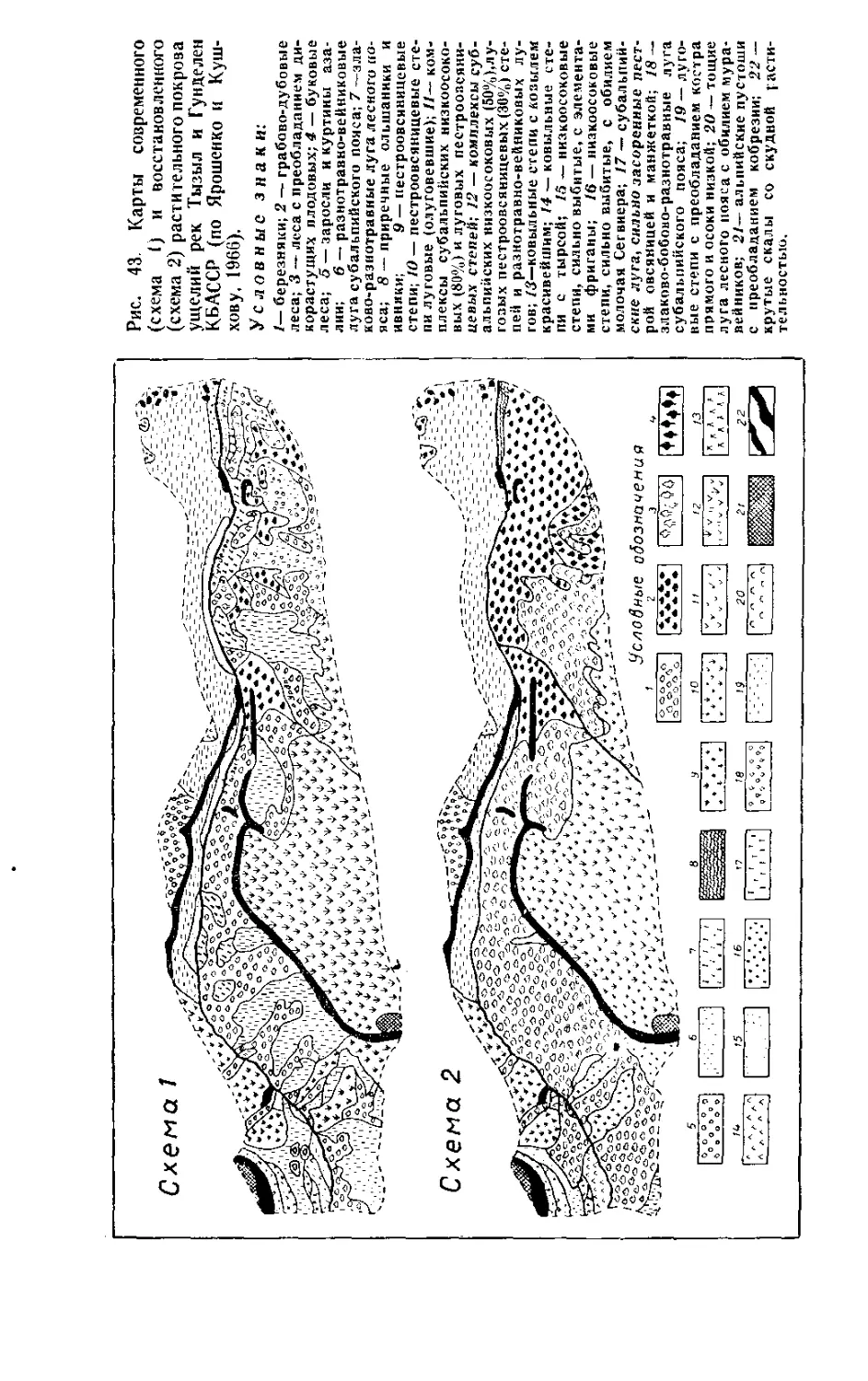
/












































































































































































































Text
П. Д. Ярошенко
ГЕ О Б О ТА НИ КА
Пособие для студентов
педагогических вузов
УТВЕРЖДЕНО МИНИСТЕРСТВОхЧ ПРОСВЕЩЕНИЯ
РСФСР
ИЗДАТЕЛЬСТВО «ПРОСВЕЩЕНИЕ» МОСКВА 19 6 9
Ярошенко П. Д.
Геоботаника. Пособие для студентов педвузов.
М., «Просвещение», 1969.
200 с. с илл.
Пособие предназначается для студентов биолого-химических, геог-
рафо-биологических и географических факультетов педагогических ин-
ститутов. В пособии изложены основы геоботаники, молодой современ-
ной науки, изучающей растительный покров, его состав, распределение,
видоизменения и многообразные связи его с другими элементами жи-
вой природы, с почвой и подстилающими ее породами.
2-10-5
23-69
581.5
П редисловие
Настоящий курс отличается от имеющихся
руководств, пособий и монографий по геоботани-
ке большей доступностью для студентов, впер-
вые изучающих этот предмет и не слушавших
лекций по экологии растений и ботанической гео-
графии.
В конце каждой главы приведены примерные
темы рефератов для семинаров, научных круж-
ков или курсовых работ. Последние большей
частью рассчитаны на сбор материалов в поле-
вых условиях, и многие из них могут служить в
дальнейшем темами дипломных работ. В ряде
случаев в пособии дана методика выполнения
курсовых работ, либо приведены списки литера-
туры в расчете на самостоятельную работу сту-
дентов.
Темы рефератов и курсовых работ подобра-
ны так, -хтобы расширить знания студентов, оз-
накомить их с более подробными руководствами
по геоботанике и с другой ботанической литера-
турой.
Автор
ЗАДАЧИ ГЕОБОТАНИКИ
И КРАТКИЙ ОБЗОР
ЕЕ ИСТОРИИ
Слово геоботаника означает в переводе земельная ботаника
(от греческого ге — земля). Специфика геоботаники состоит в
том, что объектами ее изучения являются не отдельные растения
в их отношениях к почве, климату и другим факторам среды,
а образуемые растениями группировки, или сообщества. По-
скольку сообщества растений называют также фитоценозами (от
греческих фитан — растение и койнос— общий), то геоботанику
называют также фитоценологией. Геоботаника изучает фитоце-
нозы в их связи с факторами среды: климатом, почвой, другими
фито ценозами, деятельностью животных, влиянием человека.
Геоботаника изучает не только отдельные растительные сообще-
ства, но и образуемый их сочетаниями растительный покров.
\ Фитоценоз, или растительное сообщество, — это совокупность
'растений, занимающих определенный, относительно однородный
участок и объединенных взаимодействиями со средой, а через
посредство среды и между собой. Сообщество само создает спе-
цифическую среду, которая является его неотъемлемой принад-
лежностью. Среда сообщества все время изменяется в процессе
его жизнедеятельности и вместе с тем, изменяясь, вызывает из-
менения в составе и распределении образующих сообщество
растений.
В. В. Алехин (1935; относил к фитоценозам лишь достаточно
устойчивые сочетания растений, обладающие способностью са-
мовосстановления, например степные фитоценозы, самовосста-
навливающиеся на старых залежах степной зоны. Все недоста-
точно устойчивые сочетания, еще не приобретшие способности к
самовосстановлению, В. В. Алехин называл не фитоценозами, а
группировками. Так, он не относил к фитоценозам заросли, обра-
зующиеся на песках в первых фазах их зарастания, культурные
посевы и т. д. Однако, следуя приведенному выше определению,
большинство геоботаников считает понятия «фитоценоз», «расти-
тельная группировка», а также «растительное сообщество» сино-
нимами. В том числе к фитоценозам, или сообществам
относятся, как это подчеркивал академик В. Н. Сукачев, даже
чистые от сорняков посевы полевых культур.^
Все взаимоотношения между растениями в сообществе осу-
ществляются через среду. Даже чисто механические взаимоот-
ношения, при которых растения теснят друг друга ветвями, ли-
ствой, корнями, неминуемо вызывают определенные изменения
в обмене веществ этих растений с окружающей их воздушной
и почвенной средой. Поэтому даже если механическое стеснение
не связано с затенением, оно обязательно в той или иной мере
отражается на фотосинтезе, дыхании, транспирации, поступлении
веществ из почвы, а также на росте и развитии растений, что, в
свою очередь, тесно связано с обменом веществ.
Еще более наглядна взаимозависимость растений при их взаи-
моотношениях с окружающей средой в таких явлениях, как за-
тененность одними растениями других, микориза (сожительство
высшего растения с грибом), бактериориза (сожительство высше-
го растения с бактериями) и другие.
Сообщество не простая сумма особей, оно обладает своими
специфическими качествами. Вместе с тем нельзя уподоблять
растительное сообщество животному организму или человече-
скому обществу. Растительное сообщество является чрезвычайно
своеобразной совокупностью: это и не организм, и не механизм,
и не социальная единица. По определению В. Н. Сукачева (1957)
«фитоценоз представляет собой лабораторию, в которой проис-
ходит аккумуляция и превращение вещества и энергии». Расти-
тельное сообщество живет и развивается согласно своим специ-
фическим закономерностям. Всестороннее изучение взаимодейст-
вий сообщества со средой, а также вмешательства человека в
эти взаимодействия, активное их изменение для пользы народ-
ного хозяйства и являются главными задачами геоботаники.
Можно различать внутреннюю среду сообщества и среду
внешнюю. Внутренняя среда сообщества является его неотдели-
мой частью, поскольку она в значительной мере создается самим
сообществом в процессе его жизнедеятельности: это прежде всего
свойственные .ему почва и внутренний климат (фитоклимат).
К факторам внешней среды относятся, например, приходящие
издалека холодные или теплые воздушные течения, речные раз-
ливы, массовые переселения насекомых, грызунов и других
животных, а также деятельность человека. Необходимо подчерк-
нуть, что провести резкую границу между внутренней и внешней
средой невозможно, и в процессе взаимосвязи сообщества с
внешней средой отдельные элементы последней переходят во
внутреннюю среду сообщества либо, наоборот, отдельные эле-
менты внутренней среды сообщества постоянно переходят во
внешнюю среду. Так, речной разлив, периодически переувлажняя
почву, изменяет внутреннюю среду сообщества. С другой сторо-
ны, само сообщество не может не влиять на скорость течения
5
воды, па специфику отложения наносимых ею частиц мелкозема
и т. д. Если же взять совокупность многих сообществ, т. е. весь
растительный покров данной местности в целом, то он может
оказывать значительное влияние и на само возникновение реч-
ных разливов, сдерживать их стремительность и т. д.
Растительное сообщество является частью так называемого
биоценоза — сообщества живых организмов, в котором взаимо-
действуют растения, животные и микроорганизмы. Вместе с тем
растительное сообщество нельзя представить себе иначе, чем в
его взаимоотношениях с окружающей средой, а также в его не-
отъемлемой связи с создаваемой им самим местной средой. Это
же положение целиком применимо и к биоценозу.
В. Н. Сукачев в целом ряде своих работ применяет термин
биогеоценоз, в котором частичка гео как раз и указывает на
взаимосвязь организмов биоценоза с неживой частью среды.
В. Н. Сукачев поясняет, что при выделении биогеоценозов в при-
роде целесообразно пользоваться фитоценозом (растительным
сообществом), иными словами, границы каждого в отдельности
биогеоценоза определяются границами фитоценоза. Это зависит,
в сущности, от того, что среди компонентов биогеоценоза фито-
ценозу принадлежит наибольшая, если так можно выразиться,
биогеоценозообразующая роль.
у Биогеоцеиологические исследования, естественно, должны
быть комплексными в полном смысле этого слова, в то время как
изучение биоценозов проводится нередко лишь силами зоологов
и ботаников либо одними из них и далеко не всегда сочетается
с разносторонним и глубоким исследованием среды. В изучении
биогеоценозов должны участвовать также и почвоведы, и кли-
матологи, и микробиологи, и гидрологи.
Опыт такого комплексного подхода к биогеоценозам пред-
ставляет собой вышедший в 1964 г. под редакцией В. Н. Сукаче-
ва и Н. В. Дылиса коллективный труд «Основы лесной биогеоце-
нологии».
» Будущее геоботаники, очевидно, в ее все более тесном кон-
такте с биогеоценологией. Правда, геоботаники всегда в той или
иной мере изучают среду фитоценоза, но при вовлечении геобо-
таников в комплексные биогеоценотические исследования изуче-
ние среды становится гораздо более разносторонним и глубоким,
а вместе с тем и еще более эффективным для народного хо-
зяйства.
Обратимся теперь к краткому обзору истории геоботаники.
Геоботаника зародилась в недрах ботанической географии,
изучающей закономерности географического распространения
отдельных видов растений и растительных сообществ. Отдельные
положения, впоследствии оказавшие немалое влияние на разви-
тие как ботанической географии в целом, так и геоботаники, мы
находим уже у ученых XVIII в. Сюда относятся высказывания
а
М: В. Ломоносова о происхождении чернозема вследствие гние-
ния растительных остатков, Бюффона о сменах пород в лесу и о
влиянии на этот процесс освещения и затенения, Вильденова,
который ввел термин ассоциация для естественных групп расте-
ний, и др. Однако работы с применением специальных геобота-
нических методов стали появляться лишь в первой половине
XIX в. Одним из таких первых исследований считают описание
растительности кантона Глярус в Швейцарии, опубликованное
Хеером в 1835 г. В этой работе уже были выделены и описаны
различные растительные сообщества.
В России еще более детальным исследованием такого рода
явилась работа Тициана, управляющего имением Аскания-Нова,
выполненная им в 1837 г. Изучая степную растительность, Тиц-
иан не только описывал пробные площадки, но и зарисовывал
покрытие растениями почвы, а также впервые применил зари-
совку профилей фитоценозов. Другой исследователь, колонист
Мелитопольского уезда Корнисс, использовав методику Тициана
при изучении степи, впервые применил весовой анализ траво-
стоя. Работы Тицмана и Корнисса были опубликованы Россий-
ской Академией наук, но долгое время оставались неизвестными.
Однако это были еще только эпизодические исследования.
Зарождение геоботаники как науки относится в России к
60-м годам прошлого столетия. Тогда в России бурно развивался
капитализм, ломая старые феодально-крепостнические устои.
Эти же годы ознаменовались началом деятельности таких круп-
ных ученых, как Д. И. Менделеев, К. А. Тимирязев, И. М. Сече-
нов, братья В. О. и А. О. Ковалевские, И. И. Мечников и др.
Значительное влияние на развитие науки в России оказывали
русские философы-материалисты: А. И. Герцен, Н. Г. Чернышев-
ский, Д. И. Писарев и др., а также опубликованная в 1859 г.
книга Ч. Дарвина «Происхождение видов».
К числу первых русских геоботаников того времени нужно
отнести акад. Ф. И. Рупрехта и бывшего некоторое время его
сотрудником, а впоследствии ставшего профессором Киевского
университета И. Г. Борщова.
Развитие капитализма стимулировало организацию широких
научных экспедиций для поисковгновых запасов сырья и рынков
сбыта. Борщов юыл участником одной из таких экспедиций, орга-
низованной Академией наук в 1857—1858 гг. на берег Араль-
ского моря и в бассейн Сырдарьи. Собранные материалы дали
возможность Борщову опубликовать в 1865 г. «Материалы для
ботанической географии Арало-Каспийского края». В этом пер-
вом капитальном труде по геоботанике мы еще не находим ни
самого термина «геоботаника», ни даже терминов «растительное
сообщество», «растительная формация» и т. д. Однако, описывая
частные, или областные, «флоры», на которые разделяется вся
флора Арало-Каспийской низменности, Борщов понимает под
7
этими областными «флорами» не что иное, как единицы расти-
тельного покрова; он различает области: ковыльной степи, гли-
нистых пустынь, соленых пустынь, бугристых песков, В пределах
области ковыльной степи Борщов выделяет флоры: травянистой
степи, каменистой степи, глинисто-гальковатой степи. Эти флоры
Борщова представляют собой, как отметил его биограф и попу-
ляризатор В. Б. Сочава, «зачаток современного представления о
растительной формации», т. е. об определенном типе раститель-
ных группировок, Борщов характеризует эти флоры в их тесном
взаимодействии с климатическими и почвенными условиями и,
что особенно ценно, рассматривает их в процессе исторического
формирования.
Через год после появления труда Борщова вышла знамени-
тая работа Рупрехта «Геоботанические исследования о черно-
земе». В ней Рупрехт впервые ввел термин геоботаника, понимая
под этим учение о взаимосвязи растительного покрова со средой,
Рупрехт еще не останавливается на мелких единицах раститель-
ного покрова — на отдельных растительных сообществах, пли
фитоценозах; он рассматривает целые растительные зоны и ланд-
шафты, или, как он их называет, флоры, в первую очередь черно-
земные степи в целом.
Рупрехт связывал историю степей не только с историей фор-
мирования чернозема как специфически степной почвы, но и с
геологической историей страны, понимая геоботанику как геоло-
гическую ботанику. Он установил связь распространения степной
растительности русской равнины не только с распространением
чернозема, но и с южной границей валунов, предполагая, что
валуны эти нанесены морем, а не ледниковым покровом.
Работы Борщова и Рупрехта явились в России первыми гео-
ботаническими исследованиями, выросшими на почве ботаниче-
ской географии как науки о закономерностях географического
распределения растений.
За границей выделение геоботаники как особой дисциплины
из ботанической географии стало намечаться, как и в России,
в середине XIX в. Шведский гляциолог Пост опубликовал в
1851 —1856 гг. работы, в которых обратил внимание на то, что
одни и те же виды растений могут расти в местах, довольно
сильно различающихся по почвенным и климатическим услови-
ям, Однако анализ их распространения показывает, что на раз-
ных местах они встречаются в неодинаковых количественных
соотношениях. Отсюда Пост пришел к признанию существования
групп растений как совокупностей, обладающих чертами опреде-
ленной целостности.
В числе первых геоботанических исследований методологи-
ческого значения следует отметить работу австрийского ботаника
Лоренца (1858) о высокогорных (предальпийских) болотах
Зальцбурга, Автор описывает не только болотную раститель-
8
ность в целом, но и мелкие группировки, на которые она расчле-
няется и которые соответствуют принятому нами понятию ми-
крогруппировок, или микрофитоценозов.
Существенным вкладом в развитие геоботаники явилась кни-
га другого австрийского ученого — Кернера (1863) «Жизнь
растений дунайских стран». Здесь автор, используя выделенные
Гумбольдтом жизненные формы (био-физиономические типы)
растений, подразделяет растительность на формации и дает
каждой из них довольно подробное описание.
В 1863 г. русский профессор агроном А. М. Бажанов опубли-
ковал в журнале министерства государственных имуществ
статью «Об искусственно возделываемых лугах», которая содер-
жит целый ряд положений геоботанической теории. Для разра-
ботки наилучших приемов создания искусственных лугов Бажа-
нов рекомендует предварительно рассмотреть «законы, по
которым действует в подобных случаях природа», и первая часть
его труда посвящена поэтому природе естественных лугов. Здесь
он рассматривает такие вопросы, как сложение естественных
луговых травостоев не одной, а несколькими группами растений,
главным образом злаками и бобовыми. Это разнообразие и яв-
ляется, по мнению Бажанова, причиной «продолжительной, почти
бесконечной производительности лугов». Еще не употребляя
терминов надземной и подземной ярусности, он образно сравни-
вает распределение растений лугов по высоте их роста с этажер-
ками, на которых так размещают горшечные растения, чтобы
все они равномерно обеспечивались светом, теплом, воздухом.
Рассматривая смены естественной растительности лугов, Бажа-
нов обосновывает этим необходимость плодосмена в земледелии.
С. П. Корелыциков опубликовал в 1865 г. в журнале «Сель-
ское хозяйство и лесоводство» статью «Луговые и сорные тра-
вы», в которой рассмотрены взаимоотношения между луговыми
травами, смены растительных сообществ и другие в полном
смысле слова геоботанические вопросы.
Таким образом, работники сельского хозяйства не только
ставили некоторые геоботанические проблемы, но и участвовали
в их разработке.
Значительное развитие получила геоботаника в России в 80-х
и особенно 90-х годах XIX и начале XX в. К этому периоду
относятся работы С. И. Коржинского, И. К. Пачоского,
П. Н. Крылова, А. Н. Краснова, Д. И. Литвинова, Г. И. Тан-
фильева, Г. Ф. Морозова, первые работы А. Я. Гордягина,
В. Н. Сукачева, Б. А. Келлера и др.
Большинство геоботанических работ этого периода, наряду
с ошибочным признанием в мире растений социальных явлений,
имело положительное значение. Именно в этих работах впервые
растительное сообщество рассматривалось не как простая сумма
особей, а как нечто большее, целое, обладающее . некоторыми
Рис. !. Долинный лиственный лес
в Приморском крас (Дальний Восток)
новыми качествами, присущими только всей совокупности осо-
бей. Эта точка зрения была развита с особенной полнотой в кни-
ге Г. Ф. Морозова «Учение о лесе» и отразилась также в знаме-
нитом эпиграфе к этой книге, заимствованном из «Оснований
лесоводства» Котта. В эпиграфе .лес сравнивается с часами, а
отдельные, составляющие ого растения, с пружинами и другими
деталями часового механизма. Правда, аналогия здесь весьма
грубая, так как лес, как и всякая другая группировка растении,
не механизм, но ценность этой аналогии для того времени была
в подчеркивании особых качеств леса как целого. На рисунке 1
изображен участок лиственного леса — определенная группиров-
ка растений, т. е. растительное сообщество.
Наряду с развитием прогрессивной идеи о сообществе как о
совокупности идет обособление учения о растительных сообще-
ствах в особую отрасль ботаники. Так, И. К. Пачоский в статье
«Стадии развития флоры» (1891) предложил выделить учение
о растительных сообществах в особую науку под названием
флорология, которая, в его понимании, есть паука о генезисе,
жизни, развитии и распространении растительных ассоциаций
(формаций). В этой же статье Пачоский, рассматривая взаимо-
отношения между пустыней, степью и лесом, высказывает мысль,
что генезис растительного покрова приводит к смене пустыни
IQ
степью, а степи лесом. Впоследствии Пачоский отказался от тер-
мина «флорология» и предложил в 1896 г. термин фитосоциоло-
гия, который затем применил и П. Н. Крылов (1898) в своем
«Очерке растительности Томской губернии». Последний термин
продержался в нашей науке недолго и был заменен термином
фитоценология, предложенным Гамсом (1918). Этот термин те-
перь употребляется часто как синоним термина геоботаника.
В. Н. Сукачев в своих первых геоботанических работах, как,
например, «О ботанико-географических исследованиях в Бузу-
лукском бору Самарской губернии» (1904), в очерке «Лесные
формации и их взаимоотношения в Брянских лесах» (1908), а
позднее в своей выдержавшей четыре издания книге «Раститель-
ные сообщества» также высказался за обособление этого учения
в качестве отдельной дисциплины, новой отрасли знания.
В некоторых геоботанических работах того времени чрезмер-
но переоценивалось значение борьбы за существование между
растениями и неодооценивалась взаимосвязь растений и их груп-
пировок с почвенно-климатическими условиями. К числу таких
работ относится монография С. И. Коржинского «Северная гра-
ница черноземностепной области восточной полосы европейской
России в ботанико-географическом и почвенном отношении»,
первый том которой вышел в 1888 г., а второй в 1891 г.
После Коржинского и Пачоского вопросом взаимоотношения
леса и степи на Русской равнине занимался Г. И. Танфильев,
изложивший свою точку зрения в работе «Пределы лесов на юге
России» (1894), где он отмечает надвигание леса на степь, но
связывает это в основном с выщелачивающим действием ку-
старниковой лесной опушки на богатую карбонатами степную
почву. Карбонатность степных почв, по мнению Танфильева,
является основной причиной безлесья наших степей. Позднее
Танфильев опубликовал работу «Пределы лесов в полярной
России по исследованиям в тундре тиманских самоедов» (1911),
где надвигание тундры на лес он объясняет опять-таки лишь
изменением почвенных условий, не учитывая изменений климата.
Работы Танфильева, и особенно его труд о взаимоотношении
леса и степи, вызвали оживленные дискуссии и появление цело-
го ряда других работ на эту же тему. Вообще нужно отметить,
что проблема леса и степи явилась одной из ведущих в развитии
русской геоботаники. Тогда в обсуждении этой проблемы прини-
мали горячее участие В. И. Талиев, П. Н. Крылов, Г. Н. Высоц-
кий и ряд других исследователей, а впоследствии ею занимались
В. Р. Вильямс, Л. С. Берг и многие другие ученые. К этому важ-
ному вопросу мы еще вернемся ниже, при изложении общих, или
вековых, смен растительного покрова.
Кроме оригинальных исследований, за описываемый период
стали появляться и некоторые научно-популярные работы по гео-
ботанике. К числу первых таких работ нужно отнести «Пособие
II
к изучению растительных сообществ Средней России» А. Флеро-
ва и Б. Федченко (1902). Содержание этой работы было шире,
чем значилось в ее названии, так как она, по существу, пред-
ставляла не только пособие к изучению сообществ, но и краткую
сводку основных положений, уже начавших к тому времени
укрепляться в геоботанике. Так, в этой книге развито положение
о теснейшей взаимосвязи растений сообщества с почвенно-кли-
матической средой и о том, что ни одно растительное сообщество
не может существовать, не изменяясь и не сменяясь другим сооб-
ществом. Кратко, но очень ярко описаны смены сообществ при
зарастании озер, заболачивании лесов, рассмотрены смены лес-
ных сообществ под влиянием деятельности человека, затронут
вопрос о взаимоотношении степи и леса и о возникновении лу-
гов на месте сведенного человеком леса. В работе показано, что
длительное существование лугов без смены их лесом обычно под-
держивается сенокошением или выпасом. С прекращением же
культуры луговые сообщества шаг за шагом уничтожаются и вы-
тесняются надвигающимися на них более мощными лесными со-
обществами. Последняя глава посвящена группе сообществ, свя-
занных с культурной деятельностью человека. Авторы совершен-
но правильно также относят культурные посевы к сообществам.
Ими последовательно проведена мысль о том, что жизнь расти-
тельного сообщества определяется в значительной степени кли-
матическими, почвенными и метеорологическими условиями.
В книге помещены программы исследования водных, болотных,
лесных сообществ и их смен, степных сообществ, растительности
обнажений, луговой растительности и, наконец, сорной расти-
тельности. В конце приложена классификация растительных со-
обществ.
Почти забытая и редко упоминаемая работа А. Флерова и
Б. Федченко тем не менее имеет историческое значение как пер-
вое не только в России, но и за ее предела-
ми учебное пособие по геоботанике.
Огромное значение для развития отечественной геоботаники
имели работы В. В. Докучаева, основоположника генетического
почвоведения. Докучаеву принадлежат такие классические тру-
ды, как «Русский чернозем» (1885), «Наши степи прежде и те-
перь» (1892) и многие другие. Докучаев разработал учение
о почве как продукте совокупной деятельности грунта, климата,
растительных и животных организмов, возраста страны и отча-
сти рельефа местности. Он считал это положение почвенной акси-
омой, краеугольным камнем и вернейшим залогом будущности
почвоведения как науки. Это докучаевское положение оказалось
очень важным для развития геоботаники, так как, помимо пра-
вильного представления о почве как неотъемлемой части фито-
ценоза, геоботаники стали рассматривать и сам фитоценоз как
результат совокупной деятельности природных факторов. И это
12
представление оказалось единственно правильным, В статье
«К учению о зонах природы» (1899) Докучаев рассматривает
зональность как следствие совокупного действия тех же основ-
ных природных факторов в их закономерном пространственном
изменении. Он намечает пять почвенных и вместе с тем естест-
венно-географических зон: тундровую, таежную, черноземную,
аэральную (лессовую и солонцовую) и красноземную (или лате-
ритную). Позднее ученик и последователь Докучаева Н. М. Си-
бирцев выделил еще черноземно-лесную зону, называемую те-
перь лесостепной, в то время как аэральная зона теперь разде-
ляется на полупустынную и пустынную зоны.
В этой же статье Докучаев указывает, что, несмотря на
гигантские успехи естествознания в XIX в., «нельзя- не заметить
одного весьма существенного и важного недочета... Изучались
главным образом отдельные тела — минералы, горные породы,
растения и животные — и явления, отдельные стихии — огонь
(вулканизм), вода, земля, воздух, в чем, повторяем, наука и до-
стигла удивительных результатов, но не их соотношения, не та
генетическая, вековечная и всегда закономерная связь, какая су-
ществует между силами, телами и явлениями, между мертвой и
живой природой, между растительными, животными и минераль-
ными царствами, с одной стороны, и человеком, его бытом и да-
же духовным миром — с другой. А между тем именно эти соот-
ношеная, эти закономерные взаимодействия и составляют сущ-
ность познания естества, ядро истинной натурфилософии — луч-
шую и высшую прелесть естествознания».
Изучая взаимодействия между почвой, климатом и раститель-
ностью, Докучаев пришел к мысли о возможности коренного
улучшения водного режима степной зоны. В книге «Наши степи
прежде и теперь» он посвятил специальную главу способам упо-
рядочения водного хозяйства в степях России, предлагая регули-
рование рек путем сужения и выпрямления их течений с целью
уменьшения весенних разливов; закрепление оврагов и балок
путем простых заградительных сооружений и древесных поса-
док; закладку на водораздельных степных пространствах прудов;
устройство рядов снегозадерживающих живых изгородей; лесо-
насаждение и др.
Докучаев рекомендовал организовать сеть опытных станций.
В 1892 г. после постигших страну тяжелых засух и неурожаев он
по предложению лесного департамента организовал экспедицию,
которая заложила несколько опытных участков в Каменной сте-
пи (теперь Таловский район Воронежской области), Старобель-
ской степи (теперь Луганская область) и Велико-Анадольской
степи (теперь Донецкая область). На всех трех участках были
уже в конце XIX в. заложены лесные полезащитные полосы. Ве-
лнко-Анадольский и Каменностепной участки существуют и в на-
стоящее время, причем в Каменной степи после Октябрьской ре-
13
Болюцип работы значительно расширились, и теперь там
создан Институт земледелия центрально-черноземной полосы
им. В. В. Докучаева.
На основе докучаевского представления о взаимодействии
всех природных факторов стало развиваться учение о ландшафте
как совокупности физико-географических процессов (Б. Б. Полы-
нов, 1925). Под ландшафтом при этом понимали естественно бо-
лее или менее обособленную часть природной зоны.
Из русских геоботаников едва ли не первым применившим
ландшафтный подход при изучении растительности был
Р. И. Аболин. Исследуя растительность болот, Аболин стал трак-
товать болото как определенный ландшафт или, как он называл,
эпитип. Болото, по Аболину, характеризуется пресыщенностью
в течение большей части года поверхностных слоев земной коры
водой. Это обилие влаги вызывает появление определенных ра-
стительных сообществ и особого почвенного процесса, а также
может приводить к накоплению неразложившихся растительных
(отчасти животных) остатков в виде торфа. Все это, вместе взя-
тое, влечет за собой и особый режим температуры в почве и ниж-
них слоях атмосферы, а также особый режим влажности возду-
ха. Вместе с тем и сама возможность обильного увлажнения суб-
страта определяется сочетанием природных факторов: климата,
рельефа, геологического строения местности и др. Указанная
трактовка болота как ландшафта изложена Аболиным в его по-
лучившей широкую известность работе «Опыт эпигенологической
классификации болот», опубликованной в журнале «Болотоведе-
ние» за 1914 г. В этой же работе Аболин развивает представле-
ние о так называемой эпигенеме как о совокупности всех ланд-
шафтов, образующих на поверхности земли одно сложное комп-
лексное целое, подразделяющееся на отдельные ландшафты, или
эпитипы.
В Советском Союзе геоботанические исследования разверну-
лись очень широко. Если в царской России эти работы проводи-
лись (да и то в ограниченных масштабах) преимущественно в
центральных районах Русской равнины и отчасти в Средней Азии
и Западной Сибири, то в советское время ими были охвачены все
уголки от Крайнего Севера и тихоокеанского побережья до Па-
мира и субтропиков Закавказья, потому что этого требовала не-
обходимость хозяйственного освоения и преобразования многих
территорий. Органи ация столь широких геоботанических иссле-
дований вызвала к жизни как применение целого ряда новых
методов, так и глубокую разработку теоретических проблем.
Запросы практики, неоднородные в различных частях обшир-
ной территории СССР, способствовали тому, что в советской гео-
ботанической школе складывались отдельные направления. Ча-
стично эти направления локализовались географически, как, на-
пример, закавказское и казанское. Другие направления опреде-
14
лялись в значительной мере тем, что их первые исследования
были посвящены какому-нибудь одному типу растительного по-
крова. Таково возглавляемое В. Н. Сукачевым направление, на-
зывавшееся прежде ленинградской школой, которое первона-
чально возникло при исследовании лесов, в частности, в работах
Г. Ф. Морозова. В Московском Университете оформилось направ-
ление, возглавлявшееся В. В. Алехиным и получившее название
московской школы. Свои первые шаги эта школа начала с иссле-
дования степей Европейской части СССР. Третье направление,
возглавлявшееся Л. Г. Раменским, возникло в Воронеже как чи-
сто луговое, а затем развивалось как широкогеоботаническое
в Институте кормов под Москвой, где впоследствии работал Ра-
менский.
Особенности каждого из этих трех направлений в сильной
мере определяются спецификой самого изучаемого объекта. Так,
основные единицы растительности — ассоциации — оказались бо-
лее крупными у ленинградского направления и более мелкими
у московского, где к тому же нередко выделялись и заимствован-
ные у шведов еще более мелкие — социации. Вместе с тем, рабо-
тая в степях, исследователи московской школы столкнулись с те-
ми весьма резкими различиями, которые наблюдаются в степной
зоне между сообществами: пионерами песчаных и глинистых об-
нажений и только что брошенных пашен, с одной стороны, и
сформированными степными сообществами — с другой. Отсюда
возникло представление о том, что не всякую комбинацию сов-
местно произрастающих растений можно назвать фитоценозом.
Как было упомянуто выше, В. В. Алехин предложил различать
группировки растений как менее выработавшиеся и фитоценозы
как более выработавшиеся, способные восстанавливаться на ме-
стах, где они были нарушены или уничтожены. Этот взгляд не
поддержали В. Н. Сукачев и другие представители ленинград-
ской школы, причисляющие к фитоценозам любые комбинации
совместно произрастающих растений, включая и культурные по-
севы. Этому последнему представлению следует, как уже упоми-
налось, большинство геоботаников. Ему будем следовать и мы
в нашем курсе.
Направление Л. Г. Раменского характеризуется перенесением
основного внимания при изучении сообществ на распределение
отдельных видов с широким использованием массового учета и
его статистической обработки. Такое направление могло возник-
нуть именно при изучении лугов, где сообщества менее резко от-
граничены и менее устойчивы, во-первых, в связи с неустойчи-
востью самой среды (в речных поймах) и, во-вторых, в связи
с вторичным происхождением многих луговых сообществ, возни-
кающих на местах вырубленного леса, не могущего снова восста-
новиться из-за ежегодного сенокошения или выпаса. Тщательное
изучение закономерностей в распределении на лугах отдельных
15
видов растений привело к разработке методики точных экологи-
ческих характеристик и даже экологических формул для каждого
вида растений, а также к изучению роли каждого вида в том или
ином сообществе с помощью проективного метода.
В лесах относительно легко обнаружить явление смены по-
род. Поэтому учение о сменах растительных сообществ получило
наиболее детальную разработку в ленинградской школе. Можно
сказать, что, Ито время как московская школа уделяла первенст-
вующее внимание географическим и топографическим измене-
ниям растительных сообществ, ленинградская школа имела
в центре внимания смены их во времени. Вместе с тем, сочетая
выявление такого рода смен с исследованием экологии сооб-
ществ, ленинградская школа предложила метод эколого-ценоти-
ческих рядов, использованный при изучении как лесов (В. Н. Су-
качев, С. Я. Соколов и др.), так и лугов (А. П. Шенников),
В. Н. Сукачев разработал детальную классификацию типов смен,
основанную главным образом на соотношениях «внутренних»
и «внешних» факторов в жизни растительных сообществ.
В московской школе исследование динамики сообществ ве-
лось в отношении не столько смен одних сообществ другими,
сколько сезонных изменений сообщества (смены аспектов). Так,
у В. В. Алехина мы находим превосходные красочные описания
смен аспектов Стрелецкой степи под Курском и других степных
участков.
В Казанском университете еще в конце прошлого столетия
оформилась геоботаническая школа, начало которой было поло-
жено работами С. И. Коржинского. Долгое время этой школой
руководил А. Я. Гордягин, а в настоящее время ее возглавляет
М. В. Марков, под руководством которого ведутся исследования
лесов, лугов и агрофитоценозов. Со времени А. Я. Гордягина
в работах этой школы выработались здоровые традиции: тща-
тельное и полное изучение видового состава сообществ и их
местообитаний, в особенности же режима увлажнения и связан-
ных с ним почвенных условий. В своей известной работе «Мате-
риалы к познанию почв и растительности Западной Сибири»
(1900) Гордягин, подчеркивая необходимость изучения раститель-
ного покрова в тесной связи с почвами, писал, что растительная
ассоциация, как все живое, довольно непостоянна и может под
влиянием изменившихся условий довольно быстро исчезнуть; но
если она существует на данной территории достаточно долго, то
она оставляет себе памятник, который в течение ряда веков
будет свидетельствовать о минувших условиях: памятник этот —
почва. Изучение почв помогает лучше понять не только совре-
менное распределение растительных сообществ, по и их рас-
пределение в прошлом, понять смены одних сообществ другими.
В отделе геоботаники Ботанического института им. В. Л. Ко-
марова Академии наук СССР (Ленинград) сформировалась шко-
16
ла геоботаников-картографов, возглавляемая в настоящее время
Е. М. Лавренко и В. Б. Сочавой. Заметим, что составление геобо-
танической карты нашей страны явилось одной из главных задач
отдела геоботаники уже с его возникновения, т. е. с 1922 г. Пер-
вым руководителем этой большой работы был Н. И. Кузнецов,
организовавший в 1923 г. составление геоботанической карты
Европейской части СССР в масштабе I : 1 050 000. Восемь листов
этой карты было издано в 1930—1932 гг., еще при жизни
Н. И. Кузнецова. Это издание явилось значительным событием
не только в советской, но и в мировой ботанической науке. В то
время за границей еще не было ни одной геоботанической кар-
ты такого масштаба, охватывающей столь обширную терри-
торию. Методическое значение карты возрастало потому, что
на ней были показаны и восстановленный (т. е. такой, каким он
был до влияния на него человека), и современный раститель-
ный покров.
Одновременно в те же годы разные учреждения начали со-
ставление большого числа преимущественно среднемасштабных
геоботанических карт. Все это дало возможность опубликовать
впоследствии ряд сводных карт, таких, как «Карта растительно-
сти Европейской части СССР» в масштабе 1 : 2 500 000 (1948),
«Геоботаническая карта СССР» в масштабе I ‘.4 000 000 (1954)
и др. Эти и другие геоботанические карты были положены в осно-
ву геоботанического и в целом природного районирования нашей
страны.
Первый широкий обмен мнениями между представителями
направлений советской геоботаники произошел на III Всесоюзном
съезде ботаников, состоявшемся в 1928 г. в Ленинграде. К этому
времени учение о растительных сообществах уже настолько опре-
делилось, что на съезде работала отдельная секция фитосоцио-
логии и экологии растений под председательством В. Н. Сука-
чева. Общим установкам этой новой науки было посвящено на
съезде и несколько специальных докладов. Наряду с этим были
высказаны мнения, отвергающие фитосоциологию как отдельную
дисциплину. Так, с очень резкой критикой фитосоциологии вы-
ступил В. И. Талиев. Он утверждал, что фитосоциология исполь-
зует обычные экологические методы исследования, что никакой
«социальной жизни» у растений не существует и что нет никакой
нужды выделять учение о растительных сообществах в отдель-
ную, обособленную от экологии науку.
После выступления Талиева сразу же определились мнения
по этому вопросу многих из членов съезда. Преобладающим,
если не единодушным, мнением было: если и можно говорить
о социальной жизни у растений, то лишь в переносном смысле,
никоим образом не отождествляя эти явления с социальной жиз-
нью людей. По вопросу же о выделении учения о сообществах
в отдельную науку мнения разделились, но все же это выделение
17
было принято. Был сохранен и термин фитосоциология, хотя
Л. Г. Раменский уже на съезде предложил заменить его терми-
ном фитоценология, который хотя и был предложен в 1918 г. Гам-
сом, но в Западной Европе почти не получил распространения.
Но термин «фитосоциология» у нас долго не продержался.
В советской науке в те годы уже развернулось широкое и реши-
тельное движение за овладение диалектическим методом, а для
этого требовалось в первую очередь критическое отношение ко
всем течениям, которые могли этот метод извратить. Одним из
таких извращений было стремление найти общие законы в раз-
витии человеческого общества, с одной стороны, и общества ра-
стений — с другой. Развернувшаяся в последующие годы борьба
с такого рода неправильными взглядами потребовала упраздне-
ния термина «фитосоциология». Вот почему уже в 30-х годах
у нас в Советском Союзе вошел в употребление термин фитоце-
нология. В настоящее время вместо этого термина чаще приме-
няют термин геоботаника, подчеркивая этим, что объектом иссле-
дования являются не только отдельные фитоценозы, но и расти-
тельный покров в целом в его взаимоотношениях со средой.
В 1934 г. Ботаническим институтом Академии наук СССР
была организована дискуссия на тему «Что такое фитоценоз?».
Итоги ее помещены в журнале «Советская ботаника» за 1935 г.
главным образом в статьях В. В. Алехина, В. Н. Сукачева и
А. П. Шенникова. Можно сказать, что эта дискуссия ознамено-
вала начало нового этапа в развитии советской геоботаники.
Дискуссия в значительной мере сблизила различные школы,
уточнила некоторые основные понятия и терминологию. Как
III Всесоюзный съезд ботаников, так и дискуссия о фитоценозе
были вызваны назревшей необходимостью объединения и обоб-
щения тех больших научных материалов, которые накопились
в связи с разработкой поставленных перед страной народнохо-
зяйственных проблем. Вопросы земледелия в новых, еще не осво-
енных районах Средней Азии, Закавказья, Крайнего Севера,
вопросы поднятия производительности сенокосов и пастбищ, упо-
рядочения лесного хозяйства стимулировали организацию широ-
ких геоботанических исследований.
Отдел геоботаники Ботанического института Академии наук
СССР руководит многочисленными геоботаническими исследова-
ниями. В ботанических институтах академий наук союзных рес-
публик имеются отделы и лаборатории геоботаники, а в крупней-
ших университетах — кафедры геоботаники. В настоящее время
наряду с опубликованием крупных сводных работ по геоботани-
ке шире развиваются исследования экспериментального характе-
ра, которые приводят к более тесному контакту геоботаники
с физиологией растений, биохимией и биофизикой.
Значительную роль приобретают биогеоценологические иссле-
дования, в которых геоботаники работают вместе с зоологами,
18
почвоведами, микробиологами, биофизиками и т. д. Комплексные
биогеоценологические исследования дают основу для наиболее
всестороннего и глубокого решения многих народнохозяйствен-
ных задач.
♦ Первой и основной задачей геоботаники на современном
этапе ее развития является определение значения растительного
покрова как важнейшего природного ресурса, изучение и доку-
ментация на геоботанических картах его современного состоя-
ния и потенциальных возможностей с целью его правильного
использования и преобразования. Е. М. Лавренко во вводной
главе к методическому руководству «Полевая геоботаника»,
первый том которой вышел в 1959 г., охарактеризовал народно-
хозяйственное значение геоботаники в следующих словах: «Ра-
стения, как дикие, так и культурные, почти всегда встречаются
в сообществах. Геоботаника — наука о растительном покрове —
наряду с другими ботаническими дисциплинами является биоло-
гической основой лесоводства, луговодства и полеводства. Ра-
стительный покров принадлежит к основным природным ресур-
сам нашей страны. Отсюда выясняется большое народнохозяй-
ственное значение геоботанических исследований» (стр. 68).
* Геоботанические исследования играют значительную роль
при выборе земель для распашки и при планировании работы по
их освоению. Типы растительных сообществ являются показате-
лями (индикаторами) качества земель, степени их пригодности
для вспашки и освоения под те или иные культуры. По составу
и характеру естественного растительного покрова можно судить
о механическом составе почв, степени их солопчаковатости, со-
лонцеватости или заболоченности. Поэтому участие геоботаников
при освоении целинных и залежных земель может значительно
ускорить и облегчить работу почвоведов.
* В проблеме осушения и освоения болот значение геоботани-
ческих исследований также очень велико. По распределению раз-
личных растительных группировок геоботаник устанавливает типы
болот, каждый из которых требует своих, специфических мер по
осушению, освоению под те или иные сельскохозяйственные куль-
туры или же по использованию запасов торфа. Геоботаническая
карта болотного массива является поэтому необходимой основой
для его освоения.
* В лесном хозяйстве все большее значение приобретает выде-
ление типов леса. Это дает возможность разработать хозяйствен-
ный план эксплуатации не суммарно для всего лесного массива
в целом, а для каждого типа леса или по крайней мере для каж-
дой группы близких типов в отдельности. Хотя история лесной ти-
пологии показывает, что первые шаги в ней были сделаны не
геоботаниками, а лесоводами, в дальнейшем при характеристике
типов леса и их классификации оказались совершенно необходи-
мыми геоботанические методы.
2*
19
В сущности одним из важнейших стимулов к развитию гео-
ботаники как особой научной дисциплины явилась именно по-
требность народного хозяйства в разработке классификации ле-
сов, лугов, болот, пустынь, тундр и других типов растительного
покрова с целью их правильного освоения и использования.
Другой областью хозяйственного применения геоботаники
является создание искусственных фитоценозов, а также измене-
ние естественных путем того или иного нарушения их состава
и структуры. Подсев кормовых трав на лугах, подбор травосме-
сей для сеяных лугов, удобрение сенокосов и пастбищ, осушение
заболоченных лугов и уничтожение кочек, создание искусствен-
ных лесонасаждений, и в том числе полезащитных лесных по-
лос,— все эти мероприятия требуют геоботанических методов при
своей разработке.
Сельскохозяйственное районирование наряду с районирова-
нием почвенным и климатическим, с изучением современного
распределения и урожайности отраслей растениеводства и про-
дуктивности животноводства имеет в качестве необходимого зве-
на геоботаническое картирование. Как это будет подробно пока-
зано ниже, более надежными показателями тех или иных поч-
венно-климатических условий, а следовательно, и хозяйственных
возможностей являются не отдельные виды растений, а их соче-
тания (сообщества), изучать которые можно лишь с примене-
нием геоботанических методов.
Особую ценность геоботанические исследования имеют при
микрорайонировании. Например, во многих южных областях
нашей страны при выделении субтропических микрорайонов
геоботанические исследования уже сыграли весьма значитель-
ную роль.
Здесь перечислены лишь некоторые из тех проблем народ-
ного хозяйства, которые тесно связаны с геоботаникой. Добавим,
что в нашу эпоху максимального освоения природных богатств
сильно возрастает значение мероприятий по охране природы.
Можно ли предполагать, что от природной растительности
в будущем, когда человек еще больше освоит поверхность нашей
планеты, останутся лишь сравнительно небольшие участки, пре-
имущественно в заповедниках? Очевидно, такое предположение
не имеет оснований. В. Б. Сочава (1963), например, полагает,
что при значительных урожаях, к которым стремится сельскохо-
зяйственная наука, не будет необходимости чрезмерно увеличи-
вать площадь культурных земель. Основываясь на подсчетах
академика Н. Н. Семенова (1962), можно принять, что около
70% суши даже и в отдаленном будущем останутся покрытыми
растительностью, в той или иной мере близкой к природной. Ко-
нечно, эта растительность не останется без воздействия человека,
который, пользуясь техникой будущего, будет ее многократно
трансформировать, но тем не менее это будут огромные зеленые
20
массивы, обеспечивающие необходимое для жизни человека
и животных содержание кислорода в атмосфере. Отсюда мы при-
ходим К выводу, что геоботаника не только не утратит никогда
своего практического значения, но, наоборот, значение ее будет
в известной степени возрастать и вместе с тем приобретать но-
вые, пока для нас еще неизвестные формы.
Темы рефератов на семинаре
или в кружке
Что такое фитоценоз
Литература: Ш е н и и к о в А. П. Введение в геоботанику.
Изд-во ЛГУ, 1964, стр. 9—25.
Сукачев В. Н. Общие принципы и программа
изучения типов леса. В кн. «Методические указа-
ния к изучению типов леса». М,—Л., Изд-во АН
СССР, 1957 — 1-е изд., 1962 —2-е изд.
Г. И. Танфильев и его главнейшие работы
Литература: Танфильев Г. И. Географические работы.
М., Географгиз, 1953.
И. К. Пачоский и его главнейшие работы
Литература: Пузанов И. И. и Гольд Т. М. Выдаю-
щийся натуралист И. К. Пачоский. М., Изд-во
АН СССР, 1965.
Воронов А. Г. И. К. Пачоский (к 100-летию со
дня рождения). «Бюлл. Моск, о-ва исп. природы».
Отд. биол. 1964, № 6.
Темы курсовых работ
Взаимоотношения леса и степи по работам
русских ученых
Литература: Б е р г Л. С. Климат и жизнь. М., Географгиз,
1947, стр. 4—87.
Вильямс В. Р. Почвоведение. Земледелие с
основами почвоведения. М., Сельхозгиз, 1939.
Комаров Н. Ф. Этапы и факторы эволюции
растительного покрова черноземных степей. М.,
Географгиз, 1951.
Мильков Ф. Н, Лесостепь Русской равнины.
М., Географгиз, 1950, стр. 91 — 132 и 260—263.
Значение работ В. В. Докучаева для геоботаники
Литература: Докучаев В. В. Избранные труды. М.—Л.,
Изд-во АН СССР, 1949.
21
2
ОСНОВНЫЕ ЧЕРТЫ
ВЗАИМОДЕЙСТВИЙ
МЕЖДУ РАСТЕНИЯМИ
В СООБЩЕСТВАХ
Взаимодействия в сообществе между растениями и средой
очень сложны. Чтобы лучше представить эти взаимодействия,
необходимо подойти к сообществу с исторической точки зрения.
Сформировавшееся сообщество представляет не случайное
сочетание растений, а такое, которое складывалось в процессе
эволюции веками и тысячелетиями и в котором растения в зна-
чительной мере приспособлены к совместному произрастанию в
естественных условиях.
Известно, например, что травы, обитающие в тенистом лесу,
настолько приспособлены к тени, что многие из них страдают,
будучи искусственно осветлены. Эта приспособленность к про-
израстанию в тени создавалась тысячелетиями в результате
естественного отбора.
Однако только одной приспособленностью к тени нельзя,
конечно, объяснить определенный состав травяной флоры того
или иного леса. Тенистые леса севера и юга часто бывают резко
различны по своей травяной флоре. Следовательно, кроме фак-
тора тени, тут играют роль климат, почва, история расселения
того или иного вида растения и т. д.
Кроме того, сообщество непрерывно изменяется. К сравни-
тельно более быстрым его изменениям относятся сезонные из-
mjrhphuh Так, например, состав и структура травяного покрова
в том же тенистом лесу весной и летом бывают различны. Вес-
ной цветут одни виды трав, летом—другие. Если лес листвен-
ный, тени в лесу значительно больше летом, чем весной, и лучше
приспособленные к тени травы развиваются поэтому именно
летом.
Бывают еще и другие изменения фитоценозов, более медлен-
ные, чем сезонные изменения, выражающиеся в смене одного
сообщества другим. Очень наглядно смены одного сообщества
22
другим бывают выражены при заселении растениями новых
местообитаний, например при зарастании песка, нанесенного
рекой. Сначала на песке появляется негустая заросль трав,
состоящая нередко из одного только вида, например из
хвоща (Equisetum). С течением времени в эту заросль посте-
пенно внедряются другие травянистые виды: кипрей (Epilobium),
вейник (Calamagrostis) и др., так что образуется смешанная
заросль. Далее начинают появляться всходы древесных пород,
например ив (Salix), и благодаря их быстрому росту лет через
десять на данном месте уже образуется ивовая заросль (рис. 2),
в которой травянистые растения занимают второй ярус, при-
чем и состав травяного покрова к тому времени также изме-
няется.
В этом примере изменения фитоценоза хотя и были непре-
рывными, тем не менее в них легко можно выделить три основ-
ные стадии, сменившие друг друга во времени: негустую за-
росль, часто из одного травянистого вида; густую смешанную
заросль из нескольких травянистых видов; смешанную двухъ-
ярусную заросль с ивами в верхнем ярусе и травами в
нижнем.
Как же менялись при этой смене взаимоотношения видов
внутри сообщества и взаимоотношения всего сообщества со
средой?
В негустой заросли хвоща влияние растений друг на друга
еще довольно слабое. В густой смешанной заросли растения
уже сильнее влияют друг на друга, в связи с чем виды, не спо-
собные к произрастанию в густой заросли, постепенно заменяют-
ся другими, более приспособленными.
Меняется и характер грунта. Под зарослью хвоща был еще
рыхлый, незакрепленный песок. По мере накопления в песке
перегнивших растительных остатков намечается верхний пере-
гнойный, или гумусовый, горизонт почвы. Наносимые ветром и
водой частички глины и пыли смешиваются с поверхностным
слоем песка и, склеиваясь, образуют комочки почвы. Так по-
степенно песок закрепляется, обогащается перегноем, глини-
стыми и пылеватыми частицами и превращается в почву. Под
ивняком имеется обыкновенно почва уже с хорошо выражен-
ным профилем. Почва по сравнению с песком обеспечивает воз-
можность произрастания более богатой видами травянистой
растительности. Таким образом, в процессе формирования та-
кого сравнительно сложного сообщества, как ивовая заросль,
создается почва, которая в свою очередь способствует усложне-
нию растительного сообщества.
