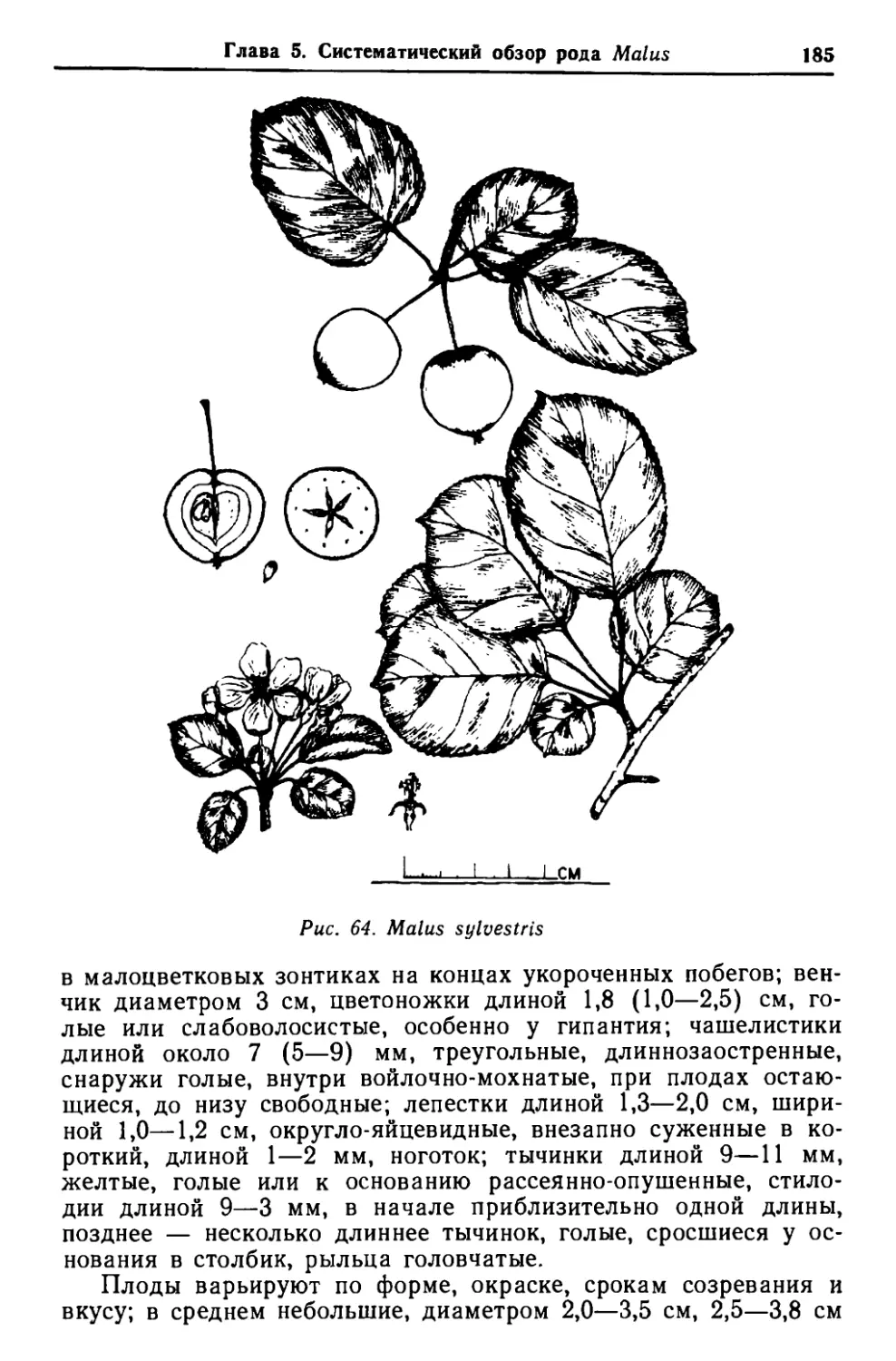
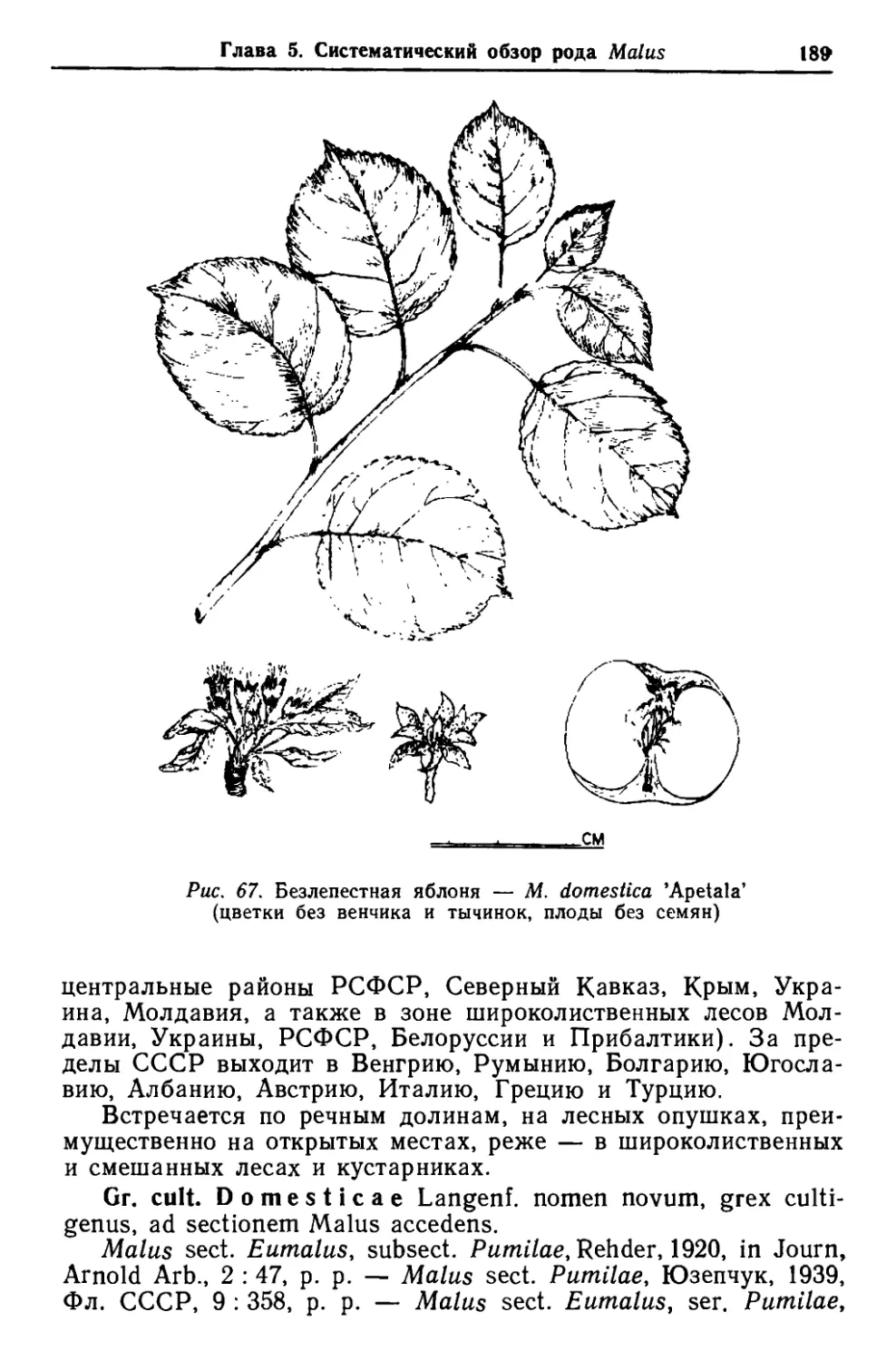
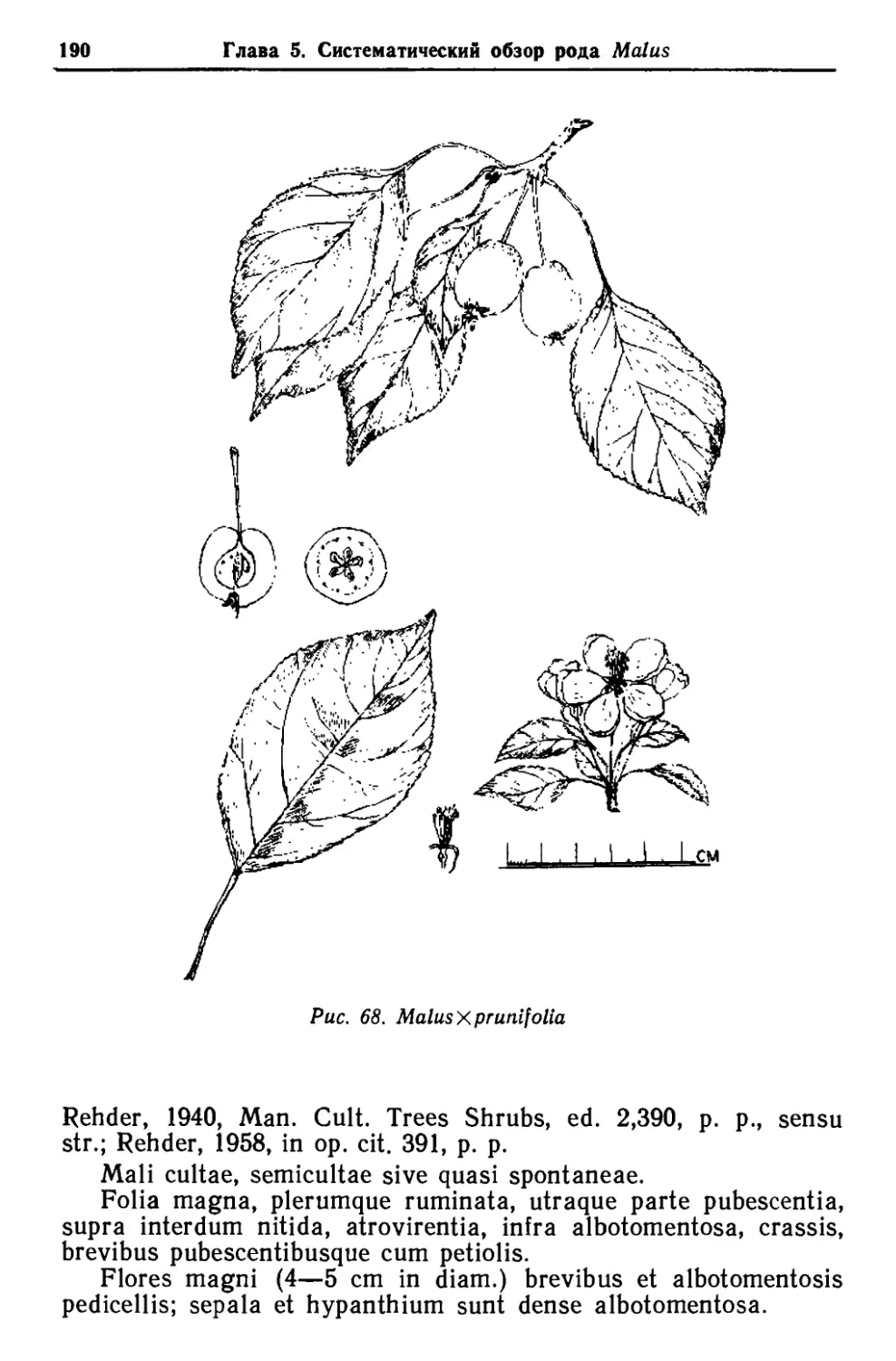
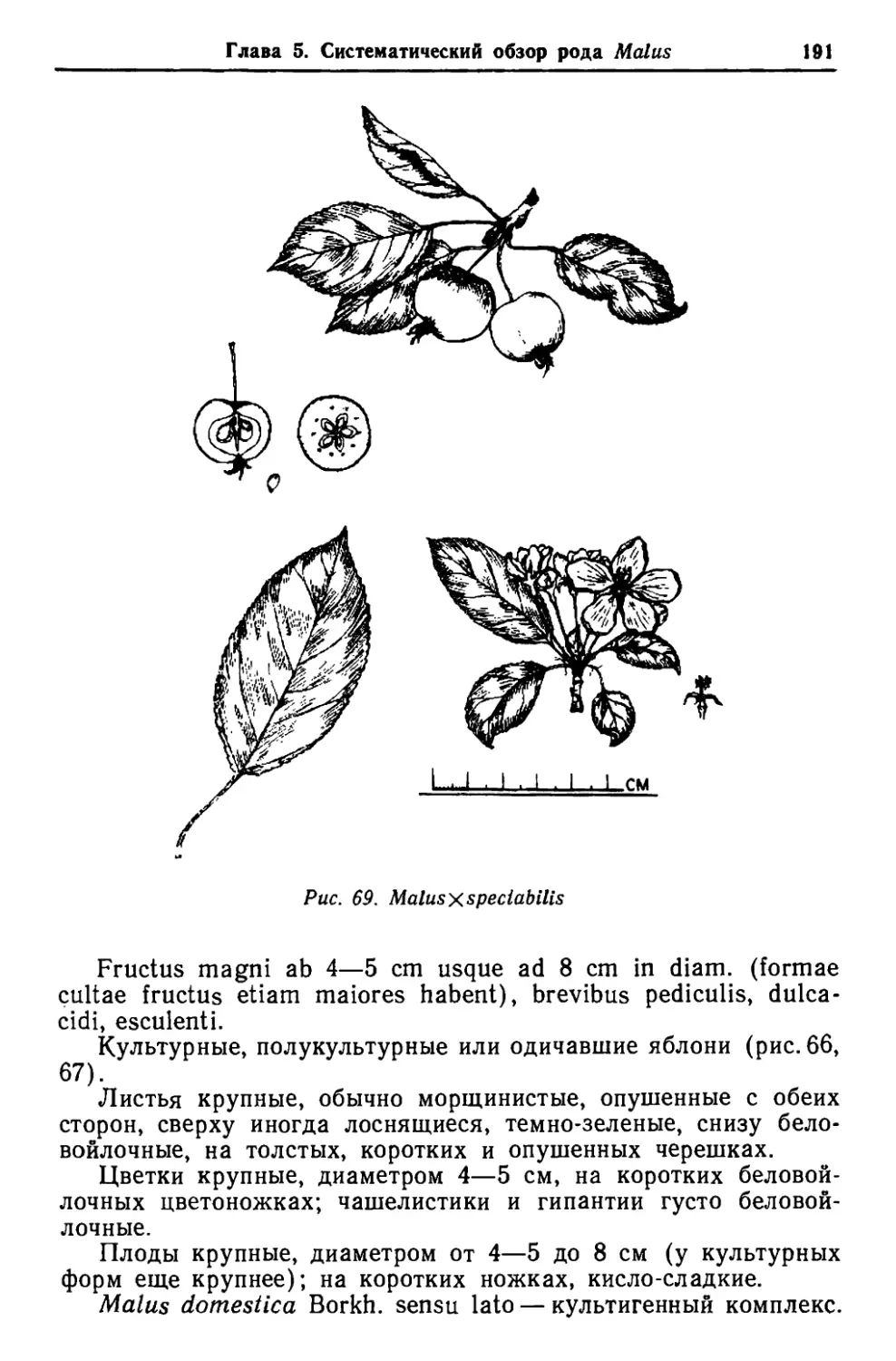
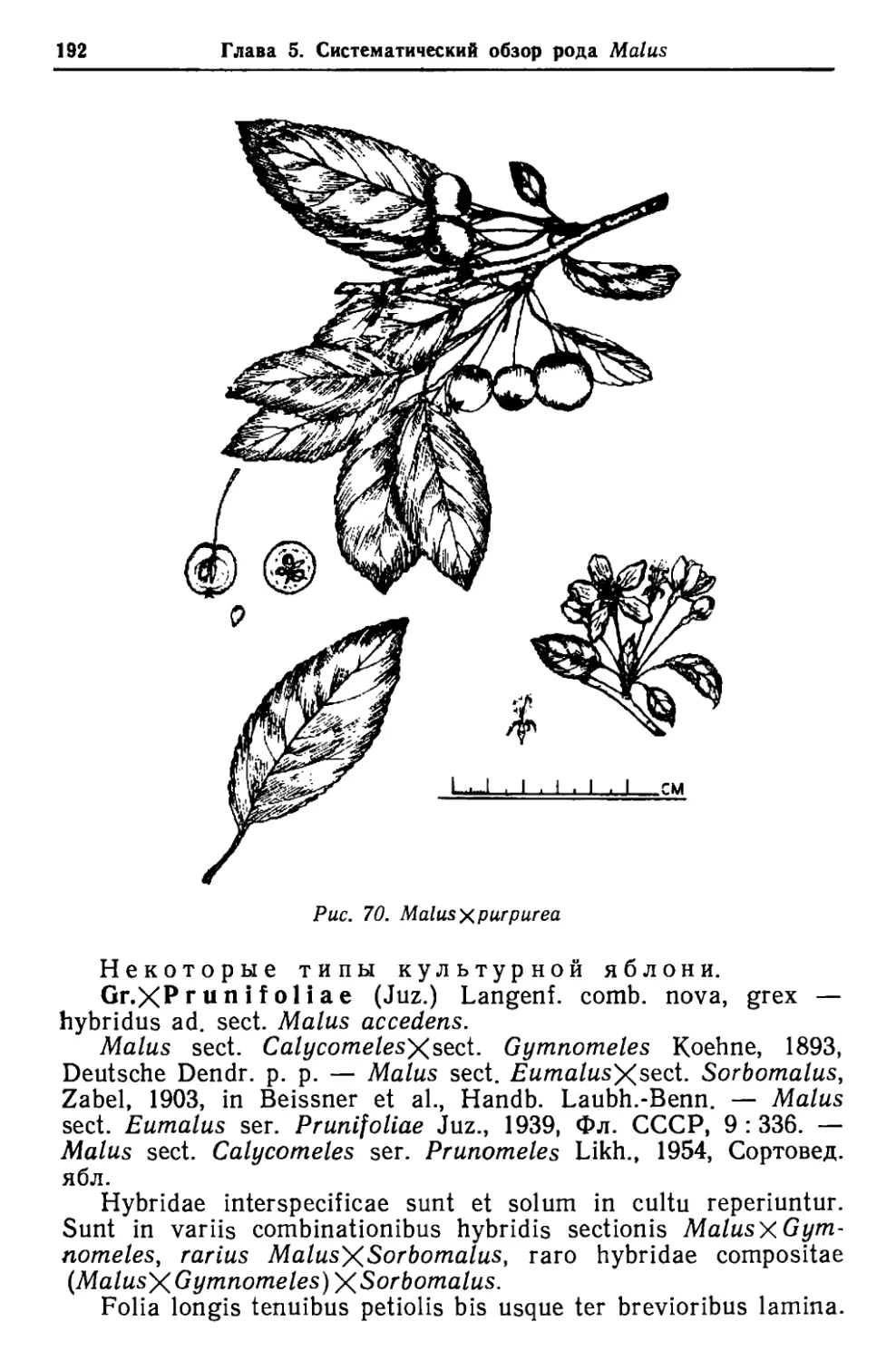
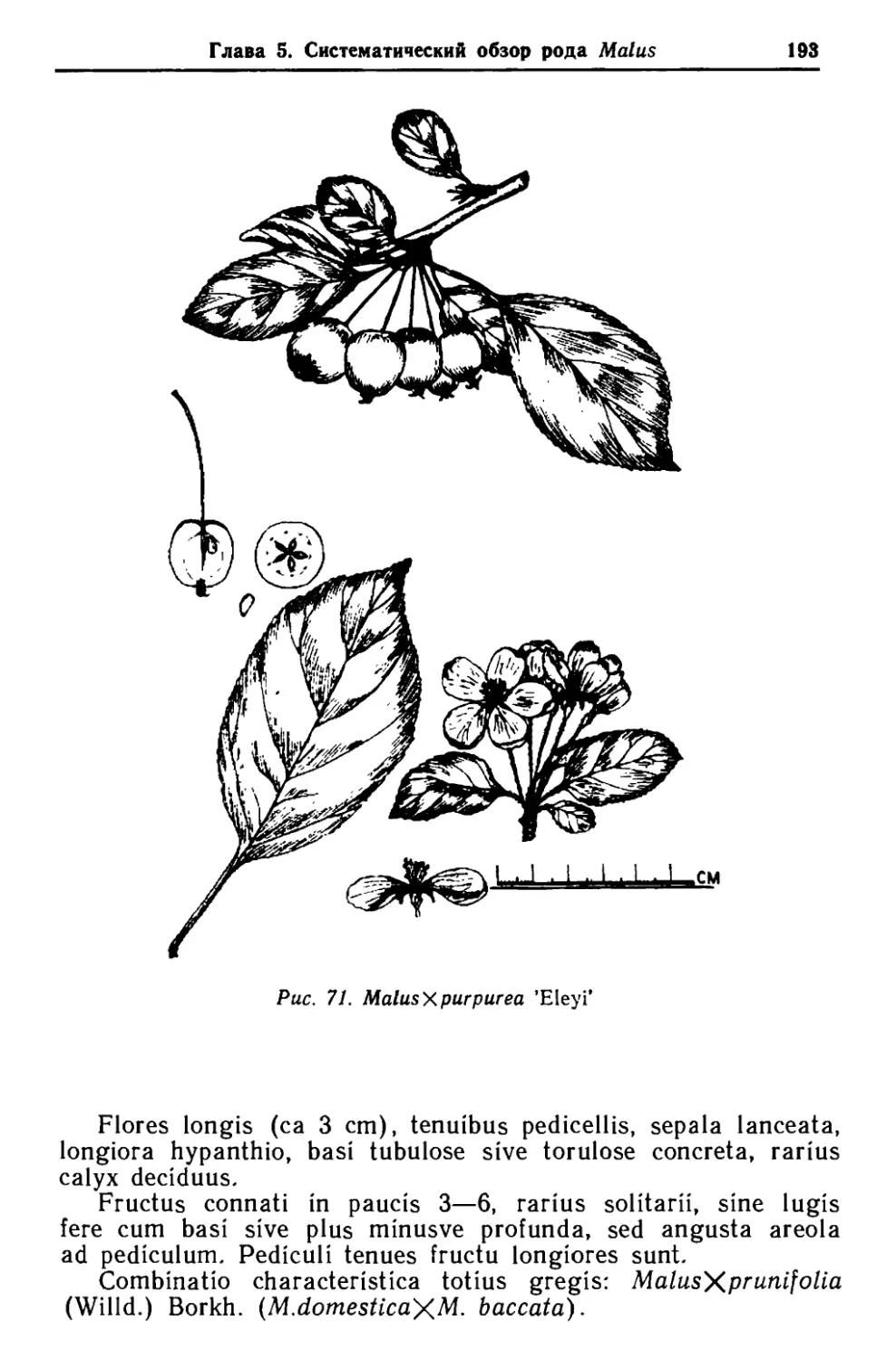
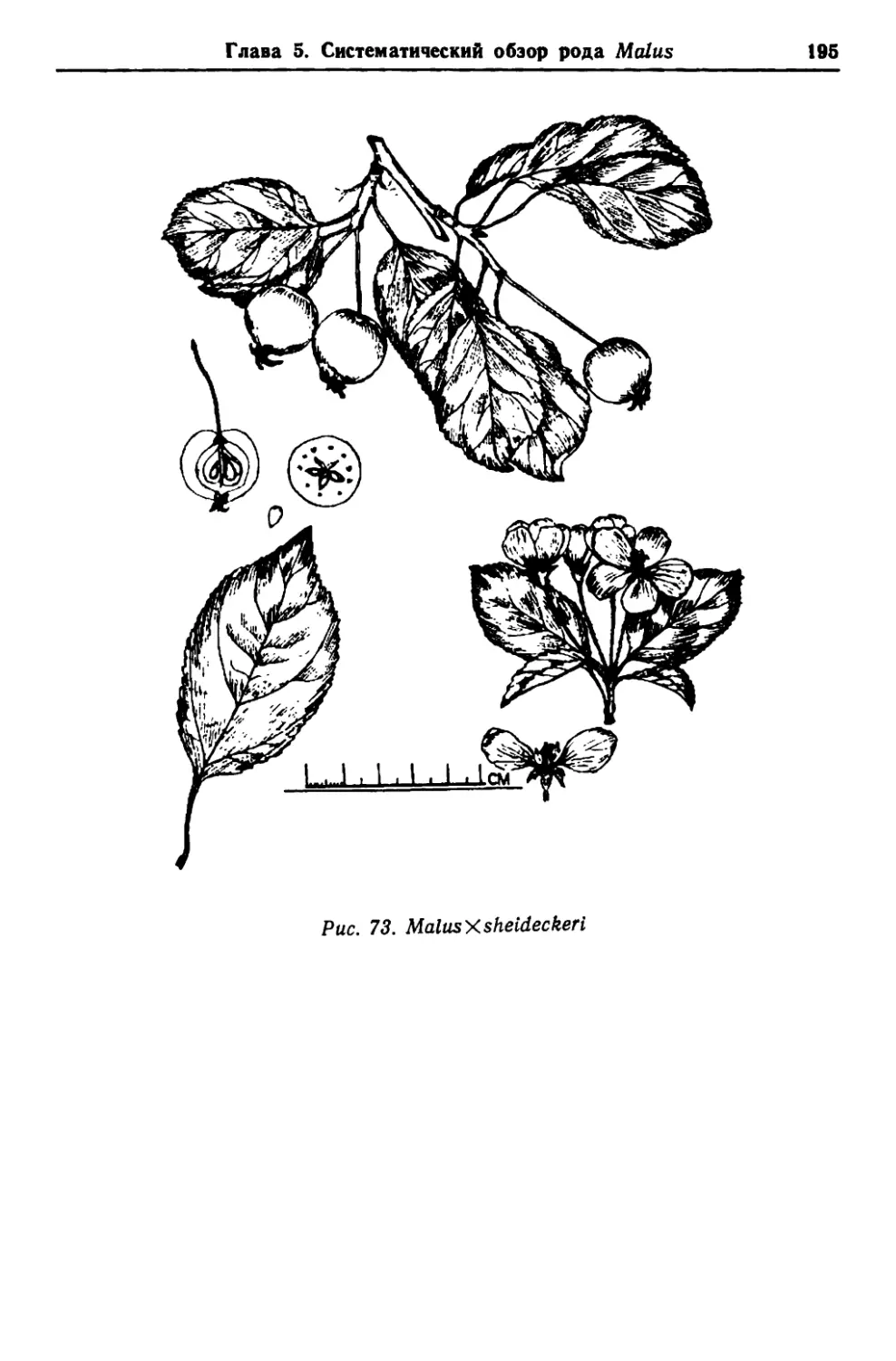
/











































































































































































































Author: Лангенфельд В.Т.
