнмакса нежного.
Обозначения как на рис. 14.
При жизни слизни обычно одноцветные. Верхняя сто-
рона окрашена в желтый, зеленовато- или серовато-желтый
цвет, реже оранжевато-желтая, вокруг пневмостома более
светлая; голова и щупальца черные или темно-бурые.
Подошва светло-желтая или кремовая. Молодь обычно
окрашена светлее — кремовая или зеленовато-кремовая.
Слизь желтая, жидкая.
После фиксации нередко на мантии проявляется лиро-
образный темный рисунок и темное пятно в центре, а на
спине пара продольных полос. При этом желтый пигмент
быстро исчезает и тело становится беловато-кремовым.
При движении длина тела до 50 мм.
Вторая петля кишечника заметно короче третьей; сле-
пой кишки нет.
Для гениталий особенно характерно следующее. Семя-
провод перед впадением в пенис плавно расширяется,
срастается с ним, образуя ряд небольших пигментиро-
13(5
Ванных вздутий. Пенис коротко-цилиндрический или меш-
ковидный, у заднего конца вздутый, иногда неглубокой
перетяжкой разделен на короткий передний и более длин-
ный задний отделы.
В СССР населяет северо-
западные, западные и цен-
тральные области европей-
ской части. Общее распрост-
ранение — почти вся Европа.
Обитает только в лесах,
как на равнинах, так и в го-
рах. Изредка встречается
в старых парках. Явно пред-
почитает лиственные леса,
однако вместе с арионом бу-
рым является одним из не-
многих видов слизней, живу-
щих и в хвойных лесах,
особенно в сосняках. Скры-
вается в подстилке, под ва-
лежинами и в гнилых пнях.
Главной пищей служат
шляпочные грибы, а также
и некоторые лишайники. Мо-
лодь питается мицелием раз-
личных грибов. Безвреден
для культурных растений.
Рис. 54. Леманниа каемчатая.
Леманниа каемчатая — Lehmannia marginata (Muller, 1774)
(рис. 54, 55)
Очень мягкие и скользкие слизни. Тело стройное,
сзади заостренное. Киль очень короткий и слабо выражен;
от него к мантии простирается узкая светлая киле-
вая полоска. Мантия занимает около уз длины тела,
сзади с отчетливым углом.
Окраска фона и особенно рисунок сильно варьируют.
Фон чаще всего светло-кремовый или беловатый, а рису-
нок от темно-серого до почти черного. Мантия с двумя
боковыми полосами, которые сзади смыкаются, образуя
137
Лирообразный рисунок, внутри которого видна еще одна
нечеткая полоса или ряд пятен. На спине ниже светлой
килевой полоски имеются 1 или 2 пары темных полос;
нижняя пара нередко распадается на ряды пятен. Голова
и щупальца черноватые. Подошва и бока светлые.
При движении длина тела до 75 мм.
Кишечник не перекручен, трехпетлевый; имеется длин-
ная слепая кишка (рис. 12, IV).
Рис. 55. Гениталии леманнин каемчатой.
б •—бич; остальные обозначения как на рис. 14.
Пенис относительно крупный, к заднему концу кони-
чески расширяется. Здесь в него с одной стороны впадает
семяпровод, с противоположной — короткий роговидный
выступ (бич).
В СССР обитает в Прибалтике, на Карельском пере-
шейке, в западных областях Белоруссии и Украины.
Вне СССР — средняя и северная Европа.
Живет в смешанных и лиственных лесах. Питается пре-
имущественно лишайниками. В силу такого образа жи-
зни для культурных растений не опасен.
Леманниа валенсийская — Lehmannia valentiana
(Ferussac, 1823) (рис. 56, 57)
Тело умеренно стройное, сзади ясно заостренное.
Мантия занимает менее уз длины тела. Щупальца узкие
и длинные.
Окраска сильно варьирует. Фон чаще всего кремовый,
но бывает коричневатым и грязно-оранжевым. Рисунок
красновато- или темно-коричневый; на мантии всегда
2 боковые полосы, между которыми лежит нечеткая
средняя полоса или ряд пятен. На спине имеется пара
138
полос, доходящих до заднего конца тела, ниже которой
может быть еще пара, но менее четких полос или рядов
Рис. 56. Леманниа валенсий-
ская.
пятен. Задний конец тела
без рисунка или покрыт
неясной темной сетью из
штрихов или пятен. Щу-
пальца бурые. Подошва
одноцветная, кремовая,
слегка просвечивающая.
При движении длина
тела до 60 мм.
Кишечник, как у преды-
дущего вида.
Пенис конический,
с крупным бичом, имею-
щим форму широкой ло-
пасти. Бич впадает в пе-
нис рядом с ретрактором
с брюшной стороны (у ле-
маннии каемчатой он впа-
Рис. 57. Передняя часть
гениталий леманнии ва-
ленсийской.
б — бич; остальные обо-
значения как на рис. 14.
дает в пенис с противоположной стороны от ретрак-
тора).
В СССР как антропохорный вид обнаружен в садовод-
ствах Ленинграда, Ростова-на-Дону и Ташкента. Природ-
139
ним ареалом этого вида является Пиренейский полуостров
и, частично, Франция. Завезен на другие континенты
и острова. В Европе, вне природного ареала, известен
почти во всех странах, но здесь живет только в оранже-
реях и садоводствах, где питается листьями, плодами
и клубнями овощных и декоративных культур. При этом
он может наносить существенный ущерб.
БОРЬБА СО СЛИЗНЯМИ
Для эффективной борьбы со слизнями па сельскохо-
зяйственных землях необходимо, во-первых, иметь пред-
ставление об общих закономерностях стадиального рас-
пределения и динамики численности этих моллюсков
в агроландшафте; во-вторых, располагать оперативной
информацией о состоянии их популяций, получаемой
в результате постоянных наблюдений; в-третьих, пра-
вильно сочетать основные методы регулирования их чис-
ленности.
Тактика борьбы со слизнями изменяется в зависимости
от фактического состояния их популяций. Так, в фазу
депрессии численности основные усилия должны быть
нацелены на выявление и (если это возможно) ликвида-
цию природных резерваций слизней, которые, как было
показано выше, играют важную роль. Если это своевре-
менно сделать не удалось, то необходимо предотвратить
или, по крайней мере, уменьшить интенсивность рассе-
ления моллюсков из резерваций на поля при наступлении
благоприятных для них погодных условий. В том случае,
если борьба со слизнями начинается в фазу массового
размножения и широкого расселения вредителей, необ-
ходим строго дифференцированный подход к снижению
численности моллюсков в пределах конкретного угодья
с учетом особенностей защищаемой культуры, популяции
вредителя, микроклиматических и других условий.
В свете вышеизложенного предупредительные меро-
приятия по борьбе со слизнями в их резервациях могут
показаться неоправданными с хозяйственно-экономиче-
ских позиций в силу неокупаемости затрат на борьбу
прибавками урожая текущего или даже следующего се-
зона. Однако многолетние исследования показывают, что
невнимание к превентивным контролирующим мероприя*
140
тиям значительно усложняет борьбу с вредителями в годы
их массового размножения, когда возможность для при-
нятия профилактических мер уже упущена.
Борьба со слизнями в настоящее время слагается из
рационального применения агротехнического, механиче-
ского и химического методов. Химический метод борьбы —
наиболее эффективный, однако его целесообразно приме-
нять лишь в тех случаях, когда с помощью двух других
методов не удается снизить численность вредителей до
хозяйственно неощутимого уровня. Рассмотрим назван-
ные методы.
Агротехнический и механический методы
Агротехническая борьба слагается из хозяйственных
мероприятий, осуществляемых в процессе возделывания
культурных растений и приводящих к ограничению вред-
ной деятельности слизней. Эти мероприятия преследуют
следующие цели: изменить условия внешней среды в сто-
рону, неблагоприятную для моллюсков; увеличить по-
левую устойчивость защищаемых растений к слизням;
механически уничтожить вредителей. Механический ме-
тод борьбы — применение специальных приемов для вы-
полнения последнего пункта.
Основные агротехнические мероприятия по борьбе
со слизнями описаны ниже.
Мелиорация земель. Проведение мелиора-
тивных работ в Нечерноземье имеет исключительно важ-
ное значение для дальнейшей интенсификации сельскохо-
зяйственного производства. Это объясняется тем, что зна-
чительная часть потенциальных сельскохозяйственных
угодий здесь практически выпадает из хозяйственного
оборота. В зоне почти пятая часть земель заболочена;
многие участки покрыты кустарником и мелколесьем,
закочкарены и засорены камнями; размеры отдельных
полей, как правило, небольшие. Такие условия создают
благоприятные возможности для вредной деятельности
слизней, а мелиорация содействует ограничению числен-
ности вредителей. Особенно велико значение этих меро-
приятий на Северо-Западе СССР, климатические условия
которого наиболее благоприятны для развития моллюсков.
Основные культурнотехнические работы по коренному
улучшению земель, снижающие в то же время численность
слизней, это: уничтожение кочек на лугах и пастбищах;
141
гидротехническая мелиорация избыточно увлажненных
земель; приведение в пахотное состояние поверхности уго-
дий, имеющих выраженный микрорельеф; уборка камней.
Подробные сведения об этих и других мероприятиях со-
держатся в специальной литературе (Преображенский,
1983, и др.).
Обработка почвы. Наибольший эффект
в борьбе со слизнями дает такая обработка, при которой
в почве сокращается количество щелей и углублений,
служащих убежищами для моллюсков, и создается мелко-
комковатая, лишенная глыб почвенная структура. На-
пример, разрыхление пахотного слоя почвы трехкратным
боронованием может привести к сильному (на 25—30 %)
сокращению численности слизней (Runham, Hunter, 1970).
Особенно большое значение имеет предпосевная обра-
ботка почвы под культуры сплошного сева, прежде всего
под озимую рожь и пшеницу. В северо-западных и цен-
тральных областях Нечерноземной зоны РСФСР и в При-
балтике предшественниками озимых в севообороте часто
служат занятые пары с вико-овсяной смесью, многолет-
ними травами, вспаханными после первого укоса, дру-
гими рано убираемыми культурами. В этих угодьях слизни
нередко накапливаются в большом количестве и осенью
могут серьезно повредить всходы озимых. Очень важно
поэтому своевременно (за 3 недели до посева озимых)
убрать парозанимающую культуру, провести лущение
стерни, вспашку на полную глубину пахотного горизонта
с одновременным боронованием, а в дальнейшем, до по-
сева озимых, проводить систематическое уничтожение сор-
няков.
Уничтожение сорной растительности не только ухуд-
шает микроклимат местообитания для слизней, но и, при
отсутствии культурных растений, лишает вредителей
важного источника питания.
Создание рыхлой, мелкокомковатой почвенной струк-
туры и борьба с сорняками имеют важное значение и на
участках с пропашными культурами; это достигается
своевременной и качественной культивацией посевов в те-
чение сезона.
Как показали наши исследования в хозяйствах Ле-
нинградской и Новгородской областей в 1975—1980 гг.,
проведенная осенью качественная вспашка почвы явля-
ется препятствием для перезимовки на этом участке мо-
лодых и взрослых слизней. Поэтому подъем зяби после
№
многолетних трав или длительного задерненпя существен-
но снижает плотность популяции вредителей. Кроме того,
при этом сильно снижается возрастное разнообразие
поселений моллюсков, что заметно упрощает и повышает
эффективность химической борьбы с ними (см. ниже).
Посев и посадка растений. Ранние ве-
сенние сроки посадки рассады и посева семян яровых
культур всегда предпочтительны, поскольку в этом слу-
чае растения успевают пройти наиболее уязвимые по от-
ношению к моллюскам начальные стадии развития до-
массового вылупления из перезимовавших яиц главных
вредящих видов слизней. Если не ожидается чрезмерно
теплая и влажная осень и опасность перерастания ози-
мых невелика, то ранние сроки посева можно рекомендо-
вать и для этих культур: до наступления «пика» размно-
жения и вредоносности слизней растения успеют пройти
наиболее уязвимые стадии — прорастания семян и всхо-
дов.
Для снижения вредоносности слизней важное значение
имеет предпосевная подготовка и проверка качества поса-
дочного материала: дружные всходы семян, имеющих вы-
сокую энергию прорастания, в меньшей степени повреж-
даются моллюсками (табл. 8).
Высадку рассады овощных культур, сильно повреж-
даемых слизнями, также нужно проводить в наиболее
ранние агротехнические сроки. Если высадка рассады
на северо-западе зоны продолжается до 15—20 июня,
то заметно возрастает вероятность ее повреждения мо-
лодью моллюсков, отрождающейся в это время из пере-
зимовавших яиц.
Таблица 8
Зависимость повреждаемости пшеницы сорта Мироновская
808 сетчатым слизнем от продолжительности прорастания
семян (оригинальные данные)
Продолжитель- ность прораста- ния семян (сут) Число повторностей Повреждаемость (%)
прорастаю- щие семена всходы
8 6 5.2 7.0
10 6 9.4 13.7
13 6 12.6 18.5
15 6 14.3 22.7
18 6 17.5 25.8
143
Установлено, что прикатывание почвы, проведенное
после посева семян, снижает вредоносность слизней (Ste-
phenson, 1975). Так, при давлении катка на почву 620 кг/м2
снижается поврежденность семян озимой пшеницы слиз-
нями с 10 до 5 %, а всходов — с 46.6 до 32 %. Увеличе-
ние глубины посева семян снижало их поврежденность
слизнями с 50 до 25 %. Значение этих агроприемов со-
стоит в том, что они затрудняют доступ вредителей к про-
растающим семенам.
Во всех случаях следует избегать создания загущенных
посевов и тем самым не допускать ухудшения аэрации
и повышения влажности почвы и воздуха среди рас-
тений.
Уборка урожая. Пожнивные остатки следует
как можно скорее вывезти с поля или запахать в почву
на достаточную глубину, поскольку они служат источни-
ком питания слизней в период их размножения и поэтому
существенно влияют на плодовитость моллюсков. Кроме
того, под ними вредители находят спасение от хищников
и чрезмерного обезвоживания, а в заморозки — и от низ-
ких температур.
На клеверных полях в первый год следует возможно
раньше провести уборку покровной культуры и удалить
ее с поля. В противном случае скапливающиеся под ней
слизни могут серьезно повредить молодой клевер. В по-
следующем необходимо систематически удалять с поля
отмершие органические остатки конными граблями. При
высокой численности слизней на ликвидируемых клеве-
рищах заготовку сена следует проводить только по уско-
ренной технологии, которая предусматривает сокращение
процесса уборки до 1—2 дней и последующее досушивание
сена активным вентилированием. Сразу же после уборки
клеверной массы необходимо провести тщательную обра-
ботку почвы.