Взаимозависимость сообщества и среды, отчетливо прояв-
ляющаяся при сменах одного сообщества другим, особенно на-
глядна при зарастании озер. По мере накопления растительных
остатков на дне озера оно мелеет у берегов и превращается в
23
болото. При этом создаются совершенно иные условия среды, в
связи с чем резко меняется и видовой состав растительности.
Если в озере была растительность из водорослей, кувшинок,
рдеста и др., то на болоте мы уже застаем заросли тростника
(Phragmites), а затем по мере накопления полуразложнвшпхея
растительных остатков торфа тростник обычно вытесняется раз-
личными осоковыми (Cyperus, Carex и др.). Здесь ясно видна
обоюдная зависимость изменяющегося сообщества и изменяю-
щейся среды, причем изменение среды (обмеление озера и на-
копление торфа в болоте) в значительной мерс вызвано жизне-
деятельностью самих растений, всего фитоценоза.
Кроме зарастания приречных и приморских песков и озер
(рис. 3), известен и целый ряд других случаев заселения расте-
ниями новых местообитаний. Это сопровождается формирова-
нием и сменами растительных сообществ с постепенным их услож-
нением, например зарастание скал, неподвижных каменных рос-
сыпей и подвижных осыпей, застывших лавовых потоков, глини-
стых обнажений и др.
Весьма часто пионерами на таких новых местообитаниях
бывают низшие растения: водоросли и лишайники, нс говоря
уже о бактериях, по-видимому, являющихся пионерами всегда.
Затем чаще всего появляются травянистые растения и уже
впоследствии деревянистые. В ряде случаев, например на мно-
гих местообитаниях речных пойм или в зоне степи, деревья и
кустарники не появляются вовсе. Тем не менее даже и в сменя-
Рис. 2. Приречные заросли ив на Амуре.
Рис. 3. Мертензия (Mertens:a maritima) — один из пио-
неров зарастания приморских песков на Дальнем Востоке.
ющих друг друга травянистых сообществах легко подмечается
постепенное усложнение структуры: увеличение количества ви-
дов, расчленение на ярусы, микрофитоценозы и т. д.
Каковы же те движущие силы, которые ведут к усложнению
сообществ? Это разнообразные противоречия, неизбежно воз-
никающие между растениями при их сколько-нибудь густом
произрастании. Сюда относятся противоречия как между осо-
бями разных видов растений, так и между особями одного вида,
выражающиеся во взаимном стеснении в виде затенения, меха-
нического вытеснения, вредного влияния корневых и других вы-
делений и т. д. Сюда относятся и явления конкуренции из-за
влаги и питательных веществ почвы, а также противоречия
между растениями и микроорганизмами и между растениями и
факторами неживой природы. Все эти противоречия объединя-
ются обычно понятием «борьба за существование».
Взаимоотношения между растениями бывают крайне проти-
воречивы, сочетая в себе элементы и конкуренции, и взаимо-
помощи. Так, в густых насаждениях из одного вида древесной
породы имеется некоторое взаимное угнетение, но в то же время
в последнее десятилетие вскрыты и довольно многочисленные
факты срастания корневых систем у совместно растущих де-
ревьев одного и того же вида (об этом мы расскажем подроб'
нее ниже). Известно, что всходы даже такой теневыносливой
породы, как европейская ель (Picea abies), не выдерживают
чрезмерно сильного затенения материнским пологом, по моло-
дые ели погибают также и при полном удалении последнего.
Поэтому применять по отношению к растительному сообществу
2о
термины «борьба за существование», «конкуренция», «взаимо-
помощь» можно лишь весьма условно, помня, что эти термины
весьма схематичны и отражают в большинстве случаев лишь
одну из сторон более сложных явлений. Правильнее говорить о
противоречиях и благоприятствованиях между образующими
сообщество особями растений и вместе с тем необходимо тща-
тельно анализировать как те, так и другие, разрабатывать их
классификации, что чрезвычайно поможет дальнейшему изуче-
нию растительных сообществ.
Ботаниками предложено несколько классификаций взаимо-
отношений растений в сообществах или так называемых копк-
ций. Остановимся на классификации А. А. Корчагина (1956), в
которой взаимоотношения между растениями в сообществах
подразделяются на следующие категории.
Прямые, или непосредственные, взаимоотношения:
паразитические; симбиотические; физиологические; биохимиче-
ские; механические; эпифитные.
Косвенные взаимоотношения: средообразующие; конку-
рентные.
Эти восемь типов коакций не всегда можно четко отделить
друг от друга. Например, охлестывание кроны одного дерева
ветвями другого может иметь не только механический, но в
известной мере и физиологический эффект, эпифитные взаимо-
отношения в ряде случаев трудно отделить от средообразующих
и т. д.
Рассмотрим примеры перечисленных типов взаимоотно-
шений.
Паразитические взаимоотношения достаточно общеизвестны.
Сюда относятся случаи присасывания к корням растений таких
паразитов, как заразиха, петров крест, оплетания побегов и
присасывания к ним повилики. К полупаразитам относятся
растения, ко^рые, в отличие от настоящих паразитов, имеют
зеленые листья, например, погремок, очанка и некоторые дру-
гие. Полупаразиты питаются как все зеленые растения, а пара-
зитизм служит для них дополнительным источником питания.
Все вредные грибы (ржавчинные, головневые, мучнисторос-
ные и др.) являются паразитами высших растений, так же как и
вредящие высшим растениям бактерии.
В то время как при паразитических взаимоотношениях вы-
году получает лишь паразит, а растение-хозяин терпит ущерб,
при взаимоотношениях симбиотических (т. е. при симбиозе) вы-
году получают оба симбионта, сожителя. Пример дают микори-
зы, при которых гриб сожительствует с высшим растением,
поставляя ему из почвы в легкоусваиваемой форме азотистые
соединения, а от высшего растения беря продукты фотосинтеза.
Высшие растения всегда имеют бактериоризу, т. е. скопления
бактерий вокруг корневых окончаний, причем это бактерии в
26
большинстве полезные, а без многих из них растения, по-види-
мому, не могут нормально существовать. В свою очередь бакте-
рии эти находят необходимые для своей жизнедеятельности
вещества в корневых выделениях. Широко известен симбиоз бобо-
вых с бактериями, образующими на их корнях клубеньки и усваи-
вающими свободный азот атмосферы. Известны и случаи симбио-
за высших растений с водорослями. Лишайники представляют
симбиотические организмы, составленные из гриба и водоросли,
причем в этом симбиозе наблюдаются и элементы паразитизма
гриба за счет водоросли.
Физиологические взаимоотношения растений имеют место
при любых других взаимоотношениях, но в наиболее чистом
виде проявляются при срастании корней разных особей. Боль-
шой интерес представляют случаи срастания корней разных де-
ревьев одного и того же вида в сомкнутых древостоях. В посвя-
щенной этому явлению интересной статье Н. И. Рубцов (1950)
отмечает, что оно было описано ранее целым рядом других авто-
ров и распространено в природе гораздо шире, чем это принято
думать. Рубцов считает, что срастание корней повышает ветро-
устойчивость деревьев, а при вырубке одного из сросшихся де-
ревьев оставшиеся могут использовать его корневую систему.
Срастание корней может охватывать значительное количество
деревьев. По И. И. Шишкову, в густом древостое обыкновенной
ели срастается корнями не менее 30% всех деревьев. По данным
С. В. Шевченко, наблюдается срастание корневых систем у евро-
пейской пихты в лесах Прикарпатья.
По данным А. П. Юновидова, в дендрологическом саду Бо-
ровского сельскохозяйственного техникума наблюдается сраста-
ние корней у сибирского кедра, сибирской лиственницы, евро-
пейской ели, сибирской ели, сибирской пихты. Возраст деревьев
перечисле_лых видов 25—30 лет. Срастание корнями особей од-
ного вида обнаруживалось автором главным образом по нахож-
дению «живых» пней, т. е. таких пней, которые, не имея ни одно-
го побега с хвоей, продолжают жить и расти в толщину. Ясно,
что эти пни получают органические вещества от соседних де-
ревьев через сросшиеся корни. По количеству живых пней автор
подсчитал, что в культурах кедра сросшиеся корнями деревья
составляют 26%, сибирской лиственницы—30%, сибирской
ели—34%, сибирской пихты—45%.
Оказалось, кроме того, что с увеличением густоты посадки
возрастает и процент деревьев, сросшихся корнями. Е. Г. Ор-
ленке (1§55) наблюдал сросшиеся корневые системы в гнездо-
вых посевах трехлетних дубков в Белоруссии. Однако в таком
возрасте срастание корней бывает еще неполным, так как в
местах срастания иногда еще сохраняются клетки коры разных
корней. Но у 20-летних дубков на поперечных срезах сросшихся
корней уже почти не видно границ срастания, и при насасыва-
27
нии цветного раствора через сосуды одного корня возникает
окрашивание и в сосудах другого корня.
Явление срастания корней у разных особей растений заслу-
живает тщательного дальнейшего изучения.
Биохимические взаимоотношения состоят во взаимном или
одностороннем влиянии корневых, листовых и других выделений
у совместно произрастающих растений. Взаимное биохимиче-
ское влияние высших растений называют аллелопатией. Был
предложен термин колики для тех продуктов обмена вешеств
высших растений, которые воздействуют на другие высшие ра-
стения. Колины следует отличать от фитонцидов, выделяемых
высшими растениями и действующих на микроорганизмы, а так-
же от антибиотиков, выделяемых микроорганизмами и дейст-
вующих на другие микроорганизмы. Наконец, маразмины выде-
ляются микроорганизмами и действуют на высшие растения.
В ряде случаев одно и то же вещество, очевидно, может совме-
щать в себе свойства двух или даже нескольких из перечислен-
ных групп выделений.
По Н. А. Красильникову (1958), из корней различных расте-
ний выделяются: углекислая, фосфорная, уксусная, муравьиная,
щавелевая кислоты, сахара, альдегиды, этиловый спирт, амино-
кислоты, танины, алкалоиды, витамины и другие, в том числе
еще не изученные, вещества.
Известны многие случаи как благоприятного, так и отрица-
тельного биохимического воздействия растений. Так, выращи-
вание лука в междурядьях картофеля или томатов намного
снижает поражаемость их грибным паразитом фитофторой.
В данном случае действует фитонцид, выделяемый луком. С дру-
гой стороны, еще Петр Кресценций в XIV в. отмечал, что ви-
ноград страдает в соседстве с капустой, и это было недавно
подтверждено опытами С. И. Чернобривенко. В данном случае,
очевидно, действовали калины, выделяемые капустой.
В дикой природе взаимное или одностороннее биохимическое
влияние растений также играет заметную роль. Так, по сообще-
нию Грюммера (1964), в дикорастущих рощах из грецкого оре-
ха на Ближнем Востоке и в Средней Азии однолетние растения
почти совершенно отсутствуют, что, вероятно, является резуль-
татом медленного выделения юглона, специфического колина из
листьев, как находящихся на дереве, так и уже опавших.
В наших лабораторных опытах по взаимному влиянию про-
растающих семян луговых злаков и бобовых было показано
благоприятное влияние соседства проростков тимофеевки или
луговой овсяницы на процент здоровых проростков красного и
розового клеверов.
В то же время проростки этих клеверов почти не оказывают
благоприятного влияния на проростки названных злаков, но и не
угнетают их.
28
Очевидно, благоприятное влияние клеверов на растущие
с ними злаки начинается позднее, когда на корнях клеверов обра-
зуются бактериальные клубеньки.
К механическим взаимоотношениям относится, например,
схлестывание кроны одного дерева ветвями другого, а также
взаимодействие между деревом и лианами. Всходы какого-либо
растения, растущие густой щеткой, тоже испытывают взаимное
механическое стеснение, но в этом случае его можно назвать и
конкуренцией за место, к которой к тому же примешивается
конкуренция за свет, за почвенную влагу и т. д.
Эпифитные взаимоотношения состоят в том, что растение-
эпифит поселяется на другом растении, обычно на древесном,
по не паразитирует на нем, а лишь использует его в качестве
субстрата. Одни из эпифитов прикрепляются своими корнями к
трещинам в коре дерева или к дуплам, где скопляется хотя бы
небольшое количество мелкозема (например, многие тропиче-
ские орхидные и бромелиевые), другие довольствуются уже и
незначительными неровностями, трещинками коры, как лишай-
ники и мхи.
Средообразующие и конкурентные взаимоотношения Корча-
гин назвал косвенными, но этого можно было и не делать.
Средообразующие взаимоотношения есть во всех фитоценозах,
особенно наглядно проявляясь в лесах, где деревья создают за-
тенение для кустарников, трав, мхов и лишайников. Одним зате-
нением средообразующес влияние деревьев, понятно, не ограни-
чивается. В лесу всегда выражен свой специфический фитокли-
мат (климат фитоценоза), отличный, например, от фитоклимата
луга, степи или болота, которые, в свою очередь, различаются
своими фитоклиматами и между собой.
Велико влияние на почву опадающей листвы или хвои, обра-
зующей на поверхности почвы так называемую подстилку. Тра-
вяной и мохово-лишайниковый покров, а также кустарниковый
ярус (подлесок), конечно, тоже оказывают воздействие и на
почву, и на фитоклимат леса.
Если сообщество достаточно устойчиво, т. е. если виды расте-
ний в нем хорошо уживаются друг с другом, то средообразую-
щие взаимоотношения видов в нем характеризуются заметной
ролью взаимного благоприятствования. Последнее проявляется,
например, в особенностях питания различных видов, которые в
этом отношении могут нередко дополнять друг друга.
В. Р. Вильямс (1939) отметил, что на рыхлокустовой стадии
развития луговой растительности бобовые, развиваясь на не
занятых злаками пространствах и усваивая свободный азот
атмосферы, разложением своих однолетних надземных органов
дополняют пищевой режим злаков усвояемой азотной пищей.
На лугу устанавливается известное нормальное количественное
соотношение между злаками и бобовыми.
29
Глубоко проникающие в почву (часто до глубины 2—4 ле)
корни бобовых не только устраняют соперничество из-за мине-
ральной пищи с неглубоко коренящимися злаками, ио и способ-
ствуют лучшему развитию злаков. Дело в том, что бобовые пере-
носят элементы зольного питания растений из более глубоких
почвенных слоев в верхние, причем особенно важно отметить
легкую усвояемость корнями бобовых растений солей кальция.
Все органы бобовых, как правило, содержат много кальция, и
при отмирании бобовых растений поверхностные слои почвы
обогащаются этим элементом. Обогащение почвы кальцием ве-
дет к улучшению почвенной структуры, так как, поглощая ионы
кальция, перегной приобретает способность прочно склеивать
частички почвы, образуя хорошо выраженные и достаточно
стойкие комочки. Бобовые и злаки при совместном произраста-
нии наилучшим образом содействуют созданию комковатой
структуры почвы. При отсутствии же бобовых преобладающие
в золе злаков одновалентные попы других металлов постепенно
вытесняют из перегноя поглощенный двухвалентный ион каль-
ция, что приводит к утрате комковатой структуры. Почва стано-
вится распыленной, и в ней, как говорит Вильямс, получают
преобладание волосные свойства и связность при полной утрате
прочности.
Благоприятное влияние одних видов сообщества на другие
проявляется в самых разнообразных формах: от приспособлен-
ности одних растений к затенению другими до взаимного допол-
нения по требовательности к химизму почвы, взаимного влияния
через ризосферы, корневые и другие выделения и т. д. Иногда
средообразующие взаимоотношения видов сообщества трудно
бывает отделить от симбиотических, биохимических и других.
Конкурентные взаимоотношения состоят в том, что совместно
живущие растения испытывают именно от этого совместного
произрастания взаимный ущерб в световом питании пли в поч-
венной влаге и растворенных в ней веществах пли, наконец,
просто в месте, что в то же время является механическим сте-
снением. При этом одни растения могут выйти как бы победи-
телями в этой конкуренции и достигнуть мощного роста, тогда
как другие могут оказаться угнетенными Нередко конкуренцию
отождествляют с борьбой за существование, но, следуя Дарви-
ну, нужно понимать борьбу за существование более широко, от-
нося сюда не только борьбу за жизненные потребности с дру-
гими организмами, но и борьбу со «слепыми силами природы»,
например с климатическими невзгодами. Таким образом, поня-
тие конкуренции менее широко, чем понятие борьбы за сущест-
вование.
Конкуренция бывает как между особями разных видов, так
и между особями одного вида растений. Внутривидовая конку-
ренция проявляется с наибольшей силой в определенные перио-
30
ды жизни сообщества, а именно при появлении густой щетки
всходов какого-либо вида. Внешне эта конкуренция выражается
в процессах дифференциации всходов по размеру и по степени
угнетенности: от более крупных и здоровых до мелких и угне-
тенных. За дифференциацией следует отпад, т. е. гибель многих
всходов. Эта дифференциация и отпад продолжаются и в даль-
нейшем, когда растения уже выходят из фазы всходов, хотя ин-
тенсивность этих процессов постепенно ослабевает.
Немецкий лесовод Крафт подразделил все деревья одновоз-
растного древостоя, образованного одной древесной породой, на
пять классов по высоте стволов и степени развития крон. Это
иллюстрирует явления дифференциации л отяада особей одной
породы в одновозрастных древостоях. Процесс дифференциации
и отпада начинается уже с того времени, как у древесной поро-
ды образуется густая щетка всходов, и продолжается в течение
всей жизни древостоя. Это явление наблюдается и при густом
росте особей какого-либо травянистого вида растений. Диффе-
ренциация предшествует отпаду; сначала особи разделяются на
классы в смысле Крафта, а уже затем из числа оказавшихся уг-
нетенными происходит отпад.
Следует отметить, что в противоположность диким растениям
у культурных растений часто не наблюдается дифференциации
особей даже в чрезвычайно густых посевах одного вида. В этих
случаях все особи испытывают угнетение. Это бывает связано с
тем, что в культурных посевах особи одного вида находятся
обычно в весьма сходных условиях в смысле глубины посева,
расстояний между особями, увлажнения и т. д.
В. Н. Сукачев (1952) приводит примеры, когда даже у лес-
ных деревьев в условиях культуры не наблюдалось дифферен-
циации и отпада. Он отмечает, что в тех случаях, когда густые
посевы и посадки древесных пород были проведены особенно
тщательно, с максимально равномерным их распределением по
площади и при отсутствии почвенной пестроты, все особи были
сильно подавлены и обычной, резко выраженной дифференциа-
ции их не наблюдалось. Это, например, имело место в опытах с
вязом, дубом и другими древесными породами в питомниках
научно-учебной станции Ленинградского университета «Лес на
Вбрскле» в Курской области, а также известно для слишком за-
гущенных посадок сосны в Бузулукском бору.
Бывают и такие случаи, когда нет ни угнетения всех особей,
ни сколько-нибудь заметных дифференциаций и отпада. Это
свойственно культурным посевам, выполненным с соблюдением
агротехнических правил (норма высева и его равномерность, глу-
бина заделки и т. д.), и при отсутствии почвенной пестроты. То-
гда наблюдается достаточно густое стояние и дружный рост куль-
турных растений, например хлебных злаков. Такие собрания
.особей одного вида образуют совокупности, способные хорошо
31
противостоять сорным растениям, не допуская или сильно огра-
ничивая их внедрение, а иногда и неблагоприятным климатиче-
ским влияниям, например суховеям. В этих случаях внутриви-
довая конкуренция бывает сильно ослаблена, но все же она
имеется и здесь, так как те же растения, но выращиваемые не
в заросли, а порознь, на грядках или в сосудах, достигают
обыкновенно значительно более мощного развития.
Межвидовая конкуренция может иметь различную интенсив-
ность и различный характер в зависимости от того, какие виды
произрастают в сообществе. В малоустойчивых, кратковремен-
ных фитоценозах межвидовая конкуренция сильнее, в более
устойчивых—слабее. В таких относительно устойчивых сообще-
ствах могут преобладать явления взаимопомощи, выражающие-
ся и в средообразующих, и в симбиотических, и в биохимических
взаимоотношениях между видами растений. В малоустойчивых
сообществах или в таких, в которые внедрились чуждые им
виды растений, межвидовая конкуренция может носить крайне
обостренный характер, приводя иногда к полному вытеснению
определенных видов.
Межвидовая, как и внутривидовая, конкуренция приводит к
дифференциации особей конкурирующих видов, которая пред-
шествует отпаду части или всех особей данного вида. Конкурен-
ция может иметь и немалое средообразующее значение, так как
в связи с гибелью части растений в ходе конкуренции в почву
при их разложении поступает немало органических веществ.
Совместное произрастание в сообществах всегда наклады-
вает на растения определенный отпечаток. У дерева, выросшего
на открытом месте, ветвление начинается гораздо ниже и крона
имеет раскидистую или пирамидальную форму. Наоборот, у де-
рева, выросшего в лесу, ветвление начинается на стволе высоко
и крона более компактна. Различия наблюдаются не только
в форме и величине кроны, но и в форме стволов. У дерева, вы-
росшего на открытом месте, ствол толще, короче и более кону-
совидной формы, т. е. суживается кверху резче, чем у дерева
того же вида и возраста, выросшего в лесу. Характер роста тра-
вянистых растений также различен в заросли и в разрозненном
состоянии. В первом случае растение стройнее, выше, боковые
ветви короче, во втором случае габитус более коренастый, рас-
тения чаще кустятся, боковые ветви длиннее. Взаимное влияние
связано не только с недостатком пространства, как такового, но
и с недостатком света, влаги, минеральных веществ почвы. Так,
форма кроны дерева, выросшего в лесу, обусловливается не
только механическим стеснением ее кронами соседних деревьев,
но и затенением ими, в связи с чем довольно быстро отмирают
нижние ветки.
Приближающаяся к конусовидной, или, как говорят лесово-
ды, «сбежистая» форма ствола у одиноко выросших деревьев
32
тоже представляет собой результат воздействия очень многих
факторов лесной среды,
В, Н. Сукачев (1959) считает, что сообщество, как правило,
в той или иной мере подавляет рост и развитие растений, отни-
мая у них условия для выявления всех их потенциальных воз-
можностей, Это относится не только к видам растений, играю-
щим в том или ином сообществе второстепенную роль, но и к
видам преобладающим. Сукачев приводит интересные цифры.
Бедренец (Pimpinella saxifraga), произрастая на лугу, имеет
среднюю высоту 50—60 см, причем каждое растение образует
обычно лишь один стебель с 5 сложными зонтиками, содержа-
щими в сумме до 1200 цветков. Этот же вид при произрастании
вне сообщества на глинистой неразвитой почве отвала канавы
достигает высоты более метра, причем одно растение образует
около 20 стеблей, содержащих 147 сложных зонтиков и 75 000
цветков.
С целью выяснить, не вызывает ли буйное вегетативное раз-
витие растений, выросших вне сообщества, снижения их способ-
ности к семенному размножению, В. Н. Сукачев поставил опыт
параллельной культуры кульбабы (Leontodon autumnale) из се-
мян, собранных с растений, которые хорошо развивались в сво-
бодном произрастании на грядках, и с растений па лугу. В куль-
туре те и другие семена дали хорошие всходы, причем первые
семена были даже лучшего качества, а выросшие из тех и дру-
гих семян растения не отличались заметно по своему росту и раз-
витию.
В. Н. Сукачев полагает, что при произрастании вне сообще-
ства растения обнаруживают такие морфологические, физиоло-
гические и биологические особенности, которые в сообществе
могут вовсе не проявляться. Он пишет, что постоянная угнетен-
ность растений в сообществах должна отражаться на направле-
нии и темпах процесса видообразования в природе, а это долж-
но учитываться при изучении этого процесса.
Т. А. Работнов (1950) ввел понятие о так называемом девст-
венном периоде в жизни растений. Каждое растение, развива-
ясь из семени, проходит последовательно состояния всхода, юно-
шеское и взрослое девственное. Последнее продолжается до тех
пор, пока растение не достигнет половой зрелости. Исследуя в
течение ряда лет жизненные циклы растений в луговых сообще-
ствах, Т. А. Работнов пришел к выводу, что на лугах многолет-
ние травы обычно развиваются медленнее, чем в питомниках,
имея более длительный девственный период. Некоторые расте-
ния в луговых сообществах зацветают не ранее чем в 20-летнем
возрасте, а в отдельных случаях длительность девственного пе-
риода достигает 40—50 лет. Хотя жизненные циклы луговых
трав зависят не только от естественных причин, но и от влияния
человека (сенокошение, выпас), при устранении этого влияния
3 П. Д. Ярошенко 33
основные черты этих циклов сохраняются. Таким образом, про-
израстание в луговых сообществах определенно задерживает
наступление половой зрелости растений.
В лесоводстве известны семенные годы, т. е. годы обильного
плодоношения, повторяющиеся для каждой древесной породы
через определенные промежутки времени, а в среднем через 2—
5 лет. В промежутках между семенными годами лесные деревья
плодоносят мало или даже вовсе не плодоносят, тогда как де-
ревья, растущие вне леса — в открытых местах, могут обильно
плодоносить каждый год. Это тоже, следовательно, влияние
фитоценоза на составляющие его растения.
Темы рефератов на семинаре
или в кружке
Взаимное биохимическое влияние высших расте-
ний (аллелопатия)
Литература: Чернобривенко С. И. Биологиче-
ская роль растительных выделений и межвидо-
вые взаимоотношения в смешанных посевах. М.,
Сельхозгиз, 1956.
Г р ю м м е р Г. Взаимное влияние высших рас-
тений— аллелопатия, пер. с нем. М., Изд-во
иностр, лит., 1957.
Гродзинский А. М. Аллелопатия в жизни
растений и их сообществ. Киев, Изд-во
АН УССР, 1965.
Дарвин о борьбе за существование у растений
Литература: Дарвин Ч. Происхождение видов. М.,
Сельхозгиз, 1962.
Микориза
Литература: Келли А. Микотрофия у растений, пер.
с англ. М., Изд-во иностр, лит., 1952.
Г. Ф. Морозов и его «Учение о лесе»
Литература: Морозов Г. Ф. Учение о лесе, изд. 7
(посмертное). М.—Л., Сельхозгиз, 1949.
34
Темы курсовых, работ
Взаимное биохимическое влияние прорастающих
семян и всходов разных растений
Тема проводится путем постановки лабораторного опыта.
Семена высеваются в чашки Петри на промытый и прокаленный
речной песок. Делаются как чистые, так и смешанные посевы.
Семена каждого вида подбираются однородные по окраске и
размерам. Высеваются они в строго определенном количестве и
так, чтобы прорастающие всходы не касались друг друга. По-
вторность шестикратная. Продолжительность всего опыта около
двух недель. Учет количества здоровых и угнетенных всходов
каждого вида производится через каждые три дня. Поддержи-
вается умеренная влажность песка поливом лучше дождевой
или дистиллированной водой, вносимой в одинаковом количест-
ве через маленькую лейку с тонким ситечком или путем обрыз-
гивания рукой.
В таких опытах всходы не испытывают недостатка влаги и
не могут конкурировать между собой из-за пищи, поскольку
питание их осуществляется только за счет фотосинтеза и запа-
сов в семенах, причем всходы не затеняют друг друга. Поэтому
взаимное благоприятное или неблагоприятное влияние всходов
можно объяснить лишь аллелопатией. Для изучения действую-
щих веществ можно проводить эту работу с участием студента-
химика, который тоже может оформить специальную курсовую
работу на эту тему.
Таблица 1
Образец бланка для учета всходов при смешанном посеве из двух видов
№ чашки Петри Дата Посеяно семян Число всходов клевера Число всходов тимофеевки
клевера красного тимофеевки луговой здоровых угнетенных здоровых угнетенных
Число здоровых и угнетенных всходов выражается затем в
процентах к числу посеянных семян.
Контролем служат чистые посевы каждого вида.
Литература: Ярошенко П. Д. Геоботаника. Основ-
ные понятия, направления и методы. М.—Л,,
Изд-во АН СССР, 1961, стр. 346—351.
3*
35
Изучение возобновления древесных пород в лесу
На однородном лесном участке площадью до 1 га (100Х
ХЮОл/) равномерно по всей площади закладывается 20—
25 метровых квадратов, обозначаемых вбитыми в углах колыш-
ками. На каждом из них еженедельно, с мая по сентябрь включи-
тельно, производится подсчет всходов, мелкого и крупного под-
роста одной или нескольких древесных пород. Условно за всхо-
ды принимаются особи не выше 10—15 см, за мелкий подрост —
от 15 до 30 см, за крупный подрост — выше 30 слг. Учитывается
также степень освещенности (затененности) каждого квадрата
на глаз либо с помощью экспонометра или люксметра. При гла-
зомерной оценке применяются такие градации:
1,0 — полная затененность;
0,9 — световые блики составляют в сумме около 0,1 площади
квадрата
и т. д. до 0,0 — полная освещенность.
Учитывается степень покрытия (на глаз в %) отдельно: кустар-
никового подлеска, кустарничково-травяного и мохово-лишайни-
кового покровов, а также степень покрытия (в %), мощность
(в см) и приблизительный состав подстилки. На каждом квад-
рате отмечаются преобладающие виды растений.
Для всего лесного участка отмечается состав древесных по-
род по принятому у лесоводов способу, а именно целыми чис-
лами, составляющими в сумме 10. Например, ель 5, сосна 3, бе-
реза 2 + единично осина. Выводы по теме должны содержать:
цифры среднего количества всходов, мелкого и крупного
подроста в пересчете на гектар;
данные о ходе отпада всходов с мая по сентябрь;
сопоставление количества всходов на каждом квадрате со
степенью освещенности, покрытием других растений и развитием
подстилки.
Литература: Ткаченко М. Е. Общее лесоводство.
М., Сельхозгиз, 1952.
Погребняк П. С. Общее лесоводство. М.,,
«Колос», 1964.
Изучение влияния сорняков на культурные рас-
тения
Эту тему лучше всего проводить на опытном участке. На двух
грядках производится посев какого-либо культурного растения
умеренной густоты. На половине каждой грядки ведется регу-
лярная прополка сорняков вручную, а на второй половине про-
полка не ведется. Полив (если он требуется) проводится на обе-
их половинах грядок. Опыт может продолжаться с апреля—мая
по июнь включительно. Нужно не допускать обсеменения сорня-
36
ков, чтобы они не распространились на другие участки, а по
окончании опыта грядки перекопать и все остатки сорняков
сжечь.
Опыт должен дать ответы на следующие вопросы:
Каковы средняя высота, средний вес (в воздушносухом со-
стоянии) и среднее число листьев одной особи культурного рас-
тения на засоренных и незасоренных частях грядок?
Каковы среднее число культурных растений на площадке
50x50 см и средний суммарный воздушносухой вес их на такой
же площадке с засоренных и незасоренных частей грядок?
На каждой части грядок берут по 4 таких площадки.
Каков видовой состав сорняков и какие виды их преобла-
дают?
Литература: Котт С. А. Сорные растения и борьба с
ними. М., Сельхозгиз, 1961.
Прянишников Д. Н. и Я к у ш к и н И. В.
Растения полевой культуры. М., Сельхозгиз, 1936.
Эту тему можно проводить и несколько иначе, а именно без
закладки специальных грядок, а путем наблюдения на засорен-
ных и незасоренных участках посевных полей.
3
ОТЛИЧИТЕЛЬНЫЕ
ПРИЗНАКИ
ФИТОЦЕНОЗА
Важнейшими категориями признаков, позволяющих отличать
один фитоценоз от другого,, являются следующие: видовой со-
став; количественные и качественные соотношения между вида-
ми; ярусность, мозаичность; физиономичность; периодичность,
синузиальность; характер местообитания.
Рассмотрим каждую из этих категорий признаков.
ВИДОВОЙ СОСТАВ
Видовой состав —очень важный признак сообщества. Глав-
ное значение имеют виды, преобладающие по числу особей,
или по массе, или по степени покрытия ими поверхности почвы,
но все же при описании сообществ большей частью желательно
учитывать все виды, как преобладающие (доминирующие), так
и непреобладающие. Дело в том. что среди видов, количественно
играющих в фитоценозе небольшую роль, могут оказаться такие,
которые своим присутствием указывают на определенные усло-
вия среды, например почвенные особенности, а это бывает очень
важно для полной характеристики фитоценоза. Например, при-
сутствие на лугу даже и в малом количестве лапчатки узика
(Potentilla erecta) всегда указывает на несколько пониженное
плодородие почвы. Такие растения Л. Г. Раменский назвал
детерминантами (определяющими). Преобладающие же в фито-
ценозе виды обычно находят в нем оптимальные для себя усло-
вия и поэтому являются менее чувствительными к умеренным
различиям среды, хотя определенное детерминирующее значение
имеют, конечно, и такие виды.
Видовой состав в той или иной мере отражает историю фор-
мирования фитоценоза, его прошлое. Так. в еловых лесах Мос-
ковской области встречается ветреница дубравная (Anemone
nemorosa), что свидетельствует о том, что раньше в этих местах
были дубовые леса. Позднее произошла смена дуба елью, но
3S
ветреница осталась, приспособившись к новым условиям. В дан-
ном случае ветреница является детерминантом условий среды,
имевших место в прошлом, но вместе с тем она показывает, что
смена дуба елью, очевидно, произошла здесь не очень давно
и еще не привела к очень сильному изменению почвенных и дру-
гих условий.
Существенным признаком фитоценоза является количество
видов в нем. Обычно подсчитывают среднее количество видов
растений на единице площади, например на квадрате размером
ЮхЮ (100 м2). При этом обнаруживаются немалые различия
у разных типов фитоценозов. В фитоценозах так называемых
луговых степей среднее количество видов на такой площадке
обычно около 50 и даже немного больше, тогда как на сухо-
дольных лугах лесной зоны оно обычно не менее 40, а на влаж-
ных пойменных (заливных) лугах не более 30. В темных еловых
или буковых лесах на стометровую площадку попадает не более
10 видов, а в дубравах более 30. ^Количество видов на единице
площади называют видовой насыщенностью фитоценоза. Наи-
большей видовой насыщенностью обладают тропические (дож-
девые) леса — гилеи.
При составлении полного видового списка фитоценоза мож-
но наряду с определением видовой насыщенности на площадке
определенных размеров охватывать этим списком участок фи-
тоценоза в пределах его естественных границ.
В полный видовой список должны быть, попятно, включены
как виды, находящиеся в цветении, колошении или спороноше-
нии, так и виды, находящиеся в вегетативном состоянии, в том
числе мхи, лишайники и водоросли. Неизвестные растения обо-
значаются пока условными названиями или номерами и берутся
в гербарий, с тем чтобы впоследствии их определить.
КОЛИЧЕСТВЕННЫЕ И КАЧЕСТВЕННЫЕ
ОТНОШЕНИЯ МЕЖДУ ВИДАМИ
Проще всего количественные отношения между видами мо-
гут быть выражены цифровыми или словесными баллами, кото-
рыми мы оцениваем обилие (число особей) или же преоблада-
ние (площадь, вес или объем) каждого вида в сообществе.
Существуют и другие, более точные методы определения оби-
лия и преобладания (доминирования); они будут рассмотрены
ниже, в главе, посвященной методике геоботанических исследо-
ваний.
Различные авторы применяли разные понятия и термины
для обозначения роли различных видов в растительных сообще-
ствах. Так, И. К. Пачоский различал компоненты, или главные
виды, образующие основу сообщества, и ингредиенты — виды,
39
играющие роль примеси и даже не всегда присутствующие в
сообществе.
Шведские геоботаники (Дю-Рие и др.) разработали понятие
о доминантах, как о видах, преобладающих в каждом ярусе
сообщества (о ярусности см. ниже). Сколько в сообществе яру-
сов, столько и видов доминантов. Например, в трехъярусном
лесном сообществе с преобладанием в первом ярусе сосны, во
втором — черники, в третьем — одного из видов зеленых мхов
три доминанта — сосна, черника и зеленый мох.
О степени преобладания доминантов чаще всего судят по
занимаемой ими в сообществе площади, или, как говорят, по
покрытию. Из советских геоботаников выявление доминантов для
каждого яруса применил, следуя Дю-Рие, В. В. Алехин (1936),
однако ранее и независимо от Дю-Рие понятие «доминант ассо-
циации» применил А. Я. Гордягин (1922), который вовсе не ука-
зывал на необходимость выделения доминантов в каждом ярусе
отдельно. По А. Я. Гордягину, доминанты — это виды, господ-
ствующие в сообществе, причем в последнем может быть один
доминант или несколько.
f У американских геоботаников чаще принято считать доми-
нантами преобладающие виды не всех ярусов, а лишь главного
яруса сообщества. В таком случае в вышеприведенном примере
соснового леса с черникой и мхом доминантою приходится счи-
тать только сосну. Виды растений, преобладающие в других
(не главных) ярусах сообщества, или же в имеющихся внутри
сообщества отдельных куртинах, в более или менее крупных
группах растений или же, наконец, имеющие преобладание се-
зонного характера, американские геоботаники называют субдо-
минантами (Клементс и Шелфорд, 1946).
В советской геоботанике в настоящее время обычно приня-
то называть доминантами виды, преобладающие в сообществе
независимо от его расчленения на ярусы. Если, например, в со-
обществе два яруса, но первый очень* редкий (в редколесьях),
то доминантою будет лишь преобладающий вид второго яруса.
Если первый ярус сообщества густой, а второй или 'очень ред-
кий, или настолько пестрый по своему составу, что преоблада-
ющие виды в нем выделить трудно, то один или несколько до-
минантов выделяются лишь в первом ярусе..|Те из доминантов,
которые в основном определяют специфическую среду всего
сообщества, принято, по предложению Г. И. Поплавской (1924)
и В. Н. Сукачева (1928), называть эдификаторами (строите-
лями).
Эдификаторы, например деревья в лесу, не обязательно
должны быть обильны по количеству экземпляров, но тем не ме-
нее они определяют структуру и специфические условия среды
внутри сообщества. Одинокое дерево на открытом месте также
является эдификатором для трав, приютившихся в его тени.
40
Рис. 4. Субальпийский луг на Кавказе.
В лесах, образованных двумя или несколькими главными дре-
весными породами, например в сложных многопородлых лесах
юга Дальнего Востока или в еще более сложных лесах влажных
тропических областей, будет несколько или даже много эдифи-
каторов./Виды, не являющиеся доминантами ни в главном, ни
в каком-либо другом ярусе сообщества, можно, следуя Г. И. По-
плавской и В. Н. Сукачеву, называть ассектаторами (соучаст-
никами).
По своему жизненному состоянию и своей роли в сообщест-
ве могут различаться не только разные виды, но и разные по
возрасту особи каждого вида. Так, изучая субальпийские луга
(рис. 4) Северного Кавказа, Т. А. Работнов (1945) пришел к вы-
воду, что каждый из слагающих луговые ценозы видов пред-
ставлен популяцией (совокупностью) особей, различных по воз-
расту (от всходов до старых растений) и по жизненному со-
стоянию (от нормально развитых до угнетенных в различной
степени /-'"Отдельные особи одного и того же вида сильно разли-
чаются между собой по мощности, конкурентной способности по
отношению к другим видам, а также по реакции на внешние
условия, в частности па деятельность человека. Так, у отдельных
особей субальпийской анемоны (Anemone fasciculate) число ли-
стьев варьировало от 1 до 11, а вес надземной части в сухом со-
стоянии— от 20 до 5130 мг. Лишь 77 особей из 136 отросли
после срезания и т. д. Изучив таким же образом и многие дру-
гие виды растений субальпийских лугов, Работнов пришел к вы-
воду, что роль отдельных особей одного и того же вида в жиз-
ни сообщества, а также их хозяйственное значение не равноцен-
ны. Следовательно, изучение состава популяций имеет большое
значение.
41
ЯРУСНОСТЬ
Ярусность, или вертикальное расчленение фитоценоза, луч-
ше всего выражена в лесных сообществах (рис. 5), где имеется
один или несколько древесных ярусов, часто один кустарнико-
вый ярус (подлесок), один или несколько травяных ярусов и
иногда еще непочвенный ярус мхов и лишайников. Эпифиты,
т. е. растущие па древесных стволах пепаразитные грибы, ли-
шайники, мхи, некоторые папоротники, а иногда и некоторые
цветковые растения, должны быть отнесены к впеярусным рас-
тениям, так же как и лианы.
Менее отчетливо бывают выражены ярусы в травяных сооб-
ществах, где нередко приходится испытывать затруднения при
отнесении особей какого-либо вида к тому или иному ярусу. Тем
не менее ярусность некоторых травяных ценозов представляется
сложной. Так, по В. В. Алехину (1936), па северных разнотрав-
ных степях Русской равнины можно насчитать до 7 ярусов, при-
чем они неясно отграничены друг от друга (рис. 6).
Ярусы обозначаются римскими цифрами, при этом самый
верхний ярус обозначается цифрой I.
Рис. 5. Светлый дубовый лес на Кавказе.
Рис. 6. Вертикальная проекция на Стрелецкой степи. Длина 1 м. Цифры ука-
зывают различные виды (по В. В. Алехину, 1936). Видны не вполне отграни-
ченные ярусы.
В. Н. Сукачев в ряде своих работ предложил различать
ярусы и пологи. Он приводит такой пример: древесные породы
в самом молодом возрасте не возвышаются в лесу над травами.
Следует ли их отнести к ярусу трав или к ярусу деревьев? Види-
мо, древесный молодняк, какой бы высоты он ни был, нужно
отнести к ярусу деревьев, но в данном случае к нижнему пологу
фитоценоза. Таким образом, следуя В. Н. Сукачеву, яруса.мц.
нужно называть элементы вертикального расчлрнрния сообще-
ства в соответствии с жизненной ролью тех или иных видов^ а
полога вертикального^расчлененця1гопбгпества в
каждый данный момент в соответствии лишь с относительными
высотами растений.
Кроме надземной ярусности, существует и подземная, но изу-
чение ее требует значительно больше труда и времени, и поэто-
43
му критерием для быстрого отличия одного сообщества от дру-
гого она может служить далеко не всегда. Наибольшей сложно-
сти ярусность достигает во влажных тропических лесах.
Тхай Ван Трунг (1962) в своей диссертации о лесах Вьетна-
ма отмечает в таком лесу чаще всего три древесных яруса, из
которых первый, т. е. самый верхний, прерывистый (т. е. не-
сомкнутый) образован как вечнозелеными, так и листопадными
видами деревьев.
Второй древесный ярус — сомкнутый и образован вечнозеле-
ными видами. Далее, кроме третьего древесного яруса, выраже-
ны еще кустарниковый и травянистый ярусы, а также внеярус-
ная растительность в виде лиан и эпифитов. Необходимо к этому
описанию добавить еще то, что и среди кустарников и трав в
таком лесу бывает выражено не по одному, а обычно по не-
скольку ярусов.
МОЗАИЧНОСТЬ И ЕЕ ОТНОШЕНИЕ
К КОМПЛЕКСНОСТИ
Представим себе участок степи. Если на его протяжении на-
блюдается один и тот же видовой состав, одни и те же количе-
ственные и качественные соотношения между видами, одна и та
же ярусность, одна и та же физиономичность (внешний вид)
и т. д., то мы относим этот участок к одному растительному сооб-
ществу. Однако если мы подойдем к нему с несколько иной мер-
кой, то без труда заставим себя усомниться в том, что перед
нами действительно лишь одно сообщество. Мы видим, напри-
мер, в степи многочисленные плотные дерновины ковыля. Дер-
новины эти не образуют сомкнутого покрова, а разбросаны одна
от другой на расстоянии в среднем около 30—40 см. Промежут-
ки между кустами ковыля заняты различными менее крупными
злаками, а также двудольным разнотравьем с примесью луко-
вичных. Некоторые из этих растений встречаются и внутри плот-
ных дерновин ковыля. Представляет ли каждая дерновина ко-
выля с сопровождающими ее немногочисленными другими расте-
ниями отдельное сообщество, отличное от густой заросли этих
других растений в промежутках между дерновинами ковыля?
Если мы будем руководствоваться перечисленными выше основ-
ными отличительными признаками сообщества, то придем к вы-
воду, что каждая дерновина ковыля вместе с поселившимися
внутри нее другими растениями представляет как бы отдельное
сообщество. Здесь совершенно иная комбинация основных при-
знаков сообщества, чем в промежутках между дерновинами:
иные количественные и качественные соотношения между вида-
ми, иная физиономичность и т. д. Следовательно, дерновины ко-
44
выля, с одной стороны, и растительность промежутков между
ними — с другой, представляют как бы сообщества внутри более
крупного сообщества всего степного участка.
Ясно, что такие мелкие сочетания растений имеют не только
количественные, но и качественные отличия от того сообщества,
составными частями которого они являются. Даже если каждое
из этих сочетаний и обладает формально признаками сообщест-
ва, все же качественное отличие проявляется уже в том, что все
эти признаки в таких мелких сочетаниях выражены не столь
ярко и в то же время в значительно менее устойчивой форме.
Такие мелкие сочетания растений внутри сообщества можно на-
звать микрогруппировками или микрофитоценозами. Они в гой
или иной степени бывают выражены в самых различных сооб-
ществах. Например, в лесу более затененные места представле-
ны одними микрогруппировками, осветленные — другими и т. д.
Естественно, что в различные микрогруппировки лесного сооб-
щества входят не только травы, мхи и лишайники, но и древес-
ные всходы, молодняк, кустарники, а также деревья.
Таким образом, в каждую микрогруппировку входит хотя и
небольшая часть всего сообщества, но включающая все его яру-
сы на данном месте.
v Наличие в сообществе горизонтального расчленения на мик-
рофитоценозы обусловливает то, что называют мозаичностью.
Рис. 7. Фра'мент березово-лиственничного леса близ Джалинды (верхнее При-
амурье) с тремя типами микрогруппировок (микроассоциациями):
а - михроасс. La^fx dahurica Betula platyphylla (затенение 0,6), Saussurea umbrosa 4- Lin-
naea borealis; b — микроасс. Larix dahurica (затенение 0,4) — Paccinium vitis idaea + Pirola
incarnata; c— микроасс. Betula platyphylla (затенение 0,3) — Calamagrostis langsdorfii+
Sangiiisorba oHicinalis + Sanguisorba parvihora. \
43
П, Д, Ярошенко (1956) в окрестностях Джалннды (верхняя
часть бассейна Амура) описал микрогруппировки в лесу из плос-
колистной березы (Betula platyphylla) и даурской лиственницы
(Larix dahurica). Здесь отмечены следующие типы микрогруп-
пировок {микроассоциации, рис. 7).
1. В древесном ярусе лиственница с примесью березы.
Сомкнутость полога 0,6. Кустарников нет. В травяном по-
крове соссюрея теневая (Saussurea umbrosa) с примесью лин-
ией (Linnaea borealis).
2. В древесном ярусе лиственница с сомкнутостью полога 0,4.
В травяно-кустарничковом ярусе брусника (Vaccinium vitis-ida-
еа) с примесью грушанки мясо-красной (Pirola incarnata).
3. В древесном ярусе лиственница с сомкнутостью полога
0,4. В травяно-кустарничковом ярусе брусника с примесью чины
низкой (Lathyrus humilis).
4. В верхнем ярусе невысокие кусты ольхи маньчжурской
(Alnus manshurica) с сомкнутостью полога 0,6. Кустарничков
нет. В травяном покрове майник (Majanthemum bifolium), сми-
лацина (Smilacina dahurica), вейник Лангсдорфа (Calamagros-
tis Langsdorffii).
5. В верхнем ярусе невысокая ольха маньчжурская с сом-
кнутостью полога 0,4. Под нею шиповник иглистый (Rosa acicu-
laris) и осока (Carex sp.).
6. В верхнем ярусе невысокая ольха волосистая (Alnus hir-
suta). Под нею грушанка мясо-красная. Имеются скопления
трухлявого валежа лиственницы, с чем, вероятно, и связано на-
личие грушанки.
7. Окна, иногда с редкостойной березой. Густой покров из
вейника Лангсдорфа, видов кровохлебки (Sanguisorba officina-
lis, S. parviflora) и других луговых трав.
Перечисленные типы микрогруппировок, перемежаясь друг
с другом, образуют мозаику небольших пятен, каждое по 4—
10 м2 (2x2; 2x3; 2X5 м и т. п.). В целом вся эта мозаика и
представляет одно из сообществ лиственнично-березового леса
с примесью ольхи. Лес этот является производным от листвен-
ничной тайги, возникшим на ее месте после выжиганий и выру-
бок. Повсюду рассеяны крупные обгорелые пни лиственницы.
Лиственница входит в состав первых трех микрогруппировок, по
нижним ярусам которых можно судить о былом составе травя-
но-кустарничкового яруса тайги. Микрогруппировки с ольхой
связаны лишь с повышенным увлажнением почвы в определен-
ных локальных условиях. Что касается микрогруппировок про-
галин и окон, то их состав указывает на то, какого типа луг
может возникнуть в случае полного уничтожения древостоя
вырубкой и выжиганием.
Мы видим, что изучение микрогруппировок дает возмож-
ность судить о прошлом и возможном будущем всего сообщест-
ве
ва в целом. Вместе с гем оно выявляет некоторые очень инте-
ресные закономерности, например то, что в условиях описыва-
емого леса одни травяные и кустарничковые виды оказываются
связанными с лиственницей (линнея, грушанка, брусника), а
другие — с ольхой (майник, смилацина).
А. А. Попов (1963, 1964), продолжая эти исследования, изу-
чал микрогруппировки в бруснично-долгомошном лиственнични-
ке Среднеамурской низменности, выделив здесь три типа микро-
группировок: 1) лиственница с брусникой, 2) лиственница с
мхом кукушкиным льном и 3) лиственница со сфагновым мхом.
Анализ хода роста лиственницы показал, что она лучше растет
в первой микроассоциации, так как корнеобитаемый слой почвы
там оттаивает раньше, чем во второй микроассоциации. Также
и возобновление лиственницы, т. е. появление и развитие ее мо-
лодняка, в первой микроассоциации значительно лучше, чем во
второй. Микроассоциация лиственницы со сфагновым мхом
представляет очаг заболачивания, так как сфагновый мох удер-
живает атмосферную влагу и долго сохраняет на небольшой
глубине сезонную мерзлоту почвы.
4 Мозаичность — это наличие горизонтального расчленения
внутри одного сообщества, а комплексность — это явление, при
Рис, 8, Схема мозаичности и комплексности:
ia — мозаичность плавная с микроассоциациями 1,2, 3, 4; б—мозаичность контурная
к минроассоциацнями 5, 6, 7,8; 11 — комплексность: комплекс двух фнтоценозов, одно-
го с мозаикой микроассоциаций 9, 10, Ц, а другого с мозаикой микроассоциа-
ций 12, 13, 14.
47
Рис. 9. Кочковатый болотистый луг на Дальнем Востоке.
котором перемежаются небольшие участки или пятна разных
сообществ.
Встает вопрос, как отличить мозаичность от комплексности,
в особенности если при последней слагающие ее пятна разных
сообществ невелики по размерам. Вопрос этот должен решаться
в том смысле, что при мозаичности каждое пятно представляет
одну единственную микрогруппировку, тогда как при комплекс-
ности каждое пятно слагается из пятен разных микрогруппиро-
вок (рис. 8). Ф. Я. Левина (1958) справедливо отмечает, что
комплексность — явление более сложное, чем мозаичность.
* Во многих случаях при заселении растительностью голых
мест ее мозаичность развивается одновременно с формировани-
ем всего сообщества. В других случаях оформление микрогруп-
пировок несколько запаздывает по сравнению с выработкой все-
го сообщества. Иногда, как это бывает, например в горах, при
зарастании скал и каменных россыпей, сначала возникают'от-
дельные разобщенные микрогруппировки. Затем, смыкаясь, они
образуют сообщество, в котором продолжается их дальнейшее
оформление вместе с развитием всего сообщества.
Причины возникновения мозаичности могут быть разные.
Следует различать мозаичность, обусловленную иеоднород-
43
ноетью среды внутри сообщества, и мозаичность, обусловлен-
ную разрастанием и взаимодействием отдельных особей и
куртин.
На кочковатых болотистых лугах (рис. 9) мозаичность в силь-
ной мере обусловлена существованием кочек, на вершинах ко-
торых формируются одни микрогруппировки, на склончиках и
шлейфиках — другие, а в промежутках — третьи. Но бывают
и такие сообщества, в которых мозаичность хотя и выражена
достаточно резко, но отчетливо не связана ни с микрорельефом
(который может быть и вовсе не выраженным), ни с пятнами
света и тени, ни с какой-либо иной неоднородностью среды. Та-
кова, например, мозаичность некоторых степных сообществ, вос-
станавливающихся на старых залежах с супесчаными и пылева-
тыми почвами на Приханкайской равнине (Дальний Восток).
Здесь на площадке в 1 м2 можно видеть целый ряд микрогруп-
пировок. В одних из них преобладает сибирская пижма (Тапа-
cetum sibiricum) с примесью змеевки (Cleistogenes sinensis),
в других — змеевка с примесью лапчатки земляничной (Poten-
tilla fragarioides), келерии (Koeleria gracilis) и др., в третьих —
володушка (Bupleurum scorzonerifoHum) и т. д. Есть и отдель-
ные кустики келерии, бледно-зеленой осоки (Carex leucochlora),
ломоноса шестилепестного (Clematis hexapetala), не входящие ни
в одну из микрогруппировок, или, другими словами, не сопро-
вождаемые другими сопутствующими видами растений. Но при
всей этой пестрой мозаике микрорельеф участка совершенно не
выражен: это просто ровная поверхность супесчаной или пыле-
ватой почвы, в промежутках между растениями частично при-
крытая тонким слоем сухих остатков (ветоши). Очевидно, что
эта зарослевость обусловлена только разрастанием и взаимодей-
ствием поселявшихся на залежи растений. Однако каждая из
возникших микрогруппировок постепенно создает свою специ-
фическую среду. Например, внутри густой куртины ломоноса
условия освещения, температуры, влажности и накопления ве-
тоши отличаются от таковых в негустом травостое володушки с
примесью единичных побегов змеевки.- Таким образом, мозаич-
ность растительности сама создает и мозаику микросред.
За последние 10—15 лет изучение мозаичности все больше
привлекает внима_ни£ исследователей. Таковы работы М. И. Са-
харова (1950, 1951), И. Т. Ивановой, П. Д. Ярошенко и
К. П. Берстюковой (1963), Н. В. Дылиса, А. И. Уткина и
И. М. Успенской (1964) и др.
