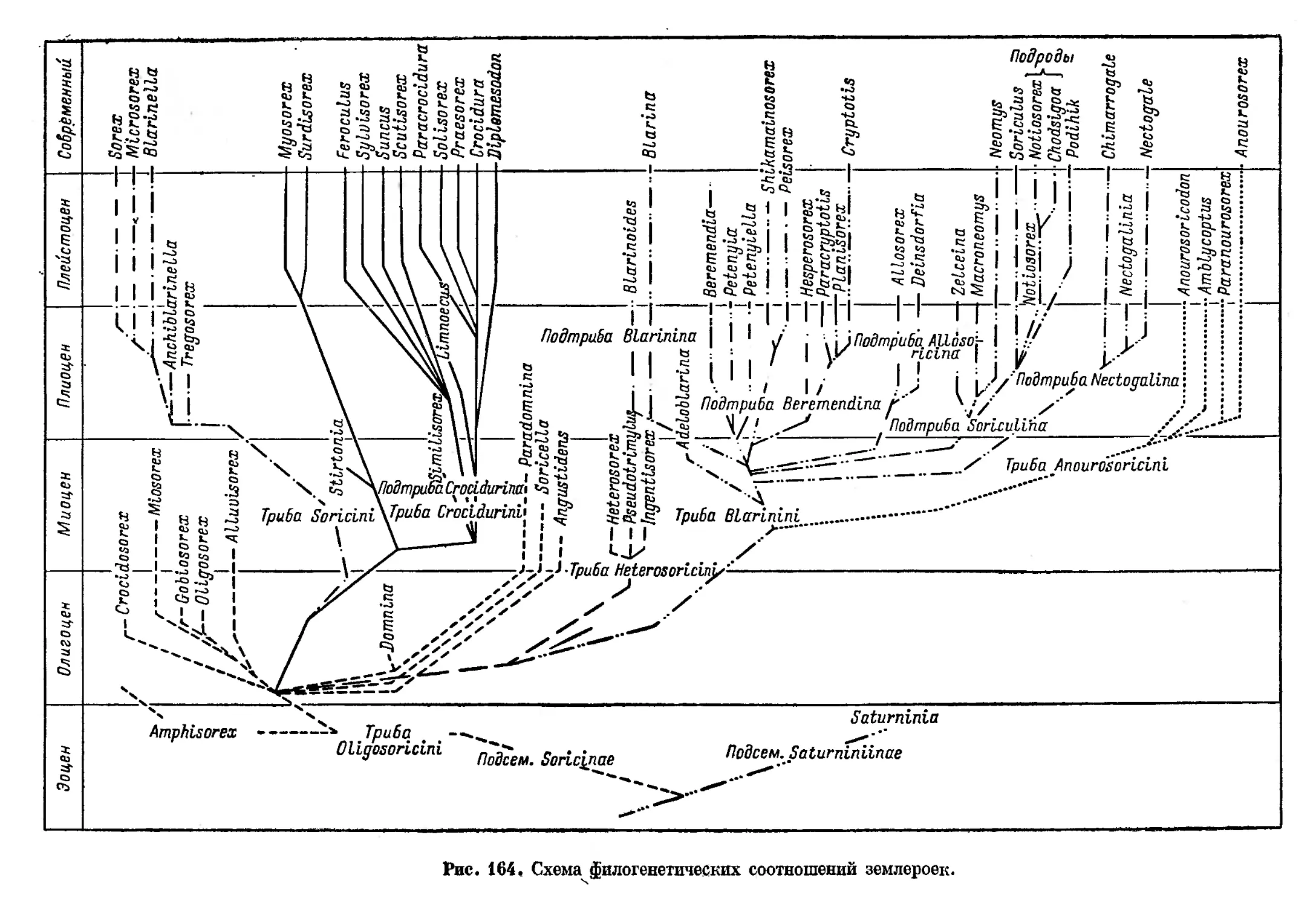
/








































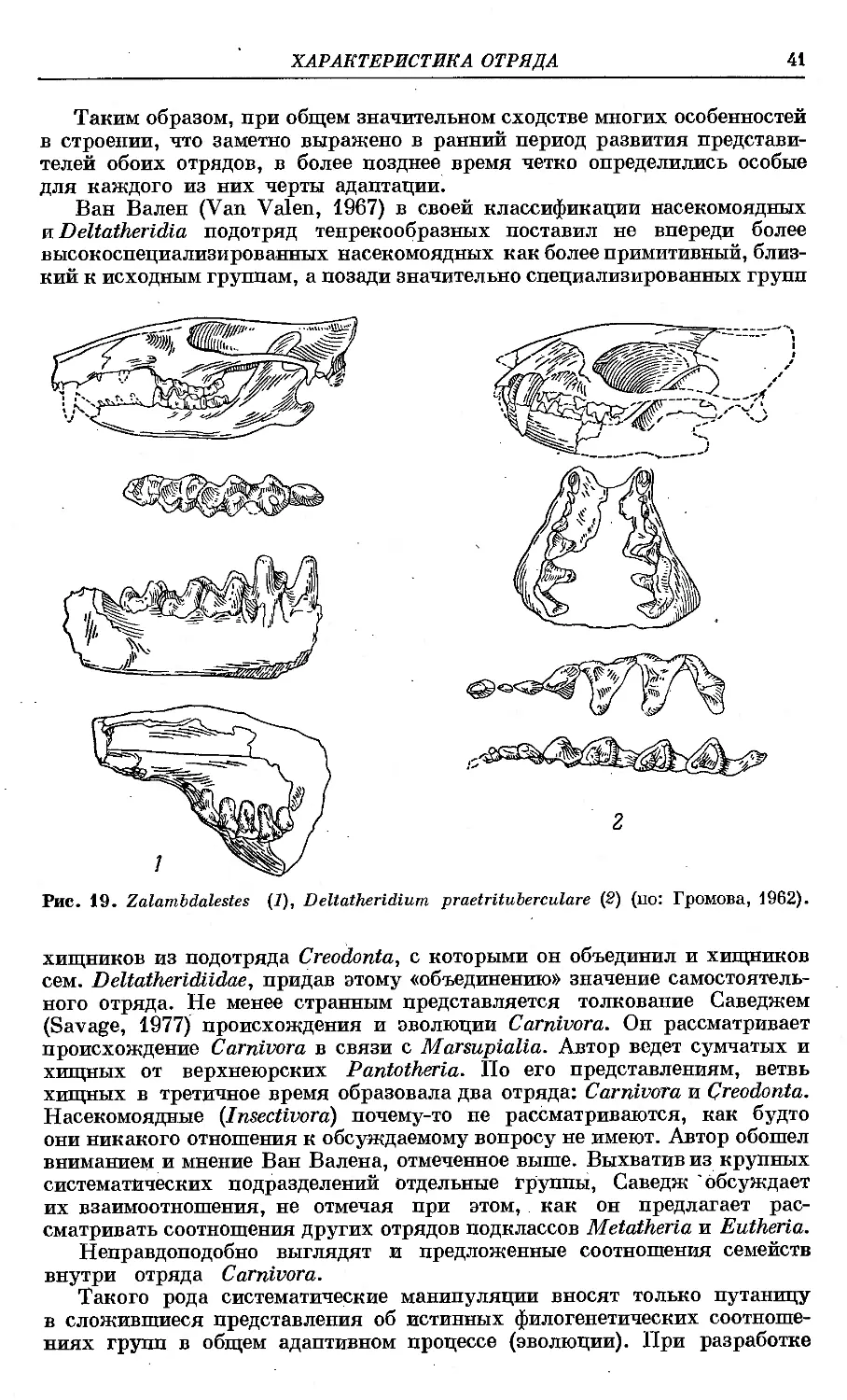




























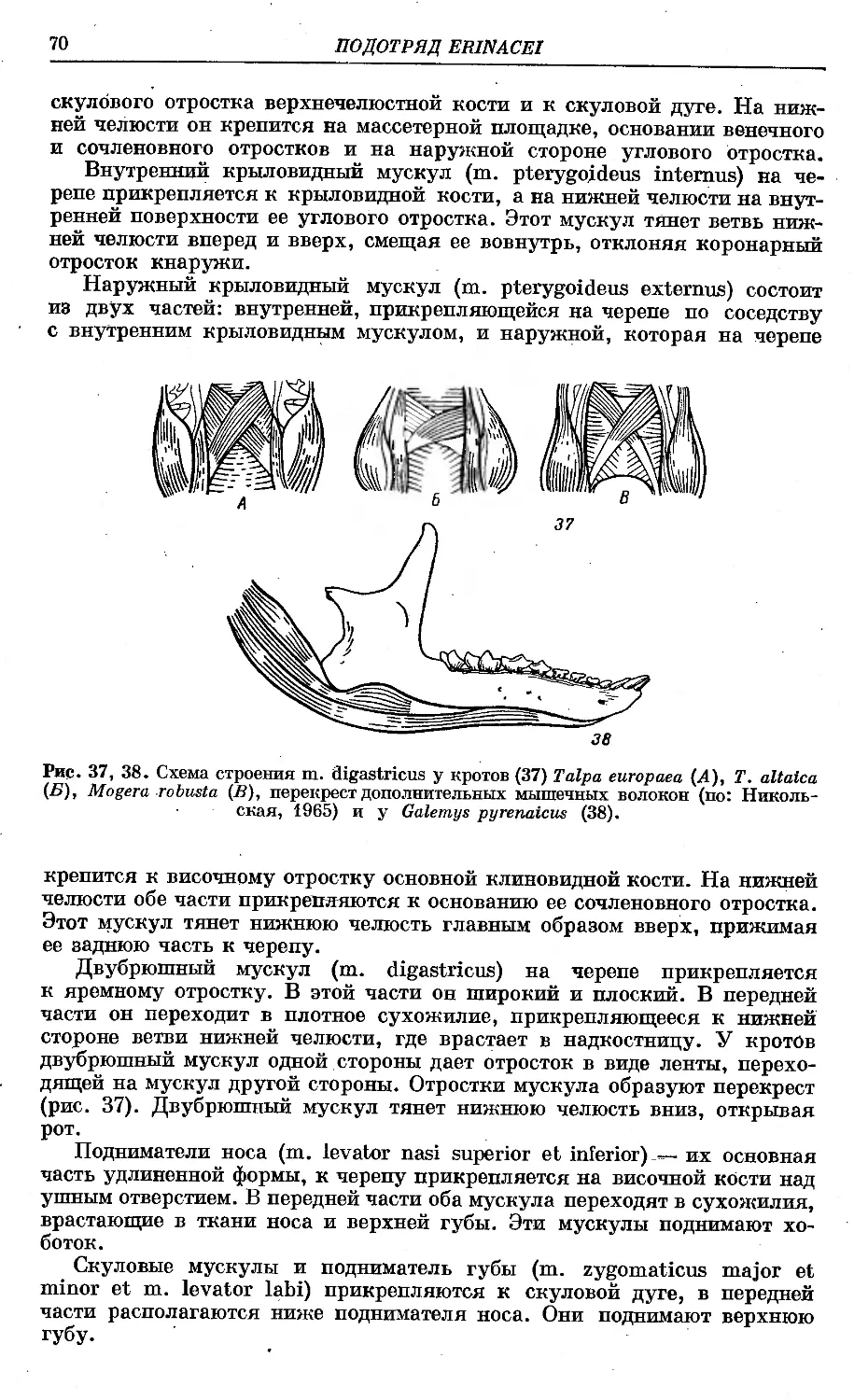













































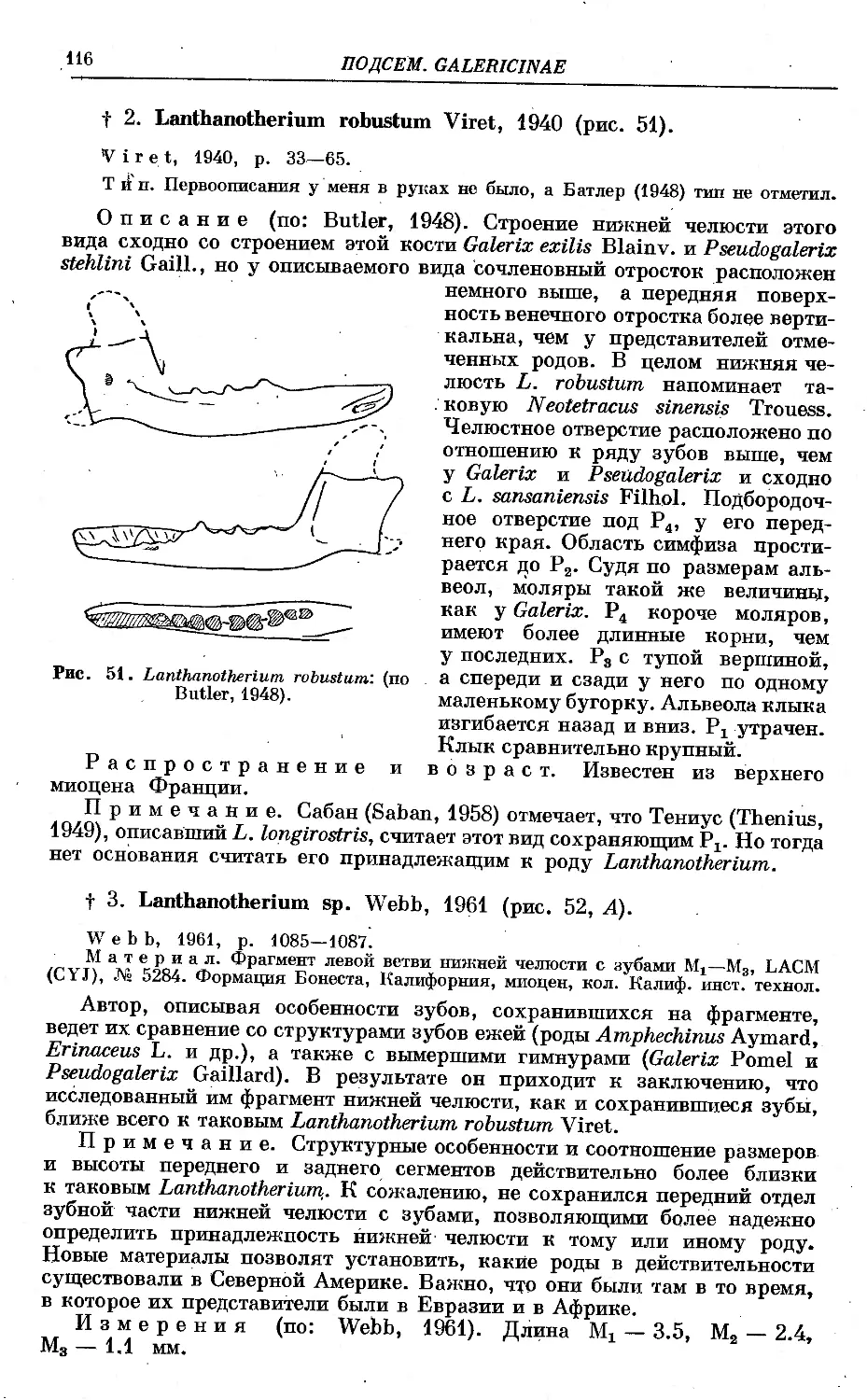






















































































































































































































































































































































































Text
ФАуНА
СССР
МЛЕКОПИТАЮЩИЕ
IV
выпуск
2
АКАДЕМИ Я НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ, № 120
ФАУНА СССР
МЛЕКОПИТАЮЩИЕ
Том IV, вып. 2
А. А. ГУРЕЕВ
НАСЕКОМОЯДНЫЕ (MAMMALIA, INSECTIVORAJ
ЕЖИ, КРОТЫ И ЗЕМЛЕРОЙКИ
(ERINACEIDAE, TALPIDAE, SORICIDAE)
ЛЕНИНГРАД
«НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1979
УДК 599.36
Насекомоядные. Ежи, кроты и землеройки (Erinaceidae, Talpidae,
Soricidae). Гуреев А. А. В серии: Фауна СССР. Млекопитающие.
Т. IV, вып. 2. Л., «Наука», 1979. 503 с.
Монография посвящена насекомоядным — полезным животным, пред-
ставляющим и большой научный интерес как группа древних млеко-
питающих. Во вводной части приведены: описание строения, адаптивные
особенности, распространение и система крупных таксонов отряда. В си-
стематической части даны характеристики семейств, родов и видов со-
временных и вымерших ежовых, кротовых и землеройковых, история
их развития и филогенетические связи, на которых основана их система-
тика^ Лит. — 281 назв., ил. — 242.
Главный редактор
директор Зоологического института О. А. С к а р л а т о
Редакционная к'оллегйя
А. А. Стрелков (ответственный редактор серии), И. М. Лихарев (зам. отв. редактора),
И. М. Громов, В. Ф. Зайцев, О. Л. Крыжановский, Л. А. Кутикова,
Г, С. Медведев, М. Е. Тер-Минасян
Редакторы тома
А. А. Стрелкову, И. М. Громов
21008-625
Ф——-—™ 656-79 2005000000
055(02)-79
© Издательство «Наука», 1979 г.
ПРЕДИСЛОВИЕ
Основная задача работы — представить сводку о насекомоядных
фауны СССР с разработанной филогенией и систематикой групп. Такая
сводка может служить основой для региональных фаунистических и част-
ных систематических исследований, зоогеографических построений
и учебных целей.
До этого было Подробно изучено большое по объему составляющих
видов сем. Soricidae. Итоги исследования в виде отдельной монографии
опубликованы в 1971 г. Целесообразность объединения в одном месте
материалов по насекомоядным фауны СССР послужила основанием для
включения в настоящую работу и материалов из монографии о земле-
ройках.
Текст книги там, где я счел нужным, переделан, дополнен новыми
данными и новоописаниями родов и видов. До упомянутой моногра-
фии, мною был написан раздел «Насекомоядные» для определителя
«Млекопитающие фауны СССР», опубликованного в 1963 г. Таким образом,
настоящая монография — третья в серии моих работ, посвященных насеко-
моядным отечественной фауны.
Она представляет результат многолетнего изучения мною насекомо-
ядных, а в последнее время и хищных, т. е. группы млекопитающих,
имеющих с насекомоядными общие древние корни в происхождении
(с. 37).
Монография выполнена в плане, предусмотренном для изданий серии
«Фауна СССР», применительно к ее современному статусу. В соответствие
с этим она представляет сводку в объеме фауны мира, в которой семейства
Erinaceidae, Talpidae и Soricidae описаны до вида (современные и вымер-
шие). В связи с тем, что в работе приводятся три наибольших по числу
видов семейства, естественно было представить читателю их положение
в системе насекомоядных. Это обстоятельство обусловило необходимость
дать характеристику отряда и подотрядов, а также включаемых ими сов-
ременных семейств и привести список их родов, без чего система носила бы
формальный характер. Эти данные позволят составить представление
о всех крупных группах современных насекомоядных животных фауны
мира.
Отряд насекомоядных объединяет наиболее примитивных плацентар-
ных млекопитающих. Тем не менее многие их группы достигли значи-
тельной специализации к передвижению по земле (Macroscelididae), рытью
(Chrysochloridae, Talpidae), плаванью \Potamogalidae, Desmaninae (Tal-
pidae)] и лазанью (Tupaiidae). Представляются своеобразными физиоло-
гические особенности отдельных семейств отряда, представители которых
сохраняют круглогодичную активность в резко различных климатических
зонах.
Среди этих млекопитающих самые мелкие виды класса и с самым
высоким темпом обмена веществ. Древность насекомоядных, связи с ними
1*
4
ПРЕДИСЛОВИЕ
в происхождении многих отрядов, в том числе обезьян и человека, осо-
бенности физиологии, отражающие высокую специализацию при общей
примитивности их организации, обусловливают повышенный интерес
зоологов многих стран к этой группе животных, особенно в последние
годы. Многие виды служат прокормителями ряда паразитов — перенос-
чиков возбудителей опасных инфекций и сами бывают спонтанными но-
сителями инфекций и с этой стороны представляют интерес для паразито-
логов и медицинской службы.
Насекомоядные как потребители огромного количества беспозвоноч-
ных животных, особенно насекомых, играют важную роль в биогеоцено-
зах. Поедание ими многих вредных видов имеет существенное практи-
ческое значение в экономике человека. Кроты служат предметом пушного
промысла. Их шкурки заготавливаются ежегодно в значительном коли-
честве. Польза, приносимая насекомоядными, с лихвой покрывает неко-
торый их вред. Они заслуживают охраны, тем более что отдельные виды
находятся под угрозой вымирания, например русская выхухоль, зане-
сенная в Красную книгу СССР.
Использованный материал позволил, как мне кажется, разобраться
в систематике насекомоядных, описанных в настоящей работе. Виды
подсем. Erinaceinae в коллекциях Зоологического института (ЗИН) АН СССР
представлены сериями почти со всей территории европейской части
Союза, Казахстана, Западной Сибири, Средней Азии, Монголии, Китая,
Дальнего Востока СССР. Меньше материала из Ирана, северной части
Аравии и Северной Африки, а также с территории Западной Европы,
немного пополненный коллекцией, полученной нами для обработки
из ГДР.
Подсем. Galericinae в нашей коллекции представлено монотипичными
родами Hylomys и Echinosorex\ материал по ним получен и из ГДР. Другие
монотипичные роды этого подсемейства (Podogymnura, Neotetracus и Neo-
hylomys) в коллекции отсутствуют, но их систематический статус ни у кого
не вызывает сомнений. Из семейства кротовых (Talpidae) в ЗИН имеется
материал по всем родам и видам из Северной Америки и Европы. Из Азии
в коллекции не представлены кроты родов Rhynchonax, Nasillus и Scapto-
пух. Они морфологически хорошо очерчены, и их систематическое поло-
жение достаточно определено. Кроты названных родов очень редки и от-
сутствуют во многих музеях мира. Из ГДР получен материал по р'Сдам
Scaptochirus и Urotrichus.
Материал по землеройкам с территории СССР в коллекции ЗИН пред-
ставлен достаточно полно. В дополнение к нему мною был исследован
материал, присланный ныне покойному профессору Б. С. Виноградову
из Института зоологии АН УССР, институтов зоологии АН АрмССР,
АН АзербССР, АН ТаджССР, Ташкентского университета. Просмотрены
коллекции МГУ.
Палеонтологический материал по отдельным группам насекомоядных
был у меня в руках из Франции, Венгрии и Монголии. С территории СССР
остатки вымерших представителей ряда семейств получены из Палеонто-
логического института АН СССР, Геологического института АН СССР,
Института геологических наук Башкирского филиала АН СССР, Инсти-
тута зоологии АН УССР, Московского и Свердловского университетов.
Изучены палеонтологические коллекции ЗИН. Кроме того, использован
значительный литературный материал.
За отсутствием достаточного материала по землеройкам родов Croci-
dura, Suncus и Sylvisorex из Африки и Cryptotis и Sorex из Северной Аме-
рики, требующих ревизии, их виды представлены списком, в котором
указаны работы с первоописанием, и дано их распространение. Они от-
ПРЕДИСЛОВИЕ , 5
мечены и в указателе видов. Это позволит составить представление об объ-
еме родов и их распространении на разных континентах. Список видов
африканских землероек рода Crocidura составлен по Дольману (Dollman,
1915, 1916), а списки африканских видов родов Suncus и Sylvisorex —
по Аллену (С. Allen, 1939). Списки американских видов родов Sorex
и Cryptotis даны по Холлу и Келсону (Hall, Kelson, 1959).
К заимствованным описаниям и диагнозам поставлены фамилии их ав-
торов. В отдельных случаях диагноз составлялся мной на основании чу-
жого описания, если он у автора не содержал важных признаков или
не был составлен.
Внутривидовую систематику я рассматриваю как особую область ис-
следования. Подвидовая дифференциация вида должна исследоваться
на фоне ландшафта и на всем его ареале. Такого рода исследование не вхо-
дило в круг задач, стоявших передо мной при изучении материала для
настоящей работы. Подвиды, приведенные для видов отечественной фа-
уны, сохраняются в их старом значении и понимании.
Очерки по экологии видов взяты из написанного мной ранее раздела
определителя (Млекопитающие фауны СССР, 1963, с. 54—121) и моно-
графии (Гуреев, 1971). Со времени их выхода ничего не прибавилось но-
вого или существенного к знаниям об экологии видов насекомоядных
нашей фауны, для которых только и составлены эти очерки. Они кратки,
поскольку содержат данные, касающиеся только конкретного вида. Об-
щие особенности вынесены в экологические характеристики крупных
таксонов.
В список литературы помещены сводки по насекомоядным, работы
общего характера, которые могут представить интерес для читателя, ра-
боты, содержащие системы разных авторов, по экологии и др. Большой
список литературы составляют издания, приведенные в синонимии.
Значительный объем информации о литературе по насекомоядным содер-
жится в помещенных в список монографиях и более или менее крупных
работах (Долгова, Юдина, 1975).
В отличие от других выпусков серии «Фауна СССР» литература при-
ведена в конце работы.
В настоящей работе верхняя граница плиоцена (верхний плиоцен)
сохраняется в ее старом значении. Это не исключает условности и пере-
ходности ее по отношению к плейстоцену. Сохраняется также и старое
обозначение времени и пространства: нижний, средний и верхний. Ясно,
что нижний плиоцен — это слой, образовавшийся ранее среднего и верх-
него.
В оценке времени формирования отдельных представителей фауны
млекопитающих Зайсанской котловины я придерживался стратиграфии
ее палеоген—неогена, разработанной Б. А. Борисовым (1963).
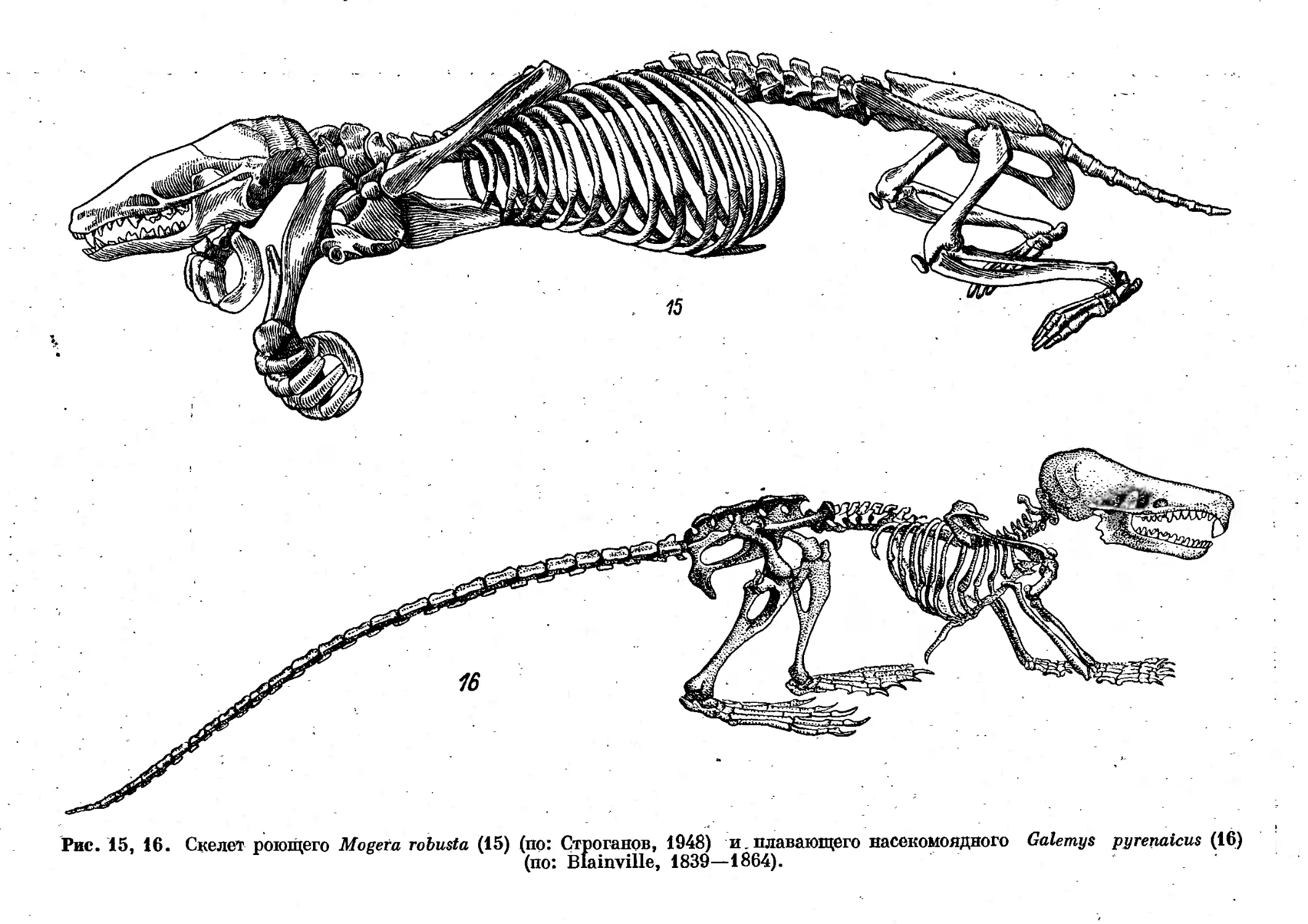
Большинство иллюстраций к введению выполнено художником
Н. Н. Фузеевой. Ею сделаны и многие рисунки к сем. Erinaceidae, Tal-
pidae.
Для сем. Soricidae использдваны рисунки из моей монографии «Земле-
ройки фауны мира» (1971), выполненные художниками М. А. Масловой,
Г. Е. Поздеевой, С. Л. Шмуйлович. Иллюстрации к сем. Talpidae (подсем.
Uropsilinae, Talpinae) сделаны художником Н. Е. Кондаковым к моно-
графии о кротах (Строганов, 1948).
В заключение следует отметить, что в последние годы отечественными
палеонтологами собраны интереснейшие материалы по многим группам
вымерших насекомоядных, значительно дополнившие наши знания о их
,6 ПРЕДИСЛОВИЕ
развитии во времени и пространстве. Эти материалы были предоставлены
мне для обработки Н. С. Шевыревой, А. Г. Малеевой, Р. С. Адаменко,
В. С. Зажигиным, А. Г. Агаджаняном, В. П. Суховым и В. А. Топачев-
ским, за что приношу им глубокую благодарность. Приношу глубокую
благодарность также за предоставление материала по современным на-
секомоядным Ангерман (Angermann, ГДР). За помощь в подготовке ру-
кописи к печати благодарю Г. И. Баранову и М. Н. Наумову.
Профессор А. А. Стрелков вложил огромный труд в редактирование
рукописи этой книги и постоянно помогал мне ценными советами и ука-
заниями. Когда рукопись была подготовлена к печати и все задуманное
с Александром Александровичем выполнено, он внезапно ушел из жизни,
оставив по себе светлую память.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
Отряд INSECTIVORA
Подотряд TEN RECH
Сем. ТЕ NRECIDAE
Подсем. TENRECINAE
1. Род Tenrec Lacepede
2. Род Setifer Frorier
3. Род Hejnicentetes Mivart
4. Род Dasogale Grandidier
5. Род Echinops Martin
Подсем. ORYZORICTINAE
6. Род Oryzorictes Grandidier
7. Род Microgale Thomas
8. Род Cryptogale Grandidier
9. Род Limnogale Major
10. Род Geogale Milne-Edwards
Сем. POTAMOGALIDAE
1. Род Potamogale Du Chaillu
2. Род Micropotamogale Heim de Balsac
Сем. CHRYSOCHLORIDAE
1. Род Chrysochloris Lacepede
2. Род Amblysomus Pomel
3. Род Chrysospalax Gill
4. Род Cryptochloris Shortridge et Carter
Сем. SOLE NODO NTIDAE
1. Род Solenodon Brandt
Додотряд ERINACEI......
Сем. M A C R О S-C E L I D I D A E
1. Род Macroscelides Smith
2. Род Nasilio Thomas et Schwann
3. Род Elephantulus Thomas et Schwann
4. Род Petrodromus Peters
5. Род Rhynchocyon Peters
Сем. TUPAIIDAE
Подсем. TUPAIINAE
1. Род Tupaia Raffles
8 СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
2. Род Anathana Lyon
3. .Род Dendrogale Gray
Подсем. PTILOCERCINAE
4. Род Ptilocercus Gray
Сем. E RINA С E I D A E
1. Подсем. GALERICINAE
f 1. Род Tetracus Aymard
fl. T. nanus Aymard.......................................................... 108
f 2. T. bouti Lavocat ...................................................... 108
f 2. Род Galerix Pome!
• • 1. G. exilis Blainville.................................................. 110
• 2. G. minor Filhol..................................................... Ill
• • 3. G. afritanus Butler .................................................. Ill
4. G. paraexilis sp. nov................................................... 112
• 5. G. tadzhikistanicus sp. nov......................................... 113
f 3. Род Pseudogalerix Gaillard
j- 1. P. stehlini Gaillard.................................................. 114
f 4. Род Lanthanotherium Filhol
• 1. L. sansaniensis Filhol ............................................... 115
• 2. L. robustum Viret...................................................... 116
3. L. sp. Webb............................................................. 116
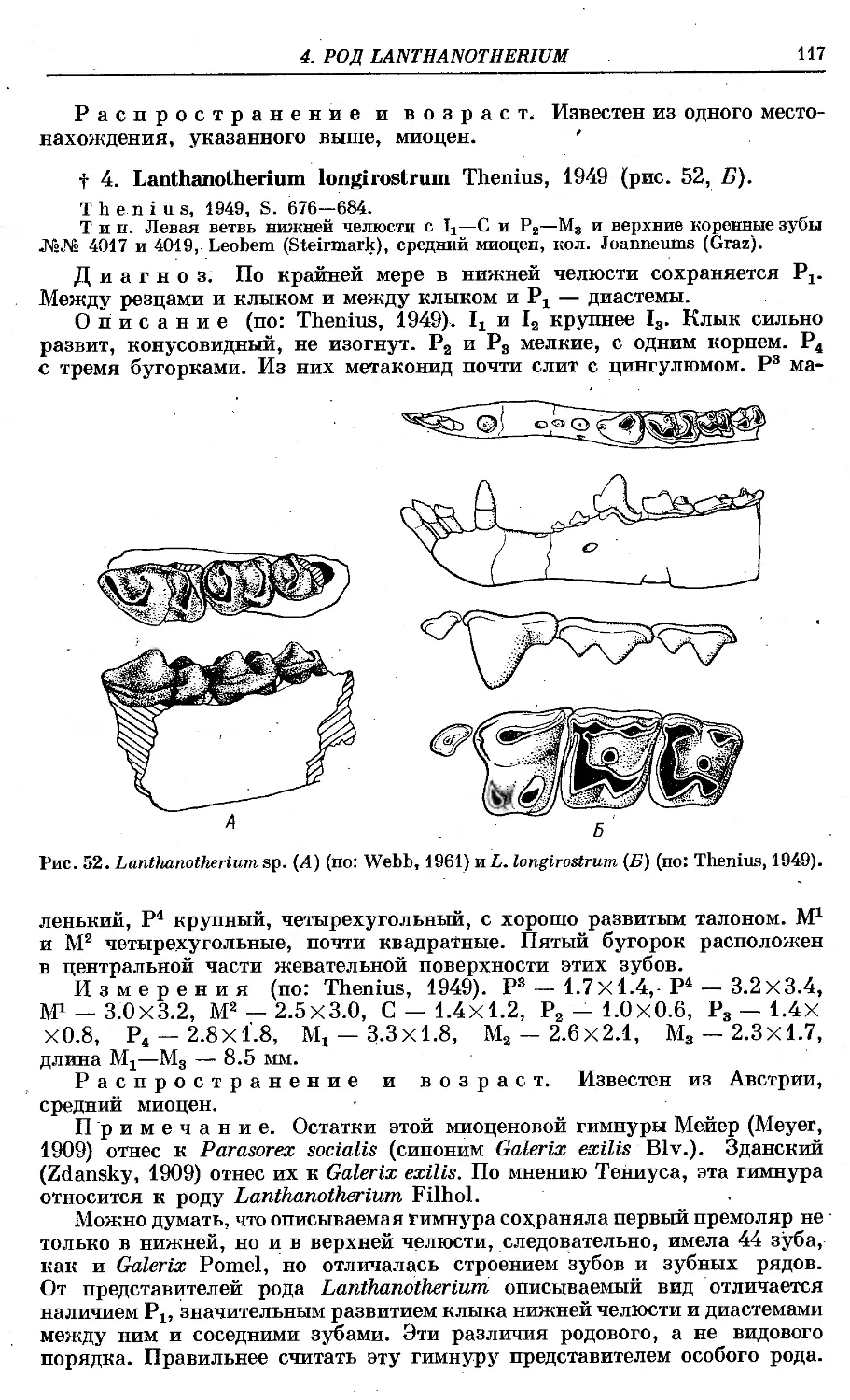
4. L. longirostrum Thenius ............................................... 117
t 5. Род Deinogalerix Freudental
f 1. D. koenigsuraldl Freudental . . . "..................................... 119
f 6. Род Ocajila Macdonald
1. O. makpiyahe Macdonald ............................................... 119
f 7. Род Neurogymnurus Filhol
1. N. cayluxi Filhol..........'............................... ........... 122
2. N. mediterraneus Viret.................................................. 123
f 8. Род Pseudeneurogymnurus gen. nov.
1. P. schevirevi sp. nov................................................... 123
2. P. zhchikuadzei sp. npv................................................ 124
9. Род Echinosorex Blainville
1. E. gymnurus Raffles.................................................... 126
10. Род Hylomys Muller et Schlegel
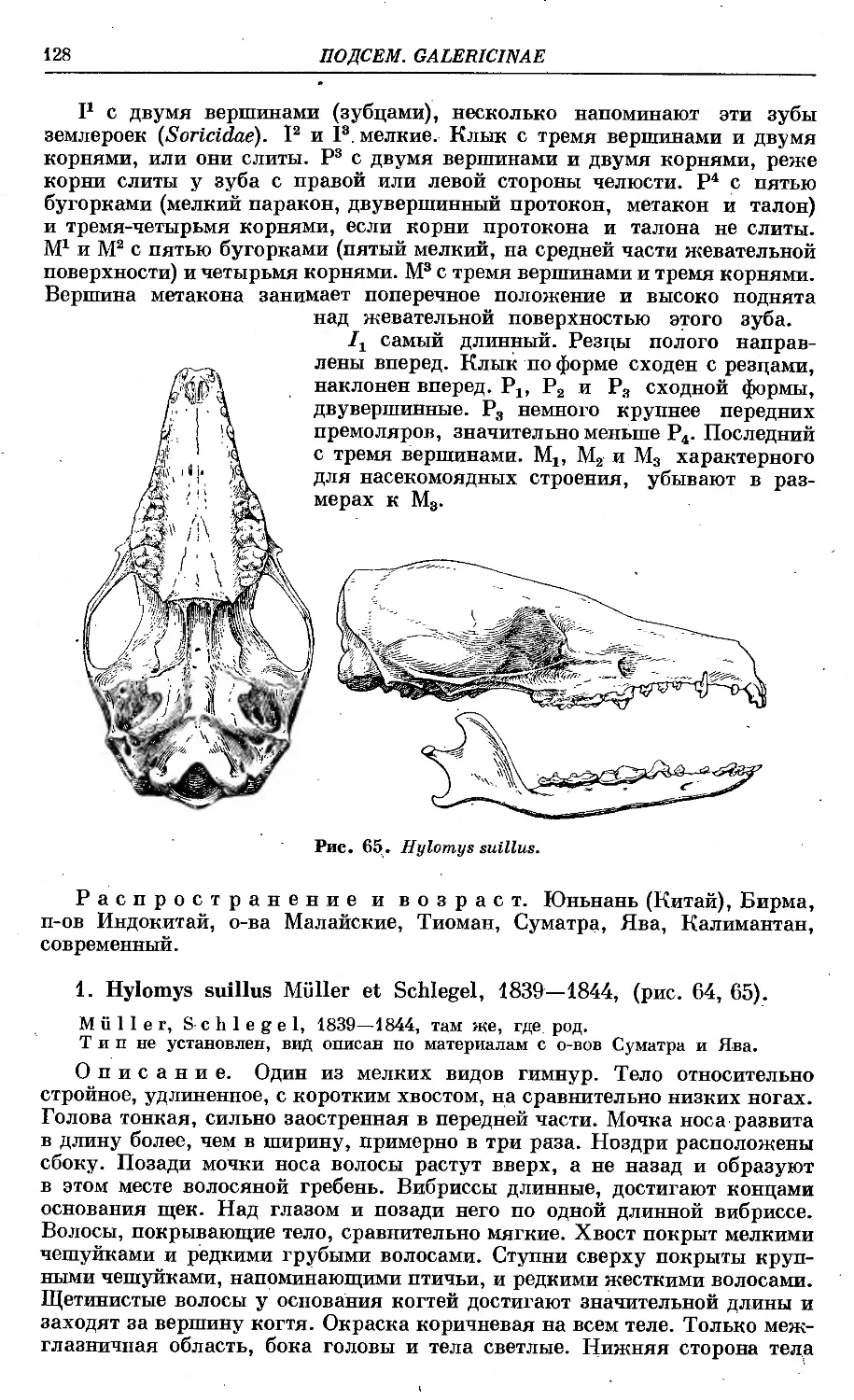
1. H. suillus Miilier et Schlegel......................................... 128
11. Род Podogymnura Mearns
1. P. truei Mearns........................................................ 130
12. Род Neotetracus Trouessart
1. N. sinensis Trouessart.............................................. 131
13. Род Neoliylomys Shaw et Wong
1. N. hainanensis Shaw et Wong............................................ 132
t 1. Триба PROTERICINI /
f 14. Род Tupaiodon Matthew et Granger
fl. T. morrisi Matthew et Granger........... 136
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
9
f 15. Род Proterix Matthew
f 1. Р. loomsi Matthew........................................................... _ 137
f 2. P. bicuspis Macdonald ................................................. 138
f 16. Род Palaeoscaptor Matthew et Granger 1
f 1. P. acridens Matthew et Granger . ....................................... . 139
t 2. Триба В К A С H Y Ё R I С I N I
f 17. Род Brachyerix Matthew
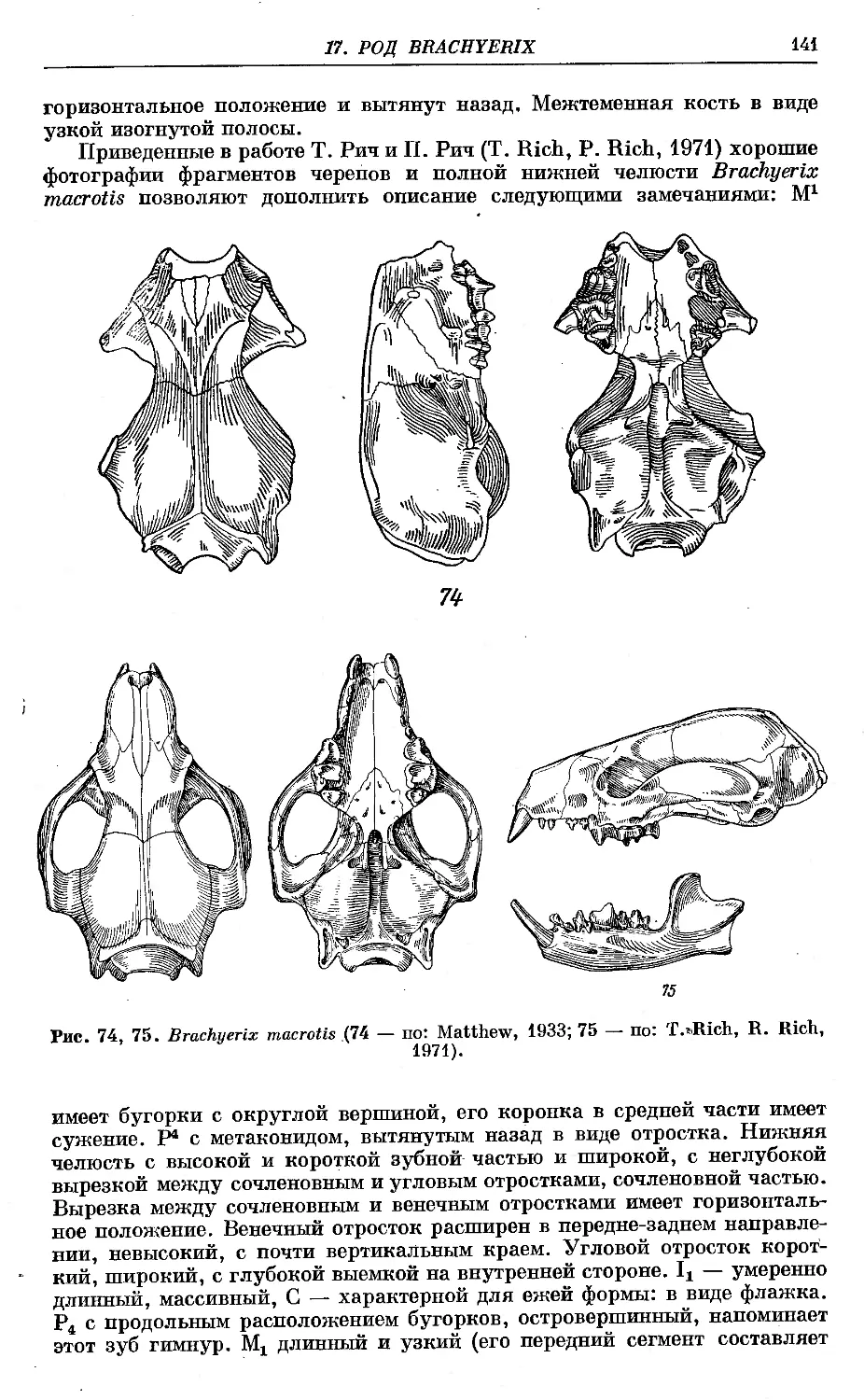
f 1. B. macrotis Matthew......................................................... 142
f 2. B. insertis Matthew ....................................................... 142
f 18. Род Metechinus Matthew
j- 1. M. nevadensis Matthew.................................................... 143 ,
f 2. M. marslandensis Meade....................................................... 144 "
f 19. Род Exallerix McKenna et Holton
f 1. E. hsandagolensis McKenna et Holton........................................ 144
f 20. Род Dimylechinus Hurzeler
fl. D. bernoullii Hurzeler . . ............................................. . 146
t 3. Триба AMPHECHINI
f 21. Род Amphechinus Aymard
fl. A. arvernensis Blain ville.................................................
f 2. A. edwardsi Filhol .......................................................
• • 3. A. intermedium Gaillard............................................. ;
• 4. A. rectus Matthew et Granger..................................
• •5. A. kansuensis Bohlin.....................................................
• 6. A. minimus Bohlin.......................................................
•7. A. rusingensis Butler ............................................ . .
8. A- homcloudi Macdonald ............................................ . .
•9. A. montanum Koerner......................................................
9. A. sp......................................................................
148
148
149
149
150
151
151
152
152
153
f 22. Род Gymnurechenus Butler
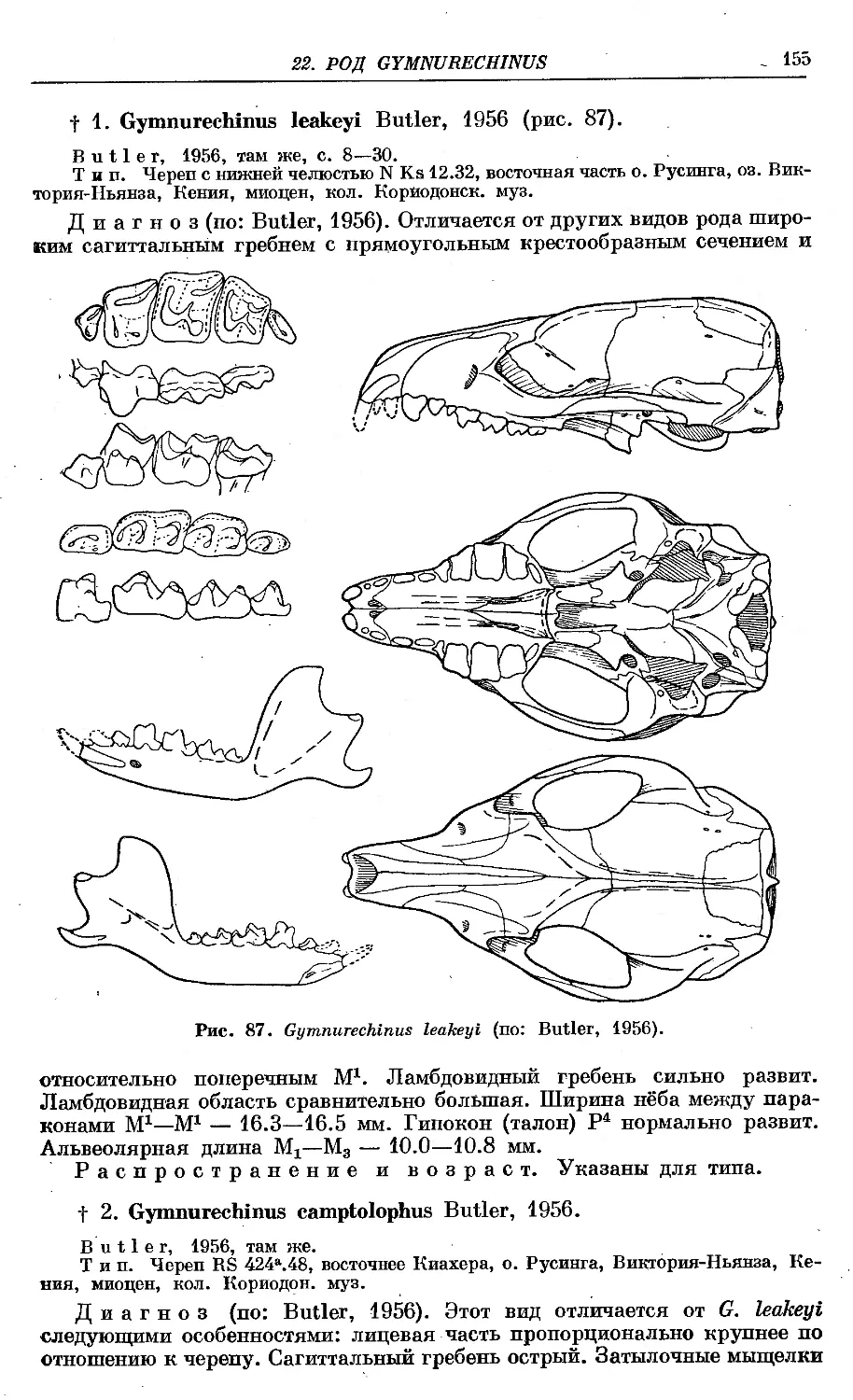
f 1. G. leakeyi Butler ................................................... 155
f 2. G. camptolophus Butler ................................................. 155
f 3. G. songhorensis Butler............<.' . ................................ 156
f 23. Род Stenoechinus Rich et Rasmussen
f 1. S. tantalus Rich et Rasmussen............................................ 156
4. Триба E R I N А С E J N .I
f 247'Род Mioechinus Butler
f 1. M. oeningensis Lydekker ........................................ 158
f 2. M. sansaniensis Lartet .......................................... 158
f 25. Род Pfotechinus Lavocat
f 1. P. salis Lavocat ...............,................................ 159
26. Род EHnaxxu&J^nnaeus^
f la. E. vireti Crusafont-Pairoet Villalta Camella..................... 162
< f l. Jamraet--. . . ........................................., ; 163j
T~ ^2. E? europaeus Linnaeus^........................................... 164
• ЗГТ?. frontalis Smith. Г .............................................. 167
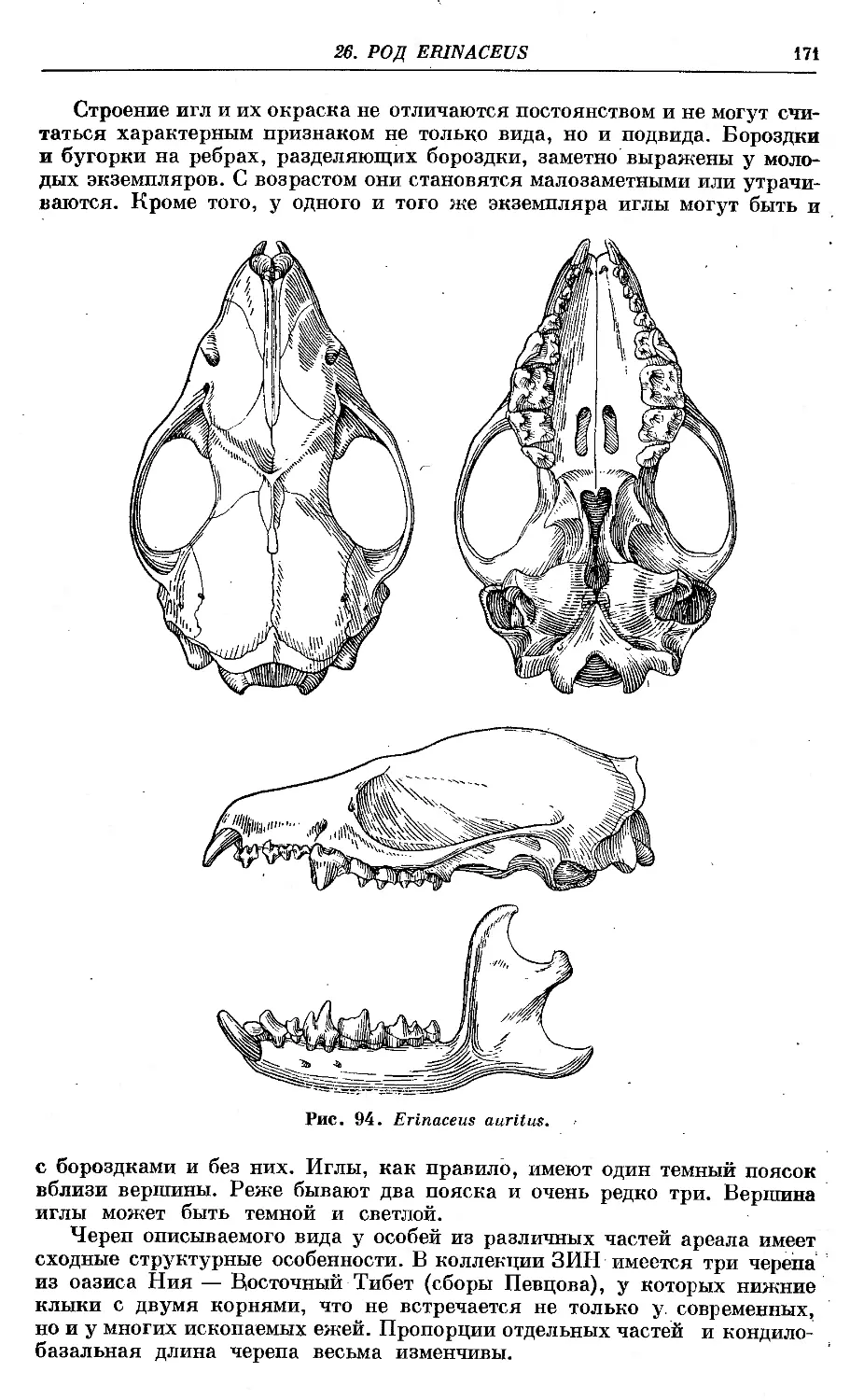
4. E. auritus Gmeliji^ ...................................... .. . . '. .169
E. dauricus Suiiclevall..........................................1-74;
6. E.. aethyopicus Ehrenberg. ...................................... 176
10 ! СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
j , . . —
Сем. TALPIDAE
1. Подсем. UROPSILINAE
1. Род Rhynchonax Thomas
1. К. andersoni Thomas........................................... 201
2. Род Nasillus Thomas
1. N. gracilis Thomas ............................................... 202
2. H. investigator Thomas ...................................... 202
3. Род Uropsilus Milne-Edwards (
1. U. soricipes Milne-Edwards...................................... 202
2. Подсем. DESMA NINAE
11?Триба G aTlLA^dFn I
f 4. Род Gaillardia Matthew
f 1. G. thomsoni Matthew 207
f 5. Род Hydroscapheus Shotwell
f 1. H. americanus Shotwell......................................... 207
2. Триба DES MANIN I
f 6. Род Mygalea Schreuder
fl. Af. antiqua Pomel 211
f 2. Af. asiatica Gureev ............................................. 212
f 7. Род Mygalinia Schreuder
fl. Af. hungarica Kormos . ............................................ 212
f 2. Af. zazhigini Gureev............................................ 213
8. Род Galemys Каир
11. G. ponticus Schreuder............................................ 214
f .2, G. semsey Kormos................................................ 214
.3. G. pyrenaicus Geoffroy Saint Hilaire............................. 214
9. Род Desmana Guldenstaedt
• 1. D. verestchagini Topatshevsky.................................. 217
2. D. nehringi Kormos ........................................... 218
• 3. D. .kormosi Schreuder...................................... 219
• 4. D. thermalis Kormos . . ..................................... . 220
5- wuwcfaito Linnaeus................................................ 220
3. Подсем. TALPINAE
3. Триба UROTRICHINI Stroganov
f 10. Род Myxomygale Filhol
f 1. A. antiqua Filhol................................................. 229
f 11. Род Paratalpa Lavocat
f 1. P. micheli Lavocat .............................................. 229
f 12. Род Mygatalpa Sehreuder
fl. Af. arvernensis' Pomel . .......................................... 231
f 13. Род Scalopoides Wilson
f 1. 8. isodens Wilson................. . ............................ 231
f 14. Род Mydecodon Wilson
ti. al martini Wilson.......................... . . .......... . 232
15. Род Scaptonix Milne-Edwards
f f. 8. ejhaardsi Gaillard .......................................... 233
f 2. 8. dolichockir Gaillard . ...................................... 233
3. S. fusieaudatus Milne-Edwards ............... . . . ............ 234
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ 11
f 16. Род Domninoides Green 1
fl. D. riparensis Green.................................. ............ 234,
f 2. D. valentinensis K. Reed.......... . ............................ 235
17. Род Urotrichus Temminck
1. U. talpoides Temminck .............................. . 236
18. Род Scapanulus Thomas
1. S. oweni Thomas . . ............................................ 238
19. Род Neurotrichus Giinter
1. N. gibbsi Baird .................................... 238
4. Триба CONDYLURINI Thomas
20. Род Condylura liliger
1. C. cristata Linnaeus . . .' 240
t 5. Триба PROSGALOPINI 1
f 21. Род Cryptoryctes C. Reed
f 1. C. kayi C. Reed.................................................. 244
f 22. Род Proscalops Matthew
fl. P. miocaenus Matthew..............................................i 244
f 2. P. tertius K. Reed................................................. 245
f 3. P. secundus Matthew ....................................... 245
f 23. Род Arctoryctes Matthew
f 1. A. terrenus Matthew ......................... 246
f 24. Род Oligosealops K. Reed
f 1. O. whitmanensis K. Reed.......................................... 247
f 25. Род Mesosealops K. Reed
f 1. M. scopelotemos K. Reed ......................................... 247
6. Триба SCALOPINI
f 26. Род Proscapanus Gaillard
fl. P. sansaniensis Lartet ........................................... 248
27. Род Parascalops True
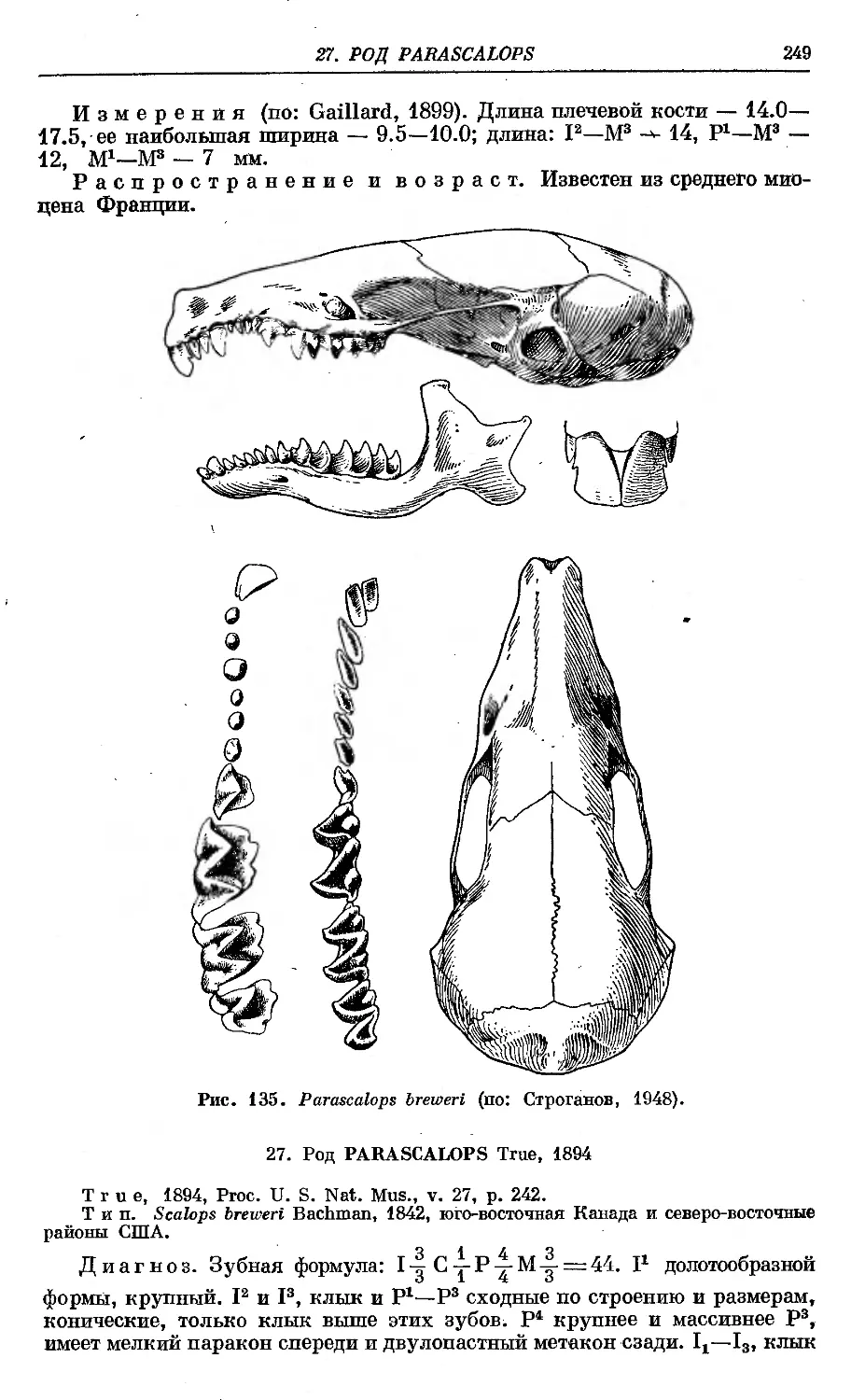
1. P. breveri Bachman.............................................. 250
28. Род Scapanus Pomel
1. S. ' townsendii Bachman.......................................... 251
2. S. orarius True................................................. 252
3. S. latimanus Bachman.......................... '. . ........... 252
t 29. Р°Д Hesperoscalops Hibbard
f 1. H. rexroadi Habbard ............................................. 252
f 2. H. sewardensis K. Reed........................................ . 253
30. Род Scalopus Geoffroy Saint Hilaire
1. S. aquaticus Linhaeus............................................ 254
2. S. aereus Bangs................................................. 254
3. S. inflatus Jackson ............................... ;......... 256
7, Триба T A L P I N I
t 31. Род Amphidozotherium Filhol
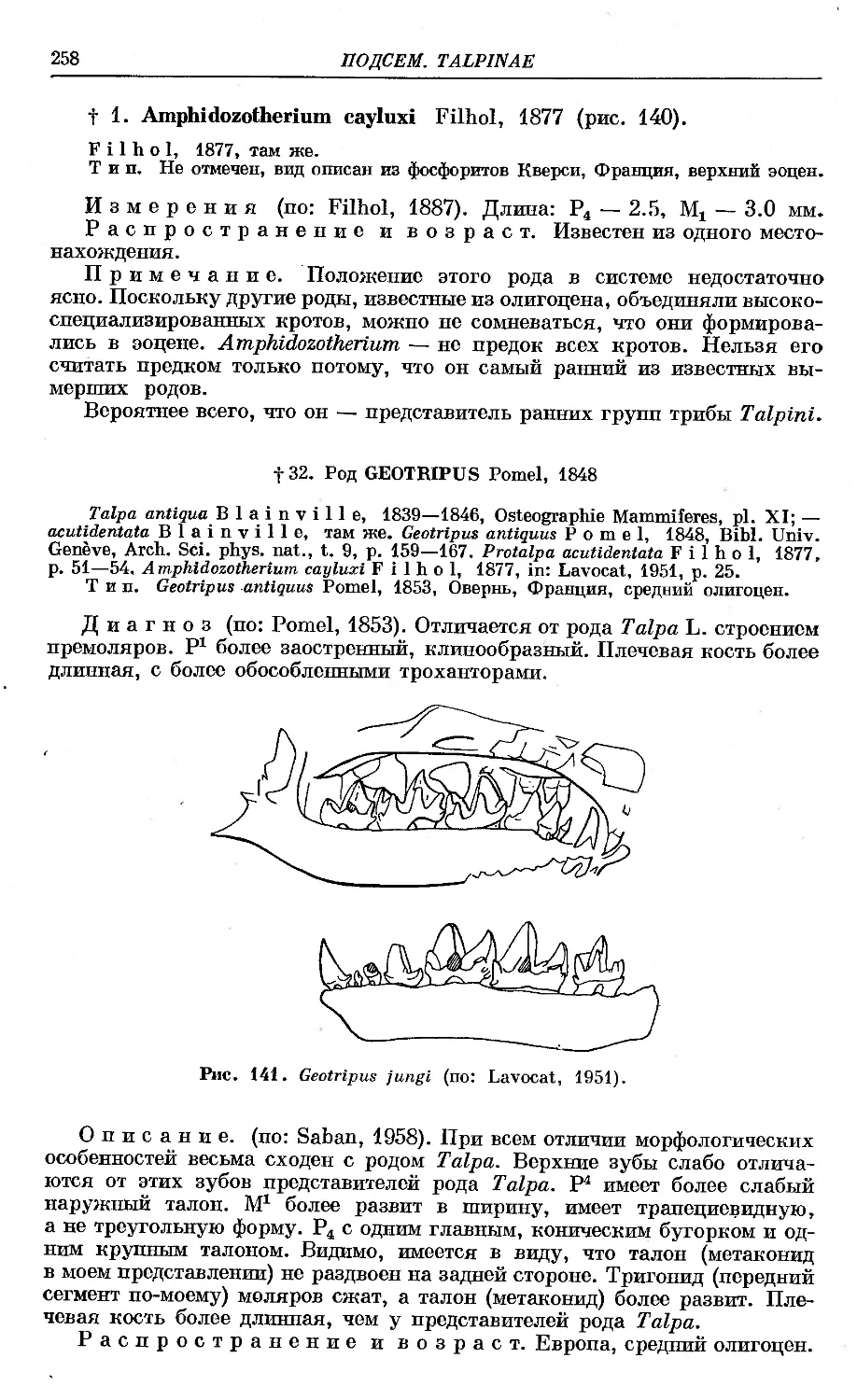
f 1. A. cayluxi Filhol............................. ................. 258
f 32. Род Geptripus Pomel
f 1. G. antlquus Pomel . . . . . . . . . ......\ ................. . 259
12
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЕ НАСЕКОМОЯДНЫХ
33. Род Talpa Linnaeus
fl. Т. minuta Blainville y ................... 260
_1^2. ТГ europaea Linnaeus /.................. 260
3. 7. altaicq.
ktiwiEi
4. T, cancasica Satunin ......................................... 267
5. 7. remoaa Thomas~~................'.'........................... 268
6. T. соеед‘Savi . .7................. .............. 269
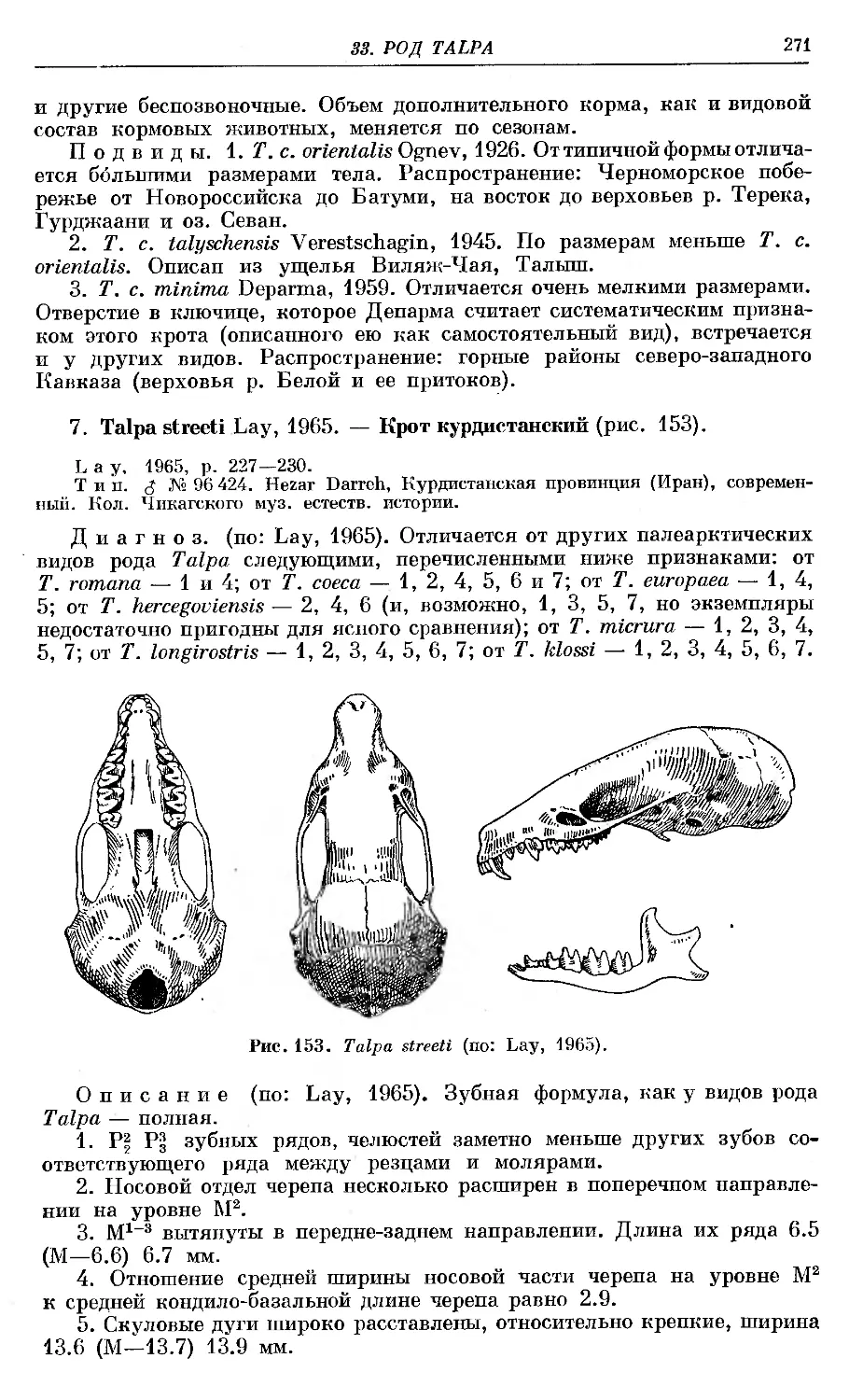
ТТ^Г7 streeti Lay.................’...........................'. . 271
8. T. klossi Thomas............................................. 273
9. T. grandis Miller.............................................. 273
10. T. parvidens Miller ......................................... 274
11. 7. longirostris Milne^Edwards ................................ 275
12. 7. micrura Hodgson . ...................................... . 276
34. Род Mogera Pomel
1. M. wogura Temmjpck ................... .'........................ 276
2, M, robusta Neliriiu? ~~~....................................... 277
3. M. latouchei Thomas ............................................. 279
35. Род Parascaptor Gill
1. P. leucura Blyth.................................................. 281
36. Род Scaptochirus Milne-Edwards
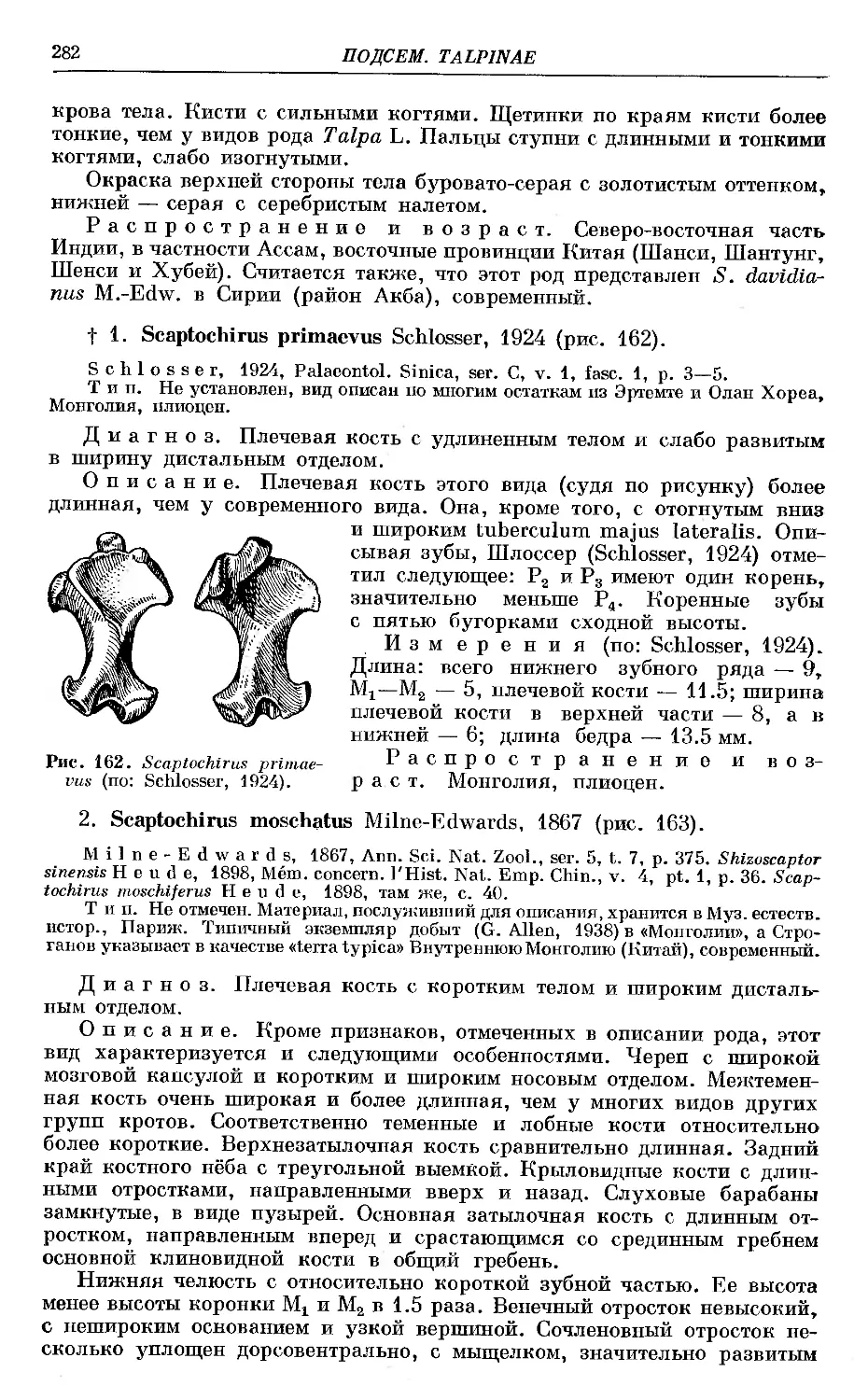
f 1. S. primaevus Schlosser ........................................... 282
2. S. moschatus Millie-Edwards.................................... 282
Сем. SORICIDAE
f 1- Подсем. SATURNINIINAE
t 1. Род Saturninia Stehlin
f 1. S. gracilis Stehlin .............................................. 316
2. Подсем. SORICINAE___
t 1. Триба OLIGOSORICINI
f 2. Род Amphisorex Filhol
j- 1. A. primaevus Filhol........................................... 319
f 3. Род Oligosorex Kretzoi
f 1. O. antiquus Pomel ................................................ 320
f 2. O. dehtni Viret et Zapfe ........................................ 320
f 4. Род Gobiosorex Sulimski
f 1. G. kingae Sulimski ............................................. 321
f 5. Род Croeidosorex Lavocat
f 1. C. piveteaui Lavocat ........................................... 323
f 6. Род Alluvisorex Hutchison
f 7. Род Miosorex Kretzoi
j- 1. M. grivensis Deperet .............................................324
f 8. Род Domnina Cope
f 9. Род Paradomnina Hutchison
f 10. Род Soricella Doben-Florin
f 11. Род Angustidens Repenning
f 1. A. vireti Wilson ............................................... 325
f 2. Триба HETERO SORICIN I
t 12. Род Heterosorex Gaillard
f 1. H. sansaniensis Lartet...................................... 327
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
13
f 13. Род Pseudotrimylus Gureev
•f 1. Р. roperi Wilson.................................................... ‘327
f 14. Род Ingentisorex Hutchison
3. Триба SORI С I N I
15. Род Sorex Linnaeus ,
t 1-
-• 2.
-• 3.
-• 4.
•• 5.
5. gracilidens Viret et Zapfe
5. compressus Wilson . .
falax Heller . . . . .
8. falax Heller . . .
S. prealpinus Heller
S. araneoides Heller
subaraneus Heller
pachyodon Разд . .
runtonensis Hinton
praearaneus Kormos
kretzoi Sulimski
S. subminutus Sulimski
S. hibbardi Sulimski
S. cudahyensis Hibbard
8. lacustris Hibbard
<S. taylori Hibbard .
S’. rexroadensis Hibbard
- 6. 8.
7. S.
8.
-• 9.
-• 10.
11.
12.
- • 13.
• • 14.
- 15.
16.
f 17. 8. hagefmanensis Hibbard et Bjork
• 18. S. powersi Hibbard et Bjork . .
f 19. 5. meltoni Hibbard et Bjork . . .
- sandersi Hibbard .............
8araneus Linnaeus} ...............
22. A. g^per 1 nomas . . ;...........
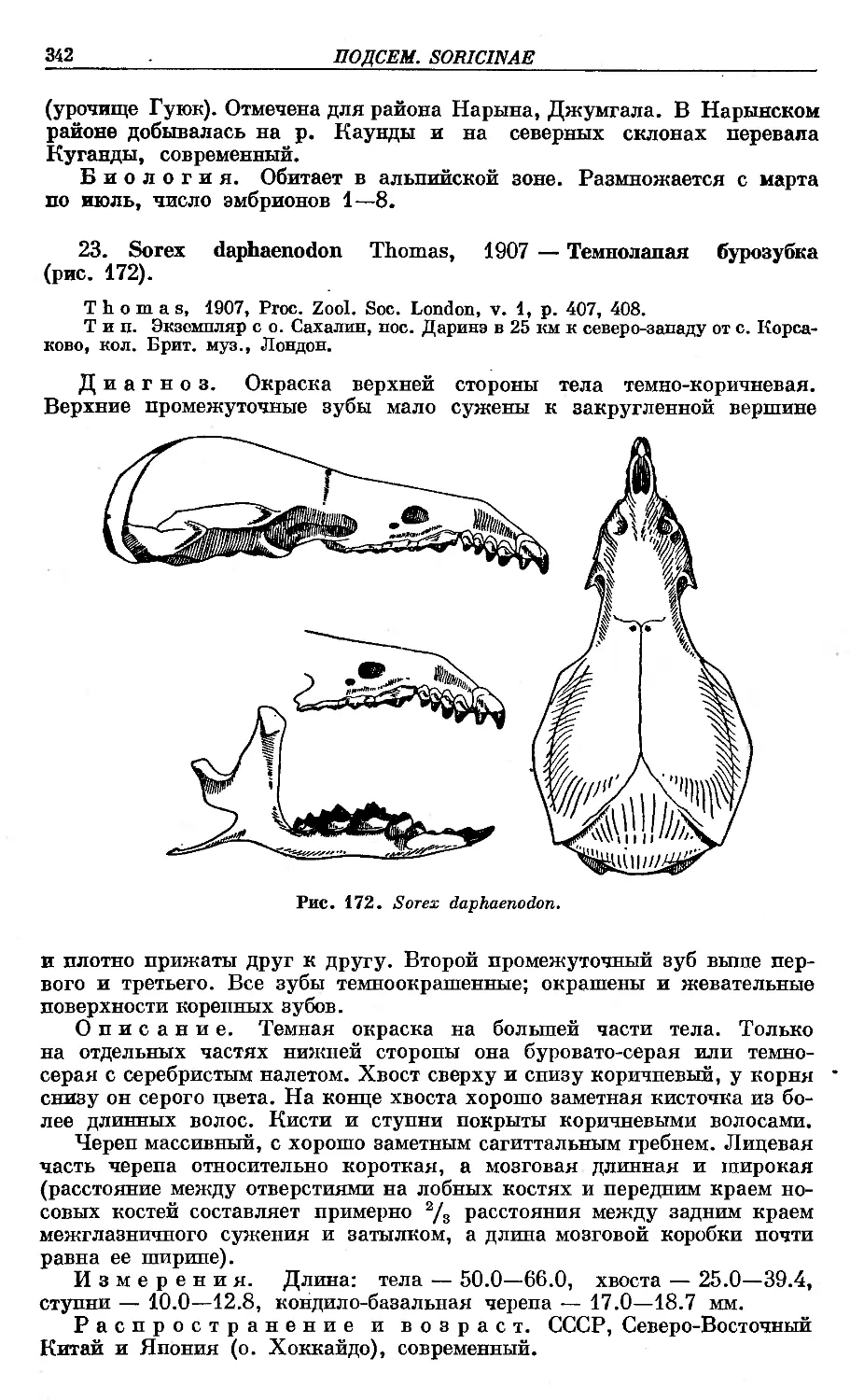
23. 8. daphaenodon Thomas ; . .. . . .
24. 8- anticus Kerr-. . .< . . . .
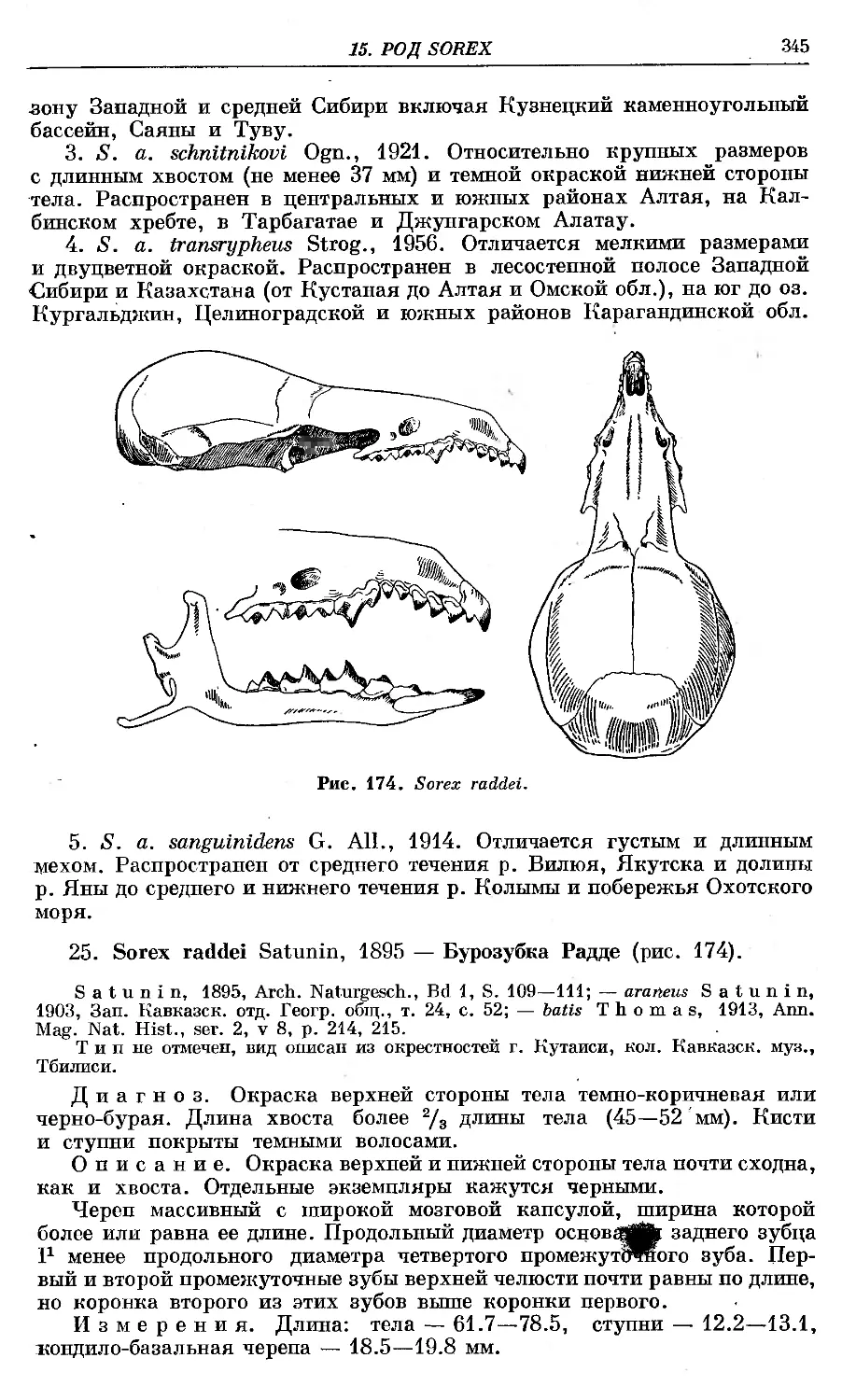
'25L A. raddfi SatunTn ..............
26. 5. gZpinas Schinz................
2/. Л'? mirabilis Ognev..............
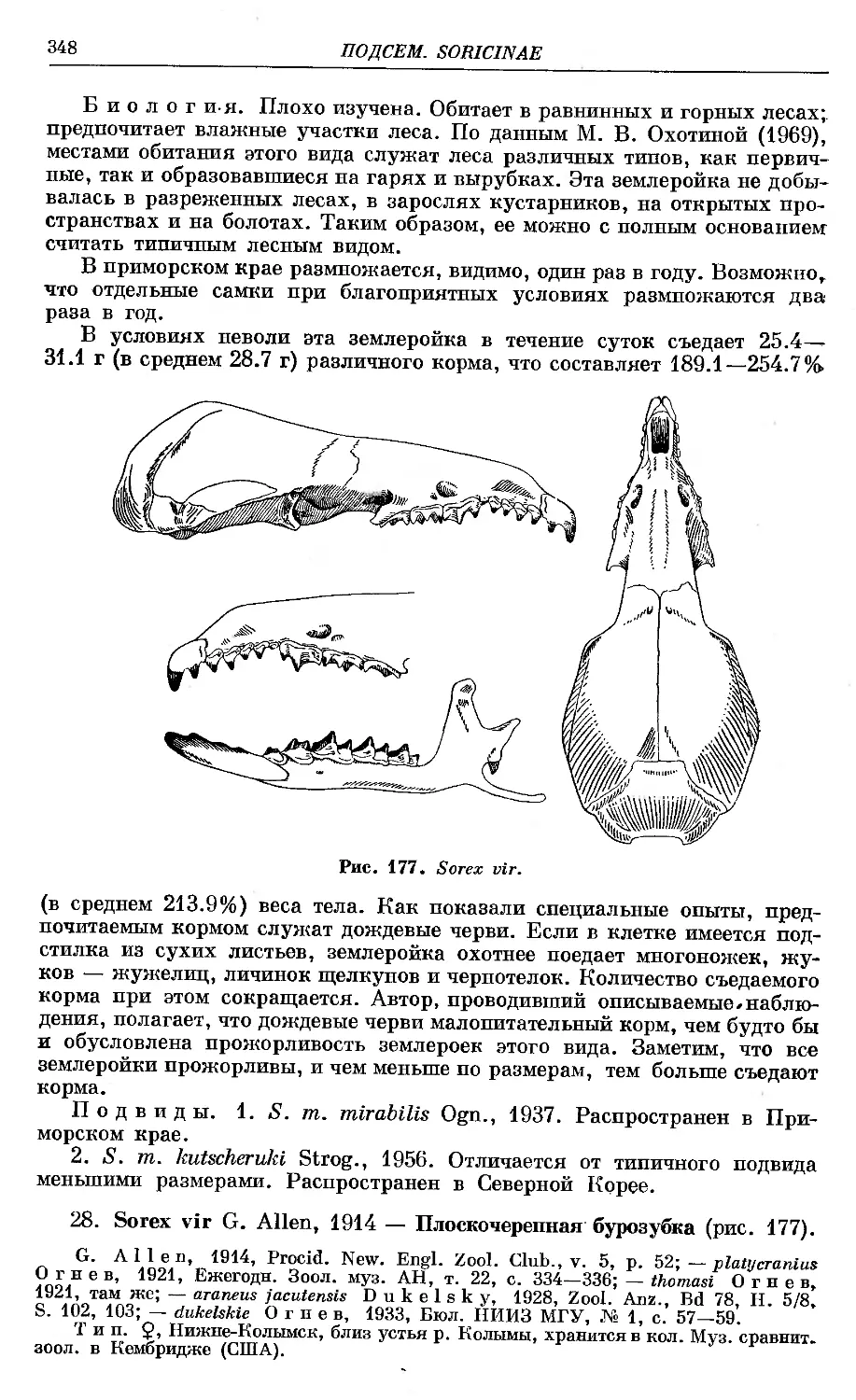
ЖХ^'^.^-АПеп-Т". ...........
29? beringtantis "Yudin <............
30. X unguZcaTaZiS TTobsoh...........
L. f'gl . 2>. catch tk'na TaxmanT........
r .......................................
S.
8.
s.
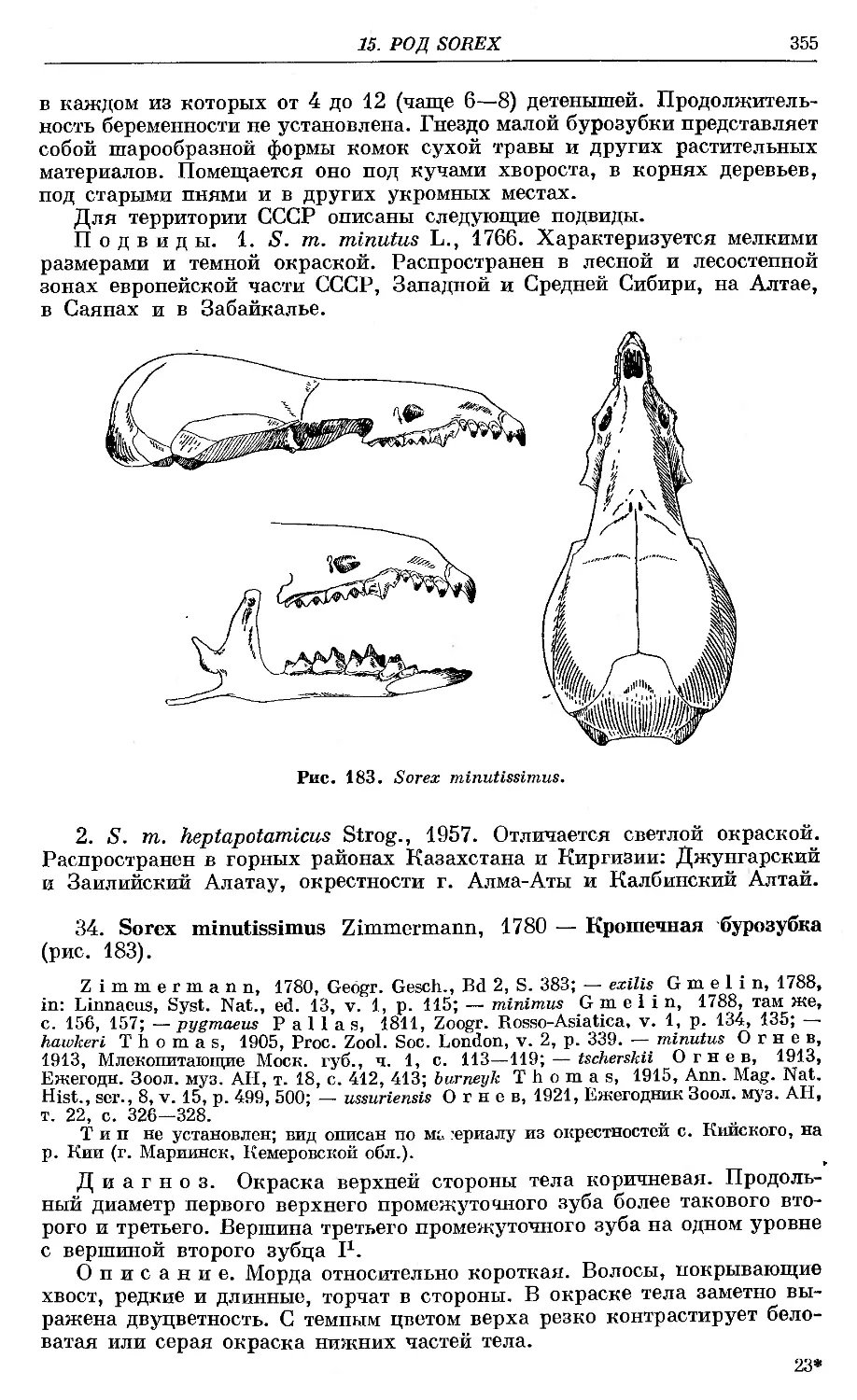
-U Q53. ,s. rginufus Liun^eus }. . . . ,
—‘"'^Tminu:iss: "i Zlinmiruian'/
4-t35. У tsodonTurov 'V . . . . . .
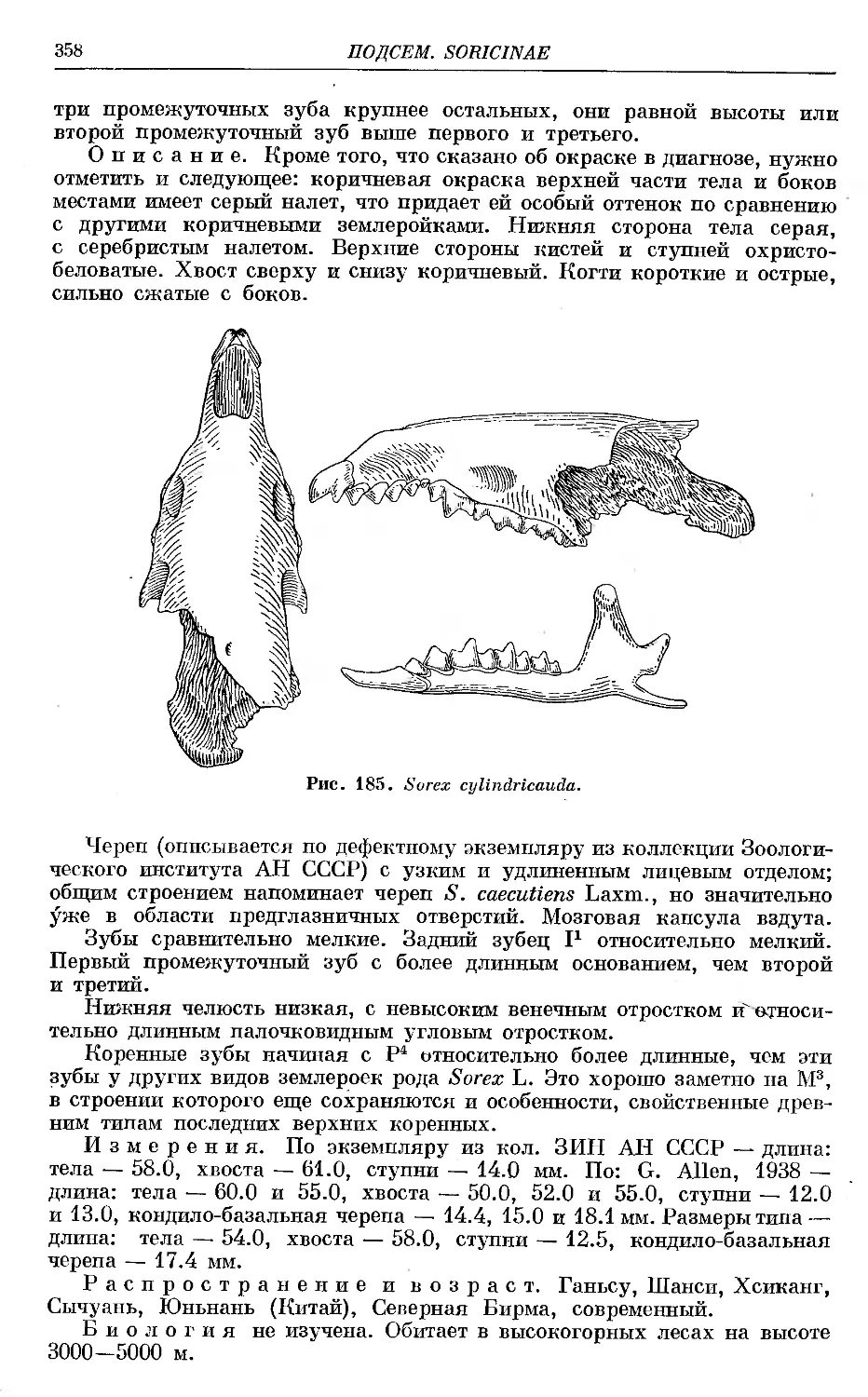
8. ct/ZinJficauda'Milne-Edwards
37. 8. cinereus Kgrr ...... .
5». S-. bucharensjsOgnev ....
39.~iS. kozZoai "Stroganov ....
8. excelsus G. Allen . >. . /
8. sinalis Thomas . . ... .
S. lyelli Merriam..................
8. preblei Jackson...............
8. milleri Jackson ......
8. longirostris Bachman . . . ,
8. vagrans Baird..................
8. ornatus Merriam................
8. tenellus Merriam .....
8. trigonirostris Jackson . . .
. 50.' S. nanus Merriam
5. juncensis Nelson et <
8. willeti von Blecker ,
S. sinuosus Grinnell
veraepacis Alston
macrodon Merriam
palustris Richardson
alascanus Merriam
bendirii Merriam
S. pribilofensis Merriam
60. S. fumeus Miller . . .
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
51.
52.
53.
54.
55.
56.
57.
• 58.
59.
s.
s.
s.
s.
s.
Goldman
333
333
334
334
334
334
334
334
335
335
335
336
336
336
337
337
337
337
338
338
, 339
341
342
343
345
346
347
348
349
350
351
353
353
355
356
357
359
361
361
362
363
363
363
363
363
363
363
364
364
364
364
364
364
364
364
364
" 365
365
365
365
14 СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
61. S. jacksoni. Hall et Gilmore . .... i .......... 365
63. S. hydrodromus Dobson....................................................... 365
63. S. gaspensis Antony et Goodwin . ........................................... 365
64. S. dispar Batchelder . . . . k .............................. . 366
65. S. trowbridgii Baird !.....................................................• 366
66. S. merriami Dobson ........................................................ 366.
67. S. saussurei Merriam .....'................................................. 366
68. S. oreopolus Merriam...................................................... 366
69. S. sclateri Merriam......................................................... 366
70. S. stizodon Merriam ....................................................... 366
16. Род Microsorex Cones
f 1. M. pfatensis Hibbard . . ....................... . ................. ... . . 367
2. M. hoyt Baird............................................................... 367
f 17. Род Tregosorex Hibbard et Jammot
fl. T. holmani Hibbard et Jammot ............................................... . 367
t 18. Род Anchiblarinella Hibbard et Jammot
f 2. A. wakeeneyensis Hibbard et Jammot............................................. 369
19. Род Blarinella Thomas
1. B. quadraticauda Milne-Adwards................................................ 370
^Триба C,R OCJ^JLRJJi-1
1. Подтриба MYOSORICINA
f 20. Род Stirtonia Gureev gen. nov.
f 1, S. niobrarensis Macdonald.................................................. 373
21. Род Myosorex Gray
, 1. M. varius Smuts ............................................................ 374
2. M. safer Sundevall . J ..................................................... 374
3. M. blarina Thomas . ............................................. 374
4. M. babaulti Heim de Balsac.............................................. . 375
5. M. preussi Mchiaste........................................................ 375
6. M. polli Heim de Balsac . . . .............................................. 375
22. Род Surdisorex Thomas
1. S. norae Thomas.......................................................... 375
2.^IIogTpH6a C^QCIDUftlNA
f 23. Род SimMisorex Stogov et Savinov
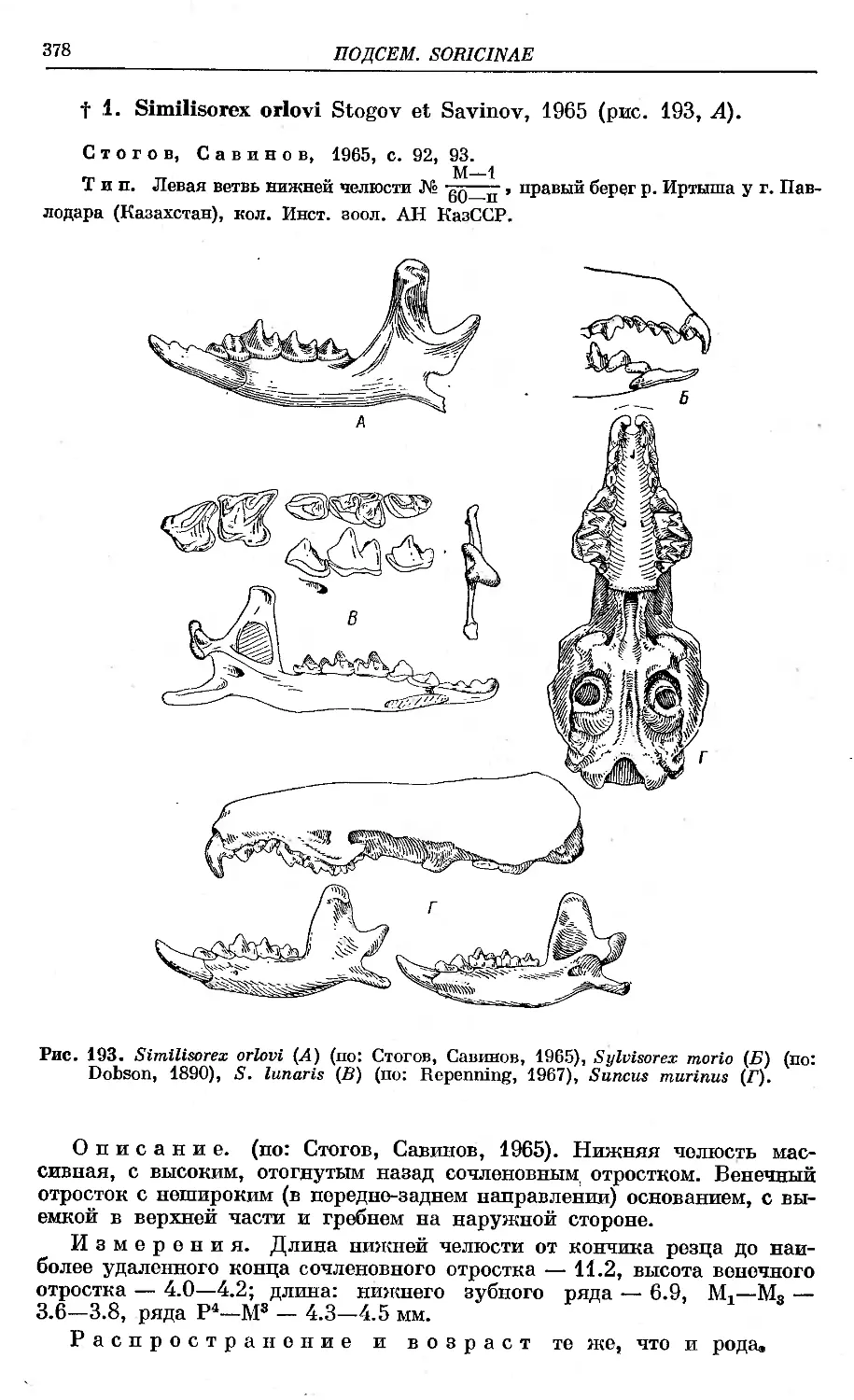
fl. S. orlovi Stogov et Savinov.................... . .................... 378
24. Род Feroculus Kelaart
1. F. macropus Bly th ..................................................... 379
25. Род Sylvisorex Thomas
1. S. morio Gray.......................................................... 380
2. S. jonstoni Dobson....................................................... 380
3. S. granti Thomas .................................................. 3j8O
4. S. lunaris Thomas ...................................................... 380
5. S. olula Thomas ...........................T. . 7 . 7 ."................. 380
6. S. oriundus Hollister ......................................................... 380
7. S. ruande Lonnberg et Gildenstolpe ............................................ 381
8. S. megalura Jentink.......................................................r . 381
9. S. sorella 'fhomas........................................................... 381
10. 5. suncoides Osgood . ......................................................... 381
26. Род Suncus Ehrenberg
1. S. murinus Linnaeus..................................................... 381
2. S. stoliczkanus Anderson................................................. 382
3. S. dayi Dobson . . . ................................................ 382,
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ 15
4. S. chriseos Kershaw ......................................................... 383
5. S. coquerellii Trouessart..................................................... 383
6. S. gracilis Blainville .................................................... 383
7. S. gratulus Thomas et Schwann ............................................... 383
8. S. infinitesimus Heller..................................................... 383
9. S. leucura Matschie ....................................................... 383
10. S. lixus Thomas .......................................................... 383
11. S. madagascarensis Coquerel . . . ......................................... . 383
12. S. orangie Roberts ........................................ 383
13. S. varillus Thomas...........................'............................... 383
14. S. warreni Roberts........................................................... 383
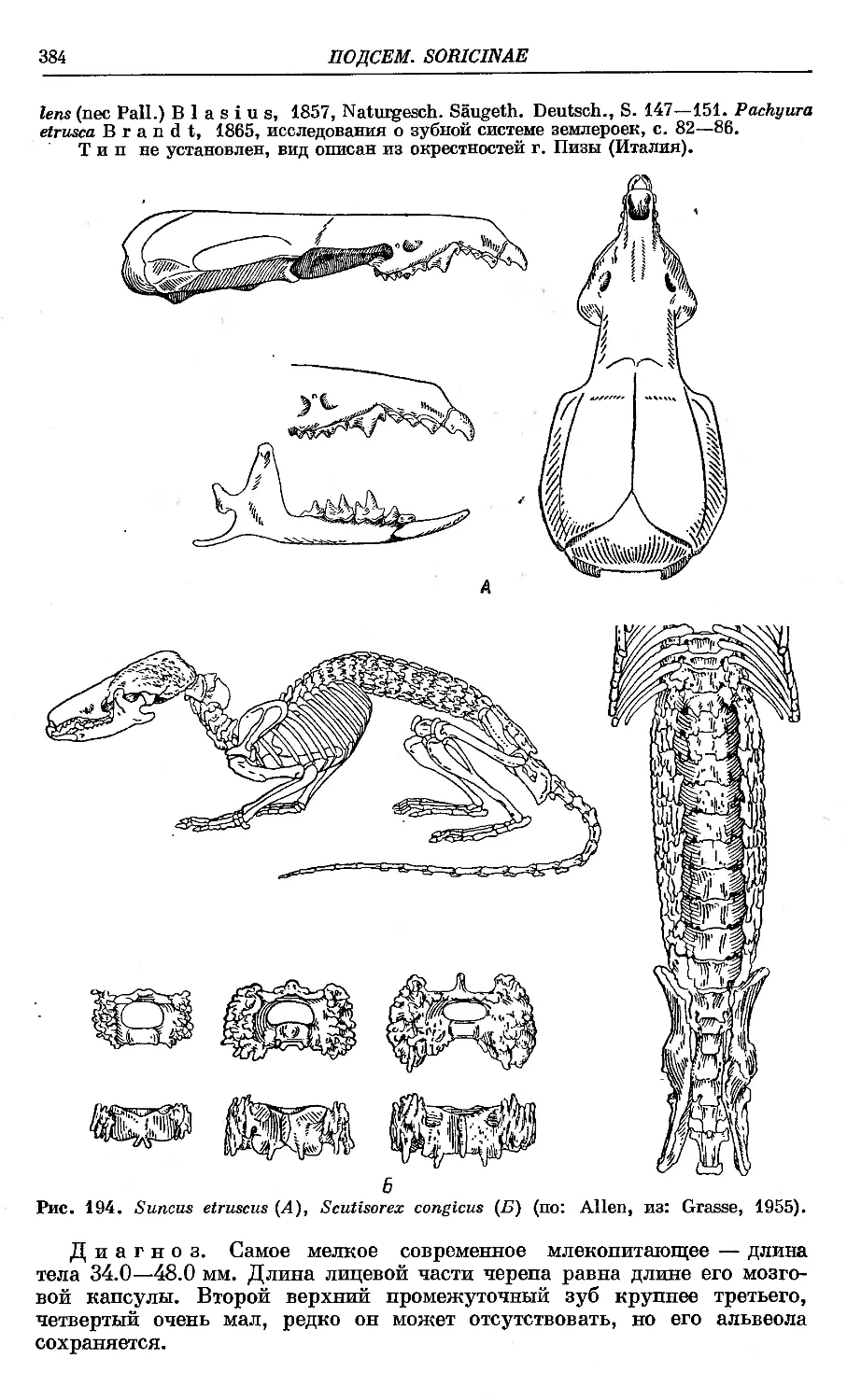
15. S. etruscus Savi...........' ........................... 383
27. Род Scutisorex Thomas
1. S. somerini Thomas................................................. . . • . . 385
2. S. congicus Thomas ......................................................... 386
f 28. Род Limnoecus Stirton
fl. L. tricuspis Stirton . . . ................................................... 386
29. Род Paracrocidura Heim de Balsac
1. P. schoutedeni Heim de Balsac •. ........................................... 387
30. Род Solisorex Thomas
1. S. pearsoni Thomas.......................................................... 388
31. Род Praesorex Thomas
1. P. goliath Thomas...................................................... . 388
32. Род Crocidura Wagler
fl. C. pavlodarica Stogov et Savinov............................................ 390
--2. C. cornfeldi Kormos........................................................ 391
•3. C. zorzii Pasa . . . ...................................................... 391
4. C. guuvevlens Pallas ............... . 7 . . ................................ 392
э. С. лугсцрс Goodwin . . . . .............................................. .t 393
6. C. pcr^ica Tnpmas ......................................................... 394
- Гоьtusa "Kretzoi ............................................................... 395
8. C. Jtucwfon Hermann......................................................... 395
C. russula Hermann . . . . ................................................ 396
IQ. C\ st&iricfl Unkebsky ........................................................ 397
11. g.l.toiura. РдЬзрр ......................................................... 398.
12. C. giildenstaedti Pallas ................................................... 399
IB. c. pamirensis Ognev........................................................ 400
'13. c. arrtienTca Gureev . .................................................... 401
, - 1!>. t'. iunnljfi JJjgnaSZ.....................................'................ 402
167 C. '.'iarudnyi Ognev........................................................ 403
17. C. calldata Miller ......................................................... 403
18. C. pergrisea Miller ........................................................ 403
19. C. attenuate Milne-Edwards.................................................. 403
20. C. dracula Thomas............................................................ 403
21. C. vorax G. Allen . . . .......................... . . .................... 404
22. C. rapax G. Allen....................................................... 404
23. C. dsinezumi Temminck....................................................... 404
24. C. horsfieldi Thomas................................................... 404
25. C. fuliginosa Blyth ................... ". . ....... Г....................... 404
26. C. hispida Thomas.................................... . ................. 404
27. C. monticola Peters ...................................................... 404
28. C. doriae Peters ................................................. 405
29. C. tenuis Muller .......................................................... 405
30. C. elongata Miller .......................................................... 405
31. C. nigripes Miller et Hollister............................................. 405
32. C. rhoditis Miller et Hollister ............................................ 405
33. C. lea Miller et Hollister................................................. 405
34. C. levicula Miller et Hollister ............................................ 405
35. C. giffardi ed Winton....................................................... 405
36. C. manni Peters ........................................................... 406
16 z" СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
37. С. odorata Leconte ........................................................ 406
38. С. biittikoferi Jentink.................................................... 406
39. С. anchietae Bocage . . . . ............................ . . ............. 406
40. C. hedenborgiana Sundevall................................................. 406
41. C. occidentalis Pucheran ................................................. 406
42. C. schweitzeri Peters ................................................... 406
43. C. martiensseni Neuman..................................................... 406
44. C. fuscosa Thomas ......................................................... 406
45. C. nyansae Neumann......................................................... 407
46. C. sururae Heller.......................................................... 407
47. C. doriana Dobson.......................................................... 407
48. C. flavescens Geoffroy..................................................... 407
49. C. zaphiri Dollman......................................................... 407
50. C. olivieri Lesson......................................................... 407
51. C. beirae Dollman........................................................ 407
52. C. hirta Peters............................................................ 407
53. C. sacralis Peters..................... . ............................ 407
54. C. mariguensis Smith ...................................................... 408
55. C. hindei Thomas ........................................................ 408
56. C. sericea Sundevall ..................................................... 408
57. C. beta Dollman.......................................................... 408
58. C. vetulina Thomas........................................................ 408
59. C. lutrella Heller ...................................................... 408
60. C. deserti Schwann......................................................... 408
61. C. butleri Thomas........................'................................. 408
62. C. percivali Doliman..................................................... 408
63. C. fischeri Pegenstecher . . . .......................................... 409
64. C. vol Osgood.......................................................... 409
65. C. suahelae Heller ........................................................ 409
66. C. smithi Thomas.......................................................... 409
67. C. turba Dollman .......................................................... 409
68. C. niloti^a Heller ....................................................... 409
69. C. zena Dollman ................................ 409
70. C. ansorgei Dollman ....................................................... 409
71. C. poensis Fraser..................................'....................... 409
72. C. batesi Dollman ......................................................... 409
73. C. foxi Dollman ........................................................... 410
74. C. arethusa Dollman ...................................................... 410
75. C. erica Dollman.....................................................' . . 410
76. C. boydi Dollman.......................................................... 410
77. C. nigricans Bocage....................................................... 410
78. C. nigrofusca Matschie ............................................. , . 410
79. C. luna Dollman........................................................... 410
80. C. ibeana Dollman........................................................ 410
81. C. argentata Sundevall.................................................... 410
82. C. cyanea Duvernoy ........................................................ 411
83. C. electa Dollman'......................................................... 411
84. C. martensi Dobson ........................................................ 411
85. C. sllacea Thomas ......................................................... 411
86. C. bovei Dobson............................................................ 411
87. C. capensoides Smith.................................................. . 411
88. C. pilosa Dobson.......................................................... 411
89. C. fumosa Thomas.......................................................... 411
90. C. raineyi Heller......................................................... 411
91. C. parvipes Osgood . .................................................. 412
92. C. sansibarica Neumann .................................................. 412
93. C. Xanthippe Osgood .......... . . . . . ................ 412
94. C. jacksoni Thomas....................................................... 412
95. C. macowi Dollman....................................... .,........... . 412
96. C. gracilipes Peters...................................................... 412
97. C. lutreola Heller .................................................... 412
98. C. maanjae Heller........................................................ 412
99. C. hildegardae Thomas .................................................. 412
100. C. planiceps Heller..................................................... 412
101. C. soinalica Thomas ...................................................... 413
102'. C. crossei Thomas .......................... у............................. 413
103. C. allex Osgood ........................................................ 4.13
104. C. alpina Heller ........................................................... 413
105. C. bicolor Bocage......................................................... 413
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ 17
106. С. Cinderella Thomas......................................................... 413
107. С. floveri Dollman.......................................................... 413
108. С. папа Dobson . . . . . ............................. 413
109. С. nanilla Thomas.......................................................... 413
110. С. pasha Dollman ........................................................... 413
111. C. glebula Dollman .......................................................... 414
112. C. religiosa Geoffroy...................................................... 414
113. C. lusitania Dollman............................................... • • . 414
114. C. dolichura Peters..................................................... . 414
115. C. maurisca Thomas ................................................... . 414
116. C. niobe Thomas............................................................. 414
117. C. bottegi Thomas...................................................... • . 414
118. С. топах Thomas ............................................................. 414
119. C. littoralis Heller....................................................... 414
120. C, ultima Dollman........................................................... 415
121. C. neavei Wroughton.......................................................... 415
122. C. silvia Thomas . .................................................. . 415
33. Род Diplomesodon Brandt
f 1. D. fossorius Repenning....................................................................... 415
2. D. pttZcfeeZZttm............................................................................ 417
5. Триба В L A R I N I M
1. Подтриба BLARININA
f 34. Род Adeloblarina Repenning
fl. A. berklandi Repenning......................................................... 421
f 35. Род Blarinoides Sulimski
f 1. B. mariae Sulimski............................................................ 422
36. Род Blarina Gray
1. B. gidleyi Gasin ........................................................ 423
• • 2. B. adamsi Hibbard ....................................................... 423
•3. B, simplicidens Cope....................................................... 424
4 . B. brevicauda Say . . .,................................................... 424
5 .' B. telmalestes Merriam............................................... . . 425
2. Подтриба BEREMENDINA
f 37. Род Beremendia Kormos
f 1. B. fissidens Peteneyi.......................................................... 426
f 38. Род Petenyia Kormos
1. P. hungarica Kormos........................................................... 427
2. P. concisa Wilson............................................................. 427 .
3. P. sp.................i...................................................... 428
f 39. Род Petenyiella Kretzoi
f 1. P. gracilis Petenyi........................................... 429
f 40. Род Shikamainosorex Hasegawa
f 1. S. densicingulata Hasegawa .. . . . . . ....................... 429
f 41. Род Peisorex Kowalski et Li-Chuan-Kuei
f 1. P. pohaiensis Kowalski et Li-Chuan-Kuei .....'.................. 430
f 1. P. sp........................................................... 430
f 42. Род Hesperosorex Hibbard
f 1. H. lovei Hibbard............................................ 431
f 43. Род Paracryptotis Hibbard
f 1. P. rex Hibbard.................................................. 431
2 А. А. Гуреев
J
18 СИСТЕМАТИЧЕСКИЕ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ
f 44. Род Planisorex Hibbard
fl. P. dixonensis Hibbard........................................................... 432
45. Род Cryptotis Pomel
fl. C. meadensis Hibbard........................................................... 433
f 2. C. kansasensis Hibbard ......................................................... 433
3. C. parva Say..........................i . .................................... 434
4. C. celatus Goodwin ...... ................................................... 434
5. C. pergracialis Elliot..................................................... 435
6. C. mericana Cones .................................................... 435
7. C. alticola Merriam ...................................................... 435
8. C. endersi Setzer...................... . . . ............................ 435
9. C. frontalis Miller........................................................ 435
10. C. griseoventris Jackson................................................... 435
11. C. guerrerensis Jackson ..................................................... 435
12. C. goodwini Jackson......................................................... 435
13. C. gracilis Miller .......................................................... 435
14. C. jacksoni Goodwin............................................................ 435
15. C. mayensis Merriam ................................................... 436
16. C. mera Goldman.......................................................... . . 436
17. C. nelsoni Merriam ........................................................... 436
18. C. obscura Merriam ............................................................. 436 '
19. C. olivacea J. Allen .......................................................... 436
20. C. zeteki Setzer............................................................. 436
21. C. fossor Merriam.............................................................. 436
22. C. magna Merriam ....................................................... . 436
23. C. mierura Tomes.............................................................. 436
24. C. soricina Merriam . . . ..................................................... 437
25. C. orophila J. Allen ....................................................... 437
26. C. nigrescens J. Allen........................................................ 437
27. C. tersus Goodwin ........................................................... 437
З.Подтриба SORICULINA
f 46. Род Zelceina Sulimski
f 1. Z. soriculoides Sulimski .....................................'........... 438
f 47. Род Macroneomys Fejfar
f 1. M. brachygnathus Fejfar............................................... 439
48. Род Neomys Каир
f . . ..................................... -. . 440
<2. IV. Mtm Bennant / ....................................................... 440
a, A. schelkoynikoyi Satunin ............................................. 442
, 4. JV. anomalus Cabrera ............................................... 443
49. Род Soriculus Blyth
f 1. 5. kubinyi Kormos........................................................... 445
2. 5. nigrescens Gray . . . .................................................. 446
3. S. caudatus Hodgson......................................................... 446
4. S. leucops Horsfield................................. . . . 447
50. Род Notiosorex Baird
1. Подрод Notiosorex Baird
f 1. N. jacksoni Hibbard .......................................... 448
2. N. crawfordi Baird ........................... . . ................. 449
3. N. gigas Merriam....................»....,.............................. 449
2. Подрод Chodsigoa Kastschenko
4. N. hybsibius de Winton et Styan........................................ 450
5. N. salenskii Kastschenko ............................................... 451
6. N. lowei Osgood............. . . ....................................... 451
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ НАСЕКОМОЯДНЫХ 19
51. Род Podihik Deramyagala
1. Р. kura Deraniyagala................................................. 452
f 4. Подтриба ALLOSORICINA
f 52. Род Allosorex Fejfar
f 1. A. stenodus Fejfar ................................................. 454
f 53. Род Deinsdorfia Heller
f 1. D. franconica Heller................................................ 455
5. Подтриба NECTOGALINA
54. Род Chimarrogale Anderson
1. Ch. himalayicus Gray................................................. 456
2. Ch. styani de Winton................................................ 457
3. Ch. leander Thomas.................................................. 457
4. Ch. platy cephala Temmink....................................., . . . 458
5. Ch. sumatrana Thomas ............................................... 458
6. Ch. phacura Thomas .....'........................................... 458
7. Ch. hantu Harrison ................................................ 458
8. Ch. varennei Thomas ............................................. . 458
f 55. Род Neetogalinia Gureev gen. nov.
f 1. N~ altaica Gureev sp. nov............................................. 458
56. Род Nectogale Milne-Edwards
1. N. elegans Milne-Edwards ............................................ 460
6. Триба ANOUROSORICINI * •
f 57. Род Anourosoricodon Topatshevsky
f 1. A. pidoplitschkoi Topatshevsky................................ . . . 461
f 58. Род Amblyeoptus Kormos
f 1. A. oligodon Kormos...........'....................................... 462
f 2. A. topali Janossi . ................................................ 463
f 59. Род Paranourosorex Rzebik-Kowalska
fl. P. gigas Rzebik-Kowalska............................................. 464
60. Род Anourosorex Milne-Edwards
f 1, 2. A. inexpectatus Schlosser . . .................................... 465
f 3. A. japonicus Shikama et Hasegawa..................................... 466
»4. A. squamipes Milne-Edwards........................................... 466
2*
Отряд INSECTIVORA
Диагноз. От очень мелких до средних размеров (длина тела 50—
450 мм) животные, с мордой, вытянутой в относительно тонкий, подвиж-
ный хоботок. Резцы 1Х^2 всегда сильно развиты и служат основным ор-
ганом захвата пищи.
Характеристика. Размеры тела небольшие (50—450 мм). В пределах
отряда и самые мелкие из современных млекопитающих — землеройки
Suncus etruscus Savi и Sorex minutissimus Zimm., длина тела которых не пре-
вышает 50—53 мм.
Внешний облик насекомоядных разнообразен, что обусловлено раз-
ными формами адаптаций. Нос вытянут в виде хоботка, который укреп-
ляется у носового отверстия с помощью хрящевой трубки, перегороженной
вдоль хрящевой пластинкой, что соответствует двум ноздревым каналам.
У кротов часть хрящевой трубки окостеневает или окостеневает ее пере-
городка. У щелезуба хрящевая трубка носа поддерживается костной
пластинкой, развивающейся у нижнего края носового отверстия. Хоботок
приводится в движение носо-губным мускулом, соединенным с носом
сухожильными тяжами.
Ушные раковины хорошо развиты или рудиментарны. Глаза крупные,
мелкие, малозаметные или закрыты сросшимися веками. Хвост хорошо
развит или очень короткий, реже снаружи незаметен. Конечности пре-
имущественно стопоходящие, редко пальцеходящие, пятипалые, у Огу-
zorictes tetradactylus M.-Edw. et Grand, четырехпалые.
Волосяной покров насекомоядных весьма разнообразен, что обуслов-
лено многообразием его функций у отдельных групп и видов. Волосы,
покрывающие отдельные части тела, не одинаковы. Например, у боль-
шинства насекомоядных конец морды, йисти и ступни, а также хвост
покрыты более жесткими или щетинистыми волосами. Все разнообразие
покровов насекомоядных можно отнести к трем основным типам: 1) во-
лосяной покров мягкий, бархатистый, образован короткими тонкими
волосами и густым подшерстком, создающими относительно однородный
слой; 2) волосы, покрывающие тело, жесткие, щетинистые, в разной сте-
пени длинные, с мягким подшерстком; 3) верхнюю сторону тела и бока
покрывают короткие иглы, а переднюю часть головы, ее боковые стороны
и все нижние части тела покрывают жесткие или мягкйё волосы.
Каждый из этих типов волосяного покрова характерен для определен-
ных групп насекомоядных и обусловлен влиянием среды, в которой на-
ходится тот или иной вид. Так, все полуводные и роющие формы имеют
мягкий бархатистый волосяной покров, который у полуводных видов
не смачивается водой, а у роющих не загрязняется почвой. Обитатели
зарослей кустарников и каменистых россыпей имеют жесткий волосяной
покров, а виды, населяющие открытые пространства, покрыты иглами
или имеют мягкий волосяной покров, что в каждом отдельном случае
объясняется особенностями их образа жизни.
ХАРАКТЕРИСТИКА ОТРЯДА 21
Как уже отмечалось, конец морды насекомоядных покрыт жесткими
волосами, расположенными правильными рядами. В каждом ряду перед-
ние волосы короче задних. Эти волосы несомненно служат защитой
кожи от повреждений, которые могут нанести животные, схватываемые
челюстями, а также от механических повреждений во время рытья, пе-
редвижения в мертвой подстилке леса и зарослей кустарников. Между
упомянутыми волосами располагаются более толстые и длинные — виб-
риссы. По-видимому, защитную роль выполняют и жесткие волоски, рас-
положенные кольцом вокруг глаз, например у землероек.
Кисти и ступни насекомоядных сверху покрыты чешуйками и жесткими
волосами. У плавающих видов боковые стороны кистей и ступней, а также
пальцев покрыты плоскими длинными волосками в виде бахромы. У рою-
щих форм боковые стороны кистей усажены жесткими щетинками. Ниж-
ние поверхности кистей и ступней покрыты щитками и мозолистыми
подушечками. У ряда видов щитки, покрывающие поверхности пальцев,
напоминают щитки на ногах птиц.
Покровы хвоста весьма различны. У мелких видов хвост покрыт
мелкими чешуйками, располагающимися в виде колец, между которыми
помещаются короткие жесткие волосы, густо покрывающие поверхность
хвоста. У многих видов к коротким волоскам подмешиваются редкие
длинные щетинки, а у ряда видов длинные щетинки на нижней стороне
хвоста образуют киль. Хвост относительно крупных наземных видов
покрыт более крупными чешуйками, расположенными в виде колец или
прирастающими плотно к коже, между которыми располагаются сравни-
тельно редкие разной длины жесткие волосы. У специализированных полу-
водных форм хвост покрыт крупными чешуйками, прирастающими к коже,
и редкими или многочисленными жесткими волосками.
Следует упомянуть и о коротких волосках, расположенных на особых
бугорках кожи внутренней поверхности ушной раковины, значение кото-
рых недостаточно ясно.
Когти у всех насекомоядных хорошо развиты. Кроме защиты когтевых
фланг от повреждений, они у этих млекопитающих несут самые разно-
образные функции. У многих видов они служат для рытья и приобретают
крупные размеры и форму, соответствующую способу, которым произво-
дится эта работа. Лазающие виды имеют изогнутые и острые когти. Они
крупные и также изогнутые у видов, раздирающих свою добычу когтями.
Кожные железы у насекомоядных располагаются на боках тела и у ос-
нования хвоста. Они выделяют мускусный экскрет, который служит одним
из способов пассивной защиты, а также, видимо, и средством нахожде-
ния друг друга в период размножения, что особенно важно в годы пони-
женной численности особей в популяциях отдельных видов.
Окраска волосяного покрова не отличается яркостью, что имеет су-
щественное значение, поскольку большинство насекомоядных более
активно ночью. Контрастное сочетание темных и светлых тонов, даже
тусклых, и при' слабом освещении делает ее заметной. Основными окрас-
ками насекомоядных бывают серая, коричневая и бурая разных оттенков.
Их разнообразное сочетание с белым й черным цветами или собственными
светлыми оттенками придает общему тону известную пестроту. Разную
тональность и пестроту придают ей и разноцветность самих волос, а также
разная интенсивность окраски отдельных частей тела.
Череп у многих видов с вытянутым лицевым отделом и часто с узкой
и небольшой мозговой капсулой. Глазница незамкнутая (исключение
сем. Tupaiidae). У ряда групп насекомоядных отсутствует скуловая дуга.
Засочленовный отросток у насекомоядных выполняет разнообразную
функцию. У ежей, обитающих на открытых пространствах, он широкий,
22 ОТРЯД INSECTIVORA
с глубокой выемкой, соединяющейся со слуховой полостью. Этот отро-
сток у отдельных подродов ежей развит в разной степени также и у гим-
нур. У землероек он в виде пластинки выдается вперед и служит опорой
для нижней поверхности сочленовного мыщелка нижней челюсти, как
и у хищных.
. Барабанная кость у многих видов не образует слуховых пузырей
и представлена в виде кольца. У ряда трупп насекомоядных кости, окру-
жающие область уха, а также крыловидные кости и крыловидные от-
ростки основной клиновидной кости, вздуты, полые или имеют ячеистое
строение. У тенреков и ежей рода Erinaceus на основной клиновидной
кости, позади хоан, глубокая впадина, которая, по-видимому, служит
резервуаром для осаждения пыли из вдыхаемого воздуха.
Нижняя челюсть различной формы, с сильно развитым у большинства
венечным отростком, у Многих с длинным угловым отростком. Мыщелок
сочленовного отростка более развит в поперечном направлении и имеет
форму валика. У землероек нижцяя челюсть сочленяется с черепом двумя
суставами.
Зубная формула: 7 3^2 С ^ 4—1 М |—2 — —26. Зубы насекомо-
ядных, особенно более примитивных групп, в своей структуре еще со-
храняют многие черты строения 3-бугорчатых зубов, например, премо-
ляры. Но и в строении более сложных 4-бугорчатых коренных (моляров)
сохраняется 3-бугорчатая основа. Если рассматривать верхние премоляры
от клыка в сторону моляров у представителей отдельных групп насекомо-
ядных, имеющих наибольшее количество зубов, то можно заметить', как
постепенно усложняются структуры этих зубов и за счет каких элементов
увеличивается их жевательная поверхность при моляризации, например
у Р4. Следует отметить, что премоляры у разных групп насекомоядных
неодйнакового строения, что отражает разную степень специализации.
Так, встречаются премоляры, сохраняющие три хорошо развитые вер-
шины, два корня и крупные размеры, а также и с сильно упрощенной
структурой — мелкие, с одной вершиной и одним корнем. Только Р4
у всех насекомоядных остается крупным, а у отдельных групп он сильно
моляризован. Разная степень моляризации Р4 у ряда групп насекомоядных,
по-разному приспособленных к пережевыванию пищи, отражает ряд
переходов от простых 3-бугорчатых зубов к специализированным моля-
рам и особенности их формирования.
Анализ структурных особенностей премоляров и моляров у разных
насекомоядных позволяет представить себе картину формирования 4-
бугорчатых зубов, с W-образной жевательной поверхностью, из 3-бугор-
чатых — с V-образной. Премоляры Plj—Р3 у многих насекомоядных опи-
раются на два корня — минимальную, но достаточную опору при их про-
дольном расположении в челюсти. Корень протокона у них подвергается
редукции. Реже Р3 сохраняет протокон и его корень. При развитии верх-
них зубов в поперечном направлении при их моляризации происходит
перемещение протокона на внутреннюю сторону челюсти; его корень
сохраняется. Таким путем увеличивается жевательная поверхность зуба.
Увеличение площади жевательной поверхности на внутренней части зуба
происходит и за счет развития дополнительного бугорка — талона.
Зачаток такого бугорка хорошо заметен на верхних молярах крупных
тенреков Tenrec ecaudatus Schreb. (рис. 1). Он расположен у основания
протокона. При образовании талона удлиняется корень протокона вдоль
внутреннего края коронки зуба. Значительное развитие талона сопро-
вождается обособлением его корня. Зуб становится 4-бугорчатым с 4 кор-
нями. W-образная эмалевая структура на его жевательной поверхности
Рис. 1. Верхние зубы тенрека Tenrec ecaudatus.
Рис. 2. Верхние зубы щелезуба Solenodon paradoxus с наружной стороны (а), с же-
вательной поверхности ^б) и внутренней стороны (в).
Рис. 3. Нижние зубы щелезуба.
Обозначения как на рис. 2.
24
ОТРЯД INSECTIVORA
есть результат углубления наружных сурепок зуба в области бугров пара-
кона и метакона в виде треугольных складок, с гребнями на боковых сто-
ронах. Совокупность этих гребней и создает W-образную фигуру. Сходно
образовались W-образные складки и на жевательной поверхности ниж*,
них моляров, но в отличие от верхних складки у них входили не с наруж-
ной, а с внутренней стороны между параконидом и протоконидом и между
протоконидом и метаконидом. Направление их гребней коррелировалось
расположением гребней на жевательной поверхности верхних моляров,
что и обусловило их наиболее выгодное в функциональном отношении
направление. Этим объясняется утрата нижними молярами четких границ
их основных бугров.
Нижние моляры, не развивавшиеся в ширину, не приобрели 4-уголь-
ной формы за счет смещения протоконида на внешнюю сторону и образо-
вания талонида. Протоконид у них утратил самостоятельность, а его
корень редуцировался. Третий верхний моляр в процессе эволюции
зубной системы насекомоядных подвергся значительной редукции и сильно
изменил свою форму. У отдельных вымерших групп он не развивался,
как и нижний.
В характеристике отряда Insectivora постоянно отмечается сходство
по строению резцов, клыков и передних премоляров. У землероек все
передние зубы, кроме первого резца (Р) и четвертого премоляра (Р4),
сходные по строению и имеющие коническую форму, называют «Промежу-
точными» зубами.
Верхние зубы. Прежде всего следует отметить, что резцы
у большинства видов насекомоядных расположены на боковой стороне
межчелюстной кости и первые резцы удалены друг от друга. Реже круп-
ные первые резцы сближены внутренними сторонами и замыкают зубные
ряды спереди, что наблюдается у многих видов кротов (Talpidae), а у мно-
гих представителей трибы Talpini все резцы на передней стороне меж-
челюстной кости.1
Резцы (I1), расположенные на передней стороне межчелюстной кости,
чаще всего имеют форму долота (широкие и плоские спереди и сзади)
или 3-гранной призмы, например у выхухоли. Первые резцы, стоящие
на боковой стороне межчелюстной кости, удаленные друг от друга, дости-
гают часто значительных размеров, саблевидной формы или расширенные
спереди-назад, с одним зубцом позади главной вершины или с несколь-
кими зубцами. У землероек эти резцы с двумя корнями. I2- 3 варьируют
в размерах и по форме. У некоторых групп Is бывает с двумя корнями.
Как уже отмечалось, отдельные виды насекомоядных имеют крупные
клыки, не выступающие в стороны, как это обычно для хищных (Carni-
vora). Клыки с одним или двумя корнями. У большинства видов клык
имеет форму, сходную с резцом или передними премолярами. Передние
премоляры удлиненной формы с двумя корнями и тремя вершинами, рас-
положенными в одну линию. Реже эти зубы имеют широкоовальную
форму, а бугорки паракона и метакона смещены на наружную или внут-
реннюю сторону или отсутствуют. Это зубы с одним-двумя корнями.
У многих видов Р1, Р2, Р3 сходной формы и размеров, конические, сходные
с клыком и третьим резцом. Изредка Р3 по строению сходен с Р4, но мень-
ших размеров. Р4 всегда крупный, с тремя корнями и тремя-четырьмя
1 Можно не сомневаться, что захват пищи первыми крупными резцами, сближен-
ными на передней стороне межчел!остной кости, с переходом отдельных групп на-
секомоядных на смешанную пищу (беспозвоночные, семена растений) привел к обра-
зованию грызущего аппарата, а вместе с этим и к развитию отрядов зайцеобразных
и грызунов.
ХАРАКТЕРИСТИКА ОТРЯДА
25
вершинами (бугорками), в разной степени моляризован, чаще сохраняет
форму и функцию хищнического зуба, как у Carnivora.
, Коренные зубы двух типов: с V- или W-образными гребнями, ограни-
чивающими складки на жевательной поверхности, входящие снаружи.
V-образные верхние коренные, как показывает изучение зубов древних
групп млекопитающих и современных примитивных насекомоядных,
могут с полным основанием считаться исходными для развития коренных
зубов всех плацентарных млекопитающих, а тенрекообразные насекомо-
ядные сохраняют их до сих пор. Эти зубы у многих видов и групп при-
обретали (в прошлом) на внешнем крае зубчатую оторочку, что хорошо
заметно и у ряда современных видов насекомоядных, особенно у Sole-
nodon Brandt и других тенрекообразных.
Последний коренной (М3) часто расположен в поперечном к продоль-
ной оси челюсти направлении и служит, видимо, ограничителем движения
нижней челюсти назад, задерживая М3. Возможно также, что он задержи-
вает и направляет пищу в задний отдел ротовой полости.
Нижние зубы. Первый резец наиболее крупный, реже крупный
второй, а первый и третий мелкие. У тупай все резцы крупные, у многих
кротов все резцы мелкие. При всем этом нижние резцы принимают участие
в захвате пищи. У большинства насекомоядных нижний клык мелкий,
сходный с соседними зубами, реже крупный, как у хищных, и при сомкну-
тых челюстях входит в специальную выемку на межчелюстной кости
впереди верхнего клыка. У кротов трибы Talpini мелний клык стоит
рядом с третьим резцом и имеет сходную с ним форму. У этих кротов функ-
цию нижнего клыка выполняет Р1Т который при сомкнутых челюстях по-
мещается позади верхнего клыка. У кротов рода Mogera нижний клык
отсутствует. Р15 Р2 и Ра чаще сходной формы, с двумя или одним корнем,
с тремя, двумя или одной вершиной. Р4 всегда крупный, с двумя корнями
и почти всегда трехвершинный. Реже он значительно моляризован.
Моляры почти у всех насекомоядных имеют сходное строение и разли-
чаются частными особенностями у отдельных групп. Эти зубы состоят
из двух сегментов, сходных по форме, образующих в совокупности W-об-
разную фигуру. У отдельных групп передний сегмент имеет форму буквы
V, а задний четырехугольный или округлый. Часто М3 имеет отличное
от описанного строение, значительно меньше по размерам, чем М4 и М2,
или очень мелкий, с округлой коронкой.
Разная высота переднего и заднего сегментов нижних коренных, что
наблюдается у современных тенрекообразных и имело место у всех мело-
вых плацентарных млекопитающих, послужила основанием для выделе-
ния переднего сегмента в собственно зуб — его основную часть (тригонид),
а задний сегмент стали считать придатком — талонидом. В действитель-
ности, передний сегмент представляет собой в разной степени обособлен-
ные или слитые параконид и протоконид, а задний сегмент — метаконид,
разросшийся в связи с особенностями расположения верхних и нижних
зубов-антагонистов и эмалевых структур на их жевательной поверхности
при сомкнутых челюстях. Этим же объясняется и разная высота сегментов
у отдельных трупп насекомоядных и их предков. Достаточно посмотреть
на соотношение верхних и нижних зубов при сомкнутых челюстях
у Solenodon или Тепгес, чтобы убедиться в сказанном. Нижние коренные
зубы видов сем. Мacroscelididae представляют пример слияния переднего
и заднего сегментов с образованием общей жевательной поверхности,
на которой отдельные кониды слабо обособлены.
Можно предполагать; что у насекомоядных относительно рано опре-
делились два основных направления в развитии жевательного аппарата:
приспособление к захвату сравнительно малоактивной добычи и пере-
26 -ОТРЯД INSECTIVORA
жевыванию грубой пищи, в том числе и растительной, и приспособление
к быстрому схватыванию очень подвижных животных. Второй тип жева-
тельного аппарата приобрел в пределах отряда ведущее значение. Его
формирование сопровождалось утратой или ослаблением скуловой дуги,
а в связи с этим изменением места крепления верхней части Массетера,
сокращением размеров угловой лопасти нижней челюсти и удлинением ее
углового отростка. Всё это привело к ускорению движения нижней че-
люсти вперед и вверх при закрывании рта мощным височным мускулом
и в разной степени мощным, косо поставленным массетеро^.
Органом захвата пищи у подавляющего большинства насекомоядных
служат резцы 1} или I*. Исключение составляют тенрек (Тепгес есаи-
Рис. 4. Черепа Solenodon paradoxus (A), Tenrec ecaudatus (Б) (по: Dobson, 1883).
datus Schreb.), гимнура (Echinosorex gymnurus Raffles) и кроты трибы
Тalp ini.
Структурные особенности и функции жевательного аппарата насекомо-
ядных отражают приспособления к элементарной обработке легко пере-
вариваемой пищи (беспозвоночные, главным образом насекомые, а также
мелкие позвоночные, чаще используемые в пищу более крупными формами).
Разного рода модификации в его строении определяются частными особен-
ностями пищи отдельных групп, размерами животных потребителей и
потребляемых в пищу, а также степенью подвижности tex и других.
Можно думать, что исходным типом челюстного аппарата насекомо-
ядных был такой, верхняя и нижняя челюсти которого имели по 22 зуба,
достаточно отчетливо дифференцированных на резцы, клыки, премоляры
и моляры. Последние, в верхней челюсти, были с тремя бугорками, рас-
положенными в форме треугольника,. а в нижней — последовательно.
Верхние моляры опирались на три йорня, а нижние — на два, поскольку
корень протокона подвергся редукции. Верхние и нижние челюсти были
удлинены. Угловые лопасти нижней челюсти не были расширены в передне-
заднем направлении и имели короткий угловой отросток. Венечный
отросток не был сильно развитым. Скуловые дуги имелись. Все жеватель-
ные мышцы были одинаково хорошо развитыми. Примерно такого типа
жевательный аппарат сохранился у примитивных кротов, но только
зубы у них более специализированные.
Потребность в быстром схватывании подвижной добычи передними
концами челюстей обусловила утрату скуловой дуги, что связано, видимо,
с необходимостью преодоления инерции массетера. Переход на питание
относительно крупными животными сопровождался у форм, утративших
скуловую дугу, увеличением размеров угловой лопасти нижней челюсти
и венечного отростка, а также увеличением массы массетера и височного
мускула. Виды, питавшиеся разной по механическому составу пищей,
ПОДОТРЯД TENRECII
27
сохранили скуловую дугу, количество премоляров у них уменьшилось,
челюсти сократились в длину, а жевательная функция усилилась.
По общим структурным особенностям все жевательные аппараты на-
секомоядных могут быть отнесены к следующим двум основным типам и
их подтипам.
I тип. Зубы трехбугорчатые (реже Р4—М3 с дополнительным бугорком-
талоном), с V-образной формой жевательной поверхности. Нижние корен-
ные зубы трехбугорчатые, протоконид двухвершинный. Угловая лопасть
нижней челюсти и венечный отросток сильно развиты. Скуловых дуг нет
(исключение Chrysochloridae) (рис. 2—4).
Подтип А. Скуловой дуги нет. Первые (/}) или вторые (/|) резцы сильно
развиты и напоминают клыки или все резцы мелкие, а клыки сильно раз-
Рис. 5. Череп Chrysochloris trivelyani
(по: Dobson, 1883).
Рис. 6. Череп Erinaceus (Hemiechinus)
dauricus.
виты в верхней и нижней челюстях. Челюсти удлинены. Массетер и височ-
ный мускулы сильно развиты. Жевательный аппарат приспособлен к бы-
строму и сильному схватыванию добычи, а также к жеванию (сем. Теп-
recidae; Pot.amogalidae, Solenodontidae).
Подтип Б. Скуловая дуга сильно развита. Первые резцы в верхней
и вторые в нижней челюсти сильно развиты. Челюсти укорочены. Массетер
и височный мускулы очень сильно развиты. Жевательный аппарат при-
способлен главным образом к обработке пищи на коренных зубах (сем.
Chrysochloridae) (рис. 5).
II тип. Коренные зубы 4-бугорчатые, с W-образной жевательной по-
верхностью, часто с дополнительными бугорками. Нижняя челюсть с угло-
вой лопастью, венечным и угловым отростками, развитыми в разной
степени. Скуловые дуги имеются, за исключением Soricidae (рис. 6—8,
10).
Подтип А. Скуловая дуга имеется. Первые резцы (/]) или верхние пер-
вые резцы и нижние клыки сильно развиты. Челюсти удлинены или укоро-
чены. Массетер и височный мускулы сильно развиты или височный более
развит, чем массетер. Преобладает функция жевания (сем. Erinaceidae,
Talpidae, Tupaiidae, Мacroscelididae).
Подтип Б. Скуловая дуга отсутствует. Только первые резцы 1\ сильно
развиты. Челюсти удлинены. Развит главным образом височный мускул.
Преобладает функция схватывания и дробления добычи (сем. Soricidae)
(рис. 9, 10).
Посткраниальный скелет насекомоядных имеет разные структурные .
особенности, что обусловлено разным характером адаптаций. Общим
для всех насекомоядных следует Считать наличие ключицы (исключение —
! Рис. 7. Зубы Erinaceus europaeus верхние (Л), нижние (Б), с жевательной и’наружной
(а, б) сторон.
Рис. 8. Зубы Echinosorex gymnurus.
ХАРАКТЕРИСТИКА ОТРЯДА
29
представители сем. Potamogalidae) и сочленение лучевой кости с плечевой
шарнирным суставом, обеспечивающее пронацию и супинацию.. У ряда
групп малая берцовая кость не срастается с большой берцовой (свободна).
Движение насекомоядных разнообразно. Среди них есть бегающие,
прыгающие, лазающие, роющие и плавающие (рис. 11—16). Бег по поверх-
ности почвы осуществляется у насекомоядных, как и у грызунов, двумя
разными формами движений: перемещение с малой амплитудой волны
движения, что обусловлено невысокой траекторией прыжков (рысь,
тяжелый галоп), характерное для стопоходящих, коротконогих животных
с массивным телом (большинство насекомоядных), и перемещение с боль-
шой амплитудой волны движе-
ния при высокой траектории
прыжков (рикошетирующий га-
лоп), свойственное животным,
передвигающимся только на
двух ногах, без опоры на перед-
ние (одно сем. Macrosceltdidae,
в котором структурные особен-
ности конечностей напоминают
конечности тушканчиков сем.
Dipodidae).
Следует отметить, что у ряда
видов передвижение, например
галопом, сочетается с необходи-
мостью рыть, что накладывает
известный отпечаток на струк-
турные особенности кисти и
всей передней конечности и не
Рис. 9. Череп землеройки Myosorex
babaulti (по: Heim de Balsac, 1956).
может не оказывать влияния на характер бега. Некоторое разнообра-
зие в биомеханике движений близких видов в пределах одного типа движе-
ний обусловлено их разной степенью приспособленности к другим формам
движений (рытье, плаванье и пр.).
Приспособленйе к рытью в отдельных группах насекомоядных стоит
на разном уровне специализации. Наряду с высокоспециализированными
землероями, какими являются кроты (Talpidae) и златокроты (Chrysoch-
loridae), существуют виды, принадлежащие разным семействам, приспособ-
ления которых к рытью находятся на разной стадии совершенства в раз-
витии этой формы движения. Такие виды есть среди кротов, землероек
и др. У одних из этих видов приспособление к рытью выражается в увели-
чении длины когтей и слабом расширении кисти, у других — в укорочении
костей передних конечностей, их сильном расширении и образовании
на них сильных гребней и бугров для прикрепления мышц. Для всех
этих видов рытье не является средством передвижения в почве для добыва-
ния пищи, как у специализированных землероев, а служит для приготовле-
ния убежищ, нор и гнездовых камер для выведения потомства или пере-
живания неблагоприятных условий. Степень сложности нор, особенности
грунта и частота рытья у менее специализированных землероев опреде-
ляют уровень развития локомоторных функций в этом направлении.
При значительной специализации способ рытья у кротов и злато-
кротов различный. Кроты роют тольцо широкой кистью с сильно раз-
30
ОТРЯД INSECTIVORA
витыми пятью пальцами, с широкими плоскими когтями. Увеличению
площади ладони служит и серповидная косточка, характерная для кротов.
Златокроты роют крупными вторым и третьим пальцами с сильно раз-
росшимся когтем. Остальные пальцы мелкие и в рытье участия почти не
принимают. Златокроты роются близко от поверхности почвы. Подрыв
немного почву, златокрот просовывает голову в образовавшееся отверстие,
раздвигает комки почвы носом, покрытым толстой кожей, и поднимает
головой свод хода так, что на поверхности почвы образуется валик, по
которому можно судить о присутствии зверька и направление его движения
в почве. В связи с большой силовой нагрузкой на голову затылочная часть
черепа этих насекомоядных сильно развита. К широкому затылку при-
крепляется мощная мускулатура, поднимающая голову.
Рис. 10. Зубы землероек рода Sorex.
А верхние, Б — пюкние; те — метакон, ms — мезостиль, mts — метастиль, ра — паракон,
ps — парастиль, hy — гипокон, рг — протокон, end — энтоконид, pad — параконид, hyd — гипо-
конид, prd — протоконид, med — метаконид (по: Строганов, 1957 — Г); 1 — паракон, 2 —• метакон,
г — протокон, 4 — талон, 1а — параконид, 2а — метаконид, За — протоконид (по: Гуреев, 1971 — II).
Приспособление к плаванью, как и к рытью, находится не на одинако-
вом уровне специализации в разных группах насекомоядных. Наиболее
высокая специализация в этом направлении выражается в приобретении
несмачивающегося в воде волосяного покрова, в редукции ушных рако-
вин, укорочении трубчатых костей и в увеличении поверхностей кистей
и ступней с образованием перепонок между пальцами. Моторными орга-
нами становятся задние ноги, передние служат для подъема и погружения,
а хвост — рулевым управлением. Описанные здесь особенности строения
частей тела характерны для представителей подсемейства выхухолей
(Desmaninae). Выдровая кутора (сем. Potamogalidae) плавает с помощью
хвоста, так как у нее нет плавательных перепонок между пальцами ног.
Подъем и опускание в воде осуществляется передними ногами, двигаю-
щимися в одной плоскости, в связи с чем утрачена ключица. По-видимому,
рулевым приспособлением у нее служат задние ноги.
Виды, приспособленные к плаванью, встречаются среди тенреков,
а также и в семействе землероек (Soricidae), отдельные представители
которых достигли в этом отношении значительной специализации.
ХАРАКТЕРИСТИКА ОТРЯДА
31
Приспособление к лазанью не получило широкого распространения
среди насекомоядных. Единственное семейство, все виды которого лазают —
тупайи (Tupaiidae). Они не достигли такой специализации в лазаньи,
Рис. 11, 12. Скелеты бегающих насекомоядных Tenrec ecaudatus (11) (по: Blainville,
1839—4864) и Erinaceusaurttus (12).
как белки или приматы, ведут полудревесный, полуназемный образ жизни
и по общему характеру приспособлений напоминают бурундуков (Tamias).
Тупайи быстро передвигаются по земле прыжками и хорошо лазают,
спускаясь с деревьев вниз головой. Приспособление к лазанью в скелете
этих насекомоядных выражается в следующих особенностях. Поясничный
и тазовый отдел позвоночника сильно развит. Шея короткая. Остистые
отростки грудных позвонков высокие. Грудная клетка короткая и широ-
кая. Лопатка широкая и короткая, с сильно развитой остью. Головка
Рис. 13, 14. Скелет прыгающего Elephantulus rozeti (13) и лазающего насекомоядного Tupaia ferruginea (14) по: Blain-
ville, 1839-1864).
Рис. 15, 16. Скелет роющего Mogeta robusta (15) (по: Строганов, 1948) и.плавающего насекомоядного Galemys pyrenaicus (16)
(по: Blainville, 1839—1864).
34
ОТРЯД INSECTIVORA
длинной плечевой кости округлая. Локтевая и лучевая кости сходной
толщины, длинные. Кисть длинная, эластичная. Таз широкий, но не-
высокий, с широкими подвздошными костями. Длинное бедро с округлой
сочленовной головкой и хорошо развитыми вертелами. Голень длинная,
большая и малая берцовые кости не срастаются (свободные). Ступня
длинная. Когти на пальцах передних и задних конечностей острые, изо-
гнутые; хвост длинный.
Следует отметить, что большинство семейств насекомоядных объединяет
виды, приспособленные к одной какой-нибудь форме движения (прыжкам,
плаванью или лазанью) и достигают значительной специализации. Только-
в составе трех семейств имеются виды, приспособленные к разнообразным
движениям: Tenrecidae, Talpidae и Soricidae. Из них первое представляет
собой древнюю группу насекомоядных с ограниченным ареало'м (Мада-
гаскар и прилежащие острова), а два других наиболее специализирован-
ные, широко распространенные.
В заключение следует отметить еще некоторые особенности насекомо-
ядных, отражающие и их сравнительно низкий уровень развития. Полу-
шария головного мозга у них небольшие, с гладкой поверхностью, матка
двурогая, кость полового члена отсутствует. У отдельных групп насекомо-
ядных, в частности у тенрекообразных, анальное и половое отверстия
открываются в углублении, напоминающем клоаку однопроходных.
Обзор своеобразных вымерших семейств насекомоядных и их родов
позволяет обнаружить значительное разнообразие форм частных адапта-
ций в пределах этого отряда. В отличие от однопроходных и сумчатых,,
сохранивших многие архаичные особенности, плацентарные уже на ранних
этапах своего развития получили большие возможности для повышения
активности. Они шире расселялись по земному шару, занимая ландшафты
в различных климатических зонах. Это способствовало интенсивному
видообразованию в различных направлениях адаптации и формированию
многих специализированных групп. Интенсивное формообразование этих
примитивных плацентарных, начавшееся, вероятно, уже в середине мело-
вого времени, продолжалось до конца палеоцена, после чего, судя по-
сохранившимся остаткам, начало затухать, переходя к формированию
более устойчивых к колеблющимся условиям видов. В это время формиро-
вались предки устойчивых к условиям умеренной зоны групп насекомо-
ядных — ежей, кротов и землероек. Каждая из них по-своему приспосо-
билась к возникавшим условиям в пределах занимаемой ими территории.
Потомки групп, формировавшихся в благоприятных условиях, сохрани-
лись до настоящего времени только в тропиках, а в умеренной зоне вымерли
в миоцене.
Общая низкая организация насекомоядных ограничивала многим их
группам возможности противостоять влиянию изменения обстановки
на земле после верхнемелового времени. В кайнозое понизилась темпера-
тура на поверхности земли. В связи с этим значительно изменился харак-
тер вегетации растений и усложнились жизненные циклы кормовых
животных по сезонам. К этому времени прибавились и потребители бес-
позвоночных, прежде всего насекомых, за счет Интенсивно развивающихся
птиц, что значительно усилило давление среды на эволюцию насекомо-
.ядных млекопитающих. Для многих из них наступило время смены кормо-
вых спектров.
Вымирание отдельных групп насекомоядных началось еще в меловое-
время. Много групп вымерло в палеоцене—миоцене. Плиоцен был по-
следним относительно теплым периодом в умеренной зоне и пределом
существования многих групп и видов из вымерших и современных семейств.
Обеднение состава современных семейств происходило и в плейстоцене
ХАРАКТЕРИСТИКА ОТРЯДА 35
и продолжалось в четвертичное время. Многие современные семейства
объединяют роды с ограниченным числом видов или монОтипичные.
Из общего числа известных 25 семейств насекомоядных 16 вымерли, а коли-
чество вымерших родов и особенно видов весьма значительно.
Изучение эволюции ряда групп насекомоядных позволяет отметить
известную пульсацию во время процветания их отдельных ветвей. При
этом подъемы и спады количества форм, отражающих то или иное на-
правление специализации в пределах определенных групп, занимает
значительный отрезок времени; графически они могут быть представлен-
ными волнообразной линией, а не плавной равносторонней кривой.
Насекомоядные распространены в Европе, Азии, Африке, Северной
Америке. Их нет в Австралии и на большей части Южной Америки (в по-
следней встречаются только на крайнем северо-западе). Можно пред-
полагать, что в Южную Америку насекомоядные — землеройки (Sorici-
dae) проникли из Северной Америки не ранее плиоцена, поскольку там
встречаются представители только рода Cryptotis Pomel, известного
-с плиоцена. Наиболее широко распространено сем. Soricidae, представи-
тели которого не встречаются только в Австралии. Сем. Tenrecidae рас-
пространено на Мадагаскаре и Коморских островах; отдельные виды
завезены на Сейшельские острова. Только в Африке распространены
сем. Potamogalidae, Chrysochloridae и Мdcroscelididae, только в Азии-— сем.
Tupaiidae. Сем. Erinaceidae распространено в Европе, Азиц и Африке,
Talpidae,— в Европе, Азии и Северной Америке. На Вест-Индских остро-
звах (Куба, Гаити) распространено сем. Solenodontidae.
Геологический возраст семейств и распространение их в Прошлые
геологические периоды приводятся в описании этих семейств.
В экологическом отношении насекомоядные делятся на три основные
группы: наземные, роющие и полуводные. Наземные в свою очередь
делятся на бегающих (разными аллюрами), прыгающих и лазающих.
Отдельные их группы способны рыть. Роющие живут под землей, очень
редко выходя на ее поверхность. Полуводные значительную часть времени
проводят в воде, добывая корм, особенно те, которые поедают его в боль-
шом количестве.
Локомоторные системы перечисленных групп насекомоядных имеют
приспособления, соответствующие функциям, выполняемым конечностями
при том или ином образе жизни.
Насекомоядные населяют все типы ландшафтов земного шара, пред-
почитая леса и кустарниковые заросли, а на открытых пространствах
места с богатой травяной растительностью, иными словами, выбирают
участки с надежным укрытием от врагов и обеспечивающие этих прожор-
ливых млекопитающих разнообразным кормом.
Пищу насекомоядных составляют различные беспозвоночные, особенно
насекомые, а также мелкие позвоночные, которых они могут одолеть.
В кормовом рацирне отдельных групп преобладает растительная пища.
Гнезда устраиваются в надежно замаскированных естественных укры-
тиях или в вырытых в грунте норах. Многие виды роют сложные, много-
ярусные норы, отдельные виды делают норы неглубокие, в виде простых
углублений. Гнезда строятся из естественного материала, окружающего
пору, чаще всего из мягких растительных остатков.
Размножаются насекомоядные один-два, реже три раза в году. Коли-
чество детенышей от 3 до 10, у тенрека (Tenrec ecaudatus Schreb.) их бывает
до 25.
Насекомоядные имеют важное хозяйственное значение как истреби-
тели вредных беспозвоночных, особенно насекомых, а также мелких
позвоночных (грызунов). Некоторый вред они приносят в качестве про-
3*
36
ОТРЯД INSECTIVORA :
кормителей различных эктопаразитов — хранителей и переносчиков опас-
ных инфекций, но это не исключает необходимости их охраны, тем более
что многие виды стоят на грани вымирания.
Все современные семейства насекомоядных имеют общие корни в про-
исхождении и развитии. Значительное различие морфологии представи-
телей отдельных семейств объясняется разностью направлений частных
Рис. 17. Схема филогенетических соотношений современных подотрядов и их семейств.
адаптаций. При этом многие группы развивались в одном каком-нибудь
направлении, например, приспосабливались только к передвижению
прыжками, только к лазанью или плаванью. Специфика функций отдель-
ных частей тела в конкретных условиях среды наложила отпечаток на
внешний облик представителей разных семейств, однако не настолько,
чтобы любого насекомоядного нельзя было бы признать за ^такового.
Частная специализация насекомоядных менее всего сказалась на строе-
нии их морды и жевательного аппарата. В этом нетрудно убедиться при
анализе строения зубов и их систем у представителей крупных групп.
Особенно сходны у насекомоядных нижние зубы. Верхние коренные
ХАРАКТЕРИСТИКА ОТРЯДА
‘37
зубы двух типов, связаны переходами друг с другом. Сходно и строение
нижних челюстей, в частности сочленовного отдела с его тремя отростками:
венечным, сочленовным и угловым, образующими в совокупности фигуру,
напоминающую трилистник.
Таким образом, насекомоядные представляют целостную группу,
подразделения в которой объединяют семейства с близким типом адапта-
ций жевательного аппарата. В пределах отряда естественно выделяются
две крупные группы насекомоядных: с V-образными зубами — подотряд
Тенрекообразные (Tenrecii), сем. Tenrecidae, Potamogalidae, Chrysohcloridae,
Solenodontidae, и с W-образными зубами — подотряд Ежеобразные
(Erinacei), надсем. Soricoidea, сем. Talpidae, Soricidae', надсем Erinacoidea,
сем. Erinaceidae', надсем. Macroscelidoidea, сём. Macroscelididae', надсем.
Tupaioidea, сем. Tupaiidae.
Это кажущееся формальным подразделение насекомоядных при изу-
чении их вымерших представителей убеждает в правильности и естествен-
ности группировок. Известная контрастность морфологии семейств в си-
стеме по отношению друг к другу объясняется отсутствием промежуточных
звеньев между ними (вымершие формы, близкие к общим корням). Все
семейства тенрекообразных, как и ежеобразных, имеют сходные по струк-
туре зубы.
При общей для всех структуре зубы видов отдельных семейств еже-
образных приобрели больше частных различий (за счет большего или
меньшего развития отдельных элементов коронок и новообразований,
а также в результате слияния или уплощения основных бугорков зубов),
чем представители семейств тенрекообразных. Например, у представи-
телей сем. Macroscelididae происходит слияние бугорков пара- и протокона,
метакона и талона с образованием продолготоватых лакун, расположен-
ных в поперечном к продольной оси направлении. На нижних коренных
сливаются бугорки пара-, прото- и метаконида Q образованием одной про-
дольной лакуны, с перехватом на границах конидов. Это же происходит
с Р4, нередко и с Р3.
Мои представления о филогенетических соотношениях семейств со-
временных насекомоядных графически выражены на рис. 17. Связи
семейств с предполагаемыми общими для них корнями в происхождении
изображены сплошными линиями. Это не исключает предположительности
связей и не устраняет боковых ветвей вымерших в разное время групп,
которые предполагаются или известны, но не включены в предлагаемую
схему. Утолщенная часть линии отмечает геологический отрезок времени,
в течение которого существуют представители того или иного семейства,
а также время, с которого они известны.
К СИСТЕМЕ ВЫСШИХ ТАКСОНОВ ПЛАЦЕНТАРНЫХ
МЛЕКОПИТАЮЩИХ
Сравнительное морфобиологическое изучение насекомоядных и хищных
разного таксономического ранга, особенно их крупных групп, дает основа-
ние для обсуждения вероятных связей в происхождении и формировании
высших таксонов плацентарных млекопитающих. Филогенетические со-
отношения этих последних с другими подклассами все еще остаются не-
решенными, а положение отдельных из них в системе — спорным. Этот
вопрос имеет принципиальное значение и привлекает постоянное внимание
зоологов. В предлагаемом здесь кратком обзоре подклассов млеко-
питающих свои представления о их филогенетических соотношениях
я основываю на следующих теоретических позициях: на признании моно-
филетического преемственного развития млекопитающих. На этом фоне
38
ОТРЯД INSECTIVORA
эволюция рассматривается как адаптивный процесс, филогения — как
преемственность форм адаптаций, выраженных в особенностях последова-
тельного ряда эволюирующих видов и их групп, а система — как от-
ражение адаптивного процесса в их соподчинении.
Судя по остаткам млекопитающих мезозойского времени, их развитие
после окончательного обособления от предков пошло в двух основных
направлениях — по пути приспособления к питанию животной пищей
(подкл. Triconodonta, Pantotheria) и по пути приспособления к питанию
растительной пищей (Л llotheria). На это указывают структуры зубов и
зубных рядов представителей этих подклассов.
Triconodonta и Pantotheria в верхнем триасе еще сохраняли двойное
сочленение нижней челюсти с черепом, как у рептилий (articulare-guadra-
tum), сменившееся в юре на dentale-squamosum. Оба подкл. объединяли
мелких хищников. Представители подкл. Triconodonta нарезали пищу
линейно-бугорчатыми зубами, а виды подкл. Pantotheria нарезали й
разминали ее треугольно расположенными бугорками и гранями жева-
тельной поверхности зубов. Захват пищи осуществлялся клыками.
Представители подкл. Allotheria захватывали пищу первыми резцами.
Задние резцы, клыки и премоляры у них сильно Видоизменялись или были
утрачены. Моляры значительно разрослись, приобрели многобугорчатую
или ребристую жевательную поверхность, что способствовало улучшению
и ускорению обработки пищи этими зубами.
Все три подкласса произошли от общих корней, связанных в развитии
с тероморфными рептилиями, но развивались независимо друг от друга на
большом отрезке времени.
Можно думать, что структурные особенности зубов Triconodonta стоят
в основе развития зубов всех других подклассов млекопитающих. Резцы,
клыки и премоляры даже современных насекомоядных и хищных во
многом сохраняют тот же тип строения, что и у Triconodonta. Следова-
тельно, зубы последних представляют исходную структуру для этих
зубов всех групп млекопитающих. Изменение характерных для Tri-
conodonta премоляров за счет расширения бугорков и изменения положения
протокона верхних и протоконида нижних зубов относительно пара- и
метаконов и конидов, а также образование дополнительных бугорков
привело к развитию моляров с разной степенью сложности их строе-
ния.
Происхождение современных подклассов, как представляется, может
быть связанным общими корнями с Pantotheria. При этом Prototheria
отделились очень рано. От общих корней с последними произошли Meta-
theria. Остатки рептильных признаков у Prototheria не исключают воз-
можности признания их достаточно специализированной группой, обла-
дающий всеми особенностями млекопитающих. Ведь и среди представи-
телей подкласса тероморфных рептилий были как крайне примитивные,
так и высокоспециализированные группы, развивавшие параллельно с мле-
копитающими многие прогрессивные особенности сходного адаптивного
значения. С примитивными Metatheria, обладавшими зачаточной плацентой,
связаны в происхождении плацентарные — Eutheria.
Рождение и ранние стадии развития детенышей у трех современных
подклассов млекопитающих представляют последовательный ряд усложне-
ния связи с материнским организмом. У представителей подкл. Proto-
theria зародыш развивается из относительно крупного яйца, которое
задерживается некоторое время в половых путях самки, а затем откладыва-
ется в сумку или гнездо. Зародыш видов подкл. Metatheria развивается
из яйца, задерживающегося в половых путях самки (редко связан с за-
чаточной плацентой), затем слабо развитый детеныш при выходе яйца
ХАРАКТЕРИСТИКА ОТРЯДА 39
из половых путей самки покидает оболочку яйца и забирается в сумку
матери, где и донашивается. И у Metatheria, и у Prototheria зародыш
до выхода из яйца не связан в питании с материанским организмом.
Эмбриональное развитие плацентарных (Eutheria) проходит внутри
тела матери, 'в матке, его питание осуществляется за счет материнского
организма с помощью плаценты.
Общими предками насекомоядных и хищных — отрядов, стоящих
в основе развития всех других плацентарных, вероятно, были мелкие
формы, питавшиеся беспозвоночными, главным образом насекомыми,
наиболее многочисленными на поверхности почвы, а также мелкими позво-
ночными и их трупами. Коренные зубы этих плацентарных должны были
быть сходными с таковыми тенрекообразных (Tenrecii), но Р4 у них не были
моляризованы, а не имели строения, сходного с этими зубами тенреко-
образных. Первый сегмент (пара-протоконид — тригонид других авторов)
нижних коренных зубов мало отличался по высоте и форме от метаконида
(талонид других авторов). Резцы и клыки были крупными.
Эти полунасекомоядные-полухищные или хищные полифаги несом-
ненно имели многие примитивные особенности в отдельных органах и их
системах. Возможно, что они сохраняли клоакальное углубление или
сходное образование и еще несовершенную плаценту.
На рис. 23 дана схема филогенетических соотношений подклассов
и объединяемых современными подклассами современных отрядов млеко-
питающих.
Надотряд CARNIVORAMORPHA
Gureev, superordo nov.
Диагноз. Клыки, за очень редким исключением, имеются и у боль-
шей части видов, хорошо развиты. Р4 у подавляющего большинства видов
крупный, с режущим краем. Ключица имеется или утрачена в поздний
период филогенеза. У большей части видов малая берцовая кость свободна
(не прирастает к большой). Пальцы оканчиваются когтями, ногтевые
фаланги часто раздвоены на конце. Сочленовный мыщелок нижней челю-
сти развит в поперечном направлении. Пахучие железы развиты^
Характеристика. Сравнительное изучение черепов, скелетов
и особенно зубов современных и вымерших насекомоядных и хищных
позволило мне в свое время высказать мнение о том, что в меловое время
плацентарные млекопитающие были представлены двумя группами
хищников (насекомоядными и србственно хищными), развивавшимися
от общих корней (Гуреев, 1971). Обе группы к верхненемеловому времени
морфологически значительно обособились. Тем не менее многие их пред-
ставители еще сохраняли общие особенности в структуре зубов, которые
в пределах отдельных групп насекомоядных сохранились и до настоящего
времени. Например, у всех тенрекообразных (Tenrecii) верхние и нижние
коренные зубы сохраняют структуру, близкую плацентарным мелового
времени. Отдельные виды некоторых семейств насекомоядных еще со-
храняют крупные клыки как орган захвата пищи, хотя другие зубы и зуб-
ные ряды у них построены по плану, общему для насекомоядных.
До настоящего времени в качестве общего признака для насекомоядных
и хищных сохранилось относительно сходное строение и функциональное
значение Р4. Много общих особенностей в строении черепа и скелета
нетрудно обнаружить при сравнении таковых Insectivora и Creodonta.
Дивергенция насекомоядных и хищных происходила на основе разного
способа захвата кормовых животных и их преследования. При этом эволю-
40
ОТРЯД INSECTIVORA
ция насекомоядных шла по пути приспособления к быстрому схватыванию
мелкой добычи (беспозвоночных, преимущественно насекомых) удлинен-
ными резцами, как пинцетом, с короткой дистанции. Хшцные приспосаб-
ливались к захвату крупных, теплокровных животных клыками, на
бегу, часто после длительного преследования. Отмеченные направления
в эволюции обеих групп отразились в специфических для каждой из них
Рис. 18. Зубы низших млекопитающих Triconodonta (/), Simmetrodonta (2), Trituber-
culata (3).
особенностях строения соответствующих органов. Эти особенности могут
быть сформулированы в следующем виде.
Насекомоядные
IJ или 1.’ сильно развиты и несут
функцию органа захвата пищи. Резцы
достигают значительных размеров и у ви-
дов, еще сохраняющих крупные клыки.
Последние не выступают в стороны от
челюстей, а нижний клык входит в специ-
альную выемку на предчелюстной кости.
Ключица всегда имеется (исключение —
сем. Potamogalidae). Лучевая кость со-
членяется с плечевой костью шарнирным
суставом, пронация и супинация сохра-
няются. ' Движение передней конечности
и кисти — во многих плоскостях.
Хищные
1} и другие резцы развиты значи-
тельно менее клыков, которые выполняют
функцию органа захвата пищи. Клыки
выступают в стороны от челюстей, а ниж-
ний клык входит в пространство между
третьим резцом и верхним клыком. Клю-
чица сохранялась в ранний период раз-
вития, а затем утрачена. Лучевая кость
сочленяется с плечевой костью блоко-
видным суставом, пронация и супина-
ция утрачены. Движение передней ко-
нечности и кисти — в одной плоскости.
ХАРАКТЕРИСТИКА ОТРЯДА
41
Таким образом, при общем значительном сходстве многих особенностей
в строении, что заметно выражено в ранний период развития представи-
телей обоих отрядов, в более позднее время четко определились особые
для каждого из них черты адаптации.
Ван Вален (Van Valen, 1967) в своей классификации насекомоядных
и Deltatheridia подотряд тенрекообразных поставил не впереди более
высокоспециализированных насекомоядных как более примитивный, близ-
кий к исходным группам, а позади значительно специализированных групп
Рис. 19. Zalambdalestes (7), Deltatheridium praetrituberculare (2) (по: Громова, 1962).
хищников из подотряда Creodonta, с которыми он объединил и хищников
сем. Deltatheridiidae, придав этому «объединению» значение самостоятель-
ного отряда. Не менее странным представляется толкование Саведжем
(Savage, 1977) происхождения и эволюции Carnivora. Он рассматривает
происхождение Carnivora в связи с Marsupialia. Автор ведет сумчатых и
хищных от верхнеюрских Pantotheria. По его представлениям, ветвь
хищных в третичное время образовала два отряда: Carnivora и Creodonta.
Насекомоядные (Insectivora) почему-то не рассматриваются, как будто
они никакого отношения к обсуждаемому вопросу не имеют. Автор обошел
вниманием и мнение Ван Валена, отмеченное выше. Выхватив из крупных
систематических подразделений отдельные группы, Саведж обсуждает
их взаимоотношения, не отмечая при этом, как он предлагает рас-
сматривать соотношения других отрядов подклассов Metatheria и Eutheria.
Неправдоподобно выглядят и предложенные соотношения семейств
внутри отряда Carnivora.
Такого рода систематические манипуляции вносят только путаницу
в сложившиеся представления об истинных филогенетических соотноше-
ниях групп в общем адаптивном процессе (эволюции). При разработке
Рис. 20. Microgale longicaudata (7), Amblysomus gottentotus (2).
ХАРАКТЕРИСТИКА ОТРЯДА
43
филогении и систематики той или иной группы животных всегда следует
иметь в виду, что каждая из них имеет в своем составе виды, еще сохраняю-
щие в морфологии элементы общности с группами, с которыми они связаны
в своем происхождении. Короче говоря, морфология каждой системати-
Рис. 21. Mesonix obtusidens (У), Tshelkaria rostrata (2), Hyracolestes ermineus (3) (по: Гро-
мова, 1962).
Рис. 22. Didymoconus colgageti (7), Ardynictis furunculus (2) (по: Громова, 1962).
ческой группы начинается с морфологии предков. Нужно уметь различать
предков разных групп и их общих предков.
Общие структурные особенности зубов, из которых развились более
сложные зубы насекомоядных и хищных, можно установить путем их срав-
нения с зубами представителей этих отрядов мелового и более позднего
времени, а также современных тенрекообразных, сохраняющих древние
структуры зубов. При этом также можно установить, что ранний тип адап-
тации жевательного аппарата хищных, еще близкий к общим предкам с на-
44
ОТРЯД-INSECTIVORA
секомоядными, сохранялся в ряду семейств до олигоцена (рис. 18—22).
Эти семейства сохраняли зубы, близкие по структурным особенностям к та-
ковым насекомоядных мелового времени и современных тенрекообразных,
хотя у последних они все же более специализированные, чем у предков.
Представители этих семейств еще сохраняли ключицу, но во всем осталь-
ном были хищными в общепринятом понимании.
Предки Triconodonta
Рис. 23. Схема филогенетических соотношений отрядов Insectivora и Carnivora и отрядов,
связанных с ними в происхождении.
Насекомоядные и хищные представляют две исходные ветви, с кото-
рыми связаны в происхождении два комплекса отрядов высших млеко-
питающих. Представители отрядов каждого комплекса сохраняют в ске-
лете особенности своих предков — насекомоядных или хищных, смотря
по тому, от кого они произошли (рис. 23).
Таким образом, отмеченные здесь морфологические особенности насе-
комоядных и хищных имеют важное значение и в эволюции многих групп
млекопитающих, связанных с ними в происхождении.
СИСТЕМА КРУПНЫХ ПОДРАЗДЕЛЕНИЙ СОВРЕМЕННЫХ НАСЕКОМОЯДНЫХ
И ХАРАКТЕРИСТИКА ПОДОТРЯДОВ
Отряд Insectivora Bowich, 1821.
Подотряд Tenrecii nom. nov.
Сем. Tenrecidae Gray, 1821.
Сем. Potamogalidae Allman, 1865.
Сем. Chrysochloridae Mivart, 1868.
Сем. Solenodontidae Dobson, 1882.
Подотряд Erinacei nom. nov.
Надсем. Erinacoidea Fischer von Waldheim, 1817.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
45
Сем. Erinaceidae Fischer von Waldheim, 1817.
Надсем. Macroscelidoidea Gill, 1872.
Сем. Macroscelididae Bonaparte, 1838.
Надсем. Tupaioidea Gray, 1825.
Сем. Tupaiidae Gray, 1825.
Надсем. Soricoidea Fischer von Waldheim, 1817.
Сем. Talpidae Fischer von Waldheim, 1817.
Сем. Soricidae Fischer von Waldheim. 1817.
Примечание. Вымершие группы, стоящие в основе развития семейств,
а также группы, связывающие^ отдельные семейства в качестве общих предков,
и группы, представляющие боковые ветви в развитии семейств, требуют тщательной
ревизии. По этой причине они не включены в предлагаемую систему. Самой ранней
группой представляются тенрекообразные, известные с середины мела, имеющие
более просто устроенные зубы, во многом сходные с таковыми ранних хищных. Се-
мейства, объединяемые подотрядами, формировались в разное время, с разных уров-
ней специализации исходных и промежуточных групп.
Подотряд TENREGII nom. nov.
Zalambdodonta Gill, 1884, Boston, S. E. Cassino and Co, v. 5, p. 120.
3 13 3
Диагноз. Зубная формула: I С -т- P -5- M -5- = 40. P* сходны c mo-
лярами. Моляры имеют V-образную форму. Бугорки пара- й метаконов
без гребней. Второй сегмент Мх и М2 в 3—4 раза ниже их первого сегмента.
Талонид на верхних коренных (молярах) отсутствует. Скуловых дуг нет
(исключение Chrysochloridae).
Описание. Длина тела 40—400, хвоста — 30—290 мм. Нос в раз-
ной степени значительно вытянут в хоботок, реже последний слабо выра-
жен. Глаза хорошо развиты или очень маленькие, скрыты в волосяном
покрове головы. Ушные раковины крупные или небольшие, мало высту-
пающие из волосяного покрова. Тело массивное, ноги короткие. Кисти
и ступни пятипалые, реже четырехпалые. Когти развиты в разной сте-
пени в зависимости от функций конечностей. У специализироваиных
к рытью сильно развит коготь 3-го или 2—3-го пальцев кисти. Виды,
раздирающие добычу когтями, имеют сильные,, изогнутые когти. У пла-
вающих видов между пальцами имеются кожные перепонки, реже пере-
понка развивается только между двумя пальцами ступни. Хвост разной
длины или очень короткий и снаружи не заметен.
Волосяной покров мягкий бархатистый, грубый и щетинистый или верх-
нюю сторону тела и заднюю часть головы покрывают иглы. В окраске тела
преобладают серые, бурые или охристо-бурые цвета разных оттенков,
а также белый. Различные сочетания этих цветов и обусловливают все
разнообразие окрасок представителей подотряда. У златокротов волосяной
покров имеет металлический блеск.
Череп клиновидной формы в разной степени вытянут в длину. Скуло-
вые дуги отсутствуют (исключение Chrysohloridae)'. У тенреков основная
клиновидная кость имеет углубление, расположенное позади хоан.
I1 и 12 у большинства видов крупные и служат органом захвата пищи.
У ряда видов резцы мелкие, и пищу они захватывают клыками. При этом
нижние клыки входят в углубление на межчелюстной кости позади
третьего резца. Премоляры имеют разную форму и размеры, но преобла-
дают 3-бугорчатые, особенно верхние. Pf, реже и Р|, сильно моляризовяны
и сходны с молярами. Верхние моляры V-образной формы.
46
ПОДОТРЯД TENRECII
Распространение и возраст. Современные представи-
тели подотряда распространены в Африке, на о-вах Мадагаскар и приле-
жащих, Куба и Гаити. Вымершие группы известны из среднего мела Азии,
олигоцена Сев. Америки, из миоцена и плейстоцена Африки.
Сем. TENRECIDAE Gray, 1821
Gray, 1821, London Med. Reposit., v. 15, pt. 1, p. 296—310.
Тип. Tenrec Lacepede, 1799, Мадагаскар, современный.
Диагноз. Зубная формула: I —3— С уРМ= 40 — 38. Скуловых
дуг нет. Косточки у нижнего края переднего носового отверстия нет.
Клыки крупнее и выше, равны по высоте или ниже I1 и 12. Последние без
бороздки на их внутренней стороне. Ключица имеется. Большая и малая
берцовые кости у большинства не срастаются (свободны).
Описание. Длина тела — 40—400, хвоста — 30—160 мм. Нос вы-
тянут в хоботок. Ноздри спереди и сбоку. Глаза небольшие, ушные рако-
вины разной величины и формы. Ноги стопоходящие, кисти и ступни
5-, реже 4-палые. В пределах семейства имеются группы, приспособленные
к плаванью. Их виды имеют кисти и ступни с перепонками между паль-
цами. Хвост очень короткий и снаружи не заметен или очень длинный,
значительно превышающий длину тела.
Волосяной покров мягкий или очень жесткий, щетинистый. У ряда
видов верхняя сторона тела и задняя часть головы покрыты иглами.
Окраска волосяного покрова варьирует от серой и охристой до черно-бу-
рой разных оттенков. Встречается и пестрая окраска, образованная свет-
лыми и темными полосками или пятнами.
Череп удлиненный, сильно суженный в носовой части или носовой от-
дел сравнительно широкий. У тенреков на основной клиновидной кости
имеется углубление, расположенное позади хоан. Г1 и 12 развиты в разной
степени. Клыки сильно развиты или мало отличаются по форме и величине
от соседних зубов. Передние премоляры клиновидные. PJ сходны по строе-
нию с молярами. Верхние моляры V-образной формы. Большая и малаа
берцовые кости свободны или срастаются в нижней части.
Распространение и возраст. Мадагаскар и Коморские
острова, завезены на Сейшельские острова. Известны из миоцена Африки.
По характеру адаптаций сем. Tenrecidae представляет неоднородную
группу, в пределах которой одни виды типичные наземные, способные рыть,
норы, другие приспособлены к плаванью и ведут полуводный образ жизни.
Кормовые связи представителей семейства в значительной степени обус-
ловлены составом животных в соответствующих средах. Более крупные
виды питаются различными животными, которых могут одолеть, — мел-
кими позвоночными, а также беспозвоночными (червями, моллюсками,
насекомыми). Поедают и растительную пищу. Мелкие виды питаются пре-
имущественно насекомыми, добавляя к ним и других беспозвоночных. По-
луводные тенреки питаются в основном водными растениями, отчасти
рыбой.
Объединяет подсем. Tenrecinae Cabrera, 1925 — роды Tenrec
Lacepede, 1799, Setifer Frorier, 1806, Hemicentetes Mivart, 1871, Dasogale
Grandidier, 1928, Echinops Martin, 1838, и подсем. Oryzorictinae,
Dobson, 1882 — роды Oryzorlctes Grandidier, 1870, Microgale Thomas, 1882,
Cryptogale Grandidier, 1928, Limnogale Major, 1896, Geogale Milne-Edwards,
1872.
ХАРАКТЕРИСТИКА Пр ДОТРИ ДА 47
Сем. POTAMOGALIDAE Allman, 1865
Allman, 1865, Proc. Zool. Soc. London, p. 467.
Тип. Potamogale Du Chaillu, 1860, западная и центральная части Экваториальной
Африки, современный.
3 13 3
Диагноз. Зубная формула: /-5-Ст-Р-5-М-5- = 40. Скуловых дуг
нет. Косточка у нижнего края переднего носового отверстия отсутствует.
Клыки значительно меньше Iх и /2. Последние без бороздки на их внутрен-
ней стороне. Ключица отсутствует. Большая и малая берцовые кости сра-
стаются в нижней части на уз их длины.
Описание. Длина тела — 147—350, хвоста — 111—290 мм. Нос
относительно мало вытянут вперед, несколько притуплен на конце.
Ноздри расположены на боковой стороне носа, замыкаются краями ноздри,
действующими как клапан во время погружения животного в воду. Глаза
небольшие. Ушные раковины небольшие, мало выступают из волосяного
покрова. Ноги невысокие, стопоходящие, 5-палые. Кожной перепонкой
соединены только два пальца ступни — второй и третий. Хвост длинный,
в сечении округлый или уплощен на большей части.
Волосяной покров образован мягкими волосами, густо покрывающими
чело. Окраска верхней стороны тела темно-коричневая или черноватая,
нижние части тела окрашены в белый, серый или желтоватый цвета.
Череп с высоким, сжатым с боков носовым отделом и уплощенной моз-
говой капсулой. Скуловые дуги отсутствуют. Iх и /а очень крупные, клино-
образные. Клыки, особенно верхний, сходны по форме и размерам с со-
седними зубами. Р| и Р| особенно последний, сходны с молярами. Верхние
моляры Р-образной формы. Ключицы нет. Большая и малая берцовые
кости срастаются в нижней части на 1/3 их длины.
Распространение и возраст. Западная и центральная
части Экваториальной Африки, современный.
Представители семейства обитают в лесных ландшафтах по берегам
ручьев и мелких стоячих водоемов. Поднимаются в горы до 1800 м.
Пища — пресноводные крабы, рыбы и амфибии.
Объединяет два рода — Potamogale Du Chaillu, 1860 и Micropotamogale
Heim de Balsac, 1954.
Сем. CHRYSOCHLORIDAE Mivart, 1868
Mi vart, 1868, J. Anat, Physiol. London, v. 2, p. 150.
Тип. Chrysochloris Lacepede, 1799, южная часть Африки, современный.
3_2 13 3
Диагноз. Зубная формула: I -д—С -j- Р -g- М -д- = 40—36. Скуловые
дуги имеются. Косточка у нижнего края переднего носового отверстия
отсутствует. Клыки меньше Iх и 12. Последние без бороздки на их внутрен-
ней стороне. Ключица имеется. Большая и малая берцовые кости сраста-
ются в нижней части на х/3 их длины.
Описание. Длина тела — 76—235 мм. Нос не вытянут в хоботок,
покрыт на конце толстой ороговевшей кожей. Ноздри расположены под
складкой кожи на передней стороне носа. Глаза слабо развиты, скрыты
в волосяном покрове или закрыты кожей. Ушные раковины маленькие, не
выступают из волосяного покрова. Ноги короткие. Кисти 4-, а ступни 5-
цалые. Коготь 3-го пальца сильно развит. Пальцы ступни соединены ко-
жей. Хвост снаружи не заметен.
48 . . ПОДОТРЯД TEHRECH
Волосяной покров мягкий и густой. Окраска верхней стороны тела ко-
ричневая, буровато-оливковая или сероватая разных оттенков, с метал-
лическим блеском — красноватым, желтоватым, зеленоватым, бронзовым
и фиолетовым. Нижняя сторона тела окрашена сходно с верхней, но свет-
лее, желтоватая или рыжая.
Череп конусовидный, с короткой носовой частью и широкой, округлой
мозговой капсулой. Межчелюстные кости у нижнего края носового отвер-
стия утолщены и образуют здесь направленный вперед выступ, нависаю-
щий над 1Х. Скуловая дуга широко соединяется с височной костью и только
нижним краем с очень коротким ее скуловым отростком, располагаясь над
засочленовным отростком, на уровне слухового отверстия или за ним.!
Засочленовный отросток имеет впадину, куда входит сочленовный мыщелок
нижней челюсти. Слуховой барабан замкнутый. I1 и /2 сильно развиты..
Клыки сходны с Р|. Р| с тенденцией к моляризации, а Р| близки по строе-
нию к молярам. Моляры Т-образной формы. Большая и малая берцовые
кости соединены вблизи лодыжки.
Распространение и возраст. Камерун, Конго, Уганда;
Танзания, Танганьика, миоцен—современный.
Представители семейства обитают на открытых равнинах, в лесах и на
культурных землях. Питаются различными беспозвоночными. Отдельные
виды, кроме беспозвоночных, поедают и ящериц. Роющие виды.
Объединяет роды Chrysochloris Lacepede, 1799, Amblysomus Pomel;
1848, Chrysospalax Gill, 1883, Cryptochloris Shortridge et Carter, 1938.
Сем. SOLENODONTIDAE Dobson, 1882
Dobson 1882, p. 1—96.
Тип. Solenodon Brandt, 1833, Куба, Гаити, современный.
Диагноз. Зубная формула: /-|-С-|-Р-|-М-|-==40. Скуловых дуг нет.'
Косточка у нижнего края переднего носового отверстия имеется. Клыки
значительно меньше I1 и /2, имеющих бороздки на их внутренней стороне.
Ключица имеется. Большая и малая берцовые кости свободные, но у ста-
рых особей могут сростаться в области лодыжки.
Описание. Длина тела — 280—325, хвоста — 175—255 мм. Нос
вытянут в длинный хоботок. Ноздри на боковой стороне носа. Глаза не-
большие. Ушные раковины относительно крупные. Ноги стопоходящие,
кисти и ступни 5-палые. Когти на пальцах кисти крупные, серповидные.
Хвост длинный, покрыт чешуйками и короткими жесткими волосками.
Волосяной покров образован сравнительно жесткими волосами.
Окраска тела пятнисто-черная, бурая, красновато-коричневая и черно-
коричневая с белыми отметинами.
Череп удлиненный, сильно сужен в носовом отделе. К нижнему краю’
носового отверстия прикрепляется маленькая косточка, поддерживающая’
хоботок. Скуловые дуги отсутствуют. I1 и 72 сильно развиты, с бороздкой
на внутренней стороне. Клыки мало отличаются от соседних зубов.'
Р| сходны по строению с молярами. Верхние моляры Т-образной формы.
Большая и малая берцовые кости свободны, но у старых особей они'
могут и срастаться.
Распространение и возраст. О-ва Куба и Гаити, совре-
менный. Представители семейства на континенте Северной Америки изве-;
стны с олигоцена.
В настоящее время включает один род — Solenodon Brandt, 1833.'
ХАРАКТЕРИСТИКА ПОДОТРЯДА &
Подотряд ERINACEI nom. nov.
Ertnaceota Van Valen, 1967, p. 261.
Диагноз. Зубная формула: 1|=| С Р М |—| = 44—26. Р4 отли-
чаются от моляров. Моляры не У-об разной формы, имеют на же-
вательной поверхности Жуобразный гребень, образованный соединением
F-образных гребней пара- и метанонов. Второй сегмент нижних моляров
равен по высоте первому или ниже первого в 1.5—2 раза. Талонид на
верхних молярах, как правило, развит. Скуловые дуги имеются (исключе-
ние Soricidae).
Описание. Длина тела — 35—450, хвоста — 11—285 мм. Нос
в разной степени значительно вытянут и хоботок хорошо выражен. Глаза,
крупные или маленькие, малозаметные, реже закрыты кожей. Ушные
раковины хорошо развиты или редуцированы и скрыты в волосяном по-
крове. Тело разного общего строения.
Кисти и ступни 5- или 4-палые. Когти развиты в разной степени, имеют
разную форму. Виды, приспособленные к плаванью, имеют кожные пере-
понки между пальцами кисти и ступни. Хвост разной длины. У плаваю-
щих видов он сжат с боков.
Волосяной покров мягкий и бархатистый, жесткий и щетинистый или
верхняя сторона тела и головы покрыта иглами. Окраска тела разнооб-
разная. От черной, бурой и серой до коричневой и охристой,' с примесью
белого цвета. Различные сочетания этих цветов определяют окраску раз-
ных видов, среди которых встречаются и пестроокращенные.