Tags: систематика растений плодоводство растения биология флора растениеводство
ISBN: 5-7966-0379-5
Year: 1991




Text
>mодщ
В. Т. ЛАНГЕНФЕЛЬД ЯБЛОНЯМОРФОЛОГИЧ ЕСКАЯЭВОЛЮЦИЯ,ФИЛОГЕНИЯ,ГЕОГРАФИЯ,С И СТЕМАТ И КА
V. LANGENFELDS APPLE-TREES МОЯ',РНОЬ061САЬ EVOLUTION, PHYLOGENY, GEOGRAPHY, SYSTEMATICS RIGA «ZlNATNE» 1991
ЛАТВИЙСКИЙ УНИВЕРСИТЕТ В. Т. ЛАНГЕНФЕЛЬД ЯБЛОНЯ МОРФОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ, ФИЛОГЕНИЯ, ГЕОГРАФИЯ, СИСТЕМАТИКА РИГА «ЗИНАТНЕ» 1991
28.592.72 Л221 УДК 582.001.4+ 634.12 Рецензенты: канд. биол. наук В. В. ПОНОМАРЕНКО, канд. биол. наук Р. Я. ЦИНОВСКИС Л 1906000000 — 007 55 91 М811(11) — 91 � В. Т. Лангенфельд, 1991 1Я В N 5-7966-0379-5 Л а н ген фельд В. Т. Яблоня: Морфологическая эволюция, филогения. география, систематика. — Рига: Зинатне, 1991. — 234 с. — 1SBN 5-7966-0379-5. В монографии обобщены многолетние исследования автора в области морфологической эволюции, географии, экологии и систематики дикорасту- щих (автохтонных) видов яблони. Приведены результаты изучения структур- ных особенностей и выявлены морфологические черты эволюционного уровня различных таксонов яблони, сопоставлены закономерности географического распространения видов яблони и их экологии в различных частях земного шара. На основе этих исследований определены основные филогенетические взаимоотношения таксономии и проведено построение эволюционной системы рода Malus. Охарактеризована хозяйственная ценность различных яблонь. Даны рекомендации по их дальнейшему использованию. Табл. 1, ил. 73, библиогр..98 назв.
Памяти моего учителя Александра Александровича Федорова посвящаю ПРЕДИСЛОВИЕ Среди плодовых деревьев в СССР яблоня занимает первое место по своему потребительскому, хозяйственному и промыш- ленно-экономическому значению. Однако яблоня (род Malus) несмотря на широкое распространение до сих пор пользуется репутацией трудного для систематики рода. Существующие системы классификации видов рода разработаны с различной степенью совершенства. Нередко они противоречивы и неясны во всех своих частях. В них недостаточно четко отражены род- ственные взаимоотношения видов, так как не всегда учитыва- ются все их признаки, указывающие на эволюционный уро- вень того или иного таксона. Наблюдаются существенные раз- ночтения в части количества видов, их объема и наименования. Некоторые автохтонные виды, преимущественно наиболее примитивные и произрастающие в Восточной Азии, пропущены, а явные гибриды или географически неясные популяции не- редко описаны в качестве самостоятельных видов. В связи с этим не всегда соблюдена географическая обособленность их ареалов, а также экологическая определенность таксонов. Особые трудности для объективной систематики рода созда- лись в результате гибридизации (иногда межвидовой) . Ряд видов яблони очень давно был взят в культуру. Отбирая наи- более пригодные для себя формы, человек создал громадное число разнообразных сортов. Сеянцы этих сортов, полученных главным образом путем гибридизации, частично одичавшие, внедрялись в естественную растительность в местах древней культуры яблонь, многократно смешивались с дикими видами и дали целый ряд форм, происхождение которых определить сейчас весьма трудно. Описывая дикие яблони СССР, С. В. Юзепчук [1939, с. ЗБ8] отмечал: «Изучение рода представляет громадные (в настоящее время покуда непреодолимые) трудности». Эти трудности, по мнению С. В. Юзепчука, таковы: 1) невозможность всегда строго различить дикие и одичав- шие формы яблонь, не говоря уже об их гибридах;
Предисловие 2) существование значительного полиморфизма, даже у не- сомненно диких форм, затрудняющих применение географичес- кого метода; 3) существование молодых, недостаточно отграниченных ви- дов, самостоятельность которых может оспариваться. В труде С. В. Юзепчука были обобщены имеющиеся к тому времени сведения о диких яблонях. Почти все авторы более поздних работ не выходили за рамки исследований этого ав- тора либо вносили некоторые небольшие изменения, главным образом по линии описания новых видов. Однако после опубликования труда С. В. Юзепчука прошло уже более 50 лет. За это время значительно расширилось наше понимание сущности и критериев вида. Появилось немало новой литературы, описаны новые виды яблонь. Накопился богатый материал, возрос объем гербарных фондов, особенно из Сред- ней Азии (Ташкент, Алма-Ата, душанбе, Ашхабад) и с Кавказа (Баку, Тбилиси, Ереван), отчасти и из европейской части СССР (Киев). Все это следует критически оценить и обобщить. На- зрела также необходимость современной критической обработки систематики рода MaLus, так как существующие системы рода разработаны недостаточно. Задача настоящей работы — дать критический пересмотр и обзор видового состава яблонь, произрастающих в СССР и за рубежом, и на этой основе разработать систематику рода MaLus. За большую помощь, без которой завершить данную работу в намеченном объеме было бы невозможно, автор глубоко бла- годарен своему учителю — члену-корреспонденту АН СССР, доктору биологических наук, профессору ~Александру Алексан- дровичу Федорову). Автор искренне признателен также курато- рам Гербария и сотрудникам Ботанического института им. В. Л. Комарова АН СССР, особенно И. Т. Васильченко, О. И. Рожковой), )В. Г. Беру~ и многим коллегам в нашей стране и за рубежом за предоставление необходимого мате- риала. Автор
Глава 1МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ Работа с яблонями была начата в 1957 г. с изучения видов, представленных на территории Латвии, и продолжалась до 1960 г. В последующие годы были изучены яблони Прибалтики, затем — европейской части СССР, Кавказа и Средней Азии, Сибири и дальнего Востока. Однако ограничиваясь материа- лом, собранным только в нашей стране, нельзя получить ре- альную картину всего разнообразия и видового состава рода Malus и полное представление о его географическом распро- странении, без чего невозможно проследить родственные взаи- моотношения видов и пути их эволюционного становления, нельзя создать объективную систематику всего рода. Поэтому рамки исследования пришлось раздвинуть и критически прора- ботать систематику всего рода Malus. Изучение яблонь проводилось с учетом наблюдений за рас- тениями в природе и привлечением обширного гербарного ма- териала. Проводились также наблюдения в культуре. Наблюдения в природе осуществлялись в ходе экспедиций по местам, где произрастают крупнейшие в СССР заросли ди- ких яблонь. В 1967 и 1968 гг. нам удалось совершить поездки но территории ГЯР. В отдельных, наиболее крупных зарослях диких яблонь проводились стационарные наблюдения. Материалами для исследований, помимо наших собственных сборов и наблюдений, послужили многочисленные фонды круп- нейших гербариев СССР, а также за рубежом. Были изучены следующие фонды основных гербариев. Советские: Ботанический институт им. В. Л. Комарова АН СССР — общий гербарий, гербарий европейской части СССР. Кавказа, Средней Азии, Сибири и дальнего Востока, Китая и Восточной Азии; гербарии Института ботаники АН КазССР, Института ботаники АН ТССР и Ботаннческого сада АН ТССР. Института ботаники АН АЗССР, Вильнюсского университета и Института ботаники АН ЛитССР, Воронежского университета Ботанического института АН ТаджССР, Института ботаники АН АрмССР, Института ботаники АН УССР, Центрального бо- танического сада АН УССР и Киевского университета, Ботани- ческого сада АН МССР, Всесоюзного института растениеводства
Глава 1. Материал и методы исследоваиий ВАСХНИЛ (Ленинград); Львовского университета и его Бота- нического сада; Института экспериментальной ботаники и мик- робиологии АН БССР; Московского университета, Латвийского университета, Ботанического сада АН ЛатвССР, Института зоо- логии и ботаники АН ЭССР и Тартуского университета, Инсти- тута ботаники АН УЗССР и Ташкентского университета, Инсти- тута ботаники АН ГССР, Государственного музея Грузии и Тбилисского университета; Института биологии АН КиргССР; Никитского ботанического сада. Зарубежные*: Naturhistorisches Museum, Wien; Ботаничес- кий институт АН Болгарии; The Herbarium and Library Royal Botanik und Pflanzengeographie, Halle; Botanisches Institut, Ros- tanicum Academiae Horti-et Viniculturae, Budapest; Institut fiir gu1turpf1anzenforzchung, Gatersleben; Institut fiir systematische Botanic Gardens, Кебич» British Мизиет, London; Institutum Bo- tock; Botanisches Institut, Greifswald; Forest Research Institute, Dehra Рип; Central National Herbarium, Calcutta; Herbarium Universitatis Florentinse, Firenze; Arnold Arboretum, Jamaica Plain; Мизее National d'Histoire Naturelle, Paris; Conservatoire et Jardin Botaniques, Geneve; Botanical Departament Naturhisto- riska Riksmuseum, Stockholm; Institute of Forest Botany, Tokyo. По ряду объективных причин мы не могли воспользоваться гербарными сборами, полученными непосредственно из Китая, Вьетнама, Лаоса, а также Ирака, Ирана и Афганистана, где яблони встречаются автохтонно и в культуре. Однако нами были исследованы значительные китайские и восточно-азиат- ские сборы, а также из Ирана, Афганистана и многих других стран, хранящиеся в гербарии Ботанического института им. В. Л. Комарова АН СССР. Все помещенные в работе карты ареалов различных видов рода Malus являются оригинальными. Они составлены на ос- новании обширного гербарного материала СССР, а также дан- ных, полученных из разных стран. Ареалы видов, представлен- ных на территории СССР и стран центральной и Западной Европы, нанесены на карты точечным методом в соответствии с имевшимися в нашем распоряжении гербарными экземпля- рами, а также собственными данными, полученными при обследовании различных районов страны. Кроме того, были ис- пользованы литературные данные. В тех случаях, когда материала было недостаточно, а также при вычерчивании обобщающих карт ареалов вида и секции был использован контурный метод. Там, где контуры ареала нельзя указать точно из-за недостатка материала (особенно в Китае, а также в некоторых слабоизученных районах Юго-Вос- точной Азии), сплошная линия ареала заменена пунктиром. * Зарубежные материалы были получены при содействии Ботанического института им. В. Л. Комарова АН СССР.
Глава 2КРАТКИЙ ОБЗОРИСТОРИИ ИЗУЧЕНИЯ РОДА MALUS С давних времен человек, собирая плоды растений для сво- его пропитания, отбирал наиболее интересные для него формы, отличающиеся величиной, наилучшими вкусовыми и другими качествами. Такой стихийный отбор явился основой для буду- щего процесса сознательной селекции яблони и научного изу- чения ее видового и внутривидового разнообразия. Кроме ос- татков плодов яблони и их схематических изображений, най- денных при раскопках древнейших поселений человека, этот период больше ничего не оставил. Можно только приблизи- тельно определить возраст таких находок — 10 — 15 тысяче- летий. Виды рода Ма/из, известные первобытному человеку, веро- ятно, были распространены достаточно широко. Это подтверж- дается тем, что яблоня получила у различных этнических групп населения земли свои наименования, свести которые в единую строгую языковую систему не представляется возможным. Од- нако некоторую ясность можно внести, особенно если учесть вероятный ареал распространения рода, а также миграцию населения земного шара. Так, у народов Европы яблоня назы- вается совершенно иначе, чем в Азии. В то же время в евро- пейских языках названия яблони часто схожи. Попытка анализа и генезиса названий многих растений, в том числе и разнооб- разных плодовых, предпринята в известной работе Хана, кото- рая сохраняет свое значение и в настоящее время. Наиболее древние письменные свидетельства о яблоне отно- сятся к IV в. до н. э. Это труды греческого философа и естествоиспытателя Теофраста, в которых описаны главнейшие признаки яблони, особенности ее хозяйственного использова- ния, дана характеристика некоторых сортов яблони и приемов их возделывания. Однако, судя по этим данным, Теофрасту были известны яблони, встречавшиеся лишь в культуре. Слово Malus (яблоня), применяющееся ныне в качестве родового названия яблони, впервые встречается у Плиния и других древнеримских писателей. Оно и вошло в науку как родовой эпитет яблони. древние римляне, видимо, были хорошими садоводами. Об этом
10 Глава 2. Краткий обзор истории изучения рода Жа1ия можно судить по трудам Катона, Варрона, Колумеллы и Пал- ладия, где говорится о многих сортах яблони того времени, а также сообщаются сведения о способах их возделывания. Интересно отметить, что на юге Европы (в Греции и Риме) в период с 111 в. до н. э. до IV в. н. э. было известно довольно много сортов культурной яблони. В Древней Греции, по данным Теофраста, 2257 лет назад было два сорта яблони; в древнем Риме, по данным Катона, 2117 лет назад — семь сортов, по данным Плиния, 1887 лет назад — уже тридцать шесть сортов, по данным Палладия, 1577 лет назад — тридцать семь сортов. Таким образом, путем отбора лучших форм дикой яблони за 7 — 8 столетий человек создал почти четыре десятка сортов. В настоящее время их число возросло более чем в 250 раз. Первое научное описание рода Malus содержится в трудах выдающегося французского ботаника Турнефора [Tournefort, 1700]. Основным признаком для выделения яблони в самостоя- тельный род он считал вдавленное основание и верхушку пло- дов. Эта особенность в строении плодов яблони является доста- точно характерным признаком, позволяющим отграничить ее от таких близких родов, как груша (Pyrus), айва (Cydonia) и др. Однако в труде Турнефора род Malus охарактеризован довольно формально, так как в нем отсутствуют перечень и точное опи- сание диких видов, поскольку Турнефор имел дело лишь с культурной яблоней. В дальнейшем в номенклатуре рода Malus возникла боль- шая путаница, чему способствовали труды Линнея, который объединил яблоню в общий род с грушей и айвой под назва- нием Pyrus. С этого времени название Malus стало видовым эпитетом для культурной яблони, которая стала именоваться Pyrus Malus 1. Кроме культурной яблони Линней описал не- которые другие виды, но рассматривал их в качестве ее разно- видностей: Pyrus Malus sylvestris 1 . и Pyrus Malus paradisi'- aca L. Он знал также сибирскую ягодную яблоню (Pyrus Ьас- са/a 1 .) и североамериканскую венечную яблоню (Pyrus corona- па 1.). Почти одновременно с Линнеем яблоню изучал английский садовод Миллер. В 1754 г. он восстановил самостоятельность рода Malus, а в 1768 г. уточнил его видовой состав. Так, евро- пейская лесная яблоня получила у него самостоятельный видо- вой эпитет Malus sylvestris (L.) Mill., культурная яблоня стала именоваться Malus хоггulana Mill., а венечная — Afalus corona- ria (L.) Mill. Будучи хорошим садоводом, Миллер заметил, что низкорослые яблони полукультурного облика являются ценным подвоем для культурных сортов. Линней их называл «райскими яблонями», а Миллер обозначил их Malus pumila Mill., что в дал~-нейшем стало совершенно неопределенным эпитетогл.
Глава 2. Краткий обзор истории изучения рода Маарив Таким образом, в изучении яблони наметились два направ- ления: первое — созданное Линнеем (объединение яблонь с другими близкими родами) и второе — определенное Турнефо- ром и Миллером (разделение яблони, груши и айвы на самостоятельные роды). Первое привело к дальнейшей путанице, а второе способствовало более детальному изучению яблони. Многие видные ботаники XVIII, XIX и даже ХХ в. — Уэс- тон, Яекандоль, Фоке, Бэйли, — подчиняясь авторитету Линнея, приняли его трактовку рода. Так, даже в «Index I(ewensis» числится 318 наименований рода Pyrus, из которых не менее 70 относятся к яблоням. Тем не менее большинство системати- ков, изучавших яблоню, не приняли линнеевскую концепцию- Яекэнь, Мёих, 1(аррьер, Бриттон и Броун, 1(ёне, Цабель, Шней- дер, Редер, С. В. Юзепчук и другие. Наряду с упомянутыми двумя направлениями в изучении яблони следует отметить еще одно, характеризующееся стрем- лением к переименованию рода. Так, Неккер род Malus пере- именовал в А pirophorum, 1(лервиль — Pyrenia, а Декандоль, следуя Линиею, признал род Pyrus, но выделил в нем секцию Malus, объединившую все яблони. Несмотря на достаточное обоснование самостоятельности рода Malus, смешение его с дру- гими родственными родами просуществовало до конца прошлого столетия и доставило систематикам множество затруднений. Немецкий ботаник Боркгаузен [Borkhausen, 1803], призна- вая самостоятельность рода Malus преимущественно по призна- кам строения плодов, значительно уточнил его объем и описал несколько новых видов. Он пытался доказать, что существую- щее разнообразие видов — результат деятельности человека. Наряду с Malus pumila, описанной еще Миллером, Боркгаузен установил опушенную яблоню — М. dasy phylla Borkh. — и собственно культурную, которую он назвал М. domestica Borkh. Кроме того, он перевел из рода Pyrus ранее описанные виды яблонь: М. baccata (L ) Borkh., М. praecox (Pall ) Borkh., M. prunijolia (Willd) Borkh., М. spectabilis (Aii.) Borkh. Ламарк, отметив включение Линнеем яблони в общий с гру- шей род Pyrus, указал на признаки, препятствующие этому смешению. Таковыми он считал наличие сросшихся у основания пяти столбиков; шаровидную форму плода, вдавленного у осно- вания и верхушки; более короткую, чем у груши, плодоножку; зубчатость и опущенность листьев, а также особенности вкуса плодов. Яля того времени это было правильно, так как Ламарк не имел сведений о всех видах рода. культурные и полукуль- турные яблони он объединил в один вид под названием Malus communis Lam. 1(аррьер, располагая более полными сведениями о роде яб- лони, определил следующие признаки, разделяющие роды Ma- lus и Pyrus:
Глава 2. Краткий обзор истории изучения рода Malus 1) желтая окраска пыльников у яблони и фиолетовая у груши; 2) отсутствие каменистых клеток в плоде яблони и наличие их в плоде груши; 3) присутствие яблочной кислоты в плоде яблони и почти полное отсутствие ее у груши. Автор обращал внимание на относительное значение этих признаков, так как у некоторых видов яблонь пыльники даже более интенсивно окрашены, чем у груши, а в плодах яблонь из- редка встречаются каменистые клетки. На основании перечис- ленных особенностей Каррьер предложил следующее деление рода Malus: виды с желтыми пыльниками и виды с красными пыльниками. Эти группы яблонь, в свою очередь, были разде- лены на крупноплодные и мелкоплодные. Дальнейшее разделение видов рода осуществлено по при- знаку консистенции плодов и особенностям их опадения: 1) плоды сочные, опадающие; 2) плоды мясистые, неопадающие; 3) плоды размягчающиеся; 4) плоды мучнистые. Система Каррьера не получила широкого распространения, так как в ней не были учтены все особенности, свойственные роду Malus. Американские систематики Бриттон и Броун также признали яблоню самостоятельным родом, указывая главным образом на отсутствие каменистых клеток в плоде. Фундаментальный вклад в систематику рода Malus внесли немецкие ботаники Кёне и Цабель. В основу деления рода они положили следующие признаки: остающаяся или опадающая чашечка при плодах (Кёне) и цельнокрайность или лопастность листьев (Цабель). В системе Кене род Malus подразделен на две секции: Calycomeles Коейпе (виды с остающимися при плоде чашелис- тиками) и Gymnomeles goehne (виды с опадающими после цветения чашелистиками). Кроме того, была выделена промежу- точная секция между Calycomeles и Gymnomeles, куда отнесены виды гибридного происхождения. Всего в системе Кене 23 вида, из них 10 — гибридного происхождения. Цабель делил род Malus также на две крупные секции: Eumalus Zabel (виды с цельными листьями) и Sorbomalus Zabel (виды с лопастными листьями). Кроме двух основных секций в данной системе также установлена промежуточная секция. в которую включены гибридные виды. Общее число видов у Ца- беля — 30. Недостатком указанных систем рода Malus следует считать крайне односторонний подход к определению ценности диагнос- тических признаков Секции Кене и Цабеля объединяют раз-
Глава 2. Краткий обзор истории изучения рода Malus личные по происхождению и признакам виды, что нарушает естественность самих секций и единство системы в целом. Тем не менее системы рода Malus, созданные этими ботаниками, принесли известную пользу другим исследователям, так как послужили основой для иных попыток классифицировать виды яблони. Наиболее детальные обработки всего рода Malus приведены в трудах Шнейдера [Schneider, 1906], Коидзуми, Редера и Эзелтина [Ezeltine, 1933], а применительно к видам, встреча- ющимся на территории СССР, — С. В. Юзепчука [1939], а также Ал. А. Федорова и О. М. Полетика [1954]. В основе большинства работ данных авторов лежит система Цабеля. В обзоре же рода яблони японского систематика Коидзуми ос- новой классификационной схемы явилась система Кёне. В 1906 г. были опубликованы две интересные работы не- мецкого садовода и ботаника Шнейдера [Schneider, 1906а,b], в которых он привел оригинальную систему рода Malus и впер- вые описал две новые секции и ряд видов. В данной системе своеобразно синтезированы классификации рода Malus, пред- ложенные Кёне и Цабелем. в качестве основных подразделений рода использованы секции Цабеля, а признаки секции Кёне учтены внутри секций. Шнейдер исключил промежуточные сек- ции, а гибридные виды поместил вместе с основными. Он зна- чительно дополнил данные систематики культурных и полукуль- турных яблонь, относимых ранее к М. pumila Mill. Как садовод и ботаник Шнейдер не признавал существование сборных куль- тигенных видов типа М. horfulana Mill. и М. domesfica Borkh., а пытался все культурные яблони отнести к автохтонным видам в качестве разновидностей последних (например, М. sylvesfris var. hortulana С. К. Schneid., М. pumila var. domesfica С. К. Schneid., М. pumila var. paradisiaca С. К. Schneid., М. pu- mila var. fransluscens С. К. Schneid.). Однако проблема клас- сификации всех культурных и полукультурных яблонь им не была решена. Шнейдер установил принадлежность к роду Malus некото- рых малоизученных видов, которые ранее относили к родам Pyrus, Sorbus и Crafaegus: М. kansuensis (Batal.) С. К. Schneid., М. yunnanensis (Franch.) С. К. Schneid., М. florenfina (Zuc- cagni) С. К. Schneid., М. fusca (Raf.) С. К. Schneid., М. ргайа (Hemsl.) С. К. Schneid., М. frilobata (Labill.) С. К. Schneid., М. fschonoskii (Maxim.) С. К. Schneid. По системе Шнейдера род Malus делится следующим об- разом. Sect. 1 Eumalus Zabel I. Виды с остающимися при плоде чашелистиками.
