/




























































































































































Author: Смирнов О.А.
Tags: pinophyta , coniferae (conifers) pinopsida экология растений этология растение и окружающая среда биология экология
Year: 1978




Text
РЕДКИЕ
и ИСЧЕЗАЮЩИЕ
ДРЕВЕСНЫЕ
РАСТЕНИЯ
ЮГА
ДАЛЬНЕГО
ВОСТОКА
АКАДЕМИЯ НАУК СССР
ДАЛЬНЕВОСТОЧНЫЙ НАУЧНЫЙ ЦЕНТР
ботанический сад
РЕДКИЕ И ИСЧЕЗАЮЩИЕ
ДРЕВЕСНЫЕ РАСТЕНИЯ
ЮГА ДАЛЬНЕГО ВОСТОКА
(БИОЛОГИЯ, экология,
КАРИОЛОГИЯ)
Владивосток .--------
1 У / с I _ .
УДК 582.47:581.5(571.6)
В сборник включены материалы о результатах изучения биологии, эко-
логии, кариологии и анатомии древесных видов флоры Дальнего Востока,
нуждающихся в охране и представляющих интерес для народного хозяйства
(лианы, лиственницы, сосны, можжевельники, тис). Обсуждаются програм-
ма изучения биологии редких и исчезающих видов и возможности их охра-
ны и воспроизводства. Статьи содержат новые сведения по внутривидовой
изменчивости, экологии и ритмам развития хвойных Дальнего Востока. Со-
общаются результаты их кариологического и морфолого-анатомического
анализа.
Сборник рассчитан на ботаников, преподавателей биологии, озеленителей
и лесоводов.
Ответственный редакто1р
г. н. О. А. СМИРНОВА
Издано по решению редакционно-издательского совета
Дальневосточного научного центра АН СССР
40508 282_ ^ез объявления
055(02)6—78
© ДВНЦ АН СССР, 1978
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ОСНОВНЫЕ ЗАДАЧИ ИЗУЧЕНИЯ ИСЧЕЗАЮЩИХ,
РЕДКИХ И СОКРАЩАЮЩИХСЯ ВИДОВ
ДЕНДРОФЛОРЫ ПРИМОРЬЯ
И ВОЗМОЖНЫЕ ПУТИ ИХ КУЛЬТИВИРОВАНИЯ
Л. Н. СЛИЗИК, О. А. СМИРНОВА
«...Необходимо соединить
знание и исследование видов в
природе с экспериментальным
их изучением в лаборатории и
на опытных делянках ботани-
ческих садов».
М. Г ПОПОВ
Планомерная инвентаризация флоры Дальнего Востока, вы-
полненная советскими ботаниками (Комаров, Клобукова-Али-
сова, 1931 —1932; Воробьев и др., 1966; Ворошилов, 1966; Во-
робьев, 1968; Воробьев и др., 1974; Никитин, Бондаренко, 1975),
в значительной мере выявила ее богатства и своеобразие. Од-
новременно многолетние флористические исследования на
Дальнем Востоке определили очередные конкретные задачи з
деле изучения, использования и охраны растительных ресурсов
региона. Одной из таких задач, имеющих государственное зна-
чение, является разработка научных основ охраны и использо-
вания исчезающих, редких и сокращающихся вадов флоры
Дальнего Востока. Значение этой работы определяется рядом
законодательных актов (Емельянова, 1975; Проект Конститу-
ции СССР, 1977), а также появлением Красной книги (1975).
Продолжительные наблюдения и коллекционирование растений
в Ботаническом саду ДВНЦ АН СССР позволили накопить
опыт и материалы по культуре, использованию и охране ред-
ких и ценных видов дендрофлоры Приморья. Совершенно оче-
видно, что эта группа видов может оказаться значительно бо-
лее многочисленной, чем указывается для Дальнего Востока
в первом издании Красной книги (1975). Поэтому в настоящее
время представляется необходимым уточнение региональных
перечней видов растений, нуждающихся в охране. В результате
анализа литературных и гербарных материалов по флоре При-
морья нами Определялся состав и объем группы древесных ви-
дов с указанием принятых в Красной книге категорий (1—4).
По нашим данным, в целом к этой группе следует относить бо-
лее 50 видов дендрофлоры Приморья (см. таблицу).
Количественные соотношения объемов подгрупп (категорий)
3
свидетельствуют о действии ряда факторов, определяющих
состояние региональной флоры. Виды 1 и 3 категорий (исче-
зающие и сокращающиеся) имеют в разной степени и по раз-
ным причинам тенденцию к сокращению ареала и численности.
Так, известны островные местообитания Ilex rugosa Fr,
Schmidt, Rhododendron fauriei Franch.; местообитания на се-
верных пределах ареалов — Parthenocissus tricuspidata (Sieb.
et Zucc.) Planch., Rhododendron schlippenbachii Maxim., Pinus
densiflora Sieb. et Zucc. и лр. Все это говорит о длительном и
своеобразном флорогенезе региона. Виды категории 2 (редкие)
достаточно многочисленны и включают как эндемы, так и виды
корейско-китайской и бореальной флор. Значительное число ви-
дов 1, 2 и 3 категорий представлены в Приморье своими погра-
ничными популяциями, т. е. обитают здесь в экстремальных
условиях. Состав видов категории 4 (неопределенные) в ден-
дрофлоре Приморья свидетельствует о степени изученности
таксономически сложных групп (Laris Mill; Tilia L ); не иск-
лючено, что ряд критических видов являются лишь формами
(Actinidia giraldii Diels, Deutzia glabrata Korn.). В целом число
видов указанных категорий в регионе — достаточно подвижный
показатель, так как может меняться по мере накопления и
уточнения фактического материала, вследствие таксономиче-
ских обработок, при организации эффективной охраны рас-
тений.
Вслед за уточнением порайонных перечней исчезающих,
редких и сокращающихся видов следует создавать коллекцион-
ные фонды этих растений в ботанических садах и дендрариях
Дальнего Востока. В Ботаническом саду ДВНЦ АН СССР
такая работа проводилась с конца пятидесятых годов. Здесь
собрана значительная часть древесных видов дендрофлоры
Приморья, в том числе и занесенных в Красную книгу (1975):
лианы, хвойные, лиственные породы, что является одним из
итогов практической работы интродукторов В. К. Василюка,
В. Т. Зориковой, Т. П. Ильченко, Е. Н. Литвинцева, М. А. Скрип-
ки, Л. Н. Слизик, Т. И. Таранковой, В. М. Урусова, А. А. Ча-
щухиной. В последние годы выполнялись работы по изу-
чению особенностей биологии и размножения ряда наиболее
ценных видов из этой группы (Грушвицкий, 1961; Нечаев, 1963;
Слизик, 1970; Богданова, 1971; Зорикова, 1973; Ильченко, 1975:
Слизик, Древецкая, 1975; Урусов, 1975; Гурьев, 1977). Но, как
видно из прилагаемого списка, ряд видов еще не представлен
в коллекциях Сада, поэтому поиски их, изучение и создание
коллекционных фондов на юге Приморья являются неотлож-
ными задачами ботаников-интродукторов.
Опыт нашей работы в Приморье показал, что привлечение
в коллекции растений редких и исчезающих видов из сопре-
дельных районов может быть лишь попутной задачей. Основное
внимание ботаников-интродукторов должно быть обращено на
4
представителей местной флоры. Так. интродукция в южное
Приморье ряда видов Сахалина и Курил выявила слабую толе-
рантность их относительно новых условий существования. Как
правило, древесные виды, переметенные в соседние области за
пределы родового ареала, плохо развиваются Skimmia repens
Nakai, Magnolia obovata Thunb., виды Sasa Makino et Shibata
и др. При переселении видов дендрофлоры, родовые ареалы
которых охватывают островную и материковую части Дальнего
Востока, наблюдаются как случаи вполне благополучного роста
и развития, так и постепенной гибели растений разных видов.
Так, при длительных наблюдениях не отмечаются нарушения в
сезонных циклах у интродуцированных с Сахалина Juniperus
sargentii (Henry) Takeda, Vitis kaempferi Koch., Celastrus stri-
gillosa Nakai. Но сахалинско-курильские гортензии ведут себя
по-разному: Hydrangea paniculata Sieb. et Zucc. нормально
вегетирует и обильно цветет; Н. petiolaris Sieb. et Zucc. в ок-
рестностях Владивостока имеет вид небольшой куртинки сла-
боветвящихся побегов, с незначительным годичным приростом
(10—12 см). Viburnum wrightii Miq. и V- furcatum Blume сов-
сем не приживаются в коллекциях Сада. Общеизвестно, что
поведение растений при переселении определяется их генези-
сом, условиями формирования данного вида. В еще большей
степени это относится к исчезающим и редким видам, так как
специфичность растений, причисляемых к категориям 1, 2 и 3,
состоит именно в теснейшей связи с узкими экологическими
нишами. Поэтому для успешного изучения, использования и
охраны исчезающих, редких и сокращающихся видов необхо-
димо накапливать региональные коллекционные фонды и куль-
тивировать наиболее ценные из указанных видов прежде всего
в районах их естественного произрастания, где биология этих
растений обычно не нарушается непривычными условиями оби-
тания.
Следует отметить, что до настоящего времени значительная
часть редких и исчезающих видов дендрофлоры Приморья
остается практически неизученной — неизвестны их сезонная и
возрастная ритмика (большой и малый циклы), особенности
цветения, плодоношения и прорастания, оптимальные и экстре-
мальные условия существования, степень изменчивости. Для
правильных оценок состояния видов в природных местообита-
ниях на Дальнем Востоке и сравнительного изучения их био
логических особенностей необходимы планомерные эколого-био-
логические исследования вилов названных категорий по единой
программе как в природе, так и в культуре (в пределах и за
пределами ареалов).
Для видов 1 и 2 категорий важно выяснить приспособитель-
ные структуры и механизмы, обеспечивающие существование
вида в данных эколого-географических условиях, найти крити-
ческие точки жизненных циклов и определить лимитирующие
5
факторы среды. В какой-то мере это относится и к видам 3 ка-
тегории, но, по-видимому, эти виды сокращаются в основном
под действием антропогенных факторов; в этом случае особен-
но важно выявить районы и причины сокращения. К числу сок-
ращающихся относится немало хозяйственно ценных видов,
испытывающих на себе действие давнего потребительского
интереса населения: лекарственные, пищевые, декоративные и
д. Поэтому биология этих групп растений исследовалась в
разных планах: аралиевые (Грушвицкий, 1961; Богданова,
1971), ягодные лианы (Титлянов, 1969). В целом программное
сравнительное изучение биологии видов всех трех категорий
даст, несомненно, материал большого прикладного и теорети-
ческого значения.
Сравнительные физиолого-морфологические исследования
академика В. Ф. Купревича (1947) определенного ряда видов
позволяют установить пути приспособительных изменений в
процессе эволюции, определить ее направленность; в случае
достоверно известной истории расселения вида и климатиче-
ских колебаний на территории региона можно выявить факто-
ры эволюции и механизм их действия. При эколого-биологиче
ском исследовании редких и исчезающих видов представляется
наиболее важным и перспективным именно сравнительное изу-
чение приспособительных и характерных структур и механиз-
мов. Для этого в какой-то мере возможно использование схем
описания объектов, принятых в ряде работ (Бобров, 1947; Био-
логическая Флора Московской области, приложение 1, 1974).
По нашему мнению, единая программа должна предусмат-
ривать изучение биологии редких, исчезающих и сокращаю-
щихся видов по нескольким основным разделам:
I. Жизненные циклы и сезонные ритмы развития
1. Морфология и темпы развития проростков. 2. Ювениль-
ный период и его особенности. 3. Особенности развития кроны,
корневой системы. 4. Вступление в плодоношение: возраст,
динамика.
II. Вегетативная сфера
1. Возрастные изменения размеров. 2. Характер и зоны по-
чек возобновления. 3. Периоды префлоральной и постфлораль-
ной вегетации, температурный фон. 4. Характер и темп роста
годичных побегов. 5. Сроки и особенности одревеснения. 6. Спо-
собность к укоренению. 7. Способность к перезимовке.
III. Генеративная сфера
А. Г е н е р а т и в н ы е п о б е г и. 1. Размеры, сроки форми-
рования. 2. Характеристика цветков и соцветий (шишек—у
хвойных). 3. Антэкологические наблюдения. 4. Соотношение
6
общего числа бутонов и цветков — на модельных побегах; при-
чины их осыпания.
Б. Семена и плоды. 1. Соотношение общего числа
цветков и плодов — на модельных побегах, причины осыпания.
2. Характеристика плодов и семян: размеры, вес. 3. Сроки и
динамика созревания, температурный фон. 4. Особенности- и
температурные режимы прорастания семян. (Морфо-биологи
ческие особенности хвойных требуют несколько иного подхода
к наблюдениям за их вегетативной и генеративной сферами —
Шкутко и др., 1974).
IV. Популяции
1. Характеристика местообитаний; оптимальные и экстре-
мальные условия; факторы, угнетающие вегетативную и (или)
генеративную сферу вида. 2. Общая характеристика популя-
ции. 3. Темп и характер возобновления в пределах популяции.
4. Оценка возможности использования популяции как источни-
ка ценных форм и посадочного материала.
В зависимости от объема и степени изученности региональ
ных групп видов возможны разные варианты комплексного
подхода к их исследованию, но результатом должно стать
выявление биологических особенностей, связей
данного вида со средой обитания, что поможет объяснить усло-
вия становления его в данной флоре, т. е. особенности гене-
зиса.
Выявление особенностей поведения — наиболее важный мо-
мент при изучении видов 1 и 2 категорий, а также видов 3 ка-
тегории, обитающих в Приморье на границах естественных
ареалов. Систематизированный фактический материал позволит
проводить сравнительный анализ поведения и состояния раз-
личных видов в разных аспектах — таксономическом (напри-
мер, рододендроны в пределах СССР), географическом (по-
пуляции тиса остроконечного на территории Дальнего Восто-
ка), интродукционном (поведение вида в пределах ареала и
вне его). Особенности прохождения жизненных циклов (боль-
шого и малого) в разных эколого-географических условиях
вскрывают природу вида, лимитирующие факторы среды и
экологический оптимум — элементы, которые по матричному
принципу «проявляют» природный фон (в частности, темпера-
турный), сформировавший данный вид.
Жизненный цикл с определенной сезонной ритмикой — это
комплекс приспособительных реакций растения к гем условиям
среды, в которых вид как таковой формировался; это неотъем-
лемый признак вида, важнейший морфобиологический показа-
тель, сложившийся при становлении вида (Ворошилов. 1960,
Сергеева, 1971; Лапин, 1974). Сходное значение имеют ритми-
ка и режимы прорастания семени. Общеизвестно, что нормаль-
ное, успешное сезонное развитие и прорастание проходят на оп-
7
ределенном температурном фоне. Таким образом, в онтогенезе
реликтовых видов разных эпох заложена соответствующая ин-
формация, которая может быть извлечена при анализе условий
и степени успешности прохождения основных фаз сезонного
развития и роста. При учете толерантности вида она можег
быть использована для достаточно конкретной характеристики
той отдаленной эпохи, в условиях которой сложилась эколого-
биологическая сущность данного вида. Особенный интерес
представляет сравнительное изучение поведения реликтовых
видов, а также видов на границе их ареалов, которые в экс-
тремальных условиях наиболее ярко проявляют структуры и
механизмы, позволяющие виду в том или ином состоянии удер-
живаться в составе данной флоры. Сравнение поведения вида
в оптимальных и экстремальных условиях выявляет также сте-
пень его толерантности — «терпимость», способность выдержи-
вать колебания факторов среды той или иной амплитуды. Эти
данные важны для понимания флорогенеза региона, генезиса
отдельных видов или родовых комплексов. Кроме того, функ-
циональное состояние вегетативной и генеративной сфер вида
в конкретных местообитаниях отражает степень соответствия
его природы условиям среды и тем самым определяет сущ-
ность мер по его охране и воспроизводству.
Целенаправленное программное изучение биологических и
эколого-физиологических особенностей хозяйственно ценных ви-
дов (лекарственных, пищевых, декоративных) из числа сокра-
щающихся под действием антропогенных факторов позволит
выявить в пределах ареалов формы, представляющие наиболь-
ший практический интерес, — крупноплодные, сахаристые, зи-
мостойкие, быстрорастущие и т. д. Это является необходимым
в планомерном познании и использовании генофонда ценных
видов.
Для понимания природы видов, уникальных по своим гене-
тическим связям, представляется крайне интересным их срав-
нительное эколог о-физиологическое исследование.
Такне работы дадут характеристики оптимумов и предельных
значений основных факторов среды для ряда редких и исче-
зающих видов. Именно эти данные очень существенны для объ-
яснения состояния видов и степени соответствия их природы
условиям конкретного местообитания.
Другое важнейшее направление изучения таких видов —
ценотические исследования: оценка состояния популяций
и степени способности к воспроизводству в разных частях ареа-
ла, выявление связей с другими компонентами ценоза — в пер-
вую очередь с опылителями и вредителями. Данные ценотиче-
ских наблюдений также необходимы для разработки мер по
охране и культуре редких и исчезающих видов растений.
Сохранение отдельных видов растений нельзя рассматривать
вне связи с природными комплексами всех рангов — от мини-
8
мального (фации), до географического ландшафта. Исчезнове-
ние или сокращение отдельных видов растений, а также искус-
ственное увеличение их численности приводит к изменениям во
флористическом составе растительности — одного из компонен-
тов ландшафтных комплексов. Любые изменения в структуре
этого компонента проявляются во внешнем облике природных
комплексов быстрее, чем нарушения во всех прочих компонен-
тах. Постепенное накопление даже незначительных изменений
может привести к появлению измененных ландшафтов — куль-
турных, если эти изменения положительные, и акультурных,
если изменения отрицательные.
В связи с этим проблему охраны отдельных видов следует
решать в рамках проблемы охраны ландшафтов в целом. Сле-
довательно, в изучение видов 1—3 категорий необходимо, по-
мимо ценотических, включить и ландшафтные исследова-
ния. Они помогут определить приемлемые объемы и формы
восстановления исчезающих и сокращающихся видов растений,
а также избежать отрицательных изменений в природных комп-
лексах и наметить возможные пути совершенствования их внут-
ренней структуры и внешнего облика.
Ландшафтные исследования необходимо проводить в типич-
ных, «эталонных» местообитаниях изучаемых видов, которые
комплексно описываются с детальным рассмотрением флори-
стического состава и доли участия изучаемых видов. Описания
должны сопровождаться характеристикой внешних качеств эта-
лонного местообитания (мы умышленно не называем это эсте-
тической оценкой, которая выражается в баллах и не отражает
индивидуальных особенностей конкретной ландшафтной едини-
цы). Для целей создания или дальнейшего формирования и
обогащения ландшафтов необходимо детальное описание
внешних качеств эталона: 1) горизонтальное членение изучае-
мой территории с зарисовкой в масштабе (схема плана).
2) вертикальное членение растительности (ярусность); 3) сомк-
нутость по отдельным ярусам; 4) объемная характеристика
наиболее типичных и декоративных группировок; указываются
объемные соотношения отдельных растений или их групп друг
с другом; сопровождается зарисовкой; 5) цветовая характери-
стика (даются общее колоритное впечатление, а также цветовые
сочетания и соотношения); 6) краткая характеристика эмоцио-
нального воздействия. Опираясь на сведения, полученные при
ландшафтном изучении эталонных местообитаний, можно опре-
делить те характерные черты и особенности, которые присущи
сочетаниям, ьключающим изучаемые виды.
Опыт изучения в природе и в культуре исчезающих и ред-
ких древесных видов флоры Приморья и практическая работа
по их коллекционированию позволяют уверенно судить о воз-
можных путях сохранения наиболее ценных представителей
этой группы. Традиционные — заповедание и введение в куль-
9
туру — недостаточны. Представляется возможным дополнить
их такими, как полукультура (термин, применяемый иногда в
ботанической литературе — Толмачев, 1974) и восстановление
природных популяций. Полукультура — это создание насажде-
ний в условиях, близких к естественным местообитаниям. В от-
личие от культивирования в городских парковых посадках —
на определенном агрофоне и в искусственно созданных агро-
ценозах — полукультура представляет собой введение растений
в естественные ценозы. Наличие зеленых зон, созданных на
базе естественных лесов, национальные и природные парки
позволяют вводить в их насаждения виды из числа редких и
исчезающих. Такие посадки при их точном учете и наблюдении
могут служить научным целям, являясь одновременно коллек-
ционным фондом. Подобная работа выполняется в Ботаниче-
ском саду ДВНЦ АН СССР, где на территории дендроучастка
природной флоры высажен ряд видов, занесенных в Красную
книгу (1975). Так, на участке хвойно-широколиственного леса
(чефнопихтарник липово-кленовый с грабом, разнокустарниково-
папоротниковый) на северный склон высажены Aristolochia
manshuriensis Кот., Hydrangea petiolaris Sieb. et Zucc. — под
полог, Parthenocissus tricuspidata (Sieb. et Zucc.) Planch.— на
осветленные каменистые места, Taxus cuspidata Sieb. et Zucc. и
Pinus funebris Kom.— на осветленные разрубкой места в ниж-
ней и верхней частях склона северо-западной экспозиции. Из-
других редких видов высажены также Ampelopsis japonica
(Thunb.j Makino, Atragene ochotensis Pall.
В некоторых случаях слелует проводить восстановление при-
родных популяций, т. е. посев семян или посадку выращенного
материала в пределах угасающей или исчезнувшей популя*
ции с целью ее восстановления. Такие работы возможны только
с хорошо изученными видами; на юге Приморья могут быть
восстановлены в природных местообитаниях (peинтpoдyциpoвai
ны) некоторые виды древесных лиан и хвойных. Научной ос-
новой для таких практических работ являются данные по био-
логии и размножению этих растений, много лет наблюдаемых
в дендрологических коллекциях Ботанического сада и Горно
таежной станции ДВНЦ АН СССР. Все такие посадки должны
либо соответствовать структуре существующих ландшафтов,
либо быть органической частью вновь создаваемых. Очевидно,
что такая работа, как восстановление природных популяций
наиболее ценных видов растений на территории Приморского
края возможна лишь при совместном участии в ней соответст-
вующих научных учреждений и лесхозов и поэтому должна
координироваться. Опыт культивирования редких и исчезаю-
щих растений в нашей стране ограничивается пока в основном
выращиванием их в коллекциях, т. е. первичной интродукцией.
Однако полукультура и реинтродукция — реальные пути к со-
хранению и восстановлению таких видов.
10
Список видов дендрофлоры Приморья 1—3 категорий; собранных или
подлежащих сбору для коллекций Ботанического сада ДВНЦ АН СССР
(1977 г.)
Категория, название вида Наличие в коллекции вег. цв., пл. Примечание
Категория 1— исчезающие виды
1. Aristolochia manshuriensis Korn. + +
2. Ilex rugosa Fr. Schmidt —> —
3. Parthenocissus tricuspidata (Sieb. et Zucc.)
Planch. + +
Категория 2 — редкие виды
1. Juniperus rigida Sieb. et Zucc. + —
2. Ephedra monosperma С. A. M. — —
3. Myrica tomentosa (DC.) Aschers. et Graebn. —
4. Alnus japonica Sieb. et Zucc. — —
5. Ulmus macrocarpa Hance + —
6. Deutzia glabrata Korn. —
7. Grossularia burejensis (Fr. Schmidt) Berger — — ценный
8. Sorbaria rhoifolia Korn. генофонд эндем
9. Cotoneaster melanocarpa Lodd. +
10. Pyrus ussuriensis Maxim. — + ценный
11. Malus manshurica (Maxim.) Korn. —♦ + тенофонд то же
12. Micromeles alnifolia (Sieb. et Zucc.) Koehne — +
13. Dasiphora mandshurica (Maxim.) Juz. — +
14. D davurica (Nestl.) Korn. — +
15. Rosa maximowicziana Rgl. + +
16. Prunus ussuriensis Koval, et Kost. + ценный генофонд
17. Prinsepia sinensis (Oliv.) Korn. — + то же
18. Armeniaca sibirica (L.) Lam.
19. Lespedeza cytrobotrya Miq. — —
20. Desmodium oldhami Oliv. —
21, Pueraria lobata (Willd.) Ohwi 4- —
22. Phellodendron amurense Rupr.* + _L
23. Securinega suffruticosa (Pall.) Rehd. + —
24. Celastrus orbiculata Thunb. + +
25. Acer komarovii Pojark. +
26. Ampelopsis brevipedunculata (Maxim.) Trautv. + +
27. A. hctcrophylla (Thunb.) Sieb. et Zucc. + —
28. A. japonica (Thunb.) Makino + +
29. Actinidia polygama (Sieb. et Zucc.) Maxim.
30. Daphne kamtschatica Maxim. +
31. Rhododendron sjchotense Pojark. + + эндем
32. Rh. shlippenbachii Maxim. + 4-
33. Rh. fauriei Franch. +
34. Syringa wolfii Schneid. + +
35. Abelia corcana Nakai — +
3G. Sambucus latipjnna Nakai —
Категория 3 — сокращающиеся виды
1. Taxus cuspidata Sieb. et Zucc. * + +
2. Pinus densiflora Sieb. et Zucc. + +
Продолжение табл.
Категория, название вида Наличие в коллекции вег. цв., пл. Примечание
3. Р. funebris Кот. + +
4. Abies holophylla Maxim. + +
5. Lanx olgensis Henry +
6. L. lubarskii Sukacz. +
7. Microbiota decussata Korn. + + эндем
8. Carpinus cordata Blume + +
9. Betula schmidtii Rgl. *
10. Quercus dentata Thunb. — —
11. Ribes fontinale Bochkar. — — ценный генофонд
12. Cerasus glandulosa (Thunb.) Lois. — + то же
13. C. sachalinensis (Fr. Schmidt) Korn, et Aliss. — + »
14. Armeniaca mandshurica (Maxim.) Skvortz. + + »
15. Acer mandschuricum Maxim. +
16. A. pseudosiboldianum (Pax) Kom. + +
17. Actinidia arguta (Sieb. et Zucc.) Planch. + + ценный генофонд
18. Echinopanax elatus Nakai + —
19. Kalopanax septemlobum (Thunb.) Koidz.* + +
20. Rhododendron mucronulatum Turcz. + +
21. Weigela praecox (Lemoine) Bail. + +
* Рубки запрещены циркулярами Министерства лесного хозяйства
РСФСР.
ЛИТЕРАТУРА
Биологическая флора Московской области. В. 1. Изд-во МГУ, 1974, 238 с.
Бобров Е. Г. Биологическая флора Британских островов (реферат). «Бот.
ж.», т. 32, 1947, №4, с. 181—182.
Богданова В. М. Условия прорастания семян аралиевых Дальнего Востока
и пути его ускорения. Автореф. канд. дис. Л., 1971, 23 с.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 277 с.
Воробьев Д. П., Ворошилов В. Н., Горовой П. Г., Шретер А. И. Опреде-
литель растений Приморья и Приамурья. М.—Л., «Наука». 1966. ,
Воробьев Д. П., Ворошилов В. Н., Гурзенков Н. Н., Доронина Ю. А., Его-
рова Е. М., Нечаева Т. И., Пробатова Н. С., Толмачев А. И., Чер-
няева А. М. Определитель высших растений Сахалина и Курильских
островов. Л., «Наука», 1974, 372 с.
Ворошилов В. Н. Ритм развития у растений. М., Изд-во АН СССР. 1960,
136 с.
Ворошилов В. Н. Флора советского Дальнего Востока. М., сНаука», 1966,
478 с.
Грушвицкий И. В. Женьшень. Л.. Изд-во АН СССР, 1961, 344 с.
Гурьев А. Д. Береза Шмидта в лесах южного Приморья, перспективы ее
использования и охраны. Автореф. канд. дис. Владивосток, 1977,23 с.
[ мельямова В. Г. Охрана заповедников, заказников, памятников Природы.
М., изд-во «Юридическая литература», 1976, 64 с.
Зорикова В. Т. Биологические особенности дальневосточных рододендронов
12
и введение их в культуру в условиях Приморского края. Автореф.
канд. дис. Владивосток, 1973, 24 с.
Ильченко Т. П. Сравнительно-кариологическое исследование лиственниц
Приморья. Автореф. канд. дис. Красноярск, 1975, 34 с.
Комаров В. Л., Клобукова-Алисова Е. Н. Определитель растений Дальне-
восточного края. Л., Изд-во АН СССР, 1931 — 1932, *1175 с.
Красная книга. Дикорастущие виды флоры СССР, нуждающиеся в охране.
М., «Наука». 1975, 204 с.
Купревич В. Ф. О задачах и возможных путях экспериментального изучения
растений в естественных условиях.— «Бот. ж.», т. 32, 1947/ №5,
с. 183—193.
Лапин П. И. Значение изучения жизненных циклов при интродукции.— Бюл.
ГБС АН СССР, т. 91, 1974, с. 3—7.
Проект Конституции (Основного Закона) Союза Советских Социалистиче-
ских Республик. Гл. 7, ст. 67. «Правда», 4 июня 1977 г.
Сергеева К. А. Физиологические и биохимические основы зимостойкости дре-
весных растений. М., «Наука». 1971, 174 с.
Слизик Л. Н. Краткий обзор декоративных деревянистых лиан юга Дальне-
го Востока.— Деревья, кустарники, многолетники для зеленого строи-
тельства на юге Дальнего Востока. Владивосток, 1970, с. 101 —108.
Слизик Л. Н., Древецкая Р. К. Опыты по зеленому черенкованию дальне-
восточных деревянистых лиан (в условиях юга Приморья).— Экспе-
риментальная экология и акклиматизация растений на Дальнем Вос-
токе. Владивосток, 1975, с. 35—42.
Титлянов А. А. Актинидии и лимонник. Дальневост, книжн. изд-во. 1969,
175 с.
Толмачев А. И. Введение в гео.ргфию растений. Изд-во ЛГУ, 1974, 165 с.
Урусов В. М. Эколого-биологические особенности и внутривидовая изменчи-
вость сосны погребальной (Pinus funebris Кот.) в Приморском крае.
Автореф. канд. дис. М., 1975, 31 с.
Шкугко Н. В., Александрова М. С., Фролова Л. А. К методике фенологиче-
ских наблюдений над хвойными растениями в ботанических садах.—
Бюл. ГБС АН СССР, т. 91, 1974, с. 8—14.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ОСНОВНЫЕ ПРОГРАММНО-МЕТОДИЧЕСКИЕ
НАПРАВЛЕНИЯ ЭКОЛОГО-ФИЗИОЛОГИЧЕСКОГО
ИЗУЧЕНИЯ РЕДКИХ И ИСЧЕЗАЮЩИХ ВИДОВ
ДЕРЕВЬЕВ И КУСТАРНИКОВ ДАЛЬНЕГО ВОСТОКА
к. в. мигин
Одной из важнейших особенностей большинства высших
древесных и кустарниковых растений, в том числе и предста-
вителей флоры Дальнего Востока, является весьма широкий
полиморфизм видов, на что неоднократно указывал Н. И. Ва-
вилов (Vavilov, 1922). При этом не всегда можно в естествен-
ных условиях уверенно выявить и отличить, когда полиморфизм
определяется различием экологических условий, а когда он ге-
нотипически присущ виду в одинаковых условиях произраста-
ния. Поэтому подлежат срочной и обязательной охране не
только исчезающие виды (возможно, уже поздно сейчас гово-
рить об изучении их полиморфизма и об отборе особо ценных
форм), но и те безусловно малочисленные или даже единичные
формы или растения внутри широко распространенных пока
видов, которые могут исчезнуть под влиянием различных ан-
тропогенных или даже случайных природных факторов.
Прежде чем перейти к непосредственному рассмотрению
программных и методических вопросов эколого-физиологиче-
ского изучения редких и исчезающих видов флоры Дальнего
Востока, хочется указать на то, что группировать объекты ис-
следования следует не только по признаку редкости и возмож-
ности исчезновения (как это сделано в Красной книге, 1975),
но также выделять цели изучения.
а) растения малочисленные, требующие на данном этапе
только сохранения либо в естественных условиях, либо путем
экстренного перенесения в культуру и в коллекции. При изуче-
нии таких растений важнейшими направлениями будут биоло-
гия семенного воспроизводства и физиология вегетативного
размножения.
б) растения, явно интересные с точки зрения выявления и
сохранения ценных форм. Здесь первостепенной задачей явля-
ется выделение локальных малых центров полиморфизма, в
которых будет проводиться изучение и дальнейший отбор форм.
в) растения, перспективные для «ближайшего использования,
широко полиморфные и в связи с этим нуждающиеся в уско-
ренном углубленном изучении для выделения и сохранения
перспективных генотипов для использования в народном хозяй-
стве.
U
При необходимости ускоренного и в то же время достаточно
надежного отбора наиболее ценных форм деревьев и кустарни-
ков выращивание большого формового разнообразия на- кол-
лекционных участках в течение 10—20 лет, пользуясь традици-
онной методикой, не только трудоемко, но и продолжительно
по срокам. Некоторые разработанные нами современные ме-
тоды экспресс-диагностики различных характеристик генотипов
на принципах моделирования внешних условий представляются
более прогрессивными.
В предлагаемой работе рассматриваются основные методи-
ческие направления эколого-физиологического изучения полез-
ных растений. Большая сложность применения фитотроники,
импульсного метода ядерного магнитного резонанса, математи-
ческого моделирования и др. еще не позволяет нам широко
рекомендовать такие методы, но для экстренного изучения, от-
бора и сохранения ценных редких и исчезающих генотипов,
находящихся под угрозой исчезновения, оправдываются затра-
ты, связанные с (применением методов экспресс-диагностики.
В задачи экспериментально-экологического изучения расте-
ний в естественной обстановке входят: выяснение форм и путей
функциональных приспособлений к определенным физическим
и биотипическим условиям; анализ структурных или морфоло-
гических приспособлений; изучение степени или амплитуды
физиологической или морфологической пластичности как нас-
ледственно закрепленной реакции на действие внешней среды
(Купревич, 1947).
Главнейшими физиологическими показателями для изуче-
ния и отбора ценных редких и исчезающих форм растений,
а также для введения их в культуру следует считать оценку
пределов устойчивости к факторам среды и физиологические
особенности вегетативного размножения. Последнее особенно
важно для сохранения оригинальных в хозяйственном отноше-
нии генотипов (Основные перспективные направления исследо-
ваний по физиологии п биохимии растений, 1969).
При изучении и введении в культуру ценных редких и исче-
зающих растений флоры Дальнего Востока нам представляется
наиболее целесообразной следующая программа эколого-физио-
логических характеристик генотипа.
Изучение форм в естественных условиях
1. Особенности местоположения данной группы или отдель-
ного растения. Рельеф, микрорельеф, почва, ее генезис, меха-
нический состав, признаки засушливости или заболачивания,
режим увлажнения, наличие засухоустойчивых или влаголюби-
вых растений.
2. Особенности и типы зимних повреждений растений: сол-
нечные ожоги; вымерзание древесины ствола и ветвей разных
15
возрастов; повреждения сердцевины однолетнего прироста,
переходной зоны от почки к побегу и проводящей системы реп-
родуктивных образований; вымерзание цветков в почках. Ха-
рактеристики должны быть по возможности количественными
(например, в процентах от общего числа учетных единиц, а
вымерзание древесины — по годам).
3. Характеристика возрастных особенностей годичного при-
роста для общей оценки силы роста в конкретных условиях
микро- и фитоклимата, особенно в данных условиях освещения.
В ряде случаев но данному признаку могут быть отобраны кар-
ликовые, а если необходимо — особо сильнорослые формы.
4. Долговечность репродуктивных органов. При этом опре-
деляется возраст репродуктивных образований и проводится
определение соотношения на растении образований разного
возраста. Такая характеристика позволит определить способ
формирования кроны для получения максимального урожая.
Эколого-физиологическое изучение форм в культуре
1. Оценка способности растений к вегетативному размно-
жению и разработка способов ускоренного размножения цен-
ных клонов. Любые случайные или микроклиматические при-
чины могут неожиданно привести к значительным повреждени-
ям или гибели части растений. Поэтому необходимо иметь
некоторый минимальный запас растений для восстановления
коллекции или для специальных экспериментальных целей.
В связи с этим особый интерес представляет изучение спо-
собности данного вида и формы к вегетативному размножению.
Отбор форм с наивысшим процентом укоренения зелеными че-
ренками даст возможность быстрейшего размножения и произ-
водственного испытания ценного материала. Существенное
значение имеет также отбор форм, способных размножаться
прививками черенком или окулировкой. Прибегать к пересадке
маточных экземпляров из природной обстановки в культуру
нужно лишь в крайнем случае, так как при этом возможна ги-
бель ценной формы при транспортировке или плохой прижи-
ваемости.
При изучении растений в естественных условиях необходи-
мо также учесть способность данного вида, формы или экотипа
к семенному размножению, ибо в случаях трудного укоренения
или неудовлетворительной приживаемости прививок единст-
венной возможностью сохранения ценных признаков генотипа
остается отбор среди его сеянцев. В таких случаях весьма не-
надежно переносить единственное маточное растение в куль-
туру с риском потерять его, проще провести отбор среди сеян-
цев этой формы.
2. Изучение физиологии покоя и прорастания семян. Это
особенно важно для растений, характеризующихся резко выра-
16
женной периодичностью плодоношения, и растений, не размно-
жающихся вегетативно. При этом необходимо изучать не толь-
ко сезонную ритмику прорастания семян, но и особенности ус-
ловий (включая термические режимы) их стратификации.
3. Создание методами вегетативного размножения клопово-
го потомства перспективных форм для углубленного исследо-
вания. Большое значение в этой работе приобретает вегетаци-
онный метод. В частности, для ускоренного изучения пределов
морозостойкости корневой системы нами разработана специ-
альная методика на основе вегетационного метода (Мигин,
заявка, на изобретение №2443521/15). Испытание зимостойко-
сти надземной системы при выращивании растений в вегетаци-
онных сосудах с использованием принципа прогрессирующих
термовоздействий (аналогично работам I. Thiele, 1957) и на-
шей методики (Мигин, 1975) мы также считаем весьма пер-
спективным.
4. Оценка пределов устойчивости перспективных форм дол-
жна проводиться ио трем параллельным направлениям: термо-
устойчивости, засухоустойчивости и устойчивости к переувлаж-
нению почвы.
Оценку термоустойчивости и зимостойкости растений флоры
Дальнего Востока целесообразно проводить по методикам, раз-
работанным нами для плодовых и ягодных культур, раздель-
но — по оценке пределов морозостойкости корневой системы
(Мигин, заявка на изобретение №2443521/15) и по определе-
нию пределов устойчивости надземных органов и тканей в
режимах переменных температур (Мигин, 1975). Эта методика
была нами проверена и получила подтверждение на других
древесных растениях (Петухова, Мигин, 1975), Для массовой
экспресс-диагностики пределов термоустойчивости надземных
органов перспективным является использование импульсного
метода ядерного магнитного резонанса (спиновое эхо) в соче-
тании с воздействием переменными температурами (Мигин и
лр., 1973).
При изучении физиологии засухоустойчивости и устойчиво-
сти к переувлажнению почвы главным мы считаем вегетацион-
ный метод, позволяющий решать поставленные вопросы вне
зависимости от конкретных метеорологических условий года.
Ускоренный отбор ценных форм и сортов по уровням их
термоустойчивости, на наш взгляд, необязательно проводить в
универсальных дорогостоящих фитотронах. Практически мы по-
лучали достаточно надежные результаты в термокамере с эле-
ментарным регулированием длины светового дня и программ-
ным регулированием суточного хода температуры, даже без
регулирования влажности воздуха. Такая камера была скон-
струирована нами и прошла многолетние успешные испытания
на базе Приморской плодово-ягодной опытной станции (i. Ар-
тем). Для зимы в южных районах Дальнего Востока характер-
2 заказ 488 Г* Li 2 17 I О«танмчоск^й т
на довольно низкая относительная влажность воздуха, которая
самопроизвольно создается и в термокамерах при отрицатель-
ных температурах. Именно поэтому в автоматике регулирова-
ния параметров в термокамере мы не придаем ведущего зна-
чения регулированию влажности воздуха.
5. Определение биологических температурных порогов нор-
мального жизнеобеспечения на различных фазах годичного цик-
ла роста и развития. Этому вопросу до сего времени уделяется
крайне мало внимания, хотя от термических порогов нормаль-
ного функционирования той или иной живой системы растения
в решающей степени зависит возможность его культуры в дан-
ных макро- и микроклиматических условиях и перспективы его
дальнейшего распространения. В этой связи весьма важной мы
считаем концепцию о роли суточного хода температуры воздуха
в развитии растений, разработанную 3. А. Мищенко (1962) и
особенно введенное ею впервые понятие о градусо-часах (нам
представляется удобнее оперировать как единицей измерения
«часо-градусом») для характеристики сумм действительно эф-
фективных температур на той или иной фазе годичного цикла
роста и развития растений.
Изучение термических порогов и амплитуд, а также опти-
мумов нормального жизнеобеспечения целесообразно проводить
по следующим основным этапам годичного цикла роста и раз-
вития:
а) начало вегетации (например, массовое появление зеле-
ного конуса, начало появления первого развернутого листа);
б) цветение (начало и массовое);
в) интенсивный рост побегов и прекращение роста побегов:
г) естественный листопад и продолжительность вегетацион
ного периода (от появления зеленого конуса до массового ли-
стопада);
д) термические пределы, динамика и сроки прохождения
зимнего развития растений.
Для изучения ритмики зимнего развития приемлема наша
методика определения процента распустившихся почек в раз-
ные сроки зимы относительно максимального процента почек,
способных распуститься (у данного вида или формы) в естест-
венных условиях весной. Общеизвестные методы отращивания
(изучение сроков выхода растений из «глубокого покоя»), при
которых начало распускания некоторого числа почек принима-
ется за дату «окончания глубокого покоя», мы считаем биоло-
гически недостаточно обоснованными, так как известно, что в
разные сроки зимы может распускаться разное число почек
(в процентном отношении) (Мороз, 1948)
По результатам наших исследований, постепенное, генотипи-
чески и фенотипически характерное нарастание процента почек,
способных к распусканию в определенные сроки зимы при ис-
кусственном отращивании, и особенно энергия распускания
18
почек в разные сроки зимы (выражается процентом распустив-
шихся почек при отращивании через определенные промежутки
времени, например через каждые 3—5 дней) являются наибо-
лее характерными показателями (и притом интегральными)
не «покоя», не сроков его окончания, а именно ритмики и энер-
гии зимнего развития растений. Под последним термином, пред-
ложенным еще К. М. Поплавским (1953), мы понимаем такое
динамичное нарастание качественных и количественных физио-
лого-биохимических изменений в клетках и тканях растений,
которое приводит к нормальному и дружному началу вегета-
ции.
Изучение ритмики и энергии зимнего развития весьма важ-
но для оценки уровня приспособленности данной формы к кон-
кретным климатическим и микроклиматическим условиям. Если
по основным фазам вегетации можно судить о приспособлен-
ности растения в данных экологических условиях только к ве-
гетационному периоду, то по ритмике зимнего развития можно
в сочетании с изучением динамики термоустойчивоеги судить о
зимостойкости растения в данной зоне, о типе зимостойкости
и отсюда — о возможных границах распространения растения
в другие зоны по пределам его термоустойчивости.
В процессе изучения ритмики зимнего развития равноценное
значение имеет как само исследование сроков и энергии подго-
товки растений к новой вегетации, так и определение темпера-
турных границ и режимов оптимального и замедленного про-
хождения зимнего развития у данного генотипа. Это позволит
также обосновать экологические направления введения в куль-
туру ценных видов или форм.
6. Основой эколого-физиологического изучения и оценки
редких видов и форм полезных растений является принцип мо-
делирования внешних условий. Это моделирование должно
приводиться по следующим основным 'параметрам:
а) вегетационный период (длина светового дня, интенсив-
ность освещения, спектральный состав света, термический ре-
жим воздуха и почвы, влажность почвы, относительная влаж-
ность воздуха);
б) период зимовки (длина светового дня, интенсивность ос-
вещения, спектральный состав света, термический режим возду-
ха и почвы, относительная влажность воздуха).
Моделирование должно осуществляться с применением ве-
гетационных и зимних камер искусственного климата. Напри-
мер, в серии вегетационных климакамер моделируются клима-
тические условия Владивостока, Хабаровска, Магадана и т. д.,
после чего подопытные растения из каждого варианта вегета-
ционного режима могут быть перемещены в климакамеры зим-
него периода соответствующих или инородных климатических
характеристик. Испытание зимостойкости растений, подготов-
ленных в вегетационном режиме влажных южных районов При-
2*
19
морского края, на режимах зимы суровых северных районов,
может дать характеристики потенциальной пластичности данно-
го генотипа для целей прогноза интродукции этого генотипа.
Более полные характеристики можно получить из разных соче-
таний условий вегетационных и зимних периодов, что и явля-
ется экспериментальной эколого-физиологической основой на-
учного прогноза путей и направлений введения наиболее цен-
ных растений в культуру.
7. Анализ географии распространения ценного материала и
прогноз введения его в культуру должен проводиться в следую-
щих основных направлениях:
а) оценка пределов устойчивости генотипа в основных вари-
антах климатических характеристик;
б) оценка генотипа по пределам экологической пластично-
сти (в условиях моделирования внешних факторов)* по основ
ным биологическим и хозяйственно ценным признакам (зимо-
стойкость, продуктивность, иммунитет, способы и интенсивность
размножения, продуцирование биологически активных полез»
ных веществ и т. д.);
в) разработка математических моделей оптимальных усло-
вий культивирования или воспроизводства в естественных ус-
ловиях;
г) разработка на принципах математического моделирова-
ния географических и микроклиматических границ возможного
и перспективного культивирования или распространения цен-
нейших отборных форм (генотипов).