Севооборот. Следует избегать посева озимой
ржи и пшеницы, а также других сильно повреждаемых
слизнями культур в низинных и сырых местах, в непо-
средственной близости от кустарниковых зарослей, лесных
и лесопарковых массивов, пустошей и других естествен-
ных резерваций вредителей. Эти культуры следует раз-
мещать и вдали от тех участков, с которых возможно
расселение слизней (посевов клевера, капусты, карто-
феля и др.). Сильно повреждаемые культуры не следует
выращивать по пласту многолетних трав, а также на вновь
144
вводимых в севооборот целинных и залежных участках -
здесь слизни часто скапливаются в большом количестве.
Удобрения. Некоторые удобрения при опре-
деленном способе их внесения могут попутно служить
контактными моллюскоцидами. Эту роль отчасти могут
выполнить вносимые при удобрении пастбищ пылевидные
формы суперфосфата, известковой и доломитовой муки,
отходы промышленности — сланцевая и цементная пыль
и другие. Их вносят автомобильными разбрасывателями
АРУП-8 и прицепными разбрасывателями РУП-8, авто-
цементовозами (С-571, С-853), тракторами-цементовозами
(ТПР-7, ТПР-8).
Механический метод. Его рационально
практиковать при наличии свободной рабочей силы на не-
больших территориях (в садах, огородах, парниках,
теплицах, на селекционных участках и т. п.). Здесь можно
применять ручной сбор слизней, создание защитных ка-
навок и ловушек. Защитные канавки в почве вокруг
участков с возделываемыми растениями делают на глу-
бину 15—30 см и наполняют материалом, затрудняющим
передвижение моллюсков (хвоей, опилками, песком).
Ловушками могут служить мешковина, доски, фанера,
кучи растительных остатков. Их размещают на расстоя-
нии не более 3—5 м, а собравшихся под ними слизней
периодически уничтожают. Механический метод борьбы
можно сочетать с химическим, размещая в укрытиях
отравленные приманки (см. ниже).
Биологический метод
Биологический метод борьбы со слизнями до настоя-
щего времени не разработан. Использование естественных
врагов слизней из числа членистоногих или позвоночных
малоперспективно из-за отсутствия среди них в Нечерно-
земной зоне специализированных малакофагов, способных
эффективно регулировать численность вредных видов.
Из этого, однако, не следует, что нужно вообще отказаться
от использования врагов слизней. На небольших участках
полезно привлекать и охранять птиц, земноводных, хищ-
ных жужелиц и стафилинид, питающихся слизнями или
их яйцами (с. 58). Значение таких мер возрастает в годы
высокой численности моллюсков, когда их доля в пищевом
рационе хищников заметно увеличивается.
Ю И. М. Лихарев и Я. С. Шапиро
145
Разработку биологической борьбы со слизнями сле-
дует вести в направлении изучения паразитов и болезней
этих моллюсков. Весьма перспективно исследование роли
паразитических инфузорий-тетрахименид (Ciliophora, Те-
trahymenidae) в регулировании численности слизней
(Brooks, 1968). Как показало исследование, если инфузо-
рию Tetrahymena limacis (Warren) следует считать слабо-
патогенным облигатным высокоспецифичным паразитом
слизней, то другой вид — Т. rostrate, (Kohl) является
Таблица 9
Распространение инфузории Tetrahymena rostrata в некоторых
местообитаниях ее хозяина — сетчатого слизня (Ленинградская обл.,
1981—1982 гг., оригинальные данные)
Стация Экстенсивность инвазии (%) Встречаемость инфузории в почвенных пробах (%)
слизни яйца
Капустные поля 0.3 0.8 1.0
Картофельные поля 0.2 0.9 1.5
Пастбища и сенокосы 0.9 3.0 5.8
Луга-пустыри в населенных 1.2 5.2 15.3
пунктах Злаково-разнотравные луга 0.9 5.7 16.8
Лесопарки 7.0 16.4 22.5
Залежные земли с древесно-ку- 11.4 18. 9 28.4
старниковой растительностью Заброшенные сады 10.3 17.5 26.0
факультативным паразитом моллюсков, весьма патоген-
ным для сетчатого слизня и, возможно, для других видов.
Упомянутое исследование пока не нашло продолжателей,
хотя к нему обращаются при разработке теории парази-
тизма инфузорий (Corliss, 1972). Современные публика-
ции, в которых упоминается Т. rostrata, посвящены в ос-
новном методам культивирования, цитологической обра-
ботке и анализу жизненного цикла инфузорий (Lynn,
1975; McArdle, 1978; Dobra et al., 1980, и др.).
Несколько расширить представление о роли инфузо-
рии-паразита в регулировании численности сетчатого
слизня и о возможных путях ее использования для борьбы
с вредителями позволяют исследования, проведенные
Я. С. Шапиро. Результаты обследования сельскохозяй-
ственных земель и природных местообитаний слизней,
проведенного в Ленинградской области в 1981—1982 гг.,
146
представлены в табл. 9. Во всех обследованных резерват-
ных стациях экстенсивность инвазии сетчатого слизня
весьма высокая. Напротив, в стациях расселения, где
моллюски появляются обычно лишь в периоды с благо-
приятными для них погодными условиями, зараженность
слизней очень низкая. Анализ почвенных проб на присут-
ствие цист паразита выявил такую же закономерность.
Если учесть, что в 1979—1982 гг. наблюдалось весьма
широкое распространение сетчатого слизня на сельско-
%
Рис. 58. Влияние паразитирования инфузорип-тетрахимены на питание и мо-
бильность зараженных сетчатых слизней.
По оси абсцисс — время после заражения, по оси ординат — суточное потребле-
ние пищи (I) и суточная активность (2) зараженных моллюсков в процентах
к контролю (незараженные моллюски).
хозяйственных землях области, то результаты оценки
указывают на пространственно-временное отставание рас-
селения инфузории-паразита от расселения слизня-хо-
зяина.
Такое отставание отчасти объяснимо различиями в воз-
растной структуре популяций слизня. Дело в том, что
в активной инвазионной стадии инфузории способны за-
ражать слизня лишь в период его постэмбрионального
развития (в яйца моллюска паразит проникает только
во время их формирования в теле хозяина). В резерва-
циях моллюсков их постэмбриональные стадии присут-
ствуют в течение всего года, тогда как в стациях рассе-
ления — лишь в течение 5—6 мес (см. с. 72). Поэтому
вероятность заражения слизней в разных стациях раз-
лична.
10*
147
Другая важная причина низкой встречаемости инфу-
зории в стациях расселения слизней, по-видимому,
заключается в поведенческих реакциях зараженных мол-
люсков. Как показали исследования, интенсивность
питания и передвижения слизней после их заражения инфу-
зорией заметно снижается (рис. 58). Поэтому и тенден-
ция к активному расселению из резерваций, которые по
существу являются очагами протозойного заболевания,
у зараженных слизней также снижается. Следствие это-
Рис. 59. Экстенсивность инвазии сетчатого слизня инфузорией-тетрахимеиоп
в 15 резерватных популяциях (1) и среди расселяющихся на них моллюсков (£)•
По оси ординат — экстенсивность инвазии.
го — низкая встречаемость инвазированных моллюсков
среди эмигрантов из резерваций (рис. 59).
Оценка последствий паразитирования инфузории Т. го-
strata показывает вполне реальное регулирующее влия-
ние паразита на численность хозяина. Исход заражения
наиболее четко прослеживается в период эмбрионального
развития слизня: погибает либо зараженный эмбрион,
либо проникший в яйцо паразит. На рис. 60 приведены
результаты учета смертности эмбрионов в зараженных
и не зараженных инфузорией яйцах сетчатого слизня
в 15 популяциях (инкубация проведена в лаборатории).
В полевых условиях труднее оценить регулирующее
влияние паразита в постэмбриогенезе хозяина: трупы по-
гибших слизней очень быстро разлагаются при участии
бактерий, являющихся антагонистами инфузорий. Вос-
148
питание слизней, зараженных инфузорией, в лаборатории
показало, что 70—90 %-нал смертность при температуре
18—22 °C наблюдается у взрослых моллюсков через 30—
40 сут, у молоди — через 10—15 сут после заражения.
Таким образом, в природных популяциях слизней
Т. rostrata является вполне реальным регулирующим
агентом. Однако это регулирующее действие проявляется
лишь в очагах протозооза — резерватных стациях мол-
люсков. При широком распространении слизней в агро-
ландшафте регулирующее влияние инфузории не проявля-
ло
W
32
24
16
8
О
Рис. 60. Смертность эмбрионов сетчатого слизня в яйцах, зараженных (1) и не
зараженных (2) инфузорлей-тетрахимсной.
По оси ординат — смертность эмбрионов в выборках из 15 популяций мол-
люска.
ется из-за низкой экстенсивности инвазии в стациях
расселения моллюсков. В связи с этим можно наметить
некоторые пути использования паразита для ограничения
численности слизней. Во-первых, это увеличение инва-
зии в резервациях слизней с целью снижения интенсив-
ности расселения моллюсков на культивируемые земли.
Во-вторых, — сезонная колонизация инфузории в местах
высокой численности моллюсков в период их широкого
распространения на сельскохозяйственных землях. Пер-
вый путь представляется более реальным, поскольку
площадь резерваций слизней в агроландшафте всегда
меньше потенциально заселяемой вредителями террито-
рии. К тому же известная вялость протекания заболева-
ния, особенно при низких температурах, не будет иметь
большого отрицательного значения в случае предупреди-
тельных мероприятий ио заражению моллюсков в их
резерватных популяциях.
149
В заключение отметим, что большой практический
интерес представляет изучение болезней слизней, вызывае-
мых вирусами и бактериями. Методические рекоменда-
ции по изучению различных патогенных для моллюсков
организмов содержатся в работах Горохова (1980, 1983).
Химический метод
Основные этапы развития химической борьбы со слиз-
нями отчасти отражают эволюцию этого метода уничто-
жения вредных организмов вообще. Однако морфофизио-
логические особенности наземных моллюсков, резко отли-
чающие их от прочих вредителей сельского хозяйства
(насекомых, клещей, фитогельминтов), определяют специ-
фику ограничения численности слизней с помощью пести-
цидов. Одна из таких особенностей, сильно затрудняющая
борьбу со слизнями, заключается в высокой устойчивости
этих моллюсков практически ко всем известным хлорорга-
ническим и фосфорорганическим биоцидам, используемым
для защиты культурных растений. Поэтому значительный
прогресс в химической защите растений, связанный с ши-
роким использованием синтетических препаратов этих
двух групп, практически не отразился на эффективности
борьбы со слизнями.
В конце прошлого века моллюскоцидные свойства
были обнаружены у ряда неорганических соединений —
медного и железного купороса, извести, суперфосфата,
сульфата аммония, соединений мышьяка и других (Лиха-
рев, 1954). Однако эти соединения не получили широкого
распространения ввиду больших норм расхода, высокой
трудоемкости применения и низкой эффективности.
Органические препараты контактного действия — дуст
этилового эфира диметилдитиокарбаминовой кислоты (Ва-
сина, 1953) и этиленхлоргидрин (Пайкин, 1949) — также
оказались малоэффективными, поскольку их применение
предусматривает обязательный контакт с телом моллюска.
Не нашел применения и метод наземной фумигации аммиа-
ком (Страховская, 1949) — пары аммиака оставляют на
растениях ожоги и вредны для человека.
В 30—40-х годах этого столетия сначала в Англии,
а затем и в^других странах для борьбы со слизнями стали
применять препараты полимера уксусного альдегида —
метальдегида, которые удачно сочетают в себе высокую
контактную и кишечную токсичность для моллюсков
1.50
(• высокой селективностью действия. Кроме того, эти пре-
параты не относятся к числу высокотоксичных соедине-
ний для человека и теплокровных животных.
Поиски эффективных моллюскоцидов привели к вы-
явлению соединений, оказывающих на слизней сильное
отпугивающее действие. Репеллентными свойствами обла-
дают ак-пвированный креолин (Разауска, 1963), диметил-
фталат ] аибутилфталат (Stephenson, 1962), тирам (Sou-
llion, 19cJ).
Высокотоксичными для слизней оказались препараты
па группы производных карбаминовых кислот — кар-
баматы, тио- и дитиокарбаматы, которые в настоящее
время довольно широко применяются за рубежом для
борьбы ( моллюсками — вредителями сельского хозяй-
ства, а в СССР — в основном как инсектициды.
И< пытания названных выше соединений в различных
агроклиматических условиях показали как положитель-
ные, так и отрицательные свойства, которые необходимо
учитывать при выборе моллюскоцида. Рассмотрим некото-
рые из препаратов, представляющие, по нашему мнению,
наибольший интерес для борьбы со слизнями.
1. Метальдегид. Полимер ацетальдегида со
строением (СН8—СН = О)П, где п=4—6. Это бесцветные
кристаллы, плохо растворимые в ксилоле, эфире, хлоро-
форме, бензоле, практически не растворимые в воде.
Метальдегид подвергается деполимеризации, поэтому его
хранят в позапаянпой таро. Метальдегид среднетоксичен
для человека и теплокровных животных. Он не обладает
к у м ул яти в п ы м, эмбриотропным, бластомогенным дейст-
вием, является слабым аллергеном. Через кожу прони-
кает плохо, не раздражает ее, но хорошо всасывается
через дыхательные пути. Концентрация метальдегида
в воздухе не более 0.05 мг/м3 является безвредной для
человека; такая концентрация устанавливается в полевых
условиях через одни сутки после проведения химической
обработки (Конашевич и др., 1972).
В СССР промышленность выпускает два препарата
метальдегида, синтезированных в ВНИИХСЗР.1 5%-ный
гранулированный препарат, приготовленный на пшенич-
ных отрубях, применяется как отравленная пищевая при-
манка. Норма расхода препарата на зерновых культурах
1 Всесоюзный научно-исследовательский институт химических
средств защиты растений.
151
15—20 кг/га, на овощных, ягодных, технических и про-
чих культурах — до 30—40 кг/га. 50%-ный смачиваю-
щийся порошок метальдегида предназначен для опрыски-
вания растений и почвы (концентрация суспензии 0.5 —
2 %). Он содержит 50—52 % технического метальдегида,
препарат ОП-7, концентрат сульфитно-спиртовой барды
и каолина. Норма расхода этого препарата 4—8 кг/га
на всех культурах. Оба препарата метальдегида можно
применять не позднее чем за 20 дней до уборки урожая
защищаемой культуры. Выпас скота на участках, обрабо-
танных метальдегидом, можно проводить не ранее чем
через 25 дней после обработки.
При соблюдении регламента применения метальдегид
не угнетает защищаемые растения и в них не накапли-
вается. Допустимые остатки метальдегида в сельскохо-
зяйственной продукции 0.7 мг/кг.
Препараты метальдегида, в первую очередь гранули-
рованные, отличаются высокой избирательностью токси-
ческого действия и не оказывают заметного отрицатель-
ного влияния на безвредную фауну.
