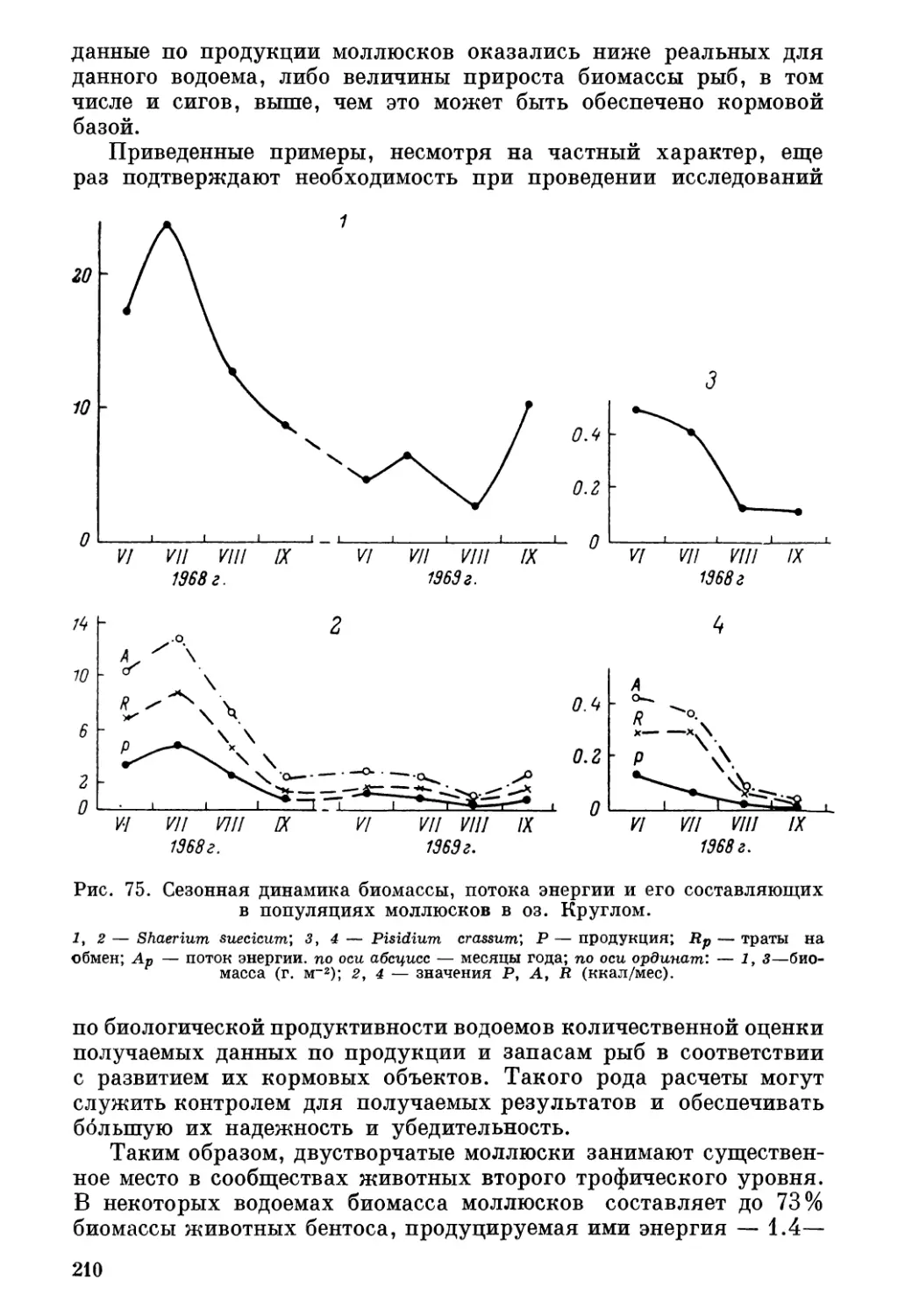
/



























































































































































































































































Text
А. Ф. АЛИМОВ
ФУНКЦИОНАЛЬНАЯ
ЭНОЛОГИЯ
ПРЕСНОВОДНЫХ
ДВУСТВОРЧАТЫХ
моллюсков
АКАДЕМИЯ НАУК СССР
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА
Т о м 96
А. Ф. АЛИМОВ
ФУНКЦИОНАЛЬНАЯ
ЭКОЛОГИЯ
ПРЕСНОВОДНЫХ
ДВУСТВОРЧАТЫХ
моллюсков
е
ЛЕНИНГРАД
«НАУК А»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1981
УДК 577.472
Функциональная экология пресноводных двустворчатых моллюсков.
Алимов А. Ф. Л., Наука, 1981. 248 с. (Тр. Зоол. ин-та АН СССР.
Монография представляет собой сводку по экологии пресноводных дву-
створчатых моллюсков в ее функциональном аспекте. Изложены и проанализи-
рованы материалы количественных полевых и экспериментальных исследо-
ваний по росту, интенсивности обмена, питания, размножения отдельных
видов моллюсков, динамике численности, возрастного состава, продукции
их популяций, определены балансы энергии особей в процессе их роста,
а также популяций массовых видов. Установлены общие для пресноводных
и морских моллюсков зависимости скорости обмена и фильтрации от веса
животных, а также влияние факторов внешней среды на процессы роста,
обмена, фильтрации у моллюсков. Приводятся количественные оценки свя-
зей между ростом, плодовитостью особей и скоростями увеличения числен-
ности их популяций. На основе эколого-физиологических и популяционных
характеристик отдельных видов оценено функциональное значение популя-
ций двустворчатых моллюсков в водных экологических системах, процессах
самоочищения водоемов, питании рыб. Обращено внимание на важное зна-
чение зоологических и фаунистических исследований в решении экологи-
ческих вопросов. Лит.—510 назв., ил. — 79, табл. 62.
Ответственный редактор
Г. Г. Винберг
Александр Федорович Алимов
ФУНКЦИОНАЛЬНАЯ ЭКОЛОГИЯ
ПРЕСНОВОДНЫХ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
(Труды Зоологического института АН СССР. Том 96)
Утверждено к печати
Зоологическим институтом Академии наук СССР
Редактор издательства Е. И. Васьковская
Художник Л. А. Яценко
Технический редактор М. Э. Карлайтис
Корректоры С. В. Добрянская и С. И. Сем и глазова
ИБ № 20220
Сдано в набор 23.07.80. Подписано к печати 2.12.80. М-21067.
Формат 60x90Vie. Бумага № 1. Гарнитура обыкновенная.
Печать высокая. Печ. л. 1572 = 15.50 усл. печ. л. Уч.-изд. л. 17.29.
Тираж 1100. Изд. № 7720. Тип. зак. № 1631. Цена 2 р. 70 к.
Ленинградское отделение издательства «Наука»
199164, Ленинград, В-164, Менделеевская лин., 1
Ордена Трудового Красного Знамени
Первая типография издательства «Наука»
199034, Ленинград, В-34, 9 линия, 12
21009-501
А___________ 585-81 2001050100
055 (02)-81
© Издательство «Наука», 1981 г.
ПРЕДИСЛОВИЕ
Экология в наши дни популярна. Актуальность и. ответствен-
ность ее задач общеизвестны. Экология, опираясь на весь комп-
лекс биологических и смежных с ними наук, создает научную
базу для гармоничного сочетания возрастающей мощи челове-
ческого общества с незыблемыми законами природы, управляю-
щими биосферой, в которой мы обитаем, вернее, в которую входим
как ее неотъемлемый компонент. Конечные цели экологии ясны,
по путь к ее вершинам тяжел и требует большой затраты сил на
его отдельных этапах. Изучение главного, точнее, специфического
объекта экологии — экосистемы — должно быть обеспечено эколо-
гически направленными исследованиями на всех уровнях органи-
зации жизни, без которых невозможно успешное продвижение
к решению главнейших экологических проблем.
Экология молода и с бесстрашием молодости берется за реше-
ние своих грандиозных задач. В начале своего жизненного пути
она нуждается в помощи со стороны породивших ее биологиче-
ских наук старшего поколения, таких как зоология и ботаника,
вернее, имеет с ними обширную область общих интересов. С раз-
витием экологических представлений обогащается и зоология,
получая возможность рассматривать и оценивать экологические
особенности животных на строго объективной основе экологи-
ческих закономерностей.
Область общих интересов экологии и зоологии прежде всего
охватывает аутоэкологические вопросы, понимаемые как эколо-
гия вида на уровне особи и популяции. При выяснении аутоэко-
логических закономерностей встречаются не только с видоспеци-
фическими явлениями, но и с общими свойствами, присущими
всем видам, входящим в соответствующую таксономическую еди-
ницу более высокого ранга. Благодаря этому тесное взаимодей-
ствие аутоэкологических и зоологических исследований оказы-
вается естественным, неизбежным и взаимополезным. Это хорошо
показано на примере пресноводных двустворчатых моллюсков
в монографии А. Ф. Алимова, содержащей результаты долголет-
3
них целенаправленных исследований зоолога и гидробиолога,
или эколога водных сообществ.
В области общих интересов, в пограничной зоне двух смеж-
ных наук, где зарождаются новые направления исследований, нет
и не может быть проторенных путей. Приходится отыскивать
верную дорогу к цели, поиски которой не могут быть свободны
от блужданий и ошибок. А. Ф. Алимов устранил эти трудности,
ограничив тематику своих работ вполне определенным комплек-
сом важнейших взаимосвязанных вопросов, отраженных в назва-
ниях глав книги. Умело пользуясь уже известными общебиоло-
гическими закономерностями и заметно пополнив их благодаря
результатам своих исследований, А. Ф. Алимов на основании соб-
ственных и литературных данных количественно охарактеризо-
вал интенсивность обмена веществ, скорость и другие особенности
питания, тип и скорость роста, т. е. основные функции пресно-
водных двустворчатых моллюсков, через которые осуществляется
их участие в процессах биотической трансформации вещества и
энергии. Тем самым он создал необходимую основу для понима-
ния роли двустворчатых моллюсков в биотическом круговороте
веществ и продуктивности пресных вод.
Несмотря на то что объектом исследования служили виды,
принадлежащие к определенному таксону, монографию А. Ф. Али-
мова по ее содержанию нельзя отнести к экологии так называе-
мых таксоценов. В абстрактно-экологическом течении научной
мысли, представленном главным образом в США, распространены
построения, базирующиеся на подсчетах числа видов и особей
в рассматриваемом тацсоцене, т. е. в совокупности популяций
видов, принадлежащих к некоторому определенному таксону.
Даже если будет показано, что соотношения между числом видов
и особей в отдельных таксоценах могут передавать существенные
свойства экосистемы и заслуживают изучения, построения аб-
страктной экологии в лучшем случае могут претендовать только
на отражение некоторых особенностей биотической (по термино-
логии Ю. Одума), но не функциональной структуры экосистемы
и, быть может, некоторых сторон взаимоотношения видов в пре-
делах таксоцена, но не экосистемы в целом.
А. Ф. Алимов идет по другому пути, оставаясь на твердой
почве конкретной экологии водных экосистем (гидробиологии),
в которой на наших глазах успешно создается действенная тео-
рия экологических явлений, столь нужная практике приложения
экологических знаний к решению кардинальных задач современ-
ного естествознания. В исследованиях А. Ф. Алимова получило
творческое развитие то направление изучения водных экосистем,
в котором видную роль играют работы советских авторов, во мно-
гом опережающие зарубежную науку. Эти работы направлены на
выяснение закономерностей, управляющих функционированием
водных экосистем, их продуктивностью и биотическим круго-
воротом веществ.
4
Для достижения этих целей надо располагать возможностью
количественно выражать основные функции животных в их вза-
имосвязи и зависимости от условий среды. Необходимые для этого
снедения получают путем последовательного применения в поле-
вых и лабораторных исследованиях энергетического принципа
количественной трактовки жизненных явлений, широко исполь-
зованного А. Ф. Алимовым. Это дает возможность накапливать
знания, нужные для понимания значения и места изучаемого вида
в механизме протекающих в экосистеме процессов. Особенно
существенно, что именно это и следует знать для решения любых
задач, выдвигаемых перед биологией практикой использования
биологических ресурсов природы и охраны окружающей среды,
как это убедительно показано А. Ф. Алимовым на ряде примеров
но результатам его работ.
Естественно, что в пределах конкретной экологии есть разные
направления изучения структуры и функций экосистем и других
объектов экологических исследований. А. Ф. Алимов подчеркнул
принадлежность своих работ к очерченному выше направлению,
удачно назвав его функциональной экологией.
Несомненно, что зоологи, экологи, гидробиологи, заинтере-
сованные в основной тематике книги, единодушно признают,
что она представляет собой не только важный итог эколого-зоо-
логического изучения двустворчатых моллюсков, но и основу
для последующих исследований и высоко оценят научный уровень,
своевременность и полезность монографии А. Ф. Алимова. Од-
нако ее содержание имеет и более широкое значение. Принципи-
ально важно, что дан пример плодотворного исследования в об-
ласти общих интересов зоологии и экологии. Это открывает путь
выполнению подобных же высокоактуальных работ по отношению
к животным самого разного систематического положения.
Г. Г. В и н б е р г
ВВЕДЕНИЕ
ОБЩИЕ ПОЛОЖЕНИЯ
Прогрессирующее развитие экологических исследований в по-
следние десятилетия обусловлено насущными потребностями чело-
вечества в использовании биологических ресурсов Земли, со-
стояние которых в связи с усиливающейся урбанизацией, ростом
промышленности и сельского хозяйства принимает в ряде слу-
чаев угрожающий характер. При этом обращает на себя внимание
современное и грядущее состояние водоемов и водотоков, под-
вергающихся все более интенсивной эксплуатации и загрязне-
нию. Поэтому охрана и сохранение природных ресурсов Земли
приобретают не только государственное, но и общечеловеческое
значение.
Разумное использование водоемов и рациональное ведение
водного хозяйства возможно только на основе фундаментальных
теоретических положений о взаимосвязи явлений и процессов
в них и применении этих положений для практики. Именно это
и рассматривается как одна из главных целей гидробиологии.
Гидробиология — раздел экологии со своим специфическим
объектом исследований — водными экологическими системами.
Она изучает население водной среды, его взаимоотношения с ус-
ловиями обитания, значение отдельных организмов в процессах
трансформации энергии и вещества и биологической продуктив-
ности гидросферы. Задачи гидробиологии определяются понима-
нием становления, развития, структуры и функций экологиче-
ских систем с целью управления ими (Винберг, 1972).
Экология, и гидробиология в том числе, исследует системы
надорганизменного уровня, для познания которых необходимы
надежные сведения по экологии отдельных видов животных.
Учение об экологических системах объединяет неразрывно
связаппые между собой ауто- и синэкологические вопросы.
«Каждое взятое в отдельности биологическое исследование
лишь постольку принадлежит к экологии, поскольку помогает
iKHuni. значение изучаемого явления в экосистеме или
а
экосистемах, определяющих среду обитания вида» (Винберг,
1972, с. 77).
В начале своего возникновения экология трактовалась как
учение об образе жизни и приспособительных реакциях организ-
мов в связи с их средой обитания, т. е. ограничивалась изучением
аутоэкологических проблем. Такое понимание экологии, нередко
встречающееся в наше время, не согласуется с современным ее
толкованием и стоящими перед ней задачами. Более расширен-
ное по сравнению с традиционным понимание экологии требует
и более широкого подхода к аутоэкологии как частной экологии.
Аутоэкология водных животных должна рассматриваться как
общая область интересов зоологии и гидробиологии. Она как част-
ная экология животных представляется составной частью зооло-
гии. В тех же случаях, когда аутоэкологические исследования
направлены на получение данных, способствующих пониманию
места и роли отдельных видов в водных экологических систе-
мах, они естественно входят в гидробиологию. Поэтому для эколо-
гии особое значение приобретают зоологические исследования,
имеющие вполне определенную направленность. Становятся не-
обходимыми количественные представления о роли популяций
отдельных видов и их сообществ в процессах биологической про-
дуктивности, превращения веществ и трансформации энергии
в экосистемах. Поэтому следует говорить о функциональной эко-
логии отдельных видов животных, понимая под этим количествен-
ные аутоэкологические исследования, направленные на выясне-
ние функционального значения популяций в экологических си-
стемах. Такое расширенное понимание аутоэкологии не всту-
пает в противоречие с ее традиционным толкованием. Изучение
образа жизни животных в определенных условиях (в обычном
понимании этого термина) должно рассматриваться как обязатель-
ная часть аутоэкологических работ. Вместе с тем в понятие «об-
раз жизни» включаются такие важнейшие количественные харак-
теристики организмов и популяций, как интенсивность их обмена,
роста, питания, размножения и их зависимость от меняющихся
условий внешней среды. Получение такого рода сведений об об-
разе жизни животных, необходимых для решения общеэкологи-
чгских проблем, должно стать одной из обязательных задач
зоологических исследований.
Начиная с XIX в. и в подавляющем большинстве случаев
до сего времени зоологи направляют свои усилия на получение
материалов, используемых для эволюционных построений. Все
।((‘обходимое в этой связи тщательно ими изучается, и соответ-
ствующие материалы в обязательном порядке содержатся в их
работах. Наряду с этим приводятся фрагментарные сведения по от-
дельным моментам аутоэкологии животных, не объединяемые по-
ниманием общеэкологических закономерностей. Теперь, когда
экология становится одной из ведущих биологических дисцип-
лин, призванной решать важнейшие вопросы, в том числе связан-
7
ные с воздействием людей на природные условия, естественно
ожидать большей экологизации зоологических работ. Это не-
сомненно обогатит и зоологию, которая тем самым получит воз-
можность в большей мере, чем сейчас, оперировать количествен-
ными понятиями, что представляется естественным и закономер-
ным процессом ее развития, результатом достигнутого биологи-
ческими науками уровня и требованием времени.
Следует специально рассмотреть правомочность использова-
ния предложенного автором понятия «функциональная эко-
логия».
В биологии, особенно в зоологии, сложились и успешно раз-
виваются такие разделы, как, например, функциональная морфо-
логия или эволюционная морфология и т. п. При этом, конечно
же, никто не имеет в виду, что можно обнаружить какие-либо
не функционирующие и не возникшие в процессе эволюции морфо-
логические структуры. Задачи этих разделов морфологии сводятся
к исследованиям функционирования или к исследованиям раз-
вития в историческом плане определенных морфологических
структур на основе знания их строения. Такая постановка науч-
ной задачи вполне обоснована, имеет цель и решается с помощью
специфических методов.
Экология — весьма обширная область биологических знаний —
исследует, как уже было сказано, системы надорганйзмен-
ного уровня. Естественно, что при этом необходимо изучать,
например, пространственное распределение, структуру популя-
ций, биоценозов, экосистем, демографию популяций, реакции
организмов на изменения факторов внешней среды и т. д. и т. п.
Эти экологические задачи успешно решаются многими исследо-
вателями на разных объектах. Однако они еще не дают возмож-
ности определения места и роли популяций или их сообществ
в общих процессах круговорота веществ и трансформации энер-
гии в экологических системах. Количественные исследования
на уровне популяции, биоценоза, способствующие пониманию
их роли и значения в функционировании экосистем с учетом фи-
зиологических особенностей организмов, правомочно отнести
к функциональной экологии. Этот раздел экологии имеет вполне
определенную цель и специфические методы исследования, среди
которых, в частности, можно указать балансовый метод, успешно
использующий энергетический принцип, эколого-физиологиче-
ские методы и т. д. На современном уровне развития экологии
и в связи со стоящими перед нею научными и практическими за-
дачами выделение функциональной экологии отдельных групп
организмов представляется полезным, способствующим разви-
тию теории экологии.
С изложенных позиций автор и подошел к изучению экологии
пресноводных двустворчатых моллюсков.
X
ЗНАЧЕНИЕ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
В КОНТИНЕНТАЛЬНЫХ ВОДОЕМАХ
Двустворчатые моллюски населяют все типы континентальных
водоемов — от крупных рек и озер до временных, пересыхающих.
Они обитают на всех глубинах, достигая профундали глубоких
озер. В ряде водоемов биомасса этих животных составляет до 70—
80% биомассы всех донных животных, а создаваемая ими про-
дукция оказывается сопоставимой с продукцией рыб.
Двустворчатые моллюски как облигатные фильтраторы в водо-
емах выступают в роли природных биофильтров, активно уча-
ствуя в процессах самоочищения вод. Это было убедительно по-
казано, в частности, экспериментальными работами К. А. Воск-
ресенского (1948) и Э. А. Бервальда (1964, 1965) и др.
Большинство мелких и молодь крупных моллюсков входят
как постоянный компонент в рацион многих видов рыб, в том:
числе имеющих важное промысловое значение, охотно поедаются
утками и другими водными животными. В питании рыб особенно
велико значение дрейссен. Они составляют до 42.2% (по весу)
в нище воблы, до 17 — сазана, до 13% — бычков, постоянно
встречаются в рационе леща и осетра Каспийского моря (Шо-
ры гип, 1952). В питании воблы в Северном Каспии в разные годы
в период с 1935 по 1971 г. удельный вес дрейссен колебался от 2.1
до 52.2%, составляя в среднем около 28.7% (Белова, 1974). В Вол-
гоградском водохранилище эти моллюски составляют 98.1%
в нище густеры и до 80.3% — плотвы (Небольсина, 1965). Среди
рыб Куйбышевского водохранилища дрейссенами питаются лещ
(92.5%), густера, плотва, язь (до 100%) (Михеев, 1963), сазан
(17-100%) (Егерева, 1971). Дрейссены входят в состав пищи
густеры (74%), карпа, воблы Каховского водохранилища (Bip-
rep, Маляревська, 1964). Их поедает тарань (49.7—71.5%) в водо-
хранилищах Приднепровья и Крыма (Чаплина и др., 1976), лещ,
рыбец (до 74.3%), густера, плотва (до 84%) в Куршском заливе
(Кублицкас, 1959) и т. д.
Рыбы охотно потребляют разные виды сфериид, которые со-
ставляют от 10 до 40% в пище плотвы в Куйбышевском водохра-
нилище (Платонова, 1966). В Куршском заливе важное значение
в питании леща, рыбца, густеры, плотвы (весной до 64.4%) при-
надлежит моллюскам Pisidium и Sphaerium (Кублицкас, 1959).
В составе пищи плотвы постоянно присутствует Sphaeriastrum
rlricola (Небольсина, 1965). Особое место занимают представи-
тели сфериид в питании рыб северных водоемов. Например, они
доминируют в рационе чира и сига в оз. Большой Харбей (Коми
АССР), составляя в разные сезоны от 15.5 до 68% их рациона
(Сидоров, 1976).
Рыбы предпочитают поедать мелких моллюсков, длина кото-
рых не превышает 2—3 мм. Поэтому наиболее активно они по-
i ребляют моллюсков в летние месяцы, когда происходит их ин-
9
тенсивное размножение. Более крупные по размерам рыбы могут
питаться и более крупными моллюсками. Так, например, сазаны
длиной в 65—120 мм поедали моллюсков, длина раковины которых
составляла 5—8.5 мм (Михеев, 1963).
Велико значение двустворчатых моллюсков в образовании
паразитарных связей в экосистемах континентальных водоемов.
Достаточно сказать, что личиночные стадии унионид паразитируют
на пресноводных рыбах, многие виды сфериид служат промежу-
точными хозяевами трематод.
Некоторые виды пресноводных двустворчатых моллюсков
имеют промысловое значение., Перловицы относятся к немногим
представителям фауны пресных вод, заготовка которых произ-
водится для нужд легкой промышленности, в частности для пуго-
вичного производства и изготовления декоративных поделок.
Добыча моллюсков ведется в бассейнах Дона, Днепра, Волги,
где еще в конце 60-х годов этого столетия ежегодно добывалось
до 2300 тонн створок перловиц, а мясо их использовалось на корм
домашней птице (Негробов, 1971). В пресноводных жемчужницах
образуется драгоценный жемчуг, добываемый с давних времен
в России, Западной Европе, Северной Америке. В России насчи-
тывалось до 77 рек, в которых обитала наиболее ценная в этом
отношении обыкновенная жемчужница. Особенно много жем-
чуга добывали в районе г. Кеми (Зорина, 1972). Однако нерегу-
лируемый промысел, массовая гибель моллюсков от загрязнения
рек сточными водами и сплава леса привели к резкому сокраще-
нию запасов моллюсков и добычи жемчуга. Восстановление за-
пасов жемчужниц, равно как и разработка методов искусствен-
ного выращивания жемчуга, столь удачно применяемых в Япо-
нии, где созданы специальные фермы морских жемчужниц, могут
иметь важное хозяйственное значение. Интересно отметить, что
еще в начале XX в. в России были предприняты первые удач-
ные попытки выращивания жемчуга в пресноводных мол-
люсках.
Развернувшееся в СССР гидростроительство, создание новых
судоходных каналов и водохранилищ ускорило процесс расселе-
ния моллюсков дрейссен, которые способны прикрепляться к дни-
щам судов. Их интенсивное развитие приводит в ряде случаев
к серьезным помехам в работе промышленных установок. Об-
растание дрейссенами трубопроводов систем технического водо-
снабжения, водозаборов и водоводов, питающихся водой из рек
и водохранилищ, в которых распространены эти моллюски,
может вызвать полную остановку в подаче воды. Массовое раз-
витие дрейссен на решетках гидростанций приводит к снижению
мощности турбин. Поселяясь внутри водопроводных труб, они
не только затрудняют водоснабжение, но при отмирании, раз-
лагаясь, портят питьевую воду.
Итак, с одной стороны, двустворчатые моллюски служат кор-
мовыми объектами для рыб, очищают воду, вступают в парази-
10
тарные связи, с другой — массовое развитие некоторых из них
может привести к серьезным затруднениям в водопользовании.
Повышение кормовой базы рыбохозяйственных водоемов,
увеличение запасов промысловых моллюсков, борьба с вред-
ными последствиями массового развития отдельных представи-
телей этих животных невозможны без точного знания их экологии
и функционального значения в водных экологических системах.
Поэтому исследование экологии пресноводных двустворчатых
моллюсков помимо теоретического имеет важное практическое
значение. В то же время отдельные аспекты экологии этих живот-
ных до сих пор остаются изученными недостаточно полно или
не изученными вовсе. До сих пор не было исследования, в кото-
ром были бы критически рассмотрены и обобщены имеющиеся
материалы по экологии двустворчатых моллюсков и выяснена их
роль в водных экосистемах.
ЦЕЛИ И МАТЕРИАЛЫ ИССЛЕДОВАНИЯ
Настоящая работа посвящена изучению экологии пресноводных
двустворчатых моллюсков в ее функциональном аспекте. В од-
ном исследовании невозможно охватить весь круг экологиче-
ских проблем, поэтому основное внимание было уделено количе-
ственному изучению обмена, роста, питания, размножения у раз-
личных видов моллюсков. Эти данные, рассматриваемые как
основа для определения динамики численности, биомассы, воз-
растного состава, продукции, энергетического баланса популя-
ций, позволили количественно выразить их функциональное зна-
чение в сообществах гидробионтов и экосистемах пресноводных
водоемов различного типа.
Поставленные задачи решались с позиций энергетического
принципа понимания биотического баланса, который занял гос-
подствующее положение в современных гидробиологических ис-
следованиях, особенно в связи с исследованиями, выполненными
по Международной биологической программе. Основу этого прин-
ципа составляет положение о возможности лишь однократного
использования потенциальной энергии для совершения работы,
что нашло отражение в концепции о потоке энергии в экологи-
ческих системах.
Материалом для работы послужили полевые наблюдения ав-
тора над популяциями разных видов моллюсков в водоемах раз-
личных частей Советского Союза, в том числе Карельской АССР,
Мурманской, Ленинградской, Калининградской, Московской об-
ластей, сборы моллюсков на оз. Байкал, в бассейне р. Амур,
результаты экспериментов, выполненных в полевых и лаборатор-
ных условиях. При этом наиболее подробно были изучены широко
распространенные массовые виды двустворчатых моллюсков,
а также наиболее многочисленные их представители в озерах,
исследованных Лабораторией пресноводной и экспериментальной
11
гидробиологии Зоологического института АН СССР по Между-
народной биологической программе: Кривое, Круглое (побережье
Белого моря), Зеленецкое и Акулькино (побережье Баренцева
моря). В этих озерах двустворчатые моллюски представлены
только сфериидами, среди которых большей численности дости-
гали Sphaerium suecicum Clessin, Pisidium crassum Stelfox, P. till-
jeborgi Clessin, широко распространенные в северных водоемах,
озерах северо-запада и запада европейской части СССР. Кроме
того, широко привлекались литературные сведения.
Наряду с изучением эколого-физиологических характеристик
отдельных видов выявлялись закономерности роста, обмена, пита-
ния, общие для Bivalvia. Такого рода закономерности для пресно-
водных моллюсков до сих пор не были установлены. Достаточно
привести следующий пример. Несмотря на множество работ по
росту пресноводных двустворчатых моллюсков, особенно имею-
щих промысловое значение, не было работы, в которой бы были
обобщены полученные данные. В обширной статье Вилбура и
Овена (Wilbur, Owen, 1964), представляющей собой главу двух-
томного издания «Physiology of mollusca», некоторые закономер-
ности роста Bivalvia были установлены только для морских ви-
дов этих животных. Возможные отклонения эколого-физиологи-
ческих характеристик отдельных видов от общих закономерностей
для моллюсков представляют особый интерес и требуют специ-
ального пристального изучения.
Для характеристики условий в местах обитания моллюсков
использованы сведения о физико-химических особенностях от-
дельных водоемов, взятые* из различных изданий справочного
характера, таких как «Справочник по водным ресурсам СССР»,
сводка О. А. Алекина (1948) по гидрохимии рек. Привлечены не-
которые другие источники (Бадер, 1934; Жадин, 1940; Францев,
Лебедева, 1941; Соловьева, 1950; Ярошенко, 1957; Лукин, 1960;
Максимова, 1963; Баранов, 1965; Буторин, 1970; Бульон, 1975а,
19756; Озерецковская, 1975; Озерецковская, Умнова, 1975; Али-
мов и др., 1976, и др.).
Результаты полевых и экспериментальных исследований об-
работаны в соответствии с методами, рекомендованными в различ-
ных руководствах по биологической статистике, и способами об-
работки данных и анализа экспериментов, в том числе изложен-
ными в работах Л. 3. Румшиского (1971) и А. А. Умнова (1976).
Используемые в работе обозначения, единицы измерений и экви-
валенты приведены в соответствии с рекомендациями, изданными
в 1972 г. Советским Национальным комитетом по Международ-
ной биологической программе.
Систематика пресноводных двустворчатых моллюсков во мно-
гих своих аспектах (особенно на видовом уровне) разработана
еще недостаточно полно. Однако основные положения системы
этих животных изложены Я. И. Старобогатовым (1970, 1977),
а в отношении Unionidae европейской части СССР уточнены
12
О. П. Кодоловой (1977). Все это принято автором во вни-
мание.
Автор не считает, что ему удалось рассмотреть все аспекты
функциональной экологии пресноводных двустворчатых моллю-
сков. Несомненно, необходимы дальнейшие исследования в этом
направлении. Поэтому он с благодарностью примет все замеча-
ния и советы, которые будут способствовать развитию функцио-
нальной экологии пресноводных животных, среди которых
не последнее место принадлежит двустворчатым моллюскам.1
1 В процессе выполнения данной работы автор чувствовал постоянную
помощь и поддержку со стороны сотрудников Лаборатории пресноводной
и экспериментальной гидробиологии ЗИН АН СССР. Это в первую очередь
относится к ее руководителю чл.-корр. АН СССР, проф. Г. Г. Винбергу,
который уделял работе большое внимание и сделал ряд весьма ценных за-
мечаний и уточнений. Отдельные разделы рукописи неоднократно обсужда-
лись в дружеских дискуссиях с А. Н. Голиковым, Я. И. Стар’обогатовым,
А. И. Кафановым. Крайне полезным для автора было критическое прочте-
ние рукописи Л. М. Сущеней, В. Е. Заикой, В. В. Хлебовичем, А. В. Мона-
ковым, М. Б. Ивановой. Неоценимую пользу принесли консультации
А. А. Умнова по вопросам статистики и математической обработки данных.
Большая помощь при оформлении и подготовке рукописи была оказана
В. Г. Власовой. Автор считает своим приятным долгом выразить всем искрен-
нюю признательность и благодарность.
Глава I
СКОРОСТЬ ОБМЕНА У МОЛЛЮСКОВ
Ассимиляция пищи и рост организмов тесно связаны с процес-
сами обмена. Имеется огромное количество работ, в которых
интенсивность обмена разнообразных животных исследовалась
как интенсивность их газообмена или дыхания. Интенсивность
обмена определялась через потребность животных в кислороде,
которая рассматривалась как наиболее содержательный в этом
плане показатель. Разумеется, все эти данные имеют несомнен-
ный экологический смысл только в том случае, если они отражают
уровень обмена в условиях, свойственных определенному виду
животных. Поскольку, потребление кислорода животными зави-
сит от самых разных факторов, таких, например, как темпера-
тура, время года, активность, размер животного и т. п., есте-
ственно учитывать их влияние и находить количественные вы-
ражения этих зависимостей в экспериментальных условиях.
Задавшись целью определить уровень обмена веществ у пресно-
водных двустворчатых моллюсков и выявить его закономерности,
автор сконцентрировал внимание на изучении потребления кисло-
рода этими животными.
При оценке только количественной стороны обмена не рас-
сматриваются механизмы процессов, что, естественно, сужает
круг вопросов, подлежащих исследованию. Однако такой под-
ход оправдан самой постановкой задачи. Расширение исследова-
ний неизбежно привело бы к необходимости выяснения интимных
механизмов обмена, применению специальных знаний и методов
из области физиологии и биохимии. Эти несомненно важные
физиологические вопросы находятся далеко в стороне от цели
и методов данной работы. Вместе с тем сведения по скорости и
интенсивности обмена у пойкилотермных животных позволяют
выделить вопросы, изучение которых необходимо для понимания
физиологических механизмов, определяющих тот или иной уро-
вень обмена. _
14
Скорость потребления кислорода животными — наиболее до-
ступный показатель интенсивности обмена. По отношению к гидро-
бионтам это особенно справедливо, поскольку содержание кисло-
рода в воде измеряется сравнительно просто, а получаемые ре-
зультаты достаточно точны и надежны, в то время как, напри-
мер, содержание в воде углекислоты с помощью общепринятых
методов определяется со значительно меньшей точностью.
Весьма важно, что по скорости потребления кислорода живот-
ными можно рассчитать траты энергии на обменные процессы
помощью оксикалорийного коэффициента, показывающего коли-
чество калорий, выделяющееся при полном окислении белков,
жиров и углеводов, поскольку 1 мг кислорода эквивалентен
3.4 кал.
Интенсивность обмена двустворчатых моллюсков, в особен-
ности пресноводных, в сравнении, например, с рыбами или рако-
образными исследована недостаточно полно. Приведенные в лите-
ратуре данные не всегда равноценны, получены различными мето-
дами, не во всех случаях строго выдерживалось сходство усло-
вий экспериментов. Все это заставляет критически подходить
к использованию литературных материалов. Тем не менее уже
накопилось достаточное количество сведений, позволяющих об-
общить материалы по скорости потребления кислорода пресновод-
ными двустворчатыми моллюсками и прийти к вполне определен-
ным выводам.
В данной работе применены термины, принятые в настоящее
время в физиологии водных животных.
Различают «стандартный обмен», под которым понимается
обмен у животных, находящихся в малоподвижном состоянии,
измеренный в стандартных условиях. Однако понятие «стандарт-
ные» условия относительно и зависит как от особенностей жи-
вотного, так и от использованной методики. По отношению к гидро-
бионтам, например, условия чаще всего стандартизируются
но температуре. В данной работе стандартный обмен не исследо-
вался. Термины «обмен покоя» и «основной обмен» употребляются
для определения дыхания животных в состоянии полной непо-
движности. Определение этих величин обычно производится на
наркотизированных объектах. Общее потребление кислорода
животными в их обычном состоянии относительного покоя или
движения со скоростью, не превышающей нормальную для дан-
ного вида, мы называем «общим обменом». Термин «активный об-
мен» относится только к той части обмена, которая непосредст-
венно связана с затратами энергии животными на движение.
В гаком случае активный обмен представляет собой разницу между
общим и основным обменом. Большая часть имеющихся в лите-
ра гуре сведений по газообмену пресноводных двустворчатых
моллюсков (за исключением немногочисленных специальных ис-
ледований) получена в результате изучения у них общего об-
мена.
15
Потребление кислорода животными обычно выражается
в величинах, отнесенных либо ко всему животному в целом, либо
к единице его веса. В первом случае рассматривается скорость,
во втором — интенсивность обмена.
МЕТОДЫ ОПРЕДЕЛЕНИЯ ПОТРЕБЛЕНИЯ
КИСЛОРОДА ГИДРОБИОНТАМИ
Изучение потребления кислорода водными животными обычно
проводится с помощью двух основных методов: проточной и не-
проточной воды. В первом случае применяются респирометры,
в которых осуществляется постоянный проток воды с заданным
содержанием кислорода. Второй метод предполагает измерение
потребления кислорода животными в замкнутых сосудах без про-
тока воды (метод замкнутых сосудов). В этом случае подопытные
животные помещаются в специальный сосуд (респирометр) строго
определенного объема, который заполняется водой с известным
содержанием растворенного в ней кислорода. Вода в респиро-
метре тем или иным способом защищается от соприкосновения
с атмосферным воздухом для устранения проникновения кисло-
рода из атмосферы. Через определенное время экспозиции из-
меряется количество растворенного в воде кислорода. По раз-
нице между содержанием его в начале и конце опыта рассчиты-
вается скорость потребления кислорода животными с учетом объ-
ема респирометра и времени экспозиции. Более подробно об этих
методах можно узнать из различных руководств, в том числе
из работы Е. А. Веселова (1959).
Метод замкнутых сосудов, впервые примененный в 1809 г.
(Humoldt, Provencal, 1809, цит. по: Винберг, 1956), остается
до сих пор наиболее распространенным. Это связано с его про-
стотой, достаточной точностью, возможностью использования
в полевых условиях. Однако в отличие от метода проточной
воды он не дает возможности проследить за изменениями интен-
сивности дыхания животных в процессе опыта и исключить яв-
ления, связанные с так называемым «эффектом сосуда». Под этим
понимается увеличение скорости потребления кислорода живот-
ными в первое время после помещения их в респирометр как след-
ствие их возбуждения и повышения двигательной активности.
Например, увеличение потребления кислорода некоторыми ли-
чинками насекомых в этот период может превышать уровень
их стандартного обмена на 250—300%. Благодаря этому даже
после установления постоянного (стандартного) уровня дыхания
на него продолжают накладываться результаты адаптационного
периода и возникает как бы «видимый» уровень обмена, постоянно
снижающийся до стандартного. В связи с этим существенное зна-
чение приобретает время экспозиции. При короткой экспозиции,
когда измеряется потребление кислорода животными в адапта-
ционный период, получаемые результаты могут оказаться не-
16
оправданно завышенными. Это позволило некоторым исследова-
телям (Kamler, 1969) поставить под сомнение пригодность метода
замкнутых сосудов для изучения интенсивности дыхания водных
животных.
Специально проведенные С. М. Голубковым (1979) опыты по-
казали, что интенсивность дыхания у личинок поденок С1оёоп
dipterum L. через час после начала опыта превышает уровень
стандартного обмена на 125%. Далее по мере увеличения экспо-
зиции происходит быстрое снижение скорости потребления кис-
лорода животными, и после 3-часовой экспозиции устанавливается
стандартный уровень дыхания. Аналогичные результаты были
получены В. С. Ивлевым (19386) и Л. В. Камлюк (1964).
Наиболее точные результаты могут быть получены методом
замкнутых сосудов только в тех случаях, когда содержание кис-
лорода в воде опытных сосудов в конце эксперимента не умень-
шается по сравнению с исходным более чем на 10—20%. В про-
тивном случае снижение потребления кислорода животными может
быть, в частности, обусловлено их кислородным голоданием.
Кроме того, при использовании этого метода важное значение
особенно для малоподвижных животных приобретают условия
перемешивания воды в респирометрах. Однако для двустворча-
тых моллюсков, постоянно фильтрующих воду, это не имеет суще-
ственного значения.
При соблюдении указанных условий результаты, получаемые
методом замкнутых сосудов, не отличаются от данных, получен-
ных с помощью проточного метода. Метод замкнутых сосудов
дает достаточно надежные результаты. В частности, это наиболее
распространенный метод изучения скорости обмена у двуствор-
чатых моллюсков.
В зависимости от способа количественного определения кисло-
рода методы определения его содержания в воде могут быть раз-
биты на три группы.
К первой группе относятся методы с применением
объемного (титровального) анализа растворенного в воде кисло-
рода с помощью обычных гидрохимических методик. Среди них
для рассматриваемых целей всеобщее признание получил метод
Винклера, применяемый в микро- и макромодификациях.
Методы объемного анализа достаточно просты, не требуют
сложной аппаратуры и дают надежные результаты. Они широко
используются при изучении скорости газообмена водных живот-
ных. При исследованиях интенсивности обмена двустворчатых
моллюсков они применяются наиболее часто (Weinland, 1919;
(iartkiewicz, 1922; Hiestand, 1938; Hers, 1943; Ludwig, Kryewien-
zyk, 1950; Hiscock, 1953; Иванчик, 1956; Алиев, 1960; Botnariuc
el al., 1961; Woynarovich, 1961; Михеев, 1964a; Hagashi, 1965;
(Jrnatowski, 1967; Pikoij, Cucerzan, 1967; Арабика, Рубинова, 1968;
Tudorancea, Florescu, 1968a, 1968b, 1970; Draghici, Picoij, 1969;
Ковалева, 1970; Lomte, Nagabhushanam, 1971, и др.).
2 А. Ф. Алимов
17
Изучая потребление кислорода моллюсками сем. Sphaeriidae,
автор также применял метод замкнутых сосудов, определение
содержания кислорода производилось методом объемного ана-
лиза (Алимов, 1975). В качестве респирометров использовались
сосуды объемом 60—100 мл. В сосудах такого объема неболь-
шие по размерам сферииды могли свободно перемещаться, и, сле-
довательно, полученные величины интенсивности обмена отра-
жали общий обмен моллюсков в их обычном состоянии. Время
экспозиции составляло 4 ч. За этот период уменьшение содержа-
ния кислорода в воде опытных сосудов не превышало 15—20%.
Опыты обычно проводились при температуре 20°. Дыхание мол-
люсков Pisidium crassum в экспедиционных условиях изучалось
при температуре 15°. При этом в качестве «термостата» исполь-
зовалось озеро, в котором обитали эти моллюски. Во всех случаях
животные акклимировались к температуре опыта в течение суток.
Поскольку эксперименты ставились обычно в летний период,
когда температура в природных местообитаниях моллюсков
близка к 20°, то столь малый срок акклимации не мог отразиться
на результатах.
Вторая группа объединяет манометрические методы.
В основу их положена следующая схема. При выделении (или
поглощении) газа в опытном сосуде давление этого газа внутри
сосуда меняется пропорционально изменению количества газа.
Поскольку температура и атмосферное давление в течение опыта
постоянны, а меняется только объем газа в сосуде, то, зная ем-
кость манометра и отношение его объема к объему опытного со-
суда, можно рассчитать изменение объема газа в сосуде за опре-
деленный отрезок времени.
Основное преимущество манометрических методов заключается
в том, что нет необходимости в химическом анализе растворен-
ного в воде кислорода. Однако при изучении газообмена относи-
тельно крупных гидробионтов преимущество метода перекрыва-
ется теми недостатками, которые он в себе несет. Основной из них
заключается в резком изменении в манометрических респиромет-
рах условий газообмена между жидкой и газовой фазами. Это
особенно сильно сказывается при изучении потребления кисло-
рода малоподвижными животными. Однако даже при условии,
когда животные интенсивно перемешивают воду, парциальное
давление кислорода в водной фазе будет в большей или меньшей
степени сниженным (Винберг, Беляцкая-Потаенко, 1963). По-види-
мому, в силу сказанного эти методы очень редко используются
для исследований скорости обмена у моллюсков, в том числе
и двустворчатых. Интенсивность обмена последних манометри-
ческими методами изучалась Веземейером (Schwarzkopf, Wese-
meier, 1959; Wesemeier, 1960).
К третьей группе могут быть отнесены методы,
основанные на измерении диффузионных токов при электролизе
воды (электрохимические методы). Они стали применяться сравни-
18
тельно недавно и позволяют в отличие от других методов исполь-
зовать любые по размерам сосуды для опытных животных, быстро
проводить в течение всего опыта определения содержания кисло-
рода в воде, не нарушая условий эксперимента. Существенное
преимущество этих методов заключается в возможности примене-
ния автоматической регистрирующей аппаратуры. К их недостат-
кам следует отнести не всегда стабильные результаты, обусловлен-
ные конструктивными особенностями аппаратуры, особенно кисло-
родных датчиков. В некоторых модификациях получаемые ре-
зультаты по содержанию кислорода в воде связаны не столько
со скоростью газообмена животных, сколько с влиянием на элек-
троды химического состава воды, что может привести к значитель-
ным погрешностям в определении потребления кислорода живот-
ными.
Эти методы вследствие их преимуществ нашли довольно широ-
кое распространение особенно в зарубежных исследованиях
при изучении дыхания различных гидробионтов. В различных
модификациях с их помощью изучалась интенсивность обмена
брюхоногих (Berg, 1952, 1961; Berg et al., 1958; Berg, Ockelmann,
1959) и двустворчатых (Berg et al., 1962; Akerlund, 1969; Спири-
донов, 1972, 1973) пресноводных моллюсков.
Разные исследователи использовали неодинаковые методы
изучения дыхания моллюсков и проводили эксперименты в раз-
личных условиях. Тем не менее, как будет показано далее, на ос-
новании уже имеющихся сведений по потреблению кислорода
пресноводными двустворчатыми моллюсками возможно устано-
вить общие закономерности, которым подчиняется количествен-
ная сторона обмена у этих животных, и охарактеризовать его
уровень вполне конкретными величинами.
Для получения по возможности более строгих результатов
в дальнейшем почти не принимались во внимание результаты тех
исследований, которые получены с применением манометрических
методов. Некоторое исключение было сделано лишь для данных
Веземейера (Wesemeier, 1960) по дыханию отдельных видов мол-
люсков, поскольку они не противоречили общим закономерно-
стям для всех двустворчатых моллюсков.
ЗАВИСИМОСТЬ СКОРОСТИ ОБМЕНА У МОЛЛЮСКОВ
ОТ ТЕМПЕРАТУРЫ
Температура внешней среды представляет собой, пожалуй, один
из наиболее важных факторов, в значительной степени определя-
ющий скорость протекания основных жизненных процессов
у пойкилотермных животных. Поэтому неудивительно, что имеется
огромное количество работ, выполненных на различных их пред-
ставителях, в которых предпринимались попытки найти количе-
ственную сторону зависимости скорости обмена от температуры
окружающей среды. Многочисленными тщательными экспери-
2*
19
ментами на объектах разного систематического положения было
убедительно показано, что зависимость скорости основного об-
мена от температуры подчиняется единой закономерности. Эта
закономерность была установлена А. Крогом (Krogh, 1914) и
имела вид кривой, которая в дальнейшем была названа «нормаль-
ной кривой Крога». Она нашла широкое применение и до сего
времени используется большинством исследователей как универ-
сальная закономерность. Например, обобщение огромного экс-
периментального материала позволило установить ее справед-
ливость для всего класса рыб (Винберг, 1956). Результаты этого
серьезного исследования сразу были взяты на вооружение по-
давляющим большинством по крайней мере советских гидробио-
логов и зачастую без достаточной проверки переносились на вод-
ных животных других групп, хотя «. „нормальная44 кривая
должна рассматриваться только как эмпирически полученная
общая закономерность, которая в каждом отдельном случае
в большей или меньшей мере перекрывается более частными за-
висимостями, отражающими особенности объектов и условий.
Есть основания думать, что в некоторых случаях они (отклоне-
ния от нормальной кривой, — А. А.) имеют закономерный харак-
тер и отражают существенные особенности объекта и условия,
при которых производятся измерения» (Винберг, 1956, с. 33).
Действительно, рядом работ на гидробионтах было показано,
что очень часто наблюдаются отличия от «нормальной» кривой.
Такого рода отклонения были, например, обнаружены для мор-
ских (Bruce, 1926) и пресноводных моллюсков (Berg, 1952; Berg,
Ockelmann, 1959). Они наиболее четко проявляются в области
низких и высоких температур и рассматриваются как физиоло-
гическое приспособление, компенсирующее неблагоприятное воз-
действие условий внешней среды (Belehradek, 1930; Scholander
et al., 1953; Rao, Bullock, 1954; Bullock, 1955, и др.).
Тщательными исследованиями И. В. Ивлевой (1973, 1976)
и И. В. Ивлевой и М. И. Попенкиной (1968) на большом факти-
ческом материале было убедительно показано, что гипотеза
«метаболической компенсации» (способность пойкилотермных жи-
вотных проявлять определенную температурную независимость
функционирования повышением или снижением обмена в неблаго-
приятных условиях) не может считаться обоснованной. Получае-
мые в экспериментах с различными животными существенные
изменения обмена при неблагоприятных для них температурах
представляют собой первичную их реакцию на изменения темпера-
турных условий. Существенный вывод этих работ заключается
также и в том, что при соответствующей акклимации к экспери-
ментальным температурам уровни обмена арктических и тропи-
ческих по происхождению животных при одинаковых температу-
рах практически неотличимы.
Большинство исследователей рассматривают «нормальную»
кривую не более как эмпирическую зависимость, которая пока
20
нс имеет теоретического обоснования. В то же время она пред-
с гавляется удобной для практического применения и в ряде слу-
чаев хорошо описывает экспериментальные данные.
Имеющиеся в литературе сведения по интенсивности обмена
у пресноводных двустворчатых моллюсков при разных темпера-
турах позволяют количественно оценить зависимость скорости и
интенсивности обмена у этих животных от температуры. Следует
разу отметить, что в большинстве работ животные акклимирова-
лись к экспериментальным температурам не менее суток. Те ис-
следования, в которых нет сведений о времени акклимации, нами
в дальнейшем во внимание не принимались. Все данные были
обработаны с помощью обычного способа, использованного, на-
пример, Г. Г. Винбергом для рыб, а для ракообразных Л. М. Су-
।цепей (1972). Скорость обмена при 20° принималась за 100%,
а обмен при всех других температурах рассчитывался в долях
от обмена при 20° Такой прием дает возможность сопоставления
измерений, выполненных на различных объектах, различаю-
щихся по уровню обмена.
Сравнение величин потребления кислорода моллюсками
Unionidae, Sphaeriidae, Dreissenidae (рис. 1) при разных темпера-
турах свидетельствует о сходном влиянии температуры на скорость
обмена у этих животных. Формы кривых, отражающие зависи-
мость обмена от температуры, как видно из рис. 1, у них одинаковы
и заметно отличаются от «нормальной» кривой. Особенно резко
эти отличия проявляются при температурах выше и ниже 15—20°.
Для сравнения изменения скоростей биологических процес-
сов с изменением температуры широко используется коэффи-
циент Вант-Гоффа, или (210. Он показывает, во сколько раз увели-
чивается скорость реакции при возрастании температуры на 10°.
Величина этого коэффициента может быть определена из простого
соотношения
где Vr и Т2 — скорости реакции при температурах Т\ и Т2. Для
большинства химических реакций величина ()10 находится в пре-
делах от 2 до 3.
Для биологических явлений правило Вант-Гоффа оказывается
не всегда справедливо, и величина ()10 закономерно изменяется
с температурой, так как одна и та же температура различно воз-
действует на разные органы или организмы. Непостоянство
<210 при различных температурах обусловлено и тем, что биологи-
ческие процессы, протекающие в организме, основаны на серии
химических и физических явлений, каждое из которых характери-
зуется своим температурным коэффициентом (Belehradek, 1930).
Тем не менее коэффициент ()10 представляет собой удобную форму
сравнения скоростей протекания биологических процессов
при различных температурах. В частности, с его помощью можно
21
Рис. 1. Скорость обмена у моллюсков при разных температурах, выраженная в процентах от обмена при 20° (из: Али-
мов, 1975).
А — Dreissena polymorpha по данным: 1 — Ковалевой, 1970; 2 — Woynarowich, 1961; 3 — Михеева, 1964; 4 — Спиридонова, 1971.
Б — Sphaeriidae (по данным Ковалевой, 1970): 1 — S. rivicola", 2 — S. согпеит', 3 — S. solidum. В — Unionidae: 1, 2 — U. tumidus; 3,4 —
U. pictorum; 5 — Anodonta piscinalis (1, 2 — по данным Ковалевой, 1970; 3—5 — по данным Tudorancea, Florescu, 1970). Прерывистая линия
соответствует «нормальной» кривой. По оси абсцисс — температура (°C); по оси ординат — относительная скорость обмена (%).
легко и надежно приводить результаты экспериментов к опреде-
ленной температуре.
Рассчитанные величины ()10 для пресноводных двустворчатых
моллюсков достоверно отличаются от величины ()10 «нормальной»
кривой в области низких и высоких температур (рис. 2). Значе-
ния (?10 в интервале температур 5—10° оказываются выше, чем
Рис. 3. Зависимость логарифма от-
носительной скорости обмена у дву-
створчатых моллюсков от обратной
величины абсолютной температуры.
По оси абсцисс — обратные величины
абсолютной температуры (1 / Т° К • 10“3к
по оси ординат — логарифм относительной
скорости обмена.
Рис. 2. Зависимость значений С?!0
от температуры (из: Алимов, 1975).
/ — «нормальная» кривая; 2 — пресно-
водные двустворчатые моллюски. По оси
абсцисс — температура (°C); по оси орди-
нат — значения Q10.
для «нормальной» кривой, для всех других температурных ин-
тервалов они, наоборот, ниже (табл. 1). Иными словами, при низ-
ких температурах интенсивность обмена увеличивалась с боль-
шей скоростью, а в остальных случаях — с меньшей, чем этого
следовало бы ожидать в соответствии с «нормальной» кривой.
Таблица 1
Значения Qlo для разных интервалов температур (°C)
Название моллюсков 5—10° 10—15° 15-20° 20—25° 25—30°
Unionidae 3.99 2.44 2.51 1.78 1.57
Sphaeriidae 3.72 2.04 1.68 1.23 1.19
Dreissena polymorpha . . . 3.74 3.47 2.23 1.59 1.46
Bivalvia (пресноводные) . . 3.65 2.65 2.26 1.60 1.40
По кривой Крога
3.50 I 2.90 I 2.50 I 2.30 I 2.20
23
Зависимость обмена у пойкилотермных животных, как изве-
стно, может быть описана не только «нормальной» кривой Крога.
Возможные функции, с помощью которых можно выразить эту
зависимость, подробно рассмотрены Г. Г. Винбергом (1968).
Одна из них представляет собой известное уравнение Вант-Гоффа—
Аррениуса, разработанное для химических реакций. И. В. Ив-
лева (1973, 1976) на большом числе морских пойкилотермных
животных показала высокое соответствие температурных измене-
ний скоростей обмена у них, предусмотренных законом Вант-
Гоффа —Аррениуса .
Применительно к обмену у животных уравнение этого закона
может быть записано в виде
R = RQe~* (2)
где R1 — газовая постоянная, равная 1.986 кал/(моль-град);
Т — температура (°К); &F/R1 — безразмерный коэффициент, опре-
деляющий ускорение процесса с возрастанием температуры.
В физико-химическом понимании AF — энергия активации
рассматриваемой реакции. При изучении биологических явлений
эту величину принято называть «температурной характеристикой»
и обозначать р..
Исходя из уравнения (2), отношение скоростей обмена при тем-
пературах Т± и Т2 находят как
(3)
^2
В полулогарифмическом масштабе, когда на оси абсцисс отклады-
вается обратная величина абсолютной температуры, а на оси
ординат — логарифм скорости процесса, график функции (2)
представляет собой так называемую «прямую Аррениуса» (рис. 3).
Значения ДГ=р и ()10 находятся между собой в прямом отноше-
нии:
^^„ = 2.187^. (4)
Как видно из рис. 3 и табл. 2 в интервале температур от 5
до 20° изменения интенсивности обмена у пресноводных Bivalvia
с определенным приближением могут быть описаны прямой Ар-
рениуса. При этом среднее значение р. для этого интервала темпе-
ратур составляет 16140 кал/(моль-град). Эта величина выше,
чем установленная для пойкилотермных животных (14 000;
Ивлева, 1973), в том числе и для всего класса ракообразных
(12 186; Сущеня, 1972).
В соответствии с законом Аррениуса скорость изменения об-
мена у пресноводных моллюсков при низких температурах ока-
зывается ниже, а при высоких — выше, чем это следует из экспе-
риментальных данных (табл. 2). Таким образом, зависимость ин-
24
генсивности обмена у этих животных от температуры не может
быть достаточно строго описана как с помощью «нормальной»
кривой, так и законом Вант-Гоффа—Аррениуса. Однако, учиты-
вая небольшое количество данных, нет оснований приписывать
какой-то особый характер этой зависимости для пресноводных
Bivalvia.
Таблица 2
Значения Qlo для двустворчатых моллюсков,
рассчитанные разными способами
Интервалы температур (°C) По данным *
5—10 10-15 15—20
11 ресноводные Bivalvia: по экспериментальным дан- ным 3.65 2.65 2.26 Автора
то же в соответствии с зако- ном Аррениуса . . 2.81 2.71 2.62 То же
Modiola adriatica . 2.75 2.66 2.57 Ивлевой, 1973
Но кривой Крога 3.50 2.90 2.50 То же
* Здесь и далее в графе «По данным» приведены работы авторов, данные которых
были использованы в качестве исходного материала для расчета необходимых пара-
метров.
Представленные результаты убедительно указывают на необ-
ходимость проведения тщательных исследований, которые по-
зволят установить более строгую зависимость интенсивности об-
мена пресноводных двустворчатых моллюсков от температуры.
Полученная зависимость может рассматриваться как первое при-
ближение, достаточно точно описывающее известные к настоя-
щему времени экспериментальные данные. Она представляется
удобной и может быть использована для практических целей.
Подтверждением тому могут служить количественные закономер-
ности обмена у моллюсков, которые нам удалось установить. Они
будут рассмотрены в дальнейшем.
Используя установленную для пресноводных моллюсков за-
висимость обмена от температуры, нетрудно определить скорость
обмена у них при любой температуре или привести эксперимен-
тальные данные к одной температуре расчетным путем по урав-
нению (1), из которого
V — V О 10
Для облегчения расчетов при приведении данных к опре-
деленной температуре вычислена вспомогательная табл. 3. В этой
таблице скорость обмена при 20° принята за единицу, а значения
25
множителя определены для каждой из температур
в интервале от 5 до 30°. Соответствующие множители для про-
межуточных дробных значений температуры легко находятся
линейной интерполяцией.
Таблица 3
Температурные поправки q для приведения скорости обмена
у пресноводных моллюсков к 20°
т т т q
5 9.30 14 1.69 23 0.869
6 8.00 15 1.540 24 0.850
7 5.68 16 1.400 25 0.845
8 4.65 17 1.270 26 0.838
9 3.85 18 1.160 27 0.832
10 3.19 19 1.070 28 0.811
11 2.69 20 1.000 29 0.819
12 2.28 21 0.944 30 0.834
13 1.95 22 0.902
Примечание, q = q№ ~~ г)/10 , где Т — температура (°C).
Уровень обмена у пойкилотермных животных при разных тем-
пературах следует рассматривать с учетом приспособлений отдель-
ных видов к термическим условиям среды, при которых они фор-
мировались в процессе эволюции. Максимум скорости обмена
у тропических-яо происхождению видов находится при темпера-
турах, близких к 30, для арктических — ниже 15° (Bullock,
1955; Prosser, 1955; Vernberg, Vernberg, 1969, и др.).
На рис. 4 показаны кривые изменения обмена в зависимости
от температуры у пресноводных моллюсков умеренной (Unio
tumidus Retz, U. pictorum L., Anodonta cygnea L.), субтропи-
ческой (Hyriopsis schlegeli Martens, U. biwae (Kob.)) и тропиче-
ской (Parreysia corrugata (Mull.)) зон. Видно, что максимум по-
требления кислорода тропическими моллюсками находится при
температурах, близких к 35°, субтропическими — около 30 и
умеренной зоны — около 25°. Существенно, что максимум по-
требления кислорода моллюсками Dreissena polymorpha (Pall.)
наблюдается при температурах около 27°, т. е. близких к таковым
для субтропических моллюсков. В целом это совпадает с пред-
ставлениями о формировании рода в водоемах южной Европы
(Старобогатов, 1970).
Сказанное позволяет надеяться, что дальнейшее более тща-
тельное изучение скорости метаболизма у моллюсков при разных
температурах окажется несомненно полезным для определения
центров происхождения животных различных видов. Так, можно
предполагать сдвиг оптимума по скорости обмена в область более
26
низких температур у моллюсков Margarittfera margaritifera (L.),
населяющих водоемы севера Европы и Америки, по сравнению,
например, с унионидами умеренной зоны.
Рис. 4. Зависимость скорости обмена у моллюсков от температуры.
Л — по данным Ковалевой, 1970: 1 — Unio tumidus (4 г); 2 — U. pictorum (4 г); 3 — Апо-
donta piscinalis (3 г). Б — Dreissena polymorpha, по данным: 1 — Woynarowich, 1961;
2 — Михеева, 1964. В : 1 — Hyriopsis schlegeli, 2 — U. biwae (по данным Hagashi, 1965).
Г — Parreysia corrugata (по данным Lomte, Negabhushanam, 1971). По оси абсцисс — тем-
пература (°C); по оси ординат — скорость обмена (мгО/ч).
СЕЗОННЫЕ ИЗМЕНЕНИЯ МЕТАБОЛИЗМА
Скорость обмена у моллюсков в разные сезоны года определяется
не только сезонными изменениями температуры окружающей
среды, но также и особенностями их жизненных циклов, в част-
ности степенью физиологической или репродуктивной активности
животных (Weinland, 1919; Gartkiewicz, 1922; Hopkins, 1946;
Дрегольская, 1961; Botnariuc et al., 1961; Lukacsovicz, 1966;
27
Pico?, Cucerzan, 1967; Pamatmat, 1969; Kinne, 1970, и др.), био-
химическими изменениями в составе их тканей (Berg et al., 1958;
Kruger, 1960; Ghiretti, 1966).
У половозрелых моллюсков Unio tumidus в течение года отме-
чаются два максимума потребления кислорода. Первый прихо-
дится на ранневесенние месяцы, второй — на июль. Для Anodonta
piscinalis Nills. характерны три максимума интенсивности обмена:
в апреле, июне и ноябре (рис. 5). Существенно, что у обоих этих
видов наибольшие величины потребления кислорода весной и
—।—।—।—।—।—।—।—।—।—1_ о —।—।—।—।—t—।—।—।—।—।—l
/ /// V VU IX XI I III V VI! IX XI
Рис. 5. Сезонные изменения потребления кислорода моллюсками.
А — Uniot umidus (W=20—30 г); Б — Anodonta piscinalis (W=20—30 г). I — температура
воды (°C); 2 — скорость потребления кислорода; 3 — скорость потребления кислорода
с учетом температурной зависимости (по данным Tudorancea, Florescu, 1970). По оси абс-
цисс— месяцы года; по оси ординат: слева —потребление кислорода (мг-г-1 *ч-1);
справа — температура (°C).
осенью приходятся на периоды, когда температура воды доста-
точно низкая и не превышает весной 15, а осенью 5°.
Из приведенного рисунка видно, что изменения скорости
обмена у этих животных обусловлены не только сезонным ходом
температуры в водоеме. Повышение температуры весной несом-
ненно увеличивает скорость обмена у моллюсков, но уровень
обмена оказывается значительно выше, чем можно было бы
ожидать только за счет воздействия температуры. В то же время
характер изменений потребления кислорода моллюсками в летние
месяцы достаточно хорошо согласуется с температурными усло-
виями, и, по-видимому, наибольшие величины скорости обмена
в этот период связаны с повышением температуры воды.
Увеличение скорости метаболизма у моллюсков весной со-
впадает по времени с началом их репродуктивного цикла, с уве-
личением скорости роста и, возможно, с пробуждением животных
от состояния зимнего покоя. Последнее может быть подтверждено
тем, что после анабиоза двустворчатые моллюски способны уве-
личивать скорость обмена до 300% (Picos, Cucerzan, 1967). Все
28
это вместе взятое определяет нарастание обмена у моллюсков
в весенние месяцы. Однако одним из ведущих факторов, влияю-
щих на сезонные изменения метаболизма у моллюсков, кроме
температуры, следует считать их репродуктивную активность.
Четкая связь изменений скорости обмена со степенью репродук-
тивной активности была показана при исследовании популяции
моллюсков Pisidium walkeri Sterki (Burky, Burky, 1976). У этих
животных, имеющих две генерации в течение года, отмечены три
периода размножения: в июне, июле и сентябре—октябре. В это же
время происходило увеличение скорости обмена у моллюсков,
обусловленное не только повышением температуры. Характерно,
что в осенние месяцы при общем понижении температуры скорость
обмена у моллюсков оставалась на достаточно высоком уровне.
Усиление обмена у A. piscinalis в ноябре, когда температура воды
снижалась до 5° (рис. 5), может быть объяснено тем, что в ок-
тябре—ноябре у этих моллюсков начинается развитие глохидиев,
которые покидают материнские раковины с конца марта следую-
щего года. Увеличение обмена при понижении температуры воды,
совпадающее с периодом размножения, отмечено у Mytilus edu-
Us L. (Bruce, 1926; Vooys, 1976). Потребление кислорода моллю-
сками Macoma baltica (L.) в 4—5, a Cerastoderma edule (L.) почти
в 2 раза выше в периоды размножения, чем при тех же темпера-
турах в другое время (Sparck, 1936).
Таким образом, при изучении потребления кислорода моллю-
сками в различные сезоны года специальное внимание должно
быть обращено на сроки репродуктивной активности отдельных
видов.
ВЛИЯНИЕ НАСЫЩЕНИЯ ВОДЫ
КИСЛОРОДОМ
В пределах своих ареалов моллюски могут обитать в водоемах,
различающихся по степени насыщения воды кислородом. Со-
держание кислорода в воде одного и того же водоема подвержено
сезонным и суточным колебаниям.
При снижении содержания кислорода в воде гидробионты спо-
собны поддерживать интенсивность дыхания на неизменном уровне
до определенных (критических) величин давления кислорода.
Ниже значений критических давлений, которые меньше, чем
парциальное давление насыщенного воздуха, происходит резкое
уменьшение потребления кислорода животными. Одновременно
снижается их активность (Бидл, 1964). Это явление характерно
для животных с так называемым «независимым» типом дыхания.
Животные с «зависимым» дыханием снижают потребление кисло-
рода пропорционально понижению его содержания в воде. Между
этими крайними выражениями зависимости обмена у животных
от количества кислорода в среде отмечены переходы. Для неко-
торых гидробионтов свойственно зависимое дыхание до опреде-
29
ленных величин насыщения кислородом воды и независимое при
более высоких значениях. Значения критического давления за-
висят от различных факторов, в частности от температуры,
уровня обмена и т. п. Критическое давление кислорода неодина-
ково у животных разного возраста — молодые особи двустворча-
тых моллюсков в ряде случаев оказываются более чувствитель-
ными к содержанию кислорода в среде (Bayne, 1971а, 1971b).
Поэтому разделение на зависимый и независимый типы дыхания
в какой-то мере условно. Условность типов дыхания по отношению
к содержанию кислорода в воде была, например, показана на
ракообразных (Сущеня, 1972).
В то же время независимость интенсивности обмена от содер-
жания в воде кислорода была установлена для рыб (Винберг,
1956) и некоторых ракообразных (Сущеня, 1972). Большинство
морских двустворчатых моллюсков способны поддерживать ды-
хание независимым в достаточно широком диапазоне изменений
парциального давления кислорода в воде (Ghiretti, 1966; Проссер,
Браун, 1967).
Результаты измерений скорости обмена у нескольких видов
пресноводных моллюсков показывают, что они способны поддер-
живать независимость обмена в широком интервале изменений
насыщения воды кислородом (рис. 6). Скорость обмена, как видно
из рис. 6, у Anodonta остается практически неизменной при пар-
циальном давлении кислорода примерно до 100 мм рт. ст.,
у Utterbackia imbecillis Say.—до 70—80, у Pisidium casertanum
(Poli.) — до 30 мм рт. ст. Вероятно, можно считать, что крити-
ческое давление кислорода в воде для этих моллюсков находится
соответственно около 61, 24 и 9—10% нормального насыщения
воды кислородом. Близкие к этим величины критического давле-
ния были получены также для моллюсков Congeria sallei (Manga-
pathi et al., 1974) и Anodonta cygnea (Hers, 1943). При более низких
значениях насыщения воды кислородом скорость обмена у живот-
ных оказывается в разной степени зависимой от парциального
давления кислорода. Среди изученных моллюсков наиболее
четко эта зависимость (до величины критического давления
кислорода) проявляется у анодонт и Р. casertanum. Более резко
по сравнению с ними уменьшается потребление кислорода (при
давлениях ниже критического) моллюсками Unio imbecillis.
Кривая потребления кислорода моллюсками Pareysia corrugata
не имеет четко выраженного перегиба, и определение вели-
чины критического давления кислорода для них затрудни-
тельно.
Приведенные для пресноводных моллюсков значения крити-
ческого давления кислорода оказываются близкими к характер-
ным и для морских представителей этих животных. Для Муа
arenaria L. критическое давление близко к 25—30% (Bruce,
1926), Mytilus perna L. — 56—69, M. edulis — 50, Ostrea edu-
lis L. — 65 % насыщения воды кислородом (Bayne, 1967).
30
Различия в величинах критического давления у разных видов
обусловлены скорее всего особенностями их экологии. Например,
моллюски Р. casertanum, для которых отмечено низкое значение
критического давления, могут нормально расти и размножаться
в небольших, мелководных, часто даже пересыхающих водоемах,
где насыщение воды кислородом может падать до весьма низких
величин, в то время как беззубки (более высокое значение кри-
тического давления) обитают в реках и озерах с достаточно бла-:
Рис. 6. Зависимость скорости обмена у моллюсков от парциального дав-
ления кислорода в воде.
А — Anodonta sp. (по данным Dakin, Dakin, 1925); Б — Pisidium casertanum (по данным
Berg et al., 1962); В — Parreysia corrugata (по данным Lomte, Negabhushanam, 1971);
Г — Utterbackia imbecillis (по данным Heistand, 1938). По оси абсцисс — парциальное-
давление кислорода (мм рт. ст.); по оси ординат— скорость обмена (мгО/ч).
гоприятными кислородными условиями. В озерах они населяют
прибрежные части, подверженные постоянному прибойному пере-
мешиванию воды. Существенно, что у моллюсков Муа arenaria,
зарывающихся глубоко в грунт, величина критического давления
кислорода оказывается ниже, чем у живущих на открытой лито-
рали или сублиторали мидий и устриц. В целом на самых разных
беспозвоночных было показано, что животные, обитающие в ус-
ловиях с низким содержанием кислорода, способны связывать
его при меньшем давлении по сравнению с живущими в условиях
с высоким содержанием кислорода (Коштоянц, 1950).