В итоге предлагаемых системных разработок можно опре-
делить перспективы использования, культивирования и геогра-
фических границ расселения новых полезных растений любого
народнохозяйственного назначениям В биологическом и природо-
охранном отношении именно такая постановка работы позволит
за сравнительно короткий отрезок времени сконцентрировать
ценнейший генофонд исходного материала на ближайшую и
дальнюю перспективу его изучения и использования, а также
сохранить этот генофонд от случайной гибели при прогресси-
рующем освоении новых территорий природных ресуросов.
ЛИТЕРАТУРА
Вавилов Н. И. Дикие родичи плодовых деревьев Азиатской части СССР и
Кавказа и проблема происхождения плодовых деревьев (Докл. IX
Международному конгрессу по садоводству в Лондоне. 1930 г.)—Изб-
ранные произведения в 2-х томах. Л., «Наука», 1967, т. 1, с. 245.
Красная книга. Дикорастущие виды флоры СССР, нуждающиеся в охране
Л., «Наука», 1975, с. 9.
Купревич В. Ф О задачах и возможных путях экспериментального изуче-
ния растений в естественных условиях.—«Бот. ж.», т. 32, 1947, №5,
с. 185—193.
Мигин К. В. Способ определения морозостойкости корней древесных и ку-
20
старниковых растений. Заявка на изобретение № 2443521/15, 1977,
М.КЛ.2А01 Н 1/04.
Мигни К. В. Диагностика зимостойкости сортов плодовых и ягодных куль-
тур в режимах переменных температур.— Информационный листок
ДВ ЦНТИ №287—75. Владивосток, 1975, 5 с.
Мигин К. В., Кулеш Ю. Г., Беликов И. Ф., Черницын А. И. Способ лабо-
раторного диагностирования зимостойкости сортообразцов плодовых и
ягодных растений. Описание изобретения к авт. свид. № 388719,
СССР, 1971, М. Кл. 2 А 01 Н 1/04.
Мищенко 3. А. Суточный ход температуры воздуха и его агроклиматическое
значение. Л., Гидрометеонздат, 1962, с. 67.
Мороз Е. С. Экспериментально-экологические исследования периода покоя
у древесных растений.— «Экспериментальная ботаника», сер. IV, 1948,
в. 6.
Основные перспективные направления исследований по физиологии и биохи-
мии растений. М.. «Наука», 1969, 58 с.
Поплавский К. М. Период зимнего развития у яблони.— Труды Плодоовощ-
ного института им. И. В. Мичурина, т. VII, 1953, с. 43—78.
Петухова И. П., Мигин К. В. Диагностика зимостойкости древесных и кус-
тарниковых растений на Дальнем Востоке (некоторые вопросы мето-
дики).— Озеленение городов Дальнего Востока. Владивосток, 1975,
с. 157—167.
Thiele I. Progressive Frostversuche mit keimenden und einjarigen Kemobst-
samlingen.— «Zuchter», Bd. 27, 1957, S. 161—172.
Vavilov N. I. The low of homologous series in variation.— «Journ. Gene-
tics», XII, No. 1, 1922, p. 7-61.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
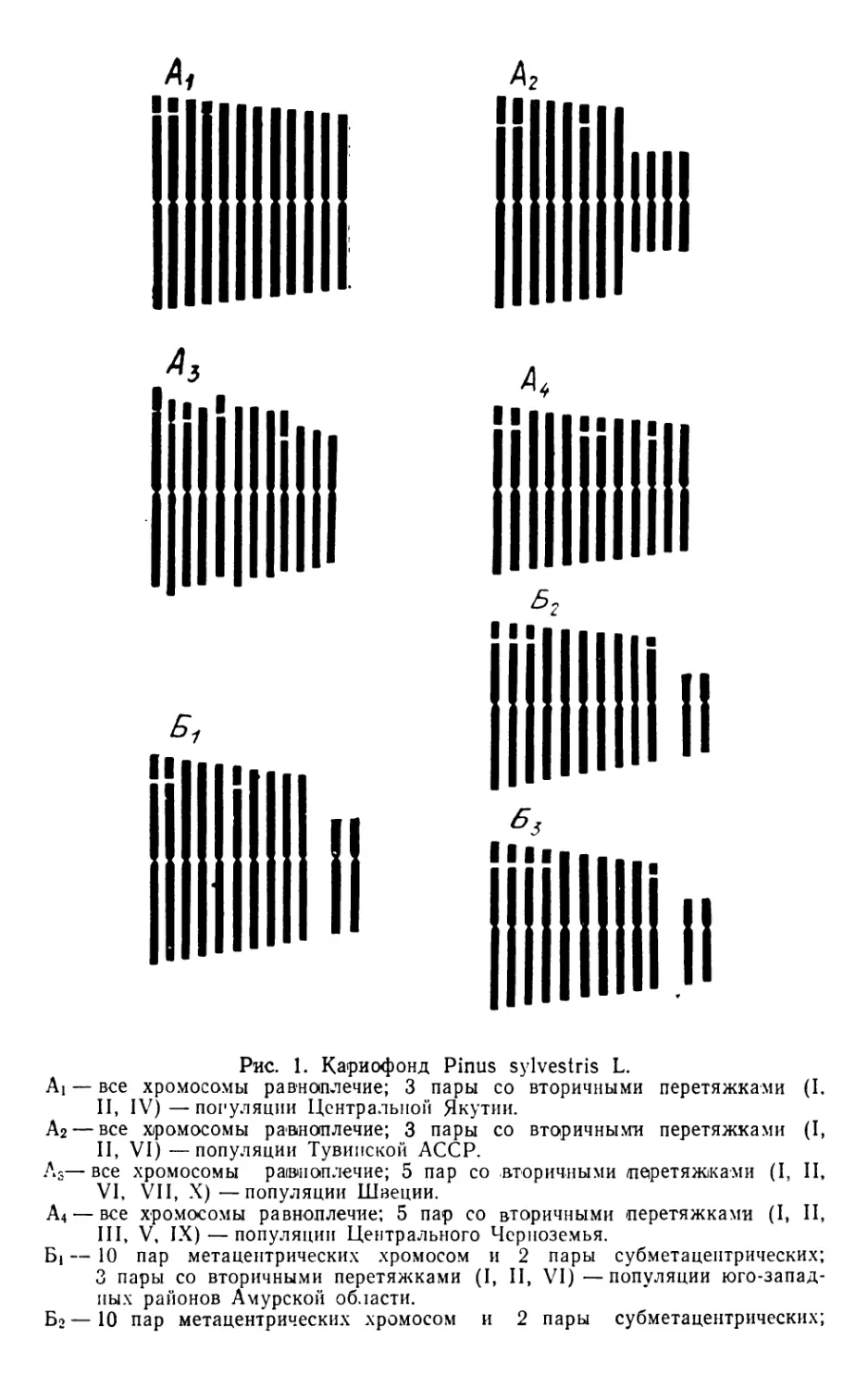
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА КАРИОТИПОВ
НЕКОТОРЫХ ВИДОВ РОДА LARIX Miller
Т. П. ИЛЬЧЕНКО
Долгое время существовало мнение о стабильности карио-
типа хвойных (Sax, Sax, 1933). Однако за последние десятиле-
тия результаты многих работ в области кариологии (Будара-
гин, 1973; Круклис, 1970, 1974а, б; и др.) позволили авторам
сделать обоснованный вывод о кариотипической изменчи-
вости этой группы растений. Сейчас уделяется особое внима-
ние хромосомному полиморфизму, что имеет немаловажное
значение в познании процессов эволюции и видообразования.
Исследования кариотипа на видовом и популяционном
уровнях позволяют выяснить кариотипическое разнообразие и
провести сравительный их анализ. Примером могут служить
некоторые виды рода Larix Miller.
К настоящему моменту изучены кариотипы Larix sibirica
Ledb., L. sukaczewii Dyl., L. griffithiana Carr., L. decidua Mill.
(Simak, 1962, 1964, 1966), L. dehurica Turcz., L. czekanowskii
Sz., L- leptolepis Gord. (Круклис, 1970, 1974a, 6), L. olgensis
A. Henry, L. lubarskii Sukacz. L- komarovii B- Kolesn. (Ильчен-
ко, 1973). В результате цитологического анализа авторы при-
шли к единому мнению о стабильности и сходстве кариотипов
изученных видов лиственниц. Причем единство заключалось не
только в количестве хромосом диплоидного набора (2п = 24).
что характерно в целом для семейства, но и в положении цент-
ромеры, локализации вторичной перетяжки, абсолютной и от-
носительной длине хромосом (табл. 1). Особенностью кариоти-
па лиственниц является и стабильное отношение морфологи-
чески различных хромосом (из 24 хромосом диплоидного
набора — 12 изобрахиальных и 12 гетеробрахиальных). У дру-
гих представителей семейства (сосна, ель, пихта) этот признак
варьирует. Близость морфологии хромосом указывает на гене-
тическую совместимость различных видов рода Larix, что спо-
собствует массовой гибридизации между ними и в результате
усложняет таксономию.
Однако авторы указанных выше работ неоднократно отме-
чали и различия в кариотипах лиственниц, которые необходимо
учитывать при характеристике видов. Эти различия выражают-
ся в суммарной абсолютной длине хромосом в наборе, в нали-
22
Таблица 1
Относительная длина набора хромосом различных лиственниц
Вид Порядковый номер хромосом Автор
1 3|4 5 6 7 8 9 10 И 12
Л. даур- Круклис
ская 133 128 123 120 114 106 87 86 82 79 75 72 (1970)
Л.европей- Сакс
ская 128 121 121 118 118 100 97 90 77 74 74 72 (1933)
Л. европей- Шимак
ская 144 128 124 118 116 106 88 86 78 74 71 67 (1962)
Л.ольгин- по нашим
ская 133 125 117 112 ПО 99 90 84 79 74 68 65 данным
Л. Любар-
ского 136 127 120 117 111 102 89 84 80 73 69 66 то же
Л. Кома-
рова 136 127 120 120 112 106 86 83 79 80 80 68 то же
чии хромосом со вторичными перетяжками и во встречаемости
их в популяции. Так, М. Шимак (Simak, 1964), учитывая эти
признаки и придавая им таксономическое значение, выделил
две кариотипические группы лиственниц — европейскую и си-
бирскую. В первую включена L. decidua, во вторую — L. sibi-
rica и L. sukaczewii. На основании кариологических данных
М. В. Круклис (19746) расширила и дополнила эту классифи-
кацию, объединив лиственницы в три группы:
1. L. griffithiana, в кариотипе которой только одна пара
хромосом со вторичной перетяжкой (III или IV).
2. L. leptolepis, L. sibirica, L. sukaczewii, в кариотипе две
пары хромосом со вторичной перетяжкой (III и IV).
3. L- dahurica и L. decidua — вторичные перетяжки наблюда-
ются в трех парах хромосом (III, IV и VII).
L . czekanowskii как естественный гибрид не имеет своего
характерного кариотипа и поэтому не включена в эти группы.
Кариологическое исследование лиственниц Приморья (Иль-
ченко, 1973) позволило в свою очередь дополнить кариотипи-
ческую классификацию лиственниц и распределить их по ка-
риотипу следующим образом:
I — L. griffithiana (рис. А),
II — кариотипическая группа сибирских лиственниц (рис. Б).
L. sibirica, L. sukaczewii. L. leptolepis,
III — кариотипическая группа европейских лиственниц
(рис. В): L. decidua,
IV — кариотипическая группа дальневосточных лиственниц
(рис. Г); L. dahurica,
а) подгруппа приморских лиственниц: L. komarovii, L. оЬ
gensis, L. lubarskii.
Лиственницы Приморья объединены с L. dahurica в одну
23
Illllllll
liiiiiiiiii
liiiiiiiiii
6
Идиограммы некоторых видов рода Larix Miller:
Л — L. griffithiana Carr. По данным M. Ши-
мака (Simak, 1966);
Б — кариотипическая группа сибирских ли-
ственниц. По данным М. Шимака (Simak, 1964),
L. sibirica Ledb., L. sukaczewii Dvl., L. leptole-
pis Gord. (Круклис, 1974a);
В - кариотипическая группа евоопенских лист-
венниц. По данным М. Шимака (Simak, 1962)
L. decidua Miller;
Г — кариотипическая группа дальневосточ-
ных лиственниц. L. dahurica Turcz. (М. В. Крук-
лис, 1970). Подгруппа приморских лиственниц.
По данным Т. П. Ильченко (1973) L. komarovii
В. Kolesn., L. olgensis Henry, L. lubarskii Su-
kacz
группу по наличию трех пар хромосом со вторичными перетяж-
ками (III, IV и VII), что указывает на филогенетическую бли-
зость, общность происхождения и генетическое родство этих ви-
дов. Но по встречаемости вторичной перетяжки приморские
виды выделены в самостоятельную подгруппу. По данным
Ч. В. Круклис (1970), вторичная перетяжка в VII паре хро-
мосом L. dahurica встречалась регулярно, чего не было
отмечено у лиственниц Приморья. Чаще всего она наблюда-
лась у L. komarovii и реже у L. lubarskii (табл. 2). Эти разли-
чия, видимо, можно объяснить не только изолированным поло-
жением ареалов, но и гибридным влиянием L. dahurica. Инте-
ресно отметить, что у приморских лиственниц (L. komarovii,
L. olgensis, L. lubarskii) вторичная перетяжка VII пары -тасто
встречалась только у одного гомолога. Это указывает, по край-
ней мере, на относительную молодость видов, так как при ви-
дообразовании хромосомные перестройки первоначально вы-
ражаются в появлении гетерозиготных хромосомных наборов,
где изменен только один гомолог.
24
Таблица 2
Встречаемость хромосом со вторичными перетяжками, %
Вид лиственниц . Хромосомы
3-я изобрахи- альная 4-я изобра- хиальная 7-я гетеро- брахиаль- ная
Larix olgensis 35 52 12
Larix lubarskii 34 55 8
Larix komarovii 30 48 20
Контакт ареалов Larix olgensis и L. komarovii 41 26 6
Если по встречаемости вторичной перетяжки в VII паре у
приморских лиственниц наблюдались различия, то по III и IV
парам получены близкие показатели (табл. 2), что позволяет
судить об общности происхождения видов. Возможно, они име-
ли общего предка, занимавшего в геологическом прошлом
сплошной ареал и имевшего кариотип, идентичный лиственни-
цам сибирской кариотипической группы (L. sibirica, L. sukacze-
wii, L. leptolepis). Следовательно, на основании анализа карио
типа просматривается генетическое родство двух кариотипи-
ческих групп рода Larix — сибирской и дальневосточной.
Отличия приморских лиственниц отмечались и в суммарной
абсолютной длине хромосом (L. komarovii — 217 мк, L. olgen-
sis— 238 мк, L. lubarskii— 240 мк).
Хромосомный полиморфизм наблюдается не только на ви-
довом, но и на популяционном уровне. Популяции кариотипи-
чески неоднородны и характеризуются определенным карио-
фондом, что четко просматривается на лиственницах Приморья.
М. В. Круклис (19746) рассматривает наличие вторичной
перетяжки в кариотипе рода Larix как прогрессивный струк-
турный признак хромосом и по данному признаку к филогене-
тически молодым видам относит L. dahurica и L. decidua.
К филогенетически более специализированным — L. leptolepis,
L. sibirica, L. sukaczewii- Самым древним видом она считает
L griffithiana, что согласуется с гипотезой В. Н. Сукачева
(1924) о древности лиственниц, произрастающих в Китае, в том
числе и L. griffithiana.
Для приморских лиственниц характерно наличие как при-
митивных (с одной парой хромосом в кариотипе со вторичной
перетяжкой или все хромосомы без вторичной перетяжки), так
и более организованных кариотипов (три пары хромосом со
вторичными перетяжками). По встречаемости этих кариотипов
и отличаются кариофонды популяций. Так, только популяции
контакта ареалов L. olgensis и L. komarovii содержали боль-
шой процент менее организованных кариотипов (до 20% пла-
стинок, где не обнаружены вторичные перетяжки). В других
25
же популяциях число таких пластинок не превышало 2%.
Популяции отличались и по встречаемости более организован-
ного кариотипа. Это явление, видймо, можно объяснить не
только влиянием процесса гибридизации, который постоянно
действует в природе, но и дивергентной эволюцией кариотипа,
в основу которой легла географическая изоляция популяций.
Очевидно, наличие менее специализированных кариотипов у
приморских лиственниц говорит о сохранении здесь примитив-
ных форм (предков), и в то же время наличие продвинутых
форм указывает на процесс эволюции. Видимо, Приморье яв-
ляется центром формообразования, а возможно, и центром
происхождения рода, по заслуживает в дальнейшем присталь-
ного детального изучения.
Таким образом, на основании исследования кариотипа в
сочетании с морфолого-анатомическими, физиологическими и
другими признаками можно судить о генетической ценности
отдельных популяций. Популяции, в которых наблюдается наи-
большее разнообразие кариотипов, имея обогащенный карио-
фонд, представляют, по нашему мнению, высшую степень при-
спосабливаемое™ к условиям среды. Эти популяции представ-
ляют большой интерес и в селекции, так как содержат ценный
генетический материал с огромным кариотипическим разнооб-
разием. Следовательно, генетическая ценность отдельных по-
пуляций лиственниц Приморья неоспорима. Но, к сожалению,
эти уникальные популяции в настоящее время находятся на
стадии гибели (Гуков, 1971), что ведет к выпадению из эволю
ционного процесса определенного цикла развития органиче-
ского мира. Поэтому необходимо сегодня решать вопрос об
охране таких популяций.
ЛИТЕРАТУРА
Бударагин В. А. Анализ кариотипов изолированных популяций сосны обык-
новенной (Pinus silvestris L.) в Северном и Центральном Казах-
стане— «Генетика», т. IX, 1973, №9, с. 41—52.
Гуков Г. В. Лиственница Любарского (Larix lubarskii Suk.) в Приморском
крае.— «Лесоведение», 1971, №6, с. 23—32.
Ильченко Т. П. Сравнительно-кари о логический анализ лиственниц Примо-
рья.— «Лесоведение», 197J, №6, с. 69—72.
Круклис М. В. Кариологическое исследование лиственницы даурской (Larix
dahurica Turcz.).— Лесная генетика, селекция и семеноводство. Пет-
розаводск, 1970.
Круклис М. В. Кариологические особенности лиственницы Чекановского
(Larix czekanowskii Sz.).— Изменчивость древесных растений Сибири.
Красноярск. 1974а, с. 11—19.
Круклис М. В. Характеристика кариотипов некоторых хвойных растений в
связи с их эволюцией.— Труды Ин-та экол. раст. и животн., в. 90.
Свепдловск. 19746, с. 95—10 i.
Сукачев В. Н. К истории развития лиственниц.— Лесное дело. М.—Л., 1924.
Sax К. and Sax Н. J. Chromosome numbers and morphology in the Coni-
fers.— J. Arnold Arboretum, 1933, v. 14.
26
V
Simak M. Karyotype analysis of Larix decidua Mill, from different provenan-
ces.— Medd fran Statens Skogsforskningsinstitut, 1962.
V
Simak M. Karyotype analysis of Siberian Larch (Larix sibirica Land, and
Larix Sukaczewii Dyl.).— Studia Forestalia Suecica, 1964.
V
Simak M. Karyotype analysis of Larix griffithiana Carr. Hereditas, 1966,
v. 56.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ИЗМЕНЧИВОСТЬ СОСНЫ ПОГРЕБАЛЬНОЙ
В ПРИМОРЬЕ В СВЯЗИ С ОСОБЕННОСТЯМИ КЛИМАТА
Т. П. ИЛЬЧЕНКО, Л. С. ЛАУВЕ, В. М. УРУСОВ
Элементы климата в отдельных частях ареала сосны погре-
бальной, определяемые в значительной мере режимом атмо
сферной циркуляции, не претерпевшие коренных изменений по
крайней мере с конца плейстоцена, различны. В связи с
этим логично было предположить наличие определенной при-
способительной дивергенции географических групп популяций,
которая и выявлена нашими исследователями. Она настолько
значительна, что не объясняется сложностью становления вида
в целом (Урусов, 1973а) и, на наш взгляд, иллюстрирует один
из возможных путей вычленения более продвинутых таксонов.
Современный ареал вида состоит из островов, локализован-
ных в отрогах Восточно-Маньчжурских гор и Сихотэ-Алиня
{рис. 1), новейшее и окончательное разъединение которых
представляется явлением последних столетий и обусловлено
антропогенными воздействиями (к западу от бассейна р. Или-
стая) и климатогенными сменами. Приморская часть ареала
заключена между 4ОГ'ЗО' и 46°20' с. ш. и 130°15' и 135°00' в. д.
К югу и востоку идет увеличение осадков при одновременном
снижении испаренця и некоторой стабилизации снежного пок-
рова, причем гидротермический коэффициент возрастает от ве-
личины, близкой 1 (Ханкайский район), до 2 и более (влажное
побережье моря, п-ов Гамова), сумма активных температур
от 2500 до 2900°, а годовая сумма осадков — от 500 до 900 мм.
В условиях наиболее жесткого климатического режима (мак-
симальные перепады температуры и влажности воздуха и поч-
вы, периодические суховеи) находятся ханкайокие популяции
(Приханкайский округ Маньчжурской лесостепной области).
Прибрежно-морские популяции (Северо-Корейская провинция
области хвойно-широколиственных лесов) находятся в условиях
относительно ровного климатического режима (Колесников,
1956). Сихотэ-алинские сосняки (Маньчжурско-Южно-Примор-
ский округ Дальневосточной хвойно-широколиственной лесной
области) по климату ближе к ханкайским, но увлажняются
равномерней, снежный покров в них мощней.
В морфолого-анатомическом аспекте изучены главные по-
пуляции по всему приморскому ареалу. Кариологическим ис-
28
Рис. 1. Остаточные насаждения из сосны погребальной в Приморье. Раз-
мещение исследованных популяций и их групп: 1—популяция запанной
террасы оз. Ханка, 2 — о-ва Соснового, 3 — верховий р. Комиссаровка, 4 —
п-ова Гамова, 5 — верховий р. Илистая; группы популяций К — Кировского
н Ш — Шкотовского районов, II — верховий ip. Пойма и Б — р. Барабашев-
ьа Хасанского района. 1—современные сосновые рощи и их скопления; 2 —
сосна за пределами основного ареала; 3 — граница приморской части ценоти-
ческого ареала вида; 4 — районы со среднегодовым количеством осадков
в мм
следованием охвачено пять популяций, для четырех из них при-
веден сравительно-кариологический анализ в данной статье.
Пятая — сихотэ-алинская (одна из популяций бассейна р. Или-
стая) — по этому ряду признаков оказалась близкой третьей
29
популяции (верховья р. Комиссарова, Пограничный район).
Имеющиеся в литературе сведения о морфолого-анатомиче-
ских особенностях сосны погребальной представляют собой
анализ небольшого количества гербарных листов и так же,
как представления о ее таксономическом ранге, противоречивы.
Однако уже они отчасти характеризуют изменчивость цвета,
формы и величины шишек, формы апофизов семенных чешуй,
длины хвои (Комаров, 1901, 1917. 1934; Федченко, Флеров,
1909; Вольф, 1925; Строгий, 1934; Каппер, 1954; Солодухин,
1962; Усенко, 1966; Бородина, Плотникова, 1967; Воробьев,
1968). Исследованная нами изменчивость этих и многих других
признаков оказалась неодинаково широкой в разных геогра-
фических группах популяций и в целом очень высока. В по-
пуляциях обнаружены более или менее многочисленные группы
деревьев с четко выраженной морфологической однородностью,
а группы популяций различаются не только участием отдель-
ных форм, но и их набором.
Вариации генеративных органов. Обследование свыше
4 тыс. семяносящих сосен позволило заключить, что плоская и
выпуклая формы апофизов, указанные в предшествующих ра-
ботах 1как единственные, лишь более распространены. У оз.
Ханка до Уз сосен имеют шишки с бугорчатыми апофизами
(рис. 2). Участие вариантов этой формы в данной группе по-
пуляций выше, чем в других, хотя преобладающей, как и пов-
семестно в западных и центральных районах, остается форма
с выпуклыми апофизами. Только на террасе Ханки и о-ве Сос-
новом (в озере) найден вариант с крючковатыми апофизами.
В южных популяциях существенна роль формы со щитками,
выпуклыми в верхней половине. Популяциям правых притоков
р. Раздольная характерно горизонтальное ребро на щитках
семенных чешуй. Деревьев с абсолютно плоскими апофизами
больше на юге края (до 36%) и около 1% на севере ареала.
Участие формы с плоскими, но переходящими в бугорчатые у
ножек шишек апофизами снижается к северу только до 12—
14% общего количества обследованных сосен (около 1,5 тыс.
экземпляров из Ханкайского и Кировского районов). В Сихо-
тэ-Алине появляются растения с короткими лопатчатыми чешуя-
ми и особенно многочешуйными шишками (2—5% сосен)
Как и у других сосен секции Eupiitys Spach, семенные че-
шуи образуют легко просчитывающиеся спирали, одно число
которых всегда 8, другое 5 (не преобладают лишь у ханкай-
ских) или 13. Последнее число в Шкотовском районе и на
юге Хасанского встречается редко (рис. 3), а в некоторых по-
пуляциях (бассейн р. Пойма) вообще отсутствует. Шишки ми-
нимальной величины свойственны сосне хасанских и шкотов-
ских популяций (соответственно 35,23±0,18 и 35,25±0,32 см
длины и 21,25±0,9 и 20,98±0,18 см в диаметре), максималь-
ной— популяциям хр. Пограничного (длина 44,94±0,14, диа-
30
Рис. 2. Распространение
сосен с апофизами раз-
ных форм в группах по-
пуляций. Географиче-
ские группы популяций:
X — ханкайская, П —
пограничная (хр. Погра-
ничный, Пограничный
район), С — сихотэ-алин-
ская (верховья р. Илис-
тая). К — кировская,
Ш — шкотовская, Б —
барабашевская, Г — га-
мовская. Апофизы: 1 —
бугорчатые, 2—» выпук-
лые, 3 — плоские
Рис. 3. Участие в по-
пуляциях сосны некото-
рых форм (группы по-
пуляций обозначены как
на рис. 2): 1—форма с
8 и 13 линиями парастих,
2—с красными мужски-
ми стробилами, 3—с за-
смоленными почками
метр 24,68±0,07 см): зес и размеры их возрастают к западу
и северо-западу. Однако с экологией, весьма разнообразной по
типам местопроизрастаний повсюду, это не может быть связа-
31
но. Климатические и почвенные условия местопроизрастаний
оптимальны если не на юге Хасанского района, то, по крайней
мере, в верховьях впадающих в море рек. Довольно обычная в
западно- и центральноприморских популяциях крупношишеч-
ная форма (средняя длина шишек 60—80 мм, 0,2—0,5% дере-
вьев) южнее крайне редка. В южных популяциях ножки шишек
наиболее короткие (в среднем около 3 мм); уже в верховьях
Илистой они вдвое длиннее. По величине шишек и ножек юж-
ные популяции достоверно не различаются, форма шишек и их
окраска в 1 и 2 годы жизни здесь однородны, и наоборот, раз-
нообразны северней. В ханкайских популяциях цвет шишек
1 года в конце июня — начале июля варьирует от светло-зеле-
ного до фиолетово-черного, в других пунктах — темно-фиоле-
товый. В Шкотовском и Хасанском районах встречены только
фиолетовые шишки 1 года. Перед раскрыванием (сосна погре-
бальная высыпает семена на юге со второй половины октября
по начало ноября, севернее — весь октябрь) шишки в западных
и центральных районах — от сизо-зеленых до коричневых,
вскрывшиеся — от оливковых до глянцевато-коричневых, юж-
ных соответственно зелено-бурые и песочные.
Еще более специфичны наборы цветосеменных форм.
У оз. Ханка учтены деревья с черными, темно-серыми, кофей-
ными, желтовато-белыми, пестрыми и другой окраски полно-
зернистыми семенами. На п-ове Гамова кроме пестрых изредка
встречаются однотонные кофейные семена, а в Шкотовском
районе еще и черно-пестрые. Крылатки семян тоже пигменти-
рованы различно: оранжеватые и темно-коричневые встреча-
ются в центральных и западных районах, пергаментные свет-
лые повсеместно редки. В среднем длина семени с крылом до-
стигает максимальной величины по р. Комиссарова (18,34 ±
±0,11 мм), здесь же максимальна и величина шишек, затем
по р. Илистая и минимальна на п-ове Гамова (14,3±0,11). По
величине крылаток и ширине семян первые 2 группы популя-
ций достоверно не различаются. И вообще в них в основном
совпадают варианты генеративных форм.
В южных популяциях мужские стробилы перед пылением
только зеленые, в западных и центральных — от зеленых до
пурпурных. К северо-западу сосен с красными колосками боль-
ше. Форма чешуй на ножках женских стробилов и приколоско-
вых чешуй, на которых специально останавливался В. Л. Ко-
маров, позднее считавший, что в основном сосна погребальная
отличается от ее южного соседа — сосны густоцветковой (Pi-
nus densiflora Sieb. et Zucc.) — прицветными чешуями мужских
колосков (1934), меняется от узкоклиновидной до почковидной.
В центральных районах преобладает ширококлиновидная фор-
ма, в Хасанском и Шкотовском прицветники мужских колос-
ков зачастую с оттянутыми остроконечиями. Средняя ширина
чешуй возрастает к северу. Непостоянна она и у сосны густо-
32
цветковой собственно в Японии (анализ материалов гербария
БИНа).
По особенностям генеративных органов приморские популя-
ции распадаются по крайней мере на 3 группы: 1) высокопо-
лиморфные в районе наиболее суровых зим и жаркого нерав-
номерно влажного лета — ханкайские, где выявлено 8 форм
по особенностям апофизов шишек из 9 обнаруженных в крае,
8 цветосеменных форм и 6 по окраске крылаток (учтенные);
2) однородные в тяготеющих к морю районах со смягченным
ходом температур и влажности в течение всего года (форм по
апофизам шишек 5, цвету семян 2—4, крылаток 3, сосен с
красными колосками нет); 3) умеренно полиморфные в услови-
ях менее напряженного, чем у оз. Ханка, экологического режи-
ма, но порой (Сихотэ-Алинь, вторая популяция) сложных
фитоценотических отношений — идут климатогенные (гологене-
тические) смены доминант (форм апофизов 6—7, окраски се-
мян 3, окраски крылаток 6). Наиболее резко различны 1 и
2 группы.
Морфолого-анатомические особенности хвои. По данным ря:
да авторов, длина хвои колеблется от 45 до 80 мм (Комаров,
1901; Федченко, Флеров, 1909; Вольф, 1925; Ворошилов, 1966),
от 50 до 100 мм (Комаров, 1934; Каппер, 1954; Лыпа, 1955;
Усенко, 1966; Новиков, 1967; Воробьев, 1968; Dallimore, Jack-
son, 1954) и более 100 мм (Бородина, Плотникова, 1967).
Анатомическое строение хвои рассматривалось только
В. Л. Комаровым (1901, 1917) и (установлено по материалам
БИНа) на очень небольшом числе срезов, в основном из Кореи.
Сравнительное изучение хвои проведено методом частного
обследования популяций, хотя предварительно на массовом
материале вскрыты основные изменения ее в онтогенезе. Рас-
смотрены особенности покровных, проводящих, механических,
ассимиляционных тканей и смоляных каналов. В 1—4 популя-
циях образцы хвои второго года взяты в десятикратной повтор-
ности от 152 деревьев в возрасте 27—70 (100) лет с западных
ветвей в средних частях крон. Параметры внутренней струк-
туры замерены на 1520 срезах при помощи окуляр-микрометра.
В 5 популяции изучено около 600 срезов с 59 сосен. Всего же
по Приморскому краю просмотрено 4320 срезов хвои с 432 де-
ревьев типичного онтогенеза в возрасте 30 (45)—80 (120) лет —
в этом возрастном интервале величина хвои сосны погребаль-
ной наиболее стабильна (рис. 4). Изменчивость хвои изучалась
также на материале, привлеченном из экстремальных условий.
Независимо от географии местонахождения наблюдается
наличие однослойной гиподермы. Клетки эпидермиса с сильно
утолщенными оболочками. Ассимиляционная ткань двухрядная
с внутренней стороны и трехрядная с наружной. Механическая
ткань (склеренхима) над проводящими пучками однослойная,
реже двухслойная, причем второй слой может состоять всего из
3 Заказ 48S
33
Рис. 4. Изменчивость средней и экстре-
мальной длины хвои сосны погребальной
в онтогенезе (популяции центральных и
западных районов края). 1—деревья опти-
мального онтогенеза, выросшие без угне-
тения, 2—деревья, выросшие в лесу
нескольких клеток. Наибольшая длина хвои на о-ве Сосновом
(в среднем 74 мм), наименьшая в бассейне р. Комиссарова
(52,2 мм). Максимально длинная хвоя встречена на террасе
Ханки (117 мем). Отношение толщины хвои к ширине умень-
шается к северу и максимально в сосняках по р. Пойма, где
оно равно 0,56. Довольно близки по толщине хвои, величине
проводящих пучков и смоляных каналов, а также по расстоя-
нию между проводящими пучками популяции западной терра-
сы оз. Ханка, о-ва Соснового, верховий р. Комиссарова и
п-ова Гамова. Существенны различия ширины хвои, величины
жилки, количества и расположения смоляных каналов. Наи-
более четко по анатомическим признакам различаются сосны
центрального и южного Приморья с соснами западных районов,
где все параметры анатомической структуры хвои значительно
выше.
Толщина слоя эпидермиса и гиподермы повсеместно сходна
и различается мало; ее экстремальные величины колеблются
соответственно от 0,01 до 0,04 мм и от 0,01 до 0,03 мм, а сред-
ние равны 0,018—0,030 и 0,016—0,020 мм. Исключение состав-
ляет о-в Сосновый, где средняя велична этих признаков мини-
мальная (соответственно 0,016 и 0,012 мм), в то время как в
бассейне р. Комиесаровка, на п-ове Гамова и оз. Ханка сред-
няя величина эпидермиса 0,03, гиподермы 0,02 мм. Необходимо
отметить значительное развитие клеточных оболочек гиподермы
и механических элементов в западных и центральных популя-
циях.
Число смоляных каналов в хвое сосны погребальной варьи-
рует от 2 до 14 и лишь в 4 популяции их не свыше 11. На
террасе Ханки, по р. Илистая, на п-ове Гамова наиболее часто
встречается хвоя с 6 каналами, на о-ве Сосновом и по р. Ко-
миссаровка — с 8—9. Срезы с крайними числами смоляных
каналов немногочисленны: хвои с 2 каналами на террасе Хан-
ки 1,8%, о-ве Сосновом 1%, в бассейне Илистой 2,9% и на
п-ове Гамова 13% от общего количества. Малоканальность
увеличивается к югу. Максимальное среднее число смоляных
34
каналов 8 (бассейн р. Комиссарова), 7 (о-в Сосновый); в бас-
сейне р. Илистая и на п-ове Гамова их не более 6. Среднее
число смоляных каналов на срезе существенно разнится по
краю — от 4 у сосен в системе р. Пойма до 8 на хр. Погранич-
ном. С увеличением количества каналов наблюдается и уве-
личение всех параметров структуры хвои.
По расположению смоляных каналов выделено 3 типа хвои
(Лауве, Урусов, 1973). Третий тип — все каналы лежат в па-
ренхиме— характерен только некоторым южным популяциям.
Хвоя с 1—2 паренхиматическими каналами (второй тип) име-
ется повсеместно, но больше всего деревьев и срезов с такой
хвоей в бассейне р. Комиссарова и на о-ве Сосновом (соот-
ветственно 88,0 и 26,4% и 88,9 и 36,9%). В целом по примор-
ской части ареала встречаемость срезов второго типа выше у
экземпляров с большим числом каналов. Встречаемость же 3 и
более каналов в паренхиме с их общим числом не связана и
обусловлена особенностями генотипа. Обычно смоляные кана-
лы окружены одним рядом клеток склерохимы с сильно утол-
щенными оболочками. Каналы, соединенные с гиподермой толь-
ко одной клеткой, — переходные между периферическими и па-
ренхиматическими (Правдин, 1964) — чаще встречаются в юж-
ных популяциях, где отмечено наличие третьего типа хвои.
Популяции сосны погребальной по морфологии и анатомии
ассимилирующего аппарата различны. По образцам хвои воз
можно идентифицирование гербарного и посадочного материа-
ла. Существенны различия между особями западных (о-в Сос-
новый, берег Ханки и бассейн р. Комиссаровка) и южных (га-
мовская и другие) популяций. Западные популяции различа-
ются мало. Сходны по анатомическому строению и величине
структур также южные и сихотэ-алинские популяции, но раз-
витием механических тканей последние ближе западным.
С продвижением к северо-западу увеличиваются длина хвои и
все параметры анатомических структур, а также число смоля-
ных каналов. В популяциях южного Приморья появляется хвоя
с исключительно паренхиматическими каналами и слабо вы-
раженной механической тканью.
Кариологическая характеристика сосны погребальной. Ка-
риологические исследования рода Pinus L. неоднократно про-
водились как в нашей стране (Правдин, 1964; Шишниашвили,
1968; Бударагин, 1973; и др.), так и за рубежом (Sax К.,
Sax Н., 1933; Sax К., I960; Mehra, Khoshoo, 1956; Santamour.
1960; и др.). В настоящее время известна морфология хромо-
сом более 40 видов сосен (Р. silvestris L., Р. nigra Агп., Р. ра-
lustris Mill, и целого ряда других), но кариотип сосны погре-
бальной до настоящего времени не изучен. Кариологический
анализ данного вида проведен по общепринятой методике
(Правдин и др., 1972). При этом определяли следующие пара-
метры хромосом: число хромосом в диплоидном наборе; абсо-
3*
35
лютную длину хромосом, мк; относительную длину хромосом,
%; центромерный индекс, %: локализацию вторичной перетяж-
ки, %; индекс спирализации, %.
Учитывали и частоту встречаемости вторичной перетяжки.
Диплоидный набор хромосом — 24 (2п=24). Чис-
ло хромосом у сосны погребальной в метафазных пластинках
меристемы кончиков корешков установлено впервые. По дан-
ному признаку сосна погребальная не отличается от других
видов. Причем это число хромосом характерно не только для
сосен, но и для других представителей семейства сосновых
(лиственниц, пихт, елей).
Ниже приводятся данные кариотипического анализа сосны
погребальной из разных популяций Приморского края.
Абсолютная длина хромосом. Во всех парах хро-
мосом равноплечие явно нарушено, и верхнее плечо несколько
короче нижнего. Однако хромосомы I—X пар близки к равно-
плечим. У хромосом XI—XII пар западных популяций верхнее
плечо значительно короче нижнего (табл. 1). Кариотип южной
популяции не стабилен, особенно хромосомы VIII—XII пар:
в одних пластинках они ведут себя ближе к равноплечим, в
других же явно разноплечие. Во втором случае в кариотипе
отмечено пять пар разноплечих хромосом. В VIII—XII парах
хромосом южной популяции особенно варьирует верхнее плечо.
Эта популяция отличается и по суммарной абсолютной длине
хромосом гаплоидного набора. Так, на п-ове Гамова последняя
составляет 141,3 (136,9) мк, на о-ве Сосновом 169 мк, по р. Ко-
миссаровка 186,7 мк и на террасе Ханки 193 мк; хромосомы с
п-ова Гамова значительно короче, чем в других популяциях.
Ошибка опыта m колеблется в пределах ±1 мк.
Центромерный индекс в основном лежит в преде-
лах 41—49%. В 4 популяции он колеблется от 33 до 47%
(табл. 2). Значительное колебание центромерного индекса от-
мечено в основном в VIII—XII парах южной популяции, что
подтверждает наличие здесь двух типов хромосом VIII—XII
пары.
Относительная длина хромосом западных, цент-
ральных и южных популяций варьирует в пределах 3—5%.
Значительных различий по данному параметру между популя-
циями не обнаружено (табл. 3). Точность опыта 3—5%.
Вторичные перетяжки. По этому признаку карио-
тип сосны погребальной разных популяций имеет некоторые
отличия. Так, в ханкайских популяциях мы наблюдали непо-
стоянное число хромосом со вторичными перетяжками. В от-
дельных пластинках таких встречалось 4 пары, в других же
1—2 пары. Но наиболее часто их было 3. У сосны по р. Ко-
миссарова и Илистая более 1 пары хромосом со вторичными
перетяжками в пластинке не найдено. Большой процент прихо-
дится здесь на пластинки, где все хромосомы без вторичных
36
Таблица I
Абсолютная длина хромосом сосны погребальной
Пары хромосом оз. Ханка о-в Сос- новый р. Комисса- рова п-ов Га- мова п-ов Га- мова
а La а L а La а La а La
b b b b b
I 9,1 - 19,1 8,1 - 17,0
10,0 8,9
II 8,8 - 18,5 7,7 - 16,1
9,7 8,4
III 8,5 - 18,0 7,5 - 15,6
9,5 8,1
IV 8,4 - 17,8 7,3 - 15,1
9,4 7,8
V 8,1 - 17,1 7,1 - 14,8
9,0 7,7
VI 8,0 17,0 6,9 - 14,6
9,0 7,7
VII 8,0 - 16,5 69 - 14,3
8,5 7,4
VIII 7,6 - 15,8 6.6 - 14,0
8,2 7,4
IX 7,2 - 15,2 6.1 - 13,2
8,0 7,1
X 6,7 - 14,4 5,9 12,8
7,7 6,9
XI 5,5 - 12,9 5,1 - 11,6
7,4 6,5
XII 4,4 - 10,7 4,2 - 10,7
6,3 5,9
Суммарная 193,0 169,4
длина хромосом
Примечание. L — абсолютная
плеча; b — длина нижнего плеча.
8,6 9,5 8,6 9,2 8,4 9,0 8,2 8,7 7,9 8,7 7,8 8,4 7,4 8,4 7,5 . 18,1 - 17,8 . 17.4 . 16,9 . 16,6 16,2 , 15,8 7,1 7,8 6,5 7,4 6,5 7,0 6,1 6,9 6,0 6,7 5,9 6,9 5,3 6,3 5,3 14,9 13,9 13,5 13,0 12,7 12,8 11,6 11,6 6,8 7,6 6,5 7,0 6,5 7,0 6,2 6,8 6,0 6,8 5,3 6,7 5,0 6,0 4,2 — 144 — 13,5 - 13.5 — 13,0 — 12,8 — 12,0 — 12,0 ИЛ
15,5 6,3 5,3 68 11 ,и
8,0 7,0 . 15,0 11,5 3,2 6,3 — 9,5
8,0 6.3 . 13,5 6,2 5,3 11,4 3,5 6,0 9,5
7,3 5,3 . 12,7 6,1 4,5 9,5 3.3 5,3 2,7 5,4 8,3
7,4 4,7 . 11,2 5,0 4,3 9,6 “ 8,1
6,5 186,7 5,3 141,3 135,9
длина хромосом; а — длина верхнего
Таблица 2
Центромерный индекс хромосом сосны погребальной, %
Место произраста- ния Пары хромосом
I II III IV V VI VII VIII IX X XI XII
Оз. Ханка 47 48 47 48 48 47 49 48 44 47 42 41
О-в Сосновый Р. Комисса- 1 48 48 48 47 48 48 48 47 46 46 44 42
ровка 47 49 49 49 48 48 47 49 46 45 42 42
П-ов Гамова 47 47 4^ 47 47 45 46 45 46 46 45 43
П-ов Гамова 47 47 47 46 46 46 47 36 34 36 34 33
Таблица 3
Относительная длина хромосом сосны погребальной, %
Место произраста- ния Пары хромое о м
I II III IV V VI VII VIII IX X XI XII
Оз. Ханка 4,8 4,6 4,6 4,5 4,4 4,4 4,3 4,1 3,9 3,7 3,3 2,9 О-в Сосно- вый 5,0 4,7 4,6 4,4 4,4 4,3 4,2 4,1 3,9 3,7 3,4 3,2 Р. Комисса- ровка 4,8 4,7 4,6 4,5 4,4 4,3 4,2 4,1 3,9 3,6 3,4 3,3 П-ов Гамова 5,0 4,9 4,8 4,6 4,5 4,1 4,1 3,9 3,6 3,5 3,2 3,1
перетяжек (табл. 4), чего не наблюдалось в других популяци-
ях. Популяции с п-ова Гамова характеризуются наличием двух
пар хромосом.
Таблица 4
Встречаемость хромосом со вторичными перетяжками в различных
популяциях сосны погребальной, %
Пары хромосом
первая вторая третья четвертая без вторич- ных перетя- жек
Место
произрастания
Оз. Ханка 57 40 4 * 5 —
О-в Сосновый 60 48 46 8 —
Р. Комиссаровка — 50 — 50
П-ов Гамова 30 50 65 —
Л о к а л и з а ц и я вторичной пер етяжки. Во всех
популяциях вторичные перетяжки встречаются в группе рав-
ноплечих (метацентрических) хромосом. Расположены они на
верхнем плече ближе к проксимальному концу, реже встре-
чаются на дистальном. Иногда кроме вторичной перетяжки
одновременно наблюдалась третичная, расположенная на ниж-
нем плече. Частота встречаемости таких хромосом не превы-
шала 1%.
Итак, в Приморье отдельные популяции сосны погребальной
различаются и по абсолютной длине и по морфологии хромо-
сом: между западно- и центральноприморокими эти различия
незначительны, резко отличима южная популяция. Причем
различия есть и внутри популяции. Как отмечалось выше,
в популяции п-ова Гамова обнаружено два типа хромосом
VIII—XII пар и соответственно выделено два кариотипа—в од-
ном все пары равноплечие (метацентрические), в другом 7 пар
равноплечих и 5 разноплечих (субметацентрических). В запад-
ных и центральных популяциях установлен один кариотип —
10 пар равноплечих и 2 разноплечих. Разнообразие кариотипов
38
сосны погребальной возможно объяснимо явлением дивергент
ной эволюции, а на п-ове Гамова произрастанием здесь двух
видов сосен кариотипически различных и их гибридизацией,
о чем будет сказано ниже. Но для решения данного вопроса
необходимы дальнейшие кариологические исследования сосен
не только Дальнего Востока, а и сопредельных стран.