Череп клиновидной или овальной формы. Скуловые дуги имеются
(исключение Soricidae). У ежей (Erinaceinae) основная клиновидная кость
имеет углубление позади хоан, как у тенреков (Tenrecinae). ZJ или 72
у большинства видов крупные и служат органом захвата пищи. У видов
некоторых групп резцы мелкие, и органом захвата пищи у них служат
клыки. Нижние клыки могут отсутствовать. Функционирующий крупный
клык нижней челюсти входит в специальную выемку на межчелюстной
кости. Передние премоляры имеют разное строение и размеры. Р£ отли-
чаются по строению от моляров. Моляры с большой жевательной поверх-
ностью и с ГУ-образными гребнями. Талонид у большинства развит.
Распространение и возраст. Европа, Азия, Африка,
Северная Америка и крайний северо-запад Южной Америки. Вымершие
представители подотряда известны с верхнего мела.
Сем. MACROSCELIDIDAE Bonaparte, 1838
Macroscelidina Bonaparte, 1838, Nuovi Ann. Sci. Nat. Bologna, t. 2, anno 1,.
p. 105—133.
Тип. Macroscelides Smith, 1829, Африка, современный.
Диагноз. Зубная формула: 7^-g-^C уР ~ м|—|=42—36. Ску-
ловые дуги без отверстия и без отростка, замыкающего орбиту. I] крупнее
Клыков (исключение Rhynchocyon Pet.), Верхние и нижние резцы располо-
жены вертикально. Коронки верхних моляров округло-четырехугольной
формы, состоят из двух сегментов, разграниченных складками, входя-
щими с внутренней и наружной сторон. ‘
Описание. Длина тела — 95—315, длина хвоста — 80—265 мм..
Нос вытянут в длинный хоботок. Ноздри открываются на передней сто-
4 А. А. Гуреев
50
ПОДОТРЯД ERIN АСЕТ
роне носа. Глаза и ушные раковины большие. Задние конечности длиннее
передних. Кисть с 5 пальцами. Ступня длинная с 4—5 пальцами.
Волосяной покров образован мягкими или жесткими волосами.
Окраска верхней стороны тела варьирует от светло-серой до бурой и ко-
ричневой разных оттенков. Нижняя сторона тела окрашена светлее верх-
ней, серая или белая. На отдельных частях тела имеются светлые или.тем-
ные пятна, реже они образуют пестрый рисунок в виде пятен или крупной
и мелкой штриховки.
Череп с вздутой мозговой капсулой и узким и относительно коротким
носовым отделом. Скуловые дуги хорошо развиты. Слуховые барабаны
в разной степени замкнуты. Костное нёбо сильно перфорировано. Z},
особенно Z2, крупные, остальные резцы мелкие. Клыки у большинства
видов сходны с передними премолярами. Pg. | в большей или меньшей
степени моляризованы. Коренные зубы (моляры) со слитыми бугорками,
разделены складками на два (верхние) или три (нижние) сегмента. Ниж-
няя чёлюсть с высоко поднятым сочленовным отростком. Венечный отро-
сток развит слабо.
Большая и малая берцовые кости срастаются на 2/3 своей длины.
Распространение и возраст. Южная и восточная части
Африки и о. Занзибар. Один вид распространен в Марокко и Алжире.
Вымершие представители известны из нижнего олигоцена, миоцена, и
плейстоцена.
Виды семейства населяют молодые леса, кустарниковые заросли, рав-
нины, покрытые травяной растительностью, скальные выходы. Питаются
муравьями, возможно, и термитами (мелкие виды), дополняя этот корм
мягкими почками, корешками и ягодами растений. Крупные виды пред-
почитают жуков.
Объединяет роды Macroscelides Smith, 1829, Nasilio Thomas et Schwann,
1906, Elephantulus Thomas et Schwann, 1906, Petrodromus Peters, 1846,
Rhynchocyon Peters 1847.
Сем. TUPAIIDAE Gray, 1825
Gray, 1825, Ann. Philos., nev. scr., v. 10, p. 337—344.
Тип. Tupaia Raffles, 1822, Юго-За .адный Китай, Индия, юго-восточная часть
Азии, о-ва Суматра, Ява, Калимантан (Борнео), Фялиипнпы.
2 13 3
Диагноз. Зубная формула: I С -т- Р М -д- = 38. Скуловые дуги
имеют отверстие, орбита замкнута соединенными скуловым и заглазнич-
ным отростками. Верхние и нижние передние резцы немного крупнее
клыков. Zx, 2 расположены почти горизонтально. Верхние моляры тре-
угольной формы с W-образной складкой, образованной гребнями пара- и
метаконов.
Описание. Длина тела — 100—220, длина хвоста — 90—225 мм.
Нос мало вытянут вперед и хоботок слабо выражен. Ноздри открываются
на передне-боковой стороне носа. Глаза крупные. Ушные раковины хо-
рошо развиты и у отдельных видов достигают значительных размеров.
Кисти и ступни 5-палые, с изогнутыми когтями. Хвост длинный, весь
покрыт длинными волосами или длинные волосы развиты только на конце
хвоста в виде плоской кисТочки, похожей на перо птиц.
Волосяной покров относительно мягкий. Окраска тела рыжая, бурая,
коричневая или серая разных оттенков. Отдельные части тела имеют бо-
лее темную или светлую окраску или темная окраска располагается
в виде пятен.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
5t
Череп овального контура, если рассматривать его сверху. Мозговая
капсула вздута, межглазничное пространство широкое. Носовой отдел
постепенно суживается к переднему носовому отверстию. Скуловая дуга
с отверстием, орбита замкнута. Костное нёбо перфорировано. Слуховые
барабаны замкнуты. Пары I1 и Z2 отодвинуты одна от другой.
Большая и малая берцовые кости свободны (не срастаются).
Распространение и возраст. Юго-Западный Китай,
Индия, юго-восточная часть Азии, п-ов Индокитай, о-ва Хайнань, Суматра,
Ява, Калимантан (Борнео), Бали, мелкие острова западной части Филип-
пин и Минданао; современный. Геологическая история семейства не ясна,
но несомненно, что это одна из древних групп насекомоядных, связан-
ная общими предками с приматами.
Представители семейства обитают в тропических и дождевых лесах
и зарослях кустарников. Живут на деревьях й на земле. Питаются пло-
дами растений и насекомыми, поедают яйца и птенцов птиц. Отдельные
виды едят и ящериц.
Объединяет подсем. Tupaiinae Lyon, 1913 — роды Tupaia Raf-
fles, 1822, Anathana Lyon, 1913, Dendrogale Gray, 1848, Urogale Mearns,
1905 и подсем. P t i l о c er cinae Lyon, 1913 — род Ptilocercus Gray,
1848.
Остальные семейства этого подотряда подробно описаны далее.
ОБЩИЕ АДАПТИВНЫЕ ОСОБЕННОСТИ СЕМЕЙСТВ
ПОДОТРЯДА ERINACEI ФАУНЫ СССР
В настоящий раздел выделены системы органов, имеющие определяю-
щее значение в характеристике адаптивного типа той или иной группы мле-
копитающих, особенно жевательный аппарат и органы движения. Дивер-
генция и эволюция крупных групп млекопитающих и их подразделений
шли по пути освоения разных кормов и специализации к их обработке
во рту, а также приспособления локомоторной системы к разным формам
движения в разных средах ландшафтов. В связи с этим представилось
целесообразным описать эти системы в одном месте.
Для удобства материал приведен в виде описаний для отдельных се-
мейств, связанных общими замечаниями. Такое расположение материала,
сохраняя сравнительный аспект, исключает чересполосицу и путаницу
в представлениях об особенностях строения тех или иных органов ежей,
кротов и землероек при чтении.
Адаптивные особенности кожи
и ее дериватов
Ежовые. Кожа у представителей сем. Erinaceidae толстая, особенна
у ежей, тело которых покрыто иглами. Она сравнительно плотно при-
растает к их телу.
Представители семейства имеют неодинаковый покров. Поэтому era
описание следует вести по подсемействам, каждое из которых характери-
зуется в этом отношении своими особенностями.
Виды более древней группы ежовых — гимнуры (подсем. Galericlnae}
имеют на всем теле только волосяной покров. Он состоит из трех групп
волос: длинных и жестких, особенно на задней части тела, сравнительно
редких — направляющих, густо растущих — остевых и подшерстка.
Как и у других млекопитающих, степень развития волосяного покрова
стоит в прямой зависимости от климатических условий в местах обитания
животных.
4* ’
52
ПОДОТРЯД ERINACET
Виды, населяющие влажные леса высокогорий, имеют сравнительно
густой и высокий волосяной покров, а обитатели равнин и окраин лесов
тропической и субтропической зон в разной степени густой и невысокий
волосяной покров. Особенности волосяного покрова меняются и по сезонам.
При всем этом направляющие и остевые волосы наощупь у всех гимнур
кажутся жесткими. Несомненно, что эта общая тенденция к образованию
жестких волос, наблюдающаяся в пределах семейства ежовых, послужила
основой для развития игл при освоении ими степных и пустынных ланд-
шафтов.
Тело ежей из подсем. Erinaceinae сверху и с боков покрыто иглами.
Иглы покрывают заднюю часть головы. Ранние стадии развития иглистого
покрова, что можно наблюдать у новорожденных ежей, показывают, что
Рис. 24. Иглистый покров ежат (Erinaceus attritus).
первоначально он слагается из продольных рядов игл, между которыми
располагаются полоски голой кожи. Полоска голой кожи вдоль позво-
ночника сохраняется длительное время и заходит на заднюю часть го-
ловы. Остаток этой полосы на голове, разделяющей иглы на два пучка,
сохраняется у взрослых ежей из подродов Erinaceus и Paraechinus и носит
название «лысины» (рис. 24, 25).
Общее строение игл у всех ежей сходно. Отличие заключается в раз-
ной степени развития бороздчатой и бугорчатой структуры на их поверх-
ности, а также в степени их постоянства у взрослых особей. Относительно
постоянно сохраняют внешние структуры иглы представителей подрода
Paraechinus, у которых они, кроме того, наиболее длинные и толстые.
Сравнительно рано исчезают бороздки на иглах ежей подрода Erinaceus.
Строение игл у видов подрода Hemiechinus варьирует в значительных пре-
делах. Наряду с типичной для этих ежей структурой игл (наличие бороз-
док и бугорков) встречаются особи, имеющие иглы только с бороздками
(без бугорков). Попадаются и экземпляры, у которых иглы имеют раз-
ную структуру. Варьирует и количество темных поясков (1—4, даже 5) на
тонких иглах, образующих пучки по бокам от полоски голой кожи на го-
лове отдельных представителей подрода Paraechinus. Чаще всего встре-
чаются экземпляры с двумя темными поясками на иглах, реже с тремя,
но есть экземпляры, у которых имеются иглы с двумя и тремя поясками.
Нижняя сторона тела ежей покрыта волосами, мягкими или грубыми.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
53
Представители семейства не отличаются яркостью окраски, преобла-
дают черно-бурая, бурая, серая и белая, реже охристая и белая. Различ-
ные сочетания этих цветов и создают разнообразие их окраски. Среди них
встречаются однотонно окрашенные сверху и снизу или пестро окрашен-
ные, а также белого цвета (гимнуры). Иглистый покров всегда имеет пе-
стрый рисунок разных оттенков, что обусловлено пестрой окраской от-
дельных игл. Нижняя сторона тела ежей окрашена в бурый, серый или
белый цвета. Морда ежей всегда окрашена темнее, чем другие части тела.
При этом часто темные волосы образуют на лицевой части головы своеоб-
разную маску (рис. 26).
Как и другие млекопитающие, ежи — обитатели аридных зон имеют
светлую окраску, а обитатели территорий с влажным климатом — в раз-
ной степени темную до почти черной. Исключение составляют ежи подрода
Рис. 25. Erinaceus europaeus (У), Е. auritus (2) (а — полоска голой кожи).
Paraechinus, среди которых встречаются особи с почти черными иглами,
обитающие в пустынях.
В связи с развитием иглистого покрова как средства пассивной защиты
от врагов стоит развитие у ежей подкожной кольцевой мускулатуры,
сворачивающей их тело в шар, а также утолщение кожи в зоне основа-
ния игл, что предохраняет мышцы тела от уколов и повреждений основа-
ниями игл при надавливании на них сверху.
Кротовые. Кожа кротов сравнительно толстая, плотная, слабо при-
растает к телу; прочно прикрепляется она только в области кистей и ступ-
ней, а также переднего отдела головы. Это позволяет кротам относительно
легко менять объем той или иной части тела в кожном «мешке», пролезая
в узких ходах вперед или пятясь назад. На кистях и ступнях, а также на
хоботке кожа значительно утолщена. Верхняя поверхность кисти и ступни
покрыта щитками и чешуйками из утолщенной кожи и рогового вещества.
Нижняя поверхность ннмей и ступней, особенно ладоней, покрыта круп-
ными щитками и бугорками из ороговевшей кожи. Все это помогает за-
щищать кожу от повреждений во время рытья и выбрасывания грунта из
ходов. Волосяной покров мягкий, бархатистый, образован слабо дифферен-
цированными волосами ости и подшерстка, имеющими извитую форму и
отогнутую вершину. В совокупности они образуют плотно сомкнутый
покров, сходный на всем теле, кроме головы, кистей и ступней.
Голова представителей триб Talpini и Scalopini покрыта более гру-
быми волосами, а у видов Urotrichini голова сверху покрыта жесткими ще-
54
ПОДОТРЯД ERINACEI
тинистыми волосами, напоминающими щетинки на хоботке. У одних кро-
тов верхняя сторона хоботка, края губ, нижняя губа и подбородок по-
крыты жесткими иглистыми щетинками, у других сверху хоботок покрыт
короткими и жесткими волосами. Вибриссы развиваются при основании
хоботка, они разной длины и толщины. Встречаются кроты, у которых
вибриссы развиваются на голове между глазами, а также на нижней губе.
Рис. 26. Темные пятна (маска) на лицевой части головы ежей, (а —кожные образова-
ния по краям ноздрей, характерные для всех ежовыХ).
Как и у других, они служат органом осязания. Возможно, что они в сово-
купности позволяют определить ширину и высоту хода, а также степень
его проходимости.
Щетинки на морде кроме защитной функции несут, возможно, и функ-
цию очистки от грунта, добываемого в земле корма, особенно дождевых
червей, извивающихся при их схватывании. Это тем более важно, что
кишечник дождевых червей часто содержит почву.
Толстые щетинки покрывают края кистей, защищая кожу от повреж-
дений, а также увеличивая ее поверхность. Они, кроме того, удерживают
грунт на кисти. Менее толстые щетинки покрывают дистальные отделы
голеней, в небольшом количестве одевают ступню сверху и более густо рас-
полагаются вокруг пяточного бугра.
Когти на пальцах кистей широкие в разной степени и плоские, реже
сжатые с боков, изогнутые. На пальцах ступней когти всегда сжатые
с боков, в разной степени изогнутые.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
55-
Окраска волосяного покрова однотонная, черно-бурая, черная и ко-
фейно-серая, серая и коричневая. У отдельных видов имеются белые от-
метины. Нижняя сторона тела немного светлее верхней.
Линька волосяного покрова происходит три раза в году, что обуслов-
лено, по-видимому, значительным стиранием волосяного покрова грун-
том при постоянном рытье.
Волосяной покров кротов, как явствует из его краткого описания, не-
сет разнообразную функцию в зависимости от места расположения и функ-
ции той части тела, которую он покрывает. Отогнутые назад вершины
остевых волос, отчасти и подшерстка образуют покровную зону над вер-
тикально стоящими, волнистыми частями волос, мешающую проникнове-
нию почвенных частиц в гущу волос, а кроме того, уменьшающую трение
волос о грунт, что облегчает передвижение кротов в подземных ходах.
У кротов, приспособленных к плаванью, волосяной покров слабо смачи-
вается водой. Можно думать, что вертикально и горизонтально направлен-
ные части волос служат улучшению термоизоляции. Видимо, теплоотдача
при перегревании тела во время работы осуществляется за счет голой кожи
на голове, (хоботок) кистях и ступнях, где покров состоит из редких гру-
бых щетинок.
Окраска кротов почти одноцветная на всем теле. Серая разных оттенков,
черная, реже коричневая. У отдельных видов бывают белые пятна на раз-
ных частях тела.
Землеройковые. Как й у кротов, кожа у землероек относительно
толстая и плотная. Прирастает ц телу лишь в отдельных местах, главным
образом на голове, дистальных отделах конечностей и хвосте. Это обстоя-
тельство позволяет телу относительно свободно перемещаться в кожном
мешке, что весьма выгодно при передвижении в узких пространствах или
при пролезании в мелкие отверстия. Кожа при этом свободно сдвигается,
позволяя уменьшать объем тела то в одном, то в другом месте. Этому же
отчасти служит и короткий, плотный, бархатистый волосяной покров.
На отдельных частях тела он несет различные функции. Необходимостью
постоянного передвижения в приземном слое растительности, в мертвой
подстилке или в норах грызунов, где приходится соприкасаться с почвой
или остатками отмерших растений, а также и в воде обусловлено развитие
плотного волосяного покрова, почти одинакового по высоте на всем теле
и относительно однообразного по составу. В связи с этим волосы ости и
подшерстка морфологически слабо дифференцированы. К волосяному
покрову землероек не пристает снег, а у водных форм он не смачивается
водой, к нему не пристает почва, а в волосах не задерживается сухой му-
сор. Он хорошо защищает тело от сильного охлаждения.
Конец морды у землероек усажен толстыми иглистыми щетинками, их
размер увеличивается кзади. Они, видимо, несут функцию защиты конца
морды от укусов животных, служащих кормом, а также при разрыхлении
почвы в поисках корма или укрытия. Эти щетинки приводятся в движе-
ние специальной мускулатурой. Осязательную функцию несут вибриссы,
достигающие у многих видов значительной длины. Короткие жесткие ще-
тинки также окружают глаза. Они, видимо, не только служат для защиты
глаз, но и мешают мягким волосам покрова головы свешиваться на глаза.
Мягкие короткие волосы покрывают края ушной раковины и верхние
края кожных выростов на ее внутренней части. Очень короткие волоски,
сидящие на особых бугорках кожи, покрывают внутреннюю поверхность
ушной раковины. Возможно, что они несут осязательную функцию.
Неяркая монотонная окраска землероек имеет важное защитное зна-
чение. Коричневая окраска характерна для северных лесных видов. Этот
цвет мало заметен на коричневом фоне опавшей листвы, сухой хвои и
56
ПОДОТРЯД ERINACEI
сучьев. Серая и бурая окраска южных форм сливается с серым фоном почв,
а черная — с почвами южных лесов и бурой окраской их мертвого покрова.
К этому нужно добавить, что темные окраски преобладают у видов, оби-
тающих во влажном климате, а светлая — у видов, распространенных
в относительно сухих климатических зонах.
Существуют возрастная сезонная и географическая изменчивости
окраски. Землеройки линяют дважды в году — осенью и весной. Осенняя
линька, видимо, представляет собой переход от ювенильной во взрослую
стадию волосяного покрова (у поздних генераций несомненно), а весенняя,
как обычно, из зимнего в летний волосяной покров.
В разных частях ареала вида каждая из линек начинается в разное
время, что обусловлено разными сроками наступления условий, обеспе-
чивающих физиологические предпосылки для этого процесса.
Движение
Движение животных из сем. Talpidae и Soricidae весьма разнообразно,
чего нельзя сказать о движении зверьков из сем. Erinaceidae.
Ежовые. Передвижение ежей по земле, как это приходилось наблю-
дать в пустынях Средней Азии, в пересеченной местности в Закавказье
и на лесных лужайках в разреженных лесах Ленинградской обл. пред-
ставляет собой ускоренный шаг или рысь. Передвигается еж сравнительна
быстро, если чует опасность. Если не слышит каких-либо пугающих его
звуков в воздухе или отраженных почвой и не видит опасного предмета,
он не спеша перемещается, придерживаясь избранного направления.
Общий характер движения всех ежей сходен. При внезапном испуге еж
свертывается в шар, остается некоторое время в таком состоянии, а затем
медленно разворачивается, прислушиваясь и нюхая воздух. Развернув-
шись, он медленно движется, по временам останавливается, оценивая
обстановку, или снова свертывается на короткое время, не доверяя
окружающим его предметам. После испуга осторожность его долго не
покидает. Мне не приходилось видеть, чтобы сильно испуганный еж
убегал галопом. Возможность быстро свернуться в колючий шар и таким
образом избежать опасность исключила необходимость развития при-
способлений к быстрому бегу. Длительная активность ежей, наблюдае-
мая в неволе, позволяет думать, что ежи приспособлены передвигаться
в поисках корма в течение продожительного времени. В пустыне, судя
по следам, ежи уходят от своих убежищ на значительные расстояния.
Все ежи в большей или меньшей степени способны рыть, устраивая себе
убежища и камеры для выведения потомства, особенно обитатели откры-
тых пространств. Норы они строят простые и неглубокие, тем не менее
достаточные для переживания зимы в состоянии глубокой спячки.
Гимнуры имеют тело более стройное, чем ежи. Их скелет образован
сравнительно тонкими костями, что позволяет предполагать у них более
быстрый бег и не только рысью, но и тяжелым галопом. Известно, чта
большая гимнура (Echinosorex gymnurus Raffles) способна плавать,
a Neohylomys hainanensis Shaw et Wong приспособился к рытью и ведет по-
луподземный образ жизни, на что указывает его внешность (см. рис. 68)
и что установлено наблюдениями. Таким образом, гимнуры в отношении
движения представляют более универсальную группу, хотя и не достиг-
шую значительной специализации в отдельных формах движения.
При всем этом бег для них имеет ведущее значение. Об этом свидетель-,
ствуёт строение дистальных отделов их конечностей, пальцев и когтей.
Строение остатков скелетов вымерших ежевых сходно во многом
с таковым современных й позволяет считать, что эти млекопитающие
ХАРАКТЕРИСТИКА ПОДОТРЯДА
57
на протяжении всего пути их развития были и остаются малоактивными
животными, не приспособленными к быстрому бегу.
Кротовые. Естественно представить себе, что кроты произошли от
бегающих форм, тем более что в составе семейства и в настоящее время
существуют три рода с четырьмя видами наземных бегающих кротов
(с. 200). Приспособление кротов к подземному образу жизни и постоянному
рытью началось; видимо, очень рано, поскольку уже в нижнем олиго-
цене обитали специализированные землерои, о чем свидетельствуют от-
носящиеся к этому времени находки плечевых костей, близких по строе-
нию к таковым современных
видов.
Можно думать, что спе-
циализация кротов к рытью
началась в эоцене, а может
быть, и еще ранее. Далее я
вернусь к этому вопросу при
обсуждении филогенетичес-
ких связей кротов, а пока
следует отметить, что о вы-
мерших кротах мы еще мало
знаем.
Обзор морфологических
особенностей кротов позво-
ляет отметить, что в пре-
Рис. 27. Предплечье и кисть бе-
гающего крота Uropsilus soricipes.
Рис. 28. Предплечье, кисть и ступ-
ня специализированного землероя
Talpa еигораеа (По: Строганов,
1948).
делах сем. Talpidae существует, с одной стороны, известное разно-
образие форм движения, с другой — эти формы движения представ-
ляют ряд ступеней или степеней специализации от передвижения гало-
пом по земле до высокого уровня специализации к рытью или плаванью.
При этом и рытье, и плаванье осуществляются гребным движением перед-
них конечностей ц отталкиванием или гребным движением (особенно
в воде) задних конечностей. Следует также отметить, что строение пояса
передних конечностей у роющих и плавающих кротов во многом сходно,
как и дистальных отделов задних ног, например у видов, приспособлен-
ных и к рытью, и плаванью (рис. 29).
Таким образом, при всем разнообразии направленности специализа-
ции роющих и плавающих видов кротов она имеет не только общие ис-
ходные морфологические позиции, но и общий характер движения ко-
нечностей (особенно передних), а также общие морфологические струк-
туры и их функции, хотя и уклоняющиеся в отдельных деталях в связи
•со спецификой среды, в которой они функционируют.
Нужно отметить также, что разная специалйзация к рытью и плаванью
имеет общую основу и в том, что одни кроты приспосабливались к греб-
ным движениям в почве в поисках относительно постоянных запасов
корма при значительных изменениях среды по сезонам, другие, при-
спосабливаясь к плаванью, нашли пригодные кормовые усдовия в воде.
58
ПОДОТРЯД ERINACEI
При этом роющие кроты могут плавать, а плавающие хорошо роют
(звездорыл) или роют сложные норы (выхухоль).
Всех кротов по характеру движения можно разбить на следующие
группы: 1) бегающие наземные кроты; 2) полуроющие, со свободными
пальцами на кистях; 3) специализированные землерои, с пальцами, соеди-
ненными кожей до когтей; 4) роющие и плавающие, имеющие приспо-
собления к достаточно специализированному рытью, а также и к пла-
ванью (вплоть до кожных придатков на носу для взмучивания ила при
добывании корма); 5) специализированные пловцы со всеми атрибутами
для этой локомоторной функции.
Среди этих подразделений мож-
но выделить три основные формы
движения, отражающие высокую
специализацию: бег, рытье и пла-
ванье (рис. 27—30).
Специализированные землерои
по земле передвигаются крайне
неуклюже, наподобие пингвинов,,
отталкиваясь задними ногами'
скользят на груди и брюхе, помо-
гая движению передними лапами.
Кроты, имеющие кисти со свобод-
ными пальцами, передвигаются,
видимо, быстрее, а отдельные виды
лазают. Что же касается наземных
Рис. 29. Ступня роющего и плавающего
крота Condylura cristata.
кротов, конституция тела которых сходна с таковой землероек, то они,
вероятно, и бегают сходно, но точных данных в этом отношении нет.
Передвижение по поверхности почвы выхухолей также неуклюжее, хотя
и относительно более быстрое, чем кротов. Нормально передвигаться
выхухоли мешают крупные и широкие ступни с изогнутыми пальцами.
Отсутствие необходимости, за редким исключением, в передвижении по
поверхности земли и высокая специализация к рытью и к плаванью, не
способствовали у этих групп кротов сохранению морфологического
обеспечения этой возможности.
Бегающих и плавающих кротов значительно меньше, чем роющих.
Следовательно, главным направлением в специализации локомоторной
системы кротов было приспособление к передвижению в толще почвы.
Сходные условия и сходные функции роющих кротов обусловили извест-
ную стабилизацию их адаптивных особенностей и ограничили возмож-
ности новообразований, что привело к унификации морфо-функциональ-
ного типа и вместе с этим затормозило процесс видообразования. Боль-
шинство родов роющих кротов имеют ограниченное число видов или
монотипичные. (
Начавшаяся рано эволюция кротов в сторону приспособления к жизни
под землей, сопровождавшаяся высокой специализацией к рытью, при-
вела к изменению скелета от универсального типа в такой степени, что
изменение условий жизни, несомненно, уже не сможет изменить строения
их тела в ином направлении. Высокая специализация к рытью или пла-
ванью, сопровождающаяся значительными изменениями скелета и дру-
гих органов тела, — тупик в эволюции.
Для кротов тупиком оказался и наземный образ жизни с использо-
ванием для передвижения универсально приспособленных конечностей.
Эти кроты сохранились в небольшом числе видов на ограниченном про-
странстве в Юго-Восточной Азии.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
59
Зсмлеройконые. Локомоторный аппарат землероек представляет собой
универсальную систему, обеспечивающую преодоление пространства при
разнообразных условиях и ситуациях. Это обстоятельство затрудняет
определение общих особенностей их бега.
По разнообразию форм движения землеройки стоят на одном из пер-
вых мест среди других групп насекомоядных (рис. 31—34). Среди них
есть виды, приспособленные к рытью, плаванью, быстрому бегу и пере-
движению прыжками. Отдельные виды способны лазать. Тем не менее
основной формой движения им служит бег галопом. Отдельные виды,
рызыскивая корм, способны проходить в течение одной ночи значитель-
Рис. 30. Кисть
(Л) и ступня (Б) Desmana moschata.
иые расстояния, например, путорак — Diplomesodon pulchellum Licht,
до 5—8 км (Стальмакова, 1955).
Необходимость потребления пищи в значительных количествах вы-
нуждает землероек большую часть врейени расходовать на добывание
кормовых животных, особенно в зимнее время в умеренной зоне, и, сле-
довательно, много находиться в движении, что само по себе обеспечи-
вает поддержание тепла в организме за счет мышечной энергии.
Следует отметить значительное развитие бугров для прикрепления
мышц на трубчатых костях передних конечностей землероек, что на пер-
вый взгляд кажется странным, учитывая ничем не примечательный их
бег. К этому надо добавить, что у них' удлиненная лопатка с хорошо
развитой остью и акромиальным отростком с вильчатым концом, а также
и широкая рукоятка грудины, к которой прикрепляется длинная клю-
чица. Однако все это находит объяснение, если проанализировать об-
становку, в которой приходится действовать землеройкам, например
в ландшафтах умеренной зоны.
С уменьшением размеров животного увеличиваются затраты энергии
на преодоление пространства в единицу времени во много раз. Там, где
крупное животное делает несколько шагов, преодолевая лужи, прими-
ная кусты и траву, например в лесу, землеройка должна пробежать,
перелезть через препятствие, подрыться или подлезть, обежать даже самые
мелкие кустарники и травы, переплыть лужи. Все это в совокупности
требует от землероек способности бегать, лазать, ползать, прыгать и
плавать, что определяет универсальность локомоторной системы этих
насекомоядных. Но не только этим обусловлено значительное развитие
60
ПОДОТРЯД ERIN AC El
и укрепление переднего пояса и свободных конечностей и усиление их
мускулатуры. Землеройкам часто приходится рыть, приспосабливая
естественные убежища для своих нужд. Добывание корма также часто
сопряжено с опорой на передние конечности, когда землеройка, добывая
насекомых или червей в лесной подстилке, под слоями листьев или мха,
поднимает их головой или вынуждена вести борьбу с крупными насеко-
мыми и другими животными.
Видимо, частое передвижение в низких норах и в пространствах
между слоями мертвой подстилки и валежника обусловило п развитие
Рис. 31. Скелеты землероек.
А — Suncus murinus универсального типа; В — Diplomesodon pulchellum, приспособленная к дли-
тельному передвижению.
у них бега, напоминающего рысь. Так бегают землеройки и на открытых
местах, плотно прижимаясь к почве. Среди землероек встречаются от-
дельные виды с сильно удлиненными задними конечностями (за счет удли-
нения голени и ступни), бег которых представляет собой длинные прыжки,
напоминающие таковые лесных и желтогорлых мышей. Возможно, что-
этот способ передвижения стоит в связи с добыванием быстро бегающих
животных.
При всем том, что все землеройки в большей или меньшей степени роют,,
приспособление к этой локомоторной функции конечностей у них выра-
жено слабо. У видов, роющих чаще других, как например у Sorex ungui-
culatus Dobs., передние конечности массивные, с расширенными кистями
и длинными когтями. Наиболее приспособленными к рытью следует счи-
тать землероек из рода Anourosorex M.-Edw., в морфологии которых
выражены особенности, свойственные типичным землероям. Из внешних
признаков следует отметить отсутствие ушных раковин, короткий хвост,,
кисти с острыми когтями.
Приспособление к плаванью у землероек стоит на разном уровне спе-
циализации. Так, ряд представителей рода Sorex L. в Северной Америке
ХАРАКТЕРИСТИКА ПОДОТРЯДА
61
ведет полуводный образ жизни и напоминает виды более специализиро-
ванных в этом отношении родов землероек, но приспособления к пла-
ванью у них не выражены. Приспособления к плаванью у представите-
лей родов Neomys Каир и Chimarrogale Anders, выражаются в следующем.
Рис. 32. Скелет конечностей землероек.
А — Neomys fodiens, приспособленная к плаванью, Б — Sarex unguieulatus много роющая землеройка.
Волосяной покров густой, не смачивающийся водой. На боковых сторо-
нах кистей и ступней, а также пальцев длинные щетинки образуют бах-
рому, увеличивающую их поверхность при гребных движениях. На
' нижней стороне хвоста длинные волосы образуют киль, что служит
увеличению боковых поверхностей при его функционировании как органа
движения и руля. Гораздо более приспособлены к плаванию землеройки
рода Nectogale M.-Edw. У них, кроме бахромы из щетинок на боковых
сторонах кистей и ступней, как у видов предыдущих родов, имеются
между пальцами плавательные перепонки. Хвост в концевой части упло-
62
ПОДОТРЯД ERINACEI
щен. Ушные раковины редуцированы. Мозолистые бугорки на нижней
поверхности кистей и ступней выполняют функцию своеобразных при-
сосок, устраняющих скольжение на мокрых камнях водных потоков.
Рис. 33. Кисть и ступня Neomys fodiens сверху и снизу.
Морфофункциональные особенности
жевательного аппарата
Жевательный аппарат представителей семейств Erinaceidae, Talpidae
и Soricidae при общем сходном типе зубов с W-образной складкой на
жевательной поверхности имеет много существенных отличий, что обус-
ловлено строением черепа и нижней челюсти, особенностями сочленения
Рис. 34. Лазающая землеройка Sylvisorex megalura (по: Vogel, 1974, с фот.).
нижней челюсти с черепом, количеством зубов и многими другими
особенностями, характерными для каждого из этих семейств.
Ежовые. Их жевательный аппарат сложился, видимо, в эоцене, так
как в олигоцене он уже имел все особенности, характерные для такового
этих млекопитающих. Каких-либо существенных отличий в его стро-
ении у гимнур (подсем. Galericinae) и ежей (подсем. Erinaceinae) отметить
не удается. Отличия заключаются в большей или меньшей удлиненности
ХАРАКТЕРИСТИКА ПОДОТРЯДА 63
мышц или сухожильных тяжей, что обусловлено разной длиной отдель-
ных частей черепа, соответственно и челюстей, при разном количестве
зубов.
Жевательный аппарат ежеобразных приспособлен к захвату добычи,
преимущественно резцами (исключение Echinosorex и Podogymnura) и
пережевыванию разнообразной, сравнительно грубой по механическому
составу пищи.
Верхние зубы. Р крупные, удлиненной формы. Следующие за
ними мелкие, разной формы, с 1:—2 корнями. Реже первый резец невы-
сокий, расширен в передне-заднем направлении, с двумя вершинами
(зубцами), напоминает этот зуб землероек. Все резцы расположены на
боковой стороне межчелюстной кости и между первыми резцами остается
широкий промежуток. Клык у большинства ежеобразных мелкий, напо-
минает строением задний резец или передние премоляры, с 1—2 корнями.
У Echinosorex gymnurus он крупный, хищнического вида. Между ним
и последним резцом глубокая впадина на межчелюстной кости, куда
входит нижний клык. Первый, второй и третий премоляры сходной
формы и величины или разной формы и размеров, особенно Р3, который
бывает крупным, с тремя, а не с двумя или одним корнем, сходный с Р4.
Чаще всего названные зубы простые пО строению, с 1—2 корнями.
Из них первый и второй могут отсутствовать. Как отмечалось, резцы или
резцы и клык служат для захвата пищи, а передние премоляры ее сми-
нают и дробят. Р4 многобугорчатый, в разной степени моляризован,
у большинства видов с острыми краями бугров, которые нарезают пищу
или сминают ее. М1 самый крупный зуб в верхней челюсти, четырех-
угольный, с пятью буграми на жевательной поверхности. М2 сходен
с первым по форме и строению, но меньших размеров. М3 отличается фор-
мой и мелкими размерами, у ежей расположен в поперечном направлении.
Нижние зубы. Первые.нижние резцы длиннее других, полого
(за небольшим исключением) направлены вверх и вперед. 12 и 13 значи-
тельно меньше 14, сходно с ним расположены в челюсти. В совокупности
они образуют ложкообразной формы лезвие. Несколько выше и позади
них, с таким же наклоном вперед расположен клык, формой напомина-
ющий задние резцы. Передние премоляры (Рх, Р2 и Р3), если есть, имеют
коническую форму, с 1—2 корнями, образуют ряд острых лезвий.
Нередко они имеют коронку, отогнутую вперед в виде флага и несут
скорее опорную функцию для более острых верхних антагонистов,
чем режущих зубов. Р4 с вильчатым (двувершинным) протоконидом,
более высоким, чем коронки других зубов, или с одновершинным, мало
поднятым вверх. Коренные зубы сходной (W-образной) формы убывают
в размерах спереди назад.
Соотношение зубов верхней и нижней челюстей, когда они сомкнуты,
у видов разных подсемейств имеет некоторое различие, поэтому целе-
сообразно описать их отдельно. Более других в этом отношении отли-
чается большая гимнура Echinosorex gymnurus. Взаиморасположение
верхних и нижних зубов при сомкнутых челюстях у этого вида сле-
дующее. Между двумя I1 располагаются мелкие 1Х. Более крупные 12
расположены между вторыми верхними резцами. 13 располагаются
между I2 и I3, отчасти упираясь во второй верхний резец. Нижний клык
входит во впадину на межчелюстных костях между I3 и верхним клыком.
Мелкие Р4 и Р2 не соприкасаются ни с какими зубами. Они часто отсут-
ствуют на той или иной стороне челюсти, особенно у старых особей.
Р3 расположен впереди Р3. Р4 помещается между Р3 и Р4. Передний сег-
мент Mj расположен на задней части Р4, а его задний сегмент помещается
на первом сегменте М1. Первый сегмент М2 опирается на задний сегмент М1.
64
ПОДОТРЯД ERINACEI
Его второй сегмент упирается в первый сегмент М2. М3 упирается во вто-
рой сегмент М2 и первый, сегмент М3. Задний сегмент М3 (с высоким зад-
ним краем) служит ограничителем движения пищи по зубному ряду
назад,. поскольку он заходит за задний край коронки М3.
Расстояние между нижними зубными рядами меньше такового между
верхними почти на ширину зубного ряда (коренных). Это обстоятельство,
я также особенности стирания зубов, направление гребней и бугров
на жевательных поверхностях и строение сочленовных мыщелков нижней
челюсти позволяют считать, что во время жевания пищи происходит сме-
щение нижней чел,гости в ту или иную сторону, без чего невозможно раз-
мельчение пищи. Однако боковое смещение нижней челюсти весьма огра-
ниченно, не более 2/4 ширины коронки моляра. Это представление под-
тверждается сравнительно малым стиранием бугорков на внешнем крае,
Ограничивающих боковое движение нижних зубов к внешнему краю же-
вательной поверхности верхних коренных зубов.
Взаиморасположение зубов у мелких гимнур сходно с описанным
для большой гимнуры. Отличия заключаются в том, что мелкий ниж-
ний клык имеет наклонное положение и сближен с резцами, напоминая
д этом отношении клык ежей. Премоляры более сближены друг с другом,
будучи относительно крупнее, чем у большой гимнуры. Соотношение
задних премоляров (Р3 и Р4) и моляров сходно с описанным для большой
гимнуры.
У ежей в связи с меньшим числом зубов, особенно в нижней челюсти,
соотношение верхних и нижних зубов заметно отличается от такового гим-
нур. 11 У ежей более крупные, они занимают все пространство между
I1—I1. 12 и нижний клык располагаются между I2 и I3 или клык только
упирается в I3. Р3 помещается между верхним клыком и Р2 ближе к клыку
(у ежей нет Р1 и нет 13, Р4 и Р2). Р4 высокий, помещается между Р3 и Р4.
Взаиморасположение М| и М| как у гимнур, М3 мелкий, весь помещается
на заднем сегменте М2. М3 расположен под углом к зубному ряду, он
с высоким протоковом, заходящим при сомкнутых челюстях за задний
край коронки М3 и служит ограничителем движения нижней челюсти
назад.
Жевательная и лицевая мускулатура ёжеобразных
характеризуется общностью для всех групп семейства. Она не отлича-
ется разнообразием мест прикрепления отдельных мышц. Различия
в частных особенностях заключаются в удлинении или укорочении мышц,
что обусловлено разной длиной отдельных частей черепа и челюстей
при разном количестве зубов, а также в степени развития отдельных
мышц, что связано с преобладанием жевательной или хватательной
функции в жевательном аппарате, когда усиливается то одна, то другая
мышца. Наблюдаемые различия в мышцах характеризуются количест-
венными показателями: больше—меньше, что определяет в каждом от-
дельном случае их качественное значение при функционировании.
Прп общем сходстве жевательного аппарата у ежовых гимнуры за-
метно отличаются удлиненной мордой, что обусловлено большим числом
зубов у них и что в свою очередь связано с добыванием относительно мел-
кой и подвижной добычи. При этом важное значение имеет ее быстрое
охватывание, осуществляемое с помощью крупных первых резцов.
В связи с этим у мелких гимнур сильно развит височный мускул. У круп-
ных гимнур, питающихся соответственно крупными животными с более
грубыми тканями, требующими значительной их обработки на зубах,
сильно развит и жевательный мускул.
С переходом на многоядность,. когда пищей служит все, что оказыва-
ется съедобным, вплоть до плодов растений, что особенно характерно
ХАРАКТЕРИСТИКА ПОДОТРЯДА
65
для ежей, развился более универсальный жевательный аппарат.
Его формирование сопровождалось уменьшением количества зубов,
особенно в нижней челюсти и соответственно уменьшением длины че-
люстей, а вместе с этим усилением жевательного давления 'на зубы
с помощью жевательных мускулов. Захват и удержание добычи челю-
стями осуществляется сильным височным мускулом и жевательным,
развивающими значительное давление нижних резцов на верхние.
Здесь описывается мускулатура жевательного аппарата ежей
(рис. 35, 36, А). Височный мускул (m. temporalis) у ежей разделен на
три части — переднемедиальную, основную и верхнескуловую. Деление
основной части на наружный и внутренний слои выражено слабо. Передне-
медиальная часть височного мускула прикрепляется к переднему краю
сагиттального гребня широкой плотной связкой, а сама мышца, сужаясь,
переходит на переднюю сторону венечного отростка нижней челюсти
и прикрепляется у его основания. Основная часть этого мускула прикреп-
ляется к сагиттальному, височному и затылочному гребням, покрывая
значительную поверхность мозговой капсулы. Передняя часть этого
мускула крепится к венечному отростку нижней челюсти. Отдельные
пучки волокон глубокого слоя этой части височного мускула крепятся
к сочленовному отростку на его внутренней стороне. Верхнескуловая
часть височного мускула (m. temporalis pars suprazygomaticus) хорошо
развита. Эта часть представляет собой два мускульных тяжа, из которых
один прикрепляется к верхней стороне скуловой дуги в ее задней части,
а другой к основанию венечного отростка. Височный мускул приводит
нижнюю челюсть к черепу, закрывая рот. Он служит также укреплению
нижней челюсти у черепа, поскольку его сочленовный отросток только
соприкасается с височной костью, не имеющей сочленовной впадины.
Особое значение как подвеска нижней челюсти имеет его верхнескуловая
часть. Эту же функцию выполняет и переднемедиальная часть этого
мускула, а также и основная часть.
Жевательный мускул (m. masseter) разделен на наружную и внутрен-
нюю части. Разделяющая их фасция слабо развита. Внутренняя часть
в свою очередь делится на переднюю и заднюю доли. У ежей этот мускул
массивный и широкий, прикрепляется к скуловой дуге на ее наружной,
нижней и внутренней сторонах. При этом на наружной и нижней сторонах
скуловой дуги прикрепляется верхний конец наружной части этого мус-
кула, а на внутренней стороне скуловой дуги крепится внутренняя часть
жевательного мускула. Нижний конец этого мускула прикрепляется
к наружной стороне нижней челюсти на массетерной площадке и наруж-
ной поверхности углового отростка. Этот мускул несет основную, жева-
тельную функцию, оказывая давление на коренные зубы.
Внутренний крыловидный мускул (m. pterygoideus internus) у ежей
сильно развит. Он прикрепляется к крыловидной кости и внутренней
поверхности сочленовного отдела нижней челюсти и углового отростка.
Этот мускул двигает нижнюю челюсть вперед и вверх, прижимая ее
к черепу.
Наружный крыловидный мускул (m. pterygoideus externus) состоит
из двух частей — массивной внутренней и более тонкой наружной.
Внутренняя часть этого мускула прикрепляется к крыловидному отростку
клиновидной кости латеральное внутреннего крыловидного мускула,
а наружная часть к височным отросткам клиновидной кости. На нижней
челюсти обе части этого мускула прикрепляются на внутренней стороне
сочленовного отростка вблизи мыщелка. Оба крыловидных мускула обес-
печивают смещение нижней челюсти в боковом направлении.
5 А. А, Гуреев
66
ПОДОТРЯД ERINACEI
Двубрюшный мускул (m. digastricus) прикрепляется к яремному
отростку затылочной кости, а передней частью — к нижней стороне ветви
нижней челюсти на уровне М2. Этот мускул открывает рот.
Кротовые. При общем сравнительно сходном строении зубов отдельные
группы кротов имеют ряд специфических особенностей в их структуре,
что обусловлено частными приспособлениями к захвату и обработке пищи
на зубах (см. рис. 99, 126, 146). Известное разнообразие наблюдается
в строении отдельных зубов, а также в строении зубных рядов. Общее
число зубов 44—34. Уменьшение их общего числа в ряду происходит
за счет редукции передних премоляров, реже за счет резцов и нижних
клыков, что приводит к укорочению длины челюстей. Последнее может
происходить и в результате уменьшения длины зубов или изменения их
строения (приобретение узкоцилиндрической, узкоклиновидной формы).
Удлинение челюстей связано с редким расположением зубов впереди
от моляров или обусловлено образованием небольших диастем между
отдельными группами зубов.
Уменьшение длины челюстей, как представляется, служит сокращению
длины рычагов и соответственно увеличению давления зубов антагонистов
при обработке пищи на коренных зубах или в резцовом отделе при захвате
пищи. Удлинение челюстей, как можно думать, связано с захватом че-
люстями мелкой добычи, как пинцетом.
Более близкой к исходной для кротов как по строению зубов й зубных
рядов, так и по численному составу зубов представляется зубная система
кротов трибы Talpini, в частности обыкновенного крота — Talpa еиго-
раеа.
Верхние зубы. Резцы уплощены в передне-заднем направлении.
I1 крупнее I2 и I3. Все резцы с одним корнем, расположены на передней
стороне межчелюстной кости, по дуге. Клык кинжаловидный, сравни-
тельно плоский, с двумя корнями, с заметной продольной впадиной на
наружной стороне. С внутренней стороны клык имеет продольный гре-
бень и глубокую выемку у передней стороны. При основании коронки
клыка, на его задней стороне, имеется конический выступ или зубец.
Следует отметить, что у кротов имеет место образование дополнительных
зубцов при основании коронки не только на клыках, но и резцах.
Р1, Р2 и Р3 сходны по внешнему виду, мелкие, с двумя корнями; па-
ракон у них не развит, а метакон в виде мелкого бугорка. Р1 выше Р2
и Р3, с отогнутой назад вершиной протокона. Р2 ниже Р1 и Р3. Р3 наиболее
длинный и массивный.
Р4 развит в высоту более чем в длину, остроконечный, с тремя кор-
нями. Протоков этого зуба с продольной бороздой на передней стороне.
Пара- и метакон — мелкие бугорки при основании протокона. Все три
бугра имеют последовательное расположение, сходное с таковым передних
премоляров. Корень протокона мало выступает на внутреннюю сторону
челюсти.
Моляры треугольной формы, с тремя корнями. М1 более развит в длину,
чем в ширину. Его паракон на наружной стороне имеет слабо выражен-
ную впадину. Метакон с более глубокой впадиной и длинным отростком,
направленным назад. В средней части коронки он образует остроконеч-
ный зубец. Протокой низкий, треугольный, напоминает, носовую часть
лодки. М2 в ширину развит более, чем в длину. Его пара- и метакон имеют
глубокие треугольные выемки, в совокупности они образуют на жева-
тельной поверхности этого зуба W-образную фигуру. Протокой М2 сходен
с таковым М1, но с менее широким основанием. М3 несколько напоминает
строение М2, но метакон у него подвергся редукции, наружный край
ХАРАКТЕРИСТИКА ПОДОТРЯДА 67
зуба скошен и повернут к задней стороне черепа. Протокон этого зуба
сходен с протоконом М2. М3 самый мелкий из моляров.
Нижние зубы. Резцы лопаточковидной формы, с одним корнем.
Расположены на передней стороне нижней челюсти, задние резцы от-
части и на передне-боковой стороне ее ветви. Клык по форме сходен
с резцами, с одним корнем', расположен позади 13 и прижат к нему, до-
полняя резцовый отдел зубного ряда. Рх самый высокий из нижних пре-
моляров, с двумя корнями, имеет внешнее сходство с клыком. Его пара-
конид не развит, а метаконид в виде гребнистого выступа. Между этим
зубом и клыком небольшой промежуток. Р2, Р3 и Р4 сходной формы,
увеличиваются в размерах спереди назад, с двумя корнями. Параконид
у этих зубов представлен небольшим бугорком, а метаконид значительно
развит, в виде выступа.
Моляры W-образной формы, с мелкими отростками, похожими на
ушки, на переднем и заднем краях внутренней стороны коронки зуба.
Соотношение верхних и нижних зубов при сомкнутых челюстях сле-
дующее. Нижние резцы и клык располагаются позади верхних резцов.
Верхний клык располагается между нижним клыком и Рх, последний
помещается между верхним клыком и Р1. Р2 помещается между Р1 и Р2,
а Р8 между Р2 и Р3, Р4 между Р3 и Р4. Первый сегмент Мг между Р4 и М1,
его второй сегмент проходит между пара- и метаноном. Мх опирается
на протокон М1. Первый сегмент М2 помещается между М1 и М2, а его
второй сегмент между пара- и метаконом Ма, опирается на протокон М2.
Первый сегмент М3 — между М2,и М3, а второй сегмент на протоконе М3.
Нижняя челюсть кротов низкая и длинная или сравнительно корот-
кая с тремя лопастями в сочленовном отделе, представляющими собой
венечный, сочленовный и угловой отростки, расположенные по радиусам,
с вершинами, почти равно удаленными от основания. Только у выхухо-
лей сильно развитый венечный отросток резко выделяется своими круп-
ными размерами. Степень развития того или иного отростка отражает
степень развития жевательной мышцы или мышц, прикрепляющихся
к нему, и соответственно функциональное значение этих мышц. При по-
вышении значения массетера увеличивается в размерах угловой отросток
или расширяется его поверхность и на ней развиваются гребни. При уси-
лении функции височного мускула увеличивается в размерах венечный
отросток.
Удлинение венечного и углового отростков, а также увеличение угла
расхождения их с сочленбвиым отростком и точкой его опоры на черепе
определяют выгодное направление тяги мышц для наибольшего давле-
ния нижних зубов на верхние в области резцов, премоляров или моляров,
а также связано с ускорением вращения челюсти на сочленовном мыщелке
для захвата добычи. Можно думать, что глубокие и широкие вырезки
между отростками служат уменьшению веса челюсти при сохранении
его общей большой поверхности.
Подавляющее большинство кротов захватывает пищу резцами. Только
кроты трибы Talpini сохраняют крупные клыки и пользуются ими для
захвата пищи. Однако нижние клыки у них мелкие, прижаты к резцам
и при сомкнутых челюстях не соприкасаются с верхними. Функцию ниж-
него клыка выполняет у этих кротов крупный Р15 но при сомкнутых
челюстях он располагается позади верхнего клыка. Таким образом,
функционирование клыков у кротов отличается от такового хи: ;ных
млекопитающих (Carnivora), у которых нижний клык входит между Is
и С. Верхний клык и Pi кротов, видимо, не только захватывают добычу,
но и разрезают ее острыми встречными ребрами. Подавляющее боль-
шинство специализированных землероев — длиннорылые. Одни из них
5*
68
ПОДОТРЯД ERINACEI
имеют крупные клыки (верхние и удлиненные, треугольной формы пре-
моляры, другие имеют мелкие верхние и нижние клыки, не отличающиеся
по форме от премоляров, а последние из них в виде столбиков или узких
клиньев образуют ряд, напоминающий гребенку. У всех кротов пре-
моляры дробят пищу (хитин насекомых, панцирь ракообразных и др.).
При всем разнообразии зубов и зубных систем верхние коренные
зубы у всех кротов —• 3-бугорчатые, что резко отличает их от многих
групп насекомоядных. Нужно отметить также, что их верхние и нижние
коренные имеют на жевательной поверхности острые грани и бугры, что
Рис. 35. Схема расположения жевательных и лицевых мускулов.
А — Ermaceus europaeus, Б — Desmana moschata, В — Talpa еигораеа, Г — Mogera robusta; 1 —
m. levator nasi inferior, 2 — m. levator nasi superior, 3 — m. levator labi, 4 — m. zigomaticus major,
5 — m. zigomaticus minor, 6 — m. masseter, 7 — m. temporalis, 3 — m. temporalis pars suprazigoma-
tlcus, S — m. digastrlcus (по: Никольская, 1965).
позволяет предполагать нарезание и раздробление пищи, а не ее пере-
тирание. При этом передний сегмент нижнего коренного, располагаясь
между смежными коренными верхней челюсти, режет или дробит пищу,
а его задний сегмент, опираясь на протокон заднего из двух смежных
верхних коренных зубов, сминает ее. Такое впечатление возникает,
когда рассматриваешь отпечатки зубов антагонистов на пластилине.
Кроты, менее приспособленные к рытью и наземные — бегающие,
имеют укороченный лицевой отдел черепа, массивную скуловую дугу,
укороченные зубные ряды и более чем у специализированных землероев
мощную нижнюю челюсть. Таким образом, жевательный аппарат этих
кротов приспособлен к обработке сравнительно грубых по механическому
составу кормов, что очень важно при малых размерах их тела.
Хорошо роющие кроты, приспособленные и к плаванью, имеют зубы,
позволяющие схватывать водных животных со скользкой поверхностью
тела. Первые резцы у них полого направлены вперед, имеют вогнутую
поверхность, обращенную навстречу: верхних к нижним, а нижних
к верхним. Их клыки и премоляры с заостренными вершинами, отогну-
тыми назад (щучьи зубы). Эти приспособления зубов наряду со складкой
кожи, расположенной вдоль щечной стороны зубных рядов верхней и
нижней челюстей, служащие своего рода изоляцией ротовой полости
от водной среды (рис. 130) во время захвата добычи (иного толкования
ХАРАКТЕРИСТИКА ПОДОТРЯДА
69
значению этих образований дать нельзя), позволяют предполагать веду-
щую роль водных животных в пищевом рационе этих кротов.
Выхухоли, приспособленные к питанию водными организмами, осо-
бенно моллюсками, имеют сильно развитые I1 и значительно развитые 12.
Их клыки и премоляры с широким основанием и тупой вершиной служат
для раздробления раковин моллюсков, панцирей раков и хищных водных
жуков. Кожные складки в полости рта у выхухолей развиты сильно
(рис. 115).
Жевательная мускулатура (рис. 35—38)-1 Височный
мускул (m. temporalis). У мускула слабо разграничены, внутренний
и наружный слои. Его передняя часть разделена на три фракции, из
' Рис. 36. Схема расположения крыловидных мускулов.
А — Erinaceus auritus, Б — Desmana moschata, В — Talpa еигораеа, Г — Mogera robusta', 1 — m. pte-
rygoideus externus pars lateralis, 2 — m. pterygoideus externus pars medialis, з — m. pterygoideus
internus (по: Никольская, 1965).
которых передневнутренняя слабо отделена от основной части мускула
и в виде узкого тяжа располагается на передней стороне венечного от-
ростка нижней челюсти, прикрепляясь к его основанию. Вторая фрак-
ция прикрепляется к верхней части венечного отростка, а третья, в виде
двух пучков волокон, прикрепляется одним пучком на внутренней сто-
роне скулового отростка височной кости, другие на наружной стороне
венечного отростка нижней челюсти.
Основная часть височного мускула прикрепляется к сагиттальному
гребню, а если он не развит, — широкой фасцией вдоль средней линии моз-
говой капсулы к костям черепа в области затылка и вдоль височно-темен-
ного шва, покрывая значительную площадь верхней стороны мозговой
капсулы. Поднимая нижнюю челюсть, этот мускул развивает значитель-
ное давление в переднем отделе зубо-челюстного аппарата при захвате
пищи. Вместе с массетером он выполняет и жевательную функцию.
Жевательный мускул (m. masseter). Слабо дифференцирован на две
части — наружную и внутреннюю, между которыми отсутствует заметно
развитая фасция.
Внутренняя часть этого мускула делится на переднюю и заднюю
доли. У кротов массетер имеет форму веретена, в разной степени удли-
ненного и утолщенного в средней части. Особенно он укорочен и утолщен
у короткомордых кротов. На черепе этот мускул прикрепляется вдоль
1 Описание жевательной мускулатуры сделано по таковой Talpa еигораеа L.,
Urotrichus talpoides Теппп., Galemys pyrenaicus Geoff, и Desmana moschata L.
70
ПОДОТРЯД ERINACEI
скулового отростка верхнечелюстной кости и к скуловой дуге. На ниж-
ней челюсти он крепится на массетерной площадке, основании венечного
и сочленовного отростков и на наружной стороне углового отростка.
Внутренний крыловидный мускул (m. pterygojdeus intemus) на че-
репе прикрепляется к крыловидной кости, а на нижней челюсти на внут-
ренней поверхности ее углового отростка. Этот мускул тянет ветвь ниж-
ней челюсти вперед и вверх, смещая ее вовнутрь, отклоняя коронарный
отросток кнаружи.
Наружный крыловидный мускул (m. pterygoideus externus) состоит
из двух частей: внутренней, прикрепляющейся на черепе по соседству
с внутренним крыловидным мускулом, и наружной, которая на черепе
Рис. 37, 38. Схема строения m. digastricus у кротов (37) Talpa еигораеа (4), Т. altaica
(Б), Mogera robusta (В), перекрест дополнительных мышечных волокон (по: Николь-
ская, 1965) и у Galemys pyrenaicus (38).
крепится к височному отростку основной клиновидной кости. На нижней
челюсти обе части прикрепляются к основанию ее сочленовного отростка.
Этот мускул тянет нижнюю челюсть главным образом вверх, прижимая
ее заднюю часть к черепу.
Двубрюшный мускул (m. digastricus) на черепе прикрепляется
к яремному отростку. В этой части он широкий и плоский. В передней
части он переходит в плотное сухожилие, прикрепляющееся к нижней
стороне ветви нижней челюсти, где врастает в надкостницу. У кротов
двубрюшный мускул одной стороны дает отросток в виде ленты, перехо-
дящей на мускул другой стороны. Отростки мускула образуют перекрест
(рис. 37). Двубрюшный мускул тянет нижнюю челюсть вниз, открывая
рот.
Подниматели носа (m. levator nasi superior et inferior) — их основная
часть удлиненной формы, к черепу прикрепляется на височной кости над
ушным отверстием. В передней части оба мускула переходят в сухожилия,
врастающие в ткани носа и верхней губы. Эти мускулы поднимают хо-
боток.
Скуловые мускулы и подниматель губы (m. zygomaticus major et
minor et m. levator labi) прикрепляются к скуловой дуге, в передней
части располагаются ниже поднимателя носа. Они поднимают верхнюю
губу.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
71
У выхухолей в жевательной мускулатуре, да и в поднимателях носа
и губы имеются некоторые отличия и особенности, которые целесообразно
описать отдельно.
Например, внутренний крыловидный мускул в отличие от кротов на
черепе прикрепляется не только к крыловидной кости, но и к крыловид-
ному отростку основной клиновидной кости, а наружный крыловидный
мускул одной частью крепится рядом с внутренним, а другой частью
к височному отростку основной клиновидной кости.
Двубрюшный мускул у выхухолей в передней части делится на две
ветви, из которых одна, более короткая, прикрепляется к угловому
отростку нижней челюсти, а другая — к нижней стороне ее зубной части
на уровне Mj. У выхухолей, как и у кротов, имеется перекрест ленто-
видных отростков мышцы, переходящих с одной стороны на другую.
Прикрепление двубрюшной мышцы к угловому отростку и зубной части
ветви нижней челюсти связано, видимо, с необходимостью отодвигать ниж-
нюю челюсть назад при открывании рта, поскольку при сомкнутых че-
люстях нижние резцы плотно прижаты к верхним. В этом нетрудно убе-
диться, взяв в руки череп выхухоли.
Подниматели носа у выхухолей крепятся во впадине, образованной
гребнями теменной и височной костей, в височно-затылочной области че-
репа. По соседству с этими мышцами прикрепляются скуловые мышцы
и подниматель губы.
Землеройковые. Зубная система землероек представляется наиболее
специализированной среди насекомоядных, как и их весь жевательный
аппарат. Его эволюция шла по пути усиления хватательной функции,
что обусловлено питанием преимущественно мелкими, очень подвижными
животными, главным образом насекомыми. В связи с этим формировались
своеобразные его особенности, выраженные в строении зубов, челюстей
и мускулатуры и соответственно и в их функциях. Можно считать, что
строением жевательного аппарата и особенностями терморегуляционной
системы землероек определились возможности для их широкого рассе-
ления и приспособления к условиям разнообразных ландшафтов и
климатов.
Описываемые ниже структурные и функциональные особенности же-
вательного аппарата землероек сложились относительно рано — в конце
эоцена, так как у олигоценовых видов он был уже значительно специали-
зирован.
Несмотря на кажущееся однообразие зубных систем землероек, обус-
ловившее представление об однообразии видовых адаптаций, имеют место
значительные различия в характере их функций, чем и определяется разно-
образие частных приспособлений видов и более крупных таксономиче-
ских подразделений этой группы насекомоядных.
Различие в общем количестве зубов у землероек разных родов (44—24),
обусловленное разным числом верхних и нижних промежуточных зубов,
породило значительную путаницу в их наименовании, а также и в пред-
ставлении о числе резцов, премоляров и моляров. Выяснение этого
вопроса затруднено тем, что швы на черепе, особенно в лицевой части,
у землероек рано зарастают и граница между межчелюстной и верхне-
челюстной костями не видна, а это мешает установлению границы между
клыком и резцами. Брандт (1878), специально занимавшийся этим
вопросом, пришел к выводу, что у представителей рода Sorex L. в верх-
ней челюсти 8 резцов, а у видов рода Crocidura Wagl. их 6. Он считал
также, что коренных (моляров) в верхней челюсти у всех землероек по 4.
Добсон (Dobson, 1882—1890) тоже признавал по 8 резцов в верхней че-
люсти у видов некоторых родов, но моляров у всех насчитывал только 3.
72
ПОДОТРЯД ERIN АСЕ I
Сравнением наиболее полных зубных систем землероек с зубными си-
стемами представителей других семейств насекомоядных и последних
между собой удалось установить, что резцов в верхней челюсти у всех
насекомоядных только 6, по 3 с каждой стороны. Землеройки не могут
быть исключением из этого общего правила. Когда было установлено
количество резцов и моляров, которых тоже по три с каждой стороны
у всех насекомоядных, а также постоянное присутствие последнего пре-
моляра, сходного по строению у всех групп землероек, оставалось опре-
делить название трех промежуточных зубов в наиболее полном зубном
ряду у видов рода Sorex. Редукция промежуточных зубов у землероек
Рис. 39. Типы сочленения нижней челюсти с черепом у землероек, например у видов
рода Sorex (4) п у видов рода Blarina (В).
а — верхняя, б — нижняя сочленовные фасетки.
6
начинается с Рз, что хорошо заметно у видов многих современных родов,
например Crocidura, Diplomesodon и др. (рис. 40). Обусловлено это уси-
лением передней части зубного ряда в связи с развитием хватательной
функции. У современных землероек с наиболее полным зубным рядом Р§
нет. Как шла редукция верхних промежуточных зубов у представителей
разных родов, хорошо иллюстрируют приведенные их зубные формулы:
I с р
Предки........................... 3 1
Sorex araneus L.................. 3 1
Neomys fodiens Penn. .... 3 1
Crocidura suaveolens Pall. ... 3 1
Diplomesodon pulchellum Licht. 3 0
4
3
2
1
1
M
3
3
3
3
3
В нижней челюсти постоянно 3 моляра. Зуб, примыкающий к первому
моляру, — последний премоляр. Рядом с внутренним резцом располага-
ется второй резец. Следовательно, в нижнем зубном ряду, как и в верхнем,
редукции подверглись первый, второй и третий премоляры, клык,
а также и третий резец, и только у видов родов Myosorex Gray и Scuti-
sorex Thos. сохраняется клык в виде мелкого вставочного зуба, а у оли-
гоценовых и миоценовых форм сохранялся и Pv
Строение отдельных зубов землероек весьма своеобразно и специфично
для них, хотя многие из коренных имеют сходство с коренными зубами
кротов, что несомненно связано с их общим происхождением, а также
со сходством функций.
Верхние зубы. Внутренний резец (I1) с широким основанием
и двумя зубцами (вершинами), из которых передний крупнее заднего,
серповидно изогнут, расположен на боковой стороне межчелюстной кости.
I2 и I3 одновершинные, в виде конусов с широким основанием, как и сле-
дующий за ними клык, а также Р1 и Ра. На нижней части задней стороны
ХАРАКТЕРИСТИКА ПОДОТРЯДА
73
этих зубов имеется выемка, в которую входит передний край основания
следующего за ним зуба. I2, Is, С, Р1 и Р2 сходны по форме, но разнятся
по величине. У видов, питающихся не только мелкими, но и относительно
крупными животными (мелкие позвоночные), I2 бывает очень массивным
и высоким, а I3 и клык мелкими. У многих видов клык и премоляры,
кроме Р4, полностью редуцированы. Воротнички (цингулюмы) у проме-
жуточных зубов, как, впрочем, и у других зубов, сильно развиты, они
прикрывают десну и служат, видимо, для ее защиты от поранений. Кроме
того, они несут опорную функцию, увеличивая основание зуба. Проме-
жуточные зубы служат для раздробления хитина насекомых, раковин
моллюсков, оболочек семян и др. Этим, возможно, и объясняется их ко-
ническая форма и сходство в строении. Изогнутый внутренний резец
служит для схватывания добычи. У видов, питающихся животными
со скользкой поверхностью тела (рыба и ее икра, водяные жуки и др.),
промежуточные зубы имеют глубокую выемку на задней стороне, что
придает им форму крючков, концами направленных назад.
Последний премоляр сильно моляризован. Он 4-бугорчатый с 4 кор-
нями. Наружная стенка этого зуба укреплена эмалевыми валиками, окан-
чивающимися острыми зубцами. Протокон его выступает вперед далее
паракона, его корень массивный. Талон со скошенным в разной степени
задним краем. Вершины протокона и талона нижи вершин пара- и мета-
кона, с вогнутой верхней поверхностью и эмалевым гребнем на внутрен-
нем крае коронки зуба.
М1 и М2 почти четырехугольной формы, с W-образной выемкой на на-
ружном крае. Протокон и талон в совокупности образуют вогнутую же-
вательную поверхность, ограниченную на внутренней стороне коронки
эмалевым гребнем. М3 с тремя вершинами и тремя или двумя корнями,
сильно редуцирован, что обусловлено его ограниченной жевательной
функцией. Вследствие низкого расположения протокона и талона по от-
ношению к высоте пара- и метакона жевательные поверхности Р4, М1
и М2 скошены на внутреннюю сторону челюсти. Эти зубы разрезают
и раздавливают пищу. Перетирать ее они не могут, так как нижняя че-
люсть способна двигаться только сверху вниз. Вперед и в стороны она
двигаться не может из-за сцепления острых граней и бугров на жеватель-
ных поверхностях верхних и нижних зубов-антагонистов.
Нижние зубы. Ijj длинные, направлены вперед и слабо изо-
гнуты. Их верхняя сторона ровная или с 1—3 выступами или бугорками.
Эти бугорки у землероек с большим числом промежуточных зубов в верх-
ней челюсти выполняют функцию утраченных нижних промежуточных
зубов. 12 имеет удлиненную в продольном направлении форму. Р4 также
удлинен, с 1—2 вершинками. Все моляры сходного строения, с W-образно
расположенными эмалевыми гребнями на жевательной поверхности.
Взаиморасположение верхних и нижних зубов, а также эмалевых
образований на их жевательных поверхностях при сомкнутых челюстях
может быть представлено в следующем виде (зубная система Sorex ага-
neus L.). Передняя часть режущего края 14 упирается в заднюю поверх-
ность переднего зубца I1. Передний зубец на верхней поверхности 14 рас-
полагается между задним зубцом I1 и I2. 2-й — между I2 и Is; 3-й — между
I3 и клыком. 12 входит в промежуток между верхним клыком и Р1.
Р4 передней вершиной упирается в задний край Р2, а задней — в прото-
кон Р4. Следует отметить, что все конические части перечисленных ниж-
них зубов не располагаются строго в промежутках между верхними
зубами, а сдвинуты несколько вперед и своими передними краями йсодят
в выемки на задней стороне верхних промежуточных зубов. Передняя
часть Мг опирается на талон Р4, а задняя часть Мх — на протокон М1.
74
ПОДОТРЯД ERINACEI
М2 и М3 также одной частью опираются на талон предыдущего верхнего
моляра и паракон последующего. Эмалевые гребни жевательных поверх-
ностей верхних и нижних моляров при закрывании рта проходят друг
возле друга под разными углами, что способствует лучшей обработке
пищи на этих зубах.
Переход к питанию не только жесткой пищей, но и волокнистой, на-
пример мелкими позвоночными, сопровождался следующими изменениями
в строении жевательной поверхности моляров: гребни входящих складок
на верхних молярах отодвигаются к наружному краю коронок и стано-
вятся 1иенее высокими, что приводит к увеличению площади, на которую
и опираются притупленные гребни нижних моляров. При этих условиях
пища не только нарезается, но и раздавливается.
Рис. 40. Число и форма верхних промежуточных зубов у землероек.
I — Sorex агапеив, 2 — Neomys fodiens, 3 — Suncus etruscus, 4 — Crocidura leucodon, 5 — Diplomesodon
pulchellum (из: Dobson, 1890).
В связи с приспособлением жевательного аппарата землероек к бы-
строму схватыванию добычи находится строение нижней челюсти. Важ-
ное значение верхних и нижних внутренних резцов, схватывающих
как пинцетом мелкую подвижную добычу в виде насекомых, привело
к значительному увеличению их размеров. Сильное удлинение нижних
резцов обусловило сокращение длины нижней челюсти впереди от сочле-
новного отростка, что повлекло за собой редукцию части премоляров
И клыка, утрату которых компенсировали бугорки, развившиеся на верх-
ней стороне нижних внутренних резцов. Ёыстрое схватывание требует
ускорения движения нижней челюсти и усиления ее давления в резцовой
части. Этим и обусловлено удлинение плечей рычагов, вращающих че-
люсть в сочленовной впадине при закрывании рта, — венечного и угло-
вого отростков. Сильное давление сверху вниз, которое испытывает сочле-
новный отросток нижней челюсти при закрывании рта мощным височным
мускулом, крепящимся на длинном венечном отростке, обусловило разви-
тие на первом нижней сочленовной фасетки и соответствующей опоры
для нее, которой служит разросшийся засочленовный отросток височной
кости. Последний, как и нижняя сочленовная фасетка, у представителей
триб Heterosoricini, Blarinini и Anourosoricini, смещены далеко вперед.
Это позволяет им при переводе сочленовного отростка в крайнее заднее
положение и при незначительном напряжении жевательных мышц дер-
жать челюсти сомкнутыми. Механизм такого замыкания челюстей заклю-
чается в том, что расположенная далеко впереди нижняя фасетка сочле-
новного отростка нижней челюсти не может вращаться на засочленовном
отростке височной кости до тех пор, пока верхняя сочленовная фасетка
ХАРАКТЕРИСТИКА ПОДОТРЯДА
75
сочленовного отростка не перейдет в крайнее переднее положение и пере-
станет упираться в верхнюю стенку сочленовной впадины на височной
кости (рис. 39).
Описываемое приспособление имеет важное значение при схватывании
крупной добычи. С ускорением движения нижней челюсти и усилением
ее давления в резцовой части связано уменьшение площади прикрепле-
ния верхней части массетера, что уменьшило его инерцию и соответственно
вызвало редукцию скуловой дуги, к которой он прикреплялся значитель-
ной частью своей поверхности. Прикрепление массетера только в двух
точках — к скуловому отростку верхнечелюстной кости и к длинному
42
Рис. 41. Схема расположения жевательных и лицевых мускулов у землероек.
1 — m. masseter, е — tn. temporalis, 8 — m. levator nasi, 3a — pars superior, 36 — pars Inferior, 4 —
m. temporalis pars suprazigomaticus, 5 — m. zigomatlcus, 6 — m. digastricus.
Рис. 42. Крыловидные мускулы: 1 — m. pterygoideus: pars lateralis (а) и pars medialis
(6), 2 — pterygoideus internus.
угловому отростку нижней челюсти — обусловило его расположение
под меныпим углом к продольной оси черепа, что позволило ему при со-
кращении усилить давление нижних резцов вперед и вверх, а также уско-
рить вращение нижней челюсти при закрывании рта.
Жевательная мускулатура. Височный мускул (m. tem-
poralis, рис. 41) — наиболее крупный и сильный из всех жевательных
мышц землероек. Он состоит из нескольких слоев и основной частью рас-
полагается на всей поверхности теменной и межтеменной костей. В перед-
ней части он прикрепляется к венечному отростку нижней челюсти на его
верхней и внутренней сторонах. Особая часть этого мускула, которой
часто дают название suprazygomaticus, обособлена от его основной массы
и соединена с ним лишь в основании. Она имеет форму изогнутой ленты,
расположенной на нижнем крае височного мускула, боковой стороне
венечного отростка и при основании последнего прикрепляется к нижней
челюсти. Эта порция височного мускула у землероек очень слабая и предч
ставляет собой, видимо, лишь прослойку между основной частью височ-
ного мускула и скуловым мускулом, исключающую их взаимное тормозное
действие. Височный мускул служит основным закрывателем рта. Его
сильное развитие связано с ускорением движения нижней челюсти и ее
76
ПОДОТРЯД ERINACEI
значительным давлением на верхнюю челюсть в передней части при схва-
тывании добычи и на моляры при обработке пищи на зубах.
Массетер, или собственно жевательный мускул (m. masseter, рис. 41),
двуслойный, реже однослойный, передней частью прикрепляется к ску-
ловому отростку верхнечелюстной кости, задней — к палочковидному
угловому отростку нижней челюсти. Сокращаясь, этот мускул поднимает
нижнюю челюсть вверх и вперед. Он помогает височному мускулу быстро
поднимать нижнюю челюсть вверх, ускоряя ее вращение, и усиливает
ее давление в резцовой части.
Наружный крыловидный мускул (m. pterygoideus externus, рис. 42)
состоит из двух частей — внутренней и наружной. Обе части передними
концами прикрепляются к крыловидной кости, а задними — к сочленов-
ному отростку нижней челюсти. При сокращении этот мускул тянет
нижнюю челюсть вперед и вверх.
Внутренний крыловидный мускул (m. pt. internus, рис. 42) не диффе-
ренцирован на отдельные части (или слои) и представляет собой единую
мышцу. Его передний конец прикрепляется к крыловидной кости, а зад-
ний — к угловому отростку нижней челюсти. При сокращении этот
мускул тянет нижнюю челюсть вперед и вверх, а нижние края ее ветвей
к средней линии черепа, отклоняя при этом верхние части, особенно ве-
нечный отросток. Последнее весьма важно в связи с сильным притяже-
нием венечного отростка к черепу височным мускулом.
Двубрюшный мускул (m. digastricus, рис. 41) с толстыми брюшками
прикрепляется к яремным отросткам и к нижней челюсти в области сим-
физа ее ветвей. Двубрюшный мускул опускает нижнюю челюсть, откры-
вая рот.
МОРФОЭКОЛОГИЧЕСКИЕ ТИПЫ КАК ФОРМЫ ЖИЗНИ
Исследование всякой систематической группы, ее филогении на фоне
геологической истории и морфо-функциональных особенностей ее совре-
менных представителей неизменно приводит к вопросу: чем обусловлено
появление этой группы на земле? Естественно представить себе, что нет
побочных или случайных групп в эволюции. Каждая из них появляется
в том или ином отрезке времени в качестве плеяды специализированных
для своего времени видов, выполняющих в эволюции функцию времен-
щиков — носителей и накопителей нового или в роли своего рода инжек-
торов — конкурентов для анцестральных групп, определяющих главное
направление развития того или иного таксона. В течение какого-то
времени они находятся на подъеме, а затем приходят в упадок, оставляя
ограниченное количество видов, существующих разное по продолжи-
тельности время. Таким образом, эволюция представляет собой поток
преемственно-последовательных приспособительных изменений, отражен-
ных в адаптациях сменяющихся во времени видов. — появляющихся
и исчезающих групп. Следовательно, в эволюции существует закономер-
ная цикличность.
Ежовые появились в компании других семейств как группа «чистиль-
щиков», подбирающих с земли все, что они могут разжевать и переварить
из животной и растительной среды. Это главное, что определило возмож-
ность их существования в разнообразных условиях различных лянд-
шафтов на занимаемых ими территориях в значительном отрезке времени.
К верхнему эоцену определились две их группы — гимнуры (Galeri-
cinae) и ежи (Erinaceinae). К этому времени они уже имели все особенности,
свойственные каждой из них и в современный период.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
77
Внешний вид животного в значительной степени отражает особенности
его адаптации. Представители сем. Erinaceidae, особенно ежи, харак-
теризуются относительно массивным телом на коротких ногах. Этому
соответствует и их сравнительно малая активность. Последнее обстоя-
тельство обусловило развитие у ежей иглистого панциря как средства
пассивной защиты. Гимнуры имеют более стройный скелет, но и они
не отличаются, видимо, большой подвижностью. Возможность питания
разнообразной пищей, сравнительно легко добываемой, не Способствовала
развитию быстрого бега и соответствующих приспособлений в локомотор-
ном аппарате.
Отдельные виды приспосабливались к рытью, и в современный период
существует роющая гимнура Neohylomys hainanensis Shaw et Wong, но ее
приспособления носят характер начальных стадий формирования этой
формы локомоций. Большая гимнура Echinosorex gymnurus Raff, способна
плавать и добывать пищу в воде, но она не приобрела специальных при-
способлений к плаванию.
Остальные виды современных гимнур не имеют и этих ограниченных
приспособлений. Представители подсемейства гимнур передвигаются
рысцой или тяжелым галопом, как и другие млекопитающие со сходным
складом тела. Видимо, все гимнуры способны вырыть себе нору.
Современные ежи передвигаются шагом или рысью. Почуяв опасность,
сворачиваются в шар и перестают двигаться. Это приспособление у них
вырабатывалось, вероятно, параллельно с развитием открытых ланд-
шафтов еще в миоцене и оказалось полезным и в лесной зоне, где отдель-
ные виды приспособились к существованию. У видов, обитающих в пу-
стыне, крупные ушные раковины и более развитые слуховые барабаны.
Ряд костей, окружающих барабанную кость, вздуты, а у одного вида
вздуты и крыловидные кости. Укрытые соединительной тканью, эти
вздутые кости в совокупности образуют систему полостей, соединяющихся
со слуховой полостью, что, видимо, повышает акустические свойства
слухового аппарата.
Жевательный аппарат ежовых описан выше и здесь следует отметить
только некоторые его общие особенности. Разнообразие пищи, разной
по механическому составу, обусловило развитие коренных зубов жева-
тельного типа, значительно отличающихся по строению от таковых боль-
шинства групп насекомоядных. Жевательная поверхность этих зубов имеет
пять бугорков с притупленной вершиной. Нижняя челюсть массивная,
приводится к черепу сильной мускулатурой, особенно мощным Массете-
ром, прикрепляющимся к толстой скуловой дуге и широкой массетерной
площадке нижней челюсти.
В заключение следует отметить, что известный универсализм жеватель-
ного аппарата, видимо, в более позднее время затормозил тенденцию
к его изменению соответственно и в других системах, что привело к суже-
нию круга частной специализации и ограничению видообразования. Все
роды современных гимнур монотипичны, а в пределах рода ежей —
небольшое число видов при значительном ареале рода.
Появление кротов на земле как особой формы жизни было обуслов-
лено необходимостью ухода от условий, неблагоприятных в кормовом
отношении, возникавших, видимо, в отдельные сезоны года. Сем. Talpidae
несомненно формировалось в период, когда поверхность земли испытала
значительное охлаждение в послемеловое время. К этому времени многие
группы насекомоядных были широко распространены и подвергались
воздействию условий, которые складывались в разных климатических
поясах и вертикальных зонах гор, поднимавшихся в результате горо-
образовательных процессов. Многие группы кротов и сейчас распростра.
78
ПОДОТРЯД ERINACEI
йены в горных местностях. Уход под землю для многих групп млеко-
питающих — закономерное явление, а приспособление к рытью разное,
что обусловлено его разным значением для отдельных видов в их общем
жизненном цикле. У многих видов рытье связано с необходимостью укры-
тия от неблагоприятных условий на поверхности земли или для размно-
жения, реже они большую часть жизни проводят под землей, находя
в ней и пищу. Большинство кротов относятся к животным, которые прово-
дят всю жизнь под землей. Приспособление кротов к рытью началось,
видимо, с рытья в лесной подстилке для добывания пищи, что характерно
и для многих современных кротов трибы Urotrichini и подсем. Uropsilinae,
а также многих групп землероек. Рытье в лесной подстилке, в которой,
кроме насекомых и других беспозвоночных, в большом количестве встре-
чаются дождевые черви, привело к использованию их в пищу многими
насекомоядными млекопитающими. В отличие от других беспозвоночных
дождевые черви при высыхании или охлаждении лесной подстилки и верх-
них слоев почвы уходят в глубину почвенных слоев. Изменение количества
пищи в подстилке и верхних слоях почвы в зонах умеренного климата
в отдельные сезоны вынуждало кротов следовать за ними в почву. Это
и привело к необходимости рыть землю, а вместе с этим — к специализации
локомоторной системы к рытью как средству добывания корма, а в даль-
нейшем — к передвижению и расселению. Разная степень необходимости
в рытье в разных климатических зонах обусловила разную специализацию
кротов к рытью, а представители подсем. Uropsilinae pt не приспосабли-
вались к нему, сохранив до настоящего времени наземный образ жизни —
они бегают как землеройки (Soricidae).
Отдельные группы роющих кротов приспособились к плаванию,
например представители трибы Condylurini. Высокой специализации
к плаванию достигли выхухоли — подсем. Desmaninae.
Основным направлением эволюции представителей Сем. Talpidae
следует считать специализацию к рытью, наиболее высокую среди млеко-
питающих. Это направление сохранило ведущее значение и в современный
период. Быстрое передвижение в почве для добывания корма — необхо-
димое условие существования кротов. Роют кроты кистью и при прокла-
дывании глубоких ходов вращают тело по спирали, как будто ввинчиваясь
в прорываемый ход, обминая его стенки и выталкивая разрыхленную
почву задними ногами.
Короткая плечевая кость и короткие кости предплечья служат опорой
для кисти. Стабилизирует положение плеча в выгодном для работы ра-
курсе мощная мускулатура. Плечевой пояс у специализированных земле-
роев выдвинут вперед, в область шеи, что позволяет сближать передние
конечности, а вместе с этим увеличивать размах кистей в стороны, что
было бы невозможно при положении конечностей по бокам тела в области
грудной клетки.
Вынос вперед передних конечностей позволяет кротам убирать голову
между плечевыми костями и костями предплечья, освобождая пространство
для работы кистей впереди морды. Возможность выдвижения передних
конечностей в область шеи обусловлена значительным развитием ру-
коятки грудины (manubrium) вперед и приобретением ею значительной
прочности за счет развития верхнего и нижнего продольных гребней.
Короткая массивная ключица причленяется непосредственно к плечевой
кости. Она несет функцию опоры, удерживает плечевые кости в одной,
общей системе.
Возможность сближения кистей и неширокая расстановка задних
конечностей, при том что конечности, кроме кистей и ступней, погружены
в кожный мешок (кожный покров тела), способствуют сужению переднего
: ХАРАКТЕРИСТИКА ПОДОТРЯДА
79
и заднего отделов тела, а это позволяет кротам свободно передвигаться
в узких ходах вперед и назад. Этому же служат короткие волосы, по-
крывающие тело крота.
Орудием рытья кротам служит кисть. Она образована короткими
и широкими уплощенными костями. Пальцы соединены толстой кожей
до когтей. Когтевые фаланги раздвоены на конце и несут плоские, широ-
кие когти. Увеличению ширины кисти служит серповидная кость (falci-
forme), а также и жесткие волосы, покрывающие края кисти. Кисти
повернуты ладонями наружу. Ступни узкие. Вероятно, для некоторого
увеличения их поверхности, что весьма существенно для отбрасывания
почвы при рытье, служит косточка, сходная по значению, возможно
и происхождению, с серповидной костью, расположенная с внутренней
стороны ступни. Когти ступни, сжатые с боков, заостренные.
Скелет и мускулатура роющих кротов обладают значительной мощ-
ностью, что позволяет кроту выталкивать землю на поверхность почвы,
поднимать свод хода, прорываемого близко от поверхности почвы, в виде
валика и рыть большую часть суток, добывая корм.
Приспособление к плаванью у кротов развилось, видимо, из гребного
движения кистей в.почве во время рытья, но еще мало специализирован-
ного. Возможность использования роющей кисти для плавания наблю-
дается у всех кротов, особенно это выражено у Condylura cristata L., спе-
циализированного землероя и хорошо плавающего вида, добывающего
пищу в воде и в зимнее время подо льдом, покрывающим водоем.
Существенным подтверждением мнения о происхождении плавания
у кротов от роющей деятельности может служить, как мне кажется,
сходство в строении плечевого пояса, особенно лопаток, у специализиро-
ванных к плаванию выхухолей и роющих кротов. Например, форма
плечевых костей выхухолей сходна с таковой кротов, мало специализи-
рованных к рытью, но с более широким проксимальным и дистальным
отделами, а лопатка по строению напоминает таковую кротов, специали-
зированных к рытью, в чем нетрудно убедиться. Существенным отличием
служит сравнительно длинная ключица у выхухолей, причленяющаяся
не только к плечевой кости, но и к лопатке, а также ряд особенностей
в строении кисти и ступни, специфичных для плавающих форм. Из ска-
занного не следует, что выхухоли произошли от специализированных
землероев, а С. cristata не прообраз кротов, от которых развились вы-
хухоли. Можно, однако, с уверенностью сказать, что предки выхухолей
рыли.
Приспособление к плаванью у выхухолей выражено в обтекаемой
форме тела, покрытого волосяным покровом, не смачивающимся водой.
Между пальцами кисти и ступни у них развиты плавательные перепонки.
Пальцы ступни наклонены наружу, а очертания ступни напоминают лапу
птиц из отряда Cyconiformes, Хвост уплощен на большей части длины
или только на конце. Функцию движителя выполняют задние ноги, хвост
выполняет функцю рулевого управления, а передние конечности ускоряют
движение вверх и вниз и служат для поворотов по оси тела. Естественно,
что хвост может служить двигателем на короткой дистанции, при схваты-
вании добычи со дна, при прижатых к телу кистях и ступнях и при пас-
сивном погружении тела по наклонной линии.
При погружении в воду ноздри у выхухолей замыкаются. По мнению
Е. И. Ивановой (1961), в замыкании ноздрей принимают участие m. m. le-
vator labii superior et superior proprius, а также depressor nasis. По моим
представлениям, замыкателем служит мускул, который проходит вдоль
носовой перегородки в носовом канале и который хорошо виден на по-
перечном разрезе хоботка.
80
ПОДОТРЯД ERINACEI
Приспособление к пребыванию длительное время в воде и под землей
потребовало ряда приспособлений в дыхательной и кровеносной системах,
а также усиления функций кроветворных органов. Усложнилась и си-
стема терморегуляции.
Подробное освещение этих вопросов в настоящее время затруднено,
поскольку сведения в этой области все еще носят отрывочный характер.
Причиной служит ограниченное число исследований в затронутых здесь
областях физиологии, проведенных на малом количестве видов, часто
без сравнительного анализа результатов.
Жевательный аппарат у всех групп кротовых имеет сходное строение
и функции. Он более специализированный у выхухолей. У большинства
видов в структуре коренных зубов сохраняются древние особенности.
Частичной или значительной модернизации, начавшейся очень рано
(эоцен—нижний олигоцен), подверглись зубы с молочной сменой (резцы,
клыки и премоляры), выполняющие функцию захвата пищи и ее предва-
рительного раздробления. У отдельных видов и групп сократилось число
резцов, или премоляров (реже исчезают и нижние клыки). Сокращение
числа зубов привело к сокращению длины челюстей (рычагов) или со-
провождалось удлинением коренных зубов и Р*. Это позволило усилить
жевательное давление челюстей или улучшить обработку пищи на корен-
ных зубах, что очень важно, особенно для мелких кротов-, в частности
из подсем. Uropsilinae, в пределах которого это явление особенно распро-
странено.
У крупных кротов изменено положение премоляров в верхней и ниж-
ней челюстях. Они располагаются под углом к продольной оси челюсти,
что, возможно, обусловлено удлинением коренных зубов, а также служит
улучшению предварительной обработки пищи на премолярах или луч-
шему размельчению и раздроблению хитина и других покровов бес-
позвоночных.
По-видимому, сходство пищи по механическому составу на всем про-
тяжении развития кротов не способствовало значительному изменению
структурных особенностей их зубов. Они сохраняют многие архаичные
черты, например, верхние коренные — треугольной формы, с тремя
корнями (рис. 146). Вероятно, облик современных землероек мало отли-
чается от внешнего облика их давних предков. Это представление под-
тверждается сходством строения частей черепа и других частей скелета
ископаемых и современных видов и близкими размерами, что позволяет
предполагать, что землеройки всегда были мелкими животными. Одно-
образие внешнего облика мало вяжется с разнообразием ландшафтов
и условий жизни в пределах их широкого ареала, охватывающего все
зоны жизни от тундр до тропических лесов и саванн. Это можно объяснить
только относительным сходством микроусловий, выбираемых землерой-
ками в различных ландшафтах, например приземный слой раститель-
ности или мертвый покров почвы. Возможно, что и обособление землероек
от общих корней с кротами шло по линии освоения животного корма
в этих ярусах растительных формаций разных климатических поясов,
а кротов — под землей.
Можно считать, что как общий адаптивный процесс, так и частная
специализация в сем. Soricidae шли не по линии морфологического про-
гресса и внешнего разнообразия, а по пути расширения диапазона физио-
логических реакций на разнообразные условия среды. Это позволило
землеройкам выработать сложную систему функциональных отношений
между органами, связанными с обменом веществ, и органами термической
регуляции. Высокий темп обмена веществ, свойственный землеройкам
(Тупикова, 1949), позволил им нивелировать вредное влияние низких
ХАРАКТЕРИСТИКА ПОДОТРЯДА
81
температур в условиях высоких широт и освоить территории у крайних
пределов жизни.
Круг адаптаций землероек определялся не только обстановкой в ме-
стах обитания, но и экологическими особенностями их кормовых живот-
ных. Все перечисленные особенности землероек позволили им широко
расселиться во многих климатических зонах и сохранить круглогодичную
активность при самых неблагоприятных условиях.
Круглогодичная активность землероек — необходимое условие их су-
ществования. Обладая высоким темпом обмена веществ, землеройки не-
способны накапливать жир, а без него не могут впадать в спячку при
наступлении неблагоприятных условий.
Землеройки весьма подвижны, что связано с высоким энергетическим
обменом, необходимым для поддержания жизненного тонуса на уровне,
соответствующем условиям среды. Но высокая активность и повышенный
тонус жизненных процессов приводят к быстрой морфо-функциональной
депрессии организма, к его быстрому старению. Можно думать, что этим
и объясняется короткий период жизни землероек и быстрая смена особей
в популяциях за счет новых генераций. Это нельзя не рассматривать
как приспособление к колеблющимся условиям среды, как своего рода
выравнивание, если можно так выразиться, физиологии особей отдельных
популяций. Различие условий, в которых появляются и развиваются
особи отдельных генераций, обусловливает известную разность их физио-
логических особенностей и соответственно возможность лучшего при-
способления в то или иное время.
Ускорению течения жизненных процессов подчинено, видимо, уско-
рение морфогенеза (формирование еще в период эмбрионального развития
дефинитивной зубной системы, раннее слияние костей лицевой части
черепа и др.), что способствует ускорению развития и быстрому приобре-
тению самостоятельности особей в постэмбриональный период. При
всем этом, землеройки сохраняют много ювенильных черт в структурах
черепа и скелета. Развитие заканчивается только весной и летом следую-
щего года, когда наступает их половая зрелость. Это явление можно рас-
сматривать как приспособление к регулированию развития отдельных
систем и органов в соответствии с условиями среды. В связи с этим половое
созревание землероек представляет довольно пеструю картину. У одних
групп землероек, например у видов рода Neomys Каир, оно происходит
в год рождения, у видов рода Sorex L. в разное время. Отдельные самки
ранних генераций созревают в год рождения, а большинство самок
и самцы — на следующий год. Но в годы пониженной численности полово-
зрелыми становится большинство самцов и самок в год рождения. Таким
образом, степень общего развития и половое созревание у землероек стоят
в прямой зависимости от условий развития и физиологического состояния
родившихся особей, а также от времени их рождения (весна, лето, осень),
определяющих период, необходимый для окончательного развития. Все
периоды онтогенеза землероек подчинены задаче ускорения жизненных
функций организма как средству существования в разнообразных и ме-
няющихся условиях среды, а само существование видов с коротким жиз-
ненным циклом представляет поток быстро сменяющихся генераций.
В связи с этим стоят и известная простота внешнего строения тела, дери-
ватов кожи, и однообразие окраски, а также мелкие размеры. Все это
объясняется экономией времени, материала и энергии на развитие, но
за этим стоит значительный биологический прогресс, которому соответ-
ствует широкое распространение, разнообразие частных адаптаций и боль-
шое число особей видов.
6 А. А, Гуреев
82
ПОДОТРЯД ERINACEI
Существенным в общем комплексе адаптаций землероек можно считать
колебания численности и значительную амплитуду других форм измен-
чивости. Последние отражают не только случайные по отношению к на-
следственным уклонения в морфогенезе, но и влияние ряда условий среды
на ход развития. Все это создает гамму морфологической и соответственно
физиологической изменчивости, что значительно расширяет адаптивный
диапазон отдельных генераций и популяций и создает предпосылки для
нарастания численности особей. Массовые размножения, сопровождаю-
щиеся разнообразной изменчивостью, позволяют ускорить расселение
и освоить новые территории и соответственно расширить ареал. Так ве-
дут себя политипические виды землероек, например Sorex araneus L.
После массового размножения наступает период длительной депрессии,
во время которой происходит отбор особей по физиологическим пара-
метрам, более соответствующим жизни при наименее благоприятных усло-
виях. Видимо, отбираются особи с высокой устойчивостью и высокой
энергией развития. Они дают начало новой разнообразной изменчивости
внутри популяции вида. Таким образом, виды землероек с широким
адаптивным диапазоном имеют два цикла функционирования: существо-
вание одних популяций в период депрессии, а других — в период массо-
вых размножений. Именно разных популяций, потому что землеройки
живут один, максимум два года.
Нетрудно представить себе, что этими особенностями землероек, опре-
деляющими состояние их популяции в каждый отдельный период, регули-
руется их численность и обеспечивается сохранение вида при самых
разнообразных условиях обстановки и при жестком отборе.
В связи с особенностями питания землероек находится строение их же-
вательного аппарата. Он прцспособлен к быстрому схватыванию мелких
и очень подвижных животных. Остробугорчатые зубы служат для раз-
дробления твердых покровов животных, которые служат им пищей,
и только острые грани входящих складок на жевательной поверхности
моляров нарезают и раздавливают их мягкие ткани. Особенности строе-
ния нижней челюсти и ее сочленения с черепом (с. 74), а также строение
мускулатуры обусловлены приспособлением к быстрому закрыванию
рта и сильному сжатию челюстей с усилением давления в области перед-
него края зубного ряда. С этим связано значительное развитие височного
мускула и частичная редукция массетера, а также потеря его связи со ску-
ловой дугой, чем, по-видимому, и обусловлена ее утрата землеройками.
Массетер стал служить преимущественно для ускорения вращения че-
люсти при закрывании рта и усиления давления нижних резцов на верх-
ние при схватывании добычи.
Общие структурные особенности зубов землероек сложилось относи-
тельно рано (конец эоцена—олигоцен) и в основных чертах сохранились
до настоящего времени, что позволяет считать их достаточно универ-
сально приспособленными. Кроме того, можно думать, что пища, во вся-
ком случае ее механический состав, мало изменилась с того времени.
Адаптивный процесс затронул не столько структуру зубов, сколько соотно-
шение размеров отдельных зубов в ряду, а также их общее число. При
сокращении числа зубов или при уменьшении размеров отдельных зубов про-
исходит укорочение длины зубного ряда и соответственно длины челюстей,
что сопровождается укорочением длины рычагов и усилением жеватель-
ного давления челюстей. Разное число промежуточных зубов в верхней
челюсти и разнообразные соотношения размеров промежуточных зубов
отражают разнообразие в приспособлениях отдельных видов к схватыва-
нию и размельчению разных по форме, величине и консистенции кормовых
животных в наименее благоприятный для их жизни период.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
83
Органы ориентации у землероек развиты неодинаково хорошо, но
в совокупности они обеспечивают их жизнедеятельность в разнообразных
условиях среды. Зрительный рецептор не отличается совершенством и слу-
жит, видимо, для ориентации на небольшом расстоянии и только в днев-
ное время. Обонятельный рецептор, насколько можно судить по сложности
макроструктур, развит достаточно, чтобы воспринимать и дифференци-
ровать запахи и находить друг друга по следам и выделениям мускусных
желез, а также разыскивать корм. Можно думать, что у землероек хорошо
развита осязательная рецепторная система. Однако в условиях леса
в ночное время во всех ландшафтах, а также в приземном слое раститель-
ности и разного рода укрытиях лучшим способом ориентации служит
эхолокация, установленная у землероек экспериментальным путем. Так,
по данным Гоулда с соавторами (Gould et al., 1964), землеройки (Sorex
palustris Rich., S. vagrans Baird и S. cinereus Kerr) находили заданный
в опыте участок в специальном аппарате, не пользуясь осязательными,
зрительными и другими рецепторами. Зверьки с закрытыми ушными от-
верстиями его не находили. При поиске они издавали звуковые импульсы
частотой 30—60 кгц, продолжительностью 4.9—33 м/с, интенсивностью
0.02—0.4 дн/см2. Такие же импульсы землеройки издают при обследова-
нии незнакомого места или предмета. Возможно, с эхолокационным при-
способлением уха землероек связано своеобразное устройство их слухо-
вого аппарата.
Однообразие внешности землероек породило представление о том, что
в пределах сем. Soricidae адаптивный процесс ограничился образованием
узкого круга видов, различающихся отдельными мелкими признаками
и размерами. Кажущееся сходство в образе жизни, обусловленное не-
достаточной изученностью биологии землероек, также служит основа-
нием для суждения о слабой дифференцировке частных адаптаций внутри
семейства. 1 Однако ближайшее знакомство с морфологией и образом жизни
землероек позволяет обнаружить значительное разнообразие форм при-
способлений, специфичных для отдельных видов, специализация кото-
рых на общем адаптивном фоне землероек шла по пути приспособления
к захвату различных по форме и величине животных, использованию
разного типа укрытий, передвижению по разным грунтам с разной ско-
ростью и передвижению в воде. Структурные особенности зубов, харак-
терные для отдельных видов землероек, отличаются постоянством. Это
обстоятельство делает строение их зубных систем важной морфологи-
ческой основой для филогенетических и систематических исследований
и построений.
Своеобразные адаптивные особенности отдельных видов землероек отра-
жаются не только в экологии, но и в их распространении. Например,
виды, приспособленные к обитанию только в хорошо освещенных, сухих
участках леса, где они собирают корм прямо с поверхности земли, распро-
странены в пределах своего, нередко обширного, ареала спорадически.
Это объясняется пятнистым распределением биотопов, занимаемых этими
видами. Виды, пластичные в экологическом отношении, распространены
в пределах своего ареала в самых разнообразных биотопах. При этом
они весьма однообразны по внешности.
Подавляющее большинство землероек относительно быстро бегает,
хотя ноги у них короткие. Высота конечностей определяется необходи-
мостью быстро передвигаться в низких укрытиях и узких ходах, что свя-
зано с добыванием мелких подвижных животных, которыми они питаются.
По временам землеройкам приходится рыть как передними конечностями,
так и носом, а особям отдельных популяций наземных видов переплывать
мелкие водоемы. Кроме того, землеройкам приходится бороться со сравни-
6*
84
ПОДОТРЯД ERINACEI
тельно крупной добычей. Все это требует устойчивости и значительной
подвижности тела. Этим и объясняется известная универсальность локо-
моторных функций их конечностей. В связи с особенностями адаптации
землероек узкая специализация в какой-либо одной форме движения
среди них не получила широкого распространения. Тем не менее в пре-
делах семейства землероек имеются формы, более других специализиро-
ванные к рытью, плаванью, быстрому бегу или лазанью, что дает им пре-
имущество при определенных условиях (рис. 31—34).
О КАРИОСИСТЕМАТИКЕ
В настоящее время наблюдается повышенный интерес к использованию
для целей систематики данных цитогенетики, в частности числа и особен-
ностей строения хромосом. Это направление исследований получило
специальное обозначение — кариосистематика. Следует отметить, что
это название не соответствует содержанию понятия «систематика», при-
нятому в биологии. Кариосистематика занимается структурой хромосом
и их числом, т. е. морфологией хромосомных наборов видов и обслужи-
вает диагностику последних, представляя один из морфологических
методов.
Он получил широкое распространение, но нужно сказать, что, как
показал опыт, и этот метод определения видоспецифичности не может
считаться достаточно надежным. Причиной этого служит ряд обстоя-
тельств, и прежде всего наличие хромосомного полиморфизма, широко
распространенного во многих группах млекопитающих. Кроме того, суще-
ствуют роды, виды которых имеют одинаковое число хромосом, и роды,
объединяющие виды с разным числом хромосом. Объективных данных
о причинах возникновения отмеченных особенностей хромосомных на-
боров разных родов и видов нет. Объяснения, которые даются по этому
поводу, — умозрительные предположения.
Для установления вида по хромосомному набору необходимо знать,
к какому роду он относится: к объединяющему виды с одинаковым числом
хромосом или с разным, да и сам вид может оказаться полиморфным в этом
отношении. Нужно также знать систематическое положение вида, опре-
деляемого по хромосомам, чтобы не получилось так, как это вышло у Бор-
гаонкар (Borgaonkar, 1969, цит. по: Орлов, 1974), поместившего один и
32
тот же вид в два разных рода: Suncus murinus L. (gg—40 хромосом) и
Crocidura murina L. (40 хромосом) — синоним первого.
Определение вида должно сочетаться с определением его и по другим
морфологическим признакам. Существуют виды, хорошо дифференциро-
ванные морфологически, с одинаковым числом хромосом, и виды, сход-
ные по внешним признакам, с разным числом хромосом. Последних на-
зывают видами-двойниками. Их систематическое положение не ясно.
Ажиотаж вокруг теперь уже не нового кариологического метода все
еще продолжается. Это мешает спокойной критической оценке его реаль-
ных возможностей, порождает необъективных толкователей отмеченных
выше явлений неустойчивости в хромосомных наборах видов и не способ-
ствует поискам решения противоречий в самом хромосомном методе.
В заключение привожу хромосомные числа насекомоядных млекопита-
упо^янутой книги Орлова.
ющих,. заимствованные из
Insectivora
Solenodontidae
Solenodon paradoxus Cabr. 34.
Tenrecidae
Centetes ecaudatus Schreb. 38.
Hemicentetes semispinosus Cuv. 38.
H. nigriceps Giint. 38.
Echinops telfairi Martin 40.
Setifer setosus Fror. 40.
Microgale dobsoni Thos. 30.
M. talazaei Major. 30.
M. cowanii Thos.'54.
ХАРАКТЕРИСТИКА ПОДОТРЯДА
85
Erinaceidae
Erinaceus europaeus L. 48.
- E. rumanicus Barr.-Ham. 48.
E. amurensis Schrenk 48.
Hemiechinus hypomelas Brandt 48.
H. megalotis Blyth 48.
H. auritus Gmel. 48.
Macroscelididae
Elephantulus rupestris Smith. 30.
E. rozeti Duv. 28.
E. rufescens Peters 34.
E. brachyrhynchus Thos. et
Schwan 26.
Soricidae
Sorex cinereus Kerr 66.
«S. fumeus Mill. 66.
S. alpinus Schinz 58?
S. minutissimus Zimm. 42.
S. minutus L. 42.
S. volnuchini Ogn. 40*.
S. shinto Thos. 42*.
S. caecutiens Laxm. 42.
S. unguiculatus Dobs. 42.
S. isodon Turov 42.
S. vir Allen 42.
S. raddei Satun. 36.
S. arcticus Kerr 28—29.
S. arcticus Kerr 33.
S. arcticus Kerr 36—37.
S. araneus L. 20—32
21—33.
5. gemellus Ott 22—23*.
•S’. caucasicus Satun. 25a.
Microso rex hoyi Baird 62?
Notiosorex crawfordi Coues 62, 68.
Neomys fodiens Penn. 52.
N. anomalus Cabr. 52.
Blarina brevicauda Say 49—50.
Crocidura russula Herm. 42.
C. murina L. 40**.
C. suaveolens Pall. 40.
C. leucodon Herm. 28.
C. occidentalis Puch. 50.
C. bottegi Thos. 40.
C. jouvenetae Bals. 52.
C. theresae Bals. 50.
C. lamottei Bals. 52.
C. horsfieldi Thos. 26.
Suncus murinus L. 32
38-40.
S. etruscus Savi 42.
Cryptotis parva Say 52.
Talpidae
Neurotrichus gibbsi Baird 38.
Galemys pyrenaicus Geoff. 42.
Talpa europaea L. 34.
T. coeca Savi 36.
T. caucastca Satun. 38.
Mogera insularis Swinhoe 32.
Condylura cristata L. 34.
Parascalops breweri Bach. 34.
Scapanus latimanus Bach. 34.
Примечание. Одной звездочкой мною отмечены виды, требующие тщатель-
ной проверки; двумя звездочками отмечен вид — синоним Suncus murinus L. (А. Г.).
СИСТЕМАТИЧЕСКИЙ ОБЗОР
Сем. ERINACEIDAE Fischer von Waldheim, 1817
Erinaceini Fischer von Waldheim, 1817, p. 368—428.
Диагноз. Зубная формула: I JL СP т22! M = 44—28. Длина
тела от 105 до 450 мм. Глаза и ушные раковины крупные. Тело по-
крыто жесткими волосами или спина и бока имеют иглистый покров.
Череп с коротким лицевым отделом. Скуловые дуги массивные, широко
расставлены в стороны. I1 саблевидной формы, длинные или короткие,
с уступом на задней стороне. М1 и М2 четырехугольные, с 5 бугорками
на жевательной поверхности и 4 корнями.
Описание. Представители семейства имеют относительно разнооб-
разную внешность и форму тела, разные размеры (105—450 мм) и вес
(50—1400 г). Одни виды обладают относительно стройным телом на сра-
внительно высоких ногах, другие массивным телом с короткими ногами.
Все виды стопоходящие.
Голова клиновидная, с носовой частью, вытянутой в хоботок. Ноздри
расположены спереди или немного смещены на боковую сторону хоботка.
Края ноздрей с зубчатым гребнем, характерным для всех видов семей-
ства. Глаза всегда хорошо развиты. У отдельных видов они крупные,
округлой формы, выпуклые. Уши преимущественно крупные, клиновид-
ные. Хвост длинный или очень короткий (10—210 мм или 7—50% длины
тела). Кисти и ступни пятипалые. Первый палец кисти развит в разной
степени, изредка он отсутствует.
Тело сверху и снизу покрыто волосами (часто грубыми) или только
снизу, а сверху и с боков — иглами. Хвост покрыт чешуйками, образую-
щими кольца, расположенными одно за другим от корня хвоста до его
вершины. Между рядами чешуек располагаются отдельные грубые ще-
тинки. У многих видов кисти и ступни сверху покрыты крупными щит-
ками, щетинками и волосами или преимущественно волосами. Нижняя
сторона кистей и ступней покрыта толстой кожей, образующей на опор-
ных частях кожные утолщения — мозоли и подушки. Когти прямые или
в разной степени изогнуты.
Окраска от однотонной коричневой, бурой разных оттенков на верх-
ней и светлой на нижней стороне тела до серой, охристо-бурой и черно-
бурой. У отдельных видов имеются светлые пятна на голове, шее, загривке.
У видов, тело которых сверху и с боков покрыто иглами, окраска зависит
от особенностей окраски игл. Последние бывают серые, палевые или
черно-бурые, и тогда верхняя сторона тела соответствующего цвета.
Если светлые иглы имеют темные пояски, то окраска приобретает мелкий
пестрый рисунок. Иногда она бывает пятнистой, что обусловлено разной
окраской отдельных игл, например черной и белой. Нижняя сторона
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
87
тела ежей окрашена в бурый, черный или белый цвета. Кожа нижней
стороны кистей и ступней имеет бурую или черно-бурую окраску разных
оттенков, когти темнее или светлее пальцев.
Череп узкий или широкий, особенно в скулах, с узкой или сравнительно
широкой и вздутой мозговой капсулой. Межчелюстные кости короткие,
с отростком, направленным назад, или без него. Носовые кости в перед-
ней части более широкие, чем в остальной, или значительно уже. Лоб-
ные кости в области общего шва, а также и носовые кости вдавлены в виде
желобка, или желобок заметен только на лобных костях, реже совсем
отсутствует. Сагиттальный гребень развит на всем протяжении или
только в задней трети мозговой капсулы, реже совсем отсутствует. Лам-
бдовидный гребень развит в разной степени. Скуловые дуги массивные
или сравнительно тонкие, более или менее широко расставлены. Костное
нёбо только с мелкими отверстиями в передней части или с редкими мел-
кими и крупными отверстиями на нёбной кости. Крыловидные отростки
невысокие, у представителей подрода Paraechinus вздутые, полые внутри,
соединяются с полостью слуховых камер. Барабанная кость не сраста-
ется с соседними костями и не образует замкнутого барабана. Клино-
видная кость позади хоан с гладкой поверхностью или с глубокой выем-
кой. Засочценовный отросток широкий, с чашевидной впадиной, или
слабо развитый, с неглубоким вдавлением. Сочленовная впадина на ви-
сочной кости неглубокая. Нижняя линия профиля нижней челюсти в раз-
ной степени изогнута. Подбородочное отверстие у одних видов под Р3,
у других под Р4. Сочленовная область нижней челюсти образована
лопастью с тремя отростками (венечным, сочленовным и угловым), раз-
граниченными сходными по форме выемками. Венечный отросток сильно
уплощен в боковом направлении. У отдельных видов он утолщен спереди
и на вершине. Мыщелок сочленовного отростка более развит в попереч-
ном направлении и имеет форму валика. Угловой отросток в разной сте-
пени развит, разной толщины и массивности.
I1 крупные саблевидные или средних размеров конические, реже
с двумя вершинами, напоминающие таковые землероек {Soricidae). I2 и
I3 значительно меньше I1. I2 у отдельных видов подрода Erinaceus и всех
видов подродов Hemiechinus и Paraechinus с двумя корнями. Клык с одним
или двумя корнями, мелкий. Редко он очень крупный и входит в специаль-
ную выемку на межчелюстной Кости, сходнук? с этим образованием у тен-
река Tenrec ecaudatus L. Между резцами и клыком у всех ежей имеется
небольшой промежуток кости, лишенный зубов (диастема). Возможно,
что это остаток впадины для нижнего клыка, изменившего свое поло-
жение (из вертикального в наклонное вперед). Р1 и Р2 мелкие или отсут-
ствуют. Они бывают с одним или двумя корнями, Р3 мелкий, с двумя
или тремя корнями. Р4 с тремя корнями, крупный, часто более высокий,
чем моляры. Моляры М1 и М2 с четырьмя корнями, округло-четырехуголь-
ной формы, с пятью бугорками. Из них пятый мелкий, в центральной
части жевательной поверхности зуба. М3 мелкий, округлой формы или
вытянутый спереди назад, состоящий из двух сегментов, с двумя или
тремя корнями и с тремя в разной степени выраженными бугорками.
В нижней челюсти два или три резца, наиболее развит Р, реже I2.
Клык наклонен вперед, не входит в выемку перед верхним клыком,
с одним, редко с двумя корнями. Сравнительно редко клык сильно, раз-
вит, направлен вверх и при сомкнутых челюстях соприкасается с верхним,
как у хищных млекопитающих. Премоляры Рх и Р2 (если есть) мелкие,
Р3 и Р4 крупные, или крупный только Р4. Мх и М2 состоят из двух сег-
ментов. Из цих первый образован параконидом и протоконидом (слив-
шимися основаниями), а второй — метаконидом. М3 из двух сегментов
88
СЕМ. ERINACEIDAE
с двумя корнями, значительно меньший, чем предыдущие зубы, или мел-
кий, округлый, с одним корнем.
Следует отметить, что зубы, подверженные молочной смене, как и
других насекомоядных, нередко образуют на своей коронке дополнитель-
ные бугорки или зубцы, а постоянные верхние моляры — зубчатый или
волнистый наружный край.
Скелет образован массивными костями у одних групп ежей или сра-
внительно тонкими у других. В связи с этим стоит и размерное соотно-
шение элементов, составляющих скелет конечностей. У видов, имеющих
скелет более тонкого сложения, предплечье, голень и ступня относительно
длиннее, чем у видов, имеющих массивный скелет.
Шейные позвонки короткие. За относительно широким атлантом сле-
дует более узкий эпистрофей с высоким и длинным остистым отростком.
Следующие шейные позвонки тесно прижаты друг к другу. 6-й позвонок
с пластинчатыми отростками, направленными вниз. В совокупности по-
звонки образуют короткую шею. Грудные и поясничные позвонки с не-
высокими остистыми отростками и неширокими поперечными. Они одно-
образны по внешнему облику. Грудные позвонки с более короткими
телами. Длина поясничных позвонков возрастает от первого к последнему.
Крестец образован тремя сросшимися позвонками. 4 первых хвостовых
позвонка еще сохраняют в разной степени развитые поперечные и ости-
тый отростки.
В пределах сем. Erinaceidae число позвонков варьирует только в хво-
стовом отделе. У всех представителей семейства шейных позвонков — 7,
грудных — 15, поясничных — 6, крестцовых — 3, хвостовых — 9—27.
У ежей, тело которых покрыто иглами, грудной и поясничный отделы
позвоночника выгнуты вверх. Это связано, видимо, с быстрым свертыва-
нием тела этих животных в шар кольцевой мускулатурой.
Ребер 14—15 пар. Они образуют объемистую грудную клетку. У пред-
ставителей подсем. Galericinae она несколько сужена в передней части,
более длинная, чем у видов подсем. Erinaceinae. Грудина образована
сравнительно узкими и тонкими костями. Лопатка умеренной ширины
(ширина почти в 2 раза менее длины). Ость лопатки хорошо развита,
простирается на 2/3 ее длины, с длинным вильчатым акромиальным
отростком, к медиальному концу которого причленяется длинная клю-
чица.
Плечевая кость разной длины и массивности. В дистальном отделе
она значительно уплощена и расширена в области сочленовных мыщел-
ков. Медиальный надмыщелок более развит, чем латеральный, особенно
у видов, приспособленных к рытью. Надмыщелковое отверстие отсут-
ствует или небольшое. Локтевая кость более широкая, чем лучевая. Обе
кости в дистальном отделе значительно расширены и сочленяются с от-
носительно широкой кистью, особенно у роющих норы видов.
Кисть сравнительно короткая и широкая или относительно длинная
и узкая. Образована короткими метакарпальными костями, узкими при
основании и расширенными на конце, и широкими фалангами пальцев
или узкими на всем протяжении метакарпальными костями и фалангами
пальцев.
Таз образован узкими костями. Подвздошная кость узкая, у одних
короткая, у других длинная. В связи с этим вертлужная впадина рас-
положена ближе к середине таза или отнесена назад. Вертлужная впа-
дина глубокая. Ее края образуют почти правильный круг. Запиратель-
ное отверстие большое. Симфиз лобковых костей образован связкой,
нередко окостеневающей. Бедро массивное. Отношение его длины к длине
голени разное. Большой и малый вертелы развиты в разной степени. Голень
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
89
массивная и сравнительно короткая или тонкая и Длинная. Малая бер-
цовая кость срастается с большой берцовой костью на 1/2—2/3 своей длины.
Ступня у всех видов уже кисти. У одних она лишь немного превышает
длину кисти, у других значительно длиннее ее. У отдельных видов длина
ступни равна длине бедра или немного более. Первый палец (hal-
lux) у некоторых видов слабо развит, а у отдельных особей может отсут-
ствовать.
Распространение. В современный период Erinaceidae мо-
жет считаться одним из широко распространенных семейств насекомояд-
ных. Его ареал охватывает большую часть Европы, Азии и Африки.
В третичное время его ареал менялся во времени. В эоцене Европы
было распространено подсем. Galericinae (род Neurogymnurus Filhol).
Можно думать, что в это же время Европу населяли и представители под-
сем. Erinaceinae, и оба подсемейства в эоцене были и в Азии. Это пред-
положение основывается на том, что олигоценовые представители обоих
подсемейств были значительно специализированными в направлении, ха-
рактерном для каждого из них и в современный период.
В олигоцене подсем. Galericinae было представлено в Европе родами
Neurogymnurus, Tetracus Aymard и Galerix Pom el, а в Азии — Galerix,
Pseudoneurogymnurus. Из подсем. Erinaceinae в это время обитали предста-
вители трибы Amphechini (род Amphechinus Aymard), а Азию населяли
виды трибы Protericini (роды Tupaiodon Matt, et Grang., Palaeoscaptor Matt,
et Grang.), трибы Brachyericini (род Exallerix McKenna et Holt.) и Amphechini
(род Amphechinus). В Северной Америке в олигоцене существовали виды
трибы Protericini (род Proterix Matt.).
В миоцене гимнуры и ежи широко распространились. Миоцен был
временем расцвета сем. Erinaceidae. Из подсем. Galericinae в Европе су-
ществовали виды родов Galerix, Pseudogalerix Gaill., Lanthanotherium
Filhol, Deinogalerix Freudent., в Азии — родов Galerix и Lanthanotherium,
в Африке — рода Galerix, а в Северной Америке — родов Ocajila Macdon.,
Lanthanotherium?
Ежи в миоцене Европы были представлены видами трибы Amphe-
chini (род Amphechinus), трибы Brachyericini (род Dimylechinus Hiirz.)
и трибы Ertnaceini (роды Mioechinus Butler и Erinaceus L.); в Азии — ви-
дами трибы Amphechini (род Amphechinus) и трибы Erinaceini (род Eri-
naceus L.); в Африке — представителями трибы Amphechini (роды Amp-
hechinus и Gymnurechinus Butler), а в Северной Америке — трибы Bra-
chyericini (роды Brachyerix Matt., Metechinus Matt.). Последний род в Аме-
рике несомненно был в миоцене, поскольку в нижнем плиоцене этот род
вымер, будучи уже высокоспециализированным. Для этого нужно было
достаточно времени.
В плиоцене окончательно сложились современные роды и подроды
сем. Erinaceidae, а в плейстоцене формировались современные их пред-
ставители.
В настоящее время подсем. Galericinae распространено в Юго-Вос-
точной Азии, где его представители занимают часть материка и терри-
тории крупных островов и полуостровов. Все роды подсемейства энде-
мичны для Азии. Род Echinosorex Blainv. распространен на Тенассериме,
в Таиланде, на островах Малайского архипелага, на Суматре и Кали-
мантане. Предствители рода Hylomys Mull, et Schleg. встречаются в Юнь-
нани (Китай), на п-ве Индокитай и на островах Малайского архипелага,
на Тиомане, Суматре, Яве и Калимантане. Род Podogymnura Meams
известен только с о-ва Минданао (Филиппины). Род Neotetracus Trouess.
распространен в Сечуани и Юньнани (Китай), Северной Бирме и на п-ве
90
СЕМ. ERINACEIDAE
Индокитай. Род Neohylomys Shaw et Wong известен с о-ва Хайнань (Юж-
ный Китай).
Распространение подсем. Erinaceinae, имеющего в своем составе в со-
временный период один род — Erinaceus L., значительно шире, чем под-
сем. Galericinae. Со времени наивысшего расцвета семейства (миоцен)
гимнуры сократили ареал за счет Европы, большей части Азии, Африки
и Северной Америки, а ареал ежей сократился только за счет Северной
Америки.
Род Erinaceus распространен в Европе, южной части Сибири, Средней
и Центральной Азии, Индии, Аравии, Малой Азии, в Африке, на остро-
вах Средиземного моря и Канарских. В пределах этой территории под-
род Erinaceus L. распространен в Европе (от Англии, Ирландии, Швеции,
Норвегии, Финляндии, северных окраин европейской части СССР на
юг до побережья Средиземного моря и его островов, Крыма, Кавказа,
на Южном Урале, в северо-западном Казахстане, Западной Сибири до
Оби, в Приморском крае, в Северо-Восточном Китае, на п-ве Корея,
в Иране, северной части Аравии, Малой Азии, Африке и на Канарских
островах.
Подрод Hemiechinus Fitz, распространен на юго-востоке европей-
ской части СССР включая Кавказ, в Казахстане, Средней Азии, на юге
Западной Сибири и Алтае, в Центральной Азии, на северо-западе Индии,
в Афганистане, Иране, в Малой Азии, Аравии и в Северной части
Африки.
Подрод Paraechinus Trouess. в пределах рода имеет наименьший ареал.
Он распространен в Средней Азии, в северо-западной, западной и южной
части п-ва Индостан, в Аравии и в северной части Африки.
В заключение следует отметить, что некогда широкий общий ареал
гимнур и ежей к современному периоду оказался разорванным. В конце
миоцена гимнуры вымерли в Европе, Африке, Северной Америке и на
большей части территории Азии, сохранившись на крайнем ее юго-вос-
токе и прилежащих к ней островах. В миоцене вымерло большинство
родов ежей, от которых сохранился один род Erinaceus L. Вымирание
гимнур носит почти глобальный характер. Объяснение этому трудно найти
без подробного анализа обстановки, сложившейся к концу миоцена
в местах нахождения гимнур, а также древних групп ежей на континен-
тах, где они вымерли в это время.
Можно думать, что родиной сем. Erinaceidae была Азия, где предста-
вители его сохранили большее разнообразие форм адаптации и более
древние морфологические особенности и где только и остались виды
подсем. Galericinae.
ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ СЕМЕЙСТВА ЕЖОВЫХ
И ЗНАЧЕНИЕ ИХ ДЛЯ ЧЕЛОВЕКА
Представители сем. Erinaceidae в пределах своего широкого ареала
населяют разнообразные ландшафты в лесах, степях, пустынях и горных
местностях. Гимнуры распространены только в Юго-Восточной Азии и
на крупных островах прилежащих к ней архипелагов. Они населяют
преимущественно лесные ландшафты, поднимаясь в горы на высоту до
2800 м. Биология гимнур изучена очень плохо. Известно, что они питаются
разнообразной пищей, состоящей из насекомых, червей и других беспоз-
воночных, а также из мелких позвоночных и плодов растений. Большая
гимнура — Echinosorex gymnurus Raff., способная плавать, добывает себе
пищу и в воде: водных беспозвоночных и рыбу. Роющий вид — Neohy-
lomys hainanensis Shaw et Wong — ведет полуподземный образ жизни.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА 91
Hylomys suillus Mull, и Neotetracus sinensis Trouess. часто встречаются
в полосе культурных земель, где они селятся по берегам канав, заросших
кустарниками и деревьями.
Ежи населяют в зоне их распространения в Евразии и в Африке все
типы ландшафтов, придерживаясь и в зоне пустынь зарослей травы и
кустарников, обеспечивающих укрытие.
Ежи ночные и сумеречные животные. В дневное время в пустынях
и степях прячутся в вырытых ими неглубоких норах, а в лесных ланд-
шафтах в различных естественных укрытиях — в корнях деревьев, под
пнями, в кучах валежника и других укромных местах. Там, где ежей
не беспокоят, они покидают свои укрытия задолго до наступления тем-
ноты, что приходилось наблюдать в Ленинградской области у обыкновен-
ного ежа, а в Каракумах — у ушастого ежа. В окрестностях г. Кутаиси
я добыл несколько ежей днем в виноградниках и под инжирами, где они
поедали спелые плоды.
Видимо, глаза ежей способны различать предметы ночью и хорошо
видеть днем. Мне неоднократно приходилось держать ежей (обыкновен-
ного, ушастого и длинноиглого) в неволе. Они хорошо различали пред-
меты в комнате ночью и днем. Можно думать, что при добывании пищи
ежам больше, чем глаза, помогает орган обоняния. Ноздри у всех пред-
ставителей семейства имеют зубчатый край, отогнутый на внешнюю сто-
рону. Эти своеобразные фестоны образованы выростами кожи и служат
как поверхности, улавливающие и направляющие в ноздри токи воздуха.
Но это только предположение.
Вероятно, для ежей обонятельный рецептор имеет важное дистантное
значение. Например, в Ленинградской области, где даже в юго-западной
ее части ежей не так много, они в течение недели ежедневно попадались
в живоловки в одном и том же месте, причем разных возрастов. Приманкой,
которая их привлекала, служил хлеб, поджаренный с луком. Видимо,
запах привлекает ежей и на помойки, где они поедают кухонные остатки,
что наблюдалось в ряде мест Средней Азии.
Пищу ежей составляют насекомые и другие беспозвоночные, а также
мелкие позвоночные, которых они могут одолеть, а также мягкие сочные
плоды растений. В каждой местности и в каждом ландшафте состав корма
ежей зависит от обитающих в них видов животных, употребляемых ими
в пищу.
Естественно, что состав корма разный у обитателей пустынь, степей
и лесов, а в пределах каждой из названных зон, он разный в отдельных
биотопах. Состав корма разнится и по сезонам, а также в зависимости
от численности того или иного вида, употребляемого ежами в пищу.
Суточная и сезонная активность у одного и того же вида разная в раз-
ных географических пунктах и климатических поясах к югу и к северу
от экватора.
При наступлении неблагоприятных условий ежи впадают в спячку,
которая в разных климатических зонах продолжается неодинаковое
время. Спячку проводят в относительно глубоких норах (до 1.5 м), в ко-
торых устраивают зимовочные гнезда. Так проводят спячку ежи, обитаю-
щие в степях и пустынях. В лесной зоне ежи устраивают зимовочные
гнезда в различного рода естественных укрытиях. Изредка роют не-
глубокие норы.
Размножаются ежи один-два раза в году в зависимости от широты
местности и соответственно длины теплого периода года. На севере,
как правило, бывает один помет, на юге — чаще два. Для выведения
потомства в норе обитатели открытых пространств устраивают специаль-
92
СЕМ. ERINACEIDAE
ную выводковую камеру, а в лесных ландшафтах ежи строят гнезда
в тех же укромных местах, где и зимуют.
Ежи приносят большую пользу поеданием вредных насекомых и мы-
шевидных грызунов — вредителей сельского хозяйства. Этим же они
существенную пользу приносят и лесному хозяйству. Некоторый вред
ежи приносят разорением гнезд птиц, в том числе и промысловых. Кроме
того, существенный вред ежи приносят как прокормители ряда парази-
тов, особенно клещей — переносчиков возбудителей болезней человека
и домашних животных.
Однако польза, приносимая этими животными, их существенная роль
как своеобразных санитаров в биоценозах и их значение в общей эко-
номике природы, превышают приносимый вред. Ежей нужно охранять,
тем более что у них и так достаточно врагов, из которых главнейшими
считаются лисица и филин.
ФИЛОГЕНЕТИЧЕСКИЕ СООТНОШЕНИЯ ОТДЕЛЬНЫХ ГРУПП ЕЖОВЫХ
И ЭВОЛЮЦИЯ ИХ ЗУБОВ
Ежовые (Erinaceidae) — одна из древних групп насекомоядных, до-
живших до настоящего времени, сохраняющая в строении зубов архаи-
ческие особенности (передние премоляры конической формы, нередко
со слабо развитыми пара- и метаконами и конидами; часто I3 и верхний
клык, редко и нижний клык, имеют два корня. Их коренные зубы очень
рано (в эоцене) приобрели форму, а жевательные поверхности структуру,
пригодную для перетирания пищи. Возможность обработки на коренных
зубах разнообразной пищи обусловила и возможность существования
ежовых в самых разнообразных ландшафтах и привела к относительному
однообразию форм адаптаций и внешности. Возможность использования
всякого корма не способствовала возникновению в пределах семейства
разных по значению приспособлений ни в жевательном аппарате, ни
в органах движения.
В сем. Erinaceidae очень рано обособились две группы: гимнуры (Gale-
ricinae) и ежи (Erinaceinae). Обе группы давно развиваются независимо
друг от друга.
История развития ежовых ранее верхнего эоцена не ясна. Из верхнего
эоцена известен род Neurogymnurus Filhol, представители которого уже
обладали основными особенностями гимнур и во многих отношениях были
специализированными. Существенным отличием этого рода от других
вымерших и современных родов гимнур служит ряд своеобразных особен-
ностей в строении зубов и черепа, отдельные из которых повторились
у ряда миоценовых ежей, а некоторые из них сохранились и у современ-
ных видов. Премоляры еще имели крупные размеры, особенно Р4 —
наибольший зуб в верхнем зубном ряду. Височная кость имела направлен-
ный назад отросток, достигающий значительного развития. Сходное
образование имело место и у миоценовых ежей рода Brachyerix Matt,
из Северной Америки. Кроме того, у представителей рода Neurogymnu-
rus костное нёбо было перфорировано, что характерно и для ежей родов
Amphechinus Aymard (олигоцен-миоцен) и современного Erinaceus L.
У всех других гимнур эти образования отсутствуют.
Таким образом, наряду с признаками, общими для всех гимнур, гим-
нуры описываемого рода имели особенности, характерные и для отдель-
ных групп ежей, но более позднего времени. Можно думать, что признаки,
близкие ежам и характерные для описываемого рода гимнур, развивав-
шиеся у них в разное время, представляют своеобразный параллелизм.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
93
Это явление можно рассматривать и как частное для отдельных групп
ежовых, не имеющее филогенетического значения.
Гимнуры рода Neurogymnurus не могут считаться предками других
гимнур, поскольку их особенности не встречаются ни у одного другого
рода подсем. Galericinae; Это несомненно особая группа гимнур, рано
сложившаяся и к верхнему эоцену достигшая высокой специализации.
Ее можно считать особой ветвью с характерным для нее направлением
адаптации, произошедшей от общих корней с другими гимнурами, давно
обособившейся и развивавшейся независимо от других. Представители
этого рода дожили до нижнего олигоцена.
Из олигоцена известны виды родов Tetracus Aymard и Galerix Pomel.
Род Tetracus представлен ограниченным числом остатков, которые от-
носят к двум видам, различающимся сравнительно небольшим числом
признаков в структурах зубов. Основным отличием этого рода следует
признать остробугорчатость зубов и мелкие размеры их коронок при зна-
чительной массивности корней, кроме того, массивность нижней челю-
сти. Создается представление, что мощность жевательного давления
у гимнур рода Tetracus создавалась массой нижней челюсти, при том
что мускулатура затрачивала энергию главным образом на приведение
челюсти. Судя по строению зубов, гимнуры этого рода питались преиму-
щественно мелким кормом, скорее всего насекомыми и червями, поскольку
их шиповатые зубы мало годились для жевания.
Род Galerix был более широко распространен и в олигоцене обитал
в Европе и Азии, а в миоцене был и в Африке. Эта группа гимнур и в оли-
гоцене, и в миоцене представляла, видимо, основное ядро этих млеко-
питающих, была широко распространена и универсально адаптирована.
Зубная формула и структура зубов приближались к таковым современ-
ного рода Hylomys, но премоляры были крупнее. Отдельные виды этого
рода отличаются деталями строения зубов и нижней челюсти.
Гимнуры олигоценового времени представляют потомков эоценовых
гимнур, развивающихся по пути уменьшения размеров резцов и клыков,
образованием на резцах дополнительных зубцов, как это имеет место и
у мелких современных гимнур. Можно считать это направление главным
в развитии гимнур, сохранившим свое значение и в современный период.
Наряду с этими должны были существовать в олигоцене и гимнуры,
которых мы пока не знаем, сохранявшие крупные клыки, и с ними не-
сомненно связаны в происхождении и развитии роды гимнур миоцено-
вого времени, имеющие крупные клыки, и современные роды, объеди-
няющие виды с крупными клыками. Отсюда можно сделать заключение,
что в олигоцене гимнуры представлены двумя группами с более и менее
развитыми клыками.. Описываемых вымерших (как и современных) гим-
нур нельзя поделить на отдельные группы, поскольку их основные осо-
бенности были и остаются общими.
Таким образом, уже в олигоцене гимнуры разнообразны по своим
частным особенностям: одни с крупными, другие с мелкими клыками,
но все имели 44 зуба — максимальное для плацентарных млекопитаю-
щих число.
В миоцене продолжали существовать виды рода Galerix. Они были
широко распространены, встречались в Европе, Азии и Африке. Суще-
ствование этой группы гимнур ограничено миоценом. За пределами этого
времени они не обнаружены. Причины вымирания этого рода, как, впро-
чем, и других, в миоцене Европы, Азии и Африки не ясны. Если их исчез-
новение в Европе можно объяснить с некоторой долей вероятности по-
холоданием уже в верхнем миоцене, когда они и вымерли, то их гибель
в Африке или Азии, где и сейчас обитают гимнуры других родов, правда
94
СЕМ. ERINACEIDAE
на островах или в горных местностях, объяснить весьма трудно. До
появления человека на земле вымерло огромное количество млекопитаю-
щих, вероятно, во много раз больше, чем существует ныне. Группы по-
являлись и исчезали. Создается впечатление, что многие из них существо-
вали разное время, т. е. обладали разным циклом развития от возникно-
вения до становления и от расцвета до упадка и вымирания. Возможно,
что для каждой из групп имело значение соотношение химического со-
става тела их представителей и химического состава их пищи и неор-
ганической среды. Нарушение благоприятных соотношений приводило
к вымиранию одних групп и развитию других, с другим сочетанием со-
отношений. Это не больше чем предположение.
Кроме рода Galerix, в миоцене существовало еще несколько родов,
также вымерших к концу этого времени, Из них прежде всего нужно
отметить род Pseudogalerix Gaill. Этот род весьма близок к роду Galerix,
отличается размерами зубов. Так, у него более крупный клык и Р2 и мел-
кие Р|. При том что отдельные зубы у представителей этого рода мелки,
их нижняя челюсть была крупнее и, следовательно, общие размеры этих
гимнур были больше, чем видов рода Galerix. Описание этого рода весьма
схематично, чтобы можно было сказать что-либо определенное о харак-
тере его адаптаций. Создается впечатление, что этот род более близок
к роду Galerix, чем это думают, и, возможно, представляют собой его
подрод или даже вид.
Среднемиоценовый род Lanthanotherium Filhol значительно отлича-
ется от плеяды родов из олигоцена и миоцена главным образом тенден-
цией к сокращению числа зубов и соответственно длины лицевого отде-
ла черепа. В связи с укорочением длины рычагов усиливается жеватель-
ное давление зубов-антагонистов, что очень важно при переходе на бо-
лее грубую по механическому составу пищу. На развитие жевательной
функции зубо-челюстного аппарата у видов этого рода указывают и
крупные, квадратно-четырехугольные верхнекоренные зубы и укорочен-
ные и широкие нижние.
Сокращение общего числа зубов за счет утраты Р] или только Рх ха-
рактерно и для современных гимнур. Однако менее 40 зубов ни в прош-
лом, ни в настоящее время у гимнур не было. Будучи более активными,
чем ежи, гимпуры захватывают в пищу более подвижных животных,
а для этого удобнее иметь и более длинную морду.
Особняком среди миоценовых гимнур стоит род Deinogalerix Freuden-
tal, недавно описанный из Италии. Прежде всего нужно отметить огром-
ные размеры известного представителя этого рода — D. koenigswaldi
Freudental. Наибольшая длина черепа этого вида достигает 21 см — в два
раза более длины черепа современного Echinosorex gymnurus Raffles.
Резцы, клыки и премоляры впереди от Р| отделены диастемой, Р| и Pf
крупные. В верхней челюсти самый крупный зуб Р4, а в нижней М,.
1} крупные и массивные, развитые в равной степени. Коренные зубы
остробугорчатые. М| и М| мелкие. Строение жевательного аппарата соз-
дает впечатление что он приспособлен для схватывания крупной и сколь-
зкой добычи и пережевывания пищи разной консистенции.
Корни современных родов гимнур пока не ясны. Возможно, что они
миоценового времени.
Нахождение в эоценовых и олигоценовых отложениях Евразии и
Северной Америки представителей сем. Erinaceidae со всеми основными
особенностями, характерными и для видов более позднего времени,
позволяет считать, что оно формировалось еще в раннем эоцене, а может
быть, и еще ранее. Можно считать также, что ежи из нижнего олигоцена
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
95
сохраняли в морфологии многие особенности, сходные с таковыми ежей
эоценового времени.
Сток (Stock, 1935) описал из верхнего эоцена Калифорнии Proterixotdes
davisi, которого он считает предком Proterix Matt. Батлер (Butler, 1948)
отметил некоторое сходство этого рода с древними ежами и гимнурами,
но в свою систему ежеобразных его не поместил, оставив лишь в графи-
ческой схеме филогении ежей в качестве боковой ветви от предковых
форм Proterix. Сабан (Saban, 1958) поместил род Proterixotdes в подсемейство
гимнур. Ван Вален (Van Valen, 1967) отнес этот род к сем. Adapisoricidae,
которое би в своей системе насекомоядных поставил сопряженно с сем.
Erinaceidae в выделенном им надСем. Erinacoidea. В пределах сем. Adapi-
soricidae находится и подсем. Geoldbinae, описанное Маккенна (McKenna,
1960), основанное им па роде Geolabis Соре. Последний по морфологическим
особенностям (форма черепа, строение зубов, отсутствие скуловых дуг
и др.) сходен с представителями тенрекообразных. Таким образом, сем.
Adapisoricidae, поставленное в системе Ван Валена в качестве предкового
или связанного в развитии, с Erinaceidae, представляет собрание разно-
родных групп, часть которых имеет сомнительное отношение к ежовым.
Фотографии верхних и нижних челюстей Proterixotdes с соответствую-
щими им зубами, приведенные Стоком, позволяют отметить, что при не-
котором отдаленном сходстве верхних коренных зубов Proterixotdes с тако-
выми Proterix его нижние коренные зубы, особенно М3, не похожи на
эти зубы ежовых. Кроме того, нижняя челюсть Proterixotdes- не имеет
углового отростка, что не свойственно ежовым. Можно думать, что этот
род имеет отдаленное отношение к Erinaceidae.
Надо полагать, что к верхнему эоцену ежи сложились столь же близко
к их современному типу, как и гимнуры. Это представление подтвержда-
ется морфологическими особенностями олигоценовых представителей
подсем. Erinaceinae. Наиболее примитивные ежи известны из олигоцена
Азии и Северной Америки. Они были малых размеров. Отмечено (Mat-
thew, Granger, 1924), что представители описанных родов Tupatodon
и Palaeoscaptor достигали размеров крота или крупной землеройки —
Blarina brevicauda Say. Немногим крупнее были виды североамериканского
олигоценового рода Proterix Matt. Все названные роды относятся к трибе
Protericini.
Можно думать, что их эоценовые предки также не были крупных раз-
меров. Известные виды этой группы родов отличались формой черепа
и общим числом зубов, что указывает на имевшуюся в то время разно-
направленную адаптацию. Одни виды имели удлиненный череп легкого
сложения, другие массивный, с короткой лицевой частью. Из этого можно
заключить, что одни из них были с легким телом, более активные, а другие
с тяжелым складом тела и менее активные. Одни известные виды сохра-
няли наибольшее для плацентарных количество зубов — 44, другие —40,
что обусловлено утратой PJ. На коренных зубах еще сохраняются за-
остренные бугорки у многих видов. Общим для всей группы родов призна-
ком следует считать треугольной формы М3, сохраняющий метакон.
Начавшееся у отдельных родов олигоцена сокращение общего числа
зубов носит начальный характер и происходит за счет Р|, как и в пределах
других семейств насекомоядных.
Тенденция к сокращению числа зубов и укорочению в связи с этим
длины рычагов, каковые представляют собой челюсти, более значительное
выражение получила в пределах трибы Brachyericini, развившейся не-
сомненно от общих корней с трибой Protericini.
Наиболее ранний представитель трибы Brachyericini известен из Азии,
откуда описан (McKenna, Holton, 1967) род Exallerix. К сожалению, он
96
СЕМ. ERINACEIDAE
описан по правой ветви нижней челюсти с отломанным сочленовным от-
делом. Как и у других представителей этой трибы, у вида описываемого
рода имело место увеличение размеров нижнего резца и значительное
уменьшение размеров I2, С, Р3, и Р4. Все перечисленные зубы, кроме Р4,
имеют один корень. РЛ одновершинный, с укороченным основанием коронки.
Значительных размеров и длины достигает М15 который вместе с М2 вы-
полняет жевательную функцию. Укорочение переднего отдела нижнего
зубного ряда и увеличение размеров моляров указывают на усиление
жевательной функции коренных зубов. Это может служить основанием
для предположения о специализации представителей трибы Brachyericini
к обработке преимущественно грубой по механическому составу пищи.
Описываемое направление адаптаций более резко выражено у предста-
вителей отдельных родов этой трибы из миоцена.
Третью олигоценовую группу ежей представляли виды рода Amphe-
chinus Aymard (триба Amphechini). Эта группа ежей в олигоцене Европы
и Азии представлена значительным числом видов, особенно в Азии, и,
вероятно, была в то время достаточно процветающей. Основные особен-
ности ежей этого рода заключаются в том, что их череп и нижняя челюсть
имеют строение, сходное с таковым более поздних и современных видов
ежей. Однако они имели много специфичных для них особенностей, а глав-
ное, еще сохраняли признаки, общие с древними ежами, как и более при-
митивные ежи трибы Protericini.
Короче говоря, эта группа ежей уже в олигоцене располагала особен-
ностями, позволяющими считать ее как развивавшуюся в направлении,
близком к специализации ежей более позднего времени, но на многих
еще примитивных основах.
Общие контуры нижней челюсти, и особенно черепа, напоминают
таковые ежей позднего времени. Зубная формула полностью совпадает
с зубной формулой ежей рода Erinaceus L. Следовательно, и жевательный
аппарат имел значительное сходство в морфологическом и функцио-
нальном отношениях. Это представление подтверждается и тем, что струк-
туры зубов представителей рода Amphechinus напоминают, а отдельные
зубы имеют значительное сходство с зубами поздних специализированных
ежей.
Видимо, ежи трибы Amphechini в фауне олигоцена играли роль, сход-
ную с ролью поздних ежей, но преимущественно в закрытых ландшафтах,
как и отдельные современные виды. На это указывает отсутствие у них
многих особенностей, характерных для ежей, обитающих на открытых
пространствах. В то же время они уже обладали особенностями, характер-
ными для адаптаций и современных видов, иными словами, у них были
многие формы адаптаций, свойственные группам, универсальным в адап-
тивном отношении.
Если сопоставить морфологические особенности представителей трех
названных олигоценовых триб, то нетрудно заметить бросающуюся в глаза
некоторую общность в структурах их черепа и зубов, но на разном уровне
эволюционного развития и специализации, что объясняется разным вре-
менем возникновения групп и разным характером адаптации. Есть основа-
ния считать, что все олигоценовые группы ежей имеют общие корни в проис-
хождении. Тенденция к сокращению общего числа зубов в каждой челюсти,
начавшаяся за счет сокращения Pj у Proterix в трибе Protericini, привела
к значительному сокращению числа зубов в верхней и нижней челюстях
путем сокращения числа премоляров и последнего моляра у представи-
телей трибы Brachyericini, что можно рассматривать как крайнее выраже-
ние этой общей в пределах подсем. Erinaceinae тенденции. Сокращение
числа зубов до характерного для видов трибы Amphechini, сохранившегося
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
97
и у современных видов, оказалось биологически более выгодным, доста-
точно универсально обеспечивающим потребности организмов разного
качества в разно адаптированных группах поздних ежей, сменявшихся
во времени.
Таким образом, в олигоцене ежи были представлены тремя трибами,
отражавшими три разных направления специализации. Более древнего
типа, преемственно развивавшегося с давнего времени, сохранившего
большое число зубов — триба Protericini (роды Tupaiodon, Palaeoscaptor,
Proterixp, специализированные к пережевыванию сравнительно грубой
пищи, с крупными резцами для ее захвата, с укороченными челюстями
и зубными рядами — ежи трибы Brachyericini (род ExaUerix), и ежи трибы
Amphechini (род Amphechinus) с направлением специализации универсаль-
ного характера, которое преемственно развивалось в более позднее время
и с рядом изменений и усовершенствований частного порядка сохрани-
лось у развивавшейся от общих корней с нею трибы Erinaceini.
Ежи всех трех описанных здесь триб имели следующие общие при-
знаки: слезное отверстие открывается вовнутрь орбиты, нёбо не перфориро-
вано (исключение триба Amphechini), базисфеноидное углубление от-
сутствует (углубление на основной клиновидной кости позади хоан).
Эти же признаки присущи гимнурам (подсем Galericinae), а следовательно,
и общим предкам всех ежеобразных.
При всей примитивности отдельных групп ежей олигоцена, особенно
трибы Protericini, виды которой сохраняли большое число зубов, все
они были высокоспециализированными для своего времени, в частности
триба Amphechini. Это позволяет считать их развивавшимися давно, зна-
чительно ранее верхнего эоцена. Ведь ежеобразпым нужно было обосо-
биться от других групп насекомоядных, направиться по пути формиро-
вания специфичной для них морфологии и ее специализации, чтобы к верх-
нему эоцену образовать род Neurogymnurus Filhol, сравнимый с родами
значительно более поздних и современных гимнур, а также и специализиро-
ванные роды ежей, которые уже в нижнем олигоцене имели облик, близ-
кий к современным ежам.
Олигоцён следует считать временем доживания архаичных групп
и временем формирования новых направлений специализации, получив-
ших максимальное развитие в миоцене.
Миоцен был периодом расцвета ежей. Представители трибы Protericini
к этому времени, вероятно, вымерли. Их остатки нигде не обнаружены.
Триба Brachyericini в Северной Америке была представлена, видимо,
несколькими родами, поскольку эта триба известна оттуда и из плиоцена,
и одним родом в Европе. В миоценовой фауне Азии они до сих пор не
обнаружены. Возможно, что к началу миоцена они там вымерли. Триба
Amphechini распространилась в Африку, где сформировался род Gymnu-
rechinus Butler и в Северную Америку (роды Amphechinus Aymard и Steno-
echinus Rich et Rasmussen). При сходстве отдельных морфологических
особенностей и при наличии общих признаков, отмеченных выше для
о лигоценовых триб, представители американских родов трибы Brachyericini
значительно отличались от видов этой трибы из Европы. Следовательно,
в миоцене обе группы представителей трибы Brachyericini развивались
своими путями, независимо друг от друга. При этом европейские пред-
ставители трибы не зашли так далеко в сокращении зубов в верхней и
нижней челюстях, как это имело место среди американских видов, от-
дельные группы которых сохранили только 28 зубов, в то время как
у европейских их было 32.
При всем отличии американских и европейских представителей трибы
Brachyericini у них сохранились общие признаки, позволяющие установить
7 Д. А. Гуреев
98
СЕМ. ERINACEIDAE
их родственные отношения. Из них нужно отметить вытянутый назад отрос-
ток метаконида у Р4, суженный в средней части М1 и сравнительно мелкий,
со скошенным назад внешним краем М2. Таким образом, у них имеется
общий принцип строения зубов и зубного ряда при общем отсутствии М|
и утрате отдельных премоляров в верхней и нижней челюстях. Кроме того,
у всех сохраняются и другие общие признаки в строении черепа и его
отдельных частей. При этом нужно отметить, что представители европей-
ского рода этой трибы (Dimylechinus Hiirz.) имели некоторые внешние
особенности, сходные с таковыми рода Amphechinus.
Все представители трибы Brachyericini в морфологии черепа и зубов,
а также в построении всего жевательного аппарата отражают приспособле-
ние к питанию грубой по механическому составу пищей. Вероятно, их
основную пищу составляли малоактивные животные, которых можно было
хватать короткими челюстями с крупными первыми резцами.
Триба Brachyericini в миоцене была представлена следующими родами:
Dimylechinus Hurz, Европа; Brachyerix Matth., Северная Америка.
В миоцене продолжала развиваться и шире распространилась триба
Amphechini. Миоценовые группы этой трибы сохраняли свои основные
особенности, совершенствуя частные приспособления, отраженнные в спе-
циализации отдельных родов, отчасти и видов. Тем не менее на протяжении
этого времени резких изменений в основных структурах зубов и всего
жевательного аппарата даже у развившегося в миоцене Африки рода
Gymnurechinus отметить не удается. Род Amphechinus распространился
в Африку и в Северную Америку, значительно расширив ареал, и был
в то время господствующей группой ежей. Распространение же рода
Gymnurechinus ограничено Африкой, за пределами которой он пока не
обнаружен и, возможно, представляет эндемичную африканскую группу
ежей из трибы Amphechini.
За пределами миоцена триба Amphechini неизвестна; триба Brachyeri-
cini еще сохранялась в плиоцене Северной Америки. Можно думать, что
миоцен был пределом существования отдельных групп ежей, как и многих
групп млекопитающих из других отрядов. Возможно, что причиной этого
служит значительное изменение ландшафтов, начавшееся еще в олигоцене.
В развитии ряда мне известных групп млекопитающих, и в частности
ежовых (Erinaceidae), существует некоторая своеобразная законо-
мерность, в которой приходится все больше убеждаться. Группы,
возникшие в эоцене, исчезают из фауны в олигоцене; возникшие
в олигоцене вымирают в миоцене; развившиеся в миоцене вымирают
в плиоцене и т. д. Однако есть роды, развившиеся в миоцене, которые,
как правило, сохраняются до современности, утрачивая из своего состава
отдельные виды в плиоцене и плейстоцене. Многие из сохранившихся
миоценовых родов к современности становятся монотипичными или со-
держат ограниченное число видов, преимущественно близких по характеру
адаптаций, а вся группа несет черты депрессии в развитии и формировании
чего-либо нового. Она кажется остановившейся во времени (например,
ежи).
Миоцен был периодом становления и развития трибы Erinaceini,
представители которой дожили доныне. Можно не сомневаться, что она
происходит от общих корней с трибой Amphechini, но ее исходные позиции
недостаточно ясны. С трибой Amphechini ее сближает ряд общих признаков,
из которых нужно отметить сходство контуров черепа и нижней челюсти,
одинаковое число зубов и близкое их строение. Сходство и в том, что нёбо
перфорировано, хотя и в разной степени значительно. Но только у пред-
ставителей трибы Erinaceini слезное отверстие открываетс^впе орбиты,
на предорбитальном гребне, и только у них имеется углубление на основ-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
99
ной клиновидной кости позади хоан (сфеноидное углубление), которое
встречается из млекопитающих только у отдельных групп тенрекообраз-
ных. Пока не ясно функциональное значение отмеченных образований,
но, видимо, оно весьма существенно, особенно сфеноидного углубления,
Рис. 43. Схема филогенетических соотношений ежовых.
I — Amphechinus, 2 — Gymnur echinus, 3 — Stenoechinus, 4 — Mioechinus, 5 — Protechinus.
встречающегося у очень поздних групп ежей и их далеких предков —
тенрекообразных.
Морфологические особенности представителей трибы Erinaceini практи-
чески исчерпываются приведенным сравнением с трибой Amphechini,
более подробно они приведены в характеристике ее родов.
Миоцен был периодом расцвета ежовых вообще и ежей в частности.
В это время существовали ежи трибы Brachyericini, трибы Amphechini
и трибы Erinaceini — роды Mioechinus Butler, Protechinus^ Lavoc., Erina-
7»
100
СЕМ. ERINACEIDAE
ceiis L. Представители трибы Amphechini вымерли к концу миоцена.
К этому времени вымерли многие виды триб Brachyericini и Erinaceini.
Плиоцен был временем становления и развития подродов рода
Erinaceus (триба Erinaceini) и временем доживания ежей в Новом Свете
Только до нижнего плиоцена там просуществовали потомки трибы Brachye-
ricini — род Metechinus Matth. Его представители сохраняли все особен-
ности, присущие ежам этой трибы, и, пожалуй, отличались более укорочен-
ным лицевым отделом черепа и более широкой расстановкой скуловых дуг,
что указывает на значительное развитие жевательной мышцы (musculus
masseter). Верхний зубной ряд впереди от Р4 состоял из мелких, редко
Эоцен I Олигоцен
Миоцен
Рис. 44. Схема филогенетических соотношений ежовых Батлера (Butler, 1948). i
Собрем.
Echinosorex
Podoffymnura
Hylomys
Neotetracus
стоящих зубов. Жевательную функцию выполняли два зуба — Р4 и М1,
остальные зубы мелкие, одновершинные впереди и с ограниченной жева-
тельной поверхностью М2 позади них.
Таким образом, укорочение лицевого отдела черепа в пределах трибы
Brachyericini первоначально шло по пути сокращения общего числа
зубов, а затем и по пути уменьшения их размеров, что сопровождалось,
по-видимому, ослаблением функции зубов, особенно премоляров, и пере-
ходом на питание легко обрабатываемой пищей с помощью сильных жева-
тельных мышц. При этих условиях малейшее изменение в кормовых спек-
трах должно было приводить особей к гибели, а в целом к вымиранию
группы.
Видимо, в плиоцене окончательно сформировался род Erinaceus L.
(триба Erinaceini). В это время он расширял ареал, а в разных по своему
характеру ландшафтах формировались его подроды, окончательно сложив-
шиеся в плейстоцене. Его ранние подроды к верхнему плиоцену вымирали,
например Postpalerinaceus Crusaf. et VilL Приспособление отдельных
подродов к открытым ландшафтам сопровождалось развитием специфиче-
ческих особенностей, описанных в их характеристиках. До настоящего
времени дожил только один описываемый род с тремя подродами, занимаю-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
101
щими в совокупности значительную территорию в Европе, Азии и Африке.
В Северной Америке ежи вымерли в нижнем плиоцене.
Мои представления о филогенетических соотношениях в пределах
семейства ежовых приведены на рис. 43. Схема филогенетических со-
отношений ежовых Батлера (Butler, 1948) изображена на рис. 44.
КРАТКАЯ ИСТОРИЯ ИССЛЕДОВАНИЯ СИСТЕМАТИКИ ERINACEIDAE
Исследование систематики семейства началось с описания Линнеем
(Linnaeus, 1758) рода Erinaceus. В 1817 г. Фишер (Fischer von Waldheim)
выделил ежей в трибу Erinaceini, а Грей (Gray) в 1821 г. установил
сем. Erinaceidae. Помель (Pomel) в 1848 г. в пределах сем. Erinaceidae выделил
подсем. Galericinae, куда отнес вымерших гимнур. В 1872 г. Гил (Gill)
выделил гимнур в подсем. Cymnurinae, а ежей в подсем. Erinaceinae.
Кабрера (Cabrera, 1925) назвал подсем. гимнур Echinosoricinae. Добсон
(Dobson, 1882) в своей монографии о насекомоядных, сохраняя под-
семейства гимнур и ежей, первых объединял в один род Gymnura,
а вторых — в род Erinaceus.
Метью (Matthew, 1929) в своей таблице, иллюстрирующей геологи-
ческий возраст и географическое распространение родов ежовых, роды
Tetracus Aymard и Neotetracus Trouess. отнес в подсем. Erinaceinae. К этому
же подсемейству род Tetracus отнесли Симпсон (Simpson, 1945) и Сабан
(Saban, 1958).
В 1945 г. Симпсон предложил свою систему ежеобразных: •
Сем. Erinaceidae Fischer von Waldheim, 1817 (=Erinaceidae Gray, 1821, p. 300); Erinaceini
Fischer, 1817. Верхний эоцен—современный, Европа; олигоцен—современный,
Азия; нижний эоцен—нижний плиоцен, Сев. Америка; современный, Африка.
Подсем. Echinosoricinae Cabrera, 1925, (—Gymnurinae Gill, 1872, p. 19). Верхний
эоцен—верхний миоцен, Европа; нижний эоцен—нижний плиоцен, Сев. Аме-
рика; современный, Азия.
f Entomolestes Matthew, 1909. Нижний эоцен, Сев., Америка.
f Proterixoides Stock, 1935. Нижний эоцен, Сев. Америка.
f Metacodon Clark, 1936. Нижний—средний олигоцен, Сев. Америка.
f Ankylodon Patterson et McGrew, 1937. Средний олигоцен, Сев. Америка,
f Proterix Matthew, 1903. Средний олигоцен. Сев. Америка.
f Brachyerix Matthew, 1933.. Верхний миоцен, Сев. Америка.
f Metechinus Matthew, 1929. Нижний миоцен—нижний плиоцен. Сев. Америка,
f Meterix Hall., 1929. Нижний плиоцен. Сев. Америка.
f Neurogymnurus Filhol, 1877 (=Necrogymnurus Lyddeker, 1891, и большинство
авторов). Верхний эоцен, Европа, Сев. Америка.
t Lantanotherium Filhol, 1888 (f Lanthanotherium большинства последующих
авторов). Средний миоцен, Европа.
f Galerix Pomel, 1848 (включает f Parasorex Meyer, 1865). Верхний миоцен,
Европа.
f Pseudogalerix Gaillard, 1929. Верхний миоцен, Европа.
Echinosorex De Blainville, 1838 (=Gymnura Lesson, 1827, не Kuhl, 1824).
Современный, южн. Азия.
Hylomys Muller, 1839. Современный, Южн. Азия.
Podogymnura Mearns, 1905. Современный, Филиппинские острова.
Neotetracus Trouessart, 1909. Современный, Вост. Азия.
Подсем. Erinaceinae Gill, 1872, р. 18. Олпгоцен—современный, Европа, Азия;
верхний миоцен, Сев. Америка?; современный, Африка.
j- Tetracus Aymard, 1850. Олигоцен, Европа.
f Palaeoerinaceus Filhol, 1879. Верхний олигоцен—нижний миоцен, Европа,
f Tupaiodon Matthew et Granger, 1924. Олигоцен, Азия.
f Palaeoscaptor Matthew et Granger, 1924. Олигоцен, Азия.
f Parvericius Koerner, 1940. Верхний миоцен, Сев. Америка.
Aetechinus Thomas, 1918. Современный, Африка.
Erinaceus Linnaeus, 1758. Миоцен—современный, Европа; плейстоцен—со-
временный, Азия.
102
СЕМ. ERINACEIDAE
Atelerix Pomel, 1840. Современный, Африка.
Hemiechinus Fitzinger, 1866 (=Ericius Sundevall, 1841, не Tilesius, 1813).
Современный, Азия и Сев. Африка.
Paraechinus Trouessart, 1879 (включает Macroechinus Satunin, 1907). Современ-
ный, Южн. Азия и Сев. Африка.
В системе Симпсона, естественно, отсутствует ряд родов, описанных
после выхода его книги. Ряд родов ежей автор отнес в подсемейство
гимнур, а одну из гимнур поместил в подсемейство ежей. Несколько родов
позднее были переведены в другие семейства.
В 1948 г. Батлер (Butler) на основании исследований морфологии
современных и вымерших ежеобразных представил в своей работе их
систему. Как и Симпсон, Батлер одни близкие друг к другу роды
поместил среди гимнур, другие — среди ежей. Получилась смесь, не
отражающая истинных соотношений групп. Кроме того, вкрались и
некоторые неточности в отношении ареалов отдельных родов.
Система (по: Butler, 1948)
Сем. Erinaceidae
Подсем. Echinosoricinae. Примитивные Erinaceidae.
Триба Galericini. Примитивные формы с нормальным для Insectivora М3.
f Galerix. Олигоцен—миоцен, Европа.
f Pseudogalerix. Миоцен, Европа.
f ? Tetracus Олигоцен, Европа.
f Tupaiodon. Олигоцен, Монголия.
Триба Echinosoricini. Со специфичным типом М3.
f Lanthanotherium. Миоцен, Европа.
Podogymnura. Современный, Филиппинские острова.
Echinosorex. Современный, Индо-Малайская область.
Hylomys. Современный, Индо-Малайская область.
Neotetracus. Современный, Китай.
-f Подсем. Neurogymnurinae. Со специфичной мастоидной областью черепа.
f Триба Neurogymnurini. Примитивные формы.
f Neurogymnurus. Олигоцен, Европа.
t Триба Brachyericini. Короткочерепные формы.
f Brachyerix. Миоцен, Сев. Америка.
f Metechinus. Миоцен, Сев. Америка.
f ? Parvericius. Миоцен, Сев. Америка.
Подсем. Erinaceinae.
f Триба Protericini. Примитивные формы с нормальным для Insectivora М3.
f Proterix. Олигоцен, Сев. Америка.
Триба Erinaceini. Со специализированными зубами.
f A mphechinus. Олигоцен—миоцен, Европа, Азия.
ф Dimylechinus. Миоцен, Европа.
f Mioedhinus. Миоцен, Европа.
Hemiechinus. Современный, главным образом Южн. Азия.
Paraechinus. Современный, Сев. Африка, Южн. Азия.
Atelerix, Плейстоцен—современный, Африка.
Aethechinus. Современный, Южн. Европа, Африка.
Erinaceus. Плейстоцен—современный, Палеарктика.
В 1967 г. Ван Вален (Van Valen) опубликовал свою систему Erinaceidae.
Сем. Erinaceidae Fischer von Waldheim, 1817 (=Acanthionidae Schulze, 1900). Поздний
эоцен—современный, Европа; средний олигоцен—ранний плиоцен, Сев. Америка;
поздний олигоцен—современный, Азия; миоцен—современный, Африка.
Подсем. Galericinae Pomel, 1848. Поздний эоцен—поздний миоцен, Европа; средний
миоцен—ранний плиоцен, Сев. Америка; миоцен, Африка; современный,
Азия.
f Триба Galericini Pomel, 1848 (=Parasoricidae Schlosser, 1887). Ранний олиго-
цен—поздний миоцен, Европа; миоцен, Африка.
f Tetracus Aymard, 1850. Ранний олигоцен, Европа.
f Galerix Pomel, 1848 (=Parasorex Meyer, 1865). Поздний миоцен, Европа;
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
103
миоцен, Африка.
f Pseudogalerix Gaillard, 1929. Поздний миоцен, Европа.
-[• Триба Neurogymnurini Butler, 1948 (=Cayluxotheriini Winge, 1917). Поздник
эоцен—средний олигоцен, Европа.
f Neurogymnurus (не Necrogymnurus) Filhol, 1877 (=Cayluxptherium Filhol’
1880). Поздний эоцен—средний олигоцен, Европа.
Триба Echinosoricini (Cabrera, 1925) Gill, 1872 (=Gymnurinae Gill, 1872;'
Hylomyidae J. Anderson, 1879). Средне-поздний миоцен, Европа; среди и й.
миоцен—поздний плиоцен, Сев. Америка; современный, Азия.
f Lanthanotherium (не Lantanotherium) Filhol, 1888. Средне-верхний миоцен,
Европа; поздний миоцен—ранний плиоцен, Сев. Америка.
f Ocajila MacDonald, 1963. Ранний миоцен, Сев. Америка.
Podogymnura Mearns, 1905. Современный, Филиппины.
Echinosorex Blainville, 1838 (Gymnura Lesson, 1827). Современный, Азия..
Hylomys Miller, 1829 (включает Neotetracus Trouessart, 1909; Neohylomys
Shaw et Wong, 1959). Плейстоцен—современный, Азия.
Подсем. Erinaceinae Fischer von Waldheim, 1817. Ранний олигоцен—современный,
Европа; средний олигоцен—ранний плиоцен, Сев. Америка; поздний олиго-
цен—современный, Азия; миоцен—современный, Африка.
f Триба Protericini Butler, 1948 (включает Brachyericini Butler, 1948). Ранний
олигоцен—поздний миоцен, Европа; средний олигоцен—ранний плиоцен,
Сев. Америка; поздний олигоцен—ранний миоцен, Азия; миоцен, Африка.
f Proterix Matthew, 1903. Средний олигоцен, Сев. Америка.
f Brachyerix Matthew, 1933. Поздний миоцен, Сев, Америка.
f Amphechinus Aymard, 1850 (включает Рalaeoerinqceus (не Paleoerinaceus^
Palaerinaceus или Palerinaceus) Filhol, 1879; Palaeoscaptor Matthew et Gran-
ger, 1924; Parvericius Koerner, 1940). Ранний олигоцен—поздний миоцен,
Европа; поздний олигоцен—ранний миоцен, Азия; миоцен, Африка;
средне-поздний миоцен, Сев. Америка.
f Dimylechinus Hurzeler, 1944. Ранний миоцен, Европа.
f Metechinus Matthew, 1929. Поздний миоцен—ранний плиоцен, Сев. Аме-
рика.
Триба Erinaceini Fischer von Waldheim, 1817. Ранний миоцен—современный,
Европа; миоцен—современный, Африка; плейстоцен—современный, Азия,
f Gymnurechinus Butler, 1956. Миоцен, Африка.
f Postpalerinaceus Crusafont-Pairo et Villalta—Cornelia, 1947. Ранний плио-
цен, Европа.
f Protechinus Lavocat, 1961. Поздний миоцен, Африка.
f Mioechinus Butler, 1948. Ранний—поздний миоцен, Европа.
Erinaceus Linnaeus, 1758 (=или включает Setiger Е. Geoffroy Saint Hi-
laire, 1803; Atelerix Pomel, 1848; Peroechinus Fitzinger, 1866; Herinaceus
Mina Palumbo, 1868; Aethechinus Thomas, 1918; Mesechinus Ognev, 1951).
Поздний миоцен—современный, Европа; плейстоцен—современный,
Азия, Африка.
Hemiechinus Fitzinger, 1866 (включает Ericius Sundevall, 1842; Erinaceulus
Ognev, 1928). Современный, Азия, Африка.
Paraechinus Trouessart, 1879 (включает Macroechinus Satunin, 1907).
Современный, Азия, Сев. Африка.
Ван Вален, как и другие авторы, занимавшиеся до него систематикой
ежовых, поместил роды с укороченным зубным рядом в трибу более
примитивных ежей, разделив при этом близкие роды между двумя трибами.
Примечание. В сводке по насекомоядным С. И. Огнев (1928)
сохранил роды Erinaceus L., Hemiechinus Fitz, и Paraechinus Trouess.,
а также все ранее описанные виды этих родов, перечислив их названия.
С. У. Строганов (1957а), в своей книге о насекомоядных Сибири сохраняет
распространенных там Erinaceus и Hemiechinus в качестве самостоятельных
родов. При этом даурского ежа он отнес к роду Hemiechinus. Попутно
отмечу, что Огнев (1954), сохраняя этот вид в составе рода Erinaceus^
выделил его в подрод Mesechinus.
В 1975 г. В. А. Долгов и Б. С. Юдин приводят для фауны Советского
Союза следующую систему ежей
104
СЕМ. ERINACEIDAE
Erinaceidae
Род Erinaceus Род Hemiechinus
E. europaeus L. Il. auritus Gmelin
E. (Ilemiechinus?) dauricus Sun- H. hypomelas Brandt
devall II. megalotis Blyth (syn. H. choro-
sanicus Laptev)
II. (Erinaceus?) dauricus Sundevall
КЛАССИФИКАЦИЯ
При построении системы ежовых мною использованы структурные
особенности черепа, зубной системы и отдельных зубов в ряду, а для
современных представителей и строение других частей тела.
При изучении морфологии черепа и зубов ежовых выяснилось, что
для многих их групп как современных, так и вымерших характерно
сочетание особенностей строения черепа и структуры зубов или числа
последних. Выяснилась также общая тенденция в эволюции гимнур
к сохранению большого числа зубов и к уменьшению его в эволюции ежей.
Внешние структуры тела отражают адаптации к жизни в определенном
типе ландшафтов. Они также представляют сочетание структур, при-
способленных к ускорению движения, защите от факторов среды или об-
наружению опасности при пассивной защите. Каждое из сочетаний от-
ражает то или иное направление частностной специализации отдельных
групп в общей ходе эволюции сем. Erinaceidae.
Сходство структурных особенностей однозначных органов, имеющих
определенное адаптивное значение, позволяет устанавливать родственные
отношения и преемственность в развитии между вымершими и современ-
ными группами.
Сем. Erinaceidae Fischer von Waldheim, 1817. Верхний эоцен —современный, Европа;
олигоцен—современный, Азия; олигоцен—нижний миоцен, Сев. Америка; мио-
цен—современный, Африка.
I. Подсем. Galericinae Pomel, 1848. Верхний эоцен — верхний миоцен, Европа;
олигоцен—современный, Азия; миоцен—нижний плиоцен, Сев. Америка;
миоцен, Африка.
f 1. Tetracus Aymard, 1850. Нижний олигоцен, Европа.
f 2. Galerix Pomel, 1848. Олигоцен—миоцен, Азия; миоцен, Европа и Африка.
3. Pseudogalerix Gaillard, 1929. Верхний миоцен, Европа.
f 4. Lanthanotherium Filhol, 1888. Средний—верхний миоцен, Европа; миоцен—
нижний плиоцен, Сев. Америка.
t 5. Deinogalerix Freudental, 1972. Миоцен, Европа.
f 6. Ocajila Macdonald, 1963. Нижний миоцен, Сев. Америка.
f 7. Neurogymnurus Filhol, 1877. Верхний эоцен—нижний олигоцен, Европа.
f 8. Pseudoneurogymnurus gen. nov. Нижний олигоцен, Азия.
9. Echinosorex Blainville, 1833. Современный, Азия.
10. ' Hylomys Muller et Schlegel, 1839—1844. Современный, Азия.
11. Podogymnura Mearns, 1905. Современный, Азия.
12. Neotetracus Trouessart, 1909. Современный, Азия.
13. Neohylomys Shaw et Wong, 1959. Современный, Азия.
II. Подсем. Erinaceinae Fischer von Waldheim, 1817. Нижний олигоцен—совре-
менный, Европа; олигоцен—современный, Азия; средний олигоцен—нижний
плиоцен, Сев. Америка; миоцен—современный, Африка.
f 1. Триба Protericini Butler, 1948. Олигоцен—верхний миоцен, Европа;
олигоцен, Азия; средний олигоцен—нижний плиоцен, Сев. Америка.
14. Tupaiodon Matthew et Granger, 1924. Олигоцен, Азия.
•• 15. Proterix Matthew, 1903. Средний олигоцен, Сев, Америка.
16. Palaeoscaptor Matthew et Granger, 1924. Олигоцен, Азия.
• 2. Триба Brachyericini Butler, 1948. Средний олигоцен, Азия; верхний мио-
цен, Европа; средний миоцен—нижний плиоцен, Сев. Америка.,
| 17. Brachyerix Matthew, 1933. Верхний миоцен, Сев. Америка.
t 18. Metechinus Matthew, 1929. Верхний миоцен—верхний плиоцен, Сев.
Америка.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
105
f 19. Exallerix McKenna et Holton, 1967. Олигоцен, Азия.
•f- 20. Dimylechinus Hurzeler, 1944. Нижний миоцен, Европа.
f 3. Триба Amphechini Gureev triba nova. Олигоцен—миоцен, Европа
и Азия; миоцен, Африка; средний—верхний миоцен, Сев. Америка,
f 21. Amphechinus Aymard, 1850. Олигоцен—миоцен, Европа и Азия; миоцен,
Африка; средне-поздний миоцен, Сев. Америка.
f 22. Gymnurechinus Butler, 1956. Миоцен, Афюика.
f 23. Stenoechinus Rich et Rasmussen, 1973. Нижний миоцен, Сев. Америка.
4. Триба Erinaceini Fischer von Waldheim, 1817. Миоцен — современный,
Европа, Азия и Африка.
f 24. Mioechinus Butler, 1948. Миоцен, Европа.
| 25. Prothechinus Lavocat, 1961. Верхний миоцен, Африка.
26. Erinaceus Linnaeus, 1758. Верхний миоцен—современный, Европа, Азия
и Африка.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ
1 (2). По крайней мере у современных тело сверху покрыто иглами.
Зубов 36—28 ........................ 2. Erinaceinae, с. 133.
2 (1). Все тело покрыто волосами. Зубов 44—40 ......................
..........................................1. Galericinae, с. 105.
1, Подсем. GALERICINAE Pomel, 1848
Pomel, 1848, Arch. Sciet. de Geneve, t. IX, p. 1—164. Gymnurinae Gill, 1872,
p. 19. Parasoricidae Schlosser, 1887, S. 1—224. Echinosoricinae Cabrera, 1925,
Mus. Nation, de Natur., p. 57—58.
Тип Galerix Pomel, 1848, Европа, Азия, олигоцен—миоцен, Африка, множен.
Д и а г но з.< Зубная формула: IСР^—| М-5. = 44—40. Тело сверху
и снизу покрыто волосами. Позади хоан, между крыльями основной
клиновидной кости, образующими стенку барабанной камеры, нет округ-
лой впадины. М3 с тремя конами и тремя корнями. М3 имеет три конида
(средний — протоконид может быть слит с первым — параконидом)
и два корня.
Описание. Размеры тела варьируют (105—445 мм). Тело от-
носительно стройное, удлиненное. Ноги разной длины, стопоходящие.
Кисти и ступни пятипалые. Хвост разной длины (12—210 мм). Голова
удлиненноклиновидная. Носовой отдел вытянут в хоботок. Глаза хорошо
развиты, у отдельных видов относительно крупные. Уши округлой или
овальной формы. Все тело покрыто жесткими щетинистыми волосами.
Подшерсток мягкий, шелковистый. Уши покрыты короткими редкими
волосами и кажутся голыми. Хвост покрыт чешуйками, расположенными
кольцами, и редкими щетинками или волосами. Ступни и кисти покрыты
сравнительно крупными роговыми щитками, напоминающими таковые
птиц, редкими щетинками и волосами или только редкими, грубыми воло-
сами. Подошвенные стороны кистей и ступней покрыты толстой кожей,
образующей на их опорных частях разной формы бугорки (мозоли). Когти
тонкие, изогнутые в разной степени сильно. Окраска не отличается
яркостью. Основной цвет темно-коричневый или бурый. Встречаются
светло окрашенные или почти белые особи, а также с пятнистой окраской.
Мочка носа и глаза окрашены темнее тела. Когти светлые или темные.
Череп с удлиненным, клиновидной формы лицевым отделом у большин-
ства видов. Особенно длинный лицевой отдел черепа у представителей
рода Podogymnura Mearns, а более короткий у Neotetracus Trouessart.
Мозговая капсула сильно сужена или умеренно широкая. Межглаз-
ничное пространство сужено в разной степени. Межчелюстные кости от-
106
ПОДСЕМ. GALERICINAE
яосительно длинные, с отростком, направленным вперед. Носовые кости
в передней части у большинства видов значительно расширены. Они здесь
более широкие, чем в средней части. Лобные кости не имеют вздутий.
Носовые и лобные кости не вдавлены по среднему шву, и здесь не образу-
ется продольный желобок, реже желобок имеется вдоль общего шва лоб-
ных костей. Предорбитальный гребень (передний край орбиты) хорошо
развит, особенно у мелких видов. . Он прикрывает слезное отверстие,
жоторое у гимнур открывается в орбиту. Этот гребень соединяется с над-
ербитальным гребнем, ограничивающим орбиту сверху, а внизу переходит
в гребень скуловой дуги. Слезная кость срастается с окружающими ее
костями, и ее границы не заметны. Скуловая дуга почти горизонтальна.
Скуловая кость короткая, помещается в центральной части дуги. В перед-
ней части скуловой дуги глубокая выемка для прикрепления мускула,
поднимающего верхнюю губу и нос (musculus nasi-labialis). Сагиттальный
гребень развит в разной степени. Ламбдовидный гребень хорошо развит.
Мыщелковое отверстие помещается в выемке мыщелка. Костное нёбо
-только с мелкими отверстиями. Задний край нёба заканчивается костным
валиком. Клиновидная кость позади хоан гладкая, без впадины или вдав-
ления. Засочленовный отросток неширокий, с небольшим углублением
или без него.
Нижняя челюсть, как у всех представителей сем. Erinaceidae, с подня-
той сочленовной частью. Вырезки между сочленовным и угловым отрост-
ками сходной формы и размеров. Подбородочное отверстие под Р3 или Р4.
Нижнечелюстное отверстие расположено ближе к сочленовному мы-
щелку.
I1 крупные, саблевидные или более мелкие, конические, реже с двумя
зубцами, похожие на эти зубы землероек. I2 и I3 мелкие. Клык маленький,
сходный по форме с задними резцами, реже крупный, как у хищных
млекопитающих. В последнем случае между клыком и последним резцом
имеется выемка в межчелюстной кости, куда входит вершина нижнего
клыка. Между резцами и клыком имеется короткая диастема.
Все резцы с одним корнем, как и клык, если он мелкий. Крупный клык
с двумя корнями. Р1, Р2, часто и Р3 с одним корнем, или Р3 с двумя или
тремя корнями. Р4всегда с тремя корнями. Р1 и Р2 конической формы или
трехвершинные, если сохраняются бугорки пара- и метакона. Р3 простой,
напоминающий по форме предшествующие премоляры, или сложной струк-
туры, крупный, с тремя корнями, напоминающий высоким бугром
паракона Р4. Р4 с высоким и вытянутым назад или смещенным кнаружи
метаконом. Протокой и талонид сходной высоты, ниже других бугорков.
М1 и М2 сходны с этими зубами видов подсем. Erinaceinae. М3 треугольной
формы, широкий, с бугорками, расположенными треугольником, а не
в одну линию, как у Erinaceinae, с тремя корнями (а не с двумя, как
у Erinaceinae).
Нижний зубной ряд начинается длинным, слабо изогнутым I1, реже
этрт зуб мелкий или сходный по размерам с 12.13 мелкий. Клык у большин-
ства видов небольшой, вершиной направлен вперед, реже крупный, сходный
с клыком хищных. Резцы, клык и Рх с одним корнем, Р2 редко с одним
корнем. Как правило, P2i 3> 4 с двумя корнями, хотя у Р2 они могут быть
соединены. Рх и Р2 сходны по величине и форме, с вершиной, направлен-
ной вперед. Р3 крупнее предыдущих, с тремя бугорками и представляет
упрощенную структуру Р4. Последний с хорошо обособленными буграми
пара-, прото- и метаконидов. М4, М2 и М3 сходны по общему строению,
убывают в размерах к М3, который в отличие от этого зуба у представи-
телей Erinaceinae не только сходен с передними молярами, но имеет также
два корня.
1. РОД TETRACUS ЮТ
Скелет представителей Gelericinae более тонкого сложения, чеяг
представителей Erinaceinae. Он образован сравнительно тонкими костями,,
что особенно заметно в строении передних и задних конечностей и со-
ответствующих поясов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВЫМЕРШИХ РОДОВ
1 (2). Длина черепа более 150 мм.................5. Deinogalerix.
2 (1). Длина черепа менее 150 мм.
3 (8). Общее число зубов 44, Pj имеются.
4 (5). Р4 самый крупный зуб верхней челюсти. Костное нёбо перфориро-
вано. Теменные кости с морщинистой поверхностью.................. .
...........................................7. Neurogymnurus.
5 (4). Признаки иные.
6 (7). Верхний клык и Р2 крупные . . . . . . . . .3. Pseudogalerix.
7 (6). Верхний клык и Р2 не выделяются размерами . . . . 2. Galerix.
8 (3). Общее количество зубов 40, Р{ отсутствуют . .............
....................................... 10. Lanthanotherium.
Примечание. Роды 6. Ocajila и 1. Tetracus не вошли в определительнук»
таблицу за отсутствием сравнимых признаков.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СОВРЕМЕННЫХ РОДОВ
1 (4). Общее число зубов 44.
2 (3). Размеры тела 260—450 мм. Нижний клык крупный, при сомкнутых
челюстях входит в специальную выемку в межчелюстной кости:
............................................. 9. Echinosorex.
3 (2). Размеры тела менее 260 мм. Нижний клык мелкий, выемки для
него на межчелюстной кости нет.......................10. Hylomys.
4 (1). Общее число зубов менее 44.
5 (6). Общее число зубов 42. В нижней7 челюсти отсутствует Р4 . . _
......................................... 13. Neohylomys.
6 (5). Общее число зубов 40.
7 (8). Верхний и нижний клыки сильно развиты...................
...........................................11. Podogymnura.
8 (7). Верхние и нижние клыки сходны с соседними зубами ....
.............................................12. Neotetracus.
fl. Род TETRACUS Aymard, 1850
Aymard, 1850, Ann. Soc. d’Agricult. Sci. et du Puy, t. 14, p. 80—86.
Тип. Tetracus nanus Aymard, 1846. Франция, нижний олигоцеп.
Диагноз, (по: Aymard, in Gervais, 1848, из: Lavocat, 1951).
Зубная формула: I? Сх P4 M3. Животное, близкое к ежам, у которого Ms
с четырьмя бугорками. Передняя часть нижней челюсти удлиненная.
Промежуток нижней челюсти, занимаемый премолярами, таков, что можно-
предполагать их большее число, чем у ежей. Отличается от Plesiosorex:
Pomel и Soricidae, имеющих Р4 простого строения (близкий к одновершин-
ному, низкий), более высоким, трехвершинным Р4.
Нижняя челюсть длинная, круто поднимается от клыка к Р4. Одио>
подбородочное отверстие под Р4. Задняя часть нижней челюсти, как
у ежеобразных.
Примечание. Можно думать, что у представителей этого рода
зубная формула такая же, как у Neurogymnurus Filhol, Galerix Pomel
или Hylomys Mull, et Schl. (l-у С у P у M = 44^.
108
ПОДСЕМ. GALERICINAE
Распространение и возраст. Европа, нижний оли-
гоцен.
Ниже я привожу описания видов рода Tetracus, сделанные мной по
фотографиям их нижних челюстей, помещенным Лавока в его книге,
посвященной ревизии остатков олигоценовой фауны млекопитающих
D’Auvergne et du Velay (Франция).
t 1. Tetracus nanus Aymard, 1846.
Aymard, in Gervais, 1848, Zool. Palaeont. Fr. 1 edit., p. 11, note 2.
Тип. Фрагмент левой ветви нижней челюсти с тремя сохранившимися зубами
Р3—Мх, № 1890-5-1, Ronzon, Франция, кол. Парижского музея.
45
Рис. 45. Левая ветвь нижней челюсти
Tetracus bouti.
46
Рис. 46. Фрагменты ветвей нижней че-
люсти Galerix exllis (по: Lavocat, 1951,
с фот.).
Диагноз (по: Lavocat, 1951). Вид того же склада, что и Tetracus
bouti. Р4 более близкий таковому Lanthanotherium. Мх немного более
островершинный, его кониды более обособлены.
Описание (по фотографии с типа, отмеченного выше). ;У челюсти
отсутствует ее передняя часть до Р2. Повреждена наружная стенка в обла-
сти корневой системы Мх и видны его корни. Отломлены все отростки сочле-
новного отдела челюсти. Оставшися части челюсти по очертаниям и
структуре близки к типу, общему и для современных гимнур. Р2 и Р3
мелкие, с двумя корнями. Р3 по строению похож на Р4, но значительно
мельче, с низким протоконидом. Р4 с хорошо развитым и обособленным
параконидом, с высоким, удлиненным в основании протоконидом, вершина
которого отогнута назад. Мх с выступающим вперед параконидом и двух-
вершинным протоконидом. Метаконид этого зуба массивный, с двумя
заостренными бугорками на заднем крае. Корни Мх высокие и массивные,
особенно метаконида. При сравнительной массивности нижней челюсти
зубы относительно мелкие, в частности премоляры.
Измерения (по: Lavocat, 1951). Длина Р3—Мх — 5.5, Мх —
2.50, ширина — 1.35 мм (тип).
Распространение и возраст. Франция, нижний оли-
гоцен.
t 2. Tetracus bouti Lavocat, 1951; рис. 45.
Tetracus nanus Aymard, pars, in Gervais, 1848, Zoologie et Paleontologie fran-
$aise, 1 edit., p. 11, note 2. Erinaceus nanus A у m a r d, in Pomel, 1853, Catalogui me-
thodique et descriptif des Vertebres fossils decouverts dans le basin hydrographique su-
perieur de la Loir et surtout dans la vallee de son affluent principal 1’Allier. — Paris
2. РОД'GALERIX
109
J. В. Balliere. Tetracus bouti L a v о c a t, 1951, Revision de la Faune des mammiferes
oligocenes D’Auvergne et du Velay, 12—14.
Тип. Фрагмент правой ветви нижней челюсти с С—Р2—М8 № 1890—5—3,
Ronzon, Франция, олигоцен, кол. Парижского музея. >
Диагноз (по: Pomel, 1853, из: Layocat, 1951). «Вид почти наполо-
вину крупнее живущего в нашей местности (по-видимому, имеется в виду
Erinaceus europaeus L.). Нижние моляры с четырьмя бугорками. Премоля-
ров, вероятно, много».
Описание (по фотографиям с тица, отмеченного выше). Нижняя
челюсть низкая, почти сходной высоты от Р3 до основания венечного от-
ростка. Венечный отросток, судя по его соотношению с положением с со-
членовным отростком, не расширен в передне-заднем направлении. К со-
жалению, вершина венечного отростка отломлена. Сочленовный отросток
высоко поднят и сближен с венечным. По отношению к нижней челюсти
зубы крупные, менее остробугорчатые, чем у Т. nanus. Р3 с менее развитыми
бугорками пара- и метаконидов. Параконид Р4 слабо обособлен, а мето-
конид более развит; чем у Т. nanus. Протоконид менее высокий. Р4 и мо-
ляры менее развиты в длину, чем у Т. nanus, с притупленными вершинами
бугорков. Крупное подбородочное отверстие расположено между Р2 и Р3.
Измерения (по: Lavocat, 1951). Длина С—М3 — 15.75, Р2—М3 —
11.55, Р2 - 1.35, Р3 - 1.35, Р4 - 2.70, М2 - 2.20, М3 - 1.70 мм.
Распространенне й возраст. Известен из Франции,
олигоцен.
f 2. Род GALERIX Pomel, 1848
Viverra exilis Blainville, 1839, Osteograph, Insect, t. I, H, p. 109, Galerix
viverroides Pomel, 1848, Suppl. bibl. Geneve, t. IX, p. 164, Parasorex socialis v. Meyer,
1855, Neues Jahrb., S. 844 .Galerix exilis Filhol, 1891, Ann. Sci. Geol. Franc., t. 21, p. 22.
Тип. Galerix exilis Brainville, 1839, Европа, нижний олигоцен—верхний миоцен.
Диагноз. Зубная формула: 7-|-СуРуЛ/-|-== 44. Р2 с двумя корнями.
Р3 сходен по строению с Р4, но менее его, хотя этот зуб более крупный,
чем у других гимнур. Метаконид (талонид других авторов) Р3 более высо-
кий, чем у Р4. Р4 имеет высокий параконид. Параконид Мх высокий, не
выступающий вперед. М2 сходен с Мх, но меньше по размерам.
Описание (по: Viret, 1938). Зубная формула такая же, как у Neu-
rogymnurus Filhol. Верхние и нижние моляры сохраняют строение, близ-
кое Neurogymnurus. Galerix обладает косыми молярами с двумя рядами
бугорков. Резцы и клыки сходны с таковыми Neurogymnurus. Р3 сходен по
строению с Р4. (по: Butler, 1948). Форма черепа Galerix сходна с таковой
Hylomys, однако лицевой отдел у Galerix несколько длиннее. Форма
лицевой части черепа и строение костного нёба сходны с этими частями
черепа Hylomys. Теменные кости не образуют выступа, вдающегося в лоб-
ные кости, как у Hylomys и Neotetracus Trouess. Височные кости имеют
направленный назад выступ. Имеется надглазничный выступ.
Нижняя челюсть сходна с этой костью Hylomys, но сочленовный от-
росток и его мыщелок расположены ниже, а передняя поверхность венеч-
ного отростка более отклонена назад.
За исключением сильно развитых премоляров зубная система Galerix
напоминает зубную систему Hylomys, но более примитивная. Клыки
больше. У Р4, М1 и М2 корень протокона и талонида слиты в один общий
корень, расширенный в сагиттальном направлении. М1 находится под
орбитой. М1 и М2 более вытянуты в поперечном направлении, чем у Hylo-
mys. Метаконуль (пятый бугорок) этих зубов имеет V-образную форму.
Р3 сравнительно большой. Метаконид (внутренний бугорок протоконида
110
ПОДСЕМ. GALERICINAE
в нашем понимании) Р4 сильно развит (по фот. Lavocat, 1951). Нижняя
челюсть с относительно высокой зубной частью. Венечный отросток имеет
отклоненную назад переднюю поверхность. Р4 с высоко поднятым пара-
конидом, отчего вершина протоконида кажется мало обособленной от
него. Вершина протоконида не отклонена назад, как у Tetracus Aymard.
Метаконид Р4 мало развит в Длину. Мг с выступающим вперед паракони-
дом, длинный. М2 и М3 мало отличаются по длине и высоте. Протоконид
Рис. 47. Передняя часть черепа Galerix exilis (по: Butler, 1948).
Рис. 48. Фрагмент левой ветви нижней челюсти и зубы G. exilis (по: Thenius, 1949).
и метаконид моляров, как и у других, двувершинные (у протоконида гре-
бень, а у метаконида задний край разделены выемкой на два бугорка).
Распространение и возраст. Европа, олигоцен—мио-
цен; Азия, олигоцен—миоцен; Африка, миоцен.
t 1. Galerix exilis Blainville H. M. D., 1839 (рис. 46—48).
Blainville, 1839—1864, Osteographie, t. 1, Osteographie mammiferes in-
sectivores. Paris.
T и п не установлен, вид описан по остаткам, собранным в La Grive St. Alban,
Франция, верхний миоцен.
Диагноз (по: Butler, 1948). Р4 одновершинный. Р2 и Р3 разных
размеров.
Описание (по: Butler, 1948). Нижние челюсти (длиной 27—29 мм)
напоминают таковые Hylomys, но отличаются слабо изогнутым и менее
вертикальным венечным отростком, ниже расположенным сочленовным
отростком. Угловой отросток с крепким выступом на внутренней стороне.
Подбородочное отверстие чаще расположено под Р3. Передний край орбиты
расположен на уровне переднего края М1. Подглазничное отверстие рас-
положено впереди Р4, а скуловая дуга поднимается против М1 и М2.
На нёбе продолговатая борозда для нёбного нерва, сходная с таковой
Hylomys. М1 отличается от всех современных Erinaceidae тем, что он более
развит в ширину. М2 сходен с предыдущим, но отличается меньшими раз-
мерами. Общие структурные особенности этих зубов имеют сходство
с современными. Оба зуба имеют по три корня. М3 отличается от современ-
ных треугольной формой, с тремя почти равными корнями. Состоит из
пара-, прото- и метаконов, без вторичных образований. Его внешняя сто-
рона косая, а задняя короткая. Р4 сходен с этим зубом современных видов,
отличается от современных поперечным расположением коронки и только
2. РОД GALERIX
111
одним язычным корнем. Р3 по форме сходен с Р4, но меньших размеров
и этим отличается от современных. Р2 и Р1 сравнительно крупные, оваль-
ной формы, с двумя корнями или у Р1 корни могут быть слитыми в один.
Нижние моляры мало отличаются от этих зубов Hylomys и Neotetracus.
Р4 имеет хорошо развитый метаконид. Передний бугорок этого зуба не
приподнят. Р3 крупный, как у современных видов. Его метаконид в виде
выступа (автор за метаконид принимает внутреннюю часть протоконида).
Р2 имеет два корня (редко один). Рг широкоовальной формы, одновершин-
ный, с одним корнем.
Верхний клык крупный с двумя корнями, нижний более мелкий с од-
ним корнем. 13 мельче 12 и 14. Его корень отогнут лингвально (по: Gail-
lard, 1899, 13 крупнее 12, а 1Х крупнее 13).
Измерения (по: Gaillard, 1899). Длина нижнего зубного
ряда — 13.0—16.0, Мх—М3 — 7.0—8.5, Рх—Р4 — 6.0—7.5, Мх — 2.8—3.3,
высота нижней челюсти на уровне Мх — 4.0—5.0 мм.
Распространение и возраст. Франция, верхний миоцен.
t 2. Galerix minor Filhol, 1880.
Filhol, 1880b, p. 120.
Тип. Нижняя челюсть № M2388g, фосфориты Caylux, Франция,- нижний оли-
гоцен.
Диагноз (по: Butler, 1948). Рх с несколькими бугорками. Р2 и Р3
сходных размеров. .