Зависимость скорости метаболизма от парциального давления
кислорода в среде может быть описана уравнением
jP, , (5)
—F ^2.Р '
31
где R — потребление кислорода животным; р — парциальное
давление кислорода; к1ч к2 — константы уравнения.
Переписав уравнение (5) как
p/R=k1-\-k.ip, (6)
видим, что функция p[R=f (р) представляет собой прямую линию,
определяющуюся параметрами кг и к2. Значения параметров легко
находятся графически. Прямая линия пересекает ось ординат
в точке, соответствующей значению /сх, а ее наклон по отношению
к оси абсцисс отражает величину /с2.
Несмотря на то что уравнение (5) выражает гиперболическую
функцию, с его помощью могут быть описаны все известные виды
зависимости потребления кислорода животными от давления его
в среде. Когда значение к± оказывается больше, чем к2р, то вели-
чина R примерно равна р!кг. В этом случае скорость потребления
кислорода животными прямо пропорциональна насыщению среды
кислородом. При ki^k^p величина R может оставаться в значи-
тельной степени постоянной, что и наблюдается в случае незави-
симого дыхания.
Тенг (Tang, 1933), который впервые предложил аналитическое
выражение зависимости скорости обмена от парциального дав-
ления кислорода в среде в виде уравнения (5) и провел исследо-
вания его параметров, указывал на сходство этого уравнения
с уравнением абсорбции газов на твердую поверхность. В этом
случае скорость потребления кислорода должна быть пропорцио-
нальна количеству кислорода, абсорбированного респирационной
поверхностью путем оседания на нее молекул кислорода. Однако
это лишь механическая сторона явления. Эффективность связыва-
ния кислорода и переноса его к различным органам животных
в значительной степени обусловлена специфичностью дыхательных
пигментов. Кроме того, количество потребленного кислорода,
как это было показано на различных животных и в дальнейшем
будет продемонстрировано на примере моллюсков, пропорцио-
нально массе организма. Поэтому пока нет достаточных данных
и оснований для биологического толкования значений констант
уравнения (5). Можно лишь отметить, что коэффициент к2 в при-
веденном уравнении связан с температурой таким образом, что
при увеличении температуры его величина должна уменьшаться
(Tang, 1933).
Рассмотренные выше эмпирические данные по скорости обмена
у нескольких видов пресноводных двустворчатых моллюсков
достаточно хорошо могут быть описаны уравнением (5), что под-
тверждается расположением экспериментальных точек на графике
p/R=f (р) (рис. 7) вдоль прямых линий. Параметры уравнений
для отдельных видов, рассчитанные графически, приведены
в табл. 4. Уравнение (5) хорошо описывает зависимость потребле-
ния кислорода моллюсками Mytilus регпа от насыщения воды
32
Таблица 4
Значения параметров к19 к2 в уравнениях зависимости
потребления кислорода моллюсками от его парциального
давления
Название моллюска
Utterbackia imbecillis............................
Anodonta sp.......................................
Pisidium casertanum...............................
Pareysia corrugata................................
10
20
80
240
4.45
1.43
7.70
5.40
кислородом. Для этих животных параметры уравнения имели
значения: ^=96.12, /с2=2.00 (рассчитано по: Bayne, 1967).
Таким образом, экспериментальные данные, отражающие за-
висимость потребления кислорода моллюсками от содержания
его в воде, могут быть достаточно хорошо аппроксимированы
уравнением гиперболической функции. В то же время необхо-
димы дальнейшие более подробные и тщательные исследования
Рис. 7. Зависимость отношения p!R от парциального давления кислорода р
в воде.
А — Parreysia corrugata (по данным Lomte, Negabhushanam, 1971); Б — Pisidium caserta-
пит (по данным Berg et al., 1962). По оси абсцисс — парциальное давление кислорода
(мм рт. ст.); по оси ординат — отношение p/R (объяснения в тексте).
с целью уточнения аналитического вида и теоретического обосно-
вания связи между интенсивностью обмена у моллюсков и степенью
насыщения воды кислородом. Работы в этом направлении поз-
воляют количественно оценить степень оксифильности отдельных
животных и помогут в определенной степени объяснить их про-
странственное расселение.
Животные с фильтрационным типом питания поглощают из
потока фильтруемой ими воды небольшие количества растворен-
ного в ней кислорода. Губки утилизируют в среднем около 20%,
асцидии не более 3—10% кислорода из воды (Проссер, Браун,
1967). Морские двустворчатые моллюски способны потреблять
от 2.5 до 15% кислорода, содержащегося в фильтруемом ими
3 А. Ф. Алимов 33
объеме воды (Ghiretti, 1966; Bayne, 1967). В отношении пресно-
водных моллюсков таких данных почти нет. Известно лишь, что
Anodonta в обычном состоянии утилизируют 5—10% растворенного
кислорода, после анаэробиоза — до 20% (Van Dam, 1937). Нет
оснований ожидать, что пресноводные моллюски могут утилизи-
ровать большие по сравнению с морскими количества кислорода
из воды, хотя и имеются сведения (Bruin, Davids, 1970) о более
эффективной утилизации кислорода жабрами у этих животных.
СКОРОСТЬ ОБМЕНА
И ОБЩИЕ РАЗМЕРЫ МОЛЛЮСКОВ
Многочисленными экспериментальными исследованиями на раз-
ных представителях животного мира с полной очевидностью
установлена тесная зависимость между скоростью, интенсивностью
обмена и размерами организмов. Скорость обмена у животных
нарастает, а его интенсивность соответственно падает с увеличе-
нием их веса. Нет нужды перечислять огромную литературу,
в которой эта теперь уже тривиальная истина была неоднократно
подтверждена. По-видимому, достаточно сослаться на серьезную
сводку Хеммингсена (Hemmingsen, 1960), в которой на огромном
фактическом материале показано, что общий обмен R пойкило-
термных и гомойотермных животных и вес их тела W связаны
между собой степенной зависимостью
R = aWb, (7)
откуда интенсивность обмена
R/W = aWb-1, (8)
где а и Ъ — константы уравнения.
Использование степенного уравнения применительно к соот-
ношению между обменом и весом животных было специально
рассмотрено и обсуждено Г. Г. Винбергом (1976а). На большом
и разнородном материале он убедительно показал правомочность
его использования для этой цели как эмпирического приема обра-
ботки, аппроксимации и усреднения результатов наблюдений.
При этом специально подчеркивалось, что аппроксимация данных
в виде степенной функции еще не указывает на какой-либо опре-
деленный механизм явления.
Разными исследователями показана пригодность степенного
уравнения для выражения связи между R и W и сравнения между
собой уровней обмена особей одного вида или при межвидовых
сопоставлениях. Впрочем, была предпринята попытка опроверг-
нуть справедливость использования степенной функции для выра-
жения рассматриваемой зависимости (Каширская, Константинов,
1973). Несостоятельность такой попытки, основанной на рассмот-
рении эмпирических данных в узком интервале весов, была с оче-
34
видностью продемонстрирована в ряде работ (Алимов и др., 1974;
Заика, 19746; Сущеня, 1974; Винберг, 1976а). Поэтому нет ни-
каких серьезных оснований описывать зависимость между обме-
ном и весом животных какой-либо другой функцией, не имеющей
реальных преимуществ перед степенной. Это лишь затруднило бы
( равнение материалов, полученных разными авторами в различ-
ных условиях и привело бы к невозможности обоснования общих
выводов.
В приведенных уравнениях (7, 8) размеры животного выражены
в весовых единицах. Вес организмов — более удобный и оправ-
данный по сравнению с линейными размерами показатель, так
как отражает массу живого вещества, участвующую в про-
цессах метаболизма. Кроме того, вес может быть легко с помощью
соответствующих коэффициентов переведен в энергетические или
другие эквивалентные единицы, что несомненно придает указан-
ной зависимости определенную пластичность.
Равенство (7) представляет собой уравнение аллометрии.
Первая его константа а отражает величину обмена у животного,
вес которого равен единице. Постоянная уравнения Ъ показывает
скорость изменения обмена при увеличении веса организма.
Значения этой константы для разных животных находятся обычно
в пределах 0.6—1.0. В последнем случае не происходит повышения
обмена с увеличением веса тела. Для большинства животных
Это означает, что величина обмена возрастает с меньшей
скоростью, чем масса тела животного в процессе его роста.
В литературе уже имеется значительное количество рассеян-
ных по разным изданиям данных по интенсивности обмена у раз-
личных видов пресноводных двустворчатых моллюсков. Наличие
этих сведений дает возможность сделать обобщения и количе-
ственно определить зависимость скорости обмена от размеров
моллюсков.
Исследователи проводили эксперименты при разных темпера-
турных условиях. Поэтому все литературные и собственные данные
пами приведены к температуре 20° с помощью ранее описанной
зависимости скорости обмена от температуры. Температуры,
близкие к 20°, — наиболее обычные летние температуры в пре-
делах ареалов большинства изученных нами видов. Величины
потребления кислорода моллюсками обычно выражаются в милли-
граммах (мг), миллилитрах (мл) либо в кубических сантиметрах
(см3) и т. п. кислорода в единицу времени (чаще всего в час).
Используя соответствующие переводные коэффициенты, мы пред-
ставили все данные в миллиграммах кислорода в час. Параметры
искомых зависимостей рассчитаны на ЭВМ. Во всех случаях вес
животных выражался в граммах сырого веса вместе с их раковиной.
Сырой вес моллюсков сильно колеблется в зависимости от сте-
пени обсушивания животных перед взвешиванием, от их физио-
логического состояния и т. д. Тем не менее представляется целе-
сообразным пользоваться сырым весом моллюсков с раковиной,
3*
35
0.01
1, 6—8 — U. tumidus', 2 — U. biwae', 3—5 — U. pictorum’, 9 — C. crassus (no
данным: 1 — Tudorancea, Florescu, 1968; 2 — Hagashi, 1965; 3—6 — Кондратьева,
1970; 4—9 — Ковалевой, 1970; Wesemeier, 1960; Ornatowski, 1967). Масштаб
логарифмический. По оси абсцисс — вес моллюсков (г); по оси ординат —
скорость обмена (мгО/ч).
гак как он соответствует их естественному состоянию. Раковина
секретируется специальными железами моллюсков в течение
всей жизни, на образование ее животными постоянно затрачива-
ется определенное количество энергии. Раковина — продукт их
жизнедеятельности и в этом отношении коренным образом отли-
чается, например, от домиков ручейников иличехликов хирономид.
Кроме того, при определении сырого веса целого моллюска не
Рис. 9. Зависимость скорости обмена от веса у моллюсков рода Anodonta.
1,3,6 — A. piscinalis', 2 — A. cygnea\ 4 — A. zellensis’, 5 — A. anatina; 1 — A. woodiana
(по данным: 1 — Tudorancea, Florescu, 1970; 2 — Ковалевой, 1970; 3 — Кондратьева,
1970; 4,5 — Ornatowski, 1967; 6 — Botnariuc et al., 1961; 7 — измерения автора). Осталь-
ные обозначения как на рис. 8.
происходит уничтожения живого материала, необходимого для
дальнейших эколого-физиологических или других исследований.
Потребление кислорода моллюсками сем. Unionidae изучалось
главным образом на наиболее широко распространенных предста-
вителях подсемейств Unioninae (род Unto) и Anodontinae (роды
Anodonta, Cristaria, Hyrtopsis). Полученные результаты приведены
на рис. 8—10, на основании которых зависимость скорости
обмена от веса для моллюсков Unto может быть аппроксимирована
уравнением
R = (0.074 ± 0.013) И/(°-б4±0 ()4) мгО/ч,1 (9)
для Anodonta —
R = (0.0861 + 0.0193) W-73±0-07) мгО/ч. (10)
1 Здесь и в последующих уравнениях приводятся ошибки средних зна-
чений параметров.
37
В уравнениях (9—10) относительная ошибка константы а
составляет 17.4% для Unto и 22.4% для Anodonta, а константы Ъ
соответственно 6.2 и 9.1%. Между уравнениями (9—10) не обна-
руживается статистически достоверных различий при принятом
уровне достоверности.
По нескольким (19) измерениям графически определены пара-
метры уравнения для моллюсков Cristaria plicata Leach., оби-
тающих в водоемах Дальнего Востока, и Hyriopsis schlegeli (во-
доемы Индокитая), кото-
рые оказались по своим
значениям близкими к по-
лученным для Unio и
Anodonta:
Я = 0.070И70-73 мгО/ч. (И)
Все это дает возмож-
ность и достаточные осно-
вания вывести единое для
всего сем. Unionidae урав-
нение, определяющее функ-
циональную связь скоро-
сти обмена и веса этих
животных:
Я = (0.076 ± 0.016) X
0.01*—__________________I__________________L- х W-713±0-055) мгО/ч. (12)
7 10 100
Рис. 10. Зависимость скорости обмена
от веса у Cristaria plicata (7) и Hyriopsis
schlegelii (2) (по данным Hagashi, 1965)
(из: Алимов, 1975).
.Остальные обозначения как на рис. 8.
Важно отметить, что близ-
кие значения параметра Ъ
были получены Г. П. Кон-
дратьевым (1970а, 1970в)
для U. tumidus (0.7) и
A. piscinalis (0.73). Везе-
мейер (Wesemeier, 1960) приводит величины этого параметра
для U. pictorum — 1.1, а для A. cygnea— 0.93. Однако, как
уже отмечалось, примененный им манометрический способ
измерения дыхания моллюсков таит в себе возможность су-
щественных ошибок, чем, по-видимому, и обусловлены такие
высокие значения этого параметра. Близкие к единице
(С7. tumidus — 0.8, U. pictorum — 0.93, A. anatina L. —0.82,
A. zellensis Gmelin — 0.96) значения этого параметра получены
Орнатовским (Ornatowski, 1967). Однако этот автор проводил
свои эксперименты только с молодыми моллюсками (вес подопыт-
ных животных находился в пределах от 1 до 15 г). Естественно,
что в столь узком для таких крупных животных, как униониды,
диапазоне весов (дефинитивный вес некоторых — сотни граммов)
ошибка в определении параметров уравнений возрастает в значи-
тельной степени.
38
Как можно видеть на рис. 8 и 9, наши уравнения выведены
для диапазона весов от 0.2 до 100 г (Unto) и от 1 до 160 г (Anodontd).
Кроме того, количество измерений, сделанных нами (84 — Unio
и 76 — Anodontd), значительно превышает число измерений,
па основании которых были выведены уравнения Орнатовским
и Веземейером. Это дает основание считать, что уравнения (9—12)
представляют более строгое выражение исследуемой зависи-
мости по сравнению с ранее пред-
ложенными в отношении Unioni-
(I ае.
Отдельные измерения интен-
с явности обмена индийских и
американских моллюсков сем. Ат-
blemidae, близкого в системати-
ческом отношении к Unionidae,
приведены на рис. 11. Эти данные
могут быть аппроксимированы
уравнением
Д = 0.096И™-76 мгО/ч. (13)
Полученные значения параметров
уравнения (13) близки к таковым
для унионид.
Результаты измерений скоро-
сти обмена моллюсков сем. Sphae-
riidae (роды Sphaerium, Musculium,
Plsidium), сведенные на рис. 12,
показывают, что среди животных
этого семейства не наблюдается родовой или видовой специфич-
ности обмена. Это позволяет вывести единое для данного семей-
ства уравнение
Рис. 11. Зависимость скорости
обмена от веса у моллюсков
Amblemidae (из: Алимов, 1975).
1 — Parreysia corrugata (по данным
Lomte, Negabhushanam, 1971); 2 —
Amblema perplicata\ 3 — Quadrula quad-
rula (по данным Read, 1954). Осталь-
ные обозначения как на рис. 8.
В = (0.129 + 0.057) W-895±0-151) мгО/ч. (14)
Обращают на себя внимание довольно большие ошибки сред-
них значений параметров уравнения (14) (44% для а и 17 — для
'О-
Значительные отклонения экспериментальных данных от сред-
них значений могут быть связаны в первую очередь с живорожде-
нием у сфериид. Разные исследователи проводили эксперименты
с моллюсками, собранными в различные сезоны года. При этом
по обращалось специального внимания на количество и степень
развития молоди в материнских раковинах. Неодинаковое коли-
чество и размеры развивающейся молоди у половозрелых живот-
ных сходного веса, естественно, могли обусловить разницу в по-
треблении ими кислорода. Скорее всего этим могут объясняться
и довольно высокие значения параметров уравнения (14). Мы
проводили измерения дыхания сфериид обычно с середины июля
39
0.001
дй-
0.01
0. 1
Рис. 12. Зависимость скорости
обмена от веса у моллюсков
Sphaeriidae.
1— S. scaldianum; 2, 5, 6 — S. ri-
vicola\ 3 — Pisidium nitidum; 4, 7, 8 —
S. corneum; 9 — S. solidum; 10 — Mu-
sculium lacustre (по данным: I, 2, 3, 8_
измерения автора; 4 — Арабикой, Ру-
биновой, 1971; 5 — Кондратьева, 1970;
б» 7, 9 — Ковалевой, 1970; Wesemeier,
196,0). Остальные обозначения как
на рис. 8.
до середины августа и, как можно видеть из рис. 12, потребле-
ние кислорода моллюсками одинакового веса в наших опытах
отличалось незначительно. Веземейер (Wesemeier, 1960) приводит
высокое значение параметра Ъ (0.94) для моллюсков Musculium
lacustre Muller, но мы уже обсуждали возможность возникновения
в его экспериментах ошибок определений количества потреб-
ленного животными кислорода. Высокие значения параметра Ъ
(0.816 и 0.834) были получены и при исследованиях количества
Рис. 13. Зависимость скорости обмена от веса у моллюсков Corbicula sandai
(по данным Hagashi, 1965) (из: Алимов, 1975).
Остальные обозначения как на рис. 8.
углекислоты, выделяемой при дыхании двумя видами Pisidium
(Р. casertanum и Р. henslowanum) при температуре 20° в зависи-
мости от их веса (Holopainen, Ranta, 1977). Поэтому можно пред-
положить, что для сфериид вообще характерны более высокие
но сравнению с другими видами пресноводных Bivalvia значения
параметра Ъ в уравнениях связи скорости обмена и веса животных.
Известно лишь одно исследование по скорости обмена у моллю-
сков сем. Corbiculidae. По пяти измерениям потребления кисло-
рода моллюсками Corbicula sandai Reinhardt (рис. 13) нами были
в первом приближении графически рассчитаны параметры урав-
нения скорости обмена от веса животных
R = 0.045Ж0-73 мгО/ч. (15)
Низкие значения константы а в уравнении (15) , вероятно, могут
быть объяснены тем, что использование для этих тропических по
41
происхождению моллюсков обсуждавшейся ранее зависимости
скорости обмена от температуры с целью приведения эксперимен-
тальных данных к 20° приводит к занижению у них обмена.
Скорость обмена у моллюсков Dreissenidae изучена в основном
у Dreissena polymorpha. Как видно из рис. 14, на котором приве-
дены все известные данные по дыханию этих животных, резуль-
таты экспериментов позволяют вывести уравнение
Я=(0.1396 + 0.0084) W-63±0-04> мгО/ч. (16)
Рис. 14. Зависимость скорости обмена от веса у моллюсков Dreissena poly-
morpha (по данным: 1 — Спиридонова, 1972; 2 — Ludwig, Krywienzyk,
1950; 3 — Кондратьева, 1970; 4 — Ковалевой, 1970).
Остальные обозначения как на рис. 8.
Ошибки средних значений параметров уравнения (16) не пре-
вышают 6 и 6.3%. Близкая к полученной нами для этих моллюсков
величина параметра Ъ приведена Г. П. Кондратьевым (1970а,
1970в) и неоправданно высокая (1.06) Веземейером (Wesemeier,
1960). Более низкое, чем в уравнении (16), значение этого параметра
(0.5845) известно для тропических представителей семейства —
Congeria sallei (Mangapathi et al., 1974). Однако следует отметить,
что параметры уравнения для этих моллюсков были рассчитаны
только на основании 19 измерений потребления ими кислорода.
Анализируя рис. 8—14 и уравнения (9—16), можно прийти
к важным заключениям. Рассмотрены все имеющиеся к настоя-
щему времени данные по интенсивности обмена у представителей
по сути всех семейств пресноводных двустворчатых моллюсков,
42
распространенных на территории СССР. Результаты свидетель-
ствуют о том, что в пределах этих семейств скорость обмена
у животных — функция их веса (массы) и родовой или видовой
ее специфичности не обнаруживается. Более того, доверительные
интервалы для найденных уравнений взаимно перекрываются
(рис. 15). Следовательно, даже между животными одного размера
из разных семейств нет достоверных отличий в величинах скорости
обмена. Отметим, что к аналогичному выводу пришел и Г. Г. Вин-
берг в отношении брюхоногих моллюсков (Винберг, Беляцкая,
1958, 1959). Все это дает возможность вывести единое уравнение
для всех исследованных моллюсков (250 измерений потребления
кислорода 20 видами):
R = (0.0938 + 0.0061) W-721±0-026) мгО/ч. (17)
При этом в уравнении (17) относительная ошибка параметра а
составляет 6.5, b — 3.6%. Максимальная величина первой кон-
43
станты не превышает 0.0999, минимальная — 0.0877, второй
соответственно 0.747 «0.75 и 0.695 «0.7.
В первой попытке обобщить сведения по скорости обмена
у пресноводных и некоторых морских двустворчатых моллюсков
(Алимов, 1969) было получено уравнение
R = (0.1075 + 0.024) ТУ0-73 мгО/ч, (18)
которое по своим параметрам очень близко к уравнению (17).
Значения константы Ь, только незначительно отличающиеся
от ее величины в уравнении (17), были получены при изучении
некоторых морских моллюсков: Scrobicularia plana (Da Costa) —
0.75 (Hughes, 1970), Marisa cornuarietis L. — 0.78 (Akerlund,
1969), Mytilus galloprovincialis Lamarck — 0.73, Mactra subtrun-
cata (Da Costa) — 0.73, Venus gallina Linne — 0.77, Mytilaster
linneatus (Gmel.) — 0.69—0.71 (Турпаева и др., 1970), несколь-
ких видов каспийских моллюсков — 0.76, 0.79 (Иванова, 1973)
и др. По мнению Е. П. Турпаевой, величина константы Ъ для
черноморских моллюсков может быть принята равной 0.75.
Заметим, что для ракообразных ее значение 0.756 (Сущеня, 1972),
пресноводных гастропод — 0.75 (Винберг, Беляцкая, 1959), 0.73
(Berg, Ockelmann, 1959), олигохет — 0.75 (Камлюк, 1974),
фитофильных личинок хирономид — 0.73 (Касаткина, I960),
реснитчатых инфузорий — 0.76 (Хлебович, 1974). Для пресно-
водных двустворчатых моллюсков величина этой константы,
как видно из уравнения (17), также достоверно не отличается
от 0.75. Существенно отметить, что, по мнению Хеммингсена
(Hemmingsen, 1960), такое значение константы справедливо для
всего животного мира.
Уровень обмена, который может быть охарактеризован кон-
стантой а, среди пресноводных двустворчатых моллюсков ока-
зывается наибольшим у представителей сем. Dreissenidae
(0.1396 мгО/(ч-г)), наименьшим — у Corbiculidae (0.045 мгО/(ч-г)),
в среднем — около 0.076. Среди черноморских моллюсков уро-
вень обмена изменялся от 0.052 до 0.148 мгО/ч на грамм сырого
веса тела. Приняв, что вес раковины у пресноводных моллюсков
в среднем составляет 30% их сырого веса, получаем среднее зна-
чение константы а для этих животных (в пересчете на сырой вес
тела) 0.122 мгО/(ч-г) или 0.085 млО/(ч.г). Эта величина прак-
тически неотличима от уровня обмена черноморских и каспий-
ских моллюсков (0.127 мгО/(ч-г); Иванова, 1973). Следовательно,
морские и пресноводные двустворчатые моллюски по уровню
обмена не различаются сколько-нибудь достоверно, на что ранее
уже было обращено внимание (Алимов, 1969).
Полученные для отдельных представителей животного мира
обобщенные количественные зависимости скорости обмена от их
размеров следует рассматривать не более как статистические,
отражающие средние для них уровни обмена и закон изменения
44
скорости обмена по мере увеличения веса животных. Поэтому
было бы ошибочным надеяться на совпадение величин обмена
в каждом конкретном опыте со средними статистическими зна-
чениями.
Непосредственно по значениям параметра а невозможно при
разных значениях константы Ъ проводить сравнение уровней
обмена у различных животных. В свое время В. С. Ивлев (1963)
предложил формулу, в которой принимались во внимание оба
параметра уравнения и определялся показатель уровня обмена.
Однако этот показатель, как отмечал и сам автор, оказался
функционально связан с максимальным весом животных, что
несомненно затрудняет его использование для сравнения обмена
у животных, различающихся по весу.
Сравнение интенсивности обмена у различных животных воз-
можно лишь при равенстве констант Ъ или по величине RIW
у животных одинакового веса. Естественно, что при этом все
составляющие сравниваемых уравнений должны быть выражены
в одинаковых единицах. Например, нельзя сравнивать значения а
в двух уравнениях, если в одном из них а выражено в милли-
литрах, в другом — в миллиграммах кислорода и потребление
кислорода животным в миллиграммах или миллилитрах отнесено
либо к его весу в 1 г, либо в 1 мг. В последнем случае необходимо
помнить, что в уравнениях зависимости скорости обмена от веса
животных их вес находится под знаком степени. Следовательно,
если в двух уравнениях показатели степени различны, то коэффи-
циенты а сравнивать между собой нельзя.
Для приведения в сравниваемых уравнениях показателей
степени к единому значению следует учитывать средний вес
особей в диапазоне весов, для которых рассчитано каждое урав-
нение. При расположении экспериментальных данных по скорости
обмена в зависимости от веса животных в логарифмической си-
стеме координат, т. е. когда
lg R = lg а + Ъ lg W,
lg W представляет собой как бы «центр тяжести» прямой линии,
относительно которого ее наклон меняется, если изменяется
значение константы Ь. Величина константы а определяется зна-
чениями угла наклона прямой линии на графике lg R=f (lg W)
и логарифма среднего веса животных lg W. При этом зависимость а
от lg W проявляется тем в большей степени, чем больше изменя-
ется угол наклона.
С целью сравнения уровней обмена животных, различающихся
по своему систематическому положению, учитывая сказанное
и рассмотренные выше данные, можно принять для них значение
константы Ъ равным 0.75. Сравнение рассчитанных при таком
условии величин а показывает, что уровень обмена оказывается
наибольшим у рыб, наименьшим (среди изученных животных) —
у двустворчатых моллюсков (табл. 5).
45
Таблица 5
Значения параметра а при Ъ== 0.75 = const в уравнениях
зависимости скорости обмена от веса у различных
водных пойкилотермных животных
Название животных а (мгО/ч на грамм сырого веса) По данным
Pisces 0.488 Винберга, 1956
Chironomidae 0.298 Касаткиной, 1960
Crustacea . 0.180 Сущени, 1972
Gastropoda Около 0.120 * Винберга, Беляцкой, 1959; Berg, Ockelmann, 1959
Oligochaeta 0.105 Камлюк, 1974
Bivalvia пресноводные . . 0.089 Автора
* Рассчитано как среднее из данных указанных авторов при допущении, что вес
раковины составляет около 30% общего веса моллюсков.
Анализируя данные табл. 5, можно заметить, что закономерное
уменьшение величины а может быть связано с уменьшением
подвижности животных. Это согласуется с предположением, ко-
торое было высказано Г. Г. Винбергом (1976а) о том, что интенсив-
ность обмена выше у высокоподвижных видов: «. . .активные виды
имеют высокую интенсивность обмена не потому, что они высоко-
подвижны, а скорее, наоборот, они высокоподвижны потому, что
имеют высокую интенсивность обмена» (с. 62). Последнее положе-
ние, подтверждаемое данными табл. 5, весьма существенно, так
как из него следует, что уровень обмена у животных не определя-
ется увеличением или уменьшением доли активного обмена от ос-
новного.
Уровень обмена, выраженный в рассматриваемых уравнениях
в единицах кислорода, с помощью соответствующих коэффициентов
может быть представлен в единицах энергии (калории или джоули).
Из физики известно, что энергия может быть определена как спо-
собность совершать работу. Более высокий уровень энергетичес-
кого обмена у животных обеспечивает им возможность совершать
большую работу за единицу времени, т. е. повышается их мощ-
ность.
Действительно, большинство рыб — прекрасные пловцы, по-
стоянно находящиеся в движении. Даже самые медлительные из
них способны в короткие отрезки времени развивать высокие ско-
рости. С другой стороны, двустворчатые моллюски или черви —
в большинстве своем малоподвижные, закапывающиеся животные.
Существенно, что у более подвижных гастропод (среди которых,
кстати, Prosobranchia и Pulmonata не различаются между собой по
уровню обмена) уровень обмена выше, чем у малоподвижных,
в ряде случаев прикрепляющихся двустворчатых моллюсков.
Уровень обмена остается неизменным в пределах таксонов
высокого ранга, например класса (Дольник, 1968). Таким образом,
46
его можно рассматривать как показатель организации общего
плана строения в самом широком понимании этого термина. Обмен
есть интегрированное отражение основных и важнейших элементов
организации животных, которые не меняются при изменениях их
экологии и морфологической дифференцировки.
КАЛОРИЙНОСТЬ МОЛЛЮСКОВ
В процессе энергетического обмена (метаболизма) в результате
дыхания происходит разрушение органических соединений с ос-
вобождением энергии, заключенной в их молекулах. Траты ве-
щества или энергии на обменные процессы в течение развития жи-
вотных могут быть определены на основе соотношения между ин-
тенсивностью обмена и их индивидуальным весом. В этом случае
скорость или интенсивность обмена должна быть выражена в тех же
единицах, что и вес животного, например в энергетических.
По отношению к жирым организмам следует различать энерге-
тическую ценность, или энергетический эквивалент веса организма,
и энергетическую ценность организма, отнесенную к единице его
массы. Размерность первой величины — единицы энергии (кало-
рии), приходящиеся на особь, второй — калории на 1 г сырого,
сухого либо беззольного вещества ее тела. Различие двух по-
следних понятий было специально подчеркнуто (Остапеня, 1968),
но еще до сих пор их иногда путают.
Основные материалы по калорийности пресноводных двуствор-
чатых моллюсков нами были получены на унионидах Unio tumidus,
£7. pictorum, Anodonta piscinalis, а также на Sphaerium suecicum
(Алимов, Шадрин, 1977).
Сырой вес моллюсков измерялся после подсушивания их на
фильтровальной бумаге, сухой — после высушивания до постоян-
ного веса при температуре 75—82° в течение 5—6 ч. Зольный ос-
таток определялся в навеске 80—150 мг высушенных до постоян-
ного веса и тщательно измельченных животных после сжигания
в течение 5—6 ч в муфельной печи при температуре 550—600°.
Калорийность определялась методом бихроматного окисления
(Остапеня, 1968) в 2—4 повторностях. Пересчет в калории прово-
дился с помощью оксикалорийного коэффициента — 3.4 кал/мгО
(Остапеня, 1965).
Соотношение между сухим (Жс) и сырым (FT) весом моллюсков
Unio и Anodonta (рис. 16) может быть выражено уравнениями:
для Unio
И/с=(0.431 + 0.041) W-992±0-018) (19)
и Anodonta
Жс=(0.224 + 0.050) Ж(ь070±0-015). (20)
Коэффициенты корреляции г в этих уравнениях соответственно
равны 0.996 и 0.901, вес — в граммах.
47
Сухой вес моллюсков S. suecicum составлял 19—34, в среднем
27.2% их сырого веса.
Содержание воды в раковинах и теле унионид в среднем близко
к 3.3% от веса раковины у перловиц и 4.7% — у беззубок. Тело
перловиц содержит в среднем 84.2, беззубок — 85.6% воды. Сухой
остаток полостной жидкости перловиц составляет 0.6, беззубок —
Рис. 16. Соотношение между логарифмами сухого и сырого весов у моллю-
сков.
1 — Unio‘t 2 — Anodonta. По оси абсцисс — логарифм сырого веса (г); по оси ординат —
логарифм сухого веса (г).
0.7% веса жидкости. Количество полостной жидкости изме-
няется с возрастом моллюсков, и зависимость его от их веса мо-
жет быть аппроксимирована степенными уравнениями:
для перловиц
Жж=(0.204 ± 0.049) W-101±°-042\ (21)
г=0.993,
для беззубок
И7в=(0.303 ± 0.051) ^(1.06910.040), (22)
г=0.984, вес — в граммах.
48
Из уравнений (19—22) следует, что количество полостной жид-
кости и сухой вес моллюсков в весьма слабой степени зависят от
их размера. Значения показателей степени в этих уравнениях до-
стоверно не отличаются от единицы. Приняв в уравнениях (19—22)
величину показателя степени равной единице, получаем, что сухой
вес у Unto составляет около 43, у Anodonta — 22%, а количество
полостной жидкости соответственно около 20 и 30% сырого веса
моллюсков.
Отношение веса раковины к весу моллюсков с возрастом ме-
няется незначительно, и у перловиц оно в два раза выше (около
32%), чем у беззубок (около 16%).
Зола в раковинах перловиц составляет 85.5%, беззубок — 49,
сфериид — 53.9% их веса. В сухом остатке полостной жидкости без-
зубок на золу приходится 17.1 %. Из данных табл. 6 видно, что
у разных видов унионид в диапазоне их весов от 5 до 30 г содержа-
ние золы возрастает с увеличением веса животных от 6 до 15%.
По мере роста беззубок зольность их тела продолжает увеличи-
ваться и, например, у моллюсков весом 30—35 г оказывается около
22—23%, у перловиц такого же веса зольность остается практи-
чески на одном уровне (14—16.5%). Количество золы у моллюсков
Sphaerium suecicum в процессе их роста убывает, т. е. происходит
относительное увеличение содержания органических веществ, что,,
вероятно, связано с живорождением:
Сухой вес (мг) Содержание золы (%)
4.5 74.4
5.5 74.0
26.5 51.7
29.0 55.0
37.0 47.4
Среди исследованных моллюсков по мере увеличения их веса
не было обнаружено сколько-нибудь закономерного изменения ка-
лорийности раковины. Калорийность раковин перловиц в среднем
не превышала 0.22, беззубок — 0.16, шаровок — 0.19 кал/мг
сухого веса раковины. Калорийность тела у перловиц оказалась
несколько более высокой, чем у беззубок (табл. 6). У животных
весом от 6 до 30 г калорийность снижалась с 4.6—4.8 до 3.6 кал/мг
сухого веса. У перловиц, вес которых превышал 30 г, она остава-
лась в пределах 3.8—4.0 кал/мг сухого веса.
При расчете калорийности сухого вещества целого моллюска
исходили из равенства
/гри/р + *ти\
4 А. Ф. Алимов
(23)
49
Таблица 6
Содержание золы и калорийность моллюсков сем. Unionidae
Вес сырой (г) Зола (%) Кал/мг сухого веса тела Кал/мг органи- ческого вещества
Unto tumidus
9.810 6.10 5.10 5.43
13.180 8.03 4.22 4.63
15.070 6.70 4.81 5.16
16.835 6.38 4.85 5.18
25.620 15.16 3.81 4.51
30.505 11.50 4.07 4.57
35.505 15.92 3.70 4.48
41.485 14.19 3.91 4.56
45.460 15.91 4.17 4.96
52.900 14.43 3.85 4.50
59.040 16.50 4.00 4.96
Unio pictorum
8.650 6.85 4.81 5.17
6.845 7.00 4.60 4.95
8.535 6.85 4.80 5.17
8.600 8.39 4.39 4.79
9.900 6.85 4.48 4.81
44.465 12.20 4.65 5.30
14.950 9.10 5.10 5.60
18.715 11.10 5.31 5.95
20.305 8.47 5.10 5.57
32.860 12.50 4.50 5.19
33.930 16.12 4.40 5.25
Anodonta piscinalis
4.770 9.57 4.70 5.20
5.095 9.99 5.05 5.60
8.640 6.31 4.25 4.54
11.010 9.86 4.29 4.76
11.365 11.70 4.72 5.32
12.460 7.89 4.59 4.98
15.020 15.70 4.17 4.95
15.075 9.49 4.83 5.34
15.400 11.65 4.30 4.87
15.390 6.77 4.68 5.02
16.950 8.49 4.50 4.92
18.710 13.52 3.88 4.49
18.960 12.50 4.45 5.05
19.020 10.68 4.15 4.64
19.710 10.50 4.69 5.24
22.155 12.97 4.46 5.12
22.305 13.07 3.94 4.54
22.610 24.50 3.57 4.73
23.210 14.96 3.56 4.20
23.970 17.40 3.88 4.70
32.570 20.90 — —
33.045 23.40 4.17 5.32
50
где К — калорийность сухого вещества моллюска; Кр, Кт — ка-
лорийность раковины и тела соответственно; Wp, WT, W — вес
раковины, тела и целого моллюска соответственно.
Расчеты показали, что калорийность целых моллюсков раз-
личных видов сем. Unionidae практически одинакова и по мере их
роста уменьшается (рис. 17):
у = (1.136 ± 0.041) —(0.01 + 0.002)я, (24)
где у — калорийность (кал/мг сухого веса); х — сырой вес моллю-
сков (г); г=0.7.
Рис. 17. Зависимость калорийности моллюсков Unionidae от их веса.
1 — Unio tumidus', 2 — U. pictorum', 3 — Anodonta piscinalis. По оси абсцисс — сырой
вес моллюсков (г); по оси ординат — калорийность моллюсков (кал/мг сух. веса).
Для более точного определения калорийности были рассчитаны
уравнения отдельно для перловиц (25) и беззубок (26):
у = (0.926 ± 0.022) — (0.006 + 0.001) х, (25)
у = (1.329 + 0.169) —(0.015 ± 0.005) х. (26)
Калорийность сфериид слабо зависит от их размеров (табл. 7).
Она, как видно из таблицы, незакономерно колеблется от 1.29 до
0.66 кал/мг сухого веса животного.
Как и следовало ожидать, по мере роста животных доля рако-
вины в их энергетическом эквиваленте нарастает (рис. 18). При
этом, как видно из уравнений (27, 28), у перловиц она выше, чем
у беззубок:
для перловиц
у = (19.18 ± 2.53)+ (0.253 ± 0.028) .г, (27)
4*
51
Таблица 7
Калорийность моллюсков Sphaerium suecicum
(определено вместе с раковиной)
Вес сухой (мг) Кал/мг сухого веса Вес сухой (мг) Кал/мг сухого веса
2.5 1.02 23.0 0.81
2.5 1.09 23.0 1.02
3.5 1.19 26.5 1.05
4.5 0.82 27.0 1.00
4.5 1.29 29.0 0.73
4.5 1.14 29.0 0.67
4.5 0.90 36.0 0.83
5.5 1.05 37.0 0.76
5.5 1.00 40.0 0.75
5.5 0.87 42.0 0.78
7.0 0.83 47.5 0.77
9.5 0.88 52.5 0.96
11.0 0.96 62.5 0.74
17.5 0.90 191.1 0.66
19.0 1.01 — —
19.5 0.92 — —
для беззубок
у = (8.51 ± 2.38) 4- (0.205 ± 0.069) х, (28)
где у — доля энергетического эквивалента раковины (%); х —
сырой вес моллюска (г).
Рис. 18. Зависимость между отношением калорийности раковины и всего
моллюска Unio К от веса животного (из: Алимов, Шадрин, 1977).
По оси абсцисс — сырой вес моллюсков (г); по оси ординат — отношение Кр/К (%) (объяс-
нения в тексте).
Калорийность органического вещества перловиц и беззубок
практически одинакова и не меняется с возрастом (табл. 6). Ве-
52
личина ее находится в пределах 4.2—5.6, в среднем 5.0 кал/мг
органического вещества. Калорийность тела также изменяется
незакономерно — от 3.7 до 5.1 кал/мг сухого веса тела.
Аналогичные значения калорийности тела получены для Unio-
nidae — 3.22—4.78, Dreissenidae — 3.96—5.1, Sphaeriidae — около
3.77 кал/мг сухого веса тела (Biprep, 1961; Биргер, Маляревская,
1965; Tudorancea, Florescu, 1968а, 1968b; Шерстюк, 1971; Stanczy-
kowska, Lawacz, 1976, и др.) и ряда морских моллюсков (Geng,
1925; Ивлев, 1939; Biprep, 1961; Kuenzler, 1961; Hughes, 1970).
Это дает основание считать, что калорийность тела двустворчатых
моллюсков не меняется с возрастом и находится в пределах 3.22—
5.1 кал/мг сухого веса. Калорийность раковины весьма низка и,
например, для Unto не превышает 1.5, Anodonta —0.29, Sphaerium —
0.41, в среднем 0.74 кал/мг органического вещества.
Описанные зависимости калорийности моллюсков от их веса
в виде прямолинейной функции следует рассматривать просто как
наиболее удобную аппроксимацию экспериментальных данных,
а функциональную связь этих двух величин — как эмпирическую,
которая вряд ли имеет какие-либо теоретические обоснования.
Аппроксимация эмпирических материалов прямой дднией в дан-
ном случае имеет смысл только в строго определенных пределах
весов. Для изученных нами моллюсков уравнения (24—26), отра-
жающие зависимость калорийности от их веса, могут быть исполь-
зованы лишь в пределах весов от 0.8 до 80 г.
Приведенного вида зависимости между двумя рассматриваемыми
величинами весьма огрубленно представляют изучаемую связь.
Действительно, известно, что калорийность животных зависит
от времени наступления половозрелости, физиологического со-
стояния, условий питания, температуры, времени года и т. п.
(Biprep, 1961; Остапеня и др., 1969; Шерстюк, Зимбалевская,
1973; Tyler, 1973, и др.). Однако, несмотря на это, для получения
средних значений калорийности животных эти уравнения вполне
могут быть использованы. При более точных исследованиях необ-
ходимо учитывать возможные отклонения от средних значений,
обусловленные влиянием различных факторов. Однако для многих
экологических работ в ряде случаев оказывается достаточным ис-
пользование средних величин калорийности животных разного
возраста, рассчитанных в соответствии с приведенными выше
уравнениями.
Количество кислорода, потребляемое животными определен-
ного веса за единицу времени, рассчитанное по приведенным ранее
уравнениям связи обмена и веса животных с помощью оксикало-
рийных коэффициентов (3.4 кал/мгО или 4.86 кал/млО), может быть
выражено в единицах энергии. В этом случае получаем скорость
рассеяния энергии в процессе обмена, а траты на обмен выражаются
в тех же единицах, что и масса животных. Их можно представить
в единицах энергии, используя сведения по калорийности живот-
ных. Например, моллюски рода Unio весом в 1 г в соответствии
53
с уравнением (9) потребляют за сутки 1.78 мгО или рассеивают
эквивалентно 6.04 кал энергии. Калорийность 1 г сырого веса
перловиц близка к 475 кал. Нетрудно рассчитать, что траты на
обмен у моллюска весом в 1 г составят за сутки около 1.3% веса.
Суточные траты на обмен у моллюсков весом 10 г не превышают
0.6, весом 50 г — 0.31%.
Рассчитанные аналогичным образом величины трат на обмен
за сутки у Sphaeriidae весом 1 г (330 кал) составляют 3.2%, весом
0.01 г — 3.59, весом 0.1 г — 2.82% от веса животных. Для срав-
нения, например, суточные энергетические траты на обмен у С1а-
docera весом 1 мг (0.5 кал), согласно уравнению Л. М. Сущени (1972),
оказываются близкими к 22.8, а весом 10 мг — к 12.8% веса рачков.
Таким образом, у моллюсков, как и других животных, в про-
цессе их развития относительные величины трат энергии на об-
менные процессы уменьшаются.
УЧАСТИЕ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
В ДЕСТРУКЦИИ ОРГАНИЧЕСКИХ ВЕЩЕСТВ В ВОДОЕМАХ
В процессе дыхания моллюски рассеивают в окружающее про-
странство энергию, количество которой эквивалентно потреблен-
ному ими кислороду или деструкции определенного количества
органических веществ. Таким путем, зная скорость или интенсив-
ность обмена отдельных особей и их численность, можно оценить
минерализационную работу моллюсков в сообществах водных жи-
вотных или экологических системах водоемов.
Рассмотрим на некоторых примерах роль двустворчатых мол-
люсков в деструкции органических веществ в водоемах. В оз.
Круглом численность моллюсков Sphaerium suecicum в июне
1968 г. составляла 255 экз./м2, средний вес одной особи в популя-
ции не превышал 0.067 г. В соответствии с уравнением (14) особь
такого веса при температуре 20° потребляет в среднем 0.276 мгО/сут
(0.937 кал/сут), а вся популяция на площади дна в 1 м2 зоны оби-
тания моллюсков в озере — 238.7 кал/сут или 7.17 ккал за месяц.
Учитывая, что средняя температура в озере в июне не превышала 16°,
энергия, рассеиваемая популяцией моллюсков, близка к 5.12 ккал
за месяц. Проведенные аналогичным образом расчеты показали,
что траты энергии популяцией моллюсков на обменные процессы
составляли в августе и сентябре соответственно 2.07 и 0.84 ккал
за месяц. В течение вегетационного сезона (июнь—сентябрь)
особи этой популяции рассеивали на процессы обмена 14.97 ккал
энергии, что эквивалентно деструкции 2.67 г беззольного органи-
ческого вещества. Однако в этом озере моллюски населяют в ос-
новном прибрежную мелководную часть, площадь которой не пре-
вышает 19% всей площади озера. В таком случае в пересчете на
площадь всего озера они рассеивали в течение вегетационного
сезона около 2.9 ккал/м2, т. е. 15.4% от трат на обменные процессы
54
за тот же период всеми донными животными (19.47 ккал/м2) в оз.
Круглом.
В соседнем оз. Кривом биомасса двустворчатых моллюсков
была в 3.7 раза ниже, чем в оз. Круглом, и их траты на обменные
процессы за вегетационный сезон не превышали 3.5% от общих
трат всеми животными бентоса в озере в течение того же отрезка
времени.
В отдельных биоценозах донных животных роль двустворчатых
моллюсков в деструкции органических веществ может быть зна-
чительно более высокой. Так, например, в биоценозе Sphaerium
suectcum—Ephemera vulgata в оз. Круглом биомасса моллюсков
составляла около 79% общей биомассы животных биоценоза, и на
их долю приходилось около 65 % энергии, рассеиваемой всеми жи-
вотными этого сообщества в течение вегетационного сезона
(Алимов, Финогенова, 1975а).
Первичная продукция в оз. Круглом за вегетационный сезон
1968 г. не превышала 68 ккал/м2. Напомним, что двустворчатые
моллюски за это время рассеивали 2.9 ккал/м2, т. е. способны были
окислить около 4.5% органических веществ первичной продукции.
В оз. Зеленецком за тот же период двустворчатые моллюски окис-
ляли 5.9% первичной продукции.
Исследования минерализационной способности двустворчатых
моллюсков Волгоградского водохранилища (Кондратьев, 1970в)
показали, что в разных его зонах эти животные тратили на обменные
процессы от 0.323 до 0.712 ккал/м2 в сутки. С учетом площадей
отдельных зон это составляет около 100.3 ккал за вегетационный
сезон или около 9.7% от величины первичной продукции в этом
водоеме. Энергетические траты моллюсков на обменные процессы
в Северном Каспии достигают 5.1% от первичной продукции водо-
ема (Иванова Л. М., 1973а, 19736). Более низкие значения отно-
шения метаболизированной моллюсками энергии к первичной
продукции (около 1%) отмечены для оз. Дривяты (рассчитано по
данным Винберга, 1970). Однако в этом озере было учтено дыхание
лишь крупных моллюсков (униониды), которым свойственна низкая
интенсивность обмена.
Приведенные расчеты показывают, что траты энергии дву-
створчатыми моллюсками на обменные процессы в различных водо-
емах не превышают 1—10, в среднем около 5% энергии, заключен-
ной в первичной продукции в этих водоемах. В то же время траты
на обмен всеми личинками водных насекомых за вегетационный
сезон в оз. Кривом не превышали 4.4%, в оз. Круглом 15.9, в оз.
Зеленецком 12.3% от энергии первичной продукции в этих водоемах
за тот же период (Алимов, Финогенова, 1975а, 19756).
Значение двустворчатых моллюсков в деструкции органических
веществ по сравнению с личинками насекомых в оз. Круглом оказы-
вается только в 3.5, а в оз. Зеленецком в 2.1 раза ниже. Это позво-
ляет заключить, что по крайней мере в малопродуктивных озерах,
в состав фауны которых входят мелкие виды двустворчатых мол-
55
люсков с высокой интенсивностью обмена, их роль в процессах
деструкции оказывается сопоставимой с участием в этих процессах
других наиболее массовых представителей донных животных. Для
сравнения можно сказать, что траты на обмен животными зоопланк-
тона за вегетационный сезон в эвтрофном оз. Дривяты достигали
16%, в олиготрофных озерах Кривое и Зеленецкое соответственно
36 и 38, в дистрофном оз. Круглом 19% от энергии первичной
продукции в этих водоемах.
Таким образом, скорость, интенсивность обмена и калорийность
двустворчатых моллюсков, как и других животных, находятся
в строгой зависимости от их веса и изменения факторов внешней
среды. Среди последних наиболее сильное влияние оказывают
термические условия и физиологическое состояние животных.
Использование сведений и полученных на их основе закономер-
ностей по интенсивности обмена животных дает возможность коли-
чественно оценить роль и значение их популяций в процессах
деструкции органических веществ в экологических системах. Уча-
стие пресноводных двустворчатых моллюсков в процессах деструк-
ции в водоемах разных типов вполне сравнимо с ролью, которую
играют в этих процессах другие донные животные.
Глава II
ИНТЕНСИВНОСТЬ ПИТАНИЯ моллюсков
Двустворчатые моллюски получают пищевой материал и необхо-
димый для дыхания кислород из воды, которая засасывается ими
внутрь раковины вододвижущим аппаратом. Вододвижущий ап-
парат моллюсков состоит из вводного и выводного (клоакального)
сифонов, мантийной полости, жабр, околоротовых лопастей и от-
носится к типу засасывающего мерцательно-гидрокинетического
(Беклемишев, 1964).
Основная сосущая сила вододвижущего аппарата моллюсков
создается в результате работы мерцательного эпителия, который
выстилает жабры и околоротовые лопасти. У некоторых сфериид
в этот механизм включается также и эпителий ноги (Митрополь-
ский, 1966). Движением ресничек эпителия на огромной поверх-
ности аппарата создается значительная сосущая сила, под дей-
ствием которой вода засасывается внутрь мантийной полости.
Проходя через систему отверстий жаберных полостей, стенки ко-
торых выстланы мерцательным эпителием, она постепенно осво-
бождается от взвешенных частиц. Затем через наджаберный канал
вода выносится в мантийную полость и оттуда наружу.
Взвешенные в воде частички гонятся мерцательными ресничками
внутренних полужабр к их нижнему краю, где имеется желобок,
образованный филаментами восходящей и нисходящей пластинок.
По этому желобку частички движением мерцательных щетинок
направляются к околоротовым лопастям. По наружным полужаб-
рам частички движутся вверх. Реснички внутренней поверхности
перемещают их на внутренние полужабры. Частички, движу-
щиеся по наружной поверхности жабр, достигая их верхнего края,
направляются по мантии непосредственно к околоротовым лопастям.
По мере продвижения по жабрам частички склеиваются в жгутики
слизью, которая секретируется специальными железами эпителия.
В конечном счете все частицы, собираемые жабрами, попадают или
на передний край внутренней жабры, или на участок мантии над
околоротовыми лопастями. Отсюда склеенные слизью комочки
57
частиц попадают или на околоротовые лопасти, или, если они
достигают значительных размеров, под действием силы собствен-
ной тяжести падают в нижнюю часть мантийной полости. Частички,
которые попадают на околоротовые лопасти, по желобку лопастей,
выстланному мерцательным эпителием, направляются в ротовое
отверстие, а частички, упавшие в нижнюю часть мантийной по-
лости, выносятся наружу потоком воды через мантийную щель.
Околоротовые лопасти, помимо транспортной, выполняют также
функцию сортирующего аппарата. Их роль как сортирующего ме-
ханизма была изучена Алленом (Allen, 1921). Он показал, что
околоротовые лопасти могут отсортировывать пищевые частички
от минеральных (в качестве последних он использовал в экспери-
ментах частички карборунда). Сортировка частичек лопастями про-
изводится пассивно. Реснички нижней их половины гонят материал
вниз и назад, а верхней — вверх и вперед. Пищевые частички, как
более легкие, направляются по верхнему краю лопастей вверх и
вперед — к ротовому отверстию. Минеральные, как более тя-
желые и грубые, под силой собственной тяжести падают с околоро-
товых лопастей вниз в мантийную полость. Экспериментами Аллена
было установлено, что на околоротовых лопастях возможно разде-
ление, например, частичек карборунда и кармина. Первые падали
в мантийную полость, вторые направлялись к ротовому отверстию
моллюсков. Моллюски, как показало вскрытие их органов пище-
варения, почти не заглатывают неорганических частичек, не имею-
щих пищевой ценности. В пищеварительной системе моллюсков
Sphaerium corneum (L.) нам не удалось обнаружить минеральных
частиц. Их желудки и кишечники были наполнены планктонными
водорослями, частицами детрита, переваренными в разной степени.
Содержимое кишечников у 10 экз. этих моллюсков состояло из про-
стейших (главным образом инфузорий — около 75%), диатомовых
водорослей (6%), протококковых водорослей (около 10%), обрыв-
ков нитчаток (около 8%), а также большого количества
бактерий.
В литературе часто встречается упоминание о способности
моллюсков не только к механической сортировке частиц, но и к се-
лективному их отбору. Для доказательства этого положения при-
водились факты различия состава фитопланктона в морской воде
и кишечном тракте моллюсков (Grave, 1916). В кишечниках Mytilus
californianus Conrad динофлагелляты составляли 97.4%, диатомо-
вые — только 2.6%, в то время как в морском планктоне, наоборот,,
количество первых не превышало 2.4, вторых — 97.6% (Buley,
1936). В экспериментальных условиях устрицы оказывались спо-
собными отбирать бактерии из культуры водорослей (Loosanoff,
1949). Однако, учитывая особенности морфологии двустворчатых
моллюсков и строение сортирующих механизмов их вододвижущего
аппарата, такого рода селекцию скорее всего следует рассматри-
вать не более как механический отбор моллюсками частиц по их
размерам. Тем более что ни в одной из упомянутых работ не при-
58
водятся достаточно подробные сведения о количестве и размере
частиц, находившихся во взвешенном состоянии в воде.
Минеральные и органические частицы, падающие в нижнюю
часть мантийной полости моллюсков в виде агглютинированных
слизью комочков (псевдофекалий), выталкиваются животными
наружу. Псевдофекалии образуются в значительных количествах
особенно при высоких концентрациях взвешенных в воде частиц.
Вододвижущий аппарат Lamellibranchia представляет наиболее
совершенный тип фильтрационного аппарата среди беспозвоночных
животных и позволяет практически полностью очистить воду от
взвешенных в ней частиц. Фильтрационный аппарат двуствор-
чатых моллюсков обладает высокой улавливающей способностью.
Эти животные способны удалять 92—100% всех частиц, взвешен-
ных в фильтруемом ими объеме воды (Кондратьев, 1970в), и за-
держивать единичные клетки крупных бактерий (Михеев, Сорокин,
1966). Дрейссены могут улавливать частички размером 1—3 мкм
(Михеев, 1967а), униониды — 2—3 мкм (Кондратьев, 1970в),
Mytilus edulis — менее 2 мкм (Vahl, 1972а), Chlamys opercula-
ris (L.) — до 7—10 мкм (Vahl, 1972b), Brachidontes exustus (Wieg-
mann) — 1—5 мкм (Paul, 1971) и т. д. Опытами А. Г. Родиной (1948)
было установлено, что моллюски Sphaerium, Musculium, Pisidium
нормально растут на чисто бактериальной пище. При этом размер
клеток в культуре бактерий, использованной для питания моллю-
сков, составлял около 1 мкм.
В качестве количественной характеристики фильтрационных
способностей двустворчатых моллюсков обычно используют ско-
рость фильтрации воды животными. Скорость фильтрации выра-
жают как объем воды, который моллюски способны пропустить
через свой вододвижущий аппарат за определенный отрезок вре-
мени. В иностранной литературе для этих же целей используют
термин «скорость прокачивания» («pumping rate»), что в принципе
эквивалентно скорости фильтрации. В ряде случаев, особенно
при изучении фильтрационной способности планктонных ракооб-
разных, скоростью фильтрации называют обловленный, или ос-
ветленный, животными объем воды за единицу времени. По отно-
шению к моллюскам обычно говорят о скорости фильтрации, или
прокачивания воды. Можно сказать, что скорость фильтрации —
условный осветляемый объем, но применительно к двустворчатым
моллюскам, обладающим высокой степенью улавливания частиц,
эта условность практически исчезает.
МЕТОДЫ ОПРЕДЕЛЕНИЯ СКОРОСТИ ФИЛЬТРАЦИИ
Существующие методы изучения скорости фильтрации воды мол-
люсками сводятся к двум основным типам — так называемым пря-
мым и непрямым.
Прямые методы основаны на раздельном измерении
потоков воды, проходящих через вводной или выводной сифоны
59
моллюсков. Обычно измеряют объем воды, вытекающий из клоа-
кального сифона, для чего в сифон вводится специальная трубка.
Вода, вытекающая из трубки, собирается и измеряется ее объем
(Allen, 1914; Galtsoff, 1928; Tammes, Dral, 1955, и др.). При этом
принимается, что при неизменных условиях размеры отверстий
сифонов и скорость прохождения воды через них остаются
постоянными. Такие допущения позволили определять объем
протекающей воды с помощью специально изготовленных моде-
лей сифонов (Кондратьев, 1963а, 19636, 1967). Модель копирует
форму и размеры клоакального сифона моллюсков определен-
ной величины, и через нее пропускается струя воды с такой же
скоростью, какую имеет струя воды, вытекающая из сифона
моллюска.
Основной недостаток прямых методов заключается в необхо-
димости введения в организм моллюсков чужеродных предметов
или препарирования животных, что может оказать существенное
влияние на точность и качество получаемых результатов. Исполь-
зование же моделей полностью исключает животное как таковое
с его сложными регуляторными механизмами. В этом случае все
сводится по сути дела к изучению чисто гидродинамических харак-
теристик вододвижущего аппарата моллюсков. Применение прямых
методов становится затруднительным или даже практически не-
возможным при изучении фильтрационных способностей мелких
по размерам животных, таких как сферииды. Прямые методы по-
зволяют исследовать скорость прокачивания воды моллюсками.
Оценка фильтрационных способностей этих животных прямыми
методами возможна лишь при допущении, что они полностью улав-
ливают все частицы, находящиеся в потоке воды. Такое допуще-
ние, как было сказано ранее, в отношении пресноводных моллю-
сков может быть вполне оправданным.
Прямые методы не находят широкого применения и использу-
ются обычно со специальными целями.
Наибольшее применение для оценки фильтрационной способ-
ности моллюсков нашли непрямые методы. В этом случае
объем воды, пропускаемый моллюском, определяется расчетным
способом как функция изменения концентрации взвеси за опреде-
ленное время в результате фильтрации воды животным. В ка-
честве взвесей в экспериментах могут быть использованы сус-
пензии минеральных веществ, например глинистого грунта, гра-
фита, некоторых красителей и т. п. В большинстве случаев при-
меняют взвеси из планктонных водорослей, бактерий, дрожжей,
детрита разной степени разложения, органических красителей
и т. д. Расчеты скорости фильтрации воды моллюсками основы-
ваются на результатах измерения концентрации взвешенных в воде
частиц в начале и конце эксперимента. При этом допускается, что
моллюски из прокачиваемого ими объема воды полностью удаляют
взвешенные частицы, которые равномерно распределены во всей
толще воды экспериментального сосуда, и процесс фильтрации
60
воды животными между двумя измерениями концентрации взвеси
протекает непрерывно и с постоянной скоростью.
Концентрация частиц в воде определяется их прямым подсче-
том в определенном объеме, измерением мутности воды с помощью
нефелометров, использованием клеток, меченных радиоактивными
препаратами.
Скорость фильтрации воды рассчитывается как объем воды,
прокачиваемый животными за исследуемый отрезок времени.
Поскольку допускается, что за время эксперимента процесс фильт-
рации воды животными протекает с неизменной скоростью, то по
мере удаления частичек концентрация их в воде уменьшается,
и она становится функцией удаления частичек, т. е.
(29)
где с — концентрация взвеси; V — объем воды; t — время; F —
скорость фильтрации воды одной особью; п1 — число животных
в опыте; а — скорость небиологического осаждения взвешенных
частичек. Решая уравнение (29), получаем
или
откуда
(30)
гдес0 — концентрация взвеси в начале, a ct — в конце э ксперимента.
Скорость осаждения взвешенных частиц а может быть определена
с учетом изменения концентрации их в контрольных сосудах без
животных за тот же отрезок времени:
In сп — In
л -----У_____L
Разными исследователями (Fuller, 1937; Jorgensen, 1943;
Gauld, 1951; Theede, 1963) были предложены уравнения расчета
скорости фильтрации воды моллюсками. Однако анализ показал,
что они все могут быть сведены к приведенному выше уравнению
(30) (Coughlan, 1969), которое известно как уравнение Голда.
В тех случаях, когда уменьшение концентрации взвеси в течение
эксперимента не превышает 10—20% от начальной ее величины,
результаты, полученные с использованием разных уравнений,
не отличаются от рассчитанных по уравнению (30).
61
Применение непрямых методов для определения скорости
фильтрации воды моллюсками исключает какие-либо серьезные
нарушения функционирования вододвижущего аппарата или пове-
дения подопытных животных. Этими методами могут быть полу-
чены экологически значимые характеристики: объем воды, прока-
чиваемый вододвижущим аппаратом моллюсков, количество от-
фильтрованных и осажденных ими частиц, взвешенных в воде,
а также зависимость этих характеристик от размеров животных,
их экологических особенностей и изменений факторов внешней
среды.
Непрямой метод был использован автором при изучении филь-
трационных способностей сфериид (Алимов, 1965, 1967а, 1969;
Алимов, Бульон, 1972). Опыты проводились с минеральными или
органическими взвесями. В первом случае взвесь приготовлялась
из мелкорастертого глинистого грунта. Приготовленная взвесь
до начала опытов отстаивалась в течение суток. За это время наи-
более тяжелые фракции оседали на дно сосуда, что уменьшало
возможность небиологического осаждения частиц в эксперименте.
В качестве органических взвесей использовались культура хло-
реллы, пекарские дрожжи или природный грунт, состоящий глав-
ным образом из частиц детрита. Изменение количества взвешенных
частиц в опытных сосудах определялось по изменению оптической
плотности воды. Для расчета концентрации взвеси предварительно
были получены градуировочные графики, показывающие связь
концентрации данной взвеси и оптической плотности воды. В неко-
торых случаях концентрация взвеси определялась по величине
бихроматной окисляемости взвешенных в воде органических ве-
ществ (Алимов, Бульон, 1972). Объемы экспериментальных со-
судов и длительность опытов рассчитывались таким образом,
чтобы уменьшение концентрации взвешенных в воде частичек
в конце эксперимента не превышало 20—30% от начальной их кон-
центрации. Обычно опыты продолжались в течение 3—4 ч. Тем-
пература поддерживалась постоянной и близкой к 20° или соот-
ветствовала температуре в водоеме, из которого были взяты для
опытов моллюски. До начала эксперимента животные в течение
суток акклимировались к этим температурам.
При изучении интенсивности питания животных-фильтраторов
следует различать скорость фильтрации воды животными (облов-
ленный, или осветленный объем) и скорость питания, т. е. коли-
чество пищи, потребленной ими за единицу времени (рацион).
Рационом принято считать количество пищи, которое поступает
за единицу времени в пищеварительный тракт животного. Пища
претерпевает там определенные изменения, в результате которых
часть пищевого материала усваивается и расходуется на процессы
обмена и роста, а часть в виде неусвоенных продуктов выводится
наружу. У фильтраторов, а у двустворчатых моллюсков в особен-
ности, через организм проходит огромное количество пищевых
частиц. У моллюсков только часть из них попадает в пищеваритель-
62
ный тракт, остальные же выводятся из организма в виде псевдо-
фекальных масс.
Если количество пищевого материала, удаленного из фильтру-
емого моллюсками объема воды, рассматривать как их рацион,
то необходимо различать физиологический (С) и «экологический»
(С1) рацион (Алимов, 1969). Первый представляет собой количество
пищи, поступившее в его пищеварительную систему, второй — все
количество пищи, удаленное из отфильтрованного им объема воды.
Заметим, что понятие экологического рациона и соответственно
экологической усвояемости было предложено В. С. Теном (1967}
при рассмотрении трофических взаимодействий пары хищник—
жертва у водных организмов.
Соотношение между физиологическим и «экологическим» ра-
ционами представляется следующим:
C1 = qV, (31)
где V — объем воды, профильтрованной моллюском; q — вес или
эквивалентная ему величина веществ, находящихся во взвешен-
ном состоянии в этом объеме воды.
Физиологический рацион можно выразить как
С = Л + /. (32).
Тогда «экологический» рацион представляет собой
^ = 67 + /, (33>
или, согласно уравнениям (32) и (33),
С1 = Л+/ + /р. (34)
В равенствах (32—34) А — количество пищи, ассимилированной
моллюском; / и /р — соответственно количество фекальных и псев-
дофекальных масс.
По отношению к двустворчатым моллюскам введение двух оп-
ределений рациона необходимо с экологических позиций в силу
их неодинакового значения в процессах превращения веществ и
трансформации энергии в водоемах. При определенных условиях:
количество псевдофекальных масс, выводимых моллюсками, мо-
жет в несколько раз превышать количество органических веществ,
выделяемых ими в виде фекалий. И те и другие отлагаются на дно
водоема, входят в состав иловых отложений и служат источником
пищи для различных видов донных животных. Естественно,
благодаря различной степени переработки пищевое значение
фекальных и псевдофекальных масс неодинаково. Для минерали-
зации их также требуется разное количество кислорода.
63>
ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ НА СКОРОСТЬ
ФИЛЬТРАЦИИ ВОДЫ МОЛЛЮСКАМИ
Температура
Температура воды оказывает существенное влияние на работу
вододвижущего аппарата двустворчатых моллюсков. Скорость
фильтрации воды моллюсками Unio tumidus, U. pictorum, Dreissena
polymorpha, Sphaeriastrum rivicola Lamark, широко распростра-
ненными в водоемах умеренной зоны, нарастает с увеличением
температуры до ее значений близких к 30° (рис. 19). Аналогичным
образом, как видно из приведенного рисунка, изменяется скорость
фильтрации при увеличении температуры и у субтропического
моллюска Hyriopsis schlegelii. Опытами Г. П. Кондратьева (1970а,
1970в) было показано, что при температурах близких к 30° процесс
фильтрации воды у унионид становится неравномерным: полная
нагрузка сменяется полным прекращением фильтрации воды. Это
связывается с нарушениями нормальной активности ресничек мер-
цательного эпителия (Lagerspetz, Terri, 1968). Нарушения фильтра-
ционной способности при температурах около 30° более резко про-
являются у молодых животных (Hopkins, 1935; Rao, 1953; Конд-
ратьев, 1963а; Morton, 1971).
Повышение температуры до 33—34° вызывало прекращение
фильтрации воды моллюсками в водоемах умеренной и субтро-
пической зон (Hagashi, 1966; Михеев, 1967а, 19676; Кондратьев,
1970а, 1970в). В ряде случаев при таких температурах отмечалась
даже гибель дрейссен (Михеев, 19676; Morton, 1971). Прекращение
процесса фильтрации при этих же температурах, связанное с нару-
шением активности ресничек эпителия, наблюдалось у большинства
бореальных и субтропических видов морских двустворчатых мол-
люсков (Galtsoff, 1928; Gray, 1928; Hopkins, 1935; Jorgensen, 1955;
Loosanoff, 1958; Nagabhushanam, Sarojini, 1965a, 1965b; Hughes,
1969; Arakawa, 1970).
Данные, полученные на пресноводных двустворчатых моллю-
сках, позволяют предполагать, что прекращение ими фильтрации
воды при температурах выше 30° (по крайней мере в отношении
видов умеренной и субтропической зон) обусловлено тем, что верх-
ний температурный порог физиологической активности мерцатель-
ного эпителия у этих животных находится при температурах близ-
ких к 33°.
Температура влияет не только на активность мерцательного
эпителия, но определяет степень раскрытости створок раковин
у моллюсков. Верхний температурный порог для мускулов аддук-
тора оказывается ниже, чем для ресничек эпителия (Hopkins,
1933, 1935). По наблюдениям этого автора при температурах
выше 25° отмечалась неравномерность сокращения мускулов аддук-
тора. В связи с этим створки раковины то полностью замыкались,
то полностью раскрывались. Вероятно, благодаря различиям
64
верхних температурных порогов для ресничек эпителия и мускулов
аддуктора из-за более низкого температурного порога последнего
при температурах выше 25° возникает как бы своеобразный тормоз
фильтрационной способности моллюсков. Поэтому нарушения
процесса фильтрации у них уже наступают при температурах, ко-
торые ниже физиологического температурного порога активности
мерцательного эпителия.
30 - А
Рис. 19. Скорость фильтрации у моллюсков при разных температурах воды.
А: 1 — Unio pictorum (вес 5 г); 2 — U. tumidus (вес 4 г) (по данным Кондратьева, 1963а).
Б: 1 — Dreissena polymorpha; 2 — Sphaeriastrum rivicola (вес. 0.8г) (по данным Кондратьева,
1969а). В — Hyriopsis schlegelii (по данным Hagashi, 1966). По оси абсцисс — температура
(°C); по оси ординат — скорость фильтрации (л/сут).
Рассмотренные данные позволяют сделать предположение, что
верхний температурный порог для фильтрационной активности
двустворчатых моллюсков умеренной и субтропической зон на-
ходится при температурах близких к 30°. В этой связи ста-
новится понятным резкое снижение скорости фильтрации воды
унионидами при температурах выше 30° (рис. 19).
Снижение температуры приводит к замедлению фильтрации
воды моллюсками, и при температурах 1—3° большинство из на-
селяющих водоемы умеренных широт видов полностью прекращают
фильтровать. Очень слабая фильтрационная активность отмеча-
лась у унионид при 4°, у дрейссен при 4.2 и у сфериид при 4.6—
5.9° (Кондратьев, 1960в). Вероятно, эти температуры (4—5.9°)
можно рассматривать как нижний температурный порог для
фильтрационной активности изученных моллюсков. Следует за-
метить, что при этих температурах кишечники у Musculium trans-
versum Say. оставались пустыми (Gale, Lowe, 1971).
5 а. Ф, Алимов 65
Оптимальные температуры находятся ниже верхнего темпера-
турного порога и могут отличаться у разных по происхождению
и местообитанию животных. В этом отношении показательно,
что, например, оптимум по фильтрации у Ostrea gigas Thunberg,
как и у Crassostrea virginica Gmelin, находится при температурах
27—28° (Hopkins, 1935), у О. eduliswpn 15—28 (Newell et al., 1977),
у Scrobicularia plana около 24° (Hughes, 1969), a у Hiatella arctica
Lam. при 15—17° (Ali, 1970). Для
пресноводных моллюсков таких
сведений пока не имеется. Можно
лишь предполагать, что оптималь-
ные температуры для фильтраци-
онной активности Margaritifera
margaritifera будут ниже, чем
Рис. 20. Зависимость относительной
скорости фильтрации (% от значе-
ний при 20°) от температуры у раз-
личных пресноводных моллюсков (по
данным рис. 19).