Анализ морфологии хромосом позволяет сделать вывод о
различии популяций сосны погребальной по кариотипу. Поэтому
возникла необходимость в составлении карты распространения
кариотипов (рис. 5). В дальнейшем, на наш взгляд, при состав-
лении таких карт необходимо учитывать кариофонд не только
вида, но и каждой популяции и отмечать на карте все карио-
типическое разнообразие популяции, а не наиболее часто встре-
чающиеся. Эти карты значительно облегчат работу селекцио-
неров. Учитывая кариотипическое разнообразие популяции,
следует обращать внимание и на полиморфизм других призна-
ков растений (морфолого-анатомических, физиологических и
эколого-биологических) Так, в западных популяциях нами от-
мечен высокий полиморфизм не только кариотипов, но и гене-
ративных органов.
Различие кариотипов по представляющим географические
группы популяциям можно объяснить и давней изоляцией
этих популяций, значительно более ранней, чем изоляция мест-
ных лиственниц, при изучении кариотипов которых существен-
ных расхождений не обнаружено (Ильченко, 1973).
Обсуждение результатов. Изучение 32 морфолого-анатоми-
ческих признаков генеративных органов и хвои сосны погре-
бальной, из которых 10, выраженных линейными мерами, обна-
руживают тесную связь в трех группах, позволило заключить,
что описанный В. Л. Комаровым (1901) вид полиморфен и
представлен рядом форм по строению апофизов, числу пара-
стих, цвету и форме генеративных органов, типам хвои. В раз-
ных группах популяций вариабельность вида неодинаково ши-
рока и гомологична изменчивости сосны обыкновенной только
в Ханкайском районе. По ареалу меняется также связь между
величиной генеративных органов и их формами и собственно
морфологическими формами, например шишек и семян, т. е.
они неравноценны. Морфолого-анатомические формы, разнооб-
разие которых частично освещено,— внешнее выражение гене-
тического, физиологического и в целом биологического несход-
ства (неодновременны цветение и высыпание семян, различны
энергия прорастания и интенсивность роста и т. д.), с возра-
станием которого увеличивается устойчивость популяций и их
систем, а экологическая амплитуда вида расширяется в про-
странстве и времени.
По набору форм, анатомии листа и особенно четко по ка-
риотипу выделяются 3 круга популяций, характеризующихся
вполне конкретно и в качественном аспекте: кроме макрокли-
39
Рис. 5. Кариофонд Pinus funebris Кот. (распространение кариотипов): А —
Ханкайский р-он (оз. Ханка, о-в Сосновый), Б — Пограничный и Михайлов-
ский районы (бассейны рек Комиссаровки и Илистая), ВГ — п-ов Гамова
матической и географической им присуща морфолого-анатоми-
ческая и кариотипическая определенность. Кариотипически
наиболее продвинутой оказалась группа популяций в условиях,
приближающихся к таковым в сосняках из лесной сосны на
южном (Юнатов, 1950; Правдин, 1964) и юго-восточном (Ко-
лесников, 1945) пределах ее распространения. Здесь в окрест-
ностях оз. Ханка и прежде всего на его террасе впервые в крае
сообщества оказываются организованными и по типу настоя-
щих боров. Видимо, многообразие местопроизрастаний, широ-
чайшая амплитуда сезонных и флюктуационных колебаний
элементов климата, не свойственные побережью моря, ряду
территорий восточного мегасклона Сихотэ-Алиня и в целом
горным его склонам (Грушвицкий, 1940), покровительствовали
накоплению форм и сложению их набора, обеспечившего вы-
сокую, возможно, наивысшую в пределах вида жизненность
этих популяций. Таким образом, можно констатировать более
энергичный темп стабилизирующего отбора именно в популя-
циях Ханкайского района, приближающегося по климатическим
характеристикам к континентальным районам. Расширение
ареала приханкайских популяций внутри общего ареала вида
и дальнейшая перестройка последнего, а это одно из главных
условий становления нового таксона, невозможны вследствие
давления антропогенных факторов. Но при искусственном вос-
становлении сосны погребальной, необходимость которого бес-
спорна (Куренцова, 1952; Розенберг, Колесников, 1958), следу-
ет привлечь посадочный материал прежде всего из ханкайских
популяций, которые необходимо особенно тщательно охранять
и восстанавливать.
Явлением этого же ряда — аналогичной кариоморфологи-
ческой дивергенцией при изоляции в стабильно различных ус-
ловиях — мы склонны объяснять и разнообразие кариотипов,
недавно обнаруженное у меловой сосны (Машкин и др., 1974).
Эволюция кариотипа сосны погребальной, а также лиственниц,
видимо, идет по пути уменьшения ДНК и нарастания асиммет-
рии хромосом. В результате увеличивается число хромосом
разноплечих и со вторичными перетяжками. Разноплечесть воз-
никает в основном за счет редукции верхнего плеча (Ильчен-
ко, 1975).
Однако в южных популяциях сосны погребальной просле-
живается несколько иной путь развития кариотипа как в связи
с приспособлением растений, так и специализацией вида — на-
растание асимметрии хромосом здесь происходит за счет поте-
ри ДНК без образования вторичной перетяжки. А!орфологиче-
ское разнообразие хромосом VIII—XII пар в данной популя-
ции подтверждает принадлежность ее сосне густоцветковой на
северо-восточном пределе материкового распространения пос-
ледней (Урусов, 1973а, б). И тогда в принципе допустимо пред-
положение о перекрывании ареалов двух видов секции на этом
41
пространстве. Совместное произрастание особей двух видов, в
связи с наложением их ареалов, могло привести и к гибридным
перестройкам, сказываясь и на морфологии хромосом. К тому
же морское побережье Приморья являлось местом перекрыва-
ния ареалов сосен секции Eupitys Spach в плейстоцене и в бо-
лее ранние эпохи (Гитерман и др., 1968; Короткий, Караулова.
1975). Однако гибридогенный процесс и его результаты в на-
стоящей статье мы не затрагиваем, как и происхождение кон-
кретных популяций и популяционных комплексов: вид как си-
стема в данном случае интересует нас в системе климатических
районов.
Морфолого-анатомические особенности оказавшихся в более
блаюприятных условиях популяций хр. Пограничного и Сихо-
тэ-Алиня близки к особенностям на первоначальных стадиях
формирования вида. Здесь логично допустить сходный с райо-
ном Ханки, но замедленный процесс, на что указывают, во-пер-
вых, аналогия наиболее распространенных морфологических
форм (по крайней мере пограничных), во-вторых, достаточно
продолжительный период времени — возраст сосны погребаль-
ной определяется нами как миоцен-среднеплиоценовый.
Необходимо отметить, что резкие различия анатомии хвои
обусловлены в большей мере происхождением определенных
популяций, чем их последующей дивергенцией: уже на ранних
этапах филогенеза вида лист должен обладать особенностями,
позволяющими оптимально функционировать в большой кли-
матической амплитуде, чего нельзя сказать, например, о гене-
ративных органах. Но и здесь налицо достоверное увеличение
числа смоляных каналов, механических клеток, размеров струк-
тур с нарастанием континентальности. Аналогично варьируют
среднепопуляционные величины хвои и продолжительность ее
жизни. Типы срезов, выделенные по особенностям смоляных
ходов, тоже определяются конкретным генезисом популяций.
Но сохранение особей с хвоей паренхиматического типа, ха-
рактерной, как известно, соснам низкоширотных достаточно
теплых районов (Япония, Центральный и Юго-Восточный Ки-
тай, Кавказ, юг Европы), обусловлено рельефно более опти-
мальной внешней средой ряда территорий Приморского края.
Таким образом, в результате дивергенции в длительно различ-
ных условиях сосна погребальная представлена в Приморье
системой экотипов, или рас, вычленение которых ускорялось
в районах нараставшей континентальности. Наоборот, в более
стабильных условиях с относительно большим или менее под-
верженным сезонным и годичным колебаниям количеством
тепла и влаги фенотипический состав популяций достаточно
однороден даже несмотря на присутствие особей с сохранив-
шимися архаичными чертами внутренних и внешних структур
(колючие апофизы, паренхиматические смоляные ходы, свет-
лая окраска генеративных органов и др.). Повышенная встре-
42
чаемость последних форм сосны погребальной, по нашему мне-
нию, может служить показателем благоприятности почвенно-
климатических условий и должна учитываться при наборе ас-
сортимента садовых и лесных культур.
ЛИТЕРАТУРА
Бородина Н. А., Плотникова Л. С. Определитель родов и видов семейства
Pinaceae Lindl.— Бюл. ГБС. 1967, в. 66.
Бударагин В. А. Анализ кариотипов изолированных популяций сосны обык-
новенной (Pinus silvestris L.) в Северном и Центральном Казахстане —
«Генетика», 1973, №9.
Вольф Э. Л. Хвойные деревья и кустарники европейской и азиатской ча-
сти СССР. Л., изд. Лесного ин-та, 1925.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока.
Л., «Наука», 1968, с. 29—33.
Ворошилов В. Н. Флора советского Дальнего Востока. М., «Наука», 1966,
478 с.
Гитерман Р. Е., Голубева Л. В., Заклинская Е. Д., Коренева Е. В.,
Матвеева О. В., Скиба Л. А. Основные этапы развития растительно-
сти Северной Азии в аитропогене.— Труды Геологического ин-та
АН СССР. в. 177. М., «Наука», 1968, 271 с.
Грушвицкий И. В. Явление инверсии растительности в Уссурийском крае.—
«Бот. ж », т. 25, 1940, № 1, с. 52—67.
Ильченко Т. П. Сравнительный криологический анализ лиственниц Примо-
рья.— «Лесоведение», 1973, №6, с. 69—72.
Ильченко Т. П. Об эволюции кариотипов хвойных.— Материалы XII Меж-
дунар. бот. конгр. 1975.
Каппер О. Г. Хвойные породы. М.—Л., Гослесбумиздат, 1954.
Колесников Б. П. Обыкновенная сосна (Pinus silvestris L. s. 1.) на юго-вос-
точной границе своего ареала.— Бюлл. МОИП, огд. биол., нов. сер.,
1945, в. 50, №5-6, с. 112-125.
Колесников Б. П. Природное районирование Приморского края. — Вопр.
сельск. и лесн. хоз-ва Дальнего Востока, з. 1, 1956.
Комаров В. Л. Флора Маньчжурии. I. СПб., 1901. с. 177—178.
Комаров В. Л. Типы растительности Южно-Уссурийского края.— Труды
почв.-бот. экспед. по исслед. колониз. районов. Аз. России, ч. 2. Бот.
исслед. 1913 г., в. 2. Пгр., 1917.
Комаров В. Л. Флора СССР. 1. Л, Изд-во АН СССР, 1934, с. 160—170.
Короткий А. М., Караулова Л. П. Новые данные о стратиграфии четвер-
тичных отложений Приморья.— Вопросы геоморфологии и четвертичной
геологии юга Дальнего Востока СССР. Владивосток, 1975, с. 79—110.
Круклис М. В. Характеристики кариотипов некоторых хвойных растений
в связи с и\ эволюцией.— Труды Института экологии растен. и жи-
вотн., в. 90. Свердловск, 1974. с. 95—101.
Курснцова Г Э. Порослевые дрсвссно кустарниковые заросли юго-западной
части Приморского края.— Труды ДВФ АН СССР сер. растениевод-
ческая, в. 1, 1952, с. 141—,147.
Лауве Л. С., Урусов В. М. К морфолого-анатомическим особенностям хвои
сосны погоебальной (Pinus funebris Кот.) в Приморском крае.— Бюл.
ГБС, 1973, в. 89, с. 49—55.
Лыпа А. Л. Определитель деревьев и кустарников. 1. Киев, Изд-во Киевск.
гос. ун-та им. Т. Г Шевченко, 1955.
Машкин С. И., Буторина А. К., Белозерова М. М., Пожидаева И. М., Му-
рая Л. С., Катунцева Л. Н., Доронин Ю. А. Сравнительное изучение
кариотипов сосны меловой (Р. crctaceae Kalen.) и сосны обыкновенной
(Р. silvestris L.) в популяциях Центрального Черноземья.— Генетика,
43
селекция и интродукция лесных пород, в. 1. Воронеж, изд. ЦНИИГиС,
1974. с. 10—31.
Новиков А. Л. Определитель хвойных деревьев и кустарников. Минск, «Вы-
шэйш. школа», 1967, с. 145—176.
Правдин Л. Ф. Сосна обыкновенная. М., «Наука», 1964, 190 с.
Правдин Л. Ф., Бударагин В. А., Круклис М. В., Шершукова О. П. Мето
дика кариологического изучения хвойных пород.— «Лесоведение», 1972,
№2.
Розенберг В. А., Колесников Б. П. Порослевые древесно-кустарниковые за-
росли малолесных районов Приморского края.— Труды ДВФ СО АН
СССР, сер. бот., в. 4(6), 1958, с. 5—45.
Солодухин Е. Д. Деревья, кустарники и лианы советского Дальнего Востока.
Уссурийск, изд. Прим. с.-х. ин-та, 1962.
Строгий А. А. Деревья и кустарники Дальнего Востока. Москва — Хаба-
ровск, Дальгиз, 1934.
Урусов В. М. Результаты и перспективы культуры хвойных в Приморье.—
Природа и человек. Владивосток, Дальневост. кн. изд-во, 1973а,
с. 280—315.
Урусов В. М. Семеношение соснь густоцветковой и погребальной в Южном
Приморье.— Половая репродукция хвойных. 2. Новосибирск, «Наука»,
19736, с. 111—113.
Усенко Н. В. Хвойные деревья и кустарники Дальнего Востока. Хабаровск,
кн. изд-во, 1966, с. 7—18.
Федченко Б. А., Флеров А. Ф. Иллюстрированный определитель растений
Сибири. В II. Голосеменные. СПб.. 1909.
Шишниашвили Р. М. Внутривидовая изменчивость сосны Сосновского. Ав-
тореф. канд. дис. Тбилиси, 1968.
Юнатов А. А. Основные черты растительного покрова Монгольской Народ-
ной Республики.— Труды Монг. ком. АН СССР, в. 39, 1950.
Dallimore W., Jackson А. В. A handbook of Coniferae including Ginkgoasceae.
London, 1954.
Mehra P. N., Khoshoo T. N. Cytology of conifers. — «J. Genet.», 54, № 1.
Santamour F. S. New chromosome counts in Pinus and Picea.— «Silvae Ge-
net ica», 1960, 9, №3.
Sax K. Meiosis in interspecific pine hybrids.— «Forest Sci.», 1960, 6, №2.
Sax K., Sax H. Chromosome number and morphology in the Conifers.—
«J. Arnold Arboretum», 1933, 14.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
СОСНОВЫЕ ЛЕСА ПОЛУОСТРОВА ГАМОВА
И ОСНОВНЫЕ ЧЕРТЫ ИХ ДИНАМИКИ
В. М. УРУСОВ
В южной части Хасанского района Приморского края на
побережье Японского моря в значительной мере распростране-
ны коренные сосняки и их дериваты. Леса из сосны, которую
специальные исследования позволили считать северными по-
пуляциями густоцветковой Pinus densiflora Sieb. et Zucc. (Уру-
сов, 1973, 1975; Бобров, 1975), отнесены Г. Э. Куренцовой к
соснякам с северокорейскими флористическими элементами и
неоднократно описаны (Куренцова, 1956, 1968а, б). Отдельные
группы сосны и ее небольшие выделы разбросаны в горах по
всему Хасанскому району. К 1972 г. они занимали около
0,5 тыс. га и были окружены громадными площадями дерива-
тов. Но лучше всего сохранились фитоценозы с сосной на п-ове
Гамова, где на 170 га сосна преобладает. Для коренных сос-
няков характерны прежде всего береза Шмидта в древостое,
рододендрон Шлиппенбаха и вейгела ранняя — в подлеске.
Вместе с широко распространенной в Приморье леспедецей
двуцветной здесь встречается и леспедеца плотная. Эти фи-
тоценозы являются естественным продолжением сосновых лесов
п-ова Корея, однако флористически значительно обедненным.
Тем не менее флора сосняков п-ова Гамова богаче других сос-
новых лесов Приморья и насыщена декоративными и относи
тельно теплолюбивыми видами кустарников и трав.
Вследствие разнообразных антропогенных влияний, прежде
всего палов, отмечавшихся в описываемом районе на протяже-
нии по крайней мере полутора тысячелетий, сосновые леса и
насаждения с сосной в значительной степени нарушены и пред-
ставлены на полуострове фитоценозами на разных этапах
разрушения. О прошлом лесов морского побережья можно су-
дить по растительности малодоступных участков — скалистых
островков (кекуров) у побережья, узких, отгороженных скаль-
ными гребнями распадков разных экспозиций.
Фитоценозы с сосной густоцветковой в процессе исследова-
ния разделены нами на группы: сосняки кекуров, береговых
обрывов и материкового склона. Первые две группы фитоце-
нозов сохранились лучше. Только в них наблюдается достаточ-
ное крупномерное возобновление сосны. Для них, как и для
45
насаждений из сосны погребальной Р. funebris Кот., характер-
но мозаичное сложение микрогруппировок и биогрупп, обус-
ловливающее ступенчатость полога. Соседствуют обычно наи-
более далекие в возрастном отношении микрогруппировки,
например: 1 (микрогруппировка по ходовой линии)—деревья
в возрасте 40—60 лет, 2 — деревья в возрасте 140—180 лет,
3 — подрост и 4 — экземпляры сосны старше 100 лет. Таким
образом, древостой на кекурах (рис. 1) разновозрастный, вклю-
Рис. 1. Участок разновозрастного сосняка на кекуре
чающий деревья в возрасте от 30—40 до 180 лет. Распределе-
ние стволов по естественным ступеням толщины здесь шире
нормального для сосны обыкновенной Р- sylvestris L. (табл. 1).
К 120—180 годам на кекурах с дифференцированным поч
венным профилем сосны достигают высоты 12—13 м и диа-
метра 40—52 см. т. е. их можно отнести к V классу бонитета
(рис. 2). В местах, где перегнойно-аккумулятивный горизонт
только формируется (относительно недавно заселенные осыпи),
рост еще более замедлен — Va бонитет. На шлейфе кекура,
находящемся на высоте 10 м над ур. м., в насаждении с сомк-
нутостью крон 0,6 деревья в возрасте 75—80 лет имеют высоту
4—7 м и средний диаметр 10,5 см. Сосна на островках медлен-
но растет первые 20—25 лет. Начальная задержка в росте про-
слеживается при наблюдении над подростом и при анализе
46
модельных деревьев. С возрастом прирост по высоте и диамет-
ру улучшается, но к 70—80 годам ослабевает. Очевидно, это
связано здесь с очень значительной влажностью воздуха и по-
стоянными туманами.
Запасы древесины сосны установлены по модельным дере-
вьям и абсолютной полноте соснового элемента древостоев, а
также по наиболее близким для сосны густоцветковой объем-
ным таблицам сосны обыкновенной Среднего Поволжья (Тю-
рин и др., 1956). Запасы лиственных пород определены по
местным таблицам объемов, в том числе и маломерных деревь-
ев (Ефимов, 1955; Петров, 1967).
Малые высоты стволов обусловливают и небольшие запасы
древостоев кекуров: в разновозрастном сосняке при полноте
3,6 учтено 260 м3 древесины. На вновь развивающихся почвах
запасы еще меньше и едва превышают 100 м3 на га, хотя
густота деревьев здесь велика — до *3000 стволов на 1 га
(табл. 2) — и соответствует густоте жердняка сосны обыкно-
венной. Кроме сосны в древостоях кекуров участвуют дуб мон-
гольский, липа корейская и береза даурская. Обычно они не-
многочисленны, но по северным склонам крупных островков
на них может приходиться несколько единиц состава древо-
стоя. В подлеске на осыпях отмечен только шиповник Макси-
мовича, а вообще на кекурах в роли подлеска выступают кус-
тарниковые формы маакии и мелкоплодника, не превышающие
1 м в возрасте 30 лет, но нормально цветущие и плодоносящие.
Рододендроны Шлиппенбаха и остроконечный малочисленны,
заросли их встречаются в разрушающихся сосняках материко-
вого склона.
Обильное возобновление, вполне обеспечивающее лесовос-
становление на кекурах, по лесоводственным шкалам (Несте-
ров, 1954) может быть оценено как хорошее. Имеющееся в ли-
тературе мнение о близости сосновых насаждений Приморья
борам ряда областей Советского Союза (Куренцова, 19686)
подтверждается и нашими данными: прослеживаются некото-
рые черты аналогичности местопроизрастаний, состава, строе-
ния и роста одновозрастных сосновых биогрупп, возобновитель-
ных процессов, связанные как с общностью происхождения
настоящих евразиатских сосняков, прежде всего формаций ки-
тайской Р. tabulaeformis Carr., густоцветковой и обыкновенной,
гак и — на современном этапе это самое главное — развитием
под влиянием одних и тех же лимитирующих почвенно-клима-
тических факторов. Возобновление — молодое поколение ле-
са— нами разделялось, согласно В. Г. Нестерову (1954), на
следующие качественные категории: всходы — растения до
I года, самосев — молодые особи до ветвления, которые в усло-
виях морского побережья обычно не выше 10 см и не старше
10 лет; подрост — растения с момента начала формирования
кроны до диаметра, соответствующего минимальной ступени
47
Распределение живых сосен по естественным ступеням толщины в
Местопро- израстание № про- бы Фаза дегра- дации Высо- та над ур. м., м Естествен ные
0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9
Кекур 3 — 30 _1 2 3 5 6 7 7 __7 „ , , 1 3 6 11 17 24 31 38 Южный склон 1 I 100 1 3 6 10 11 11 14 „ „ 1 4 То~ '20 “31 ~42 “56 Южный склон 2 I 150 1 3 5_ 7 11 14 17 Юго-западный 1 4 9 "ЙГ ~27 41 58 склон 5 I 300 _1 1 _3 4 4 7 13 15 1 2 5 9 13 20 33 48 Юго-восточный I 350 6 6 9 14 22 склон 6 — — — 6 "12 1? 35 37 Западный склон 8 II 450 9 12 26 — — — 9 21 47 Юго-западный III 250 5 6 24 склон 10 _ _ _ 5 11 35 * Цифры под чертой — нарастающий итог.
Некоторые таксационные характеристики насаждений с сосной
Местопроизрастание Высота над ур. м., м Сомкнутость крон Абсолютная полно- та, м2/га
сосны дуба и бе- рез по сосне по сопут- ствующим породам
Кекур:
осыпь (уступ) 10 0,6 — 20,8 —
склоны и вершина 30 0,6 0,05 25,4 1,4
Гора Туманная:
южный склон 100 0,6 0,3 22,4 8,1
южный склон 150 0,5 0,3 19,6 6,7
юго-западный склон 300 0,6 0,2 23,0 8,1
юго-восточный склон 350 0,7 0,2 42,9 6,1
западный склон 400 0,2 0,4 6,1 7.0
юго-западный склон 450 0,3 0,4 8,5 12,0
перечета. Таким же образом отделяет крупный подрост от дре-
востоя Ю. И. Манько (1967). Подрост еще не имеет самостоя-
тельного места в пологе крон древостоя, в то время как дере-
вья замедленного роста уже утратили его и, следовательно, к
подросту нами не относились. Всходов, самосева и подроста
на одном из кекуров оказалось соответственно 3, 1 и 4 тыс.
особей на 1 га (средняя оценка, табл. 3). Возобновление сосны
48
Табл II I» а
прибрежно-морской фации сосны густоцветковой (п-ов Гамова), %
61 73 82 84 86 89 92 94 96 97 98 98 99 100
42 12_ 6_ 5____ ________ _______
77 89 95 100
Таблица 2
густоцветковой на кекуре и материковом склоне п-ова Гамова
Возраст, сосны, лет Средняя высота сосны, м Средний диаметр сосны, см Число стволов на га Запас на га, м3 Класс бони- тета по сосне
сос- ны ДРУ- ГИХ по род всего сосны ДРУ- ГИХ пород все- го
80 (30—120) 4,7 9,4 3000 — 3000 120 .— 120 Va
100 (30—180) 10,5 20,6 780 30 810 200 16 216 V
80 (50—130) 12,5 25,0 450 740 1190 170 60 230 IV
80 (50—130) 13.3 25,6 380 670 1050 140 50 190 IV
70 (50—120) 10,5 28,2 370 660 1030 210 50 260 IV
70 (50—120) 11,2 30,2 600 460 1060 320 40 360 IV
80 (50—110) 10,0 23,5 140 740 880 50 40 90 Va
80 (60—120) 9,0 22,2 220 1300 1520 80 90 /170 Va
распределено неравномерно: оптимальными для него являются
более сухие и менее задерненные южные склоны; слабее, хотя и
удовлетворительно, восстанавливаются западные, северные и
восточные, а также вершины кекуров. По северным и восточ-
ным склонам основным препятствием для восстановления сосны
является 100%-ное задерненне почвы, а на вершинах — целый
комплекс неблагоприятных факторов: сдувание семян, выбива-
4 Заказ 488
49
Рис. 2. Ход роста сосны густоцветковой по высоте в разных местопроизра-
станиях. Рост сосны: 1 — в древостоях материкового склона; 2 — на кеку-
рах; 3 — на арунденелловых горных лугах; 4 — на скалах (скальная модифи-
кация)
ние всходов дождем и ветром, толстый слой подстилки в мик-
ропонижениях. Подтверждается довольно тесная зависимость
количества всходов и самосева от степени развития травяного
покрова и мощности подстилки — с их нарастанием возобнов-
ление сосны ухудшается. При сплошном слое подстилки тол-
щиной до 4 см количество всходов и самосева составляет лишь
треть среднего (шлейф кекура, табл. 3) Из лиственных пород
в возобновлении участвуют маакия, диморфант, мелкоплодник
(соответственно 500, 400 и 200 растений на 1 га), иногда бере-
зы Шмидта и даурская, ясень носолистный (горный).
Всходов и самосева на кекурах меньше, чем на материко-
вых склонах (табл. 4), что связано с высокой степенью задер-
нения. типичной для мест с обильным и равномерным увлажне-
нием и достаточной освещенностью. По нашим подсчетам, в
1967, 1969 и 1970 годах средне развитое дерево давало до
400 шишек, содержащих не менее 5 тыс. всхожих семян. Если
учесть ослабленное семяношение мелких и наиболее крупных
(старше 160 лет) особей, то на 1 м2 поверхности островков
выпадает до 100 жизнеспособных семян, лишь ничтожная часть
50
которых прорастает. В то же время в фитоценозах с сосной на
кекурах отпад всходов и самосева минимальный — соответст-
венно 10—20 и 30%.
Возобновление сосны растет медленно: средний возраст под-
роста равен 14 годам, а высота —46,4±3,8 см. Приросты вы-
соты за последние 5 лет возросли незначительно — с 43 в
1966 г. до 55 мм в 1970, хотя подрост в этих условиях жизне-
способен, а освещенность составляет 14—19% от полной. В сос-
няках центральных районов Приморья такие приросты отмече-
ны у отмирающего возобновления сосны при относительной
освещенности менее 8%. Еще хуже на кекурах растет затенен-
ный подрост сосны (освещенность 7—10%), прирастающий в
год на 15—20 мм. Крону он начинает образовывать с 10 лет, а
к 19—20 годам при высоте менее 0,5 м и слабом охвоении ча-
сто отмирает.
Среди трав наиболее распространены осока низенькая, мят-
лики ложнодубравный и кистевидный, арундинелла, мискантус
краснеющий, полыни Кейске, Гмелина и маньчжурская, очитки
живучий, Максимовича, Сельского, скабиоза, соссюрея Мак-
симовича, лилии поникшая и двурядная, лук стареющий,
поводник лучистый, ландыш, подмаренник настоящий, гвоздика
амурская, ясколка мелкоцветная, лапчатка земляниковидная,
патринии скабиозолистная и скальная, ширококолокольчик, го-
ричники прибрежный и тайваньский, фальката японская, вика
приятная, камнеломка маньчжурская. На шлейфе кекура к
ним добавляются вейник наземный, гетеропаппус скально-при-
морский, гления прибрежная, вербейник густоцветный, майник
двулистный. Травяной покров достаточно разнообразен и пестр,
что обусловлено контрастностью условий местопроизрастаний,
которые на очень небольшой площади меняются от влажных
до периодически сухих (скальные обнажения на южных скло-
нах). Поэтому наряду с типичными ксерофитами (осока низень-
кая, арундинелла) здесь в наличии мезофильные и мезогигро-
фильные виду (ландыш, майник, лилия двурядная, камнелом-
ка). Последние две группы трав также отмечены по обрывам
восточных и северных экспозиций и в широколиственных лесах
с отдельными деревьями сосны по северным склонам распадков,
где они становятся более многочисленными.
Надо особо отметить способность сосны во влажных усло-
виях побережья заселять осыпи и скалы, где при слабовыра-
женном гумусовом слое сосна образует экологическую форму,
характеризующуюся крайне медленным ростом, но большой
продолжительностью жизни и хорошим семяношением. К воз-
расту 100—250 лет деревья здесь редко достигают высоты 3 м
при весьма значительном диаметре у основания (36 см и бо-
лее) (рис. 3).
На скальных островах и выступах экологические формы
представлены стелющимися кустарниками или же (если они
4* 51
Возобновление сосны на кекурах
Местопроизрастание Покрытие, % Всходы, шт. Самосев, шт.
тра- вами под- стил- кой живые мерт- вые живой мерт- вый
Вершины 47 62 2000 400 200 300
Южные склоны 40 80 10000 2500 2400 2800
Западные склоны 50 70 2600 900 1800 200
Северные и восточные
склоны 70 80 1250 500 400 600
Шлейф кекура 25 95 900 300 200 200
В среднем по кекуру 44 70 2800 200 700 300
Рис. 3. Сосны скальной модификации в возрасте 70—100 лет
защищены от ветров) деревьями с вертикальными стволами
обычно с зонтичными кронами. На 100 м2 скальной поверхности
насчитывается 15—60 особей старше 30 лет и 10—30 более
молодых растений. Густота размещения сосен обусловлена на-
личием заполненных мелкоземом трещин. Их корневые системы
особенно мощны и по объему равны почти всей надземной ча-
сти стволиков. Судя по размерам хвои и шишек, здесь имеется
как короткохвойное и мелкошишечное потомство собственно
скальной модификации сосны, так и особи из семян нормаль-
ных деревьев. Хвоя и шишки у последних почти нормальной
52
в расчете на 1 га
Таблица 3
Подрост, шт.; высота, м
Всего
0—0,25 0,25—0,5 0,5—1,0 более 1,0 живого и сом- нитель- ного под- роста, шт.
ЖИ- ВОЙ сомни- тсльи. мерт- I вый 1 живой сомни- тельн. мерт- вый-' жи- вой сомпи- тельн. мерт- вый ' жи- вон сом- иит. мерт- вый
600 350 500 — 50 200 — — — г— — 1650
4000 60 90 1600 — 10 700 — — 50 — — 6410
2800 200 — 1450 — — 300 — 100 50 — — 4800
1100 200 1100
— — 50 — 20 450 — 30 30 — — 4180
2400 200 —
длины (соответственно 40—60 и 30—40 мм), рост в первые
годы относительно быстрый.
Сосняки прибрежной полосы вытянуты вдоль обрывов вос-
точной кромки полуострова. Их протяженность около 20 км,
а площадь — 40 га, ширина большей частью не превышает
20 м. Это опушка сплошного соснового массива, занимавшего
ранее весь полуостров, исключая нижние части северных скло-
нов. Сосна сохранилась на вполне развитых почвах склонов,
частично подверженных обвалам, и на осыпях камней и мел-
козема под скалами берега. Эти участки наименее подвержены
палам. Деревья типично разновозрастны (30—200 лет), разно-
высотны, но крупный подрост есть только на недоступных огню
обнажениях и осыпях, где восстановление сосны идет вполне
нормально: до 3 тыс. экз/га подроста высотой от 25 см до 2 м.
Таблица 4
Возобновление сосны густоцветковой на материковом склоне в зависимости
от высоты над уровнем моря
(средние данные учетов в августе — сентябре 1969—1970 гг.)
Экспозиция склона Высота над ур. м., м Возобновление в пересчете на га
Всходы, шт. Самосев, шт Под- рост (жи- вой),
живые мерт- вые жи- вой мерт- вый
Южная 100 45000 6000 18400 400
Южная 150 30400 1700 20800 1700 500
Юго-западная 1 300 4000 10000 9000 3000 100
Юго-восточная 350 1000 2000 9000 2000 100
Западная 400 2500 3000 5000 2200 200
Юго-западная 450 1500 2000 8500 1000 100
53
Па развитых почвах даже у береговой кромки рост сосны
соответствует IV классу бонитета. Наибольшая высота деревь-
ев у обрывов— 15 м при диаметре 80 см. Кроны старых сосен
здесь особенно мощны и приобретают флагообразную или зон-
тиковидную форму. Последняя вообще типична для сосны гу-
стоцветковой в разреженных насаждениях. На осыпях под об-
рывами сосны Va бонитета по своему росту приближаются к
экземплярам скальной формы. На склонах вдоль обрывов сос-
на возобновляется слабо — менее 2000 всходов и самосева на
1 га. Возобновление здесь не старше 7 лет (рис. -1). Рост луч-
ше, чем у подроста сосны на кекурах. Восстановлению сосны
препятствует задерненне и толстый слой опада, накапливаю-
щийся благодаря обилию кустарников, но главная причина
отсутствия подроста — проникающие и сюда весенние палы.
Рис. 4. Самосев сосны густоцветковой в сложном береговом сосняке со
свежими почвами. В подлеске преобладает рододендрон Шлиппепбаха
54
Состав трав более разнообразен. К видам, отмеченным для
кекуров, добавляются осока ржавопятнистая, ариземы, качим
тихоокеанский, марьянник розовый, фиалки бутеневидная и
маньчжурская. По осыпям у моря наряду с камнеломкой по-
являются осока Гмелина и повой солданелловый. По перифе-
рии проекций сосновых крон разнотравье образует сплошную
дернину. Непосредственно под кронами изредка встречаются
осока низенькая и марьяник. Местами сохранились копытень
Зибольда, аралия материковая, волжанка азиатская.
Кустарники в сосняках по обрывам и осыпям многочислен-
ны: у пляжей преобладают розы Максимовича и морщинистая,
выше идут спирея средняя, вейгела ранняя, барбарис амурский
и смородина маньчжурская. По обрывам и в полосе над ними
наиболее обилен рододендрон Шлиппенбаха, которому сопутст-
вуют рододендрон остроконечный, леспедецы, тимьян Комаро-
ва. Сомкнутость кустарников — 0,4—0,8. Таким образом, под-
лесок береговых сосняков богат декоративными видами, цве-
тение которых продолжается с конца апреля (рододендрон ост-
роконечный) по сентябрь (леспедецы).
По южным экспозициям гребней сложные береговые сосня-
ки соединяются с лесами и редколесьями из сосны материко-
вого склона, поднимающимися до 450 м над ур. м. (высшая
точка полуострова — сопка Туманная — 508 м). На примере
фитоценозов с сосной материкового склона представляется воз-
можным проследить основные этапы деградации соснового леса
и даже восстановления или образования его вновь на некото-
рых участках: имеются сомкнутые сосняки и производные от
них редколесья, кустарниковые заросли и луга с отдельными
соснами на склонах всех экспозиций. В отличие от сосняков
кекуров и береговой полосы местопроизрастания сосны на мате-
риковом склоне носят черты большей сухости и в целом могут
быть отнесены к сухим и свежим.
Для выявления связи возобновления и роста сосны с высо-
той над морем пробные площади на материковом склоне зак-
ладывались так, чтобы по возможности разница высот равня-
лась 50 м. Высотные отметки и экспозиция очень влияют на
перераспределение влаги и тепла. Прохождение фенологиче-
ских фаз сдвинуто: рост побегов сосны и распускание почек
лиственных пород на высоте 450 м начинается на неделю рань-
ше, чем у берега моря. Количество выпадающих осадков, осо-
бенно в лесу, с подъемом уменьшается, так как заметная доля
их обеспечивается за счет конденсации тумана, а туманы плот-
нее и держатся более продолжительное время на небольшой
высоте над морем. Только за счет горизонтальных осадков,
сконденсированных кронами, в июне — августе под пологом
леса воды выпадает в дни с туманом на 1—5 мм больше, чем
на открытых пространствах. Если учесть, что, по данным ме
теостанции на мысе Гамова, находящейся в полукилометре от
55
ближайшего сосняка, в июне и июле в среднем соответственно
17 и 19 дней с туманом (Справочник по климату.., 1969), ко-
личество добавочной влаги довольно велико. Особенности рас-
пределения горизонтальных осадков — важный фактор, опреде-
ляющий, по нашему мнению, прорастание семян сосны. Видимо,
в связи с этим ее семена здесь прорастают в мае, в то время,
как в центральной части края — в июне. Разрушение подстил-
ки многочисленными стадами пятнистых оленей и часто слабое
задернение поверхности почвы (25—40%) способствуют попа-
данию семян в основном в землю, а довольно ровный ход су-
точных температур и достаточная влажность приводят к их
обильному прорастанию. В насаждениях с сомкнутостью крон
0,5—0,6 на высоте 100—200 м над ур. м. в 1970 г. отмечено от
50 до 30 тыс. всходов на га и 18—21 тыс. самосева (табл. 4,
рис. 5). Распределяется возобновление здесь исключительно
равномерно, но подрост представлен очень небольшими расте-
ниями, отрастающими заново после повреждений оленями.
И все-таки главная причина отсутствия крупномерного подро-
ста — периодические палы, которые, судя по подсушинам на
модельных деревьях, за последние 70 лет случались в насажде-
ниях не менее 8 раз
Выше 200 м над ур. м. количество всходов сосны резко
уменьшается. На высоте 300 м и выше к августу остается
4000 живых всходов на 1 га. Их отпад составляет 60—80% из-за
периодического пересыхания поверхностного слоя почвы, кото-
рый в прогалинах и на полянах в солнечные дни нагревается
до 60°, а под пологом леса — до 40° Отпад среди самосева
менее значителен: даже на южных склонах, приподнятых над
морем на 400—450 м, учтено 5—8 тыс. растений самосева на
1 га. В целом возобновление сосны в данных условиях идет
удовлетворительно. В возобновлении на материковом склоне
участвуют также дуб, диморфант, ясень горный, березы даур-
ская и Шмидта, липа корейская, но самосева и 1незд поросли
каждой из этих пород в сосняках не более 500 на 1 га.
Сосняки материкового склона имеют в основном двухъярус-
ное строение; в первом ярусе — сосна, второй образован дубом
монгольским и березами с единичной примесью липы. В связи
с затенением сосной, относительной бедностью почв и палами
эти породы невелики по размерам и редко достигают 10—11 м
высоты. Наблюдается характерное для фитоценозов с сосной
мозаичное размещение сосновых, дубовых и железноберезовых
микрогруппировок. Непосредственно под пологом сосны порос-
левые дуб и береза искривлены, суховершинны и отличаются
медленным ростом по диаметру. Они часто старше сосны и, как
и она, носят следы неоднократного подпаливания. Дуб, липа и
береза даурская повреждены сердцевинными гнилями. Во мно-
гих случаях они восстановились на гарях одновременно с сос-
ной. Сомкнутость второго яруса в среднем не превышает 0,4,
56
Рис. 5. Изменение возобновительных потенций сосны густоцветковой и лист-
венных пород (в основном дуба монгольского, берез и кленов) в связи с
увеличением абсолютной высоты над морем и понижением интенсивности
горизонтальных осадков: 1 — всходы и 2 — самосев сосны густоцветковой;
3 — порослевое и 4 — семенное (всех категорий крупности) возобновление
лиственных пород; 5 — степень развития трав; 6 — интенсивность горизон-
тальных осадков
57
хотя густота его значительна — от 400 до 700 живых стволов
на га (табл. 2). Доля участия березы Шмидта в нем по числу
и запасу колеблется от единичного до 50%.
В случае уничтожения сосны здесь образуются дубовые и
железноберезовые редколесья.
Сосновый ярус материкового склона почти исключительно
состоит из экземпляров старше 50 лет. В то же время макси-
мальный возраст их не превышает 140 лет. Отсутствие подро-
ста и более молодых деревьев свидетельствует о длительном
воздействии огня на эти насаждения, в результате чего моло-
дые особи усохли, а наиболее мощные сосны, выгорая у осно-
ваний со стороны склонов, уничтожены буреломом. Тем не ме-
нее изменчивость стволов по диаметрам в насаждениях все
еще велика и шире нормальной для сосны обыкновенной. Зна-
чительно изменяются и высоты: минимальная высота состав-
ляет 0,4—0,6 средней, максимальная—1,2. Такая же изменчи-
вость высот выявлена нами в Приморье в семидесятилетней
посадке сосны обыкновенной (Урусов, 1970), где, однако, варь-
ирование диаметров было нормальным. В отличие от насажде-
ний кекуров сосняки непосредственно полуострова не имеют
длительной начальной задержки роста (рис. 2), но и они лишь
в лучшем случае соответствуют III, а с возрастом приближа-
ются к IV классу бонитета. Энергичный рост по высоте н
диаметру наблюдается до 30—40 лет. В это время деревья в
одновозрастных микрогруппировках дифференцируются так,
что вполне могут быть разделены по шкале Крафта. Большин-
ство отставших в росте деревьев развивают флагообразные
кроны, обеспечивающие им в условиях крутых склонов доступ
к прямому свету. Таких деревьев в насаждениях больше 20%,
что вместе с широкой изменчивостью диаметров (от 8 до
56 см) и высот свидетельствует о существенном своеобразии
строения описываемых сосняков. Однако чем больше они раз-
рушены, тем уже габитуальная изменчивость стволов в них
(табл. 1), в рединах она даже меньше, чем в насаждениях сос-
ны обыкновенной. В древостоях материкового склона диаметры
и высоты сосен ближе к средним, чем на кекурах. Плотность
средней естественной ступени толщины в нормальных сосняках
п-ова Гамова меньше установленной для сосны обыкновенной —
10% вместо 1-8. В деградирующих насаждениях она растет и
может превысить 20%.
Древостои к настоящему моменту образованы соснами вы-
сотой 8—16 м с куполовидными или плоскими вершинами
(рис. 6), свидетельствующими о замедлении роста. Прирост по
высоте у модельных деревьев в возрасте 50—100 лет не превы-
шает за последние 5 лет 20 см в год. Очищение от сучьев до-
вольно слабое и зависит от густоты как сосен, так и листвен-
ного яруса. В наименее разрушенных насаждениях с сомкну-
тостью крон 0,5—0,7 насчитывается 400—600 живых сосен на
58
Рис. 6. Сосняк на материковом склоне, опушка. На неродном плане фрагмент
вторичного горного арундинеллового луга
га. Наибольший запас стволовой массы сосны — 320 м3 на 1 га —
учтен в сухом сосново-дубовом лесу на склоне юго-восточной
экспозиции крутизной 25° Подлесок образован рододендрона-
ми, леспсдецами, всйгслой (Валова, 1967, Куренцова, 1968а).
По понижениям и более влажным местам отдельными пятнами
встречается тимьян Комарова. В широколиственно-сосновых
группировках найдены виноград амурский и bhhoiрадовннк ко-
роткоцветоножковый. Сомкнутость подлеска зачастую незна-
чительна (0,1—0,3), он хорошо развит только в рощах пони
!шоч горимости, например на выступах скал. Здесь в же-
лезпоберезовых и дубовых микрогруппировках, а также в про-
галинах наблюдаются плотные куртины рододендронов высо-
той более 1 м. Накапливающийся под кустарником опад пре-
пятствует возобновлению древесных пород. Рододендроны в
сосновых и широколиственных лесах полуострова хорошо вос-
станавливаются семенами, но обилие их самосева тоже опре-
деляется влажностью почвы, а следовательно, и высотой над
уровнем моря. В сомкнутых сосняках на 1 га насчитывается от
4—5 тыс. (100—150 м над ур м.) до 1,5 тыс. (400 м над ур. м.)
особей самосева рододендрона Шлиппенбаха и соответственно
1—0;5 тыс. самосева рододендрона остроконечного.
59
Общему более ксерофитному облику растительности соот-
ветствует и травяной покров, проективное покрытие почвы ко-
торым здесь не превышает 20%. Обычно преобладает осока
низенькая, распространены также арундинеллы, мискантус, по
лыни Кейске и Гмелина, атрактилодес, очиток Сельского, ка-
сатик одноцветковый, ширококолокольчик, патриния скальная,
гвоздика амурская. Мятлики, лапчатка земляниковидная, бу-
бенчик широковатый, володушка козелецелистная, лилия по-
никшая встречаются редко. Из папоротников отмечена только
многоножка уссурийская — на северной стороне комля сосны.
Весной на склонах аспектирует фиалка восточная (1—10 ра-
стений на 1 м2), что свидетельствует о большей влажности
этих сосняков по сравнению с произрастающими в центральной
части края, где фиалки в сосняках почти нет. На крупных
полянах формируются синузии арундинеллово-серобороднико-
вого луга с характерным для них составом. На данном этапе
они внедряются в дубовые редколесья и в сосняки и являются
поэтому прогрессирующими.
Занимаемые сосняками почвы — ближе всего к бурым лес-
ным оподзоленным (Иванов, 1964). За темно-серым перегной-
но-аккумулятивным горизонтом толщиной 7—12 см до глубины
30 см идут более светлые слои, переходящие в красновато-бу
рый или желто-бурый суглинок, наполненный дресвой и облом-
ками гранита. Судя по размывам и оползням, эта порода
залегает на глубине 1,5—2 м, препятствуя дальнейшему углуб-
лению корней. Основная их масса заключена в слое 16—60 см.