В США и Англии мозаичностью занимаются мало, больше
обращая” внимание на закономерности распределения отдельных
видов внутри сообщества. Понятие узора (pattern) относится там
чаще всего именно к такого рода закономерностям.
4 П. Д. Ярошенко
49
ФИЗИОНОМИЧНОСТЬ, ЖИЗНЕННЫЕ ФОРМЫ
И ПЕРИОДИЧНОСТЬ
Каждое растительное сообщество характеризуется опреде-
ленным внешним видом, или физиономичностью. Эю обусловле-
но тем, что в состав сообщества входят растения того или иного
габитуса, облика. Классификация растений по их внешнему виду
ведет свое начало еще от Аристотеля и Теофраста, а затем раз-
рабатывалась рядом ученых. Основоположник ботанической
географии Гумбольдт в 1805 г. предложил классификацию расте-
ний по внешнему виду, установив 19, как он называл, раститель-
ных форм. Среди них он выделял, например, злаковидные, папо-
ротники, лианы, кактусовые, банановые, лавровые и т. д. После
Гумбольдта подобного рода классификации разрабатывались
целым рядом исследователей, причем с течением времени в них
все более и более учитывались признаки не только внешнего об-
лика, но и всей организации растения, отражающей определен-
ные условия существования. Так создалось представление о жиз-
ненных формах растений. Этот термин был предложен Вармип-
гом в 1901 г.
Наибольшее распространение получила система жизненных
форм датского бстаника Раункиера (1934). Этот автор положил
в основу своей классификации расположение на растении почек
возобновления, различая следующие типы:
Фанерофиты. Почки возобновления находятся более или ме-
нее высоко над поверхностью земли. Сюда относятся деревья
и кустарники.
Хамефиты. Почки возобновления находятся невысоко над по-
верхностью земли. Сюда относятся полукустарники (с деревяни-
стым основанием стебля), низкорослые стелющиеся кустарнич-
ки (например, брусника и линнея), растения-подушки, наземные
мхи и лишайники.
Гемикриптофиты. Почки возобновления расположены на са-
мой поверхности почвы. Сюда относится большинство травяни-
стых многолетников.
Криптофиты. Почки возобновления находятся под землей
(геофиты, к которым принадлежат луковичные, клубневые и кор-
невищные травы) или на дне водоема.
Терофиты. Неблагоприятный сезон переживают только семе-
на. Сюда относятся однолетники.
Под почками возобновления нужно понимать почки, зимую-
щие в условиях холодных и умеренных климатов или же пере-
живающие летнюю засуху в условиях сухих тропических и суб-
тропических климатов.
Перечисленные пять основных типов жизненных форм Раун-
киер подразделяет еще на группы, всего различая более 30 групп
жизненных форм.
SO
Проблема жизненных форм растений, отчасти соприкасаясь
с физиономичностью сообществ, в то же время является гораздо
более широкой и разносторонней. Некоторые жизненные формы
растений могут, например, различаться только по подземным
органам, и тогда они не будут иметь значения для физиономич-
ности. Могут быть и жизненные формы, различающиеся физио-
логически, но сходные по внешнему виду.
Классификация жизненных форм Раункиера является в ос-
новном биологической, т, е. основанной на таких особенностях
строения и образа жизни растений, которые представляют собой
результат совокупного воздействия многих факторов среды.
Сюда относятся особенности общего габитуса растений, их
перезимовки, размножения и т. д. В то же время имеются и эко-
логические классификации жизненных форм, основанные на
отношении растений к строго определенным факторам среды. Из-
вестно, например, подразделение на ксерофиты, мезофиты,
гигрофиты и гидрофиты либо подразделение на мегатермы,
мезотермы и микротермы и т. д.
Классификация жизненных форм, предложенная И. Г. Сереб-
ряковым (1962, 1964), является более экологической, чем класси-
фикация Раункиера, но вместе с тем учитывает и такие, напри-
мер, признаки, как поликарпия и монокарпия, т. е. многократное
и однократное плодоношение на протяжении жизни растения.
Классификация Серебрякова, включающая жизненные формы
покрытосеменных и хвойных, имеет следующие основные подраз-
деления:
Отдел А. Наземные и эпифитные древесные растения.
I тип. Деревья.
1-й класс. Кронообразующие деревья с полностью одревес-
невшими удлиненными побегами.
1-й подкласс. Наземные кронообразующие деревья.
2-й подкласс. Г емиэпифитные (полуэпифитные) кронообра-
зующие деревья (в тропиках).
2-й класс. Розеточные деревья (образуют на вершине укоро-
ченные побеги с розетками крупных листьев; тропики и субтро-
пики).
3-й класс. Суккулентно-стеблевые безлистные деревья (на-
пример, кактусы).
II тип. Кустарники (с подразделениями).
III тип. Кустарнички (с подразделениями).
Отдел Б. Полудревесные растения.
IV тип. Полукустарники и полукустарнички (с подразделе-
ниями).
Отдел В. Наземные травянистые растения.
V тип. Травянистые поликарпики.
1-й класс. Травянистые поликарпики с ассимилирующими по-
4!
51
бегами несуккулентного типа (многолетние травы обычного
типа).
1-й подкласс. Стержнекорневые.
2-й подкласс. Кистекорневые и короткокорневищные.
3-й подкласс. Дерновинные.
4-й подкласс. Столонообразующие и ползучие.
5-й подкласс. Клубнеобразующие.
6-й подкласс. Луковичные.
7-й подкласс. Корнеотпрысковые.
2-й класс. Травянистые поликарпики с ассимилирующими по-
бегами суккулентного типа.
1-й подкласс. Суккулентностеблевые.
2-й подкласс. Суккулентнолистные.
3-й класс. Сапрофитные и паразитные травянистые поликар-
пики (с подразделениями).
4-й класс. Эпифитные травянистые поликарпики (с подразде-
лениями) .
5-й класс. Лиановидные травянистые поликарпики (много-
летние травянистые лианы).
VI тип. Травянистые монокарпики (с подразделениями).
В эту классификацию не включены водные и земноводные
травы.
В классификации И. Г. Серебрякова приведены еще более
мелкие, чем подклассы, подразделения: группы, подгруппы, сек-
ции жизненных форм и собственно жизненные формы. Для более
подробного ознакомления с этой и другими классификациями
жизненных форм рекомендуем статью И. Г. Серебрякова в кни-
ге «Полевая геоботаника», т. 3 (М.—Л., Изд-во АН СССР, 1964).
Используя классификации жизненных форм при характери-
стике сообществ, мы, с одной стороны, устанавливаем внешний
облик последних, а с другой — даем основу для выяснения спе-
цифических черт условий их существования. Физиономичность
сообществ неразрывно связана с их экологией.
На протяжении сезонов года внешний вид сообщества ме-
няется. Это называют сезонной периодичностью или сменой ас-
пектов, а под аспектом понимают физиономичность в каждое
данное время. При общей характеристике аспекта обычно ука-
зывают образующие его виды растений, различая фон и на нем
отдельные выделяющиеся особи или группы и отмечая окраску
как фоновых, так и других растений. Кроме того, отмечают фе-
нологическую фразу каждого вида растений.
Сезонные смены аспектов тесно связаны и с сезонными изме-
нениями структуры и жизнедеятельности сообщества. Наряду
с некоторыми другими признаками их кладут в основу для выде-
ления, например, таких крупных типов фигоценозов, как луго-
вой, степной и т. д. Внешне сезонная периодичность жизни ра-
стительного сообщества проявляется в смене аспектов, т. е. в
52
Рис. 10. Цветущая сольданелла (Soldanetla hungarica)
в Карпатах.
смене сезонных фаз, каждая из которых характеризуется своей
определенной физиономией (рис. 10). Однако для более глубо-
кого познания сезонной периодичности в жизни сообществ нель-
зя ограничиваться этой внешней стороной, необходимо изучать
также и такие вопросы, как сезонные изменения в корневых си-
стемах, в почвенной микрофлоре и обусловленные этим измене-
ния физико-химических процессов, протекающих в почве. Боль-
шое значение имеет также исследование по сезонам количества
и состояния всходов и подрастающего молодняка различных
компонентов сообщества.
СИНУЗИАЛЬНОСТЬ
В числе существенных признаков растительного сообщества
приводятся еще и синузии, являющиеся его составными частями.
Под синузиями чаще всего понимают за кие структурные едини-
цы внутри сообщества, которые объединяют растения одной жиз-
ненной формы или по крайней мере близких жизненных форм.
Следовательно, синузия соответствует или ярусу (если он обра-
зован одной или близкими жизненными формами), или части
яруса, или же группе внеярусных эпифитов или лиан внутри со-
общества. Можно, конечно, выделять синузии и среди почвенной
микрофлоры. Примером, когда синузия соответствует части
яруса, может служить синузия куртин осоки лесной (Carex silva-
53
iica) в травяном покрове леса. В этом же травяном ярусе могут
присутствовать (в зависимости от типа лесного фитоценоза) и
другие синузии, например синузия куртин мужского папоротни-
ка (Dryopteris filis mas) -и синузия промежутков между курти-
нами осоки и куртинами папоротника, представленная негустым
покровом некоторых двудольных с небольшой примесью злаков.
Что касается микрогруппировки, то она может совпадать, да
и то не всегда, лишь с синузией одноярусного сообщества. Го-
раздо чаще в одну микрогруппировку входит несколько или
даже много синузий.
Интересные и ценные в лесоводственном отношении данные
могут быть получены при одновременном изучении как синузий,
так и микрогруппировок в лесных сообществах. Такое исследо-
вание выполнил С. В. Лысенко (1968) для И типов лесных со-
обществ Владимирской области. Этот автор изучал в каждом
типе лесных сообществ (лесной ассоциации) синузии древесного
яруса и синузии напочвенного покрова и затем устанавливал со-
вместную встречаемость гех и других, используя несложные ста-
тистико-математические методы. Так, в типе «сосняк лишайни-
ковый» в древесном ярусе имеются две синузии — сосновая и
березовая, а в напочвенном покрове — шесть синузий: мертвопо-
кровная, лишайниковая, вейниковая изреженная, вейниково-по-
левицевая, зеленомошная и вересковая. При этом к мертвопо-
кровной синузии автор относит пятна, в которых нет ни лишай-
ников, ни мхов, ни высших растений, а имеются, следовательно,
только микроорганизмы и почвенные грибы.
У сосновой синузии оказалась наибольшая совместная встре-
чаемость с мертвопокровной синузией (20%), а наименьшая —
с вересковой синузией (!%) У березовой синузии наибольшая
совместная встречаемость (6%) с вейниково-полевицевой сину-
зией, наименьшая (1%) с вейниковой изреженной синузией, то-
гда как совместной встречаемости березовой синузии с синузия-
ми мертвопокровной, зеленомошной и вересковой нет вовсе. Что
касается прогалин, то они заняты чаще всего лишайниковой си-
нузией (31%), реже всего (1%) —зеленомошной синузией и н*
них вовсе не встречены синузии мертвопокровная и вейниковая
изреженная. Каждое сочетание сосновой или березовой синузии
или прогалины с той или иной синузией напочвенного покрова
представляет определенный тип микрогруппировки. Автор спра-
ведливо полагает, что такого рода данные о внутренней струк-
туре лесных сообществ, как имеющие непосредственное значение
при ведении лесного хозяйства, должны отражаться и при круп-
номасштабном картировании лесов путем приложения к лесным
картам планов наиболее важных типов лесных сообществ с на-
несением контуров как микрогруппировок, так и синузий.
К синузиям относятся и те группировки внутри сообщества,
которые имеют сезонный характер, обусловливая тог или иной
аспект, как, например, синузия эфемеров в пустыне или полупу-
стыне; впрочем, эта синузия одновременно является и особым
ярусом (сезонный ярус).
ХАРАКТЕР МЕСТООБИТАНИЯ
Неотъемлемым признаком растительного сообщества являет-
ся .характер местообитания, т. е. условия среды данного сообще-
ства. При этом необходимо подчеркнуть, что растительное сооб-
щество невозможно резко отграничить от местообитания. Так,
древесный полог представляет собой элемент самого сообщества
и в то же время элемент среды не только для подлеска и тра-
вяного покрова, но и для составляющих его деревьев. Подсы-
хающий лист, пока он находится еще на дереве, представляет
собой часть растения, но стоит ему упасть на землю, как он ста-
новится частью лесной подстилки, т. е. по отношению к выра-
стившему его дереву становится уже элементом среды. Некото-
рые авторы различают местообитание и экотоп, понимая под
первым все условия среды, а под вторым — лишь совокупность
факторов неживой природы, т. е. климатических условий, а так-
же водного и солевого режима почвы и отчасти характера релье-
фа. Но такое подразделение совершенно искусственно, так как
на климатические условия влияет и сам фитоценоз, образуя так
называемый фитоклимат, тогда как водно-солевой режим почвы
определяется в значительной мере характером разложения в ней
органических веществ.
Практически характеризуя местообитание того или иного со-
общества, отмечают: географическое местоположение, высоту
над уровнем моря, характер рельефа (экспозиция склона, мик-
рорельеф и т. п.), почвенные условия, характер воздействия жи-
вотных и человека. Все это в совокупности — весьма существен-
ный признак фитоценоза.
Таковы основные признаки, позволяющие исследователю вы-
делять на местности растительные сообщества и этим самым
подходить вплотную к пониманию сложения растительного по-
крова. Хотя изучение видового состава растений, т. е. флоры ка-
кой-либо территории, является задачей флориста, а изучение со-
обществ и образуемой ими растительности, или растительного
покрова, относится к деятельности геоботаника, тем не менее
геоботаник не должен оставлять в стороне и изучение флоры —
видового состава как описываемых им сообществ, так и всей ис-
следуемой местности. Он должен интересоваться и вопросами
истории флоры данного района, связывая их с изучением исто-
рии растительности. Каждый геоботаник должен быть в то же
время и флористом-систематиком. Только при этом условии
изучение растительного покрова будет действительно полноцен-
ным.
55
Тема реферата на семинаре
ила в кружке
Строение фитоценозов
Реферат главы III учебника А. П. Щенникова «Введение в
геоботанику». Изд-во ЛГУ, 1964.
О мозаичности лесных фитоценозов
Реферат может быть разделен между двумя докладчиками.
Литература: Иванова И. Т., Ярошенко П. Д. и
Берстюкова К- И. Микрофитоценозы неко-
торых сообществ хвойно-широколиственпых лесов
Приморья, «Комаровские чтения», выи. XL Вла-
дивосток, Изд-во АН СССР, 1963.
Д ы л и с Н. В., Уткин А. И., У с п е н-
с к а я И. М. О горизонтальной структуре лесных
биогеоценозов. Бюлл. МОИП, т. 69, 1964, № 4.
Темы курсовых работ
Сравнение разных методов определения степени
преобладания (доминирования) видов в фитоце-
нозах (по покрытию, объему и весу)
На участке какого-либо фитоценоза (лесного, лугового
и т. д.) намечают 5—6 видов растений и определяют степень их
доминирования разными методами. В выводах дается сравнение
этих методов.
Литература: Шенн и ков А. П. Введение в геоботанику.
Изд-во ЛГУ, 1964, стр. 96—108.
Воронов А. Г. Геоботаника. М., Изд-во «Выс-
шая школа», 1963, стр. 238—252.
Микрогруппировки и синузии одного из фито-
ценозов
Описываются и наносятся на план микрогруппировки и глав-
нейшие синузии на участке лесного (0,25—0,5 га'} или лугового
(ЮХЮл) фитоценоза.
Литература: Ярошенко П. Д. Геоботаника. М.—Л.,
Изд-во АН СССР, 1961, стр. 56—71,
Лысенко С. В. О методах изучения внутрен-
ней структуры лесных сообществ на примере ле-
сов Владимирской области. «Лесоведение», 1968,
№ 1.
56
Рис. 11. Сенокосные луга в долине р. Колымы.
Определение урожайности и группового
состава травостоя на лугу или степи
На участке луга (рис. 11) или степи площадью до 1 га, ис-
пользуемом как сенокос, срезают на высоте около 2 см перед на-
чалом сенокошения до 10 укосов, каждый с площадки 50X50 см.
Площадки эти равномерно распределяются по всему участку.
Трава с каждой площадки взвешивается сначала в свежем виде,
а затем в воздушносухом состоянии, т, е. в виде сена. Вес сена
с 10 площадок, умноженный на 4000, дает урожайность сена с I га.
Одни из наиболее типичных укосов (с площади 50x50 см) раз-
бирается в сухом виде па группы или фракции, а именно: злаки,
бобовые, осоковые, ситниковые и хвощи; разнотравье; вредные
травы, труха. Каждая группа взвешивается, и вес дается в про-
центах к общему весу сена площадки.
Урожайность сена с 1 га можно определить и более точно
с указанием и степени достоверности полученной цифры. Для
этого находят не вес сена со всех 10 площадок, а средний вес
сена с одной площадки 50X50 см. Помножив его на 40 000, нахо-
дят соответственно средний вес сена с 1 га. Затем вычисляют
среднюю ошибку (т) по формуле: /и = ± |/ где s — знак
суммирования, А — разность между весом каждого укоса (в цент-
нерах с гектара) и средним арифметическим весом, п — число
наблюдений, т. е. в данном случае 10. Допустим, что у нас от-
дельные величины, полученные с каждого из десяти укосов, бу-
дут: 22, 18, 20, 19, 20, 18, 23, 19, 19, 22 ц[га. Тогда среднее ариф-
метическое будет 20 ц/га, а общая сумма квадратов разностей
57
между весом каждого укоса и средним арифметическим соста-
вит 4 + 4 + 1+4 + 9 + 1 + 1+4 = 28. Подставив в формулу цифры,
/ 2g"
получим: m = dz у = ±5,3 : 9,5 = 0,6 (с округлением). Следо-
вательно, урожайность сена с нашего луга будет 20±0,6 ц[га.
Литература: Важенин И. Г. Применение метода ва-
риационной статистики в почвенно-агрохимиче-
ских исследованиях. «Почвоведение», 1963, № 2.
Воронов А. Г. Геоботаника. М., «Высшая
школа», 1963, стр. 262—267.
У рано в А. А. Наблюдения на летней практи-
ке по ботанике. М., «Просвещение», 1964.
Ярошенко П. Д. Геоботаника. М.—Л.,
Изд-во АН СССР, 1961, стр. 371—373.
4
экология
РАСТИТЕЛЬНЫХ
СООБЩЕСТВ
Предложенное еще в 1869 г. Геккелем название «экология»
употребляется для обозначения науки о взаимоотношениях жи-
вых организмов со средой. Существует экология растений и эко-
логия животных.
Равным образом можно говорить о взаимоотношениях со сре-
дой не только отдельных индивидов и видов, но и сообществ.
Изучение экологии растительных сообществ представляет
собой один из важнейших разделов геоботанического исследова-
ния. В нашу задачу не входит подробное изложение теории и
практики изучения экологии фитоценозов, тесно связанного с изу-
чением экологии составляющих их растений. Но некоторые
главнейшие понятия мы все же рассмотрим.
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ И ИХ ВЛИЯНИЕ
НА РАСТИТЕЛЬНЫЕ СООБЩЕСТВА.
ВЗАИМОДЕЙСТВИЕ, ВЗАИМНАЯ
ЗАМЕЩАЕМОСТЬ
И ПРЕДЕЛЫ ЗАМЕЩАЕМОСТИ
ЭКОЛОГИЧЕСКИХ ФАКТОРОВ
Среда представляет собой сложный комплекс экологических
факторов, действующих на растения. Среди них следует отме-
тить четыре основные группы, а именно: климатогенные, или
климатические, факторы (обусловленные в основном атмосфер-
ными явлениями), эдафогенные, или эдафические (обусловленные
субстратом, почвой), биогенные, или биотические (обусловлен-
ные другими растениями или их сообществами и животными),
и антропогенные (обусловленные человеком).
Каждая из перечисленных групп состоит из многих факторов,
например в климатические факторы включаются: свет, тепло, ве-
тер, влажность воздуха, осадки и т. п. В свою очередь каждый
59*
из этих элементарных факторов также является, в сущности, це-
лой группой их. Например, фактор света может быть подразде-
лен на силу света, состав света (в смысле преобладания тех или
других лучей солнечного спектра), и продолжительность дневно-
го освещения. Фактор тепла может быть подразделен на не-
сколько факторов в зависимости от суммы годовых температур,
от зимних минимумов, от летних максимумов и других особенно-
стей годового и суточного хода температуры и т. д. Отсюда вид-
но, какое сложное явление представляют собой экологические
факторы; сложность их действия значительно увеличивается еще
от того, что в природе они никогда не действуют порознь, но все-
гда совместно.
Вопросу совместного действия экологических факторов, а так-
же замещаемости одних факторов другими посвятил интересное
аналитическое исследование швейцарский эколог и геоботаник
Рюбель (1922, 1935). Он подчеркивает, что при изучении сов-
местного действия экологических факторов необходимо всегда
иметь в виду то, что каждый экологический фактор может толь-
ко тогда полностью проявить свое действие, когда другие факто-
ры действуют в достаточных размерах. Например, если почва
богата питательными веществами, то эти вещества не проявляют
своего полезного действия при отсутствии необходимой влажно-
сти почвы или, наоборот, вода и солнце не могут быть полезны
растению без питательных веществ почвы.
Более четко это правило было сформулировано В. Р. Вильям-
сом, который отметил равнозначимость или незаменимость четы-
рех факторов, или четырех условий жизни растений: света, тепла,
пищи и воды. Таким образом, если и можно- говорить о заме-
щаемости экологических факторов, то лишь в определенных пре-
делах. Замещаемость факторов может иметь место лишь при
обеспечении этих условий жизни растений. Например, один вод-
ный фактор (осадки) может быть иногда заменен другим вод-
ным же фактором (влажность воздуха, искусственный полив),
но не может быть заменен фактором света, тепла или почвенно-
го питания. Таким образом, замещаемость экологических факто-
ров может быть принята лишь с определенными ограничениями,
вытекающими из правила Вильямса.
Рюбель включает деятельность человека в число биотических
факторов наряду с воздействием растений и животных. В дей-
ствительности же антропогенные факторы (деятельность челове-
ка) принципиально глубоко отличны от биотических. В то время
как биотические факторы обусловлены в первую очередь причи-
нами биологического порядка, антропогенные факторы и харак-
тер их влияния на растительный покров зависят прежде всего от
социальных условий человеческого общества.
Примером замещаемости одного эдафического фактора дру-
гим, тоже эдафическим, может служить замещение почвы легко-
60
го механического состава почвой глинистой, содержащей известь.
Это объясняется коагулирующим действием иона кальция, что
обусловливает возникновение в глинистой почве структурных аг-
регатов.
Интересны случаи замещения климатических факторов эда-
фическими. В средней Европе сухие известняковые холмы быва-
ют покрыты в особенности на южных склонах растительностью,
характерной для районов Средиземноморья. Это следствие сухо-
сти и теплоты известковой почвы, замещающих южный среди-
земноморский климат, который, как говорит Рюбель, «репроду-
цируется эдафически на сухой теплой почве». Наоборот, хо-
лодность и влажность почвы могут способствовать поселению
северной растительности в странах с жарким южным климатом.
В районах Средиземноморья и далее на юг до пустынь Африки
лесная растительность по берегам рек и ручьев близка к средне-
европейским летнезеленым лесам (Aestisilvae), как, например,
тополевые заросли в ливийских оазисах. Вообще вода эдафиче-
ски замещает некоторые факторы климата. В жарких районах
вода действует как охладитель, заменяя низкие температуры
воздуха, в холодных — как утеплитель.
С первого взгляда может показаться, что случаи замещения
климатических факторов эдафическими противоречат правилу
Вильямса. Однако в действительности это не так. Например,
если средиземноморская растительность встречается на южных
склонах известняковых холмов средней Европы, то в этом слу-
чае атмосферное тепло замещено теплом же, но только поч-
венным, а вовсе не фактором почвенного питания или каким-
либо иным.
Ю. Н. Куражсковский (1957) приводит данные, которые, по
его мнению, противоречат правилу Вильямса. Так, он указывает,
что в поселке Саранпауль (на крайнем севере Ханты-Мансий-
ского национального округа, в зоне лесотундры) колхозник
Г. П. Семяшкин неоднократно успешно выращивал в открытом
грунте теплолюбивую культуру тыквы. Это достигалось замеще-
нием климатических факторов эдафическими: дренажем, созда-
нием высоких гряд с хорошо аэрируемой почвой, большими до-
зами навозного удобрения и т. д. Но и здесь все эти мероприя-
тия сводились в основном к утеплению почвы, т. е. фактор тепла
опять-таки не был замещен ни фактором света, ни фактором
влаги, ни фактором почвенного питания. К тому же Куражсков-
скип отмечает, что, помимо всех перечисленных мер, для тыквы
выбирались и наименее морозобойные участки.
Все же можно привести и такие факты, которые свидетель-
ствуют о возможности частичной взаимной замены четырех отме-
ченных Вильямсом условий жизни растений. Так, например, из-
вестно, что красный клевер является растением недолговечным,
причем гибель его наступает в условиях культуры чаще всего на
61
третий или четвертый год, обычно выражаясь в зимнем вымерза-
нии. Но при условиях улучшенного почвенного питания (что до-
стигается удобрениями) клевер становится более долговечным
(Калинкевич, 1955). В этом примере мы видим частичную (но
не полную) замену фактора тепла фактором почвенного пита-
ния. Приведенный случай характеризует поведение лишь одного
вида растения, но не сообщества из нескольких видов. Для со-
обществ же факты хотя бы частичного замещения одного из че-
тырех условий жизни растений нам неизвестны.
Вопросу совместного действия экологических факторов и их
взаимной замещаемости уделяет значительное внимание и швед-
ский эколог Г. Люндегорд (1937). Он отмечает, что «относитель-
ное действие фактора тем значительнее, чем более он находится
в минимуме по сравнению с другими факторами. С возрастаю-
щей интенсивностью фактора относительное действие его посто-
янно падает, и в области максимума действия величина его при-
ближается к нулю». Этим так называемым законом относитель-
ности объясняется, например, то, что в сообществах мезофитов,
у которых все факторы среды находятся в максимуме или в оп-
тимуме их действия, даже значительное изменение того или ино-
го фактора, если только оно не приведет его к минимуму, не
дает существенного эффекта. Поэтому одинаковые или сходные
сообщества мезофитов могут встречаться в неодинаковых кли-
матических и почвенных условиях.
Значительную роль в распределении растений играют биоти-
ческие факторы. Не говоря уже о воздействии животных, очень
важно глубоко уяснить себе роль так называемых фитогенных
факторов, т. е. взаимоотношений одних видов растений с други-
ми. Не принимая этого во внимание, легко впасть в ошибки при
истолковании тех пли иных закономерностей в распределении
различных видов растений и образуемых ими сообществ по опре-
деленным типам местообитаний. Существуют растения весьма
неприхотливые к почвенным условиям и в то же время на лучших
почвах легко уступающие свое место другим видам. К таким ра-
стениям относится сосна, которая в лесной области Закавказья
растет чаще всего на сухих каменистых склонах южной экспози-
ции. Однако изучение истории расселения здесь сосны показы-
вает, что раньше она была распространена и на склонах север-
ной экспозиции, отличающихся хорошей глубокой влажной поч-
вой, но впоследствии была вытеснена оттуда буком, пихтой
и некоторыми другими породами. Не зная об этом, можно поду-
мать, что сосна предпочитает сухие каменистые почвы, тогда как
в действительности она селится там лишь потому, что это един-
ственные места, где она не встречает соперничества со стороны
других древесных пород.
62
ГЛАВНЫЕ И ВТОРОСТЕПЕННЫЕ
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Не все экологические факторы имеют одинаковое значение
для распределения растений и сообществ. Например, из эдафи-
ческих факторов наиболее важны влажность почвы и ее плодо-
родие, т. е. богатство усвояемыми питательными веществами.
Такие эдафические факторы, как почвенная кислотность и т. п.,
имеют обычно более местное значение. Например, если вид при-
способлен к произрастанию на влажных почвах, то он в той или
иной мере сохраняет эту особенность во всех, даже самых разоб-
щенных частях своего ареала. И наоборот, приспособленность
вида к той или иной кислотности почвы чаще оказывается свя-
занной с нею лишь косвенно, на самом деле зависящей от дру-
гих свойств почвы, сопряженных с кислотностью только при опре-
деленных условиях.
Таким свойством часто является почвенная влажность: напри-
мер, на севере нашей страны влажные почвы обычно бывают
кислыми. Часто с кислотностью бывает сопряжена и бедность
почвы питательными веществами: так, почвы верховых болот
Крайнего Севера кислы и очень бедны.
Многие авторы составляют списки растений, будто бы обяза-
тельно приуроченных к почве с большим содержанием извести,
но большинство этих видов было впоследствии найдено (при
иных условиях климата, почвенной структуры и т. п.) и на бед-
ных известью почвах.
Исследования индийских ученых Бхаруча и Сатьянарайян
(1954) показали, что проблема растений, приуроченных к извест-
ковым почвам, равно как и неприспособленных к произраста-
нию на них, значительно сложнее, чем это могло бы показаться
при поверхностном подходе. Отмечая, что кальцефильные расте-
ния в ряде случаев хорошо чувствуют себя и на кислых почвах,
исследователи пришли к выводу, что эти растения поглощают
большие количества кальция независимо от его содержания в
почве. Вместе с тем оказалось, что существование кальцефиль-
ных растений на более или менее нейтральных почвах зависит
от общего содержания в почве обменных оснований. Поэтому под
кальцефильными растениями (кальцефитами) следует понимать
как растения щелочных почв, имеющих pH от 7,0 до 8,6 и содер-
жащих, кроме того, карбонат кальция или обменный кальций,
так и растения нейтральных или слабокислых почв, имеющих
pH от 6,2 до 7,0, но обладающих высоким содержанием обмен-
ных оснований. Попутно Бхаруча и Сатьянарайян выяснили, что
присутствие или отсутствие в почве свободных карбонатов не
всегда коррелируется с величиной ее pH и что возрастание со-
держания карбоната кальция не обязательно влечет возрастание
щелочности.
63
Не все факторы способны взаимно замещаться, поскольку
для любого растения и любого сообщества необходимы опреде-
ленные нормы влажности, тепла, света и почвенного питания.
Эти экологические факторы следует считать главными, а осталь-
ные — второстепенными.
Постоянство отношения того или иного вида растения к поч-
венной влажности всегда в достаточной мере оценивалось иссле-
дователями, и на нем основано известноечподразделение расте-
ний на ксерофиты, мезофиты и гигрофиты. Менее популярно
подразделение растений по их отношению к плодородию почвы.
По этому признаку различают эутрофы, т. е. растения плодород-
ных почв, и олиготрофы— растения бедных, тощих почв. Более
детально такое подразделение разработал шведский ботаник
Гард а Зегерштад (1924), различающий следующие группы:
А. Стенотрофы — виды, предъявляющие в том или ином отно-
шении ярко выраженные требования к почвенному бонитету.
1. Эутрофы или мегатрофы — виды, достигающие наилучше-
го развития на плодородных почвах.
2. Мезотрофы — виды, достигающие наилучшего развития на
посредственных почвах.
3. Олиготрофы— виды, достигающие наилучшего развития
на тощих почвах.
Б. Эвритрофы — виды, не предъявляющие ясно выраженных
требований к почвенному бонитету.
Гард а Зегерштад приводит списки эутрофов, мезотрофов,
олиготрофов и эвритрофов для Швеции. Изучая эти списки и со-
поставляя их с соответствующими данными по Закавказью, я убе-
дился в том, что из видов, общих для Швеции и Закавказья,
все, ведущие себя в Швеции как эу-, мезо-, олиго- или эвритро-
фы, так же ведут себя и в Закавказье. И это несмотря на силь-
ную удаленность друг от друга этих областей и значительную
разницу всех прочих почвенно-климатических условий.
Причины эутрофности тех или иных видов растений вполне
понятны. Плодородие почвы действует как благоприятный фак-
тор. Поэтому все наши культурные растения, за немногим исклю-
чением, эутрофы. Также не требует особых пояснений эвритроф-
ность каких-либо видов: она связана с широтой их эдафической
амплитуды. Однако та трактовка, какую дает Гард а Зегерштад
мезотрофности, не вполне ясна. В особенности непонятно, почему
олиготрофы достигают наилучшего развития именно на тощих
почвах.
Сущность этого кроется в том, что олиготрофы представ-
ляют собой неоднородную группу. Одни из них лучше развивают-
ся на бедных почвах потому, что имеют эндотрофную микоризу,
т. е. находятся в симбиозе с грибами, гифы которых проникают
в их клетки. Грибы, образующие эндотрофную микоризу, нередко
64
приспособлены в своем развитии лучше к кислым почвам, а бед-
ные почвы бывают часто кислы, тогда как богатые чаще ней-
тральны либо щелочны. Отсюда и появляется олиготрофность
у многих растений, имеющих эндотрофную микоризу.
Если та или иная степень кислотности или щелочности суб-
страта является для многих высших растений второстепенным
фактором, то для низших — грибов и бактерий — она играет
часто решающую роль. Поэтому и высшие растения, имеющие
эндотрофную микоризу, становятся чувствительными к кислот-
ности, Они часто бывают лучше приспособлены к кислым почвам,
в связи с чем и являются олиготрофами. Это одна категория
олиготрофов. К ней относятся многие представители семейства
вересковых (Ericaceae), как-то: обыкновенный вереск (Calluna
vulgaris), многие виды рододендронов, черника, брусника и др.
К олиготрофам с эндотрофной микоризой относится также злак
белоус (Nardus stricta), который обилен на некоторых типах
наших пастбищ, но не представляет никакой ценности в кормо-
вом отношении.
Наилучшей мерой борьбы с белоусом является органическое
удобрение почвы, увеличивающее ее аэрацию и понижающее
кислотность.
Другую категорию олиготрофов представляют сфагновые мхи.
У них нет ни корней, ни даже ризоидов (последние бывают лишь
в начальной стадии их развития) и вовсе отсутствует микориза.
Тем не менее сфагновые мхи явно тяготеют к бедным почвам
с кислой реакцией. Известны опыты Ольсена с культурами сфаг-
новых мхов на почвах различной степени кислотности. Опыты
эти показали, что рост сфагнумов протекает лучше всего на кис-
лых почвах с pH 3,5—4,5, тогда как на почвах нейтральных
и щелочных сфагнум испытывает явное угнетение.
Сфагновые мхи являются примером растений, ярко выражен-
ная олиготрофность которых не связана с наличием микоризы.
Вместе с тем вся организация сфагновых мхов, в особенности их
способность впитывать огромные количества воды, представляет
собой приспособление к произрастанию в условиях недостаточно-
го почвенного питания. По мнению В. Р. Вильямса, поглощае-
мая сфагнумом вода служит для него одним из важных источни-
ков зольного питания.
Те растения, которые к плодородию почвы, в сущности, нераз-
борчивы, но на лучших почвах не выдерживают конкуренции
многих других видов и поэтому вынуждены селиться на тощих
почвах, следует относить уже не к олиготрофам, а к эвритрофам.
Таковы, например, обыкновенная сосна и некоторые виды мож-
жевельников. При выращивании их на плодородных почвах они
повышают свою жизненность, чего нельзя сказать про настоящих
олиготрофов.
5 П. Д. Ярошенко 65
К эвритрофам же следует относить, очевидно, и многие расте-
ния, находящиеся в симбиозе с бактериями-азотособирателями,
образующими на их корнях мелкие клубневидные вздутия. Сюда
относятся, кроме бобовых, также облепиха (Hippophae), лох
(Elaeagnus) и некоторые другие.
ФИЗИЧЕСКАЯ И ФИЗИОЛОГИЧЕСКАЯ
СУЩНОСТЬ ЭКОЛОГИЧЕСКИХ
ФАКТОРОВ
При исследовании экологических свойств местообитаний нуж-
но отличать две стороны действия того или иного экологического
фактора. Например, болотные почвы физически часто очень
влажны, но влага эта может быть малодоступной для растений
или вследствие, сильнокислой реакции, или ввиду связанности
воды с почвенными коллоидами, или же по причине своей низкой
температуры. Про такие почвы говорят, что они физиологически
сухие. В таком же смысле можно говорить о почвах богатых, но
в то же время неплодородных, т. е. физиологически бедных. К по-
следним относятся некоторые болотные почвы, например некото-
рые почвы низинных болот, нередко содержащие много зольных
элементов, но в такой форме, которая слабо усваивается расте-
ниями.
Холодный или прохладный климат северных и высокогорных
местностей способствует неполному разложению органических
веществ, что и вызывает накопление в почвах торфа. Характер-
ные для таких почв растения обычно имеют черты ксероморфно-
го строения: мелкие, прижатые к стеблю листья, опушение,
восковой налет, развитие кутикулы и т. д. Однако в отличие от
настоящих ксерофитов, свойственных грунтам, которые сухи не
только физиологически, но и физически, эти растения называют
психрофитами. Очень многие (если не все) типичные психрофиты
являются в то же время и олиготрофами, так как физиологически
сухие почвы обычно в то же время бедны и усвояемыми питатель-
ными веществами. Но если все или почти все психрофиты вместе
с тем и олиготрофы, то далеко не все олиготрофы являются
психрофитами. Есть немало олиготрофов, свойственных физиче-
ски сухим и теплым, но в то же время тощим почвам, например,
многим каменистым сухим почвам юга. Произрастающие на них
олиготрофы являются в то же время ксерофитами, но нс психро-
фитами.
Примеры олиготрофов-психрофитов дают нам болотные и
тундровые растения. Примерами олиготрофов-ксерофитов могут
служить некоторые растения сухих каменистых склонов, доволь-
ствующиеся незначительными скоплениями мелкозема между об-
ломками камней.
.66
Таковы, например, акантолимоны, чебрецы и многие другие
ксерофильные кустарнички и полукустарники.
Климатические факторы тоже могут различаться по своему
физическому и физиологическому действию. Например, две мест-
ности с равным количеством годовых осадков, но с неодинако-
вым их распределением по временам года всегда значительно
отличаются и по условиям произрастания растений. Страны
Средиземноморья имеют среди своей флоры гораздо больший
процент ксерофитов, чем страны средней Европы, хотя годовое
количество осадков в Средиземноморье обычно выше. Это объяс-
няется тем, что основная масса осадков здесь приходится на
осенне-зимние месяцы, тогда как в средней Европе осадки рас-
пределены по временам года более равномерно.
УЧЕНИЕ ОБ ИНДИКАТОРАХ В СВЯЗИ
С ЗАМЕЩАЕМОСТЫО ЭКОЛОГИЧЕСКИХ
ФАКТОРОВ
Использование дикой растительности в качестве индикатора
(показателя) тех или иных почвенно-климатических условий при-
менялось издавна и особенное развитие имело первоначально
в Северной Америке, где быстрое распространение земледелия
при заселении этой страны европейцами требовало ускоренного
диагноза природных условий. А дикая растительность, если толь-
ко умело использовать знание ее особенностей, действительно
может явиться хорошим индикатором природных условий, а от-
сюда и хозяйственных достоинств местности.
Но если при этом не принимать во внимание замещаемости
экологических факторов, то можно впасть в грубые ошибки. На-
пример, увлечение индикаторами доходило у некоторых исследо-
вателей до того, что они стремились найти такие виды растений,
которые в любых частях своего ареала являлись бы показателя-
ми одних и тех же условий среды. Но на деле такие виды найти
трудно, а часто и вовсе невозможно, потому что если ареал дан-
ного вида достаточно обширен, то в разных его частях этот вид
обычно свойствен неодинаковым местообитаниям, и в частности
различным почвам. Например, европейский бук (Fagiis silvatica)
в Англии распространен на известковых почвах, а в Средней
Европе — на обыкновенных лесных почвах, имеющих реакцию,
близкую к нейтральной. Это объясняется тем, что в умеренном
климате средней Европы для бука более благоприятны обыкно-
венные лесные почвы, а в более влажном приморском климате
Британских островов он предпочитает почвы более сухие. Здесь
мы имеем пример замещения климатического фактора эдафиче-
ским. Впрочем, возможно, что вопрос этот еще сложнее, так как
по всей вероятности сюда примешивается еще и влияние конку-
5*
67
ренции других видов, вытеснивших бук в северо-западной Англии
с обыкновенных (неизвестковых) лесных почв. На этом примере
мы видим, что какой-либо вид растений, особенно если он имеет
широкий ареал, захватывающий области с различным климатом
(или различной историей взаимоотношений среди растений), не
может явиться во всех частях этого ареала надежным индикато-
ром одних и тех же почвенных условий. Вообще виды растений
как индикаторы любых условий среды могут иметь лишь ограни-
ченное применение, не выходящее за пределы одной природной
провинции.
Более универсальны как индикаторы не отдельные виды, а со-
общества. Например, сосна (Pinus silvestris) встречается в Евро-
пейской части СССР в пределах лесной области на самых разно-
образных почвах, от скал и довольно сухих песков до заболочен-
ных почв, а черника, малоразборчивая к климату, встречается
от лесной области до области тундры включительно, причем рас-
тет как на увлажненных почвах, так и на каменисто-скалистых
грунтах. Но такой тип леса, как Pinetum myrtillosum (бор-чернич-
ник) свойствен лишь строго определенному типу почвенных усло-
вий и встречается только в лесной области.
А. Я. Гордягин в уже упоминавшейся нами известной работе
«Материалы к познанию почв и растительности Западной Сиби-
ри» (1900) писал, что отдельные растительные формы в боль-
шинстве случаев не являются почвопостоянными: это свойство
присуще обыкновенно лишь немногим видам в данной местности,
большей частью редким и потому малохарактерным. По-видимо-
му, более почвопостоянными следует считать целые комбинации
форм, растительные сообщества, но и тут это постоянство явля-
ется делом довольно условным. Если посреди степи истреблен
лес, то на его месте развивается травянистая растительность,
более или менее сходная с той, которая процветает в степи; отли-
чия ее от типичной степной в некоторых случаях улавливаются
опытным глазом, но и то, может быть, не всегда; во всяком слу-
чае они не такого сорта, чтобы их можно было выразить в точной
форме.
В Закавказье многопородный лиственный колхидский лес
служит хорошим индикатором влажносубтропического климата.
Он встречается в Колхиде (западное Закавказье) не выше 600—
700 м над уровнем моря. Древесные породы, образующие основу
этого леса, а именно: бук, каштан, граб, дуб и ольха, в отдельно-
сти поднимаются в горы значительно выше. Но весь фитоценоз
колхидского леса, состоящий в верхнем ярусе из смеси всех этих
пород, ограничен лишь указанным вертикальным пределом.
В то же время почвенные условия, в которых развивается этот
лес, бывают довольно различными. Достаточно указать, что он
встречается как на маломощных скелетных почвах, так и на поч-
вах огромной мощности.
68
Использование растительных сообществ в качестве индикато-
ров затруднено в тех случаях, когда в одном сообществе встреча-
ются виды, очень сильно различающиеся по своей экологии. Так,
недалеко от предгорий Алтая Б. А. Келлер (1911) наблюдал со-
общество, в котором в смеси со степным ковылем произрастал
тростник. По этому поводу Келлер писал: «Чем объяснить это
парадоксальное на первый взгляд совместное нахождение болот-
ного растения-—тростника со степным растением — ковылем?
Нет сомнения, что тростник приурочен здесь своей корневой си-
стемой к более глубоким горизонтам грунта, богатым водой,
а ковыль к сравнительно сухим поверхностным слоям». «Возни-
кает вопрос,— пишет далее Келлер,— как поселился тростник на
сухом в поверхностных слоях субстрате? Однако при некотором
анализе всех сопутствующих условий этот вопрос разрешает-
ся без труда. Именно, мы вправе допустить, что каждый год
в известное время вегетационного периода (например, весной)
грунтовые воды на данном местообитании подходят близко к по-
верхности или даже выпотевают наружу, или, может быть, такое
явление происходило в какие-нибудь исключительные годы, и то-
гда здесь поселился тростник».
Использование растительных группировок в качестве индика-
торов имеет широкое применение в лугопастбищном хозяйстве,
лесоводстве, при сельскохозяйственном районировании и микро-
районировании.
Не менее велико значение индикаторов и при различного
рода мелиоративных работах, а также при общей бонитировке
земель и в том числе при выборе целинных и залежных массивов
для распашки.
Особой отраслью является использование растительного по-
крова в качестве индикатора более или менее глубинного залега-
ния тех или иных руд или вообще горных пород, а также подзем-
ных вод.
Интересная и важная работа по этой проблеме была проведе-
на С. В. Викторовым (1955) и другими исследователями в раз-
личных частях Средней Азии. Автор предупреждает, что нет таких
растений или растительных сообществ, которые указывали бы на
определенную горную породу в любых географических условиях.
Он подчеркивает, что как литологическая, так и гидрогеологиче-
ская специфика одних и тех же видов растений может оказаться
неодинаковой в разных природных зонах и районах. Это ограни-
чение в полной мере относится к индикаторам и почв и климата.
В пустынях и полупустынях (рис. 12) очень часто более вла-
голюбивые сообщества располагаются вдоль линии контакта
различных пород, где обыкновенно имеются хотя бы незначитель-
ные различия в водопроницаемости соприкасающихся слоев. Это
доказывают и наблюдения Е. А. Востоковой (1953) в западном
Казахстане. При таких контактах, когда верхний горизонт был
69
Рис. 12. В Ногайской
полупустыне.
более водопроницаемым, чем нижний, обозначались полосы та-
ких влаголюбивых группировок, как заросли чия (Lasiagrostis
splendens) и тростника (Phragmites communis) с участием гре-
бенщика (Tamarix) и ивы. Нередко наблюдаются также и линей-
ные участки специфической растительности вдоль линий тектони-
ческих нарушений.
Использованию растительности при поисках рудных место-
рождений также уделяется немало внимания.
Существуют растения, указывающие на повышение содержа-
ния в почвах или горных породах тех или иных металлов. Напри-
мер, в Европе на богатство пород цинком безошибочно указыва-
ют галмейная фиалка (Viola calaminaria) и галмейная ярутка
(Thlaspi calarmnarium), а в штате Монтана (США) залежи сереб-
ряной руды были открыты по распространению растения Eryogo-
num ovalifolium.
При использовании геоботанического метода для поисков тех
иди иных металлов большое значение приобретает исследование
химического состава золы растений. При обычных условиях хи-
мический состав золы каждого вида и даже рода растений явля-
ется более или менее постоянным. Но близ рудных месторожде-
70
ний этот состав меняется, показывая повышенное содержание
определенных металлов.
В 1956 и в последующие годы состоялись совещания геобота-
ников и геологов, показавшие значительную помощь и экономиче-
скую выгодность геоботанического метода как вспомогательного
в целом ряде полевых исследований по геологии и гидрогеологии.
Было отмечено, что индикационные, свойства растительности
с успехом используются также при дешифровке аэрофотосним-
ков, когда приходится распознавать многие геоморфологические,
почвенные, гидрогеологические и геологические особенности
местности. Вместе с тем совещания обсудили и некоторые труд-
ности, нередко возникающие при использовании растений-инди-
каторов и их сообществ при геологических исследованиях. Не
всегда удается установить достаточно отчетливые индикационные
признаки, что объясняется как сложностью окружающей расте-
ния обстановки, так и разнообразием, а подчас трудной уловимо-
стью приспособительной реакции растений. Совещания подчерк-
нули необходимость дальнейшей работы в этом направлении.
В книге Б. В. Виноградова «Растительные индикаторы и их
использование при изучении природных ресурсов» (1964) имеют-
ся главы о растительных индикаторах природных вод, горных
пород, полезных ископаемых и геологического строения. Интерес-
ны сводная работа Н. Г. Несветайловой «Ботанические показате-
ли полезных ископаемых» (1965) и сборник статей под общим
заглавием «Растительные индикаторы почв, горных пород и под-
земных вод» (1964), явившийся результатом совещания 1961 г.
ВЛИЯНИЕ РАСТИТЕЛЬНЫХ СООБЩЕСТВ
НА ПОЧВУ И КЛИМАТ
Зависимость фитоценозов от почвенных и климатических
условий была показана выше на целом ряде примеров. Однако
фитоценоз не только испытывает те или иные воздействия со сто-
роны внешней среды, но и сам непрерывно изменяет среду. Эти
изменения не всегда уловимы. При их изучении иногда нельзя
четко отграничить влияние фитоценоза на почву от его влияния
на климат. Например, создавая затенение, лесной или другой
ценоз изменяет этим до некоторой степени как почвенный режим,
так и степень влажности воздуха, а эта последняя в свою очередь
влияет на испарение верхнего слоя почвы и покрывающих ее
трав или мхов. Тем не менее в методических целях все же можно
условно представить их в относительно изолированном виде.
Можно различать влияние растительных сообществ на солнечную
радиацию, влажность почвы, температуру почвы, температуру
воздуха, влажность воздуха, осадки, ветер, испарение, химиче-
ский состав почвы и ее физическое строение.
71
ВЛИЯНИЕ НА СОЛНЕЧНУЮ РАДИАЦИЮ
Лучи солнца, падая на растительный покров, частично им от-
ражаются (главным образом листьями), частично поглощаются
и частично пропускаются. При этом многое зависит от цвета
листьев, от содержания в них воды, от их консистенции и т. д.
Характер листвы влияет не только количественно, но и качествен-
но па состав света внутри фитоценоза. Листва больше поглощает
одни лучи солнечного спектра и меньше — другие. Возможно, что
в этом состоит одна из причин наблюдаемого иногда различия
в составе травяного покрова иод кронами разных древесных по-
род в тени леса.
Необходимо, однако, иметь в виду, что под пологом леса мы
имеем два вида радиации: прямую и рассеянную. Прямая радиа-
ция — это та, которая не отражается и не пропускается частями
растений, а проникает под полог леса через его просветы в виде
бликов. В прямой радиации спектральный состав солнечных
лучей остается неизменным, т. е. полным, но интенсивность света,
естественно, ослаблена. Рассеянная радиация слагается из света,
падающего под полог леса через просветы, находящиеся ближе
к зениту, а также света, отраженного растениями и прошедше-
го сквозь их листву. Кроме того, часть солнечных лучей падает
не на растения, а на другие поверхности: обнаженную почву,
камни и т. д. Если эти поверхности обладают способностью отра-
жать заметную часть радиации, то такие отраженные лучи, пада-
ющие затем на растения, могут вновь ими отражаться.
Л. С. Берг (1947) приводит пример, когда отражаемая снегом
в лесу радиация нагревает стволы деревьев, а последние, в свою
очередь, излучая тепло, способствуют таянию снега у основания
стволов. Получаются всем известные воронки в снежном покро-
ве у основания деревьев, хорошо заметные, если в течение
известного времени стояла ясная морозная погода. Эти прота-
линки, очевидно, могут содействовать более раннему прораста-
нию весенних трав в непосредственном соседстве со стволами
деревьев.
В. Р. Вильямс (1939) дал более полный анализ этого явления.
Он отметил, что если бы причиной образования воронок было
только отражение тепла от стволов, то воронки были бы эксцент-
ричны по отношению к стволу, что мы и замечаем вокруг столбов
цлк неживых стволов. Но стволы живых деревьев всегда зани-
мают центр воронки, и Вильямс объясняет это тем, что ранней
весной кроны и стволы деревьев днем нагреваются, и в течение
круглых суток поглощенное тепло легко передается вниз к осно-
ванию ствола, так как живое влажное дерево представляет собой
прекрасный проводник тепла. Здесь тепло, излучаемое живым
растением, беспрерывно расходуется на согревание почвы и от-
таивание снега вокруг стволов деревьев.
72
Рис, 13. Ветреница нарциссоцветная (Anemone narcissi Пота)
на лесной поляне в Карпатах.
Отражаемая часть солнечной радиации, выраженная в процен-
тах от общего количества се, называется альбедо. Разные фито-
ценозы могут быть охарактеризованы различными величинами
альбедо, которые, однако, меняются как по сезонам года, так
и в течение суток. Имеются, например, данные, что, в то время
как в лиственном лесу в летнее время альбедо составляет в сред-
нем 13—17, в травяном покрове поляны (рис. 13) оно равно 25,
а в травяном покрове, выгоревшем на солнце,— 22. Об отноше-
нии между отражательной, поглощательной и пропускатсльиоп
способностью отдельных листьев некоторых растений говорят
наблюдения Н. Н. Калитина (1938), показавшие, что, в то тре-
мя как альбедо листа составляет около 25—35%, поглощается
и расходуется на транспирацию около 20—25% всей падающей па
поверхность листа лучистой энергии и до 45--50% пропускается
через его пластинку.
I
ВЛИЯНИЕ НА ВЛАЖНОСТЬ ПОЧВЫ
Влияние растительности на влажность почвы очень сложно.
В этом отношении лучше изучена лесная растительность. Обстоя-
тельные данные мы находим у Вильямса. Он прежде всего отме-
тил, что в лесу ранней весной, еще задолго до обнажения поверх-
ности почвы кольцом вокруг стволов, можно увидеть талую
почву вокруг всех толстых корней деревьев, вследствие чего ве-
сенняя вода впитывается в лесную подстилку и отчасти в талую
почву леса.
73
Лесная подстилка, состоящая из опавших листьев, хвои, побе-
гов и коры и представляющая рыхлый слой мертвого органиче-
ского вещества, обладает упругостью, так как главную часть
подстилки составляют отмершие побеги. По Вильямсу, 90% всех
побегов, образовавшихся на дереве весной, отмирает в то же лето.
Накопляясь в подстилке, упругая древесина побегов сообщает
и ей значительную упругость. В результате такого упруго-рых-
лого расположения элементов лесная подстилка обладает влаго-
емкостью в высшей степени ее проявления и одновременно отли-
чается высокой степенью проницаемости. Вода в лесной подстил-
ке содержится поэтому как в состоянии волосной воды, так и в
состоянии капельножидком, в связи с чем устанавливаются
одновременно два типа водного режима.
- Влагопроницаемость лесной подстилки очень велика. В связи
с этим образование стремительных делювиальных потоков на
покрытой лесной подстилкой поверхности неосуществимо. На
этом основано защитное влияние лесных группировок, даже на
крутых горных склонах. Только лесное насаждение в состоянии
предотвратить эрозию, т. е. размыв и снесение почвы с горных
склонов. Из этого указания Вильямса мы видим, что защитное
действие леса на горных склонах связано не столько со скрепле-
нием почвы древесными корнями, сколько с предотвращением
размыва почвы, энергично впитывающей воду лесной подстилкой.