Глава 2. Краткий обзор истории изучения рода Malus 1. М. sylvestris (L.) Mill. var. typica С. К. Schneid. var. hortulana С. К. Schneid. гибриды: М. sylvestris>(M. pumila М. sylvestris>(M. baccata 2. М. pumila Mill. var. domestica С. К. Schneid. var. niedzwetzkyana (Dieck.) С. К. Schneid. var. paradisiaca (Medic.) С. К. Schneid. f. apetala С. К. Schneid. f. pendula (R. Мй11ег) С. К. Schneid. var. praecox (Pa11.) С. К. Schneid. var. transluscens С. K. Schneid. гибриды: М. pumila>(M. spectabilis М. pumila>(M. baccata а) М. astracanica Dum.-Cours. Ь) М. prunifolia Borkh. с) М. cerasifera Borkh. 3. М. spectabilis (Ait.) Borkh. II. Виды с опадающими на плоде чашелистиками. 4. М. halliana goehne 5. М. baccata (L.) Borkh. var. himalaica (Maxim.) С. К. Schneid. var. mandshurica (Maxim.) С. К. Schneid. Sect. II Sorbomalus Zabel 1. Виды с опадающими на плоде чашелистиками. 6. М. ~ит1 Rehd. 7. М. ьагдеп1а Rehd. 8. М. toringo Sieb. 9. М. fusca (Raf.) С. К. Schneid. 10. М. kansuensis (Batal.) С. К. Schneid. П. Виды с остающимися при плоде чашелистиками. 11. М. florentina (Zuccagni) С. К. Schneid. 12. М. coronaria (L.) Mill. 13. М. angustifolia Michx. 14. М. yunnanensis (Franch.) С. К. Schneid. 15. М. ргаПа (Hemsl.) С. К. Schneid. Sect. III Eriolobus С. К. Schneid. 16. М. trilobata (Labill.) С. К. Schneid. Sect. IV Docyniopsis С. К. Schneid. 17. М. tschonoskii (Maxim.) С. К. Schneid. Систематика рода Malus, предложенная Шнейдером, по сравнению с классификациями предыдущих авторов явилась
Глава 2. Краткий обзор истории изучения рода Malus крупным шагом вперед. Однако она не отражает естественных отношений между видами, так как в ее основу были положены чисто формальные признаки. Несмотря на недостатки данная работа долгое время служила руководством для дальнейшего изучения рода Ма1ия. Следующая, более полная обработка рода предложена не- мецким дендрологом Редером. Его система является усовершен- ствованной классификацией Шнейдера и в последнем своем ва- рианте имеет следующий вид. Sect. I Eumalus Zabel $ег. 1. Pumilae Rehd. 1. М. pumila Mill. 2. М. prunifolia (Willd.) Borkh. 3. М. spectabilis (Ait.) Borkh. 4. М. micromalus Rehd. $ег. 2. Baccatae Rehd. 5. М. baccata (L.) Borkh. 6. М. г'пеi era Rehd. 7. М. hal iana Koehne. Sect. II Sorbomalus Zabel $ег. 3. Sieboldinae Rehd. 8. М. floribunda Sieb. 9. М. гитi (Mats.) Rehd. 10. М. sieboldii (Regel.) Rehd. 11. М. sargentii Rehd. $ег. 4. Florentinae Rehd. 12. М. florentina (Zuccagni) С. K. Schneid. $ег. 5. Капяиепяея Rehd. 13. М. fusca (Rat.) С. К. Schneid. 14. М. toringoides (Rehd.) Hughes. 15. М. kansuensis (Batal.) С. К. Schneid. $ег. 6. Yunnanenses Rehd. 16. М. prattii (Hemsel.) С. K. Schneid. 17. М. yunnanensis (Franch.) С. К. Schneid. Sect. Ш Chloromeles (Реапе) Rehd. 18. М. platycarpa Rehd. 19. М. glaucescens Rehd.
Глава 2. Краткий обзор истории изучения рода Malus 16 coronaria (L.) Mill. t,oensis Britt. lancifolia Rehd. angustifolia (Ait.) Michx. 20. М. 21. М. 22. М. 23. М. Sect. IV Eriolobus С. К. Schneid. 24. М. trilobata (Labill.) С. К. Schneid. Sect. V Docyniopsis С. К. Schneid. 25. М. tschonoskii (Maxim.) С. К. Schneid. При построении своей системы Редер использовал все важ- нейшие признаки яблони: положение листьев в почкосложении (свернутые или сложенные), цельность или лопастность листьев, строение частей цветка (количество столбиков, окраска пыль- ников), положение чашечки при плодах (остающаяся или опа- дающая), наличие или отсутствие каменистых клеток в плодах. Редер впервые ввел в систематику рода Malus серии (или ряды) викарных видов. Однако данная система оказалась не выдержанной для всех видов, поэтому в одни и те же серии попали как настоящие виды, так и гибриды, описанные в ка- честве самостоятельных видов. В этом смысле крайне пестрой и неопределенной оказалась серия Pumilae, где с М. pumila объединены явные гибриды — М. >(prunifolia, М.)<ярес1а1и- lis и M>(micromalus. Нечто подобное можно сказать и о се- рии 3. Sieboldinae, в которой виды М. sieboldii и М. sargentii числятся вместе с гибридными — M>(floribunda и М.)<гит1. Кроме того, М. >( floribunda не имеет лопастных листьев, что характерно для других видов этой серии и секции Sorbomalus в целом. В серии 5. Kansuenses оказались географически отда- ленные виды, встречающиеся лишь в Северной Америке и Цен- тральной Азии (Центральный и Юго-Западный Китай). По неизвестным причинам Редер игнорировал основной европейский вид М. sylvestris (L.) Mill., а также виды, встре- чающиеся в пределах Средней Азии и Кавказа. В его системе пропущены некоторые реликтовые виды яблонь, обладающие ограниченными ареалами: М. laosensis А. Cheval., М. formo- sana gawak et КоЫк., М. sikkimensis (Wenz.) goehne. Но все эти недостатки данной системы не лишают ее ценности, и она продолжает сохранять свое значение и поныне. Некоторые изменения в систему Редера внес Эзелтин. Сек- ции, принятые Редером, в системе Эзелтина остались без изменений, но в секции Sorbomalus cepuu Kansuenses и Yunna- nenses объединены в одну большую серию под общим названием Капяиепвея. Из нее была выделена новая монотипная серия— Fuscae Eselt., представленная лишь одним оригинальным севе-
Глава 2. Краткий обзор истории изучения рода Malus 17 роамериканским видом — М. fusca (Raf.) С. К. Schneid. Если выделение серии Fuscae можно принять, то объединение серий Капяиепзея и Yunnanenses нельзя признать целесообразным. Дальнейшее усовершенствование системы Кёне провел япон- ский систематик Коидзуми. В основе этой системы — секции Кене, но в секцию Calycomeles Qoehne включена в виде серии Chloromeles, а в секцию Gymnomeles Коеппе — серии Baccatae и Sorbomalus. Полностью исключены секции Eriolobus С. К. Schneid. и Docyniopsis С. К. Schneid., из которых образо- ван новый род Sinomalus goidz. Все виды американского мате- рика подчинены М. coronaria (L.) Mill., и значительно расши- рено число видов, близких к M. prunifolia (Willd.) Borkh., кото- рые японским ботаником Ассами рассматривались в качестве разновидностей сливолистной яблони. Общее число видов в этой системе — 36. Подразделение рода Malus Коидзуми не получило широкого распространения. Нельзя согласиться с подчинением всех аме- риканских яблонь одному виду — М. coronaria. Недостатком системы следует считать и то, что виды секций Lriolobus и Docyniopsis он не считал яблонями. На самом деле их следует считать реликтами и тем первичным типом яблонь, от которых в далеком прошлом произошли и постепенно эволюциониро- вали современные виды рода. В 1937 г. свою систематику яблонь предложил американский садовод Бэйли. Его труд представляет большой интерес, так как в основу подразделения яблонь положено сочетание при- знаков структуры генеративных и вегетативных органов, их хозяйственное значение (культурные сорта, декоративные виды и формы), а также географическое распространение. Однако в своей работе Бэйли решает главным образом чисто садовод- ческие проблемы и допускает ряд серьезных ошибок, прежде всего номенклатурного характера. В группе «Помологические сорта, культивируемые в садах ради съедобных плодов» при- водится архаическое наименование «Pyrus Malys 1 .», к кото- рому в качестве основных разновидностей отнесены var. sylvestris, var. pumila, var. niedzwetzkyana, var. apetala и са- довые формы — var. аигеа, ~аг. plana, var. pendula. Единствен- ный самостоятельный вид, по Бэйли, это Pyrus soulardi Bailey, который на самом деле является гибридом — М. ioensis>(M. pu- mila, что было установлено Редером. К культурным сортам автор относит также группу яблонь с опадающей при плодах чашечкой, куда он включает единст- венный вид — М. baccata (L.) Borkh. и его разновидность— var. sibirica С. К. Schneider. Под названием «Декоративные яб- лони» числится несколько групп, различаемых по признакам листьев, типам их почкосложения, состоянию чашечки при пло- дах, а также по некоторым признакам строения цветка~ ~сте-
18 Глава 2. Краткий обзор истории изучения рода Mglus пень опушения столбиков, сочетание раскраски венчика) и гео- графического распространения. Система Бэйли наглядна и удобна с практической точки зрения, но виды в ней разбросаны по разным группам без учета истинного значения признаков. В 1938 г. появилась работа В. В. Пашкевича «Яблоня». Ав- тор делит род Malus на два подрода — Eumalus (Zabel) Pash. и Sorbomalus (Zabel) Pash. В подроде Eumalus (Zabel) Pash. серию Pumilae Rehd. предлагается переименовать в отдел Sgl- vestres Pash., а серию Baccatae Rehd. оставить с прежним на- званием, повышая ее лишь до ранга отдела. В подрод Sorbo- malus (Zabel) Pash. включены отдел, характеризующийся чашечкой, остающейся при плоде, и отдел, в котором виды имеют опадающую чашечку. В первый отдел Са!дх persistens Pash. введены подотделы — Sclerocellulatae Pash. (плоды с ка- менистыми клетками) и Mollscellulatae Pash. (плоды без ка- менистых клеток). Все эти подразделения (отделы и подотделы) не соответствуют правилам международной ботанической но- менклатуры и не могут быть приняты во внимание. Следующая попытка классифицировать яблони была пред- принята Г. Г. Тарасенко [1941]. В его работе яблони делятся по географическому принципу и признакам строения органов. Автор выделяет следующие группы. 1. Европейские виды, . для которых характерны признаки культурной яблони, т. е. цельные, обычно опушенные листья и довольно крупные плоды. Это «родоначальники» культурной яблони — М. pumila Mill., М. dasgphglla Borkh., M. sglvestris (L.) Mill. 2. Восточно-азиатские виды, которые отличаются от видов других групп главным образом мелкими (ягодовидными) пло- дами, опадающей чашечкой и цельными или лопастными листь- ями. К цельнолистным яблоням отнесены следующие группы ягодных яблонь: М. baccata Borkh., M. himalaica Кот., M. sik- kimensis (Wenzig), КоеЬпе, M. flonbunda Sieb., M. halliana КоеЬпе, М. theifera Rehd. К видам с лопастными листьями при- числены М. sargentii Rehd., M. sieboldii Rehd., а также М. for- mosana I(awak. et I(oidz. и М. spectabilis (Ait.) Borkh. По наличию чашечки при плоде и по его величине, а также по ряду других признаков последние два вида (особенно формозская яблоня) сильно отличаются от остальных видов этой группы. 3. K североамериканским видам, обладающим ароматными плодами (с неопадающей чашечкой) и обычно несущим ло- пастные, сильноопушенные листья, отнесены М. coronaria (L.) Mill., M. ioensis (Wood.) Britt., M. angustifolia Michx., M. glau- cescens Rehd. и М. fusca (Raf.) С. К. Schneid. Последний вид отличается мелкоплодностью, опадающей при плодах чашечкой, в связи с чем его нельзя относить к данной группе яблонь.
Глава 2. Краткий обзор истории изучения рода Afalus вселение яблонь на географические группы получилось у Г. Г. Тарасенко формальным. Он, во-первых, не учел всего разнообразия видов, характерного для указанных географичес- ких зон, а, во-вторых, разделил азиатские яблони лишь по признаку структуры края листа, что нельзя признать достаточ- ным. Данная система рода совершенно искусственна, и ее едва ли следует принимать во внимание. Систематика яблонь, произрастающих в СССР в диком виде, дана С. В. Юзепчуком '[1939] во «Флоре СССР». В ее основу положена система Редера с учетом некоторых соображений В. В. Пашкевича. Автор описал всего 12 видов, относящихся только к секции Eumalus Zabel и характеризующихся цельными, свернутыми в почкосложении листьями. Род Malus Mill. в пре- делах СССР разделен на подрод Eumalus (Zabel) Juz., в котором серии, установленные Редером, повышены до ранга секций (по- добно тому, как секция Eumalus возведена в ранг подрода) . К секции Pumilae (Rehd.) Juz. отнесено два ряда — Silvestres Juz. и Prunifoliae Juz. Ряд 1. Silvestres Juz. характеризуется тем, что чашелистики при плодах несросшиеся, плоды более или менее вдавленные при основании. Сюда отнесено 7 видов: M. sylvestris (L.) Mill., M. praecox (Pall.) Borkh., М. orientalis (Uglitz.) Juz., M. sie- versii (Ledeb.) М. Roem., M. niedzwetzkyana Dieck., M. turkme- погит Juz. et Рор. и М. domestica Borkh. Ряд 2. Prunifoliae Juz. характеризуется чашелистиками при плодах, сросшимися в трубку, и плодами с округленным осно- ванием. Сюда отнесен вид М. prunifolia (Willd.) Borkh. Секция Васса1ае (Rehd.) Juz. (у Редера — серия) содержит 4 вида: М. Ьасса1а (L.) Borkh., М. pallasiana Juz., М. mand- shurica (Maxim.) Кот. и M. sachalinensis (Кот.) Juz. Обработка С. В. Юзепчука — первая полная сводка о видах яблонь, произрастающих на территории СССР. Она имеет боль- шую ценность, так как здесь описаны новые виды, в частности M. orientalis — для Кавказа, M. 1urkmenorum — для Средней Азии; восстановлен М. praecox и в новом, более полном смысле,. с указанием разнообразия и уточнения ареала — М. иеоегис. Однако описание сибирских яблонь в качестве нового вида— М. pallasiana Juz. — едва ли следует считать удачным. Линней [Linnaeus, 1767] описал свою Pyrus baccata, происходящую из центра ареала этого вида — Забайкалья (р. Шилка). Поэтому нет никаких оснований отказываться от приоритетного назва- ния Линнея и заменять его другим, более поздним. Наличие ряда популяций М. baccata в культуре, встречающихся в раз- личных зонах земного шара, не может служить достаточным основанием для рассмотрения их в качестве самостоятельного вида, отделенного от сибирских яблонь. Недостаточно четко
20 Глава 2. Краткий обзор истории изучения рода Malus обосновано в системе С. В. Юзепчука отнесение к самостоятель- ному виду М. п~ейглюе zkyana. В 1947 г. была издана работа немецкого садовода-исследо- вателя и селекционера Хеннинга [Henning, 1947]. В результате опытов по скрещиванию различных видов яблони ученый при- шел к следующим выводам: 1) при семенном размножении многие «виды» яблони, бу- дучи гибридными по происхождению, обычно не соответствуют описаниям, в связи с чем Нх видовой ранг является сомнитель- ным; 2) большинство видов группы, используемых в селекционной работе, является крупноплодными формами ягодных яблонь, от- носимых разными авторами к М. baccata, М. cerasifera или М. prunifolia; 3) число североамериканских видов яблонь должно быть сокращено, так как некоторые H3 ННх описаны без достаточных оснований; 4) в создании крупноплодных европейских и американских сортов культурной яблони участие М. baccata и М. prunifolia (а также подобных им гибридов) нельзя считать доказанным. Работа Хеннинга осталась незаконченной, так как автор по- гиб во время войны. Однако его соображения, изложенные выше, представляются интересными и должны быть приняты к сведению. В послевоенный период появился ряд работ Ал. А. Федо- рова, в которых содержатся сведения о яблонях, произрастаю- щих в Средней Азии. В результате тщательных исследований, проведенных в Южной Киргизии, Ал. А. Федоров и Ан. А. Федоров [ 949] описали новый вид яблони, названный ими яблоней киргизов (М. kirghisorum А1. et An. Tkeod.), которая распро- странена в пределах Ферганского, Таласского и Чаткальского хребтов, а также в Заилийском и джунгарском Алатау. М. Ыг- ghisorum — яблоня мезофильного облика, характерная для горных лесов. Она размножается не только семенным путем, но главным образом вегетативно (корневыми отпрысками и отводками ветвей) и имеет целый ряд интересных биологичес- ких особенностей. Ей свойственна большая изменчивость признаков, часть которых описана. Ал. А. Федоров и Ан. А. Фе- доров [1949] поясняют, что М. Ыгдйыогит имеет связь с ябло- ней Сиверса — М. sieversii (Ledeb.) М. Roem. — ксероморфным видом, также встречающимся в пределах упомянутых выше горных хребтов и в Тарбагатае. В 1948 г. опубликована работа Г. П. Сумневича «Новые виды яблони Западного Тянь-Шаня», в которой описаны 4 но- вых вида яблони Западного Тянь-Шаня (из бассейна р. Ан- грен): М. kudrjaschevii Sumn., М. tianschanica Sumn., М. ani- sophylla Sumn. и М. persicifolia Sumn. В 1949 г. П.П.Поляков
Глава 2. Краткий обзор истории изучения рода Malus описал еще 3 новых вида яблони: М. schischkinii P. Pol., М. jar- molenkovii P. Pol. и М. linczevskii P. Pol. Все новоописанные Г. П. Сумневичем и П. П. Поляковым виды яблонь, видимо, являются локальными формами среднеазиатских яблонь. Новоописанные виды Г. П. Сумневича отвергает В. И. За- прягаева [1964], так как по своим морфологическим признакам они не отличаются от М. sieversii и являются отдельными попу- ляциями этого очень полиморфного вида. Б. А. Быков во «Флоре 1~азахстана» дал подробную схему классификации яблонь 1~азахстана. Она имеет следующий вид. М. иеоелй (Ledeb.) М. Roem. ло~1Кц д о0тэо~зц4з явл- с шаровидными плодами — f. jarmolenkovii (P. Ppl.) В. Bykov. с зелеными плода ми — f. schischkinii (P. Pol.) В. Bykov. с розоватыми или красными плодами — f. рао1оой В. Bykov. с янтарно-желтыми плодами М. sieversii (Ledeb.) М. Roem — var. longicarpa В. Bykov. с продолговато-яйцевидными плодами (типа 1~андиль синан) — f. роЦайоой В. Bykov. с зелеными плодами — f. linczevskii (P. Pol.) В. Bykov. с красными или красноватыми плодами — f. dragavcevii В. Bykov. с янтарно-желтыми плодами. Таким образом, Б. А. Быков не считает яблоню киргизов особым видом, но, по-видимому, отождествляет ее с М. иеоегвй. В 1950 г. С. Н. 1~удряшовым был описан еще один вид из числа среднеазиатских яблонь — М. hissarica из бассейна р. 1~ашкадарья, с верхней части ее притоков (Южный Узбеки- стан, район Шахрисабза). Существование этого вида также оспаривается В. И. Запрягаевой [1964]. Известный вклад в изучение яблони внес и И. Т. Василь- ченко. Он описал не только среднеазиатские, европейские, кав- казские и сибирские яблони [Васильченко, 1963], но и восточно- азиатские [Васильченко, 1959]. Опубликованный им в 1963 г. труд «Новые для культуры виды яблони» по существу является справочником, включающим сведения о видовом составе, гео- графии, экологии, биологии, ресурсах и практическом значении яблони. В нем есть необходимые для практики материалы, а также научные данные. И. Т. Васильченко описаны некоторые новые виды яблони: для Средней Азии — М. juzepczukii Vass. (бассейн р. 1~угарт,
Глава 2. Краткий обзор истории изучения рода Malus 22 Южная Киргизия) и для Восточной Азии — М. seto' Vass (Камское нагорье, бассейны рр. Меконг и Бар-Чю) и М. cen- tralasiatica Vass. (высокогорные районы Центральной Азии, Китай) [Васильченко, 1959]. Здесь приводятся сведения о серии 1(ansuenses Rehd. секции Sorbomalus Zabel, восстановлен вид М. himalaica (Maxim.) Vass. Ал. А. Федоров и Q. М. Полетика [1954] описали 37 видов,. 3 разновидности и 25 форм яблони, встречающихся в СССР как в диком виде, так и в культуре. Авторы этой обработки hfalus придерживались, в основном, системы Редера. Яблони СССР они делят на следующие секции: 1 — Eumalus Zabel, II— Sorbomalus Zabel, I I I — Chloromeles (Decne.) Rehd. В 1954 г. новую систематику рода разработал помолог Ф. Д. Лихонос. В ее основе — дополненные и усовершенство- ванные системы Кёне и Коидзуми. Она имеет следующий вид. Секция 1. Calycomeles КоеЬпе: — ряд 1. Eumalus (Zabel) Likh. (8 видов крупноплодных яб- лонь среднеазиатского, кавказского и европейского очагов); — ряд 2. Prunomeles Likh. (3 вида, 6 разновидностей и 8 форм гибридных яблонь); — ряд 3. Chloromeles (Decne.) Rehd. (4 вида и 10 разновид- ностей североамериканских яблонь); — ряд 4. Eriomeles Likh. (2 вида). Секция II. Gymnomeles Koehne: — ряд 5. Baccatae Rehd. (наиболее крупный; 9 видов, 2 раз- новидности и 5 видов гибридного происхождения); — ряд 6. Sorbomalus (Zaibel) Likh. (6 видов). gJaHHaH система не получила широкого признания, так как выделенные в ней секции объединяют весьма различные как по происхождению, так и по морфологическим признакам и био- логическим свойствам яблони. Кроме того, здесь не учтен весь видовой состав яблонь, а также целиком отсутствуют виды, при- веденные Редером в составе серий E(ansuenses и Yunnanenses. Ф. ц. Лихонос, следуя Коидзуми, исключил из рода hfalus видьг M. tschonoshii и М. trilobata, ряд Eriomeles Likh. дал в ином понимании, куда включил М. formosana и М. prattii. В своих более поздних трудах Ф. gI. Лихонос [1963, 1964] высказал интересные соображения о происхождении некоторых яблонь. Относительно М. pumila он писал: «Название «низкая яблоня», которое было установлено Миллером д~я дусена и парадизки — широко известных карликовых подвоев яблони, потеряло со временем свое первоначальное значение и стало общим и совершенно неопределенным обозначением низкой яб- лони вообще» [Лихонос, 1963, с. 60].