1 — Unio pictorum, 2 — U. tumidus", 3 —
Sphaeriastrum rivicola; 4 — Dreissena po-
lymorpha. По оси абсцисс — температура
(°C); по оси ординат — относительная
скорость фильтрации (%).
Рис. 21. Значения <210 для скорости
фильтрации воды моллюсками при
разных температурах.
Прерывистая линия соответствует «нор-
мальной» кривой. По оси абсцисс — тем-
пература (°C); по оси ординат — значе-
ния Qio-
у Unio tumidus, равно как и у обитателей профундали глубоких
озер Pisidium conventus Cl ess. по сравнению, например, с моллю-
сками Sphaerium согпеит, населяющими в большинстве случаев
прибрежную зону мелководных, хорошо прогреваемых водоемов.
Сходный характер воздействия температуры на фильтрацию
воды моллюсками Unio pictorum, U- tumidus, Dreissena polymorpha*
Sphaeriastrum rivicola (рис. 19) позволил вывести общую для них
зависимость скорости фильтрации от температуры, для установле-
ния которой был использован тот же прием, что и при изучении
аналогичной зависимости скорости обмена у моллюсков. В соот-
ветствии с данными, приведенными на рис. 19, скорость фильтрации
воды изученными видами моллюсков при температуре 20° была
66
принята равной единице, ее значения при других температурах рас-
считывались в процентах от скорости фильтрации при 20° (рис. 20).
По преобразованным таким образом данным определялись значе-
ния ()10 для скорости фильтрации
в разных интервалах исследованных
температур.
Сравнение <210, полученных для
процесса фильтрации и по кривой
Крога, показало почти полное сов-
падение их во всем диапазоне рас-
смотренных температур (рис. 21).
Поэтому при изучении скорости филь-
трации у широко распространенных
моллюсков водоемов умеренной зоны
в случае необходимости могут быть
внесены температурные поправ-
ки в соответствии с «нормальной»
кривой Крога. С этой целью можно
с успехом использовать таблицу тем-
пературных поправок, рассчитанную
в свое время Г. Г. Винбергом (1956)
и перепечатанную В. С. Ивлевым
(1962).
Имеющиеся очень немногочислен-
ные данные по скорости выделения
моллюсками фекальных и псевдофе-
кальных масс при разных температу-
рах позволяют сделать предвари-
тельное заключение о том, что ско-
рость этих процессов при разных
температурах также изменяется в со-
ответствии с «нормальной» кривой
(рис. 22). Эти данные лишь под-
тверждают заключение о характере
изменения скорости фильтрации под
Рис. 22. Относительная ско-
рость (% от значений при 20°)
выделения фекальных и псев-
дофекальных масс моллю-
сками при разных температу-
рах.
влиянием температуры.
Сравнение зависимостей скорости
обмена и скорости фильтрации у мол-
люсков от температуры показывает
существенные различия ее влияния
на эти процессы. Наиболее явно эти
1 — Hyriopsis schlegelii (по данным
Hagashi, 1966); 2 — Dreissena po-
lymorpha (по данным Михеева,
1967а); 3 — соответствует «нор-
мальной» кривой. По оси абсцисс —
температура (°C); по оси ординат —
относительная скорость выделения
фекальных и псевдофекальных масс
моллюсками.
различия проявляются при темпера-
турах, превышающих 20° (табл. 8). Наибольшая скорость обмена
у изученных пресноводных моллюсков оказывается при темпера-
турах около 20°, а фильтрации — около 30°.
Различие в скоростях этих двух протекающих одновременно
процессов при разных температурах, вероятно, может быть объяс-
нено следующим. Известно, что потребление кислорода двуствор-
5*
67
Таблица 8
Средние значения Qlo для скоростей фильтрации и обмена
у двустворчатых моллюсков при различных интервалах
температур
Показатель 5—10° 10—15° 20—25° 25-30° 30-35°
Скорость фильтрации . . . 3.66 3.11 2.78 2.28 1.57
Скорость обмена 3.65 2.65 2.26 1.60 1.40
чатыми моллюсками не связано прямо с двигательной активностью
цилий жаберного аппарата (Gray, 1924). У пресноводных двуствор-
чатых моллюсков в связи с особенностями их размножения жабры
у ряда видов утрачивают значение органов дыхания, что наиболее
резко проявляется у живородящих форм, в жаберных карманах
которых вынашивается молодь. Дыхательная функция у них
выполняется отдельными участками мантии. Кроме того, имеются
данные о том, что окислительная способность митохондрий оказы-
вается наибольшей при температурах близких к 25°; дальнейшее
увеличение температуры приводит к уменьшению их окислитель-
ной способности (Newell, 1967). В то же время, как было показано
ранее, механическая, двигательная активность цилий оказывается
максимальной при температурах около 30°.
Таким образом, изменение скорости фильтрации воды широко
распространенными моллюсками умеренной зоны при увеличении
температуры в пределах температур от 5 до 30° в отличие от изме-
нений скорости обмена происходит в соответствии с температурной
кривой Крога.
Поскольку сведения по влиянию температуры на скорость филь-
трации у пресноводных моллюсков немногочисленны, то к сделан-
ным на их основе выводам следует относиться с определенной
осторожностью и рассматривать их как предварительные. Однако
несомненно, что при обычных в летний период температурах
в водоемах умеренных широт изменения температуры влияют
на скорость обмена и фильтрации у моллюсков, населяющих
их, одинаковым образом.
Концентрация взвешенных
в воде частиц
В опытах с моллюсками Sphaerium suecicum (Алимов, Бульон,
1972) было показано, что скорость фильтрации воды моллюсками
и концентрация взвешенных в воде частиц связаны между собой
обратной зависимостью почти во всем диапазоне использованных
в экспериментах концентраций. В области низких концентраций
близких к природным скорость фильтрации у моллюсков оказы-
68
вается максимальной. Аналогичная зависимость была получена
при изучении фильтрации у ракообразных (Сущеня, 1958, 1975;
Иванова, 1970).
Рассмотрим отдельно участки АВ и ВС кривой, отражающей
зависимость скорости фильтрации у моллюсков S. suecicum от
концентрации взвесей в воде (рис. 23). На участке А В увеличение
концентрации взвеси вызывает нарастание скорости фильтрации.
На участке ВС, наоборот, с увеличением концентрации происхо-
дит уменьшение скорости фильтрации.
Рис. 23. Зависимость скорости фильтрации воды моллюсками Sphaerium
suecicum от концентрации взвешенных частиц (из: Алимов, Бульон, 1972).
По оси абсцисс — концентрация взвеси (мгО/л); по оси ординат — скорость фильтрации
(мл/ч).
Увеличение скорости фильтрации при малых концентрациях
частиц взвесей связано с повышением эффективности улавливания
аппаратом животных этих частиц по мере нарастания их плотности
в воде до определенных пределов, близких по значениям к наблю-
даемым в природе (участок АВ). Поскольку расчет скорости
фильтрации производится по количеству удаленных из воды ча-
стиц, при низких концентрациях, когда моллюски улавливают
их с меньшей эффективностью, рассчитанные значения скорости
фильтрации могут быть значительно занижены. Скорее всего при
таких концентрациях скорость фильтрации воды моллюсками
остается постоянной и близкой к максимальной. Величина ее опре-
деляется наибольшей физиологической активностью вододвижу-
щего аппарата животных при определенной его морфологии.
На участке ВС нарастание концентрации взвеси представляет
собой фактор, тормозящий фильтрационную активность животных.
Многочисленными исследованиями с моллюсками и ракообразными
установлено снижение скорости фильтрации воды животными
с увеличением концентрации взвешенных в ней частиц (Касталь-
ская-Корзинкина, 1942; Loosanoff, Engle, 1947; Сущеня, 1958,
69
1964; Монаков, Сорокин, 1961; Богатова, 1967; Михеев, 1967а,
19676; Иванова, 1970; Walz, 1978а, и др.). При достаточно высоких
концентрациях, превышающих природные в несколько раз, затруд-
няется работа фильтрационного аппарата животных. У моллюсков
при таких концентрациях наблюдаются нарушения ритма работы
аддуктора, сокращение сифона л ьных отверстий, уменьшение про-
должительности фильтрации (Кондратьев, 1967). Кроме того,
при очень высоких концентрациях происходит механическое за-
бивание фильтрационного аппарата моллюсков. Уменьшение ско-
рости фильтрации обычно наблюдается уже при концентрациях,
которые превышают природные примерно вдвое (Allen, 1921;
Winter, 1969; Ali, 1970; Алимов, Бульон, 1972). Важно отметить,
что у моллюсков S. suecicum, как показали наши эксперименты,
наибольшие скорости фильтрации были при концентрациях взве-
шенных частиц около 2 мгО/л. Эта величина соответствует среднему
значению концентрации взвешенных органических веществ в лет-
ний период в воде оз. Круглого, из которого собирались животные
для опытов.
Другой фактор, который может угнетать фильтрационную спо-
собность моллюсков при высоких концентрациях взвешенных
веществ, — уменьшение количества кислорода, получаемого жи-
вотными по мере сокращения фильтруемого объема воды. Рас-
смотрим пример. Моллюску S. suecicum весом 0.038 г (средний вес
животных в опытах) на нужды энергетического обмена при темпе-
ратуре 15° требуется 0.038 мгО/сут. Моллюски, как было показано
ранее, при нормальном насыщении воды кислородом способны
утилизировать до 10% его количества. Вместе с тем при резком
дефиците кислорода они кратковременно могут значительно увели-
чивать поглощение его из воды (в ряде случаев до 80%). Из табл. 9
видно, что при минимальных (в изученных пределах концентраций)
скоростях фильтрации траты на обмен у моллюсков не превышали
25 % от имеющегося в фильтруемом животными объеме воды кисло-
Таблица 9
Количество кислорода, получаемое моллюском Sphaerium
suecicum (И7 = 0.038 г) из фильтруемого им объема воды
при разных концентрациях взвешенных в воде веществ
(температура 15°)
с (мгО/л) V (мл/сут) О (мг) к/о (о/о)
1.20 131.04 1.15 3.3
2.49 91.11 0.80 4.8
3.40 38.40 0.34 11.0
8.10 28.80 0.25 15.2
15.99 24.74 0.22 17.2
33.96 17.52 0.15 25.2
Примечание, с — концентрация взвеси в воде; V — объем профильтрованной
моллюском воды; О — содержание кислорода в этом объеме воды; R — потребление
кислорода моллюском (мгО/сут).
70
рода. Это дает основание предполагать, что даже при самых высоких
концентрациях взвесей, которые были в опытных сосудах, потреб-
ности животных в кислороде могли быть удовлетворены полностью.
Существенно, что минимальные скорости фильтрации в экспери-
ментах были получены при концентрациях взвешенных в воде
веществ, которые превышали природные в десятки раз. Таким об-
разом, в природных условиях при нормальном насыщении воды
кислородом, количество его в объеме воды, фильтруемом мол-
люсками, не может оказывать тормозящего влияния на их фильтра-
ционную активность.
Влияние концентрации взвешенных частиц на скорость филь-
трации изучалось у моллюсков, резко отличавшихся по своим
фильтрационным способностям. Поэтому для количественного
выражения единой для разных видов зависимости скорости филь-
трации от концентрации взвеси лучше использовать относительные
величины. Выразим концентрацию взвеси в виде отношения к ее
минимальному значению, использованному в эксперименте, —
c/cmln=.r, скорость фильтрации — в долях от максимальной
величины — Отметим, что обычно нижние величины
концентраций, используемые в опытах, близки или соответствуют
концентрациям, характерным для природных водоемов, где живут
моллюски.
Преобразованные таким образом данные, полученные разными
исследователями, приведены на рис. 24, из которого видно, что
у всех изученных животных скорость фильтрации начинает умень-
шаться при значениях концентраций более высоких, чем природ-
ные. Но когда экспериментальные концентрации превышают при-
родные более чем в 10 раз, скорость фильтрации с увеличением
концентрации меняется мало, и значения ее остаются крайне низ-
кими. Полученные результаты дают возможность аппроксими-
ровать зависимость между скоростью фильтрации у различных
моллюсков и концентрацией взвешенных в воде веществ в виде еди-
ного уравнения:
1
т (1 — е~пх) ’
т%е y=F!Fmx3i — относительная скорость фильтрации; х=с)ст^ —
относительная концентрация; е — основание натуральных лога-
рифмов; тп, п — константы (в данном случае ?п=8.85, тг=0.062).
Знаменатель уравнения (35) аналогичен зависимости рациона
животных от концентрации корма, которая была получена для
рыб и ракообразных (Ивлев, 1955; Сущеня, 1964, 1975). Это го-
ворит о том, что скорость фильтрации воды моллюсками в зави-
симости от концентрации пищевых частиц в воде представляет собой
величину, обратную изменению их физиологического рациона при
нарастании концентрации пищи в среде.
«Экологический» рацион у моллюсков и количество неусвоенной
ими пищи (H=f-\-fp) находятся в прямой зависимости от концентра-
71
ции взвесей в воде (Алимов, Бульон, 1972). Расположение экспери-
ментальных данных на двойной логарифмической шкале указывает
на степенную зависимость «экологического» рациона моллюсков и
неусвоенной пищи от концентрации пищи в среде (рис. 25). При
этом по мере нарастания концентрации «экологический» рацион
увеличивается не так быстро, как рассчитанная величина неусво-
енной пищи. Расчеты показали, что при некоторой концентрации,
Рис. 24. Зависимость относительной скорости фильтрации у от относительной
концентрации х взвесей в воде для моллюсков.
1—3— Dreissena polymorpha; 4 — Unio pictorum; 5 — U. tumidus: 6 — Anodonta piscinalis;
7 — Sphaeriastrum rivicola; 8, 10 — Sphaerium corneum, 9 — S. suecicum (по данным:
1 — Михеева, 19676; 2 — Кондратьева, 19696; 3—7 — Кондратьева, 1970; 8,9 — изме-
рения автора; 10 — Митропольского, 1966). По оси абсцисс — значения х; по оси орди-
нат — значения у (объяснения в тексте).
значительно превышающей наибольшую из использованных в опы-
тах, рацион и неусвоенная пища оказываются равными (прямые
на рис. 25 пересекаются). При такой ситуации усвояемость пищи
равна нулю, т. е. животные перестают питаться. В рассматривае-
мом случае с моллюсками S. suecicum такое явление могло теорети-
чески наступить, когда концентрация взвешенных веществ превы-
сила бы природную примерно в 60 раз. Вместе с тем при концентра-
циях взвесей, существенно превышающих природные, физиологиче-
ский рацион, согласно уравнению (35), с нарастанием концентрации
увеличивается, хотя и с замедляющейся скоростью. В связи с этим
при таких концентрациях, когда неусвоенная пища оказывается
равной рациону, создается маловероятная ситуация — прекращение
потребления пищи животными при продолжающейся фильтрации
ими воды. Скорее всего при столь высоких концентрациях взвеси
из-за механического забивания частичками фильтрационного ап-
72
парата моллюски просто перестают фильтровать воду. Поэтому
связь между величинами «экологического» рациона, неусвоенной
пищи и концентрацией взвеси в виде степенной зависимости можно
рассматривать лишь до определенных пределов концентрации.
В опытах с моллюсками S. suecicum зависимость такого вида
оказывалась справедливой при концентрациях, не превышающих
129—136 кал/л (38—40 мгО/л). Однако в природных местообита-
ниях моллюсков столь высокие концентрации взвешенных орга-
нических веществ вряд ли когда-либо могут быть достигнуты.
Рис. 25. Зависимость «экологиче-
ского» рациона 1g С1 и неусвоенной
пищи 1g Я у моллюсков Sphaerium
suecicum при разных концентрациях
взвеси 1g С (из: Алимов, Бульон,
1972).
По оси абсцисс — значения 1g С; по оси
ординат — значения 1g С1 и 1g Н (объяс-
нения в тексте).
Рис. 26. Скорость выделения псевдо-
фекалий fp моллюсками Sphaerium
suecicum при разных концентрациях
взвеси (из: Алимов, Бульон, 1972).
По оси абсцисс — концентрация взвеси
(кал/л); по оси ординат — значения fp
(кал/сут).
При увеличении концентрации взвеси нарастает масса выделя-
емых моллюсками псевдофекалий (Yonge, 1926; Михеев, 1967а,
19676; Кондратьев, 1970b; Walz, 1978а). Аналогичные данные были
получены для моллюсков Mytilus edulis, Mercinaria mercinaria
(L.), Crassostrea virginica (Tenore, Dunstan, 1973), Cerastoderma
lamarki (Reeve) (Санина, 1976). Выделение псевдофекалий мол-
люсками Cerastoderma edule, Venerupis pullastra Montegu было
отмечено при концентрациях планктонных водорослей, превыша-
ющих 75—250-103 клеток/мл, что составляло 9.4—31 % наибольшей
концентрации, использованной в опытах (Foster-Smith, 1975).
Моллюсками Dreissena polymorpha выделялись псевдофекальные
массы, когда концентрация пищи достигала примерно 2% от
максимальной в воде опытных сосудов (Walz, 1978а). В наших
опытах моллюски Sphaerium suecicum выделяли псевдофекалии
начиная с концентрации около 10 кал/л (примерно 26% от макси-
мальной в опытах), затем с увеличением концентрации их количе-
ство нарастало (рис. 26).
73
Расчеты показали, что при наибольших концентрациях, исполь-
зованных в опытах, количество выделяемых моллюсками S. sueci-
сит псевдофекалий составляло около 437% суточных трат на об-
мен и прирост животных в условиях опытов, что можно рассматри-
вать как избыточную величину рациона по сравнению с физиологи-
ческим. Поскольку при высоких концентрациях пищи по мере
их увеличения физиологический рацион возрастает столь незначи-
тельно, что его можно считать практически неизменным, нет осно-
ваний говорить о так называемом «избыточном» питании животных,
так как псевдофекалии никакого отношения к процессу питания не
имеют. Понятие «избыточного» питания тем более неприменимо
к питанию животных в природных условиях.
При постоянной концентрации взвесей скорость фильтрации
воды моллюсками может изменяться в зависимости от размера
частиц или качества взвеси. Например, моллюски Unio pictorum,
Anodonta piscinalis, Dreissena polumorpha фильтровали с большей
скоростью воду с взвесью из более крупных частиц, однако взвеси
с частицами больше 30 мкм фильтровались с одинаковой скоростью
(Кондратьев, 1970b). Аналогичные данные были получены для не-
которых морских моллюсков (Rice, Smith, 1958). Мортон (Morton,
1971) пришел к выводу, что чем меньше размер частиц, тем боль-
шей должна быть их концентрация, с тем, чтобы скорость фильтра-
ции воды у дрейссен была наибольшей.
Скорость фильтрации воды моллюсками, кроме того, зависит от
качественного состава взвесей. Так, например, в наших опытах
моллюски Sphaerium согпеит с большей скоростью фильтровали
воду, в которой находились минеральные частички (глина), по
сравнению с водой, содержащей взвеси из хлореллы или пекар-
ских дрожжей. Такого же рода данные были получены Г. П. Конд-
ратьевым (1970в) при изучении фильтрации воды унионидами, сфе-
риидами и дрейссенами. Возможно, что такие различия связаны
с разной питательной ценностью взвесей.
Таким образом, экспериментальные исследования показали,
что скорость фильтрации воды двустворчатыми моллюсками и
интенсивность их питания в существенной степени определяются
концентрацией взвешенных в воде частиц, их размерами и каче-
ственным составом.
Активная реакция среды
Влияние активной реакции среды (pH) на скорость фильтрации изу-
чалось в опытах с Sphaerium согпеит (Алимов, 1967а). При измене-
нии величины pH в опытных сосудах в пределах 3.1—9.1 путем до-
бавления в воду 10% H2SO4 или 20% NaHCO3 были получены сле-
дующие значения скорости фильтрации воды моллюсками:
pH.......... 3.1 4.5 5.0 6.0 7.0 8.0 9.0
F, мл/ч .... 0 1.07 2.01 6.4 4.4 1.65 0
74
Величины pH 3 и 9 можно рассматривать как крайние, при кото-
рых процесс фильтрации у S. corneum прекращается. В опытных
сосудах с такими значениями pH животные на протяжении всего
опыта неподвижно лежали на дне с плотно закрытыми створками.
В сосудах с другими значениями pH воды они активно перемеща-
лись по дну и стенкам. При перенесении животных из сосудов
срНЗи9 в воду с pH 6—7 они уже в течение первого часа рас-
крывали створки и начинали активно перемещаться. Следова-
тельно, пребывание моллюсков в воде со значениями pH 3 и 9
(по крайней мере в течение 5 ч — продолжительность опыта) не
приводит к их гибели.
Увеличение скорости фильтрации наблюдалось при изменениях
значений pH от 3 до 6 (при pH 6 она была наибольшей). Дальней-
шее подщелачивание приводило к уменьшению скорости филь-
трации, и при pH 9 моллюски практически прекращали фильтро-
вать.
Интервал значений pH, в пределах которого скорость фильтра-
ции воды моллюсками сокращалась не более чем на 50 % от макси-
мальной величины, зарегистрированной в опытах, ограничивался
значениями pH 5.5—7.5. По-видимому, эти значения pH можно
рассматривать как зону оптимума фильтрации по этому фактору
среды. Поскольку моллюски для опытов были взяты из неболь-
ших прудов, где активно идут процессы разложения органиче-
ских веществ мощных донных отложений, влекущих за собой под-
кисление придонных слоев воды, сдвиг зоны оптимума в сторону
значений pH, характерных для более кислых вод, становится
объяснимым. Уменьшение скорости фильтрации при понижении
pH от 7 до 6.5 и почти полное прекращение фильтрации при pH
4.25 отмечалось Лузановым и Томмерсом в опытах с морскими мол-
люсками Crassostrea virginica (Jorgensen, 1966). Несколько иные
результаты были получены Мортоном (Morton, 1971) в опытах
с Dreissena polymorpha. Скорость фильтрации у этих животных была
максимальной при pH 8 и 9.
Приведенные данные показывают, что активная реакция
среды, представляя собой специфический фактор, в достаточной
степени сильно влияет на активность молдюсков. Незначительные
отклонения ее от оптимальных значений для конкретных видов
приводят к ослаблению фильтрационного процесса у животных.
РИТМИЧНОСТЬ ФИЛЬТРАЦИИ
Все двустворчатые моллюски не постоянно фильтруют воду.
Различают суточные и сезонные изменения фильтрационной ак-
тивности у моллюсков. В течение суток периоды активности сме-
няются периодами покоя. В периоды активности створки раковин
открыты, в периоды покоя — закрыты. Периодичность активности
животных связана с периодическими изменениями обмена веществ
и находится в зависимости от условий среды.
75
Барнес (Barnes, 1955,1962) описал два эндогенных ритма муску-
лов аддуктора у Anodonta cygnea*. медленный и быстрый. Медленный
ритм характеризуется сменой периодов активности и покоя с часто-
той от 3 до 40 раз в неделю. В течение активного периода медленного
ритма наблюдается быстрый ритм, при котором периоды покоя и
активности чередуются с частотой до 20 раз в час. Фильтрация
воды моллюсками осуществляется только во время активной фазы.
В этот период моллюски потребляют кислорода в 5—10 раз больше,
чем во время покоя (Коштоянц, Шаланки, 1958; Salanki, 1964;
Salanki, Vero, 1969).
Суточные колебания активности, кроме того, могут быть свя-
заны с изменениями температуры воды, освещенности, степени
насыщения кислородом и т. д. Так, например, падение концентра-
ции растворенного в воде кислорода вызывает увеличение ритмич-
ности работы аддуктора, укорочение во времени периодов активно-
сти, увеличение респираторной активности моллюсков (Barnes,
1962; Salanki, 1965; Salanki, Vero, 1969; Morton, 1969). Чаще всего
такое явление можно наблюдать в ночные часы. Однако в опытах
с Actinonaias carinata Barnes, Elliptio dilatatus (Rafinesguc) и
Pleurobema coccineum Conrad было установлено, что при гипоксии
скорость фильтрации воды этими моллюсками снижается (Badman,
1975). На этом основании автор отвергает гипотезу об усилении
снабжения тканей кислородом при гипоксии благодаря большей
интенсивности движения створок раковины. По данным Мортона
(Morton, 1971), дрейссены фильтруют главным образом в ночное
время, и наиболее активная фильтрация у них отмечается в тече-
ние первой половины суток. У анодонт и дрейссен процесс филь-
трации в течение суток занимает примерно 70% времени (Bruin,
Davids, 1970; Walz, 1978а). Более высокая фильтрационная ак-
тивность в ночное время отмечена у морских моллюсков Crassostrea
virginica и Mytilus galloprovincialis (Arakawa, 1970). В то же время
Г. П. Кондратьев (1969в, 1970в) отмечает, что сферииды и дрейс-
сены наиболее активно фильтруют в дневные часы, что, по мнению
автора, обусловлено фототаксисом у этих животных. Пока нет
единого мнения о том, в какие периоды суток моллюски фильтруют
наиболее интенсивно. Однако общий результат этих работ со-
стоит в том, что продолжительность фильтрации у моллюсков в те-
чение суток составляет 50—70% времени. При этом в лабораторных
условиях периоды активности оказываются короче, чем в природе
(Jorgensen, 1966). Эти заключения следует принимать во внимание
при количественной оценке роли моллюсков в процессах превра-
щения веществ в водоемах.
Скорость фильтрации у моллюсков не остается постоянной в те-
чение года. Наибольшая фильтрационная активность и интенсив-
ность питания у них наблюдается в летние месяцы. Осенью, зимой,
ранней весной моллюски резко уменьшают или вовсе прекращают
фильтрацию (Arakawa, 1970; Gale, Lowe, 1971). Усиление филь-
трационной активности унионид совпадает со сроками их жабер-
76
ной беременности, у шаровой и дрейссен — с периодами интенсив-
ного размножения. Увеличение фильтрации в это время связыва-
ется с большими потребностями моллюсков в кислороде (Кон-
дратьев, 1973). В этом отношении показательно, что у неполово-
зрелых животных, по наблюдениям Г. П. Кондратьева, сезонная
ритмичность фильтрации выражена значительно слабее, чем у по-
ловозрелых, размножающихся особей.
СКОРОСТЬ ФИЛЬТРАЦИИ
И ОБЩИЕ РАЗМЕРЫ МОЛЛЮСКОВ
В процессе индивидуального развития двустворчатых моллюсков
скорость фильтрации воды ими возрастает с увеличением их раз-
меров (Fox et al., 1937; Воскресенский, 1948; Willemsen, 1952;
Кондратьев, 1962, 19636; Jorgensen, 1955, 1960, 1966, и др.).
Впервые количественное выражение зависимости величины скоро-
сти фильтрации от размеров животных было получено в резуль-
тате экспериментов с несколькими видами сфериид (Алимов, 1965).
Оказалось, что скорость фильтрации у этих моллюсков F и их
вес W связаны между собой зависимостью, которая, так же как
аналогичная зависимость для скорости обмена, может быть ап-
проксимирована в виде степенной функции
F = mWn. (36)
Справедливость выражения этой зависимости степенной функ-
цией была впоследствии подтверждена на ряде морских моллюсков
(Ali, 1970; Vahl, 1973а, 1973b; Санина, 1976).
В отдельных немногочисленных работах скорость фильтрации
у моллюсков рассматривается в связи с их линейными размерами.
Так, например, Хьюз (Hughes, 1969) установил, что у Scrobicularia
plana скорость фильтрации у изменяется в зависимости от длины
моллюска х, согласно степенному уравнению
y = qxn, (37)
где q и п — константы уравнения (в данном случае п=2.21).
Однако известно, что вес и длина животных связаны между собой
зависимостью
W = axLbl. (38)
Используя равенство (38), можно уравнение (37) выразить в весовых
единицах:
у = /(W) или y — iW”1, (39)
r!*e i = -^ibT’ h=nlb.
Выражение скорости фильтрации воды животными в зависимости
от их веса более значимо, так как в процессы обмена, питания,
ассимиляции пищи вступает масса развивающегося организма.
77
1000 -
ф 1
• 2
°3
*4
05
*6
*7
т 8
v 9
ф 10
▲ 11
♦ 12
о 13
А Н
10
Рис. 27. Зависимость скорости фильтрации воды
от веса моллюсков.
I, 8—11, 14 —1 Dreissena polymorpha; 2, 3 — Unio pictorum;
4,5 — U. tumidus; 6,7 — Anodonta piscinalis; 12 — Sphaeri-
astrum rivicola; 13 — Anodonta woodiana (по данным: 1, 4,
6,8 — Кондратьева, 19636; 9 — Львовой-Качановой,
1971; 11 — Михеева, 1966, 1967а, 19676; 14 — Ми-
хеева, Сорокина, 1966; 3, 5, 7, 10, 12, 13 — измерения ав-
тора). Масштаб логарифмический. Прерывистые линии —
доверительный интервал для уровня вероятности 0.95.
По оси абсцисс — вес моллюсков (г); по оси ординат — ско-
рость фильтрации (мл/ч).
Для количественного выражения зависимости скорости фильтра-
ции у различных представителей двустворчатых моллюсков от их
веса результаты экспериментов автора и литературные материалы
были приведены в соответствии с установленной ранее зависимостью
к температуре 20°. Во всех случаях скорость фильтрации выра-
жалась в миллилитрах воды за час.
На рис. 27 приведены все известные к настоящему времени экс-
периментальные данные по скорости фильтрации воды представи-
Рис. 28. Зависимость скорости фильтрации воды от веса моллюсков рода
Sphaerium.
1,6 — Sphaerium corneum’, 2 — S. scaldianum’, 3 — S. solidum; 4 — S. korotnevi; 5 — S. sue-
cicum (I—5 — измерения автора; 6 — по данным Hinz, Scheil, 1972). Остальные обо-
значения как на рис. 27.
телями семейств Unionidae и Dreissenidae. Из рисунка видно,
что животные одинакового веса, относящиеся к разным родам
этих семейств, в пределах изученных весов от 0.03 до 150 г досто-
верно не различаются между собой по скорости фильтрации.
Среди моллюсков сем. Sphaeriidae по величине скорости филь-
трации выделяются три группы: представители родов Sphaerium
(рис. 28), Pisidium (рис. 29) и Sphaeriastrum (рис. 27), что соответ-
ствует разделению семейства на роды и по морфологическим при-
знакам (Алимов, 1965, 1969).
Моллюски, относящиеся к роду Sphaeriastrum, как следует
из рис. 27, по своим фильтрационным способностям: не отличаются
достоверно от унионид и дрейссен, что дает основание рассчитать
для них единое уравнение скорости фильтрации от веса животных
79
(табл. 10). В этой же таблице приведены аналогичные уравнения
для моллюсков родов Sphaerium и Pisidium.
Несомненно важно, что эндемики такого своеобразного водоема,
как оз. Байкал, моллюски Sphaerium korotnevi (W. Dyb.) и Pisidium
Рис. 29. Зависимость скорости фильтрации воды от веса моллюсков рода
Pisidium.
1 — Р. amnicum', 2 — Р. supinum', 3 — Р. casertanum', 4 — Р. granum‘, 5 — Р. crassum*
6 — Р. nitidum. Остальные обозначения как на рис. 27.
granum (W. Dyb) или водоемов бассейна р. Амур — Anodonta wo-
odiana Lea. не отличаются по величине скорости фильтрации воды
от животных одинакового размера, но относящихся к другим родам
Таблица 10
Параметры т, п и ошибки их средних значений в уравнениях
зависимости скорости фильтрации от веса у пресноводных
двустворчатых моллюсков
Название моллюсков Пределы весов (Г) m (мл/ч • г) п
Unionidae, Dreissenidae, Sphaeriastrum .... Sphaerium Pisidium Musculium 0.0430—156 0.0070—0.403 0.0016—0.261 0.0320—0.153 85.5 ±0.54 12.8 ±0.36 24.1 ±2.50 7.45 0.605 ±0.066 0.625 ±0.040 0.598 ±0.149 0.58
80
семейства и живущим, например, в водоемах европейской части
СССР. Это говорит о широком значении связи скорости фильтра-
ции воды животными с их весом.
Статистическая обработка приведенных выше материалов по-
казала достоверность (с вероятностью 0.95) различия между рас-
считанными параметрами уравнений скорости фильтрации у мол-
люсков Sphaerium, Pisidium и совместного уравнения для Unioni-
Рис. 30. Линии регрессий зависимостей скорости фильтрации от веса мол-
люсков.
1 — Musculium; 2 — Sphaerium', 3 — Pisidium', 4 — Unionidae; Dreissenidae, Sphaeri*
astrum. Остальные обозначения как на рис. 27.
dae, Dreissenidae, Sphaeriastrum (рис. 30). Небольшое количество
данных по скорости фильтрации у моллюсков рода Musculium
приводит к слишком большой ошибке при их статистической обра-
ботке. Тем не менее расположение экспериментальных точек
(рис. 30) дает основание предполагать, что эти моллюски отлича-
ются по фильтрационным способностям от представителей рода
S phaerium.
Среди пресноводных моллюсков наиболее активные фильтра-
торы — виды семейств Unionidae, Dreissenidae и рода Sphaeriastrum,
самые низкие скорости фильтрации из изученных животных —
у представителей рода Musculium (рис. 30, табл. 10). Приведенные
в табл. 11 значения параметров уравнений зависимости скорости
фильтрации от веса у некоторых морских моллюсков показывают,
что для них характерны более высокие скорости фильтрации по
6 А. Ф. Алимов
81
Таблица И
Параметры ш, п уравнений зависимости скорости фильтрации
воды от сырого веса у морских двустворчатых моллюсков
Вид m (мл/ч на грамм сырого веса) п По данным
Pecten irradians 1250 0.62 Chipman, Hopkins, 1954
Mytilus edulis ........ 650 0.60 Willemsen, 1952
M. edulis . . 386 0.66 Воскресенского, 1948
M. edulis .... 500 0.52 Ali, 1970
M. californianus 406 0.43 То же
Mercinaria mercinaria 370 0.42 »
Hiatella arctica — 0.67 »
Mercinaria mercinaria 334.2 0.66 Coughlan, Ancell, 1964
Scrobicularia plana 270 0.66 Hughes, 1969
Arctica islandica . .... 180 0.70 Winter, 1969
Modiolus modiolus 240 0.61 Winter, 1969
Cardium edule 225 0.60 Нейферт, 1967
Mytilus edulis 3900 * 0.60 Vahl, 1973a
Cardium edule 2000* 0.58 Vahl, 1973b
* Рассчитано на 1 г сухого веса тела моллюска.
сравнению с пресноводными моллюсками. На эти различия не-
однократно обращалось внимание (Кондратьев, 19636; Jorgensen,
1966; Bruin, Davids, 1970, и др.). Наиболее активные фильтраторы
«реди пресноводных моллюсков — униониды и дрейссены — филь-
труют в среднем около 85.5 мл/ч на грамм сырого веса, в то время
как Arctica islandica (L.) — около 180, a Pecten irradians Lam. —
1250 мл/ч на грамм сырого веса.
Обращает на себя внимание близость значений константы п
в уравнениях F=f(W) для пресноводных и морских моллюсков,
параметры которых были рассчитаны независимо разными авто-
рами. Степенная зависимость между физиологическим рационом
и весом животных была установлена в экспериментах с двумя
видами черноморских двустворчатых моллюсков (Цихон-Лука-
нина, 1976). Для нас важно, что величины показателей степени
б уравнениях, аппроксимирующих эту зависимость, находились
б пределах от 0.55 до 0.66 и оказались близкими к значениям пока-
зателей степени в рассмотренных нами уравнениях связи скорости
фильтрации и веса моллюсков. Сходство показателей степени
в уравнениях для фильтрации и питания у морских и пресновод-
ных двустворчатых моллюсков указывает на общую для этих жи-
вотных закономерность изменения скорости фильтрации и питания
в процессе их роста.
Различия фильтрационных способностей пресноводных и мор-
ских моллюсков могут быть обусловлены разными причинами.
Одна из них — количество пищевого материала в толще воды.
Скорость фильтрации воды моллюсками, населяющими водоемы
>82
с малым количеством взвешенных в воде частиц, оказывается при
прочих равных условиях выше, чем у тех, которые живут в водоемах
с большим количеством взвесей (Алимов, 1969; Hughes, 1969).
Обитателям рек или литорали озер (Unio, Anodonta) свойственны
более высокие по сравнению с обитателями стоячих водоемов
(Sphaerium, Pisidium, Musculium) скорости фильтрации воды.
Моллюски Mytilus edulis, Mercinaria mercinaria, живущие в усло^
виях, где воды несут мало взвешенных веществ, характеризуются
более высокими фильтрационными способностями, чем Scrobicularia
plana или Му a arenaria, обитающие в зонах с большим количеством
взвешенных в воде частиц (Hughes, 1970).
Усиление фильтрационной функции вододвижущего аппарата
в эволюции Bivalvia совпадает с прогрессивным развитием их
жаберного аппарата в направлении его усложнения от примитив-
ного у Protobranchia до сложного у Eulamellibranchia. Наиболее
сложное строение и мощное развитие гидрокинетического аппарата
последних привело к переходу их к питанию только фильтрацион-
ным способом.
Изменение скорости фильтрации у моллюсков с одинаковым
морфологическим строением жаберного аппарата может быть
обусловлено изменениями либо скорости биения цилий мерца-
тельного эпителия, либо площади фильтрационного аппарата.
Увеличение скорости фильтрации за счет ускорения движения ци-
лий, казалось бы, должно было повлечь за собой повышение ско-
рости активного, и в конечном счете общего, обмена у животных.
Однако усложнение вододвижущего аппарата, колоссальное раз-
витие фильтрационной функции у двустворчатых моллюсков, равно
как и увеличение фильтрационных способностей в пределах одного
отряда, не сопровождалось в процессе эволюции этих животных
изменением уровня их метаболизма.
Наиболее простым способом интенсификации фильтрационной
функции у животных с одинаковым морфологическим строением
гидрокинетического аппарата представляется изменение площади
жаберного аппарата. Этот путь не ведет к сложным изменениям
в морфологии животных. В обширной сводке по фильтрационному
питанию беспозвоночных Йоргенсен (Jorgensen, 1966) уделяет спе-
циальное внимание этому вопросу. Он подчеркивает, что скорость
транспорта воды моллюсками, имеющими одинаковую структуру
органов питания, в значительной степени зависит от размеров
этих органов и должна быть в определенной мере генетически
фиксированной.
Развитие органов вододвижущего аппарата моллюсков, в том
числе и их жабр, обусловлено количеством взвешенных в воде ча-
стиц в местах обитания животных. Для животных, обитающих в во-
дах с высокой мутностью, типично сильное развитие сортирующих
механизмов (Ansell, 1961) и наличие меньших по размерам жабр
(Yonge, 1951) по сравнению с обитателями более прозрачных вод.
Скорость фильтрации воды некоторыми морскими моллюсками
6*
83
оказалась тесно скоррелированной с количеством воды, прокачива-
емой ими в расчете на единицу зоны жабр (Hughes, 1969; Foster-
Smith, 1975), что в свою очередь связано с количеством взвешенных
в воде частиц.
В качестве показателя степени развития жаберного аппарата
моллюсков можно использовать его относительную площадь
(Алимов, 1969). Под относительной площадью жаберного аппарата
понимается площадь жабры, отнесенная к площади проекции створ-
ки раковины моллюска. Это отношение показывает, какую долю от
площади раковины составляет жабра. Технически площадь жабры
и проекции створки раковины могут быть определены как площади
фигур, близких к ним по форме, таких, например, как эллипс, тра-
пеция, треугольник и т. п. Измерения показали, что относительная
площадь жаберного аппарата остается достаточно постоянной
в процессе развития животных определенного вида, в то время как
абсолютная площадь жабр моллюсков увеличивается пропорцио-
нально нарастанию массы животного (Vahl, 1973а).
Среди пресноводных моллюсков наибольшими величинами отно-
сительной площади жаберного аппарата характеризуются унио-
ниды и дрейссены, самые низкие ее значения — у представителей
рода Musculium. Значения относительной площади жаберного
аппарата морских моллюсков (Mercinaria mercinaria, Mytilus
edulis) оказались больше, чем у унионид и дрейссен: Musculium
lacustre — 0.140, Sphaerium согпеит — 0.210, S. solidum — 0.210,
S. korotnevi — 0.203, Pisidium casertanum — 0.203, P. amnicum —
0.240, Sphaeriastrum rivicola — 0.360, Dreissena polymorpha —
0.390, Unio pictorum — 0.420, Mercinaria mercinaria — 0.470, Mytilus
edulis — 0.540. Самая малая величина относительной площади
жаберного аппарата — у обитателей небольших, в ряде случаев
временных водоемов, в воде которых достаточно много взвешенных
веществ. Самые большие ее значения среди пресноводных моллюс-
сков — у унионид и дрейссен, населяющих прибрежные участки
рек или прибойную зону озер, т. е. места с постоянным течением
воды или волновым перемешиванием. Несомненно, что в таких
условиях получение необходимого количества пищевого материала
может быть обеспечено путем увеличения сосущей силы вододвижу-
щего аппарата, что и достигается расширением площади жаберного
аппарата.
Относительная площадь жаберного аппарата моллюсков и
интенсивность фильтрации воды ими в расчете на единицу веса жи-
вотных (константа т в уравнении (36)) находятся между собой
в функциональной зависимости (рис. 31), которая может быть
описана в виде степенного уравнения
m=KS\ (40)
где S — относительная площадь жаберного аппарата; К и а —
константы уравнения.
84
Константы уравнения (40), рассчитанные графически по данным
рис. 31, для изученных пресноводных моллюсков имели следующие
значения: К=2150 мл/ч на грамм сы- .
рого веса, при 5 = 1; а=3.
Поскольку F=mWn, а иг в свою
очередь функционально связано с от-
носительной площадью жаберного ап-
парата, скорость фильтрации воды
моллюсками может быть представлена
как функция двух переменных — веса
животного и относительной площади
его жаберного аппарата: 100
F = KS*Wn. (41)
С помощью уравнения (41) возможно
описать все рассмотренные выше дан-
ные по скорости фильтрации воды прес-
новодными и морскими моллюсками.
Как уже отмечалось, вода, фильтруе-
мая моллюсками, служит одновременно
источником пищи и кислорода для дыха- Ю
ния. Поэтому определенный интерес
может представлять величина, показы-
вающая объем воды, профильтрованный
моллюском весом в 1 г, при потребле-
нии им 1 мг кислорода, которую мо-
жно рассматривать как коэффициент
фильтрации
Q = FjR, (42)
где Q — коэффициент фильтрации
(мл/мгО); F, R — соответственно скоро-
сти фильтрации и потребления кисло-
рода моллюсками как функция их
веса.
С помощью коэффициента фильтра-
ции можно сравнивать количество пищи,
получаемой моллюсками разного систе-
матического положения в различных
экологических условиях. Значения
этого коэффициента для отдельных
групп пресноводных моллюсков, при-
нимая, что зависимость обмена от их
веса выражается уравнением (17), при-
ведены в табл. 12. Данные этой таб-
1 ------------------------L-
0.1 1-П
Рис. 31. Зависимость ин-
тенсивности фильтрации
(константа т в уравнениях
зависимости скорости филь-
трации от веса) от относи-
тельной площади жабер-
ного аппарата моллюсков
(из: Алимов, 1969).
1 — Musculium', 2 — Sphaerium',
3 — Pisidium', 4 — Unionidae,
Dreissenidae, Sphaeriastrum', 5 —
Mercinaria mercinaria', 6 —
Mytilus edulis. Масштаб лога-
рифмический. По оси абсцисс —
относительная площадь жабры;
по оси ординат — интенсив-
ность фильтрации (мл-ч-1 «г-1)-
лицы показывают, что наибольшие величины коэффициента филь-
трации среди изученных животных у Unionidae, Dreissenidae
и Sphaeriastrum, которые профильтровывают около 910 мл воды
на каждый миллиграмм потребленного кислорода. Значительно
выше эта величина у морских двустворчатых моллюсков. В сред-
нем они фильтруют 14—16 л воды на миллилитр потребленного
кислорода (Colin, 1960; Jorgensen, 1960, 1966). Для некоторых
из них эта величина не превышает 2—4 л (Jorgensen, 1966;
Hamwi, Haskin, 1969).
Таблица 12
Коэффициенты фильтрации Q для пресноводных
двустворчатых моллюсков
Название моллюсков Диапазон весов (г) Q (мл/мгО)
Musculium 0.0320-0.153 7.98
Sphaerium . . 0.0070-0.403 136.00
Pisidium 0.0016—0.261 272.00
Unionidae, Dreissenidae, Sphaeriastrum 0.0430-156 910.00
Различия в коэффициентах фильтрации между морскими и
пресноводными двустворчатыми моллюсками определяются адап-
тациями этих животных к жизни в водоемах, сильно различаю-
щихся по содержанию в воде взвешенных органических веществ.
Среднее количество взвешенных органических веществ в эвтроф-
ных озерах достигает 13.3 мг/л, в мезотрофных — около 2.1, в оли-
готрофных не превышает 0.5 мг/л (Бульон, 1976), в некоторых реках
средней полосы европейской части СССР — около 2—3 (Алимов,
1969), в морских водоемах — не более 0.2—0.5 мг/л (Armstrong,
Atkins, 1950).
Используя приведенные данные по содержанию взвешенных
веществ в водоемах разного типа и коэффициенты фильтрации
для соответствующих групп моллюсков, проведем ориентировочные
расчеты количества пищи, которое они могут получить. Для про-
стоты расчетов примем, как и ранее, что зависимость скорости
обмена от веса животных для пресноводных и морских двуствор-
чатых моллюсков описывается уравнением (17), а величины по-
казателей степени в уравнениях 7? =/(W) и F=f(W) одинаковы.
В таком случае для моллюсков рода Sphaerium^ населяющих,
например, эвтрофные водоемы, среднее значение коэффициента
фильтрации не превышает 136 мл/мгО. Поскольку в воде таких
водоемов в среднем содержится примерно 20 мг/л органических
взвесей, очевидно, моллюски вместе с фильтруемой водой могут
получить около 1.8 мг пищевых частиц при потреблении ими 1 мг
кислорода. Аналогичным образом можно рассчитать, что оби-
татели рек (например, Unio) получат 1.82—2.7 мг, а морские,
например, Мytilusedulis (в соответствии с табл. 11), — 1.1—2.73 мг
органических веществ.
При принятых условиях, согласно уравнению (17), легко
определить, что моллюски весом в 1 г потребляют 1 мг кислорода
86
примерно за 11 ч и при этом получают от 1.1 до 2.73 мг (в сред-
нем около 1.9 мг) пищевого материала, т. е. суточный рацион
животных в среднем составляет около 4.2 мг, или около 0.4%
от их веса. В соответствии с уравнением связи физиологического
рациона и веса моллюсков, приведенном Е. А. Цихон-Луканиной
(1976), суточный рацион, например, митилястера весом в 1 г не пре-
вышает 8.04 мг, или 0.8% его веса. Близкие величины суточных
рационов, полученные нами расчетным путем и установленные
Е. А. Цихон-Луканиной в результате экспериментов по усвояемости
пищи, лишний раз подтверждают справедливость приведенных
выше закономерностей обмена и фильтрации у моллюсков, а также
выполненных на их основе вычислений.
Обращают на себя внимание сходные количества пищи, по-
лучаемой в результате фильтрации воды моллюсками различного
систематического положения, происхождения и резко отличаю-
щимися по характеру местообитания. Это дает основание для
заключения об адаптации их фильтрационных аппаратов к со-
держанию взвешенных веществ в воде водоемов, в которых они
живут.
Рассмотрим этот вопрос по отношению к пресноводным мол-
люскам в историческом плане. По классификации Я. И. Старо-
богатова (1970), сферииды — наиболее ранние вселенцы в прес-
ные воды (палеолимнические), Unionidae и Dreissenidae — более
поздние (мезолимнические). При этом Driessenidae позже всех
осваивали пресные водоемы.
Палеолимнические моллюски, приспособленные ко всему раз-
нообразию условий жизни в континентальных водоемах, харак-
теризуются низкими значениями коэффициентов фильтрации.
Обычно они населяют водоемы с высоким содержанием взвешен-
ных органических веществ (эвтрофные, мезотрофные озера, пруды,
небольшие пересыхающие водоемы, вплоть до крупных луж).
Большинство видов рода Musculium, характеризующиеся самыми
низкими для изученных моллюсков коэффициентами фильтра-
ции, — наиболее частые обитатели водоемов с очень высоким
содержанием взвешенных органических веществ, таких как пруды,
крупные пересыхающие лужи, мелководные участки рек или
озер, с сильно развитой водной растительностью и т. п. Пред-
ставители рода Sphaerium живут в прудах, прибрежных участках
рек, заводях, старицах или в литоральной зоне эвтрофных и ме-
зотрофных, реже олиготрофных озер, поселяясь чаще всего на
участках с развитой подводной растительностью. В различных
озерах они концентрируются в основном в литоральной и суб-
литоральной зонах. Моллюски Pisidium, у которых более вы-
сокие по сравнению с Musculium и Sphaerium коэффициенты филь-
трации, заселяют различные по характеру водоемы. Однако мно-
гие из них более обычны в олиготрофных или мезотрофных озе-
рах, где обитают в суб литорали или профундали, доходя до зна-
чительных глубин. В реках они живут на участках с медленным
Ь7
течением на песчаных или слабозаиленных грунтах. Большинство
видов, которые обычно населяют большие глубины, более глубо-
ководны по сравнению со Sphaerium. На самых больших глу-
бинах (до 120 м и более) крупных олиготрофных озер обычен
Р. conventus. Бблыпая глубоководность видов рода Pisidium не-
однократно отмечалась различными авторами (Berg, 1938; Масап,
1953; Rawson, 1953; Саватеева, 1964; Solem, 1973; Sivertsen,
1973, и др.). По нашим наблюдениям, моллюски S. suecicum
в озерах Карелии (Кривое, Круглое) и Кольского п-ова (Зеленец-
кое) обитают только в литоральной зоне, не опускаясь глубже
4—6 м, в то время как Р. crassum и Р. lilljeborgi распространяются
до максимальных в этих озерах глубин (около 30 м). Существенно,
что количество взвешенных веществ в прибрежных зонах этих озер
примерно вдвое выше, чем в более глубоководных участках. Все
это говорит о том, что моллюски Pisidium могут жить в водоемах
или на их отдельных участках, которые характеризуются не-
высоким содержанием взвешенных в воде органических веществ.
Одно из косвенных подтверждений тому — более далекое про-
никновение на север этих животных. Так, например, распро-
странение большинства немногочисленных в Карелии видов рода
Sphaerium ограничено преимущественно озерами ее южной части.
Представители рода Pisidium расселяются далеко на север,
а Р. conventus — единственный вид, двустворчатых моллюсков,
который был найден на Новой Земле (Саватеева, 1964; Александ-
ров, 1965). Фильтрационные способности моллюсков этого рода
позволяют им получать необходимое количество пищи в условиях
олиготрофных северных водоемов. Можно предположить, что
эти их особенности наряду с прочими обусловливают по сравне-
нию с видами рода Sphaerium возможность обитания в более се-
верных водоемах или на больших глубинах.
Представители мезолимнических моллюсков, имеющих более
высокий по сравнению с палеолимническими коэффициент филь-
трации, населяют главным образом реки или прибрежные, под-
верженные волновому перемешиванию зоны крупных озер. Наи-
более плотные поселения унионид в озерах отмечаются на глу-
бинах не более 1.5—2 м, лишь отдельные экземпляры встречаются
на бблыпих (до 8—9 м) глубинах. Показательно, что численность
их резко уменьшается на участках с развитой водной раститель-
ностью (Soszka, 1968; Cvancara, 1972; Haukioja, Hakala, 1974) или
с увеличением мутности воды (Cvancara, 1970). Несомненно, что
благодаря высокой скорости фильтрации мезолимнические мол-
люски могут обитать в условиях постоянно движущейся воды даже
при достаточно высоком содержании в ней взвешенных пищевых
частиц.
По мере проникновения моллюсков из морских водоемов в кон-
тинентальные, т. е. в процессе заселения ими водоемов с повышен-
ным содержанием взвешенных веществ, происходило уменьшение
относительной площади их жаберного аппарата и ослабление
88
скорости фильтрации воды. Параллельно усиливалась роль жабр
как органа для вынашивания молоди. У мезолимнических Dreis-
senidae, как и у морских моллюсков, личинка пелагическая, у унио-
нид — паразитирующая, палеолимнические моллюски живоро-
дящие. Последние — наиболее древние все ленцы в пресные воды —
в связи со способностью жить в водоемах с высоким содержанием
взвешенных в воде веществ и благодаря переходу к живорождению
получили возможность заселять также наиболее мелкие, изоли-
рованные водоемы, включая крупные пересыхающие лужи и бо-
лота.
Бесспорно, нельзя всю сложную эволюцию и историю рассе-
ления пресноводных моллюсков объяснить лишь изменениями их
фильтрационных способностей, отбросив все прочие факторы.
В то же время следует учитывать значение важнейшего пище-
вого фактора при изучении и анализе адаптаций животных к усло-
виям существования, их филогении и эволюции.
ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ
КАК ПРИРОДНЫЕ БИОФИЛЬТРЫ
Двустворчатые моллюски в водоемах выступают как природные
биофильтры, очищающие воду от находящихся в ней во взвешен-
ном состоянии веществ. В ряде водоемов эффективность удаления
моллюсками из воды взвесей достигает 92—100% (Кондратьев,
19706). Одновременно ими создается биогенная циркуляция воды
(Воскресенский, 1968), способствующая перемешиванию водных
масс.
Для количественной оценки роли моллюсков в очищении воды
водоемов от взвешенных веществ могут быть использованы
установленные зависимости скорости фильтрации воды животными
от их размеров.
Рассмотрим простой пример. По нашим наблюдениям, чис-
ленность моллюсков Sphaerium согпеит в одном из небольших
прудов Калининградской об л. (площадь пруда не более 0,-01 га,
объем воды 51 м3) составляла 565 экз./м2. Средний вес одного
животного в популяции не превышал 0.096 г. Согласно урав-
нению F=f (РИ) (см. табл. 10), моллюск такого размера филь-
трует воду со скоростью 69.6 мл/сут. Следовательно, все моллюски
на площади в 1 м2 отфильтруют за сутки 39.1 л воды, а вся попу-
ляция животных — 4.2 м3, или 8.2% объема воды в пруду. При-
нимая, что моллюски фильтруют воду в течение 12 ч в сутки,
получаем, что за сутки популяция этих животных способна
профильтровать около 4.1% объема пруда, т. е. весь его объем —
примерно за 24 дня. Бихроматная окисляемость веществ, нахо-
дившихся во взвешенном состоянии в воде этого водоема, была
около 12.8 мгО/л, что эквивалентно содержанию примерно 8 мг
органических веществ. Учитывая это, определяем, что за сутки
вся популяция моллюсков может удалить из воды около 16.8 г
89
органических веществ, или около 3% общего их содержания в воде
пруда.
Аналогичные подсчеты, выполненные различными исследо-
вателями, показывают, что в водоемах разного типа двустворча-
тые моллюски в период наибольшей их активности (вегетационный
сезон) способны прокачивать огромные массы воды. Например,
дрейссены в Учинском водохранилище с мая по октябрь профиль-
тровывают объем воды, равный двум объемам водохранилища
(Львова-Качанова, 1971; Извекова, Львова-Качанова, 1972).
Образование в Учинском водохранилище широкого пояса моллю-
сков-фильтраторов увеличило более чем в три раза скорость осаж-
дения взвеси в зоне обитания дрейссен, в результате чего умень-
шилась мутность и улучшились питьевые качества воды за время
ее отстоя в водохранилище (Львова, 1977). В Пяловском водохра-
нилище эти животные за сутки пропускают 1/20 часть всего объема
воды (Михеев, 19676), они способны профильтровать весь объем
Штеттинского залива за 36 дней (Wiktor, 1963). Популяция дрейс-
сен на разных участках канала Днепр—Кривой Рог на площади
1 м2 за сутки способна отфильтровывать от 38 до 4317 л воды (Каф-
танникова, Шевцова, 1972). Принимая длительность вегетацион-
ного сезона равной 6 мес, получаем, что за это время животные
на площади в 1 м2 прокачивают от 6.84 до 777.1 м3 воды. Дрейс-
сены оз. Миколайского за вегетационный сезон отфильтровывают
53-106 м3 воды, что составляет около 97% объема воды в озере
(Stanczykowska et al., 1976).
Приведенные примеры наглядно показывают, сколь велика
фильтрационная способность популяций двустворчатых моллю-
сков. В то же время в водоемах разной продуктивности роль этих
моллюсков в фильтрации воды оказывается неодинаковой.
С увеличением продуктивности водоемов нарастает и значение
фильтрационной деятельности моллюсков, что, однако, не всегда
сопровождается увеличением их биомассы (табл. 13). Такое по-
ложение связано, вероятно, с тем, что эвтрофные и мезотрофные
водоемы, как уже говорилось, населяют моллюски с большей
фильтрационной способностью (дрейссены, некоторые униониды).
В олиготрофных же водоемах фауна двустворчатых моллюсков
бедна, и представлены они главным образом мелкими видами,
преимущественно шаровками и горошинками.
Важно знать не только объем воды, фильтруемой моллюсками,
но также и количество взвешенных веществ, удаляемых ими из этого
объема. Проведенные разными исследователями расчеты пока-
зали, что, например, в оз. Миколайском все двустворчатые мол-
люски за сезон осаждают из фильтруемой ими воды 164.5 т сухого
веса сестона, из них 160 т приходится на дрейссен (Stanczykowska
et al., 1976). В Штеттинском заливе они ежемесячно отлагают на
дно в виде фекальных и псевдофекальных масс до 51.5 т сырого
веса органического вещества (Wiktor, 1963). Дрейссены в Учин-
ском водохранилище в разные годы осаждали от 1610 до 4700 г
90
Таблица 13
Фильтрационная деятельность двустворчатых моллюсков
в водоемах разной продуктивности
Водоем Первичная продукция (ккал/м2 за сезон) В (г/м2) VnlVb По данным
Озеро: Андронниковой, Драбковой, 1976: Кузьмен- ко, 1976 Lewandowski, Stanczykowska, 1975
Красное 1486 1315.00 0.30
Миколайское 3100 3102.50 1.10
Дривяты 1770 61.0 0.80 Гаврилова, 1970
Круглое 55 15.80 0.21 Автора
Зеленецкое 28 0.12 0.01 То же
Водохранилище: Львовой-Качано- вой, 1971 Кондратьева, 1970а
Учинское ... — 500.00 2.00
Волгоградское .... 1127 50.85 0.32
Примечание. V — объем воды, отфильтрованной за вегетационный сезон мол-
люсками; Vb — объем воды в водоеме; В — средняя за вегетационный сезон биомасса
моллюсков.
органического вещества (Львова-Качанова, 1971). Осаждаемые
моллюсками органические вещества в виде агглютинатов сно-
сятся на большие глубины и входят в состав донных отложений.
Специальными экспериментальными исследованиями установ-
лено, что агглютинированные дрейссенами в процессе фильтра-
ции органические вещества оказались наиболее питательным
кормом для личинок хирономид (Извекова, Львова-Качанова,
1972). В связи с этим существенно знать, в какой степени пищевые
потребности личинок хирономид могут быть удовлетворены за счет
агглютинатов моллюсков. Проведем несложный расчет на при-
мере оз. Миколайского.
В профундальной зоне этого озера средняя биомасса личинок
хирономид составляет около 2.4 г/м2 (Kajak et al., 1975). Вы-
полненные нами расчеты показали, что в различных озерах ко-
личество потребляемой личинками хирономид пищи в течение
сезона превышает среднюю их биомассу за тот же период примерно
в 23 раза. Исходя из этого, рацион хирономид в оз. Миколайском
за сезон должен быть около 55.2 г/м2, или 138.6 т сырого органи-
ческого вещества в расчете на площадь профундали озера (251 га).
Примем, что сухой вес сестона составляет около 10% его сырого
веса. В таком случае рацион хирономид в профундальной зоне
озера за сезон не превышает 13.9 т органических веществ (в сухом
весе). За этот же период, как уже было сказано, моллюски осаж-
91
дают 164.5 т сестона. По данным А. П. Остапени (1976а), в сестоне
содержится около 36% минеральных веществ. Учитывая это,
получаем, что моллюски в течение сезона осаждают на дно около
105.3 т сухого органического вещества. Это количество превышает
пищевые потребности личинок хирономид в озере примерно
в 7.7 раза. Аналогичным образом нетрудно подсчитать, что все
нехищные животные бентоса в профундали этого озера, биомасса
которых около 7.97 г/м2, а рацион превышает их биомассу примерно
в 13 раз, должны потребить за сезон около 16.7 т органических
веществ, или примерно 16% всего количества веществ, осажденных
моллюсками за тот же период. Таким образом, пищевые потреб-
ности донных животных в этом озере могут быть полностью удов-
летворены за счет фильтрационной деятельности моллюсков.
Неутилизированные донными животными органические ве-
щества, которые были осаждены моллюсками, подвергаются де-
струкции, например, бактериями или окисляются за счет раство-
ренного в воде кислорода. В незагрязняемых водоемах удельное
потребление кислорода сестоном находится в пределах 0.01 —
0.1 мгО/мг органического вещества за сутки (Остапеня, 19766),
т. е. в среднем около 0.06 мгО/мг органического вещества. Отсюда
нетрудно определить, что для деструкции осаждаемых моллюсками
и не использованных донными животными органических веществ
в течение сезона потребуется около 6.4 мгО/м2. Последняя вели-
чина сопоставима с потреблением кислорода в единице объема
воды олиготрофного озера.
Приведенные данные и расчеты, несмотря на всю их прибли-
женность, демонстрируют важное значение двустворчатых мол-
люсков в процессах очищения воды, образования донных отло-
жений органического происхождения и обеспечении тем самым
пищевых потребностей бентосных животных.
Заключая главу, следует подчеркнуть, что, используя уста-
новленные количественные закономерности фильтрационного пи-
тания пресноводных двустворчатых моллюсков, удается объяснить
некоторые особенности их расселения, происхождения и оценить
их роль в процессах самоочищения природных водоемов.
Глава III
ЗАКОНОМЕРНОСТИ РОСТА
И ОСОБЕННОСТИ ФОРМЫ РАКОВИН МОЛЛЮСКОВ
В процессе роста происходит увеличение линейных размеров и
массы развивающегося организма. Различают линейный и весо-
вой рост, скорость которых определяется степенью развития орга-
низма, его физиологическим состоянием, условиями внешней среды.
При этом соотношение между массой и линейными размерами,
так же как и относительные скорости изменения линейных разме-
ров в разных измерениях, определяются законом аллометрического
роста.
При изучении роста моллюсков основное внимание обычно
концентрируется на исследовании их постэмбрионального роста.
Конечно, это сужает рамки исследований и возможность обоб-
щений. Впрочем, длительность эмбрионального развития у боль-
шинства видов несоизмеримо мала по сравнению с общей про-
должительностью жизни животных. Поэтому в результате иссле-
дования только постэмбрионального периода развития моллюсков
мы получаем достаточно полное представление о закономерностях
их роста*
ЛИНЕЙНЫЙ РОСТ
Линейные размеры раковин моллюсков и изменения их в онто-
генезе могут быть определены с помощью простейших инструмен-
тов и приемов с достаточной точностью.
Изучение роста моллюсков обычно проводится четырьмя основ-
ными методами: 1) непосредственным наблюдением за ростом,
2) мечением животных, 3) подсчетом годовых колец на их ракови-
нах, 4) путем анализа размерной структуры популяций отдельных
видов.
Непосредственные наблюдения за ростом
подопытных животных производятся в специально изготовленных
садках, устанавливаемых в естественных местах обитания моллю-
93
сков, или в условиях аквариума. Периодически проводятся про-
меры животных, на основании которых ‘ определяется скорость
их роста. Результаты наиболее точно отражают картину роста,
но для этого требуются длительные наблюдения. В аквариуме
получение надежных данных осложняется трудностью поддер-
жания в течение продолжительных отрезков времени стабильных
условий. Время наблюдений и экспериментов может быть сокра-
щено, если одновременно прослеживать рост нескольких возра-
стных групп моллюсков одного вида. Изучение роста методом
непосредственных наблюдений используется в основном при иссле-
дованиях животных с быстрым темпом роста, например сфериид
и дрейссен (Thiel, 1926; Касымов, 1957; Качанова, 1963; Wiktor,
1963; Михеев, 19646; Thomas, 1965; Арабина, 1966; Алимов,
1970; Little, Gentner, 1970, и др.).
Рост раковин моллюсков может быть изучен с помощью ме-
чения животных красками, специальными химическими
реактивами (например, тетрациклином), радиоактивными препа-
ратами, нанесением царапин, надпилов на раковинах |и т. п.
Меченые животные периодически отлавливаются и измеряются.
При изучении пресноводных моллюсков этот метод использовался
редко. Можно отметить лишь работу по изучению роста Маг-
garitifera margaritifera (Rubbel, 1913) и незаконченное исследование
дальневосточных моллюсков Cristaria pUcata, Anodonta woodiana
Lea, Nodularia douglasiae Griff et Pidg (Булдовский, 1935).
Наиболее широко для определения возраста двустворчатых
моллюсков применяется метод подсчета «годовых» к о-
л е ц. Кольца на раковинах моллюсков образуются в резуль-
тате длительных остановок роста в периоды неблагоприятных
условий. На раковинах четко выражены кольца, образовавшиеся
в результате замедления и остановки роста в зимний период
(зимние кольца — их называют годовыми). У ряда тропических
моллюсков такие же кольца образуются в неблагоприятные сухие
периоды года.
Привлекательность этого метода состоит в том, что при нали-
чии хорошо сохранившихся раковин без специальных экспери-
ментов, приборов и приспособлений и при минимальных затра-
тах времени можно проследить изменения размеров животных в те-
чение их жизни. Неудивительно, что он широко используется
для изучения роста моллюсков, имеющих большую продолжи-
тельность жизни (Rubbel, 1913; Altnoder, 1926; Chamberlain,
1931; Бластов, 1933, 1934, 1935; Троицкий, 1934; Безходарная,
1937; Лукин, 1937а, 19376, 1937в; Муратова, 1937; Сальдау,
1939; Граевский, Баранов, 1949; Карпевич, 1952, 1964; Crowley,
4957; Hendelberg, 1960; Okland, 1963; Stanczykowska, 1963; Ляхов,
Михеев, 1964; Петров, 1964; Haranghy et al., 1965; Heard, 1965;
Negus, 1966; Tudorancea, Florescu, 1968a, 1968b, 1969; Haukioja,
Hakala, 1978, й др.). В ряде случаев возраст животного опреде-
ляется подсчетом колец на лигаменте (Hendelberg, 1960; Bjork,
94
1962), которые эквивалентны кольцам на створках раковины
(Strober, 1972).
Определение возраста моллюсков по годовым кольцам имеет
определенные трудности, связанные с тем, что на раковинах кроме
годовых колец зимней остановки роста имеются также кольца,
образовавшиеся в результате менее продолжительных остано-
вок роста, обусловленных резкими, кратковременными измене-
ниями условий внешней среды. В периоды размножения рост
моллюсков резко замедляется или даже может прекратиться,
что также отражается на скульптуре раковины в виде четкого
кольца, которое подчас трудно отличить от зимних колец. На-
пример, на раковинах некоторых сфериид можно различить одно
или два кольца. Длительность же жизни большинства из них редко
превышает один год. Одно кольцо отделяет часть раковины,
соответствующей размерам моллюска, впервые приступившего
к размножению, второе — закладывается в результате зимней
остановки роста. Поэтому у моллюсков, родившихся весной и
погибших осенью того же года, на раковинах имеется только одно
кольцо, а у особей, рожденных в конце лета и доживших до осени
следующего года, можно различить два кольца (Алимов, 19676).
Аналогичные данные были получены и для дрейссен (Morton,
1969). Таким образом, при оценке возраста моллюсков по годо-
вым кольцам следует учитывать, что после достижения животными
половозрелости в течение года на раковинах образуются два четко
различимых кольца, превышающих по ширине прочие кольца.
Дополнительные кольца как результат кратковременных остано-
вок роста порой настолько осложняют определение возраста
животных, что, как считают некоторые исследователи, этот метод
может привести к серьезным ошибкам. Вместе с тем многочисленные
данные подтверждают правомочность и надежность такого способа
определения возраста моллюсков. Изучая структуру раковин
унионид, Б. В. Бластов (1935) пришел к выводу, что основной
признак годовых колец — их правильная цикличность, а коли-
чество их отражает реальный возраст животных. Это было пока-
зано также и на морских представителях (Haskin, 1954; Seed,
1969; Садыхова, 1972; Золотарев, 1974; Coutts, 1974; Lutz, 1976,
и др.).
В некоторых работах кривые роста моллюсков определяются
в результате анализа динамики размерной
структуры популяций, в частности по гистограммам
частотно-размерного распределения животных. Достоверные ре-
зультаты могут быть получены, когда каждый возрастной класс
может быть уверенно выделен в любые периоды времени. Воз-
можные ошибки при использовании этого метода заключаются
в трудности определения возраста группы животных, выделяемых
как возрастной класс, в оценке возраста самых молодых живот-
ных при отсутствии дополнительной информации. Растянутый
во времени нерест осложняет выделение возрастных групп. Обычно
95-
этот способ используется в сочетании с другими (Журавель, 1951;
Михеев, 19646; Митропольский, 1965; Morton, 1969; Алимов,
1970; Алимов, Финогенова, 1975а).
Один из методов выделения возрастных групп по гистограммам
размерного распределения особей в популяции был предложен
для морских моллюсков А. Н. Голиковым (1970) и затем исполь-
зован для пресноводных моллюсков Н. Ф. Смирновой (1975а).
В популяциях видов с прерывистым размножением молодь, по-
явившаяся за некоторый промежуток времени, «... составляет
единую размерно-весовую группу, отделенную от родителей раз-
мерным промежутком, превышающим индивидуальные вариации
в размерах внутри каждой группы. . . Испытывая единовремен-
ное воздействие факторов, тормозящих и стимулирующих рост
и размножение, популяция сохраняет дискретность своей раз-
мерно-весовой структуры на всех возможных уровнях» (Смирнова,
1975а, с. 201). Иными словами, в популяции отдельные возрастные
группы по своим размерам не трансгрессируют и наблюдается
так называемая генеративная дискретность. Это позволяет, по
мнению авторов, различать и подсчитывать количество возрастных
групп в популяции, изучать групповую и межгрупповую измен-
чивость роста особей.
Естественно, что отрожденная молодь по своим размерам отли-
чается от родителей, признать же отсутствие трансгрессии по раз-
мерам между всеми последующими возрастными группами весьма
трудно. В этом случае придется допустить, что все особи каждой
размерно-возрастной группы растут с равными скоростями, их
рост продолжается в течение абсолютно одинаковых отрезков
времени, все возможные изменения внешних условий действуют
на всех особей в популяции, независимо от стадии их развития
и состояния, строго однозначно. Весьма сомнительна возможность
строгого соблюдения этих условий в природной обстановке. Не-
соблюдение любого из них неизбежно приведет к разной степени
трансгрессии размеров возрастных групп, что обычно и наблю-
дается в природных популяциях. На этот счет можно привести
множество примеров популяций животных самого разного си-
стематического положения. Тем не менее утверждается: «Метод
определения группового роста и возраста по генеративной дискрет-
ности в структуре локальных популяций оказывается универ-
сальным» (Голиков, 1976, с. 106). Однако какого-либо точного ме-
тода выделения возрастных групп, кроме различения «на глаз»,
не предлагается. Несомненно, что такая оценка произвольна и
в значительной степени основана на интуиции исследователя, что,
естественно, может в ряде случаев приводить к серьезным и не-
учитываемым ошибкам.