Самый старый, т. е. прикасающийся к почве, слой подстилки на-
зывается лесным войлоком и состоит из уже в значительной сте-
пени разложившихся органических остатков, почти потерявших
свою упругость, слежавшихся в более плотную массу и связанных
между собой бесчисленным множеством нитей грибницы. Гори-
зонтальное движение воды по массе этого войлока происходит
крайне медленно, так что значительная ее часть успевает впиты-
ваться в почву и накапливается в ней.
На ровных или слабонаклонных местоположениях большое
значение приобретает вопрос о влиянии леса на уровень грунто-
вых вод. Исследования, проводившиеся в конце прошлого и нача-
ле нынешнего столетия Г. Н. Высоцким и П. В. Отоцким, привели
их к выводу, что по сравнению с пахотным полем или поляной лес
высушивает почву, снижая уровень грунтовых вод (Высоцкий,
1938). Сравнивая изменение уровня грунтовых вод в Велико-
Анадоле под лесом и под поляной в течение 1901 —1902 гт.,
Г. Н. Высоцкий отмечал, что, в то время как весной 1901 г.
уровни воды на этих двух участках были одинаковы, с конца
мая — начала июня началось снижение уровня вод, более силь-
ное под лесом. В начале сентября под лесом началось было под-
нятие воды, но новые сухие дни его снизили, и лишь со второй
половины сентября, с началом листопада, начался новый подъем
уровня воды, снова приведший к выравниванию уровней в течение
зимы и весны 1902 г. После этого наступило новое расхождение.
74
Даже и временный подъем грунтовых вод под лесом
Г. Н. Высоцкий объяснил тем, что наблюдения производились
в «выпотной низине». В целом же Высоцкий приходит к выводу,
что лес усиливает расход почвенной влаги, тогда как вырубка
леса вызывает, наоборот, повышение влажности почвы, за ис-
ключением лишь ее самых верхних слоев.
Однако данные Всесоюзного научно-исследовательского ин-
ститута лесного хозяйства (ВНИИЛХ) не подтвердили вывода
Высоцкого о том, что лес сушит равнины. Сотрудник этого инсти-
тута В. И. Рутковский (1954) указывает, что как Г. Н. Высоцкий,
так и П. В. Отоцкий проводили свои основные исследования
в период длительных засух, и именно это привело их к выводу,
что уровень грунтовых вод под лесом понижается. В действитель-
ности же уровень грунтовых вод сильно зависит от погодных
условий. Во влажные периоды уровень грунтовых вод под лесами
поднимается выше, чем в поле, а в засушливые годы — падает
ниже. Но когда после засухи наступает дождливая погода, то
в лесу почва поглощает значительно больше влаги, чем на от-
крытом месте.
Следует также отметить, что низкий уровень грунтовых вод
далеко не всегда связан с недостатком влаги в лесной почве. До-
бавим к этому, что огромное накопление влаги в почве под лесом
в условиях влажной погоды приводит к тому, что впоследствии
значительное расходование ее (путем прямого испарения и тран-
спирации) в засушливое время хотя и создает значительный де-
фицит влаги в почве, но умеряет сухость атмосферы, а этим са-
мым уменьшает иссушение и соседних открытых мест. Таким
образом, мы приходим к выводу, что общая гидрологическая
роль леса должна оцениваться не отдельно по его влиянию на
влажность почвы, а по его влиянию на режим влаги и в почве,
и в воздухе. Тогда для нас станет еще очевиднее не иссушающее,
а водорегулирующее значение леса. К этому вопросу мы вернемся
еще и ниже при рассмотрении прямого испарения и транспирации.
По сравнению с безлесными участками лес уменьшает сток,
а это, в свою очередь, создает условия для повышения уровня
грунтовых вод как непосредственно под лесом, так и на ближай-
ших к нему открытых местах.
По исследованиям Г. А. Харитонова (1949), проводившимся
в 1938—1941 гг. на юге лесостепной зоны, в районе Шипова леса,
за время зимних оттепелей сток в поле составлял 10,3 т, тогда
как в лесу стекало всего 2,4 т с гектара. Тот же автор приводит
Исследования Г. Ф. Басова в Каменной степи (Воронежская об-
ласть), где при наличии лесных полос, занимающих всего лишь
6% сельскохозяйственной площади, стекал только 31% снеговых
вод, тогда как на безлесной территории их стекало 54%. В связи
с этим в Каменной степи за 50 лет существования лесных полос
уровень грунтовых вод повысился на 1,7 м.
75
ВЛИЯНИЕ НА ТЕМПЕРАТУРУ ПОЧВЫ
Л. С. Берг приводит чрезвычайно интересные данные иссле-
дований В. Н. Оболенского, Э. Лоске и др., свидетельствующие
о различиях теплового режима почвы под лугом и под степью.
Оказывается, что, в то время как почва под лугом летом бывает
холоднее, чем оголенная, под степью она, напротив, теплее.
Значительное влияние на температурный режим почвы оказы-
вает способность растительного покрова задерживать и накап-г
ливать снег. Эта способность неодинакова у разных типов фито-
ценозов. О нагревании почвы живыми стволами и корнями де-
ревьев было упомянуто выше. Несомненно, что и травы нагрева-
ют почву непосредственным излучением тепла своими корнями и
подземными побегами.
Д. П. Буриапкий, И. К. Винокурова и др. (1952) сообщают
интересные данные о температурном режиме почвы в открытой
степи и на участках вблизи лесных полос (между ними) на тер-
ритории Института земледелия центрально-черноземной полосы
им. В. В. Докучаева (Каменная степь). Оказалось, что соотноше-
ние температуры почвы до глубины 20 см на полях среди лесных
полос и в открытой степи меняется в течение вегетационного
периода. Весной температура почвы среди лесных полос выше,
чем в открытой степи, а в начале лета более высокая температу-
ра почвы, наоборот, свойственна открытой степи. На глубине от
50 см и ниже на межполосном участке летом наблюдается более
сильное прогревание, чем в открытой степи. Указанные особен-
ности теплового режима почвы межполосных полей весьма бла-
\ гоприятмы для сельскохозяйственных культур.
ВЛИЯНИЕ НА ТЕМПЕРАТУРУ ВОЗДУХА
Шестилетние наблюдения А. П. Тольского (1918) в Бузулук-
ском бору (Чкаловская область) показали, что в лесу бывает
холоднее, чем на прилегающих полянах, причем в среднем за год
эта разница температуры воздуха составляет 0,5°. Кроме того,
выяснилось, что годовая амплитуда в лесу меньше, чем на поля-
не. Меньше в лесу и суточные амплитуды.
Сходные соотношения были получены А. И. Палечек (1927—
1928) при сравнительном изучении температуры воздуха в иво-
вых зарослях и на лугу в окрестностях г. Перми в течение трех
летних дней. Ежедневно температура измерялась с И до 19 часов
через каждые 30 минут. Измерения производились во всех слу-
чаях на высоте 13 см от поверхности почвы как в ивняках, так
и на лугу внутри травяного покрова. В результате оказалось, что
в ивняках средняя температура была всегда ниже, чем на лугу,
а именно: 25 июля на 0 2°, 26 июля на 1,5° и 31 июля на 2°, а сред-
76
няя амплитуда в ивняках оказалась в 1,2 раза меньше, чем
на лугу.
Полезное свойство леса умерять колебания температуры воз-
духа сочетается, таким образом, с некоторым снижением средних
температур внутри леса по сравнению с открытым местом. Вме-
сте с тем стена леса действует, как плотина, задерживая движе-
ние холодных приземных течений воздуха. Поэтому близ лесной
опушки могут при определенных условиях возникать морозобой-
ные ямы. Г. Н. Высоцкий (1938) обращал внимание на то, что
самые резкие морозобойные ямы образуются на пониженных
местах полян, защищенных от ветров, но достаточных для мест-
ного стекания нижних охлажденных слоев.
С. И. Костин (1954) отмечает, что по картам распределения
средних температур воздуха за период от мая до сентября для
центральной части лесостепной зоны (Воронежская область)
можно видеть, что в районах лесных массивов находятся как бы
острова пониженных температур. При этом становится очевид-
ным, что лесные массивы в лесостепной полосе понижают темпе-
ратуру воздуха не только на территории, которую они занимают,
но и в местности, которая к ним прилегает.
ВЛИЯНИЕ НА ВЛАЖНОСТЬ ВОЗДУХА
В упомянутой работе Д. П. Бурнацкого с соавторами было
исследовано влияние на влажность воздуха полезащитных полос
на участках Института им. В. В. Докучаева. Оказалось, что на
межполосном участке увеличение минимальной относительной
влажности воздуха доходит по сравнению с таковой в открытой
степи в течение отдельных суток до 10%. Наряду с этим было
показано, что влажность воздуха над полями с растительным
покровом выше, чем над паровыми полями; это указывает на
увлажнение воздуха за счет транспирации растений.
J ВЛИЯНИЕ НА ОСАДКИ
Следует различать способность различных типов раститель-
ности задерживать осадки и’влияние различных типов раститель-
ности на образование дождевых туч. Было замечено, что дожде-
мер, установленный в лесу, дает большую цифру осадков, чем
установленный на открытом месте. Однако некоторые авторы
указывали, что это увеличение количества осадков оказывается
мнимым, так как на лесной поляне, где ветры слабы, осадки,
особенно снег, не так сильно выдуваются из дождемера, как
в поле.
Г. А. Харитонов (1949) сообщает, что, по исследованиям
А. В. Шипчинского в Каменной степи Воронежской области,
77
было установлено, что над лесом осадков выпадает на 26—35 мм
больше, чем над полем. О влиянии леса на увеличение количест-
ва осадков писали Г. Н. Высоцкий, Н. С. Нестеров, И. И. Касат-
кин и многие другие авторы.
По данным С. И. Костина (1954), в районах Хреновского,
Усманского и Шипова лесных массивов Воронежской области
и Цнинского лесного массива Тамбовской области отмечаются
пятна повышенных осадков. Особенно надежные материалы име-
ются для Хреновского бора, где еще в конце прошлого века
В. В. Докучаев организовал дождемерную сеть из четырех пунк-
тов в степи и шести — в бору и на его опушке. Данные этих пунк-
тов показывают, что над лесом от мая до сентября осадков выпа-
дает на 10—12% больше, чем над безлесными районами.
Сходная цифра (12%) получена для Усманского лесного мас-
сива. Костин упоминает о двух факторах, повышающих коли-
чество осадков на территориях, занятых лесом. Это, во-первых,
повышенная влажность воздуха и пониженная температура и,
во-вторых, так называемая турбулентность. Последняя вызыва-
ется тем, что движущиеся массы воздуха, встречая на своем пути
среди открытой местности лесной массив, обтекают его со всех
сторон, и в том числе сверху. Но так как верхняя поверхность
лесного полога бывает неровной (из-за различной высоты деревь-
ев, наличия окон, прогалин и т. д.), то при ее обтекании образу-
ются многочисленные маленькие вихри. Это и есть турбулент-
ность. Она приводит к добавочной конденсации водяного пара
и к некоторому перераспределению осадков. Автор сообщает, что
подъем воздушных масс и увеличение турбулентности над Шипо-
вым лесом и лесными полосами Каменной степи были в 1950—
1951 гг. подтверждены исследованиями экспедиции Главной
геофизической обсерватории, работавшей со специально обору-
дованным самолетом.
По всей вероятности, влияние леса на возникновение атмос-
ферных осадков тесно переплетается с действием леса, задержи-
вающим осадки. Кроны деревьев задерживают значительную
часть выпадающих осадков и затем возвращают их в атмосферу
при испарении.
Стационарные наблюдения над задержанием осадков поло-
гом леса были организованы еще в 1906 г. проф. Н. С. Нестеро-
вым в Лесной опытной даче Петровской (теперь им. К. А. Тими-
рязева) сельскохозяйственной академии и продолжались
непрерывно до 1941 г. (после 1926 г. ими руководил уже
проф. Г. Р. Эйтинген, который и обработал все материалы). На-
блюдения велись на четырех лесных участках, относящихся к раз-
ным типам насаждений, причем в каждом типе было установлено
по пяти дождемеров. Сравнение многолетних данных показало,
что наименьшее количество осадков задерживается пологом спе-
лого березняка, несколько большее — сосновым насаждением и
78
больше, всего — пологом чистого ельника. Средневозрастный
густой ельник задерживает на своих кронах в среднем за год
одну треть (32%) осадков, выпадающих в открытом месте. При
этом зимой полог ельника задерживает в среднем 16% осадков,
а весной, летом и осенью 31—38% осадков. Однако наблюдают-
ся значительные колебания в зависимости от силы выпадающих
дождей. Очень слабые дожди целиком поглощаются кронами.
Слабые дожди задерживаются пологом елового леса более чем
на 60%, почти не достигая почвы.
Увеличение количества осадков над лесом может объяснять-
ся и действием деревьев как преграды. Когда в горах туча дви-
жется через лес, ветки, сучья и листья деревьев препятствуют ее
движению. Вследствие этого она приходит в такое состояние, при
котором влага не может оставаться в воздухе взвешенной
и осаждается на листьях и ветвях деревьев в форме росы, капель
или кристаллов инея, перехватываясь лесом подобно тому, как
это бывает с речными отложениями, осаждающимися из потока
при замедлении стремительности его течения.
В отношении влияния травяной растительности на увеличение
образования осадков исчерпывающих данных еще нет, но, оче-
видно, оно значительно меньше, чем влияние леса. Влияние же
различных травяных сообществ на задержание осадков очень
велико, причем здесь сказывается специфическое значение и со-
стояние самих осадков (дождь, роса, снег, иней и т. д.).
ВЛИЯНИЕ НА ВЕТЕР
На силу ветра особенно сильно влияют лесные сообщества.
По данным В. А. Бодрова (1950), в Партизанском опытном пунк-
те Генического района Херсонской области в 1946 г. посевы
озимой пшеницы на полях, защищенных лесными полосами, пол-
ностью сохранились, в то время как незащищенные посевы по-
страдали на 50—90% от пыльной бури.
Ветрозащитная роль как естественных лесов, так и искус-
ственных лесных полос сильно зависит от их густоты и строения.
Г. Н. Высоцкий в упомянутой выше итоговой работе (1938), по-
священной гидрологическому и метеорологическому влиянию
лесов, подробно разъяснил механизм влияния леса на движение
воздуха. Проникая внутрь леса, воздушные потоки испытывают
трение о стволы и кроны деревьев. Это трение их тормозит. Набе-
гающие новые воздушные потоки наталкиваются на сопротивле-
ние уже замедленных и в свою очередь замедляются. Поэтому
воздушный поток уменьшает скорость, еще не доходя до лесной
опушки. Далее Высоцкий обратил внимание на то, что воздуш-
ные потоки тем менее проникают в глубь леса, чем гуще древо-
стой. При этом на проникновение воздушного течения в глубь
леса влияет также густота опушки. Если она закрытая, т. е. за-
79
щищена густыми и высокими кустарниками, или же если ветви
деревьев (например, у елей) сбегают до самой земли, то ветер
проникает в лес в меньшей степени, чем при открытой опушке.
Поэтому если опушка плотно закрытая и низбегакицая, то поток
ветра разбивается слабо. Воздушный поток обтекает лесной
полог, сначала поднимаясь кверху, затем плывя над пологом
и, наконец, лишь несколько разреживаясь, стекает снова вниз па
заветренной стороне. Высоцкий отмечал, что влияние такой
опушки на прилегающие окраины полей, по-видимому, невелико,
в противоположность влиянию высокой и воздухопроницаемой
(продуваемой) опушки, которая сильно рассеивает поток нижних
слоев воздуха. Это нашло полное подтверждение в практике
защитного лесоразведения.
В настоящее время различают три основных типа конструк-
ции лесных полос: непродуваемый, ажурный и продуваемый. Осо-
бенности непродуваемых полос ясны из приведенных выше дан-
ных Высоцкого. Ажурные полосы имеют сквозные мелкие просве-
ты, более или менее равномерно распределенные сверху донизу.
Воздушный поток средней силы проходит сквозь такую полосу,
не меняя своего основного направления. Продуваемые полосы
имеют крупные сквозные просветы и чаще всего лишены как ку-
старникового подлеска, так и древесного молодняка (подроста).
Г. И. Матякин в книге «Лесные полезащитные полосы и мик-
роклимат» (1952), представляющей сжатую сводку результатов
исследований Всесоюзного научно-исследовательского института
агролесомелиорации (ВНИАЛМИ), отдает предпочтение ажур-
ным, по возможности многоярусным полосам. Вместе с тем автор
предостерегает против увлечения широкими полезащитными
(40—60-метровыми) полосами, так как они не оправдывают себя
прежде всего с экономической стороны, требуя много труда и
средств и отнимая большую площадь у пахотных земель. Важна
не столько ширина полос, сколько их правильная конструкция.
Поэтому следует остановиться на ширине полезащитных лесных
полос от 10 до 20 м. Добавим, что речь идет именно о полезащит-
ных полосах колхозов и совхозов, тогда как полосы более круп-
ного значения могут быть более широкими.
Интересны наблюдения С. В. Зонна (1959) о ветрозащитной
роли влажных тропических лесов (гилей) южного Китая. Хотя
здесь деревья имеют большей частью неглубокую и не очень
мощную корневую систему, тем не менее гилеи очень ветроустой-
чивы, Это объясняется необычайной густотой древостоя, обили-
ем переплетающих деревьев лиан, большой сомкнутостью и плот-
ностью крон. Поэтому даже сильнейшие ураганы, достигая плот-
ной стены такого леса, теряют свою скорость, а над поверхностью
крон ветер как бы скользит, не будучи в состоянии сильно раска-
чивать деревья.
80
ВЛИЯНИЕ НА ИСПАРЕНИЕ. ТРАНСПИРАЦИЯ
РАСТИТЕЛЬНОГО ПОКРОВА
Влияние растительной группировки на испарение (испа-
ряемость) следует рассматривать прежде всего как результат
действия растительности на ветер, температуру и влажность воз-
духа. Г. М. Тумип (1926) привел результаты наблюдений над
изменением испарения под влиянием защитных лесных полос.
Наблюдения проводились в Каменностепной опытной станции
(теперь Институт земледелия центрально-черноземной полосы
им. В. В. Докучаева). Оказалось, что в теплый период испарение
па участках между лесными полосами при общей облесенности
в 17% уменьшилось по сравнению с открытым местом в среднем
на 39%.
Прямое испарение, или просто испарение, не следует смеши-
вать с транспирационным испарением, или транспирацией. В то
время как прямое испарение представляет собой процесс физиче-
ский, транспирация является очень сложным физиологическим
процессом. Хотя транспирация растений и зависит от тех же са-
мых факторов, что и прямое физическое испарение, т. е. от ветра,
температуры и влажности воздуха и почвы, она зависит еще и от
физиологического состояния растений. Поэтому судить о тран-
спирации только по прямому испарению, учитываемому просто
по убыли воды в испарителях, нельзя.
Л. А. Иванов (1956) приводит очень важные и интересные
данные о соотношении между прямым испарением и транспира-
цией. Эти материалы получены в исследованиях Института леса
Академии наук СССР и других учреждений как на юге степной
зоны, так и в лесостепной зоне и в подзонах смешанных и хвой-
ных лесов. Кроме того, использованы гидрологические карты, со-
ставленные на основании многолетних наблюдений осадков
и стока в речных бассейнах (Троицкий, 1948). Как поясняет ав-
тор, полный расход влаги лесом слагается из: транспирации дре-
востоя; испарения осадков, задержанных кронами: испарения
почвы или лесной подстилки; транспирации травяного покрова.
Хотя две последние величины, т. е. испарение почвы и транспи-
рация травяного покрова, не всегда учтены с достаточной полно-
той и точностью, при использовании «балансного метода»
Г. Н. Высоцкого это и не требовалось, так как величины эти вхо-
дят в расход почвенной влаги. Иванов отмечает большие рас-
хождения между полным расходом влаги лесом и суммарным
испарением поверхности в том же лесу. В то время как в степной
зоне полный расход влаги лесом значительно меньше, чем сум-
марное испарение (на 22—33%), в зоне лесостепи эта разность
уменьшается уже до 8—10%, а в лесной зоне полный расход
влаги лесом уже превышает суммарное испарение на 7—16%
6 П. Д. Ярошенко
81
в подзоне смешанных лесов и на 38—47% в подзоне хвойных
лесов.
Эти расхождения обусловлены в основном тем, что лесной
древостой в степной зоне транспирирует значительно меньше, чем
в зоне лесной, тогда как в тех же самых лесах суммарное испаре-
ние поверхности, наоборот, больше в степной зоне и меньше
в лесной зоне. Чем объяснить, что, хотя в степной зоне условия
температуры и влажности воздуха способствуют более сильному
испарению, транспирация древостоев там меньше, чем в лесной
зоне? Это явление Л. А. Иванов (1952) объясняет экономностью
транспирации, проявляющейся в условиях недостатка влаги.
У разных пород экономность транспирации сильно разли-
чается, причем она зависит от влажности почвы. При более вы-
соком стоянии грунтовых вод и при искусственном поливе эко-
номность транспирации у многих пород уменьшается. Хотя на
транспирацию влияют влажность воздуха, температура и осве-
щение, главным фактором, ограничивающим транспирацию
в условиях степной зоны, является недостаток почвенной влаги.
Это может быть объяснено, как отмечает Л. А. Иванов, извест-
ным положением К. А. Тимирязева о том, что растение автомати-
чески обращает в свою пользу действие тех самых сил, с кото-
рыми оно ведет борьбу. В упомянутой работе Иванова с сотруд-
никами (1952) указано, что вследствие недостатка почвенной
влажности в степной зоне интенсивность транспирации древесных
пород усиливается от воздействйя метеорологических факторов
только до известного предела. Каждый фактор, усиливающий
испарение, за этим пределом автоматически вызывает в растении
сопротивление этому воздействию. Так, повышение температуры
воздуха, ускоряющее отдачу воды, выше известного предела
начинает уже автоматически тормозить транспирацию, так как
часть воды при обмене веществ переходит в связанное состояние,
снижающее упругость паров, а следовательно, и транспирацию.
Поскольку же перегрев тормозит транспирацию, то сходным
образом влияют и возрастающие выше определенного предела
другие, косвенно связанные с перегревом факторы, например,
сухость воздуха и освещение.
Учитывая все это, Л. А. Иванов делает важный вывод, что
увеличение площади леса в степи ведет не к иссушению ее, как
предполагал Г. Н. Высоцкий, а, наоборот, к более экономному
и равномерному расходованию влаги и в то же время оказыва-
ет положительное влияние на водный режим и урожай полевых
культур в степи. Что касается лесной зоны, то там, наоборот,
леса (в особенности темнохвойные) играют осушающую роль,
и поэтому сокращение площадей еловых лесов севера может
в условиях равнины вызвать расширение существующих и возни-
кновение новых болот.
82
ВЛИЯНИЕ РАСТИТЕЛЬНЫХ СООБЩЕСТВ НА КЛИМАТ
В ЦЕЛОМ
Это влияние выражается, во-первых, в том, что каждое расти-
тельное сообщество имеет свой фитоклимат, и, во-вторых, в том,
что оно в той или иной мере воздействует и на климат окружаю-
щей местности, в том числе и на фитоклиматы других фитоце-
нозов.
Под фитоклиматом понимают климат.рпределенного фитоце-
ноза, зависящий от состава и строения сообщества. Так, в густом
еловом лесу проникает под полог ели не более 1 % всей радиации,
воздух в таком лесу безветренный и влажный. Летом почва
в лесу холоднее, а зимой теплее, чем в открытом поле. Амплиту-
да суточных температур воздуха в еловом лесу мало меняется
в течение года. Снежный покров в лесу, особенно темнохвойном,
стаивает значительно позднее, чем на соседнем открытом поле,
так как лес в значительной мере задерживает проникновение
в него теплых масс воздуха. Повышенная влажность и понижен-
ная температура воздуха в лесу и над ним, а также вихревые
движения воздуха над лесом приводят к увеличению количества
осадков на территории, занятой лесом (Б. П. Кароль, 1959).
Значительную специфику имеют и фитоклиматы травяных
фитоценозов. Например, выше было упомянуто о том, что почва
под лугом летом бывает холоднее, чем под степью. Чем гуще
травостой, тем ниже температура и среди растений. В луговом
травостое, особенно состоящем из широколиственных трав,
днем больше всего нагревается поверхность травяного покрова;
отсюда температура снижается как вверх, так и вниз к почве.
Ночью самая низкая температура бывает немного ниже верхней
поверхности травяного покрова (Кароль).
Травостой способен задерживать немало осадков, а также
ливневый и весенний сток. Сток более задерживается теми фито-
ценозами, где сильнее развит слой ветоши (подстилки). Из тра-
вяных сообществ это чаще свойственно степям, особенно целин-
ным.
На сфагновых торфяниках фитоклимат характеризуется, по
А. П. Шенникову (1964), повышенной влажностью воздуха, силь-
ной дневной нагреваемостью приповерхностного слоя воздуха,
сменяемой сильным ночным охлаждением, частыми туманами,
почти полной освещенностью и доступностью ветру. Сфагновый
торф имеет плохую теплопроводность: он медленно оттаивает
весной, мало прогревается летом. Для сфагновых торфяников
характерны ранние осенние заморозки.
Фитоклиматическое влияние сообщества на прилегающую
местность особенно хорошо изучено для лесных сообществ.
Г. Н. Высоцкий назвал.это «лесной пертиненцией», т. е. распро-
странением влияния леса на окружающую территорию. Влияние
6*
83
это видно уже из практики защитных лесонасаждений. На приле-
гающих к полезащитным полосам полях это влияние проявляется
е значительном увеличении урожаев полевых культур. Отмечая,
что влияние крупных лесных массивов простирается на далекие
расстояния, А. П. Шенников (1964) пишет, что современный эко-
логический режим степной зоны в некоторой степени зависит от
состояния лесов и сохранности их на далеком лесном севере.
ВЛИЯНИЕ РАСТИТЕЛЬНЫХ СООБЩЕСТВ НА ПОЧВУ
В ЦЕЛОМ
Каждому крупному типу растительного покрова свойствен и
свой специфический тип почвы. Так, степям свойственны черно-
земы и каштановые почвы; лесам — подзолистые почвы, подзо-
лы, серые лесные почвы, лесные буроземы и некоторые другие
типы; полупустыням — светло-каштановые и бурые почвы; пу-
стыням — бурые почвы и сероземы; высокогорным лугам — гор-
нолуговые почвы и т. д. Все эти типы почв отличаются один от
другого целым рядом признаков. Например, для серых лесных
почв характерна ореховатая структура второго почвенного го-
ризонта, несвойственная ни степным, ни луговым почвам. Черно-
зему свойственна зернистая структура верхнего горизонта, зна-
чительная мощность почвы, нередко наличие в нижних слоях
твердых включений углекислой извести в виде так называемой
белоглазки и т. д. Типы почв сильно отличаются также по своей
микрофлоре и химическому составу.
В настоящее время, особенно в Советском Союзе, по различ-
ным вопросам воздействия растительности на почву накопилась
обширная литература.
Весьма интересные исследования проведены Институтом ле-
са Академии наук СССР (С. В. Зонн, А. Я. Орлов, В. Н. Мина
и др.) по влиянию на почву лесных сообществ.
Так, в горах Северо-Западного Кавказа довольно часто
встречаются смешанные одновозрастные древостои из восточной
ели (Picea orientals) и кавказской пихты (Abies Nordmanniana).
Также обычны смешанные одновозрастные древостои из кав-
казской пихты и восточного бука, тогда как одновозрастные сме-
шения бука с елью совсем нехарактерны. По данным С. В. Зонна
(1950), это объясняется тем, что пихта и ель, а в значительной
мере также пихта и бук по влиянию на почву дополняют друг
друга. Ель по сравнению с пихтой извлекает из почвы гораздо
больше кальция и гораздо меньше железа и, возможно, алюми-
ния. Подстилка из еловой хвои разлагается медленно, и поэтому
запас извлеченного из почвы кальция не успевает пополняться.
Пихтовая же подстилка минерализуется гораздо быстрее. По-
этому под ельником почва приобретает кислую реакцию и сильно
оподзоливается, а под пихтарниками бывает нейтральной или
84
слабо кислой, интенсивно накопляя кальций в поверхностных
горизонтах. Под ельниками формируются подзолистые почвы,
а под пихтарниками типичные бурые лесные почвы. Что касается
бука, то он по своему влиянию на химизм почвы занимает про-
межуточное положение между елью и пихтой.
Воздействие древостоев одной и той же породы на почву
может быть совершенно неодинаковым в зависимости от характе-
ра самой почвы. Еловые древостои, например, во многих случаях
вызывают оподзоливание, содействуя выщелачиванию зольных
элементов. Но, как сообщает С. В. Зонн (1955), исследования
И. М. Розановой в урочище Моховом Орловской области показа-
ли, что еловые древостои, растущие на выщелоченном черноземе,
не только не оподзоливали его, но, наоборот, создавая условия
более активной деятельности грибной и бактериальной флоры,
способствовали обогащению почвы питательными веществами.
Также и дубовые леса в одних случаях увеличивали запасы
питательных веществ в темно-серых почвах и на черноземах,
в других же случаях, при условии обеспечения хотя бы сезонного
внутрипочвенного оттока части влаги атмосферных осадков, они
вызывали оподзоливание серых лесных почв (Зонн, 1955).
Целый ряд исследований касается воздействия защитных лес-
ных насаждений на степные почвы. По данным Института земле-
делия центрально-черноземной полосы им. В. В. Докучаева,
особенно быстрое преобразование степной почвы в лесную
наблюдается внутри широких лесных полос, где создаются луч-
шие условия для накопления подстилки.
В. Р. Вильямс отметил, что для лесных почв, в отличие от
степных и луговых, характерно наличие в верхних слоях не бакте-
риального, а грибного аэробного разложения органического ве-
щества, что объясняется большим количеством дубильных ве-
ществ, попадающих в почву при гниении частей древесных расте-
ний и препятствующих развитию бактерий.
Опыты И. Н. Гоголева (1952) показали, что внесение в почву
определенных количеств дубильных веществ резко подавляет
бактериальный процесс нитрификации. В то же время энергично
размножается грибная флора, обусловливая максимальное на-
копление аммиака. Уменьшение дозы дубильных веществ вызы-
вает обратный эффект: увеличение количества нитратов и умень-
шение аммиака.
Изменения, вызываемые в почве растительным сообществом,
неизбежно ведут к тому, что сообщество само же создает небла-
гоприятные для себя условия и подготовляет свою смену другим
сообществом. Например, растительность солонцов, действуя на
почву в сторону ее рассолонцевания, при определенных клима-
тических условиях подготовляет свою смену растительностью со-
лодей, а эта последняя, еще более рассолонцовывая почву, при-
водит к появлению растительных группировок незаселенных почв.
8о
Довольно сильное оподзоливание почв, наблюдающееся в неко-
торых влажных типах буковых лесов Карпат, вызывается не чем
иным, как жизнедеятельностью самого букового леса. Однако
такое же оподзоливание приводит в определенных случаях к пре-
кращению возобновления бука и к смене букового леса ело-
вым лесом.
Конечно, кроме растительного покрова, большое влияние
на формирование различных типов почв оказывает также климат
зон и подзон. Но непосредственными создателями того или иного
типа почвы являются все же растительность и связанные с ней
почвенная фауна и микрофлора, начинающие превращение ма-
теринской породы в почву с накоплением и разложением в ней
органических остатков. Являясь непосредственным образовате-
лен почвы, растительный покров выполняет в то же время роль
неизбежного и постоянного посредника при воздействии на почву
климатических факторов.
Темы рефератов на семинаре
или в кружке
Книга В. В. Докучаева
«Наши степи прежде и теперь» *
Литература: Докучаев В. В. Наши степи прежде и те-
перь. Избр. труды. М., Изд-во АН СССР, 1949.
Влияние на растительность физико-географиче-
ских условий
Литература: Шенников А. П. Введение в геоботанику.
Изд-во ЛГУ, 1964, стр. 153—198.
Влияние растительности на среду
Литература: Шенников А. И. Введение в геоботанику.
Изд-во ЛГУ, 1964, стр. 198—239.
Индикация растений
Литература: Статьи С. В. Викторова и др. (стр. 7—12) и
Б. В. Виноградова (стр. 210—220) из сб. «Расти-
тельные индикаторы почв, горных пород и под-
земных вод», под ред. А. Г. Чикишева. Изд-во
МГУ, 1964.
86
Темы курсовых работ
Материалы к изучению фитоклимата некоторых
растительных сообществ
Работа выполняется двумя студентами с помощью двух пси-
хрометров Ассмана. Проводятся единовременные определения
в разные часы температуры и относительной влажности воздуха
в двух различных фитоценозах, лучше всего в лесу и на поляне.
Литература: Кароль Б. П. Микроклимат и методы его
изучения при геоботанических исследованиях.
В кн.: «Полевая геоботаника», под ред. Е. М. Лав-
ренко и А. А. Корчагина, т. 1. М., Изд-во
АН СССР, 1959.
Сравнение почв двух различных фитоценозов
Литература: Садовников И. Ф. Почвенные исследова-
ния и составление почвенных карт. М., Сельхоз-
гиз, 1954.
Захаров С. А. Краткий курс практических за-
нятий по почвоведению. М.—Л., Сельхозгиз, 1930.
Рожнова Т. А, О методике полевого изучения
почв при геоботанических исследованиях. В кн.:
«Полевая геоботаника», под ред. Е. М. Лавренко
и А. А. Корчагина, т. 1. М., Изд-во АН СССР,
1959.
5
АССОЦИАЦИЯ
И ДРУГИЕ
ЕДИНИЦЫ
РАСТИТЕЛЬНОГО
ПОКРОВА
АССОЦИАЦИЯ В ТРАКТОВКЕ РАЗЛИЧНЫХ
ГЕОБОТАНИЧЕСКИХ ШКОЛ
Основной единицей растительного покрова принято еще
с Брюссельского Международного ботанического конгресса
1910 г. считать ассоциацию.
Впоследствии начавшие оформляться после Брюссельского
конгресса геоботанические школы разных стран развивали
и уточняли определение ассоциации, сохраняя в то же время ее
значение как основной единицы растительного покрова. Лишь
упсальская школа выдвинула в качестве основной единицы более
мелкую — сопиацию, но в то же время признала, что сходные
социации объединяются в ассоциацию,
СОВЕТСКАЯ ШКОЛА
Советскими геоботаниками было предложено несколько
определений ассоциации. Наиболее популярно определение
В, Н. Сукачева (1938), согласно которому «растительная ассо-
циация, или тип фитоценоза, объединяет фитоценозы, характери-
зующиеся однородным составом, строением и в основном одина-
ковым сложением составляющих их синузий и имеющие оди-
наковый характер взаимоотношений как между растениями, так
и между ними и средой»,
В 1957 г. В. Н. Сукачев предложил еще одно, более полное,
определение ассоциации, которое, однако, не противоречит пер-
вому, а лишь углубляет его. Согласно этой новой формулировке
«растительная ассоциация (тип фитоценоза), являясь основной
таксономической единицей в фитоценологии, объединяет все фп-
тоценозы, однородно участвующие в аккумуляции и в трансфор-
мации вещества и энергии на поверхности земли, или, точнее, ее
фитогеосферы, В соответствии с этим опа характеризуется
в основном однородным видовым составом, однородной синузи-
альной структурой, отражающей соответствующий состав эколо-
гических типов растений, и однородным составом факторов
среды, влияющим на фитзценотический процесс».
88
В обоих определениях подчеркивается, что ассоциация — это
прежде всего тип фитоценоза. Это подразумевает при характе-
ристике ассоциации некоторую неизбежную схематизацию отно-
шений, наблюдающихся в природе, а именно исключение край-
них, редко встречающихся признаков и выпячивание признаков
средних, наиболее обычных. В своем известном «Руководстве
к исследованию типов лесов» В. Н. Сукачев (1930) писал, что
всякая заросль растений представляет собой сообщество. Таким
образом, в любом лесу мы имеем очень большое количество сооб-
ществ. Например, в сосновом лесу на дюнных всхолмлениях, вер-
шины которых заняты лишайниковым бором, на любой вершине
мы имеем особое сообщество, совершенно нетождественное вся-
кому другому, даже на соседней дюне. Чтобы разобраться в этом
разнообразии сообществ, необходимо свести это разнообразие
к определенным типам сообществ, которые объединяли бы одно-
родные по своим существенным чертам сообщества. Эти типы
сообществ и получили название растительных ассоциаций.
Понятие ассоциации в некоторой мере абстрактно как тип фи-
тоценозов и в то же время конкретно как их совокупность. Ряд
признаков случайного характера может быть присущ отдельным
фитоценозам данной ассоциации, но при характеристике ассо-
циации в целом мы эти нехарактерные признаки отбрасываем,
и именно этим ассоциация приобретает некоторую абстрактность.
В связи со всем сказанным необходимо подчеркнуть, что и сам
фитоценоз, или растительное сообщество, также представляет
единственно конкретного (и в то же время частного) и абстракт-
ного (и в то же время общего). Как конкретное понятие сущест-
вует каждый данный фитоценоз, т. е. конкретный «участок ассо-
циации», но вместе с тем о фитоценозе мы говорим и в общем,
отвлеченном смысле, подобно тому как в ботанике говорим о ра-
стении вообще (Сукачев, 1950).
В основу выделения ассоциаций В. Н. Сукачев кладет преж-
де всего наличие тех пли иных преобладающих видов и в их чи-
сле видов-эдификаторов, определяющих основные черты строе-
ния сообщества и создающих в нем некоторые специфические
условия среды.
В качестве примера ассоциации назовем ассоциацию Pinetiim
myrtillosum (бор-черничник), для которой характерно наличие
трех ярусов: первого, образованного сосной (Pinus silvestris),
второго — с преобладанием черники (Vaccinium niyrtillus) и
третьего — в виде сплошного покрова мхов, чаще всего Hypnum
Schreberi или же Hylocomiutn splendens. Ассоциация эта встре-
чается в центральных частях лесной равнинной области СССР,
в местах с более или менее развитым рельефом и небогатыми
почвами, где она занимает более влажные участки с малопро-
точной водой. По соседству с участками бора-черничника неред-
ко встречаются участки другой ассоциации — бора-брусничника
89
Рис. 14. Бор-брусничник (Владимирская обл.).
(Pineluni vacciniosum), приуроченные к несколько более сухим
и бедным почвам (рис. 14).
Л. Г. Раменский принимает ассоциацию лишь как продукт
субъективного выделения из будто бы совершенно непрерыв-
ного ряда растительных сообществ. Последняя ошибка совер-
шенно аналогична ошибке тех систематиков, которые не признают
реальности вида. В обоих случаях методологический дефект со-
стоит в том, что не осознается единство непрерывности и преры-
вистости в природе.
По степени выработанности, или устойчивости, ассоциаций
предложено несколько подразделений. В. II. Сукачев различает
ассоциации коренные и производные. Коренные ассоциации раз-
виваются в природе без влияния человека и таких необычных
факторов, как катастрофическое нападение вреди гелей или
сплошной ветровал, но обязаны своим характером только клима-
ту и почвенно-грунтовым условиям. Производные же ассоциа-
ции — это те, в образовании которых участвуют указанные выше
91)
факторы. Но как те, так и другие ассоциации могут быть в раз-
ной степени постоянства и временности. Например, некоторые
коренные ассоциации при быстро развивающемся заболачивании
могут носить временный характер, а производные могут по-раз-
ному отличаться от тех коренных, из которых они возникли.
Я предложил (П. Д. Ярошенко, 1953) различать узловые
и кратковременные ассоциации. Первые более устойчивы и мо-
гут длительно существовать на каждом занимаемом ими участке
без существенных изменений. Вторые существуют на каждом
конкретном месте относительно недолго. Впоследствии (1961)
мною было добавлено еще подразделение этих узловых и кратко-
временных ассоциаций на первичные, т. е. не вызванные к су-
ществованию деятельностью человека (вырубка, выпас, выжига-
ние, распашка, осушение, орошение), и вторичные, т. е. вызван-
ные к жизни воздействием человека. Таким образом, мое
подразделение выглядит так:
Узловые ассоциации:
первичные, 'вторичные.
Кратковременные ассоциации:
первичные, вторичные.
Узловые первичные ассоциации соответствуют коренным
ассоциациям В. Н. Сукачева. Примером может служить ельник-
кисличник. После вырубки на его месте возникает вторичная
кратковременная ассоциация березняка, соответствующая произ-
водной ассоциации в смысле Сукачева.
Узловые вторичные ассоциации, по Сукачеву, также будут
и производными, причем такие длительно существующие на каж-
дом конкретном месте производные ассоциации Сукачев назы-
вает устойчиво-производными. Например, на месте коренной
(узловой) ассоциации сосны со вторым ярусом дуба после вы-
рубки сосны возникает дубовый лес, в котором обычно сосна ужо
вовсе не появляется. Это будет, по Сукачеву, устойчиво-произ-
водная ассоциация, а по моей терминологии — узловая вторич-
ная. Узловые вторичные ассоциации весьма обычны также среди
травянистой растительности, например на пастбищах.
Что касается кратковременных первичных ассоциаций, то в
подразделении В. Н. Сукачева они места не находят. Между тем
сюда должны быть отнесены все те ассоциации, которые пред-
шествуют коренным или узловым при зарастании так называе-
мых молодых или новых местообитаний: песков, скал, глинистых
обнажений, галечников и т. д. В. Б. Сочава (1957) называет та-
кие ассоциации серийными, заимствуя этот термин у англо-аме-
риканских геоботаников, а среди производных различает дли-
тельнопроизводные и кратковременнопроизводные. Таким обра-
зом, развивая подразделение В. Н. Сукачева, Сочава различает:
коренные, серийные, длительнопроизводные и кратковременно-
производные ассоциации.
91
Рис. 15. Сосново-Березовый лес (Владимирская обл.).
В. В. Алехин отличает фитоценозы как более устойчивые об-
разования от менее устойчивых группировок. Ассоциации он счи-
тает лишь единицами фитоценозов. Этому следует и А. П. Шеи-
инков. Итак, мы видим, что у отдельных советских исследовате-
лей имеются некоторые различия в понимании основной единицы
растительности — ассоциации. Вместе с тем для советских геобо-
таников общим является признание единства растительного сооб-
щества (а следовательно, и его ассоциации) со средой, а также
признание того, что в развитии растительных сообществ’пе может
быть остановки, что развитие это постоянно. Выделение корен-
ных и узловых ассоциации означает признание нс их неизменяе-
мости, а лишь их отиоснтелыю большей устойчивости по сравне-
нию с кратковременными ассоциациями. Одна из таких узловых
вторичных ассоциаций изображена на рис. 15.
Аналогично ассоциациям выделяются микроассоциадии, т. е.
типы микрогрунпировок (микрофитоценозов). Подобно ассоциа-
циям можно различать микроассоциации: коренные и производ-
ные, первичные и вторичные, узловые if кратковременные.
92
ШВЕДСКАЯ ШКОЛА
Упсальская (шведская) школа, наиболее ярким выразителем
которой является Дю-Рие, при установлении и характеристике
единиц растительных сообществ придает первостепенное значе-
ние видам, доминирующим в каждом ярусе. Вместе с тем э':а
школа придает большое значение подразделению ассоциаций на
более мелкие единицы-—социации. По Дю-Рие (Du-Rietz, 1930),
социация объединяет устойчивые фитоценозы с одинаковыми до-
минантами в каждом ярусе. Так, среднеевропейские заросли
брусники с покровом лишайника Cetraria islandica относятся
к одной социации, тогда как заросли брусники с различными
покровами составляют ассоциацию.
В. В. Алехин, заимствовавший у шведов понятие социации,
подразделяет ассоциацию бора-черничника Pinetum myrtillosurn
на социацию Pin. silv. — Vacc. myrt. — Hypnum Schreberi и co-
циацию Pin. silv. — Vacc. myrt.— Hylocomium splendens.
Шведская школа прежде выделяла ассоциации в основном по
доминантам, но впоследствии стала чаще выделять их по так
называемым характерным видам, заимствуя этот принцип у фло-
ристической школы (см. ниже).
Большое значение эта школа придает при характеристике со-
циаций и ассоциаций также константам, т. е. видам, которые мо-
гут и не доминировать, но встречаются на всех или почти на всех
участках данной социации или ассоциации.
Как ассоциацию, так и социацию Дю-Рие считает единицами
так называемых стабильных растительных сообществ, т. е. таких,
которые находятся в относительном равновесии с местообита-
нием, являясь более или менее выработавшимися. Для лабиль-
ных же, т. е. невыработавшихся и потому чрезвычайно изменчи-
вых, растительных сообществ следует, по мнению Дю-Рие, разра-
ботать другие классификационные единицы. К лабильным сооб-
ществам он относит неустойчивые заросли на молодых место-
обитаниях, заросли сорняков и т. д.
ФЛОРИСТИЧЕСКАЯ (ФРАНЦУЗСКАЯ] ШКОЛА
Флористическую школу можно назвать также французской,
поскольку ее главой является французский геоботаник Браун-
Бланке, но в настоящее время ей следуют также многие немец-
кие, венгерские, польские, румынские и некоторые другие геобо-
таники. Принимая в качестве основной единицы ассоциацию, эта
школа устанавливает ее по так называемым характерным ви-
дам растений. Это те виды, которые имеют определенную вер-
ность данной ассоциации, т. е. свойственны только ей или глав-
ным образом ей. Отсюда и крупный объем ассоциаций этой шко-
лы. Характерные для какой-либо ассоциации виды не всегда
93
являются ее доминантами, но могут произрастать в ней и как
примесь. Так, в ассоциации рододендрона волосистого и сосно-
вого стланика (Mugeto— Rhodoretum hirsuti) характерными ви-
дами являются не только рододендрон и сосновый стланик, но и
горная смородина, и силезская ива, и некоторые другие виды.
В ассоциации дубово-березового леса с Quercus robur и Betula
pendula (Querceto — Betuletum) характерными видами оказы-
ваются не дуб и не береза (хотя они и доминируют), а осина,
майник двулистный, седмичник и некоторые другие.
Весьма крупная по объему ассоциация флористической шко-
лы подразделяется на более мелкие единицы — субассоциации.
В то время как ассоциация устанавливается по характерным ви-
дам, субассоциации устанавливаются по так называемым диффе-
ренциальным видам, т. е. по видам, которые в ряду близких со-
обществ встречаются не во всех из них и по которым можно
близкие сообщества легко отличать одно от другого.
В качестве примера ассоциации, установленной по описывае-
мому принципу, назовем Lathyreto — Fagetum submontanum,
описанную в 1938 г. Гартманном и представляющую богатые ви-
дами буковые леса нижних горных местоположений на субстра-
те, богатом основаниями. Характерными видами этой ассоциации
являются лесной ячмень (Hordeum europaeum), орхидные (Cepha-
lanthera rubra, C. grandiflora) и др. Ассоциация разделяется на
четыре субассоциации, различающиеся по дифференциальным ви-
дам: медвежий лук (Allium ursinum) и хохлатка (Corydalis
cava); лесной ячмень (Hordeum europaeum); типичная субассо-
циация без дифференциальных видов; ландыш (Convallaria
majalis). Таким образом, сам бук (Fagus silvatica), являющий-
ся эдификатором этой ассоциации, не входит ни в число ее ха-
рактерных видов, ни в число ее дифференциальных видов. Сле-
дует отметить, что в третьей субассоциации дифференциальных
видов нет вовсе, в связи с чем она и названа типичной. Отметим
также, что, в то время как характерные виды действительно
свойственны в пределах средней Европы главным образом толь-
ко этой одной ассоциации, дифференциальные виды ее субассо-
циаций встречаются и в других ассоциациях, например ландыш
встречается не только в некоторых других ассоциациях буковых
лесов, но и в ассоциациях дубовых и грабовых лесов.
В последнее время Браун-Бланке (1964) признает в качестве
одного из подразделений ассоциации также и социацию, понимая
последнюю приблизительно в том же объеме, который принят
в шведской школе.
Недооценка роли местообитания и упор, который делается
исключительно на флористический состав, придают пониманию
ассоциации флористической школы односторонность и затруд-
няют использование такого рода материалов в луговодстве, ле-
соводстве и т. д. Однако в методике флористической школы есть
94
и положительные черты. Так, придавая большое значение харак-
терным видам, эта школа устанавливает ассоциации не только
по преобладающим растениям, но и по тем видам, которые хотя
и представлены небольшим количеством особей, но за пределы
данной ассоциации не выходят или почти не выходят. Этот под-
ход имеет свои положительные стороны. Из советских геоботани-
ков против одностороннего упора только на доминанты возражал
Л. Г. Раменский, а в последнее время также X. X. Трасс (1965).
Вместе с тем и принцип выделения ассоциаций по характерным
видам вызвал целый ряд возражений, особенно участившихся за
последние десять — пятнадцать лет. Так, анализируя ассоциации,
выделенные по этому принципу Тюксеном (1937) в Северо-За-
падной Германии, Айхингер (1954) отмечает, что такого рода
ассоциации часто не соответствуют наблюдаемым в природе отно-
шениям и не всегда могут служить основой для хозяйственного
использования. Например, описанная Тюксеном ассоциация Epi-
lobium angustifolium — Senecio silvaticus (заросли иван-чая в
смеси с лесным крестовником) формируется на сплошных вы-
рубках трех совершенно различных лесных ассоциации, а имен-
но: березово-дубовой, еловой и буковой. Поэтому для лесовода
такая объединенная ассоциация не имеет никакого значения, и
при картировании с лесоводственными целями нужно все три
типа ее развития наносить отдельно. Добавим к этому, что поч-
венные условия в каждом из трех типов формирования этой
«ассоциации», конечно, неодинаковы. Это по меньшей мере три
различные ассоциации.
Айхингер отмечает, что некоторые сообщества вообще нельзя
различать по характерным видам, как, например, лесные сооб-
щества, возникшие на лесосеках сплошной рубки и используемые
для сбора подстилки. Однако Айхингер упоминает и такие сооб-
щества, которые можно различать по характерным видам, но
нельзя различать по доминантам. К ним он относит богатые мно-
голетними травами буково-пихтово-еловые смешанные леса.
А. П. Шенников (1956) подверг критике систему Браун-Блан-
ке. Отмечая переоценку в ней значения верных видов и недооцен-
ку других признаков сообщества, Шенников утверждает, что для
отнесения нескольких фитоценозов к одному типу (к одной ассо-
циации) необходимо сходство их по большому количеству при-
знаков, которые в совокупности и составляют диагноз ассоциа-
ции, а именно: необходимо сходство флористического состава,
жизненности общих видов растений, истории и тенденции разви-
тия фитоценозов, количественных соотношений между видами
растений и других признаков строения фитоценозов, отражаю-
щих взаимоотношения между растениями и между фитоценозом
и средой. Однако наряду с недостатками Шенников отмечает
в качестве одной из положительных черт системы Браун-Бланке
требование ею многочисленных детальных описаний сообществ
95
как исходного материала при установлении ассоциаций и других
единиц растительности.
Браун-Бланке (1964) склонен признать неизбежным выде-
ление ассоциаций по характерным видам лишь потому, что их
невозможно объективно выделять сразу по многим признакам.
Многомерное выделение ассоциаций и других единиц раститель-
ности нужно, по Браун-Бланке, разрабатывать биоматемати-
чески, но это, по его мнению, — дело будущего.
ФИЗИОНОМИКО-ЭКОЛОГИЧЕСКАЯ {ШВЕЙЦАРСКАЯ,
ИЛИ ЦЮРИХСКАЯ) ШКОЛА
Эта школа настаивает, чтобы установление как ассоциаций,
так и более крупных единиц растительности было не «линейным»,
как у Браун-Бланке, т. е. основанным не на одном признаке, а
«многомерным», основанным на многих признаках. Этим цюрих-
ская школа более чем какая-либо другая приближается к совет-
ской геоботанической школе. Один из представителей этой школы,
Гаме (1954) писал, что многомерное разнообразие, представляе-
мое сообществами даже в пределах небольшой флористической
области, не дает возможности уложить их в линейную систему.
Цель исследования — осветить это разнообразие различными
способами, не отдавая предпочтения ни одному из них.
Мейзель (1954) довольно полно выразил требования цюрих-
ской школы в следующих трех тезисах.
Нужно иметь в виду, что любое растительное сообщество не
изолировано; оно всегда должно рассматриваться только в об-
щем комплексе растительности определенного естественного
ландшафта или части ландшафта, в котором является локальным
пространственным выражением.
Нужно рассматривать каждое растительное сообщество в гео-
графическом аспекте, т. е. определять его региональное место.
При этом немаловажную роль играет рассмотрение ареалогиче-
ских спектров для образующих сообщество видов растений.
Нужно развиваемую школой Браун-Бланке систему расти-
тельных сообществ Южной и средней Европы в наибольшей мере
приспособить для более широкого использования.
Один из современных руководителей физиономико-экологиче-
ской школы — Э. Айхингер (1966) хотя и критикует флористиче-
скую школу, но находит возможным использование и ее принци-
пов в числе других. Изучение многомерного сообщества требует
охвата разнообразных признаков, в том числе и характерных ви-
дов. Но главное, по мнению Айхингера, это развитие сообщества
во времени с учетом не только его состояния в настоящее время,
но его прошлого и будущего. Основной единицей должен быть, по
Айхингеру, тип развития растительности.
96
Иногда в геоботанических характеристиках какого-либо рай-
она представители цюрихской школы вовсе не применяют поня-
тия «ассоциация».
Э. Шмид при изучении растительности Швейцарских Альп
выделяет так называемые модельные фитоценозы, представле-
ние о которых создается путем синтеза большого количества опи-
саний с учетом видового состава, жизненных форм и особенно-
стей структуры. По крайней мере в некоторых случаях модель-
ные фитоценозы соответствуют ассоциациям. Они характеризуют
растительность каждого горного пояса. В отдельных случаях
моделью служит не синтезированный тип, а конкретное сообще-
ство, но чаще модель выводится синтетически.
Значительное внимание уделяет цюрихская школа синузиаль-
ному сложению сообществ, причем в некоторых работах даже
сквозит тенденция к признанию основной единицей не ассоциа-
ции, а синузии, в особенности при описании травянистых и ку-
старничковых, а иногда и лесных сообществ. Так, Липпмаа
(1939) описал синузию Galeobdolon-Asperula-Asarum (зеленчук-
ясменник-копытень), характерную для европейских широколист-
венных лесов и повторяющуюся в различных их ассоциациях.