При более близком залегании коренных пород изреженные сос-
няки ветровальны. Гумусовый горизонт менее развит в средней
части склона. Сверху он носит черты прогорания и последую-
щего выветривания. В этом случае на поверхности 1—3 см слой
дресвы гранита, под которым почвенные горизонты, начиная с
остатков перегнойно-аккумулятивного, располагаются нормаль-
но. На очень крутых склонах почвы могут иметь фрагментар-
ный характер — полный профиль выражен в западинах между
скалами. Именно таковы почвы в рединах и дубово-сосновых
лесах выше 400 м над ур. м.
С нарастанием сухости местопроизрастаний низовые пожа-
ры становятся более частыми и сильными. Поэтому на южных
и юго-западных склонах сопки Туманной с высоты 300 м все
большие площади занимают дубово-сосновые насаждения и
дубняки с единичными соснами, а также сосновые редины. Из
300 га сосняков и насаждений с сосной материкового склона
полуострова на редины и дубняки с остатками сосен прихо-
дится около половины площади. В дубовых лесах с сосной
сосредоточены толстомерные, часто усыхающие или усохшие,
сосны, росшие в начале жизни как на просторе, так и в лесу.
В первом случае нм характерны особенно мощные зонтиковид-
ные кроны со средним диаметром от 7 до 12 м. В древостое
60
дубу сопутствуют березы даурская и Шмидта. Возобновление
представлено всходами и самосевом сосны, гнездами поросли
дуба, редким самосевом берез. Восстановить преобладание
сосна не может из-за палов и сбоя. Если не исключить эти влия-
ния, дубово-сосновые насаждения сменяются чистыми дубня-
ками.
Особняком стоят сосновые редины на горных арундинелло-
вых лугах с серобородником сибирским, мискантусом краснею-
щим — тоже одна из стадий разрушения сосновых фитоцено-
зов. Частые палы в этом случае изреживают еосняки и полно-
стью уничтожают широколиственные породы, порослевая спо-
собность которых истощается при жизни одного поколения
сосны. Судя по очищению от сучьев и ходу роста, сосны редин
первые 10—20 лет находились в лесу и росли по III бонитету
(рис. 2), а позднее, на просторе, по IV, V, в то время как при-
рост по диаметру замедлялся только после 40 лет. В 1970 г.
средние высоты сосен в рединах составляли 8—12 м, диамет-
ры — 32—36 см, кроны по форме близки к зонтиковидным.
Преобладают деревья старше 100 лет. На 1 га редин насчиты-
вается 50—100 сосен и небольшое количество дубов монголь-
ского и зубчатого и липы. Весьма многочисленны выгоревшие
до основания пни крупных сосен (до 200 на 1 га).
Разрушение предшествовавших рединам сосняков началось
в конце прошлого столетия. По данным анализа модельных де-
ревьев, первый большой пожар отмечен в 1889 г. Теперь здесь
полностью сформировался горный луг с элементами остепнения.
В травяном покрове его наряду с арундинеллой, серобород-
ником, мискантусом участвуют мятлики охотский, ложнодуб-
равный, однолетний, овсяница ложнобороздчатая, келерия ази-
атская, касатик одноцветковый, лук стареющий, горичник тай-
ваньский, ширококолокольчик, гвоздика амурская, полыни,
дендрантема нактонгенская, анафалис посьетский, горечавка
Цоллингера, фиалки восточная и маньчжурская, прострел ки-
тайский. На выходах скальных обнажений встречаются патри-
ния скальная и горноколосник хрящеватый. Для арундинелло-
вых лугов с отдельными соснами можно отметить количествен-
ное преобладание растений остепненных лугов над боровыми
видами, число которых пока значительно. Объясняется это
относительной молодостью лугов, которые устойчивы только
благодаря пожарам и объеданию древесно-кустарниковой по-
росли оленями. При снятии указанных факторов возможно
формирование лещинно-рододендроновых зарослей с дубом и
сосной; а по днищам тальвегов — молодняков из ольхи япон-
ской, ясеня носолистного и липы. Сомкнутость кустарников по
средним частям склонов в настоящее время 0,2—0,4, но высота
их не более 30 см. На 1 гектар луга в среднем насчитывается
1000 гнезд поросли дубов (800 дуба монгольского и 200 зубча-
того) и 10—13 тыс. кустов лещины разнолистной (северные
61
экспозиции). На лугах южных экспозиций среди кустарников
преобладает рододендрон Шлиппенбаха (2—4 тыс. на 1 га), ро-
додендрон остроконечный и лещина малочисленны — по 500 —
1000 экземпляров на 1 га. Много подроста сосны, часто кустя-
щегося из-за многократного объедания (2000 растений на 1 га).
Количество возобновления сосны изменчиво и определяется за-
дернением почвы и наличием семян, так как в наиболее старых
и ослабленных палами и размывом рединах сосен мало, и они
почти не семяносят. На лугах, удаленных от деревьев сосны
ие более чем на 60 м, возможно успешное восстановление этой
породы; здесь насчитывается в среднем 500 всходов, 1400 са-
мосева, 300 экземпляров подроста .на 1 га. На размытых
участках самосев сосны обильнее.
На южных склонах разрушение коренного соснового леса
идет следующим образом: разновозрастные сосняки со вторым
ярусом из березы Шмидта и дуба под влиянием пожаров и
палов преобразуются в сосново-дубовые редколесья, дубняки,
редины, которые в свою очередь сменяются арундинелловыми
лугами. При интенсивных палах луговая растительность на
склонах круче 25° подвержена размыву, что иногда ведет к об
разованию каменных россыпей. На россыпях восстановление
лесной растительности начинается через формирование родо-
дендроново-лещинных зарослей, в которых снова может посе-
ляться сосна. Видимо, уже сейчас сосняки материкового склона
в значительной мере представлены именно вторичными сосня-
ками. С прекращением палов и вытаптывания можно ожидать
(при наличии семян) восстановления сосны на любом этапе
деградации (схема 1). Площади, на которых возможно естест-
венное возобновление сосны густоцветковой, не превышают
2000 га, поэтому активная оптимизация возобновительного ре-
жима необходима уже сейчас.
На северном склоне сопки Туманной, как и вообще в Ха-
санском районе, в свежих и влажных местопроизрастаниях сос-
новые насаждения естественно сменились хвойно-широколист-
венными лесами маньчжурского типа еще в доагрикультурный
период. Сосна была вытеснена более теневыносливыми поро-
дами в процессе вековых смен, благоприятствующих мезофит-
ным формациям (Куренцова, 1971). По нижним частям скло-
нов и обращенным к северу тальвегам сохранились исключи-
тельно многопородные широколиственные насаждения с верти-
кальной сомкнутостью крон. В древостое здесь дуб монголь-
ский, ясень носолистный, диморфант, клены мелколистный,
зеленокорый, ложнозибольдов, бородатый, вишня сахалинская,
березы Шмидта и даурская, бархат, граб, липа, ивы цельно-
листная и козья, мелкоплодник, бересклет Маака. Подлесок
образован лещиной маньчжурской, рододендронами, акантопа-
наксом, вейгелой, рябинником, таволгой средней, жимолостью
раннецветущей, дейцией, крыжовником буреинским. Присутст-
62
Схема 1. Деградация под влиянием пожаров и сбоя и восстановление
сосняков на южных склонах
вуют виноград, актинидии, лимонник. Судя по составу расти-
тельности, здесь произрастали пихта цельнолистная и кедр,
уничтоженные пожарами. Насаждения с сосной занимали не-
давно верхние трети северных склонов, но к настоящему пе-
риоду они и здесь уничтожены практически невозвратно. О них
напоминают только отдельные сосны среди дубового леса. Под
влиянием пожаров сосновый ярус распался, все более замет-
ную роль играли широколиственные породы второго яруса, осо-
бенно дуб. В дальнейшем дубово-широколиственные насажде-
ния частично превращались в редины с формирующимся под
их пологом разнотравным лугом (схема 2). С ослаблением
палов в данном случае лишь возможно восстановление широко-
лиственных насаждений, которые будут все более обогащаться
по составу растительности всех ярусов. Такие длительно устой-
чивые насаждения в сущности представляют смену коренного
соснового типа леса новым коренным типом.
63
Схема 2. Разрушение сосновых лесов под действием периодических па-
лов на северном склоне
Выводы
1. Наиболее сохранившимися на полуострове являются сос-
новые насаждения кекуров, характеризующиеся в основном
нормальным строением разновозрастных древостоев. Чистые
дубняки и дубово-черноберезовые насаждения являются новей'
шими производными сосняками.
2. Сосняки на обрывах представляют опушку соснового
массива, в недавнем прошлом расчлененного и деградировав-
шего.
3 Достаточное возобновление сосны имеется только на ке-
курах и в сосняках береговых обрывов. Только эти сосняки в
настоящее время устойчивы. Сосняки материкового склона в
условиях палов и вытаптывания лишены подроста и обречены
на уничтожение.
4. Возобновление сосны, а также и других древесных и кус-
тарниковых пород определяется увлажнением почвы и степенью
ее доступности для семян. С увеличением высоты над морем
количество всходов и самосева падает прежде всего из-за
уменьшения горизонтальных осадков, доля которых может со-
ставлять 30% и более от общего количества выпадающих в ле-
су в мае — августе осадков.
5. Древостои сосны густоцветковой представляют простран-
ственное чередование микрогруппировок, внутри которых де-
ревья относительно одновозрастны. Поэтому при работе с не-
большими участками насаждений всегда выделяется 1—2 поко-
64
ления сосны. С увеличением исследованной площади распре-
деление стволов по возрастам становится более равномерным.
6. Изменчивость высот и диаметров в древостоях сосны гу-
стоцветковой (в том числе и в одной микрогруппировке) в силу
биологических особенностей вида значительно шире, чем от-
мечаемая для сосны обыкновенной. Однако в деградирующих
под воздействием палов насаждениях ее амплитуда резко су-
жается.
7. На высоте 50 м и менее сосна в береговых насаждениях
я на островках растет особенно медленно. У подроста наблю-
дается длительная задержка в росте, вызванная неблагоприят-
ным микроклиматом. Однокачественными являются деревья
в возрастном интервале, равном 40 годам. С подъемом, как и
с удалением от моря, рост сосны улучшается.
8. Низовые пожары в фитоценозах с сосной в текущем сто-
летии случались в среднем через 8—9 лет. По модельным де-
ревьям усиление палов прослеживается с конца прошлого сто-
летия.
9. При устранении палов и вытаптывания в дубово-сосновых
лесах и редколесьях и на арундинелловых лугах с отдельными
соснами даже без применения активных мер содействия воз-
можно восстановление сосны.
10. Арундинелловые луга устойчивы в результате разнооб-
разных антропогенных влияний, при исключении которых
восстановятся древесные и кустарниковые заросли.
11. На северных склонах сосновые леса в основном замене-
ны широколиственными (вначале чернопихтово-широколиствен-
ными) под влиянием естественных причин. Их восстановление
здесь невозможно.
12. Богатые теплолюбивыми видами фитоценозы с сосной
на полуострове уникальны для страны и исключительно ценны
как источник обогащения хозяйственных и декоративных на-
саждений и интересны для выявления флорогенеза Приморья.
13. Разработка мер по сохранению и восстановлению сосно-
вых лесов п-ова Гамова совершенно необходима.
ЛИТЕРАТУРА
Бобров Е. Г. Что такое Pinus funebris Korn.— «Бот. ж.». Вып. 60, 1975, №5,
с. 698—702.
Валова 3. Г. Флора и растительность юга Хасанского района (Приморский
край). Автореф. канд. дис. Владивосток, 1967, 20 с.
Ефимов Н. В. Справочник таксатора. Хабаровск, изд. ДВ треста «Леспро-
ект», 1955, 133 с.
Иванов Г. И. Почвы Приморского край. Владивосток, Дальневост кн. изд-во,
1964, 107 с. е u ч
Куренцова Г. Э. Формация сосны могильной (Pinus funebris Кот.) в При-
морском крае. — Труды ДВФ АН СССР, сер. бот., в. 3. М. Л., 1956,
с. 93—104.
5 Заказ 488 65
Куренцова Г. Э. Реликтовые растения Приморья. Л., «Наука», 1968а,
с. 50—58.
Куренцова Г. Э. Растительность Приморского края. Владивосток, Дальне-
вост. кн. изд-во, 19686, с. 102—111.
Куренцова Г. Э. Смены растительности Приморья и южного Приамурья в
связи с естественной динамикой ландшафтов и деятельностью чело-
века. Автореф. докт. дис. Новосибирск, 1971, с. 47—57.
Манько Ю. И. Пихтово-еловые леса Северного Сихотэ-Алиня. Л., «Наука»,
1967, 244 с
Нестеров В. Г. Общее лесоводство. М. — Л. Гослесбумиздат., 1954, 656 с.
Справочник по климату СССР, в. 26, ч. V. Л., Гидрометеоиздат. 1969,
с. 135—153.
Петров П. Г. Временные таблицы для вычисления объема маломерных де-
ревьев.— Рубки ухода в лесах Приморья. Владивосток Дальневост,
кн. изд-во. 1967, с. 131—138.
Тюрин А. В., Науменко И. М., Воропанов П. В. Лесная вспомогательная
книжка. Изд. 2-е, доп. М.—Л., Гослесбумиздат. 1956, с. 106—107.
Урусов В. М. Опыт использования сосны обыкновенной в лесопосадках юга
Приморья.— Наука — сельскому и лесному хозяйству. Владивосток.
Дальневост. кн. изд-во, 1970, с. 29—31.
Урусов В. М. Семяношение сосны густоцветковой Pinus densiflora Sieb. et
Zucc. и погреба чьной Р. funebris Кот. в Южном Приморье.— Поло-
вая репродукция хвойных. II. Новосибирск, «Наука», 1973, с. 111—114.
Урусов В. М. Эколого-биологические особенности и внутривидовая 'изменчи-
вость сосны погребальной (Pinus funebris Кош.) в Приморском крае.
Автореф. канд. дис. М., 1975, 30 с.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
КАРИОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
PINUS SYLVESTRIS L.
Т. П. ИЛЬЧЕНКО
Сосна обыкновенная (Pinus sylvestris L.) на материке Ев-
разии занимает обширную территорию — от 70 до 37° с. ш. и от
7° з. д. до 126° в. д. Этот вид в разных частях ареала неодно-
роден по морфологическим, биологическим и эколого-физиоло-
гическим особенностям (Правдин, 1964),
В кариологическом отношении сосна обыкновенная иссле-
дована далеко не достаточно. Изучение же кариотипа на про-
тяжении всего ареала важно для селекции, внутривидовой
систематики, познания закономерностей формо- и видообразо-
вания. Результаты кариолсгического анализа помогут пред-
ставить возможные пути дифференциации таксонов в процессе
эволюции. Для экспериментальных селекционных работ необ-
ходимы также точные данные о морфологии хромосом.
Изучение кариологической изменчивости — один из аспек-
тов исследования фенотипической и, более того, генотипической
изменчивости природных популяций.
Кариотип Pinus sylvestris L. в настоящее время изучен в
Норвегии (Aass, 1957), Швеции (Natarajan, Ohba, Simak,
1961), США (Saylor, 1964) и в СССР - Тувинской АССР
(Правдин, 1964), Северном и Центральном Казахстане (Буда-
рагин, 1973), Центральном Черноземье (Машкин и др., 1974).
В данной работе представлены результаты кариолргическо-
го анализа сосны обыкновенной, произрастающей на юго-запа-
де Амурской области (Свободненский район) и районах Цент-
ральной Якутии (Покровский и Якутский). Материал обраба-
тывался по методике, разработанной группой сотрудников
Лаборатории лесоведения АН СССР (Правдин и др., 1972).
Работа проводилась на давленных препаратах из меристема
тических тканей кончиков корешков проросших семян. Семена
проращивались при температуре 25±1° и обрабатывались
1%-ным раствором колхицина. Затем фиксировались в спирто-
во-уксусной смеси и окрашивались ацето-кармином. Для маце-
рации и просветления клеток использовалась 40%-ная уксусная
кислота и хлорал-гидрат. Препараты просматривались и фото-
графировались при помощи микроскопа МБИ-6 (увелич.
60X10). С одного корешка (препарата) отбирали только одну
5*
67
Рис. 1. Кариофонд Pinus sylvestris L.
Ai — все хромосомы равноплечие; 3 пары со вторичными перетяжками (I.
II, IV)—популяции Центральной Якутии.
Аг — все хромосомы равноплечие; 3 пары со вторичными перетяжками (I,
II, VI)—популяции Тувинской АССР.
As—все хромосомы равноплечие; 5 пар со вторичными /перетяжками (I, II,
VI, VII, X) —популяции Швеции.
А4 — все хромосомы равноплечие; 5 пар со вторичными перетяжками (I, II,
III, V, IX) — популяции Центрального Черноземья.
Б1 — 10 пар метацентрических хромосом и 2 пары субметацентрических;
3 пары со вторичными перетяжками (I, II, VI) —популяции юго-запад-
ных районов Амурской области.
Бо — 10 пар метацентрических хромосом и 2 пары субметацентрических;
пластинку. Всего же отобрано 90 пластинок. Хромосомы изме-
рялись на микрофотографиях.
Определяли абсолютную длину хромосом (и суммарную аб-
солютную длину хромосом набора), мк; относительную длину
хромосом: отношение абсолютной длины хромосом к суммарной
длине диплоидного или гаплоидного набора, %; центромерный
индекс: отношение абсолютной длины короткого плеча хромо-
сомы к абсолютной длине длинного плеча, %; локализацию
вторичной перетяжки: отношение расстояния от вторичной пе-
ретяжки до центромеры к общей длине данного плеча, %.
Большое значение придавалось также соотношению морфо-
логически различных хромосом в кариотипе и наличию вто-
ричных перетяжек, с учетом их встречаемости, так как эти приз-
наки характерны для кариотипа хвойных. Конечным результа-
том кариологического анализа является построение идиограммы,
которая строилась для кариотипа каждой популяции отдельно.
Учитывалось и число хромосом в диплоидном наборе.
В исследованных хромосомных наборах существует разно-
образие по степени митотипической спирализации хромосом.
Поэтому для отбора сравнительно однородных метафазных
пластинок по степени спирализации хромосом в качестве кри-
терия использовался индекс спирализации I4 s —отношение
суммарной длины в абсолютных единицах двух самых малень-
ких хромосом к суммарной длине двух больших хромосом
данного набора (%).
По числу хромосом отклонений не обнаружено. Сосна
обыкновенная в диплоидном наборе имеет 24 хромосомы (2п--'
24), что характерно в целом для рода Pinus L. и семейства
Pinaceae Lindl.
Абсолютная длина хромосом от 16,7 мк (I пара) до
9 мк (XII пара). Несколько длиннее оказались хромосомы
сосны обыкновенной из Амурской области. Первая пара —
16,7 мк, последняя — 9,9 мк, в то время как сосна обыкновен-
ная из Якутии имела первую пару 15,6 мк, последнюю 9,6 мк
(табл. 1). В результате эти кариотипы отличались и по сум-
марной абсолютной длине хромосом гаплоидного набора
(Амурская обл.— 63,4 мк, Якутия — 57,12 мк).
Показатели по локализации вторичной перетяжки, центро-
мерному индексу и относительной длине хромосом сосны обык-
новенной из Амурской обл. и Якутии близки (табл. 2).
4 пары со вторичными перетяжками (I, II, III, X). X пара с добавоч-
ной вторичной перетяжкой. Популяции Северного и Центрального Ка-
захстана (Кокчетау — Мунчактинская).
Ег—10 пар метацентрических хромосом и 2 пары субметацентрических;
5 пар со вторичными перетяжками (I, II, III, IV, X)—X пара с доба-
вочной вторичной перетяжкой, Северный и Центральный Казахстан
(западные и южные популяции).
69
Таблица 1
Абсолютная длина хромосом сосны обыкновенной, мк
Место
произрастания
Пары а
I II III IV V VI VII VIII IX X XI
XII
Амурская обл.
(юго-запад-
ные районы) 16,7 16,0 15,5 15,3 14,9 14,6 14,2 13,8 12,7 12,3 11,2 9,9
Центральная
Якутия 15,6 14,7 14,1 13,9 13,5 13,2 13,2 12,5 12,0 11,1 10,8 9,0
Таблица 2
Относительная длина хромосом сосны обыкновенной, %
Место произрастания Пары хромосом
1 । П III IV V VI VII VIII IX X XI XII
Амурская обл.
(юго-запад-
ные районы) 4,8 4,8 4,6 4,5 4,4 4,4 4,3 4,1 3,7 3,6 3,3 2.9
Центральная
Якутия 4,8 4,5 4,5 4,4 4,4 4,2 4,1 4,0 3,8 3,6 4,2 3,0
Различия кариотипов сосны наблюдались в соотношении
морфологически различных хромосом и в количестве вторичных
перетяжек. Причем эти различия были отмечены и ранее
(Правдин, 1964; Бударагин, 1973; Машкин и др., 1974). Поль-
зуясь нашими данными и результатами других авторов, мы
провели сравнительный анализ кариотипов сосны обыкновен-
ной, произрастающей:
I) в Швеции (Natarajan и др., 1961) —все хромосомы рав-
ноплечие; со вторичнымй перетяжками 5 пар (I, II, VI, VII, X).
Рис. I, А3;
2) в Норвегии (Aass, 1957)—одна пара хромосом со вто-
ричной перетяжкой (I);
3) в США (Saylor, 1964)—три пары хромосом со вторич-
ными перетяжками (I, II, X).
К сожалению, мы не имеем данных по соотношению различ-
ных типов хромосом в указанных выше кариотипах.
4) В СССР: v
а) в Тувинской АССР, Балгазикском бору в пределах
51° с. ш. и 96° в. д. (Правдин, 1964) —все хромосомы равно-
плечие, но из 12 пар последние 4 пары значительно короче
других; со вторичными перетяжками 3 пары (I, II, VI).
Рис. 1, А2;
70
б) в Северном и Центральном Казахстане (Бударагин,
1973) — 10 пар разноплечих (метацентрических) и 2 пары раз-
ноплечих (субметацентрических). Автор обследовал несколько
популяций и отметил различия кариотипов этих популяции
по числу вторичных перетяжек. Так, в Кокчетау-Мунчактин-
Рис. 2. Распространение кариотипов Pinus sylvestris L. в пределах ареала
71
ской популяции — 4 пары таких хромосом (I, II, III, X) в за-
падных и южных популяциях (Челябинская степная, Ар-Кара-
гайская, Аман-Карагайская, Наурзумская, Баян-Аульская,
Каркаралинская, Кзыл-Райская)—5 пар (I, II, III, IV, X).
Рис. 1 Б2, Б3;
в) в Центральном Черноземье, Левобережном лесничестве
Воронежской Л. Т. И. (Машкин и др., 1974) —5 пар со вто-
ричными перетяжками (I, II, III, V IX). Число равно- и раз-
ноплечих хромосом авторы не указывают. (Рис. 1 А4);
г) в Амурской области (по нашим данным) — 100 пар рав-
ноплечих и 2 пары разноплечих; 3 пары хромосом со вторич-
ными перетяжками (I, И, VI). Рис. 1 Бг,
д) в Центральной Якутии (по нашим данным) — все хромо-
сомы равноплечие; 3 пары хромосом со вторичмыми перетяж-
ками (I, II, IV). Рис. 1 Ai.
На рис. 1 представлен кариофонд сосны обыкновенной в
пределах ареала. На рис. 2 показано распространение карио-
типов сосны обыкновенной в пределах ареала.
Как отмечалось выше, изменчивость кариотипов в основном
наблюдается в отношении морфологически различных хромо-
сом и числе вторичных перетяжек. Эти признаки варьируют и
в пределах популяции. Возможно, кариотипический полимор
физм есть результат генетической адаптации растений к эко-
логическому разнообразию, что ведет в свою очередь к дивер-
гентной эволюции кариотипа. Но механизм, вызывающий эти
различия, пока не ясен, и поэтому мы их лишь констатируем.
Дальнейшее исследование кариотипического полиморфиз-
ма, особенно внутрипопуляционного, необходимо для вскрытия
механизма формо- и видообразования.
ЛИТЕРАТУРА
Бударагин В. А Анализ кариотипов изолированных популяций сосны обык-
новенной (Pinus sylvestris L.) в Северном и Центральном Казахста-
не. — «Генетика», 1973, № 9.
Машкин С. И., Буторина А. К., Белозерова М. М., Пожидаева И. М., Му-
рая Л. С., Хатунцева Л. Н., Доронин Ю. А. Сравнительное изучение
кариотипов сосны меловой (Р. cretaceae Kalen) и сосны обыкновенной
(Р. sylvestris L.) в популяциях Центрального Черноземья. — Генетика,
селекция и интродукция лесных пород. Темат. сборн. научи, трудов,
в. 1. Воронеж, изд. ЦНИИЛГИС, 1974, с. 10—31.
Правдин Л. Ф. Сосна обыкновенная. М., «Наука», 19G4.
Правдин Л. Ф., Бударагин В. А., Круклис М. В., Шершукова О. П. Мето-
дика кариологического изучения хвойных пород.— «Лесоведение»,
1972, №2.
Aass J. A. Cytological analysis of Scots Pine (Pinus silvestris L.) from
Skjak, Norway.— Medd, Norske Skogforsoksvesen, 1957, Bd. 14.
Natarajan A. T.. Ohba K., Simiak M. Karyotype analysis of Pinus silves-
tris. Hereditas, 1961, v. 47.
Saylor L. C. Karvotype analysis of Pinus Group. Lariciones. Silvae genetica.
1964, Bd. 1'3, 6.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ЦЕНОТИЧЕСКАЯ РОЛЬ, СОСТОЯНИЕ И ПУТИ
СОХРАНЕНИЯ МОЖЖЕВЕЛЬНИКОВ И ДРУГИХ
ХВОЙНЫХ КУСТАРНИКОВ ПРИМОРЬЯ
В. м. УРУСОВ
На юге Дальнего Востока разнообразие хвойных кустарни-
ков нарастает, так как к видам с евразиатскими и восточноси-
бирскими ареалами присоединяются эндемы более или менее
широких регионов востока и юго-востока Азии. Этот процесс
ощутим уже в Приморском крае, где естественно произрастает
5 кустарниковых хвойных, принадлежащих к 3 родам семейств
Cupressaceae F Y. Neger и Pinaceae Lindl. Следует отметить,
что родовое разнообразие хвойных в целом приближается к
максимальному именно здесь, хотя видовое разнообразие, на-
пример можжевельников, более значительно в Центральной
Азии, где, по нашему мнению, уточнение его осложнено нали-
чием признаваемых за виды географических рас и гибридов.
Выявление географического и ценотического распространения
можжевельников Приморья позволило заключить, что один из
них, твердый, Juniperus rigida Sieb. et Zucc., является элемен-
том сообществ гемиксерического облика, относящихся к немо-
ральным лесам на переходе к лесостепи. Можжевельники си-
бирский J. sibirica Burgsd. и даурский J. dahurica Pall, тяготе-
ют к бореальным растительным комплексам, а их современное
распространение в значительной мере определяется послелед-
никовым изменением климата, крупной морской трансгрессией
и вызванным ими вертикальным смещением растительных поя-
сов в антропогене. Можно предположить, что ценозы этих
можжевельников представляют собой фрагменты подняв-
шихся зон; иногда они сохраняются в виде синузий поглотивших
их генетически чуждых растительных комплексов. То же отно-
сится к сообществам кедрового стланика Pinus pumila (Pall.)
PgL и микробиоты перекрестнопарной Microbiota decussata
Кот. в низкогорьях.
К настоящему времени наиболее известны состав ассоциа-
ций с микробиотой (Шишкин, 1935; Воробьев, 1935; Кабанов,
1937; и др.) и некоторые черты их динамики в отдельных
пунктах ареала (Пономаренко, 1961; Куренцова, 1968а, б).
В литературе имеются и сведения о кедровое гл аниковой фор-
мации, а также распространении и составе фитоценозов мож-
жевельников (Колесников, 1969; Куренцова, 1968а, б). Однако
73
ценодинамизм сообществ с хвойными кустарниками, их био-
логические свойства, а для можжевельников и распростране-
ние и состояние возобновления требуют порой значительных
уточнений.
Здесь мы коснемся некоторых вопросов, таких как приро«
да и перспективы ценозов с хвойными кустарниками для всей
территории края, их место и роль в растительном покрове и
меры по сохранению.
Можжевельник твердый — вмкарирующий по отно-
шению к обыкновенному J. communis L- вид, являющийся юго-
восточным звеном ряда Communies Kondr (Кондратюк, 1960)
и распространенный от Внутреннего Китая до севера Приморья,
а также по всей Корее, в Средней и Южной Маньчжурии, Япо-
нии. В Приморском крае вид расселен в низкогорьях Восточно-
Маньчжурских гор и Сихотэ-Алиня (рис. 1), причем в послед-
нем случае сообщества с можжевельником тяготеют к освещен-
Рис. 1. Распространение можжевельников в Приморском крае и состояние
их популяций.
Местонахождение можжевельников: 1—твердого, 2 — даурского, 3 — сибир-
ского; исчезающие популяции можжевельников: 4—твердого, 5— даурского.
6 — наиболее интересные и перспективные для расширения популяции
74
ным обрывистым склонам и скалам долин, впадающих в
Японское море, и местностям, открытым к Ханкайской равнине.
Можжевельник твердый, изредка поднимается до высоты
300 м над ур. м. (хр. Пограничный), обычно же он не прохо-
дит выше гребней береговых скал и не углубляется в свежие
и влажные леса. Кроме обычных древовидных встречены кус-
тарниковая, или распростертая (m. prostrata т,), и плоская
(т. depressa in.) модификации. В целом насаждения с можже-
вельником твердым занимают в крае около 1 тыс. га, но лишь
на 300 га его участие значительно: в бассейнах рек Нестеров-
ка, Артемовна, Арсеньевна, Партизанская, Черная, Самарга,
на обрывах мысов Островного и Туманного, на уступе террасы
западного берега оз. Ханка. Островной ареал обусловлен в ос-
новном флородинамическими причинами, в частности затопле-
нием морем ряда связующих популяций и ускоряющейся рег-
рессией в результате интенсивного освоения территории.
По крайней мере 80% площадей фитоценозов с этим мож-
жевельником приурочено к известняковым и приморским ска-
лам, крутым и обрывистым южным и западным склонам.
В удаленных от моря районах на гранитах, кварцевых порфи-
рах, глинистых сланцах вид встречается редко, плотность попу-
ляций ниже, а участки ассоциаций фрагментарны. Наблюдаемая
в настоящее время узость экологической амплитуды — след-
ствие сохранения можжевельника твердого в менее подвержен-
ных низовым пожарам местах. Такая вторичная однородность
эдатопов свойственна можжевельнику даурскому, некоторым
другим относимым к ксерофитам растениям, а в еще большей
степени — соснам густоцветковой и погребальной.
Выделено 6 групп ассоциаций с можжевельником твердым
(схема 1). Можжевельник твердый входит в состав и может
доминировать в ксерофитных группировках. Имеются можже>
велово-полынные, можжевелово-абрикосовые с полынью, мож-
жевелово-широколиственные с полынью и тимьяном ценозы,
которые можно объединить в можжевелово-тимьянниково-полын-
ную группу ассоциаций, довольно характерную для скал н наи
более распространенную. Видимо, еще в недавнем прошлом и
в Восточно-Маньчжурских горах существовала самостоятельная
можжевеловая формация, представленная теперь участками
ассоциаций дубово-можжевелово-злаковой группы. Насаждения
формации занимали крутые южные и западные склоны с диф-
ференцированными почвами и граничили с лесами из погре-
бальной сосны. Можжевельник твердый доминирует и на пес-
чаных валах морского берега. Псаммофитные можжевелово-
лишайниковые ассоциации, существуя в устьях рек, являются
самостоятельными в течение длительного времени или возни-
кают на скалах ниже зарослей.
Состав следующих 3 групп ассоциаций наиболее сложен,
а доминирование можжевельника твердого в них возможно в
75
_____________________ 1
-----2
-----3
Схема 1. Группы ассоциаций с можжевельником твердым в связи с их сук-
цессионными и генетическими связями. Двойной линией обведены группы ас-
социаций, наиболее распространенные и устойчивые в настоящее время. Ну-
мерация групп дана в направлении снижения ценотической роли можжевель-
ника твердого:
1—направление основных смен ассоциаций в связи с почвообразованием,
2 — смены в процессе почвообразования,
3 — генетические связи групп ассоциаций
лучшем случае лишь в микрогруппировках. При отсутствии
пожаров вид является постоянным участником определенных
синузий сосново-можжевелово-злаковой, сосново-можжевелово-
разнокустарниковой и широколиственно-разнокустарниковой с
можжевельником групп ассоциаций. Причем для сухих зла-
ковых, свежих и влажных разнокустарниковых сосняков мож-
жевельник твердый характерен. Здесь он нормально растет и
плодоносит., выдерживая значительное затенение. В целом
ассоциации с сосной и можжевельником территориально отно-
сительно велики и занимают около 200 га. Группа ассоциаций
широколиственно-разнокустарниковых лесов с можжевельни-
ком твердым встречена только в бассейне р. Партизанская.
Эта группа наиболее богата по составу дендрофлоры, но зани-
мает лишь несколько га и, пожалуй, наиболее обособлена.
Можно предполагать, что в определенные периоды восстанов-
ление можжевельника в ней идет за счет семян, заносимых со
скал по бортам речных долин.
На обрывистых склонах и скалах можжевеловые рощи мо-
76
гут занимать десятки гектаров, в древостое насчитывается
иногда свыше 1000 экземпляров можжевельника твердого на
1 га (табл. 1). Густота можжевеловых рощ больше <в бассей-
нах впадающих в море рек. В центральных районах Приморья
встречено не более 600 стволов можжевельника в пересчете
на 1 га. Его сравнительно крупные куртины приурочены к осо-
бенно мощным скалам. Однако несмотря на иногда очень зна-
чительное численное участие в древостое, запасы древесины
можжевельника невелики и в группе скальных ассоциаций
обычно менее 10 м3 на 1 га. При диаметре у основания ство-
лов до 50 и на высоте груди до 24 см экземпляры можжевель-
ника твердого достигают здесь высоты 7, редко 10 м, но сред-
ние диаметры и высоты соответственно не более чем 17 см и
4 м. Со второго столетия жизни деревья суховершинят, но
продолжают плодоносить. Типичным для можжевеловых по-
пуляций скал является значительное преобладание мужских
особей: на горе Змеиной Шкотовского района к ним относится
70% экземпляров древостоя, в верховьях р. Илистая — еще
больше, х
Изменчивость семяношения в связи с флюктуациями кли-
мата наиболее четка у старых суховершинящих экземпляров
можжевельника (Шмидт, Урусов, 1973). Дерево продуцирует
до 4 тыс. шишкоягод в год, но в среднем их не менее 1 тыс.
На 1 га можжевеловых рощ на скалах в урожайные годы все
еще можно собирать до 12—15 кг шишкоягод с 400 тыс. выпол-
ненных семян (вес 1 тыс. шишкоягод 102 г). Некоторое коли-
чество семян образуется ежегодно.
Можжевельнику сопутствуют ясень носолистный, яблоня
маньчжурская, осина, низкорослый кедр корейский (гора Змеи-
ная в Шкотовском районе), боярышник перистонадрезанный,
ильм крупноплодный, акатник (бассейны рек Арсеньевна и
Партизанская), абрикос маньчжурский, клен мелколистный
(Пограничный и Партизанский районы), дуб зубчатый (Парти-
занский район). В группировках участвуют секуринега, ши-
повник тонконожковый, карагана уссурийская, пузыреплодник
амурский, тимьяны, а по р. Партизанская также и курильский
чай кустарниковый, вишня железистая. В верховьях Артемовки
и на горе Чандалаз в подлеске встречен и можжевельник даур-
ский. В центральных районах края, а частично и в Шкотовском
районе в скальные можжевеловые рощи проникают настоящие
сосны.
На склонах и скалах морского побережья полынь Гмелина
с древеснеющими стеблями и кустарниковая модификация
можжевельника изредка содоминируют в одном ярусе, хотя
чаще на скалах преобладает полынь или можжевельник даур-
ский. Древовидная форма можжевельника твердого на обры-
вах Японского побережья не встречена: здесь представлена
только распростертая модификация. Его проективное покрытие
77
Некоторые таксационные характеристики древостоев с можжевельником
Местонахождение пробной площади Ярус Состав древо- стоя Сомк- ну- тость крон Поро- да Средние и возраст, лет
Сухие скалы в бассейне I 8См 2Д+Мт, 0,8 См 80(20—130)
р. Арсеньевна ед К, Л па Мт 140(90—260)
Скалы в верховьях р. Ар- I 5Мт 4К1Яг 0,4 Мт 80(40—240)
темовна К 60(40—120)
Южный склон крутизной Яг 35(30—50)
I 7Д2Мт 1Бч 0,55 Д 50(40—65)
20° в верховьях р. Не- стерова Мт 70(40—100)
Терраса оз. Ханка 1-П 7См 2Д 1Мт 0,5 См 40(20—90)
ед Мк, Бч, Аа, Км, Лпп Мт 60(40—130)
Уступ- террасы оз. Хан- I—II 9Д 1Мк+Мт 0,5 Д 30(20—40)
ка, северо-восточный склон крутизной 35° Мт 50(40—90)
Уступ террасы оз. Ханка, I—II 6См 2Мт 2Д 0,7 См 50(20—110)
юго-западный склон крутизной 35° с сьгоы- ми почвами ед. Ол Мт 80(40—160)
Условные обозначения пород: См — сосна могильная. Мт —
Аа— акатник амурский, Бч — береза даурская, Д — дуб монгольский, Км —
ская и пекинская, Мк — мелкоплодник, Ол— ольха волосистая, Яг — ясень
обрывов морского берега составляет 0,01—5%. На 100 м2 скал
учтено до 11 растений старше 20 лет и 14 более молодых. На
мысе Островном на 1 км берега приходится не более чем по
250 экз. возобновления и взрослого можжевельника. Отдельные
синузии вида занимают до 15 м2, диаметры кустов достигают
2—3,5 м, корневой шейки — 12 см, приросты ветвей составляют
2—15 см в год, на 1 м2 зарослей насчитывается от нескольких
до 100—150 шишкоягод, которые на растениях прибрежных
песков могут быть несколько обильней (до 200—500). На из-
вестняковых скалах возобновление можжевельника многочис-
ленней, чем других пород (табл. 2). Так, на горе Змеиной на
нижней трети обрыва в можжевеловой роще в 1972 г. было уч-
тено по 30—100 экз/100 м2 всходов и самосева можжевельника
твердого. Подроста можжевельника в рощах по 100—800экз/га.
На выходах других горных пород самосева и подроста мож-
жевельника до 100—200 экз/га (бассейны Арсеньевки и Ко-
миссаровки) и 300—500 экз/га (в верховьях Илистой), лучше
восстанавливаются дуб и акатник. Возобновление можжевель-
ника твердого даже здесь страдает от пожаров и сбоя. Вслед-
ствие этого в ассоциациях группы идет упрощение состава
78
Таблица J
твердым в разных районах Приморского края
предельные Число ство- лов на 1 га Запас дре- весины, м3 га Класс бони- тета
высота, м диаметр, см жи- вые мерт- вые жи- вые мерт- вые
10,3(3—17) 28,6(8—52) 367 178 V 4,0(2—9) 17,0(8—24) 65 5,3 Va 3,5(2-5,5) 6,0(2—10) 1120 150 7,3 1,0 Va 4,5(2—8) 9,0(6—16) 275 5,5 Va 2,5(2—3,5) 4,0(2—6) 450 1,4 Va 4,0 (3,5—6) 12,0(8—16) 1400 60 Va 3,0(2—4) 8,0(2—14) 917 15 Va 7,6(2-13) 12,4(3—32) 717 30 40 1,0 IV 3,8(2—8) 6,2(4—14) 170 106 2 1,0 Va всех пород 2117 140 53 2,0 4,0(2,5—4,5) 6,0(4—8) 2000 14 IV 4,5(3,0-5,5) 4,0(2—8) 20 0,1 Va 10,0(7,0—11,0) 16,0(12—28) 450 — 59 IV 6,0(2,5—8,0) 9,0(4—16) 400 50 8,5 1,0 Va
можжевельник твердый, К — кедр корейский,
клен мелколистный, Лпа и Лпп — липы амур-
горный.
(схема 2) с последующей сменой их полынными зарослями.
Под влиянием естественных причин, связанных с особенностями
современного климата, в можжевеловые рощи скал внедряются
кедр, сосна и мезофильные широколиственные породы. Однако
ядро ксерофитов, образовавшееся здесь до начала климатиче-
ских смен, и сейчас четко выделяется. К характерным для
можжевеловых рощ следует отнести абрикос, ильм крупноплод-
ный, вишню железистую, пузыреплодник амурский, секуринегу,
тимьяны.
В травяном покрове доминирует полынь Гмелина, очень
редко — кругонос язычный (бассейны Комиссаровки и Черной),
участвуют вудсии, полынь Кейске, некоторые осоки и злаки,
лилии поникшая и тонколистная, луки, смолевки, гвоздика
амурская, лапчатки, горноколосники, очитки, патринии, ломо-
нос шестилепестковый, василистник вонючий, рапонтикум одно-
цветковый и др. Синузии трав, как и кустарников, составлены
преимущественно ксерофитами разных флористических комп-
лексов, в том числе и тяготеющими к бореальным. Смешение
разнородных ценоэлементов указывает на то, что и в этих ме-
стопроизрастаниях сменились фитоценозы, принадлежащие
79
Таблица 2
Возобновление древесных пород в ассоциациях с можжевельником твердым
в Приморском крае, по данным учета летом и осенью 1972 г.
Местонахождение пробной площади Состав подроста Порода Количество возобновления в пересчете на 1 га
всходы самосев подрост
живые мертвые живые ме₽т- вые живые выТ
Сухие скалы в бассейне р. Ар- 5Аа2См1Мт1Д1Ик ед. Бч См 500 сеньевка Все лиственные породы Мт 100 1250 Скалы в верховьях р. Арте- Мт 775 мовка 8Мт1К1Яг К 150 Яг 125 g Южный склон крутизной 20° 6Д4Мт Мт 500 в верховьях р. Нестеровка Д 1500 Терраса оз. Ханка ЗМт2См2Мк2Д Мт 140 133 574 105 1Аа+Ос, ед. Км, Лпп См 414 269 325 28 Все лиственные породы 30 100 1036 Уступ террасы оз. Ханка, се- 4Мт2СмЗМк1Км+Д, Бп Мт 1250 4000 4000 веро-восточный склон кру- См 3250 6750 3000 тизной 35° Все лиственные породы Мт 2000 3000 4500 Уступ террасы оз. Ханка, юго- 3570 6970 6430 западный склон крутизной 4Мт4См1Мк1Д См 16250 7860 5000 6400 35° ед. Км, Ол, Ос, Лпп Все лиственные породы 3500 4000 500 5000
Условные обозначения пород: См — сосна могильная, Мт —можжевельник твердый, К — кедр корейский,
Аа — акатник амурский, Бч — береза даурская, Д — дуб монгольский, Км— клен мелколистный, Лпп — липа пекинская,
Мк — мелкоплодник, Ол — ольха волосистая, Яг — ясень горный, Ик — ильм крупноплодный, Ос — осина, Бп — боярыш-
ник перистонадрезанный.
5 Заказ 488
I
II
Сосново-можжевелово-злако-
вая по обрывистым склонам
IV
V
Ильмово-широколиствен-
ная с можжевельником
I в /речных долинах
Схема 2. Коренные (двойная обводка) и производные
дым. Указаны направления характерных или на современном
1'ип. Замещающие друг друга на экологических профилях
ассоциации и группы ассоциаций с можжевельником тзер-
этапе уже нетипичных (пунктирные линии) смен ассоциа-
ассоциации на схеме соединены плюсами
к разным зонам. Обращают на себя внимание зикарирование
видов и форм определенных родов и довольно существенные
различия полных флористических списков. Это, по нашему
мнению, связано с изоляцией, достаточно продолжительной для
дивергентной эволюции (Ильченко, 1975), популяций в скаль-
ных местопроизрастаниях, например тимьянов, очитков, овся-
ниц. Изоляция, видимо, сказалась и на можжевеловых (не
только можжевельника твердого) популяциях. По крайней мере,
экземпляры морского побережья и более континентальных рай-
онов различимы.
Наиболее разнообразны травы на участках скальных ассо-
циаций побережья и повсеместно у подножий скал. Здесь резко
возрастает участие более влаголюбивых видов, являющихся
для рассматриваемой группы ценозов случайными и второсте-
пенными.
Дубово-можжевелово-злаковые ассоциации крутых и пока-
тых склонов — вторая группа ассоциаций, остатки собственно
можжевеловой формации, еще недавно значительной в Восточ-
но-Маньчжурских горах. Сейчас ее участки, в отдельности по
превышающие 1—2 га, занимают всего 50 га (в верховьях
р. Нестеровка — 40 га и до 10 га по окраинам скальных мож-
жевеловых рощ в других районах края). В можжевелово-дубо-
вых и дубовых с можжевельником рощах хр. Пограничного на
1 га сохранилось до 900 живых растений можжевельника твер-
дого в возрасте 50—130 лет высотой 5 м и ниже. Плодоносит
менее 20% деревьев. По запасу и почти всегда численно в дре-
востоях преобладает дуб монгольский, хотя максимальный за-
пас можжевельника твердого учтен именно здесь. Есть пни
крупных стволов можжевельника с диаметром у шейки корня
до 60 см (свыше 200, а иногда до 1000 на 1га), доживавших
до 300 лет. Эти сообщества довольно распространены в Мань-
чжурии (Лукашкин, 1933). Ассоциации можжевелово-дубового
леса со злаково-разнотравным покровом в результате клима-
тогенных смен голоцена в значительной мере поглощены фор-
мацией сосны погребальной или уничтожены пожарами. Не-
многочисленный подрост можжевельника—до 500 экз. на
1 га — уничтожается периодическими пожарами и сейчас. По-
жары, сдерживая восстановление сосны, способствуют разви-
тию редкостойных изреженных дубняков.