В Англии метальдегид выпускается в форме мелких
гранул, содержащих 6 % технического метальдегида,
и в форме 50%-ного смачивающегося порошка. Небольшой
размер гранул позволяет снизить норму расхода препа-
рата в 1.5—2 раза и вдвое повысить эффективность его
использования. Во Франции выпускаются 5%-ные гра-
нулы метальдегида (ариотокс).
Сравнительные испытания показали, что препараты
метальдегида, выпускаемые в нашей стране, не уступают
зарубежным. В некоторых случаях отмечена более высо-
кая эффективность отечественного гранулированного пре-
парата (Дмитриева, 1963). В арсенале отечественных
средств защиты растений от слизней метальдегид яв-
ляется пока единственным моллюскоцидом. Его примене-
ние разрешено как в производственных условиях, так
и на приусадебных участках рабочих и служащих (на по-
следних разрешен только 5%-ный препарат).
Действие метальдегида на слизней.
Метальдегид может проникать в организм слизня через
рот и пищеварительный тракт при поедании отравленной
приманки или растений, обработанных моллюскоцидом.
В этом случае он становится кишечным ядом. Кроме того,
после попадания на тело слизня метальдегид легко про-
никает в его организм через кожу, т. е. действует как
152
контактный яд. Действие метальдегида на слизней до-
вольно сложно. С одной стороны, он обладает хорошо
выраженными аттрактивными свойствами: 5%-ные гра-
нулы оказывают на слизней привлекающее действие на
расстоянии до 0.5 м — моллюски их активно отыскивают
и поедают.
С другой стороны, выявлены основные аспекты токси-
ческого действия метальдегида на моллюсков: на пище-
варительный тракт при поедании отравленного субстрата;
нервно-паралитическое действие; раздражающее действие
на кожные покровы, которое сопровождается обильным
выделением слизи (Barnes, Weil, 1942; Thomas, 1948).
Поведенческие реакции отравленных метальдегидом
слизней и некоторые физиологические изменения в их
организме исследованы Дмитриевой и Шапиро (19756).
Внешние признаки отравления метальдегидом появляются
у слизней уже через 1—2 мин после поедания отравлен-
ной приманки. Вначале наблюдается явление гиперакти-
вации, которое проявляется в усиленной двигательной
активности («реакция бегства») и некоординированных
судорожных движениях. Продолжительность гиперакти-
вации колеблется в пределах 5—20 мин, причем она нахо-
дится в обратной зависимости от принятой дозы моллюско-
цида. Гиперактивация сопровождается повышением ин-
тенсивности дыхания — при отравлении сублетальными
дозами в 3.5—4 раза, летальными дозами — в 2 раза,
а также катаболизмом запасных веществ.
После гиперактивации наступает депрессия, которая
выражается в утрате слизнями двигательной активности,
резком снижении интенсивности дыхания и дальнейшем
катаболизме гликогена и липидов. Продолжительность
этого периода у моллюсков, отравленных сублетальными
дозами, составляет 35—50 мин. Если в течение периода
депрессии животные находятся в оптимальных гидротер-
мических условиях, то после него наблюдается возобнов-
ление активности, сопровождающееся возвращением по-
движности и повышением интенсивности дыхания. При
отравлении слизней летальными дозами депрессия про-
должается 1—1.5 ч, после чего моллюски погибают.
Таким образом, гиперактивация более значительна
при отравлении животных сублетальными дозами, причем
в благоприятных метеорологических условиях сопут-
ствующие ей физиологические изменения обратимы. Де-
прессия более глубока в случае летальных доз, а насту-
153
пающий при этом паралич необратим и приводит к гибели
слизня.
Помимо механизма токсического действия метальде-
гида на слизней, при организации защитных мероприятий
необходимо также учитывать те факторы, которые опре-
деляют устойчивость вредителей к моллюскоциду. Весьма
полезными здесь могут оказаться сведения о влиянии
моллюскоцида на отдельные органы отравленного жи-
вотного.
В исследованиях Дмитриевой и Шапиро (1979) чув-
ствительность некоторых органов сетчатого слизня к ме-
тальдегиду оценивалась посредством их прижизненного
окрашивания нейтральным красным. Этот метод основан
на изменении поглощения (сорбции) красителя тканью
при ее повреждении. Количество поглощенного красителя
прямо зависит от степени повреждения органа и служит
объективной характеристикой его физиологического со-
стояния (Граменецкий, 1963). Сравнение данных, полу-
ченных при разных способах введения яда в организм
слизня, позволяет судить о тех физиологических барьерах,
которые препятствуют проникновению пестицида к жиз-
ненно важным центрам моллюска.
Данные табл. 10 характеризуют сравнительную повреж-
даемость некоторых органов слизня при введении метальде-
гида в полость тела. При этом обеспечивается одинаково
свободный доступ яда ко всем органам отравленного мол-
люска. Оказалось, что метальдегид в концентрациях
не более 20 °/00 вызывает лишь обратимые нарушения
жизнедеятельности органов. Причем в этом случае разли-
чия в поглощении красителя разными органами незначи-
тельны. В концентрации же 40 °/00 метальдегид необратимо
повреждает органы слизня.
Поступая в организм моллюска естественным путем
(с пищей с отравленной приманкой), моллюскоцид оказы-
вает резко иное воздействие (табл. И). Наиболее глубо-
кие нарушения происходят в тканях печени слизня,
тогда как другие органы в начале отравления повре-
ждаются слабее. Эти различия особенно заметны при
отравлении сублетальными дозами метальдегида, не при-
водящими к гибели слизня. В случае же смертельных доз
яда наступает весьма быстрое (в течение 1 ч) необратимое
повреждение всех органов.
Эти исследования позволили заключить, что на ран-
них стадиях отравления моллюска метальдегидом наибо-
154
Таблица 10
Изменение сорбции нейтрального красного органами сетчатого слизня
при введении метальдегида в полость тела (из: Дмитриева, Шапиро, 1979)
Орган моллюска Изменение сорбции красителя (% к контролю) при разных концентрациях метальдегида (%о)
5 10 20 40
Печень + 10.0 + 105.5 + 180.3 4-260.3
Ганглии + 15.3 +100.2 +125.0 4-183.6
Гонада +7.8 +82.4 +140.7 4-192.0
Пенис +19.0 +128.6 +194.4 4-287.3
Таблица 11
Изменение сорбции нейтрального красного органами сетчатого слизня при
отравлении метальдегидом через рот (из: Дмитриева, Шапиро, 1979)
Орган моллюска Доза металь- дегида Изменение сорбции (% к контролю) в разные сроки после начала отравления
5 мин 30 мин 1 ч 12 ч 24 ч 48 ч
Печень а +30.5 +97.7 + 186.6 + 167.5 +84.6 +58.7
б +20.1 +105.3 + 190.9 Гибель слизня
Ганглии а +11.5 +24.3 +32.7 4-54.8 I I 4-39.4 +35.0
б +10.2 +46.1 +95.0 Гибель слизня
Гонада а + 17.6 +26.1 +44.2 4-58.0 I I +62.5 +50.7
б +20.0 + 108.8 +120.5 Гибель слизня
Пенис а +43.0 +56.2 +89.0 +90.6 I +80.4 +43.2
б +52.3 +130.2 +145.1 Гибель < 1 злизня 1
Примечание, а — летальная, б — сублетальная.
лее сильной токсикации подвергается печень. Относи-
тельно слабое повреждение других органов связано с ча-
стичным обезвреживанием яда в печени. Такой вывод
представляется вполне правомерным, если учесть особо
важную роль печени в метаболизме у слизней. Работа
мощного ферментативного аппарата печени резко умень-
шает количество яда, достигающего другие органы живот-
ного, и это, вероятно, наиболее важное проявление за-
щитной функции печени. В случае разрушения этого
барьера, при отравлении смертельными дозами моллюско-
цида, яду открывается доступ к жизненно важным орга-
нам и моллюск быстро погибает.
Подтверждением этих выводов служат результаты ис-
следований названных авторов, показавшие, что возраст-
155
Таблица 12
Динамика некоторых физиолого-биохимических показателей
в постэмбриогенезе сетчатого слизня (по: Дмитриева, Шапиро, 1976,
с дополнениями)
Стадия развития слизня Интенсивность дыхания, мл О2/г • ч Содержание гликогена, мг% Содержание липи- дов, мг% Средняя летальная ; доза метальдегида, мг/г
в целом организме в печени в белковой железе в целом организме в печени в белковой железе
Ювенильные осо- 1.2 564 685 Следы 180 219 Следы 0.06
би 0.3 1180 1832 117 630 1380 56 0.15
Интенсивно раз- 0.3 1180 1832 117 630 1380 56 0.15
множающиеся особи 0.6 723 895 860 620 1100 380 0.0S
0.6 723 895 860 620 1100 380 0.08
Стареющие особи о.оэ 53 107 170 87 98 106 0.02
Примечание. В числителе — данные на начало стадии развития
слизня, в знаменателе — на ее конец.
пая устойчивость сетчатого слизня к метальдегиду су-
щественно зависит от физиологического состояния печени,
в частности, от ее обеспеченности гликогеном и липидами
(табл. 12).
В начале постэмбрионального развития устойчивость
слизня к метальдегиду возрастает в 2.5 раза. Это связы-
вается как с постепенным снижением уровня обмена ве-
ществ (потребление О2 снижается в 4 раза), так и с интен-
сивным накоплением запасных веществ в печени мол-
люска.
С наступлением половой зрелости и периода размно-
жения уровень обмена веществ (метаболизма) у слизня
заметно возрастает. Одновременно наблюдается как общая
утилизация запасных веществ, так и их отток от печени
к органам, обеспечивающим яйца слизня питательными
веществами, в первую очередь к белковой железе. В ре-
зультате перераспределения этих веществ защитная функ-
ция печени ослабляется, а устойчивость моллюска к ме-
тальдегиду снижается почти вдвое.
В конце жизни слизня в его печени остается минималь-
ное количество гликогена и липидов за весь постэмбрио-
156
нальный период. Этому соответствует и Минимальная
устойчивость моллюска к яду.
Итак, на последствия отравления слизней метальдеги-
дом существенно влияют возраст и физиологическое со-
стояние моллюсков. Последнее, в свою очередь, зависит
как от особенностей развития (онтогенеза), так и от раз-
личных экологических факторов, например от обеспечен-
ности моллюсков доступным кормом. Поэтому учет фи-
зиолого-биохимических показателей состояния вредителей
важен как при лабораторных испытаниях моллюскоцидов,
так и в практической защите растений (например, для
определения оптимального расхода моллюскоцида в пре-
делах установленных регламентом норм).
Важно учитывать, что в полевых условиях на устой-
чивость популяции слизней к обработкам метальдегидом,
кроме отмеченных выше факторов, влияют пищевая спе-
циализация и пространственное распределение разных
возрастных групп моллюсков в биотопе (Шапиро, 1979).
В табл. 13 показано изменение возрастной структуры
популяции сетчатого слизня в результате обработки
участка двумя препаратами метальдегида. Относительно
высокая полевая устойчивость самых молодых моллюсков
1-го и 2-го классов при проведении опрыскивания
50%-ным препаратом метальдегида объясняется тем, что
молодь обитает и питается в ярусах растительности,
менее доступных для пестицида.
При рассеве метальдегидной приманки (5%-ных гра-
нул) происходит еще более резкое омолаживание популя-
ции, потому что гранулы метальдегида, приготовленные
на пшеничных отрубях, привлекают значительно больше
взрослых слизней, чем молодых, которые питаются пре-
имущественно мицелием грибов и гниющими частями
растений.
Особенности действия метальдегида на слизней в поле-
вых условиях необходимо учитывать при планировании
и проведении мероприятий по химической защите расте-
ний от этих моллюсков и борьбе с ними на пастбищах.
На эффективность химической борьбы со слизнями
влияет комплекс различных факторов, которые необхо-
димо учитывать в целях достижения наилучшего резуль-
тата. Среди этих факторов — правильный выбор формы
препарата моллюскоцида, состояние защищаемых расте-
ний и популяций вредителей, погодные и другие условия.
Оба рассмотренных выше препарата метальдегида
157
Таблица 1#
Изменение Возрастной структуры популяции сетчатого слизня в результате
обработки клеверного поля двумя препаратами метальдегида (оригинальные
данные)
Весовой класс слизней Пределы весового класса, мг Доля весового класса, %
до обработки после обработки
50 %-ный смачиваю- щийся порошок 5 %-ные гранулы
1-й 50 35.5 38.5 46.5
2-й 51—100 31.0 37.0 44.0
3-й 101—250 22.5 17.0 5.0
4-й 251—500 11.0 7.5 4.5
Таблица 14
Восстановление популяции сетчатого слизня на краевой полосе капустного
поля шириной 10 м после обработки метальдегидом (оригинальные данные)
Препарат и норма расхода Плотность популяции (экз./м2) после максималь- ного снижения численности в результате обработки, по дням учета
1-й 3-й 6-й 9-й 12-й 15-й 18-й 21-й
50 %-ный смачива- ющийся порошок, 8 кг/га М 1.0 1.5 1.5 3.0 8.5 12.5 18.5 22.0_
5 %-ные гранулы, 40 кг/га 1.5 1.5 1.5 3.0 5.5 6.0 7.5 7.5
имеют как положительные свойства, так и недостатки.
Многочисленные исследования, проведенные в СССР и
за рубежом, показали, что на больших и достаточно одно-
родных массивах рациональнее опрыскивать растения
и почву суспензией 50 %-него порошка метальдегида.
Преимущество опрыскивания состоит прежде всего в бы-
стром и резком снижении численности вредителей, по-
гибаюп-ь?г в результате питания обработанных моллюско-
цидом растений, а также контакта с ними и с почвой.
При опрыскивании, проведенном в часы наибольшей
активности слизней, отравление моллюсков может насту-
пить и в результате непосредственного попадания яда
на тело животного.
Другое преимущество опрыскивания заключается в его
универсальности: опрыскивать можно растения в любых
158
фазах их развития при условии соблюдения регламенти-
рованного срока последней обработки. Рассев же грану-
лированного препарата на участках с густым и высоким
травостоем затруднен, а его эффективность как пищевой
приманки резко снижается. Универсальность опрыскива-
ния проявляется и в том, что этот способ эффективен
против всех стадий роста слизней. Эффективность же
приманки против молоди моллюсков, как отмечалось
выше, очень низка.
Опрыскивание, наконец, — это более производитель-
ный технологический процесс, чем рассев отравленных
приманок, и это особенно заметно проявляется при обра-
ботке метальдегидом больших сельскохозяйственных мас-
сивов.
Однако применение 50%-ного препарата имеет и свои
недостатки, главный из которых заключается в малой
продолжительности защитного действия метальдегида, на-
несенного на растения и почву в форме суспензии. Мол-
люскоцид при этом легко смывается и быстро теряет свою
токсичность для слизней в результате деполимеризации.