Описание (по: Butler, 1948). Нижняя челюсть этого вида отлича-
ется от таковой G. exilis Blainville менее приподнятым сочленовным мы-
щелком, слабее развитым угловым отростком с менее выдающимся на вну-
треннюю сторону выступом на его внутренней поверхности. Фильхол
отмечает отсутствие метаконида на Р4, но, как пишет Батлер, зуб сильно
изношен и это может служить причиной его отсутствия. Судя по альвео-
лам, Р3 и Р2 были с двумя корнями и более близки по общим размерам, чем
у G. exilis. От последнего описываемый вид отличается и разделением
коронки на бугорки у Р4, имеющего один корень. Диастема между клыком
и Pj у G. minor длиннее, чем у G. exilis. Клык более крупный, а 13 очень
мелкий, смещен на внешнюю сторону, как у G. exilis.
Примечание. Сабан (Saban, 1958, р. 852) этот вид отметил как
синоним Тetracus Aymard и отнес его вместе с последним к подсем. Erinacei-
nae. При этом не упоминаются виды Tetracus nanus Aymard (тип рода) и
Т. bouti Lavocat. Большинством систематиков-палеонтологов род Tetracus
совершенно правильно считается принадлежащим подсем. Galericinae,
в котором он сохраняется и в настоящей работе.
Распространение и возраст. Франция, нижний оли-
гоцен.
t 3. Galerix africanus Butler, 1956 (рис. 50).
Butler, 1956a, p. 62—64.
Тип. Фрагмент правой ветви нижней челюсти с зубами Р3, Р4, Sgr № 315-49,
Сонгхор, о. Русинга на оз. Виктория-Нианца, Кения, миоцен.
Диагноз (по: Butler, 1956). Вид рода Galerix, сходный с G. exilis
Blainv. скорее, чем с Pseudogalerix stehlini Gaill. в относительных размерах
Р3 и Р4 и в развитии метаконида Р4 (внутреннего бугорка протоконида
в моем представлении), но отличается от G. exilis более обособленным пара-
конидом Р4. Более крупный, чем G. exilis (Р3—М2=11.0 мм).
Описание (по: Butler, 1956, с сокращениями). Нижний край
ветви нижней челюсти узкий, возвышающийся к М3. Угловой отросток,
112
ПОДСЕМ. GALERICINAE
вероятно, был более высокий, чем у G. exilis, только слегка ниже уровня
зубов. Передний край венечного отростка возвышался более круто, чем
у сравниваемого вида. Гребень на лингвальной стороне челюсти проходит
назад и вверх от основания венечного отростка, как у G. exilis. У типа
конец симфиза расположен под Р3, а подбородочное отверстие открыва-
ется под передним краем Р4. Р2, от которого сохранилась только задняя
альвеола, вероятно, был такой же длины, как и Р3. Р3 сравнительно шире,
Рис. 49. Galerix paraexilis (Л) и G. tadzhikistanicus (Б) sp. nov.
Рис. 50. Galerix africanus (по: Butler, 1956а).
чем у G. exilis и Р. stehlini. Его задняя часть шире, чем передняя. Р4 выше
и длиннее Р3.
Распространение и воз р а с т. Известен из одного место-
нахождения, указанного для типа.
f 4. Galerix paraexilis sp. nov. (рис. 49, A).
Тип. Фрагмент правой ветви нижней челюсти с зубами P4hMj, № 2977 (13—25)—1,
Зайсанская котловина, правый берег р. Сарыбулак, Казахстан, верхний миоцен, кол.
Палеонтол. инет. АН СССР.
Материал. Серия зубов и фрагментов нижних челюстей № 2977 (13—25),
(26—91), (111—181), Зайсанская котловина, р. Сарыбулак, Казахстан, верхний мио-
цен, сборы Н. С. Шевыревой, кол. Палеонтол. инет. АН СССР.
Диагноз. Верхние коренные зубы широкие, массивные. М3 оваль-
ной формы, не треугольный. Нижние моляры со сложной эмалевой скульп-
турой на жевательной поверхности.
Описание. Р4 характерной для гимнур формы. Его паракон высо-
кий и массивный. Метакон вытянут назад, гребнистый. От средней части
зуб резко сужен к внутреннему краю. Протокон и талон узкие, прижаты
друг к другу, разделены узкой бороздкой. Талон на задней стороне ко-
ронки зуба отделен бороздкой от третьего бугорка, представляющего
утолщение задней стенки зуба. М1 самый широкий и крупный зуб верхней
челюсти. Наружный край этого зуба скошен, что обусловлено значитель-
ным удлинением метакона на внешнюю сторону и назад. Передний край
2. РОД GALERIX
ИЗ
этого зуба выгнут вперед, а задний вогнут. Паракон развит менее, чем
метакон. У нестертого зуба вершина переднего срединного бугорка (пара-
стиль) прижата к паракону, а у стертого приобретает треугольную выемку
и принимает карманообразную форму. Такую же форму приобретает и
задний срединный бугорок (метастиль). Протокой развит значительно,
выступает на внутренней стороне коронки далее более мелкого и более
уплощенного талона. Цингулюм не развит только на внутренней стороне
зуба.
Очертания коронки М2 близки к прямоугольнику, но этот зуб, так же
как и М1, имеет выпуклую наружную сторону и вогнутую заднюю. Цин-
гулюм развит так же, как и у М1. Паракон и метакон у этого зуба имеют
сходные размеры. Передний срединный бугорок развит более, чем у М1.
Протокон пропорционально крупнее протокона М1. М3 овальной формы,
со скошенным назад наружным краем. Паракон, парастиль и протокон
развиты хорошо, метакон и метастиль развиты слабо, а талон отсутствует.
Цингулюм имеется на передней и задней сторонах. Мх удлиненной формы.
Выступающий вперед высокий паракон соединен гребнем с наружным
бугорком протокона. Внутренний бугорок протокона обособлен. Метакон
(талон других авторов) удлинен. Его внутренне-задний бугорок высокий,
со сложным эмалевым рельефом, а наружно-задний бугорок низкий,
обособленный. На наружной стороне коронки между передним (пара—
протокон) и задним (метакон) сегментами зуба впадина. Цингулюм не
развит только на задней стороне коронки. М2 сходен по строению с Мх,
его паракон не выступает вперед. Внутренний бугорок протокона крупнее,
чем у Мх, прижат к паракону, выше последнего. М3 сходен с М2 по строе-
нию, но самый мелкий из нижних коренных зубов. Неповрежденная сти-
ранием жевательная поверхность нижних коренных зубов имеет сложную
эмалевую скульптуру.
Измерения. Длина по наружному краю М1 — 2.26, 2.27, 2.28,
3.0, 3.0; М2 - 2.25, 2.35, 2.52; М3 - 1.35, 1.5 (в средней части); Р4 - 1.5
(в средней части); наибольшая ширина Р4 — 3.5; М1 — 3.5, 3.9; М2 —
2.5, 3.25, 3.35, 3.5; М3 — 2.3, 2.5; длина Р4‘— 1.8; Мх — 3.0, 3.25, 3.3, 3.5;
М2 — 2.5, 2.35, 2.7 мм.
Распространение и возраст. Казахстан, верхний
миоцен.
Примечание. Описание этого и следующего видов проведено
в сравнении с экземпляром G. exilis Blainville (Grive St. Alban, верхний
миоцен), полученным из Франции.
+ 5. Galerix tadzhikistanicus sp. nov. (рис. 49, Б).
Тип. Фрагмент левой ветви нижней челюсти с зубами Р3, Р4, Мх и М2,
№ 3848
' 1000-106 ’ Кулябская область, Таджикистан, миоцен, кол. Геология, инет.
АН СССР.
Описываемый вид резко отличается от G. paraexilis структурными
особенностями зубов нижней челюсти, которые по строению ближе к тако-
вым G. exilis. Тем не менее они хорошо отличаются и от зубов последнего,
что видно из их сравнения.
Galerix tadzhikistanicus
Р3. Параконид развит более. Метако-
нид сужен.
Р4. Параконид относительно высокий.
Наружный бугорок протоконида немас-
сивный, невысокий, а внутренний буго-
8 А. А. Гуреев
Galerix exilis
Р3. Параконид развит менее. Метако-
нид расширен, более развит.
Р4. Параконид невысокий. Наружный
бугорок протоконида массивный, а его
внутренний бугорок слабо развит, мало
114
ПОДСЕМ. GALERICINAE
рок хорошо развит, обособлен. Метако-
нид хорошо развит и вытянут на внутрен-
ней стороне назад.
Mj. Наружный край внешнего бугорка
почти вертикальный, мало скошен к ос-
нованию. Зуб слабо расширен кзади.
Метаконид удлинен.
М2. Не широкий, с косым задним
краем. Цингулюм Мг и М2 слабо развит.
обособлен. Метаконид развит слабо, не
вытянут, а в виде ступеньки.
Mi. Наружный край внешнего бугорка
протоконида сильно скошен от вершины
к его основанию. Зуб сильно расширен
кзади. Метаконид укорочен.
М2. Широкий, с прямым задним краем.
.Цингулюм на Мх и М2 хорошо развит.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
t 3. Род PSEUDOGALERIX Gaillard, 1929
Gaillard, 1929, р. 7.
Тип. Pseudogalerix stehlinii Gaillard, 1929, Griv St. Ablan (Isere), Франция,
верхний миоцен.
Диагноз (по: Butler, 1948; Viret, 1938). Нижняя челюсть крупнее
таковой Galerix Pomel (около 32 мм). Ее угловой отросток менее развит,
чем у Galerix, также и выступ на его внутренней стороне. Клык выше
и менее уплощен, чем у Galerix. Рх с одним корнем. Р2 сравнительно
крупный, Р| мелкие, простые по строению. М| с тремя корнями.
Зубная формула у представителей рода судя по литературным источ-
никам: I^C^-P^-M^- — 44.
3. 14 3
Описание (по: Butler 1948). Кроме признаков, указанных в диаг-
нозе, нужно отметить и следующие: подбородочное отверстие располо-
жено под Р3 или между Р2 и Р3. Нижние моляры такие же, как у Gale-
rix. Внутренний зубец протоконида Р4 (метаконида по: Butler, 1948, и др.)
развит слабо или отсутствует.
Примечание. Первоописание этого рода (Gaillard, 1929) мне
получить не удалось. По замечанию Вире (Viret, 1938), род и вид описаны
по двум фрагментам: по левой половине верхней челюсти с пятью зубами
(С—Р4) и правой половине верхней челюсти с тремя молярами. Последний
фрагмент, по его мнению, скорее принадлежит представителю рода Gale-
rix. Следовательно, Pseudogalerix более точно может определяться по верх-
ним зубам С—Р4. В 1948 г. Батлер описывает нижнюю челюсть Pseudo-
galerix в сравнении с таковой Galerix, хотя Вире, ссылаясь на заметку
(Gaillard, 1929) с описанием Pseudogalerix, не упоминает про нижнюю
челюсть. Были ли подобраны нижние челюсти к верхним и соответ-
ствуют ли они друг другу и этому роду не ясно. Тем не менее Батлер
(Butler, 1956) при описании Galerix africanus сравнивает его особенности
с особенностями представителей родов Galerix и Pseudogalerix. То же
делают и другие авторы.
Распространение и возраст. Европа, верхний
миоцен.
+ 1. Pseudogalerix stehlini Gaillard, 1929.
Описан там же, где и род.
Тип для меня остался не известен.
Диагноз. Совпадает с таковым рода.
Распространение и возраст. Отмечены в описании
рода.
4. РОД LANTHANOTHERIUM 115
f 4. Род LANTHANOTHERIUM Filhol, 1888
Filhol, 1888b, p. 24, 25.
Тип. Lanthanotherium sansaniensis Filhol, 1888, San-San (Gers), Франция, средний
миоцен.
3 13 3
Диагноз. Зубная формула: I4т-С-т-РМ-=- = 40, Р] отсутствуют.
О 1 О О
В отличие от Galerix Pomel и Pseudogalerix Gaill. верхний клык с двумя
корнями.
Описание (по: Saban, 1958). Нижняя челюсть удлиненная, с изо-
гнутым нижним краем. Подбородочное отверстие под Р3. Три верхних
резца клыкообразной формы, с одним корнем, убывают в размерах от
первого к третьему. Клык массивный, с двумя корнями, Р1 отсутствует.
Р2 мелкий, с двумя корнями. Р3 небольшой, с двумя корнями и имеет мел-
кий бугорок (дополнительный) на внутренней стороне коронки. Внешне
этот зуб сходен с Р4. Р4 трапециевидной формы, с тремя корнями. М1 и М2
квадратно-четырехугольной формы, с тремя корнями. Лингвальный
корень самый крупный (образовался в результате слияния корня прото-
кона и талона, что имеет место у представителей рода Galerix, материал
по которым был у меня в руках). Метаконуль (пятый бугорок) занимает
центральное положение на жевательной поверхности зуба. М3 треугольной
формы, с тремя корнями. Нижние резцы лопаточковидной формы, накло-
нены вперед, убывают в размерах от первого к третьему. Нижний клык
с двумя корнями, объемистый, слабо отогнут назад. Р2 и Р3 сравнительно
мелкие, с двумя корнями, имеют небольшой талон. Р4 высокий и массив-
ный, приближается по строению к молярам. Мх и М2 массивные. Их триго-
нид более высокий, чем талонид (передний сегмент (параконид и прото-
конид) выше второго сегмента (метаконида), по моим представлениям).
Распространение и возраст. Европа, средний—верх-
ний миоцен.
t 1. Lanthanotherium sansaniensis Filhol, 1888.
Описан там же, где и род.
Тип. Не отмечен, вид описан из San-San (Gers), Франция, средний миоцен.
Описание (по: Filhol, 1888b). Вид описан по фрагменту верхней
челюсти с зубами (три резца, клык, три премоляра и один моляр). Резцы
направлены наклонно, они удлиненные, убывающие в размерах от первого
к третьему. Эта редукция гораздо менее выражена, чем у тупай, у которых
коронка третьего резца значительно редуцирована по сравнению с пред-
шествующими резцами. Клык опирается на один корень. Его коронка кони-
ческая с выпуклым передним и вогнутым задним краями. Первый премоляр
(Р2 в действительности) с одним корнем. Его коронка треугольной формы,
массивная и низкая. Второй премоляр (Р3) имеет сходную с Р2 форму,
такой же низкий. Его передний край выпуклый, а задний вогнутый.
Р4 сходного с предшествующими строения. М1 описан так, что нельзя по-
нять, какого он строения.
Примечание. Автор описания этого вида отмечает, что клык и Р2
имеют по одному корню, а Сабан (Saban, 1958) для этих зубов отмечает
два корня. Думаю, что это расхождение не имеет принципиального значе-
ния, поскольку у ежеобразных корни резцов, клыков и первых двух
премоляров часто сливаются. Например, у Hylomys suillus Р2 чаще
бывает с корнями, слитыми в один, но может быть и с двумя свободными
корнями.
Распространение и возраст. San-San (Gers), Франция,
средний миоцен.
8*
116
ПОДСЕМ. GALERICINAE
t 2. Lanthanotherium robustum Viret, 1940 (рис. 51).
Viret, 1940, p. 33—65.
Тип. Первоописания у меня в руках не было, а Батлер (1948) тип не отметил.
Описание (по: Butler, 1948). Строение нижней челюсти этого
вида сходно со строением этой кости Galerix exilis Blainv. и Pseudogalerix
stehlini Gaill., но у описываемого вида сочленовный отросток расположен
Рис. 51. Lanthanotherium robustum'. (по
Butler, 1948).
немного выше, а передняя поверх-
ность венечного отростка более верти-
кальна, чем у представителей отме-
ченных родов. В целом нижняя че-
люсть L. robustum напоминает та-
ковую Neotetracus sinensis Trouess.
Челюстное отверстие расположено по
отношению к ряду зубов выше, чем
у Galerix и Pseudogalerix и сходно
с L. sansaniensis Filhol. Подбородоч-
ное отверстие под Р4, у его перед-
него края. Область симфиза прости-
рается до Р2. Судя по размерам аль-
веол, моляры такой же величину,
как у Galerix. Р4 короче моляров,
имеют более длинные корни, чем
у последних. Р3 с тупой вершиной,
а спереди и сзади у него по одному
маленькому бугорку. Альвеола клыка
изгибается назад и вниз. Рх утрачен.
Клык сравнительно крупный.
возраст. Известен из верхнего
Распространение и
миоцена Франции.
Примечание. Сабан (Saban, 1958) отмечает, что Тениус (Thenius,
1949), описавший L. longirostris, считает этот вид сохраняющим Рх. Но тогда
нет основания считать его принадлежащим к роду Lanthanotherium.
t 3. Lanthanotherium sp. Webb, 1961 (рис. 52, A).
Webb, 1961, p. 1085—1087.
Материал. Фрагмент левой ветви нижней челюсти с зубами М,—М3, LACM
(CYJ), № 5284. Формация Бонеста, Калифорния, миоцен, кол. Калиф, инет, технол.
Автор, описывая особенности зубов, сохранившихся на фрагменте,
ведет их сравнение со структурами зубов ежей (роды Amphechinus Aymard,
Erinaceus L. и др.), а также с вымершими гимнурами (Galerix Pomel и
Pseudogalerix Gaillard). В результате он приходит к заключению, что
исследованный им фрагмент нижней челюсти, как и сохранившиеся зубы,
ближе всего к таковым Lanthanotherium robustum Viret.
Примечание. Структурные особенности и соотношение размеров
и высоты переднего и заднего сегментов действительно более близки
к таковым Lanthanotherium.. К сожалению, не сохранился передний отдел
зубной части нижней челюсти с зубами, позволяющими более надежно
определить принадлежность нижней челюсти к тому или иному роду.
Новые материалы позволят установить, какие роды в действительности
существовали в Северной Америке. Важно, что они были там в то время,
в которое их представители были в Евразии и в Африке.
Измерения (по: Webb, 1961). Длина Мг — 3.5, М2 — 2.4,
Мд -- 1.1 ММ.
4. РОД LANTHANOTHERIUM
117
Распространение и возраст. Известен из одного место-
нахождения, указанного выше, миоцен. '
t 4. Lanthanotherium longirostrum Thenius, 1949 (рис. 52, Б).
Thenius, 1949, S. 676-684.
Тип. Левая ветвь нижней челюсти с h—С и Р2—М3 и верхние коренные зубы
4017 и 4019, Leobem (Steirmark), средний миоцен, кол. Joanneums (Graz).
Диагноз. По крайней мере в нижней челюсти сохраняется Р4.
Между резцами и клыком и между клыком и Р, — диастемы.
Описание (по: Thenius, 1949). Д и 12 крупнее 13. Клык сильно
развит, конусовидный, не изогнут. Р2 и Р3 мелкие, с одним корнем. Р4
с тремя бугорками. Из них метаконид почти слит с цингулюмом. Р3 ма-
Рис. 52. Lanthanotherium sp. (А) (по: Webb, 1961) и L. longirostrum (Б) (по: Thenius, 1949).
ленький, Р4 крупный, четырехугольный, с хорошо развитым талоном. М1
и М2 четырехугольные, почти квадратные. Пятый бугорок расположен
в центральной части жевательной поверхности этих зубов.
Измерения (по: Thenius, 1949). Р3 — 1.7x1.4,- Р4 — 3.2x3.4,
М1 — 3.0x3.2, М2 — 2.5x3.0, С — 1.4x1.2, Р2 - 1.0х0.6, Р3 — 1.4Х
Х0.8, Р4 — 2.8Х1.8, М7 — З.ЗХ1.8, М2 — 2.6х2.1, М3 — 2.3Х1.7,
длина М4—М3 — 8.5 мм.
Распространение и возраст. Известен из Австрии,
средний миоцен.
Примечание. Остатки этой миоценовой гимнуры Мейер (Меуег,
1909) отнес к Parasorex socialis (синоним Galerix exilis Blv.). Зданский
(Zdansky, 1909) отнес их к Galerix exilis. По мнению Тениуса, эта гимнура
относится к роду Lanthanotherium Filhol.
Можно думать, что описываемая гимнура сохраняла первый премоляр не
только в нижней, но и в верхней челюсти, следовательно, имела 44 зуба,
как и Galerix Pomel, но отличалась строением зубов и зубных рядов.
От представителей рода Lanthanotherium описываемый вид отличается
наличием Р1? значительным развитием клыка нижней челюсти и диастемами
между ним и соседними зубами. Эти различия родового, а не видового
порядка. Правильнее считать эту гимнуру представителем особого рода.
US
ПОДСЕМ. GALERICINAE
f5. Род DEINOGALERIX Freudental, 1972
Freudental, 1972, p. 1—11.
Тип. Deinogalerix koenigswaldi Freudental, 1972, Италия, миоцен.
Диагноз (no: Freudental, 1972). Зубная формула: I С у Р М у =
=40—42. Гимнура гигантских размеров. Премоляры разделены диасте-
мами. Лицевая часть черепа удлинена. Череп очень длинный по сравнению
с телом;
Описание (по: Freudental, 1972). Череп сверху плоский, его
наибольшая длина 210 мм. Длина костного нёба от заднего гребня до
Рис. 53. Deinogalerix koenigswaldi (по: Freudental, 1972).
переднего края черепа 129 мм. Носовой отдел черепа по сравнению с обыч-
ными Erinaceidae очень длинный. Морда плоская и длинная. Подглазнич-
ное отверстие впереди переднего края Р4. Сагиттальный гребень высотой
в 1 см, узкий. Венечный отросток нижней челюсти очень низкий. Сочленов-
ный отросток на уровне жевательной поверхности зубов. Угловой отросток
длинный, массивный, с внутренней стороны плоский. Подбородочное
отверстие под передним краем Р3.
I1 очень крупный, толстый, слабо изогнут. I2 и I3 одновершинные,
с одним корнем. Клык, Р1 и Р2 сходные по внешнему виду, высокие,
с двумя корнями. Р3 треугольной формы, с одной крупной вершиной на
внешней стороне и двумя мелкими на внутренней. Р4 имеет крупную вер-
шину на наружной части коронки и три мелких на внутренней. Структур-
ные особенности этого зуба напоминают таковые у хищных. Этот зуб соче-
тается с крупным Mlt как у хищных. Коренные зубы уменьшаются в раз-
6. РОД OCAJILA
119
мерах от М1 к М3. Последний треугольной формы, с тремя бугорками.
Верхние зубы разделены четырьмя диастемами: большой диастемой между
Is и С, меньшей между С и Р1 и двумя мелкими между Р1 и Р2, и Р2 и Р3.
Р4 более похож на этот зуб Lanthanotherium Filhol, чем Galerix Pomel,
Р2 одновершинный, с двумя корнями. Впереди Р2 длинная диастема.
Распространение и возраст. Италия, миоцен.
t 1. Deinogalerix koenigswaldi Freudental, 1972 (рис. 53—56).
Freudental, 1972, там же.
Тип. Большая часть скелета от № RGM 177777 до № 177884. Череп № RGM
177777. San Giovanino, провинция Foggia, Италия, миоцен, кол. Rijksmuseum Geol.
and Mineral.
Измерения (no: Freudental, 1972). Длина плечевой кости —
103, локтевой — 125, лучевой — 102, бедра — 114, голени — 136,
ступни — 105 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
f 6. Род OCAJILA Macdonald, 1963
Macdonald, 1963, р. 166.
Тип. Ocajila makpiyahe Macdonald, 1963, Южная Дакота, США, нижний миоцен.
Диагноз (по: Macdonald, 1963). Тригонид сжат в передне-заднем
направлении. Передне-задний диаметр тригонида сходен с таковым его
талонида. Параконид редуцирован до гребня.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
t 1. Ocajila makpiyahe Makdonald, 1963 (рис. 57).
Macdonald, 1963, p. 167, 168.
T и n. SDSM № 56105, фрагмент левой ветви нижней челюсти со слегка стертыми
М2, М3, формация Sharps, Южная Дакота, США, нижний миоцен, кол. Муз. г. Лос-
Анджелеса.
Описание. Передний сегмент (тригонид других авторов) М2 сжат
спереди-назад. Параконид слабо обособлен от протоконида и представлен
в виде гребня, соединенного на внешнем крае коронки с наружным бугор-
ком протоконида. Протоконид с двумя бугорками (наружным и внутрен-
ним). Метаконид этого зуба (талонид других авторов) широкий, с двумя
бугорками. Из них внешний менее высокий и менее массивный. М3 сходен
по строению с М2, но меньше по размерам.
Измерения (по: Macdonald, 1936). М2 — 1.78x1.45, М3 — 1.42х
Х1.14 мм.
Распространение и возраст. Известен из одного место-
нахождения, отмеченного выше для типа.
Примечание. Структурные особенности сохранившихся зубов и
нижней челюсти (фрагмент ее задней части) схо’дны с таковыми гимнур.
Это основательный документ, подтверждающий существование представи-
телей подсем. Galericinae в Северной Америке в конце палеогена—начале
неогена. Нет оснований отрицать вероятность их существования на этом
континенте.
120
ПОДСЕМ. GALERICINAE
t 7. Род NEUROGYMNURUS Filhol, 1877
Filhol 1877, p. 51—54. Cayluxotherium, Filhol, 1880b, p. 120—125.
Тип. Neurogymnur cayluxi. Filhol, 1877, Quersi, Франция, верхний эоцен.
55
Рис. 54—56. Deinogalerix koenigswaldi (no: Freudental, 1972).
Диагноз. Зубная формула: 1-|-С-|-Р-|-М-|- = 44. Теменные кости
в отличие от всех других ежеобразных с морщинистой поверхностью Заты-
лочный выступ сильно развит. Костное нёбо перфорировано. Длина ряда
премоляров более ряда моляров. Р4 самый крупный зуб в верхней челюсти.
7. РОД NEUROGYMNURUS
121
Описание (по: Butler, 1948). Верхняя линия профиля черепа пря-
мая. Основание скуловой дуги имеет выемку, как и у современных гимнур.
Слезное отверстие в орбите. Подглазничное отверстие перед Р4. М1 пол-
ностью под орбитой. Имеется надглазничный выступ, оканчивающийся
Рис. 54—56 (продолжение).
заглазничным отростком. Барабанная область сходна с таковой современ-
ных гимнур. Барабанные камеры крупные, а промежуток между ними
узкий. Чешуйчатая и клиновидная (крылья) кости участвуют в покрытии
57
Рис. 57. Ocajlla ma.kpiya.he (по:
Macdonald, 1963).
58
Рис. 58. Neurogymnurus cayluxi (по: Viret, 1947).
(образовании) барабанных камер. Базисфеноидная выемка отсутствует.
Межтеменная кость треугольной формы. Рх с одним бороздчатым корнем
(при слиянии двух корней) или с Двумя корнями. Р2 с двумя корнями. Ниж-
няя челюсть такая же, как у Galerix Pomel.
Распространение и возраст. Европа, верхний эоцен—
нижний олигоцен.
122
ПОДСЕМ. GALERICINAE
t 1. Neurogymnurus cayluxi Filhol, 1877 (рис. 58, 59).
Filhol, 1877, p. 51—54, Cayluxotherium elegans Filhol. 1880b, p. 120—125.
Тип. Установить не удалось. Описан из Франции (Quersi), верхний эоцен.
Рис. 59. Neurogymnurus cayluxi (по: Dechascaux, 1964).
Рис. 60. Neurogymnurus mediterraneus (по: Viret, 1947).
Описание. Особенности, отмеченные в характеристике рода, для
вида могут быть дополнены следующими (по: Viret, 1938): резцы в полном
наборе простые. I2 и I8 с одним корнем. Премоляры слабо редуцированы.
8. РОД PSEUDONEUROGYMNURUS
123
Р4 очень крупный (самый крупный в верхнем зубном ряду). Верхние
моляры сложной структуры. М3 треугольной формы. М3 сходен с М2-
Распространение и возраст. Известен (Friant, 1971)
из верхнего эоцена Франции и Швейцарии.
t 2. Neurogymnurus mediterraneus Viret, 1947 (рис. 60).
Viret, 1947, Eclog. Geol. Helvet., v. 40, N 2, p. 341—343.
Тип. Фрагмент левой ветви нижней челюсти с зубами (С—М3) № Маг—221,
добытой в глине вблизи Марселя (Франция), нижний олигоцеи, кол. Муз. естест. истор.
в г. Бале.
Вире (Viret, 1947) отмечает следующие особенности этого вида: раз-
меры составляют около 1.5 размеров N. cayluxi Filhol, горизонтальная
ветвь имеет две диастемы, одна между Р3 и Р2, другая между Р2 и Р2; буго-
рок метаконида очень мелкий или отсутствует; параконид Мх обособлен
в виде массивного бугорка с широким основанием.
И з м е р е н и я (по: Viret, 1947). Длина фрагмента от сочленовного
кондилюса до альвеол резцов — 53, длина ряда сохранившихся зубов
(С—М3) — 2.7, длина четырех премоляров — 12.7, высота фрагмента на
уровне М3 — 7.9 мм.
Распространение и возраст. Франция, нижний оли-
гоцен.
|8. Род PSEUDONEUROGYMNURUS gen. nov.
Тип. Pseudoneurogymnurus schevyrevii sp. nov., Казахстан, олигоцей.
Диагноз. P4 не массивный, по размерам меньше М1 и М2. Послед-
ние имеют прямоугольную форму, и только у М1 наружный край немного
скошен. Задний срединный бугорок (метастиль) пирамидальной или
карманообразной формы.
Описание. Бугорки паракона и ме-
такона М1 и М2 конической формы, обособ-
лены друг от друга (не соединены на на-
ружной стороне гребнем). Задний срединный
бугорок часто соединен с протоконом или
с протоконом и талоном.
Рис. 61. Pseudoneurogymnurus schevyrevii (А),
Р. zhchikvadzei (Б) sp. sp. nov.
Распространение и возраст. Казахстан, олигоцен.
t 1. Pseudoneurogymnurus scheyyrevi sp. nov. (рис. 61, A).
T и п. M1 левой стороны верхней челюсти № 2979 (134—305) (1), Зайсанский район,
Конуркура, Казахстан, олигоцен.
Материал. Р4—1 экз., М1— 2 экз., М2 — 1 экз., Зайсанский район, Конур-
кура, Казахстан, М1 и М2 без номера. Хонгуркульджа, Казахстан, олигоцен.
Диагноз. Передняя и задняя стороны коронки М2 не параллельны.
Бугорок паракона меньше бугорка метакона. Метастиль мелкий, пира-
мидальный. ,
Описание. Р4 вытянут в поперечном к продольной оси челюсти
направлении. Его паракон высокий, а метакон низкий, гребнистый, вытя-
нут назад. Впереди паракона низко расположенный массивный выступ.
С этим выступом соединен гребнем протокон. Последний достигает при-
мерно половины высоты паракона. Талон в виде обособленного бугорка,
задний край которого переходит в цингулюм. М1 со скошенным немного
наружным краем, бугорок паракона развит менее бугорка метакона.
124
ПОДСЕМ. GALERICINAE
Задний срединный бугорок (метастиль) в виде трехгранной пирамидки,
а передний срединный бугорок (парас,тиль) представлен утолщением
задней стенки зуба. Бугорок талона обособлен. М2 отличается от М1
меньшей величиной, слабым развитием протокона и пропорционально
более крупным талоном, а также наличием цингулюма не только на задней,
но и на передней сторонах коронки зуба. М2 с массивным передним сег-
ментом (паракопид-протоконид) и коротким задним (метаконид).
Измерения. Длина Р4 — 1.75, М1 — 1.75, 1.76, М2 — 1.5, М2 —
2.0; ширина Р4 — 2.4, М1 — 2.6, 2.75, М2 — 2.1, М2 — 1.5 мм.
Распространение и возраст. Казахстан, олигоцен.
t 2. Pseudoneurogymnurus zhchikvadzei sp. nov. (рис. 61, Б).
T ип. M2 правой стороны верхней челюсти, 2979 (134—305), (2), Зайсанский район,
Конуркура, Казахстан, олигоцен.
Материал. Р4— 1 экз., М2 — 2 экз. (один поврежден), М2— 1 экз.
Диагноз. Передний и задний края М2 почти параллельны. Буго-
рок паракона сходен по размерам с бугорком метакона. Метастиль круп-
ный, карманообразный.
Описание. Паракон Р4 в виде высокого массивного бугорка,
соединенного гребнем с вытянутым назад метаконом. Протокон его низ-
кий, малых размеров, талон стерт, он располагался на широком, лопасте-
видном основании. Паракон и метакон М2 сходных размеров, широкие и
массивные. Протокон широкий в основной части, с высокой вершиной.
Метастиль соединен гребнем с протоконом. Небольшой талон в виде от-
дельного бугорка, тонким гребнем соединен с основанием метастиля.
М2 с высоким передним сегментом (параконидом—протоконидом). Бугорки
пара- и протоконида сильно сближены и образуют узкий треугольник.
Метаконид (талонид других авторов) низкий, бугорок на задне-внутрен-
ней стороне отломан, на передне-наружной невысокий, узкий. Цингулюм
развит на передней и задней сторонах коронки зуба.
Измерения. Длина Р4 — 1.5; М2 — 1.6; М2 — 1.65; ширина Р4 —
1.7; М2 — 2.0; М2 — 1.25 мм.
Распространение и возраст. Казахстан, олигоцен.
f Genus et sp. nov.
Фрагмент M2 левой стороны, № 2979 (134—305) (1), 19 км к С-3 от г. Зайсан;
Конуркура, Казахстан, олигоцен. У фрагмента сохранились часть внутренней сто-
роны паракона' метастиль со стертой вершиной, парастиль,бугорки протокона и талона.
В отличие от М2 других гимнур описываемый зуб имеет следующие
особенности: протокон не крупнее талона, парастиль в виде хорошо обособ-
ленного бугорка, протокон окружен широким цингулюмом. Длина зуба —
1.0 мм, что немного менее длины М2 землеройки Sorex araneus L.
9. Род ECHINOSOREX Blainville, 1838
Vlverra gymnura Raffles, 1821, Trans. Linn. Soc. London, v. 13, p. 272.
Gymnura Lesson, 1827, .Man. Mamm., p. 171; Horsfield, Vigors, 1828,
Zool. J., v. 3, p. 246. Echinosorex Blainville, 1838, p. 142.'Echinosorex Blain-
ville, 1839—1864, Osteogr., Insectiv., p. 109.
Тип. Echinosorex gymnurus Raffles, 1821, Тенассерим, Сиам, о-ва Малайские,
Суматра, Калимантан, современный.
Диагноз. Зубная формула: /~С ~ Р-^- м'^- = 44. Длина тела — 262—
450 мм, длина хвоста — более 50% длины тела. Верхний и нижний клыки
крупные. В отличие от других современных гимнур нижний клык входит
в глубокую выемку на межчелюстной кости.
9. РОД ECHINOSOREX
125
Описание. Тело удлиненное, на сравнительно коротких ногах.
Голова вытянутой формы, с заостренным носовым отделом. Ноздри имеют
передне-боковое расположение. Глаза относительно небольшие. Ушные
раковины хорошо развиты, почти голые. Хвост длинный — 200—210 мм.
Кисти и ступни 5-палые. Когти пальцев короткие, сжатые с боков. Волосяг
ной покров грубый, щетинистый. Окраска от бурой и черно-бурой со свет-
лыми пятнами на голове и плечах до одинаковой на всем теле — белой или
серовато-белой.
Рис. 62. Echinosorex gymnurus.
Череп удлиненной формы, с вытянутой лицевой частью, массивный.
Переднее носовое отверстие широкое. Подглазничное отверстие крупное.
Носовые кости расширены спереди и сужены сзади. Заглазничных отрост-
ков в виде бугорков нет. Мозговая капсула неширокая. Нижняя челюсть
массивная, с низкой зубной частью и широкой лопастью,, образованной
основаниями венечного, сочленовного и углового отростков.
I1 длинные, изогнутые, I2, и особенно Is, мелкие. Верхние клыки круп-
ные, с двумя корнями, нижние также крупные, с одним корнем. Р1 и Р2
мелкие, чайто не все развиты. Р3 крупный и высокий, но менее массивный,
чем Р4. Оба зуба с тремя корнями. М1 и М2 крупные, четырехугольной
формы, с четырьмя крупными и одним мелким (в центре жевательной по-
верхности) бугорками. М3 треугольной формы, вытянут назад, с тремя
бугорками и тремя корнями. 1х длиннее остальных. Размеры резцов убы-
вают к I3. Pj и Р2 мелкие, одновершинные, с одним корнем. Р3 трехвершин-
ный с двумя корнями. Р4 многовершинный с двумя корнями. Мх—М3
характерного для насекомоядных строения, убывают в размерах спереди
назад.
Распространение и возраст. Тенассерим и Сиам, Ма-
лайский архипелаг, Суматра и Калимантан, современный.
126
ПОДСЕМ. GALERICINAE
1. Echinosorex gymnurus Raffles, 1821 (рис. 62, 63).
Viverra gymnura Raffles, 1821, Trans. Linn. Soc. London, v. 13, p. 272—273.
Gymnura birmanica Trouessart, 1879, Rev. Zool. Paris, p. 240, G. rafflesi Bland-
ford, 1888, p. 220. G. gymnura minor Lyon, 1909, Proc. U. S. Nat. Mus. v. 36, p. 453.
Типне установлен, вид описан по материалам с о. Суматра. Автором описания
Рис. 63. Echinosorex gymnurus (по экз. из музея Унив. им. Гумбольдта, Берлин, ГДР).
вида отмечено, что несколько лет назад животное этого вида было доставлено из лесов
внутренней Малакки. Следовательно, ранее он был известен из Тенассерима, но не был
описан.
Описание. Кроме особенностей, приведенных в характеристике
рода, отмечены следующие. На большей части ареала особи описываемого
вида окрашены в темно-бурый цвет, со светлыми пятнами на голове,
плечах и загривке, а также на горле и шее. Эти пятна могут сливаться
в своеобразный светлый рисунок. Через глаз проходит темная полоса,
сливающаяся с темной окраской лба и темени. Хвост покрыт мелкими
10. РОД HYLOMYS
127
чешуйками и короткими и редкими волосами, кажется голым. На 1/3
от основания он бурый, остальная часть белая. Кисти и ступни сверху
густо покрыты жесткими коричневыми волосами и только кожа когтевых
фаланг одета светлыми волосами.
На о-ве Калимантан встречаются только белые особи этой гимнуры,
которых некоторые авторы выделяли в отдельный вид.
Отдельные вибриссы, расположенные на верхней губе и по бокам носа,
достигают значительной длины, прижатые к голове, заходят до основания
уха. Две вибриссы расположены на щеке и одна за глазом.
Измерения (по: Lyon, 1909). Длина тела — 311—445, хвоста —
190—278, ступни — 58—74, основная длина черепа — 61—87 мм (по:
Dobson, 1882, 60—87 мм).
Распространение в тех же границах, что и рода.
10. Род HYLOMYS Muller et Schlegel, 1839—1844
Muller, Schlegel, 1839—1844, Verhandelungen ovar de natuurlijke Ge-
schiedenis der- Nederlansche overseesche bezittungen Leiden, S. 153—158.
Тип. Hylomys suillus Miller, 1839, Юньнань, Бирма, п-ов Индокитай, о-ва Ма-
лайские, Тиоман, Суматра, Ява, Калимантан.
Диагноз. Зубная формула I -|- С у Р у М -|- = 44. Длина тела менее
260 мм. Длина хвоста не более 15°/0 длины тела. Верхний и нижний
клыки мелкие.
Описание. Тело удлиненное, на коротких ногах. Голова вытянута,
с заостренным носовым отделом (в виде хоботка). Ноздри расположены
сбоку, глаза крупные. Ухо небольшое, с округлой вершиной. Хвост
короткий, 12—30 мм. Кисти и ступни 5-палые. Когти короткие, неширо-
Рис. 64. Hylomys suillus (по: Temminck, 1839—1844).
кие. Волосяной покров сравнительно мягкий. Окраска однотонная на
всем теле, коричневая.
Череп с удлиненным лицевым отделом и сравнительно выпуклой
мозговой капсулой, не массивный. Переднее носовое отверстие неширокое.
Посдлазничное отверстие небольшое. Носовые кости более широкие в пе-
редней части. Заглазничные отростки в виде очень мелких бугорков.
Мозговая капсула неширокая. Нижняя челюсть тонкая, не массивная,
с низкой зубной частью и неширокой лопастью, образованной основаниями
венечного, сочленовного и углового отростков, разграниченных глубокими
выемками.
128
ПОДСЕМ. GALERICINAE
I1 с двумя вершинами (зубцами), несколько напоминают эти зубы
землероек (Soricidae). I2 и I3 мелкие. Клык с тремя вершинами и двумя
корнями, или они слиты. Р3 с двумя вершинами и двумя корнями, реже
корни слиты у зуба с правой или левой стороны челюсти. Р4 с пятью
бугорками (мелкий паракон, двувершинный протокон, метакон и талон)
и тремя-четырьмя корнями, если корни протокона и талона не слиты.
М1 и М2 с пятью бугорками (пятый мелкий, на средней части жевательной
поверхности) и четырьмя корнями. М3 с тремя вершинами и тремя корнями.
Вершина метакона
занимает поперечное положение и высоко поднята
над жевательной поверхностью этого зуба.
Д самый длинный. Резцы полого направ-
лены вперед. Клык по форме сходен с резцами,
наклонен вперед. Рь Р2 и Р3 сходной формы,
двувершинные. Р3 немного крупнее передних
премоляров, значительно меньше Р4. Последний
с тремя вершинами. Мп М2 и М3 характерного
для насекомоядных строения, убывают в раз-
мерах к М3.
Рис. 65. Hylomys suillus.
Распространение и возраст. Юньнань (Китай), Бирма,
п-ов Индокитай, о-ва Малайские, Тиоман, Суматра, Ява, Калимантан,
современный.
1. Hylomys suillus Muller et Schlegel, 1839—1844, (рис. 64, 65).
Muller, Schlegel, 1839—1844, там же, где род.
Т и п не установлен, вид описан по материалам с о-вов Суматра и Ява.
Описание. Один из мелких видов гимнур. Тело относительно
стройное, удлиненное, с коротким хвостом, на сравнительно низких ногах.
Голова тонкая, сильно заостренная в передней части. Мочка носа развита
в длину более, чем в ширину, примерно в три раза. Ноздри расположены
сбоку. Позади мочки носа волосы растут вверх, а не назад и образуют
в этом месте волосяной гребень. Вибриссы длинные, достигают концами
основания щек. Над глазом и позади него по одной длинной вибриссе.
Волосы, покрывающие тело, сравнительно мягкие. Хвост покрыт мелкими
чешуйками и редкими грубыми волосами. Ступни сверху покрыты круп-
ными чешуйками, напоминающими птичьи, и редкими жесткими волосами.
Щетинистые волосы у основания когтей достигают значительной длины и
заходят за вершину когтя. Окраска коричневая на всем теле. Только меж-
глазничная область, бока головы и тела светлые. Нижняя сторона тела
11. РОД PODOGYMNURA
129
имеет охристый или сероватый оттенок. Верхняя сторона кистей и ступней
бурого или черно-бурого цвета. Вдоль средней части головы и тела про-
ходит черно-бурая полоса, малозаметная на темном фоне спины. Когти
светлее окраски пальцев, с тонкими изогнутыми концами.
Измерения (по: Miiller, Schlegel, 1839—1844). Длина тела —
136, хвоста — 12, высота уха — 12 мм.
Распространение и. возраст. Юньнань (Китай), Бирма,
п-ов Идокитай, Сиам, о-ва Малайские, Тиоман, Суматра, Ява и Калиман-
тан, современный.
11. Род PODOGYMNURA Mearns, 1905
М е ar ns, 1905, р. 436—437.
Тип. Podogymnura truei Mearns, 1905, о. Минданао (Филиппины), современный.
Диагноз (по: Mearns, 1905). Зубная формула: 1-|- СуРуМ^- =40.
Is мелкий. Хвост толстый, умеренно обволосен, более 1/3 длины тела.
Ступни голые, длиной 3/4 длины головы. «Бакенбарды» длинные, протяги-
ваются до плечей. Диаметр глаз — 1/5 длины головы. Длина головы менее
чем J /4. общей длины тела. Уши крупные, полуголые, высотой от основания
более х/3 длины головы. Кисти полуголые сверху, совершенно голые снизу,
с четырьмя бугорками. Ступни
с пятью бугорками.
Диагноз, составленный мной,
сопоставим с таковыми других
родов: длина тела менее 260 мм.
Длина хвоста 35—45% длины
тела. Верхний и нижний клыки
крупные.
Описание (по: Mearns,
1905). Череп по сравнению с че-
репом Hylomys suillus Mull, et Sch-
leg. имеет более удлиненную рост-
ральную часть, а нижний край
верхнечелюстной кости выпрямлен-
ный (не дугообразный). Носовые
Рис. 66. Podogymnura truei (по: Walker
et al., 1964).
отверстия более наклоненные. Носовые кости узкие. Ветви нижней
челюсти более широкие. Первый (второй в действительности) верхний
резец крупный, но много меньше, чем клык. Второй (третий) верхний
резец мелкий. Р2 крупнее, чем I2 или Is, одновершинный, клыко-
образный. Р3 с двумя корнями, плоский, с выступающим цингулюмом.
Третий (четвертый) верхний премоляр внешне сходен со вторым, только
с более выступающим цингулюмом и выступом с двумя бугорками,
придающими зубу с внешней стороны квадратную моляровидную форму.
М1 и М2 квадратные, с четырьмя выступающими буграми и мелким цен-
тральным бугорком. М3 мелкий, треугольный, имеет три выступающих
бугорка и передне-наружный выступ. Нижпие резцы как Echinosorex
gymnurus Raff. Они уменьшаются в размерах спереди назад, с увеличен-
ными наружными остриями, чем хорошо отличаются от этих зубов верх-
ней челюсти. Нижний клык с одним корнем. Премоляры плоские, с двумя
корнями и выступающими цингулюмами. Моляры убывают в размерах
спереди назад, с четырьмя бугорками.
Распространение и возраст. О-ва Минданао, Филип-
пинские, современный.
9 А. А. Гуреев
130
ПОДСЕМ. GALERICINAE
1. Podogymnura truei Mearns, 1905 (рис. 66).
M e a r n s, 1905, p. 437—438.
Тип. Взрослая самка (череп с останками в спирте) № 125286, гора Апо, около
1050 м, южная часть Минданао, Филиппинские острова, 1904, кол. Нац. муз., США.
Описание (по: Mearns, 1905). Окраска свежего экземпляра.
Нос и пальцы мясного цве1а. Волосы, покрывающие верхние части тела,
аспидно-серые, смешанные с грубыми красновато-коричневыми волосами.
Нижние части тела седые, с небольшой примесью коричневатого тона,
образованного редкими коричневыми волосами. Когти с красновато-
коричневыми пятнами в середине.
Уолкер (Walker et al., 1964) отмечает, что волосяной покров у этого
вида мягкий и густой. Хвост умеренно обволосенный, от коричневого
до пурпурно-мясного цвета.
Измерения (тип). Длина: тела (вероятно, с хвостом) 210, хвоста
по позвонкам — 62, ступни — 36, кисти — 23.5, высота уха — 19 мм.
По: Walker et al., 1964 длина: тела — 130—150, хвоста около 40—70 мм.
Этот вид известен только с о. Минданао, где он найден в горах Апо
на высоте 1700—2100 м, на восточном склоне горы Мкинлей на высоте
1800—2300 м и на горе Катанглад на высоте 1600 м. Одна из нор, около
которой добыты 3 особи этого вида, помещалась под крупным деревом
в сплетении корней деревьев в моховом покрове, в густом лесу с папорот-
ником, вблизи реки.
Р а с п р ос транение и возраст. О. Минданао (Филип-
пины), современный.
12. Род NEOTETRACUS Trouessart, 1909
Trouessart, 1909, р. 389—390.
Тип. Neotetracus sinensis Trouessart, 1909, Сычуань, Юньнань (Китай), Северная
Бирма и п-ов Индокитай, современный.
Диагноз (по: Trouessart, 1909). Зубная формула: 1-|-СуР-|-М-|- — 40.
По внешнему виду напоминает Podogymnura Mearns или Мacroscelides,
но с зубами Erinaceus. Двукорневой клык в каждой челюсти не выше,
чем мелкие резцы, которые предшествуют ему, и мелких премоляров,
следующих за ним. Костное нёбо не заканчивается окостенением, сходным
с таковым у Erinajceus. Волосы мягкие, не колючие, хвост очень развит.
Диагноз, сопоставимый с диагнозом других родов. Длина тела
менее 260 мм, длина хвоста не менее 50% длины тела. Верхний и нижний
клыки мелкие.
Описание (по: Trouessart, 1909). Р высокий, конический, обычной
для семейства формы. I2 отделен от I1 широким промежутком, сходен
с I1 по ширине, но меньше по высоте. I3 очень мелкий. Клык с двумя кор-
нями и треугольной вершиной, не выше резцов. Р2 и Р3 имеют сходную
с клыком форму, но ниже его. Паракон и метакон (передний и задний
бугорки по: Trouessart, 1909) у них отсутствуют. Р4 очень крупный,
с четырьмя бугорками, один из которых на передне-наружной стороне
сильно развит. Моляры уменьшаются по высоте от М1 к М3. М1 четырех-
угольный, с 5 бугорками, как у других представителей семейства. М3
треугольный, имеет три бугорка: два спереди и один сзади.
В нижней челюсти Ij очень крупный, наклонен, лопатковидный,
с ложкообразной впадиной на верхне-внутренней стороне. 12 и 13 очень
мелкие, их вершины ниже первого, как у Erinaceus. Клык мелкий, сходен
с резцами. Р2 и Р3 мелкие, одновершинные. Р4 очень крупный, трехбугор-
чатый. Средний бугорок очень крупный, выступающий. Моляры посте-
12. РОД NEOTETRACUS
131
пенно уменьшаются в размерах от М2 к М3. имеет пять бугорков, рас-
положенных 1—2—2. Два бугорка последней пары объединяются в по-
перечный гребень. М2 имеет четыре бугорка — 2 и 2. М3 мелкий, сходен
с М2, немного сужен сзади. Все зубы обеих челюстей имеют острые вершины
и не сходны с Erinaceus.
Распространение и возраст. Сычуань, Юпьнань (Ки-
тай), Бирма, п-ов Индокитай, современный.
i
1. Neotetracus sinensis Trouessart, 1909 (рис. 67).
Trouessart, p. 390—391.
T и п не выделен, вид описан по 7 экз., добытым в Та-Тшен-Лу (Ta-tsien-lou),
Китай.
Описание (по: Trouessart, 1909). Голова коническая, удлиненная,
с мордой (носовым отделом), выдающейся над нижней губой. Глаза
округлые, очень крупные. Кисти и ступни 5-палые, стройные, особенно
Рис. 67. Neotetracus sinensis (по: Walker et al., 1964).
ступни. Все нижние поверхности ног покрыты очень короткой шерстью
так же, как и хвост. Волосяной покров тела сходен с таковым Hylomus
suillus Muller et Schlegel, состоит из двух сортов волос. Основная окраска
тела красновато-желтая. Основания волос серого цвета. Длинные жесткие
черные волосы подмешиваются к остевым волосам на спине и боках.
Нижняя сторона тела серо-белая. Уши покрыты очень короткой шерстью,
кисти и ступни окрашены в сравнительно темный коричневый цвет. Кисти
с красноватым оттенком на передней части верхней стороны. Нижние
поверхности кистей и ступней с 6 бугорками, расположенными попарно.
Уолкер (Walker et al., 1964) приводит следующие данные об этой
гимнуре: шерсть мягкая, густая и длинная. Верхние части тела оливково-
коричневые, охристо-коричневые или их окраска смешанная из кремового
и черного цветов. На боках головы, шеи окраска иногда имеет рыжий
оттенок. Изредка на теле бывает черная штриховка. Нижние части тела
красноватые, буро-серые или кремового цвета, образованного светлыми
концами волос с черными основаниями.
Измерения (по: Trouessart, 1909, вероятно, по 1 экз.). Длина
тела — 115, головы — 35, хвоста — 55, ступни — 26, высота уха —
15 мм. (По: Walker et al., 1964). Длина тела — 105—148, хвоста —
60—82 мм.
Распространение и возраст. В границах рода, со-
временный.
Биология. Уолкер приводит некоторые данные по биологии этого
малоизученного вида. В горы поднимаются на высоту 2100—2800 м.
Эти животные, несомненно, строго наземные и ночные. Обыкновенны
в местах их обитания. Занимают участки леса, покрытые мхом и папорот-
9*
132
ПОДСЕМ. GALERICINAE
ником. Здесь встречаются их тропинки и норы, которые располагаются
под упавшими деревьями и под камнями. Строят ли норы эти животные
сами или используют норы других млекопитающих, не известно. Из двух
вскрытых желудков один содержал земляных червей, другой — только
растительные остатки. Можно думать, что пища этой гимнуры смешанная.
Самка с 4 эмбрионами добыта в апреле и две самки (одна с 4, другая
с 5 эмбрионами) добыты в Китае в августе. По этим данным можно считать
размножение растянутым или ограниченным двумя генерациями.
13. Род NEOHYLOMYS Shaw et Wong, 1959
S haw, Wong, 1959, Acta Zool. Sinica, v. 11, N 3, p. 422—428.
Тип. Neohylomys hainanensis Shaw et Wong, 1959, о. Хайнань (Пай-са-Хенан),
Китай, современный.
Диагноз. Зубная формула: IСР-f-М=42. Размеры тела
О 1 О О
менее 260 мм. Длина хвоста не менее 28°/0 длины тела. Верхний и ниж-
ний клыки мелкие.
Описание (по: Shaw, Wong, 1959, с некоторыми сокращениями).
Череп овальной формы, крупнее, чем у Hylomys Miller из Юньнани, широ-
кий. Скуловая ширина в задней части более х/2 длины черепа. Носовой
Рис. 68. Neohylomys hainanensis (по: Shaw a. Wong, 1959).
отдел удлиненный, плоский на верхней стороне. Носовые кости узкие,
лишь немного расширены в средней части. Межчелюстные кости короткие,
а верхнечелюстные сравнительно длинные. Лобные кости не слиты, ромбо-
видной формы. Межглазничное пространство неширокое. Заглазничные
отростки развиты, они небольшие, но хорошо заметные. Теменные кости
крупные. Швы между ними и соседними костями у взрослых не явствен-
ные. Ламбдовидный гребень хорошо развит. Мозговая капсула сверху
плоская. Костное нёбо позади поперечного гребня имеет узкий пластин-
чатый выступ, отсутствующий у Hylomys. Нижняя челюсть удлиненная,
невысокая. Наибольшая высота зубной части ее ветви менее х/3 зубного
ряда. Венечный и сочленовный отростки крупные. Угловой отросток не-
большой, серповидный. Подбородочное отверстие крупное, расположено
под Р2 и Р3.
Распространение и возраст О. Хайнань, Китай,
современный.
1. Neohylomys hainanensis Shaw et Wong, 1959 (рис. 68).
Shaw, Wong, 1959, там же, где и род.
Тип. Взрослая самка (шкура и череп), № 70886, Pai-sa-Hsian, о. Хайнань,
Китай, кол. Муз. Инет, зоол., Акад. Наук Китая.
Описание. Внешний вид полевкообразный. Окраска головы
черно-серая, с примесью коричневого тона. Спина ржаво-коричневая,
с примесью черно-серого тона. По середине спины проходит черная полоса,
13. РОД NEOHYLOMYS
133
соединяющаяся с темной окраской головы. Эта полоса на задней части
тела становится светлее. Бока тела размытого светлого оливково-желтого
цвета. Нижние части тела светло-серые или желто-белые. Вибриссы черные,
длиной не более 35 мм.
Измерения (по: Shaw, Wong, 1959). Длина тела — 120—147,
хвоста — 36—43, ступни — 24—29, высота уха — 16—19 мм. Кондило-
базальная длина черепа — 33.2—39.9 мм, вес тела — 51—69 г.
Распространение и возраст. В границах рода, со-
временный.
Биология. Об образе жизни этого интересного вида описавшие
его авторы сделали следующие краткие замечания: «Этот вид предпочел
древесному образу жизни полуподземный и трату большого количества
времени на рытье подземных ходов. Цилиндрическая форма тела, короткий
квост и слабо развитые когти есть приспособление к роющей деятельности».
2. Подсем. ERINACEINAE Fischer von Waldheim, 1817
Erinacihi Fischer von Waldheim, 1817, p. 368—428.
Тип. Erinaceus Linnaeus, 1758, Европа, Азия, Африка, миоцен—современный.
Диагноз. Зубная формула: I -|- С Р |—| М |—| = 36—28. Тело сверху
покрыто иглами, а снизу волосами. Позади хоан, между крыльями основ-
ной клиновидной кости, образующими стенку барабанной камеры, име-
ется округлая впадина. М3 с двумя конами и двумя корнями (исключение
триба Protericini). М3 имеет два конида или они слиты и один корень,
часто разделенный бороздками с внутренней и наружной сторон.
Описание. Средние по размерам животные (135—300 мм) с мас-
сивным телом, на сравнительно коротких ногах, стопоходящие. Кисти
и ступни 5-палые, изредка первый палец ступни отсутствует. Хвост корот-
кий (16—37 мм), снаружи малозаметен. Голова клиновидная. Носовой от-
дел вытянут в хоботок. Глаза хорошо развитые, округлые, выпуклые. Уши
с широким основанием, с округлой или с заостренной вершиной, разной
высоты (26—57 мм). Тело сверху и с боков или только сверху покрыто
иглами. Последние у разных видов отличаются размерами и разной сте-
пенью постоянства в строении. Иглы на задней части головы располага-
ются равномерно по всей поверхности или двумя группами, разделен-
ными на средней части головы полосой голой кожи (пробором). При этом
по бокам от переднего края полоски голой кожи высокие и тонкие иглы
образуют пучки, заметно выступающие на фоне остальных более коротких
игл. Большая часть головы, нижняя сторона тела и конечности покрыты
грубыми или сравнительно мягкими волосами.
Окраска верхней стороны тела беловатая, палевая, охристо-серая,
серая, бурая, коричневая до почти черной с мелким, в разной степени
светлым, легким пестрым рисунком, образованным пестрой окраской
игл (чередованием светлых и темных поясков на иглах и светлой и темной
окраской их вершин). Изредка встречаются особи с пятнистой окраской,
обусловленной пятнистым расположением темно и светло окрашенных
игл. Окраска волосяного покрова часто темнее, чем иглистого, светлее
его или белая. Мочка носа и глаза окрашены темнее тела. Когти светлые
или темные.
Межчелюстные кости короткие, с коротким отростком, направленным
назад. Носовые кости сужены в передней части, реже немного шире, чем
в их средней части. Лобные кости в разной степени значительно вздуты
в передней части. Лобные и носовые кости по средней линии вдавлены
134 ПОДСЕМ. ERINACEINAE
и образуют желобок, или вдавлена только вся передняя часть лобных
:крстей. Предорбитальный гребень развит слабо. Его продолжение —
надорбитальный гребень — заметен только у видов подрода Hemiechinus
Fitzinger. Слезная кость сливается с окружающими костями, и ее границы
незаметны. Слезное отверстие открывается на предорбитальном гребне,
на лице (у ряда вымерших групп — в орбиту). Скуловая дуга в передней
части опущена вниз, а в задней Поднята вверх, в совокупности образует
угол. Скуловой отросток верхнечелюстной кости с желобком на внешней
стороне или с гладкой поверхностью, без глубокой выемки, как у гимнур.
1 Скуловая кость короткая, помещается в центральной части дуги. Сагит-
тальный гребень невысокий. Ламбдовидный гребень хорошо развит.
Мыщелковое отверстие расположено под мыщелком. Оно может быть
двойным. Костное нёбо с мелкими отверстиями и крупными на нёбной
кости (у ряда вымерших групп крупных отверстий нет). Костное нёбо
заканчивается сзади валиком или продолжается за ним в виде костной
пластинки. Клиновидная кость позади хоан с глубокой округлой впадиной.
Засочленовный отросток широкий, с чашевидным углублением или узкий,
со слабым вдавлением.
Нижйяя челюсть характерной для сем. Erinaceidae формы.' Вырезки,
разграничивающие венечный, сочленовный и угловой отростки, сходной
формы и величины. Подбородочное отверстие под Р4. Нижнечелюстное
•отверстие расположено ближе к зубной части ветви челюсти.
I1 крупный, разной высоты, и по-разному сильно изогнут. I2 мелкий,
•с одним корнем. Is крупнее I2, с двумя вершинами, у большинства видов
с двумя корнями. Клык по строению напоминает Is и передние премоляры.
•Он конусовидный, с одним корнем или с тремя бугорками и двумя корнями.
.Между клыком и третьим резцом короткая диастема. Р2 мелкий, с одним
корнем или сравнительно, крупный ,с тремя бугорками и двумя корнями.
-Р® бывает* мелким с одним корнем или сравнительно крупным с двумя
корнями или с тремя бугорками, расположенными не вдоль коронки,
а треугольником, и с тремя корнями. Р4 крупный, с высоким, хорошо
обособленным протоконом, с выступающим назад метаноном, и широким
талоном. Этот зуб всегда с тремя корнями. .. ,
М1 самый крупный зуб, он четырехугольный, с пятью бугорками.
Паракон и метакон на внешней стороне коронки зуба, протокон и талон
-на ее внутренней стороне, а мелкий пятый бугорок — на центральной
, части жевательной поверхности. М2 четырехугольный, но меньших раз-
меров, со скошенным назад внешним краем. Он также с пятью бугорками,
расположенными как на М1. М3 мелкий, вытянутый вдоль заднего края
.верхнечелюстной кости и расположен под углом к зубному ряду. Этот зуб
с двумя бугорками и двумя корнями.
Нижний зубной ряд начинается длинным, направленным вершиной
вперед и слегка вверх 1х. За ним следует мелкий, со скошенной вперед
вершиной 12, а за последним клык, также с направленной вперед вершиной,
мелкий, редко сравнительно крупный, с двумя корнями. Ра с 2—3 в раз-
ной степени хорошо выраженными бугорками, с одним корнем. Р4 с двумя
корнями, характерной для этого подсемейства формы: с высоким пара-
конидом и протокоцидом. Последний выемкой разделен на два зубца.
Метаконид этого зуба низкий. Mi — самый крупный зуб, расширяющийся
спереди назад более, чем . у гимнур. Параконид этого зуба смещен на
внутреннюю сторону коронки. Протоконид высокий с раздвоенной верши-
ной- Метаконид широкий с высоким наружным и внутренним краем и
гребнем на задней стороне. Ма .меньший, чем Мц с более широким пара-
конидом, сходен по строению с предыдущим зубом. М3 мелкий, с редуциро-
ванным метаконидом. Параконид этого зуба в разной степени Хорошо раз-
14. РОД TUPAIODON 135.
вит. Протоконид, как и у предыдущих зубов, с двумя зубцами. Реже Ма
со слитыми бугорками, округлой формы.
Скелет отличается массивностью всех составляющих его костей.
Все его сложение несет черты приспособления к значительной весовой
нагрузке, а также и к свертыванию тела в шар. Более выгнутый вверх
позвоночник, опущенный вниз таз, вертикально расположенные лопатки,
широкие кисти и короткие ступни. Вес телй создается толстой кожей,
несущей иглы, подкожной мускулатурой и кольцевой мышцей, сворачиваю-
щей тело в шар.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ТРИБ
1 (6). Слезное отверстие открывается в орбиту. Базисфеноидного углубле-
ния нет.
2 (5). М| имеются.
3 (4). М3 треугольной формы (не расширен в поперечном к зубному ряду
направлении). Нёбо не перфорировано ... 1. Триба Protericini.
4 (3). М3 расширен в поперечном зубному ряду направлении, не треуголь-
ной формы. Нёбо перфорировано.............3. Триба Amphechini.
5 (2). М| отсутствуют...................2. Триба Brachyericini.
6 (1). Слезное отверстие открывается вне орбиты. Базисфеноидное углуб-
ление имеется . ......................4. Триба Erinaceini.
f 1. Триба PROTERICINI Butler, 1948
Butler, 1948, p. 488.
Тип. Proterix Matthew, 1903, Северная Америка, олигоцен.
Диагноз. Слезное отверстие открывается в орбиту. Нёбо не перфо-
рировано. Базисфеноидного углубления нет. В верхней челюсти 3—4 пре-
моляра. Мз имеются. М3 треугольной формы, в одном ряду с другими
молярами (не вытянут в поперечном к зубному ряду направлений). М3
сходен по строению с М2, но меньше его по размерам.
Группа наиболее примитивных ежей, сохраняющих ряд особенностей,
встречающихся у представителей подсем. Galericinae — хранителей мно-
гих архаичных признаков, свойственных общим предкам с ежами.
Распространение и возраст. Азия, олигоцен; Северная
Америка, олигоцен—нижний миоцен.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). В верхней челюсти четыре прембляра...........14. Tupaiodon.
2 (1). В верхней челюсти три премоляра ...............15. Proterix.
Примечание. За отсутствием сравнимых признаков, в таблицу не вклю-
чен Palaeoscaptor.
fl4. Род TUPAIODON Matthew et Granger, 1924
Matthew, Granger, 1924, p. 1, 2.
Тип. Tupaiodon morrisi Matthew et Granger, 1924, Монголия, олигоцен.
Диагноз (no: Matthew, Granger, 1924). Верхние моляры и пре-
моляры до некоторой степени похожи на таковые Ptilocercus (тупайя),
но отличаются более крупным и более обособленным гипоконом (тало-
ном) на М1 и М2. Резцы не известны. Клык, Р1 и Р2 мелкие, с двумя корнями,
с невысокой массивной вершиной и рудиментарной пяткой (метаконидом).
Р3 и Р4 очень крупные, с хорошо развитыми внутренними бугорками.
Р4 с крупным метастилем (пятым бугорком) и рудиментарным тетраконом.
М1 и М2 в сечении почти квадратные, с высокими, несколько угловатыми
136
ПОДСЕМ., ERINACEINAE
бугорками. М3 треугольный, без метастиля или гипокона, пропорциями
напоминает (близок) этот зуб Ptilocercus.
Нижние моляры с несколько широким и коротким тригонидом (перед-
ним сегментом), с тремя угловатыми вершинами. Протоконид слегка
возвышающийся. Талонид (метаконид) такой же ширины, как тригонид,
с глубокой чашей, с бугорками у задцих углов. Эти бугорки не так вы-
соки, как у тригонида.
Подглазничное отверстие над Р3. Корень скуловой дуги над М2.
Размеры тела близки к таковым европейского крота или блярины
(Blarina brevicauda Say). Длина С—М3 — 13 мм.
Рис. 69. Tupaiodon morrisi (по: Matthew, Granger, 1924).
Распространение и возраст. Азия, олигоцен.
f 1. Tupaiodon morrisi Matthew et Granger, 1924 (рис. 69).
Matthew, Granger, 1924, p. 1 —2.
Тип. Верхняя челюсть и часть черепа, фрагмент ветви нижней челюсти с М2^М3,
№ 19134, формация' Хзанда-Гол, красные слои, Ло, Монголия, олигоцен, Амер. муз.
естест. истор., Нью-Йорк.
Распространение и возраст. Известен из одного место-
нахождения.
Примечание. К этому же роду с вопросом авторы отнесли и еще
один вид, названный ими Т. ? minutus (тип: фрагмент правой ветви нижней
челюсти с Р3—М1? № 19135, из одного с Т. morrisi местонахождения).
Этот вид меньше, коренные зубы близки по размерам, но более узкие,
чем у типичного вида, и с заметным цингулюмом на внешней стороне
коронки. Передняя часть нижней челюсти значительно тоньше и слабее.
Р3 с двумя корнями, простой, с одним острым не прижатым бугорком,
мелким передним основным бугорком и задним цингулюмом. Р4 полу-
моляризован, мельче, чем моляры, имеет два хорошо обособленных
бугорка, низкий передний бугорок и широкий низкий цингулюм на
округлом заднем конце зуба. Длина Р3—М2 — 9 мм.
Положение рода Tupaiodon в системе ежевых недостаточно ясно.
Батлер (Butler, 1948) сближает его с гимнурами, в частности с родом
Galerix Pomel, а Ван Вален (Van Valen, 1967) поставил этот род в сем.
Adapisoricidae. Структурные особенности зубов представителей этого
рода близки к таковым ежовых, особенно их древних групп. Главное от-
личие от ежей заключается в том, что Tupaidon имеет большее количество
премоляров в верхней челюсти (вероятно, и в иижней), характерное для
гимнур. Однако это не дает основания для исключения его из состава
подсем. Erinaceinae, развившегося от общих корней с гимнурами, но
имеющего значительно меньшее количество зубов, чем гимнуры. Сокраще-
ние количества зубов характерно для всех ежеобразных, но происходило
оно в разных их группах в разное время и с разной интенсивностью.
В одном отрезке геологического времени, например в олигоцене, существо-
вали ежи и гимнуры с разным числом зубов.
По строению зубов род Tupaiodon ближе всего к роду Proterix
Matthew.
15. РОД PROTERIX
137
f 15. Род PROTERIX Matthew, 1903
Matthew, 1903, p. 227—229.
Тип. Proterix loomsi Matthew, 1903, Южная Дакота (США), олигоцен.
Диагноз. Зубная формула: I С Р М -f- =40. Подглазничное
отверстие не закрыто костной стенкой и видно сбоку. Слезное отверстие-
открывается в орбиту. Нёбная кость не перфорирована. Базисфеноидного’
углубления нет. крупные. С крупный, с двумя корнями.
Распространение и возраст. Северная Америка, олигоцен.
1. f Proterix loomsi Matthew, 1903 (рис. 70).
Matthew, 1903, p. 228—229.
Тип. Лицевой отдел черепа № 9756, Индийский волок, р. Шейенн, горы Ореодон,
Южная Дакота (США), олигоцен, кол. Амер. муз. естеств. истории, Нью-Йорк.
Диагноз Р2 и 13 имеются.
Описание (по: Matthew, 1903). I1 широкий. Р2 мелкий, с одним
корнем. Р3 мелкий, с тремя корнями и сильно развитым протоконом. Р4 очень
Рис. 70. Proterix loomsi (по: Matthew, 1903).
крупный, сильно моляризованный, с мелким протоконом (талоном).
М1 более широкий, чем длинный, М2 почти квадратный в сечении. Оба
зуба с пятью бугорками — 4 по углам коронки и 1 в центре жеватель-
ной поверхности (метастиль). М3 треугольный, более развит в ширину,
не расположен в поперечном направлении По отношению к зубному
ряду. 12 и 13 массивные, также и клык, имеющий округлую форму
в сечении и вершину, не отогнутую вперед в виде флага. Р2 округлый
в сечении, с одним корнем. Р3 с продольно треугольной коронкой и
слитыми корнями (в один). Р4 с одновершинным протоконидом, как у гим-
нур, высокий. Мх и М2 сходной формы, но разных размеров, с широким
метаконидом (талонид других авторов). М3 с хорошо развитым и высоким
метаконидом, с двумя корнями, похож на этот зуб гимнур, отогнут на
внутреннюю сторону челюсти. Поперечный гребень нёбной кости далеко
отодвинут назад от М8.
138
ПОДСЕМ. ERINACEINAE
Измерения (по: Matthew, 1903). Ширина нёба между молярами —
17.6, длина С—М3 — 18.4, М1 — 3.4, М2 — 2.9, М3 — 2.0, ширина М1 —
4.8, М2 — 3.9, М3 — 2.8 мм.
Распространение и возраст. Известен из Южной Да-
коты и Небраски (США), олигоцен.
t 2. Proterix bicuspis Macdonald, 1951 (рис. 71).
Apternodus bicuspis M a c d о n a 1 d, 1951, Journ. Palaeont., v. 25, p. 257—265. —
Proterix bicuspis Gawne, 1968, p. 11—25.
Тип. Apternodus bicuspis Macdonald, 1951, череп S. D. S. M. № 4048, семь миль
к востоку от Рокифорда, область Шаннон, Южная Дакота (США), верхний олигоцен,
кол. муз., Лос-Анджелес.
Диагноз (по: Gawne, 1968). Р2 и 13 отсутствуют.
Описание. Р неизвестен. На Р3 протокона нет. Р3 около половины
высоты Р4. Гипокон (талон) Р4 меньше, чем у Р. loomsi. Гребень параконида
и талонид (метаконид) Р4 неболь-
шие. М1 длинный' (длина равна
ширине). Размеры М2 и М3 меньше,
чем М1. М2 несколько треуголь-
ной формы. М3 расположен вблизи
заднелобного гребня. Скуловая
Рис. 71. Proterix bicuspis (по: Gawne, 1968).
дуга поднимается над М1. Она широкая по вертикали. Орбитосфеноидная
•область длинная. Барабанные камеры очень крупные.
Примечание. Судя по фотографиям и рисункам (Gawne, 1968),
описываемый вид имел удлиненный череп со сравнительно короткими
зубными рядами и в отличие от Р. loomsi имел более широкий М3, не ото-
гнутый на внутреннюю сторону нижней челюсти. Существенным отличием
следует считать и большую остробугорчатость зубов этого вида.
Измерения (по: Gawne, 1968). Длина: черепа — 55, Р4 — 3.5—
4.0, М1 - 4.3-4.9, Р4 - 3.2, М4 — 3.9-4.0, М2 - 3.5, М3 - 2.6 мм;
ширина: Р4 - 4.3-5.3, М1 — 4.7-5.1, Р4 - 2.6-2.8, Мх — 3.1, М2 —
3.1, М3 — 2.2 мм.
Распространение и возраст. Известен из Небраски
и Южной Дакоты (США), верхний олигоцен.
Примечание. Отсутствие Р| и 13, а также структурные особен-
ности черепа позволяют считать, что этот вид принадлежит особому роду,
‘более специализированному, чем род Proterix.
f 16. Род PALAEOSCAPTOR Matthew et Granger, 1924
Matthew, Granger, 1924, p. 2.
Тип. Palaeoscaptor acridens Matthew et Granger, 1924, Азия, средний олигоцен.
Диагноз (по фотографии верхней челюсти AMNH 22 080, приведен-
ной Rich, Rasmussen, 1973, и описанию авторов рода). Р3 сходен по форме
«с Р4, но значительно меньших размеров. Р4 с сильно развитым метакони-
16. РОД PALAEOSCAPTOR
139
дом, направленным назад в виде отростка. М2 с обособленным талонидом
в виде бугорка. М3 треугольной формы, сходен по внешнему виду с М2.
М3 по строению сходен с М2, но значительно меньше по длине и ширине.
Описание (диагноз по: Matthew a. Granger, 1924); Первый резец,
(IJ широкий, мало удлиненный, слабо наклонен. Второй резец не со-
хранился, его альвеола маленькая, округлоовальной формы. Клык не-
большой, его корни слиты и разграничены только углублением. Коронка'
клыка сходна с коронкой премоляра, вытянута вперед. Премоляр тре-
тий меньше клыка, также со слитыми корнями, мало вытянут вперед..
Р4 с двумя корнями, небольшой, моляризован. М4 очень крупный, с триго—
нидом (передний сегмент, образованный пара- и протоконидом, по моим
Рис. 72, 73. Рaleoscaptor acridens. (72, А — по: Rich, Rassmnsen, 1973, с фот., Б'—
по: Matthew, Granger, 1924, 73 — по: Sulimski, 1970).
представлениям), более длинным, чем широким. М2 значительно меньше-
Мх, со сходным тригонидом и почти так же развитым талонидом-(мета-
конидом, — А. Г.), менее развит в длину. М3 сильно редуцирован, с низким
талонидом (метаконидом), значительно сужен в боковом направлении.
Распространение и в оз р а с т. Азия, олигоцен.
f 1. Palaeoscaptor acridens Matthew et Granger, 1924 (рис. 72, 73).
Matthew, Granger, 1924, p. 2. — Palaeoerinaceus acridens M a t t h e w,
Granger, Bohlin, 1942, Palaeontologia Sinica, new ser., C, N 8a, pt 1, p. 18.
Тип. Фрагмент левой ветви нижней челюсти с зубами (Р4—М3) № 19138, Хзанда-
Гол, красные пески в 15 милях от Ло, Монголия, олигоцен.
Диагноз (по: Трофимов, 1960). Размеры мелкие, параконид Р4
ниже протоконида, талонид (метаконид) у М3 имеется.
Описание (по: Трофимов, I960). Череп мелкий, лицевая часть
впереди Р4 узкая, нёбо длинное (позади доходит до М3), в задней части
значительно расширено. Мозговая коробка крупная, уплощенная, без
стреловидного гребня. Скуловые дуги тонкие. Нижняя челюсть невысокая,
узкая и довольно толстая (не очень ясное сочетание). Подбородочное от-
верстие расположено на уровне передней части Р4 (на 3 экз.). Массетерная
впадина крупная, глубокая. Сочленовный отросток расположен низко,
на уровне альвеолярного края челюсти. Венечный отросток (полностью ‘
не сохранился) направлен почти вертикально вверх и, видимо, довольно- '
узкий и высокий.
140
ПОДСЕМ. ERINACEINAE
Измерения (по: Трофимов, 1960). Длина черепа — около 40;
высота на уровне заднего края нёба — 10; скуловая ширина — около 20;
длина Р4 — 1,5, 1.2; Мх — 3.5, 2.0; М2 — 2.0; М3 — 1.0; ширина Р4 —
1.0, 0.9; Мх — 2.0, 1.5; М2 — 1.2; М3 — 1.0 мм.
Распространение и возраст. Известен из Хзанда-Гол
и Татал-Гол (Монголия) из юго-восточной части Гоби и Сан-Тао-Хо
(Китай), олигоцен.
f 2. Триба BRACHYERICINI Butler, 1948
Butler, 1948, р. 488.
Тип. Brachyerix Matthew, 1933, Северная Америка, средне-верхний миоцен.
Диагноз. Слезное отверстие открывается в орбиту. Нёбо не перфо-
рировано. Базисфеноидного отверстия нет. В верхней челюсти 2—3 пре-
моляра. М| отсутствуют.
Мелкие примитивные ежи с укороченным лицевым отделом черепа.
Зубная система, насколько можно судить по длине зубного ряда и строе-
нию зубов, приспособлена к обработке грубой пищи, возможно и расти-
тельной. По-видимому, в связи с укорочением морды, в качестве компенса-
ции, увеличились размеры резцов 1} как органа захвата пищи.
Распространение и возраст. Азия, средний олигоцен;
Европа, верхний миоцен; Северная Америка, средний миоцен—нижний
плиоцен.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). Р4 одновершинный.
2 (3). Длина Р4 менее длины Мх в 4—5 раз...............18. Metecbinus.
3 (2). Длина Р4 менее длины Мх в 2—2.5 раза............19. Exallerix.
4 (1). Р4 двувершинный.
5 (6). В нижней челюсти два премоляра...............20. Dimyllechinus.
6 (5). В нижней челюсти один премоляр.................17. Brachyerix.
f 17. Род RRACHYERIX Matthew, 1933
Brachyerix Matthew, 1933, in Matthew, Mook, 1933, Amer. Mus. Novitates,
N 601, p. 1—4.
Тип. Brachyerix macrotis Matthew, 1933, западная часть Северной Америки,
средне-верхний миоцен.
Диагноз. Зубная формула (по: Т. Rich, Р. Rich, 1971) 1уС-р
2 2
Р т М — 26 или 28. Подглазничное отверстие закрыто костной тканью
и сбоку мало заметно. Слезное отверстие открывается в орбиту. Нёбная
кость не перфорирована. Базисфеноидного углубления нет.
О п и с а н и е (по: Matthew, 1933). М1 округло-четырехугольной
формы, М2 треугольный, значительно редуцирован. Р4 с крупной, не-
сколько треугольной формы, близкой к конической, передне-наружной
вершиной (наружной вершиной проток она), с большой, гребнистой вну-
тренней пяткой (талоном) и задне-наружным гребнем (метаконом).
Р3 сильно редуцирован, мелкий, простой, с двумя корнями и продольной
гребнистой коронкой. Две альвеолы впереди Р3 позволяют предполагать,
что впереди стоящий зуб также мелкий, с двумя корнями. Заднелобная
часть узкая и короткая. Барабанные крылья основной клиновидной кости
образуют большую часть барабанных камер, а в передней части вместе
с крыловидными костями образуют дополнительную слуховую полость,
соединяющуюся с барабанной полостью. Мастоидный отросток имеет
17. РОД BRACHYERIX
141
горизонтальное положение и вытянут назад. Межтеменная кость в виде
узкой изогнутой полосы.
Приведенные в работе Т. Рич и П. Рич (Т. Rich, Р. Rich, 1971) хорошие
фотографии фрагментов черепов и полной нижней челюсти Brachyerix
maxrotis позволяют дополнить описание следующими замечаниями: М1
Рис. 74, 75. Brachyerix macrotis (74 — по: Matthew, 1933; 75 —- по: T.»Rich, R. Rich,
1971).
имеет бугорки с округлой вершиной, его коронка в средней части имеет
сужение. Р4 с метаконидом, вытянутым назад в виде отростка. Нижняя
челюсть с высокой и короткой зубной частью и широкой, с неглубокой
вырезкой между сочленовным и угловым отростками, сочленовной частью.
Вырезка между сочленовным и венечным отростками имеет горизонталь-
ное положение. Венечный отросток расширен в передне-заднем направле-
нии, невысокий, с почти вертикальным краем. Угловой отросток корот-
кий, широкий, с глубокой выемкой на внутренней стороне. Ij — умеренно
длинный, массивный, С — характерной для ежей формы: в виде флажка.
Р4 с продольным расположением бугорков, островершинный, напоминает
этот зуб гимнур. Мх длинный и узкий (его передний сегмент составляет
142
ПОДСЕМ. ERINACEINAE
длины зуба), островершинный, с удлиненным параконидом. М2 —
значительно ниже и короче, отогнут на внутреннюю сторону.
Распростране ни е и возраст. Северная Америка,
средне-верхний миоцен.
f 1. Brachyerix macrotis Matthew, 1933 (рис. 74, 75).
Matthew, 1933, in Matthew, Mook, 1933, там же.
Тип. Череп без носового отдела, № 21335, р. Глубокая, вблизи форта Логап,
Монтана (США), средне-верхний миоцен, кол. Амер. муз. естеств. истории, Нью-Йорк.
Диагноз этого вида сходен с таковым рода и не сопоставим с диагнозом
другого вида.
Описание. Морфологические особенности этого вида приведены
в характеристике рода.
Измерения (по: Т. Rich, Р. Rich, 1971). Скуловая ширина —
20.3—20.5; ширина нёба между М1 — 11.4—14.1; ширина межглазничного
сужения — 6.2—8.3; максимальная ширина мозговой части черепа
12.0—16.3; длина Р4—М2 — 7.2—8.0 мм.
Распространение и возраст. Известен из Калифорнии,
Вайоминга, Монтаны и Небраски (США), средне-верхний миоЦен.
f 2. Brachyerix insertis Matthew, 1924.
Talpa incerta Matthew, 1924, Bull. Amer. Mus. Nat. Hist., v. 50, art. 2, p. 74.
Metechinus fergusoni Henshaw, 1924, Publ. Carnegie Inst. Washington, N 530, p. 101.
Тип; Фрагмент правой ветви нижней челюсти № 18891, с сохранившимися го-
ризонтальной ветвью до альвеолы Mj, восходящей ветвью и угловым отростком. Из зу-
бов сохранился только М2. Средний—верхний миоцен, кол. Амер. муз. естеств.
истории, Нью-Йорк.
Диагноз (по: Т. Rich, Р. Rich, 1971). Мелкий вид рода. Слабый
цингулюм на Р3 проходит на лингвальной стороне коронки от парастиля
до мелкого протокона на заднюю часть паракона.
Примечание. Более важными отличительными признаками
этого вида следует считать значительно более крупные, округлые и выпук-
лые барабанные камеры, менее массивную нижнюю челюсть с крупными
и широкими зубами.
Распространение и возраст. Известен из Калифорнии,
Невады и Небраски (США), средний—верхний миоцен.
f 18. Род METECHINUS Matthew, 1929
Matthew, 1929, р. 93—102.
Тип. Metechinus nevadensis Matthew, 1929, Невада (США), нижний плиоцен.
Диагноз (по: Matthew, 1929). Зубная формула: 1уС-|-РрМ-|-.
Череп короткий и широкий, строение зубов, как у Erinaceus L., только
премоляры и задние моляры сильно редуцированы, а первый моляр широ-
кий и специализированный. М2 более мелкие, чем М|._М| отсутствуют.
Р4 очень мелкий и проще, чем у Erinaceus. Р3 выпадающий. Мозговая
капсула очень короткая и широкая. Нёбная кость не перфорирована,
вытянута назад до М1 (к этому нужно добавить, что слезное отверстие
открывается в орбиту и базисфеноидной впадины, видимо, нет).
Описание (по: Matthew, 1929). Ширина черепа приблизительно
только на х/6 менее длины черепа (видимо, скуловая ширина). Верхняя
ветвь межчелюстной кости широкая и короткая. Морда укорочена и орбита
передним краем располагается над серединой Р4. Подглазничное отверстие
18. РОД METECHINUS
143
широкое. Скуловая дуга короткая, широкая, выгнутая кнаружи и изо-
гнутая под орбитой и в задней части. Межглазничного сужения черепа нет.
Сагиттальный гребень высокий (автор почему-то не отмечает гребни на
боковых сторонах лобных и теменных костей, хорошо заметные на приве-
денной им фотографии). Сочленовная впадина на височной кости неглубо-
кая, треугольной формы. Засочленовный отросток неширокий. Сосцевид-
ный отросток, кажется, значительно редуцирован вследствие значительного
расширения чешуи височной кости позади засочленовного отростка, а зад-
няя часть ламбдовидного гребня позади сосцевидного отростка вздута.
Рис. 76, 77. Metechinus nevadensis (по: Matthew, 1929).
Костное нёбо короткое и широкое. Заднелобный гребень хорошо развит.
Сочленовный отросток нижней челюсти незначительно выступает назад
и не возвышается над зубным рядом. Венечный отросток короткий и ши-
рокий, уплощенный; его передний край почти вертикальный.
Распространение и возраст. Северная Америка, верх-
ний миоцен—нижний плиоцен.
f 1. Metechinus nevadensis Matthew, 1929 (рис. 76, 77).
Matthew, 1929, p. 93—102.
Тип. Неполный череп и фрагмент нижней челюсти № 29600, Рыбье озеро, Не-
вада (США), нижний плиоцен, кол. Палеонтол. муз. Калифорн. унив., Беркли.
Диагноз (по: Meade, 1941). Крупнее М. marslandensis Meade.
Тригонид и талопид (передний и задний сегменты) Мх и М2 разной ширины
у каждого зуба. Гипоконид у Mi удлинен в передне-заднем направлении
и ниже, чем у М. marslandensis. М2 более крупный, с укороченным
тригонидом.
'Описание. Морфологические особенности этого вида, приведенные
в характеристике рода, известны только для этого вида.
144
ПОДСЕМ. ERINACEINAE
Измерения (по: Matthew, 1929). Длина от переднего премакси-
лярного края до парокципитального отростка — 36.7, от переднего
премаксилярного края до задненёбного гребня — 18.5, верхнего ряда
зубов (I3—М2) — 15.95, крыши черепа от переднего премаксилярного
края до верхнезатылочного гребня — 37.3, ширина между наружными
краями скул — 30.2, костного нёба — 17.2, заглазничного сужения —
10.9, между орбитами — 13.2, морды между подглазничными отвер-
стиями — 10.7, мозговой части черепа на уровне верхней стороны скуловых
дуг — 18.1 мм.
Распространение и возраст. Известен из Невады
(США), нижний плиоцен.
t 2. Metechinus marslandensis Meade, 1941.
M e а д e, 1941, p. 43—47.
Тип. Левая ветвь нижней челюсти с зубами М, и М2, F. М. № Р. 26399, около
7 миль к северо-западу от Марсланда, Небраска (США), верхний миоцен?, кол. Област-
ного муз., Чикаго.
Диагноз (по: Meade, 1941). Меньше, чем М. nevadensis. Тригонид
и талонид М| и М2 сходной ширины. У М. nevadensis талонид М4 шире,
чем тригонид, а тригонид М2 шире, чем талонид. Гипоконид М4 менее
удлинен в передне-заднем направлении и выше, чем у М. nevadensis.
М2 пропорционально меньше и со слегка удлиненным тригонидом.
Распространение и возраст. Известен из одного место-
нахождения.
119. Род EXALLERIX McKenna et Holton, 1967
McKenna, Holton, 1967, p. 2—3.
T и n. Exallerix hsandagolensis McKenna et Holton, 1967, Хзанда-Гол, Монголия,
средний олигоцеи.
Диагноз (по: McKenna, Holton, 1967). Зубы редуцированы. Пер-
вый резец крупный. Р4 с двумя корнями. Его коронка редуцирована до
одной вершины, параконид и метаконид им утрачены. М4 очень крупный,
с сильно развитым параконидом. Долинка параконида неглубокая. Мета-
конид сближен с протоконидом, его вершина немного крупнее, чем вер-
шина протоконида (по моим представлениям, обе вершинки принадлежат
протокониду, а метаконид — задний сегмент зуба — талонид других
авторов). Челюсть высокая, с выпуклым, сильно складчатым гребнем,
идущим непрерывно из-под Р4 до нижнего края ветви нижней челюсти и
до передней части ее углового отростка.
Распространение и возраст. Монголия, средний
олигоцен.
t 1. Exallerix hsandagolensis McKenna et Holton, 1967 (рис. 78).
McKenna, Holton, 1967, p. 3—9.
Тип. Правая ветвь нижней челюсти с сохранившейся областью симфиза и зуб-
ной частью, А. М. N. Н. № 22 083, Тсаган-Нур, Хзанда-Гол, Монголия, средний оли-
гоцен. Амер. муз. естеств. истории, Нью-Йорк.
Диагноз (по: McKenna, Holton, 1967). Только один вид рода.
По размерам приблизительно на 50% крупнее, чем Brachyerix macrotis
Matthew, 1933.
Описание (по: McKenna, Holton, 1967). Вероятная зубная фор-
мула: I—С— Р—М—?. 14 с отломанной вершиной, длинный, сжатый
с боков. Часть лабиальной стороны этого зуба имеет ясный скульптурный
рельеф в виде небольших, округлой формы шишечек. Сходный, но более
грубый скульптурный рельеф располагаемся и вдоль основания коронки
19. РОД EXALLERIX. 20. РОД DIMYLECHINUS
145
P4 и Mx. Шероховатость симфиза ветви нижней челюсти простирается от
резцового отдела до Р4. Три альвеолы между Р4 и 1х принадлежат одно-
корневым зубам. Из них передняя — 12, средняя — С, задняя — Р3.
Р4 имеет два корня, он ниже Мх.
Приведенные авторами фотографии и рисунки позволили познако-
миться с некоторыми деталями строения сохранившихся в челюсти зубов.
Параконид Р4 в виде выступа при основании протоконида на его передней
стороне, а метаконид в виде ступеньки или ската на задней стороне прото-
конида.
Рис. 78. Exallerix hsandagolensis (по: McKenna, Holton, 1967).
Длина ряда зубов (по альвеолам) между 1Х и Мх примерно равна длине
основания коронки Мх. Последний — самый крупный зуб в челюсти. Судя
по альвеолам, длина М2 составляет примерно 2/3 длины Мх. Параконид Мх
удлиненный, протоконид со сближенными бугорками, разделенными узкой
бороздкой, метаконид (талонид других авторов) не более х/3 длины Мх,
с глубокой выемкой, ограниченной снаружи и с внутренней стороны эма-
левыми стенками, с бугорком на задне-наружном крае. Крупное под-
бородочное отверстие — под передним корнем Мх.
Измерения (по: McKenna, Holton, 1967). Максимальный диа-
метр 1х — 2.4, общая длина по альвеолам 1х—Р4 — 3.0, Р4 — 2.5 X 2.0,
Mi — 5.0 X 2.6, высота нижней челюсти у переднего края альвеолы Мх —
7.2, у заднего края альвеолы Мх — 5.9 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
f 20. Род DIMYLECHINUS Hurzeler,. 1944
Hiirzeler, 1944, S. 460—467.
Т и п. Dimylechinus bernoullii Hiirzeler, 1944, Европа, верхний миоцен.
Диагноз (по: Hurzeler, 1944). Череп значительно меньше такового
Palaeoerinaceus edwardsi Filhol, кроме того, очень похож на .него.
Зубная формула (по: Hurzeler, 1944): •
Примечаниек диагнозу. Хюрцелер (Hurzeler, 1944) сравнивает
описываемый им род с родом Palaeoerinaceus Filhol. Последний, по совре-
менным воззрениям, должен быть отнесен в синонимы рода Amphechinus
Aymard. В дальнейшем описании будет фигурировать не Palaeoerinaceus
edwardsi Filhol, a Amphechinus edwardsi Filhol. Зубная формула, пред-
10 А. А. Гуреев
146
ПОДСЕМ. ERINACEINAE
ставленная автором, может быть изображена в следующем виде:
3 13 2
I уСТРТ "2 ~32' Можно не сомневаться, что зубная формула Dimy-
lechinus отличается от таковой Erinaceus только меньшим числом моляров*
Описание. По мнению Хюрцелера, структурные особенности
черепа Dimylechinus сходны с таковыми черепа Amphechinus edwardsi
Filhol. Сравнение приведенных им изображений черепов и нижних челю-
стей убеждает в этом. Тем не менее имеются существенные различия.
У Dimylechinus более суженная в передней части орбита. Менее развиты
орбитальные гребни. Ниже расположено слезное отверстие, более развита
Рис. 79. Dimylechnus bernoullii (по: Iliirzeler, 1944).
впадина впереди подглазничного отверстия и другие отличия. Однако
степень постоянства этих отличий может быть подтверждена лишь на боль-
шем материале.
Строение верхних зубов, как справедливо отмечает автор, в типе сходно
со строением зубов других Erinaceidae, в частности представителей рода
Amphechinus, но более узкие в средней и внутренней частях и более широ-
кие в наружной части. В связи с этим их жевательные поверхности имеют
меньшую площадь.
Нижняя челюсть Dimylechinus с более расширенным спереди-назад
венечным отростком, более обособленным сочленовным отростком и более
тонким и длинным угловым отростком (рис. 79, 81).
Нижние моляры отличаются от этих зубов других представителей
Erinaceidae, в частности Amphechinus, большим развитием пара- и прото-
конида и меньшим — метаконида. Это же заметно выражено и у Р4
(рис. 79, 81).
Распространение и возраст. Европа, верхний миоцен.
t 1. Dimylechinus bernoullii Hiirzeler, 1944 (рис. 79).
Hiirzeler, 1944, S. 460—467.
Тип. Фрагмент черепа с сохранившимися зубами: на правой стороне М2 и С
и альвеолы Р2—Р4 и М2—Р4 и С, а также альвеолы Р2, Р3 на левой стороне, № 1692,
Saulcet — Allier, Франция, верхний миоцен, кол. Базельского музея.
Распространение и возраст. Saulcet, Yaligny, Chav-
roche — Allier, Франция, верхний миоцен.
21. РОД AMPHECHINUS
147
Примечание. Автором описываемых здесь рода и вида дан схема-
тичный диагноз и в сравнении с видом, которого может не оказаться при
определении. Палеонтологи и зоологи чаще всего имеют дело с фрагмен-
тами черепа и нижней челюсти с неполным рядом зубов. Поэтому опреде-
ление следует вести по описанию и рисункам, позволяющим достаточно
ясно представить себе морфологию Dimylechinus bernoullii.
f 3. Триба AMPHECHI N I Gureev triba nova
T и ii. Amphechinus Aymard, 1850, Европа, Азия, олигоцен—миоцен; Африка,
миоцен.
Диагноз. Слезное отверстие открывается в орбиту. Нёбо перфо-
рировано. Базисфеноидного углубления нет. В верхней челюсти 3 пре-
моляра. М| имеются. М3 узкий, расположен в поперечном к зубному ряду
направлении. М3 без метаконида (талонида других авторов).
Мелкие и средних размеров ежи, сохраняющие многие примитивные
особенности, характерные для общих предков с гимнурами, поскольку они
встречаются и у последних. Тем не менее их специализация характери-
зуется направлением, близким к таковому поздних групп ежей, в част-
ности рода Erinaceus (перфорированное нёбо, сходное количество зубов
и др.).
Распространение и возраст. Европа и Азия, олиго-
цен—миоцен; Северная Америка и Африка, миоцен.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). 1} значительных размеров. Корень Д под Р4. Р4 расположен под
орбитой........................................21. Amphechinus.
2 (1). I] менее крупные. Корень Д под Р3. Р4 расположен впереди орбиты
............................................ 22. Gymnurechinus.
Примечание. Род Stenoechinus Rich et Rasmussen не включен в таблицу за
отсутствием сравнимых признаков.
-f-21. Род AMPHECHINUS Aymard, 1850
Aymard, 1850, Ann. Soc. Agricult. Sci. du Puy, v. 14, p. 80—86; Palaeoerina-
ceus Filhol, 1879, p. 1—252; Parvericius Koerner, 1940, p. 837.
Тип. Erinaceus arvernensis Blainville, 1839—1864, Auvergne, Франция, верхний
олигоцен.
3 13 3
Диагноз. Зубная формула: I у С у Р -у М у — 36. Подглазничное от-
верстие не закрыто костной стенкой и видно сбоку. Слезное отверстие
открывается в орбиту. Нёбная кость перфорирована. Базисфеноидного
углубления нет. 1} значительных размеров. Корень Д под Р4. I2 имеет два
корня.
Описание (по: Viret, 1938; Baller, 1948). Кроме признаков, отме-
ченных в диагнозе, представители рода характеризуются и следующими.
Череп общим обликом напоминает таковой современных ежей рода Eri-
naceus L. В отличие от последних скуловая дуга менее поднята вверх
в задней части (более горизонтальна). Межчелюстная кость более длин-
ная. V-образный гребень на лобных костях и соединяющийся с ним сагит-
тальный гребень сильно развиты, также ламбдовидный гребень. Мозго-
вая капсула более вздута, особенно в задней части. Засочленовный от-
росток, как и у отдельных современных видов рода Erinaceus, сильно
развит, с глубокой выемкой.
Нижняя челюсть в разной степени массивная, с расширенным в пе-
редне-заднем направлении венечным отростком.
10*
148
ПОДСЕМ. ERINACEINAE
I* очень крупные, массивные. Расстояние между первым и вторым рез-
цами больше, чем у современных ежей. Верхний клык небольшой, с двумя
корнями. Р4 с вытянутым назад метаконидом. Верхние коренные мелкие
и Р4 помещаются под орбитой. Талон верхних коренных более обособлен,
чем у современных. Р3 с тремя, а Р3 с двумя корнями.
Распространение и возраст. Европа, Азия, олигоцен—
миоцен; Африка, миоцен; Северная Америка, средний миоцен.
Рис. 80. Amphechinus edwardsi (по: Viret, 1938).
t 1. Amphechinus arvernensis Blainville, 1839—1864.
Blainville, 1839—1864, Osteographie Mammiferes Insectivores, 1.1, p. 102—105.
T и п не отмечен. Вид описан из Auvergne, Франция, верхний олигоцен.
Характерной особенностью этого вида служит значительное развитие
переднего сегмента на Мх и М2, а также сильно выступающий назад мета-
конид Р4.
Измерения (по: Viret, 1929), Общая длина нижней челюсти —
32.0—33.5, нижнего зубного ряда'— 17.0—18.8, зубного ряда позади 1Х —
15.0-16.0, Р2-Р3 - 3.8-4.3, Мх—М3 - 8.0-8.5 мм.
Распространение и возраст. Известен из верхнего
олигоцена Франции.
t 2. Amphechinus edwardsi Filhol, 1879 (рис. 80, 81).
Erinaceus priscus Meyer, 1846, Noues Yahrb. fiir Min., S. 474; Palaeoerinaceus
edwardsi Filhol, 1879, p. 1—252.
Тип. He отмечен. Вид описан из St.-Gerard le Puy, Франция, нижний миоцен.
Хорошо сохранившийся череп и другие остатки скелета этого вида
послужили материалом для подробной характеристики не только вида, но
и рода Amphechinus (см. выше). Этот вид отличается короткой лицевой
частью черепа и расширенной в передне-заднем направлении массетерной
площадкой, ограниченной сзади неглубокими вырезками между слабо
выступающими венечным, сочленовным и угловым отростками.
Измерения (по: Viret, 1929). Длина составленного ряда верхних
зубов (С—М3) — 16, Р3 — 2.0, Р4— 3.7; ширина М3 — 2.0; длина Р4 —
2.7, Мх — 4.0, М2 — 3.2, М3 — 1.5 мм.
Распространение и возраст. Известен из миоцена
Франции и ФРГ.
21. РОД AMPHECHINUS
149
t 3. Amphechinus intermedius Gaillard, 1899 (рис. 82).
Palaeoerinaceus intermedius G a i-11 a r d, 1899, p. 15—17.
Тип. He установлен. Вид описан по фрагментам нижней и верхней челюстей из
Grive-Saint-Alban (Isere), Франция, средний миоцен.
Рис. 81. Amphechinus edwardsi (по: Hurzeler, 1944).
Структурные особенности верхних и нижних зубов сходны с таковыми
других представителей рода Amphechinus, также и строение нижней че-
люсти этого вида отличается главным образом крупными размерами,
приближающимися к размерам видов
рода Erinticeus L.
Измерения (по: Gaillard 1899).
Общая длина зубного ряда (по альвео-
лам) — 22.5, М1—М3 — 11.5. М1 —
5.5 X 3.5, длина Р4 - 3.5, С-Р4 (по
альвеолам) — 13 мм.
Распространение и возр
Рис. 82. Amphechinus intermedius
(по: Viret, 1938).
а с т. Известен из одного место-
нахождения, отмеченного для типа.
t 4. Amphechinus rectus Matthew et Granger, 1924 (рис. 83, 84).
Palaeoscaptor rectus Matthew, Granger, 1924, p. 3; Palaeoerinaceus rectus
Matthew, Granger, Bohlin, 1942, Palaeontologia Sinica. New ser., C, N 8“,
pt 1, p. 18.
Тип. Фрагмент нижней челюсти с зубами (М2 и М3) № 19146, Хзанда-Гол, Ло,
Монголия.
150
ПОДСЕМ. ERINACEINAE
Диагноз. По: Matthew Granger, 1924. Зубы отдаленно напоминают
таковые A. acridens Matth. et Grang. но более крупные. М3 у этого вида
редуцирован, талонид (метаконид) отсутствует. По: Трофимов, 1960.
Размеры сравнительно крупные. Параконид Р4 выше протоконида, М3 без
талонида (метаконида).
Описание (по: Bohlin, 1942). 14 слабо наклонен. Его основание
расположено под передним корнем Р4. Нижний клык крупнее 12 и Р3.
Р4 несколько изношен, но более или менее ясно, что его внутренний буго-
а
8k
Рис. 83, 84. Amphechinus rectus (a), A. kansuensis (б), A. minimus (в) (83 — по: Bohlin,
1942; 84 — по: Sulimski, 1970).
рок был массивным, высоким, не обособленным в такой степени, как
у Erinaceus europaeus L. Тригонид (передний сегмент пара-протокониды —
А. Г.) более развит, чем талонид (метаконид). М2 значительно меньше Mt
(2/3 его длины). Его параконид короткий. М3 состоит из тригопида (мета-
конид отсутствует). Венечный отросток более наклонен назад чем у Eri-
naceus. Судя по длине нижней челюсти, череп A. rectus длиннее, чем череп
A. edwardsi.
Распространение и возраст. Известен из Хзанда-Гол
и Татал-Гол, Монголия, средний олигоцен.
t 5. Amphechinus kansuensis Bohlin, 1942 (рис. 83).
Palaeoerinaceus kansuensis Bohlin, 1942, Palaeontologia Sinica, New ser., C,
N 8% pt 1, p. 23.
Тип. Фрагмент левой ветви нижней челюсти с Р4 и альвеолами других зубов,
Т. Ь. 593а, Табен-Булук, Китай, верхний олигоцен.
21. РОД AMPHECHINUS
151
Диагноз (по: Bohlin, 1942). Вид, средний по размерам. Зубы между
Р4 и 1Х мелкие. Судя по альвеолам, 12 > С > Р3. Р3 отделен от Р4 хорошо
выраженной диастемой.
Описание (по: Bohlin, 1942). Длина Р4—М3 около 6.2 мм, или
около 60% длины этих зубов сравниваемого вида. Подбородочное отвер-
стие впереди Р4 (под диастемой между Р3 и Р4). Угол между передней по-
верхностью венечного отростка и альвеолярным краем нижней челюсти
больше, чем у А. rectus. Судя по мелкой округлой альвеоле, М3 у этого
вида очень мелкий.
Распространение и возраст. Известен из одного место-
нахождения, отмеченного для типа.
Рис. 85. Amphechinus rusingensis (по: Butler, 1956).
t 6. Amphechinus minimus Bohlin, 1942 (рис. 83, 84).
Palaeoerinaceus minimus Bohlin, 1942, там же, с. 23—24.
Тип. Почти полная правая ветвь нижней челюсти с зубами (Р4—М3) и альвео-
лами С и Р3, Т. Ь. № 235, Табен-Булук, Китай, верхний олигоцен. Типы описанных
Болиным видов, хранятся, видимо, в г. Упсала (Швеция).
Диагноз (по: Bohlin, 1942). Мелкий вид с тонкой нижней челюстью.
Горизонтальная ветвь низкая, ее нижний край ровный под молярами и
премолярами и параллелен альвеолярному краю. Зубы, типичные для рода
Palaeoerinaceus (Amphechinus), крупные по отношению к размерам нижней
челюсти. Судя по альвеолам, 12 С Р3.
Описание (по: Bohlin, 1942). Размеры зубов сходны с таковыми
A. kansuensis (Р4—М3 — 6.0 мм), но величина нижней челюсти много
меньше и ее форма также отличается. Размеры Мх и М2 у этого вида более
сходны, чем у A. rectus Matth. et Grang. M3 с одним корнем, треугольный.
Подбородочное отверстие под передней частью Р4. Венечный отросток
отодвинут назад, и между его основанием и М3 имеется промежуток.
Распространение и возраст. Известен из одного место-
нахождения, отмеченного для типа.
t 7. Amphechinus rusingensis Butler, 1956 (рис. 85).
Butler, 1956, p. 54,—62.
Тип. Лицевая часть черепа с нижней челюстью N RS723. 47, 47, Hiwegi, место-
нахождение Rx, о. Русинга, на оз. Виктория-Ньянза, Кения; миоцен.
Диагноз (по: Butler, 1956). Отличается от A. edwardsi Filhol сле-
дующими особенностями. Размеры меньше (С—М3 — 13.2—13.5 мм). С и
Р^ менее редуцированы (в действительности С, Р2 и Р3). Р2 (Р3) с двумя
корнями. Щечный край М1 слегка длиннее, чем у Р4. М2 пропорционально
крупнее Мх. Отношение длины М2 к длине"Мх составляет 0.82.
152
ПОДСЕМ. ERINACEINAE
Описание (по: Butler, 1956). Крупнее Л. acridens Matth. etGrang.,
но меньше, чем A. arvernensis Blainv., A. edwardsi и A. rectus Matth. et
Grang. Размеры нёба и зубов составляют 75—85% таковых A. edwardsi.
Носовые кости с почти параллельными краями, задними концами далеко
заходят За передний край орбиты. Подглазничное отверстие над Р3, а Р1
под орбитой. Скуловая дуга поднимается над М1, она более тонкая, чем
у Gymnurechinus Butler. Посторбитальные отростки и лобные гребни более
развитые, чем у Gymnurechinus. Нёбо слабо перфорировано. Костное нёбо
лишь немного заходит за поперечный гребень. Слезное отверстие открыва-
ется внутрь орбиты. Нижняя челюсть сходна с этой костью других видов
рода, а также рода Gymnurechinus.
Измерения (по: Butler, 1956). Длина по наружному краю (тип)
Р3 — 1.8, Р4 — 2.9, М1 — 3.0, М2 — 2.2, М3 — 1.6; (котип) Р4 — 1.9x1.5,
Мх — 3.3X 2.0, М2 — 2.7 X 1.6; ширина нёба между М1 — 12.2 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
f 8. Amphechinus horncloudi Macdonald, 1970
Palaeoerinaceus horncloudi Macdonald, 1970, Bull. Los Angeles Co. Mus. Nat. Hist.,
v. 8, p. 1—82.
Тип. Фрагмент правой ветви нижней челюсти с зубами Р2—Mt, поврежденным
С, корнями h-g и передним корнем М2 N SDSM 62.113, Южная Дакота (США), средний
миоцен; кол. Геология, музея, Школы шахтеров и технологов, Лос-Анджелес.
Диагноз (по: Bich a. Rasmussen, 1973). Отличается от других видов
рода Amphechinus Aymard более крупным и широким, чем тригонид, тало-
нидом.
Распространение и возраст. Известен из Южной
Дакоты, средний миоцен.
f 9. Amphechinus montanus Koerner, 1940.
Parvericius montanus Koerner, 1940, p. 837—862.
Тип. Фрагмент правой части верхней челюсти с зубами — Р3—М3, № YPM
19356, Montana (США), средний миоцен, кол. Yale Peabody Mus.
Диагноз (по: Rich a. Rasmussen, 1973). М1 крупный и широкий.
Метаконид М| отсутствует. Д крупный.
Примечание. Этими признаками авторы диагноза отличают вид
от видов всех других родов и считают его принадлежащим к особому
роду — Parvericius Koerner.
Сравнение перечисленных в диагнозе признаков с таковыми Amphechi-
nus не оставляет сомнения в принадлежности описываемого вида к роду
Amphechinus. Поскольку, как пишут авторы, диагнозы рода и вида совпа-
дают, то и род Parvericius должен быть отнесен в синонимы рода Amphe-
chinus, что и сделано в свое время (Van Valen, 1967).
Распространение и возраст. Известен из Монтаны,
Вайоминга и Небраски (США), средний миоцен.
ПРЕДСТАВИТЕЛИ РОДА AMPHECHINUS AYMARD
С ТЕРРИТОРИИ СССР
Материал по вымершим ежам из олигоцена и миоцена Казахстана
(сборы Н. С. Шевыревой в Зайсанской котловине), полученный из Палеон-
тологического института АН СССР для обработки, позволил установить,
что составляющие его остатки относятся к разным видам рода Amphechinus.
Видовую принадлежность установить не удалось за отсутствием
сравнимого материала по видам, описанным из других частей ареала рода,
расположенных за пределами Союза.
21. РОД AMPHECHINUS
153
Amphechinus sp.
Фрагмент левой ветви нижней челюсти со стертым М3 № 2979 (134—305)—1, 19 км
к С-3 от г. Зайсана, Конуркура, Казахстан, олигоцен.
Очень мелкий вид: высота нижней челюсти у основания М3 — 1.5 мм.
Коронка этого зуба удлиненной формы с более, чем у других, развитым
параконидом. Длина М3 — 0.45, ширина — 0.35 мм.
Amphechinus sp. (рис. 86).
Два зуба — М1 и М2 левой стороны К» 2979 (134—305), 19 км к С-3 от г. Зайсана,
Конуркура, Казахстан, олигоцен.
Рис. 86. Amphechinus sp. а — верхний эоцен? —нижний олигоцен, б — олигоцен,
в — миоцен.
М1 с сильно скошенным наружным краем. Бугорки пара- и метаконов
крупные. Срединный бугорок (мезостиль) сильно развит и менее гребнист,
чем у более поздних видов. Слаборазвитый цингулюм отсутствует только
на внутренней стороне коронки. На задней стороне последней глубокая
выемка. М2 с коротким параконидом. Передний сегмент зуба (пара-, прото-
кониды) по длине примерно равен заднему — метакониду (талонид других
авторов). Цингулюм у М2 развит на наружной и задней сторонах, он утол-
щен в выемке между сегментами. Длина М1 по его наружному краю —
1.5, по внутреннему — 1.0, наибольшая ширина — 2.0; длина М2 — 1.65,
ширина — 1.1 мм.
Amphechinus sp. (рис. 86).
М1 — правой стороны, Хонгуркульджа, Зайсанский район, Казахстан, олигоцен.
Бугорки пара- и метакона М1 сильно развиты. Метакон имеет длин-
ный выступ, направленный назад. На задней стороне коронки глубокая
выемка, по краю которой проходит слаборазвитый цингулюм. На других
сторонах зуба цингулюм отсутствует. Срединный бугорок (мезостиль)
в виде небольшого гребнистого возвышения. Наибольшая длина зуба — 1.9,
наибольшая ширина—-2.5 мм.
Amphechinus sp. (рис. 86).
Обломки нижних челюстей и зубы № 3462 (1—45) (22), гора Ашутас, I терраса,
Казахстан, нижний миоцен.
Р4 трапециевидный. Его пара- и метакон островершинные, плоские,
в виде двузубой вилки. Протокон с конической вершиной. Талон лопасте-
видный с двумя бугорками.
154
ПОДСЕМ. ERINACEINAE
М1 почти четырехугольный, с глубокой выемкой на задней стороне.
Пара- и метакон в виде островершинных конусовидных бугорков. Протокон
с треугольной жевательной поверхностью, соединен гребнем с параконом.
Талон в форме конусовидного бугорка, расположенного на наклонном,
допастевидном широком основании. Срединный бугорок (мезостиль) греб-
нистый, соединен гребнями с протоконом и талоном. Цингулюм не развит
только на внутренней стороне коронки. М2 сходен по общим структурным
особенностям с М1, но более узкий, а гребень срединного бугорка чаще
соединен только с протоконом. М3 удлиненно-овальный. Его жевательная
поверхность поднята в виде гребня, расположенного в поперечном к про-
дольной оси челюсти направлении, окаймленного цингулюмом.
Р4 напоминает строением моляры, но метаконид (талонид других авто-
ров) у него низкий, короткий. Мх с удлиненным, треугольной формы,
со скошенным внутренним краем параконидом. Его протоконид двувер-
шинный, а удлиненный метаконид с двумя пирамидальными бугорками
на задней стороне. Цингулюм развит на внешней и задней сторонах. М2
в общих чертах сходен с Мх, но более короткий. М3 мелкий, округлый, без
метаконида, а бугорки протоконида слиты в гребень.
Высота ветви нижней челюсти у основания М3 — 2.6, 2.65; длина
Р4 — 1.7, 2.0; М1 — 2.1, 2.1; М2 — 2.0, 2.0; М3 — 0.5; Р4 — 1.0, 1.0;
Мх — 2.5, 2.5; М2 — 1.9; М3 — 0.9, 1.0; ширина Р4 — 2.2, 2.25; М1 —
2.35, 2.9; М2 - 2.45, 2.5; М3 — 1.25; Р4 - 1.05, 1.1; Мх - 1.5, 1.6; М2 -
1.5; М3 — 0.6, 0.85 мм.
Примечание. В этом же местонахождении обнаружен М1 очень
мелкого вида (длина 1.55 мм). По строению он сходен с М1 предыдущей
формы, но все грани и бугорки у него очень острые.
Amphechinus sp.
М1 — 3 экз. с дефектными отдельными частями и М2 — 2 экз. Из них один в пол-
ной сохранности, № 2977 (lit—181), р. Сарыбулак, Зайсан, Казахстан, верхний мио-
цен. Отткуда же М1 — 1 экз. и М2 — 2 экз., № 2977 (26—91).
М1 при общем сходстве с этим зубом нижнемиоценового представителя
этого рода из местонахождения гора Ашутас № 3462 (1—45) отличается
значительно меньшим развитием лопастевидного основания талона, менее
вытянутым на наружную сторону и назад метаконом и менее гребнистым
срединным бугорком. Слабо развитый цингулюм имеется и на внутрен-
ней стороне этого зуба. М2 также сходен с таковым упомянутой нижнемио-
ценовой формы. Отличается более обособленным талоном и наличием
цингулюма на внутренней стороне коронки.
f 22. Род GYMNURECHINUS Butler, 1956
Butler, 1956, р. 11—62.
Тип. Gymnurechinus leakeyi Butler, 1956, восточная часть о. Русинга, Виктория-
Ньянза, Кения, миоцен.
Диагноз (по Butler, 1956). Ежи, сходные по многим особенностям
черепа с Amphechinus Aymard, отличаются: менее длинным I1 и располо-
жением корня I1 под Р3; более крупными коренными зубами так, что Р4
расположен впереди орбиты. Р3 имеет два корня. Отверстия костного нёба
небольшие. Базисфеноидной впадины нет. Слезное отверстие открывается
внутрь орбиты.
Распространение и возраст. Кения, Африка, миоцен.
22. РОД GYMNURECHINUS
_ 155
f 1. Gymnurechinus leakeyi Butler, 1956 (рис. 87).
Butler, 1956, там же, с. 8—30.
Тип. Череп с нижней челюстью N Ks 12.32, восточная часть о. Русинга, оз. Вик-
тория-Ньянза, Кения, миоцен, кол. Кориодонск. муз.
Диагноз (по: Butler, 1956). Отличается от других видов рода широ-
ким сагиттальным гребнем с прямоугольным крестообразным сечением и
Рис. 87. Gymnurechinus leakeyi (по: Butler, 1956).
относительно поперечным М1. Ламбдовидный гребень сильно развит.
Ламбдовидная область сравнительно большая. Ширина нёба между пара-
конами М1—М1 — 16.3—16.5 мм. Гипокон (талон) Р4 нормально развит.
Альвеолярная длина Мх—М3 — 10.0—10.8 мм.
Распространение и возраст. Указаны для типа.
f 2. Gymnurechinus camptolophus Butler, 1956.
Butler, 1956, там же.
Тип. Череп RS 424а.48, восточнее Киахера, о. Русинга, Виктория-Ньянза, Ке-
ния, миоцен, кол. Кориодон. муз.
Диагноз (по: Butler, 1956). Этот вид отличается от G. leakeyi
следующими особенностями: лицевая часть пропорционально крупнее по
отношению к черепу. Сагиттальный гребень острый. Затылочные мыщелки
156
ПОДСЕМ. ERINACEINAE
широкие в дорсальной части. М1 и М2 менее поперечные. Р4 с редуцирован-
ным гипоконом (талоном). Ширина костного нёба между параконами
М1—М1 — 18.7 мм. Альвеолярная длина Мх—М3 — 11.0—11;4 мм.
Распространение и возраст. Указаны для типа.
t 3. Gymnurechinus songhorensis Butler, 1956.
Butler, 1956, там же.
Тип. Задняя часть черепа NSgr 513.49, Сонгхор (Songhor), Кения, миоцен.
Д и а г н о з (по: Butler, 1956). Отличается от других видов этого рода
менее высоким затылком, меньшей ламбдовидной областью, слабым ламбо-
видным гребнем, мелким параокципитальным отростком и слабым раз-
витием кондилюсов в дорсальной части. Сагиттальный гребень острый.
М1 сходной длины и ширины. Гипокон (талон) Р4 очень развит. Ши-
рина нёба между параконами М1—М1 — 18.7 мм. Альвеолярная длина
Мх—М3 — 11.3 мм.
Распространение и возраст. Отмечены для типа.
ф 23. Род STENOECHINUS Rich et Rasmussen, 1973
Rich, Rasmussen, 1973, p. 1—54.
T и n. Stenoechinus tantalus Rich et Rasmussen, 1973. Северная Америка, ранний
миоцен.
Диагноз (по: Bich, Bassmussen, 1973). Отличается от Aethechinus,
Amphechinus, Atelerix, Dimylechinus, Erinaceus, Gymnurechinus, Hemiechi-
nus, Mioechinus, Paraechinus, Parvericius и Postpalerinaceus большей шири-
ной Mx относительно его длины; от Amphechinus, Dimylechinus, Mioechinus,
Palaeoscaptor и Postpalerinacius, сжатым в передне-заднем направлении
тригонидом Мх; от Aethechinus, Amphechinus, Atelerix, Erinaceus, Hemiechi-
nus, Mioechinus, Paraechinus, Parvericius и Poctpalerinaceus наличием вы-
ступающего заднего цингулюма на М3; от Palaeoscaptor талонидом, редуци-
рованным до выступа цингулюма и от Dimylechinus наличием М3.
Примечание. Из этого странного диагноза видно, что описан-
ный авторами род отличается от всех родов относительно большей шириной
Мх. Кроме того, от одной части родов отличается укороченным первым
сегментом (тригонидом авторов), от другой — укороченным до степени
воротничка (цингулюм) метаконидом (талонидом авторов), наличием М3
от рода Dimylechinus Hiirz.
Авторы описываемого здесь рода отличают его от рода Palaeoscaptor
следующими признаками: 1) талонид (метаконид) М3 значительно редуци-
рован у Stenoechinus-, 2) тригонид (передний сегмент — пара-протоконид)
сжат в передне-заднем направлении; 3) параконид Р4 заметно ниже, чем
протоконид; 4) высота протоконида менее длины этого зуба; 5) корень 1х
не простирается далеко назад.
Все перечисленные признаки по своему значению не могут быть
признанными за родовые. Можно думать, что более важные, если они есть,
окажутся в строении черепа и в строении и общем количестве зубов.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
t 1. Stenoechinus tantalus Rich et Rasmussen, 1973.
Rich, Rasmussen, 1973, там же.
Тип. КИ 18001, фрагмент правой ветви нижней челюсти с зубами —Мх—М3
Powell Со, Монтана, нижний миоцен, кол. Муз. Канзас, унив., Лоренс (США).
Описание. Весьма расплывчатое описание в сравнении с призна-
ками других родов, крайне многословное, не прибавляет ничего особенно
существенного к описанию рода Stenoechinus.
23. РОД STENOECHINUS
157
Судя по фотографии, М1 и М2 угловатой формы, гребнистые, со слабо
развитым бугорком в центре жевательной поверхности. Нижние коренные
зубы характерные для ранних групп ежей. М3 с воротничком (цингулюмом)
на месте метаконида (талонида авторов). Этот зуб сходен с этим же зубом
Amphechinus rectus Matt, et Grang.
Измерения (no: Rich, Rasmussen, 1973). Передне-задний диаметр
M1 — 1.7; М2 — 1.2; поперечный диаметр (ширина) М1 — 2.2, М2 — 1.9 мм.
Распространенней возраст. Известен из одного место-
нахождения, указанного для типа.
Примечание. Общие структурные особенности зубов на
представленных авторами фотографиях позволяют отметить, что никаких
специфичных для этого рода признаков, отражающих особый путь разви-
тия, обнаружить не удается. Можно думать, что этот род при накоплении
достаточного материала будет объединен с родом Amphechinus.
Триба Е R I NACEI NI Fischer von Waldheim, 1817
Fisher von W aldheim, 1817, p. 368—428.
Тип. Erinaceus Linnaeus, 1758, Европа, Азия, Африка, миоцен — современный.
Диагноз. Слезное отверстие открывается вне орбиты, на пред-
орбитальном гребне. Нёбо перфорировано. Базисфеноидное углубление
имеется. В верхней челюсти 3 премоляра. М| сохраняются. М3 узкий, рас-
положен по отношению к зубному ряду в поперечном направлении.
Самые крупные ежи в пределах подсем. Erinaceinae. Приспособлены
к существованию в различных условиях -открытых ландшафтов и в лесной
зоне. Основные особенности этой группы ежей приведены в характери-
стике подсемейства и родов.
Распространение и возраст. Европа, Азия и Африка,
миоцен—современный.
ТА БЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ПОДРОДОВ
1 (4). На коренных зубах М1 и М2 в центре жевательной поверхности
гребень.
2 (3). Р3 и Р4 крупные............................24. Mioechinus.
3 (2). Р4 крупный, а Р3 очень мелкий..............25. Protechinus.
4 (1). На коренных зубах М1 и Ма в центре жевательной поверхности
имеется бугорок, а не гребень......................26. Erinaceus.
5 (6). Засочленовный отросток без полости или она очень маленькая.
Барабанные отростки основной клиновидной кости закрывают
слуховую полость только с внутренней стороны. Длина уха не более
половины длины головы...........................Подрод Erinaceus.
6 (5). Засочленовный отросток с полостью (углублением). Барабанные
отростки основной клиновидной кости закрывают слуховую полость
с внутренней и наружной стороны. Длина уха более половины длины
головы, а если короче, то на голове нет полоски голой кожи, разде-
ляющей иглы на два пучка.
7 (8). Крыловидные кости и крыловидные отростки основной клиновид-
ной кости не вздуты, в них нет полостей, сообщающихся со слуховой
полостью ......................................Подрод Hemiechinus.
8 (7). Крыловидные кости и крыловидные отростки основной клиновид-
ной кости вздуты и их полость сообщается со слуховой полостью
............................................ . Подрод Paraechinus.
Примечание. В таблицу не вошел подрод Postpalerinaceus Crusafont-
Pairo et Villalta-Camella за отсутствием сравнимых признаков
158
ПОДСЕМ. ERINACEINAE
f 24. Род MIOECHINUS Butler, 1948
Butler, 1948, p. 478—482.
Тип. Mioechinus oeningensis Lydekker 1886 (= Erinaceus oeningensis Lydekker,
1886), Европа, верхний миоцен.
судя по верхнему
как у видов рода
Диагноз (по: Butler, 1948). Зубная формула,
зубному ряду, приведенному Батлером (Butler, 1948),
Erinaceus L. Базисфеноидное углубление имеется. Нёбо перфорировано.
В отличие от рода Erinaceus метаконуль (бугорок на жевательной поверх-
ности) М1 в виде складки. Р3 и Р4 крупные. Р2 с двумя корнями. Можно
думать, что слезное отверстие открывалось на предорбитальном гребне.
Распространение и возраст.
Европа, плиоцен.
t 1. Mioechinus oeningensis Lydekker, 1886
(рис. 88).
Erinaceus oeningensis Lydekker, 1886, Quart.
J. Geol. Soc., v. 42, -p. 23. Mioechinus oeningensis Lydek-
ker, Butler, 1948, p. 478—482.
Тип. He установлен.
Описание (no: Butler, 1948). Мыщел-
ковое отверстие расположено в углублении на '
границе мыщелка, как у гимнур.
Задне-наружный выступ у М1 менее развит,
чем у Erinaceus, а углубление на наружном
крае менее значительно. М1 имеет почти квад-
ратную форму. Его язычный корень сходен
’ с таковым Erinaceus. М2 меньше М1. При этом
Рис. 88. Mioechinus oenin- разница в их размерах более, чем у Erinaceus.
gensis (по: Butler, 1948). Клык лишь немного крупнее Р2, с двумя кор-
• нями. I3 крупнее клыка, от которого отделен
небольшим пространством, он также с двумя корнями. I2 мелкий, с высту-
пом спереди и сзади, с одним корнем. I1 удлиненной формы. Его корень
проходит над резцами и клыком. Этот зуб более развит, чем у Erinaceus.
Распространение и возраст. Европа, верхний миоцен.
f 2. Mioechinus sansaniensis Lartet, 1851.
Erinaceus sansaniensis Lartet, 1851, Notice sur la colline de Sansan, p. 12. Mioe-
chinus sansaniensis Lartet, Butler, 1948, p. 478—482.
Этот вид, описанный по нижней челюсти, Батлер (Butler, 1948) отнес
к роду Mioechinus провизорно, не желая, как он выразился, вводить в си-
стему новый род. Его филогенетическое и систематическое положение
остаются неясными. Как отмечает Батлер, сравнение М. sansaniensis
с М. oeningensis Lidek. невозможно, поскольку первый описан по нижней
челюсти, а второй — по черепу. Сравнение обломков нижней челюсти
М. sansaniensis с представителем другого рода, как это сделал Батлер,
или описание обломков при значительной вариабельности и сходстве
нижней челюсти у разных групп ежей из триб Amphechini и Erinaceini
ничего положительного дать не могут.
Примечание. Род Mioechinus, как и род Protechinus Lavoc.,
представляет, видимо, миоценовую группу ежей, близкую по характеру
адаптации к роду Erinaceus. Можно также рассматривать род Mioechinus
как подрод рода Erinaceus, сохранивший в миоцене более архаичные
25. РОД PROTECHINUS
159
особенности. Миоцен был периодом расцвета и формирования окончательно-
го адаптивного типа ежей, отраженного в особенностях приспособлений
представителей современных групп рода Erinaceus L.
f 25. Род PROTECHINUS Lavocat, 1961
Lavocat, 1961, Notes et Memoires du service Geologique, N 155, p. 68—75.
Тип. Protechinus salts Lavocat, 1961, Бени-Меллаль (Марокко), миоцен.
Диагноз, (по: Lavocat, 1961).'Нижнеглазничное отверстие впереди
переднего края Р4. Слезное отверстие расположено на тонком возвышении
(на предглазничном гребне) вне орбиты. Р4 развит пропорционально
по отношению к М1. Он переходного типа между этим зубом ранних и
поздних видов Palaeoerinaceus (синоним Amphechinus Aymard) и современ-
ных ежей.
Гипоконы верхних моляров высокие.
Кондилюс нижней челюсти низкий. Первый резец клыкообразный,
с одним корнем, короткий, типичен для представителей рода Erinaceus L.
и не типичен для рода Palaeoerinaceus.
Р4 удлиненный, его тригонид узкий, параконид низкий, обособлен,
метаконид заметный, но не обособлен.
Р а с п р о стр анение и возраст. Африка, миоцен.
t 1. Protechinus salis Lavocat, 1961.
Lavocat, 1961, там же, с. 68—75.
Тип. Фрагмент правой стороны верхней челюсти с зубами (Р4 и М1), № 1501,
Бени-Меллаль (Марокко), миоцен.
Описание. Изучение приведенных автором рода и вида фото-
графий костных остатков, послуживших материалом для их описания,
позволяет отметить некоторые особенности строения зубов этого вида.
I1 с несколько стертой вершиной, округлый в сечении, невысокий, напоми-
нает этот зуб видов рода Erinaceus L. I3 с двумя корнями, конической
формы, с заметным сзади метаконом. Клык одновершинный, с двумя кор-
нями. Р3 с тремя корнями и тремя бугорками, очень мелкий по сравнению
с Р4. Последний имеет вытянутый назад отросток метакона, он более
широкий, чем у представителей рода Erinaceus. М1 и М2 сходны с этими
зубами современных ежей, но у М1 метакон более массивный и менее
остробугорчатый. Кроме того, на месте бугорка в центре жевательной
поверхности имеется гребень, соединяющий метакон и протокон, более
заметно выраженный на М2. М3 по строению сходен с этим зубом современ-
ных представителей рода Erinaceus.
1Х близок по строению к резцу видов рода Erinaceus. 12 одновершин-
ный, с коронкой, мало отогнутой вперед. Клык также менее наклонен
вперед. Р3 напоминает этот зуб современных ежей, только более округлой
формы. Р4 напоминает таковой многих других родов ежей. Его параконид
вытянут вперед и обособлен от протоконида. Протоконид двувершинный.
Его внутренний бугорок (метаконид других авторов) выдвинут вперед.
Метаконид более развит (талонид других авторов). Мх и М2 сходного строе-
ния, но М2 меньше Мх, с более широким, но низким протоконом и с более
широко расставленными бугорками переднего сегмента. М3 округлой
формы, относительно менее развит, чем у видов рода Erinaceus.
Измерения (по: Lavocat, 1961). Длина: с наружной стороны
Р4 — 3.6, М1 — 3.8, М2 — 3.0, с внутренней стороны — Р4 — 2.2, М1 —
3.2, М2 — 2.0; Р4 — 2.8, Мх — 4.0, М2 — 3.84 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
160
ПОДСЕМ. ERINACEINAE
Примечание. При том, что материал сравнительно ограничен-
ный и многие важные детали в строении черепа описываемых рода и
вида непосредственно видеть нельзя, можно все-таки представить их вероят-
ное сочетание с теми, которые известны. Прежде всего можно с уверен-
ностью сказать, что представители рода Protechinus, слезное отверстие
у которых находится за пределами орбиты, имеют впадину на базисфено-
иде, а нёбо у них с крупными отверстиями. 'Это заключение основывается
на том, что наличие впадины на базисфеноиде и крупные отверстия на нёбе
сочетаются с расположением слезного отверстия на предорбитальном гре-
бне.
Из всех известных групп ежей такое сочетание отмеченных признаков
характерно только для видов рода Erinaceus L. В связи с этим есть
основания считать, что род Protechinus очень близок к роду Erinaceus,
если это не его ранний подрод.
26. Род ERINACEUS Linnaeus, 1758
Linnaeus, 1758, р. 52. Ericius Sundeva 1, 1842, Kongl. Svenska Vetensk.
Akad. Handl., p. 225—237. Atelerix Pomel, 1848, Bibl. Univ. Geneve, Arch. Sci.
phys. nat., v. 9, p. 251. Peroechinus Fitzinger, 1866. Sitz. Ber. Ost. Akad. Wiss.
Math.-naturw. KI. Wien, Bd 1, S. 565. Hemiechinus Fitzinger, 1866, там же. He-
rinaceus Mina-Palumbo, 1868, Ann. Agric. SiciL, ser. 2, v. 12, p. 37. Paraechi-
nus Trouessart, 1879, Revue et Magaz. de ZooL, Paris, ser. 3, t. VII, p. 242. Ma-
croehinus S a t u n i n, 1906, Annuaire du Musee ZooL de 1’Academie Imper. de Scien-
ces de St. Petersb., t. XI, p. 23. Aethechinus Thomas, 1918, p. 194. Erinaceulus О r-
н e в, 1928, c. 168.
Тип. Erinaceus europaeus Linnaeus, 1758, Европа, Азия, плиоцен—современный.
Диагноз. Слезное отверстие открывается на предорбитальном
гребне. Нёбо перфорировано. Основная клиновидная кость с углублением
позади хоан.
Описание. Представители рода характеризуются массивным
телом и коротким малозаметным хвостом. Длина тела от 135 до 300 мм.
Ноги короткие, пятипалые, стопоходящие. Редко первый палец ступни
(hallux) у отдельных особей может отсутствовать. Кисти массивные,
с широко расставленными пальцами. Голова клиновидной формы, с вытя-
нутой в короткий хоботок носовой частью. Затылок, верхняя сторона шеи,
спина и в разной степени бока тела покрыты иглами. Иглы разной длины
и толщины, с разной скульптурой на поверхности (бороздки, бугорки). Они
имеют светлые и темные зоны (пояски), число которых варьирует. Нижняя
сторона тела и конечности покрыты волосами грубыми или мягкими. Общий
тон окраски варьирует от охристо-беловатого, охристого, серого и бурова-
того до темного, черно-бурого. Различные сочетания перечисленных цветов
создают разнообразную окраску иглистого панциря и волосяного покрова.
Нижняя сторона тела может быть белого цвета. Встречаются светло
окрашенные особи, а также с пятнистой окраской.
У подавляющего большинства особей видов, объединяемых родом,
темные волосы образуют на лицевой части головы своеобразный рисунок
(маску) (рис. 26).
Череп сравнительно широкий, массивный, с широко расставленными
скуловыми дугами. Его носовой отдел сравнительно широкий или в раз-
ной степени сужен. Подглазничное отверстие закрыто сбоку костным вы-
ростом. У видов с крупными слуховыми барабанами, засочленовый от-
росток или последний и крыловидные кости вздуты и имеют полость,
сообщающуюся со слуховой полостью. Барабанная кость не срастается
с окружающими ее костями черепа и не образует замкнутого барабана.
Скелет образован массивными костями. Построен по общему для всего
рода плану, со слабым различием структур костей даже у видов разных
26. РОД ERINACEUS 161
подродов. Строение скелета соответствует нагрузке, которую создает
массивное тело и иглистый панцирь с подкожрой кольцевой мускулатурой,
сворачивающей тело в шар. В соответствии с общим устройством тела
стоит и характер передвижения ежей: медленная и быстрая рысь. Ежи
роют себе норы, используя для этого широкие сильные кисти с хорошо
развитыми когтями. Брошенный в воду еж выплывает, двигая ногами
, сходно с их движением на земле.
Представители рода известны из нижнего миоцена Европы, плейстоцена
Азии и Африки. Можно думать, что они были в Азии и в Африке в то же
время, что и в Европе, т. е. еще в миоцене.
Специализация видов рода шла по пути приспособления к жизни
в условиях открытых ландшафтов, на что указывает развитие иглистого
панциря и способность сворачивать тело в шар, устройство слухового
аппарата, крупные для насекомоядных глаза и др., а также способность
питаться разнообразной животной пищей разной консистенции.
Распространение и возраст. Европа, Средняя и Цен-
тральная Азия, северная Аравия, Африка, миоцен—современный.
Примечание. Исследованный значительный материал по раз-
личным видам современных ежей позволяет отметить, что большая часть
приписываемых им отличительных признаков варьирует в значительных
пределах и многие из них не могут считаться таковыми.
Отличия (считаемых самостоятельными родами) Erinaceus L., Hemiechi-
nus Fitzinger и Paraechinus Trouessart имеют переходный характер и могут
быть оценены показателями: больше—меньше. Например, величина
ушей, степень развития засочленовного отростка и его полости, степень
развития барабанного отростка основной клиновидной кости, закрывающей
слуховую полость, крыловидного отростка основной клиновидной кости,
крыловидной кости, а также образования в них полости, сообщающейся
со слуховой полостью. Варьирует число корней у зубов и строение игл.
К этому следует добавить, что все виды этих родов имеют общую зуб-
ную формулу и общую структуру зубов.
Все вышесказанное позволяет рассматривать перечисленные роды
только в качестве подродов описанного рода Erinaceus. Что же касается
родов Atelerix Pomel и Aethechinus Thomas, то они давно сведены в сино-
нимы друг к другу и к роду Erinaceus. Правда, в последнее время некоторые
авторы их вновь возродили, но это уже проявление анахронизма.
Объединение названных родов в один род Erinaceus не нововведение
автора настоящей работы. Добсон (Dobson, 1882) в своей монографии
о насекомоядных, не обсуждая, отнес все виды ежей к роду Erinaceus.
Андерсон (Anderson, 1902) сделал то же самое с африканскими видами,
относимыми к трем разным родам.
Морфологический анализ структурных особенностей видов перечислен-
ных родов не оставляет сомнения в том, что современные ежи представ-
ляют собой целостную группу, приспособленную к жизни в открытых
ландшафтах.
Подродовая дифференциация отражает разную степень приспособлен-
ности представителей отдельных подродов к жизни в степях и пустынях,
что оставляет возможность существования отдельных видов в условиях
лесостепи, а также в лесной зоне, на удобных для них местах.
f 1. Подрод POSTPALEOERINACEUS Crusafont-Pairo et Villalta Camella, 1947.
Palerinaceus (Postp alerinaceus) ^Crusafont-Pairo, Villalta Ca-
mella, 1947, p. 320—333.
Тип. Palaeoerinaceus (Postpalaeoerinaceus) vireti Crusafont-Pairo et Villalta Ca-
mella, 1947, Барселона, Испания, верхний плиоцен.
11 А. А. Гуреев
162
ПОДСЕМ. ERINACEINAE
Диагноз, (по: Crusafont-Pairo, Villalta Camella, 1947). Нижняя
челюсть с более длинной, чем у современных представителей рода Erina-
ceus L. зубной частью. Предкоренные и особенно нижние коренные с удли-
ненной коронкой и более остробугорчатые. Нижний клык крупнее. К этому
нужно добавить, что все гребни на черепе рельефнее и выше (рис. 89).
Примечание. Приведенные авторами признаки не дают основа-
ния для' сближения описанного ими подрода с родом Palaeoerinaceus
Рис. 89. Erinaceus vireti (по: Crusafont-Pairo, Villalta Camella, 1947).
Filhol (синоним рода Amphechinus Aymard). Напротив, есть все основания
считать, что они ближе к роду Erinaceus.
К описываемому подроду относится один вид, отмеченный как тип.
Распространение и возраст. Пиренейский полуостров
(Европа), верхний плиоцен.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЕЖЕЙ
ФА УНЫ СССР
1 (4). Ушная раковина, будучи отогнута вперед, достигает только зад-
него края глаза. Волосяной покров жесткий, щетинистый.
2 (3). На темени имеется полоска голой кожи, лишенная волос и игл.
I1 и клык имеют один корень. Очень редко клык имеет два слабо
обособленных корня....................2. Erinaceus europaeus L.
3 (2). На темени полоски голой кожи нет. Ни клык всегда имеют два
корня .....................................5. Е. dauricus Sundevall.
4 (1). Ушная раковина, будучи отогнута вперед, заходит за передний
край глаза. Волосяной покров мягкий.
26. РОД ERINACEUS
163
5 (6). На темени имеется полоска голой кожи. Крыловидные отростки
вздуты и полые .............................6. Е. aethyopicus Ehrenb.
6 (5). Полоски голой кожи нет. Крыловидные отростки не вздуты, не
полые .........................................4. Е. auritus Gmelin.
2. Подрод ERINACEUS s. str. Linnaeus, 1758 — Обыкновенные ежи.
Linnaeus, 1758, p. 52. Atelerix Pomel, 1842, Bibl. Univ. Geneve, Arch.
Sci. Phys, nat., v. 9, p. 257. Aethechinus Thomas, 1918, p. 194.
Тип. Erinaceus europaeus Linnaeus, 1758, Европа, Азия, миоцен—современный.
Диагноз. Иглистый панцирь спускается на бока тела. Барабан-
ные отростки основной клиновидной кости образуют внутреннюю стенку
слухового барабана (у отдельных особей узким краем заходит на его верх-
нюю сторону). Засочленовный отросток не шире сосцевидного, с неглубо-
ким углублением или без него.
Описание. Уши небольшие, прижатые к голове не заходят за
глаза. Волосяной покров жесткий. На голове полоска голой кожи раз-
деляет иглы на две группы. Иглы в этом месте более длинные, чем в области
шеи. Иглы со слабо выраженными бороздками или без них.
Череп крупный или средних для ежей размеров. Скулы широко рас-
ставлены. Носовой отдел черепа широкий и высокий. Предглазничный
гребень развит слабее, чем у других подродов. V-образный гребень,
начинающийся от предорбитального гребня и заканчивающийся углом на
сагиттальном гребне в области лобно-теменного шва, слабо развит
й мало заметен или отсутствует. Череп не имеет вдавления в области лобно-
носового шва. Межглазничное сужение менее выражено, чем у других
подродов. Сагиттальный гребень невысокий. Ламбдовидный гребень сильно
развит. Затылочное отверстие развито в высоту и в ширину примерно
одинаково, имеет сердцевидную форму или в виде трилистника. Крыло-
видные отростки основной клиновидной кости слабо вздуты. Крыловидные
кости не вздуты или слабо вздуты, не сообщаются со слуховой полостью.
I3 и С с одним или с двумя корнями.
Распространение и возраст. Европа, Передняя Азия,
Западная Сибирь, южная часть Амурской обл., Приморский край (СССР),
Северный и Северо-Восточный Китай, п-ов Корея, Африка, верхний мио-
цен—современный.
f 1. Erinaceus s. str. davidi Jammot, 1973 (рис. 90).
J a m m о t, 1973, p. 42—44.
Тип. Правая ветвь нижней челюсти LF (1) 5019, Aven la Fage (Correze) A 6 couche
5, — 686 cm (Франция), средний плейстоцен. Для типа автор указал № 5019, а в таб-
лицах и в подписи к рисунку указан № 5012. Какой из них истинный, знает только
автор.
Диагноз (по: Jammot, 1973). Самый крупный еж из описанных
с территории Европы. Нижняя челюсть характеризуется широким (пра-
вильнее удлиненным) апофизом венечного отростка в его верхней части.
Горизонтальная ветвь между Р4 и М3 сходной высоты. Сочленовный от-
росток высоко поднят. Строение моляров сходно с таковым современных
видов. Верхние премоляры более широкие, а нижние премоляры при
крупных размерах с редуцированным талонидом (метаконидом).
Описание. Нижняя челюсть описываемого вида хорошо отлича-
ется от таковой других европейских видов строением сочленовного отдела.
Вот что явствует из описания автора вида. Другие приведенные им при-
знаки не отличаются постоянством.
11*
164
ПОДСЕМ. ERINACEINAE
Измерения (по: Jammot, 1973). Длина вершины венечного
отростка у Е. davidi — 6.1, у алжирского ежа — 3.4, у обыкновенного —
3.2—5.3 мм.
Примечание. Описанные в разное время: Е. praeglacialist
Е. lechei, Е. stramosi, Е. samsonovici в настоящей работе не приводятся.
Эти виды требуют тщательной ревизии на большом материале.
2. Erinaceus s. str. europaeus Linnaeus, 1758. — Обыкновенный еж
(рис. 91, 92).
Рис. 90. Erinaceus davidi (а), Е. frontalis (б) (по: Jainmot, 1973).
Linnaeus, 1758, р. 52; — sibiricus Erxleben, 1777, Syst. Regn. Anim.,
p. 172. Hystrix erinaceus Blumenbach, 1779, Handbuch Naturgesch., S. 72. Eri-
naecus concolorM a r t i n, 1838, Proc. Zool. Soc. London, p. 103; — caniceps H.Sini th,
1845, Naturalist’s Libr. (Jardines), ed. 2, v. 15, p. 148; — amurensis Schrenk, 1858,
в кп.: Огнев, 1928, с. 97—100; — dealbatus S winhoe, 1870, Proc. Zool. Soc. Lon-
don, p. 450; — echinus Schulze, 1897, Helios, Berlin, Bd 14, S. 91; — rumanicus
В arrett-Hamilton, 1900, в кн.: Огнев, 1928, с. 104—107; — danubicus М а-
t s с h i e, 1901, Sitzungsber. Ges. Naturf. Freunde, Berlin, Bd 9, S. 229; — orientalis J.
A lien, 1903, Bull. Amer. Mus. Nat. Hist., v. 19, p. 179; — ussuriensis С а т у н и н,
1906, Ежегодн. Зоол. муз. Акад, наук, т. XI, с. 170; — chinensis С атунин, 1906,
там же, с. 173; — krayenbergi М at sc hi е, 1907, Exped. Filchner Mamin., S. 135; —
tschifuensis M a t s c h i e, 1907, там же, с. 137; — hanensis Matschie, 1907,
там же, с. 138; — unghi Thomas, 1908, Abstr. Proc. Zool. Soc. London, p. 44; —
suillus M i 11 e r, 1912, Cat. Mamm. Western Europe, p. 120. — caninus Miller,
1912, там же; — koreanus L 6 n b e г g, 1922, Ann. Mag. Nat. Hist., v. 9, p. 624; —
Hemiechinus mandschuricus Mori, 1926, Anuot. Zool. Jap., v. 11, p. 108.
Тип. He установлен, вид описан из южной части о. Готланд (Wamlingbo),
Швеция.
Диагноз. Костное нёбо не заходит за нёбный поперечный гребень
в виде пластинки. Третий резец и клык с одним корнем. Редко клык имеет
два слабо обособленных корня или они разделены бороздкой.
26. РОД ERINACEUS 165
Описание. Просмотр значительного материала из, различных
ландшафтов обширной территории ареала обыкновенного ежа (из по-
давляющего большинства областей европейской части СССР, включая
Прибалтийские республики, Белоруссию, Украину, Крым, Кавказ и
Башкирию; из Казахстана, Западной Сибири и Дальнего Востока, а также
из Англии, ГДР, Болгарии, Венгрии, Италии и островов Средиземного
моря, Ирана и Турции) позволяет отметить значительную изменчивость
морфологии этого вида, в частности, окраски тела.
Экземпляры из Ленинградской обл. имеют иглистый панцирь, окрашен-
ный в охристо-желтоватый или охристо-серый цвета. Волосяной покров
боков тела бурый разных оттенков, а нижней стороны — бурый с сереб-
ристым налетом, охристо-палевым или пятнами бурый и палевый. При
Рис. 91. Erinaceus europaeus (по: Walker et al., 1964).
общем охристом оттенке покровов у ежей из северных окраин евро-
пейской части СССР этот цвет более развит у экземпляров из Калинин-
ской обл., в меньшей из Вологодской. Окраску, близкую к северным, имеют
экземпляры из Венгрии. Два однотипных экземпляра из Англии имеют
темную бурую окраску, более близкую к окраске южных экземпляров.
От Белоруссии и Рязанской обл. до Крыма и Кавказа, Южного Урала и
Казахстана в окраске волосяного покрова преобладают темно-бурые
тона, достигающие черно-бурого цвета в Закавказье, Иране и Турции.
Окраска иглистого панциря варьирует от светло-бурого и серого до черно-
бурого. Для южных представителей этого вида характерны белые пятна
на горле и груди, часто и на брюхе. Следует отметить, что молодые экзем-
пляры светло окрашенных ежей из Западной Сибири и Дальнего Востока
имеют темную окраску и белое пятно на нижней стороне тела, например,
Erinaceus europaeus amurensis Schrenk, молодые экземпляры которого
представлены в коллекции Зоологического института АН СССР большой
серией. Экземпляры из Западной Сибири имеют светлую охристо-беловатую
окраску иглистого и волосяного покровов. Сходную окраску имеют и
дальневосточные ежи.
Череп по сравнению с Erinaceus frontalis Smith более сужен в передней
части, особенно на уровне слезных отверстий. Скуловые дуги не сужены
в заднем отделе. Сочленовная поверхность для сочленения с нижней
челюстью на скуловом отростке височной кости менее длинная, чем
у сравниваемого вида. Зубы менее массивны. Зубная часть нижней челюсти
более высокая.
166
ПОДСЕМ. ERINACEINAE
Измерения. Длина: тела — 195—300, хвоста — 23—37, задней
ступни — 34—47, кондило-базальная черепа — 43.0—63.0 мм.
Рас пр остранение и во з р а с т. Западная Европа, Перед-
няя Азия, Северный и Северо-Восточный Китай, п-ов Корея. В СССР
от северный берегов Лддожского озера на юг до Крыма и Кавказа включи -
тельно; западные районы Северного Казахстана, Западная Сибирь, южная
чаСть Амурской обл. и Приморский край, верхний плиоцен—современный.
Рис. 92. Erinaceus europaeus.
Биология. Населяет разнообразные ландшафты, избегая за-
болоченных мест и больших массивов высокоствольных лесов.
В лесных ландшафтах предпочитает опушки лесов, вырубки и заросли
кустарников. В открытых ландшафтах селится среди мелких кустарников,
высоких травянистых растений и в других укромных местах, предпочитая
всюду участки под укрытием древесно-кустарниковой растительности.
В отдельных местах он селится и на песчаной почве, например, в Волго-
Уральской степи и Прикумских степях северо-восточного Предкавказья.
Пищей ему служат разнообразные беспозвоночные и мелкие позво-
ночные животные, а также сочные плоды растений. Из позвоночных он
поедает мелких грызунов, ящериц, лягушек и змей, яд которых ему, по-
видимому, не вредит, а кроме того, еж умеет защищаться от укусов змей.
Иногда поедает яйца и птенцов птиц, гнездящихся на земле. Активен
в сумеречные и ночные часы. Летнее гнездо устраивает в углублении почвы,
под укрытием из нависшей травы, в основании куста и в других защищен-
ных местах. Гнездо строится из сухих растений, пбднятых над неглубокой
ямкой в виде свода, с выстилкой пола из мелкой сухой травы. Залегает
26. РОД ERINACEUS 167
в спячку в октябре—ноябре. На зиму устраивает наземное гнездо, выби-
рая для этого укромное место под корнями деревьев, в пространстве под
пнем, расположенном в укрытом месте, под кучами валежника и др.
Гнездо крупное, строится из сухой травы, листьев и разнообразных мяг-
ких растительных остатков. Отмечены случаи, когда еж для устройства
зимнего гнезда выкапывал нору на глубину в 40 см, с длиной хода до
70—80 см. Весной првбуждается от зимней спячки в разное время, что свя-
зано с различием климата в отдельных частях его ареала и погодными ус-
ловиями в разные годы. Спаривание начинается вскоре после пробуждения
от зимней спячки. В Закавказье это происходит в марте. Отмечены драки
между самцами из-за самок. В течение года у ежа бывает один помет.
Период размножения может быть растянут с весны до осени. Возможно,
что на юге ареала их бывает два. Продолжительность беременности около
7 недель. Следовательно, второй помет может быть, но при очень благо-
приятных условиях. Количество детенышей в помете 3—6, реже бывает
8—9. Половозрелость наступает на втором году жизни.
Из врагов ежа главнейшими считаются лисица и филин, отмечается и
барсук.
Подвиды. I. Е. е. europaeus L., 1758. Отличается преобладанием
серой окраски в волосяном покрове. Кондило-базальная длина черепа —
50.0—58.5 мм. Распространение: северные и северо-западные области
европейской части СССР, включая Эстонию, Латвию и Литву, северная и
западная часть Польши, северная и западная части Чехословакии, ГДР,
ФРГ, Франция, Италия, Испания, острова Средиземного моря, на севере
Ирландия, Англия, Швеция, Норвегия и Финляндия. Не ясно взаимо-
отношение этого подвида и Е. е. concolor Martin на севере Италии, в Австрии,
Швейцарии и Венгрии.
2. Е. е. concolor Martin, 1837. Характеризуется темно-бурой до почти
черной окраской волосяного покрова. Кондило-базальная длина черепа —
56.0—62.0 мм. Распространение: к югу от северных границ Белоруссии,
Костромской и Кировской областей до Крыма, Кавказа, Ирана и Турции.
Западная часть ареала охватывает южную и восточную Польшу, юго-
восточную часть Чехословакии, Румынию, Югославию и Болгарию.
На востоке распространен на Южном Урале и в северо-западном Казах-
стане.
3. Е. е. pallidus Ognev, 1928. Волосяной покров светлый, преобладает
песчано-серая или охристо-беловатая окраска. Кондило-базальная длина
черепа — 55.0—56.0 мм. Распространение: Южная часть Западной Сибири
к востоку до бассейна р. Оби.
4. Е. е. amurensis Schrenk, 1858. Отличается очень светлой окраской
иглистого панциря, в котором до ЗО’/о игл без темных поясков (беловатых).
Окраска волосяного покрова варьирует от светло-серой до желто-серой и
буровато-серой. Кондило-базальная длина черепа — 54.0—59.0 мм.
Окраска волосяного покрова молодых (1/3—г/2 длины взрослого)
резко отличается от таковой взрослых темно-бурым цветом и наличием
белого пятна на горле, груди и брюхе и совершенно совпадает с таковой
Е. е. concolor. Распространение: Приморский край и южные районы Амур-
ской обл.
3. Erinaceus s. str. frontalis Smith, 1831 — Африканский еж (рис. 906).
Smith, 1831, South-African Quarterly J., v. 1, N 5, p. 10; — capensis Smith,
1831, Philos Mag., v. 9, p. 61 (nomen nudum); — albiventris Wagner, 1841, Suppl.
Schreb. Saugeth., Bd 2, S. 22; — pruneri W a g n e r, 1841, там же, с. 23; — heterodac-
tylus Sundevall, 1841, Kongl. Swenska Vetensk.-Akad. Handl., Stockholm,
S. 227; — algirus Duvernoy et Lere boullet, 1842, Mem. Soc. Sci. Nat.
168
ПОДСЕМ. ERINACEINAE
Strasbourg, v. 3, N 2, p. 4; — dimidiatus Fitzinger, .1867, Sitzungsb. K. Akad.
WiSs. Wien, v. 56, Sect; 1, p. 853; — krugi Peters, 1877, Sitzungsb. Gesellsch. na-
turf. Ereunde, Berlin, S. 78; — fractilis Peters, 1877, там же, с. 79; — f allax Dob-
son, 1882, Monograph Insectivora, system a. anatomie, part. 1, p. 9; — diadematus
Dobson, 1882, там же, с. 10', — algirus vagans Thomas, 1901, Proc. ZooL Soc.,
v. 1, p. 38; — hindei Thomas, Wroughton, 1907, Ann. Mag. Nat. Hist., ser. 7,
v. 19, p. 370; — spiculus Thomas, Wroughton, 1907, там же, с. 371; — al-
girus cuniculus Thomas, 1915, Ann. Mag. Nat. Hist., v. 16, p. 152; — angolae Tho-
mas, 1918, p. 230; — spinifex Thomas, 1918, p. 231; — kilimanus Thomas, 1918,
p. 232; — jaradjius J. Allen,’ 1922, p. 13—14; langi J. A llen, 1922, p. 15—17; —
algirus lavandeni Cabrera, 1928, BoL Soc. Esp. Hist. Nat., t. 28, p. 454; — (Atelerix)
pruneri mreni Setzer, 1953, J. Washingt. Acad. Sci., v. 43, N 7, p. 237. Atelerix pru-
neri lovei Setzer, 1956, Proc. U. S. Nat. Mur., v. 106, N 3377, p. 453.
Тип. He установлен. Вид описан из Южной Африки.
Д и а г н о з. Костное нёбо в виде пластинки заходит за небный
поперечный гребень. I3 и клык с двумя корнями.
Описание. Общий тип окраски иглистого панциря светлый,
охристый или охристо-серый, у отдельных экземпляров серый или бурый.
На морде темный рисунок, характерный для представителей рода Erina-
ceus L. Нижняя сторона тела серо-буроватая с белым или беловатым пятном
на брюхе, охристо-серая или белая. Пояски на иглах темно- или светло-
бурые. Верхние стороны кистей и ступней бурого цвета разных оттенков.
Кисти и ступни крупные, что особенно заметно при сравнении с другими
видами рода. Первый палец ступни короткий, у отдельных особей одного
из подвидов может отсутствовать.
Череп сходен по внешнему виду с таковым Е. europaeus L., но с более
широким и массивным лицевым отделом. Лобные кости впереди вздуты.
Скуловые дуги сужены в заднем отделе. Сочленовная поверхность для
сочленения с нижней челюстью на скуловом отростке височной кости
удлиненной формы. Нижняя челюсть с относительно низкой зубной частью.
Зубы крупные, массивные.
Измерения. Длина: тела — 168—230, хвоста — 19.0—50.0,
ступни — 25.0—34.0; высота уха — 20.0—30.0; кондило-базальная чере-
па — 42.0—50.5 мм.
Распространение и возраст. От Северной до Южной
Африки, Канарские острова, юго-восточная Франция, Испания, Балеарские
острова, плейстоцен—современный.
Подвиды. Не изучены.
Примечание. В связи с тем что у меня был только 1 экз. этого
вида, притом молодой особи, пришлось описание его окраски и измерения
дополнить из описаний Аллена (Allen, 1922) и Робертса (Roberts, 1954).
3. Подрод HEMIECHINUS Fitzinger, 1866 — УШАСТЫЕ ЕЖИ
Ericius Sundeva 1, 1842, Kongl. Svenska Vetensk. Akad. Handl., S. 225—227.
Hemiechinus Fitzinger, 1866, Sitzungsber. Ost. Akad. Wis< Math.-naturwiss. KI.
Wien, Bd 1, S. 565. Erinaceulus Огнев, 1928, c. 168. Mesechinus Огнев, 1951,
Бюл. МОИП, Нов. сер., отд. биол., т. LVI, вып. 1, с. 8—14.
Тип. Hemtechinus auritus G m е 1 i n, 1770, юго-восточная Европа, Азия, Се-
верная Африка, плейстоцен—современный.
Диагноз. Иглистый панцирь не спускается на бока тела. Барабан-
ные отростки основной клиновидной кости образуют внутреннюю и верх-
нюю стенки слухового барабана. Засочленовный отросток шире сосцевид-
ного, с глубоким вдавлением.
Описание. Уши большие, прижатые к голове заходят за глаза
(исключение Erinaceus {Hemiechinus) dauricus Sundev., у которого они
небольшие). Волосяной покров мягкий или жесткий. На голове нет по-
лоски голой кожи, разделяющей иглы на две группы. Иглы с заметно
26. РОД ERINACEUS
169
выраженными бороздками, с бороздками и бугорками на валиках между
бороздками, или гладкие, без всех отмеченных образований.
Череп варьирует в размерах, относительно широкий. Скуловые дугй
широко расставлены. Носовой отдел черепа заметно суживается к перед-
нему носовому отверстию, низкий, что хорошо заметно при взгляде на
череп сбоку. Предглазничный гребень хорошо развит, как и V-образный
гребень на лобных костях. Вдавление в области лобно-носового шва за-
метно выражено, так же и межглазничное сужение. Сагиттальный гребень
невысокий. Ламбдовидный гребень хорошо развит. Затылочное отверстие
более развито в ширину, эллипсовидной формы. Крыловидные кости не
вздуты, не сообщаются со слуховой 'полостью. С последней сообщаются
вздутые крыловидные отростки основной клиновидной кости, образую-
щие полость. I3 и С всегда с двумя корнями.
Распространение и возраст. Юго-восточная Европа,
Передняя, Средняя Азия, Забайкалье, Центральная и Южная Азия,
Индия, Северная Африка, плейстоцен-современный.
4. Erinaceus (Hemiechinus) auritus Gmelin, 1770 — Ушастый еж
(рис. 93, 94).
Erinaceus auritus Gmelin, 1770, Nov. Comment. Acad. Sci. Petrop., t. 14,
p. 519; — aegyptius Fischer, 1829, Syn. Mamm., p. 262; — collaris Gray, 1830,
Illustr. Indian Zool., v. 1, pl. 8; — spatangus Bennett, 1832, Proc. Zool. Soc. Lon-
don, p. 123; — grayi Bennet, 1832, там же, с. 124; — indicus Boyle, 1833, Il-
lustr. Bot. Himalaija, p. 6; — libicus Ehrenberg, Hemprich, Ehren-
berg, 1833, Symb. Phys. Mamm., dec. 2; — auritus caspicus Sundevall, 1842,
Kongl. Svenska Vetensk. Akad. Handl., p. 237; — (Ericius) platyotis Sundevall,
1842, там же, с. 232; — megalotis В 1 у t h, 1845, J. Asiat. Soc. Bengal, v. 14, p. 353; —
(Hemiechinus) albulus Stoliczka, 1872, там же, т. 41, № 2, с. 226; — syriacus
Wood, 1876, Bible Animals, p. 83; — calligoni С а т у н и н, 1901. Прилож. к про-
ток. Об-ва Естеств. при Казанск. унив., № 191, с. 1—2; — albulus turanicus С а т у-
н и н, 1905, Зап. Кавк. отд. Имп. геогр. Об-ва (nomen nudum); 1905—1906, Извест.
Кавк. Муз., т. 2, с. 45, 46. Hemiechinus albulus minor С а т у и и и, 1906, Ann. Mus.
Zool. 1'Acad. St. Petersb., v. 11, p. 180; — albulus alaschanicus", С а т у н и и, 1906,
там же, с. 181; — persicus С а т у в и н, 1906, там же, с. 184; — russowi С а т у и и н,
1906, там же, с. 177; — holdereri Matschie, 1922, Sitzungsb. Ges. Naturf. Freunde,
Berlin, S. 73; — microtis Лаптев, 1925, Бюл. Средне-Азиат. унив., № 8, с. 66; —
chorassanicus Лаптев, 1926, там же, № 13, с. 115, 116; — brachyotis Огнев, 1928,
с. 130—132; — albulus major Огнев, 1928, с. 135—138; — homalocanthus Стро-
ганов, 1944, ДАН СССР, т. 44,№ 3, с. 120.
Тип. Не установлен, вид описан с низовьев р. Волги.
Диагноз. Окраска мягкого волосяного покрова светлая. От светло-
бурого до чисто белого цветов. Ухо, отогнутое вперед, заходит вершиной
за глаз. Засочленовный отросток широкий, с глубокой впадиной. Барабан-
ные отростки основной клиновидной кости крупные, подняты вверх.
Описание. Окраска волосяного и иглистого покровов варьирует
в значительных пределах. Голова всегда окрашена темнее боков и нижней
стороны тела, даже в том случае, когда все части тела имеют темную
окраску преобладающей. Темный цвет головы чаще всего обусловлен
примесью бурых волос, разных оттенков, к более светлым или белым
волосам. От их количества в примеси зависит степень потемнения головы.
Реже вся голова равномерно покрыта темными, охристо-бурыми волосами.
Часто голова имеет двуцветную окраску: от глаз к ушам более светлую,
а от глаз к носу — темную. В такой же мере часто темные волосы на голове
образуют своеобразный рисунок, как и у других видов ежей. У основания
ушей всегда в разной степени развитые кисточки из светлых или белых
волос, переходящие и на передний край уха. Мочка носа черная или черно-
бурая. Вибрисы бурые или смешанной окраски. Большая часть поверх-
ности ушей покрыта короткими, светлыми, охристыми или белыми, реже
170
ПОДСЕМ. ERINACEINAE
буроватыми волосами. Волосяной покров тела от • белого до бурого со
всеми степенями переходов одного цвета в другой, что обусловлено раз-
ным количеством темных или светлых волос в примеси. Буроватая окраска
волос бывает разной интенсивности. На теле темная окраска развивается
чаще всего на боках, задней части, на бедрах и плечах.
Окраска этого вида представляет довольно пеструю картину, если ее
рассматривать на всем ареале. Так, экземпляры из Ростовской, Волго-
градской и других областей и частей европейской территории СССР —
темные, сероватые, бурые с темной головой, иглы с сероватыми концами.
Юго-восточное Предкавказье, Закавказье, Центральный и Западный Иран,
Переднюю Азию и Северную Африку населяют светлые особи, но встреча-
ются и особи с темным оттенком волосяного покрова. Экземпляры из
Рис. 93. Erinaceus auritus (по: Walker et al., 1964).
Туркмении имеют общую буроватую окраску что объясняется кремовым
и охристым оттенком в окраске светлых частей игл и волосяного покрова,
но встречаются и светлые экземпляры. Экземпляры из Кушки имеют
охристой окраски иглистый покров, волосяной покров головы бурый,
а на остальных частях тела, кроме кистей и ступней, белый. Сходно окра-
шены экземпляры из Тахта-Базара, Кизыл-Имама. Экземпляры из Тер-
меза, Репетека и Бухары имеют темную и светлую окраску иглистого
и волосяного покрова. Ежи северной части Казахстана более темные,
чем из южной его части, а в восточной части не светлее, чем в западной.
Все монгольские экземпляры (11 шт.) довольно однотипны, светлые сверху
(концы игл беловатые) и белые снизу. Темные только конец морды и верх-
ние поверхности кистей и ступней — буровато-серые. Близки к мон-
гольским по окраске и экземпляры из Китая. В северной части Таджики-
стана преобладают светлоокрашенные особи, а в южном темные. Еще более
темные экземпляры преобладают в Афганистане и Индии. Темные экзем-
пляры попадаются и в восточной части Ирана. Если проанализировать
общую тенденцию в особенностях окраски этого вида по отдельным зонам,
то можно отметить следующее: северная полоса ареала занята особями
с преобладанием серого или буровато-серого цвета, средняя зона занята
особями с преобладанием белого или охристо-белого цвета. Эта зона про-
стирается далеко на восток, а на западе переходит в северную часть
Африки. Третья зона населена преимущественно темно-бурыми особями
Эта зона включает юго-восточную Туркмению, восточную часть Ирана,
южную часть Таджикистана, Афганистана и Индию.
26. РОД ERINACEUS
171
Строение игл и их окраска не отличаются постоянством и не могут счи-
таться характерным признаком не только вида, но и подвида. Бороздки
и бугорки на ребрах, разделяющих бороздки, заметно выражены у моло-
дых экземпляров. С возрастом они становятся малозаметными или утрачи-
ваются. Кроме того, у одного и того же экземпляра иглы могут быть и
Рис. 94. Erinaceus auritus.
с бороздками и без них. Иглы, как правило, имеют один темный поясок
вблизи вершины. Реже бывают два пояска и очень редко три. Вершина
иглы может быть темной и светлой.
Череп описываемого вида у особей из различных частей ареала имеет
сходные структурные особенности. В коллекции ЗИН имеется три черепа
из оазиса Ния — Восточный Тибет (сборы Певцова), у которых нижние
клыки с двумя корнями, что не встречается не только у современных,
но и у многих ископаемых ежей. Пропорции отдельных частей и кондило-
базальная длина черепа весьма изменчивы.
172
ПОДСЕМ. ERINACEINAE
Основные особенности черепа Е. auritus выражаются в следующем:
носовой отдел сужен и уплощен. Костная стенка подглазничного отверстия
не отогнута кнаружи в виде чешуйки. Слезное отверстие мелкое. Скуловая
дуга в задней части полого поднимается вверх. Засочленовный отросток
широкий, с глубокой выемкой. Слуховые пузыри крупные, вздутые.
Сочленовный отросток нижней челюсти расположен ближе к венечному
отростку.
Измерения. Длина: тела — 142—300, хвоста — 17.0—31.2,
ступни — 29—43; высота уха — 29—51; кондило-базальная длина черепа
41.2—60.0 мм.
Распространение и возраст. Обитает в границах,
указанных для подрода. В СССР от западных границ Ростовской обл.,
восточных частей Предкавказья и Закавказья на западе, через всю Сред-
нюю Азию до Алтая и Тувинской АССР на востоке. Известен с плейсто-
цена.
Биология. Ушастый еж населяет открытые пространства —
степи, полупустыни и пустыни. При этом в каждой из названных зон
он встречается в разнообразных растительных формациях. В полосе
степей избегает участков с густым травостоем и распаханных земель.
В горы поднимается до высоты 2400 м. Пищу этого вида составляют бес-
позвоночные, особенно насекомые, и мелкие позвоночные животные яще-
рицы, мелкие грызуны, а также генеративные органы растений. Состав
кормов в пределах ареала определяется видовым составом фауны той или
иной географической области, а также плотностью поселений кормовых
животных, колебанием их численности и сменой видов беспозвоночных
по сезонам. Таким образом, видовой состав кормовых животных ушастого
ежа представляет довольно пеструю картину. Гнездо ушастого ежа
помещается, как правило, в норе. Нору еж выкапывает сам или при-
спосабливает для жилья чужие заброшенные норы. В зависимости от
механического состава грунта глубина норы бывает от 1 до 1.5 м. Широкое
и низкое входное отверстие ведет в круто спускающийся ход. Гнездовая
камера помещается в боковом отнорке, расположенном недалеко от вход-
ного отверстия. С целью сохранения потомства ежей этого вида в неволе
А. И. Колоденко (1969) специально исследовал строение выводковых нор
ушастого ежа. Устройство помещений, имитирующих форму норы в естест-
венных условиях, позволило названному автору сохранять ежей, рождав-
шихся в условиях неволи.
По данным Колоденко, ушастые ежи в Туркмении размножаются
дважды в году. Первое спаривание происходит во второй половине марта.
Молодые разного возраста и веса встречаются с апреля по ноябрь включи-
тельно. Такая растянутость размножения объясняется разновременным
половым созреванием ежей, родившихся в разные месяцы предыдущего
года. Этик) же автором установлено, что половое созревание ежей наступает
в возрасте 230—240 дней. Число детенышей у самок, рожавших в неволе,
4—5, а в естественных условиях их бывает от 3 до 7.
Врагами ежа считается лисица, волк, бродячие собаки; из птиц — филин
коршун, болотный лунь и др.
Подвиды. 1. Е. a. auritus Gmelin, 1770. Размеры относительно
мелкие. Кандило-базальная длина черепа до 50 мм. К белой окраске
волосяного покрова в значительной степени подмешаны охристо-серые
и охристо-буроватые тона. Иглистый покров с сероватым оттенком. Рас-
пространение: северная часть ареала (юго-восточные области европейской
части СССР, Северный Казахстан, южная часть Западной Сибири, север-
ная часть Алтая).
26. РОД ERINACEUS
173
2. Е. a. aegyptius Fischer von Waldheim, 1829. Мелкая форма. Кондило-
базальная длина черепа до 46 мм. Окраска волосяного покрова белая,
только на голове примесь буроватых волос. Распространение: восточная
часть Предкавказья, Закавказье, Западный Иран, Передняя Азия, Малая
Азия, Северная Африка.
3. Е. a. turanicus Sat., 1905. Средних размеров, сильно варьирующая
форма. Кондило-базальная длина черепа до 55 мм. Окраска волосяного
покрова белая, белая с примесью бурых волос или местами светло-бурая
(на боках и задней части тела). Распространение: от восточного берега
Каспийского моря через всю Среднюю Азию и большую часть ареала
в Центральной Азии, включая Монголию и Туву.
4. Е. a. megalotis Blyth. 1845. Размеры крупные, кондило-базальная
длина черепа до 60 мм. Окраска волосяного покрова темно-бурая или бу-
рая, светлеющая на западе ареала. Распространение: южный Гиндукуш,
Афганистан, Южный Таджикистан, Восточный Иран, Туркмения (Копет-
даг, юго-западные районы).
5. Е. a. collaris Gray, 1830. Мелкий подвид. Кондило-базальная длина
черепа до 50 мм. Окраска волосяного покрова темно-бурая. Бурый от-
тенок имеет и иглистый покров. Распространение: Северо-западная Индия,
западная часть Пакистана. Западная граница не ясна.
6. Е. a. albulus Stoliczka, 1872. Мелкая форма. Столичка отмечает, что
единственный измеренный им экземпляр имел длину тела — 7 дюймов
(175 мм), а высота его уха достигала одного дюйма (25 мм).
Краткая характеристика этого подвида из описания названного автора.
Морда очень длинная. Уши умеренной величины, овальные на вершине.
На верхней стороне тела преобладает белый цвет. Голова рыжеватая сверху
и с боков, снизу белая. Основания ушей белые. Нижняя часть тела белая.
Уши, нижняя часть лба, задние ноги и хвост темно-бурые. К сожалению,
автор не приводит описание черепа. О местонахождении экземпляра,
послужившего для описания, Столичка указывает следующее: Гендерсон
(Henderson) добыл этого ежа в Langur near Sanju (Яркенд, Китай).
Сходно с описанным Столичкой экземпляром окрашены экземпляры
из оазиса Ния, что примерно в 150 км к западу от г. Карасая, добытые
Певцовым и экземпляр с р. Хотана, пересекающей пустыню Такла-Макан,
из сборов Б. Л. Громбчевского (кол. ЗИН АН СССР).
Черепа этцх экземпляров отличаются от черепов других подвидов
Е. auritus Gmelin менее] угловатой и менее отведенной в сторону скуловой
дугой, менее развитым барабанным отростком основной клиновидной
кости. Кроме того, нижний клык у описываемых черепов имеет два корня
и отличается формой коронки. Два корня у нижнего клыка сохранились
только у гимнур. Недостаток материала не позволяет в настоящее время
решить вопрос о таксономическом ранге описываемой здесь, вероятно,
реликтовой формы.
Можно думать, что ареал Е. a. albulus ограничен территорией йустыни
Такла-Макан и прилежащих к ней пространств, окруженных горными
системами.
Попутно следует отметить, что из яркенда Гюнтер (Gunther, 1875)
списал реликтового зайца — Caprolagus yarkandensis, добытого затем
Пржевальским в системе р. Тарима, на оз. Лобнор и на р. Хотане. Воз-
можно, что эта территория хранит древние виды.
Примечание. Нитхамер (Niethamer, 1969), выясняя системати-
ческое положение афганского ежа — Hemiechinus megalotis Blyth, пришел
к заключению, что он, как и другие виды рода — Hemiechinus (подрод
в моем понимании), описанные разными авторами в разное время, пред-
174
ПОДСЕМ. ERINACEINAE
ставляет собой подвид Н. auritus Gmelin. При этом он сохраняет в составе
вида следующие подвиды: 1) Н. a. aegyptius Fischer, 1829; 2) Н. a. auritus
Gmelin; 3) Н. a. albulus Stoliczka, 1872 (в старом значении среднеазиатского
подвида); 4) Н. a. chorosanicus Laptev, 1926; 5) Н. a. megalotis Blyth,
1845; 6) Н. a. collaris Gray, 1830.
Описанного Строгановым в 1944 г. Н. homalocanthus он не упоминает.
Очевидно, этот вид остался ему неизвестным.
Рис. 95. Erinaceus dauricus.
5. Erinaceus (Hemiechinus) dauricus Sundevall, 1841 — Даурский еж
(рис. 95).
Erinaceus auritus Pallas, 1811, Zoographia Rosso-Asiatica, t. 1, p. 138—139; —•
dauricus Sundevall, 1841, Kongl. Vetensk. Acad. Handl., Stockholm, p. 237; —
europaeus R a d d e, 1862, Reisen im Suden von Ost-Sibirien, Bd 1, S. 117—124. Hemie-
chinus przewalskii С а т у н и н, 1906. Ежегодн. Зоол. муз. Акад [наук, т. XI, с. 181; —
(?) dauricus С а т у н и н, 1906, там же, с. 185; — dauricus С а т у н и н, 1914, Опре-
делитель млекопитающих Российской империи Тифлис, с. 54. Erinaceus (Mesechinus')
dauricus Sundevall, по: Огнев, 1951, с. 8—14.
Тип. Не установлен, вид описан из Даурии•
26. РОД ERINACEUS
175
Диагноз. Окраска грубого волосяного покрова темная, бурая.
Ухо отогнутое вперед не заходит за глаз. Засочленовный отросток слабо
расширен, с неглубокой впадиной. Барабанные отростки основной клино-
видной кости небольшие, не подняты вверх, а полого направлены в сто-
роны.
Описание. Общий тон окраски розовато-серый, однообразный.
На всей верхней стороне тела. Только на голове и особенно на верхней
стороне кистей и ступней окраска более темная, бурая. Нижняя сторона
тела буровато-белесая с серебристым налетом. Несколько серее и светлее
окрашены 2 экз. ежей этого вида, добытые в Хо-Ней, полученные из Китая
в 1956 г. На голове нет полоски голой кожи, разделяющей иглы на два
пучка.
Череп мало сужен в носовой части и слабо уплощен. Костная стенка
подглазничного отверстия отогнута кнаружи. Слезное отверстие сравни-
тельно крупное. Скуловая дуга в задней части круто поднята вверх.
Слуховые капсулы небольшие, уплощенные. Сочленовный отросток нижней
челюсти расположен посредине между височным и угловым отростками
или ближе к угловому отростку.
Измерения. Длина: тела — 233—270, хвоста — 24—38, ступни—
33—45, уха — 29—34; кондило-базальная черепа —50.0—57.5 мм.
Распространение и возраст. Недостаточно изучены.
Он обитает в ряде районов южного и юго-восточного Забайкалья, в северо-
западной Монголии и в Северо-Восточном Китае, современный.
Биология. Образ жизни этого вида во многом напоминает образ
жизни других ежей. Это типичный степной вид, но не избегающий участков
с редким древостоем (березово-осиновые колки). В степи занимает разно-
образные участки, предпочитая места с хорошими ремизными условиями:
заросли кустарников и высоких трав под пологом ивняков и черемухи.
В горы поднимается до высоты 1000 м. Питается, как и другие ежи, разно-
образными беспозвоночными и позвоночными. Из беспозвоночных основ-
ной корм составляют многочисленные в степях прямокрылые и крупные
виды жуков. Из позвоночных питается грызунами, ящерицами, змеями
и бесхвостыми амфибиями. Поедает ягоды степных растений.
Жилища устраивает в заброшенных норах даурских сусликов и сурков,
приспосабливая их и для зимовки. О размножении имеются скудные дан-
ные. Молодые появляются в июне—июле. Число детенышей 1—7. Даурс-
кий еж не строго ночное животное, кормиться выходит еще до наступления
темноты.
Подвиды 1. Е. (Hemiechinus) d. dauricus Sundevall, 1841. Относи-
тельно крупный, темноокрашенный еж. Распространение: южная и юго-
восточная части Забайкалья. Северо-западная часть Монголии.
2. Е. (Н.) d. mandschuricus Mori, 1927. Отличается от номинальной фор-
мы меньшими размерами и более светлой окраской. Распространение:
Северо-Восточный Китай (его западные пустынные районы).
Примечание. Систематическое положение этого вида длительное
время считалось неопределенным. Одни авторы относили его к роду
Erinaceus L., другие — к роду Hemiechinus Fitz. Изучение мной черепов
и шкур этого вида позволило установить его несомненную близость
к представителям подрода Hemiechinus. У этого вида на голове нет полоски
голой кожи, разделяющей иглы на два пучка. I3 и С с двумя корнями.
Сравнительно широкий засочленовный отросток. Отличается он от видов
подрода Hemiechinus небольшим размером ушей, жестким волосяным
покровом, а также меньшим развитием барабанных отростков основной
клиновидной кости. Отмеченные отличительные признаки могут быть
объяснены приспособлением этого вида к более суровым условиям кли-
176
ПОДСЕМ. ERINACEINAE
мата в пределах его ареала, а также обстановкой, отличной от таковой
общего ареала подрода. Hemiechinus, приведших к утрате особенностей,
характерных для пустынных видов.
4. Подрод PARAECHINUS Trouessart, 1879
Trouessart, 1879, Revue et Magaz. de Zool., Paris, ser. 3, t. 7, p. 242. Macroe-
chinus S a t u n i n, 1906, Annuaire du Musee Zool. de I’Academie Imper. de Sciences de
St.-Petersb., t. 11, p. 23.
Тип. Erinaceus micropus Blyth, 1846, Индия, современный.
Диагноз. Иглистый панцирь не спускается на бока тела. Барабан-
ные отростки основной клиновидной кости образуют внутреннюю, верх-
нюю и наружную стенки слухового барабанад Засочленовный отросток
шире сосцевидного, с глубоким вдавлением. В отличие от других подродов
рода Erinaceus L. у представителей этого подрода крыловидная кость
вздута и сообщается со слуховой полостью.
Описание. Уши крупные, прижатые к голове, заходят за глаза.
Волосяной покров относительно мягкий. На голове полоска голой кожи
разделяет тонкие и длинные иглы на две группы. Иглы всегда с хорошо
выраженными бороздками и бугорками на валиках между ними.
Череп варьирует в размерах. Относительно узкий. Скулы расставлены
менее широко, чем у других подродов. Носовой отдел черепа сужен,
низкий. Предглазничный гребень хорошо развит, как и V-образный
гребень на лобных костях. Вдавление в области лобно-носового шва
хорошо заметно. Межглазничное сужение резко выражено. Сагиттальный
гребень невысокий, ламбдовидный гребень хорошо развит. Затылочное
отверстие более развито в ширину, эллипсовидной формы. Крыловидные
кости вздуты и через вздутые крыловидные отростки основной клино-
видной кости сообщаются со слуховой полостью. I3 и С всегда с двумя
корнями.
Распространение и возраст. Северная Африка, Север-
ная Аравия, восточная часть Ирана, Средняя Азия, Афганистан, северо-
западная, западная, южная и юго-восточная части Индии.
6. Erinaceus (Paraechinus) aethyopicus Ehrenberg, 1833 — Длинноиг-
лый еж (рис. 96, 97). р -j Л (и е <( ’ •
Erinaceus aethyopicu Ehrenberg (in: Hemprich, Ehrenberg),
1833, Symb. Phys. Mamm., dec. 2; —hypomelas Brandt, 1836, Bull. Sci. St.-Petersb.,
т. 1, p.32; — sennaarensis Hedenborg, 1839, Isis, N 32, p. 8 (nomen nudum); —
brachydactylusVv a g n e r, 1840, Schreber Saugeth., suppl. 2, p. 24; — micropus Blyth,
1846, J. Asiat. Soc. Bengal, v. 15, p. 170; —nudiventris Horsfield, 1851, Cat.
Mamm. Mus. E. India Co., p. 136; — deserti L о c h e, 1858, Cat. Mamm. Oiseaux Al-
gerie, p. 20. Hemiechinus pectoralis Heugliu, 1861, Nov. Acta Phys. Med. Acad.
Caes. Leop. Carol., v. 29, p. 22; — pallidus Fitzinger, 1867, Sitzungsber. Akad.
Wiss. Wien, Bd 56, N 1, S. 866; — mentalis Fi tzinger, 1867, c. 874. Erinaceus
(Hemiechinus) pictus Stoliczka, 1872, J. Asiat, Soc. Bengal, v. 41, N 2, p. 223.
Erinaceus macracantus Blanford, 1875, Ann. Mag. Nat. Hist., v. 16, p. 310; —
blanfordii Anderson, 1878, J. Asiat. Soc. Bengal, v. 47, N 2, p. 208; — jerdoni A n-
d e r s о n, 1878, там же, с. 209; — niger В 1 a n f о r d, 1878, там же, с. 212; — dor-
salis Anderson, de Winton, 1901, Ann. Mag. Nat. Hist., v. 7, p. 42. Paraechi-
nus amir Thomas, 1918, p. 232; — ludlowi Thomas, 1919, J. Bombay, Nat. Hist.
Soc., v. 26, p. 748; — deserti blancalis Thomas, 1921, Ann. Mag. Nat. Hist., v. 8,
p. 570; — niger seniculus Thomas, 1922, там же, с. 142; — niger sabaeus Thomas,
1922, там же, с. 143; — dorsalis albatus Thomas, 1922, там же, т. 9, с. 144; — anis-
cus Thomas, 1922, там же, т. 10, с. 307; — hypomelas eoersmannii Ognev, 1927,
Zool. Anz., Bd 69, S. 218; — dorsalis albior P о с о c k, 1934, Ann. Mag. Nat. Hist.,
v. 14, p. 636.
T и п. He установлен, вид описан из пустыни Дагола, Судан.
Диагноз. Совпадает с таковым подрода. В качестве дополнения
следует отметить, что у Е. (Paraechinus) aethyopicus иглы длинные и толстые.
t
26. РОД ERINACEUS
177
На голове имеется полоска голой кожи, разделяющая иглы на два пучка.
Описание. Крупный еж, покрытый длинными толстыми иглами,
с крупными ушами, сравнительно высоконогий. Голова, бока и нижняя
часть тела покрыты мягкими волосами. У светлых экземпляров окраска
волосяного покрова белая и только на голове имеются темные пятна.
У темных особей примесь черных волос вызывает потемнение окраски
на боках, конечностях и голове. Местами черные волосы почти полностью
вытесняют белые и окраска становится черной. Окраска темных поясков
и концов игл черно-бурая или черная у темноокрашенных особей придает
их иглистому покрову темную окраску.
Череп варьирует в размерах. Он сравнительно узкий, особенно в носо-
вой части и вдавлен в области лобно-носового шва. Впереди от орбит оц
Рис. 96. Erinaceus aethyopicus (по: Walker et al., 1964).
ниже, чем в затылочной части. Межглазничное сужение хорошо выражено.
Барабанные отростки основной клиновидной кости образуют внутреннюю
верхнюю и отчасти наружную стенку слуховой капсулы. I3 и С с двумя
корнями.
Измерения. Длина: тела — 140—272, хвоста — 10—42; ступни—
35.9—40.0, уха — 44—57, кондило-базальная черепа — 45.8—60.1 мм.
Распространение и возраст. Северная Африка, Аравий-
ский полуостров, Малая Азия, Иран, Афганистан, Индия. В СССР п-ова
Мангышлак, Бузачи, плато Устюрт и прилежащие к ним участки пустыни
(Казахстан), в Туркмении и Узбекистане к северу до Самаркандской обл.,
современный.
Биология. Обитает в глинистых и песчаных пустынях, пред-
горьях и горных местностях, поднимаясь в горы до зоны арчи. Встреча-
ется и в полосе культурных земель, например, в садах. Ведет ночной образ
жизни. День проводит в норах, достигающих, как указывают некоторые
авторы, 1 м в длину. Основную пищу составляют насекомые и другие
беспозвоночные, а также мелкие позвоночные. По имеющимся данным,
размножение этого вида происходит в мае—июне. Детенышей бывает
от 3 до 6. Интересные данные о развитии ежат в постэмбриональный период
приводит П. М. Ланкин (1967).' Ему удалось поймать двух беременных
самок с шестью эмбрионами каждая. Одна добыта 28 мая 1965 г. у сква-
жины Кыск-Кудук, другая в первых числах июня на п-ове Бузачи
в районе поселка Акшимрад. Обе самки содержались в неволе и принесли
помет. Судя по описанию, молодые ежата этого вида рождаются со свет-
лыми мягкими иглами слепые, с крупным носом и большими, чем задние,
передними ногами. Молодые быстро растут и прибавляют в весе.
12 А- А. Гуреев
178
ПОДСЕМ. ERINACEINAE
К сорокадневному возрасту они все же не достигают, как пишет автор,
размеров взрослого ушастого ежа.
Подвиды 1. Erinaceus (Paraechinus) aethyopicus aethyopicus Ehren-
berg, 1833. Голова впереди глаз буровато-коричневая, лоб, нижняя губа
и нижняя сторона тела белые. Конечности окрашены в буровато-коричне-
Рис. 97. Erinaceus aethyopicus.
вый цвет. Р3 с одним корнем. Распространение: Марокко, Алжир, Тунис,
Египет, Судан, Аравийский полуостров, Аден, Ирак.
2. Е. (Р). a. hypomelas Brandt, 1836. Голова окрашена в светлые тона
с примесью темных волос, образующих местами темные пятна, на осталь-
ных частях тела волосяной покров белый (светлая форма). У темных осо-
бей примесь черных волос обусловливает темную окраску головы, боков
и конечностей, а также задней части тела. Р3 с двумя-тремя корнями.
Распространение: Малая Азия, северная часть Аравии, Иран, Афганистан,
Средняя Азия, северо-западная Индия. В СССР от п-овов Мангышлак
ВЫМЕРШИЕ ЕЖОВЫЕ С ТЕРРИТОРИИ СССР
179
и Бузачи, плато Устюрт и прилежащих к ним участков пустыни (Казах-
стан), в Туркмении и Узбекистане к северу до Самаркандской обл.
3. Е. (Р.) a. micropus Blyth, 1846. Передняя часть головы и полоски,
идущие от глаз назад, дистальные части передних ног, задняя часть тела
включая заднюю часть живота, хвост и задние ноги коричневого цвета.
На остальных частях тела волосяной покров белый. Р3 с одним или двумя
сросшимися корнями. Распространение: южная и юго-восточная, западная
и северо-западная части Индии.
Вымершие ежовые с территории СССР
На протяжении значительного отрезка времени, особенно в последние
годы, в различных местонахождениях на территории СССР вместе с остат-
ками разных групп млекопитающих попадались и остатки ежей (Erina-
ceinae). Большинство из них позднего времени (плиоцен, плейстоцен и еще
ближе к современному периоду). Остатки относятся к отдельным видам
рода Erinaceus L., населяющим различные ландшафты европейской и азиат-
ской частей СССР и в настоящее время, или близким к ним формам, таксо-
номический ранг которых установить пока невозможно вследствие фраг-
ментарности материала: отдельные зубы, фрагменты нижних челюстей
с альвеолами зубов и др. Эти материалы собраны Н. К. Верещагиным,
И. М. Громовым (кол. ЗИН АН СССР), В. С. Зажигиным (кол.
ГИН АН СССР), В. А. Топачевским (кол. Инет. зоол. АН УССР) и др.
Отмеченный материал позволил установить, что современные виды
ежей давно занимают присущие им ареалы и что более ранние их представи-
тели относились к современным скорее как подвиды, чем виды. Кроме того,
и на этом материале можно прийти к выводу, что в плиоцене, плейстоцене
и позднее в Западной Европе не могли одновременно существовать пять
описанных оттуда вымерших видов (см. с. 164) и два современных Erina-
ceus frontalis Smith и Е. europaeus L. Из последних ареал первого охва-
тывает всю Африку и только часть Западной Европы, а второго — уме-
менную зону Евразии. Трудно себе представить, чтобы на ограниченной
территории Западной Европы имело место такое разнообразие ландшаф-
тов, а в них столь различные условия, что неизбежно должны были сформи-
роваться в каждом из них отдельные виды. Напротив, поразительная
экологическая пластичность ежей позволяет предполагать замедленное
формообразование на всех этапах их эволюционного развития. Все извест-
ные роды ежей существовали и существуют в значительном отрезке времени,
образуя в каждый из геологических периодов ряд форм, существование
которых разграничено во времени, как например у рода Amphechinus
Aymard.
Большой интерес представляют остатки олигоценовых и миоценовых
ежовых, найденных в Зайсанской котловине (Казахстан) Н. С. Шевыре-
вой и В. М. Чхиквадзе. В олигоценовых отложениях обнаружены зубы
гимнуры, по общему характеру строения близкие к таковым Neurogymnurus
Filhol,.но отличающиеся большей шириной и более заостренными в виде
шипов бугорками, а также мелким в отличие от крупного у названного
рода Р4. На этом основании описан новый род Pseudoneuro gymmirus
с двумя видами, различающимися размерами и строением зубов.
Из этого же местонахождения и того же времени описаны без названия
три формы вымерших ежей рода Amphechinus. Степень их обособленности
друг от друга не вызывает сомнений, но отличие их от видов, описанных
ранее из других частей ареала рода, за отсутствием материала для сравне-
ния установить не удалось.
12*
180
СЕМ. TALPIDAE
Из миоцена Казахстана стала известной гимнура, близкая к Galerix
exilis Blainv., названная мной G. paraexilis, хорошо отличающаяся от
сравниваемого вида структурными особенностями зубов. Новая гимнура
описана и из миоцена Таджикистана (сборы В. С. Зажигина), названная
G. tadzhikistanicus.
В заключение следует отметить, что перечисленные данные о представи-
телях сем. Erinaceidae, найденных в палеогеновых и неогеновых отложениях
Казахстана и Таджикистана, позволяют составить представление о двух
главных группах ежовых, игравших, видимо, важную роль в соответствую-
щих биогеоценозах в. разное время. Гимнуры на территории азиатской
части СССР существовали возможно дольше, чем в Европе, Африке и
Северной Америке, где они вымерли в конце миоцена и медленно отступали
в юго-восточную часть Азии, где сохранились до настоящего времени.
Сем. TALPIDAE Fischer von Waldheim, 1817
Talpini Fischer von Waldheim, 1817, p. 368—428.
3__2 1 4_2 3
Диагноз. Зубная формула: I g—। C P g M у = 44—34. Длина
тела 60—230 мм. Глаза мелкие, малозаметные, часто закрыты кожной пере-
понкой. Ушные раковины рудиментарные в виде кожного валика, очень
редко они развиты, небольшие, выступают из волосяного покрова. Тело
покрыто густым бархатистым покровом.