В кустарниковом ярусе преобладают характерные для сос-
няков и дубняков леспедеца двуцветная и рододендрон остро-
конечный, встречаются секуринега и некоторые розы, в травя-
ном — обычные для сосняков виды: налицо выровненный по
составу в результате пожаров растительный покров, одинаково
свойственный дубовым и сосновым группировкам на сухих и
частично свежих почвах, а также горным лугам. Абсолютное
большинство учтенных видов относится к дубравным и лугово-
степным элементам. Бореальных видов, в отличие от ассоциа-
82
ций скал, почти нет. Появляются мезофиты, не свойственные
рощам на скалах, но их доля в составе синузий возрастает за
счет общего обеднения тревяного покрова. Можно предполагать,
что коренной состав можжевеловых рощ на склонах с нормаль-
ным почвенным профилем был близок составу рощ на скалах.
Остатки ассоциаций можжевелово-лишайниковой группы
сохранились на значительном участке морского берега между
устьями рек Черная и Киевка, причем на речных наносах, от-
стоящих от можжевеловых зарослей скал на многие километ-
ры, эти сообщества существуют изолированно по крайней мере с
конца прошлого тысячелетия. Можжевельник плоской модифика
ции рассеянно встречается на песках у моря вместе с розой мор-
щинистой, травянистыми ксерофитами, некоторыми литоральны-
ми видами, а в устье р. Киевка в непосредственной близости
с синузиями эфедры. Южнее с. Глазковка на береговом валу
сохранилась интереснейшая можжевелово-лишайниково-злако-
вая группировка, занимающая около 5 га. Можжевельники
даурский (от моря не удаляется) и твердый покрывают 5—
70% вала. Диаметр кустов можжевельника твердого (плоская
модификация) там достигает 3,5 м, высота 0,7 м, диаметр
стволов у основания достигает 10—16 см. Плодоношение мож-
жевельника твердого протекает успешно. На 100 м2 насчиты-
вается 1—7 взрослых кустов и 1 —10 экз. возобновления.
Распростертая модификация в можжевелово-яблоневой микро-
группировке прибрежных кустарниковых зарослей характеризу-
ется высотой кустов можжевельника твердого 1,8 м. Под кро-
нами женских особей на 1 м2 там учтено по 10—20 экз. всхо-
дов, самосева и подроста высотой до 0,3 м. К сожалению, по-
жары и здесь изреживают и уничтожают можжевеловые за-
росли.
Сосне погребальной можжевельник твердый сопутствует в
ее горноколосниковом скальном и арундинелловом горных ти-
пах леса в западных и центральных районах Приморья, а так-
же в осоково-разнотравном и широкотравно-таволговом слож-
ных равнинных на уступе ханкайской террасы, сосне густоцвет-
ковой— в осоковом и марьянниково-разнотравном типах в от
рогах горы Змеиной. В сосняках можжевельник твердый может
рассматриваться как характерный вид, способный слагать соб-
ственные микрогруппировки или синузии, в то время как в
кленово-лещинном грабовом кедровнике с пихтой целыюлист-
ной (гора Змеиная) и его дериватах присутствие можжевель-
ника объясняется заносом семян со скал или поглощением
можжевеловых ассоциаций. В таких лесах можжевельник
твердый не семяносит.
В древостоях сухих сосновых типов живых стволов можже-
вельника от 30 до 100, они достигают 9 м в высоту, запас мож-
жевеловой древесины менее 6—7 м3 на 1 га. Невелико и уча-
стие можжевельника в возобновлении— 100—200, в верховьях
6’
83
р. Нестеровка. где сосна надвигается на собственно можжеве-
ловую формацию, 700 растений на 1 га. В возобновлении пре-
обладают сосны и широколиственные породы.
В свежих и влажных сосняках ханкайской террасы — ас-
социации с можжевельником занимают около 50 га — можже-
вельник твердый отличается лучшим ростом (рис. 2), семяно-
шением и возобновлением. В древостоях уступов южной экс-
позиции наибольшее учтенное количество можжевельника —
400 экз./га, северной— 130 экз./га, однако запас можжевеловой
древесины в связи с небольшими средними диаметрами ство-
лов не превышает 10 м3/га. К НО—150 годам некоторые особи
можжевельника достигают высоты 12 м при диаметре 32 см.
На слоистых песках уступа юго-западной экспозиции — влаж-
ный сосняк — учтено 3,5 тыс. всходов, 6,9 тыс. самосева и
6,4 тыс. подроста можжевельника на 1 га. Возобновление его
приурочено к западинам, тальвегам, микрогруппировкам дуба,
где освещенность на поверхности почвы колеблется от 9 до 11 —
14% полной и на высоте 2 м не превышает 16%. При наличии
семян можжевельник твердый хорошо возобновляется и и ч
северных склонах, но на свежих, порой пересыхающих почвах
Рис 2. На уступе
ханкайской террасы
фрагмент древостоя
свежего осоково-раз-
нотравного сосняка с
хорошим возобновле-
нием можжевельника
твердого (высота
лее 8 м). Насажде-
ние находится север-
нее с. Новокачалрз-
ское
поверхности террасы самосева и подроста менее 1 тыс. экз./га.
Отчасти это объясняется и прошедшими здесь пожарами.
В древостоях ханкайской террасы V2 особей можжевель-
ника— женские. Здесь из 10 лет урожайными являются 3—4,
в течение еще двух лет урожаи шишкоягод близки средним
многолетним. В семенной год 1 особь продуцирует в среднем
0,15 кг шишкоягод (от 0,3 до 8 тыс. шт.}, 1 га фитоценозов —
20 кг и более. Около трети этого количества шишкоягод мо-
жет быть собрано в средние по урожайности годы. Нормальный
и непрерывный возобновительный процесс при исключении по-
жаров привел бы к увеличению роли можжевельника твердого
и расширению фитоценозов с ним. Но возобновление и взрос-
лые деревья (до 30—50%) периодически гибнут: при обследо-
ваниях учтено до 150 мертвых стволов можжевельника на 1 га.
Во всех ярусах сосново-можжевеловых фитоценозов хан-
кайской террасы многочисленны мезофиты, мезогигрофиты, а
на влажных почвах и гигрофиты. Там же в значительной сте-
пени сохраняются деревья, кустарники и травы, типичные для
скальных можжевеловых рощ.
Антропогенные смены пород ведут к замене сосново-мож-
жевеловых насаждений дубово-широколиственными, хотя, судя
по характеру естественного возобновления, они вполне соот-
ветствуют здесь современным почвенно-климатическим услови-
ям, наиболее сложны и продуктивны и могут считаться кли-
максовыми. Климаксовыми можно считать также можжевело-
во-широколиственные заросли скал, в которых до известных
пределов будет возрастать участие темнохвойных пород, а так-
же можжевеловые группировки прибрежных песков. На скалах
и береговых валах синузии можжевельника твердого выступа-
ют как почвообразователи и сами становятся более мощными
по мере формирования почв. Вместе с можжевельником твер-
дым пионерные ценозы образуют можжевельник даурский, ку-
рильский чай, горноколосники, очитки, и видимо, луки и другие
ксерофиты. Большинство перечисленных растений генетически
связано с бореальными формациями. Несмотря на это, можно
предполагать, что можжевельнику твердому до вертикальных
перемещений высотных растительных поясов, особенно на ска-
лах нижнего и среднего пояса гор, сопутствовали растения,
относимые к ценоэлементам лесов неморального и близкого к
ним гемпксерического ряда: кругонос язычный, гвоздика амур-
ская, некоторые лилейные и сложноцветные.
На влажных почвах можжевельник твердый (по 10—20
стволов на 1 га) встречен также к востоку от с. Новицкое (бас-
сейн р. Партизанская) в многопородном широколиственном
долинном лесу с дубом зубчатым. Его высота до 5 м и диаметр
до 12 см, приросты по высоте в год не превышают 15 см. Как
и на уступе ханкайской террасы, при исключении пожаров вид
вполне устойчив.
85
Обобщая сведения о возобновлении можжевельника твер-
дого в фрагментах различных ассоциаций, мы пришли к заклю-
чению, что достаточно интенсивно вид возобновляется только
на влажных почвах (рис. 3). На легких почвах влажных и сы-
рых местопроизрастаний возобновление можжевельника может
быть более многочисленным, чем других пород. На более сухих
лучше возобновляются настоящие сосны и даже широколист-
венные породы. Здесь в последние 4—5 десятилетий прослежи-
вается не связанное с ухудшением плодоношения падение во
Рис. 3. Возобновление неко-
торых пород в различных
по увлажнению почвы ас-
социациях с можжевельни-
ком твердым. При питы сле-
дующие градации местопро-
израстаний: 0 — очень су-
хие скалы, I — сухие ска-
лы, II—свежие почвы
склонов и западин, III —
свежие почвы террасы оз.
Ханка и обрывов, IV —
влажные почвы уступа
террасы и подножий обры-
вов, V — сырые почвы ус-
тупа ханкайской террасы.
1 — всходы, 2 — самосев, 3—
подрост можжевельника
твердого; 4— всходы, 5—
самосев, 6— подрост сосны
погребальной. Возобновле-
ние: 7— дуба монгольского,
8—акатника амурскою или
мелкоплодника; 9—осве-
щенность подроста можже-
вельника в процентах от
полной
зобновления можжевельника, обусловленное, возможно, его
естественной периодичностью. Поэтому надо обратить особое
внимание на сохранение вида на достаточно влажных почвах:
прежде всего на террасе оз. Ханка и ее уступе. Вполне возмож-
но и введение можжевельника под полог дубняков у скал и на
влажных фрагментарных почвах нижних частей обрывов. Со-
вершенно необходимо отграничивание участков с можжевель-
ником твердым хотя бы в перспективных для восстановления
его популяциях минерализованными полосами.
Можжевельник сибирский в Приморье не только
образует группировки в верхних частях сухих и щебнистых
склонов предгольцового пояса и по каменным россыпям среди
елово-пихтовых лесов (Колесников, 1969), но и участвует в
кедровостланиково-лишайниковых сообществах (покрывает до
10—30% поверхности), а также входит в подлесок дубово-ка-
86
тленноберезовых лесов с рябиной амурской, кедровым стлани-
ком, брусникой (южные склоны хр. Брусничного). Ветви мож-
жевельника сибирского приподнимаются на 0,3—0,5 м над
почвой и умеренно плодоносят: в зарослях с кедровником
образуется от нескольких до 50 шишкоягод на 1 м2 можже-
веловых синузий, в дубово-каменноберезовом лесу — до 20, но
при сомкнутости крон 0,8 и выше их может не быть. Под поло-
гом леса можжевельник занимает до 40% поверхности почвы,
на 100 м2 учтено до 50 его растений. Обычно же на 1 га камен-
ноберезняков или каменноберезово-темнохвойных лесов с мож-
жевельником сибирским насчитывается от 100 до 3000 его
взрослых растений и до 700—800 экз. возобновления. В отли-
чие от можжевельника даурского, доминирующего иногда на
нескольких га, площади синузий сибирского не превышают 1 —
2 аров и не образуют таких мощных зарослей, как, например,
в Якутии. Однако в более сухие периоды антропогена заросли
можжевельника сибирского могли составлять самостоятельный
пояс на верхней границе леса, объединявший также и высоко-
горные ксерофитные кустарнички и травы, как это считает
Б. П. Колесников (1969:66). Характерные виды можжевеловой
формации и ее фрагменты, сохранившие можжевельник сибир-
ский, в Среднем Сихотэ-Алине встречаются уже в низкогорьях.
поднимаясьхдо высоты 1730 м над ур. м.
Группы ассоциаций с можжевельником сибирским, в отли-
чие от групп другого вида этого же видового ряда — можже-
вельника твердого, представляют единый комплекс, связанный
со стадиями почвообразовательного процесса. Можжевельник
сибирский доминирует единственно в группе ассоциаций мож-
жевелово-кустарниковых зарослей каменных россыпей на юж-
ных склонах у вершин, хотя в настоящее время типичней участ-
ки кедровостланиковых ассоциаций с можжевельником сибир-
ским, другими кустарничками и некоторыми травами (рис. 4).
По мере формирования почв ассоциации группы сменяются
каменноберезняками или дубово-каменнобсрезовыми лесами с
можжевельником сибирским в подлеске. На данном этапе в под-
леске может быть более значительной роль кедрового стланика.
Третья группа ассоциаций с видом — подгольцовые пихтово-
еловые леса, при развитии которых хвойные кустарники посте-
пенно исчезают.
Большее единство групп ассоциаций с можжевельником си-
бирским, а по существу и кедровостланиковых и микробиото-
вых, по сравнению с ассоциациями можжевельника твердого
стоит в связи с лучшей сохранностью таежного и субальпий-
ского поясов в целом, молодостью и однотипностью их расчле-
нения и деградации.
Можжевельник даурский, принадлежащий к ряду
Sabinae polyspermae Кот., квазибореальный ценоэлемент,
покрывает значительные площади прибрежных скал, каменных
87
Рис, 4. Участок кедровостланиковой (с можжевельником сибирским и рододендроном снхотинским)
южном склоне одной из вершин хр. Брусничного. Средний Сихотэ-Алинь
циации
россыпей, галечников и в меньших количествах встречаются на
скалах и россыпях среди лесов в отрогах Восточно-Маньчжур-
ских гор и Сихотэ-Алине. Заселяя скалы, он преобразует мик-
роклимат приземного слоя воздуха и способствует почвообра-
зованию. Его пенотическая и ценодинамическая роль анало-
гична таковой микробиоты и кедрового стланика. Можжевель-
ник даурский встречается от уровня моря до высоты около
’300 м в следующих семи группах ассоциаций: в кустарниково-
можжевеловой субальпийской на каменных россыпях высоко-
горий и водоразделов; изреженных таежных, как правило под-
гольцовых, лесов с можжевельником в подлеске;, вторичных
березовых или широколиственных лесов, сменяющих тайгу; за-
растающих галечников речных долин; зарослей приморских
береговых валов; кустарничково-травяных зарослей скал; мож-
жевелово-разнотравных группировок каменистых склонов
(схема 3). Видимо, исходными следует считать подгольцовые
Схема 3. Группы ассоциаций с можжевельником даурским и их генети-
ческие связи. Более устойчивые' в настоящее время группы выделены двой-
ной обводкой
ассоциации, фрагменты которых теперь территориально незна-
чительны. Ассоциации других групп являются их недавними
(фитоиенозы подгольцовых лесов с участием вида) или очень
древними (заросли скал в низкогорьях) производными. Имен-
но на скалах низкогорий можжевельник даурский нашел новый
экологический оптимум и способен к ограниченному расселению
отсюда в прибрежные дубняки и лиственничники, а также со-
общества литоральных видов.
Можжевельник даурский покрывает несколько сотен
гектаров скал Японского побережья, где наиболее сохранился
в составе сообществ можжевеловой полынно-тимьянниковой
ассоциации. Вид здесь может доминировать и покрывает иног-
89
да 40% поверхности скал. Отдельные синузии занимают до
40 м2, длина приподнимающихся не более чем на 30 см ветвей
достигает 5 м, диаметр — 5—7 см, возраст — 90—130 лет, при-
росты по длине (у старых и вегетативного происхождения
растений) —7—И см в год, по диаметру — 0,6—2,5 мм. Ветви
укореняются через 1 м и более, но с увеличением влажности —
через 0,5—0,7 м.
Изредка можжевельник даурский присутствует вместе с
можжевельником твердым на известняковых скалах — в кус-
тарниковом ярусе (горы Змеиная, хр. Чандалаз).
С береговых обрывов можжевельник даурский заходит в
дубняки и другие насаждения. На песках берегового вала
его синузии достигают 60 м2, а ветви укореняются через 0,2—
1,2 м. На участке можжевелово-лишайниковой ассоциации с
можжевельником твердым отмечены укоренившиеся в 7—10
местах ветви можжевельника даурского длиной до 12 м, а об-
разованный одним материнским растением покров распадается
на ряд самостоятельных синузий, чья былая связь прослежива-
ется на площади 300—400 м2 и более. Семенного возобновления
можжевельника даурского не более 100 экз./га ценозов и их
гарей. Еще малочисленней оно на скалах и у их подножий.
Можжевельник даурский семенного происхождения несколько
десятилетий имеет почти исключительно игольчатую хвою, ею
ветви прирастают на 11—24 см в год, проекции ковров звездо-
образны.
На скалах можжевельник даурский продуцирует шишко-
ягод больше, чем твердый, — до 800—1400 экз./м2, на песках
примерно столько же, сколько и твердый — 100—400 экз./м2.
Но выполненных семян у можжевельника даурского до 3% от
общего количества или нет вовсе.
Фрагменты подгольцовых ассоциаций с можжевельником
даурским с типичными для высокогорий горичниками, сложно-
цветными и другими растениями встречаются уже с высоты
200 м (окрестности залива Ольги), в Хасанском районе — с
400 м над ур. м. На западном склоне горы Крестовой в зал.
Ольги кроме участка скальной можжевелово-рододендроново-
полынной с лишайниками ассоциации, в которой доминирует
можжевельник, присутствуют фрагменты можжевелово-шипов-
никовой (розы уссурийская и тонконожковая) с тимьяном не-
равным, можжевелово-разнотравной и кедровостланиковой с
можжевельником даурским ассоциаций. В зарослях кедрового
стланика, достигающего высоты 3—4 м, можжевельник даур-
ский образует сплошной покров под пологом стланика и по ок-
раинам его биогрупп. Участки такой ассоциации на каменных
россыпях горы Крестовой распространены весьма широко.
В них присутствуют почти все виды — спутники можжевело-
вых зарослей на каменистых развалах высокогорий.
На каменных россыпях с фрагментами белопихтарников и
90
каменноберезняков в Восточно-Маньчжурских горах можже-
вельник даурский теперь исчезает. Севернее присутствие его
под пологом леса в верхнем поясе растительности —-в камен-
ноберезняках, лиственничниках, белоберезняках — так же, как
микробиоты, кедровника, можжевельника сибирского, свиде-
тельствует, по нашим v данным, о завершении первых этапов
почвообразовательного процесса и надвигании леса на субаль-
пийский пояс или связано с характером филоценогенеза (мож-
жевельник даурский в широколиственных насаждениях с пих-
тами белокорой и цельнолистной). Именно характер филоцено-
генеза — особенности смещения вертикальных растительных
поясов и сохранения их фрагментов — обусловил совместное
произрастание и содоминирование ценотически чуждых видов,
в том числе и можжевельников даурского и твердого.
Следует особенно подчеркнуть, что можжевельник даур-
ский на побережье Приморья по форме и цвету хвои, цвету
шишкоягод, длине их ножек, цвету коры существенно отличен
от экземпляров вида из континентальных районов Дальнего
Востока. По этим признакам можжевельник даурский прибли-
жается к можжевельнику Саржента Juniperus sargenfii (Hen-
ry) Takeda и может рассматриваться как замещающий его на
побережье материка таксон. По нашему мнению, которое
поддержано и В. Н. Ворошиловым, можжевельник даурский из
Восточного Сихотэ-Алиня целесообразно выделить в самостоя-
тельный подвид. Происхождение и значение этого подвида мо-
гут быть поняты при специальном изучении сабнновых мож-
жевельников Азии, путей их расселения и взаимоотношений.
Микробиота перекрестнопарная, эндемичная,
для Сихотэ-Алиня, приурочена в основном к каменным россыпям
верхних частей склонов и вершин среднегорий и более распро-
странена в Южном и Среднем Сихотэ-Алине, где, очевидно, и
возникла на окраине лесов с участием древовидного предста-
вителя рода во время регрессии лесов. Становление ее пред-
ставляется совершенно аналогичным вычленению кедрового
стланика и других стелющихся видов хвойных, например гор-
ных стланиковых сосен Европы, и представляет собой уцелев-
шее звено экологического ряда видов, существовавшего в близ-
ких современным условиях относительно широких амплитуд
температуры и влажности. Ксероморфность, стойкость к суро-
вым зимам и дневному перегреву летом — даже на северо-вос-
точных склонах поверхность каменных россыпей среди зарос
лей ми1кробиоты нагревается до 60° и более — позволяют от-
нести время ее наибольшего распространения к периодам
превращения окраинных морей в замкнутые водоемы, регрес-
сии океана, сухости климата, которая вследствие особенностей
атмосферной циркуляции становилась наибольшей на юге и
юго-востоке горной страны. Микробиота перекрестнопарная,
если учесть закономерности ее распространения и расселения,
91
появилась не позже олигоцена и должна быть отнесена к тре-
тичным видам. Она моложе представителей родственных родов,
например биоты восточной, а также можжевельников, но рас-
пространилась в Сихотэ-Алине скорее всего до появления здесь
кедрового стланика. Микробиота однодомна. Сообщения о ее
двудомности (Шкутко, 1970:244), как и о том, что она является
ювенильной формой Biota orientalis Endl. (Rehder, 1949:55),
не подтвердились нашими исследованиями.
Фитоценозы с микробиотой, преимущественно с ее домини-
рованием и содоминированнем, занимают в Сихотэ-Алине ты-
сячи гектаров. Многочисленные ассоциации с микробиотой —
нами описана 21 такая ассоциация — генетически тесно связа-
ны и составляют 3 группы: 1) микробиотовые заросли вершин
и водоразделов с незначительным участием других видов, ха-
рактерные Южному и отчасти Среднему Сихотэ-Алиню; 2) кус-
тарниково-микробиотовые заросли, уже со Среднего Сихотэ-
Алиня становящиеся преобладающими ассоциациями с микро-
биотой; 3) ассоциации каменноберезовых и таежных лесов и их
дериватов с микробиотой в подлеске, встречающиеся более
равномерно, но занимающие меньшие площади.
Монодоминантные сообщества> микробиоты представляют
сплошные стелющиеся по каменным россыпям массивы пло-
щадью— на хр. Ливадийском — до 1—3 га (рис. 5). Водой
микробиота обеспечивается здесь за счет конденсации влаги в
камнях, а почвы наиболее молоды и проходят первые этапы
формирования. По химизму они близки к бурым горно-лесным
неоподзоленным почвам, «подвешены и аэрируются сверху и
снизу» (Богатырев, 1954:39). В условиях периодической летней
и погодичной ксеричности сообщества микробиоты являются
мощнейшими почвообразователями и превосходят по этим ка-
чествам другие кустарники и кустарнички, лишайники и травы.
В заселении находящегося необычно низко гольцового пояса и
его фрагментов им предшествуют здесь только лишайники.
Другие растения идут за микробиотой и поселяются под ее по-
логом. Однако уже с центральной части Среднего Сихотэ-Али-
ня условия горных каменных россыпей оказываются более
благоприятными для кедрового стланика: перепады темпера-
туры и влажности менее резки.
Россыпи с отдельными участками микробиоты, рябинника
рябинолистного (в западинах), малины Комарова, жимолостей
Максимовича и съедобной только на хр. Ливадийском занима-
ют около 1000 га преимущественно крутых южных и западных
склонов и могут быть окружены каменноберезово-елово-пихто-
выми лесами. На контакте с россыпями в подлеске этих лесов
присутствует узкий пояс микробиоты, растущей в сторону рос-
сыпей и укореняющейся на них. Под пологом леса вдоль пояса
микробиоты иногда сохраняются синузии кедрового стланика,
семяносящего в прогалинах (хр. Ливадийский), остатки кото-
92
Рис. 5. В субальпийском поясе Южного Сихотэ-Алиня формация микробиоты
абсолютно преобладает. Хр. Ливадийский, Шкотовский район
рого встречены и среди россыпей. В целом микробиота здесь
устойчивей, а россыпи склонов образовались на гарях ее и
стланика. Только у вершин возможно первичное надвигание
микробиоты на горные тундры и пустыни.
Наряду с заманихой высокой, сиренью Вольфа, жимоло-
стью съедобной микробиота входит в подлесок мелкотравно-
зеленомошного пихтово-елового леса, в том числе и на север-
ных склонах. Состояние и семяношение микробиоты здесь опре-
деляются освещенностью: при сомкнутости древесного яруса
0,7 и выше хвоя ее живет 2—3 года (на россыпях обычно свы-
ше 10 лет), семяношение отсутствует, хотя годичные приросты
ветвей достигают 3—7 см. На пологих участках вершин отме-
чены и каменноберезняки-микробиотники (гора Лысая Спас-
ская), где сомкнутость микробиоты 0,7—1,0, а ветви ее при-
поднимаются на высоту 1 м и успешно семяносят.
Ценодинамизм микробиотников и сообществ с ними весьма
прост: микробиота надвигается на реликтовые холодные камен-
ные пустыни и горную лишайниковую тундру, формируя поч-
ву; под пологом микробиоты селятся зеленые мхи, брусника,
бадан тихоокеанский, другие кустарнички; кустарники (родо-
дендроны, багульники, жимолости, сирень Вольфа и т. д.), тра-
вы; в сообщества с доминированием микробиоты внедряются
каменная береза, пихта белокорая, ель аянская, рябина амур-
ская, иногда кедр корейский и дуб монгольский; формируются
каменноберезняки с микробиотой, в которых возрастает роль
темнохвойных пород, и, наконец, таежные насаждения со
сплошным слоем дифференцированной почвы и постепенно от-
мирающими синузиями микробиоты, чьи масса ветвей и семя-
ношение в 10—40 раз меньше, чем на открытых пространствах.
А1икробиота может снова появиться только после сильных по-
жаров с россыпей и гарей или за счет сохранивших всхожесть
семян. Здесь вместе с ней в большей или меньшей степени вос-
станавливаются и идущие за ней виды, формирование пока
вновь низкопроизводительной тайги ускоряется, иногда возни-
кают лиственничники с микробиотой в подлеске (Васильев, Ку-
ренцова, 1960). Правомерен вывод, что в современном климате
существует тенденция не только к сокращению микробиотни
ков, но и — в исторической перспективе — к исчезновению мик-
робиоты вообще.
Созревающие и высыпающиеся в сентябре года цветения
семена микробиоты не имеют распространителей и остаются в
пределах ее зарослей, прорастая только после пожаров. Запа-
сы семян в гумусовом слое могут быть очень значительными:
в монодоминантных синузиях ежегодно на 1 м2 образуется от
150 до 700 выполненных семян (высота над ур. м. 900—1200 м)
и от 50 до 700 (высота 500—700 м). После уничтожения за-
рослей слабыми пожарами (остаются отмершие ветви диамет-
ром 0,3 см и более) на западных и юго-западных склонах
94
появляется до 300 экз. возобновления на 100 м2, а на значи-
тельных площадях вновь обнажившихся россыпей — до 10—
12 тыс. экз. на 1 га. Микробиота здесь быстро смыкается, но
даже слабое семяношение ее начинается в возрасте 14—17 лет.
Повторные пожары вызывают новые волны возобновления,
постепенно истощая запас семян: на горе Лысой Спасской пос-
ле второго пожара появилось до 6 тыс. экз. возобновления,
после третьего—1,52 тыс. на 1 га. Устойчивые пожары, после
которых* сохраняются только ветви с диаметром 5 см, форми-
руют слабо восстанавливающиеся гари — имеется 0,3—1,2 тыс.
экз. возобновления микробиоты на 1 га — гумус и семена в
нем в основном выгорают. На южных склонах, круче 35°, гари
восстанавливаются слабо, но остаются участки не уничтожен-
ных старых зарослей. Интенсивность пожаров определяется
конкретной влажностью микробиотников и гумуса, поэтому
микробиота может сохраняться или лучше возобновляться у
стен леса. Малочисленно возобновление микробиоты и на га-
рях лесов с ее участием в подлеске. В целом вид успешно
возобновляется лишь на 71 своих современных гарей: для
заселения вторичных каменных россыпей ему в основном пот-
ребуются столетия — микробиота распространяет свои семена
только в процессе роста.
В условиях «жесткой погоды» гольцов (Сочава, 1944)
микробиота создает микроклимат, в котором возможно почво-
образование. Как и кедровый стланик (Гроссет, 1959; и др.),
она обладает биологическими приспособлениями к экстремаль-
ным условиям, в том числе и осенним пониканием ветвей, ло-
жащихся на россыпи и перезимовывающих под снегом.
Кедровый стланик не только образует особый пояс
стелющихся темнохвойных лесов (Колесников, 1969), но и
вкраплен в дубовую и таежные формации. На нижнем пределе
пояса стволы его достигают 11 м длины, 4—5 м высоты и при
густоте до 4 тыс. на 1 га запас древесины в зарослях около
100 м3. Как и микробиотники, кедровостланиковые заросли, фор-
мируя почву и особый микроклимат, создают условия для бо-
лее сложных сообществ, а потом и биогеоценозов с преоблада-
нием деревьев. Кедровостланиковые группировки и формацион-
ные реликты в пространстве и времени образуют ряды фито-
ценозов, все более мощных на последующих этапах почвооб-
разования. На современном этапе формация кедрового стлани-
ка, видимо, не соответствует климату и, как и ряд других фор-
маций Сихотэ-Алиня и Восточно-Маньчжурских гор, утрачивает
стабильность.
Ассоциации с кедровником относятся к группам: 1) моно-
доминантных стланиковых лесов, 2) субальпийских кустарни-
ковых зарослей (содоминирует несколько видов), 3) подголь-
цовых каменноберезовых и 1) елово-пихтовых лесов и их дери-
ватов (в том числе дубняки) с кедровником в подлеске.
95
Ассоциации двух первых групп подробно описаны в Среднем
Сихотэ-Алине: Б. П. Колесников (1969) выделил здесь 12 ас-
социаций кедровниковых лесов. Группировки кедрового стла-
ника с можжевельником сибирским и микробиотой — субаль-
пийские кустарниковые заросли — встречены и нами. Выше в
настоящей статье приведена характеристика ранее не описы-
вавшейся группировки кедровника с можжевельником даур-
ским. С продвижением к югу состав всех ярусов зарослей
стланика становится более разнообразным за счет присоеди-
нения эндемичных высокогорных видов и растений иных
высотных поясов. Преобладание от. кедровника может перехо-
дить к сирени Вольфа, ольхе маньчжурской, жимолостям,
спиреям. В микрогруппировках горного луга доминирование
от бореальных трав (вейники) переходит к шнрокотравью,
свойственному лесам из кедра корейского. В то же время даже
на юге Сихотэ-Алиня еще сохранились достаточно мощные
синузии кедрового стланика. Однако, как уже говорилось, сов-
ременный климат Среднего Сихотэ-Алиня благоприятней для
микробиоты, и синузии стланика фрагментарны.
Синузии кедрового стланика под пологом каменноберезня-
ков и их вариантов и особенно подгольцовой тайги являются
угасающими, хотя после пожаров восстановление кедровостла-
никовых лесов за счет орнигохории возможно. В этих группах
ассоциаций позиции кедровника несколько прочней в каменно-
березняках и лиственничниках.
К интереснейшим ассоциациям с кедровником в Приморье
относятся группировки рододендроновых, леспедецевых и дру-
гих дубняков с его синузиями, с брусникой и другими субаль-
пийскими кустарниками и кустарничками, а также сгланиково-
дубовые леса, в которых высота стланика (до 5 м) и дуба мон-
гольского (4—6 м) практически равны. Нахождение осколка
кедровостланиковой формации и массы субальпийских видов
на небольших абсолютных высотах позволяет предположить,
что в конце плейстоцена — начале голоцена на востоке Сихотэ-
Алиня в гораздо большей, чем на западе, степени была развита
высокогорная растительность. Возможно также, чго опускание
восточного склона было сравнительно недавним.
Возобновление стланика в дубняке с его участием — высота
над ур м. 30—70 м — происходит из спрятанных птицами семян.
Ba 1 га фитоценоза насчитывается 3000 экз. всходов, самосева
и подроста кедровника. Взрослые растения на южном склоне
усыхают в связи с летним перегревом. Климатогенная смена
стланика дубом реализуется здесь через пирогенные, что для
Приморья типично.
Обсуждение результатов исследования. Формация можже-
вельника твердого могла существовать в районах на переходе
к аридному климату и в горной местности аридной зоны. Поэ-
тому закономерно нахождение ее фрагментов именно на древ-
96
нем рубеже особенно сухой части бассейна Амура (Николь-
ская, 1972), проходившем по Восточно-Маньчжурским и Буре-
инским горам и, видимо, в конце плейстоцена, смещавшемся
к восточному склону Сихотэ-Алиня. В поясе ксерофитизованных
формаций можжевеловые леса — первоначально стелющиеся —
могли выступать почвообразователями. Ландшафты с можже-
веловыми и можжевелово-широколиственными насаждениями
тогда, очевидно, были характерными, их сменяли на более влаж-
ных почвах настоящие сосняки. Однако особенности возобнов-
ления вида в современный период заставляют предположить,
что изменение климата в голоцене шло в сторону выравнивания
влажности: в стабильно сухом климате можжевельник твердый
возобновляется очень слабо и не может образовать сколько-ни-
будь густых рощ. В современном климате имеющаяся густота
можжевеловых рощ на скалах не сохранится вследствие того,
что возобновление не обеспечивает достаточного пополнения
древостоев. В этом и заключена причина столь явного старения
и разрушения последних. Поэтому в прошлом, несомненно,
имели место особенно дождливые годы, в которые и прораста-
ли семена вида. В настоящее время периодичность семяноше-
ния и ритмы влажных лет не совпадают.
Интереснейшим явлением можно считать изменение можже-
вельником твердым жизненной формы в изолированных по-
пуляциях побережья, потому что именно в них обусловленные
специфичностью естественного отбора популяционные волны
могут благоприятствовать накоплению и дальнейшему размно-
жению мутантных форм. Поэтому возможно, что внутривидо-
вые отдельности, описанные нами как модификационные видо-
изменения в условиях особых почв и микроклимата, на самом
деле наследственны и должны рассматриваться как экотип.
Тогда можно предположить, что и можжевельник прибрежный
Juniperus conferta Pari- возник из подобных экотипов. Этот
всеми признаваемый хорошим видом можжевельник совершен-
но сходен с плоской модификацией твердого по жизненной фор-
ме, густоте и особенностям хвои и шишкоягод.
Современное распределение хвойных кустарников и других
растений, связанных с субальпийским поясом и бореальными
формациями, позволяет сделать вывод, что в периоды похоло-
дания, синхронные последним оледенениям антропогена и рег-
рессии океана, территория края в ее нынешних границах была
занята поясом субальпийских кустарников и тайгой, фрагменты
которых еще встречаются в различных формациях от уровня
моря. Изменение климата вызвало прогрессивность верхних
границ растительности в целом и ее вертикальных поясов в
частности. Надвигание субальпийских кустарников на гольцы
и лесов на стланиковые заросли продолжается, а скорость
этого процесса определяется темпом почвообразования. Мик-
робиота и кедровник — сугубо пионерные растения. Их станов-
7 Заказ 488 97
ление, как и появление можжевельников сибирского и даур’
ского, мыслимо в олигоценовую и более ранние эпохи в
верхнем поясе хвойных лесов при обусловленной изменением
климата регрессии и деградации лесной растительности. Ви-
димо, большинство горных почв Сихотэ-Алиня прошло первые
этапы формирования в их зарослях, а частично и в сообщест-
вах можжевельников сибирского и даурского. Стелющиеся
можжевельники и ныне имеют вспомогательное значение в кед-
ровостланиковых и некоторых других пионерных ценозах.
Уничтожение хвойных кустарников и особенно^ микробиоты
при интенсивных пожарах отбрасывают почвообразовательный
процесс по крайней мере на столетия.
Являясь доминантами самостоятельных пионерных форма-
ций, микробиота, кедровый стланик и можжевельник сибир-
ский постепенно сокращают свои площади: на протяжении
всего голоцена их ареалы регрессировали по естественно-исто-
рическим причинам, уступая место тайге и кедрово-широко-
лиственному лесу, но в настоящее время их крупные массивы
гибнут в результате проникновения палов и в субальпийский
пояс. Антропогенные смены дают двойственный результат: во-
первых, убыстряют выпадение хвойных кустарников из слож-
ных фитоценозов с ними, во-вторых, они частично консервируют
стелющиеся заросли. В любом случае продуктивность биогео-
ценозов снижается. В естественно-исторической перспективе
микробиоту, стланик, можжевельник сибирский? в Приморье
можно рассматривать как исчезающие. Только можжевельники
твердый и даурский нашли новые экологические оптимумы —
первый в сосновой формации, второй — на скалах и песках по-
бережья.
Хотя ветви можжевельника сибирского и стелющихся моди-
фикаций твердого укореняются, вегетативное размножение у
них существенной роли не играет. Вследствие пассивной миг-
рации можжевельник даурский оказался в несвойственных для
него условиях и почти не размножается семенами. Прибрежные
заросли этого красивого быстрорастущего кустарника могут
полностью уничтожаться палами. Можжевельник сибирский —
один из древнейших элементов флоры восточной окраины
Азии — в настоящее время поглощается кедровым стлаником
и субальпийскими высокоствольными лесами. Мероприятия по
сохранению стелющихся хвойных предполагаются в общей си-
стеме мер по защите высокогорной растительности и ценозов
береговых скал. В среднем и южном Приморье появилось уже
много мест, лишившихся главных пионерных растений. Мож-
жевельники и микробиоту можно вернуть на них только посад-
кой по периферии и в нижних неперегревающихся частях как
россыпей, так и обрывов с минимально выраженными почва-
ми. Особую задачу должно составить возвращение стелющих-
ся можжевельников на прибрежные скалы. Большой природо-
96
охранительный эффект даст введение хвойных кустарников в де
коративные посадки. Это особенно важно для можжевельника
твердого, который, видимо, утратил распространителя, перено-
сившего семена вверх по склонам при трансгрессии v моря.
К тому же на скалах преобладают мужские особи. Некоторый
опыт введения вида в декоративные посадки имеется в Погра-
ничном, Партизанском и Лазовском районах. Для сохранения
видов необходимо как можно шире культивировать хвойные
кустарники.
ЛИТЕРАТУРА
Богатырев К. П. Почвы зарослей микробиоты. — Сообщ. ДВФ АН СССР,
в. 6. Владивосток, 1954, с. 37—42.
Васильев Н. Г., Куренцова Г. Э. Поясность растительного покрова на горе
Ко в Среднем Сихотэ-Алине.— Комаровские чтения ДВФ СО АН
СССР, в. 8. Владивосток, 1960. с. 21—40.
Воробьев Д. П. Растительный покров Южного Сихотэ-Алиня и дикорасту-
щие плодово-ягодные растения в нем.— Труды ДВФ АН СССР, сер
бот. Т. 1. М.—Л., Изд-во АН СССР, 1935, с. 287—374.
Гроссет Г. Э. Кедровый стланик.— Матер, к познанию флоры и фауны
СССР, издаваемые МОИП, нов. сер., отд. ботанический, в. 12(20),
1959, 140 с.
Ильченко Т. П. Об эволюции кариотипов хвойных.— Тезисы докладов XII
Международного ботанического конгресса. Л., «Наука», 1975, с. 25.
Кабанов Н. Е. Типы растительности южной оконечности Сихотэ-Алиня.—
Труды ДВФ АН СССР, сер. бот. Т. 2, М—Л., Изд-во АН СССР,
1937, с. 273—332.
Колесников Б. П. Высокогорная растительность Среднего Сихотэ-Алиня. Вла-
дивосток, Дальневост. кн. изд-во, 1969, 106 с.
Кондратюк Е. Н. Дикорастущие хвойные Украины. Киев, Изд-во АН УССР,
1960 (на укр. яз.), 120 с.
Куренцова Г. Э. Растительность Приморского края. Владивосток, Дальне-
вост. кн. изд-во, 1968а, 192 с.
Куренцова Г. Э. Реликтовые растения Приморья. Л., «Наука», 19686, 72 с.
Лукашкин А. С. Озеро Дзиньбоху и его бассейн.— Вести. Маньчжурии,
1933, №7, с. 42—54.
Никольская В. В. Морфоскульптзра бассейна Амура. М., «Наука», 1972,
295 с.
Пономаренко В. М. О динамике верхней границы леса в горах Южного
Сихотэ-Алиня.—Изв. СО АН СССР, 1961, №5, с. 100—109.
Сочава В. Б. Причины безлесья ольцов Восточной Сибири и в Приамурье.
«Природа», 1944, №2, с. 63—65.
Шишкин И. К. Microbiota decussata Кош. как элемент растительного покро-
ва Уссурийского края.— Труды ДВФ АН СССР, сер. бот. Т. 1. Л.—М.,
Изд-во АН СССР, 1935, с. 227—243.
Шкутко Н. В. Хвойные экзоты Белоруссии и их хозяйственное значение.
Минск, «Наука и техника», 1970, с. 244.
Шмидт И. Н., Урусов В. М. Семяношение можжевельника твердого в При-
морском крае.— Половая репродукция хвойных. II (материалы 1-го
Всесоюзного симпозиума). Новосибирск, «Наука», 1973.
Rehder A. Manual of cultivated trees and shrubs hardy in North America.
Exclusive the subtropical and warmer temperate regions. N. Y., The Mac-
millan company, 1949, p. 55.
7'
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
МЕЛКОПЛОДНИК ОЛЬХОЛИСТНЫЙ
MICROMELES ALNIFOLIA (Sieb. et Zucc.) Koehne
(распространение, возобновление, использование, охрана)
В. к. ВАСИЛЮК
Формирование растительности Приморья, его разнообразной
флоры вообще и дендрологической в особенности неотделимо
от геологической истории края. Современное положение на
окраине крупнейшего материка и крупнейшего океана и гори-
стый рельеф формируют массу оригинальных микроклимати-
ческих участков и приводят к чрезвычайной пестроте экологи-
ческих условий. Перечисленными факторами и объясняется
концентрация большого флористического разнообразия в дан-
ном регионе.
Только на территории Приморья и Приамурья насчитывает-
ся 322 вида с деревенеющими стеблями. По данным исследо-
вателей-ботаников, 32 рода, произрастающие на Дальнем
Востоке, нигде больше в СССР не имеют своих представителей
(Воробьев, 1958).
В их число входит род Micromeles Decne. — мелкоплодник,
сем. Rosaceae Juss.
Род Micromeles, по данным одних исследователей, включает
10 видов (Пояркова, 1939), по данным других—15, распро-
страненных на Дальнем Востоке и в Юго-Восточной Азии (Ко-
новалов, 1954; Качалов, 1970).
На территории советского Дальнего Востока мелкоплодник
представлен видом Micromeles alnifolia (Sieb et Zucc.)
Koehne — мелкоплодник ольхолистный.
Систематическое положение рода до настоящего времени
четко не определено и является объектом споров системати-
ков. Часть исследователей (Rheder, 1949; Колесников, 1974;
Ворошилов, 1966) считают его принадлежащим к роду Sor-
bus L. Другие (Воробьев, 1958, 1968; Пояркова, 1939; Конова-
лов, 1954; Качалов, 1970; Комаров, Клобукова-Алисова, 1932;
Строгий, 1934; Цымек, 1950; Толмачев, 1956; Усенко, 1969;
Лучник, 1970; Плотникова, 1971; Василюк, 1975) признают его
самостоятелвным в ранге рода Micromeles.
В настоящей работе мы не ставили целью сделать критиче-
ский обзор рода и, придерживаясь мнения большинства иссле-
дователей, принимаем описываемое растение как Micromeles
alnifolia (Sieb. et Zucc.) Koehne.
100
Мелкоплодник — одна из наиболее южных пород для При-
морья. Северная граница его ареала проходит в бассейне
э. Киевка, в Чугуевском и Анучинском районах, по ключу Бе-
эезовому, выше с. Таежка и Пихтовое. Растет на островах
<унашир и Итуруп. Общее распространение: Япония, п-ов Ко-
рея, КНР (Воробьев, 1968).
Мелкоплодник ольхолистный — дерево первой величины, до-
стигающее 20 м в высоту и 28 см в диаметре. Ствол прямой,
стройный, покрыт светло-серой, реже коричнево- или темно-
серой гладкой корой, хорошо очищенный от сучьев. Крона вы-
сокоприкрепленная, овальной, округлой или рваной формы,
достаточно плотная.
Листья от широкоовальных до округлых с клиновидным или
округленным основанием, равнобокие, неравномерно туподвоя-
козазубренные. 5—10 см длины, 5—8 см ширины, сверху изум-
рудно-зеленые со слабым опушением по жилкам, снизу более
светлые с рельефным жилкованием.
Цветки белые, до 1 см в диаметре, собраны в многоцветко-
вые щитковидные соцветия. Очень редко — одиночные. Цветет
в конце мая — начале июня. В отдельные годы продолжитель-
ность цветения достигает 15 дней.
Плоды — яблочки, округлой, чаще продолговатой формы,
6—7 мм длины и 8—12 мм ширины с 2—4 семенами, кирпично-
красные или красно-оранжевые с голубоватым налетом, съе-
добные, приятно кисловатые.
Широко используются местным населением для приготовле-
ния компотов. Содержат сахар, каротин, яблочную и лимонную
кислоты и дубильные вещества. Виноделами-любителями ис-
пользуются в виде добавки при изготовлении вин (особенно
из актинидий). Созревают в сентябре и длительное время ос-
таются на деревьях. Охотно поедаются птицами, особенно ряб-
чиками.
Осенняя окраска — в конце сентября, листопад — в третьей
декаде октября.
Древесина красновато-бурая с лимонно-желтой заболонью,
тяжелая, твердая, крепкая, ровного строения, не коробится,
пригодна для токарных изделий. Ранее использовалась для из-
готовления чертежных принадлежностей. В настоящее время,
ввиду незначительных запасов, в промышленности не приме-
няется. Лесозаготовители используют стволы молодых растений
при работе с бревнами и кряжами как ваги или так называе-
мые «слеги», или «стежки». Особую прочность им придают
путем поверхностного обжига в кострах.