Поэтому на краевых полосах обработанного таким
способом посева популяция слизней может частично вос-
становиться уже через 2—3 недели (табл. 14). Восста-
новление же популяции вредителей на небольших участ-
ках в результате иммиграции слизней с соседних терри-
торий может потребовать проведения повторной обработки
моллюскоцидом.
Преимущество отравленных приманок метальдегида
заключается прежде всего в высокой продолжительности
их защитного действия. В опытах Увалиевой (1979) этот
период составлял 20 дней; в наших опытах, в зависимости
от погодных условий, — от 15 до 24 дней. Одноразовое
внесение гранул защищает рассаду табака в течение всего
периода ее выращивания в парниках (Акрамовский,
Бабаян, 1976). Даже в условиях Черноморского побе-
режья Кавказа, где высокая температура и влажность
воздуха и почвы снижают устойчивость метальдегида,
токсичность 5%-ной приманки сохранялась более 2 недель
(Алан, 1972; Вашадзе, 1972; Мурусидзе, 1972; Загайный,
1978).
Не менее важно, что применение отравленных металь-
дегидных приманок в большей степени, чем опрыскивание
суспензией препарата, создает щадящий режим для без-
вредной фауны агробиоценоэов и предпочтительнее с по-
159
зиций охраны труда работников, осуществляющих обра-
ботку моллюскоцидом.
На эффективность применения препаратов метальде-
гида сильное влияние оказывает активность слизней,
которая, в свою очередь, тесно связана с погодными усло-
виями (см. с. 83). Прежде всего это касается использова-
ния метальдегидных приманок, действие которых основано
на активном привлечении слизней. Именно поэтому при
низкой температуре воздуха эффективность приманочного
метода борьбы со слизнями резко снижается. При темпе-
ратуре воздуха ниже 15 °C его применение неоправданно.
Химическая защита различных сельскохозяйственных
культур имеет свои особенности, связанные со специфи-
кой их выращивания, фенологией и уязвимостью со сто-
роны вредителей.
Применение препаратов метальде-
гида.
Озимые зерновые. На долю Северо-Запад-
ного и Центрального экономических районов приходится
около половины всей площади, занятой зерновыми куль-
турами в Нечерноземье; 30 % приходится на озимые
рожь и пшеницу. Из всех зерновых культур именно ози-
мые хлебные злаки повреждаются слизнями особенно
часто. Это связано прежде всего с фенологией озимых —
наиболее уязвимые для слизней фазы развития (прораста-
ние семян и всходы) эти растения проходят в конце лета—
начале осени, когда численность и вредоносность мол-
люсков на пашне наибольшие.
В этот период популяции слизней представлены здесь
преимущественно половозрелыми и размножающимися
особями, в борьбе с которыми эффективны 5%-ные гра-
нулы метальдегида. Их применение целесообразно здесь
еще и потому, что осенние миграции слизней из резерва-
ций на поля с озимыми культурами определяют необхо-
димость продолжительного защитного действия хими-
ческих обработок.
В случае теплой затяжной осени в северо-западных
и центральных областях зоны возможно появление вто-
рого (осеннего) поколения вредителей, отродившихся
из отложенных моллюсками в конце лета яиц. Молодые
слизни перед уходом на зимовку активно питаются неж-
ными всходами озимых и наряду со взрослыми моллю-
сками могут нанести серьезный ущерб. В случае массо-
вого осеннего отрождения молоди слизней для их у ни что-
160
жения следует провести опрыскивание всходов растений
и почвы суспензией 50%-ного порошка метальдегида.
Такую же обработку проводят и в борьбе со взрослыми
моллюсками при низкой температуре воздуха и почвы,
когда приманки метальдегида малоэффективны.
Овощные культуры открытого
грунта. В структуре посевных площадей овощных
культур Северо-Западного и Центрального районов,
а также Калининградской области наибольший удельный
вес занимает капуста (соответственно 60, 44 и 53%)
и морковь (18, 17 и 15%). Эти культуры наряду с сала-
том, свеклой, огурцами, помидорами и другими овощами
сильно повреждаются слизнями.
Особенно большой вред моллюски наносят прорастаю-
щим семенам, всходам и рассаде овощных растений.
Повреждение молодых растений часто приводит к их
гибели и большой изреженности посева.
Для защиты от слизней овощных культур с коротким
периодом вегетации (салата, редиса и др.) следует приме-
нять гранулированный метальдегид, который вносят одно-
временно с посевом семян. В этом случае будет соблюден
временной регламент применения моллюскоцида и, бла-
годаря высокой продолжительности защитного действия
5%-ных гранул, обеспечена защита растений от слизней.
В конце весны—начале лета овощам вредит молодь
слизней, отрождающаяся из перезимовавших яиц.
В борьбе с ней на среднеспелых и поздних по срокам
созревания культурах эффективно опрыскивание рассады
и всходов суспензией 50%-ного препарата метальдегида.
Следует учитывать, что во время затяжной весны и про-
хладного лета отрождение слизней из яиц может растя-
нуться на 1—1.5 мес. Это порой определяет необходимость
проведения повторной химической обработки.
В конце лета—начале осени в результате миграций
слизней из диких стаций на поля со средними и поздними
овощами возможно повреждение растений на краевых
полосах. С целью их предупреждения не позднее чем
за 20 дней до уборки урожая можно провести краевые
обработки полей: на культурах с большой вегетативной
массой — опрыскивание, на прочих — рассев отравлен-
ной приманки. Ширина обработанной полосы должна
быть не менее 50 м.
Многолетние кормовые травы. Боль-
шой вред слизни наносят различным видам клевера, ко-
11 И. М. Лихарев и Я. С. Шапиро
161
торый является основной многолетней бобовой культурой
в травосмесях, распространенных в северо-западных и
центральных областях Нечерноземья. Причина высокой
вредоносности обусловлена не только тем, что клевер —
одно из предпочитаемых слизнями кормовых растений,
но и благоприятным для моллюсков микроклиматом кле-
верных полей, а также продолжительным их существова-
нием (до 3 лет).
Популяции слизней на клеверных полях отличаются
высокой возрастной неоднородностью. Зимуют здесь на-
ряду с яйцами и молодые слизни, вредоносность которых
на этой культуре проявляется уже рано весной в период
отрастания перезимовавших растений.
Для борьбы со слизнями в начале сезона, до укоса,
возможно применение любой формы препарата металь-
дегида. Однако в случае невысокого удельного веса в по-
пуляции отрождающейся из яиц молоди эффективнее
использовать 5%-ные гранулы моллюскоцида. Во второй
половине лета и осенью в популяциях доминируют взрос-
лые слизни. Поэтому в случае высокой численности вре-
дителей следует провести рассев гранул. Этот прием будет
предохранять посев и от мигрирующих с соседних участ-
ков моллюсков, вредная деятельность которых в против-
ном случае может ослабить растения и резко снизить их
зимостойкость.
Необходимо учитывать, что клеверные поля, особенно
двух-, трехлетние, служат резервациями слизней, по-
этому борьба с ними в этих угодьях существенно влияет
и на численность вредителей на соседних участках.
Земляника. Эта культура относится к числу
сильно повреждаемых слизнями, и в годы массового раз-
множения вредителей ее потери составляют 60% и более.
Значительная часть потребляемых населением ягод зем-
ляники (до 80%) в северо-западных и центральных
областях выращивается в коллективных садах и на при-
усадебных участках, где численность и ' вредоносность
слизней обычно высокие. Поэтому химическая защита
земляники, особенно в годы высокой численности вреди-
телей, становится необходимой для получения устойчивых
и качественных урожаев этой ценной культуры.
Между тем борьба со слизнями на земляничных план-
тациях часто сопряжена с рядом трудностей. Главная
из них в том, что вредность слизней начинает прояв-
ляться обычно лишь в период созревания ягод, когда
162
применение моллюскоцида в соответствии с регламентом
уже невозможно. Предупредительные обработки земля-
ничных посадок оказываются малоэффективными, по-
скольку небольшие по площади плантации быстро вновь
заселяются на всю глубину слизнями, мигрирующими
сюда с соседних участков в период созревания ягод.
Поэтому в целях предотвращения потерь урожая необ-
ходима предупреждающая обработка соседних с земля-
ничными плантациями участков в радиусе до 100—150 М;
В зависимости от характера растительности и состава
популяции вредителей эту обработку можно провести
либо путем опрыскивания территории суспензией
50%-ного порошка метальдегида, либо рассевом отравлен-
ных приманок. Непосредственно на посадках земляники
предпочтительнее применять 5%-ные гранулы как более
устойчивую форму моллюскоцида. Как правило, погод-
ные условия в это время благоприятны для использования
приманки.
Метальдегид в условиях защищен-
ного грунта.2 В парниках, оранжереях и теплицах
слизни нередко серьезно вредят рассаде овощей и декора-
тивных растений, а также салату, огурцам, томатам
и другим культурам. Испытание в условиях защищенного
грунта двух стандартных форм метальдегида, проведенное
Косоглазовым (1973), показало большую эффективность
5%-ных гранул, которые вносили из расчета 2—3 г/м2.
Для предотвращения преждевременного размокания гра-
нул и потери ими токсичности во влажной атмосфере
теплиц и оранжерей автор предлагает размещать при-
манку под небольшими переносными зонтиками, рас-
ставляя их на грунте и стеллажах на расстоянии до 1 м.
Высокая эффективность гранулированного метальде-
гида выявлена и при защите рассады табака в парниках.
Минимальная норма расхода препарата, обеспечивающая
100%-ную гибель слизней, 1 г на 1 погонный метр пар-
ника (Акрамовский, Бабаян, 1976, 1977).
Метальдегид на пастбищах. Если в юж-
ных районах страны роль слизней как промежуточных
хозяев гельминтов домашних животных невелика, то
в более влажных центральных и особенно северо-запад-
ных областях Нечерноземья эти моллюски становятся
2 На период до 1985 г. применение метальдегида было разрешено
лишь в условиях открытого грунта (Список. . ., 1982).
весьма существенным компонентом биоценозов пастбищ.
Борьба с ними может стать необходимой для предотвра-
щения распространения паразитарных заболеваний среди
сельскохозяйственных животных.
На пастбищах Калининской области в борьбе с мол-
люсками были испытаны оба препарата метальдегида:
50%-ный смачивающийся порошок и 5%-ные гранулы
(Горохов, Осетров, 1978). Наиболее целесообразно вно-
сить препарат в мае—начале июня, в дни с пасмурной
погодой, но без дождя. Обработкам предшествует выявле-
ние инвазированных моллюсков и картирование террито-
рии пастбища, а также определение площади, подлежащей
обработке моллюскоцидом. Нормы расхода препаратов;
50 %-ного порошка — 8—16 кг/га, разведенных в 600 л
воды; 5 %-ных гранул — 40—60 кг/га. Расход препаратов
повышается при обработке густой и высокой (более 20 см)
растительности. Температура воздуха при внесении гра-
нулированного препарата должна быть не ниже 15 °C,
влажность — достаточно высокой, но не избыточной.
Если после обработки прошел дождь, ее следует по-
вторить.
По нашим наблюдениям, популяции слизней пастбищ
и лугов отличаются высокой возрастной неоднородностью
(с. 77). Периоды размножения и вылупления молоди
из яиц часто растягиваются до 1.5—2 мес. Поскольку
метальдегид не оказывает токсического действия на яйца
моллюсков, а 5%-ная приманка малоэффективна против
отрождающихся из яиц молодых слизней, то в этих био-
топах более целесообразно обрабатывать растения и почву
суспензией 50%-ного препарата метальдегида. Сроки про-
ведения опрыскивания по возможности должны совпадать
с тем периодом, когда в популяциях отсутствуют яйца
слизней (например, в Ленинградской обл. это конец
июня—начало июля). Если в период обработки значи-
тельная часть популяции представлена яйцами, то ее сле-
дует повторить после вылупления слизней.
Выпас скота на обработанных метальдегидом пастби-
щах разрешено проводить не ранее чем через 25 дней
после обработки.
Рекомендации по борьбе со слизнями на пастбищах
можно использовать и для уничтожения этих моллюсков
в окрестностях птицеферм. Такая необходимость нередко
возникает в связи с ролью слизней в передаче ленточного
червя давайнеи — паразита куриных (с. 59).
164
Из всего вышеизложенного можно сделать вывод, что
в производственных условиях следует использовать оба
препарата метальдегида. Подходящую форму препарата и
способ применения моллюскоцида необходимо выбирать
в зависимости от вида и состояния растений, популяции
вредителя, погодных условий и преследуемых целей. От-
метим, что препараты метальдегида относятся к числу де-
шевых пестицидов (цена 1 кг гранул 40 к.). В случае пра-
вильного их применения обеспечивается не только высо-
кая биологическая и хозяйственная (урожайная) эффек-
тивность, но и хороший экономический результат.
Так, в Новгородской области при защите белокочан-
ной капусты в совхозе «Прогресс» эффективным оказался
50%-ный смачивающийся порошок метальдегида: биоло-
гическая эффективность составила 97 %, а полученный
чистый доход — 960 р. с 1 га. В Ленинградской области
применение 5%-ных гранул метальдегида на культуре
салата из расчета 40 кг на 1 га обеспечило снижение
численности сетчатого слизня на 98 % уже на 7-е сутки.
Продолжительность защитного действия составила 22—
24 сут, дополнительная прибавка урожая 43 ц/га, полу-
ченный чистый доход — 788 р. с 1 га (Дмитриева, Ша-
пиро, 1974).
По данным Акрамовского и Бабаяна (1976), дополни-
тельный урожай табачного листа, полученный в резуль-
тате защиты рассады от слизней с помощью 5%-ного пре-
парата метальдегида, составил 6 ц/га, а затраты на борьбу
с вредителями оказались почти в 100 раз меньше тех
расходов, которые приходилось нести из-за повреждения
рассады.
За рубежом за полвека интенсивного применения пре-
паратов метальдегида в борьбе со слизнями были выяв-
лены не только достоинства этого моллюскоцида, но
и недостатки. Например, было установлено, что при высо-
кой влажности окружающей среды отравление слизней
метальдегидом не всегда заканчивается их гибелью. Пред-
полагается, что отравление метальдегидом, вызывая серь-
езное нарушение водного обмена моллюска, не приводит
к смертельному исходу в условиях свободного пополне-
ния запаса воды (Stringer, 1949; Hermann, 1971; Mallet,
Bougaran, 1971).
Поэтому параллельно с разработкой новых препаратов
па основе метальдегида велись активные поиски других
соединений с моллюскоцидными свойствами. Такие соеди-
165
нения были обнаружены среди производных карбамино-
вых кислот — карбаматов. Наибольшее распространение
в борьбе с моллюсками получили метиокарб, севин,
зектран и другие.