Трудность выделения отдельных возрастных групп может
быть преодолена применением, например, метода «вероятностной
бумаги», предложенного Хардингом (Harding, 1949) и развитым
Касси (Cassie, 1950, 1954). В его основе лежит вполне реальное
96
допущение, что каждая возрастная группа имеет нормальное рас-
пределение по размерам особей. Использование «вероятностной
бумаги», позволяющей спрямить кривые нормального распределе-
ния, предоставляет возможность аналитической дифференцировки
размерных (возрастных) групп в результате анализа полимо-
дальных частотно-размерных распределений животных в попу-
ляции. Этот метод был рекомендован для определения возраста
двустворчатых моллюсков (Садыхова, 1972) и апробировался на
некоторых популяциях этих животных (Табилова, Бергман, 1975).
К сожалению, по-видимому, из-за кажущейся сложности он
не используется пока исследователями пресноводных моллюсков.
Перечисленные выше методы определения возраста и изуче-
ния роста моллюсков каждый в отдельности или в сочетании друг
с другом дают достаточно надежные результаты. В этом отноше-
нии особый интерес представляет исследование роста Scrobicu-
laria plana (Hughes, 1970). Рост моллюсков изучался с помощью
маркировки отдельных особей, подсчетом годовых колец, а также
определялся в результате анализа возрастной динамики попу-
ляции с использованием вероятностной бумаги. На основании
тщательно проведенных исследований и статистической обработки
полученных данных автор пришел к выводу, что все три способа
изучения роста дают очень сходные результаты и каждым из них
можно пользоваться в равной степени. Этот вывод еще раз под-
тверждает правомочность определения возраста моллюсков под-
счетом годовых колец.
В литературе накопилось много сведений по размерам и про-
должительности жизни самых разных видов пресноводных дву-
створчатых моллюсков, которые, однако, еще ждут осмысливания
и обобщения. Данные же о росте целого ряда представителей этих
животных крайне скудны или полностью отсутствуют. Поэтому
важно обобщить уже имеющиеся сведения, восполнить по воз-
можности недостающие и попытаться представить результаты
в аналитическом выражении.
Собственные и литературные данные по росту пресноводных
Bivalvia были обработаны единым образом. С этой целью исполь-
зовался способ описания кривых изменения линейных размеров
в онтогейезе животных, предложенный Валфордом (Walford, 1946)
и расширенный Риффенбургом (Riffenburgh, 1966). Способ Вал-
форда представляет собой графический вариант определения
коэффициентов уравнения роста Берталанфи
Lt = Lm(\-e~kt}, (43)
где Lt — длина животного ко времени Z; — теоретическая
максимальная длина животного, представляющая собой асимптоту
роста; к — константа роста; е — основание натуральных ло-
гарифмов.
С помощью уравнения (43) могут быть описаны кривые
линейного роста всех моллюсков (Wilbur, Owen, 1964).
7 А. Ф. Алимов 97
Чтобы выразить уравнение (43) в линейной форме, запишем
его относительно La+1} и Lt:
L(t+v=Lme~^, Lt = Lme~kt.
Поделив эти выражения и сделав некоторые преобразования г
получаем]
= = _к
^со L^e~kt
откуда:
L(i+1) —— е~к)Lte~k, (44)
где Lt — длина животного ко времени t; L(f+1) — длина животного
ко времени £+1-
Выражение (44) представляет собой линейную форму уравне-
ния (43). Используя уравнение (44), строится график, выражаю-
щий зависимость Lt=f [£(/+1J. В том случае, когда изменения
линейных размеров в течение роста моллюсков могут быть аппрок-
симированы уравнением (43), экспериментальные точки на гра-
фике располагаются вдоль прямой линии, которая характеризуется
начальным значением L{t+1} при Lt=Q и проходит под углом ме-
нее 45° к оси абсцисс (рис. 32).
Опустив перпендикуляр на ось абсцисс из точки пересечения
прямой, проведенной через экспериментальные точки, и прямой,
проходящей под углом 45° через начало координат, находим один
из параметров уравнения роста — теоретический максимальный
размер Л а,, который потенциально может быть достигнут живот-
ным, обладающим данным темпом роста. Второй параметр урав-
нения — константа роста к — определяется через тангенс угла
наклона прямой, проходящей через экспериментальные точки.
Тангенс угла наклона этой прямой tg а и параметр уравнения
к связаны между собой равенством
tg а — е~к,
откуда —к = I n tg а или
jfc= tga
Ige ‘
Параметры и к можно рассчитать аналитически из уравне-
ния (44) как уравнения прямой линии, в котором Ьт(1—е~к)
представляет собой свободный член, а е~к — угловой коэффициент.
Определив е~\ нетрудно рассчитать Lm:
т ___-k(?+i) — Lte~k
00 ’ (l-e-fc) *
Таким образом, определив численные значения параметров
уравнения для каждого конкретного случая, можно построить
98
теоретическую кривую роста, сравнить ее с опытными дан-
ными и, что весьма важно, исследовать параметры уравнения.
Рис. 32. Зависимость длины раковины моллюсков ко времени Н“1 (Л^+1) от
длины ко времени t (Lt).
А — Unio tumidus, р.Ока; Б — Crassiana crassus, р. Ока; В — Anodonta zellensis, р. Волга;
Г — Corbicula flumenalis (1 — реки Закавказья; 2 — р. Мургаб: 3 — р. Кумбашни).
По оси абсцисс — значения Lft по оси ординат — значения L^+i (объяснения в тексте).
Способ Валфорда предоставляет возможность определить мак-
симальный размер моллюсков и по уравнению (43) рассчитать
продолжительность их жизни. Существенно, что для решения урав-
7*
99
нений при помощи этого способа не обязателен, хотя и желателен
подсчет абсолютно всех годовых колец на раковинах моллюсков.
Кстати, при изучении роста Cerastoderma edule наряду с подсче-
том всех годовых колец на раковинах этого моллюска для сравне-
ния подсчитывались лишь некоторые из них, образованные через
равные промежутки времени. В обоих случаях результаты полу-
чились идентичные (Hancock, 1965). Вообще говоря, когда тип
постэмбрионального роста животных не меняется с возрастом,
для построения теоретической кривой роста достаточно под-
считать несколько четко различимых, следующих друг за другом
годовых колец, зная при этом, в каком возрасте было образо-
вано некоторое определенное кольцо. Это приобретает особое зна-
чение при определении возраста медленнорастущих моллюсков,
например, таких, как Margarittfera margarittfera. У этих животных
последние кольца столь плотно прилегают друг к другу, что раз-
личить их чрезвычайно трудно, а иногда и просто невозможно.
Например, М. П. Сальдау (1939) приводит данные по росту жем-
чужниц только до 13-летнего возраста. Подсчет последующих
колец оказался невозможным из-за их большой плотности. Однако,
используя только эти данные по размерам моллюсков (до 13-
летнего возраста) и рассчитав по ним параметры уравнения роста,
возможно построить кривую их линейного роста и определить
продолжительность жизни животных.
Благодаря столь очевидным возможностям метод Валфорда
получает все более широкое применение в практике малаколо-
гических исследований. Его использовали для изучения роста
Dreissena polymorpha (Morton, 1969; Львова, 1977), мидий (Theisen,
1967—1969), Mercinaria mercinaria (Ansell, 1968), Scrobicularia
plana (Hughes, 1970), Cerastoderma edule (Hancock, 1965), Cor-
biculamanilensis Phillip., C. fluminalis (Mull.) (Morton, 1973,
1977) и др.
При изучении скорости роста, продолжительности жизни и
изменчивости формы по коллекциям Зоологического института
и собственным сборам автор измерил раковины моллюсков раз-
личного возраста главным образом широко распространенных ви-
дов. Всего было промерено около 1500 раковин представителей
сем. Unionidae, 250 экз. — Corbiculidae, около 900 экз. — Sphae-
riidae. Кроме того, были использованы результаты измерения
раковин отдельных видов Sphaeriidae из рек Сухона, Вологда,
Кама, Печора, выполненные в свое время В. И. Жадиным и любезна
переданные автору для обработки.
По полученным результатам с привлечением литературных ма-
териалов для наиболее широко распространенных видов семейств
моллюсков, населяющих территорию Советского Союза, методом
Волфорда рассчитывались параметры уравнений линейного роста,
которые даны в табл. 14—27. В этих же таблицах приводятся зна-
чения максимальной из измеренных в каждом водоеме длины ра-
ковины Лтах и соответствующая наибольшая продолжительность
100
жизни животных £max, рассчитанные в соответствии с уравнениями
(43, 44):
± __ In Lqq In (Z>oo ^max)
^max- fa •
Представленные таким образом материалы дают возможность
исследования внутри- и межвидовой изменчивости роста моллюсков.
Особенности роста жемчужниц (Margaritiferidae) были рассмо-
трены по литературным данным на примере Margaritifera marga-
ritifera. К сожалению, рост этих важнейших промысловых мол-
люсков до сих пор исследовался неоправданно мало. Это, вероятно,
связано с большими трудностями определения их возраста по ра-
ковинам, сложностью поддержания условий, приближающихся
к природным, при содержании этих стенобионтных животных в ла-
бораторной обстановке. Величины параметров уравнений роста
для жемчужниц из разных водоемов показывают (табл. 14), что
Таблица 14
Параметры уравнений роста моллюсков Margaritifera
margaritifera
Река h (год-1) Длина (мм) 2 max (годы) По данным
Г оо Lmax
Сайда . .... 0.050 140 140 41 Граевского, Бара- нова, 1949
Акулинина . . . 0.023 140 140 90 То же
Ташечная . ... 0.030 120 120 86 »
Кыма . . ... 0.035 140 140 61 »
Емешь . ... 0.030 140 140 61 »
Кереть 0.041 110 99 56 Властова, 1934
Варзуга . ... 0.030 100 90 76 Сальдау, 1939
Муна . . ... 0.051 110 100 84 То же
Ламукса 0.030 120 108 76 »
Пана 0.030 140 99 76 »
Реки Германии . . 0.041 120 108 76 Rubbel, 1913
Реки Швеции ... 0.021 145 130 105 Hendelberg (1960)
Реки Скандинавии . . . 0.084 120 108 58 Altnoder (1926)
константа роста у них колеблется от 0.023 до 0.084 год”1, а про-
должительность жизни соответственно от 41 до 105 лет. Средние
значения константы роста и продолжительности жизни М. mar-
garitifera из изученных водоемов оказались близкими со-
ответственно к 0.038 год-1 и 73 годам. Величины констант роста
и продолжительности жизни моллюсков, вероятно, отражают
реальные их значения в природной обстановке, так как они вполне
согласуются с общими представлениями о росте пресноводных
двустворчатых моллюсков, которые будут рассмотрены в даль-
нейшем.
101
Константа роста у моллюсков рода Unto находится в пределах
от 0.1 до 0.347 для U. tumidus и от 0.183 до 0.500 год-1 — для
U. pictorum, продолжительность жизни соответственно от 6.5
до 22.5 и от 4.7 до 11.5 лет (табл. 15, 16). Средние значения этих
величин не превышают 0.244 год-1 (8.7 лет) (СЛ tumidus) и
0.282 год-1 (7.7 лет) (U. pictorum). Близкие величины констант
роста — от 0.16 до 0.357 (средняя около 0.222) год"1 и продол-
жительности жизни — 4—12 (средняя 7.9) лет отмечены для мол-
люсков Crassiana crassus (табл. 17).
Анодонты и псевданодонты характеризуются более быстрым
темпом роста по сравнению с Unio и Margaritiferidae. Величины
констант роста для A. anatina близки к 0.187—0.437 (средняя
около 0.236) год"1, A. ponderosa — 0.235—0.490 (0.369), A. zel-
lensis — 0.223—0.485 (0.350), A. seisanensis Kob. — 0.212—
0.382 (0.269), A. piscinalis — 0.282—0.415 (0.369), Pseudanodonta
complanata Resm. — 0.324—0.420 (0.370) год"1, продолжитель-
ность их жизни — в пределах от 3.3 до 13.4 года (табл. 18—23).
В табл. 24 приведены параметры уравнений роста для неко-
торых дальневосточных унионид и жемчужниц. В литературе
до сих пор отсутствовали сведения о темпе роста и продолжи-
тельности жизни этих животных. Поэтому выделение в особую
таблицу сведений о росте этих моллюсков представляется оп-
равданным. Поскольку количество изученных животных незна-
чительно, полученные данные следует рассматривать как пред-
варительные. Тем не менее они дают представление о скорости
роста этих моллюсков. Наиболее медленно (так же, как и евро-
пейские жемчужницы) растут Margaritifera dahurica Midd.
(&=0.122 год"1) и Cristaria plicata (fc=0.127 год"1). В среднем
скорость их роста оказывается ниже, чем у моллюсков рода
Unio, но выше, чем у европейских жемчужниц.
Сравнение темпа роста у представителей семейств Unionidae
и Margaritiferidae показывает, что среди Unionidae наибольшей
скоростью роста характеризуются виды родов Anodonta и Pseud-
anodonta. При этом самый быстрый рост отмечен у A. ponde-
rosa С. Pf., A. piscinalis и Р. complanata. Наиболее медленно ра-
стут М. margaritifera. Промежуточное положение занимают пред-
ставители рода Unio, среди которых быстрее всего растут
U. pictorum, медленнее — U. tumidus.
При внимательном рассмотрении приведенных данных по росту
унионид и жемчужниц можно заметить важную закономерность.
Максимальные и минимальные значения констант роста у
М. margaritifera различаются в 3.6, а продолжительность жизни
в 2.6 раза. Аналогичные величины у видов рода Unio отличаются
соответственно в 2.5 и 3.5, в среднем примерно в 3 раза. У Ano-
donta — по константам роста в 1.5—2.3, по продолжительности
жизни — 1.7—3, т. е. в среднем приблизительно в 2 раза. Такие
в общем-то сходные различия наибольших и наименьших значе-
ний констант роста и продолжительности жизни у видов, отличаю-
102
Таблица 15
Параметры уравнений роста моллюсков Unio tiimidus
Водоем h (год-1) Длина (мм) fmax (годы) По данным
Lqo Ещах
Река:
Оскол 0.200 НО 99 8.7 Сальдау, 1939
Проня 0.100 но 99 22.5 То же
Упа 0.210 70 63 11.0 »
Воронеж 0.200 70 63 11.6 »
Ока (Башкирия) .... 0.149 115 103 14.9 Властова, 1935
Дон 0.343 87 78 6.5 Троицкого, 1934
Кичуй 0.146 90 81 15.9 Безходарной, 1937
Свияга .... 0.315 105 95 7.5 Лукина, 1937а
Карла 0.322 100 90 7.4 Лукина, 19376
Волга 0.268 75 67 8.4 Лукина, 1937в
Казанка 0.268 70 60 7.3 Муратовой, 1937
Казанка (пруд) .... 0.242 85 76 9.3 То же
Темза 0.235 90 81 9.7 Negus, 1966
Ока (у Пущина) .... 0.248 100 78 6.1 Автора
Ока (у Льгова) .... 0.260 100 71 4.8 То же
Ока (у Шилова) .... 0.282 85 75 7.6 »
Теша 0.118 103 74. 10.8 »
Ока (Велетьминский за-
тон) 0.229 92 75 7.3 »
Ока (Липинский затон) 0.296 73 66 7.9 »
Ока (затон Култук) . . 0.209 93 73 7.3 »
Сура 0.186 103 79 9.0 »
Ветлуга 0.235 100 68 4.9 »
Днестр 0.184 98 75 7.9 »
Днепр 0.237 95 82 8.4 »
Вента 0.268 71 49 4.4 »
Сев. Двина 0.131 73 37 5.4 »
Рейн 0.221 104 97 12.2 »
Луара 0.264 108 74 4.4 »
Велетьма 0.296 83 76 8.3 »
Нижний Ик 0.268 129 105 7.8 »
Дон 0.160 100 71 7.7 »
Озеро:
Чудское 0.160 81 47 5.2 »
Палеостом 0.271 100 77 5.4 »
Студенец (пойма) . . . 0.235 94 68 5.6 »
Крапина 0.163 90 81 13.8 Tudorancea, Flo-
rescu, 1968b
Залив:
Финский 0.138 95 67 8.9 Автора
103
Таблица 16
Параметры уравнений роста моллюсков Unto pictorum
Водоем k (год-1) Длина (мм) ^тах (годы) По данным
Lqo Lmax
Река:
Ока (Башкирия) . . 0.336 120 108 6.7 Властова, 1935
Дон 0.313 95 86 7.4 Троицкого, 1934
Оскол 0.320 НО 99 7.1 Сальдау, 1939
Проня 0.256 НО 99 8.8 То же
Упа 0.223 70 63 10.1 »
Воронеж .... 0.200 70 63 11.5 »
Темза 0.210 85 77 11.0 Negus, 1966
Ока (нижнее течение) Ока (Велетьминский за- 0.362 96 93 9.6 Автора
тон) 0.302 87 71 10.2 То же
Теша . ... 0.206 90 76 9.1 »
Ветлуга .... 0.256 100 77 5.8
Вента 0.183 80 60 7.6 »
Казанка . 0.284 90 80 7.8
Карла 0.350 128 115 6.5 »
Кичуй .... 0.218 92 82 10.1 »
Свияга 0.392 96 85 5.5 »
Сура 0.301 87 79 8.0 »
Рейн . 0.376 83 75 6.2 »
Кубань 0.250 100 83 7.1 »
Волга . 0.500 83 76 5.6 »
Крынка . . 0.392 124 116 7.0 »
Чусовая .... 0.256 93 65 4.7
Озеро:
Барские Кулики . . 0.210 НО НО 10.2 Сальдау, 1939
Крапина 0.223 90 81 10.2 Tudorancea, Flo- rescu, 1968а
Студенец 0.228 198 83 6.4 Автора
Великое 0.376 83 80 8.8 »
Таблица 17
Параметры уравнений роста моллюсков Crassiana crassus
Водоем к (год-1) Длина (мм) 2 max (годы) По данным
^00 Lmax
Река: Ока (Башкирия) .... 0.256 85 77 9.1 Властова, 1935
Оскол 0.357 70 63 6.4 Сальдау, 1939
Воронеж 0.187 60 54 12.0 То же
Свияга 0.272 77 62 5.9 Лукина, 1937а
Кичуй ........ 0.172 72 58 9.5 Безходарной, 1937
Ока (нижнее течение) . 0.219 80 50 6.2 Автора
Ока (среднее течение) 0.219 73 55 6.4 То же
104
Таблица 17 (продолжение)
Водоем к (ГОД"1) Длина (мм) /max (годы) По данным
Lqo Lmax
Крынка . . . 0.310 88 79 7.3 Автора
Южный Буг 0.160 80 50 6.2 »
Чусовая . . 0.218 80 63 7.1 »
Урал .... ... 0.244 90 57 5.6 »
Днестр ... ... 0.187 80 50 5.2 »
Теша .... 0.302 81 68 4.0 »
Москва 0.282 72 51 4.4 »
Ока (Липинский затон) 0.322 75 57 4.4 »
Финский залив .... 0.134 84 58 8.8 »
Таблица 18
Параметры уравнений роста моллюсков Anodonta anatina
Водоем к (год-1) Длина (мм) /шах (годы) По данным
^СО Lmax
Река, озеро: Англии 0.229 90 81 9.3 Crowley, 1957
Швеции 0.248 110 99 9.1 Okland, 1963
Река: Дон 0.385 85 77 5.7 Троицкого, 1934
Онега 0.070 104 48 10.0 Автора
Сев. Двина 0.302 100" 76 4.7 То же
Ветлуга 0.350 85 68 4.6 »
Теша 0.254 100 85 7.5 »
Ока (среднее течение) 0.394 97 71 3.3 »
Ока (верхнее течение) 0.437 85 70 4.0 »
Озеро: Чудское 0.187 100 65 5.7 »
Колодозеро 0.316 93 82 6.8 »
Погозеро 0.316 93 84 7.4 »
Онежское 0.187 105 69 5.8 »
Ушкозеро 0.187 105 79 7.6 »
Таблица 19
Параметры уравнений роста моллюсков Anodonta ponderosa
(по измерениям автора)
Река к (год-1) Длина (мм) /max (годы)
^ОО Lmax
Клязьма 0.235 220 190 7.9
Ока 0.490 98 95 7.1
Ингулец . .... 0.262 130 117 9.9
Днепр . .... 0.420 110 99 5.5
Волга 0.239 110 100 7.2
105
Таблица 20
Параметры уравнений роста моллюсков Anodonta zellensis
(по измерениям автора)
Река к (год-1) Длина (мм) *max (годы)
Ьсо Lmax
Клязьма 0.223 246 200 7.3
Молочная 0.390 152 134 5.5
Уда 0.336 154 135 6.2
Кубань 0.485 149 142 9.9
Волга 0.315 115 102 6.8
Таблица 21
Параметры уравнений роста моллюсков Anodonta seisanensis
(по измерениям автора)
Река к (год-1) Длина (мм) ^тах (годы)
•^00 Lmax
Чусовая 0.392 86 80 6.8
Обь (верхнее течение) 0.237 86 67 6.4
Лена (верхнее течение) 0.303 62 58 13.4
Лена (среднее течение) 0.212 85 76 10.6
Енисей (среднее течение) 0.234 78 61 6.5
Енисей (нижнее течение) 0.237 84 76 9.9
Подкаменная Тунгуска 0.302 70 57 5.6
Нижняя Тунгуска 0.218 80 62 6.8
Чона 0.282 67 55 6.2
Таблица 22
Параметры уравнений роста моллюсков Anodonta piscinalis
(по измерениям автора)
Река к (год-1) Длина (мм) (годы)
Lqo Lmax
Днепр (у Киева) 0.362 112 90 4.5
Днепр (у Херсона) 0.350 115 101 5.8
Ока (верхнее течение) 0.392 104 97 7.5
Ока (оз. Студенец) 0.415 89 76 4.6
Ока (Велетьминский затон) 0.392 104 86 4.5
Теша ... 0.364 104 85 4.6
Вента 0.282 93 75 5.9
Волга (дельта) 0.387 106 90 4.9
106
Таблица 23
Параметры уравнений роста моллюсков Pseudanodonta
complanata (по измерениям автора)
Водоем А (год *) Длина (мм) 2 max (годы)
^ОО Ьшах
Река:
Днестр 0.420 108 90 3.2
Днепр 0.392 97 79 4.2
Дон 0.342 115 36 5.3
Днепровское водохранилище 0.324 105 80 4.4
Таблица 24
Параметры уравнений роста некоторых дальневосточных
моллюсков (по измерениям автора)
Вид k (год-1) Длина (мм) J max (годы)
•Loo Ьтах
Cristaria plicata ... 0.127 262 246 22.0
Lanceolaria grayana Lea 0.194 112 54 3.8
L. decurtata Shrenck 0.194 163 131 6.7
L, cylindrica (Simps.) 0.230 128 107 8.6
Margaritifera dahurica 0.122 '204 158 12.2
щихся как по темпу роста, таки по особенностям мест обитания,
могут свидетельствовать о реальности полученных параметров
их роста.
Особенности роста Dreissenidae были изучены по литературным
Данным главным образом на моллюсках D. polymorpha (табл. 25).
Как видно из этой таблицы, константа роста дрейссен в разных
водоемах изменялась от 0.122 до 0.492, не превышая в среднем
0.278 год"1, а продолжительность жизни от 4.2 до 11 лет (в сред-
нем около 6.5 года). Кроме того, мы рассчитали параметры урав-
нения роста для моллюсков D. caspia Eichwald из Аральского моря
(по данным Карпович, 1952), которые оказались следующими:
к=0.228 год"1,1/00=18.5 мм, Lmax=15.5 мм, £тах=8 лет. Рост дру-
гих представителей этого семейства пока остается неизученным.
Сведения по росту Corbiculidae очень скудны. Можно при-
вести лишь работы по исследованию скорости роста у Corbicula
manilensis (Morton, 1973), С. fluminalis (Morton, 1977) и C. japo-
nica Prime (Fuji, 1957). Константа роста для первого вида близка
к 0.093 (продолжительность' жизни 8 лет), второго — 0.34,
С. japonica живет около 3—4 лет. Слабая изученность корбику-
107
Таблица 25
Параметры уравнений линейного роста моллюсков
Dreissena poly morpha
Водоем к (год-1) Длина (мм) 2 max (годы) По данным
-Ьоо Lmax
Река Волга 0.248 32.5 26.0 6.6 Карлович, 1952
Аральское море .... Водохранилище: 0.234 29.0 23.2 6.6 То же
Куйбышевское .... 0.492 32.0 23.7 4.2 Михеева, 19646
Пяловское 0.122 45.0 35.1 11.0 То же
Рыбинское 0.235 32.0 24.9 6.4 »
Учинское 0.242 38.0 36.0 8.0 Качановой, 1963
Водоемы Англии .... 0.370 51.0 42.0 4.6 Morton, 1969
Щецинский залив . . . 0.235 35.0 29.4 7.6 Wictor, 1963
Мазурские озера .... 0.343 32.5 28.0 5.4 Stanczykowska, 1963
лид, вероятно, связана с трудностью различения годовых колец
на раковинах моллюсков, которые испещрены линиями кольце-
вой скульптуры. Однако при внимательном изучении скульптуры
раковин этих моллюсков оказывается возможным достаточно на-
дежно различать годовые кольца. Таким способом Мортоном
(Morton, 1973) был изучен рост С. manilensis.
В результате измерений 30 раковин С. purpurea Prime, 25 ра-
ковин С. tibetensis Prash. и 36 раковин С. fluminalis из некоторых
водоемов Средней Азии и Закавказья нами были рассчитаны па-
раметры уравнений роста для этих животных, величины которых,
как видно из табл. 26, не расходятся с литературными данными.
Наибольшая продолжительность жизни отмечена у С. fluminalis,
которые характеризуются низкой скоростью роста.
Рост Sphaeriidae изучался различными методами. Исследова-
ние роста моллюсков Sphaerium suecicum из оз. Круглого прово-
дилось в специально изготовленных садках, установленных не-
посредственно в озере. Опыты длились с 15 июля по 15 августа
1968 г. при средней за этот период температуре воды в озере 15°.
Прослеживался одновременно рост отдельных размерных групп
моллюсков, которые помещались в разные садки. Каждые десять
дней проводились измерения длины раковин, а затем рассчиты-
вались их средние значения за эти отрезки времени. В результате
была получена кривая изменения длины раковин моллюсков
в зависимости от времени наблюдения (рис. 33). При этом учиты-
валось, что средняя длина новорожденного моллюска составляла
2.4 мм, а максимальная длина взрослого животного в данном во-
доеме не превышала 7.2 мм.
Кривая линейного роста моллюсков этого вида показывает,
что если бы они росли при постоянной температуре 15°, то средний
(из наибольших) размер взрослого животного (принято как 90%
108
Таблица 26
Параметры уравнений линейного роста моллюсков
сем. Corbiculidae (по измерениям автора)
Водоем k (год-1) Длина (мм) *тах (годы)
•^00 Ьтах
С. purpurea
Река: Сырдарья 0.394 28.4 22.0 3.7
Амударья 0.336 32.5 27.0 5.7
Зеравшан 0.429 30.6 23.5 3.0
С. tibetensis
Река: Амударья 0.580 28.5 17.0 4.1
Ануй, Чарыша .... 0.580 29.0 27.0 4.5
Аральское море .... 0.429 29.0 25.0 3.0
С. fluminalis
Река: Мургаб 0.320 35.5 31.0 6.4
Араке 0.223 32.0 29.0 6.3
пт максимального для данного водоема) был бы ими достигнут
через 150 сут. Однако весной, осенью и зимой температура воды
в озере значительно ниже, и рост животных в эти периоды резко
замедляется или прекращается. Следовательно, общая продол-
жительность жизни моллюсков S. suecicum при коротком северном
лете и довольно длительном в течение года периоде низких тем-
ператур превышает 12 мес, в то время как, например, в водоемах
Запада и Северо-Запада европейской части СССР длительность
жизни Sphaerium не превышает года (Алимов, 19676). Константа
роста для S. suecicum составляла 0.504 за мес, у S. corneum в од-
ном из прудов г. Ленинграда (Алимов, 19676) не превышала
0.450 за мес, а длительность жизни была около года (табл. 27).
Кривая линейного роста моллюсков Pisidium crassum (рис. 33)
была построена в результате анализа гистограмм динамики раз-
мерно-возрастного состава популяции этих животных в оз. Круг-
лом (рис. 34). При этом были установлены следующие размерные
группы (по длине раковины): 1.2—2.0 мм, 2.1—2.7 мм и 2.8—
3.0 мм. Максимальная длина взрослого животного в этом озере
не превышала 3.0 мм, длина раковины новорожденного — 1.2 мм.
Константа роста моллюсков составляла 0.360 в месяц, а время,
в течение которого они достигали среднего максимального для
данного водоема размера при температуре 15°, равнялось 100 сут.
Параметры уравнений роста Pisidium dubium (Say), Musculium
109
Таблица 27
Параметры уравнений линейного роста моллюсков
сем. Sphaeriidae
Вид h (мест1) Длина (мм) lmax (годы) По данным
Boo Lmax
Sphaerium suecicum . . 0.504 7.5 6.8 1 Автора
S. согпеит 0.450 11.0 9.9 1 То же
Pisidium crassum .... 0.360 3.0 2.7 1 Автора
P. dubium 0.110 9.5 8.55 2 Heard, 1965
Musculium lacustre . 0.486 7.5 6.8 0.3 Митропольского, 1965
M. partumeium 0.357 9.0 8.1 1 Thomas, 1959
lacustre и М. partumeium (Say) рассчитаны в результате анализа
динамики размерного состава популяций этих моллюсков
(табл. 27). Из таблицы видно, что наименьшие значения кон-
станты роста характерны для моллюсков М. partumeium и Р. du-
Ыит.
Рис. 33. Кривые линейного роста моллюсков Sphaerium suecicum (Л), Pi-
sidium crassum (Б) из оз. Круглого.
По оси абсцисс — время (сут); по оси ординат — длина раковины (мм).
У сфериид, животных с наиболее коротким среди двуствор-
чатых моллюсков жизненным циклом, при определении величины
скорости роста за год важно учитывать длительность периода
реального роста в течение года. Так, например, у моллюсков
S. suecicum и Р. crassum, населяющих северные озера, длитель-
ность периода роста в течение года не превышает в среднем 3 мес,
у S. согпеит из средней полосы Советского Союза период роста
около 5 мес, а у М. partumeium — около 6 мес. Если учесть дли-
тельность периода роста в течение года, то величины констант
110
роста для этих животных получатся следующие: S. suecicum —
1.512 год-1; 5. согпеит — 2.250, М. partumeium — 2.142,
М. lacustre — Р. crassum — 1.080, Р. dubium — 0.333 год-1.
Эти величины говорят о том, что среди изученных видов сфериид
наибольшей скоростью роста характеризуются S. согпеит, а наи-
меньшей — М. lacustre и Р. dubium.
Естественно, что продолжительность жизни моллюсков свя-
зана со скоростью их роста. Определение наибольшей продолжи-
3VII
200
ЮО
0
22 VII
II, Г ПТ ттТ
60
о
22 VIII
Рис. 34. Гистограмма размерного состава популяции моллюсков Pisidium
crassum в оз. Круглом (1968 г.).
По оси абсцисс — длина (мм); по оси ординат — численность моллюсков (экз./м2).
тельности жизни пресноводных двустворчатых моллюсков кроме
теоретического интереса имеет практическое значение. Особенно
важно знать предельный возраст для промысловых видов (раз-
личные униониды). Поэтому не случайно ряд исследователей,
изучая этот вопрос, особое внимание уделяли крупным униони-
дам. По мнению, например, Раббла (Rubbel, 1913), длительность
жизни унионид не превышает 10—12 лет, Фишер (Fischer, 1950)
определяет их наибольший возраст в 12—14 лет. Предельный
возраст беззубок составляет 7—10 лет (Wesenberg-Lund, 1937).
В сводке Комфорта (Comfort, 1957), в которой приведены данные
о наибольшей длительности жизни различных двустворчатых
моллюсков, в частности, длительность жизни перловиц определя-
ется в 10 лет, беззубок — 10—15, жемчужниц — в 100 лет.
В табл. 28 приведены наши и литературные данные по продол-
111
Таблица 28
Продолжительность жизни моллюсков (годы)
Вид Средняя Максималь- ная Источник
Unio tumidus U. pictorum .... A nodonta anatina A. piscinalis A. piscinalis { А. cygnea Pseudanodonta complanata . . Margaritifera margaritijera . . j Dreissena polymorpha { Sphaeriastrum rivicola .... Sphaerium corneum S. suecicum S. corneum S. occidental# S. transversum (Say) Musculium lacustre Pisidium dubium P. fallax P. compressum P. nitidum P. variabile P. walkeri P. casertanum P. henslowanum P. hibernicum P. lilljeborgi P. milium P. pulchellum P. subtruncatum P. casertanum P. conventus Neopisidium montessierianum Pa- ladilhe Около 9 Около 9 Около 7 Около 9 Около 6 12 11 8—11 7-8 8-11 12 10—15 6—8 8—11 7—10 11—12 11—13 7 7 70-80 116 Около 60 Около 100 5 5 Около 2 1 1.5 1 Около 2 Около 1 Около 1 3 1 1 1 1 1 1 1 1 1 1 1 1 3 3 2 Петров, 1964 Negus, 1966 Tudorancea, 1969 Сальдау, 1939 Троицкий, 1934 Negus, 1966 Бластов, 1933 Сальдау, 1939 Троицкий, 1934 Negus, 1966 Okland, 1963 Franz, 1939, 1941 Agre]l, 1949 Троицкий, 1934 Rubbel, 1913 Heidelberg, 1960 Altnoder, 1926 Comfort, 1957 Morton, 1969a Wiktor, 1963 Groenwegen, 1926 Алимов, 19676 Алимов, 1970 Thiel, 1926 Herrington, 1948 Dietz, Stem, 1977 Митропольский, 1965 Heard, 1965 Там же » » » » » » » » » » » Holopainen, 1978 » »
112
жительности жизни различных моллюсков. Из таблицы нетрудно
видеть, что разные исследователи приводят то максимальную,
то среднюю продолжительность жизни животных. По-видимому,
более рационально определять какую-то условную среднюю
продолжительность жизни моллюсков, приняв ее как возраст
животного, имеющего длину раковины, которая представляет
собой среднюю из максимальных величин для данного вида в оп-
ределенных условиях (Lmax), понимая под этим наиболее часто
встречающийся в популяции максимальный размер моллюска.
Возраст животного, имеющего такую длину раковины, можно
считать средним максимальным и наиболее характерным для дан-
ного вида в условиях определенного водоема (£тах). Его величина
легко может быть определена из основного уравнения роста (43)
Lt = La)(l—ё~ы).
Приняв Lt=Lm^ получаем
f ___________________ In T'oo In (Z/qo ^max)
^max- ft •
Рассчитанная таким образом продолжительность жизни изучен-
ных двустворчатых моллюсков представлена в табл. 29. По дан-
ным этой таблицы можно видеть, что отношение Zmax к для
самых разных представителей колеблется в узких пределах.
Обычно Zmax составляет у различных животных 72—98%, в сред-
нем 80% от теоретической максимальной длины. Это весьма по-
лезное соотношение. Принимая его во внимание, можно более
строго интерпретировать результаты определения параметров
уравнения линейного роста, а по рассчитанному уравнению ана-
литически определить среднюю максимальную длину раковины
и соответствующий ей возраст животного.
Сравнивая табл. 28 и 29, легко заметить, что продолжитель-
ность жизни моллюсков, определенная разными авторами с ис-
пользованием различных методов, имеет достаточно близкие ве-
личины. Наши данные почти не отличаются от литературных.
Однако определение среднего максимального возраста имеет,
на наш взгляд, преимущество — единообразие способа опреде-
ления. Кроме того, поскольку в уравнении роста представ-
ляет собой теоретический максимальный размер (асимптота на
графике), к которому стремится животное в процессе роста,
время жизни моллюска при приближении к этому размеру ста-
новится неопределенным. Поэтому, ограничивая определение наи-
большего возраста животного размерами его раковины, соответ-
ствующими 80% теоретического максимального размера, мы
тем самым отражаем наиболее реальную и экологически более
значимую величину длительности его жизни. В дальнейшем
будем называть ее реальной продолжительностью жизни fmax.
8 А. Ф. Алимов
ИЗ
Таблица 29
Параметры уравнений линейного роста для разных видов моллюсков
Вид Число измерений Zco (мм) (У lmax (mm) (У /max (годы) <r к (год-1) lmax /Leo (%)
Margaritifera margaritifera . . 13 124.20 3.48 117.80 4.35 73.60 5.570 0.038 0.003 95
Unio tumidus 36 95.97 1.60 75.60 2.72 8.65 0.480 0.244 0.009 79
U. pictorum 31 95.96 2.43 82.85 2.46 7.70 0.326 0.282 0.009 87
Crassiana crassus 13 78.30 2.70 61.38 2.52 7.85 0.224 0.222 0.020 79
A nodonta anatina 14 96.57 । 2.05 75.25 4.18 6.50 0.549 0.236 0.030 79
A. ponderosa 5 133.60 25.62 120.00 19.95 7.50 0.924 0.369 0.055 91
A. zellensis 5 163.20 27.50 142.60 20.60 7.20 0.924 0.350 0.056 88
A. seisanensis 9 77.56 2.86 66.22 3.00 8.00 0.960 0.269 0.021 85
A. piscinalis 8 103.40 3.12 87.50 3.38 5.30 1.040 0.369 0.018 85
Pseudanodonta complanata . . . 4 107.00 5.04 86.30 4.76 4.30 0.588 0.370 0.027 81
Cristaria plicata 3 262.0 — 246.00 — 22.00 — 0.127 — 94
Dreissena polymorpha 10 35.5 2.38 29.10 1.50 6.50 0.704 0.278 0.040 82
Corbiculidae 9 29.6 2.58 25.60 1.68 5ДЗ 0.715 0.402 0.039 87
Lanceolaria grayana 1 112.0 — 54.00 — 3.80 — 0.194 — —
L. decurtata 2 163.0 — 131.00 — 6.70 — 0.194 — 80
L. cylindrica 1 128.0 — 107.00 — 8.60 — 0.230 — 84
Примечание. В графе «число измерений» приводится количество водоемов, в которых исследовался рост моллюсков; — средняя
квадратичная ошибка.
Из данных табл. 29 следует, что виды одного рода отличаются
по величине константы роста. Например, нетрудно рассчитать,
что моллюски Unto tumidus, U. pictorum, а также Anodonta ana-
tina. A, seisanensis, A. piscinalis имеют статистически достоверные
(с вероятностью 0.95) отличия между собой по этому признаку.
Рассмотренные выше особенности роста некоторых предста-
вителей пресноводных двустворчатых моллюсков позволяют
прийти к заключению о том, что каждый вид моллюсков кроме
морфологических признаков характеризуется свойственными ему
дефинитивными размерами, определенной длительностью жизни
и специфической скоростью роста. Так же как и вся организа-
ция животного, эти признаки представляют собой результат исто-
рического развития данного вида и закреплены наследственно
(Алимов, 1974).
До сих пор мы рассматривали постэмбриональный рост мол-
люсков, полагая в уравнении роста, что ко времени t0 длина мол-
люска равна нулю, т. е. £о=О. Необходимо определить теорети-
ческий возраст tQ, при котором длину моллюска можно считать
равной нулю. В таком случае мы будем как бы принимать во вни-
мание личиночный период роста животных. Уравнение роста
может быть переписано в виде
—е-м (45)
или с учетом Ло соответственно
Л0 = Л00(1-^о),
откуда
£°°-£» =е-^. (46)
Ьоо
Поделив (45) на (46), получаем
Lt = Лоо - (Лоо - ло) (47)
Определение теоретического возраста t0 производится следую-
щим образом. При вычерчивании зависимости 1g (Л т—Lt)=f (t)
на графике получается прямая линия с отрицательным наклоном.
По графику находится значение 1g (Лт—Lt) при £=0, а из урав-
нения роста — значение tQ (Бивертон, Холт, 1969). В таком слу-
чае кривая роста охватывает весь период развития животного.
Величина же tQ будет показывать рассчитанную длительность
личиночного развития, которая может не совпадать с фактиче-
ской, когда закономерности роста эмбрионов и личинок иные,
чем в постэмбриональный период. Рассчитанные значения
(годы) показывают, что наибольшее время личиночного разви-
8*
115
тия следует ожидать у моллюсков Margaritifera margaritifera*.
М. margaritifera — 0.94, Unio tumidus — 0.30, U. pictorum —
0.25, Crassiana crassus — 0.54, Anodonta piscinalis — 0.43,
A. ponderosa — 0.16, Pseudanodonta complanata — 0.20, Cristaria
plicata — 0.40, Dreissena polymorpha — 0.41, D. caspia — 0.16.
Разумно поставить вопрос, в какой степени полученные нами
расчетные величины tQ соответствуют реальному времени личи-
ночного развития отдельных видов. Экспериментами по росту
отдельных личиночных стадий моллюсков Dreissena polymorpha
б Куйбышевском водохранилище было установлено, что этот пе-
риод жизни животных в среднем близок к 130 дням (около 0.36
года) (Кирпиченко, 1)62). Вынашивание глохидиев в жабрах
Unio tumidus в оз. Хепо-Ярви составляло 20—40 дней, а их пара-
зитирование на рыбах 22—35 дней (в зависимости от температуры).
Весь же период развития личинок с момента оплодотворения у мол-
люсков этого вида занимает около 3 мес (0.25 года), а у моллюсков
U. pictorum около 2.5 мес (0.2 года) (Жадин, 1938). Сравнение
этих данных с ранее приведенными показывает, что расчеты t^
дают достаточно точное представление о длительности личиноч-
ного развития моллюсков. С другой стороны, отношение tjtm^
у исследованных моллюсков не превышает 0.08 (8%): Margari-
tifera margaritifera — 0.013, Unio tumidus — 0.034, U. pictorum—
0.032, Crassiana crassus — 0.068, Anodonta piscinalis — 0.080,
A. ponderosa — 0.021, Pseudanodonta complanata — 0.050, Cri-
staria plicata — 0.020, Dreissena polymorpha — 0.060. Столь ма-
лые величины отношения tQltm^ позволяют не принимать во вни-
мание tQ при анализе особенностей линейного роста моллюсков.
Интересно, что у моллюсков, согласно данным табл. 29, длитель-
ность жизни различается в 17 раз, в то время как различия
в продолжительности их личиночного развития не превышают
6 раз.
Конечные размеры глохидиев унионид или велигеров дрейссен
весьма различны (табл. 30). Наиболее крупные личинки у Ano-
donta и Unio, наиболее мелкие — у Margaritifera, глохидии
Nodularia и велигеры Dreissena по своим размерам занимают сред-
нее положение.
Сравнивая между собой данные табл. 29 и 30, нетрудно за-
метить, что длина личинок различных видов оказывается несоиз-
меримо малой по отношению к размерам раковин взрослых жи-
вотных, составляя не более 0.2—0.4% длины последних. Поэтому
при расчетах кривых роста моллюсков мы можем без больших
погрешностей пренебречь и величиной Ло. В таком случае исполь-
зование для наших целей уравнения роста
особенно для долгоживущих моллюсков, вполне оправданно.
Важным оказалось сопоставление средних для различных
пресноводных моллюсков значений констант роста с размерами
116
Таблица 30
Средняя длина личинок (глохидиев и велигеров)
различных моллюсков
Вид
Средняя длина
(мкм)
Источник
Margaritifera margaritifera . .
Unio pictorum........................
U. gibbosus Raf......................
U. tumidus...........................
Crassiana crassus....................
Nodularia douglasiae.................
Anodonta cygnea .....................
A, piscinalis........................
Pseudanodonta complanata ......
Cristaria plicata....................
Anodonta grandis Say.................
A. corpulenta Cooper.................
Tritogonia verrucosa (Raf.)..........
Lampsilis anodontoides (Lea).........
Dreissena polymorpha.................
48
220
200
162
145
140—180
340
330
285
264—320
410—420
350
85
210
100—150
Жадин, 1938
Там же
Surber, 1912
Жадин, 1938
Там же
»
»
»
»
»
Surber, 1912
Там же •
»
»
Кирпиченко, 1964
личинок (глохидиев, великонхов), с которых начинается пост-
эмбриональный рост (рис. 35). Там же приведены данные и для
некоторых морских видов.1 Из рис. 35 следует, что у видов, ха-
рактеризующихся быстрым ростом, личинки оказываются боль-
шего размера, чем у более медленно растущих животных.
Среди изученных моллюсков можно наметить три группы.
К первой относятся перловицы, беззубки, дрейссены (пресновод-
ные) и морской моллюск Mytilus edulis, ко второй — главным
образом морские виды — Cerastoderma edule, Mytilaster linneatus,
Adacna minima Ostr., Mezuchopecten yessoensis (ley), Spisula sa-
chalinensis (Schrenck), а также пресноводный моллюск Lampsilis
seliquides (Barnes), к третьей — Margaritifera margaritifera, Cri-
staria plicata, Crenomytilus gray anus (Dunker). Между констан-
той роста и длиной личинок I скорее всего имеется зависимость,
которая может быть аппроксимирована степенной функцией
k = clm. (48)
Существенно, что такого же рода связь скорости роста и размера
личинок (как можно видеть на примере двух видов азовских
моллюсков) наблюдается и в период эмбрионального роста.
По данным А. Ф. Карпевич (1964), моллюски Syndesmya ovata
(Phil.) растут более интенсивно по сравнению с Corbulomya moetica
1 Значения констант роста для морских представителей были рассчитаны
по данным А. Ф. Карпевич (1964), А. Н. Голикова, О. А. Скарлато (Golikov,
Scarlato, 1970). Размеры личинок соответствующих видов взяты из работ
Торсона (Thorson, 1946), А. Ф. Карпевич (1962, 1964), К. А. Захваткиноп
(1959).
117
(Milachewich). Заметим, что у первых длина личинок близка
к 150 мкм, у последних — не превышает 72 мкм. По-видимому,
указанная выше зависимость константы роста и размера личинок
вполне реальна. Она имеет статистический характер и дальней-
шие более подробные ис-
следования позволят найти
ее количественное выра-
жение.
Максимальная длина
животных £тах по урав-
нению (43)
^тах--^оо (1 ^тах)*
Произведя необходимые
преобразования и поделив
обе части равенства на
цолучаем
^тах
^оо
Рис. 35. Зависимость константы роста от
длины личинки у различных двустворчатых
моллюсков (из: Алимов, 1977).
1 — Anodonta ponderosa; 2 — A. piscinalis; 3 —
Pseudanodonta complanata; 4 — Cristaria plicata;
5 — Lampsilis seliquides; 6 — Margiritifera marga-
ritifera; 7 — Unio tumidus; 8 — U.* pictorum;
9 — Crassiana crassus; 10 — Dreissena polymorpha;
11 — Tritogonia verrucosa; 12 — Cerastoderma edule;
13 — Mytilaster linneatus; 14 — Adacna minima;
15 — Mytilus edulis; 16 — Macoma baltica', 17 —
Mezuchopecten yessoensis; 18 — Spisula sachalinensis;
19 — Crenomytilus grayanus. Масштаб логарифми-
ческий. По оси абсцисс — длина личинки, (мм);
по оси ординат — константа роста (год'1).
Подставив в это уравне-
ние значения константы
роста из уравнения (48),
находим
f —_____L \/
‘'max с]т
Х1п(1—^-). (49)
Используя уравнение (49),
можно рассчитать реаль-
ную продолжительность
жизни животных, зная
Lmax и длину личинки.
Уравнение (49) показывает, что чем меньше размер личинки
(или скорость роста), тем дольше живут моллюски. Поскольку
пока еще недостаточно данных для количественного выражения
параметров зависимости константы роста и длины личинок, она
представлена только графически, а средние значения констант
роста как функции длины личинок лежат на проведенных пункти-
ром линиях. Тем не менее, используя их, можно попытаться оце-
нить по уравнению (49) реальную продолжительность жизни мол-
люсков, имеющих определенную длину личинки при разных
значениях отношения L^IL^. Пример такого расчета при
£тах/£оо=0.9 приведен в табл. 31, из которой видно, что теоре-
тически (с учетом ранее высказанных допущений) реальная про-
должительность жизни изученных моллюсков не превышает
118
60 лет, что согласуется с литературными данными. Такого рода
расчеты имеют важное значение, так как могут служить свое-
образным контролем при изучении роста моллюсков разными
методами.
Таблица 31
Продолжительность жизни моллюсков (годы)
при Z/max/Z/oo = 0.9 (из: Алимов, 1977)
Группа моллюсков (согласно рис. 35) Длина личинки (мм)
0.05 0.1 0.2 0.3 0.4
I 19.2 12.1 8.1 7.0 5.8
II 27.0 16.4 12.0 10.0 8.2
III 57.5 33.0 23.0 18.0 14.4
Изучение скорости линейного роста и продолжительности
жизни различных видов основных семейств пресноводных дву-
створчатых моллюсков, населяющих водоемы территории СССР,
показало, что у каждого вида наблюдается изменчивость скорости
роста. Пределы внутривидовых изменений скорости роста у близ-
ких видов достоверно различаются между собой, и их можно рас-
сматривать наравне с морфологическими как видовые признаки.
Скорость роста, связанная также с размерами личинок отдель-
ных видов, в значительной степени определяет продолжитель-
ность жизни моллюсков. Наиболее важным с экологических по-
зиций представляется определение реальной продолжительности
жизни животных, которая отражает наибольший возраст, до ко-
торого доживают пусть 5—10% особей одного поколения, а не
отдельные долгожители. Параметры уравнения роста животных
имеют статистический характер. В связи с этим для получения
их достоверных значений необходимо проводить исследования
на достаточно больших выборках.
ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ
НА СКОРОСТЬ РОСТА МОЛЛЮСКОВ
Многочисленными исследованиями на разных животных было
установлено, что скорость их роста, конечные размеры и длитель-
ность жизни в значительной степени определяются условиями
окружающей среды. Можно привести много примеров, показы-
вающих справедливость этого. Ограничимся лишь одним. На-
пример, дрейссены в Волгоградском водохранилище быстрее
растут в тех его частях, где более высокая температура, нет рез-
ких суточных ее колебаний, нет прибоя, меньше минеральных
взвесей в воде (Спиридонов, 1969).
119
Значения параметров уравнений линейного роста моллюсков
одного вида, как уже было показано, различны в разных водоемах.
Важным и почти не разработанным, по крайней мере для пресно-
водных двустворчатых моллюсков, остается вопрос о степени и
характере влияния факторов среды на их рост. Не менее суще-
ственно выяснить, при каких условиях определенный вид будет
характеризоваться наибольшими величинами параметров роста.
Поскольку невозможно охватить все разнообразие факторов среды
и проследить их воздействие на рост моллюсков, ограничимся
лишь некоторыми из них. Рассмотрим, в какой степени измене-
ния окисляемости и жесткости воды, скорости течения и темпе-
ратуры влияют на величину константы роста у наиболее широко
распространенных моллюсков в пределах их ареалов.
Окисляемость воды
Окисляемость воды по отношению к животным с фильтрационным
способом питания может рассматриваться как фактор, находя-
щийся в прямой связи с пищевыми условиями, так как в неокра-
шенных водах она в известной степени зависит от количества
сестона. Конечно, окисляемость воды, а не одних взвешенных
в воде органических частиц, не может рассматриваться как по-
казатель пищевых условий в чистом виде, но в значительной мере
их характеризует.
Обеспеченность пищей — один из факторов, влияющих на
рост животных. Для морских моллюсков это было показано экс-
периментальными и полевыми исследованиями на ряде видов
(SegerstrAle, 1960; Stickney, 1964; Walne, 1965, 1970; Theisen,
1967—1969; Seed, 1969, и др.). Пресноводные моллюски в этом
отношении оказались почти не изученными. Можно отметить
лишь экспериментальные исследования роста трех видов сфериид
при питании разными видами взвешенных частиц (бактериально-
инфузорная взвесь, Chlorella, Ankistrodesmus} (Митропольский,
1970) и Dreissena polymorpha (Walz, 1978b) при различных кон-
центрациях пищи.
Оценим влияние окисляемости воды на рост различных пред-
ставителей пресноводных моллюсков на основе полученных ранее
данных линейного роста и сведений по окисляемости воды в ме-
стах их обитаний.
Моллюски Margaritifera margaritifera населяют водоемы с окис-
ляемостью воды от 4.5 до 14 мгО/л, однако наиболее интенсивный
рост у них наблюдается в пределах окисляемости от 5.5 до
8.5 мгО/л (рис. 36). Известно, что окисляемость воды — один
из важнейших факторов, ограничивающих распространение этих
моллюсков (Жадин, 1939).
Моллюски родов Unio и Crassiana населяют водоемы с окисля-
емостью воды от 3.5 до 14 мгО/л (рис. 37). Из приведенного ри-
сунка видно, что наиболее быстрый рост у моллюсков U. tumidus
120
наблюдается при окисляемости воды 6—9 мгО/л, U. pictorum
и С. crassus быстрее всего растут при более низких величинах
окисляемости — 5—8 и 5—7 мгО/л соответственно. Кривые
изменения константы роста у этих моллюсков в зависимости
Рис. 36. Зависимость константы роста у Crassiana crassus (X), Unio pictorum
(Б) и U. tumidus (В) от окисляемости воды (из: Алимов, 1974).
По оси абсцисс — окисляемость воды (мгО/л); по оси ординат — константа роста (год-1).
от окисляемости воды имеют достаточно четко выраженные мак-
симумы. У наиболее широко распространенных видов U. tumidus
и Z7. pictorum значения констант роста не так резко варьируют
—I_______________________I_______I_______1_______1—
2 6 8 Ю 12 /4
Рис. 37. Зависимость константы роста у моллюсков Margaritifera margariti-
fera от окисляемости воды (из: Алимов, 1974).
Обозначения как на рис. 36.
при изменениях окисляемости, как у более специализированных
к местообитанию С. crassus.
Виды рода Anodonta обитают в водоемах с окисляемостью воды
от 4.0 до 17 мгО/л (рис. 38). Для широко распространенных
A. piscinalis и A. anatina наибольшие скорости роста отмечаются
в более широком диапазоне значений окисляемости: около 5—
121
9 мгО/л у A. anatina и 5—11 мгО/л у A. piscinalis. У моллю-
сков A. seisanensis, населяющих преимущественно реки Сибири
со своеобразным режимом, оптимальные величины окисляемости
оказываются в пределах 10—14 мгО/л. Кривая зависимости кон-
стант роста от окисляемости воды для этих животных имеет чет-
кий максимум, сдвинутый в область более высоких, чем для дру-
гих изученных видов беззубок, значений окисляемости.
Рис. 38. Зависимость константы роста у Anodonta piscinalis (Л), A. seisa-
nensis (Б), A. anatina (В) от окисляемости воды (из: Алимов, 1974).
Обозначения как на рис. 30.
Моллюски Dreissena polymorpha живут в водоемах с окисляе-
мостью воды от 8 до 14 мгО/л (рис. 39). Наиболее интенсивный
рост их отмечен при окисляемости воды 9—11 мгО/л.
Относительно представителей семейств Sphaeriidae и Corbi-
culidae пока не имеется сколько-нибудь надежных данных, позво-
ляющих сделать какие-либо заключения о влиянии окисляемости
воды на скорость их роста.
Таким образом, представители семейств Margaritiferidae,
Unionidae, Dreissenidae живут в водоемах с окисляемостью от
3.5 до 17 мгО/л. Для всех изученных видов верхняя и нижняя
границы ее значений практически одинаковы. В то же время от-
дельным видам свойственны в разной степени отчетливо выражен-
ные зоны окисляемости, в которых их рост протекает с наиболее
высокими скоростями. Их можно рассматривать как зоны опти-
мума по этому фактору, различающиеся у видов одного рода.
Для некоторых пресноводных моллюсков в свое время были
определены так называемые экологические спектры (Жадин,
122
1938, 1952). Рассматривая наши данные с этих позиций, можно
заметить, что, хотя униониды и населяют водоемы, относящиеся
к олиготипу или мезотипу по окисляемости воды, зоны оптималь-
ных значений окисляемости для
0.0
0.2
них лежат в олиготипе по этому
фактору. Исключение состав-
ляют A nodonta seisanensis — наи-
большие скорости роста этих
моллюсков приходятся на мезо-
ОА -
0.2
____।____।___।___।____।___।____1_
О 2 4 6 8 10 12 14
Рис. 39. Зависимость константы
роста у моллюсков Dreissena poly-
morpha от окисляемости воды (из:
Алимов, 1974).
_______j_______।______।_______।_______1_
0 г 4 6 8 10
Рис. 40. Зависимость константы
роста у моллюсков Margaritifera
margaritifera от содержания каль-
ция в воде (из: Алимов, 1974).
Обозначения как на рис. 36.
По оси абсцисс — содержание Са** (мг/л);
по оси ординат — константа роста (год-1).
тип по окисляемости. Дрейссены также наиболее активно растут
в мезотипе окисляемости. Эти данные достаточно хорошо, согла-
суются с ранее сделанными нами заключениями о том, что унио-
ниды и дрейссены — наиболее активные фильтраторы среди прес-
новодных моллюсков — могут получать необходимое количество
пищи в водоемах с невысоким содержанием взвешенных в воде
органических веществ.
Содержание кальция в воде
Кальций составляет около 52—53% веса раковины двустворча-
тых моллюсков, которая на 98% состоит из СаСО3, остальное
приходится на фосфорнокислый кальций, магний, полисиликат
кальция, кремневокислый магний и азотсодержащие органиче-
ские вещества (Овчинников, 1932, 1933).
Наиболее подходящие условия для роста большинства мол-
люсков оказываются в водоемах с умеренно жесткой водой с со-
держанием кальция от 20 до 100 мг/л, лишь Margaritifera marga-
ritifera предпочитает воды с малым содержанием кальция 5 —
20 мг/л. Наиболее интенсивный рост у них отмечен при содержа-
нии кальция около 6—7 мг/л (рис. 40).
Несмотря на то что моллюски могут населять водоемы с раз-
личной концентрацией кальция, оптимальные его количества,
при которых их рост происходит быстрее всего, имеют значительно
более узкий диапазон, отличающийся у разных видов.
123
Среди моллюсков Unio и Crassiana оптимальные условия
роста С. crassus находятся при концентрациях кальция близких
к 100 мг/л, у широко распространенных U. tumidus и U. pictorum
соответственно при 60—70 и 50—60 мг/л (рис. 41). У беззубок
оптимум роста по сравнению с перловицами несколько сдвинут
в сторону меньших концентраций кальция (рис. 42): для A nodonta
Рис. 41. Зависимость константы роста у моллюсков Crassiana crassus (Л),
Unio pictorum (J5), U. tumidus (В) от содержания кальция в воде (из: Алимов,
1974).
Обозначения как на рис. 40.
anatina он наблюдается при 35—60 мгСа/л, для A. piscinalis
при 35—50, для A. zellensis при 40—50, для A. seisanensis —
при концентрациях кальция около 30 мг/л. Пожалуй, только
A. ponderosa растут интенсивнее в более жестких водах с содержа-
нием кальция 50—70 мг/л. У моллюсков Dreissena polymorpha.
по-видимому, наиболее быстрый рост при концентрациях каль-
ция близких к 70 мг/л (рис. 43).
Приведенные на рис. 40—43 кривые изменения константы
роста моллюсков в зависимости от содержания кальция в воде
показывают, что большинство унионид растут более интенсивно
в мезотипе кальция. При этом перловицы предпочитают более
жесткие по сравнению с беззубками воды.
124
8
Рис. 42. Зависимость константы роста у моллюсков Anodonta zellensis (Л),
А. ponderosa (Б), Л. piscinalis (В), Л. seisanensis (Г), Л. anatina (Д) от содержа-
ния кальция в воде (из: Алимов, 1974).
Обозначения как на рис. 40.
Таблица 32
Вес створок J₽c (г) и длина раковины L (мм) трех видов унионид в разных реках (из: Жадин, 1938)
Река
Ока Ока (затоны) Сура Варнава Теша Сережа Ветл у га Верхний Ик Нижний Ик Ока (Башкирия)
Unto pictorum
L 68 76.24 ±1.1 93.16 ±0.38 87.9 ±1.4 69.5 ±1.54 69.08 ±1.7 61.5±2.9 108.75 ±0.7 109.5 ±0.6 —
wc 6.25 7.37 ±0.3 14.25 ±0.73 13.31 ±0.7 5.88 ±0.38 U. tumidus 6.8 ±0.55 4.08 ±0.29 24.5 ±0.5 26.5 ±0.1 —
L 62.5 ±0.47 70.56 ±0.92 84.4±1.1 87.2±1.2 65.9 ±2.1 — 51.98 ±0.55 86.16 ±1.5 101.9 ±0.8 103.8 ±1.03
Wc 5.97 ±0.47 8.08 ±0.33 15.79 ±0.74 16.09 ±0.4 6.38 ±0.46 Crassiana crc ISSUS 4.12 ±0.15 19.16 ±0.9 37.0±l.l 42.5±1.5
L 53.11 — 76.18 51.92 63.36 60.02 — 75.0 84.2 —
6.11 — 14.50 5.21 7.68 7.64 — 15.7 22.92 —
Существует вполне определенное мнение о том, что увеличение
концентрации солей кальция в воде до определенных пределов
влечет за собой утолщение створок раковин моллюсков (Wil-
bur, Owen, 1964). Изучая изменчивость унионид, В. И. Жадин
(1938) пришел к выводу о большой внутривидовой изменчивости
среди этих моллюсков и выделял различные морфы. Среди перло-
виц, например, выделялись морфы рек с мягкой, умеренно жест-
кой, жесткой и ультражесткой водой, морфы запруд с водой раз-
личной жесткости и т. д. У мол-
люсков С7. tumidus было отмечено
уменьшение длины животного по
мере падения жесткости воды.
Иными словами, подразумевалось,
что жесткость воды в значитель-
ной степени определяет особен-
ности внешней морфологии рако-
вины моллюсков.
Попытаемся рассмотреть это
положение в количественном ас-
пекте. В качестве показателя тол-
щины раковины можно исполь-
зовать ее вес. В табл. 32, взятой
из монографии В. И. Жадина,
в каждом случае приведен вес однс
Рис. 43. Зависимость константы
роста у Dreissena polymorpha
от содержания кальция в воде.
Обозначения как на рис. 40.
створки раковины и ее длина
у трех видов унионид из водоемов, различающихся по содержанию
кальция. Данные этой таблицы были нанесены на графики, на
которых, кроме того, приведены веса створок раковин у этих же
моллюсков, определенные другими исследователями (рис. 44) г
и измеренные автором веса створок у трех видов Anodonta из раз-
ных водоемов (рис. 45). Все эти реки различаются по содержанию
кальция в воде. В р. Оке, например, его концентрация составляет
около 60 мг/л, в р. Суре — 80.3, в р. Ветлуге — 24.1 мг/л и т. д.
Тем не менее, как следует из приведенных рисунков, вес раковины
находится в прямой зависимости от ее длины. С другой стороны,
не выявляется четкой корреляции между длиной раковины мол-
люсков и содержанием кальция в воде различных водоемов
(табл. 33).
Таблица 33
Длина раковин моллюсков Unio tumidus в водоемах
с разным содержанием кальция в воде
Река Са-- (мг/л) Ьоо (мм) ^тах (мм) Река Са-- (мг/л) Ьоо (мм) tmax (мм)
Оскол 72.8 110 99 Сура 80.3 103 79
Днестр 34.4 98 75 Дон 90.0 100 71
Проня 88.1 НО 99 Ока 60.0 100 71
Упа 76.7 70 63 Вента 59.5 71 49
127
A
Рис. 44. Зависимость веса створки раковины от ее длины у моллюсков
А — Unio tumidus: 1 — разные водоемы (по данным Жадина, 1938); 2 — р. Темза (Negus
1966); 3 — р. Волга (Лукин, 1937в), Б — U. pictorum: 1 — разные водоемы (Жадин, 1938)
2 — р. Свияга (Лукин, 1937а); 3 — р. Темза (Negus, 1966). В — Crassiana crassus:
1 — разные водоемы (Жадин, 1938); 2 — р. Свияга (Лукин, 1937а). По оси абсцисс —
длина раковины (мм); по оси ординат — вес створки (г).
Рис. 45. Зависимость веса створки раковины от ее длины у моллюсков.
А — Anodonta piscinalis: 1 — р. Ока; 2 — р. Теша; 3 — р. Днепр; 4 — Куршский залив.
Б — A. anatina: 1 — р. Теша; 2 — р. Онега; 3 — оз. Чудское; 4 — р. Ока. В — A. sei-
sanensis: 1 — р. Нижняя Тунгуска; 2 — р. Енисей (измерения автора). Остальные обо-
значения как на рис. 44.
Все это вместе взятое позволяет заключить, что содержание
кальция в воде не влияет прямо на толщину створок раковин
моллюсков. Количество его в воде в значительной степени опре-
деляет скорость роста этих животных, с которой в свою очередь
связаны размеры и толщина створок их раковин.
Скорость течения воды
Скорость течения воды накладывает определенный отпечаток
на темп роста моллюсков. Так, например, константа роста для
Unto tumidus в мельничном пруду р. Свияги не превышает (в ра-
счете за год) 0.267, в самой реке — 0.135, для U. pictorum соот-
Рис. 46. Зависимость константы роста у моллюсков Crassiana crassus (А),
Unio pictorum (Б), U. tumidus (В) от скорости течения воды.
По оси абсцисс — скорость течения (м/с); по оси ординат — константа роста (год-1).
ветственно 0.268 и 0.392 (рассчитано по данным Лукина, 1937а).
Оба вида растут быстрее на течении в реке, по сравнению с мель-
ничным прудом.
Моллюски Unio и Crassiana населяют реки, в которых ско-
рость течения составляет 0.4—1.3 м/с, но для каждого вида
характерны свои оптимальные скорости течения, при которых
их f рост происходит более интенсивно (рис. 46). Моллюски
U. tumidus растут с большей скоростью при течении воды около
0.7 м/с, U. pictorum — 0.5—0.55, С. crassus — около 0.8 м/с.
130
Беззубки населяют реки со скоростями течения от 0.2 до 2.0 м/с.
Однако европейские широко распространенные A nodonta anatina
и A. piscinalis живут в реках, скорость течения воды в которых
не превышает 1.0 м/с. Наиболее реофильными оказываются
A. seisanensis, живущие в реках со скоростью течения от 0.6 до
2.0 м/с. Так же как перловицам, каждому виду беззубок свой-
ственны свои оптимальные скорости течения, при которых их
рост протекает быстрее всего: 0.6—0.7 м/с для A. anatina, 0.8 для
A. piscinalis, около 1 м/с для A. seinsanensis.
Анализируя рассмотренные данные, можно сказать, что зоны
оптимума роста унионид находятся в мезотипе течения. Фильтра-
ционные способности унионид, которым свойственны высокие коэф-
фициенты фильтрации, обеспечивают возможность получения
моллюсками необходимых для роста количеств пищевого мате-
риала в условиях постоянно движущейся с достаточно высокими
скоростями воды.
ТЕМПЕРАТУРНЫЕ УСЛОВИЯ
Влияние температуры, этого важнейшего фактора среды, на рост
двустворчатых моллюсков изучалось главным образом на мор-
ских представителях этих животных (Сое, Fox, 1944; Сое, 1948;
Taylor, 1959; Segerstr^le, 1960; Stickney, 1964; Walne, 1965;
Ansell, 1968; Theisen, 1967—1969; Quayle, 1969, и др.).
Этими работами, в частности, было показано возрастание скоро-
сти роста моллюсков при увеличении температуры до определенных
пределов, превышающих оптимальные для конкретных видов.
Дальнейшее повышение температуры вызывало уменьшение
темпа роста. К сожалению, в отношении пресноводных моллюсков
такие работы крайне немногочисленны. Можно привести лишь
отдельные исследования влияния температуры на рост несколь-
ких видов (Howard, 1922; Morton; 1969; Митропольский, 1970).
Изучение роста моллюсков при разных температурах этими авто-
рами проводилось либо экспериментально, либо такого рода
связь устанавливалась в результате обработки полевых наблю-
дений. Эксперименты проводились с видами, имеющими короткий
•жизненный цикл, например Pisidium obtusdle (Pf.) (Митрополь-
ский, 1970), или с личинками видов с длительным жизненным
циклом (Кирпиченко, 1971а, 19716). Полученные сведения по
росту пресноводных моллюсков неполны, отрывочны и не дают
количественной оценки явления.
Влияние температуры на рост моллюсков можно рассматри-
вать в сезонном и широтном аспектах. В каждом биотопе неза-
висимо от его географического положения температура воды
в течение года претерпевает значительные колебания. Результаты
обобщения немногочисленных литературных материалов показали,
что скорость роста (абсолютный прирост — &L/&t) и темпера-
тура Т в пределах изученных значений связаны между собой экс-
поненциальной зависимостью (экспериментальные точки доста-
9*
131
точно хорошо укладываются на прямую в полулогарифмическом
масштабе; рис. 47), которая может быть выражена в виде
^L/M = ce^T, (50)
где с и ср — константы уравнения.
Существенно, что такого же вида зависимость между констан-
той роста и температурой была установлена для моллюсков
o.i\-
Рис. 47. Зависимость линейного прироста у моллюсков от температуры.
А — Pisidium obtusale (Митропольский, 1970); Б — Lampsilis radiata (Howard, 1922).
Масштаб полулогарифмический. По оси абсцисс — температура (°C); по оси ординат: А —
линейный прирост (мм/сут); Б —то же (мм/год).
Siliqua patela (Taylor, 1959). Эта работа представляет специаль-
ный интерес, так как в ней изучался рост моллюсков от северной
до южной границы их ареала (средние температуры на севере
не превышали 5°, на юге были около 14°).
Все указанные исследования проводились в интервалах тем-
ператур, которые характерны для природных биотопов. Это важно
отметить, поскольку в данном случае не превышался температур-
ный оптимум для вида, и поэтому не наблюдалось снижения ско-
рости роста при температурах выше оптимальных.
132
Кривая с максимумом в области оптимальных температур
и уменьшением скорости процесса при температурах выше опти-
мальных была теоретически обоснована Янишем (Janisch, 1927)
и впоследствии использована для введения температурной зави-
симости в уравнение роста (Ursin, 1963; Theisen, 1967—1969).
Однако увеличение продолжительности развития, или уменьше-
ние скорости роста при наивысших температурах, наблюдаемое
в лаборатории, не имеет места в природных условиях, поскольку
такие температуры просто не свойственны природным местообита-
ниям моллюсков. Поэтому не имеет смысла рассматривать пра-
вую нисходящую ветвь кривой с экологической точки зрения.
По-видимому, экспоненциальная зависимость скорости роста
от температуры воды достаточно правдоподобно описывает это
явление в природных условиях. Поскольку
7 dL 1
*= dFT’
7 dL
следует ожидать, что величина к, как и —, и температура
среды также связаны между собой экспоненциальной зависимостью
к = схе^т. (51)
Обозначив
k = cre^T
через т] или т]=/ (Г), получаем
£, = ^[1-6-^] Или = (52)
В таком виде уравнение линейного роста будет отражать сезонные
изменения роста конкретного вида моллюсков, связанные с дина-
микой температуры в определенном биотопе.