Глава 2. Краткий обзор истории изучения рода Malus 23 М. praecox, по Ф. Д. Лихоносу, можно рассматривать как подвид, или даже разновидность М. sylvestris, а М. sieversii и М. niedzwetzkyana представляют собой лишь подвиды основного вида М. kirghisorum. Автор отрицает существование в природе новоописанных ви- дов: Г. П. Сумневича — М. kudrjaschevii, М. tianschanica, M. anisophyIIa, M. persicifolia; П. П. Полякова — М. schischkinii, М. jarmolenkovii, М. Iinczevskii; С. Н. Кудряшова — М. hissa- r<ca. И. Т. Васильченко — М. juzepczukii. М. sargentii Rehd. (из Японии) он считает карликовой разновидностью М. sieboldii (Regel.) Rehd. Дальневосточные виды ягодных яблонь М. mandshurica (Maxim.) Кот. и M. sachalinensis (Кот.) JUz. — предлагает рассматривать в качестве разновидностей основного вида ягодных яблонь — M. baccata (L.) Borkh. Коли- чество североамериканских видов рекомендует уменьшить с шести [Rehder, 1958] до двух. Предложения Ф. Д. Лихоноса по сокращению видового со- става яблонь заслуживают внимания. Он прав, что критический пересмотр видового состава позволит «более четко представить содержание рода Malus и более полно использовать морфолого- биологические особенности видов яблони в работах по систе- матике как дикорастущей, так и культурной яблони и в селек- ции» [Лихонос, 1964, с. 12]. В принципе соглашаясь с этими предложениями, все же необходимо подчеркнуть, что к ревизии видов следует подходить с большой осторожностью, не меняя их объема произвольно и не впадая в крайности исключительно :из соображений удобства. На основании результатов исследований Г. Г. Тарасенко и собственных опытов по скрещиванию яблонь Ф. Д. Лихонос [1963, 1964] пришел к следующим выводам: 1) при скрещивании сибирской яблони с культурными сор- тами в потомстве наблюдается доминирование признаков М. baccata; 2) от скрещивания сибирской яблони с культурными сор- тами получаются формы вишнеплодной яблони; 3) при последующих скрещиваниях вишнеплодной яблони с .культурными сортами в потомстве появляются китайки; 4) многие виды, близкие к М. prunifolia, следует рассмат- ривать в качестве разновидностей и форм, так как они сущест- вуют только в культуре; 5) при скрещивании культурных сортов между собой дико- растущие виды в потомстве не появляются; 6) новые «виды» яблони, описанные на основании изучения единичных экземпляров, т. е. «лишенные ареала», нельзя при- знать видами. Китайскими авторами Чжи-Те-всунем и Чжень-Чень-Лунгом приведено для Китая 20 видов яблони, из них 14 автохтонных,
24 Глава 2. Краткий обзор иИофии изучения фода Ма1иь' остальные в культуре. В их работе даны оригинальные рисунки китайских яблонь, а также данные об их распространении во всех 23 провинциях Китая. Однако вопросы систематики авторы не решают. Польский дендролог К. Брович [Browicz, 1969] исключил из рода Malus яблоню трехлопастную — М. trilobata, повышая секцию Eriolobus в ранг рода. Таким образом, реликтовый наи- более древний вид яблони, по Бровичу, получил прежнее назва- ние — Егсolobus trilobata (Poir.) Roem. Он же [Browicz, 1970] показал возможность гибридного происхождения яблони фло- рентийской — М. florentina (Zucc.) С. К. Schneid. По его убеж- дению, флорентийская яблоня является межродовым гибридом [Malus sylvestris (L.) Mill.>(Sorbus torminalis (L.) Сгапк] .. В результате им описан новый гибридный род Malosorbus Bro- wicz с единственным видом гибридного происхождения Malo- sorbus Х florentina (Zucc.) Browicz. В ряде работ В. В. Пономаренко [1972 — 1976, 1988] сделана попытка уточнения видового и внутривидового статуса дикорас- тущих яблонь Сибири, Дальнего Востока, Сахалина, Средней Азии и Кавказа. Им изучены некоторые культурные, одичавшие и интродуцированные яблони, описан новый подвид маньчжур- ской яблони — яблоня Жуковского. Труды В. В. Пономаренко, безусловно, являются существенным вкладом в распознавание видов и способствуют более детальному изучению рода яблони
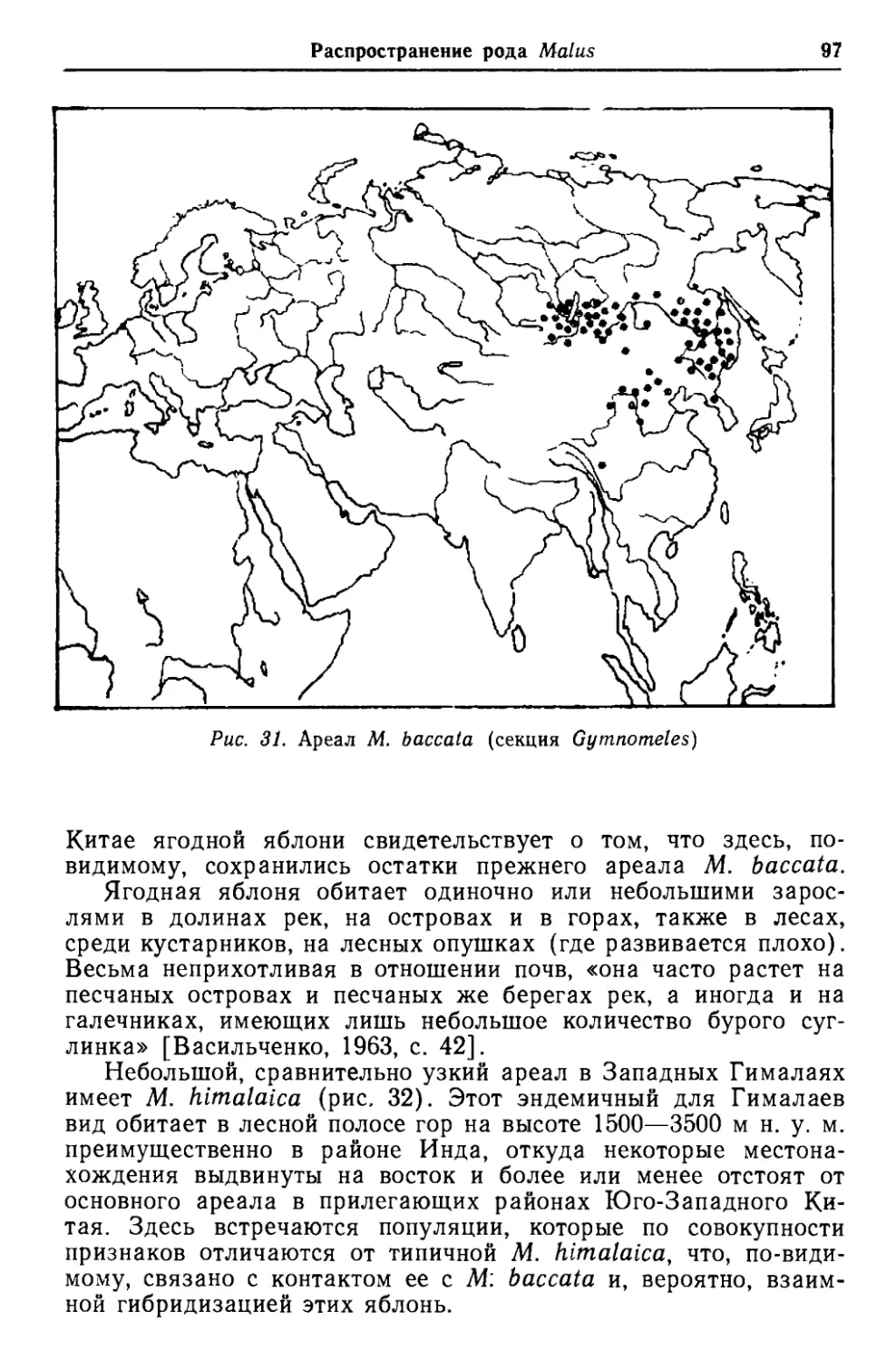
Глава 3СТРУКТУРНЫЕ ОСОБЕННОСТИ,ЭВОЛЮЦИЯ И ФИЛОГЕНИЯ ЯБЛОНИ Организм, достигший определенного развития, характеризу- ется целой системой признаков, которые в известной степени отражают путь его эволюционного становления. Признаки, свой- ственные любому организму, находятся на разных уровнях раз- вития в зависимости от его генетической основы и способности реагировать на воздействие факторов среды. Для описания лю- бого вида и определения его положения в системе, а также для выяснения уровня его филогенетического развития необходимо тщательное изучение признаков, а также оценка их примитив- ности или продвинутости в эволюционном отношении. Основными признаками, определяющими филогенетические отношения видов рода Malus, являются общая структура расте- ний, их облик (или габитус), строение листьев, цветков и пло- дов, а также другие особенности, поддающиеся различению. Изучение и сопоставление признаков в пределах рода приобре- тает первостепенное значение и позволяет выявить филогенез рода. Однако при выявлении основных признаков и установлении путей их эволюции возникают значительные трудности. Дело в том, что род Malus представлен сложной, диффузной системой популяций, состоящих нередко из элементов случайной комби- нации генов (мутации, гибридизация и т. п.). Оценку примитив- ности или продвинутости таксона нельзя проводить, учитывая только сложность или простоту структуры признаков, так как простота в строении не всегда является признаком примитив- ности. Наоборот, целесообразное упрощение структурных эле- ментов таксона, приобретенное в процессе пройденного филоге- нетического пути, должно оцениваться как признак продвину- тости. У рода Malus это ярко выражено в эволюции структуры листовой пластинки, а также других элементов. В качестве основы построения исходных позиций для выяв- ления основных направлений эволюции рода Malus мы исполь- зовали сравнение амплитуд изменчивости признаков внутри рода в связи с географическим распространением видов, осо- бенно при их продвижении с юга на север, а также трудно
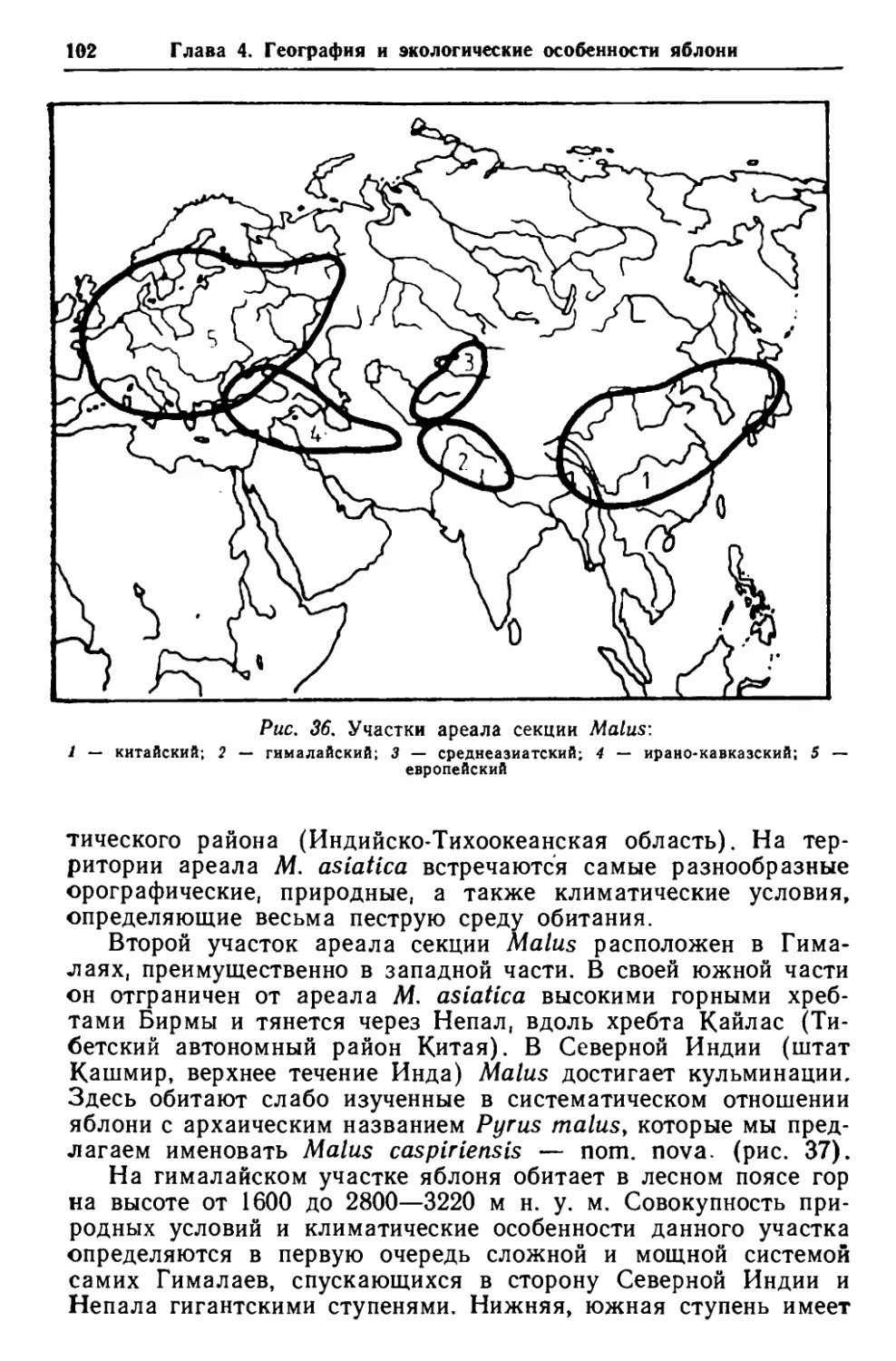
Глава 3. Структурные особенности, эволюция и филогения 26 Прогрессивные признаки Примитивные признакиМестообитание субтропическое Местообитание в умеренных широтах северного полушария деревца (деревья) с менее вы- раженными стволами или кус- тарники Сильноразветвленные деревца (деревья) или кустарники, иногда ксероморфного облика, с симподиальным ветвлением Листья вторично упрощенные, цельные, по краю зубчатые, пильчатые, городчатые, в поч- ках свернутые Крупные, прямые деревья со стволами Слаборазветвленные деревья иезофильного облика с неопре- деленным типом моно- и сим- подиального ветвления Листья лопастные, до раздель- ных, на укороченных побегах, переходящие в цельные, иногда цельнокрайные, в почках сло- женные вдоль центральной жилки )Килкование листа перисто- краебежное, слаборазветвлен- ное )Килкование листа петлевид- ное или сетчатое, хорошо раз- ветвленное опровергаемых признаков связи Malus с другими, как несом- ненно примитивными, так и более продвинутыми родственными родами. Если в области определения происхождения как всех розо- вых в целом, так и рода Malus в частности из-за отсутствия ясных палеоботанических свидетельств ситуация пока малобла- гоприятна и сводится лишь к некоторым гипотетическим сооб- ражениям, то основные направления эволюции рода Malus изучены несколько лучше. Естественность рода Malus и тесное родство между яблонями и такими более древними родами, как Docynia, Pyrus и частично Cydonia, — вне сомнений. Следова- тельно, мы имеем опору для суждения о том, какие группы внутри рода Malus являются наиболее примитивными. По-види- мому, это группы, которые ближе всего стоят к доциниям и наиболее примитивным представителям родов Pyrus и Cydonia— обитателям Юго-Восточной Азии, ныне входят в секции Eriolo- bus и Docyni opsis. Наиболее прогрессивные признаки выявляются при сравне- нии развития структурной изменчивости внутри рода (в сторону продвинутости) в связи с географическим распространением ви- дов, т. е. в результате продвижения видов от более благоприят- ных условий местообитания к менее благоприятным. Основыва- ясь на этих позициях, основные направления эволюции внутрк рода Malus можно представить следующие образом:
27 Корневая система и побег Цветки в малоцветковых, ре- дуцированных, щитка- или зонтиковидных соцветиях Цветки в многоцветковых (ци- мозных) соцветиях неопреде- ленного типа Стилодии, сросшиеся до поло- вины и выше Плоды крупные, с остающейся чашечкой и свободными до ос- нования чашелистиками (ли- ния крупноплодности) или мелкие с опадающей чашечкой (линия мелкоплодности) Стилодии, сросшиеся у самого основания Плоды мелкие, с грубоватой мякотью, содержащей каме- нистые клетки, и с остающейся при плоде у основания мясис- той чашечкой, состоящей из сросшихся у основания чаше- листиков Семенные камеры открыты, сливаются в общую централь- ную полость плода, края пло- долистиков не срослись пол- ностью Семенные камеры полузакры- ты или закрыты, срастание плодолистиков неполное (верх- ние концы не срослись) или полное КОРНЕВАЯ СИСТЕМА И ПОБЕГ Корень. У яблони, так же как и у большинства растений, различаются 2 типа корней: первичные (или настоящие) и вто- ричные (или придаточные). Первичные корни развиваются из первичного корешка зародыша. В результате дальнейшей диф- ференциации из него образуется главный корень, ветвление которого дает боковые корни. Главный корень нулевого порядка обычно имеет вертикальное положение и углубляется в нижние горизонты почвы. При усиленном развитии главного корня об- разуется стержневая корневая система (когда главный корень развивается сильнее боковых) . Стержневая, относительно малоразветвленная корневая сис- тема характерна для яблонь, произрастающих на сухих скло- нах, и яблонь на ранних этапах жизни. В естественных условиях (Гиссарский хребет) яблоня Сиверса, по данным В. И. Запрягаевой [1964], в первый год жизни развивает стерж- невой корень, проникающий в глубину до 30 см. Такой верти- кальный стержневой корень либо прекращает свой рост в глу- бину на втором году жизни дерева, либо сохраняется в течение первых 4 — 5 лет и достигает глубины до 2,5 м. Таким образом, для молодых яблонь характерны стержневые корни, которые, однако, существуют недолго. Вскоре происхо- дит редукция главного корня, в результате чего возникает
Глава 3. Структурные особенности, вволюция и филогения 28 Рис. 1. Корневая система 12 — 15-летней яблони Сиверса на Гиссарскомхребте (по В. И. Запрягаевой [1964]).Цифрами обозначена глубина (см) система боковых корней. По данным В. А. Колесникова [1966], у 50 — 80~/а сеянцев (в условиях культуры) концы главных кор- ней усыхают в длину на 5 — 20 см уже на первом году жизни. То же можно наблюдать и в природных условиях на участках, не обеспеченных влагой. Здесь уже с первой недели само- стоятельного существования сеянца происходит замена главного корня боковым. После редукции главного корня образуются боковые корни первого порядка, которые дают начало боковым корням второго порядка, из которых, в свою очередь, возникают боковые корни третьего порядка и т. д. К 7 — 10 годам корневая система яблони Сиверса (Гиссарский хребет, по данным В. И. Запря- гаевой [1964] ) становится сильноразветвленной, распространя- ется в стороны до 2 м и проникает в глубину до 1,5 — 2,0 м. В 12 — 15-летнем возрасте проекция корневой системы превы- шает проекцию кроны более чем в 10 раз ~[Запрягаева, 1964] (рис. 1). В этом возрасте наиболее интенсивное разрастание корней происходит в горизонтальном направлении. Основная масса корней у деревьев в возрасте от 5 до 42 лет в Заилийском Алатау, по данным А. П. Ярагавцева [1956], располагается в горизонте от 17 до 35 см, а в возрасте более 55 лет — на глу- бине 34 — 55 см. Глубина проникновения вертикальных корней яблони Си- верса не превышает 2 — 3 м. Горизонтальные же корни разраста- ются от стволов вниз по склону на 12 — 20 м, поперек склона- на 8 — 10 м и вверх по склону — на 5 — 8 м. В отличие от вер- тикальных, у горизонтальных корней на поверхности коры име- ется большое число чечевицей и спящие почки,
Корневая система и побег p~~S ' Ф Рис. 2. Корневая система 100-летней яблони Сиверса (по В. И. Запрягае- вой [1964]). Пунктиром обозначена проекция кроны Корневая система старой яблони Сиверса уже не столь значительно превосходит крону по величине. У 100-летней яб- лони Сиверса проекция корневой системы примерно в 2 — 3 раза больше проекции кроны ~[Запрягаева, 1964] (рис. 2). В культуре при создании благоприятных условий для жизне- деятельности корней проекция корневой системы лишь в 1,5— 2,0 раза превышает проекцию кроны, причем активная (всасы- вающая) часть корней находится за пределами кроны. Так, у культурной яблони Сары синан в Крыму в возрасте 45 лет ветви кроны отходили от ствола на 3 — 5 м, а корни — на 11 м Из них под кроной было 3~о~о всех корней (в Казахстане— 30%, в Грузии — 38,9%). Общая длина всех корней крымской яблони Ренет шампанский составила 543 м, из них в проекции кроны было 216 м и за ее пределами — 327 м. При этом из всех 543 м корней данной яблони только 77 м были направлены вертикально — в глубину до 5 м, а остальная масса распро- странялась горизонтально, охватывая более поверхностные го- ризонты почвы до 50 — 65 см [Колесников и др., 1966, с. 75]. Интенсивность проникновения корней яблони в толщу суб- страта и горизонтального их распространения зависит от многих
Глава 3. Структурные особенности, эволюция и филогения ЗО Рис. 8. Размножение яблони киргизов корневыми отпрысками (по Ал, А. Федорову и Аи. А. Фе- дорову [19491) природных факторов; рельефа местности, плотности грунта, влажности и аэрации почвы, глубины залегания грунтовых вод, а также от наследственной природы вида и др. Большое влия- ние на расположение корневой системы яблонь оказывает место произрастания: горные склоны, различные возвышения, речные террасы, равнинные участки. В пределах данных элементов рельефа создаются различные условия для жизни корней. Поло- жительное влияние на развитие корней яблони оказывают раз- личные почвенные бактерии, грибы, водоросли, а также сопут- ствующие яблоне высшие растения. На горизонтально расположенных боковых корнях яблони нередко развиваются адвентивные (придаточные), а также спя- щие почки, из которых образуются надземные побеги — корне- вые отпрыски. Яблоня киргизов, в изобилии произрастающая в горах Средней Азии, отличается такой энергией образования корневых отпрысков, что крупные естественные насаждения этого вида, насчитывающие до нескольких сотен стволов, ока- зываются связанными общей корневой системой. А среди них находится вполне жизнеспособное исходное материнское дерево. Образование корневых отпрысков у этой яблони происходит на всем протяжении горизонтальных корней, но только при ус- ловии залегания последних на глубине от 30 до 40 см. Много- численные и хорошие отпрыски возникают на значительном расстоянии от дерева (от 15 до 20 м) и растут рядами, ориен- тированными вдоль горизонтального корня (рис. 3). Количество отпрысков у одного дерева достигает 500. 1~орневые системы яблонь по пластичности и ее зависимости от различных природных факторов не имеют прямого диагности- ческого значения.