Череп с удлиненным лицевым отделом. Скуловые дуги тонкие, не
выступают за боковые стороны черепа. I11 плоские, долотообразные или
трех-четырехгранные. М1 и М2 треугольные, с W-образной складкой на
жевательной поверхности, с тремя корнями.
Описание. Относительно мелкие и средних размеров для насекомо-
ядных — млекопитающие (длина тела — 60—230 мм). Подавляющее боль-
шинство видов семейства сходно по внешнему облику. Имеют массивное
тело на коротких ногах. Голова широкая в затылочной области, значи-
тельно сужена в лицевой части, заканчивается длинным хоботком. Шей-
ное сужение очень редко выражено. Ноздри направлены вперед или имеют
боковое расположение. Глаза мелкие, спрятаны в шерсти или закрыты
кожной перепонкой. Ушные раковины не развиты, реже в виде кожного
валика, ограничивающего сзади слуховое отверстие очень редко хорошо
развиты. У специализированных к рытью видов кисти широкие, с отороч-
кой из щетинистых волос, ладонями повернуты наружу. Пальцы соедине-
ны кожей до когтей. Когти широкие, плоские. У менее специализирован-
ных к рытью кисти удлиненные, пальцы свободные. У бегающих кротов
кисти обычные, когти сжатые с боков, напоминают таковые землероек.
Кроты, приспособленные к плаванью — выхухоли, имеют на кистях
и ступнях оторочку из жестких волос, пальцы соединены кожной перепон-
кой, более развитой между пальцами ступни. Ступня у роющих видов
узкая, пальцы свободные, когти сжатые с боков. Хвост очень короткий или
длинный, достигающий у наземных и плавающих видов длины тела.
При этом он у плавающих почти весь или только на конце уплощен.
Череп в типичном случае удлиненный, клиновидной формы. Его наи-
большая ширина в затылочной области менее длины в 2—2.5 раза. Кости
черепа рано сливаются, особенно межчелюстные и челюстные, а также
носовые и прилежащие к ним. Скуловые дуги образованы тонкими костями,
прижаты к черепу, реже они сравнительно толстые, выступающие в сто-
роны. Межтеменная кость широкая, у выхухолей может быть узкая,
в виде чешуйки, налегающей на соседние кости. Только у последних
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
181
хорошо, у других малозаметно развиты затылочный и сагиттальный
гребни. Продолжением окостеневающей носовой перегородки служит
особая предносовая косточка, поддерживающая хоботок. Нёбные кости
узкие, простираются до последних моляров или заходят за них. Меж-
жрыловидное пространство относительно глубокое. Крыловидные кости
вздуты, часто образованы губчатой костной тканью, также и крылья основ-
ной клиновидной кости. Барабанные камеры сравнительно плоские,
образованы слуховыми костями и отростками основной клиновидной кости.
Положение и размеры слуховых отверстий варьируют. Реже барабанные
камеры отсутствуют, а барабанная кость в виде кольца, не прирастающего
к соседним костям.
Нижняя челюсть удлиненная, у большинства низкая с небольшим
поперечным сочленовным мыщелком. Венечный отросток разной высоты
и формы. Его передняя поверхность расположена под разным углом по
отношению к продольной оси зубной части ветви нижней челюсти. Угловой
отросток развит в разной степени, широкий или палочковидный.
Верхние зубы. I1-1 уплощены в передне-заднем направлении,
клиновидной формы или в виде трехгранной призмы. Они крупнее I2 и I3
или очень крупные, превышающие размерами все другие зубы. Занимают
отвесное положение или направлены режущим краем вперед. Часто I1
имеет на наружном крае основания коронки в разной степени развитый
дополнительный зубец. I2 и I3 варьируют по форме и размерам. Клыки
хорошо развиты только у видов трибы Talpi.nl, у которых они вынесены
вперед, как у хищников (Carnivora). У представителей трибы Scalopini
они отодвинуты назад, что объясняется расположением сравнительно
крупных резцов (I2 и I3) на боковой стороне межчелюстной кости, а не на
ее передней стороне, как у Talpini. У других кротов клыки мелкие, сход-
ные по форме и размерам с соседними зубами. Р13, как правило, сходной
формы, конические, со слабо развитыми бугорками пара- и метакона при
©сновании коронки, разных размеров. Р4 всегда крупный, характерной для
насекомоядных формы, реже он напоминает строением другие премоляры.
Моляры чаще всего с W-образной, гребнистой жевательной поверхностью
и низким протоколом, с треугольной формы верхней поверхностью.
Их талонид не развит или в виде небольшого выступа протокона. У одних
все моляры сходной формы, но разной величины, у других они отличаются
не только размерами, но и строением отдельных частей коронки.
Резцы всегда с одним корнем. Клыки и Р1-3 с двумя или одним корнем,
Р4 и моляры с тремя корнями.
Нижние з у б ы. Нижние резцы мельче верхних, уплощены
спереди назад или конической формы; реже лопаточковидные, направлены
вперед. Клыки напоминают строением резцы и прижаты к ним (Talpini),
конусовидной и цилиндрической формы или сходны по строению с перед-
ними премолярами. Р2—Р3 сходной формы, часто и по размерам. Только
у видов трибы Talpini Р4 по форме и размерам похож на клык и выполняет
его функцию, поскольку истинный клык прижат к резцам. Р4 крупнее
остальных премоляров и более сложного строения. Моляры W-образной
формы, ребристы, убывают в размерах от первого к третьему. На переднем
и заднем крае их внутренней стороны имеется ушковидный отросток.
Скелет не отличается разнообразием строения, что обусловлено
сходством локомоторных функций, и в целом отражает общий для семейства
адаптивный тип.
Позвоночник состоит из 7 шейных, 13—14 грудных, 5—6 поясничных,
4 крестцовых и 8—27 хвостовых позвонков. Сравнительно тонкие, широко
расставленные друг от друга ребра образуют суженную спереди и широкую
сзади грудную клетку. Грудина составлена шестью сегментами. Рукоятка
182
СЕМ. TALPIDAE
грудины выдвинута вперед, в шейную область. На нижней и верхней сторо-
нах грудины гребни. Степень развития переднего выступа рукоятки гру-
дины, ее нижнего киля (гребня) и боковых лопастевидных отростков
обусловлена уровнем приспособления к рытью, большим или меныпим выно-
сом пояса передних конечностей в шейный отдел. У менее приспособлен-
ных к рытью кротов рукоятка грудины менее выдвинута вперед, нижний
гребень развит слабее, а боковые отростки более развиты. Лопатка удли-
ненной формы, ее наибольшая длина более ширины в 2.5—5 раз. Ость
лопатки в задней части развита в высокий гребень с отростком, направлен-
ным вверх или в сторону позвоночника. Акромиальный отросток развит
в разной степени. Он мейее развит у специализированных землероев.
Лопатки расположены параллельно продольной оси тела, их дистальные
отделы скреплены сухожильным тяжем.
Ключица короткая и толстая у специализированных землероев. У них
ее длина равна ширине или ширина более длины. Менее специализирован-
ные землерои имеют более длинную ключицу, ширина которой менее длины
в четыре раза. У бегающих кротов ключица длинная и тонкая.
У кротов ключица сочленяется непосредственно с плечевой костью
и только у выхухолей она отчасти причленяется и к лопатке.
Плечевая кость кротов своей формой сильно отличается от этой кости
других млекопитающих. Она короткая и широкая, с многими отростками,
к которым прикрепляются мышцы. У менее специализированных к рытью
кротов она удлиненная и узкая, с менее длинными отростками. Головка
плеча удлиненно овальная, у бегающих кротов она округлой формы,
а у выхухолей по форме средняя между отмеченными двумя. Надмыщел-
ковое отверстие отсутствует.
Локтевая кость у типичных землероев с гребнем на нижней стороне
и сильно развитым локтевым отростком. В дистальной части она значи-
тельно расширена. Лучевая кость более короткая и массивная, широкая
в дистальном отделе, с округлой сочленовной поверхностью для сочленения
с плечевой костью. Последнее позволяет предплечью вращаться на со-
членовном отростке плечевой кости на значительный угол по отношению
к оси плеча.
Кисть широкая, с короткой пястной и запястной частями и крупными
фалангами пальцев. Когтевые фаланги у специализированных к рытью
кротов раздвоены на конце. У них же хорошо развита дополнительная
кость — серповидная, расположенная на внутренней стороне кисти.
Дополнительная кость слабо развита у кротов, менее приспособленных
к рытью, а у бегающих она отсутствует. Бегающие кроты имеют кисть
удлиненной формы.
Таз длинный, образован двумя крестцовыми и тремя хвостовыми
позвонками. Лобковые кости не образуют симфиза (исключение составляют
выхухоли). Бедро с крупной головкой, цилиндрической формы. Малая
берцовая кость срастается с большой берцовой почти на половину ее длины.
Ступня по форме сходна с таковой землероек. Она узкая, образована длин-
ными костями. У кротов, приспособленных к плаванью, особенно у выху-
холей, длинные пальцы изогнуты, отклонены наружу. Расправленные
пальцы и перепонки между ними в совокупности придают ступне форму
лапы баклана. Os penis отсутствует.
Распространение
Ареал семейства охватывает значительные территории в Евразии и
в Северной Америке. Видимо, кроты никогда не населяли Африку и
Южную Америку, не говоря уже об Австралии, где обитают немногие
виды плацентарных млекопитающих.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
183
Роющие кроты сформировались очень рано (эоцен) и с того времени
предъявляют к ландшафтам и почвам определенные требования, чем и
объясняется их отсутствие не только на отдельных континентах, куда
они не смогли проникнуть по многим обстоятельствам, но и во многих
местах их сложившегося ареала. По-видимому, главной преградой при
расселении кротов служат широкие водные пространства и пустынные
ландшафты, а также зона вечной мерзлоты. Можно думать, что ареал
кротов был значительным по своей площади уже в третичное время,
по крайней мере с эоцена. Отдельные подсемейства и роды сокращали
или увеличивали свой ареал, но общие границы ареала семейства сохра-
нились до настоящего времени.
Кроты эоценового времени известны только из Европы, откуда опи-
саны Му хоту gale и Amphidozotherium Filhol. Единственный известный
вид последнего рода A. cayluxi Filhol, судя по описанию его зубов, об-
ладал особенностями, близкими к олигоценовым кротам.
В олигоцене кроты были широко распространены, о чем можно су-
дить по остаткам разнообразных групп этого семейства, найденным на
разных континентах. В Европе в это время существовали виды рода
Geotripus Pomel (триба Talpini); Paratalpa Lav. и Mygatalpa Schr. (триба
Urotrichini). В Северной Америке в олигоцене были распространены
кроты трибы Proscalopini (роды: Proscalops Matt., Cryptoryctes С. Reed,
Arctoryctes Matt, и Oligoscalops К. Reed). Олигоценовые кроты из Азии
известны по фрагментам нижней челюсти и зубам — Мг и М2, по строе-
нию близких к таковым представителей подсем. Uropsilinae и Talpinae.
Можно не сомневаться, что они там были задолго до олигоцена, поскольку
этот континент — родина сем. Talpidae. Отсутствие находок древних
вымерших кротов в Азии объясняется, видимо, тем, что раскопки ведутся
в ограниченных масштабах, и, возможно, в местах, где их не было.
' Судя по тому, что в эоцене выхухоли (подсем. Desmaninae) были пред-
ставлены группами, специализированными близко к уровню современных
видов, на что указывают находки их зубов, они развивались еще ранее и
были широко распространены в Азии. В олигоцене они были еще более
специализированными и более разнообразными, обусловив развитие це-
лой плеяды миоценовых выхухолей.
В миоцене Европу населяли многие представители подсем. Desmaninae
(из родов Му galea, Mygalinia Schr. и Galemys Капр). Виды этих же родов,
а также рода Desmana Guld. были распространены и в Азии. Роющие
кроты (подсем. Talpinae) были представлены видами трех триб: Urotrichini
(род Scaptonyx M.-Edw.), Scalopini (род Proscapanus Gaill.) и Talpini
(род Talpa L.). В Северной Америке в это время существовали виды триб:
Proscalopini (род Mesoscalops К. Reed), Urotrichini (роды Scalopoides Wils,
и Midecodon Wils.), а также и трибы Scalopini, типичной для Северной
Америки группы кротов, сложившейся несомненно в миоцене, хорошо
известной с нижнего плиоцена.
Обзор миоценовых кротов позволяет отметить следующее: в миоцене
сложились все основные направления специализации кротовых — при-
способление к рытью, плаванью и бегу. Приспособление к бегу сохраняют
виды подсем. Uropsilinae, представляющие и ныне исходный тип кротов
и их локомоторной системы.
Плиоцен был пределом существования многих групп кротовых, раз-
вившихся ранее этого времени, а также периодом развития новых групп,
особенно видов. R Европе в плиоцене существовали представители родов
Mygalinia Schr.?, Desmana Giild. и Galemys Капр (подсем. Desmaninae).
Подсем. Talpinae было представлено видами рода Talpa L. (триба Tal-
pini), а в Азии — родом Scaptochirus M.-Edw. В Северной Америке под-
184
СЕМ. TALPIDAE
сем. Desmaninae было представлено родами Gaillardia Matt, и Hydrosca-
pheus Schotv., а подсем. Talpinae — родами Domninoides Green (триба
Urotrichini), Scapanus Pomel и Scalopus Geoff, (триба Scalopini).
В конце плиоцена в Европе вымерли виды рода Mygalinia (Desmaninae)
и ряд видов рода Talpa (Talpinae), а в Северной Америке вымерли вы-
хухоли родов Gaillardia и Hydroscapheus.
В плейстоцене сложилась современная фауна.
В современный период Европу населяют виды монотипичных ныне
родов Galemys Капр, и Desmana Giild. (подсем. Desmaninae) и 4 вида рода
Talpa L. (подсем. Talpinae). Наиболее богата представителями сем.
Talpidae Азия. На ее территории обитают виды подсем. Uropsilinae (роды
Rhynchonax Thos. — 1 вид, Nasillus Thos. — 2 вида, Uropsilus M.-Edw. —
1 вид) и подсем. Talpinae — триба Urotrichini (роды Scaptonyx M.-Edw. —
1 вид, Urotrichus Ternm. — 1 вид и Scapanulus Thos. — 1 вид), триба
Talpini (роды Talpa — 7 видов, Mogera Pomel — 3 вида, Parascaptor
Gill. — 1 вид и Scaptochirus M.-Edw. — 1 вид).
В Северной Америке обитают только представители подсем. Talpinae —
триба Urotrichini (род Neurotrichus Glint. — 1 вид), триба Scalopini (роды
Parascalops True — 1 вид, Scapanus Pomel — 4 вида, Scalopus Geoff. —
3 вида).
Видимо, подсем. Uropsilinae на протяжении всего времени развития
сем. Talpidae было эндемичным для Азии.
В современный период подсем. Desmaninae (роды Desmana и Galemys)
эндемично для Европы. Подсем. Talpinae — общее для Европы, Азии и
Северной Америки. При этом триба Urotrichini общая для Азии и Север-
ной Америки, триба Scalopini эндемична для Северной Америки, а триба
Talpini общая для Европы и Азии.
Из трибы Urotrichini эндемичны для Азии роды Scaptonyx, Urotrichus
и Scapanulus, а для Северной Америки — род Neurotrichus. Из трибы
Talpini — род Talpa общий для Европы и Азии, а роды Mogera, Paras-
captor и Scaptochirus эндемичны для Азию
Экологические особенности семейства кротов
и значение их для человека
Привычное представление о кротах всегда связано с роющими видами,
обитающими в Европе, Азии и Северной Америке. Значительно менее
известны плавающие кроты — выхухоли, сохранившиеся в Португалии,
Испании, Франции (Galemys Капр) и в СССР, в системе бассейна Волги
(Desmana Guld). «Полуроющие—полуназемные» кроты трибы Urotrichini.,
распространенные в Юго-Восточной Азии, Японии и на западе Северной
Америки, а также наземные — бегающие кроты подсем. Uropsilinae,
населяющие ограниченные территории Юго-Восточной Азии, еще менее
известны.
Уже это перечисление групп кротов с разными локомоторными функ-
циями их двигательных систем показывает, насколько разнообразны
приспособления в пределах сем. Talpidae. В связи с этим трудно дать и
их общую экологическую характеристику. Общей экологической особен-
ностью кротовых следует считать их привязанность к древесно-кустар-
никовой растительности и близко к ним расположенным территориям
(кроты) или к поросшим лесом и кустарниками берегам равнинных и
горных водоемов (выхухоли). Связь кротов с лесными и кустарниковыми
формациями обусловлена многими благоприятными для них факторами,
особенно характерными для лесных ландшафтов. Из них прежде всего
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
185
нужно отметить менее резкие, чем на открытых пространствах, колебания
температуры и влажности, а также относительное постоянство запасов
разнообразных кормов во всех ярусах растительности, в подстилке леса
и его почве. Имеются в виду различные группы беспозвоночных, особенно
насекомых, зимующих на разных стадиях развития в подстилке и верх-
них слоях почвы, многоножки и многочисленные дождевые черви, оби-
тающие в различных горизонтах почвы и в лесной подстилке.
Основным кормом кротам служат земляные черви, хотя в отдельные
сезоны года они поедают и насекомых, а также представителей других
групп беспозвоночных, а в отдельных случаях и мелких позвоночных,
которых могут одолеть. Но благополучие кротов основывается на пита-
нии дождевыми червями, многочисленными в почве и широко распрост-
раненными. Можно с уверенностью утверждать, что достаток и широкое
распространение дождевых червей определили и широкое распростране-
ние большинства групп семейства, объединяющих специализированных
землероев.
Вероятно, приспособление кротов к подземному образу жизни нача-
лось с питания дождевыми червями и шло параллельно с приспособле-
нием к их экологии. Расположение охотничьих ходов кротов в большей
или меньшей степени согласуется с зонами скопления дождевых червей
ж отдельные сезоны года.
Норы для гнезда кроты строят под землей, помещая его под защитой
стоящего сверху пня, камня или дерева. На открытых пространствах
кроты над местом, где помещается гнездо, насыпают высокий и широкий
холмик земли. Гнездо представляет собой расширенное углубление
в почве, соединяющееся короткими ходами с кормовыми ходами. Под-
стилкой гнезда служат остатки сухих растений, подстилки может и не
быть. Русская выхухоль строит гнездо в высоком участке берега, с си-
стемой ходов, соединяющихся с ходом под воду. Пиренейская выхухоль
для устройства гнезда довольствуется укрытиями в каменных осыпях,
расселинами в скалах или норами водяной полевки (Arvicola terrestris L.).
Размножаются кроты 1—2 раза в году в зависимости от погодных
условий и широты местности, в Которой они обитают. От этих же условий
зависит и время наступления и окончания размножения, а также и число
эмбрионов, отдельные из которых могут подвергнуться резорбции в слу-
чае недостатка корма при изменении режима условий, необходимых
животным, служащим кормом. При малой подвижности самок в период
беременности достаток корма имеет особенно важное значение.
За пределами сельскохозяйственных угодий кроты приносят большую
пользу, разрыхляя землю и открывая доступ воздуху в почвенные слои.
На пастбищах они приносят некоторый вред, выбрасывая на поверхность
менее плодородную почву из ее глубоких слоев, на которой поселяется
рудеральная растительность, худшая по питательным свойствам, чем
луговая, а чаще непригодная для кормления скота. Кроме того, кучки
земли мешают низкой косьбе машинами, что ведет к потерям сена,
а часто и к порче машин.
Б садах и огородах кроты, подрывая корни, вызывают снижение
урожая и даже гибель растений, что приходилось наблюдать при обсле-
довании сельскохозяйственных угодий.
Кроты считаются хранителями некоторых опасных инфекций в период
депрессии численности спонтанных носителей инфекции из числа грызу-
нов. Но в целом вред от кротов не настолько ощутим, чтобы было осно-
нание причислить их к вредным видам. Миллионы их шкурок, добывае-
мых ежегодно, с лихвой покрывают приносимый ими вред в отдельных
местах. Кроме того, кроты поедают много насекомых, особенно личинок
186
СЕМ. TALPIDAE
жуков-щелкунов и пластинчатоусых — майских и июньских хрущей,
а также других насекомых, среди которых много вредителей зерновых
культур, плодовых деревьев и лесных питомников. Кротов нужно охра-
нять. Нет животных, которые приносят только пользу. Вред в одном
случае может оказаться пользой в другом.
Выхухоль — ценный пушной зверек, но настолько сократилась в чи-
сле, что можно говорить лишь об ее охране.
Филогенетические соотношения
отдельных групп кротовых и эволюция
их жевательного аппарата
Местом формирования кротовых следует считать Азию, где они и
в современный период представлены разнообразно адаптированными груп-
пами, в том числе и примитивными, сохранившими исходный для эволю-
ции этих млекопитающих тип локомоторной системы, приспособленной
к бегу галопом. Слабая изученность Центральной Азии в палеонтологи-
ческом отношении — причина того, что мы не знаем ее древних вымер-
ших кротов.
Происхождение кротовых, как, впрочем, и многих других групп насе-
комоядных, остается неясным. Одно несомненно, что их ранние предки
связаны общими корнями с тенрекообразными (Tenrecii). Трехбугорча-
тое строение верхних коренных зубов, с тремя корнями, характерное
для всех групп кротовых, отражает архаичную структуру ранних типов
зубов с W-образной складкой на жевательной поверхности. Некоторые
изменения деталей жевательной поверхности верхних коренных зубов
в процессе эволюции отдельных групп не повлияли на их общую при-
митивную структуру.
Значительному изменению в процессе развития сем. Talpidae под-
верглись конечности ряда его групп, причем очень рано. Свидетельством
раннего становления и обособления трех адаптивных типов кротовых
(подсем. Uropsilinae, Desmaninae, Talpinae} служит нахождение в эоцене
и нижнем—среднем олигоцене плечевых костей, близких по строению
к таковым современных видов. Это также доказывает, что кротовые раз-
виваются давно.
Исходным типом локомоторной системы кротовых следует считать
эту систему наземных бегающих кротов подсем. Uropsilinae, сходных
по внешнему облику с землеройками. Предки этого подсемейства не най-
дены. Несомненно, что его современные виды значительно уклонились
от исходного типа и представляют собой достаточно специализированную
группу, для видов которой характерно движение галопом по различным
субстратам и грунтам поверхности земли. Зубная система описываемых
кротов образована наименьшим среди кротов количеством зубов, что
привело к сокращению длины челюстей и несомненно к усилению жева-
тельного давления на коренные зубы. Это, надо думать^ позволило мел-
ким кротам подсемейства питаться разнообразной пищей, в том числе
и грубой консистенции. Мелкие общие размеры описываемых кротов
позволяют предполагать потребление ими большого количества корма
по отношению к весу их тела и поддерживать высокий темп обмена ве-
ществ, что очень важно для терморегуляции при значительных перепа-
дах температуры в среде обитания.
Этими общими замечаниями и приходится ограничиться в характе-
ристике кротов подсем. Uropsilinae. Можно только отметить, что при
всей специализации его представителей к настоящему времени, их внеш-
ний облик, строение конечностей и экстерорецептеров позволяют пред-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
187
ставить себе достаточно достоверный структурный портрет предков кро-
товых современных подсемейств.
Для представителей подсем. Desmaninae бегающие кроты не могли
служить непосредственными предками. Пояс передних конечностей выху-
холей близок по строению к таковому роющих кротов, но мало специали-
зированных. Наиболее близок по строению пояс передних конечностей
и плечевая кость выхухолей к этим частям скелета некоторых предста-
вителей подсем. Talpinae, например Urotrichus talpoides Temm. (триба
Urotrichini). Можно думать, что выхухоли произошли от предков, общих
с трибой Urotrichini. Наиболее ранняя выхухоль известна из эоцена—
нижнего олигоцена Казахстана, кратко описанная в настоящей работе,
но оставленная без названия за отсутствием достаточного материала.
Коренные зубы описанных здесь (с. 209) представителей подсем. Desma-
ninae из олигоцена сходны по строению с этими зубами поздних и совре-
менных выхухолей, но имеют и свои особенности. Значительная специа-
лизация зубов древних выхухолей в направлении, характерном и для
современных их представителей, отражает давнее развитие этой группы
кротовых. Есть все основания предполагать, что становление видов
подсем. Desmaninae происходило еще в начале эоцена.
Судя по большому количеству остатков вымерших групп выхухолей
из разных местонахождений, в миоцене они были широко распространены.
Они обитали в Европе (роды Му galea Schr., Mygalinia Schr., Galemys
Каир, Desmana Guld) и в Азии {родьъ Mygalinia, Му galea Schr., Desmana).
Несомненно, что там будут найдены и другие роды. О миоценовых выху-
холях Северной Америки ничего не известно, но они должны были там
быть, поскольку они найдены на этом континенте в плиоцене и состав-
ляли особую группу.
Миоценовые выхухоли отличались размерами: от зверьков величиной
с Neomys fodiens Penn, до размеров современной Desmana moschata L.
Различная длина и форма плечевых костей позволяют предполагать и
разный характер плаванья. Некоторые различия в деталях строения
зубов и их размеров отражают, видимо, различия в характере пищи и
ее обработки во рту, но основные структуры зубов сходны у всех групп.
У видов отдельных родов, при общих особенностях строения нижней
челюсти, венечный отросток был строго вертикальным, наклоненным
вперед или отклоненным назад, что, можно думать, связано с широтой
открывания рта и усилением давления зубов-антагонистов в резцовом,
среднем и заднем отделах зубного ряда.
Разная ширина дистальных отделов плечевой кости и ее разная тол-
щина и высота отражают разную степень приспособленности к рытью,
передвижению по земле и плаванью. Значительное число родов, существо-
вавших почти одновременно, позволяет предполагать значительное разно-
образие частных форм адаптации и широкую избирательность в местах
обитания. Сохранявшийся в миоцене теплый климат способствовал раз-
витию разнообразной растительности, а вместе с этим разнообразным
беспозвоночным, что в совокупности обеспечивало возможность существо-
вания разно адаптированным к обстановке выхухолям. Миоцен, судя
по всему, был периодом расцвета этих кротовых.
По-видимому, значительное изменение обстановки в плиоцене послу-
жило причиной вымирания выхухолей с более древним типом адапта-
ции, например Му galea Schr., Mygalinia Schr. Их остатки в послемиоце-
яовое время не обнаружены. Плиоцен был периодом расцвета представи-
телей рода Desmana Guld., разнообразные виды которого были широко
распространены в Европе и Азии. Несомненно, что в то же время в Европе
происходило сужение ареала выхухолей рода Galemys Каир. Как пред-
188
СЕМ. TALPIDAE
ставляется, род Galemys относится к плеяде родов Му galea и Mygaliniar
объединявших виды мелких выхухолей, развивавшихся длительное время
и в тропических и субтропических условиях, утративших потенции к фор-
мообразованию, а затем и к преодолению влияния условий менявшейся
среды. Позднее появившийся род Desmana объединял виды, обладавшие-
значительным биологическим потенциалом. Его представители в миоцене
и особенно в плиоцене были широко распространены, что объясняется
их разнообразным приспособлением к среде, о чем можно судить по
строению челюстей и скелетов их конечностей. Большинство видов рода
Desmana относительно крупных размеров, что, возможно, помогало
в борьбе за места обитания.
В конце плиоцена представители рода Desmana начали сокращать
ареал сначала за счет Западной Европы, а затем и за счет его азиатской
части, закончив этот процесс к концу плейстоцена.
В плиоцене Северной Америки обнаружены более примитивные, чем
в Старом Свете, выхухоли (роды Gaillardia Matt., Hydroscapheus Shotw.).
У вида рода Gaillardia нижняя челюсть напоминает эту кость крота, а зубы
сходны с таковыми выхухоли. Представитель рода Hydroscahpeus имел
плечевую кость с округлой сочленовной головкой, что, вероятно, позво-
ляло двигать переднюю конечность во многих плоскостях. Позднее плио-
цена выхухоли в Северной Америке не обнаружены.
Несходство в строений известных остатков американских выхухолей
с таковыми выхухолей из Евразии позволяет предполагать, что обе
группы рано разошлись и развивались, независимо в значительном от-
резке времени. Об особенностях истории развития американских выху-
холей мы ничего не знаем. Что же касается евроазиатских выхухолей,
то их развитие шло сопряженно в одном, общем направлении, что и вьь-
разилось в сходстве их многих морфологических особенностей. Сходства
механического состава пищи, о чем можно судить по сходству состава
фауны водоемов, обусловливало сохранение общего типа в строении зубов
и зубной системы, а также и всего жевательного аппарата. Различия
в жевательном аппарате выхухолей разных родов разного времени столь
малы, что все они могут быть сведены в один род, если бы не было других
признаков, важных и существенных.
Зубы выхухолей образовались из таковых примитивных кротов. Верх-
ние коренные, особенно олигоценовых видов, более близки к этим зубам
кротов трибы Urotrichini (подсем. Talpinae), но с большим числом бугор-
ков на протоконе. Они менее длинные и более широкие. Премоляры
более массивные и низкие, верхние резцы трехгранные и более высокие,
чем у других кротовых, что придает всей зубной системе выхухолей
своеобразный облик.
Обособление подсем. Desmaninae, как уже отмечалось, происходило
относительно рано по отношению ко времени общего развития кротовых,
а параллельное развитие его групп протекало сравнительно долго, так
и не дав сколь-нибудь значительных уклонений от общего адаптивного
типа выхухолей. Какой род с каким родом связан в происхождении,
пока сказать трудно, но одно несомненно, что мелкие формы, с отклоненным
назад венечным отростком нижней челюсти, с более простой эмалевой
структурой на жевательной поверхности коренных зубов и с большим
числом корней у премоляров — более древние. Изменение перечисленных
признаков от рода к роду во времени и представляет преемственную
иерархию форм в адаптивном процессе эволюции выхухолей. Это и при-
нято в предлагаемой мной схеме филогенетических соотношений отдель-
ных групп выхухолей.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
189
По сравнению с другими подсемействами подсем. Talpinae представ-
ляет весьма разнородную группу, причем разнородность сохраняется
и в пределах его подразделений. Прежде всего следует отметить, что
кроты делятся на две группы: менее специализированных, относительно
универсально приспособленных землероев, со свободными пальцами на
кистях и высокоспециализированных роющих кротов, с пальцами, соеди-
ненными кожей до когтей.
Первые объединяются трибой Urotrichini и представляют собой сме-
шанную группу, разно приспособленных к рытью кротов, особенно из
числа вымерших форм. Высокоспециализированные землерои делятся на
четыре трибы, объединяющие роды и виды с разно приспособленной
зубной системой.
До недавнего времени наиболее ранним представителем подсем. Tal-
pinae считался род Amphidozotherium Filhol. В системе Симпсона (Rimpson,
1945) этот род стоит в качестве предкового в подсем. Talpinae, в старом
понимании этого подсемейства. СтроганЬв (1948) в своей схеме филоге-
нетических соотношений родов сем. Talpidae отметил его в качестве
боковой ветви, сопряженной в развитии с подсем. Talpinae. Лявока
(Lavocat, 1951) отнес его в синонпмы рода Geotripus Pomel. Вире (Viret,
1955) приводит этот род как самостоятельный и дает рисунок его плече-
вой кости (с. 1704). Ван Вален (Van Valen, 1967) в своей системе насеко-
моядных поставил его в сем. Adapisoricidae, а Кроше (Crochet, 1974) отнес
этот род к неизвестному семейству надсем. Erinacoidea. В настоящей ра-
боте представители рода Amphidozotherium считаются кротами — ран-
ней группой трибы Talpini, но боковой, архаичной ветвью.
Представители рассматриваемого здесь рода имели крупные первый
и второй резцы (рис. 140), что наблюдается и среди современных кротовых,
у отдельных видов которых особенно развит второй резец. У большинства
кротовых, как и у большинства других групп насекомоядных, органом
захвата пищи служат резцы. Судя по числу зубов в одной ветви нижней
челюсти (11), виды описываемого рода имели в верхней и нижней челюстях
44 зуба. Резцы и клык имели в поперечном сечении округлую форму,
а первые 3 премоляра имели строение, близкое таковому современных
представителей подсем. Uropsilinae или трибы Urotrichini (подсем. Tal-
pinae). Коренные зубы были менее вытянуты спереди назад, чем у совре-
менных кротовых, с более широким метаконидом (талонид других авто-
ров). Плечевая кость приближалась по форме к таковой современных
роющих кротов. Таким образом, по общим морфологическим особенностям
представители рода Amphidozotherium приближались к адаптивному типу
землероев, но еще сохраняли многие примитивные черты (округлой формы
резцы и клыки, широкие и короткие моляры и др.). Вряд ли этот род
можно считать предковой группой всех кротовых или какой-либо их
группы. Несмотря на отмеченное выше сходство в отдельных структурах
с некоторыми видами вымерших и современных групп кротовых, он все
же стоит особняком и представляет особую группу эоценовых кротов,
специализированных для своего времени.
Сходство Amphidozotherium и Geotripus, усмотренное Лявока, позво-
лившее ему отнести первого в синонимы второго, дает основание пред-
полагать развитие последнего от общих корней с Amphidozotherium или
близких к нему групп эоценовых кротов. Судя по значительной специа-
лизации кротов к верхнему эоцену—нижнему олигоцену, развитие этого
подсемейства началось значительно ранее верхнего эоцена.Описываемые
здесь роды представляют, как мне кажется, направление развития кротов
трибы Talpini.
190
СЕМ. TALPIDAE
Из верхнего эоцена Европы известен и еще один род — Myxomygale
Filhol. По особенностям строения нижней челюсти и зубов этот род дол-
жен быть отнесен к трибе Urotrichini. Несмотря на ранний геологический
возраст, представители Myxomygale имели черты известной специали-
зации, что выражено в вертикально стоящем венечном отростке и редук-
ции или слиянии корней до одного на передних премолярах. Последнее
несомненно служило укорочению длины челюстей. Эта тенденция сохра-
нила свое значение во все последующее время развития группы кротов.
Но сокращение длины челюстей происходило не только за счет сокраще-
ния числа корней, айв результате уменьшения количества зубов. Наряду
с отмеченными особенностями специализации представители рода Myxo-
mygale сохраняли и признаки, которые можно рассматривать как архаич-
ные по отношению к специализации трибы в более позднее время: крупный,
судя по альвеоле, клык и большое количество зубов — 44.
Отмеченные находки эоценовых кротов позволяют с уверенностью
сказать, что трибы подсем. Talpinae формировались еще в эоцене и уже
тогда, в большей или меньшей степени, отражали в морфологии общее
для каждой из них направление развития при всех частных особенностях
их эоценовых видов.
В олигоцене кроты были представлены разнообразными группами.
По-видимому, все современные трибы подсем. Talpinae окончательно
складывались в олигоцене, поскольку в миоцене они были представлены
высоко и разнообразно специализированными группами.
Триба Urotrichini в олигоцене была представлена родами Paratalpa
Lavoc. и Mygatalpa Schr. Как отмечают Лявока (Lavocat, 1951),
Шрейдер (Schreuder, 1940), кроты этих родов совмещают признаки
крота и выхухоли. При этом отмечается, что для представителей первого
рода характерна менее выгнутая нижняя челюсть, а представители вто-
рого имели трубчатую плечевую кость. Виды того и другого рода имели
нижние коренные зубы, напоминающие таковые выхухолей. Следует
отметить, что плечевая кость с трубчатой средней частью характерна
и для современных видов трибы Urotrichini, как и несколько выпрямлен-
ный нижний край нижней челюсти. Что же касается сходства нижних
коренных зубов с этими зубами выхухолей, то в этом нет ничего удиви-
тельного: зубы отдельных групп кротовых в прошлом могли быть сход-
ными, как это имеет место и среди современных кротов. Зубы предста-
вителей подсем. Desmaninae развились из. таковых кротов, степень
их сходства или различия — результат частных адаптаций.
Своеобразную группу олигоценовых кротов представляла триба Рго-
scalopini. Черепа представителей этой трибы формой напоминали тако-
вой златокротов (Chrysochloridae'). Зубы, типичные для кротов. Плечевые
кости имели разнообразную структуру. Например, роды Cryptoryctes
С. Reed и Arctoryctes Matt, отличались — первый перехватом средней
части этой кости при широких проксимальном и дистальном ее отделах,
а другой имел широкую в средней части плечевую кость с суженным
проксимальным и широким дистальным отделами. При всей фрагментар-
ности данных о морфологических особенностях представителей описываемой
трибы известный фактический материал отражает своеобразие форм их
адаптации и значительную разнонаправленность частной специализации.
В олигоцене эта триба была представлена следующими родами: Prosca-
lops Matt., двумя уже упомянутыми и Oligoscalops К. Reed. Адаптивные
особенности представителей трибы Proscalopini можно рассматривать
как частный путь развития кротов, не повторяющийся в других трибах.
Существование этой трибы на континенте Северной Америки закончилось
в миоцене. Однако особый путь развития этой трибы не исключает ее
ХАРАКТЕРИСТИКА СЕМЕЙСТВ^
191
связи в происхождении с другими группами кротов, в частности с трибой
Scalopini.
Как уже отмечалось, триба Talpini в олигоцене была представлена
родом Geotripus Pomel. Известный представитель этого рода G. antiqaus
Pomel, судя по описанию ряда авторов, имел много сходства с видами
рода Talpa L. Основные отличия заключаются в островершинности пре-
моляров, особенно Р1, имеющего клиновидную форму, Р4 с одной кони-
ческой вершиной. М1 более широкий, чем у представителей рода Talpa,
а плечевая кость более длинная. Перечисленные признаки рода Geotri-
pus отражают архаичность олигоценовых представителей трибы Talpini.
В заключение обзора олигоценовых кротов следует отметить, что
они в то время составляли разнообразные по характеру адаптаций группы.
Представители подсем. Talpinae, как и ныне, были по-разному приспо-
соблены к рытью. Из них наиболее универсально приспособленными
были виды трибы Urotrichini. Возможно, что они, как и теперь, населяли
верхние слои почвы, лесную подстилку, а отдельные виды могли плавать,
сочетая особенности кротов и выхухолей. Виды триб Proscalopini и Tal-
pini были по-разному специализированными к рытью, но достаточно
высоко. Можно не сомневаться, что в это время развивались и предста-
вители трибы Scalopini.
Миоценовая фауна кротовых несет отпечаток высокой специализации
ее видов в основных направлениях адаптации. В это время окончательно
сложились все основные крупные группы кротов, большинство которых
сохранилось до настоящего времени.
Триба Urotrichini была представлена разно специализированными груп-
пами. Из Северной Америки известны два рода этой трибы: Scalopoides
Wils, и Mydecodon Wils. Первый, судя по числу зубов в одной ветви ниж-
ней челюсти (11), имел полную зубную систему, у другого она была не-
полной. Строение плечевой кости представителей рода Scalopoides сходно
с таковым Condylura cristata L., что позволяет предполагать их значитель-
ную специализацию к рытью. Сокращенная зубная система Mydecodon
отражает специализацию жевательного аппарата. Из миоцена Европы
известны остатки современного рода Scaptonyx M.-Edw., представители
которого в то время были менее специализированы к рытью, чем совре-
менные виды, на что указывает более удлиненная плечевая кость, с ме-
нее развитыми надмыщелками. Таким образом, миоценовые роды трибы
Urotrichini, как и ныне, объединяли кротов, в разной степени приспо-
собленных к рытью и с разно специализированной зубной системой.
При изучении этой группы кротов все больше приходится убеждаться
в том, что разность форм адаптации ее видов определила возможность
существования ее представителей в значительном отрезке времени, пе-
реходя с помощью разно и универсально приспособленных форм, с одной
адаптивной позиции на другую.
Из трибы Proscalopini в миоцене существовали представители двух
родов: Proscalops Matt, и Mesoscalops К. Reed. Виды этих родов сохраняли
особенности, близкие их олигоценовым предкам. По-видимому, общий
характер адаптаций видов этой трибы ограничил возможности их приспо-
соблений к условиям, быстро менявшимся в ландшафтах миоцена.
Триба Scalopini была представлена в это время европейским родом
Proscapanus Gaill. Зубная система представителей этого рода по числу
зубов и их структуре близка к таковой современного рода Scapanus Po-
mel, а плечевая кость более удлиненная, чем у видов последнего. Род
Proscapanus стоит особняком среди кротов Старого Света, что можно объяс-
нить отсутствием материала по другим группам трибы Scalopini из Евразии,
вымершим в разное время.
Talpa Mogera Parascalops Scalops
i I t r.j____________________
Scaptochjrus
| Paras captor
Hesperoscalops
I
Myxomygalt\
Триба Proscalopini'--
Триба Talpini.
Geotripus \
\ \ Oligoscalops\ Proscalops \
\ \ ...'-•Arrtoryctes
\ Cryptoryctes
Amphidosotherium
Mesoscalops
j Триба Scalopini y
: \ \ / Paratalpa
Scapanulus Scaptonix
*. Urotrichus I
i I :
Galemys
Desmana Uropsilus
\Nasillus\ Rynchonax
V/
Gaillardia Hydroscapheus
|| Domninoides
.Scalopoides^ Midecodon
Proscapanus I
у gen. et sp. (3y6w)j
• gen.etsp. (зуб)
\ : r gen.et sp.(3y6t>t)/
Подсем. Talpinae '-JПодсем. Desmaninae /
/ Подсем. Uropsilinae
Mygalinia
Рис. 98. Схема филогенетических соотношений кротовых.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
193
Представители трибы Talp ini из миоцена известны по плечевым костям,
сходным по строению с современным их состоянием у видов рода Talpa.
Это мешает достоверному определению их систематического положения.
При общем сходстве плечевых костей современные роды трибы Talpini
отличаются составом зубных систем, строением зубов и черепа, отдель-
ными внешними признаками. Для трибы Talpini в ее современном состоя-
нии характерно уменьшение размеров нижнего клыка и сближение его
с резцами. Функцию нижнего клыка выполняет Pv Эти особенности имеют
все ее роды и виды. Такое своеобразное строение нижнего зубного ряда
позволяет предполагать длительный путь его развития. С другой стороны,
у кротов этой трибы в верхней челюсти сохраняется крупный клык —
признак явно архаичный и не только для кротов.
Это, по-видимому, и служило поводом для некоторых систематиков,
например Симпсона (Simpson, 1945), считать эту группу кротов более
примитивной, чем триба Scalopini. Но у кротов трибы Talpini верхние
резцы расположены по широкой дуге, с чем связано расположение ниж-
него клыка вблизи нижних резцов, а также близкое к ним строение.
Это среди насекомоядных признак прогрессивный, близкий по располо-
жению резцов к хищным (Carnivora). Все вместе взятое позволяет видеть
в этом особый путь развития плеяды родов кротов трибы Talpini, укло-
нившийся от общего направления развития зубных систем кротовых,
а также предполагать, что эта триба с миоцена развивается в направлении,
характерном для нее и ныне.
Плиоцен был периодом окончательного формирования родов кротов,
сложившихся в миоцене. В это время появились отдельные новые роды,
угасшие к концу этого периода. Формировались и новые виды, но все
это не меняло общего облика плиоценовой фауны кротовых, еще сохра-
нявшей многие давно развивавшиеся группы.
В плиоцене существовали все современные роды подсем. Talpinae.
Плиоценовый род Domninoides Green (Северная Америка), вымерший
к концу этого периода, представлял древние группы трибы Urotrichini.
В конце плиоцена и в плейстоцене окончательно сложилась совре-
менная фауна сем. Talpidae.
Свои представления о филогенетических соотношениях различных
групп кротовых я привел на рис. 98.
Краткая история исследования
систематики Talpidae
Изучение кротов, как и многих других групп млекопитающих, нача-
лось с описания Линнеем в 1758 г. Европейского крота Talpa еигораеа
и американских кротов Condylura cristata и Scalopus aquaticus. Гюльден-
штедт (Giildenstaedt, 1777) описал род Desmana. С начала XIX в. началось
более обстоятельное изучение кротовых. Жофруа Сент Илер (Geoffray
Saint Hilaire, 1803) описал, род Scalopus, Иллигер (liliger, 1811) описал
род Condylura, Фишер (Fischer von Waldheim, 1817) объединил известных
к тому времени кротов в трибу Talpini, Маррей (Murray, 1866) в пределах
этого семейства выделил подсем. Talpinae, только Томас (Thomas, 1912)
выделил подсем. Uropsilinae, Desmaninae, Scalopinae, Condylurinae, Стро-
ганов (1948) установил подсем. Urotrichinae, Рид (Beed, 1961) описала
подсем. Proscalopinae. Если современные кроты начали изучаться с сере-
дины XVIII в., то их вымершие виды — с середины XIX в. А в общей
сложности кротовые изучаются более 2 веков. За это время описано много
родов и видов вымерших и современных кротов. В последние 30—40 лет
13 А. А. Гуреев
194
СЕМ. TALPIDAE
предложено несколько систем кротовых, как для всего семейства, так
и для отдельных подсемейств.
Добсон (Dobson, 1883) в своей известной монографии при описании
особенностей мускулатуры и скелета представителей сем. Talpidae рас-
положил роды в следующей последовательности:
1. Myogale (ныне Desmana) moschata. 7. Parascaptor
Myogale (ныне Galemys) pyrenaica. 8. Scaptochirus
2. Condylura 9. Scaptonyx
3. Scapanus 10. Neurotrichus
4. Scalopus 11. Urotrichus
5. Talpa 12. Uropsilus
6. Mogera
Вряд ли такое соотношение родов было принято ради удобства изло-
жения. Скорее всего расположение родов отражает их систематические
соотношения, какими они представлялись автору.
Симпсон (Simpson, 1945) представил следующую систему кротов:
Сем. Talpidae Gray, 1825 (—Talpini Fischer, 1817) (включает Scalopidae Cope, 1889. —
Myogalidae Milne-Edwards, 1868—1874—Myoladae Gray, 1821). Верхний эоцен—
современный, Европа; верхний олигоцен—современный, Сев. Америка; современ-
ный — Азия.
Подсем. Uropsilinae Thomas, 1912. Современный, Азия.
Upopsilus Milne-Edwards, 1871 (включает Rhynchonax Thomas, 1911). Сов-
ременный, Азия.
Nasillus Thomas, 1911. Современный, Азия.
Подсем. Desmaninae Thomas, 1912 (=Myogalina Bonaparte, 1845, Myolade Gray,
1821). Верхний миоцен—современный, Европа; современный, Азия.
- \-Mygalea Schreuder, 1940. Верхний миоцен, Европа.
^Mygalinia Schreuder, 1940. Нижний плиоцен, Европа.
Desmana Guldenstaedt, 1777 (—Му gale Cuvier, 1800; Myogale Brandt,.
1836). Нижний плиоцен—современный, Европа; современный, Азия.
Galemys Каир, 1829. Современный, Европа.
Подсем. Talpinae Murray, 1866. Верхний эоцен—современный, Европа; современ-
ный, Азия, средне-верхний миоцен? Сев. Америка?
fAmphidozotherium Filhol, 1876. Верхний эоцен, Европа.
fGeotripus Pomel, 1848. Средний олигоцен, Европа.
- \-Prose ар anus Gaillard, 1899. Миоцен, Европа.
Talpa Linnaeus, 1758. Миоцен—современный, Европа; средний—верхний
миоцен? Сев. Америка? Современный, Азия.
Mogera Pomel, 1848. Современный, Азия.
Parascaptor Gill, 1875. Современный, Азия.
Scaptochirus Milne-Edwards, 1867. Современный, Азия.
Подсем. Scalopinae Thomas, 1912 (—Scalopidae Соре, 1889). Верхний олигоцен—
современный, Сев. Америка, Вост. Азия; средний миоцен? Европа?
• \Proscalops Matthew, 1901. Верхний олигоцен—нижний миоцен, Сев.
Америка.
Scaptonyx Milne-Edwards, 1871. Средний миоцен? Европа? современный,
Вост. Азия.
Dymecodon True, 1886. Современный, Япония.
Neurotrichus Gunter, 1880. Современный, Сев. Америка.
Urotrichus Temminck, 1838—1839. Современный, Япония.
Scapanulus Thomas, 1912. Современный, Вост. Азия.
Parascalops True, 1894. Плейстоцен—современный, Сев. Америка.
• \Hesperoscalops Hibbard, 1941. Верхний плиоцен, Сев. Америка.
Scapanus Pomel, 1848. Плейстоцен—современный, Сев. Америка.
Scalopus Geoffroy, 1803 (—Schleps liliger, 1811). Плиоцен—современный,
Сев. Америка.
Подсем. Condylurinae Thomas, 1912. Современный, Сев. Америка.
Condylura liliger, 1811. Современный, Сев. Америка.
В системе Симпсона, естественно, отсутствуют многие роды, описанные
позднее. Трудно согласиться с объединением Симпсона родов Scaptonyx,
Urotrichus, Scapanulus и др. с родами Scalopus, Scapanus и др. в одно подсем.
Scalopinae. Обе названные группы родов эволюировали в разных направ-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
195
лениях. У одних более специализированный жевательный аппарат, у дру-
гих локомоторная система, высокоспециализированная к рытью. Кроме
того, род Dymecodon синоним рода Urotrichus.
В 1948 г. С. У. Строганов опубликовал свою систему современных
кротов. Подсем. Desmaninae он
в отдельное семейство.
Сем. Talpidae
Подсем. Uropsilinae
Rhynchonax Thomas
Nasillus Thomas
Uropsilus Milne-Edwards
Подсем. Urotrichinae
Condylura liliger
Parascalpps True
Scaptonyx Milne-Edwards
Urotrichus Temminck
Scapanulus Thomas
Neurotrichus Gunter
выделил из состава сем. Talpidae
Подсем. Scalopinae
Scapanus Pomel
Scalopus Geoffroy
Подсем. Talpinae
Talpa Linnaeus
Подрод Talpa s. str
Подрод A sioscalops Stroganov
Euroscaptor Miller
Eoscalops Stroganov
Mogera Pomel
Parascaptor Gill
Scaptochirus Milne-Edwards
Включение Строгановым родов Parascalops и Condylura в подсем.
Urotrichinae — группу, объединяющую менее специализированных к ры-
тью, чем упомянутые роды, кротов, но с более специализированным
в другом направлении жевательным аппаратом, представляется искус-
ственным.
Нельзя не согласиться со Строгановым в том, что Rhynchonax, Nasil-
lus и Uropsilus — хорошо дифференцированные роды, объединение кото-
рых в один род Uropsilus [как это сделал с родом Rhynchonax Симпсон
(Simpson, 1945), Эллерман и Моррисон-Скотт (Ellerman, Morrison-Scott,
1951), объединившие с родом Uropsilus роды Rhynchonax и Nasillus, а за-
тем Ван Вален (Van Valen, 1967) и В. Е. Соколов (1973)], лишено основа-
ний. Все кроты — кроты, но при этом разные.
Шварц (Schwarz, 1948) свел роды: Asioscalops Strog., Mogera Pomel,
Scaptochirus M.-Edw., Chiroscaptor Heude (синоним Scaptochirus), Euro-
scaptor Miller, Eoscalops Strog. и Parascaptor Gill в синонимы рода Talpa L.
Можно согласиться co Шварцем только в том, что роды: Asioscalops,
Euroscaptor и Eoscalops должны быть объединены с родом Talpa, по-
скольку морфологические особенности их видов не отражают отличного
от представителей рода Talpa направления адаптации. Нелепость объеди-
нения других, перечисленных здесь родов с родом Talpa, подчеркивается
сведением в синонимы всех их видов к одному виду Тalpa micrura Hodgs.
Избежал этого только Talpa altaica Nik. Вряд ли можно согласиться
со Шварцем, что разные зубные формулы и другие отличные особенности
этих родов представляют собой лишь вариации.
Штейн (Stein, 1960), применив аллометрический метод исследования
структур черепа, предложил свою систему видов и родов кротов в пре-
делах подсем. Talpinae. По представлениям этого автора, в Евразии
существуют следующие роды и виды кротов подсем. Talpinae.
Talpa europaea Linnaeus
T. coeca Savi
T. hercegovinensis Bolkay
T. romana Thomas
T. longirostris M.-Edw.
T. klossi Miller
T. micrura Hodgson
Parascaptor leucura Blyth
Mogera robusta Nehring
M. wogura Temminck
M. latouchei Thomas
Scaptochirus moschatus M.-Edw.
Совершенно справедливо Штейн объединяет роды Euroscaptor Miller,
Eoscalops и Asioscalops Stroganov с родом Talpa L. Все они обладают
четко выраженными общими признаками. Представители ни одного из них
13*
196
СЕМ. TALPIDAE
не отражают в морфологии признаков особого направления в развитии,
что собственно и служит основой для выделения в отдельный род.
Ван Вален (Van Valen, 1967) в своей общей системе насекомоядных
дал свою систему Talpidae.
Сем. Talpidae (Fischer von Waldheim, 1817) Vicq d’Azur, 1792 (Myaladae Gray, 1822;
Oryeteri Blainville, 1834). Поздний эоцен—современный, Европа; ранний олиго-
цен—современный, Сев. Америка; Средний плиоцен—современный, Азия (McDo-
well, 1958; Строганов, 1948).
j-Подсем. Proscalopinae К. М. Reed, 1961 (Arctoryctinae С. A. Reed, Turnbull, 1965).
Ранний олигоцен—средний миоцен, Сев. Америка (Reed, 1961; Reed, Turn-
bull, 19 >5)
^Cryptoryctes C. A. Reed, 1954. Ранний олигоцен, Сев. Америка.
\Oligoscalops К. A. Reed, 1961. Средний олигоцен, Сев. Америка.
\-Proscalops Matthew, 1901 (включает Arctoryctes Matthew, 1907).
Средний олигоцен—средний миоцен, Сев. Америка.
-[Mesoscalops К. М. Reed, 1960. Средний миоцен, Сев. Америка.
Подсем. Uropsilinae Dobson, 1883. Современный, Азия (Ellerman, Morrison-Scott,
1951).
Uropsilus A. Milne-Edwards, 1872 (=Nasillus Thomas, 1911; Rhynchonax
Thomas, 1912). Современный, Азия.
Подсем. Desmaninae (Thomas, 1912), Mivart, 1871 (—Myogalina Mivart, 1871).
Средний олигоцен—современный, Европа; средний плиоцен, Сев. Америка.
fParatalpa Lavocat, 1951. Средний олигоцен, Европа.
\Му galea Schreuder, 1940. Поздний миоцен, Европа.
уMygalinia Schreuder, 1940. Ранний олигоцен—плейстоцен. Европа.
Desmana Guldenstaedt, 1777 (=включает Desman Lacepede, 1799;
Mygale G. Cuvier, 1800, Desmagnus Rafinesque, 1815; Myogalea, J. B.
Fischer, 1829; Caprios Wagler, 1830; Myogala Brandt, 1836; Palaeospa-
lax Owen, 1846). Ранний плиоцен—современный, Европа. Galemys
Каир, 1829 (=Mygalina I. Geoffrey Saint Hilaire, 1835; Galomys
Agassiz, 1846). Плиоцен—современный, Европа.
-\Gaillardia Matthew, 1932 (включает Hydroscapheus Shotwell, 1956).
Средний плиоцен, Сев. Америка.
-\-Geomana Brunner, 1937. Плейстоцен, Европа.
\Desmagale Kretzoi, 1954. Плейстоцен, Европа.
Подсем. Talpinae Fischer von Waldheim, 1917. Поздний эоцен—современный,
Европа; поздний миоцен—современный, Сев. Америка; средний плиоцен—
современный, Азия.
Триба Scaptonychini Van Valen, 1967. Ранний эоцен—ранний миоцен, Европа;
современный, Азия.
\Myxomygale Filhol, 1890. Верхний эоцен, Европа.
\-Mygatalpa Schreuder, 1940. Средне-поздний олигоцен, Европа.
Scaptonyx A. Milne-Edwards, 1872. Средне-поздний миоцен, Европа;
современный, Азия.
Триба Urotrichini Dobson, 1883. Ранний миоцен—современный, Сев. Америка;
миоцен, Европа; средний плиоцен—современный, Азия.
\Domninoides Green, 1958. Ранний миоцен—ранний плиоцен, Сев.
Америка.
Parascalops True, 1894. Плейстоцен—современный, Сев. Америка.
• -\Mydecodon Wilson, 1960. Средний миоцен, Сев. Америка.
yScalopoides Wilson, 1960. Средний миоцен, Сев. Америка.
Proscapanus Gaillard, 1899 (=Proscapaneus Winge, 1917). Миоцен, Европа.
Neurotrichus Gunther, 1880. Современный, Сев. Америка.
Urotrichus Temmink, 1841 (включает Dymecodon True, 1886). Современ-
ный, Япония.
Scapanulus Thomas, 1912. Средний плиоцен—современный, Азия.
Триба Talpini Fischer von Waldheim, 1817. Поздний миоцен—современный,
Европа; плейстоцен—современный, Азия (см. Schwarz, 1948; fmazumi,
1955; Stein, I960).
Talpa Linnaeus, 1758 (включает Mogera Pomel, 1848; Meterotalpa Peters,
1863; Taipops Gervais, 1868; Parascaptor Gill, 1875; Euroscaptor Miller,
1940; Eoscalops Stroganov, 1941; Asioscalops Stroganov, 1941). Поздний
миоцен—современный, Европа; плейстоцен—современный, Азия.
Scaptochirus A. Milne-Edwards, 1867 (=Chiroscaptor Heude, 1898). Совре-
менный, Азия.
Триба Condylurini Trouessart, 1879? Поздний миоцен, плейстоцен—современ-
ный, Сев. Америка.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
197
Condylura liliger, 1811 (=Talpasorix Shinz, 1821; Astromycter Har-
ris, 1825; Rhinaster Wagler, 1830). Поздний? миоцен, плейстоцен—совре-
менный, Сев. Америка.
Триба Scalopini Trouessart, 1879 (включает Scapanei Winge, 1917). Нижний
плиоцен—современный, Сев. Америка.
Scapanus Pomel, 1848 (—Scapanei Winge, 1817). Ранний плиоцен—совре-
менный, Сев. Америка.
Scalopus Е. Geoffroy Saint Hilaire, 1803 (включает Scalops Hibbard,
1941). Ранний плиоцен—современный, Сев. Америка.
В отличие от других систематиков Ван Вален объединил подсем.
Scalopinae и Condylurinae с подсем. Talpinae, выделив в пределах послед-
него три соответствующих трибы и включив в него трибу Urotrichini
(подсем. Urotrichinae — Строганов, 1948). С этим нельзя не согласиться,
поскольку такое систематическое подразделение правильно отражает
уровень адаптивного соотношения подсем. Uropsilinae, Desmaninae и Tal-
pinae, а также отдельных групп внутри последнего.
В чем трудно согласиться с Ван Валеном, так это с выведением из со-
става трибы Urotrichini рода Scoptonyx и выделением его вместе с родом
Mygatalpa Schr. в особую трибу Scaptonychini. То, что известно о морфо-
логии представителей этих родов, не дает оснований для представления
об их общем и особом направлении специализации.
То же можно сказать и о подсем. Proscalopinae — группе специализи-
рованных землероев, близкой к Talpini, особенно трибе Scalopini, объе-
диняющей кротов с общим для всех роющих видов направлением специали-
зации, но с частными особенностями приспособления отдельных частей
локомоторной системы и жевательного аппарата, близкого по общему
характеру к таковому Scalopini. Соотношение уровня обособленности,
направления специализации этой группы кротов не выше такового других
триб, объединяемых подсем. Talpinae.
Здесь не упомянуты еще несколько авторов, из которых одни придер-
живаются системы, сложившейся в итоге работы многих поколений си-
стематиков от Линнея (Linnaeus, 1758) до середины текущего столетия,
полно и естественно представляющей соотношение групп и многообразие
форм частных адаптаций, другие придерживаются упрощенной системы
вроде инвентарного списка — чем короче, тем лучше, — от которой
веет духом прагматизма.
В 1975 г. В. А. Долгов и Б. С. Юдин представили для Фауны СССР
следующую систему кротов.
Род Talpa Род Adoscalops,? Род Mogera
Т. europaea L. As. (?) altaica Nikolsky M. robusta Nehring
T. caucasica Satunin M. w ogura Tcmminck
T. coeca Savi
Авторы «системы» не отметили, к какому виду рода Talpa они отнесли
в качестве синонимов описанного С. У. Строгановым (1944) Т. romana
ognevi и описанного Депарма (Deparmat, 1959) Т. minima. Последнего
Штейн (Stein, I960) отнес в подвиды Т. hercegovinensis Bolkay. Тогда
синонимом какого вида нужно считать последний вид? Как эти, сущест-
вующие в литературе виды, нужно теперь называть?
Под вопросом, без оговорки, ставить в систему ничего нельзя. Прежде
чем ставить в систему, нужно решить: существует ли в природе род Asio-
scalops или нет. Он сведен в синонимы рода Talpa давно (Schwarz, 1948;
Ellerman; Morrison-Scott, 1951; Stein, 1960; Van Valen, 1967; Соколов,
1973).
198 СЕМ. TALPIDAE
Классификация
Предлагаемая система сем. Talpidae основана на подробном сравни-
тельном изучении особенностей морфологии его современных и вымерших
представителей.
Как и в других группах, которые я изучал, и, как принято в систе-
матике млекопитающих, основным материалом, позволяющим сравни-
тельное изучение морфологии современных и вымерших групп, послужил
жевательный аппарат (челюсти и зубы) и отдельные части скелета. Опыт
показал, что определенному типу жевательного аппарата конкретной
группы млекопитающих соответствует комплекс признаков в структуре
скелета и внешней морфологии видов, объединяемых этой группой.
При всех частных сомнениях в системе важно, что в ней отражены
главные направления специализации и преемственность основных групп
в адаптивном процессе, выраженная в их соподчинении, а также после-
довательность в смене форм адаптаций во времени и пространстве.
Сем. Talpidae Fischer von Waldheim, 1817. Европа, верхний эоцен—современный;
Азия, эоцен—современный; Сев. Америка, нижний олигоцен—современный.
1. Подсем. Uropsilinae Thomas, 1912. Азия, эоцен—современный.1
1. Rhynchonax Thomas, 1911. Азия, современный.
2. Nasillus Thomas, 1911. Азия, современный.
3. Uropsilus Milne-Edwards, 1871. Азия, современный.
2. Подсем. Desmaninae Thomas, 1912. Европа, миоцен—современный; Азия, оли-
гоцен—плейстоцен; Сев. Америка, плиоцен.
t 1. Триба Gaillardini nov. Сев. Америка, плиоцен.
f4. Gaillardia Matthew, 1932. Сев. Америка, средний плиоцен,
f5. Hydroscapheus Shotwell, 1956. Сев. Америка, средний плиоцен.
2. Триба Desmanini Thomas, 1912. Европа, миоцен—современный; Азия,
олигоцен—плейстоцен.
f6. Mygalea Schreuder, 1940. Европа, средний миоцен; Азия,
средний—верхний миоцен.
f7. Mygalinia Schreuder, 1940. Европа, верхний миоцен—плиоцен;
Азия, миоцен—плиоцен.
8. Galemys Каир, 1829. Европа, верхний миоцен—современный.
9. Desmana . Giildenstaedt, 1777. Европа, миоцен—современный;
Азия, миоцен — плейстоцен.
3. Подсем. Talpinae Murray, 1866. Европа, верхний эоцен—современный; Азия,
эоцен—современный; Сев. Америка, нижний олигоцен—современный.
3. Триба Urotrichini Stroganov, 1948. Европа, верхний эоцен—миоцен;
Азия, олигоцен—современный, Сев. Америка, миоцен—современный.
flO. Myxomygale Filhol, 1890. Европа, верхний эоцен.
fll. Paratalpa Lavocat, 1951. Европа, олигоцен.
|12. Mygatalpa Schreuder, 1940. Европа, средний—верхний олигоцен.
•|‘13. Scalopoides Wilson, 1960. Сев. Америка, миоцен.
f!4. Midecodon Wilson, 1960. Сев. Америка, миоцен.
15. Scaptonyx Milne-Edwards, 1872. Европа, миоцен; Азия, мио-
цен—современный.
fl6. Domninoides Green, 1956. Сев. Америка, нижний плиоцен.
17. Urotrichus Temminck, 1841. Азия, современный.
18. Scapanulus Thomas, 1912. Азия, современный.
19. Neurotrichus Gunther, 1880. Сев. Америка, современный.
4. Триба Condylurini Thomas, 1912. Сев. Америка, верхний миоцен?—совре-
менный.
20. Condylura liliger, 1811. Сев. Америка, современный.
5. f Триба Proscalopini К. Reed, 1961. Сев. Америка, олигоцен—средний мио-
цен.
+21. Cryptoryctes С. Reed, 1954. Сев. Америка, нижний олигоцен.
f22. Proscalops Matthew, 1901. Сев. Америка, средний олигоцен.
+23. Arctoryetes Matthew, 1907. Сев. Америка, средний олигоцен.
+24. Oligoscalops К.' Reed, 1961. Сев. Америка, средний олигоцен.
+25. Mesoscalops К. Reed, 1960. Сев. Америка, средний миоцен.
6. Триба Scalopini Thomas, 1912. Европа, средний миоцен; Азия? Сев. Аме-
рика, плиоцен—современный.
+26. Proscapanus Gaillard, 1899. Европа, средний миоцен.
КЛАССИФИКАЦИЯ 199'
27. Parascalops True, 1894. Сев. Америка, плейстоцен—современный..
28. Scapanus Pomel, 1848. Сев. Америка, плиоцен—современный.
+29. Hesperoscalops Hibbard, 1941. Сев. Америка, верхний плиоцен.
30. Scalopus Geoffroy Saint Hilaire, 1803. Сев. Америка, плиоцен—
современный.
7. Триба Talpini Fischer von Waldheim, 1817. Европа, верхний эоцен—со-
временный; Азия, эоцен—современный.
+31. Amphidozotherium Filhol, 1876. Европа, верхний эоцен.
+32. Geotripus Pomel, 1848. Европа, верхний эоцен?—олигоцен.
33. Talpa Linnaeus, 1758. Европа, Азия, миоцен—современный.
34. Mogera Pomel, 1848. Азия, современный.
35. Parascaptor Gill, 1875. Азия, современный.
36. Scaptochirus Milne-Edwards, 1867. Азия, современный.
Примечание. Положение в системе родов Scaptogale Trouessartr
1897 и Camphotherium Filhol, 1884 не ясно. Род Scaptogale под вопросом
в системе Симпсона (Simpson, 1945) стоит среди кротов, также и у Ван
Валена (Van Valen, 1967), а род Camphotherium никем не упоминается.
Возможночто последний род, описанный из верхнего эоцена Европы,
вместе с родом Amphidozotherium (Filhol, 1876) представляет плеяду
эоценовых кротов, близких к роду Geotripus или его синонимов.
В систему не помещен род Geomana Brunner (1957), описанный по пле-
чевой кости с явными признаками уродства (рис. 101, Л), а также Desma-
gale Kretzoi (1954), требующий более обстоятельного описания.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ И ТРИБ
1 (2). Ушная раковина хорошо развита. Передние конечности не при-
способлены к рытью или плаванью. Кисть узкая, когти сжатые
с боков. Когтевые фаланги не раздвоены на конце. Хвост длинный
и тонкий................................Подсем. Uropsilinae (с. 200).
2 (1). Ушная раковина не развита. Передние конечности приспособлены
для рытья или плаванья. Кисть расширена и повернута ладонями
наружу, с широкими плоскими когтями или не повернута ладонями
наружу, с обычными когтями, с кожными перепонками между
пальцами. Когтевые фаланги раздвоены или нет. Хвост короткий
тонкий или длинный, в разной степени толстый.
3 (4). Кисти и ступни с перепонками между пальцами. Длина хвоста
не менее 75—80% длины тела. Он утолщен у основания, а далее
уплощен с боков или округлый в сечении, уплощен на конце и в этом
месте с гребнем из жестких волос сверху и снизу, напоминает фор-
мой перо. Когти сжатые с боков. I1 очень крупные, в виде трех-
гранной пирамиды........................Подсем. Desmaninae (с. 204).
4 (3). Кисти и ступни без перепонок между пальцами, а если они есть,
то кисть в разной степени расширена, повернута ладонями наружу
и приспособлена к рытью. Когти на ее пальцах широкие, плоские.
Длина хвоста менее 50% длины тела. Резцы плоские или кониче-
ские ......................................Подсем. Talpinae (с. 226).
5 (6). На конце морды розетка со щупальцами, образованная выро-
стами кожи. Премоляры мелкие, редко стоящие, остробугорчатые,
с вершинами, отогнутыми назад .... Триба Condylurini (с. 239).
6 (5). Признаки иные.
7 (8). Мозговая капсула сильно вздута. Череп по форме напоминает
таковой представителей сем. Chrysochloridae (рис. 133) или плечевая
кость сильно расширена в проксимальном и дистальном отделах,
а сравнительно узкое ее тело изогнуто или ширина тела этой кости
сходна с шириной ее проксимальной части (рис. 132).....................
....................................Триба Proscalopini (с. 242).
200
ПОДСЕМ. UROPSILINAE
к
8 (7). Мозговая капсула не вздута, а плечевая кость иной формы
(рис. 127).
9 (10). Мелкие кроты с резко суженной к носу головой и заметным
шейным перехватом. Кисти с относительно длинными пальцами,
не соединенными кожей (свободными). Серповидная кость развита
слабо. Плечевая кость удлиненная. Слуховые барабаны не полные,
барабанная кость не срастается с соседними костями................
.....................................Триба Urotrichini (с. 227).
10 (9). Крупные кроты с головой, не отделенной от тела шейным пере-
хватом. Кисти с укороченными пальцами, соединенными кожей
до когтей. Серповидная кость развита сильно. Длина плечевой кости
почти равна ее ширине (отношение длины к ширине 60—70%).
Слуховые барабаны полные, барабанная кость у подавляющего
большинства видов срастается с окружающими костями.
11 (12). Верхние клыки хищнической формы, крупные. Нижние клыки
мелкие, прижаты к задним резцам. Рх крупный, выполняет функцию
нижнего клыка...............................Триба Talpini (с. 256).
12 (11). Верхний клык мелкий, конический, сходен с соседними зубами.
Нижний клык также мелкий, стоит на обычном для него месте.
Рх мелкий...................................Триба Scalopini (с. 247).
Подсем. UROPSILINAE Thomas, 1912j
Thomas, 1912, Ann. Mag. Nat. Hist., ser. 8, v. 10, p. 397.
Тип. Uropsilys Milne-Edwards, 1868—1874, Азия, современный.
2 1 4—3 з
Диагноз. Зубная формула: I-р С у Р^—М у = 38—34. Ушная рако-
вина хорошо развита. Передние конечности не приспособлены к рытью
или плаванью. Кисть узкая, когти сжатые с боков. Когтевые фаланги
не раздвоены на конце. Хвост длинный и тонкий.
Описание. Небольшие (длина тела 60—88 мм) наземные бегаю-
щие кроты. Внешним обликом напоминают землероек {Soricidae). Тело
стройное на относительно высоких ногах. Хвост достигает длины тела.
Морда заостренная, ушные раковины хорошо развиты. Глаза мелкие,
скрыты в волосяном покрове головы. Кисти и ступни напоминают таковые
землероек.
Волосы, густо покрывающие тело, короткие, образуют бархатистый
покров. Окраска тела от темно-коричневой до темно-серой.
Череп относительно широкий, особенно мозговая капсула. Скуловые
дуги относительно массивные, вогнуты в орбиту. Барабанная кость в виде
кольца. Количество зубов варьирует. I1 крупные. Кости скелета тонкие,
напоминают соответствующие кости землероек. Ключица длинная, тон-
кая. В скелете кисти отсутствует серповидная кость.
Наземные кроты, составляющие своеобразную по характеру адаптаций
группу, сохраняющую исходный тип локомоторного аппарата, и с этой
точки зрения представляются наименее специализированными в общем
направлении развития семейства. С другой стороны, кроты этого под-
семейства имеют весьма специализированный жевательный аппарат
и в этом отношении могут рассматриваться весьма прогрессивной группой.
Однако наличие ряда архаических особенностей, таких как неполное раз-
витие барабанных камер, что обусловлено несрастанием барабанной
кости с соседними костями, массивные скуловые дуги и нижняя челюсть
и другие, ставит их в особое положение в системе кротовых.
ПОДСЕМ. UROPSILINAE
201
Это не переходная группа между кротами и землеройками, как считал
Строганов (1948), а до него Гайляр (Gaillard, 1915).
Насколько известно, кроты этого подсемейства обитают в лесной
и альпийской зонах гор, например, Uropsilus soricipes M.-Edw. на вы-
соте от 1250 до 4500 м. Следовательно, они подвергаются влиянию смены
условий среды по сезонам. Вряд ли эти мелкие, несомненно подвижные
животные способны накапливать жир и впадать в зимнюю спячку. Ско-
рее всего они активны в течение круглого года, как землеройки, близкие
к ним по размерам тела. Возможно также, что приспособление к жизни
в высокогорьях способствовало их сохранению до настоящего времени.
Распространение и возраст. Азия, эоцен—совре-
менный.
+ Gen. et sp.?
М2 правой стороны верхней челюсти № 2979 (1), 19 км к С-3 от г. Зайсана (Казах-
стан), верхний эоцен? сб. Н. С. Шевыревой и В. М. Чхиквадзе, кол. ПИН АН СССР,
Москва.
Сравнение этого зуба с таковым известных мне кротов позволяет отме-
тить его значительное сходство только с М2 представителя подсем. Uro-
psilinae — Uropsilus soricipes M.-Edw., которое выражается в четырех-
угольной форме коронки зуба, в значительном развитии талона и в
наличии лопасти на месте разросшегося цингулюма на задней стороне ко-
ронки зуба, с впадиной на ее средней части. Отличия заключаются в сле-
дующем: у вымершего вида треугольный бугор метакона развит более,
чем у современного вида; протокон более широкий, талон отделен от него
складкой; задний край коронки почти прямой (у современного на нем
глубокая выемка); цингулюм на передней стороне зуба отсутствует.
Длина описываемого зуба — 1.8, современного — 1.6 мм.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). Общее количество зубов 38.
2 (3). В верхней челюсти отсутствует I3, а в нижней — 13 и Рх ...
.......................'.................. 1. Rhynchonax.
3 (2). В верхней челюсти отсутствует I3, а в нижней — 12 и 13 ...
........................................... 2. Nasillus.
1 (4). Общее количество зубов 34 (в верхней челюсти отсутствуют I9
и Р1, а в нижней — 1213 и Рт)....................3. Uropsilus.
1. Род RHYNCHONAX Thomas, 1911 1
Thomas, 1911, Abstr. Proc. Zool. Soc. London, p. 49.
Тип. Rhynchonax andersoni Thomas, 1911. Сычуань, Юпьнань(Китай), современный.
Диагноз. Зубная формула IС4-РМ= 38. Верхние и нижние
резцы крупные.
Распространение и возраст. Сычуань, Юньнань (Ки-
тай), современный.
1. Rhynchonax andersoni Thomas, 1911.
Thomas, 1911, там же.
Т и п. № 12.2.1.25, М. Р. А. 25ОН, Omei-Shan, Сычуань (Китай), кол. Брит,
муз., Лондон
1 Диагноз этого и следующего родов, а также описания их видов по: Allen, 1938.
202 ПОДСЕМ. UROPSILINAE
Описание. Окраска верхней стороны тела темно-коричневая,
нижней темно-серая.
Измерения (по: G. Allen, 1838). Длина: тела — 67—70, хвоста —
65—68, ступни — 14.5—15.5, общая черепа — 19.9—20.0 мм.
Распространение и возраст. В границах рода.
2. Род NASILLUS Thomas, 1911
Thomas, 1911, Abstr. Proc. Zool. Soc. London, p. 49.
T и n. Nasillus gracilis Thomas, 1911, Сычуань (Китай), современный.
Диагноз. Зубная формула: 1-^-С-|-Р-^-М-|- =38.
Распространение и возраст. Сычуань, Юньнань (Ки-
тай), современный.
1. Nasillus gracilis Thomas, 1911.
Thomas, 1911, там же.
Тип. Ks 11.9.1.13, Shinfu-Shan, юго-восточная часть Сычуани (Китай), кол.
'Брит, муз., Лондон.
Диагноз. Длина тела менее 70 мм. Окраска верхней стороны тела
шоколадно-коричневая, нижней серая.
Измерения (по: G. Allen, 1938). Длина: тела — 66, хвоста — 55,
ступни — 13.5, ушной раковины — 9, общая черепа — 21.3 мм.
Распространение и возраст. Известен из одного
местонахождения, указанного для типа. Добыт на высоте 2000 м.
2. Nasillus investigator Thomas, 1922.
Thomas, 1922, Ann. Mag. Nat. Hist., ser. 9, v. 10, p. 393.
Тип. №22.9.1.16. Kinkiang-Salween, северо-западная часть Юньнани, кол.
Брит, муз., Лондон.
Диагноз. Длина тела более 70 мм. Окраска верхней стороны
тела черно-коричневая, нижней темно-серая.
Измерения (по: G. Allen, 1938). Длина: тела — 70—88, хвоста —
54—75, ступни — 14—16, ушной раковины — 8—10, общая черепа —
21.2—21.8 мм.
Распространение и возраст. Юньнань (Китай). Встре-
чается на высоте 1100—1400 м, современный.
3. Род UROPSILUS Milne-Edwards, 1871
Milne-Edwards, 1871, in David. Nouv. Arch. Mus. d’Hist. Nat., Paris,
1. 7, Bull., p. 92.
Тип. Uropsilus soricipes Milne-Edwards, 1871, Сычуань, Юньнань (Китай), се-
рверная Бирма, современный.
2 13 3
Диагноз. Зубная формула: I-t-C-t-P-^-M —= 34. Р крупные. I2 очень
1 1 О. О
мелкие.
Распространение и возраст. Юго-Восточная Азия,
•современный.
1. Uropsilus soricipes Milne-Edwards, 1868—1874 (рис. 99).
Milne-Edwards, 1871, там же.
Тип. Не отмечен, вид описан из Muping, Сычуань (Китай), кол. Парижск. муз.
Описание. Верхняя сторона тела темно-коричневая, нижняя
темно-серая.
3. РОД UROPSILUS
203
Измерения (по: Walker et al. 1964). Длина: тела — 63—88,
хвоста — 54—75 мм. (По: G. Allen, 1938). Длина: тела — 63—72, хвоста —-
64—65, ступни — 14.5—15.5, общая черепа — 21.0—21.2 мм.
Рис. 99. Uropsilus soricipes (по: Строганов, 1948).
Распространение и возраст. Сычуань, Юньнань (Ки-
тай), северная Бирма, современный. Экземпляр, хранящийся в коллек-
ции ЗИН АН СССР, № 9514, добыт в окрестностях г. Фу-чао.
Примечание. Виды подсем. Uropsilinae представлены в кол-
лекциях отдельных музеев ограниченным числом экземпляров. Объясня-
ется это, по-видимому, не только сравнительно редким посещением райо-
нов Азии, где они обнаружены, но и условиями их местообитаний —
высокогорья от 1000 до 4500 м. Этим же объясняется и слабая изучен-
ность их биологии, а также отсутствие точных данных об их ареале.
204
ПОДСЕМ. DESMANINAE
В последнее время роды Rhynchonax Thos и Nasillus Thos объединяют
с родом Uropsilus M.-Edw. в качестве его синонимов. Такого рода объеди-
нение представляется весьма странным. Хорошо известно, что упомянутые
роды отличаются разным составом зубных рядов верхней и нижней че-
люсти. Разве это не отражает разное направление специализации объеди-
няемых ими кротов? В настоящей работе они сохраняются в своем перво-
начальном статусе.
2. Подсем. DESMANINAE Thomas, 1912 — Выхухоли
Thomas, 1912. Ann. Mag. Nat. Hist., ser. 8, v. 9, p. 397.
Тип. Desmana Giildenstaedt, 1777, Европа, плиоцен, современный; Азия, эоцен—
плейстоцен.
Диагноз. Зубная формула: 1-|-С-|-Р-|-М-|-=:44. Кисти и ступни
с широкими перепонками между пальцами (приспособление к плаванью).
Хвост длинный, не менее 75—80% длины тела, уплощен с боков
на 2/3 длины и покрыт крупными щитками и редкими щетинками или уп-
лощен только на конце, с бахромой из волос по краям.
Череп характерной для кротов формы: с удлиненным лицевым отде-
лом и уплощенной мозговой капсулой, расширенной в слуховой области.
Межтеменная кость хорошо развита или в виде чешуйки. I1 очень круп-
ные, в виде трехгранной пирамиды. Моляры сходны по строению с этими
зубами кротов, но с более развитыми эмалевыми гребнями и большим
числом бугорков по периферии коронки зуба.
Описание. Тело выхухолей толстое, массивное, на коротких
ногах. Голова клиновидная, с длинным, свисающим, расширенным
на конце хоботком. Ноздри расположены спереди или сбоку. Глаза мел-
кие, скрыты в волосяном покрове. Ушные раковины не развиты. Шейный
перехват не выражен. Кисти и ступни широкие. Между их пальцами
натянута кожная плавательная перепонка. По наружному краю кисти
и ступни бахрома из жестких волос. Хвост равен или длиннее тела, сжа-
тый с боков или в сечении округлый, плоский на конце, покрыт крупными
плоскими или мелкими чешуйками, а на конце волосами в виде бахромы.
Тело покрыто мягкими волосами, образующими густой, бархатистый
покров. Впереди глаз, морда и хоботок покрыты разной длины жесткими
щетинками. Вибриссы длинные.
Череп удлиненной формы, с узкими межглазничной областью и носо-
вым отделом. Мозговая капсула в задней части широкая, суживающаяся
в передней. Скуловые дуги тонкие, в средней части параллельны. За-
тылочный отдел черепа с сильно или слабо развитыми гребнями. Боковые
затылочные кости с крупными или мелкими отверстиями. Передние нёб-
ные отверстия большие. Крыловидные и барабанные отростки основной
клиновидной кости, срастаясь и соединяясь с барабанной костью, обра-
зуют удлиненный формы слуховые барабаны, незамкнутые полностью.
У Galemys pyrenaicus Geoff, барабанная кость более свободна, а слуховые
барабаны более плоские. Ветви нижней челюсти с удлиненной зубной
частью. Венечный отросток высокий. Его передний край почти перпенди-
кулярен к линии альвеолярной части ветви нижней челюсти, наклонен
вперед или назад. Сочленовный отросток короткий с цилиндрической,
реже с округлой, сочленовной головкой. Угловой отросток разной длины
и массивности, с многими или небольшим числом, в разной степени раз-
витыми гребнями.
I1 очень крупный, трехгранный, с широким основанием. Задние резцы
мелкие. Клык развит в разной степени, сходен с задним резцом или перед-
ним премоляром, но крупнее. Передние премоляры низкие, массивные,
ПОДСЕМ. DESMANINAE
205
убывают в размерах спереди назад или крупные и мелкие зубы череду-
ются в ряду. Моляры широкие, многобугорчатые. Первый нижний резец
небольпюй, а второй крупный или первый и второй резцы сходных раз-
меров. Третий резец мелкий. Клык сходен по форме с резцом или с пре-
моляром, его размеры изменчивы. Р15 Р2 и Р3 сходной формы, но разных
размеров. Pf отличаются от других премоляров строением и крупными
размерами. Нижние моляры сходного строения, убывают в размерах
спереди назад.
Шейный отдел позвоночника короткий. Ключицы удлиненной формы
направлены вперед и вверх, причленяются к плечевой кости, частью —
к лопатке. Лопатки длинные. Их наибольшая ширина в 4 раза менее
длины. Они расположены параллельно, сближены и соединены поперечной
связкой, а их ости направлены вверх. Акромиальный отросток лопатки
длинный, заостренный. Передние края лопаток находятся на уровне
второго шейного позвонка.
Плечевая кость относительно длинная, с массивным проксимальным
отделом и широким дистальным. Последнее обусловлено главным образом
значительным развитием медиального и латерального надмыщелков.
Олекранон с широким поперечным отростком, соединенным на внешней
стороне с гребнем, цроходящим по нижне-боковому краю олекранона
и локтевой кости. Седалищные кости не соединены перемычками с позво-
ночником. Симфиз лобковых костей имеется хотя бы в виде хрящевой
связки. Бедро сильно расширено в проксимальной части, что обусловлено
значительным развитием троханторов, в дистальном отделе оно утол-
щено. Гребень большой берцовой кости сильно развит и несет направлен-
ный на внутреннюю сторону длинный отросток.
Распространение и возраст. Европа, миоцен—совре-
менный, Азия, эоцен—плейстоцен, Северная Америка, плиоцен.
Основное направление адаптаций представителей подсемейства —
приспособление к плаванью для добывания корма. Специализация к пла-
ванью наложила отпечаток на всю организацию выхухолей, в частности
на локомоторную систему, ограничив возможности использования конеч-
ностей в других формах движения. Из современных выхухолей Desmana
moschata L. еще сохранила способность рыть и роет довольно сложные
норы, a Galemys pyrenaicus Geoff, эту способность, по-видимому, в значи-
тельной степени утратила, поскольку обитает на берегах горных ручьев,
где пользуется укрытиями в скальных обломках, трещинах скал или
норами Arvicola terrestris L.
Оба названных вида выхухолей плохо передвигаются по поверхности
почвы, что объясняется строением их конечностей, особенно кисти и ступни.
Следует отметить, что евроазиатские выхухоли с миоцена имеют сход-
ное с современными видами строение жевательного аппарата и локомотор-
ной системы. Американские выхухоли в строении отмеченных систем
и в плиоцене сохраняли черты начальных стадий их формирования
в направлении, характерном для типичных представителей подсемейства.
fl.Триба GAILLARDINI nov.—АМЕРИКАНСКИЕ ВЫХУХОЛИ
Тип. Gaillardia Matthew, 1932, Северная Америка, средний плиоцен.
Диагноз. Вероятная зубная формула: I С-1- Р М = 44. Триба
включает два рода, описанных по разным остаткам. Поэтому диагноз отра-
жает особенности того и другого. Нижняя челюсть длинная, тонкая,
формой напоминает таковую кротов. Венечный отросток длинный, отогнут
назад. Сочленовный мыщелок округлой, а не цилиндрической формы.
206
ПОДСЕМ. DESMANINAE
Угловой отросток короткий, плоский. Плечевая кость с округлой (а не
овальной формы) сочленовной головкой. Ps и Р4 сходной формы и разме-
ров, конусовидные, с двумя корнями, простые, напоминают эти зубы
представителей рода Desmana Guld.
Примечание. Отмеченные морфологические особенности пред-
ставителей трибы отражают не только ранний этап в формировании при-
способлений, характерных для сем. Desmaninae, но и своеобразный путь
развития американских выхухолей. По-видимому, движение передних
конечностей у видов отдельных групп трибы осуществлялось во многих
плоскостях, что расширяло возможности их использования для работы
разного назначения. Сходное с современными выхухолями строение бедра
указывает и на сходное движение задних ног в воде, а сходное строение
задних премоляров на сходство пищи. Шарнирное, а не блоковидное
сочленение нижней челюсти с черепом (округлый, а не цилиндрической
формы сочленовный мыщелок), позволяет предполагать бблыпую сво-
боду движения нижней челюсти в стороны, что, возможно, связано с пи-
танием и растительной пищей.
Распространение и возраст. Северная Америка, сред-
ний плиоцен.
f 4. Род GAILLARDIA Matthew, 1932
Matthew, 1932, р. 5, 6.
Тип. Gaillardia thomsoni Matthew, 1932. Северная Америка, средний плиоцен.
Диагноз (по: Matthew, 1932). Приблизительная зубная формула
1.1.4.3. Ветви нижней челюсти длинные, тонкие, пропорции, как у Talpa.
Сочленовный мыщелок в виде нароста, округлый, а не вытянут в попереч-
ном направлении. Угловой отросток короткий, расширен в поперечном
направлении и уплощен.
2^ Венечный отросток длинный,
отогнут назад от линии зуб-
ного ряда под углом в 45°.
Зубная часть под премоля-
рами низкая, как у Talpa.
Заднее «подбородочное» от-
верстие под Мх. Зубной ряд
сплошной без диастем, зубы
также без промежутков, тесно
сближены. Последний моляр
брахиодонтный, трехбугорча-
тый, тригонид (пара-прото-
Рис. 100. Gaillardia thompsoni (по: Matthew, 1932). КОНИД, по-моему) более ко-
роткий и широкий, чем у
Talpa или Condylura. Его
талонид (метаконид, по-моему) ниже, чем тригонид и немного шире.
Ра и Р4 сходных размеров, с двумя корнями, простые, массивные, сходны
с этими зубами Desmana Guld. Впереди этих зубов две альвеолы Р2,
более длинного, чем Ря или Р4. Впереди альвеол Р2 одна альвеола Р4,
за которой следуют две альвеолы клыка и одна пологая, крупная, разру-
шенная альвеола.
Примечание. Автор этого рода (Matthew, 1932) в приведенной им
приблизительной зубной формуле отмечает один резец. Сломанный резцо-
вый отдел не может служить основанием для предположения о том, что
этот род выхухолей имеет один нижний резец. Можно думать, что их
было три, как и у современных выхухолей.
4. РОД GAILLARDIA. 5. РОД HYDROSCAPHAEUS
207
Нижняя челюсть этого рода выхухолей отличается от таковых выхухо-
лей Старого Света значительным ее сходством с нижней челюстью кротов
в частности представителей рода Condylura liliger. Но зубы, особенно
премоляры, весьма сходны с зубами выхухолей Старого Света. Можно
предполагать, что американские выхухоли рано отделились от общего
ствола выхухолей, давно развивались независимо от других, и до конца
своего существования на американском континенте сохраняли многие
структуры, близкие кротам. До настоящего времени на американском
континенте сохранился крот, приспособленный к рытью и плаванью
Condylura cristata L., который, возможно, может служить прототипом
кротов, от общих корней с которыми развились выхухоли. На то, что они
произошли от роющих кротов, указывает строение их плечевого пояса
и ряд других общих особенностей.
Рис. 101. Части скелета
А— уродливая плечевая
передней и задней конечностей Hydroscapheus americanus
(по: Shotwell, 1956).
кость, по которой описан род Geomana Brunner, (по:
Brunner, 1957).
Распространение и возраст. Северная Америка, сред-
ний плиоцен.
f 1. Gaillardia thomsoni Matthew, 1932 (рис. 100).
Matthew, 1932, там же.
Тип. Левая ветвь нижней челюсти, с отломанным резцовым отделом, с зубами
Р3—Р4, М3 и сохранившимися альвеолами других зубов, № 20508, горы в верховьях
р. Змеиной (Snake Creek), кол. Амер. муз. естеств. истории, Нью-Йорк.
Измерения. Длина нижней челюсти приблизительно 35 мм.
f 5. Род HYDROSCAPHEUS Shotwell, 1956
Shotwell, 1956, р. 724—726.
Тип. Hydroscapheus americanus Shotwell, 1956, Северная Америка, средний
плиоцен.
Диагноз (по: Shotwell, 1956). Размеры около 4/5 современной
Desmana moschata L. Плечевая кость без двуглавого тоннеля. Головка
плечевой кости округлой формы. На ней нет углубления или ямки для
прикрепления большой связки мускула flexor digitorum profundus. Бедро
имеет овальную головку и сильно развитые сесамовидные кости на эпи-
кондилюсах (лучше сказать: сильно развитые надмыщелковые отростки —
медиальный и латеральный).
Распространение и возраст. Северная Америка, сред-
ний плиоцен.
t 1. Hydroscapheus americanus Shotwell, 1956 (рис. 101).
Shotwell, 1956, там же.
Тип. Плечевая кость NF — 4101, местонахождение McKay, Орегон, кол. Муз.
естеств. истории. Орегопского унив. (США).
208
ПОДСЕМ. DESMANINAE
Описание (по: Shotwell, 1956). Плечевая кость приблизительно
на х/з больше этой кости Mygalinia^ или Galemys и около 4/6 размеров пле-
чевой кости Desmana. Ее головка округлой формы, а не удлиненнооваль-
ная, как у других вихухолей. Она не отделена от малой бугристости,
как у Desmana и Galemys.
Малая бугристость меньше, чем у Desmana. Двуглавый желобок
не закрытый, в форме тоннеля^ как у современных выхухолей, неглубо-
кий. Ориентация грудного гребня кости, как у Desmana. Выступ дельто-
видного отростка заходит дальше назад, чем у Desmana. Мелкий гребень
от дельтовидного отростка проходит по спинной стороне головки плечевой
кости. Дистальный отдел расширен и уплощен, как у других выхухолей.
Отверстие внутреннего надмыщелка открыто в глубокую ямку на передней
стороне, как у Desmana. Неясная ямка олекранона менее глубокая,
чем у Desmana. Нет ямки для прикрепления большой связки мускула
flexor digitorum profundus. Эта ямка хорошо развита у Desmana. У Hydro-
scapheus отмеченная связка прикрепляется к плоской поверхности.
Ключица в основном сходна с этой костью Desmana, отличается мел-
кими деталями, в частности более широким и плоским проксимальным
отделом и более длинным и узким, изогнутым дистальным отделом.
Бедро по внешнему виду сходно с бедром других выхухолей. Отли-
чается главным образом более тонкой трубчатой частью, более крупным
третьим вертелом, расположенным более проксимально, чем у Desmana,
и менее обособленным от большого трохантора. Третий вертел, кроме
того, удлиненный и узкий, морщинистый. Головка бедра овальная,
а не округлая, как у Desmana или Galemys, направлена более вперед,
чем у Desmana.
Измерения (по: Shotwell, 1956). Длина плечевой кости —
18.1, ширина ее проксимальной части — 4.8, дистальной — 10.2, тол-
щина головки — 4.1, ширина трубчатой части кости — 2.8, ее толщина —
2.2; длина ключицы — 8.4 мм, ширина — 1.0, толщина — 0.8; длина
бедра — 19.7 мм, ширина трубчатой части — 3.0, ширина проксималь-
ной части — 7.9, дистальной — 6.4 мм.
Распространение и возраст. Известна из Орегона
(США), средний плиоцен.
2. Триба DESMANINI Thomas, 1912 —
ЕВРОАЗИАТСКИЕ ВЫХУХОЛИ
Desmaninae Thomas, 1912, Ann. Mag. Nat. Hist., ser. 8, v. 9, p. 397.
Тип. Desmana Giildenstaedt, 1777. Европа, плиоцен—современный; Азия, мио-
цен-плейстоцен.
Диагноз. Зубная формула: 1уСуР —М-|- = 44. Нижняя челюсть
длинная, массивная, с вертикально стоящим венечным отростком, реже
отклонен назад или вперед. Сочленовный мыщелок цилиндрический.
Угловой отросток разной длины, массивный, не уплощен. Плечевая
кость с овальной сочленовной головкой. Р2 и Р4 сходной формы и разме-
ров или их форма и размеры разные, конусовидные, Р3 с одним или двумя
корнями, а Р4 с двумя, простые или с бугорками при основании коронки.
Эта группа выхухолей, по-видимому, уже в эоцене обладала многими
особенностями, свойственными их миоценовым представителям и совре-
менным видам. Это представление подтверждается значительной специали-
зацией описываемых млекопитающих в приспособлении к плаванью,
достигнутой в миоцене и практически сохранившейся на том же уровне
у современных родов и видов. Для этого потребовался значительный
ПОДСЕМ. DESMANINAE
209
отрезок времени. Можно думать, что приспособление локомоторной
системы к плаванью шло параллельно с приспособлением жевательного
аппарата, в частности структуры зубов, к питанию преимущественно
водными животными, поскольку обе системы достигли высокого развития
к одному времени и у всех известных групп в равной степени. Они специ-
фичны и сопряжены друг с другом.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). Передний край венечного отростка отогнут назад и образует с ли-
нией альвеолярного края ветви нижней челюсти тупой угол.
2 (3). Наибольшая длина Р2 меньше таковой Р3..........6. Mygalea.
3 (2). Наибольшая длина Рй более таковой Р3 или почти равна . . . .
...............................................7. Mygalinia.
1 (4). Передний край венечного отростка вертикальный, образует с ли-
нией альвеолярного края ветви нижней челюсти прямой угол или
близкий к нему.
5 (6). I2 меньше I3. Р| повернут поперек продольной оси челюсти. Р3
с двумя корнями. 12 спереди вогнут................... . 8. Galemys.
6 (5). I2 больше I3. PJ не повернут поперек продольной оси челюсти.
Р3 с одним корнем. 12 спереди не вогнут..........9. Desmana.
Ниже перечисляются и кратко описываются зубы выхухолей, найденные
в эоцене?—олигоцепе Казахстана. Можно не сомневаться, что они при-
надлежали видам особых родов, отличных от более поздних и современных.
По отдельным зубам не принято описывать формы. Они имеют значе-
ние как фактический материал, позволяющий установить преемственность
между систематическими группами и время существования их представи-
телей. Независимо от того, какими представляются выхухоли эоцена
и олигоцепа по внешности и особенностям адаптаций, факт существова-
ния этой группы кротовых в отмеченные геологические периоды имеет
огромное значение для понимания общего хода развития семейства кро-
товых во времени, а также и для установления вероятного времени фор-
мирования его отдельных подсемейств как особых направлений специали-
зации.
f Gen. et sp.?
Mj левой ветви нижней челюсти с поврежденным параконидом, беловатого цвета,
№ 2979—1005 (1), Конуркура, Зайсанская котловина (Казахстан), эоцен—олигоцен
(сб. Н. С. Шевыревой), кол. ПИН АН СССР, Москва.
Зуб очень мелкой выхухоли (наибольшая ширина — 1 мм). Можно
думать, что размеры выхухоли, которой он принадлежал, были меньше,
чем представителей рода Mygalinia Schr.
При общем сходстве этого зуба с таковым других выхухолей трибы
Desmanini он отличается следующими особенностями: гребень, пересекаю-
щий межсегментную впадину (долинку), не доходит до вершины линг-
вального бугорка протокона (метакона других авторов). У более поздних
и современных видов он всегда достигает вершины этого бугорка. Слабо
развитый цингулюм имеется только на задней стороне коронки зуба.
В том же местонахождении обнаружены два фрагмента ветвей нижней
челюсти с альвеолами зубов, № 2979—134—305 (1—2). Из них один левой
ветви с альвеолами Мх и М2, по размерам (высота и ширина) близок к та-
ковым этой кости Galemys pyrenaicus Geoff., другой — правой ветви
с альвеолами М2 и М3, по размерам средний между таковой Galemys py-
renaicus Geoff, и представителей рода Mygalinia Schr.
Если принять, что описанный выше моляр принадлежал выхухоли,
достигавшей размеров видов рода Mygalinia, а два описываемых фраг-
14 А. А. Гуреев
210
ПОДСЕМ. DESMANINAE
мента нижней челюсти — двум более крупным выхухолям (варианты
размеров исключаются), то можно считать, что имеются документы о трех
формах выхухолей, существовавших в эоцене—нижнем олигоцене
в Казахстане (рис. 107).
t Gen. et sp.?
Mj левой ветви нижней челюсти, коричневого цвета, № 2435—5 (1), Кусто-Кызыл-
кан, Зайсанская котловина (Казахстан), олигоцен, кол. ПИН АН СССР, Москва.
Зуб сходен по размерам и строению с таковыми Galemys pyrenaicus
Geoff., но отличается наличием узкого цингулюма на передней стороне,
в межсегментной впадине, и более широкого — на задней стороне ко-
ронки; задняя сторона коронки не вогнута, как у сравниваемого вида.
Длина коронки описываемого зуба — 2 мм.
t Gen. et sp.?
Мх правой ветви нижней челюсти, черного цвета, № 2976—35—45 (1), р. Аксыир,
Голубые пески, Зайсанская котловина (Казахстан), нижний олигоцен, кол. ПИН
АН СССР, Москва.
Близок по размерам к этому зубу Galemys pyrenaicus. Отличается зна-
чительным развитием цингулюма на передней и задней сторонах коронки,
а также наличием эмалевого наплыва в основании межсегментной выемки.
Последняя более узкая. Задняя сторона этого зуба вогнута сильнее.
Длина по наиболее выступающим частям коронки — 2 мм.
t Gen. et sp.?
Mx левой ветви нижней челюсти, темно-коричневого цвета, № 2076—35—45 (1),
р. Аксыир, Голубые пески, Зайсанская котловина (Казахстан), нйжний олигоцен, кол.
ПИН АН СССР, Москва.
Зуб по длине сходен с таковым Galemys pyrenaicus Geoff. Межсегмент-
ная впадина узкая, в виде борозды. У ее нижнего края поперечная бо-
роздка. Параконид несколько вытянут вперед, а лингвальный и лабиаль-
ный бугорки протоконида (метаконид и протоконид других авторов)
сближены. Длина зуба — 2.3 мм.
t Gen. et sp.?
M1 левой половины верхней челюсти, коричневый, без №, Конуркура (буранская
свита), Зайсанская котловина (Казахстан), олигоцен, кол. ПИН АН СССР, Москва.
Зуб принадлежал мелкой выхухоли (наибольшая длина по внешнему
краю — 1.7 мм, у Galemys pyrenaicus Geoff. — 3.3 мм). Метаконид сильно
вытянут назад. Задний край коронки с глубокой выемкой. Цингулюм
задней стороны коронки образует у нижне-заднего края протокона хорошо
заметный бугорок. Бугорки по краям протокона характерные для выху-
холей, небольшие, слаборазвитые.
f 6. Род MYGALEA Schreuder, 1940
Mygale Pomel, 1848, Bibl. Univ. Geneve, Arch. Sci. phys. nat., t. 9, p. 161.
Mygalea Schreuder, 1940, p. 318—320.
Тип. Mygale antiqua Pomel, 1848, Франция, средний миоцен.
Диагноз. Размеры зубов увеличиваются от передних к задним.
Р1 и Р2 с двумя корнями, а Р3 и Р4 с тремя корнями. Корни Р3 расположены
вдоль челюсти.
Распространение и возраст. Западная Европа, сред-
ний миоцен. Можно не сомневаться, что представители этого рода были
6. РОД MYGALEA
211
распространены и в восточной части Европы, так как в Азии (Казахстан)
они найдены в отложениях средне-верхнего миоцена.
t 1. Mygalea antique Pomel, 1848 (рис. 102).
Mygale pyrenaicus Blainville, 1838, Osteogr. Insect., p. 99—115, pl. XI; —
antiquaP о m e 1, 1848, Bibl. univ. Geneve, Arch. Sci. phys. nat., t. 9, p. 161; Pictet,
Рис. 102. Mygalea antiqua (no: Schreuder, 1940).
Рис. 103. M. asiatica sp. nov.
1848, Paleont., v. 1, p. 175; — sansaniensis L a r t e t, 1851, Notice sur la colline de
Sansan, Paris, p. 18. Mygalea antiqua Schreuder, 1940, p. 318—320.
, Тип. He указан, вид описан из Сансан, Франция.
Диагноз. Очертания верхних моляров близки к квадрату. Цин-
гулюм на нижних молярах отсутствует.
Рис. 104. Mygalinia hungarica Рис. 105. М» zazhigini sp. nov.
(по: Schreuder, 1940).
Описание (по: Schreuder, 1940). Размеры сходны с таковыми
Galemys pyrenaicus Geoff., но кости массивнее. Р1 и Р2 с двумя корнями,
а Р3 и Р4 с тремя корнями. Очертания верхних моляров близки к квадрату.
Подбородочных отверстий два. Ц не очень удлинен. 13 тоньше клыка.
Клык не похож на резец, с одним корнем, сходен с Рх. Р2 с двумя корнями,
Р3 с двумя корнями, стоящими вдоль зубного ряда. Цингулюм на ниж-
них молярах отсутствует. Плечевая кость по длине сходна с этой костью
Galemys Капр, а по пропорциям с мелкими видами рода Desmana Guld.
Измерения (по: Schreuder, 1940). Длина плечевой кости — 12.7;
высота нижней челюсти у Мх — 3.2; длина: Р2 — 1.2; Р3 — 1.3; Р4 — 1.95;
М1 —3.15; М2 —2.3; М3 — 2.0; Р3 — 1.2; Р4 — 1.7; М, — 2.55; М2—
2.5; М3 — 2.1 мм.
Распространение и возраст. Западная Европа (Фран-
ция), средний миоцен.
14*
212
ПОДСЕМ. DESMANINAE
f 2. Mygalea asiatica sp. nov. (рис. 103).
Тип. Фрагмент ветви нижней челюсти с зубами Р8 и Р4, № 952—1, правый
берег р. Ишима, Петропавловск (Казахстан), средний—верхний миоцен, сборы
В. С. Зажигина, 1964, кол. Геологического института АН СССР, Москва.
Диагноз. Очертания верхних моляров близки к треугольнику.
Цингулюм на нижних молярах имеется.
Описание. Средние размеры костей близки к таковым Galemys
pyrenaicus Geoff., отдельные экземпляры крупнее. Кости массивнее.
Рис. 106. Galemys ponticus (по:
Schreuder, 1940).
Рис. 107. Desmaninae gen. et sp.? (a — эо-
цен? — олигоцен, б — олигоцен).
Передний край венечного отростка наклонен назад и образует с линией
альвеолярного края ветви нижней челюсти тупой угол. Сочленовный
отдел челюсти широкий. Заднемандибулярное отверстие расположено
на его заднем крае, как у Desmana verestchagini Topatsh. Р3 и Р4 с тремя
корнями. Подбородочных отверстий два. Р2 с двумя корнями, Р3 с двумя
корнями, расположенными вдоль зубного ряда.
Измерения. Высота зубной части нижней челюсти у Мх — 3.3,
3.5, 3.8; длина: Р4 — 2.5; М1 — 3.3; М2 — 2.3, 2.5; М3 — 1.8; Р2 — 1.8;
Р3 — 2.0; Р4 — 2.3; Мх — 2.8, 2.8, 2.9, 3.0; М2 — 2.5, 2.5 мм.
Распространение и возраст. Известен из одного ме-
стонахождения, указанного для типа.
f 7. Род MYGALINIA Schreuder, 1940
Galemys К о г ш о s, 1913, Ann. Mus. Nation. Hung., v. 11, p. 192—194. Mygali-
nia Schreuder, 1940, p. 316, 317.
Тип. Galemys hungaricus Kormos, 1913, Polgardi, Венгрия, верхний миоцен.
Диагноз. Передний край венечного отростка отогнут назад и
образует с линией альвеолярного края ветви нижней челюсти тупой угол.
Наибольшая длина Р2 более таковой Р3 или почти равна ей.
Распространение и возраст. Западная Европа, верх-
ний миоцен; Азия, миоцен—нижний плиоцен.
f 1. Mygalinia hungarica Kormos, 1913 (рис. 104).
Galemys hungaricus Kormos, 1913, Ann. Mus. Nation, Hung., v. 11, p. 132—134;
1938, Festschr. Embrik Strand, v. 4, p. 163—180.
Тип. He установлен, вид описан из Венгрии.
Диагноз. Рхс одним корнем, не повернут под углом к продоль-
ной оси ветви челюсти.
7. РОД MYGALINIA. 8. РОД GALEMYS
213
Описание. Мелкая выхухоль. Судя по размерам челюстей и
зубов, раза в полтора крупнее Neomys fodiens Penn. Зубы низкие. Подбо-
родочных отверстий два. Р4 по форме напоминает передний сегмент ниж-
них моляров. Рх с одним корнем, а все остальные нижние премоляры
с двумя корнями, расположенными один за другим. Цингулюм развит
на всех зубах. Плечевая кость очень тонкая с цилиндрической средней
частью.
Измерения (по: Schreuder, 1940). Длина плечевой кости —
11.2 мм.
Распространение и возраст. Западная Европа (Вен-
грия), верхний миоцен.
Примечание. Диагноз и описание составлены по: Schreuder,
1940.
f 2. Mygalinia zazhigini sp. nov. (рис. 105).
Тип. Фрагмент левой ветви нижней челюсти с Р4 и альвеолами С—Р3, а также
альвеолами 1х—13 с поврежденными верхними краями, № 952-2, правый берег
р. Ишима, Петропавловск, Казахстан, верхний миоцен—нижний плиоцен, сборы
В. С. Зажигина, 1964 г., кол. Геологического института АН СССР, Москва.
Кроме того, фрагмент левой ветви нижней челюсти с Р2; фрагмент правой ветви
нижней челюсти с альвеолами Р4—М2 и отдельные зубы — Мх (2 экз.). Все из одного
места с типом.
Диагноз. В отличие от Mygalinia hungarica Kormos у этого вида
Р] имеет два корпя и он повернут под углом к продольной оси ветви че-
люсти.
Описание. Первое подбородочное отверстие расположено впереди
Pv а второе — у переднего края Мх. Судя по расположению и величине
альвеол резцов, 1Х у этой выхухоли был тонким и невысоким, 12 был самым
крупным, а 13 немного уступал по размерам 12. Клык был крупным.
Рх с двумя корнями, расположен под углом к оси ветви челюсти. Р2 круп-
нее Р3, имеет три бугорка, Р4 с крупным параконидом. Его метаконид
слит с сильно развитым цингулюмом на задней стороне этого зуба. Мх
относительно крупный, с острыми бугорками характерного для выхухо-
лей строения. Цингулюм хорошо развит, прерывается только на внутрен-
ней стороне метаконида (талонида других авторов). Передний сегмент
(тригонид других авторов) со сближенными бугорками. Ушковидный
отросток на передне-наружной стороне этого зуба слабо развит. Такой же
отросток на его задней стороне хорошо развит.
Судя по размерам челюстей и зубов, описываемый вид имел длину
тела равной или немного более длины тела современной Neomys fodiens
Penn.
Измерения. Высота зубной части ветви нижней челюсти на
уровне Р2 — 1.85—2.5; длина (по альвеолам): С—Р4 — 4.5, Р4—М2 —
4.1; длина: Р2 — 1.25, Р4 — 1.4, Мх — 1.5—1.6 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
8. Род GALEMYS Каир — ГОРНЫЕ ВЫХУХОЛИ
Каир, 1829, Skizz. Еигор. Tierwelt, Bd 1, S. 119. Mygalina Geoffroy
Saint Hilaire in: Gervais, 1835, Resume des Logons de Mamm.; p. 45. Galomys
Agassiz, 1846, Nom. Zool. Index Univ., p. 159.
Тип. My gale pyrenaica Geoffroy Saint Hilaire, 1811, Португалия, Испания, юго-
западная часть Французских Пиренеев, плейстоцен—современный.
214
ПОДСЕМ. DESMANINAE
Диагноз. Передний край венечного отростка вертикальный,
а задний наклонен вперед. I2 меньше I3. PJ повернуты поперек продольной
оси челюсти. Р2 и Р3 с двумя-тремя корнями. 12 спереди вогнут.
Описание. Череп в ростральной части не расширен в области
первых верхних резцов. Шов между предчелюстной и верхнечелюстной
кбстями проходит между I1 и I3. Гребни, особенно затылочные, слабо
развиты. Межтеменная кость широкая, трапециевидная. Гребни на вну-
тренней стороне углового отростка слабо выступают.
Распространение и возраст. Европа, верхний мио-
цен—современный.
f 1. Galemys ponticus Schreuder, 1940 (рис. 106).
Galemys hungaricus Kormos, 1913, Ann. Mus. Nation. Hung., v. 11, p. 132—134.
Desmana pontica Schreuder, 1940, p. 317—318; Topatshevsky, 1962, c. 14.
Тип. В качестве типа Шрейдер (Schreuder, 1940) взят фрагмент верхней челюсти
с зубами С—Р2 (01615074), кол. Венгерск. геол, управл., Будапешт, (Polgardi) Поль-
гарди, верхний миоцен.
Диагноз. Р2 и Р3 с тремя корнями.
Описание (по: Schreuder, 1940). Размеры, близкие к Galemys
pyrenaicus Geoff. Зубы невысокие. Р1 не прижат к клыку. Симфизная
площадка нижней челюсти заканчивается сзади у Рх. Подбородочных
отверстий два. 12 не более мощный, чем 1Х. 13 мелкий. Нижний клык не
похож на резец. Рх и Р3 сильно прижаты к крупному Р2, так что корни
Рх повернуты поперек продольной оси челюсти, но у Р3 корни стоят по-
следовательно.
Распространение и возраст. Европа (Венгрия, Ав-
стрия), верхний миоцен.
t 2. Galemys semseyi Kormos, 1913.
Kormos, 1913, Ann. Mus. Nation. Hung., v. 11, p. 131—132.
Тип. He установлен, вид описан из Piispokfurdo (Венгрия), верхний плиоцен.
Описание (по: Schreuder, 1940). По размерам тела близка к Ga-
lemys pyrenaicus Geoff.
Цингулюмы сильно редуцированы. Р4 удлиненный, со значительным
метаконидом и низким параконидом. Долинка между передним и задним
сегментами нижних моляров узкая, неширокая, как у современной Ga-
lemys pyrenaicus.
Примечание. В. А. Топачевский (1962) считает этот вид ближе
к видам рода Desmana Guld. Пока нет достаточного материала, чтобы ре-
шить этот вопрос более основательно.
Распространение и возраст. Известен из одного место-
нахождения.
3. Galemys pyrenaicus Geoffroy Saint Hilaire,* 1811 — Пиренейская
выхухоль (рис. 108—110).
Mygale pyrenaica Geoffrey Saint Hilaire, 1811, Ann. Mus. Hist.
Nat., t. 17, p. 193. Myogalea rufula G r a e 11 s, 1897, Mem. R. Acad. Madrid, t. 17,
p. 460.
Тип. He установлен, вид описан из Французских Пиренеев.
Диагноз. Р2 и Р3 с двумя корнями.
Описание. Небольшая выхухоль с массивным телом. Голова
крупная, не отделена от тела шейным перехватом. Хоботок относительно
длинный, ноздри на его переднебоковой стороне. Глаза мелкие, окружены
длинными волосами, но хорошо заметные. Ушные раковины не развиты.
Кисти и ступни с плавательными перепонками. Хвост длинный, в сече-
8. РОД GALEMYS
215
нии округлый, и только на конце уплощен. В этом месте его края покрыты
щетинками. Жесткие щетинки покрывают голову впереди глаз и хоботок,
а также края кисти и ступни. Голова и тело покрыты мягкими волосами,
не смачивающимися водой. Окраска тела коричневая сверху, беловатая
снизу. Волосы имеют металлический блеск, что характерно для этой вы-
хухоли.
Череп сравнительно крупный. Носовой отдел с почти параллельными
сторонами. Сагиттальный гребень невысокий, ламбдовидный гребень не
развит. Мозговая капсула более выпуклая, чем у Desmana moschata L.
Нижняя челюсть тонкого сложения, со слабо расширенным (в передне-
Рис. 108. Galemys pyrenaicus.
заднем направлении) венечным отростком и с менее массивными, чем
у D. moschata сочленовным и угловым отростками.
Верхние зубы. I1 с полулунной бороздой на боковой стороне.
I2 и I3 мелкие, цилиндрической формы. I2 немного более I3, оба отклонены
назад. С — с двумя корнями, по строению и размерам сходен с Р2. Оба
эти зуба имеют овальную форму, они вытянуты в передне-заднем направле-
нии. У клыка реже, а у Р2 чаще бывают бугорки — один на передней,
другой на задней стороне коронки. Передний бугорок — паракон, а зад-
ний — метакон. Р1 мелкий, с одним корнем, поставлен поперек челюсти
так, что бугорок паракона расположен на внутренней стороне зубного
ряда, а метакона — на наружной. Р3 меньше Р2, округлых очертаний,
с двумя корнями. Р4 с тремя корнями. Основная вершина протокона этого
зуба смещена к наружной стенке коронки и стоит на одной линии с пара-
и метаконом. Бугорок при основании протокона смещен на задне-внутрен-
нюю сторону коронки зуба и представляет собой талон. Р4 имеет треуголь-
ное очертание, если смотреть на него сверху. М1 с удлиненным наружным
краем, что обусловлено удлинением назад паракона и особенно метакона.
Паракон с тремя бугорками, расположенными треугольником. Из них
передний отделен от других впадиной- Метакон с треугольно располо-
женными гребнями. Протокон не рассечен спереди и с внутренней стороны
глубокими складками, а имеет впадины на передней и задней сторонах,
как у кротов. М2 с параконом и метаконом треугольной формы, разви-
тыми в поперечном направлении, с глубокими выемками между гребнями,
образованными складками, входящими с наружной стороны коронки.
По внешнему краю этого зуба расположены 4 бугорка. Из них два сред-
них соединены в основании. Протокон треугольной формы, с двумя выем-
216
ПОДСЕМ. DESMANINAE
ками: одной спереди, другой сзади. Паракон М3 сходен с таковым М2.
Метакон в виде лопасти, ограниченной сзади высоким зубчатым гребнем.
Протокон треугольной формы, без выемок на передней и задней сторонах.
Рис. 109. Galemys pyrenaicus.
Нижние зубы. 1Х и 12 с вогнутой передней поверхностью. 1Х
с гребнем на средней части коронки. 12 немного крупнее 1Х. На 1/3 от осно-
вания уплощен, с бугорком на внешнем крае. 13 ниже 12 в 2 раза. С и Рх
сходной формы и размеров, с одним корнем. Рх повернут поперек челюсти.
Р2 короче Р3, без бугорка метакона, с двумя корнями. Р3 со всеми тремя
бугорками с двумя корнями. Р4 крупный, с двумя корнями и хорошо
развитыми тремя бугорками. Уплощен в боковом направлении. Мх, М2
и М3 сходной формы, убывают в размерах спереди назад. М3 с более длин-
9. РОД DESMANA
217
ным и менее широким метаконидом (талонидом других авторов) и без бу
горка на его задне-внутреннем крае.
Измерения (по: Niethammer, 1970). Длина: тела — 109—130
хвоста — 123—155, ступни — 31—36, кондило-базальная черепа — 32.5—
34.8 мм.
Распространение
ивозр а с т. Северная часть
Пиренейского полуострова и юго-
западная часть Франции, плей-
стоцен? — современный.
Рис. НО. Расположение зубов в ниж-
нем зубном ряду (а), форма шва между
предчелюстной и челюстной костями (б),
форма нижних резцов (в) у Galemys
pyrenaicus (по: Schreuder, 1940).
Биология. Обитает по берегам горных потоков, поднимаясь
в горы на высоту до 1800 м. Сумеречный и ночной зверек. День проводит
в укромных местах в скальных обломках, древесном хламе, выброшенном
потоками на берег или в норах Arvicola terrestris L. Гнездо устраивает
из травы и сухих отмерших частей растений. Питается водными насеко-
мыми, мелкой рыбой, ракообразными и моллюсками. В неволе (Nietham-
mer, 1970) поедал мясо, рыбу, мышцы моллюсков (Anodonta), земляных й
мучных червей. Ловил рыбу, пускаемую в аква-террариум. В лаборатор-
ных условиях съедал в день от 14 до 21 г мяса.
9. Род DESMANA Goldenstaedt, 1777 — ВЫХУХОЛЬ
Guldenstaedt, 1777, Naturforsch. Freunde, Bd 3, S. 108. Desman Lacepede,
1799, Tabl. divis. Mammifer., Paris, p. 7. Mygale Cuvier, 1800, Tabl. elment. clas-
sific. anim., p. 109. Desmanus Rafinesque, 1815, Analyse de la Nature, p. 59.
Myegalea Fischer von Waldheim, 1829, Synopsis Mammalium, Stuttgart,
p. 250. Caprios W aglet, 1830? Nat. System. Amphib. mit Vorang. Classific. der Sau-
geth. und Vogel, S. 17. Myogale Brandt, 1834, Wiegmann’s Archiv fur Naturg., p. 176.
Тип. Castor moschatus Linnaeus, 1758 (==Desmana moschata L.), Восточная Ев-
ропа, современный.
Диагноз. Передний край венечного отростка вертикальный,
образует с линией альвеолярного края ветви нижней челюсти прямой
угол или близкий к нему. I2 больше I3. Р{ не повернуты поперек продоль-
ной оси челюсти. Р3 с одним корнем, а если их два, то они слиты. 1г спереди
не вогнут.
Описание. Череп в ростральной части расширен в области пер-
вых резцов. Шов между межчелюстной и верхнечелюстной костями про-
ходит между I8 и С. Гребни на мозговой капсуле сильно развиты. Меж-
теменная кость узкая, в виде чешуйки налегает на теменные части.
Гребни на внутренней стороне углового отростка хорошо развиты, как и
на наружной стороне.
Распространение и возраст. Европа, верхний миоцен —
современный; Азия, миоцен—плейстоцен.
Примечание. В Западной Европе известен из плиоцена.
f 1. Desmana verestchaginii Topatshevsky, 1961 (рис. 111).
Desmana sp. В e p e щ а г и н, 1954, Матер, по изуч. Ставроп. края, вып. 6,
с. 172; — verestchagini Topatshevsky, 1961, Палеонтол. журн., № 4, с. 131—137.
218
ПОДСЕМ. DESMANINAE
Тип. Правая ветвь нижней челюсти с зубами Р2—М3, без резцовой части, ве-
нечного сочленовного и углового отростков, № 26967, Косякинский карьер, у Став-
рополя Кавказского, шикпий плиоцен, кол. Зоол. инет. АН СССР.
Диагноз. Размеры ветви нижней челюсти меньше, чем у совре-
менной выхухоли в 1.5 раза. Задний край симфизной площадки доходит
до заднего края альвеолы Р2. Премоляры с удлиненными коронками,,
передний край которых приподнят. Рх и последующие премоляры с двумя
Рис. 111. Desmana verestchagini (по: Топачевский, 1962).
корнями. Р2, Р3 и Р4 с тремя бугорками (пара-, прото- и метаконидом)
Параконид высоко поднят, гребнистый.
Описание. Нижний край зубной части ветви нижней челюсти
более поднят, чем у современной D. moschata L. Передний край венечного-
отростка немного отогнут назад. Угловой отросток, видимо, не был рас-
ширен в дорсо-вентральном направлении, а венечный отросток, судя по
строению сочленовного отдела челюсти, не был высоким. Мандибулярное
отверстие приближено к основанию сочленовного отростка. Заднее под-
Рис. 112. Desmana nehringi (по: Rzebik-Kowalska, 1971).
бородочное отверстие расположено под передним корнем Мх. Рх с двумя
корнями, судя по размерам альвеолы, меньше Р2 и равен Р3. Р2 крупнее
Р3, сходный с последним по форме. Р4 с высоким, островершинным прото-
конидом. Моляры характерной для выхухолей формы. Уменьшаются
в размерах от Мх к М3. Межсегментная долинка у Мх более узкая, чем у М2.
Измерения (по: Топачевский, 1962). Длина: Рх—М3 (по аль-
веолам) — 12.3, Мх—М3 — 6.6 мм.
Распространение и возраст. Известен из одного место-
нахождения.
t 2. Desmana nehringi Kormos, 1913 (рис. 112).
Kormos, 1913, Ann. Mus. Nation. Hung., v. 11, p. 138; 1930, там же, т. 37,
с. 3.
9. РОД DESMANA
219
Тип. Не отмечен, вид описан по нижней челюсти с зубами Р4—М4. Венгрия,
верхний плиоцен.
Диагноз (по: Schreuder, 1940). Размеры средние между современ-
ными видами рода Desmana Giild. и Galemys Каир. Зубы низкие. Моляры
по сравнению с премолярами мелкие. Носовой отдел черепа несколько
сужен, не расширен в области первых резцов. Клык не крупнее, чем Р2,
но выше, не сходен по форме с премолярами. Премоляры удлиненные.
Р2, Р3 и Р4 с тремя корнями. Р3 много меньше, чем Р2, треугольный, с округ-
лыми углами, трехвершинный. Зубная часть ветви нижней челюсти отно-
сительно низкая. Коронарный отросток по высоте средний между таковым
современных выхухолей. На угловом отростке только один гребень. 12
не сильно удлинен (не очень высокий). Р2 сравнительно крупный, со слабо
Рис. 113. Desmana ther mails (a), D. kormosi (б, в) (по: Schreuder, 1940).
развитым параконидом. Р3 удлиненный. Параконид на Р3 и Р4 более или
менее явственный. Метаконид на Р4 может быть выражен. Цингулюм
имеется на Мх и М2. М3 относительно мелкий.
Измерения (по: Schreuder, 1940). Длина: плечевой кости —
16.4—17.7, бедра — 18.2—18.4, Рх—М3 (по альвеолам) — 13.8—14.4,
Mj-—М3 — 7.4—8.3 мм.
Распространение и возраст. Известен из Венгрии,
ФРГ, Польши и СССР, верхний плиоцен.
t 3. Desmana kormosi Schreuder, 1940 (рис. 113, б, в).
Galemys semseyi Kormos, Heller, 1936, Neues Jahrb. Mineral, etc. Beil.,
Bd 76, S. 99—160; Kormos, 1938, Festschrift zum 60 Geburtstage von Profess. Dr.
Embrik Strand, v. 4, p. 163—180. Desmana Kormosi Schreuder, 1940, p. 314—316.
Тип. За тип принят фрагмент верхней челюсти с зубами Р4—М3, Венгрия, верх-
ний плиоцен, кол. Национ. муз., Будапешт.
Диагноз (по: Schreuder, 1940). Размеры сходны с размерами
Galemys semseyi Kormos. Наружная стенка подглазничного отверстия
тоньше и более наклонена, чем у Galemys pyrenaicus Geoff. Зубы относи-
тельно выше, чем у D. nehringi Kormos. Цингулюмы более расширены,
чем у других выхухолей. Премоляры не удлиненные. Р2, Р3 и Р4 с тремя
корнями. Р4 короткий, округлый. Ветви нижней челюсти повышаются
спереди назад. Гребни на внутренней стороне углового отростка не удли-
ненные, а массивные, один около другого. Р3 несколько редуцирован.
Р2 удлинен немного. Метаконид Р4 представлен гребнем или выступом.
Таким образом его тригонид не явственный. На Мх и М2 цингулюм пред-
ставлен на заднем крае зуба. Плечевая кость с более округлым телом,
чем у D. nehringi Kormos.
И з м е р е н и я (по: Schreuder, 1940). Длина: плечевой кости —
14.3—14.4, бедра — 14.4—15.3, Рх—М3 (по альвеолам) — 11.5—11.9,
Mj—М3 (по альвеолам) — 6.6—6.9 мм.
Распространение и возраст, Известен из Венгрии,
ФРГ, Польши и СССР, поздний плиоцен.
220
ПОДСЕМ. DESMANINAE
t 4. Desmana thermalis Kormos, 1930 (рис. 113, a).
Kormos, 1930, Ann. Mus. Nation. Hung., v. 37, p. 5—19. Myogale moschata
П i д о п л i ч к o, 1954, XI наукова cecia КДУ, присвячена 300-p ччю возз еднання
Украши з Pociero, тези доп. с. 94; — thermalis Kormos, Топачевський, 1956,
Тр. Ун-ту зоологИ АН УРСР, т. 13, с. 93.
Тип.- Не отмечен, вид описан по фрагменту верхней челюсти с зубами—Р2—М5 * * 8,
Piispokfiirdo, Венгрия, верхний плиоцен, кол. Национ. муз., Будапешт.
Диагноз (по: Schreuder, 1940). Размеры крупные. Наружная
стенка подглазничного отверстия широкая, вертикальная. Зубы высокие.
Р1 и Р2 с одним, Р3 с двумя корнями, его коронка четырехугольная. Ниж-
няя челюсть более тонкая и низкая, чем у D. moschata L.
Рис. 114. Desmana moschata.
Измерения (по: Schreuder, 1940). Длина: плечевой кости —
18.9—19.7, бедра — 21.3, Рх—М3 (по альвеолам) — 15.8—16.1, Мх—
М3 — 8.9 мм.
Распространение и возраст. Известен из Венгрии,
Нидерландов и СССР, верхний плиоцен.
5. Desmana moschata Linnaeus, 1758 — Русская выхухоль (рис. 114—117).
Castor moschatus Linnaeus, 1758, р. 59. Mygale moscovitica Geoffrey
Saint Hilaire, 1811, Ann. Mus. Hist. Nat., Paris, v, 17, p. 192; — moschatum
Двигубский, 1829, Опыт естественной истории всех животных Российской им-
перии, с. 10. Sorex moschatus Pallas, 1831, Zoographia Rosso-Asiatica, p. 128. Myo-
gale moschata Brandt, 1836, Wiegmanns Archiv fur Naturg., Bd 1, S. 176; — mos-
covitica С и м а ш к o, 1851, Русская фауна, т. 2, с. 101—132.
Тип. Не установлен. При описании вида Линней в качестве «terra typica», от-
метил: «Habitat in Russiae aquosis» (c. 59). Эту цитату привел С. И. Огнев (1928).
Диагноз. Отличается от других видов крупными размерами
узкой наружной стенкой подглазничного отверстия и редукцией корней
на отдельных верхних и нижних премолярах, что легко установить при
сравнении этих зубов с таковыми D. nehringi Kormos, D. kormosi Schreu-
der, D. verestchagini Toparshevsky. Кроме того, D. moschata у нижнего
9. РОД DESMANA
221
края долинки между сегментами нижних моляров имеет бугорок или
морщинистую площадку.
Описание. Размеры тела крупные (длина — 180—226 мм). Тело
массивное, ноги короткие. Голова клиновидная, с длинным массивным
хоботком. Ноздри на переднем крае хоботка. Глаза мелкие, но хорошо
заметные. Ушные раковины не развиты. Шейный перехват отсутствует,
и голова кажется втянутой в тело. Хвост длинный, у основания сужен,
в первой трети утолщен и округлый в сечении, а далее уплощен в боковом
направлении. В утолщении хвоста расположена мускусная железа.
Рис. 115. Кожные складки на губах у Desmana moschata и ее эмбрион.
Кисти и особенно ступни с удлиненными пальцами, между которыми
натянута плавательная кожная перепонка.
Волосяной покров головы и тела мягкий, бархатистый, не смачиваю-
щийся водой. Переднюю часть головы покрывают более жесткие волосы
и разной длины щетинки. Щетинки в разной степени густо покрывают по-
верхность хоботка сверху и снизу. Вибриссы длинные. Жесткие щетинки
покрывают края кистей и стухшей. Хвост покрыт крупными плоскими
чешуйками, плотно прирастающими к его поверхности и редкими щетин-
ками.
Окраска тела этой выхухоли серовато- или темно-коричневая, с се-
ребристым налетом, образованным светлыми концами остевых волос.
Нижняя сторона тела и бока окрашены в серебристо-белый или серебристо-
серый цвета. Встречаются белые выхухоли (альбиносы).
Череп удлиненный, сильно суженный в лицевой части. Задний отдел
мозговой капсулы широкий, с сильно развитыми гребнями. Носовой отдел
в передней части, на уровне первых резцов, расширен. Межглазничная
область сужена. Скуловые дуги тонкие, в средней части параллельны.
Межтеменная кость маленькая, в виде чешуйки, налегает на теменные
кости.
Нижняя челюсть длинная, массивная, с высоким венечным отростком,
передний край которого вертикальный, образует с линией альвеолярной
части ветви нижней челюсти почти прямой угол. Сочленовный отросток
222
ПОДСЕМ. DESMANINAE
короткий, массивный. Угловой отросток высокий, массивный, с сильно
развитыми гребнями на наружной и внутренней сторонах.
Верхние зубы. I1 крупный, трехгранный, с гладкой боковой
поверхностью, I2 крупнее I3, удлиненной формы. I3 мелкий, с конусовид-
Рис. 116. Desmana moschata.
ной коронкой. С с двумя корнями, овально-конусовидный, с одним мел-
ким бугорком на передне-внутренней стороне (паракон) и одним на задне-
наружной (метакон). Центральная вершина протокон. Р1 с одним корнем,
мелкий, с мелкими бугорками у основания протокона, одним на передней,
одним на задней и одним на наружной сторонах. Они могут быть слабо
развитыми и малозаметными. Р2 с двумя корнями, более других вытянут
' в продольном направлении, с тремя бугорками при основании протокона,
на передне-внутренней, задней и наружной сторонах. Р3 мелкий, с одним
9. РОД DESMANA
223
корнем, округлой формы, с тремя бугорками при основании протокона,
расположенными там же, где и у Р2, но более развитыми. Р4 с тремя кор-
нями. Вершина протокона выше вершины Р3, а сходные бугорки при ос-
новании протокона более развитые, чем на Р3.
Р4 имеет округлое очертание. М1 с сильно удлиненной наружной сто-
роной коронки, с тремя корнями. Метакон сильно развит в поперечном
направлении. Протокон широкий, с йногими бугорками. Протокон рас-
сечен на две части складкой, входящей с внутренней стороны. Талон
расположен против метакона в виде выступа. Корень протокона слит
с корнем талона. В месте их слияния сохраняется бороздка или впадина.
М2 с тремя корнями. Корень протокона и талона слиты, но хорошо за-
метны их трубки. Метакон й паракон вытянуты в поперечном направлении
в виде треугольников с впадиной между их гребнями. Протокон рассечен
с внутренней стороны складкой. Та-
лон в виде выступа на одной линии
с метаконом. М3 с тремя корнями,
его паракон широкий, треугольной
Рис. 117. Форма шва между предчелюстной
и челюстной костями (а), форма нижних
резцов (б)у Desmana moschata (по: Schreuder,
1940).
формы, короче, чем эта часть коронки на М2. Метакон на задней стороне
зуба, вытянут в поперечном направлении. Протокон по форме сходен
с таковым М2, но относительно большей величины. Талон отсутствует.
Нижние зубы. 1х сходной формы с 12, но меньше последнего
в 1.5—2 раза. 12 с плоской передней, и задней поверхностями. 13 очень
мелкий, с округлым в сечении основанием и гребневидной вершиной,
направленной вперед. С и Рх сходны по форме с 13, с одним корнем. Р2
с двумя корнями, крупнее Р3, удлиненный. Р3 с двумя корнями, округлый.
Р4 с двумя корнями, высоким протоконом. Метакон расположен на зад-
ней, а паракон, в разной степени хорошо заметный, — на передней сто-
роне основания протокона. Мх, М2 и М3 сходной формы, разной величины.
В складке между сегментами (долинке), у ее нижнего края, бугорок или
морщинистая площадка.
Измерения. Длина: тела — 180—226, хвоста — 170—215,
ступни — 51—58, кондило-базальная черепа — 50.0—55.5 мм.
Распространение и возраст. В настоящее время со-
хранилась в центральных частях бассейнов Волги, Дона и Урала. Успешно
реакклиматизирована на юге Горьковской обл. и по р. Сейму в Курской
(Барабаш-Никифоров, Шапошников, 1974). Выпущена в реки Таган и
Тартас (бассейн Оби) в 1958 и 1968 гг. и пока там сохранилась (Юдин,
1971). Восточная часть Европы, плейстоцен—современный, Азия, плей-
стоцен.
Биология. Местами обитания выхухоли служат различные
пойменные старицы, реже она селится по берегам небольших речек,
предпочитая места с тихим течением или заводи. Для ее существования
необходимо открытое зеркало воды, а также наличие прибрежной и вод-
ной растительности. Для рытья нор выхухоль выбирает высокие, но не
крутые берега у мест, где глубина водоема бывает не менее 2 м. Норы
роет в берегах, а на болотистых участках устраивает их в кочках и дру-
гих более или менее сухих местах. Гнездовая камера с подстилкой устраи-
вается близко от поверхности земли. В местах с сильно меняющимся
уровнем воды выхухоль роет многоярусные норы. В высоких берегах
224
ПОДСЕМ. DESMANINAE
норы не имеют ярусов и идут наклонно к поверхности воды. Выход из нор
всегда располагается под водой. В зависимости от характера грунта
входные отверстия могут быть от 12x15 до 30 x 20 см. Средние размеры
гнезд — около 20x25x17 см. Выхухоль активна в сумерки и ночью.
При переселении из водоема в водоем, при наступлении неблагоприятных
условий, чаще связанных с разливом рек, выхухоль может пройти до 5 км.
Пищей выхухоли служат различные животные: из позвоночных —
мелкая рыба, лягушки и их икра и головастики, из беспозвоночных —
различные моллюски, черви, насекомые и их личинки, ракообразные.
Выхухоль поедает также стрелолист, тростник, камыш, рогоз и другие
прибрежные растения (их корневища и клубни). В неволе выхухоль по-
едает разнообразные корма, животные и растительные.
Размножение изучено недостаточно. Массовое появление родивших
самок в конце июня и в октябре позволяет предполагать два периода раз-
множения: весенний (конец апреля—май) и осенний (конец августа—
сентябрь). Известны случаи размножения выхухоли в зимнее время. Срок
беременности 40—45 дней (по: Юдин, 1971 — 45—50 дней). В помете
1—5 детенышей, в среднем 3—4; осенние пометы меньше весенних. Моло-
дые рождаются голыми и слепыми; двухмесячные весят около 200 г,
а в возрасте одного года достигают размера взрослых.
Врагами выхухоли считаются лисица, горностай, хорь и норка, а также
бродячие собаки, которые при встрече давят этих зверьков. Остатки вы-
хухоли найдены в погадках филина и в желудке щуки.
Выхухоль — ценный пушной зверек, но настолько малочисленный,
что добывание его запрещено. С каждым годом его количество умень-
шается. На многих водоемах он полностью вымер. Причиной этого служат
разного рода хозяйственные мероприятия, ловля рыбы орудиями, в кото-
рые попадают и выхухоли, браконьерство. Во многих местах благоприят-
ному существованию выхухоли мешает сильно размножившаяся ондатра.
Охранные мероприятия пока не дали ощутимых результатов.
Вымершие выхухоли с территории СССР. До недавнего времени наибо-
лее ранние вымершие выхухоли были известны из Западной Европы
(роды Му galea Schr. и Mygalinia Schr. из среднего и верхнего миоцена),
а с территории СССР только плиоценовые и плейстоценовые представи-
тели рода Desmana Guld., характерные и для Западной Европы. Исклю-
чение составлял нижнеплиоценовыйD. verestchagini Topatsh., известный
пока только из СССР (Северный Кавказ).
В 1966—1974 и в последующие годы в Зайсанской котловине (Ка-
захстан) Н. С. Шевыревой и В. М. Чхиквадзе найдены остатки выхухо-
лей разного геологического возраста (кол. ПИН АН СССР). Наиболее
ранние из них относятся к эоцену—олигоцену. Остатки этих выхухолей
описаны на с. 209.
Сходство в строении зубов ранних по геологическому возрасту и позд-
них выхухолей не только подтверждает представление о раннем разви-
тии этой группы кротовых, но и позволяет предполагать, что выху-
холи в эоцене были достаточно хорошо приспособлены к плаванью и,
как и теперь, питались водными организмами. Известно, что сходные
зубы формируются в сходном зубном ряду при сходной функции жева-
тельного аппарата, а сходному жевательному аппарату соответствуют
сходные по строению конечности, обеспечивающие добычу сходного корма.
В 1964 г. на берегу р. Ишима вблизи Петропавловска (Казахстан)
В. С. Зажигиным в миоценовых отложениях собраны остатки выхухолей
родов Mygalea Schr. и Mygalinia Schr. (кол. Геол. инет. АН СССР). До-
статочный для определения и сравнения материал (несколько фрагментов
ветвей нижней челюсти и серия зубов верхней и нижней челюстей) позво-
3. РОД DESMANA
225
.------------—------------------------------------------------------
лил установить, что оба названных рода в Азии были представлены ви-
дами, отличными от европейских (с. 212, 213).
Остатки выхухолей из плиоцена и более позднего времени обнаружены
во многих областях европейской и азиатской частей СССР. В. А. Топа-
чевский (1962) в своей монографии о выхухолях отмечает их нахождение
в следующих областях Украины: Одесской, Херсонской, Запорожской,
Днепропетровской, Черкасской, Киевской и Тернопольской. Таким
Рис. 118. Схема распространения вымерших выхухолей в Евразии и на западе Сев.
Америки по данным находок.
образом, в плиоцене—плейстоцене выхухоли были широко распростра-
нены на территории УССР.
В коллекции Зоологического института АН СССР хранятся остатки
вымерших выхухолей (Desmana nehringi Kormos, D. moschata L., D. kor-
mosi Schr.) из юго-западной Украины (окр. оз. Ялпух, Котловина), верх-
ний плиоцен; из западной части Одесской области (Этулия), средне-
верхний плиоцен, те же виды; из Воронежской области (Урыв, правый
берег р. Дона), средний плиоцен, остатки мелкой выхухоли с узкими верх-
ними коренными зубами, близкой по размерам к D. nehringi-, из Ростов-
ской области (Левенцовка), верхний плиоцен, остатки (зубы), по размерам
близкие к D. kormosi и D. nehringi или D. thermalis-, из Тамани, верх-
ний плиоцен, отдельные зубы, близкие по размерам к D. moschata и D. kor-
mosi. Все материалы из сборов И. М. Громова (1962, 1965, 1972, 1976 гг.).
Там же хранятся материалы по вымершим выхухолям из Ставрополя
Кавказского (Косякин карьер), Северный Кавказ, нижний плиоцен, пра-
вая ветвь нижней челюсти D. verestschagtni Topatsh., сборы Н. К. Вере-
15 А, А, Гуреев
226
ПОДСЕМ. TALPINAE
щагина и др. (1952—1958 гг.); из При-Дон раннечетвертичные остатки
D. thermalis и D. moschata, сборы JL М. Громова и II. К. Верещагина
(1945, 1946 гг.) и из Иркутской обл. (верховья р. Лены, Малые Голы,
250 км от. Байкала), верхний плиоцен, сборы Р. С. Адаменко (1972 г.).
В коллекции Института геологических наук Башкирского филиала
АН СССР хранятся остатки .(плечевые кости и зубы) верхнеплиоценовых
выхухолей, найденных В. П. Суховым в окрестностях дер. Симбугино
(Башкирия) в 1973, 1974 гг. Кости и зубы принадлежат двум видам —
D. kormosi и более мелкой, близкой по размерам и строению плечевой
кости к Galemys pyrenaicus Geoff., но, несомненно, другого вида.
В палеонтологической коллекции Московского университета хранятся
остатки D. kormosi, D. nehringi или D. thermalis (точно определить трудно
по размерам коренных зубов), собранные Р. В. Красненковым в средне-
плиоценовых отложениях Воронежской обл. (Урыв II, Чугуновка, Анги-
новка, 1972, 1973 гг.), предоставленные мне для изучения А. К. Агад-
жаняном.
Среди материалов по палеонтологии насекомоядных коллекции ка-
федры зоологии Свердловского университета мной обнаружен зуб выхухоли,
близкой к D. moschata L., найденный А. Г. Малеевой в Челябинской обл.
(Батурине). Возраст этой находки — верхний плиоцен.
Кроме перечисленных ранее, в коллекции Геологического института
АН СССР хранятся материалы по крупным выхухолям плиоценового
времени, близким к D. moschata из Омской обл. (правый берег р. Иртыша,
Карташево) и с Алтая. Там же хранится фрагмент левой ветви нижней
челюсти, найденный в Туве (р. Холу), средний плиоцен, размерами,
расположением первого и второго подбородочных отверстий, а также
строением альвеолы Р2 ближе всего к Galemys Каир. Все материалы соб-
раны В. С. Зажигиным в 1964—1966 гг.
В заключение следует отметить одну особенность: почти во всех ме-
стонахождениях крупные и мелкие выхухоли встречаются вместе. Вряд
ли это можно объяснить только смешением слоев разного геологического
времени. Современные выхухоли разных родов живут одновременно.
Перечисленные материалы и их местонахождения позволяют составить
представление о разнообразии видов выхухолей, населявших обширную
территорию в европейской и азиатской частях Союза, а также о времени
существования этих кротовых на земле (рис. 118).
3. Подсем. TALPINAE Thomas, 1912
Thomas, 1912, Ann. Mag. Natur. Hist., ser. 8, v. 10, p. 396—397.
Тип. Talpa Linnaeus 1758, Европа и Азия, миоцен—современный.
Диагноз. Зубная формула. -Р :p-f Af-|-=44—36.
Ушные раковины не развиты. Кисти широкие, без перепонок между паль-
цами, повернуты ладонями наружу. Когтевые фаланги раздвоены на
конце. Когти широкие, плоские. Первые верхние резцы плоские, а если
трехгранные, то небольшие.
Описание. Представители семейства имеют вальковатую форму
тела, мелких и средних для насекомоядных размеров. Голова относительно
крупная, клиновидная. Шейный перехват хорошо выражен или отсут-
ствует. Глаза маленькие или закрыты кожей. Ушные раковины не раз-
виты. Кисти широкие, лопатовидные, с крупными плоскими когтями,
с пальцами, соединенными кожей до когтей или свободными, ладонями
ПОДСЕМ. TALPINAE
227
повернуты наружу. Ступни обычной для стопоходящих насекомоядных
формы. Длина хвоста варьирует.
В скелете этой группы кротов следующие особенности: позвонки со слабо-
развитыми остистыми отростками; шейный отдел укорочен; рукоятка
грудины выступает далеко вперед; ключицы короткие и широкие; также
и плечевые кости; олекранон локтевой кости сильно развит; лопатки
длинные и узкие; пояс передних конечностей далеко продвинут вперед
так, что голова помещается между передними конечностями.
Волосяной покров образован короткими, густо растущими волосами —
бархатистый. Преобладает темная окраска, сходная на всем теле —
темно-,черно-серая или темно-бурая, реже коричневая. У отдельных
видов бывают белые пятна на голове, груди и хвосте или хвост весь белый.
Распространение и возраст. Европа, Азия и Северная
Америка, эоцен—современный.
Подсем. Talpinae объединяет землероев, специализированных к рытью
в разной степени. Например, триба Urotrichini объединяет кротов как
малоприспособленных к рытью, могущих передвигаться по поверхности
земли и даже забираться на деревья, так и значительно приспособленных
к роющей деятельности. Эта триба представляет универсальную по ха-
рактеру адаптаций группу, сохранившую до настоящего времени многие
особенности, характерные и для ее олигоценовых и миоценовых пред-
ставителей. С предками этой трибы связывают происхождений предста-
вителей подсем. Desmaninae.
Разнообразие форм адаптации позволило видам подсем. Talpinae ши-
роко расселиться в Европе, Азии и Северной Америке и занять там самые
разнообразные ландшафты. Эта группа кротов населяет преимущественно
лесную зону континентов, выбирая участки с умеренно влажной почвой,
богатой перегноем. В таких местах обитают разнообразные беспозвоноч-
ные, особенно дождевые черви, составляющие основной корм кротов.
Кроты этого подсемейства роют сложнйе норы с многоярусной систе-
мой ходов и большую часть жизни проводят под землей.
3. Триба UROTRICHINI Stroganov, 1948
Urotrichinae Stroganov, 1948, с. 334—335. Urotrichini Van Valen, 1967, p. 264.
Тип. Urotrichus Temminck, 1842, Япония, современный.
Диагноз. Зубная формула: 1СРМ— 42—36.
Мелкие кроты с длинным хвостом (25—100% длины тела). Кисти с отно-
сительно длинными, разделенными пальцами (не соединены кожей).
Серповидная кость слабо развита. Слуховые барабаны незамкнутые (не
в виде пузыря). Барабанная кость не срастается с окружающими костями.
Верхний и нижний клыки мало отличаются от соседних зубов. Рх не круп-
нее соседних зубов или отсутствует.
Описание. В современном состоянии внешним обликом напоми-
нают роющих кротов, но отличаются мейьшими размерами и длинным
хвостом. Голова резко суживается кпереди в хоботок, оканчивающийся
мочкой носа с овальной плоской передней поверхностью. Глаза очень
маленькие, у отдельных видов скрыты под тонкой кожей. Ушные раковины
не развиты. Передние конечности роющие, но менее приспособленные
к рытью, чем у специализированных землероев из других триб. Пальцы
кисти разделены (не соединены кожей), относительно длинные, с силь-
ными когтями, не сжатыми с боков. Длина хвоста варьирует, достигая
у отдельных видов длины тела. Барабанная кость кольцеобразная, не
сросшаяся с соседними костями, не образует замкнутого барабана. Ко-
15*
228
ПОДСЕМ. TALPINAE
личество зубов варьирует. Резцы, клыки и премоляры слабо дифферен-
цированы.
I1 долотообразные или конические, у большинства поставлены вер-
тикально. Серповидная кость развита слабо. Когтевые фаланги раздвоены
на конце.
Триба объединяет кротов, по-разному приспособленных к рытью.
Как отмечают некоторые авторы, отдельные виды способны свободно
передвигаться по земле и даже лазать на деревья. Роются эти кроты под
поверхностью почвы или в лесной подстилке и моховом покрове, возможно,
используя при этом хоботок с голым пятачком на конце, как это делают
златокроты (сем. Chrysochloridae), раздвигающие толщу почвы головой,
просунув предварительно в небольшое отверстие нос, покрытый толстой
голой кожей. Все виды трибы обитатели лесов и зарослей кустарников.
Отдельные виды поднимаются высоко в горы, до 2150—4500 м. Питаются
насекомыми, дождевыми червями, многоножками, пауками и другими
беспозвоночными, отчасти мелкими позвоночными.
Распространение и возраст. Европа, олигоцен—миоцен;
Азия, олигоцен—современный; Северная Америка, нижний миоцен—совре-
менный.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВЫМЕРШИХ РОДОВ
1 (4). Р2, Р3, Р4 с двумя корнями каждый.
2 (3). Корни Ра и Р3 расположены вдоль продольной оси нижней челюсти
................................................11. Paratalpa.
3 (2). Корни Р2 и Р3 расположены поперек продольной оси нижней че-
люсти ......................................16. Domninoides.
4 (1). Р2 с одним, а Р3.с одним или двумя корнями.
5 (6). Р2 с одним, а Р3 и Р4 с двумя корнями........14. Mydecodon.
6 (5). Р2 и Р3 с одним корнем, а Р4 с двумя корнями......
.................................... 10. Myxomygale.
Примечание. Роды Mygatalpa и Scalopoides не включены в таблицу за от-
сутствием сравнимых признаков.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СОВРЕМЕННЫХ РОДОВ
1 (2). Общее количество зубов — 42 .... ..............15. Scaptonyx.
2 (1). Общее количество зубов — 36.
з
3 (4). Резцов -= ..................................19. Neurotrichus.
О
4 (3). Количество резцов иное.
5 (6). Резцов 4 ....................................17. Urotrichus.
2
6 (5). Резцов .........................................18. Scapanulus.
110. Род MYXOMYGALE Filhol, 1890
Filhol, 1890, Bull. Soc. Philomatique, Paris, v. 2, p, 170.
Тип. Myxomygale antiqua Filhol, 1890, Франция, верхний эоцен.
Диагноз, (по: Filhol, 1890, р. 170, из: Crochet, 1974, р. 114). Резцы
уменьшаются спереди назад. Альвеола клыка крупнее таковой 13 и Pv
Pj-з с одним корнем, увеличиваются спереди назад. Р4 с двумя корнями.
Моляры уменьшены в задней части. Талоны этих зубов (метакониды по-
моему) сжаты спереди назад. Восходящая ветвь нижней челюсти сходна
с таковой выхухолей.
10. РОД MYXOMIGALE
229
К этому диагнозу Кроше (Crochet, 1974) добавляет: впереди Р4 семь
альвеол. Моляры с цингулюмом. Метаконид впереди протоконида. Та-
лонид высокий. Параконид заметно выдвинут вперед. Восходящая ветвь
нижней челюсти типична для трибы.
П римечание. Кроше в описании вида сравнивает род Myxo-
mygale с рядом родов трибы Urotrichini, что, мне кажется, не может дать
достаточных основании для каких-
либо определенных заключений о его
близких родственных связях с какой-
нибудь из сравниваемых групп.
Описываемый род самый древний из
известных, объединяемых этой три-
бой. Основные морфологические осо-
бенности челюсти и зубов представ-
лены в диагнозе рода и на рисун-
ках. В отличие от более поздних и
современных родов известный пред-
ставитель рода Myxomygale обладал
полной зубной системой и более вер-
тикально направленным венечным
отростком нижней челюсти, напоми-
нающим в этом отношении выхухо-
лей.
Распространение и
возраст. Европа, верхний эоцен.
f 1. Myxomygale antiqua Filhol,
1890 (рис. 119).
Рис. 119. Myxomygale antiqua (no: Cro-
chet, 1974).
Filhol, 1890, там же.
Тип. QU 8682, правая ветвь нижней челюсти с зубами Р4—М2 и альвеолами
остальных зубов полного ряда; с повреждс.г’ ымл венечным и угловым отростками,
фосфориты Кверси, Франция, верхний эоцен, кол. Инет, палеонтол. Национ. муз.
естеств. истории, Париж.
Всё, отмеченное в характеристике рода, относится и к виду. Следует
только отметить, что уже в эоцене представители рода Myxomygale Filh.
имели относительно специализированные зубы, в частности премоляры,
напоминавшие формой эти зубы более поздних и современных видов
кротов.
Измерения. Р4: длина — 1.33Хширина — 0.84 мм; М4 — 1.91 X
X 1.24; М2 — 1.91X1.18.
Распространение и возраст. Франция, верхний эоцен.
I
f 11. Род PARATALPA Lavocat, 1951
Lavocat, 1951, р. 30.
Тип. Paratalpa micheli Lavocat, 1951, Овернь, Франция, олигоцен.
Диагноз, (по: Lavocat, 1951). Нижняя челюсть кротовидная.
Три задних премоляра с двумя корнями. Р2 и Р4 массивные, простые. Ра
мелкий. Моляры по строению сходны с этими зубами Galemys Каир.
Талонид (метаконид) этих зубов крупнее тригонида (переднего сегмента).
Распространение и возраст. Европа, олигоцен.
f 1. Paratalpa micheli Lavocat, 1951 (рис. 120).
Lavocat, 1951, p. 32.
Тип. Фрагмент ветви нижней челюсти с.вубами Р2—М2. Ju 19, Овернь, Франция,
олигоцен.
230
ПОДСЕМ. TALPINAE
Описание, (по: Saban, 1958). Нижняя челюсть менее выгнута,
чем у Talpa L. Премоляры с двумя корнями. Р3 меньше соседних премо-
Рис. 120. Paratalpa micheli (по: Lavocat, 1951).
Рис. 121. Scalopoides isadens (по: Wilson, 1960).
Рис. 122. Midecodon martini (no: Wilson, 1960).
ляров (P2 и P4), более массивных. Строение сохранившихся моляров не-
сколько напоминает строение этих зубов Desmaninae, но отличается
наличием одного цингулюма (передне-наружного), со значительно ре-
дуцированным его соединительным образованием между гипоконидом и
метаконидом, что характерно для Talpinae.
Измерения (по: Lavocat, 1951). Длина: Мх—М3 — 3.3, Р2—Мх —
4.8 мм.
Распространение и возраст. Франция, олигоцен.
f 12. Род MYGATALPA Schreuder, 1940
Schreuder, 1940, р. 320—324.
Т и п. Mygatalpa arvernensis Pomel, 1848, Овернь, Франция, верхний олигоцен.
13. РОД SCALOPOIDES
231
Д и а г но в (по:. Lavocat, 1951). Насекомоядное, представляющее сме-
шение признаков Talpinae и Desmaninae.
Описание (по: Saban, 1958). Структурные особенности скелета и
зубов представителей этого рода сочетают признаки кротов и выхухолей.
Р4 имеет три корня и круговой цингулюм, задне-внутренний талон, на
котором два отдельных бугорка. На верхних молярах острия (бугорков)
менее развиты, чем у выхухолей. Паракон и метакон образуют букву W,
что характерно для кротов. Нижние моляры имеют соединенные прото-
и метаконид. Плечевая кость с цилиндрическим телом как у выхухолей.
Распространение и возраст. Европа, верхний олигоцен.
t 1. Mygatalpa arvernensis Pomel, 1848.
Desman arvernien Pomel, 1844. Bull. Soc. Geol. France, p. 592. — Mygale naya-
dum et arvernensis Pomel, 1848, Bibl. Univ. Geneve, Arch. Sci. pliys. nat., t. 9,
p. 162. — Talpa cfr. brachychir Viret, 1929, p. 44.
Тип. He отмечен, вид описан по фрагментам верхней и нижней челюсти и пле-
чевой кости, Овернь, Франция, верхний олигоцен.
Измерения (по: Schreuder, 1940). Длина: Р4 — 1.35, М1 — 2.1,
М2 — 1.55, М3 — 1.55, Р4 — 1.0, Мх — 1.5, М2 — 1.65, М3 — 1.25 мм.
Распространение и возраст. Франция, верхний оли-
гоцен.
Примечание. Своеобразные особенности представителей этого
рода создают впечатление, что род основан на костных остатках не одного
рода. С другой стороны, не вызывает сомнения возможность существо-
вания в пределах отдельных групп кротов тенденции к развитию при-
способлений для плавания и использования животных водной среды как
дополнительной кормовой базы. Эта тенденция могла в конце концов
выразиться в разной форме приспособлений, часть которых отражается
в структурах отдельных частей скелета. Возможно, что с такими груп-
пами кротов имеют общие корни и выхухоли.
f 13. Род SCALOPOIDES Wilson, 1960
Wilson, 1960, р. 42—46.
Тип. Scalopoides isodens Wilson, 1960, северо-восточная часть Колорадо (США),
нижний миоцен.
Диагноз (по: Wilson, 1960). Крот из группы, близкой к Scalo-
pus Geoff. S. Hilaire.
Зубная формула: I—С—P—M—. Резцы и клык мелкие. Рх мелкий,
сходный по форме с резцами. Р2 с одним корнем. Р3—Р4 с двумя корнями.
Коронки моляров более или менее мезодонтные. Прото- и гипоконид
угловатые. Выступающий метастиль имеется на фланге метаконида М2 и
М3. Передний выступ гипоконидов простирается до оснований метакони-
дов. Мх и М2 почти равны. М3 меньше, но массивнее. Нижняя челюсть
по форме кротовидная. Имеются два подбородочных отверстия. Одно
расположено позади заднего корня Р4 или, что более часто, между Р4 и
Мх, а другое позади второго корня Р3.
Верхние моляры трехбугорчатые. Гипокон (талон, по-моему) и бу-
горок на передней стороне протокона малозаметны или отсутствуют.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
t 1. Scalopoides isodens Wilson, 1960 (рис. 121).
Wilson, 1960, p. 42—46.
Тип. Левая ветвь нижней челюсти с зубами 13—Рх, Р4—М2, № 10067 область Ло-
ган, Колорадо (США), нижний миоцен, кол. Муз. естеств. истории Канзас, унив.,
Лоренс.
232
ПОДСЕМ. TALPINAE
Примечание. Сравнение плечевой кости S. isodens и Condylura
cristata L. позволяет отметить их значительное сходство. Можно думать,
что имело место известное сходство и уровня приспособления к рытью.
Некоторое сходство имеет и нижняя челюсть этих видов, но строение
зубов и зубных рядов резко отлично.
Измерения (по: Wilson, 1960). Альвеолярная длина Р2—М3 —
8.4, М4—М3 — 5.3—5.8 мм.
Распространение и возраст. Северо-восточная часть
Колорадо (США), нижний миоцен.
f 14. Род MYDECODON Wilson, 1960
Wilson, 1960, р. 38—42.
Тип. Mydecodon martini Wilson, 1960, северо-восточная часть Колорадо (США),
нияший миоцен.
Диагноз (по: Wilson, 1960). Мелкий, землеройкообразный крот.
Зубная формула: I—С—Р—М—? Длина Мх—М3 — 4.2 мм. Клык медкий.
Рх отсутствует? Р2 с одним корнем. Р3—Р4 с двумя корнями. Р2—Р4 по-
степенно увеличиваются в размерах спереди назад. Моляры с умеренно
брахиодонтными коронками. Наружный цингулюм у них имеется. Перед-
ние отростки гипоконидов простираются до основания метаконидов (не
ясно, что за образования имеются в виду). Бугорки тригонида (переднего
сегмента) Мх тесно сближены. Второе подбородочное отверстие располо-
жено под передним корнем Мх, а первое — под передним корнем Р4.
Распространение и возр ас т. Северная Америка, ниж-
ний миоцен.
t 1. Mydecodon martini Wilson, 1960 (рис. 122)
Wilson, 1960, p. 38—42.
Тип. Неполная правая ветвь нижней челюсти с зубами Р4—М3 № 10058, об-
ласть Логан, Колорадо (США), нижний миоцен, кол. Муз. естеств. истории Канзас,
унив., Лоренс.
Примечание. Автор, описавший этот вид, отмечает, что его
размеры меньше таковых Urotrichus talpoides Temm. Далее он сравнивает
структурные особенности зубов этого вида с зубами многих родов, находя
то сходство, то различие. Отсутствие многих деталей на фрагментах
ветвей нижней челюсти, послуживших автору для описания вида, а также
отсутствие полного ряда зубов не позволяет решить вопрос о его система-
тической принадлежности. Предположительно он должен быть отнесен
к трибе Urotrichini.
Измерения (по: Wilson, 1960). Длина: С—М3 — 6.6, Р2—М3
(по альвеолам) — 5.8, М4—М3 — 4.2 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
15. Род SCAPTONYX Milne-Edwards, 1868—1874
Milne-Edwards, 1868—1874, р. 278.
Тип. Scaptonyx fasicaudatus Milne-Edwards, 1868—1874, Известен из северной
части Юньнани, Сычуани, Тибета и оз. Кукунор (Китай), современный.
Диагноз. Зубная формула: I С Р М -j- = 42. I1 долотообразные.
О 1 .о о
I1 немного шире I2 и Is, сходной с ними высоты, не ниже клыка.
Описание. Глаза открытые (не покрыты кожной перепонкой).
Пальцы кисти короткие. Верхние и нижние клыки сходны с премолярами
и лишь немного крупнее их. Р2 и Р3 конусовидные. Р4 более высокий
15. РОД SCAPTONYX
233
в зубном ряду, имеет сзади добавочный бугор. Р2 меньший в нижнем
зубном ряду. Р3 сходен по размерам с клыком, а Р4 крупнее других пре-
моляров, с бугорком на задней стороне. Верхние и нижние молярыпо-
хожи на эти зубы представителей рода Talpa L.
Ключица удлиненной формы. Плечевая кость менее укорочена, чем
у других, приспособленных к рытью, кротов.
Распространение и возраст. Европа, средний миоцен,
Азия (Китай, Северная Бирма), средне-поздний миоцен—современный.
Рис. 123. Нижвяя челюсть (Л) и плечевая кость (В) Scaptonyx edwardsi; S. dolichochir
(В) (no: Zapfe, 1951).
f 1. Scaptonyx edwardsi Gaillard, 1899 (рис. 123, А, Б).
Gaillard, 1899, p. 29—30.
Тип. He отмечен. Вид описан из Grive-Saint-Alban (Isere), Франция, средний
миоцен.
Описан по фрагменту правой ветви нижней челюсти с зубами Р3—М2
и плечевой кости. Зубы имеют известное сходство с этими зубами Scapto-
nyx fusicaudatus М. Edw., а плечевая кость настолько значительно отли-
чается от плечевой кости сравниваемого вида, что не вызывает сомнения
в принадлежности ее другому роду.
Распространение и возраст. Известен из одного место-
нахождения.
t 2. Scaptonyx dolichochir Gaillard, 1899 (рис. 123, В).
Gaillard, 1899, р. 30, 31.
Тип. Не отмечен, вид описан из Grive-Saint-Alban (Isere), Франция, средний
миоцен.
Этот вид описан по плечевой кости. Сравнение ее с плечевой костью
Scaptonix fusicaudatus позволяет отметить известное сходство. Отличие
заключается в большей длине, меньшем развитии медиального и лате-
234
подсем; talpinae
рального надмыщелков, а также в меньшем развитии гребня на месте
малого вертела. Кроме того, сочленовная головка плеча с меньшим пере-
хватом у основания (шейки).
Распространение и возраст. Известен из одного место-
нахождения.
3. Scaptonix fusicaudatus Milne-Edwards, 1868—1874 (рис. 124).
Milne-Edwards, 1868—1874, р. 278.
Тип. Не установлен, вид описан из окрестностей оз. Кукунор (Китай).
Рис. 124. Scaptonyx fusicaudatus (по: Milne-Edwards, 1868—1874).
Диагноз. Тот же, что и рода, поскольку род описан по морфо-
логическим особенностям этого вида. Другие виды рода требуют проверки
их отношения к роду Scaptonyx.
Описание. Волосяной покров мягкий, густой, бархатистый.
Волосы аспидно-серые, с коричневыми концами, которые придают окраске
при сером общем тоне буроватый оттенок.
Измерения. Длина: тела — 65—90, хвоста — 26—31, ступни —
12.0—15.5, наибольшая черепа — 22.4—24.3 мм.
Распространение и возраст. Сычуань, Юньнань (Ки-
тай), Северная Бирма, современный.
Биология. Не изучена. Поднимается в горы до 2150—4500 м.
1 экз. в долине р. Меконга пойман в моховом покрове елового леса, днем.
Примечание. Род Scaptonyx M.-Edw. и его виды описаны по:
Gaillard, 1899; G. Allen, 1938; Walker et al., 1964.
f!6. Род DOMNINOIDES Green, 1956
Green, 1956, J. Paleont., v. 30, N 1, p. 152—153.
Тип. Domninoides riparensis Green, 1956, Южная Дакота (США), нижний плио-
цен.
Диагноз, (по: Green, 1956). Гипоконид соединен с метаконидом
гребнем. Ложбина талонида М2 и М3 открыта на лингвальной стороне.
Талонид М3 не редуцирован.
Распространение и возраст. Северная Америка, верх-
ний миоцен, нижний плиоцен.
f 1. Domninoides riparensis Green, 1956 (рис. 125).
Green, 1956, там же.
Тип. Левая ветвь нижней челюсти с обломками первого резца, корнями Р, и
Р2 и сохранившимися зубами Р3—М3, SDSN № 53107, Южная Дакота (США), нижний
плиоцен, кол. Муз. Канзас, унив., Лоренс.
16. РОД DOMNINOIDES
235
Д и а г н оз. (по: Green, 1956). Передние цингулюмы М2 и М3 сильно
развиты.
Описание, (по: Green, 1956, с сокращениями). 14 немного накло-
нен вперед. Р4 имеет наклон. Его единственный корень сходен с таковым
клыка, он крупнее, чем Р2 и Р3. Р2 с двумя корнями, его корни расположены
в поперечном направлении по отношению к продольной оси челюсти.
Р3 с двумя корнями, его корни также расположены в поперечном направ-
лении. Вершина этого зуба немного отогнута назад. Между Р3 и Р4 имеется
короткая диастема. Р4 с двумя корнями, расположенными по продольной
оси челюсти. Вершина этого зуба немного изогнута. Задний сегмент М4
много шире переднего. Задний сегмент М2 относительно менее широкий.
М3 — наименьший из моляров, его задний сегмент уже переднего.
Рис. 125. Domninoides ripariens (по: М. Green, 1956).
Распространенней возраст. Известен из одного место-
нахождения.
t 2. Domninoides valentinensis К. Reed, 1962.
К. Reed, 1962, р. 1—3.
Тип. Фрагмент правой ветви нижней челюсти с зубами: Рх, Р2—Р3, М2 и М3,
№ ИСМР 33152, формация Валентина, форт Ниобрага, область Чарри, Небраска
(США), верхний миоцен, кол. Палеонт. муз. Калифорй. унив., Беркли.
Диагноз (по: К. Reed, 1962). Более крупный и более массивный,
чем D. riparensis Green. Диастемы между Р3 и Р4 нет. Бугорок на мета-
кониде М2 (метастиль других авторов) ясно выражен.
Измерения (по: К. Reed, 1962). Длина: М2 — 3.1—3.2, (при-
близительная) М3 — 2.4; высота нижней челюсти в области М4 — 3.7 мм.
Распространение и возраст. Известен из верхнего
миоцена Небраски (США).
t Gen. et sp.?
M2 левой стороны верхней челюсти, № не установлен, Зайсан, Хонгуркульджа
(Казахстан), средний олигоцен, кол. ПИН АН СССР, Москва.
Сравнение описываемого зуба с таковым разных групп кротовых
позволило установить его наибольшую близость к М2 представителей
трибы Urotrichini. Из двух сравниваемых видов трибы — Neurotrichus
gibbsi Baird и Urotrichus talpoides Тепла., он ближе к последнему. При
236
ПОДСЕМ. TALPINAE
общем сходстве зуб олигоценового вида отличается следующими особен-
ностями: большей шириной и меньшей длиной коронки зуба; эмалевый
треугольник паракона более развит, чем у современного вида; впадина между
параконом и метаконом менее широкая; протокон крупнее; бугорок талона
менее развит, а бугорок на передней стороне протокона отсутствует, под
бугорком талона выемка, которой нет у современного.
17. Род UROTRICHUS Temminck, 1842
Temminck, 1842, Fauna Japonica, v. 1, p. 22. Dymecodon True, 1886, Proc.
U. S., Nat.-Mus., v. 9, p. 97.
Тип. Urotrichus talpoides Temminck, 1842, Япония (о-ва Кюсю и Сикоку).
3 13 3
Диагноз. Зубная формула: I -к- С -тг Р М — — 36. Р конические,
Z и о о
трехгранные. Значительно превышают высоту I2 и Is, а также высоту
клыка.
Описание. Череп внешне напоминает череп Galemys, что обус-
ловлено сходным строением I1 и формой носового отдела черепа, а также
строением одновершинных клыков и премоляров. Кроме того, формой
скуловой дуги и межтеменной кости.
I2 и I3 более или менее округлой формы в поперечном сечении. I2
с отогнутой наружу вершиной. Верхние клыки и премоляры сходной
формы, конические, увеличиваются в размерах спереди назад.
по форме сходны с верхними, но меньше их по размерам. 12 и 13
имеют наклонное положение, меньше Iv Нижние премоляры одновер-
шинные, конические. Верхние и нижние моляры общей для кротов
формы.
Распространение и в оз р а с т. Азия (Япония), совре-
менный.
1. Urotrichus talpoides Temmink, 1842 (рис. 126, 127).
Т е m m i n k, 1842, там же.
, Тип. Не отмечен, вид описан с Kiu-shiu (Кюсю, Япония).
Описание. Волосяной покров мягкий, но не бархатистый как
у других кротов (по: Walker et al., 1964). Ушные раковины имеются,
небольшие и скрыты в шерсти. Окраска темно-коричневая до черной,
с металлическим блеском. Экземпляр из Зоологического музея Берлин-
ского университета, бывший у меня в руках, имел одинаковую окраску
верхней и нижней сторон, сверху она имела охристый оттенок, снизу
серебристо-охристый. Так же окрашены экземпляры из коллекции ЗИН
АН СССР.
Измерения (по: Walker et al., 1964). Длина: тела — 64—102,
хвоста — 24—41 мм (по: Строганов, 1948). Длина: тела — 80—101,
хвоста — 26—34, ступни — 14—16, общая черепа 24.7—26.5 мм.
Распространение и возраст. Хондо и другие острова
Южной Японии, современный.
Биология (по: Walker et al., 1964). Этот вид обитает в лесах
от основания гор до высоты 2000 м, роется под поверхностью почвы,
активен и на ее поверхности. Забирается на кустарники и деревья. Зимой
Часто устраивается в пустых гнездах птиц. При этом он карабкается по
дереву на высоту 2—4 м.
126
121 ।
Рис. 126, 127. Urotrtchus talpoides (по: Строганов, 1948).
238
ПОДСЕМ. TALPINAE
18. Род SCAPANULUS Thomas, 1912
Thomas, 1912, Ann. Mag. Nat. Hist., ser. 8, v. 10, p. 396—397.
Тип. Scapanulus oweni; Thomas, 1912, Шэньси, Ганьсу, Сычуань (Китай), со-
временный.
Диагноз. Зубная формула: Г-|-СуРуМу=36. I1 долотообразные,
крупнее I2, Is и клыка.
Описание. Морда длинная и тонкая, с желобком на нижней
стороне хоботка. Кисти у кротов этого, рода более широкие, а, когти длин-
ные и слабые. Хвост относительно толстый, густо • покрыт волосами.
Р4 такой же высоты, как и клык, его внешняя сторона треугольной формы.
Распространение и возраст. Азия (Китай), плиоцен—
современный.
1. Scapanulus oweni Thomas, 1912.
Thomas, 1912, там же, с. 396—398.
Тип. N 12.8.5.2. Tao-chou, Ганьсу (Китай), кол. Брит, муз., Лондон.
Описание. Общим обликом напоминает представителей рода
Scaptonix M.-Edw. Окраска темно-серая с серебристым налетом.
Измерения, (по: Строганов, 1948). Длина: тела — 108—136,
хвоста — 37—38, ступни — 13.8—14.0, общая черепа — 27.0—28.3 мм.
(по: Walker et al., 1964). Длина: тела — 98—108, хвоста — 35—38 мм.
Распространение и возраст. Китай (Ганьсу, Сычуань,
Шэньси), плиоцен—современный.
Биология. Не изучена. Добывались в моховом покрове елового
леса. Поднимаются в горы на высоту 2000—3000 м.
Примечание. Описание рода и вида по: G. Allen, 1938; Walker
et al., 1964.
19. Род NEUROTRICHUS Gunther, 1880
Gunther, 1880, p. 441.
Тип. Urotrichus gibbsi Baird, 1857, Тихоокеанское побережье Северной Америки
(юго-зап. часть Британской Колумбии, Канада), западная часть штатов Вашингтон и
Орегон, и северо-западная часть Калифорнии (США), современный.
3 1 2 "3
Диагноз. Зубная формула: I-д-С у РуМу = 36. I1 долотообразные.
I1 шире и в два раза выше I2 и I3, но значительно ниже клыка.
Описание. Ключица длинная и узкая, длина в 2 раза превы-
шает ширину. Плечо удлиненное, его ширина составляет 3/4 длины. Таз
узкий, соединен с крестцовыми позвонками подвижно (места соединения
не окостеневают).
Распространение и возраст. Северная Америка (за-
падные части штатов, примыкающих к тихоокеанскому побережью),
современный;
1. Neurotrichus gibbsi Baird, 1857 (рис. 128).
Urotrichus gibbsi В a i r d, 1857, Rep. Pacific R. R. Survey, v. 8, p. 1, Mammals,
p. 76. Neurotrichus gibbsi Gunther, 1880, p. 441.
Тип. He отмечен, вид описан c White River Pass, штат Вашингтон (США).
Описание. Внешностью напоминает других кротов этой трибы.
Ноздри расположены по бокам мочки носа. Волосяной покров густой,
мягкий, с металлическим блеском. Окраска тела варьирует от темно-
серого, голубовато-черного до черного цветов.
19: РОД NEUROTRICHUS
239
Распространение и возраст. Влажное тихоокеанское
побережье Северной Америки (от южной части Британской Колумбии
до средней части Калифорнии). Современный.
Биология. Не изучена. Известно, что он обитает в лесах и ку-
старниковых зарослях. Поднимается в горы до высоты 2500 м. Питается
теми же группами животных, что и другие кроты.
4. Триба CONDYLURINI Thomas, 1912
Thomas, 1912, р. 357.
Тип. Condylura liliger, 1811, Северная Америка, современный.
Диагноз. Зубная формула: I-|-C-i-P-|-=44. Средних размеров
кроты с длинным хвостом. Пальцы соединены кожей до когтей. Серпо-
видная кость слабо развита. Слуховые барабаны полностью не замкнуты.
Барабанная кость срастается не со всеми окружающими костями. Верх-
ний клык слабо развит, нижний клык высокий, с отогнутой назад верши-
ной. Рх немного ниже клыка. В отличие от других триб, представители
240
ПОДСЕМ. TALPINAE
этой трибы имеют на конце морды розетку с 22 щупальцами, или отрост-
ками, представляющую собой видоизмененную мочку носа.
Описание. Первые верхние резцы повернуты задней стороной
внутрь и направлены вперед. I2 — мелкий. I3 клыкообразный, с бу-
горком на задней стороне основания коронки. Верхний клык мелкий,
Р1, Р2 и Р3 трехбугорчатые, возрастают в размерах спереди назад. Их
протокон отогнут назад («щучьи» зубы). Р4 сходен по строению с преды-
дущими, но значительно крупнее их. 1х долотообразные, с ложкообразной
выемкой, направлены вперед. 12 и 13 мелкие. Нижный клык высокий,
тонкий, трехбугорчатый, с отогнутой назад вершиной протокона. Строе-
ние нижних премоляров сходно со строением верхних. Верхние и нижние
моляры сходного строения, сжатые с боков и вытянуты спереди назад.
Клыки и премоляры отделены друг от друга большими промежутками.
При значительной специализации к рытью кроты этой трибы хорошо
плавают. Если приспособления к рытью и жизни под землей у этих кротов
хорошо выражены в структуре тела и передних конечностей, а также
в степени развития зрительного и слухового аппаратов, то приспособле-
ние к плаванью малозаметно, будучи нивелировано большей специали-
зацией к рытью и жизни под землей. Тем не менее они достаточно опре-
деленно выражены и становятся более заметными при сравнении со спе-
циализированными к плаванью кротами — выхухолями.
Специализация кротов к плаванью берет начало от роющей деятель-
ности, гребных движений при рытье. «Плаванье» в почве перешло с до-
полнительными приспособлениями, развившимися на конечностях в пла-
ванье в воде. Многие кроты хорошо плавают, если попадают в воду.
У представителей этой трибы черты приспособления к передвижению
в воде и пребыванию в ней во внешних органах выразились в следующем.
Ступня длинная, способна поворачиваться подошвенной стороной внутрь,
как у выхухолей, но не в такой степени, как у последних, имеющих
к тому же ряд специфичных особенностей в строении ступни.
Хвост длинный, заметно сжат с боков. Имеется складка кожи, сви-
сающая с верхней губы, достигающая угла рта (губ). Складка кожи име-
ется и на нижней губе. В совокупности они образуют шторки, способству-
ющие, по-видимому, лучшей изоляции полости рта от водной среды при
сомкнутых челюстях. Такие же складки имеются и у выхухолей, но более
развитые. Зубы этих кротов с отогнутыми назад вершинами служат для
захвата скользких водных животных: жуков-плавунцов, водолюбов, мел-
ких рыбок и др.
Распространение и возраст. Северная Америка, со-
временный.
20. Род CONDYLURA liliger, 1811
liliger, 1811, Prod. Syst. Mamm. et Avium, p. 125. Talpasorex Sehina,
1821, Cuvier’s Thierreich, Bd 1, S. 191. Astromycter Harris, 1825, Amer. J. Sci. a.
Arts, v. 9, p. 400. Rhinaster W a g 1 e r, 1830, Nat. Syst. Amphib., p. 14. Astromycter
Gray, 1843, List Spec. Mamm. Brit. Mus., p. 76. Astromydes В 1 у t h, 1863, Cat.
Mamm. Asiat. Soc. Mus., p. 87. Condylus V an Binning, 1913, Science, n. s., v. 38,
p. 243.
Тип. Sorex cristatus Linnaeus, 1758. Юго-восточная Канада, северо-восточная
часть США, современный.
Диагноз. Полностью совпадает с таковым трибы.
Распространение и возраст. Те же, что и трибы.
1. Condylura cristate Linnaeus, 1758 (рис. 129—131).
Sorex cristatus Linnaeus, 1758, p. 53. Talpa longicaudata Erxleben, 1777, Reg.
Anim., p. 118; — cristata Zimmermann, 1777, Specimen Zool. Geogr., p. 496; —
2а. РОД CONDYLURA
241
caudata Zimmerman (71- caudata L. и др.), там же, с. 497; Talpa? canadensis La
Faile, 1778, Naturgesch. des Maulwurtes, Bd 1, S. 3; Talpa radiata Shaw, 1800,
Gen. ZooL, Mamm., v. 1, p. 523. Sorex radiatus S haw, там же, с. 531. Scalopus cri-
status G e о f f r ay, 1803, Gat. Mamm. Mus. Nat. Hist., p. 77; F i s c h e r, 1814, Zoo-
gnosia, v. 3, p. 156. Condylura longicaudata Desmarest, 1819, J. de Physique,
de Chimie, d’Hist. Nat. et de Arts, t. 89, p. 230. Tai (pasorex) cristata S c h i n z, 1821,
Рис. 129. Condylura cristata (по: Строганов, 1948).
Cuviers Thierreich, Bd 1, S. 191. Condylura fissipes S chi n z, 1821, там же. Talpa flava
S c h i n z, 1821, там же; — purpurascens S c h i n z, 1821, там же. Condylura ma-
croura Harla n, 1825, Fauna Amer., p. 39. Astromycter prasinatus Harris, 1825,
Amer. J. Sci. a. Arts, v. 9, p. 400. Condylura prasinata Harris, 1825, Boston J. Phi-
los. a. Arts, v. 2, p. 582. Rhinaster cristatus, Rh. macrurus, Rh. longicaudatus W a g n e r,
1841, Suppl. Schreber’s Saugetiere, Bd 2, S. 114—117. Astromydes cristatus В 1 у t h,
Cat. Mamm. Asiat. Soc. Mus., p. 87.
T и и. He установлен, вид опи-
сан из восточной части Пенсильва-
нии (США).
Описание. Волосяной
покров густой, грубоватый, нес-
мачивающийся водой. Верхняя
сторона тела черно-коричневая,
почти черная. Нижняя сторона
светлее, более коричневая.
Череп удлиненный, узкий.
Нижняя челюсть низкая и тон-
кая, с сильно развитым сочле-
новным отростком, низким, сла-
боразвитым венечным и палоч-
ковидным угловым отростками.
Измерения (по: Jack-
son, 1915). Длина: тела —
-189 211, Хвоста —- 71 84, рис. 130. Складки пагубах Condylura cristata.
ступни — 26—30, наибольшая
черепа — 13.5—14.0 мм.
Распространение и возраст. Юго-восточная Канада
и юго-восточные штаты США от южной части Лабрадора, центральной
части Квебека и Онтарио и юго-восточной части Манитобы, от юга до
северо-востока Иллинойса и севера Индианы и Охио; в Атлантической
части ареала к югу до Виргинии (Dismal swamp)- и Георгии (Marlow)
и в Аппалачских горах до западной части Северной Каролины.
Биология. Обитает на лугах и болотах, но предпочитает дрениро-
ванные почвы, в лесистой местности. Пищей служат водные беспозвоноч-
ные и рыба, а также наземные беспозвоночные, особенно насекомые и
черви. Размножается от середины апреля до середины июня.
16 А. А. Гуреев
242
ПОДСЕМ. TALPINAE
15. Триба PROSCALOPINI К. Reed, 1961
Proscalopinae К. Reed, 1961, p. 473—474.
Тип. Proscalops Matthew, 1901, Северная Америка, олигоцен.
’Рис. 131. Condylura cristata
(по: Строганов, 1948).
Диагноз (по: К. Reed, 1961). Зубная формула 1уС-|-РуМу.
Череп по форме напоминает таковой златокротов (Chrysochloridae). Зубы
типичные для кротов. Череп широкий тт высокий в задней части. Максил-
лярный изгиб между Р3 и Р4 уже, чем у Neurotrichus Giint. Сагиттальный
и затылочный гребни слабые. Нёбо относительно длинное. Передне-
орбитальный гребень верхнечелюстной кости не соединяется с основанием
21. РОД CRYPTORYCTES
243
скуловой дуги. Ширина лингвального выступа верхних моляров и гипо-
кона варьирует.
Распространение и возраст. Северная Америка, сред-
ний олигоцен—средний миоцен.
Таблицу для определения родов этой трибы составить не представля-
ется возможным вследствие отсутствия сравнимых признаков. Одни из
них описаны по верхним зубам, другие по нижним или по плечевым костям,
типичный род — и по черепу.
f 21. Род CRYPTORYCTES С. Reed, 1954
С. Reed, 1954, р. 102—111.
Тип. Cryptorictes kayi С. Reed, 1954, Монтана (США), нижний олигоцен.
Диагноз (по: С. Reed, 1954). Плечевая кость уплощена в передне-
заднем направлении и расширена в боковом. Сочленовная головка удли-
Рис. 132. Cryptoryctes kayi (плечевая кость) (по: С. Reed, 1954).
Рис. 133. ProsCalops miocenus (по: Matthew,' 1901).
ненной формы. Малая бугристость удлиненной формы. Большая бугри-
стость небольшая и не специализированной формы в месте соединения
с кцючицей. Круглый бугорок образует медиальный край кости. Дисталь-
ный отдел гребня грудного отростка поднят в отдельный отросток. Го-
16*
244
ПОДСЕМ. TALPINAE
ловка луковицеобразная. Надмыщелковое отверстие имеется, удлиненной
формы.
Распространение и возраст. Северная Америка, ниж-
ний олигоцен.
f 1. Cryptoryctes kayi С. Reed, 1954 (рис. 132).
С. Reed, 1954, там же.
Т и п. Левая плечевая кость, № РМ 1009. Монтана (США), нижний олигоцен
кол. Муз. естеств. истор., Чикаго.
Рис. 134. Arctorycies terrenus (по: С. Reed, 1954).
Примечание. Более полное представление об особенностях
строения плечевой кости этого своеобразного крота дает приведенный
рисунок. Укорочение плечевой кости достигнуто изгибом ее тела и значи-
тельным развитием круглого бугорка (по: Reed, 1954), а также сокраще-
нием длины внутреннего края кости.
Измерения (по: С. Reed, 1954). Плечевая кость (голотип):
длина — 10.9 мм, ширина проксимальной части — 6.0, ширина дисталь-
ной части — 10.9 мм. Плечевая кость № 9777 из кол. Американского
национ. муз.: длина — 13.9 мм.
Распространение и возраст. Монтана (США), нижний
олигоцен.
f 22. Род PROSCALOPS Matthew, 1901
Matthew, 1901, Mem. Amer. Mus. Nat. Hist., v. 1, pt. 7, p. 355—447.
Тип. Proscalops miocaenus Matthew, 1901, область Логан, Колорадо (США),
средний олигоцен.
Диагноз (по: К. Reed, 1961). Внешний выступ предчелюстной
кости сжат дорсо-вентральпо. Между Р3 и Р4 имеется диастема. Р3 мел-
кий, овальный, с одним корнем. Р4 с неразвитой парастилярной областью,
с бугорком на лингвальной стороне, расположенном впереди от средней
линии зуба. Верхние моляры с в разной степени хорошо развитым гипо-
коном (талоном). Парастиль слабо развит на М1, отчетливый на Ма.
Примечание. Здесь приведен исправленный, как отмечает Рид
(К. Reed), диагноз. Метью (Matthew, 1901) практически не описал этот
род, ограничившись несколькими замечаниями. К сожалению, и ис-
правленный диагноз, как представляется, не содержит многих важных
особенностей этого рода.
Распространение и возраст. Северная Америка, сред-
ний олигоцен—нижний миоцен.
1- 1. Proscalops miocaenus Matthew, 1901 (рис. 133).
Matthew, 1901, там же.
Тип. Поврежденный череп и нижняя челюсть, № 8949а, область Логан, Коло-
радо (США), средний олигоцен, кол. Амер. муз. естеств. истор., Нью-Йорк.
23. РОД ARCTORYCTES
245
Диагноз (по: К. Reed, 1961). Мелкий из известных вид рода.
Р4 заметно уже, чем у Р. tertius К. Reed. Его лингвальный бугорок рас-
полагается позади. Лингвальные части коронок верхних моляров уже,
чем у других видов этого рода, с гипоконами (талонами) и протостилями
менее развитыми. М3 сжат в передне-заднем направлении. Его лингвальная
часть узкая, гипокон рудиментарный.
Измерения (по: К. Reed). Длина: Р4 — 2.0, М1 — 2.2, М3 —
1.5; ширина: Р4 — 1.7—1.8, М1 — 2.5, М2 — 2.7, М3 — 1.8, нёба у М2 —
9.4 мм.
Распространение и возраст. Известен из Колорадо
(США), средний олигоцен.
f 2. Proscalops tertius К. Reed, 1961.
К. Reed, 1961, там же.
Тип. Часть черепа и левая ветвь нижней челюсти с зубами — М, — 3, № 19420
«Бесплодные земли», Южная Дакота (США), олигоцен, кол. Амер. муз. естеств. истор.,
Нью-Йорк.
Диагноз (по: К. Reed, 1961). Зубы немного крупнее, а гипокон
более развит, чем у Р. secundus Matt. Лингвальный бугорок Р4 конический,
крупнее, чем у Р. miocaenus Matt., не доходит передней частью до лабиаль-
ного бугорка. Метаконид М2 с мелким метастилидом, с широкой долинкой
талонида (метаконида) внутри, как у Mogera wogura Temm.
Измерения (по: К. Reed, 1961). Длина: Р4 — 2.1, М1 — 3.2,
М2 — 2.8, М3 — 2.4; ширина: Р4 — 2.1, М1 — 2.7, М2 — 2.65, М3 — 2.1;
длина: Мх — 2.85, М2 — 2.7, М3 — 2.2; ширина черепа на уровне М2 —
12.0 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
f 3. Proscalops secundus Matthew, 1909.
Matthew, 1909, Mem. Amer. Mus. Nat. Hist., v. 9, pt. 6, p. 291—567.
Тип. Череп (немного деформированный в поперечном направлении) № 13798,
Южная Дакота, верхний миоцен, кол. Амер. муз. естеств. истории., Нью-Йорк.
Диагноз (по: К. Reed, 1961). Крупнее, чем Р. miocaenus. Линг-
вальный бугорок Р4 широкий и простирается далее вперед, чем у Р. mio-
caenus, как у Mesoscalops scopelotemos К. Reed. Лингвальная часть М1—
М3 широкая. Гипоконы (талоны) у М 1 и М2 более развиты, чем у Р. mio-
caenus. Задняя часть скуловой дуги (основание) поднята высоко — на
боковой стороне черепа.
Измерения (по: К. Reed, 1961). Длина: Р4 — 1.85, М1 — 3.1,
М2 — 2.55, М3 — 2.3; ширина: Р4 — 1.6, М1 — 2.6, М2 — 2.8, М3 — 1.9,
черепа на уровне М2 — 10.7 мм.
Распространение и возраст. Известен из одного место-
нахождения.
f 23. Род ARCTORYCTES Matthew, 1907
Matthew, 1907, р. 169—219.
Тип. Arctoryctes terrenus Matthew, 1907, нижний миоцен, Южная Дакота (США).
Диагноз. Тело плечевой кости сильно расширено в средней части
и в дистальном отделе. Грудной отросток смещен на среднюю часть кости.
Дельтовидный отросток округлой формы, короткий. Надмыщелковое
отверстие открывается в длинную бороздку на внешней стороне надмы-
щелка.
Описание. В дополнение к диагнозу следует отметить и другие
особенности описываемой плечевой кости. Большая бугристость смещена
246
ПОДСЕМ. TALPINAE
на медиальную часть кости и сильно развита. Малая бугристость на верх-
ней стороне имеет большую площадь, чем у современных специализиро-
ванных землероев. Дельтовидный отросток направлен не вниз, а в сторону.
По-видимому, меньшее, чем у других кротов, развитие проксимальной
части плечевой кости у Arctoryctes, обусловило формирование структур-
ных особенностей, характерных для плечевых костей представителей
этого рода. Детали строения описываемой кости хорошо заметны на
рис. 134.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
+ 1. Arctoryctes terrcnus Matthew, 1907 (рис. 134).
Matthew, 1907, р. 169—219.
Тип. Автором не отмечен. Рид (С. Reed, 1954) в подписи под рисунком привел
№ 12864 кол. Амер. муз. естеств. истор., Нью-Йорк. Возможно, что это номер типа.
Распространение и возраст. Южная Дакота, нижний
миоцен.
Примечание. Симпсон (Simpson, 1945) в своей системе млеко-
питающих отнес род Arctoryctes к насекомоядным под вопросом, а Ван
Вален (Van Valen, 1967) в предложенной им системе насекомоядных свел
его в синонимы рода Proscalops Matt. (Talpidae), Рид высказал мнение,
что роды Cryptoryctes Reed и Arctoryctes и форма (плечевая кость без опре-
деления систематической принадлежности № 9777, из среднего палеоцена
Северной Америки (Sweetgrass County, Монтана) кол. Амер. нац. муз.,
Вашингтон), могут быть отнесены к насекомоядным или неполнозубым
(Edentata), но не могут считаться предками Talpidae (Insectivora).
Сравнение рисунков плечевых костей различных групп вымерших
Edentata и современных Chrysochloris Lacepede и Talpa L. (Insectivora),
а также изображений плечевых костей Cryptoryctes и Arctoryctes, при-
веденных Сабан (Saban, 1958, р. 529), и сравнение последних с плечевыми
костями современных Insectivora и Edentata позволяет отметить их сходство
только с кротами. Плечевые кости обсуждаемых здесь родов имеют тот же
план строения и те же структурные особенности, что и соответствующие
кости кротов. Сокращение длины плечевых костей кротов в пределах сем.
Talpidae происходило, видимо двумя путями: в результате изгиба кости
(Cryptoryctes) и путем сокращения ее длины (Arctoryctes и др.).
Примечание. Описание рода и вида сделаны мной путем
сравнения рисунков и костей кротовых. К сожалению, описания не сделал
ни автор рода и вида (Matthew, 1907), ни Рид (Reed, 1954).
f 24. Род OLIGOSCALOPS К. Reed, 1961
К. Reed, 1961, р. 474, 475.
Тип. Oligoscalops whitmanensis К. Reed, 1961, Вайоминг и Колорадо, средний
олигоцен.