Положительным свойством карбаматов с моллюско-
цидной активностью является стабильность их токсиче-
ского действия на слизней как в сухой, так и во влажной
среде. Карбаматы не вызывают заметного нарушения вод-
ного баланса слизней, но они обладают значительно более
сильным парализующим действием, чем препараты ме-
тальдегида.
Таблица 15
Сравнительная эффективность метальдегида и метиокарба против еетчат^о
слизня при разной температуре воздуха (но: Mallet, Bougaran, 1971)
Препарат Снижение численности слизней (%) при температуре воздуха (°C)
6 8 10 16 20 22 25
Метальдегид, 5 %-пая приманка 0 15 15 40 80 100 100
Метиокарб, 4 %-пая при- манка 90 95 90 85 90 95 85
Так, при сравнительной оценке биологической эффек-
тивности метиокарба (при норме расхода 4.5 кг/га) и ме-
тальдегида (9.7 кг/га) при влажности воздуха 98 % в пер-
вом варианте погибло 72 % слизней, а во втором — лишь
55 % (Hermann, 1971).
Другое ценное свойство карбаматов — высокая ста-
бильность их моллюскоцидного действия в широком диа-
пазоне температур (табл. 15). Дело в том, что при низкой
температуре воздуха и почвы слизни малоподвижны и пи-
таются менее активно. В этих условиях эффективность
моллюскоцида во многом зависит от его контактной ток-
сичности и скорости действия, а эти показатели значи-
тельно выше у карбаматов, чем у метальдегида.
Недостатком некоторых карбаматов является их более
высокая токсичность для безвредных беспозвоночных,
а также позвоночных животных и значительно меньшая,
чем у метальдегида, селективность биоцидного действия.
Рассмотрим некоторые из этих препаратов.
2. Метиокарб, мезурол, меркаптоди-
мет у р, Б а й е р - 37 344 (4-метил-меркапто-3,5-диме-
16G
тилфенил-1\т-метилкарбамат).3 Кристаллическое вещество
без запаха, в воде почти нерастворимо, но хорошо раство-
ряется в большинстве органических растворителей. Вы-
пускается фирмой «Байер» (ФРГ) в форме 50 %-ного сма-
чивающегося порошка, 3%-ных и 5%-ных гранул. Это
контактный и кишечный пестицид (кроме моллюсков эф-
фективен против гусениц плодожорок, листоверток, коло-
радского жука). Для человека и теплокровных животных
высокотоксичен. При норме расхода порошка 3 кг/га
эффективен против слизней при высокой влажности воз-
духа и почвы. Применение 4%-ных гранул в борьбе
со слизнями снижает поврежденность клубней картофеля
на 38—71 % (Hunter, Symonds, 1970). При опрыскивании
растений и почвы суспензией метиокарба гибнет до 68 %
слизней, а от применения гранулированного препарата —
до 90 % вредителей; 3%-ный и 4%-ный метиокарб в ко-
личестве 11—13 кг/га оказался более эффективен, чем
метальдегид (Getzin, 1965).
В Англии для защиты от слизней картофеля, овощных
и зерновых культур наряду с метальдегидом успешно
применяется метиокарб. При внесении 4%-ных гранул
метиокарба в количестве 11 кг/га в период цветения
гороха поврежденность культуры слизнями снизилась
на 90 % (Hunter, Symonds, 1970). Биологическая эффек-
тивность 50%-ного препарата метиокарба на 12-е сутки
после обработки достигла 100%.
От смачивающегося порошка метиокарба на влажных,
поросших кустарником пастбищах Калининской области
погибло до 60 % моллюсков разных видов, в том числе
и слизней (Горохов, Осетров, 1978).
Не отмечено токсического действия метиокарба на по-
лезных насекомых агробиоценозов — жужелиц и стафи-
линид, но имеются данные о губительном влиянии мол-
люскоцида на дождевых червей и многоножек.
3. Зектран, цектран (4-диметиламино-3,5-ди-
метилфенил-1Ч-метилкарбамат). Белое кристаллическое ве-
щество, плохо растворимое в воде, хорошо — в боль-
шинстве органических растворителей. Высокотоксичен
для человека и теплокровных животных. Обладает высо-
кой моллюскоцидной активностью; при норме расхода
2%-ного гранулированного зектрана 1 г на 1 м2 биологи-
3 В скобках указано действующее вещество препарата.
167
ческая эффективность против слизней составляет до 80 %
(Godan, 1965; Mallet, Bougaran, 1971).
4. Пиримор, пиримикарб (2-диметил-
амино-5,6-диметилпиримидинил-4-1\т, N-диметилкарбамат).
Бесцветное кристаллическое вещество, слабо растворимое
в воде, хорошо растворимое в органических растворите-
лях. Пиримор, инсектицид, применяемый в СССР для
борьбы с различными видами тлей, выпускается фирмой
«Си-Ай-Си» (Англия) в форме 50%-ного смачивающегося
порошка, 20%-ного эмульгирующегося концентрата,
5%-ных гранул и дымовых шашек. Отмечено контактное,
системное и фумигационное действие на насекомых. Мало-
токсичен для пчел и энтомофагов тлей. Для человека
и теплокровных животных высокотоксичен. Для борьбы
со слизнями представляет интерес смесь пиримора и ме-
тальдегида (соответственно 2% и 3%). Предпола-
гается усиление действия этих соединений на слизней
при их совместном применении (Mallet, Bougaran,
1971).
5. Севин, карбарил, нафтилкарба-
м а т, к а р п о л и н (а-нафтил-метил-карбамат). Белое
кристаллическое вещество, плохо растворимое в воде,
хорошо в органических растворителях. Севин не обладает
избирательным действием и вызывает гибель полезных
насекомых (в том числе пчел, жужелиц, стафилинид),
дождевых червей, многоножек. В почве сохраняется
до 1—2 лет и может распространяться с водой по дренаж-
ной системе. В СССР применяется 85%-ный смачиваю-
щийся порошок севина против вредителей яблони, хлоп-
чатника, кукурузы. Для человека и теплокровных живот-
ных среднетоксичен. Отмечены высокие моллюскоцидные
свойства севина в борьбе с раковинными формами и слиз-
нями. Используется для опрыскивания мест скопления
моллюсков и в приманках (1.25 и 2.5%-ной). Биологи-
ческая эффективность 2.5%-ной приманки севина, приго-
товленной на зерновых отрубях, составила на 5-е сутки
100 %, а 1.25%-ной приманки — 76 %. Отмечен синер-
гизм смеси севина с метальдегидом (Ruppel, 1959).
6. П и р о л а н [(3-метил-1-фенилпиразолил-5) -N,N-
диметил-карбамат]. Бесцветное кристаллическое вещество,
слабо растворимое в воде, хорошо — в спирте, ацетоне,
ароматических углеводородах. Высокотоксичен для чело-
века и теплокровных животных. Биологическая эффектив-
ность 2.5 %-ной нримацки пиролана, приготовленной на
158
пшеничных отрубях, составила в борьбе со слизнями
92 % на 5-е сутки (Горохов, Осетров, 1978).
7. Букстен [смесь из 1-метилбутилфенил-1Ч-ме-
тилкарбамата (75 %) и 1-этилпропилфенил-1У-метилкар-
бамата (25 %)]. Выпускается фирмой «Хеврон» (США)
в форме 2%-ных гранул и применяется как почвенный
пестицид. Один из наименее токсичных карбаматов для
человека и теплокровных животных. Биологическая эф-
фективность букстена против слизней достигает 100 %
(Mallet, Bougaran, 1971).
8. Карбатион, вапам (N-метилдитиокарбамат
натрия). Белое кристаллическое вещество, хорошо раство-
римое в воде, по плохо в органических растворителях.
Карбатион действует как стерилизатор почвы, уничто-
жает нематод, возбудителей болезней растений, сорняки.
Промышленность выпускает препарат карбатиона, пред-
ставляющий собой 40%-ный водный раствор действую-
щего вещества. В СССР карбатион применяется для сте-
рилизации грунта парников и теплиц при норме расхода
1 л на 1 м2. Стерилизацию проводят за 30 дней до исполь-
зования грунтовой смеси. Кроме того, препарат исполь-
зуют и на открытом грунте — в борьбе с возбудителями
заболеваний капусты, тыквенных, пасленовых, табака
п махорки, декоративных культур. Норма расхода от 750
до 2000 л/га, обработку проводят не позднее чем за 30 дней
до высадки рассады или посева семян. Карбатион исполь-
зуют также для уничтожения фитонематод, паразити-
рующих на картофеле, овощных культурах (огурце, тома-
тах, моркови, перцах, баклажанах, редисе, салате, луке,
чесноке), землянике, декоративных культурах. Норма
расхода в зависимости от культуры составляет от 1000
до 2000 л/га. Карбатион среднетоксичен для человека
и теплокровных животных.
Моллюскоцидные свойства карбатиона довольно хо-
рошо изучены. При норме расхода 15 кг действующего
вещества на 1 га карбатион вызывает массовую гибель
слизней (Пишнамазов, Бахышов, 1967). В опытах Увалие-
вой (1979) 2%-ный раствор карбатиона при норме расхода
200 мл на 1 м2 вызывал 100%-ную гибель слизней.
Таким образом, согласно регламенту, карбатион при-
меняется лишь на участках, свободных от защищаемых
растений. Численность слизней на таких участках редко
бывает высокой. Поэтому представляет интерес изучить
169
действие карбатиона на яйца моллюсков, находящиеся
в почве.
Rv Кроме метальдегида и карбаматов, моллюскоцидными
свойствами обладают и другие органические пестициды
различного химического строения. Некоторые из них
описаны ниже.
9. ТМТД, тирам [бис-(диметилтиокарбамоил)-
дисульфид]. Мелкокристаллический порошок желтовато-
серого цвета, практически нерастворим в воде, но раство-
рим в большинстве органических растворителей. Выпу-
скается промышленностью в форме 80%-ного препарата
для предпосевной обработки семян и обеззараживания
посадочного материала. В СССР ТМТД используется для
сухого или с увлажнением протравливания семян куку-
рузы, пшеницы, ржи, сорго, гречихи, многолетних кор-
мовых трав, огурца, свеклы, моркови, капусты и других
культур, а также для обработки семенных клубней кар-
тофеля и корнеплодов. В парниках ТМТД применяется
для обработки рассады капусты против комплекса возбу-
дителей заболеваний.
ТМТД среднетоксичен для человека и теплокровных
животных. Способен накапливаться в организме, а в боль-
ших дозах обладает мутагенным и канцерогенным дей-
ствием. В организме сохраняет токсичность.
Отмечено отпугивающее действие ТМТД на птиц
и слизней: семена пшеницы, обработанные препаратом
и высеянные в почву, не повреждались слизнями и не вы-
клевывались птицами (Southon, 1959).
Как показали наши лабораторные опыты, слизни не
повреждали предварительно обработанные ТМТД семена
капусты, огурца, клевера, пшеницы, овса, ржи, ячменя
в период их прорастания в почве. Однако продолжи-
тельность защитного действия препарата после появления
всходов была незначительной и составляла от 5 до 12 дней.
По нашему мнению, необходима широкая проверка репел-
лентного действия ТМТД на слизней и изучение возмож-
ности сочетания этого препарата с метальдегидом.
10. П р е п а р а т № 19 [N, (З-хлор-4-метилфенил)-
бутирамид]. Разработан в СССР для борьбы с наземными
моллюсками — промежуточными хозяевами гельминтов.
Малотоксичен для человека и теплокровных животных.
Препарат в количестве 6 г на 1 м2 губителен для назем-
ных моллюсков, обитающих на пастбищах в средней по-
лосе европейской части СССР, в том числе для слизней
170
(Горохов, Осетров, 1978). В качестве химического средства
защиты растений не испытан.
11. W L 21 959 [8-2-цианоэтил-М-(метилкарбамои-
локси)-тиоацетимидат]. Экспериментальный моллюскоцид
(Англия). Выпускается в форме 50%-ного смачивающегося
порошка и 5%-ных гранул с желатиной. В количестве
15 кг/га оказался более эффективным против сетчатого
слизня, чем метальдегид и метиокарб (Symonds, 1975).
12. Р Е 9 6 8 3 [З-метил-Ы-метил-К-(карбамоилокси)-
тиоацетимидат]. Моллюскоцид испытан в форме 5 %-ной
приманки в борьбе со слизнями. Эффективность препарата
оказалась намного выше, чем 5%-ного метальдегида
и 4%-ного метиокарба. Отмечено особенно сильное дей-
ствие смеси, содержащей РЕ 9683 (2%) и метальдегид
(3 %). Выпускается фирмой «Дюпон» (США) (Mallet,
Bougaran, 1971; Moens, 1971).
В заключение обзора отметим, что применение мол-
люскоцидов как в производственных условиях, так и
в хозяйствах частного сектора должно находиться в стро-
гом соответствии с перечнем химических средств защиты
растений, утвержденным Министерством сельского хо-
зяйства и Министерством здравоохранения СССР, а также
с установленным регламентом их использования (Спи-
сок . . ., 1982).
Между тем наличие в арсенале моллюскоцидов лишь
одного метальдегида нельзя считать удовлетворительным.
Необходимы испытания и других химических средств
защиты растений, в частности описанных выше.
Аппаратура для внесения моллю-
скоцидов. Для внесения моллюскоцидов пригодна
различная аппаратура, выбор которой зависит от формы
препарата и способа применения пестицида, а также
от размеров обрабатываемого участка. При обработке
небольших площадей можно использовать ручные и ран-
цевые опрыскиватели, а рассев отравленных приманок
осуществлять вручную. Для внесения препаратов на
большие территории необходимы более совершенная тех-
нология и аппаратура.
Лаборатория механизации ВНИИ защиты растений
рекомендует использовать для разбрасывания гранулиро-
ванных препаратов разбрасыватель удобрений НРУ-0.5.
При этом равномерность распределения гранул на по-
верхности почвы (коэффициент вариации) составляет 55 %
при норме расхода препарата 36 кг/га. Удовлетвори-
171
тельно выполняет тот же технологический процесс туко-
вая сеялка СТН-2.8 (равномерность распределения 28—
35 %). Однако эта машина имеет небольшую ширину
захвата и, следовательно, невысокую производительность.
Поэтому ее целесообразно использовать на небольших
участках (Белецкий и др., 1977).
На небольших участках может найти применение ран-
цевый разбрасыватель отравленной приманки РП, разра-
ботанный в Дальневосточном НИИ лесного хозяйства
и выпускаемый серийно опытным заводом института.
Бункер разбрасывателя рассчитан на 10 кг приманки
(емкость бункера 16 л). Дозатор обеспечивает как порцион-
ный выброс приманки (по 7—700 см3), так и беспрерывный.
Аппарат размещается на спине рабочего, его вес без прима-
нки 5 кг, производительность до 2 га/ч (Тимченко, 1976).