Рассчитав по данным В. И. Митропольского (1970) (>10 для
скорости роста моллюсков Pisidium obtusale, получаем, что в ин-
тервале температур 10—20° его величина составляет 2.51; при
20—25° — 2.04, т. е. оба значения практически не отличаются
от значений ^10 для кривой Крога (соответственно 2.7 и 2.3).
Надо отметить, что невероятно высокие значения (>10 (при 5—10° —
9.25, при 15—20° — 8.17) получаются для моллюсков Lampsilis
radiata Lam. при расчете по данным Ховарда (Howard, 1922).
Это может свидетельствовать лишь о недоброкачественности
исходных данных.
Конечно, столь отрывочные сведения не позволяют делать
какие-либо выводы. Можно лишь предполагать, что зависимость
скорости роста, так же как и скорости фильтрации, от температуры
скорее всего соответствует кривой Крога.
Рассматривая вид в его ареале, мы, естественно, обнаружи-
ваем, что в разных участках ареала животные живут в условиях,
отличающихся по среднегодовым или среднесезонным значениям
133
температуры. Но не обязательно самая низкая температура будет
наблюдаться в северных частях ареала. Например, нижнее течение
р. Сев. Двины расположено на 62—64° с. ш. Средняя за летний
период температура воды на этом участке реки около 14.7° С.
В нижнем течении р. Чусовой (56—57° с. ш.) средние темпера-
туры за тот же период не превышают 10°. Поскольку дрейссены
и униониды прекращают рост при температурах ниже 10°, а мак-
симальные температуры даже в южных частях их ареалов не
превышают 22—23°, вероятно, использование температурной
шкалы для выявления влияния температурных условий на рост
моллюсков в столь узких границах температур малоэффективно.
В ряде случаев моллюски живут в водоемах, характеризую-
щихся довольно близкими по абсолютным величинам средними
температурами. Те же водоемы достаточно сильно различаются
по сумме температур в период активного роста животных. Сумму
температур можно выразить в виде градусодней, учитывая при
этом лишь те периоды, когда температура воды равна или выше 10°.
В таком случае для Unio tumidus самой холодной рекой оказы-
вается р. Сев. Двина, в которой средняя температура воды в период
роста составляет 14.7°, сумма температур за тот же период —
1764 градусодня, а самой теплой — р. Днестр (18°, 3780 градусод-
ней); для U. pictorum соответственно р. Чусовая (10°, 1438
градусодней) и р. Кубань (16.9°, 3335 градусодней), для Anodonta
anatina — р. Онега (14.6°, 1634 градусодней) и р. Ока (14.6°,
2643 градусодня), для A. seisanensis — р. Чусовая и р. Чона
(13.7°, 1671 градусодень). В связи с этим для сравнения скорости
роста моллюсков в разных частях ареала, отличающихся по клима-
тическим особенностям, представляется удобным использование
сумм температур, учитывающих среднюю температуру и продол-
жительность периода роста в конкретных местообитаниях.
На рис. 48 показана связь между константами роста у трех
видов унионид, значения которых взяты из табл. 15, 16, 18, и
суммами температур за период роста моллюсков в соответствующих
водоемах (из разных участков ареалов), рассчитанными по спра-
вочникам. Из приведенного рисунка видно, что величины констант
роста животных находятся в экспоненциальной зависимости от
суммы температур (разброс точек может быть отнесен за счет
влияния прочих неучтенных факторов). Отдельные виды неодина-
ково реагируют на изменения температурных условий. Наиболее
резкая реакция у A. anatina, в меньшей степени сумма темпера-
тур влияет на рост U. pictorum. Вероятно, такие различия свя-
заны с особенностями происхождения видов. В какой-то степени
на их происхождение могут указывать пределы или средние
величины сумм температур за период роста, определенные по всему
ареалу вида. Наиболее холодостойкими из перловиц оказываются
U. pictorum, а из беззубок — А. seisanensis, наиболее теплолюбивы—
A. zellensis и A. piscinalis. В целом моллюски рода Anodonta
несколько более теплолюбивы по сравнению с родом Unio. В то же
134
время можно предполагать, что у этих животных были до-
вольно близкие по климатическим особенностям центры про-
исхождения, так как средние значения сумм температур за период
роста в пределах их ареалов оказываются довольно близкими: для
Unio tumidus — 2707, U. pictorum — 2144, Crassiana crassus —
2550, Anodonta seisanensis — 1553, A, ponderosa — 2589, A. ana-
tina — 2604, A. piscinalis — 2906, A. zellensis — 3021.
Рис. 48. Зависимость константы роста у моллюсков Unio tumidus (А),
U. pictorum (Б), Anodonta anatina (В) от суммы температур (из: Алимов,
1974). Масштаб полулогарифмический.
По оси абсцисс — сумма температур (градусодни); по оси ординат — константа роста
(год-»).
Изученные моллюски довольно стенотермны, даже у самых
широко распространенных видов разница средних температур
в наиболее холодных и теплых частях ареала не превышает 4° .
Поэтому, вероятно, оправданно, что зависимость скорости роста
моллюсков от температурных условии в местах их обитания,
характеризуемых суммой температур за период активного роста,
не имеет вида кривой с зоной оптимума, как это было показано,
например, для Mercinaria mercinaria (Ansell, 1968). Автор поль-
зовался простой температурной шкалой для характеристики
разных частей ареала, средние температуры в которых изменя-
лись от 5 до 31°.
По-видимому, в первом приближении можно считать, что для
изученных моллюсков константа роста и суммы температур (£Г)
135
связаны между собой экспоненциальной зависимостью
к = те6^т^
Обозначив
к = тес^т^ = р,
введем эту зависимость в уравнение роста
£, = £„(1-^). (53)
В таком виде уравнение роста становится более содержатель-
ным и позволяет проследить изменения кривой роста моллюсков
в разных точках видовых ареалов в зависимости от сумм темпе-
ратур, которыми они могут быть охарактеризованы.
Согласно известному правилу Бергмана (Рубнера), животные
одного вида в более северных (холодных) частях ареала достигают
больших размеров. Это правило было установлено для гомойотерм-
ных животных. Впоследствии без достаточного критического
подхода оно было перенесено на пойкилотермных животных,
и в отдельных случаях была показана его справедливость (Кон-
стантинов, 1967; Мина, Клевезаль, 1976).
Не обсуждая правомочность распространения этого правила
вообще на всех пойкилотермных животных (хотя физиологический
механизм его действия у них неясен), попытаемся оценить его
только применительно к изучаемым моллюскам.
Приведенные на рис. 49 данные указывают на прямолинейную
зависимость длины раковины моллюсков от суммы температур
в период их роста в различных областях ареалов, т. е.
Дпах = т + ^7’. (54)
где у и | — константы уравнения.
Следует отметить, что зависимость аналогичного характера
была получена, например, для мидий (Bdetius, 1962). Отмечено
увеличение размеров дрейссен с повышением температуры (Спи-
ридонов, 1969; Кирпиченко, 1971а, 19716). Это означает, по-
видимому, что по крайней мере для пресноводных моллюсков
правило Бергмана вряд ли может быть распространено.
Ранее было показано, что для изученных моллюсков отноше-
ние Zmax/^co довольно постоянная величина. Если
^оо--TO Z/qq----^max/0^
НО
bml,x = /(2?1) ИЛИ £niax = 'r + £2
т. е.
г — Г / _? +
^оо---^тах/^- п
136
Подставив выражение для Lm в уравнение (53), получаем
(55)
В приведенном виде уравнение роста отражает особенности роста
моллюсков в зависимости от температурных условий местообита-
ний с учетом сезонных и широтных изменений температуры.
Рис. 49. Зависимость средней из максимальных длины раковины £тах
у моллюсков Unio tumidus (Л), U. pictorum (Б), A nodonta anatina (В) от суммы
температур.
По оси абсцисс — сумма температур (градусодни); по оси ординат — Lxa&x. (мм).
АЛЛОМЕТРИЧЕСКИЙ РОСТ
В процессе онтогенетического развития животных отдельные
части их тела растут с разными скоростями относительно друг
друга. Изучая закономерности роста отдельных частей организмов,
Гексли (Huxley, 1932) установил закон, называемый законом
гетерогонии, или законом неравномерного, или аллометрического
роста. Аналитически он записывается в виде простого уравнения:
у = ах₽, (56)
где х, у — размеры изучаемых частей (органов) тела; а, Р — кон-
станты уравнения.
137
В этом равенстве не имеется явной зависимости от времени.
Время в нем заменяется неким функционалом (я), который начи-
нает играть роль «физиологического» времени, т. е. скорость изме-
нения другого функционала у измеряется уже не по отношению
к абсолютному времени, а по отношению к другому функционалу,
выступающему в роли «физиологических часов». Поскольку же
каждый функционал в процессе роста животных изменяется от-
носительно реального времени, то в рассматриваемом уравнении
отсутствие зависимости от времени кажущееся, а не действитель-
ное. Параметр а уравнения (56) определяется только минималь-
ными значениями х и у, (3 характеризуется соотношениями скоро-
стей их изменения.
Различают положительную (Р > 1), отрицательную (Р<1)
аллометрию и изометрию (Р=1). При изометрии в процессе роста
сохраняется геометрическое подобие формы растущего организма.
Рост раковины моллюсков в трех ее основных измерениях
(длина, высота, толщина) происходит по законам аллометрии.
С удивлением следует признать, что изучению особенностей алло-
метрического роста пресноводных двустворчатых моллюсков,
способствующего пониманию адаптивного характера формы их
раковины, уделялось незаслуженно мало внимания. Во многих
зоологических или экологических работах приводятся данные изме-
рений линейных размеров раковины, но каких-либо сопоставле-
ний и выводов чаще всего не делается. В этом отношении несом-
ненный интерес представляют исследования аллометрического
роста сфериид (Nomura, 1926а, 1926Ь; Алимов, 19676, 1976) и
унионид (Agrell, 1949). В неявном виде аллометрический рост
раковин моллюсков постоянно используется специалистами по
систематике моллюсков при составлении определительных таб-
лиц и описании видов, в которых, в частности, приводятся вели-
чины индексов соотношений основных линейных параметров
раковин.
Результаты выполненных автором по общепринятой схеме
измерений длины L, высоты Н и толщины S 1 раковин отдельных
представителей двустворчатых моллюсков из различных биотопов
в пределах ареалов видов (рис. 50 , 51) и рассчитанные величины
параметров уравнений аллометрического роста (табл. 34) пока-
зали, что среди изученных животных наблюдаются все типы алло-
метрического роста. У перловиц, некоторых беззубок и сфериид
раковина растет изометрически во всех направлениях, у других
видов изометрия роста наблюдается только в одном направлении.
В связи с этим у первых индексы отношений параметров раковин
(HjL, S/L)He меняются с возрастом, у вторых они не остаются
постоянными в процессе роста. Например, для некоторых видов
1 У моллюсков Anodonta, Pseudanodonta, Cristaria, кроме того, измеря-
лась высота раковины h у лигамента.
138
Unio индексы могут рассматриваться как видовой признак,
так как их раковины с высокой степенью достоверности (р=0.01)
различаются по их значениям (табл. 35).
Рис. 50. Зависимость толщины и высоты
от длины раковины моллюсков.
A, Ai — Unio tumidus; Б, Б, — U. pictorum; В, Вг — Crassiana crassus: 1 — р. Волга;
2 — р. Днепр; 3 — р. Рейн; 4 — Финский залив; 5 — р. Ока; 6 — р. Ветлуга; 7 —
р. Вента; 8 — р. Теша; 9 — р. Елатьма; 10 — оз. Студенец; 11 — Таганрогский залив;
12 — оз. Великое; 13 — р. Москва; 14 — р. Крынка; 15 — р. Ю. Буг (I—15 — данные
автора); 16 — разные водоемы (по данным Жадина, 1938); 17 — по данным Schnitter, 1922.
По оси абсцисс — длина раковины (мм); по оси ординат: А, Б, В — толщина раковины
(мм); Alt Бп Bi — высота раковины (мм).
Сложнее использовать для целей диагностики индексы для
видов, у которых величина их закономерно изменяется по мере
роста моллюсков.
139
Рис. 51. Зависимость толщины и высоты от длины раковины у моллюсков.
A, Ai — Anodonta piscinalis: 1 — р. Днепр; 2 — р. Теша; 3 — р. Ока; 4 — р. Вента.
Б, Б1 — A. anatina'. 1 — р. Онега; 2 — р. Ветлуга; 3 — р. Теша; 4 — р. Ока; 5 —
р. С. Двина; 6 — оз. Чудское; 7 — оз. Колдозеро; 8 — оз. Погозеро; 9 — Онежский залив;
10 — оз. Укшозеро (измерения автора). Остальные обозначения как на рис. 50.
Таблица 34
Уравнения аллометрического роста для различных моллюсков
Вид Число измерений н = f (L) h = f(L) S = /(L)
Unio tumidus 107 0.5L 0.362;
U. pictorum 97 0.442/ — 0.322/
Crassiana crassus . 80 0.562/ — 0.392;
Anodonta zellensis 25 0.205L1-12 0.752/0-9 0.0162?-84
A, seisanensis . 67 0.5252; 1.302Л8 0.2102?-18
A. anatina 40 1.1002/0-86 1.142A86 0.3502;
A. piscinalis 50 0.552/ 0.0942Л92 0.3002;
Sphaerium corneum 171 0.5782Л18 — 0.4652?-18
S. solidum 60 0.6242? •80 — 0.5862;
S. suecicum 45 1.1222A86 — 1.1352A79
S. scaldianum 40 1.3122A7* — 1.2452A66
Musculium creplini 30 0.8132/ — 0.4932;
Sphaeriastrum rivicola 90 1.1352/0-88 — 0.7622A86
Всего . . . . 917 1
Таблица 35
Сравнение моллюсков Unio и Crassiana по индексам их раковин
Вид Число измере- ний HIL SIL t
пределы м n пределы м n № п/п HIL SIL
Unio tumidus 107 0.54-0.47 0.50 0.0016 0.38—0.34 0.36 0.016 1-2 25 11.80
U. pictorum 97 0.41-0.46 0.44 0.0027 0.30-0.36 0.32 0.0030 2-3 40 13.04
Crassiana crassus 80 0.53-0.59 0.56 0.0045 0.30-0.41 0.38 0.0035
Примечание. t = (М2 — Mj/ (nl 4- ni), где Ми М2 — средние значения, пп п2 — ошибки средних. Различия статистически досто-
верны, если выдерживается неравенство t > 3.
Исследования линейных размеров раковин у нескольких ви-
дов сфериид (Алимов, 1976) показали бблыпую индивидуальную
изменчивость индекса S/L по сравнению с HfL. Разные виды
достоверно не различались по величине индекса Н/L и различа-
лись по индексу S/L, что позволяет рассматривать индекс H/L
как признак более высокого ранга, чем видовой.
При изучении моллюсков всегда отмечалась незначительная
изменчивость величин индексов раковин одного вида при значи-
тельных различиях животных по абсолютным размерам их раковин
0.8
0.4
О —।------1----l_
0.2 0.6 1.0
Рис. 52. Зависимость индексов hlL и Н/L от скорости течения воды для мол-
люсков.
А — Anodonta: 1 — A. anatina; 2 — A. piscinalis; 3 — A. seisanensis. Б— Unio и Cras-
siana: 1 — U. pictorum; 2— U. tumidus, 3 — С. crassus (измерения автора). По оси абсцисс—
скорость течения (м/с); по оси ординат: А — значения h/L, Б — значения Н/L (объясне-
ния в тексте).
(Schnitter, 1922; Жадин, 1938; Станьчиковска, 1977). Внутри-
и межвидовая изменчивость формы раковин оказывается обуслов-
ленной в первую очередь физическими и химическими особенностями
условий мест обитаний (Ortmann, 1920; Ball, 1922; Brown et al.,
1938; Agrell, 1949; Алимов, Богатов, 1975; Алимов и др., 1976).
Наиболее низкие и плоские раковины у унионид характерны
для стагнофильных видов (U. pictorum, U. tumidus), наиболее
высокие и толстые — для реофилов {Crassiana crassus). Высота
раковины оказывается положительно скоррелированной с опти-
мальными для каждого вида скоростями течения воды (рис. 52).
Такая же тенденция отмечается и среди анодонт. С возрастанием
реофилии их раковина, как правило, становится более высокой,
что достигается главным образом за счет увеличения высоты
у лигамента. У сфериид наблюдается обратная картина — рео-
фильные моллюски имеют более приземистые и плоские по срав-
нению со стагнофильными раковины (Алимов, 19676, 1976).
Условия в местах обитания определяют особенности аллометри-
ческого роста моллюсков, в результате которого создается форма
раковин, адаптированная к этим условиям. Униониды, как из-
вестно, закапываются в грунт почти на 2/3 длины, выставляя
142
наружу лишь часть заднего конца раковины. Естественно, для
того чтобы удержаться в грунте на течении, им необходимо иметь
большую площадь соприкосновения с грунтом. В противном
случае они будут опрокинуты потоком воды. Поэтому неудиви-
тельно, что реофильные представители этого семейства обладают
более широкой раковиной. У анодонт, живущих в водоемах со
стоячей водой на мягких илистых грунтах, увеличение высоты,
особенно у лигамента, делает их раковины более широкими. При
такой форме увеличивается площадь соприкосновения раковины
с грунтом, что препятствует погружению достаточно тяжелых
моллюсков в более глубокие слои ила. Сферииды же, напротив,
редко закапываются в грунт. Они преимущественно располага-
ются на его поверхности, а наиболее стагнофильные из них ак-
тивно ползают по поверхности грунта и водным растениям. Отсюда
становится понятным, почему для реофильных видов этих моллю-
сков характерна более низкая и плоская, т. е. более обтекаемая
форма раковины.
Перечисленные примеры не могут полностью объяснить все
многообразие форм раковин двустворчатых моллюсков. В то же
время они показывают основные направления формообразования
раковин, связанные с адаптациями к условиям существования.
Из сказанного можно сделать вывод о том, что форма раковин
моллюсков, обусловливаемая особенностями аллометрического
роста животных в различных условиях, может быть охаракте-
ризована индексами отношений основных линейных размеров
раковин, пределы изменчивости которых в ряде случаев могут
быть использованы как один из видовых признаков.
ВЕСОВОЙ РОСТ МОЛЛЮСКОВ
Уравнение аллометрического роста в подавляющем большинстве
случаев используется для описания соотношения между линей-
ными размерами и весом самых разных водных животных: рыб,
моллюсков, ракообразных, личинок насекомых и т. п. (Винберг,
1968, 1971). Соотношение между линейными размерами L и
весом W различных животных может быть аппроксимировано
степенной функцией
W=a1Lb'. (57)
Изменения сырого веса пресноводных двустворчатых моллю-
сков в процессе их роста показаны на рис. 53—55, рассчитанные
величины параметров соответствующих уравнений — в табл. 36.
Из данных этой таблицы отчетливо видно, что параметры уравнений
для всех изученных моллюсков не имеют статистических различий
с высокой степенью достоверности (/7=0.01).
Важно отметить, что параметры уравнений для изученных
представителей различных семейств пресноводных и морских
143
Таблица 36
Параметры ах, &х и доверительные интервалы средних их значений
(р = 0.01) уравнений зависимости сырого веса (г) от длины
раковины (мм) у различных моллюсков
Название моллюсков «1 bl
Unionidae
Unioninae 0.00023 ±0.000021 2.85 ±0.16
Anodontinae 0.00005 ±0.000020 3.10 ±0.18
Lampsilidae 0.00019 2.70
Dreissenidae 0.00018 ±0.000072 2.87 ±0.16
Sphaeriidae 0.00031 ±0.000200 3.02 ±0.34
Margaritiferidae 0.00013 3.00
Примечание. Доверительные интервалы для Lampsilidae и Margaritiferidae
из-за небольшого количества измерений не рассчитывались.
моллюсков достоверно не различаются между собой (Алимов,
Голиков, 1974). Кроме того, полученные для пресноводных мол-
люсков значения показателя степени Ъ± достоверно не отличаются
от 3 (при /7=0.01).
При принятой степени достоверности крайние величины пока-
зателя степени для изученных моллюсков находятся в пределах
2.691—3.357. Полученные рядом исследователей значения этого
показателя полностью укладываются в эти пределы. Например,
у Unio tumidus и U. pictorum из оз. Крапина его величина равна
соответственно 2.99 и 3.22 (Tudoranceae, Florescu, 1968а, 1968b),
U. tumidus из оз. Крапина и Жижила — 2.76 (Tudorancea, 1969),
Grassiana crassus из р. Нера — 3.03 (Tudorancea, Gruia, 1968).
Для моллюсков белорусских озер значения этого параметра были
следующие: Sphaerium согпеит — 2.6, Anodonta anatina — 3.0,
U. tumidus — 2.8 (Арабина, Гаврилов, 1967). В тех же озерах:
A. anatina — 3.6—2.4 (среднее 3.06), S. согпеит 2.5—3.4 (2.99),
U. pictorum — 2.8, U. tumidus 2.5—3.0 (2.78) (Десятик, 1968).
Для Dreissena polymorpha — 2.7, D. bugensis — 2.69 (Днепровское
водохранилище, Шевцова, 1971а), U. tumidus, U. pictorum,
A, piscinalis, D. polymorpha, S. rivicola — 3.0 (Волгоградское
водохранилище, Кондратьев, 19706), у семи видов рода Pisidium
в одном из озер Южной Финляндии от 2.3 до 3.1 (Holopainen,
1979).
Рис. 53. Зависимость логарифма сырого веса 1g W от логарифма длины ра-
ковины lg L у моллюсков.
А — Unio: U. pictorum по данным: 1 — Кондратьев, 1970; 2 — Троицкий, 1934; 3 — изме-
рения автора; U. tumidus', 4 — Кондратьев, 1970; 5 — Троицкий, 1934; 6 — измерения
автора; 7 — Арабина, Гаврилов, 1967. Б — Anodonta: 1 — A. woodiana (измерения ав-
тора); A. piscinalis: 2 —„измерения автора (Куршский залив); по данным: 3 — Кондра-
тьева, 1967, 1970; 4 — Okland, 1963. В — Lampsilidae (по данным Brown et al., 1938):
1 — Ptychobrancus fasciolaris', 2 — Proptera alba\ 3 — Leptodea fragilis', 4 — Lampsilis
seliquodea', 5 — Elliptio dilatatus. Г — Margaritifera margaritifera (p. Кереть, измерения
автора). По оси абсцисс — значения lg L (мм); по оси ординат — значения lg W (г).
144
-/ Н
Рассчитанные разными авторами значения параметра 6^ для
моллюсков одного вида из одного и того же водоема могут зна-
чительно отличаться. Такого рода различия, как и отличия его
Рис. 54. Зависимость логарифма сырого веса 1g W от логарифма длины ра-
ковины lg L у моллюсков Sphaeriidae.
1 — Sphaeriastrum rivicola; (Кондратьев, 1970); 2 — S. rivicola (измерения автора); 3 —
Sphaerium согпеит (измерения автора); 4 — S. согпеит (Десятин, 1968); 5 — S. scaldianum
-(измерения автора); 6 — Pisidium casertanum (измерения автора); 7 — Р. атпгсит (Деся-
тин, 1968). Прерывистые линии — доверительный интервал для уровня вероятности 0.95.
Остальные обозначения как на рис. 53.
величины от 3, обусловлены разными причинами: степенью обсу-
шивания животных перед взвешиванием, точностью измерений,
физиологическим состоянием моллюсков, диапазоном линейных
размеров, использованных для расчетов параметров уравнений
146
и т. п. Вес моллюсков одинакового размера может иметь закономер-
ные сезонные колебания (Скиркявичене, 1970; Барыбина, Санина,
1975). Вместе с тем точность расчета параметров уравнений степей-
Рис. 55. Зависимость логарифма веса lg W от логарифма длины раковины
lg L у моллюсков Dreissena polymorpha.
1 — Штеттинский залив (Wictor, 1969); 2 — Куршский залив (измерения автора); 3 —
Куйбышевское водохранилище (Ляхов, Михеев, 1964); 4 — Волгоградское водохранилище
(Кондратьев, 1970); 5 — канал Днепр—Кривой Рог (Шевцова, 1971); 6 — Каспий (Кар-
певич, 1964); 7 — Аральское море (Карпевич, 1964); 8 — Таганрогский залив Азовского
моря (Некрасова, 1971). Остальные обозначения как на рис. 53.
ной функции возрастает при увеличении диапазона рассматривае-
мых значений функции.
Известно, что параметр в таких уравнениях отражает
степень аллометрии изменения формы животных в процессе их
10* 147
роста (Thompson, 1959; Винберг, 1971). Когда рост протекает
с сохранением геометрического подобия, т. е. без изменения формы,
то должно быть равно 3. Однако, несмотря на отсутствие для
изученных моллюсков достоверных отличий значений от 3,
как было показано выше, не для всех видов характерен изометри-
ческий рост в трех измерениях.
Приведенные уравнения и их параметры отражают лишь общую
закономерность связи между длиной раковины и весом моллюсков
включающую в себя все возможные варианты. Они не выражают
особенностей этой зависимости для конкретных видов в опреде-
ленных условиях. В то же время для отдельных видов, у которых
не наблюдается изометрического роста, величина показателя сте-
пени в уравнении зависимости веса от линейных размеров
оказывается отличной от 3. Сказанное можно проиллюстриро-
вать на примере моллюсков S. согпеит из некоторых прудов
г. Ленинграда. Для них характерно закономерное относитель-
ное увеличение раковины по высоте и особенно толщине по мере
роста моллюсков. При этом показатель степени в уравнении W=
=/ (L) равен 3.23 (Алимов, 1965).
При изометрическом росте с сохранением подобия формы
генеративная линия створки раковины моллюсков нарастает
по закону логарифмической спирали (Thompson, 1959; Stasek,
1963; Кафанов, 1975), а в качестве меры выпуклости раковины
может быть использован постоянный угол этой спирали или индекс
отношения линейных размеров S/Н. Например, среди изученных
моллюсков, согласно данным табл. 34, следует ожидать роста
с сохранением формы в постэмбриональный период развития
у некоторых видов унионид и сфериид (Unio tumidus, U. pictorum,
Anodonta piscinalis, Sphaerium solidum (Normand), Musculium crep-
lini (Dunker)). Для моллюсков, рост раковины которых под-
чиняется закону логарифмической спирали, постоянный угол ее
может быть использован в качестве видового признака (Алимов,
19676; Кафанов, 1975).
Вес раковины и тела моллюсков связаны также степенной за-
висимостью с линейными размерами животных (рис. 56, 57,
табл. 37). Сравнение данных табл. 36 и 37 показывает, что по мере
роста моллюсков относительный вес их раковин увеличивается,
а тела — уменьшается. При этом изменение относительного веса
раковины у анодонт происходит с меньшей, а тела — с большей
скоростью по сравнению с перловицами.
Наиболее тяжелые раковины оказываются у тех моллюсков,
для которых свойственны более высокие величины оптимальных
для их роста концентраций Са** (рис. 58). Из приведенного рисунка
видно, что вес створок и оптимальные значения концентраций
Са,е для разных видов связаны между собой прямолинейной зави-
симостью. Экстраполяция данных рис. 58 показывает, что тео-
ретический нижний предел содержания Са” в воде, при котором
эти животные уже не могут жить, должен быть у Unio вдвое
148
Т а б л и ц а 37
Параметры а19 &х уравнений зависимости веса раковины И7р (г)
и тела (г) от длины раковины L (мм) и их отношения
к сырому весу W унионид
Название моллюсков Wp = f (L) WT = f (L) b
«1 bi bi WplW WTIW
U nio 0.000012 3.2 0.000450 2.53 0.35 —0.32
Anodonta .... 0.000006 3.3 0.000018 2.30 0.20 —0.80
выше (около 20 мг/л), чем у Anodonta (10 мг/л). Действительно,
в целом беззубки по сравнению с перловицами предпочитают
Рис. 56. Зависимость логарифма веса раковины lg Wp от логарифма ее длины
lg L у моллюсков.
А — Unio: U. pictorum по данным: 1 — Жадин, 1938; 2 — Лукин, 1937а; 19376, 1937в;
3 — Negus, 1966; U. tumidus: 4 — Лукин, 1937а, 19376, 1937в; 5 — Negus, 1966; 6 — Жа-
дин, 1938. Б — Anodonta (измерения автора): 1 — A. anatina; 2 — A. seisanensis. По оси
абсцисс — значения lg L (мм); по оси ординат — значения lg (г).
более мягкие воды и обладают более легкими раковинами. Можно
предполагать существование различий в механизмах кальцификации
у этих животных.
149
Утяжеление раковин перловиц по сравнению с анодонтами
в сочетании с особенностями их форм и условий обитания способ-
ствует более прочному закреплению перловиц в грунте. Если
учесть, что, как у первых, так и у вторых, более тяжелым оказы-
вается передний, погруженный в грунт конец раковин, то адап-
тивный характер формы и веса створок раковин, связанный с осо-
бенностями роста животных, становится более понятным и объяс-
нимым. Уместно напомнить, что, например, среди сфериид самыми
Рис. 57. Зависимость логарифма веса тела lg WT от логарифма длины ра-
ковины lg L у моллюсков.
А — Unio (по данным Кондратьева, 1967); 1 — U. pictorum', 2 — U. tumidus. Б—Ano-
donta piscinalis (измерения автора). По оси абсцисс — значения lg L (мм); по оси ординат—
значения lg WT (г).
тяжелыми и прочными раковинами обладают реофильные виды
S. solidum и Р. amnicum (Muller), а более тонкими и хрупкими —
стагнофильные S. corneum, S. suecicum и виды рода Musculium.
У последних, особенно живущих во временных, пересыхающих,
мелководных водоемах, раковина может быть настолько тонкой,
что через нее просвечивает тело животного.
Для некоторых видов моллюсков возможно определить средние
величины отношений веса их тела, раковины и мантийной жид-
кости к общему весу животных. У унионид вес тела составляет
51—79%, дрейссен и сфериид — только 35—44% от их общего
веса (табл. 38). Вес раковины у моллюсков S. corneum не превы-
шает 22% веса животного (рассчитано по данным Винберга и
Беляцкой, 1959). По данным А. Ф. Карпевич (1964), у дрейссен
на раковину приходится 63%, на тело — 35% веса моллюсков.
150
Таблица 38
Отношение веса раковины Жр, тела JFT и мантийной жидкости
Wn к сырому весу W (г) моллюсков (%)
Вид WplW wTiw wxiw
Unio tumidus 34.0 60.0 6.0
U. pictorum 31.2 60.0 8.8
A nodonta anatina 16.0 79.0 5.0
A. piscinalis 35.0 51.0 14.0
Dreissena polymorpha — 35.0 —
Sphaeriastrum rivicola — 44.0 —
Примечание. WM = W — (Wp + WT).
Вес тела у кристарий составляет не более 38% их обЬцего веса
(Булдовский, 1935). Легко рассчитать, что 25% веса моллюска
приходится на мантийную жидкость. У остальных моллюсков
вес мантиинои жидкости ко-
леблется в пределах 5—14%
от общего веса животных.
С помощью зависимости веса
от линейных размеров моллю-
сков (уравнение 57) можно пе-
рейти от линейного к весовому
росту этих животных. Для этого
достаточно в уравнении линей-
ного роста (43) подставить зна-
чения веса, выраженные через
линейный размер:
Lt = е~м), но W = arLь',
откуда
L = (W/a1)1lb^
Рис. 58. Зависимость веса рако-
вины (при длине 10 мм) от содержа-
ния кальция в воде для моллюсков.
1 — Unio; 2 — Anodonta. По оси абсцисс —
содержание Са** (мг/л); по оси ординат —
вес раковины (г).
Подставляя в (43), получаем
— W1*1 (1 — e~ki), (58)
где Wt — вес ко времени
FTqo — максимальный теоретиче-
ский вес животных.
Равенство (58) представляет собой уравнение 5-образной
кривой. Такого вида кривая изменения веса в процессе развития
особей свойственна подавляющему большинству моллюсков (Wil-
bur, Owen, 1964). Естественно, что все основные закономерности
изменения параметров этого уравнения, обусловленные влиянием
151
факторов внешней среды и установленные для линейного роста,
остаются справедливыми и для весового роста.
Для продукционных исследований весьма важны закономер-
ности именно весового роста, а также характер изменения с воз-
растом величин абсолютного и относительного прироста веса
животных.
В данной работе, как и во многих работах других авторов,
абсолютный прирост определяется как прирост веса животного
ДЖ=И/2 — W± за исследуемый отрезок времени &t=t2 — tx.
При Az -> 0 получаем абсолютную скорость весового роста
dWfdt. Относительный, или удельный прирост представляет собой
отношение абсолютного прироста к среднему весу животного за
рассматриваемый отрезок времени Az, т. е. AJT/AMF. При
Az -> 0 удельная скорость весового роста Cw равна
£„ = 1/^^. (59)
Средняя величина удельной скорости роста Cw за рассматри-
ваемый период времени Z2 — может быть определена из равен-
ства
г _1пЖ2-1пТГ1 _ 1g РГ2 — lg
w t2 — t! (t2 — 0.4343 ’
В том случае, если разность W2 — W± не превышает 10% от
начального веса JV19 значение С№ можно с достаточной точностью
найти как
_ {w2-W1)2
w (W2 - И\) (t2 - •
Характер изменения относительной и абсолютной скорости
роста в процессе развития животного проследим на примере мол-
люсков Unio tumides. Удельная скорость роста Cw, как это видно
на рис. 59, уменьшается с возрастом животного. Абсолютная
скорость роста dW[dt сначала нарастает, затем, достигнув своего
максимального значения, снижается. Такой характер изменения
удельной и абсолютной скорости роста следует непосредственно
из основного уравнения весового роста. Точка перегиба кривой
роста определяется соотношением
^ = 0.296^00,
где Wn — вес животного, соответствующий точке перегиба кривой
весового роста.
По времени точка перегиба совпадает с достижением максималь-
ного значения абсолютной скорости роста. Однако замедление
нарастания абсолютной скорости наступает несколько раньше
точки перегиба кривой весового роста.
152
Моллюски U. tumidus достигают половозрелости к 3—4-му
году жизни (Бластов, 1933; Лукин, 1937а, 19376; Tudorancea,
1969, 1972). Примерно с этого времени замедляется скорость
их весового роста. Замедление скорости роста с наступлением
половозрелости отмечено у различных видов пресноводных дву-
створчатых моллюсков (Карпевич, 1964; Botnariuc, Tudorancea,
1967). Вес моллюсков U. tumidus, как можно видеть из рис. 59,
при наступлении половозрелости составляет около 17% от FKqo»
Рис. 59. Кривые весового роста моллюска Unio tumidus.
А — изменения веса в процессе роста моллюска: 1 — средний вес при достижении поло-
возрелости; 2 — вес в точке перегиба кривой; з — средний максимальный вес и7тах. Б —
изменения относительного (1) и абсолютного (2) приростов. По оси абсцисс — время (годы);
по оси ординат: А — вес моллюска (г);*5Б: слева — относительный прирост (г-год-1),
справа — абсолютный прирост (год-1).
25 % от РИшах или 59 % веса в точке перегиба кривой весового роста
для этих животных. Такого же порядка соотношения отмечены
и для других моллюсков (табл. 39).
Уменьшение прироста веса моллюсков после наступления
половозрелости связано с расходом энергии потребленной пищи
не только на соматический, но и генеративный рост.
Необходимо уточнить, что понимать под временем наступле-
ния половозрелости применительно к двустворчатым моллюскам.
Моллюски с перекрестным оплодотворением считаются полово-
зрелыми в возрасте, когда у них впервые созревают половые
продукты, и они способны нереститься. Сложнее обстоит дело
с живородящими видами, особенно со сфериидами, у которых
наблюдается внутреннее самооплодотворение (Odhner, 1921; Okada,
1935; Thomas, 1959). Принято считать их половозрелыми в воз-
расте, когда они впервые выметывают молодь. К этому времени
длина раковин у них составляет 50—60% дефинитивного размера
(Алимов, 19676). Истинная же половозрелость, т. е. возраст, при
котором созревают половые продукты, у этих животных наступает
153
Таблица 39
Соотношение между весом при наступлении половозрелости
(FFnoJ, в точке перегиба (JFn), максимальным (FFmax)
и теоретически максимальным (Wra) весом для различных
моллюсков
Вид Woo, г Wmax, г ^пол, г WJ10a wJ10J wJt0J Wn
Woo Wmax
Unio tumidus 183.7 126.0 32.00 0.17 0.25 0.59
U, pictorum 158.9 79.8 25.40 0.16 0.32 0.54
Anodonta piscinalis .... 54.3 32.8 8.60 0.16 0.26 0.54
Dreissena polymorpha . . . 4.9 2.7 0.78 0.16 0.29 0.53
Sphaeriastrum rivicola . . . — 2.5 0.54 — 0.22 —
Примечание. Половозрел ость U. pictorum наступает на 3-й год (Бластов, 1933;
Лукин, 1937а, 19376), A. piscinalis — около 2 лет (Botnariuc, Tudorancea, 1967), D. poly-
morpha — к концу 1-го года жизни (Карпевич, 1964), S. rivicola — при длине раковины
12 мм (Groenwegen, 1926).
значительно раньше, и пока нет надежных данных, позволяющих
его определить.
РАЗМНОЖЕНИЕ
И ПЛОДОВИТОСТЬ МОЛЛЮСКОВ
По способу размножения пресноводные двустворчатые моллюски
могут быть разделены на три группы. Представители сем. Dreis-
senidae размножаются с помощью свободно плавающей личинки,
Unionidae — паразитирующей на рыбах личинки (глохидий),
Sphaeriidae и некоторые Corbiculidae — живородящие. У послед-
них наряду с живорождением отмечено и яйцерождение (Fuzi-
wara, 1975).
Оплодотворение может быть наружное и внутреннее (само-
оплодотворение). Среди двустворчатых моллюсков широко рас-
пространен гермафродитизм. Он бывает постоянный (сферииды),
частичный и случайный. Постоянный гермафродитизм отмечен
у некоторых североамериканских унионид, частичный и случай-
ный — у британских анодонт, жемчужниц из Германии, амери-
канских унионид (Schalie, 1970). Различная степень гермафроди-
тизма в разных частях видовых ареалов рассматривается как
адаптация животных к определенным условиям существования.
Молодь развивается в обеих полужабрах (Margaritiferidae,
Amblemidae) или только в наружных (Unionidae, Lampsilidae).
Развитие зародышей у сфериид протекает в специальных
выводковых карманах (марсупиальные карманы). В соответствии
со степенью развития зародышей различают первичные, вторичные
и третичные марсупиальные карманы. Последние содержат за-
родышей наиболее поздних стадий развития, имеющих уже вид
взрослого животного.
154
После наступления половозрелости процесс размножения
у моллюсков повторяется ежегодно. Однако известно, что в не-
которые годы при очень неблагоприятных условиях, главным обра-
зом температурных, униониды не приступают к размножению
(Tudorancea, 1969).
Большинство двустворчатых моллюсков пресных вод размно-
жаются в летние месяцы. Так, дрейссены приступают к размноже-
нию, когда температура воды поднимается выше 10—15° (Кача-
нова, 1961; Hillbricht-Ilkowska, Stanczykowska, 1969; Wiktor,
1963; Гальперина, 1976; Лубянов, Золотарева, 1976). В связи
с этим в северных частях ареала период размножения у них короче,
чем в южных (Гальперина, 1976).
Представители подсемейств Unioninae и Ambleminae размно-
жаются в летние месяцы с мая по август (Жадин, 1938; Троицкий,
1939; Властов, 1962; Tudorancea, 1972; Clarke, 1973) при темпера-
турах воды выше 15°, виды подсем. Anodontinae — при более
низких температурах. Период их размножения начинается
в конце лета—осенью и заканчивается весной следующего года
(Tudorancea, 1972; Clarke, 1973).
Нерест большинства видов сфериид растянут во времени, и
молодь встречается в популяциях этих моллюсков с конца весны
до осени. Для некоторых видов характерно наличие одного или
двух отчетливо выраженных пиков рождения молоди. Два помета
в течение года, или вегетационного сезона, — у Pisidium henslo-
wanum (Sheppard), P. hibernicum Wstl., P. lilljeborgi, P. milium
Held, P. nitidum Jenyns, P. pulchellum Jenyns (Heard, 1965),
Sphaerium corneum (Thiel, 1926; Митропольский, 1965; Алимов,
19676), S. simile (Say) (Zemoff, 1973), P. sybtruncatum, P. casertanum
(Holopainen, 1979). Один раз в году размножаются Р. compres-
sum Prime, Р. fallax Sterki, P. variabile Prime, P. valkeri (Sterki)
(Heard, 1965), P. henslowanum (Holopainen, 1979), P. amnicum
(Miiller) (Danneel, Hinz, 1976), Musculium sucuris (Say) (Mackie
et al., 1976a, 1976b) и др. Моллюски, живущие при постоянных
температурах, например Р. conventus, в профундали глубоких
озер, или тропические виды размножаются в течение всего года
(Meier-Brook, 1970). Периоды размножения Р. obtusale, населяю-
щих пересыхающие водоемы, приурочены к тем летним и осенним
месяцам, когда водоемы заполнены водой (Митропольский,
1969).
Вымет половых продуктов у унионид происходит в достаточно
короткие отрезки времени: у Unio pictorum, U. tumidus, Crassiana
crassus — в течение июля, Anodonta piscinalis — в мае, Pseudano-
donta complanata — в июне (Жадин, 1938). У перловиц наблюда-
ется двукратный, у беззубок — однократный нерест за этот период
(Властов, 1962). Б. В. Властов различал четыре фазы процесса
размножения у этих моллюсков, каждая из которых характери-
зуется определенным количеством беременных самок в популяциях,
которое достигает 100% к середине—концу мая, после чего наблю-
155
дается постепенное уменьшение их численности, и к концу июля
почти у всех животных жабры оказываются пустыми.
Рождение молоди у сфериид происходит порционно. Моллюски
Sphaerium securis нерестятся один-два, редко три раза в течение
жизни, рождая за один помет от 2 до 7 потомков (Mackie et al.,
1976а, 1976b), S. simile — два-три раза (6—8 потомков) (Zumoff,
1973), Pisidium amnicum — один, редко два раза (Danneel,
Hinz, 1976). У большинства шаровок новые зародыши начинают
развиваться раньше, чем происходит вымет готовых к рождению
(Алимов, 19676; Meier-Brook, 1970). Лишь у S. solidulum Prime,
так же как и у большинства горошинок, новые яйца появляются
и начинают развиваться только после рождения молоди предыду-
щей порции (Harley et al., 1947; Митропольский, 1969; Meier-
Brook, 1970).
В тесной связи с особенностями размножения находится инди-
видуальная плодовитость моллюсков, т. е. количество яиц или
молоди, приходящееся на одну самку. Поскольку среди двуствор-
чатых моллюсков, населяющих пресноводные водоемы, широко
распространен гермафродитизм, более верно рассматривать их
индивидуальную плодовитость по отношению к особи, а не только
к самке.
Индивидуальная плодовитость двустворчатых моллюсков изу-
чена еще недостаточно полно. Обычно определяется среднее или
наибольшее количество яиц или молоди у разных видов. Некото-
рые морские представители этих животных одновременно мечут до
нескольких десятков миллионов яиц на одну особь. Например,
число яиц у Ostrea edulis достигает 1.5-IO6, Mytilus edulis — 1-107
(Fretter, Graham, 1964). По сравнению с морскими моллюсками
моллюски пресных и солоноватых вод продуцируют в среднем
меньшее количество яиц, но более крупных по величине (Bondesen,
1940). Среди пресноводных двустворчатых моллюсков наиболее
плодовитыми оказываются жемчужницы: Margaritifera dahurica
мечет от 2-106 до 22-106 яиц (Микулич, 1954), М. margaritifera —
в среднем около 3«106яиц (Жадин, 1938). Несколько менее
плодовиты беззубки — от 288-103 до 694-103 (в среднем около
396-103) и перловицы — от 7-103 до 260-103 яиц (Троицкий,
1939). Среди последних наиболее плодовиты Unio pictorum —
от 360-103 до 430-103, менее — Crassiana crassus, которые мечут
около 133-103 яиц (Жадин, 1938).
Значительно меньшая плодовитость у живородящих сфериид.
Наибольшее количество молоди у представителей Pisidiinae и
Eupirinae, меньше — у Sphaeriastrinae (табл. 40).
Обычно у более крупных, т. е. более старших по возрасту
особей, оказывается и большее количество яиц (большая инди-
видуальная плодовитость). Это неоднократно отмечалось для
различных пойкилотермных животных. Было показано, что между
линейными размерами родительских организмов и количеством
яиц или молоди существует прямая зависимость, которая может
156
Таблица 40
Количество молоди у некоторых сфериид
Вид Среднее Максималь- ное Источник
Sphaerium corneum 8—10 28 Наши данные
S. solidulum — 8-18 Harley et al., 1947
5. simile 6—8 12 Zumoff, 1973
S. transversum — 38 Dietz, Stem, 1977
S. rivicola — 10—16 Groenwegen, 1926
Musculium lacustre 8—12 — Митропольский, 1970
M. heterodon — 16 Okada, 1935
M. securis — 5—7 Mackie et al., 1976b
M. partumeium 10 — Clarke, 1973
Pisidium milium ... — 12 Ladle, Baron, 1969
P. casertanum 12.4 53 Heard, 1965
P. nitidum 5.4 13 Там же
P. punctatum Sterki . . . 6.0 12 »
P. compressum 13.0 42 »
P. fallax 8.6 24 »
P. conventus 6.0 — »
P. dubium — 26 »
P. adamsi 26.0 66 Heard, 1964
P. variabile Prime 28.0 34 Там же
P. henslowanum — 40 Митропольский, 1970
P. obtusale 5—10 20 Там же
P. hibernicum — 8 Meier-Brook, 1970
P. lilljeborgi — 13 Там же
P. forense Meier-Brook . . 5—10 30 Meier-Brook, 1970
P. amnicum — 12 Holopainen, 1979
P. casertanum ... — 27 Там же
P. henslowanum . . . — 25 »
P. lilljeborgi — 9 »
P, hibernicum — 4 »
P. conventus — 5 »
Neopisidium moUessiarianum ... — 5 »
Eupera cubensis (Prime) .... — 25-35 Heard, 1964
E. plafensis — 66 Там же
быть аппроксимирована степенной функцией (Юровицкий, 1958;
Зотина, Зотин, 1967; Хмелева, 1969; Walz, 1978b; Holopainen,
1979, и др.)
I = mLn, (60)
где I — число яиц (молоди); L — длина взрослого животного;
т, п — константы уравнения.
Такого же вида зависимость (60) справедлива для различных
видов сфериид (рис. 60). На примере Sphaerium corneum из водо-
емов окрестностей г. Ленинграда видно, что характер этой зави-
симости сохраняется в различные сезоны (рис. 61). Однако зна-
чения параметров уравнения не остаются постоянными. Наиболь-
шая скорость увеличения числа молоди у моллюсков одинаковых
размеров наблюдается в летние месяцы, меньшая — весной и осенью.
157
Среднее за рассматриваемый период значение параметра п ока-
зывается близким к 2.7. Приняв эту величину постоянной для
всего периода, получаем наибольшие значения параметра т
Рис. 60. Зависимость логарифма индивидуальной плодовитости 1g I от
логарифма отношения 1g (L/Zmax) у различных видов сфериид.
1 — Pisidium lilljeborgi; 2 — Р. nitidum; 3 — Р. conventus; 4 — Р. hibernicum; 5 — Ми-
sculium heterodon; 6 — М. partumeum; 7 — М. lacustre; 8 — М. transversum; 9 — Р. fo-
rense. По оси абсцисс — значения отношения 1g L/Zmax *, по оси ординат — значения 1g I.
весной и осенью (табл. 41), т. е. когда вымет молоди моллюсками
резко сокращается или полностью прекращается. В летние же
Таблица 41
Значения параметров т и п в уравнениях Зависимости
количества молоди от длины раковин для моллюсков
Sphaerium согпеит в разные сезоны года
Месяц 7И п т (при п = 2.7)
Май 16.8 2.00 27.3
Июнь ... .... 10.0 3.15 7.3
Июль 9.8 3.00 7.9
Август 12.0 3.00 9.7
Сентябрь 10.8 2.37 13.6
CJp е д н е е . . . . 11.88 2.70 13.6
II р и м е ч а н и е. В табл. 41, 42 значение m рассчитано при длине моллюска,
равной 10 мм.
158
Рис, 61, Зависимость индивидуальной плодовитости I от длины раковины моллюсков Sphaerium согпеит из водоемов
окрестностей г. Ленинграда в разные месяцы. Масштаб логарифмический.
По оси абсцисс — длина раковины (мм); по оси ординат — значения I (экз./особь),
месяцы, когда животные размножаются наиболее интенсивно,
количество молоди у моллюсков одинакового возраста уменьша-
ется. Это совпадает с представлениями о том, что количество
молоди у сфериид изменяется в зависимости от сезона года (Zumoff,
1973) и условий внешней среды (Mackie et al., 1976b).
Значения параметров уравнения (60) для различных сфериид
(табл. 42) показывают, что наиболее плодовиты моллюски Muscu-
lium и некоторые виды Pisidium ( Р. forense, Р. lilljeborgi,
Р. nitidum). Высокая плодовитость этих животных неоднократно
отмечалась (Harley et al., 1947; Thomas, 1959; Heard, 1964),
и она в значительной степени обусловлена особенностями место-
обитаний и жизненных циклов этих животных. Например, мол-
люски Musculium, обычно населяющие временные или пересыхаю-
щие водоемы, нерестятся после достаточно длительного периода
покоя в короткие периоды, когда водоемы заполнены водой.
Несомненно, что только высокая плодовитость отдельных особей
позволяет в этих условиях поддерживать оптимальную численность
популяции.
Таблица 42
Значения параметров т и п в уравнениях зависимости
количества молоди от отношения Z//Zmaz для некоторых сфериид
(рассчитано по данным рис. 50)
Название моллюсков m n Пределы Z/Zmax
Sphaerium 11.88 2.70 0.40-0.96
Musculium 33.00 2.56 0.45-0.85
Pisidium conventus . . . 6.31 1.96 0.50—0.92
P. lilljeborgi 19.50 2.67 0.50—0.90
P. forense 41.78 2.77 0.44—0.88
Значения константы п, рассчитанные для различных видов
сфериид, оказываются близкими к таковым для других водных
беспозвоночных. У моллюсков, ракообразных и хирономид вели-
чины этого параметра находятся в пределах 1.96—3.0 (табл. 43),
средняя величина, вероятно, близка к 2.6. В этом отношении
интересно отметить, что для рыб значение константы п чаще всего
превышает 3 (Юровицкий, 1958; Бруенко, 1967).
Однако биологический смысл и значение имеет зависимость
плодовитости животных от их веса, а не от линейных размеров.
Основные функции и процессы развивающегося организма, как
это было показано на примере обмена и питания, связаны именно
с его массой. Рассмотренные выше зависимости плодовитости
животных от их линейных размеров могут иметь количественные
выражения только вследствие наличия функциональной связи
между весом и размерами животных, которая описывается алло-
метрическими уравнениями.
160
Таблица 43
Значения константы п в уравнениях зависимости количества яиц
(молоди) от длины для некоторых водных животных
Название животных n По данным
Mollusca
Sphaerium 2.70 Автора
Musculium 2.56 То же
Pisidium conventus, P, hibernicum 1.96 »
P. lilljeborgi, P. nitidum 2.67 »
P. forense 2.77 »
Margaritifera dahurica 2.86 Микулич, 1954
Patella coerulea 3.00 Чухчина, 1972
Crustacea
Crangon septemspinosa 2.50 Микулич, Козак, 1971
Idotea baltica 2.32 Хмелевой, 1969
I. ochotensis 2.53 Стрельниковой, 1971
I. chelipes 2.81 Семенченко, Романова, 1976
Oithona папа ........... 2.78 Ковалева, 1968
Четыре вида черноморских крабов 2.56 Аболмасовой, 1973
Chironomidae
Chironomus dorsalis 3.00 Константинова, 1958
Обсуждению зависимости плодовитости от линейных размеров
животных было уделено внимание лишь потому, что до сих пор
в практике исследований она широко используется и обсуждается
в литературе, правда, без необходимых оговорок и критического
осмысливания. Поскольку для целого ряда животных еще не
установлены количественные выражения зависимостей их веса
от размеров, соотношения между их плодовитостью и линейными
размерами особей пока остаются единственным материалом,
который может быть использован для сравнительных целей.
Кроме того, по крайней мере по отношению к пресноводным дву-
створчатым моллюскам в полевых условиях в ряде случаев бывает
проще получить сведения по плодовитости в соответствии с их
линейными размерами. Поэтому, не придавая биологического
значения этой связи, мы посчитали возможным и необходимым
привести ее количественные выражения.
Используя полученные ранее количественные зависимости
между весом и линейными размерами пресноводных двуствор-
чатых моллюсков, нетрудно рассчитать, что величина показателя
степени в уравнениях, отражающих связь между плодовитостью
животных и их весом, находится в пределах 0.7—0.96. Существенно,
что значения этой константы для различных ракообразных
колеблются от 0.7 до 0.88 (Хмелева, 1969). Столь незначитель-
11 А. Ф. Алимов
161
ные различия величин этой константы в уравнениях для пред-
ставителей разных водных беспозвоночных несомненно отражают
важное общебиологическое значение функциональной связи между
плодовитостью животных и их весом. И в этом смысле она стано-
вится в ряд установленных и достаточно хорошо изученных важ-
нейших соотношений скорости обмена, роста питания и массы
растущих организмов. Значения показателей степени в уравне-
ниях, отражающих количественные связи между весом и раз-
мерами у различных животных, как известно, колеблются в до-
статочно узких пределах. Учитывая это, можно предполагать,
что приведенные величины этого параметра в уравнениях «пло-
довитость—вес животных» вряд ли должны претерпеть в даль-
нейшем значительные изменения.
Необходимо особо отметить, что полученные степенные зави-
симости для плодовитости справедливы в пределах определенного
размерно-возрастного диапазона животных. Нижняя граница
этого диапазона определяется размерами (весом), которые дости-
гаются животными ко времени наступления половозрелости.
По достижении взрослыми организмами некоторого размера,
количество потомков у них уменьшается. Это связано с ослабле-
нием воспроизводительной способности у особей старших воз-
растов. Размеры животных, при которых сокращается их плодо-
витость, можно рассматривать как верхнюю границу размерного
диапазона. В частности, для сфериид размерный диапазон, в пре-
делах которого справедлива степенная зависимость числа молоди
от размеров взрослого животного, ограничен значениями L/Lmax
от 0.4 до 0.96 (табл. 42). Вместе с тем в природных популяциях
в отличие от лабораторных большинство водных животных
просто не доживают до такого возраста, когда у них сокращается
плодовитость, или доживают лишь единичные особи. В связи
с этим для экологических исследований, при которых учитыва-
ется реальная, а не возможная продолжительность жизни живот-
ных, использование зависимости плодовитости от их веса (раз-
меров) в виде степенной функции может считаться вполне прием-
лемым.
Несмотря на то что по мере роста моллюсков до определенного
размера (возраста) количество яиц (молоди) увеличивается, число
беременных особей с их возрастом в популяциях сокращается.
Наибольшее количество беременных самок наблюдается в попу-
ляциях унионид среди 4—5-летних моллюсков (Асташкина,
1965), больше всего молоди рождают моллюски Pisidium amnicum,
имеющие длину раковины 4—5.95 мм (Danneel, Hinz, 1976).
Среди особей более старших возрастов число беременных особей
уменьшается.
Количественное выражение линейного или весового роста
моллюсков, зависимости количества потомков от размеров мате-
ринского организма и продолжительности эмбрионального раз-
вития позволяют определить абсолютную плодовитость животных,
162
под которой в данном случае понимается общее количество по-
томков, рожденное одной самкой (особью) в течение ее жизни.
Рассмотрим это на примере моллюсков Sphaerium согпеит. Кривая
линейного роста моллюсков этого вида, как было показано ранее,
при температуре 16° (близкая к средней температуре за вегета-
ционный сезон) может быть описана уравнением
Zz/ = 12[1 —
При этом Л=0.342 мес"1, £0=27.9 сут. Зависимость количества
молоди от длины моллюска описывается уравнением 1=11.88 L?-7
(см. табл. 42). Исходя из этих данных, нетрудно рассчитать, что
за 200 сут, т. е. за время достижения животным размера £шах=
=0.9£оо, моллюск сможет в среднем выметать около 35 потомков.
Аналогичным образом рассчитана абсолютная плодовитость
для других видов сфериид и Unio tumidus. При определении плодо-
витости U. tumidus принималось, что эти моллюски становятся
половозрелыми в возрасте 3—4 лет, а каждая особь ежегодно
мечет в среднем около 146-103 яиц (Троицкий, 1939).
Абсолютная плодовитость моллюсков (количество молоди или
яиц), размножающихся с помощью паразитирующей личинки,
примерно в 40*103 раз выше, чем у живородящих, что несомненно
компенсирует высокую смертность на ранних стадиях развития
у животных, размножающихся с помощью личинки: Sphaerium
согпеит — 35, S. suecicum — 48, Pisidium crassum — 19, Р. lill-
jeborgi — 30, Unio tumidus — 1168-103.
Изучение роста наиболее распространенных и массовых видов
пресноводных двустворчатых моллюсков с использованием коли-
чественных методов позволило определить некоторые важнейшие
положения.
Каждый вид моллюсков может быть охарактеризован разме-
рами, весом, продолжительностью жизни (роста), связанной со
скоростью роста, пределы внутривидовой изменчивости которой
могут рассматриваться в качестве одного из видовых признаков.
Форма раковин моллюсков, образующаяся в соответствии с осо-
бенностями аллометрического роста, зависящими от условий
в водоемах, может быть охарактеризована индексами отношений
линейных размеров раковин, которые с учетом их внутри- и
межвидовой изменчивости применимы для видовой диагностики.
Необходимы специальные экспериментальные исследования для
выяснения отдельных моментов роста моллюсков и, в частности,
влияния различных факторов среды (особенно температуры) на
его скорость. Совершенно не исследованы воздействия пищевых
условий на скорость роста и особенно плодовитость моллюсков.
Глава IV
ФУНКЦИОНАЛЬНО-ЭКОЛОГИЧЕСКИЕ
ХАРАКТЕРИСТИКИ ПОПУЛЯЦИЙ
ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
Перенос энергии в экосистемах осуществляется через трофичес-
кие цепи, возникающие в результате пищевых взаимоотношений
организмов. Поэтому функциональное значение в экологических
системах популяций отдельных видов, входящих в состав сооб-
ществ организмов, может рассматриваться только с позиций коли-
чественного выражения доли их участия в общих процессах
превращения веществ и трансформации энергии. В связи с этим ста-
новятся необходимыми количественно представленные закономер-
ности питания, обмена и роста организмов, соотнесенные с популя-
ционными характеристиками динамики численности, возрастного
состава, биомассы, воспроизводства и смертности.
Естественно, прежде чем рассматривать трансформацию энер-
гии в популяции, следует рассмотреть этот процесс применительно
к особи в процессе ее развития. Уместно вспомнить, что в экологии
основные положения балансовых равенств и следствия из них были
впервые разработаны именно для особи, а затем перенесены на
популяционный уровень. Кроме того, сравнение степени эффек-
тивности использования энергии развивающимися особями и по-
пуляциями того же или других видов животных несомненно пред-
ставляет общеэкологический интерес.
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС ОСОБИ
Пища, потребленная животными, используется ими на прирост
массы тела, покрывает траты энергии в процессах обмена. Неус-
военная часть пищи выделяется в виде фекальных масс.
Скорость потребления пищи (рацион) обозначим С, скорости
изменения массы тела, разрушения органических веществ в про-
цессах обмена, выделения неусвоенной части пищи соответственно
Р, R и /. Скорость ассимиляции пищи, равную сумме Р и 7?, обо-
164
значим А. Сказанное принято выражать в виде балансового ра-
венства
C = P + # + f = A + f. (61)
Когда известна величина усвояемости пищи U=A{C, рацион
животного может быть выражен как
C = 1IU (P-\-R) (62)
или
C — A/U.
Скорость изменения массы животного равна сумме скоростей
соматического (Р8), генеративного (Рд) роста и скорости отчуж-
дения веществ в процессе развития (Ре), т. е.
+ + (63)
Эту величину, по балансовому равенству равную Л—7?,В. Е. Заика
(1972) назвал продукцией особи в отличие от прироста Р — Ре,
т. е. продукция особи определяется равенством
Р = А — R. (64)
Для многих животных экспериментально показано, что их
рацион, так же как и траты на обмен, находится в степенной за-
висимости от веса:
C = aWi‘. (65)
Принимая в уравнении (65) C7=const и обозначив aU=ax, полу-
чаем
A = UC = aJVb'. (66)
В таком случае уравнение (64), или модель продукции особи, пред-
ставляет собой следующее выражение:
P = aiwb' — aWb, (67)
где
aWb = R.
Такой подход к моделированию роста и продукции особи ак-
тивно разрабатывается В. Е. Заикой и Н. П. Макаровой (Заика,
Макарова, 1971; Макарова, Заика, 1971; Заика, 1975; Макарова,
1976). Используя его, необходимо иметь возможность экспери-
ментально определять величину ассимилированной пищи, так же
как, например, скорость обмена, как функцию веса животных.
Однако для подавляющего большинства водных животных это
пока не представляется возможным.
По отношению к двустворчатым моллюскам, как уже было
сказано, определение величины физиологического рациона в экспе-
165
риментальных условиях в большинстве случаев встречает непре-
одолимые пока методические трудности. Поэтому применительно
к этим, как и ко многим другим, животным более надежно коли-
чество ассимилированной ими пищи или эквивалентное количество
энергии находить в виде суммарных величин прироста и трат в про-
цессах обмена (A=P+R), а их рацион — в соответствии с урав-
нением (62). В связи с этим в дальнейшем мы будем использовать
термин «продукция» применительно к скорости образования орга-
нического вещества в популяции в отличие от «прироста», относи-
мого к скорости роста особи.
Балансовые равенства (61, 62) представляют собой закон со-
хранения вещества и энергии. В случае выражения их составляю-
щих в единицах энергии роль популяций и сообществ животных
в экологических системах рассматривается с позиций энергети-
ческого принципа.
Величины составляющих энергетического баланса для отдель-
ных отрезков времени развития и роста моллюсков могут быть оп-
ределены либо экспериментальным путем, либо рассчитаны в со-
ответствии с приведенными выше закономерностями их роста, об-
мена и питания с использованием необходимых для этого урав-
нений. Для расчета величины физиологического рациона моллюсков
необходимо знать усвояемость ими пищи.
Усвояемость пищи у моллюсков разных видов находится в пре-
делах 0.5—0.9 (Алимов, 1970). Близкие к ним значения были
также получены для Anodonta cygnea — 0.62 (Burlacu et al., 1968)
и Helixpomatia — 0.73 (Grossu et al., 1968). Вероятнее всего усвоя-
емость пищи моллюсками в среднем близка к 0.6, т. е. к средней
величине, характерной для водных животных, питающихся расти-
тельной пищей и детритом (Ивлев, 1966), на что указывают резуль-
таты экспериментов с различными видами моллюсков (Цихон-
Луканина, Солдатова, 1973).
Экологический рацион, представляющий собой скорость по-
ступления взвешенных органических веществ в фильтрационный
аппарат животного, может значительно превышать их физиологи-
ческий рацион. В то же время при концентрациях взвешенных
веществ, близких к наблюдаемым в природных водоемах, псевдо-
фекалии у моллюсков еще не образуются или их количество по
сравнению с фекальными массами столь мало, что при расчетах
им можно пренебречь. При таких концентрациях величина эко-
логического рациона становится практически неотличимой от
физиологического. Это дает возможность использовать закономер-
ности интенсивности фильтрации воды моллюсками для определе-
ния их физиологического рациона в условиях природных водоемов.
Пользуясь уравнениями для весового роста, скорости обмена
и фильтрации и сделав соответствующие температурные поправки,
автор по собственным данным рассчитал величины и построил
кривые изменения составляющих энергетического баланса Р, R,
С в процессе индивидуального развития моллюсков Sphaerium
166
suecicum (оз. Круглое), Unio tumidus (p. Луга), Pisidium lillje-
borgi (оз. Зеленецкое) и P. crassum (оз. Круглое) (рис. 62). 1
Абсолютные значения составляющих энергетического баланса*
С, Р, R, как и их динамика в процессе роста животных, представ-
ляют несомненный интерес, но практически не пригодны для срав-
нительных целей. Для этих целей более удобны их относительные
величины.
Соотношения составляющих баланса для этих моллюсков по-
казывают, что по мере роста, т. е. старения, животных у них
увеличивается доля ассимилированной энергии, идущая на энер-
гетический обмен. Ко времени достижения половозрелости, или
первого размножения, у живородящих моллюсков траты энергии
па обмен составляют 45—54% от ассимилированной энергии.
Когда они достигают возраста, соответствующего fmax, величина R
в среднем оказывается близкой к 0.844. Вероятно, соотношение
между тратами на обмен и ассимиляцией (R)A =1 — K2)f тесней-
шим образом связанное с коэффициентом использования пищи
на рост (К2=Р/А), можно рассматривать в качестве количест-
венного показателя степени старения организма (Алимов,
1970).
Величины коэффициента К2, рассчитанные для отдельных от-
резков времени развития, у моллюсков изменяются сходным обра-
зом (рис. 62). Характер их изменений, как видно из рис. 63,
может быть удовлетворительно выражен с помощью известного
уравнения (Винберг, 1968)
^2 = [l/Vma^ + ir1, (68)
где Vrmax=P/7?max; п=1 — а/b, но а/b — показатель степени
в уравнении зависимости скорости обмена от веса животных;
к — константа роста; t — время. При этом среднее значение Х2тах
для изученных моллюсков может быть принято равным 0.8 (7? =0.24),
т. е. близким к теоретическому для пойкилотермных животных
(Заика, 1974а). Показательно, что у личинок Ostrea edulis A2max
составляло 0.7—0.8 (Walne, 1965).
Таким образом, изменения величин коэффициента К2 в про-
цессе развития и роста у различных по систематическому положе-
нию двустворчатых моллюсков находятся в пределах от 0.8 до
0.2-0.16 (7? - от 0.2 до 0.844).
Особое значение имеют соотношения суммарных за весь период
реальной продолжительности жизни животных величин составля-
ющих баланса энергии, или так называемого кумулятивного ба-
ланса, понятие и определение которого было предложено Кле-
ковским (Klekowski, 1970; Klekowski et al., 1972).
1 Все значения составляющих баланса определены для средней темпе-
ратуры воды в водоемах за время роста моллюсков; для S. suecicum,
Р. crassum, U, tumidus она равнялась 15°, для Р. lilljeborgi — 9°.
167
п
Б
&С * о • R ° F * к2
Рис. 62. Динамика составляющих энергетического баланса в течение жизни
моллюсков (из: Алимов, 1977).
Значения составляющих кумулятивного баланса могут быть
вычислены интегрированием соответствующих функций: C=f (£)
или F=/2(0; Р=/4(0-
Отношение интегральных величин прироста Рс к тратам на
обмен Rc за жизненный цикл животных можно рассматривать как
своеобразный коэффициент полезного действия для особи; отно-
шение объема воды, отфильтрованного моллюсками за время их
жизни Fc, к тратам на обмен за тот же период Rc — как коэффи-
циент фильтрации.
Используя значения составляющих кумулятивного баланса,
введем величину К*:
K* = PJAC = PJ(PC^RC). (69)
На важность близкого по смыслу показателя в свое время ука-
зывал В. С. Ивлев (19386).
Величины составляющих кумулятивного баланса и их отно-
шения для некоторых видов моллюсков приведены в табл. 44.
Из данных этой таблицы видно, что отношения прироста (калорий-
ность особи дефинитивного размера) к тратам на обмен Rc у изу-
ченных моллюсков практически одинаковы — от 0.35 до 0.39.
Следовательно, значения К* также весьма близки (0.259—0.279).
Коэффициент фильтрации воды оказался наибольшим у моллюсков
Pisidium и Unio, которые характеризуются бблыпими величинами
относительной площади жаберного аппарата.
Таблица 44
Составляющие кумулятивного баланса энергии для некоторых
видов моллюсков (обозначения в тексте)
Вид Pc (ккал) Rc (ккал) Fc (л) Cc (ккал) PcIRc PcICc FcIRc
Unio tumidus . . 26.500 68.500 13864.6 158.700 0.30 0.167 202.8 0.279
Sphaerium suecicum 0.170 0.457 29.2 1.047 0.37 0.163 63.8 0.272
Pisidium crassum 0.0021 0.0055 1.692 0.013 0.38 0.162 307.6 0.278
P. lilljeborgi . . . 0.0034 0.0098 — 0.022 0.35 0.155 — 0.259
Отношение PcfCc обозначим как К*. Величины К* для изу-
ченных моллюсков оказываются практически одинаковыми (0.155—
0.167), в среднем близки к 0.162. Иными словами, как у мелких
сфериид, так и у крупных унионид энергия пищи, потребленной
А — Sphaerium suecicum', Б — Unio tumidus; В — Pisidium lilljeborgi; Г — P. crassum
c — физиологический рацион (усвояемость пищи 0.6); Р — прирост; R — траты на обмен
/<’ — скорость фильтрации воды; К2 — коэффициент использования пищи на рост. По оси
абсцисс: А, Г — время (сут), Б — время (годы), В — время (мес); по оси ординат: слева:
\, Г — значения С, Р, R (кал-сут-1); Б: I — значения К2, II — значения С, Р, R
(ккал -год-1); В — значения С, Р, R (ккал «мес-1). Справа: А, Г: I — значения К2, II —
значения F (мл^сут"1); Б — значения F (л-год-1); В — значения К2-
169
ими в течение жизни (физиологический рацион), превышает энер-
гию, запасенную в телах особей дефинитивных размеров (разли-
чия по Рс составляют 78 раз) в 6.2 раза.
Значения К* соответствуют величине К2 особи, когда возраст
ее составляет около 0.7—0.8 реальной продолжительности жизни.
Рис. 63. Изменения величины К2 в про-
цессе роста моллюсков (из: Алимов, 1977).
1 — Sphaerium suecicum] 2 — Pisidium lilljeborgi]
3 — P. crassum] 4 — Unio tumidus. Масштаб полу-
логарифмический. По оси абсцисс — значения
отношения ///max’, по оси ординат — значения К2
(объяснения в тексте).
При этом существенно,
что возраст моллюсков,
впервые приступающих
к размножению, и время
достижения ими возраста,
при котором К2 соответ-
ствует К2, находятся
в определенном соотно-
шении. У сфериид это от-
ношение составляет 0.12—
0.18, у перловиц 0.37.
Возраст первого размно-
жения у всех изученных
животных в среднем близок
к 0.17 реальной продол-
жительности их жизни.
Когда они достигают воз-
раста , соответствующего
fmax, значения К 2 ока-
зываются близкими к
0.2 (7?=0.8Л). Как уже
было показано, в таком
возрасте воспроизводи-
тельная способность жи-
вотных сводится к мини-
муму.
Таким образом, особи
старших возрастных групп,
возраст которых превы-
шает реальную продолжи-
тельность жизни особей
в популяциях, оказыва-
ются выполнившими функ-
цию размножения, обеспечивающую поддержание уровня числен-
ности, необходимого для существования популяции и вида
в целом. Большая часть энергии потребленной ими пищи расхо-
дуется на обменные процессы, т. е. рассеивается. Учитывая низ-
кую численность в популяции особей такого возраста, можно счи-
тать, что их экологическое значение в сообществах животных
и экологических системах крайне незначительно. _
Обращают на себя внимание довольно высокие значения К*.