Диагноз (по: К. Reed, 1961). Корни Р3 частично разделены.
Зуб снаружи сжатый, прилегающий к Р4. Р4 с большой парастилярной
областью и лингвальным бугорком, расположенным на средней линии.
Протоконы моляров занимают передне-внутреннее положение. Ширина
М2 более его длины. Метастиль М8 почти параллелен линии, соединяю-
щей протокон и метакон. Его гипокон (талон) рудиментарный.
Примечание. Признаки, установленные автором для рода,
могут оказаться (на большем материале) недостаточными и для вида.
Но вопрос этот может быть решен на более обильном фактическом мате-
риале.
25. РОД MESOSCALOPS
247
t 1- Oligoscalops whitmanensis K. Reed, 1961.
K. Reed, 1961, там же.
Тип. Неполный череп и челюсти, NP. 25800, из Brule fm. Whitman, область
Ниобрара, Вайоминг, средний олигоцен, кол. Муз. естеств. истор. в Чикаго (США).
Диагноз. Совпадает с родовым. Можно только отметить, что этот
вид наименьший в пределах трибы Proscalopini.
Измерения (по: К. Reed, 1961). Длина М1 — 2.2, М2 — 1.7—
2.1, М3 — 1.4—1.5, ширина М1 — 1.7, М2 — 2.0, М3 — 1.4 мм.
Распространение и возраст. Известен из Вайоминга
и Колорадо (США), средний олигоцен.
f 25. Род MESOSCALOPS К. Reed, 1960
К. Reed, 1960, р. 2—3.
Тип. Mesoscalops scopelotemos К. Reed, 1960, область Фремонт, Вайоминг (США),
средний миоцен.
Диагноз (по: К. Reed, 1960). В основном близок к Proscalops
Matt. Отличается следующим: протокон верхних моляров более округлый
и несколько более крупный, чем гипокон (талон), расположен непосредст-
венно у внутреннего края паракона и придает контуру зубов квадратную
форму. Р4 на лингвальной стороне имеет выемку и широкий выступ.
Нижние моляры с небольшим медиальным цингулюмом, имеющим основа-
ние выше линии десны между гипоконидом (талонидом) и протоконидом.
Талонид (метаконид в моем представлении) Мх шире, чем тригонид (перед-
ний сегмент зуба).
Распространение и возраст. Северная Америка, сред-
ний миоцен.
t 1. Mesoscalops scopelotemos К. Reed, 1960.
К. Reed, 1960, там же, с. 24.
Тип. Изолированный М1 № АСМ 10 461, область Фремопт, Вайоминг (США),
средний миоцен, кол. Муз. Amherst.
Описание (из: К. Reed, 1960). Кроме признаков, отмеченных
в диагнозе, характерны и следующие;' М2 более квадратный, чем М1,
а М3 по очертаниям приближается к треугольнику.
Примечание. Ограниченность и фрагментарность материала,
которым располагал автор первого описания этого рода и вида, обусловли-
вает известные сомнения в их качественной определенности.
Распространение и возраст. Известен из среднего
миоцена Вайоминга (США).
6. Триба SCALOPINI Thomas, 1912
Scalopidae Соре, 1889, Amer. Nat., v. 23, p. 849—877. Подсем. Scalopinae Tho-
mas, 1912, Ann. Mag. Nat. Hist. ser. 8, v. 10, p. 397.
Тип. Scalopus Geoffroy Saint Hilaire, 1833, Северная Америка, плиоцен—со-
временный.
Диагноз. Зубная формула: I СРМ= 44—36. Крупные
кроты с хвостом разной длины, преимущественно коротким. Пальцы
соединены кожей до когтей, по крайней мере у современных видов. Серпо-
видная кость сильно развита. Барабанная кость слита с соседними костями
или слита не полностью и барабанные камеры замкнутые и вздуты или
не замкнуты полностью. Верхний клык мелкий, не отличается по форме
от соседних зубов, также и нижний клык. Рх мелкий или отсутствует.
248
ПОДСЕМ. TALPINAE
Распространение и возраст. Европа, средний миоцен:
Азия (можно думать ранее, чем в Европе), Северная Америка, плиоцен—
современный.
Триба объединяет сравнительно крупных кротов, высоко специализиро-
ванных к рытью. Зубная система преимущественно полная (44 зуба),
в значительной степени специализированная, приспособленная в большей
степени для нарезания и раздробления пищи острыми вершинами и гра-
нями зубов. Все современные представители трибы — обитатели окраин
лесов и открытых участков в лесной зоне, а также открытых ландшафтов
с кустарниковыми зарослями — равнин и склонов гор. Основную пищу,
как и кротов других триб, составляют земляные черви, насекомые и дру-
гие беспозвоночные.
. ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Мг и М3 с добавочными бугорками на передней стороне первого
сегмента зуба (тригонид других авторов) . . .29. Hesperoscalops.
2 (1). М2 и М3 без добавочного бугорка на передней стороне переднего
сегмента зуба.
3 (8). Общее количество зубов 44.
4 (5). Верхний клык, Р2 и Р3 с двумя корнями .... 26. Proscapanus.
5 (4). Верхний клык, Р2 и Р3 с одним корнем.
6 (7). Верхний клык крупнее соседних зубов..........27. Parascalops.
7 (6). Верхний клык не крупнее соседних зубов..........28. Scapanus.
8 (3). Общее количество зубов 36 ..................... 30. Sealopus.
f 26. Род PROSCAPANUS Gaillard, 1899
Gaillard, 1899, p. 23—27.
Тип. Proscapanus sansaniensis Lartet, 1851. Sansan (Gers) et Grive-Saint Alban
(Isere), Франция, средний миоцен.
Диагноз. Зубная формула: I -|- С у Р М -|- = 44. Плечевая кость от-
носительно длиннее, чем у Talpa europaea L. Отношение длины к наиболь-
шей ширине у Т. europaea — 72—73%, у Proscapanus sansaniensis Lart. —
66—69%. I1-1 (судя по альвеоле) трехгранные. Рх не крупнее нижнего
клыка и соседнего премоляра.
Распространение и возраст. Европа, средний миоцен.
f 1. Proscapanus sansaniensis Lartet, 1851.
Talpa sansaniensis Lartet, 1851, Notice sur la colline de Sansan, p. 14; — vul-
garis Blainville, 1839—1864, Osteographie, p. 96, 97; — telluris Gervais,
1852, Zool. et Paleont. fran?. p. 58.
Тип. He установлен, вид описан по верхней и нижней челюстям и плечевой
кости из Сансан (Gers), Франция, средний миоцен.
Описание (по: Saban, 1958). I2 и I3 мелкие. Верхний клык с двумя
корнями. Первые три премоляра крупные, сходной формы. Р1 с одним,
Р2 и Р3 с двумя корнями. Р4 крупный, с тремя корнями, как и моляры.
1Х крупнее других, наклонен вперед. Задние резцы и передние премоляры
одновершинные. Р4 крупный, с двумя корнями. Моляры с одним цингу-
люмом, развиты в передне-заднем направлении. Нижняя челюсть сходна
с таковой Talpa europaea L.
27. РОД PARASCALOPS
249
Измерения (по: Gaillard, 1899). Длина плечевой кости — 14.0—
17.5, ее наибольшая ширина — 9.5—10.0; длина: I2—М3 -*• 14, Р1—М3 —
12, М1—М3 — 7 мм.
Распространение и возраст. Известен из среднего мио-
цена Франции.
Рис. 135. Parascalops breweri (по: Строганов, 1948).
27. Род PARASCALOPS True, 1894
True, 1894, Proc. U. S. Nat. Mus., v. 27, p. 242.
Тип. Scalops breweri Bachman, 1842, юго-восточная Канада и северо-восточные
районы США.
Диагноз. Зубная формула: I-| С уР-|-М-|- —44. I1 долотообразной
формы, крупный. I2 и I3, клык и Р1—Р3 сходные по строению и размерам,
конические, только клык выше этих зубов. Р4 крупнее и массивнее Р3,
имеет мелкий паракон спереди и двулопастный метакон сзади. 1х—13, клык
250
ПОДСЕМ. TALPINAE
и Рг—Р3 однотипные, конические, мало разнятся по величине. Р4. на 1/3
крупнее Р3, строением напоминает Р4, но пара- и метаконид у него слабее
развиты. Верхние и нижние коренные зубы сходны по форме с этими
зубами других кротов.
Описание (по: Jackson, 1915). Тело массивное, слабо уплощено.
Голова коническая, уплощенная. Носовой отдел удлинен, но короче,
чем у представителей рода Scalopus Geoffr. и Scapanus Pomel, с продоль-
ной бороздкой по средней части его верхней стороны. Ноздри направлены
в стороны с полулунной выемкой. Глаза маленькие, спрятаны в волося-
ном покрове. Длина кисти равна ее ширине. Пальцы кистей и ступней
без перепонок. Когти на пальцах кисти массивные, широкие и плоские,
на пальцах ступней короткие, слабые. Хвост короткий, но длиннее, чем
у Scalopus. Он густо покрыт жесткими волосами.
Волосяной покров тела более жесткий, чем у видов других родов.
Окраска тела черно-серая или коричневато-черная. Бывают белые пятна
на груди, животе, голове, носу и хвосте.
Ключица относительно длинная и массивнее, чем у других родов трибы.
Ее длина близка к ширине. Имеется сквозное отверстие вблизи верхнего
края этой кости. Плечевая кость массивная. Ее ширина около 3/4 длины.
Распространение и возраст. Северная Америка, плей-
стоцен—современный.
1. Parascalops breweri Bachman, 1842 (рис. 135).
Bachman, 1842, Boston J. Nat. Hist., v. 4, p. .32. Scapanus breweri Pomel,
1848, Arch. Sci. Physique et Nat., t. 9, p. 247. Talpa reposta L e Conte, 1853, Proc.
Acad. Nat. Sci. Philadelphia, v. 6, p. 327; — breweri LeConte, 1853, там же. Scapa-
nus americanus Cones, 1879, Amer. Nat., v. 13, p. 190. — Scaphanus breweri Her-
rick, 1892, Geol. et Nat. Hist. Surv. Minnesota, bul. 7, p. 55. Parascalops breweri J a-
c k s о n, 1915, North Amer. Fauna, N 38, p. 79—82.
Тип. He известен, вид описан из Marthas Vineyard, Массачусетс (США).
Измерения, (по: Walker et al., 1964). Длина: тела — 116—140,
хвоста — 23—36 мм. (По: Hall, Kelson, 1959). Длина: тела — 139—153,
хвоста — 23—36, ступни — 18—20, наибольшая черепа — 31..0—33.8 мм.
Распространение и возраст. Юго-восточная Канада
и северо-восточные штаты США, плейстоцен—современный.
28. Род SCAPANUS Pomel, 1848
Pom e l, 1848, Bibl. Univ. Geneve, Arch. Sci. phys. nat., t. 9, p. 247. Scaphanus
Herrick, 1892, Geol. Nat. Hist. Surv. Minnesota, bull. 7, p. 55. Scapasius В e d-
d a r d, Cambridge Nat. Hist., v. 10, p. 518.
Тип. Scalops townsendii Bachman, 1839, Северная Америка (западная часть),
современный.
Диагноз. Зубная формула: С у РМ-|-= 44.
I1 долотообразный крупный. Задние резцы, клык и Р1—Р2 сходной
формы и размеров, конические. На клыке и Р1—Р3 спереди и сзади имеются
бугорки (остатки пара- и метаконов). Р4 с хорошо развитым метаконом.
Его паракон очень слабо развит или отсутствует. 12 крупнее 14 и 13. Клык
и Рх—Р3 мелкие, конические, сходные по форме. Р4 с пара- и метаконидом.
Верхние и нижние коренные зубы сходны с таковыми других кротов.
Голова коническая, уплощенная. Ноздри верхние, полулунные,
с углублением, повернутым латерально. Глаза мелкие, спрятанные в шер-
сти. Слуховое отверстие небольшое. Ширина кисти равна ее длине. Когти
широкие, плоские, массивные. Ступни большие, мясистые, сверху покрыты
волосами, снизу голые. Когти короткие, слабые. Пальцы передних и зад-
них ног без перепонок.
28. РОД SCAPANUS
251
Описание (по: Jackson, 1915). Тело крепкое, низкое, хвост корот-
кий, округлый, толстый и мясистый, стянутый при основании и суживаю-
щийся к концу. Его покров образован грубыми волосами.
Волосы, покрывающие тело, мягкие, сходные по высоте, образуют
бархатистый покров черно-серого, коричнево-черноватого или темно-
серого цвета.
Ключица короткая, массивная. Ее ширина составляет 3/4 длины.
Плечевая кость мощная. Ее ширина равна 3/4 длины. Таз узкий.
Распространение и возраст. Северная Америка, ниж-
ний плиоцен—современный.
1. Scapanus townsendii Bachman, 1839.
Scalops canadensis Richardson, 1829, Fauna Boreali — Amer., pt 1, p. 9; —
townsendii Bachman, 1839, J. Acad. Nat. Sci., Philadelphia, v. 8, pt 1, p. 58. Sca-
panus townsendii Pomel, 1848, Arch. Sci. Physiques et Nat., v. 9, p. 247. Scalops me-
tallescens C a s s i n, 1853, Proc. Acad. Nat. Sci. Philadelphia, v. 6, p. 242, — aeneus
C a s s i n, 1853, там же, с. 299. Talpa aenea Le С о n t e, 1853, там же, с. 327; — town-
sendii Le С о n t e, 1853, там же, с. 327; — taeniata Le С о n t e, 1853, там же, p. 327.
Scapanus townsendii Peters, 1863, Monatsber. Konig. Preuss. Akad. Wissensch.,
Berlin, p. 656.
Тип. Котип № 449, форт Ванкувер, кол. Акад, естеств. наук в Филадельфии.
Диагноз (по: Jackson, 1915, с сокращениями). Самый крупный
вид в пределах рода. Череп крупный, с удлиненным носовым отделом.
Задне-наружные углы теменных костей без отростков, вклинивающихся
между височной и межтеменной костями.
Измерения (по: Hall, Kelson, 1959). Длина: тела — 195—237,
хвоста — 34—51, ступни — 24—28, наибольшая черепа — 41.2—44.6 мм.
Распростране н»и е и возраст. Западная часть штатов
Вашингтон и Орегон, северо-западная Калифорния (США), современный.
252
ПОДСЕМ. TALPINAE
2. Scapanus orarius True, 1896.
True, 1896. Proc. U. S. Nat. Mus., v. 19, p. 52.
Тип. § (шкура и череп) № 1381/37434 мелководный залив, Вашингтон, кол.
Амер. нац. муз., Вашингтон.
Диагноз (по: Jackson, 1915). Размеры средние. Череп по строению
сходен с таковым S. townsendii, но с более узкой ростральной частью.
Измерения (по: Hall, Kelson, 1959). Длина: тела — 162—175,
хвоста — 26—37, ступни — 21.5—23.0, наибольшая черепа — 32.8—
39.0 мм.
Распространение и возраст. Юго-западная часть Бри-
танской Колумбии, штаты Вашингтон и Орегон и северо-западная часть
Калифорнии, современный.
3. Scapanus latimanus Bachman, 1842 (рис. 136).
Bachman, 1842, Boston J. Nat. Hist., v. -4, p. 34. Scalops californicus Ayres,
1856, Proc. California Acad. Sci., v. 1, p. 54; True, 1896, Proc. U. S. Nat Mus., v. 19,
p. 52. Scapanus latimanus Osgood, 1907, Proc. Biol. Soc. Washington, v. 20, p. 52.
Тип. Джексон указал, что тип находится в Берлинском музее. Вид описан из
Санта-Клара, Калифорния.
Диагноз (по: Jackson, 1915). Череп уплощен в максиллярной
области. Носовой отдел черепа относительно короткий и высокий. Задне-
наружные углы теменных костей с отростком, вклинивающимся между
височной и межтеменной костями.
Измерения (по: Hall, Kelson, 1959). Длина: тела — 132—192,
хвоста — 21—45, ступни — 18—25, наибольшая черепа — 29.7—37.4 мм.
Распространение и возраст. Калифорния, Невада,
Орегон (США), современный.
f 29. Род HESPEROSCALOPS Hibbard, 1941
Hibbard, 1941, Amer. Midi. Nat., v. 26, p. 337—339.
Тип. Hesperoscalops rexroadi Hibbard, 1941, Meade County, Канзас (США), верх-
ний плиоцен.
Диагноз (составлен по описанию: Hibbard, 1953). Зубная формула
I -х- С -о- Р -д- М -к- • Плечевая кость и дистальный отдел ключицы не крупнее
Л о о о
и не выше развиты, чем эти кости Scalopus Geoff. М2 и М3 имеют дополни-
тельный бугорок на передней стороне переднего сегмента коронки. Нижние
коренные высокие и массивные. Р4 с хорошо развитым метаконидом.
Распространение и возраст. Северная Америка, верх-
ний плиоцен.
t 1. Hesperoscalops rexroadi Hibbard, 1941 (рис. 137, A).
Hibbard, 1941, там же.
Тип. Остался мне неизвестным, поскольку у меня не было в руках работы с пер-
воописанием этого вида.
Диагноз. Мельче II. sewardensis. Бугорок на передней стороне
М2 и М3 менее развит.
Измерения (по: Hibbard, 1953). Передне-задняя длина Mt—М3
по альвеолам 3 экз. соответственно 7.8, 7.15, 7.6 мм.
Распространение и возраст. Канзас (США), верхний
плиоцен.
Примечание. Род Hesperoscalops Ван Вален (Van Valen, 1967)
отнес в синонимы рода Scalopus. Хиббард, описавший этот род, достаточно
30. РОД SCALOPUS
253
опытный палеонтолог и систематик, чтобы не считаться с его мнением
о качественной определенности обсуждаемого рода. Это представление
подтверждается также и описанием Рид (Reed, 1962) другого вида этого
рода, более позднего времени.
Л
Б
Рис. 137. Hesperoscalops rexroadi (Л) (по: Hibbard, 1953), Н. sevardensis (Б) (по:
К. Reed, 1962).
t 2. Hesperoscalops sewardensis К. Reed, 1962 (рис. 137, Б).
Reed, 1962, р. 4—5.
Тип. Фрагмент правой ветви нижней челюсти с зубами: М2 и М3, фрагмент ле-
вой ветви, передняя часть левой ветви со стертым М,, локтевая кость, часть лопатки и
бедро, № ИММР 27276, из Saw Rock, область Seward, Канзас (США), верхний плио-
цен или нижний плейстоцен, кол. Палеонт. муз. Мичиган, унив., Анн Арбор.
Диагноз (по: Reed, 1962). Заметно крупнее Н. rexroadi Hibb.
и с более развитым основанием добавочного бугорка (имеется в виду перед-
ний бугорок на М2 и М3).
Измерения (по: Reed, 1962). Длина: М2 — 3.0, М3 — 2.8, М2—
М3 — 5.8, высота нижней челюсти перед М2 на лингвальной стороне —
4.5 мм.
Распространение и возраст. Известен из Канзаса
(США), верхний плиоцен или нижний плейстоцен.
30. Род SCALOPUS Geoffrey Saint Hilaire, 1803
Scalops Cuvier, 1800, Lecons d’Anat. Compar., t. 1, pl. 1 (nomen nudum). Sca-
lopus Geoffroy Saint-Hilaire,- 1803, Cat. Mamm. Mus. Nat. Hist. Nat.,
p. 77. Talpasorex Lesson, 1827, Manuel de Mamm., p. 124. Scalops Brooks, 1911,
Rept. W. V. Stat Board Agric. for Quarter ending dec. 10, 1910, p. 28.
Тип. Sorex aquaticus Linnaeus, 1758 (= Scalopus aquaticus Geoffroy, 1803).
254
ПОДСЕМ. TALPINAE
Диагноз. Зубная формула I-|-C-q-P-|-М-| =36.
I1 широкий, долотообразный, I2 и I3 мелкие, конические, часто вы-
падают. 'Клык крупный, выше Р1—Р3, почти равен по высоте Р4. Послед-
ний с конической вершиной, имеет пара-, метакон и талонид на лингваль-
ной стороне.
1Х некрупный, конический. 12 вдвое крупнее 1х, клыкообразный.
Р2 и Р3 конические, с бугорками на задней стороне (остатки метаконида).
Р4 сходен по форме с Р3, но крупнее и с более развитым метаконидом.
Верхние коренные зубы в отличие от других с менее удлиненным наружным
краем и менее приближены друг к другу.
Описание (по: Jackson, 1915, с сокращениями). Тело массивное,
низкое. Голова коническая, уплощенная. Нос удлинен, узкий, ноздри
сверху полулунные, с углублением, повернутым латерально. Глаза ма-
ленькие, скрыты в шерсти. Ушное отверстие небольшое. Ноги короткие,
сильные. Кисти большие. Их ширина более длины. Пальцы кисти и ступни
соединены перепонкой. Когти на кистях широкие, плоские, поднятые,
на ступнях заостренные, узкие, слабые. Хвост короткий. Волосяной покров
густой, мягкий, бархатистый. Его окраска варьирует от темно- и черно-
серого до черно-коричневого.
Череп конусовидный, плоский, с укороченным и узким носовым отделом
и относительно широкой мозговой капсулой. Межглазничная область
сильно сжата. Передняя часть межчелюстной кости утолщена. Выступает
вперед за носовые кости. Межтеменная кость короткая и узкая, неправиль-
ных очертаний с выступом в передней части. Скуловая дуга слабая, впереди
вогнута в орбиту. Слуховые барабаны замкнутые. Ключица короткая,
массивная, ее ширина более 2/3 длины. Плечевая кость широкая (ширина
составляет 3/4 длины). Таз узкий.
Распространение и возраст. Северная Америка, ниж-
ний плиоцен—современный.
1. Scalopus aquaticus Linnaeus, 1758 (рис. 138, 139).
Sorex aquaticus Linnaeus, 1758, p. 53. Talpa еигораеа flavescens Erxleben,
1777, Syst. Regn. Anim., p. 118; —flava Zimmermann, 1777, Specim. Zool. Geogr.,
p. 496; — fusca Zimmermann, 1777, там же, с. 497. Scalopus virginianus Geof-
frey Saint-Hilaire, 1803. Cat. Mamm. Mus. Nat., p. 78. Talpa cupreata R a-
finesque, 1814, Precis des decouv. et travaux somiologiques, p. 14. Scalops cana-
densis Desmarest, 1820, Mammalogie, 1 partie, p. 155; — pensylvanica Harlan,
1825, Fauna Amer., p. 33. Talpasorex pensylvanica Lesson, 1827, Manuel de Mamm.,
p. 124. Talpa virginiana Blainville, 1838, Annales Franc, et Etrang. d’Anat. et
de Physiol., t. 2, p. 219; — sorex pensylvanicus Blainville, 1838, там же; — (Sca-
lops) virginiana Blainville, 1839—1864, Osteographie, Atlas 1, pl. 5, 9; — aqua-
tica L e Conte, 1853, Proc. Acad. Nat. Sci. Philadelphia, v. 6, p. 327. Scalopus aqua-
ticus Oberholser, 1905, Mammals and Summer Birds of Western North Caro-
lina, p. 3.
Тип. He установлен, вид описан из Филадельфии (США).
Диагноз. Череп крупный массивный, со слуховыми барабанами,
не выступающими за контуры черепа.
Описание. Приведено в характеристике рода.
Измерения (по: Jackson, 1915). Длина: общая тела — 134—208,
хвоста — 19.7—38.0, ступни — 15.5—24.0, черепа — 31.4—39.1 мм.
Распространение и возраст. Восточная часть США,
плиоцен—современный.
2. Scalopus aereus Bangs, 1896.
Scalops texanus aereus Bangs, 1896, Proc. Biol. Soc. Washington, v. 10, p. 138; —
aereus Elliot, 1901, Field Columb. Mus., publ. 45, Zool. ser., v. 2, p. 390. Scalopus
aereus Elliot, 1905, там же, публ. 105, т. 6, с. 471.
Рис« 138„ Scalopus aquaticus (по: Строганов, 1948).
Рис. 139. Scalopus aquaticus (по: Строганов, 1948).
256
ПОДСЕМ. TALPINAE
Тип. j, шкура и череп. № 5475, Stilwell, Оклахома (США), кол. Муз. сравнит,
зоол. Гарвард, унив.
Диагноз. Отличается от других видов рода коричневато-бронзовой
окраской летнего меха, слабым и узким черепом с удлиненным носовым
отделом.
Измерения (по: Jackson, 1915). Длина: общая тела — 154,
хвоста — 24, ступни — 19, черепа — 33.2 мм.
Распространение ,и возраст. Известен по 1 экз.
3. Scalopus inflatus Jackson, 1914.
J a c k s о n, 1914, Proc. Biol. Soc. Washington, v. 27, p. 21.
Тип. Шкура и череп № 52709, Тамалиупас, Мексика, кол. Нац. муз. США,
Вашингтон.
Диагноз. Череп мелкий, с крупными слуховыми барабанами,
выступающими за контуры задней части черепа.
Измерения (по: Jackson, 1915). Длина: хвоста — 18, ступни —
16.5 мм.
Распространение и возраст. Известен из одного место-
нахождения.
7. Триба Т A L Р I N I Fischer von Waldheim, 1817
Fischer von W aldheim, 1817, p. 368—428.
Тип. Talpa Linnaeus, 1758, Европа, Азия, миоцен—современный.
3 । 4_з з
Диагноз. Зубная формула IС—gР—3М у = 44—40. Пальцы
соединены кожей до когтей. Серповидная кость сильно развита. Слухо-
вые барабаны замкнутые, в виде пузырей. Барабанная кость срастается
с окружающими ее костями. Верхний клык крупный, напоминает этот зуб
хищных млекопитающих. Нижний клык мелкий, прижат к последнему
резцу. Рх или Р2 крупные, выполняют функцию клыка.
Описание. Размеры тела варьируют. Все представители имеют
форму тела, характерную для кротов, специализированных к рытью.
Голова округло-клиновидная, внешне не отделена от тела шейным пере-
хватом. Носовой отдел головы вытянут h хоботок. Глаза маленькие,
заметные или закрыты кожной перепонкой. Ушные раковины не развиты.
Пальцы плотно соединены кожей, с плоскими когтями. Ладони повернуты
наружу. Хвост короткий. Волосяной покров мягкий, бархатистый.
Окраска тела от темно-серой до черно-коричневой и почти черной. Нижняя
сторона тела окрашена светлее.
Череп удлиненной формы, с более или менее широкой мозговой капсу-
лой, относительно плоский. Слуховые барабаны развиты.
Зубы дифференцированы на резцы, клыки, премоляры и моляры.
Верхние резцы имеют плоскую переднюю поверхность, расположены
на передней стороне межчелюстной кости полукругом или в виде тре-
угольника. I1 крупнее остальных или только шире. Верхние клыки
крупные, кинжаловидные. Нижние резцы менее крупные и менее широкие,
чем верхние, немного наклонены вперед. Они расположены на передне-
боковой стороне ветви нижней челюсти. Нижние клыки мелкие, прижаты
к задним резцам. Нижний клык может отсутствовать, как и Р{. Общие
структурные особенности зубов близки таковым кротов других триб
подсем. Т alpinae.
Плечевая кость широкая и невысокая (ширина 74—83% длины),
уплощена спереди назад. Ключица короткая (длина около 50—85% ши-
рины).
31. РОД AMPHIDOZOTHERIUM
257
В пределах семейства — это одна из наиболее однообразных, как по
внешнему виду, так и по характеру адаптаций, групп кротов. Все виды
этой трибы высокоспециализированные землерои. Многие ее представители
широко распространены и обитают в разнообразных ландшафтах.
Распространение и возраст. Европа, эоцен—современ-
ный; Азия, миоцен—современный.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Зубная формула полная для плацентарных млекопитающих: общее
количество зубов 44 .................................. 33. Talpa.
2 (1). Общее количество зубов 42—40.
3 (6). Общее количество зубов 42.
4 (5). В верхней челюсти отсутствует Р1.................35. Parascaptor.
5 (4). В нижней челюсти отсутствует клык...................34. Mogera.
6 (3). Общее количество зубов 40. Отсутствуют Р} 36. Scaptochirus.
Примечание. В определительную таблицу не вошли роды: Amphidozotherium
Filhol и Geotripus Pomel за отсутствием сравнимых признаков.
f 31. Род AMPHIDOZOTHERIUM Filhol, 1877
Filhol, 1877, Bull. Soc. Philomath., ser. 7, t. 1, p. 51—54.
T и n. A mphidozotherium cayluxi Filhol, 1877, фосфориты Кверси, Франция, верх-
ний эоцен.
Диагноз. По: Filhol, 1877 — зубная формула: I? С—М-§-=? Первые
три премоляра очень мелкие. Р4 весьма развит, с конической коронкой
и остроконечной вершиной. По: Saban, 1958 — клык некрупный. Первые
три премоляра уменьшаются в размерах спереди назад. Р4 не моляризо-
ванный, крупнее, чем Mv имеет два бугорка слаборазвитого талона. Мо-
ляры с одним невысоким тригонидом и большим талонидом (метаконидом).
Рис. 140. Amphidosotherium cayluxi (по: Crochet, 1974).
Примечание. Оба приведенных диагноза весьма схематичны.
Следовало бы отметить строение клыка и премоляров. Сколько корней
имеют клык и премоляры. Этот род — самый ранний из известных, и его
систематическое положение представляет значительный интерес. Остается
неясным, почему Лавока (Lavocat, 1951) свел этот род в синонимы рода
Geotripus Pomel, а Ван Вален (Van Valen, 1967) поставил его в сем. Ada-
pisoricidae.
Распространение и возраст. Европа, верхний эоцен.
17 А. А. Гуреев
258
ПОДСЕМ. TALPINAE
+ 1. Amphidozotherium cayluxi Filhol, 1877 (рис. 140).
Filhol, 1877, там же.
Тип. He отмечен, вид описан из фосфоритов Кверси, Франция, верхний эоцен.
Измерения (по: Filhol, 1887). Длина: Р4 — 2.5, Мх — 3.0 мм.
Распространение и возраст. Известен из одного место-
нахождения.
Примечание. Положение этого рода в системе недостаточно
ясно. Поскольку другие роды, известные из олигоцена, объединяли высоко-
специализированных кротов, можно не сомневаться, что они формирова-
лись в эоцене. Amphidozotherium — не предок всех кротов. Нельзя его
считать предком только потому, что он самый ранний из известных вы-
мерших родов.
Вероятнее всего, что он — представитель ранних групп трибы Talpini.
+ 32. Род GEOTRIPUS Pomel, 1848
Talpa antiqua Blainville, 1839—1846, Osteographie Mammiferes, pl. XI; —
acutidentata Blainville, там же. Geotripus antiquus Pomel, 1848, Bibl. Univ.
Geneve, Arch. Sci. phys. nat., t. 9, p. 159—167. Protalpa acutidentata Filhol, 1877,
p. 51—54. Amphidozotherium cayluxi Filhol, 1877, in: Lavocat, 1951, p. 25.
Тип. Geotripus antiquus Pomel, 1853, Овернь, Франция, средний олигоцен.
Диагноз (по: Pomel, 1853). Отличается от рода Talpa L. строением
премоляров. Р1 более заостренный, клинообразный. Плечевая кость более
длинная, с более обособленными троханторами.
Рис. 141. Geotripus jungi (по: Lavocat, 1931).
Описание, (по: Saban, 1958). При всем отличии морфологических
особенностей весьма сходен с родом Talpa. Верхние зубы слабо отлича-
ются от этих зубов представителей рода Talpa. Р4 имеет более слабый
наружный талон. М1 более развит в ширину, имеет трапециевидную,
а не треугольную форму. Р4 с одним главным, коническим бугорком и од-
ним крупным талоном. Видимо, имеется в виду, что талон (метаконид
в моем представлении) не раздвоен на задней стороне. Тригонид (передний
сегмент по-моему) моляров сжат, а талон (метаконид) более развит. Пле-
чевая кость более длинная, чем у представителей рода Talpa.
Распространение и возраст. Европа, средний олигоцен.
33. РОД TALPA
259
f 1. Geotripus antiquus Pomel, 1848.
Talpa antiqua Blainviile, 1839—1864, pl. XI. Geotripus antiquus Pomel,
1848, Bibl. Univ. Geneve, Arch. Sci. phys. etnat., t. IX, p. 159—167. Talpa acutidentata
Blainville, 1839—1864, pl. XI. Protalpa codurcensis Filhol, 1877, p. 51—54.
Тип. Плечевай кость, номер и место хранения не отмечены. Лавока (Lavocat,
1951) указал, что тип (плечевая кость) — изображение на табл. XI в Osteographie de
Blainville, 1839—1864. Вид описан по фрагментам верхней и нижней челюсти и плече-
вой кости из Оверни, Франция, олигоцен.
Распространение и возраст. Франция, олигоцен.
Примечание. К роду Geotripus Лавока отнес описанные им виды:
G. jungi и G. arambourdi. Кроме того, он приводит «забытый» вид — G. аси-
tidens, описанный Помелем (Pomel, 1848, 1853) и сравнивает его с предста-
вителями 5 родов. К настоящему времени еще очень мало материала,
чтобы можно было разобраться в истинном систематическом положении
этих видов. Пока их реальность весьма сомнительна.
За отсутствием у меня изображения G. antiquus Pomel привожу рисунок
G. jungi Lavocat (рис. 141).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КРОТОВ ФАУНЫ СССР
1 (Ю). Клык в нижней челюсти имеется.
2 (5). Глаза открыты и снабжены подвижными веками. Ряд верхних
резцов образует более или менее правильную дугу. Все верхние
резцы почти равны по высоте.
3 (4). Наибольшая ширина М2 более 2 мм . . . .2. Talpa europaea L.
4 (3). Наибольшая ширина М2 менее 2 мм........3. Т. altaica Nik.
5 (1). Глаза закрыты кожей и снаружи не видны. Ряд верхних резцов
образует угол. I1 заметно выше остальных.
6 (7). Длина тела меньше НО мм. Ширина когтя третьего пальца кисти
1.5— 2.0 мм ................................6. Т. соеса Savi.
7 (6). Длина тела не меньше 110 мм. Ширина когтя третьего пальца
кисти более 2 мм.
8 (9). Ширина М2 не превышает 2.5 мм .... 4. Т. caucasica Satun.
9 (8). ШиринаМ2 2.8—3.2 мм...................5. Т. romana Thom.
10 (1). Клык в нижней челюсти отсутствует.
11 (12). Длина ступни 22.4—25.5 мм, кондило-базальная длина черепа
40.0—47.4 мм, длина максиллярного зубного ряда 15.6—17.2 мм
...........................................2. Mogera robusta Nehr.
12 (11). Длина ступни 18.3—20.5 мм, кондило-базальная длина черепа
30.0—38.7 мм, длина максилл:ярного зубного ряда 10.6—14.9 мм
. . . .......................................1. М. wogura Temm.,
33. Род TALPA Linnaeus, 1758
Linnaeus, 1758, p. 52.
Тип. Talpa europaea Linnaeus, 1758, Европа и Азия, миоцен—современный.
Диагноз. Зубная формула: I ф С Р |-М ~ = 44.
Р1 по величине равен Р2 или немного выше. Р2 меньше Р3. Р4 сравни-
тельно крупный. Р1 Р2 и Р3 стоят по оси челюсти. Р2 и Р3 или только Р3
стоят под углом к продольной оси ветви нижней челюсти. Рх крупный,
клинообразный.
Описание. Размеры тела 90—234 мм. Длина хвоста не менее
длины ступни. Глаза открыты и снабжены подвижными веками или за-
крыты кожей. Окраска тела черная, черно-бурая или черно-серая. Ниж-
няя сторона тела светлее, нередко с коричневым налетом.
17*
260
ПОДСЕМ. TALPINAE
Седалищные кости соединены с позвоночником костным выступом,
соединены только на одной стороне или не соединены совсем (рис. 143).
Распространение и возраст. Европа и Азия, мио-
цен—современный.
Обитатели преимущественно открытых участков горных и равнинных
ландшафтов.
t 1. «Talpa» minuta Blainville 1839—1864 (рис. 142).
Blainville, 1839—1864, p. 97. Talpa (?) minuta Blainv. Gaillard, 1899,
p. 27, 28.
Тип. He отмечен, вид описан по плечевой кости, найденной в миоцене Сансан
(Франция), изображение которой помещено на табл. XI (Blainville, 1839—1864).
диагноз. оа таковой
Бляинвиля: плечевая кость
Рис. 142. «Talpa» minuta (по:
Zapfe, 1951).
здесь условно принято краткое описание
Т. minuta менее таковой современного
Т. europaea L. примерно на х/3 общей
длины. Тело кости пропорционально более
удлиненное, что позволяет считать этого
крота менее приспособленным к рытью.
Описание (по: Gaillard, 1899).
Гайллярд составил описание по большему
числу экземпляров плечевых костей, добы-
тых в миоценовых отложениях Grive-Saint-
Alban (Франция). Этот автор отмечает
следующие особенности строения плечевой
кости описываемого вида: проксимальный
отдел расширен; фасетка для сочленения
с ключицей большая; гребень для прикреп-
ления сухожилия musculus pectoralis на
верхней и нижней сторонах кости сходной длины; апофизы для прикрепле-
ния большого дорсального мускула не удлинены, хорошо заметны; дисталь-
ный отдел кости не имеет специфических особенностей и мало отличается
от этой части плечевой кости других видов кротов; справа и слева над
кондилюсами имеются бугорки, которые часто повреждены; характер-
ной для дистального отдела плечевой кости описываемого вида следует
считать глубокую впадину для локтевого отростка локтевой кости.
Примечание. Нельзя не согласиться с мнением названных здесь
авторов, что описанные особенности плечевой кости Т. minuta не позволяют
установить его систематическое положение. Для этого необходимо иметь
хотя бы его челюсти.
Измерения (по: Gaillard, 1899). Общая длина плечевой кости —
9.5 и 10.5 ширина ее проксимальной части — 7.0 и 7.5, ширина ее дисталь-
ной части — 5.5 и 6.0, отношение ширины проксимальной части к длине
кости — 70 и 73%.
Распространение и возраст. Франция, миоцен.
В заключение следует отметить, что мелкие плечевые кости кротов
известны и из других частей Западной Европы, а также из СССР. Можно
думать, что часть из них принадлежит к описанному здесь виду. Возможно
также, что они, как и этот вид, принадлежит роду Talpa L., а может
быть, все принадлежат разным родам. Пока ничего точно установить нельзя.
2. Talpa europaea Linnaeus; 1758 — Европейский крот (рис. 143—
148).
Linnaeus, 1758, р. 52; — frisius М ii 11 е г, 1766, Natursystem. Suppl. und
Register-Bd 36 (Австрия). {Talpa) caudata Boddaert, 1772, Kortbegrip van het za-
menstel der Natur, Bd 1, S. 50, (Talpa) vulgaris Boddaert, 1785, Elenchus Anim.,
33. РОД TALPA
261
Bd 1, S. 126. Talpa еигораеа L. Д в и г у б с к и й, 1829, Опыт естеств. истории Всех
животных Российской империи, с. И; — scalops Schultze, 1897,- Abh. und Vbrtr.
Gesammtb. Naturw., ser. 4, Bd 10, S. 19.
'Тип. He установлен, вид описан из Упсалы (Швеция).
Рис. 143. Тазовые кости кротов Talpa еигораеа (/), Т. ronama (?), Mogera robusta (5)
Т. caucasica (по: Строганов, 1948).
Диагноз. Длина тела — 120—165 мм. Глаза открыты и снабжены
подвижными веками. Резцы расположены в верхней челюсти по дуге
(рис. 144). I1 такой же высоты, как и I2 и I3 или немного выше. Ширина
М2 более 2 мм. Ширина когтя третьего пальца более 2 мм.
Описание. Окраска варьирует от черно-серой до черно-бурой
и почти черной. Верхнечелюстные кости слабо выступают в стороны, и
предглазничные отверстия при
взгляде на череп сверху мало-
заметны (рис. 148). Зубы крупные.
Костная перемычка, соединяющая
седалищные кости с позвоночником,
имеется (рис. 143).
И з мерение. Длина: хвоста
— 28.0—42.0, ступни — 17.0—22.2,
кондило-базальная черепа — 32.0
—37.5 мм.
Распространение
и возраст. Лесная и лесостеп-
ная зоны Европы и Азии, от север-
ной части Пиренейского полуост-
рова на западе до Западной Сибири
на востоке и от Шотландии, Юж-
ной Швеции и Западной Финлян-
Рис. 144. Соотношение размеров и распо-
ложение резцов у кротов Talpa еигораеа (/),
Т. caucasica (2) (по: Строганов, 1948).
дии на севере до побережья Средиземного моря на юге. В СССР
распространен от западной государственной границы на юг до линии:
Запорожье, Изюм, Лиски, Пенза, Вольск, через Оренбург до Тюмени
(по р. Днепру до Черного моря). На севере граница распространения
проходит через район Кандалакши, устье р. Онеги, восточный берег
Онежской губы и Белого моря на восток до низовьев рек Кулоя и Мезени,
долины р. Пезы и верховьев р. Пеши; далее северная граница идет по
рекам Цыльме, Сосьве и долине р. Печоры до Нарьян-Мара, проходит
262
ПОДСЕМ. TALPINAE
к низовьям рек Лаи, Колвы и Усы, пересекает Северный Урал около
64° с. ш., проходит на Березов и спускается к Ирбиту и Тюмени. Восточная
граница неясна, плейстоцен—современный.
Биология. Обитает в лесной и лесостепной зонах. В лесостепной
зоне он не выходит на широкие, открытые пространства, а придерживается
участков с лесной или кустарниковой растительностью. Объясняется
Рис. 145. Крот из среднего плейстоцена Франции — Talpa ci. europaea (Jammot, 1973).
это континентальностью климата южных окраин лесостепной и степной
зон, обусловливающей значительную сухость верхних слоев почв, а в связи
с этим ограниченное количество дождевых червей, составляющих основ-
ную пищу крота. Главным условием существования этого вида, всю жизнь
проводящего под землей, служат достаточно увлажненные почвы, богатые
перегноем, в которых он находит в достатке дождевых червей и других
беспозвоночных животных и в их числе насекомых. Крот избегает участков
с сильно заболоченной и песчаной почвой. При сильном переувлажнении
почвы талыми водами весной кроты прокладывают жировочные ходы
близко от поверхности почвы, часто настолько, что земля поднята над
33. РОД TALPA
263
ходом в виде валика. Это мне приходилось наблюдать в окрестностях
поселка Ропша Ленинградской обл. в мае 1968 г.
Место обитания крота можно узнать по кучам земли, выброшенной им
на поверхность почвы. Диаметр отдельных куч может достигать 70—100,
в среднем 30—40 см; они располагаются чаще всего группами. Прорывая
ходы в сильно уплотненном грунте, например под дорогой или тропинкой,
крот роет ход близко от поверхности земли, отчего почва в этом месте
Рис. 146. Talpa еигораеа (7), Т. соеса (2), Т. altaica (3) (по: Строганов, 1948).
поднимается в форме грядки. Ходы крота по своему значению делятся
на две основных категории: кормовые, располагающиеся близко от поверх-
ности почвы, другие — подходы к гнезду, водопою и кормовым ходам.
В общей сложности они образуют многоярусную систему и располагаются
на глубине от 5 до 60 см. Стенки ходов гладкие, уплотненные. Путем
кольцевания установлено, что ходы простираются на значительные пло-
щади и крот передвигается по ним на значительные расстояния (до 4 км).
Передвижение кротов в земляных ходах происходит относительно быстро.
Окольцованный молодой самец через 20 ч оказался на расстоянии около
700 м по прямой от места выпуска, а самка через 6 дней — в 2 км. В случае
необходимости крот хорошо плавает.
Гнездо крота помещается под корнями деревьев, под пнями или кам-
нями, реже на открытом месте. Гнездо в виде небольшого углубления вы-
264
ПОДСЕМ. TALPINAE
стилается сухими растениями. Оно располагается в почве на глубине 20—
70 см.
Пищу крота, как уже отмечалось, составляют различные беспозвоночные,
но основным кормом, определяющим повсеместно его численность, служат
земляные черви (Oligocheta). На фоне этого основного корма в разных
биотопах и в разные сезоны года меняется состав беспозвоночных, главным
образом насекомых, составляющих небольшое дополнение к основной пище.
По данным О. С. Русакова (1965), насекомые составляют 3—12% от об-
щего количества корма, поедаемого кротом в сутки. Этот же автор при-
водит данные о количестве поедаемого корма кротом в течение суток.
За 10 сут крот съел 960 г дождевых червей и 63 г насекомых, что составляет
в среднем за сутки 125% веса его тела. Максимальное количество корма,
Рис. 147. Talpa europaea (по: Строганов, 1948).
съеденное этим кротом за сутки — 150 г, или 183% веса его тела (вес
крота 82 г). Недостаток пищи кроты переносят плохо, а при полном голода-
нии быстро погибают. Они быстро погибают и от ожирения в условиях
неволи при недостатке работы. Высокий темп обмена веществ у кротов
скоррелирован с большими затратами энергии на работу по прокладке
ходов, а также с затратами энергии на согревание при понижении темпера-
туры в отдельные сезоны года. При круглогодичной активности крота
затраты энергии на его нормальную жизнедеятельность и размножение
стоят в прямой зависимости от конкретных условий в годичном цикле
природы, они то увеличиваются, то уменьшаются.
Половозрелыми и самцы, и самки становятся на следующий год после
рождения. Размножается крот один раз в году. Спаривание в зависимости
от температурных условий происходит в марте—апреле. Беременность
длится около 40 дней. Число детенышей 1—9, в среднем 5 или несколько
более. В отдельных случаях отмечается 12 детенышей. В период эмбрио-
нального развития, отдельные зародыши могут подвергнуться резорбции.
Период размножения растянут в относительно небольшом отрезке времени.
Колебание сроков появления молодых обусловлено погодными условиями
каждого года, скоростью прогревания почвы в разных биотопах, временем
схода талых вод и многими другими. Возможно также, что может меняться
длительность эмбрионального развития: указывается беременность в 3—
4 недели, 28—32 дня и 35—45 дней.
33. РОД TALPA
265
Линяет крот три раза в году: весной, летом и осенью. Летняя линька
неполная.
Подвиды. 1. Т. е. europaea L., 1758. Отличается высоким и вы-
пуклым спереди I1. Распространение: северные и средние области евро-
пейской части СССР, на восток до Северного Урала, Соликамска, по
р. Каме до р. Волги и по Волге до Куйбышева.
2. Т. е. brauneri Satunin, 1908. Отличается менее широкими и плоскими
спереди Р. Распространение: южные области европейской части СССР.
Границы ареала этого подвида неясны.
3. Т. е. transuralensis Stroganov, 1956. Отли-
чается наличием талона и зачаточного гипоконуса
у Р4. Распространение: восточные склоны Южного
и Среднего Урала, бассейны рек Конды и Тавды, ни-
зовья рек Тобола, Ишима, Иртыша и Демянки.
Рис. 148. Talpa europaea (по: Строганов, 1948).
4. Т. е. obensisScaAmt et Rajevsky (1940). Отличается мелкими размерами
и формой I1. Он низкий, невыпуклый спереди и мало отличается по ширине
от I2 и I3. Распространение: бассейн р. Малой Сосьвы, долина р. Ляпин,
район г. Березова.
3. Talpa altaica Nikolsky, 1884 — Крот сибирский, или алтайский
(рис. 149).
Nikolsky, 1884, Тр. СПб. о-ва естествоисп., т. 11, с. 165—170. Talpa (Asio-
scalops) altaica Nikolsky Строганов, 1941. Зоол. жури., т. 20, вып. 3,
с. 371—381. Asioscalops altaica Nik. Строганов, 1957.
Тип. Не установлен, вид описан из окрестностей дер. Тоурак на Алтае.
Диагноз. Длина тела — 150—232 мм. Глаза открыты и снабжены
подвижными веками. Резцы расположены в верхней челюсти по дуге
(рис. 144). I1 такой же высоты, как и I2 и I3 или немного выше. Ширина
М2 менее 2 мм. Ширина когтя третьего пальца более 2.5 мм.
Описание. Окраска варьирует от черно-серой и черно-бурой
до бурой разных оттенков. Верхнечелюстные кости слабо выступают
в стороны, и предглазничные отверстия при взгляде на череп сверху
незаметны или почти незаметны (рис. 149). Зубы мелкие. Костная пере-
266
ПОДСЕМ. TALPINAE
мычка, соединяющая седалищные кости с позвоночником, отсутствует
(рис. 143).
Измерения. Длина: хвоста — 21—33, ступни — 15—25, кон-
дило-базальная черепа — 34.5—41.0 мм.
Распространение и возраст. Западная Сибирь, запад-
ная часть Восточной Сибири и южное Забайкалье. Западная граница про-
ходит приблизительно по линии г. Сургута, Тюмени, по системе р. Иртыша
до Семипалатинска и оз. Зайсан и по р. Иртышу уходит в северо-западную
часть Монголии. К северу распространен приблизительно до 68° с. ш.
Восточная граница неясна, плейстоцен?—современный.
Биология. Обитает в разнообразных усло-
виях, от лесостепи на юге до зоны вечной мерзлоты
на севере и от низменных местностей Западной Си-
бири до субальпийского пояса Саян и Алтая, подни-
маясь в горы до 2000—3500 м над ур. м. Населяет
разнообразные леса, но избегает понижений с высоким
уровнем грунтовых вод и болот. Предпочитает площади
Рис. 149. Talpa altaica (но: Строганов, 1948).
с рыхлой и богатой гумусом почвой. В наибольшем количестве
встречается в смешанных лесах с редко стоящими деревьями, с подлеском
из различных кустарников и густым травяным покровом. В ряде районов
отмечены сезонные перекочевки крота. Последние обусловлены изменением
режима грунтовых вод, разливом рек, таянием снега весной и другими
обстоятельствами.
Многоярусная система кротовых ходов имеет значительную протяжен-
ность и пронизывает почву во многих направлениях. Гнездо устраивается
в хорошо аэрируемой почве, в местах, где весной рано сходит снег. Гнездо
представляет собой расширенное углубление с выстилкой из сухих расте-
ний, соединенное с системой кормовых ходов кольцевыми и радиальными
ходами.
Основную пищу сибирского крота, как и европейского, составляют
дождевые черви. Насекомые и другие беспозвоночные служат небольшим
дополнением к основному корму. Количество и состав этого дополнитель-
ного корма меняется по сезонам.
Самцы сибирского крота достигают половозрелости на втором году
жизни, а самки уже на 4—5-м мес. способны к размножению. Спариваются
летом, с июня по август (Строганов, 1957а). По данным Т. Л. Бородулиной
33. РОД TALPA
267
(1953), беременность длится около 9 мес. (видимо, развитие зародыша
задерживается). Рождение молодых происходит весной. Беременные
самки под Томском добывались в апреле. Новорожденные добывались
в мае, а в июне молодые ведут уже самостоятельный образ жизни. Остается
неясным, когда рождаются и спариваются самки-сеголетки, способные
размножаться в 1.5—3-месячном возрасте (Юдин, 1971), 4—5 мес. от-
роду (Строганов, 1957; Бородулина, 1953). По данным Бородулиной,
спаривание самок-сеголеток происходит в июне—июле. Количество дете-
нышей в помете бывает 3—6.
Линяет крот три раза в году.
Подвиды. 1. Т. a. altaica Nikolsky, 1883. Отличается темной
окраской летнего волосяного покрова с заметным коричневым оттенком.
Распространение: Алтай и приалтайские ленточные боры.
2. Т. a. suschkini Kastschenko, 1905. Имеет относительно светлую окраску
и более крупные размеры (длина тела — 165—232 мм). Распространение:
Западный и Восточный Саяны, к-северу до Красноярска, на восток до
Байкальского хребта.
3. Т. a. gusevi Fetisov, 1956. Распространение: Тува и юго-западная
часть Забайкалья.
Подвиды Т. a. salairica Egorin, 1936 (Салаир, Кузнецкий Алатау),
Т. a. tymensis Egorin, 1937 (бассейн рек Васюгана и Тыма) и Т. a. sibirica
Egorin, 1937 (Мариинская тайга, бассейн р. Енисея) слабо дифференциро-
ваны. Требуется более тщательный сравнительно-морфологический их
анализ.
4. Talpa caucasica Satunin, 1908 — Крот кавказский (рис. 143,
144, 150).
Talpa europaea L. М ёп ё tries, 1832. Cat. raisonne des objets Zoologie rec. dans
voyage au Caucase etc. St. Petersbourg, p. 17; — coeca Savi Satunin, 1896, Zool.
Jahrb., Bd 9, System., S. 288; — coeca caucasica, С а т у н и н, 1908, Изв. Кавк, муз.,
т. 4, с. 5; — caucasica Огнев, 1926, с. 45.
Тип. Не установлен, вид описан из Ставрополя (Кавказ).
Диагноз. Длина тела — 110—136 мм. Глаза закрыты кожей.
Резцы расположены в верхней челюсти под углом (рис. 144). I1 значи-
тельно выше I2 и I3. Ширина М2 не более 2.5 мм. Ширина когтя третьего
пальца более 2 мм.
Описание. Окраска черно-серая, черно-бурая или черная. Верхне-
челюстные кости слабо выступают в стороны, и предглазничные отверстия
при взгляде на череп сверху малозаметны (рис. 150). Зубы относительно
крупные. Костная перемычка, соединяющая седалищные кости с позво-
ночником, как правило, отсутствует, но встречаются отдельные особи,
у которых она имеется на одной стороне таза.
Измерения. Длина: хвоста — 24—37, ступни — 17.5—19.2, кон-
дило-базальная черепа — 33.1—35.7 мм.
Распространение и возраст. Северный Кавказ от
Азовского моря на западе до Каспийского на востоке, Черноморское по-
бережье; южная граница к востоку от г. Батуми неясна, плейстоцен?—
современный.
Примечание. Путаница в систематике кавказских видов кро-
тов не способствовала выяснению их ареалов, они все еще весьма схе-
матичны.
Биология. Обитает в лесостепи предгорий, в лесах разных
типов, в субальпийской и альпийской зонах высокогорий на высоте до
2100 м. Наибольшее количество кротов встречается в лесостепи и широко-
268
ПОДСЕМ. TALPINAE
лиственных лесах, меньшее — в альпийской зоне и наименьшее в темно-
хвойных лесах. Предпочитает участки с достаточно влажной почвой,
что связано с большим содержанием в ней различных беспозвоночных
животных, особенно дождевых червей.
Роет многоярусные ходы, которые делятся на ряд категорий: основные-
ходы поверхностные и глубинные (жировочные' и гнездовые). Глубина
залегания ходов зависит от характера грунта, уровня
а также от степени промерзания почвы в зимнее
время. Глубина охотничьих, или жировочных ходов
от 3 до 20 см. В связи с отмеченными факторами изме-
няется и глубина расположения гнездовой камеры.
Последняя может быть двойной. Число ответвлений
ходов от гнездовой камеры бывает различным, как и
грунтовых вод,
Рис. 150. Т. caucasica (по: Строганов, 1948),
количество изгибов и поворотов, чем и объясняется разная степень
сложности строения гнездовой камеры. Повышение уровня грунтовых
вод часто вынуждает крота строить новую гнездовую камеру, выше ста-
рой. Подстилка гнезда пз сухих растений.
Пищей кроту служат различные беспозвоночные животные: насекомые,
многоножки и другие, но основной корм составляют дождевые черви.
В желудках находят и остатки растений. Количество корма, необходимого
кроту в сутки, равно примерно половине веса его тела.
Размножается кавказский крот один раз в году. К размножению
приступает в феврале. В разные годы период размножения охватывает
разные сроки. Беременные самки попадаются с середины февраля до конца
марта. Наибольшее количество беременных самок добыто в марте.
Запоздавшие с размножением самки встречаются в конце мая. Продолжи-
тельность беременности около 35—40 дн. Эмбрионов — 2—5, в среднем —
3. Молодые растут быстро и приблизительно через 30—40 дн. после
рождения начинают попадаться в ловушки.
Линяет кавказский крот три раза в году — весной, летом и осенью.
Подвиды. Не изучены.
5. Talpa romana Thomas, 1902 — Крот крупнозубный (рис. 143, 151).
Thomas, 1902, Ann. Mag. Nat. Hist., set. 7, v. 10, p. 517.
T и п. He установлен, вид описан из окрестностей Рима (Италия).
33. РОД TALPA
269
Диагноз. Длина тела — 130—145 мм. Глаза закрыты кожей.
Резцы расположены в верхней челюсти под углом (рис. 144, 2). I1 выше и
почти в два раза шире I2 и I3. Ширина когтя третьего пальца более 2 мм.
Описание. Окраска черно-бурая или черно-серая. Верхне-
челюстные кости сильно выступают в стороны, и предглазничные отверстия
при рассматривании черепа сверху хорошо видны (рис. 151). Зубы крупные,
например, наибольший диаметр М1 — 3.7—4.3 мм. Костная перемычка,
соединяющая седалищные кости с позвоночником, отсутствует.
Измерения. Длина: хвоста — 27—30, ступни — 19—20, кондило-
базальная черепа — 35.9—37.6 мм.
Распространение и возраст. Из-
вестен из Италии (окрестности Рима), Югославии
(Македония) и с Кавказа (Бакуриани, Кутаиси),
современный.
Биология. На территории Кавказа не изу-
чена.
Рис. 151. Т. romana. (по: Строганов, 1948).
И о д в и д ы. Т. г. ognevi Stroganov (1944). Отличается от других
подвидов крупными размерами зубов. Распространение: найден в окрест-
ностях Бакуриани и Кутаиси (Кавказ).
Примечание. Пока у меня нет никаких оснований исключать
из фауны СССР крупнозубого крота — Т. romana ognevi Stroganov.
Сравнение экземпляров кротов из Бакуриани и Кутаиси, послуживших
С. У. Строганову материалом для описания этой формы, с экземплярами
Т. romana из Югославии позволило обнаружить их значительное сходство.
В то же время и те и другие отличаются от Т. caucasica и Т. соеса общими
для них признаками.
Ареал Т. romana разделен на три разобщенных участка: Италия,
Югославия, Кавказ. Из Ирана и Ирака описаны кроты, не сходные ни
с европейскими, ни с азиатскими видами, а также и с кавказскими,
ближайшими к ним. Разрыв ареала ряда видов кротов, населяющих Среди-
земноморскую подобласть Палеарктики, в результате горообразовательных
процессов обусловил развитие отдельных видов и подвидов.
6. Talpa соеса Savi, 1822 — Крот слепой, или малый (рис. 152).
S a v i, 1822, Nuova Giorn. de Latterati, Pisa, t. 1, p. 265; — olympica C h a-
wor th-Must er s, 1932, Ann, Mag. Nat. Hist., ser. 10, v. 9, p. 166, 167; — her-
270
ПОДСЕМ. TALPINAE
cegovinensis В о Ik ay, 1925. Novit. Mus. sarajev, v. 1, p. 1—16; — minima Д e-
парма, 1959, Бюл. МОИП, отд. биол., т. 64, вып. 6, с. 31—36.
Тип. Не установлен, вид описан из окрестностей Пизы (Италия).
Диагноз. Длина тела — 90—106 мм. Глаза закрыты кожей.
Резцы расположены в верхней челюсти под углом (рис. 144). I1 значи-
тельно выше и шире I2 и I3. Ширина М2 не превышает 2 мм. Ширина когтя
третьего пальца — 1.5—2 мм.
Описание. Окраска от черно-серой до почти черной с буроватым
налетом. Верхнечелюстные кости выступают в стороны настолько, что
предглазничные отверстия при рассматривании черепа сверху хорошо
видны (рис. 152). Зубы относительно мелкие. Костная перемычка между
седалищными костями и позвоночником, как правило, отсутствует, но
может быть на одной стороне, реже на двух в виде
слабых костных анастомозов.
Измерения. Длина: хвоста — 27.0—
ступни — 16.0—17.9 кандило-базальная че-
— 29.8—32.6 мм.
Рис. 152. Talpa coeca (по: Строганов, 1948).
30.2,
репа
Распространение и возраст. Португалия, Испания,
Швейцария, Италия, о. Сардиния, Балканы, Турция. В СССР на Кавказе,
современный.
Биология. Обитает в разнообразных биотопах преимущественно
в лесных ландшафтах. Предпочитает мягкие почвы. Ходы роет двух типов.
На открытых местах роет ходы на глубине 10—40 см. Кучки выброшенной
при рытье земли располагает равномерно друг от друга. Из гнездовых
ходов на землю выбрасывает в нескольких местах, до четырех на 1 м2.
В лесу кротовые ходы располагаются близко от поверхности почвы и
хорошо заметны по валикообразным возвышениям. Почва в лесу рыхлая,
поэтому крот не столько роет, сколько уминает почву по бокам хода и
поднимает его свод. В связи с этим он выбрасывает на поверхность мало
земли. Встречающиеся в лесу относительно крупные кучки земли рас-
полагаются над ходом, идущим на глубину до 70 см и заканчивающимся
гнездовой камерой — небольшим расширенным тупичком. Подстилка
в гнездах отсутствует. Кормовые ходы в лесу крот роет от дерева к дереву,
между кустами по краям и скал и под камнями, а также прорывает ходы
в гнилых, поваленных деревьях и под слоями моховой подстилки.
Размножение не изучено. Как и для других кротов, основной пищей
этому виду служат дождевые черви, к которым примешиваются насекомые
33. РОД TALPA
271
и другие беспозвоночные. Объем дополнительного корма, как и видовой
состав кормовых животных, меняется по сезонам.
Подвиды. 1. Т. с. orientalis Ognev, 1926. От типичной формы отлича-
ется большими размерами тела. Распространение: Черноморское побе-
режье от Новороссийска до Батуми, на восток до верховьев р. Терека,
Гурджаани и оз. Севан.
2. Т. с. talyschensis Verestschagin, 1945. По размерам меньше Т. с.
orientalis. Описан из ущелья Виляж-Чая, Талыш.
3. Т. с. minima Deparma, 1959. Отличается очень мелкими размерами.
Отверстие в ключице, которое Депарма считает систематическим призна-
ком этого крота (описанного ею как самостоятельный вид), встречается
п у других видов. Распространение: горные районы северо-западного
Кавказа (верховья р. Белой и ее притоков).
7. Talpa streeti Lay, 1965. — Крот курдистанский (рис. 153).
Lay, 1965, р. 227—230.
Т и п. № 96 424. Hezar Darrell, Курдистанская провинция (Иран), современ-
ный. Кол. Чикагского муз. естеств. истории.
Диагноз, (по: Lay, 1965). Отличается от других палеарктических
видов рода Talpa следующими, перечисленными ниже признаками: от
Т. romana — 1 и 4; от Т. соеса — 1, 2, 4, 5, 6 и 7; от Т. еигораеа — 1, 4,
5; от Т. hercegoviensis — 2, 4, 6 (и, возможно, 1, 3, 5, 7, но экземпляры
недостаточно пригодны для ясного сравнения); от Т. micrura — 1, 2, 3, 4,
5, 7; от Т. longirostris — 1, 2, 3, 4, 5, 6, 7; от Т. klossi — 1, 2, 3, 4, 5, 6, 7.
Рис. 153. Talpa streeti (по: Lay, 1965).
Описание (по: Lay, 1965). Зубная формула, как у видов рода
Talpa — полная.
1. Р2 Р| зубных рядов, челюстей заметно меньше других зубов со-
ответствующего ряда между резцами и молярами.
2. Носовой отдел черепа несколько расширен в поперечном направле-
нии на уровне М2.
3. М1-3 вытянуты в передне-заднем направлении. Длина их ряда 6.5
(М—6.6) 6.7 мм.
4. Отношение средней ширины носовой части черепа на уровне М2
к средней кондило-базальной длине черепа равно 2.9.
5. Скуловые дуги широко расставлены, относительно крепкие, ширина
13.6 (М—13.7) 13.9 мм.
272
ПОДСЕМ. TALPINAE
6. Ширина наиболее выступающих в стороны частей теменных и височ-
ных костей (наибольшая ширина мозговой капсулы) — 16.5 (М — 16.8)
17.1 мм.
7. Ладони значительно расширены. Когти всех пяти пальцев широкие
и округлые на конце, сохраняют в поперечнике овальную форму, снизу
плоские.
Измерения (по: Lay, 1965). Длина: кондило-базальная черепа —
30.6—31.7, М1—М3 — 6.5—6.7 мм.
Распространение и возраст. Иран, современный.
Примечание. Отсутствие материала, хороших фотографий или
рисунков не позволяет составить нормальный диагноз. Судя по описанию,
значение отдельных признаков Т. streeti автором преувеличено. По-
видимому, череп описываемого вида по общему строению близок
к таковому Т. romana Thos. Наиболее существенным отличием этого вида
от других служат мелкие Р| и Р|. От кротов, территориально близких
к Т. streeti, — Т. caucasica Sat. и Т. соеса Savi, он отличается резцами,
расположенными полукругом, как у Т. europaea L., а не под углом, как
у названных видов. Полукругом резцы расположены и у представителей
рода Mogera Pomel. Можно думать, что такое расположение резцов у от-
дельных видов рода Talpa L. и у всех видов рода Mogera возникло парал-
лельно и обусловлено определенным функциональным значением в кон-
кретных условиях.
Примечание. Следующие пять видов рода Talpa L. (описаны
ниже) требуют дополнительного описания на достаточном материале
и в сравнении с другими видами этого рода. Они несомненно принадлежат
к этому роду, а не представляют какое-то особое направление специализа-
ции, чтобы их можно было выделить в особые роды, как это сделал Мил-
лер (Miller, 1940), выделив виды Т. klossi Thos., Т. parvidens Miller и
Т. grandis Miller в род Euroscaptor, или С. У. Строганов (1941), .объеди-
нивший в род Eoscalops виды Т. longirostris M.-Edw. и Т. micrura Hodgs.
Нелепым представляется объединение отдельных из названных видов
с видами родов Mogera Pomel, Scaptochirus M.-Edw., Parascaptor Gill,
и ранее описанных в один вид — Talpa micrura Hodgs. в качестве под-
видов последнего Шварц (Schwarz, 1948).
Географическое распространение обсуждаемых видов таково, что может
только подтверждать, а не вызывать сомнений в их качественной опре-
деленности. В самом деле, эти виды населяют Кашмир, Тибет, Западный,
Центральный и Юго-Восточный Китай, Непал, Сикким, Ассам, Индо-
китай. Вряд ли эта разноликая по физико-географическим условиям
территория, даже при том, что кроты живут в сходных условиях под
землей, населена одним видом, а не несколькими.
Описание этих интересных видов заимствовано из монографии
С. У. Строганова (1948) и работы Миллера. Материал из коллекции
ЗИН АН СССР позволил немного дополнить описание отдельных зубов
Т. micrura и Т. longirostris. Сравнительные диагнозы, к сожалению, не
получились.
В заключение следует отметить, что сравнение черепов Т. micrura
и Т. longirostris с таковыми Т. europaea L., Т. caucasica Sat., Т. соеса Savi
и Т. romana Thos. позволило обнаружить значительное различие между
ними и Т. europaea и Т. romana и сходство черепов этих кротов с черепами
Т. caucasica и Т. соеса. Сходство выражается в суженности носового отдела,
выступающих вперед резцах Р~2, в изменчивости числа корней (1 или 2)
у Р1 и Р2, особенно у последнего, в их общих размерах и другом.
Возможно, что сравниваемые группы видов кротов имели близкую
территориальную связь в прошлом и были распространены по южным
33. РОД TALPA
273
окраинам Азии от Кашмира до северной части Аравии и Малой Азии,
откуда проникли на Кавказ и Балканы, а затем и в южную часть Европы.
В более позднее время связь отдельных видов кротов между Ираном и
Кашмиром оборвалась. Но пока это только предположение, его подтвердить
могут лишь находки вымерших кротов в промежуточной зоне.
8. Talpa klossi Thomas, 1929.
Thomas, 1929, Ann. Mag. Nat. Hist., ser. 10, v. 3, N 13, p. 206—207. Euroscap-
tor klossi Miller, 1940, J. Mammal., v. 21, N 4, p. 442—444.
Тип. N 28.5.3.1, окрестности Hahend, западная часть Сиама (Таиланд), кол.
Брит, муз., Лондон.
Описание (по: Miller, 1940). Череп мало удлинен, с широким нёбом
(ширина нёба составляет 65% его длины), зубы крупные, особенно £корен-
ные. Р4 не имеет поперечного бугорка.
/54
Рис. 154. Talpa longirostris (по: Строганов, 1948).
Измерения (по: Miller, 1940). Длина тела — 138, хвоста —
11.9, ступни — 16, кондило-базальная черепа — 30—32 мм.
Распространение и возраст. От северной и западной
части Таиланда к востоку до Чапа и Бакбо, Северный Вьетнам, Китай,
современный.
9. Talpa grand is Miller, 1940.
Euroscaptor grandis Miller, 1940, J. Mammal., v. 21, N 4, p. 444.
Тип. j ad N 255 370, Mount Omei, Сычуань, Китай, кол. Амер. нац. муз., Ва-
шингтон .
Описание (по: Miller, 1940). По внешнему облику сходен
с Т. klossi Thos, отличается более крупными размерами. От европейских
видов рода Talpa L. и от Т. altaica Nik. отличается расположением ноздрей
на боковой стороне мочки носа, коротким булавовидным хвостом и корич-
невой окраской волосяного покрова.
Измерения (по: Miller, 1940). Длина: тела — 150, хвоста —
9.6, ступни — 18, наибольшая черепа около 37 мм.
Распространение и возраст. Известен от южной до
северной части Бакбо, Чапа, Северный Вьетнам, современный.
18 А. А. Гуреев
274
ПОДСЕМ. TALPINAE
10. Talpa parvidens Miller, 1940.
Miller, 1940, J. Mammal., v. 21, N 2, p. 203, 204. Euroscaptor parvidens M i 1-
I e r, 1940, там же, N 4, c. 442—444.
Тип. $ ad. N 258 342, Blao, верховья p. Donai, Трунбо, Северный Вьетнам, кол.
Амер. нац. муз., Вашингтон.
Рис. 155. Talpa longirostris (по: Строганов, 1948).
Описание (по: Miller, 1940). По внешнему виду сходен с Т. klossi
Thos. Череп удлиненный с относительно узким небом и мелкими зубами.
Р4 с хорошо развитым передним бугорком.
Измерения (по: Miller, 1940). Длина: тела — 140, хвоста — 6,
ступни — 14.4, наибольшая черепа — 34 мм.
Распространение и возраст. Кроме местонахождения
типа, известен из окрестностей Pakho на границе с Китаем, современный.
33. РОД TALPA
275
11. Talpa longirostris Milne-Edwards, 1870 (рис. 154, 155).
Milne-Edwards, 1870, Compt. Rend. Acad. Sci., Paris, t. 70, p. 70. Eos-
calops longirostris Строганов, 1941, c. 270—272.
T и п. He установлен. Аллен (Allen, 1938) предложил считать за terra typica
горы Muping, Сычуань, Китай.
Описание (по: Строганов, 1948, с некоторыми дополнениями).
Мелкий крот. Хвост булавовидный, покрыт редкими беловатыми волосами.
Рис. 156. Talpa micrura (по: Строганов, 1948).
Череп с удлиненным и тонким носовым отделом. Лобные кости в межглаз-
ничной области выпуклые. Предглазничные отверстия не широкие,
но хорошо заметные при взгляде на череп сверху. Слуховое отверстие очень
маленькое. I1 шире и выше остальных. На внутренней стороне клыка
имеется продольная бороздка. Наружная сторона коронки М1 длиннее
этой части коронки М2. Талон на М2 не обособлен в виде бугорка.
Измерения (по: Строганов, 1948). Длина: тела — 100—105,
хвоста — 17—20, ступни — 14.5—14.8, общая черепа — 32.1—32.5 мм.
Распространение и возраст. Кашмир, Тибет, Западный
Китай (Сычуань), современный.
18*
276
ПОДСЕМ. TALPINAE
12. Talpa micrura Hodgson, 1840 (рис. 156).
Hodgson, 1840, J. Asiat. Soc. Bengal., v. 19, N 1850, p. 215;—cryptura
Blyth, 1843, там же, т. 12, с. 928. Eoscalops micrura Строганов, 1941, с. 270—
272; 1941, Зоол. журн., т. 20, вып. 3, с. 371—380.
Т н п. Не установлен, вид описан из Непала.
Описание (по: Строганов, 1948, с некоторыми дополнениями).
Крупнее Т. longirostris M.-Edw. Хвост очень короткий. Предглазничные
отверстия очень узкие и малозаметны при рассматривании черепа сверху.
Слуховое отверстие относительно крупное. Размеры верхних резцов
мало разнятся от первого к третьему. На внутренней стороне клыка нет
продольной канавки. Наружная сторона коронки Ма длиннее этой части
М1. Талон на Ма в виде хорошо обособленного бугорка.
Измерения (по: Строганов, 1948). Длина: тела — ИЗ—120,
хвоста — 4.6—5.0, ступни — 12.6—15.1, общая черепа — 32.9 мм.
Распространение и возраст. Найден в Ассаме, Непале
и Сиккиме, современный.
34. Род MOGERA Pomel, 1848
Pomel, 1848, Bull. Univ. Geneve, Arch. Sci. phys. nat., v. 9, p. 247.
T и n. Talpa wogura Temminck, 1833, Япония, современный.
Диагноз. Зубная формула: I -|- С Р у М у = 42. Р1 больше Р2,
а Р2 меньше Р3. Р1 и Р2, часто и Р3 стоят под углом к продольной оси
челюсти. Р4 очень крупный. Рх крупный, клыкообразный. Р2 меньше Р3.
Описание. Размеры тела 90—210 мм. Длина хвоста менее длины
ступни или немного длиннее. Глаза закрыты кожной перепонкой и снаружи
не видимы. Окраска тела буровато- или черновато-коричневая, разных
оттенков. Седалищные кости таза срастаются с позвоночником двумя
костными перемычками. Нижнего С нет (рис. 143, 3).
Распространение и возраст. Япония (от Иокогамы
до Нагасаки и Якусимы), Китай (о-ва Тайвань и Хайнань, на континенте —
в Сычуани, Гуанчжоу, Гуаней, Хунани, Цзяньси, Гуандуне, Хубее,
Фуцзяни, на северо-востоке), п-ов Корея. В СССР в Приморье, на север
до устья р. Самарчи и г. Хабаровска, современный.
1. Mogera wogura Temminck, 1833 — Крот японский (рис. 157).
Talpa wogura Temminck, 1833, Fauna Japonica, v. 1, p. 19. Mogera wogura
Dobson, 1883, p. 140.
T и п. He установлен, вид описан из Японии.
Диагноз. Длина тела — 120—156, ступни — 18.3—20.5 мм.
Р1 с двумя корнями, выше Р2 или равен ему по высоте.
Описание. Окраска буровато-коричневая разных оттенков.
На горле и брюхе охристые пятна. Сходна с таковой М. robusta Nehr. Задний
край предглазничного отверстия на линии, проходящей между Ма и М3.
I1-1 выступают вперед и ряд верхних резцов образует выпуклую дугу.
Верхние резцы мало разнятся по величине. М. В. Охотина (1966) ука-
зывает следующие признаки, характерные для этого вида: penis притуп-
лен на конце, прианальные железы отсутствуют.
Измерения. Длина: хвоста — 20.3—24.0, кондило-базальная
черепа — 30.0—38.7 мм.
Распространение и возраст. Япония, п-ов Корея,
о-ва Тайвань, Хайнань, Фуцзянь, Ляонин (Восточный Китай). В СССР
в Приморье (Хасанский район).
34. РОД MOGERA
277
Биология. Мало изучена. По данным Охотиной (1963, 1966)
и других авторов, сведения которых она приводит в своих работах, япон-
ские кроты предпочитают безлесные пространства и придерживаются
заброшенных лугов, сельскохозяйственных угодий, возвышенностей
с редким кустарником. Реже они поселяются в горных лесах с преобла-
данием лиственных деревьев.
Ходы диаметром 3—4 см хорошо уплотнены и прокладываются на
глубине 50—70 см от поверхности почвы. Второй ярус ходов располага-
•ется на глубине 1.0—1.5 м. Верхний и нижний ярусы ходов связыва-
Рис. 157. Mogera wogura (по: Юдин, 1971).
ются вертикальными ходами. Близко от поверхности этот крот прокла-
дывает ходы редко.
Можно думать, что основные жизненные функции этого вида сходны
или близки к таковым Mogera robusta Nehr.
Подвиды. В СССР, по-видимому, М. w. coreana Thomas (1907).
2. Mogera robusta Nehring, 1891 — Крот уссурийский (рис. 158—160).
Talpa wogura R a d d е, 1862, Reisen im Siiden u. Ost-Sibirien, S. 115, 116. Mogera
robusta Nehring, 1891, Sitzungs-Ber. Gesell. Naturforsch. Freunde, Berlin, S. 95—
103. Talpa (Mogera) robusta Бобринский, 1935, Определитель охотничьих и про-
мысловых зверей СССР, с. 76.
Тип. Не установлен, вид описан из Владивостока.
Диагноз. Длина: тела — 160—210, ступни — 22.4—28.0 мм.
Р1 с двумя корнями, выше Р2.
Описание. Окраска буровато-коричневая, разных оттенков, мало
отличается от окраски М. wogura Ternm. Задний край предглазничного
отверстия — расположен впереди линии, разделяющей М2 и М3. I1 выше
и шире I2 и I3.
В дополнение к этим признакам Охотина (1966) отмечает и следующие:
penis заострен на конце. Прианальные железы имеются.
Измерения. Длина: хвоста — 26—37, кондило-базальная черепа—
40.1—47.4 мм.
Распространение и возраст. Лесные ландшафты Севе-
ро-Восточного Китая, п-ова Корея и крайнего юго-востока СССР (южная
часть Приморского края).
278
ПОДСЕМ. TALPINAE
Биология. Наиболее характерные местообитания крота — ши-
роколиственные леса равнин и горных склонов с густым травостоем и
Рис. 158—160. Mogera robusta (по: Строганов, 1948).
рыхлой богатой перегноем почвой. Наиболее многочислен он в лесах
долин рек и ручьев. Реже встречается в хвойных лесах, а широких откры-
тых пространств избегает.
34. РОД MOGERA
279
Ходы располагаются на незначительной глубине от 3—5 до 7—10 см
-от поверхности земли и тянутся на многие сотни метров. Рыхлая почва
в местах обитания уссурийского крота позволяет ему не только рыть,
но и сминать почву, не выбрасывая ее на поверхность. Только в твердом
грунте крот вынужден рыться глубоко и выбрасывать почву на поверх-
ность земли. Основную пищу этого вида, как и других кротов, составляют
.дождевые черви. В отдельные сезоны к этому корму в значительном ко-
личестве примешиваются другие беспозвоночные животные, особенно
насекомые. Видовой состав дополнительного корма и его количество
зависят от видового состава беспозвоночных того или иного биотопа
и сезона года. Общее количеств.© корма, которое может съесть крот за
-одни сутки, весьма значительно. По весу оно равно весу крота. При
интенсивной работе и понижении температуры этот крот, как и другие
виды, несомненно съедает больше, чем то количество, которое регистри-
руется при содержании кротов в неволе, при пониженной их актив-
ности.
К размножению этот вид приступает в марте—апреле, а при небла-
гоприятных условиях значительно позднее, в мае. В конце мая появля-
ются новорожденные. Выкармливание молодых занимает 28—32 дня.
Незадолго до окончания выкармливания детенышей, самка спаривается
второй раз и в июле рождает детенышей второго помета. В отличие от
других видов кротов уссурийский крот может размножаться 1—2 раза
в году в зависимости от погодных условий каждого года. Число дете-
нышей 2—10. В августе размножение этого крота заканчивается.
Подвиды. Не изучены.
3. Mogera latouchei Thomas, 1907 — Крот китайский.
Thomas, 1907, Proc. Zool. Soc., London, p. 467; — mogera Shin, 1930, Bull.
Dept. Biol. Sun Yatsen Univ. Canton, v. 4, p. 3.
T и п. Шкура и череп, N 98.8.17.1, Kuatun, северо-западная часть пров. Фуц-
зянь (Китай), кол. Брит, муз., Лондон.
Диагноз. Длина: тела — 93—115, ступни — 13.5—14.0 мм.
Р1 с одним корнем, ниже Р2.
Описание. Окраска верхней стороны тела аспидно-серая, с рас-
плывчатым темно-коричневым налетом. Бока тела, кисти, ступни и хвост
покрыты редкой грязно-белой шерстью, более длинной на хвосте.
Измерения. Длина: хвоста — 14—20, наибольшая черепа —
27.7—29.5 мм.
Распространение и возраст. Встречается в восточной
части Сычуани, в Гуанчжоу, Гуаней, Хунани, Изанси, Фуцзяни, Гуан-
дуне (Китай), современный.
t Gen. et sp.?
Фрагмент правой ветви нижней челюсти с зубами Мх и М2, № 2976 (1), правый бе-
рег р. Аксыира, Зайсанская Котловина (Казахстан), олигоцен, кол. ПИН АН СССР,
Москва.
Сравнение зубов упомянутого фрагмента нижней челюсти с таковыми
представителей различных групп кротов позволило установить их наи-
большее сходство только с зубами видов рода Parascaptor Gill.
При этом зубы олигоценового крота отличаются более высоким греб-
нем между передним и задним сегментами зуба, на его задней стороне
отсутствием ушковидного выступа на передне-внутренней и низко рас-
положенным ушковидным выступом на задне-внутренней сторонах ко-
ронки зуба.
280
ПОДСЕМ. TALPINAE
35. Род PARASCAPTOR Gill. 1875
Gill, 1875, p. 110.
Тип. Talpa leucura В 1 у t h, 1850, Cherra Punji, Ассам (Индия), современный.
Диагноз. Зубная формула; 1-|-СуР-|-Му = 42.
Рис. 161. Parascaptor leucura (по: Строганов, 1948).
Р2 выше или крупнее Р3. Р3 стоит под углом к оси челюсти. Р4 очень
крупный и высокий. Pj крупный, клыкообразный. Р2 значительно меньше
Р3. Р2 и Р3 стоят по оси ветви нижней челюсти.
Описание (по: G. Allen, 1938). Внешним обликом напоминает
обыкновенного крота Talpa europaea L., отличается очень коротким
хвостом булавовидной формы, покрытым белыми волосами и скрытым
35. РОД PARASCAPTOR. 36. РОД SCAPTOCHIRUS
281
в волосяном покрове задней части тела. Окраска мягкого волосяного
покрова от коричневой до почти черной.
Череп с относительно коротким носовым отделом. Межчелюстные
кости сужены в области носового отверстия.
Распространение и возраст. Юго-восточная часть
Азии от восточных районов Бенгалии и Северной Бирмы на северо-вос-
ток до южной части Хингана, современный.
1. Parascaptor leucura Blyth, 1850 (рис. 161).
В 1 у t h, 1850, J. Asiat. Soc. Bengal, v. 19, p. 215.
Тип. He установлен, вид описан из Cherra Punji, Ассам (Индия).
Описание (по: G. Allen, 1932). Кроме признаков, перечисленных
в описании рода, характеризуется и следующими: череп тонкого сложе-
ния, с суживающимся к концу носовым отделом. Зубы варьируют в раз-
мерах. В верхней челюсти только один мелкий одновершинный премоляр
(Р2), расположенный между крупным клыком и крупным Р3. Р1 утрачен
Измерения (по: G. Allen, 1938). Длина: тела —• 105—НО
хвоста — 10.0—15.5. ступни — 15, копдило-базальная черепа — 27.8—
28.8, верхнего зубного ряда — 10.5—11.6 мм.
Распространение и возраст. Ассам (Индия), Юньнань
(Китай), Бирма.
Примечание. Томас (Thomas, 1881) описал по 1 экз., добытому
в окрестностях Пекина, особый вид — Talpa leptura (Ann. Mag. Nat.
Hist., ser. 5, v. 7, p. 470), который Добсон (Dobson, 1883) отнес к роду
Parascaptor, а Томас (Ann. Mag. Nat. Hist., 1910, ser. 8, v. 5, p. 350)
перевел в род Scaptochirus M.-Edw. и он стал называться Scaptochirus
leptura. Аллен (G. Allen, The Mammals of China and Mongolia, 1938,
pt. 1, p. 73—75) свел его в синонимы Scaptochirus moschatus M.-Edw.
С. У. Строганов (1941, c. 270—272) по 1 экз. из Гунцзя-Инцзя (южный
Хинган) из кол. ЗИН АН СССР описал Parascaptor grandidens. Из опыта
известна значительная вариабельность структурных особенностей черепа
и зубов кротов. Признаки, положенные в основу разграничения Р. lep-
tura и Р. grandidens друг от друга и от Р. leucura, недостаточно основа-
тельны: длина зубного ряда — 14.0 (по 1 экз. Р. leptura), 12.5 (по 1 экз.
Р. grandidens) и 11.5—11.6 мм (по 2 экз. Р. leucura).Разный угол между
продольной осью отдельных зубов и ветви нижней челюсти варьирует
в широких пределах, также ширина носового отдела, длина хвоста, его
форма и другие.
До получения достаточного материала, на котором можно будет ра-
зобраться в видоспецифичных особенностях Р. leptura и Р. grandidens,
вряд ли нужно считать их самостоятельными видами.
36. Род SCAPTOCHIRUS Milne-Edwards, 1867
Milne-Edwards, 1967, Ann. Sci. Nat. Zool., ser. 5, t. 7, p. 375. Chiroscap-
tor H eude, 1898, Mem. concern. 1'Hist. Nat. Emp. Chin., v. 4, pt 1, p. 36.
Тип. Scaptochirus moschatus Milne-Edwards, 1867, Внутренняя Монголия (Ки-
тай), современный.
Диагноз. Зубная формула: 1С ^РМ = 40. Р2 крупнее Р3. Р3
О 1 О О
стоит под углом к продольной оси челюсти. Р4 очень крупный. Р2 очень
крупный, клыкообразный. Р3 очень мелкий, с одним корнем, а не двумя,
как у представителей других родов трибы Talpini.
Описание. Размеры тела, сходные с таковыми Talpa еигораеа L.
Морда относительно короткая. Хвост мало выступает из волосяного по-
282
ПОДСЕМ. TALPINAE
крова тела. Кисти с сильными когтями. Щетинки по краям кисти более
тонкие, чем у видов рода Talpa L. Пальцы ступни с длинными и тонкими
когтями, слабо изогнутыми.
Окраска верхней стороны тела буровато-серая с золотистым оттенком,
нижней — серая с серебристым налетом.
Распространение и возраст. Северо-восточная часть
Индии, в частности Ассам, восточные провинции Китая (Шанси, Шантунг,
Шенси и Хубей). Считается также, что этот род представлен 5. davidia-
nus M.-Edw. в Сирии (район Акба), современный.
f 1. Scaptochirus primaevus Schlosser, 1924 (рис. 162).
Schlosser, 1924, Palaeontol. Sinica, ser. C, v. 1, fasc. 1, p. 3—5.
T и п. He установлен, вид описан по многим остаткам из Эртемте и Олан Хореа,
Монголия, плиоцен.
Диагноз. Плечевая кость с удлиненным телом и слабо развитым
в ширину дистальным отделом.
Описание. Плечевая кость этого вида (судя по рисунку) более
длинная, чем у современного вида. Она, кроме того, с отогнутым вниз
Рис. 162. Scaptochirus primae-
vus (по: Schlosser, 1924).
и широким tuberculum majus lateralis. Опи-
сывая зубы, Шлоссер (Schlosser, 1924) отме-
тил следующее: Р2 и Р3 имеют один корень,
значительно меньше Р4. Коренные зубы
с пятью бугорками сходной высоты.
Измерения (по: Schlosser, 1924).
Длина: всего нижнего зубного ряда — 9,
Мх—М2 — 5, плечевой кости — 11.5; ширина
плечевой кости в верхней части — 8, а в
нижней — 6; длина бедра — 13.5 мм.
Распространение и воз-
раст. Монголия, плиоцен.
2. Scaptochirus moschatus Milne-Edwards, 1867 (рис. 163).
Milne-Edwards, 1867, Ann. Sci. Nat. Zool., ser. 5, t. 7, p. 375. Shizoscaptor
sinensis H e u d e, 1898, Mem. concern. I'Hist. Nat. Emp. Chin., v. 4, pt. 1, p. 36. Scap-
tochirus moschiferus Hen d e, 1898, там же, с. 40.
T и п. Не отмечен. Материал, послуживший для описания, хранится в Муз. естеств.
истор., Париж. Типичный экземпляр добыт (G. Allen, 1938) в «Монголии», а Стро-
ганов указывает в качестве «terra typica» Внутреннюю Монголию (Китай), современный.
Диагноз. Плечевая кость с коротким телом и широким дисталь-
ным отделом.
Описание. Кроме признаков, отмеченных в описании рода, этот
вид характеризуется и следующими особенностями. Череп с широкой
мозговой капсулой и коротким и широким носовым отделом. Межтемен-
ная кость очень широкая и более длинная, чем у многих видов других
групп кротов. Соответственно теменные и лобные кости относительно
более короткие. Верхнезатылочная кость сравнительно длинная. Задний
край костного нёба с треугольной выемкой. Крыловидные кости с длин-
ными отростками, направленными вверх и назад. Слуховые барабаны
замкнутые, в виде пузырей. Основная затылочная кость с длинным от-
ростком, направленным вперед и срастающимся со срединным гребнем
основной клиновидной кости в общий гребень.
Нижняя челюсть с относительно короткой зубной частью. Ее высота
менее высоты коронки Мг и М2 в 1.5 раза. Венечный отросток невысокий,
с нешироким основанием и узкой вершиной. Сочленовный отросток не-
сколько уплощен дорсовентрально, с мыщелком, значительно развитым
36. РОД SCAPTOCHIRUS
283
в поперечном направлении. Основание углового отростка почти такой же
ширины, как основание венечного отростка, он с широкой вогнутой по-
верхностью, отогнут внутрь.
Верхние зубы. Верхние резцы расположены на переднем крае
межчелюстных костей. Первый резец лишь немного крупнее второго и
третьего. Клык с двумя корнями, сильно развит. Р2, Р3 с двумя кор-
Рис. 163. Scaptochirus moschatus.
нями. М1—М3 сходны по общим структурным особенностям. Гребни,
образованные складками, входящими с внешней стороны, имеют разную
высоту, М3 самый мелкий, повернут в косом направлении по отношению
к продольной оси зубного ряда.
Нижние зубы. Нижние резцы сходной формы и размеров.
Нижний клык располагается в одном ряду с резцами, он более длинный
и менее плоский, чем резцы, имеет небольшой выступ при основании
коронки. Его корень немного смещен к средней линии челюсти, а вершина
коронки отклонена кнаружи. Р2 сильно развит, клыкообразный, выпол-
няет функцию клыка. Мх—М3 сходного строения, убывают в размерах
к М3.
284
ПОДСЕМ. TALPINAE
Измерения. По 25 из кол. Зоологического музея Берлинского
университета (ГДР) — длина: тела — 115 и 125, ступни — 18 и 18,
хвоста — 24 и 25, кондило-базальная черепа (1 экз.) — 31.5; ширина:
наибольшая мозговой капсулы — 17.2 и 18.3, когтя второго пальца —
2.4 и 2.5, третьего — 2.4 и 2.8, четвертого — 2.4 и 2.4 мм. По: Строганов,
1948, — длина: ступни — 19.5—20.0, хвоста — 15.0—16.0, общая че-
репа — 30.2—34.3; его наибольшая ширина — 16.9—18.1; длина верх-
него зубного ряда — 12.5—14.6 мм.
Распространение и возраст. Внутренняя Монголия,
Шэньси, Шанси, Шаньдунь, Цзяньсу, Аньхой, Heisui (Китай), Ассам
(Индия).
Примечание. Р. У. Строганов (1948) привел в своей монографии
о кротах в качестве отдельного вида S. davidianus Milne-Edwards, 1884
(Compt. Rend. Acad. Sci. Paris, t. 99, p. 1141), описанного по 1 экз. из
Сирии (Акба), хранящемуся в Парижском музее. Строганов отметил, что
S. davidianus отличается от S. moschatus M.-Edw. более удлиненной мор-
дой и сравнительно мелкими зубами, а также мелким Р3. Строганов,,
по-видимому, имел в виду Р4, который он в упомянутой работе часто на-
зывает Р3, если в зубном ряду всего три премоляра, третий по счету.
Это замечание справедливо и потому, что Строганов знал зубы S. mos-
chatus, у которого Р3 мелкий, а Р4 очень крупный. Следовательно, отли-
чать названные виды можно только по разным размерам Р4.
Лей (Lay, 1965) описал из Восточного Ирана (Курдистанская пров.)
Talpa streeti, у которого морда длиннее, чем у Scaptochirus moschatus, и
короче, чем у Talpa еигораеа L., а зубы мелкие, меньше, чем у этих двух
видов. Возможно, что S. davidianus M.-Edw. и Talpa streeti Lay — одно
и то же. К этому нужно добавить, что Эллерман и Моррисон-Скотт (Е1-
lerman, Morrison-Scott, 1951) отнесли S. davidianus в синонимы Talpa
соеса Savi. Разобраться в этом вопросе можно только путем сравнения
материала по соответствующим видам.
Ко всему сказанному здесь следует добавить, что Суинго (Swinhoe,
Proc. Zool. Soc. London, 1870, p. 620) описал по 1 экз. из окрестностей
Пекина (Китай) Scaptochirus davidianus, а Томас (Thomas, 1881) назвал
его Talpa leptura. Через два года Добсон (Dobson, 1883) отнес Talpa
leptura к роду Parascaptor и он стал называться Parascaptor leptura. Томас
(Thomas, 1910) отнес последний к роду Scaptochirus и он стал называться
S. leptura, но Аллен (G. Allen, 1938) свел этот вид в синонимы S. mos-
chatus M.-Edw., а через 10 лет Строганов (1948) вернул его в лоно рода
Parascaptor. Единственный экземпляр крота, доставленный консулом
Суинго в 1860 г. в Британский музей, служил предметом систематических
упражнений пяти зоологов из трех стран в течение 88 лет. А он, между
прочим, относится к Parascaptor leucura Blyth, 1850. Добсон был прав,
отнеся Talpa leptura Thomas к роду Parascaptor, но нужно было отнести
его в синонимы Р. leucura.
О вымерших кротах с территории СССР
Вымершие кроты известны из ряда районов европейской и азиатской
территории СССР. Они представлены преимущественно остатками пред-
ставителей трибы Talpini (подсем. Talpinae), близких к роду Talpa L,
Наиболее ранний представитель сем. Talpidae найден в слоях олигоцена
Казахстана (окрестности г. Зайсана). М2 правой стороны верхней челюсти,
близкий по строению к таковому представителей подсем. Uropsilinae.
Из этого же местонахождения, из олигоценовых слоев, по зубам известны
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
285
два крота, из которых один близок к видам трибы Urotrichini, а другой
к видам азиатских кротов трибы Talpini.
Миоценовые остатки кротов (плечевые кости) найдены на о. Ольхон
(оз. Байкал). По строению они близки к этим костям представителей
рода Parascaptor Gill. Плиоценовые кроты известны с Алтая, из Казах-
стана и ряда местонахождений на территории европейской части СССР.
От них сохранились только плечевые кости и единичные зубы. Плечевые
кости разных размеров, но не крупнее таковых обыкновенного крота.
По строению они близки к этим костям представителей рода Talpa. То же
можно сказать и о плечевых костях плейстоценовых кротов.
В заключение следует отметить, что пока еще нет такого материала
по вымершим кротам, который позволил бы разобраться в составе и
истории развития подсемейства.
Места нахождения кротов на территории СССР по сборам А. К. Агад-
жаняна, кол. МГУ; Н. К. Верещагина и И. М. Громова, кол. ЗИН АН
СССР; В. С. Зажигина, кол. ГИН АН СССР; В. А. Топачевского, кол.
Инет. зоол. АН УССР; Н. С. Шевыревой и М. В. Чхиквадзе кол. ПИН
АН СССР. Казахстан, эоцен?—олигоцен; о. Ольхон (оз. Байкал), миоцен—
нижний плиоцен; Тамань, плиоцен—нижний плейстоцен; Урыв (Воро-
нежская обл.), плиоцен; Котловина (оз. Ялпух, юго-зап. Украина),
верхний плиоцен; Этулия (западная часть Одесской обл.), средне-верх-
ний плиоцен; Новопетровское (Украина), средний плиоцен; Алтай (Бар-
наул, р. Чумыш), нижний плейстоцен.
Сем. SORICIDAE Fischer von Waldheim, 1817
Soricini Fischer von Waldheim, 1817, p. 368—428.
Диагноз. Зубная формула: 13^2 СP M |—| = 44—24. Длина
тела 34—180 мм. Глаза маленькие, малозаметные. Ушные раковины круп-
ные или небольшие, скрытые в волосяном покрове. Тело покрыто густым
бархатистым волосяным покровом. Череп с удлиненным лицевым отделом.
Скуловые дуги с миоцена отсутствуют. Первые верхние резцы серповид-
ные, с зубцом на задней стороне. Верхние коренные зубы (Мх и М2) че-
тырехугольные, с W-образными гребнями на жевательной поверхности
и четырьмя корнями.
Описание. Преимущественно мелкие животные (длина тела
34—180 мм).
Тело удлиненное, вальковатой формы, на коротких ногах, редко
задние ноги сравнительно длинные. Голова относительно крупная, с вы-
тянутым в хоботок носовым отделом. Глаза очень маленькие, малозамет-
ные в волосяном покрове. Изредка они закрыты сросшимися веками.
Ушная раковина крупная, выступающая из волосяного покрова или
скрытая в нем, небольшая, очень редко она не развита. На ее внутренней
поверхности кожные выросты образуют своего рода карманы. Волосы,
покрывающие тело, короткие, мягкие, бархатистые, реже длинные, же-
сткие, щетинистые. Конец морды густо покрывают в разной степени тол-
стые и жесткие щетинки. Вибриссы относительно длинные, нередко до-
стигают длины головы. Хвост разной длины, покрыт только короткими
волосами или короткими волосами и редкими длинными щетинками. У ряда
видов на нижней стороне хвоста имеется киль из длинных щетинистых
волос. Кисти и ступни сверху покрыты мелкими чешуйками и корот-
кими жесткими волосами. Пальцы нередко покрыты крупными чешуй-
ками, похожими на роговые щитки птиц. Нижние стороны кистей и
286
СЕМ. SORICIDAE
ступней покрыты мелкими щитками и разной формы и величины подо-
швенными бугорками — «мозолями». У видов, приспособленных к пла-
ванию, боковые стороны кистей и ступней, а также и пальцев, покрыты
плоскими щетинками в виде бахромы. Редко между пальцами имеются
кожные плавательные перепонки. На боках тела у самцов расположены
мускусные железы, функционирующие в период размножения. У самок
они слабо развиты или отсутствуют. Окраска тела различная: коричне-
вая, серая, черно-бурая и бурая разных оттенков, местами белая. Раз-
ные сочетания этих цветов обусловливают все разнообразие расцветок
волосяного покрова землероек.
Череп в разной степени удлиненной формы. Швы между от-
дельными костями его лицевой части зарастают очень рано, между ко-
стями же мозгового отдела долго не зарастают, особенно венечный
шов. Мозговая капсула выпуклая, округлая или плоская, овальная.
Лицевая часть черепа суживается к передней части. Нижние части бо-
ковых сторон верхнечелюстных костей расширены в стороны и упло-
щены сверху вниз. Лобные кости в межглазничной области с двумя от-
верстиями или без них. Скуловые дуги (с конца миоцена) отсутствуют.
Крыловидные отростки не развиты. Барабанная кость имеет форму кольца,
не прирастает к черепу. Нижняя челюсть с относительно низкой зубной
частью и удлиненным палочковидным угловым отростком. Венечный от-
росток сильно развит и отогнут на внешнюю сторону. Сочленение с че-
репом осуществляется с помощью двух суставов. Сочленовный отросток
нижней челюсти имеет две сочленовные фасетки (рис. 39).
Зубная формула: I С Р М ^-5 — 44—24.
о—/j 1—U 4—1 о—Z
Зубы окрашены в красный, коричневый цвет или белые, а также и
смешанной окраски: резцы, клыки и премоляры с желто-коричневыми
вершинами, моляры белые. Уменьшение количества зубов происходит за
счет редукции верхних и нижних промежуточных зубов, реже и за счет
последних моляров.1 У современных землероек верхних промежуточных
зубов 5—2, а нижних 3—2. Верхние внутренние резцы имеют два зубца.
Верхние промежуточные зубы одновершинные, варьируют в числе, разме-
рах и форме. Последний премоляр в большей или меньшей степени моля-
ризован. Его пара-и метакон гребнистые. Бугорок протокона разной формы,
часто уплощен в передне-заднем направлении, островершинный. Талон ши-
рокий, плоский. Его корень нередко слит с корнем протокона. М1 и М2 округ-
лочетырехугольной формы. Пара- и метакон этих зубов имеют обособленные
корни. Их бугорки с V-образными гребнями в совокупности образуют
фигуру, как буква W. Протокон и талон Р4, М1 и М2 расположены ниже
пара- и метакона. Они с вогнутой жевательной поверхностью, а на внут-
реннем крае имеют гребень. М3 трехвершинный, с тремя или двумя кор-
нями, вытянут в поперечном направлении. Ij длинный, прямой или слабо
изогнутый вверх, с гладкой верхней поверхностью или с 1—3 зубцами.
Первый нижний промежуточный зуб удлиненный, одновершинный, реже
двухвершинный. Второй промежуточный зуб с одной или двумя верши-
нами, относительно короткий и высокий. Мь М2 и М3 с W-образной же-
вательной поверхностью и с двумя корнями. Бугорок протокона этих
зубов вошел в состав эмалевого образования на их жевательной поверх-
ности, а его корень подвергся редукции.
1 Промежуточными зубами у землероек принято называть специфичные для нихв
сходной формы — резцы (I2 и I3), клык и премоляры, кроме Р4.
Характеристика семейства
287
Расстояние между зубными рядами верхней челюсти больше, чем
между нижними примерно на ширину зубного ряда. Вследствие этого
при сомкнутых челюстях коронки нижних коренных зубов располага-
ются ближе к внутренним краям коронок верхних.
Скелет. Шейные позвонки с узкими дугами, без остистых отростков.
Атлант с широкими боковыми крыльями, охватывающими затылочную
кость, с отростком на нижней стороне, направленным вперед и опираю-
щимся на основание затылочной кости. Пятый шейный позвонок имеет
с каждой стороны по одному отростку, направленному назад. Грудные
позвонки с короткими остистыми отростками, сильно наклоненными назад,
несут 14 пар ребер. Крестец образован 4 сросшимися позвонками. Ко-
личество хвостовых позвонков варьирует. (У представителен рода Scu-
tisorex Thos. тела позвонков, их дуги и отростки разрастаются в виде
губчатых образований, и только хвостовые позвонки более или менее
обычной формы). Ключица длинная, причленяется к внутреннему от-
ростку вилочки акромиального отростка лопатки и к крупной рукоятке
грудины. Лопатка удлиненная, ее ширина менее длины в 2—3 раза.
Ость лопатки сильно развита, на переднем крае несет длинный акро-
миальный отросток, раздвоенный на конце.
Плечевая кость короткая (в 1.5—1.25 раза короче костей предплечья),
с относительно крупными буграми в проксимальной и дистальной ее
частях. Гребень большого бугра плечевой кости сильно развит. Ниже
проксимального эпифиза на наружной и внутренней сторонах плечевой
кости имеются гребни, из которых гребень на внутренней стороне больше
наружного. Локтевая кость часто толще лучевой, с крупным олекраноном.
Кисть тонкая и длинная или короткая и широкая.
Таз узкий, образован относительно тонкими костями. Запирательное
отверстие крупное. Подвздошная кость в передней части слабо расши-
рена. Симфиз лобковых костей отсутствует (исключение Anourosorex
squamipes M.-Edw.). Небольшая вертлужная впадина расположена по-
средине боковой стороны тазовой кости. Бедренная кость с хорошо
развитыми вертлугами. Наиболее развит большой вертел. Соотношение
длины бедра и голени варьирует. Длина этих отделов почти равна у рою-
щих форм, у прыгающих видов голень значительно длиннее бедра. Сте-
пень развития гребней и бугров для прикрепления мышц на большой
берцовой и бедренной костях зависит от особенностей локомоторных
функций конечностей отдельных видов. Малая берцовая прирастает
к большой берцовой кости на г/2—г/3 в ее нижнем отделе. Ступня узкая
и длинная или короткая и широкая. Пяточная кость относительно ко-
роткая.
Из внутренних органов особенно бросается в глаза крупная вели-
чина печени и почек. Последние достигают таких размеров, что передней
частью вдаются в задне-нижнюю часть печени. Значительное развитие
1 этих органов — печени как депо крови, «кладовой» питательных веществ
и пищеварительной железы, а почек как органа выделения, а возможно,
и резервуара для избыточной воды (как нагретого балласта для сохра-
нения тепла зимой) обусловлено высоким обменом у землероек.
Os penis отсутствует. Половое и анальное отверстия окрываются
в углублении, напоминающем клоаку, что особенно хорошо заметно у самок.
Малая величина тела землероек, как уже отмечалось, есть комплек-
сное приспособление: к передвижению в естественных укрытиях (лесная
подстилка, приземный слой растительности, норы и др.), к питанию
мелкими подвижными животными и к существованию в самых разно-
образных условиях в состоянии круглогодичной активности при потреб-
лении огромного количества корма по отношению к весу тела животного.
288
СЕМ. SORICIDAE
Распространение
Ареал семейства охватывает территории Европы, Азии, Африки, Се-
верной и Центральной Америки и крайний северо-запад Южной Аме-
рики. Их нет в Австралии, на о. Мадагаскар они, возможно, завезены.
Если семейство в целом заселяет на упомянутых континентах все
зоны от тундр до тропических лесов, то распространение отдельных
триб не столь широкое. Сопоставление времени появления и вероятной
обстановки, в которой складывались отдельные трибы, с одной стороны,
и их современных ареалов — с другой, позволяет отметить, что ареалы
охватывают либо территории с условиями среды, близкими к тем, в ко-
торых развивались предки той или иной трибы, или с условиями, к ко-
торым приспосабливались их более поздние роды и виды. При этом трибы,
объединяющие более специализированных землероек, распространены
в южной части ареала семейства, а менее специализированные в се-
верной.
Развитие одних землероек шло по линии приспособления к питанию
преимущественно мелкими животными, при сохранении примитивных
особенностей в зубной системе (Soricini), а в других к питанию относи-
тельно крупными животными и при специализации зубной системы
(Crocidurini и Blarinini).
По-видимому, в условиях северных широт питание мелкими живот-
ными, преимущественно насекомыми, представляет больше возможностей
для выживания, чем питание более крупными (мелкие позвоночные),
одни из которых зимой впадают в спячку, а другие бывают в малом ко-
личестве вследствие колебания численности. Вероятно, этим и объясня-
ется вымирание в умеренной зоне в послеплиоценовое время многих
родов землероек из трибы Blarinini. виды которых питались крупными
животными, и сокращение ареалов родов, сохранившихся до настоящего
времени. Только в условиях субтропиков и тропиков питание сравни-
тельно крупными животными привело к значительному разнообразию
частных адаптаций жевательного аппарата и широкому расселению
многих видов.
Распространение землероек шло сопряженно с формированием лес-
ных ландшафтов, как наиболее обеспечивающих постоянным кормом,
а на севере и в соответствии с приспособлениями к резким колебаниям
температуры. Последнее обусловливало особенности добывания корма,
а также и особенности годового цикла энергетического обмена. В связи
с этим со стороны отдельных групп землероек потребовалась высокая
активность и приспособление к добыванию мелких животных в самых
разнообразных условиях, особенно в зимнее время. По-видимому, пи-
танием мелкими животными обусловлено сохранение удлиненных челю-
стей с большим числом мелких промежуточных зубов в верхней челюсти,
сохранение зубцов на нижних внутренних резцах, а также значительной
подвижности нижней челюсти.
Расселение землероек к северу сопровождалось стабилизацией при-
знаков, характерных для примитивных групп раннего времени развития
семейства. Можно предполагать, что исходные группы землероек также
питались мелкими беспозвоночными животными, преимущественно на-
секомыми — наиболее многочисленными на поверхности земли.
Важное значение отдельных элементов рельефа, в частности горных
систем и водных бассейнов, в распространении землероек не вызывает
сомнения. Распространение мелких животных имеет своеобразные осо-
бенности и представляет сложный процесс, оно требует длительного вре-
мени и соответствующих условий. Возможно, что широкие ареалы мел-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
289
ких млекопитающих указывают не только на процветание вида, но и
на его эвритопность, а также на длительную историю развития.
Землеройки, по-видимому, мало связаны с определенными местами
обитания. Они, насколько известно, не пользуются длительное время
одними и теми же норами, довольствуясь чаще разнообразными есте-
ственными укрытиями. В поисках корма постоянно перемещаются с од-
ного места на другое, особенно вне периода размножения. Такие формы
несомненно скорее и шире распространяются, чем типичные норники
или колониальные виды.
Следует отметить, что распространение одних видов семейства идет
сопряженно с распространением других как того же, так и других се-
мейств и отрядов. В результате распространяющийся вид не оказывается
на новом месте в совершенно иных по составу биоценозах.
Изучение распространения землероек в отдельные периоды их исто-
рии и в настоящее время позволяет предположить, что расселение их
в местах с подходящими условиями происходило диффузно, а в аридных
зонах — по относительно узким пространствам вблизи побережий рек
или по горным хребтам. Совокупность рек, речек и других мелких водое-
мов, густой сетью покрывающих огромные пространства, благоприят-
ствует расселению. Два основных стока вод на север и на юг всегда слу-
жили средством как активного, так и пассивного расселения землероек
в двух противоположных направлениях, равно как и сохранения многих
видов в прибрежных зонах.
Речные долины, как правило, населяет более разнообразная расти-
тельность, чем водораздельные территории, что объясняется многими
благоприятными для нее условиями: достатком влаги и света (последнее
очень важно в ландшафтах), а также менее низкими температурами
почвы и воздуха вблизи воды, оказывающей на них смягчающее влияние.
Не случайно по долинам рек леса уходят далеко к северу — в тундру,
или к югу — в степь. Водные системы рассекают леса на отдельные уча-
стки, по краям которых (на берегах водоемов) развивается богатая и
относительно разнообразная растительность, содержащая нередко виды,
чуждые флористическим комплексам, сложившимся на той или иной
территории. Естественно, что такие места очень богаты различными
мелкими животными, которые служат кормом многим мелким хищникам.
Реки, кроме того, регулируют режим воды в почвах окружающих пло-
щадей, покрытые льдом, служат мостами в зимнее время. Все это поз-
воляет считать, что речные системы имеют важное значение в жизни
и распространении животных, в частности землероек.
Горные системы служат местом обитания разнообразных групп земле-
роек, а также главнейшими путями их расселения. Наиболее важными
в этом отношении следует считать горные системы средиземноморской
геосинклинали и тихоокеанское горное кольцо. Пульсация горообразо-
вательных процессов во времени, сопровождавшаяся многократными из-
менениями обстановки, служила также причиной вымирания не только
отдельных видов землероек, но и их групп, а также и целых фаунисти-
ческих комплексов, характерных для того или иного геологического
периода.
Предками землероек считаются виды сем. Nyctitheriidae (Simpson,
1945), еще сохраняющие особенности строения, близкие тенрекообразным.
Пока остатки его представителей найдены лишь в палеоцене—эоцене
Северной Америки. Можно, однако, не сомневаться в том, что они были
в это время распространены и в Азии, еще слабо изученной в этом отно-
шении.
19 А. А. Гуреев
290
СЕМ. SORICIDAE
Областью формирования сем. Soricidae следует считать Азию, в ко-
торой в настоящее время обитает наибольшее количество разнообразных
групп землероек, в том числе с особенностями переходного характера,
например роды Soriculus Blyth, Podihik Deraniyag. и др. Кроме того,
близкое по происхождению к землеройкам семейство кротов представ-
лено в Азии большим числом примитивных групп. В Северной Америке
настоящие землеройки известны только с олигоцена, а в Африке и Азии —
с миоцена. В Европе они найдены в верхнем эоцене. Можно предполагать,
что роды с примитивными признаками (Myosorex Gray и Surdisorex Thos.),
обитающие в Южной Африке, также азиатского происхождения.
Наиболее древняя группа землероек — подсем. Saturniniinae — изве-
стна из верхнего эоцена Европы. Позднее этого времени и за пределами
Европы остатки их пока не обнаружены. Возможно, что ее распростра-
нение было ограничено Евразией.
Верхнеэоценовые представители подсем. Soricinae (род Amphisorex
Filh.) также известны только из Европы. Можно думать, что это были
типичные тропические группы, существование которых ограничено эоце-
ном. Олигоценовые группы из того же подсемейства известны по наход-
кам из Европы, Азии и Северной Америки, это роды Oligosorex Kretz.,
Crocidnsorex Lavoc. (Европа), Gobiosorex Sulims. (Азия), Domnina Cope
(Северная Америка), просуществовавшие, видимо, до среднего олигоцена.
Есть основание предполагать, что более примитивные землеройки трибы
Oligosoricini были широко распространены в Европе, Азии и в Северной
Америке, послужив основой для развития миоценовых групп землероек.
Их потомки получили широкое распространение в миоцене Европы, Азии,
Африки и Северной Америки. К тому времени уже сформировались со-
временные трибы: Soricini — Европа, Азия, Северная Америка; Blari-
nini — Европа, Азия и Северная Америка; Anourosoricini — Европа и
Азия; Crocidurini — Европа, Азия, Африка и Северная Америка, а также
вымершая в конце этого периода триба Heterosoricini, известная по остат-
кам из Европы и Северной Америки.
Таким образом, миоцен был периодом широкого распространения
одной из специализированных групп землероек трибы Crocidurini. Резкое
сокращение ее ареала за счет Северной Америки и большей части Европы
в конце миоцена—плиоцена позволяет считать, что она в то время, как
и теперь, представляла собой типичную тропическую группу. Только ее
отдельные представители в послемиоценовое время проникли в умерен-
ную зону Евразии.
Широко распространенные роды трибы Blarinini в конце плиоцена
в Северной Америке стали отступать к югу, в субтропическую и тропи-
ческую зоны. Для многих групп этой трибы подобная картина наблюдалась
в Европе и Азии. Виды трибы Crocidurini отступили к югу широким
фронтом. Напротив, представители трибы Soricini начали распростране-
ние преимущественно к северу, приспосабливаясь к резким колебаниям
климата. Давнее известное однообразие условий жизни в северных ра-
стительных формациях не способствовало образованию многих родов
в пределах этой трибы. В ее составе сформировавшийся, видимо, в ран-
нем миоцене циркумполярный род Sorex L., Blarinella Thos. (Азия);
Tregosorex Hibb. et Jamm., Anchiblarinella Hibb. et Jamm., Microsorex
Coues (Северная Америка). Распространение отдельных представителей
Soricini к югу происходило по горным системам Европы, Средней и Цент-
ральной Азии, где впоследствии сформировалось несколько своеобразных
видов.
Общие данные о распространении землероек в третичную эпоху сле-
дующие. В эоцене в Европе были распространены подсем. Saturniniinae
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
291
(род Saturninia Stelil.) п Soricinae (род Amphisorex Filh.). Об эоценовых
землеройках из других частей света ничего не известно. Б олигоцене
в Европе существовали и представители трибы Oligosoricini (роды OU-
gosorex Kretz. и Crocidosorex Lavoc.), в Азии Gobiosorex Sulimski, в Север-
ной Америке виды рода Domnina Соре. Об олигоценовой фауне Африки
ничего не известно.
В миоцене в Европе были распространены три трибы: Oligosoricini
(роды Soricella Dob.-Flor. и Miosorex Kretz.), Heterosoricini (род Hetero-
sorex Gaill.) и Soricini (род Sorex L.). В Азии в это время существовали
представители триб Crocidurini (роды Crocidura Wagl. и Similisorex Stog.
et Sav.) и Anourosoricini (род Anourosorex M.-Edw.). Можно не сомневаться,
что в это время в Азии существовали представители трибы Oligosoricini
и др. В нижнем миоцене в Африке уже существовали представители трибы
Crocidurini (роды Crocidura Wagl. и Myosorex Gray). Из миоцена Северной
Америки известны представители четырех триб: Oligosoricini (род Angu-
stidens Repenn.), Heterosoricini (род Pseudotrimylus Gur.), Crocidurini
(роды Limnoecus Stirt. и Stirtonia gen. nov.) и Soricini (род Sorex L.).
В конце этого периода окончательно сформировались представители
трибы Blarinini (род Adeloblarina Repenn.). В конце миоцена вымерли
виды триб Oligosoricini и Heterosoricini.
В плиоцене в Европе были распространены четыре трибы: Soricini
.(род Sorex L.), Blarinini (роды Blarinoides Sulims., Beremendia Korm.,
Petenyia Korm., Petenyiella Kretz., Soriculus Blyth, Zelceina Sulims.,
Allosorex Feif.), Anourosoricini (роды Anourosoricodon Topatsli., Amblycop-
tus Korm., Paranourosorex Rzeb.-Kow.), Crocidurini (роды Crocidura
Wagl., Suncus Ehrenb.). В Азии в это время были распространены те же
трибы: Soricini (род Sorex L.), Blarinini (роды Soriculus Blyth, Shikamai-
nosorex Haseg., Peisorex Kow. et Li-Chuan-Kuei), подтриба Nectogalina
(род Nectogalinia Gur.), Anourosoricini (Anourosorex M.-Edw., более ши-
роко распространенный, чем в современный период) и триба Crocidurini
(роды Crocidura и Suncus). ''
В Африке в плиоцене существовали представители разнообразных
групп трибы Crocidurini. Вероятно, все африканские роды этой трибы
сформировались окончательно в плиоцене. Можно думать, что предста-
вители других триб в Африку не проникали.
В плиоцене в Северной Америке широко распространились виды
трибы Blarinini (роды Blarina Gray, Hesperosorex Hibb., Paracryptotis
Hibb., Planisorex Hibb., Notiosorex Baird, а также Cryptotis Pom.). Про-
должали существовать виды трибы Crocidurini (род Limnoecus Stirt.),
вымершие в конце этого периода.
В конце плиоцена сформировался эндемичный род Microsorex Coues.
В это время формировались многочисленные американские виды рода
Sorex L. Представители родов Blarina Gray и Cryptotis Pom. отступили
к югу. При этом виды последнего интенсивно распространялись в суб-
тропической и тропической зонах американских континентов. По-види-
мому, в это время уже широко распространились в Северной Америке и
представители рода Microsorex Coues.
Таким образом, плиоценовая фауна землероек была весьма богата
видами и родами, о чем можно судить по остаткам этих млекопитающих
из Европы, Азии и Северной Америки. Можно считать, что плиоцен
был временем расцвета представителей трибы Blarinini, широко распро-
страненной в это время и разнообразной по видовому составу.
В плейстоцене на всех континентах были распространены трибы и роды,
характерные для них и в современный период. В это время вымерли пред-
ставители ряда древних родов в Европе, Азии и Северной Америке,
19*
292
СЕМ. SORICIDAE
а также ряд видов современных родов. Значительно сократился видовой
состав многих триб, особенно трибы Blarinini.
В современный период землеройки распространены значительно шире,
чем представители других семейств насекомоядных. Они представлены
большим числом различно адаптированных групп в тропической и субтро-
пической зонах. В умеренной зоне обосновались немногие роды с большим
числом видов.
В Европе распространены четыре рода землероек, но ни один из них
не эндемичен для нее. В самой северной ее части обитают виды двух родов:
Sorex L. и Neomys Каир, в средней части Европы встречаются, кроме ука-
занных, также и виды рода Crocidura Wagl., и только на крайнем юге этого
континента — один широко распространенный в Южной Азии и Африке
вид рода Suncus Ehrenb. (Suncus etruscus Savi). Из 16 видов перечисленных
родов бурозубых землероек к эндемичным для Европы нужно отнести
Sorex alpinus Schinz, S. raddei, Neomys anomalus Cabr., N. schelkovnikovi
Sat., из белозубых землероек — Crocidura russula Herm., C. giildenstaedti
Pall., C. dinniki Ogn., C. armenica Gur.
Азиатский материк и в настоящее время богат родами. На его террито-
рии обитают представители 13 родов — это более чем в 2.5 раза превышает
количество родов на каждом из остальных материков, 8 из них эндемичны
для Азии. Без ревизии на достаточно обильном материале нельзя уста-
новить истинное количество видов, обитающих в Азии, но их несомненно
более 60.
Sorex L.
Blarinella Thos
Suncus Ehrenb.
Feroculus Kelaart
Solisorex Thos
Crocidura Wagl.
— 17 видов, из них 9 эндемичных, 5 общих с Европой и
3 общих с Северной Америкой.
— 1 эндемичный вид.
— 3 эндемичных вида и 1 общий с Европой и Африкой.
— 1 эндемичный вид.
— 1 эндемичный вид.
•— около 25 видов, 1 общий с Европой и Африкой и 1 общий
с Европой.
Diplomesodon Brandt — 1 эндемичный вид (плейстоцен, Африка).
Neomys Каир ' "
Soriculus Blyth
Notiosorex Baird
Podihik Deraiyag.
Chimarrogale Anders. — 8 эндемичных видов.
Nectogale M.-Edw. — 1 эндемичный вид.
Anourosorex M.-Edw. — 1 эндемичный вид.
— 1 вид, общий с Европой.
— 3 эндемичных вида.
— (подрод Chodsigoa Kastscht.) — 3 эндемичных вида.
— 1 эндемичный вид.
Большинство родов распространено в южной части Азии. Как и в Ев-
ропе, на севере Азии распространены виды двух родов: Sorex L. и Neomys
Каир. В средней части этого континента к названным двум присоединя-
ются роды Suncus Ehrenb., Crocidura Wagl. та Diplomesodon Brandt, а в гор-
ной части, к востоку, отдельные виды родов Notiosorex Baird, Soriculus
Blyth, а также рода Nectogale M.-Edw., Chimarrogale Anders, и Anouro-
sorex M.-Edw.
По данным Г. Аллена (G. Allen, 1939), в Африке обитает около 150
видов землероек. Только принадлежащих роду Crocidura Wagl. около 112,
а по другим данным (Dollman, 1915, 1916) — еще более.
Африканские землеройки относятся к родам Myosorex Gray, Surdi-
sorex Thos, Scutisorex Thos, Suncus Ehrenb., Sylvisorex Thos, Praesorex
Thos, Paracrocidura H. de Bals. Все роды, кроме Suncus Ehrenb. и Croci-
dura Wagl., эндемичны для Африки. Подавляющее большинство видов
родов Crocidura Wagl. и Suncus Ehrenb. эндемичны. Эндемичные роды рас-
пространены преимущественно в южной части Африки. Из них роды Myo-
sorex Gray и Surdisorex Thomas объединяют виды с древними особенно-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
293
стями, характерными для землероек олигоценового и нижнемиоценового
времени. Их распространение ограничено небольшой территорией. Виды
рода Suncus Ehrenb. и особенно рода Crocidura Wagl. широко распростра-
нены. Формирование фауны землероек Африки началось, по-видимому,
еще в олигоцене, так как в миоцене Кении уже обнаружены представители
трибы Crocidurini. Потомками ранних поселенцев Африки следует считать
виды родов Myosorex Gray и Surdisorex Thos, а несомненно более поздними
представителей рода Scutisorex Thos, сохранившихся к настоящему вре-
мени в популяциях двух видов. Видимо, уже в миоцене роды Suncus
Ehrenb и Crocidura Wagl. были общими для Азии и Африки. С этого же
времени африканские землеройки развивались независимо.
В Северной Америке еще с миоцена развиваются виды триб Blarinini
и Soricini. Триба Soricini представлена двумя родами: Sorex L. (29 видов)
и Microsorex Cone's (1 вид); триба Blarinini — тремя родами: Blarina Gray
(2 вида), Cryptotis Pom. (25 видов) и Notiosorex Baird (2 вида).
Виды трибы Soricini населяют северную часть Северной Америки,
а трибы Blarinini — ее южную часть, откуда они проникли в Центральную
Америку и на крайний северо-запад Южной Америки. Обе трибы в Аме-
рике представлены большим числом видов, что вряд ли объясняется
только видодробительством, свойственным многим териологам. Так, в Се-
верной Америке обитают 30 видов двух родов трибы Soricini, в Азии их
18 из двух родов, а в Европе — 6 одного рода. Триба Blarinini представ-
лена 29 видами трех родов, а в Азии — 16 из шести родов. В Европе —
3 вида одного рода.
Из пяти американских родов три эндемичных: Microsorex Coues, Blarina
Gray, Cryptotis Pom. Род Notiosorex Baird, общий с Азией. Отметим, что
из отряда зайцеобразных древний род Caprolagus Blyth имеет своих пред-
ставителей, как и род Notiosorex Baird, в Сонорской подобласти и также
может считаться общим для Азии и Северной Америки. Таким образом,
Сонорская подобласть имеет древние связи с Азией.
Изучение распространения землероек в прошлые геологические пе-
риоды и в настоящее время позволяет установить, что Европа во все этапы
развития этих млекопитающих, да и других, только заселялась пришель-
цами из Азии (частично из Африки). Она никогда, по-видимому, не слу-
жила местом формирования особых групп животных, а только отдельных,
слабодифференцированных видов, и ее роль в этом отношении ничтожна.
Ее ранние и временные связи с другими материками не имели особого зна-
чения в расселении землероек. Поскольку Азия была местом формирова-
ния млекопитающих, для их расселения особенно важное значение имело
тихоокеанское геосинклинальное кольцо и средиземноморская геосин-
клиналь.
Экологические особенности семейства землероек
и значение их для человека
Несмотря на кажущееся внешнее однообразие землероек, приспособ-
ления многочисленных видов, объединяемых семейством, весьма разнооб-
разны. Эти частные адаптации видов и их групп обусловлены неодинако-
выми условиями среды в пределах широкого ареала Soricidae, охватываю-
щего зоны жизни от тундр до саванн и тропических лесов, на четырех из
пяти континентов земного шара.
Местом постоянного пребывания землероек служит поверхность почвы
с приземным слоем растительности, а в качестве укрытий используется
лесная подстилка или всякого рода углубления в почве и норы грызунов.
Отдельные виды роют норы, особенно в период размножения, а один вид
294
СЕМ. SORICIDAE
из рода Anourosorex M.=Edw. ведет полуподземный образ жизни. Подав-
ляющее большинство землероек обитает в равнинных и горных лесах или
зарослях кустарников, реже в высоких травах луговых или прибрежных
биотопов. Сравнительно немного видов живет в степях, на альпийских
лугах, в россыпях и нагромождениях камней или в пустынях.
Пищу землероек составляют преимущественно насекомые на разных
стадиях их развития. Можно сказать, что из всех представителей отряда
насекомоядных землеройки самые насекомоядные. Поедают они и других
беспозвоночных, а также мелких позвоночных животных. Преобладание
того или иного корма зависит от наличия его в то или иное время. Кормо-
вые спектры меняются на протяжении всего года. У северных видов земле-
роек насекомые преобладают в пище в теплый период года. Многие север-
1 ные виды в зимнее время поедают семена травянистых и древесных расте-
ний, особенно хвойных.
Возможно, что с питанием мелкими животными и семенами связано
сохранение видами трибы Soricini большего, чем у видов других триб, ко-
личества мелких верхних промежуточных зубов и наличие функционально
соответствующих им лопастей на режущем крае нижних первых резцов.
Землеройки размножаются в период, благоприятный в кормовом отно-
шении. В различных местах их обширного ареала эти периоды приходятся
на разное время года и имеют разную продолжительность. Гнезда, в кото-
рых рождаются молодые, строятся зверьками самостоятельно в различных
укрытиях, в большинстве случаев на земле, между корнями деревьев, под
старыми пнями; альпийская бурозубка строит свои гнезда на стеблях ра-
стений. Многие виды роют специальные норы или используют чужие,
заброшенные. Форма гнезд изменчива, что обусловлено в каждом случае
индивидуальными особенностями строителей, а также характером окру-
жения гнезда. Чаще всего оно полушаровидное. Материалом служат раз-
личные сухие части растений: стебли, листья, мелкие корешки, раститель-
ные волокна п пух, а также мхи и лишайники.
- В разных широтах число генераций у одного и того же вида разное.
’Можно считать, что зверьки размножаются не более трех раз в году при
благоприятных условиях на юге и один раз на севере. Продолжительность
беременности, установленная в лабораторных условиях, 25—30 дней.
.Число детенышей в помете в среднем 5—10, но может быть меньше или не-
сколько больше. Детеныши рождаются голыми, слепыми и беспомощными.
Период выкармливания молоком около 21 дня. Подросшие детеныши вы-
ходят из гнезда сначала на небольшие расстояния. По наблюдениям
В. А. Стальмаковой (1955), при переселении семьи пегого путорака па но-
вое место молодые цепляются зубами за хвост впереди стоящего, а первый
за хвост матери, и такой цепочкой отправляются в путь. Развитие у земле-
роек длится 50—58 дней.
Среди землероек имеются виды, подверженные массовым размноже-
ниям, например, обыкновенная землеройка, а также виды, широко распро-
страненные. по особи которых встречаются крайне редко — Sorex minutis-
simus Zimm. Продолжительность жизни землероек крайне невелика, не
более 1.5—2 лет, отчего они с полным правом могут быть названы эфеме-
рами среди млекопитающих.
Линька происходит 2 раза в жизни, первая связана со сменой юве-
нильного волосяного покрова на взрослый — он же зимний на севере;
вторая, весенняя, — с обычной сменой зимних волос на летние, послед-
няя в жизни для большинства особей.
Считается, что врагов у землероек немного, так как их почти никто не
ест из7за мускусного запаха; нередко встречаются трупы задавленных, по
не съеденных зверьков. Изредка землероек поедают хищные птицы, от-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
295
части лисицы. Остатки землероек встречаются в погадках мелких видов
сов (болотная и ушастая сова, сычи). Но в основном регулятором их числен-
ности служит отбор. Каннибализм в природных условиях не доказан, од-
нако в условиях неволи при недостатке пищи землеройки нередко поедают
друг друга. В естественных условиях иногда приходится видеть трупы
объеденных землероек, но вряд ли можно быть уверенными, что они заживо
загрызены своими собратьями, вернее, что поедены были уже погибшие
экземпляры.
Землероек следует считать полезными животными. Как уже отмеча-
лось, основной корм их составляют насекомые, многие виды которых так
или иначе приносят вред сельскохозяйственным культурам, лесам, лугам
и пастбищам, а также продовольственным запасам в местах их храпения.
При этом насекомые добываются там, где многие другие животные достать
их не могут, например в верхних слоях почвы, в лесной подстилке и дру-
гих малодоступных местах (отчасти им в этом помогают кроты). Поедают
землеройки и других беспозвоночных, среди которых много вредных видов.
Многие землеройки, особенно крупные виды, поедая мелких позвоночных,
в частности грызунов, сокращают число вредителей зерновых и овощных
культур. Поедают землеройки и трупы, тем самым выполняя санитарную
функцию. Вред, который приписывают землеройкам, выражающийся
в объедании промысловых зверей, попавших в капкан, в порче продуктов
или в поедании мальков в рыборазводнях (куторы), в общем незначителен.
В последнее время отмечается вредное значение землероек как носи-
телей разного рода инфекций, опасных для человека: на землеройках
кормятся личинки иксодовых клещей, гамазовые клещи и другие экто-
паразиты — переносчики возбудителей трансмиссивных заболеваний.
Однако эпидемиологическое значение землероек пока еще изучено недо-
статочно .
В заключение следует отметить существенное значение землероек
в экономике природы. В совокупности с другими массовыми видами они
поедают огромное количество растительноядных беспозвоночных, сокра-
щая, таким образом, число потребителей растительной массы.
Филогенетически? взаимоотношения
отдельных групп землероек
и эволюция их жевательного аппарата
Представители сем. Soricidae известны с верхнего эоцена. Они рано
сформировались как особая группа насекомоядных с характерными только
для нее особенностями, сохранившимися в значительной степени и в морфо-
логии современных видов. Морфо-физиологические особенности, определив-
шие возможности широкого приспособления землероек к разнообразным
условиям среды, формировались несомненно давно и в связи с процес-
сами адаптаций их кормовых животных, а также сопряженно с эволю-
цией климата и ландшафта.
Общая тенденция в эволюции млекопитающих к увеличению актив-
ности п в связи с этим к усилению энергетического обмена получила в пре-
делах класса наибольшее выражение у землероек, наименьших по разме-
рам. тела. Эволюция последних шла по линии максимального ускорения
жизненных функций организма, и в этом отношении они не сравнимы ни
с какими другими млекопитающими.
Вопрос о происхождении и родственных отношениях землероек, как и
других семейств, весьма сложен, что объясняется многими обстоятель-
ствами, но прежде всего недостатком палеонтологического материала.
296
СЕМ. SORICIDAE
Однако и то, что имеется в этом отношении, позволяет составить общее
представление об их эволюции и высказать некоторые соображения по
этому поводу.
В палеоцене землероек еще не было, но уже существовали группы, с ко-
торыми они связаны в происхождении: тенрекообразные (Tenrecii) и еще
близкие к ним, но и имеющие особенности, позволяющие сближать их
с землеройками, представители сем. Nyctitheriidae. Эта группа насекомо-
ядных известна пока только из Северной Америки, но можно не сомне-
ваться, что она будет найдена и в Азии. Таким образом, филогенетические
связи землероек с другими семействами насекомоядных удается просле-
дить еще в палеоцене.
Землеройки эоценового времени были представлены разными группами.
Одни еще сохраняли особенности, свойственные и кротам, например пред-
ставители рода Saturninia Stehl. (подсем. Saturniniinae), другие были по-
хожи на землероек из подсем. Soricinae. Филхол (Filhol, 1884), описывая
нижнюю челюсть землеройки Amphisorex primaevus из фосфоритов Кверси
(Франция), отмечает, что ее зубная формула нормальная. Она все же отли-
чалась от таковой современных видов числом промежуточных зубов,
а зубы — строением.
Возможно, что в эоцене (его верхнем отделе) окончательно сложился
адаптивный тип землероек, характерный и для современных видов. Это
представление подтверждается структурными особенностями челюстей и
зубов землероек из нижнего олигоцена, представленных рядом своеобраз-
ных групп.
Представители упомянутого выше подсем. Saturniniinae еще сохра-
няли жевательный аппарат, мало приспособленный к быстрому схватыва-
нию добычи, которой было много. Тем не менее ряд особенностей в строении
отдельных зубов показывает, что изменение в сторону развития быст-
рого схватывания и удержания добычи уже началось и у видов этого под-
семейства. Оно выражалось в увеличении высоты верхних первых резцов
и образовании на них второй вершины или зубца, а также в удлинении
второго, а не первого, как у поздних Soricinae нижнего резца ц возникно-
вении на нем бугорков. Жевательные поверхности коренных зубов, начи-
ная с Р4, приобрели еще слабозаметные складки, входящие с наружной сто-
роны коронок, а бугорки пара-, прото- и метаконов уменьшились в разме-
рах, коронки этих зубов стали широкими.
Структуры зубов представителей подсем. Saturniniinae были исходными
для зубов, характерных для видов трибы Oligosoricini (олигоцен) и ряда
миоценовых групп землероек, на основе которых преемственно развились
структуры зубов и весь жевательный аппарат современных видов.
Подсем. Saturniniinae известно только из Европы и позднее эоцена не
обнаружено. Открытие его Штелином (Stehlin, 1940) позволило устано-
вить связь в развитии землероек не только с близкими к ним кротами, но
и с более древними группами насекомоядных — Nyctitheriidae и Tenrecii.
Как уже отмечалось, в эоцене существовали землеройки, близкие по
своим морфологическим особенностям к видам подсем. Soricinae. За отсут-
ствием достаточно обстоятельных описаний, а также общности в представ-
лениях о границах между эоценом и олигоценом приходится ограничиться
указанием на то, что описываемые далее группы примитивных землероек
из нижнего олигоцена я отношу и к верхнему эоцену и думаю, что с до-
статочным основанием.
К олигоцену окончательно сложился адаптивный тип землероек.
Благоприятные условия существования позволили землеройкам в олиго-
цене широко распространиться. Они известны из олигоцена Европы, Азии
п Северной Америки.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
297
Частные адаптации у олигоценовых землероек достаточно разнообразны,
что отражено в морфологии отдельных групп того времени. Их жеватель-
ный аппарат еще сохранял многие примитивные особенности. Верхние
коренные зубы были с неглубокими входящими складками на наружной
стороне коронки и слаборазвитым рельефом из гребней на жевательной по-
верхности. Сохранялось значительное количество промежуточных зубов
не только в верхней, но и в нижней челюсти (в нижней до 5). Строение зу-
брв, как и их число, у олигоценовых землероек представляет промежуточ-
ную стадию в развитии зубной системы между современным, более специа-
лизированным типом, характерным, например, для трибы Soricini,
и древним типом, свойственным землеройкам подсем. Saturniniinae. Преем-
ственность между зубами Saturniniinae и в разной степени специализи-
рованными зубами триб подсем. Soricinae наглядно выступает при их срав-
нении (рис. 165).
Из олигоцена известны землеройки трибы Oligosoricini, объединявшей
роды Crocidosorex Lavoc. Oligosorex Kretz. (Европа). Gobiosorex Sulims. (Азия)
и Domnina Cope (Северная Америка). Эта триба была основной группой
в развитии всех послеолигоценовых землероек. Сложившийся в ее пределах
тип зубов послужил основой для развития зубов всех последующих групп
семейства.
Вероятно, у многих представителей трибы Oligosoricini были скуловые
дуги или сухожильные тяжи на месте скуловой кости, соединявшие скуло-
вые отростки. Судя по строению йпжней челюсти представителей родов
Domnina Соре и Oligosorex Kretz., олигоценовые землеройки питались мел-
кими подвижными беспозвоночными животными, в основном насекомыми.
Нижние челюсти были относительно тонкими, с небольшим сочленовным
отростком, что позволяет предполагать возможность их быстрого приве-
дения к черепу прп схватывании добычи. Представители трибы Oligoso-
ricini просуществовали до среднего, местами верхнего миоцена.
В миоцене в пределах трибы Oligosoricini сформировалось значительное
количество родов, адаптации которых на фоне общих еще сохранявшихся
примитивных особенностей, присущих этой трибе, сильно разошлись.
Возникли группы землероек, приспособленных к питанию не только мел-
кими, но и относительно крупными животными, например, представители
рода Angustidens Repenn.
Одни, видимо, еще сохраняли скуловые дуги, другие их утратили.
У видов большинства родов сократилось общее количество прдмежуточных
зубов в верхней и нижней челюстях. Изменилась окраска зубов. В мио-
цене триба Oligosoricini стала по своим адаптациям в значительной степени
универсальной. Тем не менее в основных адаптивных позициях она сохра-
няла древние особенности, что и привело большую часть ее состава к выми-
ранию в конце этого периода. В миоцене триба Oligosoricini была представ-
лена следующими родами: Miosorex Kretz., Soricella Dob .-Flor. (Европа;
вероятно, и Азия), Angustidens Repenn., Alluvisorex и Paradomnina Hutchis.
(Северная Америка).
Начавшаяся в пределах, трибы Oligosoricini еще в верхнем олигоцене
дивергенция привела к обособлению в миоцене ряда групп, определивших
отдельные направления в эволюции землероек, получившие конкретное
выражение в более позднее время. Еще в миоцене складывались предста-
вители триб Soricini (род. Sorex L., Европа, Азия, Северная Америка),
Crocidurini (роды Similisorex Stog. et Sav., Азия; Crocidura Wagl,
Европа, Азия, Африка; Limnoecus Stirl., Stirtonia Gur., Северная Америка),
Deterosoricini (роды: Heterosorex Gaill., Европа; Pseudotrimylus Gur. и
Ingentisorex Hutchis, Северная Америка; вероятно, и Азия); Blarinini (род
298
СЕМ. SORICIDAE
Adeloblarina Repenh., Северная Америка), Anourosoricini (род Anouroso-
rex M.=Edw., Азия).
По-видимому, большинство из названных триб сложилось уже к верх-
нему миоцену, так как в плиоцене они были представлены разнообразно
специализированными группами.
Вероятно, к концу миоцена окончательно сложился тип зубов, харак-
терный и для современных землероек. В дальнейшем изменениям подверга-
лась лишь степень развития отдельных эмалевых образований на жеватель-
ной поверхности зубов, что характеризовало большее или меньшее приспо-
собление к жеванию. Кроме того, изменениям подвергалась величина коро-
нок, их форма и величина цингулюма. Качество обработки пищи зубами
у землероек во многом зависит от массивности нижней челюсти, угла ее
смещения в сторону и степени развития отдельных жевательных мышц, оп-
ределяющих величину жевательного давления зубов.
Как показало изучение современных видов землероек, для них особое
значение имеет структура всего зубного ряда и степень подвижности ниж-
ней челюсти, что обеспечивает скорость и надежность захвата пищи. В этом
направлении и шло в основном развитие жевательного аппарата земле-
роек, определившее возможности их широкого распространения до край-
них пределов жизни.
Землеройки — одни из самых мелких млекопитающих, среди них есть
и самые мелкие, поэтому величина животных, которые служат им пищей, их
форма, консистенция и активность имеют для них важное значение. Раз-
мерные соотношения тела землеройки и добываемых ею животных нередко
превышают соотношение размеров тигра и оленя.
Таким образом, добывание пищи землеройками сопряжено со значи-
тельными затратами энергии, требует многих специальных приспособле-
ний со стороны зубо-челюстного аппарата. С этим и связано значительное
разнообразие в структурах зубных рядов и верхних и особенно нижних
внутренних резцов (образование на последних зубцов, острых граней или
гребней). В зубных рядах имеют существенное значение величина от-
дельных промежуточных зубов, соотношение размеров отдельных зубов
в ряду (чередование крупных и мелких), количество промежуточных зубов
в верхней и нижней челюстях. Совокупностью всех этих особенностей зуб-
ных систем землероек и определяются кормовые связи отдельных видов и
их групп.
Становление отдельных типов жевательных аппаратов и их элементов
имеет известную протяженность во времени, поскольку их структурные
особенности изменяются относительно медленно, что объясняется уни-
версальностью их функций. В связи с этим становление групп в эволюции
не сопровождается перестройкой их жевательного аппарата в короткий
срок. В строении жевательного аппарата объединяемых ими видов до поры,
до времени сохраняются черты, свойственные предшественникам, что и
позволяет устанавливать преемственность в адаптивном процессе между
отдельными группами во времени при филогенетических построениях.
Видимо, еще в самом начале миоцена в развитии жевательного аппарата
землероек определились два основных направления: приспособление к схва-
тыванию мелких, подвижных, мягкой консистенции животных и приспо-
собление к схватыванию и удержанию в челюстях относительно крупных
животных. В конечном счете развились два типа жевательных аппаратов.
Из них первый (при всех модификациях в более позднее время) сохраняет
особенности, свойственные и древним группам землероек (относительно
тонкие и легкие ветви нижней челюсти, некрупный сочленовный отросток
с небольшими сочленовными фасетками и др.); второй значительно спе-
циализированный (все части нижней челюсти массивные, нижняя сочленов-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
299
ная фасетка с направленной вперед внутренней частью). Первый тип ха-
рактерен для триб Soricini и Crocidurini, а второй для триб Heterosoricini,
Blarinini и Anourosoricini.
Сравнение структурных особенностей жевательных аппаратов совре-
менных и ископаемых землероек позволило установить, что происхожде-
ние триб Soricini и Crocidurini связано непосредственно с древними груп-
пами сорицид из трибы Oligosoricini, дериватами которых представляются
роды Oligosorex Kretz., Gobiosorex Sulim, и Crocidosorex Lavoc. У последних,
как и у представителей родов, объединяемых трибами Soricini и Crocidu-
rini, жевательный аппарат с очень подвижной нижней челюстью, на что
указывают ее тонкие ветви с небольшим сочленовным отростком. Отличия .
заключаются главным образом в общем числе промежуточных зубов, ,
а также в разной степени развития эмалевых образований на коренных
зубах. Обособление триб Soricini и Crocidurini от общих корней с древними
землеройками происходило еще в верхнем олигоцене, а с нижнего миоцена
в ряду групп они развиваются независимо друг от друга.
Что же касается триб Heterosoricini, Blarinini и Anourosoricini, то их
связи с древними группами землероек представляются более сложными,
опосредованными через ряд промежуточных групп с боковыми ветвями,
каковыми можно считать роды Domnina Соре, Soricella Dob.-Flor.,
Angustidens Repenn. и др. Эти роды появились в разное время. У них.в раз-
ной степени выражено приспособление к схватыванию и удержанию отно-
сительно крупной добычи. Наиболее подчеркнуто это в жевательном ап-
парате представителей рода Angustidens Repenn., у которых массивная ,
нижняя челюсть с крупным сочленовным отростком имеет выдвицутую ,
вперед нижнюю сочленовную фасетку. Жевательный аппарат представи- ,
телей триб Blarinini и Anourosoricini весьма сходен с таковым видов трибы
Heterosoricini, вымершей в миоцене. Это позволяет считать, что они проис-
ходят от общих с нею корней и развивались некоторое время параллельно,.,
а затем резко разошлись.
Представители трибы Heterosoricini при всей высокой специализации
для своего времени (жевательный аппарат образован мощными челю-
стями и зубами, сочленовные отростки с выдвинутой вперед нижней сочле- .
новной фасеткой, в верхней челюсти четыре промежуточных зуба) еще со- ,
хранили скуловые дуги (возможно лишь тяжи на месте скуловой кости),
а в нижней челюсти у них было три, а не два промежуточных зуба, как у бо-
лее поздних групп землероек.
Виды триб Blarinini и Anourosoricini обособлялись от общих корней
с трибой Heterosoricini, по-видимому, в нижнем—среднем миоцене, так как
в верхнем они уже окончательно обособились и сформировались и разви-
ваются с этого времени независимо. При общем сходстве жевательного
аппарата, приспособленного к схватыванию и удержанию относительно
крупной добычи, последний у представителей Anourosoricini более приспо-
соблен к выполнению жевательной функции. У них более широкие и менее
остробугорчатые зубы, более мощные нижние челюсти, соответственно й
жевательная мускулатура.
Представление о том, что трибы Blarinini и Anourosoricini окончательно ,
сложились к верхнему миоцену, подтверждается находками их родов
начиная с верхнего миоцена. При этом виды, объединяемые родами, уже
обладали всеми особенностями соответствующей трибы.
Если эоцен ' был временем формирования землероек, олигоцен —
периодом становления их основного типа и периодом расцвета примитив-
ных видов, доживших до среднего миоцена, а в миоцене складывались от-
дельные направления развития новых крупных групп землероек, то плио-
цен можно считать периодом формирования частных адаптаций в пределах
300
СЕМ. SORICIDAE
сложившихся триб. В плиоцене сильно изменилась обстановка в большей
части ареала землероек. Значительная пятнистость в распределении ланд-
шафтов и соответственно условий жизни стимулировали возникновение
многих своеобразных приспособлений. Известная монотонность в адап-
тациях олигоценовых и миоценовых землероек, характерная для давно
сложившихся групп, в плиоцене сменилась разнообразием частных приспо-
соблений.
Возникшие еще в миоцене отдельные направления в эволюции земле-
роек в плиоцене получили более определенное выражение. В это время раз-
вивались две группы триб с разным типом жевательного аппарата: трибы
Soricini и Crocidurini с жевательным аппаратом, приспособленным для
быстрого схватывания мелкой добычи, и трибы Blarinini и Anourosoricini
с жевательным аппаратом для схватывания относительно крупных
животных.
Специализация зубной системы представителей трибы Soricini шла по
линии приспособления к схватыванию преимущественно мелких беспоз-
воночных разной формы и консистенции, что отразилось на структуре
ряда промежуточных зубов. У одних видов промежуточные зубы при сход-
ной форме убывают в размерах к заднему краю их ряда, у других эти зубы,
разные по величине, отчасти и по форме комбинируются в разных соче-
таниях. Виды трибы и до настоящего времени сохранили наибольшее ко-
личество промежуточных зубов в верхней челюсти. На нижнем первом
резце у них три бугорка. В пределах этой трибы к концу плиоцена обо-
собились от общих корней с родом Sorex L. роды Microsorex Baird (Север-
ная Америка) и Blarinella Thos (Азия). Представители рода Sorex L.
к концу плиоцена распространились циркумполярно, а роды Tregosorex,
Anchiblarinella, Hibb. et Jamm. вымерли в нижнем плиоцене.
В трибе Crocidurini специализация зубной системы шла по пути сокра-
щения общего количества промежуточных зубов и уменьшения их разме-
ров в задней части ряда. Только первый промежуточный зуб сильно уве-
личился. Это привело к сокращению длины зубного ряда и соответственно
укорочению длины челюстей (и плечей рычагов), что усилило давление
челюстей в передней части и позволило захватывать относительно круп-
ных животных.
Наряду со значительной специализацией, как у видов родов Crocidura
Wag], или Diplomesodon Brandt, в пределах трибы сохранились роды с мно-
гими примитивными особенностями в зубной системе (бугорки на режущем
крае нижнего первого резца, 4 промежуточных зуба в верхней челюсти и
др.) — роды Feroculus Kelaart, Sylvisorex Thos, Paracrocidura H. de Bals.,
а, например, у видов рода Myosorex Gray и до настоящего времени в нижней
челюсти сохранился третий промежуточный зуб.
Триба Blarinini в плиоцене достигла наибольшего расцвета. Частные
приспособления ее представителей в это время были весьма разнообразны.
При том, что строение жевательного аппарата было общим для всех пред-
ставителей трибы, а различия заключаются главным образом в размерных
соотношениях составляющих его элементов, эта группа в плиоцене была
представлена разнообразно приспособленными формами, объединявшимися
многими родами. Многие роды уже в плиоцене составляли группы с об-
щим направлением в развитии. Так, в начале этого периода обособилась
подтриба Вlarinina, объединившая роды Blarinoides Sulims, и Blarina Gray.
Их представители еще сохраняли 5 промежуточных зубов в верхней че-
люсти, но уже тогда у них началась редукция предпоследних премоляров.
Это несомненно наиболее примитивная группа в пределах трибы, хотя виды
рода Blarina Gray и сохранились до настоящего времени. В это же время
формировалась и подтриба Beremendina, объединяющая роды, виды которых
Рис. 164, Схема филогенетических соотношений землероек.
302
СЕМ. SORICIDAE
имеют 4—3 промежуточных зуба в верхней челюсти. У них относительно
короткие и массивные нижние челюсти, что представляет более высокую
специализацию землероек к питанию относительно крупными животными,
получившую наибольшее выражение у представителей подтрибы Allosori-
cina. По-видимому, еще в плиоцене виды подтрибы Soriculina специализи-
ровались в двух основных направлениях: к быстрому бегу — род Sori-
culus Blyth, отчасти Notiosorex Baird (подрод Chodsigoa), и полуводному
образу жизни — роды Zelceina Sulims, и Neomys Каир. Наибольшей спе-
циализации к водной среде достигли представители подтрибы Nectogalina—
роды Chimarrogale Anders, и Nectogale M.-Edw. особенно последний.
Начавшееся в конце миоцена и особенно в плиоцене быстрое развитие
трибы Вlarinini к концу плиоцена резко замедлилось. Многие ее роды вы-
мерли. Из подтрибы Blarinina остался один род Blarina Gray. Только род
Cryptotis Pom. сохранился из подтрибы Beremendina. Полностью вымерли
представители подтрибы Allosoricina. Вымерли многие роды и из других
подтриб. Сохранились в основном роды, менее специализированные в глав-
ном направлении специализации трибы Вlarinini. Это виды, ведущие полу-
водный образ жизни, отступившие в южные части ареала, а также форми-
ровавшиеся на территориях с безморозными зимами. Отсюда можно сде-
лать вывод, что землеройки, приспособленные к питанию относительно
крупными животными, при изменении климатических условий столкну-
лись в умеренной зоне с затруднениями в добывании пищи. Видимо,
со сменой кормовых спектров связано также вымирание в плиоцене и родов
трибы Anourosoricini. До настоящего времени из этой трибы дожил один
вид рода — Anourosorex M.-Edw., приспособленный к рытью.
Таким образом, более перспективным для Soricidae в северной части
ареала оказался более универсальный жевательный аппарат, характер-
ный для трибы Soricini, которая объединяет мелких землероек. Крупные
виды семейства в современный период распространены в южной части аре-
ала. В плейстоцене, в его нижнем отделе, существовали отдельные ранние
виды современных родов и триб, вымершие в раннечетвертичное время.
К началу четвертичного периода вымерли в Европе, Азпп и в Северной
Америке многие роды из трибы Blarinini. В Европе из ее состава сохра-
нился только род Neomys Каир. Подавляющее большинство представите-
лей этой трибы в современный период обитает в местах с короткой зимой и
незамерзающими водоемами или на территориях с безморозными зимами.
По-видимому, триба Soricini на протяжении плейстоцена пополнялась
новыми видами и подвидами и расширяла ареал к северу, виды же трибы
Crocidurini продолжали в это время сокращать свой ареал за счет север-
ных окраин, например: на Украине и в других местах Средней Европы
к этому времени вымерли представители рода Suncus Ehrenb. К концу плей-
стоцена окончательно сложилась современная фауна.
Процветающими группами в современный период следует считать две
группы землероек — трибу Soricini, род Sorex L., и трибу Crocidurini, род
Crocidura Wagl. Первая широко распространена в умеренной зоне Пале-
арктики и Неарктики, а вторая — в субтропиках и тропиках Старого Света.
Мои представления о филогенетических взаимоотношениях различных
групп землероек приведены на рис. 164.
Краткая история
исследования систематики
Описанием в 1758 г. землеройки Sorex araneus Линией положил начало
изучению этой группы насекомоядных. В последующее время все новые
виды землероек относили к роду Sorex L. Только через 70 лет Каупом
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
303
(Каир, 1829) был описан род Neomys, а на протяжении следующих 40 лет
разными авторами — еще 12 родов. В этот же период были описаны 2 под-
сем. — Soricinae (Murray, 1866 и Crocidurinae (Milne-Edwards, 1868—
1874), а также надсем. Soricoidea (Gill, 1872). В первой половине XX в.
были описаны еще 7 родов современных землероек и подсем. Scutisoricinae J.
Allen, 1917.
Исследование ископаемых землероек началось только во второй поло-
вине XIX в., а наиболее обстоятельное их изучение — в текущем столетии.
Большое значение для познания землероек отечественной фауны имела
и сохранила важное значение и в настоящее время капитальная работа по
насекомоядным фауны СССР С. И. Огнева (1928). Существенный вклад
в изучение землероек сделал своими работами и С. У. Строганов. К сожа-
лению, ни тот, ни другой автор не занимались разработкой общей си-
стемы этой важной во многих отношениях группы насекомоядных.
Стиртон (Stirton, 1930), основываясь на изучении современных и иско-
паемых землероек, пришел к выводу, что сем. Soricidae представляет собой
целостную группу и выделение внутри его подсемейств нецелесообразно.
Он предложил объединить современные роды в следующие группы.
Группа — Crocidura
Роды: Crocidura, Pachyura, Surdisorex, Sylvisorex, Myosorex
Группа — Sorex
Роды: Sorex, Microsorex
Группа — Blarina
Роды: Neomys, Soriculus, Notiosorex, Chimarrogale, Cryptotis, Blarina
Группа — Blarinella
Роды: Blarinella
Группа — Anourosorex
Роды: Anourosorex
В основу подразделений современных землероек Стиртоном положено
строение отдельных частей черепа и зубов, отчасти и окраска последних.
Это правильное в основном, как мне кажется, подразделение современных
землероек в более поздних систематических работах по землеройкам не
отмечено и не комментировано. Даже Кретцой (Kretzoi, 1965), предло-
живший свое подразделение современных и ископаемых землероек, не упо-
минает работу Стиртона.
Симпсон (Simpson, 1945) предложил следующую классификацию сов-
ременных и ископаемых групп землероек.
Сем. Soricidae Gray, 1821. Нижний олигоцен—современный, Европа; средний олиго-
цеп—современный, Северная Америка; нижний плиоцен—современный, Азия;
современный, Африка.
Подсем. Soricinae Murray, 1866. Нижний миоцен—современный, Северная Аме-
рика; нижний плиоцен—современный, Азия.
t Saturninia Stehlin, 1940. Верхний эоцен, Европа.
f Necrosorex Filhol, 1889. Нижний олигоцен, Европа.
f Domnina Соре, 1873. Нижний—средний олигоцен, Северная
Америка.
f Heterosorex Gaillard, 1915. Средний миоцен, Европа.
t Mystipterus Hall, 1930. Нижний плиоцен, Северная Америка.
Sorex Linnaeus, 1758. Нижний олигоцен?, нижний миоцен—со-
временный, Европа; верхний плиоцен—современный, Северная
Америка; плиоцен—современный, Азия.
Microsorex Coues, 1877. Плейстоцен—современный, Северная
Америка.
Soriculus Blyth, 1855. Современный, Азия.
Neomys Каир, 1829. Современный, Европа, Азия.
Blarina Gray, 1838. Верхний плиоцеп—современный, Северная
Америка.
Blarinella Thomas, 1911. Нижний плиоцен—современный, Ев-
ропа, Азия.
304
СЕМ. SORICIDAE
Cryptotis Pomel, 1848. Современный, Северная Америка, север-
ная часть Южной Америки.
Notioserex Baird, 1877. Плейстоцен—современный, Северная
Америка.
Подсем. Crocidurinae Milne-Edwards, 1868—1874. Верхний миоцен, Северная Аме-
рика; верхний миоцен—современный, Европа; нижний плиоцен—
современный, Азия; современный, Африка.
f Limnoecus Stirton, 1930. Верхний миоцен, Северная Америка.
Crocidura Wagler, 1832 (включает Heliosorex Heller, 1910). Ниж-
ний миоцен—современный, Европа; нижний плиоцен—современ-
ный, Азия; современный, Африка.
Praesorex Thomas, 1913. Современный, Западная Африка.
Suncus Ehrenberg, 1832 b=Pachyura Selys-Longchamps, 1839).
Современный, Европа, Азия, Африка.
Feroculus Kelaart, 1852. Современный, о. Шри Ланка (Цейлон).
Solisorex Thomas, 1924. Современный, о. Шри Ланка.
Sylvisorex Thomas, 1904. Современный, Африка.
Myosorex Gray, 1838. Современный, Южная Африка.
Surdisorex Thomas, 1906. Современный, Африка.
Diplomesodon Brandt, 1853. Современный, Азия.
Anourosorex Milne-Edwards, 1870. Нижний плиоцен—современ-
ный, Европа; миоцен—современный, Азия.
Chimarrogale Anderson, 1877 (включает Crossogale Thomas, 1921).
Современный, Азия.
Nectogale Milne-Edwards, 1870. Современный, Азия.
Подсем. Scutisoricinae J. Allen, 1917. Современный, Африка.
Scutisorex Thomas, 1913. Современный, Африка.
Кретцой (Kretzoi, 1965) предложил следующие группировки в пределах
сем. Soricidae.
f Подсем. Trimylinae (= Нeterosoricinae Gaillard, 1915)1 с родом Trimylus Roger,
1885 {—Heterosorex Gaillard, 1915).
Подсем. Soricinae. В пределах последнего он выделяет следующие группы.
Soricini: Sorex-gmppe (Atophyrax, Sorex s. str., Neosorex, Oti-
sorex, Eurosorex), Microsorex, Oligosorex, Drepanosorex.
Soriculini: S oricuZus-gruppe (Asoriculus, Nesiotites, Soriculus,
Episoriculus, Chodsigoa), Petenyiella, Zelzeina, Neomys.
Crocidurini: Crocidura—Suncus—Pachyura-gruppe, Praesorex,
Sylvisorex.
Blarinini: Blarina, Blarinella, Beremendia, Paracryptotis, Cryp-
totis, Shikamainosorex, Petenyia, Peisorex.
Близкими к Soriculini Кретцой считает роды Notiosorex и Megasorex.
Роды Myosorex и Surdisorex он выделил в трибу Myosoricini. Для рода
Diplomesodon и другого азиатского рода (по-видимому, Anourosorex M.-
Edw.) автор не определил места. Род Scutisorex он считает особой трибой,
близкой к трибе Crocidurini, а род Amblycoptus—трибой Amblycoptini.
Роды Crocidosorex, Limnoecus и Hesperosorex никуда им не определены.
Следует отметить, что трибы Soricini, Crocidurini и Blarinini совпадают
с группами Sorex, Crocidura и Blarina Stirton (1930). Триба Soriculini
в моей системе фигурирует как подтриба трибы Blarinini, а триба Myoso-
ricini — как подтриба трибы Crociduruni. Род Scutisorex ни в какие особые
группы я не выделял, а оставил родом в трибе Crocidurini. Род Amblycoptus
я отнес в трпбу Anourosoricini, выделенную в качестве особой группы еще
Стиртоном (Stirton, 1930).
В 1967 г. Репеннинг (Repenning) предложил следующую систему.
1 Подсем. IIeterosoricinae Viret et Zapfe, 1967
Domnina Cope, 1873
Paradomnina Cope, 1873
1 Подсем. Heterosoricinae выделили Вире и Запфе (Viret, Zapfe) в 1951 г.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
305
Trimylus Roger, 1885
Ingentisorex Hutchison, 1966
Подсем. Crocidurinae Milne-Edwards, 1868—1874
Crocidura Wagler, 1832
Diplomisodon Brandt, 1853
Feroculus Kelaart, 1852
f Miosorex Kretzoi, 1959
Myosorex Gray, 1838
Paracrocidura Balsac, 1956
Praesorex Thomas, 1913
Scutisorex Thomas, 1913
Solisorex Thomas, 1924
f Soricella Doben-Florin, 1964
Suncus Ehrenberg, 1832
Surdisorex Thomas, 1906
Sylvisorex Thomas, 1905
f Подсем. Limnoecinae Repenning, 1967
f A ngustidens Repenning, 1967
f Limnoecus Stirton, 1930
Подсем. Soricinae Fischer von Waldheim, 1817
Триба Soricini Fischer von Waldheim, 1817
f Crocidosorex Lavocat, 1951
f Antesorex Repenning, 1967
Sorex Linnaeus, 1758
f Drepanosorex Kretzoi, 1941
Microsorex Cones, 1877
f Alluvisorex Hutchison, 1966
f Petenyia Kormos, 1934
Blarinella Thomas, 1911
t Zelceina Sulimski, 1962
Триба Blarinini Stirton, 1930
f Adeloblarina Repenning, 1967
Cryptotis Pomel, 1848
f Paracryptotis Hibbard, 1950
f Shikamainosorex Hasegawa, 1957
Blarina Gray, 1838
f Blarinoides Sulimski, 1959
Blarinini? incertae sedis Peisorex Kowalski et Li-Chuan-Kuei, 1963
Триба Neomyini Repenning, 1967
Neomys Каир, 1829
f Petenyiella Kretzoi, 1956
Episoriculus Ellermann et Morrison-Scott, 1951
Chodsigoa Kastschenko, 1905
f Beremendia Kormos, 1934
f Nesiotites Bate, 1945
Nectogale Milne-Edwards, 1870
Soriculus Blyth, 1854
Chimarrogale Anderson, 1877
Anourosorex Milne-Edwards, 1870
f Amblycoptus Kormos, 1926
f Hesperosorex Hibbard, 1957
Notiosorex Baird, 1877
f Megasorex Hibbard, 1950
Neomyini incertae sedis f Deinsdorfia Heller, 1963
t Подсем. Allosoricinae Fejfar, 1966
f Allosorex Fejfar, 1966
Soricidae incertae sedis Podihik Deraniyagala, 1958
Признаки, использованные Репеннингом (Repenning, 1967) для построе-
ния системы, крайне изменчивы, а потому непригодны. Роды, близкие
по происхождению и характеру адаптаций, отнесены автором к разным
подсемействам или трибам.
Выбор нижних челюстей и их зубов в качестве основного материала
для построения системы и филогенетических соотношений групп землероек
вряд ли можно считать удачным. Опыт изучения последних показывает,
что структурные особенности нижних челюстей этих млекопитающих
20 А. Л. Гуреев
306
СЕМ. SORICIDAE
поразительно однообразны. Это можно объяснить не только сходством
основных функций, но и универсальностью. Если внимательно присмотре-
ться к нижним челюстям землероек, то можно заметить, что по строению они
могут быть разделены на два основных типа: нижняя челюсть с сочленовным
отростком, имеющим неширокие верхнюю и нижнюю фасетку, и нижняя
челюсть с сочленовным отростком, имеющим неширокую верхнюю фасетку
и широкую нижнюю, внутренний край которой продвинут вперед на линг-
вальной стороне ветви нижней челюсти.
Нижние челюсти ископаемых групп землероек как первого, так и
второго типов были или с карманообразной птеригоидной (темпоральной)
ямкой или с неглубокой впадиной на месте ее. Это позволяет считать,
что приспособление к быстрому схватыванию добычи в пределах семейства
уже давно идет в двух основных направлениях: по пути приспособления
к быстрому захвату относительно мягких и мелких животных (I тип) и
по пути приспособления к захвату и удержанию относительно крупных
жестких животных (II тип).
Усиление функции быстрого сжатия челюстей для схватывания добычи
обусловило значительное развитие височного мускула и сопровождалось
образованием нижней фасетки на сочленовном отростке нижней челюсти,
а также значительным давлением последней во время сжатия челюстей на
свою опору — засочленовный отросток. В связи с этим потребовалось ук-
репление основания сочленовного отростка по его горизонтальной оси,
что и выразилось в костном утолщении при основании венечного отростка
с образованием карманообразной ямки. В этой ситуации роль массетера
стала сводиться к функции ускорителя закрывания рта. Сила его тяги
направилась в основном по линии угловой отросток—скуловой отросток
верхнечелюстной кости. Ослабла его связь со скуловой дугой, а вместе
с этим уменьшилась и его инерция. Остается неясным вопрос, на какой
стадии формирования жевательного аппарата в целом и развития эмале-
вых гребней на жевательной поверхности зубов у землероек полностью
исчезла скуловая дуга. Последняя более или менее достоверно известна
только у видов рода Heterosorex Gaill. {-Trimylus Roger). Что же касается
других родов, как более ранних, так и того же времени, что и Heterosorex
Gaill., то о них в этом отношении ничего неизвестно. Судить о наличии
или отсутствии скуловой дуги на основании степени развития птеригоид-
ной (темпоральной) ямки вряд ли можно.
В связи с изложенным мне представляется малоубедительным объеди-
нение в одну группу, хотя бы и на уровне подсемейства, родов Domnina
Соре и Heterosorex Gaill.
Перетасовка родов в системе Soricidae на основании строения бугорков
и ямочек на жевательной поверхности зубов при полном пренебрежении
к такому важному признаку, как общее количество зубов в верхней и
нижней челюсти, вызывает по меньшей мере удивление.
При всех сделанных здесь замечаниях в отношении систематики земле-
роек работа Репеннинга имеет ряд несомненных достоинств. Она пред-
ставляет наиболее полную сводку по ископаемым и современным родам
землероек, содержит богатый иллюстративный материал и достаточно
подробные описания структурных особенностей зубов и других элементов
нижней челюсти.
Ван Вален (Van Valen, 1967) опубликовал систему насекомоядных,
в которой представлена и система землероек.
Сем. Soricidae Ficher von Walheim, 1817. РВерхний эоцен, нижний олигоцен—совре-
менный, Северная Америка; средний олигоцен—современный, Европа; нижний
миоцен—современный, Азия; миоцен—современный, Африка; современный,
Южная Америка.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
307
f Подсем. Heterosoricinae Viret et Zapfe, 1951. ’Верхний эоцен, нижний олиго-
цен—верхний миоцен, Северная Америка; средний олигоцен—нижний плио-
цен, Европа.
f Domnina Соре, 1873 (Miothen Соре, 1873; Protosorex Scott, 1895).
?Верхний эоцен, нижний олигоцен—нижний миоцен, Северная
Америка.
f Paradomnina Hutchison, 1966. Верхний миоцен, Северная Аме-
рика.
f Ingentisorex Hutchison, 1966. Верхний миоцен, Северная Аме-
рика.
f Trimylus Boger, 1885 (= Heterosorex Gaillard, 1915).
Средний олигоцен—верхний миоцен, Северная Америка; сред-
ний олигоцен—пижний плиоцен, Европа.
Подсем. Crocidurinae Milne-Edwards, 1872. Нижний миоцен—современный, Ев-
ропа; миоцен—современный, Африка; плейстоцен—современный, Азия.
Триба Crocidurini Milne-Edwards, 1872. Нижний миоцен—современный,
Европа; миоцен—современный, Африка; плейстоцен—современный,
Азия.
j- Miosorex Kretzoi, 1959. Верхний миоцен, Европа.
Crocidura Wagler, 1832 (=Hhinomus Murrey, 1861; Leucodon Fa-
tio, 1869; Paurodus Schulze, 1897; f Heliosorex Heller, 1910).
’Миоцен, плейстоцен—современный, Африка; плейстоцен—со-
временный, Азия, Европа.
Diplomesodon Brandt, 1853. Плейстоцен, Африка; современный,
Азия.
Feroculus Kelaart, 1852. Современный; Шри Ланка (Цейлон).
Myosorex Gray, 1838 (включает Congosorex Heim de Balsac et
Lamotte, 1956). Плейстоцен—современный, Африка.
Paracrocidura Heim de Balsac, 1956. Современный, Африка.
Praesorex Thomas, 1913. Современный, Африка.
Solisorex Thomas, 1924. Современный, Шри Ланка (Цейлон),
f Soricella Doben-Florin, 1964. Нижний миоцен, Европа.
Suncus Ehrenberg, 1833 (=Pachyura Selys Longchamps, 1839;
Suncus Sundevall, 1843; Paradoxodon Wagler, 1855; Pleurodus
Schulze, 1897). Нижний плиоцен—современный, Европа; плей-
стоцен—современный, Африка; современный, Азия.
Surdisorex Thomas, 1906. Современный, Африка.
Sylvisorex Thomas, 1904. Современный, Африка.
Триба Scutisoricini J. A. Allen, 1917. Современный, Африка.
Scutisorex Thomas, 1913. Современный, Африка.
f Подсем. Limnoecinae Bepenning, 1967. Средний миоцен—средний плиоцен, Се-
верная Америка.
t A ngustidens Bepenning, 1967. Средний миоцен, Северная Аме-
рика.
f Limnoecus Stirton, 1930. Верхний миоцен—средний плиоцен,
Северная Америка.
Подсем. Soricinae Ficher von Waldheim, 1817. Средний олигоцен—современный,
Европа; средний миоцен—современный, Северная Америка; средний плио-
цен—современный, Азия; современный, Южная Америка.
Триба Soricini Ficher von Waldheim, 1817. Средний олигоцен—современ-
ный, Европа; средний миоцен—современный, Северная Америка;
средний плиоцен—современный, Азия.
f Crocidosorex Lavocat (включает Oligosorex Kretzoi, 1959). Сред-
ний олигоцен—нижний миоцен, Европа.
у Antesorex Repenning, 1967. Средний миоцен, Северная Аме-
рика.
Sorex Linnaeus, 1758 (=Musaraneus Brisson, 1762; Oxirhin Каир,
1829; Amphisorex Duvernoy, 1835; Corsira Gray, 1838; Otisorex
De Kay, 1842; Hydrogale Pomel, 1848; Neosorex Baird, 1858;
Atophyrax С. H. Merriam, 1884; Homalurus Schulze, 1890; Sorici-
dus Altobello, 1927). Верхний плиоцен—современный, Европа;
Северная Америка; плейстоцен—современный, Азия.
f Drepanosorex Kretzoi, 1941. Плейстоцен, Европа.
Microsorex Baird, 1877. Плейстоцен—современный, Северная
Америка.
t A lluvisorex Hutchison, 1966. Верхний миоцен—нижний плио-
цен, Северная Америка.
Blarinella Thomas, 1911. Средний плиоцен—современный, Азия.
20*
308
СЕМ. SORICIDAE
f Petenyia Kormos, 1934. Плиоцен—плейстоцен, Европа.
f Zelceina Sulimski, 1962. Плиоцен, Европа.
Триба Blarinini Stirton, 1930. Верхний миоцен—современный, Северная
Америка; плиоцен—плейстоцен, Европа; плейстоцен, Азия; совре-
менный, Южная Америка.
f Adeloblarina Repenning, 1966. Верхний миоцен, Сев. Америка.
Cryptotis Pomel, 1848 (=Brachysorex Duvernoy, 1842, Soriciscus
Cones, 1877). Средний плиоцен—современный, Северная Аме-
рика; современный, Южная Америка.
f Paracryptotis Hibbard, 1950. Средне-верхний плиоцен, Северная
Америка.
t Shikamainosorex Hasegawa, 1957. Плейстоцен, Япония.
f «Sorex» dehneli Kowalski, 1956. Плейстоцен, Европа.
Blarina Gray, 1838 (Talpasorex Pomel, 1848, A notus Wagner, 1855,
Mamblarinaus Herrera, 1899). Верхний плиоцен—современный,
Северная Америка.
f Blarinoides Sulimski, 1959. Плиоцен, Европа.
Триба Neomyini Repenning, 1966 (=IIydrosoridae аноним, 1838; Crossopinae
Milne-Edwards, 1872, Nectogalinae J. Anderson, 1879; Anourosoricinae
J. Anderson, 1879, Soriculi Wingo, 1917, Amblycoptinae Kormos,
1926). Нижний плиоцен—современный, Европа; средний плиоцен—
современный, Азия, Северная Америка.
Neomys Каир, 1829 (=Leucorrhynchus Каир, 1829, Hydrogale
Каир, 1829, Crossopus Wagler, 1832, Hydrosorex Duvernoy, 1835,
Amphisorex Duvemoi, 1835; Pinalia Gray, 1838, Myosictis Pomel,
1854). Плиоцен—современный, Европа; современный, Азия,
f Petenyiella Kretzoi, 1956 (=Allop achy ига Kormos, 1934). Плио-
цен-плейстоцен, Европа.
Episoriculus Ellerman et Morrison-Scott, 1951 (=Asoriculus
Kretzoi, 1959). Верхний плиоцен—плейстоцен, Европа; современ-
ный, Азия.
Chodsigoa Kastschenko, 1905. Плейстоцен—современный, Азия,
f Beremendia Kormos, 1934. Плиоцен—плейстоцен, Европа; плей-
стоцен, Азия.
f Nesiotites Bate, 1945. Плейстоцен, острова Средиземного моря.
Nectogale Milne-Edwards, 1870. Современный, Азия.
Soriculus Blyth, 1854. Современный, Азия.
Chimarrogale J. Anderson, 1877 (=Crossogale Thomas, 1921).
Современный, Азия.
Anourosorex Milne-Edwards, 1870 (=Pygmura J. Anderson, 1875;
Anurosorex J. Anderson, 1875). Средний плиоцен—современный,
Азия.
f Amblycoptus Kormos, 1926. Нижний плиоцен, Европа.
f Hesperosorex Hibbard, 1957. Средний плиоцен, Северная Америка.
Notiosorex Baird, 1877. Верхний плиоцен—современный, Северная
Америка.
f Deinsdorfia Heller, 1963. Верхний плиоцен—плейстоцен, Европа.
Megasorex Hibbard, 1950. Современный, Северная Америка.
Триба Allosoricini Fejfar, 1966. Средний миоцен—верхний плиоцен,
Европа;
f «Sorex» gracilidens Viret et Zapfe, 1951. Средний миоцен, Европа,
f A llosorex Fejfar, 1966. Верхний плиоцен, Европа.
(Подсемейство не выяснено).
f (изолированный моляр) Bohlin, 1942. Нижний миоцен, Азия.
Podihik Deraniyagala, 1958. Современный, Шри Ланка (Цейлон).
Система Ван Валена (Van Valen, 1967) мало отличается от таковой
Репеннинга (Repenning, 1967). Отличие заключается в том, что добавлены
некоторые трибы и изменен порядок чередования родов в трибах и под-
семействах. Обе системы представляют произвольную смесь родов.
В 1975 г. В. А. Долгов и Б. С. Юдин опубликовали следующую систему
землероек фауны СССР.
В систему рода Sorex L. авторы ввели как самостоятельный вид S. саи-
casicus Satunin, не имеющий отличительных признаков от S. araneus L.,
в чем нетрудно убедиться, просмотрев серии черепов и шкур того и другого
вида. Прав С. И. Огнев (1928), выделяя последний в подвид S. araneus.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
309
Плоскочерепная бурозубка — S. vir Allen сведена в синонимы S. roboratus
Hollist. Первая описана Алленом (Allen, 1914) из окрестностей Нижне-
Колымска (тип хранится в Музее сравнительной зоологии в Кембридже,
США), вторая описана Холлистером (Hollister, 1913) с Алтая по одному
дефектному экземпляру (тип хранится в Национальном музее в Нью-Йорке,
США). Не ясно, что послужило Юдину (1969) основанием для переимено-
ния A. vir в S. roboratus. Последняя считается подвидом A. araneus. В «Фауне
Сибирй» (1975, выл. 23) в статье Лукьяновой (с. 70) плоскочерепная буро-
зубка носит название S. vir, а в статье Швецова, Литвинова и Моложни-
кова (с. 106) она приведена под названием A. roboratus. Кому должен верить
читатель? В действительности S. vir Allen — морфологически хорошо
очерченный вид, близкий к американским группам бурозубок, a S. robora-
tus Hollist. подвид A. araneus L. с более мелкими зубами.
В систему рода Crocidura Wagler авторы ввели, а следовательно, и посе-
лили на территории СССР С. russula Негш. Этот вид распространен только
в Западной Европе. Он близок к С. leucodon Негш., а не к С. guldenstaedti
Pall., как думал Огнев (1928). Эллерман и Моррисон-Скотт (Ellerman,
Marrison-Scott, 1951), ничтоже сумняшись, расселили С. russula через
всю Евразию до Японских островов.
Сем. Soricidae
Род Sorex
S. araneus L.
S. caucasicus Satunin
S. raddei Satunin
S. arcticus Kerr
S. asper Thomas
S. isodon Turov
S. unguiculatus Dobson
5. alpinus Schinz
S. bucharensis Ognev
5. daphaenodon Thomas
S. roboratus Holister (syn. S.
vir. Allen)
S. mirabilis Ognev
S. caecutiens Laxmann
S. minutus L.
S. gracillimus Thomas
S. minutissimus Zimmermann
S. cinereus Kerr
5. beringianus Judin
Род Neomys
N. anomalus Cabrera
N. fodiens Pennant
N. schelkovnikovi Satunin
Род 5 uncus
S. etruscus Savi
Род Crocidura
C. suaveolens Pallas
C. russula Hermann
C. giildenstaedti Pallas
C. leucodon Hermann
C. lasia Thomas
C. lasiura Dobson
C. sibirica Dukelsky
C. persica Thomas
C. hyrcana Goodwin
C. gmelini Pallas
C. pergrisea Miller (syn.
C. serezkyensis, C. armenica)
Род Diplomesodon
D. pulchellum Lichtenstein
C. lasia Thomas введена в фауну СССР Огневым (1926). В качестве
отличительного признака этого вида от С. leucodon Herm. отмечена боль-
шая величина С. lasia (Огнев, 1928; Виноградов, 1958), никаких других
отличий у этих видов нет. Зачем вводить в систему вид, отсутствующий
в природе.
В рассматриваемую систему помещена С. gmelini Pallas — вид с неизве-
стным систематическим положением. Паллас в своей книге (Zoographia
Rossoasiatica, 1811, т. 1, с. 134) описал землеройку под названием Sorex
gmelini. В диагнозе отмечено: A. inauritus rufescens, cauda concolore,
tereti, setosa — уши невыступающие, рыжеватая, хвост одноцветный,
округлый, щетинистый. Хвост у всех землероек покрыт щетинками —
только короткими или короткими и длинными. И те, и другие отличаются
от волос, покрывающих тело. За диагнозом следует замечание Палласа
о том, что он получил от Гмелипа-младшего экземпляр землеройки,
пойманной в Гиркании (старое название области в северо-восточной части
Иранского плоскогорья к северу от Парфии), который изображен и без-
310
СЕМ. SORICIDAE
условно отличается от предыдущего вида (Crocidura suaveolens, описанной
им на с. 133 той же книги). Затем приведено описание, в котором отмечено
следующее: размеры и пропорции сходны с Sorex pygmacus, но крупнее,
нос менее удлиненный, чем у S. araneus. Уши выступают незначительно,
окраска серо-рыжеватая, снизу желтоватая, хвост рыжевато-желтый,
округлый, покрыт щетинками, лапки простые. Из этого описания не ясно,
что эта землеройка из рода Crocidura. В атласе рисунков, приложенном
к трехтомнику «Zoographia Rossoasiatica», на двух таблицах изображены
четыре землеройки: на одной Crocidura giildenstaelti и С. suaveolens, а на
другой Sorex pygmaeus и S. gmelini. У двух первых нарисованы крупные
уши, а на хвостах — короткие и длинные щетинки, особенно заметные
у С. suaveolens, а вторые нарисованы с короткими щетинками на хвосте
и с маловыступающими ушами. По характеру рисунков можно заключить,
что Паллас хорошо различал особенности покрова хвостов и строения
ушей землероек и поместил на таблицах близкие по морфологии виды
попарно.
Поскольку тип S. gmelini Pall, не сохранился, нельзя восстановить
истинное систематическое положение этого вида. Какие основания были
у Огнева (1928) считать этот вид подвидом S. minutus L., а местом нахожде-
ния типа Крым? неясно.
Неясность систематического положения S. gmelini не помешала И. И.
Стогову и Е. П. Бондарю (1966) восстановить этот вид в качестве реального,
объединив под этим названием еще Два вида: Crocidura hyrcana Gudw.
(северо-восточный Иран, юго-западная Туркмения) и С. dinniki Ognev,
которую Огнев распространил от Ставропольского края до северо-восточ-
ного Ирана. Получился триумвират с одним неизвестным. Насколько
можно судить, поводом для этой мыслительной находки послужили сле-
дующие обстоятельства: обитание С. dinniki в северо-восточном Иране
(Огнев, 1928), введение этого вида в состав фауны Туркмении М. К. Лапте-
вым (1936) и описание С. hyrcana из юго-западной Туркмении Б. С. Вино-
градовым (1958), а также получение землеройки из Гиркании Палласом
в 1811 г. Сопоставление географии видов, описаний перечисленных видов
разными авторами и собственные заблуждения привели не к восстановле-
нию приоритета S. gmelini Pall., а к путанице в систематике землероек
рода Crocidura.
В системе Долгова и Юдина в отличие от взглядов Стогова и Бондаря
С. gmelini (=S. gmelini) не включает С. hyrcana, помещенную в систему
в качестве самостоятельного вида, а только, по-видимому, С. dinniki,
не отмеченную как ее синоним. Последним видом в системе рода Crocidura
стоит С. pergrisea Miller, синонимами которой авторы сочли С. serezkyensis
и С. armenica, а С. pamirensis Ognev не па шла места в системе.
Нельзя не согласиться с Долговым и Юдиным, что род Crocidura —
весьма трудная группа землероек, но из этого не следует, что можно нару-
шать правила систематики.
В заключение следует специально остановиться на некоторых работах,
внесших путаницу не только в систематику, но на этой «основе» ив геогра-
фическое распространение видов.
Чаще всего виды сводятся в синонимы не па основании анализа матери-
ала или сопоставления и анализа описаний одного и того же вида не-
сколькими авторами, а на основании чужого мнения. Можно и должно
доверять мнению других авторов о статусе видов. Всего сам не посмот-
ришь, когда не хватает материала, для этого и пишутся книги, но нужно
хорошо знать морфологию большей части видов исследуемой группы.
К сожалению, этому не всегда следуют.
КЛАССИФИКАЦИЯ
311
Например, на каком основании Эллерман и Моррисон-Скотт (Ellerman,
Morrison-Scott, 1951) свели в синонимы Crocidura suaveolens Pall. — С. hyr-
сапа Goodw; в синонимы С. russula Herm. — С. giildenstaedti Pall.,
С. dsi-nezumi Ternm., C. pamirensis Ogn., C. rapax, C. vorax Allen и др.
К C. leucodon Herm. сведена в синонимы С. sibirica Dukelsky, к С. pergrisea
Miller, С. zarudnyi Ogn. В синонимы С. lasiura Dobs, сведен подвид С. leuco-
don lasia Thomas. С другой стороны, сохранены виды, требующие проверки.
Основанием для положения отдельных видов в «системе» названных
авторов послужило сведение А. П. Кузякиным (1944, с. 56, 57) всех земле-
роек-белозубок фауны СССР к четырем видам: С. suaveolens, С. russula,
С. leucodon и С. lasiura. С. russula объединяет С. giildenstaedti и С. pami-
rensis’, синонимом С. lasiura стала С. leucodon lasia. Это послужило для
Эллермана и Моррисон-Скотта основанием для расселения С. russula
через всю Евразию до Японии, а С. lasiura оказалась не только на Даль-
нем Востоке, но и на Кавказе и в Малой Азии, по мнению Харрисона
(Harrison, 1964), и в Аравии.
Ошибки названных авторов были восприняты как последнее слово
в систематике землероек Стоговым и Бондарем (1966), а также Рихтером
(Richter, 1970), который свел всех землероек в группы подвидов двух
слабо различающихся видов — С. russula и С. leucodon.
Классификация
В основу классификации землероек мною положено строение жеватель-
ного аппарата, что позволило установить известную преемственность между
современными и ископаемыми группами, поскольку в руках оказались
сравнимые и важные части их тела. Но строение одного жевательного
аппарата не может быть признано в качестве абсолютного критерия (как
•и любой другой орган) для оценки правильности выводов при системати-
ческих и филогенетических построениях. Только после того, как удалось
установить, что каждому типу жевательного аппарата соответствуют
определенные структурные особенности и в других частях тела, можно было
использовать его для целей систематики. При этом можно предполагать,
что и у ископаемых форм имели место адекватные коррелятивные связи.
Сравнительное изучение зубных систем и черепов, а также жевательной
мускулатуры, скелетов и других частей тела современных землероек
палеарктических родов и параллельно представителей других семейств
отряда Insectivora убедительно показало, что жевательные аппараты,
характерные для определенных крупных групп семейства, специфичны
также и для более дробных систематических подразделений включая
п виды, и коррелятивно связаны с другими особенностями строения.
Например, с определенным соотношением разных по величине промежу-
точных зубов у отдельных видов совпадают: специфичная структура
сочленовного отростка нижней челюсти, длина и форма углового отростка,
а также и форма нижней челюсти. С этими особенностями связано специ-
фическое расположение жевательных мышц, определяющее особенности
движения нижней челюсти и силу, и место ее наибольшего жевательного
давления.
При том, что виды отдельных родов отличаются специфическими
особенностями структур зубных рядов и частными приспособлениями
в характере движения нижней челюсти, для всех них характерно и общее
в строении жевательного аппарата, например, общее число верхних про-
межуточных зубов, относительное постоянство крупных по величине
первых трех промежуточных зубов, значительная длина углового отростка,
большая высота венечного отростка, длинная носовая часть черепа и др.
312
СЕМ. SORICIDAE
При резком различии частных особенностей отдельных родов легко уда-
ется установить общие признаки в их жевательном аппарате, характерные
для трибы, к которой они принадлежат. Например, у всех родов трибы
Soricini имеются три лопасти (бугорка) на режущем крае нижнего внутрен-
него резца, длинный угловой отросток, а массетер расположен под малым
углом к продольной оси черепа.
Различие в строении penis у самцов отдельных видов установлено
Строгановым (1957) и Виноградовым (1958). Но для диагностики строение
пениса имеет малое значение: определяются только самцы. Изменчивость
этого органа не изучена.
Зубные системы давно служат важнейшей основой классификации
млекопитающих, в чем нетрудно убедиться путем знакомства с системами,
предложенными многими авторамп, начиная с К. Линнея.
Известно, что способ захвата и обработки пищи, ее механический
состав имеют ведущее значение в эволюции класса. Это, однако, не озна-
чает, что при построении предложенной мной системы не учитывались
и другие морфологические особенности видов и их групп.
Поскольку, как мне представляется, филогения отражает преемствен-
ность в адаптивном процессе (основанную на родственных связях), а си-
стема — ее последовательность во времени, я старался отразить это
в предлагаемой мной системе.
Сем. Soricidae Ficher, von Waldheim, 1817, верхний эоцен—современный, Европа (не-
сомненно и Азия); нижний олигоцен—современный, Сев. Америка; нижний
миоцен—современный, Африка.
f I. Подсем. Saturniniinae Gureev, 1971, верхний эоцен, Европа.
f 1. Saturninia Stehlin, 1940, верхний эоцен, Европа.
II. Подсем. Soricinae Fischer von Waldheim, 1817, верхний эоцен—современный, Ев-
ропа (несомненно и Азия); нижний олигоцен—современный, Сев. Америка; плио-
цен—современный, Центр, и Южн. Америка; миоцен—современный, Африка.
f 1. Триба Oligosoricini Gureev, 1971, верхний эоцен—нижний миоцен,
Европа (а также и Азия); нижний олигоцен—нижний миоцен, Сев. Аме-
рика (вероятно, и Африка).
2. Amphisorex Filhol, 1884, верхний эоцен, Европа.
• 3. Oligosorex Kretzoi, 1959, олигоцен, Европа.
4. Gobiosorex Sulimski, 1970, нижний олигоцен, Азия.
5. Crocidosorex Lavocat, 1951, олигоцен, Европа.
6. Alluvisorex Hutchison, 1966, верхний миоцен, Сев. Америка.
7. Miosorex Kretzoi, 1959, миоцен, Европа.
8. Domnina Соре, 1873, олигоцен, Сев. Америка.
9. Paradomnina Hutchison, 1966, верхний миоцен, Сев. Америка.
- 10. Soricella Doben-Florin, 1964, средний миоцен, Европа.
11. Angustidens Repenning, 1967, нижний миоцен, Сев. Америка.
t 2. Триба Heterosoricini Viret et Zapfe, 1951, нижний миоцен, Европа (веро-
ятно, и Азия), Сев. Америка.
t 12. Heterosorex Gaillard, 1915, нижний миоцен, Европа.
f 13. Pseudotrimylus Gureev, 1971, миоцен, Сев. Америка.
t 14. Ingentisorex Hutchison, 1966, верхний миоцен, Сев. Америка.
3. Триба Soricini Fischer von Waldheim, 1817, нижний миоцен—современный,
Европа, Азия, Сев. Америка.
15. Sorex Linnaeus, 1758, нижний миоцен—современный, Европа,
Азия, Сев. Америка.
16. Microsorex Cones, 1877, плейстоцен—современный, Сев. Америка,
f 17. Tregosorex Hibbard et Jammot, 1971, нижний плиоцен, Сев.
Америка.
t 18. Anchiblarinella Hibbard et Jammot, 1971, нижний плиоцен, Сев.
Америка.
19. Blarinella Thomas, 1911, современный, Азия.
4. Триба Crocidurini Stirton, 1930, миоцен—современный, Азия, Африка;
миоцен—плиоцен, Сев. Америка; плиоцен—современный, Европа.
1. Подтриба Myosoricina Kretzoi, 1965, миоцен—современный, Африка;
миоцен, Сев. Америка.
t 20. Stirtonia gen. nov., миоцен, Сев. Америка.
КЛАССИФИКАЦИЯ
313
21. Myosorex Gray, 1837, миоцен—современный, Африка.
22. Surdisorex Thomas, 1906, современный, Африка.
2. Подтриба Crocidurina s. str. Stirton, 1930, миоцен—современный,
Азия и Африка; миоцен—плиоцен, Сев. Америка; плиоцен—совре-
менный, Европа.
f 23. Similisorex Stogov et Savinov, 1965, миоцен, Азия.
24. Feroculus Kelaart, 1852, современный, Азия (Шри Ланка).
25. Sylvisorex Thomas, 1904, современный, Африка.
26. Suncus Ehrenberg, 1833, плиоцен—современный, Европа, Азия,
Африка.
27. Scutisorex Thomas, 1913, современный, Африка.
f 28. Limnoecus Stirton, 1930, миоцен—плиоцен, Сев. Америка.
29. Paracrocidura Heim de Balsac, 1956, современный, Африка.
30. Solisorex Thomas, 1924, современный, Азия (Шри Ланка).
31. Praesorex Thomas, 1913, современный, Африка.
32. Crocidura Wagler, 1832, миоцен—современный, Азия и Африка;
плиоцен—современный, Европа.
33. Diplomesodon Brandt, 1852, плейстоцен, Африка; современный,
Азия.
5. Триба Blarinini Stirton, 1930, плиоцен—современный, Европа, Азия,
Сев., Центр, и Южн. Америка.
1. Подтриба Blarinina s. str. Stirton, 1930, плиоцен, Европа; плиоцен—
современный, Сев. Америка. ,
f 34. Adeloblarina Repenning, 1967, миоцен, Сев. Америка.
f 35. Blarinoides Sulimski, 1959, плиоцен, Европа.
36. Blarina Gray, 1838, плиоцен—современный, Сев. Америка.
2. Подтриба Beremendina Gureev, 1971, плиоцен, Европа; плиоцен-
плейстоцен, Азия; плиоцен—современный, Сев, Центр, и Южн.