АНГЛО-АМЕРИКАНСКАЯ ШКОЛА
Большинство исследователей этой школы проводит резкую
грань между так называемыми кли макс-сообществ а ми и серий-
ными сообществами, или стадиями формирования климакс-со-
обществ.
При этом, хотя климакс-сообществами и являются конкретно
как раз те самые сообщества, которые В. Н. Сукачев называет
коренными (а я называю узловыми), трактовка этих понятий у
англо-американских и советских исследователей неодинакова.
Совпадает лишь то, что как те, так и другие считают необходи-
мым различать, во-первых, сообщества достаточно сформировав-
шиеся, стойкие и сообщества недостаточно сформированные, не-
стойкие. Трактовка же самого понятия стойкости растительных
сообществ совершенно различна.
Дело в том, что климакс-сообщества понимаются многими
представителями англо-американской школы как сообщества
стабилизировавшиеся, достигшие окончания своего развития и
могущие в дальнейшем изменяться лишь под влиянием сугубо
внешних причин, а не в результате своей жизнедеятельности.
Советские же исследователи понимают стойкость коренных или
узловых сообществ как стойкость относительную, как замедление
развития, а не полную его остановку и считают, что изменения
сообщества происходят всегда, осуществляясь под влиянием
как внешних, так и внутренних причин в их теснейшем взаимо-
проникновении.
7 П. Д. Ярошенко
97
По определению американского ботаника Клементса, ассо-
циация представляет единицу климакс-сообществ и объединяет
те из них, которые сходны по составу своих доминант и по поло-
жению этих доминант в структуре сообщества. Определение
ассоциации здесь приближается к упсальскому определению со-
циации. Но по своему объему ассоциация англо-американской
школы гораздо крупнее не только социации, но и ассоциации
упсальской школы.
Для серийных сообществ англо-американские геоботаники
предлагают особую систему единиц. В этой системе единицей,
параллельной ассоциации, является ассоция (associes).
Нужно еще упомянуть о том направлении англо-американ-
ской школы, которое в некоторой степени приближается к взгля-
дам Л. Г. Раменского. Сюда относятся, например, такие иссле-
дователи, как Глизон (1939) с его концепцией «индивидуально-
сти ассоциации». Сущность этой концепции состоит в том, что
каждый конкретный фитоценоз есть в то же время и особая ас-
социация, так как вполне сходных между собой фитоценозов в
природе не бывает. Сюда же примыкают Картис (1955) и дру-
гие, развивающие идею «непрерывности» (континуум) расти-
тельного покрова. Эта непрерывность создается тем, что различ-
ные сообщества связаны между собой постепенными перехода-
ми, затрудняющими установление четких границ. Виттэкер
(1953, 1956) придает первостепенное значение в сложении ра-
стительного покрова не сообществам, а популяциям, т. е. сово-
купностям особей отдельных видов.
По мнению этих исследователей, нельзя уложить необычай-
ную пестроту растительного покрова со множеством переходных
группировок в ограниченное число хорошо выраженных типов
или ассоциаций: такая пестрота свойственна не только серийным
или стадийным, но и климакс-группировкам, которые представ-
ляют динамическое равновесие популяций (Виттэкер, 1953).
Многие сторонники этого направления не устанавливают ас-
социаций, заменяя их «скоплениями», или «роями», видов, выво-
димыми статистическим путем. Это направление ярко выражено
в недавно вышедшей во втором издании книге П. Грейг Смита
«Количественная экология растений», переведенной и на русский
язык (1967).
Более умеренную позицию занял Кэйн (1947), отвергая объ-
ективную реальность ассоциации, но вместе с тем отмечая, что
принятие ассоциации как типологической единицы может ока-
заться полезным.
Кершоу (1964), хотя и отмечает, что выбор площадей для
описания ассоциаций целиком субъективен, в то же время скло-
няется к тому, что ассоциации соответствуют сообществам, наи-
более часто встречающимся в каждом данном районе. Он счи-
тает удовлетворяющим применение к ассоциациям понятия иода
98
(nodal—центральный, узловой), введенного Пури. С нашей точки
зрения, нода соответствует не всяким ассоциациям, а лишь уз-
ловым.
Если взгляды Глизона и Картиса больше подходят к взгля-
дам Раменского в его более ранних работах, то взгляды Кэйна и
Кершоу приближаются к позднейшим высказываниям Рамен-
ского, когда он стал признавать возможность выделения типоло-
гических единиц, приближающихся к ассоциациям.
Общей ошибкой всего этого направления, делающего упор на
непрерывность растительного покрова, является, как уже упоми-
налось, недопонимание диалектики единства непрерывности
(континуума) и прерывистости (дискретности). Если же глубоко
осмыслить такое единство, то даже постепенные переходы между
сообществами в пространстве и времени не помешают выделе-
нию типологических единиц, т. е. ассоциаций, и признанию их
реальности. Вместе с тем это не помешает и признанию свой-
ственного ассоциациям, как и любым другим типологическим
единицам, единства отдельного и общего, конкретного и аб-
страктного.
НАИМЕНОВАНИЕ АССОЦИАЦИИ
В советской геоботанической школе чаще всего применяются
два способа наименования ассоциаций. Первый из них состоит
в использовании двойных латинских названий, например Pine-
turn hylocomiosum (бор-зеленомошник), Fagetum asperulosum
(бучина ясменниковая) и т. д. Для правильного составления
двойного латинского названия нужно окончания «etum» и «osum»
приставлять к корням слов, а корни слов определять по их роди-
тельному падежу, отбрасывая окончание, образованное паде-
жом. В некоторых случаях, если двойное название не может дать
достаточное представление о той или иной ассоциации, приме-
няют трех-, а иногда и четырехчленные названия, например:
Betuletum mixto-herbosum (березняк со смешанным травяным
покровом), Piceetum sphagno-herbosum (ельник сфагново-тра-
вяной) .
Второй способ наименования ассоциаций состоит в перечисле-
нии главнейших доминирующих видов, чаще всего по ярусам,
например: Pinus silvestris— Vaccinium myrtillus — Hylocomium
splcndens (сосновый бор-черничник с моховым покровом). Если
же в сообществе ярусы выражены неясно, можно на первое
место поставить доминирующий вид, а на второе субдоминирую-
щий, например Stipa capillata + Artemisia campestris, или же со-
ставить наименование из названий двух равноправных доминан-
тов, например Stipa sareptana — Artemisia lessingiana.
Если перед названием растения стоит знак «плюс», это озна-
чает, что участие данного вида в составе сообщества меньше, чем
т
99
вида, название которого стоит впереди. Если же названия двух
видов соединены знаком «тире», это значит, что участие обоих
видов в составе сообщества приблизительно одинаково. Послед-
нее относится чаще всего к доминантам разных ярусов сообще-
ства, что возможно при условии, когда ярусы выражены доста-
точно отчетливо.
При составлении названий ассоциаций и других единиц
растительного покрова следует стремиться к тому, чтобы эти
названия не были слишком громоздкими и были доходчивы для
производственников: агрономов, лесоводов, землеустроителей
и т. д. Поэтому наряду с латинскими названиями необходимо
давать и более популярные. В ряде случаев можно с успехом
использовать и местные народные названия растительных груп-
пировок, например: боры, субори, груды и др.
Советские геоботаники часто отражают в названии ассоциа-
ции преобладание в ней тех или иных хозяйственных и в то же
время биологических групп растений, а в скобках дают перечис-
ление латинских названий основных видов, например: ассоциа-
ция кустарниково-разнотравно-злаковая (Bromus inermis + Agro-
pyrum repens + Festuca sulcata + Galatella glabra + Artemisia
austriaca — Spiraea hypericifolia). Здесь в русском названии пре-
обладающая группа, т. е. злаки, стоит на последнем месте.
Принцип помещения в русском названии ассоциации преоб-
ладающей группы или преобладающего растения на последнем
месте принят Ботаническим институтом им. В. Л. Комарова Ака-
демии наук СССР и рекомендуется выпущенным этим институ-
том совместно с Институтом леса и Комплексной научной экспе-
дицией по вопросам полезащитного лесоразведения «Кратким
руководством для геоботанических исследований» (1952). Так,
например, название ассоциации «ковыльно-типчаковая» показы-
вает, что в ней преобладает не ковыль, а типчак.
ВЫСШИЕ ЕДИНИЦЫ СИСТЕМАТИКИ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
В геоботанике применяется определенная система таксоно-
мических единиц, в некоторой степени (ио, конечно, далеко не
полностью) аналогичных видам, родам, семействам и другим
единицам систематики. Сходные ассоциации объединяются
в группы ассоциаций, далее следуют формации, группы форма-
ций, классы формаций и, наконец, типы растительности.
В качестве примера приведем подразделение лесного типа
растительности. Этот тип делится на несколько классов форма-
ций, в числе которых есть, например, класс формаций летнезеле-
ных лесов, сбрасывающих листву на холодное время. К этому
классу относятся группы формаций буковых лесов, дубовых ле-
сов, полидоминантных (многодоминантных) лиственных лесов
100
и др. В группе формаций дубовых лесов одной из формаций
являются леса из черешчатого дуба (Quercus robur). В этой фор-
мации можно выделить группу ассоциаций сухих дубрав, а в по-
следней одной из ассоциаций будет осоковая дубрава (Quercus
robur-Carex pilosa).
Кроме лесного (точнее, кустарниково-лесного) типа расти-
тельности, различают еще типы растительности: степной, луго-
вой, пустынный, саванновый и др., различающиеся по жизненным
формам, по сезонной ритмике и другим признакам.
Тема реферата на семинаре
или в кружке
О геоботанических школах и направлениях
Литература: Воронов А. Г. Геоботаника. М., «Высшая
школа», 1963, стр. 352—365.
Ярошенко П. Д. Геоботаника. М.—Л., Изд-во
АН СССР, 1961, стр. 10—22, 26—36, 106—126.
Темы курсовых работ
Экологические ряды растительных ассоциаций
Растительный покров окрестностей населенного
пункта
Литература по этим темам: У р а н о в А. А. Наблюдения
на летней практике по ботанике. Изд. 2-е. М.,
«Просвещение», 1964, стр. 169—198.
6
ОСОБЕННОСТИ
КЛАССИФИКАЦИИ
ФИТОЦЕНОЗОВ
В РАЗЛИЧНЫХ ТИПАХ
РАСТИТЕЛЬНОГО
ПОКРОВА
Классификация растительных сообществ должна основывать-
ся на понимании сообщества как единства организмов и среды.
Взяв этот принцип за основу, мы в то же время должны учитывать
некоторые специфические черты, характеризующие различные
типы растительности: леса, степи, луга и т. д. В каждом из этих
типов классификация сообществ будет иметь некоторые характер-
ные особенности. Основы такой классификации лучше всего раз-
работаны для лесных сообществ, и поэтому на них мы остановим-
ся более подробно,
ЭКОЛОГО-ФИТОЦЕНОТИЧЕСКИЕ РЯДЫ
В. Н. СУКАЧЕВА
Если каждая ассоциация, или (в лесоводстве) тип леса, харак-
теризует определенные условия среды в более или менее узких
пределах, то несколько близких ассоциаций будут характеризо-
вать условия среды уже в более широких пределах, В связи с этим
можно по группам ассоциаций и по ассоциациям (типам леса)
классифицировать типы местообитаний.
Для северных лесов, в особенности для еловых и сосновых,
такие классификации подробно разработаны В, Н. Сукачевым.
Еловые леса северной половины Европейской части СССР Сука-
чев подразделяет на пять групп ассоциаций, соответствующих
пяти типам условий произрастания (рис. 16).
1. Ельники-зеленомошники— Piceeta hylocomiosa.
Рельеф большей частью достаточно развит, места хорошо дре-
нированы, почвы — более или менее богатые суглинки, глинистые
или супесчаные, не заболочены.
2. Ельники-долгомошникн — Piceeta polytrichosa. Рельеф ме-
нее развит, места слабо дренированы, почвы такие же, но уже
несколько заболочены.
102
3. Сфагновые ельники — Piceeta sphagnosa. Рельеф равнин-
ный или представляет собой дно котловины. Места не дренирова-
ны, почвы заболочены.
4. Болотно-травяные ельники — Piceeta uliginoso-herbosa. Дно
логов с заболоченными почвами, но с проточной водой.
5. Сложные ельники — Piceeta composita. Места с богатыми,
хорошо дренированными почвами, большей частью с близким
залеганием известняков.
Каждая из этих групп слагается из ряда ассоциаций, или типов
леса. Например, группа ельников-зеленомошников, характеризую-
щаяся покровом из так называемых блестящих мхов (Hylocomium
proliferum, Pleurozium Schreberi, Rhytidiadelphus triquetrus и др.),
подразделяется на следующие три основные ассоциации:
1) Ельник-кисличник — Piceetum oxalidosum. Характерно зна-
чительное участие в травяном покрове кислицы (Oxalis acetosel-
la), майника (Majanthemum bifolium) и др. Занимает обычно луч-
шие, хорошо дренированные почвы. Древостой высокий, полнодре-
весный, высоко очищающийся от сучьев. Из мхов в покрове часто
преобладает Rhytidiadelphus triquetrus, но обилен также Pleuro-
zium Scheberi, реже Hylocomium proliferum и др.
2) Ельник-черничник — Piceetum myrtillosum. Характеризует-
ся преобладанием в покрове черники и отсутствием или слабым
распространением кислицы и других показателей лучших почв.
Занимает более влажные, с худшей аэрацией почвы. Древостой
ниже и дает меньшие запасы древесины. Из мхов чаще всего
Hylocomium proliferum, реже Pleurozium Schreberi, Ptilium crista
castrensis и др.
Этот тип очень распространен по более ровным местам.
Р. hylocomiosa
Рис. 16. Система эколого-фнтоценотических рядов типов еловых лесов (по-
Сукачеву, 1928);
Р. ох. — ельник-кисличник; р.гп. — ельничек-черничник; Р. V. — ельник-брусничник;
Р. pl. — елькик-долгомошнкк; Р. sph. — ельник сфагновый; Р. с. —sph. — ельник осоково-
сфагновый; Р. tl. — ельник лиловый; Р. gr. — ельник дубовый; Р. f. — ельник лриручье-
вой; Р. sph. — h. — ельник сфагновотравяной.
103
3) Ельник-брусничник — Piceetum vacciniosum. Этот еще
мало изученный тип распространен местами на севере, но отмечен
также и в средних областях Европейской части СССР; приурочен
к несколько более сухим и бедным, часто более песчанистым поч-
вам, чем почва типа ельников-кисличников. Древостой меньше ро-
стом и дает меньшие запасы древесины. Часто наблюдается, хотя
и небольшая в естественных условиях, примесь сосны. Второй
ярус и подлесок отсутствуют, В покрове преобладает брусника;
кислица и майник мало распространены или вовсе отсутствуют.
В моховом покрове преобладает Pleurozium Scheberi.
В каждой из установленных Сукачевым групп типов леса
имеется один как бы стержневой тип, наиболее полно выражаю-
щий свойства данной группы. Остальные типы каждой данной
группы то более, то менее близки к стержневому и в то же время
обнаруживают связи и с типами другой группы, образуя переход
от одной группы к другой. Таким образом, все группы типов ело-
вых лесов могут быть изображены в виде, как их назвал Сукачев,
эколого-фитоценотических рядов, отражающих степень близости
между собой как отдельных типов, так и групп типов, т. е. групп
ассоциаций.
Система таких рядов ельников, составленная Сукачевым
(1928), показана на рисунке 16. В этой системе принято, что
стержневой группой еловых типов является группа Picceta hyloco-
miosa, а стержневым типом этой группы служит тип Piceetum
oxalidosum, наиболее полно выражающий все вообще свойства
елового леса. Здесь, как говорит автор, роль ели как основного
строителя (эдификатора) ассоциации выражена наиболее сильно.
От этого типа, стержневого для всех еловых лесов севера, идут
четыре ряда: А, В, С, D.
Ряд А идет к Р. vacciniosum и характеризуется увеличением
сухости почвы, но без улучшения ее минерального состава, а, на-
оборот, даже с некоторым его обеднением.
Ряд В идет через Р. myrtillosum и Р. polytrichosum к Р. sphag-
nosum и характеризуется увеличением увлажнения и ухудшением
почвенной аэрации.
Ряд С идет через Р. tiliosum к Р. quercetosum и характеризует-
ся увеличением почвенного богатства и аэрации.
Ряд D идет к Р. fontinale и выражает увеличение увлажнения
проточной водой. Р. fontinale — это так называемый приручьевой
ельник.
Наконец, имеется еще и пятый ряд Е, представленный типами
Р. sphagnoso-herbosum и Р. caricoso-sphagnosum. Типы эти обра-
зуют переходы от приручьевого ельника к сфагновом}-' и характе-
ризуются соответственно переходными местообитаниями.
Сукачев указывает, что по такому принципу могут быть по-
строены аналогичные системы рядов ассоциаций и для других
(т. е. не лесных) типов растительности.
104
Рис. 17. Лишайни-
ковый бор (Влади-
мирская обл,).
Во многих случаях (по не всегда) такие ряды будут, хотя бы
частично, отражать и взаимоотношения ассоциаций в процессах
смен.
Приведенные выше ряды ельников являются эдафо-ценотиче-
скими, так как связаны с изменением почвенно-грунтовых условий
в пределах (в данном случае) Европейской части СССР. Но
могут быть, как указывает Сукачев, и географически-ценотиче-
скис ряды, или, точнее, климатически-цепотические ряды ассо-
циаций.
Сукачев подчеркивает отличие его эколого-фигоцснотических
рядов от экологических рядов в представлении Б. А. Келлера.
105
Экологический ряд, по Келлеру,— это конкретный ряд последова-
тельно сменяющих друг друга в пространстве ценозов, могущих
очень сильно между собой различаться. Например, в поймах рек
южной части Русской равнины можно, переходя из понижения на
бугор, видеть сначала лугово-болотные, затем луговые, лугово-
степные и наконец степные ассоциации. Это экологический ряд,
в котором участвуют ценозы трех различных типов: лугово-болот-
ного, лугового и степного. Эколого-фитоценотические ряды Сука-
чева, напротив, связывают родственные ассоциации, причем связь
эта предполагает не обязательно пространственное их чередова-
ние (сообщества эти в пространстве могут и не прилегать друг
к другу), а просто произведенное нами самими размещение их
в определенной последовательности согласно изменению известно-
го экологического фактора, в данном случае почвенно-грунтовых
условий. Ряды Сукачева учитывают не только условия местооби-
тания, но весь комплекс признаков фитоценоза. В числе этих при-
знаков мы видим характер как нижних ярусов (не только кустар-
ничково-травяного, но и мохового), так и верхних ярусов, по-
скольку для каждой, преобладающей в верхнем ярусе древесной
породы ряды составляются отдельно (для ели, сосны и т. д.).
Впоследствии В. Н. Сукачев разработал аналогичную схему
и для сосновых лесов (один из типов сосновых лесов изо-
бражен на рисунке 17): Другие исследователи предложили схе-
мы такого рода для дубовых лесов, ольшаников и др.
ЭДАФИЧЕСКАЯ СЕТКА П. С. ПОГРЕБНЯКА
П. С. Погребняк (1955) критически рассматривает эколого-
фитоценотические ряды Сукачева. Погребняк считает, что стерж-
невой, или центральный, тип леса — это искусственно выхвачен-
ный из природы отрезок. Центральное положение стержневого
типа в системе мотивируется, по Сукачеву, тем, что он наиболее
полно выражает эколого-ценотические свойства данной группы
лесных типов. Но дело в том, что и сама группа еловых лесов есть
группа не вполне естественная, ибо у Сукачева в ней нс отражены
смешанные леса с участием ели, а приняты во внимание только
чистые еловые леса. В природе же наряду с чистыми ельниками не
менее значительное место занимают и всевозможные промежуточ-
ные типы леса. Хозяйственное значение этих типов иногда бывает
более велико, чем значение чистых типов.
Свою классификационную схему типов леса Погребняк (192.9,
1951) строит иначе. В основу ее он кладет только две координаты:
изменение почвенного богатства, точнее, плодородия (трофиости)
и изменения почвенной влажности, причем условно различает
шесть степеней влажности и четыре степени трофности Все воз-
можные сочетания каждой степени влажности с каждой степенью
106
^Свсн° остролисты^ 6елеза ф Члыа
<3> diji кустарники ^сересрииьнь.'е и мезофильные
Рис, 18. Эдафическая сетка типов леса Полесья и лесостепи (по Погреб-
няку, 1955).
трофности и иллюстрируют (конечно, схематично) разнообразие
лесных типов, которые для наглядности на этой же схеме охарак-
теризованы соответственным составом древесного, травяного и ли-
шайниково-мохового ярусов. Следут отметить, что эдафическая
сетка Погребняка пользуется большой популярностью среди укра-
инских лесоводов как чрезвычайно наглядная и поэтому легко
используемая в массовой практике.
На рисунке 18 изображена эдафическая сетка Погребняка,
составленная для лесной и лесостепной областей Европейской
части СССР.
По плодородию почвы в схеме Погребняка различаются: боры,
субори, сложные субори и дубравы, т. е. четыре группы типов ле-
107
са, характеризующие различные степени почвенного плодородия
в лесах Европейской части СССР. Боры — это леса на крайне
бедных почвах, субори — леса на переходных, относительно бед-
ных почвах, сложные субори характеризуются относительно бога
тыми почвами и, наконец, дубравы — это леса на плодородных
почвах. При этом указано, что последнюю группу можно лишь
условно назвать дубравной, так как в нее (в зависимости уже от
климатических условий) приходится помещать и буковые, и пих-
товые, и кедровые, и даже некоторые еловые и лиственничные
леса. Дубравы в узком смысле слова, т. е. леса с преобладанием
дуба и его широколиственных спутников (клена, липы, ясеня,
ильмовых, граба и др.), наиболее распространены в лесостепной
зоне Европейской части СССР.
Д. В. Воробьев (1953) называет группу типов, соответствую-
щую по степени плодородия почвы сложным суборям, сугрудка-
ми, а соответствующую по этому же признаку дубравам — гру-
дами.
В пределах каждого типа леса Погребняк выделяет коренные
и производные сообщества, понимая их в том же смысле, что
и Сукачев. Например, в типе леса «сырой бор (Л4)» коренными
будут сосняки, а производными — березняки, так как вырубки
заселяются березой. В суборях (группа типов леса В) коренными
сообществами являются (в зависимости от типа леса) в северной
части лесной зоны сосново-еловые, в южной — сосново-дубовые,
а в переходной полосе — сосново-дубово-еловые, причем во всех
случаях дуб и ель образуют второй ярус под пологом сосны. Про-
изводными формами суборей являются березняки, осинники, ель-
ники, дубовые низкостволышки или чистые сосняки. Такие произ-
водные формы образуются в результате либо уничтожения вто-
рого яруса, либо сплошных рубок, либо при поселении сосны на
старых пахотных землях (залежах).
Мы видим, что Погребняк понимает тип леса очень широко,
включая в него всегда более одной ассоциации.
КЛАССИФИКАЦИИ ЛУГОВОЙ
РАСТИТЕЛЬНОСТИ
Одним из простейших путей систематизации луговых сооб-
ществ какой-либо конкретной местности является выявление
и описание экологических рядов, т. е. пространственных смен
ассоциаций в связи с изменением какого-либо одного фактора
(а не двух, как в схеме Погребняка), например фактора почвен-
ной влажности.
В. В. Алехин (1925) приводит экологический ряд ассоциаций
для центральной части поймы р. Циы (в бывш. Тамбовской губ.).
108
Существование этого экологического ряда обусловлено простран-
ственной сменой степени почвенного*увлажнения. Алехин выде-
ляет здесь 10 ассоциаций, из которых первая (с наименьшей поч-
венной влажностью) является, по существу, не луговой, а степной,
а последние (наиболее влажные) являются лугово-болотными,
тогда как основная часть ряда представлена типично луговыми
ассоциациями. Весь ряд состоит из следующих ассоциаций:
Ассоциация типчака (Festuca sulcata). Заливается весной,
по-видимому, лишь на очень короткое время.
Ассоциация собачьей полевицы (Agrostis canina).
Ассоциация разнотравно-бобовая.
Ассоциация лисохвоста (Alopecurus pratensis).
Ассоциация едкого лютика, красной овсяницы и погремка
(Ranunculus асег4-Festuca rubra + Alectorolophus major).
Ассоциация лисохвоста и болотного мятлика (Alopecurus рга-
tensis + Poa palustris).
Ассоциация болотного мятлика (Роа palustris). Это уже луг
низкого уровня (сильного увлажнения).
Ассоциация бекманнии (Beckmannia eruciformis). Обычно
располагается по дну западин или на занесенных старицах.
Ассоциация болотного манника (Glyceria fluitans). Это уже
ассоциация луговых болот, которая развивается, если западины
еще более углубляются.
Ассоциация водного манника (Glyceria aquatica). Развивается
еще ниже, уже в самой воде.
Если же водоем достаточно глубок, то в его относительно глу-
боководной части развивается ассоциация Phragmites communis
(тростник), а в еще более глубоководной части — ассоциация
Scirpus lacustris (камыш), после чего следует уже открытое вод-
ное пространство. Алехин оговаривается, что в приведенном эко-
логическом ряду им указаны только те ассоциации, которые
встречаются наиболее часто, являясь поэтому более характерны-
ми. Второстепенные и различные замещающие типы сюда не
включены. Вместе с тем Алехин отмечает, что этот ряд имеет зна-
чение, выходящее за пределы поймы р. Цны. Так, им обнаружен
совершенно такой же ряд на лугах р. Вороны (в той же бывш.
Тамбовской губ.).
На лугах выделять ассоциации обыкновенно несколько труд-
нее, чем в лесах. Объясняется это тем, что на лугах нередко сла-
бее выражено доминирование тех или иных видов растений, на
лугах чаще, чем в лесах, имеет место пространственное чередова-
ние сообществ на сравнительно небольших протяжениях, а также
наблюдаются и более плавные, чем в лесах, переходы между сооб-
ществами разных ассоциаций. Поэтому в луговой типологии,
в особенности при более кратковременных маршрутных исследо-
ваниях, в большей мере, чем в лесной типологии, приобретают
значение единицы крупнее ассоциации.
109
Мохово -лишайниковые
пустоши
психрофитно
оксилофитные
Травянистые пустоши
психрофитная олиготрофная
растительность
Остепненные пустоши
психрофитно -
зуксерофитные
неостро фные
Промежуточные психро- Пустошные луга Промежуточ-
оксиломезофитные фор- психромезофитные ные психрозу-
моции олиготрофные олиготрофные ксеромезофит-
ные формации
Ксероморфная тра-
вянистая ц мохово- /
кустарничковая
ра стительность
оксилофитоё
/Ц\\11Ж
"оксипомезофитные зумезофитные луга зуксеро-т^
олиготрофные мезотрофные мезофитные
\ X. аназрофитные зутрофные / jf
Промежуточные болотистые луга Промежуточные
оксилогидрс-мезо гидромезофитные гидро мезо фит-
мезотрофные
азр&фитнь/е
олиготрофные мезотрофные мезофитные
оксилогидромезо -
фитные формации
елиготрофнь/е
ные зутрофные
формации
Водная растительность
олиготрофных водоемов
Водная растительность
(гида тофитная)
мезотрофных водоемов
Степи зуксерофитныа
зутрофные формации
травянистых много*
летников
Водная растительность
зутртрных водоемо$~
Рис. 19. Схема экологических рядов луговой растительности и смежных типов
(по Шенникову, 1941).
Классификация всей луговой растительности СССР приведена
А. П. Шенниковым в его сводной монографии «Луговая расти-
тельность СССР» (1938), а впоследствии в несколько доработан-
ном виде в книге «Луговедение» (1941). Луга в целом Шенников
считает не классом формаций (вопреки А. П. Ильинскому
и В, В. Алехину), а типом травянистой растительности наряду
с такими типами, как травянистые степи, травянистые пустоши,
травянистая водная растительность, травянистая оксилофильная
растительность и растительность из травянистых однолетников
(Ephemeretum).
Характерной чертой лугового типа растительности (Prata или
Pratoherbosa) Шенников считает то, что к этому типу относятся
ассоциации травянистых многолетних мезофитов. При этом Шен-
ников (1938) поясняет, что мезофитами называются растения,
нормально произрастающие в средних условиях увлажнения,
аэрации, тепла и минерального богатства почвы.
Луговой тип растительности Шенников подразделяет на сле-
дующие пять классов формаций:
настоящие, или эумезофитные, луга;
110
остепненные, или эуксеромезофитные, луга;
пустошные луга;
гидромезофитные, или болотистые, луга; -
оксиломезофитные, или торфянистые, луга.
Далее Шенников (1941) приводит интересную схему экологи-
ческих рядов, показывающую экологические соотношения между
этими классами формаций и их место среди соседних, не луговых.
Схема эта представлена на рисунке 19. Автор приводит и подраз-
деления внутри каждого из этих классов, указывая также наибо-
лее распространенные формации. Так, внутри класса формаций
настоящих лугов „выделяются такие группы формаций: крупно-
злаковые луга, или крупнозлачники; мелкозлаковые луга, или
мелкозлачники; низкозлаковые луга, или низкозлачники; крупно-
разнотравники; мелкоразнотравники; низкотравники; крупнозла-
ково-разнотравные настоящие луга; мелкозлаково-разнотравные
настоящие луга; злаково-разнотравные низкотравники.
В качестве примера приведем подразделение одной из этих
групп формаций на формации. Так, группа мелкозлаковых лугов,
или мелкозлачников, включает такие формации: красноовсянич-
ники; белополевичники; обыкновенные полевичники; душистоко-
лосковые луга; лугомятличники; болотномятличники; трясунко-
вые луга; гребенниковые луга; бескильницевые луга, представ-
ляющие галофитный вариант группы.
Мною (Ярошенко, 1961) в основу подразделения лугового типа
растительности взяты прежде всего признаки структуры траво-
стоя. Важным признаком луговых сообществ, отражающим не
только их структуру в статическом понимании, но и их генезис,
может служить наличие или отсутствие отчетливой мозаичности.
По этому признаку можно разделить все луга как тип раститель-
ности на два подтипа: простые и сложные луга.
Простые луга. Микрогруппировки высших растений лишь едва
намечаются, так что травостой в горизонтальном направлении
почти не расчленен. Если ?ке такое расчленение в некоторой мерс
и выражено, то микрогруппировки в своем большинстве слабо
оформленные, невыработавшиеся. Сюда относятся, во-первых,
многие луга, образованные злаками и осоками, имеющими длин-
ные ползучие корневища и образующие неплотные травостои. Эго,
например, луга из ползучего пырея или из вейника наземного. Во-
вторых, к этому же подтипу мы относим луга из вейника Лангс-
дорфа (рис. 20) и вейника узколистного на Дальнем Востоке,
луга из императы в Юго-Восточном Закавказье и Средней Азии.
К простым лугам следует отнести и большинство искусственных
сеяных лугов.
Сложные луга. Микрогруппировки выражены более или менее
отчетливо, образуя или контурную, или хотя бы плавную, но ясно
заметную мозаику. К сложным лугам относится значительное
большинство луговых сообществ, причем дернообразователями
111
Рис. 20. Луг с преобладанием вейника (Calamagrostis langs-
dorfii) на Дальнем Востоке.
в них могут быть как корневищные, так и рыхло- и плотнокусто-
вые злаковндныс травы. Микрогруппировки образованы сочета-
ниями злаковидных и других трав. При контурной мозаичности
м икр огру ппировки чаще всего распределены по элементам микро-
рельефа. При плавной же мозаичности микрорельеф может быть
почти или вовсе не выражен, и распределение микрогруппировок
обусловливается тогда в большей мере особенностями разраста-
ния и взаимодействия особей в условиях богатого видами сооб-
щества.
Необходимо отметить, что подразделение лугов на простые
и сложные по степени выраженности относительно сформирован-
ных микрогруппировок, будучи легкодоступным для геоботаника,
может оказаться затруднительным для тех работников сельского
хозяйства, которые не приобрели достаточного навыка в геобо-
танических исследованиях.
Однако практически распознавание простых и сложных лугов
облегчается еще и тем, что первые значительно беднее видами,
чем вторые, что отражено в таблице 2.
Данные таблицы 2 показывают, что общее количество видов
значительно больше па сложных лугах, чем па простых, и что еще
большая разница получается, если сравнивать не общие видовые
112
списки, а списки видов, играющих в сообществах сколько-нибудь
заметную роль. Количество таких видов на сложных лугах При-
морского края превышает в два и более раз количество их на
простых лугах. В то время как на простых лугах здесь на метро-
вую площадку попадает обыкновенно не более 5—6 видов, на
сложных лугах на такую же площадку попадает 10—15 видов.
Далее подтип простые луга я подразделяю на два класса фор-
маций: простые луга с преобладанием злаковидных однодольных
и простые луга с преобладанием двудольных.
Подтип сложные луга подразделяется на классы формаций:
сложные рыхлозадерненные луга и сложные луга с сочетанием
рыхлого и плотного задернення.
Классы формаций делятся непосредственно на формации (без
нагромождения промежуточных единиц в виде порядков форма-
ций и групп формаций), формации — на группы ассоциаций и эти
последние—на ассоциации. По такому принципу мною и была
разработана классификация лугов Приморского края на Даль-
нем Востоке, Однако эта классификация пока не доведена до
ассоциаций и заканчивается формациями, а в ряде случаев груп-
пами ассоциаций. Формации и группы ассоциаций как раз и явля-
ются единицами, наиболее удобными для хозяйственной оценки
и разработки тех или иных мер улучшения лугов. Термин «луга»
принят в нашей классификации не в хозяйственном, а в геобота-
ническом смысле, т, е. не как синоним сенокосных угодий, а как
определенный тип растительности. Луга как тип растительности
могут использоваться и как сенокосы, и как пастбища, отдельные
Таблица 2
Количество видов в луговых сообществах Приморского края
Группа ассоциации Общее к о ли че ство Ко л и че с т во видов
видов травянистых растений трав, играющих заметную роль
Простые луга Горные луга из вейника Лангсдорфа .... 19 9
Низинные луга из вейника Лангсдорфа . . . 34 13
Приречные заросли мискантуса 32 15
Пастбищные луга из белого клевера с мят- ликом 16 12
Сложные луга Влажные разнотравные луга с купальницей, осоками и вейником Лангсдорфа 57 45
Суховатые злаково-разнотравные луга с по- левицей Триниуса 48 30
Кочковатые болотистые луга с осоками и вейником Лангсдорфа 46 24
8 П, Д. Ярошенко
113
же типы лугов могут иметь или только сенокосное, или только
пастбищное значение.
Мы привели примеры различных классификаций двух типов
растительности: лесного и лугового. Разрабатывались и класси-
фикации других типов растительности: степного, болотного и т, д..
но мы на них здесь останавливаться не будем.
КЛАССИФИКАЦИЯ И ОРДИНАЦИЯ
В последнее время в геоботанике более или менее четко отли-
чают классификацию растительности от ее ординации. По опреде-
лению английского исследователя П. Грейс-Смита (1967) класси-
фикация состоит в объединении насаждений (сообществ.— П. fl.)
в классы, члены каждого из которых имеют некоторое количество
общих характеристик, благодаря чему они оказываются отграни-
ченными от членов других классов...
Ординация насаждений (сообществ) состоит в расположении
каждого из них в некоторой системе координат (образованной
одной или большим числом осей) таким образом, чтобы иметь
возможность получить максимальную информацию о его составе.
Простейшей ординацией будет, например, расположение сооб-
ществ по одной оси в виде экологического ряда. Несколько более
сложной ординацией будет эдафическая сетка, построенная по
двум координатам.
Классификациями в рассматриваемом узком смысле этого сло-
ва являются системы Браун-Бланке и скандинавских геоботани-
ков. Ординациями в последнее время особенно увлекаются
в США. Эта позиция американских исследователей весьма отчет-
ливо изложена в новой работе Виттакера (1967), посвященной так
называемому градиентному анализу растительности, под которым
обычно понимается соотношение между участием в сообществе
тех или иных видов растений и одним или несколькими факто-
рами.
Эколого-фитоценотические ряды В. Н. Сукачева, как правиль-
но отмечают Т. А. Работнов и А. А. Уранов 1, совмещают принци-
пы ординации и классификации. Это относится и к схеме эколо-
гических рядов луговой растительности А. П. Шенникова, по-
строенной по тому же методу В. И. Сукачева.
Эдафическая сетка П. С. Погребняка построена уже в основ-
ном по принципу ординации, тогда как наше подразделение луго-
вого типа растительности является, наоборот, в большей мере
классификационным.
Одностороннее проведение принципа ординации связано
с представлением о растительном покрове как континууме (не-
1 См. предисловие Т. А. Работнова и А. А. Уранова к русскому изданию
книги П. Грейг-Смита (1967).
114
прерывности). Одностороннее проведение принципа классифика-
ции связано с представлением о дискретности (прерывистости)
растительного покрова. Но так как растительный покров следует
рассматривать как единство непрерывности и прерывистости, то,
очевидно, наиболее правильным будет совмещение принципов
классификации и ординации, что, как указывают Т. А. Работнов
и А. А, Уранов, широко вошло в обиход советских ботаников бла-
годаря талантливой разработке сходной идеи В. Н. Сукачевым на
примере еловых и сосновых лесов.
Темы рефератов на семинаре
или в кружке
О разных направлениях лесной типологии
Литература: Сукачев В. Н. Руководство
к исследованию типов лесов. М., Сельхозгиз, 1931.
Ярошенко П. Д. Геоботаника. М.—Л., Изд-во
АН СССР, 1961, 137 -161.
Классификация степной растительности
Литература: Л а в р е н к о Е. М. Степи
и сельскохозяйственные земли на месте степей.
В кн.: «Растительный покров СССР», пояснитель-
ный текст к геоботанической карте СССР, т. 2.
М.— Л., Изд-во АН СССР, 1956.
Ярошенко П. Д. Геоботаника. М.—Л., Изд-во
АН СССР, 1961, стр. 178—188.
Тема курсовой работы
Ассоциации лесного (или лугового) массива
Выделить ассоциации, описать их и расположить их в эколо-
го-фитоценотические ряды и в эдафическую сетку. Дать сравни-
тельную оценку обоим методам.
Литература: Алехин В. В. Методика по-
левого изучения растительности и флоры. М.,
Изд-во Наркомпроса, 1938.
Ярошенко П. Д. Геоботаника. М.—Л., Изд-во
АН СССР, 1961, стр. 137—150 и 161 — 178.
8*
7
ДИНАМИКА
РАСТИТЕЛЬНЫХ
СООБЩЕСТВ
Как мы видели, растительное сообщество представляет собой
не застывшее в неподвижности, а непрерывно изменяющееся об-
разование. Основные типы этих изменений следующие: сезонные;
разногодние, или флюктуации, т. е. изменения сообщества по го-
дам в зависимости от метеорологических факторов и других усло-
вий данного года; частные смены, или сукцессии; общие смены;
эволюция ценозов.
В любой момент своего существования каждое сообщество
находится в изменении, и это изменение всегда включает элемен-
ты перечисленных пяти типов. Действительно, сообщество в каж-
дый момент своей жизни проходит одну из фаз своего сезонного
развития; отражает на себе влияние условий данного года; нахо-
дится на пути к смене каким-либо другим сообществом; находит-
ся под влиянием общей смены растительного покрова всей мест-
ности и, наконец, проходит одну из стадий своей эволюции. Отсю-
да видно, что все эти изменения есть не что иное, как различные
стороны единого общего процесса — развития растительного по-
крова.
Частные смены, или сукцессии,— это смены, осуществляющие-
ся на каждом отдельном участке, занятом растительным покро-
вом. Другими словами, это смены определенных фитоценозов.
Естественно, что время, необходимое для осуществления какой-
либо частной смены, обыкновенно бывает значительно меньшим,
чем время, необходимое для вековой, или так называемой общей,
смены. Общая смена — это смена, которую претерпевает каждая
растительная зона или крупная область, представленная значи-
тельным числом различных фитоценозов.
Все перечисленные типы изменений тесно связаны между со-
бой и в значительной мере обусловливают друг друга. Так, напри-
мер, частные смены подготовляются разногодними и даже сезон-
ными изменениями. Это можно проследить в тех случаях, когда
нормальный для данного сообщества ход сезонных смен сильно
замедляется под воздействием каких-либо причин и в особенности
116
под влиянием климата данной местности. Например, в субальпий-
ской зоне Кавказа встречаются своеобразные сообщества высоко-
травий. Они характеризуются преобладанием широколиственного
разнотравья гигантского размера и весьма незначительным уча-
стием злаков, почти не образующих задернения. Однако, по моим
наблюдениям (1948), в Северной Армении, где высокотравья
находятся уже на окраине своего ареала, ранневесенняя фаза
высокотравий сближает их с луговыми сообществами. В это вре-
мя года травостой едва достигает высоты 50—60 см и в нем зна-
чительную роль играют дерновины злаков: Dactylis glornerata,
Роа iberica, Festuca gigantea и др.
К лету широколиственное разнотравье, вырастающее выше
человеческого роста, заглушает и почти нацело вытесняет злако-
вый дерн. Если же мы представим себе, что ранневесенняя фаза
высокотравий в силу климатических причин настолько «задержа-
лась», что не сменилась летней, то этим самым мы примем, что
произошла смена высокотравных сообществ луговыми. И такие
смены в Северной Армении действительно наблюдаются, причем
причиной их служит как деятельность человека, (сенокошение
высокотравий), так и (при отсутствии заметного вмешательства
человека) естественные причины, скорее всего изменение климата,
действующее и на почвообразовательный процесс. В обоих слу-
чаях такие смены, несомненно, подготовляются ранневесенней
фазой высокотравных сообществ.
Различия, но в то же время тесная связь и взаимная обуслов-
ленность сезонных и разногодних изменений сообществ, с одной
стороны, и их частных смен — с другой, достаточно ясны. В свою
очередь имеются различия и в то же время тесная связь и взаим-
ная обусловленность между этими категориями, с одной стороны,
и общими (вековыми) сменами и эволюцией сообществ —
с другой.
Вместе с тем нужно подчеркнуть, что при сезонных и разно-
годних изменениях сообщества последнее остается самим собой,
т. е. еще не сменяется другим сообществом, хотя эта смена шаг за
шагом и подготовляется. При сукцессиях же и общих сменах,
а также при эволюции сообществ происходят не только изменения
в рамках одного и того же сообщества, но и смены одних сооб-
ществ другими.
ЧАСТНЫЕ СМЕНЫ, ИЛИ СУКЦЕССИИ
В своих статьях «Идея развития в фитоценологии» (1942)
и «О некоторых основных вопросах фитоценологии» (1950)
В. Н. Сукачев, говоря о сменах растительных сообществ, указы-
вает, что в них находят выражение три следующих процесса.
1. Сингенетические смены, т. е. процессы заселения растения-
ми молодых местообитаний, например песков, каменных осыпей,
117
свежих речных наносов и т. д. При этом основную роль играет
пока лишь подбор растений друг к другу и к почвенно-климатиче-
ской среде, тогда как взаимодействие со средой всего сообщества
еще только начинает намечаться. Это потому, что элементы среды,
создаваемые самим сообществом (почва и фитоклимат), еще на-
ходятся в зачаточной стадии образования.
2. Эндоэкогенетические смены следуют за сингенетическими
уже после того, как сообщество создало свою определенную сре-
ду и в дальнейшем изменяется вместе с дальнейшим изменением
среды самою же жизнедеятельностью сообщества.
3. Экзогенные смены вызываются причинами, лежащими не
в самом сообществе и не в той части среды, которая тесно и по-
стоянно с ним связана, а вне ее, и поэтому являющимися по отно-
шению к сообществу чисто внешними и случайными, например
смены, вызванные нашествием насекомых, оползнями, обвалами
и т. д. К экзогенным сменам относятся, по Сукачеву, также смены,
вызванные деятельностью человека (пожар, вырубка, выпас).
Впоследствии Сукачев (1954) выделил еще одну категорию
смен, а именно:
4. Гологенетические смены. Это процесс изменения раститель-
ного сообщества вследствие изменения более общего крупного
единства, в состав которого входит данное сообщество. Эти сук-
цессии, как говорит Сукачев, раньше не отделялись им от эндо-
экогенетических. Примером их могут, очевидно, служить смены
сообщества под влиянием изменений климата всей местности,
смены, вызванные развитием речной долины с постепенным
углублением русла реки и отодвиганием его при соответственном
расширении прирусловой террасы и т. д.
Можно сказать, что гологенетическая смена — это частная
смена сообщества, вызванная общей сменой растительного покро-
ва целого массива или страны.
Мною (Ярошенко, 1961) было предложено несколько иное
подразделение частных смен, или сукцессий.
А. Природные смены.
1. Последовательные: а) эндоэкогенетические, б) гологенети-
ческие.
2. Внезапные.
Б. Антропогенные смены.
1. Последовательные.
2. Внезапные.
В этом подразделении противопоставляются природные сме-
ны антропогенным, т. е. вызванным воздействиями человека (та-
ковы вырубка леса, выпас, распашка, осушение, орошение
и т. д.). Кроме того, последовательные смены противопоставляют-
118
ся внезапным. Примерами внезапных природных смен могут слу-
жить смены, происходящие в результате оползней и обвалов
в горах, в результате налета саранчи и других вредных насекомых
и т. д. Внезапные антропогенные смены бывают, например, при
вырубке леса, после распашки, выжигания и т. д. Последователь-
ные антропогенные смены — при сенокошении, умеренном выпа-
се, умеренном регулярном осушении или орошении, повторяю-
щемся из года в год.
Сенокошение, выпас, осушение, орошение, если они проводят-
ся в данном месте впервые, вызывают в первое время внезапную
смену, но затем, если то или иное из этих воздействий из года
в год регулярно повторяется, оно может вызвать уже последова-
тельные смены.
Среди антропогенных последовательных смен можно (как
и среди природных последовательных смен) выделять антропо-
генные гологенетические смены. Последние имеют место, на-
пример, при осушении крупных болотных массивов. При этом
осушение болот может снижать уровень грунтовых вод и на при-
легающих суходолах, в результате чего на последних также
происходит смена одних растительных сообществ другими
(И. Д. Юркевич и Л. П. Смоляк, 1968).
В. Д. Александрова (1964) вместо термина «внезапные сме-
ны» применяет термин «катастрофические смены». Однако вне-
запные смены не всегда носят катастрофический характер. Напри-
мер, при несплошной, выборочной рубке леса смена может ока-
заться внезапной, но не катастрофической; также внезапной, по
не катастрофической будет смена при умеренном и кратковре-
менном заливании речным разливом.
Каждая смена слагается из ряда последовательных этапов,
или стадий. Так, при смене с образованием на приречных песках
ивовых зарослей можно различать: стадию редкостойных зарос-
лей одного или нескольких травянистых видов; стадию более
густых смешанных зарослей хвоща, кипрея, вейника и других
травянистых многолетников; стадию смешанных зарослей много-
летних трав с ивовым молодняком; стадию ивовых зарослей.
Наблюдения показывают, что если ивняки не вырубать и не вы-
пасать в них скот, то в условиях приречных (тугайных) лесов
пустынной и полупустынной зон Закавказья начинается постепен-
ное внедрение в ивовую заросль тополя и с течением времени
ивняк сменяется тополевым лесом, в котором светолюбивая ива
остается лишь иа прогалинах и-по опушкам. Тополевый лес
при отсутствии резкого вмешательства человека может существо-
вать в таких местах очень долгое время без существенных изме-
нений. Эта стадия в жизни сообщества, когда оно изменяется
весьма медленно и внешне недостаточно заметно, соответствует
образованию коренных или узловых ассоциаций. Предшествуют
же им серийные или кратковременные ассоциации.
119
ОБЩИЕ СМЕНЫ И ИСТОРИЯ
РАСТИТЕЛЬНОГО ПОКРОВА СТРАНЫ
Общая смена растительных сообществ какой-либо страны по
своему направлению вовсе не обязательно соответствует общему
процессу эволюции этих сообществ, так как более древние сооб-
щества могут в отдельных случаях приходить на смену более
новым их типам. Так, если климат местности претерпевает иссу-
шение, то леса могут смениться пустынями, хотя в процессе эво-
люции лесная растительность не предшествовала пустынной.
И тем не менее при общих сменах типы сообществ непрерывно
проходят и свою эволюционную выработку. Также и какая-либо
частная смена вовсе не обязательно соответствует по своему на-
правлению общей смене сообществ или их эволюции, но тем не
менее всегда является одним из мелких этапов того и другого
процесса.
Можно говорить не только об общих сменах, но и шире — об
истории фитоценозов. Эту историю можно изучать различными
методами. К ним относится, например, метод спорово-пыльцевого
анализа слоев торфа или почвы, дающий представление о чередо-
вании растительности в данной местности за ряд прошедших
эпох. Сюда относится и восстановление истории растительности
путем изучения ископаемых отпечатков и окаменелостей (палео-
ботанический метод).
Кроме спорово-пыльцевого и палеоботанического методов, на
историю растительности какой-либо страны проливают свет и та-
кие данные, как уцелевшие остатки былой растительности, харак-
тер рельефа и почвы, сведения по истории народов, местные
названия тех или иных урочищ, нередко происшедшие от названий
растений, и т. д.
Некоторые вопросы исторических смен растительности явились
предметом оживленных дискуссий, например вопрос о взаимоот-
ношении между лесом и степью на Русской равнине. Научные
споры на эту тему начались еще в прошлом столетии.
По мнению С. И. Коржинского, лес надвигается на степь
и после отступания великого ледника на месте современных лесов
на Русской равнине сначала были развиты степи, которые уже
впоследствии сменились лесами. Процесс смен степи лесом, по
Коржинскому, продолжается и в настоящее время, но этому часто
препятствует человек, вызывающий -своей деятельностью нередко
даже расширение степных участков за счет лесных. Концепцию
Коржинского поддерживал ряд авторов, в том числе Г. И. Тан-
фильев и Г. Н. Высоцкий.
С совершенно другой точкой зрения выступал В. И. Талиев.
Он считал, что большинство степных пространств Русской рав-
нины целиком обязано своим возникновением населявшим ранее
эти места кочевым племенам. Степи развились па месте лесов,
120
уничтоженных главным образом массовыми пожарами. Одним из
доказательств существования лесов на месте современных степей
Талиев считал встречаемые местами в степной почве следы ходов
древесных корней. Однако другие авторы указывают, что эти
ходы в действительности чаще всего являются кротовыми хода-
ми •— кротовинами.
Есть и другие доказательства былого существования степей
па некоторых современных лесных участках, например старые
курганы, обнаруженные среди лесов. Обычай насыпать курганы
имелся, как известно, у кочевых племен, населявших степи.
Ф. Н. Мильков (1950) разработал теорию, согласно которой
ведущим фактором соотношения леса и степи на равнинах СССР
является рельеф. Поскольку же рельеф подчинен определенным
закономерностям своего развития и ими в значительной мере
определяются и условия почвообразования, то эти причины явля-
ются главными и в распределении леса и степи. Рельеф — не
застывшее образование: он подвержен то более быстрым, то более
медленным изменениям, в его развитии играют роль не только
эрозионные явления, в том числе деятельность рек, но и эпейроге-
нетические движения (медленные вековые опускания или подня-
тия участков земной коры). Все это определяет и динамику рас-
пределения леса и степи. Лесные и степные участки постоянно
перераспределяются, хотя это перераспределение и является
очень медленным процессом, не могущим быть наглядно просле-
женным на протяжении жизни одного человека.
Ф. Н. Мильков полагает, что перераспределение лесных и степ-
ных участков, хотя и может иметь внешние черты циклического
процесса, в действительности не циклично. Если, например, какой-
либо участок был занят лесом, а затем по прошествии тысячеле-
тий в силу изменения рельефа его заняла степь, а затем снова
повторно лес, то это второе наступание леса уже не могло проис-
ходить в условиях, тождественных предыдущим, так как за столь
большой промежуток времени и почвы и климат не могли не
измениться. Вместе с тем Ф. Н. Мильков (1947, 1950, 1953) выска-
зывает мысль, что лесостепной ландшафт, который в настоящее
время представлен на территории СССР в виде сплошной полосы
от Карпат до Алтая, является древним ландшафтом, существо-
вавшим на данной территории еще с доледникового периода.
Кстати, современная точка зрения на происхождение северо-
американских прерий также приписывает им значительную древ-
ность. Известный американский эколог и геоботаник Вивер
в своей книге о прериях Северной Америки (1954) пишет,
что прерии ведут свое начало с третичного времени, имея, таким
образом, возраст около 25 млн. лет. Правда, впоследствии прерии
частью распространились и на места, освобожденные от леднико-
вого покрова. Но крупные участки их современной территории на
юго-западе никогда и не были подо льдом.
121
О надвигании степи на лес высказывался П. Н. Крылов. В из-
вестной мере такие же взгляды имел и В. Л. Комаров, а позднее
Н. В. Павлов, основываясь на материалах изучения растительно-
сти Средней Азии. Решительным противником гипотезы надвига-
ния степи на лес выступал до конца своих дней Л. С. Берг,
подробно обосновавший свою точку зрения в книге «Климат
и жизнь» (1947). Основываясь главным образом на данных повы-
шения уровня озер Средней Азии за последние столетия и сопо-
ставляя это с многолетними данными европейских метеорологиче-
ских станций, Берг приходит к выводу, что все северное полуша-
рие испытывает за последние столетия потепление и одновременно
увлажнение климата, что, по Бергу, совершенно исключает не
только гипотезу надвигания степи на лес, но и гипотезу усыхания
Средней Азии.
М. С. Цыганов (1952) критикует теорию Берга, но отмечает,
что широкое развитие мероприятий по обводнению и лесонасаж-
дениям в степной зоне способно не только полностью предотвра-
тить ее дальнейшее иссушение, но и направить изменение кли-
мата в сторону увлажнения.
В. В. Докучаев был убежден в иссушении Восточно-Ев-
ропейской равнины, рассматривая его как постоянный и последо-
вательный (а не колеблющийся) процесс, упорный, хотя и мед-
ленный, длящийся столетиями и тысячелетиями и неуклонно про-
грессирующий.
Причины этого иссушения Докучаев подробно рассматривает
в своей книге «Наши степи прежде и теперь».
В специальной главе «Способы упорядочения водного хозяй-
ства в степях России» Докучаев наметил ряд мероприятий по
предотвращению дальнейшего усыхания степной зоны.