у изученных моллюсков. Однако, рассчитанные по данным И. П. Ара-
биной и Л. А. Рубиновой (1971), величины этого показателя для
170
моллюсков Sphaerium согпеит оказались близкими к 0.35, для
Fissurella barbadensis Gmelin (рассчитано по данным Hughes,
1971а) — 0.33, у слизня Arion rufus (L.) — 0.27 (Stern, 1969).
Более низкие величины К* отмечены для некоторых ракооб-
разных. Амфипода Gammarus lacustris Sars. из оз. Кривого тратит
в течение жизни на процессы обмена 242.2 ккал, а ее прирост
составляет 37.44 ккал (Алимов, Финогенова, 1975а), т. е. значе-
ние К*2 для этих животных не превышает 0.14. Столь же низкие
величины этого коэффициента известны для Asellus aquaticus L. —
0.18 (Klekowski et al., 1972) и для Idotea baltica basteri (And.) —
0.22 (рассчитано по данным Хмелевой, 1973).
Величины К* свидетельствуют об эффективном использовании
пищи моллюсками на рост. В то время как, например, ракообраз-
ные используют пищу с меньшей эффективностью.
Уместно задать вопрос, насколько полученные эксперименталь-
ным путем величины скоростей обмена и роста у моллюсков со-
ответствуют таковым в условиях природных местообитаний?
Очевидно, положительный ответ может быть получен в том случае,
когда пищевые потребности животных, определенные по эмпири-
ческим данным скорости роста и обмена, будут удовлетворяться
при известной скорости фильтрации количеством пищевого ма-
териала в виде органических частиц, находящихся во взвешенном
состоянии в воде природных водоемов.
Ожидаемую (исходя из закономерностей роста и обмена) ве-
личину рациона моллюсков можно определить, зная количество
ассимилированной ими энергии при определенной усвояемости
пищи. Рассмотрим примеры.
В оз. Круглом моллюски S. suecicum населяют только прибреж-
ную его часть до глубины 1.5 м. На бблыпих глубинах (максималь-
ная глубина озера 4 м) они или вовсе не встречаются, или встре-
чаются лишь отдельные экземпляры. На глубинах 2—4 м в этом
озере многочисленны моллюски Р. crassum. В прибрежной части
озера содержание взвешенных органических веществ в воде в лет-
ний период не бывает ниже 1.2 мгО/л (4.1 кал/л), в центральной
части — близко к 0.6 мгО/л (около 2 кал/л). Естественно, что
при волнении количество взвешенных частиц особенно в прибрежных
частях озера может быть в несколько раз выше за счет волнового
перемешивания. Поэтому приведенные ранее цифры можно рас-
сматривать как минимальные.
В условиях спокойной воды моллюски S. suecicum (средний вес
особи в популяции 0.038 г) при температуре 15° могут отфильтро-
вать за сутки 131 мл. При этом они получат в виде пищевого мате-
риала 0.537 кал. На нужды энергетического обмена им необходимо
в сутки 0.218 кал, а суточный их прирост — 0.185 кал, т. е. их
пищевые потребности составляют 0.403 кал/сут. Из приведенных
данных видно, что даже минимального количества взвешенных
веществ оказывается достаточно для удовлетворения пищевых
171
потребностей моллюсков, и усвояемость пищи при этом не превы-
шает 0.75.
В центральной части озера количество взвешенных частиц
вдвое ниже, чем у берегов. Легко рассчитать, что такого количества
недостаточно для нормальной жизнедеятельности 5. suecicum.
У моллюсков Р. crassum, которые живут в центральной части озера,
скорость фильтрации примерно в два раза выше, чем у 5. suecicum.
Поэтому им оказывается вполне достаточно имеющегося в воде
центральной части озера количества органических веществ. Таким
образом, распределение в этом озере популяций двух видов мол-
люсков обусловлено кроме других причин особенностями кормовых
условий в различных его частях.
Наши наблюдения в августе 1973 г. за популяцией Unio tumi-
dus в р. Луге показали, что средний вес особи был близким к 27.6 г
(возраст около 6 лет). Температура воды составляла 15°, содержа-
ние органических веществ в сестоне достигало 3.8 мгО/л. Траты на
обмен у моллюсков такого веса оказываются близкими к 41 кал/сут,
а их прирост — 26 кал/сут. При усвояемости пищи равной 60%,
физиологический рацион животных не превышает 112 кал/сут.
Животные весом 27.6 г способны отфильтровать за сутки около
11.5 л воды, т. е. их экологический рацион составляет 148 кал/сут.
Это показывает, что пищевые потребности животных удовлетворя-
ются, при этом они выделяют около 81 кал/сут в виде фекальных
и псевдофекальных масс.
Приведенные примеры показывают, что экспериментально уста-
новленные закономерности роста, обмена и питания у моллюсков
отражают аналогичные процессы в природных условиях и с успе-
хом могут быть использованы при анализе функционирования
экосистем водоемов.
Генеративный прирост за время реальной продолжительности
жизни моллюсков может быть определен как суммарный вес яиц
или молоди, выметанных ими в течение жизни с момента наступле-
ния половозрелости (абсолютная плодовитость). Так как специ-
альных исследований по определению калорийности яиц унионид
не проводилось, для дальнейших расчетов их калорийность была
принята равной 5 ккал/г сухого веса (сухой вес составляет 30%
сырого). Калорийность новорожденных сфериид принималась
такой же, как у взрослых животных.
Суммарный вес выметанных яиц у унионид составляет 1.9%
веса взрослого животного (табл. 45). Для живородящих сфериид
это отношение значительно выше (от 65 до 133%). Из этой же
таблицы видно, что калорийность выметанных яиц или молоди
колеблется от 2.4% (униониды) до 50% (горошинки) энергии, рас-
ходуемой моллюсками в течение жизни на обменные процессы.
У живородящих сфериид энергетические траты на репродукцию,
как и следовало ожидать, оказались выше, чем у унионид. В этой
связи существенно, что, например, у моллюсков Neritacea и Fissu-
rella barbadensis (размножаются откладкой яиц) энергия половых
172
Таблица 45
Соотношение индивидуальной плодовитости Тр и трат на обмен
Rc у некоторых моллюсков
Вид Ip (экз.) q (кал) ZP (кал) Rc (кал) Wp Wd Rc
Unio tumidus 1168-103 0.0014 1.635 68.5 0.019 0.024
Sphaerium suecicum . . 48 2.3100 0.111 0.457 0.650 0.243
Pisidium lilljeborgi . . . 30 0.1300 0.004 0.010 — 0.400
P. crassum 19 0.1300 0.008 0.006 1.330 0.500
Примечание. Wp — суммарный вес ниц или молоди; Wd — вес взрослого жи-
вотного; q — калорийность одного потомка. Остальные обозначения в тексте.
продуктов составляет 1.8—3.9% энергетических трат на дыхание
(Hughes, 1971а, 1971b). Еще более низкие величины (0.04—1.42%)
известны для травяного и мраморного крабов (Аболмасова, 1973).
Таким образом, при переходе к живорождению у животных
уменьшилась индивидуальная и абсолютная плодовитость и воз-
росла доля энергии, заключенной в молоди, от общих затрат на
ее обменные процессы. Благодаря этому появилась возможность
вымета вполне сформированных молодых животных, достаточно
защищенных с самого начала самостоятельной жизни от неблаго-
приятных воздействий внешней среды.
Двустворчатые моллюски более эффективно по сравнению с ра-
кообразными используют энергию пищи на прирост своей массы.
Уровень обмена, как было показано ранее, остается неизменным
в пределах класса животных. Эффективность использования
особями энергии пищи на рост с учетом скорости роста и энергети-
ческих трат на обменные процессы можно рассматривать как одну
из важнейших интегральных характеристик класса животных,
обусловленную спецификой морфологического строения и физио-
логией животных, относимых к конкретному классу.
ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
МОЛЛЮСКОВ
С экологических позиций популяцию животных можно рассмат-
ривать как систему особей одного вида, различающихся по полу
и возрасту и находящихся в отношениях размножения (Алимов,
Финогенова, 1975а). Определяя популяцию как систему, а не
группу особей, мы тем самым хотим подчеркнуть, во-первых,
наличие популяционного уровня организации и, во-вторых, что
популяция, занимающая определенное место в экосистеме, может
быть охарактеризована некоторыми энергетическими парамет-
рами, в частности потоком энергии.
173
Поток энергии в популяции (часть энергетического ее баланса)
может быть рассчитан как суммарное количество энергии, асси-
милированной особями различных возрастов. Абсолютные вели-
чины составляющих потока энергии в значительной степени опре-
деляются особенностями динамики численности и возрастного
состава популяции, обусловленными скоростями роста числен-
ности и смертности в конкретных условиях.
Известно, что численность особей в популяции при отсутствии
факторов, тормозящих ее увеличение, нарастает согласно экспо-
ненциальному закону:
NtINQ = erm\ (70)
где No — начальная численность; — численность ко вре-
мени £; гт — максимальная мгновенная удельная скорость роста
численности или «врожденная скорость естественного увеличения»
(Andrewartha, Birch, 1954).
Величина гт взаимосвязана с плодовитостью, длительностью
жизни животных и скоростью их развития. Для популяций не-
которых видов животных в условиях лабораторных культур были
определены значения rm (Lotka, 1925; Birch, 1948, 1953; Howe,
1953, и др,). Эти данные позволили Фенчилу (Fenchel, 1974) по-
казать, что величина гт уменьшается с увеличением веса живот-
ных W, согласно степенному уравнению
гт = ^-”, (71)
где р и п — параметры уравнения.
Для пойкилотермных животных им было рассчитано уравнение
lg rm = —1.6391 — 0.2738 lg W,
или
гт = 0.02291 Ж"0-2738. (72)
Врожденная скорость увеличения численности популяции
и интенсивность обмена у животных могут быть переданы одинако-
вой по форме зависимостью от их веса. Действительно, согласно
уравнению Фенчила,
rm = P-W-”,
в то время как по уравнению (8) интенсивность обмена и вес живот-
ных связаны между собой степенной зависимостью:
R!W = aW^1}.
Из этих уравнений видно, что
Обозначив
р/а = 2,
174
получаем
rm __
R/W — ™
откуда
гт = 2 4жС””'Ь_1)1- (74)
Согласно уравнению (72) значение константы п может быть
принято равным 0.27, константа Ъ — 1 по уравнению (17) состав-
ляет 0.28, а для всех пойкилотермных животных, как показал
Хемингсен, — 0.25. Столь близкие величины констант в рассмат-
риваемых уравнениях позволяют принять п^Ъ — 1. В таком
случае
= (75)
т. е. врожденная скорость увеличения численности в популяциях
пойкилотермных животных находится в прямой зависимости от
интенсивности обмена у них. Следовательно, у животных с высокой
интенсивностью обмена можно ожидать большей скорости увели-
чения численности и большей популяционной плодовитости (ко-
личества потомков, приходящегося на одну особь в популяции
в единицу времени).
Врожденная скорость естественного увеличения численности
популяций гт, кроме того, зависит от изменения различных фак-
торов среды, таких, как, например температура (Birch, 1948;
Shift, 1964а; Shiff, Garnett, 1966; Fenchel, 1974, и др.), а для
моллюсков — также от химического состава воды (Harrison,
Shiff, 1966), степени осушения водоема (Shiff, 1964b). Графически
характер изменения гт под влиянием отдельных факторов среды
может быть представлен в виде выпуклой кривой с одним макси-
мумом в области значений, оптимальных по данному фактору.
В природных условиях всегда имеется значительное количество
факторов (хищники, конкуренция, изменение абиотических усло-
вий и т. п.), тормозящих нарастание численности животных.
Поэтому определение гт для природных популяций не представ-
ляется возможным, кроме редких, исключительных ситуаций,
которые могут наблюдаться, например, в начальный период раз-
вития популяции при вселении животных в новые, благоприятные
для них условия.
В качестве примера роста численности популяции по экспо-
ненциальному закону можно привести данные, полученные Стань-
чиковской (Stanczykowska, 1975) для популяции моллюсков
Dreissena polymorpha в одном из Мазурских озер. В этом озере
численность моллюсков в 1959 г. достигала 2200 экз./м2, в 1960 г.
в силу неясных причин численность их упала практически до нуля,
а с 1962 г. до 1972 г. наблюдалось нарастание численности от 30
до 700 экз./м2 (рис. 64).
175
Рис. 64. Изменение численности (1)
и логарифма численности lg N (2) в по-
пуляции моллюсков Dreissena polymor-
pha из оз. Миколайского в период
1962—1972 гг. (по данным Stanczykow-
ska, 1975).
По оси абсцисс — годы; по оси ординат:
слева — численность моллюсков (экз*м“2),
справа — значения lg N.
Выполненные по данным Станьчиковской расчеты показали,
что численность популяции моллюсков в озере в течение 10 лет
увеличивалась согласно экспоненциальному закону, и ее измене-
ние может быть описано уравнением
7V/ = ZVoe°-315f,
где No — численность в 1962 г.; t — время (годы). Принимая,
что рост дрейссен в водоеме продолжается около 100 сут, получаем
Nt = NQe^15t,
где t — время (сутки).
Средний вес моллюсков
в популяции в этот период
изменялся незначительно и
составлял 2.1—2.45 г (вес
без раковины — 0.83—0.835 г,
вес раковины — около 70 %
от общего веса моллюска).
Нетрудно определить, что
значение гт, согласно урав-
нению (71), для животных
такого веса должно быть
0.0187-0.0180, т. е. в 5.9-
5.7 раз выше, чем рассчитан-
ное по данным Станьчиков-
ской. Этот пример показы-
вает, что даже при условии
экспоненциального роста чис-
ленности популяции в при-
родной обстановке величина мгновенной скорости роста оказы-
вается ниже теоретически возможной.
Численность особей в природных популяциях в отдельные пе-
риоды года или вегетационного сезона нарастает или уменьшается,
что связано с различиями в скоростях рождения и смертности
особей разных возрастов. Однако в достаточно короткие по отно-
шению к году или сезону периоды изменение численности популя-
ций может быть описано без больших ошибок экспоненциальной
функцией. Это было, в частности, показано на примере популяций
Sphaerium suecicum и Pisidium crassum (Алимов, 1970). В этом слу-
чае уравнение экспоненциального роста численности особей в по-
пуляции (70) может быть записано как
Nt = N^, (76)
где Nq — численность популяции в момент времени, предшествую-
щий Nt; г — действительная мгновенная скорость увеличения
численности популяции; t — время.
176
В таком виде уравнение используется довольно широко и вошло
в методическую литературу (A manual on Methods. . 1971).
Величина г представляет собой разность между мгновенными
удельными скоростями рождаемости Ъ и смертности d особей в по-
пуляции:
Г = Ъ — d = (In Nt — In N0)/t.
При d=0 r=rm=b; когда b > d, численность популяции уве-
личивается и г положительно; при b < d г отрицательно, числен-
Рис. 65. Динамика численности популяций моллюсков.
Л — S. suecicum (оз. Круглое); Б — Р. crassum (оз. Круглое, 1968 г.); В—Р. nitidum
(оз. Зеленецкое, 1971 г.); Г — Р. lilljeborgi (оз. Зеленецкое, 1971 г.). По оси абсцисс —
месяцы года; по оси ординат — численность моллюсков (экз«м~2).
ность уменьшается. По уравнению (76) могут быть определены зна-
чения и знак г для отдельных отрезков времени развития попу-
ляции.
Изучение динамики численности популяций некоторых видов
моллюсков (рис. 65) позволило рассчитать значения г для отдель-
ных отрезков времени в течение вегетационного сезона (табл. 46).
Из таблицы видно, что г может быть положительной и отрицательной
и в некоторые моменты, совпадающие с наиболее интенсивным раз-
множением, ее величина положительна и достигает наибольших
значений. В эти периоды численность особей в популяции нара-
стает с наибольшей удельной скоростью, которую назовем макси-
мальной удельной скоростью увеличения численности в природ-
ных популяциях гтр.
В ряде случаев необходимо оценить направление развития по-
пуляции. Наиболее строго это можно выполнить, изучив скорости
рождаемости и смертности для различных возрастных групп в по-
12 А. Ф. Алимов
177
Таблица 46
Значения г для популяций некоторых моллюсков
Вид Дата г Водоем
1968 г.
^Sphaerium suecicum VI-VII VII-VIII VIII-IX 0.0222 —0.0167 -0.0103 Оз. Круглое
1969 r. VI-VII VII-VIII VIII-IX 1968 r. —0.0132 —0.0151 0.0470
JPisidium crassum VI-VII VII-VIII VIII-IX 1969 r. VI-VII VII-VIII VIII-IX 1971 r. 0.0120 —0.0210 —0.0081 0.0130 —0.0210 —0.0230 Оз. Круглое
P. lilljeborgi V-VI VI-VII , VII-VIII 1971 г —0.0330 0.0030 0.0220 Оз. Зеленецкое
P. nitidum V-VI VI-VIII VII-VIII -0.0530 0.0370 —0.0070 Оз. Зеленецкое
пуляции. Однако для большинства представителей водных живот-
ных исследования в этом направлении только начинают разви-
ваться, а для многих из них сведения о популяционных характери-
стиках рождаемости и смертности просто отсутствуют. По отноше-
нию к двустворчатым моллюскам их определение сопряжено с ря-
дом методических трудностей, особенно для видов с личиночной
стадией развития. Поэтому неудивительно, что такие данные только
начинают появляться и на их основании пока рано делать какие-
либо заключения. Вместе с тем можно использовать в качестве
показателя направления развития популяций этих животных от-
ношение суммарных значений положительных и отрицательных
величин г, рассчитанных для отдельных одинаковых отрезков вре-
мени, на которые разбито время развития популяции, обозначив
478
его 7s. Знак г указывает на преобладание в популяциях рождае-
мости или смертности, независимо от формы кривой изменения
численности особей.
Расчеты значений г для популяций некоторых видов моллюсков
и ракообразных (табл. 47) показали, что в отдельные годы г может
быть положительной или отрицательной, и это соответствует осо-
бенностям жизненных циклов животных в конкретных усло-
виях. Конечно, такие расчеты представляют собой весьма грубую
оценку направления развития популяции, но в ряде случаев она
может оказаться полезной, тем более что, как будет показано в даль-
нейшем, г связано со средним возрастом особей в популяциях.
Таблица 47
Величины т для популяций некоторых моллюсков и ракообразных
Вид r Водоем ‘ Год
Sphaerium suecicum . . . . j -0.82 + 1.66 Оз. Круглое Тот же 1968 1969
РШйштп crassum -0.43 » » 1968
Р. lilljeborgi +0.69 Оз. Зеленецкое 1971
Р. nitidum -0.60 Тот же 1971
Gammarus lacustris { + 1.64 + 1.02 Оз. Кривое Тот же 1968 1969
Pontoporeia affinis +3.68 » » 1969
Несомненно важно сопоставить наибольшие значения г в при-
родных популяциях (гтр) с максимальной мгновенной удельной
скоростью увеличения численности (rm), рассчитанной по уравне-
нию (72) для животных того же веса. Отношение rmpfrm показывает,
в какой мере в природных популяциях реализуются потенции уве-
личения их численности. В этом смысле его можно рассматривать
как количественную оценку степени давления природных факто-
ров на популяцию конкретного вида животных.
Величина этого отношения оказывается наибольшей у более
крупных животных (табл. 48). Она не имеет таксономической спе-
цифичности и у разных видов зависит от веса животных с дефини-
тивными размерами (рис. 66):
гтр1гт = (0.607 + 0.122) И^ООЗ), (77)
где Жшах — дефинитивный вес животного определенного вида,
выраженный в граммах.
Уравнение (77) отражает важную закономерность (справедли-
вую по крайней мере для рассмотренных представителей водной
фауны), подтверждающую тот несомненный факт, что популяции
12*
179
более мелких по размерам животных подвержены воздействиям
внешней среды в большей степени, чем крупных. Один из воз-
можных механизмов поддержания численности, необходимой для
существования популяций мелких животных, обладающих высо-
кими ростовыми потенциями, — большая их популяционная пло-
довитость по сравнению с более крупными животными.
В пользу такого заключения можно указать на обратную за-
висимость удельной продукции от среднего веса животных (Заика,
Рис. 66. Зависимость логарифма отношения lg (rmplr„^ от логарифма сред-
него веса lg W особей в популяциях различных животных (обозначения
см. табл. 48).
По оси абсцисс — значения lg по оси ординат — значения отношения lg {ттр/тт).
1972). Заметим, кстати, что по скорости роста численности и опре-
деляют продукцию популяций бактерий, коловраток и некоторых
рачков.
Запишем уравнение (77) в общем виде:
rmplrm = qW^,
где q и М — коэффициенты уравнения.
Когда нарастание численности популяции подчиняется закону
экспоненциального роста и мгновенная удельная скорость увели-
чения численности максимальна, смертность низка и ею можно
пренебречь. Согласно В. Е. Заике, гтя&Съ, где Сь — удельная про-
дукция популяции.
Приняв во внимание сказанное, получаем
rmplCb — qW^,
откуда
rmp = qCbW^.
(78)
180
Таблица 48
Значения гтр и их отношения в популяциях различных
водных беспозвоночных животных
Группа и вид животных Bee (г . IO-3) I'm, (сут-1) rmp (сут-1) Гтр/Гщ
Rotatoria *
Keratella cochlearis 0.0003 1.3995 0.0359 0.025
Polyarthra sp 0.0006 1.1576 0.0709 0.061
Asplanchna priodonta 0.0120 0.5097 0.0810 0.159
Cldadocera *
Bosmina longirostris 0.0120 0.5097 0.0830 0.163
Daphnia cristata 0.0200 0.4432 0.0750 0.169
Ceriodaphnia pulchella 0.0150 0.4795 0.0680 0.142
Insecta
Ephemera vulgata 42.15 0.0550 0.0230 0.420
Sialis flavilatera 34.50 0.0570 0.0210 0.369
T any tarsus mancus 0.23 0.2270 0.0320 0.141
Sergentia coracina 1.60 0.1340 0.0280 0.209
Amphipoda
Gammarus lacustris 11.58 0.0780 0.2050 0.320
Pontoporeia affinis . 5.60 0.0950 0.2060 0.274
Bivalvia
Sphaerium suecicum 55.40 0.0320 0.0222 0.430
Pisidium crassum 3.50 0.1080 0.0130 0.121
P. nitidum 0.96 0.1536 0.0380 0.248
P. compessum ** 0.16- 0.17 0.2510— 0.2470 0.0279— 0.0313 0.110— 0.113
Примечание. Одной звездочкой обозначены данные, любезно сообщенные
М. Б. Ивановой, двумя — данные, рассчитанные по материалам Гиллиспи (Gillespie,
19G9). Материалы по остальным видам получены автором на озерах Кривое, Круглое,
Зеленецкое. тт рассчитано цо уравнению (71).
В то же время, как показал В. Е. Заика, удельная продукция
популяции связана со средним весом W особей в ней
Cb = f(W) и = (79)
где риг — коэффициенты уравнения.
Средний вес животных в популяции представляет собой один
из показателей возрастного состава и может быть легко определен
но величине биомассы В и численности животных N:
W = B/N.
Подставляя уравнение (79) в (78), получаем
181
обозначив
?Р = &>
получаем
W*1
г^=^- (8°)
Последнее равенство показывает, что при принятых допущениях
максимальная скорость увеличения численности в природных по-
пуляциях животных определяется соотношением дефинитивного
веса животных конкретного вида и средним весом особи в популя-
ции того же вида. Поскольку в условиях обитания популяции
дефинитивный вес животных данного вида можно считать постоян-
ным, максимальная скорость увеличения численности, так же как
и удельная продукция популяции, находится в обратной зависи-
мости от среднего веса особей в ней.
Однако, как уже было показано, животные одного вида в разных
условиях различаются по размерам. Время достижения ими дефи-
нитивных размеров определяется характером и скоростью изме-
нения их размеров в процессе роста (типом роста).
Как известно, в случае 5-образного весового роста
Подставляя это выражение в уравнение (80), получаем
или, обозначив
mb = cp,
(81)
Однако W а, и для моллюсков находятся в определенном
соотношении между собой:
откуда
Подставляя это выражение в уравнение (81), получаем
а обозначив
&
и-71’
182
получаем
r^ = 7l-^(1-e’fc/max)?- <82)
Из уравнения (82) следует, что величина гтр для видовой по-
пуляции характеризуется соотношением дефинитивного веса живот-
ных (при определенной продолжительности жизни) и среднего
веса особей в популяции, а также константой роста. Поскольку
уравнение (82) остается открытым для учета влияния внешних
условий на скорость роста животных, имеется возможность внут-
ривидовых сопоставлений значений гтр.
Константа роста и продолжительность развития животных на-
ходятся между собой в обратной зависимости. Поэтому в стацио-
нарной популяции, когда возрастной состав остается постоянным
(И7=const), максимальная скорость увеличения численности опре-
деляется константой роста особи конкретного вида. Когда сред-
ний вес особи в популяции не остается постоянным, т. е. популяция
находится в нестационарном состоянии, величина гтр, кроме того,
будет зависеть и от среднего веса особей. Поскольку величина кон-
станты роста определяется также и изменениями факторов внешней
среды (температура, окисляемость воды и т. п.), следует ожидать
различий в максимальных скоростях увеличения численности
природных популяций данного вида животных в разных условиях,
в том числе, например, в разных участках ареала. Иными словами,
максимальная скорость увеличения численности популяции в при-
родных местообитаниях определяется возрастным составом попу-
ляции, закономерностями роста животных данного вида и специ-
фикой условий их обитания.
Попытаемся проанализировать возможные изменения величины
г для популяций моллюсков, приняв в уравнении (82) для про-
стоты расчетов tq=1,7И=0.16 (из уравнения 77), i=1.6, т. е. в 10 раз
больше М.
Исследования популяций некоторых видов моллюсков показали,
что средний вес особи в популяции Sphaerium suecicum в разные
годы колебался от 0.26 до 0.28, у Pisidium crassum достигал 0.6,
у Р. lilljeborgi не превышал 0.23, у Unio tumidus (р. Луга) был бли-
зок к 0.54 от максимального веса в конкретном водоеме, т. е. ве-
личина этого отношения в изученных популяциях находилась
в пределах 0.23—0.45 (2.6 раза).
Рассмотрим возможные изменения максимальной скорости уве-
личения численности популяции на примере моллюсков U. tumidus
из водоемов разных частей ареала вида, допустив при этом колеба-
ния соотношения между средним весом особи в популяции и мак-
симальным весом животного (приняв его равным 100 г) в указанных
выше пределах. Константа роста у этих моллюсков в зависимости
от условий меняется от0.1 до 0.343 год-1, а связанная с ней продол-
жительность жизни — от 6.5 до 22.5 года (см. табл. 15). Заметим,
183
что для всех унионид изменения константы роста не превышают
3—3.5 раза.
Рассчитаем крайние варианты при принятых нами условиях.
Допустим, что рост моллюсков характеризуется &=0.1=const.
Когда Й^/РИтах=0.23, гтр оказывается равной 0.0102, приЙ7РКшах=
=0.6 — соответственно 0.0025 год"1. Второй вариант — популя-
ция стационарна, WfW^=0.6=const, величина к меняется от 0.1
до 0.34 год"1. В этом случае значения гтр ограничиваются преде-
лами 0.0025—0.0028 год"1. Сравнивая два крайних варианта, легко
заметить, что при изменении константы роста в 3.4 раза гтр меня-
ется всего в 1.1 раза, в то время как увеличение отношения W^Wmax
в 2.6 раза приводит к уменьшению гтр в 4 раза.
Приведенный условный и приближенный пример показал, что
максимальная скорость увеличения численности в природных по-
пуляциях в большей степени определяется возрастным их соста-
вом, чем параметрами роста особи. В стационарных популяциях
возможны изменения гтр в небольших пределах в связи с меняю-
щимися условиями обитания.
Наши построения, касающиеся возможной связи скорости увели-
чения численности популяций с ростовыми характеристиками от-
дельных видов в разных условиях, несомненно схематичны, прибли-
женны и несут в себе ряд условностей и допущений. Тем не менее
они представляются полезными и необходимыми, так как помогают
обратить внимание на возможность установления определенных
количественных связей. Намечаются пути поиска строгих законо-
мерностей общебиологического значения между скоростью увели-
чения численности, возрастной структурой популяции и характе-
ристиками роста особей отдельных видов.
ПРОДУКЦИЯ ПОПУЛЯЦИЙ моллюсков
Продукция популяции животных представляет собой скорость
образования ее биомассы. Существуют несколько способов рас-
чета величины продукции. Однако по отношению к популяциям
пресноводных моллюсков чаще всего используют два из них.
Некоторые исследователи рассчитывают продукцию популяций
моллюсков Р, используя метод Бойсен-Йенсена, согласно кото-
рому она определяется как сумма прироста биомассы в на-
чале (5J и конце (Р2) года и биомассы элиминированных за
тот же период особей (5J:
Р = В< + (52-51). (83)
При этом молчаливо принимается, что в уравнении (83) время
равно единице (году).
Метод был разработан и использовался его автором для вы-
числения продукции популяций животных с длительностью
184
жизни в несколько лет и размножающихся ежегодно в течение
краткого периода. В этих случаях величины продукции, рас-
считанные с его помощью, оказывались достаточно близкими
к действительным. Для видов с относительно коротким жизнен-
ным циклом и более или менее растянутым по времени или мно-
гократным в течение года размножением этот метод может дать
лишь приближенное представление о действительных величинах
продукции популяций.
Используя различные модификации этого метода, рассчиты-
вали продукцию популяций дрейссен (Wiktor, 1963; Львова-
Качанова, 1970; Львова, 1976). Поскольку эти моллюски живут
более одного года, полученные авторами величины скорее всего
близки к действительным.
Наиболее широкое применение для определения продукции
популяций двустворчатых моллюсков нашел метод расчета,
при котором учитываются скорость роста особей отдельных воз-
растных групп, динамика численности и возрастного состава
популяции. Такой метод более строго соответствует понятию
продукции как скорости образования биомассы в популяции.
Продукция j-й возрастной группы рассчитывается следую-
щим образом:
<84>
при -> 0
где — прирост особей j-й возрастной группы; — сред-
няя численность данной возрастной группы за отрезок вре-
мени Д£.
Продукция всей популяции в рассматриваемый момент вре-
мени t представляет собой сумму продукций отдельных возраст-
ных групп:
к
(86)
t=0
Количество вещества, продуцируемого в популяции за длитель-
ный период, например вегетационный сезон или год, определя-
ется площадью, ограниченной на графике осью времени и линией,
соединяющей значения продукции в отдельные моменты времени t.
Эта площадь может быть вычислена путем интегрирования
соответствующей функции
Уп к
Р= ( 2 Pi(t)dt
/ ,=о
с использованием методов численного интегрирования и, в част-
ности формулы трапеций.
185
Такой метод вычисления продукции использовался при
расчете продукции популяций унионид в р. Темзе (Negus, 1966),
оз. Крапина (Tudorancea, Florescu, 1968а, 1968b, 1969), в во-
доемах окрестностей Монреаля (Magnin, Stanczykowska, 1971),
в оз. Миколайском (Lewandowski, Stanczykowska, 1975), сфериид
в белорусских озерах (Арабина, 1973), в озерах Кольского п-ова
и Карелии (Алимов, 1970; Алимов, Финогенова, 1975а, 19756,
1976; Алимов и др., 1977; Алимов, Умнов, 1978), планктонных
личинок дрейссен (Hillbricht-Ilkowska, Stanczykowska, 1969).
В последнее время Н. Ф. Смирнова (1975а, 19756) предло-
жила применить для расчетов продукции популяций пресновод-
ных двустворчатых моллюсков метод А. Н. Голикова (1970),
разработанный им на некоторых видах морских брюхоногих
моллюсков. Суть его сводится к тому, что в популяциях разли-
чаются ростовая и поддерживающая продукции, которые рас-
считываются для отрезка времени равного году. При этом при-
нимается, что популяция стационарна.
Ростовая продукция Рд равна весовому приросту всех особей
за год ко времени анализа ее состояния:
Р.= 2 N-(И^+1 — W‘) = 2
i=l i=l
где N\ — численность особей i-й возрастной группы ко вре-
мени £; W*+1—W\ — весовой прирост особи этой группы за год.
Поддерживающая продукция Ps представляет собой ту
«. . .часть вещества, которая образуется за определенный отрезок
времени (в данном случае год) и остается в популяции» (Смир-
нова, 1975а, с. 204), т. е.
Р, - Во + 1 N‘* - И")=Во + 2 А^ДИ"
1=1 i=i
где BQ — биомасса молоди и особей до одного года включи-
тельно.
Предлагаемый метод определяется как новый и рекомендуется
к широкому использованию, так как с его помощью, по мнению
Н. Ф. Смирновой, преодолеваются методические трудности,
свойственные ранее разработанным методам расчета продукции
популяций различных животных, в том числе моллюсков.
Тщательный критический анализ этого метода проведен
Г. Г. Винбергом (19766) и В. В. Сухановым (1978). Поэтому не
станем здесь повторять уже ими изложенное. Остановимся лишь
на некоторых моментах.
Существенное отличие расчетов Рд и Р8 от общепринятой
величины Р заключается в следующем. В уравнениях, исполь-
зуемых для определения этих видов продукции, принимается,
что численность особей каждого возраста в течение года остается
186
неизменной и равной N*. или N*+\ Нетрудно показать, что при
сложении Рд и Р8 и делении этой суммы пополам получается
выражение, которое неотличимо от выражения для продукции
в общепринятом понимании Р:
р , р 2 +в« + 2
__ >=1 »=1
2 2 •
Поскольку
JV? + W,-+1 Л-
* 1 1- — /у
2
ТО
^=2^+4».
1=1
Следовательно, Рд оценивает состояние популяции с учетом
ее начальной, а Р8 — конечной численности за исследуемый
отрезок времени — год, тогда как при расчете Р учитывается
средняя численность популяции за то же время. Поэтому при-
ходится удивляться утверждению, что ростовая продукция Рд
оказывается несмещенной оценкой по отношению к Р (Голиков,
Меншуткин, 1971). Это противоречит уравнению для Рд и пока-
зывает некорректность расчета продукции в использованной
А. Н. Голиковым и В. В. Меншуткиным модели. Определение
ростовой продукции не имеет никаких преимуществ перед обычно
рассчитываемой продукцией Р и ни в коей мере не устраняет
каких-либо методических затруднений.
Сомнителен и биологических смысл, который вкладывается
в понятие поддерживающей продукции: та часть вещества, ко-
торая образуется за год и остается в популяции. Напомним,
что продукция — скорость образования биомассы. Трудно со-
гласиться с тем, что скорость или ее часть (?) остается в популя-
ции. В ней реально может оставаться только биомасса особей,
выживших к определенному (любому) отрезку времени, которая
представляет собой результат продукционного процесса.
Следовательно, разделение продукции на Рд и Р8 в предло-
женном понимании и их расчет не имеет смысла для изучения
продукционных свойств популяций животных. Кроме того,
символ Рд был уже применен Н. Н. Хмелевой (1967) для обозна-
чения продукции выметанных яиц. Использование его и для
обозначения ростовой продукции может привести лишь к не-
нужной путанице.
Продемонстрируем расчет продукции Р с учетом прироста
особей отдельных возрастных групп, динамики численности
и возрастного состава популяции на примере ’популяции моллю-
187
ков Spaerium suecicum из оз. Круглого. В популяции были вы-
делены три размерные группы животных (по длине раковин):
2.4—4.0 (I), 4.0-6.0 (II), 6.0-7.0 мм (III) (табл. 49). Кривая
Таблица 49
Абсолютный (dWjdt) и относительный (Cw) приросты
для отдельных размерных групп, численность (N) и продукция
(Р) популяции моллюсков Sphaerium suecicum в оз. Круглом
(июнь 1968 г., средняя температура воды 15°)
Размерная группа моллюсков W (г) Cw (сут-1) dWIdt (г/сут) jV (экз./м2) Р (г/м2 сут)
I 0.018 0.0280 0.0005 231 0.116
II 0.063 0.0220 0.0014 203 0.284
III 0.144 0.0021 0.0003 62 0.019
весового роста моллюсков рассчитывалась на основе их линей-
ного роста (см. рис. 33) с учетом связи между весом W (г)
и длиной L (мм) у моллюсков этого вида:
Ж = 0.0005Л2-94.
В соответствии с кривой весового роста определялись относитель-
ные и абсолютный приросты для особей каждой размерной группы
(табл. 49). При расчете продукции отдельных размерных групп
и популяции в целом (по уравнению 86) использовались гисто-
граммы размерного состава популяции (рис. 67).
В июле 1968 г. продукция популяции в среднем за сутки
оказалась равной 0.419 г/м2 или 12.97 г/м2 за мес. Аналогичным
образом с учетом изменения температуры воды в озере вычис-
лялись величины продукции популяции в другие сроки (табл. 50).
В табл. 51 приведены величины прироста массы животных
в популяциях некоторых видов сфериид за вегетационный сезон
в исследованных нами северных озерах. В литературе обычно
приводятся сведения о приросте в популяциях моллюсков в рас-
чете за год, но, так как все виды этих животных в континенталь-
ных водоемах умеренных широт в зимние месяцы практически
прекращают расти, естественно рассматривать прирост, рассчи-
танный за вегетационный сезон как годовой.
Важнейший и существенный показатель продукционного про-
цесса — так называемый Р//?-коэффициент. Чаще всего Р)В-
коэффициент находят как отношение количества образованной
в популяции за длительный период (месяц, сезон, год) массы
животных или накопленной в ней энергии к средней за это же
время биомассе популяции. Получается безразмерная величина,
показывающая число оборотов биомассы или энергии за изу-
чаемый интервал времени. В тех случаях, когда Р/5-коэффициент
188
Рис. 67. Гистограмма размерного состава популяции моллюсков Sphaerium
suecicum в оз. Круглом (1968 г.) (из: Алимов, Умнов, 1978).
По оси абсцисс — длина раковины (мм); по оси ординат — численность моллюсков (экз«м~2).
Таблица 50
Количество продуцируемого вещества (Р) и биомасси (В)
в популяции моллюсков Sphaerium suecicum в оз. Круглом
в разные месяцы 1968 и 1969 гг.
Дата Ё (г/м2) Р (г/м2) Р/5
1968 г.
VI 17.2 8.77 0.51
VII 24.0 12.97 0.54
VIII 13.1 6.69 0.51
IX 8.8 2.54 0.29
За сезон 13.3 20.07 1.53
1969 г.
VI 4.4 3.23 0.73
VII 6.65 2.10 0.32
VIII 2.5 0.96 0.39
IX 10.5 2.31 0.22
За сезон 5.98 8.60 1.44
Таблица 51
Количество продуцируемого за сезон вещества (Р)
в популяциях некоторых сфериид
Вид Водоем Р (г/м2) Р/5
Pisidium crassum . . Оз. Круглое 0.618 2.16
Р. lilljeborgi Оз. Зеленецкое 0.219 1.19
Р. nitidum Оз. Зеленецкое 0.055 0.80
определяется как отношение продукции (т. е. скорости образо-
вания биомассы в некоторый момент времени) к биомассе, он
представляет собой удельную продукцию, понятие которой было
предложено В. Е. Заикой (1972) и имеет размерность 1/время.
Поэтому в дальнейшем будем употреблять термин /^-коэффи-
циент по отношению к длительному периоду времени в отличие
от удельной продукции, относимой к моменту времени. Величина
Р/Б-коэффициента, как и удельной продукции, в значительной
степени определяется скоростью прироста биомассы, которая
связана со скоростью роста животных конкретного вида, воз-
растной структурой популяции и численностью особей в ней.
С помощью Р/Б-коэффициента и удельной продукции можно
сравнивать продукционные способности популяций разных видов
или одного вида животных в различных условиях.
190
Скорость роста животного зависит от ряда абиотических
факторов, важнейшее место среди которых принадлежит темпе-
ратуре. Поэтому величина Р/5-коэффициента также находится
в зависимости от факторов среды. Изменение Р/5-коэффициента
под влиянием абиотических и биотических факторов достаточно
подробно было рассмотрено М. Б. Ивановой (1973) на примере
популяций планктонных ракообразных. Полученные ею законо-
мерности справедливы для широкого круга водных беспозвоноч-
ных, в том числе и моллюсков.
Величины Р/5-коэффициентов, рассчитанные как отношение
годового прироста органического вещества к средней за год
биомассе популяции моллюсков, как и для других животных,
различны у коротко- и долгоживущих организмов. Например,
для популяций моллюсков Sphaerium suecicum из оз. Круглого
и 5. согпеит из оз. Нарочь (Арабина, 1966) значения этих коэф-
фициентов оказались весьма близкими и в разные годы состав-
ляли 1.44—1.53, в популяциях Pisidium casertanum и Р. con-
ventus в оз. Пяярви — 1.3, 1.0 (Holopainen, 1979). У более
долгоживущих унионид они не бывают больше единицы. Зна-
чения P/jB в популяциях животных изменяются в течение года
в зависимости от возрастной структуры популяций. В периоды
активного размножения моллюсков Р[В будет выше, чем в другие
периоды (табл. 50).
В популяциях животных одного вида, населяющих водоемы,
которые отличаются по физико-химическим условиям, значе-
ния Р/Р-коэффициентов могут существенно различаться. Так,
например, для популяций моллюсков Elliptic dilatatus в двух
водоемах Канады величины этого коэффициента составляли
0.17 и 0.094, для Lampsilis radiata в тех же водоемах — 0.19
и 0.32 (Magnin, Stanczykowska, 1971). У моллюсков Unio tumi-
dus в р. Темзе годовая величина Р/Р не превышала 0.13 (Negus,
1966), в р. Дунай — 0.26 (Tudorancea, Florescu, 1968b),
в оз. Миколайском — 0.28 (Lewandowski, Stanczykowska, 1976),
у U. mancus (Lam.) из двух водоемов Ломбардии — 0.25 и 0.31
(Nardi, 1978) и т. д.
Все это говорит о том, что формальное применение Р/7?-коэф-
фициентов, установленных в определенных условиях, для вычис-
ления массы или энергии, в ней накопленной, продуцируемой
популяцией того же вида животных в других условиях без учета
сказанного и внесения соответствующих корректив может при-
вести к существенным погрешностям. Особенно значительны
могут быть ошибки при такого рода расчетах для популяций
с кратким жизненным циклом.
Удельная продукция популяций связана с продолжитель-
ностью жизни животных. В частности, по отношению к моллю-
скам, в том числе и двустворчатым, В. Е. Заика (1972) показал^
191
что величина удельной продукции Сь находится Ц в обратной
степенной зависимости от их продолжительности жизни. Однако
он не привел количественного выражения этой зависимости.
Используя его данные по двустворчатым моллюскам и добавив
собственные определения Съ (сут"1) для популяций S. suecicum
(0.0164), Р. crassum (0.018), Р. nitidum (0.0112), Р. lilljeborgi
(0.0132), мы рассчитали параметры уравнения этой зависимости
(рис. 68):
Сь = (0.0362 ± 0.018) т(1-б9±о.зб2) cyT-i, (87)
Рис. 68. Зависимость логарифма удельной продукции 1g Съ от логарифма
продолжительности жизни моллюсков 1g т.
1 — Pisidium crassum} 2 — Sphaerium suecicum; 3 — Pisidium lilljeborgi; 4 —‘P. nitidum;
5 —‘ Elliptic complanata; 6 — E. delatatus; 7 — Lampsilis radiata; 8 — Anodonta piscinalis
(по данным: 1—4 — автора; 5—7 — Magnin, Stanczykowska, 1971; 8 — Lewandowski,
Stanczykowska, 1976; 9—15 — Заики, 1972). По оси абсцисс — значения 1g т, по оси
ординат — значения lg С*.
где т — продолжительность жизни моллюсков (год). Относи-
тельная ошибка функции в данном случае составляет 32 %.
Теперь, зная продолжительность жизни моллюсков с опреде-
ленной вероятностью (в нашем случае 0.95), по уравнению (87)
можно определить удельную продукцию их популяций.
Удельная продукция представляется удобной для сравни-
тельных целей, так как отражает мгновенную по отношению
к сезону или году скорость продуцирования. С ее помощью
удобно сравнивать не только популяции одного вида в различ-
ных условиях, но также разных, далеко отстоящих друг от друга
по систематическому положению видов (например, долгоживу-
щих моллюсков и личинок насекомых или планктонных рако-
образных и т. п.).
Величина удельной продукции зависит от изменения факто-
ров внешней среды и, в частности температуры (Заика, 1972).
При внесении соответствующих поправок, например, на влияние
температуры на скорость роста животных, использование удель-
192
ной продукции для определения количества органического веще-
ства, продуцируемого популяциями за продолжительные периоды
времени, может быть вполне допустимым. В этом случае масса
вещества, продуцируемого в популяции за отрезок времени i2—
может быть определена, исходя из величины средней за этот же
период биомассы популяции:
^2 ^2
Р= j Cb(t)dtB = B j Cb(t)dt.
Несомненно, получаемые в этом случае величины продукции
окажутся менее точными, чем при расчетах, основанных на не-
посредственном изучении скорости роста особей и анализе ди-
намики численности и возрастного состава популяций. Тем не
менее при предварительных, ориентировочных оценках может
быть рекомендован такой способ расчета с использованием ве-
личин удельной продукции.
МОДЕЛЬ ПОПУЛЯЦИИ моллюсков
При исследовании динамики численности, возрастного состава,
биомассы, продукции популяций, механизмов, управляющих ею,
особое значение приобретают математические модели. В процессе
математического моделирования, кроме того, проверяется как
справедливость и надежность полевых исследований структуры
популяций, результатов изучения роста и размножения живот-
ных, так и сделанных на их основе выводов.
С этими целями была создана и проанализирована математи-
ческая модель популяции моллюсков 5. suecicum из оз. Круг-
лого, для построения которой использовались уже обсуждав-
шиеся кривые линейного и весового роста животных и гис-
тограммы ее размерно-возрастного состава (рис. 67) (Алимов,
Умнов, 1978). Анализ динамики размерно-возрастного состава
популяции показал, что моллюски размножаются наиболее актив-
но в июле и августе.
Для построения кривой выживаемости моллюсков возраст-
ной ряд особей в популяции был разделен на отдельные, отли-
чающиеся по возрасту (на 20 сут) группы. Для каждой из них
в соответствии с рис. 69 определялось среднее количество эмб-
рионов. Выживаемость особей отдельных размерных групп lf
находилась как отношение средней численности особей каждой
группы за изученный период к среднему количеству молоди,
выметанной в популяции за это же время 2VM, т. е.
(88)
Построенная таким образом кривая выживаемости особей
различных возрастных групп (рис. 70) показала, что наибольшая
’/а 13 А. Ф- Алимов
193
смертность, как и следовало ожидать, наблюдается на самых
ранних стадиях развития животных. Выживаемость моллюсков
в возрасте от 23 до 90 сут, имеющих длину раковины 2.5—5.5 мм,
оставалась практически на одном уровне и затем возрастала
у более старших особей.
В соответствии с кривой роста продолжительность развития
моллюсков от момента рождения до теоретического максималь-
ного возраста разделялась на К стадий. Продолжительность
каждой стадии зависит от температуры. Предполагалось, что она
Рис. 69. Среднее количество молоди
в. жаберных карманах моллюсков
Sphaerium suecicum разного размера
(из: Алимов, Умнов, 1978).
По оси абсцисс — длина раковины (мм);
по оси ординат — количество молоди (экз.).
определяется среднесезоннои тем-
пературой воды и постоянна внутри
каждого сезона. Популяция рас-
сматривалась в дискретные мо-
менты времени с шагом равным
длительности стадии для каждого
сезона S.
Рис. 70. Кривая выживаемости мол-
люсков разных возрастов в оз. Круг-
лом (из: Алимов, Умнов, 1978)
По оси абсцисс — возраст моллюсков (сут)
по оси ординат — выживаемость.
Диапазон возможных весов -особей разбивался на т групп.
Таким образом, все особи популяции распределялись на Кт
групп в соответствии с тем, на какой стадии развития, они
находились и в какую весовую группу попадали. Образовав-
шиеся группы особей нумеровались двойным индексом i, /.
Первый индекс i обозначает стадию развития, второй j — весо-
вую группу.
Пусть Nt (i, 7), W (г, /) — соответственно численность и сред-
ний вес особей (i, ;)-й группы в момент времени t. Для каждой
группы вводилась смертность fg (i, /) как доля особей, умерших
за время и плодовитость Р8 (i, 7) как среднее число рожден-
ных особью потомков за то же время.
В рассмотрение вводились три матрицы (элемент этих мат-
риц с номером (г, 7; £♦, 7*) показывает, какая доля особей из
(Z, /)-й группы перешла в (i*, /*)-ю группу):
194
1) выживаемости Fg, диагональные элементы которой Fg
(j, /; h j)=l—Л (*, /), остальные равны нулю;
2) плодовитости Р8, элементы первого столбца которой
равны P9(i, j; 1, l)=P8(i, j), остальные равны нулю;
3) роста гв, образованной элементами (j, 7; &♦, /*), показываю-
щими долю особей (г, /)-й группы, переходящих в (j*, /*)-ю
группу. Они определялись по скорости роста особи с учетом
температуры. Остальные элементы матрицы равны нулю.
Введенные соотношения позволяют записать уравнения, опре-
деляющие:
1) динамику численности каждой группы особей в популяции:
A^,=A^(^ + g), (89)
где Nt — вектор строка, (г, /)-я компонента которого равна
численности (j, /)-й группы особей в популяции соответственно
в + t — моменты времени;
2) средний прирост веса особи (i, /)-й группы за время
bW,(i, j)= 2 [(РГ(Г, Г)-Ж(ь 7))r,(i, j, i*, ;*)]; (90)
i*t j*
3) средний вес потомков, рожденных особью (f, /)-й группы
за время 5:
/)=W(1, 1)РД/, /; 1, 1); (91)
4) средний прирост особи (Z, /)-й группы за время S\
j) = (bW8(i, + 7)); (92)
5) продукцию популяции за время
Р(Ъ) = (ДГ(ВД). (93)
где ДТУ — вектор строка, (Z, /)-я компонента которого равна сред-
нему приросту особи (Z, 7‘)-й группы;
6) биомассу популяции:
(94)
где W — вектор строка, (Z, /)-я компонента которого равна среднему
весу особи (Z, 7’)-й группы.
При моделировании распределение особей по группам про-
водилось, исходя из экспериментальной кривой весового роста
моллюсков. Продолжительность развития особей делилась на
20 стадий длительностью 20 сут каждая при температуре 15°.
Год подразделялся лишь на два сезона: зиму и лето. Средняя
температура сезонов была принята равной летом 15°, зимой 6°.
С понижением температуры зимой продолжительность развития
моллюсков увеличивалась в соответствии с законом Крога
до 40 сут.
13*
195
Диапазон возможных весов особи подразделялся на 20 групп.
Поскольку темп роста особи с возрастом меняется, удобнее про-
водить подразделение на весовые группы в соответствии с при-
ростом за одну стадию развития. При этом средний вес особи
в (j, /)-й группе — W (i, j) — находился по кривой весового
роста. Смертность особей (i, /)-й группы определялась по кривой
выживаемости Поскольку экспериментальные исследования
не обнаружили зависимости выживания особи от веса, для всех
весовых групп одного возраста смертность принималась одина-
ковой:
/з(ь 7) = /2(«, Л; if =1, 2, 3 ... 20.
Предполагалось, что смертность в зимний период, так же как
и длительность развития, увеличивается вдвое по сравнению
с летом. Используя кривую выживаемости, смертность определяли
из равенства
/2^ ])— •
Плодовитость особей одинакового возраста принималась сход-
ной, а также вводилось допущение, что вся молодь, имеющаяся
у одной особи, рождается ею в течение одной стадии. В таком
случае, плодовитость особей (г, /)-й группы Р8 (Z, /) равна коли-
честву молоди, поделенному на долю половозрелых особей
в (i, /)-й группе а (г, /):
P2(i, i) = —-—-
2V " »0u)
В зимний период моллюски не размножаются, поэтому для всех
групп Р± (г, f)=0.
На основе полевых наблюдений сделано предположение,
что независимо от возраста все особи определенного веса за
время т2 переходят в следующую весовую группу, т. е. элементы
матрицы r2(i, j; i+1, /+1) равны единице, остальные — нулю.
В зимний период рост отсутствует, и элементы т\ (j, 7; i+1, 7)
равны единице, остальные — нулю.
Построенные на основе сделанных предположений и исходных
данных матрицы F8, Р8 и г8 позволили рассчитать динамику
численности каждой возрастной группы особей в популяции
и ее продукцию.
Численность и биомасса модельной популяции при принятых
условиях плавно нарастают с начала лета, достигают максималь-
ных величин и затем постепенно уменьшаются до исходных
уровней к началу лета следующего года (рис. 71). Кривая дина-
мики продукции имеет два максимума: в начале и конце лета.
После достижения второго из них продукция резко уменьшается
и к осени оказывается равной нулю. На следующий год картина
изменения численности, биомассы и продукции популяции
196
повторяется. Это позволяет рассматривать популяцию стацио-
нарной за год.
Кривая выживаемости характеризуется наличием плато
с одинаковым уровнем для животных, возраст которых составлял
23—90 сут. Первая величина соответствует возрасту новорож-
денных, вторая — наиболее активно размножающимся моллю-
скам, возраст которых близок к среднему возрасту особей в данной
популяции, так как реальная продолжительность жизни при
принятой температуре не превышает 200 сут.
Рис. 71. Динамика численности (7), продукции (2) и биомассы (3) моллюсков
в модельной популяции (из: Алимов, Умнов, 1978).
По оси абсцисс — сезон; по оси ординат’, слева — биомасса, продукция (г-м“2), справа —
численность моллюсков (экз./м"2).
Изменение размера плато на кривой выживаемости («плато
выживаемости») вызывает существенные колебания численности
популяции. Увеличение «плато выживаемости» на одну стадию
(20 сут) приводит к возрастанию численности особей в популяции
почти вдвое, сокращение его на одну стадию, наоборот, влечет
за собой резкое падение численности (рис. 72).
Таким образом, для поддержания необходимой численности
в популяции существенно сохранение неизменного уровня вы-
живаемости особей среднего возраста, которые способны активно
размножаться. Они оказываются как бы заблокированными от
воздействий внешней среды, и незначительное увеличение или
уменьшение их выживаемости приводит к резким и значительным
колебаниям численности особей в популяции. Эти колебания
более существенны по сравнению с теми, которые могли бы быть
обусловлены изменениями выживаемости особей более младших
и старших возрастных групп. Заметим, что наиболее низкая
197
смертность была отмечена для животных среднего возраста
в популяциях моллюсков Sphaerium striatinum (Lam.) и S. si-
mile из двух американских озер (Avolizi, 1976).
Рис. 72. Численность особей в модельной популяции при увеличении (I)
и сокращении (2) «плато выживаемости» (из: Алимов, Умнов, 1978).
По оси абсцисс — сезон; по оси ординат — численность моллюсков (экз./м.“2).
Несомненно важно сравнить динамику численности, био-
массы, продукции модельной и природной популяций моллю-
сков. Характер изменения и уровень численности обеих популя-
ций в 1968 и 1969 г. оказались весьма сходными (рис. 73).
Рис. 73. Динамика численности в модельной (7) и природной (5) популяциях
моллюсков (из: Алимов, Умнов, 1978).
По оси абсцисс — месяцы года; по оси ординат — численность моллюсков (экз./м~2).
В то же время такое совпадение оказывается возможным только
при условии, что в 1969 г. «плато выживаемости» особей в при-
родной популяции было короче по сравнению с 1968 г. на одну
стадию модельной популяции. Иными словами, следует предпо-
198
дожить, что отмирание некоторой части особей среднего воз-
раста в 1969 г. происходило с большей скоростью, чем в 1968 г.
Такая ситуация вполне возможна и могла быть, в частности,
связана с более высокой температурой воды в озере в 1969 г.
Динамика биомассы модельной и природной популяций ока-
залась также весьма сходной. Несмотря на то что, например,
в 1968 г. в природной популяции наибольшая биомасса
(23.998 г/м2) оказалась выше, чем должна была бы быть в соот-
ветствии с моделью (20.4 г/м2), средние значения биомасс за лет-
ний период модельной (14.02 г/м2) и природной (15.7 г/м2) по-
пуляций различались в меньшей степени. Суммарные величины
продукции модельной и природной популяций за этот же период
оказались также близкими — соответственно 38.81 и 31.23 г/м2.
Сравнение озерной и модельной популяций в период актив-
ного размножения моллюсков (июль 1968 г.), когда продукция
популяций была максимальна, показало * достаточно хорошее
сходство их размерно-возрастных структур (табл. 52).
Таблица 52 Таблица 53
Размерно-возрастная структура природной (П) и модельной (М) популяций (июль 1968 г.) Соотношение между генеративной (Рд)>. суммарной (Р) (за 20 сут) и удельной (Сь) продукциями в модельной популяции Моллюсков
Размерная Экз./м3 Р (г/м2) Рд (г/м2).' Рд1Р сь
группа моллюсков (мм)
п м - ‘
_1 5— 6.651 4.811 0.723 0.043
2.4—4.0 4.0-6.0 6.0^-7.5 231 . 203 62 290 180 84 7.920 7.'841 ,7.564 9.836 ' 4.480 4.010 3.711 4.507 0.570 0.510 0.490 0.458 0.034 0.026 0.023 0.028
В большинства ; работ . определение . продукции популяций
двустворчатых моллюсков проводится с учетом только их весо-
вого прироста. В силу методических трудностей особенно для
живородящих видов обычно не учитывается генеративная со-
ставляющая продукции. Построенная модель позволяет оценить
соотношение между продукцией генеративной и создаваемой
за счет соматического роста животных в популяции. Генератив-
ная продукция, как можно видеть из табл. 53, изменяется летом
на отдельных отрезках времени от 46% до 72%, составляя
в среднем 55% общей продукции популяции. Продукция генера-
тивного роста в популяциях Р. casertanum и Р. conventus дости-
гала 17—34% продукции соматического роста (Holopainenr
1978). Известно, что продукция глохидиев унионид не превы-
шает 10% их продукции за счет роста мягких тканей моллюсков
(Negus, 1966).
199
Эти данные дают основание сделать вывод, что генеративная
продукция живородящих моллюсков в значительной степени
определяет величину общей продукции.
При обычных методах определения продукции популяций
живородящих моллюсков генеративная ее составляющая косвен-
ным образом учитывается в виде увеличения веса взрослых
животных за счет развивающейся в них молоди и выживших
новорожденных особей. Однако без специальных исследований
невозможно установить, какую часть веса моллюсков состав-
ляет развивающаяся молодь. Поэтому без учета скорости раз-
множения, плодовитости и выживаемости животных нельзя
получить реальные величины генеративной продукции их попу-
ляций. Подтверждение этому мы находим в различиях абсолют-
ных величин продукции и Р/5-коэффициентов модельной и при-
родной популяций. Величина ^/^-коэффициента за вегетацион-
ный сезон в природной популяции, продукция которой была
определена главным образом с учетом соматического роста,
составляла 1.98. За тот же период для модельной популяции
Р/5-коэффициент оказался равным 2.84. Существенно, что
удельная продукция находится в прямой зависимости от отно-
шения генеративной части к общей продукции популяции
(табл. 53). Это указывает на необходимость при расчетах удель-
ной продукции популяций живородящих моллюсков принимать
во внимание генеративную составляющую.
Таким образом, с помощью математической модели удалось
даже при приближенных исходных данных описать особенности
динамики численности, размерно-возрастного состава, биомассы
и продукции природной популяции. Следовательно, исходные
условия для построения мбдели, полученные в результате поле-
вых наблюдений и использования установленных закономерно-
стей роста и плодовитости моллюсков, оказались достаточно
надежными.
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС ПОПУЛЯЦИЙ
Популяции животных, занимая вполне конкретное место в цепи
трофических связей, могут быть отнесены к определенному
уровню использования энергии первичной продукции в экоси-
стемах. Изучение энергетического баланса популяций позволяет
рассмотреть энергетические соотношения внутри них и коли-
чественно выразить их взаимоотношения с другими компонентами
экологической системы.
Энергетический баланс популяций пресноводных двуствор-
чатых моллюсков исследован мало. Известно лишь несколько
работ такого плана (Negus, 1966; Tudorancea, Florescu, 1968а,
1968b, 1969; Алимов, 1970; Tudorancea, 1972). Поступление
энергии в популяцию животных за единицу времени определя-
ется величинами их рационов. Продукция Р и траты Rp на об-
200
менные процессы в популяции животных представляют собой
составляющие потока энергии Ар (ассимилированная энергия)
за то же время.1 Баланс энергии в популяциях моллюсков рас-
считывался на основе сведений об их динамике численности,
возрастного состава, продукции и составляющих энергетического
баланса в процессе развития особей.
Таблица 54
Энергетический баланс популяции моллюсков Sphaerium
suecicum за вегетационный сезон 1968 г. (ккал/м2)
(из: Алимов, Финогенова, 1975а)
Время р Rp •Ар ср
VI 3.07 6.83 9.90 . 16.50
VII 4.39 8.98 13.37 22.30
VIII 2.33 4.56 6.89 11.50
IX 0.86 1.49 2.35 3.92
За сезон 10.65 21.86 32.51 54.60
Примечание. Здесь и далее рацион рассчитан при условии усвоения живот-
ными 60% пищи. Обозначения в тексте.
В качестве примера в табл. 54 приведены величины состав-
ляющих энергетического баланса популяции моллюсков Sphae-
rium' suecicum из оз. Круглого. Аналогичным образом были
рассчитаны балансы энергии популяций Pisidium crassum из
того же озера и Р. lilljeborgi из оз. Зеленецкого, суммарные
за вегетационный сезон значения составляющих которых в рас-
чете на 1 м2 площади дна зоны обитания животных сведены
в табл. 55. Однако площадь этой зоны, например, в оз. Круг-
лом для моллюсков S. suecicum составляет около 19%, Р. cras-
sum — 29% площади озера (около 0.1 км2). С учетом этого об-
стоятельства и данных табл. 55 суммарное (за сезон 1968 г.)
потребление пищи моллюсками S. suecicum не превышало 10.4,
а Р. crassum — 0.44 ккал/м2 площади озера, т. е. рацион Bivalvia
в озере был близок к 22% первичной продукции фитопланктона
за тот же период (49 ккал/м2). Энергия неусвоенной пищи
fp=Cp—Ар моллюсками S. suecicum не превышала 4.2, Р. cras-
sum — 0.18 ккал на 1 м2 площади озера за сезон. В результате
жизнедеятельности популяций двустворчатых моллюсков в те-
чение сезона на дне озера отлагались органические вещества,
количество которых эквивалентно 4.4 ккал/м2 (около 9% пер-
вичной продукции).
1 Для отличия обозначений соответствующих понятий по отношению
к особи и популяции в дальнейшем составляющие энергетического баланса
популяций будут обозначаться индексом р: Rp, Ар, Ср, К2р и т. д.
14 А. Ф. Алимов
201
Таблица 55
Суммарные за вегетационный сезон величины составляющих
энергетического баланса популяций некоторых моллюсков
(ккал/м2)
Вид В Р Rp А.р Ср —* К<2.р к1р ip
1968 г.
Sphaerium suecicum 1 6.10 | 10.70 | 21.90 | 32.60 | 54.50 | 0.33 | 0.20 | 21.90
1969 г.
2.10 3.00 5.40 8.40 14.00 0.36 0.21 5.60
Pisidium crassum P. lilljeborgi 1 0.11 0.06 1 0.23 0.07 0.69 0.17 0.92 0.24 1.53 0.40 0.25 0.29 0.15 0.18 0.61 0.16
Примечание. Баланс энергии рассчитан на 1 м2 зоны обитания моллюсков;
/ — энергия неусвоенной пищи. Обозначения в тексте.
Все нехищные донные животные в оз. Круглом потребили
за вегетационный сезон в виде пищи 26.7 ккал/м2, из которых
4 ккал приходились на личинок хирономид (Алимов, Финоге-
нова, 1975а). Нетрудно определить, что энергия неусвоенной
двустворчатыми моллюсками пищи составляла около 15.6% ра-
циона донных животных и оказалась равной пищевым потреб-
ностям личинок хирономид. Напомним, что в оз. Миколайском
количество органических веществ, осаждаемых дрейссенами, пре-
вышало рацион хирономид в 7.7 раза (см. гл. II). В оз. Зеленец-
ком количество неусвоенной двустворчатыми моллюсками пищи
составляло не более 1.2% рациона нехищных животных бентоса,
около 6 % рациона личинок хирономид. Но, как видно из
табл. 55, биомасса моллюсков в этом озере была значительно
ниже, чем в оз. Круглом. В оз. Круглом она составляла 29%
биомассы донных животных (3.38 ккал/м2), в оз. Зеленецком —
лишь 11.6%.
Биомасса и продукция двустворчатых моллюсков в некото-
рых водоемах могут достигать значительных величин (табл. 56).
В р. Темзе, например, биомасса этих моллюсков вдвое выше,
а создаваемая ими продукция лишь в два раза ниже, чем у рыб
в этой реке (Mann, 1964). Биомасса моллюсков в различных
водоемах колеблется от 6 до 73% биомассы животных бентоса,
продукция — от 2 до 59% продукции нехищных животных бен-
тоса (табл. 57). Величины продукции популяций моллюсков за
год обычно составляют десятые или сотые доли процента, не пре-
вышая 1.4—2.4% от первичной продукции в водоемах, рассчи-
танной за тот же период. Правда, в некоторых водоемах про-
дукция моллюсков достигает 10% от первичной продукции
202
Таблица 56
Среднегодовая биомасса (В) и прирост биомассы в расчете за год
(Р) популяций моллюсков
Группа моллюсков Водоем Я (г/м2) Р (г/м2) Источник
Униониды Р. Темза 292.20 45.20 Negus, 1966
То же Оз. Миколайское . . . 6.56 2.24 Lewandowski, Stanczykowska, 1976
» Оз. Монтагнес .... 85.80 16.02 Magnin, Stanczy- kowska, 1971
» Оз. Красное 940.00 564.00 Кузьменко, 1976
Дрейссены Учинское в-ще .... 1050.00 2430.00 Львова, 1976
(Gillespie, 1969). Вместе с тем намечается определенная тенденция
к увеличению продукции популяций моллюсков по мере нара-
стания продуктивности водоемов (рис. 74). Это связано скорее
всего с большим видовым разнообразием, большей численностью
и биомассой этих животных в эвтрофных водоемах по сравнению
с менее продуктивными. Так, в олиготрофных и мезотрофных
озерах Швеции численность двустворчатых моллюсков, в частно-
сти унионид, не превышает 1.8, в эвтрофных — 8.2, достигая
в высокоэвтрофных 20—30 экз./м2 (по данным Agrell, 1949, цит.
по: Lewandowski, Stanczykowska, 1976).
Таблица 57
Соотношение биомассы (Вм) и продукции (Рм) моллюсков,
биомассы животных бентоса (Вб), продукции нехищных
животных бентоса (Рн.х.) и первичной продукции (Рп) в водоеме
*м/*б Р м/^н.х. По данным
Озеро:
Зеленецкое 0.0050 0.116 0.059 Автора
Кривое . . . 0.0012 0.064 0.092 То же
Круглое . ... 0.0240 0.392 0.348 »
Дривяты 0.0069 0.730 0.587 Гаврилова, 1970
Река:
Темза ... 0.014— 0.0134 — 0.483 0.194 Mann et al., 1972
В о дохранилище:
Киевское . . 0.0019 0.080 0.035 Гак и др., 1972
Рыбинское 0.0005 0.070 0.023 Сорокина, 1972
14*
203
Траты на обменные процессы популяциями двустворчатых
моллюсков, как уже было показано, в течение вегетационного
сезона близки к 1—10% (в среднем 5%) энергии первичной про-
дукции за тот же период. В частности, в оз. Круглом эта ве-
личина составляла 4.5, в оз. Зеленецком — 5.9%.
Рассмотрим еще несколько примеров, демонстрирующих воз-
можности применения балансового подхода для оценки функцио-
нального значения популяций от-
дельных видов в экологических
системах.
На одном из участков р. Пре-
голи (выше Калининграда непо-
средственно у его городской черты)
в августе 1974 г. была изучена
популяция моллюсков Dreissena
polymorpha. В этом месте реки
вода характеризовалась следую-
-2 -
Рис. 74. Зависимость логарифма
годовой продукции популяций
моллюсков 1g Рм от логарифма
первичной продукции 1g Рп в раз-
личных водоемах.
1 — оз. Кривое; 2 — оз. Круглое;
3 — оз. Дривяты; 4 — р. Темза; 5 —
оз. Зеленецкое; 6 — Киевское водо-
хранилище; 7 — Рыбинское водохра-
нилище (поданным: 1, 2, 5 — автора;
3 — Гаврилова, 1970; 4 — Mann et al.,
1972; 6 — Гак и др., 1972; 7 — Соро-
кина, 1972). По оси абсцисс — значения
1g Рп; по оси ординат — значения 1g Рм.
щими показателями: скорость тече-
ния — 0.15 м/с, прозрачность —
0.75 м, температура — 16.3°, на-
сыщение кислородом — 68.1%,
бихроматная окисляемость воды —
41.8, сестона — 2.16 мгО/л (Али-
мов и др., 1976). На глубине около
1.5 м численность дрейссен дости-
гала 1750 экз./м2, биомасса —
1170 г/м2. Средний вес моллюсков
в популяции составлял 0.668 г.
Принимая удельную продукцию
равной 0.0014 сут-1 и калорийность
моллюсков 0.24 ккал/г сырого
веса (Биргер и др., 1967 с поправкой по: Остапеня, 1968), полу-
чаем, что продукция популяции составляет 1.64 г/м2, или
0.384 ккал/м2 в сут, траты энергии моллюсками на обменные
процессы — около 8.4 ккал/м2, поток энергии в популяции —
8.78 ккал/м2 в сут. В течение суток популяция на площади 1 м2
способна отфильтровать до 1.96 м3 воды, в которой содержится
7.48 ккал/л сестона, т. е. их экологический рацион составляет
14.66 ккал/сут. Допуская, что моллюски фильтруют не полные
сутки, а только 20 ч, величина их рациона равна 12.21 ккал/сут.
В столбе воды над площадью дна 1 м2 содержится 11.22 ккал
сестона. Вода на этом участке реки за 1 ч проходит 0.54 км
(1 км — за 1.85 ч). За время пробегания водой 1 км (т. е. над
популяцией, располагающейся на площади 1000 м2) моллюски
потребят 0.941 ккал сестона, потратят на обменные процессы
0.648 ккал, что составляет соответственно 12.6 и 8.7% количества
сестона в протекающей воде. Физиологический их рацион (при
усвояемости 0.6) близок к 0.475 ккал, или 4.2 % количества сестона.
204
Аналогичные расчеты для популяций двустворчатых моллю-
сков на других участках р. Преголи показали, что наиболее
значима роль этих животных на участках вблизи Калининграда
(табл. 58). Степень загрязнения реки постепенно нарастает по
направлению к городу. Так, если вода в реке ниже городов Зна-
менска и Гвардейска может быть оценена как умеренно загряз-
ненная, вблизи Калининграда — как загрязненная. В черте
города река принимает наибольшее количество загрязнений,
здесь двустворчатые моллюски практически отсутствуют.
Таблица 58
Отношение величины потребленного популяциями моллюсков
сестона (Сс) и трат энергии на обмен (Rp) к количеству сестона
под 1 м2 (Es) на разных участках р. Преголи и р. Лавы
за время прохождения воды над 1 м2 площади дна
Река Вид моллюска N (экз./м2) В (г/м2) о GO <u о Э ь? "а.