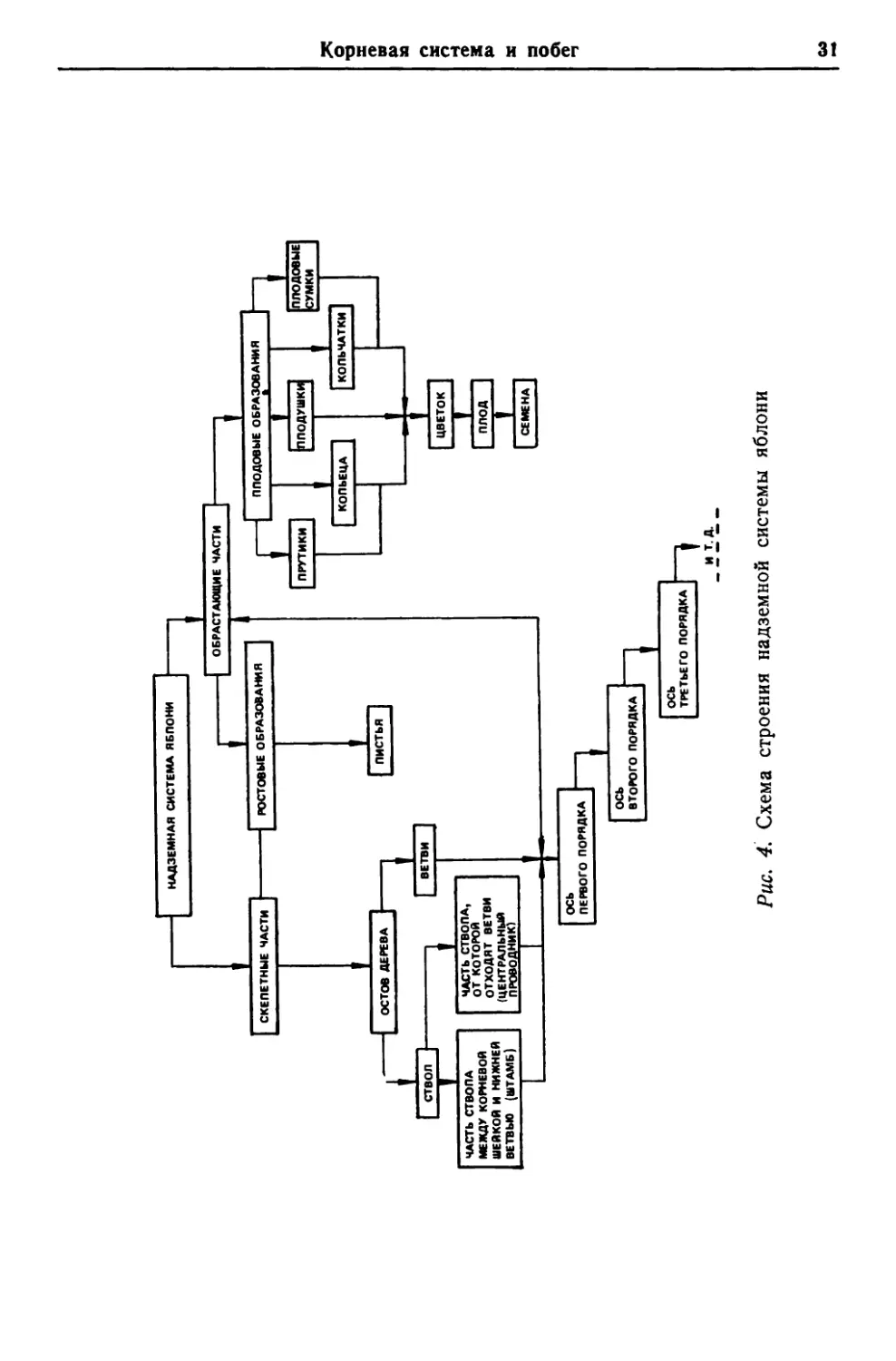
СХЕПЕТННЕ ЧАСТИ ОСТОВ ДЕРЕВА QCb ПЕРВОГО ПОРЯДКА РР~ИЗЕМН*Я СИСТЕМА ЯБПОНИ УЦ~ЕБМЕ ОБР~АИИЯ ЕИЕ ВТОРОГО ПОРЯДКА ПИСТЬЯ ОСЬ ТРЕТЬЕ ГО ПОРЯДКА ОБРАСТРАБЕИЕ ЧАСТИ ПМ'ЪИКИ ППОИЕИЫЕ ОБРЕ:И~АНКИ ЦВЕТОК YH3N30 I I I Корневая систеыа и побег 3l
32 Глава 3. Структурные особенности, эволюция и филогения Побег. Строение побега (надземной системы) яблони до- вольно сложно. Учитывая данные общей морфологии растений, а также принятую в плодоводстве терминологию, мы предла- гаем следующую схему структуры надземной системы яблони (рис. 4) . Побег, т. е. ствол и крона яблони, состоит из скелетных и обрастающих частей. Основная часть дерева состоит из ствола и самых крупных скелетных ветвей, а также менее круп- ных — полускелетных, дающих оси первого, второго, третьего ;и последующих порядков. Часть ствола между корневой шейкой (местом соединения ствола и корня) и нижней ветвью обычно называется штамбом, а часть ствола, от которой отходят ветви, в плодоводстве принято называть центральным проводником. Вместе с ветвями он составляет крону дерева. Обрастающие части надземной системы яблонь состоят из ростовых и плодо- вых образований. Ростовые образования — это побеги одно- .летнего возраста. Типичными ростовыми побегами следует считать однолетние верхушечные или боковые приросты продолжения, характери- зующиеся удлиненными междоузлиями. Они образуются из терминальной (верхушечной) почки или из аксиллярных (боко- вых) почек прошлогоднего прироста. Растут они вертикально и достигают значительной длины (от 40 — 60 до 80 — 100 см) . Спустя некоторое время верхушечный рост постепенно затухает, что вызывает интенсивное распускание боковых почек и способ- ствует заложению новых. В результате образуются укороченные побеги. Они весьма многочисленны и имеют короткие междо- узлия и горизонтальное направление роста. Укороченные росто- вые побеги обеспечивают развитие в кроне всей массы листьев, т. е. весь фотосинтезирующий аппарат. Таким образом, яблони имеют два типа ростовых побегов: удлиненные (долихобласты) и укороченные (брахибласты) . Первые преобладают в период усиленного роста молодых яб- лонь и осуществляют вегетативный прирост особи. Вторые появ- ляются позже и обеспечивают вступление дерева в пору плодо- ношения. У яблонь различают несколько типов плодовых образований, которые берут свое начало от укороченных ростовых побегов (см. рис. 4). 1. Плодовые прутики — самые длинные (10 — 15 см) одно- летние приросты, оканчивающиеся цветочной почкой. Они обычно изогнуты и отходят от центральной оси под большим углом. 2. Копьеца — побеги средней длины (5 — 15 см), располо- женные под прямым углом к центральнои оси, имеющие укоро- ченные междоузлия и сильно сближенные боковые почки. Иногда они оканчиваются колючкои.
Корневая система и побег 33 3. Кольчатки — самые короткие (3 — 5 cM) годичные побеги, обладающие кольцеобразными рубцами и хорошо развитой ко- нечной почкой. 4. Плодушки — многолетние приросты, состоящие из коль- чаток, копьец и прутиков. Плодухи — разновидность плодушек, т. е. многолетние приросты, состоящие из разветвлений пло- душек. 5. Плодовые сумки — вздутые на конце плодовые образо- вания, образующиеся в результате утолщения луба, где откла- дываются запасы питательных веществ и где ранее прикрепля- лись плоды. Рядом с плодовыми образованиями, а иногда и вместе с ними, у некоторых диких видов яблонь появляются колючки— образования стеблевого происхождения. Колючки представляют собой укороченные побеги длиной до 5, реже 10 см. Иногда они несут у основания нормальные или более мелкие листья и, видимо, защищают растения от объедания животными. Яблони с колючками характерны для аридных условий. В условиях сильного увлажнения или в культуре колючки могут исчезать и превращаться в нормальные облиственные побеги. Наличие колючек довольно характерно для многих видов груши; они также встречаются у многих плодовых растений, относящихся к семейству Posaceae. П о ч к и, расположенные на побегах, являются зачаточными побегами, находящимися в состоянии относительного покоя. Они имеют конусообразную форму, состоят из очень короткой оси, на конце которой расположен конус нарастания, прикры- тый спирально расположенными листовыми, а также цветоч- ными зачатками. Снаружи почки покрыты 2 — 6 спирально рас- положенными почечными чешуями, которые у яблонь образу- ются из прилистников, срастающихся с основанием листового черешка. Почки у различных видов рода Malus имеют различную окраску (оливково-зеленые, буроватые, каштаново-бурые, руби- ново-красные и др.). Они различаются по величине, в меньшей мере — по форме. Яблони имеют как вегетативные (ростовые) почки, обеспе- чивающие рост надземной части, так и репродуктивные (гене- ративные, или цветочные), из которых развиваются цветки, а позднее плоды. Вегетативные почки структурно отличаются от генеративных своей формой, величиной и расположением на рос- товых побегах. Репродуктивные почки обычно крупнее, более округлой формы и расположены на концах плодовых образо- ваний. У яблонь нередко встречаются смешанные почки, из ко- торых после их прорастания развиваются как укороченные по- беги (листовые розетки), так и соцветия (цветки).
Глава 3. Струнтурные особенности, эволюции и филогении 34 Почки у яблонь одиночные. По расположению на побеге они бывают терминальными (верхушечными, конечными) или аксиллярными (пазушными, боковыми). Развитие терминальных почек обеспечивает рост побега в длину, а аксиллярных — его ветвление. В случае утраты терминальной почки ее заменяют аксиллярные. Из общего числа закладывающихся аксиллярных почек раз- виваются лишь немногие. Часть аксиллярных почек отмирает, часть остается в покое, превращаясь в так называемые спящие почки. Они отличаются малыми размерами, небольшим числом зачатков листьев, располагаются в пазухах листьев и вокруг оснований побегов. Спящие почки у яблонь сохраняют жизне- способность десятки лет. На побегах яблонь иногда образуются адвентивные (или придаточные) почки, обычно слабо дифференцированные. Они закладываются вне пазух листьев. Эти почки также способны к развитию. Морфологически они почти не отличаются от спя- щих, однако последние, как правило, образуются в пазухах листьев, а адвентивные — вне их. Как спящие, так и адвен- тивные почки в жизни яблонь выполняют сходную функцию. Они являются как бы запасными зачатками побегов. Способ- ность многих яблонь образовывать адвентивные почки имеет важное значение при вегетативном размножении. Почки различных видов яблонь в общих чертах по своей структуре одинаковы, однако имеются некоторые отличия, слу- жащие систематическими признаками при диагностике некото- рых таксонов рода Malus. Это форма и величина, структура, окраска и опущенность покровных чешуй почек, расположение зачаточных листьев в почкосложении (свернутые или сложен- ные). ГАБИТУС И ~ВЕТВЛЕНИЕ В процессе длительного эволюционного пути у яблони выра- ботались различные приспособительные свойства, служащие признаками отличия одного вида от другого. Эти различия прежде всего можно связать с наличием в роде Malus следую- щих форм габитуса (рис. 5). 1. Крупные деревья с хорошо выраженными мощными ство- лами — макрофанерофиты. K ним относятся Malus laosensis, M. formosana, M. tschonoskii, M. sieversii var. kirghisorum, M. sylvestris, M. mandshurica и др. 2. Менее крупные деревья с небольшими или несколькими стволами — мезофанерофиты. К, ним относятся M. sikkimensis, M. baccata, M. sieversii, М. sieversii var. hissarica, M. orientalis, M. toringo и др.
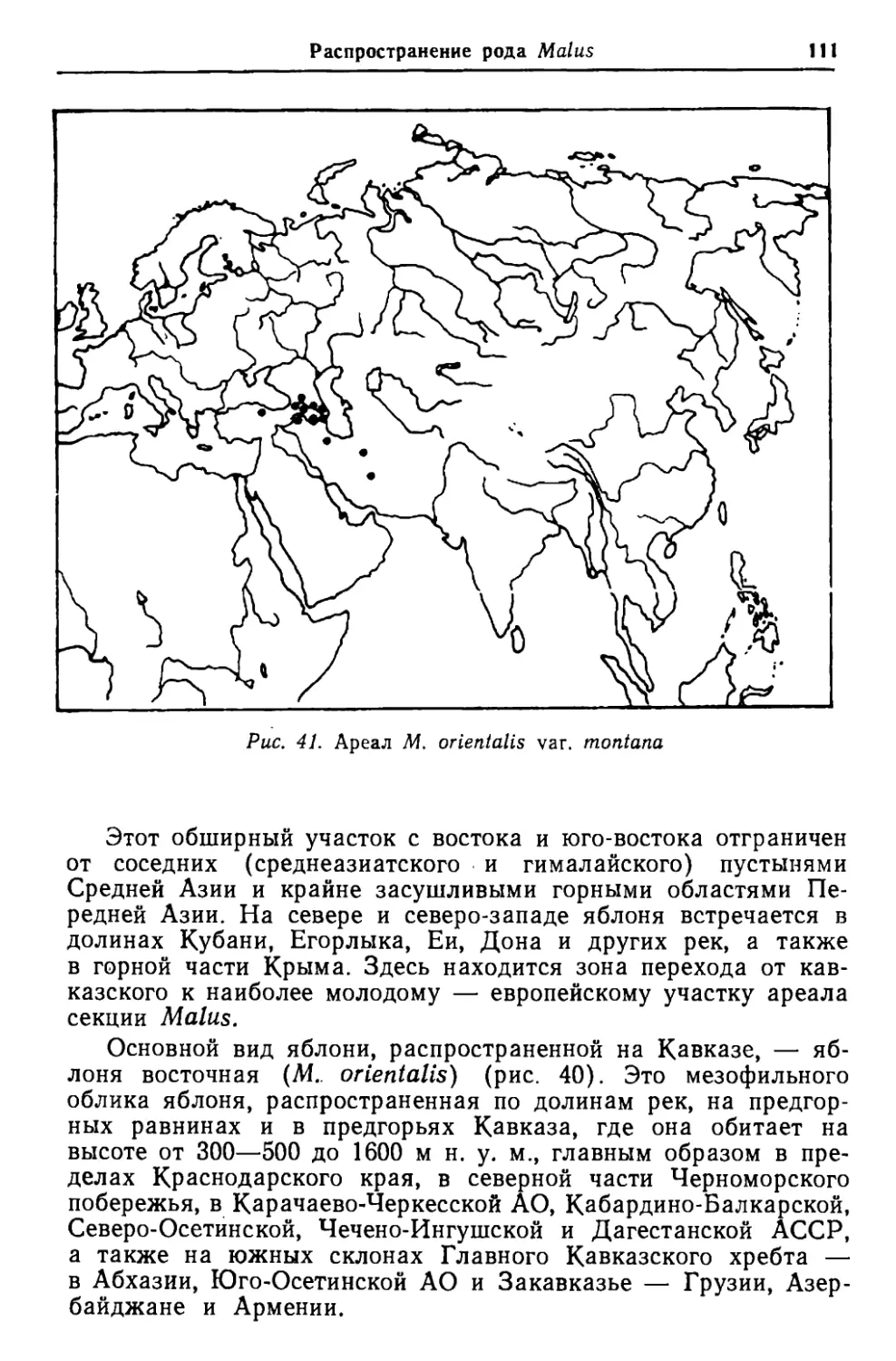
Габитус и ветвление Рис. 5. Основные формы габитуса яблони: а, б — макрофанерофиты; в, г — мезофанерофиты; д, е — нанофанерофиты 3. Низкие кустообразные формы с несколькими стволами- нанофанерофиты. К. ним относятся М. turkmenorum, М, Orienta- lis subsp. montana, М. praecox, а также описанные И. Т. Ва- сильченко [1959] виды для Пакистана — М. chitralensis и для Афганистана — М. nuristanica и др. Обособление основных форм габитуса яблонь объясняется их возникновением в онтогенезе в результате сочетания роста и развития в определенных условиях среды. Таким образом, жизненные формы габитуса возникли исторически, как приспо- сабливание к наиболее полному использованию всего комплекса условий местообитания, т. е. жизненная форма является выра- жением способности растений к расселению и закреплению их на определенной территории. Kate сообщает В. И. Запрягаева [1964], М. sieversii в Таджи- кистане (Памиро-Алай) в засушливых условиях приобретает форму куста при наличии многоствольности. Сходные законо- мерности наблюдаются и на Тянь-Шане, Кавказе и особенно в Передней и Малой Азии — в Турции, а также в европейской части СССР, где местные дикие яблони под воздействием за- сушливых условий приобрели форму небольших деревьев или кустарников. Следовательно, эти типы габитуса яблонь следует рассматривать в эколого-морфологическом аспекте как жизнен- ные формы.
36 Глава 3. Структурные особенности, эволюция н фнлогення Вместе с тем условия обитания, воздействующие на форми- рование внешнего облика растения, определяют и длительность жизни последнего. Продолжительность жизни яблони не пре- вышает 100 — 150 лет, редко достигает 200 лет. Это относится к небольшому числу наиболее крупных по размерам яблонь, преимущественно мезофильного облика. Другие яблони, обита- ющие в засушливых районах, проходят свой жизненный цикл в течение 30 — 80 лет. Основная масса видов яблонь в среднем живет 60 — 80 лет, реже — более 100 лет. Яблони с хорошо выраженными стволами во взрослом со- стоянии достигают в высоту 15 — 20 м, с менее выраженными— 8 — 12 м, а кустовидные яблони не превышают 4 — 8 м. Встреча- ются и карликовые яблони, едва достигающие 0,5 — 2,0 м в вы- соту. Диаметр кроны в различных условиях не превышает 3 — 5 (реже 6 — 8) м. Внешний облик, или габитус, и связанные с ним продолжи- тельность жизни и размеры яблонь определяются характером их ветвления. Эволюция габитуса и особенностей ветвления яблонь прохо- дила по пути трансформации деревьев в небольшие деревья и кустарники. Переход от неопределенного моноподиально-симпо- диального ветвления к строго симподиальному можно наблю- дать у большинства современных видов рода Malus. Основные ветви кроны многих яблонь и особенно удлиненные побеги со- храняют черты моноподиального ветвления. Наоборот, укоро- ченные побеги, несущие цветки, развиваются как симподии. На более ранних этапах развития у подавляющего большинства яблонь хорошо выражено моноподиальное ветвление, впоследст- вии переходящее в симподиальное. Черты моноподиального ветвления лучше всего сохранились у современных реликтовых видов доциниевидных яблонь. Влаж- ный субтропический климат, в котором обитают указанные яб- лони (в частности, M. laosensis, произрастающая в Лаосе), долгое время сохраняет верхушечные меристемы, в связи с чем они обладают почти непрерывно функционирующим конусом роста, что приводит к сильному развитию главной оси и подав- лению боковых ответвлений. Виды доциниевидных яблонь, про- израстающих в более умеренных зонах (М. 1зсйопояйгг' — в Юж- ной Японии и М. sikkimensis — в Гималаях), отражают посте- пенный переход к симподиальному ветвлению. Яблони, расту- щие в еще более суровых климатических условиях, имеют почти исключительно симподиальное ветвление. Большое значение в эволюции габитуса и ветвления, свойст- венных яблоням, сыграло их распространение в широты с уме- ренным климатом, в условиях которого они приобрели период глубокого зимнего покоя. Это затормозило развитие их вер- хушечных почек и неизбежно стимулировало появление боль-
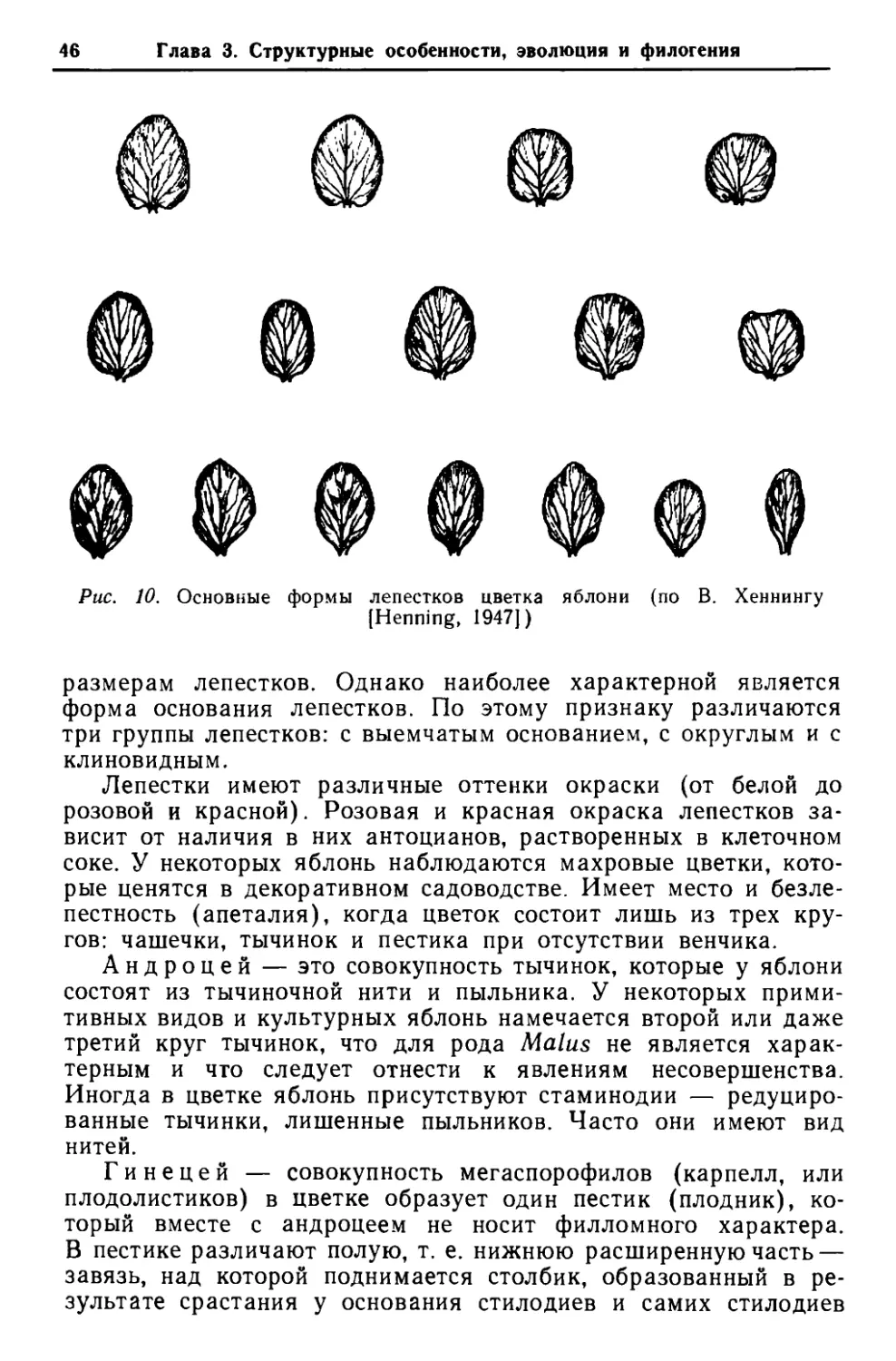
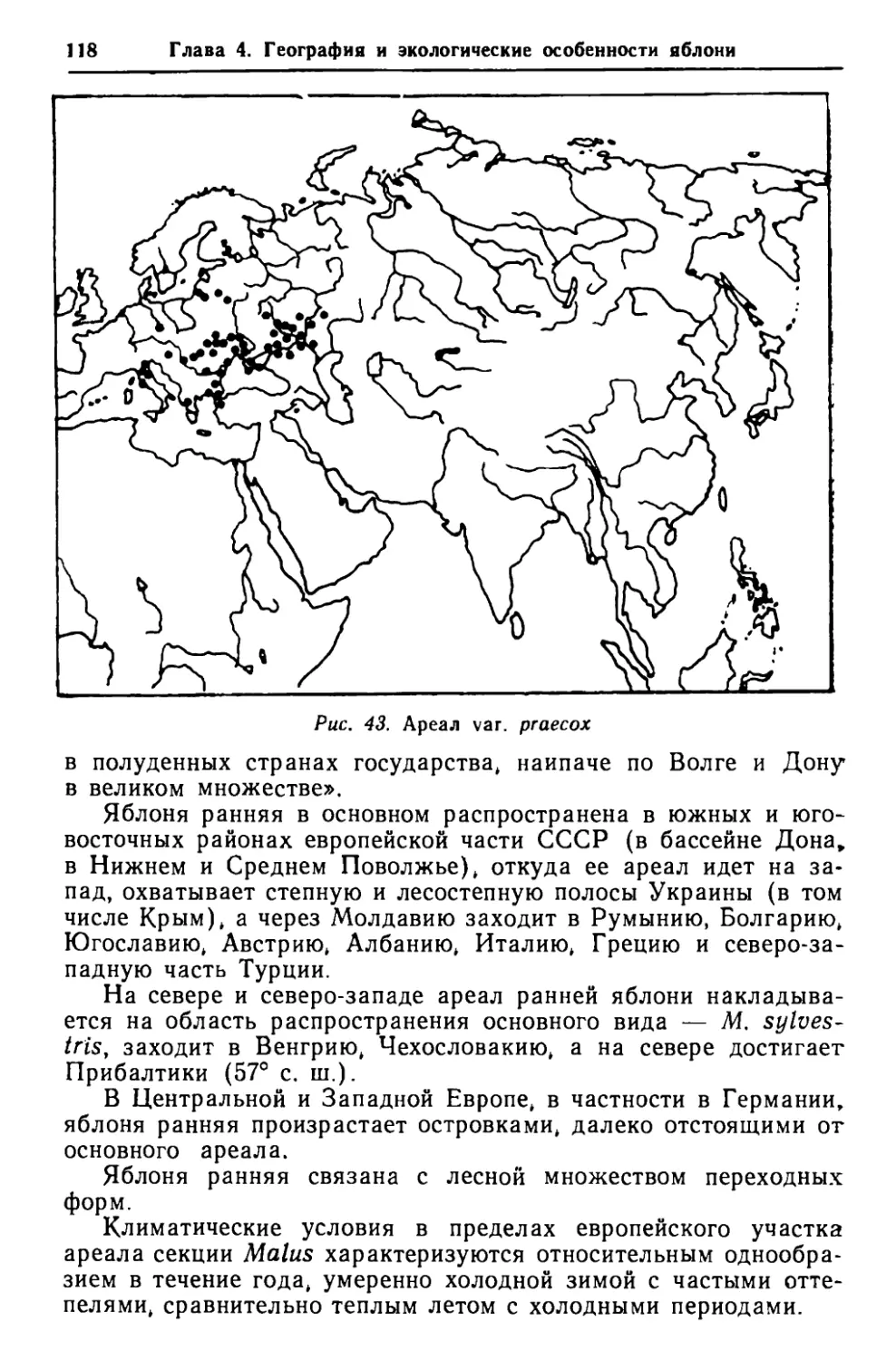
Лист шего числа боковых ответвлений, что, в свою очередь, способ- ствовало переходу от моноподиальной системы ветвления к симподиальной. Такой процесс перехода от одного типа ветвле- ния к другому привел к формированию всего облика кроны яблони. Следует также указать, что параллельно с формированием видов яблонь, приспособившихся к жизни в условиях климата с резкими температурными перепадами в течение года, проис- ходил процесс ксероморфогенеза, так как виды рода Malus (при их продвижении с юга на север) попали под действие за- сушливых условий. Последнее обстоятельство привело к форми- рованию видов ксероморфного облика и способствовало замене крупных древесных форм формами более низкими и даже кус- тообразными. ЛИСТ Листья яблони весьма полиморфны и различаются по форме, величине, очертанию, опушению, жилкованию и другим призна- кам (рис. 6). Однако несмотря на разнообразие и сильную пластичность структура листьев в роде Malus является важней- шим показателем для выяснения эволюции и филогенеза рода. Листья яблони укреплены на стеблевых узлах поочередно, листорасположение спиральное, приблизительно 2/5, под углом 144'. На удлиненных и укороченных побегах яблонь различа- ются три категории листьев: 1) низовые (folia т~гепога ч. рпт~депа) — первые листья побега, недоразвитые, не вполне принявшие характерную для вида форму и не достигшие нормальных размеров; 2) серединные, или ординарные (folia ordinaria), — наибо- лее полно развитые, с характерной для вида формой, размерами и другими показателями, расположенные не только посередине, но и на большей части побега; 3) верховые (folia altima) — один-два самых верхних ли- ста, которые не только недоразвиты, но и имеют несколько уклоняющуюся форму. Для оценки систематического взаимоотношения видов по признакам структуры листьев крайне важно различать перечис- ленные категории. Форма листовой пластинки яблонь определяется соотноше- нием ее длины и ширины. При этом длина листа измеряется от основания пластинки до ее вершины, а ширина — в самом широком месте пластинки (выше или ниже середины ее длины). По степени рассеченности листья яблони могут быть цель- ными (краевые вырезы листовой пластинки не доходят до одной
Глава 3. Структурные особенности, эволюция и филогения Ъ % Ъ I 1 Ф I Р I t Р I 1 Ъ Ъ I % \ 1 1 Ъ Ъ Ъ Ъ 1 1 ! I 1 1IРtIРР/ % Ъ \ Ъ 1 1 1 I ю Ю Ю Ф \ с с Рис. б. Разнообразие листьев яблони: а — М. sylveslris; б — М. domeslica; в — М. fusca; г — М. Xzumi; д — М.Xcerasi- fera; е — М. X floribunda; ж. з — М. X purpurea; и — М. Xsargenlii; к — М. loringoides; л — прилистники четверти ее ширины), лопастными (вырезы доходят до одной четверти ширины пластинки) и раздельными (вырезы превы- шают четверть пластинки). Разнообразие форм листовых плас- тинок у различных видов Malus в большой степени зависит от условий обитания. Рассеченность листьев или тенденция к этому в молодом возрасте или на удлиненных побегах проявляется почти у всех видов яблони. Подобные явления довольно хорошо изучены у родственного рода Pyrus ~[Васильченко, 1965]. В ходе исследований выяснено, что рассеченность листьев груши в большой мере связана с ухудшенными условиями произраста- ния (засушливые районы, маломощные каменистые почвы и т. п,). По этому поводу И. Т. Васильченко '[1965 с. 19 — 20] писал: «В условиях особенно неблагоприятных, например, на сухих каменистых склонах, скалах и т. п., у Ругила гедеИ на- блюдается особенно интенсивное расчленение листов». Вся поверхность листовой пластинки армирована жилками, которые составляют ее скелет. Листья яблони обычно имеют перистое, сетчатое и петлевидное жилкование.