Для опрыскивания растений и почвы суспензией ме-
тальдегида можно использовать, например, универсаль-
ный опрыскиватель ОН-400, который навешивается на
тракторы Т-25, Т-54 и МТЗ. Ширина захвата в полевом
варианте 14 м, рабочая скорость до 10 км/ч, емкость
полиэтиленового резервуара 400 л, расход рабочей
жидкости регулируется в пределах 150—1000 л на 1 га.
Этот опрыскиватель может быть использован в садах
и на полях. Необходимо учитывать, что опрыскивание
растений при помощи наземной аппаратуры возможно
при скорости ветра до 3 м/с.
Моллюскоциды можно вносить и с помощью другой
аппаратуры, сведения о которой содержатся в специаль-
ной литературе.
Методы оценки моллюскоцидов. Со-
вершенствование химической борьбы со слизнями преду-
сматривает, в частности, поиск и испытание новых соеди-
нений с моллюскоцидными свойствами, а также апроби-
рование новых способов применения уже известных пре-
паратов. В силу вышеизложенных причин такой поиск
особенно актуален для отечественной химической защиты
растений от моллюсков.
Методы оценки моллюскоцидов включают лаборатор-
ные испытания, которые позволяют оценить токсичность
соединения для моллюсков, и полевые, в результате кото-
рых выясняется эффективность препарата (или способа
его применения). Под эффективностью понимают конеч-
ный результат химической обработки. Рассмотрим наибо-
лее важные из этих методов.
П2
Для определения токсичности кишечных моллюскоци-
дов используют метод «сэндвичей». На поверхность лис-
товой пластинки растения наносят известное количество
пылевидного моллюскоцида или его суспензию известной
концентрации. Эту пластинку покрывают другой пластин-
кой такой же формы, предварительно смоченной крахма-
лом (для закрепления). «Сэндвичи», наколотые на энтомо-
логические булавки, предлагают отдельно рассаженным
слизням. После питания моллюска таким «сэндвичем» его
переносят в другой садок, где он продолжает питаться
обычной (не отравленной) пищей.
За отравленными слизнями ведутся наблюдения, в про-
цессе которых регистрируют их поведенческие реакции.
Для выяснения влияния водного баланса слизней на по-
следствия отравления модельных животных помещают
в садки с субстратом разной влажности. В тех случаях,
когда в результате отравления наступает гибель слизня,
с помощью миллиметровой бумаги определяют площадь
съеденной им части листовой пластинки. Затем, зная рас-
ход моллюскоцида на единицу поверхности листа и массу
моллюска, нетрудно рассчитать летальную дозу для ка-
ждого моллюска.
Существуют и другие методы оценки кишечной ток-
сичности моллюскоцидов, в частности, основанные на
принудительном скармливании моллюскам отравленного
субстрата (Henderson, 1969).
Токсичность контактных моллюскоцидов ориентиро-
вочно можно установить при групповой обработке слиз-
ней, а более точно — способом индивидуального нанесе-
ния. В первом случае 50—100 слизней одного возраста,
содержащихся до опыта в одинаковых условиях, поме-
щают в эксикатор и обрабатывают (опыливают или опры-
скивают) моллюскоцидом с помощью лабораторного опры-
скивателя или опыливателя. Обработанных слизней пере-
носят в чистые садки с кормом и ведут за ними наблюде-
ния. Процент смертности определяют в сравнении с кон-
тролем, которым служат необработанные слизни из той же
лабораторной популяции.
Токсичность контактных моллюскоцидов более точно
определяется индивидуальным нанесением яда на тело
слизня. Наиболее простой способ индивидуального дози-
рования заключается в нанесении раствора моллюскоцида
известной концентрации на тело моллюска проволочной
петлей. Один конец петли следует закрепить в держателе
173
с низкой теплопроводностью, а другой — свернуть в ко-
лечко с внутренним диаметром 0.5—1 мм. Для калибровки
петли ее погружают в сосуд с раствором моллюскоцида
и набранную каплю жидкости переносят в предварительно
взвешенный на аналитических весах бюкс. Таким образохм
в бюкс из сосуда переносят 20—30 капель, взвешивают
его и определяют вес одной капли. Раствор моллюскоцида
наносят на определенный участок тела каждого слизня
прикосновением петли с набранной каплей. Зная концен-
трацию испытываемого раствора, среднюю массу одной
капли и одного слизня, можно установить дозу яда (в мг
на 1 г живой массы), полученную моллюском.
При нанесении моллюскоцида на тело слизня необхо-
димо отмечать реакцию животного (выделение слизи,
изменение двигательной активности и т. п.). Полезно
проследить различия в реакции моллюска на контакт
с растворами различной концентрации.
Для сравнения токсичности двух или более моллюско-
цидов, а также токсичности одного и того же моллюско-
цида для слизней разного возраста или физиологического
состояния сопоставляют средние летальные дозы яда.
Их определяют методом пробит-анализа.
При оценке токсичности моллюскоцидов для слизней
нужно учитывать, что чувствительность модельных экзем-
пляров определяется не только свойствами моллюскоцида,
но и физиолого-биохимическими характеристиками самих
животных (с. 157). Поэтому для получения корректных
результатов оценки такие характеристики желательно
знать.
Определение биологической эффективности моллюско-
цида — это установление процента смертности, скорости
гибели слизней в результате проведенной химической
обработки, а также процента снижения поврежденности
ими растений. При испытании новых моллюскоцидов
определение биологической эффективности проводится на
опытном участке, который должен быть однородным
по заселенности моллюсками, по типу почвы, рельефу,
агротехнике защищаемой культуры и другим показа-
телям.
Следует избегать чрезмерного насыщения опыта вари-
антами, ибо для обеспечения достоверности его результа-
тов химическая обработка участка и последующие учеты
должны проводиться в возможно короткий срок. Размер
делянок при мелкоделяночном опыте может составлять
174
от 5 до 200 м2, при крупйоделяночном — до 1 га. При
оптимальном размере делянок число повторностей можно
ограничить 4—6.
Испытание новых моллюскоцидов, так же как и нового
способа применения уже известного препарата, должно
включать эталонный вариант, т. е. обработку моллюско-
цидом с установленной ранее эффективностью. Так, при
испытаниях препаратов метальдегида отечественного про-
изводства эталонами служили импортные препараты этого
пестицида и суперфосфат, который ранее широко приме-
нялся как контактный яд. Теперь отечественный металь-
дегид может служить эталоном при оценке эффективности
других препаратов с моллюскоцидными свойствами (на-
пример, карбаматов и др.). Поскольку смертность слиз-
ней может определяться не только действием яда, но
и другими причинами (болезнями, деятельностью парази-
тов, неблагоприятными погодными условиями), для про-
ведения корректного опыта необходим контрольный ва-
риант — на не подвергавшихся химической обработке
делянках.
После обработки участка моллюскоцидом проводят
определение ее эффективности путем подсчета живых
и мертвых слизпей по каждой повторности варианта,
вычисляют процент смертности по каждой повторности,
а также средний процент по каждому варианту. Эффек-
тивность определяют по формуле:
с _ с с _ 100Б 1006
1 2 А а 9
где С — смертность моллюсков с поправкой на контроль
(в %), Сг — смертность в варианте с обработкой, С2 —
смертность в контрольной! варианте, А — общее число
моллюсков в варианте с обработкой, Б — число погибших
моллюсков в варианте с обработкой, а — общее число
моллюсков в контрольном варианте, б — число погибших
моллюсков в контрольном варианте.
Во влажной атмосфере и при высокой температуре
воздуха погибшие слизни очень быстро подвергаются
разложению. Кроме того, они могут быть уничтожены
естественными врагами. Поэтому для определения в этих
условиях биологической эффективности моллюскоцида
можно проводить учет не погибших слизней, а оставшихся
в живых. Снижение численности вычисляют по разности
между средней численностью до обработки (Л) и после
175
обработки (Б), Процент снижения численности (С”) опре-
деляют по формуле
г, —ЛМ(А-Б)
С — А
Изменение численности с поправкой на контроль будет
равно
= 100 (Л — Б) 100 (д — 6) 100 (Лб- Ба)
А а Аа ’
где а — численность слизней в контроле до обработки,
б — численность в контроле после обработки.
При испытании новых моллюскоцидов или известных
препаратов новым способом важно определить скорость
их действия в полевых условиях. Для этого после обра-
ботки проводят несколько учетов (через каждые сутки)
до получения стабильных результатов. В дальнейшем
учеты можно продолжить для определения продолжи-
тельности токсического действия препарата. Для этого
следует проводить подсадки слизней на обработанные де-
лянки в садках или изоляторах.
В качестве дополнительного показателя эффективности
моллюскоцида может быть использована сравнительная
поврежденность слизнями защищаемой культуры (см. ме-
тоды учета поврежденности растений слизнями — с. 97).
Этот показатель позволяет установить своевременность
проведения обработки.
Хозяйственную (урожайную) эффективность химиче-
ских обработок, так же как и других методов борьбы
со слизнями, определяют путем сравнения урожая с обра-
ботанного и необработанного (контрольного) участков.
Учитывать следует не только бункерный и амбарный, но
и биологический урожай с пробных площадок, поскольку
при уборке и транспортировке возможны потери про-
дукции.
Хозяйственную эффективность выражают в виде при-
бавки урожая. Необходимо, кроме того, учитывать и то-
варный вид, качество продукции (ее сортность) в целях
получения более объективного представления об эффек-
тивности борьбы с вредителями.
Экономическую эффективность химических обработок
рассчитывают сопоставлением стоимости прибавки уро-
жая, полученной в результате проведения защитных
мероприятий, с затратами на их проведение.
176
При определении сравнительной экономической эф-
фективности химических обработок для ускорения и упро-
щения расчетов можно ограничиться подсчетом только
прямых затрат, без накладных расходов. К прямым за-
тратам относятся: расходы денежных средств (оплата
пестицидов, плата за работы специализированных отря-
дов, средства на приобретение различных материалов,
оплата наемного транспорта); затраты труда и живой
тягловой силы; собственно транспортные расходы (авто-
транспорт, тракторы); аммортизационные отчисления; за-
траты на уборку и транспортировку прибавки урожая,
полученной в результате проведения мероприятий по за-
щите растений.
При определении стоимости прибавки урожая необхо-
димо учитывать доход от улучшения качества продукции
вследствие успешной борьбы с вредителями; он опреде-
ляется разницей в цене продукции разных сортов.
Для выражения экономической эффективности хими-
ческих обработок можно принять следующие показатели:
1) полученный дополнительный чистый доход (разность
между стоимостью сохраненного урожая и величиной
затрат по его защите); 2) норма рентабельности (отноше-
ние чистого дохода к величине затрат по защите); 3) оку-
паемость затрат (отношение стоимости дополнительной
прибавки урожая к величине затрат по защите).
В заключение отметим, что затраты на химическую
борьбу со слизнями будут оправданы лишь в случае ре-
альной угрозы снижения урожая защищаемой культуры.
Для высокоурожайных культур в качестве статистически
достоверного и хозяйственно ощутимого снижения урожая
принят уровень свыше 3% (Танский, 1977).
Ожидаемые потери урожая от вредителей при опреде-
ленной плотности их популяций можно рассчитать в том
случае, если известен коэффициент их вредоносности.
Последний выражается в процентах снижения биологи-
ческого урожая. К сожалению, до настоящего времени
коэффициенты вредоносности слизней не определены ни
для одной из повреждаемой ими культуры, а, следова-
тельно, и не рассчитаны пороговые значения численности
этих вредителей, выше которых химическая борьба стано-
вится экономически оправданной. Поэтому установление
критериев экономической целесообразности применения
моллюскоцидов следует считать одной из актуальных
задач защиты растений от слизней.
12 И. М. Лихарев и Я. С. Шапиро
177
Меры безопасности при работе
с моллюскоцидами. Все работы, связанные
с применением моллюскоцидов, а также с их перевоз-
ками, приемом, отпуском, хранением и уничтожением,
необходимо проводить в строгом соответствии с действую-
щими правилами охраны труда и производственной сани-
тарии (Инструкция. . ., 1976).
Применение моллюскоцидов проводится под руковод-
ством агрономов по защите растений или соответствующих
специалистов. К этим работам допускаются лица, достиг-
шие 18 лет, прошедшие медосмотр и инструктаж о мерах
предосторожности при работе с ядохимикатами. К работе
с пестицидами не допускаются подростки, кормящие
и беременные женщины, лица, страдающие заболеваниями
желудочно-кишечного тракта, сердечно-сосудистой си-
стемы и некоторыми другими заболеваниями. Кроме того,
мужчины старше 55 лет и женщины старше 50 лет не до-
пускаются к приготовлению рабочих смесей, протравли-
ванию семян и к фумигации.
Общая продолжительность рабочего дня на операциях,
непосредственно связанных с метальдегидом, — не более
6 ч, а с высокотоксичными моллюскоцидами — пе более
4 ч. Во время работы с пестицидами запрещается прием
пищи, курение, питье, а также присутствие посторон-
них лиц.
Помещение для хранения моллюскоцидов, а также
площадки для заправки ими аппаратуры, приготовления
отравленных приманок должны находиться не ближе
200 м от жилых и животноводческих помещений, источни-
ков водоснабжения, мест концентрации полезных живот-
ных и птиц и не менее 2000 м от рыбохозяйственных
водоемов.
Склады для хранения пестицидов должны иметь хоро-
шую вентиляцию и надежные запоры. Здание склада
должно быть бесчердачной конструкции и не ниже III сте-
пени огнестойкости. В каждом складе должно быть два
отделения — для хранения ядохимикатов и для хранения
спецодежды, воды и аптечки. Тара, в которой хранятся
моллюскоциды, должна иметь этикетки. В складе выве-
шивается инструкция по противопожарной безопасности,
а снаружи возле входа устанавливаются бочка с водой,
ящик с песком, огнетушитель и щит с противопожарным
инвентарем. Транспорт, предназначенный для перевозки
пестицидов, обеспечивается огнетушителем марки ОП-5.
178
При приготовлении ядовитых рабочих смесей или
отравленных приманок необходимо остерегаться попада-
ния пестицида в глаза, на губы и другие места тела,
особенно влажные от пота. Не следует допускать скот
близко к месту приготовления рабочих смесей и оставлять
последние без охраны в поле. На местах работы и осо-
бенно на дорогах, проходящих через обработанные места,
за 300 м от границы обработанного участка выставляются
предупредительные надписи единого образца. Выпас
скота, косьба травы, сбор грибов и ягод и т. п. на местах,
обработанных моллюскоцидами и в соседстве с ними,
не допускаются в течение срока, соответствующего стой-
кости пестицида.
Остающиеся после окончания работы неизрасходован-
ные пестициды должны быть учтены и немедленно отправ-
лены к месту постоянного хранения. Места приготовления
рабочих смесей по окончании работ должны быть пере-
паханы или перекопаны.