Теневыносливая порода, предпочитает свежие, хорошо дре-
нированные, богатые гумусом почвы, но нередко встречается и
на маломощных, обедненных гумусом. На почвах, сформиро-
ванных тяжелыми суглинками, растет хуже. Встречается еди-
нично или небольшими группами как примесь в чернопихтово-
101
широколиственных, кедрово-широколиственных или широко-
лиственных лесах, массивов не образует. Встречается и в корен-
ных дубняках. Фитоценотические связи с какой-либо формаци-
ей, по нашему мнению, отсутствуют. Это дает основание , счи-
тать мелкоплодник ольхолистный реликтовым видом, развив-
шимся в третичных листопадных лесах.
Специальных работ, освещающих участие мелкоплодника в
сложении фитоценозов, его естественное возобновление, найти
не удалось; для выяснения этих вопросов было предпринято
обследование лесной площади Ботанического сада ДВНЦ в
пригородном районе г. Владивостока. Почти вся его террито-
рия—178 га — покрыта смешанным лесом южноуссурийского
типа, представленным двумя формациями: чернопихтово-широ-
колиственной с кедром корейским на востоке и дубовой — на
западе.
Лес полностью сохранился и находится в удовлетворитель-
ном состоянии. С 1949 года установлен усиленный режим ох-
раны, тем не менее ни пирогенный, ни другие отрицательные
антропогенные факторы исключить полностью не удалось.
Вся территория разделена на 12 кварталов. Детальному
обследованию подверглись три — 6, 8 и II, представляющие
значительный интерес. Суммарная площадь их 53 га.
Квартал 6. Площадь 14 га.
Преобладают пихтарники IV кл. возраста, III кл. бонитета.
Почвы бурые, лесные, глеевато-оподзоленные, средней мощно-
сти, скелетные, увлажненные. Северная, северо-западная экс-
позиция, крутизна до 15°
Распределение покрытой лесом площади и запасов насаж-
дений по преобладающим породам и группам возраста: пихта
цельнолистная — 230, дуб монгольский— НО, липа амурская —
100, орех маньчжурский — S0 м3 в пересчете на 1 га.
Пробная площадь № 1 — 100 м2 Взрослое одно дерево мел-
коплодника, высота 12 м, диаметр на высоте груди — 16 см.
Учтено подроста:
высотой до 1,5 м — 1,
высотой до 2,0 м — 2.
В пересчете на 1 га приходится 100 взрослых экземпляров и
300 экземпляров подроста мелкоплодника.
Квартал 8. Площадь 20 га.
Расположен в центральной части сада, представляет наи’
больший интерес во флористическом отношении и поэтому
наиболее детально изучен. Преобладают пихтарники VI—
VII кл. возраста, II, III кл. бонитета. Почвы бурые, лесные,
слабо глеевато-оподзоленные, влажноватые, средней мощности.
Склоны северной экспозиции.
Распределение по преобладающим породам: пихта цельно-
листная— 190, дуб монгольский—130, липа амурская—135,
береза — ПО м3 в перечете на 1 га.
102
Пробная площадь №2 —100 м2. Взрослых экземпляров
мелкоплодника на площади не учтено. Подроста в пересчете на
1 га оказалось:
в возрасте 6—10 лет—100 экз. жизнеспособного,
в возрасте 11 —15 лет— 100 экз. жизнеспособного и 50 экз.
поврежденного,
в возрасте 16—20 лет — 80 экз. жизнеспособного, 20 экз.
угнетенного и 50 поврежденного до стадии прекращения роста.
Всего на гектаре жизнеспособного подроста мелкоплодни-
ка— 280 экз., угнетенного — 20, сильно поврежденного—100.
Квартал 11. Площадь 19 га.
Преобладают чернопихгарники VII кл. возраста, III кл.
бонитета и липняки IV кл. возраста, III IV кл. бонитета. Поч-
вы бурые, лесные, глеевато-оподзоленные, сырые, средней мощ-
ности. Склоны северной, северо-западной экспозиций.
Распределение по преобладающим породам: пихта цельно-
листная— 245, кедр корейский — 200, липа амурская—115,
дуб монгольский—ПО м3 в пересчете на 1 га.
Пробная площадь №3—100 м2. Крупных деревьев мелко-
плодника на площади не учтено. Подрост высотой:
до 1,5—2,0 м на 1 га — 200 экз.,
до 2,0—2,5 м на 1 га — 200 экз.
В квартале на 1 га приходится 400 экз. жизнеспособного
подроста.
Как видно, возобновление мелкоплодника на обследован-
ной территории вполне удовлетворительное. Однако эти дан-
ные справедливы для территории с повышенным режимом ох-
раны.
Мелкоплодник очень устойчив к неблагоприятным для про-
израстания условиям крупного промышленного города.
За несколькими деревьями, растущими в селитебной части,
автор проводил наблюдения на протяжении двух десятилетий.
Первый экземпляр — на Океанском проспекте г. Владивос-
тока; второй — на мысе Чуркина; третий — на станции Сана-
торная. Судя по всему, эти деревья — уцелевшие представите-
ли некогда росших здесь лесов — крупные, хорошо развитые
растения. Все экземпляры находятся в условиях уплотнения
почвы и запыленности кроны, однако у них хороший рост,
нормальное развитие и через 3—4 года обильное цветение и
плодоношение. Следов угнетения не обнаружено.
Таким образом, мелкоплодник ольхолистный следует реко-
мендовать для широкого использования в озеленении населен-
ных мест южного Приморья, южного Сахалина и части остро-
вов южных Курил. Он может быть использован в качестве со-
литера и для создания куртин и мелких декоративных групп.
По нашим наблюдениям, это лучшее парковое дерево, очень
эффектное в течение всего вегетационного периода. Красив
103
весной распускающимися бронзовыми листочками на фоне
нежно-розовых чешуи, в пору цветения — щитками белых цве-
тов, в пору созревания плодов — сизовато-красными яблочка-
ми. Очень эффектен осенью, когда листья окрашены в золотис-
то-бронзовый или бронзово-багряный цвета.
Может быть использован как подвой для семечковых в пло-
доводстве и как исходный материал для селекции.
Хорошая мелиоративная порода. Следует создавать посад-
ки для использования технических свойств древесины. Размно-
жать лучше всего свежесобранными семенами осенью. Сле-
дующей весной появляются хорошие дружные всходы. При
весеннем посеве требуется длительная стратификация при пе-
ременных температурах в течение 2—4 месяцев. Даже при та-
кой обработке всходы получаются изреженные. Неплохо раз-
множается путем черенкования и возобновляется пневой по-
рослью.
Для охраны и воспроизводства этого ценного вида следует
установить твердый лицензионный режим.
ЛИТЕРАТУРА
Василюк В. К. Ассортимент деревьев и кустарников для озеленения г Вла-
дивостока.— Озеленение городов Дальнего Востока. Владивосток, 1975,
с. 114—120.
Воробьев Д. П. Определитель деревьев и кустарников Приморья и Приаму-
рья. Благовещенск, Амурское кн. изд-во, 1958, 184 с.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 277 с.
Ворошилов В. Н. Флора советского Дальнего Востока. М., «Наука», 1966.
Древесные растения Главного ботанического сада АН СССР. М., «Наука»,
1975, 577 с.
Качалов А. А. Деревья и кустарники. Справочник. М., «Лесная промышлен-
ность», 1970. 408 с.
Колесников А. И. Декоративная дендрология. М., «Лесная промышлен-
ность», 1974. 704 с.
Комаров В. Л., Клобукова-Алисова Е. Н. Определитель растений Дальне-
восточного края. Т. 1—2. Л., Изд-во АН СССР, 1931—1932, 1175 с.
Коновалов И. Н. Мелкоплодник — Micromeles Decne. Деревья и кустарники
СССР. М—Л., 1954, т. III, с. 487—488.
Лучник 3. И. Интродукция деревьев и кустарников в Алтайском крае. М.,
«Колос», 1970 656 с.
Плотникова Л С. Интродукция древесных растений Китайско-Японской
флористической подобласти в Москве. М., «Наука», 1971. 136 с.
Пояркова А. И. Мелкоплодник — Micromeles Decne.— Флора СССР. М.—Л.,
1939, т. IX, с. 406—407.
Строгий А. А. Деревья и кустарники Дальнего Востока, их лесоводственные
свойства, использование и техническое применение. Москва — Хаба-
ровск, Дальнезост. кн. изд-во, 1934, 235 с.
Толмачев А. И. Деревья, кустарники и деревянистые лианы острова Саха-
лина. Краткий определитель. М. — Л., Изд-во АН СССР, 1956, 172 с.
Усенко Н. В. Деревья, кустарники и лианы Дальнего Востока. Хабаровск,
кн. изд-во. 1969, 416 с.
Цымек А. А. Главнейшие лиственные породы Дальнего Востока. Хабаровск,
Дальгиз 1950 197 с
Rheder A. Manual of cultivated trees and shrubs hardy in North America.
N —Y. Macmillan C°. 1949, 996 p
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ОСОБЕННОСТИ СЕЗОННОЙ РИТМИКИ РАЗВИТИЯ
НЕКОТОРЫХ РЕЛИКТОВЫХ ДРЕВЕСНЫХ ЛИАН
ФЛОРЫ ПРИМОРЬЯ
Л. н. слизик
Региональные исследования по выявлению редких и исче-
зающих видов лиан флоры Приморья представляют собой пер-
вый этап познания этой интереснейшей группы растений. Ис-
следованиями дальневосточных ботаников уточнены ареалы и
состояние этих видов на юге Дальнего Востока (Воробьев,
1968; Куренцова, 1968; Валова, Куренцова, 1974; Красная кни-
га, 1975). Изучение их биологии, особенностей сезонного раз-
вития и жизненных циклов составляет следующий этап иссле-
дования дальневосточных видов деревянистых лиан.
Сезонная ритмика, механизмы семенного и вегетативного
размножения, сложившиеся в определенных естественно исто-
рических условиях, в известной мере отражают особенности
генезиса вида. Так, по определению К. А. Сергеевой (1971),
«годичная морфофизиологическая периодичность древесных ра-
стений является наследственно закрепленным свойством, фор-
мировавшимся в процессе эволюционной адаптации к опреде-
ленному климату». ।
Наблюдения за сезонной ритмикой редких и исчезающих
видов помогают выявить особенности и различия их поведения
в разных условиях, найти критические точки жизненных цик-
лов в данном природном регионе и на этой основе четко опре-
делить комплекс охранных мер. В этом аспекте изучались и
сравнивались два вида древесных лиан — аристолохия маньч-
журская и пуэрария волосистая — в природных местообитани-
ях (юг Приморского края) и в первичной культуре (окр. Вла-
дивостока— Ботанический сад ДВНЦ АН СССР). Определе-
ние и моделирование условий, необходимых для нормального
завершения сезонных циклов изучаемых видов, уточняет фи-
логенез этих видов и позволяет прогнозировать пределы и пу-
ти успешной интродукции этих растений.
1. Аристолохия маньчжурская, кирказон
маньчжурский — Aristolochia manshuriensis Кот., Hocqu-
artia manchuriensis (Кот.) Nakai.
Типичная деревянистая лиана, обитающая в отдельных ло-
кусах на юге Приморья,— в широколиственных лесах северной
части Хасанского района. Известные местонахождения в пре-
105
делах Приморья являются островными относительно основного
ареала вида: четкое выделение из окружающих растительных
группировок, приуроченность к определенным микроклиматиче-
ским условиям, монодоминантный характер зарослей свойствен-
ны этому виду в данном регионе. Основной ареал — Северная
Корея, северо-восток Китая (Воробьев, 1968).
Листья аристолохин маньчжурской служат специфическим
субстратом для одного из реликтовых видов насекомых (Ку-
р’енцова, 1968) .Таким образом, и ценотические связи вида свое-
образны и свидетельствуют о его реликтовости.
Аристолохия маньчжурская растет по распадкам, подножи-
ям склонов, встречаясь отдельными плотными пятнами по кра-
ям опушек или на лесных полянах. Наиболее крупные и силь-
ные заросли аристолохин маньчжурской известны из окр.
с. Горное (Хасанский район). Здесь аристолохия растет у под-
ножия северного склона вдоль ручья и представлена крупными
плодоносящими экземплярами, поднимающимися до вершин
деревьев — более 10—15 метров. Это лесной вид, избегающий
склонов с ярким солнечным освещением и нагреванием почвы;
в условиях коллекции значительно лучше себя чувствует буду-
чи посаженным в тени — дает больший прирост, образует более
крупные листья и более обильные цветки и плоды.
По нашим наблюдениям, аристолохия маньчжурская начи-
нает свою вегетацию в конце апреля — начале мая: набухание
почек наблюдается в течение II и III декад апреля; разверты-
вание листьев отмечалось с 20.IV по 5—8.V. Почки аристолохин
маньчжурской интересны своими особенностями — они густо
покрыты серебристыми волосками, расположены сериально (по
2—3) и полупогружены в ткани побега; в зимнем состоянии
довольно мелки.
Развитие листьев идет быстро, и к 15.V листья достигают
15 см в длину; завершение облиствения — не позднее 20.V.
Побеги текущего года 2 типов: укороченные, на которых раз-
виваются цветки, и ростовые, обвивающие опору. Кора молодых
побегов ярко-зеленая и сохраняет такой вид на следующий
год; более старые части стволов покрыты светло-серой, глубо-
коморщинистой коркой. Молодые побеги почти не страдают от
зимних морозов — обмерзание составляет 10—40 см. Годичный
прирост побегов (в коллекции) на солнце — до 2, в полуте-
ни — до 3 м.
Завершение облиствения совпадает с началом бутонизации.
Наиболее ранние сроки цветения — 20.V. На солнечном откры-
том месте в коллекции отмечено запаздывание этой фазы на
3—5 дней.
Цветки аристолохин маньчжурской известны оригинальной
формой; они одиночные, со слабым неприятным ароматом. Вен-
чик их в виде изогнутой трубки с цельным волнистым отгибом
не закрывается на ночь. Опылители — различные мухи. В кол-
106
лекции наблюдается осыпание цветков и завязей. Массовое
цветение отмечается в конце мая — начале июня; последние
цветки формируются не позднее середины июня. На коллекци-
онных экземплярах обычно отмечалось небольшое число цвет-
ков: на открытой перголе — 2—5; в полутени, на более старом
экземпляре—10—15 цветков. Однако замечено, что после не-
скольких неблагоприятных — холодных и дождливых — вегета-
ционных сезонов наблюдается обильное цветение в первое же
теплое лего. В этих случаях число цветков даже на открытых
перголах достигает 20—25 штук, хотя завязывание плодов идет
не столь интенсивно — 5—8 завязей на тех же экземплярах.
Такое обильное, цветение объясняется тем, что формирование
цветочных почек в неблагоприятные периоды сдерживается
низкими температурами и происходит их «накопление».
Формирование плодов — с середины июня до начала сен-
тября; плоды созревают к середине октября, растрескиваются
и частично высыпают семена в октябре — ноябре.
В природных местообитаниях обнаруживаются самосев и
разновозрастные экземпляры в пределах одной пробной пло-
щади; порослевое отрастание или укоренение узлов ни в при-
роде. ни в культуре не наблюдались. Таким образом, вид обес-
печивает самовозобновление путем семенного размножения;
однако даже в наиболее крупной и благополучной популяции
аристолохин маньчжурской (окр. с. Горное) оно недостаточно
интенсивно — преобладают крупные плодоносящие экземпляры.
Здесь вид сохраняет определенное равновесие, по совершенно
очевидно отрицательное действие антропогенного фактора, на-
рушающего сбалансированное состояние популяции (сбор пло-
дов, срезание побегов). В других обнаруженных нами местах
произрастания аристолохин маньчжурской (в 20—30 км к югу
от с. Горное) имеются значительно более слабые, почти угне-
тенные группы растений этого вида, включающие, однако, осо-
би разного возраста (не более 6—8 м длиной) и редкий, ослаб-
ленный подрост.
В целом представляется несомненным реликтовое, остров-
ное состояние ариссолохии маньчжурской на юге Приморья.
Но при этом она обладает сезонным ритмом развития, который
совпадает с общей ритмикой окружающих растительных груп-
пировок, и проходит все фенологические фазы в числе первых.
Об этом говорит раннее набухание и развертывание почек, цве-
тение в первой половине лета (до 20 нюня), успешное и свое-
временное созревание плодов, которые высыпают семена в ос-
новном под снег.
Таким образом, и вегетативная, и генеративная сферы у
аристолохин маньчжурской относительно нормально развивают-
ся в указанных местообитаниях. Лимитирующее действие раз-
нообразных факторов среды сказывается в умеренной степени
самовоспроизводства вида — отсутствие обильного семяноше-
107
нйя и подроста; нет признаков расширения границ известных
популяций и процветания их. Но тем не менее аристолохия
маньчжурская обладает сложившимся устойчивым ритмом се-
зонного развития, соответствующим феноклиматическим ха-
рактеристикам региона.
Аристолохия маньчжурская совсем не растет на южных
склонах, где обычны резкие перепады дневных и ночных тем-
ператур; это указывает на то, что она нуждается не в макси-
муме температур, а в минимуме их колебаний, и говорит об
узком спектре экологических факторов, благоприятных для
данного вида в экстремальных условиях на пределе ареала.
Стабильность жизненной формы, отсутствие порослевого возоб-
новления свидетельствуют о консервативности его вегетативной
сферы и значительной филогенетической древности аристоло-
хин маньчжурской как компонента флоры южного Приморья.
2. Пуэр ария волосистая— Pueraria hirs-uta (Thunb.)
С. К. Schn..
Известна в одичавшем состоянии на юге СССР — в Крыму,
на Кавказе, где отмечается ее агрессивность (Борисова, 1933;
Петров, 1971), а также в культуре на Кубани (Медведев,
1948). Сравнительно недавно она была найдена в дикорасту-
щем состоянии на юге Приморья (Воробьев и Валова, 1962).
Основной ареал — Япония (не севернее о-ва Хонсю), Северная
Корея, Китай. Пуэрария волосистая имеется в коллекциях Бо-
танического сада ДВНЦ АН СССР и на Горнотаежной станции
(окр. г. Уссурийска); в течение 14 лет здесь наблюдаются осо-
бенности ее развития, роста и размножения.
В Северной Корее пуэрария волосистая довольно обычное
растение и имеет жизненную форму, которую можно опреде-
лить как лианоидный кустарник (Серебряков, 1962). Скелетные
оси в виде прочных деревенеющих стелющихся и вьющихся
побегов с диаметром 2,5—4 см густо переплетаются, образуя
плотные заросли, чему способствует легкая укореняемость пуэ-
рарии в узлах. Пуэрария волосистая в Северной Корее ежегод-
но цветет и дает семена; семяношение необильное.
В пределах Приморского края пуэрария волосистая встре-
чается только в окрестностях оз. Тальми (Хасанский р-н) в ви-
де плотных островковых зарослей. Эти популяции являются,
по-видимому, самыми северными, обитающими на пределах
ареала вида, где он заселяет соответствующие экологические
ниши — подножия южных открытых склонов.
В этих условиях пуэрария волосистая приобретает свойства
полукустарника, несколько утрачивая лианоидный характер.
Вегетативная сфера пуэрарии волосистой представлена здесь
главным образом в виде однолетних слегка древеснеющих
побегов, ежегодно обмерзающих или выгорающих почти до' ос-
нования. Наибольший условный возраст живых одревесневших
частей побегов (по Смирновой, 1967) в районе оз. Тальми —
108
3 года. Побеги текущего года формируются начиная с первой
декады мая из придаточных почек, развившихся у основания
перезимовавших побегов, и из нижних пазушных почек. Разви-
тие первых листьев — II—III декады мая.
Генеративные органы развиваются только на побегах теку-
щего года. Цветение начинается в первой декаде августа и
длится до конца сентября вместе с продолжающимся ростом
побегов Ось густой цветочной кисти направлена вертикально
вверх; в зрелом состоянии она имеет длину 9—16 (20) см и
несет от 15 до 50 цветков (чаще 20—40), плотно сидящих в
пучках по 2—3. Цветки раскрываются в базипетальном поряд-
ке; одновременно на оси соцветия наблюдаются все стадии —
от неокрашенных бутонов до сформировавшихся завязей —
бобов. Довольно обычным является опадание цветков, особенно
нижних — 5—10 штук в одной кисти.
Цветение пуэрарии волосистой на юге Приморья начинается
8—15.VIII, максимум фазы — III декада августа.
Развитие соцветий продолжается в течение двух месяцев
вместе с ростом побегов текущего года; последние кисти ос-
таются недоразвитыми, засыхая с бутонами. Такая растяну-
тость продуцирования генеративных органов определяется в
какой-то мере степенью зрелости побегов, развитие которых
заметно задерживается весной до наступления устойчивого
тепла. Характер цветения пуэрарии волосистой в указанном
районе можно определить как затяжной (Аврорин, 1956).
Этот тип обычно свойствен переселенным растениям, которые,
по определению Н. А. Аврорина, в новых условиях существова-
ния находятся в состоянии «биологического тупика». Продол-
жительное позднелетне-осеннее цветение пуэрарии, заканчиваю-
щееся гибелью последних соцветий, свидетельствует о том, что
сезонный цикл пуэрарии волосистой выпадает из общего ритма
сезонного развития сопутствующих видов и не укладывается в
сроки вегетационного периода на юге Приморья. При этом
затяжной тип цветения пуэрарии волосистой в данных клима-
тических условиях сочетается с активным состоянием ее веге-
тативной сферы. По темпу и энергии роста пуэрария волоси-
стая первенствует даже среди лиан. В природных местообита-
ниях ее побеги за сезон отрастают на 6—И м; в культуре
(ГТС) — 10—16 (22) метров. Растущие побеги имеют обычно
в пазухах по 3 коллатеральных почки, из которых, начиная с
июня, развиваются боковые ветви. При повреждении верхушеч-
ной почки рост боковых побегов активизируется, и чаще всего
годичный побег состоит из подиумов разного порядка.
Цветоносные побеги — III, IV порядков; кисти пазушные.
Нередко в зоне узлов на стелющихся побегах возникают
2—3 прямых и толстых стеблевых корешка, быстро проникаю-
щих в почву. Как и основной корень, стеблевые корешки у
пуэрарии волосистой стержневые. Они довольно хрупкие, легко
109
повреждаются, поэтому пересадка укорененных узлов или
черенков у этого вида требует особенной осторожности.
Формирование завязей наблюдается с конца августа, при
этом часть завязавшихся бобов, так же как цветков и бутонов,
опадает. Судя по положению на оси соцветия, все бобы с семе-
нами формируются из нижних цветков, развивающихся в нача-
ле цветения. Наибольшее число цветков в кисти — 51; наиболь-
шее число бобов (в фенофазе цвз—плг) —8—13, что составляет
7s от количества цветков в соцветии. Это свидетельствует о
том, что потенциальные возможности семяношения, заложен-
ные в многоцветковых кистях пуэрарии волосистой, в данном
районе реализуются слабо. В какой-то мере оно определяется
условиями сезона; так, в отдельные годы цветение и плодоноше-
ние пуэрарии волосистой выражены столь слабо, что обнару-
живаются лишь при продолжительном и тщательном осмотре
ее зарослей. Вегетирование и продуцирование завязей с семя-
зачатками обрывают отрицательные температуры — первые
заморозки на почве в середине октября.
Вызревшие бобы содержат зрелые семена обычно в числе
1—2 шт. Бобы не растрескиваются и нс опадают, сохраняясь
на оси соцветия всю зиму.
Самосев у пуэрарии волосистой в условиях юга Приморья не
наблюдается ни в природных местообитаниях, ни в условиях
коллекции. В лабораторных опытах прорастание семян этого
вида отмечалось при следующих условиях: скарификация,
длительное набухание, температура не ниже 115°; энергия
прорастания слабая. Осенне-весенний сезон на юге Приморья
(окр. оз. Тальми) весьма неблагоприятен для набухания и про-
растания семян пуэрарии из-за сухой осени и бесснежной зимы.
11о данным метеослужбы, зимний температурный минимум от-
мечается в январе: воздуха — до —28°, поверхности почвы —
до —36°; промерзание почвы от 29 до 127 см (Справочник,
1966). Кроме того, наблюдаются частые пожары в осенне весен-
нее время, которые уничтожают растения.
На коллекционных участках (Ботанический сад, ГТС ДВНЦ
АН СССР) пуэрария волосистая представлена недревеснеющи-
ми или полудревеснеющими экземплярами. Здесь она быстро
разрастается на солнечных открытых местах и угнетается в
тени. Цветет (ГТС), но отмечается заметное опадание цветков.
Семена — редкость, самосев отсутствует. Стелющиеся плети
легко укореняются в узлах и образуют плотные куртины.
Таким образом, проведенный анализ состояния вегетатив-
ной сферы, ритма сезонного развития и основных климатиче-
ских факторов среды в местах произрастания пуэрарии воло-
систой на юге Приморья дает основание достаточно уверенно
судить об особенностях поведения вида на северном пределе
его ареала. Пуэрария волосистая, будучи представителем суб-
тропических флор, находит условия для существования на
110
крайнем юге Приморья лишь в определенных локусах. Актив-
ность вегетативной сферы вида в тальминских популяциях и
клонах значительна и обеспечивает присутствие и даже некото-
рое распространение вида при его низкой реальной урожайно-
сти и отсутствии заметного самосева.
По-видимому, в условиях юга Приморья для нормального
завершения сезоннного цикла пуэрарии волосистой в природе
и в культуре пороговое значение приобретают следующие фак-
торы. 1) недостаточные температуры в начале вегетации
(май) и во второй половине периода цветения (сентябрь), а
также во время плодоношения (октябрь); 2) отсутствие актив-
ных опылителей; 3) малоблагоприятные условия зимовки для
семян и почек возобновления. Именно эти факторы определяют
состояние и пределы распространения пуэрарии волосистой на
северной окраине ее ареала в южном Приморье.
Создается впечатление, что этот вид в значительной мере
является чужеродным элементом во флоре южного Приморья.
Не исключено, что он является заносным, так как некоторые
черты поведения его на юге Приморья напоминают инорайон-
ное растение; кроме того, сходным образом ведет себя пуэра-
рия волосистая в Крыму и на Кавказе, где она одичала. Су-
щественным следует считать указание А. Г. Борисовой (1933)
на то, что пуэрария волосистая культивируется в Маньчжурии.
Во всяком случае, ритм сезонного развития этого вид^ на юге
Приморья в значительной степени не соответствует общей рит-
мике природных явлений и развития основной массы сопутст-
вующих видов. При этом монодоминантные островные заросли
пуэрарии удалены на незначительное расстояние — не более
100—150 км от мест, где она обычна (Северная Корея).
Тем не менее реликтовый характер пуэрарии волосистой
как компонента флоры Приморья следует признать. Не исклю-
чено, что она культивировалась прежде на самом юге При-
морья. Но значительно существеннее те факты, что пуэрария
найдена в третичных отложениях на территории Китая (Криш-
тофович, 1957) и оптимум условий обитания этого вида в на-
стоящее время наблюдается на юге п-ова Корея, где он пред-
ставлен крупными деревянистыми лианами. По-видимому,
пуэрария волосистая, будучи пластичной и весьма жизнеспособ-
ной, является автохтонным видом флоры Юго-Восточной Азии,
обладающим ареалом с пульсирующими границами. На север-
ных пределах ареала пуэрария волосистая становится стелю-
щимся полукустарником, сохраняя некоторую лиановидность и
удерживаясь в экстремальных условиях благодаря активному
состоянию вегетативной сферы. Крайне интересными могут
быть палеоботанические находки этого вида на территории
Приморья.
Таким образом, изучение ритмики сезонного развития
вскрывает существенные моменты поведения, характерные осо-
111
бенности жизненных и сезонных циклов, степень приспособлен^
ности у разных видов. Это особенно интересно и важно выявить
у редких и реликтовых растений, поскольку «изучение жизнен-
ных циклов — коренной вопрос для познания сущности видов»
(Алехин, 1938).
ЛИТЕРАТУРА
Аьрорин Н. А. Переселение растений на Полярный север (эколого-геогра-
фический анализ). М.—Л., Изд-во АН СССР, 1956, 286 с.
Алехин В. География растений. М., «Сов. книга», 1938, 283 с.
Борисова А. Г. Pueraria hirsuta (Thunb.) С. К. Schneid.— новое прядильное
растение в пределах СССР.—«Сов. ботаника», 1933, №3—4, с. 76—84.
Валова 3. Г., Куренцова Г. Э. Репиктовые лианы в юго-западном Приморье.
Комаровские чтения, XXI. Владивосток, 1974, с. 43—60.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 275 с.
Воробьев Д. П., Валова 3. Г. Новая для флоры СССР деревянистая лиана
Pueraria hirsuta (Thunb.) С. К. Schneid.— «Бот. ж.», 1962, 47, 3,
с. 1194-1196.
Красная книга. Редкие и исчезающие виды природной флоры СССР. М.,
«Наука», 1975, 203 с.
Криштофович А. Н. Палеоботаника. Л., Гостехнефттопиздат, 1957, 650 с.
Куренцова Г. Э. Реликтовые растения Приморья. М., «Наука», 1968, 72 с
Медведев П. Ф. Пуэрария,—Новые кормовые культуры СССР. М., Сельхоз-
гиз, 1948, с. 210—215.
Петров В. В. В мире субтропических (растений. М., «Наука», 1971, 175 с.
Сергеева К. А. Физиологические и биохимические основы зимостойкости
древесных растений. М., «Наука», 1971, 174 с.
Серебряков И. Г. Экологическая морфология растений. М., «Высш, школа»,
1962, 378 с.
Смирнова О. В. Жизненный цикл пролески сибирской (Scilla sibirica
Andr.) Науч. докл. высш, шк., биол. науки, 19о7, № 9.
Справочник по климату СССР. Приморский край, в. 26, II. Л., Гидро-
метеоиздат, 1966, 220 с.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ ПО РАЗМНОЖЕНИЮ
ДРЕВЕСНЫХ ЛИАН ПРИМОРЬЯ
л. н. слизик
В Ботаническом саду ДВНЦ АН СССР (окр. г. Владивосто-
ка) продолжительное время изучалась коллекция древесных
лиан, которая включала инорайонные и местные виды, с целью
исследования в природе и в культуре особенностей поведения
лиан флоры Приморья и обоснования рекомендаций для их ис-
пользования и размножения.
Дальневосточные лианы как материал для садовой культу-
ры или озеленения довольно известны за пределами своих аре-
алов (Мичурин, 1955; Шульгина, 1955; Деревья и кустарники,
1959; Бибиков, 1962; Павленко, 1965; Кравченко, 1966; Головач,
1973; Писаный, 1974). В указанной литературе имеются раз-
розненные или общие сведения о приемах размножения расте-
ний этой группы в разных природных зонах. Более подробно
данные приводятся лишь в работах, посвященных изучению не-
которых ягодных лиан в природе и в культуре в Приморье —
виноград амурский (Лебедева, Лебедев, 1962), актинидии и
лимонник (Титлянов, 1969).
В целом группа древесных дальневосточных лиан насчиты-
вает более 20 видов (Воробьев, 1968; Слизик, 1970). Они рас-
пространены в основном в южной половине Дальнего Востока,
более всего в Приморье, но при этом значительно отличаются
по своему состоянию во флоре региона. Три приморских вида —
аристолохия маньчжурская, девичий виноград триостренный,
пуэрария волосистая — занесены в Красную книгу (1975). Од-
нако анализ литературных данных и продолжительные наблю-
дения автора за лианами в природных местообитаниях дают
основания считать почти все древесные лианы Дальнего Восто-
ка редкими И сокращающимися видами, которые нуждаются в
охране или бережном отношении.
К редким (категория 2), по-видимому, следует отнести при-
морские виды лиан, которые отличаются узкой экологической
приуроченностью и незначительной численностью — виноградов-
ники, краснопузырники, шиповник Максимовича, княжик охот-
ский, некоторые ломоносы. Группа сокращающихся видов (ка-
тегория 3) включает все ягодные лианы — актинидии, виноград
амурский, лимонник китайский. Численность их заметно сок-
8 Заказ 488
113
ращается в Приморье, особенно в местах строительства и ле-
соразработок, в населенных районах края. Генофонд этих цен-
нейших видов обедняется из-за давнего нерегулируемого сбора
ягод населением в естественных зарослях лиан.
Совершенно очевидна актуальность задач по сохранению
и воспроизводству растительных ресурсов — в первую очередь
редких и ценных видов отечественной флоры. С этими целями
в Ботаническом саду изучалось семенное и вегетативное раз-
множение древесных лиан Приморья (Слизик, Древецкая,
1975). Предлагаемые рекомендации касаются 18 видов (см.
таблицу). Для некоторых из них приемы размножения в усло-
виях Приморья предлагаются впервые (виноградовник япон-
ский, княжик охотский, краснопузырник плетистый, пуэрария
волосистая). По ряду видов (актинидия, аристолохия маньч-
журская) результаты наших опытов отличаются от известных
в литературе (Звиргзд, 1967; Титлянов, 1969).
Способность к возобновлению древесных лиан Приморья и рекомендуемые
приемы по их размножению
Категория, вид Возобновление в природе Рекомендуемые приемы
семей. вегет.
1 2 3 4
Категория 1 — исчезающие виды Девичий виноград три- слабое значит. Черенкование— 10.VII,
остренный Parthenocissus tricuspi data (Sieb. et Zucc.) Planch. Аристолохия маньчжур- ская значит. не набл. гетероауксин, песок;срок укоренения 20 дн.; посев семян без страти- фикации— III, IV Черенкование — 15. VI, гетероауксин, песок с
Aristolochia manshurien- sis Korn. Категория 2 — редкие виды Пуэрария волосистая Pueraria hirsuta (Thunb.) С. К. Schneid. не набл. значит. перегноем; срок укорене- ния 45 дн. (65%); посев семян без страти- фикации — IX, IV Черенкование — 5—15. VIII, гетероауксин, песок с пе-
регноем; срок укорене- ния 20 дн. (60%); посев скарифицирован- ных семян при темпера- туре не ниже 15е (гор- шки, регулярный по- лив)
Продолжение таблицы
1
2 3
4
Виноградовник японский Ampelopsis japonica (Thunb.) Makino слабое не набл.
В. коротконожковый A. brevipeduculata (Ma- xim.) Trautv. значит. слабое
В. разнолистный незначительное
A. heterophylla (Thunb.)
Sieb. et Zucc.
Княжик охотский незначит. слабое
Atragene ochotensis Pall.
Краснопузырник кругло-
листный значит.
Celastrus orbiculata
очень
слабое
Thunb.
К. плетистый не
С. flagellaris Rupr. значит. набл.
Шиповник Максимовича не
Rosa maximdwicziana значит. набл.
Rgl.
Ломонос короткохвостый не
Clematis brevicaudata значит. набл.
DC.
Л. пильчатый
С. serratifolia Rehd.
не
значит.
набл.
Черенкование — 15.VII,
гетероауксин, песок;
срок — 30 дн. (60 %);
посев семян без стра-
тификации — IX, IV
Черенкование— 15.VII,
гетероауксин, песок;
срок —30 дн.; посев се-
мян после стратифика-
ции в течение 1 месяца
при температуре 2—8°
Черенкование — 15. VII,
гетероауксин, песок с
перегноем; срок — 30 дн.
(60%)
Черенкование— 10.VII,
гетероауксин, песок с
перегноем; срок —30 дн.
(менее 50%); посев се-
мян по мере созрева-
ния, регулярный полив
Черенкование— 10.VII,
гетероауксин, песок с пе-
регноем; срок — 35 дн.;
посев семян весной в
грунт после предвари-
тельного выдерживания
при температуре 18 —
23° в течение 15 дн.
Черенкование— I.VII,
гетероауксин, песок с пе-
регноем; срок—35 дн.;
посев семян после стра-
тификации при темпера-
туре 18—23° в течение
15 дн.
Черенкование — 1 .VII,
гетероауксин песок с
перегноем; срок—30 дн.;
посев семян после стра-
тификации при темпера-
туре + 4° в течение 3 ме-
сяцев — IV; возможен
осенний посев — IX
Черенкование— 15.VI,
гетероауксин, песок с
перегноем; срок—30 дн.;
посев семян после стра-
тификации при темпера-
туре 2—5° в течение
1 месяца
То же
115
Окончание таблицы
Категория 3 —
сокращающиеся виды
Лимонник китайский значит.
Schizandra chinensis
Bail.
Виноград амурский значит.
Vitis amurensis Rupr.
Актинидия коломикта значит.
Actinidia kolomicta Ma-
xim.
А. многодомная A. polygama (Sieb. et значит.
Zucc.) Planch.
А. острая значит.
А. arguta (Sieb. Zucc.) Planch. et
А. Жиральда не изв.
А. giraldii Diels.
не изв. »
не Черенкование—15.VI,
набл. гетероауксин, песок с
перегноем; срок—25 дн.;
посев семян весной пос-
лё стратификации при
переменном режиме:
1 месяц 18—23°, 2 ме-
сяца 2—5°
слабое Черенкование—15.VI,
гетероауксин, песок с
перегноем; срок—30 дн.;
посев семян весной (IV)
или осенью (IX—X) —
без стратификации, но
после суточного намачи-
вания
слабое Черенкование—1.VII,
гетероауксин, песок;
срок — 25 дн.; посев се-
мян после стратифика-
ции при переменном ре-
жиме: 1—2 месяца 18 —
23°, 1—2 месяца 2'—5°
слабое То же
слабое >
Разработанные рекомендации следует использовать при соз-
дании маточных и производственных питомников лиан, а также
для полукультурных посадок в зеленых зонах и заказниках на
юге Дальнего Востока.
ЛИТЕРАТУРА
Бибиков Ю. А. Интродуцированные вьющиеся древесные растения для вер-
тикального озеленения в БССР. Автореф. канд. дис. Минск, 1962, 18 с.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 275 с.
Головач А. Г. Лианы, их биология и использование. Л., «Наука», 1973,
191 с.
Деревья и кустарники. Краткие итоги интродукции в ГБС АН СССР. М.,
Изд-во АН СССР, 1959, 190 с.
116
Звиргзд А. В. Предварительная схема подготовки и посева семян деревьев п
кустарников при интродукции. — Бюл. ГБС, в. 65, 1967, с. 18—23.
Кравченко Л. К. Дикие винограды, интродуцированные Ботаническим садом
АН УзССР, Ташкент, изд-во «Фан», 1966, 80 с.
Красная книга. Дикорастущие виды флоры СССР, нуждающиеся в охране.
М., «Наука»,1975, 203 с.
Лебедева Л. Я., Лебедев А. И. Виноград в Приморском крае. Владивосток,
1962, 159 с.
Мичурин И. В. Актинидия.— Избранные сочинения. М., Гос. изд-во с.-х. лит.,
1955, с. 472—500.
Павленко Ф. А. Размножение вьющихся растений. М., Стройиздат., 1965,
с. 60.
Писаный Г. Г. Эколого-биологические особенности деревянистых лиан, инт-
родуцированных в Донбассе. Автореф. канд. дис. Донецк, 1974, 24 с.
Слизик Л. Н. Краткий обзор группы декоративных деревянистых лиан юга
Дальнего Востока.— Деревья, кустарники и многолетники для озеле-
нения юга Дальнего Востока. Владивосток, Дальневост. кн. изд-во,
1970, с. 101—108.
Слизик Л. Н., Древецкая Р. К. Опыты по зеленому черенкованию дальне-
восточных деревянистых лиан (в условиях юга Приморья).— Экспери-
ментальная экология и акклиматизация растений на Дальнем Востоке.
Владивосток, 1975, с. 35—42.
Титлянов А. А. Актинидии и лимонник. Владивосток, Дальневост. кн. изд-во,
1969, 175 с.
Шульгина В. В. Древесные лианы и их культура в Ленинграде. Труды БИН
АН СССР, сер. VI, в. 4, 1955, с. 157—194.
9 Зак. 488
Н7
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
МОРФОЛОГО-АНАТОМИЧЕСКИЕ ХАРАКТЕРИСТИКИ
ХВОИ СОСНЫ ПОГРЕБАЛЬНОЙ
(PINUS FUNEBRIS Кот) В ПРИМОРСКОМ КРАЕ
Л. С. ЛАУВЕ
Популяции сосны секции Eupitys Spach, объединенные под
именем погребальной (Комаров, 1901), в Приморском крае ока-
зались неодинаково полиморфными, причем по набору наслед-
ственных морфологических форм вскрыто два круга общности:
а) тяготеющие к Японскому морю комплексы популяций сгла-
/кенного полиморфизма; б) западно- и центральноприморские
комплексы с большей изменчивостью признаков. Различие этих
двух макрокомплексов прослеживается и на анатомическом ма-
териале. Последний позволил идентифицировать популяции и
обосновать причины изменчивости некоторых из них.
Существует немало исследований, где анатомия хвои прив-
лекалась в качестве одного из признаков при разделении сосен:
В. П. Малеев (1925) разграничил виды сосен в том числе и по
расположению смоляных каналов; по ряду анатомических приз-
наков П. Я. Соколов (1928) различает географические расы
сосны обыкновенной, а М. Моисеева (1937)—разновидности
и формы некоторых видов сосен. В. Л. Комаров (1901) при
выделении сосны погребальной в самостоятельный вид тоже
использовал признаки расположения смоляных каналов и ме-
ханических тканей в хвое в качестве диагностических и указал
на их существенные особенности. Однако небольшой объем
просмотренного В. Л. Комаровым материала даже схематично
не характеризует строения хвои сосны погребальной. А так
как ее систематическое положение не вполне ясно, нами про-
веден анатомический анализ, охватывающий все значительные
популяции, результаты которого частично опубликованы (Лау
ве, Урусов, 1973). Работы велись в соответствии с программой
исследований В. М. Урусова.
Изучены морфолого-анатомические особенности хвои второ-
го года с западных ветвей в середине кроны. Брали по 10 хвои-
нок из средней части побегов первого порядка у деревьев в
возрасте от 27—40 до 45—120 лет (табл 1). Собранный мате-
риал представляет наиболее распространенные поколения осо-
бей однородных групп популяций Ханкайского, Пограничного и
Михайловского районов и отдельные популяции Октябрьского,
Шкотовского и Хасанского. Поперечные срезы в средней части
118
хвоинок делались от руки и заливались желатин-глицерином.
На них измерены ширина и толщина хвои, жилки проводяших
пучков, расстояние между пучками, вертикальные и горизон-
тальные диаметры смоляных каналов, толщина эпидермиса и
гиподермы. Специально рассмотрены особенности покровных,
проводящих и механических тканей и смоляных каналов. По
каждому срезу учитывалось общее количество каналов и число
каналов, расположенных в паренхиме. Параметры внутренней
структуры хвои измерены с помощью окуляр-микрометра на
2670 срезах с 245 сосен. Полученные результаты обработаны
статистическим методом (Рокицкий, 1961).
Необходимо отметить, что ряд структурных анатомических
характеристик хвои сосны погребальной не подвержен геогра-
фической изменчивости. Так, эпидермис и гиподерма, преры-
вающиеся устьицами на обеих сторонах листа, повсеместно
однослойны Форма клеток эпидермиса округлоконическая,
стенки их сильно утолщены. Вытянутые в продольном направ-
лении клетки гиподермы в углах хвои располагаются в 2,
реже в 3 ряда. Степень развития гиподермальных клеточных
оболочек в различных районах неодинакова: в Ханкайском и
Пограничном районах, где отмечена самая значительная в крае
сухость воздуха и почвы, они более утолщенные. Толщина ги-
подермы почти не меняется (табл. 1), в то время как толщина
слоя эпидермиса возрастает к югу, достигая максимума в вер-
ховьях р. Илистая (Михайловский район). Ассимиляционная
паренхима расположена в 3 слоя с верхней стороны листа и в
2 — с нижней. Околопучковая склеренхима обычно находится
рядом с флоэмной частью проводящих пучков и в большинстве
случаев однослойна, реже двуслойна, но второй слой склерен
химы уже. Форма поперечного среза хвои плосковыпуклая,
реже выпукло-вогнутая, очертания прямые, иногда слабоизви-
листые. Различия в длине и ширине хвои по популяциям
определяются не только их наследственной и экологической ин-
дивидуальностью, но и физиологической ослабленностыо дере-
вьев в результате палов. Наиболее короткая хвоя отмечена у
деревьев в Октябрьском районе, длинная — в Ханкайском и
Хасанском. Исключив изолированные от палов популяции,
можно сделать вывод, что в целом длина хвои к югу возраста-
ет при одновременном уменьшении ширины и довольно посто-
янной толщине. Минимальные ширина и толщина ее жилки в
Михайловском районе, к северо-западу и югу они возрастают,
достигая максимума в окрестностях оз. Ханка.
Два биколлатеральных пучка (три отмечено не более чем
на 0,1% срезов) располагаются по краям жилки. В них хорошо
выражены флоэма и ксилема. Ширина и толщина пучков в
среднем равны. Резких различий в размерах пучков нет, хотя
на о-ве Сосновом (Ханкайский район) и в верховьях р. Арте-
мовна (Шкотовский) они крупнее. Наименьшее среднее рас-
9*
119
Варьирование морфолого-анатомических показателей
Районы Возраст деревьев, лет Хвоя Жилка
длина шири- на толщина ширина
Ханкайский 27—40 65,0 ± 1,07 ± 0,51± 0,54 ±
(хр. Синий, о-в Сое- 0,5 0,005 0,005 0,004
новый)
Пограничный 35—80 54,2 ± 1.05 ± 0,54 ± 0.51 ±
(хр. Пограничный) 0,5 0,02 0,001 0,001
Октябрьский 27—40 52,0 ± 0,84 ± 0,57 ± 0,45 ±
(бас. р. Казачка) 0,1 0,017 0,004 0,0009
Михайловский 35—80 58,0 ± 0,99 ± 0,51± 0,43 ±
(хр. Пржевальского) 0,3 0,005 0,005 0,004
Шкотовский 30—50 55,0± 0,97± 0,52 ± 0,48 ±
(верховья р. Артемов- на) Хасанский 2,0 0,013 0,005 0,0024
45—120 66,0 ± 1,02± 0,51 ± 0,48 ±
(п-ов Гамова) 0,5 0,02 0,005 0,003
стояние между проводящими пучками отмечено в Октябрьском
и Шкотовском районах, наибольшее — в Ханкайском и Хасан-
ском, причем аналогично этим величинам изменяется и мощ-
ность механических тканей. В Ханкайском, Михайловском и
Шкотовском районах соответственно на 1,5; 3 и 9,5% срезов
пучки примыкают непосредственно.