Лист Одновременно с пластинкой листа образуются парные при- листники, первоначально покрывающие весь зачаток, а позже имеющие вид маленьких листочков. Наличие прилистников у яблонь рассматривается как примитивный признак, так как продвинутые в эволюционном отношении яблони прилистников не имеют или же последние сильно редуцированы. Составной частью листа яблони является черешок. Черешки у различных видов, как правило, желобчатые, с верхней вогну- той и нижней выпуклой сторонами. Разнообразие черешков у различных видов может быть сведено к варьированию их раз- меров (длина, толщина) и характеру поверхности (степень опущенности). Соотношение длины черешка и листовой плас- тинки в среднем для видов секции Eriolobus — 3: 4; Docyniop- sis — 3: 8; Sorbomalus — 2: 5; Chloromeles — 2: 6; Gymnome- les — 3: 7; Malus — 2: 6. Черешки бывают голыми и опушенными (реже войлочными) Однако полной корреляции в опушении черешка и листьев почти не наблюдается. При наличии опушенного черешка листовая пластинка может быть голой, но при опушенной пластинке го- лый черешок наблюдается редко. Морфологическая эволюция листьев яблони Отсутствие палеоботанических данных о строении листьев у бо- лее примитивных яблонь, равно как и эволюционная пластич- ность структуры листа, не позволяют установить их первона- чальный облик. Однако сравнительное изучение современных яблонь дает нам право заключить, что приведенная ниже схема в основном правильно отражает ход эволюционных преобразо- ваний структуры листовой пластинки. Основываясь на изучении морфологии листовой пластинки различных видов рода Malus, а также учитывая имеющиеся данные специальной литературы по эволюции листьев, мы счи- таем возможным принять сложные, раздельные или лопастные листья с перисто- или пальчато-краебежным жилкованием за исходные. В дальнейшем ходе эволюции выявилась тенденция к редукции, и в результате образовались наиболее продвинутые формы редуцированных цельных листьев с более густым петле- видным или сетчатым жилкованием. Таким образом, прослеживая эволюционный путь становле- ния структуры листа в пределах рода Malus, можно выделить три ступени эволюции: ступень сложных, раздельных или ло- пастных листьев, ступень редукции сложных листьев и переход к упрощенным структурам и ступень вторично (целесообразно) упрощенных простых листьев. Следует, однако, заметить, что расположенные на укороченных и удлиненных побегах одного дерева листья при различном онтогенетическом возрасте имеют значительные структурные различия. Сильное влияние на струк- турные особенности имеют различные факторы экологии. Все
Глава 3. Структурные особенности, эволюция и филогения 40 Рис. 7. Схема эволюции листьев в роде Ма1иь.Римскими цифрами обозначены типы, арабскими — линии это придает различиям указанных ступеней эволюции листьев яблони общий, иногда относительный и условный характер. Учитывая то, что эволюция проходит по спирали, мы пред- лагаем следующую схему развития структуры листьев рода Ма!из (рис. 7). Т и п 1. Eriolobus. Трехраздельные листья. 1 линия. Trilobatae. Весьма оригинальные, округло-яйцевид- ные (в очертании), голые сверху и пушистые снизу, кожистые с сердцевидным основанием, глубоко трехраздельные листья с боковыми отстоящими, двухлопастными долями и трехлопаст- ной верхушечной долей. ~Килкование перисто- или пальчато- краебежное. На укороченных побегах листья иногда цельные. Эту линию характеризует М. trilobata. Т и п П. Sorbomalus. Раздельные и лопастные, переходящие к простым цельным листья. Наиболее сложный и разнообраз- ный тип, в пределах которого можно проследить все переходы. 2 линия. l(ansuenses. Пяти-трехраздельные, глубоко пяти- трехлопастные, неглубоко трехлопастные, в очертании яйцевид- ные листья. Линию характеризует постепенный переход формы листьев от пяти- к трехраздельным (М. centralasiatica~M. tran- sitoria), от пятираздельных к пятилопастным (M. centralasia- fica~M. toringoides), от пяти- к трехлопастным (M. toringo- ides~M. kansuensis) или от трехраздельных к трехлопастным (M. trilobata~M. komarovii) .
Лист 43 3 линия. Toringonae. Гетерофилия, листья на удлиненных побегах пяти-трехлопастные, на укороченных — цельные, цель- нокрайные, в очертании яйцевидные, продолговато-яйцевидные до продолговатых. Линию характеризует M. toringo, представ- ляющий незначительное ответвление от типа Sorbomalus. 4 линия. Florentinae. Своеобразные, четырех-шестилопаст- ные, яйцевидные или почти округлые в очертании, остроконеч- ные, с пильчато-зубчатыми треугольными лопастями, при осно- вании усеченные листья. Линия представлена единственным, по всей вероятности гибридным, видом — M.>(florentina, распро- страненным далеко от основного ядра ареала секции Sorboma- l us — в Итал и и. 5 линия. Fuscae. Продолговато-яйцевидные, остропильчатые, едва трехлопастные или цельные листья с сердцевидным осно- ванием. Линия представлена видом M. fusca в Северной Америке. 6 линия. Yunnanenses. Листья по форме едва лопастные, преимущественно цельные, от широко- до продолговато-яйце- видных или овально-продолговатых. Эта линия явно характери- зует редуцированный переход от слаболопастных (М. honanen- sis) к цельным листьям (М. ombrophila~M. prattii~M. yunna- nensis), что связано с постепенно увеличивающейся мезофильностью этих яблонь, возрастающим порядком и укруп- нением размеров цельной листовой пластинки. В отличие от всех предыдущих линий, Yunnanenses вместе с линиямк Tschonoskianae и Coronariae находится на второй ступени раз- вития, т. е. в стадии редукции сложных и перехода к упрощен- ным структурам эволюции листьев рода Malus. T и п 111. Docyniopsis. Крупные цельные, на удлиненных по- бегах, едва лопастные листья кожистой консистенции. 7 линия. Tschonoskianae. Листья с клиновидным, неравносто- ронним или округло-клиновидным основанием, заостренной, реже притупленной верхушкой, на более молодых побегах листья едва лопастные. Линия представлена видами M. laosen- sis, M. formosana, M. tschonoskii и M. sikkimensis, обитающими во влажном климате Юго-Восточной Азии. Здесь осуществляется переход от лопастных к цельным листьям. T и п IV. Chloromeles. Продолговато-яйцевидные, зубчато- пильчатые, городчатые или почти цельнокрайные, на длинны~с побегах слаболопастные листья. 8 линия. Coronariae. Слаболопастные листья, переходят к цельным яйцевидным — эллиптическим — продолговатым с сердцевидным или клиновидным основанием, острой или за- кругленной верхушкой и опушенной, преимущественно по ниж- ней стороне, пластинкой: M. coronaria — М. ioensis — М. angusti- folia. На этом заканчиваются первые две ступени эволюционного
Глава 3. Структурные особенности, эволюция и филогения 42 СОЦВЕТИЕ Соцветия различных видов яблони обычно бывают щитко- видными или зонтиковидными (рис. 8). Если боковые оси, несу- щие цветки, отходят от укороченной, но еще заметной главной развития листьев рода Ма1из, характерным признаком которых, кроме описанных выше структурных особенностей, является еще один важный систематический признак — сложенность листьев в почкосложении. Следующие два типа и две их линии, достигшие наиболее продвинутого в эволюционном отношении положения, характе- ризуются свернутыми в почкосложении листьями. Т и п Ч. Gymnomeles. Характерной чертой эволюции этого типа следует считать его приспособленность к условиям холод- ного климата, типичным для него длительным периодом глубо- кого покоя. Эволюция листьев относящихся к нему яблонь начинается от простого цельного листа типа М. sikkimensis, у которого в связи с применением к более суровым зимним усло- виям приобретается новое качество — свернутость листьев в почкосложении. 9 линия. Baccatae. Опушенные в молодом возрасте, позже— голые овальные или яйцевидные, а также продолговатые, почти цельнокрайные, городчато-пильчатые или острозубчатые листья, с округлым или клиновидно-суженным основанием и умеренно заостренной или удлиненной верхушкой. Наиболее примитивным типом листа этой линии обладают М. mandshurica и M. hima- laica, а более продвинутым — M. baccata. Среднее положение занимают М. hupehensis и М. halliana. Ти п VI. Malus. Цельные вторично упрощенные листья на- стоящих яблонь, сформировались в суровых условиях на об- ширной территории в Гималаях, Средней и Передней Азии, на кавказе и в Европе. 10 линия. Malus. Простые, цельные, в почкосложении свер- нутые, различной формы и консистенции листья, чаще всего продолговатые или почти округлые; по краю пильчатые, зубча- гые, городчатые, имеющие клиновидное или округлое основание и заостренную или тупую верхушку, голые травянистые или бумажистые — у мезофильных яблонь — и опушенные снизу (или с обеих сторон), плотные — у ксероморфных. В пределах этой линии более примитивным типом мы счи- гаем крупные и тонкие листья, встречающиеся у крупных де- ревьев мезофильного облика М. sieversii var. kirghisorum, а наиболее продвинутым — более плотные, опушенные листья, характерные для ксероморфных яблонь, например, М. turkme- norum, М. orientalis var. montana, М. sylvestrisvar.ргаесохидр.
Соцветие Рис. 8. Схема редукционного ряда образования соцветий в роде 34alus а — сложное цимозное соцветие — ~л (исходная форма); б — редуциро- ванная форма — ложный щиток; в — редуцированная промежуточная форма ложный зонтик; г — щитковидное соцветие; б — зонтиковидное соцветие; е малоцветковое щитковидное соцветие; ж — малоцветковое зонтиковидное соцветие» з — одиночный цветок оси цветочного побега, то образуются щитковидные соцветия- тип 1. В щитковидных соцветиях нижние боковые цветки имеют более длинные цветоножки, а верхние — короткие, вследствие чего все цветки располагаются почти в одной горизонтальной плоскости. Такие соцветия встречаются у видов секций Sorbo- malus и Chloromeles. Их следует рассматривать как более при- митивные по отношению к типу II. Зонтиковидные соцветия (тип II) образуются в том случае, когда боковые оси выходят будто из одной точки и главная ось цветоносного побега полностью редуцирована. Цветки рас- полагаются на цветоножках равной длины, но не в одной плос- кости, а куполообразно. Такие соцветия характерны для видов секций Malus и Gymnomeles. Характерной особенностью соцветий яблони является бази- петальная последовательность распускания цветков. Первым расцветает средний, т. е. самый верхний цветок главной оси,. который иногда развивается лучше, так как является первичным по отношению к боковым. Это подтверждается ходом эволюции соцветий яблони (и других розоцветных) . Исходным типом соцветия, свойственным видам Malus, сле- дует считать плейохазий. В результате редукции главной и бо-
Глава 3. Структурные особенности, эволюция и филогения 44 ЦВЕТОК Цветки яблонь расположены на метаморфизированных уко- роченных побегах, обладающих ограниченным ростом. Такой побег представляет собой ось (главную или боковую), к кото- рой прикреплены видоизмененные в различной степени образо- вания, составляющие цветок. Ось метаморфизированного побега ковых осей образовались щитковидные или зонтиковидные соцветия. Терминальный и наиболее крупный цветок, находя- щийся на вершине главной оси соцветия и расцветающий пер- вым (базипетальная последовательность распускания цветков в соцветиях), является характерной особенностью цимозных соцветий. Рассматривая форму соцветий современных видов яблони, следует прежде всего указать на неопределенность описатель- ных терминов «зонтиковидный» или «щитковидный», так как в природе встречаются более сложные и, по-видимому, менее ре- дуцированные формы, представляющие собой либо ложный щиток (преимущественно более примитивные яблони из секций Sorbomalus и Docyniopsis), либо ложный зонтик (примитивные яблони из секции Eriolobus). Встречаются также малоцветковые и, по-видимому, сильно редуцированные щитка- и зонтиковид- ные соцветия (первые из секции Chloromeles, вторые — пред- ставители крупноплодных яблонь из секции Malus). Таким образом, основываясь на изложенных представлениях о формировании соцветий в роде Malus на основе редукции от цимозных к щитковидным и зонтиковидным, можно наметить две линии эволюции. Одна из них шла по схеме: ложный щи- ток»-щитковидное соцветие — ~-малоцветковый щиток (Docyniop- sis~Sorbomalus~Chloromeles); другая — ложный зонтик — э- зонтиковидное соцветие- малоцветковый зонтик (Eriolobus~ — ~-б утпоте1ея-~-Ма1 из) . Есть и иная точка зрения на происхождение соцветия яб- лонь, поддерживаемая А. Л. Тахтаджяном '[1964] и некоторыми другими исследователями. Согласно ей, эволюционный путь становления соцветий рода Malus, мог осуществляться по дру- гой схеме. Ось цветоносного побега ограничивается в росте и заканчивается одиночным цветком. У его основания появляются боковые оси, быстро сравнивающиеся по длине с главной. Об- разуется либо мало-, либо многоцветковое соцветие типа щитка или зонтика. В отличие от этих представлений, В. Тролль ,'[Troll, 1957] главное внимание уделяет не появлению боковых осей, а их редукции. Нам представляется, что Тролль прав, так как утрату любых органов легче себе представить, чем их возникно- вение.
ЦветОк Рис. 9. Основные формы отгиба чашечки цветка яблони (по В. Хен- нингу [Henning, 1947] ) (цветок яблони) имеет расширенную часть — гипантий, или тор (цветоложе). К нему циклически (по направлению от перифе- рии к центру) прикреплены основные его части: чашечка, венчик, андроцей и гинецей. Ч а ше ч к а у яблони всегда имеет филломный характер, что подтверждается наличием хлоропласта и способностью к фото- синтезу (подобно листьям). Чашелистики у различных яблонь морфологически отличаются по форме, ширине и длине (рис.9). По форме они либо широко-, либо узкотреугольные или тре- угольно-ланцетные, опушенные, иногда беловойлочные (осо- бенно у ксероморфных яблонь), или голые (преимущественно у мезофильных яблонь). У всех яблонь чашелистики сросшиеся у основания. Виды рода Malus значительно разнятся по степени развития свободной, эпигинной части чашечки, которая вместе с отгибом участвует в образовании на верхушке плода углубле- ния (umbilicus), имеющего форму довольно длинной трубки (более примитивные виды секции Эосупьорз1з, например, М. formosana) или же более или менее глубокой воронки (виды секции Malus). Чашечка у яблонь может быть остающейся при плодах (крупноплодные яблони), частично опадающей (некото- рые рябиновидные яблони) или же опадающей после цветения вместе с верхней частью гипантия (мелкоплодные ягодные, а также некоторые рябиновидные яблони). У многих видов яб- лони чашечка вместе с завязью и гипантием принимает участие в образовании плода. В е н ч и к состоит из лепестков. Лепестки у яблонь свобод- ные. Каждый лепесток состоит из расширенной части (плас- тинки) и нижней, суженной части (ноготка), при помощи кото- рой лепесток прикрепляется к основанию цветоложа. По форме лепестки цветка яблони бывают округлые, эллиптические или продолговатые, Они имеют закругленную или несколько заос- тренную, а также выемчатую верхушку, сердцевидное, закруг- ленное или клиновидное основание. Жилкование лепестков ди- хотомическое. Последний признак свидетельствует о тычиночной природе лепестков. Лепестки у яблонь весьма неоднородны (рис. 10). Даже цветки одного дерева могут различаться как по форме, так и по
Глава 3. Структурные особенности, эволюция и филогения 46 Рис. 10. Основные формы лепестков цветка яблони (по В. ХеннингуtHenning, 1947)) размерам лепестков. Однако наиболее характерной является форма основания лепестков. По этому признаку различаются три группы лепестков: с выемчатым основанием, с округлым и с клиновидным. Лепестки имеют различные оттенки окраски (от белой до розовой н красной). Розовая и красная окраска лепестков за- висит от наличия в них антоцианов, растворенных в клеточном соке. У некоторых яблонь наблюдаются махровые цветки, кото- рые ценятся в декоративном садоводстве. Имеет место и безле- пестность (апеталия), когда цветок состоит лишь из трех кру- гов: чашечки, тычинок и пестика при отсутствии венчика. А н д р о ц е й — это совокупность тычинок, которые у яблони состоят из тычиночной нити и пыльника. У некоторых прими- тивных видов и культурных яблонь намечается второй или даже третий круг тычинок, что для рода Malus не является харак- терным и что следует отнести к явлениям несовершенства. Иногда в цветке яблонь присутствуют стаминодии — редуциро- ванные тычинки, лишенные пыльников. Часто они имеют вид нитей. Г и н е ц е й — совокупность мегаспорофилов (карпелл, или плодолистиков) в цветке образует один пестик (плодник), ко- торый вместе с андроцеем не носит филломного характера. В пестике различают полую, т. е. нижнюю расширенную часть— завязь, над которой поднимается столбик, образованный в ре- зультате срастания у основания стилодиев и самих стилодиев
47 Цветок Рис. 11. Степени срастания стилодиев в цветке яблони: а — стилодии, сросшиеся у самого основания (I стадия); б стилодии, сросшиеся до половины Д1 стадия); e — стилодии, сросшиеся до 2/3 длины (III стадия) «Термин «приросшая завязь» предложен А. Л. Тахтаджяном [ 1964] вместо неудачного понятия «нижняя завязь». (отдельные, свободные на конце ветви столбика), несущих на своих вершинах рыльце. Завязь — самая главная часть пестика. У яблонь она ниж- няя или приросшая*, синкарпная, состоящая из пяти сросшихся в различной степени между собою карпелл (плодолистиков) и гипантия в цветочной трубке. Завязь имеет пять гнезд, в каж- дом из которых располагается от двух (секции Eriolobus и Docyniopsis), четырех (секции Sorbomalus и Chloromeles) до шести (секции Afalus и Gymnomeles) семяпочек. Число семя- почек — важный признак эволюционного уровня таксонов, при- чем более примитивные яблони характеризуются меньшим чис- лом семяпочек (от двух до четырех), а наиболее продвинутые— большим числом (до шести) . В пределах рода МШиз у различных видов столбик, образо- ванный пятью стилодиями, находится еще в стадии образова- ния. Степень срастания стилодиев, а также форма, опушенность столбика, характер свободных концов столбика — весьма тон- кие и неустойчивые признаки у рода яблони, имеющие значение в сортоведении (помологии) ~[Лихонос, 1963, 1964]. По степени срастания стилодиев можно отметить три стадии эволюционного становления цветка яблони: стилодии, сросшиеся у самого основания (рис. 11,a), стилодии, сросшиеся до поло- вины (рис. 11 6), и стилодии, сросшиеся до 2/3 длины (рис. 11,в). Срастание стилодиев и образование столбиков— бесспорный признак продвинутости видов. По числу стилодиев исходным типом у яблони следует счи- тать цветки с пятью стилодиями, которые образовались в ре- зультате неполного срастания верхних концов пяти карпелл.
Глава 3. Структурные особенности, эволюция и филогения Рис. 12. Типы цветков яблони в зависимости от соотношения длины столбиков и тычинок (венчик опушен) (по В. Хен- нннгу [Henning, 1947], дополнено): а — столбики и тычинки одной длины (I тип); б — столби.<и ко- роче тычинок (11 тип); в — столбики длиннее тычинлс (1~1 тип) В цветках некоторых яблонь наблюдаются три стилодия. Это несомненный результат редукции, т. е. признак продвинутости. Степень срастания карпелл — важный показатель эволюци- онного уровня цветка яблони — хорошо выражена в структуре стилодиев и образовании столбика. Не менее важным, но не- явно выраженным в цветке признаком следует считать степень срастания карпелл, образующих завязь. Этот признак можно хорошо проследить в структуре эндокарпа плодов. При определении эволюционного уровня признаков цветка яблони, так же как у соцветия, следует учитывать степень ре- дукции его частей по пути экономии и специализации цветка к опылению. В ходе развития по линии энтомофилии у цветка яблони сложился довольно сложный механизм для привлечения насекомых-опылителей: яркий венчик, более или менее сильный запах, а также нектарники и т. д. Однако такие приспособления цветков различных видов яблонь к энтомофилии не .вполне совершенны. Это выражается прежде всего в наличии сравни- тельно большого количества пыльцы, остающейся в цветке и поедаемой разнообразными жуками (кантарофилия), хотя яб- лони опыляются по преимуществу пчелами, прилетающими за нектаром. Примитивным признаком цветка яблони также сле- дует считать и постоянное число тычинок.