Преголя:
ниже г. Знаменска Р. amnicum . . 580 141.50 2.30 0.60
ниже г. Гвардейска Р. amnicum . . 200 2.68 0.42 0.33
ниже г. Калининграда D. polymorpha 1750 1170.00 12.60 8.70
Лава
у г. Знаменска Р. henslowanum 1380 220.80 1.00 0.50
Примечание. N — численность, В — биомасса моллюсков.
Сказанное позволяет рассматривать роль двустворчатых мол-
люсков в процессах самоочищения в водоемах как существенную.
Наиболее ощутимо их участие в этих процессах и умеренно за-
грязненных водах. Оно повышается по мере нарастания содержа-
ния органических веществ в воде до определенных пределов,
выше которых моллюски просто не могут жить. Роль этих живот-
ных в процессах трансформации энергии оказывается наиболь-
шей в эвтрофных, незагрязняемых водоемах.
В состав планктонных сообществ некоторых водоемов как
непременный компонент входят личинки (велигеры) дрейссен.
Их количество в ряде случаев доходит до сотен тысяч в одном
кубометре воды, составляя до 60% численности животных зооплан-
ктона (Вьюшкова, 1965). Средняя биомасса личинок за время их
пребывания в планктоне в различных водоемах колеблется от 0.7 до
20% биомассы зоопланктона за тот же отрезок времени (табл. 59),
достигая наибольших величин в периоды массового выхода личинок
к планктон. Например, биомасса личинок дрейссен в Каховском
205
водохранилище в июне 1959 г. составляла 10%, в июле 13.3, в ав-
густе 42% биомассы зоопланктона (рассчитано по данным Цееба,
1964).
Таблица 59
Отношение биомассы личинок (Вл) дрейссен к общей биомассе
животных (В8) в разных водоемах
Водоем БЛ1В3 (%) По данным
Каховское водохранилище 20 Волгоградское водохранилище ... 5 Цимлянское водохранилище .... 0.9 Мазурские озера . .' 0.7 Цееба, 1964 Вьюшковой, 1965 Дзюбана, 1958 Hillbricht-Ilkowska, Stanczykowska, 1969; Kajak et al., 1970
Несмотря на значительную численность и биомассу велигеров
дрейссен, до сих пор не было сделано попытки количественного
выражения значения их в составе планктонных сообществ и в эко-
системе водоемов. Попытаемся это сделать в первом приближении.
Немногочисленные исследования по продукции личинок дрейс-
сен, выполненные на Мазурских озерах (Hillbricht-Ilkowska,
Stanczykowska, 1969) и в канале Днепр—Кривой Рог (Шевцова,
19716), показали, что удельная продукция личинок в планктонной
стадии составляет около 0.23—0.29 сут'1. Она близка к величинам,
известным для ракообразных зоопланктона. Для осевших ли-
чинок значения удельной продукции значительно ниже и не превы-
шают 0.017—0.082 сут"1. Прирост массы личинок в двух исследован-
ных Мазурских озерах в разные годы составлял 2—4% прироста
массы животных зоопланктона.
Используя эти данные, определим продукцию планктонных ли-
чинок дрейссен в Куршском заливе. Средняя численность велиге-
ров в планктоне этого залива достигала 84«103 экз./м3 (Киселите,
1959), или (при средней глубине 3.7 м) 311-103 экз. под 1 м2.
Поскольку средний вес одной личинки не превышает 6*10"4 мг
(Hillbricht-Ilkowska, Stanczykowska, 1969), биомасса велигеров
в Куршском заливе составляет около 0.187 г/м2. Средняя за сезон
биомасса зоопланктона в заливе достигает 10.83 г/м2, из которой,
как легко определить, 1.7% приходится на велигеры дрейссен.
Нетрудно рассчитать, что продукция личинок (при удельной про-
дукции 0.26) составляет 0.049 г/м2 в сут. За время пребывания
их в планктоне (около 120 сут) они продуцируют 5.85 г/м2.
Животные зоопланктона в заливе за вегетационный сезон,
по расчетам Т. Киселите, продуцируют 216 кг/га, или 21.6 г/м2.
Скорее всего это заниженная величина, так как, принимая ее,
получим неоправданно низкий Р/7?-коэффициент для зоопланк-
206
тона — около 2. Приняв более реальную величину этого коэф-
фициента, например равную 10, получаем, что животные зоо-
планктона за вегетационный сезон продуцируют примерно 108 г/м2.
В таком случае, прирост массы велигеров дрейссен за тот же
период составит около 5.4% от прироста биомассы зоопланктона.
Эта величина близка к полученной польскими исследователями
на Мазурских озерах.
Приведенные данные и расчеты позволяют заключить, что
продукция личинок дрейссен в различных водоемах составляет
2—5% продукции животных зоопланктона.
Для определения пищевых потребностей планктонных личи-
нок дрейссен, потребляющих в значительных количествах водо-
росли фитопланктона, произведем несложный расчет, используя
сведения Хилбрихт-Илковской и Станьчиковской по численности
и продукции велигеров в оз. Талтовистко (одно из Мазурских
озер). Средняя численность велигеров дрейссен в этом озере за
86 сут пребывания их в планктоне была 462-103 экз./м2.
Согласно уравнению (16) нетрудно определить, что личинка
со средним весом 6-10"4 мг должна потреблять на нужды обмена
5-10'4 мгО/сут. Все личинки в озере за время их планктонной
жизни потратят на нужды обмена около 6.75 ккал/м2. При кало-
рийности личинок 0.5 ккал/г сырого веса и удельной продукции
0.26 сут'1 прирост массы этих животных за тот же период составит
3.29 ккал/м2. Величина их рациона при усвояемости 0.6 не превысит
16.77 ккал/м2 (около 2% от первичной продукции в озере за время
пребывания личинок в планктоне).
Это несомненно интересный и важный результат, показываю-
щий сопоставимость пищевых потребностей велигеров и нехищных
животных бентоса. Как можно видеть из данных табл. 60, отноше-
ние количества пищи, потребляемой нехищными животными бен-
тоса, к первичной продукции в большинстве изученных водоемов
Таблица 60
Соотношение между величинами рационов за вегетационный
сезон нехищных животных зоопланктона (О3) и бентоса (Сб) и
первичной продукцией (Рл) в различных водоемах
Водоем Сб/Ри По данным
Озеро:
Красное 0.08 0.015 Андрондиковой и др.,
Миколайское . — 0.040 Kajak et al., 1972
Снярдви ... — 0.022 То же
Каракуль . . . . 0.22 0.012 Хусаиновой и др., 1973
Мястро ... . . — 0.013 Умнова, 1973
Дривяты 0.27 0.019 Винберга, 1970
Кривое 0.56 0.114 Автора
Киевское водохранилище . . . 0.64 0.019 Цееба и др., 1973
207
изменяется от 0.01 до 0.04. Более высокие значения отмечены,
в оз. Кривом (0.114). В то же время рацион нехищных животных
зоопланктона составляет большую по сравнению с бентосными
организмами долю от первичной продукции. Пищевые потребности
планктонных личинок моллюсков оказываются значительно ниже,
чем у животных зоопланктона и близкими к таковым у донных
животных.
Приведенные расчеты дают основание говорить о существенном
значении личинок дрейссен в планктонных сообществах. При ко-
личественных исследованиях процессов трансформации вещества
и энергии в пресноводных водоемах необходимо учитывать участие
в них личинок моллюсков. Тем более что в периоды массового
размножения этих животных количество их планктонных личинок
может достигать значительных величин.
Двустворчатые моллюски составляют заметную долю в питании
рыб. Особенно охотно потребляют рыбы мелких моллюсков, длина
раковин которых не превышает 2—3 мм. Поэтому наиболее активно
рыбы питаются моллюсками в летние месяцы в периоды интенсив-
ного размножения последних. Более крупные по размерам рыбы
могут поедать и более крупных моллюсков. Оценим зна-
чение моллюсков в питании рыб на примере некоторых водо-
емов.
В пищевых рационах бентосоядных рыб Куршского залива зна-
чительная доля принадлежит дрейссене. Средняя биомасса дрейс-
сен в наиболее благоприятных для них участках этого водоема
колеблется от 300 до 3500 г/м2 (Аристова, 1968), в среднем около
1900 г/м2. Величина годового Р /5-коэффициента для этих моллюс-
сков близка к 0.4. Приняв это», получаем продукцию дрейссен за год,
равную 760 г/м2, или 7.6 т/га. Вылов рыб, питающихся дрейссеной
(плотва, густера, лещ), в заливе может достигать 15.1 кг/га
(рассчитано по данным Манюкаса, 1959). С достаточным основанием
можно принять, что вылов составляет около 30% запаса рыб в водо-
еме. В таком случае величина последнего оказывается близкой
к 50 кг/га. Допустим, годовой рацион рыб при питании моллюсками
превышает собственный вес рыб в 20 раз, исходя из того, что при
таком же корме годовой рацион воблы больше ее веса в 24 раза (Bip-
rep, Маляревська, 1964). Следовательно, рацион рыб за год, если
они питаются только дрейссенами, составит около 1 т/га. В то же
время на долю дрейссен в этом водоеме приходится около 30 % ра-
циона рыб, т. е. количество потребленных рыбами моллюсков за год
не должно превышать 300 кг/га. Однако, по данным Г. И. Аристо-
вой, рыбы поедают дрейссен, длина раковины которых не более
15 мм. Моллюски такого размера составляют 13% биомассы дрейс-
сен в заливе. Учитывая это, легко определить доступную для рыб
часть продукции моллюсков, которая при принятых допущениях
оказывается близкой к 968 кг/га. Таким образом, величина продук-
ции мелких дрейссен примерно в 3.2 раза выше по сравнению
с количеством моллюсков, необходимых для питания бентосоядных
208
рыб в этом водоеме. Иными словами, основные потребители дрейс-
сен в Куршском заливе для удовлетворения своих пищевых потреб-
ностей должны использовать примерно третью часть веществ, про-
дуцируемых моллюсками за год. Поскольку рыбы питаются глав-
ным образом молодью дрейссен, сделанный расчет позволяет пред-
полагать, что количество молоди, не элиминированной рыбами,
в популяции моллюсков оказывается достаточным для сохранения
ее численности на уровне, необходимом для существования вида.
Это заключение дает основание предполагать соответствие между
количеством бентосоядных рыб и их кормовой базой в Куршском
заливе.
Другой пример. В оз. Большой Харбей в пище леща около
39% (по весу) приходится на моллюсков сфериид (Сидоров, 1976).
За два исследованных летних месяца продуцируемая популяцией
моллюсков в этом озере энергия была близкой к 0.33 ккал/м2
(Попова, 1976), или около 0.5 ккал за вегетационный сезон (3 мес).
В озере кроме сфериид обитают также два вида гастропод. Сделаем
в нашем примере допущение, что все моллюски в озере представле-
ны только сфериидами. Кроме того, введем еще одно крайне неверо-
ятное допущение — в течение сезона сиги потребят целиком все
органическое вещество, продуцируемое популяцией моллюсков за
тот же период. Известно, что суточный рацион рыб при питании
моллюсками может достигать 6% от их веса, а при потреблении
иного корма лишь 2.5 %. Учитывая, что калорийность сфериид вмес-
те с раковиной выше, чем, например, у дрейссен, примем величину
рациона рыб при питании сфериидами за сутки близкой к 4 % их
веса.
Приняв эти допущения, нетрудно рассчитать, что рацион сигов
(сферииды составляют в нем 39%) за вегетационный сезон должен
быть близким к 1.26 ккал/м2. Эта величина превышает известный
для озера прирост биомассы моллюсков в 2.5 раза. Биомасса
сигов за этот же период, исходя из сказанного, оказывается близкой
к 0.14 ккал/м2. Биомасса всех рыб в этом озере была около
5.9 ккал/м2. Рассчитанная нами величина биомассы сигов состав-
ляет около 2.5% общей биомассы рыб в озере. По данным Г. И. Си-
дорова (1976), значение Р/2?-коэффициента рыб в озере не превы-
шало 0.29. Примем для сигов величину этого коэффициента такой же.
В этом случае при средней биомассе 0.14 ккал/м2 сиги продуцируют
около 0.041 ккал/м2 за сезон, что составляет примерно 2.4% энер-
гии, продуцируемой всеми рыбами в озере (1.7 ккал/м2 за сезон),
тогда как, согласно Г. И. Сидорову, прирост биомассы сигов
(7.1 кг/га) за сезон достигал 40% прироста биомассы всех рыб
(17 кг/га).
Рассчитанные, исходя из продуктивности кормовых организмов,
величины возможной биомассы сигов и ее прироста за сезон в оз.
Большой Харбей оказались даже при невероятно благоприятных
для питания рыб допущениях ниже известных для этого озера более
чем в 10 раз. Это позволяет предполагать, что либо опубликованные
209
данные по продукции моллюсков оказались ниже реальных для
данного водоема, либо величины прироста биомассы рыб, в том
числе и сигов, выше, чем это может быть обеспечено кормовой
базой.
Приведенные примеры, несмотря на частный характер, еще
раз подтверждают необходимость при проведении исследований
Рис. 75. Сезонная динамика биомассы, потока энергии и его составляющих
в популяциях моллюсков в оз. Круглом.
I, 2 — Shaerium suecicum] 3, 4 — Pisidium crassum', P — продукция; Rp — траты на
обмен; Ар — поток энергии, по оси абсцисс — месяцы года; по оси ординат: — 1, 3—био-
масса (г. м-2); 2,4 — значения Р, A, R (ккал/мес).
по биологической продуктивности водоемов количественной оценки
получаемых данных по продукции и запасам рыб в соответствии
с развитием их кормовых объектов. Такого рода расчеты могут
служить контролем для получаемых результатов и обеспечивать
большую их надежность и убедительность.
Таким образом, двустворчатые моллюски занимают существен-
ное место в сообществах животных второго трофического уровня.
В некоторых водоемах биомасса моллюсков составляет до 73%
биомассы животных бентоса, продуцируемая ими энергия — 1.4—
210
2.4%, энергия, рассеиваемая на процессы обмена — около 5%
от энергии, заключенной в первичной продукции. Энергия, по-
требляемая популяциями моллюсков в виде пищи, в различных
водоемах достигает 4—21 % от первичной продукции. При этом от
2 до 12% энергии потребленной пищи ассимилируется популяци-
ями в виде потока энергии.
Рис. 76. Зависимость логарифма потока энергии 1g Ар от логарифма био-
массы 1g В в озерах Кривое, Круглое, Зеленецкое для популяций животных.
А — Bivalvia: 1 — Sphaerium suecicum', 2 — Pisidium crassum', 3 — P. lilljeborgi. Б —
Amphipoda: 1 — Gammarus lacustris', 2 — Pontoporeia affinis. В — Chironomidae: 1 — Ta-
nytarsus mancus: 2 — Procladius nigriventris', 3 — Sergentia coracina. По оси абсцисс — зна-
чения 1g В (ккал/м2 за мес); по оси ординат — значения 1g Ар (ккал/м2 за мес).
Поток энергии, или темп ее ассимиляции в популяциях мол-
люсков, имеет сходный с биомассой характер сезонных изменений
(рис. 75). Наибольшие его величины совпадают по времени с пе-
риодами активного размножения животных. В популяциях мол-
люсков, амфипод и личинок хирономид из озер Кривое, Круглое
и Зеленецкое изменения потока энергии в течение вегетационного
сезона находились в прямой зависимости от величин биомассы, и
эта зависимость может быть выражена в виде степенной функ-
ции (рис. 76):
Ар = уВл ккал/м2 за месяц, (95)
где Ар — поток энергии в популяции; В — ее биомасса; у и а—
параметры уравнения.
211
Значения параметров соответствующих уравнений для популя-
ций моллюсков, амфипод и хирономид рассчитаны графически.
Для всех изученных представителей донных животных величина
показателя степени а оказалась практически неотличимой от еди-
ницы. Поэтому параметр ^=Ар)В представляет собой удельный
поток энергии в популяциях, и его значения в популяциях хироно-
мид выше (3.98), чем двустворчатых моллюсков (1.51) и амфипод
(1.35). Аналогичным образом поток энергии в расчете за сезон или
год (сумма его значений за отдельные отрезки времени) связан
со средней за этот же период биомассой животных в популяции
(рис. 77):
Ар — ^В ккал/м2 за сезон, при этом
На примере популяций моллюсков можно показать, что вели-
чина удельного за сезон потока энергии находится в обратной
зависимости от среднего веса особи в популяциях разных видов
(табл. 61). Аналогичный вывод был сделан Г. И. Аболмасовой (1976)
в результате изучения популяций нескольких видов морских
ракообразных.
Таблица 61
Соотношение между средним весом особей (JFj и ЛР1В
в популяциях некоторых видов моллюсков
Вид W (r) Apia По данным
Sphaerium suecicum . . 0.0500 * 6.50 * Алимова, 1975a
0.0550 ** 4.40 ** To же
Pisidium crassum .... 0.0025 9.55 »
Unio tumidus 4.0000 3.80 Tudorancea, 1972
U. pictorum 7.6000 2.56 Tudorancea, Florescu,
1968a
* Данные за 1968 г.
** Данные за 1969 г.
Несомненно, зависимость потока энергии от биомассы особей
в популяциях не представляется простой функцией, поскольку
удельный поток энергии в свою очередь связан со средним весом
особей. Дальнейшие исследования и количественные выражения
этих функций позволят создать своеобразные номограммы, с по-
мощью которых станет возможным с определенной вероятностью
находить величины потоков энергии в популяциях отдельных видов
животных по их биомассам и среднему весу особей.
Ранее было показано, что отношение гтр1гт, так же как Ар)В, свя-
зано со средним весом особей в популяциях, т. е.
= (Рй),
212
по в свою очередь
Л/В = /2(Ж),
поэтому
= (96)
Это означает, что в популяциях более мелких по размерам животных
подверженных в большей степени воздействиям внешней среды,
Рис. 77. Зависимость логарифма потока энергии 1g Ар от логарифма средней
биомассы 1g В в популяциях животных.
I — Chironomidae: 1 — Tanytarsus mancus; 2 — Procladius nigriventris; 3 — то же; 4 —
Sergentia coracina; 5 — Chironomus plumosus; 6 — Ch. antracinus. II — Bivalvia и Amphi-
poda: 7 — Sphaerium suecicum', 8 — Pisidium crassum; 9 — Unio pictorum; 10 — Anodonta
piscinalis; 11 — P. lilljeborgi; 12 — U. tumidus; 13 — Gammarus lacustris; 14 — Candona
Candida; 15 — Pontoporeia affinis (по данным: 1, 2, 4, 7, 8, 11, 13—15 — автора; 3, 5, 6 —
Андронниковой и др., 1973); 9, 12, 16 — Tudorancea, 1972). По оси [абсцисс — значения
lg S (ккал/м2 за сезон); по оси ординат — значения 1g Ар (ккал/м2 за сезон).
следует ожидать более высоких величин удельного потока энергии.
Анализ данных табл. 55 показывает, что, несмотря на значитель-
ные различия абсолютных величин составляющих энергетического
баланса в популяциях разных видов моллюсков, относительные
их значения колеблются в очень узких пределах. Так, количество
потребленной животными пищи и поток энергии в популяциях
отличались в 136 раз, в то время как коэффициенты К1р=Р 1Ср и
K2p=PjAp — не более чем в 1.4 раза. Поэтому мы можем рассматри-
213
вать эти коэффициенты как показатели, общие для изученных по-
пуляций, или как популяционные характеристики.
Незначительные различия коэффициентов К2р указывают на на-
личие общей зависимости между составляющими потока энергии
в популяциях разных видов.
Многочисленные данные различных исследователей по величи-
нам продуцируемой энергии Р и суммарным тратам на обмен Rp за
год или вегетационный сезон в популяциях животных, отличающих-
ся по систематическому положению, позволили установить на-
личие функциональной связи между этими параметрами популя-
ций. Впервые такого рода связь в виде степенной функции была
выявлена Энгеллманном (Engellmann, 1966) для почвенной фауны:
7?р = 4.17Р0-86 ккал/м2 за год. (97)
Несколько позже аналогичная зависимость была получена для са-
мых разных животных МакНейлом и Лаутоном (McNeill, Lawton,
1970). Они привели уравнения для гомойотермных и пойкило-
термных животных, а среди последних — отдельно для животных
с продолжительностью жизни не более и более двух лет. Для всех
пойкилотермных животных авторы рассчитали следующее урав-
нение:
7?р = 2.371Р1,073 ккал/м2 за год. (98)
Наконец, Левек (Leveque, 1973), не согласившись на разделение ко-
ротко- и догложивущих особей, для ряда популяций пойкилотерм-
ных животных рассчитал общее уравнение
Rp — 2.730Р1-0816 ккал/м2 за год. (99)
Для изученных популяций Bivalvia, некоторых видов
Chironomidae и Amphipoda в озерах Кривое, Круглое и Зеленец-
кое и по литературным материалам автором были определены их
суммарные траты на обмен, продуцируемая энергия в популяциях
(рис. 78) и рассчитано соответствующее уравнение
Rp = (2.879 + 0.046) Р(1<006±0-045) ккал/м2 за год. (100)
Уравнение (100) не имеет статистически достоверных отличий
от уравнений (97—99). Показатель степени в этом, как, впрочем,
и в других приведенных уравнениях, достоверно не отличается
от единицы. Это свидетельствует о прямопропорциональной
зависимости энергетических трат на обмен от продукции попу-
ляции.
Из уравнения (100) легко определить, что продуцируемая в те-
чение года в популяциях энергия составляет примерно третью часть
энергии, рассеиваемой животными за тот же период в виде трат
на обмен:
Р = Др/2.879 ккал/м2 за год. (101)
214
Из уравнения (101) получаем:
PjRp = 1/2.879 = 0.347,
р//?р=л:2/(1 + л’2),
или
К2р = 0.347 :1.347 = 0.258 0.26.
Рис. 78. Зависимость логарифма трат на обмен lg Rp и логарифма продук-
ции lg Р в популяциях различных животных (из: Алимов, 1978).
1 — Sphaerium suecicum, 1968, 1969 гг.; 2 — Pisidium crassum', 3 — Gammarus lacustris;
4 — Tanytarsus mancus; 5 — Procladius nigriventris; 6 — Sergentia coracina; 7 — Ponto-
poreia affinis', 8 — Pisidium lilljeborgi; 9 — Schrobicularia plana; 10 — Littorina irrorata;
11 — Modiolus demisus; 12 — Pisidium verginicum; 13 — Physa integra; 14 — Fissurella
barbadensis; 15 — Nerita pelorata; 16 — N. versicolor; 17 — N. testelata (по данным: 1—8 —
автора; 9—10 — Hughes, 1970; 11 — Kuenzler, 1961; 12 — Teal, 1957; 13 — Tilly, 1968;
14 — Hughes, 1971a; 15—17 — Hughes, 1971b). По оси абсцисс — значения lg Р (ккал/м2
за год); по оси ординат — значения lg Rp (ккал/м2 за год).
Следовательно, согласно уравнениям (100, 101), возможные значе-
ния К2р в расчете за год при уровне вероятности 0.999 не выходят
за пределы 0.224—0.304.
Разными авторами были получены в принципе одинаковые коли-
чественные выражения для зависимости трат энергии на обмен
популяциями животных от энергии, продуцируемой ими в течение
года, что говорит о ее важном биологическом значении. Действи-
тельно, в популяциях животных, далеко отстоящих по системати-
ческому положению, соотношения между тратами на обмен и проду-
цируемой энергией оказываются весьма сходными.
Сказанное дает основание рассматривать поток энергии как
одно из фундаментальных свойств популяции как системы особей
определенного вида животных, которое может быть охарактери-
зовано коэффициентом К2 или связанным с ним отношением
Р/Я,.
215
Зависимость P=f (Rp) может быть использована для определе-
ния количества энергии, продуцируемой популяцией за длительный
отрезок времени, по величинам трат на обмен у животных, ее со-
ставляющих. Траты на обмен могут быть с достаточной надеж-
ностью рассчитаны по установленным для различных представи-
телей животного мира зависимостям от их веса. При этом может
быть учтено влияние факторов внешней среды на скорость обмена
у животных. Сложнее обстоит дело с изучением продукционных
способностей популяций. Определение их связано с исследовани-
ями роста животных, которые требуют длительных экспериментов,
сопряженных для некоторых видов с большими методическими
трудностями. Поэтому возможность расчета годовых величин
энергии или вещества, продуцируемого популяциями, минуя изуче-
ние роста животных, несомненно заслуживает внимания. Опреде-
ление энергии, продуцируемой в популяции за год, с использова-
нием уравнений (100, 101) представляет собой известный физиоло-
гический способ расчета продукции популяций (Шушкина, 1968),
при котором учитываются интенсивность обмена животных и зна-
чения коэффициента К2. Для популяций двустворчатых моллюсков
величина этого коэффициента может быть принята рав-
ной 0.26.
Как уже отмечалось, МакНейл и Лаутон привели отдельные
уравнения для животных, имеющих продолжительность жизни более
и менее двух лет, считая последних короткоживущими. Вниматель-
ное рассмотрение рис. 78 показывает, что популяции конкретного
вида животных, для которых характерна определенная продолжи-
тельность жизни, в разные годы или при разных условиях проду-
цируя сходные количества энергии, различались по величинам
трат на обмен. Так, например, отношение PIRp в популяции
моллюсков Sphaerium suecicum в 1968 г. было 0.49, в 1969 — 0.56;
амфипод Gammarus lacustris соответственно 0.163 и 0.178; для по-
пуляции Pontoporeia affinis Lindstr. величина этого отношения на
разных глубинах озера составляла 0.54, 0.3 и 0.25. Аналогичные
изменения отношения Р /R были отмечены для популяции мол-
люсков Scrobicularia plana на разных горизонтах литорали —
0.26, 0.32 и 0.27 (Hughes, 1970).
Во всех случаях, как и следовало ожидать, отличия величин
отношения PIRp совпадали с различиями возрастного состава
популяций. В популяции S. suecicum средний возраст особей
в 1968 г. был меньшим по сравнению с 1969 г. В популяции G. la-
custris, наоборот, в 1969 г. преобладали более молодые по возрасту
особи. Поэтому особое значение приобретают определения среднего
возраста особей в популяциях, который обусловлен динамикой
численности отдельных возрастных групп, соотношением рождае-
мости и смертности. Вероятно, в связи с этим более правильно гово-
рить, обсуждая связь продукционных возможностей популяции
с тратами ими энергии на обмен, не о различиях в продолжительно-
го
сти жизни определенного вида животных, а о разном относительном
среднем возрасте особей в популяции.
Значения К*р, рассчитанные для популяций моллюсков разных
видов (табл. 62), находятся в пределах величин этого коэффициента,
определенных согласно уравнению 101. При этом крайние величины
коэффициентов соответствуют величинам К*, для растущих особей
(см. табл. 44), т. е. относительный средний возраст животных
Таблица 62
Значения коэффициентов К*р (за год или сезон)
в популяциях моллюсков
Вид Источник
Sphaerium suecicum . . . 0.33—0.36 Нашц данные
Pisidium crassum . . .... 0.25 Там же
P. lilljeborgi ... . . 0.29 » »
P. nitidum ... .... 0.20 » »
Bellamya unicolor . . . 0.32 Leveque, 1973
Modiolus modiolus 0.30 Kuenzler, 1961
Scrobicularia plana 0.24 Hughes, 1970
в популяциях адекватен среднему возрасту особи конкретного
вида. Преобладание в популяциях возрастной группы животных,
способных активно размножаться, несомненно существенно,
так как тем самым поддерживается необходимая численность
особей и жизнеспособность популяции. Для подавляющего боль-
шинства популяций конкретных видов моллюсков характерна вы-
сокая смертность на ранних стадиях развития (вогнутая кривая
выживания). В случаях, когда в популяциях этих животных будут
доминировать особи, возраст которых отличается от среднего,
свойственного данному виду в данных условиях, следует ожидать
или омоложения (высокая рождаемость при одновременной повы-
шенной смертности старших возрастов), или, наоборот, старения
(уменьшение рождаемости при высокой смертности) популяции.
В популяциях унионид обычно преобладают особи старших возраст-
ных групп, и в связи с этим величины К*2р для них не превышают
0.058—0.098 (рассчитано по данным Tudorancea, 1972); столь же
низкие значения этого коэффициента (0.098) отмечены, например
для Corbicula africana (Кг.) (Leveque, 1973). Наоборот, высокие
значения К*р характерны для популяций, в которых большое ко-
личество молодых особей, как это наблюдалось у моллюсков
Pisidium verginicum Gmelin, где значение этого коэффициента со-
ставляло 0.47 (Teal, 1957).
В литературе часто отмечается преобладание в популяциях
унионид животных, возраст которых более 4—6 лет, при этом
15 А. Ф. Алимов
217
1—2-летние особи обычно единичны. Такое явление требует
специального изучения, так как вряд ли можно поверить в дли-
тельное существование популяции при условии почти полного
отсутствия в ней молоди. Вероятно, дело в том, что популяции
этих моллюсков занимают значительно большие территории, чем те,
где производится сбор животных для анализа их состава. Пара-
зитирующие на рыбах глохидии этих моллюсков могут рассеиваться
по мере созревания активно плавающими рыбами на значительном
удалении от места обитания их родителей. Выход глохидиев
Рис. 79. Соотношение между К$р за
сезон и г за тот же период в популя-
циях моллюсков.
1 — Sphaerium suecicum’, 2 — Pisidium cras-
sum; 3 — P. lilljeborgi; 4 — P. nitidum.
По оси абсцисс — значения г; по оси орди-
нат — значения К* (объяснения в тексте).
2р
из жабр вовсе не обязательно дол-
жен происходить в тот момент,
когда рыбы подходят к берегу.
В то же время массовые поселе-
ния этих моллюсков известны
вблизи берегов на глубинах,
редко превышающих 1.5—2 м.
Можно предположить, что рас-
сеиваемые рыбами на значитель-
ных площадях глохидии мол-
люсков мигрируют на прибреж-
ные мелководья. Поскольку эти
животные малоактивны, их ми-
грации могут продолжаться
длительное время, в течение
которого они успевают сущест-
венно подрасти. Косвенным под-
тверждением такого предполо-
жения может служить тот факт,
что обычно на глубинах более 2—3 м обнаруживаются лишь отдель-
ные экземпляры унионид, притом обычно только молодые моллюски,
возраст которых чаще всего не превышает 2 лет. Скорее всего
площадь, занимаемая популяциями этих моллюсков в водоемах,
охватывает значительную часть дна, а не ограничивается прибреж-
ными мелководными участками. Приняв это во внимание при
сборе и обработке материалов количественного учета унионид, мы,
вероятно, должны получить вполне обычное, как и для других
видов, соотношение численности особей различных возрастов в их
популяциях.
Величины для популяций разных видов моллюсков оказыва-
ются скоррелированными со средней скоростью изменения их
численности г в течение сезона или года (рис. 79). При небольшом
количестве имеющихся пока данных можно говорить лишь о тен-
денции увеличения К*р с нарастанием г. Однако такого вида связь
между этими двумя характеристиками популяций вполне воз-
можна, так как изменения каждой из рассматриваемых величин
определяются соотношениями рождаемости и смертности особей
разных возрастных групп.
218
Из данных рис. 79 видно, что величины К*2р в изученных по-
пуляциях моллюсков изменялись незначительно — от 0.2 до
0.36; в то же время значения г колебались в пределах от +1.65 до
—1.6. Минимальные из рассмотренных значения К*р наблюдались
в популяции моллюсков Р. nitidum, когда смертность животных
была довольно высокой (г=—1.6), но при этом популяция не
деградировала. Учитывая достаточно высокий относительный
средний возраст особей (что нашло отражение в величине К*р) и пре-
обладание в этой популяции смертности над рождаемостью, можно
рассматривать отмеченные низкие значения К*р и г как близкие
к критическим. Возможно, что дальнейшее увеличение смертности
может привести к резкому нарастанию численности особей старших
возрастов, репродукционные способности которых не смогут ком-
пенсировать смертность, и популяция потеряет конкурентно-
способность.
Из уравнения (95), учитывая, что К2=Р1А, получаем
Р = ЪВК*р,
откуда
Р/В = иК*р. (102)
Последнее равенство показывает, что Р/2?-коэффициент для по-
пуляции животных определенного вида находится в прямой связи
с коэффициентом К*р и удельным потоком энергии, рассчитанными
за тот же период. Поскольку ранее было показано, что значения
удельного потока энергии, например для популяций хирономид и
двустворчатых моллюсков, различаются между собой, коэффици-
енты Р{В и К*р у них связаны прямой зависимостью, и оба отра-
жают возрастную структуру популяций. Удельные потоки
энергии представляют собой коэффициенты пропорциональности,
различающиеся для популяций этих групп животных по абсолют-
ным величинам.
Принимая во внимание все изложенное, популяцию животных
с экологических позиций можно определить как локализованную
в пространстве и времени, сбалансированную по энергии систему
особей одного вида животных, различающихся по полу и возрасту
и находящихся в отношениях размножения. Популяция характе-
ризуется возрастным составом, скоростью изменения ее численно-
сти и потоком энергии. Последний может рассматриваться как
одно из фундаментальных свойств популяции.
Изучение энергетического баланса популяций разных видов
моллюсков позволило найти соотношения между показателями из-
менения их численности, биомассой, продукцией и потоками энер-
гии. Это открывает перспективы установления связей между
статическим, динамическим и функциональным аспектами иссле-
дований популяций животных. Рассматривая полученные ре-
15’
219
зультаты как первую попытку, требующую дальнейшей проверки
и уточнения, кажется несомненным, что работы в этом направле-
нии могут привести к широким обобщениям общебиологического
значения.
Популяции конкретных видов животных как составные части
биоценозов занимают вполне определенное место в экологических
системах водоемов. Составляющие их особи, вступая в отношения,
среди которых важнейшее место принадлежит трофическим
связям, с особями популяций других видов, тем самым включаются
в общие процессы круговорота веществ и трансформации энергии
в экосистемах.
ЗАКЛЮЧЕНИЕ
Многие закономерности, установленные при изучении функци-
ональной экологии двустворчатых моллюсков, справедливы для
большинства водных животных. Поэтому данное исследование
в значительной мере может рассматриваться как модельное в отно-
шении количественной оценки участия гидробионтов в процессах,
протекающих в водных экологических системах.
Большая часть количественных зависимостей от внешних и
внутренних факторов отдельных функций моллюсков, как и других
животных, могут быть выражены в виде простых уравнений,
чаще всего степенных или показательных. Степенная зависимость,
как показал опыт многих исследований, представляется универ-
сальной при описании связи с массой животных таких важнейших
функций, как скорость обмена, питания, плодовитость. В виде
степенных или показательных уравнений выражаются продукцион-
ные или энергетические характеристики популяций животных.
Показатели степени в уравнениях, аппроксимирующих зависимо-
сти скорости обмена, питания и плодовитости от массы моллюсков,
меньше единицы (0.72 — для обмена, 0.6 — для фильтрации).
Это означает, что при увеличении веса животных в 10 раз скорость
их обмена возрастает лишь в 100*72, т. е. в 5.3 раза, а скорость филь-
трации воды — примерно в 4 раза. Особи одного вида моллюсков
в процессе развития увеличивают свою массу более чем в 100 раз.
При увеличении веса животных в 100 раз скорость обмена у них во-
зрастает в 28, а скорость фильтрации воды — почти в 16 раз.
Изменение температуры на 10° вызывает изменение скоростей об-
мена и фильтрации не более чем в 2—3 раза, возрастание концен-
трации взвеси в воде в 30 раз (в природных местообитаниях ко-
лебания концентрации значительно ниже) приводит к уменьшению
скорости фильтрации только в 7 раз, сезонные колебания скорости
обмена не превышают 4—5 раз и т. д. Таким образом, ни один
из факторов внешней среды не оказывает такого сильного влияния
на скорости этих функций у животных, как изменение их веса.
Уравнения, описывающие вышеуказанные зависимости, отражают
средние величины скоростей обмена или питания и их количе-
221
ственные выражения для всех моллюсков или отдельных их групп,
и с их помощью невозможно обнаружить различия, имеющие таксо-
номическую значимость. Видовая специфичность интенсивности
обмена или питания у моллюсков может быть обнаружена при
рассмотрении влияния факторов внешней среды на скорости этих
процессов у животных. Как правило, в большинстве случаев у ви-
дов, отличающихся по происхождению, зоны оптимальных значе-
ний по определенному фактору различны. Поэтому особое значение
для работ по систематике и фаунистике приобретают исследований,
позволяющие количественно оценить влияние отдельных факторов
в пределах их природных колебаний на важнейшие физиологиче-
ские функции животных. Так, например, изучение влияния темпе-
ратуры на скорость метаболизма, питания или роста моллюсков
позволяет выявлять зоны оптимальных для каждого вида темпе-
ратур, что несомненно может быть использовано для более строгого
определения центров их происхождения.
Анализ роста моллюсков показал, что отдельные виды могут
быть охарактеризованы не только морфологическими признаками,
но и пределами изменчивости их линейных размеров, веса, скорости
роста и связанной с ней продолжительности жизни. При уста-
новлении видового ранга для конкретных групп особей привле-
чение сведений по росту животных будет способствовать, особенно
в спорных случаях, объективной оценке их систематического по-
ложения. Поскольку важнейшие функции животных, такие как
питание, обмен, плодовитость, скорость роста, зависят от веса
особей, особое значение приобретает исследование именно весо-
вого роста животных, выявление его специфичности в различных
условиях обитания видов.
В процессе выполнения данной работы выяснились наиболее
слабо изученные стороны экологии двустворчатых моллюсков.
Недостаточно полно изучено влияние температуры на темп роста,
плодовитость, скорость фильтрации воды моллюсками. Почти не ис-
следована зависимость плодовитости от качественного состава
пищи и пищевых условий в местах обитания моллюсков. Необходимы
работы по установлению количественных закономерностей ди-
намики численности, возрастного состава, рождаемости и смертно-
сти в популяциях отдельных видов.
Аутоэкологические исследования — необходимый этап изу-
чения функциональной экологии животных. Количественные све-
дения по росту, обмену веществ, питанию, размножению особей
различных видов предствляют собой основу определений про-
дукции, потоков энергии в популяциях, сообществах животных и
экологических системах. Важное значение приобретает выяснение
количественных закономерностей, определяющих динамику чис-
ленности и возрастного состава популяций конкретных видов.
В настоящее время наметилась диспропорция между успешно
развивающимися исследованиями баланса веществ, и особенно
потоков энергии, в популяциях, сообществах животных в водных
222
экосистемах, при которых обязательно учитываются изменения чи-
сленности и возрастного состава популяций, и неоправданно слабо
разрабатываемыми проблемами так называемой популяционной
экологии, выясняющей механизмы, управляющие динамикой чис-
ленности и возрастного состава. Предпринятая в данной работе
попытка установления зависимостей между скоростью увеличения
численности в популяциях, их удельной продукцией и интенсив-
ностью обмена особей, составляющих популяцию, привела к обна-
деживающим результатам. Они показали, что есть возможность
выяснения количественных связей между динамическим и функ-
циональным аспектами изучения популяции. Нужны поиски
закономерностей в этом направлении. Существенную роль в ис-
следованиях такого рода может играть математическое моделиро-
вание. Моделирование служит необходимым инструментом, с по-
мощью которого проверяются установленные связи и закономер-
ности, выясняются внутренние механизмы явлений и круг вопросов,
требующих специального изучения.
Весьма важно, что изучение балансов энергии особей в процессе
их развития и популяций разных видов моллюсков привело к за-
ключению о том, что экспериментальные данные и установленные
на их основе закономерности роста, питания, обмена, размножения
могут быть успешно применены по отношению к природным попу-
ляциям животных и использованы для количественной оценки их
значения и роли в экологических системах. По отношению к дву-
створчатым моллюскам применение экспериментальных данных
позволило установить, что фильтруемые ими органические вещества
стабилизируются в экосистемах в виде накопленной живой массы.
Одновременно происходит окисление органических веществ
со скоростью, эквивалентной интенсивности обмена у животных.
Неусвоенная ими часть потребленных органических веществ по-
ступает в донные отложения и впоследствии активно используется
в пищу организмами инфауны. Осветляя воду и окисляя органи-
ческие вещества, двустворчатые моллюски выступают как активные
агенты в процессах самоочищения природных водоемов. Недооценка
функционального значения двустворчатых моллюсков в экологи-
ческих системах континентальных водоемов в ряде случаев может
привести к существенным погрешностям и неточностям при изуче-
нии баланса систем, их моделировании и прогнозировании на-
правления их развития или определения норм эксплуатации.
Материалы работы и сделанные выводы показывают, что для
решения экологических задач необходимы исследования, выпол-
ненные зоологами, в сфере внимания которых находятся вопросы
аутоэкологии животных. Важнейшие сведения по весовому,
линейному росту и их соотношению, продолжительности развития,
плодовитости особей, равно как и количественные характеристики
динамики численности, возрастного состава, плодовитости в попу-
ляциях конкретных видов в различных условиях и участках
ареалов, могут быть получены в процессе сбора и обработки мате-
223
риалов зоологических и фаунистических исследований. Существен-
но, что многие важные для экологии и зоологии закономерности
могут быть установлены при изучении массовых видов. В тех же
случаях, когда необходимо решать вопросы, значимые для понима-
ния происхождения и эволюции отдельных, часто не массовых
весьма специфичных видов, знание биологии которых существенно
для разработки проблем происхождения и эволюции фаунисти-
ческих комплексов или фаун или эволюции отдельных групп
животных, следует проводить детальные (в том числе эксперимен-
тальные) исследования экологии этих видов.
Разумеется, неверно ожидать, что все необходимые материалы
экологи могут и должны получать самостоятельно. Более есте-
ственно, если аутоэкологические данные будут черпаться эколо-
гами из работ, выполненных зоологами — специалистами по от-
дельным систематическим группам животного мира. Успешное,
надежное и по возможности быстрое, что немаловажно, решение
важнейших экологических проблем, связанных с охраной природы
и сохранением природных ресурсов при рациональной их эксплуа-
тации, возможно в результате совместных, целенаправленных
усилий специалистов экологов и зоологов.
ЛИТЕРАТУРА
Аболмасова Г. И. Плодовитость и расход энергии на образование яиц у неко-
торых черноморских крабов. — Тр. Всесоюз. гидробиол. о-ва, 1973, т. 18,
с. 50—62.
Аболмасова Г. И. Поток энергии в популяциях некоторых видов беспозвоноч-
ных. — В кн.: Экспериментальная экология морских беспозвоночных.
Матер, симп. Владивосток, 1976, с. 9—12.
Алекин О. А. Гидрохимия рек СССР. Л., 1948, вып. 3. 160 с.
Александров Б. М. Двустворчатые моллюски озер Карелии. — В кн.: Фауна
озер Карелии. М.; Л., 1965, с. 96—110.
Алиев А. А. К вопросу потребления кислорода беззубкой. — ДАН АзССР,
1960, т. 16, № 8, с. 793—795.
Алимов А. Ф. Фильтрационная способность моллюсков рода Sphaerium
(Scopoli). — ДАН СССР, 1965, т. 164, № 1/3, с. 195—197.
Алимов А. Ф. Влияние некоторых факторов среды на фильтрационные спо-
собности моллюсков Sphaerium corneum (L.). — Гидробиол. журн., 1967а,
т. 3, № 2, с. 26—32.
Алимов А. Ф. Особенности жизненного цикла и роста пресноводного мол-
люска Sphaerium corneum (L.). — Зоол. журн., 19676, т. 46, вып. 2, с. 192—
199.
Алимов А. Ф. Некоторые общие закономерности фильтрации у двустворча-
тых моллюсков. — Журн. общ. биол., 1969, т. 30, № 5, с. 621—631.
Алимов А. Ф. Поток энергии через популяцию моллюсков (на примере Sphaeri-
idae). — Гидробиол. журн., 1970, т. 6, № 2, с. 63—71.
Алимов А. Ф. Закономерности роста пресноводных двустворчатых моллю-
сков. — Журн. общ. биол., 1974, т. 35, № 4, с. 576—589.
Алимов А. Ф. Интенсивность обмена у пресноводных двустворчатых моллю-
сков. — Экология, 1975, № 1, с. 10—20.
Алимов А. Ф. Заметки по изменчивости некоторых моллюсков сем. Sphaeri-
idae. — В кн.: Гидробиологические исследования самоочищения водое-
мов. Л., 1976, с. 119—128.
Алимов А. Ф. Использование энергетического принципа при исследовании
популяций (на примере донных животных). — Тр. Всесоюз. гидробиол.
о-ва, 1977, т. 21, с. 231—249.
Алимов А. Ф., Богатов В. В. Рост беззубки Anodonta piscinalis в водохрани-
лищах Калининской области. — Зоол. журн., 1975, т. 54, вып. 1, с. 27—31.
Алимов А. Ф., Бульон В. В. Фильтрационная активность моллюсков Sphae-
rium suecicum Clessin при разных концентрациях взвешенных веществ. —
Журн. общ. биол., 1972, т. 33, № 1, с. 97—104.
Алимов А. Ф., Бульон В. В., Гутельмахер Б. Л., Иванова М. Б. Методы изу-
чения участия гидробионтов в процессах самоочищения водоемов. — В
кн.: Роль гидробионтов в очистке сточных вод. Фрунзе, 1977, с. 3—42.
Алимов А. Ф., Бульон В. В,, Озерецковская Н. Г,, Умнова Л. П. Общая харак-
теристика исследованных участков некоторых рек Ленинградской, Кали-
225
нинградской и Московской областей. — В кн.: Методы биологического
анализа пресных вод. Л., 1976, с. 5—15.
Алимов А, Ф., Голиков А. Н. Некоторые закономерности соотношений
между размерами и весом у моллюсков. — Зоол. журн., 1974, т. 53, вып. 4,
с. 517—530.
Алимов А. Ф., Иванова М. Б., Лебедев Ю. М., Максимова Л. П. Рец. на кн.:
Трофология водных животных. Итоги и задачи. М., Наука, 1973. — Зоол.
журн., 1974, т. 53, вып. И, с. 1743—1745.
Алимов А. Ф., Умнов А. А. Использование моделирования при изучении
продуктивности популяций моллюсков. — Журн. общ. биол., 1978,
т. 39, № 6, с. 886—894.
Алимов А. Ф., Финогенова Н. П. Биоценозы и продуктивность бентоса. —
Тр. Зоол. ин-та АН СССР. Л., 1975а, т. 56, с. 156—195.
Алимов А. Ф., Финогенова Н. П. Продуктивность бентоса. — Тр. Зоол. ин-та
АН СССР. Л., 19756, т. 57, с. 150—167.
Алимов А. Ф., Финогенова Н. П. Количественная оценка роли сообществ дон-
ных животных в процессах самоочищения пресноводных водоемов. —
В кн.: Гидробиологические основы самоочищения вод. Л., 1976, с. 5—14.
Алимов А. Ф., Шадрин Н, В. Калорийность некоторых представителей пресно-
водного бентоса. — Гидробиол. журн., 1977, т. 13, № 3, с. 80—86.
Андронникова И. Н., Драбкова В. Г. Биотический баланс озера Красного. —
В кн.: Биологическая продуктивность оз. Красного и условия ее форми-
рования. Л., 1976, с. 192—198.
Андронникова И. Н., Драбкова В. Г., Кузьменко К. Н. и др. Продукция основ-
ных сообществ оз. Красного и его биотический баланс. — В кн.: Продук-
ционно-биологические исследования экосистем пресных вод. Минск, 1973,
с. 5—19.
Арабина И. П. Рост пресноводного моллюска Sphaerium согпеит. — ДАН
БССР, 1966, т. 10, Яг 12, с. 978—981.
Арабина И. И., Гаврилов С. И. Соотношение веса и линейных размеров у пред-
ставителей пресноводного бентоса. — Гидробиол. журн., 1967, т. 3,
№ 2, с. 71—73.
Арабина И. П., Рубинова Л. А. Зависимость между скоростью роста и ин-
тенсивностью обмена у пресноводного моллюска. — В кн.: Биопродуктив-
ность озер Белоруссии. Минск, 1971, с. 141 — 145.
Аристова Г. И. Дрейссена Куршского залива и перспективы ее использова-
ния. — Тр. Калинингр. техн, ин-та рыб. пом-сти и хоз-ва, 1968, вып. 20,
с. 85-92.
Асташкина А. Н. О течении беременности донских перловиц (Unionidae)
в связи с их возрастом (по данным 1964 г.). — В кн.: Сб. науч, работ аспи-
рантов Воронеж, ун-та. Воронеж, 1965, вып. 2, с. 134—142.
Бадер Ф. Ф. Состав воды Аральского моря. — В кн.: Исследования озер
СССР. Л., 1934, вып. 6, с. 5—12.
Баранов И. В. Гидрохимический режим и грунты Горьковского водохрани-
лища по данным 1960—1961 гг. — Изв. НИИ озер, и реч. рыб. хоз-ва.
Л., 1965, т. 59, с. 19—42.
Барыбина И. А., Санина Л. В. Исследования зависимостей между размерно-
весовыми характеристиками моллюсков методами корреляционно-ре-
грессионного анализа. — Гидробиол. журн., 1975, т. 2, № 6, с. 34—40.
Беюодарная Т. Ф. К биологии перловицы р. Кичуй. — Тр. о-ва естество-
испыт. при Казан, ун-те, 1937, т. 55, вып. 1—2, с. 93—114.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. М.,
1964, т. 2. 444 с.
Белова Л. Н. Питание воблы в Северном Каспии. — В кн.: Биологическая
продуктивность Каспийского моря. М., 1974, с. 98—107.
Бервалъд Э. А. Определение интенсивности биологической очистки воды
методом фильтрационных контейнеров. — В кн.: Радиоактивные изотопы
в гидробиологии и методы санитарной гидробиологии. Л., 1964, с. 101 — 107.
226
Бервалъд Э. А Биологическая очистка природных вод с помощью фильтра-
ционных контейнеров. — В кн.: I съезд Всесоюз. гидробиол. о-ва. Тез.
докл. М., 1965, с. 31—32.
Бивертон Р., Холт С. Динамика численности промысловых рыб. М., 1969,
248 с.
Бидл Л. Адаптации некоторых водных животных к недостатку кислорода и
к анаэробным условиям. — В кн.: Механизмы биологической конкурен-
ции. М., 1964, с. 153—168.
Бгргер Т. I. Кормова цшшсть для риб массовых форм безхребетних Дншра
и Дшпровсько-Бузького лиману. Кшв, 1961. 110 с.
Бъргер Т. 1., Маляревсъка О. Я. Б1ох1м1чный склад i кормова цшшсть массо-
вих форм безхребетних Каховського водоймища. — В кн.: Каховсько
водоймище. Кшв, 1964, с. 200—213.
Биргер Т. И., Маляревская А. Я. Биохимический состав некоторых видов
пресноводных моллюсков и их использование рыбами. — В кн.: Моллюски.
Вопросы теоретической и прикладной малакологии. Тез. докл. М.; Л.,
1965, с. 40—42.
Биргер Т. И., Маляревская А. Я., Оливари Т. А. Кормовая ценность бентоса
Днепра, Каховского и Кременчугского водохранилищ и ер изменение
под влиянием зарегулирования стока. — В кн.: Гидробиологический
режим Днепра в условиях зарегулированного стока. Киев, 1967, с. 331—
350.
Богатова И. Б. К вопросу о питании Moina rectirostris (Leydig). — Тр.
ВНИИ пруд. рыб. хоз-ва, 1967, т. 15, с. 106—116.
Бруенко В. П. Плодовитость сома низовьев Дуная. — Гидробиол. жури.,
1967, т. 3, № 1, с. 83—85.
Булдовский А. Т. О промысловых пресноводных моллюсках Дальнего Вос-
тока СССР. — Вести. Дальневост. фил. АН СССР, 1935, № 12, с. 39—63.
Бульон В. В. Первичная продукци озер.я — Тр. Зоол. ин-та АН СССР. Л.,
1975а, т. 56, с. 32—41.
Бульон В. В. Первичная продукция тундровых озер. — Тр. Зоол. ин-та
АН СССР. Л., 19756, т. 57, с. 19—36.
БулЬон В. В. Содержание органического вещества и фотосинтетических
пигментов в планктоне озера Байкал. — В кн.: Гидробиологические
основы самоочищения вод. Л., 1976, с. 60—68.
Буторин Н. В. О комплексных исследованиях Иваньковского водохранилища
в районе Конаковской ГЭС. — Информ, бюл. Ин-та биол. внутр, вод
АН СССР, 1970, № 5, с. 81—83.
Веселов Е. А. Методы изучения газообмена рыб и водных беспозвоночных. —
В кн.: Жизнь пресных вод СССР. М.; Л., 1959, т. 4, ч. 2, с. 79—124.
Винберг Г. Г. Интенсивность обмена и пищевые потребности рыб. Минск,
1956. 253 с.
Винберг Г. Г. Рост, скорость развития и плодовитость в зависимости от усло-
вий среды. — В кн.: Методы определения продукции водных животных.
Минск, 1968, с. 45—78.
Винберг Г. Г. Общие особенности экологической системы оз. Дривяты. — Тр.
Всесоюз. гидробиол. о-ва, 1970, т. 13, с. 185—196.
Винберг Г. Г. Линейные размеры и масса тела животных. — Журн. общ.
биол., 1971, т. 32, № 6, с. 714-723.
Винберг Г. Г. Цели и задачи гидробиологии пресных вод при комплексном
использовании водных ресурсов. — Водные ресурсы, 1972, № 3, с. 76—102.
Винберг Г. Г. Зависимость энергетического обмена от массы тела у водных
пойкилотермных животных. — Журн. общ. биол., 1976а, т. 37, № 1,
с. 56-70.
Винберг Г. Г. Соотношение между продукцией морских животных и «росто-
вой» и «поддерживающей» продукциями. — Океанология, 19766, т. 16,
вып. 5, с. 854—867.
Винберг Г. Г., Беляцкая Ю. С. Зависимость интенсивности обмена от общих
размеров тела у пресноводных брюхоногих моллюсков. — Бюл. Ин-та
биол. АН БССР, 1958, вып. 3, с. 273—276.
227
Винберг Г. Г., Беляцкая Ю. С. Соотношение интенсивности обмена и веса
тела у пресноводных брюхоногих моллюсков. — Зоол. журн.,7Р5Р, т. 38,
вып. 8, с. 1146—1151.
Винберг Г. Г., Беляцкая-Потаенко Ю. С. Суточный ход интенсивности газо-
обмена у пойкилотермных животных. — Зоол. журн., 1963, т. 42, вып. И,
с. 1593—1600.
Властов Б. В. Unionidae (перловицы) р. Оки (Башреспублика). — Бюл. Моск,
о-ва испыт. природы. Отд-ние биол., 1933, т. 72, № 3/4, с. 195—234.
Властов Б. В. Биология жемчужницы Margaritana margaritifera L. и про-
блема использования ее раковин как перламутрового сырья. — Тр. Боро-
динск биол. ст. в Карелии. Петрозаводск, 1934, т. 7, вып. 2, с. 5—36.
Властов Б. В. Методы определения возраста Unionidae по раковине и их
значение в научно-промысловых исследованиях. — В кн.: Зап. Болшев-
ской биол. ст. М., 1935, вып. 7—8, с. 133—148.
Властов Б. В. Порционность яйцекладок, фазы размножения и продолжи-
тельность эмбриогенеза при разных температурах у видов перловиц
(Unio). — В кн.: Вопросы общей зоологии и медицинской паразитологии.
М., 1962, с. 63—85.
Воскресенский Я. А, Пояс фильтраторов как биогидрологическая система
моря. — Тр. Гос. океаногр. ин-та, 1948, вып. 6 (18), с. 55—120.
Въюшкова В. П. Зоопланктон Волгоградского водохранилища по материалам
1962—1964 гг. — Тр. Саратов, отд-ния НИИ озер, и реч. рыб. хоз-ва,
1965, т. 8, с. 42—54.
Гаврилов С. И. Бентос оз. Дривяты и его продуктивность. — Тр. Всесоюз.
гидробиол. о-ва, 1970, т. 15, с. 106—125.
(Гак Д. 3., Гурвич В. В., Корелякова И. Л. и др.) Gak D. Z., Gurvich V. V.,
Korelyakova I. L. et al. Productivity of aquatic organism communities of
different trophic levels in Kiev reservoir. — In: Proceedings of the IBP—
UNESCO symposium of productivity problems of freshwater. Kazimierz
Dolny, Poland, May 6—12, 1970. Warszawa; Krakow, 1972, p. 447—455.
Гальперина Г. E. Размножение двустворчатых моллюсков (Bivalvia) Север-
ного Каспия. Автореф. канд. дис. М., 1976. 29 с.
Голиков А. Н. Метод определения продукционных свойств популяций по
размерной структуре и численности. — ДАН СССР, 1970, т. 193, № 3,
с. 730—733.
Голиков А. Н. Некоторые закономерности роста и изменчивости на примере
моллюсков. — В кн.: Гидробиологические исследования самоочищения
водоемов. Л., 1976, с. 97—118.
Голиков А. Н., Меншуткин В. В. Модельное исследование продукционного
процесса популяции брюхоногого моллюска Epheria turrita (A. Adams). —
ДАН СССР, 1971, т. 197, № 4, с. 944—947.
(Голиков А. Н., Меншуткин В. В.) Golikow A. N., Menshutkin V. V. Estima-
tion of production properties of mollusk populations. — Mar. Biol., 1973,
vol. 20, N 1, p. 6—13.
(Голиков A. H., Скарлато О. A.) Golikov A. N., Skarlato O. A. Abundance,
dinamics and production properties of populations of edible mollusks Mizu-
chopecten yessoensis (Zag) and Spisula sachalinensis (Schrenk) related to
problem of organization of controllable submarine ferms at the Western
shore of the Sea of Japan. — Helg. Wiss. Meers, 1970, vol. 20, p. 498—513.
Голубков С. M. Рост и обмен личинок Cloeon dipterum L. (Ephemeroptera)
при различных температурах. — В кн.: Экспериментальные и полевые
исследования биологических основ продуктивности озер. Л., 1979,
с. 147-156.
Граевский Э. Я., Баранов И. В. Промыслово-экологические исследования
жемчужницы бассейна оз. Вадозера. — Вести. ЛГУ. Сер. биол., 1949,
№ 8, с. 14—31.
Десятик И. И. Соотношение веса и линейных размеров у некоторых видов
пресноводных моллюсков. — ДАН БССР, 1968, т. 12, № 9, с. 845—848.
Дзюбан Н. А. Зоопланктон Цимлянского водохранилища. — Изв. НИИ
озер, и реч. рыб. хоз-ва. Л., 1958, т. 45, с. 51—74.
228
Дольник В. Р. Энергетический обмен и эволюция животных. — Успехи
соврем, биол., т. 66, вып. 2 (5), 1968, с. 276—293.
Дрегольская И, Н. Отношение некоторых видов двустворчатых моллюсков
к изменению температуры, солености и газового режима. — Тр. Карадаг.
биол. ст. АН УССР. Севастополь, 1961, вып. 17, с. 52—61.
Егерева И, В. Питание и пищевые отношения рыб в Куйбышевском водохра-
нилище. — В кн.: Волга-1. Матер. Первой конф, по изучению водоемов
бассейна Волги. Куйбышев, 1971, с. 268—273.
Жадин В. И, Моллюски семейства Unionidae. М.; Л., 1938, 170 с. (Фауна
СССР; Т. 4, ч. I).
Жадин В. И. К экологии жемчужницы (Margaritana margaritifera L.). —
Изв. НИИ озер, и реч. рыб. хоз-ва. Л., 1939, т. 21, с. 351—358.
Жадин В. И. Фауна рек и водохранилищ. — Тр. Зоол. ин-та АН СССР.
М.; Л., 1940, т. 5, вып. 3—4, с. 519—591.
Жадин В. И. Моллюски пресных и солоноватых вод СССР. М.; Л., 1952. 374 с.
Журавель П. А. О Dreissena bugensis (Mollusca) из системы Днепра и недавнем
ее появлении в Днепровском водохранилище. — Зоол. журн., 1951,
т. 30, вып. 2, с. 186—188.
Заика В. Е. Удельная продукция водных беспозвоночных. Киев, 1972. 143 с.
Заика В. Е. Соотношение продукции и метаболизма популяции. — Вкн.:
Биологическая продуктивность южных морей. Киев, 1974а, с. 100—111.
Заика В. Е. Рец. на кн.: Трофология водных животных. Итоги и задачи. М.,
Наука, 1973. — Экология, 19746, № 6, с. 98—100.
Заика В. Е. Балансовые уравнения роста животных. — В кн.: Количествен-
ные аспекты роста организмов. М., 1975, с. 25—36.
Заика В. Е., Макарова Н. П. Биологический смысл параметров, входящих
в уравнение роста Берталанфи. — ДАН СССР, 1971, т. 199, № 1, с. 242—
244.
Золотарев В. Н. Изучение закономерностей роста морских моллюсков по
структурам и вещественному составу их раковин. Автореф. канд. дис.
Владивосток, 1974. 23 с.
Зорина И. П. Жемчуг. М., 1972. 46 с. (Новое в жизни, науке, технике. Сер.
биол.; № 6).
Зотина Р. С., Зотин А. И. Количественные соотношения между весом, дли-
ной, возрастом, размерами яиц и плодовитостью у животных. — Журн.
общ. биол., 1967, т. 28, № 1, с. 82—92.
Иванова Л. М. Роль массовых форм северокаспийского бентоса в деструкции
органического вещества. — Тр. ВНИИ мор. рыб. хоз-ва и океанографии,
1973а, т. 80, с. 173—181.
Иванова Л. М. Скорость потребления кислорода донными животными Север-
ного Каспия. — Тр. ВНИИ мор. рыб. хоз-ва и океанографии, 19736,
т. 80, с. 159—172.
Иванова М. Б. Влияние концентрации пищи на скорость фильтрации у Clado-
сега. — Журн. общ. биол., 1970, т. 31, № 6, с. 721—731.
Иванова М. Б. Продукция популяций планктонных животных в пресных
водах СССР. — Экология, 1973, № 3, с. 52—62.
Иванчик Г. С. Материалы к изучению потребления кислорода перловицей
обыкновенной. — Науч, ежегодник Черновиц. ун-та, 1956, т. 1, № 2,
с. 139—141.
Ивлев В. С. Влияние температуры на дыхание рыб. — Зоол. журн., 1938а,
т. 17, вып. 4, с. 645—661.
Ивлев В. С. О превращении энергии при росте беспозвоночных. — Бюл.
Моск, о-ва испыт. природы. Отд-ние биол., т. 67, № 4, 19386, с. 267—277.
Ивлев В. С. Метод определения калорийности гидробиологических проб. —
В кн.: Научно-исследовательские записки Главного управления по за-
поведникам. М., 1939, № 5, с. 65—72.
Ивлев В. С. Экспериментальная экология питания рыб. М., 1955. 252 с.
Ивлев В. С. Метод вычисления количества пищи, потребляемой растущей
рыбой. — В кн.: Тр. 7-й науч. конф, по изучению внутр, водоемов При-
балтики. Петрозаводск, 1959 г. М.; Л., 1962, с. 132—137.
229
Ивлев В, С.О параметрах, характеризующих уровни энергетического обмена
животных. — В кн.: Применение математических методов в биологии. Л.,
1963, кн. 2, с. 146—151.
Ивлев В. С. Элементы физиологической гидробиологии. — В кн.: Физиология
морских животных. М., 1966, с. 3—45.
{Ивлева И. В.) Ivleva I. V. Quantitative correlation of temperature and respira-
tory rate in poikilothermic animals. — Pol. arch, hydrobiol., 1973, t. 20,
N 2, p. 283—300.
Ивлева И. В. Изменение уровней энергетического обмена у морских беспозво-
ночных животных при адаптации к разным температурам. — В кн.:
III съезд Всесоюз. гидробиол. о-ва. Тез. докл. Рига, 1976, с. 128—130.
Ивлева И. В., Попенкина М. И. О температурной зависимости обмена у пой
килотермных животных. — В кн.: Биология моря. Киев, 1968, вып. 15,
с. 29—51.
{Извекова Э. И., Львова-Качанова A. A.) Isvekova Е. I., Lvova-К at chanova А. А.
Sedimentation of suspended matter by Dreissena polymorpha Pallas and its
subsequent utilization by Ghironomidae larvae. — Pol. arch, hydrobiol.,
1972, vol. 19, N 2, p. 203—210.
, Камлюк Л. В. Энергетический обмен у св ободножив ущих плоских и кольча-
тых червей и факторы, его определяющие. — Журн. общ. биол., 1974,
т. 35, № 6, с. 874—885.
Карпевич А. Ф. Рост некоторых моллюсков Каспийского и Аральского мо-
рей. — Тр. ВНИИ мор. рыб. хоз-ва и океанографии, 1952, вып. 1, с. 116—
120.
Карпевич А. Ф. Влияние вод с увеличенным количеством кальция на азов
ских моллюсков; — Тр. Всесоюз. гидробиол. о-ва, 1962, т. 12, с. 5—42.
Карпевич А. Ф. Особенности размножения и роста двустворчатых моллюсков
солоноватоводных морей СССР. — В кн.: Экология беспозвоночных юж-
ных морей СССР. М., 1964, с. 3—60.
Касаткина М. Н. Зависимость между весом и интенсивностью газообмена
у экологически различных видов хирономид. — ДАН СССР, 1960, т. 135,
№ 1, с. 182—184.
Касталъская-Корзинкина М. А. Материалы по питанию дафний. — Зоол.
журн., 1942, т. 21, вып. 4, с. 153—164.
Касымов А. Г. Изучениё жизненного цикла моллюска Sphaerium (Muscu-
lium) lacustre (Mull.). — ДАН АзССР, 1957, т. 13, № 3, с. 325—327.
Кафанов А. И. Об интерпретации логарифмической спирали в связи с анали-
зом изменчивости и роста двустворчатых моллюсков. — Зоол. журн.,
1975, т. 54, вып. 10, с. 1457—1467.
Кафтанникова О. Г., Шевцова Л. В. Дрейссена в водоснабжающих каналах
и участие ее в формировании качества воды. — В кн.: Гидробиология ка-
налов и биологические помехи в их эксплуатации. Тез. докл. Киев, 1972,
с. 44—46.
Качанова А. А. Некоторые данные о размножении Dreissena polymorpha
Pallas, в Учинском водохранилище. — Тр. Всесоюз. гидробиол. о-ва,
1961, т. 2, с. 117—121.
Качанова А. А. О росте Dreissena polymorpha Pallas в Учинском водохрани-
лище и каналах Мосводопровода. — В кн.: Учинское и Можайское водо-
хранилища. М., 1963, с. 226—234.
Каширская Е. В., Константинов А. С. О возможностях оценки роли гидро-
бионтов в трансформации энергии по величине потребления кислорода. —
В кн.: Трофология водных животных. М., 1973, с. 149—163.
Кирпиченко М. Я. Изучение биологии моллюска Dreissena polymorpha Pallas
в Куйбышевском водохранилище. — В кн.: Тр. зональн. совещ. по типо-
логии, биологии, основам рыбохозяйств. использования внутр, (пресно-
водных) водоемов степной зоны СССР. Кишинев, 1962, с. 139—143.
Кирпиченко М. Я. Фенология, динамика численности и рост личинок дрейс-
сены в Куйбышевском водохранилище. — Тр. Ин-та биол. внутр, вод
АН СССР, 1964, вып. 7, с. 19—30.
230
Кирпиченко М. Я. К экологии Dreissena polymorpha Pallas в Цимлянском
водохранилище. — Тр. Ин-та биол. внутр, вод АН СССР, 1971а, вып. 21
(24), с. 142—154.
Кирпиченко М. Я. Речная дрейссена на северо-восточной окраине ее ареала.—
Тр. Ин-та биол. внутр, вод АН СССР, 19716, вып. 21 (24), с. 130—141.
Киселите Т. С. Зоопланктон залива Куршю Марес. — В кн.: Куршю Марес.
Вильнюс, 1959, с. 169—189.
Ковалев А. В. Зависимость между размером самки Oithona папа Giesbr;
(Crustacea, Copepoda) и числом яиц в кладке. — Гидробиол. журн., 1968,
т. 4, № 3, с. 76—80.
Ковалева А, А. Уровень газообмена у некоторых двустворчатых и брюхоногих
моллюсков. — В кн.: Вопросы физиологии и популяционной экологии.
Саратов, 1970, вып. I, с. 31—37.
Кодолова О. П. Сравнительный анализ моллюсков семейства Unionidae Евро-
пейской части СССР по системе миогенов и морфологии раковины. Автореф.
канд. дис. М., 1977. 22 с.
Кондратьев Г. П. О некоторых особенностях фильтрации у Dreissena poly-
morpha Pallas. — Тр. Саратов, отд-ния НИИ озер, и реч. рыб. хоз-ва,
1962, т. 7, с. 280—283.
Кондратьев Г. П. Влияние температуры на фильтрацию пресноводных раку-
шек. — Науч. докл. высш, школы. Биол. науки, 1963а, № 3, с. 17—20.
Кондратьев Г. П. О некоторых особенностях фильтрации у пресноводных мол-
люсков. — Науч. докл. высш, школы. Биол. науки, 19636, № 1, с. 13—16.
Кондратьев Г. П. К методике изучения фильтрации у моллюсков. — В кн.:
Фауна Волгоградского водохранилища и влияние на нее загрязнения.
Саратов, 1967, с. 100—106.
Кондратьев Г. П. Величина фильтрационной работы у некоторых двустворок
Волгоградского водохранилища. — В кн.: Влияние хозяйственной дея-
тельности человека на животный мир Саратовского Поволжья. Саратов,
1969а, с. 32—36.
Кондратьев Г. П. Влияние концентрации взвесей на длительность фильтра-
ционной активности у некоторых пресноводных ракушек. — В кн.: Видо-
вой состав, экология и продуктивность гидробионтов Волгоградского
водохранилища. Саратов, 19696, с. 37—40.
Кондратьев Г. П. Влияние температуры воды на длительность фильтрацион-
ной активности у некоторых пресноводных ракушек. — В кн.: Видовой
состав, экология и продуктивность гидробионтов Волгоградского водо-
хранилища. Саратов, 1969в, с. 31—36.
Кондратьев Г. П. О масштабах фильтрационной и минерализационной ра-
боты некоторых двустворок в Волгоградском водохранилище. — В кн.:
Вопросы физиологии и популяционной экологии. Саратов, 1970а, вып. 1,
с. 38-40.
Кондратьев Г. П. О соотношении между весом и линейными размерами тела
у некоторых моллюсков. — В кн.: Вопросы физиологии и популяционной
экологии. Саратов, 19706, вып. 1, с. 56—60.
Кондратьев Г. Я. Фильтрационная и минерализационная работы двустворча-
тых моллюсков Волгоградского водохранилища. Автореф. канд. дис.
Саратов, 1970в. 23 с.
Кондратьев Г. П. Сезонность изменения фильтрационной активности у дву-
створчатых моллюсков Волгоградского водохранилища. — В кн.: Тр.
комплекс, экспедиции Саратов, ун-та по изуч. Волгоград, и Саратов,
водохранилищ. Саратов, 1973, вып. 3, с. 127—130.
Константинов А. С. Биология хирономид и их разведение. — Тр. Саратов,
отд-ния НИИ озер, и реч. рыб. хоз-ва, 1958, т. 5. 359 с.
Константинов А. С. Общая гидробиология. М., 1967. 431 с.
Коштоянц X. С. Основы сравнительной физиологии. М.; Л., 1950, т. I. 524 с.