И. К. Пачоский (1904, 1908, 1917) отмечал еще один очень
существенный фактор, способствующий усыханию степей,— вы-
бивание скотом тех участков, которые остаются нераспаханными.
Большое значение для выяснения вопроса о взаимоотношениях
леса и степи имеют палеоботанические и палеозоологические
данные, а также данные спорово-пыльцевых анализов, сводка
которых приводится в работе К. К. Маркова, В. П. Гричука,
Н. С. Чеботаревой и др. (1950). Авторы приходят к выводу, в не-
которой степени сходному с выводом Ф. Н. Милькова, о том, что
лесостепной ландшафт является очень древним. Но в то время как
/Мильков считает непосредственным предшественником этого
ландшафта ландшафт саванн, указанные исследователи пола-
гают, что современная лесостепь развилась из плейстоценовой
«холодной лесостепи», в которой вместо наблюдающихся теперь
дуба, вяза и некоторых других широколиственных пород участ-
вовали береза и сосна. Северная граница этой «холодной лесосте-
пи» шла на север несколько дальше границы современной лесо-
степи.
122
Таким образом, мы видим, что вопрос о природных взаимоот-
ношениях леса и степи на Русской равнине все еще не может счи-
таться окончательно решенным.
В то время как выяснение природных общих смен, происхо-
дивших в историческое время и в современности, бывает сопря-
жено с большими трудностями из-за искажающего их влияния
человека, спорово-пыльцевой анализ дает нам довольно отчетли-
вую картину общих смен не только растительности, но и климатов
в доисторическое время. Для средней полосы Европейской части
СССР эти доисторические, послеледниковые смены растительно-
сти и климатов схематически показаны на таблице 3. По имею-
щимся данным, ледниковый период, или плейстоцен, а именно его
последняя, вюрмская ледниковая эпоха закончилась здесь около
13 000 лет назад, после чего и происходили все эти смены. После-
ледниковый период, или голоцен, имел, как видно из таблицы 3,
вплоть до наших дней пять основных климатических эпох, каж-
дой из которых соответствовал и определенный состав раститель-
ности.
Таблица 3
Послеледниковые смены растительности и климата в средней полосе Европей-
ской части СССР (по М. И. Нейштадту, 1952, и Л, С. Бергу, 1947)
Хронологизация от 1900 Климатическая эпоха Растительность
ОТ 0 ДО — 2000 Субатлантическая (климат бо- лее влажный и менее теплый, чем в предыдущую эпоху) Максимум ели, много сосны, меньше березы, мало широко- лиственных пород
от — 2000 до — 5000 Суббореальная(сухая и теплая) Березово - елово - сосновые леса. Много широколиственных по- род
от — 5000 до — 7000 Атлантическая (умеренно теп- лая и влажная) Максимум широколиственных лесов, немного березовых и сосновых, мало ели
от — 7000 до _ Ю 000 Бореальная (сухая и теплая) Безусловное господство сосново- березовых лесов. Других по- род ничтожное количество
от — 10 000 до — 12 000 Субарктическая (менее холод- ная, чем предыдущая) Нижний максимум ели и полы- ни. Сосны и березы немного
от — 12 000 до — 13 000 Арктическая (холодная) Господство березы и полыни. Мало ели и сосны
123
ЭВОЛЮЦИЯ РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Эволюция растительности, в отличие от ее смен (как частных,
так и общих), представляет собой процесс выработки новых, ра-
нее не существовавших типов растительных сообществ. Все со-
временные типы фитоценозов — результат эволюционной выра-
ботки, что особенно отчетливо выражено у относительно более
устойчивых, узловых сообществ. Вместе с тем все даже наиболее
оформившиеся узловые сообщества продолжают эволюциониро-
вать, хотя изучение этого процесса, обычно весьма длительного,
значительно труднее, чем изучение смен.
Возьмем к примеру ассоциацию Pinetum myrtillosum (рис. 21),
1. е. бор-черничник с сосной в первом ярусе, черникой во вто-
ром и гипновыми мхами в третьем. Эта ассоциация — не случай-
ный набор растений, т. к. она занимает целый ряд участков, мно-
гие из которых хотя и бывают разбросаны далеко один от дру-
гого, но на каждом состав и структура сообщества все же очень
сходны. Кроме того, известны многочисленные случаи восстанов-
ления какого-либо узлового сообщества на том самом месте, где
оно было уничтожено человеком. Так, на пожарищах еловых
лесов севера России временно возникают сосновые леса, но за-
тем они при отсутствии вмешательства человека, как прави-
ло, рано или поздно сменяются елью. На залежах (заброшен-
ных пашнях), если их предоставить самим себе, через некоторое
(длительное) время в результате ряда смен стадий их зара-
стания возникает та же самая растительность, какая была
до распашки.
Этот процесс восстановления сообществ Г. Н. Высоцкий наз-
вал демутацией, а И. К. Пачоский — регенерацией. Процесс де-
мутации, или регенерации, свидетельствует, что узловое сооб-
щество представляет собой стойкое образование, в котором
виды растений приспособлены друг к другу, а все сообщество со-
ответствует определенным почвенно-климатическим условиям.
В процессе эволюции создались все типы растительного по-
крова: лесной, степной, луговой и т. д. Возникает вопрос: в ка-
ком же родственном отношении находятся между собой эти типы,
какие из них более древние, какие моложе, произошел ли лес от
луга, или луг от леса, или степь от луга и т. д.?
Еще в конце прошлого века к разрешению этой задачи пы-
тался подойти один из основоположников геоботаники И. К. Па-
чоский. В уже упоминавшейся статье «Стадии развития флоры»
(1891) Пачоский выдвинул мысль, что растительность каждой
страны проходит в своем естественном развитии три последова-
тельные стадии: пустыни, степи и леса. По Пачоскому, следова-
тельно, лес произошел из степи и степь из пустыни. Впоследствии
(1925) Пачоский продолжил разработку своей первоначальной
схемы и дал такой последовательный ряд, отражающий уже не
124
Рис. 21. Бор-черничник (Владимирская обл.).
только «прогресс», но и «регресс» в эволюции растительного по-
крова:
пустыня (временная);
прерия-степь (прогрессивный тип);
лес (прогрессивный тип, регрессивный тип);
саванна (регрессивный тип), частично тундра;
пустыня (регрессивный тип).
125
Как видим, Пачоский и здесь не изменил своего представле-
ния о последовательном ходе прогрессивной эволюции раститель-
ных сообществ: степь развивается из пустыни и лес из степи.
Однако и схема Пачоского не отражает процессов эволюци-
онной выработки типов растительного покрова. Нужно иметь в
виду, что прототипами современных растительных формаций яви-
лись скорее всего не другие современные формации той же стра-
ны, а формации, которые в настоящее время частью уже вовсе
не существуют, а частью встречаются как угасающие реликты
или распространены лишь в тех странах, где уцелели более
древние растительные типы. Другими словами, если даже в со-
временную эпоху в данной местности и происходит смена пусты-
ни лесом, то это вовсе не значит, что лесной тип растительности
в процессе своей эволюции произошел из пустынного.
В одной из своих работ (Ярошенко, 1946) я сделал попытку
построить схему именно эволюции (а не частных смен) основ-
ных типов растительности. Эта схема исходила из того, что
«предком» современных лесов были так называемые гилеи (Ну-
laeion), формации, которые чаще называют тропическими и суб-
тропическими «дождевыми лесами»; они были четко выделены
в особый тип растительности А. П. Ильинским, хотя само на-
звание «гилея» было введено в науку значительно раньше, еще
Гумбольдтом. Ильинский (1935) подчеркнул, что гилеи отлича-
ются от лесов: преобладанием индивидуальных и видовых рит-
мов над сезонными (т. е. в них каждый растительный вид или
даже индивид имеет свой ритм развития); пестротой и мощ-
ностью верхнего древесного полога с участием лиан; обилием
эпифитов, в том числе цветковых; отсутствием у большинства
древесных пород почечных чешуи; крайним разнообразием и бо-
гатством жизненных форм. Путем обеднения гилей и приспособ-
ления их к резкому чередованию времен года могли образовать-
ся леса.
Другим чрезвычайно древним типом растительности были
тропические пустыни, а также саванны, занимавшие области, пе-
реходные от тропических пустынь к гилеям. Саванны в результа-
те полного исчезновения из их состава древесных пород, а также
общего обеднения и приспособления к иной сезонной ритмике мо-
гли дать начало степям умеренного климата. В связи с этим и су-
ществуют формации, явно переходные от саванн к степям, как,
например, описанные в Средней Азии и Закавказье полустепи
и описанные в Средней Азии полусаванпы. Уже после образова-
ния из саванн степей в местах контакта степей с пустынями могли
возникнуть в качестве переходных формаций и полупустыни.
Луговой тип растительности развился, с одной стороны, из
травянистого покрова лесов в местах их изреживания по тем
или другие причинам. С другой стороны, «предком» лугов яви-
лось субальпийское выбокотравье, древний третичный тип ра-
126
стительности и сейчас еще уцелевший местами на Кавказе, Кам-
чатке и т. д. И наконец, «предком» горных лугов можно считать
открытые сообщества высокогорных скал и осыпей. Образование
лугов из этих открытых сообществ путем частных смен, наблю-
дающихся и в настоящее время, является тем случаем, когда част-
ная смена отражает один из путей эволюции сообществ. Приве-
денная схема эволюционных связей типов растительности
(рис. 22) представляет собой лишь первый набросок. Последую-
щие исследования должны дать материал для построения более
совершенной и уточненной схемы эволюции типов растительно-
сти. В качестве предпосылок к такой будущей схеме можно при-
вести следующие соображения.
По-видимому, лишь в сравнительно редких случаях можно
допустить, что какой-либо тип растительности является в полном
смысле слова эволюционным предшественником, или «пред-
ком», другого типа растительности. С достоверностью такие слу-
чаи не установлены, но зато не вызывают сомнения факты дру-
гой категории, когда одна крупная единица растительного по-
крова имеет не один, а несколько исходных типов, принявших
участие в ее эволюционной выработке.
На рисунке 23, например, показано, что высокогорные луга
Кавказа имеют три исходных типа сообществ: травяной покров
лесов; высокотравье и открытые группировки высокогорных скал
и осыпей. Возможно, и многие другие типы растительности так-
же имеют не один, а несколько типов «сообществ-предков». На-
зывать эти последние предками можно, однако, лишь в перенос-
Рис. 22. Схема эволюционных связей основных типов растительности.
127
Рис. 23. Схема эволюционной выработ-
ки высокогорных лугов Кавказа.
ном значении этого слова.
Если какой-нибудь эволюци-
онно более молодой тип рас-
тительности имеет несколько
таких «предков», то это мо-
жет означать, что он впер-
вые возник в местах их вза-
имного контакта, причем
каждый из них вложил свою
долю в образование этого
молодого типа. Такие исход-
ные сообщества нужно на-
зывать не предками, а вклад-
чиками или участникам и
(партиципнентами). Этим
наименованием подчеркива-
ется их специфическая роль
в эволюционной выработке
производных типов расти-
тельного покрова, роль, со-
стоящая как во «вложении»
типы соооществ определенных
плексов, так одновременно и
структур сообществ. При этом
от сообществ-вкладчиков,
в новые, формирующиеся
видов растений и видовых ком-
в их участии в выработке новых
виды и их комплексы, полученные
попадая в эволюционно молодые,
формирующиеся сообщества, получают новые стимулы к своей
дальнейшей эволюции в связи с развитием приспособлений к
изменившейся экологической обстановке. Предпосылкой же к
созданию таких эволюционно молодых сообществ может явиться
контакт нескольких различных типов растительного покрова,
создающих новые сочетания, сначала неустойчивые, но затем
оформляющиеся в новый, более устойчивый тип.
Дальнейшая работа по изучению эволюции сообществ должна
иметь в виду установление вкладчиков (участников) и специфи-
ческой роли каждого из них в формировании того или иного типа
сообществ. При этом нужно учитывать, что эволюция внешних
признаков сообществ, их «формы», имеет две стороны, слагается
из двух взаимно проникающих процессов, а именно: структуро-
генеза, или эволюционной выработки той или иной структуры це-
нозов, и флорогенеза, или видообразования в связи с эволюцией
ценозов.
Процесс образования новых типов сообществ в местах кон-
такта вкладчиков представляет один из путей эволюции сооб-
ществ, который можно назвать эволюцией путем вбирания. Дру-
гим путем является противоположный процесс расчленения (се-
грегации) одного типа сообществ на несколько производных.
128
Пример такого расчленения описан для смешанных лиственных
лесов южных штатов Северной Америки (Браун, 1935, и др.),
а также для смешанного колхидского леса (Синская, 1933; Со-
чава, 1946). Кроме того, сходные процессы описаны Ричардсом
(1945) для тропических гилей. Во всех этих случаях наблюдает-
ся расчленение очень сложной макрогруппировки со многими
преобладающими древесными видами на ассоциации с преобла-
данием в каждой из них одного-двух видов. Например, колхид-
ские леса нижнего горного пояса (до 600—700 м над ур. м.), обра-
зованные в основном пятью породами: буком, каштаном, дубом,
грабом и ольхой,— расчленяются на ассоциации с господством
одной-двух из этих древесных пород. Эти процессы наблюдаются
в пространстве при продвижении к северным и восточным окраи-
нам Колхиды, и они же, несомненно, происходили и происходят
во времени под воздействием начавшегося еще с конца плиоцена
некоторого усиления континентальности климата Кавказа.
Расчленение в некоторой степени противоположно вбиранию,
но тем не менее всегда сочетается с этим последним, так как во
всех случаях исходные полидоминантные типы, а также и про-
дукты их расчленения испытывают влияние окружающих типов
растительного покрова, в той или иной мере играющих по отно-
шению к ним роль вкладчиков.
В. Б. Сочава (1944) предложил понятие фратрия для сово-
купности фитоценозов, связанных территориальным соседством
и общностью происхождения в результате расчленения какого-то
ранее более однородного растительного покрова. При этом он
отмечает, что в одну фратрию могут входить фитоценозы и лес-
ные, и кустарниковые, и травяные.
Темы рефератов на семинаре
или в кружке
Способы образования болот
Литература: Сукачев В. Н. Болота, их
образование, развитие и свойства. М., Сельхозгиз,
1926.
Алехин В. В. Растительность СССР в основ-
ных зонах. М., «Советская наука», 1951,
стр. 387—402.
Кац Н. Я. Типы болот СССР и Западной Евро-
пы и их географическое распространение. М., Гео-
графгиз, 1948.
История спорово-пыльцевого метода в СССР
Литература: НейштадтМ. И. Спорово-
пыльцевой метод в СССР. М.—Л., Изд-во
АН СССР, 1952, стр. 3—48.
9 П, Д. Ярощенко
12)
Темы курсовых работ
Наблюдения над сменой пород в лесу
Литература: У ранов А. А. Наблюдения на летней прак-
тике по ботанике. М., «Просвещение», 1964,
стр. 198—205.
Ткаченко М. Е. Общее лесоводство, ДЕ, Сель-
хозгиз, 1952.
Морозов Г. Ф. Учение о лесе, изд. 7. М„ Сель-
хозгиз, 1949.
Влияние выпаса на травостой луга (или степи)
Подыскивается участок луга, защищенный от выпаса (на-
пример, на окраине огороженного сада), и рядом участок сход-
ного луга, но выпасаемый. За вегетационный период делают не-
сколько параллельных описаний и берут пробные укосы для ве-
сового анализа с обоих участков. Если площадь каждого участ-
ка невелика (менее 1 га), то достаточно взять в каждый из сро-
ков по 4 пробных укоса с площадок 50X50 см или по 16 пробных
укосов с площадок 25x25 см, выбираемых в типичных местах.
Кроме того, учитывают видовой состав, обилие и размеры осо-
бей каждого вида, а также покрытие (густоту) и среднюю высоту
всего травостоя.
Литература: Алехин В. В. Методика полевого изучения
растительности и флоры, М., Изд-во Наркомпро-
са, 1938.
Ярошенко П. Д. Геоботаника. М.—Л.,
Изд-во АН СССР, 1961, стр. 426—429 и 433—436,
8
ЗАКОНОМЕРНОСТИ
ТЕРРИТОРИАЛЬНОГО
РАСПРЕДЕЛЕНИЯ
РАСТИТЕЛЬНОГО
ПОКРОВА
Геоботаническое изучение какой-либо одной ассоциации (ли-
бо нескольких ассоциаций) не должно носить узко ограни-
ченного, описательного характера.t Исследователя закономерно-
стей территориального распределения растительных группиро-
вок должны интересовать и такие вопросы, как географическое
распространение ассоциации, приуроченность ее к той или иной
зоне, родственные ей другие ассоциации, комплексирование дан-
ной ассоциации с другими в одну макрогруппировку и многое
другое. Объектом геоботаники является и само по себе изучение
крупных комплексов растительного покрова — фитоландшаф-
тов, зон. В связи со всем этим становится ясной исключительная
важность для геоботаники изучения закономерностей в террито-
риальном распределении растительного покрова.
ЗОНЫ И ПОЯСА РАСТИТЕЛЬНОСТИ
Уже давно было подмечено, что растительный покров земного
шара распределяется по следующим друг за другом с севера на
юг или с юга на север широтным зонам. На континенте Евразии
различают начиная с севера следующие зоны растительности:
холодные (арктические) пустыни; тундры; хвойные леса; листвен-
ные леса; степи; пустыни; субтропические леса; тропические леса.
Кроме этих основных зон, существуют еще переходные: лесотунд-
ры, лесостепи, полупустыни.
Можно говорить как о зонах растительности, так и о зонах
природы в целом, причем правильнее считать (хотя с этим сог-
ласны не все географы), что зоны растительности и зоны всей
природы совпадают. Элементы несовпадения могут в той или
иной мере проявляться лишь в местах перехода одной зоны
в другую, поскольку, как правильно заметил Ф. Н. Мильков
(1956), границы зон нельзя представить в виде линий.
Мысль о существовании зон растительности и климата была
высказана еще Гумбольдтом, однако идея о зональности всей
9*
131
90'
Океан
Климаты
Континент
Морской Средний Континентальный
- л. * л л « Арктические пустыни*- * •*• •» *.» » .«.
3
£
6
60
Е
<3
3
<ч
60'
Зкдитор О'
«ш Вечнозеленые ...iinfint,",",'1,<|,li|‘l ♦ t х ± i ± ± ♦ t
HlUruill II |Щ,| IIHIHHI'IllinHtuH * £ £
:=ж1tj < e 11 < t м t111
V'.a"a "h Q Q.Q ^o*o*ot 11 t til
Хф 9 лист Bod летом Q ty Q I* t | | $ £ 1 £
9 9 $ 9 9 9 9 9 9 9 9 9 9 9 < Mil t H
••••• ф ф ф ф Q Q Q Q Q ЛIA Q
ШЛ 'n1^,'
—“—|C^ Жестколистные I I 1 ।
——— < «i’i’i ( 'леса I’l’i'j*, |,Л,1
p^---^',' 7.:,::.; л1
-Mipaim^Ux','.' a a a a Q ।,',’
^£-€-£=5.' qVq qVq".1!
9 9 9 Q 9odi
Q 9 9 9 9 9:;:
Ъ' Тропические jr-9 ^eca сдетые о * о
.QL.9—9£2-° g.
Г сырыёТлеса'-Q 9 9 9 9 9 ° * ° e"
—'—9 листВой зимой r oC^e
о 'J
о
о
о
е
о
о
о
о
о
о
Q ф ф Q 9 ффо-о-
Гф ф Q ф $ 9 0$ф°: -
Г 99999999 9'^
^QQQQQQQQQQ
о
о
о
Рис. 24. Схема распределения растительности на идеальном континенте (по
Брокман-Ерошу из Щенникова, 1950).
природы в целом была впервые глубоко и всесторонне обосно-
вана В. В. Докучаевым. Рассматривая каждую зону как резуль-
тат взаимодействия всех факторов живой и неживой природы,
Докучаев вместе с тем первый разработал понятие о зоне как
исторически формирующемся явлении.
В статье «О почвенных зонах вообще и вертикальных зонах
в особенности» Докучаев (1898) писал, что почвенные зоны в то
же время являются и зонами природными, что в них проявляется
теснейшая генетическая связь климата, почвы, животных и ра-
стительных организмов.
132
Кроме изменений растительности в широтном направлении
(север — юг), известны и изменения ее в долготном направлении
(запад — восток), причем последние связаны прежде всего со
степенью близости или отдаленности от океана.
Расположение в пределах Евразии наиболее сухих областей
в центральной части (Сибирь, Центральная Азия) дало повод
П. Н. Савицкому (1927), а позднее В. В. Алехину (1936) говорить
о симметричности системы зон на территории СССР. Так, на
крайнем западе и крайнем востоке СССР хвойные, более север-
ные леса переходят к югу в леса широколиственные, тогда как
в центре, т, е. в Сибири, широколиственные леса отсутствуют.
Симметричность системы зон сказывается не только в западно-
восточном направлении, но и в северо-южном. Безлесные про-
странства располагаются и на севере СССР (тундры) и на юге
(степи и пустыни), а между ними находится широкая зона лесов.
Интересно обратить внимание еще на то, что как южная, так
Рис. 25. Схема распределения типов растительности на континенте Евразии
от экватора до полюса (по Шенникову, 1950).
10 П. Д. Ярошенко
133
и северная граница лесной зоны представлена породами с опа-
дающей на зиму листвой: на южной границе это дуб, осина и бе-
реза, а на северной — лиственница (а на Кольском полуостро-
ве— береза), тогда как вечнозеленые хвойные играют там вто-
ростепенную роль или вовсе отсутствуют.
Брокман-Ерош и Рюбель составили схему распределения ра-
стительности на так называемом идеальном континенте (рис. 24).
Эта схема отражает закономерные пространственные изменения
растительного покрова в связи с изменениями режима темпера-
туры и влажности. Схема основана на условном предположении,
что океан находится лишь к западу от идеального континента,
и поэтому континентальность климата неуклонно возрастает по
направлению с запада на восток. Однако в действительности это-
го ни на одном материке не наблюдается, так как океан всегда
находится и с запада и с востока, а наиболее континентальные
области тяготеют поэтому к самым внутренним частям мате-
риков.
В «Экологии растений» А. П. Шенникова (1950) приведена
схема распределения типов растительного покрова на материке
Евразии, более соответствующая действительному положению
вещей (рис. 25). Как видим, на этой схеме океанические хвойные
леса есть и на северо-западе, и на северо-востоке Евразии,
лиственные леса — и на западе, и на востоке, тогда как сухие
пустыни располагаются в центральной и западной частях конти-
нента, а на востоке отсутствуют вовсе. На этой же схеме показа-
ны широтные зоны, причем видно, что степная зона выражена
на материке Евразии лишь в его центральной части. Не по всему
материку протянута и лесостепная зона, а в Сибири выпадает
зона лиственных лесов. Все эти различия связаны с изменения-
ми климата в долготном направлении.
В связи с этим в физической географии разработано подраз-
деление земного шара на крупные страны или области, каждая
из которых характеризуется своим набором широтных зон. По-
этому характеристика той или иной природной зоны не будет
полной, если не указать, к какой физико-географической стране
она относится. Например, Ф. Н. Мильков (1956) дает следующее
определение природной, или географической, зоны: «Зона пред-
ставляет собой широтно вытянутую часть географической стра-
ны, характеризующуюся определенным сочетанием тепла и влаги,
господством одного или своеобразной комбинацией нескольких
типов почв и растительности». Что касается физико-геогра-
фических стран, то примерами их могут служить: восточ-
ноевропейская страна, западносибирская страна, восточно-
сибирская страна, дальневосточная страна и др. Под географи-
ческой (физико-географической) страной нужно, следуя
Ф. Н. Милькову, понимать крупную географическую область,
характеризующуюся определенной госструктурой, определенным
134
Рис. 26. Пояс можжевелового
стом склоне в Закавказье.
редколесья на сухом камени-
макроклогатическим режимом и определенным
пенью выраженное™ Широтных „ в“"хзон " " СТ<>’
выраХ77Ж
называют, в. отличие от равнинных, поясами (рис.^б)30™ Ч^Т°
10*
да-
система океанического типа,
III. Лесной пояс: 7 — темнохвойные еловые леса бореального
типа; 8 темнохвойные елово-пихтовые леса средиземноморского
типа; 9 — широколиственные леса (дубовые, буковые) бореально-
го типа: 10 — широколиственные (дубовые, буковые) леса среди-
земноморского типа.
Рис. 27. Горнопоясная
I, Альпийский пояс;/ — каменистая горная тундра (голь-
цы); 2 — лишайниковая и кустарничковая горная тундра; 3 — пус-
тоши и аустошные луга. JL Субальпийский пояс: 4 — кри-
волесье, вересковые кустарники и луга; 5 — лугово-парковые кед-
ровники и лиственничники сибирского типа; 6 — сосновые леса сре-
дня емком орского, отчасти бореального типа и остепненные луга.
40'
система континентального типа.
га. IV. Горно-лесостепной пояс:// — остепненные лист-
венничные леса (на фоне лугов, луговых кустарничковых и типчако-
вых степей); 12 — островные еловые леса (на фоне лугов, луговых
кустарничковых и типчаковых степей). V. Горно-степной
п о я с: /3 — разнотравно-ковыльные и типчаковые степи, остров-
ные широколиственные (кленовые) леса и фрагменты арчевников;
14 — типчаковые степи, арчевники и фрагменты нагорных ксеро-
фитов; /5 — мезофильный клик атлантического климатического
воздействия.
Рис. 28. Горнопоясная
1. Альпийский пояс:/ - каменистая горная тундра (гольцы);
2 — мохово-лишайниковая горная тундра; 3 — высокогорные пусто-
ши и пустошные луга; 4 — высокогорная пустыня. П. Субаль-
пийский пояс: б — лишайниковое и кустарничковое листвен-
ничное криволесье; б—кедровый стланик, ерники (Алтай, Саяны)
и арчевники (Тянь-Шань); 7—лугово-парковые лиственничные и кед-
рово-лиственничные лесаи луга; 8 — лугово-парковые островные
еловые леса и луга. III. Горно-лесной пояс: 9 — листвен-
ничная горная тайга: 10 — пихтовая и кедрово-пихтовая горная тай-
Рис. 29. Верхняя граница леса в Карпатах.
Естественно, что характер вертикальной зональности или по-
ясности всегда зависит от широтной зональности, а также от рас-
положения в отношении морей и оксанов.
Еще Гумбольдт в своих «Идеях ботанической географии»
различал типы вертикальной зональности.
Интересные схемы горной поясности в связи с широтой
и океаничностью или континентальностью климата составлены
Р. А. Еленевским (1940).
Схемы эти показаны на рисунках 27, 28.
Каждая зона, или пояс,— это целая мозаика (макромозаика)
фитоценозов. Образующие эту мозаику типы растительных сооб-
ществ часто бывают свойственны только данной зоне и отсутст-
вуют в других. Например, в Карпатах (рис. 29) поясу темно-
хвойных лесов свойственны растительные сообщества из ели
(Picea excelsa); на Дальнем Востоке (рис. 30) субальпийскому
поясу свойственны густые заросли кедрового стланика (Piiius
pumila), а па Кавказе тип альпийских ковров свойствен только
альпийскому поясу гор и в других поясах не встречается. Типы
ценозов, которые наиболее полно отражают условия лишь одной
определенной зоны или пояса (на равнине или в горах), назы-
вают зональными.
Но бывают и такие типы ценозов, которые встречаются в не-
скольких зонах. Например, солончаковая растительность встре-
138
чается в зонах пустынь, полупустынь и степей. Такая раститель-
ность называется интразональной. Травяные болота встречают-
ся во всех зонах и поясах от пустынь до альпийских высот
и тундры. Про такие типы ценозов говорят, что они азональны.
Однако азональными и интразональными могут быть лишь круп-
ные единицы растительности, совершенно же одинаковых ассо-
циаций в различных зонах обычно не бывает.
Иногда в связи с какими-либо местными условиями в горных
областях наблюдаются очень сильные нарушения обычной пояс-
ности. Например, полупустыни, которые в восточном Закавказье
свойственны высотам не более 500—600 м над уровнем моря,
в окрестностях Еревана распространены и на высотах
1500—1600 м. Это объясняется тем, что близ Еревана прости-
раются обширные лавовые плато, вследствие чего местность
открыта постоянному доступу иссушающих ветров и очень бедна
осадками. Такого рода местности имеют «климат высоких пла-
то» (Берг, 1938), характеризующийся также и большими суточ-
ными амплитудами температуры.
Более мелкие нарушения зональности наблюдаются иногда
в специфических элементах рельефа. Так, дубовые колки по ов-
рагам проникают на равнине в степную зону; высокотравно
в горах Кавказа спускается по оврагам далеко вниз, в лесной
пояс, и т. д. В этих случаях про дубовые колки и высокотравье
Рис. 30. Заросли кедрового стланика в горах Сихотэ-Алиня (Дальний Восток).
можно сказать, что они являются экстразональными сообщест-
вами по отношению к той зоне, с которой связано их основное
местонахождение.
ПРАВИЛО ПРЕДВАРЕНИЯ
Изучая экстразональные сообщества на Русской равнине,
Б. А. Келлер, а позднее В, В. Алехин отметили интересные за-
кономерности. Келлеру принадлежит заслуга первого точного
описания экологических рядов растительности, т. е. закономер-
ного чередования в пространстве сообществ в связи с простран-
ственным изменением экологических факторов в определенном
направлении.
Изучая в зоне южнорусской полупустыни экологические ряды
в понижениях мезорельефа, Келлер (Димо и Келлер, 1907) обна-
ружил там постепенные переходы от полупустынных до травяно-
степных группировок, причем эти переходы были вполне сходны
с теми, какие наблюдаются на громадном пространстве равнины
при движении из зоны полупустыни в зону степи. Близкая к от-
меченной Келлером, но несколько иная закономерность, связан-
ная уже с экспозицией склона, выражена Алехиным в так назы-
ваемом правиле предварения. Алехин (1950) указывает, что
экстразональные явления противоположного значения, т. е. пред-
ставляющие отображение и более северных плакорных условий
и более южных, могут быть наблюдаемы в непосредственной бли-
зости. Например, если балка прорезает степь в направлении
с запада на восток, то на северных и южных ее склонах мы на-
блюдаем эти экстразональные явления, иногда с необычайной
отчетливостью и резкой контрастностью.
Таким образом, на самом небольшом пространстве мы видим
и зональный тип данного места (на нлакорной степи), и более
северный зональный тип (на северных склонах), и более южный
зональный тип (на южных склонах).
Следовательно, экстразональная растительность, с одной сто-
роны, как бы отображает более северные или более южные зо-
нальные типы, но с другой — она их и предваряет. Такие законо-
мерные отношения дают возможность говорить о правиле пред-
варения, которое дает возможность предсказывать раститель-
ность еще не исследованных мест; восстанавливать ее былой
характер там, где она уже уничтожена; изучать растительность
в свете широких географических представлений.
По мнению Н. Ф. Комарова (1938), это правило дает слиш-
ком упрощенную трактовку зависимости между растительностью
и рельефом. Он пишет, что в работах самого В. Алехина можно
найти немало указаний на то, что наряду с экспозицией склона и
его гидротермическими условиями в той мере, в какой они могут
зависеть непосредственно от экспозиции, существенное значение
140
Рис. 31. В горах Казахстана: на северных склонах — леса
из тяныианьской ели, южные склоны безлесны.
имеет также характер материнских пород, высотное положение
по склону и многое другое.
Добавим к этому, что если все же в условиях обширных рав-
нин правило предварения имеет большое значение, то в горных
областях его значение меньше, так как здесь в распределении
фитоценозов играют роль более сложные совокупности экологи-
ческих факторов, чем на равнине. Правда, экспозиция склона
в горах играет громадную роль, по к ее влиянию примешиваются
еще такие факторы, как степень крутизны склона и высота над
уровнем моря.
ВЛИЯНИЕ ГОРНОГО РЕЛЬЕФА
НА РАСПРЕДЕЛЕНИЕ РАСТИТЕЛЬНЫХ
СООБЩЕСТВ
Влияние на распределение растительных сообществ экспози-
ции склона в сочетании с его крутизной и высотой над уровнем
моря в горах очень велико. Так, в северном полушарии склоны,
обращенные на север, обычно холоднее и лучше увлажнены, поч-
ва на них более мощная; склоны, обращенные на юг, при прочих
равных условиях, наоборот, более теплые и сухие, с менее мощ-
ным почвенным покровом (рис. 31).
Склоны западной и восточной экспозиций занимают по своему
экологическому режиму переходное /место. При этом можно при-
нять, что склоны западной экспозиции при равенстве остальных
условий теплее и суше склонов восточной экспозиции, так как
141
Рис. 32. В горах Кабардино-Балкарии (Сев. Кавказ): на северных склонах —
березняки, на южных —ковыльная степь.
солнце нагревает их во вторую половину дня, т. е. когда действие
этого нагрева сильнее. На крутых склонах влияние экспозиции
обыкновенно сказывается сильнее, чем на пологих.
В приморских местностях прямое влияние экспозиции неред-
ко сводится на нет действием влажных морских ветров и тума-
нов; влажнее здесь бывают (независимо от экспозиции) склоны,
обращенные к морю. Обратное действие может иметь близость
обширных сухих пустынь. Влияние высоты над уровнем моря ска-
зывается в том, что с высотой климат делается в общем холод-
нее. Что же касается изменений влажности климата, то она также
увеличивается обычно с подъемом, но до известного предела,
после чего с дальнейшим поднятием опять несколько уменьшает-
ся. Самым влажным поясом в горах Кавказа является субаль-
пийский пояс, и особенно его нижняя часть, у верхней границы
леса. Это полоса наибольшей облачности, сильных туманов и
чрезвычайно обильных рос. Существование в указанной полосе
высокотравья, этого «леса без деревьев», связано как раз с силь-
ной влажностью почвы и воздуха. В альпийском же поясе влаж-
142
ность снова уменьшается, а кроме того, там влияние влажности
несколько парализуется холодностью климата, вызывающей в те-
чение значительной части года физиологическую сухость почв.
Поэтому типичные растения-альпийцы всегда несут те или
иные черты ксероморфной организации.
Кроме высоты над уровнем моря, экспозиции и крутизны
склона, большое значение имеет нередко едва заметная выпук-
лость или вогнутость склона.
КОМПЛЕКСЫ
Как уже упоминалось, комплексами в геоботанике называют
пространственные чередования небольших участков различных
ассоциаций.
Различают следующие геоботанические комплексы:
поясные комплексы, когда участки ассоциаций располагаются
узкими полосами, представляя экологические ряды;
мозаичные комплексы, когда чередование участков разных
ассоциаций носит пятнистый характер, напоминая шахматную
доску. Мозаичные комплексы ассоциаций не следует путать с мо-
заичностью каждой ассоциации в отдельности; о различиях меж-
ду комплексностью и мозаичностью было упомянуто при рас-
смотрении мозаичности (стр. 47—48);
диффузные комплексы, когда одна из ассоциаций образует
фон, в который вкраплены мелкие участки другой или других
ассоциаций. Бывают и комплексы переходные между этими тре-
мя основными типами.
Иногда различают комплексы и сочетания. Некоторые авто-
ры, например болгарский исследователь X. Кочев (1967),
считают, что в комплексах составляющие их сообщества связаны
сукцессионными взаимоотношениями, а в сочетаниях этого нет.
Однако такое подразделение не имеет всеобщего признания, так
как находящиеся рядом различные фитоценозы не могут в той
или иной степени не влиять друг на друга, а эти влияния в той
или иной мере не могут не отражаться на сменах (сукцессиях).
Кроме комплексов ассоциаций, называемых также мезокомп-
лексами, или иногда микрокомплексами, отличают еще макро-
комплексы, т. е. комплексы комплексов. Различные исследовате-
ли называют их разными терминами (объединение, макроассо-
циация, катена и др.).
Темы рефератов на семинаре
или в кружке
О зонах и поясах
Литература: Гумбольдт А. География растений.
М.—Л., Сельхозгиз, 1936.
143
Докучаев В. В. Учение о зонах природы,
Избр. труды. М., Изд-во АН СССР, 1949.
К истории растительности Русской равнины
и ближайших к ней горных стран
Литература: Мильков Ф. Н. Воздействие рельефа на
растительность и животный мир. М., Географгиз,
1953, стр. 124—150.
Темы курсовых работ
О вековых смещениях природных зон и поясов
Литература: Берг Л. С. Климат и жизнь, М., Географ-
гиз, 1947.
Мильков Ф. Н. Воздействие рельефа на расти-
тельность и животный мир. М., Географгиз, 1953,
стр. 112—120.
Шенников А. П. Введение в геоботанику.
Изд-во ЛГУ, 1964, стр. 348—356.
Цыганов М. С. К вопросу об усыхании сте-
пей. «Почвоведение», 1952, № 4.
Ярошенко П. Д. Геоботаника. М.—Л.,
Изд-во АН СССР, 1961, стр. 221—225 и 264—270.
Растительность .....................республики
(области, района)
По литературным данным. Литературу подобрать самостоя-
тельно.
9
ГЛАВНЕЙШИЕ МЕТОДЫ
ГЕОБОТАНИЧЕСКИХ
ИССЛЕДОВАНИЙ
ЗАКЛАДКА И ОПИСАНИЕ ПРОБНЫХ
ПЛОЩАДЕЙ И УЧЕТНЫХ ПЛОЩАДОК
Для описания отдельных сообществ приходится выделять
пробные площади. Эти площади должны дать достаточно полное
представление о данном сообществе, и потому размеры их не
должны быть очень малы. Для травяных сообществ выделяют
площади от 1 до 100 я2 в зависимости от размеров естественных
участков отдельных сообществ, для лесов — от 100 до 1000 и
5000 я2. На равнине можно закладывать более крупные площа-
ди, в горах нередко необходимы более мелкие, в связи с тем что
здесь пестрота экологических условий создает и быструю смену
растительных сообществ в пространстве.
При очень крупных (в четверть гектара и более) площадях,
закладываемых в лесах, целесообразно древесную и кустарнико-
вую растительность описывать на всей этой площади, а для тра-
вянистой выделять в ее пределах несколько площадок менее
крупных размеров и описывать только их.
Пробные площади могут быть как строго определенных, так
и менее определенных размеров и очертаний. В первом случае
площадь имеет обыкновенно квадратную или прямоугольную
форму и границы ее, хотя бы в углах, обозначаются колышками
или другими знаками. Во втором случае, если размер описывае-
мого ценоза невелик (что часто бывает в горах), можно огра-
ничить пробную площадь естественными границами данного со-
общества, не задевая полос, переходных к другим сообществам.
Если же естественный размер сообщества (участка ассоциации)
более крупный, то в его пределах можно выделить площадь
меньших размеров.
На пробной площади производится общее описание расти-
тельности по программе, которая приводится ниже. При исследо-
вании лесных сообществ пробная площадь используется также
для пересчета деревьев, количество которых затем пересчиты-
вается на гектар. В этих случаях следует выделять пробные
площади точных размеров, привязывать к каким-либо ориенти-
рам, например к просекам, и наносить на план.
145
Если имеется необходимость точного подсчета более мелких
объектов, например: древесных всходов, побегов или кустов трав,
покрытия отдельных видов травяного, мохового или лишайнико-
вого ярусов, проективной съемки границ микрогруппировок
и т. д.,— то в пределах пробной площади закладывают еще учет-
ные площадки. Эти последние имеют значительно меньшие раз-
меры, чем пробная площадь, а именно чаще всего от 1 до 4 .ч2.
Для весовых анализов травостоя нередко применяют квадраты
размером.0,25 м2 (со стороной, равной 50 см).
Описание пробной площади может быть произведено с различ-
ной степенью подробности. Чаще всего учитываются важнейшие
признаки сообщества, которые были перечислены выше, и к ним
добавляется еще описание местообитания, т. е. рельефа, почвы и
т. д. Перейдем к рассмотрению методов учета всех этих признаков.
УЧЕТ ВИДОВОГО СОСТАВА РАСТИТЕЛЬНОГО
СООБЩЕСТВА
Необходимо очень тщательно составлять список растений, с
тем чтобы в него попали все виды, находящиеся как в зрелом
состоянии, так и в виде молодняка и даже всходов. Рекоменду-
ется располагать растения в списке по жизненным формам,
например: сначала деревья, затем кустарники, кустарнички, по-
лукустарники, многолетние травы, однолетние травы, мхи, ли-
шайники, грибы, водоросли. Можно использовать и более деталь-
ные классификации жизненных форм. При составлении списков
на сенокосах и пастбищах с целью их кормовой оценки можно
располагать виды по хозяйственным группам: злаки, осоковые
и ситниковые, бобовые, разнотравье, вредные и ядовитые расте-
ния. В пределах каждой из групп виды растений перечисляются
или в систематическом, или в алфавитном порядке.
ОБИЛИЕ
Обилием называется количество экземпляров какого-либо ви-
да в пределах пробной площади. При учете обилия приходится
нередко сталкиваться с трудностью разграничения отдельных
индивидов, В таких случаях подсчет индивидов можно заменять
подсчетом стеблей. И то и другое представляет собой объектив-
ную оценку обилия. Такого рода оценка бывает нередко очень
громоздкой и кропотливой, и поэтому в особенности для травя-
ных ценозов чаще применяют субъективную оценку с помощью
какой-либо шкалы. Были предложены различные шкалы обилия:
одни из них цифровые, чаще всего пяти- или десятибалльные,
другие же — словесные.
Из цифровых шкал самая простая — пятибалльная шкала
Хульта, цифровые баллы которой следует понимать приблизи-
тельно так: 5 — очень обильно, 4 — обильно, 3 — не обильно, 2 —
мало, 1 — очень мало.
146
В шестибалльной шкале Друде даны не цифры, а следующие
словесные обозначения:
soc. (sociales) — растение смыкается надземными частями,
образуя фон;
сорз (copiosae5) — растение встречается очень обильно;
сор2 (сорюэаег) — особей много;
сор] (copiosaei) —особей довольно много;
sp. (sparsae) — растение встречается в небольшом количест-
ве, рассеянно;
sol. (solitariae) — растение встречается в очень малом коли-
честве, редкими экземплярами.
Часто употребляют еще обозначение un. (unicum), когда вид
встречен на площадке в единственном экземпляре.
При пользовании шкалой Друде неизбежно приходится соче-
тать представление о количестве экземпляров каждого вида с
представлением о покрытии, т. е. о занимаемой им площади.
Приблизительно можно принять, что во многих случаях отметка
soc. будет соответствовать покрытию особями данного вида бо-
лее 90% площади участка, отметка сорз — 90—70%, сор2 — 70—
50%, сор] — 50—30%, sp. — 30—10%, sol. — менее 10%. В тех
случаях, когда особей какого-либо вида очень много, но они
очень мелкого размера (например, некоторые весенние эфемеры,
как крупка и др.), будет иметь место большое расхождение меж-
ду обилием и покрытием. В этих случаях при пользовании шка-
лой Друде в основном принимают во внимание именно количе-
ство особей, а не площадь покрытия. Могут быть и разные пере-
ходные случаи.
ПРЕОБЛАДАНИЕ, ИЛИ ДОМИНИРОВАНИЕ
Этот признак характеризуется, во-первых, покрытием, т. е.
площадью, покрываемой надземными частями того или иного
вида в сообществе, во-вторых, занимаемым ими объемом и,
в-третьих, их весом. Определять покрытие растений в сообществе
можно различными методами.
Определение проективного покрытия. В травостоях разли-
чают так называемое истинное покрытие и проективное покры-
тие. Истинное покрытие —это процент площади, занятой основа-
ниями побегов растений, а проективное покрытие — это процент
площади, покрываемой верхними частями растений. При изуче-
нии этих двух показателей следует сначала определить проектив-
ное покрытие, а затем, срезав травостой, определить истинное
покрытие. Методика определения проективного покрытия наибо-
лее детально разработана Л. Г. Раменским. При этом определя-
ются три показателя, а именно: общее проективное покрытие
(так называемая проективная полнота травостоя); ярусное по-
крытие, т. е. степень покрытия нижних ярусов травостоя верхни-
147
Рис. 33. Сеточки для учета проективного покрытия (по Раменскому, 1938):
Д —малая сеточка; Б — зеркальная сеточка.
ми, и проективное обилие, т. е, проективное покрытие отдельных
видов или их групп. Для хозяйственных целей особенно важны
первый и третий показатели.
Для их более точного (не глазомерного) определения приме-
няется небольшая пластинка (картонная, деревянная, из пласт-
массы), в которой вырезано прямоугольное отверстие размером
2x5 см или 3X7,5 см. Отверстие это разделено белой ниткой или
тонкой проволокой на 10 квадратных клеток (ячеек) по 1 или
по 1,5 см2 каждая. Рассматривая травостой сверху через эту ма-
лую сеточку (рис. 33, Л), определяют, сколько ячеек приходится
на проекцию растительности и сколько на не прикрытую травой
поверхность почвы. Повторяя такой учет в разных местах участ-
ка, получают среднюю величину проективного покрытия с точ-
Рис. 34. Масштабные вилочки для определения проективного покрытия (по Ра-
менскому, 1938):
слева — для учета на квадратах размером 100 х 100 см с площадью в 4% от I мг‘, спра-
ва— для учета на квадратах размером 50Х50сл с площадью в 10% от 0,25 м*.
148
ностью до 5% и большей. Оно выражается как для всего траво-
стоя, так и для отдельных видов в процентах. Следует иметь в
виду, что проективное покрытие всего травостоя не является
простой суммой проективных покрытий отдельных видов, так как
последние отчасти перекрывают друг друга.
Кроме упомянутой малой сеточки, Раменским предложены
еще так называемая зеркальная сеточка, квадрат-сетка и мас-
штабные вилочки. Зеркальная сеточка (рис. 33, Б) может быть
легко приготовлена из карманного зеркальца с откидной крыш-
кой, в которой нужно прорезать оконце размером лсчше не
менее 4X10 см и разделить его проволокой на квадратные
ячейки. Крышка с такой сеточкой должна быть скреплена с зер-
кальцем подвижно и иметь приспособление для установки к пло-
скости зеркальца под углом в 45°. Весь прибор устанавлпваюг
таким образом, чтобы в зеркальце отражалась и проекция тра-
востоя и ячейки оконца. Зеркальная сеточка пригодна не только
при определении покрытия травостоев обычного типа, но и
очень высоких (например, зарослей тростника), а также при
определении (снизу вверх) сомкнутости крон деревьев в лесу.
Квадрат-сетка — это просто квадратная рамка (чаше всего
метровая), подразделенная внутри проволокой или шпагатом на
мелкие квадратики, позволяющие прикидывать площадь проек-
тивного покрытия отдельных видов и всего травостоя. Более
точные результаты дают масштабные вилочки (рис. 34), изго-
Рис. 35. Эталоны градаций проективного покрытия, по Раменскому (1938).
11 П. Д. Ярошенко
149
товляемые из крепкой проволоки и позволяющие учитывать
площадь проекции отдельных частей травостоя и отдельных ра-
стений. Для облегчения учета Раменский приводит иллюстра-
ции — эталоны, наглядно показывающие разные градации про-
ективного покрытия (рис. 35).
Опытные исследователи определяют проективное покрытие
прямо на глаз с достаточной степенью точности, выражая его
также в процентах от всей описываемой площадки, которая при
глазомерном определении покрытия не обязательно должна
иметь определенный размер. Нередко вместо обозначений в про-
центах применяют баллы, причем каждый балл соответствует
определенным величинам процентов, например: балл I — менее
10%, балл 2 — от 10 до 20% и т. д. Некоторые исследователи
предпочитают этот метод обозначению обилия по шкале Друде,
так как он позволяет более объективно определить количест-
венную роль вида (или группы видов) в сообществе.
Определение истинного покрытия травостоя проводится с по-
мощью той же аппаратуры, что и проективное покрытие (кроме
зеркальной сеточки). Можно определять истинное покрытие
также линейным методом. А. А. Гроссгейм применял этот метод
для вычисления покрытия основаниями дерновин. Разделенную
на сантиметры метровую линейку кладут на землю и по ее длине
измеряют пространства, покрытые дерном. Полученное «линей-
ное покрытие» выражают в процентах.
Определение покрытия в лесоводстве. Для древостоя покры-
тие выражается в двух величинах; в покрытии стволами и покры-
тии кронами.
Покрытие стволами — это сумма поперечных сечений всех
стволов данного древесного вида па уровне человеческой груди,
принятом в среднем за 1,3 м от поверхности земли. Покрытие
стволами или учитывают на глаз, или вычисляют после пересче-
та стволов и измерений их диаметров мерной вилкой. Выража-
ют покрытие в десятых долях от общего покрытия стволов всего
древостоя. Например, выражение: бук ь/10, граб 4/ю, липа l/io —
представляет пример такого учета в смешанном насаждении с
преобладанием бука. Сумма дробей должна быть всегда равна 1,
независимо от того, густым или изреженным является древо-
стой в целом. Чаще, однако, обозначают это сокращенно, отме-
чая лишь числители дробей, например 5Б, 4Г, 1Л, и тогда их
сумма должна быть равна 10. Иногда такого рода обозначения
применяют для учета не относительного покрытия каждой дре-
весной породы, а просто относительного количества ее стволов.
В обоих случаях, если сумма дробей должна быть равна 1, сум-
ма числителей будет равна 10.
Покрытие стволов называют в лесоводстве полнотой древо-
стоя, применяя этот термин обычно ко всему древостою в целом,
а не к отдельным составляющим его древесным породам. Полно-
150
та древостоя либо выражается в долях от пробной площади,
либо определяется по таблицам хода роста путем сравнения с
полнотой так называемого «нормального насаждения» данного
типа, выражаясь также в десятых долях единицы от последней.
Покрытие кронами, или сомкнутость крон («световая полно-
та» насаждения), представляет собой отношение поверхности
почвы, затененной кронами деревьев (суммарно для всех видов
древесного полога), к общей поверхности почвы пробной пло-
щади, выраженное в десятых долях, например при световой пол-
ноте 0,9 затенено кронами 9/ю поверхности почвы и '/ю не зате-
нена. Световую полноту нужно определять по возможности в
полдень, так как ее величина зависит от угла падения солнечных
лучей и поэтому неодинакова в разное время дня. Кроме того,
при работе в лиственных лесах ее нужно определять в наших
широтах летом, т. е. в то время, когда листва на деревьях
уже вполне развита. При стационарных же исследованиях
световую полноту полезно определять в течение всего вегетацион-
ного периода, но в одни и те же часы, чтобы иметь сравнимые
данные.
Объемный метод определения преобладания применяется
главным образом в лесной таксации. Для определения объема
дерева, или, как говорят в лесоводстве, его запаса, нужно знать
площадь сечения ствола на высоте груди и высоту дерева. При
определении запаса площадь сечения измеряют обыкновенно
уже не на глаз, а мерной вилкой. Высоту дерева измеряют или
высотомером, или с помощью той же мерной вилки. В последнем
случае на неподвижной ножке мерной вилки, как раз против ну-
левого деления подвижной ножки укрепляют бечевку с грузом
в качестве отвеса. Затем отходят от дерева на расстояние, при-
мерно равное высоте дерева, и расстояние это точно измеряют
рулеткой. После этого раздвигают ножки вилки на столько сан-
тиметров, сколько метров от наблюдателя до дерева, и визиру-
ют по нижнему ребру неподвижной ножки на вершину дерева.
Число сантиметров на подвижной ножке в точке пересечения ее
с отвесом покажет высоту дерева в метрах. При беглых исследо-
ваниях опытный наблюдатель может определять высоты деревь-
ев, так же как и площади их сечений, глазомерно. Зная площадь
сечения и высоту дерева, определяют запас по специальным таб-
лицам или же просто считают объем дерева равным ’/г произве-
дения площади сечения на высоту. Практика показывает, что и
последний способ дает удовлетворительные результаты. Зная
запас каждого дерева или среднего дерева той или иной породы,
нетрудно вычислить запас всего древостоя пробной площади.
Для травяных сообществ объемный метод определения гос-
подства применяется довольно редко. Он основан на погружении
растений в градуированные сосуды с водой и последующем вы-
числении по объему вытесненной воды.
151
11*
Весовой метод определения преобладания применяется, на-
оборот, только для травяных ценозов. Для этого с площадки в
1 jw2 срезают или скашивают весь травостой на уровне почвы,
разбирают по видам, и каждый вид взвешивают отдельно в све-
жем или воздушносухом состоянии (смотря по заданию). Затем
вес надземных частей каждого вида выражают в процентах. Оп-
ределенный процент веса всегда отходит на так называемую
труху, т. е. мелкие обрывки, которые не поддаются определению,
а также высохшие на поле, побуревшие кусочки и т. п. Такой
анализ называется ботаническим анализом травостоя.
При агрономическом же анализе травостой разбирают не по
видам, а по хозяйственным группам. Для луговой растительно-
сти обычно принимают шесть групп: злаки; бобовые; разно-
травье; кислые травы (осоки, ситниковые, хвощи и другие пока-
затели кислых почв); вредные травы (колючие, ядовитые, слиш-
ком жесткие) и труха (мелкие обрывки, не поддающиеся опреде-
лению) . Для степной и полупустынной растительности принимают
подразделение на следующие хозяйственные группы: злаки и
осоки; бобовые; полыни; маревые; ядовитые и сильно колючие;
прочие растения; труха.
Кроме того, в группе злаков и осок, а также в группе бобо-
вых выделяют две подгруппы: а) хорошо и отлично поедаемые
и б) удовлетворительно (реже плохо) поедаемые. В остальных
группах (кроме ядовитых и сильно колючих) выделяют подгруп-
пы: а) хорошо и отлично поедаемые и б) плохо поедаемые.
Лучше всего взвесить весь укос в тот же день в свежем виде,
затем разобрать по видам или по хозяйственным группам, вы-
сушить его и уже в сухом состоянии взвесить отдельно каждую
фракцию. Срезать травостой можно острым кухонным ножом,
овечьими ножницами, обыкновенными ножницами или серпом.
Срезать следует отдельными пучками, укладывая их в снопик,
в котором все верхушки побегов направлены в одну сторону: это
облегчит впоследствии разбор по фракциям. В степных и пу-
стынных фитоценозах целесообразнее срезать сразу по видам.
Срезать нужно в период массового сенокошения. При агрономи-
ческих анализах срезают не на уровне почвы, а на высоте 2 см.