Цветок 49 а В Г рис. 73 эволюция рылец яблони (по В. Хеннингу [Непп!п~, 19471, с неко- торыми изменениями) Вместе с тем у яблони иногда наблюдается гетеростилия. В цветках отдельных растений, принадлежащих к одному виду, все рыльца длинностолбчатые, а тычинки укороченные. другие растения того же вида, наоборот, имеют в цветках короткие столбики и длинные тычинки, или же на одной особи располо- жены цветки с длинными столбиками и короткими тычинками. В этом отношении цветки яблони, согласно В, Хеннингу [Hen- ning, 1947], могут быть разделены на три типа (рис. 12): стол- бики и тычинки примерно одной длины, столбики короче тычи- нок, столбики длиннее тычинок. В эволюционном отношении исходным следует считать 1 тип (norma!), от которого позже развились 1! тип и наиболее совершенный 111 тип. В цветках яблонь нередко наблюдается дихогамия — разно- временное созревание в одном цветке тычинок и пестиков. Раз- личаются протерандрия, когда пыльники созревают раньше пестиков, и протерогиния, когда сначала созревают пестики. В пределах одного вида встречаются как протерандрические, так и протерогинические растения. Нередки случаи, когда, как при гетеростилии, на одном растении встречаются и те и другие цветки, что свойственно, по преимуществу, более примитивным яблоням. В пределах рода Ма1ия по этим признакам можно проследить пути эволюции цветка, так как у различных видов данные показатели находятся на различных уровнях развития. В целом как гетеростилия, так и дихогамия из-за своей неопределенности и неустойчивости при семенном размножении яблонь не могут использоваться как систематические признаки в диагностике видов. Однако эти признаки с успехом могут быть применены в помологии для классификации вегетативно размножающихся клонов и сортов культурной яблони, что и сделано Ф. Д. Лихоносом i[1963, 1964]. Наличие разных эволюционных уровней в строении цветка яблони выражено в структуре рыльца. По А. Л. Тахтаджяну [1964, с. 87], «рыльце развивалось из двух сблизившихся в процессе эволюции и затем сросшихся краев плодолистиков, т. е. оно произошло из шва». Следуя этим соображениям, ис- 4 — 727
Глава 3, Структурные особенности, эволюция и филогения ходным типом рыльца в роде Ма/из мы считаем двурасщеплен- иое, несросшееся рыльце (рис. 13,a). В дальнейшем развились продолговатые и расширенные, имеющие продолговатый шов посередине рыльца (рис. 13,б), от которых эволюция, видимо, шла по двум линиям. Одна из них привела к исчезновению шва и развитию продолговатых, вздутых рылец (рис. 13,в), а дру- гая — к головчатым рыльцам (рис. 13,г). ПЛОД Плоды яблонь — яблоки — обычно рассматриваются как ложные плоды. Понятие «ложный плод» бессмысленно, так как плод не может быть ложным. В строении типичного яблока различают три части: экзокарп (внеплодник), мезокарп (межплодник) и эндокарп (внутри- плодник) . Происхождение экзокарпа и особенно мезокарпа из обраста- ющей завязи цветочной трубки можно хорошо проследить у незрелых плодов. Наличие в них проводящих пучков, остатка чашелистиков, а также лепестков указывает на участие в обра- зовании мезокарпа оснований чашечки и венчика. Присутствие в верхней части плода остатков тычинок и столбиков со стило- диями подтверждает факт участия в образовании плода тычи- нок и пестика. Образование эндокарпа в результате разрастания плодолистиков (карпелл) доказывается наличием дорзальных и вентральных пучков карпелл, проходящих через звездообраз- ный эндокарп. Экзокарп, так называемая кожица плода, состоит из 1 — 2 слоев однородных живых клеток эпидермиса с придаточным об- разованием в виде кутикулы, воскового налета или волосков, бугорков и пр. Характер эпидермиса плода коррелятивно связан с аналогичными образованиями других органов. Экзокарп при- митивных видов яблонь покрыт сплошной кутиновой пленкой (М. laosensis, М. formosana), иногда бывает бугорчатым (М. tschonoskii). У более продвинутых в эволюционном отно- шении яблонь на поверхности экзокарпа имеется восковой на- лет. Среднее положение занимают яблони, обладающие воло- систым или опушенным снаружи экзокарпом. Мезокарп занимает ббльшую часть плода (80 — 90%). Парен- химатические его клетки содержат ценные питательные веще- ства, что с давних пор привлекало внимание человека. В зависимости от принадлежности к тому или иному виду, а также от комплекса условий местообитания и степени зрелости плодов химический состав мезокарпа меняется. В результате длительной селекции яблони созданы разнообразные сорта, ко- торые по химическому составу плодов значительно превосходят своих диких предков.
Плод 5! Большую часть плода яблони занимает вода (в зрелых плодах ее около 85%). Химический состав плодов характеризу- ется наличием в них растворимых веществ, содержание кото- рых, по данным К,. Гутманиса l[Gutmanis, 1961], в условиях Латвии колеблется от 9,7 до 16,0О/о. Содержание сухого веще- ства в плодах яблони зависит не только от вида или сорта, но и от расположения плода в кроне, освещения и целого ряда дру- гих факторов. Содержание сухого вещества, а следовательно, химический состав в различных частях плода также неодина- ковы. Из других наиболее существенных компонентов в химичес- кий состав яблок входят: сахара — около 8,8О/о, органические кислоты — 0,85%, минеральные вещества — 0,8О/о, пектииовые вещества — 0,43%, дубильные вещества — 0,07О/о, азотистые вещества (белки) — 0,3О/о, витамины: С — 21,4 мго/о, В2 0,015 Mr%. 1(расящие и ароматические вещества, обусловлива- ющие окраску и аромат плодов, содержатся в незначительных количествах. Важным показателем, определяющим вкусовые качества плода, является соотношение сахаров и кислот. Наилучшие вку- совые качества имеют плоды при соотношении сахаров и кислот 15/20. Примитивные виды диких яблонь характеризуются не- высоким содержанием в плодах сахаров (4 — бо/о ) и кислот (0,2 — 2,0О/о) и значительным наличием дубильных (около 0,4/ц) и пектиновых (до 1,3%) веществ. С продвижением в более су- ровые условия, в связи с повышением зимостойкости, резко возрастает общее количество сахаров (до 10,89О/о) и кислот (до 3,47О/о) — М. baccata c дальнего Востока. По мере окультури- вания, в результате отбора, у культурных яблонь (сортов) со- держание дубильных и пектиновых веществ в плодах уменьша- ется, а сахаров и кислот — увеличивается. В зависимости от химического состава у различных яблонь меняется консистенция плодов: от хрящеватой, грубой, содержащей каменистые клетки, — к- мелкозернистой, плотной, сочной — до рыхлой, мучнистой, суховатой. Эндокарп составляет так называемое сердечко яблока и является наиболее специализированной частью плода. Эндокарп яблока слагается из пяти звездообразно расположенных семен- ных камер. Семенные камеры могут быть открытыми, когда они сливаются в общую центральную полость плода (края плодо- листиков не срослись), полуоткрытыми, когда между срастаю- щимися краями плодолистиков остаются отверстия (неполиое срастание плодолистиков), и закрытыми (полиое срастание пло- долистиков). Вдоль каждой семенной камеры проходят прово- дящие пучки. Они расположены по одному в наружных и по два во внутренних камерах. Наружные пучки соответствуют дор- зальиым, а внутренние — веитральиым пучкам карпелл. Таким
Глава 3. Структурные особенности, эволюция и филогения образом, в целом эндокарп яблока имеет 5 дорзальных и 10 вентральных пучков. У некоторых видов (секции Docyniopsis и Sorbomalus юньнаньские яблони) эндокарп состоит из склереид, подобно видам Ругила, причем склереиды имеют волокнистую структуру. У других видов (секция Sorbomalus — ганьсуйские яблони, яблоня торинго) сочный слой в результате одревеснения обра- зует твердый костянковидный эндокарп, как у видов Crataegus, Cotoneaster. Происхождение эндокарпа, несомненно, карпеллярное, о чем свидетельствуют его строение и наличие дорзальнь|х и вен- тральных пучков карпелл. В продольном разрезе эндокарп бо- лее разнообразен. Продольная форма эндокарпа иногда кор- релятивно связана с формой плода. корреляция форм плода и эндокарпа указывает на первичность эндокарпа и вторичность мезокарпа. Эндокарп, непосредственно возникший из плодо- листиков и предохраняющий семена, — первичное образование, а мезокарп — вторичное. Яблони по внешнему виду плодов могут быть разделены на две большие группы: крупноплодные — плоды от 2,0 до 3,0 см в диаметре (и крупнее), имеют характерную форму яблока и приспособились к распространению семян при помощи назем- ных животных (териохория), — и мелкоплодные — плоды 0,5— 1,5 см в диаметре, напоминают ягоду, семена распространя- ются птицами (орнитохория). Около 2/3, или 65%, всех видов рода Ма1из принадлежит к мелкоплодным яблоням. Дальнейшее подразделение плодов основывается на форме, величине, окраске, длине плодоножки, наличии при плодах ча- шечки, консистенции мякоти плода и ряде других признаков, приобретенных в конкретных условиях местообитания. В эволюции плодов яблони можно отметить несколько на- правлений, приведших к большому разнообразию современных видов рода Malus (рис. 14). Участие чашечки и других частей цветка в образовании плода составило основу формирования плода у крупноплодных яблонь. Возникновение мелких плодов шло по пути разрастания одной завязи. Первично простым исходным типом плодов в пределах со- временного рода Malus мы считаем плоды, принадлежащие ви- дам секций Eriolobus и Docyniopsis. Плоды яблонь этой группы обычно средней величины (2 — 3 см в диаметре), имеют не- сколько удлиненное и хорошо заметное основание чашечки у верхушки плода, грубохрящеватую мякоть, иногда содержащую каменистые клетки. Осыпавшиеся с деревьев плоды поедают животные, обеспечивающие распространение семян. В пределах этих, по плодам наиболее примитивных, видов наблюдается и некоторая неопределенность структуры плодов. Например, у со- временного М. laosensis встречаются как популяции с характер-
53 Рис. 14. Эволюция плодов яблони: а — первично примитивный тип — Docyniopsis; б — примитивный тип — Eriolobus; в — тип Chloromeles; г — ыедкоплодный тип — So<bomalus; д — промежуточная форма типа Росуп~орз~з (М. зйЫтепз~з); e — тип настоящих яблонь — Malus; ж медкоплодн ый тип — G ymnomeles ными для описанного типа Docyniopsis плодами, так и отдель- ные популяции с более мелкими плодами, притом с частично опадающей чашечкои. В ходе дальнейшей эволюции плоды укрупнялись за счет появления сочного мезокарпа и образовывались настоящие яб- лони (тип плодов Malus). Плоды этого типа одиночные, имею- щие характерную форму яблока, обычно округлой, шаровидной, сплюснутой формы, иногда сдавленные с полюсов и слегка реб- ристые, 2 — 3 или 3 — 8 см в диаметре, на коротких толстоватых плодоножках, которые в 2 — 4 раза короче плода, и с хорошо развитым, нежным (без каменистых клеток) мезокарпом. Одновременно или несколько раньше у типа Docyniopsis появилась тенденция к уменьшению плода. В результате обра- зовались яблони с типом плодов, характерных для Eriolobus. Плоды этих яблонь шаровидные, 1,0 — 1,3 см в диаметре, на тонких длинных (около 2,5 см) плодоножках, со слабо грубо- хрящеватым мезокарпом. Плоды Eriolobus, Docyniopsis и Malus развивались самосто- ятельно, вероятно, от общих предков типа Docynia. Однако линию крунноплодности продолжает еще одна ветвь, а именно,
Глава 3. Структурные особенности, эволюция и филогения 54 * Окраска плодов как способ привлечения птиц в ходе эволюции ста- новилась все более разнообразной. виды яблонь из Северной Америки. Плоды этих яблонь очень похожи на плоды доциниевидных яблонь, в частности M. Шйо- noskii. Так образовался четвертый тип крупноплодных яблонь— Chloromeles, представленный обитателями Северной Америки. Типы плодов Docyniopsis, Malus, Eriolobus и Chloromeles являются типичными териохорами. Такой способ распростране- ния семян нельзя признать достаточно эффективным, так как животные не всегда распространяют семена на дальние расстоя- ния, кроме того, их привлекает только сочная мякоть плода. Поэтому лучшим способом распространения семян яблонь сле- дует считать орнитохорию. Орнитохория способствовала умень- шению размеров плода, опадению после цветения чашечки (вследствие чего чашечка играет незначительную роль в обра- зовании плода), удлинению плодоножек, собиранию плодов по несколько штук и более яркой их окраске. Мелкоплодность, свя- занная с орнитохорией, способствовала более широкому распро- странению таких яблонь. Склонность к мелкоплодности имеет место уже у примитив- ных доциниевидных яблонь. Так, M. sikkimensis, встречающаяся в Восточных Гималаях, имеет кубаревидные, около 1,5 см в диаметре красные плоды*; чашечка при плодах еще удержива- ется, но по мере их созревания она становится частично опада- ющей. От таких, как М. sikkimensis, и подобных ей доциниевид- ных яблонь, по-видимому, возникли мелкоплодные яблони типа О ymnomeles. Плоды Gymnomeles характеризуются следующими призна- ками: они мелкие (в среднем около 1,0 см в диаметре), широко- овальные до почти шаровидных, на тонких, длинных плодонож- ках (длина плодоножки в 4 — 5 раз превышает диаметр плода); по консистенции твердые, долго удерживающиеся (в течение всей зимы) на материнском растении. После первых заморозков они становятся мягкими и полупрозрачными, их охотно поедают птицы. Семенные гнезда на верхушке вытянутые, острые. По признакам плодов связь с другими доциниевидными яб- лонями (в частности, М. laosensis) имеет наиболее обширный и разнообразный, довольно примитивный тип мелкоплодных яб- лонь — Sorbomalus. Плоды этого типа мелкие (0,5 — 1,0 см в диаметре), несколько вытянутые или шаровидные, различной консистенции — от грубой (с каменистыми клетками до неж- ной и мучнистой мякоти, на коротких плодоножках (плодо- ножка — в 2 — 3 раза превышает диаметр плода). Чашечки при плодах остающиеся, частично или полностью опадающие. Се- менные гнезда на верхушке тупые.
55 Особснности индивидуального развития яблони Среди рябиновидных яблонь Sorbomalus встречаются яблони (серия Yunnanenses) с плодами, обладающими грубохрящева- той мякотью, содержащей каменистые клетки. При плодах оста- ются чашелистики, как у доциниевидных яблонь. Однако мелко- плодность как новый качественный признак отделяет юньнань- ские яблони от доциниевидных и позволяет выделить их в ка- честве группы Yunnanenses в составе типа плодов 5огЬота1из. Непосредственную связь по признакам плодов с группой Vunnanenses (в пределах типа Sorbomalus) имеют ганьсуйские яблони — серия Капзиепзея. Плоды этих яблонь мелкие (около 1,0 см в диаметре). Некоторые виды (М. transitoria) достигают предельно мелкой величины — 0,5 — 0,7 см в диаметре. Мякоть плодов — от грубоватой, реже с каменистыми клетками, до нежной, часто с одревесненным костянковидным эндокарпом. Чашечка при плодах опадающая, реже с остатками чашелис- тиков. Наконец, третья группа типа рябиновидных яблонь (серия Toringonae) имеет мелкие (0,6 — 1,0 см в диаметре), терпкие или кисловатые плоды с нежной, иногда мучнистой мякотью и прочным эндокарпом, с наружной стороны ярко окрашенные и с легким восковым налетом. Чашечка при плодах полностью опа- дающая. ОСОБЕН НОСТИИНДИВИДУАЛЬНОГО РАЗВИТИЯ ЯБЛОНИ На различных этапах индивидуального развития (онтоге- неза) растения постепенно меняют многие из своих признаков. Прежде всего это относится к морфологии вегетативных орга- нов, в особенности листьев. Так, у сеянцев яблонь листья значительно мельче, чем у взрослых особей, они слабее опу- шены и сильнее зазубрены, а иногда имеют даже лопастность, не свойственную взрослым растениям. Форма листьев, находя- щихся на различных по возрасту побегах, нередко сильно варь. ирует. Почки, в особенности на многочисленных и преждевре- менно возникших побегах, как правило, мелкие и слаборазви- тые. Эти побеги в течение лета нередко усыхают и превраща- ются в колючки. Меняются и другие признаки: габитус, осо- бенности ветвления и др. Поэтому при учете признаков, приме- няемых в систематике яблонь, крайне важно знать «возраст» изучаемого признака. Рассматривая важнейшие для систематики особенности онтогенетического развития яблонь, мы предлагаем схему, в ко- торой прежде всего различаются две категории изменений. Одна из них связана с периодической сменой признаков в течение года, повторяется ежегодно и носит название годичного цикла
56 Глава 3. Структурные особенности, эволюция и филогения
Особенности индивидуального развития яблони 57 развития. Другая, обусловленная изменениями на протяжении всей жизни яблони и происходящими один раз в жизни особи, называется жизненным циклом развития (рис. 15). )3(изненный цикл развития начинается с момента деления оплодотворенной яйцеклетки и кончается естественным отмира- нием особи. Этот сложный цикл развития в известной степени обусловлен всем предшествующим развитием данного вида и состоит из ступеней, различающихся физиологическими, анато- мическими и морфологическими признаками, а также биологи- ческими особенностями. В ходе развития по данным ступеням у растения постепенно совершенствуются вегетативные органы, обеспечивающие нормальное питание организма, и постепенно накапливается потенциал для создания органов воспроизведе- ния. В определенный момент наступает цветение, заканчиваю- щееся образованием плодов и семян. Интенсивное плодоноше- ние вызывает постепенное старение организма, и, наконец, растение отмирает. )Кизненный цикл яблонь, как и многих дру- гих деревьев, делится на четыре этапа: эмбриональный, юве- нильный, взрослый (продуктивный) и сенильный (старческий). Эмбриональный этап раз вити я. Началом эмбрио- нального этапа развития принято считать образование зиготы. Однако до этого решающим фактором оплодотворения и после- дующего развития являются количество и качество пыльцы. Вся пыльца, попавшая на рыльце и прорастающая на нем, образует пыльцевые трубки, проникающие через ткани пестика и слива- ющие свое содержание с яйцеклеткой, образуя зародыш и эндо- сперм — главные части семени и плода в целом. Эти процессы следует считать наиболее существенными моментами данного этапа. Оплодотворение является сложным физиологическим процес- сом, основанным на обмене веществ. Чем больше пыльцы попа- дает на рыльце пестика и чем больше образуется пыльцевых трубок (содержание которых вливается в завязь и зародыше- вый мешок), тем лучше создаются условия для оплодотворения и развития полноценных семян и плодов. Закономерности развития яблони на эмбриональном этапе следующие: 1) решающая роль наследственной природы расте- ния (его генетической основы), носителями которой являются половые элементы цветка; 2) течение биохимических процессов, проходящих в период формирования семян и плодов; 3) условия среды, в которых находится растение. Для систематики рода Malus познание особенностей эмбрио- нального этапа, свойственных разным видам рода, является весьма важным. Еще более ценным материалом для выяснения филогенеза рода Malus следует считать данные о биохимичес- ких процессах, проходящих при образовании, дозревании и прорастании семян на эмбриональном этапе развития. Однако
Глава 3. Структурные особенности, эволюция и филогения подобных сведений почти не имеется, и это составляет задачу для будущих исследователей. Ювенильный (юношеский) этап р а зв ития начина- ется с развертывания семядолей и появления первых настоящих листьев и завершается появлением первых цветков. В течение этого этапа усиливается рост вегетативных частей растения, но они остаются стерильными, бесплодными, что обусловлено дей- ствием гормонов. В процессе развития вегетативных органов можно установить ряд различий морфологического и анатоми- ческого строения, химического состава органов, а также физио- логического поведения яблони. Вместе с тем ткани растений, образующие побег, имеют различный календарный возраст. Весьма важной особенностью ювенильного этапа развития сле- дует считать способность побегов к укоренению. Вегетативное размножение яблонь укоренением частей стебля на ювенильном этапе развития не представляет особых трудностей. Знание признаков, свойственных растению на ювенильном этапе развития, в сравнении с признаками взрослого растения позволяет в некоторой степени определить направление их раз- вития, решить вопрос об их первичности или вторичности. Прежде всего это относится к характеру ветвления побега, обус- ловливающего габитус растения. Ювенильный тип ветвления у яблони в большинстве случаев характеризуется горизонтальным расположением боковых ветвей по отношению к вертикальному основному побегу. Многие горизонтальные ювенильные ответ- вления оказываются недолговечными. На протяжении лет они усыхают и превращаются в колючки. Почки обычно мелкие, раз- виваются медленно и нередко остаются спящими. Ювенильные листья обычно более мелкие и, как правило, менее опушенные, но имеют более острую зазубренность, нередко появляется и лопастность. Изучение строения первых цветков и плодов, еще полностью не развившихся, представляет особый интерес для понимания процесса становления признаков генеративных органов. Однако при сравнении признаков на различных этапах раз- вития необходимо принимать во внимание широкую амплитуду их изменчивости как у индивидуума, так и в пределах одного вида. П р оду к т и в н ы й э т а п, или этап возмужалости растения, характеризуется преобладанием в нем процессов плодоношения. Данный этап является наиболее длительным в жизни яблони. Несмотря на то что в этот период растение имеет характерные и вполне установившиеся признаки, могут возникать клоновые изменения, когда имеют место значительные отклонения от типа материнских растений. Плодоводы давно подметили, что они возникают в конусах роста листовых или цветочных почек и проявляются в структуре побегов, цветов и плодов. Их обычно
Особенности индивидуального развития яблони 59 называют почковыми вариациями, спортивными уклонениями, или спартами. По существу клановые изменения являются поч- ковыми мутациями, носят наследственный характер и могут быть закреплены в потомстве. Клановая изменчивость имеет практическое значение, так как почковые мутации используются для создания многих новых сортов яблони. Этап возмужалости со свойственным ему плодоношением имеет свои особенности. Так, в начале его наблюдается силь- ный прирост вегетативной массы, стабилизирующейся к сере- дине этапа и сокращающейся в конце. Меняется облик побегов и корней. Крона яблони становится сначала более компактной, потом раскидистой, а в конце этапа с отмиранием мелких ске- летных побегов приобретает неправильные очертания. Изменения габитуса в период возмужалости обусловлива- ются возр астными особенностями анатомического строения. Значительно увеличиваются покровные ткани, происходит одре- веснение, образуются широкие сосуды, а в ситовидных трубках флоэмы появляются разрывы, нарушающие доступ органических веществ к периферийным побегам. В точках роста периферий- ных побегов создаются неблагоприятные для роста условия. Дерево еще продолжает плодоносить, а у основания скелетных ветвей появляются вертикальные побеги — волчки, иногда спо- собствующие восстановлению кроны. Засыхание и отмирание периферийных побегов вызывает наступление последнего — се- нильного этапа жизненного цикла растения. С е н и л ь н ы й э т а п начинается с усыхания наиболее крупных скелетных ветвей. Старая крона дерева отмирает и сменяется новой, возникающей из водяных побегов. У основа- ния ствола и скелетных ветвей из спящих почек появляются сильные побеги, которые в скором времени образуют нижний этаж кроны. В результате дерево приобретает вид, показанный на рис. 16. Дерево еще может произрастать несколько лет, но плодоношение отсутствует. Так, сенильные М. sgluestris харак- теризуются полным отсутствием плодоношения. Возобновления, возникающие на сенильном этапе из пневых пророслей, оказы- ваются нежизнеспособными и недолговечными. Они характери- зуются резкой неустойчивостью, у них проявляется тенденция к удлинению вегетационного периода, в связи с чем они чаще подвергаются обмерзанию. Они часто и легко повреждаются вредителями и болезнями. Годичный цикл — это совокупность изменений в разви- тии яблонь, проходящих в течение одного года. Данный цикл начинается распусканием почек и заканчивается их новой за- кладкой, повторяется ежегодно и характеризуется определен- ными периодами и фазами. Переход от одного периода к дру- гому совершается последовательно. Однако некоторые периоды годичного развития яблони проходят одновременно. Особенно-
Глава 3. Структурные особенности, эволюция и филогения 60 Рис. 1б. Лесная яблоня на сенильном этапе жизненного цикла сти годичного цикла зависят от природных условий, и прежде всего от времени года. Здесь наблюдаются два состояния вегетации и покоя. Состояние вегетации наступает рано весной и характеризу- ется набуханием почек. Яалее следуют распускание листьев, цветение, образование и созревание плодов и семян, заложение и дифференциация новых почек. Все это сопровождается акти- визацией многих жизненных процессов. Состояние вегетации завершается с осенним листопадом, после которого растения входят в состояние относительного покоя. В состоянии покоя можно различать два периода — авто- номный и вынужденный (или относительный). Автономный период является обязательной составной частью годичного цикла, без которого невозможен переход к новой вегетации, так как в течение его завершаются необходимые для дальнейшего развития растения изменения. Вынужденный период покоя на- ступает зимой и продолжается до тех пор, пока не наступят условия для новой вегетации. Изменения развития в автономном периоде проходят при положительных температурах (от 0 до 10'С). С этим связана продолжительность автономного периода у различных по про- исхождению видов яблони. Восточно-азиатские виды яблонь характеризуются относительно коротким периодом покоя, по- скольку в условиях Восточной Азии оптимальная температура для прохождения данного периода бывает кратковременной (осенью и весной). В странах Западной Европы, наоборот, про-
Особенности индивидуального развития яблони 63 должительная осень и теплая зима вызывают более длительный автономный период покоя. Это важно учитывать при интродук- ции яблонь. Так, восточно-азиатские яблони в условиях Запад- ной Европы обычно плохо зимуют, так как теплые оттепельные зимы здесь способствуют преждевременному наступлению новой вегетации. Различные части растения в данный период покоя ведут себя по-р азному. Продолжительный автономный период хар а ктерен для боковых почек однолетних побегов. Вегетативные (листо- вые) почки имеют более длительный автономный период, не- жели цветочные. Краткий автономный период свойствен ство- лам, у корней он вовсе отсутствует. В зависимости от характера изменений, проходящих в со- стоянии вегетации, можно наметить пять его периодов: 1) рас- пускание почек и цветение; 2) вегетативный этап; 3) заложение и дифференциация цветочных почек; 4) образование и созрева- ние плодов и 5) листопад. Период р аспускания по чек и цветения насту- пает рано весной и состоит из восьми фаз: 1) набухание почек, раздвижение почечных чешуй и появление кончиков листьев; 2) распускание почек, появление листьев и зачатков соцветий; 3) обособление соцветий, отделение и образование листовых ро- зеток; 4) появление соцветий; 5) обособление бутонов, отделе- ние их друг от друга; 6) появление свернутых лепестков, рас- крытие чашелистиков и рост лепестков; 7) раскрытие лепестков и бутонов соцветий; 8) опадение лепестков (рис. 17). Продолжительность цветения яблони, а также долговечность цветка в значительной степени зависят от природных условий. В жаркую и сухую весну цветение яблонь может закончиться в течение недели, тогда как в холодную и сырую погоду цветение длится более двух недель. Таким образом, продолжительность жизни одного цветка яблони может колебаться от 5 — 6 до 12— 15 дней. Поэтому целесообразно в процессе цветения яблонь различать его начало (появление первых распустившихся цвет- ков), массовое цветение (когда на растении распускается более 50Я всех цветков) и конец (опадение лепестков с нормально развитых цветков). Далее наступает период вегетативного роста. Рост побегов и корней в длину осуществляется конусами роста, а рост в толщину — за счет деятельности камбия. В периоде ве- гетативного роста яблонь различают три фазы развития: 1) де- ление цветков, 2) их растяжение и 3) дифференциация. Из них визуально более заметной является последняя, когда образу- ются ткани и отдельные части побегов и корней. В процессе вегетативного роста наблюдается известная пе- риодичность. Весной после периода покоя возобновляется рост и наступает весенний максимум роста, продолжительность ко-
Глава 3. Структурные особенности, эволюция и филогения 62 Рис. 17. Развитие цветочной почки лесной яблони (по В. А. Колесникову[1966], с некоторыми изменениями) торого составляет от 20 до 90 дней (в среднем 30 — 40 дней). После небольшого спада наступает новый максимум роста летний. Он обычно бывает в конце июня и длится 15 — 20 дней. В конце лета иногда наступает еще и третий максимум вегета- тивного роста — осенний, особенно когда после летней засуш- ливой погоды выпадают обильные осадки. Осеннему росту не- редко сопутствует осеннее цветение. Период заложения и диффере нци ации цветоч- н ы х п о ч е к начинается с формирования покровных чешуй молодых почек и заканчивается образованием внутренних час- тей цветка. Он проходит в шесть фаз: 1) развитие покровных чешуй, покровных листьев и прицветников; 2) появление в ко- нусе роста зачатков цветков; 3) формирование чашечки; 4) об- разование венчика; 5) формирование тычинок и пыльников и 6) образование пестика. длительность дифференциации цветочных почек у яблони зависит также от многих условий и составляет 21 — 89 дней. К осени в цветочной почке яблони имеются вполне сформиро- ванные цветки, в которых отсутствуют плодовые клетки. Разви- тие последних происходит рано весной после прекращения периода покоя. Развитие цветочных почек яблони наступает после первого (весеннего) максимума вегетативного роста, а также после первых, наиболее интенсивных фаз образования плодов, в среднем с 15 июля по 20 — 30 августа.