Америка.
•37. Beremendia Kormos, 1930, плиоцен, Европа.
• 38. Petenyia Kormos, 1930, плиоцен, Европа, Азия, Сев. Америка.
• •39. Petenyiella Kretzoi, 1956, плиоцен, Европа.
• 40. Shikamainosorex Hasegawa, 1957, плейстоцен, Азия.
- •41. Peisorex Kowalski et Li-Chuan-Kuei, 1963, плейстоцен, Азия.
• 42. Hesperosorex Hibbard, 1957, плиоцен, Сев. Америка.
43. Paracryptotis Hibbard, 1957, плиоцен, Сев. Америка.
• 44. Planisorex Hibbard, 1972, нижний плейстоцен, Сев. Америка.
4 5. Cryptotis Pomel, 1848, плиоцен—современный, Сев., Центр, и
Южн. Америка.
3. Подтриба Soriculina Kretzoi, 1965, плиоцен—современный, Европа,
Азия, Сев. Америка.
t 46. Zelceina Sulimski, 1962, плиоцен, Европа.
f 47. Macroneomys Fejfar, 1966, нижний плейстоцен, Европа.
48. Neomys Каир, 1829, плиоцен—современный, Европа, Азия.
49. Soriculus Blyth, 1854, плиоцен, Европа; плиоцен—современный,
Азия.
50. Notiosorex Baird, 1877, плиоцен—современный, Азия и Сев.
Америка.
1. Подрод Notiosorex s. str. Baird, 1877, плиоцен—современный,
Сев. Америка.
2. Подрод Chodsigoa Kastschenko, 1905, плиоцен—современный,
Азия.
51. Podihik Deraniyagala, 1958, современный, Азия (Шри Ланка).
f Подтриба Allosoricina Gureev, 1971, плиоцен—плейстоцен, Европа.
| 52. Allosorex Fejfar, 1966, плиоцен—плейстоцен, Европа.
f 53. Deinsdorfia Heller, 1963, нижний плейстоцен, Европа.
5. Подтриба Nectogalina Gureev, 1971, современный, Азия.
54. Chimarrogale Anderson, 1877, современный, Азия.
f 55. Nectogalinia Gureev gen. nov., плиоцен, Азия.
56. Nectogale Milne-Edwards, 1870, современный, Азия.
6. Триба Anourosoricini Stirton, 1930, миоцен—современный, Азия; плиоцен,
Европа.
f 57. A nourosoricodon Topatshevski, 1965, плиоцен, Европа.
t 58. Amblycoptus Kormos, 1926, плиоцен, Европа.
-j- 59. Рaraanourosorex Rzeb.-Kow., 1975, плиоцен, Европа.
60. Anourosorex Milne-Edwards, 1870, миоцен—современный, Азия.
314
СЕМ. SORICIDAE
Таким образом, в отличие от предлагаемой Симпсоном (Simpson, 1945)
классификации землероек я сохраняю в системе не три подсемейства,
а только два, при этом одно из них новое, выделенное мной — подсем.
Saturniniinae.
Мне кажется более естественным выделение двух крупных групп
землероек: древних — кротовидных с большим числом примитивных
зубов — подсем. Saturniniinae и собственно землероек — со специализи-
рованными зубами и их системой — подсем. Soricinae. Выделение в особое
подсемейство белозубых землероек (Crocidurinae), а тем более представи-
телей рода Scutisorex Thomas (подсем. Scutisoricinae) с гиперостозом
позвонков, по моим представлениям, лишено достаточных оснований.
Специализация отдельных групп современных и близких к ним ископае-
мых землероек не зашла столь далеко, чтобы можно было говорить о прин-
ципиально иных в каждом случае или особых направлениях адаптивного
процесса. Роды с общим направлением развития выделены мной в трибы
или подтрибы.
Цвет зубов оказался малопригодным систематическим признаком.
Существуют два типа бурозубок и два типа белозубок. Можно думать,
что существуют две пары параллельных групп, но все группы развивались
своими особыми путями и появились в разное время: белозубые землеройки
родов Crocidura Wagl. и Suncus Ehrenb. более древние, а родов Nectogale
M.-Edw. или Chimarrogale Anders, более позднего происхождения, и
те и другие связаны в развитии с разными группами бурозубых землероек.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ, ТРИБ И ПОДТРИБ
1 (2). Общее количество зубов более 36. Жевательная поверхность
М1 и М1 2 3 4 5 6 7 8 без W-образно расположенных гребней, а с бугорками на
месте пара- и метакона (рис. 165, Б).........................
................. Подсем. Saturniniinae (с. 315).
2 (1). Общее количество зубов не более 36. Жевательная поверхность
М1 и М2 с развитыми в разной степени W-образно расположенными
гребнями, образованными краями складок, входящих с наружной
стороны их коронок в области пара- и метакона................
.................................Подсем. Soricinae (с. 316).
3 (4). Скуловые дуги или хотя бы их остатки в виде сильноразвитых
скуловых отростков верхнечелюстной и височной костей имеются.
На внутренней стороне венечного отростка птеригоидной ямки нет
................................... Триба Heterosoricini (с. 326).
4 (3). Скуловых дуг нет, как и их остатков. Птеригоидпая ямка на
внутренней стороне венечного отростка имеется.
5 (6). В нижней челюсти 4 промежуточных зуба, а если 3, то в верхней
челюсти 5 промежуточных зубов, или складки, входящие с наруж-
ной стороны коронки верхних моляров (М1 и М2) неглубокие . . .
.....................................Триба Oligosoricini (с. 318).
6 (5). В нижней челюсти 2 промежуточных зуба, а если 3, то в верхней
челюсти не более 4 промежуточных зубов. Складки, входящие с на-
ружной стороны коронки М1 и М2, глубокие, с хорошо развитыми
гребнями на боковых сторонах.
7 (12). Нижняя сочленовная фасетка на сочленовном отростке нижней
челюсти не выдвинута вперед, как и засочленовный отросток височ-
ной кости, на который она опирается.
8 (9). Все зубы с пигментированными вершинами................
.................................. Триба Soricini (с. 329).
КЛАССИФИКАЦИЯ
315
9 (8). Все зубы белые .................Триба Crocidurini (с. 371).
10 (И). В нижней челюсти 3 промежуточных, зуба..................
.................................Подтриба Myosoricina (с. 372).
11 (10). В нижней челюсти 2 промежуточных зуба..................
.................................Подтриба Crocidurina (с. 376).
12 (7). Нижняя сочленовная фасетка на сочленовном отростке нижней
челюсти выдвинута вперед, как и засочленовный отросток височной
кости, на который она опирается.
13 (14). В верхней челюсти 2 промежуточных зуба, а если 3, то М3 и М3
отсутствуют.........................Триба Anourosoricini (с. 460).
14 (13). В верхней челюсти не менее 3 промежуточных зубов; М3 и М3
всегда имеются.............................Триба Blarinini (с. 418).
15 (18). Все зубы пигментированы.
16 (17). В верхней челюсти 5 промежуточных зубов ...............
.................................Подтриба Blarinina (с. 420).
17 (16). В верхней челюсти 4 промежуточных зуба.................
...............................Подтриба Beremendina (с. 425).
18 (15). Все зубы белые или часть зубов пигментирована, а если пигмент
заметен на всех зубах, то кисти и ступни с длинными щетинками
по краям.
19 (20). Зубы белые и пигментированные. Вершины резцов, клыков и
премоляров пигментированы, а моляры белые, если и моляры с пиг-
ментом, то имеются плавательные щетинки, расположенные по краям
кистей и ступней......................Подтриба Soriculina (с. 437).
20 (19). Все зубы белые.
21 (22). Венечный и сочленовный отростки слабо обособлены . . . .
.................................Подтриба Allosoricina (с. 453).
22 (21). Венечный и сочленовный отростки хорошо обособлены . . .
.................................Подтриба Nectogalina (с. 455).
Примечание. Количество зубов указывается для половины верхней или
нижней челюсти.
t 1. Подсем. SATURNINIINAE Gureev, 1971
Т и и. Saturninia Stehlin, 1940, Западная Европа, верхний эоцен (рис. 165, Б).
Диагноз. Зубная формула (по комбинированным челюстям, но,
по-видимому, достоверна): 1-|-С-|-Р-^-М-|- = 44.
Пара- и метакон моляров — первого и второго — без складок, входя-
щих с наружной стороны, отчего на их жевательной поверхности нет
W-образно расположенных эмалевых гребней. Р4 характерной для Sorici-
dae формы, также и I1.
Описание. Общие структурные особенности коронок верхних
и нижних моляров сходны с таковыми Soricinae, но зубы сохраняют при-
митивные признаки, что выражаеся и в строении жевательной поверх-
ности: она бугорчатая, а не складчатая, как у более поздних вымерших
и современных землероек. Судя по числу альвеол, у Saturninia Stehl.
в верхней челюсти 6 промежуточных зубов. Второй зубец I1 слабо развит.
В нижней челюсти крупный и длинный 12. Форма нижней челюсти напоми-
нает эту кость кротов.
Подсем. Saturniniinae отличается от подсем. Soricinae количеством
зубов и их структурой. В состав подсемейства входит один извест-
ный монотипичный род.
Распространение и возраст. Западная Европа, верхний
эоцен.
316
ПОДСЕМ. SATURNINIINAE, SORICINAE
fl. Род SATURNINIA Stehlin, 1940
Stehlin, 1940, p. 301—306.
Тип. Saturninia gracilis Stehlin, 1940, Швейцария, верхний эоцен.
Диагноз совпадает с диагнозом подсемейства. Род включает один
вид, отмеченный как тип рода.
Длина зубного ряда Р4—М3=7.5 мм.
Примечание. Saturninia StehL, как пишет автор этого рода,
обладает многими признаками, близкими представителям Soricidae. Однако,
как он сам замечает, ему трудно назвать семейство насекомоядных, к кото-
рому следует отнести этот род. Рисунки, приведенные в работе Штелина,
сделаны с комбинированных челюстей, поскольку автор располагал только
мелкими обломками костей и отдельными зубами. Тем не менее нет никаких
оснований сомневаться в правильности представлений Штелина о строении
зубной системы Saturninia. Структурные особенности челюстей и особенно
зубов служат достаточным основанием для признания существования
в прошлом своеобразной группы насекомоядных, близких, с одной стороны,
к кротам, с другой — к землеройкам, особенно к последним.
Положение Saturninia в качестве рода в основании системы землероек
(Simpson, 1945) не может считаться правильным. Если нет достаточных
оснований считать его принадлежащим к особому семейству, то выделение
этого рода в особое подсемейство примитивных землероек вполне оправды-
вается его особенностями, отмеченными выше.
3. Подсем. SORICINAE Fischer von Waldheim, 1817
Soricidae Gray, 1821, London Med, Reposit., v. 20, p. 300. Soricini Fischer,
1817, Advers. Zool. Mem. Soc. Imp. Nat. Moskow, v. 5, p. 368—428. Подсем. Soricinae
Murray, 1866, The geographic distribution of mammals, v. 16, p. 231.
Тип. Sorex Linnaeus, 1758, Европа, Азия, Северная Америка, миоцен—совре-
менный.
Диагноз. Зубная формула I С—1=36—24. На же-
вательной поверхности коренных зубов (моляров) на месте пара- и ме-
такона имеется W-образный рисунок, образованный эмалевыми краями
складок, входящих с наружной стороны коронок верхних коренных, и
с внутренней — на нижних.
Описание. Жевательная поверхность верхних и нижних моляров
складчатая, а не бугорчатая, как у представителей подсем. Saturniniinae.
В верхней челюсти не более 5 промежуточных зубов. Второй зубец I1
сильно развит. В нижней челюсти крупный и длинный It. Форма нижней
челюсти типичная для землероек.
Подсемейство объединяет 4 современные и 2 вымершие трибы и большое
число современных и ископаемых родов, включающих более 300 видов.
Распространение и возраст. Европа, эоцен—совре-
менный; Азия, Африка, миоцен—современный (? эоцен—современный);
Северная Америка, олигоцен—современный, и крайний северо-запад
Южной Америки, верхний плейстоцен—современный.
Примечание. Подсемейство составляют внешне однообразные,
но весьма различные по морфофизиологическим особенностям насекомояд-
ные, что позволило им существовать в разнообразных условиях ланд-
шафта и климата. Однако основная масса видов землероек приспособлена
к жизни в условиях леса и зарослей кустарников. В лесу, в мертвой под-
стилке и приземном слое растительности эти млекопитающие постоянно
находят корм и достаточно надежные убежища. Все виды при всех условиях
активны в течение круглого года.
Рис. 165. Челюсти и зубы древних землероек Oligosorex antiqus (Л), Saturninia gra-
cilis (Б) (по: Stehlin, 1940), Ol. dehmi (В) (no: Viret, Zapfe, 1951).
318
ПОДСЕМ. SORICINAE
11. Триба OLIGOSORICINI Gureev, 1971
Тип. Oligosorex Kretzoi, 1959, Европа, олигоцен.
Диагноз. Зубная формула I-|-Су РМ-|-=: 36—34.
Верхних промежуточных зубов 5. В нижней челюсти 4 или 3 промежу-
точных зуба. Зубы пигментированные или белые. Верхняя и нижняя
сочленовные фасетки на сочленовном отростке нижней челюсти распо-
ложены одна под другой — нижняя не продвинута вперед (исключение —
род Angustidens Repenn.).
'Структурные особенности зубов и их общее количество (большее,
чем у других) хорошо отличают представителей этой трибы от видов
других триб.
Распространение и возраст. Известны из Европы
и Северной Америки, верхний эоцен?—олигоцен—нижний миоцен.
Примечание. Представители этой трибы отличались значитель-
ной для своего времени специализацией жевательного аппарата в направле-
нии, характерном и для более поздних и современных групп землероек.
Прежде всего следует отметить, что виды трибы Oligosoricini так же специа-
лизировались в быстром захвате подвижной добычи, как и современные
землеройки, но еще сохраняли ряд примитивных особенностей, унаследо-
ванных от более древних групп Soricidae. Этими примитивными особен-
ностями следует считать еще слабое углубление складок входящих с на-
ружной стороны коронок верхних коренных зубов в области пара- и
метакона, а также наличие 3—4 промежуточных зубов (а не 2) в нижней
челюсти. Можно думать, что вместе с этими архаическими чертами в строе-
нии зубной системы видов трибы Oligosoricini у них сохранялись и другие
древние особенности, не способствовавшие их прогрессу в более позднее
время.
Эта группа сыграла важную роль в эволюции землероек, поскольку
с нею связаны в развитии более поздние и специализированные группы.
Можно допустить, что вымирание землероек этой трибы обусловлено
относительно быстрым изменением и исчезновением ландшафтов и кормо-
вых животных в конце олигоцена, к которым они были приспособлены.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (4). В нижней челюсти 4 промежуточных зуба.
2 (3). В верхней челюсти 5 промежуточных зубов ... 3. Oligosorex.
3 (2). В верхней челюсти 4 промежуточных зуба .... 8. Domnina.
4 (1). В нижней челюсти 3 промежуточных зуба.
5 (6). Все три нижних промежуточных зуба крупные . . 10. Soricella.
6 (5). Из 3 нижних промежуточных зубов второй очень мелкий.
7 (8). Сочленовный отросток нижней челюсти массивный. Его нижняя
сочленовная фасетка продвинута вперед на внутренней стороне
ветви челюсти .................................11. Angustidens.
8 (7). Сочленовный отросток нижней челюсти не массивный. Его нижняя
сочленовная фасетка не продвинута вперед.
9 (10). Первый нижний резец с тремя бугорками на режущем крае
............................................... 6. Alluvisorex.
10 (9). Первый нижний резец с двумя бугорками на режущем крае . . .
................................................... 7. Miosorex.
Примечание. В таблицу не вошли роды Crocidosorex Lavocat, Amphisorex
Filhol, Gobiosorex Sulimski, Paradomnina Hutchison за отсутствием сравнимых при-
знаков.
2. РОД AMPHISOREX. 3. РОД OLIGOSOREX
319
f 2. Род AMPHISOREX Filhol, 1884
Filhol, 1884, Bull. Soc. Philomathiq., ser. 7, t. 8, N 2, p. 63, 64.
Тип. Amphisorex primaevus Filhol, 1884, Франция, верхний эоцен.
Диагноз (по: Filhol, 1884; Crochet, 1974). Предполагаемая формула
ряда зубов нижней челюсти: lyCyPyMy. Сочленовный отдел нижней
челюсти с массеторной впадиной. 1х с тремя бугорками на его верхней
стороне. Между 1Х и Мх 4 альвеолы. Это позволяет предполагать, что у пред-
ставителей описываемого рода было 3 промежуточных зуба в нижней
челюсти, если Р4 был с двумя корнями.
Описание. Приведено для вида.
Рис. 166. Amphisorex primaevus (по: Crochet, 1974).
Распространение и возраст. Европа, верхний эоцен.
t 1. Amphisorex primaevus Filhol, 1884 (рис. 166).
Filhol, 1884, там же.
Тип. Правая ветвь нижней челюсти с зубами — 1х и Мх и М2, № QU 8681 (по:
Filhol — de Lamondine Haute, dans le Tarnet Garonne, a no: Crochet—Lamandine-
Basse?), Франция, верхний эоцен.
Описание (по: Filhol, 1884). Зубная формула нормальная. Гх с тремя
лопастями на верхнем крае, крупнее, чем у современных видов. Моляры
образованы двумя сегментами, из которых передний выше заднего. Они
убывают в размерах от первого к последнему. Из описания, приведенного
для этого вида Кроше (Crochet, 1974), здесь помещены наиболее существен-
ные: нижняя челюсть массивная, с неглубокими массетерной и темпораль-
ной впадинами.
Измерения (по: Crochet, 1974). Длина: 1х — 3.4, серии альвеол
между 1Х и Мх —6.5, Мх —2.09, М2 — 1.92 мм.
Распространение и возраст. Франция, верхний эоцен.
Примечание. В 1974 г. Кроше свел род Amphisorex в синонимы
рода Domnina Соре. Для этого пока нет никаких оснований. В настоящей
работе оба рода остаются в прежнем систематическом положении.
f 3. Род OLIGOSOREX Kretzoi, 1959
Sorex Pomel, 1853, Cat. method, vertebr. foss. decouv. dans bassin hydrogr.
superieur et de Loire et d’Allier; Stehlin, 1940, p. 298—306. Oligosorex Kretzoi,
1959, p. 237—239; 1965, p. 123.
Тип. Sorex antiquus Pomel, 1853, Западная Европа, олигоцен.
Диагноз. Зубная формула: I С — Р ~ М 4 = 36. В нижней челюсти 4,
olio
а в верхней 5 промежуточных зубов. Структурные особенности нижней
челюсти близки к таковым видов рода Sorex L., но зубная часть ее ветвей
320
ПОДСЕМ. SORICINAE
относительно длиннее, что обусловлено большим числом промежуточных
зубов.
Распространение и возраст. Западная Европа, нижний
олигоцен—нижний миоцен.
t 1. Oligosorex antiquus Pomel, 1853 (рис. 165, A).
Sorex antiquus Pomel, 1853, там же, где и род.
Тип. Фрагмент нижней челюсти из Phryganidenkalk von Landy (Allier), Франция,
олигоцен.
Диагноз. Венечный отросток резко суживается к вершине, без
выступа, направленного назад. Нижняя сочленовная фасетка слабо
развита (мало отличается от верхней по величине). Сочленовный и угловой
отростки не массивные. Зубная часть нижней челюсти низкая.
Описание. Судя по рисунку, приведенному Штелиным (Stehlin,
1940), сочленовный отросток нижней челюсти значительно меньше, чем у
современных видов рода Sorex L. Подбородочное отверстие, как указывает
Штелин, расположено впереди Р4, а не под Мх, как у Sorex araneus L.
Нижний внутренний резец меньше, чем у современных видов, с двумя
зубцами. Форма и строение жевательных поверхностей нижних моляров
близки S. araneus L.
Распространение и возраст. Западная Европа, олиго-
цен.
Примечание. При всей еще значительной примитивности в струк-
туре зубов описываемый вид уже весьма близок к современным землерой-
кам. Известное сходство он имеет с видами современных родов Myosorex
Gray и Surdisorex Thos., с которыми его сближает большее, чем у других
групп, число промежуточных зубов в нижней челюсти: у Oligosorex anti-
quus Pom. их 4, у видов рода Myosorex Gray и Surdisorex Thos. — 3, а у всех
прочих — 2.
К роду Oligosorex Кретцой (Kretzoi, 1959, 1965) относит и вид, опи-
санный Вире и Запфе (Viret, Zapfe, 1951) как Sorex dehmi, а также
Oligosorex meyeri Kretz., 1959 (=pusillus Meyer, 1846).
t 2. Oligosorex dehmi Viret et Zapfe, 1951.
Sorex pusillus Zapfe, 1949, Anz. Osterr. Akad. Wiss. Math.-naturwiss., KI.,
S. 177; — dehmi Viret, Zapfe, 1951, Eclogae geol. helv., t. 44, N 2, p. 415—417.
T и п. He установлен, вид описан по остаткам из Нейдорфа (Чехословакия) и из
Франции.
Распространение и возраст. Западная Европа, ниж-
ний миоцен.
Примечание. Отличается от типичного вида рода более мас-
сивным венечным отростком, более длинными ветвями нижней челюсти
и крупными зубами.
|4. Род GOBIOSOREX Sulimski, 1970
Sulimski, 1970, р. 66—69.
Тип. Gobiosorex kingae Sulimski, 1970, Татал-Гол, северная часть Гоби, Монго-
лия, нижний олигоцен.
Диагноз (по: Sulimski, 1970, с сокращениями). Зубная формула
? ? ? ? 3
Ъэ л оз Г» а в обычном изображении: I^C^P^M^-. Можно думать,
Zr 1г Zr о Zr 1г Zr о
что общее количество зубов 36. 1Х (у Сулимского 12) крупный, с явствен-
ным цингулюмом. Между 1Х и Мх 5 альвеол. Это позволяет предполагать,
4. РОД GOBIOSOREX
321
что у представителей этого рода в нижней челюсти было 4 промежуточных
зуба. Мх, М2, М3 сходны по длине, но М3 более узкий.
Описание. Талонид (задний сегмент — метаконид, по-моему)
Мх шире, чем тригонид (передний сегмент). Наружный цингулюм этого
зуба развит сильно. Метаконид М3 короткий и узкий. Подбородочное
отверстие под передним корнем Мх. Ветвь нижней челюсти тонкая, удли-
ненная в передней части, ее наружная сторона снизу с выемкой или вы-
пуклая.
Рис. 167. Gobiosorex kingae (Л) (по: Sulimski, 1970), Domnina thompsoni (Б) (по: Simpson,
1941), D. gradata (Б) (no: Repenning, 1967).
Распространение и возраст. Азия, нижний олигоцен.
t 1. Gobiosorex kingae Sulimski, 1970 (рис. 167).
Sulimski, 1970, с. 66—67.
Тип. Фрагмент левой ветви нижней челюсти с 1х и Мх—М8, Z. Pal. N MgM —
111/27, нижний олигоцен, кол. Палеозоол. инет. Польск. АН, Варшава.
Примечание. Наличие пяти хорошо сохранившихся альвеол
между резцом и первым моляром на фрагменте нижней челюсти, послу-
жившем типом для описания нового рода и вида, позволяет предполагать,
что представители рода Gobiosorex имели в каждой половине нижней
челюсти 3 резца, клык и Р4. Вымершие землеройки с древним типом
зубной системы были известны только из Европы и Северной Америки.
Их существование в Азии в олигоцене—миоцене предполагалось, теперь
подтвердилось.
21 А. А. Гуреев
322
ПОДСЕМ. SORICINAE
Измерения (по: Sulimski, 1970). Длина: 12—М3 — 6.3; М,—М3 —
4.6, 4.7, Мх—М2,—3.5, 3.6, М2-М3 —2.7, 2.8, Мх — 2.0, 2,1, М2 -1.7, 1.8,
М3 — 1.3, 1.3; высота ветвей нижней челюсти под Мх — 1.7, 1.7 мм.
Распространение и возраст. Монголия, нижний оли-
гоцен.
Рис. 168. Miosorex grivensis (Л) (по: Viret, Zapfe, 1951), Soricella discrepans (Б) (no: Do-
ben-Florin, 1964).
|5. Род CROCIDOSOREX Lavocat, 1951
Sorex sp. Blainville, 1839, Ann. Anat., t. 3; araneus auvergne Blain-
ville, 1839? Osteograph. Insectiv., pl. 11. Crocidosorex Lavocat, 1951, Revis.
Faune Mammifer, Oligoc. Auvergne et du Velay, p. 23, 24.
Тип. Crocidosorex piveteaui Lovocat, 1951, Франция, олигоцен.
6. РОД ALLUVISOREX
323
Диагноз (по: Lavocat, 1951). Землеройка обладает зубами, лишен-
ными пигмента. Р4 простой, как у Crocidura Wagl.; талон М3 двухбугор-
чатый, как у Sorex L.
Распространение и возраст. Франция, олигоцен.
1. Crocidosorex piveteaui Lavocat, 1951.
Lavocat, 1951, там же, где и род.
Тип. Фрагмент левой ветви нижней челюсти с зубами Р4—М3, мергель Marcoin
вблизи Вольвик (Volvic), олигоцен, кол. Croizet Mus., Париж.
Диагноз. Тот же, что и рода, также и распространение.
Рис. 168 (продолжение). Angustidens vireti (В) (по: Wilson, 1960).
f 6. Род ALLUVISOREX Hutchison, 1966
Hutchison, 1966, Geol. Surv. Profess. Paper, N 565, p. 33, 34.
Тип. Alluvisorex arcadensis Hutchison, 1966, Орегон (США), верхний миоцен.
Описание (по:- Repenning, 1967, с сокращениями). Землеройка
с примитивным сочленевным отростком нижней челюсти, сравнимым
по контурам с Antesorex Repenn. {=Sorex L.), но с меньшей межартику-
лярной (межфасеточной) областью, разграничивающей верхнюю и ниж-
нюю сочленовные фасетки. Зубная формула нижней челюсти 1—3—3.
Щечные зубы крепкие и низкие. Нижняя челюсть высокая. 1Х с тремя
низкими бугорками на режущем крае. Р3 мелкий, когда имеется.
Род включает один вид, отмеченный как тип рода.
Распространение и возраст. Орегон (США), верхний
миоцен.
Примечание. К этому роду Репеннинг отнес и вид A. chasseae
Tedford, 1961 из нижнего плиоцена Невады (США), с двумя, а не тремя
промежуточными зубами в нижней челюсти, что, как мне кажется, неверно.
21*
324
ПОДСЕМ. SORICINAE
f 7. Род MIOSOREX Kretzoi, 1959
Sorex pusillus (race) grivensis D e p e г e t, 1892, Arch. Mus. Lyon, t. 5; — grivensis
Viret, Zapfe, 1951, Eclogae geol. helv., t. 44, N 2, p. 411, 415. Miosorex Kret-
zoi, 1959, p. 237—239; 1965, p. 123.
Тип. Sorex pusillus (race) grivensis Deperet, 1892, Франция, нижний миоцен.
Диагноз. В нижней челюсти 3 промежуточных зуба. 1Х с двумя
бугорками (лопастями) на верхнем крае, как и у представителей рода
Oligosorex Kretz. М1 и М2 с более развитыми, чем у них, складками, входя-
щими с наружной стороны.
Распространение и возраст. Европа, миоцен.
t 1. Miosorex grivensis Deperet, 1892 (рис. 168, Л).
Sorex pusillus (race) grivensis Deperet, 1892, Arch. Mus. Lyon, t. 5.
Тип. He отмечен, вид описан из Франции (Grive-Staint-Alban).
Диагноз совпадает с таковым рода.
Распространение и возраст. Известен из Франции,.
ГДР и ФРГ, миоцен.
|8. Род DOMNINA Соре, 1873
Соре, 1873, Paleontol: Bull., N 16, р. 1—8.
Тип. Domnina gradata Соре, 1873, Северная Америка, олигоцен.
Диагноз (по: Repenning, 1967, с сокращениями). Зубная формулам
3 1 2 3
I -5- С -т- Р -т- М = 34. В верхней и нижней челюсти по 4 промежу-
о 1 1 <5
точных зуба.
Описание (по: Repenning, 1967). Подбородочное отверстие под
серединой Мх. Массетерная ямка хорошо развита. Птеригоидная (темпо-
ральная) ямка неглубокая, не карманообразная. К этому нужно доба-
вить, что судя по рисунку нижней челюсти, приведенному Симпсоном
(Simpson, 1941), структурные особенности нижней челюсти представи-
телей этого рода близки таковым видов рода Oligosorex Kretz.
Описано 4 вида: Domnina gradata Соре, 1873 — тип рода; D. thomp-
soni Simps., 1941, Amer. Mus. Novitates, N 1150, описан из Монтаны:
D. compressa Galbr., 1953, Univ. Kansas Paleontol. Contrib., Vertebrate,,
art. 13, p. 46—48 описан из Колорадо; D, greeni Macdon., 1963, Bull.
Amer. Mus. Nat. Hist., v.125, art. 3 p. 168, описан из Южной Дакоты.
Примечание. Степень обособленности видов рода Domnina Соре
требует проверки на достаточном материале. Возможно, что многие из
них будут объединены.
Виды рода Domnina (кроме D. thompsoni) описаны по мелким облом-
кам нижних челюстей, главным образом по нескольким сохранившимся
зубам. В связи с этим относительное постоянство отмеченных для видон
признаков, как и степень обособленности, недостаточно ясно.
f 9. Род PARADOMNINA Hutchison, 1966
Hutchison, 1966, Oregon Univ. Mus. Nat. Hist. Bull., N 2, p. 23.
Тип. Paradomnina relictus Hutchison, 1966, Орегон (США), верхний миоцен.
Описание (по: Repenning, 1967, с сокращениями). В нижней
челюсти 5 промежуточных зубов с корнями, направленными назад.
Щечные зубы удлиненные, не луковицеобразные. Нижняя челюсть удли-
ненная.
Распространение и возраст. Орегон (США), верхний
миоцен.
10. РОД SORICELLA. 11. РОД ANGUSTIDENS
325
Примечание. Вряд ли в верхнем миоцене у землероек сохра-
нилось 5 промежуточных зубов в нижней челюсти. По-видимому, автор,
описавший этот род, ошибся.
f 10. Род SORICELLA Doben-Florin, 1964
Doben-Florin, 1964, Bayer. Acad. Wissenschaft. math.-naturwiss. KI.,
Abb., Neue-Folge, H. 117, S. 7—81.
Тип. Soricella discrepans Doben-Florin, 1964 (Sorex aff. neumarianus Schlosser,
1937), Европа, средний миоцен.
3 12 3
Диагноз. Зубная формула I-^-C-t-Pt-M-^-^:32. В верхней че-
Z 1 L О
люсти из четырех промежуточных зубов самый крупный первый, второй
меньше первого и третьего, четвертый меньше третьего. В нижней че-
люсти все 3 промежуточных зуба крупные. 1Х с двумя бугорками (лопа-
стями).
Описание, (по: Doben-Florin, 1964). Резцовый отдел в носовой
части черепа массивный и широкий, что хорошо заметно при взгляде
на череп с нижней или верхней стороны. Нижняя челюсть относительно
длинная, массивная в резцовой части, напоминает строением таковую
Saturninia Stehl. 1Х сравнительно короткий с круто загнутой вверх кон-
цевой частью. Род включает один вид, отмеченный как тип рода';
Распространение и возраст. Европа, средний миоцен
(возможно, и олигоцен).
Примечание. В структуре нижней челюсти представителей
этого рода еще сохранились архаичные особенности, свойственные земле-
ройкам эоценового времени. Можно предполагать, что их пищу составляли
относительно крупные, малоактивные животные. По-видимому, не только
в олигоцене, но и в миоцене уровень специализации отдельных групп
землероек был весьма различным. Наряду со специализированными,
появившимися уже в олигоцене, в миоцене продолжали существовать и
примитивные землеройки, как например из описываемого здесь рода
или рода Heterosorex Gaill. Представители последнего еще сохраняли
скуловую дугу.
f 11. Род ANGUSTIDENS Repenning, 1967
Sorex vireti Wilson, 1960, p. 33—35. Angustidens Repenning, 1967,
p. 25, 26.
Тип. Sorex vireti Wilson, 1960, Северная Америка, нижний миоцен.
3 13 3
Диагноз. Вероятная зубная формула I-^C-p Р-р Му = 34.
В нижней челюсти, судя по числу альвеол и сохранившимся зубам, 7 зу-
бов. 1Х с одним бугорком на верхнем крае, массивный. Нижняя челюсть
массивная. Сочленовный отросток крупный, его нижняя фасетка выдви-
нута вперед (сходство с представителями трибы Blarinini).
Представители рода стоят ближе к более специализированным груп-
пам миоценового времени. &
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
t 1. Angustidens vireti Wilson, 1960.
Sorex vireti Wilson, 1960, p. 33—35.
Тип. Правая ветвь нижней челюсти с зубами Мх—М3, № 10037, Колорадо (США),
нижний миоцен, кол. Муз. Канзас, унив., Лоренс.
326
ПОДСЕМ. SORICINAE
Диагноз (по: Wilson, 1960, с сокращениями). Подбородочное
отверстие под передней частью Мх. Зубное отверстие ниже средней линии
межтемпоральной впадины (странное название ложбины под венечным
отростком), которая относительно низкая и вытянута спереди назад.
Р4 однокорневой. Пятка М3 одновершинная с небольшим основанием.
Зубная формула, возможно, 2 3 •
Распространение в границах рода.
Примечание. Судя по рисункам и описанию особенностей
строения, этот вид никакого отношения к роду Sorex L. не имеет. По числу
зубов в нижней челюсти он стоит близко к представителям трибы OUgo-
soricini. Структурные особенности нижней челюсти, особенно ее сочленов-
ного отростка, позволяют выделить этот вид в особый род с чертами спе-
циализации в направлении, близком к трибе Blarinini.
f 2. Триба HETEROSORICINI Viret et Zapfe, 1951
Подсем. Heterosoricinae Viret, Zapfe, 1951, Eclogae geol. helv., t. 44, N 2,
p. 419-426.
Тип. Heterosorex Gaillard, 1915, Европа, Северная Америка (вероятно, и Азия),
миоцен.
3 113
Диагноз. Зубная формула Ij СР у М j = 28. В верхней че-
люсти 3 промежуточных зуба. Скуловая дуга сохранялась. Ямки на внут-
ренней стороне венечного отростка (птеригоидная ямка) нет.
Распространение и возраст. Европа, Северная Аме-
рика, нижний миоцен.
Примечание. По-видимому, основную пищу этих землероек
составляли животные с волокнистыми или упругими тканями, обработка
которых на зубах требовала значительных усилий. Это несомненно и
обусловило необходимость сохранения значительно развитого массе-
тера и соответственно места для его широкого прикрепления — скуловой
дуги. При этом височный мускул сохранял свое важное значение для
быстрого закрывания рта и сильного сжатия челюстей при схватывании
добычи. Его внутренняя порция прикреплялась на большей части внут-
ренней поверхности венечного отростка. У этих землероек, как и у пред-
ставителей триб Blarinini и Anourosoricini, нижняя фасетка на сочленов-
ном отростке нижней челюсти и засочленовный отросток височной кости,
на который она опирается, продвинуты вперед. Можно думать, что их
пищу составляли относительно крупные и сильные животные.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). На режущем крае 1х имеется хорошо заметный бугорок ....
............................................. 14. Ingentisorex.
2 (1). На режущем крае 1х бугорка нет.
3 (4). В верхней и нижней челюсти по 3 промежуточных зуба ....
......................................... 12. Heterosorex.
4 (3). В верхней и нижней челюсти по 4 промежуточных зуба . . .
........................................... 13. Pseudotrimylus.
112. Род HETEROSOREX Gaillard, 1965
Sorex sansaniensis Lartet, 1851, Notice sur la collie de Sansan. Trimylus schlos-
seri Roger, 1885, Palaont. Mitt., p. 106, Sorex (Crocidura) schlosseri Roger,
in: Schlosser, 1887—1888, schlosseri Roger, in: Filhol 1891, Ann. Sci. Geol., p. 21.
Heterosorex Gaillard, 1915, Ann. Soc. Linn., t. 62, p. 83—98.
13. РОД PSEUDOTRIMYLUS
327
Тип. Heterosorex delphinensis Gaillard, 1915 (=sansaniensis Lartet, 1851), Grive-
Saint-Alban, Франция, миоцен.
Диагноз. В верхней и нижней челюсти по 3 промежуточных
зуба.
Распространение и возраст. ^Франция, миоцен.
+ 1. Heterosorex sansaniensis Lartet, 1851 (рис. 169).
Sorex sansaniensis Lartet, 1851, Notice sur la colline de Sansan. Heterosorex
delphinensis Gaillard, 1915, Ann. Soc. Linn., t. 62, p. 83—98.
T и п не отмечен, вид описан из Франции, миоцен.
Описание. Отмеченные в диагнозе трибы морфологические осо-
бенности, характерные и для рода, заимствованы из описания этого вида.
К общей характеристике можно добавить следующее: череп массивный
и высокий, с относительно короткой лицевой частью. Сочленовная впадина
для сочленовного отростка нижней челюсти образована скуловым и за-
сочленовным отростками височной кости, глубокая. I1 с выступом на
внутренней стороне и слабо развитым задним зубцом. Судя по величине
альвеол, I2 крупнее остальных, второй крупнее третьего. Р4, М1 и Ма
без выемки на задней стороне.
Распространение и возраст. Франция, миоцен.
Примечание. К роду Heterosorex Gaill. относится и описанный
в 1887 г. Шлоссером (Schlosser) из Германии Н. nemayrianus.
Соотношения этих видов с Pseudotrimylus ropert Wils, недостаточно
ясны, хотя структурные особенности черепа последнего не вызывают
сомнения в его принадлежности к этой трибе.
|13. Род PSEUDOTRIMYLUS Gureev, 1971
Heterosorex Wilson 1960, p. 27—33. Trimylus Repenning, 1967, p. 13.
T и n. Heterosorex roperi Wilson, 1960, Колорадо (США), миоцен.
Диагноз. В верхней и нижней челюсти 4 промежуточных зуба.
Распространение и возраст. Северная Америка,
миоцен.
f 1. Pseudotrimylus roperi Wilson, 1960.
Heterosorex roperi Wilson, 1960, p. 27—33.
Тип. Левая ветвь нижпей челюсти с зубами Мх—Мя. № 10008, Колорадо (США),
нижний миоцен, кол. Муз. Канзас, унив., Лоренс.
Диагноз (по: Wilson, 1960, с сокращениями). Поперечные от-
ростки костного нёба лучше развиты, чем у Н. sansaniensis Lart. Верхние
моляры относительно шире, чем у Н. sansaniensis, с протоконом и гипоко-
ном более островершинными. Передний край восходящей ветви нижней
челюсти, возможно, более близок к вертикали, чем у Н. sansaniensis.
Описание (по: Wilson, 1960, с сокращениями). Череп этого
вида практически неизвестен; то, что сохранилось от него, вероятно,
соответствует таковому типа рода Н. sansaniensis ( в отношении обладания
полной скуловой дугой). Передняя часть скуловой дуги сохранилась
у всех экземпляров как хорошо развитые гребни, большие, чем у боль-
шинства существующих землероек. Нёбо частично сохранилось и имеет
хорошо развитый поперечный гребень, которым заканчивается сзади.
Этот гребень более развит, чем у Н. sansaniensis, и это не пример индиви-
дуальной изменчивости. Зубы не демонстрируют убедительно заметного
пигмента.
Рис. 169. Heterosorex sansaniensis (Л) (no: Gaillard, 1915), Pseudotrimylus roperl (Б)
(no: Wilson, 1960).
14. РОД INGENTISOREX
329
Зубная формула Р. roperi Wils, точно не известна; можно написать
3.0.3.3 2.1.3.3 „ -
ее так: ь „ или Нижняя челюсть характеризуется особен-
1.0.4.3 1.1.3.3 1 J
ностями, диагностичными для рода: положением подбородочного отвер-
стия под передним корнем М2 и оканчивающимся сзади значительным
углублением в боковой стороне челюсти; крупным резцом, корень кото-
рого располагается под М2; высоким венечным отростком, имеющим глу-
бокую массетерную ямку, но неглубокую межтемпоральную ямку на
внутренней стороне челюсти (птеригоидная ямка — у других авторов);
общей массивностью нижней челюсти. 14 более массивен, чем у европей-
ских видов.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
114. Род INGENTISOREX Hutchison, 1966
Hutchison, 1966, Oregon Univ. Mus. Nat. Hist. Bull., N 2, p. 23.
Тип Ingentisorex tumididens Hutchison, 1966, Орегон (США), верхний миоцен.
Описание (по: Repenning, 1967, с сокращениями). Землеройка
с луковицеобразными зубами, сходными с таковыми видов рода Hetero-
sorex Gaill. (Trimylus Roger), исключая отсутствующие верхние и ниж-
ние моляры. Р4 очень крупный, крупнее первого нижнего промежуточ-
ного зуба. 1Х не толстый в основании, с одним сильно выступающим бу-
горком на его режущем крае.
Род включает один известный вид, отмеченный как тип рода.
Распространение и возраст. Орегон (США), верхний
миоцен.
3. Триба SORICI NI Fischer von Waldheim, 1817
Fischer von Waldheim, 1817, p. 368—428. Soricinae Murray, 1866,
p. 231.
Тип Sorex Linnaeus, 1758, Европа, Азия, Северная Америка, миоцен—современный.
3 13 3
Диагноз. Зубная формула: 1-2-С-^-Р-рМ-д- = 32. Верхних про-
межуточных зубов пять. Все зубы пигментированы. Верхняя и нижняя
сочленовные фасетки на сочленовном отростке нижней челюсти находятся
на одном уровне: одна под другой. Нижняя фасетка, как и засочленов-
ный отросток височной кости, на который она опирается при сочленении
с черепом, не продвинуты вперед. Надглазничные отверстия имеются.
Описание. В морфологии представителей трибы Soricini должны
быть отмечены следующие особенности: малые размеры тела и относительно
короткие ноги. Морда узкая, удлиненная. Уши всегда спрятаны в во-
лосяном покрове и выступает только их верхний край, покрытый волосами.
Глаза мелкие, скрытые в шерсти, но хорошо заметные. Волосяной покров
густой, мягкий бархатистый. Волосы, покрывающие кисти и ступни сверху
и отчасти снизу, жесткие, плоские, часто с бороздкой вдоль средней
части. У видов, обитающих вблизи болот и рек и способных плавать,
волосы на боковых сторонах кистей и ступней, а также пальцев дости-
гают значительной длины. Нижняя сторона кистей и ступней покрыта
крупными и мелкими щетинками и чешуйками, а в опорной части — круп-
ными мозолями. Хвост покрыт короткими жесткими волосами.
Распространение и возраст. Европа, Азия, Северная
Америка, миоцен—современный.
Эта группа землероек сравнительно рано (миоцен) обособилась от
общего ствола Soricidae. Среди других групп землероек, доживших до
современного периода, виды этой трибы сохраняют наиболее архаичные
330
ПОДСЕМ. SORICINAE
особенности, не помешавшие им, однако, занять обширную территорию
в Евразии и Северной Америке.
По-видимому, с самого начала представители Soricini приспосабли-
вались к существованию в умеренном климате. Основное их местооби-
тание — леса и заросли кустарников, значительно реже они встречаются
вдали от больших массивов лесов или кустарниковой растительности,
в полосе ивняков и травянистых растений на открытых местах или
в степной зоне, куда проникают по водным системам. В горных ландшаф-
тах они оказались, видимо, в результате расселения по горным хребтам,
а также в результате подъемов земной коры.
Обитание отдельных видов на небольших площадях, изолированных
от основного ареала трибы, может рассматриваться как сохранение ре-
ликтовых участков некогда большого ареала трибы.
Приспособление видов шло по линии освоения кормов в виде разно-
образных беспозвоночных, особенно насекомых, добываемых в призем-
ном слое растительности, в лесной подстилке и различных укромных
местах на земле, а также в норах мелких животных. Сохранение большого
числа промежуточных зубов обусловлено у них питанием преимущественно
мелкими подвижными животными.
Основным способом передвижения в пределах этой трибы служит
бег. Последний не отличается ни особой скоростью, ни разнообразием,
поскольку совершается в сходных для большинства видов условиях:
приземная растительность, мертвая подстилка леса, моховой покров и
другие субстраты и низкие укрытия. Только отдельные виды приспособ-
лены и к другим формам движения, таким как плавание (например,
Sorex palustris Rich.) или рытье (S. unguiculatus Dobs.). Однако приспо-
собления к последним носят характер начальных стадий их формирования.
В связи с тем что бег служит основным способом передвижения, он
сохранил ведущее значение в эволюции этой группы землероек, и при-
способление к нему нивелирует приспособления к другим локомоциям.
В скелете видов этой трибы отражается приспособление к выпол-
нению разнообразных функций, т. е. известная универсальность локо-
моторного аппарата. Ни передний, ни задний пояса конечностей в своем
строении не обнаруживают специальных приспособлений к рытью или
быстрому бегу прыжками, а также каким-либо другим формам движения.
Соотношения длины отдельных частей конечностей соответствуют при-
способлению к передвижению шагом, рысью и тяжелым галопом — ши-
роко распространенным формам движений неспециализированных локо-
моторных систем. Передними конечностями при некотором удлинении
когтей животные могут рыть, а при развитии длинных щетинок по краям
ступней, кистей и пальцев — грести в воде. В жевательном аппарате
кроме перечисленных уже особенностей имеются и следующие характер-
ные черты. Угловой отросток длинный; в связи с этим массетер имеет бо-
лее наклонное положений, чем у землероек других триб. Это позволяет
при его сокращении ускорять вращение нижней челюсти при закрывании
рта височным мускулом.
Сохранение длинной морды (соответственно и челюстей) обусловлено
необходимостью быстро схватывать насекомых как пинцетом.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СОВРЕМЕННЫХ РОДОВ
1 (2). Первые три верхних промежуточных зуба крупные, а если третий
меньше четвертого, то он не дисковидной формы, как у Microsorex
• Coues ............................................... . 15. Sorex.
2 (1). Крупные только два верхних промежуточных зуба.
15. РОД SOREX
331
3 (4). Три задних верхних промежуточных зуба мелкие................
.............................................. 19. Blarinella.
4 (3). Из трех задних верхних промежуточных зубов мелкие третий
и пятый ...........................................16. Microsorex.
15. Род SOREX Linnaeus, 1759 — ЗЕМЛЕРОЙКИ БУРОЗУБКИ
Linnaeus, 1758, р. 53. Mus araneus Brisson, 1762, Regnum Animate,
p. 126. Oxirin Каир, 1829, Entwickl. Gesch. und Natiirl. Sistem. europ. Thierwelt,
Bd 1, S. 119. Amphisorex Duvernoy, 1835, Mem. Soc. Mus. Hist. Nat. Strasb.,
t. 2, p. 23. Hydro-Sorex Duvernoy, 1835, там же, с. 24. Corsira Gray, 1837—
1838, p. 123. Hydrogale Pomel, 1848, Bill. Univ. Geneva, Arch. Sci. phys. nat., t. 9,
p. 248. Otisorex De Kay, 1842, Zoology, N. Y., pt 1, Mammalia, p. 22. Neosorex
Baird, 1858, Mammals, in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1, p. 11. Homa-
lurus Schultze, 1890, Schriften Nat. Vereins Harzes, in: Wernigerode, Ed. 5,
S. 28. Soricidus Al tobello, 1927, Rev. France Mammif., t. 1, p. 6.
По-видимому, в синонимы этого рода должен быть отнесен и род Antesorex R е-
penning, 1967, р. 1—74.
Тип. Sorex araneus Linnaeus, 1758, Европа, Азия, плиоцен—современный.
Диагноз. Первые три верхних промежуточных зуба крупные,
а если третий меньше четвертого, то он не дисковидный, как у Microso-
rex Cones.
Описание. Уши мало выступают из волосяного покрова. Хвост
разной длины, но не короче половины длины тела, покрыт короткими
.жесткими волосами. Волосяной покров тела мягкий, бархатистый.
Окраска верхней части тела коричневая, бурая или почти черная. У So-
rex cylindricauda M.-Edw. вдоль средней части спины проходит темная
полоса. Окраска нижней стороны тела белая, серая или бурая, сходная
с окраской спины.
Череп с широкой, угловатой, округлой или овальной формы мозговой
капсулой. Лицевая часть черепа сильно суживается к переднему носовому
отверстию, и только боковые стороны верхнечелюстных костей расширены,
а их нижние края несколько уплощены в дорсо-вентральном направлении.
На лобных костях в межглазничном пространстве имеются два надглаз-
ничных отверстия.
Скелет несет черты приспособления к бегу в различных условиях. Отно-
сительно тонкие и длинные кости передних и задних конечностей сооответ-
ствуют быстрому передвижению разными аллюрами по поверхности почвы.
Относительно короткие кисти и ступни несут черты приспособления
к передвижению в различных низких укрытиях и по стволам деревьев,
а также рытью. Разнообразие в выборе мест обитания у землероек этого
рода обусловило разные приспособления скелета, в частности конечностей.
При этом каждый вид имеет свои особенности в строении скелета.
Представители рода известны с нижнего миоцена в Европе, видимо,
с этого же времени в Азии и Северной Америке.
Специализация видов рода шла по пути приспособления к жизни
в условиях главным образом лесных ландшафтов с различными почвен-
ными условиями в разных климатических поясах и к питанию мелкими
беспозвоночными животными, преимущественно насекомыми (отдельные
виды в период бескормицы питаются и семенами растений). Некоторые
виды приспособлены к плаванию.
Распространение и возраст. Европа, Азия до северо-
западного Памира, Тибета и Центрального Китая, Северная Америка;
нижний миоцен—современный.
Примечание. В пределах рода Sorex L. американские авторы
выделяют два подрода: Otisorex De Kay, объединяющий виды, у боль-
шинства из которых нет заднемандибулярного отверстия, а пигмент на
332
ПОДСЕМ. SORICINAE
зубах имеется только на их вершинах, и Sorex L., виды которого имеют
' заднемандибулярное отверстие, а пигмент на зубах занимает всю поверх-
ность до цингулюма. Автором настоящей работы эти подроды не призна-
ются.
В 1941 г. Кретцой (Kretzoi) в пределах рода Sorex L. выделил под-
род Drepanosorex, поставив около этого названия вопросительный знак.
Типом подрода он поставил новый вид X. tasnadii и привел рисунок,
из которого видно, что новый вид, а следовательно, и подрод отличаются
только размерами. В дальнейшем (Kretzoi, 1965) подрод Drepanosorex
возведен Кретцоем в ранг рода. При этом типом его оставлен вид S. tas-
nadii, хотя, как он сам пишет: «Genoholotypus (Drepanosorex) tasnadii
Kretzoi, 1941 (=Sorex savini austriacus Kormos, 1937 =? Sorex savini
Hinton, 1911», он должен считаться синонимом S. savini. В настоящей
работе этот род не сохраняется за отсутствием принципиальных отличий
от рода Sorex L.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЗЕМЛЕРОЕК
РОДА SOREX L. ФАУНЫ СССР
1 (4). Длина хвоста не менее 55 мм.
2 (3). Кондило-базальная длина черепа более 22 мм...............
..................................... 27. S. mirabilis Ognev.
3 (2). Кондило-базальная длина черепа менее 22 мм...............
......................................26. S. alpinus Schinz.
4 (1). Длина хвоста менее 55 мм.
5 (14). Второй верхний промежуточный зуб выше первого и третьего.
6 (13). Третий верхний промежуточный зуб выше четвертого, верхние
промежуточные зубы без выемки на задней стороне.
7 (10). Череп с узкой и удлиненной лицевой частью.
8 (9). Первые три промежуточных зуба очень крупные, с округлой вер-
шиной. Второй промежуточный зуб равен или крупнее первого
зубца I1, измеренного от выемки до его вершины. Окраска тела
сверху светло-бурая, снизу беловато-серая......................
........................................ 22. S. asper Thom.
9 (8). Первые три промежуточных зуба не очень крупные, островер-
шинные. Второй промежуточный зуб мельче переднего зубца I1.
Окраска тела темная сверху и снизу (бурая или черно-бурая) . . .
.......................................... 25. S. raddei Satunin.
10 (7). Череп со сравнительно короткой лицевой частью.
11 (12). Верхние промежуточные зубы мало суживаются к притуплен-
ным вершинам. Жевательные поверхности коренных зубов окра-
шены. Лапы и хвост темно-коричневые ............................
...................................23. S. daphaenodon Thom.
12 (11). Верхние промежуточные зубы сильно суживаются к заостренным
вершинам. Жевательные поверхности коренных зубов не окрашены.
Хвост двуцветный, лапы беловатые .... 24. S. arcticus Kerr.
13 (6). Третий верхний промежуточный зуб ниже четвертого или равен
по высоте. Верхние промежуточные зубы с выемкой на задней сто-
роне. Второй верхний промежуточный зуб выше первого, пятый
очень мелкий, смещен на внутреннюю сторону зубного ряда . . .
............................................ 28. S. vir G1. АП.
14 (5). Второй верхний промежуточный зуб не выше первого и третьего.
15 (22). Второй верхний промежуточный зуб по величине (или только
по высоте) равен первому и крупнее третьего, или первый промежу-
точный зуб крупнее второго и третьего.
15. РОД SOREX
333
16 (17). Первый верхний промежуточный зуб равен или почти равен
второму, третий крупнее четвертого ............21 .<S. araneus L.
17 (16). Первый верхний промежуточный зуб крупнее остальных, третий
и четвертый равны или почти равны по величине.
18 (19). Кондило-базальная длина черепа не менее 19 мм. Череп массив-
ный, мозговая капсула на боковых сторонах имеет хорошо заметные
выступы. Длина когтей более 3 мм...............................
...................................30. S. unguiculatus Dobs.
19 (18). Кондило-базальная длина черепа менее 19 мм. Череп не мас-
сивный, мозговая капсула без выступов. Длина когтей менее 3 мм.
20 (21). Кондило-базальная длина черепа больше 16 мм. Мозговая капсула
округлой формы, выпуклая. Лицевая часть черепа суженная к но-
совой части и удлиненная .................31. S. caecutiens Laxm.
21 (20). Кондило-базальная длина черепа меньше 16 мм. Мозговая кап-
сула овальной формы, плоская. Лицевая часть черепа не резко
суженная, не удлиненная.................34. S. minutissimus Zimm.
22 (15). Второй верхний промежуточный зуб ниже и мельче первого
и третьего, все три равны по высоте или размерные соотношения
зубов иные.
23 (24). Второй верхний промежуточный зуб ниже и мельче первого и
третьего, зубы узкие, клиновидные .............................
...................................28. S. bucharensis Ognev.
24 (23). Второй верхний промежуточный зуб ниже или равен по высоте
первому и третьему, зубы не узкие, не клиновидные, а в виде равно-
сторонних треугольников........................33. S. minutus L.
25 (26). Второй верхний промежуточный зуб равен по высоте третьему
и четвертому, пятый меньше четвертого, а первый крупнее всех
........................................ . 35. S. iso don Turov.
26 (25). Второй верхний промежуточный зуб меньше первого и четвер-
того, пятый равен четвертому или крупнее его. Все зубы с глубокой
выемкой на задней стороне...............32. S. gracillimus Thom.
Примечание. В таблицу не включена A. cinereus Kerr (37), сходная с другими
мелкими видами рода. Отличается выпуклыми наружными сторонами зубов. Первый
и второй, а также третий и четвертый верхние промежуточные зубы образуют сходные
пары. При этом второй крупнее первого, а четвертый выше третьего; длина основания
второго более длины первого, а третьего более четвертого.
t 1. Sorex gracilidens Viret et Zapfe, 1951.
Viret, Zapfe, 1951, p. 417, 418.
Тип. Фрагмент левой ветви нижней челюсти с зубами Р4—М3, Нейдорф, Чехо-
словакия, миоцен, кол. Г. Запфе (Н. Zapfe).
Распространение и возраст. Европа, миоцен.
t 2. Sorex (?) compressus Wilson, 1960.
Wilson, 1960, p. 35—38.
Тип. Фрагмент левой ветви нижней челюсти с зубами Р4—М2, № 10050, Колорадо
(США), нижний миоцен, кол. муз. Канзас, унив., Лоренс.
Диагноз (по: Wilson, 1960). Подбородочное отверстие под Р4
и передней частью Мх. Зубное отверстие под средней линией межтемпо-
ральной ямки, которая расположена относительно высоко, и короткая
•спереди назад. Эмаль окрашена? (автор, видимо, не уверен в пигмента-
ции зубов). Зубная формула неизвестна (если автор считает описываемый
вид принадлежащим к роду Sorex L., то зубная формула его известна).
Р4 с двумя корнями, коронка его сжата. Метаконид Мх малозаметен,
334
ПОДСЕМ. SORICINAE
будучи прижат к протокониду. Вершина Р4 приблизительно на одном
уровне с параконидом Мх.
Распространение и возраст. Северная Америка, ниж-
ний миоцен.
t 3. Sorex falax Heller, 1936.
Heller, 1936, Sonder-Abdruck Neuen Jahrb. Mineral, et. Beil., Bd 76, Abt. B,
S. 109, 110.
T и п не отмечен, вид описан из Рейнгисей, Германия, верхний плиоцен.
Описание. Судя по фотографии, I1-1 у этого вида широко рас-
ставлены; Р4 длиннее М1; М1 и М2 почти четырехугольной формы со слабой
выемкой на задней стороне коронки, также и Р4. Складки, входящие
с наружной стороны коронки М1 и М2, неглубокие, не доходят до середины
жевательной поверхности зуба.
Распространение и возраст. Европа, верхний плиоцен.
f 4. Sorex praealpinus Heller, 1930.
( Heller, 1930, там же, т. 63, с. 260.
Тип не отмечен, описан из Германии, плиоцен.
Распространение и возраст. Европа, плиоцен.
t 5. Sorex araneoides Heller, 1930.
Heller, 1930, там же, с. 260, 261.
Тип не отмечен, вид описан из Германии, плиоцен.
Распространение и возраст. Европа, плиоцен.
t 6. Sorex subaraneus Heller, 1958.
Heller, 1958, Neues Jahrb. Geol. PalaontoL, Bd 107, N 1, S. 15—18.
T и п не отмечен, вид описан из ФРГ, нижний плейстоцен.
Распространение и возраст. Европа, нижний плей-
стоцен.
t 7. Sorex pachyodon Pasa, 1949.
P a s a, 1949, I Mammif. di Alcune Antiche Brecce Veronesi, p. 8—11.
Тип не отмечен, вид описан из Италии.
Распространение и возраст. Европа, плиоцен.
t 8. Sorex runtonensis Hinton, 1911.
Hinton, 1911, Geol. Mag. N. S., v. 8, p. 532.
Тип не отмечен за отсутствием работы с оригинальным описанием и соответ-
ствующих ссылок других авторов. Вид описан из плиоцена Англии.
Описание, (по: Sulimski, 1959, как более обстоятельному, чем
У Других авторов). 1Х с тремя лопастями. Из них две задние более слабо
выражены. Р4 с двумя вершинами и гипоконом не столь хорошо развитым,
как у современных Soricidae. М3 с пятью вершинами. Венечный отросток
тонкий, в основании широкий. Птеригоидная ямка на венечном отростке
нижней челюсти высокая и широкая, треугольной формы. Подбородоч-
ное отверстие у переднего края Мх, или под Р4. Вершины зубов лишены
пигмента (депигментированы при фоссилизации) или окрашены.
Измерения, (по: Sulimski, 1959). Длина: общая нижней челюсти
11.2—12.4, основная нижней челюсти — 8.6—9.1, нижнего зубного ряда
(1Х—М3) — 6.9—7.3; высота венечного отростка — 3.8—4.1 мм (измере-
ния сделаны на молодых экземплярах).
15. РОД SOREX
335
Распространение и возраст. Западная Европа, плиоцен.
Примечание. По мнению Сулимского (Sulimski, 1959), распо-
лагавшего серией в 25 экз. нижних челюстей этого вида, венечный от-
росток S. runtonensis Hint, сходен с этим отростком «S', caecutiens Laxm,
а по другим признакам — близок S. araneus L.
f 9. Sorex praearaneus Kormos, 1934.
Kormos, 1934, p. 3, 4.
Тип. Фрагмент черепа № 08 (3681), Kgl. Ung. Geol. Anst., Виллань (Венгрия),
плиоцен.
Диагноз (по: Kormos, 1934). Размеры черепа относительно круп-
ные. Четвертый верхний промежуточный зуб меньше пятого. Подбородоч-
ное отверстие под коронкой Р4.
Измерения. Основная длина нижней челюсти — 9.5; ее высота
вместе с высотой венечного отростка — 4.4; высота сочленовного отростка —
2.4—2.5; длина нижнего зубного ряда (1Х—М3) — 7.4—7.6 мм.
Распространение и возраст. Европа, плиоцен.
f Ю. Sorex kretzoi Sulimski, 1962.
Sulimski, 1962, p. 469—472.
Тип. Фрагмент черепа с сохранившейся ростральной частью и правой ветвью
нижней челюсти. У нижней челюсти отсутствует угловой отросток и поврежден венеч-
ный отросток, NMZ VIII (Vm-312) 1, Венже, Польша, плиоцен.
Диагноз (здесь и далее по: Sulimski, 1962). 1Х длинный, со сла-
бым цингулюмом. Р4 со слабо развитыми двумя вершинами. Подбородоч-
ное отверстие под Мх, в углублении. Венечный отросток высокий, широкий,
лопатовидный.
Описание. 1Х крупный. I2 и I3 одинаковой величины. С и Р1
вдвое меньше предыдущих зубов. Р2 очень мелкий. Носовая часть отно-
сительно короткая. Слезное отверстие над задним корнем М2. Массетер-
ный гребень с шипом. Птеригоидная ямка на венечном отростке треуголь-
ная, глубокая.
Измерения. Длина: верхнего зубного ряда — 8.5, М1—М3 —
3.8—4.0; ширина: костного нёба впереди — 2.3, сзади — 5.3, межглазнич-
ная черепа — 4.6. Длина: общая нижней челюсти — 12.2—12.5, внутрен-
няя I — 4.2—4.3, нижнего зубного ряда (1х—М3) —8.0—8.2; высота ве-
нечного отростка — 5.0—5.4 мм.
Распространение и возраст. Европа, плиоцен.
f 11. Sorex subminutus Sulimski, 1962.
Sulimski, 1962, p. 461—465.
Тип. Фрагмент черепа с сохранившейся ростральной частью, верхним первым
резцом и М1—М3, NMZ VIII (Vm-309) 1, Венже, Польша, плиоцен.
Диагноз. 1х короткий, толстый, с тремя отчетливыми лопастями.
13 короткий, высокий, одновершинный. Р4 короткий, высокий, двувер-
шинный. Подбородочное отверстие под Мх.
Описание. Ростральная часть черепа узкая. Подглазничные от-
верстия между Р4 и М1. Верхний первый резец без бугорка. Р4 сильно
моляризован. Венечный отросток высокий, стройный, заостренный. Пте-
ригоидная ямка на венечном отростке глубокая, треугольная.
Измерения. Длина: верхнего зубного ряда (I1—М3) — 5.3—5.5,
костного нёба — 5.0, общая пижней челюсти — 8.0, ее основная — 6.4—
336
ПОДСЕМ. SORICINAE
6.6, нижнего зубного ряда (Ц—М3) — 5.4—5.6, М4—М3 — 3.0; высота
венечного отростка — 2.5—2.8; ширина костного нёба между М1-1 —
3.0 мм.
Распространение и возраст. Европа, плиоцен.
t 12. Sorex hibbardi Sulimski, 1962.
Sulimski, 1962, p. 467—469.
Тип. Левая ветвь нижней челюсти с полным рядом зубов, без углового отростка,
NMZ (Vm-311) 1, Венже, Польша, плиоцен.
Диагноз. 1г длинный, с тремя лопастями. Из них передняя слабо
развита, в передней части не обособлена от гребня вершины этого зуба.
Средняя лопасть крупная, высокая, задняя мелкая. 12 мелкий, плоский.
Р4 широкий, двувершинный. М3 мал. Подбородочное отверстие под М4
в углублении.
Описание. В дополнение к диагнозу нужно отметить следующие
особенности в строении нижней челюсти. Массетерный гребень с шипом.
Венечный отросток низкий, отклоненный кнаружи и назад. Птеригоид-
ная ямка этого отростка небольшая, трапециевидная, глубокая. Угло-
вой отросток толстый у основания.
Измерения. Общая длина нижней челюсти — 12.0—12.5 мм.
Распространение и возраст. Европа, плиоцен.
t 13. Sorex cudahyensis Hibbard, 1944.
Hibbard, 1944, Bull. Geol. Soc. Amer., v. 55, p. 720, 721.
Тип. Фрагмент правой ветви нижней челюсти с зубами (Р4—М„), № 6513, Канзас
(США), плейстоцен, кол. Муз. Канзас, унив., Лоренс.
Диагноз (по: Hibbard, 1944). По размерам приблизительно схо-
ден с Sorex merriami Dobs. Отличается от ископаемых и современных
землероек степенью развития Р4. Р4 массивный и недостаточно опущен
на лингвальной стороне; мелкий передний бугорок (вершина) на фронталь-
ной основной вершине. Цингулюм вдоль лабиального края Р4 широкий
и высокий.
Распространение и возраст. Канзас (США), плейс-
тоцен.
t 14. Sorex lacustris Hibbard, 1944.
Hibbard, 1944, там же, с. 721, 722.
Тип. Фрагмент правой ветви нижней челюсти с М4—М3, № 6630, Канзас (США),
плейстоцен, кол. Муз. Канзас, унив., Лоренс.
Диагноз (по: Hibbard, 1944). По размерам сходен с Neosorex
bendirii dlbiventer Merr. Характер зубов и ветвей нижней челюсти как
у рода Neosorex Baird. Наибольший поперечный диаметр М4, М2 и М3
пропорционально значительно менее передне-заднего диаметра моляров,
чем у современных представителей рода Neosorex Baird.
Примечание. Хиббард (Hibbard, 1944) сравнивает этот вид
с представителями так называемых длиннохвостых землероек, при-
способленных к плаванию, — S. bendirii, S. palustris Richards, и др.,
которых Байрд (Baird) в 1858 г. отнес к особому роду Neosorex, а Мерриам
(Merriam) в 1895 г. — к подроду. В настоящее время эти виды считаются
принадлежащими к роду Sorex L.
Распространение и возраст. Канзас (США), плейс-
тоцен.
15. РОД SOBEX
337
f 15. Sorex taylori Hibbard, 1938.
Hibbard, 1938, Trans. Kansas Acad. Sci., v. 40, p. 242, 243.
Тип. Фрагмент правой ветви нижней челюсти с зубами М2—М3, № 3906, Канзас
(США), Rexroad Fauna, верхний плиоцен, кол. Муз. Канзас, унив., Лоренс.
Диагноз (по: Hibbard, 1938). Протокопид подавляет степенью
развития и занимаемой площадью на коронке зуба остальные бугорки.
Метаконид на Мп М2 и М3 крупнее, чем у современных видов. На моля-
рах развиты все пять вершин. Добавочные бугорки отсутствуют. Тонкие
цингулюмы развиты. Восходящая ветвь хорошо развита. Коронарный
отросток высокий, узкий, отходит от горизонтальной ветви под прямым
углом. М3 стоит ближе к восходящей ветви, чем у современных видов.
Распространение и возраст. Канзас (США), верхний
плиоцен.
f 16. Sorex rexroadensis Hibbard, 1953.
Hibbard, 1953, J. Paleontol., v. 27, N 1, p. 23, 24.
Тип. Задний отдел левой ветви нижней челюсти с Мх—М3, № 28147, Канзас
(США), верхний плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн Арбор.
Диагноз (по: Hibbard, 1953). Мелкая землеройка, по размерам
близкая к 5. preblei Jacks., с относительно высокой ветвью нижней
челюсти. Талонид (метаконид) Мг и М2 небольшой сравнительно с триго-
нидом (передним сегментом зуба), как у современных видов. Энтоконид
относительно более явственно обособлен от метаконида у Мх и М2, чем
у современных форм Sorex L.
Распространение и возраст. Канзас (США), верхний
плиоцен.
f 17. Sorex hagermanensis Hibbard et Bjork, 1971.
Hibbard, Bjork, 1971, p. 171, 172.
T и п. Фрагмент левой ветви нижней челюсти с зубами Мх—М3 и сохранившимися
отростками сочленовной части, V 56100, Twin Falls, Айдахо (США), верхний плиоцен,
кол. Палеонтол. муз. Мичиган, унив., Анн Арбор.
Диагноз (по: Hibbard, Bjork, 1971). По размерам близка к Sorex
obscurus Merr., переднее мандибулярное отверстие расположено более
кпереди, чем у S. arcticus Kerr, S. obscurus и S. trowbridgii. Мелкое заднее
мандибулярное отверстие имеется, напоминает положением таковое
S. jumeus Miller. Зубы пигментированы. На задневнутренней стороне М3
имеется хорошо заметный бугорок (эпиконид).
Описание. Авторы вида в описании голотипа привели те же при-
знаки, что и в диагнозе. При этом указали, что описываемый вид отли-
чается от S. poverst Hibbard et Bjork крупной и высокой нижней челю-
стью, более передним расположением мандибулярного отверстия и ши-
роким талонидом (метаконидом).
Измерения (по: Hibbard, Bjork, 1971). Длина: Мх—М3 — 3.62,
Мх — 1.38, М2 — 1.30, М3 — 1.12 мм.
Распространение и возраст. Айдахо (США), верхний
плиоцен.
t 18. Sorex powersi Hibbard et Bjork, 1971.
Hibbard, Bjork, 1971, p. 172.
Тип. Фрагмент левой ветви нижней челюсти с зубами М,—М3 и сохранивши-
мися отростками сочленовной части, V 55028, Twin Fals, Айдахо (США), верхний
плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн. Арбор.
22 А. А. Гуреев
338
ПОДСЕМ. SORICINAE
Диагноз (по диагнозу и описанию: Hibbard, Bjork, 1971). Сходен
по размерам с 5. saussurei Merr. и 5. trowbridgii Baird. Ширина талонида
(метаконида) Мх—М2 близка к ширине их тригонида (переднего сегмента).
Авторы почему-то не отмечают значительного развития М3. Оба сегмента
этого зуба достигают значительных размеров, что хорошо заметно на
приведенных ими рисунках.
Описание. В описании авторы приводят следующие особенности
этого вида: Р4 явственно пигментирован как и Мх и М2; у М3 голотипа
сильно развит лабиальный цингулюм; зубы более луковицеобразные,
чем у современных видов.
Измерения (по: Hibbard, Bjork, 1971). Длина: Мх—М3 — 3.57,
Р4—М2 — 3.25 мм.
Распространение и возраст. Айдахо (США), верхний
плиоцен.
t 19. Sorex meltoni Hibbard et Bjork, 1971.
Hibbard, Bjork, 1971, p. 172—174. 4
Тип. Фрагмент правой ветви нижней челюсти с зубами Мх—М3, венечным и
сочленовным отростками V 55173, Twin Falls, Айдахо (США), верхний плиоцен, кол.
Палеонтол. муз. Мичиган, унив., Анн Арбор.
Диагноз (по: Hibbard, Bjork, 1971). Размеры сходны с таковыми
5. cinereus Kerr. На нижней челюсти хорошо заметны переднее и заднее
мандибулярные отверстия. Талонид (метаконид) Мх и М2 не шире триго-
нида (переднего сегмента), как и у современных видов. Энтоконид обо-
соблен от метаконида (бугорки на протокониде — гребне, граничащем
с задним сегментом зуба).
Примечание. Здесь приведены наиболее существенные признаки
из отмеченных авторами. Они тем не менее не могут считаться постоян-
ными и видоспецифичными. Строение нижней челюсти и структура ее
зубов столь вариабельны, что не всегда можно на них положиться. Из
этого однако не следует, что верхний зубной ряд и строение черепа не
имели специфических особенностей, резко отличных от таковых совре-
менных видов.
Измерения. Авторы привели только длину Мх—М3, равную
2.92 мм.
Распространение и возраст. Айдахо (США), верхний
плиоцен.
t 20. Sorex sandersi Hibbard, 1956.
Hibbard, 1956, Pap. Michigan Acad. Sci. Arts Letters, v. 148, p. 145—203-
1972, p. 77, 78.
Тип. Фрагмент правой ветви нижней челюсти с зубами 12—М3, ИММР V 31976,
Meade, Канзас (США), нижний плейстоцен, кол. Муз. Канзас, унив., Лоренс.
Диагноз (по: Hibbard, 1972). Верхняя сторона коронки первого
нижнего промежуточного зуба покрыта передним краем Р4. Задняя часть
ветви нижней челюсти по ширине равна ширине основания вентральной
сочленовной фасетки. Мандибулярное отверстие одно, переднее. Подбо-
родочное отверстие впереди или под передним корнем Мх.
Измерения (по: Hibbard, 1972). Длина: 1Х—М3 — 7.12, 1а—Мэ —
5.00, 5.13, Р4—М3 — 4.36, 4.57, Мх—М3 — 3.61, 3.77 мм.
Распространение и возраст. Канзас, Небраска (США),
нижний плейстоцен.
15. РОД SOREX
33»
21. Sorex araneus Linnaeus, 1758 — Обыкновенная бурозубка
(рнс. 170).
Linnaeus, 1758, p. 53; — vulgaris Linnaeus, 1764, Mus Regni Adelphi,
p. 10; — tetragonurus Hermann, In: Zimmermann, 1780, Geogr. Gesch., Bd 2,
S. 383; — quadricaudatus Kerr, 1772, Animal Kingdom, p. 208; — coronatus Mil-
let, 1828, Faune de maine et Loire, v. 1, p. 18; — personatus Millet, 1828, там же;—
concinnus W a g 1 e г, 1832, Isis, p. 54; — rhindophus W a g 1 e r, 1832, там же; —<
melanodon W a g 1 e г, 1832, там же; — hermanni Duvernoy, 1834, L’Institut,
p. 299; — labiosus Leonard Janyns, 1838, Ann. Nat. Hist. Bd 2, S. 326; —
macrotrichus de Selys Longchamps, 1839, Etudes Micromammal., p. 20.
Crossopus ou Sorex ignotus F a t i o, 1905, Arch. Sci. Phys. Nat. Geneve, v. 19, N 4,
p. 201. — Sorex santonus M о 11 a z, 1908, Bull. Soc. Zool. Geneve, v. 1, p. 118; —•
roboratus Hollister, 1913, Smiths. Misc. Coll., v. 60, № 24, p. 2; — grantii Bar.
Рис. 170. Sorex araneus (по: Строганов, 1957).
rett-Hamilton, Hinton, 1913, Abstract Proc. Zool. Soc. London, p. 18; —•
samniticus Al to bell o, 1926, Boll. Inst. Zool. Roma, v. 3, p. 102. Soricoidus mons-
vairant Al tobello, 1927, Rev. Franse Mammif., t. 1, p. 6.
Тип не сохранился (?). Вид описан из Упсалы (Швеция).
Диагноз. Окраска верхней стороны тела светло- или темно-
коричневая до почти черной (в западной части ареала). Первый и второй
верхние промежуточные зубы равны или почти равны по величине. Раз-
меры остальных постепенно убывают к заднему краю зубного ряда.
Описание. Окраска тела варьирует в значительных пределах.
Она может быть почти сходной сверху и снизу или нижние части тела
в разной степени светлее спины, особенно в зимнее время, серые или серо-
бурые. Хвост двуцветный, сверху темно-коричневый, снизу светло-корич-
невый или буро-серый. На конце хвоста кисточка из волос, более длинных,
чем на остальных частях хвоста.
Череп массивный с хорошо и рано развивающимся сагиттальным греб-
нем. Длина лицевой части черепа примерно равна длине мозговой. Верх-
ние промежуточные зубы массивные с заостренными вершинами. Первый
и второй промежуточные зубы верхней челюсти значительно выше осталь-
ных. Окраска зубов светло-красно-коричневая.
Измерения. Длина: тела — 60.0—84.0, хвоста — 30.0—50.0,
ступни — 12.0—13.7, кондило-базальная черепа — 18.0—21.7 мм.
Распростра не ни е и возраст. Населяет обширную тер-
риторию от Финляндии, Норвегии и Швеции на севере, до Франции,
22*
340
ПОДСЕМ. SORICINAE
Италии и Югославии на юге. На восток ареал простирается до Западной
Сибири, Алтая и северо-западной Монголии включительно. В европей-
ской части СССР распространена от полосы тундр на севере до зоны сте-
пей на юге, в лесах Северного Кавказа и Закавказья; известна с плиоцена.
Биология. Обыкновенная бурозубка населяет все зоны от тундры
до степи, но предпочитает леса. В лесной зоне держится в смешанных,
широколиственных и чисто хвойных лесах с хорошо развитым подлеском
или высоким травостоем. Селится в березовых колках и ольшаниках, из-
бегая сфагновых болот и сильно заболоченных участков леса, а также
очень сухих мест.
Наиболее благоприятные условия жизни для обыкновенной бурозубки
представляют леса с рыхлой и толстой мертвой подстилкой, богатой на-
секомыми и другими беспозвоночными, расположенные по берегам водое-
мов. В степных районах селится по берегам рек и других водоемов, до-
вольствуясь укрытием из высоких прибрежных травянистых растений.
Открытых мест избегает. Встречается в садах и на огородах, иногда за-
ходит в жилые помещения.
Пища — разнообразные насекомые и их личинки, пауки, земляные
черви, моллюски и другие беспозвоночные, которых зверек добывает
в лесной подстилке или из верхних слоев почвы, роя для этого мелкие ходы
и используя чужие. Насекомые с твердым хитином — навозники, мед-
ведки и пр. — поедаются менее охотно. Поедают детенышей мышей и по-
левок, мелких лягушек. Используют в небольших количествах раститель-
ные корма — семена ели, сосны, лиственницы, липы, а также вегетативные
части растений. По-видимому, существенное значение в питании земле-
роек имеют и дождевые черви, особенно в дождливые годы, когда они
в большом количестве появляются на поверхности почвы, что наблюда-
лось летом 1962 г. под Ленинградом. В это время дождевые черви состав-
ляли большую часть содержимого вскрытых желудков этого вида.
Размножение. Половозрелость наступает на следующий год после рож-
дения. Наблюдаются редкие случаи размножения самок-сеголеток ран-
них выводков. В годы пониженной численности способными к размноже-
нию оказываются и самцы-сеголетки. Размножение охватывает период
с апреля по октябрь, начинаясь и заканчиваясь раньше или позднее,
в зависимости от погодных условий и физиологического состояния особей
вида в разных частях ареала. Продолжительность беременности около
3 недель, в лабораторных условиях 20 дней. За весь период размножения
бывает 2—3 помета. В одном помете 3—11 (чаще 6—8) детенышей.
Гнездо шарообразной формы, строится из мелких растительных остат-
ков и располагается на поверхности земли в укромном месте или в чужой
заброшенной норе, приспособленной для этой цели.
По наблюдениям в лаборатории, молодые особи этого вида развива-
ются 21 день. Волосы на теле детенышей появляются на второй неделе
жизни, а глаза открываются на третьей. Характерный для землероек
облик молодые приобретают на 12-й день. Через 17 дней начинают выхо-
дить из гнезда, а на 22-й покидают его и становятся самостоятельными.
Длина тела новорожденных в среднем 20.0 мм, вес 350—500 мг.
На 25-й день после родов у самки вновь наступает течка и она готовится
к размножению.
Продолжительность жизни 1—1.5 года. В неволе отдельные экзем-
пляры живут значительно дольше.
Подвиды. 1. S. a. araneus L., 1758. Характеризуется средними
размерами тела и относительно светлой окраской. Распространен в евро-
пейской части СССР, на Северном Урале и в Западной Сибири, где встре-
чается в бассейнах рек Тавды, Туры, Исети и Тобола.
• 15. РОД SOREX
341
2. S. a. satunini Ogn. 1920. Некрупная землеройка с укороченной ли-
цевой частью черепа. Распространена на Кавказе и в Закавказье.
3. S. a. tomensis Ogn., 1921. Слабо дифференцированная от типичного
подвида форма. Распространена в лесной и лесостепной полосе равнинной
части Западной Сибири, к югу от предгорий Алтая, Салаира, Кузнецкого
Алатау и Саянов.
4. S. a. roboratus Hollist.., 1913. Характеризуется крупными разме-
рами и светлой окраской. Распространена на Алтае, Салаире, Кузнецком
Алатау и Саянах к востоку до Забайкалья.
Рис. 171. Sorex asper.
22. Sorex asper Thomas, 1914 — Тянь-шаньская бурозубка (рис. 171).
Thomas, 1914, Ann. Mag. Nat. Hist., ser. 8, v. 13, p. 565, 566; — araneus asper
Огнев, 1940, Бюл. МОИП, выл. 3 (XVIII), с. 22, 23.
Тип. № 5.4.8.2, долина р. Текеса, Тянь-Шань, кол. Брит, муз., Лондон.
Диагноз. Окраска верхней стороны тела светло-коричневая.
Верхние промежуточные зубы в отличие от таковых у других видов этого
рода очень крупные, высокие, массивные, с притупленными вершинами,
темноокрашештые. Второй промежуточный зуб верхней челюсти выше
первого и третьего.
Описание. Тело и хвост двуцветные. Коричневая окраска свет-
лых и тусклых оттенков верхней стороны тела и хвоста довольно резко
контрастирует с серой или буровато-серой окраской нижних поверхностей
тела. Кисточка на конце хвоста слабо развита или отсутствует.
Череп массивный, с хорошо заметными сагиттальным и ламбдовидным
гребнями и узкой, удлиненной лицевой частью. Второй верхний проме-
жуточный зуб равен или крупнее переднего зубца I1, измеренного от вер-
шины до основания выемки между зубцами.
Измерения. Длина: тела — 65.0—77.0, хвоста — 34.0—45.0,
ступни — 11.6—13.0, кондило-базальная черепа — 18.0—19.4 мм.
Распространение и возраст. Тянь-Шань. Известна
из долин рек Текеса и Коксу, Заилийского Алатау, верховьев р. Большой
Алмаатинки, северных склонов Терскей-Алатау, Киргизского хребта
342
ПОДСЕМ. SORICINAE
(урочище Гуюк). Отмечена для района Нарына, Джумгала. В Нарынском
районе добывалась на р. Каунды и на северных склонах перевала
Куганды, современный.
Биология. Обитает в альпийской зоне. Размножается с марта
по июль, число эмбрионов 1—8.
23. Sorex daphaenodon Thomas, 1907 — Темнолапая бурозубка
(рис. 172).
Thomas, 1907, Proc. Zool. Soc. London, v. 1, p. 407, 408.
Тип. Экземпляр с о. Сахалин, пос. Даринэ в 25 км к северо-западу от с. Корса-
кове, кол. Брит, муз., Лондон.
Диагноз. Окраска верхней стороны тела темно-коричневая.
Верхние промежуточные зубы мало сужены к закругленной вершине
Рис. 172. Sorex daphaenodon.
и плотно прижаты друг к другу. Второй промежуточный зуб выше пер-
вого и третьего. Все зубы темноокрашенные; окрашены и жевательные
поверхности коренных зубов.
Описание. Темная окраска на большей части тела. Только
на отдельных частях нижней стороны она буровато-серая или темно-
серая с серебристым налетом. Хвост сверху и снизу коричневый, у корня
снизу он серого цвета. На конце хвоста хорошо заметная кисточка из бо-
лее длинных волос. Кисти и ступни покрыты коричневыми волосами.
Череп массивный, с хорошо заметным сагиттальным гребнем. Лицевая
часть черепа относительно короткая, а мозговая длинная и широкая
(расстояние между отверстиями на лобных костях и передним краем но-
совых костей составляет примерно 2/3 расстояния между задним краем
межглазничного сужения и затылком, а длина мозговой коробки почти
равна ее ширине).
Измерения. Длина: тела — 50.0—66.0, хвоста — 25.0—39.4,
ступни — 10.0—12.8, кондило-базальная черепа — 17.0—18.7 мм.
Распространение и возраст. СССР, Северо-Восточный
Китай и Япония (о. Хоккайдо), современный.
15. РОД SOREX
343
В СССР встречается спорадически на обширной территории от бас-
сейна нижнего и среднего течения р. Оби и верховьев р. Ишима, на во-
стоке до низовьев р. Колымы, побережья Охотского моря, Камчатки,
Курильских островов, Сахалина и Приморья.
Биология. Обитает преимущественно в лесных и лесостепных
биотопах, но численность этого вида всюду невелика. Встречается в разре-
женных широколиственных лесах с кустарниковым ярусом и толстым
слоем мертвой подстилки, в смешанных лесах гор и равнин, в захламлен-
ных, относительно влажных и малозатененных хвойных лесах с преобла-
данием папоротника-орляка в травяном покрове, в березовых колках
и осиновых рощах с травяным покровом.
Рис. 173. Sorex arcticus.
Пищей служат разнообразные мелкие беспозвоночные: черви, мол
люски, насекомые. Последние составляют основной корм.
Размножение темнолапой бурозубки изучено недостаточно. Известно,
что начало размножения приходится на конец апреля—начало мая,
а заканчивается к октябрю. В период размножения бывает 2—3 помета,
по 4—9 детенышей в каждом. В Барабинской степи в течение июня—
августа попадались беременные и кормящие самки. В .Новосибирской
области беременные самки попадались до конца сентября, число эмбрио-
нов было 6—7.
Подвиды: 1. S. d. daphaenodon Thos, 1907. Характеризуется отно-
сительно мелкими размерами, длинным хвостом и темно-коричневой
окраской. Распространен в Забайкалье, Восточной Сибири, на Куриль-
ских островах, Сахалине, в Приморском крае, Амурской обл. и в низовьях
Амура.
2. S. d. scaloni Ogn., 1933. Отличается от номинального подвида более
крупными размерами, более длинным носовым отделом черепа и черно-
бурой окраской. Распространен в Западной Сибири.
24. Sorex arcticus Kerr, 1792 — Арктическая бурозубка (рис. 173).
Kerr, 1792, Animal Kingdom, р. 206; — richardsonii В a g h m a n, 1887, J.
Acad. Nat. Sci. Philadelphia, v. 7, pt 2, p. 383; — pachyurus В a i r d, 1858, Mammals,
in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1, p. 20; — sphagnicola С о u e s, 1877,
Bull. U. S. Geol. and Geogr. Surv., v. 3, p. 650; — belli Merriam, 1892, Proc. Biol.
Soc. Washington, v. 7, p. 25; — pribilofensis Merriam, 1895, p. 87; — tandrtnsi»
344
ПОДСЕМ. SORICINAE
Merriam, 1900, Proc. Wash. Acad. Sci., v. 2, p. 16; — buxtoni J. Allen, 1903,.
Bull. Amer. Mus. Nat. Hist., v. 19, p. 181, 182; — araneus borealis Кащенко, 1905,
c. 85—90; — baicalensis Огнев, 1913, Млекопитающие Моск, губ., ч. 1, с. 105—
107; — sanguinidens G. Allen, 1914, Proc. New. Engl. Zool. Club., v. 5, p. 54; —
amasari Огнев, 1921, Ежегодн. Зоол. муз. АН, т. 22, с. 316, 317; — sibiriensis
Огнев, 1921, там же, с. 328, 329; — ultimus Огнев, 1926, с. 22; — daphaenodon
sibiriensis Огнев, 1928, с. 212, 213; — baicalensis Огнев, 1928, с. 227, 228; —
jenissejensis Dukelsky, 1930, Zool. Anz., Bd 88, S. 75, 76;-—macropygmaeu.s
Ognev, Бобринский, Кузнецов, Кузякин, 1944, с. 50, 51; —
caecuttens Ellerman, Morrison-Scott, 1951, p. 48.
Тип не установлен, вид описан по экземпляру из окрестностей форта Северн,
Онтарио, Канада.
Диагноз. У северных экземпляров на верхней стороне тела тем-
ный «чепрак». Бока головы, а также вся нижняя сторона белые или серо-
белые. К югу темная окраска становится преобладающей.
Верхние промежуточные зубы остроконечные, плотно сомкнутые,
массивные. Продольный диаметр оснований коронок этих зубов менее
их высоты (отличие от Sorex caecuttens). Второй верхний промежуточный
зуб выше первого и третьего.
Описание. У особей отдельных географических популяций зим-
ний волосяной покров образован длинными мягкими волосами. В север-
ных частях ареала окраска резко двуцветная, в южных приближается
к однотонной. Хвост двуцветный, сверху темно-коричневый, снизу белый.
Череп с относительно короткой и широкой лицевой частью (длина
мозговой части черепа примерно равна лицевой).
Измерения. Длина: тела — 51.0—70.0, хвоста — 21.0—47.0,
ступни — 11.0—12.0, кондило-базальная черепа — 15.0—18.0 мм.
Распространение и возраст. Северо-восточная Европа
и Азия к югу до Северо-Восточного Китая и Монголии; Северная Америка
от Аляски, западных областей Канады и США к югу до верховьев Мисси-
сипи. В СССР от низовий Печоры на восток до Чукотского полуострова,
от полосы тундр на севере, на юг до Приморского края, южных окраин
Западной Сибири и Семиречья, современный. '
Биология. Арктическая бурозубка обитает в разнообразных
биотопах от арктических тундр, равнинной и горной тайги, широколист-
венных лесов и до лесостепных и степных пространств Сибири и Казах-
стана. В зоне тундр селится по берегам водоемов с древесной раститель-
ностью, а также в открытой тундре, но всюду отдает предпочтение участ-
кам, занятым лесом или кустарником. В лесной зоне селится в сырых
болотистых местах. В Казахстане отмечена в лесах по берегам рек и в гор-
ных лесах, а также в зарослях тростника и кустарниках вблизи водоемов
(в открытых ландшафтах). Питается преимущественно насекомыми, но пое-
дает и других беспозвоночных, в том числе и моллюсков.
Размножение не изучено. Беременные самки под Томском попадались
в июне—августе. Число эмбрионов 3—10, чаще 6—7.
Подвиды. 1. 5. a. borealis Kastsch., 1905. Характеризуется дву-
цветной окраской и коротким хвостом (до 33 мм). Распространен в зоне
тундр от долины р. Печоры, Енисейского залива и низовий р. Енисея
до Чукотского полуострова, Анадыря и тундровой полосы Камчатки.
По-видимому, этот же подвид обитает на крайнем северо-западе Северной
Америки.
2. S. a. buxtoni J. АП., 1903. Отличается слабовыраженным «чепра-
ком» (окраска не двуцветная). Распространен в лесной зоне Сибири от бас-
сейна р. Вилюя и нижнего течения р. Витима на восток до побережья
Охотского моря; на север до полосы лесотундры, к югу до лесостепной
и степной части Забайкалья. За пределами СССР — на севере Монголии
и в Северо-Восточном Китае. Видимо, этот же подвид населяет таежную
15. РОД SOREX
345
зону Западной и средней Сибири включая Кузнецкий каменноугольный
бассейн, Саяны и Туву.
3. S. a. schnitnikovi Ogn., 1921. Относительно крупных размеров
с длинным хвостом (не менее 37 мм) и темной окраской нижней стороны
тела. Распространен в центральных и южных районах Алтая, на Кал-
бинском хребте, в Тарбагатае и Джунгарском Алатау.
4. S. a. transrypheus Strog., 1956. Отличается мелкими размерами
и двуцветной окраской. Распространен в лесостепной полосе Западной
Сибири и Казахстана (от Кустаная до Алтая и Омской обл.), на юг до оз.
Кургальджин, Целиноградской и южных районов Карагандинской обл.
Рис. 174. Sorex raddei.
5. S. a. sanguinidens G. All., 1914. Отличается густым и длинным
мехом. Распространен от среднего течения р. Вилюя, Якутска и долины
р. Яны до среднего и нижнего течения р. Колымы и побережья Охотского
моря.
25. Sorex raddei Satunin, 1895 — Бурозубка Радде (рис. 174).
Satunin, 1895, Arch. Naturgesch., Bd 1, S. 109—111; — araneus Satunin,
1903, Зап. Кавказск. отд. Геогр. общ., т. 24, с. 52; — batis Thomas, 1913, Ann.
Mag. Nat. Hist., ser. 2, v 8, p. 214, 215.
T и п не отмечен, вид описан из окрестностей г. Кутаиси, кол. Кавказск. муз.,
Тбилиси.
Диагноз. Окраска верхней стороны тела темно-коричневая или
черно-бурая. Длина хвоста более 2/3 длины тела (45—52 мм). Кисти
и ступни покрыты темными волосами.
Описание. Окраска верхней и нижней стороны тела почти сходна,
как и хвоста. Отдельные экземпляры кажутся черными.
Череп массивный с широкой мозговой капсулой, ширина которой
более или равна ее длине. Продольный диаметр основгйк заднего зубца
I1 менее продольного диаметра четвертого промежуточного зуба. Пер-
вый и второй промежуточные зубы верхней челюсти почти равны по длине,
но коронка второго из этих зубов выше коронки первого.
Измерения. Длина: тела — 61.7—78.5, ступни — 12.2—13.1,
кондило-базальная черепа — 18.5—19.8 мм.
346
ПОДСЕМ. SORICINAE
Распространение и возраст. Найдена в Пятигорске,
на Черноморском побережье Кавказа, вблизи Кутаиси, в Бакуриани
и Лагодехах в Грузинской ССР, в лесах Карского плоскогорья, Шам-
шадинского, Кироваканского и Калининского районов Армянской ССР,
в лесах Карабаха Азербайджанской ССР и в Дагестанской АССР. За пре-
делами СССР известен из Турции (окрестности Трабзона), современный.
Биология. Малр изучена. Охотнее всего населяет сырые ле-
систые местности, а на открытых пространствах — берега водоемов.
Данные о размножении крайне скудные. В Армении в конце сентября
добыты беременная (6 эмбрионов, 1.5—2.0 мм) и кормящая самки.
Рис. 175. Sorex alpinus.
26. Sorex alpinus Schinz, 1837 — Альпийская бурозубка (рис. 175).
Sc h in z, 1837, Neue Denkschr, Allgem. Schweiz. Gesell. Naturwiss. Neuchatel,
Bd 1, S. 13; — antinori Bonaparte, 1840?, Iconogr, Faun. Ital., v. 1, fasc. 29; —
intermedins Cornalia, 1870?, Cat. Descr. Mammal. Ital., p. 27.
Тип не установлен, вид описан с Сен-Готардского перевала, Швейцария.
Диа г н о з. Окраска верхней стороны тела черная, черно-серая,
черно-бурая или бурая. Кисти и ступни покрыты белыми или охристо-
белыми волосами. Длина хвоста равна или немного менее длины тела
(59.6—71.0 мм).
Описание. Окраска верхней и нижней сторон тела сходная
или нижняя светлее — темпо-серая. Верхняя сторона хвоста темно-
бурая. Конец хвоста часто покрыт белыми волосами.
Череп с массивной лицевой частью, округлой и вздутой мозговой
капсулой. Продольный диаметр основания заднего зубца I1 менее тако-
вого четвертого или пятого промежуточных зубов, которые почти равны
между собой. Первый промежуточный зуб верхней челюсти равен вто-
рому или последний более высокий.
И з м е р е щи я. Длина: тела — 59.0—73.0, ступни — 13.6—16.0,
кондило-базальЙЙг черепа — 19.2—20.6 мм.
Распространение и возраст. Франция (к югу до Пи-
ренеев), ГДР, ФРГ, Италия, Швейцария, Австрия, Венгрия, Югославия,
Румыния, Чехословакия и Польша. В СССР известна из Ивано-Франков-
ской и Закарпатской обл. УССР, современный.
15. РОД SOREX
347
Биология. Обитает в разнообразных ландшафтах от лесов, реч-
ных долин, до альпийских лугов, поднимаясь в горы до 2000 м. Образ
жизни недостаточно изучен. Гнезда строит на стеблях растений на вы-
соте 10—20 см от поверхности почвы. Хорошо лазает по стеблям растений,
обвивая их при передвижении хвостом. Зимние гнезда строит на почве
или в норах. Молодых находили с июня по август включительно. Они
темнее взрослых и с более коротким хвостом.
27. Sorex mirabilis Ognev, 1937 — Гигантская бурозубка (рис. 176).
Огнев, 1937, Бюлл. МОИП, т. 46, вып. 5, с. 268—271; — pacificus Baird, Боб-
ринский, Кузнецов, Кузякин, 1944, с. 52.
Тип. j № 274, р. Кигаинка, Приморский край, кол. С. И. Огнева, хранится
в Зоол. муз. МГУ.
Рис. 176. Sorex mirabilis.
Диагноз. Самая крупная из палеарктических землероек-буро-
зубок (длина тела 74.0—97.0 мм). Окраска верхней стороны тела варьирует
в оттенках коричнево-бурого цвета. Задний зубец I1 очень низкий, не выше
или немного выше цингулюма первого верхнего промежуточного зуба.
Второй промежуточный зуб меньше первого.
Описание. Окраска сходная на всем теле или нижняя сторона
несколько светлее и тусклее верхней. На этом фоне хорошо выделяются
светло-серые длинные вибриссы (до 28 мм). Хвост сверху и снизу темный,
без кисточки на конце.
Череп крупный, с массивным лицевым отделом и неширокой плоской
мозговой капсулой. Зубы массивные, у отдельных экземпляров третий
промежуточный зуб лшпь немного крупнее пятого, а четвертый почти
равен второму.
Измерения. Длина: хвоста — 63.0—72.0, ступни — 16.0—17.5,
кондило-базальная черепа — 22.3—24.3 мм.
Распространение и возраст. Извести из ряда гео-
графических пунктов Северо-Восточного Китая и п-ова Корея, а в СССР
из Приморья (долина р. Кишинки, заповедники «Кедровяаападь», Лаза-
ревский и Супутинский), современный.
348
ПОДСЕМ. SORICINAE
Биология. Плохо изучена. Обитает в равнинных и горных лесах;
предпочитает влажные участки леса. По данным М. В. Охотиной (1969),
местами обитания этого вида служат леса различных типов, как первич-
ные, так и образовавшиеся на гарях и вырубках. Эта землеройка не добы-
валась в разреженных лесах, в зарослях кустарников, на открытых про-
странствах и на болотах. Таким образом, ее можно с полным основанием
считать типичным лесным видом.
В приморском крае размножается, видимо, один раз в году. Возможно,
что отдельные самки при благоприятных условиях размножаются два
раза в год.
В условиях неволи эта землеройка в течение суток съедает 25.4—
31.1 г (в среднем 28.7 г) различного корма, что составляет 189.1—254.7%.
Рис. 177. Sorex vir.
(в среднем 213.9%) веса тела. Как показали специальные опыты, пред-
почитаемым кормом служат дождевые черви. Если в клетке имеется под-
стилка из сухих листьев, землеройка охотнее поедает многоножек, жу-
ков — жужелиц, личинок щелкунов и чернотелок. Количество съедаемого
корма при этом сокращается. Автор, проводивший описываемые*наблю-
дения, полагает, что дождевые черви малопитательный корм, чем будто бы
и обусловлена прожорливость землероек этого вида. Заметим, что все
землеройки прожорливы, и чем меньше по размерам, тем больше съедают
корма.
Подвиды. 1. S. т. mirabilis Ogn., 1937. Распространен в При-
морском крае.
2. S. т. kutscheruki Strog., 1956. Отличается от типичного подвида
меньшими размерами. Распространен в Северной Корее.
28. Sorex vir G. Allen, 1914 — Плоскочерепная бурозубка (рис. 177).
G. Allen, 1914, Procid. New. Engl. Zool. Club., v. 5, p. 52;—platycranius
Огнев, 1921, Ежегодн. Зоол. муз. АН, т. 22, с. 334—336; — thomasi Огнев,
1921, там же; — araneus jacutensis Dukelsky, 1928, Zool. Anz., Bd 78, H. 5/8,
S. 102, 103; — dukelskie Огнев, 1933, Бюл. НИИЗ МГУ, № 1, с. 57—59.
Тип. 9, Нижне-Колымск, близ устья р. Колымы, хранится в кол. Муз. сравнит,
зоол. в Кембридже (США).
15. РОД SOREX
349
зубы невысокие (продольный
Рис. 178. Sorex beringianus (по: Юдин, 1967).
Диагноз. Окраска верхней стороны тела светло-, реже темно-
коричневая. В отличие от других палеарктических видов длина лицевой
части черепа в 1.25—1.50 раза более длины мозговой. Коронки проме-
жуточных зубов имеют сзади выемку. Второй промежуточный зуб верх-
ней челюсти выше первого, пятый очень мелкий зуб, смещен на внутрен-
нюю сторону зубного ряда.
Описание. Нижняя сторона тела светлее верхней, серо-бурая
или темно-серая. Хвост сверху темно-коричневый, снизу светло-коричне-
вый или серо-бурый.
Череп с удлиненной лицевой частью, неширокий, с уплощенной моз-
говой капсулой. Верхние
диаметр их коронок у ос-
нования более высоты зу-
бов).
Измерения. Дли-
на: тела — 60.0—80.8,
хвоста — 30.0—42.0, ступ-
ни — 12.7—14.0, кондило-
базальная черепа — 19.0—
21.5 мм.
Распрос т р ане-
ние и возраст.
Только в СССР. Алтай,
Западная и Восточная Си-
бирь до побережья Охотс-
кого моря и Приморского
края и от бассейна р. Ниж-
ней Тунгуски, устья Индигирки и Нижне-Колымска к югу до среднего
течения р. Хамнея в южном Забайкалье и окрестностей г. Уссурийска
в Приморском крае, современный.
Биология. Обитает в разнообразных условиях от болотистых
низменностей до альпийских лугов (на Алтае до 2200 м над ур. м.). Встре-
чается в лесах различных типов, зарослях кустарников и болотах. По пи-
танию этой бурозубки данных нет, но можно думать, что она в этом отно-
шении мало отличается от других видов своего рода. Размножение не изу-
чено. 11 июня в долине р. Юктали (бассейн р. Нижней Тунгуски) была
добыта беременная самка с 14 эмбрионами.
Подвиды. l.S. v. vir G. АП. 1914. Характериузется массивным
и менее уплощенным, чем у других подвидов, черепом, более коротким
хвостом и светлой окраской. Распространен в северо-восточной Сибири
(низовья Лены, бассейн рек Яны, Индигирки, Колымы и Анадыря).
2. S. V. jacutensis Dukelsky, 1928. Отличается более длинным хвостом
и темно-коричневой окраской. Распространен в Западной и Средней
Сибири, на Алтае, в Саянах и Туве, на восток до Байкала и бассейна
среднего течения р. Лены.
3. S. V. thomasi Ogn., 1921. Отличается от других подвидов меньшими
размерами и монотонной окраской тела. Распространен в Забайкалье.
4. S. V. platycranius Ogn., 1921. Отличается широко расставленными
предглазничнымй отверстиями. Известен из окрестностей г. Ворошилова
(Приморский край).
И® 29. Sorex beringianus Judin, 1967 — Берингийская бурозубка
(рис. 178).
Юдин, 1967, Изв. Сиб. отд. АН СССР, № 5, вып. 1, с. 155—157.
Тип. (j № 2341, Курильская гряда, о. Парамушир, кол. Зоол. муз. Биол. инет.
СО АН СССР, Новосибирск.
350
ПОДСЕМ. SORICINAE
Диагноз. Верхняя сторона тела коричнево-бурая, бока значи-
тельно светлее, нижняя сторона тела серая. Первый и второй верхние
промежуточные зубы крупные, сходных размеров. Третий промежуточный
зуб меньше второго и четвертого, пятый очень мелкий.
Описание. В окраске тела заметно выраженная «чепрачность».
Темная окраска спины резко отграничена от светлой окраски боков.
Длина хвоста более 70% длины тела. Окраска его верхней стороны бурая,
нижней — охристо-серая.
Череп со слабо вздутой мозговой капсулой и относительно широкой
и высокой носовой частью.
Измерения. Длина: тела — 46.0—56.0, хвоста — 44.0—47.0,
ступни — 11.0—11.2, кондило-базальная черепа — 15.0—15.8 мм.
Рис. 179. Sorex unguiculatus.
Распространение и возраст. Курильская гряда, о. Па-
рамушир, современный.
Биология не изучена.
Примечание. Диагноз и описание по: Юдин, 1967. Система-
тическое положение этого вида не ясно.
30. Sorex unguiculatus Dobson, 1890 — Когтистая бурозубка (рис. 179)
Sorex vulgaris Schrenk, 1858, Reisen und Forschung im Amur Lande, Bd 1,
S. 106, 107; — unguiculatus Dobson, 1890, Ann. Mag. Nat. Hist., v. 5, p. 115; —
araneus Linnaeus Бобринский, Кузнецов, Кузякин, 1944, с. 52.
Тип. $ № 2387, кол. ЗИН АН СССР, Ленинград.
Диагноз. Окраска верхней стороны тела коричневая или бурая.
В отличие от других палеарктических бурозубок длина когтей на пальцах
кисти более 3 мм. Первый верхний промежуточный зуб в 1.5 раза более
второго.
Описание. Окраска тела сверху и снизу сходная или нижняя сто-
рона светлее — буровато-серая. Хвост покрыт относительно короткими
волосами, не образующими кисточки на конце.
Череп массивный, с широкой и угловатой мозговой капсулой. Наиболь-
шая ширина последней примерно равна расстоянию между отверстиями
15. РОД SOREX
351
на лобных костях и передним краем межчелюстных костей. Второй про-
межуточный зуб равен третьему или немного менее его.
Тело массивное. Кисти широкие и массивные (роющие лапы). Пальцы
кистей и ступней более длинные, чем у других видов палеарктических
землероек бурозубок.
Измерения. Длина: тела — 79.0—91.0, хвоста — 44.0—47.0,
ступни — 13.0—14.0, кондило-базальная черепа — 19.0—20.7 мм.
Распространение и возраст. Северо-Восточный Китай,
п-ов Корея и Япония. В СССР — Приморье, Курильские острова и Са-
халин, современный.
Биология. Слабо изучена. Обитает в разнообразных биотопах.
В Приморье живет в горных и равнинных лесах различных типов, а также
Рис. 180. Sorex caecuttens.
в полосе культурных земель, расположенных вблизи леса или зарослей
кустарников. Встречается также в жилых постройках. Пища — различ-
ные беспозвоночные, главным образом насекомые и их личинки, поедают
и мелких позвоночных.
Размножение не изучено. Беременные самки добыты в Приморье
(Сучанский район) 16 и 28 июня. Число эмбрионов 4—7.
31. Sorex caecuttens Laxmann, 1788 — Средняя бурозубка (рис. 180).
L axmann, 1788, Nova Acta Acad. Set. Petropol., v. 3, p. 284, 286; — macro-
pygmaeus Miller, 1901, Proc. Biol. Soc. Wachington, v. 14, p. 158; — shinto Tho-
mas, 1905, Proc. Zool. Soc. London, v. 2, p. 238; — annexus Thomas, 1907, ibid.,
v. 1, p. 859; — centralis Thomas, 1911, Ann. Mag. Nat. Hist., ser. 8, v. 7, p. 758.
Тип не сохранился, вид описан по экземпляру с юго-западного побережья
оз. Байкал.
Диагноз. Верхняя сторона тела коричневая разных оттенков.
Продольный диаметр оснований коронок верхних промежуточных зубов
более их высоты. Задний край этих зубов имеет выемку, вследствие чего
они кажутся редко стоящими. Первый верхний промежуточный зуб зна-
чительно крупнее остальных, следующие за ним постепенно убывают
в размерах.
Описание. Окраска тела и хвоста двуцветная. Темно-коричневая
верхняя сторона резко контрастирует с серой или серо-белой нижней.
352
ПОДСЕМ. SORICINAE
Череп с округлой и вздутой мозговой капсулой и сильно суженной
лицевой частью. Из внешних признаков следует отметить хорошо замет-
ные, особенно на свежих и живых экземплярах, крупные уши, выступаю-
щие в стороны.
Измерения. Длина: тела — 48.7—63.0, хвоста — 35.0—45.0,
ступни — 10.0—11.0, кондило-базальная черепа — 16.0—17.0 мм.
Распространение и возраст. Лесная зона Восточной
Европы и Азии (включая Монголию, Северный Китай, п-ов Корея и Япо-
нию). В СССР от западной государственной границы на восток до Чукот-
ского полуострова, Камчатки, Курильских островов, Приморья, Сахалина
включительно и от северных окраин тундровой зоны на юг до средней части
Украины. Тамбовской обл., южной Башкирии, Северного Казахстана
Рис. 181. Sorex gracillimus (по: Юдин, 1965).
и горных областей Южной
Сибири, современный.
Биология. В преде-
лах столь обширного ареала
средняя бурозубка обитает
в весьма разнообразных био-
топах. В лесах предпочитает
более влажные участки с рых-
лой почвой и толстым слоем
мертвой подстилки. В зоне
тундр обитает на берегах во-
доемов, поросших древесной
или кустарниковой расти-
тельностью. Сухих участков,
а также мест с глубоким
моховым покровом эта земле-
ройка избегает, что объяс-
няется, по-видимому, труд-
ностью рытья на сухих почвах или бедностью корма на моховых боло-
тах. Пищей этому виду, как и другим видам землероек, служат различ-
ные беспозвоночные, обитающие в почве или лесной подстилке. Средняя
бурозубка лучше других видов приспособлена к жизни в различных ус-
ловиях среды, так как вероятно, может плавать и рыть, на что указы-
вают ее широкие кисти и сильно развитые волосяные оторочки на кистях
и ступнях, особенно у особей из северо-восточной части ареала.
Размножение, видимо, не приурочено к определенному периоду, а про-
исходит в течение всего теплового времени года. Так, под Томском и Ново-
сибирском беременные самки, как и кормящие, попадаются с мая по ок-
тябрь. В Башкирском заповеднике беременные самки добывались с се-
редины апреля до середины октября. Число эмбрионов, по данным для
Западной Сибири, колеблется от 2 до 11, чаще 7—8. Детеныши, как и у дру-
гих видов землероек, рождаются слепыми.
Подвиды. 1. S. с. caecutiens Laxm., 1788. Характеризуется сравни-
тельно крупными размерами тела и бледной окраской верхней стороны
тела. Распространен на Алтае, Тарбагатае, Сауре, Саянах, побережье
р. Енисея, к северу до бассейна р. Нижней Тунгуски и в районе оз. Байкал.
2. S. с. pleskei Ogn., 1921. Отличается средними размерами и темно-
коричневой окраской верхней стороны тела. Распространен в лесной
и лесостепной зонах европейской части СССР и Западной Сибири, на восток
до предалтайских степей и р. Енисея.
3. S. с. koreni G. All. 1914. Отличается мелкими размерами, коротким
для этого вида хвостом и преобладанием светлой окраски на всем теле,
15. РОД SOREX
353
особенно на боках. Распространен по всей тундровой полосе от р. Пе-
чоры до Анадыря.
4. S. с. macropygmaeus Mill., 1901. Отличается длинным хвостом.
Распространен на Камчатке, на востоке Амурской обл., в Приморском
крае и на Шантарских островах.
5. S. с. shinto Thos, 1905. Отличается более длинным, чем у других
форм, хвостом. Распространен на Курильских островах, Сахалине и в Япо-
нии.
32. Sorex gracillimus Thomas, 1907. — Тонконосая малая бурозубка
(рис. 181).
Sorex minutus gracillimus Thomas, 1907, Proc. Zool. Soc. London, v. 1, p. 408,
409; — gracillimus Юдин, 1965, в кн.: Новые и малоизвестные виды фауны Сибири,
с. 44—47.
Тип. $ № 7.2.5.2.3, о. Сахалин, пос. Дарпнэ в 25 км к северо-западу от с. Корса-
кове, кол. Брит, муз., Лондон.
Диагноз. Окраска верхней стороны тела коричнево-бурая разных
оттенков. Первый верхний промежуточный зуб равен четвертому или
выше его. Пятый промежуточный зуб равен четвертому или крупнее.
Все зубы с глубокой выемкой на задней стороне, отчего кажутся (особенно
первые три) редко стоящими.
Описание. Мелкая землеройка размерами с малую бурозубку,
с длинным тонким носом. Длина хвоста достигает 80% длины тела. Зим-
няя и летняя окраски тела отличаются яркостью темного «чепрака» на
спине, а также степенью посветления боков тела, которые в зимнее время
более светлые и резко отграничены от темной спины.
Измерения (по: Юдин, 1965). Длина: тела — 48.0—58.0,
хвоста — 40.0—45.5, ступни — 10.8—11.4, кондило-базальная черепа —
15.0—15.9 мм.
Распространение и возраст. Приморский край, Шан-
тарские острова, о. Сахалин, северная часть п-ова Корея, Китай, Япония,
современный.
33. Sorex minutus Linnaeus, 1766 — Малая бурозубка (рис. 182).
Linnaeus, 1766, Sist. Nat. ed. 12, t. 1, p. 73.; — pygmaeus L a x m a n n,
1769, Sibir. Briefe, S. 72; — minutissimus Zimmermann, 1780, Geogr. Gesch.,
Bd 2, S. 385; — exilis Gmelin, 1788, in: Linnaeus, Syst. Nat., ed. 13, v. 1, p. 115; —
canaliculatus L jungh, 1806, K. Svenska Vetensk. Acad. Handl., Bd 27, S. 263; —
minimus Geoffroy, 1811, Ann. Mus. Hist., Nat., Paris, t. 17, p. 186; — gmelini
Pallas, 1811, Zoogr. Rosso-Asiatica, v 1, p. 134;—pumilio W a g 1 e r, 1832,
Isis, v. 25, p. 54; — rusticus J a n у n s, 1838, Ann. Nat. Hist. Bd 11, S. 423; — pumi-
lus Nilsson, 1844, K. Svensca Vetensk. Acad. Handl., Bd 1, S. 33; — planiceps
Miller, 1911, Proc. Biol. Soc. Washington, v. 24, p. 242; — leucogaster Kuroda,
1933, Bull. Biogeogr. Soc. Jap., ser. 3, v. 3, p. 155.
T и п не сохранился, вид описан из окрестностей г. Барнаула.
Диагноз. Окраска верхней стороны тела коричневая разных от-
тенков. Первые три верхних промежуточных зуба равны по высоте или
второй меньше (ниже) первого и третьего.
Описание. Морда сильнд, удлиненная и заостренная, что осо-
бенно бросается в глаза при сравнении с другими видами бурозубок.
Относительно длинный хвост покрыт длинными, снизу очень светлыми
волосами; он резко сужен у основания и имеет хорошо заметную кисточку
на конце. Нижняя сторона тела буровато-серая или серая, реже бурая,
но и тогда она кажется значительно светлее коричневых верхних частей
тела.
23 а. А. Гуреев
354
ПОДСЕМ. SORICINAE
Череп с округлой, вздутой мозговой капсулой и узкой лицевой частью.
Наибольшая высота мозговой капсулы примерно в два раза или более
высоты лицевой части черепа в области Р4.
Измерения. Длина: тела — 40.0—59.0, хвоста — 33.0—50.0,
ступни — 9.0—11.3, кондило-базальная черепа — 14.0—16.0 мм.
Распространение и возраст. Лесные и лесостепные
ландшафты Европы, Западной Сибири, восточного берега Байкала,
Центрального и Северо-Западного Китая, Кашмира. Эллерман и Морри-
сон-Скотт (Ellerman, Morrison-Scott, 1951) указывают этот вид и для
Палестины. В пределах СССР распространен в европейской части вклю-
чая Крым и Кавказ, в Семиречье, Сибири к востоку до Енисея и Баргу-
Рис. 182. Sorex minutus.
зина. На севере найден на полуостровах Гыданском и Ямале. Известен
с плиоцена, современный.
Биология. Обитает в разнообразных растительных ассоциациях.
Предпочитает разреженные, хорошо освещенные солнцем участки в лист-
венных и смешанных лесах, молодые сосновые боры со скальными выхо-
дами и валунами, сухие освещенные склоны берегов рек и озер, поросших
редколесьем с богатым травяным покровом. Встречается также в зарослях
высоких трав на открытых местах и в полосе культурных земель, главным
образом в садах и огородах, попадается и вблизи построек. В горных
местностях поднимается до высоты 3500 м и более.
Малая бурозубка более требовательна к местам обитания, чем другие
виды бурозубок. Прихотливостью в отношении жизненных условий,
видимо, и объясняется ее невысокая численность.
Питается разнообразными мелкими беспозвоночными — насекомыми
на разных стадиях развития, а также малощетинковыми червями, мол-
люсками и др. Предпочитает мягкие корма, что обусловлено мелкими
размерами тела и зубов этой землеройки. Корм добывает преимущественно
с поверхности почвы и в верхних слоях лесной подстилки.1
Размножение — половозрелость, как полагают, наступает й возрасте
7—8 мес. Период размножения около 5 мес. В конце апреля—начале
мая добывались беременные самки, а в начале июня попадались молодые
особи. В течение теплого периода у малой бурозубки бывает 1 —3 выводка,
15. РОД SOREX
355
в каждом из которых от 4 до 12 (чаще 6—8) детенышей. Продолжитель-
ность беременности не установлена. Гнездо малой бурозубки представляет
собой шарообразной формы комок сухой травы и других растительных
материалов. Помещается оно под кучами хвороста, в корнях деревьев,
под старыми пнями и в других укромных местах.
Для территории СССР описаны следующие подвиды.
Подвиды. 1. S. т. minutus L., 1766. Характеризуется мелкими
размерами и темной окраской. Распространен в лесной и лесостепной
зонах европейской части СССР, Западной и Средней Сибири, на Алтае,
в Саянах и в Забайкалье.
Рис. 183. Sorex minutissimus.
2. S. т. heptapotamicus Strog., 1957. Отличается светлой окраской.
Распространен в горных районах Казахстана и Киргизии: Джунгарский
и Заилийский Алатау, окрестности г. Алма-Аты и Калбинский Алтай.
34. Sorex minutissimus Zimmermann, 1780 — Крошечная бурозубка
(рис. 183).
Zimmermann, 1780, Gedgr. Gesch., Bd 2, S. 383; — exilis G m e 1 i n, 1788,
in: Linnaeus, Syst. Nat., ed. 13, v. 1, p. 115; — minimus G m e 1 i n, 1788, там же,
с. 156, 157; — pygmaeus Pallas, 1811, Zoogr. Rosso-Asiatica, v. 1, p. 134, 135; —
hawkeri Thomas, 1905, Proc. Zool. Soc. London, v. 2, p. 339. — minutus Огнев,
1913, Млекопитающие Моск, губ., ч. 1, с. ИЗ—119; — tscherskii Огнев, 1913,
Ежегодн. Зоол. муз. АН, т. 18, с. 412, 413; Ьигпеук Thomas, 1915, Ann. Mag. Nat.
Hist., ser., 8, v. 15, p. 499, 500; — ussuriensis Огнев, 1921, Ежегодник Зоол. муз. АН,
т. 22, с. 326—328.
Тип не установлен; вид описан по мь :ериалу из окрестностей с. Кийского, на
р. Кии (г. Мариинск, Кемеровской обл.).
Диагноз. Окраска верхней стороны тела коричневая. Продоль-
ный диаметр первого верхнего промежуточного зуба более такового вто-
рого и третьего. Вершина третьего промежуточного зуба на одном уровне
с вершиной второго зубца I1.
Описание. Морда относительно короткая. Волосы, покрывающие
хвост, редкие и длинные, торчат в стороны. В окраске тела заметно вы-
ражена двуцветность. С темным цветом верха резко контрастирует бело-
ватая или серая окраска нижних частей тела.
23*
356
ПОДСЕМ. SORICINAE
Череп с уплощенной, овальной формы мозговой капсулой и относи-
тельно короткой лицевой частью (длина мозговой капсулы больше или
равна длине лицевой части черепа, измеренной от лобных отверстий
до переднего края межчелюстных костей).
Измерения. Длина: тела — 38.0—53.0, хвоста — 20.0—31.0,
ступни — 6.2—8.7, кондило-базальная черепа — 11.7—14.0 мм.
Распространение и возраст. Известна из северной
и средней частей Восточной Европы, более широко распространена в Азии,
включая Монголию и Японию, распространена также в Северо-Восточном
Китае и на п-ове Корея. В СССР — от западной Карелии на восток
до р. Анадыря, Камчатки, Приморья и Сахалина; на север до тундровой
зоны, к югу до Орловской обл. и Северного Казахстана.
В раннечетвертичное время была распространена в Западной Европе.
Известна из Польши, Франции и Англии, современный.
Биология. Населяет разнообразные биотопы. Встречается в раз-
личных горных и равнинных ландшафтах севера и юга, на болотах и в пой-
менных низинах, полупустынях Казахстана (Большие Басуки). Тем не ме-
нее она не населяет сплошь те или иные биотопы, а выбирает в них,
по-видимому, наиболее выгодные для себя условия. Возможно, этим объяс-
няется ее немногочисленность и спорадичность в распространении. Пи-
тается, как и все землеройки, мелкими беспозвоночными, главным обра-
зом насекомыми и их личинками.
Размножение этого вида мало изучено. Беременные самки попадались
в мае и июле. Число эмбрионов 4—5.
По двиды. 1. S. т. minutissimus Zimm., 1780. Характеризуется
светлой окраской. Распространен в центральных и восточных районах
европейской части СССР и таежной зоне Сибири, к востоку до Енисея
и Забайкалья.
2. S. т. barabensis Strog., 1956. Отличается темной окраской и почти
одноцветным хвостом. Известен пз Барабинской низменности; вероятно,
распространен и в западносибирской лесостепи.
3. S. т. tschuktschorum Strog., 1949. Отличается густым мехом, яркими
тонами окраски спины и боков тела и резко двуцветным хвостом. Распро-
странен в Якутии, на Чукотском полуострове, Анадыре и Камчатке.
4. S. т. tscherskii Ogn., 1913. Отличается преобладанием серых тонов
в окраске тела; хвост слабо двуцветный. Распространен в Приморье
и на Сахалине.
5. S. т. caudatus Judin, 1964. Отличается длинным хвостом и нали-
чием «чепрака». Распространен на Салаирском кряже, Кузнецком Ала-
тау, Западном Саяпе и северо-восточном Алтае.
6. S. т. stroganovi Judin, 1964. Отличается тусклой светлой окраской.
Распространен на Алтае (Чуйская степь). Возможно, распространение
охватывает южный Алтай и сопредельные части Тувы, Казахстана и Мон-
голии.
35. Sorex isodon Turov, 1924 — Равнозубая бурозубка (рис. 184).
Sorex araneus tomensis isodon Туров, 1924, ДАН СССР, с. 109—112; — gravest
Goodwin, 1933, Amer. Mus. Novitates, N 681, p. 3, 4; — isodon ruthenus Стро-
ганов, 1936, Зоол. журн., т. 15, вып. 1; — isodon isodon Туров, 1936, Сб. трудов
Гос. зоол. муз. (при МГУ), т. 3, с. 26, 27; — centralis Долгов, 1964, с. 898—903.
Т и п не установлен, вид описан по экземпляру из долины реки Сосновки, Баргу-
зинская тайга.
Диагноз. Окраска верхней стороны тела темно-коричневая.
Относительно мелкий верхний внутренний резец с крупным задним зуб-
цом. Первый верхний промежуточный зуб крупнее остальных, второй,
15. РОД SOREX
357
третий и четвертый почти равны по высоте, реже третий ниже второго
и четвертого. Пятый промежуточный зуб по высоте не менее 2/а четвертого.
Описание. Окраска верхней и нижней сторон тела сходная или
нижняя несколько светлее верхней — серовато-бурая.
Череп массивный с угловатой мозговой капсулой, более плоский,
чем череп Sorex araneus L., на который он несколько похож, что обус-
ловлено слабым развитием сагиттального и ламбдовидного гребней, со-
здающих впечатление высокого черепа. Зубы имеют темную окраску,
они не отличаются крупными размерами, а промежуточные мало раз-
Рис. 184. Sorex isodon.
нятся по величине. Последнее обстоятельство и послужило основанием
для названия «равнозубая».
Измерения. Длина: тела — 63.0— 75.0, хвоста — 36.0—46.0,
ступни — 12.0—13.5, кондило-базальная черепа — 18.3—19.3 мм.
Распространение и возраст. Известна из Финляндии,
КАССР и ряда районов центральной полосы северо-востока европей-
ской части СССР, из Западной и Восточной Сибири, Камчатки и Дальнего
Востока, современный.
Биология. Специально не изучалась, поскольку вид считался
до недавнего времени подвидом S. araneus L. Вероятно, образ жизни
этой бурозубки напоминает таковой обыкновенной бурозубки, но равно-
зубая бурозубка, можно думать, отличается большей экологической
пластичностью, на что указывает ее более широкий ареал.
Подвиды не изучены.
36. Sorex cylindricauda Milne-Edwards, 1868—1874 — Темнополосая
бурозубка (рис. 185).
Milne-Edwards, 1868—1874, р. 260; — bedfordiae Thomas, 1911, Abstract
Proc. Zool. Soc. London, № 90, p. 3; — wardi fumeolus Thomas, 1911, там же,
№ 100, с. 49; — bedfordiae gomphus G. A 1 1 e n, 1923, Amer. Mus. Novitates, № 100, p. 3.
Тип не установлен, вид описан из Моупина, Сычуань (Китай).
Диагноз. Окраска верхней стороны тела коричневая. Вдоль
хребта проходит темная, коричнево- или черно-бурая полоса. Первые
358
ПОДСЕМ. SORICINAE
три промежуточных зуба крупнее остальных, они равной высоты или
второй промежуточный зуб выше первого и третьего.
Описание. Кроме того, что сказано об окраске в диагнозе, нужно
отметить и следующее: коричневая окраска верхней части тела и боков
местами имеет серый налет, что придает ей особый оттенок по сравнению
с другими коричневыми землеройками. Нижняя сторона тела серая,
с серебристым налетом. Верхние стороны кистей и ступней охристо-
беловатые. Хвост сверху и снизу коричневый. Когти короткие п острые,
сильно сжатые с боков.
Рис. 185. Sorex cylindricauda.
Череп (описывается по дефектному экземпляру из коллекции Зоологи-
ческого института АН СССР) с узким и удлиненным лицевым отделом;
общим строением напоминает череп «S', caecutiens Laxm., но значительно
уже в области предглазничных отверстий. Мозговая капсула вздута.
Зубы сравнительно мелкие. Задний зубец I1 относительно мелкий.
Первый промежуточный зуб с более длинным основанием, чем второй
и третий.
Нижняя челюсть низкая, с невысоким венечным отростком и' относи-
тельно длинным палочковидным угловым отростком.
Коренные зубы начиная с Р4 относительно более длинные, чем эти
зубы у других видов землероек рода Sorex L. Это хорошо заметно па М3,
в строенпп которого еще сохраняются и особенности, свойственные древ-
ним типам последних верхних коренных.
Измерения. По экземпляру из кол. ЗИН АН СССР — длина:
тела — 58.0, хвоста — 61.0, ступни — 14.0 мм. По: G. Allen, 1938 —
длина: тела — 60.0 и 55.0, хвоста — 50.0, 52.0 и 55.0, ступни — 12.0
и 13.0, кондило-базальная черепа — 14.4, 15.0 и 18.1мм. Размеры типа —
длина: тела — 54.0, хвоста — 58.0, ступнп — 12.5, кондило-базальная
черепа — 17.4 мм.
Распространение и возраст. Ганьсу, Шанси, Хсиканг,
Сычуань, Юньнань (Китай), Северная Бирма, современный.
Биология не изучена. Обитает в высокогорных лесах на высоте
3000-5000 м.
15. РОД SOREX
359
Подвиды. 1. S. с. cylindricauda M.-Edw., 1868—1874. Описан
выше. Распространен в Шанси и Сычуани.
2. 5. с. wardi Thos, 1912. Более светлая и с более плоским черепом,
чем типичная форма. Распространена в Ганьсу.
3. S. с. gomphus G. АП., 1923. Отличается более мелкими размерами
и более темной окраской. Распространен в Юньнани (Китай) и Северной
Бирме.
37. Sorex cinereus Kerr, 1792 — Трансарктическая бурозубка
(рис. 186).
Sorex arcticus cinereus Kerr, 1792, Animal Kingdom, p. 206; —personatus Geof-
froy St. - Hilaire, 1828, .Mem. Mus. Hist. Nat., Paris, t. 15, p. 122; — for-
steri Richardson, 1828, Zool. J. v. 3, N 12, p. 516; — cooperi Bachman, 1837,
Рис. 186. Sorex cinereus.
J. Acad. Nat. Sci., Philadelphia, v. 7, pt. 2, p. 388; — jimbripes В achmdn, 1837,
там же, с. 331. Corsira forsteri Gray, 1837—1838, p. 124. Otisorex platyrhynus D e
Kay, 1842, Zool. N. Y., pt 1, Mammalia, p. 22. Siprex) platyrhynchus L i n s 1 e y,
1842, Amer. J. Sci. and Arts, v. 43, N 2, p. 346. Amphisorex lesueurii Duvernoy,
1842, Mag. Zool., ser. 2. v. 4, p. 33, 34. Hiydrogale) jimbripes Pomel, 1848, Bibl.
Univ. Geneve, Arch. Sci. Phys, nat., v. 9, p. 248. Sorex acadicus Gilpin, 1867, Proc,
and Trans. Nova Scotian Inst. Nat. Sci., v. 1, pt 2, p. 2. Crocidura cooperi Fitzinger,
1868, Sitzungsber. Osterr. Acad. Wiss. Math.-naturwiss. KL, Abt. 1, Bd. 57, S. 513; —
platyrhyncha F itzinger, 1868, там же, с. 585. Crossopus jimbripes Fitzinger,
1868, там же, с. 631. Crocidura jimbripes Fitzinger, 1868, там же, с. 631.
Тип не, установлен, вид описан из форта Северн, Онтарио, Канада.
Диагноз. Окраска верхней стороны тела коричнево-бурая раз-
ных оттенков. Первый и второй, а также третий и четвертый верхние
промежуточные зубы образуют пары сходных зубов, но второй немного
выше первого, а четвертый выше третьего. Длина основания первого
промежуточного зуба более длины второго, а третьего — более четвер-
того. Все верхние промежуточные зубы без выемки на заднем крае,
но с глубокой выемкой на передне-внутренней стороне.
Описание. К описанию окраски тела, отмеченной в диагнозе,
следует сделать следующие добавления: у экземпляров из СССР окраска
яркая и темная даже летом, с хорошо выраженным коричневатым чепра-
360
ПОДСЕМ. SORICINAE
ком на спине, резко двуцветная, у отдельных экземпляров трехцветная,
что обусловлено примесью желтоватого тона. Окраска особей этого вида
из США (по: Hall, Kelson, 1959), серая, рыжевато-коричневая или ко-
ричневатая, двух- и трехцветная. Зимой светлее, летом темнее.
Череп с относительно узкой носовой частью и высокой мозговой кап-
сулой. Зубы сравнительно крупные с выпуклыми наружными сторонами.
Последний премоляр слабо развит в длину и опорный зубец наружной
стенки этого зуба расположен почти посредине, а не ближе к переднему
краю, как у S. caecutiens Laxm. Наружные стенки коронок коренных
зубов мало пигментированы.
Измерения. По экземплярам из СССР — длина: тела — 39.0—
48.0, хвоста — 23.0—26.0, ступни — 9.0—11.0, кондило-базальная че-
Рис. 187. Sorex bucharensis.
репа — 14.9—15.2 мм. По экземплярам из США, Hall, Kelson, 1959 —
длина: тела — 71.0—111.0, хвоста — 25.0—50.0, ступни — 10.0—14.0,
кондило-базальная черепа — 14.6—16.9 мм.
Распространение и возраст. Северный Квебек и вся
Северная Канада, северная, западная и центральная части Аляски,
запад Британской Колумбии (исключая побережье), к югу распростра-
нена до Нью-Джерси, гор Северной Каролины и Тенесси, центральной
части Огайо, Южной Индианы, севера Иллинойса, северо-востока Айовы,
востока Миннесоты, северной и восточной части Манитобы, северного
Саскачевана, гор Айдахо, западной Монтаны и западной части Вайоминга
до северной части Нью-Мексико и северной и центральной части штата
Вашингтон. В СССР известна из северо-восточной Сибири и Камчатки,
современный.
Биология. Предпочитает открытые пространства вблизи водое-
мов, по нередко населяет и другие биотопы включая сухие леса низких
ярусов, например на востоке США.
Пищей служат разнообразные беспозвоночные, главным образом
насекомые, и мелкие позвоночные. Размножается с апреля по октябрь.
Число эмбрионов 4—10.
Подвиды на территории СССР не изучены. На американской
части ареала этого вида описано 13 подвидов.
15. РОД SOREX
361
38. Sorex bucharensis Ognev, 1921 — Бухарская бурозубка (рис. 187).
Огнев, 19.21, Ежегодн. Зоол. муз. АН, т. 22, с. 230, 231.
Тип. (? № 8647, ледник Ошанина, долина р. Дивансу, хр. Петра Первого, Тад-
жикистан, кол. ЗИН АН СССР, Ленинград.
Диагноз. Окраска верхней стороны тела буровато-коричневая,
с сероватым оттенком. Верхние промежуточные зубы имеют форму узких
клиньев; второй промежуточный зуб ниже первого и третьего. Задний
зубец I1 так же сильно заострен, как и промежуточные зубы.
Описание. Нижняя сторона тела и хвоста серовато-белая. Череп
с относительно короткой лицевой частью и неширокой удлиненной мозго-
Рис. 188. Sorex kozlovi (А) (по: Строганов, 1952), Microsorex hoyi (Б).
вой капсулой. Длина мозговой капсулы, измеренной от мыщелков до зад-
него края межглазничного сужения, примерно равна расстоянию между
отверстиями на лобных костях и передним краем носовых костей.
Измерения. Длина: тела — 49.0—69.5, хвоста — 42.0—50.0,
ступни — 10.4—12.3, кондило-базальная черепа — 16.0—17.9 мм.
Распространение и возраст. Таджикистан (хр. Петра
Первого, Гиссарский), современный. Ареал этого вида, по-видимому,
значительно шире, чем представляется. Близкий к нему вид (вероятно,
подвид — Sorex kozlovi Strog.) найден в Каме (Восточный Тибет).
Биология. Места обитания — березовые и тополевые, реже
арчовые леса высокогорий. Выбирает участки с влажной почвой, покры-
той толстым слоем мертвой подстилки и густой травянистой раститель-
ностью. Пищей служат различные насекомые. В июле—августе добыва-
лись молодые. В это же время были добыты кормящая и беременная самки,
у последней оказалось 7 эмбрионов. Этими краткими данными и исчер-
пываются наши сведения по биологии этой своеобразной землеройки.
39. Sorex kozlovi Stroganov, 1952 — Тибетская горная бурозубка
(рис. 188).
Строганов, 1952, с. 21, 22.
Тип. $ № 8123, Кам, р. Дзачу (Меконг), Восточный Тибет, сборы П. К. Коз-
лова, кол. ЗИН АН СССР (черепа нет), Ленинград.
362
ПОДСЕМ. SORICINAE
Диагноз (по: Строганов, 1952). Отличается от Sorex bucharensts
Ogn. строением и более мелкими размерами I1. Его вторая (задняя) вер-
шина лишь слегка ниже передней и заметно выше первого промежуточ-
ного зуба (у <9. bucharensts Ogn. задняя вершина I1 значительно ниже
как передней вершины этого зуба, так и первого промежуточного). Раз-
меры мелкие.
Описание (по: Строганов, 1952). Летний мех длинный, довольно
пушистый и шелковистый. Хвост покрыт относительно длинными и жест-
кими волосами, образующими на конце длинную кисточку. Окраска
меха, насколько можно судить по спиртовому экземпляру, относительно
светлая, шоколадно-буроватая на спинной стороне, постепенно светлеет
на боках и переходит в белую с палевым налетом на брюшной стороне.
Хвост слабо двуцветный. Верхняя сторона кистей и ступней грязно-
белая. К этому следует добавить, что мех не столь длинный и шелкови-
стый, как пишет Строганов.
Измерения (по: Строганов, 1952). Длина: тела — 40.6, хвоста —
32.8, ступни — 10.5, кондило-базальная черепа — 15.2, верхнего зуб-
ного ряда — 7.0 мм.
Распространение и возраст. Кам, Восточный Тибет,
современный.
Примечание. При отсутствии черепа трудно судить, чем в его
строении данный вид отличается от Sorex bucharensts Ogn. Можно думать,
что он все же представляет собой лишь подвид последнего. Строганов
оба вида выделил в подрод Eurosorex, типом которого ему послужил
S. bucharensts Ogn. Изучение представителей рода Sorex L. из разных
областей широкого ареала последнего не позволяет выделить и достаточно
обосновать какие-либо группировки видов с общим для них адаптивным
направлением. То же можно сказать и о подроде Eurosorex.
Виды Sorex bucharensts Ogn. и S. kozlovi Strog. В. А. Долгов и P. С. Гоф-
ман (1977) свели в синонимы >9. minutus thlbetanus Kastschenko, 1905,
добавив туда и «S', planiceps Miller, 1911. Этому новообразованию авторы
дали название >9. thibetanus Kastschenko. Не вдаваясь в обсуждение
систематического соотношения A. bucharensts и S. kozlovi с другими ви-
дами бурозубок Центральной Азии за отсутствием материала отмечу,
что оба названных вида не могут быть сведены в синонимы A. minutus
thibetanus, потому что последняя — типичная S. minutus, в чем нетрудно
убедиться, посмотрев череп типа <9. minutus thibetanus Kastschenko № 6557
(Цайдам, сборы Н. М. Пржевальского), хранящегося в коллекции ЗИН
АН СССР.
40. Sorex excelsus G. Allen, 1923.
G. Allen, 1923, Amer. Mus. Novitates, N 100, p. 4.
Тип. в (шкура и череп) № 44359, Хо-Шан, Пейтай, в 30 милях к югу от Хунг-
Тени, восточная часть Юньнани, Китай (высота 2200 м), кол. Амер. муз. естеств.
истор., Нью-Йорк.
Описание (по: G. Allen, 1938). Верхняя сторона тела серо-корич-
невая, разных оттенков, нижняя — сероватая, с коричневым оттенком.
Хвост двуцветный — коричневатый сверху, светлый снизу. Верхняя
сторона кистей и ступней серебристо-серая.
Череп с узкой лицевой частью. Соотношение размеров и форма верхних
промежуточных зубов напоминает таковые S. araneus L.
Измерения. Длина: тела — 60.0—62.0, хвоста — 50.0—51.0,
ступни — 13.0—14.0, наибольшая черепа — 18.7, нижнего зубного ряда —
7.8 мм.
15. РОД SOREX
363
Распространение и возраст. Юньнань (Китай), совре-
менный.
41. Sorex sinalis Thomas, 1912.
Thomas, 1912, Ann. Mag. Nat. Hist., ser. 8, v. 10, p. 398.
Тип. <5 № 12.8.5.3, 45 миль южнее Фенасиангфу, Шэнси, Китай, кол. Брит,
муз., Лондон.
Описание (по: G. Allen, 1938). Крупная землеройка, коричневая
сверху и снизу. Верхняя сторона кистей и ступней коричневато-белая.
Хвост сверху коричневый, снизу светлее.
Череп с удлиненной лицевой частью.
Измерения. Длина: тела — 70.0, хвоста — 35.0, ступни — 14.0,
кондило-базальная черепа — 20.3, нижнего зубного ряда — 9.0 мм.
Распространение и возраст. Шэньси, Ганьсу (Китай),
современный.
42. Sorex lyelli Merriam, 1902.1
Sorex tenellus lyelli Merriam, 1902, Proc. Biol. Soc. Washington, v. 15, p. 75.
Тип не установлен, вид добыт в Маунт-Лиель, Калифорния (США).
Распространение. Калифорния (США).
43. Sorex preblei Jackson, 1922.
J a c k s о n, 1922, J. Wash. Acad. Sci., v. 12, p. 263.
Тип из долины Джордан, Орегон (США).
Распространение. Орегон (США).
44. Sorex milleri Jackson, 1947.
J a c k s о n, 1947, Proc. Biol. Soc. Washington, v. 60, p. 131.
Тип из Madera Camp., Sierra del Carmen, Coahuila, Мексика^
Распространение. Мексика.
45. Sorex longirostris Bachman, 1837.
Bachman, 1837, J. Acad. Nat. Sci. Philadelphia, ser. 1, v. 7, № 2, p. 370.
Тип из болот Sante River, Южная Каролина (США).
Распространение. Юго-восточные штаты Северной Америки.
46. Sorex vagrans Baird, 1858.
Baird, 1858, Mammals, in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1, p. 15; —
suckley Baird, 1858, там же, с. 18—20; — pacificus Coues, 1877, Bull. U. S. Geol.
Surv. Terr., v. 3, p. 560; — dobsoni Merriam, 1895, North. Amer. Fauna, № 5, p. 33,
34; — amoenus Merriam, 1895, p. 69, 70; — nevadensis Merriam, 1895, p. 71,
72; — shastensis Merriam, 1899, там же, № 16, с. 87.
Тип с побережья мелководного залива, штат Вашингтон (США).
Распространение. Западные штаты Северной Америки.
47. Sorex ornatus Merriam, 1895.
Merriam, 1895, p. 79.
Тип из каньона San Emigdio, Калифорния США).
Распространение. Калифорния (США).
1 Этот вид и следующие за ним виды рода Sorex L., из США, требующие тщательной
ревизии, приведены без описания и указания возраста.
364
ПОДСЕМ. SORICINAE
48. Sorex tenellus Merriam, 1895.
Merriam, 1895, p. 81.
Тип из Lone Pine Crick, Калифорния, (США).
Распространение. Калифорния, Невада (США).
49. Sorex trigonirostris Jackson, 1922.
J а с к s о и, 1922, J. Wash. Acad. Sci., v. 12, p. 264.
Тип из Ashland, Орегон (США).
Распространение. Орегон (США).
50. Sorex nanus Merriam, 1895.
Sorex tenellus nanus M e r r i a m, 1895, p. 21.
Тип из Восточного парка, Колорадо (США).
Распространение. Калифорния, отдельные части Вайо-
минга. Южной Дакоты, Колорадо, Нью-Мексико, Юты и Аризоны (США).
51. Sorex juncensis Nelson et Goldman, 1909.
Sorex californicus juncensis Nelson, Goldman, 1909, Proc. Biol. Soc. Wa-
shington, v. 22, p. 27.
Тип из Soccoro, п-ов Калифорния (США).
Распространение. Известен с п-ова Калифорния (США).
52. Sorex willeti von Bloeker, 1942.
Von Bloeker, 1942, Bull. Counthern Calif. Acad. Sci., v. 40, p. 163.
Тип из каньона Авалон на о. Санта-Каталина.
Распространение. Известен из одного местонахождения.
53. Sorex sinuosus Grinnell, 1913.
Grinnell, 1913, Univ. Calif. Publ. Zool., v. 10, p. 187.
Тип co. Гризли, Калифорния (США).
Распространение. Известен только из типичного место-
нахождения.
54. Sorex veraepacis Alston, 1877.
Sorex verae-pacis Alston, 1877, Proc. Zool. Soc. London, p. 445.
Тип из Кобан, Гватемала.
Распространение. Южная часть Мексики и Гватемала.
55. Sorex macrodon Merriam, 1895.
Merriam, 1895, p. 82.
Тип из Орисаба, Веракрус (Мексика).
Распространение. Известен из типичного местонахождения.
56. Sorex palustris Richardson, 1928.
Richardson, 1928, Zool. J. v. 3, № 12, p. 517.
Тип из болотистых равнин Гудзонова залива.
Распространение. Аляска, Канада (северные окраины),
северо-западная и северо-восточная части США.
15. РОД SOREX
365
57. Sorex alascanus Merriam, 1900.
Sorex navigator alascanus Merriam, 1900, Proc. Wash. Acad. Sci., v. 2, p. 18; —
alascanus J а с к s о n, 1926, J. Mammal., v. 7, p. 58.
T п п из пункта Gustavus, зал. Ледовый, Аляска.
Распространение. Аляска.
Примечание. В заключение описания Холл и Келсон (Hall,
Kelson, 1959) высказывают предположение, что этот вид может быть
подвидом S. palustris Rich.
58. Sorex bendirii Merriam, 1884.
Merriam, 1884, Trans. Linnaean Soc., N. Y., v. 2, p. 217.
Тип добыт в 1 миле от р. Вильямсон и 18 милях от форта Klamath, Орегон (США).
Распространение. Британская Колумбия (Канада), Ва-
шингтон, Орегон, Калифорния (США).
59. Sorex pribilofensis Merriam, 1895.
Merriam, 1895, p. 87.
Тип добыт на о. Св. Павла, Аляска (США).
Распространение. Известен из типичного местонахождения.
60. Sorex furneus Miller, 1895.
Miller, 1895, p. 50.
Тип из Питерборо, Медисон, штат Нью-Йорк (США).
Распространение. Онтарио, Квебек (Канада), Висконсин,
Нью-Гэмпшир, Вермонт, Кентукки, Джорджия (США).
61. Sorex jacksoni Hall et Gilmore, 1932.
Hall, Gilmore, 1932, Univ. Calif. Publ, Zool., v. 38, p. 392.
Тип из Севунга, о. Св. Лаврентия (США).
Распространение. Известен только с о. Св. Лаврентия.
62. Sorex hydrodromus Dobson, 1889.
Dobson, 1889, Ann. Mag. Nat. Hist., ser. 6, v. 4, p. 373.
T и и № 2389 с о. Уналашка, Алеутские острова, Аляска (США), добыт И. Г. Воз-
несенским в 1842 г. Хранится в ЗИН АН СССР, Ленинград.
Примечание. До настоящего времени известны только два, пло-
хой сохранности, спиртовых экземпляра этого вида. После Вознесен-
ского нпкто на этом острове землероек не находил. Сравнение экземпля-
ров, добытых Вознесенским, с другими видами землероек из Северной
Америки и северо-восточной Азии позволило обнаружить их значитель-
ное сходство с экземплярами <9. cinereus Kerr. — вида, широко распро-
страненного в Северной Америке и северо-восточной Азии. Есть все осно-
вания считать, что S. hydrodromus Dobs, должен быть отнесен в синонимы
S. cinereus Kerr.
63. Sorex gaspensis Anthony et Goodwin, 1924.
Anthony, Goodwin, 1924, Amer. Mus, Novitates, N 109, p. 1.
Тип с п-ова Гаспе, Квебек (Канада).
Распространение. Квебек, Канада.
366
ПОДСЕМ. SORICINAE
64. Sorex dispar Batchelder, 1911.
Sorex macrurus Batchelder, 1896, Proc. Biol. Soc. Washington, v. 10, p. 133;
— dispar Batchelder, 1911, там же, т. 24, с. 97.
Тип из области Эссек, штат Нью-Йорк (США).
Распространение. Нью-Гемпшир, Нью-Йорк, Пенсильва-
ния, Нью-Джерси, Виргиния, Западная Виргиния, Северная Каролина,
Теннеси (США).
65. Sorex trowbridgii Baird, 1858.
Baird, 1858, Mammals, in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1, p. 13.
Тип добыт из Астории (устье р. Колумбии), Орегон (США).
Распространение. Британская Колумбия (Канада), Ва-
шингтон, Орегон, Невада, Калифорния (США).
66. Sorex merriami Dobson, 1890.
Dobson, 1890, pl. 23, fig. 6.
Тип ср. Малый Биг-Хорн, около мили выше форта Кастер, область Биг-Хорн,
Монтана (США).
Распространение. Вашингтон, Монтана, Северная Дакота,
Орегон, Вайоминг, Невада, Калифорния, Колорадо, Аризона, Нью-
Мексико (США).
67. Sorex saussurei Merriam, 1892.
Merriam, 1892, Proc. Biol. Soc. Washington, v. 7, p. 173; — durangae J a-
c k s о n, 1925, Proc. Biol. Soc. Washington, v. 38, p. 127.
Типе северного склона Сьерра-Невадо-де-Колима, Халиско, Мексика.
Распространение. Мексика. Гватемала.
68. Sorex oreopolus Merriam, 1892.
Merriam, 1892, Proc. Biol. Soc. Washington, v. 7, p. 173.
Тип с северного склона Сьерра-Невадо-де-Колима, Халиско, Мексика.
Распространение. Мексика.
69. Sorex sclateri Merriam,' 1897.
Merriam, 1897, Proc. Biol. Soc. Washington, v. 11, p. 228.
Тип из Тумбала, Чьяпас, Мексика.
Распространение. Мексика.
70. Sorex stizodon, Merriam, 1895.
Merriam, 1895, p. 98.
Тип из Сан-Кристобаль, Чьяпас, Мексика.
Распространение. Известен только из типичного место-
нахождения.
16. Род MICROSOREX Cones, 1877
Cones, 1877, Bull. U. S. Geol. and Geogr. Surv., v. 3, p. 646.
Тип. Sorex hoyi Baird, 1858, Канада, северная и восточная части США.
Диагноз. Третий и пятый верхние промежуточные зубы очень
мелкие, причем третий в виде диска, зажатого между соседними зубами.
Примечание. По внешнему облику представители этого рода
сходны с особями мелких видов рода Sorex L. Описываемый род хорошо
отличается от других родов трибы Soricini строением верхних промежу-
точных зубов.
16. РОД MICROSOREX. 17. РОД TREGOSOREX
367
Распространение ивозраст. Канада, северные и вос-
точные штаты США, плейстоцен—современный.
t 1. Microsorex pratensis Hibbard, 1944.
Hibbard, 1944, Bull. Geol. Soc. Amer., v. 55, p. 722.
Тип. Фрагмент левой ветви нижней челюсти с Р4—М3, № 6470, Канзас, плей-
стоцен, кол. муз. Канзас, упив., Лоренс.
Д и ат? н о з. (по: Hibbard, 1944). Размеры крупнее, чем у Microsorex
hoyi intervectus Jackson, P4 крупнее, а-ветви нижней челюсти и зубы выше.
Распространение и возраст. Канзас (США), плейсто-
цен.
2. Microsorex hoyi Baird, 1858.
Baird, 1853, Mammals, in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1, p. 32.
Тип не установлен, добыт в Рацине, Висконсин (США).
Диагноз. Череп плоский, неширокий в затылочной части. Верх-
ние внутренние резцы крупные, сильно выступают вперед. Их передний
зубец раза в два и более крупнее заднего зубца.
Описание. Верхняя сторона тела серо-коричневая или серая.
Хвост двуцветный. Череп с короткой и более или менее широкой ростраль-
ной частью. Зубы мелкие.
Измерения. Длина: тела — 50.0—67.0, хвоста — 27.0—35.0,
ступни — 8.5—12.0; кондило-базальная черепа — 13.9—15.8 мм. Холл
и Келсон (Hall, Kelson 1959) приводят длину тела 78.0—98.0 мм.
Распространение ивозраст. Северная часть Квебека,
Онтарио и северо-западные территории Канады, центральная и западные
части Аляски, северные и восточные окраины США.
f 17. Род TREGOSOREX Hibbard et Jammot, 1971
Hibbard, Jammot, 1971, Con trib. Mus. Paleontol. Univ. Michigan, v. 23,
N 24, p. 377, 378.
T и n. Tregosorex holmani Hibbard et Jammot, 1971, область Трего, Канзас (США),
нижний плиоцен.
Диагноз (по: Hibbard, Jammot, 1971). Ветви нижней челюсти
высокие. Сочленовный отросток нижней челюсти имеет выемку только
на лингвальной стороне. Его нижняя сочленовная фасетка имеет сильно
развитую бороздку, отделяющую ее основание от дорсальной части меж-
кондилярной вырезки. Крупное заднемандибулярное отверстие располо-
жено позади заднего края темпоральной ямки на венечном отростке.
Р3 и Р4 (12 и Р4) с одним корнем.
Описание (по: Hibbard, Jammot, 1971). Размеры крупнееBla-
rina brevicauda Say. Темпоральная ямка венечного отростка высокая,
треугольная, с несколько округлой вершиной. Подбородочное отверстие
расположено под Мд.
Распространение и возраст. Северная Америка, ниж-
ний плиоцен.
f 1. Tregosorex holmani Hibbard et Jammot, 1971 (рис. 189).
Hibbard, Jammot, 1971, там же, с. 377, 378.
Тип. Левая ветвь нижней челюсти с зубами 1д и Мд—М3. № V 60444, область
Трего, Канзас (США), нижний плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн
Арбор.
Описание, (по: Hibbard, Jammot, 1971). Метаконид на Мд и М2
придвинут к сильно развитому протокониду. Энтоконид соединен с мета-
368
ПОДСЕМ. SORICINAE
конидом, не отделен выемкой. Хиполофид Мх и М2 отделен от основания
метаконида. М3 сильно редуцирован за счет уменьшения размеров тало-
нида (метаконида). Мх—М3 с сильно развитыми цингулюмами. Все зубы
пигментированы.
Измерения (по: Hibbard, Jammot, 1971). Длина: Мт—М3 — 5.01,
Мх — 1.90 мм.
Распространение и возраст. Канзас (США), нижний
плиоцен.
Рис. 189. Tregosorex holniani (по: Hibbard, Jammot, 1971).
Примечание. Структурные особенности нижней челюсти пред-
ставителей этого рода (массивные ветви челюсти, также сочленовный
и венечный отростки, короткий, с утолщенным основанием первый ниж-
ний резец, без бугорков на его верхней стороне) позволяют считать эту
группу землероек приспособленной к питанию грубой пищей. Сходство
в строении сочленовного и венечного отростков с таковыми видов трибы
Soricini, а не Blarinini, может служить основанием для включения этого
рода в первую трибу. При всем этом следует отметить, что виды рода
Tregosorex были более специализированы по сравнению с видами рода
Sorex L., но, по-видимому, менее универсально приспособленными, что,
возможно, и обусловило их вымирание.
118. Род ANCHIBLARINELLA Hibbard et Jammot, 1971
Hibbard, Jammot, 1971, там же, с. 380.
Тип. Anchtblarinella wakeeneyensis Hibbard et Jammot, 1971, Канзас (США),
нижпип плиоцен.
18. РОД ANCHIBLARINELLA 19. РОД BLARINELLA
369
Диагноз (по: Hibbard, Jammot, 1971). Тригонид М3 (передний
сегмент, пара-протоконпд, по-моему) редуцирован, лопастевпдный. Ве-
нечный отросток более широкий, чем у видов рода Sorex и имеет хорошо
развитый шип. Сочленовный отросток направлен назад, сочленовные фа-
сетки широко разведены. Верхняя фасетка овальной формы, нижняя
расширена книзу, как у Blarinella Thomas (ссылка на рисунок: Repen-
ning, 1967, р. 35, fig. 24).
Описание (по тем же авторам). Немного крупнее Sorex taylori
Hibb., с пигментированными зубами. Ветви нижней челюсти строением
напоминают таковые представителей рода Sorex, сходен и Р4. Подбо-
родочное отверстие расположено под средней частью М4. Крупные пе-
Рис. 190. Anchiblarienella wakeensyensis (по: Hibbard, Jammot, 1971).
реднее и заднее мандибулярные отверстия расположены довольно низко.
Распространение и возраст. Северная Америка, ниж-
ний плиоцен.
t 1. Anchiblarinella wakeeneyensis Hibbard et Jammot, 1971 (рис. 190).
Hibbard, Jammot, 1971, там же, с. 380.
Тип. Левая ветвь нижней челюсти с зубами Р4—М3, № V 60446 область Трего,
Канзас (США), нижний плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн Арбор.
Описание. Кроме признаков, приведенных в диагнозе и описа-
нии, составленных мной по диагнозу Хиббарда и Джаммо, названные ав-
торы в описании вида привели и следующие: «Темпоральная ямка (на ве-
нечном отростке) треугольной формы, как у видов рода Sorex. Выемка
на лабиальной стороне ветви нижней челюсти расположена под М2». Дру-
гие признаки, данные авторами в сравнении с таковыми Petenyia concisa
Hibb. et Jamm., здесь не приведены.
Измерения, (по указанным выше авторам). Длина: Р4—М3 —
4.50, Мх-М3 - 3.82, М4 — 1.36, М2 — 1.31, М3 - 1.13 мм.
Распространение и возраст. Канзас (США), нижний
плиоцен.
19. Род BLARINELLA Thomas, 1911
Thomas, 1911, Proc. Zool. Soc. London, p. 166; Sorex Milne-Edwards, 1868—
1874, p. 261.
Tnn. Sorex quadraticauda M.-Edw., 1868—1874, Центральный и Южный Китай,
Северная Бирма.
Диагноз. Первые два промежуточных зуба крупные; остальные
мелкие. Четвертый и пятый смещены на внутреннюю сторону зубного
ряда и снаружи часто незаметны. i
Описание. Окраска тела одноцветная сверху и снизу, реже
нижние части значительно светлее верхних. На фоне темной окраски
кистей и ступней резко выступают светлые длинные когти, особенно
на пальцах кисти. Хвост черно-бурый сверху и снизу. Жесткие волосы
24 А. А. Гуреев
370
ПОДСЕМ. SORICINAE
на конце морды не такие длинные и толстые, как у видов рода Sorex L.
Череп относительно крупный, массивный, с узкой лицевой частью
и слабо выступающими в стороны верхнечелюстными костями. Переднее
носовое отверстие большое. Его задний край находится на одном уровне
с передним краем последнего премоляра, если их соединить линией.
Нижняя челюсть относительно массивная, с более высокой зубной частью,
чем у видов Sorex L., такого же размера. Нижние зубы сравнительно мел-
кие, что обусловлено, по-видимому, укорочением верхнего зубного
ряда и в связи с уменьшением размеров третьего, четвертого и пятого
промежуточных зубов.
Рис. 191. Blarinella quadraticauda.
Отличия этого рода от других хорошо выступают при сравнении
диагнозов.
Распространение и возраст. Центральный и Южный
Китай, Северная Бирма, современный.
Примечание. Ряд особенностей, характерных для представи-
телей рода Blarinella Thos., например массивное тело, укороченный
хвост, малые размеры ушных раковин и длинные когти, позволяет счи-
тать их приспособленными к рытью.
1. Blarinella quadraticauda Milne-Edwards, 1872 (рис. 191).
Milne-Edwards, 1868—1874, р. 261; — grtselda Thomas, 1912, Ann.
Mag. Nat. Hist., ser. 8, v. 10, p. 400; — wardi Thomas, 1915, там же, сер. 8, т. 15,
с. 336.
Тип не выделен. Экземпляр, послуживший для описания, добыт в Моупине (За-
падный Китай) и хранится в Муз. естеств. истор. в Париже.
Диагноз совпадает с таковым рода.
Единственный экземпляр этого вида, хранящийся в ЗИН АН СССР,
имеет двуцветную окраску. Верхняя сторона тела коричнево-серая с се-
ребристым, местами пепельно-серым налетом. Нижние части тела светлые,
сероватые. Череп (дефектный) с сильно стертыми зубами.
19. РОД BLARINELLA
371
Измерения. Длина: тела — 62.0—70.0, хвоста — 33.0—39.0,
ступни — 11.0—12.0, кондило-базальная черепа (тип одного из подви-
дов) — 18.6, основная черепа — 17.0—18.0 мм.
Распространение и возраст. Ареал вида имеет сравни-
тельно ограниченную протяженность в горах и горных районах Ганьсу,
Сычуани и Юньнани в Китае и Северной Бирме. Плейстоцен? —совре-
менный.
Подвиды. 1. В. q. quadraticauda M.-Edw. 1868—1874. Распро-
странен в Моупине, Сычуань (Китай).
2. В. q. griselda Thos., 1912. Отличается мелкими размерами, более
серой окраской и коротким хвостом. Распространен в Ганьсу (Китай).
3. В. q. wardi Thos., 1915. Отличается темной, резко двуцветной окрас-
кой и относительно более коротким черепом. Распространен в Юньнани
(Китай) и в Северной Бирме.
4. Триба CROCIDURINI Stirton, 1930
Crocidurinae Milne-Edwards, 1868—1874, p. 256. Подсем. Scutisoricinae
J. A I 1 e n, 1917, p. 769—784. Crocidura-group S t i r t о n, 1930, p. 224.
Тип. Crocidura Wagler, 1832, Европа, Азия и Африка, миоцен—современный.
Диагноз. Зубная формула: I-f-С Р-у-М= 32—26. Верхних
промежуточных зубов 4, 3 или 2. Все зубы белые. Верхняя и нижняя
сочленовные фасетки сочленовного отростка нижней челюсти находятся
одна под другой (нижняя сочленовная фасетка не выдвинута вперед,
как и засочленовный отросток височной кости, на который она опирается
при сочленении с черепом). Надглазничных отверстий нет (исключение —
подтриба Myosoricini). Хвост покрыт короткими волосами и редкими
длинными щетинками или только короткими волосами.
Распространение и возраст. Европа, плиоцен—совре-
менный; Азия, миоцен—современный; Африка, миоцен—современный;
Северная Америка, миоцен—плиоцен.
Примечание. Представители трибы известны с нижнего миоцена,
но можно считать, что они обособлялись еще в конце олигоцена, так как
в нижнем миоцене уже имели все характерные для этой трибы особен-
ности.
Обособление и развитие этих землероек шло по пути приспособления
к быстрому схватыванию добычи, главным образом сравнительно крупных
насекомых и мелких позвоночных, удержанию челюстями и разжевыванию
их относительно жестких покровов и волокнистых тканей.
В связи с этим приспособлением у видов трибы Crocidurini происхо-
дило сокращение числа верхних промежуточных зубов (например, у Dip-
lomesodon Brandt — до 2), а также увеличение размеров первого верхнего
промежуточного зуба, выполняющего у них функцию клыка. В связи
с изменением количества промежуточных зубов и сокращением размеров
оставшихся (кроме I2) произошло сокращение длины верхней челюсти
и изменение соотношения размеров верхних промежуточных зубов,
чем и обусловлена редукция лопастей (бугорков) на верхнем (режущем)
крае нижнего внутреннего резца у ряда групп этой трибы. Важное зна-
чение быстрого закрывания рта обусловило сохранение более древнего
типа сочленения нижней челюсти с черепом. Верхняя и нижняя сочленов-
ные фасетки на сочленовном отростке нижней челюсти расположены одна
под другой, соответственно засочленовный отросток на черепе (опора
для нижней сочленовной фасетки) не выдвинут вперед. Перечисленные
здесь особенности жевательного аппарата являются общими для всех
родов и видов, объединяемых трибой Crocidurini.
24*
372
ПОДСЕМ. SORICINAE
Вместе с этим следует отметить, что по характеру адаптаций объеди-
няемых видов описываемая триба не может считаться однородной. На-
ряду с видами со значительной специализацией жевательного аппарата,
например представители рода Diplomesodon Brandt, существуют виды
из родов Myosorex Gray и Surdisorex Thos., сохраняющие архаичные осо-
бенности, свойственные древним группам землероек — три промежуточных
зуба в нижней челюсти.
Виды, сохраняющие приспособление к питанию относительно мелкими,
очень подвижными животными, сохраняют и большое число верхних
промежуточных зубов (4), а также и относительно длинные челюсти,
удобные для быстрого схватывания мелкой добычи. Землеройки, приспо-
собленные к питанию относительно крупными и сильными животными
(крупные беспозвоночные, мелкие позвоночные, в частности ящерицы),
имеют наименьшее число верхних промежуточных зубов (2) и укорочен-
ные челюсти, что более выгодно для схватывания и удержания сильных
животных.
Подавляющее большинство видов трибы распространено в субтропи-
ческой и особенно в тропической зонах. Сравнительно небольшое их число
населяет южные окраины умеренной зоны.
1. Подтриба МYOSORICINA Kretzoi, 1965
Триба Myosoricini Kretzoi, 1965, р. 124.
Тип. Myosorex Gray, 1837, Африка, плейстоцен—современный.
Диагноз. Зубная формула: I С Р ~~ М -|- == 32—30. Верхних про-
межуточных зубов 4 или 3. В нижней челюсти 3 промежуточных зуба.
Надглазничные отверстия имеются. Хвост покрыт только короткими
волосами или короткими волосами и длинными редкими щетинками.
Распространение и возраст. Южная Африка, совре-
менный.
Примечание. Следует отметить известное сходство между со-
временными представителями родов Myosorex Gray и Surdisorex Thos.
и видами древних групп землероек, например миоценового рода Mioso-
rex Kretz., выражающееся в большем, чем у других современных родов,
числе нижних промежуточных зубов — 3, у других современных — 2.
В связи с этим представляется правильным мнение о давнем обособ-
лении представителей трибы Crocidurini от общего ствола землероек и
раннем их проникновении в Африку.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Когти пальцев кисти короткие. Верхних промежуточных зубов 4
или 3. Отверстия на лобных костях имеются . . . .21. Myosorex.
2 (1). Когти пальцев кисти длинные (5—6 мм). Верхних промежуточных
зубов 3. Отверстий на лобных костях нет..........22. Surdisorex.
Примечание. В таблицу не включен род Stirtonia nov. за отсутствием срав-
нимых признаков.
f 20. Род STIRTONIA Gureev gen. nov.
Limnoecus niobrarensis Macdonald, 1947, Amer. J. Sci., v. 245, p. 123—126;
Гуреев, 1971, c. 137.
T и n. Limnoecus niobrarensis Macdonald, 1947, Небраска (США), верхний миоцен.
Диагноз. Ij короткий, массивный, с двумя бугорками на верх-
ней стороне. Нижних промежуточных зубов 3.
20. РОД STIRTONIA. 21. РОД MYOSOREX
373
Описание. Нижняя челюсть низкая с высоким венечным отро-
стком. Ямка для прикрепления височного мускула на венечном отростке
небольшая. Сочленовный отросток крупный, массивный, особенно в ниж-
ней части. Основание углового отростка широкое.
Рис. 192. Stirtonia niobrarensis (Л) (по: Macdonald, 1947), Surdisorex погае (Б) (по: Re-
penning, 1967), Myosorex varius (В) (no: Dobson, 1890).
Распространение и возраст. Северная Америка, верх-
ний миоцен.
Примечание. Род выделен на основании описания Limnoecus
niobrarensis Macdonald, 1947.]
f 1. Stirtonia niobrarensis Macdonald, 1947 (рис. 192, A).
Limnoecus niobrarensis Macdonald, 1947, там же.
Тип. Правая ветвь нижней челюсти с полным рядом зубов № 36 171, р. Ниоб-
рара, Небраска (США), верхний миоцен, кол. Палеонтол. муз. Калифор.унив., Беркли.
Распространение и возраст. Небраска (США), верх-
ний миоцен.
21. Род MYOSOREX Gray, 1837 — МЫШИНЫЕ ЗЕМЛЕРОЙКИ
Gray, 1837—1838, р. 124. Suncus (Myosorex) Е И erm an, Morrison-
Scott, Hayman, 1953. Southern African Mammals, p. 20.
Тип. Sorex varius Smuts, 1832, Южная Африка, современный.
374
ПОДСЕМ. SORICINAE
Д и а г н о з. Зубная формула: I С Р М == 32—30. Ушные рако-
вины нормально развиты. Когти короткие. Отверстия на лобных костях
имеются. В верхней челюсти четыре илй три промежуточных зуба.
Описание (по: Gray, 1837—1838). Голова удлиненная. Уши спря-
таны в мягкой шерсти. Хвост длинный, тонкий, покрыт короткими, же-
сткими, плотно прилегающими волосами. С возрастом он становится
четырехугольным (очевидно, в сечении). Пальцы кисти и ступни не опу-
шены ресницами (имеются в виду плавательные щетинки). Зубы белые.
Режущих зубов (промежуточных ) 8. Два верхних центральных зуба
(Р) — двураздельные (с двумя вершинами). Второй наружный (I3) —
умеренный. Третий очень мелкий, рудиментарный; четвертый мелкий,
но крупнее третьего. Передний нижний режущий зуб (IJ удлиненный,
с совершенно острым верхним краем. Второй и третий наружные зубы
(промежуточные) мелкие, простые, теснятся на основании Iv
Дополнение к описанию (по: Walker et al., 1964): окраска бурая,
желто-коричневая, коричневая, серая или черноватая разных оттенков,
нередко с пятнами вследствие смешения светлых и темных волос. У одних
видов кисти и ступни сверху имеют светло серую окраску, у других они
окрашены в коричневый или черноватый цвет. Крупные мускусные же-
лезы у самцов располагаются, как и у других групп землероек, на боках
тела позади передних конечностей. В нижней челюсти семь зубов. По-
зади второго резца имеется мелкий зуб. Если он отсутствует, то его аль-
веола сохраняется.
Измерения. Длина тела — 60.0—109.0, хвоста — 29.0—56.0,
ступни — 12.0—16.0, уха — 7.0—12.0, черепа — 21.0—25.7 мм.
Распространение и возраст. Африка к югу от Сахары,
миоцен—современный.
Можно думать, что представители рода сохраняют морфологические
особенности миоценовых предков.
Биология. Образ жизни этих землероек недостаточно изучен.
Основными местами их обитания служат сырые местности с густыми
лесами или зарослями кустарников, а также берега горных потоков и
горные долины, покрытые густой растительностью.
1. Myosorex varius Smuts, 1832 (рис. 192, В).1
Sorex varius Smuts, 1832, Mammal. Capens., p. 8. Myosorex varius Gray,
1837—1838, p. 124.
Тип пе отмечен, вид описан из Капской провинции (ЮАР).
Распространение. Южная Африка.
2. Myosorex cafer Sundevall, 1848.
Sundevall, 1848, Ofvers Vetensk. Akad. Forh., v. 3, p. 119; — sclateri Tho-
mas, Schwann, 1905, Abstrakt Zool. Soc. London, N 15, p. 10; — tenuis Tho-
mas, Schwann, 1905, Proc. Zool. Soc. London, v. 1, p. 132; — swinnyi Chubb.,
1908, Ann. Transvaal Mus., v. 1, p. 2.
Распространение. Южная Африка, современный.
3. Myosorex blarina Thomas, 1906.
Thomas, 1906, Ann. Mag. Nat. Hist., ser. 8, v. 18, p. 139.
Распространение. Уганда.
1 Этот вид и другие современные виды рода Myosorex Gray, требующие ревизии,
приводятся без описания и указания возраста.
21 РОД MYOSOREX . 22. РОД SURDISOREX
375
4. Myosorex babaulti Heim de Balsac, 1956.
Heim de Balsac, 1956a, Mammalia, t. 20, N 2, p. 150—153.
Распространение. Киву, Конго.
5. Myosorex preussi Matschie, 1893.
Matschie, 1893, Sitzungsber. Ges. Naturf. Freunde, Berlin, S. 177.
Г. Аллен (G. Allen, 1893) отнес этот вид к роду Sylvisorex Thos.
Распространение. Камерун, Южная Нигерия.
6. Myosorex polli Heim de Balsac, 1956.
Heim de Balsac, 1956a, p. 155—157.
Распрост p’a некие. Конго.
Примечание. Этот вид выделен автором в подрод Congosorex.
Основанием к этому послужило отсутствие в верхней челюсти четвертого
промежуточного зуба, характерного для представителей рода Myosorex
Gray.
22. Род SURDISOREX Thomas, 1906
Thomas, 1906, Ann. Mag. Nat. Hist., ser. 7, v. 28, p. 223.
Тип. Surdisorex norae Thomas, 1906, Кения.
3 113
Диагноз. Зубная формула: 1у Су Ру М-==30. Ушные раковины
рудиментарны. Когти длинные (6.5—6.0 мм). Отверстий на лобных ко-
стях нет. В верхней челюсти 3 промежуточных зуба.
Примечание. Г. де Бальзак (Heim de Balsac, 1956), сопоставляя
особенности родов Myosorex Gray и Surdisorex Thos., отмечает для послед-
него отсутствие третьего промежуточного зуба в нижней челюсти. Этот
зуб иногда отсутствует и у представителей рода Myosorex Gray. Возможно,
что и у Surdisorex Thos. он также иногда отсутствует.
Распространение и возраст. Кения, современный.
Примечание. В заключение описания этого рода Томас отметил,
что он, очевидно, родствен роду Myosorex Gray, но объединяемые им
виды более приспособлены к рытью. Доказательством этого служат:
недоразвитые ушные раковины, длинные когти на пальцах передних
конечностей и короткий хвост, что делает их скорее похожими на крота,
чем на землеройку.
Наличие дополнительного зуба в нижней челюсти у видов рода Surdi-
sorex Thos. и Myosorex Gray, позволяет предполагать прямую связь этих
групп с олигоценовыми и миоценовыми землеройками, а также давнее их
обособление от других и давнее поселение на Африканском континенте.
1. Surdisorex norae Thomas, 1906 (рис. 192, Б).
Thomas, 1906, там же, где и род, с. 223, 224.
Тип. $ № 6.7.8.1, добыта вблизи Ниере, Кения, кол. Брит, муз., Лондон.
Описание (по: Thomas, 1906). Размеры тела скорее крупнее,
чем у известных видов рода Myosorex Cray. Шерсть близка к кротовой,
грубее чем у видов Myosorex Gray. Волосы на спине около 6 мм в длину.
Основная окраска сверху темно-бурая с зеленоватым отливом. Отдель-
ные волосы на 5/fi их длины аспидно-серые со светлым основанием и тем-
ной вершиной. Нижняя поверхность окрашена сходно с верхней, но не-
376
ПОДСЕМ. SORICINAE
сколько светлее. Верхние стороны кистей и ступней темно-коричневые.
Хвост очень короткий: менее, чем двойная длина задней ступни, покрыт
шерстью и длинными щетинками, темно-коричневый сверху и снизу.
Череп длиннее, чем у известных видов рода Myosorex Gray, но более
стройный. Зубы крупные как у Myosorex Gray, но второй верхний проме-
жуточный зуб пропорционально крупнее первого на 1/3 его поперечного
сечения. Третий более длинный и расположен вблизи (или касается) Р4.
Измерения. Длина: тела (на свежем экземпляре) — 108.0,
хвоста — 25.0, ступни — 14.0, наибольшая черепа, включая внутренние
резцы — 25.5, нижнего зубного ряда — 10.0 мм.
Распространение. Кения.
Примечание. Эта землеройка весьма интересна как пример
параллельного развития приспособлений в пределах одного семейства
и разных семейств Soricidae и Talpidae. И в том, и в другом семействе
приспособление к рытью есть средство обеспечения относительно ста-
бильными условиями жизни в сильно изменчивой среде. Кроме описан-
ного вида к этому роду относится и S. polulus Osg. с западной части
горы Кения.
2. Подтриба CROCIDURINA s. str. Stirton, 1930
Crocidura-group S t i r t о n, 1930, p. 224.
Тип. Crocidura Wagler, 1832, Европа, Азия и Африка, миоцен—современный.
Диагноз. Зубная формула: I С Р М — 30—26. Верх-
них промежуточных зубов 4, 3 или 2. В нижней челюсти 2 промежуточ-
ных зуба. Надглазничных отверстий нет. Хвост покрыт только корот-
кими волосами или короткими волосами и редкими длинными щетинками.
Распространение и возраст. Европа, плиоцен—совре-
менный; Азия и Африка, миоцен—современный; Северная Америка,
миоцен—плиоцен.
Примечание. Представители подтрибы известны из нижнего
миоцена, когда они уже обладали особенностями, близкими к современ-
ным видам. Можно думать, что формирование подтрибы Crocidurina на-
чалось еще в олигоцене.
При всем разнообразии частных приспособлений в пределах трибы
всех ее представителей объединяет общее для них строение жевательного
аппарата. Последний приспособлен к быстрому схватыванию добычи
и удержанию ее в челюстях. По-видимому, в пределах этой подтрибы,
приспособление к питанию относительно крупными животными началось
сравнительно рано. У всех землероек этой группы первый промежуточный
зуб значительно крупнее остальных. Он выполняет функции клыка и
режущего аппарата.
Основное ядро подтрибы Crocidurina составляет род Crocidura Wagl.,
объединяющий и в настоящее время значительное число видов, отчасти
и род Suncus Ehrenb. Ряд родов, например Feroculus Kelaart, Sylvisorex
Thos. и др., сохраняет архаичные черты. У объединяемых ими видов
на нижнем внутреннем резце имеются бугорки. Наибольшей специали-
зации достигли представители рода Diplomesodon Brandt, обитатели
пустынь, основную пищу которых составляют мелкие позвоночные, осо-
бенно ящерицы.
Большинство представителей подтрибы Crocidurina распространено
в субтропической и тропической зонах Европы, Азии и Африки. Сравни-
тельно небольшое число видов проникло в южные окраины Палеарктики.
23. РОД SIMILISOREX
377
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (8). В верхней челюсти 4 промежуточных зуба.
2 (5). 1Х с тремя бугорками на верхней стороне.
3 (4). Хвост покрыт короткими волосами и длинными редкими щетин-
ками ................................................24. Feroculus.
4 (3). Хвост покрыт только короткими волосами . . . 25. Sylvisorex.
5 (2). 1Х без бугорков на верхней стороне.
6 (7). Хвост покрыт короткими волосами и длинными редкими щетин-
ками ................................................. 26. Suncus.
7 (6). Хвост покрыт только короткими волосами . . .27. Scutisorex.
8 (1). В верхней челюсти 3 или 2 промежуточных зуба.
9 (16). В верхней челюсти 3 промежуточных зуба.
10 (15). Хвост покрыт только короткими волосами.
11 (12). На верхней стороне 1Х два бугорка........29. Paracrocidura.
12 (11). На верхней стороне 1Х бугорков нет.
13 (14). Длина хвоста не более половины длины тела. Волосяной покров
образован короткими мягкими волосами. Уши небольшие, скрыты
в волосяном покрове. Когти длинные....................30. Solisorex.
14 (13), Длина хвоста значительно более половины длины тела. Воло-
сяной покров образован длинными жесткими волосами. Уши не
скрыты в волосяном покрове. Когти не длинные . .31. Praesorex.
15 (10). Хвост покрыт короткими волосами и длинными редкими щетин-
ками .................................................32. Crocidura.
16 (9). В верхней челюсти 2 промежуточных зуба ....................
.............................................33. Diplomesodon.
Примечание. В таблицу не включены роды Similisorex, Limnoecus за от-
сутствием сравнимых признаков.
23, Род SIMILISOREX Stogov et Savinov, 1965
Стогов, Савинов, 1965, с. 92, 93.
Тип. Similisorex orlovi Stogov et Savinov, 1965, Казахстан, средний—верхний
миоцен.
Диагноз. Зубная формула: — 28—30 (Р).1 1Х с двумя
лопастями на режущем крае. 12 с двумя вершинами, как и Р4. Судя по
относительно тонкому основанию углового отростка, последний достигал
значительной длины.
Распространение и возраст. Казахстан, средний—
верхний миоцен.
Примечание. Основываясь на отмеченных морфологических
особенностях рода Similisorex Stog. et Sav., можно думать, что объеди-
няемые им виды имели и другие признаки, встречающиеся и у современ-
ных представителей некоторых азиатских и африканских примитивных
родов трибы Crocidurini, что позволяет предполагать многократную дав-
нюю связь азиатской и африканской фаун и их родственные отношения.
1 Судя по структурным особенностям зубов, у представителей этого рода в верх-
ней челюсти было не более четырех промежуточных зубов, как у видов рода Feroculus
Kelaart или Suncus Ehrenb.
378
ПОДСЕМ. SORICINAE
+ 1. Similisorex orlovi Stogov et Savinov, 1965 (рис. 193, A).
Стогов, Савинов, 1965, с. 92, 93.
M—1
Тип. Левая ветвь нижней челюсти № 'gQ_n > правый берег р. Иртыша у г. Пав-
лодара (Казахстан), кол. Инет. зоол. АН КазССР.
Рис. 193. Similisorex orlovi (А) (по: Стогов, Савинов, 1965), Sylvisorex morio (Б) (по:
Dobson, 1890), S. lunaris (В) (по: Repenning, 1967), Suncus murinus (Г).
Описание, (по: Стогов, Савинов, 1965). Нижняя челюсть мас-
сивная, с высоким, отогнутым назад сочленовным отростком. Венечный
отросток с нешироким (в передне-заднем направлении) основанием, с вы-
емкой в верхней части и гребнем на наружной стороне.
Измерения. Длина нижней челюсти от кончика резца до наи-
более удаленного конца сочленовного отростка — 11.2, высота венечного
отростка — 4.0—4.2; длина: нижнего зубного ряда — 6.9, Мх—Ms —
3.6—3.8, ряда Р4—М3 — 4.3—4.5 мм.
Распространение и возраст те же, что и рода.
24. РОД FEROCULUS. 25. РОД SYLVISOREX
379
24. Род FEROCULUS Kelaart, 1852
Sorex В 1 i t h, 1851, J. Asiat. Soc. Bengal., v. 20, p. 163. Corsira Kelaart,
1851, Ann. Mag. Nat. Hist., ser. 2, v. 8, p. 338. Feroculus Kelaart, 1852, Prodromus
Faunae Zeylanicae, p. 32, 33.
Тип. Sorex macropus Blith, 1851, Nuvara Ellia, Шри Ланка (о. Цейлон).
3 12 3
Диагноз. Зубная формула: 1уС-^-Р-рМ-д- = 30. В верхней челю-
сти четыре промежуточных зуба. На верхней стороне нижнего внутрен-
него резца три бугорка (лопасти). Хвост покрыт короткими волосами
и длинными редкими щетинками. В диагнозе (Kelaart, 1852) отмечены
и следующие особенности этого рода: зубы мелкие; верхние внутренние
резцы короткие (невысокие) и менее крючковатые, чем у типичных со-
рицид, с крупной задней вершиной; два средних верхних промежуточных
зуба равны по размерам; кисти с длинными когтями.
Описание (по: Blanford, 1888—1891). Волосы, покрывающие
тело, относительно длинные, мягкие, однообразно окрашенные — голу-
боватые (по-видимому, аспидно-серые), с легким рыжеватым пятном на
верхней стороне (на спине). Нижняя сторона скорее светлая. Хвост
черный, исключая концевую часть, она голая — мясного цвета; покрыт
короткими волосами и длинными редкими щетинками.
Распространение и возраст. Шри Ланка, совре-
менный.
1. Feroculus macropus Blith, 1851.
Sorex macropus Blith, 1851 (см. синонимику рода). Corsira nuvara ellia Kela-
art, 1851 (см. синонимику рода). Feroculus macropus Kelaart, 1852 (см. синони-
мику рода). Crocidura macropus Blanford, 1888—1891, p. 237, 238.
T и п не установлен. Добыта Келаартом в Nuvara Ellia, Шри Ланка (о. Цейлон).
Описание (по: Blanford, 1888—1891). Хвост суживающийся
к концу, покрыт редкими волосами и еще более редкими длинными ще-
тинками. Кисти и ступни большие. Кисти с очень длинными когтями,
в два раза более длинными, чем когти на ступнях. Мех средней длины,
мягкий. Уши небольшие, одеты волосами и хорошо заметны в редком
волосяном покрове.
Тело сверху темного аспидно-серого цвета с рыже-коричневым оттен-
ком. Нижняя сторона тела светлее. У типичного экземпляра конец хвоста
белый. Когти светлые.
Измерения (по: Blanford, 1888—1891). Длина: тела — 196.2,
хвоста — 56.0, ступни с когтями — около 22.0, когтей на кистях — около
6.0 мм.
Распространен и.е в границах рода.
Биология. Об образе жизни этой землеройки ничего не известно.
Судя по длине когтей на пальцах кистей, этот вид, по-видимому, спо-
собен рыть, как например Sorex unguiculatus Dobs., у .которого также
длинные ногти на пальцах кистей, а ее способность к рытью установлена.
25. Род SYLVISOREX Thomas, 1904
Thomas, 1904, Abstract Proc. Zool. Soc., London, N 10, p. 12. Crocidura Gray,
1862, Proc. Zool. Soc. London, p. 180. Myosorex Dobson, 1887, там же, с. 75.
Тип. Crocidura morio Gray, 1862, южная Нигерия.
3 12 3
Диагноз. Зубная формула: I у С у Р -р М у = 30. В верхней челю-
сти четыре промежуточных зуба. На верхней стороне Д три бугорка
(лопасти). Хвост покрыт только короткими волосами, длинных щети-
нок нет.
380
ПОДСЕМ. SORICINAE
Распространение и возраст. Центральная и Южная
Африка, современный (можно думать, что представители этого рода суще-
ствуют в Африке по крайней мере с плиоцена).
Примечание. Роды Sylvisorex и Myosorex Gray Эллерман, Мор-
рисон-Скотт и Гайман (Ellermann et al., 1953) объединили с родом Sun-
cus Ehrenb. в качестве подродов последнего.
1. Sylvisorex morio Gray, 1962 (рис. 193, Б).
Crocidura morio Gray, 1862, Proc. Zool. Soc. London, p. 180.
T и п не установлен, вид описан из Камеруна.
Описание (по: Dobson, 1887). Уши средних размеров, покрыты
короткой шерСтыо, также и верхние стороны кистей и ступней. Окраска
верхней стороны тела красно-коричневая, нижней — немного светлее.
Зубы относительно мелкие. Второй верхний промежуточный зуб
ниже третьего.
Измерения. Длина: тела — 64.0, ступни 13.5, кондило-базаль-
ная черепа — 16.5 мм.
Распространение и возраст. Камерун, плиоцен?—со-
временный.
2. Sylvisorex johnstoni Dobson, 1887.1
Dobson, 1887, Proc. Zool. Soc. London, p. 577, 578.
Тип. Спиртовый экземпляр, добытый Джонстоном (Johnston) в Камеруне (Rio
de Key).
Описание (по: Dobson, 1887). Немного крупнее Suncus etruscus
Savi. Пальцы кистей и ступней с короткими когтями. Окраска верхней
стороны тела темно-коричневая, нижней — светлее. Череп относительно
крупный. Второй верхний промежуточный зуб значительно меньше
третьего.
Измерения. Длина: тела — 39.0, хвоста — 25.0, ступни — 9.0,
кондило-базальная черепа — 13.0 мм.
Распространение и возраст. Камерун, Гвинея (о. Ма-
сиас-Нгема-Бийого), современный.
3. Sylvisorex granti Thomas, 1907.
Thomas, 1907, Ann. Mag. Nat. Hist., ser. 7, v. 19, p. 118; — mundus Osgood,
1910, Field Mus. Nat. Hist. (zool. ser.), v. 10, N 18, p. 7.
Распространение. Кения.
4. Sylvisorex lunaris Thomas 1906 (рис. 193, В).
Thomas, 1906, там же, т. 18, с. 139.
Распространение. Уганда.
5. Sylvisorex olula Thomas, 1913.
Thomas, 1913, там же, сер. 8, т. 11, с. 321.
Распространение. Камерун.
6. Sylvisorex oriundus Hollister, 1916.
Hollister, 1916, Bull. Amer. Mus. Nat. Hist., v. 35, p. 672.
Распространение. Конго.
1 Другие виды этого рода кроме (S. morio Gray) требуют ревизии. Они приведены
без описания и указания возраста.
26. РОД SUNCUS
381
7. Sylvisorex ruande Lonnberg et Gildenstolpe, 1925.
Lonnberg, Gildenstolpe, 1925, Arkiv. zool., 17 B, N 5, p. 1.
Распространение. Конго.
8. Sylvisorex megalura Jentink, 1888.
Pachyura megalura Jentink, 1888, Notes Lyden Mus., v. 10, p. 48. Sylvisorex
megalura Thomas, 1904, Proc. Zool. Soc. London, v. 2, p. 190.
Распространение. Либерия.
9. Sylvisorex sorella Thomas, 1898.
Myosorex (?) sorella Thomas, 1898, Proc. Zool. Soc. London, p. 930. Sylvisorex
angolensis Roberts, 1929, Ann. Transvaal Mus., v. 13, p. 84; — gemmeus Heller,
1910, Smiths. Misc. Coll., v. 56, N 15, p. 7; — sorelloides Lonnberg, 1912, Ann. Mag.
Nat. Hist., ser. 8, v. 9, p. 67.
Распространение. Судан, Эфиопия, Камерун, Уганда,
Кения, Ботсвана.
10. Sylvisorex suncoides Osgood, 1936.
Osgood, 1936, Field Mus. Nat. Hist. (Zool. ser.), v. 20, p. 217.
Распространение. Конго,
26. Род SUNCUS Ehrenberg, 1833
Ehrenberg, 1833, Symbolac Physic., Mammal., dec. 2, Folio K. Crocidura
W ag 1 er, 1832, Isis, p. 275. Pachyura (как подрод Sorex L.) de S e 1 у s Long-
champs, 1839, Etudes Micromammal., p. 32. Paradoxodon (подрод Sorex L.) W a g-
1 e r, in: Schrebers, 1855, Saugeth., Bd. 5, S. 805. Pleurodus Schulze, 1897, Helios,
Berlin, Bd 14, S. 90.
Тип. Suncus sacer Ehrenberg, 1833. Суэц, Египет (в настоящее время синоним
Suncus murinus L>. = Sorex murinus L., 1766.)
3 12 3
Диагноз. Зубная формула: IуС-^РуМj = 30. В верхней челю-
сти четыре промежуточных зуба. На верхней стороне нижнего внутрен-
него резца бугорков (лопастей) нет. Хвост покрыт короткими волосами
и редкими длинными щетинками.
Описание. Размеры тела 34.0—160.0 мм. Голова впереди глаз
резко суживается к носу. Уши не скрыты в волосяном покрове, у боль-
шинства крупные. Окраска разнообразная. Верхняя и нижняя стороны
тела мало разнятся по цвету.
Распространение и возраст. Южная Европа, Передняя,
Средняя, Центральная и Южная Азия, восточная и южная части Африки.
В Европе известен с позднего плиоцена.
Примечание. В 1953 г. Эллерман, Моррисон-Скотт и Гайман
(Ellerman, Morrison-Scott, Hayman) объединили этот род с родом Myo-
sorex Gray, сохранив за последним значение подрода. В 1958 г. Б. С. Ви-
ноградов объединил этот род с родом Crocidura. Основанием для этого
послужило сходство в строении penis.
1. Suncus murinus Linnaeus, 1766 (рис. 193, Г).
Sorex murinus Linnaeus, 1766, Syst. Nat., ed 12, t. 1, p. 74; — myosurus Pal-
las, 1781, Acta Acad. Sci. PetropoL, v. 2, p. 337; — caerulaeus Kerr, 1772, Animal
Kingdom, p. 207; — pilorides Shaw, 1796, Mus. Lever, v. 2, p. 31; — caerulescens
Shaw, 1800, Genus Zool. Mammal., v. 1, p. 533; — indicus Geoffroy, 1811, Ann.
Mus. Hist., v. 17, p. 183; •— conneratii Geoffroy, 1827, Mem. Mus. Nat. Hist.,
Paris, t. 15, p. 183; — giganteus Geoffroy, 1831, Voy. Belanger, Indies Orient.
382
ПОДСЕМ. SORICINAE
Zool., p. 117; — serpentarius Geoffroy, in: Belanger, 1831, там же, с. 119; — sa-
cer Ehrenberg, 1833, Symbolac Physic., Mammal., dec. 2, Folio K; — crassicau-
dus Hemprich, Ehrenberg, 1834; in: Lichtenstein’s Darstellung Saugeth.,
pl. 40, fig. 1; — tristrami Bodenheimer, 1835, Animal Life in Palestine, p. 95; —
soccatus Hodgson, 1845, Ann. Mag. Nat. Hist., v. 15, p. 270; — nemorivagus Hodg-
son, 1845, там же, с. 269; — montanus Kelaart, 1850, J. Ceylon Br. Asiat. Soc.,
v. 2, p. 211; — ferrugineus Kelaart, 1850, там же, с. 212; — griffithi Horsfield,
1851, Cat. Mammal. E. India Mus. Co., p. 134; — niger Horsfield, 1851, там же,
с. 135; — kandianus Kelaart, 1852, Prodr. Faunae Zeylanicae, p. 30; — heterodon
Blyth, 1855, J. Asiat. Soc. Bengal, v. 24, p. 31; — kelaarti Blyth, 1855, там же,
с. 32; — saturatior Hodgson, 1855, Ann. Mag. Nat. Hist., v. 16, p. 110; — virides-
cens Blyth, 1859, J. Asiat. Soc. Bengal, v. 28, p. 285; — swinhoei Blyth, 1859,
там же, с. 285; — albinus Blyth, 1868, там же, т. 29, с. 90. Pachyura duwernoyi F i-
t z i n g e r, 1868, Osterr. Acad. Wiss. math.-naturwiss. Kl., Abt. 1, Bd 57, S. 136. Cro-
cidura microtis Peters, 1870, Mber. Preuss, Acad. Eiss., S. 589; — (Pachyura.) walde-
marii Peters, 1870, там же, с. 590; — zeylanica Peters, 1870, там же, с. 591; — —
media Peters, 1870, там же, с. 532;-fulvovinerea Anderson, 1877, J. Asiat.
Soc. Bengal, v. 46, N 2, p. 263;-blythii Anderson, 1877, там же, с. 264;-
sindensis Anderson, 1877, там же, с. 266;-pealana Anderson, 1877,
там же, с. 267;-blanfordii Anderson, 1877, там же, с. 269; — — rubicunda
Anderson, 1877, там же, с. 277;-andersoni Trouessart, 1879, Rev. Zool.,
p. 253. Sorex beddomei Anderson, 1881?, Cat. Mammal. E. Ind. Mus. Co., p. 179.