Коштоянц X. С., Шаланки Я. Данные о физиологической основе периоди-
ческой деятельности у анодонт. — Журн. общ. биол., 1958, т. 19, № 3,
с. 212—216.
231
Кублицкас А. Питание бентосоядных рыб залива Куршю Марес. — В кн.:
Куршю Марес. Вильнюс, 1959, с. 463—519.
Кузьменко К. Н, Роль двустворчатых моллюсков в процессах самоочищения
оз. Красного (Карельский перешеек). — В кн.: Методы биологического
анализа пресных вод. Л., 1976, с. 134—135.
Лубяное И, П., Золотарева В. И. Особенности биологии Dreissena bugensis
Andrusov — важного биофильтра Запорожского водохранилища. —
В кн.: Гидробиологические исследования самоочищения водоемов. Л.,
1976, с. 129—133.
Лукин А. В. Биология и запасы промысловых ракушек бассейна р. Свияги
в пределах Татреспублики. — Тр. о-ва естествоиспыт. при Казан, ун-те,
1937а, т. 55, вып. 1—2, с. 15—55.
Лукин А. В. Отчет о работе по изучению запасов промысловых ракушек
в водоемах TAGGP. — Тр. о-ва естествоиспыт. при Казан, ун-те, 19376,
т. 55, вып. 1—2, с. 1 — 14.
Лукин А. В. Перловица из Красновидовского затона (р. Волги). — Тр.
о-ва естествоиспыт. при Казан, ун-те, 1937в, т. 56, вып. 1—2,
с. 57-74.
Лукин А. В. Краткая характеристика условий существования рыб и водных
животных в Куйбышевском водохранилище. — Тр. Татар, отд-ния НИИ
озер, и реч. рыб. хоз-ва. Казань, 1960, вып. 9, с. 5—10.
Львова А. А. К расчету продукции Dreissena polymorpha (Pall.) Учинского
водохранилища. — Информ, бюл. Ин-та биол. внутр, вод АН СССР,
1976, № 29, с. 37—41.
Львова А. А. Экология Dreissena polymorpha (Pall.) Учинского водохрани-
лища. Автореф. канд. дис. М., 1977. 22 с.
Львова-Качанова А. А. Продукция дрейссены — основного компонента бен-
тоса Учинского водохранилища. — В кн.: Биологические процессы
в морских и континентальных водоемах. Кишинев, 1970, с. 231.
Львова-Качанова А. А. О роли дрейссены (Dreissena polymorpha Pallas)
в процессах самоочищения воды Учинского водохранилища. — В кн.:
Комплексные исследования водохранилищ. М., 1971, вып. 1, с. 196—203.
Ляхов С. М., Михеев В. П. Распределение и количество дрейссены в Куй-
бышевском водохранилище на седьмом году его исследования. — Тр.
Ин-та биол. внутр, вод АН СССР, 1964, вып. 7 (10), с. 3—18.
Макарова Н. П. Балансовые модели в экологии особи и популяций. Автореф.
канд. дис. Севастополь, 1976, 23 с.
Макарова Н. П,, Заика В. Е. Связь между ростом животных и количеством
усвоенной пищи. — Гидробиол. журн., 1971, т. 7, № 3, с. 5—13.
Максимова М, П, Материалы по гидрохимической характеристике рек
Карельского побережья Белого моря. — В кн.: Проблемы использова-
ния промысл, ресурсов Белого моря и внутр, водоемов Карелии. М.;
Л., 1963, вып. 1, с. 40—49.
Манюкас И, Ихтиофауна, состояние запасов и промысел рыб в заливе Куршю
Марес. — В кн.: Куршю Марес. Вильнюс, 1959, с. 293—401.
Микулич Л. В. Плодовитость даурской жемчужницы (Margaritana dahurica
Midd.). — Изв. Тихоокеан. НИИ рыб. хоз-ва и океанографии. Влади-
восток, 1954, т. 39, с. 357—358.
Микулич Л. В., Козак Л, П. Плодовитость некоторых Decapoda залива
Петра Великого. — Гидробиол. журн., 1971, т. 7, № 1, с. 97—101.
Мина М, В., Клевезаль Г. А. Рост животных. М., 1976, 291 с.
Митропольский В. И. Наблюдения над жизненным циклом, темпом роста
и способностью к перенесению высыхания у Musculium lacustre (Mul-
ler). — Тр. Ин-та биол. внутр, вод АН СССР, 1965, вып. 8 (11), с. 118—124.
Митропольский В, И, О механизме фильтрации и о питании сфериид (Мо1-
lusca, Lamellibranchia). — Тр. Ин-та биол. внутр, вод АН СССР, 1966,
вып. 12 (15), с. 129—133.
Митропольский В. И. Жизненный цикл Pisidium obtusale Jenyns. — Информ,
бюл. ин-та биол. внутр, вод АН СССР, 1969, № 3, с. 17—21.
232
Митропольский В. И. Особенности биологии сфериид Верхневолжских водо-
хранилищ. Автореф. канд. дис. Л., 1970, 26 с.
Михеев В. П, О питании рыб дрейссеной на мелководьях Куйбышевского
водохранилища в 1961 г. — В кн.: Материалы по биологии и гидробиоло-
гии Волжских водохранилищ. М.; Л., 1963, с. 80—83.
Михеев В, П, О скорости отмирания дрейссены в анаэробных условиях. —
В кн.: Биология дрейссены и борьба с ней. М.; Л., 1964а, с. 76—81.
Михеев В, П, О линейном росте Dreissena polymorpha Pallas в некоторых,
водохранилищах Европейской части СССР. — Тр. Ин-та биол. внутр,
вод АН СССР, 19646, вып. 7, с. 55—65.
Михеев В, П. Питание дрейссены в прудах и водохранилищах в зависимости
от условий среды. — Тр. ВНИИ пруд. рыб. хоз-ва, 1966, т. 14, с. 169—
178.
Михеев В. П, Фильтрационное питание дрейссены. — Тр. ВНИИ пруд,
рыб. хоз-ва, 1967а, т. 15, с. 117—129.
Михеев В. П. Питание дрейссены (Dreissena polymorpha Pallas). Автореф.
канд. дис. М., 19676, 19 с.
Михеев В. П., Сорокин Ю. И, Количественные исследования питания дрейс-
сены радиоуглеродным методом. — Журн. общ. биол., 1966, т. 27, № 4,
с. 463—472.
Монаков А, В., Сорокин Ю, И, Количественные данные о пит&нии дафний. —
Тр. Ин-та биол. внутр, вод АН СССР, 1961, вып. 4 (7), с. 251—261.
Муратова Р. X. Перловицы реки Казанки и их запасы. — Тр. о-ва естество-
испыт. при Казан, ун-те, 1937, т. 55, вып. 1—2, с. 75—91.
Небольсина Т. И. Качественная и количественная оценка питания леща,
густеры и плотвы Волгоградского водохранилища в 1962—1964 гг. —
Тр. Саратов, отд-ния НИИ озер, и реч. рыб. хоз-ва, 1965, т. 8, с. 108—
127.
Негробов В. П. Промысел перловицы в СССР и перспективы его разви-
тия. — В кн.: Моллюски. Пути, методы и итоги их изучения. Тез. докл.
4-го совещ. по изучению моллюсков. Л., 1971, с. 96—97.
Нейферт А. В. Скорость фильтрации и прохождение пищевого комка у Car-
dium edule (L.). — В кн.: Донные биоценозы и биология бентосных орга-
низмов Черного моря. Киев, 1967, с. 113—121.
Некрасова М. Я. Эколого-биологическая характеристика доминирующих_
видов зообентоса в Таганрогском заливе Азовского моря. — Гидробиол.
журн., 1971, т. 7, № 6, с. 49—55.
Овчинников И. Ф. Unio crassus m. ater Nils, и его промысловое значение. —
Тр. Зоол. ин-та АН СССР. Л., 1932, т. 1, с. 1—24.
Овчинников И. Ф. Материалы к изучению советского перламутра и его про-
дуцентов. — Тр. Севан, озер. ст. Ереван, 1933, т. 4, вып. 1—2, с. 10—72.
Озерецковская Н, Г, Физико-географические и гидрохимические особенности
озер Кривого и Круглого. — Тр. Зоол. ин-та АН СССР. Л., 1975, т. 56,
с. 10—31.
Озерецковская Н. Г,, Умнова Л, П. Краткий гидрохимический очерк озер
Зеленецкое и Акулькино. — Тр. Зоол. ин-та АН СССР. Л., 1975, т. 57,
с. 3—18.
Остапеня А. П, Полнота окисления органического вещества водных бес-
позвоночных методом бихроматного окисления. — ДАН БССР, 1965,
т. 9, № 4, с. 273—276.
Остапеня А. П. Биомасса и способы ее выражения. — В кн.: Методы опре-
деления продукции водных животных. Минск, 1968, с. 20—44.
Остапеня А. П, Взвешенное вещество как структурный и функциональный
компонент водных экосистем. — В кн.: III съезд Всесоюз. гидробиол.
о-ва. Тез. докл. Рига, И—15 мая 1976 г. Рига, 1976а, т. 2, с. 36—38.
Остапеня А, П, Сестон и его участие в процессах самоочищения. — В кн.:
Методы биологического анализа пресных вод. Л., 19766, с. 141 — 142.
Остапеня А, П., Печень Г, А,, Бабицкий В. А., Павлютин А. П. Интенсив-
ность обмена Diaptomus graciloides (Lili.) при низкой температуре. —
Гидробиол. журн., 1969, т. 5, № 5, с. 121—124.
16 А. Ф. Алимов
233
Петров В. В. К оценке промысловой мощности популяций моллюсков се-
мейства Unionidae. — Изв. НИИ озер, и реч. рыб. хоз-ва. М.. 1964
т. 57, с. 42-46.
Платонова О. П. Питание бентосоядных рыб Куйбышевского водохрани-
лища. — Учен. зап. Казан, ун-та, 1966, т. 123, кн. 7, с. 59—102.
Попова Э. И. Бентос оз. Б. Харбей и его продукция. — В кн.: Продуктив-
ность озер восточной части Болыпеземельской тундры. Л., 1976, с. 101 —
103.
Проссер Л., Браун Ф. Сравнительная физиология животных. М., 1967. 766 с.
Родина А. Г. Бактерии как пища для пресноводных моллюсков. — Микро-
биология, 1948, т. 17, вып. 3, с. 232, 239.
Румшиский Л. 3. Математическая обработка результатов эксперимента.
М., 1971. 192 с.
Саватеева Е. Б. О распространении и экологии моллюсков семейства Sphaeri-
idae в некоторых озерах Карелии. — Изв. НИИ озер, и реч. рыб. хоз-ва.
М., 1964, т. 57, с. 199—205.
Садыхова И. А. Методика определения возраста двустворчатых моллюсков.
М., 1972. 40 с.
Сальдау М. П. Темп промысловых моллюсков некоторых районов Европей-
ской части СССР. — Изв. НИИ озер, и реч. рыб. хоз-ва. М., 1939. т. 22,
с. 244-269.
Санина Л. В. О связи скорости фильтрации и питания с весом тела при раз-
личных концентрациях корма у каспийского кардиума. — Гидробиол.
журн., 1976, т. 12, № 1, с. 108—112.
Семенченко В. П., Романова 3. А. Размерно-весовая характеристика и
плодовитость Idotea chelipes (Pallas) (Crustacea, Isopoda) из Балтий-
ского моря. — Гидробиол. журн., 1976, т. 12, № 1, с. 115—118.
Сидоров Г. П. Биологическая характеристика и продукция рыб оз. Б. Хар-
бей. — В кн.: Продуктивность озер восточной части Болыпеземельской
тундры. Л., 1976, с. 110—130.
Скиркявичене 3. Ю. Закономерности увеличения роста дрейссены. 2. Влия-
ние периода года. — Тр. АН ЛитССР. Сер. биол. науки, В, 1970, № 2
(52), с. 91-97.
Смирнова Н. Ф. Определение продукции моллюсков. — В кн.: Методика
изучения биоценозов внутренних водоемов. М., 1975а, с. 200—208.
Смирнова Н. Ф. Рост и .продукция унионид (Mollusca, Unionidae) в районе
Костромской ГРЭС. — В кн.: Экология организмов водохранилищ-
охладителей. Л., 19756, с. 272—286.
Соловьева Н. Ф. К вопросу о динамике солевого баланса Аральского моря. —
В кн.: Материалы к познанию фауны и флоры СССР. Нов. сер. Отд-ние
зоол. М., 1950, № 19 (34), с. 62—69.
{Сорокин Ю. И.) Sorokin Y. I. Biological productivity of the Rybinsk reser-
voir. — In: Proceedings of the IBP—UNESKO symposium of producti-
vity problems of freshwater. Kazimierz Dolny, Poland, May 6—12, 1970,
Warszawa; Krakow, 1972, p. 493—503.
Спиридонов Ю. И. О линейном росте дрейссен в Волгоградском водохрани-
лище близ Саратова. — В кн.: Видовой состав, экология и продуктив-
ность гидробионтов Волгоградского водохранилища. Саратов, 1969,
с. 70—74.
Спиридонов Ю. И. Исследование дыхания у дрейссен полярографическим
методом. — В кн.: Вопросы физиологической и популяционной экологии.
Саратов, 1972, вып. 2, с. 15—21.
Спиридонов Ю. И. Влияние некоторых факторов на газообмен дрейссен
по данным микрополярографического метода. — В кн.: Физиологическая
и популяционная экология животных. Саратов, 1973, вып. 1 (3), с. 5—9.
Справочник по водным ресурсам СССР. Л., 1933—1936, т. 2—17.
Станъчиковска А. Размер и вес Dreissena polymorpha (Pal.) из озер Запад-
ной Европы. — Гидробиол. журн., 1977, т. 13, № 3, с. 26—29.
Старобогатов Я. И. Фауна моллюсков и зоогеографическое районирование
континентальных водоемов земного шара. Л., 1970. 372 с.
234
Старобогатов Я. И. Класс двустворчатые моллюски Bivalvia. — В кн.:
Определитель пресноводных беспозвоночных Европейской части СССР.
Л.» 1977, с. 123—151.
Стрельникова В. М. Взаимосвязь интенсивности обмена и плодовитости
у Idotea ochotensis Brand из Японского моря. — Гидробиол. журн., 1971,
т. 7, № 2, с. 118-121.
Суханов В. В. О понятиях «ростовой» и «поддерживающей» продукции по-
пуляций. — Биология моря, 1978, № 5, с. 83—87.
Сущеня Л. М. Зависимость скорости фильтрации у планктонных ракообраз-
ных от концентрации пищевых частиц. — В кн.: Гидробиологические ис-
следования на рыбоводных прудах БССР. Минск, 1958, с. 241—260.
Сущеня Л. М. Количественные закономерности фильтрационного питания
Artemia salina L. — Тр. Севастоп. биол. ст. Севастополь, 1964, т. 15г
с. 434—445.
Сущеня Л. М. Интенсивность дыхания ракообразных. Киев, 1972. 195 с.
Сущеня Л. М. О содержании книги «Трофология водных животных. Итоги
и задачи». — Гидробиол. журн., 1974, т. 10, № 3, с. 124—130.
Сущеня Л. М. Количественные закономерности питания ракообразных.
Минск, 1975. 206 с.
Табилова Т. X., Брегман Ю. Э. Рост двустворчатого моллюска Mizuhopecten
yessoensis в бухте Троицы (залив Посьет, Японское море). — Экология,
1975, № 2, с. 65—72.
Тен В. С. О трофическом взаимодействии примитивных пар хищник—жертва
у водных организмов. — В кн.: Структура и динамика водных сообществ
и популяций. Киев, 1967, с. 16—43.
Троицкий С. К. Пресноводные ракушки Азово-Черноморского края. Ростов
н/Д, 1934. 44 с.
Троицкий С. Б. Материалы по размножению унионид в водоемах Ростов-
ской области и Краснодарского края. — Тр. Ростов, обл. биол. о-ва.
Ростов н/Д, 1939, вып. 3, с. 77—101.
Турпаева Е. П., Солдатова И. Н., Симкина Р. Г. Интенсивность обмена
у черноморских двустворчатых моллюсков. — Зоол. журн., 1970, т. 49,
вып. 10, с. 1571—1572.
Умнов А. А. Математическая модель озера Мястро. — В кн.: Продукционно-
биологические исследования экосистем пресных вод. Минск, 1973, с. 95—
109.
Умнов А. А. Применение статистических методов для оценки параметрон
эмпирических уравнений, описывающих взаимосвязь между энергети-
ческим обменом и массой тела животного. — Журн. общ. биол., 1976,.
т. 37, № 1, с. 71—86.
Францев А. В., Лебедева С. И. Химизм воды Учинского водохранилища. —
Тр. Зоол. ин-та АН СССР. Л., 1941, т. 7, вып. 1, с. 31—52.
Хлебович Т. В. Интенсивность дыхания у инфузорий разного размера. —
Цитология, 1974, т. 16, № 1, с. 103—105.
Хмелева Н. Н. Трансформация энергии у Artemia salina (L.). — ДАН СССР,.
1967, т. 175, № 4, с. 934—937.
Хмелева Н. Н. Связь плодовитости с величиной тела и энергетическим об-
меном у Idotea baltica basteri (And) и у других ракообразных. — ДАН
СССР, 1969, т. 185, № 1, с. 198—301.
Хмелева Н. Н. Биология и энергетический баланс морских равноногих рако-
образных (Idotea baltica basteri). Киев, 1973. 184 с.
Хусаинова Н. 3., Митрофанов В. П., Мамилова Р. X. и др. Биологическая
продуктивность оз. Каракуль. — В кн.: Продукционно-биологические
исследования экосистем пресных вод. Минск, 1973, с. 32—43.
Цееб Я. Я. Зоопланктон. — В кн.: Каховське водоймище. Киев, 1964, с. 60—
111.
Цееб Я. Я., Денисова А. И., Приймаченко А. Д. и др. Продуктивность,
сообществ водных организмов Киевского водохранилища. — В кн.:
Продукционно-биологические исследования экосистем пресных вод.
Минск, 1973, с. 60—71.
16*
235^
Цихон-Луканина Е. А. Питание черноморских двустворчатых моллюсков. —
Биология моря, 1976, Яг 3, с. 42—48.
Цихон-Луканина Е. А., Солдатова И. Н. Усвоение пищи водными беспоз-
воночными. — В кн.: Трофология водных животных. М., 1973, с. 108—
121.
Чаплина А. М., Булахов В. Л., Тарасенко С. П. Вселение и развитие тарани
и судака в водохранилищах степного Приднепровья и Крыма. — В кн.:
III съезд Всесоюз. гидробиол. о-ва. Тез. докл. Рига, 11—15 мая 1976 г.
Рига, 1976, т. 3, с. 121—124.
Чухчин В, Д. Биология размножения брюхоногого моллюска Patella сае-
rulea L. в Черном море. — В кн.: Биология моря. Киев, 1972, вып 26,
с. 98—109.
Шевцова Л. В. Определение веса Dreissena polymorpha Pall, и D. bugensis
Andr. по их размерам. — Гидробиол. журн., 1971а, т. 7, № 1, с. 123—
125.
Шевцова Л. В. Продукция осевших личинок дрейссен в канале Днепр—
Кривой Рог. — В кн.: Матер. 4-го совещ. по изучению моллюсков.
Автореф. докл. Л., 19716, с. 74—75.
Шерстюк В. В. Калорийность кормовых организмов Кременчугского водо-
хранилища. — Гидробиол. журн., 1971, т. 7, № 6, с. 99—103.
Шерстюк В. В., Зимбалевская Л. Н. Калорийность фитофильных беспоз-
воночных в водохранилищах Днепра. — Гидробиол. журн., 1973, т. 9,
№ 4, с. 83—87.
Шорыгин А. А. Питание и пищевые взаимоотношения рыб Каспийского моря.
М., 1952. 268 с.
Шушкина Э. А. Физиологический способ расчета продукции видовых по-
пуляций. — В кн.: Методы определения продукции водных живот-
ных. Минск, 1968, с. 126—135.
Юровицкий Ю. Г. Некоторые вопросы методики изучения плодовитости
рыб. — Вопр. ихтиологии, 1958, вып. 10, с. 12—28.
Ярошенко М. Ф. Гидрофауна Днестра. М., 1957. 169 с.
Agrell I. The shell morphology of some Swedish Unionides as affected by ecolo-
gical conditions. — Arkiv Zool., 1949, Bd 41 A, N 15, S. 1—30.
Akerlund G. Oxygen consumption of the ampullariid snail Marisa comuarie-
tis L. in relation to body weight and temperature. — Oikos, 1969, vol. 20,
N 2, p. 529—533.
AU R. M. The influence of suspension density and temperature on the filtra-
tion rate of Hiatella arctica. — Mar. Biol., 1970, vol. 6, N 4, p. 291—302.
Allen W. R. The food and feeding habits of freshwater mussels. — Biol. Bull.,
1914, vol. 27, N 3, p. 121—141.
Allen W. R. Studies of the biology of freshwater mussels. — Biol. Bull., 1921,
vol. 40, N 4, p. 210—241.
A It nd cl er K. Beobachtungen fiber die Biologie von Margaritana margaritifera
und fiber die Okologie ihres Wohnorts. — Arch. Hydrobiol., 1926, Bd 17,
H. 3, S. 423—491.
A manual on methods for the assessment of secondary productivity in fresh
waters. IBP Handbook, N 17. Oxford; Edinburgh, 1971. 358 p.
Andrewartha H. C., Birch L. C. The distribution and abundance of animals.
Chicago, 1954. 782 p.
Ansell A. D. The functional morphology of the British species of Veneracea
(Eulamelli branchi a). — J. Mar. Biol. Assoc. U. K., 1961, vol. 41, N 2,
p. 489—515.
Ansell A. D. The rate of growth of the hard clam Mercenaria mercenaria (L.)
throughout the geographical range. — J. Conseil perman. intern, explor.
mer., 1968, vol. 31, N 3, p. 364—409.
Arakawa К. I. Statological studies of the Bivalvia (Mollusca). — Adv. Mar.
Biol., 1970, vol. 8, p. 413—429.
.236
Armstrong F. A. J., Atkins W. R. G. The suspended matter of sea water. —
J. Mar. Biol. Assoc. U. K., 1950, vol. 29, N 1, p. 139—144.
Avolizi R. J. Biomass turnover in populations of viviparous sphaeriid clams:
comparisons of growth, fecundity, mortality and biomass production. —
Hydrobiologia, 1976, vol. 51, N 2. p. 163—180.
В adman D. G. Filtration of neutral red by fresh water clams in aerobic and
hypoxic conditions. — Comp. Biochem. and Physiol., 1975, vol. 51, N 4A,
p. 741—744.
Ball G. H. Variation in freshwater mussels. — Ecology, 1922, vol. 3, N 1,
p. 93—121.
Barnes G. E. The behaviour of Anodonta cygnea L. and its nuerophysiological
basis. — J. Exp. Biol., 1955, vol. 32, N 1, p. 158—174.
Barnes G. E. The behaviour of unrestrained Anodonta. — Anim. Behav., 1962,
vol. 10, N 1—2, p. 174—176.
Bayne B. L. The respiratory response of Mytilus perna L. (Mollusca; Lamel-
libranchia) to reduced environmental oxygen. — Physiol. Zool., 1967,
vol. 40, N 3, p. 307—313.
Bayne B. L. Oxygen consumption by three species of lamellibranch molluscs
in declining ambient oxygen tension. — Comp. Biochem. and Physiol.,
1971a, vol. 40, N 4, p. 955—970.
Bayne B. L. Ventilation, the heart beat and oxygen uptake by Mytilus edulis L.
declining oxygen tension. — Comp. Biochem. and Physiol., 1971b, vol. 40,
N 4, p. 1065—1085.
Belehradek J. Temperature coefficients in biology. — Biol. Rev., 1930, vol. 5,
p. 30—58.
Berg K. Studies on the bottom animals of Esrom lake. — K. danske vidensk.
Selsk. Skr., 1938, Bb 9, N 8, S. 131—255.
Berg K. On the oxygen consumption of Ancylidae (Gastropoda) from an Ecolo-
gical point of view. — Hydrobiologia, 1952, vol. 4, N 3, p. 225—267.
Berg K. On the oxygen consumption of some freshwater snails. — Verb. Intern.
Ver. theoret. und angew. Limnol., 1961, vol. 14, pt 2, p. 1019—1022.
Berg K., Jonasson P. M., Ockelmann K. W. The respiration of some animals
from the profundal zone of a lake. — Hydrobiologia, 1962, vol. 19, N 1,
p. 1—39.
Berg K., Lumbye J., Ockelmann K. W. Seasonal and experimental variation
of the oxygen consumption of the limpet Ancylus fluviatilis (O. F. Mtil-
ler). — J. Exp. Biol., 1958, vol. 35, N 1, p. 43—73.
Berg K., Ockelmann K. W. The respiration of Freshwater snails. — J. Exp.
Biol., 1959, vol. 36, N 4, p. 690—708.
Birch L. C. The intrinsic rate of natural increase of the insect population. —
J. Anim. Ecol., 1948, vol. 17, N 1, p. 15—26.
Birch L. C. Experimental background to the study of the distribution and
abundance of insects. — Ecology, 1953, vol. 34, N 4, p. 698—726.
Bjork S. Investigations on Margaritifera margaritifera and Unio crassus. —
Acta limnol., 1962, vol. 4, p. 1—109.
Bdetius L. Temperature and growth in a population of Mytilus edulis (L.) from
the N. Harbour of Copenhagen (the Sound). — Medd. Danm. fisk.-og havun-
dersog., N. S., 1962, Bd 3, N 11, S. 339—346.
Bondesen P. Preliminary investigations in to the development of Neritina
fluviatilis L. in brackish and freshwater. — Vidensk. Medd. Dansk Naturh.
Foren, 1940, vol. 104, p. 283—318.
Botnariuc N., Negrea A., Pico? C. A. Observa0i asupra Anodontelor din com-
plexul de Bal^i Crapina—Jijila. — St. cere. biol. anim., 1961, t. 13, N 1,
p. 93—102.
Botnariuc N., Tudorancea Cl. Beitrage zur Populationsdynamik bei Anodonta
piscinalis im Flachsee Jijila (Uberschwemmungsgebiet der Donau). —
Arch. Hydrobiol., 1967, Bd 30, SuppL 2, N 4, S. 400—419.
Brown C. J. D., Clark C.. Gleissner B. The size of certain Naiades from Wes-
tern lake Erie in relation to shoal, exposure. — Amer. Midland Naturalist,
1938, vol. 19, N 3, p. 682—701.
237
Bruce J. R. The respiratory exchange of the mussel Mytilus edulis. — Biochem.
J. Cambridge, 1926, vol. 20, p. 829—846.
Bruin J, P. C., Davids C. Observations on the rate of water pumping of the
freshwater mussel Anodonta cygnea zellensis (Gmelin). — Neth. J. ZooL,
1970, vol. 20, N 3, p. 380—391.
Buley H. M. Consumption of diatoms and dinoflagellates by the mussel. —
Bull. Scripps Inst. Oceanogr. Univ. Calif. Techn. set., 1936, vol. 4, N 1,
p. 19-27.
Bullock T. H. Compensation for temperature in the metabolism and activity
of poikilotherms. — Biol. Rev., 1955, vol. 30, N 3, p. 311—342.
Burky A. J., Burky K. A. Seasonal respiratory variation and acclimation in
the Pea clam, Pisidium walkeri Sterki. — Comp. Biochem. and Physiol.,
1976, vol. 55, p. 109—114.
Burlacu Gh., Nastasescu Gh., Marinescu G., Raduca C., Voiculescu T. Eficien^a
utilizarii energieri scoicilor (Anodonta cygnea L.) administrate in Hrana
Gainilor. — St. cere. biol. Ser. zool., 1968, t. 20, N 2, p. 181—191.
Cassie R. M. The analysis of Polymodal Frequency distributions by the Proba-
bility paper method. — N. Z. Sci. Rev., 1950, vol. 8, p. 89—91.
Cassie R. M. Some uses of probability paper in the analysis of size frequency
distributions. — Austr. J. Mar. and Freshw. Res., 1954, vol. 5, N 3, p. 513—
522.
Chamberlain T. K. Annual growth of freshwater mussels. — Bur. Fish. USA,
1931, vol. 46, N 1103, p. 713—739.
Chipman W. A., Hopkins J. G. Water filtration by the bay scallop, Pecten
irradians, as observed with the use radiactive plankton. — Biol. Bull.,
1954, vol. 107, N 1, p. 80—91.
Clarke A. H. The freshwater molluscs of the Canadian Interior Basin. — Mala-
cologia, 1973, vol. 13, N 1—2. 509 p.
Coe W. R. Nutration, environmental conditions and growth of marine bivalve
molluscs. — J. Mar. Res., 1948, vol. 7, p. 586—601.
Coe W. R., Fox D. L. Biologia of the California sea mussel (Mytilus california-
nus). III. Environmental conditions and rate of growth. — Biol. Bull.,
1944, vol. 87, N 1, p. 59-72.
Colin N. I. A. The biology of marine animals. London, 1960. 707 p.
Comjort A. The duration of life irt molluscs. — Proc. Malacol. Soc. London,
1957, vol. 32, N 6, p. 219—241.
Coughlan J. The estimation of filtering rate from the clearance of suspensions. —
Mar. Biol., 1969, vol. 2, N 4, p. 356—358.
Coughlan J., Ansell A. D. A direct method for determining the pumping rate
of siphonate bivalves. — J. Conseil perman. intern, explor. met., 1964,
vol. 29, N 2, p. 205—213.
Coutts P. J. F. Growth characteristics of the bivalva Chione stutchburyi. —
N. Z. J. Mar. and Freshw. Res., 1974, vol. 8, N 2, p. 333—339.
Crowley T. E. Age determination in Anodonta. — J. Conchology, 1957, vol. 24,
N 6, p. 201-207.
Cvancara A. M. Mussels (Unionidae) of the Red river valley in North Dakota
and Minnesota, U. S. A. — Malacologia, 1970, vol. 10, N 1, p. 57—92.
Cvancara A. M. Lake mussel distribution as determined with Soube. — Eco-
logy, 1972, vol. 53, N 1, p. 154—157.
Dakin W. J.. Dakin С. M. G. The oxygen requirements of certain aquatic
animals and its bearing upon the source of food supply. — Brit. J. Exp.
Biol., 1925, vol. 2, p. 293—322.
Danneel I., Hinz W. Trockenresistenz dreier Pisidium-Populationen (Bival-
via) in Abhangigkeit von der relativen Luftfeuchtigkeit. — Hydrobiolo-
gia, 1974, vol. 45, N 1, p. 39—43.
Danneel I., Hinz W. Zur Biologie von Pisidium amnicum O. F. Muller (Bival-
via). — Arch. HydrobioL, 1976, Bd 77, H. 2, S. 213-225.
Dietz T, H., Stern E. M. Seasonal change in reproductive activity and biochemi-
cal composition of fingernail clam, Sphaerium transversum. — Nautilus,
1977, vol. 94, N 4, p. 136—140.
238
Draghici О., Picos C. A. CercetSri asupra consumului de oxigen la molupte
(Unio pictorum L.). — St. cere. biol. Ser. zool., 1969, t. 21, N 12, p. 151—
156.
Engellmann M. D. Energetics terrestial field studies and animal productivity. —
Adv. Ecol. Res., 1966, vol. 3, p. 73—115.
Fenchel T. Intrinsic rate of natural increase; the relationship with body size. —
Oecologia, 1974, vol. 14, N 4, p. 317—326.
Fischer P. H. Vie et moeurs des Mollusques. Paris (Payot), 1950, 312 p.
Foster-Smith R. L. The effect of concentration of suspension on the filtration
rates and pseudofaecal production for Mytilus edulis L., Cerastoderma
edule (L.) and Venerupis pullastra. — J. Exp. Mar. Biol., Ecol., 1975,
vol. 17, N 1, p. 1—22.
Fox D. L., Sverdrup H. V., Gunningham J, P. The rate of water propulsion
in the Californian mussel. — Biol. Bull., 1937, vol. 72, N 3, p. 417—438.
Franz V. Die Unterscheidung der zwei mitteleuropaischen Anodonta-Arten
cygnea (L.) und piscinalis Nills und die Haupttypen derselben. — Z. Na-
turwiss., 1939, Bd 72, H. 2, S. 75—210.
Franz V. Anodonta — Bemerkungen in Anschluss an meine Anodontenarbeit
von 1939. — Arch. Molluskenk., 1941, Bd 73, H. 5—6, S. 201—210.
Fretter V., Graham A. Reproduction. In: Physiology of Mollusca.. New York;
London, 1964, vol. 1, p. 127—164.
Fuji A. Growth and breeding season of the brackish water bivalve, Corbicula
japonica, in Zyusan—Gate Inlet. — Bull. Fac. Fish. Hokkaido Univ.,
1957, vol. 8, N 3, p. 178-184.
Fuller J. Z, Feeding rate Calanus finmarchicus in relation to environmental
conditions. — Biol. Bull., 1937, vol. 72, N 2, p. 233—246.
Fuziwara T. On the reproduction of Corbicula leana Prime. — Venus, 1975,
vol. 34, N 1—2, p. 54—56.
Gale W. F., Lowe R. L. Phytoplankton ingestion by the fingernail clam, Spha-
erium transversum (Say), in Pool 19, Mississippi River. — Ecology, 1971,
vol. 52, N 3, p. 507-513.
Galtsoff P, S. Experimental study of the function of the oyster gills and its
bearing on the problems of Oyster culture and sanitary control of the oyster
industry. — Bull. U. S. Bur. Fish., 1928, vol. 44, p. 1—39.
Gartkiewicz S. О oddychaniu Szczezni (Anodonta sp.) w Stanie czyunosciowym
i spoezynkowym (snu). — Arch. nauk. biol. Towar. nauk, Warszawskiego,
1922, t 1, N 17, p. 1-24.
Gauld D. T. The grazing rate of planctonic Copepods. — J. Mar. Biol. Assoc.
U. K., 1951, vol. 29, N 3, p. 675—706.
Geng H. Der Futter der natiirlichen Fischnahrung. — Z. Fischerei, 1925, Bd 23,
H. 2, S. 10—280.
Ghiretti F. Respiration. In: Physiology of mollusca. New York; London, 1966,
vol. 2, p. 175—208.
Gillespie D. M. Population studies of four species of molluscs in the Madison
River. Yellowstone National Park. — Limnol. and Oceanogr., 1969, vol, 14,
N 1, p. 101—114.
Grave C. The process of feeding in the oyster. — Science, 1916, vol. 44, p. 178—
181.
Gray J. M. A. The mechanism of ciliary movement. IV. The relation of ciliary
activity to oxygen consumption. — Proc. Roy. Soc. London, 1924, ser.
B, vol. 96, p. 95—114.
Gray J. M. A, Ciliary movement. Cambridge, 1928. 162 p.
Groenwegen J. A. W. Uber den Bau und die Entwicklung der Bruttaschen von
Sphaerium rivicola Lm. — Z. Morph., Okol. Tiere, 1926, Bd 5, H. 2, S. 207—
290.
Grossu D., Burlacu Gh., Baltac M. Cercetari asupra bilan^ului energetic le mel-
cul de livada (Helix pomatia L.). — St. cere. biol. Ser. zool., 1968, vol. 20,
N 2, p. 179—184.
239
Hamwi A., Haskin H. H. Oxygen consumption and pumping rates in the hard
clam Mercenaria mercenaria: a direct method. — Science, 1969, vol. 163,
N 3869, p. 823—824.
Hancock D. A. Graphical estimation of growth parameters. — J. Conseil
perman. intern, explor. met., 1965, vol. 29, N 3, p. 340—351.
Haranghy L., Balazs A., Burg M. Determination of age and histological exami-
nation of the biomorphosis of the gonads in mussels. — Intern. Conf. Geron-
tol., Budapest, 1965, p. 117—123.
Harding J. P. The use of probability paper for the graphical analysis of poly-
modal frequency distribution. — J. Mar. Biol. Ass. U. K., 1949, vol. 28,
N 1, p. 141—153.
Harley J., Wright C. A. G., Nixon C. W. Preliminary observations on reprodu-
ction in the molluscan. — Nautilus, 1947, vol. 61, N 1, p. 6—11/
Harrison A. D., Schiff C. J. Factors influencing the distribution of some species
of squatic snails. — South Afr. J. Sci., 1966, vol. 62, N 8, p. 253—258.
Haskin H. U. Age determination in molluscs. — Trans. N. Y. Acad. Sci., 1954,
vol. 16, N 6, p. 300—304.
Haukiofa H., Hakala T. Vertical distribution of freshwater mussels (Pelecy-
poda, Unionidae) in southwestern Finland. — Ann. zool. Fenn., 1974,
vol. 11, N 2, p. 127—130.
Haukiofa E., Hakala T. Measuring growth from shell rings in population
of Anodonta piscinalis (Pelecypoda, Unionidae). — Ann. Zool. Fenn.,
1978, vol. 15, N 1, p. 60—65.
Heard W. H. Litter size the Sphaeriidae. — Nautilus, 1964, vol. 78, 'N 2,
p. 47—49.
Heard W. H. Comparative life histories of North American Pill clams (Sphaeri-
idae: Pisidium). — Malacologia, 1965, vol. 2, N 3, p. 381—411.
Hemmingsen A. M. The relation of standart (basal) energy metabolism to total
fresh weight of living organisms. — Rep. Stenool. memor. Hospital, Copench.,
1960, vol. 4, p. 7—58.
Hendelberg I. The freshwater pearl mussel Margaritifera margaritifera (L.). —
Rep. Inst. Freshw. Res. Drottningholm, 1960, vol. 41, p. 149—171.
Herrington H. B. Further proof that Sphaerium occidentale does not attain full
growth in one year. — Canad. Field-Natur., 1948, vol. 62, N 2, p. 74—75.
Hers M. J. Relation entre respiration et circulation chez Anodonta cygnea.
L. — Ann. Soc. Roy. Zool. Belg., 1943, vol. 74, p. 45—54.
Hiestand W. A. Respiration studies with freshwater molluscs. II. Oxygen con-
sumption in relation to Hydrogen-ion concentration. — Proc. Ind. Acad.
Sci., 1938, sect. B, vol. 47, p. 223—298.
Higashi S. The respiration of Principal Molluscs in Lake Biwako. — Venus,
1965, vol. 23, N 4, p. 229—237.
Higashi S. The rate of feeding and digestibility of the freshwater mussel Hyri-
opsis schlegellii determined with crude silicate as an indicator. — Venus,
1966, vol. 25, N 1, p. 43—46.
Hillbricht-Ilkowska A., Stanczykowska A. The production and standing crop
of planctonic larvae of Dreissena polymorpha Pall, in two Mazurian lakes. —
Pol. arch, hydrobiol., 1969, vol. 16 (29), N 2, p. 193—203.
Hinz W., Scheil H. G. Zur Filtrationsleistung von Dreissena, Sphaerium und
Pisidium (Eulamellibranchiata). — Oecologia, 1972, vol. 11, N 1, p. 45—54.
Hiscock I. D. Osmoregulation in australian freshwater mussels (Lamellibranchi-
ata). II. Respiration and its relation to osmoregulation in Hyridella austra-
lis (Z.). — Austr. J. Mar. and Freshw. Res., 1953, vol. 4, N 2, p. 330—332.
Holopainen I. J. Ecology of Pisidium (Bivalvia, Sphaeriidae) populations in an
oligotrophic and mesohumic lake. — Publ. Univ. Joensuu., 1978, ser. Bn,
N 9, p. 1—12.
Holopainen I. J. Population dynamics and production of Pisidium species
(Bivalvia, Sphaeriidae) in the oligotrophic and mesohumic lake Paajarvi,
240
Southern Finland. — Arch. Hydrobiol., 1979, suppl. 54, N 4, p. 1—
23.
Holopainen I. J., Ranta E. Carbon dioxide output in the respiration of three
Pisidium species (Bivalvia, Sphaeriidae). — Oecologia (Berl.), 1977, vol. 30,
N 1, p. 1-8.
Hopkins A. E. Experiments on the feeding behavior of the oyster Ostrea gigas. —
J. Exp. Zool., 1933, vol. 64, N 3, p. 469—494.
Hopkins A. E. Temperature optima in the feeding mechanism of the oyster,
O. gigas. — J. Exp. Zool., 1935, vol. 71, N 3, p. 333—344.
Hopkins H. S. The influence of season, concentration of sea water and environ-
mental temperature upon the oxygen consumption of tissues in Venus mer-
cenaria. — J. Exp. Zool., 1946, vol. 102, N 2, p. 143—158.
Howard A.D. Experiments in the culture of freshwater mussels. — Bull. Bur.
Fish., 1922, vol. 38, N 916. 26 p.
Howe R. W. The rapid determination of the intrinsic rate of increase of an insect
population. — Ann. Appl. biol., 1953, vol. 40, N 1, p. 134—151.
Hughes R. N. A study of feeding in Scrobicularia plana. — J. Mar. Biol. Assoc.
U. K., 1969, vol. 49, N 3, p. 805—823.
Hughes R. N. Population dynamics of the Bivalve Scrobicularia plana (De Costa)
an intertidal mudflat in North Wales. — J. Anim. Ecol., 1970, vol. 39, N 2,
p. 333-356.
Hughes R. N. Ecological energetics of the keyhole limpet Fissurella barbadensis
Gmelin. — J. Exp. Mar. Biol., Ecol., 1971a’, vol. 6, N 3, p. 167—178.
Hughes R. N. Ecological energetics of Nerita (Archaegastropoda, Neritacea)
populations on Barbados, West Indies. — Mar. Biol., 1971b, vol. 11, N 1,
p. 12—22.
Huxley I. S. Problems of Relative Growth. London; Methuen, 1932. 276 p.
Janisch E. Das exponetialgesetz als Grundlage einer vergleichenden Biology.
Berlin (Springer), 1927. 383 p.
Jorgensen С. B. On the water transport through the gills of bivalves. — Acta
Physiol. Scand., 1943, vol. 5, p. 297—304.
Jorgensen С. B. Quantitative aspects of filter feeding in invertebrates. — Biol.
Rev., 1955, vol. 30, N 4, p. 391—459.
Jorgensen С. B. Efficiency of particle retention and rate of water transport in
undisturbed lamellibranchs. — J. Conseil perman. intern, explor. mer.,
1960, vol. 26, N 1, p. 94—116.
J drgensen С. B. Biology of suspension feeding. Oxford, 1966. 357 p.
Kajak Z., Hillbricht-Ilkowska A., Pieczynska E. The production processes in
several Polish lakes. — In: Proceedings of the IBP—UNESKO symposium
of productivity problems of freshwater. Kazimierz Dolny, Poland, May 6—12,
1970. Warszawa; Krakow, 1972, p. 129—147.
Kajak Z., Lawacz W., Wisniewski R. J. et al. Ecosystem of the Mikolajskie lake.
The fate of organic matter of profundal zone.— Pol. arch, hydrobiol., 1975,
vol. 22, N 1, p. 89—99.
Kamler E. A comparison of the closed-bottle and flowingwater methods for
measurements of respiration in aquatic animals. —Pol. arch, hydrobiol.,
1969, vol. 16, N 1, p. 121—144.
Kinne O. Temperature. Animals Invertebrate. — Mar. Ecol., 1970, vol. 1,
pt 1, p. 407—517.
Klekowski R. Z. Bioenergetic budgets and their application for estimation of
production efficiency. — Pol. arch, hydrobiol., 1970, vol. 17, N 1—2,
p. 55—80.
Klekowski R. Z., Fischer E., Fischer Z. et al. Energy budgets and energy trans-
formation effeciencies of several animal species of different feeding types. —
In: Proceedings of the IBP—UNESCO symposium of productivity problems
of freshwater. Kazimierz Dolny, Poland, May 6—12, 1970. Warszawa;
Krakow, 1972, p. 749—763.
Krogh A. The quantitative relation between temperature and standart meta-
bolism in animals. — Intern. Z. Physikchem., Biol., 1914, vol. 1, p. 491—
508.
241
Kruger F. Zur Frage der GrossenabhSngigkeit des Sauerstoffverbrauchs von
Mytilus edulis L. — Helgoland, wiss. Meeresuntersuch., 1960, Bd 7, H. 3,
S. 125—148.
Kuenzler E. /. Structure and energy flow of a mussel population in a Georgia
salt marsh. — Limnol. and Oceanogr., 1961, vol. 6, N 2, p. 191—204.
Ladle M., Baron F. Studies on three species of Pisidium (Molluscs: Bivalvia)
from a chalk stream. — J. Anim. Ecol., 1969, vol. 38, N 2, p. 407—413.
Lagerspetz K. Y. H., Terri R. Transmitter substances and temperature acclima-
tion in Anodonta (Pelecypoda). — Ann. Zool. Fenn., 1968, vol. 5, N 4,
p. 396—400.
Leveque C. Bilans energetiques des populations naturelles de mollusques benthi-
ques du lac Tchad. — Gahiers ORSTOM. Ser. Hydrobiol., 1973, vol. 7,
N 3-4, p. 151—165.
Lewandowski K., Stanczykowska A. The occurrence and role of bivalves of the
family Unionidae in Mikolajskie Lake. — Ekologia polska, 1975 (1976),
vol. 23, N 2, p. 317—334.
Little I. W., Centner H. W. Growth of Amblema perplicata Conrad (Pelecypoda)
in Texas river. — Nautilus, 1970, vol. 84, N 1, p. 16—21.
Lomte V. S., Nagabhushanam R. Studies on the respiration of freshwater mussel
Parreysia corrugata. — Hydrobiologia, 1971, vol. 38, N 2, p. 239—246.
Loosanojj V. L. On the food selectivity of oysters. — Science, 1949, vol. 110,
p. 122—125.
Loosanojj V. L. Some aspects of behaviour of oyster at different temperatures. —
Biol. Bull., 1958, vol. 114, N 1, p. 57—70.
Loosanojj V. L., Engle J. B. Effect of different concentrations of microorganisms
on the feeding of oysters (O. virginica). — Fish. Bull. Fish, and Wildlife
Serv., 1947, vol. 51, N 42, p. 31—57.
Lotka A. J. Elements of Physical Biology. Baltimora, 1925. 460 p. [Rep. Dover
PubL, New York, 1956].
Ludwig W., Krywienczyk J. Korpergrosse, Korperzeiten und Energiebilanz. III.
Mitteilung. Der Sauerstoffverbrauch von Muscheln in Abhangigkeit von
der Korpergrosse. — Z. vergl. Physiol., 1950, Bd 32, H. 5, S. 464—467.
Lukacsoi icz F. Hypoxial examination of Anodonta cygnea L. on the O.2-consump-
tion of gill-tissues and relation between body dimensions and respiration
on the gill-tissue. — Ann. biol. Tihany, 1966, vol. 33, p. 79—94.
Lutz R. A. Annual growth patterns in the inner shell layer of Mytilus edulis L. —
J. Mar. Biol. Assoc. U. K., 1976, vol. 56, N 3, p. 723—731.
Macan T. T> Ecology of freshwater mollusca in England lake district. — J.
Anim. Ecol., 1953, vol. 19, N 1, p. 124—146.
Mackie G. L., Qadri S. U., Clarke A. H. Intraspecific variations in growth,
birth periods and longevity of Musculium securis (Bivalvia: Sphaeriide)
near Ottawa, Canada. — Malacologia, 1976a, vol. 15, N 2, p. 433—446.
Mackie C. L., Qadri S. U., Clarke A. H. Reproductive habits of four populations
of Musculium securis (Bivalvia: Sphaeriide) near Ottawa, Canada. — Nau-
tilus, 1976b, vol. 90, N 2, p. 76—86.
Magnin E., Stanczykowska A. Quelques donnees sur la croissance, la biomasse
et la production annuelle de trois mollusques Unionidae de la region de
Montreal. — Can. J. Zool., 1971, vol. 49, N 4, p. 491—497.
Mangapathi R. K., Ramachandra R. P., Canti S. S., Kalyanasundaram N.
Studies on respiration in relation to body size and oxygen tension in the
pelecypod Congeria sallei Recluz. — Proc. Ind. Acad. Sci., 1974, ser. B,
vol. 80, N 4, p. 163—171.
Mann К. H. The pattern of energy flow in the fish and invertebrate fauna of the
River Thames. — Verh. Intern. Ver. theoret. und angew. Limnol., 1964,
vol. 15, pt 1, p. 485—495.
Mann К. H., Britton R. H., Kowalczewski A., Lack T. J., Mathews Ch. P.
McDonald I. Productivity and energy flow at all trophic levels in the river
Thames, England. — In: Proceedings of the IBP—UNESKO symposium of
productivity problems of freshwaters. Kazimierz Dolny, Poland, May 6—12,
1970. Warszawa; Krakow, 1972, p. 579—596.
242
McNeill S., Lawton J. H. Annual production and respiration in animal popula-
tions. — Nature, 1970, vol. 225, N 5231, p. 472—474.
Meier-Brook C. Untersuchungen zur Biologie einiger Pisidium-Arten (Mollusca:
Eulamellibranchiata: Sphaeriidae). — Arch. Hydrobiol., 1970, Bd 39,
Suppl., N 1—2, S. 73—150.
Morton B. S. Studies on the biology of Dreissena polymorpha Pall. III. Popula-
tion dynamics. — Proc. Malacol. Sec. London, 1969 a, vol. 38, pt 6, p. 471—
482.
Morton B. S. Studies on the biology of Dreissena polymorpha Pall. II. Correla-
tion of the rhythms of adductor activity, feeding, digestion and excretion. —
Proc. Malacol. Soc. London, 1969b, vol. 38, pt 5, p. 401—414.
Morton B. S. Studies on the biology of Dreissena Polymorpha Pall. V. Some
aspects of filter-feeding and the effect of microorganisms upon the rate of
filstration. — Proc. Malacol. Soc. London, 1971, vol. 39, pt 4, p. 289—301.
Morton B, S. Analysis of a simple of Corbicula manilensis Philipi from the pearl
river. — China. Malacol. Rev., 1973, vol. 6, N 1, p. 35—37.
Morton B. S. The population dynamics of Corbicula fluminea (Bivalvia: Corbi-
culacea) in Plover Cove Reservoir, Hong Kong. — J. Zool. London, 1977,
vol. 181, pt 1, p. 21—42.
Nagabhushanam R., Sarojini R. Rate of water propulsion of Mujinia lateralis
(Say) in relation to salinity and temperature (Mollusca: Lamelli branchi-
ataj. — Sci. and Cult., 1965a, vol. 31, N 8, p. 439—439.
Nagabhushanam R., Sarojini R. The rate of water propulsion by the mussel,
Branchidontes recurvus (Mollusca: Lamellibranchiata). — Ind. J. Physiol,
and Allied Sci., 1965b, vol. 19, N 1, p. 1—4.
Nardi P. R. Dati sperimentali sulla productivity di Unio (Lamellibranchiata). —
Boll. Mus. Ven., 1978, vol. 29, suppl., p. 201—202.
Negus Ch. L. A quantitative study of growth and production of unionid mussels
in the river Thames of reading. — J. Anim. Ecol., 1966, vol. 35, N 3,
p. 513—532.
Newell R. C. Oxidative activity of poikiloterm mitochondria as a function of
temperature. — J. Zool. London, 1967, vol. 151, pt 3, p. 299—311
Newell R. C., Johnson L. G., Kofoed L. H. Adjustment of the components of
energy balance in response to temperature change in Ostrea edulis.— Oecolo-
gia (Berl.), 1977, vol. 30, N 2, p. 97-110.
Nomura E. Further studies on the applicability of a—kbx in expressing the growth
relations in Molluscan Shells. — Sci. Repts. Tohoku Univ., Biol., 1926a,
vol. 2, N 1, p. 63—85.
Nomura E. On application of a=kbx in expressing the growth relation in the
freshwater Bivalve, Sphaerium heterodon Pils. — Sci. Repts. Tohoku Univ.,
Biol., 1926b, vol. 2, N 1, p. 57—62.
Odhner N. H. On some species of Pisidium in the Swedish State Museum. — J.
Conchology, 1921, vol. 16, N 7, p. 218—223.
Okada K. Some notes on Musculium heterodon (Pirsbry) a freshwater bivalve.
III. Fertilization and segmintation. — Sci. Repts. Tohoku Univ., Biol.,
1935, vol. 9, N 4, p. 367—483.
Okland J. Notes on population density, age distribution, growth, and habitat
of Anodonta piscinalis Nilss (Mollusca; Lamellibranchiata) in a euthrophic
Norwegian Lake. — Nytt. mag. Zool., 1963, vol. 11, p. 19—43.
Ornatowski Z. Oxygen consumption several species of freshwater mussels (Bi-
valvia). — Bull. Soc. Amis Sci. et Lett. Poznan, 1967, ser. D, livr. 8,
p. 143-148.
Ortmann A. E. Correlation of shape and station in freshwater mussels (Naiades).—
Proc. Amer. Phil. Soc. Philadelphia, 1920, vol. 59, N 4, p. 269—312.
Pamatmat M. M. Seasonal respiration of Transennella tantilla Gould. — Amer.
Zoologist, 1969, vol. 9, N 2, p. 419—426.
Paul A. Z. Rate of water transport by Brachidontes exustus. — Quart. C. Fla
Acad. Sci., 1971, vol. 34, N 2, p. 125—130.
243
Pico? C, A,, Cucerzan M. Contributions to the study of the energetic metabolism
of Mollusca. — Rev. Roumaine Biol. Ser. Zool., 1967, vol. 12, N 3, p. 195—
201.
Prosser C. L. Physiological variation in animals. — Biol. Rev., 1955, vol. 30,
N 3, p. 229—262.
Quayle D. B. Pacific oyster culture in British Columbia. — J. Fish. Res. Board
Canada, 1969, vol. 169. 193 p.
Rao К. P. Rate of water propulsion in Mytilus californianus as a function of
latitude. — Biol. Bull., 1953, vol. 104, N 2, p. 171—181.
Rao К. P., Bullock T. H. Qm as function of body size and habital temperature. —
Amer. Naturalist, 1954, vol. 88, N 838, p. 33—44.
Rawson D. S. The bottom fauna of Great Slave lake. — J. Fish Res. Board
Canada, 1953, vol. 10, N 8, p. 486—520.
Read L. B. The Pelecypoda of Dallas country. — Texas. Field and Lab., 1954,
vol. 22, N 2, p. 35—52.
Rice T. R., Smith G. J. Filtering rates of the hard clam (Venus mercenaria)
determined with radioactive phytoplankton. — Fish. Bull. Fish, and Wild-
life Serv., 1958, vol. 58, N 129, p. 73—82.
Riffenburgh R. H. On growth parameter estimation for early life stages. — Bio-
metrics, 1966, vol. 22, N 1, p. 162—178.
Rubbel A. Beobachtungen iiber das Wachstum von Margaritana margaritifera. —
Zool. Anz., 1913, Bd 46, S. 156—161.
Salanki J. Contributions to the problem of daily rhythm in the activity of the
freshwater mussel Anodonta cygnea L. — Ann. Biol. Tihany, 1964, vol. 31,
p. 109—116.
Salanki J. Oxygen level as a specific regulator in the rhythmic activity of
freshwater mussel (Anodonta cygnea L.). — Acta biol. Acad. Sci. Hung.,
1965, vol. 15, N 3, 299—310.
Salanki J., Vero M. Diurnal rhythm of activity in freshwater mussel (Anodonta
cygnea) under natural conditions. — Ann. Biol. Tihany, 1969, vol. 36,
p. 95-107.
Schalie H. Hermoprhoditism among North American freshwaters mussels. —
Malacologia, 1970, vol. 10, N 1, p. 93—112.
Schiff C. J. Studies on Bulinus (Physopsis) globosus in Phodesia. I. The Influence
of temperature on the intrinsic rate pf natural increase. — Ann. Trop. Med.
and Parasitol., 1964a, vol. 58, N 1, p. 94—105.
Schiff C. J. Studies on Bulinus (Physopsis) globosus in Phodesia. III. Bionomics
of a natural population existing in a temporary habitat. — Ann. Trop. Med.
and Parasitol., 1964b, vol. 58, N 2, p. 240—255.
Schiff C. J., Garnett B. The influence of temperature on the intrinsic rate of na-
tural increase of freshwater snail Biomphalaria pfeiferi (Krauss) (Pulmonatar
Planorbidae). — Arch. Hydrobiol., 1966, Bd 62, H. 4, p. 429—438.
Schnitter H. Die Najaden der Schweiz mit besonderes Beriicksichtigung der
Umgebung Basels. — Z. Hydrologie, 1922, Bd 2, Suppl. 202 S.
Scholander P. F., Flagg V., Walters V., Irving L. Climatic adaptation in arctic
and tropical poikiloterms. — Physiol. Zool., 1953, vol. 26, N 1, p. 67—92.
Schwarzkopf J., Wesemeier H. Zur Frage der AbhSngigkeit des Sauerstoffver-
brauchs vom Korpergewicht bei Gastropoden und Bivalviern. — Natur-
wissenschaften, 1959, Bd 46, N 8, S. 272—273.
Seed R. The ecology of Mytilus edulis L. (Lamellibranchiata) on exposed rocky
shores. II. Growth and mortality.— Oecologia, 1969, vol. 3, N3—4,p.317—
350.
Segerstrdle S. C. Investigations on Baltic populations of the bivalve Macoma
baltica (L.) Pt I. Introduction. Studies on recruiment and its relation to
depth in Finnish coastal waters during the period 1925—1959. Age and
growth. — Soc. Sci. Fenn. Comment. Biol., 1960, vol. 23, N 2, p. 1—72.
Sivertsen B. The bottom fauna of Lake Huddingsvath, based on quantitative
sampling. — Norw. J. Zool., 1973, vol. 21, N 4, p. 305—321.
Solem J. O. The bottom fauna of lake Lille-Jonsvann, Trondelag, Norway. —
Norw. J. Zool., 1973, vol. 21, N 3, p. 227—261.
244
Soszka C. Selected problems of the ecology of molluscs (Mollusca) of the brackish
lake Lebzko. — Ekologia Polska, 1968, ser. A, t. 16, N 37, p. 729—753.
Sparck R. On the relation between metabolism and temperature in some marine
lamellibranches and its zoogeographical significance. — Biol. Medd., 1936,
vol. 13, N 5, p. 1—27.
Stanczykowska A. Analysis of the age of Dreissena polymorpha Pall, in the
Masurian Lakes. — Bull. Acad. Polon. Sci. Ser. sci. biol., 7003, vol. 11,
N 1, p. 29—33.
Stanczykowska A. Ecosystem of the Mikolajskie lake. Regularities of the Dreis-
sena polymorpha Pall. (Bivalvia) occurrence and its function in the lake. —
Pol. Arch. Hydrobiol., 1975, vol. 22, N 1, p. 73—78.
Stanczykowska A., Lawacz W. Caloric value of the Dreissena polymorpha (Pall)
dry body weight in some Mazurian lakes. — Pol. Arch. Hydrobiol., 1976,
vol. 23, N 2, p. 271—275.
Stanczykowska A., Lawacz W., Mattice J., Lewandowski K. Bivalves as a factor
effecting circulation of matter in lake Mikolajskie (Poland). — Limnolo-
gica, 1976, Bd 10, H. 2, p. 347—352.
Stasek C. R. Geometrical form and gnomic growth in the bivalved mollusca. —
J. Morphol., 1963, vol. 112, N 3, p. 215—234.
Stern G. Bilan energetique de la limace Arion rufus (Mollusque Pulmone) en
periode de croissance. — C. r. Acad. Sci., D, 1969, vol. 269, N 11, p. 1015—
1018.
Stickney A. P. Feeding and growth of juvenile soft-shell clams Mya arenaria. —
Fish. Bull., 1964 , vol. 63, N 3, p. 635—642.
Strober Q, J. Distribution and age of Margaritana margaritifera (L.) in a Madison
river (Montana, U. S. A.) Mussel bed. — Malacologia, 1972, vol. 11, N 2,
p. 342—350.
Surber T. Identification of the glochidiae of freshwater mussels. — Bur. Fish.,
1912, Doc. N 771, p. 1—10.
Tammes P. M. L., Drall A. D. C. Observations on the straining of suspension
by mussels. — Arch, neerl. Zool., 1955, vol. 11, N 1, p. 87—112.
Tang P. S. On the rate of oxygen consumption by tissues and lower organisms
as a function of oxygen tension. — Quart. Rev. Biol., 1933, vol. 8, N 3,
p. 260—274.
Taylor С. C. Temperature and growth — the Pacific razor clam. — J. Conseil
perman. intern, explor. mer., 1959, vol. 25, N 1, p. 93—101.
Teal J. M. Community metabolism in a temperature cold spring. — Ecol.
Monogr., 1957, vol. 27, N 3, p. 283—302.
Tenore K. R., Dunstan W. M. Comparison of feeding and biodeposition of three
bivalves at different food levels. — Mar. Biol., 1973, vol. 21, N 3, p. 190—
195.
Theede H. Experimentelle Untersuchungen fiber die Filtrarionsleistung der
Miesmuschel M. edulis(L.).—Kiel. Meeresforsch.,IPd5, Bd 19, H. 1, S. 20—41.
Theisen B. F. Growth and mortality of culture mussels in the Danish Wadden
Sea. — Medd. Damn. fisk.-og havundersog., 1967—1969, Bd 6, N 1—4,
S. 47—48.
Thiel M. E. Formwachstumsversuche an Sphaerium corneum Wilhelm Roux. —
Z. wiss. Biologie, 1926, Bd 108, H. 1, S. 87—137.
Thomas G. J. Self-fertilization and production of young in a Sphaeriid clam.—
Nautilus, 1959, vol. 72, N 4, p. 131—140.
Thomas G. J. Growth in one species of sphaeriid clam. — Nautilus, 1965, vol. 79,
N 2, p. 47—54.
Thompson D. A. W. On growth and form. Cambridge, 1959, vol. 1—2. 793 p.
Thorson G. Reproduction and larval development of Danish Marine bottom in-
vertebrates. — Medd. Danm. fisk-og havundersog. Ser. Plankton, 1946,
Bd 4, N 1. 523 S.
Tilly L. J. The structure and dynamics of Cone Spring. — Ecol. Monorg., 1968,
vol. 38, N 2, p. 169—197.
Tudorancea CL Elemente der Dynamik der Populationen von Unio pictorum und
Unio tumidus in Jijila Flachsee (Uberschwemmungsgebiet der Donau). —
245
In: Limnol. Вег. 10. JubilSumstag. Arbeitsgemeinsch. Donauforsch. Bulga-
rian, 1966. Sofia, 1968, S. 269—275.
Tudorancea Cl. Comparison of the populations of Unio tumidus Philipsson from
complex of Crapina-Jijila marshes. — Ecologia polska, 1969, ser. A, t. 17,
N 11, p. 185—204.
Tudorancea Cl. Studies on Unionidae populations from the Crapina-Jijila com-
plex of pools (Dunabe zone liable to inundation). — Hydrobiologia, 1972,
vol. 39, N 4, p. 527-531.
1 uaorancea Cl Florescu M. Cu privere la fluxul energetic al populatiei de Unio
pictorum din balta Crapina. — An. Univ. Bucuresti. Ser. stiint. natur.
Biol., 1968a, An. 17, p. 233—243.
dudorancea Cl., Florescu M. Considerations concerning the production and
energetics of Unio tumidus Philipsson population from the Crapina marsh. —
Trav. Mus. Hist. Natur. C. r. Antipa, 1968b, vol. 8, pt 1, p. 395—409.
Tudorancea Cl., Florescu M. Aspecte ale productiei si energeticii populatiei
de Anodonta piscinalis Nilsson din balta Crapina (zona inundabila a Duna-
rii)* — St. cere. biol. Ser. zool., 1969, t. 21, N 1, p. 43—55.
Tudorancea Cl., Florescu M. Aspecte ale productiei si energetiei cii populatiei
de Anodonta piscinalis Nilsson din balta Crapina (zona inundabila) a Duna-
1 rii)* — St. cere. biol. Ser. zool., 1970, t. 22, N 4, p. 377—384.
Tudorancea Cl., Gruia Z. Observations on the Unio crassus Phillipson population
from the Nera River. — Trav. Mus. Hist. Natur. C. r. Antipa, 1968, vol. 8,
pt 1, p. 381—394.
Tyler A. V. Caloric values of some North Atlantic invertebrates. — Mar. Biol.,
1973, vol. 19, N 3, p. 258—261.
Urstn E. On the incorporation of temperature in the von Bertalanffy Growth
Equation. —Medd. Damn, fisk.-og havundersog., N. S.,7Pd5,Bd 4, N 1.
S. 1—16.
Vahl 0. Efficiency of particle retention in Mytilus edulis L. — Ophelia, 1972a,
vol. 10, N 1, p. 17—25.
Vahl 0. Particle retention and relation between water transport and oxygen
uptake in Chlamys opercularis (L.) (Bivalvia). — Ophelia, 1972b, vol. 10,
N 1, p. 67—74.
Vahl O. Pumping and oxygen consumption rates of Mytilus edulis L. of diffe-
rent sizes. — Ophelia, 1973a, vol. 12, N 1—2, p. 45—52.
Vahl O. Porosity of the gill, oxygen consumption and pumping rate in Cardium
edule (L.) (Bivalvia). — Ophelia, 1973b, vol. 10, N 2, p. 109—118.
Van Dam L. Uber die Atembewegungen und das Atemvolumen von Phryganea-
Larven, Arenicola marina und Nereis virens, sowie uber die Sauerstoffaus-
nutzung bei Anodonta cygnea, Arenicola marina und Nereis virens. — Zool.
Anz., 1937, Bd 118, S. 122—128.
Vernberg F. J., Vernberg W. B. Thermal influence on invertebrate respiration. —
Chesapeake Sci., 1969, vol. 10, N 3—4, p. 234—240.
Vooys C. G. N. The influence of temperature and time of year on the oxygen
uptake of sea mussel Mytilus edulis. — Mar. Biol., 1976, vol. 36, N 1,
p. 25—30.
Walford L. A. A new graphic method of describing growth of animals. — Biol.
Bull., 1946, vol. 90, N 2, p. 141 — 147.
Walne P. R. Observations on the influence of food supply and temperature on
the feeding and growth of larvae of Ostrea edulis L. — Fish. Investig. Lon-
don, 1965, ser. 2, vol. 24, N 1, p. 1—45.
P. R. Studies on the food value of nineteen genera of algae to juvenile
bivalvies of the genera Ostrea, Crassostrea, Mercinaria and Mytilus. — Fish.
Investig. London, 1970, ser. 2, vol. 26, N 5, p. 1—62.
Walz N. The energy balance of freshwater mussel Dreissena polymorpha. Pallas
in laboratory experiments and the lake Constance. I. Pattern of activity,
feeding and assimilation efficiency. — Arch. Hydrobiol., 1978a, Bd 55,
SuppL, H. 1, S. 83—105.
246
Walz N. The energy balance of freschwater mussel Dreissena polymorph Pallas
in laboratory experiments and the lake Constance. II. Reproduction. —
Arch. Hydrobiol., 1978b, Bd 55, Suppl., H. 1, S. 106—119.
Walz N. The energy balance of freshwater mussel Dreissena polymorpha Pallas
in laboratory experiments and the lake. Constance. III. Growth under stan-
dard conditions. — Arch. Hydrobiol., 1978c, Bd 55, Suppl., H; 2, S. 121—
141.
Walz N. The energy balance of freschwater, mussel Dreissena polymorpha Pallas
in laboratory experiments and the lake Constance. IV. Growth in lake Con-
stance. — Arch. Hydrobiol., 1978d, Bd 55, Suppl., H. 2, S. 142—156.
Weinland K. Beobachtungen uber der Gaswechsel von Anodonta cygnea L. —
Z. Biol., 1919, Bd 69, H. 1—2, S. 1—86.
Wesemeier H. Untersuchungen uber die Stoffwechselreduction. Ein intra- und
interspezifische Vergleich an 17 Molluskenarten. — Z. vergl. Physiol.,
1960, Bd 43, H. 1, S. 1—28.
Wesenb erg-Lund C. Ferskvandstaunaen, Biologisk belyst. Invertebrata. Koben-
havn, 1937, Bd 1—2. 837 S.
Wiktor J. Research on the ecology of Dreissena polymorpha Pall, in the Szcze-
cin Lagoon. — Ecologia polska, 1963, ser. A, t. 11, N 9, p. 275—280.
Wilbur К. M., Owen G. Growth. — In: Physiology of Mollusca; New York;
London, 1964, vol. 1, p. 211—237.
Willemsen J. Quantities of water pumped by mussels (Mytilus edulis) and cockles
(Cardium edule). — Arch, neerl. Zool., 1952, vol. 10, livr. 2, p. 153—160.
Winter J. E. Uber den Einfluss der Nahrungskonzentration und anderer Faktoren
auf Filterierleistung und Nahrungsausnutzung der Muscheln Arctica islandica
und Modiolus modiolus. — Mar. Biol., 1969, vol. 4, N 2, p. 87—135.
Woynarovich, E. The oxygen consumption of the Dreissena polymorpha (Lamelli-
branchiata) at different temperatures. — Ann. Biol. Tihany, 1961, vol. 28r
p. 211—216.
Yonge С. M. On some aspects of digestion in ciliary feeding animals. — J.
Mar. Biol. Assoc. U. K., 1926, vol. 20, N 2, p. 341—346.
Yonge С. M. Studies on Pacific coast mollusks. I. On the structure and adapta-
tions of Cryptomya califomica (Conrad). — Univ. Calif. Publ., Zool.r
1951, vol. 55, N 6, p. 421—438.
Zumoff С. H. The reproductive cycle of Sphaerium simile. — Biol. Bull., 1973,.
vol. 144, N 1, p. 212—228.
ОГЛАВЛЕНИЕ
Стр.
Предисловие .................................................. 3
Введение ..................................................... 6
Общие положения............................................... 6
Значение двустворчатых моллюсков в континентальных водоемах ... 9
Цели и материалы исследования................................ 11
Глава I. Скорость обмена у моллюсков......................... 14
Методы определения потребления кислорода гидробионтами....... 16
Зависимость скорости обмена у моллюсков от температуры....... 19
Сезонные изменения метаболизма............................... 27
Влияние насыщения воды кислородом............................ 29
Скорость обмена и общие размеры моллюсков.................... 34
Калорийность моллюсков . . г................................. 47
Участие двустворчатых моллюсков в деструкции органических ве-
ществ в водоемах..................................*....... 54
Глава II. Интенсивность питания моллюсков..................... 57
Методы определения скорости фильтрации........................ 59
Влияние факторов среды на скорость фильтрации воды моллюсками 64
Температура............................................. 64
Концентрация взвешенных в воде частиц.................... 68
Активная реакция среды.................................. 74
Ритмичность фильтрации....................................... 75
Скорость фильтрации и общие размеры моллюсков................ 77
Двустворчатые моллюски как природные биофильтры.............. 89
Глава III. Закономерности роста и особенности формы раковин
моллюсков ................................................... 93
Линейный рост................................................ 93
Влияние факторов среды на скорость роста моллюсков.......... 119
Окисляемость воды........................................ 120
Содержание кальция в воде.................................. 123
Скорость течения воды.................................... 130
Температурные условия.................................... 131
Аллометрический рост.......................................... 137
Весовой рост моллюсков...................................... 143
Размножение и плодовитость моллюсков........................ 154
Глава IV. Функционально-экологические характеристики популя-
ций двустворчатых моллюсков................................. 164
Энергетический баланс особи................................. 161
Динамика численности популяций моллюсков................... 17.3
Продукция популяций моллюсков.............................. is'i
Модель популяции моллюсков................................ Г.»-3
Энергетический баланс популяций.......................... ’*<>()
Заключение.................................................. '-1
Литература................................................ •'*'>