Смоляные каналы наиболее многочисленны на севере ареа-
ла (табл. 2). В целом же хвоя сосны погребальной может
быть оценена как малоканальная (Правдин, 1964:9), так как
к югу число смоляных каналов уменьшается.
Анализируемая хвоя представлена двумя типами: 1) пери-
ферическим — все каналы непосредственно примыкают к гипо-
дерме, 2) смешанным — часть каналов лежит в паренхиме.
Всюду преобладают срезы периферического типа, их особенно
много в Михайловском и Хасанском районах (табл. 3), как и
деревьев с хвоей исключительно такого типа. Вообще же во
всех популяциях, а особенно в Ханкайском, Пограничном и
Шкотовском районах преобладают деревья с обоими типами
хвои. В Пограничном и Шкотовском районах доля срезов сме-
шанного типа превышает 30%, но только в последнем наблю-
дается до 10 каналов в паренхиме: найдена хвоя, у которой из
9—13 каналов паренхиматическими были 6—10. За исключе-
нием шкэтовской популяции, среди хвои смешанного типа
абсолютно преобладают срезы с 1—2 смоляными каналами в
паренхиме. Они повсеместно приурочены к достоверно более
широкой и толстой многоканальной хвое: при 6 каналах на
срезе—1—2 канала в паренхиме (9—26% случаев), при 9 на
срезе—1—3 в паренхиме (21 —100%). Большее количество па-
120
Таблица J
>вои сосны погребальной, мм
Проводящие пучки Диаметры смо- ляных каналов Толщина слоя
толщина расстоян. между пучками ширина горизон- тальный верти- кальный эпидер- миса гипо- дермы
0.26 ± 0,05 ± 0,10± 0,08 ± 0,07± 0,018± 0,016±
0,002 0,0015 0,0014 0,004 0,0006 0,0007 0,0003
0,24 ± 0,04 ± 0,08 ± 0,07 ± 0,08 ± 0,02 ± 0,02 ±
0,001 0,001 0,001 0,0004 0,0006 0,00001 0,00001
0,25 ± 0,02 ± 0,08 ± 0,06 ± 0,06 ± 0,03 ± 0,02 ±
0,0007 0,006 0,0009 0,0009 0,0007 0,0003 0,0001
0,22 ± 0,04 ± 0,09 ± 0,08 ± 0,08 ± 0,03 ± 0,02 ±
0,01 0,0017 0,0009 0,006 0,0005 0,0002 0,00006
0,23 ± 0,03 ± 0,10± 0,07 ± 0,07 ± 0,02± 0,017±
0,0019 0,0013 0,0011 0,0009 0,002 0,0004 0,0003
0,25 ± 0,05 ± 0,09 ± 0,07 ± 0,06 ± 0,03 ± 0,02 ±
0,002 0,001 0,001 0,001 0.001 0,0005 0,0002
ренхиматических каналов с увеличением структур и общего
числа каналов не коррелирует.
Изменчивость признаков по популяциям и районам неоди-
накова.
Уровень изменчивости — его градации приняты по С. А. Ма-
маеву (1970) —величины хвои, проводящих пучков, расстояния
между пучками, толщины эпидермиса и гиподермы, количества
смоляных каналов, их горизонтальных и вертикальных диа-
метров приводится в табл. 1. В целом величина и внутренние
структуры оказались более вариабельными в Шкотовском и
отчасти Пограничном районах, а весьма стабильными в Ок-
тябрьском (за исключением ширины жилки, количества и ши-
рины смоляных каналов) и Михайловском. Резкое повышение
стабильности горизонтальной величины внутренних структур в
какой-либо популяции сопровождается повышением изменчи-
вости вертикальной, и наоборот.
Изложенное позволяет заключить, что среднепопуляционные
морфолого-анатомические характеристики хвои определяются,
во-первых, происхождением и участием конкретной генетиче
ской информации (положение смоляных каналов, их число в
целом — мало-, средне-, многоканальность, непосредственное
примыкание проводящих пучков, мощность механических тка-
ней, число слоев клеток эпидермиса и гиподермы, соотношение
площадей сечений внутренних структур хвои), во-вторых, ди-
вергенцией в разных макроклиматах (общая величина и вели-
чина внутренних структур, расстояние между проводящими
пучками, толстостенность клеток покровных тканей), в-третьих,
мощными факторами воздействия, способными изменить физио-
121
Таблица 2
Изменчивость числа смоляных каналов в хвое сосны погребальной
в Приморье
Районы Число обследо- ванных Встречаемость смоляных каналов, % Среднее число смо- ляных ка- налов
особей срезов 2 3 4 5 6 7 8 9 10 11 12 13 14
Ханкайский 69 690 0,7 2,8 6,6 15,6 18,5 17,3 19,7 9,4 4,6 5,3 4,1 0,6 7,0±0,10 Пограничный 54 540 0,1 0,1 1,0 1,5 10,8 15,2 25,2 22,2 14,6 6,2 2,5 0,3 0,3 8,0 ±0,09 Октябрьский 36 360 1,6 9,9 20,7 17,3 26,3 10,1 5,5 5,2 3,4 6,0 ±0,085 Михайловский 59 590 2,9 1,4 6,4 6,2 31,1 21,1 11,1 9,9 6,6 2,2 1,1 6,5±0,05 Шкотовский 25 250 1,3 10,4 9,5 7,8 17,8 14,8 11,7 13,1 7,4 3,6 2,2 6,7±0,1 Хасанский 22 220 0,06 6,9 10,8 23,1 21,9 20,0 8,8 6,3 1,6 6,0±0,35
Таблица 3
Распределение хвои по типам
расположения смоляных каналов, %
Районы Деревья с хвоей типов Срезы типов Встречаемость срезов второго типа Наибольшее число смоля- ных каналов в паренхиме
1-го 2-го 1-го 2-го 2 3 4 5 6 7 8 9 10 11 12 13
Ханкайский 11,6 88.4 73,6 26,4 11,5 8,0 15,6 24,4 27,7 37,3 39,6 43,9 4 Пограничный 11,1 88,9 63,1 36,9 100 9,0 23,4 36,0 49,0 63,5 83,3 55,5 9 Октябрьский 33.4 66,6 80,3 19,7 14,0 44,4 20,0 26,3 40,0 83,3 70,0 50,0 3 Михайловский 44,0 56,0 87,9 12,1 6,1 9,8 16,1 17,5 20,9 38,4 62,5 100 3 Шкотовский 17,4 82,6 68,3 31,7 18,2 22,2 14,6 32,4 33,8 50,0 52,9 100 100 100 10 Хасанский 45,4 54,6 87,2 12,8 3,2 10,9 13,3 33,3 21,5 11,1 3
логическое состояние большинства растений популяции (вели-
чина хвои и ее внутренних структур). Разумеется, вызываемая
несоответствием макроклиматов дивергенция может привести
к рельефным качественным сдвигам анатомических характери-
стик, изменяя, например, количество и положение смоляных
каналов или толщину слоя гиподермы во всей популяции, но
тогда данная популяция окажется за рамками рассматривае-
мого таксона и приобретает ранг по крайней мере подвида.
ЛИТЕРАТУРА
Комаров В. Л. Флора Маньчжурии. 1. СПб, 1901.
Лауве Л. С., Урусов В. М. К морфолого-анатомическим особенностям хвои
сосны погребальной (Pinus funebris Кот.) в Приморском крае. — Бюл.
ГБС, в. 89, 1973.
Малеев В. П. Определитель и описание видов сосен (Pinus L.), дикорасту-
щих и разводимых на Черноморском побережье Кавказа. Труды по
прикладн. бот., генет. и селекц., т. 14, 4. М., Изд-во АН СССР, 1925.
Мамаев С. А. Закономерности внутривидовой изменчивости семейства Pina-
сеае на Урале. Автореф. докт. дис. Свердловск, 1970.
Моисеева М. Применение количественно-анатомического метода при подраз-
делении рода Pinus на более мелкие таксономические единицы.—
ДАН СССР, 17, 5, 1937.
Рокицкий П. Ф. Основы вариационной статистики для биологов. Минск.
«Наука», 1961.
Соколов П. Я. К вопросу о географических расах Pinus sylvestris L —
Изв. ГБС СССР, в. 27, №5—6. Л., 1928.
Р1ДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ХАРАКТЕРИСТИКА ЭПИДЕРМИСА ХВОИ
НЕКОТОРЫХ ВИДОВ СОСЕН СЕКЦИИ EUPITYS Spach
Л. С. ЛАУВЕ
Исследование эпидермальных признаков хвои сосен секции
Eupitys Spach является частью комплексного исследования
морфолого-анатомического изучения систематического положе-
ния сосны погребальной (Pinus funebris Кот.).
Строение эпидермиса весьма разнообразно у каждого вида.
Его анатомические особенности дают возможность отличать
морфологически и систематически близкие растения.
Особенности эпидермиса хвои сосны погребальной изучены
в пределах Приморского ареала. Собранный материал пред-
ставляет наиболее распространенные поколения однородных
популяционных групп. Кроме того, для сравнения исследован
материал хвои Pinus sylvestris L., Р. densiflora Sieb- et Zucc.,
P. tabulaeformis Carr., P. massoniana D. Don., P luchuensis
Мауг, P. thynbergiana Franko- Образцы хвои этих видов сосен
были предоставлены нам сотрудниками Ботанического (институ-
та им. В. Л. Комарова.
Для получения эпидермальных препаратов использован ме-
тод мацерации в азогной кислоте (Свешникова, 1966). Описа-
ние эпидермы ведется по методике Г. С. Сашиной (1975).
Pinus funebris Кот.
Лист сосны погребальной относится к амфистоматному типу
(Свешникова, 1966).
Клетки верхнего эпидермиса имеют четырехугольную, пря-
моугольную, трапециевидную форму. Редко встречается треу-
гольная форма. Продольные стенки прямые, поперечные — пря-
мые и скошенные, мелкоизвилистые. Минимальная длина
клеток верхнего эпидермиса 320 мк, ширина—20 мк, макси-
мальная длина— 1000 мк, ширина—60 мк. На 1 мм2 поверх-
ности хвои приходится от 100 до 400 клеток.
Форма клеток нижнего эпидермиса четырехугольная, прямо-
угольная, трапециевидная. Продольные стенки прямые, попе-
речные как прямые, так и скошенные. Длина клеток колеблет-
ся от 1000 до 280 мк. Ширина клеток нижнего эпидермиса
не отличается от верхнего.
121
Устьичный аппарат у сосны погребальной изолированный,
смежный, связанный, пиноидного типа. На поверхности листа
устьица располагаются полосами, а в них — тремя прямыми,
неправильными, умеренно плотными рядами. Устьичная щель
ориентирована вдоль оси листа. На 1 мм2 приходится от 70 до
90 устьиц. Околоустьичные клетки отличаются от эпидер-
мальных размерами и формой. Располагаются они в один ряд,
и количество их колеблется от 3 до 8.
Р. densiflora Sieb. et Zucc.
Лист сосны густоцветковой амфистоматный.
Форма клеток верхнего эпидермиса четырехугольная, пря-
моугольная, трапециевидная, реже встречается треугольная.
Клеточные стенки прямые, мелкоизвилистые. Максимально
длинные клетки (1000 мк) в средней части листа, минималь-
ные (202 мк) —у основания листа. Количество клеток на 1 мм2
поверхности листа колеблется от 300 до 400.
Клетки нижнего эпидермиса по форме не отличаются от
верхнего. Поперечные клеточные стенки прямые или скошенные.
Продольные прямые, реже изогнутые. Минимальная длина
клеток у вершины листа — 280 мк, максимальная — в средней
части— 1000 мк. Их ширина колеблется в пределах от 40 до
70 мк.
Устьичный аппарат листа сосны густоцветковой изолиро-
ванный, иногда встречается смежный, пиноидного типа, моно-
циклический. Устьичные ряды умеренно плотные, прямые, не-
правильные, устьичная щель ориентирована вдоль оси листа.
Количество устьиц на единицу поверхности листа колеблется
в пределах от 50 до 100 штук (нижний эпидермис) и от 30 до
100 (верхний эпидермис). Околоустьичные клетки отличаются
от покровных формой и размерами. В основном они округлой
формы. Их 3—6 в одном ряду.
Pinus thunbergiana Franco
Лист сосны Тунберга амфистоматный. Эпидермис характе-
ризуется вытянутыми вдоль листа правильными рядами кле-
ток.
Форма клеток верхнего эпидермиса четырехугольная, прямо-
угольная, трапециевидная. Стенки их прямые, мелкоизвили-
стые. Размеры покровных клеток различные. Наибольшая дли-
на клеток верхнего эпидермиса в средней части листа
(1000 мк), наименьшая — у основания (200 мк). На 1 мм2
листа приходится от 200 до 500 клеток.
Клетки нижнего эпидермиса по форме аналогичны клеткам
верхнего, с прямыми продольными и скошенными поперечными
стенками. Оболочки их мелкоизвилистые. Наиболее длинные
Ю Заказ 488
125
клетки также в средней части листа (1000 мк), минимальная
их длина у вершины (280 мк). Ширина клеток как верхнего,
так и нижнего эпидермиса варьирует от 50 до 70 мк. Количест-
во клеток на 1 мм2 до 500.
Устьичный аппарат моноциклический, изолированный, пино-
идного типа. Устьичные ряды умеренно плотные, прямые, не-
правилные.
Устьичная щель ориентирована вдоль оси листа. Околоусть-
ичные клетки овальной и полукруглой формы, количество их
от 4 до 8 штук. Расположены они в один ряд. Количество усть-
иц на 1 мм2 50—80 штук.
Pinus tabulaeformis Carr.
По расположению устьичного аппарата лист сосны китай-
ской относится к амфистоматному типу.
Клетки верхнего эпидермиса вытянуты вдоль листа пра-
вильными рядами. Они четырехугольной, прямоугольной, пяти
угольной, трапециевидной формы, с параллельными продольны-
ми и непараллельными поперечными стенками. Клеточные
стенки их мелкоизвилистые. Размеры покровных клеток в раз-
ных частях листа различны. Наибольшей длины они достигают
в средней части, более 1000 мк. Самые короткие клетки у ос-
нования листа (290 мк). На 1 мм2 приходится от 400 до 600
клеток.
По форме клетки нижнего эпидермиса не отличаются от
верхнего. Размеры их также различны. В средней части листа
длина их максимальная (690 мк), у вершины — минимальная
(310 мк). Ширина клеток верхнего и нижнего эпидермиса 10—
60 мк.
Устьичный аппарат моноциклический, изолированный, пино-
идного типа. Устьичные ряды умеренно плотные, прямые, не-
правильные. Устьичная щель ориентирована вдоль оси листа.
Околоустьичных клеток (они от короткочетырехугольных до
округлых) от 4 до 6. На 1 мм2 количество устьиц колеблется
от 40 до 100.
Pinus massoniana D. Don.
Лист сосны Массона амфистоматный. Эпидермис характе-
ризуется правильными рядами клеток, вытянутых вдоль листа.
Форма клеток верхнего эпидермиса четырехугольная, пря-
моугольная, трапециевидная. Продольные стенки параллельные,
поперечные — непараллельные. Размеры клеток различные.
Длина их максимальна в средней части листа (960 мк) и ми-
нимальна у основания (200 мк). На 1 мм2 приходится от 50
до 100 клеток.
Клетки нижнего и верхнего эпидермиса по форме одинако-
вы. В средней части листа они достигают своей максимальной
126
длины — 940 мк, у вершины — минимальной — 390 мк. Коли-
чество клеток на 1 мм2 — 400—600. Ширина клеток 40—70 мк.
Устьичный аппарат монопиклический, изолированный, пино-
идного типа. Устьица собраны в прямые, параллельные, уме-
ренно плотные ряды. Устьичная щель ориентирована вдоль
оси листа. Околоустьичные клетки от четырехугольной до ок-
руглой формы, располагаются в один ряд в количестве 4—8
(нижний эпидермис) и 4—7 (верхний эпидермис). На единицу
поверхности листа приходится 50—100 устьиц.
Pinus sylvestris L.
Лист сосны обыкновенной амфистоматный. Клетки верхнего
эпидермиса прямоугольной или трапециевидной формы, вытя-
нуты вдоль листа с параллельными продольными и непарал-
лельными поперечными стенками. Оболочки клеточных стенок
мелкоизвилистые. Максимальной длины клетки достигают у
основания, минимальной — в средней части листа. На 1 мм2
поверхности хвои приходится от 300 до 400 эпидермальных
клеток. * *
Форма клеток нижнего эпидермиса четырехугольная, пря-
моугольная, трапециевидная. Продольные стенки клеток пря-
мые, параллельные, поперечные — непараллельные, стенки их
мелкоизвилистые. Длина клеток колеблется от 440 до 560 мк,
причем максимальная ее величина у основания, минимальная
у вершины хвои. Ширина покровных клеток 50—60 мк
Устьичный аппарат монопиклический, изолированный, смеж-
ный, пиноидного типа. Устьичные ряды умеренно плотные, пря-
мые, неправильные. Устьичная щель ориентирована вдоль ли-
ста. Околоустьичные клетки прямоугольной формы, распола-
гаются в один ряд, в количестве от 3 до 5. На 1 мм2 поверх-
ности хвои приходится от 80 до 100 устьиц.
Pinus luchuensis Мауг
Лист амфистоматный. Форма клеток верхнего эпидермиса
четырехугольная, прямоугольная, трапециевидная. Клеточные
стенки прямые, оболочки их мелкоизвилистые. Максимально
длинные клетки находятся в средней части листа (670 мк), са-
мые короткие — у основания (260 мк). Количество клеток на
! мм2 поверхности листа колеблется от 300 до 600.
Клетки нижнего эпидермиса по форме не отличаются от
верхнего. Поперечные клеточные стенки прямые, параллельные,
продольные — непараллельные, мелкоизвилистые. Минимальная
длина клеток у вершины листа 330 мк, максимальная — в сред-
ней части листа — 720 мк.
Устьичный аппарат изолированный, монопиклический, пино-
идного типа. Устьичные ряды умеренно плотные, прямые, не-
10*
127
ш
X
со о S ф • 2, ►- X Ф 2 5 я х S S о. х » -
S н
О.
ф
м 1
S х еь ного голь ная
х
о ° А
ф 2 5 К
ч 5 О W
X X. и, -
X >>
о. X
о X
е четы рех- угол
°?
X
о 1
о
X ю 1
X sr 1 со
X
o' 4—7
ч
о
X о со 1
о
0
ф со 1
1
X со
о
4-8
Й9=
ф 3
S х
о
X
X
ИНН
о
-НЕЕК0Э
X
S
Д о х »=
§ О- РЗ 3
2 g д =
со
+++++++
+
+++++++
+++++ +
+
+
+
+
+
+ +
+ + + + + I
+
+ + + ++4:+
+++++++
Си
»а в
правильные. Околоустьичные
клетки округлой формы, распола-
гаются в один ряд, в количестве
от 4 до 9 штук. Количество усть-
иц на 1 мм2 колеблется от 50 до
60 (нижний эпидермис) и от 40
до 80 штук (верхний эпидермис).
Результаты настоящего иссле-
дования показали, что строение
эпидермиса хвои изученных ви-
дов сосен имеет как сходство,
так и различие (см. таблицу). У
всех видов устьичный аппарат пи-
ноидного типа (околоустьичные
клетки располагаются в один ряд
и количество их больше 4). Усть-
ица изолированные и смежные,
исключение составляют Р. thun-
bergiana и Р. luchuensis, у кото-
рых смежные устьица отсутству-
ют. Лишь у Р. funebris встречены
все три типа устьиц.
Форма околоустьичных клеток
устьичного аппарата эпидермиса
хвои. Р. funebris и Р. sylvestris
прямоугольная, у остальных ви-
дов округлая. Количество около-
устьичных клеток зависит, по-ви-
димому, от длины хвои. По это-
му признаку виды сосен можно
подразделить на длиннохвойные
(количество 4—9) и короткохвой-
ные (количество 3—6).
Размеры клеток эпидермиса и
количество устьиц на единицу
поверхности хвои находятся в
прямой зависимости от района
произрастания.
Литература
Сашина Г. С. Устьичный аппарат хвой-
ных и вопросы его филогении. —
Тезисы докл. XII Международн.
ботан. конгресса. Л., «Наука»,
1975, 263 с.
Свешникова Н. И. Исследование листь-
ев ископаемых и современных рас-
тений эпидермально-кутикулярным
методом. — «Бот. ж.», 1966, № 4,
т. 51, с. 584—590.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
МОРФОЛОГО-АНАТОМИЧЕСКОЕ СТРОЕНИЕ
ЛИСТА И ДРЕВЕСИНЫ ТИСА
TAXUS CUSPIDATA Sieb. et Zucc.
Г. И. ВОРОШИЛОВА
Тис остроконечный — один из древнейших представителей
флоры Дальнего Востока. Это дерево первой величины, дости-
гающее 20—21 м высоты и 100—120 см в диаметре (Воробьев,
1968). Встречается одиночными экземплярами или небольшими
группами в смешанных хвойно-широколиственных лесах. У се-
верной границы ареала может принимать кустарниковую и даже
стелющуюся форму (Куренцова, 1968а, б). Его следует отнести
к видам, нуждающимся в охране.
Тис является весьма ценной породой для озеленения в усло-
виях с повышенной и устойчивой влажностью воздуха. Одна-
ко в городских посадках он пока с успехом используется лишь
на Сахалине.
Для пополнения сведений по биологии тиса мы, используя
морфолого-анатомический метод, постарались дать полное опи-
сание его листа и древесины. Сбор материала производился в
1973—1975 гг. на юге Приморского края (Шкотовский район),
в окрестностях Южно-Сахалинска и на о-ве Кунашир. Обра-
ботка материала велась по общепринятой методике.
Лист тиса остроконечного ланцетный с одной жилкой. Дли-
на его колеблется от 12 до 26 мм, ширина — 2—2,7 мм, толщи-
на 0,4—0,7 мм. Вершина заострена в шипик, основание — кли-
новиднооттянутое, край листа гладкий.
Форма поперечного среза полулунная с выступом с нижней
стороны (жилка листа). На срезе обращает внимание сильное
развитие кутикулярного слоя. На верхней стороне листа он
образует сплошной слой, а на нижней — кутикула возвышает-
ся над эпидермальными клетками в виде гребней. Минималь-
ная толщина кутикулы равна 7,5 мк, максимальная — 12,5 мк.
Установлено, что ее толщина находится в прямой зависимости
от условий существования. Так, у листа тиса, произрастающего
на заболоченном участке на о-ве Кунашир, толщина кутикулы
была в два раза меньше (лист второго года жизни).
Лист тиса относится к гипостоматному типу, так как усть-
ичный аппарат располагается на его нижней стороне (Свеш-
никова, 1970). Устьичный аппарат погруженный, устьичная ям-
ка заполнена восковым веществом. По анатомическому строе-
129
нию лист дорзивентральный. Палисадная ткань прилегает
к верхней стороне листа и представлена двумя-тремя слоями
клеток округло-прямоугольной формы; остальная часть мезо-
филла представлена губчатой тканью с клетками округлой и
округло-прямоугольной формы. В жилке листа находится один
проводящий пучок эллиптической формы. От мезофилла жил-
ка отделена слоем плотно расположенных паренхимных кле-
ток (эндодерма отсутствует). Минимальный размер жилки по
вертикали поперечного среза равен 180 мк, максимальный —
280 мк, а по горизонтали соответственно 320 и 420 мк. Размер
проводящего пучка по вертикали 100—150 мк, по горизонта-
ли— 110—220 мк. Трансфузионная ткань располагается по
бокам проводящего пучка. В листе отсутствуют смоляные ка-
налы, склеренхима и гиподерма.
Форма клеток верхнего и нижнего эпидермиса листа различ-
на, иные они и по краю листа. Описание приводится по методи-
ке Г. С. Сашиной (1975).
Форма клеток верхнего эпидермиса трапециевидная, прямо-
угольная, многоугольная, квадратная, редко встречается тре
угольная с прямыми, скошенными поперечными стенками, ту
пыми и прямыми углами. Стенки клеток верхнего эпидермиса
ровные. Минимальная длина клеток верхнего эпидермиса
составляет 28 мк, максимальная — 84 мк, ширина соответст-
венно 16 и 56 мк. Количество клеток на 1 мм2 поверхности
листа колеблется от 500 до 700 штук.
Форма клеток нижнего эпидермиса прямоугольная и тра-
пециевидная с прямыми или скошенными поперечными стен-
ками и с тупыми или острыми углами. Стенки ровные. Мини-
мальная длина клеток 53 мк, максимальная — 168 мк, а ши-
рина соответственно 16 и 39 мк.
При изучении эпидермиса листа тиса в различных его ча-
стях обнаружены различия только в размерах клеток. На верх-
нем эпидермисе самые длинные клетки на вершине листа
(100 мк), а на нижнем эпидермисе — у основания листа
(184 мк). Наиболее широкие клетки эпидермиса расположены
в средней части листа. Клетки, расположенные по краю листа,
вытянуты вдоль его оси.
Непрерывность нижнего эпидермиса нарушается устьицами,
которые располагаются полосами. Устьичный аппарат изоли-
рованный, смежный и связанный. Устьичные ряды умеренно
плотные, изгибающиеся и неправильные. Устьичная щель ори-
ентирована вдоль оси листа. На 1 мм2 имеется от 100 до 250
устьиц. Устьичный аппарат амфицикличный, неполно или
полно дицикличный пинопицеоидного типа. Околоустьичные
клетки отличаются от эпидермальных формой и размерами,
особенно клетки первого ряда своей округлой или бобовидной
формой. Околоустьичные клетки располагаются в два ряда, и
их число колеблется от 11 до 17 штук. В строении эпидермиса
130
тиса остроконечного и китайских видов (Kwei Yao-Lin, Hu Yti-
shi, 1974) обнаруживаются большие сходства.
Форма устьичной ямки пяти- и шестиугольная, квадратная,
прямоугольная, в диаметре равна или чуть больше полярных
клеток. Замыкающие клетки нарушаются при мацерации.
В клетках нижнего эпидермиса присутствуют срединные и ла-
теральные папиллы.
Обработка материала показала, что строение мезофилла
листа тиса остроконечного имеет много общего с тисом ягод-
ным, структура которого дана Г. В. Куликовым и И. А. Ругу-
зовым (1973). Это обстоятельство подтверждает близость этих
видов в систематическом отношении. При изменении условий
произрастания наблюдаются количественные изменения в
строении хвои, что свидетельствует об адаптационных способ-
ностях тиса, весьма важных при разведении его в культуре.
Краткое описание древесины тиса приводят А. А. Цымек и
В. Н. Сергеева (1951), П. Грегуш (1963) и Л. В. Рыффа
(1965). Наши данные дополняют имеющиеся сведения.
Древесина тиса с желтовато-белой заболонью и красновато-
коричневым ядром, тяжелая, твердая. Годичные слои извили-
стые, узкие, хорошо выделяются на поперечном и радиальном
срезах.
Переход от ранней Древесины к поздней постепенный. Ши-
рина годичного слоя от 0,5 до 1,2 мм. Изредка встречаются
кольца шириной 0,08 мм. Число рядов трахеид в годичном
слое 25—70, иногда присутствуют кольца прироста, которые
состоят из шести рядов трахеид. Ширина ранней древесины в
2—14 раз превышает ширину поздней. Сечение ранних трахе-
ид округло-четырех-пяти-шестиугольное, поздних — сплюснутое
в радиальном направлении.
На радиальных стенках ранних трахеид располагаются
крупные окаймленные поры в один ряд, иногда встречаются
и два ряда пор. Распределены они по членику трахеид равно-
мерно. Диаметр окаймленных пор колеблется от 8 до 21 мк.
П. Грегуш (1963) отмечает больший диаметр пор — 16—18 мк.
В поздних трахеидах поры имеют косые отверстия. Спиральные
утолщения в древесине тиса имеют все трахеиды. Они отчетли-
во видны и равномерно распределены по всей трахеиде как на
радиальном, так и на тангентальном срезах.
Радиальные лучи тиса с идентурами. Горизонтальные стен-
ки луча с редкими порами, тангентальные — гладкие. На полях
перекреста располагаются 2—3—4 купрессоидные норы. Если
пор 2—3, то они расположены в один ряд по продольной оси
клетки, если их 4, то они располагаются по углам клетки.
Радиальные лучи однорядные, гомогенные. Высота их ко-
леблется от 1 до 28 клеток Однако есть сведения (Грегуш,
1963), что высота лучей доходит до 34 клеток. Длина трахеид
колеблется от 1600 до 3100 мк. Смоляные каналы, лучевые
131
трахеиды и паренхима в древесине тиса остроконечного отсут-
ствуют. При изучении срезов древесины тиса, находящегося в
различных условиях произрастания, обнаружены лишь количе-
ственные изменения.
Итак, дорзивентральное строение, отсутствие смоляных ка-
налов и эндодермы у листа тиса остроконечного указывают на
его высокую специализацию, в то время как наличие в древе-
сине спиральных трахеид и отсутствие смоляных каналов па-
ренхимы и лучевых трахеид, наоборот, характеризуют ее про-
стое строение.
ЛИТЕРАТУРА
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 278 с.
Грегуш П. Определитель древесины голосеменных по микроскопическим
признакам. М., Изд-во МГУ, 1963, 184 с.
Куликов Г. В., Ругузов И. А. Изменчивость анатомических показателей ли-
ста тиса ягодного в зависимости от условий местообитания.— «Эколо-
гия», 1973, № 1, с. 90—94.
Куренцова Г. Э. Растительность Приморского края. Владивосток, Дальне-
вост. кн. изд-во, 1968а, с. 102—111.
Куренцова Г. Э. Реликтовые растения Приморья. Л., «Наука», 19686. 71 с.
Рыффа Л. В. Сравнительная оценка основных анатомических элементов не-
которых хвойных пород юга Дальнего Востока.— Восьмая конферен-
ция молодых ученых Дальнего Востока, сер. биол. наук. Владивосток,
1965, с. 12—13.
Сашина Г. С. Устьичный аппарат хвойных и вопросы его филогении.— Тези-
сы докладов на XII Межд. бот. конгрессе, Л., «Наука», 1975, с. 263.
Свешникова И. Н. О терминологии эпидермы листа современных и ископае-
мых хвойных.— «Бот. ж.», 1970, №4, т. 55, с. 510—521.
Цымек А. А., Сергеева В. Н. Анатомические признаки древесины главней-
ших хвойных пород Дальнего Востока.— Особенности развития лес-
ного хозяйства Дальнего Востока, в. III. М.—Л., Гослесбумиздат, 1951,
с. 119—122.
Kwei Yao-Lin. Hu Yu-shi. Epidermal feature of the leaves of Taxus in relati-
on to taxonomy.— Acta phytotaxonomica sinica, 12, 1974, №3,
p. 329—335.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
СТРОЕНИЕ ЭПИДЕРМИСА ХВОИ ЛИСТВЕННИЦ
ПРИМОРСКОГО КРАЯ
Р. А. КОРОЛЕВА
Род Larix в систематическом отношении еще недостаточно
изучен. Так, на территории Дальнего Востока В. Л. Комаров
выделяет 2 вида и 3 формы лиственницы, В. Н. Сукачев — 5
видов, Б. П. Колесников — 9 видов, а Н. В. Дылис — 4 вида
(по Боброву, 1972). Эти данные свидетельствуют о том, что
вопрос систематики рода нельзя считать окончательно решен-
ным и для его доработки необходимо привлечение комплекс-
ных исследований, в том числе морфолого^анатомических.
В настоящее время установлено, что внутренние анатоми-
ческие структуры более устойчивы и более способны к удер-
жанию древних свойств, чем внешние — морфологические. Ана-
томические особенности хвои у лиственниц были признаны
В. Н. Сукачевым (1924) в качестве весьма ценных и важных
показателей не только для диагностики видов, но и для выяс-
нения филогенетических отношений в роде Larix.
Для изучения строения эпидермиса были взяты лиственни-
цы даурская (Larix dahurica Turcz.), Комарова (L. komaro-
vii Kolesn.), Любарского (L. lubarskii Sukacz.), ольгинская
(L. olgensis Henry), приморская (L. marituna Sukacz.).
Материал для работы был собран сотрудниками кафедры
ботаники ДВГУ в 1972—1974 гг.
Для изучения строения эпидермиса пользовались методикой
И. Н. Свешниковой (1966, 1970) и Г. С. Сашиной (1975). Изу-
чался эпидермис средней части хвои.
Лиственница ольгинская. Покровная ткань хвои
лиственницы представлена эпидермисом. Эпидермальные клет-
ки вытянутотрапециевидной, неправильной формы с парал-
лельными или скошенными поперечными перегородками.
Стенки эпидермальных клеток гладкие.
По расположению устьичного аппарата хвоя гипостомат-
ная — устьица на нижней стороне листа. Устьичный аппарат
моноцикличного типа — один ряд околоустьичных клеток.
Устьица на поверхности листа располагаются двумя полосами.
В полосах устьица лежат 3—6 сплошными, плотными, иногда
умеренно плотными рядами. Между устьичными рядами лежит
один, реже два ряда эпидермальных клеток, если они исчезают,
133
то устьичные ряды соприкасаются околоустьичными клетками.
Устьичные ряды неправильные, слабоизгибающиеся, парал-
лельны продольной оси листа, в этом же направлении ориенти-
рована устьичная щель.
Для лиственниц характерен лярикоидный тип устьичного
аппарата — устьица окружены четырьмя околоустьичными
клетками. Околоустьичные клетки отличаются от эпидермаль-
ных формой и размерами. Латеральные клетки неправильной
формы, вытянуты вдоль устьичной ямки. Полярные клетки про-
долговатые. Устьичный аппарат связанный, смежный, встреча-
ется единично изолированный. Устьица погруженные. Устьич-
ная ямка с поверхности листа прямоугольной формы.
Лиственница Любарского. По расположению усть-
ичного аппарата хвоя гипостоматная, моноцикличного типа.
Клетки верхнего эпидермиса удлиненно-трапециевидной, непра-
вильной формы с прямыми и скошенными поперечными стен-
ками. Клетки нижнего эпидермиса неправильной формы.
Стенки эпидермальных клеток гладкие.
Устьица на поверхности листа располагаются двуми поло-
сами, состоящими из 4—5 сплошных плотных рядов. Устьич-
ные ряды неправильные, слабоизгибающиеся, параллельны
продольной оси листа. Устьица лярикоидного типа, погружен-
ные. Околоустьичные клетки отличаются от эпидермальных
формой и размерами. Латеральные клетки неправильной фор-
мы, расширяющиеся на границе с устьичной ямкой и суживаю-
щиеся на концах. Полярные клетки округлой, продолговатой
формы. Связанный и смежный устьичный аппарат, редко встре-
чается изолированный.
Лиственница даурская. Листовая пластинка по
расположению устьичного аппарата гипостоматная. На верх-
ней стороне листа клетки эпидермиса трапециевидной, прямо-
угольной, неправильной формы с прямыми или скошенными
поперечными стенками. Клетки нижнего эпидермиса имеют не-
правильную форму. Стенки эпидермальных клеток гладкие.
Устьица моноцикличные, лярикоидного типа, погруженные.
На поверхности листа устьица располагаются двумя полосами,
состоящими из 3—4 сплошных плотных рядов. Устьичные ряды
неправильные, слабоизгибающиеся, параллельны продольной
оси листа. Между устьичными рядами не всегда находится ряд
эпидермальных клеток; когда их нет, устьичные ряды соприка-
саются околоустьичными клетками. Устьичный аппарат свя-
занный, редко встречается смежный, и нами не обнаружен изо-
лированный тип. Околоустьичные клетки отличаются от эпи-
дермальных. Полярные клетки продолговатой формы. Лате-
ральные клетки имеют неправильную форму. Наибольшая ши-
рина латеральной клетки — на границе с устьичной ямкой,
которая с поверхности листа прямоугольной формы.
134
Лиственница приморская. Эпидермальные клетки
прямоугольной, трапециевидной формы с прямыми и скошен-
ными поперечными стенками. Стенки эпидермальных стенок
гладкие.
Устьичный аппарат лярикоидного типа расположен на ниж-
ней стороне листа — хвоя гйпостоматная. Устьица на поверх-
ности листа располагаются двумя полосами, состоящими из
5 плотных устьичных рядов. Устьичные ряды неправильные
(ширина междурядий различна), прямые.
Устьичная щель расположена вдоль продольной оси листа.
Устьица погруженные, моноцикличные. Полярные клетки у
лиственницы приморской имеют бочонковидную, округлую
форму, а латеральные — продолговатую, четырехугольную.
Устьичный аппарат связанный, редко смежный, и нами не об-
наружен изолированный тип. Устьичная ямка прямоугольной
формы.
Лиственница Комарова. Покровная ткань хвои
лиственницы представлена эпидермисом. Эпидермальные клет-
ки трапециевидной, неправильной формы. Край стенок эпидер-
мальных клеток гладкий. По расположению устьичного аппа-
рата хвоя гйпостоматная. Устьичный аппарат лярикоидного
типа, т. е. устьичная щель окружена четырьмя околоустьичны-
ми клетками. Устьица погруженные, моноцикличные. Устьица
на поверхности листа располагаются двумя полосами, состоя-
щими из 3 сплошных плотных рядов. Устьичные ряды непра-
вильные — ширина междурядья различна, слабоизгибающиеся,
параллельны продольной оси листа. Устьичные ряды либо
соприкасаются, либо между ними лежит один ряд эпидермаль-
ных клеток. Околоустьичные клетки отличаются от эпидермаль-
ных формой и размером. Полярные клетки округлой формы,
латеральные — неправильной формы, вытянуты вдоль устьич-
ной ямки и имеют разную ширину. Устьичная ямка прямо-
угольной формы. Устьичная щель параллельна продольной
оси листа. Устьичный аппарат связанного типа, редко встреча-
ется смежного, изолированный тип не обнаружен.
В результате изучения строения эпидермиса хвои пяти ви-
дов лиственницы были выявлены общие черты, характерные
для рода, а также черты, присущие отдельным видам.
Покровная ткань хвои у всех видов представлена эпидер
мисом. Клетки эпидермиса бывают прямоугольной, трапецие-
видной, неправильной формы. Стенки эпидермальных клеток
гладкие. У всех видов лиственниц устьичный аппарат моно-
дикличный, лярикоидного типа. Устьица на поверхности листа
располагаются двумя полосами. В полосах устьица располага-
ются сплошными плотными, иногда умеренно плотными (лист-
венница ольгинская) рядами. Устьичные ряды слабоизгибаю-
щиеся, редко прямые (лиственница приморская). Форма около-
устьичных полярных клеток округлая (лиственницы Комарова,
135
Любарского, приморская), продолговатая (лиственницы даур-
ская, ольгинская, Любарского) и бочонковидная (лиственница
приморская). Латеральные околоустьичные клетки неправиль-
ной формы, только у лиственницы приморской они имеют про-
долговатую и четырехугольную форму. Устьица связанные и
смежные, изолированные отмечены нами только в хвое лист-
венницы ольгинской.
ЛИТЕРАТУРА
Бобров Е. Г. История и систематика лиственниц. Л., «Наука», 1972, с. 3—95
Сашина Г. С. Устьичный аппарат хвойных и вопросы его филогении.— Тези-
сы докладов XII Межд. бот конгресса. Л., «Наука», 1975, с. 263.
Свешникова И. Н. Исследование листьев ископаемых и современных расте-
ний эпидермально-кутикуляюным методом.— «Бот. ж.», т. 51, 1966,
№4, с. 584—590.
Свешникова И. Н. О терминологии эпидермы листа современных и ископае-
мых хвойных.— «Бот. ж.», т. 55, 1970, №4.
Сукачев В. Н. К истории развития лиственниц.— Лесное дело. М.—Л., 1924,
с. 12—44.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
К АНАТОМИЧЕСКОЙ ХАРАКТЕРИСТИКЕ ДРЕВЕСИНЫ
НЕКОТОРЫХ АРАЛИЕВЫХ ПРИМОРЬЯ
С. А. СНЕЖКОВА
В Приморье произрастает наибольшее в СССР количество
представителей сем. Araliaceae Juss., объединяющего в основ-
ном роды и виды, свойственные тропическим районам. Здесь
насчитывается пять родов и семь видов, большинство из них —
деревья и кустарники.
Представители сем. Araliaceae постоянно привлекали вни-
мание многих ботаников. Однако анатомия древесины видов,
произрастающих в нашей стране, недостаточно изучена. В за-
рубежной литературе имеется большое количество работ, по-
священных анатомии аралиевых (Harms, 1918; Капо, 1936; Su-
do, 1959). Однако работы этих авторов касаются в основном
видов, не произрастающих на территории СССР. -В отечествен-
ной литературе краткие сведения по анатомии древесины ара-
лиевых имеются лишь в нескольких работах (Абраменко, 1935;
Гаммерман и др., 1946; Ванин, 1949; Андреева, 1963, 1969).
Целью нашей работы было сравнительное изучение древе-
сины трех представителей этого семейства: Eleutherococcus
senticosus (Rupr. et Maxim.) Maxim. — элеутерококка колючего,
Acanthopanax sessiliflorum (Rupr. et Maxim.) Seem.— аканто-
панакса сидячецветкового и Kalopanax septemlobum (Thunb.)
Koidz. — калопанакса семилопастного.
Многие авторы, отмечая сходство представителей этих трех
родов, объединяли их в один род Acanthopanax, например
Гармс (Harms, 1918). Еще в 1916 г. Вигье (Viguier, цит. по
Капо, 1936) отметил, что изучение анатомии этого семейства
может многое дать для его классификации. Однако, исходя из
анатомических данных, он тоже объединял три этих рода в
один большой род Acanthopanax. Наиболее детальное иссле-
дование этих родов провел Кано (Капо, 1936). Исследуя виды
этих родов, он рассматривал Eleutherococcus senticosus как
Acanthopanax senticosus, a Kalopanax septemlobum — под наз-
ванием К. ricinifolium Mig. Кано отмечает, что древесина
К. ricinifolium по анатомическому строению подобна древесине
Acanthopanax, особенно в первые несколько лет жизни, и хо-
рошо отличается от других видов Kalopanax. А древесина
A. sessiliflorum и A. senticosus по строению очень схожа. К- sep-
137
temlobum долгое время был известен под синонимом К. ricini-
folium Mig., а в 1925 г. Коидзуми переименовал его в К. sep-
temlobum Koidz- (Пояркова, 1950). В 1927 г. этот вид был вто-
рично переименован в К Dictum Nakai, под этим названием он
описан в работе В. Л. Некрасовой (1933) и в определителе
А. Ф. Гаммерман и др. (1946). Но за калопанаксом сохрани-
лось видовое название К. septemlobum (Thunb.) Koidz.
Для исследования были взяты образцы древесины с 2—3
растений каждого вида, произрастающих в смешанном лесу
вблизи ст. Анисимовка Шкотовского района Приморского края.
С кустарников образцы брались в нижней части ствола, с де-
ревьев— на высоте 1,3 м. Описание анатомического строения
древесины производилось по схеме, разработанной А. А. Яцен-
ко-Хмелевским (1954). Характеристика объектов исследования
и их распространение приводятся по определителю Д. П. Во-
робьева (1968).
Акантопанакс сндячецветковый — высокий ветвистый кус-
тарник, растущий по лесистым берегам рек и речек, по опуш-
кам и лесным прогалинам, одиночно или небольшими группами.
Элеутерококк колючий — кустарник до 2, реже 4—5 м высо-
той, растущий в смешанных и хвойных горных лесах небольши-
ми зарослями или группами. Калопанакс семилопастный — де-
рево до 20—25 м высоты и 50—80 см в диаметре на высоте
груди, предельный возраст около 200 лет. Растет калопанакс
в смешанных и широколиственных лесах.
По нашим наблюдениям, для древесины A. sessiliflorum и
Е. senticosus характерны следующие признаки. В составе дре-
весины имеются сосуды, сосудистые трахеиды, волокна либри-
форма, перегородчатые волокна, тяжевая и лучевая паренхима.
Граница годичного кольца хорошо выражена благодаря нали-
чию полоски сплюснутых волокон и сосудов в 2—4 слоя. Дре-
весина рассеяннососудистая с рисунком в виде волнистых
тангентальных полос шириной в 1—2 просвета у акантопанакса
и более широких, в 2—3 просвета,— у элеутерококка. Переход
от ранней древесины к поздней постепенный, размеры просве-
тов сосудов уменьшаются от ранней древесины к поздней в 2—
4 раза. Очертания большинства просветов овальные, радиально
вытянутые, некоторые угловатые. Сосуды многочисленные, с
диаметром просвета от 20 до 60 мк. У акантопанакса отмечены
тиллы. Членики сосудов удлиненные: 320—560 мк у элеутеро-
кокка, 350—510 мк у акантопанакса, с короткими и длинными
клювами, реже без клювов. Перфорации сосудов простые, на
поперечных скошенных и боковых стенках. Межсосудистая по-
ровость очередная, поры крупные, сближенные и многочислен-
ные свободные; округлые, реже овальные; внутренние отвер-
стия пор включенные, округлые и вытянутые. Сосудистые тра-
хеиды располагаются вместе с сосудами в группах, от узких
сосудов отличаются только отсутствием перфораций.
138
Древесинная паренхима вазицентрическая, в тяже 2—6 кле-
ток. Поры между клетками паренхимы и сосудами крупные,
овальные.