Crocidura muschata H a t о r i, 1915, Taiwan Igakukia Zasshi. Pachyura caerulla riukiu-
ana Kuroda, 1924, On New Mammal, from Riu-Kiu Islands, Tokyo, p. 3. Suncus
zeylanicus Phillips, 1928, Spolia Zeylanica, v. 14, p. 313; — niger malabaricus
Lindsay, 1929, J. Bombay Nat. Hist. Soc., v.- 33, p. 334.
Тип не установлен, вид описан с о. Ява.
Диагноз (по описанию: Blanford, 1888—1891). Размеры тела
крупные. Второй верхний промежуточный зуб значительно менее
третьего. Четвертый промежуточный зуб всегда имеется, смещен на
внутреннюю сторону зубного ряда.
Описание. Волосяной покров жесткий. Окраска тела варьирует
от коричневого и бурого до охристо- и голубовато-серого цвета. Окраска
нижней стороны тела мало отличается от окраски верхней. Когти сильно
развиты.
Измерения. Длина: тела — 100.0—160.0, хвоста — 70.0—90.0,
ступни — 19.0—23.0, кондило-базальная черепа до 30.0 мм.
Распространение и возраст. Египет, Эфиопия, Ара-
вийский полуостров, Кашмир, к югу до Шри Ланка, Юго-Восточный
Китай, о. Тайвань, Южный Вьетнам, Япония, Бирма, Малайский архи-
пелаг, Бали, Ява, Суматра, Калимантан, Сулавеси, Филиппины. Завезен
в Восточную Африку и на о. Мадагаскар, современный.
2. Suncus stoliczkanus Anderson, 1877.
Crocidura (Pachyura) stoliczkana, Anderson, 1877, J. Asiat. Soc. Bengal, v. 46,
p. 270; — bidiana Anderson, 1877, там же, с. 276;— subfulva An derson,
1877, там же, с. 278. Crocidura leucogenis Dobson, 1888, Ann. Mag. Nat. Hist., v. 1,
p. 428.
Тип не установлен, вид описан из Бомбея, Индия.
Распространение и возраст. Бомбей, Центральная и
Южная Индия, современный.
3. Suncus dayi Dobson, 1888.
Crocidura dayi Dobson, 1888, Ann. Mag. Nat. Hist., v. 1, p. 428.
T и n не отмечен, вид описан из Трихур, Кахий (Индия).
Распространение и возраст. Южная часть Индии, со-
временный.
Примечание. Из Африки описано значительное число видов
рода Suncus, таксономические границы которых не ясны. В связи с этим
здесь приводятся только их названия и распространение.
26. РОД SUNCUS
383
4. Suncus chriseos Kershaw, 1921.
Kershaw, 1921, Ann. Durban Mus., v. 3, N 31, p. 31.
Распространение. Дурбан, провинция Наталь (ЮАР).
5. Suncus coquerellii Trouessart, 1880.
Trouessart, 1880, Notes Leyden Mus., y. 2, p. 86.
Распространение, о. Мадагаскар.
6. Suncus gracillis Blainville, 1838.
Blainville, 1838, Ann. Sci. Nat. Zool., ser. 2, v. 10, p. 120.
Распространение. Южная Африка.
7. Suncus gratulus Thomas et Schwann, 1907.
Thomas, Schwann, 1907, Proc. Zool. Soc. London, p. 781.
Распространение. Трансвааль (ЮАР).
8. Suncus infinitesimus Heller, 1912.
Heller, 1912, Smiths. Misc. Coll., v. 60, N 12, p. 5.
Распространение.' Кения.
9. Suncus leucura Matschie, 1894.
Crocidura albicauda N oack, 1891, Jahrb. Hamburg. Wiss. Anst., Bd. 9, S. 117.
Suncus leucura Matschie, 1894, Sitzungsber. Ges. Naturf. Freunde, Berlin, S. 205.
Распространение. О. Занзибар.
10. Suncus lixus Thomas, 1897.
Crocidura (Pachyura) lixa Thomas, 1897, Proc. Zool. Soc. London, p. 930.
Распространение. Северная Ботсвана.
11. Suncus madagascarensis Coquerel, 1848.
Coquerel, 1848, Ann. Sci. Nat. Zool., ser. 3, v. 9, p. 194.
Распространение. О. Мадагаскар.
12. Suncus orangie Roberts, 1924.
Roberts, 1924, Ann. Transvaal Mus., v. 10, p. 61.
Распространение. Южная Африка.
13. Suncus varillus Thomas, 1895.
Crocidura (Pachyura) virilla Thomas, 1895, Ann. Mag. Nat. Hist., ser. 6, v. 16,
p. 54.
Распространение. Мыс Доброй Надежды.
14. Suncus warreni Roberts, 1929.
Roberts, 1929, Ann. Transvaal Mus., v. 13, p. 84.
Распространение. Южная Африка.
15. Suncus etruscus Savi, 1822 — Многозубка-малютка (рис. 194).
Savi, 1822, Nuovo Giorn. del Letterati, Pisa, t. 1, p. 60. Crocidura etrusca W a g-
1 e r, 1832, Isis, p. 275. Sorex pachyurus К ii s t e r, 1835, Isis, p. 77. Crocidura suaveo-
384
ПОДСЕМ. SORICINAE
lens (пес Pall.) Blasius, 1857, Naturgesch. Saugeth. Deutsch., S. 147—151. Pachyura
etrusca Brandt, 1865, исследования о зубной системе землероек, с. 82—86.
Тип не установлен, вид описан из окрестностей г. Пизы (Италия).
6
Рис. 194. Suncus etruscus (Л), Scutisorex congicus (Б) (по: Allen, из: Grasse, 1955).
Диагноз. Самое мелкое современное млекопитающее — длина
тела 34.0—48.0 мм. Длина лицевой части черепа равна длине его мозго-
вой капсулы. Второй верхний промежуточный зуб крупнее третьего,
четвертый очень мал, редко он может отсутствовать, но его альвеола
сохраняется.
27. РОД SCUTISOREX
385
Описание. Конец морды сильно сужен и удлинен. Хвост густо
покрыт относительно длинными волосами и более длинными редкими
щетинками; на конце хвоста кисточка из длинных волос. Окраска верхней
стороны тела светлая, охристо-буроватая или коричневато-серая. Ниж-
няя сторона тела сероватая или серо-белесая.
Череп мелкий, тонкого строения.
Измерения. Длина: хвоста — 22.0—31.0 мм, ступни — 6.8—8.1,
кондило-базальная черепа — 12.0—12.6 мм.
Распространение и возраст. Южная Европа, Ара-
вийский полуостров, Иран, Средняя Азия, Индия, Шри Ланка, Тайвань
и Юго-Восточный Китай, Бирма, Япония, Малайский архипелаг, Фи-
липпины, восточная и южная части Африки. В СССР — Закавказье,
юго-западная Туркмения, юго-западный Таджикистан, южная часть
Узбекистана, Северное Приаралье (в погадках птиц).
Верхнеплиоцеповые остатки этого вида известны из Харьковской обл.
(Украина), современный.
Биология. Образ жизни на территории СССР не изучен.
Подвиды. Описано 7 подвидов. На территории СССР распростра-
нена типичная форма — S. е. etruscus Savi. Характеризуется очень мел-
кими размерами. Распространена в Южной Европе, Иордании, Закавказье,
Иране, Средней Азии, Алжире и Нигерии. В пределах СССР — в грани-
цах, указанных для вида.
27. Род SCUTISOREX Thomas, 1913
Thomas, 1913, Ann. Mag. Nat. Hist., ser. 8, v. 11, p. 321.
Тип. Sylvisorex somereni Thomas, 1910, Kyetyme вблизи Кампала, Уганда.
3 12 3
Диагноз. Зубная формула: I у С у Р у М у = 30. В верхней челюсти
четыре промежуточных зуба. На верхней стороне нижнего внутреннего
резца бугорков нет. Хвост покрыт только короткими волосами. В отли-
чие от других землероек у них череп с очень сильно развитыми гребнями;
позвонки образованы сильно разрастающейся губчатой костной тканью
(гиперостоз).
Распространение и возраст. Конго, Уганда, совре-
менный.
Примечание. Своеобразные особенности в строении скелета
этого рода, обусловленные ненормальным развитием составляющих его
костей, послужили основанием для выделения рода в особое подсемей-
ство.
1. Scutisorex somereni Thomas, 1910.
Sylvisorex somereni Thomas, 1910, Ann. Mag. Nat. Hist., ser. 8, v. 6, p. 113.
Scutisorex somereni Thomas, 1913, там же, т. 11, 321.
Тип. $ N 10.5.21.1, Kyetyme, вблизи Кампала, Уганда, кол. Брит, муз.,
Лондон.
Диагноз. Выступы ламдовидного гребня выдаются назад за кон-
дилюсы затылочной кости.
Описание. Шерсть длинная, на спине достигает 11—12, а ме-
стами 16 мм. Верхняя сторона тела пепельно-серая, нижняя — аспидно-
серая. Конец морды, уйти и верхняя сторона кистей и ступней черно-
коричневые. Хвост сверху коричневый.
Стреловидный гребень (особенно в задней части) и гребни на месте
височно-теменного шва сильно развиты. Слезные отверстия крупные.
25 А. А. Гуреев
386
ПОДСЕМ. SORICINAE
Измерения (по: Thomas, 1913). Длина: тела — 120.0, ступни —
23.0, кондило-базальная черепа — 33.0, кондило-резцовая черепа — 33.7,
верхнего зубного ряда — 15.3, нижнего зубного ряда — 14.0 мм.
Распространение. Уганда.
2. Scutisorex congicus Thomas, 1915 (рис. 194, Б).
Thomas, 1915, там же, т. 16, с. 470—471.
Тип. (? коллекторский № 840, Medje, верховье р. Итури, Конго, кол.
Брит, муз.?
Д и а г н о з. Выступы ламбдовидного гребня не выдаются за конди-
люсы затылочной кости.
Описание. Окраска тела серая с коричневатым налетом, сход-
' ная с таковой Scutisorex somereni Thos. В отличие от последнего у описы-
ваемого вида шерсть короче и грубее.
Измерения. Длина: тела — 143.0, хвоста — 77.0, ступни — 20.0,
кондило-базальная черепа — 30.2, кондило-резцовая черепа — 31.3, верх-
него зубного ряда — 13.6, нижнего — 12.7 мм.
Распространение. Конго.
Примечание. Диагнозы и описание видов рода Scutisorex со-
ставлены по первоописаниям Томаса (Thomas, 1913) и Уолкера (Walker
et al., 1964).
|28. Род LIMNOECUS Stirton, 1930
S t i г t о n, 1930, p. 217—219.
Тип. Limnoecus tricuspis Stirton, 1930, Калифорния, миоцен.
Диагноз (no: Stirton, 1930). М3 трехбугорчатый. Его энтоконид
(бугорок на внутренней стороне метаконида, по принятым мной обозна-
чениям) крупнее, чем энтоконид у М2. Протоконид и метаконид (внутрен-
ний и наружный бугорки протоконида) менее широко обособлены (от-
граничены друг от друга на общем гребне), чем у других родов. Венеч-
ный отросток более заостренный, чем у других родов. По общему
характеру строения род Limnoecus ближе всего к родам Suncus Ehrenb.
и Crocidura Wagl.
Распространение и возраст. Калифорния, миоцен—
плиоцен.
t 1. Limnoecus tricuspis Stirton, 1930 (рис. 195).
Stirton, 1930, там же, с. 218, 219.
Тип. Левая ветвь нижней челюсти с полным рядом зубов № 31047, Barstow,
San Bernardino, Калифорния, миоцен, кол. Палеонтол. муз. Калифорн. унив., Беркли.
Диагноз совпадает с родовым.
Описание (по: Stirton, 1930). Венечный отросток низкий, остро-
конечный. Впадина для прикрепления височного мускула (птеригоидная
ямка — у других авторов) на его внутренней стороне — треугольного
очертания. Судя по форме основания обломанного углового отростка,
последний был округлый в сечении и относительно короткий. Вся ниж-
няя челюсть более массивная, чем у современных белозубых землероек.
Нижний внутренний резец относительно короткий, слабо изогнут вверх
и без бугорка на верхней стороне (возможно, Стерт).
Распространение и возраст. Калифорния (США),
миоцен—плиоцен.
29. РОД PARACROCIDURA. 30. РОД SOLISOREX
387
29. Род PARACROCIDURA Heim de Balsac, 1956
Heim de Balsac, 1956, p. 137—146.
T и n-. Paracrocidura schoutedeni Heim de Balsac, 1956, Касаи, Конго, современный.
3 113
Диагноз.1 Зубная формула: Г С у Р у М у — 28. В верхней челюсти
3 промежуточных зуба. На верхней стороне нижнего внутреннего резца
.2 бугорка. Хвост покрыт только короткими волосами. В отличие от дру-
гих родов этой трибы на медиальной стороне верхнего внутреннего резца
имеется бугорок (выступ); Р4, М1 и М2 без выемки на их задней стороне;
,12 крупный, с двумя вершинами, а Р4 с тремя вершинами.
Описание. Уши небольшие, почти голые. Длина хвоста более
половины длины тела, он покрыт короткими волосами.
Распространение и возраст. Конго, современный.
1. Paracrocidura schoutedeni Heim de Balsac, 1956 (рис. 195, И).
Heim de Balsac, 1956, p. 137—146.
Тип. (J № 23463 (спиртовой), Тшимбули (Дибайя), Любондайре (75 км южнее
Люксембурга), Касаи, Конго, кол. Муз. Тервирен.
Описание. Тело часто покрыто редкими волосами черноватого
цвета. Окраска верхней и нижней сторон тела сходная. Кисти и ступни
«покрыты редкими серыми волосами. Хвост черноватый. Когти короткие.
Череп массивный, с плоской мозговой капсулой. Зубы крупные.
• Второй и третий промежуточные зубы не так сильно разнятся по величине
от первого промежуточного зуба, как у представителей рода Crodidura
Wagl. Нижняя челюсть массивная с коротким и толстым угловым отро-
стком. Ij с двумя слабо выраженными бугорками.
Измерения. Длина: тела — 65.0, хвоста — 34.5, ступни — 12.0
(с когтями), кондило-резцовая черепа — 22.7, верхнего зубного ряда —
11.5 мм.
Распространение и возраст. Касаи, Конго, совре-
менный,
30. Род SOLISOREX Thomas, 1924
Thomas, 1924, Spolia Zeylanica, v. 13, N 1, p. 94.
Тип. Solisorex pearsoni Thomas, 1924, Шри Ланка (Цейлон), современный.
Диагноз (по характеристике из: Walker et al., 1964). Зубная фор-
3 113
мула: I у С у Р-р М-д-= 28. В верхней челюсти три промежуточных
зуба. На верхней стороне 1Х бугорков нет. Хвост покрыт только корот-
кими волосами. В отличие от представителей рода Praesorex Thos. у них
уши небольшие, скрыты в волосяном покрове головы, волосы, покрываю-
щие тело, мягкие; кисти с длинными когтями.
Распространение и возраст. Шри Ланка, современный.
Примечание. По тому, что известно об этом роде, трудно судить,
насколько резко он дифференцирован от других родов этой трибы. Отсут-
ствие длинных щетинок на хвосте и длинные когти вряд ли могут служить
основанием для выделения вцда в особый род. Этот вопрос можно решить
лишь при исследовании соответствующего материала, которого в моих
руках не было.
1 Диагноз и описание по характеристикам рода и вида'(Н. de Balsac, 1956; Walker
et al., 1964).
25*
388
ПОДСЕМ. SORICINAE
1. Solisorex pearsoni Thomas, 1924.
Thomas, 1924, там же.
T и п не указан, вид описан из окрестностей Нувара Элин, Шри Ланка (о. Цей-
лон).
Диагноз совпадает с родовым.
Описание (по: Walker et al., 1964). Окраска верхней стороны
тела темная, серовато-коричневая со светлым налетом, образованным
светлыми концами отдельных волос. Нижние части тела окрашены сходно
с верхними, по светлее. Кисти и ступни коричневые.
Зубы крупные и высокие. I1 сильно развиты.
Измерения. Длина тела — 125.0—134.0, хвоста — 59.0—66.0 мм.
Распространение и возраст. Шри Ланка.
31. Род PRAESOREX Thomas, 1913
Thomas, 1913, Ann. Mag. Nat. Hist., ser. 8, v. 11, p. 319, 320.
Тип. Crocidura goliath Thomas, 1906, Камерун, современный.
3 113
Диагноз. Зубная формула: I у С-q-Р-р М у = 28. В верхней челюсти
3 промежуточных зуба. На верхней стороне 1г бугорков нет. Хвост покрыт
только короткими волосами. В отличие от видов рода Solisorex Thos.
у них уши большие, выступают из волосяного покрова головы; волосы,
покрывающие тело, длинные, грубые; кисти с короткими когтями.
Распространение и возраст. Камерун, современный.
Примечание. Род Praesorex слабо отличается от рода Crocidura
Wagl. Возможно, что он будет объединен с последним.
1. Praesorex goliath Thomas, 1913.
Thomas, 1906, там же, сер. 7, т. 17, с. 177.
Тип. в № 5.10.28.2, Эуфуми, Камерун, кол. Брит, муз., Лондон.
Диагноз совпадает с таковым рода.
Описание (по: Thomas, 1913). Внешним обликом, как отмечает
Томас, эти землеройки напоминают крысу, что обусловлено их относительно
крупными размерами и грубым мехом, а также длинным хвостом, покрытым
короткими грубыми волосами. Окраска верхней стороны тела коричневая
с серым налетом, образованным отдельными беловатыми волосами. Го-
лова сверху темно-коричневая. Нижняя сторона тела только слегка
светлее верхней. Окраска хвоста черноватая. Отдельные волосы, покры-
вающие хвост, с белыми вершинами.
Измерения. Длина: тела — 155.0—180.0, хвоста — до 110.0,
ступни 26.0, наибольшая черепа — 39.3 мм.
' Распространение и возраст. Камерун, современный.
32. Род CROCIDURA Wagler, 1832 — ЗЕМЛЕРОЙКИ БЕЛОЗУБКИ
W a g 1 е г, 1832, Isis р. 275. Leucodon F a t i о, 1869, Faune Vert. Suisse, v. 1,
p. 132. Paurodus Schulze, 1897, Helios, Berlin, Bd 14, S.90. Heliosorex Heller,
1910, Smiths. Misc. Coll., v. 56, N 15, p. 6.
Тип. Sorex leucodon Hermann, 1780, Европа, Азия.
3 113
Диагноз. Зубная формула: 1-2"С-рРуМу=28. В верхней челюсти
3 промежуточных зуба. На верхней стороне Д бугорков нет. Хвост покрыт
короткими волосами и длинными редкими щетинками.
32. РОД CROCIDURA
389
Описание. Уши не скрыты в волосяном покрове. Волосяной
покров густой, бархатистый, образован относительно короткими волосами,
реже жесткий или щетинистый. Преобладающая окраска — серая или
коричневая разных оттенков. Особи отдельных видов окрашены в жел-
товатый, бурый или черно-бурый цвета.
Череп относительно массивный, с удлиненной и угловатой мозго-
вой капсулой. Отверстий на лобных .костях нет. Первый верхний проме-
жуточный зуб значительно крупнее двух других (Is и С), которые мало
разнятся по величине. Внутренний верхний резец с сильно развитым
передним зубцом, круто опущенным вниз. Нижний внутренний резец
не имеет зубцов на верхней (режущей) стороне; его передний конец в раз-
ной степени круто изогнут вверх.
Посткраниальный скелет образован массивными костями, что обуслов-
лено необходимостью увеличения мощности тела для добывания относи-
тельно крупных и сильных животных из числа беспозвоночных и мелких
позвоночных, которые часто служат им пищей.
Род объединяет большое число видов (описано более 150).
Распространение и возраст. Средняя и южная части
Европы и Азии с островами, Африка и Мадагаскар; плиоцен—современ-
ный — Европа, миоцен—современный — Азия, миоцен—современный —
Африка.
Биология. В пределах столь обширного ареала виды рода оби-
тают в разнообразных условиях открытых и закрытых ландшафтов гор
и равнин, встречаются практически всюду. Отдельные виды роют норы,
но чаще используют естественные убежища или приспосабливают для
своих нужд чужие норы, например грызунов. Для европейских видов
отмечена общественная жизнь в зимнее время, когда для обогревания
несколько особей (до 8) собираются в одном гнезде. Ведут преимущественно
ночной образ жизни, активны в течение всего года.
Питаются различными беспозвоночными и мелкими позвоночными
животными. В неволе многие виды поедали мелких млекопитающих,
а также ящериц.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ЗЕМЛЕРОЕК (РОД CROCIDURA WAGLER)
ФАУНЫ СССР (СОВРЕМЕННЫЕ)
1 (2). Длина тела более 80 мм. Кондило-базальная длина черепа более
21 мм. Окраска верхней и нижней сторон кистей и ступней темная,
черно-бурая .................................11. С. lasiura Dobs.
2 (1). Длина тела не более 80 мм. Кондило-базальная длина черепа
менее 21 мм. Окраска верхней и нижней сторон кистей и ступ-
ней светлая, белая, серая или охристая.
3 (12). Длина тела не менее 70 мм. Кондило-базальная длина черепа
не менее 18 мм. Если длина тела или черепа менее указанных разме-
ров, то длина ступни не менее 12 мм.
4 (9). Длина хвоста не менее 40 мм.
5 (6). Окраска верхней стороны тела и боков однотонная, коричневая,
бурая или буровато-серая. Окраска нижней стороны тела часто
сходна с окраской боков и спины. Зимняя окраска сходна с летней
...................................... 12. С. giildenstaedti Pall.
6 (5). Окраска верхней стороны тела пепельно-серая или серая с буро-
ватым налетом, бока тела светлее спины — светло-серые или бело-
ватые. Нижняя сторона тела окрашена в белый или беловатый цвет.
Зимняя окраска спины серебристо-серая.
390
ПОДСЕМ. SORICINAE
7 (8). Второй верхний промежуточный зуб крупнее третьего . . . .
............................................ 14. С. armenica Gur.
8 (7). Второй верхний промежуточный зуб меньше третьего . . . .
.........................................13. С. pamirensis Ognev.
9 (4). Длина хвоста менее 40 мм.
10 (И). Второй верхний промежуточный зуб меньше третьего. Длина
волос, покрывающих хвост, меньше его диаметра в средней
части ........................................8. С. leucodon Henn.
11 (10). Второй верхний промежуточный зуб равен третьему. Длина
волос, покрывающих хвост, больше его диаметра в средней части
...............................................6. С. persica Thom.
12 (3).. Длина тела меньше 70 мм. Кондило-базальная длина черепа
меньше 18 мм. Если длина тела или черепа более указанных разме-
ров, то длина ступни менее 12 мм.
13 (14). Сумма продольных диаметров второго и третьего верхних про-
межуточных зубов равна или меньше продольного диаметра осно-
вания переднего зубца I1.....................10. С. sibirica Dukels.
14 (13). Сумма продольных диаметров второго и третьего верхних про-
межуточных зубов больше продольного диаметра основания перед-
него зубца I1.
15 (16). Конус заднего зубца I1 меньше конической части коронки вто-
рого верхнего промежуточного зуба или хотя бы короче его по вы-
соте в 1.25—1.5 раза. Хвост покрыт крупными чешуйками и длин-
ными волосами, длина которых больше половины диаметра хвоста
в средней его части.......................... 5. С. hyrcana Goodw.
16 (15). Конус заднего зубца I1 больше конической части коронки вто-
рого верхнего промежуточного зуба. Хвост покрыт мелкими че-
шуйками и короткими волосами, длина которых редко превышает
Vs диаметра хвоста в его средней части ...........................
.........................................4. С. suaveolens Pall.
Примечание. В таблицу не включена Crocidura dinniki Ognev. (15). Недо-
статок материала по этому виду не позволяет более четко, чем это сделано в описании,
провести границу между ним и близкими к нему видами. При всем этом качественная
определенность его не вызывает сомнений.
t 1. Crocidura pavlodarica Stogov et Savinov, 1965 (рис. 195).
Стогов, Савинов, 1965, с. 93, 94.
Тип. Фрагмент правой ветви нижней челюсти с зубами (1Х—М2), fiO-1 i • Ка'
захстан, миоцен, кол. Инет. зоол. АН КазССР, Алма-Ата. ои 11
Диагноз (по: Стогов, Савинов, 1965). Размеры относительно
крупные, 1Т массивный, короткий и круто загнут кверху. Рг (как теперь
установлено 12) очень мал и смещен кпереди. Жевательная поверхность
талонидов (метаконидов — по моему представлению) Мг и М2 почти
плоская, без ямки посредине и со слабо выступающими гребнями по их
краям.
Описание (по: Стогов, Савинов, 1965). Нижняя челюсть довольно
массивная, широкая в поперечном сечении. Массивный короткий 11т
без лопастей на режущем крае. На режущем крае Рт (12) ясно заметна
полулунная выемка, образовавшаяся в результате стачивания его про-
тивостоящим зубом верхней челюсти. Жевательная поверхность тало-
нидов (метаконидов) Мг и М2 почти плоская, без ямки между бугорками.
Пара- и метакониды на этих зубах слабо развиты. Передние зубы нижней
челюсти очень тесно стоят в зубном ряду; Рг (12) далеко смещен кпереди,
на него сзади сильно налегает своим основанием Р2 (Р4), на который в свою
очередь также сильно налегает Mv
32. РОД CROCIDURA
391
Стогов и Савинов предполагают сходство этой белозубки с представи-
телями рода Diplomesodon Brandt.
Распространение и возраст. Окрестности г. Павло-
дара (Казахстан), миоцен.
Рис. 195. Paracrocidura schoutedeni (Л) (по: Heim de Balsac, 1956), Praesorex goliath (Б)
(no: Repenning, 1967), Crocidura pavlodarica (В) (по: Стогов, Савинов, 1965), Limnoecus
tricuspis (/’) (no: Stirton, 1930).
t 2. Crocidura comfeldi Kormos, 1934.
Kormos, 1934, p. 9, 10.
Тип. Фрагмент передней части черепа с зубами № ОБ 1386, кол. Kgl. Ung.
Geol. Anst.
Распространение и возраст. Европа, плиоцен.
t 3. Crocidura zorzii Pasa, 1949.
P a s a, 1949, Mem. Mus. Civ. Storia Natur., Verona, t. 1, p. 18—20.
Тип не указан, вид описан из Италии.
392
ПОДСЕМ. SORICINAE
Распространение и возраст. Европа, плиоцен.
4. Crocidura suaveolens Pallas, 1811 — Малая белозубка (рис. 196).
Sorex suaveolens Pallas, 1811, Zoogr. Rosso-Asiatica, v. 1, p. 132, 133; — leo-
codon (Herm.) N о r d m a n, 1840, Observ. sur Faune pontique, p. 13; — etruseus Bla-
sius, 1843, in: Wiegmann’s Arch. Naturgesch., Bd. 11, N 2, S. 29; — araneus Кес-
слер, 1850, Животные губерний Киевского учебного округа, с. 17. Pachyura suaveo-
lens Fitzinger, 1868, Sitzungsber. Osterr. Acad. Wiss. math.-naturwiss. Kl., Abt.
1, Bd 57, S. 171—-196. Crocidura suaveolens Brandt, 1873, Bull. Soc. Imp. Nat. Mos-
cow, v. 46, p. 22—27; — russula (Herm.) Огнев, 1908, Дневник Зоол. отд. Общ. лю-
бит. естествозн., т. 3, № 9, с. 22; — mimula (Miller) М и г у л и н, 1917, Млекопитаю-
щие Харьковской губ., с. 42—45.
Тип. не установлен, вид описан из Крыма.
Рис. 196. Crocidura suaveolens.
Диагноз. Граница между окраской верхней стороны тела и боков
не резкая. Череп мелкий, плоский. Конус второго зубца I1 крупнее кони-
ческой части коронки второго верхнего промежуточного зуба.
Описание. Хвост относительно тонкий, покрыт короткими во-
лосами и мелкими чешуйками, а также редкими длинными щетинками.
Верхняя сторона тела охристо-серая, буровато-серая или серая. Нижняя
сторона тела и внутренние поверхности ног — белые или серо-белые.
Измерения. Длина: тела — 50.0—68.0, хвоста — 25.0—34.0,
ступни — 10.0—11.5, кондило-базальная черепа — 15.0—17.2 мм.
Распространение и возраст. Центральная и южная
части Западной Европы, юг и центр европейской части СССР, Южный
Казахстан, Средняя Азия до южных границ СССР, Иордания, Иран,
на восток до Алтая, Северной Монголии, Северо-Восточного Китая,
Приморья и п-ова Корея включительно; Северная Африка. В СССР рас-
пространена на большей части Украины, к северу до южных окраин
Белоруссии, Московской и Горьковской обл., а отсюда северная граница
спускается к городам Оренбургу и Уральску, доходит до Семиозерного
района Кустанайской обл., Прибалхашья, Алтая и южных районов
Бурятской АССР на восток до Южного Приморья. На юг распространена
до южных границ СССР.
Биология. Образ жизни мало изучен. Обитает в разнообразных
биотопах. Ее добывали в лесах, кустарниках лесных долин и в открытой
степи. Встречается в домах.
32. РОД CROCIDURA
393
Спаривание происходит в мае; беременность длится около 4 недель.
Детенышей 5—10. Молодые рождаются слепыми; самостоятельными ста-
новятся через 6 недель. Пищей служат различные мелкие беспозвоночные,
преимущественно насекомые.
Подвиды. Описано 14 подвидов. На территории СССР следующие:
1. С. s. suaveolens Pall., 1811. Характеризуется темной окраской,
относительно коротким хвостом и мелким черепом. Распространен в евро-
Рис. 197. Crocidura hyrcana.
пейской части СССР, на Кавказе и на восточных склонах Южного и Сред-
него Урала.
2. С. s. shantungensis Mill., 1901. Мелкая форма с очень светлой окрас-
кой. Распространена в южной части Советского Приморья, Северо-Восточ-
ном Китае и на п-ове Корея.
3. С. s. ilensis Mill. 1901. Отличается светлой окраской и более круп-
ными размерами тела. Распространен в Средней Азии, точнее границы
ареала не ясны, в Сибири найден близ г. Троицка Челябинской обл.
5. Crocidura hyrcana Goodwin, 1940 — Гирканская белозубка
(рис. 197).
Goodwin, 1940, Amer. Mus. Novitates, N 1082, p. 1, 2.
Тип. J № 88738. 60 км. к северо-востоку от Астрабада на р. Горгане (Иран),
кол. Амер. муз. естеств. истор., Нью-Йорк.
Диагноз. Граница между темной окраской верхней стороны тела
и боков в разной степени резкая. Череп мелкий и плоский. Конус вто-
рого зубца внутреннего верхнего резца меньше конической части ко-
ронки второго верхнего промежуточного зуба или только короче его
по высоте.
Описание. Кроме особенностей, отмеченных в диагнозе, следует
отметить и следующие: хвост толстый, покрыт крупными чешуйками
и более или менее длинными волосами (не менее половины диаметра
хвоста в его средней части). Окраска верхней стороны тела буровато-
серая, нижней — беловато-серая или беловатая.
Измерения. Длина: тела — 51.4—62.0, хвоста — 31.0—39.0,
ступни — 9.9—11.3, кондило-базальная черепа — 15.6—17.6 мм.
394
ПОДСЕМ. SORICINAE
Распространение и возраст. Добыта на р. Горгане
в 60 км от Астрабада (Иран). В СССР известна из западной части Туркме-
нии (Кара-Калинский р-н, р. Сумбар, ущелье Ай-Дере). Современный.
Биология не изучена. Обитает в полупустынной зоне пред-
горий.
6. Crocidura persica Thomas, 1907 — Персидская белозубка (рис. 198).
Crocidura leucodon persica Thomas, 1907, Ann. Mag. Nat. Hist., v. 20, p. 198,
199;--caspica Lydekker, 1808, Zool. Rec., v. 44, p. 6031; — russula giilden-
staedti (Pall.) С атунин, 1915, Млекопитающие Кавказского края, с. 109—111; —
Рис. 198. Crocidura persica.
persica (Thomas) Виноградов, 1958, Зоол. журн., т. 37, выл. 8, с. 1236—1243.
Тип. № 7.7.14.9, Эльбурс, близ Демавенда, Северный Иран, кол. Брит, муз.,
Лондон.
Диагноз. Граница между темной окраской верхней стороны
тела и светлыми боками, особенно в зимнем волосяном покрове, резкая.
Череп массивный с крупными зубами. Второй верхний промежуточный
зуб равен или почти равен третьему. Конус второго зубца I1 менее (хотя бы
по высоте) конической части коронки второго верхнего промежуточного
зуба.
Описание. Хвост немного длиннее половины длины тела, покрыт
относительно длинными волосами и отдельными редкими щетинками,
как у всех представителей рода Crocidura Wagl. Кисти и ступни массив-
ные и широкие. Верхняя сторона тела коричневая бурая или серо-бурая,
бока и нижняя сторона тела, а также кисти и ступни белые или беловатые.
Измерения. Длина: тела — 64.0—72.0, хвоста — 35.0—39.0,
ступни — 12.0—13.5, кондило-базальная черепа 18.0—20.4 мм.
Распространение и возраст. Найдена в Северном
Иране (Эльбурс близ Демавенда) на высоте около 1980 м, на берегу Астра-
бадского залива (Молла-Кала) и близ Тераскана (к востоку от Гумбет-
Кабуза) в Горгане, Иране. Кроме того, добыта там же близ г. Дашта (Бодж-
нурд, недалеко от западного Копетдага, в бассейне р. Атрека). На терри-
тории СССР добыта (по: Виноградов, 1958) в Кара-Калинском р-не Турк-
мении (долина р. Сумбара, в 20 км от г. Кара-Калы, ущелье Ай-Дере,
32. РОД CROCIDURA
395
бассейн Чендыра, близ с. Ярты-Кала, верхнее течение р. Сумбара
Кайне-Касыр), современный.
Биология не изучена.
t 7. Crocidura obtusa Kretzoi, 1937—1938.
Kretzoi, 1937—1938, Separat. Ann. Mus. Nat. Hungarici, v. 31, pars Mineral.,
Geol., Palaeontol., p. 92.
Тип. Правая ветвь нижней челюсти (Fa=16), Венгрия, нижний плейстоцен.
Описание. Как отмечает Кретцой (Kretzoi, 1937—1938), опи-
сываемый вид по размерам близок к Crocidura leucodon Herm. Отличается
Рис. 199. Crocidura leucodon.
от последнего большей длиной 12 (лучше сказать, более крупными разме-
рами) и туповершинным Р4.
Распространение и возраст. Венгрия, нижний плей-
стоцен.
8. Crocidura leucodon Hermann, 1780 — Белобрюхая белозубка
(рис. 199).
Sorex leucodon Hermann, 1870, in: Zimmermann’s, Geogr. Gesch., Bd 2, S. 382.
Crocidura leucodon (Wagler) Brandt, 1865, Исследование о зубной системе землероек,
с. 60; — leucodus Schultze, 1897, Helios, Berlin, Bd 14, S. 90; — russula leucodon
Trouessart, 1910, Faune Mammif, d’Europe, p. 44.
Тип не установлен, вид описан из Страсбурга, Франция.
Диагноз. Граница между темной окраской верхней стороны тела
и светлыми боками резкая, особенно в зимнем волосяном покрове. Череп
крупный и массивный, с относительно широкой ростральной частью.
Второй верхний промежуточный зуб заметно менее третьего. Конус
второго зубца I1 равен или почти равен ионической части коронки второго
верхнего промежуточного зуба.
Описание. Хвост относительно короткий и толстый, не длиннее
половины длины тела, покрыт сравнительно короткими волосами и длин-
ными редкими щетинками. Из 10 экз. только у одного он оказался рав-
ным половине длины тела, у остальных значительно короче. Верхняя
396
ПОДСЕМ. SORICINAE
сторона тела охристо- или буровато-серая; бока и нижняя сторона тела
белые.
Измерения. Длина: тела — 68.0—80.0, хвоста — 28.0—37.0,
ступни — 12.0—13.5, кондило-базальная черепа 17.5—19.6 мм.
Распространение и возраст. Западная часть Франции,
Швейцария, Италия и Югославия. Найден в Иране и Турции, а также
в Китае (Синьцзян) и Монголии. Эллерман и Моррисон-Скотт (Ellermann,
Morrison-Scott, 1951) указывает на распространение этого вида в Ни-
дерландах, Бельгии, Польше и на о. Сицилия. Можно не сомневаться,
что этот вид распространен в Европе значительно шире. В пределах
СССР — в лесостепной, степной и полупустынной зонах европейской
части СССР к югу до Крыма, Закавказья и до территорий, расположенных
у северной части Каспийского моря. Подробных данных о распростра-
нении этого вида в Средней Азии нет. Он добыт в южной части Таджики-
стана, в Узбекистане. Известен из верхнего плейстоцена Азербайджана
(Бинагады), современный.
Биология не изучена.
Подвиды. 1. С. I. leucodon Herm., 1780. Характеризуется сред-
ними размерами тела (длина черепа до 20.0 мм) и относительно светлой
окраской. Распространен в Западной Европе и европейской части СССР.
2. С. I. lasia Thos., 1906. Отличается более крупными размерами
и темной окраской. Длина черепа 21.0 мм. Распространен на Черноморском
побережье Кавказа и в Турции. Описан как особый вид (см.: Виноградов,
1958, с. 1240).
9. Crocidura russula Hermann, 1870 — Европейская белозубка
(рис. 200).
Sorex russulus Hermann, 1870, in: Zimmermanns, Geogr. Gesch., Bd 2, S. 383; —
araneus Schreber, 1778, Sauget., Bd 3, S. 573; — constrictus Hermann, 1870?,
in: Zimmermanns, Geogr. Gesch., Bd 2, S. 383; — unicolor, Kerr, 1792?, Animal King-
dom, p. 208;— leucurus S h д w, 1800?, Gen. Zool., v. 1, N 2, p. 538;—fimbriatus
Wagler, 1832, Isis, p. 54; — Crocidura mischata Wagler, 1832, Isis, p. 275;
— major Wagler, 1832, Isis, p. 1218; — rufa Wagler, 1832, Isis, p. 1218; — po-
liogastra Wagler, 1832, p. 1218; — Sorex thoracicus S a v i, 1832, Nupvo Giorn de
Letterati, v. 24, p. 52; — inodorus deSelys Longchamps, 1838, Etudes Micro-
mammal., p. 34; — Crocidura aranea var. albiventris deSelys Longchamps,
там же. Sorex hydruntina Costa, 1839?, Fauna del Ragna di Napoli, Mammal.,
p. 6; — chrysithorax D e h n e, 1855, Allg. Deutsche Naturhist. Zeitung, Bd 1, S. 241;
— mauritanicus R о m e 1, 1856, C. r. Acad, sci., t. 42, p. 653; Pachyura agilis L e v s i 1-
1 a n t, 1867, in: Loyhe, Explor. Sci. de 1’Algerie, Zool. pl. 4, fig. 2; — pygmaea L о c h e,
1867, там же, с. 86. Crocidura caneae Miller, 1909, p. 418; — ichnussae Festa, 1912,
Boll. Mus. Zool. Anat. Comp., v. 27, N 648, p. 1; — yebalensis Cabrera, 1913, Bol.
Soc. Esp. Hist. Nat., v. 13, p. 400; — corsicana Heim de Balzac etReynand,
1940, Bull. Soc. Zool. Franse, t. 65, p. 216; — anthony Heim de Balzac, 1940,
Bull. Mus. Hist. Nat., t. 12, p. 382; — foucauldi A g a c i n o, 1943, Bol. Soc. Esp. Hist.
Nat., v. 41, p. 37.
Тип не установлен, вид описан из окрестностей Страсбурга (Bas-Rhin.),
Франция.
Диагноз. Граница между окраской верхней стороны тела и свет-
лыми боками не резкая и в зимнем волосяном покрове. Череп с удлинен-
ной ростральной частью (по сравнению с С. leucodon Herm.). Боковая
сторона верхнечелюстной кости в отличие от упомянутого вида не обра-
зует хорошо заметного выступа между задним краем Р1 и передним М1.
Второй и третий верхние промежуточные зубы крупнее, они почти сходны
по размерам или по высоте. Конус второго зубца внутреннего верхнего
резца заметно ниже конической части коронки второго верхнего проме-
жуточного зуба, но шире ее (в передне-заднем направлении).
32. РОД CROCIDURA
397
Описание. Хвост относительно короткий, около половины длины
тела. Покрыт очень короткими волосами и длинными щетинками. Верх-
няя сторона тела бурая или коричневая, разных оттенков. Бока и нижняя
сторона тела сероватые, охристо-сероватые,
реже серо-беловатые.
Измерения. Длина: тела — 64.0—
74.0, хвоста — 31.0—40.0, ступни — 11.5—
13.5, кондило-базальная черепа —18.0—19.8мм.
' Распространение и возраст.
Нидерланды, Бельгия, Франция, ГДР, ФРГ,
Рис. 200. Crocidura russula.
Швейцария, Италия, Югославия, Тунис, Алжир, Марокко, острова Сре-
диземного моря, современный.
10. Crocidura sibirica Dukelsky, 1930 (рис. 201).
Crocidura leucodon sibirica Dukelsky, 1930, Zool. Anzeig., Bd 88, H. 1—4,
:S. 75, 76; ognevi Строганов, 1956, Tp. Биол. инет. Зап. — Сиб. фил.
АН СССР, вып. 1, Зоол., с. 9, 10; Crocidura ognevi Stroganov Виноградов, 1958,
Зоол. журн., т. 37, вып. 8, с. 1239; — sibirica Dukelsky Юдин, 1971, Насекомояд-
ные млекопитающие Сибири (определитель), с. 146—149.
Тип. 3 № М. 3343, село Означенное на р. Енисее, в 96 км к югу от г. Минусинска,
современный, кол. Зоол. муз. МГУ.
Диагноз. Граница между окраской верха и боков тела не резкая.
Череп сравнительно крупный с очень коротким носовым отделом. Сумма
продольных диаметров очень мелких второго и третьего верхних проме-
жуточных зубов равна или менее продольного диаметра переднего зубца I1
у его основания.
Описание. Длина хвоста не менее половины длины тела. Он по-
крыт короткими прижатыми волосами и редкими длинными щетинками.
Окраска верхней стороны тела темная, коричнево-бурая, на боках более
светлая, нижний край боков тела и головы светло-серые или серые. Ниж-
няя сторона тела серая или серо-беловатая.
Измерения. Длина: тела — 58.0—65.0, хвоста — 31.0—34.0,
ступни — 11.0, кондило-базальная черепа — 18.0 мм.
Распространение и возраст (по: Строганов, 1956).
Уймонская долина и побережье Телецкого озера на Алтае; район с. Майма
на р. Катуни; окрестности дер. Кунчак в среднем течении р. Бии; Верхне-
обский бор близ пос. Боровлянка между Барнаулом и Бийском; долина
р. Тайдона (правый приток р. Томи), Кемеровская обл.; окрестности
398
ПОДСЕМ. SORICINAE
с. Кожевникове и район пристани Дубровино на р. Оби; окрестности
Новосибирска, дер. Заварзино под Томском.
Биология изучена недостаточно. В Западной Сибири населяет
леса различных типов, на Алтае найдена на разнотравном лугу вбяизи
Телецкого озера, а в предгорьях — в зарослях ивняков по луговым доли-
нам ручьев, впадающих в р. Катунь. В Кузнецком Алатау найдена в тайге,
на участках с густой и высокой травяной растительностью. В Верхне-
обском бору добыта в сосновых лесах с травяным ярусом и в смешанном
сосново-лиственном участке леса, вблизи болота и йа берегу озера. Север-
Рис. 201. Crocidura sibirica.
нее Новосибирска найдена в сосновом бору на берегу р. Оби. Под Том-
ском добывалась в затененном сыром логе, заросшем мелколиственными
деревьями. В Минусинской степи найдена на берегу р. Енисея, среди
камней, в местах, где почти совершенно отсутствовала растительность.
Питается в основном различными насекомыми. Молодые встречаются
в течение всего весенне-летнего сезона. Число детенышей в помете 5—10. 11
11. Crocidura lasiura Dobson, 1890 — Большая уссурийская белозубка
(рис. 202).
Dobson, 1890, р. 31—33; — thomasi S о w е г Ь у, 1917, Ann. Mag. Nat. Hist.,
v. 20, p. 318; — lisenkani Kishi d a, 1931, Zool. Mag., Tokyo, v. 43, p. 377 (nom.
nud.); — yamashinai Kuroda, 1934, J. Mammal., v. 15, p. 237.
Тип. № 2383, p. Уссури, кол. ЗИН АН СССР, Ленинград.
Диагноз. Череп крупный, угловатый. Второй верхний промежу-
точный зуб меньше третьего. Конус заднего зубца I1 равен или немного
меньше конической части коронки второго верхнего промежуточного
зуба. Зимой и летом окраска сходная. Длина ступни не более 16% длины
тела.
Описание. Длина хвоста менее 50% длины тела. Окраска тела
бурая или черно-бурая, на нижней стороне с серым оттенком. В отличие
от землероек этого рода нашей фауны кисти и ступни у них черно-бурые,
когти белые (светлые).
Измерения. Длина: тела — 90.0—104.0, хвоста — 37.0—42.0,
ступни — 14.5—16.4, кондило-базальная черепа — 21.5—25.0 мм.
Распространение и возраст. Широко распростра-
нена в Северо-Восточном Китае на п-ове Корея. В СССР найдена в ряде
32. РОД CROCIDURA
399
пунктов Приморья; в долинах рек Уссури и Туман-Галы, окрестностях
г. Уссурийска, в заповеднике «Кедровая падь», на оз. Ханко и в других
местах, современный.
Биология. В пределах СССР обитает в широколиственных лёсах.
Попадается и в культурной полосе, на залежах, в зарослях полыни.
Питается наземными беспозвоночными, преимущественно насекомыми.
Период размножения, по-видимому, растянут. У самки, добытой 9 ав-
густа, обнаружено 10 эмбрионов.
Рис. 202. Crocidura lasiura.
12. Crocidura giildenstaedti Pallas, 1811. — Кавказская длиннохвостая
белозубка (рис. 203).
Sorex guldenstaedti Pallas, 1811, Zoogr. Rosso-Asiatica, v. 1, p. 133. S(orex)
araneus Nathusius, 1838, Arch. Naturgesch., S. 24; — leucodon Wagner, 1841,
Sauget., Bd 2, p. 66. Sorex (Crocidura) fumigatus Filippi, 1863, Arch. Zool. Anat.
Physiol. Genova, t. 2, p. 379. — Crocidura longicaudata (Dobson) Тихомиров,
Корчагин, 1889, Изв. Общ. любит, естеств., т. 6, вып. 4, № 1, с. 17. Sorex bogda-
novi (Tscherniavski) S a t u n i n, 1896, Zool. Jahrb., Bd 9, S. 288. Crocidura russulus
(Herm.) = Crocidura araneus С а т у н и н, 1899, Изв. Кавказск. муз., т. 1, с. 53;
— guldenstaedti (Pall.) С а т у н и н, 1903, Зап. Кавказск. отд. Геогр. общ., т. 24, с. 53;
— russula monacha Thomas, 1906, Ann. Mag. Nat. Hist., v. 17, p. 417;-------caspica
Thomas, 1907, Ann. Mag. Nat. Hist., v. 20, p. 197, 198;-------aralychensis G а т y-
н и h, 1914, Изв. Кавказск. муз., т. 8, вып. 1—2, с. 92;-guldenstaedti С а т у н и н,
1915, Млекопитающие Кавказского края, с. 109—111; — guldenstaedti, Виногра-
дов, 1958, Зоол. журн., т. 37, вып. 8, с. 1241.
Тип не установлен, вид описан из окрестностей г. Душети, ГрузССР.
Диагноз. Череп с относительно узкой лицевой частью, тонкого
сложения. Второй верхний промежуточный зуб меньше третьего. Конус
заднего зубца I1 равен конической части коронки второго верхнего про-
межуточного зуба. В зимнее время окраска не приобретает серебристого
цвета. Длина ступни от 16.8 до 18.8% длины тела.
Описание. Хвост толстый у основания, постепенно суживается
к концу. Его длина составляет 63—75% длины тела. Окраска верхней
стороны тела коричневая, бурая или буро-серая, разных оттенков; ниж-
ней — серая, охристо-серая или буроватая, часто сходная с окраской
верхней стороны.
400
ПОДСЕМ. SORICINAE
Измерения. Длина: тела — 65.2—80.0, хвоста — 43.0—53.0,
ступни — 12.0—13.5, кондило-базальная черепа — 17.8—19.7 мм.
Распространение и возраст. Кавказ (от северных
предгорий Главного хребта) и северные области Турции и Ирана, совре-
менный.
Биология. Обитает в разнообразных условиях: горных и равнин-
ных лесах разных типов, в кустарниковых зарослях и на участках с луго-
вой растительностью, по берегам водоемов, а также в степи; попадается
Рис. 203. Crocidura gUldenstaedti.
в жилых постройках. Пищей служат различные беспозвоночные, главным
образом насекомые. Питается также мелкими позвоночными, которых
может осилить. Размножение не изучено.
13. Crocidura pamirensis Ognev, 1928 — Памирская белозубка
(рис. 204).
Огнев, 1928, с. 366—368; — serezkyensis Лаптев, 1929, Определитель мле-
копитающих Средней Азии, с. 16.
Тип. <J № 4553, окрестность оз. Друм, южные склоны Памирского хребта, кол.
Зоол. муз. МГУ.
Диагноз. Череп с относительно узкой лицевой частью, мелкий.
Второй верхний промежуточный зуб меньше третьего. Конус заднего
зубца I1 равен или крупнее конической части коронки второго верхнего
промежуточного зуба. Зимняя окраска серебристо-серая. Длина ступни
от 19.9 до 20.0% длины тела.
Описание. Хвост тонкий, его длина не менее 75% длины тела.
В окраске резко выражен сезонный диморфизм. Летняя окраска серо-
бурая или буроватая, зимняя — серебристо-серая, иногда с буроватым
налетом. Нижняя сторона тела серовато-белая или белая. Вибриссы
длинные.
Измерения. Длина: тела — 57.1—68.0, редко до 70.0, хвоста —
40.0—49.6, ступни — 12.0—13.3, кондило-базальная черепа — 17.5—
18.0 мм.
32. РОД CROCIDURA
401
Распространение и возраст. Известна из следующих
пунктов Западного Памира: оз. Друм на южном склоне Памирского
хребта, Нимхана близ устья р. Маца, на р. Памире, несколько пунктов
на р. Пяндже, Банче, с. Шардара, оз. Сарезское; возможно, обитает
и на других хребтах Таджикистана (Виноградов, 1958). Найдена на Гис-
сарском хребте (кишлак Наджи), современный.
Биология не изучена.
Примечание. Crocidura pamirensis Ogn., С. armenica Gur.,
C. pergrisea Mill, и другие серебристо-серые длиннохвостые землеройки
составляют в пределах рода Crocidura Wagl. своеобразную группу. Общей
Рис. 204. Crocidura pamirensis.
для них особенностью служит сходная серебристая окраска в зимнее
время, значительная длина хвоста и относительно длинная ступня при
сравнительно малой длине тела, а также крупный череп. Характерно
и их распространение: высокогорья Бирмы, Китая, Индии, Пакистана,
Афганистана, Ирана и СССР; близкие к ним формы встречаются и в Аф-
рике. Возможно, что эти землеройки при изучении достаточного материала
не только из Азии, но и Африки, будут выделены в особый подрод —
альпийских белозубок.
14. Crocidura armenica Gureev, 1963 — Армянская белозубка
(рис. 205).
Гуреев, 1963, с. 118, 119.
Тип. <5 № 45277, Армения, окрестности с. Гарни (14 км вниз по р. Гарни), кол.
ЗИН АН СССР, Ленинград.
Диагноз. Череп мелкий. Второй верхний промежуточный зуб
крупнее третьего в 1.5—1.75 раза. Конус заднего зубца I1 равен кони-
ческой части коронки второго верхнего промежуточного зуба. Зимняя
окраска пепельно-серая. Длина ступни не менее 19% длипы тела.
Описание. Длина хвоста не менее 75% длины тела. Летняя
окраска верхней стороны тела буровато-серая, на боках светлее, нижней —
беловатая с охристым налетом. Зимняя окраска верхней стороны тела
пепельно-серая с серебристым налетом, более заметным на боках, ниж-
ней — белая. Вибриссы длинные.
26 А. А. Гуреев
;402
ПОДСЕМ. SORICINAE
Измерения. Длина: тела — 60.0, хвоста — 45.0, ступни — 42.0,
кондило-базальная черепа — 16.5 мм (промеры по 1 экз.).
^Распространение и возраст. Армянская ССР. Из-
вестна из двух местонахождений: из Сисианского р-на, с. Сисеван,
и с. Гарни (на р. Гарни), современный.
Биология не изучена.
Рис. 205. Crocidura armenica.
15. Crocidura dinniki Ognev, 1921 — Белозубка Динника (рис. 206)
Огнев, 1921, Ежегодн. Зоол. муз. АН, т. 22, с. 340.
Тип. № 9169, Ставрополь, кол. ЗИН АН СССР, Ленинград.
Диагноз. Череп относительно крупный. Второй верхний проме-
жуточный зуб крупнее третьего. Конус заднего зубца I1 меньше (вернее,
ниже) конической части коронки второго верхнего промежуточного зуба.
Зимняя окраска серовато-палевая. Длина ступни 17—21% длины тела.
206
Рис. 206. Crocidura dinniki.
207
Рис. 207. С. zarudnyi (по: Огнев, 1928).
Описание. Длина хвоста не более 50—60% длины тела. Судя
по описанию Огнева (1928), летняя и зимняя окраски у этого вида мало
отличаются. Верхняя часть тела окрашена в серо-коричневый цвет,
нижняя — в белый с желтоватым налетом. Верхняя сторона кистей
и ступней желто-белая. Вибриссы длинные.
Измерения. Длина: тела — 58—70, хвоста — 37.3—44.4,
ступни — 11.5—12.3, кондило-базальная черепа — 17.6—17.9 мм.
Распространение и возраст. Известна с Северного
Кавказа (Ставрополь), Закавказья (Грузия) и из Ирана (Чай — приток
р. Гюргена), современный. Распространение по: Огнев, 1928.
32. РОД CROCIDURA
403
16. Crocidura zarudnyi Ognev, 1928 — Белозубка восточноиранская
(рис. 207).
Crocidura tatianae Огнев, 1921, Ежегодн. Зоол. муз. АН, т. 22, с. 338; — za-
rudnyi Огнев, 1928, с. 341—344.
Тип. № 6506, Восточный Иран, кол. ЗИН АН СССР, Ленинград.
Д и а г и о з. Череп с относительно узкой мозговой капсулой. Второй
верхний промежуточный зуб равен третьему. Конус заднего зубца I1
крупнее конической части коронки второго верхнего промежуточного
зуба. Зимняя окраска светло-серая с охристым налетом. Длина ступни
20—21% длины тела.
Описание. Длина хвоста не менее 75% длины тела. Окраска
верхней стороны тела (по спиртовому экз.) светло-серая с охристым от-
тенком, образованным коричневыми концами отдельных волос, нижней —
белая. Хвост сверху охристо-беловатый, снизу — белый. Вибриссы длин-
ные. По-видимому, описываемый экземпляр в зимнем волосяном покрове.
Летняя окраска, как можно предполагать по аналогии с близкими видами,
охристо-сероватая.
Измерения. Длина: тела — 60.8, хвоста — 47.5, ступни — 13.0,
кондило-базальная черепа — 18.8 мм.
Распространение и возраст. Известен по 1 экз. из вос-
точной части Ирана, современный.
Примечание. Эллерман и Моррисон-Скотт (Ellermann, Mor-
rison-Scott, 1951) отнесли этот вид к С. pergrisea Mill., что как мне кажется,
требует специальной проверки на достаточном материале.1
17. Crocidura caudata Miller, 1901.
Miller, 1901, Proc. Biol. Soc. Washington, v. 14, p. 42.
Распространение. Корсика, Сицилия, Балеарские острова.
18. Crocidura pergrisea Miller, 1913.
Miller, 1913, там же, т. 25, с. 113, 114.
Тип. $ № 175918, Балтистап, кол. Амер. нац. муз., Вашингтон.
Распространение. Восточная часть Л рана, Пакистан, Ин-
дия (Кашмир).
19. Crocidura attenuata Milne-Edwards, 1872.
Milne-Edwards, 1872, p. 263; — grisea Howell, 1926, там же, т. 39,
с. 137.
Т и п не установлен, вид описан из Моупина, Сычуань (Китай).
Распространение. Киангсу, Чекианг, Хупех, Сычуань, Ту-
кан, Фукиен, западная часть Юньнани, Хайнань (Китай); северная часть
Бирмы, Ассам, Бутан Дуарс, Сикким, Пенджаб, Кашмир (Индия).
20. Crocidura dracula Thomas, 1912.
Thomas, 1912, Ann. Mag. Nat. Hist., v. 9, p. 686; — praedax Thomas, 1923,
там же, т. 11, с. 656. — grisescens Howell, 1928, J. Mammol., v. 9, p. 60.
T i n. (J (оригинальный № 34), добыт вблизи Mengtsz, Юньнань, Китай, кол. Брит,
муз., Лондон.
Распространение. Юньнань, Фукиен (Китай), Северная
Бирма, п-ов Индокитай.
1 Виды, требующие ревизии, приведены далее без описания и указания возраста.
26*
404
ПОДСЕМ. SORICINAE
21. Crocidura vorax Allen, 1923.
Allen, 1923, Amer. Mus. Novitates, N 100, p. 8.
Тип. <j № 44383, Ликианг, Юньнань (Китай), кол. Амер. муз. естеств. истор.,
Нью-Йорк.
Распространение. Юньнань, Китай.
22. Crocidura rapax Allen, 1923.
Allen, 1923, там же, с. 9.
Тип. j № 44321, Виппанкай, р. Меконг, южная часть Юньнани, Китай, кол.
Амер. муз. естеств. истор., Нью-Йорк.
Распространение. Юньнань, Китай.
23. Crocidura dsinezumi Temminck, 1842.
Temminck, 1842, in: Siebold, Fauna Japon., Mammal., p. 26.
Эллерман и Моррисон-Скотт (Ellermann, Morrison-Scott, 1951) отнесли
этот вид к С. russula Herm. в качестве подвида, с чем трудно согласиться
хотя бы потому, что до сих пор С. russula никто для Азии не отмечал.
Окраска верхней стороны тела С. dsinezumi Temm. шоколадно-коричне-
вая, нижней — с синеватым отливом, зимой и летом сходная.
Распространение. П-ов Корея, Япония.
24. Crocidura horsfieldi Thomas, 1856.
Thomas, 1856, Ann. Mag. Nat. Hist., v. 17, p. 23; — retusa Peters, 1870,
Monatsber. Konigl. Preuss. Akad. Wiss., Berlin, S. 585. Sorex (Crocidura) myoides Blan-
ford, 1875, J. Asiat. Soc. Bengal, v. 44, N 2, p. 106. Crocidura indochinensis Robin-
son, Kloss, 1922, Ann. Mag. Nat. Hist., v. 9, p. 88; — watasei Kuroda, 1924,
New Mammal, from Riu-Kiu Islands, p. 1; — tadae Tokuda, Kano, 1936, Annot.
zool. Japon., v. 15, p. 429.
T и п не установлен; вид описан из Шри Ланка (о. Цейлон).
Распространение. Шри Ланка, Кашмир (Индия), п-ов Индо-
китай, Таиланд, Северная Бирма, Япония.
25. Crocidura fuliginosa Blyth, 1855.
В 1 i t h, 1855, J. Asiat. Soc. Bengal, v. 24, N 4, p. 362; — rubricosa and kingiana
Anderson, 1877, p. 280, 281.
Тип не установлен, вид описан из Ассама (Индия).
Распространение. Восточные Гималаи, Ассам. Тенассерим,
обычна в Сиккиме (Индия).
26. Crocidura hispida Thomas, 1913.
Thomas, 1913, Ann. Mag. Nat. Hist., ser. 8, v. 21, p. 468, 469.
Тип. № 13.3.13.1. Средние Андаманские острова в Бенгальском заливе, кол.
Брит, муз., Лондон.
Распространение. Известен только с Андаманских островов.
27. Crocidura monticola Peters, 1870.
Peters, 1870, Monatsber. Konigl. Preuss. Akad. Wiss., Berlin, S. 588.
Тип не установлен, вид описан с гор Лаву (Lawu), вблизи Саракарта, о. Ява.
Распространение. О. Ява.
32. РОД CROCIDURA
405
28. Crocidura doriae Peters, 1870.
Peters, 1870, там же, с. 587; Davis, 1962, Bull. Nat. Mus. State Singapore,
NJ31, p. 27, 28.
T и п не установлен, вид описан из Саравака, северная часть Калимантана
(о. Борнео).
Распространение. О. Калимантан.
Примечание. С Калимантана Петерс (Peters, 1870) описал
еще один вид землеройки — Crocidura foetida, отличающийся от описан-
ного меныпими размерами и более яркой коричневой окраской. Можно
думать, что это не особый вид.
29. Crocidura tenuis Muller, 1839.
М ii 11 е г, in: Temmink, 1839, Vehrh. Nat. Ges. Ned. Overz. Bezitt. Zool., v. 1,
Nj26, p. 50; — macklotti J e n t i n k, 1888?, Notes Leyden Mas., v. 10, p. 164.
Тип не установлен, вид описан с о. Тимор.
Распространение. О. Тимор.
30. Crocidura elongata Miller et Hollister, 1921
Miller, Hollister, 1921, Proc. Biol. Soc. Washington, v. 34, p. 101.
Тип. в (шкура и череп), № 217534, Тембоан (к юго-западу от оз. Тондано),
северо-восточная часть Сулавеси (о. Целебес), кол. Амер. нац. муз., Вашингтон.
Распространение. О. Сулавеси.
31. Crocidura nigripes Miller et Hollister, 1921.
Miller, Hollister, 1921, там же.
Тип. (шкура и череп), № 217545, Тембоан (к юго-западу от оз. Тондано),
северо-восточная часть Сулавеси (о. Целебес), кол. Амер. нац. муз., Вашингтон.
Распространение. Сулавеси.
32. Crocidura rhoditis Miller et Hollister, 1921.
Miller, Hollister, 1921, там же, с. 102.
Тип. 3 (шкура и череп), № 217550, Тембоан, Сулавеси (о. Целебес), кол. Амер,
нац. муз., Вашингтон.
Распространение, Сулавеси.
33. Crocidura lea Miller et Hollister, 1921.
Miller, Hollister, 1921, там же, с. 102.
Тип. <$ (шкура и череп), № 217553, Тембоан, северо-восточная часть Сулавеси
(о. Целебес), кол. Амер. нац. муз., Вашингтон.
Распространение. Сулавеси.
34. Crocidura levicula Miller et Hollister, 1921.
Miller, Hollister, 1921, там же, с. 103.
Тип. $ (шкура и череп), № 219450, Пинодапа, Сулавеси (о. Целебес), кол. Амер,
нац. муз., Вашингтон.
Распространение. Сулавеси.
35. Crocidura giffardi De Winton, 1898.
De Winton, 1898, Ann. Mag. Nat. Hist., ser. 7, v. 11, p. 484.
Тип. ? № 98.10.24.5, Moshi (Mossi). Золотой берег, кол. Брит, муз., Лондон.
Распространение. Золотой берег.
406
ПОДСЕМ. SORICINAE
36. Crocidura manni Peters, 1878.
Peters, 1878, Sitzungsber, Ges. Naturf. Freunde, Berlin, S. 19.
Тип не отмечен.
Распространение. Северные и южные провинции Нигерии.
37. Crocidura odorata Leconte, 1857.
Sorex odoratus Leconte, 1857, Proc. Acad. Nat. Sci. Philadelphia, p. 11.
T и п не установлен, вид описан из Габона, Западная Африка.
Распространение. Габон, Западная Африка.
38. Crocidura biittikoferi Jentink, 1887.
J e n t i n k, 1887, Notes Leyden Mus., v. 10, p. 47.
Тип. $ (в спирту). Порт Роберта, Либерия.
Примечание. Наличие бугорков на 1х у этого вида позволяет
предполагать, что он принадлежит к роду Paracrocidura Н. de Bals, или
даже к особому роду. Но решить этот вопрос можно лишь при сравнении
.фактического материала.
Распространение. Известен из одного местонахождения.
39. Crocidura anchietae Bocage, 1889.
В о с a g е, 1889, J. Sci. Lisb., v. 1, р. 26.
Т и п не установлен, вид описан из Анголы.
Распространение. Ангола.
40. Crocidura hedenborgiana Sundevall, 1842.
Sorex hedenborgiana Sunde all, 1842, K. Svenska Vetensk. Acad. Handl.
Stock., S. 174.
Тип не отмечен, вид добыт в Сеннаре (Судан).
Распространение. Сейнар (Судан), Эфиопия.
41. Crocidura occidentalis Pucheran, 1855.
Pachyura occidentalis P u c h e r a n, 1855, Rev. Mag. Zool., ser. 2, v. 7, p. 154;
— aequatorialis Pucheran, 1855, там же. Crocidura petersi Dobson, 1890, Ann.
Mag. Nat. Hist., ser. 6, v. 6, p. 495.
Тип не установлен, вид описан, по-видимому, из Габона (Западная Африка).
Распространение. Габон, Бибинага, Азаба, Сьерра-Леоне.
42. Crocidura schweizeri Peters, 1877.
Peters, 1877, Monatsber. Konigl. Preuss. Acad. Wiss., Berlin, S. 187; — stamp-
lit J en t ink, 1877, Notes Leyden Mus., v. 10, p. 47.
Тип. $ (спиртовой), Либерия, кол. Брит, муз., Лондон.
Распространение. Либерия.
43. Crocidura martiensseni Neumann, 1900.
Neumann, 1900, Zool. Jahrb., Abt. Syst., Bd 6, S. 544.
Тип не отмечен, вид описан с Килиманджаро.
Распространение. Килиманджаро и Усамбара (Танганьика).
44. Crocidura fuscosa Thomas, 1913.
Crocidura doriana fuscosa Thomas, 1913, Ann. Mag. Nat. Hist., ser. 8, v. 12, p. 90.
Тип. № 1.8.8.17, Какка, Белый Нил, Брит, муз., Лондон.
Распространение. Какка (Эфиопия), Белый Нил.
32. РОД CROCIDURA
407
45. Crocidura nyansae Neumann, 1900.
N e u m a n n, 1900, Zool. Jahrb., Abt. Syst., Bd 6, p. 544.
T и п не отмечен, вид добыт у места выхода р. Нила из оз. Виктория-Ньянза
(Уганда).
Распространение. Уганда, Кения, Южная Родезия, Бот-
свана.
46. Crocidura sururae Heller, 1910.
Heller, 1910, Smiths. Misc. Coll., v. 56, N 15, p. 2.
Тип. № 164637, добыт, по-видимому, в Рино-Камп, Ладо Энклаве, кол. Амер,
нац. муз., Вашингтон.
Распространение. Рино-Камп, Ладо Энклаве, Уганда.
47. Crocidura doriana Dobson, 1887.
Dobson, 1887, Ann. Mus. Civ. Stor. Nat. Genova, v. 4, p. 564.
Тип не отмечен, вид описан из Шоа, Эфиопия.
Распространение. Эфиопия.
48. Crocidura flavescens Geoffroy, 1827.
Sorex flavescens Geoffroy, 1927, Diet. Classif., v. 11, p. 324; Mem. Mus. Hist.
Nat., Paris, t. 15, p. 126; — cinnamomeus Lichtenstein, 1829, Verhandl. Ges.
Naturf. Freunde, Berlin, Bd 11, S. 381; — capensis Smith, 1833, South Afric. Quart.
J., v. 11, p. 62; — rutilus Sundevall, 1846, Ofvers Vetensk. Acad. Forh., v. 3,
p. 119.
T и п не отмечен, вид описан из «1а Cafrerie et le pays des Hottentots», Южная
Африка.
Распространение. Южная Африка.
49. Crocidura zaphiri Dollman, 1915.
D о 1 1 m a n, 1915, Ann. Mag. Nat. Hist., ser. 8, v. 16, N 91, p. 66, 67
Тип. 9 №6.11.1.12, Каффа, Эфиопия, кол. Брит, муз., Лондон.
Распространение. Каффа, Эфиопия.
50. Crocidura olivieri Lesson, 1827.
Sorex olivieri Lesson, 1827, Manusc. Mammal., p. 121,
Тип не установлен, вид описан из Египта.
Распространение. Египет.
51. Crocidura beirae Dollman, 1915.
D о 11 m a n, 1915, Ann. Mag. Nat. Hist., ser. 8, v. 16, N 91, p. 69, 70.
Тип. J № 7.6.2.28, Бейра, Мозамбик, кол. Брит, муз., Лондон.
Распространение. Мозамбик, Ботсвана.
52. Crocidura hirta Peters, 1852.
Peters, 1852, Boise Mossamb. Saug., S. 78; — canescens Peters, 1852, там же,
с. 83; anneiata Peters, 1852, там же, с. 85; — flavescens flavidula Thomas,
Schwann, 1905, Proc. Zool. Soc. London, p. 264; — pondoensis Roberts, 1913,
Ann. Transvaal. Mus., v. 4, pt 2, p. 71.
Тип не установлен, вид описан из Тете, Мозамбик.
Распространение. Мозамбик, Трансвааль.
53. Crocidura sacralis Peters, 1852.
Peters, 1852, там же, с. 82,
Тип не отмечен, вид описан с о, Кабацейра, Мозамбик.
408
ПОДСЕМ. SORICINAE
Распространение. Мозамбик.
54. Crocidura mariquensis Smith, 1849.
Sorex mariquensis Smith, 1849, Illustr. Zool. South Africa, p. 1.
Тип не установлен, вид описан из Южной Африки.
Распространение. Южная Африка.
55. Crocidura hindei Thomas, 1904.
Thomas, 1904, Ann. Mag. Nat. Hist., ser. 7, v. 14, p. 237.
Тип. $> № 1.8.7.2, Кения, кол. Брит, муз., Лондон.
Распространение. Кения.
56. Crocidura sericea Sundevall, 1842.
Sorex sericeus Sundevall, 1842, K. Svenska Vetensk. Acad. Hand!., Stock.,
S. 173—177. Crocidura strauchil Dobson, 1890, Ann. Mag. Nat. Hist., ser. 6, v. 6,
p. 225.
Тип не отмечен, вид описан из окрестностей Бар-эль-Абиад, Конго.
Распространение. Конго.
57. Crocidura beta Dollman, 1915.
D о 11 m a n, 1915, Ann. Mag. Nat. Hist., ser. 8, v. 15, p. 76—79.
Тип. № 12.7.1.67, Кения, кол. Брит, муз., Лондон.
Распространение. Кения.
58. Crocidura vetulina Thomas, 1904.
Thomas, 1904, там же,, сер. 7, т. 14, с. 237.
Тип. $ № 99.6.25.1, Усамбара (Танганьика), кол. Брит, муз., Лондон.
Распространение. Оз. Танганьика.
59. Crocidura lutrella Heller, 1910.
Heller, 1910, Smiths. Misc. Coll., v. 56, N 15, p. 4.
Тип. $ № 164640, Рино-Камп, Ладо Энклаве, Уганда, кол. Амер. нац. муз.,
Вашингтон.
Распространение. Известен из Рино-Камп, Ладо Энклаве,
Уганда.
60. Crocidura deserti Schwann, 1906.
Schwann, 1906, Proc. Zool. Soc., London, p. 103.
Тип. № 4.10.1.62, p. Молопо, Ботсвана, кол. Брит, муз., Лондон.
Распространение. Ботсвана.
61. Crocidura butleri Thomas, 1911.
Thomas, 1911, Ann. Mag. Nat. Hist., ser. 8, v. 8, p. 375.
Тип. № 874.2.10, Бар-эль-Газал, Конго, кол. Брит, муз., Лондон.
Распространение. Бар-эль-Газал, Конго.
62. Crocidura percivali Dollman, 1915.
D о 1 1 m a n, 1915, там же, т. 16, с. 126, 127.
Тип. <J № 12.7.1.61, Кения, кол. Брит, муз., Лондон.
Распространение. Кения.
32. РОД CROCIDURA
409
63. Crocidura fischeri Pagenstecher, 1885.
Pagens t e c h er, 1885, J. Wiss. Anst., Hamburg, Bd 2, S. 34.
Тип не установлен, вид описан с Танганьики.
Распространение. Оз. Танганьика.
64. Crocidura voi Osgood, 1910.
Osgood, 1910, Field Mus. Nat. Hist. (zool. ser.), v. 10, N 3, p. 18.
Тип. 3 № 16942, Вой, Кения, кол. Field Mus. Nat. Hist., Чикаго.
Распространение. Кения.
65. Croicdura suahelae Heller, 1912.
Heller, Smiths. Misc. Coll., v. 60, N 12, p. 6.
Распространение. Кения.
66. Crocidura smithi Thomas, 1895.
Thomas, 1895, Ann. Mag. Nat. Hist., ser. 6, v. 16, p. 51.
Тип. 3 № 96.8.1.1, Уэви Шебели, Сомали, кол. Брит, муз., Лондон.
Распространение. Сомали.
67. Crocidura turba Dollman, 1915.
D о 11 m a n, 1915, там же, сер. 8, т. 16, с. 130, 131.
Т и п. 3 №9.12.4.17, о. Чилюи, оз. Бангвеулу (Замбия),кол. Брит.муз., Лондон.
Распространение. Замбия, Кения, Уганда, Конго и Ангола.
68. Crocidura nilotica Heller, 1910.
Heller, 1910, Smiths. Misc. Coll., v. 56, N 15, p. 3.
Тип. § № 164638, Ладо Энклаве, Уганда, кол. Амер. муз. естеств. истор.,
Нью-Йорк.
Распространение. Уганда.
69. Crocidura zena Dollman, 1915.
D о 11 m a n, 1915, Ann. Mag. Nat. Hist., ser. 8, v. 16, p. 137, 138.
T и n. 3 №9.12.4.21, о. Чилюи, оз. Бангвеулу (Замбия),кол. Брит, муз., Лондон.
Распространение. Известен из типичного местонахождения.
70. Crocidura ansorgei Dollman, 1915.
D о 11 m a n, 1915, там же, с. 138, 139.
Тип. 3 4.4.9.29, Daque Braganca, Ангола, кол. Брит, муз., Лондон.
Распространение. Ангола.
71. Crocidura poensis Fraser, 1842.
Sorex [Crocidura) poensis Fraser, 1842, Proc. Zool. Soc., London, p. 200.
Тип не отмечен, добыт в Кларенце, О. Мо Каас-Нгема-Бийого (Фернандо-По),
Нигерия.
Распространение. Золотой берег, Нигерия.
72. Crocidura batesi Dollman, 1915.
D о I 1 m a n, 1915, Ann. Mag. Nat. Hist., ser. 8, v. 16, p. 143.
Тип. 3 № 97.7.1.4, p. Комо, Габон, кол. Брит, муз., Лондон.
Распространение. Габон, Камерун.
410
ПОДСЕМ. SORICINAE
73. Crocidura foxi Dollman, 1915.
D о 11 m a n, 1915, там же, с. 143, 144.
Тип. А №11.3.24.7, Паннам, Северная Нигерия, кол. Брит, муз., Лондон.
Распространение. Северная Нигерия.
74. Crocidura arethusa Dollman, 1915.
D о 11 ш a n, 1915, там же, с. 144, 145.
Тип. $ №14.11.8.2, Кабвир, провинция Баучи, Нигерия, кол. Брит, муз.,
Лондон.
Распространение. Северная Нигерия.
75. Crocidura erica Dollman, 1915.
Dollman, 1915, там же, с. 145, 146.
Тип. А № 4.4.9.30, Панго, Айдонго, Ангола, кол. Брит, муз., Лондон.
Распространение. Ангола.
76. Crocidura boydi Dollman, 1915.
Dollman, 1915, там же, № 94, с. 357.
Тип. $ № 7.7.8.52, Титебби, р. Велле, кол. Брит, муз., Лондон.
Распространение. Известен из одного местонахождения.
77. Crocidura nigricans Bocage, 1889.
Bocage, 1889, J. Sci. Lisb., v. 1, p. 29.
Тип добыт в Квиндомбо, Ангола; хранится в музее Лиссабона.
Распространение. Ангола.
78. Crocidura nigrofusca Matschie, 1895.
Matschie, 1895, Saugethiere, in: Deutsch. Ost. Africa, Bd 3, S. 33
Тип не отмечен, добыт в Вукалала-Камп, Кенияванга, западнее Семянки,
Уганда, кол. Берлинск. муз.
Распространение. Конго, Уганда, оз. Танганьика.
79. Crocidura luna Dollman, 1910.
Dollman, 1910, Ann. Mag. Nat. Hist., ser. 8, v. 5, p. 175.
Тип. № 9.1.3.3, p. Бункея, Катанга, Южное Конго, кол. Брит, муз., Лондон.
Распространение. Конго, Кения, Эфиопия, по-видимому,
Уганда и Танганьика.
80. Crocidura ibeana Dollman, 1915.
Dollman, 1915, там же, т. 16, с. 362.
Тип. <5 № 12.7.1.68, р. Ольгерой, Кения, кол. Брит, муз., Лондон.
Распространение. Кения.
81. Crocidura argentata Sundevall, 1858.
Sorex argentatus Sundevall, 1858, K. Svenska Vetensk. Akad. Handl.
Stock., Bd 11, N 10, S. 16.
Тип не отмечен, вид добыт в Roodeval Karroo, Южная Африка.
Распространение. Южная Африка.
82. РОД CROCIDURA
411
82. Crocidura cyanea Duvernoy, 1838.
Sorex cyaneus Duvernoy, 1838, Mem. Soc. Hist. Nat. Strasb., Suppl. 2, p. 2.
Тип не отмечен, добыт, как указывает автор, с «1а riviere des Elephants, au sud
•de 1* Afrique.»
Распространение. Известен из одного местонахождения.
Примечание. Возможно, что этот вид близок к С. argentata
Sundev.
83. Crocidura electa Dollman, 1910.
D о 11 m a n, 1910, Ann. Mag. Nat. Hist., ser. 8, v. 5, p. 175.
Тип. d № 9.12.4.15, Kamtobu, южнее оз. Танганьика, кол. Брит, муз., Лондон.
Распространение. Указано одно местонахождение, приве-
денное выше для типа.
84. Crocidura martensi Dobson, 1890.
Dobson, 1890, там же, сер. 6, т. 6, с. 496.
Тип не установлен, вид описан с мыса Доброй Надежды.
Распространение. Южная Африка.
85. Crocidura silacea Thomas, 1895.
Thomas, 1895, там же, т. 16, с. 53.
Тип. $ № 93.11.26.29, Figtree Creek, Де Каар, Трансвааль, кол. Брит, муз.,
Лондон.
Распространение. Трансвааль (ЮАР).
86. Crocidura bovei Dobson, 1887.
Dobson, 1887, Ann. Mus. Cuv. Stor. Nat. Genova, ser. 2, v. 5, p. 425.
Тип. d (добыт в Vivi, Конго), хранится в Museo Civico, Genova, Женева.
Распространение. Известен из одного местонахождения.
87. Crocidura capensoides Smith, 1833.
Sorex capensoides Smith, 1833, South Afric. Quart. J., v. 11, p. 62.
Тип. d № 45.7.3.38, добыт вблизи Кейптауна, ЮАР, кол. Брит, муз., Лондон.
Распространение. Известен из одного местонахождения.
88. Crocidura pilosa Dobson, 1890.
Dobson, 1890, Ann. Mag. Nat. Hist., ser. 6, v. 6, p. 496.
Тип не отмечен, вид описан из Трансвааля.
Распространение. Трансвааль (ЮАР).
89. Crocidura fumosa Thomas, 1904.
Thomas, 1904, там же, сер. 7, т. 14, с. 238.
Тип. J № 0.2.1.8, западная часть горы Кения, кол. Брит, муз., Лондон.
Распространение, Ботсвана, Кения, Уганда.
90. Crocidura raineyi Heller, 1912.
Heller, 1912, Smiths. Misc. Coll., v. 60, № 12, p. 7.
Тип. $ № 181816, горы Gargues, Кения, кол. Амер. нац. муз., Вашингтон.
Распространение. Кения.
412
ПОДСЕМ. SORICINAE
91. Crocidura parvipes Osgood, 1910.
Osgood, 1910, Field Mus. Nat. Hist. (zool. ser.), v. 10, N 3, p. 19.
Тип. <? № 16890, Вой, Кения, кол. Field Mus. Nat. Hist., Чикаго.
Распространение. Кения.
92. Crocidura sansibarica Neumann, 1900.
N e u m a n n, 1900, Zool, Jahrb., Abt. 3, Bd 13, S. 544.
Тип не отмечен, вид описан с о. Занзибар.
Распространение. Известен из одного местонахождения.
93. Crocidura Xanthippe, Osgood, 1910.
Osgood, 1910, Field Mus. Nat. Hist. (Zool. ser.), v. 10, N 3, p. 15.
Тип. J № 16888, Вой, Кения, кол. Field Mus. Nat. Hist., Чикаго.
Распространение. Кения.
94. Crocidura jacksoni Thomas, 1904.
Thomas, 1904, Ann. Mag. Nat. Hist., ser. 8, v. 14, p. 238.
Тип. $> № 99.8.24.27, Кения, кол. Брит, муз., Лондон.
Распространение. Кения, Конго.
95. Crocidura macowi Dollman, 1915.
D о 11 m a n, 1915, там же, т. 16, с. 378.
Тип. J № 12.7.1.65, горы Нийро, к югу от оз. Рудольфа, Кения, кол. Брит, муз.»
Лондон.
Распространение. Известен из одного местонахождения.
96. Crocidura gracilipes Peters, 1870.
Peters, 1870, Monatsber. Konigl. Preuss. Acad. Wiss., Berlin, S. 584.
Тип не установлен, вид описан из Килиманджаро.
Распространение. Кения.
97. Crocidura lutreola Heller, 1912.
Heller, Smiths. Misc. Coll., v. 60, N 12, p. 8.
Тип. $ № 181818, горы Мбололо, Кения, кол. Амер. нац. муз., Вашингтон.
Распространение. Кения.
98. Crocidura maanjae Heller, 1910.
Heller, 1910, там же, т. 56, № 15, с. 4.
Тип. <J № 164639, Kabula, Мулиро, Уганда, кол. Амер. нац. муз., Вашингтон.
Распространение. Уганда.
99. Crocidura hildegardac, Thomas, 1904.
Thomas, 1904, Ann. Mag. Nat. Hist., ser. 7, v. 14, p. 240.
Тип. № 3.3.2.7, форт Хол, Кения, кол. Брит, муз., Лондон.
Распространение, Уганда, Кения.
100. Crocidura planiceps Heller, 1910.
Heller, 1910, Smith. Misc. Coll., v. 56, N 15, p. 5.
Тип. 9 № 164641, Рино-Камп, Ладо Энклаве, кол. Амер. нац. муз., Вашингтон.
Распространение. Уганда.
32. РОД CROCIDURA
413
101. Crocidura somalica Thomas, 1895.
Thomas, 1895, Ann. Mag. Nat. Hist., ser. 6, v. 16, p. 52.
Тип. 9 № 93.6.30.7, Webi Shebali, Сомали, кол. Брит» муз., Лондон.
Распространение. Известен из одного местонахождения.
102. Crocidura crossei Thomas, 1895.
Thomas, 1895, там же, с. 53.
Тип. № 95.5.3.4, Asaba, 150 миль вверх по р. Нигеру, кол. Брит, муз.,
Лондон.
Распространение не ясно, известен из бассейна р. Нигера.
103. Crocidura allex Osgood, 1910.
Osgood, 1910, Field. Mus. Nat. Hist., (Zool. ser.), vol. 10, N 3, p. 20.
Тип. <J № 16820, Найваша, Кения, кол. Field Mus. Nat. Hist., Чикаго.
Распространение. Известен из одного местонахождения.
104. Crocidura alpina Heller, 1910.
Heller, 1910, Smiths. Misc. Coll., v. 56, N 9, p. 5.
T и n. $ № 163069, западная часть горы Кения, кол. Амер, нац/муз., Вашингтон.
Распространение. Кения.
105. Crocidura bicolor Bocage, 1889.
Bocage, 1889, J. Sci. Lisb., v. 1, p. 29.
Тип не отмечен, экземпляр хранится в музее г. Лиссабона.
Распространение. Ангола, Уганда, Ботсвана.
106. Crocidura Cinderella Thomas, 1911.
Thomas, 1911, Ann. Mag. Nat. Hist., ser. 8, v. 8, p. 119.
Тип. $ №11.6.10.13, Gemenjulla, Гамбия, кол. Брит, муз., Лондон.
Распространение. Гамбия.
107. Crocidura floveri Dollman, 1916.
Dollman, 1916, там же, т. 17, с. 192, 193.
Тип. J № 10.6.18.3, Гиза, Египет, кол. Брит, муз., Лондон.
Распространение. Известен из одного местонахождения.
108. Crocidura папа Dobson, 1890.
Dobson, 1890, там же, сер. 6, т. 6, с. 225.
Тип. № 90.3.6.1, Долло, Сомали, кол. Брит, муз., Лондон.
Распространение. Сомали.
109. Crocidura nanilla Thomas, 1909.
Thomas, 1909, там же, сер. 8, т. 4, с. 19.
Тип. $ 9.7.14.1, Уганда, кол. Брит, муз., Лондон.
Распространение. Уганда.
110. Crocidura pasha Dollman, 1916.
Dollman, 1916, там же, т. 17, с. 195, 196.
Тип. $ № 8.9.22.1, р. Атбара, Судан, кол. Брит, муз., Лондон.
Распространение. Судан.
414
ПОДСЕМ. SORICINAE
111. Crocidura glebula Dollman, 1916.
Dollman, 1916, там же, с. 196, 197.
Тип. $ № 4.7.9.14, Цунгеру, Северная Нигерия, кол. Брит, муз., Лондон,
Распространение. Нигерия.
112. Crocidura religiosa Geoffroy, 1827.
Geoffroy, 1827, Mem. Mus. Hist. Nat., Paris, t. 15, p. 128.
Тип не отмечен, вид описан из Египта.
Распространение. Египет.
ИЗ. Crocidura lusitania Dollman, 1916.
Dollman, 1916, Ann. Mag. Nat. Hist., ser. 8, v. 17, p. 197, 198.
Тип. № 13.3.7.2, область Трарза, Мавритания, кол. Брит, муз., Лондон.
Распространение. Мавритания.
114. Crocidura dolichura Peters, 1876.
Peters, 1876, Monatsber. Konigl. Preuss. Acad. Wiss. Berlin, S. 475.
Тип не отмечен, описан из Bonjongo, Камерун.
Распространение. Камерун.
115. Crocidura maurisca Thomas, 1904.
Thomas, 1904, Ann. Mag. Nat. Hist., ser. 7, v. 14, p. 239.
Тип. <j> № 1.8.9.99, Энтеббе, Уганда, кол. Брит, муз., Лондон.
Распространение. Уганда.
116. Crocidura niobe Thomas, 1906.
Thomas, 1906, там же, т. 18, с. 138.
Тип. $ № 6.7.1.32, Рувензори, Уганда, кол. Брит, муз., Лондон.
Распространение. Уганда.
117. Crocidura bottegi Thomas, 1898.
Thomas, 1898, Ann. Mus. Civ. Stor. Nat., Genova, ser. 2, v. 18, p. 677.
Тип не отмечен; котип № 98.2.5.6, между Badditu и Dime вблизи оз. Рудольфа,
Эфиопия, кол. Брит, муз., Лондон.
Распространение. Эфиопия.
118. Crocidura топах Thomas, 1910.
Thomas, 1910, Ann. Mag. Nat. Hist., ser. 8, v. 4, p. 310.
Тип. $ № 10.7.2.58, Рокгоо, Килиманджаро, Танганьика, кол. Брит, муз.,
Лондон.
Распространение. Оз. Танганьика.
119. Crocidura littoralis Heller, 1910.
Heller, 1910, Smiths. Misc. Coll., v. 56, N 15, p. 5.
Тип. J № 164642, Butiaba, восточнее побережья оз. Альберт и Виктория
Ньянза, Уганда, кол. Амер. нац. муз., Вашингтон. у
Распространение. Уганда.
32. РОД CROCIDURA
415
120. Crocidura ultima Dollman, 1916.
D oilman, 1916, Ann. Mag. Nat. Hist., ser. 8, v. 17, N 98, p. 204, 205.
Тип. № 12.7.1.60, горы Jombeni, область Nyeri, Кения, кол. Брит.муз., Лондон.
Распространение. Кения.
121. Crocidura neavei Wroughton, 1907.
Wrought on, 1907, Manchester Mem., v. 51, N 5, p. 7.
Тип. $ № 7.1.11.12, p. Кафуэ, Северная Родезия, кол. Брит, муз., Лондон.
Распространение. Замбия.
122. Crocidura silvia Thomas et Schwann, 1906.
Thomas, Schwann, 1906, Proc. Zool. Soc. London, v. 39, p. 587.
Тип. № 6.4.3.10, северо-восточный Трансвааль, кол. Брит, муз., Лондон.
Распространение. Трансвааль (ЮАР).
33. Род DIPLOMESODON Brandt, 1852 — ПУТОРАКИ
Brandt, 1852, Melanges hiologiques tires du Bull, de 1’Academy, p. 452.
Тип. Sorex pulchellus Lichtenstein, 1823, юго-восточная Европа, Средняя Азия
(в границах СССР).
Диагноз. Зубная формула: 1-н-С7-Р-7-М-5- = 26. В верхней челюсти
XU U I О
два промежуточных зуба. На верхней стороне R бугорков нет. Хвост
покрыт короткими волосами и редкими длинными щетинками.
Специализация рода шла по пути приспособления к жизни в песчаных
пустынях, передвижению по очень рыхлому грунту на значительные рас-
стояния (до 5—8 км за ночь) и к питанию животными с жесткими покро-
вами из беспозвоночных и мелкими позвоночными, преимущественно
ящерицами.
Распространение и возраст. Южная Африка, плей-
стоцен; СССР, современный.
f 1. Diplomesodon fossorius Repenning, 1965 (рис. 208, 209).
В epenning, 1965, р. 189—196.
Тип. Правая ветвь нижней челюсти, BPU N MF 430, вблизи Potgieterstrust,
Трансвааль, Южная Африка, нижний плейстоцен, кол. Bernard Price Institute for Pa-
laeontological Research. University of Witwatersrand.
Диагноз (по описанию: Repenning, 1965). Скуловой отросток
верхнечелюстной кости не заходит за передний край М2. I3 крупные
(диаметр I3 менее диаметра I2 в два раза). М3 утратил задний сегмент.
Описание (по: Repenning, 1965). Размеры черепа близки к тако-
вым черепа Anourosorex M.-Edw. Внешним обликом череп напоминает
череп Diplomesodon Brandt, но с более узким межглазничным сужением.
Диаметр I3 менее диаметра I2 в два раза, а у D. pulchellum Licht, в три
раза. Р4 имеет на задней стороне менее глубокую выемку, чем у D. pul-
chellum, также и М1. М3 имеет скорее овальную форму, чем треугольную,
как у сравниваемого вида.
Измерения (по: Repenning, 1965). Длина Р4—М3 — 6,1; ши-
рина: носового отдела черепа на уровне предглазничных отверстий —
4.0, межглазничная — 4.8 мм.
Распространение и возраст. Известен из одного место-
нахождения, отмеченного для типа. Нижний плейстоцен.
209
Рис. 208. Diplomesodon fossorius.
Рис. 209. Diplomesodon pulchellum (a), D. forssorlus (6) (no: Repenning, 1965).
33. РОД DIPLOMESODON
417
Примечание. При значительном сходстве D. fossorius и D. pul-
chellum они отличаются строением сочленовного отростка нижней че-
люсти. Возможно, что эти виды представляют два подрода с разным на-
правлением кормовой адаптации, из которых представляемая D. fossorius
не получила дальнейшего развития, и не только в пределах рода Diplo-
mesodon, но и в пределах других родов африканского континента.
2. Diplomesodon pulchellum Lichtenstein, 1823 — Пегий путорак, или
Пустынная землеройка (рис. 209, 210).
Sorex pulchellus Lichtenstein, 1823, in: Eversmann’s Reise von Orenburg
nach Buchara, S. 125. Crocidura pulchella Peters, 1852, Monatsber. Konigl. Preuss.
Acad. Wiss. Berlin, S. 175. Diplomesodon pulchellus (Licht.) Brandt, 1852, Zool.
Anhang., in: Lehmann’s Reise nach Buchara und Samarkand, S. 299.
Рис. 210. Diplomesodon pulchellum.
Тип не отмечен. Экземпляр кол. проф. Эверсманна хранится в Берлинск. зоол.
муз. Описан из Киргизской степи (восточный берег р. Урала, пески).
Диагноз. Скуловой отросток верхнечелюстной кости доходит
до середины М2. 1| мелкие (диаметр I3 менее диаметра I2 в 3 раза). М3 не
утратил задний сегмент.
Описание. Тело массивное. Конечности с широкими кистями
и ступнями. Голова широкая, морда слабо вытянутая. Уши мало высту-
пают из волосяного покрова. Волосы, покрывающие тело, короткие,
мягкие, образуют густой бархатистый покров. На боковых сторонах
кистей и ступней имеются оторочки из щетинистых волос — приспособ-
ление к передвижению по сыпучим пескам пустыни.
Окраска тела резко двуцветная. Спина серая или буровато-серая с бе-
лым продолговатым пятном на средней части, нижняя сторона тела и
бока белые.
Череп массивный и широкий, с короткой лицевой частью. Ширина
мозговой капсулы не менее ее длины, измеряемой от верхнезатылочного
гребня до лобно-теменного шва. Отверстий на лобных костях нет.
Первые два коренных зуба верхней челюсти сильно выступают за боко-
вые стороны нижней челюсти, что хорошо заметно при сомкнутых челю-
стях. М1 и М2, а также Мх и М2 массивные и широкие.
27 А. А. Гуреев
418
ПОДСЕМ. SORICINAE
Измерения. Длина: тела — 65.0—75.0, хвоста — 25.0—33.0,
ступни 12.4—13.4, кондило-базальпая черепа — 17.9—19.0 мм.
Распространение и возраст. Песчаные пустыни между
Волгой и Эмбой, в их низовьях, Большие Барсуки (песчаные участки,
прилежащие с севера к Аральскому морю), па северо-западном берегу
Аральского моря, на п-ове Дарджа (юго-восточный берег Каспийского
моря), в Каракумах, Кызылкумах, в пустынной местности к югу от р. Чу,
в песчаной пустыне между реками Или и Аксу в южных пустынях вблизи
оз. Балхаш.
Биология. Пегий путорак — обитатель различных типов пес-
чаных пустынь, где встречается в самых разнообразных условиях.
В поисках пищи уходит от своего жилища на несколько километров —
до 5—8, по наблюдениям в Каракумах (Стальмакова, 1955).
При спокойном передвижении путорак проходит в минуту 45—50,
на быстром ходу — 85—90 м. Превосходно роет, но предпочитает при-
спосабливать для своего жилища старые, брошенные норы грызунов.
Выкопанные самими путораками норы просты по устройству. Общая
протяженность хода — более 300, диаметр хода — 1.8—2.0, гнездовая
камера — 18x20 см.
Пища — различные беспозвоночные и мелкие позвоночные, главным
образом ящерицы. Путорак прожорлив, в неволе самка за одну ночь съе-
дала 12 малых круглоголовок и 25 черных тараканов, что намного пре-
вышает ее собственный вес (Стальмакова, 1955).
Размножение изучено недостаточно. По наблюдениям в неволе,
беременность продолжается 30 дней. Спаривание самки, за которой велись
наблюдения, происходило в то время, когда ее 5 детенышей еще питались
только молоком. Размножается три раза в год.
5. Триба BLARININI Stirton, 1930
BZflrina-group S t i г t о n, 1930, p. 225.
T и n. Blarina Gray, 1838, Северная Америка, верхний плиоцен—современный.
Диагноз. Зубная формула: lyC-^-Р ^-j-^M| = 32—28. Верхних
промежуточных зубов 5—3. Зубы пигментированы или белые, а также
пигментированные и белые в одном зубном ряду (у одной особи). Над-
глазничные отверстия имеются. Нижняя сочленовная фасетка на сочленов-
ном отростке нижней челюсти и засочленовный отросток височной кости,
на который она опирается при сочленении с черепом, выдвинуты вперед
(рис. 39, б).
Распространение и возраст. Европа, Азия, Северная,
Центральная и Южная Америка, миоцен—современный.
Представители трибы известны с плиоцена, когда они были широко
распространены. Они известны из плиоцена Европы, Азии, Северной
Америки. Можно, однако, считать, что виды отдельных групп трибы
формировались еще в миоцене, поскольку в нижнем плиоцене они уже
обладали всеми основными особенностями, характерными и для их потом-
ков в современный период.
В настоящее время распространены в Европе (1 род), в Средней,
Центральной и Южной Азии, включая прилежащие острова и Малайский
архипелаг (6 родов), в южной части Северной Америки, в Центральной и
на северо-западе Южной Америки (3 рода).
Как видно из распространения, большинство представителей этой
трибы сохранили привязанность к субтропическому и тропическому
климату, в котором они формировались еще в миоцене.
ТРИБА BLARINI
419
Основным направлением развития представителей трибы Blarinini
следует считать приспособление к схватыванию крупных животных
и питанию относительно жесткими волокнистыми тканями, как беспозво-
ночных, так и мелких позвоночных. При этом последние составляют
значительную долю их корма. В связи с этим у них развились своеобраз-
ные особенности в строении жевательного аппарата, обеспечивающие
возможность удерживания челюстями крупной добычи без особого напря-
жения жевательной мускулатуры. Своеобразие устройства, позволяющего
удерживать сильную добычу, заключается в том, что нижняя сочленовная
фасетка сочленовного отростка нижней челюсти и засочленовный отро-
сток височной кости, на который она опирается при сочленении с че-
репом, выдвинуты вперед. При закрывании рта с добычей неболь-
шое смешение сочленовного отростка назад делает невозможным от-
крывание рта.
Питание относительно крупными животными обусловливает необхо-
димость существования этих землероек преимущественно на территориях,
где нет морозных зим, или добывать корм в воде. Вероятно, не случайно,
что все землеройки, в большей или меньшей степени приспособленные
к плаванию, относятся к этой трибе. Подавляющее большинство родов
этой трибы объединяет небольшое число видов или монотипичны. Это
обстоятельство, а также сравнение распространения представителей
трибы в прошлом и в настоящее время позволяет считать, что расцвет
этой группы землероек был в плиоцене, т. е. в период, когда в пределах
их широкого ареала имелись благоприятные условия: теплый климат,
разнообразная теплолюбивая растительность и соответственно большой
выбор разнообразных и многочисленных кормовых животных, активных
в течение круглого года. Можно думать, что приспособления видов этой
трибы имеют существенное значение только при определенных условиях. В
современный период распространение большинства представителей трибы
ограничено в основном зоной теплого, умеренного климата.
В пределах трибы В larinini подтриба Blarinina объединяет современные
и ископаемые виды с пятью промежуточными зубами в верхней челюсти.
Подтриба Beremendia объединяет виды с четырьмя промежуточными зу-
бами в верхней челюсти. Представители этой подтрибы в плиоцене дости-
гали значительного разнообразия форм адаптаций в пределах общего
для трибы типа, а виды отдельных родов отличались крупными для земле-
роек размерами. Виды подтрибы Soriculina имеют ряд особенностей пере-
ходного характера. Одни роды этой подтрибы имеют в верхней челюсти 4
промежуточных зуба, другие — 3. У видов тех и других вершины бугров
моляров не пигментированы. Эти зубы полностью белые. А у рода Ро-
dihik Deraniyag. особенности близки к таковым представителей трибы
Crocidurini (длинные щетинки на хвосте, крупные уши и др.).
Подтрибы Nectogalina и Allosoricina объединяют виды с белыми
зубами.
Таким образом, в ряду групп землероек этой трибы наглядно пред-
ставлен процесс изменения количества промежуточных зубов в верхней
челюсти и изменение или переход темной окраски зубов в чисто белую.
Это лишний раз доказывает, что темная или белая окраска зубов не скор-
релирована с другими, более важными особенностями видов отдельных
групп, и не имеет ведущего значения для систематических построений.
Можно лишь отметить, что темная окраска зубов у землероек подсемейства
Soricinae более древняя, но с нижнего олигоцена известны и белозубые
землеройки, как более, так и менее специализированные.
Белозубые землеройки из трибы Blarinini в строении черепа имеют те же
особенности, что и виды с темными или наполовину белыми зубами.
27*
420
ПОДСЕМ. SORICINAE
Среди землероек только в пределах этой трибы отдельные виды достигли
значительной специализации в приспособлениях к плаванию и бегу,
что отмечено в соответствующих местах работы.
Основное ядро трибы в настоящее время составляют несколько более
или менее широко распространенных родов: Cryptotis Pom. (подтриба
Beremendina), Neomys Капр и Soriculus Blith (подтриба Soriculina), более
богатые видами, чем другие.
Более полное представление об особенностях трибы Blarinini дают
характеристики объединяемых ею подтриб.
1. Подтриба BLAR ININA s. str. Stirton, 1930
Blarina-group S t i r t о n, 1930, p. 225.
Тип. Blarina Gray, 1838, Северная Америка, верхний плиоцен—современный.
3 13 3
Диагноз. Зубная формула: I-^-Cy-P-y Му = 32. Верхних промежу-
точных зубов пять. Из пих два или три последних мелкие, сжатые спереди
назад. Все зубы сильно пигментированы. Специальных приспособлений
к плаванию и рытью нет.
От других подтриб отличаются количеством верхних промежуточных
зубов и их формой.
Подтриба Blarinina объединяет три рода.
Распространение и возраст. Европа, плиоцен; Север-
ная Америка, верхний миоцен—современный (были, несомненно, в Азии).
Примечание. Представители подтрибы в их современном со-
стоянии — обитатели преимущественно тепло-умеренной зоны, с ланд-
шафтами, близкими по своему характеру к таковым конца неогена. Пищу
составляют не только безпозвоночные, но и мелкие позвоночные. Эти
землеройки способны рыть, хотя и не располагают типичными для земле-
роев особенностями, правда, у них относительно широкие кисти, неболь-
шие ушные раковины и короткие хвосты. Особенности, свойственные всем
группам трибы, выражены у них весьма отчетливо, что, несмотря на
большее, чем у других представителей трибы, количество зубов, не остав-
ляет сомнения в принадлежности к этой группе землероек.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Из пяти верхних промежуточных зубов три первых крупные, а два
задних мелкие ....................................35. Blarinoides.
2 (1). Два первых крупные, а три задних мелкие........36. Blarina.
Примечание. В таблицу не включен род Adeloblarina Repenn. за отсутствием
сравнимых признаков.
f 34. Род ADELOBLARINA Repenning, 1967
Repenning, 1967, Geol. Surv. Profess. Paper, N 565, p. 37, 38.
Тип. Adeloblarina berklandi Repenning, 1967, Орегон (США) верхний миоцен»
Описание (по: Bepenning, 1967, с сокращениями). Землеройка
с крупными зубами, сходными с таковыми Blarina Gray, но с нижней че-
люстью, более похожей на эту кость Sorex L. Ыижняя зубная формула:
1—2—3. Нижняя челюсть умеренно массивная. Подбородочное отверстие
под серединой Mj. Венечный отросток узкий. Его передний край направлен
под углом 90° к линии зубов горизонтальной ветви нижней челюсти. Его
вершина отклонена кнаружи («щечно» — у автора), как у Blarina Gray
или Neomys Каир. Нижний кондилюс ориентирован под углом 90° от вер-
тикальной оси горизонтальной ветви и'под углом 45° от продольной оси;
34. РОД ADELOBLARINA. 35. РОД BLARINOIDES
421
наружная артикулярная фасетка расположена вентрально, а отросток —
лингвально к плоскости нижней челюсти у нижней сигмоидной выемки.
Смещение вперед нижнего кондилюса не так значительно, как у Blarina
Gray, Cryptotis Baird или Notiosorex Baird, менее, чем у Chimarrogale Anders,
и Neomys Каир, больше, чем у Sorex L. Межартикулярная область, вероятно,
с умеренной лингвальной выемкой, не заполнена костной пластинкой. Только
передний сочленовный отросток поперечно уплощен более, чем у многих ро-
дов Neomyini. Основание сочленовного отростка у его соединения с восходя-
щей ветвью вертикально сужено по сравнению со всеми землеройками,
исключая Soricini. Верхняя птеригоидная ямка имеет очень мелкую
чашу с недостаточно развитым бугорком для прикрепления верхнего
птеригоидного мускула. Наружная птеригоидная ямка нижней челюсти
Рис. 211. Adeloblarina berklandi (по: Repenning, 1967).
не простирается к низу венечного отростка, как верхняя сигмоидная вы-
емка. Внутренняя птеригоидная ямка нижней челюсти узкотреугольная,
с хорошо развитой перекладиной.
Распространение и возраст. Орегон (США), верхний
миоцен.
t 1. Adeloblarina berklandi Repenning, 1967 (рис. 211).
Repenning, 1967, p. 37, 38.
Тип. Фрагмент правой ветви нижней челюсти с обломком сочленовного отростка
и Mj, № 23098, кол. Амер. нац. муз., Вашингтон.
Морфологические особенности вида представлены в характеристике
рода, там же и распространение.
f 35. Род BLARINOIDES Sulimski, 1959;
Sulimski, 1959, р. 144—148.
Тип. Blarinoides mariae Sulimski, 1959, Венже (Польша), плиоцен.
Диа гноз (и описание по: Sulimski, 1959). Зубная формула:
гг 3 1 3 3
IуС-у-РуМу—32. Из пяти верхних промежуточных зубов три первых
(I2, I3 и С) крупные, а два задних (Р1 и Р2) — мелкие.
422
ПОДСЕМ. SORICINAE
Описание. 1Х с тремя лопастями. 12 мелкий, плоский. Р4 с двумя
слабо выраженными вершинами. Верхняя суставная поверхность (фа-
сетка) сочленовного отростка нижней челюсти расположена под углом в 45°
по отношению к плоскости горизонтальной части ветви нижней челюсти.
Межсуставная часть сочленовного отростка (межсуставная пластинка —
Sulimski, 1959) со слабой вырезкой на внутренней (лингвальной) стороне.
Нижняя суставная поверхность этого отростка широкая, с выемкой
в средней части и отогнутым вниз внутренним краем. Подбородочное
отверстие расположено у переднего края Мх.
Распространение и возраст. Европа, плиоцен.
f 1. Blarinoides mariae Sulimski, 1959 (рис. 212).
Sulimski, 1959, p. 144—148.
Тип. Ростральная часть черепа с неполным рядом зубов, № 803, Венже
(Польша), плиоцен, кол. Палеонтол. лабор. Польской АН.
Рис. 212.
(по:
Blarinoides mariae
Sulimski, 1959).
Описание. Ростральная часть черепа удлиненная. На верхнечелю-
стной кости между М1 и Р4 имеется углубление. Слезное отверстие над
задним корнем М1. Подглазное отвер-
стие над Р4. Передние нёбные от-
верстия расположены между вторыми
резцами. Между этими отверстиями
находится короткая щель. Задние нёб-
ные отверстия расположены впереди
первых моляров. Второй резец меньше
третьего резца, реже равен ему. Клык
и первый премоляр вдвое меньше, чем
резцы сходного строения. Р2 мелкий,
округленный, расположен ближе к внут-
реннему краю челюсти и снаружи не
виден. Р4 со стороны жевательной по-
верхности трапециевидной формы. Зад-
ние края М1 и М2 со слабой вырезкой. М3
трехбугорчатый.
Измере н и я. Длина носовых
костей — 10.6, расстояние между ор-
битальной впадиной и нижнеглазнич-
ным отверстием — 3.8, 3.8, 4.3, 3.7;
ширина черепа:
области — 7.7, 8.0,
неглазничными отверстиями — 6.6, 6.5, 6.6, 6.4;
зубного ряда I1—М3 — 15.0, 15.4, I1—М2 — 14.0, 14.2, Р4—М1 — 5.7
5.7, 5.6, 5.6, Р4-М3 — 9.0, М1—М3 — 6.0, 6.0, 6.0, 5.8, М1—М2 — 5.0,
М2—М3 - 3.5, 3.3, 3.3, 3.2, М3—1.0, 1.1, 1.0 мм.
Распространение и возраст. Польша, плиоцен.
в межглазпичной
7.8, между ниж-
длипа: верхнего
36. Род BLARINA Gray, 1838 — КОРОТКОХВОСТЫЕ ЗЕМЛЕРОЙКИ
Blarina (подрод рода Sorex L.) G г а у, 1837—1838, р. 124. Blarina (род) Les-
son, 1842, Nouv. Tableau Mammif., tabl. 89. Brachysorex (подрод) Duvernoy,
1842, Mag. Zool. v. 4, p. 37—41. Talpasorex Pomel, 1848, Bibl. Univ. Geneve, Arch.
Sci. phys. nat., t. 9, p. 248. Galemys Pomel, 1848, там же, c. 248. A notus (подрод) Wag-
ner, 1855, in: Schreber’s Sauget., v. 5, p, 550, 551.
Тип. Sorex talpoides Gapper, 1830 (=Sorex brevicaudus Gapper, 1823), Северная
Америка, плиоцен—современный.
36. РОД BLARINA
423
3 13 3
Диагноз. Зубная формула: I у С Р у М у = 32. Из пяти верхних
промежуточных зубов первые два (I2 и I3) — крупные, а три задних (С, Р1,
Р2) — мелкие.
Род включает два современных вида и несколько ископаемых форм.
От других родов трибы этот род отличается количеством верхних про-
межуточных зубов и их строением.
Распространение и возраст. От самых южных окраин
Канады на севере, Северной и Южной Дакоты, Небраски, Канзаса и
восточной части Оклахомы — на западе до морских побережий на юге
и востоке США; верхний плиоцен—современный.
Примечание. Представители рода Blarina Gray сохраняют
многие особенности, характерные для более примитивных землероек:
большое число промежуточных зубов, темный пигмент на них и др. Они
обитают в ландшафтах, близких по составу растительности к ландшафтам
конца неогена.
Современные виды этого рода приспособились к рытью, что заметно
выражено как во внешнем облике землероек, так и в строении соответ-
ствующих частей их скелета.
f 1. Blarina gidleyi Gasin, 1933.
Gasin, 1933, J. Mammal., v. 14, N 2, p. 142.
Тип. № 12650, фрагмент левой ветви нижней челюсти с Мх—М3, Айдахо (США),
верхний плиоцен, кол. Амер. нац. муз. Вашингтон.
Диагноз (по: Gasin, 1933). Размеры ближе к Blarina brevicauda
Say. Подбородочное отверстие далеко впереди. Нижний край ветви нижней
челюсти менее угловатый под задней частью М3. Тригонид нижних моляров
удлинен в переднезаднем направлении (с тупым гребнем). Задняя стенка
его не перпендикулярна к лингвальному краю зубов. Талонид узкий и
его гребень соединяется с задней стенкой тригонида. Цингулюм более
выступающий на переднем и заднем краях первого и второго моляров.
Примечание. Хиббард (Hibbard, 1957) отмечает, что Blarina
gidleyi Gas. отличается от современных представителей рода Blarina
Gray наличием лишь двух бугорков на верхнем крае нижнего внутрен-
него резца. Р4 очень крупный, а цингулюмы Р4, Мх и М2 более развиты.
Распространение и возраст. Северная Америка, верх-
ний плиоцен.
t 2. Blarina adamsi Hibbard, 1953.
Sorex ? H i bb ard, Riggs, 1949, Bull. Geol. Soc. America, v. 60, p. 835. No-
tiosorex jacksoni Hibbard, 1950, Contrib. Mus. Paleontol. Univ. Michigan, v. 8, N 6,
p. 321. Blarina adamsi Hibbard, 1953, J. Paleontol., v. 27, N 1, p. 23—31.
Тип. № 27267, левая половина верхней челюсти с сохранившимися зубами Р2—
М2, Канзас (США), верхний плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн
Арбор.
Диагноз (по: Hibbard, 1953). Меньше, чем Blarina brevicauda
carolinensis Bach и Notiosorex jacksoni Hibb. B. adamsi характеризуется
мелкими размерами, менее развитыми задними лингвальными бугорками
на I2, I3 и Р1, Р2 не редуцированный, как у Blarina brevicauda carolinensis
Bach. Выемка между протоконом и гипоконом Р4—М2 не такая глубокая,
как у современных форм.
Распространение и возраст. Канзас (США), верхний
плиоцен.
424
ПОДСЕМ. SORICINAE
f 3. Blarina simplicidens Cope, 1899.
Cope, 1899, J. Acad. Nat. Sci. Philadelphia, v. 11, p. 219.
Тип. № 150, фрагмент левой ветви нижней челюсти с зубами (часть 1х, 18, Р4,
Мх, М2 и обломок М3), порт Кеннеди, Пенсильвания, кол. Акад, естеств. наук Фила-
дельфии (США).
Отличается от Б. gidleyi Gas. следующими особенностями: мелкими раз-
мерами, менее развитыми цингулюмами у Р4, Мх, М2 и мелкими входящими
складками при основании коронок Мх и М2 на губной стороне (Hibbard,
1953).
Распространение и возраст. Северная Америка, ниж-
ний плейстоцен.
4. Blarina brevicauda Say, 1823 — Большая блярина (рис. 213).
Sorex brevicaudus Say, 1823, in: Long’s Exped. to the Rocky Mountains, v. 1,
p. 164, — talpoides G a p p e r, 1830, Zool. J., v. 5, p. 202. Corsira (Blarina) talpoides
Gray, 1837—1838, p. 124. Sorex dekayi (Cooper) Bachman, 1837, J. Acad. Nat.
Sci. Philadelphia, v. 7, pt 2, p. 377—381. Galemys (Brachysorex) micrurus Pomel, 1848,
Bibl. Univ. Geneve, Arch. Sci. Physiol, Nat. t. 9, p. 249. Blarina brevicauda Baird,
1857, Mammal. North. Amer., p. 42—45; — costaricensis J. A 11 e n, 1891, Bull. Amer.
Mus. Nat. Hist., v. 3, p. 205, 206.
Тип не установлен, вид описан с западного берега р. Миссури вблизи Blair,
Небраска (США).
Диагноз. Длина тела более 100, хвоста — менее 30 мм; длина
ступни менее половины длины хвоста. Окраска тела черно-бурая, почти
сходная сверху и снизу. Череп
крупный, массивный и угловатый.
Его ширина менее длины в два
раза или немного более.
Рис. 213. Blarina brevicauda (по: Merriam, 1895).
Описание. Окраска верхней стороны тела темно-коричневая,
серовато-бурая или черноватая. Нижняя сторона окрашена сходно с верх-
ней или светлее, серее. Ушные раковины малых размеров, не выступают из
волосяного покрова. .Когти на пальцах кисти значительно длиннее когтей
на пальцах ступни. Внешним обликом эти землеройки напоминают кротов.
Череп массивный, с относительно короткой и широкой лицевой частью.
Ламбдовидный и сагиттальный гребни сильно развиты. Зубы крупные,
темноокрашенные.
Измерения. Длина: тела — 115—135, хвоста — 22—27, ступни
13—17, кондило-базальная черепа 23—25 мм.
Распространение и возраст. Южная часть Саскачевана,
Манитобы, Онтарио и Квебека (Канада) на севере, до Северной и Южной
Дакоты, Небраски, Канзаса и восточной части Оклахомы на западе,
до морских побережий на юге и востоке США; верхний плиоцен — совре-
менный.
37. РОД BEREMENDIA
425
5. Blarina telmalestes Merriam, 1895.
Merriam, 1895, p. 15.
Тип. <J № 71823, с о. Драммонд, кол. Амер. нац. муз., Вашингтон.
Описание. Слабо отличается от В. brevicauda Say более светлой
окраской и сравнительно длинными ступнями, а также менее грубым
черепом и не столь темной окраской вершин зубов (Merriam, 1895).
Измерения. Длина: тела — 118.0—119.5, хвоста — 25.4—28.0,
ступни —16, наибольшая черепа — 24 мм.
Распространение и возраст. Болота Виргинии, совре-
менный.
f2. Подтриба BEREMENDINA Gureev, 1971
Тип. Beremendia Kormos, 1930, Румыния, плиоцен.
3 1 2—1 3
Диагноз. Зубная формула: lyC-g-P-j-Му = 30—28. Промежу-
точных зубов в верхней челюсти 4 или 3, убывающих в размерах спереди
назад, или крупные и мелкие чередуются. Все зубы пигментированы.
Распространение и возраст. Европа, плиоцен—плейсто-
цен?; Азия, плиоцен—плейстоцен; Северная Америка, плиоцен—совре-
менный. В современный период представители подтрибы сохранились
только в Новом Свете.
По характеру приспособлений они очень близки к видам подтрибы
Blarinina, но, по-видимому, ранее последних приспособились к питанию
более грубой пищей и добыванию относительно крупных животных.
В современный период подтриба представлена одним родом Cryptotis
Pomel, объединяющим сравнительно мелких землероек, распространен-
ных в субтропической и тропической зонах Америки. Только представи-
тели этого рода проникли нй северо-запад Южной Америки. Среди зем-
лероек Центральной Америки они занимают господствующее положение.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Крупные землеройки (длина нижнего зубного ряда не менее 10 мм).
На верхней стороне Ц бугорков (лопастей) нет..............
........................................ 37. Beremendia.
22 (1). Размеры мелкие и средние (длина нижнего зубного ряда менее
10 мм). На верхней стороне 1х два бугорка.
3 (4). Первый верхний промежуточный зуб крупнее второго . . . .
................................................. 38. Petenyia.
4 (3). Первый верхний промежуточный зуб меньше второго.
5 (6). Ростральная часть черепа широкая, как и область подглазничного
отверстия ....................................43. Paracryptotis.
6 (5). Ростральная часть черепа и область подглазничного отверстия
не широкие .......................................45. Cryptotis.
Примечание. Остальные роды подтрибы не включены в таблицу за отсут-
ствием сравнимых признаков, что обусловлено разной степенью сохранности их кост-
ных остатков
f37. Род BEREMENDIA Kormos, 1930
Crossopus Р е t е п у i, 1864, Hatragyott munkay, p. 60. Neomys К о r m o.s, 1911,
Mitt. a. d. Jahrb. d. Kgl. Ung. Geol. R. A., Bd 19, N 4, S. 170. Beremendia gen. nov.
Kormos, 1930, AUatt. Kozlony, p. 57 (nomen nudum); 1934, p. 4—6. Blarina П и-
допличко, 1955, ДАН СССР, т. 100, вып. 5, с. 989—991.
Тип. Crossopus fissidens Petenyi, 1864, Румыния, плиоцен.
426
' ПОДСЕМ. SORICINAE
3 12 3
Диагноз. Зубная формула: IС7-Р-7-М—- = 30. Череп массивный,
крупный, что отличает представителей этого рода от видов других родов
этой же подтрибы. Длина верхнего зубного ряда 12.5 мм. Верхние про-
межуточные зубы с широким массивным основанием и сжатой с боков
узкой коронкой. Нижний внутренний резец без бугорков на режущем
крае.
Распространение и возраст. Европа, Азия, плиоцен—
плейстоцен.
Рис. 214. Beremendia fissidens (по: Kormos, 1934).
f 1. Beremendia fissidens Petenyi, 1864 (рис. 214).
Crossopus fissidens Petenyi, 1864, Hatragyott munkay, p. 60. Neomys fissiden
(Petenyi) Kormos, 1911, Mitt. Jahrb. Kgl. Ung. Geol. R. A., Bd 19, N 4, S. 170.
Beremendia gen. nov. Kormos, 1930, Allatt. Kozlony, p. 57 (nomen nudum); — fis-
sidens (Petenyi) Kormos, 1934, p. 4—6.
Тип не установлен, вид описан Петени из Румынии. Типом для описания рода
Кормошу (Kormos, 1934) послужил череп с нижней челюстью № 08/3682, Виллань, Венг-
рия, плиоцен, кол. Венгерск. геол.,инет., Будапешт.
Описание. Лицевая часть черепа короткая и широкая. Засочле-
новный отросток височной кости, который служит опорой для нижней
сочленовной фасетки сочленовного отростка нижней челюсти, выступает
далеко вперед. Нижняя челюсть массивная. Сочленовный отросток невысо-
кий; его нижняя сочленовная фасетка выдвинута далеко вперед.
Угловой отросток нижней челюсти тонкий и короткий. I1 с невысоким
задним зубцом, массивные, Р4 с длинным наружным краем, невысокий.
Измерения. Длина верхнего зубного ряда (I1—М3) — 15.4—
16.2; ширина черепа между предглазничными отверстиями — 5.8—6.6;
длина общая: нижней челюсти — 16.0—17.0, зубного ряда Мх—М3 —
10.5—11.2 мм.
Распространение и возраст. Европа, плиоцен—плей-
стоцен. В СССР найден в пижнечетвертичпых отложениях (Таманский
полуостров).
Примечание. Диагноз и описание составлены по описанию
Кормоша (Kormos, 1934), Сулимского (Sulimski, 1959, 1962) и др.
38. РОД PETENYIA
427
f 38. Род PETENYIA Kormos, 1930
. 215. Petenyia hungarica (по:
Kormos, 1934).
Kormos, 1930, Allatt. Kozlony, p. 57 (nomen nudum); 1934, p. 6—8.
Тип. Petenyia hungarica Kormos, 1930, Виллань, Венгрия, плиоцен.
3 12 3
Диагноз. Зубная формула: ГуС-фРуМ-^30. Череп менее мас-
сивный, чем у представителей рода Beremendia Korm. Передний край Р4
(его паракон) не выступает па внешнюю сторону в виде выступа, как у Be-
remendia Korm. Длина верхнего зубного ряда — 7.0—7.4 мм; верхние про-
межуточное зубы не сжаты с боков. 1х с двумя бугорками (лопастями)
на режущем крае (верхнем).
Распространение и воз-
раст. Европа, Азия, США, плиоцен.
+ 1. Petenyia hungarica Kormos,
1934 (рис. 215).
Kormos, 1930 (см. синонимику рода);
1934 (см. синонимику рода); — neglecta Kre-
tzoi, 1943, Fold. Kozlony, v. 73, p.
607, 608.
Тип. Череп № 08/3684, Виллань, Ве-
нгрия, плиоцен, кол. Венгерск. геол, инет.,
Будапешт.
Д и а г н о з. Мандибулярное от-
верстие одно. Венечный отросток узкий.
Описание (по: Kormos, 1934).
Череп с короткой лицевой частью и
широкой мозговой капсулой. Нижняя
челюсть короткая и массивная. Ее
зубная часть высокая. Судя по основа-
нию углового отростка, сохранивше-
муся на нижней челюсти, последний,
вероятно, был тонким и относительно
коротким. Верхние промежуточные
зубы убывают в размерах спереди назад. 1х толстый и относительно
короткий, что особенно бросается в глаза при сравнении его с таковым
особей других родов.
Измерения. Общая длина нижней челюсти 10.0—11.0; высота
нижней челюсти — 2.3—2.5; длина нижнего зубного ряда (1х—М3) —6.4—
6.6; высота венечного отростка — 4.2—4.5 мм.
Распространение и возраст. Известна из плиоцена
Европы и Забайкалья.
f 2. Petenyia concisa Wilson, 1968 (рис. 216).
Parydrosorex concisus Wilson, 1968, Contrib. Mus. Paleontol. Univ. Michigan,
v. 22, N 7, p. 75—126. Petenyia concisa Hibbard, Jammot, 1971, там же, т. 23,
N 24, с. 379, 380.
Тип. Левая ветвь нижней челюсти с зубами Р4—М2, № V 55724, область Трего,
Канзас (США), нижний плиоцен, кол. Палеонтол. муз. Мичиган, унив., Анн Арбор.
Диагноз (по описанию: Hibbard, Jammot, 1971). Мандибулярных
отверстий два. Венечный отросток широкий.
Описание (по тем же авторам). Сочленовный отросток сильно
развит, все его части направлены назад, как у представителей трибы
Soricini. Строение фасеток сочленовного отростка сходно с таковым
Petenyia Kormos. Нижняя челюсть под вентральной фасеткой более узкая,
а межкондилярная выемка выше и более кпереди, чем у Р. hungarica
428
ПОДСЕМ. SORICINAE
Korm. Темпоральная ямка более высокая и глубокая и менее скошена.
Нижняя челюсть у Р. concisa имеет наибольшую высоту под М2. Талонид
(метаконид) М3 редуцирован. Моляры имеют непрерывный цингулюм
(по-видимому, вокруг всей коронки зуба).
Распространение и возраст. Канзас (США), нижний
плиоцен.
t 3. Petenyia sp.
Фрагмент левой ветви нижней челюсти с поврежденными зубной частью и венеч-
ным отростком и хорошо сохранившимся сочленовным отростком, Придонье, Воронеж-
ская обл., плиоцен, сб. И. М. Громова, кол. ЗИН АН СССР, Ленинград.
Вторая находка представителя этого, рода на территории СССР. Первая
сделана М. А. Ербаевой в Забайкалье (Береговая), верхний плиоцен—
нижний плейстоцен. Остатки землеройки, по моему мнению, относятся
к Petenyia hungarica Kormos. Вероятно, и описываемая кость особи того же
вида.
|39. Род PETENYIELLA Kretzoi, 1956
Sorex Petenyi, 1864, Hatragyott munkay, p. 85—120, Pachyura (? Allopachyura
n. gen.) Kormos, 1934, p. 4—6. Petenyiella Kretzoi, 1956, Geol. Hungar., Ser.
Б
Рис. 217. Petenyiella gracilis-
(A — no:'Sulimski, 1962; Б — no:
Kretzoi, 1965).
Palaeontol., Bd 27, S. 1—264. Suncus Sulimski, 1959, Acta palaentol. polonica,
v. 7, N 3, p. 479—480.
Тип. Sorex gracilis Petenyi, 1864, Венгрия, Польша, плиоцен.
Диагноз. Нижняя челюсть мелкая, тонкая. 1х короткий, с двумя
слабыми лопастями на верхней стороне. 12 небольшой, однобугорчатый.
Р4 высокий, однобугорчатый. Пигмент на зубах светло-желтый (отличие
от других родов этой подтрибы).
Распространение и возраст. Европа, плиоцен.
40. РОД SHIKAMAINOSOREX
429
f 1. Petenyiella gracilis Petenyi, 1864 (рис. 217).
Sorex gracilis Petenyi, 1864, Hatragyott munkay, p. 85—120. Petenyiella.
{Petenyi) Kretzoi, 1956 (см. синонимику рода). Suncus zelceus Sulimski, 1959,
Acta palaentol. polonica, v. 7, N 3, p. 479, 480.
T и и не установлен, вид описан из Беременд, Венгрия, плиоцен.
Диагноз совпадает с таковым рода, также и распространение.
Описание (по: Sulimski, 1959). 12 и Р4 почти одинаковой длины.
М3 трехбугорчатый. Венечный отросток широкий (в передне-заднем направ-
лении) у основания и резко суживается к вершине. Сочленовный отро-
сток короткий, игловидный. Подбородочное отверстие между корнями
Р4 и Mv
Примечание. В 1956 г. Кретцой (Kretzoi) только назвал род
Petenyiella. В 1959 г. дал краткий диагноз этого рода. Сулимский (Sulim-
ski). в 1959 г. описал новый вид Suncus zelceus, отнесенный им впослед-
ствии к роду Petenyiella, к его типичному виду — Р. gracilis Peten.
В диагнозе рода использованы признаки, отмеченные Кретцоем,
а также из характеристики вида.
j-40. Род SHIKAMAINOSOREX Hasegawa, 1957
Hasegawa, 1957, Sci. Repts lokogama Nat. Univ., sect. 2, N 6, p. 65—69.
Тип. Shikamainosorex densicingulata Hasegawa, 1957, префектура Тотиги, Япо-
ния, средний плейстоцен.
Диагноз (по: Hasegawa, 1957). Крупнее Paracryptitis rex Hibb.
Нижняя зубная формула: 1, 1, 1, 3 (1Ъ 12, Р4 и М4_3 по принятой мной фор-
муле,— А. Г.). Зубы беловато-желто-
коричневые. Их цингулюмы сильно
развиты и гладкие. Моляры широ-
кие, квадратные. Межкопдилярная
выемка атрофирована (лучше ска-
зать, неглубокая). Нижняя сторона
кондилюса треугольная. Надконди-
лярная выемка в виде тупого угла.
По мнению автора, описываемый
род близок к Paracryptotis rex Hibb.,
1950.
Распространение и
возраст. Япония, плейстоцен.
t 4. Shikamainosorex densicingu-
lata Hasegawa, 1957 (рис. 218). _ o.o c ... . , . .
Рис. Z18. Schikamamosorex densicingu-
lata (no: Hasegava, 1957).
Hasegawa, 1957, там же.
Тип. Левая ветвь нижней челюсти с зубами (Р4—М3), формация Kuzun, пре-
фектура Тотиги, Япония, кол. Геол. инет. нац. унив. в Иокогаме.
Описание (по: Hasegawa, 1957). Альвеола Ц крупная, удлинен-
ная, ее задний конец достигает основания М3. Р4 удлиненный по наружной
стороне. Его наибольшая длина по средней линии 2.5 мм. Цингулюм
составляет удлиненное основание коронки зуба на ее внутренней и наруж-
ной сторонах. Задний край Р4 выше параконида Mv Мх самый крупный из
моляров. Его цингулюм сильно развит, особенно от середины к заднему
краю. Впадина, разграничивающая гребни параконида и протоконида,
глубже, чем у М2. Нижняя челюсть крупнее таковой видов Paracryptotis
Hibb. Подбородочное отверстие расположено между Р4 и Мх. Маидибу-
430
ПОДСЕМ. SORICINAE
лярное отверстие почти треугольное, относительно крупное, расположено
в задней части ветви челюсти под впадиной, на коронарном отростке.
Впадина на коронарном отростке небольшая.
Измерения. Суммарная длина Р4—М3 — 7.0 мм.
Распространение и возраст. Япония, плейстоц ен
|41. Род PEISOREX Kowalski et Li-Chuan-Kuei, 1963
Kowalski, Li-Chuan-Kuei, 1963, Vertebrata Palasiatica, v. 7, N 2,
p. 138—143.
Тип. Peisorex pohaiensis Kowalski et Li-Chuan-Kuei, 1963, Chiachia Shan, Китай,
плейстоцен.
Диагноз (no: Kowalski, Li-Chuan-Kuei, 1963). Нижняя челюсть
массивная. Венечный отросток относительно узкий; его вершина не разде-
лена насечками. Он отклонен в сторону от основной оси ветви челюсти.
Рис. 219. Peisorex pohaensis (по:
Птеригоидная ямка венечного от-
ростка глубокая и широкая, сооб-
щается с крупным округлым ман-
дибулярным отверстием. Между
ними располагается тонкая кост-
Kowalski, Li-Chuan-Kuei, 1963).
пая перемычка. Сочленовный отросток имеет глубокую ямку на верхней
стороне. Его нижняя сочленовная фасетка выдвинута далеко вперед.
Подбородочное отверстие под Мх.
Распространение и возраст. Китай, плейстоцен.
f 1. Peisorex pohaiensis Kowalski et Li-Chuan-Kuei, 1963 (рис. 219).
Kowalski, Li-Chuan-Kuei, 1963, описан там же, где и род.
Тип. Фрагмент левой ветви нижней челюсти с молярами (М2—М3) N 1671, Chi-
achia Shan, вблизи Tangsham, Китай, плейстоцен, кол. Инет, палеонтол. позвон.
и палеоантропол. АН Китая, Пекин.
Измерения. Высота нижней челюсти вблизи М4 — 3.0; длина:
М4 — 2.6, М2 — 2.1, М3 — 1.5; ширина: М4 — 1.7, М2 — 1.4, М3 — 0.86 мм.
t 2. Peisorex sp.
Фрагмент правой ветви нижней челюсти № 949 (1), Карташеве, правый берег
р. Иртыша, Омская обл., плио-плейстоцен, сб. В. С. Зажигина, кол. ГИН АН СССР,
Москва.
Кость темно коричневого цвета с хорошо сохранившимися венечным
и сочленовным отростками. Зубная часть отломлена на уровне заднего
корня М2. Структурные особенности описываемой кости совпадают с тако-
выми, отмеченными Ковальским и Ли-Хуан-Куей (Kowalski, Li-Chuan-
Kuei, 1963) для Р. pohaensis, описанного ими из плейстоцена Китая.
42. РОД HESPEROSOREX. 43. РОД PARACRYPTOTIS
431
|42. Род HESPEROSOREX Hibbard, 1957
Hibbard, 1957, Trans. Kansas Acad. Sci., v. 60, N 4, p. 327—336.
Тип. Hesperosorex level Hibb., 1957, область Teton, Вайоминг (США), средний
плиоцен.
Диагноз (по: Hibbard, 1957). Тип рода по размерам близок
к Blarina brevicauda carolinensis Bach.
Mx и M2 с хорошо развитым энтоконидом (бугорком на внутренней
стороне метаконида). Зона расположения энтоконида на Мх отделена от
метаконида, что не встречается ни у одной из ископаемых и современных
землероек Северной Америки.
Род включает один вид, отмеченный как тип рода.
Распространение и возраст. Северная Америка, сред-
ний плейстоцен.
f 1. Hesperosorex lovei Hibbard, 1957.
Blarina sp. L о v e, 1956, Bull. Amer. Assoc. Petrol. Geol., v. 60, N 8, p. 1910.
Hesperosorex lovei Hibbard, 1957, Trans. Kansas Acad. Sci., v. 60, N 4, p. 328, 329.
Тип. № 21384, задняя часть левой ветви нижней челюсти с зубами Мх и Мг,
кол. Амер. нац. муз. США, Вашингтон.
Описание. Подбородочное отверстие мелкое, расположено в ниж-
ней части ветви челюсти под серединой Мх. Бугорки на метаконидах Мх
и М2 (энтокониды других авторов) относительно крупные, высокие.
Вершины зубов темные. Судя по рисунку, нижняя челюсть описывае-
мого вида менее массивная, чем у представителей рода Blarina Gray.
Распространение и возраст. Известен из одного
местонахождения.
|43. Род PARACRYPTOTIS Hibbard, 1950
Hibbard, 1950, Contrib. Mus. Paleontol. Univ. Michigan, v. 8, N 6, p. 121—127.
Тип. Paracryptotis rex Hibbard, 1950, Канзас (США), верхний плиоцен.
Диагноз (по: Hibbard, 1950). Крупная землеройка с зубной
3 12 3
формулой: 1уС-р-Р-рМ-д- = 30. Верхних промежуточных зубов четыре, из
них второй крупнее первого, третий наполовину меньше первого, четвер-
тый сильно редуцирован, смещен лингвально и виден с нёбной стороны.
Р4 и моляры крупные, без выемок сзади.
Как отмечает Хиббард, род Paracryptotis легко отличается от рода
Cryptotis Pom. относительно широким рострумом и широкой областью
предглазничного отверстия.
Распространение и возраст. Канзас (США), верхний
плиоцен.
t 1. Paracryptotis rex Hibbard, 1950.
Hibbard, 1950, там же, с. 122—127.
Тип. Передняя часть черепа с полным комплектом зубов, N 25172, формация
Rex road, Канзас (США), верхний плиоцен, кол. Палеонтол. муз. Мичиганск. унив.,
Анн Арбор.
Описание (по: Hibbard, 1950, с сокращениями). Рострум широкий
в дорсальной части, граничащей с (или, лучше сказать, вблизи) Р4—М1,
что контрастирует с равномерно суженным рострумом Cryptotis magna
Merr. Рострум у ископаемого экземпляра по форме приближается к тако-
вому Blarina brevicauda Say. Передние концы носовых костей заострены
к средней линии черепа. Подглазничное отверстие крупное.
432
ПОДСЕМ. SORICINAE
Измерения. Длина нёба — 9.4; максиллярная ширина — 7.3;
длина верхнего зубного ряда — 8.5 мм.
Примечание. Признаки, по которым Хиббард отличает род
Paracryptotis от Cryptotis Pom., мне кажутся неосновательными. Думаю,
что в дальнейшем оба рода будут объединены.
|44. Род PLANISOREX Hibbard, 1972
Hibbard, 1972, р. 78—80.
Тип. Sorex dixonensis Hibbard, 1956, Канзас (США), нижний плейстоцен.
Диагноз (по: Hibbard, 1972). Передний край подглазничного
отверстия выше и позади второго-третьего корней Р4, а его задний край
между первым и вторым наружными корнями М1, М1 и М2 обладают явствен-
ным гипоконом, не наблюдающимся ни у одного другого рода землероек
Рис. 220. Planisorex dixonensis (по: Hibbard, 1972).
(гипокон или талон имеется и у представителей других родов,—А. Г.}.
Мх и М2 четырехугольные, имеют лабиальный и лингвальный цингулюмы.
Талонид (метаконид, по-моему) М3 широкий, низкий, почти такой же
по величине, как и тригонид (передний сегмент — пара-протоконид, по-
моему). Имеется только одно переднее мандибулярное отверстие.
Примечание. Признаки, принятые Гиббардом (Hibbard, 1972)
для характеристики рода Planisorex, не могут быть признаны достаточно
основательными, поскольку их нельзя оценить как отражение общего,
а не частного направления специализации. Однако без материала нет
оснований не считаться с мнением автора рода.
Распространение и возраст. Северная Америка, ниж-
ний плейстоцен.
t 1. Planisorex dixonensis Hibbard, 1956 (рис. 220).
Sorex dixonensis Hibbard, 1956, Pap. Michig. Acad. Sci. Arts. Letters, v. 41,
p. 145—203. Planisorex dixonensis Hibbard, 1972, p. 78—80.
Тип. Фрагмент левой ветви нижней челюсти с зубами Мх—М3, UMMP V 31986,
Kingman County, Канзас (США), нижний плейстоцен, кол. Мичиган, унив., Анн
Арбор.
Описание. Кроме признаков, отмеченных в диагнозе рода, для
этого вида характерны и следующие: Ij относительно короткий, массив-
ный, с двумя бугорками на верхней стороне. Зубная часть ветви нижней
челюсти сравнительно низкая, а венечный отросток высокий. Сочленов-
ный отросток массивный.
45. РОД CRYPTOTIS
433
Измерения (по: Hibbard, 1972). Длина: Ц—М2 — 6.40, Р4—
М3 — 4.95, Mj-Ms — 3.99, 4.06, Mi — 1.87, Ма — 1.68, М3 — 0.93,
1.10, Р4 — 1.34, М1 — 1.34, М2 — 1.28 мм
Распространение и возраст. Канзас, Небраска (США),
нижний плейстоцен.
45. Род CRYPTOTIS Pomel, 1848
Pomel, 1848, Bibl. Univ. Geneve, Arch. Sci. phys. nat., v. 9, p. 249. Brachyso-
rex Duvernoy, 1842, Mag. Zool. Anat. Comp. Paleontol., t. 25, p. 40. Soriculus
Coues, 1877, Bull. U. S. Geol. a. Geogr. Surv., v. 649. Blarina (подрод Cryptotis)
Merriam, 1895, p. 16.
Тип. Sorex cinereus Bachman (=Sorex parvus Say).
3 12 3
Диагноз. Зубная формула: lyС-^Р-рМ j=30. Верхние промежу-
точные зубы с закругленными основаниями без заметно выраженного
цингу люма, сжатые с боков. Задняя сторона Р4, М1 и М2 без выемки (ровная).
1Х с двумя бугорками (лопастями) не верхнем крае.
Род объединяет более 20 современных видов (по: Merriam, 1895 —
13, по: Hall, Kelson, 1959 — 25, по другим авторам — около 30 видов).
Распространение и возраст. Юго-восточная часть США,
Мексика, Центральная Америка и крайний северо-запад Южной Америки,
плиоцен—современный.
Основным местом обитания современных представителей этого рода слу-
жат леса и заросли кустарникав. Только один вид — Cryptotis parva
Say обитает и на открытых местах.
t 1. Cryptotis ? meadensis Hibbard, 1953.
Hibbard, 1953, J., Paleontol., v. 27, N 1, p. 27, 28.
Тип. № 27 266, задняя часть левой нижней челюсти с Р9—М3, Канзас (США),
верхний плиоцен, кол. Палеонтол. муз. Мичиганск. унив., Анн Арбор.
Диагноз (по: Hibbard, 1953). Эта землеройка близка по размерам
к Cryptotis parva Say и средняя по размерам между Sorex taylori и S. rexro-
adensis Hibb.
P4, если сделать проекцию с его передней части, должен был распо-
лагаться на верхней поверхности клыка (12, по моей и других авторов номен-
клатуре нижних промежуточных зубов). Зубы не обладают выемками
на лингвальной части жевательной поверхности. Энтокониды Мх и Мя
узкие впереди, где они соединяются с соответствующими тригонидами.
М3 имеет небольшой тригонид в сравнении с высоким округлым талонидом.
Распространение и возраст. Канзас (США), верхний
плиоцен.
f 2. Cryptotis kansasensis Hibbard, 1957.
Hibbard, 1957, Trans. Kansas Acad. Sci., v. 60, N 4, p. 327—336.
Тип. № 34 447, фрагмент левой стороны верхней челюсти с Р4—М2, Канзас,
в 1 миле к востоку от Норвига (область Кингман), нижний плейстоцен, кол. Палеонтол.
муз. Мичиган, унив., Анн Арбор.
Диагноз (по: Hibbard, 1957). Землеройка крупнее современной
Blarina brevicauda talpoides Gapper. Гипокопид P4 представлен как яв-
ственный лингвальный гребень, который отделен впереди от прилежащего
к губе протоконида глубокой ложбинкой и сходен по расположению с та-
ковыми Paracryptotis rex Hibbard.
Измерения. Длина Мг—М3 — 5.15 мм.
Распространение и возраст. Канзас (США), нижний
плейстоцен.
28 А. А. Гуреев
434
ПОДСЕМ. SORICINAE
3. Cryptotis parva Say, 1823 — Малая короткохвостая землеройка
(рис. 221). .
Sorex parvus Say, 1823, in: Long’s Exped. to the Rocky Mountains, v. 1, p. 164; —
cinereus В achmam, 1837, J. Acad. Nat. Sci. Philadelphia, v. 7, p. 373—376. Blarina
exilipes Baird, 1858, Mammals, in: Repts. Expl. Surv. Railr. Pacific, v. 8, pt 1,
p. 51; — eximius Baird, 1858, там же, с. 52. — floridana Merriam, 1895, p. 19; —
berlandieri Baird, 1858, Mammals, in: Repts. Expl. Surv. Railr., Pacific, v. 8, pt 1,
p. 53.
Тип не установлен, вид описан с западного берега р. Миссури вблизи Blair,
Небраска (США).
Рис. 221. Cryptotis parva.
Диагноз. I1 полого направлен вперед. Его задний зубец хорошо
развит и по высоте равен первому промежуточному зубу. Второй про-
межуточный зуб лишь немного выше первого.
Описание. Верхние части тела окрашены в коричневый, чернова-
тый или черно-серый цвета. Нижние части тела имеют сходную с верхом
окраску или светлее.
Череп с неширокой мозговой капсулой и относительно узкой лицевой
частью.
Измерения. Длина: тела — 60.0—92.0, хвоста — 12.0—42, ступ-
пи — 9.0—17.0, кондило-базальная черепа — 14.4—23.5 мм. У одного
из 2 экз. коллекции ЗИН АН СССР — длина: тела — 72.0, хвоста — 18-0,
ступни — 11.0 мм. По: Hall, Kelson, 1959 — длина тела — 75.0—134.0 мм.
Распространение и возраст. Южные части восточных
штатов Северной Америки от Техаса и восточной части Небраски до
Атлантического побережья.
4. Cryptotis celatus Goodwin, 1956.
Goodwin, 1956, Amer. Mus. Novitates, N 1791, p. 1.
T и п из Теуантепек, Оахака, Мексика.
Распространение. Известен из типичного местонахождения
и, судя по измерениям, описан по 1 экз.
45. РОД CRYPTOTIS
435
5. Cryptotis pergracialis Elliot, 1903.
Elliot, 1903, Field Columb. Mus. (Zool. ser.), v. 3, p. 149.
Ti п из Окотлан, Халиско, Мексика.
Распространение. Тепик, Халиско, Мичоакан, Пуэбла (Мек-
сика).
6. Cryptotis mexicana Coues, 1877.
Blarina (Soriciscus) mexicana Coues, 1877, Bull. U. S. Geol. a. Geog. Surv. Terr.,
v. 3, p. 652.
T и п из Ялапа, Веракрус, Мексика.
Распространение. Оахака, Тамаулипас, Веракрус (Мексика).
7. Cryptotis alticola Merriam, 1895.
Merriam, 1895, p. 27.
Типе вулкана Попокатепетль, Мексика.
Распространение. Мексика.
8. Cryptotis endersi Setzer, 1950.
Setzer, 1950, J. Wash. Acad. Sci., v. 40, p. 300.
Тип из Панамы.
Распространение. Панама.
9. Cryptotis frontalis Miller, 1911.
Miller, 1911, Proc. Biol. Soc. Waschington, v. 24, p. 222.
T ж п из окрестностей Теуантепека, Мексика.
Распространение. Мексика.
10. Cryptotis griseoventris Jackson, 1933.
Jackson, 1933 там же, т. 46, с. 80.
Т ж п из Сан-Кристобаль, Чьяпас, Мексика.
Распространение. Мексика.
11. Cryptotis guerrerensis Jackson, 1933.
J a c k s о n, 1933, там же.
T ж п из Омильтеме, Гуерреро, Мексика.
Распространение. Мексика.
12. Cryptotis goodwini Jackson, 1933.
J a c k s о n, 1933, там же.
Тип из Galel, Гватемала.
Распространение. Гватемала.
13. Cryptotis gracilis Miller, 1911.
Miller, 1911, там же, т. 24, с. 221.
Типе верховьев р. Лари вблизи основания пика Бланко Таламанка, Коста-Рика.
Распространение. Коста-Рика.
14. Cryptotis jacksoni Goodwin, 1944.
Goodwin, 1944, Amer. Mus. Novitates, N 1267, p. 1.
Типе вулкана Ирасу, Коста-Рика.
Распространение. Коста-Рика.
28*
436
ПОДСЕМ. SORICINAE
15. Cryptotis mayensis Merriam, 1901.
Merriam, 1901, Proc. Wash. Acad. Sci., v. 3, p. 559; Miller, 1912, Bull.
U. S. Nat. Mus., v. 79, p. 26.
T и п из Майя, развалины Чичен-Ица, Юкатан, Мексика.
Распространение. Юкатан, Мексика.
16. Cryptotis mera Goldman, 1912.
Cryptotis merus Goldman, 1912, Smiths. Misc. Gool., v. 60, N 2, p. 17.
Типе верховьев p. Лимона, восточная Панама.
Распространение. Панама.
17. Cryptotis nelsoni Merriam, 1895.
Merriam, 1895, p. 26; Miller, 1912, Bull. U. S. Nat. Mus., v. 79, p. 27.
Типе вулкана Тахтла, Веракрус (Мексика).
Распространение. Веракрус (Мексика).
18. Cryptotis obscura Merriam, 1895.
Merriam, 1895, p. 23.
T и п из Тулансинго, Идальго, Мексика.
Распространение. Мексика.
19. Cryptotis olivacea J. Allen, 1908.
J. Allen, 1908, Bull. Amer. Mus. Nat., Hist., v. 24, p. 669.
T и п из San Rafael del Norte, Никарагуа.
Распространение. Никарагуа.
20. Cryptotis zeteki Setzer, 1950.
Setzer, 1950, J. Wash. Acad. Sci., v. 40, p. 299.
T и п из Церро Пунта, Панама.
Распространение. Панама.
21. Cryptotis fossor Merriam, 1895.
Blarina fossor Merriam, 1895, p. 28. Cryptotis fossor Miller, 1912, Bull.
U. S. Nat. Mus., v. 79, p. 28.
T и п из Земпоалтепес, Мексика.
Распространение. Мексика.
22. Cryptotis magna Merriam, 1895.
Blarina magna Merriam, 1895, p. 28. Cryptotis magna Miller, 1912, Bull.
U. S. Nat. Mus., v. 79, p. 28.
T и п из Тотентепес, Оахака, Мексика.
Распространение. Оахака, Мексика.
23. Cryptotis micrura Tomes, 1862.
Sorex micrurus Tomes, 1862, Proc. Zool. Soc. London, p. 279. Cryptotis micrura
Miller, 1924, Bull. U. S. Nat. Mus., v. 128, p. 32.
Тип из Кобан, Гватемала.
Распространение. Мексика, Гватемала, Гондурас, Коста-
Рика.
45. РОД CRYPTOTIS
437
24. Cryptotis soricina Merriam, 1895.
Blarina soricina Merriam, 1895, p. 22. C(ryptotis) soricina Miller, 1911,
Proc. Biol. Soc. Washington, v. 24, p. 221.
Тип из Талалпам, Мексика.
Распространение. Мексика.
25. Cryptotis orophila J. Allen, 1895.
Blarina (Soriculus) orophila J. Allen, 1895, Bull. Amer. Mus. Nat. Hist., v. 7,
p. 340. C(ryptotis) orophila Miller, 1911, Proc. Biol. Soc. Washington, v. 24, p. 221,
Типе вулкана Ирасу, Коста-Рика.
Распространение. Коста-Рика.
26. Cryptotis nigrescens J. Allen, 1895.
Blarina (Soriculus) nigrescens J. Allen, 1895, Bull. Amer. Mus. Nat. Hist., v. 7,
p. 339. C (ryptotis) nigrescens M i 11 e r, 1911, Proc. Biol. Soc. Washington, v. 24, p. 222.
Тип из San Isidro, Коста-Рика.
Распространение. Гондурас, Коста-Рика.
27. Cryptotis tersus Goodwin, 1954.
Goodwin, 1954, Amer. Mus. Novitates, N 1677, p. 1.
Тип добыт в Панаме (на дороге в 15 милях от границы с Коста-Рикой).
Распространение. Панама.
3. Подтриба SORICULINA Kretzoi, 1965
Kretzoi, 1965, р. 117—128.
Тип. Soriculus Blith, 1854, Европа, плиоцен; Азия, плиоцен—современный.
Диагноз. Зубная формула: I у С Р М = 30—28. Верхних
промежуточных зубов 4 или 3, убывающих в размерах спереди назад.
Пигментированы только резцы, клыки, премоляры, моляры белые, а если
частично пигментированы все зубы, то имеются приспособления к пла-
ванию (бахрома из длинных волос на боковых сторонах кистей, ступней
и их пальцев, а также киль на нижней стороне хвоста).
Распространение и возраст. Европа, Азия и Северная
Америка, плиоцен—современный.
Примечание. Адаптивные особенности этой подтрибы весьма
разнообразны. Виды из Северной Америки приспособлены к жизни в откры-
тых ландшафтах пустынного характера. Они имеют короткий хвост и
обычные для землероек ступни, что соответствует передвижению галопом
и в норах. Из евразииских родов одни объединяют виды, приспособленные
к плаванию и имеющие соответствующие морфологические особенности,
другие объединяют обитателей влажных лесов, приспособленных к быст-
рому передвижению прыжками. У них соответственно длинные ступни
и хвост, превышающий у многих видов длину тела. Наконец, в Шри Ланка
обитает землеройка, внешне напоминающая представителей трибы Croci-
durini, — у нее крупные, выступающие из волосяного покрова уши и
длинные щетинки на хвосте. Тем не менее всех их объединяет приспособ-
ление к схватыванию и удержанию в челюстях крупной или скользкой
добычи — рыба, крупные водные беспозвоночные.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Уши большие, сильно выступают из волосяного покрова. Хвост
покрыт короткими волосами и длинными редкими щетинками,
расположенными беспорядочно ......................51. Podihik.
438
ПОДСЕМ. SORICINAE
2 (1). Уши небольшие, мало выступают из волосяного покрова. Хвост
покрыт короткими волосами или короткими волосами и длинными
щетинками, образующими на его нижней стороне бахрому (киль).
3 (6). Длина основания верхних промежуточных зубов более их высоты
в 1.25—1.5 раза. Вершины этих зубов отклонены назад.
4 (5). На верхней стороне Д два бугорка (лопасти) . . . .-46. Zelceina.
5 (4). На верхней стороне 1х один бугорок...........48. Neomys.
6 (3). Длина основания верхних промежуточных зубов менее их высоты.
Вершины этих зубов не отклонены назад.
7 (8). В верхней челюсти 4 промежуточных зуба . . . .49. Soriculus.
8 (7). В верхней челюсти 3 промежуточных зуба....50. Notiosorex.
Примечание. В таблицу не включен род Macroneomys F ejfar за отсутствием
сравнимых признаков.
f46. Род ZELCEINA Sulimski, 1962
Neomys soriculoides Sulimski, 1959, Acta palaeontol. polonica, v. 4, N 2,
p. 149—152. Zelceina Sulimski, 1962, Acta palaeontol. polonica, v. 7, N 3—4,
p. 476—479.
Тип. Neomys soriculoides Sulimski, 1959, Венже (Польша), плиоцен.
Диагноз. Зубная формула: l4-C-lP-?-M-^- = 3O.
Л U 1 о
В верхней челюсти четыре промежуточных зуба. Длина их основания
более их высоты в 1.25—1.50 раза. Вершины этих зубов отклонены назад.
На верхней стороне нижнего внутреннего резца два бугорка.
Рис. 222. Zelceina soriculoides (по: Sulimski, 1959).
Описание. Носовой отдел черепа относительно короткий, тон-
кого сложения. I2 и Is одинаковой величины; С почти вдвое меньше, чем
I2 и Is. Р1 мал, прижат к Р4. Зубная часть ветви нижней челюсти относи-
тельно короткая и высокая.
Распространение и возраст. Европа, плиоцен.
f 1. Zelceina soriculoides Sulimski, 1959 (рис. 222).
Neomys soriculoides Sulimski, 1959 (см. синонимику рода). Zelceina soricu-
loides Sulimski, 1962 (см. синонимику рода).
Тип. Ростральная часть черепа с полным рядом зубов, № 1109, Венже, Польша,
плиоцен. Сулимский (Sulimski, 1959) указывает в качестве типов и ветви нижней че-
люсти (правая № 700 и левая № 160).
Измерения. Длина общая: нижней челюсти — 9.6—10.3, основ-
ная нижней челюсти — 7.6—8.2, нижнего зубного ряда — 6.3—6.6,
Р4—М3 — 4.0—4.3; высота: нижней челюсти в области Мх — 1.3—1.6,
венечного отростка — 2.8—4.2, сочленовного отростка — 2.3—3.1 мм.
47. РОД MACRONEOMYS. 48. РОД NEOMYS
439
Примечание. Диагноз и описание рода Zelcenia Sulimski —
составлены на основании сравнения его особенностей, почерпнутых из
описаний и рисунков Сулимского (Sulimski, 1959, 1962), и особенностей
представителей современного рода — Neomys Каир. При этом взяты
наиболее характерные для этих родов признаки.
|47. Род MACRONEOMYS Feifar, 1966
Feifar, 1966, Neues Jahrb. Geol. und Palaontol. Abhandl., H. 11, S.685—691.
Тип. Macroneomys brachygnathus Feifar, 1966, Европа, нижний плейстоцен.
Диагноз (по: Feifar, 1966). Нижний резец с одним зубцом на верх-
ней стороне, массивный. Нижние моляры короткие и высокие, массивные,
с наружным цингулюмом. Нижние
промежуточные зубы с тупыми
закругленными вершинами. Ве-
нечный отросток высокий, массив-
ный. Нижняя челюсть относи-
тельно короткая.
Распространение и
возраст. Европа, нижний
плейстоцен.
Рис. 223. Macroneomys brachygnatus (по:
Fejfar, 1966).
t 1. Macroneomys brachygnathus
Feifar, 1966 (рис. 223).
Feifar, 1966, там же.
Тип. Левая ветвь пижней челюсти
с зубами (li-г, и Мх), SUUG, OF, N
64532, карстовая пещера горы Золотой
у Кунепрузи, Средняя Богемия.
Описание. Зубы пигмен-
тированы. Нижний резец относи-
тельно короче, чем у представи-
телей рода Neomys Каир; его
цингулюм слабо развит. 12 и Р4
одновершинные. Верхние и нижние
моляры с относительно высокими
и массивными коронками. Бугорки
их жевательной поверхности с при-
тупленными вершинами. Венечный
отросток с почти параллельными передней и задней сторонами; располо-
жен под прямым или острым углом по отношению к зубной части нижней
челюсти. Подбородочное отверстие под протоконидом Мх.
Примечание. Признаки, установленные автором для рода Macro-
neomys, не убеждают в его обособленности от рода Neomys Каир. Можно
думать, что они в дальнейшем будут объединены.
Распространение и возраст. Европа, нижний плейсто-
цен.
48. Род NEOMYS Каир, 1829 — КУТОРЫ, или ВОДЯНЫЕ ЗЕМЛЕРОЙКИ
Капр, 1829, Entwickl. und Naturl. Syst. europ. Thierwelt, Bd 1, S. 117. Leucor-
hynchus Каир, 1829, там же, с. 118. Hydrogale Каир, 1829, там же, с. 122, 123.
Crossopus W a g 1 ё г, 1832, Isis, р. 275. Hydrosorex Duvernoy, 1835, Mem. Soc.
Mus. Hist. Nat., Strasb., t. 2, p. 17. Amphisorex Duwernoy, 1835, там же, с. 23.
Pinalia Gray, 1837—1838, p. 226. Galemys Pom el, 1848, Bibl. Univ. Geneve, Sci.
Physiol. Nat., v. 9, p. 249. Myosictis Pomel, 1854, Catal. method, vertebr. foss. de-
ceuv. dans bassin hidrogr. superieur et de Loir et de Allier.
Tin. Sorex fodiens Pennant, 1771, Европа, Азия, плиоцен—современный.
440
ПОДСЕМ. SORICINAE
3 12 3
Диагноз. Зубная формула: I-g-C-Q-РуМу^ЗО. В верхней челюсти
4 промежуточных зуба. Длина их оснований более высоты в 1.25—1.5 раза.
Вершины этих зубов отклонены назад. На верхней стороне нижнего вну-
треннего резца один бугорок.
Описание. Глаза хорошо заметны или скрыты в густой шерсти.
За глазами небольшое светлое пятно. Уши почти не выступают из волося-
ного покрова. Хвост значительно длиннее половины длины тела. Тело
густо покрыто волосами, образующими плотный, не смачивающийся водой
бархатистый покров. Окраска верхней стороны тела черно-бурая, бурая
или коричневая. Нижняя сторона тела и бока имеют белую, серо- или
желто-белую окраску, иногда с примесью оранжевого или коричневого
цвета.
Череп крупный и массивный с широкой мозговой капсулой. Сагитталь-
ный и ламбдовидный гребни хорошо заметны. Отверстия на лобных костях
имеются. Передний зубец внутреннего верхнего резца более заднего
в 3—4 раза.
Род Neomys отличается от других родов формой верхних промежуточ-
ных зубов, а также рядом приспособлений для плавания.
Распространение и возраст. Лесная зона Европы и
Северной Азии, плиоцен—современный.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ХУТОР
(РОД NEOMYS KAUP) ФАУНЫ СССР
1 (2). Длина хвоста менее 60 мм, киль из длинных волос развит только
на его задней трети. Плавательные щетинки на кистях и ступнях
слабо развиты и не образуют густой бахромы.....................
.......................................4. N. anomalus Сайг.
2 (1). Длина хвоста не менее 60 мм, киль из длинных волос развит не
менее, чем на 2/3 его длины. Плавательные щетинки на кистях и
ступнях образуют длинную и густую бахрому.
3 (4). Глаза скрыты в густой шерсти. Верхняя сторона кистей и ступней
покрыта редкими, светлыми волосами и крупными, черно-бурыми
или черноватыми чешуйками................3. N. schelkovnikovi Sat.
4 (3). Глаза хорошо видны в негустой шерсти. Верхняя сторона кистей
и ступней густо покрыта светлыми волосами и мелкими буроватыми
чешуйками ...................................2. N. fodiens Penn.
t 1. Neomys gastellarini Pasa, 1947.
P a s a, 1947, Mem. Mus. Civ. Storis. Natur., Verona, t. 1, p. 11—14.
Распространение и возраст. Европа, плиоцен.
2. Neomys fodiens Pennant, 1771 — Кутора обыкновенная (рис. 224).
Sorex fodiens Pennant, 1771, Sinopsis Qadrupeds, p. 308; — daubentonii E r x?
1 e b e n, 1777, Syst. Animal., p. 124; — carinatus Hermann, 1780, in: Zimmer-
mann’s Geogr. Gesch., Bd 1, S. 383; — liricatus Kerr, 1792, Animal Kingdom, p. 208. —
eremitaM e у e r, 1793, Zool. Annal., Bd 1, S. 232; — jluviatilisN. e у e r, 1793, там же.—
hydrophilus Pallas, 1811, Zoogr. Rosso Asiatica, v. 1, p. 130, 131; — lineatus Geof-
froy, 1811, Ann. Mus. Hist., Nat., t. 17, p. 181; — remifer Geoffroy, 1811,
там же; — collaris Desmarest, 1818, Nouv. Diet. Hist. Nat., t. 22, p. 65; — macru-
rus Lehmann, 1822, Obsrev. Zool. Faun. Hamburg., Bd 1, S. 5; — amphibius
Brehm, 1826, Omis, v. 11, p. 38; — natans Brehm, 1826, там же, с. 44; — stag-
nalis Brehm, 1826, там же, с. 47; — rivalis Brehm, 1830, Isis, p. 1128.
Neomys fodiens Каир, 1829, Entwickl.-Gesch. und Naturl. Syst. europ.
Thierwelt, Bd 1, S. 117. Sorex musculus W a g 1 e r, 1832, Isis, p. 54; — psilurus Wag-
ler, 1832, там же. Crossopus fodiens Wagler, 1832, Isis, p. 275. Sorex nigripes, Mel-
48. РОД NEOMYS
441
с hi or, 1834, Den danske stats on Norges Pattedyr, p. 68. Hidrosorex fodiens D u v er-
n о v, 1835, Mem. Soc. Mus. Hist., Nat., Strasb., t. 2, p. 8. Amphisorex hermanni D u-
v e г n о y, 1835, там же; — constrictus Duvernoy, 1838, там же (январское допол-
нение); — limneana Gray, 1838, Ann. Mag. Nat. Hist., p. 287. Crossopus sowerbyi
Bonaparte, 1840, Fauna Italica, t. 1, p. 29. Neomys argenteus Огнев, 1921,
Ежегодн. Зоол. муз. АН, т. 22, с. 346.
Т и и не сохранился или не установлен, вид описан из окрестностей Берлина.
Диагноз. Глаза хорошо заметны в негустой шерсти, окружающей
их. Верхняя сторона кистей и ступней покрыта светлыми волосами и мел-
кими буроватыми чешуйками. Длина хвоста не менее 60.0 мм.
Рис. 224. Neomys jodiens.
Описание. Волосяной покров во все сезоны образован относи-
тельно короткими волосами, густой и бархатистый. Киль из длинных ще-
тинистых волос на нижней стороне хвоста хорошо развит и заметен
на всем его протяжении. Окраска верхней стороны тела черная, черно-
бурая или бурая, нижней — белая, серо-белая, охристая или белая с оран-
жевым или бурым налетом. За глазом крупное, хорошо заметное светлое
пытно. Хвост одноцветный, бурый или черно-бурый, или снизу до поло-
вины длины беловатый, а в концевой половине белым остается только
киль. Окраска кожи кистей и ступней бурая сверху и сцизу. Чешуйки,
покрывающие кисти и ступни, слабо заметные. Волосы густо покрывают
ступни и кисти сверху, плавательные оторочки из длинных щетинистых
волос хорошо развиты.
Череп с угловатой и уплощенной мозговой капсулой. Продольный
диаметр конической части коронки второго и третьего промежуточных
зубов в 2 раза менее продольного диаметра основной части коронки (вклю-
чая цингулюм) этих зубов (рис. 224).
Измерения. Длина: тела — 76.0—87.0, хвоста — 60.0—71.0,
ступни — 17.0—21.5, кондило-базальная черепа — 18.6—21.8 мм.
Распространение и в оз р а с т. Англия, Франция, Дания,
Бельгия, Нидерланды, Люксембург, Швейцария, Италия, Румыния, ГДР,
ФРГ, Швеция, Норвегия, Финляндия, Польша, Монголия. В пределах
СССР распространена по всей его европейской части от Северного Ледо-
442
ПОДСЕМ. SORICINAE
витого океана до Северного Кавказа; в Западной Сибири от Полярного
Урала по всему бассейну р. Оби, среднему и верхнему течению Енисея,
добыта на Таймыре; на северо-востоке Сибири кутора не попадается.
На юге известна из Волжско-Уральских степей, из Курганской, Омской
и Новосибирской обл., из Северо-Восточного Казахстана, Целиноградской
и Семипалатинской обл. и из Алтайского края. Встречается в окрестностях
оз. Байкал. К востоку ее ареал прерывается и известны лишь отдельные
местонахождения в низовьях р. Амура и на о. Сахалин, современный.
Биология. Обитает преимущественно по берегам различных водое-
мов: озер, рек, ручьев и проточных канав, а также на сырых болотах.
Крайне редко она попадается вдали от воды в сырых лесах и колках,
что связано с пересыханием водоемов. Вслед за древесной растительностью
проникает в степную зону. В горы поднимается до альпийских лугов на вы-
соту 2000—2600 м. Наиболее предпочитаемыми местами обитания куторы
служат топкие берега водоемов, поросшие деревьями, кустарниками, гу-
стой травяной растительностью и сильно захламленные валежником.
Отмечаются случаи поселения кутор в заброшенных норах водяной
полевки (Arvicola terrestris L.). Иногда роет норы и сама.
Кутора превосходно плавает; волосяной покров ее не смачивается
водой. В зимнее время перемещается в места, где водоемы не замерзают
или болотный грунт остается мягким под глубоким снегом. Гнезда кутора
размещает в норе, а также в различных укромных местах: между корнями
деревьев и кустарников, в кочках, дерновинах и др. Строится оно из сухих
травянистых растений и имеет шарообразную форму с рыхлыми стенками
и одним входным отверстием, направленным в сторону воды.
Питается кутора насекомыми, червями, моллюсками, икрой и маль-
ками рыб, мелкими рыбами, лягушками и другими мелкими позвоночными,
с которыми может справиться. Пищу поедает только на суше.
Размножение не изучено. Беременные самки добывались с конца апреля
до августа. Продолжительность беременности точно не установлена, ука-
зываются 24 и 28 дней. Число эмбрионов для Западной Сибири — 4—14,
чаще 6—8.
Ц о д в и д ы: 1. N. f. fodiens Penn., 1771. Характеризуется средними
размерами тела, крупными ушами и относительно короткой ступней. Рас-
пространен в Западной Европе, европейской части СССР, лесостепных
районах Западной Сибири к востоку до р. Оби и Приалтайских степей.
2. N. /. orientis Thos., 1914. Более крупная форма с короткими ушами.
Распространена к востоку от р. Оби и Приалтайских степей до Охотского
моря, Приморья и Сахалина включительно.
3. Neomys schelkovnikovi Satunin, 1913 — Кутора Шелковникова
(рис. 225).
С а т у нин, 1913, Тр. Общ. для исслед. Черноморск. побережья, т. 3, с. 24, 25.
Т и п не отмечен, вид описан из Сванетии (с. Ушкул, ГрузССР).
Диагноз. Глаза скрыты в густой шерсти. Верхняя сторона кистей
и ступней покрыта редкими светлыми волосами и крупными черно-бурыми
или черноватыми чешуйками. Длина хвоста не менее 60.0 мм.
Описание. Волосы, покрывающие тело, более длинные, чем у дру-
гих видов рода, особенно в зимнее время. Киль из длинных щетинистых
волос хорошо заметен только в концевой половине хвоста. Окраска верх-
ней стороны тела черная, черно-бурая, реже бурая, нижней — серая или
беловатая с бурым налетом. Светлое пятно за глазом очень мелкое, мало-
заметное или совсем отсутствует. Хвост двуцветный, светлый: серый
сверху, буроватый снизу или сверху буроватый, а снизу серо-белый или
48. РОД NEOMYS
443
белый. Окраска кожи и чешуек на верхней и нижней сторонах кистей
и ступней черная или черно-бурая. Чешуйки хорошо дифференцированы
друг от друга. Волосы на верхней стороне кистей и ступней редкие, а во-
лосяные плавательные оторочки на последних хорошо развиты.
Череп со слабо выраженной угловатостью боковых сторон мозговой
капсулы. Продольный диаметр конической части коронки второго и
третьего промежуточных зубов в 2 раза менее продольного диаметра
основной части коронки этих зубов, включая цингулюм.
Измерения. Длина: тела — 87.0—102.0, хвоста — 60.0—75.0,
ступни — 18.0—20.0, кондило-базальная черепа — 20.0—22.3 мм.
Рис. 225. Neomys schelkovnikovi.
Распространение и возраст. Известна из ряда районов
Грузинской ССР и Армянской ССР. Границы распространения этого вида
неясны. Образ жизни не изучен, по-видимому, он мало отличается от тако-
вого обыкновенной куторы. Современный.
4. Neomys anomalus Cabrera, 1907 — Малая кутора (рис. 226).
Cabrera, 1907, Ann. Mag. Nat. Hist., v. 20, p. 214. Neomys milleri M о 11 a z,
1907, Mem. Soc. Zool. Franse, t. 20, p. 22; — teres Miller, 1908, Ann. Mag. Nat.
Hist., v. 1, p. 68; — soricoides Огнев, Ежегодн. Зоол. муз. АН, т. 22, с. 347.
Т и п не установлен, вид описан из San Martin de la Vega, Испания.
Диагноз. Длина хвоста менее 60.0 мм. Плавательные щетинки
на кистях и ступнях в отличие от других видов слабо развиты и не обра-
зуют густой бахромы.
Описание. Волосы, покрывающие тело, относительно короткие.
Глаза всегда окружены короткими волосами и хорошо заметны. Киль
из длинных щетинистых волос (на нижней стороне хвоста) слабо развит
и более заметен в его концевой трети. Окраска верхней стороны тела черно-
или коричнево-бурая, нижней — серая, серо-белесая или серая с бурова-
тым налетом. Небольшое светлое пятно за глазом хорошо заметно. Хвост
444
ПОДСЕМ. SORICINAE
сверху бурый, снизу — беловатый, реже — почти одноцветный, и только
волосы киля белые. Кожа ступней и кистей, а также покрывающие их че-
шуйки на верхней и нижней сторонах светло-бурые. Границы между
чешуйками, покрывающими кожу лап, мало заметны. Плавательные ото-
рочки на кистях и ступнях слабо развиты.
Череп с округлой, несколько вздутой мозговой капсулой. Продольный
диаметр конической части коронки второго и третьего промежуточных
зубов в 1.5 раза менее продольного диаметра основной части коронки этих
зубов, включая цингулюмы.
Измерения. Длина: тела — 70.0—82.0, хвоста — 47.0—53.0,
ступни — 15.5—17.3, кондило-базальная черепа — 19.7—20.3 мм.
Рис. 226. Neomys anomalus.
Распространение и возраст. Испания, Франция, Швей-
цария, Югославия, Греция, Венгрия, Польша и Малая Азия. В СССР
найдена в Крыму, в Закарпатской и северо-западной частях Украины,
в Молдавии и южной части Белоруссии. Указывается также (Бобринский,
Кузнецов, Кузякин, 1965) для Воронежского заповедника, современный.
Близкая к этому виду форма известна из верхнего плиоцена Молдавии и
Среднего Придонья.
Биология. Малая кутора обитает в разнообразных ландшафтах —
от равнин до высокогорий. В горах встречается по берегам горных потоков,
ручьев и родников, в буковых и буково-ясеневых лесах и в зарослях
кустарников на лугах. В равнинных ландшафтах населяет леса разных
типов, кустарники, встречается на лугах и изредка на болотах. Повсюду
связана с водой. Предпочитает быстротекущие незамерзающие водоемы
с крутыми каменистыми берегами, водный режим которых приближается
к таковому горных рек. Гнезда устраивает под землей и на ее поверхности,
в густых зарослях травы, в корнях деревьев или в щелях между камнями.
Питается беспозвоночными и мелкими позвоночными животными: червями,
моллюсками, насекомыми, мелкой рыбой и икрой, а также лягушками.
Подвиды. Из Крыма описан подвид N. a. mokrzeckii Martino,
1917, отличающийся от номинальной формы более крупными размерами
и более длинным хвостом.
49. РОД SORICULUS
445
49. Род SORICULUS Blyth, 1854
Blyth, 1854, J. Asiat. Soc. Bengal., v. 23, p. 733.
Тип. Corsira nigrescens, Gray, 1848, Индия, Непал, Бирма.
3 12 3
Диагноз. Зубная формула: I-tt-C-tcP-t-M — = 30. В верхней челюсти
V 1 О
4 промежуточных зуба. Длина их основания не более высоты зуба. Вер-
шины этих зубов не отклонены назад. На верхней стороне 1Х один бугорок.
Описание. Окраска изменчива. Красно- и темно-коричневая,
серая и черно-бурая различных оттенков. Изменчива и длина ступни,
хвоста и вибрисс, но преобладают виды с длинными хвостами, ступнями
и вибриссами. Волосяной покров мягкий, бархатистый.
Череп относительно крупный, массивный, с хорошо развитыми греб-
нями. I1 с мелким и коротким задним зубцом. I2-3, а также клык близки
по размерам. Передний премоляр мелкий, смещен на внутреннюю сторону
челюсти. 1Х с одним бугорком при основании.
Распространение и возраст. Китай (Сычуань, Юнь-
нань, Тайвань), Индия (Бутан, Сикким, Ассам), Непал, северная часть
Бирмы, Бакбо (п-ов Индокитай), плиоцен—современный. Европа —
плиоцен.
Примечание. В пределах рода Soriculus Blyth Эллерман и Мор-
рисон-Скотт (Ellermann, Morrison-Scott, 1951) выделили 3 подрода: Episo-
riculus Ell. et M.-Scott, представители которого характеризуются длиной
хвоста, превышающей длину тела: Soriculus Blyth — с хвостом, равным
длине тела или короче: Chodsigoa Kastsch., объединяющий виды с тремя
верхними промежуточными зубами, а не с четырьмя, как у Soriculus и
Episoriculus.
По моему мнению, длина хвоста не может служить основанием для
выделения подрода в пределах рода длиннохвостых землероек, так же
как и длина когтей. Поэтому в этой работе не сохраняются названные
подроды. Что же касается подрода Chodsigoa Kastsch., то я совершенно
согласен с Томасом (Thomas, 1908), что его следует считать самостоятель-
ным родом, хорошо обособленным от рода Soriculus Blyth, поскольку его
представители имеют в верхней челюсти не четыре, а три промежуточных
зуба — признак весьма существенный в пределах семейства Soricidae.
Эту же точку зрения разделял и Аллен (G. Allen, 1938). Но сравнение
рода Chodsigoa Kastsch. с американским родом Notiosorex Baird позволило
обнаружить значительное сходство представителей обоих родов по целому
ряду признаков (общее число зубов, сходная их окраска, сходство в строе-
нии черепа), что позволяет их считать генетически связанными, близкими
группами. Это обстоятельство позволило объединить названные роды
в один род — в качестве его подродов, сохранив за последним название
ранее описанного рода Notiosorex Baird. Эта группа землероек близка
к роду Soriculus Blith и весьма далека от других американских родов.
Можно думать, что американская и азиатская группы землероек этого
рода уже с плиоцена развивались независимо, чем и объясняются их под-
родовые различия в современный период.
1. Soriculus kubinyi Kormos, 1934 (рис. 227).
Kormos, 1934, v. 64, p. 8, 9.
Тип. Фрагмент черепа с нижней челюстью 06/3685, Виллань, Венгрия,
плиоцен.
Описание (по: Kormos, 1934). Кормош для этого вида отмечает
следующие особенности: подглазничное (предглазничное — у других авто-
ров) отверстие округлой формы. Слезное отверстие расположено на слез-
446
ПОДСЕМ. SORICINAE
ной кости ближе к ее переднему краю. Р3 (Р1) расположен ближе к внутрен-
нему краю зубного ряда и снаружи не заметен. Бугорок на верхней стороне
1Г слабо развит.
Измерения. Длина верхнего зубного ряда — 7.7, нижнего —
6.6—6.7 мм.
Распространение и возраст. Европа, плиоцен.
2. Soriculus nigrescens Gray, 1842 — Сиккимская когтистая земле-
ройка.
Corsira nigrescens Gray, 1842, Ann. Mag. Nat. Hist., v. 10, p. 261. Sorex aterrimus
Blith, 1842, J. Asiat. Soc. Bengal, v. 12, p. 928; — sikkimensis Hodgson, 1849, Ann.
Mag. Nat. Hist., ser. 2, v. 3, p. 203. Soriculus nigrescens Blith, 1854, J. Asiat. Soc.
Рис. 227. Soriculus kubinyi (no: Kormos, 1934).
Bengal, v. 23, p. 733. Sorex aligurus Gray, Cat. Mammal. Hodgsons, 1863, Coll. Nepal
et Tibet, Brit. Mus., 2nd. ed., p. 8; — holosericous Gray, там же, с. 9. Soriculus radulus
Thomas, 1922, J. Bombay Nat. Hist. Soc., v. 28, p. 429.
T ип не установлен, вид описан из Индии (Darjeeling).
Диагноз (по: Blanford, 1888—1891, также и описание). Отличается
от других видов рода сильно развитыми, относительно широкими когтями
на пальцах кисти, крупными размерами тела (более 70 мм) и относительно
более коротким хвостом, не превышающим 70% длины тела.
Описание. Окраска верхней стороны тела коричневая; местами
почти черная или рыжевато-коричневая. Нижняя сторона тела светлее,
с примесью серого на брюшке. Кисти и ступни сверху темные — коричне-
вые. Хвост темный сверху, светлый снизу.
Измерения (по: Blanford, 1888—1891). Длина: тела — 82.0—
86.5, хвоста — 43.0—48.0, ступни — 12.5—13.0, основная черепа —
22.0 мм (в дюймах соответственно: 3.4—3.0, 1.65—1.82, 0.54—0.55, 0.8).
Распространение и возраст. Индия (Бутан, Сикким,
Ассам). Непал, северная часть Бирмы, современный.
3. Soriculus caudatus Hodgson, 1849.
Sorex caudatus Hodgson, 1849, Ann. Mag. Nat. Hist., ser. 2, v. 3, p. 203; H or s-
field, 1851, Cat. Mamm. Mus. E. India Co., p. 135. Corsira alpina Thomas, 1856,
Ann. Mag. Nat. Hist., ser. 2, v. 17, p. 27. Sorex homourus ? Gray, 1863, Cat. Mammal.
Hodgsons, Coll. Nepal et Tibet, Brit. Mus., 2nd ed., p. 8 (не описан). Soriculus gracili-
cauda Anderson, 1877, J. Asiatic Soc. Bengal, v. 46, pt. 2, p. 282; — minor Dob-
50. РОД NOTIOSOREX
447
son, 1890, Monogr. Insectivora, pt. 3, pl. 24, fig. 2—2b;—fumidus Thomas, 1913,
Ann. Mag. Nat, Hist., v. 2, p. 216; — baileyi T h о m a s, 1914, J. Bombay Nat. Hist.
Soc., v. 22, p. 683.
T и п не установлен, вид описан из Индии (указывается Darjeeling, Сикким).
Диагноз (по: Blanford, 1888—1891). Когти на пальцах кисти
короткие. Длина тела менее 70 мм; длина хвоста равна или немного более
длины тела.
Описание. Уши покрыты волосами и частично скрыты в волося-
ном покрове головы. Окраска тела коричневая, черноватая до красноватой.
I2 заметно крупнее, чем I3 * s и клык.
Измерения (по: Blanford, 1888—1891). Длина: тела — 63.0,
хвоста — 70.0, ступни — 14.0, основная черепа — 18 мм (в дюймах со-
ответственно: 2.46, 2.57, 0.52, 0.66).
Распространение и возраст. Индия, Китай (Сычуань,
Юньнань и Тайвань), северная часть Бирмы, Бакбо (п-ов Индокитай).
4. Soriculus leucops Horsfield, 1855.
Sorex leucops Horsfield, 1855, Ann. Mag. Nat. Hist., v. 16, p. Ill; — nivicola
Gray, 1863, Cat. Mammal. Hodgsons, Coll. Nepal et Tibet, Brit. Mus., 2nded.; p. 8
(не описан); — macrurus Hodgson, 1863, там же, с. 9 (не описан). Soriculus macrurus
Blanford, 1888—1891, p. 231; — irene Thomas, 1911, Abstract Proc. Zool.
Soc. London, p. 49.
T и п не установлен, вид описан из Непала.
Диагноз. Длина тела значительно менее 70.0 мм. Длина хвоста
более длины тела в 1.5 раза. Когти короткие.
Описание. Тело и конечности стройные. Хвост округлый в сече-
нии, тонкий, очень длинный; он покрыт короткими редкими волосами,
напоминает хвост мышей. Уши волосистые, частично скрытые в волосяном
покрове головы. Волосы, покрывающие тело сравнительно длинные, мяг-
кие. Бленфорд (Blanford, 1888—1891) отмечает, что окраска тела однотон-
ная — черно-коричневая, а в описании этого вида у Аллена (G. Allen,
1938) отмечено различие окраски в зимнее и летнее время и в качестве
основного тона указывается серая (зимой более светлая). По-видимому,
окраска тела у этого вида варьирует в значительных пределах. Хвост дву-
цветный, сверху светло-коричневый, снизу — серебристо-серый. Кисти
и ступни сверху беловатые и только на средних частях с примесью корич-
неватого цвета.
Измерения. Длина: тела — 53.3—58.0, хвоста — 81.0—86.3,
ступни — 14.0—15.0, основная черепа 15.0 мм.
Распространение и возраст. Непал, Индия (Сикким),
северная часть Бирмы, Китай (Сычуань, Юньань), современный.
50. Род NOTIOSOREX Baird, 1877
Notiosorex (подрод Sorex) Baird, in: Coues, 1877, Bull. Geogr. Surv., v. 3, N 3,
p. 646, 647. Notiosorex Dobson, 1890, Monogr. Insectivora, pt 3, pl. 23, fig. 20.
Soriculus (Chodsigoa) Кащенко, 1907, Ежегодн. Зоол. муз. АН, т. 10, с. 1—4.
Megasorex Hibbard, 1950, Contrib. Mus. Paleontol. Univ. Michigan, v. 8, p.
127—129.
Тип. Sorex (Notiosorex) crawfordi Baird, 1877, юго-запад США, восточная и юго-
западная часть Мексики.
3 113
Диагноз. Зубная формула: I С у Р-р М у — 28. В верхней челюсти
3 промежуточных зуба. Длина их основания не более высоты зуба. Вер-
шина этих зубов не отклонена назад. На верхней стороне Д один бугорок.
Описание. Представители этого рода имеют относительно разно-
образный вид. Одни напоминают обыкновенных землероек, другие (с длин-
448
ПОДСЕМ. SORICINAE
ними ногами и хвостом) похожи на мышей. Среди этой группы землероек
есть и роющие формы; правда, приспособления к этой форме локомоций
слабо выражены.
Окраска тела разнообразная, преимущественно темно-бурая, коричне-
вая и серая разных оттенков. Нижняя сторона тела светлее верхней или
беловатая. Хвост двуцветный. У длиннохвостых землероек он почти голый,
как у мышей.
Череп массивный, с относительно короткой лицевой частью и длинной
мозговой капсулой. Нижняя фасетка на сочленовном отростке нижней
челюсти отодвинута значительно вперед по направлению к переднему
краю челюсти, соответственно выдвинут и засочленовный отросток височ-
ной кости, на который она опирается.
Распространение и возраст. Средний и Южный Китай,
юго-запад США, восточная и южная части Мексики, плиоцен—совре-
менный. Представители рода в Азии — обитатели преимущественно влаж-
ных окраин лесов и лесных чащ, в Америке — пустынно-степных формаций.
1. Подрэд NOTIOSOREX Baird, 1877 — АМЕРИКАНСКИЕ
МЫШЕВИДНЫЕ БУРОЗУБКИ
Notiosorex (подрод Sorex) Baird, 1877, там же, где и род. Notiosorex (род) Dob-
son, 1890, Monogr. Insectivora, pt 3, pl. 23, fig. 20.
Тип. Sorex (Notiosorex) crawfordi Baird, 1877, часть нижней Соноры (зона от вос-
точного Техаса до южной Калифорнии (по: Merriam, 1895).
Диагноз. Второй зубец Г хорошо обособлен, имеет округлую
внешнюю сторону и окрашен. Верхние промежуточные зубы имеют глад-
кую и выпуклую внешнюю сторону, а выемку только на внутренней сто-
роне; их коронки не кажутся гребнистыми, как у Chodsigoa. Первый и вто-
рой промежуточные зубы равны по величине или второй выше первого.
Распространение и возраст. Юго-запад США и северо-
восточная часть Мексики, плиоцен—современный.
Примечание. Представители этого подрода в отличие от видов
подрода Chodsigoa Kastsch. в строении зубов сохраняют некоторые древние
особенности, близкие менее специализированным группам землероек,
в частности мало различающиеся по величине верхние промежуточные
зубы. Это на фоне несомненного прогресса (сокращение общего числа
промежуточных зубов) представляется более примитивной чертой по
сравнению с заметным увеличением первого промежуточного зуба и умень-
шением следующих за ним, что наблюдается у более специализированных
представителей подрода Chodsigoa.
11. Notiosorex (Notiosorex) jacksoni Hibbard, 1950.
Hibbard, 1950, Gontrib. Mus. Paleontol. Univ. Michigan, v. 8, N 6, p. 129—
133. Sorex ? sp. Hibbard, Riggs, 1949, Bull. Geol. Soc. America, v. 60, p. 835.
Тип. Левая половина верхней челюсти с зубами (Р4—М2), № 24347, Канзас,
США, верхний плиоцен, кол. Палеонтол. муз. Мичиганск. унив., Анн Арбор.
Диагноз (по: Hibbard, 1950). Крупнее, чем Notiosorex crawfordi
Baird, но меньше, чем N. gigas Merr., с общим числом зубов — 28. Р1, М1
и М2 с глубокой выемкой сзади. Если верить рисунку Хиббарда,
то у N. jacksoni на верхней стороне 1х два бугорка, а не один, как у совре-
менных видов.
Измерения. Максимальная длина I1—М2 — 8.0 мм.
Распространение и возраст. Северная Америка, верх-
ний плиоцен.
50. РОД NOTIOSOREX
449
2. Notiosorex (Notiosorex) crawfordi Baird, 1877 — Серая, или Пустын-
ная землеройка (рис. 228).
Sorex (Notiosorex) crawfordi Baird, 1877, там же, где и род, с. 651, 652.
2653
Тип. № 4437, окрестности форта Блисс (Bliss), Новая Мексика (El Paso, Техас)
кол. Амер. нац. муз., Вашингтон.
Диагноз. Размеры мелкие (длина тела 48—65 мм). Уши крупные
для землероек, заметно выступают из шерсти. Ступня и хвост короткие.
Длина хвоста примерно равна половине длины тела.
Описание. Окраска верхних частей тела оливково-серая, ниж-
них — беловатая. Хвост двуцветный.
Череп плоский, с округлой мозговой частью.
Измерения (по: Mercian, 1895). Тип (по спиртовому экзем-
пляру) — длина: тела — 48, хвоста (по позвонкам) — 28, ступни —
v
Рис. 228. Notiosorex crawfordi (по: Merriam, 1895).
10.0 мм. Размеры тела спиртовых экземпляров из Сан-Диего (Калифор-
ния) — длина: тела — 82 и 56, хвоста — 26 и 28, ступни — 10.0 и 10.5 мм.
Общая длина черепа типичного экземпляра, включая внутренние резцы, —
17.3 мм.
Этот вид отличается от N. gigas Merr. меньшими размерами тела и олив-
ково-серой окраской, а также прямым задним краем зубов Р4, М1 и М2.
Распространение и возраст. Южная часть штата Кали-
форния, южная часть Колорадо и Оклахомы, юго-запад Арканзаса, Техас,
Нью-Мексико (США), северо-восточная и юго-западная части Мексики,
современный.
3. Notiosorex (Notiosorex) gigas Merriam, 1897 — Большая серая земле-
ройка.
Notiosorex gigas Merriam, 1897, Proc. Biol. Soc. Washington, v. 11, p. 227.
T и п не установлен, вид описан из горной местности вблизи Сан-Себастьяна,
Халиско, Мексика.
Диагноз (по: Hall, Kelson, 1959). Размеры крупные: длина тела
127—130 мм; по другим данным — 83—90 мм (Walker et al., 1964). Окраска
меха свинцово-серая.
Измерения (по: Hall, Kelson, 1959). Длина: хвоста — 39—43
(по другим данным — 39—50), ступни — 16—17, кондило-базальная че-
репа — 22.2—22.8 мм.
Распространение и возраст. Халиско, Колима, Ми-
чоакан (Мексика), современный.
j/229 А- А- Гуреев
450
ПОДСЕМ. SORICINAE
2. Подрод CHODSIGOA Kastschenko, 1907 — МЫШЕВИДНЫЕ БУРОЗУБКИ
Soriculus (Chodsigoa) Кащенко, 1905, Ежегодн. Зоол. муз. АН, т. 10, с. 1, 2.
Chodsigoa Thomas, 1908, Proc. Zool. Soc. London, p. 639.
Тип. Soriculus (Chodsigoa) beresowskii Kastschenko, 1907. ( = Soriculus hypsibius
de Winton et Styan, 1899), Китай (Ганьсу, Сычуань, Юньнань, Шэньси, Чжили).
Диагноз. Второй зубец I1 не обособлен и не оформлен в виде бугра,
а имеет форму пластинчатого выступа. Верхние промежуточные зубы сжаты
с наружной и внутренней сторон, и их коронки в виде гребней поднимаются
от широких округлых оснований. Первый верхний промежуточный зуб
значительно крупнее следующих за ним. Форма и соотношение размеров
верхних промежуточных зубов напоминают таковые представителей неко-
торых родов белозубых землероек.
Рис. 229. Notiosorex hybsibius.
Распространение и возраст. Северная часть Ганьсу,
Шэньси, Чжили, Сычуань, Юньнань (Китай), северная часть Бирмы,
Бакбо (п-ов Индокитай), плейстоцен—современный; СССР — плейстоцен.
Примечание. Возможно, что представители подрода Chodsigoa,
как и подрода Notiosorex, существовали значительно ранее плейстоцена.
Можно также считать, что представители подтрибы Soriculina могут слу-
жить известным эталоном переходных групп от бурозубых к белозубым
землеройкам.
4. Notiosorex (Chodsigoa) hypsibius de Winton et Styan (рис. 229).
Soriculus hypsibius de Winton, Styan, 1899, Proc Zool. Soc. London, p. 574.
Soriculus (Chodsigoa) beresowskii Кащенко, 1907, Ежегодн. Зоол. муз. АН, т. 10,
с. 2.—Chodsigoa larvarum Т h о m a s, 1911, Abstract Proc. Zool. London, p. 49; — la-
mula Thomas, 1912, Ann. Mag. Nat. Hist., v. 10, p. 399; — hypsibia parva G. Al-
len, 1923, Amer. Mus. Novitates, N 100, p. 5; — parva G. Allen, 1938, p. 107.
Тип. 5 (шкура и череп) № 99.3.1.10, Янглиупа, северно-западная часть Сычуани
(Китай), кол. Брит, муз., Лондон. Кащенко описал Soriculus (Chodsigoa) beresowskii
по спиртовым 6 экз. № 6430—6436, Ходзигоу, Сычуань (Китай), кол. ЗИН АН СССР,
Ленинград.
Диагноз. Длина ступни не более 16 мм. Длина хвоста не превы-
шает 80% длины тела.
Описание. Окраска верхних частей тела черная или черно-бурая.
Нижняя лишь немного светлее верхней. Окружность рта и дистальные
отделы конечностей охристо- или буровато-белые. Нижняя сторона хвоста
светлее верхней или беловатая. Вибриссы длинные — до 26 мм при общей
длине тела 70.0—84.0 мм.
Череп с относительно короткой лицевой частью, плоский и широкий.
Сагиттальный и ламбдовидный гребни хорошо развиты.
Измерения. По: Кащенко, 1907, для 2 экз. — длина: тела —
70.0—74.0, хвоста — 50.0—65.0, ступни — 14.0—15.0, наибольшая черепа
(видимо, кондило-базальная — 20.0 мм. По: G. Allen, 1938 — длина:
тела — 54.0—58.0, хвоста — 43.0—65.0, ступни — 11.0—15.0, основная
черепа — 16.7—18.5 мм.
50. РОД NOTIOSOREX
451
Распространение и возраст. Ганьсу, Шаньси, Сычуань,
Юньнань (Китай), современный.