Опрыскивание и опыливание растений следует прово-
дить в утренние или вечерние часы при ослаблении вос-
ходящих и нисходящих токов воздуха. Движение агрега-
тов при обработке растений должно быть таким, чтобы
ядовитая волна не сносилась ветром на людей. При ис-
пользовании ранцевой аппаратуры рабочие должны рас-
полагаться на "поле по его диагонали и не ближе чем
на 5—6 м друг от друга. При ветре скоростью свыше 3 м/с
химические обработки запрещены.
Для защиты организма от поступления моллюскоци-
дов применяют комплекс защитных средств: спецодежда
(комбинезон или халат), резиновые сапоги, рукавицы,
защитные очки (ПО-2, ПО-3), респиратор соответствую-
щей марки (например, при работе с препаратами металь-
дегида — марки РУ-60М с патроном марки «А», работаю-
щим при концентрации моллюскоцида, превышающей
ПДК в 15 раз).
После окончания работ лицевые части респираторов
промывают в теплой воде 0.5%-ным раствором КМпО4
или спиртом, после чего промывают водой и высушивают
при температуре 30—35°С. Оканчивая рабочий день,
верхнюю одежду тщательно вычищают.
Первая помощь при отравлениях.
При отравлении моллюскоцидом пострадавшему необхо-
димо оказать первую помощь, которая включает меро-
приятия, осуществляемые самими работающими (само-
12*
179
Помощь и взаимопомощь), а также специальные меры,
которые осуществляют медицинские работники.
Для прекращения поступления яда в организм необ-
ходимо вынести пострадавшего из отравленной зоны,
при попадании пестицида на кожу смыть его струей воды
или удалить, не размазывая, куском ткани, а затем это
место обмыть водой. При желудочно-кишечном отравле-
нии — выпить несколько стаканов воды (желательно теп-
лой) или слабо-розового раствора перманганата калия
(КМпО4) и раздражением задней стенки глотки вызвать
рвоту, после этого выпить полстакана воды с 2—3 столо-
выми ложками активированного угля, а затем принять
солевое слабительное.
При ослаблении дыхания нужно дать понюхать по-
страдавшему нашатырный спирт, а в случае прекращения
дыхания немедленно делать искусственное дыхание.
Во всех случаях отравления необходимо пострадав-
шему предоставить покой и как можно скорее обратиться
к врачу.
Важную роль в профилактике отравлений пестици-
дами имеет рациональное питание, поскольку оно повы-
шает сопротивляемость организма к действию ядовитых
веществ. Пища работающих с пестицидами должна быть
богата белками; желательно, чтобы она содержала про-
дукты, обладающие обволакивающими свойствами (крах-
мал, желатин). Перед началом работы с пестицидами не-
обходим прием пищи. Ее отсутствие в желудочно-кишеч-
ном тракте способствует более интенсивному поступлению
в кровь ядовитых веществ. Следует учитывать, что дей-
ствие пестицидов на лиц, употребляющих перед работой
или во время ее алкоголь, усиливается в несколько раз,
что крайне опасно для жизни.
ЛИТЕРАТУРА
Акрамовский И. Н., Бабаян Г. С. Разработка мер борьбы с горным
слизнем — вредителем табака в парниках Армении // Изв. с.-х.
паук. 1976. № 10. С. 68—72.
Акрамовский Н. Н., Бабаян Г. С. Борьба со слизнями в парниках //
Защита растений. 1977. № 8. С. 34.
Алан М. Н. Применение метальдегида против слизней в условиях
Абхазии // Химия в сел. хоз-ве. 1972. № 5. С. 10.
Бодренков Г. Е. О вспышке массового размножения слизней в Ор-
ловской области в 1957 году // Учен. зап. Орлов, пед. ин-та.
1963. Вып. 18, № 1. С. 21—38.
Буренков М. С. Возрастная структура популяций трех видов слиз-
ней (отр. Stylommatophora, п/кл Pulmonata) // Журн. общ.
биологии. 1977. Вып. 38, № 2. С. 296—304.
Вавилов Н. Голые слизни (улитки), повреждающие поля и огороды
в Московской губ. М.: изд-во Моск. губ. земства. 1910. 55 с.
Васина А. Н. О мерах борьбы с голыми слизнями // Сад и огород.
1953. № 7. С. 12.
Вашадзе В. Н. Метальдегид защищает цветочную рассаду // За-
щита растений. 1972. № 3. С. 36.
Белецкий И. Н., Бегенеев И. Р., Касамапский А. Ш. Для внесения
гранулированных пестицидов // Защита растений. 1977. № 6.
С. 30-31.
Горохов В. В. Методические рекомендации по изучению патологии
моллюсков. М.» 1980. 72 с.
Горохов В. В. Экологические основы борьбы с вредными моллю-
сками. М., 1983. 207 с.
Горохов В. В., Осетров В. С. Моллюскоциды и их применение
в сельском хозяйстве. М., 1978. 224 с.
Граменецкий Е. М. Прижизненная окраска клеток и тканей в норме
и патологии. Л., 1963. 243 с.
Гриванов К. П. Полевой слизень и меры борьбы с ним // На защиту
урожая. 1935. № 8. С. И—13.
Гриванов К. П. Ареал распространения и зоны вредности слизней
(Agriolimax) в европейской части СССР // Итоги н.-п. работ
ВИЗР за 1935 год. Л., 1936. С. 34-37.
Дмитриева Е. Ф. Эффективность и длительность действия препа-
ратов метальдегида в борьбе с полевыми слизнями Ц Тр. Всесоюз.
НИИ защиты растений. 1963. Вып. 19. С. 260—266.
Дмитриева Е. Ф. Динамика численности, рост, питание и размноже-
ние сетчатого слизня (Deroceras reticulatum Mull.) в Ленинград-
ской области И Зоол. журн. 1969. Т. 48, вып. 6. С. 802—810.
Дмитриева Е. Ф., Шапиро Я. С. Гранулированный метальдегид —
активный токсикант кишечного действия // Науч. тр. Ленингр.
с.-х. ин-та. 1974. Т. 216. С. 110-113.
181
Дмитриева Е. Ф., Шапиро Я. С. Прогнозирование численности
сетчатого слизня // Защита растений. 1975а. № 7. С. 47—48.
Дмитриева Е. Ф., Шапиро Я. С. Некоторые функциональные изме-
нения в организме сетчатого слизня (Deroceras reticulatum
Mull.) при отравлении метальдегидом // Зап. Ленингр. с.-х.
ин-та. 19756. Т.270. С. 115—118.
Дмитриева Е. Ф., Шапиро Я. С. Физиолого-токсикологические
аспекты постзмбрпогенеза сетчатого слизня // Науч. тр. Ленингр.
с.-х. ин-та. 1976, Т. 297. С. 91-93.
Дмитриева Е. Ф., Шапиро Я. С. Изучение неспецифической устой-
чивости сетчатого слизня к метальдегиду // Науч. тр. Ленингр.
с.-х. ин-та. 1979. Т. 374. С. 59—62.
Загайпый С. А. Для защиты от слизней // Защита растений. 1978.
№ 3. С. 49.
Инструкция по технике безопасности при хранении, транспорти-
ровке и применении ядохимикатов (пестицидов) в сельском
хозяйстве. М., 1976. 40 с.
Конашевич В. А., Белоножко Г. А., Добрянский В. М. Метальде-
гид Ц Защита растений. 1972. № 7. С. 46.
Косоглазое А. А. Метальдегид в оранжереях // Защита растений.
1973. № 6. С. 47.
Лихарев И. М. Слизни — вредители сельского хозяйства. М.; Л.,
1954. 75 с.
Лихарев И. М. Некоторые факторы, определяющие распростране-
ние синантропных наземных моллюсков // 2-е Совещ. по изуче-
нию моллюсков: Тез. докл. Л., 1965. С. 48—51.
Лихарев И. М., Виктор А. Й. Слизни фауны СССР и сопредельных
стран (Gastropoda terrestria nuda). Л., 1980. 437 с. (Фауна СССР.
Нов. сер.; № 122: Моллюски; Т. 3. Вып. 5).
Мурусидзе Т. Э. Метальдегид — эффективный лимацид в цитрусовых
насаждениях // Субтроп, культуры. 1972. № 2. С. 104—105.
Пайкин Л. М. Способ борьбы с полевыми слизнями // Бюл. изобре-
тений. 1949. № 12. С. 17.
Пахорукова Л. В. Количественное исследование питания полевого
(Deroceras agreste) и сетчатого (D. reticulatum) слизней // Зоол.
журн., 1976. Т. 55, вып. 1. С. 29—31.
Пишнамазов Г. А., Бахышев Д. С. Голые слизни и борьба с ними
в условиях Азербайджана И Тр. Азерб. НИИ защиты растений.
1967. Т. 1. С. 12-23.
Поляков И. Я. Основные предпосылки теории защиты растений
от вредителей // Энтомол. обозрение. 1968. Т. 47, вып. 2.
С. 343—361.
Поляков И. Я. Ритмика явлений в природе и динамика численности
сельскохозяйственных вредителей // Чтения памяти Л. С. Берга.
XV—XVIII. Л., 1973. С. 220—233.
Преображенский К. И. Культуртехнические работы на закустарен-
ных землях Нечерноземной зоны РСФСР. Л., 1983. 119 с.
Разауска Э. Производственное испытание активированного креолина
(АК) как средства борьбы со слизнями // Краткие итоги науч,
исследований по защите растений в Прибалт, зоне СССР. Рига,
1963. С. 90.
Соколов А. М. Полевые слизни: Новое об образе жизни их. Меры
борьбы Ц Сб. Всесоюз. НИИ защиты растений. 1933. Вып.
7. С. 39—48.
182
Соколов А. М. Материалы изучения слизней и мер борьбы с ними //
Тр. Иванов, с.-х. ип-та. 1935. № 1. G. НО.
Список химических и биологических средств борьбы с вредителями,
болезнями растений и сорняками и регуляторов роста расте-
ний, разрешенных для применения в сельском хозяйстве на
1982—1985 гг. // Защита растений. 1982. № 3. С. 40—61.
Старобогатов Я. И. Фауна моллюсков и зоогеографическое райони-
рование континентальных водоемов земного шара. М., 1970.
372 с.
Страховская Е. Н. Газообразные лпмациды мгновенного действия
в борьбе со слизнями И Докл. ВАСХНИЛ. 1949. Вып. 10.
С. 34—40.
Танский В. И. Методические указания по разработке экономиче-
ских порогов вредоносности вредных насекомых. Л., 1977. 24 с.
Тимченко Л. И. Разбрасыватель приманки // Защита растений.
1976. № 10. С. 26—27.
Увалиева К. К. Слизпи — как вредители сельскохозяйственных
культур юго-востока Казахстана. Алма-Ата, 1979. 59 с.
Хохуткин И. М. Эпибиоз моллюсков и летучей мыши: Автореф.
докл. засед. секции Урал, отд-ния Моск, о-ва испыт. природы //
Бюл. МОИП. Отд. биол. 1967. № 5. С. 153—154.
Шапиро Я. С. Изучение вирофорной способности четырех видов
слизней // Науч. тр. Леиингр. с.-х. ип-та. 1978. Т. 351. С. 56—
61.
Шапиро Я. С. Популяция сетчатого слизня (Deroceras reticulatum
Milll.) ссльскохозяйствепного ландшафта Ленинградской области:
возрастная структура и устойчивость к метальдегиду Ц 6-е
Совещ. по изучению моллюсков. Л., 1979. С. 156—157.
Шапиро Я. С. Наземные моллюски агробпоцепозов Ленинградской
области. Сообщ. 2. Возрастная структура популяций слизней
Deroceras И Науч. тр. Ленингр. с.-х. ин-та. 1980. Т. 389.
С. 42-44.
Шапиро Я. С. Прогнозирование численности слизней Deroceras
(Stylommatophora, Agriolimacidae) в сельскохозяйственном
ландшафте И Биол. науки. 1981. № 10. С. 103—106.
Шиков Е. В. О расселении паземных моллюсков во время полово-
дий Ц Зоол. журн. 1977. Т. 56, вып. 3. С. 361—367.
Шилейко А. А. Некоторые аспекты изучения современных конти-
нентальных брюхоногих моллюсков: Итоги науки п техники.
Зоол. беспозвоночных. М., 1972. Т. 1. 187 с.
Abeloos М. Forme de la croissance des especes du genre Limax Ц
G. R. Acad. sci. Paris. 1943. Vol. 217. P. 159—161.
Abeloos M. Evolution par variation du nombre des phases de crois-
sance chez les mollusques Limaciens // G. R. Acad. sci. Paris.
1945. Vol. 220, N 1—5. P. 62-63.
Arias R. O., Crowell H. H. A contribution to the biology of the grey
garden slug Ц Bull. Soc. Calif. Acad. sci. 1963. Vol. 62, N 2.
P. 83—97.
Backer A. N. Factors contributing towards the initiation of slug
activity in the field // Proc. Malacol. Soc. bond. 1973. Vol. 40,
N 4. P. 329—333.
Barnes H. F., Weil J. W. Baiting slug using metaldehyde mixed
with various substances // Ann. appl. Biol. 1942. Vol. 29,
N 1. P. 11—26.
183
Barnes H. Weil J. W. Slugs in garden: their numbers, activities
and distribution. Part 1 // J. Anim. Ecol. 1944. Vol. 13. P. 140—
175.
Barnes H. F., Weil J. W. Slugs in gardens: their numbers, activities
and distribution. Part 2 // J. Anim. Ecol. 1945. Vol. 14. P. 71—105.
Bayne C. J. Histochemical studies on the egg-capsules of eight gastro-
pod molluscs // Proc. Malacol. Soc. bond. 1968. Vol. 38, N 3.
P. 199-212.
Brooks W. M. Tetrahymenid ciliates as parasites of gray garden
slug Ц Hilgardia. 1968. Vol. 39, N 8. 72 p.
Carrick R. The life-hystory and development of Agriolimax agre-
stis L., the grey field slug // Trans. Roy. Soc. Edinburgh. 1938.
Vol. 59, N 3. P. 563—597.
Chevallier H. Taxonomie et biologies des grands Arion de France
(Pulmonata : Arionidae) Ц Malacologia. 1969. Vol. 9, N 1.
P. 73—78.
Chevallier H. Cycle biologique des grands Arion en France (Gastro-
poda, Pulmonata) // Atti Soc. ital. sci. natur. 1971. Vol. 112,
N 2. P. 316-320.
Corliss J. 0. Tetrahymena and some thoughts on the evolutionary
origin of endoparasitism // Trans. Amer. Soc. 1972. N 91.
P. 566—573.
Dainton В. H. The activity of slugs: 1 The induction of activity by
changing temperatures Ц J. exp. Biol. 1954. Vol. 31. P. 165—
167, 188—197.