Рекомендуется на каждой пробной площади брать укосы не
менее чем с четырех метровых площадок и не менее чем с 5—
6 площадок в V4 м2. При значительном числе пробных площадей
и достаточно однородных травостоях можно ограничиваться дву-
мя-тремя укосами на каждой пробной площади.
Нередко пользуются площадками еще меньшего размера, чем
четверть квадратного метра, но зато закладывают их в большом
количестве. Площадки для срезания укосов закладываются в ти-
пичных местах.
Обыкновенно весовые анализы дают цифры, несколько пре-
увеличенные против хозяйственной урожайности. Это объясняет-
152
ся прежде всего тем, что при обычном сенокошении и особенно
при сенокошении механизированном срезание производится не-
сколько выше от поверхности земли, а отчасти тем, что неизбеж-
ны некоторые потери при сушке, уборке в копны и стога, пере-
возке и скармливании. По данным И. В, Ларина, при срезании
для весовых анализов урожайность бывает преувеличена про-
тив хозяйственной в среднем на 10—30% в зависимости от ха-
рактера растительного покрова, а именно:
на 10—15% для лугов с преобладанием корневищевых и
рыхлокустовых злаков;
на 15—20% для злаково-разнотравных лугов;
на 20—25% для разнотравных и крупноосоковых лугов;
на 25—30% для крупноразнотравных лугов.
Поэтому, получив путем весовых анализов «биологический
урожай», нужно затем перевести его в «хозяйственный урожай»,
уменьшив на соответствующую величину в зависимости от ха-
рактера травостоя.
ВСТРЕЧАЕМОСТЬ
Это понятие относится к характеру распределения экземпля-
ров какого-либо вида на пробной площади. Встречаемость нахо-
дится в некоторой зависимости от обилия, но не всегда.
Так, если при большом обилии всегда велика и встречаемость,
то при средних величинах обилия встречаемость может быть в
одних случаях велика, в других мала, что зависит от равномер-
ности распределения особей данного вида на площадке (рис. 36).
При очень малом обилии встречаемость также низка.
°по°00
0>0°°°
О°о О
Q О
Рис. 36. Соотношения меж-
ду обилием и встречае-
мостью:
а — большое обилие и боль-
шая встречаемость; б — зна-
чительное обилие и малая
встречаемость; в — среднее
обилие и значительная встре-
чаемость; г — малое обилие
и малая встречаемость.
153
Таким образом, встречаемость характеризует частоту стояния
особей вида на участке, которая не всегда соответствует боль-
шому обилию.
Общепринятым для определения встречаемости является ме-
тод Раункиера. В пределах одной пробной площади закладыва-
ют некоторое количество более мелких площадок, распределяя
их возможно более равномерно по всему участку. Чем меньше
размер маленьких площадок, тем большим должно быть их ко-
личество. Раункиер рекомендует соотношения между размером
мелкой площадки и количеством площадок, приведенные в таб-
лице 4.
Таблица 4
Соотношение размера и количества мелких пробных
площадок
Размер площадки (В -п2) Количество площадок в пределах одной пробной площади
10 10
1 20
710 50
7100 200
Чаще всего пользуются 50 площадками круглой формы по
7ю м2 (1000 см2). Лучше всего приготовить проволочное кольцо
диаметром 35,6 см, которое и будет ограничивать на поверхности
земли кружок площадью 7ю м2. По пути обхода участка кольцо
кидают 50 раз и составляют полные списки видов, особи которых
коренятся в пределах кольца. При этом рекомендуют остерегать-
ся субъективности при выборе места для заброски кольца: луч-
ше всего забрасывать кольцо механически через определенное
расстояние от предыдущего места. После составления всех
50 списков для каждого вида вычисляют процент его встречае-
мости в данном ценозе. Эта цифра, называемая коэффициентом
встречаемости и обозначаемая иногда буквой R (по имени Ра-
ункиера), выражает процентное отношение числа площадок, на
которых данный вид встречен, к общему числу всех площадок,
т. е. в данном случае к 50.
Метод Раункиера требует значительных затрат времени. При
маршрутных исследованиях он применяется редко. Менее тру-
доемким является другой способ определения встречаемости, так
называемый новозеландский точечный метод, разработанный Кок-
кэйном и Леви при изучении лугов Новой Зеландии, а затем по-
лучивший довольно широкое распространение главным образом
в Австралии и Америке. Здесь вместо мелких площадок метода
Раункиера берут просто точки. Это осуществляется при помощи
154
простого прибора в виде вертикальной рамы с рядом направлен-
ных вниз игл (10 игл на расстоянии 5 см одна от другой), при-
чем отмечают виды, прокалываемые иглой или прикасающиеся
к ней (рис. 37).
Так как длина прибора невелика, то его следует передвигать
по одной или обеим диагоналям пробной площади. Встречае-
мость каждого вида растений выражается затем в процентном
отношении числа особей данного вида, прикоснувшихся к игле, к
числу всех точек касания игл по пути передвижения прибора.
При работе с травостоями, в которых преобладают узколист-
ные травы с направленными вверх листьями (например, злаки
и осоковые), редко удается прокалывать иглой все попадающие
под нее растения. В таких случаях отмечают растения, прика-
сающиеся к игле. В густых травостоях к игле могут прикасаться
сразу особи нескольких видов и тогда отмечают их все.
Точечный метод был применен нами при сравнении видового
состава луговых участков, подвергавшихся улучшению, с видо-
вым составом контрольных участков. На каждой пробной площа-
ди размером около 0,5 га мы ставили стойку с иглами в 50 равно-
мерно распределенных местах, причем в 25 случаях располагали
стойку в одном направлении и в 25 случаях в другом, перпенди-
кулярном первому. Всего на каждой пробной площади было взя-
то, таким образом, по 500 точек. Полученные результаты дали
хорошее представление о встречаемости видов на разных уча-
стках.
СКУЧЕННОСТЬ, ДРУЖНОСТЬ, ИЛИ «ОБЩЕСТВЕННОСТЬ»
Этот признак касается того, растет ли тот или иной вид на
данном участке единичными особями или, если группами, на-
сколько эти группы велики, а особи в их пределах скучены.
В бланке описания пробной площади графы для скученности
Рис. 37. Стойка с иглами
для точечного метода.
155
можно не делать, а если какой-либо из видов встречается груп-
пами, то приводить соответствующую отметку в графе обилия.
Пользуются, например, обозначениями gr. (gregariae), если рас-
тение встречается более или менее плотными группами, и cum.
(cumulosae), если группы в виде редких («облачных») скопле-
ний, в которых могут быть примешаны и другие виды. Эти обо-
значения комбинируют со значками обилия, например sp.— gr.;
copi— cum. и т. д.
Браун-Бланке (1951) приводит шкалу для обозначения ску-
ченности («общественности») в баллах: 1 — растет единично;
2 — группами или кучками; 3 — мелкими пятнами; 4—мелкими
колониями или крупными пятнами или же образует ковер;
5 — большой зарослью.
Этой шкалой пользуются многие геоботаники Франции,
Швейцарии, Германии, Польши и других стран. При этом отмет-
ку скученности принято ставить после отметки обилия. Напри-
мер, Anemone nemorosa 4.3 означает, что обилие определяется
баллом 4, а скученность — баллом 3. Или: Milium effusum-b 1
означает, что растение встречается в малом количестве и еди-
ничными особями.
ЖИЗНЕННОСТЬ
Этот признак обыкновенно указывается лишь для тех видов,
которые или явно угнетены, или, наоборот, слишком буйно раз-
виты в данном сообществе. Жизненность обозначается просто
краткой характеристикой в графе «Примечание».
При более детальных исследованиях определяют жизненность
путем взвешивания сухой массы всех особей данного вида с еди-
ницы площади данного фитоценоза. Проделав это для несколь-
ких фитоценозов, можно сравнивать жизненность популяции
данного вида (т. е. совокупности его особей) в различных фито-
ценозах. По А. А. Уранову (1960) мера мощности популяции —
это количество органического вещества, производимого популя-
цией на единице территории за год. Мера эта зависит от коли-
чества особей, возрастая по мере увеличения их численности, нс
до некоторого оптимального предела. При увеличении числен-
ности особей вида выше оптимальной жизненность популяции
уменьшается. При оптимальной численности обычно наблюдает-
ся и наибольший процент особей, проходящих все фазы разви-
тия до генеративных включительно.
На основе этого Урановым предложена оценка жизненности
популяции в баллах по следующей шкале:
I — средне- и крупнокалиберные особи видовой популяции,
имеющие в сумме проективное покрытие от 8% и выше, причем
не менее */4 взрослых особей проходит полный цикл развития;
П-а — проективное покрытие меньше 8%, но больше 0,25%,
156
причем не менее взрослых особей проходит полный цикл раз-
вития;
П-б — при проективном покрытии большем или равном 8%,
менее особей популяции достигает цветения и плодоносит;
Ш-а — проективное покрытие меньше 8%, но больше 25%;
полный цикл развития проходит менее взрослых особей попу-
ляции;
Ш-б — проективное покрытие не менее 8%, но все особи
популяции утратили генеративную функцию;
IV — проективное покрытие меньше 8%, но не меньше0,25%;
все особи утратили генеративную функцию;
V — проективное покрытие меньше 0,25%; все особи утрати-
ли генеративную функцию.
В этой шкале вместо веса особей вводится их проективное
покрытие из расчета, что оно у трав, кустарников и мхов при-
близительно пропорционально их весу. Что касается нормы 8%,
то она взята по Раменскому как нижний предел покрытия вида,
образующего фон.
Н. В. Дылис, А. И. Уткин и И. М. Успенская (1964) обратили
внимание на то, что жизненность отдельных видов растений бы-
вает неодинаковой в различных микрогруппировках одного и
того же сообщества, называемых ими парцеллами. В качестве
показателей жизненности авторы брали для марьянника
(Melampyrum nemorosum) среднюю длину соцветия, число бо-
ковых ветвей на растении, число листьев, среднюю и максималь-
ную длину листовой пластинки и черешка, средний вес одного
растения (в воздушносухом состоянии). Для мужского папоротни-
ка (Dryopteris filix-mas) были взяты среднее и максимальное
число вай в кусте, средняя и максимальная длина вайи, средний
вес одного куста и процент вай с сорусами.
ЯРУСНОСТЬ
При оценке того, к какому ярусу относится тот или другой
вид растения, нужно проявлять известную осторожность, учиты-
вая сказанное выше о различии между ярусами и пологами..
Ярусы обозначаются обыкновенно римскими цифрами, причем
отметка I применяется для самого верхнего яруса, II — для сле-
дующего и т. д. Если принадлежность вида к тому или иному
ярусу не совсем ясна, то лучше просто обозначать высоту его
особей (среднюю или лучше чаще встречающуюся) в сантимет-
рах или (для деревьев) в метрах. Для облегчения отметки высо-
ты как отдельных травянистых растений, так и целых их ярусов
полезно пользоваться в качестве экрана листом картона или фа-
неры с делениями.
Для более детального изучения ярусности травостоев ставят
вертикально метровую раму, подразделенную внутри проволокой
137
или шпагатом на дециметры, и ведут зарисовки. Повторив это
для данного фитоценоза несколько раз за сезон, получают кар-
тину сезонных изменений ярусности, которые бывают значитель-
ными.
Попутно с изучением ярусности травяных сообществ жела-
тельно отмечать высоту основной массы травостоя, под каковой
следует понимать верхний уровень той части травостоя, которая
является еще более или менее густой, но выше которой густота
уже резко падает и там располагаются лишь отдельные более
высокие стебли и цветочные стрелки. Определение высоты ос-
новной массы травостоя важно при хозяйственной оценке сено-
косов и пастбищ.
МОЗАИЧНОСТЬ (МИКРОГРУППИРОВКИ]
Для изучения микрогруппировок в пределах пробной площа-
ди закладывают более мелкие, например на лугах и степях, мет-
ровые площадки или же описывают линейные или ленточные
трансекты. Метровые площадки распределяют не просто равно-
мерно по всему участку, а размещают так, чтобы охватить все
типы микрогруппировок (микроассоциации), а еще лучше так,
чтобы каждый тип попал не в одну, а в несколько площадок. На
каждой метровой площадке желательно сделать проективную
съемку распределения пятнышек всех микроассоциаций. Для
каждой микроассоциации устанавливают преобладающие в ней
виды растений, а также некоторые специфические условия сре-
ды: элементы микрорельефа, увлажнение почвы, накопление ве-
тоши. Каждую микроассоциацию обозначают по двум, трем, че-
тырем преобладающим растениям. Например, в микроасс. Са-
lamagrostis 1 angsdorf 1 i Ч- Artemisia sto]onifera +Trifolium lupi-
naster преобладает вейник Лангсдорфа, на втором месте по пре-
обладанию — полынь и на третьем — клевер.
Описание микрогруппировок по линейным трансектам имеет
особое значение в местах резко выраженного мелкого микро-
рельефа (нанорельефа). Попутно с характеристикой микрогруп-
пировок производят съемку микрорельефа следующим простым
способом: на обоих концах намеченной линии вбивают в землю
по одному колу и на них возможно ближе к поверхности земли
натягивают строго горизонтально (по ватерпасу или по транспор-
тиру с прикрепленным к нему отвесом) бечевку с делениями че-
рез каждые 5 см. Проходя микропрофиль, попутно с описанием
видового состава и других признаков микрогруппировок ставят на
каждом делении бечевки строго вертикально (по отвесу) линей-
ку или метр с делениями и записывают уровень пересечения его
с горизонтальной бечевкой. По полученным цифрам можно впо-
следствии легко и достаточно точно вычертить микрорельеф уча-
158
стка и нанести на него (пользуясь какими-либо условными зна-
ками^- все-записанные микрогруппировки. Этим путем получают
точные данные о приуроченности каждого типа микрогруппиро-
вок к той или иной форме микрорельефа, что приближает к по-
знанию экологических особенностей различных микрогрулли-
ровок.
В лесах могут быть как микрогруппировки с участием всех
ярусов, в том числе и яруса деревьев, так и микрогруппировки
прогалин уже без участия деревьев. В лесах для проективной
съемки мозаичности приходится использовать уже не учетные
площадки, а пробные площади, выделяя непосредственно в их
пределах контуры микрогруппировок, а затем группируя послед-
ние в микроассоциации. При наименовании лесных микроассо-
циаций с участием деревьев названия последних ставятся на
первом месте, причем после названия древесной породы в скоб-
ках следует отметить степень затенения, создаваемого кроной,
например: микроасс. Larix dahurica (затенение 0.4) —Vaccinium
vitis. idaea + Lathyrus humilis. В ней средообразующая роль да-
урской лиственницы и брусники оценивается как приблизительно
одинаковая (поскольку лиственница сильного затенения не об-
разует) и поэтому после скобок поставлен не знак «плюс», а
«тире». Что касается чины низкой (Lathyrus humilis), то хотя
она также входит в число преобладающих видов микроассоциа-
ции, но роль ее меньше, чем лиственницы и брусники, и поэтому
перед латинским названием чины стоит знак «плюс». Второсте-
пенные виды растений вовсе не включаются в название микро-
ассоциации.
ПЕРИОДИЧНОСТЬ
Для изучения периодичности нужно отмечать при описании
сообщества фенологическую фазу каждого вида. Для этого мож-
но употреблять сокращенные словесные обозначения: вег., зацв.,
цв., незрел, плоды, обсемен., отмир. Удобно также пользоваться
значками, предложенными Алехиным: — (тире) — вегетация до
цветения; А —начало бутонизации: } —расцветание; О —
полное цветение; С “ отцветание; + — растение отцвело, но се-
мена (плоды) еще не созрели и не опадают;#—семена
(плоды) созрели (опадают); ~ —вегетация после обсеменения.
В последнее время чаще пользуются цифровыми обозначе-
ниями фаз развития растений, а именно: 1 —вегетация; 2 — бу-
тонизация, 3 —цветение, 4 — плодоношение, 5 — окончание ве-
гетации, 6 — период покоя. Кроме того, в пределах каждой
фазы различают еще подфазы, которые также обозначают циф-
рами.
Периодичность учитывают как при стационарных, так и при
маршрутных исследованиях. В первом случае мы получаем дан-
ные о сезонном развитии одних и тех же сообществ за ряд лет,
во втором случае получаем разовые фенологические данные для
всех тех сообществ, которые описываются при прохождении мар-
шрута.
Как при маршрутных, так и особенно при стационарных ис-
следованиях встает вопрос о том, следует ли учитывать перио-
дичность составляющих сообщество видов растений на всей проб-
ной площади, или же надо делать это на закладываемых внутри
ее небольших учетных площадках. И. Н. Бейдеман (1954), со-
ставившая обстоятельное руководство по методике фенологиче-
ских наблюдений при геоботанических исследованиях, рекомен-
дует изучать фенологию сообществ на учетных площадках раз-
мером не менее 4 м2 для травянистых сообществ и 100 м2 для
лесных, причем на одном гектаре должно быть не менее 10 та-
ких площадок. Небольшие размеры учетной площадки дают воз-
можность выявить соотношение между количествами особей
каждого вида, находящихся одновременно в разных фенологиче-
ских фазах. Постоянные учетные площадки должны быть обо-
значены колышками, на одном из которых следует написать но-
мер пробной площади и номер учетной площадки.
При более детальных исследованиях недостаточно все нецве-
тущие и неплодоносящие экземпляры обозначать лишь как про-
сто вегетирующие, но желательно отдельно отмечать для каж-
дого вида соотношение числа всходов, подроста (молодняка) и
взрослых особей. Результаты подсчетов можно выражать в бал-
лах и процентах или в абсолютных числах. До сравнительно не-
давнего времени такого рода изучение естественного возобновле-
ния практиковалось лишь в лесоводстве, однако теперь уже
можно назвать целый ряд исследователей, интенсивно работаю-
щих над изучением возобновления травяных ценозов. Из ученых,
известных своими достижениями в этом направлении, укажем в
первую очередь советских ботаников Т. А, Работнова и И. Д. Бог-
дановскую-Г иенэф.
Из травяных ценозов в этом отношении изучались главным
образом луговые, причем было отмечено, что на лугах вырастаю-
щие из семян молодые растеньица большей частью погибают
уже в первые годы, а оставшиеся развиваются очень медленно,
достигая взрослого состояния лишь на 3—8-й год. Значительно
быстрее происходит возобновление луговых растений вегетатив-
ным путем, т. е. кущением и побегообразованием.
Работнов определяет и возраст травянистых растений. Мето-
дика определения возраста предложена им для субальпийской
анемоны (Anemone fasciculata) —по остаткам проводящих пуч-
ков от листовых черешков и влагалищ в виде скоплений волокон
у оснований надземных побегов, а для зонтичных (Agasyllis 1а-
tifolia и Libanotis montana) — по следам прикрепления побегов
на вертикальных корневищах и т. д. При этом им установлено,
160
что возраст особей травянистых растений на лугах бывает до-
вольно большим, достигая 10 и более, а в отдельных случаях и
нескольких десятков лет.
ФИЗИОНОМИЧНОСТЬ
Этот признак, относящийся ко всему описываемому сообще-
ству в целом, выражается словесными описаниями.
В качестве примера таких описаний можно привести выдерж-
ку из прекрасных описаний Алехина, сделанных им на «Лотарев-
ской степи» (на территории нынешней Тамбовской области), ко-
торая в настоящее время распахана.
Вот, например, описание предвесеннего и ранневесеннего
аспектов.
«Как только сойдет снег, растительность степи начинает
быстро оживать. Прежде всего зеленеет столь характерный для
всех северных степей мох — Thuidium abietinum, который покры-
вает степь сплошным покровом, прижимаясь к самой почве и за-
нимая все промежутки между надземными частями растений. Эти
последние стоят еще безжизненные в своих бурых прошлогодних
отмерших листьях. Издали степь представляет собой бурую,
унылую, безжизненную картину...
Проходит неделя-другая, и степь начинает оживать, появля-
ются уже первые цветущие растения; в случае ранней весны пер-
вые цветки появляются приблизительно в середине апреля и да-
же ранее. В это время мы встречаем на степи крупные лиловые
анемоны (Pulsatilla patens), золотые звездочки гусиного лука
(Gagea erubescens), несколько видов фиалок, кроме того, для
данного периода крайне характерно еще одно растение, именно
один вид осоки — Carex humilis. Это совсем невзрачное растение
массами покрывает степь плотными низкими дерновинами и, за
отсутствием других растений, играет заметную роль в физиономии
степи (особенно заметны светло-желтые тычинки мужских колос-
ков). Таким образом, на светло-желтых пятнах осоки в настоя-
щий период особенно выделяются лиловые анемоны, но в общем
степь имеет еще более или менее серый вид от остатков прошло-
годней растительности: цветущие и зацветающие растения не
образуют сплошного пестрого ковра.
Добавим, что в это же время цветут бледно-лиловые бран-
душки (Bulbocodium ruthenicum), однако это растение в откры-
тую степь не выходит, а образует кольцо вокруг некоторых оси-
новых кустов в условиях несколько повышенного увлажнения.
Оно, как кажется, является первым зацветающим растением на
Лотаревской степи»
1 В. В. Алехин. Растительный покров степей центрально-черноземной
области. Воронеж, 1925, стр. 14.
161
ПРИЗНАКИ МЕСТООБИТАНИЯ
Все перечисленные признаки относились к структуре сооб-
щества. Но, кроме них, при описании пробной площади нужно
учитывать и признаки местообитания.
Прежде всего указывают характер рельефа, причем, если
пробная площадь находится в горной местности, нужно отметить
высоту над уровнем моря (приблизительно или с помощью высо-
томера-анероида), затем общий характер рельефа всей местно-
сти: хребет, плато, равнина, вершина горы и т. п. Если пробная
площадь находится на склоне, нужно отметить экспозицию и
уклон склона. Экспозицию определяют с помощью компаса и ука-
зывают точно, с применением не только обозначений Ю, С, В, 3,
ЮВ, СЗ, СВ, ЮЗ, но также и ЮЮВ, ВЮВ и т. д. Уклон склона
определяют или на глаз, или, для получения навыка, с помощью
угломера, или, наконец, просто транспортиром с отвесом, уста-
навливая линейку транспортира параллельно поверхности скло-
на. В местностях со сложным горным рельефом приходится
отмечать уклон и экспозицию не только того мелкого склона
(микро- или мезосклона), на котором непосредственно располо-
жена пробная площадь, но также и макросклона.
Затем отмечают характер почвы или (при менее детальных
исследованиях) лишь ее слоя до глубины 25—30 см. Наиболее
важными для характеристики местообитания растительных сооб-
ществ являются следующие свойства почвы.
Окраска почвы имеет важное значение, так как находится
в связи с ее химическим составом. При определении окраски,
если бывает трудно охарактеризовать ее одним цветом, указы-
вают или степень его интенсивности (светло-бурый, темно-корич-
невый и т. д.), или оттенки, или переходные цвета (коричнево-
бурый). Черпая окраска или черные оттенки почвы обусловли-
ваются содержанием в ней перегноя, или гумуса, красная —
содержанием гидратов окиси железа, беловатая —содержанием
извести, гипса, а также кремнезема. Цветовой эффект, который
получается от содержания в почве различных составных частей,
С. А. Захаров (1927) изобразил в виде очень наглядной схемы,
впоследствии дополненной С. И. Соколовым (1940). Из этой схе-
мы (рис. 38) видно, как в почве возникают самые разнообразные
окраски, промежуточные между черной, красной, белой и голу-
бой. Окраска почвы зависит отчасти также от влажности, что не-
обходимо иметь в виду при описании.
Структура почвы характеризуется теми отдельностями, или
агрегатами, чаще всего в виде комочков или зерен, на которые
она распадается, если только не является бесструктурной. Основ-
ные типы структур: глыбистая (отдельности неправильной формы
размером 5—10 см и более), комковатая (размер отдельностей
менее 5 см, причем форма их неправильно закругленная), оре-
162
ховатая (отдельности размером 0,5—1 см и более, с резко выра-
женными гранями), зернистая (размер отдельностей 0,5 см и ме-
нее). Кроме того, встречаются столбчатые, плитчатые и некото-
рые другие структуры. Структурность почвы зависит от ее меха-
нического состава и от содержания в ней цементирующих кол-
лоидов, а характер структуры зависит от режима температуры
и влажности, а также от присутствия тех или иных минеральных
солей. Кроме того, структурность почвы зависит от ее механиче-
ского состава; так, песчаные и супесчаные, а также глинистые
почвы нередко лишены структуры, тогда как суглинистые почвы
(с преобладанием глины над песком) часто являются структур-
ными.
Мощность почвы и ее отдельных, горизонтов.
В развитой почве бывают выражены следующие горизонты:
Гор. А] — перегнойно-аккумулятивный, имеющий обыкновен-
но более темную окраску.
Гор. А2 — переходный или часто элювиальный (горизонт вы-
мывания).
Гор. В — иллювиальный (горизонт вмывания).
Гор. С—почти не измененная почвообразованием материн-
ская порода или подпочва.
Выделяют еще самый верхний подгоризонт До, состоящий из
опавшей сухой листвы или хвои, обломков сухих веточек и т. д.
с той или иной примесью перегноя. В лесах этот слой называют
подстилкой, а на степях — войлоком. Его называют также мерт-
вым покровом, но этот термин неудачен, так как в этом слое при-
сутствует немало мелких почвенных животных: червей, насекомых
и т. д., а также множество микроорганизмов и мелких корней.
Рис. 38. Схема почвенных окрасок (по Захарову, 1927, с изменениями Соко-
лова, 1940).
По степени обеднениости железом, или выщелоченности, почвы располагаются по схе-
ме окрасок по ребрам BD и CD с усилением выщелачивания по направленн о к вер-
шине D.
Белая (БЮги пр.)
163
Для исследования мощности почвы и ее расчленения на го-
ризонты можно воспользоваться естественным обнажением поч-
вы (обрывом, откосом у края дороги), предварительно обновив
его поверхность лопатой. В других случаях выкапывают почвен-
ный разрез, т. е. прямоугольную яму, одну из стенок которой
делают ровной и строго отвесной (для изучения горизонтов),
а на противоположной стороне устраивают ступеньки для спус-
ка в яму.
Мощность почв, уже на небольшой глубине подстилаемых
щебнем или цельной глыбой каменной породы (что нередко бы-
вает на каменистых склонах гор), можно определять также ме-
тодом, который предложил Л. Г. Раменский. Метод этот состоит
в зондировании почвы нетолстым, но достаточно твердым прутом
из стальной проволоки длиной 35—48 см, нижний конец которого
сплющен и заострен наподобие копья, а верхний согнут в виде
ручки. Зонд с силой вгоняют в почву, слегка поворачивая его для
преодоления мелких препятствий и углубляя до упора в камень
или щебенку. После этого, захватив зонд пальцами на уровне
земли, вынимают его из почвы и измеряют, на какой глубине он
уперся в камень.
Почвенный скелет. Так называются включения в почву кам-
ней и мелких камешков. Почвы бывают нескелетными, слабоске-
летными и сильноскелетными. Кроме того, имеет значение ха-
рактер скелета. Угловатая форма камней указывает на то, что
они образовались на месте или же были перемещены на неболь-
шое расстояние, округлая форма указывает на то, что камни
были окатаны действием воды или, реже, ветра. Угловатая фор-
ма скелета характерна, кроме почв, образовавшихся на месте,
также для делювиальных отложений, т. е. для наносов с гор вре-
менными потоками. Окатанная форма скелета характерна для
аллювиальных (речных) наносов, а также для морских и озерных
отложений. По размерам обломков почвенный скелет характери-
зуется показателями, приведенными в таблице 5.
Таблица 5
Характеристика почвенного скелета
Размер ы обломков Форма обломков
округлая угловатая
1—3 ММ 3 ММ—3 СМ 3—10 см более 10 см Крупный песок (гравий) ) Хрящ ( Галька Валуны Дресва Щебень Камни (крупные обломки)
164
Механический состав. Содержащиеся в почве частицы горных
пород размером менее 1 мм называют: песком (0,25—1 мм),
пылью (0,001—0,25 мм), илом или глиной (менее 0,001 льм). Все
эти частицы, от песка до глины включительно, составляют поч-
венный мелкозем, тогда как включения крупнее I мм образуют
почвенный скелет. Частицы почвенного мелкозема не следует
смешивать с мелкими структурными отдельностями, или агрега-
тами (комочками, зернами и т. д.), состоящими из таких же
частиц мелкозема, но сцементированных почвенными кол-
лоидами.
Почва, мелкозем которой представлен исключительно песком,
называется песчаной, если же песок смешан с более мелкими
частицами (пылью, глиной), то в случае преобладания песка
почва называется супесчаной, а в случае преобладания глины —
суглинистой. Если же песка в почве нет вовсе, то такая почва
может быть или пылеватой, или глинистой.
Наибольшее распространение в природе имеют суглинистые
почвы, и они же являются наиболее благоприятными для произ-
растания большинства растений. Песчаные почвы в сухую погоду
сильно пересыхают в верхнем слое, но в нижних слоях концент-
рируют влагу, легко просачивающуюся вниз вследствие значи-
тельной водопроницаемости верхних слоев. Противоположность
представляют глинистые почвы: они с трудом проницаемы для
нисходящих токов воды, но зато сильно капиллярны, вследствие
чего вода легко поднимается в них снизу вверх. Когда пересы-
щенный водой верхний слой все же пересыхает (в засуху), то
на поверхности почвы образуется весьма вредная для многих
растений плотная, растрескивающаяся корка. Суглинистые почвы
лишены тех недостатков, которые свойственны почвам песчаным
и глинистым. Механический состав почв имеет связь также и с
их плодородием.
Механический состав почв в полевой обстановке может быть
приблизительно определен путем смачивания почвы и затем
скатывания из нее «колбаски». При этом глинистые почвы дают
очень прочную, тонкую и длинную «колбаску», суглинистые дают
«колбаску», разламывающуюся при ее дальнейшем удлинении,
из супесчаных же «колбаску» скатать невозможно.
Более точное определение механического состава почв про-
изводится лабораторным путем.
Органические остатки в почве могут быть как в ее толще, так
и на поверхности. В последнем случае они, как указывалось,
образуют так называемый мертвый покров, который в лесах
назы&ают подстилкой, а в степях войлоком.
Следует указывать, образует ли подстилка или войлок сплош-
ной или же прерывистый слой, отмечать мощность этого
слоя, а также характер остатков, из которых он образован
(сухие листья, хвоя, части отмершей дернины, отмершие остат-
163
ки мхов и т. д.). Скопление не вполне разложившихся органи-
ческих остатков в почвенной толще придает ей торфянистый
(если этих остатков много) или полуторфянистый (если их мало)
характер.
В случае необходимости более детальных исследований соби-
рают лесную подстилку (или степной войлок) с площадочек
определенного размера, лучше всего 50X50 см, подсушивают,
затем разбирают на фракции и каждую из них взвешивают.
Агрономическая или лесоводственная оценка. В заключение
полевого описания почвы необходимо дать ей агрономическую
или лесоводственную оценку, а также назвать тот почвенный
тип и, если возможно, разность (разновидность), к которой она
принадлежит: чернозем, каштановая, бурая полупустынная,
серая лесная, подзол, бурая лесная, перегнойно-карбонатная,
горнолуговая, горнолуговая торфянистая почва и т. д. Не имея
возможности характеризовать здесь все почвенные типы и разно-
сти, отсылаем читателя к руководствам по почвоведению.
Все перечисленные главнейшие свойства почвы записывают
для одного-двух верхних горизонтов (при менее детальных иссле-
дованиях) или для всех горизонтов вплоть до материнской
породы, отмечая мощность каждого горизонта отдельно. Лабо-
раторный анализ образцов может быть произведен по различной
программе, но наиболее важно бывает определить: механический
состав; содержание гумуса; содержание карбоната кальция;
концентрацию водородных ионов (pH) как показатель кислотно-
сти почвы. Существует прибор и для полевого определения pH,
который называется универсальным индикатором.
Во всех случаях pH желательно определять не в водной
вытяжке, а в вытяжке 7,5%-ного раствора КС1, так как при этом
получаются цифры, относительно более близкие к гой концентра-
ции ионов, какая имеет место в почвенном растворе, всегда
содержащем минеральные соли. В водном растворе показателем
нейтральной реакции является рН = 7, кислые почвы имеют pH
меньше 7, а щелочные — больше 7. В растворе же КС1 все цифры
несколько передвигаются в сторону уменьшения, так что ней-
тральные почвы показывают в этом случае pH —6,7; 6,5, а иногда
и 6,0 (в зависимости от других свойств почвы). Определение pH
представляет наибольший интерес в тех местностях, где есть
комплексы сильно карбонатных почв с почвами сильно
кислыми, что вызывает и соответственную пестроту раститель-
ного покрова.
Для более подробной характеристики почвы следует взять ее
образцы (отдельно по каждому естественному горизонту) для
лабораторного анализа. Последний не должен ограничиваться
изучением лишь химического и механического состава. Необхо-
димо особое внимание обратить на почвенную микрофлору, а
именно на преобладание тех или иных грибов или бактерий.
166
Рис. 39. Озеро среди сырого смешанного леса в Приморском крае
(Дальний Восток).
Чрезвычайно важно выяснить, какой тип бактериального про-
цесса протекает в почве — аэробный или анаэробный, а так-
же есть ли в ней микоризы. Большой интерес представляет
определение на единицу объема почвы общего количества бак-
терий, а также представителей тех или иных групп почвенной
фауны.
Уровень грунтовых вод очень важно определять в особенно-
сти при изучении сырых, заболачивающихся лугов и лесов
(рис. 39). Определив для каждой ассоциации или группы близ-
ких ассоциаций характерный уровень грунтовых вод и сравнивая
характер травостоя при разном уровне грунтовой воды, мы
этим самым подходим к уяснению того, в каком направлении
будет изменяться растительность при искусственном снижении
уровня грунтовых вод путем дренирования. Уровень грунтовых
вод следует определять в относительно менее влажное время
года, причем необходима повторность, т. е. для каждого типа
сообществ нужно по крайней мере 3—4 определения на разных
участках изучаемой территории.
Подробнее о почвенных и гидрологических изысканиях при
геоботанических исследованиях см. в книге «Полевая геобота-
ника» (т. I, 1959, стр. 138—248; в статьях И. Н. Бсйдеман
и Р. А. Филепко, А. А. Завалишина и Т. А. Рожновой). Там же
имеется статья Т. В. Арпстовской о методах исследования поч-
венной микрофлоры (стр. 412—434).
Хозяйственная оценка. Этот показатель является обязатель-
ным и очень важным при геоботанических исследованиях. Прав-
767
да, при экстенсивных маршрутных исследованиях обыкновенно
ограничиваются краткой хозяйственной характеристикой,
заполняя соответствующую графу в бланке описания пробной
площади, но и такая краткая характеристика должна быть
ясной, четкой, содержать в себе по возможности не только
констатацию современного состояния, но и первую наметку
способов улучшения данного угодья.
Хозяйственная характеристика лесных сообществ должна
исходить главным образом из совокупного учета бонитета древо-
стоя и обеспеченности семенным возобновлением. Если бонитет
низок, нужно искать пути изменения состава древостоя за счет
введения пород, более соответствующих данным условиям. Если
семенное возобновление незначительно или отсутствует (мы
имеем в виду не только одно-, двухлетние всходы, но и подрост
старших возрастов), то нужно искать пути содействия возоб-
новлению данной породы (или пород) за счет изменения системы
рубки и проведения других мер или же за счет подсева другой
породы, более пригодной для данных почвенно-климатических
условий.
Хозяйственная характеристика сенокосов и пастбищ дается
по трем основным показателям: участие в травостое полезных
и вредных растений; состояние травостоя в отношении главным
образом степени его выбитости (на пастбищах), а также в отно-
шении его хозяйственной урожайности; степень плодородия
почвы.
Для сенокосных участков следует отмечать, имеются ли пре-
пятствия к механизированному сенокошению, т. е. кочки и
кустарники.
При хозяйственной оценке кормовых угодий важное значение
имеет знание степени поедаемости растений теми или иными
видами сельскохозяйственных животных. Методика изучения
поедаемости разработана во всех деталях И. В. Лариным и его
сотрудниками. Отмечается, что поедаемость не обязательно
соответствует питательной ценности растения, и это особенно
характерно для пустынной и полупустынной зон, где бывает мало
ценных кормовых трав и где поэтому и такие растения, как
полыни, поедаются скотом очень охотно, представляя нередко
основной вид корма.
Поедаемость одного и того же вида растений бывает неодина-
ковой в разные периоды его жизни. Обыкновенно растения
поедаются лучше в молодом состоянии. Некоторые растения, как,
например, белоус, поедаются почти исключительно в молодом
возрасте. Наконец, нередко различные части одного и того же
растения поедаются не в одинаковой степени, чаще всего лучше
поедаются листья. Все это нужно иметь в виду при изучении
поедаемости.
168
БЛАНКИ ДЛЯ ПОЛНОГО ОПИСАНИЯ ПРОБНЫХ
ПЛОЩАДЕЙ
В зависимости от типа растительного покрова бланк для
описания пробной площади имеет те или иные особенности. Так,
бланк для описания травяных фитоценозов сходен с используе-
мым для травяного покрова лесов, но для древесного яруса фор-
ма бланка несколько иная. Ниже приводим примерные образцы
двух бланков: для луговых и лесных фитоценозов.
Первый из них пригоден также для степных фитоценозов, с
той лишь разницей, что в последних желательно отмечать также
мощность (толщину) войлока (опада) и степень покрытия им
поверхности почвы. Некоторую специфику будут иметь также
бланки для описания болотной растительности, поскольку там
необходимо детальное описание характера увлажнения (аллю-
виальное, делювиальное, грунтовое, атмосферное), степени
увлажнения, а для северных болот, кроме того, указание уровня
мерзлоты в связи с микрорельефом.
Приводимые ниже бланки пригодны для общего описания
площади, соответствующей по размерам площади выявления
ассоциации или крупнее ее (см. стр. 177).
Наряду с этим, как уже упоминалось, в пределах такой более
крупной площади закладывают более мелкие, а именно раз-
мером 1 зг или 50x50 см для взятия укосов или же метровые
площадки для учета возобновления в лесах, а также для более
детального изучения микрогруппировок.
Бланк для описания лугового фитоценоза
№ __.. ..___ 19 г.
Название типа фитоценоза .
Республика и область Район
Окрестности. Название местности
Ок ру жен не_______________________________________
Высота над уровнем моря Экспозиция склона
Уклон в градусах Общий характер рельефа .
Микрорельеф и его происхождение „________________________________________
12 11. Д Ярошенко
/69
Характер почвы
Условия увлажнения и глубина грунтовых вод
Точные или приблизительные размеры пробной площади
Список растений пробной площади
№ | п/п Название растения Обилие Покрытие (в %; про- ективное) Высота (в СМ) Фенофаза Общие замечания
Общее покрытие высшими растениями: проективное и истинное
Покрытие почвы мхами и лишайниками__________________________________________
Характер задернения (плотнокустовое, рыхлокустовое, злаковое, осоковое, жела-
тельно указать также процент задерненности злаками и осоками)
Высота травостоя: максимальная высота основной массы
травостоя (см. стр. 158)
Характер ярусности __________
Аспект (физиономическая характеристика)
Микрогруппировки и их связь с теми или иными условиями
Мертвый покров_________________________________________________________
Влияние человека_______________________________________________________
Влияние диких животных___________________________________________________
Хозяйственная оценка угодья_________________________________________________
Общие замечания (типичность сообщества, его история, характер возобновления
тех или иных видов растений и т. д.)________________________________________
Подпись_______________________
170
При описании пробных площадей в лесах приведенный выше
бланк применим для травяного покрова, тогда как для древес-
ного яруса описание составляется уже в несколько ином виде,
а именно: вместо обилия отмечают покрытие стволов или (чаще)
световую полноту древостоя, а кроме того, в специальных гра-
фах— диаметр ствола на высоте груди и возраст, учитываемые
отдельно по каждой породе. Таким образом, для лесных сооб-
ществ бланк должен подразделяться на две части: для древесных
и для нижних ярусов. Кроме того, в лесных сообществах очень
важно отмечать наличие или отсутствие возобновления каждой
древесной породы, причем для его учета желательно заклады-
вать дополнительные, более мелкие (метровые) площадки или
трансекты (шириной в 1 м) и на них производить точный подсчет
числа всходов и подроста каждой древесной породы. Кроме всхо-
дов и подроста, или, как говорят лесоводы, семенного возобнов-
ления, нужно отдельно учитывать поросль от пней и корневые
отпрыски у тех пород, которые имеют способность вегетативного
размножения.
Бланк для описания лесного фитоценоза
№ 19 г.
Тип леса или ассоциация__________________________________________________
Географическое положение (республика, область, район, лесхоз, дача, урочище,
ква рта л)_______________________________________________________________
Топографическое положение (если склон, указать его экспозицию и уклон в гра-
дусах, а также указать высоту над уровнем моря),
Общий характер рельефа L
Микрорельеф и его происхождение__________________________________________
Поверхностная горная порода______________________________________________
Почва (название типа и разности, мощность, окраска, механический состав, струк-
турность, скелетность, степень увлажнения). Признаки эти отмечаются для всех
горизонтов или хотя бы для двух верхних
Подстилка (мощность, процент покрытия),
Окружение (другие лесные типы, луга, пашни и т. д.)
Влияние человека_________________________________________________________
Влияние диких животных___________________________________________________
Степень сомкнутости крон (световая полнота) древостоя
Размер пробной площади___________________________________________________
12*
171
Описание древостоя
Порода Ярус Состав (по покрытию СТВОЛОВ} Диаметр (в сж) Высота Н (в ж) Воз- раст (преоб- ладаю- щий) Фено- фаза Общие заме- чания
преоб- ладаю- щий макси- маль- ный преоб- ладаю- щая макси- маль- ная
Описание возобновления
Порода Состав (в десятых ДОЛЯХ) Обилие Высота (в СЖ) Возраст Происхожде- ние (семенное, порослевое» Состояние (жизнен- ности)
Общие замечания______________________________________________________________ __
Описание подлеска
Порода Состав (в десятых долях) Высота Н (в ж) Состояние (жизненность) Фенофаза Общие замечания
преобла- дающая максималь- ная
Сомкнутость полога подлеска______________________________________ ,__________
Описание напочвенного покрова
Общий характер_______________________________________________________
Степень покрытия в процентах ^проективное и истинное покрытие)
Разделение на ярусы, их высота и характер
Микрогруппировки, их связь с теми или иными условиями (характер микрорель-
ефа, степень освещения, развитие подстилки и т. д.) и состав всех их ярусов
(включая и древесный) *
172
Название растения Обилие Покрытие (в % или де- сятых долях, проективное) Жизненность Фен о фа за Общие замечания
Мохово-лишайниковый покров на поверхности почвы (степень покрытия в про-
центах, мощность живого и мертвого слоя)
Внеярусная растительность (лианы и эпифиты; указать состав и обилие тех и
других)_______________________________________________________________________
Аспект кустарничково-травяного и мохово-лишайникового покровов (физиономиче-
ская характеристика)________________________________________________________
Общие замечания для всего сообщества (типичность сообщества, его динамика,
узловой или кратковременный характер, хозяйственная оценка)
Подпись__________________
При работе в районе, флора которого хорошо известна,
целесообразно использовать печатные бланки, в которых зара-
нее напечатан список видов, так что придется лишь вычеркивать
отсутствующие виды и, быть может, лишь изредка добавлять
недостающие. В плохую погоду полезно пользоваться магнито-
фонной записью (диктофон). Об этих и некоторых других мето-
дах улучшения геоботанической документации см. статью
В. В, Мазинга «О научной документации в геоботанике» (сборн.
«Проблемы современной ботаники», т. I. М.—Л., «Наука», 1965).
ОПИСАНИЕ ПРОФИЛЯ
Наряду с описанием пробных площадей производят также
описание геоботанических профилей, имеющее особенное значе-
ние в горных местностях, где этот метод позволяет прекрасно
иллюстрировать связь между растительностью и рельефом.
Выбрав определенное направление, наметив его по какому-
173
нибудь видному издали предмету (скала, дерево и т. д.) и ориен-
тировав его по компасу, исследователь пускается в путь, опи-
сывая все изменения растительного покрова. При этом он непре-
рывно ведет измерение расстояния (шагами, рулеткой или
мерной лентой) и отмечает (в расстояниях от начальной точки
профиля) границы всех сообществ, встреченных по линии про-
филя. В пределах участка каждого сообщества отмечают воз-
можно точнее и уклон склона в градусах. Если съемка такого
профиля (табл. 6) является лишь дополнением к описаниям
большого количества пробных площадей, то можно сообщества
не описывать подробно, а просто указывать их общий характер
и условное название со ссылкой на соответствующую запись
пробной площади.
По цифрам расстояний и уклонов каждый профиль может
быть затем вычерчен. При описании профилей приходится опи-
сывать отдельно не только резко выраженные, узловые, но и
все кратковременные группировки, тогда как пробные площади
и площадки закладываются обычно лишь в узловых и наиболее
типичных из кратковременных сообществ.
Таблица 6
Пример описания профиля
Расстояние (в шагах) Уклон (в градусах) Фитоценоз Примечание
0—30 5—10 Залежь, постепенно превра- щающаяся в луг: Echium vulgare copy Falcaria vulgaris copL Покрытие почвы расте- ниями 80% (проектив- ное)
30—96 25 Послелесный луг (см. описа- ние пробной площади № 24)
96—110 18 Лесная опушка: Quercus macranthera сор2; Prunus spinosa sp.; Viburnum lantana sp.; Rosa canina sol. В промежутках между де- ревьями и кустарниками луговая растительность предыдущего типа Деревья дуба порослевого происхождения, корявы и низкорослы, высотой 2—3 м
ПО—... 15—20 Дубово-грабовый лес (см. описание пробной пло- щади № 29) Подстилка из сухих листь- ев, довольно мощная (10—12 си) и покры- вает почву сплошь (в отличие от записи № 29)
174
ХИМИЧЕСКИЕ И ФИЗИОЛОГИЧЕСКИЕ
МЕТОДЫ В ГЕОБОТАНИКЕ
В. Н. Сукачев (1957) справедливо отметил, что среди компо-
нентов биогеоценоза фитоценозу принадлежит наибольшая био-
геоценозообразующая роль и что фитоценоз представляет собой
лабораторию, в которой происходит аккумуляция и превращение
веществ и энергии. Отсюда ясно, насколько важным является
изучение отдельных звеньев этого превращения, т. е. другими
словами, насколько важны химические и физиологические иссле-
дования фитоценозов.
Химические исследования растительных сообществ могут про-
изводиться в различных направлениях, например: изучение на-
копления тех или иных минеральных солей в образующих сооб-
щество различных видах растений; изучение накопления у отдель-
ных видов растений одного сообщества специфических органиче-
ских веществ — белков, жиров, углеводов, алкалоидов, витаминов
и т. д.; изучение накопления тех или иных веществ у одних и тех
же видов растений, но в разных сообществах; изучение накопле-
ния тех или иных веществ в различных типах сообществ в целом
(без учета отдельных видов растений), причем можно изучать
отдельно наземную и отдельно подземную массу фитоценозов;
исследование химического влияния одних растений на другие пу-
тем выделения ими в окружающую среду тех или иных веществ;
изучение круговорота элементов питания в растительных сооб-
ществах.
Что касается чисто физиологического исследования расти-
тельных сообществ, то под этим следует понимать изучение в по-
левой обстановке физиологических процессов у растений, обра-
зующих данное сообщество, а также у всего сообщества в целом
(симфизиология). При этом следует пользоваться методами, ко-
торые разрабатываются специально для физиологических иссле-
дований в природной обстановке. Подробная сводка этой мето-
дики дана О. В. Заленским в I томе книги «Полевая геоботаника*
(1959, стр. 245—365). Физиологическое исследование полезно со-
четать с анатомической характеристикой растений по типам со-
обществ.
Исследования по физиологии лесных древостоев наиболее
широко развернуты Л. А. Ивановым и его многочисленными со-
трудниками.
ОБРАБОТКА МАТЕРИАЛОВ ПОЛЕВЫХ
ГЕОБОТАНИЧЕСКИХ ИССЛЕДОВАНИЙ
Чтобы составить подробную характеристику того или иного
типа сообщества (как узлового, так и кратковременного), недо-
статочно описать лишь одну пробную площадь, а необходимо
175
значительное число таких описаний. То же самое относится и к
профилям: чтобы выявить закономерную связь растительности
с рельефом, недостаточно одного профиля, их нужно несколько,
и тем больше, чем полнее мы хотим получить наши выводы.
ПОНЯТИЕ О КОНСТАНТАХ И МИНИМАЛЬНОМ АРЕАЛЕ
Каждая ассоциация встречается в природе в виде значитель-
ного числа участков или отдельных сообществ, сходных между
собой, но разобщенных в пространстве. Сходство между собой
этих отдельных сообществ, или, как их назвал Браун-Бланке,
индивидуумов ассоциации, хотя и бывает значительным, но пол-
ного тождества между ними нет. Поэтому среди видов растений
данной ассоциации бывают такие, которые встречаются на всех
ее участках, и такие, которые встречаются лишь на части участ-
ков. Виды, свойственные 90—100% всех участков данной ассо-
циации, Дю-Рие назвал константами. Все виды какой-либо ассо-
циации Дю-Рие подразделяет на 10 классов константности,
подобно тому как это было указано выше по отношению к встре-
чаемости. Константы X класса встречаются на 90—100% (точнее,
91 —100%) всех исследованных участков, IX класса — на 80—
90% (точнее, 81—90% )и т. д. Итак, если встречаемость вида ха-
рактеризует его распространение в пределах одного участка
ассоциации, то константность характеризует его распределение
в пределах многих участков.
При определении константности обыкновенно не пользуются
пробными площадями крупных размеров, но в пределах каждого
участка ассоциации закладывают одну небольшую площадку.
При этом было выяснено, что константность многих видов при
увеличении размеров таких площадок сначала увеличивается, но
затем при дальнейшем увеличении размеров площадок остается
уже неизменной. Следовательно, при увеличении размеров пло-
щадок до определенного предела будет возрастать и число кон-
стант высших классов. Например, если при площадках по 0,5 .и2
число констант X и IX классов вместе равно 2, то при площад-
ках в 1 м2 оно увеличивается, скажем, до 3, а при пло-
щадках в 2 л2 — до 5. Но затем сколько бы мы ни увеличива-
ли размер каждой площадки, число констант уже более не
возрастает.
Это означает, что в нашем примере площадки менее 2 л<2 каж-
дая не выявляли еще всех присущих данной ассоциации кон-
стантных видов. Но начиная с 2 м2 и выше константные виды уже
выявляются полностью. Следовательно, истинное число констант
X и IX классов в приведенном примере равно 5. Тот наименьший
размер площадки, который включает все ее константы, называет-
ся по Дю-Рие, минимальным ареалом ассоциации. В нашем при-
мере минимальный ареал равен 2 м2, Дю-Рие и его коллеги по
176
упсальской геоботанической школе вычислили минимальный
ареал для многих ассоциаций, точнее, социаций, Швеции; его раз-
меры оказались в большинстве случаев равными 2—4 я2; для за-
рослей карликовых кустарников и лишайников — 1 я2, а для
богатых видами лугов и «луговых лесов»— 16 я2. Знание мини-
мального ареала имеет некоторое значение для определения не-
обходимого размера пробной площади при изучении тех или
иных типов сообществ и синузий. Пробная площадь должна быть
не меньше минимального ареала.
Техника определения минимального ареала вытекает из изло-
женного выше: в каком-либо типе сообщества закладывают сна-
чала более мелкие площадки и в каждой из них составляют пол-
ный список видов. Затем размер каждой площадки увеличивают
и снова составляют полные списки видов. Проделав так несколь-
ко раз, наконец находят тот размер площадки, при дальнейшем
увеличении которого число констант X класса (или X и IX клас-
сов вместе) уже не возрастает. Эта площадь и будет минималь-
ным ареалом ассоциации.
Степень константности каждого вида ассоциации может быть
выражена в процентах совершенно так же, как и встречаемость.
Константность обозначают буквой К. Распределяя все виды ассо-
циации по классам константности, можно вычерчивать кривые
константности наподобие кривых встречаемости.
ПЛОЩАДЬ ВЫЯВЛЕНИЯ И ФРАГМЕНТ АССОЦИАЦИИ
Вместо минимального ареала Л. Г. Раменский предложил по-
нятие площади выявления, т. е. той наименьшей площади, на ко-
торой выявляются в полной мере все существенные черты дан-
ного ценоза. Понятие площади выявления вместо минимального
ареала принимает и В. Н. Сукачев.
Безусловно, определение площади выявления гораздо важнее,
чем определение минимального ареала, так как последний осно-
ван только на одних константных видах, здесь же учитываются
все признаки сообщества.
Несоответствие площади выявления определяемому только по
константности видов минимальному ареалу Раменский поясняет
схематическим рисунком (рис. 40). Из рисунка видно, что пло-
щадь выявления константности какого-либо вида (Л) может ока-
заться во много раз меньше, чем площадь выявления обилия
того же самого вида растений (В). Раменский подчеркивает, что
площадь выявления растительной группировки — это «минималь-
ная величина площадок, обеспечивающая однородность количест-
венного состава их покрова» (1938, стр. 293).
С точки зрения учения о микрогруппировках площадью выяв-
ления будет та наименьшая площадь, на которую попадают все
типы микрогруппировок, свойственные данной ассоциации. Та-
177
ким путем мною (Ярошенко, 1931) была определена площадь
выявления альпийской ассоциации Alchimilla caucasica + Carex
tristis, оказавшаяся между 1 и 2 м2. Для лесных ассоциаций
площадь выявления обычно не менее 2000 л.’2, а иногда и больше.
Метод определения площади выявления какой-либо ассоциа-
ции состоит в описании ряда площадок с последовательным уве-
личением их размеров. Начиная с некоторого размера дальней-
шее увеличение площадок уже не прибавляет ничего нового из
существенных признаков сообщества. Этот размер и будет пло-
щадью выявления. Опытный исследователь определяет площадь
выявления прямо на глаз, не описывая предварительно ряда по-
следовательно увеличивающихся площадок. Надо полагать, что
чем выработаннее тип сообщества, тем определеннее размер его
площади выявления (так же как и минимального ареала). По-
этому понятия о площади выявления и минимальном ареале
имеют в основном значение для сравнительно выработавшихся
узловых типов ценозов.