5. Notiosorex (Chodsigoa) salenskii Kastschenko, 1907 (рис. 230)
Soriculus (Chodsigoa) salenskii Кащенко, 1907, Ежегодн. Зоол. муз. АН, т. 10,
с. 251. Chodsigoa salenskii Thomas, 1911, Proc. Zool. Soc. London, p. 166; — smithi
Thomas, 1911, там же.
Тип. Спиртовой экземпляр, № 6388, Лунь-нгань-фу, Сычуань (Китай), кол.
ЗИН АН СССР, Ленинград.
Рис. 230. Notiosorex salenskii.
Диагноз. Длина ступни значительно более 16 мм, у типичного
экземпляра 20.5 мм. Длина хвоста более длины тела, у типа почти
в 1.5 раза.
Опис ание. Верхняя сторона тела окрашена в черный или черно-
бурый цвет. Окружность рта, дистальные отделы конечностей и нижняя
сторона хвоста буро-белые. Вибриссы очень длинные — 38 мм при длине
тела 81 мм.
Измерения. Тип — длина: тела — 81.0, хвоста — 116.0,
ступни — 20.5, наибольшая черепа — 25 мм. По.: G. Allen, 19.38, размеры
N. (Ch.) smithi Thos. = N. (Ch.) salenskii — длина: тела — 85, хвоста — 68,
ступни — 18, наибольшая черепа — 22.5—23.0 мм.
По-видимому, N. (Ch.) smithi Thos. принадлежит к особому виду.
Распространение и возраст. Шэньси, Сычуань, Юнь-
нань (Китай), северная часть Бирмы, современный.
6. Notiosorex (Chodsigoa) lowei Osgood, 1932.
Chodsigoa lowei Osgood, 1932, Puhi. Field Mus. Nat. Hist. (Zool. ser.), v. 18,
N 10, p. 249—250.
Тип. (J № 32.4.19.4, Чапа, Бакбо (п-ов Индокитай), кол. Брит, муз., Лондон,
Диагноз (по: Osgood, 1932). Размеры средние, хвост длиннее тела.
Череп с высокой и узкой мозговой капсулой — хорошо отличается от ши-
роких и плоских черепов, характерных для рода.
Описание. Окраска тела мышино-серого цвета с более коричне-
вым оттенком на горле и груди. Конец хвоста белый или грязно-белый.
Череп удлиненной формы, стройный, кнаружи от линии сагиттального
гребня слабоокруглый. Сочленовный отросток с редуцированной ямкой,
слегка заметной сверху. Зубы несколько уже, чем у Notiosorex (Ch.)
29*
452
ПОДСЕМ. SORICINAE
hypsibius de Winton et Styan, а зубной ряд несколько длиннее. Моляры
снаружи пигментированы.
Измерения. Длина: тела — 77.0, хвоста — 86.0, ступни — 15.0,
кондило-базальная черепа — 19.2, верхнего зубного ряда — 8.8, ниж-
него — 8.2 мм.
Распространение и возраст. Таиланд, п-ов Индокитай,
современный.
Примечание. Юнгом (Young, 1934) описан из плейстоцена Ки-
тая (Хоукаутиен) новый вид вымершей землеройки Neomys bohlini (Ра-
leontol. sinica, ser. C, v. 8, fasc. 3, p. 16—18). Сравнение описания и ри-
сунков нижней челюсти этого вида с нижней челюстью Notiosorex (Chodsigoa)
hypsibius de Winton et Styan позволяет считать эту землеройку принадле-
жащей к представителям рода Soriculus Blith или Notiosorex Baird.
Но к которому она ближе, без изучения соответствующего материала
(черепа ископаемого вида) решить нельзя.
Весьма возможно, что к одному из названных родов должен быть отне-
сен и вид Neomys sinensis Zdansky, 1929 (Paleontol. sinica, ser. C, v. 5,
fasc. 4, p. 12), с которым Юнг сравнивает свою землеройку. К сожалению,
описания последнего вида у меня в руках не было. К родам Soriculus
Blith и Notiosorex Baird (подрод Chodsigoa Kastsch.) должны быть отнесены
и описанные Бате (Bate, 1945, Ann. Mag. Nat. Hist., ser. 11, v. 11, № 83,
p. 741—753) f Nesiotites hidalgo и N. corsicanus, а также N. similis Hensel,
1925 (Z. Deutsch, geol. Ges., Bd 7, H. 3, S. 458—462). В описании рода f Ne-
siotites Бате отмечает, что этот род объединяет виды с двумя и одним бугор-
ком на верхней стороне 11, а также с тремя и четырьмя верхними проме-
жуточными зубами. Зубы пигментированы.
Это обстоятельство позволяет считать, что описываемый род объеди-
няет виды двух близких, но самостоятельных родов — с двумя и одним
бугорком на 1х и с тремя и четырьмя промежуточными зубами в верхней:
челюсти. Общая характеристика представителей рода Nesiotites Bate пока-
зывает, ’гго они обладают признаками, близкими родам Soriculus Blith
и Notiosorex Baird (подрод Chodsigoa Kastsch.).
В диагнозе N. hidalgo отмечаются три (известных) верхних промежуточ-
ных зуба, а для N. corsicanus — четыре. Вероятно, эти виды должны быть
отнесены к роду Soriculus Blith. Что же касается N. similis Hensel, то,
как указывает Бате, особи этого вида имеют три и четыре верхних про-
межуточных зуба. Следовательно, к этому виду отнесены особи не только
разных видов, но и родов.
51. Род PODIHIK Deraniyagala, 1958
Deraniyagala, 1958, Admin. Rep. Director Nat. Mus. Ceylon, pt 4, Ed.
Sci. Art. (E), p. 5.
Тип. Podihik kura Deraniyagala, 1958, Шри Ланка (о. Цейлон).
Диагноз (по: Walker et al., 1964). Размеры тела мелкие. На хвосте,
как у представителей рода Crocidura Wagl., кроме покрывающих его корот-
ких волос имеются длинные редкие щетинки (рис. 231). Резцы, клыки и
премоляры с красно-коричневыми вершинами, моляры — белые, как и
у других представителей подтрибы Soriculina.
Распространение и возраст. Шри Ланка, современный.
1. Podihik kura Deraniyagala, 1958 (рис. 231).
Deraniyagala, 1958, там же.
Т и и. № 183, р. Каудулю вблизи Мадиригирийя, североцентральная часть Шри
Ланка (о. Цейлон), кол. Нац. муз. Коломбо.
51. РОД PODIHIK
453
Описание (по: Walker et al., 1964).1 Верхняя сторона тела красно-
коричневая. На голове «шапочка» из более темных волос. Морда пур-
пурно-серая. Каждая ноздря выдается как короткая притупления тру-
Рис. 231. Podihik kura (по: Walker et al., 1964).
бочка. Кисти, ступни и уши желтые. Нижние части тела однообразно
голубовато-серые.
При описании автор рода и вида отметил (Walker et al., 1964), что Po-
dihik кита, как он думает, более близко родствен роду Soriculus Blith,
что подтверждается строением черепа, а также и красной окраской вершин
зубов.
Измерения. Длина: тела — 40 и 41, хвоста — 24 и 27, ступни —
6.6 и 8.0, ушей — 5 мм.
Распространение и возраст. Североцентральная часть
о. Шри Ланка, современный.
f4. Подтриба ALLOSORICINA Fejfar, 1966
A llosoricinae Fejfar, 1966, Neues Jahrb. Geol und Palaontol. AbhandL, Bd 123,
N 3, S. 221—248.
Тип. Allosorex Fejfar, 1966, Европа, плиоцен—плейстоцен.
• Диагноз. Зубная формула неизвестна. Судя по строению нижней
челюсти, в верхней челюсти должно быть не более четырех промежуточных
зубов. Венечный отросток в отличие от такового представителей других
подтриб этой же трибы невысокий, с вытянутым спереди назад основанием.
Распространение и возраст. Европа, плиоцен—плей-
стоцен.
Примечание. Подтриба Allisoricina объединяет виды, специали-
зация которых в общем для трибы Blarinini направлении получила свое-
образное выражение. Жевательный аппарат, сохраняя функцию быстрого
1 Авторское описание рода и вида мне неизвестно.
454
ПОДСЕМ. SORICINAE
Рис. 232. Allosorex stenodus (по: Fejfar,
1966).
схватывания добычи сравнительно тонкими и длинными низкими челю-
стями, приобрел способность более основательно пережевывать пищу.
На это указывает сильное развитие основания короткого венечного от-
ростка и массивного сочленовного отростка нижней челюсти с широкими
частями и поверхностями для прикрепления жевательной мускулатуры.
Это позволяет считать, что виды
подтрибы обладали более мощными
височной и внутренней крыловидной
мышцами. Сильное развитие крыло-
видных мышц обусловлено усилением
жевательной функции зубо-челюст-
ного аппарата. Возможно, что основ-
ной пищей этим землеройкам слу-
жили животные с прочными волок-
нистыми тканями. С таким представ-
лением можно связать и особенности
строения нижних коренных зубов.
Их коронки удлиненной формы,
с невысокими буграми и притуплен-
ными сверху гранями боковых сторон
входящих складок.
При всем своеобразии описывае-
мых структур жевательного аппарата
представитзлей этой группы землероек выделение их в особое подсемейство
лишено достаточных оснований. По существу строение их жевательного
аппарата не более как частное выражение общего для всей трибы Blarinini
направления развития. Тем не менее своеобразие строения представителей
рода Allosores Fejf. позволяет выделить его в особую подтрибу.
Таблицу для определения родов этой подтрибы составить не представ-
ляется возможным.
f52. Род ALLOSOREX Fejfar, 1966
F е i f а г, 1966, там же, с. 224—226.
Тип. Allosorex stenodus Fejfar, 1966, Чехословакия, плиоцен—современный.
Диагноз. Нижний внутренний резец без бугорков (лопастей)
на верхней стороне. Все зубы белые.
Описан и е. Нижняя челюсть низкая и длинная, с сильно подня-
тыми вверх основаниями углового и сочленовного отростков. Венечный
отросток невысокий, с вытянутым спереди назад основанием и широкой
поверхностью для прикрепления височного мускула на вершине. Сочле-
новный отросток массивный, с широким основанием и крупной верхней
сочленовной фасеткой. Нижняя сочленовная фасетка небольшая, далеко
выдвинута вперед.
I1 не крупный, с круто изогнутым вниз передним зубцом, на внутренней
стороне которого имеется выступ. Ij относительно длинный, не толстый,
с гладкой верхней поверхностью и круто изогнутым вверх передним кон-
цом. Нижние моляры удлиненной формы, невысокие.
Распространение и возраст. Европа, плиоцен—плей-
стоцен.
f 1. Allosorex stenodus Fejfar, 1966 (рис. 232).
Fejfar, 1966, там же, с. 227—248.
Т и и. Правая ветвь нижней челюсти с внутренним резцом, № 651273, Ивановце,
западная часть Словакии, плиоцен—плейстоцен.
53. РОД DEINSDORFIA
455
Основные особенности отмечены в диагнозе подтрибы и в описании
рода.
Распространение. Известен из одного местонахождения.
Примечание. Диагноз и описание составлены по данным работы
Фейфара (Fejfar, 1966).
f 53. Род DEINSDORFIA Heller, 1963
Heller, 1963, Neues Jahrb. Geol und Palaontol., Bd 118, N 1, S. 1—20.
Тип. Deinsdorfia franconica Heller, 1963, Деинсдорф ( ЬРГ), нижний плейстоцен.
Диагноз. Нижняя челюсть с низкой слабо изогнутой зубной
частью. Венечный отросток с косо срезанной передней стороной. Птери-
гоидная ямка перегорожена костной перемычкой, разделяющей ее на две
Рис. 233. Deinsdorfia franconica (по: Heller, 1963).
части — верхнюю и нижнюю. Сочленовный отросток массивный, крупных
размеров, с высоким основанием, далеко выдается назад. Угловой отросток
с толстым основанием, был, по-видимому, коротким.
Распространение и возраст. Европа, нижний плейстоцен.
Примечание. По общему строению нижней челюсти описывае-
мый род ближе всего к роду Allosorex Feifar. Можно думать, что он объеди-
нял виды, приспособленные к питанию грубой пищей, обработка которой
на зубах требовала значительного жевательного давления с участием
крупного крыловидного мускула, особенно его внутренней части. О сте-
пени развития внутренней части крыловидного мускула можно судить
по значительному развитию поверхности для его прикрепления на сочле-
новном отростке нижпей челюсти у типичного вида.
t 1. Deinsdorfia franconica Heller, 1963 (рис. 233).
Heller, 1963, там же, с. 6, 7.
Тип. Фрагмент правой ветви нижней челюсти без зубов. Деинсдорф (ФРГ),
нижний плейстоцен.
Измерения. Высота: венечного отростка — 3.7, нижней челюсти
в области альвеолы М2 — 1.4 мм.
Примечание. Диагноз рода составлен по описанию Геллера
(Heller, 1963).
5. Подтриба NECTOGAL1NA Gureev, 1971
Тип. Nectogale Milne-Edwards, 1870, Азия, современный.
Диагноз. Зубная формула: 1-|-С-^-Р-^М-|- = 28. Все зубы белые-
Имеются приспособления для плавания, вплоть до перепонок между
пальцами.
Подтриба объединяет три рода с одинаковыми зубными формулами.
Распространение и возраст. Азия, плейстоцен — сов-
ременный.
Примечание. Виды этой подтрибы приспособлены к передвиже-
нию в воде, особенно представители рода Nectogale M.-Edw. Особенности
456
ПОДСЕМ. SORICINAE
строения черепа и жевательного аппарата не оставляют сомнения в том,
что виды, объединяемые этой подтрибой, как и другие виды трибы Blari-
nini, приспосабливались к захвату относительно крупной добычи, но
только в водной среде и на побережьях водоемов. Тесная связь в кормовом
отношении с водными обитателями обеспечивает полуводным землеройкам
относительно стабильные условия жизни. Этим несомненно обусловлена
возможность их существования в сравнительно разнообразных условиях
климата и в разных ландшафтах. Разная степень приспособленности
к плаванию Видов отдельных родов отражает не только разную давность
специализации в этом направлении, но и разное значение его в общем
комплексе адаптаций каждой из этих групп.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СОВРЕМЕННЫХ РОДОВ
1 (2). Ушные раковины развитые. Хвост округлый в сечении на всем про-
тяжении. Перепонок между пальцами нет............................
............................................54. Chimarrogale.
2 (1). Ушные раковины не развитые, есть ушные клапаны. Хвост в кон-
цевой части плоский (сжат с боков). Между пальцами имеются
перепонки .........................................56. Nectogale.
54. Род CHIMARROGALE Anderson, 1877 —
БЕЛОЗУБЫЕ ВОДЯНЫЕ ЗЕМЛЕРОЙКИ
Sorex Temminck, 1842, Fauna Japon, v. 1, Mammal., p. 23, pl. 5, fig. 1. Cros-
sopus Gray, 1842, Ann. Mag, Nat. Hist., v. 10, p. 261. Chimarrogale Anderson,
1877, J. Asiat. Soc. Bengal, v. 46, N 2, p. 262. Crossogale Thomas, 1921, Ann. Mag.
Nat. Hist., v. 7, p. 243.
Тип. Crossopus himalayicus Gray, Кашмир, Пенджаб, Сикким, Гималаи (Индия),
северная часть Бирмы.
Диагноз. Ушные раковины небольшие. Кисти и ступни сверху
покрыты чешуйками и светлыми и темными волосами, а по краям (также
и пальцы) плавательными щетинками. Между пальцами перепонок нет.
На подошвах обычные для землероек мозоли и подушечки. Хвост длинный,
округлый в сечении, с килем из светлых волос на нижней стороне.
Во внешнем виде и в строении конечностей и хвоста представителей
этого рода много общего с видами рода Neomys Капр, но жевательный
аппарат значительно отличается.
Распространение и возраст. Кашмир, Пенджаб, Сик-
ким, Гималаи (Индия), Бирма, Сычуань, Юньнань, Юго-Восточный Ки-
тай, Япония, п-ов Индокитай, Суматра, Калимантан (Индонезия), совре-
менный.
1. Chimarrogale himalayicus Gray, 1842 — Гималайская водяная зем-
леройка.
Crossopus himalayicus Gray, 1842, Ann. Mag. Nat. Hist., v. 10, p. 261. Chimar-
rogale himalayicus Anderson, 1877, J. Asiat. Soc. Bengal, v. 46, N 2, p. 262.
T и п не установлен, вид описан из северо-восточного Пенджаба (Индия).
Диагноз. Окраска хвоста в отличие от других видов резко дву-
цветная, темная сверху, светлая снизу. Длина ступни — 23, длина че-
репа — свыше 26 мм.
Описание. Окраска верхней стороны тела темно-серая или темно-
бурая. По всей спине распространены отдельные редкие белые щетинки,
придающие окраске серебристый налет. Нижняя сторона тела светло-
серая или беловатая. Верхняя сторона кистей и ступней темно-коричневая.
Верхний внутренний резец с выступом на внутренней стороне.
54. РОД CHIMARROGALE
457
Измерения (по: Blanford, 1888—1891). Длина: тела — 4.35 дюйма
(108.75 мм), хвоста — 3.05 дюйма (76.25 мм), ступни — 0.75 дюйма
(18.75 мм), основная черепа мелкой самки — 0.94 дюйма (23.50 мм).
Отдельные особи достигают крупных размеров: длина тела 5—6 дюймов
(125—150 мм). Ни у одного из 2 экз. коллекции ЗИН АН СССР измерения
на этикетках не отмечены.
Рис. 234. Chimarrogale platycephala.
Распространение и возраст. Чамба, северо-восточная
часть Пенджаба, Кашмир, Секким, Дарьеринг (Индия), северная часть
Бирмы, Юньнань (Китай), Лаос и Бакбо (п-ов Индокитай), современный.
Для других видов этого рода Гаррисон (Harrison, 1958, р. 282—290)
приводит следующие краткие отличия.
2. Chimarrogale styani de Winton, 1899. Proc. Zool. Soc. London, p. 574.
Отличается резко двуцветной окраской тела (сверху темная, снизу свет-
лая). Распространен в северо-западной части Сычуани (Китай) и в Се-
верной Бирме.
3. Ch. leander Thos., 1902, Ann. Mag. Nat. Hist., Ser. 7, v. 10, p. 165.
Отличается малой величиной (длина ступни 22 мм и менее, длина черепа
25Амм и менее)., распространен в Юго-Восточном Китае.
30 А. А. Гуреев
458
ПОДСЕМ. SORICINAE
4. Ch. platycephala Temm., 1842 (рис. 234). Отличается плоским чере-
пом и длинными ступнями (свыше 25 мм). Распространен в Японии.
5. Ch. sumatrana Thos., 1898, Ann. Mag. Nat. Hist., ser. 7, v. 2,
p. 246 , 247. Отличается крупными размерами и короткими ступнями
(длина ступней 22 мм и менее, длина черепа более 28 мм), распространен
на Суматре.
6. Ch. phacura Thos., 1898, там же. Отличается меньшими размерами
черепа, распространен на Калимантане.
7. Ch. hantu Harr., 1958, там же, ser. 13, v. 1, р. 282—290. Отличается
широким черепом, распространен на п-ове Малакка.
8. Ch. varennei Thos., 1927, Proc. Zool. Soc. London, p. 45.
Отличается отсутствием выступов на внутренней стороне I1, распростра-
нен на п-ове Индокитай.
Примечание. Самостоятельность большинства видов сомни-
тельна, но вопрос этот можно решить лишь при исследовании соответ-
ствующего материала, которого у меня не было.
f 55. Род NECTOGALINIA Gureev gen. nov.
Тип. Nectogalinia altaica Gureev sp. nov., Алтай, верхний плиоцен—средний
плейстоцен.
Диагноз. Зубная формула: I С-ту Р -г М-^-. Верхняя фасетка
Z v 1 о
сочленовного отростка нижней челюсти соединена гребнем с лингвальным
отростком нижней фасетки. М3 очень
Рис. 235. Nectogaltnia altaica.
мал, его длина составляет половину
длины Мх.
Описание. Приведено для
вида.
Распространение и
возраст. Азия, плиоцен—плей-
стоцен.
+ 1. Nectogalinia altaica Goreev
sp. nov. (рис. 235).
Тип. Правая ветвь нижней че-
люсти с Мх, М2 и альвеолами осталь-
ных зубов, № 664/201, Алтай, Раз-
долье, р. Алей, вблизи г. Рубцовска,
верхний плиоцен—средний плейстоцен,
кол. ГИН АН СССР, Москва.
Описание. Венечный отросток отклонен на внешнюю сторону
под углом в 35—40?. Он не высокий, сужен в верхней части. Темпоральная
ямка треугольной формы с закругленными углами. Мандибулярное отвер-
стие крупное. Верхняя фасетка сочленовного отростка соединена гребнем
с лингвальным отростком нижней фасетки. Подбородочное отверстие
под задним корнем Мг.
Зубы несут следы пигмента. Цингулюм развит вокруг всей коронки
зуба. Мя, судя по альвеолам, короче Мх в два раза.
Измерения. Высота: нижней челюсти от вершины венечного
отростка до нижнего края челюсти под ним — 6.3, ветви между Мг и М2 —
2.3; длина Мх и М2 — 4.0 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
56. РОД NECTOGALE
459
56. Род NECTOGALE Milne-Edwafds, 1870
ВЫХУХОЛЕОБРАЗНЫЕ ЗЕМЛЕРОЙКИ
Milne-Edwards, 1870, Compt. rend. Acad, sci., t. 70, p. 341.
Тип. Nectogale elegans M.-Edw., 1870, Моупин, Сычуань (Китай).
Диагноз (по: Blanford, 1888—1891). Ушных раковин нет, имеются
ушные клапаны. Ступни длинные, широкие, покрыты сверху чешуйками
и белыми волосами, а по краям (также и пальцы) плавательными щетин-
ками. Между пальцами имеются кожные перепонки. На подошвах круп-
Рис. 236. Nectogale elegans (по: Milne-Edwards, 1868—1874).
ные подушки — мозоли. Хвост длинный, с белыми щетинками по краям
и на нижней поверхности, а в концевой части на верхней стороне. В кон-
цевой трети хвост сжат с боков. Все особенности строения ступней и хвоста
специфичны для этого рода.
К роду относится одни вид, описываемый ниже.
Распространение и возраст. Сикким, Вутап (Индия),
Северная Бирма, Тибет, Сычуань, Юньнань п Шэньси (Китай), современный.
Примечание. Своеобразные приспособительные особенности, ха-
рактерные для этого рода (строение ступней и хвоста, отсутствие ушных
ЗЭ*
460
ПОДСЕМ. SORICINAE
раковин и наличие клапана, замыкающего слуховой проход и др.)
сравнимы лишь со сходными структурами выхухоли.
1. Nectogale elegans Milne-Edwards, 1870 — Тибетская водяная земле-
ройка (рис. 236).
Milne-Edwards, 1870, там же.
Т и п не установлен, вид описан из Моупина, Сычуань (Китай).
Описание (по: Blanford, 1888—1891). Глаза очень мелкие. Рыльце
покрыто мехом, сходным с таковым на теле. Ступни большие и широкие,
вдвое длиннее кистей, голые сверху. Только когтевые фаланги свободны
от перепонки и бахромы из белых волос, сходных с таковыми на кистях.
Бахрому образуют разной длины грубые, плоские, белые волосы. Волосы
на кистях более короткие и не образуют бахромы на пальцах. Мозольные
диски на нижней стороне кистей сходны с этими образованиями на нижней
стороне ступней.
Шерсть на теле очень густая, мягкая и длинная. На спине и боках,
а особенно на огузке отдельные полосы достигают значительной длины.
Окраска тела от пепельно-серой до темно-коричневой. Отдельные длин-
ные волосы блестящие, белые. Окраска губ и подбородка, а также горла
и груди беловатая или белая, переходящая на бргошке и боках тела в се-
ребристую или светло-коричневую. Хвост сверху беловато-коричневый,
снизу— белый.
Измерения. Длина: тела — 90.0—128.0, хвоста — 89.0—110.0,
ступней — 25 мм (1 дюйм), черепа — 25 мм (1 дюйм).
Распространение й возраст. В границах рода, совре-
менный.
Биология. Замечание об образе жизни, сделанное Бленфордом
(Blanford, 1888—1891) на основании морфологических особенностей, сво-
дится к следующему: внешний облик доказывает ее приспособленность
к водной среде. Крупные дискообразные мозоли на нижней стороне кистей
и ступней, если верить наблюдениям Мильне-Эдвардса (Milne-Edwards,
1870), используются, возможно, землеройкой как присоски, чтобы удер-
жаться на гладкой поверхности камней или валунов в ложе ручья. Кроме
этих замечаний, ничего не известно об образе жизни этого интересного
вида.
6. Триба ANOUROSORICINI Stirton, 1930
Amblycoptinae Kormos, 1926, Ann. H.-N. Mus. Nation. Hung., v. 24, p. 352—
391. A nourosorex-group S t i r t о n, 1930, p. 225. A mblycoptini Kretzoi, 1965, p. 125.
Тип. A nourosorex Milne-Edwards, 1870, Азия, миоцен—современный.
Диагноз. Зубная формула: 1-|-С-1-р4-М|—1 = 28—24. В верхней
челюсти два или три промежуточных зуба. Все зубы белые. Надглазничные
отверстия имеются. Нижняя сочленовная фасетка сочленовного отростка
нижней челюсти и засочленовный отросток височной кости, на который
она опирается при сочленении челюсти с черепом, выдвинуты вперед.
У современных представителей ушных раковин нет.
Распространение и возраст. Европа, плиоцен; Азия,
миоцен—сов ременный.
Представители трибы в их современном состоянии, вероятно и в прош-
лом, приспособлены к роющей деятельности и жизни под землей. Они
не достигли уровня приспособления к рытью, свойственного специализи-
рованным кротам, но все же имеют особенности, характерные для типич-
ных землероев и подземных обитателей (редуцированные ушные раковины,
короткий хвост, бархатистый волосяной покров и др-)- Приспособления
57. РОД ANOUROSORICODON
461
этих землероек следует рассматривать как параллельные таковым кротов,
обусловленные известной биологической выгодой относительно стабиль-
ных условий в кормовом и других отношениях в почвенном слое.
Строение зубных систем этих землероек позволяет предполагать, что
они служат для перетирания волокнистых тканей.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (2). Ij с двумя бугорками на верхней стороне и продольной бороздкой
на внутренней стороне........................57. Anourosoricodon.
2 (1). 1х без бугорков на верхней стороне и без продольной бороздки на
внутренней.
3 (4). I1 без заднего (второго) зубца.........59. Paranourosorex.
4 (3). I1 с двумя зубцами.
5 (6). В верхней челюсти два промежуточных зуба. В верхней и нижней
челюстях по три моляра..........................60. Anourosorex.
6 (5). В верхней челюсти три промежуточных зуба. В верхней и нижней
челюстях по два моляра..........................58. Amblycoptus.
f 57. Род ANOUROSORICODON Topatshevski, 1965
Anourosorex Топачевский, 1962, Геол, журн., т. 22, вып. 1, с. 108. Anouro-
soricodon Топачевский, 1965, Насекомоядные и грызуны Ногайской позднеплио-
ценовой фауны, с. 138 (nomen nudum); 1966, Республ. м!жвщомчий зб!рник, сер.
«Проблемы зоологии», с. 90—96.
Тип. Anourosoricodon pidoplitschkoi, Topatshevski, 1965, Украина (СССР), сред-
ний плиоцен.
Диагноз. Полная зубная формула неизвестна. Нижний зубной
ряд составлен следующими зубами: I—С—Р—М—, как у современных и
Рис. 237. Anourosoricodon pidoplitschkoi (по: Топачевский, 1966).
ископаемых представителей рода Anourosorex M.-Edw. В отличие от послед-
него и рода Amblycoptus Копп, у Anourosoricodon Д с двумя бугорками
и продольной бороздкой на его внутренней стороне.
f 1. Anourosoricodon pidoplitschkoi Topatshevski, 1965 (рис. 237).
Топачевский, 1965, 1966 (там же, где и род).
Тип. Правая ветвь нижней челюсти без восходящей ветви с зубами I, и Mt,
№ 42—1, левый берег Каховского водохранилища у с. Каменского Запорожской обл.,
Украина (СССР), средний плиоцен, кол. Инет. зоол. АН УССР, Киев.
Описание (по: Топачевский, 1966, с сокращениями). Крупная
белозубая землеройка, уступающая в размерах Anourosorex squamipes
M.-Edw. Горизонтальная ветвь нижней челюсти низкая, ее высота у заднего
края Мх менее длины этого зуба. Подбородочное отверстие на уровне
середины Mv Мандибулярное отверстие у переднего края восходящей
ветви. Мх очень велик, вытянут в передне-заднем направлении. Кониды
более высокие, чем у представителей рода Anourosorex M.-Wdw. М3 очень
мал.
462
ПОДСЕМ. SORICINAE
Измерения. Длина: 1Х—Мх (по коронкам) — 7.5, 1Х (по внешней
стороне) — 5.1, Мх — 2.6, 12—Р4 (альвеолярная) ~ 1.8; высота: Мх
на уровне протоконида — 1.8, горизонтальной (зубной) ветви нижней че-
люсти 2.0 мм.
Распространение и возраст. Украина (СССР), средний
плиоцен.
Рис. 238. Amblycoptus oligodon (по: Kormos, 1926).
f 58. Род AMBLYCOPTUS Kormos, 1926
Kormos, 1926 Ann. H.-N. Mus. Nation. Hung., v. 24, p. 370—391.
Тип. Amblycoptus oligodon Kormos, 1926 (Polgradi, Komitat Fejer, Ungarn.),
Польгарди, Венгрия, нижний плиоцен.
Диагноз. Зубная формула: 1|-С£-Р}-М^- = 24. В верхней челюсти
три промежуточных зуба. В верхней и нижней челюсти по два моляра.
Распространение и возраст. Европа, нижний плиоцен.
f 1. Amblycoptus oligodon Kormos, 1926 (рис. 238).
Kormos, 1926, там же.
58. РОД AMBLYCOTUS. 59. РОД PARANOUROSOREX
463
Т и п не отмечен, вид описан по фрагменту левой половины верхней челюсти
с пятью зубами и левой ветви нижней челюсти с пятью зубами (Polgardi, Komitat
Fejer, Ungarn.), Польгарди, Венгрия.
Диагноз. В отличие от A. topali Janossy у этого вида венечный
отросток ниже и менее широкий; Мх менее длинный (2.7—3.0 мм) I, ко-
роче и толще, его основание расположено под Р4.
Описание (по: Kormos, 1926). Череп, как и нижняя челюсть,
массивный. Зубы крупные, с притупленными бугорками. Нижняя челюсть
с сильно утолщенным и широким (спереди назад) венечным отростком,
на котором хорошо заметен бугорок в его верхней части, направленный
на внешнюю сторону и назад. Ямка для прикрепления порции височного
мускула, расположенная на внутренней стороне венечного отростка
(птеригоидная ямка), очень мала и смещена кпереди. Угловой отросток
короткий и широкий, уплощен в дорсо-вентральном направлении. Сочле-
новный отросток массивный, с разобщенными верхней и нижней фасетками.
Г2 мельче I3. I3 очень крупный, с вершиной, отогнутой вперед. Клык самый
мелкий из верхних промежуточных зубов. Коронки верхних и нижних
моляров и Р4 с широкими жевательными поверхностями и крупными,
с округлой поверхностью буграми. I1 массивные, широкие, с толстыми
корнями.
Распространение и возраст. Польгарди, Венгрия, ниж-
ний плиоцен.
t 2. Amblyeoptus topali Janossy, 1972.
J a n о s s y, 1972, там же, т. 64, с. 27—52.
Т и п. Фрагмент правой ветви нижней челюсти с зубами 1Г, 12, Р4, Мх и альвеола
М2, № V.71.49, Osztramos=l (северо-восточная часть Венгрии, 50 км севернее Miskolc),
средний плиоцен, кол. Национ. муз. естеств. истор. Будапешт.
Диагноз. В отличие от A. oligodon Kormos у этого вида венечный
отросток выше и шире; Мх более длинный (3.1—3.4 мм); 1Х более длинный,
его основание под Мх.
Описание (по: Janossy, 1972). Череп близок по размерам к тако-
вому Blarina brevicauda Say, но более высокий и широкий. Прямой в перед-
ней части сагиттальный гребень сходен с таковым других землероек. Пе-
редний и задний сегменты Мх удлиненные.
Измерения. Длина ветви нижней челюсти от основания It
до заднего края сочленовного отростка — 11.4—11.8, высота: ветви под
Мх — 2.5-—2.9, венечного отростка — 6.6, 6.8, 7.2; длина М, — 3.1-—
3.4 мм.
Этот же вид обнаружен Ржебик-Ковальской (Rzebik-Kowalska, 1975)
в пещере Замкова Дольна в Ольштыне, окрестности Ченстохова, Польша.
Я
f 59. Род PARA NO URO SOREX Rzebik-Kowalska, 1975
Rzebik-Kowalska, 1975, Acta Zool. cracoviensia, t. 20, N 6, p. 169—178.
T и n. Paranourosorex gigas Rzebik-Kowalska, 1975, Подлесицы, Польша, средний
плиоцен.
।_4__3
Диагноз (по: Rzebik-Kowalska, 1975). Зубная формула = 28
(3113 \
по моим представлениям I уС-- Ру Му = 28). I1 крупный, без заднего
(второго) зубца. Р4 крупный, моляризованный, трапециевидной формы,
с сильно вытянутым назад отростком метаконида. М1 с жевательной поверх-
ностью квадратной формы и выступающими на внешнюю сторону отрост-
ками пара- и метаконов. М2 много меньше М1, с двумя очень мелкими кор-
нями. М3 мелкий, треугольной формы. 1х удлиненный, без бугорков на ре-
464
ПОДСЕМ. SORICINAE
жущем крае. Мх крупный и массивный. М2 сходен по форме с Мь но мень-
ших размеров, а М3 очень мелкий, сохраняющий передний и задний сег-
менты. Зубы пигментированы в разной степени или не окрашены.
Распространение и возраст. Европа, средний плиоцен.
Рис. 239. Paraanourosorex gigas (по: Rzebik-Kowalska, 1975).
1. Paranourosorex gigas Rzebik-Kowalska, 1975 (рис. 239 , 240).
Rzebik-Kowalska, 1975, там же.
Тип. Левая ветвь нижней челюсти с полным рядом зубов № 1345(1), Подлесицы,
Польша, средний плиоцен.
Рис. 240. Paraanourosorex gigas (по: Rzebik-Kowalski, 1975).
Описание. Кроме признаков, вынесенных в диагноз, нужно отме-
тить и следующие. Крупное подглазничное отверстие расположено над пе-
редним корнем Р4. Маленькое слезное отверстие находится над передним
корнем М1. Нижняя челюсть массивная. Подбородочное отверстие между
корнями М1. Горизонтальная (зубная) и восходящая ветви нижней че-
люсти образуют угол, близкий к тупому. Широкая верхняя поверхность
венечного отростка выступает на его внешнюю сторону в виде гребня или
60. РОД ANOUROSOREX
465
карниза. Отверстие птеригоидной ямки очень маленькое, округлой формы
или с близким к треугольному верхним краем. Сочленовный отросток
как у других представителей трибы A nouro soricini. Цингулюм на зубах
развит в разной степени. Окраска зубов кирпично-красная, коричневато-
желтая или зубы не пигментированы.
Измерения (по: Rzibik-Kowalska, 1975). Длина: нижней челюсти
с Ц — 18.80, 19.10, нижней челюсти без Ц — 15.50, 15.70, 15.90, зубного
ряда, включая Ц, — 13.00, 13.10, зубного ряда без Ц — 9.70, 9.80; АД—
М3 — 6.46; высота: восходящей ветви — 8.00, 7.90, 7.80, горизонтальной
ветви на уровне Мх — 3.04, 3.06, 3.09, 3.11, 3.13; наибольшая длина:
Р4 — 3.08, 3.10, М1 — 2.95, 3.10, 3.16, 3.21; наибольшая ширина: Р4 —
3.06, 3.16, М1 — 2.93, 2.96, 3.00, 3.09; длина: М, — 3.49, 3.59, 3.63, 3.69,
М2 — 2.15, 2.25, 2.28, 2.30, М3 — 1.08, 1.10; ширина: Р4 — 1.60, 1.62,
1.68, 1.73, 1.76; Мг — 1.82, 1.86, 1.88, 1.90, 1.91; М2 — 1.32, 1.33, 1.34,
1.38; М3 — 0.75, 0.76 мм.
Распространение и возраст. Известен из одного место-
нахождения, указанного для типа.
60. Род ANOUROSOREX Milne-Edwards, 1870
Milne-Edwards, 1870, Compt. rend. Acad, sci., t. 70, p. 341; 1872, Rech.
Hist. Nat. Mammal., p. 264. Pygmura Anderson, 1873, Proc. Zool. Soc. London,
p. 229.
T и n. A nourosorex squamipes M.-Edwards, 1872, Шэньси к югу от Хупеха, Сычуань,
Юньнань, Тайвань (Китай), Ассам (Индия), западная и северная части Бирмы, Бакбо
(п-ов Индокитай), СССР.
Диагноз. Зубная формула: I-^-С-^Р-рМ-^-=26. В верхней челюсти
два промежуточных зуба. В верхней и нижней челюстях по три моляра.
Отличия рода Anourosorex от других этой же трибы отчетливо высту-
пают при сравнении диагнозов. Род объединяет один современный и два
вымерших вида.
Распространение и возраст. Япония, Китай, Индия,
Бирма, п-ов Индокитай в границах, указанных для типа рода, миоцен—
современный.
f 1. Anourosorex inexpectatus Schlosser, 1924 (рис. 241).
Neomys (Crossopus) inexpectatus Schlosser, 1924, Palaeontol. Sinica, ser. C,
v. 1, fasc. 1, p. 6, 7.
T и п не установлен, вид описан по фрагменту правой ветви нижней челюсти
из Ertemte, Монголия, верхний миоцен.
Ни диагноза, ни описания Шлоссер не привел, а только указал некото-
рые размеры: высота нижней челюсти между ее нижним краем и венечным
отростком — 7.5, под М2 — 2.5; расстояние между угловым отростком и
вершиной резца 1.5 мм (очевидно, сантиметра).
Распространение и возраст. Известен из одного место-
нахождения, верхний миоцен.
t 2. Anourosorex conf, inexpectatus Schlosser.
Фрагмент правой ветви нижней челюсти с М,, № 948 (1), Краснокутск, Омская обл.,
нижний плиоцен, сб. В. С. Зажигина, кол. ГИН АН СССР, Москва.
Кость серо-коричневого цвета с хорошо сохранившимися сочленовным
отростком, угловым отделом и зубной частью до переднего края Мх (ко-
торый сохранился) и альвеолами М2 и М3. М3 в отличие от Anourosorex
squamipes M.-Edw. и A. japonicus Shik. et Haseg. имеет два корня,
466
ПОДСЕМ. SORICINAE
как у А. inexpectatus Schlosser, описанного из верхнего миоцена Монголии.
При общем сходстве описываемой челюсти с таковой представителей рода
Anourosorex она ближе всего к монгольскому виду.
t 3. Anourosorex japonicus Shikama et Hasegawa, 1958.
Рис. 241. Anourosorex inexp-
ctatus (по: Schlosser, 1924).
Shikama, Hasegawa, 1958, Sci. Repts lokohama Nat. Univ., sect. 2, N 7,
p. 106—112.
Тип. Фрагмент лицевой части черепа с зубами I1—М2 (I2 на левой стороне от-
сутствует), N. An. S. 5, Инаса-чун, префектура Судзуока, Япония, средний плейстоцен,
кол. Геол. инет. Иокогам. нац. унив.
Диагноз (по: Shikama, Hasegawa, 1958). В верхней челюсти два
промежуточных зуба, из которых первый в два раза крупнее второго. М1
и Мг более развиты, чем другие зубы. М3 и М3
очень мелкие. Верхнечелюстная кость вздута
(выпуклая) в области Р4 и М1. Только одно
передненёбное отверстие.
Описание (по: Shikama, Hasegawa,
1958, с сокращениями). Лицевая часть черепа
массивная, удлиненная (вернее, суженная).
Мозговая часть широкая в области надгла-
зничных отверстий. Подглазничное отвер-
стие почти треугольной формы, крупное.
Слезное отверстие треугольное, расположено
над вершиной переднего корня М1. Задние
нёбные отверстия отсутствуют. I1 крупный
с невысоким и широким задним зубцом; Р4 сильно моляризирован; М1
сильно развит в поперечном направлении; М2 по длине и ширине состав-
ляет 4/3 М1; М3, судя по величине альвеолы, очень мал. Все моляры без
глубоких выемок на заднем крае.
Угловой отросток нижней челюсти не сильно развит, отогнут на вну-
треннюю сторону. Сочленовный отросток сильно развит, разделен на две
части.
Нижняя часть с крупной четырехугольной сочленовной фасеткой,
верхняя с небольшой треугольной. Венечный отросток выше, чем у совре-
менного вида Anourosorex squamipes M.-Edw. Подбородочное отверстие
расположено под протоконидом Mv 12 крупный, массивный. Мх самый
крупный, М2 наполовину меньше Мх, а М3 очень мал.
Измерения. Длина: костного нёба — 11.5, I1—М2 — 12.5 мм.
Распространение и возраст. Япония, средний плей-
стоцен.
Примечание. Признаки, отмеченные для японского вида, в зна-
чительной степени совпадают с таковыми Anourosorex squamipes M.-Edw.,
а отмеченные в диагнозе — полностью. По-видимому, это лишь подвид
современного вида.
4. Anourosores squamipes Milne-Edwards, 1872 (рис. 242).
Milne-Edwards, 1872, там же, где и род.
Т и п не установлен, вид описан из Моупина, Сычуань (Китай).
Диагноз. Веки почти сросшиеся, но заметные. Ушные раковины
не развиты, на их месте кожный валик. Хвост очень короткий, покрыт
чешуйками и редкими грубыми волосами, кажется голым.
Описание. Окраска оливково-серая, сходная на всем теле. Голова
крупная, носовой отдел относительно короткий. Ноздри расположены
60. РОД ANOUROSOREX
467
на боковой стороне мочки носа. Щетинистые волосы на конце морды корот-
кие, располагаются лишь на ее передней части. Вибриссы относительно
тонкие и длинные.
На месте ушной раковины кожа зверька покрыта короткими редкими во-
Л0С9МИ И кажется ГОЛОЙ. По нижнему краю слухового прохода узкая скла-
дка кожи, выполняющая роль клапана, предохраняющего слуховой про-
ход от загрязнения. Когти на пальцах кистей развиты более, чем у
обычных землероек, они острые и серповидные, вполне пригодные для
рытья в мягком грунте.
Рис. 242. Anourosorex squamipes (по: Milne-Edwards, 1868—1874).
Череп массивный, угловатый, с короткой лицевой частью. Зубы массив-
ные. В отличие от других групп землероек у представителей рода Anouro-
sorex M.-Edw. (по крайней мере в современном состоянии) М2 и М3 сильно
редуцированы, а Р4 и М1 достигают очень крупных размеров. Внешние
бугорки (пара- и метакон) верхних коренных зубов сильно выступают
в стороны
Мощная нижняя челюсть с короткой зубной частью и широким
(в передне-заднем направлении) венечным отростком. Угловой отросток
короткий и тонкий. Верхняя и нижняя сочленовные фасетки сочленов-
468 ПОДСЕМ. SORICINAE
ного отростка нижней челюсти разделены широкой и глубокой вырезкой.
Птеригоидная ямка мала.
Измерения. По 2 экз. кол. ЗИН АН СССР — длина: тела —
104 и 111 , хвоста — 11 и 15, ступни — 16 и 17 мм. По 3 экз. кол. ЗИН
АН СССР — длина кондило-базальная черепа — 23.3—25.7 мм.
Распространение и возраст. Китай (от Шэньси к югу
до Хупеха, Сычуани и Юньнани, Тайвань), Бирма (северная и западная
части), Бакбо (п-ов Индокитай), Бленфорд (Blanford, 1888—1891) указал
и Ассам (Индия), современный.
Биология. Образ жизни изучен мало. Живет под землей. Пи-
тается насекомыми, их личинками и куколками, а также червями; можно
думать, что и другими мелкими животными из беспозвоночных. Подни-
мается в горы до 1500—3100 м.
ЛИТЕРАТУРА
Абеленцсв В. У., Пидопличко У. Г., Попов Б. М. Ссавць — Фауна Украйги, т. 1,
вып. 1. Киев, 1956, с. 171—803.
Александрова И. В., Фролова В. А. Запись суточной ритмики крота с использованием
магнитофона — МАГ-8М. — Сб. Науч.-тех. информ. ВНИИЖП, 1967, вып. 21,
с. 19—23.
Асписов Д. И. Выхухоль. М., 1952. 45 с.
Барабаш-Никифоров И. Н. Состояние популяции русской выхухоли и неотложные
меры к ее сохранению. — В кн.: Редкие виды млекопитающих фауны СССР и
их охрана. 1973, с. 37—38.
Барабаш-Никифоров И. Н., Шапошников JE В. Пути сохранения Desmana mos-
chata L. — Первый Международный териологический конгресс. Реф. докл.,
т. 1. М., 1974, с. 41—42.
Бибиков Д. И. Биологические наблюдения над ушастыми ежами — лептоспироноси-
телями. — Зоол. журн., 1956, т. 35, вып. 7, с. 1059—1063.
Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Определитель млекопитающих
СССР. М., 1944, 440 с.
Борисов Б. А. Стратиграфия верхнего мела и палеоген—неогена Зайсанской котло-
вины. — Мат. по геологии и полезным ископаемым, Нов. сер., 1963, т. 94,
с. 11—75.
Бородулина Т. Л. О латентном периоде в развитии эмбриона алтайского крота. —
ДАН СССР, Нов. сер., 1951, т. 80, № 4, с. 689—692.
Бородулина Т. Л. К биологии алтайского крота. — Тр. Ин-та морфол. животных,
1953, вып. 9, с. 256—279.
Брандт Э. К. О зубной формуле землеройки (Sorex Cuv.). СПб., 1878, с. 1—12.
Виноградов Б. С. О строении наружных гениталий у землероек-белозубок (род Croci-
dura, Insectivora, Mammalia) как диагностическом признаке. — Зоол. журн.,
1958, т. 37, вып. 8, с. 1236—1243.
Воронов А. Г. О землеройках Наурзума (Кустапайск. обл.). — Изв. Ест.-науч. ин-та
при Молотовском ун-те, 1952, т. 13, вып. 4—5, с. 433—445.
Воронцов Н. Н. Значение изучения хромозомных наборов для систематики млекопи-
тающих. — Бюл. МОИП, отд. биол., 1958, т. 58, вып. 2, с. 5—36.
Галанцев В. П., Русаков О. С. Некоторые черты кровеносной системы крота в связи
с экологическими особенностями вида. —• Сб. науч.-тех. информ. ВНИИЖП,
1967, вып. 17, с. 27—31.
Григорьев П. П. Размножение крота в Белоруссии. — Первое Всесоюзное совещание
по млекопитающим. Тез. докл., ч. 2, М., 1961, с. 22—24.
Громова В. И. Отряд Carnivora — Хищные. Подотряд Creodonta — Примитивные
хищники. — В кн.: Основы палеонтологии. Млекопитающие. М., 1962, с. 184—
193.
Гуреев A. A. Insectivora — Насекомоядные. — В кн.: Млекопитающие фауны СССР,
ч. 1. М,— Л., 1963, с. 54—122.
Гуреев А. А. Землеройки (Soricidae) фауны мира. Л., 1971. 254 с.
Даль С. К. Насекомоядные и рукокрылые Армянской ССР и Нахичеванской АССР
(данные по распространению, систематике и биологии). — В кн.: Зоологиче-
ский сборник, вып. 2. Ереван, 1940, с. 5—26.
Дзуев Р. И., Иванов В. Г., Темботов А. К. Новые данные по карпологии кротов
Кавказа. — Зоол. журн., 1972, т. 51, вып. 12, с. 1898, 1899.
Долгов В. А. Об изменчивости некоторых костей посткраниального скелета землероек
(Mammalia Soricidae). — Acta theriol., 1961, v. 5, № 15, p. 203—227.
Долгов В. А. Равнозубая бурозубка — Sorex centralis Thomas, 1911 (Mammalia,
Soricidae) в фауне Советского Союза. — Зоол. журн., 1964, т. 43, вып. 6, с. 898—
903.
Долгов В. А., Киселев В. Е. Изменчивость крота (Talpa europaea L., 1758) в Башки-
рии. — Науч. докл. высшей школы, Биол. науки, 1971, № 1, с. 29—37.
470
ЛИТЕРАТУРА
Долгов В. А., Юдин Б. С. Состояние и задачи исследований насекомоядных млеко-
питающих (Mammalia Insectivora) СССР. — В кн.: Фауна Сибири. Тр. Биол.
ин-та, 1975, вып. 23, с. 5—40. На с. 18—40 дана библ. 535 назв.].
Долгов В. А., Гофман Р. С. Тибетская бурозубка — S. thibetanus Kastschenko, 1905
(Soricidae Mammalia). — Зоол. жури., 1977, т. 56, вып. 11, с. 1687—1692.
Заидова С. A. Hemiechinus azerbajdzanicus sp. nov. — Изв. АН АзССР, geol. geogr.,
1961а, № 2, с. 51—56.
Заидова С. А. Остеологические отличия Обыкновенного ежа (Erinaceus europaeus L.)
от ушастого ежа (Hemiechinus brachyotis Sat.). — Изв. АН АзССР, сер. биол.,
196'16, № 12, с. 39—46.
Заблоцкая Л. В. Растаскивание семян хвойных и липы землеройками-бурозубками. —
Тр. Приокско-Террасного заповеди., 1957, вып. 1, с. 242—247.
Иванова Е. И. Строение дыхательных путей и их адаптивные особенности у русской
выхухоли. — Тр. Хоперского заповеди., 1961, вып. 5, с. 173—190.
Исаков С. И., Юдин Б. С. Систематика и географическое распространение ушастого
ежа (Hemiechinus auritus Gmelin, 1770) в Таджикистане. — Изв. АН Тадж ССР,
отд. биол. наук, 1971, № 2, с. 61—67.
Кащенко Н. Ф. Обзор млекопитающих Западной Сибири и Туркестана, вып. 1. Томск,
1905. 102 с.
Киселев В. Е. К вопросу о характеристике изменчивости морфологических признаков
обыкновенного крота на южной границе ареала. — В кн.: XXIII Герценовские
чтения. Естествознание. Л., 1970, с. 199—201.
Колоденко А. И. К распространению и экологии белозубки-малютки (Suncus etruscus
Savi) в Туркмении. — Изв. АН ТуркмССР, сер. биол., 1971, № 3, с. 60—64.
Козловский А. Н., Орлов В. Н., Пайко Н. С. Систематическое положение кавказского
(Talpa caucasica Sat.) и обыкновенного (Talpa europaea L.) кротов. — Зоол.
журн., 1972, т. 51, вып. 2, с. 312—316.
Кудряшова Н. И. К биологии обыкновенного и ушастого ежей Ставропольского
края. — Тр. науч.-иссл. ин-та Кавказа и Закавказья, 1956, вып. 1, с. 441 —
454.
Ланкин П. М. О распространении и биологии длинноиглого ежа (Paraechinus hypo-
melas Brandt). — Зоол. журн., 1967, т. 46, вып. 9, с. 1414—1416.
Ланкин П. М., Такмовцева С. И., Никитенко Г. И. Новые материалы о распростра-
нении длинноиглого ежа (Paraechinus hypomelas Brandt) в восточном При-
каспии. — Зоол. журн., 1970, т. 49, вып. 10, с. 1586—1587.
Лапинь А. М. Биология обыкновенной бурозубки (Sorex araneus L.) в лесах Лат-
вийской ССР. — Тр. Ин-та биол. АН ЛатвССР, 1961, т. 20, с. 243—248.
Лаптев М. К. Определитель млекопитающих Средней Азии (насекомоядные, руко-
крылые, хищные, копытные), вып. 1. Ташкент, 1929. 107 с.
Лаптев М. К. Определитель позвоночных животных Туркменской ССР. Млекопитаю-
щие (Mammalia), вып. 5. Ашхабад—Баку, 1936. 99 с.
Летов Г. С., Летова Г. И. Материалы к фауне насекомоядных Тувы и их эктопара-
зитов. — Докл. Иркутск, противочум. ин-та, 1962, вып. 4, с. 164—166.
Межжерин В. А. К вопросу о питании обыкновенной и малой бурозубок (Sorex ara-
neus L. и Sorex minutus L.). — Зоол. журн., 1958, т. 37, вып. 6, с. 948—953.
Назаров А. А. Бурозубки и хищники леса. — Природа, 1963, № 11, с. 111.
Никифоров Л. П. Новое местонахождение крошечной бурозубки (Sorex minutissimus.
Zimm.). — Бюл. МОИП, отд. биол., 1955, т. 60, вып. 4, с. 93.
Никольская В. Н. Сравнительно-морфологический обзор жевательной мускулатуры
насекомоядных (Mammalia, Insectivora) фауны СССР. — Зоол. журн., 1965,
т. 44, вып. 8, с. 1228—1237.
Огнев С. И. Материалы к познанию фауны Кавказа. — Уч. зап. Северо-Кавк. ин-та
краеведения, 1926, т. 1, с. 45.
Огнев С. И. Звери восточной Европы и северной Азии, т. 1. М.—Л., 1928. 631 с.
Огнев С. И. Материалы по систематике, морфологии и географическому распростра-
нению землероек. — Зоол. журн., 1933, т. 12, вып. 4, с. 8- 16.
Огнев С. И. Об ежах (Erinaceidae) Дальнего Бостока. — Бюлл. МОИП, отд. биол.,
1951, т. 56, вып. 1, с. 8—14.
Орлов В. Н. Кариоспстематика млекопитающих. М., 1974. 207 с.
Охотина М. В. Дальневосточный крот и его промысел. М., 1966. 136 с.
Павлинин В. Н. Материалы по морфо-физиологической характеристике уральского
обыкновенного крота. — Тр. МОИП, отд. биол., 1967, т. 25, с. 56—66.
Петров Б. М. О зимнем питании ушастой совы и о находке белозубки-малютки (Suncus
etruscus Savi) в предгорьях западного Тянь-Шаня. — Зоол. журн., 1965, т. 44,
вып. 10, с. 1579—1581.
Пидопличко И. Т. Новые данные о фауне позвоночных антропогеновых отложений
Тернопольской области. — ДАН СССР, 1955, т. 100, № 5, с. 989—991.
Подковыркин Б. А, Наземные млекопитающие острова Шумшу (Курилы). — Бюл.
МОИП, отд. биол., 1956, т. 63, вып. 4, с. 125—127.
ЛИТЕРАТУРА
471
Попов В. А., Воронов Н. Л., Кулаева Т. М. Очерки по экологии землероек Раифского
леса ТатАССР. — Изв. Казанск. фил. АН СССР, сер. биол. и с.-х. наук, 1950,
№ 2, с. 173—208.
Русаков О. С. Биологическое обоснование и техника промысла крота. Л., 1965. 52 с.
Снигиревекая Е. М. Материалы но биологии размножения и колебания численности
землероек в Башкирском заповеднике. — Тр. Башкирск. заповеди., 1947, вып. 1,
с. 49—68.
Соколов В. Е. Систематика млекопитающих. М., 1973. 430 с.
Стальмакова В. А. Млекопитающие Репетекского песчанопустынного заповедника и
прилегающих районов Каракумской пустыни. — Тр. Репетекской песчано-
пустынной станции. Ашхабад, 1955, т. 3, с. 313—315.
Стогов И. И., Бондарь Е. П. Обзор землероек-белозубок южной Туркмении и Тад-
жикистана. — Зоол. жури., 1966, т. 45, вып. 3, с. 414—420.
Стогов И. И., Савинов П. Ф. Новые виды Soricidae из захоронения гиппарионовой
фауны. — Вести. АН КазССР, 1965, № 11 (248), с. 91—94.
Строганов С. У. Насекомоядные млекопитающие фауны СССР. — ДАН СССР, 1941,
т. 33, № 3, с. 270—272.
Строганов С. У. Систематика кротовых (Talpidae). — Тр. Зоол. ин-та АН СССР, 1948,
т. 8, вып. 2, с. 287—405.
Строганов С. У. Обзор подвидов и географического распространения крошечной буро-
зубки (Sorex tscherskii Ogn.). — Уч. зап. Томск, ун-та, 1949, № 12, с. 183—192.
Строганов С. У. К систематике и распространению двух малоизученных видов буро-
зубок Средней и Центральной Азии. — Бюл. МОИП, отд. биол., 1952, т. 57,
вып. 5, с. 21, 22.
Строганов С. У. Звери Сибири (Насекомоядные). М., 1957а, с. 83—259.
Строганов С. У. К вопросу о происхождении фауны насекомоядных Сибири. —
Изв. Сиб. отд. АН СССР, 19576, № 2, с. 95—103.
Строганов С. У. К распространению и таксономии белозубки-малютки (Suncus etru-
scus bactrianus subsp.). — Изв. Сиб. отд. АН СССР, 1958а, № 1, с. 122—125.
Строганов С. У. О гладкоиглом еже (Hemiechinus homalocanthus Stroganov). —
Тр. Биол. ин-та, зоол., 19586, вып. 5, с. 231—233.
Строков В. В. Зимнее размножение кавказской бурозубки. — Природа, 1957, № 7,
с. 113—114.
Татаринов К. А. Звери западных областей Украины. Кшв, 1956. 138 с.
Татаринов К. А. Знахщки землеройки блярипа на Украши. — ДАН УРСР, 1958,
№ 1, с. 81—84.
Темботов А. К. К фауне насекомоядных и грызунов Кабардино-Балкарской АССР. —
Уч. зап. Кабардино-Балкарск. ун-та, 1957, вып. 2, с. 149—165.
Топачевский В. О. Викопнш вихухол! роду Desmana з пеогепових та аптропогеповпх
В1дклад1в европейськоп частпни СРСР. — В кн.: Викопш фауни Украши
1 сум!жних територш. Кшв, 1962, с. 7—90.
Трофимов Б. А. Насекомоядные рода Palaeoscaptor из олигоцена Азии. — Тр. Палеон-
тол. ин-та АН СССР, 1960, т. 77, вып. 4, с. 35—40.
Тупикова Н. В. Питание и характер суточной активности землероек средней полосы
СССР. — Зоол. жури., 1949, т. 28, с. 561—572.
Фолитарек С. С. К биологии обыкновенной землеройки (Sorex araneus L.). — Зоол.
журн., 1940, т. 19, вып. 2, с. 324, 325.
Чернышев В. И. К экологии бухарской бурозубки (Sorex bucharensts Ogn.). —
Изв. Отд. естеств. наук АН ТаджССР, 1956, вып. 14, с. 123 —138.
Шварц С. С. Биология землероек лесостепного Зауралья. — Зоол. жури., 1955,
т. 34, вып. 4, с. 915—927.
Щеглова А. И., Мальджюнайте С. А. Химическая терморегуляция у крота. —
Тр. АН ЛитССР, Биол. науки, 1968, т. 1, с. 131—140.
Эфрон К. М. К экологии куторы (Neomys jo die ns Penn.) на среднем Урале. — В кн.:
Вопросы экологии, т. 6, Киев, 1962, с. 178.
Юдин Б. С., Исаков С. И. Систематика и фауна землероек (Soricidae) Таджикистана.
Род Suncus и Sorex. — Изв. АН ТаджССР, 1975, № 4 (61), с. 47—53.
Allen G. М. The mammals of China and Mongolia. Natural History of Central Asia, v. 2,
pt. 1. New York, 1938. 620 p.
Allen G. M. Checklist of African mammals. — Bull. Mus. Compar. Zool. Cambridge,
1939, v. 82, p. 5—53.
Allen J. A. The skeletal characters of Scutisorex Thomas. — Bull. Amer. Mus. Natur.
Hist., 1917, v. 37, p. 769- 784.
Allen J. A. The American Museum Congo expedition collection of Insectivora. — Bull.
Amer. Mus. Natur. Hist., 1922, v. 47, art. 1, p. 8—16.
Anderson J. On the osteology and dentition of Hylomys. — Trans. Zool. Soc. London,
1874, v. 8, p. 453—467.
472
ЛИТЕРАТУРА
Anderson J. Reviews new or little known Asiatic shrews in the Indian Museum. —
J. Asiat. Soc. Bengal, 1877, v. 46, pt. 2, ,N 1—3, p. 261—283.
Anderson J. Zoology of Egypt. Mammalia. 1902, p. 156—165.
Bate D. M. A. Pleistocene Shrews from the larger Western Mediterranean island. — Ann.
Mag. Natur. Hist. 1944, ser. 11, v. 11, p. 738—769.
Biswas B., Chose R. Taxonomic notes on the Indian Pale hedgehogs of the genus Pa-
raechinus Trouessart, with descriptions of a new species and subspecies. — Mam-
malia, 1970, t. 34, N 3, p. 467—477.
Blainville H. M. D. Osteographiae des Mammiferes (Insectovora), v. 1. 1838, p. 102—109.
Blanford W. I. The fauna of British India, including Ceylon and Burma. Mammalia.
London, 1888—1891. 617 p.
Brink F. A. La musaraigne masques espece circum-boreale. — Mammalia, 1953, t. 17,
N 2, p. 96—125.
Broom R. Some South African Pliocene and Pleistocene mammals. — Ann. Transvaal
Mus., 1948, v. 21, p. 1—38.
Brunner G. Die Breitenberghohle bei Go|3weinstein Ofr. (Eine Mindel-Rip- und einepost-
glaciale Mediterran-Fauna). — Neues Jahrb. Geol. Palaont. Monatschrifte, 1957,
H. 7-8, S. 351—378.
Butler В. M. On the evolution of the skull and teath in the Erinaceidae, with special
reference to fossil material in the British Museum. — Proc. Zool. Soc. London,
1948, v. 118, p. 392—500.
Butler P. M. Erinaceidae from miocene of East Africa. — In: Fossil mammals of Africa,
N 11. London, 1956a. 75 p.
Butler P. M. The skull of Ictops and the classification of the Insectivora. —- Proc. Zool.
Soc. London, 1956b, v. 126, pt 3, p. 453—481.
Butler P. M. Insectivores and Bats from the miocene of East Africa new material. —
In: Fossil vertebrates of Africa, v. 1. 1969, p. 1—37.
Butler P. M. Some functional aspects of molar evolution. — Evolution, 1972, v. 26,
N 3, p. 474—485.
Butler P. M., Hopwood A. T. Insectivora and Chtroptera from the Miocene roks of Kenia
Colony. — In: Fossil Mammals of Africa, N 13. London, 1957. 35 p.
Campbell B. The shoulder anatomy of the moles. A study in phylogeny and adaptation. —
Amer. J. Anat., 1939, v. 64, p. 1—39.
Crochet I. V. Les Insectivores des Phosphorites du Quercy. — Palaeovertebrata, 1974,
v. 6, fasc. 1—2, p. 109—159.
Crusafont P. M., Villalta J. F. Sur un nouveau Palerinaceus du Pontien d’Espagne. —
Eclogue geol. Helv., 1947, v. 40, N 2, p. 320—333.
Crusafont M., de Villalta J. E., Y. J. Truyols. El Burdigaliense Continental de la Cuenca
del Valles-Penedes. — Mem. Comunicac. Inst. Geol. Barselona, 1955, N 12,
p. 3—272.
Dechaseaux C. L’encephale de Neurogymnurus cayluxi, Insectivore des Phosphorites
du Quersy. — Ann. paleontol., 1964, t. 50, fasc. 1, p. 83—100.
Dobson G. E. A monograph of the Insectivora, systematic and anatomical. Pt. 1—3.
London, 1882, 1883, 1890.
Dubale M. S., Travadi H. S. Myology of the feeding apparatus of the Hedgehog (Erina-
ceus collaris). — J. Animal Morphol. Physiol., 1971, v. 18, N 1, p. 91—105.
Eadie W. R. The dermal glands of shrews. — J. Mammal., 1938, v. 19, N 2, p. 171—174.
Ellerman J. R., Morrison-Scott T. C. S. Checklist of Palaearctic and Indian Mammals.
London, 1951. 810 p.
Ellerman J. R., Morrison-Scott T. C. S., Hayman R. W. Southern African Mammals.
London, 1953. 364 p.
Fiedler W. Kaumusculatur der Insectivora. — Acta anatom., 1953, Bd 18, S. 101—175.
Filhol H. Considerations sur la decouverte de quelques mammiferes fossiles appartenant
a I’epoque eocene superieure. — Bull. Soc. Philomat. Paris, 1877, t. 1, p. 51—54.
Filhol H. Etude des mammiferes fossiles de Saint Gerand-leluy (Allier). — Ann. Sci.
Geol., 1879, t. 10, N 3, p. 1—252. •
Filhol H. Observations relatives a des mammiferes fossiles nouveaux provenant des de-
pots de phosphate de chaux du Quersy. — Bull. Soc. Sci. Phys. Natur. Touluse,
1880a, t. 5, p. 159—201.
Filhol H. Note sur des mammiferes fossiles nouveaux provenant des phosphrorites
du Quersy. — Bull. Soc. Philomat. Paris, 1880b, ser. 7, t. 3, p. 120—125.
Filhol H. Etudes sur les mammiferes fossiles de Ronzon. — Ann. Sci. Geol., 1882, t. 12,
₽• 8‘
Filhol H. Sur un nouveau genre d’InsectivOre. — Bull. Soc. Philomat. Paris, 1888, ser. 7,
t. 12, p. 24—25.
Fischer von Waldheim G. Adversaria zoologica. — Mem. Soc. Imp. Natur. Moscow,
1817, v. 5, p. 368—428.
ЛИТЕРАТУРА
473
Fons R. Contribution a la connaissance de la musaraigne etrusque Suncus etruscus Savi,
1822 (Mammifcre, Soricidae). — Vie et Milieu, ser. C, Biol. Terrestre, 1970, t. 21,
fasc. 1-c, p. 209—218.
Freudenthal M. Deinogalerix koenigswaldi nov. gen. nov. sp. a giant Insectivore from
‘ Neogene Italy. — Scripta Geol., 1972, N 14, s. 1—19.
Friant M. Le type priniitif des molaises chez les Insectivores (Erinaceides). — Nouv.
Arch. Mus. Hist, natur. Paris, 1934a, ser. 6, t. 11, p. 126.
Friant M. Reparatition geographique et classification (d’apres les caracteres dentaires)
des Erinaceides fossiles et actuels. — Bull. Soc. zool. France, 1934b, t. 59, p. 508—
516.
Friant M. On the evolution of the teeth in the Erinaceidae. — Proc. Zool. Soc. London,
1949, vol. 119, N 4, p. 821.
Friant M. A propos molares d'un Insectivore (Erinaceidae) d’Europe, le N eurogymmurus
de 1’Ere Tertiare. — Folia morphol., 1971, t. 19, N 2, p. 198—204.
Gaillard C. Mammiferes miocenes nouveaux ou peu connus de la Grive Saint-Alban
(Isere). — Arch. Mus. d’Hist, natur. Lyon, 1899, t. 7, N 2, p. 14—34.
Gaillard C. Nouveau genre de Musaraignes dans les depots miocenes de la Grive-Saint-
Alban (Isere). — Ann. Soc. Linn. Lyon, 1915, t. 62, p. 83—98.
Gaillard C. Nouveaux mammiferes dans les depots miocenes de la Grive-Saint-Alban
(Isere). — Bull. Soc. Natur. Arch. Ain, 1929, p. 1—17.
Galbreath E. C. A contribution to the Tertiary geology and paleontology of northeastern
Colorado-Univ. — Kansas Paleontol. Contrib. (Vertebrata), 1953, art. 5, p. 1 —120.
Ganghran G. R. L. A comparative study of the osteology and myology of the cranial and
cervical regions of the Shrew Вlarina brevicauda and the mole Scalopus aquaticus. —
Misc. Pubis Mus. Zool. Univ. Michigan, 1954, N 80, p. 1—82.
Gawne C. F. The genus Proterix (Insectivora, Erinaceidae) of the Upper Oligocene of North
America. — Amer. Mus. Novitates, 1968, N 2315, p. 1—26.
Geisler M., Gropp A. Chromosome polimorphism in the European Hedgehog Erinaceus
europaeus (Insectivora). — Nature, 1967, v. 214, N 5086, p. 396—397.
Gill T. Arrangement of the families of mammals with analytical tables. — Smiths.
Misc. Coll., 1872, v. 11, art. 1, p. 1—98.
Gill T. Synopsis of Insectivorous mammals. — Bull. U. S. Geol. Geograph. Surv., 1875,
v. 1, ser. 2, p. 91—120.
Gill T. On the classification of the insectivorous mammals. — Bull. Philos. Soc. Washing-
ton, 1882, v. 5, p. 118—120.
Gould E., Negus N. C., Novik A. Evidence for echolocation in shrews. — J. Exptl.
Zool., 1964, v. 156, N 1, p. 19—37.
Gray J. E. Revision of the genus Sorex L. — Proc. Zool. Soc. London, 1837—1838, p. 124.
Grulich I. Zur Methodik der Altersbestimmung des Maulwurfs Talpa europaea L. in der
Periode Science selbstandigen Lebensweise. — Zool. listy, 1967, v. 16, N 1,
p. 41—59.
Grulich I. Ein Beitrag zur Kenntnis asmediterranen Kleinwiichsigen, blinden Maulwurfs-
formen (Talpinai). — Zool. listy, 1972, v. 21, N 1, p. 3—21.
Gunther F. A. S. Notes on some Japanese Mammalia. — Proc. Zool. Soc. London, 1880,
p. 441.
Gupta В. B. Distribution of nodes and tracts in the scull of Indian hedgehog Paraechinus
micropus micropus Blyth. — Zool. Poloniae, 1962a, v. 11, fasc. 1, p. 85—87.
Gupta В. B. The interpretation of the rolling in the Indian hedgehogs. — Zool. Poloniae,
1962b, v. 12, fasc. 1, p. 79—84.
Gupta В. B. Studies on the morphology of the head region in the Indian hedgehogs, Pa-
ra echinus micropus micropus Blyth, and Hemichinus auritus collaris Gray et Hard-
wickii. — Zool. Poloniae, 1962c, v. 12, fasc. 2, p. 107—129.
Gupta В. B. A study on the skeleton and the musculature of the neck, the thorax and the
abdomen of the Indian hedgehogs. Paraechinus micropus micropus Blyth and He-
miechinus auritus collaris Gray et Hardwickii. — Zool. Poloniae, 1963, v. 12,
fasc. 4, p. 365—422.
Gupta В. B. Locomotion and some postures in the Indian hedgehogs. — Zool. Poloniae,
1964a, v. 14, fasc. 1—2, p. 71—74.
Gupta В. B. A study on the skeleton and the musculature of the distal posterior appendage
of the Indian hedgehogs, Paraechinus micropus micropus Blyth and Hemiechinus
auritus collaris Gray et Hardwickii. — Zool. Poloniae, 1964b, v. 14, fasc. 1—2,
p. 53—70.
Hall E. R. A second new genus of hedgehog from the Pliocene of Nevada. — Univ. Ca-
lifornia Publ. Bull. Depart. Geol. Sci., 1929, v. 18, N 8, p. 227—231.
Hall E. R., Kelson K. R. The mammals of North America. V. 1. New York, 1959, p. 22—
. 78’
Harrison J. L. The mammals of Arabia. V. 1. Insectivora, Chiroptera, Primates. 1940,
p. 40.
31 А. А. Гуреев
474
ЛИТЕРАТУРА
Harrison J. L. Chimarrogale hantu a new water shrew from the Malay Peninsula, with
a note on the genera Chimarrogale and Crossogale (Insectivora, Soricidae). — Ann.
Mag. Natur. Hist., 1958, ser. 13, v. 1, N 4, p. 282—290.
Heim de Balsac H. Notions nouvelles sur 1’evolution des Soricidae (Mamiferes, Insectivo-
res). — Compt. rend. Acad. Sci. Paris, 1955, t. 241, N 25, p. 1997—1998.
Heim de Balsac H. Evolution et phylogenie des Soricides africanus. — Mammalia, 1956a,
t. 20, N 2, p. 140—167.
Heim de Balsac H. Un Soricidae inedit et aberrant du Kasai exige la creation d’un genre
nouveau (Mammalia, Insectivora). — Res. Zool. et Bot. Afriq., 1956b, t. 54,
p. 137—146.
Heim de Balsac H. Evolution et phylogenie des soricides africains II La lignee Sylviso-
rex — Suncus — Crocidura. — Mammalia, 1957a, t. 21, N 1, p. 15—49
Heim de Balsac H. Insectivores de la famille des Soricidae de 1’Afrique orientale. — Zool.
Anz., 1957b, t. 158, N 7—8, p. 143—153.
Heim de Balsac H. Persistance inattendue de certain caracteres ancestraux fans la denture
de Soricidae (Mammaiferes, Insectivores). — Compt. rend. Acad. Sci. Paris,
1958, t. 247, N 18, p. 1499—1501.
Heim de Balsac H. Premieres donnees sur la lepartition generale, 1’ecologie et la varia-
tion morphologique du genre Paracrocidura Heim de Balsac en Afrique Centrale. —
Rev. Zool. et Bot. Afriq., 1959, t. 59, p. 19—28.
Heim de Balsac H. Nouvelle contribution a 1’etude des insectivores Soricidae du Mont
Camerun. — Bonner Zool. Beitr., 1959 (1960), Bd. 10, H. 3—4, S. 198—217.
Herder K. Gefangenschaftscbeobachtungen an einem Algerischen Igel (Aethechinus algirus
Duvernoy und Lebeboullet). — Zool. Beitr., 1964, Bd 10, H. 2, S. 189—225.
Hibbard C. W, with others. Early Pleistocene Preglacial and glacial rocks and faunas
of north-central Nebraska. — Bull. Amer. Mus. Natur. Hist., 1972, v. 148,
art. 1, p. 77—80.
Hibbard C. W., Bjork P. R. The insectivores of the Hagerman local fauna, Upper Plio-
cene of Idaho. — Contrib. Mus. Paleontol. Univ. Michigan, 1971, v. 23, N 9,
p. 171—180.
Hinton M. A. C. The British fossil shrews. — Geol. Mag., N. S., 1911, v. 5, N 8, p. 529—
539.
Hoffmann R. S. Relationships of certain Holarctic shrews, genus Sorex. — Z. Saugeti-
erkunde, 1971, Bd. 36, H. 4, S. 193—200.
Hoffmann R. S., Peterson R. S. Systematics in Zoogeography of Sorex in the Bering strait
Area. — Repr. System. Zool., 1967, v. 16, N 2, p. 127—136.
Hollister N. Mammals collected by the Smithsonian. — Harvard expedition to the Altai
mountains, 1912. — Proc. U. S. Nation. Mus., 1913, v. 45, N 1990, p. 507—532.
Hollister N. Shrews collected by the Congo expedition of the American Museum. —
Bull. Amer. Mus. Natur. Hist., 1916, v. 35, p. 663—680.
Hopwood A. T. Notes on the recent and fossil mammalian faunas of Africa. — Proc.
Linnean Soc. London, 1954, v. 165, N 1, p. 46—49.
Hurzeler J. Uber einem dimyloiden Erinaceiden (Dimylechinus nov. gen.) aus dem Aqui-
tanien der Limagne. — Eclogae geol. helv., 1944, v. 37, N 2, p. 460—467.
Hutchison J. H. Fossil talpidae (Insectivora, Mammalia) from the later Tertiary of Ore-
gon. — Bull. Mus. Natur. Hist. Univ. Oregon, 1968, N 11, p. 1—117.
Imaizumi J. New species and subspecies of Sorex from Japan (Mammalia, Insectivores). —
Bull. Natur. Sci. Mus., Tokyo, 1954, N. S., v. 1, p. 94—108.
Imaizumi J. Taxonomic studies of the Japanese mountain mole (Talpa mizura) with
description of two new subst. — Bull. Natur. Sci. Mus. Japan, 1955, v. 2, N 1,
p. 26—28.
Jammot D. Les Insectivores (Mammalia) du Gesement Pleistocene moyen des Abimes
de la Fage a Noailles (Correze). — Nouv. Arch. Mus. natur. Lyon, 1973, fasc. 11,
p. 41—51.
Jackson H. H. A Review of the American moles. — North. Amer. Fauna, 1915, N 38,
p. 3—98.
Jackson H. H. A taxonomic review of the American long-tailed shrews. — North. Amer.
Fauna, 1928, N 51, p. 1—238.
Kindal M. Same aspects of the tooth development in Soricidae. — Acta Odontol. Scand.
Stockholm, 1959, v. 17, p. 203—237.
Koerner H. E. The geology and vertebrate palaeontology of the Fort Logan and Deep
River formations of Montana. Pt. 1. New vertebrates. — Amer. J. Sci., 1940,
p. 238, 837.
Kormos T. Neue insektenfresser, fledermause und nager aus dem oberpliocen der Vil-
lanyer Gegend. — Foldt. kozloni, 1934, t. 64, p. 1—26.
Kowalski K. Katalog Ssakow pleistocenu Polski. Warszawa—Wroclaw, 1959, s.7—267.
Kratochvil J. Zur Frage der Verbreitung des Igels (Erinaceus) in der CSSR. — Zool.
listy, 1966, v. 15, N 3—4, p. 291—304.
ЛИТЕРАТУРА
475
Kratochvil J. Das Stachelkleid des Ostigels (Erinaceus concolor roumanicus) — Acta Sci.
Nat. Brno, 1974, v. 11, N 8, p. 1—52.
Kretzoi M. Weitere Beitrage zur Kenntnis der Fauna von Gombaszog. — Ann. Mus.
Natur. Hung., 1941, v. 34, p. 105—139.
Kretzoi M. Insectivoren, Nagetiere und Lagomorphen der jungstpliozanen Fauna von
Csarnota im Villanyer Gebirge (Siidungarn). — Vertebrata Hungarica, 1959,
t. 1, fasc. 2, p. 237—239.
Kretzoi M. Drepanosorex — neu definiert. — Vertebrata Hungarica, 1965, t. 7, fasc. 1—2,
p. 117—129.
Lavocat R. Revision de la Faune des mammiferes Oligocenes D’auvergne et du Velay.
Paris, 1951, p. 3—153.
Lavocat R. Etude systematique de la faune de mammiferes et conclusions generales. —
Notes Mem. Serv. Geol. Maroc., 1961, v. 155, p. 29—94.
Lay D. M. A new species of mole (Genus Talpa) from Kurdistan Province, Western
Iran. — Fieldiana Zool., 1965, v. 44, N 24, p. 227—230.
Linnaeus C. Systema naturae. Vol. 1. Ed. 10, 1758. 824 p.
Lydekker R. Description of the cranium of a new species of Erinaceus from the upper
Miocene of Oeningen. — Quart. J. Geol. Soc., 1886, v. 42, p. 23.
Macdonald .1. D. The Miocene faunas from the Wounded Knee area of western South
Dakota. — Bull. Amer. Mus. Natur. Hist., 1963, v. 175, p. 139—238.
Matthew W. D. A fossil hedgehog from the American Oligocene. — Bull. Amer. Mus.
Natur. Hist., 1903, v. 19, p. 227—230.
Matthew W. D. A lower Miocene fauna from South Dakota. — Bull. Amer. Mus. Natur.
Hist., 1907, v. 23, p. 169-219.
Matthew W. D. The Carnivora and Insectivora of the Bridger basin, Middle Eocene. —
Mem. Amer. Mus. Natur, Hist., 1909, v. 9, p. 291—567.
Matthew W. D. A new and remarkable Hedgehog from the later Tertiary of Nevada. —
Univ. Calif. Publ. Bull. Depart. Geol. Sci., 1929, v. 18, N 4, p. 92—102.
Matthew W. D. New fossil Mammals from Snake Creek quarries. — Amer. Mus. Novita-
tes, 1932, N 540, p. 1—8.
Matthew W. D., Granger W. New Insectivores and Ruminants from the Tertiary of Mon-
golia, with Remarks on the Correlation. — Amer. Mus. Novitates, 1924, N 105,
p. 1—7.
Matthew W. D., Mook С. C. New fossil Mammals from the Deep River Beds of Montana. —
Amer. Mus. Novitates, 1933, N 601, p. 1—7.
McKenna M. C. The Geolabidinae. A new subfamily of early cenozoic erinaceoid insecti-
vores. — Univ. Calif. Publ. Geol. Sci., 1960, v. 37, N 2, p. 131—164.
McKenna N. C., Holton С. P. A new insectivore from the Oligocene of Mongolia and new
subfamily of hedgehogs. — Amer. Mus. Novitates, 1967, N 2311, p. 1—11.
Meade G. E. A new erinaceid from the lower Miocene. — Field Mus. Natur. Hist., 1941,
v. 8, p. 143—147.
Mearns E. A. Descriptions of new genera and species of mammals from the Philippine
Islands. — Proc. U. S. Nat. Mus., 1905, v. 28, p. 425—460.
Merriam С. H. Synopsis of the American shrews of the genus Sorex. — North Amer.
Fauna, 1895, N 10, p. 5—34, 56—100.
Meylan A. Formules chromosomiques et polyinorphisme robertsonien chez Blarina bre-
vicauda Sey (Mammalia, Insectivora). — Canad. J. Zool., 1967, v. 45, p. 1119—
1127.
Meylan A., Hasser J. Les chromasomes des Sorex du groupe araneus — arcticus (Mam-
malia, Insectivora). — Z. Saugetierkunde, 1973, Bd 38, H. 3, S. 143—158.
Miller G. S. The long-tailed shrews of the Eastern United States. — North Amer. Fauna,
1895, N 10, p. 35—56.
Miller G. S. Twelve new European mammals. — Ann. Mag. Natur. Hist., 1909, ser. 8,
v. 3, p. 415—422.
Milne-Edwards H. Recherches pour servir a 1’histoire naturelie des mammiferes comp-
renant des considerations sur la classifications de ces animaux, t. 1—2. Paris,
1868—1874, 386 p.
Morris N. Some observations on the breeding season of the Hedgehog and the rearing
and handling of the voung. — Proc. Zool. Soc. London, 1961, v. 136, pt 5, p. 201—
206.
Murray A. The geographic distribution of mammals, v. 16. London, 1866, p. 1—420.
Niethammer J. Zur Taxonomic europaischer Zwergmaulwiirfe (Talpa «mizura»). — Bonner
zool. Beirt. 1969a, Jg. 20, H. 4, S. 360—372.
Niethammer J. Zur Taxonomie der Ohrenigel in Afganistan (Gattung Hemiechinus). —
Z. Saugetierkunde, 1969b, Bd 34, H. 5, S. 257—274.
Niethammer G. Beobachtungen am Pyrenaen-Desman, Galemys pyrenaicus. — Bonner
zool. Beitr., 1970, Jg. 21, H. 3—4, S. 157—182.
Parker W. K. On the structure and development the scull in the Mammalia. 3. Insecti-
vora. — Philos. Roy. Soc. London., 1882, pt 2, p. 121—275.
f 31*
476
ЛИТЕРАТУРА
Patterson В., McGrew Р. О. A soricid and two erinaceids from the White River Oligo-
cene. — Field Mus. Natur. Hist., 1937, v. 6, p. 245—272.
Pearson О. P. Shrews. — Sci. Amer., 1954, v. 191, N 2, p. 66—68.
Peterson R. S. The Land mammals of Unalaska Island: Present status and zoogeo-
graphy. — J. Mammal., 1967, v. 48, N 1, p. 119—129.
Peyre A. Ecologie et biogeographie du Desman (Galemys pyrenaicus G.) dans les Pyre-
nees francaises. — Mammalia, 1956, t. 20, p. 405—418.
Pucek L. Die toxische Wirkung der Glandulae submaxillares bei Neomys fodiens. — Bull.
Acad, polon. sci., 1957, Cl. 2, v. 5, N 9, p. 301—306.
Reed C. A. Locomotion and appendiculare anatomy in three Soricoid Insectivores. —
Amer. Midland Naturalist, 1951, v. 45, N 3, p. 513—671.
Reed C. A. Same fossorial mammals from the Tertiary of Western North America. —
J. Paleontol., 1954, v. 28, p. 102—111.
Reed К. M. Insectivores of the Middle Miocene Split Rock local fauna. Waioming. —
Breviora, 1960, N 116, p. 1—11.
Reed К. M. The Proscalopinae new subfamily of Talpid insectivores. — Bull. Mus.
Compar. Zool., 1961, v. 125, N 14, p. 473—494.
Reed К. M. Two new species of fossil talpid insectivores. — Breviora, 1962, N 168,
p. 1—6.
Repenning C. A. An extinct shrew from the Early Pleistocene of South Africa. —
J. Mammal., 1965, v. 46, N 2, p. 189—196.
Repenning C. A. Subfamilies and genera of the Soricidae. — Geol. Surv. Profess. Paper,
1967, N 565, p. 1—74.
Rich T. H. V., Rich P. V. Brachyerix a Miocene hedgehog from Western North America,
with a description of the tympanic regions of Paraechinus and Podogymnura. —
Amer. Mus. Novitates, 1971, N 2477, p. 1—58.
Rich T. H. V., Rasmussen D. L. New North American erinaccine hedgehogs (Mammalia,
Insectivora). — Occasion, pap. Mus. Natur. Hist., Univ. Kansas, 1973, N 21,
p. 1—54.
Richter H. Zur Systematik und Verbreitung afganischer Crociduren (Mammalia, In-
sectivora, Soricidae). — Zool. Abhandl. 1970a, Bd 31, N 15, S. 269—277.
Richter H. Zum taxonomischen status der zwei Crociduren-Formen von Kreta (Mam-
malia, Insectivora, Soricidae). Zool. Abhandl., 1970b, Bd 31, N 16, S. 279—291.
Richter H. Zur Taxonomie und Verbreitung der palaearktischen Crociduren (Mammalia,
Insectivora, Soricidae). — Zool. Abhandl., 1970, Bd 31, N 17, S. 293—304.
Roberts A. The mammals of South Africa. Johannesburg, 1954. 700 p.
Rode P., Didier R. Atlas des Mammiferes de France. Paris, 1946. 218 p.
Ruprecht A. L. Correlation structure of skull dimensions in European hedgehogs. —
Acta theriol., 1972, v. 17, N 32, p. 419—442.
Rzebik B. Crocidura Wagler and other Insectivora (Mammalia) from the Quarternary
Deposits of Tornewton in England. — Acta zool. Cracov., 1968, t. 13, N 10,
p. 251—262.
Rzebik-Kowalska B. The Pliocene and Pleistocene insectivores (Mammalia) of Poland. —
Acta zool. Cracov., 1971, t. 16, N 9, p. 436—461.
Saban R. Phylogenie des Insectivores. — Bull. Mus. Nat. Hist. Natur. Paris, 1954,
ser. 2, t. 26, p. 419—432.
Saban R. Insectivora. — Traite Paleontol., 1958, t. 6, v. 2, p. 822—912.
Saint Girons M.-C. Notes sur les Mammiferes de France. S. Donnees sur la morphologie
et la repartition de Erinaceus europaeus et Erinaceus algirus. — Mammalia, 1969,
t. 33, N 2, p. 206—218.
Savage R. J. G. Evolution in Carnivorous mammals. — Palaeontology, 1977, v. 20,
pt 2, p. 238—271.
Schlosser M. Die Affen, Lemuren, Chiropteren, Insectivoren. Marsupialier, Creodonten
und Carnivoren, der europaischen Tertiars und deren Beziehungen zu ihren leben
und fossilen aussereuropaischen Verwandten. — Beitr. Palaont. Geol. Ost. Un-
garn, 1887—1888, Abt. 1, Bd 6, S. 1—224; Abt. 2, Bd 7, S. 225—386.
Schreuder A. A revision of the Fossil water-moles (Desmaninae). — Arch. Neerland.
Zool., 1940, t. 4, livr. 2—3, p. 201—333.
Schubarth H. Zur Variabilitat von Sorex araneus L. — Acta theriol., 1958, v. 2, N 9,
p. 175—202.
Schwarz E. Revision of the Old-world moles of the genus Talpa. — Proc. Zool. Soc.
London, 1948, v. 118, p. 36—48.
Shotwell J. A. Hemphilian mannualian assemblage form northestern Oregon. — Bull.
Geol. Soc. Amer., 1956, v. 67, p. 717—738.
Siivonen L. Sorex isodon Turov (1924) and Sorex unguiculatus Dobson (1890) as indepen-
dent shrew species. — Afj^Slo, 1965, v. 4, p. 3—34.
Simpson G. G. A new insectivore from Oligocene Utan Gochu Horizon of Mongolia. —
Amer. Mus. Novitates, 1931, N 505, p. 1—22.
ЛИТЕРАТУРА
477
Simpson G. G. The principles of classifications and a classification of Mammals. — Bull.
Amer. Mus. Natur. Hist., 1945, v. 85, p. 1—350.
Simpson G. G. American cretaceous insectivores. — Amer. Mus. Novitates, 1951, N 1541,
p. 1—19.
Sorenson M. W. Some aspects of water shrew bechavior. — Amer. Midland Naturalist,
1962, v. 68, N 2, p. 445—462.
Spitzenberger F. Zur Verbreitung und Systematik turkischer Soricinae (Insectivora, Mam-
malia). — Ann. Naturhistor. Mus. Wien, 1968, Bd 72, S. 272—279.
Stehlin H. G. Zur Stammesgeschichte der Soriciden. — Eclogae geol. helv., 1940, t. 33,
N 2, p. 445—462.
Stein G. H. W. Schadelallometrien und Systematik bei altweltlichen Maulwiirfen (Tal-
pinae). — Mitt. Zool. Mus. Berlin, 1960, Bd 36, H. 1, S. 1—48.
Sterba O. Developmental anatomy of the mole, Talpa europaea Linn., 1758, 3. Prenatal
development of musculature of the trunk. — Zool. listy, 1972, v. 21, N 2, p. 319—323.
Stirton R. A. A new genus of Soricidae from the Barstow Miocene of California. — Univ.
Calif. Publ. Bull. Dept. Geol. Sci., 1930, v. 19, N 8, p. 217—228.
Stock C. Insectivora from the Sespe Uppermost Eocene, California. — Proc. Natl. Acad.
Sci., 1935, v. 21, N 4, p. 214—219.
Stolizka F. Notice of the Mammals and Birds inhabiting Kachh. — J. Asiat. Soc. Bengal,
1872, v. 41, N 2, p. 211—258.
Sulimski A. Pliocene insectivores from Weze. — Acta paleontol. polon., 1959, v. 4,
N 2, p. 119-179.
Sulimski A. Supplementary studies on the insectivores from Weze (Poland). — Acta
paleontol. polon., 1962, v. 7, N 3—4, p. 458—498.
Sulimski A. On some Oligocene insectivore remains from Mongolia. — Palaeontologia
polonica, 1970, N 21, p. 61—65.
Sych B., Rzebik-Kowalska B. Fossi^ endocranial cast of Desmana kormosi Schreuder
(Insectivora, Mammalia) from the Pliocene of Poland. — Acta zool. Cracov.,
1972, v. 17, N 2, p. 37—46.
Tate G. И. H. The mammals of the Guiana region. — Bull. Amer. Mus. Natur. Hist.,
1939, v. 76, p. 151—229.
Thenius E. Zur Revision der Insectivoren des steirischen Tertiars. — Zitz. Ber. Ost.
Akad. Wiss. Math-naturw. KI. Wien, 1949, Abt. 1, Bd 158, H. 9—10, S. 671—693.
Thomas O. The generic division of the hedgehogs. — Ann. Mag. Natur. Hist., 1918,
ser. 9, v. 1, N 2, p. 193—196.
Tobien H. Insectivoren (Mammalia) aus dem Mitteleozen (Lutetium) von Messel bei
Darmstadt. — Notizbl. Hessischen Landesamtes Bodenforsch. Weisbaden, 1962,
Bd 90, S. 7—47.
Trouessart E. L. Neotetracus sinensis, a new Insectivore of the Family Erinaceidae. —
Aim. Mag. Natur. Hist., 1909, ser. 8, v. 4, N 19, p. 389—391.
Van Valen L. Deltatheridia, a new order of mammals. — Bull. Amer. Mus Natur. Hist.,
1966, v. 132, art. 1, p. 1—126.
Van Valen L. New Paleocene insectivores and insectivore classification. — Bull. Amer.
Mus. Natur. Hist., 1967, v. 135, art. 5, p. 217—284.
Verma K. On the structure of the skulls of the Indian hedgehogs. — Mammalia, 1962,
t. 26, N 3, p. 362—401.
Viret J. Les faunes de mammiferes de 1 ’Oligocene superieur de la Limagne Bourbon-
naise. — Ann. Univ. Lyon, 1929, t. 47, p. 1—328.
Viret J. Etude sur quelques erinaceides fossiles specialement sur lei genre Palaerina-
ceus. — Trav. Lab. Geol. Univ. Lyon, 1938, t. 34, p. 1—32.
Viret J. Etude sur quelques Erinaceidae fossiles (suite): genres Plesiosorex, Lanthanothe-
rium. — Trav. Lab. Geol. Univ. Lyon, 1940, t. 39, p. 33—65.
Viret J., Zapfe H. Sur quelques soricides miocenes. — Eclogae geol. helv., 1952, t. 44,
N 2, p. 411—426.
Walker E. P., Wanick F., Lange K. et al. Mammals of the World, v. 1. Baltimore,
1964. 664 p.
Webb S. D. The first American record of Lanthanotherium Filhol. — J. Paleontol., 1961,
v. 35, p. 1085—1087.
Wilson R. W. Early Miocene rodents and insectivores from northeastern Colorado. —
Unjy. Kansas Paleontol. Contr. Verteb., 1960, art. 7, p. 1—92.
Young C.rC., Lui P. T. On the mammalian fauna at Koloshan. — Bull. Geol. Soc.
Shina, 1951, v. 30, p. 43—90.
Zapfe H. Die Fauna der miozanan spalten fiillung von Neudorf an der March (C. R. S.).
Insectivora. — Sitz. Ber. Ost. Akad. Wiss. Math.-naturw. KI. Wien, 1951, B<J| 160,
H. 5, S. 449—480.
Zdansky O. Die alttertiaren Saugetiere Chinas nebst stratigraphischen Bemerkungen. —
Paleontol. Sinica, 1929, ser. C, v. 6, N 2, p. 1—87.
Ziegler A. C. Dental homologies and possible relationships of recent Talpidae. —
J. Mammal., 1971, v. 52, N 1, p. 50—68.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ *
acadicus, Sorex 359
Acanthionidae 102
acridens, Amphechinus 150, 152
— Palaeoerinaceus 139
— Palaeoscaptor 138, 139*
acutidens, Geotripus 259
acutidentata, Protalpa 258
— Talpa 258, 259
adamsi, Blarina 423
Adapisoricidae 95, 136, 189, 257
Adeloblarina 291, 298, 301*, 305, 308,
313, 420
— berklandi 420, 421*
aegyptius, Erinaceus 169
— — europaeus 173
— Hemiechinus auritus 174
aenea, Talpa 251
aeneus, Scalops 251
aequatorialis, Pachyura 406
aereus, Scalops texanus 254
— Scalopus 254
Aethechinus 100*, 101—103, 156, 160,
161, 163
aethyopicus aethyopicus, Erinaceus 178
— Erinaceus 163, 176, 177*, 178*
— — aethyopicus 178
— hypomelas, Erinaceus^ 178
— micropus, Erinaceus 179
africanus, Galerix 111, 112*, 114
agilis, Pachyura 396
alascanus, Sorex 365
— — navigator 365
alaschanicus, Hemiechinus albulus 169
albicauda, Crocidura 383
albinus, Sorex 382
albior, Erinaceus dorsalis 176
albiventer, Neosorex bendirii 336
albiventris, Crocidura aranea var. 396
— Erinaceus 167
albulus alaschanicus, Hemiechinus 169
— Erinaceus 169
— — auritus 173
— Hemiechinus auritus 174
— major, Hemiechinus 169
— minor, Hemiechinus 169
— turanicus, Erinaceus 169
algirus cuniculus, Erinaceus 168
— Erinaceus 167
— lavandeni, Erinaceus 168
— vagans, Erinaceus 168
algiurus, Sorex 446
allex, Crocidura 413
Allopachyura 308
Allosorex 291, 301*, 305, 308, 313, 453,
454, 455
— stenodus 454*
Allosoricina 301*, 302, 313, 315, 419,
453
Allosoricinae 305, 453
Allosoricini 308
Allotheria 38, 44*
Alluvisorex 297, 301*, 305, 307, 312,
318, 323
— arcadensis 323
alpina, Corsira 446
— Crocidura 413
alpinus, Sorex 85, 292, 309, 332, 346*
altaica altaica, Talpa 267
— Asioscalops 197, 265
— gusevi, Talpa 267
— Nectogalinia 458*
— salairica, Talpa 267
— sibirica, Talpa 267
— suschkini, Talpa 267
— Talpa 70*, 195, 259, 263*, 265, 266*,
273
— — altaica 267
— tymensis, Talpa 267
alticola, Cryptotis 435
amasari, Sorex 344
Amblycoptinae 308, 460
Amblvcoptini 460
Amblycoptus 291, 301*, 304, 305, 308,
31'3, 461, 462
— oligodon 462*, 463
— topali 463
Amblysomus 48
— gotten totus 42*
americanus, Hydroscapheus 207*
amoenus, Sorex 363
Amphechini 89, 92, 96—98, 99*, 100, 105,
135, 147
Amphechinus 89, 92, 96—98, 99*, 100*,
102, 103, 105, 116, 146, 147, 149,
151, 152, 154, 156, 157, 159, 162, 179
— acridens 150, 152
— arvernensis 148, 152
— edwardsi 145, 148*, 149*, 150—152
— horncloudi 152
— intermedius 149*
— kansuensis 150*, 151
— minimus 150*, 151
* Страницы с описаниями выделены жирным; страницы с рисунками отмечены
звездочками.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
— montanus 152
— rectus 149, 150*, 151, 152, 157
— rusingensis 151*
— sp. 153*, 154
amphibius, Sorex 440
Amphidozotherium 183, 189, 193*, 194,
199, 257, 258
— cayluxi 257*, 258
Amphisorex 290, 291, 301*, 307, 308,
312, 318, 319, 331, 439
— constrictis 441
— hermanni 441
— lesueurii 359
— limneana 441
— primaevus 296, 319*
amurensis, Erinaceus 85, 164
— — europaeus 165, 167
Anathana 51
Anchiblarinella 290, 300, 301*, 312, 368
— wakeeneyensis 368, 369
anchietae, Crocidura 406
andersoni, Crocidura 382
— Rhynchonax 201
angolae, Erinaceus 168
angolensis, Sylvisorex 381
Angustidens 291, 297, 299, 301*, 305,
307, 312, 318, 325
— vireti 323*, 325
aniscus, Erinaceus 176
Ankylodon 101
anneiata, Crocidura 407
annexus, Sorex 351
Anodonta 217
/anomalus, Neomys 85, 292, 309, 440,
443, 444*
— — morkzeckii 444
Anotus, подрод 308, 422
Anourosorex 60, 291, 292, 294, 298, 301*,
302—305, 308, 313, 415, 460, 461,
465, 467
— группа 303, 460
— inexpectatus 465, 466*
— — conf. 465
— japonicus 465, 466
— squamipes 287, 461, 465, 466, 467*
Anourosoricinae 308
Anourosoricini 74, 290, 291, 298—300,
301*, 302, 304, 313, 315, 326, 460,
467
Anourosoricodon 291, 301*, 313, 461
— pidoplitschkoi 461*
ansorgei, Crocidura 409
Antesorex 305, 307, 323
— chasseae 323
anthony, Crocidura 396
antinori, Sorex 346
antiqua, Mygalea 210, 211*
— Myxomygale 228, 229*
— Talpa 258, 259
antiqus, Oligosorex 317*-
— Geotripus 191, 258, 259
anur, Paraechinus 176
Anurosorex 308
Apternodus bicuspis 138
aquatica, Talpasorex 254
aquaticus, Scalopus 192, 253, 254, 225*
— Sorex 253
aralvchensis, Crocidura russula 399
arambourdi, Geotripus 259
aranea var, albiventris, Crocidura 396
araneoides, Sorex 334
araneus areneus, Sorex.340
— asper, Sorex 341
— auvergne, Sorex 322
— borealis, Sorex 344
— buxtoni, Sorex 344
— Crocidura 399
— jacutensis, Sorex 348
— Mus 331
— roboratus, Sorex 341
— sanguinidens, Sorex 345
— satunini, Sorex 341
— schnitnikovi, Sorex 345
araneus, Sorex 72, 73, 74*, 82, 85, 124,
302, 308—310, 320, 331, 333, 335,
339*, 345, 350, 357, 362, 392, 396,
399
— — araneus 340
— tomensis isodon, Sorex 356
— — Sorex 341
— transrypheus, Sorex 345
arcadensis, Alluvisorex 323
arcticus cinereus, Sorex 359
— Sorex 85, 309, 332, 337, 343*
Arctoryctes 183, 190, 193*, 198, 245, 246
— terrenus 244*, 245, 246
Arctoryctinae 196
Ardynictis furunculus 43*
arethusa, Crocidura 410
argentata, Crocidura 410
argenteus, Neomys 441
armenica, Crocidura 292, 310, 390, 401,
402*
Artiodactyla 44*
arvernensis, Amphechinus 148, 152
— Erinaceus 147
— Mygatalpa 230, 231
arvernien, Desman 231
Arvicola terrestris 185, 205, 217, 442
asiatica, Mygalea 211*, 212
Asioscalops? 197
Asioscalops, подрод 195, 196, 265
— altaica 197, 265
Asoriculus 304, 308
asper, Sorex 309, 332, 341*
— — araneus 341
Astromycter 197, 240
— prasinatus 241
Astromydes 340
___ cristatus 241
Atelerix 100*, 101—103, 156, 160, 161, 163
— подрод 168
— pruneri lowei 168
aterrimus, Sorex 446
Atophyrax 304, 307
attenuate, Crocidura 403
auritus aegyptius, Erinaceus 173
— — Hemiechinus 174
— albulus, Erinaceus 173
— — Hemiechinus 174
— auritus, Erinaceus 172
— — Hemiechinus 174
— caspicus, Erinaceus 169
— chorosanicus, Hemiechinus 174
— collaris, Erinaceus 173
— — Hemiechinus 174
— Erinaceus 31*, 52*, 53*, 69*, 163,
169, 170*, 171*—174
480
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
— — auritus 172
— Hemiechinus 85, 104, 168
— — auritus 174
— megalotis, Erinaceus 173
— — Hemiechinus 174
— turanicus, Erinaceus 173
austriacus, Sorex savini 332
auvergne, Sorex araneus 322
babaulti, Myosorex 29*, 375
baicalensis, Sorex 344
baileyi, Soriculus 446
batesi, Crocidura 409
batis, Sorex 345
beddomei, Sorex 382
bedfordiae gomphus, Sorex 357
— Sorex 357
beirae, Crocidura 407
belli, Sorex 343
bendirii alhiventer, Neosorex 336
— Sorex 336, 365
Beremendia 291, 301*, 304, 305, 308,
313, 419, 420, 425, 427
— fissidens 426*
Beremendina 300, 301*, 302, 313, 315,
425
beresowskii, Soriculus 450
beringianus, Sorex 309, 349*
berklandi, Adeloblarina 420, 421*
berlandieri, Blarina 434
bernoullii, Dimylechinus 145, 146*, 147
beta, Crocidura 408
bicolor, Crocidura 413
bicuspis, Apternodus 138
— Proterix 138*
bidiana, Crocidura 382
birmanica, Gymqura 126
blancalis, Paraechinus deserti 176
blanfordii, Crocidura 382
— Erinaceus 176
Blarina 72, 291, 293, 300, 301*, 302—305,
308, 313, 418, 420, 422, 423, 425
— подрод 424, 433
— группа 303, 418, 420
— adamsi 423
— berlandieri 434
— brevicauda 85, 95, 136, 367, 424*,
425, 431, 463
— — carolinensis 423, 431
— — talpoides 433
— costaricensis 424
— exilipes 434
— eximius 434
— floridana 434
— lessor 436
— gidleyi 423, 424
— magna 436
blarina, Myosorex 374
Blarina nigrescens 437
— orophila 437
— simplicidens 424
— soriciana 437
— telmalestes 425
Blarinella 290, 292, 300, 301*, 303—305,
307, 312, 331, 369, 370
— группа 303
— quadraticauda 370*
— — griselda 371
— — quadraticauda 371
— — wardi 371
Blarinina 300, 301*, 302, 313, 315, 418,
420, 425
Blarinini 74, 288, 290—293, 297, 299, 300,
301*, 302, 304, 305, 308, 313, 315,
325, 326, 368, 418, 419, 420, 453,
454, 456
Blarinini? 305
Blarinoides 291, 300, 301*, 305, 308,
313, 420, 421
— mariae 421, 422
blythii, Crocidura 382
hogdanovi, Sorex 399
hohlini, Neomys 452
borealis, Sorex araneus 344
bottegi, Crocidura 85, 414
bouti, Tetracus 108*, 109, 111
bovei, Crocidura 410
boydi, Crocidura 411
brachychir conf., Talpa 231
brachydactylus, Erinaceus 176
Brachyericini 89, 95—98, 99*, 100, 102—
104, 135, 140
Brachyerix 89, 92, 98, 99*, 100*, 101 —
104, 140
— insertis 142
— macrotis 140, 141*, 142, 144
brachygnatus, Macroneomys 439*
brachyotis, Hemiechinus 169
brachyrchynchus, Elephantulus 85
Brachysorex 308, 422, 433
— подрод 424
brauneri, Talpa europaea 265
brevicauda, Blarina 85, 95, 136, 367,
424*, 425, 431, 463
— carolinensis, Blarina 423, 431
— talpoides, Blarina 433
brevicaudus, Sorex 424
breweri, Parascalops 85, 249*, 250
— Scalops 249
— Scapanus 250
— Scaphanus 250
bucharensis, Sorex 309, 33, 360*, 361, 362
bumeyk, Sorex 355
butleri, Crocidura 408
biittikoferi, Crocidura 406
buxtoni, Sorex 344
— — araneus 344
caecutiens caecutiens, Sorex 352
— koreni, Sorex 352
— macropygmaeus, Sorex 353
— pleskei, Sorex 352
— shinto, Sorex 353
— Sorex 85, 309, 333, 335, 344, 351*,
358, 360
— — caecutiens 352
caerulaeus, Sorex 381
caerulescens, Sorex 381
caerulla riukiuana, Pachyura 382
cafer, Myosorex 374
californicus juncensis, Sorex 364
— Scalops 252
calligoni, Erinaceus 169
Camphotherium 199
camptolophus, Gymnurechinus 155
canadensis, Talpa 241
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
481
— Scalops 251, 254
canaliculatus, Sorex 353
caneae, Crocidura 396
canescens, Crocidura 407
caniceps, Erinaceus 164
caninus, Erinaceus 164
capensis, Erinaceus 167
— Sorex 407
capensoides, Crocidura 411
Caprios 196, 217
carinatus, Sorex 440
Carnivora 24, 25, 41, 44*, 67, 181, 192
Carnivoramorpha 39, 44*
carolinensis, Blarina brevicauda 423, 431
caspica, Crocidura leucodon 394
— — russula 399
caspicus, Erinaceus auritus 169
Castor moschatus 217, 220
caucasica, Talpa 85, 197, 259, 261*, 267,
269, 272
— — coeca 267
caucasicus, Sorex 85, 308, 309
caudata, Crocidura 403
— Talpa 241, 260
caudatus, Sorex 446
— — minutissimus 356
— Soriculus 446
cayluxi, Amphidozotherium 257*, 258
— Neurogymnurus 120, 121*, 122*, 123
Cayluxotherium 103, 120
— elegans 122
celatus, Cryptotis 434
Centetes ecaudatus 84
centralis, Sorex 351, 356
Cetacea 44*
chasseae, Antesorex 323
Chimarrogale 61, 292, 301*, 302—305,
308, 313, 314, 421, 456
— hantu 458
— himalayicus 456
— leander 457
— phacura 458
— platycerhala 457*, 458
— styani 457
— sumatrana 458
— varennei 458
cliinensis, Erinaceus 164
Chiroptera 44*
Chiroscaptor 195, 281
Chodsigoa 304, 305, 308, 313, 445
— подрод 301*, 302, 447, 448, 450, 451,
452
— hypsibia parva 450
— lamula 450
— larwarum 450
— lowei 451
— parva 450
— salenskii 451
chorosanicus, Hemiechinus 104
— — auritus 174
chriseos, Suncus 383
chrysithorax, Sorex 396
Chrysochloridae 3, 27, 29, 35, 36*, 37,
44, 45, 47, 190, 199, 227, 242
Chrysochloris 47, 48, 246
— trivelyani 27*
Chrysospalax 48
cinereus, Sorex 23, 85, 309, 333, 338,
359*, 365, 433, 434
— — arcticus 359
cinnamomeus, Sorex 407
codurcensis, Protalpa 259
coeca caucasica, Talpa 267
— minima, Talpa 271
— orientalis, Talpa 271
— Talpa 85, 195, 197, 259, 263*, 267,
269, 270*, 271, 272, 284
— talyschensis, Talpa 271
colgageti, Didymoconus 43*
collaris, Erinaceus 169
— — auritus 173
— Hemiechinus auritus 174
— Sorex 440
compressa, Domnia 324
compressus, Sorex? 333
concinnus, Sorex 339
concisa, Petenyia 369, 427, 428*
concisus, Parydosorex 427
concolor, Erinaceus 164
— — europaeus 167
Condylura 192, 194, 195, 197, 198, 206,
207, 239, 240
— cristata 58*, 79, 85, 192, 207, 232,
240, 241*, 242*
— fissipes 241
— longicaudata 241
— macroura 241
Condylurinae 192, 194, 197
Condylurini 78, 196, 198, 199, 239
Condylus 240
congicus, Scutisorex 384*, 386
Congosorex 307, 375
conneratii, Sorex 381
constrictus, Amphisorex 441
— Sorex 396
cooperi, Crocidura 359
— Sorex 359
coquerellii, Suncus 383
coreana, Mogera wogura 277
cornfeldi, Crocidura 391
coronatus, Sorex 339
corsicana, Crocidura 396
corsicanus, Nesiotites 452
Corsira 307, 331, 379
— alpina 446
— forsteri 359
— nigrescens 445, 446
— nuvara ellia 379
— talpoides 424
costaricensis, Blarina 424
cowanii, Microgale 84
crassicaudus, Sorex 382
crawfordi, Notiosorex 85, 448, 449*
— Sorex 447—449
Creodonta 39, 41, 44*
cristata, Condylura 58*, 79, 85, 192,
207, 232, 240, 241*, 242*
— Talpa 240
— Talpasorex 241
cristatus, Astromydes 241
— Rhinaster 241
— Scalopus 241
Crocidosorex 290, 291, 297, 301*, 304,
305, 307, 312, 318, 322
— piveteani 322, 323
Crocidura 4, 71, 72, 291, 292, 293, '297,
300, 301*—305, 307, 309, 310, 313,
482
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
314, 323, 371, 376, 377, 379, 381, — goliath 388
386, 388, 394, 401, 452
группа 303, 371, 376
подрод 326, 399, 404, 409
— albicauda 383
— allex 413
— alpina 413
— anchietae 406
— andersoni 382
— anneiata 407
— ansorgei 409
— anthony 396
— aranea var. albiventris 396
— araneus 399
— arethusa 410
— argentata 410
— armenica 292, 310, 390, 401, 402*
— attenuate 403
— batesi 409
— beirae 407
— beta 408
— bicolor 413
— bidiana 382
— blanfordii 382
— blythii 382
— bottegi 85, 414
— bovei 411
— boydi 410
— butleri 408
— biittikoferi 406
— caneae 396
— canescens 407
— capensoides 411
— caudata 403
— cooperi 359
— cornfeldi 391
— corsicana 396
— crossei 413
— cyanea 411
— dayi 382
— deserti 408
— dinniki 292, 310, 390, 402*
— dolichura 414
— doriae 405
— doriana 407
— — fuscosa 406
— dracula 403
— dsinezumi 404
— dsi-nezumi 311
— einderella 413
— electa 411
— elongate 405
— erica 410
— etrusca 383
— fimbripes 359
— fischeri 409
— flavescens 407
— — flavidula 407
— floveri 413
— foetida 405
— foucauldi 396
— foxi 410
— fuliginosa 404
— fulvovinerea 382
— fumosa 411
— fuscosa 406
— giffardi 405
— glebula 414
— gmelini 309, 310
— gracilipes 412
— grisescens 403
— giildenstaedti 292, 309, 310, 311, 389,
399, 400*
— hedenborgiana 406
— hildegardae 412
— hindei 408
— hirta 407
— hispida 404
— horsfieldi 85, 404
’ — hyrcana' 309, 310, 311, 390, 393*
‘ — ibeana 410
— ichnussae 396
— indochinensis 404
— jacksoni 412
— jouvenetae 85
— kingiana 404
— lamottei 85
_.. _ lasia, 309
— lasiura 309, 311, 389, 398, 399*
— lea 405
— leucodon 74, 85, 309. 311, 390, 395*,
396
— — caspica 394
— — lasia 311, 396
— — leucodon 396
— — ognevi 397
— — persica 394
— — sibirica 397
— leucodus 395
— leucogenis 382
— levicula 405
— lisenkani 398
— littoralis 414
— lixa 383
— longicaudata 399
— luna 410
• — lusitania 414
— lutrella 408
— lutreola 412
— maanjae 412
— macklotti 405
— macowi 412
— macropus 379
— major 396
— manni 406
— mariquensis 408
— martensi 411
— martiensseni 406
— maurisca 414
— media 382
— microtis 382
— mimula 392
— monticola 404
— morio 379, 380
— murina 84, 85
— muschata 382, 396
— nana 413
— nanilla 413
— neavei 415
— nigricans 410
— nigripes 405
— nigrofusca 410
— nilotica 409
— niobe 414
— nyansae 407
— obtusa 395
— occidentalis 85, 406
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
483
— odorata 406
— ognevi 397
— olivieri 407
— pamirensis 310, 311, 390, 400, 401*
— parvipes 412
— pascha 413
— pavlodarica 390, 391*
— pealana 382
— percivali 408
— pergrisea 309, 310, 311, 401, 403
— persica 309, 390, 394*
— petersi 406
• — pilosa 411
— planiceps 412
— platyrhyncha 359
— poensis 409
— poliogastra 396
— poudoensis 407
— praedax 403
— pulchella 417
— raineyi 411
— тарах 311, 404
— religiosa 414
— retusa 404
— rhoditis 405
— rubicunda 382
— rubricosa 404
— rufa 396
— russula 85, 292, 309, 311, 392, 396,
397*, 404
— — aralychensis 399
— — caspica 399
— — giildenstaedti 394, 399
— — leucodon 395
— — monacha 399
— russulus 399
— sacralis 407
— sansibarica 412
— schweizeri 406
— serezkyensis 309, 310, 400
— sericea 408
— sibirica 309, 311, 390, 397, 398*
— silacea 411
— silvia 415
— sindensis 382
— smithi 409
— somalica 413
— stoliczkana 382
— strauchii 408
— suahelae 409
— suaveolens 72, 85, 309, 310, 311, 383,
390, 392*
— — ilensis 393
— — shantungensis 393
— — suaveolens 393
— subfulva 382
— sururae 407
— tadae 404
— tatianae 403
— tenuis 405
— theresae 85
— thomasi 398
— turba 409
— ultima 415
— vetulina 408
— virilla 383
— voi 409
— vorax 311, 404
— waldemarii '382
— watasei 404
— Xanthippe 412
— yamashinai 398
— yebalensis 396
— zaphiri 407
— zarudnyi 311, 402*, 403
— zena 409
— zeylanica 382
— zorzii 391
Crocidurina 301*, 313, 315, 376
Crocidurinae 303, 304, 305, 307, 314, 371
Crocidurini 288, 290, 291, 293, 297, 299,
300, 301*, 302, 304, 307, 312, 315,
371, 372, 377, 419
crossei, Crocidura 413
Crossogale 308, 456
Crossopinae 308
Crossopus 308, 339, 425, 439, 456
— подрод 467
— fimbripes 359
— fissidens 426
— fodiens 440
— himalayicus 356
— sowerbyi 441
Cryptochloris 48
Cryptogale 46
Cryptoryctes 183, 190, 193*, 196, 198,
243, 246
— kayi 243*, 244
Cryptotis 4, 5, 35, 291, 293, 301*, 302,
303, 304, 305, 308, 313, 420,£421,
425, 431, 433
— alticola 435
— celatus 434
— endersi 435
— fossor 436
— frontalis 435
— goodwini 435
— gracilis 435
— griseoventris 435
— guerrerensis 435
— jacksoni 435
— kansasensis 433
— magna 431, 432, 436
— mayensis 436
PCryptotis meadensis 433
— mera 436
— merus 436
— mexicana 435
— micrura 436
— nelsoni 436
— nigrescens 437
— obscura 436
— olivacea 436
— orophila 437
• — parva 85, 433, 434*
— pergracialis 435
— soriciana 437
— tersus 437
— zeteki 436
cryptura, Talpa 276
cudahyensis, Sorex 336
csmiculus, Erinaceus algirus^l68
cupreata, Talpa 254
cyanea, Crocidura 411
Cyconiformes 79
cylin dricauda cylindricauda, Sorex 359
— gomphus, Sorex 359
— Sorex 331, -357, 358*
484
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
— — cylindricauda 359
— wardi, Sorex
danensis, Erinaceus 164
daphaenodon daphaenodon, Sorex 342
— sibiriensis, Sorex 344
— Sorex 309, 332, 342*
— — daphaenodon 342
— — sealoni 342
Dasogale 46
daubentonii, Sorex 440
dauricus dauricus, Erinaceus 175
— Erinaceus 27*, 104, 162, 168, 174*
— — dauricus 175
— Hemiechinus 104, 174
— Hemiechinus (?) 174
— mandschuricus, Erinaceus 175
davidianus, Scaptochirus 282, 284
davidi, Erinaceus 163, 164*
davisi, Proterixoides 95
dayi, Crocidura 382
— Suncus 382
dealbatus, Erinaceus 164
dehmi, Oligosorex 317*, 320
— Sorex 320
dehneli, «Sorex» 308
Deinogalerix 94, 99*, 104, 107, 118
— koenigswaldi 94, 118*, 119, 120*, 121*
Deinsdorfia 301*, 305, 308, 313, 455
— franconica 455*
dekayi, Sorex 424
delphinensis, Heterosorex 327
Deltatheridia 41
Deltatheridiidae 41
Deltatheridium praetrituberculare 41 *
Deltatheriidea 44*
Dendrogale 51
densicingulata, Shikamainosorex 429*
Dermoptera 44*
deserti blancalis, Paraechinus 176
— Crocidura 408
— Erinaceus 176
Desmogale 196, 199
Desmagnus 196
Desman 217
Desmana 183, 184, 187, 188, 192, 193*,
194, 196, 198, 204, 206, 208, 209,
211, 214, 217, 218, 224
— arvernien 231
— kormosi 219, 220, 225, 226
— moschata 59*, 68*, 69*, 187, 205,
207, 215, 218, 220*, 221*, 222*, 223*,
225, 226
— nehringi 218*, 219, 220, 225, 226
— pontica 214
thermalis 219*, 220, 225, 226
— verestchagini 212, 217, 218*, 220,
224, 225
Desmaninae 3, 30, 36*, 78, 183, 184, 186—
188, 190, 192, 193*, 194—199, 204,
206, 208, 212*, 227, 230, 231
Desmanini 198, 208, 209
Desmanus 217
diadematus, Erinaceus 168
Didymoconus colgageti 43*
dimidiatus, Erinaceus 168
Dimylechinus 89, 98, 99*, 100*, 102, 103,
105, 140, 145, 146, 156
— bernoullii 145, 146*, 147
dinniki, Crocidura 292, 310, 390, 402*
Diplomesodon 72, 292, 300, 301*, 304,
305, 307, 309, 313,371, 372, 376, 377,
391, 415
— fossorius 415, 416*, 417
\— pulchellum 59, 60*, 72, 74*, 309, 415,
' 416*, 417*
Dipodidae 29
discrepans, Soricella 322*, 325
dispar, Sorex 366
dixonensis, Planisorex 432*
— Sorex 432
dobsoni, Microgale 84
— Sorex 363
dolichochir, Scaptonyx 233*
dolichura, Crocidura 414
Domnina 290, 291, 297, 299, 301*, 303,
304, 307, 312, 318, 319, 324
— compressa 324
— gradata 321*, 324
— greeni 324
— thompsoni 321*, 324
Domninoides 184, 192, 193*, 196, 198,
228, 234
— riparensis 234, 235*
— valentinensis 235
doriae, Crocidura 405
doriana, — 407
— fuscosa, Crocidura 406
dorsalis albior, Erinaceus 176
— Erinaceus 176
dracula, Crocidura 403
Drepanosorex 304, 305, 307, 332
— подрод 332
driselda, Blarinella quadraticauda 371
dsi-nezumi, Crocidura 311
dsinezumi, Crocidura 404
dukelskie, Sorex 348
duwernoyi, Pachyura 382
Dymecodon 194, 195, 236
ecaudatus, Centetes 84
— Tenrec 22, 23*, 26*, 31*, 35, 87
Echinops 46
— telfairi 84
Echinosorex 4, 63, 89, 99*, 100*—104,
107, 124
Echinosorex 124
Echinosorex gymnurus 26, 28*, 56, 63,
77, 90, 94, 124, 125*, 126*, 129
Echinosoricinae 100*, 101, 102, 105
Echinosoricini 102, 103
echinus, Erinaceus 164
Edentata 44*, 246
edwardsi, Amphechinus 145, 148*, 149*—
152
— Palaeoerinaceus 145, 148
— Scaptonyx 233*
einderella, Crocidura 413
electa, Crocidura 411
elegans, Cayluxotherium 122
— Nectogale 459*, 460
Elephantulus 50
— brachyrchynchus 85
— rozeti 32*, 85
— rufescens 85
— rupestris 85
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
485
ellia, Corsira nuvara 379
elongate, Crocidura 405
endersi, Cryptotis 435
Entomolestes 101
Eoscalops 195, 196 , 272
— longirostris 275
— micrura 276
Episoriculus 304, 305, 308, 445
eremita, Sorex 440
erica, Crocidura 410
Ericius 102, 103, 160, 168
— подрод 169
Erinacei, подотряд 36*, 37, 44*, 49, 51
Erinaceidae 3, 5, 35, 36*, 37, 44, 51, 56,
62, 77, 85, 86, 88, 89, 92, 94, 95, 98,
99*, 101, 102, 104, 106, 110, 118, 134,
146, 180
Erinaceinae 4, 49, 52, 62, 76, 88, 89, 90,
92, 95, 96, 99*, 100*, 101—107, 111,
133 136 157 179
Erinaceini 86, 89, 97, 98, 99*, 100—103,
105, 135, 157
Erinaceota 49
Erinaceulus 103, 160, 168
Erinaceus 22, 89, 90, 96, 99,* 100*, 101—
105 116, 130, 131, 133, 142, 146, 147,
149, 150, 156—159, 160, 161, 162,
168, 175, 176, 179
— подрод 52, 87, 90, 99*, 104, 157, 163,
164, 167
Erinaceus aegyptius 169
— aethyopicus 163, 176, 1.77*, 178*
— — aethyopicus 178
— — hypomelas 178
— — micropus 179
— albiventris 167
— albulus 169
— albulus turanicus 169
— algirus 167
— — cuniculus 168
— — lavandeni 168
— — vagans 168
— amurensis 85, 164
— augolae 168
— aniscus 176
— arvernensis 147
— auritus 31*, 52*, 53*, 69*, 163, 169,
170* —174
— — aegyptius 173
— — albulus 173
— — auritus 172
— — caspicus 169
— — collaris 173
— — megalotis 173
— — turanicus 173
— blanfordii 176
— brachydactylus 176
— calligoni 169
— caniceps 164
— caninus 164
— capensis 167
— chinensis 164
— collaris 169
— concolor 164
— danensis 164
— danubicus 164
— dauricus 27*, 104, 162, 168, 174*
— — dauricus 175
— — mandschuricus 175
— — davidi 163, 164*
— daelbatus 164
— desrti 176
— diadematus 168
— dimidiatus 168
— dorsalis 176
— — albior 176
— echinus 164
— europaeus ,28*, 53*, 68*, 85, 104, 109,
•1'50? 160, 162, 163, 164, 165*, 166*,
168, 174, 179
— — amurensis 165, 167
— — concolor 167
— — europaeus 167
— — pallidus 167
— fallax 168
— faradjius 168
— fractilis 168
— frontalis 164*, 165, 167, 179
— grayi 169
— heterodactylus 167
— hindei 168.
— |hypomelas /176
— — eversmannii 176
erinaceus, Hystrix 164
Erinaceus indicus 169
— jerdoni 176
— kilimanus 168
— koreanus 164
— krayenbergi 164
— krugi 168
— langi 168
— lechei 164
— libicus 169
— macracantus 176
— megalotis 169
— micropus 176
— nanus 108
— niger 176
— — sabaeus 176
— — seniculus 176
— nudi ventris 176
— oeningensis 158
— orientalis 164
— pictus 176
— platyotis 169
— praeglacialis 164
— prisons 148
— pruneri 167
— — oweni 168
— rumanicus 85, 164
— sansaniensis 158
— sennaarensis 176
— sibiricus 164
— spatangus 169
— spiculus 168
— spinifex 168
— stramosi 164
— suillus 164
— syriacus 169
— tschifuensis 164
— unghi 164
— ussurlensis 164
— vireti 162*
Erinacini 133
Erinacoidea 37, 44, 95, 189
armineus, Hyracolestes 43*
etrusca, Crocidura 383
— Pachyura 383
486
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
etruscus etruscus, Suncus 385
— Sorex 392
— Suncus 20, 74*, 85, 292, 309, 380,
383, 384*
— — etruscus 385
europaea brauneri, Talpa 265
— europaea, Talpa 265
— obensis, Talpa 265
— Talpa 57*, 66, 68*—70*, 85, 192, 195,
197, 248, 259, 260, 261*, 263*, 267,
271, 272, 280, 281, 284
---cf. 262*
— — europaea 265
— — flavescens 254
— transuralensis, Talpa 265
eurepaeus amurensis, Erinaceus 165, 167
— concolor, — 167
Erinaceus 28*, 53*, 68*, 85, 104, 109,
150, 160, 162, 163, 164, 165*, 166, 168,
174, 179
— — europaeus 167
— europaeus, Erinaceus 167
— pallidus, Erinaceus 167
Euroscaptor 195, 196, 272
— grandis 273
— klossi 273
— parvidens 274
Eusorex 262
Eutheria 38, 39, 41, 44*
eversmannii, Erinaceus 176
Exallerix 89, 95, 97, 99*, 105, 140, 144
— hsandagolensis 144, 145*
excelsus, Sorex 362
exilipes, Blarina 434
exilis, Galerix 108*, 109, 110*, 111—
113, 116, 117, 180
— Sorex 353, 355
— Viverra 109
eximius, Blarina 434
falax, Erinaceus 168
fallax,. Sorex 334
faradjius, Erinaceus 168
Feroculus 292, 300, 301*, 304, 305, 307,
313, 376, 377, 379
— macropus 379
ferruginea, Tupaia 32*
ferrugineus, Sorex 382
fimbriatus, Sorex 396
fimbripes, Crocidura 359
— Crossopus 359
— Sorex 359
fischeri, Crocidura 409
fissidens, Beremendia 426*
— Crossopus 426
— Neomys 426
Fissipedia 44*
fissipes, Condylura 241
flava, Talpa 241, 254
flavescens, Crocidura 407
— flavidula, Crocidura 407
— Sorex 407
— Talpa europaea 254
flavidula, Crocidura flavescens 407
floridiana, Blarina 434
floveri, Crocidura 413
fluviatilis, Sorex 440
fodiens, Crossopus 440
— fodiens, Neomys 442
— Hydrosorex 441
— Neomys 61*, 62*, 72, 74*, 85, 187,
213, 309, 440
— — fodiens 442
— orientis, Neomys 442
— Sorex 439, 440
foetida, Crocidura 405
forsteri, Corsira 359
— Sorex 359
fossor, Blarina 436
— Cryptotis 436
fossorius, Diplomesodon 415, 416*, 417
foucauldi, Crocidura 396
foxi, Crocidura 410
fractilis, Erinaceus 168
franconica, Deinsdorfia 455*
frisius, Talpa 260
frontalis, Cryptotis 435
— Erinaceus 164*, 165, 167, 179
fuliginosa, Crocidura 404
fulvovinerea, Crocidura 382
fumeolus, Sorex wardi 357
fumeus, Sorex 85, 337, 365
fumidus, Soriculus 447
fumigatus, Sorex 399
fum.osa, Crocidura 411
furunculus, Ardynictis 43*
fusca, Talpa 254
fuscosa, Crocidura 406
— — doriana 406
fusicaudatus, Scaptonyx 232, 233, 234*
Gaillardia 184, 188, 193*, 196, 198, 206
— thomsoni 206*, 207
Gaillardini 198, 205
Galemys 183, 184, 187, 188, 193*, 194,
196, 198, 208, 209, 211, 212, 213, 218,
226, 236, 422, 439
— hungarica 212, 214
— micrurus 424
— ponticus 212*, 214
— pyrenaicus 33*, 69, 70*, 85, 204, 205,
209—212, 214, 215*, 216*, 217*, 219,
226
— semseyi 214, 219
Galericinae 4, 51, 62, 76, 88—90, 92, 93,
97, 99*, 101, 102, 104, 105, 107, 111,
119, 135
Galericini 102
Galerix 89, 93, 94, 99*, 100*, 101, 102,
104, 105, 107, 109, 111, 114—117,
119, 121, 136
— africanus 111, 112*, 114
— exilis 108*, 109, 110*, 111—113, 116,
117, 180
— minor 111
— paraexilis 112*, 113, 180
— tadzhikistanicus 112*, 113, 180
— viverroides 109
Galomys 196, 213
gaspensis, Sorex 365
gastellarini, Neomys 440
gemellus, Sorex 85
gemneus, Sylvisorex 381
Genoholotypus tasnadii 332
Geogale 46
Geolabidae 95
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
487
Geolabis 95
Geomana 196, 199, 207*
Geotripus 183, 189, 191, 193*, 194, 199,
257, 258, 259
— acutidens 259
— antiquus 191, 258, 259
— arambourdi 259
— jungi 258*, 259
gibbsi, Neurotrichus 85, 235, 238, 239*
— Urotrichus 238
gidleyi, Blarina 423, 424
giffardi, Crocidura 405
giganteus, Sorex 381
gigas, Notiosorex 448, 449
— Paranourosorex 463, 464*
glebula, Crocidura 414
Glires 44*
gmelini, Crocidura 309, 310
— Sorex 309, 310, 353
Gobiosorex 290, 291, 297, 301*, 312,
318, 320, 321
— kingae 320, 321*
goliath, Crocidura 388
— Praesorex 388, 391*
gomphys, Sorex bedfordiae 357
— — cylindricauda 359
goodwini, Cryptotis 435
gottentotus, Amblysomus 42*
gracilicauda, Soriculus 446
gracilidens, Sorex 333
— «Sorex» 308
gracilipes 412
gracilis, Cryptotis 435
— Nasillus 202
— Petenyiella 429
— Saturninia 316, 317*
— Sorex 428, 429
gracillimus, Sorex 309, 333, 352*, 353
— — minutus 353
gracillis, Suncus 383
gradata, Domnina 321*, 324
grandidens, Parascaptor 281
grandis, Euscaptor 273
— Talpa 272, 273
grantii, Sorex 339
granti, Sylvisorex 380
gratulus, Suncus 383
graves!, Sorex 356
grayi, Erinaceus 169
greeni, Domnina 324
griffithi, Sorex 382
griseo ven tris, Cryptotis 435
grisescens, Crocidura 403
grivensis, Miosorex 322*, 324
— Sorex 324
— — pusillus 324
Gryptotis 291
guerrerensis, Cryptotis 435
gusevi, Talpa altaica 267
giildenstaedti, Crocidura 292, 309—311,
389, 399, 400*
— — russula 394, 399
— Sorex 399
Gymnura 101, 103, 124
— birmanica 126
— raffles! 126
gymnura, Viverra 124, 126
Gymnurechinus 89, 97, 98, 99*, 103, 105,
147, 152, 154, 156
— camptolophus 155
— leakeyi 154, 155*
— songhorensis 156
Gymnurinae 101, 103, 105
gymnurus, Echinosorex 26, 28*, 56, 63,
77, 90, 94, 124, 125*, 126*, 129
hagermanensis, Sorex 337
hainanensis, Neohylomys 56, 77, 90, 132*
hantu, Chimarrogale 458
hawkeri, Sorex 355
hedenborgiana, Crocidura 406
Heliosorex 304, 307, 388
Hemicentetes 46
— nigriceps 84
— semispinosus 84
Hemiechinus 100, 102—104, 156, 160,
161, 168, 173, 175
— подрод 27*, 52, 87, 90, 99*, 104,
134, 157, 168, 169, 174—176
— albulus alaschanicus 169
— — major 169
— — minor 169
— auritus 85, 104, 168
— — aegyptius 174
— — albulus 174
— — auritus 174
— — chorosanicus 174
— — collaris 174
— — megalotis 174
— brachyotis 169
— chorosanicus 104
— dauricus 104, 174
— holdereri 169
— homalocanthus 169, 174
— hypomelas 85, 104
— mandschuricus 164
— megalotis 85, 104, 173
— mentalis 176
— microtis 169
— pallidus 176
— pectoralis 176
— persicus 169
— przewalskii 174
— russowi 169
heptapotamicus, Sorex minutus 355
hercegovinensis, Talpa 195, 197, 269,
271
H erinaceus 103, 160
hermanni, Amphisorex 441
— Sorex 339
Hesperoscalops 193*, 194, 199, 248, 252
— rexroadi 252, 253*
— sewardensis 252, 253*
Hesperosorex 291, 301*, 304. 305, 308,
313, 431
- - lovei 431
heterodactylus, Erinaceus 167
heterodon, Sorex 382
Heterosorex 291, 297, 301*, 303, 304,
306, 307, 312, 325, 326, 327, 329
— delphinensis 327
— nemayrianus 327
— roperi 327
— sansaniensis 327, 328*
Heterosoricinae 304, 307, 326
Heterosoricini 74, 290, 291, 299, 301*,
312, 314, 326
488
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
hibbardi, Sorex 336
bidalgo, Nesiotites 452
Hidrosorex fodiens 441
hildegardae, Crocidura 412
himalayicus, Chimarrogale 456
— Crossopus 456
hindei, Crocidura 408
— Erinaceus 168
hirta, Crocidura 407
hispida, Crocidura 404
holdereri, Hemiechinus 169
holmani, Tregosorex 367, 368*
holosericous, Sorex 446
homalocanthus, Hemiechinus 169, 174
Homalurus 307, 331
homourus, Sorex 446
horncloudi, Amphechinus 152
— Palaeoerinaceus 152
horsfieldi, Crocidura 85, 404
hoyi intervectus, Microsorex 367
— Microsorex 85, 361*, 367
— Sorex 366
hsandagolensis, Exallerix 144, 145*
hungarica, Galemys 212, 214
— Mygalinia 211*, 212, 213
— Petenyia 427*, 428
hypsibius, Notiosorex 450*
hydrodromus, Sorex 365
Hydrogale 307, 308, 331, 439
hydrophilus, Sorex 440
Hydroscapheus 184, 188, 193*, 196, 198,
207, 208
— americanus 207*
Hydrosorex 308, 439
Hydro-Sorex 331
Hydrosoricidae 308
hydruntina, Sorex 396
Hylomyidae 103
Hylomys 4, 89, 93, 99*, 100*, 101—104,
107, 109—111, 127, 132
— suillus 91, 115, 127*, 128*, 129, 131
hypomelas, Erinaceus 176
— — aethyopicus 178
— eversmannii, Erinaceus 176
— Hemiechinus 85, 104
hypsibia parva, Chodsigoa 450
hypsibius, Soriculus 450*, 452
Hyracoidea 44*
Hyracolestes ermineus 43*
hyrcana, Crocidura 309—311, 390, 393*
Hystrix erinaceus 164
ibeana, Crocidura 410
ichnussae, Crocidura 396
ignotus, Sorex 339
ilensis, Crocidura suaveolens 393
incerta, Talpa 142
indicus, Erinaceus 169
— Sorex 381
indochinensis, Crocidura 404
inexpectatus, Anourosorex 465, 466*
— — conf. 465
— Neomys 467
infinitesimus, Suncus 383
inflatus, Scalopus 256
Jngentisorex 297, 301*, 305, 307, 312,
325, 329
— tumididens 329
inodorus, Sorex 396
Insectivora 20, 24, 39, 41, 44*, 84, 102,
311
insertis, Brachyerix 142
insularis, Mogera 85
intermedius, Amphechinus 149*
— Sorex 346
intervectus, Microsorex hoyi 367
investigator, Nasillus 202
irene, Soriculus 447
isodens, Scalopoides 230*, 231, 232
isodon isodon, Sorex 356
— ruthenus, Sorex 356
— Sorex 85, 309, 333, 356, 357*
— — araneus tomensis 356
— — isodon 356
jacksoni, Crocidura 412
— Cryptotis 435
— Notisorex 423, 448
— Sorex 365
jacutensis, Sorex araneus 348
— — vir 349
japonicus, Anourosorex 465, 466
jenissejensis, Sorex 344
jerdoni, Erinaceus 176
johnstoni, Sylvisorex 380
jouvenetae, Crocidura 85
juncensis, Sorex 364
— — californicus 364
jungi, Geotripus 258*, 259
kandianus, Sorex 382
kansasensis, Cryptotis 433
kansuensis, Amphechinus 150*, 151
— Palaeoerinaceus 150
kayi, Cryptoryctes 243*, 244
kelaarti, Sorex 382
kilimanus, Erinaceus 168
kingae, Gobiosorex 320, 321*
kingiana, Crocidura 404
klossi, Euroscaptor 273
— Talpa 195, 271, 272, 273, 274
koenigswaldi, Deinogalerix 94, 118*, 119,
120*, 121*
koceanus, Erinaceus 164
koreni, Sorex caecutiens 352
kormosi, Desmana 219, 220, 225, 226
kozlovi, Sorex 361*, 362
krayenbergi, Erinaceus 164
kretzoi, Sorex 335
krugi, Erinaceus 168
kubinyi, Soriculus 445, 446*
kura, P’odihik 452, 453*
kutscheruki, Sorex mirabilis 348
labiosus, Sorex 339
lacustris, Sorex 336
Lagomorpha 44*
lamottei, Crocidura 85
lamula, Chodsigoa 450
langi, Erinaceus 168
Lantanotherium 101, 103
Lanthanotherium 89, 94, 99*, 100*, 101—
104, 107, 108, 115, 116, 117, 119
Lanthanotherium? 89
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
489
Lanthanotherium longirostrum 116, 117*
— robustum 116*
— sansaniensis 115, 116
— sp. 116, 117*
larvarum, Chodsigoa 450
lasia, Crocidura 309
— — leucodon 311, 396
lasiura, Crocidura 309, 311, 389, 398,
399*
latimanus, Scapanus 85, 252
latouchei, Mogera 195, 279
la vandeni, Erinaceus algirus 168
lea, Crocidura 405
leakeyi, Gymnurechinus 154, 155*
leander, Chimarrogale 457
lechei, Erinaceus 164
leptura, Parascaptor 281
— Scaptochirus 281, 284
— Talpa 281, 284
lesueurii, Amphisorex 359
Leucodon 307, 388
leucodon caspica, Crocidura 394
— Crocidura 74*, 85, 309, 311, 390,
395*, 396
— — leucodon 396
— — russula 395
— lasia, Crocidura 311, 396
— leucodon, Crocidura 396
— ognevi, Crocidura 397
— persica, Crocidura 394
— sihirica, Crocidura 397
— Sorex 388, 392, 395, 399
leucod us, Crocidura 395
leucogaster, Sorex 353
leucogenis, Crocidura 382
leucops, Sorex 447
— Soriculus 447
Leucorhynchus
leucura, Parascaptor 195, 280*, 281, 284
— Suncus 383
— Talpa 280
leucurus, Sorex 396
levicula, Crocidura 405
libicus, Erinaceus 169
limneana, Amphisorex 441
Limnoecinae 305, 307
Limnoecus 291, 297, 301*, 304, 305, 307,
313, 377, 386
— niobrarensis 372, 373*
— tricuspis 386, 391*
Limnogale 46
lineatus, Sorex 440
liricatus, Sorex 440
lisenkani, Crocidura 398
littoralis, Crocidura 414
lixa, Crocidura 383
lixus, Suncus 383
longicaudata, Condylura 241
— Crocidura 399
— Microgale 42*
— Talpa 240
longicaudatus, Rhinaster 241
longirostris, Eoscalops 275
— Sorex 363
— Talpa 195, 271, 272, 273*, 274*, 275,
276
longirostrum, Lanthanotherium 116, 117*
loomsi, Proterix 137*, 138
lovei, Hesperosorex 431
32 А. А. Гуреев
— Atelerix pruneri 168
— Chodsigoa 451
— Notiosorex 451
ludlowi, Paraechinus 176
lima, Crocidura 410
lunaris, Sylvisorex 378, 380
lusitania, Crocidura 414
lutrella, Crocidura 408
lutreola, Crocidura 412
lyelli, Sorex 363
maanjae, Crocidura 412
macklotti, Crocidura 405
macowi, Crocidura 412
macracantus, Erinaceus 176
macrodon, Sorex 364
Macroechinus 102, 103, 160, 176
Macroneomys 301*, 313, 439
— brachygnathus *439*
macropus, Crocidura 379
— Feroculus 379
— Sorex 379
macropygmaeus, Sorex 344, 351
— — caecutiens 353
Macroscelides 49, 50
Macroscelididae 3, 25, 27, 29, 35, 36*,
37, 45, 49, 85, 130
Macroscelidina 49
Macroscelidoidea 37, 44
macrotis, Brachyerix 140, 141*, 142, 144
macrotrichus, Sorex 339
macroura, Condylura 241
macrurus, Rhinaster 241
— Sorex 366, 440, 447
— Soriculus 447
madagascarensis, Suncus 383
major, Crocidura 396
— Hemiechinus albulus 169
makpiyahe, Ocajila 119, 121*
malabaricus, Suncus niger 382
Mamblarinaus 308
mandschuricus, Erinaceus dauricus 175
— Hemiechinus 164
manni, Crocidura 406
mariae, Blarinoides 421, 422
mariquensis, Crocidura 408
marslandensis, Metechinus 143, 144
Marsupialia 41
martensi, Crocidura 411
martiensseni, Crocidura 406
martini, Medicodon 230*, 232
maurisca, Crocidura 414
mauritanicus, Sorex 396
mayensis, Cryptotis 436
magna, Blarina 436
— Cryptotis 431
meadensis, Cryptotis? 433
media, Crocidura 382
mediterraneus, Neurogymnurus 122*, 123
megalotis, Erinaceus 169
— — auritus 173
— Hemiechinus 85, 104, 173
— — auritus 174
megalura, Pachyura 381
— Sylvisorex 62*, 381
Megasorex 304, 305, 308, 447
melanodon, Sorex 339
meltoni, Sorex 338
490
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
mentalis, Hemiechinus 176
тега, Cryptotis 436
merriami, Sorex 336, 366
merus, Cryptotis 436
Mesechinus 103, 168
— подрод 103, 174
Mesonix obtusidens 43*
Mesoscalops 183, 191, 193*, 196, 198,
247
— scopelotemos 245, 247
Metacodon 101
metallescens, Scalops 251
Metatheria 38, 39, 41, 44*
Metechinus 89, 99*, 100*, 101—104,
140, 142
— fergusoni 142
— marslandensis 143, 144
— nevadensis 142, 143*, 144
Meterix 101
Meterotalpa 196
mexicana, Cryptotis 435
meyeri, Oligosorex 320
micheli, Paratalpa 229, 230*
Microgale 46
— cowan ii 84
— dobsoni 84
— longicaudata 42*
— talazaei 84
Micropotamogale 47
micropus, Erinaceus 176
— — aethyopicus 179
Microsorex 290, 291, 293, 300, 301*,
303—305, 307, 312, 331, 366
— hoyi 85, 361*, 367
— — intervectus 367
— pratensis 367
microtis, Crocidura 382
— Hemiechinus 169
micrura, Cryptotis 436
— Eoscalops 276
— Talpa 195, 271, 272, 275*, 276
micrurus, Galemys 424
— Sorex 436
Midecodon 183, 191, 193*, 196
— martini 230*, 232
milleri, Neomys 443
— Sorex 363
mimula, Crocidura 392
minima, Talpa 197, 270
— — соеса 271
minimus, Amphechinus 150*, 151
— Palaeoerinaceus 151
— Sorex 353, 355
minor, Galerix 111
— Hemiechinus albulus 169
— Soriculus 446
minuta, «Talpa» 260*
— Talpa? 260
minutissimus saudatus, Sorex .356
— minutissimus, Sorex 356
— Sorex 20, 85, 294, 309, 333, 353, 355*
— — minutissimus 356
— tscherskii, Sorex 356
— tschuktschorum, Sorex 356
minutus gracillimus, Sorex 353
— heptapotamicus, Sorex 355
— minutus, Sorex 355
— Sorex 85, 309, 310, 333, 353, 354*,
362
— .minutus 355
— thibetanus, Sorex 362
— Tupaiodon? 136
miocenus, Proscalops 243*, 244, 245
Mioechinus 89, 99*, 100*, 102—103, 105,
156, 157, 158
— oeningensis 158*
— sansaniensis 158
Miosorex 291, 297, 301*, 305, 307, 312,
318, 324
— grivensis 322*, 324
— swinnyi 374
mirabilis kutscheruki, Sorex 348
— mirabilis, Sorex 348
— Sorex 309, 332, 347*
— — mirabilis 348
Mogera 25, 184, 193*, 194—197, 199, 257,
272, 276
— подрод 277
— insularis 85
— latouchei 195, 279
— mogera 279
mogera, Mogera 279
Mogera robusta 33*, 68*—70*, 195, 197,
259, 276, 277, 278*
— wogura 195, 197, 245, 259, 276, 277*
— — coreana 277
mokrzeckii anomalus, Neomys 444
monacha, Crocidura russula 399
monsvairant, Soricoidus 339
montanus, Amphechinus 152
— Parvericius 152
— Sorex 382
monticola, Crocidura 404
morio, Crocidura 379, 380
— Sylvisorex 378, 380
morrisi, Tupaiodon 135, 136*
moschata, Desmana 59*, 68*, 69*, 187,
205, 207, 215, 218, 220*, 221*—223*,
225, 226
— Myogale 194, 220
moschatum, Mygale 220
moschatus, Castor 217, 220
— Scaptochirus 195, 281, 282, 283*, 284
— Sorex 220
moschiferus, Scaptochirus 282
moscovitica, Mygale 220
mundus, Sylvisorex 380
murina, Crocidura 84, 85
murinus, Sorex 381
— Suncus 60*, 84, 85, 378, 381
Mus araneus 331
Musaraneus 307
muschata, Crocidura 382, 396
musculus, Sorex 440
Myaladae 196
Mydecodon 196, 228, 232
Mygale 194, 196, 210, 217
— arvernensis 231
— moschatum 220
— moscovitica 220
— nayadum 231
— pyrenaica, 213 214
— pyrenaicus 211
Mygalea 183, 187, 188, 194, 196, 198, 209,
210, 224
— antiqua 210, 211*
— asiatica 211*, 212
— sansaniensis 211
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
491
Mygalina 213
Mygalinia 183, 184, 187, 188, 193*, 194,
196, 198, 209, 212, 224
— hungarica 211*, 212, 213
— zazhigini 211*, 213
Mygatalpa 183, 190, 193*, 196—198,
230
— arvernensis 230, 231
Myogale 196, 217
— moschata 194, 220
— moscovitica 220
— pyrenaica 194
Myogalea 217
— rufula 214
Myogalidae 194
Myogalina 194
myoides, Sorex 404
Myoladae 194
Myolade 194
Myosictis 308, 439
Myosorex 72, 290—293, 300, 301*, 303—
305, 307, 313, 320, 372, 373, 374—
376, 379—381
— babaulti 29*, 375
— blarina 374
— cafer 374
— polli 375
— preussi 375
— sceateri 374
Myosorex? sorella 381
Myosorex tenuis 374
— varius 373*, 374
Myosoricina 312, 315, 372
Myosoricini 304, 371, 372
myosurus, Sorex 381
Mystipterus 303
Myxomygale 183, 190, 193*, 196, 198,
228, 229
— antiqua 228, 229*
nana, Crocidura 413
nanilla, Crocidura 413
nanus, Erinaceus 108 -
— Sorex 364
— — tenellus 364
— Tetracus 107, 108, 109, 111
Nasilio 50
Nasillus 4, 184, 193*, 194, 195, 198, 201,
202, 204
— gracilis 202
— investigator 202
natans, Sorex 440
navigator alascanus, Sorex 365
nayadum, Mygale 231
neavei, Crocidura 415
Necrogymnurus 101, 103
Necrosorex 303
Nectogale 61, 292, 301*, 302, 304, 305,
308, 313, 314, 459, 455
— elegans 459*, 460
Nectogalina 291, 301*,’302, 313, 315,
419, 455
Nectogalinae 308
Nectogalinia 291, 301*, 313, 458
— altaica 458*
neglecta, Petenyia 427
nehringi, Desmana 218*, 219, 220, 225,
226
nelsoni, Cryptotis 436
nemayarianus, Heterosorex 327
nemorivagus, Sorex 382
Neohylomys 4, 90, 99*, 103, 104, 107,
132
— hainanensis 56, 77, 90, 132*
Neomyini 305, 308, 421
Neomys 61, 81, 292, 301*, 302—305,
308, 309, 313, 420, 421, 425, 438, 439,
. 456
— anomalus>85, 292, 309, 440, 443, 444*
— — mokrzeckii 444
— argenteus 441
— bohlini 452
— fissidens 426
— fodiens 61*, 62*, 72, 74*, 85, 187,
213, 309, 440, 441*
— — fodiens 442
— — orientis 442
— gastellarini 440
— inexpectatus 467
— milleri 443
— schelkovnikovi;292, 309, 440, 442, 443*
— sinensis 452
— soricoides 443
— soriculoides 438
— teres 443
Neosorex 304, 307, 331, 336
— bendirii albiventer 336
Neotetracus 4, 89 99*, 100*, 101—105,
107, 109, 111, 130
— sinensis 91, 116, 130, 131*
Nesiotites 304, 305, 308, 452
— corsicanus 452
— hidalgo 452
— similis 452
neumarianus, Sorex aff. 325
Neurogymnurinae 100*, 102
Neurogymnurini 102, 103
Neurogymnurus 89, 92, 93, 97, 99*, 100*,
101—104, 107, 109, 120, 179
— cayluxi 120, 121*, 122*, 123
— mediterraneus 122*, 123
Neurotrichus 184, 194—196, 198, 228,
238, 242
— gibbsi 85, 235, 238, 239*
nevadensis, Metechinus 142, 143*, 144
— Sorex 363
niger, Erinaceus 176
— malabaricus, Suncus 382
— sabaeus, Erinaceus 176
— seniculus, Erinaceus 176
— Sorex 382 #
nigrescens, Blarina 437
— Corsira 445, 446
— Cryptotis 437
— Soriculus 446
nigricans, Crocidura 410
nigriceps, Hemicentetes 84
nigripes, Crocidura 405
— Sorex 440
nigrofusca, Crocidura 410
nilotica, Crocidura 409
niobe, Crocidura 414
niobrarensis, Limnoecus 372, 373*
nivicola, Sorex 447
norae, Surdisorex 373, 375
Notiosorex 291—293, 301*, 302—305, 308,
313, 421, 438, 445, 447, 448, 450, 452
32*
492
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
— подрод 301*, 313, 447, 448, 449, 450,
452
— crawfordi 85, 448, 449*
— gigas 448, 449
— hypsibius 450*, 452
— jacksoni 423, 448
— lowei 451
— salenskii 451*
— smithi 451
nudiventris, Erinaceus 176
nuvara ellia, Corsira 379
nyansae, Crocidura 407
Nyctitheriidae 289, 296
obensis, Talpa europaea 265
obscura, Cryptotis 436
— Sorex 337
obtusa, Crocidura 395
obtusidens, Mesonix 43*
Ocajila 89, 99*, 103, 104, 107, 119
— makpiyahe 119, 121*
occidentalis, Crocidura 85, 406
— Pachyura 406
odorata, Crocidura 406
odoratus, Sorex 406
oeningensis, Erinaceus 158
— Mioechinus 158*
ognevi, Crocidura 397
— — leucodon 397
— Talpa romana 197, 268
oligodon, Amblycoptus 462*, 463
Oligoscalops 183, 190, 193*, 196, 198,
246
— whitmanensis 246, 247
Oligosorex 290, 291 297, 301*, 304, 307,
312, 318, 319, 320, 324
— antique 317*, 320
— dchmi 317*, 320
— meyeri 320
— pusillus 320
Oligosoricini 290, 291, 296, 297, 301*,
312, 314, 318, 326
olivacea, Cryptotis 436
olivieri, Crocidura 407
— Sorex 407
olula, Sylvisorex 380
olympica, Talpa 269
orangie, Suncus 383
orarius, Scapanus 252
oreopolus, Sorex 366
oriental is, Erinaceus 164
— Talpa coeca 271
orient is, Neomys fodiens 442
oriundus, Sylvisorex 380
orlovi,.Similisorex 377, 378*
ornatus, Sorex 363
orophila, Blarina 437
— Cryptotis 437
Oryeteri 196
Oryzorictes 46
— tetradactylus 20
Oryzorictinae 46
Otisorex 304, 307, 331
— platyrhynus 359
oweni, Erinaceus pruneri 168
— Scapanulus 238
Oxirin 307, 331
pachyodon, Sorex 334
Pachyura 303, 304, 307
Pachyura? 428
Pachyura, группа 304
— подрод 381—383
— aequatorialis 406
— agilis 396
— caerulla riukiuana 382
— duwernoyi 382
— etrusca 383
— megalura 381
— occidentalis 406
— pygmaea 396
— suaveolens 392
(Pachyura) waldemarii 382
pachyurus, Sorex 343, 383
pacificus, Sorex 347, 363
Palaeoerinaceus 101, 103, 145, 147, 151,
159, 162
— acridens 139
— edwardsi 145, 148
— horncloudi 152
— kansuensis 150
— minimus 151
— rectus 149
— vireti 162
Palaeoscaptor 89, 95. 97, 99*, 101, 103,
104, 135, 138, 156
— acridens 138, 139*
— rectus 149
Palaeospalax 196
Palaerinaceus 103
Paleoerinaceus 103
Palerinaceus 103, 161
pallidus, Erinaceus europaeus 167
— Hemiechinus 176
palustris, Sorex 83, 330, 336, 364, 365
pamirensis, Crocidura 310, 311, 390,
400, 401*
Pantotheria 38, 41, 44*
Paracrocidura 292, 300, 301*, 305, 307,
313, 377, 387, 406
— schoutendeni 387, 391*
Paracryptotis 291, 301*, 304, 305, 308,
313, 425, 429, 431, 432
— rex 429, 431, 433
Paradomnina 297, 301*, 304, 307, 312,
318, 324
— relictus 324
Paradoxodon 307
— подрод 381
paradoxus, Solenodon 23*, 26*, 84
Paraechinus 100*, 102, 103, 156, 160,
161
— подрод 52, 87, 90, 99*, 157, 176, 178,
179
— anur 176
— deserti blancalis 176
— ludlovi 176
paraexilis, Galerix 112*, 113, 180
Paranourosorex 291, 301*, 313, 461, 463
— gigas 463, 464*
Parascalops 184, 193*, 194—196, 199,
248, 249
— breweri 85, 249*, 250
Parascaptor 184, 193*, 194—196, 199,
257, 272, 279, 280, 281, 284, 285
— grandidens 281
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
493
— leptura 281, 284
— leucura 195, 280*, 281, 284
Parasorex 101, 102
— socialis 109
Parasoricidae 102, 105
Paratalpa 183, 190, 193*, 196, 198, 228,
229
— micheli 229, 230*
parva, Chodsigoa 450
— — hypsibia 450
— Cryptotis 85, 433, 434*
Parvericius 100*, 101—103, 147, 152,
156
— montanus 152
parvidens, Euroscaptor 274
— Talpa 272. 274
parvipes, Crocidura 412
parvus, Sorex 433, 434
Parydosorex concisus 427
pascha, Crocidura 413
Paurodus 307, 388
pavlodarica, Crocidura 390, 391 *
pealana, Crocidura 382
pearsoni, Solisorex 387, 388
pectoralis, Hemiechinus 176
Peisorex 291, 301*, 304, 305, 313, 430
— pohainensis 430*
— sp. 430
pensylvanica, Scalops 254
— Talpasorex 254
pensylvanicus, Talpasorex 254
percivali, Crocidura 408
pergracialis, Cryptotis 435
pergrisea, Crocidura 309—311, 401, 403
Perissodactyla 44*
Peroechinus 103, 160
persica, Crocidura 309, 390, 394*
— — leucodon 394
persicus, Hemiechinus 169
personatus, Sorex 339, 359
Petenyia 291, 301*, 304, 305, 308, 313,
425, 427
— concisa 369, 427. 428*
— hungarica 427*, 428
— neglecta 427
— sp. 428
Petenyiella 291, 301*. 304, 305, 308, 313,
428, 429
— gracilis 429
petersi, Crocidura 406
Petrodromus 50
phacura, Chimarrogale 458
Pholid ota 44*
pictus, Erinaceus 176
pidoplitschkoi, Anourosoricodon 461*
pilorides, Sorex 381
pilosa, Crocidura 411
Pinalia 308, 439
Pinnipedia 44*
piveteaui, Crocidosorex 322, 323
planiceps, Crocidura 412
— Sorex 353, 362
Planisorex 291, 301*, 313, 432
— olixonensis 432*
platycephala, Chimarrogale 457*, 458
platycranius, Sorex 348
— — vir 349
platyotis, Erinaceus 169
platyrhyncha, Crocidura 359
platyrhynchus, Sorex 359
platyrhynus, Otisorex 359
Plesiosorex 107
pleskei, Sorex caecutiens 352
Pleurodus 307, 381
Podinik 290, 292, 301*, 305, 308, 313, 419,
437, 452
— kura 452, 453*
Podogymnura 4, 63, 89, 99*, 100*, 101—
105, 107, 129, 130
— truei 129*, 130
poensis, Crocidura 409
— Sorex 409
pohainensis, Peisorex 430*
poliogastra, Crocidura 396
polli, Myosorex 375
polulus, Surdisorex 376
pondoensis, Crocidura 407
pontica, Desmana 214
ponticus, Galemys 212*, 214
Postpaleoerinaceus, подрод 161, 152
Postpalerinaceus 100, 103, 156, 161
Potamogale 47
Potamogalidae 3, 27, 29, 30, 35, 36*, 37,
40, 44, 47
powersi, Sorex 337
praealpinus, Sorex 334
praearaneus, Sorex 335
praedax, Crocidura 403
praeglacialis, Erinaceus 164
Praesorex 292, 301*, 304, 305, 307, 313,
377, 387, 388
— goliath 388, 391*
praetrituberculare, Deltatheridium 41 *
prasinatus, Astromycter 241
pratensis, Microsorex 367
preblei, Sorex 337, 363
preussi, Myosorex 375
pribilofensis, Sorex 343, 365
primaevus, Amphisorex 296
— Scaptochirus 282*
Primates 44*
priscus, Erinaceus 148
Proboscidea 44*
Proscalopinae 192, 196, 197, 242
Proscalopini 183, 190, 191, 193*, 198,
200, 242, 247
Proscalops 183, 190, 191, 194, 196, 198,
242, 244, 246, 247
— tertius 245
— miocenus 243*, 244, 245
— secundus 245
Proscapaneus 196
Proscapanus 183, 191, 193*, 194, 196,
199, 248
— sansaniensis 248
Protalpa acutidentata 258
— codurcensis 259
Protechinus 99*, 103, 105, 157, 158, 159,
160
— salis 159
Protericini 89, 95—97, 99*, 102—104,
133 135
Proterix 89, 95—97, 99*, 100*, 101—104,
135, 136, 137, 138
— bicuspis 138*
— loomsi 137*, 138
Proterixoides 95, 100*, 101
— davisi 95
494
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Protosorex 307
Prototheria 38, 39, 44*
pruneri, Erinaceus 167
— loweli, Atelerix 168
— oweni, Erinaceus 168
przewalskii, Hemiechinus 174
Pseudogalerix 89, 94, 99*, 100*, 101—
104, 107, 114, 115, 116
— stehlini 111, 112, 114, 116
Pseudoneurogymnurus 89, 99*, 104, 123,
179
— schevyrevii 123*
— zhchikvadzei 123*, 124
Pseudotrimylus 291, 297, 301*, 312, 326,
327
— roperi 327, 328*, 329
psilurus, Sorex 440
Ptilocercinae 51
Ptilocercus 51, 135, 136
pulchella, Crocidura 417
pulchellum, Diplomesodon 59, 60*, 72,
74*, 309, 415, 416*, 417
pulchellus, Sorex 415, 417
pumilio, Sorex 353
pumilus, Sorex 353
purpurascens, Talpa 241
pusillus grivensis, Sorex 324
— Oligosorex 320
— Sorex 320
pygmaea, Pachyura 396
pygmaeus, Sorex 310, 353, 355
Pygmura 308
pyrenaica, Mygale 213, 214
— Myogale 194
pyrenaicus, Galemys 33*, 69, 70*, 85,
204, 205, 209—212, 214, 215*, 216*,
217*, 219, 226
— Mygale 211
quadraticauda, Blarinella 370*
— — quadraticauda 371
— driselda, Blarinella 371
— quadraticauda, Blarinella 371
— Sorex 369
— wardi, Blarinella 371
quadraticaudatus, Sorex 339
raddei, Sorex 85, 292, 309, 332, 345*
radiata, Talpa 241
radiatus, Sorex 241
radulus, Soriculus 446
rafflesi, Gymnura 126
raineyi, Crocidura 411
тарах, Crocidura 311, 404.
rectus, Amphechinus 149, 150*, 151, 152,
157
— Palaeoerinaceus 149
— Palaeoscaptor 149
relictus, Paradomnina 324
religiosa, Crocidura 414
remifer, Sorex 440
reposta, Talpa 250
retusa, Crocidura 404
rex, Paracryptotis 429, 431, 433
rexroadensis, Sorex 337, 433
rexroadi, Hesperoscalops 252, 253*
Rhinaster 240
— crictatus 241
— longicaudatus 241
— macrurus 241
rhindophus, Sorex 339
Rhinomus 307
rhoditis, Crocidura 405
Rhynchocyon 49, 50
Rhynchonax 4, 184, 193*, 194, 195, 198,
201, 204
— andersoni 201
richardsonii, Sorex 343
riparensis, Domninoides 234, 235*
riukiuana, Pachyura caerulla 382
rivalis, Sorex 440
roboratus, Sorex 309, 339
— — araneus 341
robusta, Mogera 33*, 68*, 69*, 70*, 195,
197, 259, 276, 277, 278*
robustum, Lanthanotherium 116*
romana ognevi, Talpa 197, 269
— Talpa 195, 259, 268, 269*, 271, 272
roperi, Heterosorex 327
— Pseudotrimylus 327, 328*, 329
rostrata, Tshelkaria 43*
rozeti, Elephantulus 32*, 85
ruande, Sylvisorex 381
rubicunda, Crocidura 382
rubricosa, Crocidura 404
rufa, Crocidura 396
rufescens, Elephantulus 85
rufula, Myogalea 214
rumanicus, Erinaceus 85, 164
runtonensis, Sorex 334, 335
rupestris, Elephantulus 85 .
rusingensis, Amphechinus 151*
russowi, Hemiechinus 169
russula aralychensis, Crocidura 399
— caspica, Crocidura 399
— Crocidura 85, 292, 309, 311, 392, 396,
397*, 404
— guldenstaedti, Crocidura 394, 399
— leucodon, Crocidura 395
— monacha, Crocidura 399
russulus, Crocidura 399
— Sorex 396
rusticus, Sorex 353
ruthenus, Sorex isodon 356
rutilus, Sorex 407
sabaeus, Erinaceus niger 176
sacer, Sorex 382
— Suncus 381
sacralis, Crocidura 407
salairica, Talpa altaica 267
salenskii, Chodsigoa 451
— Notiosorex 451*
— Soriculus 451
salis, Protechinus 159
samniticus, Sorex 339
sandersi, Sorex 338
sanguinidens, Sorex 344
— — araneus 345
sansaniensis, Erinaceus 158
— Heterosorex 327, 328*
— Lanthanotherium 115, 116
— Mioechinus 158
— Mygalea 2111
— Prosca panus 248
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
495
— Sorex 326, 327
— Talpa 248
sansibarica, Crocidura 412
santonus, Sorex 339
satunini, Sorex araneus 341
saturatior, Sorex 382
Saturninia 291, 296, 301*, 303, 312, 315,
316, 325
— gracilis 316, 317*
Saturniniinae 290, 296, 297, 301*, 312,
314, 315, 316
saussurei, Sorex 338, 366
savini, Sorex 332
— austriacus, Sorex 332
scaloni, Sorex daphaenodon 342
Scalopidae 194, 247
Scalopinae 192, 194, 195, 197, 247
Scalopini 53, 181, 183, 184, 191, 192, 193*,
197, 198, 200, 247
Scalopoides 183, 191. 193*, 194, 198, 231
— isodens 230*, 231, 232
Scalops 197, 253
— подрод 254
— aeneus 251
— breweri 249
— californicus 252
— canadensis 251, 254
— metallescens 251
— pensylvanica 254
scalops, Talpa 261
Scalops texanus aereus 254
— townsendii 250, 251
Scalopus 184, 192, 193*, 194, 195, 197,
199, 247, 248, 250, 252, 253
— aereus 254
— aquaticus 192, 253, 254, 255*
— cristatus 241
— inflatus 256
— vigrinianus 254
Scapanei 197
Scapanulus 184, 193*, 194, 195, 196, 198,
228, 238
— oweni 238
Scapanus 184, 191, 193*, 194, 195, 199,
248, 250
— breweri 250
— latimanus 85, 252
— orarius 252
— townsendii 251, 252
Scapasius 250
Scaphanus 250
— breweri 250
Scaptochirus 183, 184, 193*, 194—196,
199, 257, 272, 281, 284
— davidianus 282, 284
— leptura 281, 284
— moschatus 195, 281, 282, 283*, 284
— moschiferus 282
• — primaevus 282*
Scaptogale 199
Scaptonychini 196, 197
Scaptonyx 4, 183, 184, 191, 193*, 194—
198, 228, 232, 234
— dolichochir 233*
— edwardsi 233*
— fusicaudatus 232, 233, 234*
schelkovnikovi, Neomys 292, 309, 440,
442, 443*
schevyrevii, Pseudoneurogymnurus 123*
schlosseri, Sorex 326
schlosseri, Trimylus 326
schnitnikovi, Sorex araneus 345
schoutedeni, Paracrocidura 387, 391*
schweizeri, Crocidura 406
sclateri, Myosorex 374
— Sorex 366, 374
scopelotemos, Mesoscalops 245, 247
Scutisorex 72, 287, 292, 293, 301*, 304,
305, 307, 313, 314, 377, 385, 386
— congicus 384*, 386
— • somereni 385, 386
Scutisoricinae 303, 304, 314, 371
Scutisoricini 307
secundus, Proscalops 245
semispinosus, Hemicentetes 84
semseyi, Galemys 214, 219
seniculus, Erinaceus niger 176
sennaarensis, Erinaceus 176
serezkyensis, Crocidura 309, 310, 400
sericea, Crocidura 408
sericeus, Sorex 408
serpentarius, Sorex 382
Setifer 46
— setosus 84
Setiger 103
setosus, Setifer 84
sewardensis, Hesperoscalops 252, 253
shantungensis, Crocidura suaveolens 393
shastensis, Sorex 363
Shikamainosorex 291, 301*, 304, 305,
308, 313, 429
— densicingulata 429*
shinto, Sorex 85, 351
— — caecutiens 355
Shizoscaptor sinensis 282
sibirica, Crocidura 309, 311, 390, 397,
398*
— — leucodon 397
— Talpa altaica 267
sibiricus, Erinaceus 164
sibiriensis, Sorex 344
— — daphaenodon 344
sikkimensis, Sorex 446
silacea, Crocidura 411
silvia, Crocidura 415
similis, Nesiotites 452
Similisorex 291, 297, 301*, 313, 377
— orlovi 377, 378*
Simmetrodonta 40*
simplicidens, Blarina 424
sinalis, Sorex 363
sindensis, Crocidura 382
sinensis, Neomys 452
— Neotetracus 91, 116, 130, 131*
— Shizoscaptor 282
sinuosus, Sorex 364
Sirenia 44*
smithi, Crocidura 409
— Notiosorex 451
soccatus, Sorex 382
socialis, Parasorex 109
Solendon 25, 48
Solendon paradoxus 23*, 26*, 84
Solendontidae 27, 35, 36*, 37, 44, 48, 84
Solisorex 292, 301*, 304, 305, 307, 313,
377, 387, 388
— pearsoni 387, 388
somalica, Crocidura 413
496 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
somereni, Scutisorex 385, 386
— Sylvisorex 385
songhorensis, Gymnurechinus 156
sorella, Myosorex (?) 381
— Sylvisorex 381
sorelloides, Sylvisorex 381
Sorex 4, 5, 30*, 60, 71, 72*, 81, 290—293,
300, 301*, 302—305, 307—309, 312,
316, 319, 320, 322, 326, 329—331,
332, 333, 336, 337, 358, 362, 366,
368—370, 379, 381, 420, 421, 456
Sorex, группа 303, 304
Sorex acadicus 359
— alascanus 365
— albinus 382
— aligurus 446
— alpinus 85, 292, 309, 332, 346*
— amasari 344
— amoenus 363
— amphibius 440
— annexus 351
— antinori 346
— antiquus 319, 320
— aquaticus 253, 254
— araneoides 334
— araneus 72, 73, 74*, 82, 85, 124, 302,
308—310, 320, 331, 333, 335, 339*,
345, 350, 357, 362, 392, 396, 399
— — araneus 340
— — asper 341
— — auregne 322
— — borealis 344
— — buxtoni 344
— — jacutensis 348
— — roboratus 341
— — sanguinidens 345
— — satunini 341
— — schnitnikovi 345
— — tomensis 341
— — — isodon 356
— — transrypheus 345
— arcticus 85, 309, 332, 337, 343*
— — cinereus 359
— asper 309, 332, 341*
— aterrimus 446
— baicalensis 344
— batis 345
— beddomei 382
— bedfordiae 357
— — gomphus 357
— belli 343
— bendirii 336, 365
, — beringianus 309, 349*
— bogdanovi 399
— brevicaudus 424
— bucharensis 309, 333, 360*, 361, 362
— burneyk 355
— buxtoni 344
— ceacutiens 85, 309, 333, 335, 344, 351*,
358, 360
— — caecutiens 352
— — koreni 352
— — macropygmaeus 353
— — pleskei 352
— — shinto 353
— caerulescens 381
— caerulaeus 381
— californicus juncensis 364
— canaliculatus 353
— capensis 407
— carinatus 440
— caucasicus 85, 308, 309
— caudatus 446
— centralis 351, 356
— chrysithorax 396
— cinereus 83, 85, 309, 333, 338, 359*,
365, 433, 434
— cinnamomeus 407
— collaris 440
— (?) compressus 333
— concinnus 339
— conneratii 381
— constrictus 396
— cooperi 359
— coronatus 339
— crassicaudus 382
— crawfordi 447—449
— cudahyensis 336
— cylindricauda 331, 357, 358*
— — cylindricauda 359
— — gomphus 359
— — wardi 359
,— daphaenodon 309, 332, 342*
— — daphaenodon 342
— — scaloni 343
— — sibiriensis 344
— daubentonii 440
— dehmi 320
— dehneli 308
— dekayi 424
— dispar 366
— dixonensis 432
— dobsoni 363
— dukelskie 348
— eremita 440
— etruscus 392
— excelsus 362
— exilis 353, 355
— falax 334
— ferrugineus 382
— fimbriatus 396
— fimbripes 359
— flavescens 407
— fluviatilis 440
— fodiens 439, 440
— forsteri 359
— fumeus 85, 337, 365
— fumigatus 399
— gaspensis 365
— gemellus 85
— giganteus 381
— gmelini 309, 310, 353
— gracilidens 308, 333
— gracilis 428, 429
— gracillimus 309, 333, 352*, 353
— grantii 339
— gravesi 356
— griffithi 382
— grivensis 324
— giildenstaedti 399
— hagermanensis 337
— hawkeri 355
— hermanni 339
— heterodon 382
— hibbardi 336
— holosericous 446
— homourus 446
— hoyi 366
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
497
— hydrodromus 365
— hydrophilus 440
— hydruntina 396
— ignotus 339
— indicus 381
— inodorus 396
— intermedius 346
— isodon 85, 309, 333, 356, 357*
— — isodon 356
— — ruthenus 356
— jacksoni 365
— jenissejensis 344
— juncensis 364
— kandianus 382
— kelaarti 382
— kozlovi 361*, 362
— kretzoi 335
— labiosus 339
— lacustris 336
— leucodon 388, 392, 395, 399
— leucogaster 353
— leucops 447
— leucurus 396
— lineatus 440
— liricatus 440
— longirostris 363
— lyelli 363
— inacrodon 364
— macropus 379
— macropygmaeus 344, 351
— macrotrichus 339
— macrurus 366, 440, 447
— mauritanicus 396
— melanodon 339
— meltoni 338
— merriami 336, 366
— micrurus 436
— milleri 363
— minimus 353, 355
— iniinutissimus 20, 85, 294, 309, 333,
353, 355*
— — caudatus 356
— — minutissimus 356
— — tscherskii 356
— — tschuktschorum 356
— minutus 85, 309, 310, 333, 353, 354*,
362
— — gracillimus 353
— — heptapotamicus 355
— — minutus 355
— — thibetanus 362
— mirabilis 309, 332, 347*
— — kutscheruki 348
— — mirabilis 348
— montanus 382
— moschatus 220
— murinus 381
— musculus 440
— myoides 404
— myosurus 381
— nanus 364
— natans 440
— navigator alascanus 365
— neumarianus aff. 325
— nemorivagus 382
— nevadensis 363
— niger 382
— nigripes 440
— nivicola 447
— obscurus 337
— odoratus 406
— olivieri 407
— oreopolus 366
— ornatus 363
— pachyodon 334
— pachyurus 343, 383
— pacificus 347, 363
— palustris 83, 330, 336, 364, 365
— parvus 433, 434
— personatus 339, 359
— petenyi 428 -
— pilorides 381
— planiceps 353, 362
— platycranius 348
— platyrhynchus 359
— poensis 409
— powersi 337
— praealpinus 334
— praearaneus 335
— preblei 337, 363
— pribilofensis 343, 365
— psilurus 440
— pulchellus 415, 417
— pumilio 353
— pumilus 353
— pusillus 320
— — grivensis 324
— pygmaeus 310, 353, 355
— quadraticauda 369
— quadricaudatus 339
— raddei 85, 292, 309, 332, 345*
— radiatus 241
— remifer 440
— rexi-oadensis 337, 433
— rhindophus 339
— richardsonii 343
— rivalis 440
— roboratus 309, 339
— runtonensis 334, 335
— russulus 336
— rusticus 353
— rutilus 407
— sacer 382
— samniticus 339
— sandersi 338
— sanguinidens 344
— sansaniensis 326, 327
— santonus 339
— saturation 382
— saussurei 338, 366
— savini 332
— — austriacus 332
— schlosseri 326
— sclateri 366, 374
— sericeus 408
— serpentarius 382
— shastensis 363
— shinto 85, 351
— sibiriensis 344
— sikkimensis 446
— sinalis 363
— sinuosus 364
— soccatus 382
— sphagnicola 343
— s. str. 304
— stagnalis 440
498
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
— stizodon 366
— suaveolens 392
— subaraneus 334
— subminutus 335
— sucldey 363
— swinhoei 382
— talpoides 424
— tasnadii 332
— taylori 337, 369, 433
— tenellus 363, 364
— — nanus 364
— tetragonurus 339
— thibetanus 362
— thomasi 348
— thoracicus 396
— trigonirostris 364
— tristrami 382
— trowbridgii 337, 338, 366
— tscherskii 355
— tundrensis 343
— ultimus 344
— unguiculatus 60, 61*, 85, 309, 330,
l’“ 333, 350*, 379
— unicolor 396
— ussuriensis 355
— vagranus 83, 363
— varius 373, 374
— veraepacis 364
— verae-pacis 364
— vir 85. 309, 332, 348*
— — jacutensis 349
— — platycranius 349
— — thomasi 349
— — vir 349
— vireti 325
— viridescens 382
— volnuchini 85
— vulgaris 339, 350
— wardi fumeolus 357
— willeti 364
Sorex? sp. 423, 428
Soricella 291, 297, 301*, 305, 307, 312,
318, 325
— discrepans 322*, 325
Soriciana, Blarina 347
— Cryptotis 437
Soricidae 3, 5, 27, 30, 34—36*, 37, 45, 49,
56, 62, 78, 80, 83, 85, 87, 107, 128,
200, 285, 293, 295, 302—306, 309,
312, 316, 318, 329, 334, 376, 445
Soricidus 307, 331
Soricinae 290, 291, 296, 297, 299—301*,
302—305, 307, 312, 314—316, 329,
419
Soricini 285, 288, 290, 291, 293, 294,
297, 299—301*, 302—305, 307, 312,
314, 316, 329, 330, 366, 368, 421, 427
soriceps, Uropsilus 57*, 201, 202, 203*
Soriciscus 308
Soricoidea 37, 45, 303
soricoides, Neomys 443
Soricoidus monsvairant 339
Soriculi 308
Soriculina 301*, 302, 313, 315, 419, 420,
437, 450, 452
Soriculini 304
soriculoides, Neomys 438
soriculoides, Zelceina 438*
Soriculus 290—292, 301*, 302—305, 308,
313, 420, 433, 437, 438, 445, 450,
452, 453
— группа 304
— подрод 437
— baileyi 447
— beresowskii 450
— caudatus 446
— tumidus 447
— gracilicauda 446
— hypsibius 450
— irene 447
— kubinyi 445, 446*
— leucops 447
— macrurus 447
— minor 446
— nigrescens 446
— radulus 446
— salenskii 451
sowerbeyi, Crossopus 441
spatangus, Erinaceus 169
sphagnicola, Sorex 343
spiculus, Erinaceus 168
spinifex, Erinaceus 168
squamipes, Anourosorex 287, 461, 465,
466, 467*
stagnalis, Sorex 440
stehlini, Pseudogalerix 111, 112, 114,
116
stenodus, Allosorex 454*
Stenoechinus 97, 99*, 105, 147, 156
— tantalus 156
Stirtonia 291, 297, 301*, 312, 372
stizodon, Sorex 366
stoliczkana, Crocidura 382
stolizkanus, Suncus 382
stramosi, Erinaceus 164
strauchii, Crocidura 408
streeti, Talpa 271*, 272, 284
styani, Chimarrogale 457
suahelae, Crocidura 409
suaveolens, Crocidura 72, 85, 309—311,
383, 390, 392*
— — suaveolens 393
— ilensis, Crocidura 393
— Pachyura 392
— shantungensis, Crocidura 393
— suaveolens, Crocidura 393
— Sorex 392
subaraneus, Sorex 334
subfulva, Crocidura 382
subminutus, Sorex 335
suckley, Sorex 363
suillus, Erinaceus 164
— Hylomys 91, 115, 127*, 128*/ 129,
131
sumatrana, Chimarropale 458
suncoides, Sylvisorex! 381
Suncus 4, 291—293, 301*, 302—305, 307,
309, 313, 314, 373, 376, 377, 380, 381,
382, 386, 428
— chriseos 383
— coquerellii 383
— dayi 382
— etruscus 20, 74*, 85, 292, 309, 380.
383, 384*
— — etruscus 385
— gracillis 383
— gratulus 383
— infinitesimus 383
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
499
— leucura 383
— lixus 383
— madagascarensis 383
— murinus 60*, 84, 85, 378, 381
— niger malabaricus 382
— orangie 383
— sacer 381
— stoliczkanus 382
— varillus 383
— warreni 383
— zelceus 429
— zeylanicus 382
Surdisorex 290, 292, 293, 301*, 303—305,
313, 320, 372, 375
— norae 373, 375
— polulus 376
sururae, Crocidura 407
suschkini, Talpa altaica 267
swinhoei, Sorex 382
swinnyi, Miosorex 374
Sylvisorex 4, 292, 300, 301, 303—305,
307, 313, 375—377, 379, 380
— angolensis 381
— gemneus 381
— granti 380
— johnstoni 380
— lunaris 378, 380
— megalura 62*, 381
— morio 378, 380
— mundus 380
— ohila 380
— oriundus 380
— ruande 381
— somereni 385 \.'
— sorella 381
— sorelloides 381
— suncoides 381
syriacus, Erinaceus 169
tadae, Crocidura 404
tadzhikistanicus, Galerix 112*, 113, 180
taeniata, Talpa 251
talazaei, Microgale 84
Talpa 183, 184, 191, 192, 193*, 194—197,
099, 206, 226, 246, 256—258, 259,
•260, 271—273, 282, 284, 285, 333
— acutidentata 258, 259
— aenea 251
—’altaica 70*, 195, 259, 263*, 265, 266*,
273
— — altaica 267
— — gusevi 267
— — salairica 267
— — sibirica 267
— — suschkini 267
— — tymensis 267
— antiqua 258, 259
— brachychir conf. 231
— canadensis 241
— caucasica' 85, 197, 259, 261*, 267,
9KC1 972
— caudata 241, 260
— coeca 85, 195, 197, 259, 263*, 267,
269, 270*, 271, 272, 284
— — caucasica 267
— — minima 271
— — orientalis 271
— — talyschensis 271
— cristata 240
— cryptura 276
— cupreata 254
tlalpa europaea 57*, 66, 68*, 69*, 70*, 85,
192, 195, 197, 248, 259, 260, 261*,
263*, 267, 271, 272, 280, 281, 284
-----conf. 262*
— — brauneri 265
— — europaea 265
— — flavescens 254
— — obensis 265
— — transuralensis 265
— flava 241, 254
— frisius 260
— fusca 254
— grandis 272, 273
— hercegovinensis 195, 197, 269, 271
— incerta 142
— klossi 195, 271, 272, 273, 274
— leptura 281, 284
— leucura 280
— longicaudata 240
— longirostris 195, 271, 272, 273*, 274*,
275, 276
— micrura 195, 271, 272, 275*, 276
— minima 197, 270
«Talpa» minuta 260*
Talpa? minuta 260
Talpa olympica 269
— parvidens 272, 274
— purpurascens 241
— radiata 241
— reposta 250
— ;romana 195, 259, 268, 269*, 271, 272
— — ognevi 197, 269
— sansaniensis 248
— scalops 261
— streeti 271*, 272, 284
— taeniata 251
— telluris 248
— townsendii 251
— virginiana 254
— vulgaris 260
— wogura 276, 277
Talpasorex 197 , 240, 253, 308, 422
— aquatica 254
— cristata 241
— pensylvanica 254
— virginiana 254
Talpidae 3—5, 24, 27, 29, 34, 35, 36*,
37, 45, 56, 57, 62, 77, 78, 85, 180,
183, 184, 186, 189, 192, 194—196,
198, 246, 284, 376
Talpinae 5, 36*, 183, 184, 186—192,
193*, 194—199, 226, 227, 231, 256
Talpini 24—26, 53, 66, 67, 180, 181, 183,
189, 191, 192, 193*, 194, 196, 197,
199, 200, 256, 258, 281, 284, 285
talpoides, Blarina brevicauda 433
— Corsira 424
— Sorex 424
— Urotrichus 69, 187, 232, 235, 236,
237*
Taipops 196
talyschensis, Talpa coeca 271
tantalus, Stenoechinus 156
tasnadii, Genoholotypus 332
— Sorex 332
tatianae, Crocidura 403
500
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
taylori, Sorex 337, 369, 433
telfairi, Echinops 84
telluris, Talpa 248
telmalestes, Blarina 425
tenellus nanus, Sorex 364
— Sorex 363, 364
Tenrec 25, 46
— ecaudatus 22, 23*. 26*, 31*, 35, 87
Tenrecidae 27, 34, 35, 36*, 37, 44, 46, 84
Tenrecii 36*, 37, 39, 44*, 45, 186, 296
Tenrecinae 46, 49
tenuis, Crocidura 405
— Myosorex 374
teres, Neomys 443
terrenus, Arctoryctes 244*, 245, 246
terrestris, Arvicola 185, 205, 217, 442
tersus, Cryptotis 437
tertius, Proscalops 245
Tetracus 89, 93, 99*, 100*, 101, 102, 104,
107, 108, 110, 111
Tetracus? 102
Tetracus bouti 108*, 109, 111
— nanus 107, 108, 109, 111
tetradactylus, Oryzorictes 20
tetragonurus, Sorex 339
texanus, Scalops aereus 254
theresae, Crocidura 85
thermalis, Desmana 219*, 220, 225, 226
thibetanus, Sorex 362
— — minutus 362
thomasi, Crocidura 398
— Sorex 348
— — vir 349
thompsoni, Domnina 321*, 324
thomsoni, Gaillardia 206*, 207
thoracinus, Sorex 396
tomensis isodon, Sorex araneus 356
— Sorex araneus 341
topali, Amblycoptus 463
townsendii, Scalops 250, 251
— Scapanus 251, 252
— Talpa 261
transrypheus, Sorex araneus 345
transuralensis, Talpa еигораеа 265
Tregosorex 290, 300, 301*, 312, 367, 368
— holmani 367, 368*
trevelyani, Chrysochloris 27*
Triconodonta 38, 40*, 44*
tricuspis, Limnoecus 386, 391*
trigonirostris, Sorex 364
Trimylinae 304
Trimylus 304—307, 327. 329
— schlosseri 326
tristrami, Sorex 382
Trituberculata 40*
trowbridgii, Sorex 337, 338, 366
truei, Podogymnura 129*, 130
tscherskii, Sorex 355
— — minutissimus 356
tschifuensis, Erinaceus 164
tschuktschorum, Sorex minutissimus 356
Tshelkaria rostrata 43*
Tubulidentata 44*
tumididens, Indentisorex 329
tundrensis, Sorex 343
Tupaia 50, 51
— ferruginea 32*
Tupaiidae 3, 21, 27, 31, 35, 36*, 37, 45,
50
Tupaiinae 51
Tupaiodon 89, 95, 97, 99*, 100*, 101, 102.
104, 135, 136
Tupaiodon? minutus 136
Tupaiodon morrisi 135, 136*
Tupaioidea 37, 45
turanicus, Erinaceus albulus 169
— — auritus 173
turba, Crocidura 409
tymensis, Talpa altaica 267
ultima, Crocidura 415
ultimus, Sorex 344
unghi, Erinaceus 164
unguiculatus, Sorex 60, 61*, 85, 309, 330,
333, 350*, 379
unicolor, Sorex 396
Urogale 51
Uropsilinae 5, 78, 80, 183, 184, 186, 189,
192, 193*, 194—199, 200, 201, 203, 284
Uropsilus 184, 193*, 194—196, 198, 200„
201, 202, 204
— soricipes 57*, 201, 202, 203*
Urotrichinae 192, 195, 197, 227
Urotrichini 53, 78, 183, 184, 187—191,
193*, 196—199, 227, 229, 232, 235,
285
Urotrichus 4, 184, 193*, 194—196, 198,
227, 228, 236
— gibbsi 238
— talpoides 69, 187, 232, 235, 236, 237*
ussuriensis, Erinaceus 164
— Sorex 355
vagans, Erinaceus algirus 168
vagrans, Sorex 83, 363
valentinensis, Domninoides 235
varennei, Chimarrogale 458
varillus, Suncus 383
varius, Myosorex 373*, 374
— Sorex 373, 374
veraepacis, Sorex 364
verae7pacis, Sorex 364
verestchagini, Desmana 212, 217, 218*,
220, 224, 225
vetulina, Crocidura 408
vireti, Angustidens 323*, 325
— Erinaceus 162*
— Postpalaeoerinaceus 162
— Sorex 325
virginiana, Talpa 254
— Talpasorex 254
virginianus, Scalopus 254
viridescens, Sorex 382
virilla, Crocidura 383
vir jacutensis, Sorex 349
— platycranius, Sorex 349
— Sorex 85, 309, 332, 348*
— — vir 349
— thomasi, Sorex 349
— vir, Sorex 349
Viverra exilis 109
— gymnura 124, 126
viverroides, Galerix 109
voi, Crocidura 409
volnuchini, Sorex 85
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
501
vorax, Crocidura 311, 404
vulgaris, Sorex 339, 350
— Talpa 260
yamashinai, Crocidura 389
yebalensis, Crocidura 396
wakeeneyensis, Anchiblarinella 368, 369
waldemarii, Crocidura 382
wardi, Blarinella quadraticauda 371
— fumeolus, Sorex 357
— Sorex cylindricauda 359
warreni, Suncus 383
watasei, Crocidura 404
whitmanensis, Oligoscalops 246, 247
willeti, Sorex 364
wogura coreana, Mogera 277
— Mogera 195, 197, 245, 259, 276, 277*
— Talpa 276, 277
Xanthippe, Crocidura 412
Zalambdalestes 41*
Zalambdodonta 45
zaphiri, Crocidura 407
zazhigini, Mygalinia 211*, 213
zarudnyi, Crocidura 311, 402*, 403
Zelceiiia 291, 301*, 302, 304, 305, 308,
313, 438, 439
— soriculoides 438*
zelceus, Suncus 429
zena, Crocidura 409
zeteki, Cryptotis 436
zeylanica, Crocidura 382
zeylanicus, Suncus 382
zhchikvadzei, Pseudoneurogymnurus 123*,
124
zorzii, Crocidura 391
ОГЛАВЛЕНИЕ
Предисловие ................................................. 3
Систематический указатель насекомоядных................................. 7
Отряд Insectivora........................................... 20
К системе высших таксонов плацентарных млекопитающих.... 37
Система крупных подразделений современных насекомоядных. 44
Подотряд Tenrecii........................................... 45
Подотряд Erinacei ............................ 49
Общие адаптивные особенности семейств подотряда Erinacei фауны СССР 51
Морфоэкологпческие типы как формы жизни............................ 76
О кариосистематике................................................. 84
Систематический обзор.................................................. 86
Семейство Erinaceidae ....................... 86
Подсемейство Galericinae ................... 105
Подсемейство Erinaceinae ................... 133
Семейство Talpidae ..................................... 180
Подсемейство Vropsilinae .................. 200
Подсемейство Desmaninae ................... 204
Подсемейство Talpinae .................... 226
Семейство Soricidae ......................... 285
Подсемейство Saturniniinae ................. 315
Подсемейство Soricinae ................... 316
Литература............................................................ 469
Алфавитный указатель латинских названий............................... 478
Книги издательства «Наука»
можно предварительно заказать
в магазинах конторы «Академкнига»
Адреса и почтовые индексы магазинов:
480091 Алма-Ата, ул. Фурманова, 91/97
370005 Баку, ул. Джапаридзе, 13
320005 Днепропетровск, пр. Гагарина, 24
734001 Душанбе, пр. Ленина, 95
375009 Ереван, ул. Туманяна, 31
664033 Иркутск, ул. Лермонтова, 289
252030 Киев, ул. Ленина, 42
252142 Киев, пр. Вернадского, 79
252030 Киев, ул. Пирогова, 4
277001 Кишинев, ул. Пирогова, 28
343900 Краматорск (Донецкой обл.), ул. Марата, 1
660049 Красноярск, пр. Мира, 84
443002 Куйбышев, пр. Ленина, 2
192104 Ленинград, Д-120, Литейный пр., 57
199164 Ленинград, В-164, Таможенный пер., 2
199004 Ленинград, В-4, 9 линия, 16
220072 Минск, Ленинский пр., 72
103009 Москва, ул. Горького, 8
117312 Москва, ул. Вавилова, 55/7
630076 Новосибирск, Красный пр., 51
630090 Новосибирск, Академгородок, Морской пр., 22
142292 Пущино (Московской обл.), «Академкнига»
620151 Свердловск, ул. Мамина-Сибиряка, 137
700029 Ташкент, ул. Ленина, 73
700100 Ташкент, ул. Шота Руставели, 43
700187 Ташкент, ул. Дружбы народов, 6
634050 Томск, наб. реки Ушайки, 18
450059 Уфа, ул. Р. Зорге, 10
720001 Фрунзе, бульв. Дзержинского, 42
310003 Харьков, Уфимский пер., 4/6
Для получения книг почтой
заказы просим направлять по адресу:
117192 Москва, Мичуринский пр., 12
Магазин «Книга — почтой»
Центральной конторы «Академкнига»
197110 Ленинград, Петрозаводская ул., 7
Магазин «Книга — почтой»
Северо-Западной конторы «Академкнига»