Основная масса древесины состоит из волокон либриформа
и перегородчатых волокон, имеющих толстые стенки — 4—
6,5 мк у акантопанакса, 2,7—5,5 мк у элеутерококка. Волокна
с простыми, мелкими, редкими порами, расположенными в од-
ном вертикальном ряду на радиальных стенках. Длина волокон
у акантопанакса от 440 до 750 мк, у элеутерококка — от 600
до 900 мк. Окончания волокон гладкие, штыкообразные и за-
зубренные.
Лучи смешанно-гетерогенные, 1—4-рядные у акантопанакса
и 1—5-рядные у элеутерококка. Однорядные лучи редки, высо-
та йх от 2 до 7 клеток. Высота многорядных лучей у акантопа-
накса от 5 до 55 клеток, у элеутерококка от 3 до 220 клеток и
более, часто 70—80 клеток. На поперечном срезе узкие лучи
при встрече с сосудами изгибаются, при переходе из одного
годичного слоя в другой лучи не расширяются у элеутерококка
и незначительно расширяются у акантопанакса. Тангентальные
диаметры широких просветов сосудов превышают ширину ши-
роких лучей. Тангентальные стенки клеток лучей и прямые и
косые. На тангентальном срезе однорядные лучи линейные,
многорядные веретеновидные, некоторые с короткими оконча-
ниями — в 2—4 клетки у акантопанакса и 2—7 клеток у элеу-
терококка; у последнего же отмечены сдвоенные лучи. На
радиальном срезе лучи составлены из лежачих, стоячих и квад-
ратных клеток. Стоячие и квадратные клетки образуют отдель-
ные слои, чаще краевые (1—3 слоя) и невысокие лучи; могут
быть вкраплены среди лежачих клеток. Длина лежачих клеток
в 2—4 раза превышает их ширину. Высота стоячих клеток в
2—3 раза превышает ширину. Поры между клетками луча и
сосудами крупные, овальные, 3—5 пор по высоте стоячих кле-
ток, 1—2 — по высоте лежачих клеток; поры имеются в боль-
шинстве слоев. Стенки клеток лучей незначительной толщины.
В отличие от древесины элеутерококка и акантопанакса,
древесина калопанакса кольцесосудистая. В ее составе: сосуды,
сосудистые трахеиды, волокна либриформа, перегородчатые
волокна, тяжевая и лучевая паренхима. Граница годичного
кольца выражена отчетливо благодаря полоске сплюснутых
сосудов и волокон в 2—4 слоя. Годичные кольца имеют хорошо
выраженное кольцо просветов сосудов из одного-двух слоев,
оно довольно плотное, хотя некоторые крупные сосуды разде-
лены сосудистыми трахеидами, клетками паренхимы. Переход
от кольца просветов к поздней древесине резкий, просветы
поздней древесины располагаются в виде изогнутых танген-
тальных полос шириной в 2—5 просветов. Большинство просве-
тов имеют овальные очертания, в сосудах имеются тиллы.
Диаметр просветов сосудов ранней древесины 75—255 мк, позд-
139
ней древесины 15—65 мк. Членики сосудов ранней древесины
220—680 мк, поздней — 200—600 мк длиной, с короткими и
длинными клювами; широкие членики чаще без клювов. Пер-
форации сосудов простые на поперечных и боковых стенках.
Межсосудистая поровость очередная, поры свободные и сбли-
женные, округлые. Внутренние отверстия пор округлые или
вытянутые, включенные. Сосудистые трахеиды похожи на уз-
кие сосуды, отличаются только отсутствием перфораций. Па-
ренхима вазицентрическая, более обильная в ранней древесине;
в тяже от 2 до 6 клеток. Поры между клетками паренхимы и
сосудами крупные, овальные.
Основная часть древесины представлена волокнами либри-
форма и перегородчатыми волокнами. Волокна с простыми,
мелкими, редкими порами в одном вертикальном ряду. Обо-
лочки волокон толстые, окончания — гладкие, зазубренные,
штыкообразные.
Лучи смешанно-гетерогенные 1—5-рядные, чаще 4-рядные.
Высота однорядных лучей — 2—7 клеток, многорядных — 4—
35 клеток. На поперечном срезе все лучи уже диаметров широ-
ких просветов. Граница годичного слоя в лучах часто совпадает
с общей границей годичного слоя. Лучи при встрече с сосуда-
ми изгибаются. Тангентальные стенки клеток лучей прямые и
косые. При переходе из одного слоя в другой лучи не расширя-
ются. Все лучи на тангентальном срезе веретеновидные, одно
рядные линейные. На радиальном срезе лучи составлены из
лежачих, стоячих и квадратных клеток. Стоячие и квадратные
клетки в 1—3 слоя расположены по краям луча. Длина лежа-
чих клеток в 3—6 раз превышает ширину. Высота стоячих
клеток в 2—3 раза превышает ширину. Поры между клетками
луча и сосудами простые, овальные, 3—6 пор по высоте стоя-
чих клеток, 1—3 по высоте лежачих клеток. С генки клеток
лучей незначительной толщины.
Итак, исследования показывают, что анатомическое строе-
ние древесины этих видов может различаться по следующим
признакам:
1. Древесина калопанакса хорошо отличается от древесины
элеутерококка и акантопанакса ясно выраженной кольцесосу-
дистостью, а кроме того меньшей высотой лучей (до 35 клеток).
2. Древесина элеутерококка и акантопанакса схожа, однако
довольно значительные различия имеются в характере лучей —
более высокие и широкие у элеутерококка, 1—5-рядные, высо-
той более 220 клеток; у акантопанакса — 1—4-рядные, от 2 до
55 клеток высотой. Кроме того, у элеутерококка на попереч-
ном срезе древесины просветы сосудов располагаются в широ-
ких тангентальных полосах, у акантопанакса в более узких.
140
ЛИТЕРАТУРА
Абраменко С. М. Определитель древесины главнейших пород СССР. Л.,
Гослестехиздат, 1935.
Андреева И. С. Анатомическое строение акантопанакса скученно-цветного.—
Сообщения ДВФ СО АН СССР, в. 18. Владивосток, 1963, с. 67—72.
Андреева И. С. Анатомия вегетативных органов калопанакса семилопастного
Kalopanax septeinlobum (Thunb) Koidz.— Ученые записки (Омского
пединститута). Вопросы биологии, в. 51, 1969, с. 104—119.
Ванин С. И. Древесиноведение. М—Л., Гослесбумиздат, 1949.
Воробьев Д. П. Дикорастущие деревья и кустарники Дальнего Востока. Л.,
«Наука», 1968, 274 с.
Гаммерман А. Ф., Никитин А. А., Николаева Т. Л. Определитель древесины
по микроскопическим признакам. М.—Л., Изд-во АН СССР, 1946.
106 с.
Некрасова В. Л. Обзор аралиевых СССР, их географическое распростране-
ние и использование.— «Советская ботаника», 1933, №6, с. 82—100.
Супрунов Н. И. Анатомическое изучение растений сем. аралиевых.— Труды
Ленинградского химико-фармацевтического института, т. 17, 1964, в. 2,
с. 121—142.
Пояркова А. И. Семейство Araliaceae Vent.— Флора СССР, т. 16. М., Изд-во
АН СССР, 1950, с. 1—36.
Яценко-Хмелевский А. А. Основы и методы анатомического исследования
древесины. М.—Л., Изд-во АН СССР, 1954, 337 с.
Harms Н. Ubersicht fiber die Arten der Gattung Acanthopanax. Mittcil. de-
utch. Gesel., 27, 1918.
Kano R.Vergleichende Anatomic йЬег die Gattung Acanthopanax und die ver-
wandten Gattunen.— «Japanese jeurn. of botany, v. VIII, 1936, №3
Sudo S. Identification of Japanese hardwoods Reprinted from Bulletin of the
government forest experiment station. № 119, Tokyo, 1959.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
О ПЫЛЬЦЕ ДАЛЬНЕВОСТОЧНЫХ БЕРЕСКЛЕТОВ
Ю. С. ПОЛИЙЧУК
Современные дальневосточные бересклеты — это листопад-
ные кустарники или деревца, обитающие чаще всего под поло-
гом широколиственного или смешанного леса; они весьма деко-
ративны как во время цветения, так и в период плодоношения.
Распространены неодинаково, многие виды довольно редки. Во
флоре Дальнего Востока род Euonymus L. представлен секция-
ми Euonymus Leon., Pseudoevonymus Nakai, Melanocaria
(Turcz.) Nakai из подрода Euonymus и секцией Uniloculares
Rouy et Fouc. из подрода Kalonymus G. Beck. (Леонова, 1974).
Сведения о пыльце бересклетов исследуемого региона, как
и в целом дендрофлоры Дальнего Востока, весьма ограниченны
и касаются лишь 3 видов (Гладкова. 1950; Ikuse, 1956; Мор-
фология пыльцы растений Китая, 1960; Мячина и др., 1971).
Нами изучена пыльца 9 видов бересклета (Euonymus maa?kii
Rupr., Е. sieboldiana Blume., E. pauciflora Maxim., E. sacrosan-
c’a Koidz., E. macroptera Rupr., E. maximowicziana Prokh ,
E. planipes Koehne, E. sachalinensis Max., E. miniata Tolm.)
с помощью светового микроскопа МБИ-3 Размеры приводятся
для ацетолизированных зеосн. Фактическим материалом иссле-
дований явились личные сборы автора и гербарные образцы,
хранящиеся в БИН им. В. Л. Комарова АН СССР.
Пыльцевые зерна бересклетов трехбороздно-оровые, почти
сфероидальные, с полюса округло-трехпластные или тре-
угольно-округлые; с экватора эллиптические или округлые.
Размеры пыльцевых зерен исследованных видов бересклета
колеблются в пределах от 25,2 до 44,8 мк; самая крупная
пыльца отмечена для Е. miniata Tolm.: 42—44,8 мк (табл.).
Экзина пыльцы бересклетов состоит из трех слоев — подсти-
лающего, столбикового и перфорированного покровного. Пос-
ледний составляет простую сетчатую скульптуру поверхности
зерен. По характеру скульптуры пыльцевые зерна могут быть
крупно- и мелкосетчатые.
На основании числа борозд, толщины экзины и размера
зерен мы подразделяем пыльцу рода Euonymus на 2 морфоло-
гических типа. Это деление соответствует таксономическому
делению рода на подроды.
142
Размеры пыльцевых зерен рода Euonymus L.
Вид Полярная ось, мк Экваториаль- ный диаметр, мк Толщина эк- зины, мк Исследованный образец
Euonymus maackii 30,8—36,4 28,0—33,6 4,2 Владивосток, 25-й км, 28 июня 1970 г., Ю. С. Полийчук о-в Сахалин, 12 июля 1956 г. Т. Г. Леонова, Г. Шухтина
Е. sieboldiana 30,8—36,4 33,6-39,2 4,2
Е. pauciflora 25,2—30,8 25,2—28,0 4,2 Владивосток, 26-й км, 2 июня 1963 г. Ю. С. Полийчук
Е. sacrosancta 30,0—33,6 28,0—30,8 4,6 Владивосток, 25-й км, 1970 г. Ю. С. Полийчук
Е. macroptera 36,4—42,0 36,4—42,0 4,6—5,5 о-в Сахалин, 24 июня 1956 г., Г. Шухтина
Е. maximowicziana 33,6—42,0 30,0—39,2 4,6—5,5 Владивосток, б. Артур, 7 мая 1972 г., Ю. С. Полийчук
Е. planipes 36,4—42,0 36,4—42,0 4,6—5,8 о-в Сахалин, 27 июня 1956 г., Т. Г. Леонова, Г. Шухтина
Е. sachalinensis 33,6—39,2 33,6—39,2 5,6 о-в Сахалин, 24 июня 1953 г. А. И. Толмачев
Е. miniata 42,0—44,8 36,4—42,0 5,6 о-в Сахалин, 27 июня 1955 г., Т. Г. Леонова, Е. Толмачева
1 тип — Euonymus.
Пыльцевые зерна мелкие, полярная ось 25,2—36,4 мк, эква-
ториальный диаметр 25,2—33,6 мк, экзина 4,2 мк в центре ме-
зокольпиума; с полюса округло-трехлопастные, с экватора эл-
липтические. Столбики ясно булавовидные. Виды: Е. maackii,
Е. sieboldiana, Е. pauciflora, Е. sacrosancta.
2 тип — Kalonymus.
Пыльцевые зерна крупные, полярная ось 33,6—44,8 мк, эк-
ваториальный диаметр 30,0—42,0 мк, экзина 4,2—5,8 мк в
центре мезокольпиума; с полюса резко трехлопастные, с эква-
тора эллиптические. Столбики почти цилиндрические, головки
слабо выражены. Число борозд варьирует от 2 до 4, кроме то-
го, помимо обычных пыльцевых зерен с меридиональными бо-
роздами отмечены слитно-трехбороздные. Виды: Е. macroptera.
Е. maxiniowicziana, Е. planipes, Е. miniata, Е. sachalinensis.
Особая трудность при определении видов бересклета по
пыльце заключается в том, что их пыльцевые зерна относятся
к одному апертурному типу. Однако такие признаки спородер
мы, как скульптура, форма и очертания зерен в полярной и
экваториальной проекциях, размер зерен, дают возможность
установить и видовые признаки. В частности, среди бороздно-
оровых пыльцевых зерен бересклетов почти все виды достаточ-
но четко отличаются по размерам и форме оры. В том случае,
когда апертуры практически сходны (Е. pauciflora и Е. sacro-
sancta), на оптическом срезе спородермы отчетливо выступает
разница между этими видами: у Е. sacrosancta длинные цилин-
дрические столбики, а пыльцевые зерна Е. pauciflora с корот-
кими булавовидными столбиками.
Секционное деление подрода Euonymus отражается в раз-
мерах пыльцевых зерен исследованных видов. Наиболее мел-
кая пыльца в секции Pseudovyenomus, несколько крупнее в
секции Melanocarya, более крупные в секции Euonymus.
Таким образом, палиноморфологическое исследование бере-
склетов показало, что деление рода на 2 подрода вполне оп-
равданно, и, видимо, нет оснований для выделения подрода
Kalonymus в самостоятельный род, хотя именно здесь заметно
разнообразие апертурного аппарата. Последнее, возможно,
объясняется полиплоидией, и тем самым подтверждается мне-
ние относительно вероятной молодости подрода.
ЛИТЕРАТУРА
Гладкова А. Н. Семейство Celastraceae — береклестовые.— Пыльцевой анализ.
М., Госгеолиздат, 1950, с. 265—266.
Леонова Т. Г. Бересклеты СССР и сопредельных стран. Л., «Наука», 1974.
Мячина А. И , Казачихина Л. Л., Мамонтова И. Б., Калинина В. С. Атлас
пыльцы и спор некоторых современных растений Дальнего Востока.
Хабаровск, 1971, 85 с.
Ikuse М. Pollen grains of Japan. Tokyo, 1956.
РЕДКИЕ И ИСЧЕЗАЮЩИЕ ДРЕВЕСНЫЕ РАСТЕНИЯ ЮГА ДАЛЬНЕГО ВОСТОКА
1978
ЛАНДШАФТНОЕ ЛЕСОУСТРОЙСТВО ЛЕСОПАРКА
АКАДЕМГОРОДКА ДАЛЬНЕВОСТОЧНОГО НАУЧНОГО
ЦЕНТРА АН СССР
А. Ф. ЖУРАВКОВ
В связи с проблемой сохранения редких и исчезающих ра-
стений флоры Приморья, в частности ее древесных представи-
телей, особый интерес представляет лесоустройство пригород-
ных территорий. Пригородные леса более других посещаются
отдыхающими, на их территории ведется в той или иной мере
строительство — все это отрицательно сказывается на флори-
стическом составе; в первую очередь сокращается численность
редко встречающихся видов.
Лесоустройство зеленых зон может сыграть свою роль в
деле создания условий для обогащения растительного покро-
ва особо ценными представителями и сохранения генофонда.
Представляет интерес ландшафтное лесоустройство Академго-
родка в зеленой зоне Владивостока. Он расположен на п-овтз
Муравьева-Амурского в северо-западной части города на поло-
гих склонах хр. Центрального, обращенных к Амурскому за-
ливу Японского моря, его территория составляет 468 га, из
которой покрытая лесом — 183 га.
Леса преимущественно вторичные, сильно измененные, пар-
ковые. Среди насаждений размещены частные постройки, ве-
домственные жилые дома, пионерские лагеря и различные
предприятия. Через всю территорию проходит электрифициро-
ванная железная дорога. Близость большого города, наличие
хороших транспортных связей способствуют интенсивному по-
сещению лесопарка в любое время года; в результате там
образовалось множество троп и грунтовых дорог.
Первоначально предусматривалось размещение объектов
строительства ДВНЦ АН СССР только на свободных от леса
участках при максимально возможной плотности застройки,
а также ведение лесопаркового хозяйства с учетом использо-
вания этих мест для отдыха горожан. Осуществление этих ус-
ловий на практике потребовало создания лесопарковой опыт-
ной станции с задачами организации лесопаркового хозяйства,
зеленого строительства, благоустройтельных работ, охраны и
защиты насаждений и отдельных редких и исчезающих видов,
а также ведения научно-исследовательской и опытно-производ-
ственной работы по этим вопросам.
145
Для составления плана детальной планировки Академгород-
ка, оценки состояния насаждений и разработки мероприятий
по лесопарковому ведению хозяйства возникла необходимость
в проведении лесоустроительных работ, которые в !973—
1974 гг. выполнены Приморским филиалом Союзгипролесхоз
совместно с лабораторией зеленых зон и ландшафтной архи-
тектуры Ботанического сада ДВНЦ АН СССР.
Площадь лесопарка была разделена на 25 кварталов, гра-
ницами которых послужили естественные разделительные ли-
нии (дороги, тропы, улицы, трассы линий электропередач и
связи, водоводы и т. п.) и только в отдельных исключительных
случаях делались затесы на деревьях до луба или наносились
меты масляной краской. Квартальная сеть увязывалась с гене-
ральным планом застройки Академгородка, разработанным
‘Дальневосточным отделением ГИПРОНИИ АН СССР. Средний
размер квартала определился в 18,7 га (максимальный — 29 га
и минимальный — 3 га).
Каждому участку помимо обычной таксационной характе-
ристики давалась дополнительная: тип ландшафта и пейзажа,
эстетическая и санитарно-гигиеническая оценки, жизнеустой-
чивость насаждений, проходимость, характер сочетания с окру-
жающими типами пейзажей, строениями, сооружениями. Ука-
зывались положение в рельефе, экспозиция и крутизна склона,
форма склона и его часть, сомкнутость полога древостоя, на-
личие видовых точек и их характеристика, наличие редких и
исчезающих видов или деревьев и кустарников, обладающих
высокими декоративными качествами, наличие дорожно-тропи-
ночной сети и ее состояние.
Таксация проводилась без применения аэрофотоснимков.
Абрисы готовились на миллиметровой бумаге в масштабе
1 2000. Хозяйственные распоряжения отражались в таксаци-
онных описаниях для каждого выдела — типа пейзажа — с ука-
занием мероприятий (кроме лесохозяйственного характера)
по благоустройству и облагораживанию того или иного участка
территории Академгородка.
В результате полевых учетных работ установлено, что лес-
ная площадь составляет 256 га, из которых 177 га покрыты
лесом. Редины занимают 6 га, несомкнувшиеся лесные куль-
туры— 2,8 га, прогалины и пустыри — 64 га. Нелесной площа-
ди здесь 212 га и представлена она огородами частных лиц
(8,8 га), водоемами (0,1 га), жилой, преимущественно мало-
этажной, застройкой (164 га), дорогами (16,6 га), трассами
линий электропередач, связи и водоводов (17,6 га), пляжами
Амурского залива (4,2 га), болотами, оврагами и крутыми
склонами (0,7 га).
Древостои почти полностью естественного происхождения.
Искусственных насаждений только 6,2 га. Из древесных пород
господствующее положение принадлежит дубу монгольскому —
146
82,7 га (45,2% покрытой лесом площади). Значительная тер-
ритория занята насаждениями с преобладанием ясеня мань-
чжурского— 37,6 га (20,5%) и березы маньчжурской—19,4 га
(10,6%) Другие лиственные породы (липы маньчжурская и
амурская, березы даурская и ребристая, ильм долинный, клен
мелколистный, орех маньчжурский, бархат амурский, осина,
ольха, яблони, тополя) представлены на небольших площадях,
и их удельный вес незначителен. Средняя полнота насажде-
ний— 0,64. Средний класс бонитета — III. В прошлом в дре-
востоях присутствовали в значительном количестве пихта цель-
нолистная и кедр корейский. Сейчас эти породы следует ак-
тивно вводить в насаждения искусственным путем. То же
можно сказать и про клен ложнозибольдов и чубушники —
высокодекоративные породы, но редкие на этой территории.
Представители семейства аралиевых встречаются на тер-
ритории Академгородка, но это преимущественно взрослые ра-
стения, а возобновление их в связи с большой рекреационной
нагрузкой протекает неудовлетворительно. Совсем нет подро-
ста декоративного реликтового вида диморфанта.
Почвы преобладают бурые лесные глеевато-оподзоленные
(40,7% от общей площади) и бурые лесные оподзоленные
(20,4%). На остальной территории распространены бурые лес-
ные сильно оподзоленные (13,1%), бурые лесные неоподзолен-
ные (12,5%), почвы оврагов (11,6%), прибрежные песчано-га-
лечные отложения (0,9%) и торфянисто-перегнойно-глеевые
(0,8%).
Подлесок и кустарничково-травяной ярус под пологом боль-
шинства древостоев развит слабо. Подлесок развит хорошо в
тех местах, где древостой изрежен, однако кустарничково-
травяной ярус остается слаборазвитым из-за высокой сомкну-
тости подлеска. Среди подлесочных пород чаще всего встре-
чаются чубушник тонколистный, жимолости, спиреи, шиповни-
ки, бересклеты, леспедеца двуцветная, лещина, сирень
(трескуп), барбарис, элеутерококк, акантопанакс и другие
кустарниковые породы. Лианы (виноград, актинидии, лимон-
ник) чаще наблюдаются в ярусе подлеска и в небольшом ко-
личестве. Редко отдельные экземпляры актинидии или вино-
града достигают стадии спелости и поднимаются в верхний
полог древостоя. На участках, примыкающих к жилым масси-
вам, остановкам транспорта или интенсивно посещаемых насе-
лением (более 10 человек на га), подрост отсутствует или
встречается единично и находится в плохом состоянии. Таких
площадей в лесопарке более 23%.
По нашей (классификации типов пейзажей зеленых зон
прибрежных городов южного Приморья (Журавков. 1974), на
территории Академгородка доминирующее положение занима-
ет низкогорный ландшафт — 87% общей площади лесопарка.
Равнинный и прибрежно-морской ландшафты распространены
147
на значительно меньших площадях—10 и 3%. Господствую-
щее положение принадлежит открытым типам пейзажа — 59%.
Закрытые типы пейзажа составляют 36 и полуоткрытые—5%.
Для более точного учета особенностей кажцого участка при
практическом выделении типов пейзажей и для полного учета
всех факторов, определяющих хозяйственные распоряжения на
этом участке, давалась эстетическая оценка отдельных участ-
ков (Журавков, 1967) Выявлено, что к первому (высшему)
классу может быть отнесено 32% площади, ко второму — 54,
третьему — 7,3 и к четвертому — 6,7%. Эти данные говорят о
том, что для повышения эстетической оценки типов пейзажей
необходимо прибегнуть к их облагораживанию путем лесопар-
ковых рубок, посадки декоративных групп из древесно-кустар-
никовых пород, мелиоративных и благоустроительных работ
или иного хозяйственного вмешательства лесоводов и ланд-
шафтных архитекторов с предварительным составлением про-
ектно-сметной документации на каждый тип пейзажа.
Оценка санитарно-гигиенического состоянии лесопарка да-
валась по трем категориям с целью определения пригодности
его участков для нормального отдыха населения на открытом
воздухе. Натурные обследования показали, что к первому
(высшему) классу можно отнести 70% территории, ко второ-
му— 28 и к третьему — 2. Таким образом, значительные площа-
ди требуют проведения ландшафтных и санитарно-оздорови-
тельных рубок, ландшафтно-декоративных и реконструктивных
посадок, благоустройства и мелиоративных работ, а также
установки малых архитектурных форм.
Для комплексной оценки типов пейзажей определялась жиз-
неустойчивость существующих насаждений по разработанной
нами шкале оценок (Журавков, 1974). В ходе обследования в
натуре выявлено, что к первому (высшему) классу относится
98% покрытой лесом площади, второму—1,6 и к третьему —
6.4%. Так, например, к третьему классу жизнеустойчивости
отнесены насаждения из ясеня маньчжурского с признаками
усыхания и естественного отмирания древостоя, которые и
назначены в первоочердную рубку по санитарному состоянию.
С целью правильной оценки и контроля за эффективностью
хозяйственной деятельности, повышения ценности насаждений
в выполнении ими лесопарковых функций производилась оцен-
ка совершенствования лесных участков по общепринятой у нас
в стране классификации при лесоустройстве зеленых зон и
рекреационных лесов. Учетные работы показали, что к первому
классу отнесено 36,7% площади насаждений. Это те участки
леса, которые пригодны для организации отдыха как жителей
Академгородка, так и трудящихся г. Владивостока. Ко второму
классу — 61.8% и к третьему— 1,5%. Участки леса, отнесенные
к третьему классу, требуют проведения многогранных и объем-
ных хозяйственных работ по их облагораживанию.
148
Существенным звеном в организации лесопарка является
оценка распределения его территории по степени крутизны
склонов гор для определения доступности п проходимости.
Натурные обследования показали, что 71,6% обшей площади
относится к первой группе склонов (до 10°) как легко проходи-
мые, ко второй — средне проходимые склоны (от И до 15°) —
12.9%, к третьей — склоны затруднительно проходимые (от 16
до 20°)—8,5%, к четвертой — трудно проходимые склоны (от
21 до 25°) —2,5%, к пятой — особо трудно проходимые склоны
(от 26 до 30°) — 1,5%. к шестой — склоны ограниченной про-
ходимости (от 31 до 35°) —-2,2%, к седьмой — склоны, непри-
годные для перехода (от 362 и выше)—0,8%. Общая оценка
проходимости приближается к средней и равна 1,6, что свиде-
тельствует о доступности территории лесопарка для массового
посетителя.
Учет показал, что посещаемость лесопарка, за малым иск-
лючением, высокая (более 10 человек на га)—87,7% общей
площади и лишь 12,3% отнесено к средней посещаемости (до
10 человек на га).
Для детальной характеристики травяного покрова лабора-
торией зеленых зон и ландшафтной архитектуры Ботанического
сада ДВНЦ АН СССР были заложены пробные площади в
слабо-, средне- и интенсивно посещаемых участках леса. Ре-
зультаты обследования изложены нами в отдельной работе
(Журавков, Селедец, 1976).
Объем и характер информации о каждом участке, фикси-
ровавшиеся при данном лесоустройстве, значительно отлича-
лись от предыдущего лесоустройства 1965—1966 гг. Составлен-
ные картографические и проектно-сметные документы исполь-
зовались Дальневосточным отделением ГИПРОНИИ АН СССР
при разработке плана детальной планировки Академгородка
ДВНЦ АН СССР.
Необходимость в проведении подобных работ в пригородных
лесах городов Приморья очевидна.
ЛИТЕРАТУРА
Журавков А. Ф. Опыт классификации типов пейзажей зеленых зон поибреж-
ных городов южного Приморья.— Лесоводственные исследования на
Дальнем Востоке. 2. Рубки ухода в лесах Приморья. Владивосток.
1967, с. 117—130.
Журавков А. Ф. Основы ведения дозяйства в лесах зеленых зон прибоежных
городов (южное Приморье). Новосибирск, «Наука», 1974, 168 с.
Журавков А. Ф., Селедец В. П. Рекреационная дигрессия травяного покрова
в дубняках (на примере лесопарка Академгородка ДВНЦ АН
СССР).— Пригородные леса городов Западной Сибири и пути их
рационального использования для рекреационных целей. Новосибирск, 1976,
с. 83—85.
СОДЕРЖАНИЕ
Слизик Л. Н., Смирнова О. А. Основные задачи
изучения исчезающих, редких и сокращающихся ви-
дов дендрофлоры Приморья и возможные пути их
культивирования ......................................... 3
М и г и н К. В. Основные программно-методические направ-
ления эколого-физиологического изучения редких и
исчезающих видов деревьев и кустарников Дальне-
го Востока .... 14
Ильченко Т. П. Сравнительная характеристика карио-
типов некоторых видов рода Larix miller . . 22
Ильченко Т. П., Лауве Л. С., Урусов В. М. Из-
менчивость сосны погребальной в Приморье в свя-
зи с особенностями климата 28
Урусов В. М. Сосновые леса полуострова Гамова и ос-
новные черты их динамики . 45
Ильченко Т. П. Кариологическая изменчивость Pinus
silvestris L. 67
Урусов В. М. Ценотическая роль, состояние и пути сох-
ранения можжевельников и других хвойных кустар-
ников Приморья . . . . 73
Василюк В. К. Мелкоплодник ольхолистный Microme-
les alnifolia (Sieb. et Zucc.) Koehne (распростране-
ние, возобновление, использование, охрана) 100
Слизик Л. Н. Особенности сезонной ритмики развития
некоторых реликтовых древесных лиан флоры При-
морья ... 105
Слизи к Л. Н. Практические рекомендации по размно-
жению древесных лиан Приморья . . . 113
Лауве Л. С. Морфолого-анатомические характеристики
хвои сосны погребальной (Pinus funebris Кот.) в
Приморском крае . 118
Лауве Л. С. Характеристика эпидермиса хвои некото-
рых видов сосен секции Eupitys Spach. 124
Ворошилова Г. И. Морфолого-анатомическое строение
листа и древесины тиса Taxus cuspidata Sieb. et Zucc. 129
Королева P. А. Строение эпидермиса хвои лиственниц
Приморского края . .133
Снежкова С. А. К анатомической характеристике дре-
весины некоторых аралиевых Приморья . . 137
Полийчук Ю. С. О пыльце дальневосточных бересклетов 142
Журавков А. Ф. Ландшафтное лесоустройство лесо-
парка Академгородка Дальневосточного научного
центра АН СССР 145
УДК 507.2(571.63)
Основные задачи изучения исчезающих, редких и сокращающихся
видов дендрофлоры Приморья и возможные пути их культивирования.
С л из и к Л. Н., Смирнова О А. «Редкие и исчезающие древесные
растения юга Дальнего Востока (биология, экология, кариология)».
Владивосток, 1976, с. 3—113.
В статье обсуждаются конкретные задачи региональных исследований
группы наиболее ценных и редких видов дендрофлоры Приморья: уточ-
нение перечня видов, нуждающихся в охране; изучение их биологии;
создание коллекционных фондов. Предлагается единая программа изу-
чения их биологии; жизненных циклов, генеративной и вегетативной
сфер и природных популяций как составной части ландшафтных комп-
лексов. Возможные пути сохранения видов в пределах естественного
ареала: культура (коллекции, озеленительные посадки), полукультура
(посадки в природных парках и зеленых зонах) и реинтродукция (вос-
становление угасающих популяций). Прилагается список видов дендро-
флоры Приморья 1—3 категорий, собранных и подлежащих сбору для
коллекций Ботанического сада ДВНЦ АН СССР.
Библ. 25.
УДК 581.5.02.05(571.6) (018) (57.34)
Основные программно-ме1 одические направления эколого-физиологи-
ческого изучения редких и исчезающих видов деревьев и кустарников
Дальнего Востока. Мигин К. В. «Редкие и исчезающие древесные
растения юга Дальнего Востока (биология, экология, кариология)».
Владивосток, 1978, с. 14—211.
Обосновывается актуальность и безотлагательность изучения и
сохранения не только редких и исчезающих видов уникальной флооы
Дальнего Востока, но и редких ценнейших форм, (подверженных опас-
ности исчезновения. Для ускорения изучения и отбора наиболее цен-
ного генофонда таких растений в целях сохранения и быстрейшего на-
роднохозяйственного использования предлагается программа и основ-
ные перспективные методы исследования, включающие наряду с изве-
стными новые методы (диагностика зимостойкости с применением
ЯМР, оценка зимостойкости растений в режимах переменных темпе-
ратур, особенности применения вегетационного метода для оценки
морозостойкости корней, изучение ритмики зимнего развития и др.).
Библ. 13.
УДК 582.47:581.14
Сравнительная характеристика кариотипов некоторых видов рода
Larix Miller. Ильченко Т. П. «Редкие и исчезающие древесные ра-
стения юга Дальнего Востока (биология, экология, кариология)». Вла-
дивосток, 1978, с. 22—27.
В результате сравнительной характеристики кариотипов некоторых
видов рода Larix Miller отмечены общие черты и различия их. Боль-
шое внимание уделяется проценту встречаемости вторичных перетяжек
в различных парах хромосом, что позволяет при сравнительной харак-
теристике кариотипов установить филогенетические отношения между
отдельными видами.
Ил. 1, табл. 2, библ. 12.
151
УДК 582.475:581.845(571.63)
Изменчивость сосны погребальной в Приморье в связи с особенно-
мн климата. Ильченко Т. П., Лауве Л С., Урусов В. М.
«Редкие и исчезающие древесные растения юга Дальнего Востока
(биология, экология, кариология)*. Владивосток, 1978, с. 28—44.
Охарактеризована морфолого-анатомическая и кариологическал
вариабельность сосны погребальной (Pinus funebris Кот.) в При-
морье. Полиморфизм популяций оценен в связи с изоляцией отдель-
ных частей ареала н дивергенцией в различающихся условиях, чем
объяснены отличия кариотипа и наборов наследственных форм. Хан-
кайские популяции признаны наиболее продвинутыми филогенетически
и перспективными для искусственного расселения. Сделан вывод об
относительной стабильности анатомической структуры хвои в резуль-
тате широких приспособительных потенций и физиологической плас-
тичности. Рассмотрены некоторые возможности эволюции кариотипа
Исследования проведены впервые.
Ил. 5, табл. 4, библ. 38.
УДК 634.94:634.95(571.63)
Сосновые леса полуострова Гамова и основные черты их дина-
мики. Урусов В. М. «Редкие и исчезающие древесные растения юга
Дальнего Востока (биология, экология, кариология)*. Владивосток,
1978, с. 45-66.
Анализируется строение древостоев сосны густоцветковой, харак-
тер ее возобновления. Приводятся особенности роста породы, подроб-
но рассматривается динамика сосняков; отмечается их уникальность
и ценность. Указывается на необходимость разработки мер по сохра-
нению и восстановлению сосновых лесов п-ова Гамова.
Ил. 6, сх. 2, табл. 4, библ. 16.
УДК 582.475:581.15
Наркологическая изменчивость Pinus sylvestris L. И л ь ч е н -
к о Т. П. «Редкие и исчезающие древесные растении юга Дальнего
Востока (биология, экология, кариология)». Владивосток, 1978,
с. 67—72.
Приводится сравнительный анализ кариотипов сосны обыкновен-
лой, произрастающей в различных точках ареала. Замечена значитель-
ная кариотипическая изменчивость вида. Предполагается зависимость
изменчивости кариотипов от экологического разнообразия.
Ил. 2, табл. 2, библ. 7.
УДК 582.47:581.5(571.63)
Ценотнческая роль, состояние и пути сохранения можжевельников
и других хвойных кустарников Приморья. Урусов В. М. «Редкие и
исчезающие древесные растения юга Дальнего Востока (биология, эко-
логия, кариология)». Владивосток, 1978, с. 73—99.
Изложены результаты изучения некоторых эколого-биологических
особенностей и распространения 5 хвойных кустарников Приморья.
Сделан вывод о реликтовом характере их сообществ в низкогорьях.
Верхняя граница зарослей стелющихся хвойных определена как прог-
рессивная в связи с изменением климата, в том числе и четвертичной
трансгрессией моря. Указано на пионерный характер сообществ хвой-
ных кустарников, их уникальную почвообразовательную роль, которую,
152
если эти ценозы уничтожаются пожарами, не могут взять па себя
другие растения, и закономерность смены стелющихся зарослей
высокоствольным лесом. Освещено современное состояние, предложе-
ны меры по охране популяций и сбережению видов.
Ил. 5, сх. 3, табл. 2, библ. 18.
УДК 580:502.7(571.6)
Мелкоплодник ольхолистный Micromeles alnifolia (Sieb. et Zucc.) Koeh-
ne (распространение, возобновление, использование, охрана). Васи-
люк В. К- «Редкие и исчезающие древесные растения юга Дальнего
Востока (биология, экология, кариология)». Владивосток, 1978, с. 100—
104.
Мелкоплодник — наиболее южный для Приморья реликтовый вид.
Приводится ареал, морфологическая и экологическая характеристика,
использование. На основании детального обследования лесной терри-
тории Ботанического сада выявлено участие мелкоплодника в сложе-
нии фитоценозов с повышенным режимом охраны, отмечается удовлет-
ворительное возобновление и возможности воспроизводства этой цен-
ной породы.
Библ. 17.
УДК 581.543(571.63)
Особенности сезонной ритмики развития некоторых реликтовых дре-
весных лиан флоры Приморья. С л и з и к Л. Н. «Редкие и исчезающие
древесные растения юга Дальнего Востока (биология, экология, карио-
логия)». Владивосток, 1978, с. 105—112.
Изучены и сравниваются сезонные ритмы двух видов древесных
лиан, обитающих на юге Приморья и занесенных в Красную книгу,—
аристолохин маньчжурской к пуэрарии волосистой; анализируются их
эколого-генетические связи.
Библ. 15.
УДК 507.2(571.63)
Практические рекомендации по размножению древесных лиан При-
морья. Слизик Л. Н. «Редкие и исчезающие древесные растения юга
Дальнего Востока (биология, экология, кариология)». Владивосток,
1978, с. 113—117.
На основе изучения особенностей поведения и размножения лиан
флоры Приморья (18 видов) анализируется их способность к возоб-
новлению в природе, а также даются рекомендации по искусственному
размножению —указываются оптимальные сроки и условия для по-
сева семян и зеленого черенкования. Почти все исследуемые виды
относятся к числу исчезающих, редких или сокращающихся.
Табл. 1, библ. 15.
УДК 581.845(571.63)
Морфолого-анатомические характеристики хвои сосны погребаль-
ной (Pinus funebris Кот.), в Приморском крае. Лауве Л С «Ред-
кие и исчезающие древесные растения юга Дальнего Востока (биоло-
гия, экология, кариология)». Владивосток, 1978, с. 118—123.
Вид в крае оказался представленным двумя морфологически чет-
кими макрокомплексами популяций с разной степени широкими на-
борами форм. Макрокомплексы различны и по морфолого-анатомиче-
ским особенностям хвои: прежде всего по количеству и расположению
153
смоляных каналов, развитию механических тканей и расстоянию меж-
ду проводящими пучками. Установлено, что южноприморские популя-
ции по изученным признакам или тождественны сосне густоцветковой
(тонкая малоканальная хзоя), или являются гибридным циклом ее и
сосны Тунберга (хвоя с преимущественно и полностью паренхиматиче-
скими каналами). Западно- и центральноприморские популяции по
строению хвои едины. Сделано заключение о факторах, определяю-
щих среднепопуляционные морфолого-анатомические характеристики,
и важности установления последних при выяснении филогенеза.
Табл. 3, библ. 10.
УДК 582.475:581.821
Характеристика эпидермиса хвои некоторых видов сосен секции
Eupltys Spach. Лауве Л. С. «Редкие и исчезающие древесные расте-
ния юга Дальнего Востока (биология, экология, кариология)». Влади-
восток, 19*78, с. 124—1128.
Изучение эпидермиса хвои у семи видов сосен показало, что ли-
стовой аппарат амфистома пюго типа. Устьичный аппарат моноциклич-
ный, пиноидного типа. Изолированные устьица присутствуют в хвое
всех сосен смежных у пяти видов (Pinus funebris Кот., Р. densiflo-
гг Sieb. et Zucc., Р. tabulaeformis Carr, P. massoniana D. Don..
P. sylvestris L.), а связанный устьичный аппарат обнаружен только
в хвое Р. funebris Кот, Количество околоустьичных клеток в хвое
этих сосен различно: 4—8 —у сосен погребальной и Тунберга, 3—6 —
у сосны густоцветковой, 4—6 — у сосны китайской, 4—7 — у сосны
Массона, 3—5 — у сосны обыкновенной и у последней — 4—9. Клетки
эпидермиса хвои у всех исследуемых видов сосен имеют прямоуголь-
ную форму, а также четырехугольную и трапециевидную
Табл. 1, библ. 2.
УДК 582.471:581.14(571.63)
Морфолого-анатомическое строение листа и древесины тиса Taxus
cuspidata Sieb. et Zucc. Ворошилова Г. И. «Редкие и исчезающие
древесные растения юга Дальнего Востока (биология, экология, ка-
риология)». Владивосток, 1978, с. 129—432.
Дается анатомическое описание строения хвои и древесины тиса
остроконечного, произрастающего на юге Приморского края, па остро-
вах Сахалин и Кунашир. Отмечается дорзивентральное строение ли-
ста тиса и отсутствие в нем эндодермы, смоляных каналов, склерен-
химы и гиподермы. Устьичный аппарат листа амфицикличный. Пере-
ход от ранней древесины к поздней постепенный. Показано, что в
древесине этой породы не обнаружено смоляных каналов, лучевых
трахеид и паренхимы.
Библ. 10.
УДК 582.475:581.821(571.63)
Строение эпидермиса хвои лиственниц Приморскою края. Коро-
лева Р. А. «Редкие и исчезающие древесные растения юга Дальнего
Востока |(биологи1Я, экология, кариология)». Владивосток, 1978,
с. 133—136.
В строении эпидермиса хвои пяти видов лиственницы выявлены
общие черты, характерные для рода, а также черты, присущие отдель-
ным видам.
У всех лиственниц устьичный аппарат моноцикличный, лярикоид-
пого типа. Устьица погруженные, на поверхности листа располагаются
154
полосами. Форма околоустьичных полярных клеток округлая (лист-
венницы Комарова, Любарского, приморская), продолговатая (лист-
венницы даурская, ольгинская, Любарского) и бочонковидная (лист-
венница приморская). Латеральные околоустьичные клетки неправиль-
ной формы, а у лиственницы приморской они имеют продолговатую и
четырехугольную форму. Устьица связанные и смежные, изолирован-
ные отмечены в хвое лиственницы ольгинской.
Библ. 5.
УДК 582.89—1.844
К анатомической характеристике древесины некоторых аралиевых
Приморья. Снежкова С. А. «Редкие и исчезающие древесные расте-
ния юга Дальнего Востока (биология, экология, кариология)». Вла-
дивосток, 1978, с. 137—141.
Рассматривается анатомическое строение древесины трех предста-
вителей аралиевых: Eleutherococcus senticosus, Acanthopanax sessiliflo-
rum и Kalopanax septemlobum. Отмечается, что исследованные виды
могут быть различимы по признакам строения древесины: древесина
калопанакса хорошо отличается от древесины элеутерококка и акан-
топанакса кольцесосудистостью, небольшой высотой лучей (до 35 кле-
ток). Древесина элеутерококка и акантопанакса схожа, однако до-
вольно значительные различия имеются в характере лучей — более
высокие и широкие у элеутерококка, 1—5-рядные, высотой более 220
к четок; у акантопанакса 1—4-рядные, от 2 до 55 клеток высотой.
Кроме того, у элеутерококка на поперечном срезе древесины просветы
сосудов располагаются в более широких тангентальных полосах, чем
акантопанакса.
Библ. 13.
УДК 581.331.2:582.766
О пыльце дальневосточных бересклетов Полийчук Ю. С. «Ред-
кие и исчезающие древесные растения юга Дальнего Востока (биоло-
гия, экология, кариология)». Владивосток, 1978, с. 142—144.
В статье приводятся данные по исследованию морфологии пыль-
ны 9 видов бересклета флоры Дальнего Востока. Пыльцевые зерна
в основном трехбороздно-оровые с сетчатой скульптурой. По харак-
теру апертур, очертаниям и размерам пыльцевые зерна подразделены
па 2 типа, соответствующие таксономическим подродам. Делается вы-
вод о целесообразности выделения подрода Kalonymus в самостоя-
тельный род.
Табл. 1, библ. 4.
УДК 634.92(571.63)
Ландшафтное лесоустройство лесопарка Академгородка Дальне*
восточного научного центра АН СССР. Журавков А. Ф. «Редкие
н исчезающие древесные растения юга Дальнего Востока (биология,
экология, кариология)». Владивосток, 1978, с. 146—/149.
Проводится первый опыт ландшафтного лесоустройства террито-
рий, разнообразных во флористическом отношении, но в условиях при-
городов педвергающихся постоянной рекреационной нагрузке, а потому
стоящих перед проблемой сохранения растительного богатства. В ос-
нове лежит типизация пейзажей и классификация типов пейзажей,
разработанные автором ранее.
Библ. 3.
155
РЕДКИЕ И ИСЧЕЗАЮЩИЕ
ДРЕВЕСНЫЕ РАСТЕНИЯ
ЮГА ДАЛЬНЕГО ВОСТОКА
(БИОЛОГИЯ, ЭКОЛОГИЯ, КАРИОЛОГИЯ)
Редактор Т. /7. Дружинина
Художник Л. Ф. Литвшико
Техн, редактор Л. С. Гришина
Корректор И. П. Прокопенко
ВД 12547. Сдано в набор 28.XI 1977 г. Подписано
к печати 13. VII. 1978 г. Формат 60Х90/1в. Усл. печ.
л. 9,75. Уч.-изд. л. 10,48. Тираж 500 экз. Цена
1 р. 60 к. Заказ 488.
Редакционно-издательский отдел Дальневосточного
научного центра Академии наук СССР
690600, Владивосток, Ленинская, 50.
Полиграфический комбинат Управления издательств
полиграфии и книжной торговли Приморского
крайисполкома
Владивосток, Океанский пр., 69