Dobra K. W., McArdle E. WEhret C. F. Growth kinetics of three
species of Tetrahymena on solid agar // J. Protozool. 1980. Vol. 27,
N 2. P. 226-230.
Duval A. Le systeme circulatoire des limaces // Malacologia. 1982.
Vol. 22, N 1—2. P. 627—630.
Duval D. M. A record of slug movements in late summer // J. Con-
chol. 1972. Vol. 27, N 7. P. 505—508.
Fromming E. Biologie der mittteleuropaischen Landgastropoden.
Berlin. 1954. S. 1—404.
Getzin L. W. Control of gray garden slug with bait formulations
of a carbamate molluscicide // J. econ. Entomol. 1965. Vol. 58,
N 1. P. 158-159.
Godan B. Untersuchungen uber die molluskizide Wirkung der Car-
bamate//Z. Pflanz. 1965. Bd 72, N 7. S. 21—30.
Henderson I. F. A laboratory method for assesing the toxicity of
stomach poisons to slug // Ann. appl. Biol. 1969. Vol. 23, N 1.
P. 161-171.
Hermann G. BekSmpfung von Landschnecken mit Methiocarb Ц
Mededel. facult. landbouw. Gent. 1971. D36, N 1. S. 197—206.
Hunter P. J. Studies on slugs of arable ground: 2. Life cycles //
Malacologia. 1968. Vol. 7, N 6. P. 379—389.
Hunter P. J., Symonds В. V. The distribution of bait pellents for
slug control // Ann. appl. Biol. 1970. Vol. 65, N 1. P. 13—19.
Kalmus H. Anemotaxis in soft-skinned animals // Nature. London,
1942. Vol. 150. P. 524.
Kerkut G. A. A neurophysiological analysis of behaviour // Proc.
15-th Int. Zool. congr. 1959. P. 845—847.
Kosinska M. The life cycle of Deroceras sturanyi (Simroth, 1894)
(Pulmonata, Limacidae) // Zool. Polon. 1980. Vol. 28, N 2.
P. 113-155.
184
Kunkel К. Zur Biologic dor Lungcnschnecken. Heidelberg. 1916.
440 S.
Langlois T. H. The conjugal behaviour of the introduced european
giant slug Limax maximus as observed on S. Bass Island, Lake
Erie Ц Ohio J. Sci. 1963. Vol. 65. P. 208-304.
Laviolette P. L’evolution de la glande hermaphrodite d’Arion rufus
et ses reports avec la croissance // G. R. Soc. biol. Paris. 1950.
Vol. 144. P. 135—136.
Laviolette P. Role de gonade dans determinisme glandulaire du
tractus genital chez quelques Gasteropodes Arionidae et Lima-
cidae Ц Bull. biol. France-Belg. 1954. Vol. 88. P. 310—332.
Lynn D. H. The life cycle of the histophagus ciliate Tetrahymena
corlissi Thompson. 1955 Ц J. Protozool. 1975. Vol. 22, N 2.
P. 188—195.
Mallet C., Bougaran H. Action molluscicide de divers carbamates //
Mededel. facult. landbouw. Gent. 1971. D36, N 1. P. 207—223.
Martin A. W., Harrison F. M., Huston M. J., Stewart D. M. The
blood volumes of some representative molluscs // J. exp. Biol.
1958. Vol. 35. P. 260—279.
McArdle E. W. Gytological staining of ciliates Protozoa cultured on
membrane filters Ц Trans. Amer. Microsc. Soc. 1978. Vol. 97,
N 4. P. 582-586.
Moens R. Test d’efficacite de quelques nouvelles preparations mol-
luscicides pour la lutte contre les limaces Ц Mededel. facult.
landbouw. Gent. 1971. D36, N 1. P. 216-223.
Newell P. F., Newell G. E. The eye of the slug, Agriolimax reticulatus
(Miill.) // Invertebr. Receptors. Symposia Zool. Soc. Lond. 1968.
N 23. P. 97—111.
Pallant D. The food of the grey field slug Agriolimax reticulatus
(Muller) in woodland // J. Anim. Ecol. 1969. Vol. 38, N 2. P. 391 —
397.
Pallant D. A quantitative study of feeding in woodland by the grey
field slug Agriolimax reticulatus (Muller) // Proc. Malacol. Soc.
Lond. 1970. Vol. 39, N 1. P. 83—87.
Pelluet D., Henderson N. E. The effect of visible radiation on the
ovotestis of the slug Deroceras reticulatum // Proc. Nova Scotian
Inst. Sci. 1954. Vol. 23. P. 420—421.
Pelluet D., Lane N. J. The relation between neurosecretion and
cell differentiation in the ovotestis of slugs (Gastropoda : Pul-
monata) // Ganad. J. Zool. 1961. Vol. 39, N 6. P. 789—805.
Pinder L. С. V. The biology and behaviour of some slugs of economic
importance Agriolimax reticulatus, Arion hortensis and Milax
budapestensis // Ph. D. thesis. University of Newcastle-upon-
Tyne. 1969.
Rees W. The aerial dispersial of Mollusca // Proc. Malacol. Soc.
Lond. 1965. Vol. 36, N 5. P. 269—282.
Richter К. O. Freez-branding for individually marking the banana
slug: Agriolimax columbianus // G. Northwest Sci. 1973. Vol. 47,
N 2. P. 109—113.
Richter К. O. A method for individually marking slug 11 J. Molluscan
Stud. 1976. Vol. 42, N 1. P. 146—151.
Runham N. W., Hunter P. J. Terrestrial slugs. Hutch. Lond. 1970.
184 p.
Ruppel R. Effectiveness of Sevin against the grey field slug // J. econ.
Entomol. 1969. Vol. 53, N 3. P. 124-127.
185
Shotton F. E. Outline of the practical problem 11 Slug res. Confe-
rence. Univ, college Cardiff. 1971. P. 2—3.
Sokolove P. G., McCrone E. J. Reproductive maturation in the slug,
Limax maximus, and the effects of artificial photoperiod //
J. Comp. Physiol. 1978. A, vol. 125, N 4. P. 317—325.
Southon T. Trials with thiram as seed and ripening grain protec-
tant Ц New Zel. J. Agrical. 1959. Vol. 99, N 2. P. 21—22.
Stephenson J. W. The molluscacidal properties of three fly repel-
lents 11 Plant Pathol. 1962. Vol. 11, N 1. P. 37.
Stephenson J. W. Laboratory observation on the effect of soil com-
paction on slug damage to winter wheat // Plant Pathol. 1975.
Vol. 24, N 1 P. 9—11.
Stringer A. A note on the action of metaldehyde on slugs 11 Res.
agricult. Res. Stn. Univ. Bristol. 1949. P. 87—88.
Symonds В. V. Evalution of potential molluscicides for the control
of the field slug, Agriolimax reticulatus (Miill.) // Plant Pathol.
1975. Vol. 24, N 1. P. 8-9.
Thomas D. C. The use of metaldehyde against slugs // Ann. appl.
Biol. 1948. Vol. 35, N 2. P. 207—227.
Wattez Ch. Influence du complexe cephalique (tentacule ocularies
cerveau) sur 1’evolution, en culture in vitro, de gonades infan-
tiles et juveniles d’Arion subfuscus Drap. (Gasteropode Pulmone) И
Gen. and Comp. Endocrinol. 1978. Vol. 35, N 4. P. 360—374.
Williamson M. Studies on the colour and genetics of the black slug 11
Proc. Roy. phys. Soc. Edinburgh. 1959. Vol. 27. P. 87—93.
УКАЗАТЕЛЬ РУССКИХ
И ЛАТИНСКИХ НАЗВАНИЙ ВИДОВ СЛИЗНЕЙ
Арион бурый — Arion subfuscus 107, 111
Арион лесной — Arion silvaticus 107, 117
Арион окаймленный — Arion circumscriptus 107, 114
Арион полосатый — Arion fasciatus 107, 112
Бётгерилла светлая — Boettgerilla pallens 108, 128
Леманниа валенсийская — Lehmannia valentiana 110, 138
Леманниа каемчатая — Lehmannia marginata 110, 137
Малаколпмакс нежный — Malacolimax tcnellus 110, 136
Слизень большой — Limax maximus 110, 132
Слизень гладкий — Deroceras laeva 109, 123
Слизень желтоватый — Limax flavus 109, 133
Слизень крапчатый — Limax maculatus 110, 135
Слизень пашенный см. Слизень полевой
Слизень полевой — Deroceras agreste 109, 121
Слизень проворный см. Слизень гладкий
Слизень сетчатый — Deroceras reticulatum 108, 118
Слизень Штурания — Deroceras sturanyi 109, 126
Слизень черный — Limax cinereoniger 110, 131
УКАЗАТЕЛЬ НАЗВАНИЙ
МОЛЛЮСКОЦИДОВ
Аммиак 150 Ариотокс 152 Байер-37 344 166 Букстен 169 Вапам 169 Дибутилфталат 151 Диметилфталат 151 Дитиокарбаматы 169 Железный купорос 150 Зектран 167 Известь 150 Карбатион 169 Карбарил 168 Карполин 168 Креолин активированный 151 Медный купорос 150 Мезурол 166 Меркаптодиметур 166 Метальдегид 151 Метиокарб 166 Нафтилкарбамат 168 Пиримикарб 168 Пиримор 168 Пиролан 168 Препарат № 19 170 Севин 168 Сульфат аммония 150 Суперфосфат 150 Тиокарбаматы 151 Тирам 170 ТМТД 170 Цектран 167 Этиленхлоргидрин 150 Этиловый эфир диметилдитио- карбаминовой кислоты 150 РЕ 9683 171 WL 21959 171
ОГЛАВЛЕНЙЕ
Введение • . ........................................... 3
Значение слизней как вредителей культурных растений ... 7
Основные особенности морфологии п физиологии слизней . . 14
Внешний вод ..................................... 14
Внутреннее строение............................... 19
Покровы (22). — Мускулатура (23). — Раковина
(24) . — Легкое и мантийный комплекс органов (24).
— Кровеносная система (24). — Выделительная си-
стема (25). — Пищеварительная система (27).
— Нервная система (30). -- Органы чувств (30).
— Половая система (32).
Размножение и онтогенез................................ 36
Копуляция (36). — Оплодотворение и формиро-
вание яиц (38). — Откладка яиц (39). — Постэмбрио-
генез и рост (39). — Жизненный цикл (42).
Питание................................................ 46
Слизни и среда обитания................................ 48
1. Абиотические факторы ......................... 48
Температура (49). — Влажность (50). — Осадки
(52). — Ветер (53). — Свет (53). — Почва (54).
2. Биотические факторы .......................... 55
Растительность, основные ландшафты и биотопы
(55) . — Хищники, паразиты и болезни (58).
3. Антропогенные факторы ........................ 60
Синантропные и антропохорные виды (60).
Динамика популяций слизней в агроландшафте и ее прогно-
зирование ............................................. 64
Особенности фенологии и структуры популяций
(66). — Сезонная и многолетняя динамика числен-
ности (73). — Расселение слизней на сельскохозяй-
ственных землях (81).
Выявление, учет численности и определение вредоносности
слизней................................................ 87
Полевые исследования ............................ 88
Маршрутные исследования (88). — Учет яиц
слизней (92). — Учет слизней постэмбриональных
стадий (93). — Учет поврежденности растений 497).
189
Лабораторные исследования .......................... 99
Выделение слизней из почвенных проб (99).
— Определение живой массы и возраста слизней
(100) . — Определение жизнеспособности яиц (101).
— Содержание слизней в неволе (102). — Фиксация
материала (103).
Определение видов слизней.......................... 105
Таблица для определения родов и видов слизней Не-
черноземной зоны европейской части СССР .... 106
Семейство Ариониды (Arionidae)............... Ill
Семейство Агриолимациды (Agriolimacidae) .... 118
Семейство Бётгериллиды (Boettgerillidae) .......... 128
Семейство Лимациды (Limacidae) .................... 131
Борьба со слизнями ...................................... 140
Агротехнический и механический методы........ 141
Мелиорация земель (141). — Обработка почвы
(142). — Посев и посадка растений (143). — Уборка
урожая (144). — Севооборот (144). — Удобрения
(145). — Механический метод (145).
Биологический метод................................ 145
Химический метод................................... 150
1. Метальдегид (151). — Действие метальдегида
на слизней (152). — Применение препаратов ме-
тальдегида (160). — Озимые зерновые (160). —
Овощные культуры открытого грунта (161). — Мно-
голетние кормовые травы (161). — Земляника (162).
— Метальдегид в условиях защищенного грунта (163).
— Метальдегид на пастбищах (163). — 2. Метио-
карб, мезурол, меркаптодиметур, Байер-37 344 (166).
— 3. Зектран, цектран (167). — 4. Пиримор, пири-
микарб (168). — 5. Севин, карбарил, нафтилкарба-
мат, карполин (168). — 6. Пиролан (168). — 7. Бук-
стен (169). — 8. Карбатион, вапам (169). —
9. ТМТД, тирам (170). — 10. Препарат № 19 (170).
— И. WL 21 959 (171). — 12. РЕ 9683 (171). — Ап-
паратура для внесения моллюскоцидов (171). — Ме-
тоды оценки моллюскоцидов (172). — Меры безопас-
ности при работе с моллюскоцидами (178). — Пер-
вая помощь при отравлениях (179).
Литература ............................................. 181
Указатель русских и латинских названий видов слизней 187
Указатель названий моллюскоцидов........................ 188
Илья Михайлович Л и х а р с и, Икон Семенович 111 а и и р о
СЛИЗНИ —ВРЕДИТЕЛИ СЕЛЬСКОГО ХОЗЯЙСТВА
НЕЧЕРНОЗЕМНОЙ ЗОНЫ
(НАУКА - СЕЛЬСКОМУ ХОЗЯЙСТВУ)
Утверждено к нсч н ти
Редакционной коллегией серии научно-популярных изданий
Академии паук СССР
Редактор издательства В. Г. Федотова
Художник И. II. Кремлей
Технический редактор Ф. Л. Юлит
Корректоры И. А. Корзинина, Г. Н. Мартьянова
ИБ № 21646
Сдано в набор 3.09.86. Подписано к печати 12.02.87. М-17049.
Формат 84хЮ81/*2- Бумага офсетная № 1. Гарнитура обыкновенная. Печать
высокая. Усл. печ. л. 10.08. Усл. кр.-от. 10.3. Уч.-изд. л. 10.26.
Тираж 35.000. Тип. зак. Я» 773. Цена 65 к.
Ордена Трудового Красного Знамени издательство «Наука». Ленинградское
отделение. 199034, Ленинград, В-34, Менделеевская лин., 1.
Ордена Трудового Красного Знамени Первая типография издательства «Наука».
199034, Ленинград, В-34, 9 линия, 12.