Практическое значение знания площади выявления каждой
ассоциации то же, что и знания ее минимального ареала; пробная
площадь должна быть не меньше площади выявления. Однако,
как было упомянуто, площадь выявления предста'вляет резуль-
тат бодее всестороннего подхода к сообществу, тогда как мини-
мальный ареал определяется лишь по одному признаку.
Иногда размеры участка ассоциации в его естественных гра-
ницах настолько малы, что оказываются меньшими, чем пло-
щадь выявления. Такой клочок, не выявляющий в полной мере
всех присущих данной ассоциации характерных признаков, на-
зывается фрагментом ассоциации.
Рис. 40. Сравнение площади выявления и минимального ареала (по Рамен-
скому, 1838):
А —минимальный ареал (площадь выявления константности какого-либо вида): В — пло-
щадь выявления обилия того же вида.
178
Фрагмент может состоять из одной микрогруппировки или
из нескольких, но не включает всех типов микрогруппировок
(т. е. микроассоциаций), свойственных данной ассоциации.
ХАРАКТЕРИСТИКА СЕЗОННОЙ ДИНАМИКИ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Материалы по изучению динамики фенологических фаз раз-
личных видов какого-либо сообщества могут быть обработаны
и сведены в так называемый фенологический спектр. Способы
изображения феноспектров могут быть различны. Феноспектр
Гамса, или, как называет его сам автор, феноэкологический
спектр, менее детализирован. Принцип составления такого спект-
ра виден на рисунке 41, Л. Расширение фигуры говорит о разви-
тии вегетации, а дальнейшее расширение — о зацветании. Макси-
мальная ширина ее соответствует цветению, после чего фигура
опять сужается, что соответствует отцветанию и обсеменению.
Максимальная ширина фигуры пропорциональна обилию вида
во время разгара его цветения.
В феноспектре Шенникова (рис. 41, Б), совокупность фено-
фаз каждого вида имеет форму удлиненного прямоугольника,
общая высота которого пропорциональна встречаемости вида в
данном сообществе. Косыми линиями прямоугольник подразде-
ляется на отрезки, каждый из которых обозначает фенофазу и
окрашен в особый цвет или же покрыт особой штриховкой. Если
провести через прямоугольник вертикальную линию в каждый из
сроков, мы можем по высоте отдельных отрезков судить о встре-
чаемости экземпляров вида, находящихся в каждой из фенологи-
ческих фаз отдельно. Вместе с тем длина каждого отрезка про-
порциональна продолжительности обозначаемой им фенофазы.
Н. Г. Москаленко (1964) предложил ввести в феноспектр
Шенникова уточнение для отражения относительной встречаемо-
сти особей, вступающих в ту или иную фазу в каждый календар-
ный срок. В результате косые линии, подразделяющие прямо-
угольники спектра Шенникова, заменяются ломаными и вер-
тикальными линиями. Это позволяет точнее отражать ход
фенологических явлений и влияние на них экологических фак-
торов (рис. 42).
Недостаток феноспектра Гамса состоит главным образом
в том, что он нс дает представления о количественных соотноше-
ниях различных фаз в каждый момент жизни сообщества.
Спектр Шенникова восполняет этот пробел, но все же и он не
дает представления о сезонном изменении количественной роли
каждого вида. С этой точки зрения заслуживает внимания
спектр Браун-Бланке (1951) для средиземноморской гариги с
Quercus coccifera. В этом спектре, который автор называет схемой
периодичности, вовсе не указываются фенофазы, а изменение
179
2 .'./...Гу W v V/l
----------- ---- ~ ~1
Апрель | май.
/ I //1///1 / I if]hi
Июнь Июль I ASeycm Сентябрь | Октябрь [ Ноябрь
/ | //1 ///1 / | //1 ///] / | // |/// J / | //1 ///171 //1 m\ / I //1 ш
1 ;•- я .aiiiiiiiiiniiiiiiiiiiiwn
* И^^ж/:у>7ЖМ1111Н11111—
в з mMlMfaj У ~г / AJiiiMiiiinMiiiiiMiiiiMiiM
1Л1шпш1111Шшш1111Ш111Ш1Ш1Ш11]1П111111ВШШ11ШШП]1Й1111111111
*
» » ВТ В Ш Е23'
Рис. 41. Различные типы фенологических спектров:
А — фрагмент спектра Гаме а для ржаного поля в Валлисе. Наибольшая ширина полосы
соответствует обилию вида во время цветения. / — щетинник зеленый; 2 — рожь; 3 —
гусиный лук; 4 — гадючий лук; б — полевая фиалка. Б — фрагмент спектра Щенникова
для луга на Средней Волге. Ширина полосы соответствует степени встречаемости вида;
а — вегетативное состояние; б — бутоны; в — цветение; г — созревание; д — обсеменение;
е — отмирание; 1 — лисохвост; 2 — звезчатка злаковидная; 3 — гвоздика Фишера; 4 —
костер безостый; 5— подмаренник членистый. В — фрагмент спектра Шалита для ковыль-
но-типчаковой степи в Аскания-Нова. Ширина полосы соответствует проективному оби-
лию вида (в процентах); а — вегетативное состояние; б — бутоны; в — цветение; г — со-
зревание плодов; Ь — обсеменение; е — отмирание; I — касатик низкий; 2 — типчак; 3 —
чертополох крючковатый; 4 — солонечник мохнатый; б — полынь австрийская.
9121518 212427 3024 1 10 14 1? 21 24 27 31 4 7 1118 21 28
июнь июль аВгуст
Рис. 42. Фенологический спектр альпийской толокнянки (Acetous alpina) по
Москаленко (1964):
1 — вегетация; 2 — бутонизация; 3 — цветение; 4 — незрелые плоды; 5 — зрелые пло-
ды; б — изменение окраски листьев; 7—границы между фенофазами по Шенникову;
9— то же по Москаленко.
ширины каждой полосы соответствует сезонной динамике расти-
тельной массы каждого вида.
М. С. Шалыт (1946) объединил феноспектры Шенпикова и
Браун-Бланке и предложил более совершенный тип спектра
(см. рис. 41, В). В спектре Шалыта ширина полосы меняется в со-
ответствии с динамикой массы или же покрытия каждого вида,
но сама полоса не одноцветная, а в нее вписаны (как у Щенни-
кова) условные обозначения фенологических фаз. Автор совер-
шенно правильно указывает, что такой прием сделает спектры
более ценными как для теоретических, так и для чисто хозяйст-
венных выводов, ибо мы сможем определить по спектру, для
каждого данного момента, какие виды там преобладают и в ка-
кой фенофазе они находятся. Думается, однако, что не обяза-
тельно степень участия вида и ее сезонную динамику учитывать
непременно по весу или по площади покрытия; при менее де-
тальных исследованиях можно учитывать просто сезонное изме-
нение обилия вида, оценивая его хотя бы по способу Друде, и
тот или иной балл выражать соответственной условной шириной
полосы каждого вида в спектре.
Сезонный ритм растительного сообщества представляет весь-
ма существенный признак, который наряду с другими структур-
ными, а также экологическими особенностями может быть по-
ложен в основу эволюционной классификации фитоценозов.
ВЫЯВЛЕНИЕ ЧАСТНЫХ СМЕН
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Такое выявление достигается главным образом двумя путя-
ми: исследованием закономерностей размещения фитоценозов в
пространстве, которые часто отражают их смену во времени, и
непосредственным наблюдением изменений во времени. Первый
способ в особенности применим при изучении такого простран-
ственного ряда фитоценозов, на одном конце которого находятся
181
«молодые» местообитания, только начавшие заселяться растени-
ями, а на другом — «зрелые». Двигаясь от молодых местооби-
таний к более зрелым, например от свежих речных наносов в
глубь поймы, мы наблюдаем пространственную смену фитоце-
нозов, которая в общих чертах повторяет их смену во времени.
Это связано с тем, что формирование всего ряда приречных со-
обществ вызвано постепенным отодвиганием речного русла,
вследствие чего каждая отдельная полоса проходит здесь после-
довательно все стадии, начиная от свежих речных наносов.
Аналогичное соответствие пространственного размещения
смене во времени можно наблюдать и в других случаях, когда
также имеет место постепенный переход от молодых местообита-
ний к зрелым. Например, если в пустынной зоне Арало-Каспий-
ской низменности мы будем продвигаться от моря в глубь стра-
ны, то сначала увидим солончаки, либо пески, или же болота.
Эти три типа местообитаний молодые, потому что каждое из них
представляет собой один из вариантов первой стадии отступания
моря. С удалением от моря первые стадии переходят в следую-
щие: солончаки — в слабозасоленные почвогрунты; рыхлые, под-
вижные пески — в более уплотненные, закрепленные, с зачатка-
ми гумусообразования; болота — во влажные, но уже без явной
заболоченности болотно-луговые почвы. Еще дальше от моря все
эти стадии переходят в узловые сообщества, чаще всего в солян-
ковую пустыню.
Соответствие пространственных смен сменам во времени
можно наблюдать и в горах. Например, если двигаться в высо-
когорной области от лишенной растений каменистой осыпи к лу-
говым массивам, можно нередко наблюдать все стадии заселе-
ния осыпи растениями, протекающего одновременно с почвооб-
разовательным процессом: камни постепенно размельчаются,
превращаясь в мелкозем, в котором, чем дальше от осыпи, тем
более содержится гумуса, и наконец следуют луга с уже вполне
развитой горнолуговой почвой.
Однако подобное соответствие пространственных смен сме-
нам во времени бывает не всегда. В долинах рек оно нередко
нарушается влиянием речных разливов, а в горах наряду с про-
цессами закрепления осыпей и почвообразования сплошь и ря-
дом протекают и противоположные им эрозионные процессы, ко-
торые тоже нарушают это соответствие. Встречаются и такие
случаи, когда процесс закрепления осыпи протекает бок о бок с
дальнейшей эрозией, так что иногда приходится видеть на одном
и том же участке элементы обоих этих процессов одновременно.
Для того чтобы расшифровать все эти сложные сплетения и
установить картину смен растительности в горах, нужно уметь
наблюдать, замечать все важные моменты, нужен хороший ис-
следовательский глаз. Начинающим геоботаникам можно реко-
мендовать практиковаться сначала на более простых случаях:
182
в приречной полосе, на берегу моря, на окраинах усыхающих бо-
лот и т. д., а затем уже браться за изучение более сложных яв-
лений в горных местностях.
Приведенные примеры соответствия пространственных смен
сменам во времени относились главным образом к сменам после-
довательным. Изучение смен этого типа является вообще делом
более легким, чем изучение смен внезапных и антропогенных.
Для выявления внезапных и антропогенных смен недостаточно
изучать пространственные смены ценозов, но нужно детально
исследовать процессы возобновления в каждом из сопри-
касающихся сообществ, изучать жизненность тех или других
«строителей сообществ» (эдификаторов) и т. д. Необходимо так-
же тщательно изучать местообитания с целью уловить те их из-
менения во времени, которые вызываются не только непосред-
ственно жизнедеятельностью фитоценозов. Надо привлечь дан-
ные об изменении климата всей местности за ряд лет, архивные
материалы, которые помогут восстановить полную картину бы-
лого растительного покрова и т. д.
Переходя к методам непосредственных наблюдений разного
типа частных смен, заметим прежде всего, что такие наблюде-
ния должны осуществляться при стационарных работах. Выде-
ляют участки и либо из года в год делают общие описания их
растительности в целом, либо с целью детализации закладывают
на них постоянные площади тех или иных размеров и на этих
площадях производят регулярный, ежедекадный или ежемесяч-
ный учет различных признаков фитоценозов. При таком учете, в
отличие от маршрутных исследований, рекомендуется пользо-
ваться по возможности методами объективной оценки.
Наглядные результаты при сравнительном изучении изменя-
емости фитоценоза по годам дают ежегодное в один и тот же ка-
лендарный срок картирование пробной площади с точным на-
несением размещения отдельных растений и куртин. В типах с
хорошо выраженными микрогруппировками последние как раз
и смогут явиться единицами такого картирования.
При стационарном изучении частных смен ценные материа-
лы может дать исследование семенного и вегетативного возоб-
новления. В настоящее время разработана методика изучения
возобновления не только в лесах, но и в травяных сообществах.
Занимающиеся этими вопросами И. Д. Богдановская-Гиенэф,
Т. А. Работнов и некоторые другие авторы дают подробные ме-
тодические указания.
Являясь связующим звеном между сезонными изменениями,
разногодними изменениями и частными семенами, процессы во-
зобновления проливают свет на многие закономерности динами-
ки сообществ, приближая нас к пониманию непосредственных
причин частных смен: последовательных, внезапных, а также
антропогенных.
183
Сотрудники возглавляемой А. А. Урановым кафедры ботани-
ки Московского государственного педагогического института
имени В. И. Ленина проводят интересную и практически важную
работу по изучению возрастного состава популяций некоторых
видов на лугах и в лесах. На основании этих исследований дела-
ются выводы и о направлениях частных смен, или сукцессий
(см., напр., сборник статей «Онтогенез и возрастной состав попу-
ляций цветковых растений». М., «Наука». 1967).
В некоторых случаях изучение общего характера сезонных
изменений растительности того или иного участка дает пред-
ставление о частных и даже общих сменах, которые привели в
прошлом к ее формированию: более того, изучение современных
сезонных изменений той или иной группировки иногда может
помочь и в предугадывании тех возможных смен, какие могут
произойти в будущем. Так, в окрестностях Львова Е. И. Шишо-
ва (1952) описала болотистые луга, претерпевающие очень рез-
кие сезонные изменения. Ранней весной, когда почва пересыще-
на водой, в травостое преобладают осоки и болотное разно-
травье. К июню, когда почва сильно подсыхает, преобладание
переходит к мезофильным луговым злакам.
Как при стационарном, так и особенно при маршрутном изу-
чении частных смен нужно иметь в виду, что влияние хозяйст-
венной деятельности человека проявляется не только при антро-
погенных сменах, но, правда, в меньшей мере, и при сменах
других типов. Нужно уметь отличать коренные виды растений
каждого фитоценоза от видов, роль которых возрастает под вли-
янием человека, или же от видов, появляющихся в данном месте
целиком под влиянием человека. Необходимо учитывать влияние
человека на структуру фитоценозов, на процессы почвообразо-
вания, на окружающую растительность.
СТАТИСТИКО-МАТЕМАТИЧЕСКИЕ МЕТОДЫ В ГЕОБОТАНИКЕ
Простейших из них мы уже касались при рассмотрении
определения встречаемости, а также константности видов в расти-
тельных сообществах. В последние десятилетия статистико-ма-
тематические методы получили чрезвычайно большое распро-
странение в англо-американской школе. При этом, например,
выявляются с помощью этих методов наиболее часто встречаю-
щиеся видовые сочетания, заменяя этим глазомерное выделение
ассоциаций, которое, по мнению этих исследователей, всегда гре-
шит субъективизмом. С другой стороны, размещение особей того
или иного вида в сообществе анализируется в свете опреде-
ленных математических закономерностей, как например бино-
миальные. Методика такого рода исследований обстоятельно из-
ложена в книге английского автора П. Грейг-Смита «Количест-
184
венная экология растений», вышедшей в 1967 г. в русском пе-
реводе.
Из советских исследователей статистико-математический
подход в геоботанике последовательно отражается в работах
А. А. Уранова, В. И. Василевича, Т. Э. Фрея, Б. М. Миркина,
Ю. А. Злобина и некоторых других.
На состоявшейся в 1968 г. в г. Владимире широкой научной
конференции по проблеме динамики растительного покрова 10
докладов в той или иной степени отражали применение стати-
стико-математических методов при изучении динамики фитоцсно-
зов и в том числе при прогнозировании с помощью этих методов
частных смен (сукцессий). Для ознакомления с этими доклада-
ми мы отсылаем к сборнику «Материалы по динамике расти-
тельного покрова» (Изд. Владимирского пед. института, 1968).
В решении конференции было отмечено, что в настоящее время
ведется активный поиск новых методов, в особенности статисти-
ческих, которые позволяют давать объективную количественную
характеристику сукцессионных серий и разногодичных флюкту-
аций.
ГЕОБОТАНИЧЕСКОЕ КАРТИРОВАНИЕ
И РАЙОНИРОВАНИЕ
Важным звеном геоботанических исследований является
составление карты распределения растительных сообществ изуча-
емой местности, или геоботанической карты. Основным материа-
лом для таких карт служат дневники, которые геоботаник обыч-
но ведет одновременно с описанием пробных площадей, профи-
лей и т. д. В этих дневниках исследователь должен отмечать
все изменения растительности, какие он встречает на пути
следования. Кроме того, материалом для геоботанических карт
могут служить геоботанические профили, а также материалы
различного рода землеустроительных и лесоустроительных
съемок.
При крупномасштабном картировании растительного покро-
ва необходимо вычерчивать карту не только на основании опи-
саний профилей, а производить сплошную контурную съемку
прямо на местности, обходя каждый выделяемый на карте кон-
тур. При этом, если в распоряжении геоботаников нет соответ-
ствующей по масштабу топографической карты района, необхо-
димо организовать совместную работу с землемером-топогра-
фом.
Контурную съемку полезно сочетать со съемкой по попереч-
никам (визирам), намечаемым с помощью компаса и вешек.
Поперечники проводят в направлении меньшей ширины карти-
руемого массива; расстояние между каждыми двумя соседними
поперечниками при значительной комплексности растительного
13 Б. Д. Ярошенко
185
покрова устанавливается в 100—200 м (или шагов, с последую-
щим переводом их в метры), а для более однообразных покровов
может быть увеличено до 500 м и даже (в сравнительно немно-
гих случаях) до 1 км. Поперечники проводят строго парал-
лельно.
Проходя по поперечникам, исследователь отсчитывает рас-
стояние шагами (с последующим переводом их в метры) или же
мерной лентой и одновременно следит за изменениями раститель-
ности по обе стороны от линии поперечника. Как только одна
ассоциация сменяется другой, исследователь тотчас же делает
отметку расстояния, а затем записывает краткую характери-
стику пройденного участка ассоциации или же просто отмечает
название ассоциации. Перенося затем все эти поперечники на
миллиметровую бумагу, он производит интерполяцию наноси-
мых на карту единиц растительного покрова на пространствах
между каждыми двумя соседними поперечниками. Иногда бы-
вает и так, что, проходя по поперечнику, съемщик видит справа
или слева от него пятно какой-либо ассоциации, не заходящее
на линию ни данного, ни соседнего поперечника. В таких случа-
ях он должен отметить, в какой стороне и на каком расстоянии
от линии поперечника находится данное пятно, а также его
приблизительные размеры, с тем 'чтобы потом нанести его
на карту.
В зависимости от масштаба геоботанической карты на нее
наносят более или менее крупные единицы растительного покро-
ва. При этом имеет значение и рельеф местности. В горных стра-
нах с сильно развитым рельефом мелкие единицы — ассоциа-
ции — не удается обыкновенно наносить даже на карты доста-
точно крупного масштаба и приходится брать в качестве исход-
ных единиц группы ассоциаций или формации. Кроме того, в
горах комплексность растительности иногда бывает выражена
настолько сильно, что нередко за единицу приходится брать ком-
плекс в целом. В равнинных условиях съемка геоботанических
карт облегчается.
В некоторых случаях в дополнение к сравнительно мелко-
масштабной карте всей местности снимают в значительно более
крупном масштабе карту отдельных наиболее типичных участков.
Это так называемый метод ключей. При этом выбираемые в ка-
честве таких ключей участки должны по возможности отражать
все разнообразие картируемой местности в смысле ее геоморфо-
логии, почвенного и растительного покрова. Так, например, если
ключ выбирают в долине реки, то в него должны входить все
полосы поймы, а если ключ более крупный, то и все террасы, а
также часть водораздела. Если назначение ключа — дать пред-
ставление о размещении растительных группировок в их связи
с микрорельефом, то такой ключ может быть очень небольшого
размера. Своеобразными ключами могут быть и однометровые
186
площадки, если на них картируется размещение отдельных
растений или куртин, как это часто практикуется, в особенности
при изучении степной, полупустынной и пустынной раститель-
ности.
При картировании комплексов, характеризующихся значи-
тельной пестротой, применяют метод линейной таксации. Сущ-
ность этого метода состоит в пересечении комплекса нескольки-
ми линиями, желательно во взаимно перпендикулярных или во-
обще в различных, не параллельных между собой направлени-
ях. Частота этих линий устанавливается в зависимости от
масштаба карты и степени пестроты комплекса. На каждой
такой линии измеряют отрезки, приходящиеся на пятна различ-
ных ассоциаций, после чего вычисляют процентные соотношения
их общих площадей.
Картирование крупных комплексов производят также путем
аэросъемки, в частности аэрофотосъемки, нашедшей особо ши-
рокое применение при картировании лесных массивов и значи-
тельных пространств полупустынь и пустынь.
Применение аэросъемки для геоботанического картирова-
ния началось у нас еще в 20-х годах и сейчас широко распрост-
ранилось, в особенности в сочетании с обычной наземной
съемкой.
Следует различать карты современного растительного покро-
ва и карты восстановленного растительного покрова. На первые
наносят фактическое распределение растительности, какое имеет
место в настоящее время. Что касается карт восстановленного
растительного покрова, то на них наносят распределение расти-
тельности, имевшее место до видоизменения растительного по-
крова хозяйственной деятельностью. Основой для восстановления
былого распределения растительности служит карта современ-
ного растительного покрова. Диагрикультурную, девственную
растительность восстанавливают на основании следующих мате-
риалов: клочков девственной растительности, уцелевших среди
культурных и вообще резко измененных хозяйственной деятель-
ностью участков; сравнения почв целинных и культурных зе-
мель; характера рельефа; наименований отдельных местностей,
иногда указывающих на прошлое их растительного покрова; ар-
хивных и исторических материалов. Кроме того, нужно иметь
в виду, что такая «реставрированная карта» отличается от «кар-
ты современной» не только тем, что на ней не показан культур-
ный растительный покров, но часто и тем. что вместо некоторых
вторичных (антропогенных) типов растительного покрова напей
обозначены их предшественники (рис. 43).
Весьма наглядным примером параллельного составления гео-
ботанических карт обоих типов является работа Алехина «Гео-
ботанические карты Московской области» (рис. 44 — на вклей-
ке), доложенная им на Международном географическом кон-
13*
/87
Рис. 43. Карты современного
(схема 0 и восстановленного
(схема 2) растительного покрова
ущелий рек Тызыл и Гун де лен
КБАССР (по Ярошенко и Куш-
хову, 1966),
Условные знаки:
/— березняки; 2 — грабово-дубовые
леса; 3 — леса с преобладанием ди-
корастущих плодовых; 4 — буковые
леса; 5 — заросли и куртины аза-
лии; 6 — разнотравно-вейниковые
луга субальпийского пояса; 7 —зла-
ково-разнотравные луга лесного по-
яса; 8 — приречные ольшаники и
ивняки; 9 — пестроовсяницевые
степи; SO — пестроовсяницевые сте-
пи луговые (олуговевшие); 11— ком-
плексы субальпийских низкоосоко-
вых (80%) и луговых пестроовсяни-
цевых степей; 12 — комплексы суб-
альпийских низкоосоковых (50%),лу-
говых пестроовсяиицевых(30%) сте-
пей и разнотравно-вейниковых лу-
гов; 13—ковыльные стели с козылем
красивейшим; 14 — ковыльные сте-
пи с тырсой; /5 — ниакоосоковые
степи, сильно выбитые,с элемента-
ми фриганы; /6— низкоосоковые
степи, сильно выбитые, с обилием
молочая Сегвиера; 17 — субальпий-
ские луга, сильно засоренные пест-
рой овсяницей и манжеткой; 18 —
злаково-бобоно-разнотравные луга
субальпийского пояса; 19— луго-
вые степи с преобладанием костра
прямого и осоки низкой; 20 — тощие
луга лесного пояса с обилием мура-
вейников; 21— альпийские пустоши
с преобладанием кобрезни; 22 —
крутые скалы со скудной расти-
тельностью.
грессе в Варшаве и изданная в 1934 г. в серии «Докладов совет-
ской делегации» на этом конгрессе (Алехин, 1934). На карте со-
временной растительности Московской области значительное
место, например, занимают мелколиственные леса из осины и бе-
резы, тогда как на карте восстановленной растительности эти
леса не показаны вовсе, и на их местах мы видим главным обра-
зом дубовые и еловые леса.
Это объясняется тем, что леса из осины и березы являются
временными типами, возникающими па вырубках и пожарищах
дубовых и еловых лесов и обязанными своему широкому распро-
странению деятельности человека. Поемные луга на карте совре-
менной растительности также занимают видное место, но их
вовсе нет на карте восстановленного покрова, так как в Москов-
ской области они имеют целиком послелеснос (антропогенное)
происхождение.
Принятые для нанесения на карту единицы растительности
обозначаются различными красками, значками или штриховкой.
Общепринятых обозначений до сих пор не выработано.
Сектор географии и картографии растительности Отдела гео-
ботаники Ботанического института им. В. Л. Комарова (БИН)
Академии наук СССР закончил в 1954 г. и опубликовал в 1956 г.
«Геоботаническую карту СССР» в масштабе 1:4 000 000, со-
ставленную под руководством и редакцией Е. М. Лавренко
и В. Б. Сочавы. На этой карте, а также на изданной под той же
редакцией в 1948 г. «Карте растительности Европейской части
СССР» в масштабе 1:2 500 000 приняты следующие цвета для
основных крупных единиц растительного покрова:
Арктические пустыни и тундры — светло-серые, серые, слабо-
фиолетово-серые и серо-синие цвета (всего 8 красочных обозна-
чений).
Высокогорная растительность — синие (горные тундры), го-
лубовато-синие (субальпийские и альпийские луга), зеленовато-
синие (высокогорная подушковидная растительность), бледно-
голубые (разреженная растительность высокогорных осыпей и
скал), фиолетово-розовые (высокогорные полынно-тересксповые
пустыни) цвета.
Темнохвойные леса — серовато-фиолетовые и серовато-синие
цвета (всего 10 красочных обозначений).
Сосновые леса — желтовато-коричневые, коричневато-розо-
вые, желтовато-розово-коричневые цвета (всего 6 красочных
обозначений).
Л иственничные леса — оливковые, синевато-оливковые, зеле-
новато-оливковые, желтовато-оливковые цвета (всего 13 красоч-
ных обозначений).
Хвойно-широколиственные леса — серовато-синевато-зеленые
цвета с участием темнохвойных пород и желтовато-коричневые
с зеленым оттенком, если участвует сосна.
189
Широколиственные леса — зеленые цвета разных оттенков
и они же с желтоватым и синеватым оттенком (всего 10 красоч-
ных обозначений).
Горные плодовые леса и кустарники Средней Азии — бирю-
зовые цвета.
Луга и травяные болота — бледно-зеленые цвета (9 красоч-
ных обозначений).
Сфагновые болота — бледно-голубые цвета.
Ксерофитные редколесья, нагорные ксерофиты и субтропиче-
ские степи — розовые, красно-розовые, оранжево-розовые, сла-
бо-фиолетово-розовые и розовато-фиолетовые цвета.
Степи и сельскохозяйственные угодья на их месте — желто-
вато-светло-зеленые, медово-желтые, зеленовато-желтые, слабо-
коричневато-желтые цвета (10 красочных обозначений).
Полукустарничковые полынные и солянковые пустыни —
светло-коричневые, светло-оранжевые, голубовато-оранжевые
цвета (8 красочных обозначений).
Сочносолянковые пустынные группировки и однолетняя со-
лянковая растительность — светло-фиолетовые цвета.
Саксаульники и кустарниковые группировки пустыни— крас-
новато-коричневые, коричневато-красные, желтовато-оранжевые,
розово-оранжевые цвета.
Слоевищная растительность такыров — бледно-голубовато-
зеленоватый цвет.
Перечисленные обозначения были приведены В. Б. Сочавой
в его докладе о принципах и задачах геоботанической картогра-
фии на VIII Международном ботаническом конгрессе в Париже
в июле 1954 г. Этими обозначениями, как указал Сочава, охва-
чены растительные группировки умеренного и арктического поя-
сов и отчасти северной окраины субтропического пояса. Для
остальной, большей части субтропического пояса, а также для
тропиков Сочава рекомендовал использовать ряд других обоз-
начений, например: для субтропических и тропических лесов
темно-зеленые, темно-фиолетовые, лиловые, багровые, винно-
красные и другие цвета, для южных пустынь коричневые и т. д.
В своем докладе на VIII Международном ботаническом кон-
грессе В. Б. Сочава заострил также вопрос о желательности
составления геоботанической карты мира в масштабе
1 : 1 000 000, которая должна явиться коллективным трудом спе-
циалистов разных стран.
Применяют также комбинации цветных обозначений с теми
или иными значками или штриховкой.
Аэрофотосъемку желательно применять не только при мелко-
масштабном, но и при среднемасштабном геоботаническом карти-
ровании. Аэрофотосъемка дает точные границы лесов, залежей,
полей и других угодий. Вместе с тем нужно сочетать аэрофото-
съемку с применением там,.где это нужно, большей детализации.
190
При проведении среднемасштабного геоботанического кар-
тирования необходимо не ограничиваться одним лишь нанесени-
ем на карту контуров, но в отдельных ключевых участках давать
возможно более полную характеристику растительности, содер-
жащую разносторонний цифровой материал.
В практику геоботанического картирования все более и более
входит сочетание показа типов растительного покрова с различ-
ными другими данными. Весьма часто применяется сочетание
геоботанических единиц с показом их хозяйственной оценки и
возможностей использования, а также улучшения. Применяется
также составление крупномасштабных геоботанических карт
с нанесением на них и направлений сукцессий. Все это можно
наносить на одну и ту же карту, но тогда она становится слиш-
ком сложной и менее наглядной. Поэтому применяют и вычерчи-
вание для одной и той же территории нескольких карт, на-
при мер:
а) карта современного растительного покрова;
б) карта восстановленного растительного покрова;
в) карта основных направлений сукцессий;
г) карта хозяйственной оценки угодий и перспектив их ис-
пользования и улучшения.
Пример картосхем современного и восстановленного расти-
тельного покрова двух смежных горных ущелий Кабардино-
Балкарии (Сев. Кавказ) показан на рисунке 43 (Ярошенко и
Кушхов, 1966).
Картосхема этой же территории по хозяйственной оценке уго-
дий и перспективам их использования и улучшения показывает
территориальное распределение рекомендуемых мероприятий по
улучшению горных пастбищ и сенокосов.
Например, для луговых степей с преобладанием костра пря-
мого и осоки низкой рекомендуется пастбищеоборот с чередова-
нием пастбищного и сенокосного использования и полного от-
дыха. Для разнотравно-вейниковых лугов субальпийского пояса
рекомендуется подрезание чемерицы и аконита в конце весны,
а также пастбищеоборот; для луговых пестроовсяницевых сте-
пей •— кратковременный ранний выпас и последующее сено-
кошение, а также постановка опытов по обжиганию с целью
вытеснения пестрой овсяницы лучшими травами. На сильно выби-
тых низкоосоковых степях — запрещение пользования в тече-
ние 5—6 лет, подсев желтой люцерны и житняка, местами под-
резание молочая.
Среднемасштабные геоботанические карты приобретают
особую ценность для сельскохозяйственного районирования.
Еще лучше, если они будут при этом сочетаться с почвенными,
климатологическими картами. Кроме того, при сельскохозяйст-
венном районировании совершенно необходимо учитывать мест-
ный агропроизводственный опыт и экономику. Одной из предпо-
191
сылок к сельскохозяйственному районированию должно явиться
геоботаническое районирование, основной единицей которого бу-
дет служить геоботанический район.
Геоботанический район, как правило, совпадает в своих гра-
ницах с ландшафтным (физико-географическим) районом.
Отличие геоботанического районирования от районирования
ландшафтного состоит не в очертаниях районов, а в их харак-
теристиках. В первом случае районы характеризуются преиму-
щественно по растительности, хотя не могут не приниматься
во внимание и черты геоморфологии, климата и почв. Во втором
случае районы характеризуются в равной мере по всем этим
факторам.
Геоботанический район, а в горных странах микрорайон явля-
ются мелкими единицами геоботанического районирования. Са-
мой же крупной единицей считают геоботаническую область, со-
ответствующую физико-географической стране (см. стр. 134—135).
В каждой геоботанической области имеется свое специфическое
разделение на растительные зоны. Далее зоны подразделяются
в широтном (с севера на юг или наоборот) направлении на под-
зоны, а в долготном, или меридиональном (с востока на запад или
наоборот), направлении каждая подзона подразделяется на гео-
ботанические провинции, которые в свою очередь делятся на
геоботанические районы. Таким образом, в условиях равнины
принимают следующую иерархию (соподчинение) единиц геобо-
танического районирования:
Геоботаническая область
J
4
Зона растительности
I
I
Подзона растительности
4
Геоботаническая провинция
I
4
Геоботанический район
Например, в геоботанической области Восточно-Европейской
равнины, в ее степной зоне можно выделить по крайней мере две
подзоны: северную подзону ковыльно-разнотравных (луговых)
и ковыльных степей и южную подзону типчаково-ковыльных сте-
пей. В северной подзоне степной зоны выделяются геоботаниче-
ские провинции: Южноукраинская, Волжско-Донская, Заволж-
ская и Предкавказская. Далее провинции подразделяются на
геоботанические районы, каждый из которых характеризуется
своей спецификой растительности. Так, одним из геоботаниче-
ских районов Волжско-Донской степной провинции является Бо-
192
гучарский волнисто-балочный степной район (Ф. Н. Мильков,
1956). Таким образом, в условиях равнины геоботанический
район обязательно является частью зоны или (если таковая вы-
ражена) подзоны.
В горных условиях появляется новый фактор — вертикаль-
ная зональность, или поясность, заставляющая несколько по-
иному понимать геоботанический район. Здесь такой район не
является частью пояса, обыкновенно он включает несколько
поясов. Горная страна может располагаться целиком в одной
провинции или охватывать несколько провинций. Далее, в гор-
ных странах между геоботанической провинцией и геоботаниче-
ским районом различается еще промежуточное звено — геобо-
танический округ, далее следует геоботанический район и, на-
конец, геоботанический микрорайон, представляющий часть
лишь одного пояса. Так, в Кавказской геоботанической области
одной из геоботанических провинций является Центральная про-
винция Большого Кавказа. Одним из округов этой провин-
ции является Терский геоботанический округ, называемый
А. А. Гроссгеймом (1948) Терским флористическим округом.
Одним из районов этого округа является Приэльбрусский геобо-
танический район, в составе которого И. И. Тумаджанов (1963)
выделяет шесть, как он называет, «ландшафтных» (геоботани-
ческих) микрорайонов. Одним из этих микрорайонов является,,
например, «субальпийский лесо-луговой микрорайон с отдельны-
ми урочищами разреженных сосняков, березовым криволесьем
и рододендроново-можжевеловыми стланиками».
Темы курсовых работ
Сравнение двух методов определения встречае-
мости
План изложения:
Характеристика метода Раункиера.
Характеристика точечного метода.
Характеристика ассоциации, изученной обоими методами.
Количество колец и точек и расположение их на изучаемом,
участке. Хронометраж обоих методов.
Результаты. Сравнительная оценка обоих методов.
Литература: Браун Д. Методы исследования и учета
растительности. М., Изд-во иностр, лит., 1957.
Полевая геоботаника. Т. 3, М.—Л., Изд-во АН
СССР, 1964.
Ярошенко П. Д. Геоботаника. М.—Л.,
Изд-во АН СССР, 1961.
193
Определение жизненности некоторых видов в раз
ных микрогруппировках сообщества
План изложения:
Понятие о мозаичности и о причинах ее вызывающих.
Характеристика изученных ассоциаций и их мозаичности.
Выбор видов и признаков для изучения жизненности.
Результаты и выводы.
Литература: Д ы л и с Н. В. и др. О горизонтальной
структуре лесных биогеоценозов. Бюлл. М.ОИП,
1964, № 4.
У р а н о в А. А. Жизненное состояние вида в
растительном сообществе. Бюлл. МОИП, 1960,
т. 65, № 3.
Ярошенко П. Д. Геоботаника. М.—Л., Изд-во
АН СССР, 1961.
Фенологические спектры растительных сообществ
Полевые наблюдения проводятся ежедекадно от ранней вес-
ны до поздней осени.
Литература: Б е й д е м а н И. Н. Методика фенологических
наблюдений при геоботанических исследованиях.
М.— Л., Изд-во АН СССР, 1954.
Шенников А. П. Введение в геоботанику.
Изд-во ЛГУ, 1964.
Составление крупномасштабной геоботанической
карты участка с разнообразной растительностью
Литература: Викторов С. В., Востокова Е. А.,
В ы ш и в к и и Д. Д. Краткое руководство по
геоботаническим съемкам. М., Изд-во МГУ, 1959,
стр. 42—46.
Воронов А. Г. Геоботаника. М., «Высшая
школа», 1963, стр. 324—342.
Дополнительный список рекомендуемой
литературы1
Александрова В. Д. Изучение смен растительного покрова. В кн.: «По-
левая геоботаника», т. 3, под ред. Е. М. Лавренко и А, А, Корчагина,
М,—Л„ Изд-во АН СССР, 1964,
Алехин В. В. Геоботанические карты Московской области. М., Изд-во Науч-
но-исследовательского института Большого советского атласа мира, 1934.
Алехин В. В. Растительность СССР в основных зонах. М_, «Советская
наука», 1951.
Бей дем ан И. Н Методика фенологических наблюдений при геоботаниче-
ских исследованиях. М.— Л., Изд-во АН СССР, 1954,
Б е р г Л. С. Климат и жизнь. М., Географгиз, 1947.
Вальтер Г., Алехин В. Основы ботанической географии. М,, Учпедгиз,
1936.
Викторов С. В. Использование геоботанического метода при геологиче-
ских и гидрогеологических исследованиях. М., Изд-во АН СССР, 1955.
Вилья м с В. Р. Почвоведение. Земледелие с основами почвоведения. М„
Сельхозгиз, 1939.
Виноградов Б. В. Растительные индикаторы и их использование при
изучении природных ресурсов. М„ «Наука», 1964.
Воронов А. Г. Геоботаника. М., «Высшая школа», 1963.
Геоботаническая карта СССР, под ред. Е. М. Лавренко и В. Б. Сочавы.
At—Л., Изд-во АН СССР, 1956.
Грейг-Смит П. Количественная экология растений. Пер. с англ. М.,
«Мир», 1967.
Докучаев В. В. Избранные труды. М., Изд-во АН СССР, 1949.
Дохман Г. И. Растительность Мугоджар. М., Географгиз, 1954.
Зонн С. В. Лесные почвы северо-западного Кавказа. М.— Л., Изд-во АН
СССР, 1950.
Иванов Л. А., С и л н н а А. А., Цель ни кер Ю. Л. О транспирации поле-
защитных пород в условиях Деркульской степи. «Ботанический журнал»,
т. 37, 1952, № 2.
Кароль Б. П. Микроклимат и методы его изучения при геоботанических
исследованиях В кн.: «Полевая геоботаника», под ред. Е. М. Лавренко и
А. А. Корчагина, т. 1. М.— Л., Изд-во АН СССР, 1959.
Карта растительности Европейской части СССР, под ред. Е. М. Лавренко
и В. Б. Сочавы. при участии Б. Н. Г о р о д к о в а. М.— Л., Изд-во АН
СССР, 1948.
Комаров Н. Ф. Рельеф и топографическое распределение растений. «Тр.
Бот. ин-та им. В. Л. Комарова АН СССР», сер. 3 (Геоботаника), вып. 4.
М,—Л., Изд-во АН СССР, 1956.
1 Названия некоторых работ менее общего значения приведены в конце
каждой главы при тематике рефератов и курсовых работ.
/95
Корчагин А. А. К вопросу о характере взаимоотношений растений в сооб-
ществе. В сб.: «Академику В. Н. Сукачеву к 75-летию со дня рождения».
М.— Л, Изд-во АН СССР, 1956.
Куражсковский Ю. Н. Соотношение особенностей действия основных
групп факторов среды в экологии. «Тр. Воронежск. гос. заповедника»,
вып. 7. Воронеж, 1957.
Л а в р е н к о Е. М. Основные закономерности растительных сообществ и пути
их изучения. В ки.: «Полевая геоботаника», i. I. М.— Л., Изд-во АН СССР,
1959.
Лисенков А. Ф. К вопросу о срастании корней дуба в густых культурах.
«Ботанический журнал», 1957, т. 42, № 3.
Лысенко С. В. О методах изучения внутренней структуры лесных сооб-
ществ на примере лесов Владимирской области. «Лесоведение», 1968, № 1.
Люндегорд Г. Влияние климата и почвы на жизнь растений, пер. с нем.
М„ Учпедгиз, 1937.
Марков К. К., Г р и ч у к В. П., Чеботарева П. С. и др. Взаимоотноше-
ния леса и степи в историческом освещении. Вопр. географии, сб. 23. М.,
Географгиз, 1950.
М а з и и г В. В. О научной документации в геоботанике. Сб. «Проблемы совре-
менной ботаники». Т. I. М.— Л., «Наука», 1965.
«Методика полевых геоботанических исследований». М.— Л., Изд-во АН СССР,
1938.
Мильков Ф. Н. Лесостепь Русской равнины. Опыт ландшафтной характе-
ристики. М., Географгиз, 1950.
Мильков Ф. Н. Физико-географический район и его содержание. М., Гсо-
графгиз, 1956.
Москаленко Н. Г. О методике построения фенологических спектров.
«Научные доклады высшей школы», биол. науки, № 2. М„ 1964.
Нейштадт М. И. Спорово-пыльцевой метод в СССР. М.— Л., Изд-во АН
СССР, 1952.
Несветайлова Н. Г. Ботанические показатели полезных ископаемых
(обзор методов и анализ индикационных показателей). М„ «Наука». 1965.
«Основы лесной биогеоценологии». Сб., под ред. В. Н. Сукачева и Н. В. Дыли-
са. М., «Наука», 1964.
Погребняк П. С. Основы лесной типологии. Киев, Изд-во АН УССР, 1955.
«Полевая геоботаника», т. I, II и III. М.— Л., Изд-во АН СССР, 1959, 1960,
1964.
П о п о в А. А. О микрогруппировках в бруснично-долгомошном лиственничнике
средне-амурской низменности. «Ботанический журнал», т. 48, № 9.
Работнов Т. А. Биологические наблюдения на субальпийских лугах Север-
ного Кавказа. «Ботанический журнал», 1945, т. 30, № 4.
Работнов Т. А. Опыт определения возраста у травянистых растений. «Бо-
танический журнал», 1946, т. 31, № 5.
Раменский Л. Г. Введение в комплексное почвенно-геоботаническое иссле-
дование земель. М., Сельхозгиз, 1938.
«Растительный покров СССР». Пояснительный текст к геоботанической карте
СССР, т. 1 и 2. М —Л., Изд-во АН СССР, 1956.
Ревердатто В. В. Введение в фитоценологию, ч. I. Томск, изд-во Том-
ского универе., 1935.
«Растительные индикаторы почв, горных пород и подземных вод». Сб. статей,
под ред. А. Г. Чикишева. (Труды МОИП, т. 8). М., Изд-во МГУ, 1964.
Рокицкий П. Ф. Биологическая статистика. Минск, «Высшая школа», 1967.
Серебряков И. Г. Экологическая морфология растений. Жизненные формы
покрытосеменных и хвойных. М., «Советская наука», 1962.
Серебряков И. Г. Жизненные формы высших растений и их изучение.
В кн.: «Полевая геоботаника.», т, 3, М.— Л., «Наука», 1964.
Соколов С. И. Краткий курс почвоведения. М., Сельхозгиз, 1940.
196
Сочава В. Б. Опыт филоценогенетической систематики растительных ассо-
циаций. «Советская ботаника», 1944, № 1.
Сочава В. Б. Основные положения геоботанического районирования. «Бо-
танический журнал», 1952, т. 37, № 3.
Сочава В. Б. Перспективы геоботанического картографирования. В со.:
«Геоботаническое картографирование». М.-~ Л., Изд-во АН СССР, 1963.
Сукачев В. Н. Растительные сообщества (введение в фитосоциологию), изд.
4-е. М., «Книга», 1928.
Сукачев В. Н. Главнейшие понятия из учения о растительном покрове.
«Растительность СССР», т. 1. М.— Л., Изд-во АН СССР, 1938.
Сукачев В. Н. О некоторых основных вопросах фитоценологии. В сб.:
«Проблемы ботаники», т. I. М.— Л., Изд-во АН СССР, 1950.
Сукачев В. Н. К вопросам теории степного лесоразведения. Лес и степь,
№ 8, 1952.
Сукачев В. Н. Некоторые общие теоретические вопросы фитоценологии.
В кн.: «Вопросы ботаники», т. 1. М.—Л., Изд.-во АН СССР, 1954.
Сукачев В. И. Общие принципы и программа изучения типов леса. (В. Н. Су-
качев, С. В. Зонн, Г. П. Мотовилов. Методические указания к изуче-
нию типов леса). М., Изд-во АН СССР, 1957, изд. 2, 1962.
Сукачев В. Н. Новые данные по экспериментальному изучению взаимоотно-
шений растений. Бюлл. МОИП, нов. сер., 1959, т. 34, № 4.
Таифильев Г. И. Пределы лесов на юге России. (Географические рабо-
ты). М., Географгиз, 1953,
Тумаджанов И. И. Опыт дробного геоботанического районирования се-
верного склона Большого Кавказа. Тбилиси, Изд-во АН ГССР, 1963.
Тхай Ван Тру и г. Экология и классификация лесной растительности Вьет-
на.ма. Автореферат докт. дисс. Л., Изд-во Бот, ин-та АН СССР, 1962.
У р а и о в А. А. Количественное выражение межвидовых отношений в расти-
тельном сообществе. Бюлл. МОИП, отд. биал. 1955, т. 60, вып. 3.
У р а н о в А. А. Жизненное состояние вида в растительном сообществе. Бюлл.
МОИП, от д. биол., 1960, т. 65, вып. 3.
У р а н о в А. А. Наблюдения на летней практике по ботанике. Изд. 2-е. М„
Учпедгиз. 1964.
Цыганов М. С, К вопросу об усыхании степей. «Почвоведение», 1952, № 4.
Шал ыт М. С. О фитофенологических спектрах. «Советская ботаника», 1946,
№ 4,
Шен ни ков А. П, Фенологические спектры растительных сообществ. «Тр.
Вологодской обл. с-х. опытн. станции», т. 2. 1927.
Шенников А. П, Луговая растительность СССР. В сб. «Растительность
СССР», т. I. М.—Л., Изд-во АН СССР, 1938.
Шенников А. П. Экология растений. М., «Советская наука», 1950.
Шенников А. П. Заметки о методе классификации растительности по
Браун-Бланке. В сб.: «Академику В Н. Сукачеву к 75-летию со дня рож-
дения». М.— Л., Изд-во АН СССР, 1956.
Шенников А. П. Введение в геоботанику. Изд-во ЛГУ, 1964.
Юновидов А. П. К познанию внутривидовых взаимоотношений в лесу.
Лесное хозяйство, 1952, № 8.
Ярошенко П. Д, О сменах растительного покрова. «Ботанический жур-
нал», 1946, т. 31, № 5.
Ярошенко П. Д. Геоботаника. Основные понятия, направления и методы.
М.— Л., Изд-во АН СССР, 1961,
Ярошенко П, Д., К у ш х о в А. X, Пастбища и сенокосы ущелий рек
Тызыл и Гунделен в КБАССР (очерк растительного покрова). Нальчик,
Каб.-Балк. кн. изд-во, 1966.
Aichinger Е. Statische und dynamische Betrachtungen in der pflanzensoziolo-
giscben Forschung. Aktuelle Probleme der PSlanzensoziologie. VeroH. des
geobot. Inst. Riibel in Zurich, 29, Bern, 1954,
197
Aichinger E. Uberlegungen zur Entwicklung der botanischen und pflanzen-
sozioiogischen Forschung. Angew. Pflanzensoz. Nr. 18—19, Wien, 1966.
Bharucha F. R„ Satyanarayan Y. The problem of calcicolus plants.
Festschrift fur E. Aichinger zum. 60. Geburtstag, 1. Wien, 1954,
Braun E. L. The undifferentiated deciduous forest climax and the association
segregate. «Ecology», vol., 16, No. 3, 1935.
Braun-Bia nquetJ. Pflanzensoziologie. 3. Aufl. Wien — New-York (1. Aufl.
1928, 2. Aufl. 1951) 1964.
Clements F. E., Shelford V. E. Bio-Ecology. Second print. (1st — 1939).
New York, 1946.
Cu rtis J. T. A prairie continuum in Wisconsin. «Ecology», vol. 36, No. 4, 1955.
Dansereau P. Biogeography on ecological perspective. New York, 1957.
Du-Rietz G. E. Vegetationsforschung auf soziationsanalytischer Grundlage.
Handb. dec biolog. Arbeltsmeth. B. 11, Nr. 5, 1930.
Gleason H. A. The individualistic concept of the plant association. «American
midland naturalist», vol. 21, No. 1, 1939.
Hard av Segerstad F. Siidsvenska florans vaxtgeografiska huvudgrupper.
Malmo. 1924.
К er sc ha w K. A, Quantitative and dynamic ecology. London, 1964.
Lippmaa T. The unistratal concept of plant communities (the unions)'.
«American midland naturalist», vol. 21, No. 1, 1939.
M e u s e 1 H. Die Grapsheiden Mitteleuropas. Bot. Arch., 41, 1940.
Meusel H. Uber die umfassende Aufgabe der Pflanzengeographie. Veroff, des
geobot. Inst. Rubel in Zurich, 29, Bern, 1954.
Paczoski J, Dziela Wybrane. Warszawa, 195L
Raunkiaer C. Life forms of plants and statistical plant geography. New
York, 1934.
Richards P. W. The floristic composition of primary tropical rain forest.
«Biological review», vol. 20. No. I, 1945.
Rubel E. Geobotanische Untersuchungsmethoden. Berlin, 1922.
R lib el E. The replaceability of ecological factors and the law of the minimum.
«Biological reviews», vol. 20, No. 1, 1945.
T й x e n R. Die Pflanzengesellschaften Nordwestdeutschland, Mitt, for soc.
Arbeitsgem. in Niedersachsen. Nr. 3, Hannover.
W e a v e r J. E. North American prairie. London, 1954.
WhittakerR. H. A consideration of the climax theory; the climax as a popu-
lations and pattern. «Ecological monographs», vol. 23, No. 1, 1953.
Whittaker R. H. Gradient analysis of vegetation. «Biological reviews»,
vol. 42, № 2, 1967.
Оглавление
Предисловие ......................................................... 3
1. Задачи геоботаники и краткий обзор ее истории .................... 4
Темы рефератов на семинаре или в кружке............................21
Темы курсовых работ.................................................—
2. Основные черты взаимодействий между растениями в сообществах . 22
Темы рефератов на семинаре или в кружке............................34
Темы курсовых работ................................................35
3. Отличительные признаки фитоценоза.................................38
Видовой состав .................................................... —
Количественные и качественные отношения между видами .... 39
Ярусность..................................................... 42
Мозаичность и ее отношение к комплексности................ . ч4
Физиономичность, жизненные формы и периодичность...................50
Синузиальность.....................................................53
Характер местообитания ........................................... 55
Тема реферата на семинаре или в кружке.............................56
Темы курсовых работ.................................................—
4. Экология растительных сообществ..................................59
Экологические факторы я их влияние на растительные сообщества.
Взаимодействие, взаимная замещаемость и пределы замещаемости
экологических факторов ............................................ —
Главные и второстепенные экологические факторы.....................63
Физическая и физиологическая сущность экологических факторов . 66
Учение об индикаторах в связи с замещаемостью экологических
факторов........................................................ 67
Влияние растительных сообществ на почву и климат...................71
Темы рефератов на семинаре или в кружке............................86
Темы курсовых раббт................................................87
5. Ассоциация и другие единицы растительного покрова.................88
Ассоциация в трактовке различных геоботанических школ ... •—
Наименование ассоциаций .......................................... 99
Высшие единицы систематики растительных сообществ . 100
Тема реферата на семинаре или в кружке............................101
Темы курсовых работ.................................................—
6. Особенности классификации фитоценозов в различных типах расти-
тельного покрова....................................................102
Эколого-фитоценотические ряды В, Н, Сукачева ...... —
Эдафическая сетка П. С. Погребняка................................106
Классификации луговой растительности.......................... . 108
Классификация и ординация.........................................114
Темы рефератов на семинаре или в кружке...........................115
Тема курсовой работы................................................—
7. Динамика растительных сообществ...................................116
Частные смены, или сукцессии.......................................117
Общие смены и история растительного покрова страны . . . . 120
Эволюция растительных сообществ....................................124
Темы рефератов на семинаре или в кружке ....... 129
Темы курсовых работ................................................130
8. Закономерности территориального распределения растительного по-
крова ...............................................................131
Зоны и пояса растительности.........................................—
Правило предварения ............................................. 140
Влияние горного рельефа на распределение растительных сообществ
Комплексы.........................................................Т4о
Темы рефератов на семинаре или в кружке........................—
Темы курсовых работ.......................................... 144
9. Главнейшие методы геоботанических исследований...................145
Закладка и описание пробных площадей и учетных площадок . . —
Описание профиля..................................................173
Химические и физиологические методы в геоботанике..................175
Обработка материалов полевых геоботанических исследований . . —
Выявление частных смен растительных сообществ......................181
Статистико-математические методы в геоботанике....................184
Геоботаническое картирование и районирование ...................... 185
Темы курсовых работ................................................193
Дополнительный список рекомендуемой литературы........................195
Ярошенко Павел Дион исьевич
ГЕОБОТАНИКА
Редактор Н. В. Королева. Редактор карт С. Г. Тютюнник. Художественный редактор
И. А. Володина. Технический редактор Е. В. Богданова. Корректор К- И. Иванова.
Сдано в набор 30/ХП 1968 г. Подписано к печати 30/IV 1969 г. Формат 60 X SO’/ie- Типо-
графская № 1 Леч. л. 12,5+ вкл. 0,25. Уч.-изд. л. 12,55 4- вкл. 0,13. Тираж 32 тыс. экз.
(Тем. пл .1969 г. № 23). Л 04088. Цена без переплета 37 коп., переплет 10 коп.
Издательство «Просвещение» Комитета по печати при Совете Министров РСФСР.
Москва, 3-й проезд Марьиной рощи, 41,
Типография издательства ЦК КП Белоруссии. Минск. Заказ № 8.