Особенности индивидуального развития яблони 63 Период образования и созревания плодов и с е м я н начинается с момента оплодотворения и продолжается до созревания плодов. У некоторых видов яблони и позднезим- них сортов процесс полного созревания плодов происходит после отделения их от дерева. Здесь мы можем различать сле- дующие фазы развития и созревания плодов: 1) завязывание плода; 2) смыкание или отделение (опадение) чашелистиков; 3) образование характерной формы плода; 4) изменение ок- раски кожицы плода; 5) созревание плода и семян и 6) дозре- вание плода и семян (см. рис. 17). Продолжительность всего периода зависит от видовых и сортовых особенностей яблонь, а также от комплекса природ- ных условий. У большинства яблонь число образующихся цвет- ков настолько велико, что часть их опадает, не завязывая пло- дов. Первое опадение наблюдается еще в период цветения. Опадают цветки с недоразвитыми или ненормально развитыми пестиками или другими частями цветка. Второе опадение насту- пает через 10 — 15 дней после цветения, когда опадают неопы- ленные цветки. Третье опадение завязей следует примерно через месяц после цветения и вызывается недостаточным снабжением дерева водой, органическими и минеральными веществами, а также неблагоприятными климатическими условиями, наличием вредителей, болезнями и др. В результате лишь 15 — 20~/о всех цветков превращаются в плоды. Оставшиеся на дереве завязи постепенно образуют плоды. Сначала образуется околоплодник, или перикарпий, потом про- исходит его дифференциация и образуются экзокарп, эндокарп и мезокарп. Л и с то п ад является заключительным периодом и одновре- менно переходом к зимнему покою. Под листопадом подразуме- вается не только опадение листьев, но также изменение био- химических процессов, способствующих переходу к зимнему покою и успешной зимовке растений. Частое опадение листьев наблюдается на протяжении всей вегетации, но оно не сопро- вождается существенными изменениями биохимических процес- сов. Массовое опадение листьев в листопадный период начина- ется под влиянием климатических условий, из которых глав- ными, обуславливающими прекращение роста, являются температура и освещение. После опадения листьев происходит перераспределение и концентрация веществ, от которых зависит успешное переживание неблагоприятного для вегетации вре- мени года — зимы. Морфогенез жизненной формы изображен на рис. 18. Ниже приводятся данные наблюдений автора за развитием М. sylves- tris в 1959, 1962 — 1963 и 1967 гг. на территории совхоза «Кри- мунас» Добельского района (Латвия) в роще лесных яблонь.
Глава 3. Структурные особенности, эволюция и филогения 64 Рис. 18. Морфогенез жизненной 10венильное развитие М. sylvestris начинается с прораста- ния семян, которое происходит по надземному типу. Сначала образуется интенсивно растущий стержневой корень. Вслед за этим при помощи усиленно развивающегося гипокотиля выно- сятся на поверхность толстоватые и мясистые семядоли с точкой роста посередине. Они вскоре зеленеют, и над ними (0,5— ,0 см) на молодом стебле появляются первичные, эллиптически заостренные, по краю зазубренные, зеленые и слегка волосистые листья. Стебель продолжает интенсивно расти в вертикальном направлении и до конца года достигает 15 — 30 см в длину (в питомнике до 25 — 50 см). Нередко уже на первом году из пазух листьев (в средней части сеянца) появляются горизон- тальные боковые побеги, которые развиваются слабее главного вертикального побега. Рост сеянца на первом году обычно завершается формированием зимующей терминальной почки. Однако моноподиальность нарастания сеянца может нарушаться
Особенности индивидуального развития яблони формы лесной яблони уже на первом году его жизни. В условиях Латвии это особенно выражается в засушливые годы, когда из-за недостатка влаги (в конце июня) рост прекращается. В июле, при достаточном количестве осадков, рост сеянца обычно возобновляется, но при этом происходит подсыхание (и гибель) терминальной почки, а также верхней части вертикального побега. Этот процесс «де- капитации», происходящий в самом начале жизни сеянца, спо- собствует превращению дерева в кустарник. На втором году рост сеянца усиливается, и он достигает в высоту 50 — 60 см. Теперь происходит интенсивное развитие боковых побегов. Иногда наблюдается особенно интенсивный рост первого (верхнего) бокового побега, который принимает вертикальное положение и вскоре достигает высоты централь- ного побега и даже превосходит его. На втором году жизни сеянца начинает формироваться крона будущего дерева. Она состоит из небольшого числа боковых побегов, расположенных
Глава 3. Структурные особенности, эволюция и филогения на высоте 20 — 40 см от земли, и является первичной (не вполне сформировавшейся) . В последующие 3 — 5 лет над первичной кроной образуется крона нормальная, состоящая из 5 — 7 сильно развитых боковых ветвей. Главная ось в этом возрасте еще отчетливо прослежи- вается, хотя она рано теряется среди мощных боковых ветвей формирующейся кроны. Нижние боковые ветви, составляющие первичную крону, уже начиная со второго года жизни сеянца постепенно отстают в росте и усыхают, а более короткие из них превращаются в колючки, у основания которых располагаются небольшие, почти округлые листья, сидящие не в очередном порядке, а в виде мутовок. В условиях Латвии 5-летние деревья М. sylvestris достигают в высоту 2,0 м, а 10 — 12-летние — 3,5 — 4,0 м. Для сравнения, на территории 1~урского государственного заповедника яблоня в этом возрасте не превышает 3 м [Серебряков, 1962]. Плодоношение у особей М. sylvestris наступает в разное время. В условиях Латвии первое цветение обычно начинается в 10 — 12-летнем возрасте, однако в отдельных случаях сильно разветвленные особи начинают цвести значительно раньше (уже на 6 — 9-м году жизни). Наблюдается и более позднее вступле- ние в пору цветения — в 15 — 20-летнем возрасте. 1~роме того, встречаются и некоторые аномальные особи, растущие в небла- гоприятных условиях (на краях канав, пообочинам дорог ит.д.) .. Они имеют облик небольших кустов (0,5 — 1,0 м в высоту), как правило, не цветущих. Эти кустообразные яблони размножа- ются вегетативно, за счет придаточных почек на корнях, даю- щих корневую проросль, а не с помощью спящих почек ствола [Серебряков, 1962]. Первое цветение и плодоношение (обычно слабые) сопро- вождаются интенсивным ростом вегетативных органов дерева. После первого плодоношения меняется внешний облик кроны. Если в начале плодоношения крона лесной яблони обычно ком- пактна, то с наступлением плодоношения она под тяжестью плодов становится более раскидистой, так как боковые сучья горизонтально отклоняются. Это способствует закладке цветоч- ных почек. В результате продолжительного плодоношения в возрасте 20 — 25 лет наблюдается резкое ослабление роста де- рева, а плодоношение становится регулярным. Однако ежегодному плодоношению нередко препятствует летняя засуха, наступающая в период заложения цвсточных почек. В условиях Прибалтики это случается сравнительно редко. После такого засушливого периода в следующем году наблюдается очень слабое цветение, сопровождающееся образо- ванием малого числа плодов. При благоприятных погодных условиях в кроне снова закладывается много цветочных почек, обеспечивающих обильный урожай нового года. Следовательно,
Некоторые сведения о филогении рода Malus 67 НЕКОТОРЫЕ СВЕДЕНИЯО ФИЛОГЕНИИ РОДА MALUS Для выяснения филогении рода Malus и филогенетических отношений видов следовало бы основное внимание уделить ана- после неурожайного года наступают весьма обильные цветение и плодоношение, которые снова препятствуют заложению цве- точных почек для следующего года. Так возникает периодич- ность плодоношения, когда урожайный год чередуется с неуро- жайным. После неоднократного и продолжительного плодоношения наряду с другими возрастными изменениями прежде всего из- меняется габитус кроны. Форма ее становится менее компактной и более раскидистой и приобретает округлую или воронковид- ную вершину, что является следствием полной утраты четко выраженной лидер ной оси и преобладающего роста боковых ветвей. И. Г. Серебряков '[1962, с. 211] считает, что «ранняя потеря лидерной оси приближает форму дерева, наблюдаемую у яб- лонь, к кустарникам, у которых, как известно, главная ось рано теряет лидерное положение в онтогенезе растения». В связи с этим автор приходит к выводу что кустообразная жизненная форма яблони экологически возникла в аридных условиях как следствие ускорения развития и сокращения жизненного цикла главной оси. Уже в 20 — 30-летнем возрасте вскоре после ослабления роста у М. sylvestris наблюдается частичное подсыхание кроны. Сна- чала подсыхают мелкие ответвления, находящиеся на стволе и нижней части крупных боковых ветвей внутри кроны. В это время при интенсивном плодоношении дерево стареет, но через некоторое время появляются признаки восстановления утерян- ных частей кроны и происходит «омолаживание» дерева. «Омо- лаживание» кроны у М. sylveskris обычно осуществляется раз- вивающимися на скелетных ветвях вертикальными побегами— волчками. П. Г. Шитт [1958] этот процесс подсыхания мелких скелетных частей и появление волчков иногда называет «изре- живанием и естественным обновлением> и объясняет это явле- ние нарушением корреляции между ростом корневой системы (всасывающей части) и нарастанием надземной части (листо- вым аппаратом). Однако эти явления более характерны для последнего — сенильного этапа жизни яблони. Продолжительность продуктивного этапа у М. sylvestris весьма различна и колеблется в больших пределах: с 10 — 12- летнего возраста до 60 — 80 и даже до 100 — 20 лет, когда пло- доношение прекращается и дерево постепенно отмирает.
68 Глава 3. Структурные особенности, эволюция и филогеиия лизу палеоботанического материала. Однако от этого пришлось отказаться по следующим причинам. 1. До настоящего времени известно лишь несколько единич- ных фрагментарных палеоботанических образцов, на основании которых невозможно составить полное представление о прежнем облике видов яблони. 2. Достоверность принадлежности этих немногочисленных ископаемых фрагментов прежней растительности к роду Malus во многих случаях еще сомнительна. 3. Прежде чем приступить к палеоботаническому изучению рода Malus, необходимо более четко разработать систематику и раскрыть основные филогенетические взаимоотношения совре- менных видов данного рода и, главное, выявить постоянные и наиболее выразительные признаки рода, на основе которых можно было бы провести точный анализ палеоботанического м атер иал а. Предполагается, что яблони возникли в меловом периоде мезозойской эры на обширной территории Евразии, еще до трансгрессии океана. В настоящее время наиболее примитивные виды яблонь — реликты прежних эпох — сосредоточены в юж- ных пределах родового ареала — Юго-Восточная Азия и страны Средиземноморья. В местах распространения предков современ- ных видов рода Malus климат, видимо, был теплым и влажным и способствовал образованию соответствующих морфологичес- ких структур. Исходной жизненной формой яблонь, очевидно, являлось вечнозеленое мезофильное дерево — обитатель влажных тропи- ческих и субтропических лесов. Сравнение особенностей строе- ния некоторых современных видов Malus, находящихся на низ- ких ступенях развития (виды секции Docyniopsis), с древней- шими родами, в частности Docynia и Pyrus, позволяет составить некоторое представление об облике древних исходных форм яблонь. Итак, первичные яблони, вероятно, были высокими (до 20 — 25 м) деревьями с хорошо выраженными толстыми (до 2 м в диаметре) стволами, слаборазветвленной (3 — 5 порядков) кроной; крупными, кожистыми, полувечнозелеными, блестящими, сложными, раздельными или лопастными, в очертании широко- эллиптическими, широкоовальными, почти округлыми или про- долговатыми листьями, собранными в пучки, преимущественно на верхушках длинных побегов; довольно крупными (3 — 5 см в диаметре) цветками, собранными в пучки или многоцветко- вые соцветия неопределенного типа (ложный щиток, ложный зонтик); продолговатыми, несколько грушевидными (2 — 3 см в длину и 2 — 5 см в ширину) плодами с грубой мякотью, содер- жащей довольно много каменистых клеток; остающейся при плодах и несколько приподнятой чашечкой и сросшимися при
Некоторые сведения о филогении рода Ма/из основании, отвернутыми чашелистиками; а также плодоножкой примерно в 2 раза длиннее плода. Семенные камеры открыты, сливаются в общую центральную полость плода (края плодо- листиков не срослись}. Подобные признаки имеются у видов рода Docynia, распро- страненных в современном Юго-Западном Китае (пров. Юнь- нань), а также у некоторых примитивных видов Pyrus, проис- ходящих из горной части Юго-Западного Китая. Сходные при- знаки имеют также современные реликтовые виды яблонь из секции Docyniopsis: М. laosensis из Лаоса, М. formosana из Южного Китая (пров. Фуцзянь и о-в Тайвань}, М. tschonoskii из Южной Японии и М. sikkimensis из Восточных Гималаев, а также ископаемый вид М. obensis, описанный М. Г. Горбуновым [1959], из третичных отложений Западной Сибири (Заобский Яр на левом берегу р. Оби, 56'15' с. ш., 84' в. д.). У перечисленных доциниевидных яблонь сочетаются наибо- лее примитивные признаки из всех ныне существующих видов. На первый взгляд кажется, что эти яблони составляют началь- ное звено в эволюции рода Malus. Однако даже наиболее при- митивные их виды (М. laosensis, М. formosana) со временем значительно изменились и в настоящее время сильно отлича- ются от первоначальных форм рода Malus. Поэтому вероятными исходными формами яблони следует считать каких-то древних предков, давших начало первичным видам Docynia, Pyrus, Ma- lus и Cydonia. Секция Docyniopsis, таким образом, представляет собой боковую ветвь филогенетического дерева целого ком- плекса исходных форм, которые послужили основой для обра- зования основных ветвей эволюции рода Malus. Это, видимо, произошло в третичном периоде. В составе третичной голарктической древесной флоры третичные яблони были широко распространены, занимая в пределах азиатского материка (вероятно, в Евразии) область между 20 — 25' и 55— 60' с. ш. Третичный ареал яблонь был обширен, что подтверж- дается остатками их потомков — реликтовых видов, распро- страненных дизъюнктивно в Юго-Восточной и Восточной, а также Центральной Азии. О предполагаемых северных пределах ареала третичных яблонь свидетельствует обнаружение уже упомянутого ископаемого вида M. obensis в Западной Сибири (56'15' с. ш.). Существованием в третичном периоде огромного ареала тре- тичных яблонь можно объяснить возникновение таких весьма оригинальных эндемических видов, как M. trilobata, обитающего дизъюнктивно на скалистых склонах Ливанских гор и на Бал- канском п-ове, а также М. fusca, распространенного в Северной Америке. Это подтверждается обнаружением М. Г. Горбуновым фоссилизированных яблонь в отложениях третичного периода в Западной Сибири.
70 Глава 3. Структурные особенности, эволюция и филогения Наиболее крупные центры резервации первичных яблонь на- ходятся в Азии: 1) в северной части современного Лаоса, Вьетнама и в ки- тайской провинции Юньнань — M. laosensis; 2) в южной части современной провинции Фуцзянь и о-ва Тайвань — М. formosana; 3) на современных о-вах ~юсю, Хонсю и Хоккайдо М. tschono ski; 4) в восточной части Гималаев — М. зйЫтепив. Здесь же, по крайней мере еще в меловом периоде, началась дифференциация первичных яблонь, положившая начало даль- нейшей эволюции рода, так как, по-видимому, эти яблони были типичными горными растениями, представленными изолирован- ными популяциями, с характерным для них процессом видооб- разования и быстрыми темпами эволюции. Наряду с климатическими особенностями третичного времени на распространение яблонь оказали воздействие и другие фак- торы. Трансгрессия океана, начавшаяся еще в меловом периоде, сильно изменила очертания древнего материка. От Азии от- делилась Северная Америка, а также о-ва Тайвань, Хонсю, Хоккайдо и ряд других, некогда соединявших Азию с Северной Америкой. Таким образом до разъединения материков на тер- ритории Северной Америки оказались некоторые формы тре- тичных яблонь, положивших впоследствии начало развитию самостоятельной ветви североамериканских яблонь. В третичном периоде образовался Тетис, произошли подня- тия гор. На развитии рода Malus особенно отразилось гигант- ское поднятие, составляющее ныне горные системы Пиренеев, Альп, Балкан, кавказа, Памира, Тянь-Шаня, Гималаев и дру- гие, которые тянутся вплоть до ~итая. Некоторые формы тре- тичных яблонь еще до образования названных горных систем распространились вдоль южных берегов Тетиса в Переднюю и Малую Азию, а также на кавказ и Балканы. Подтвержде- нием этого служат примитивные и одновременно своеобразные и оригинальные яблони, встречающиеся ныне на территории некоторых стран Средиземного моря. Это М. trilobata — энде- мик скалистых склонов среднегорной полосы Ливана, а также некоторых частей Балканского п-ова. Строение цветков и пло- дов сближает указанный вид с доциниевидными яблонями. По всей вероятности, M. trilobata является реликтом третичных яб- лонь, скорее всего «беженцем», т. е. случайно сохранившимся звеном третичных яблонь. Возникновение общих форм рельефа, равно как и заметное похолодание в конце третичного периода, ускорили дифферен- циацию яблонь. Общее распространение третичных яблонь зна- чительно сократилось. Появились популяции, сформировавшиеся в условиях умеренного климата. Уступая место формам более
Некоторые сведения о филогении рода Malus 71 * Значение скрещивания и создания новых форм растений на базе ста- рых путем отдаленной гибридизации показано М. Г. Поповым [1963]. умеренного климата, третичные яблони положили начало обра- зованию и развитию нескольких самостоятельных линий рода Ма1ия. От третичных яблонь, обитавших в условиях влажного суб- тропического климата (тип I), по-видимому, в конце третичного периода произошли и эволюционировали деревья, типичные для умеренного климата (тип II), и, наконец, кустарники ксеро- морфного облика — обитатели умеренных засушливых, преиму- щественно горных районов (тип III). Не вдаваясь в детали, ограничимся констатацией лишь основных факторов, оказавших большое влияние на образование этих трех основных линий (типов) эволюции жизненных форм яблони и возникновение полиморфизма яблонь. Третичные яблони, особенно наиболее архаичные формы, были типичными горными растениями, о чем свидетельствуют их реликты (секция Docgniopsis), встречающиеся в горах тро- пического и субтропического поясов Юго-Восточной Азии. Раз- виваясь в горах, они находились в максимально благоприятных условиях, поскольку горы являются своеобразными «лаборато- риями», создающими разнообразие форм растений, в том числе плодовых [Вавилов, 1931в]. Под воздействием горных условий образовались диффузные комплексы особей, состоявшие из не- больших, более или менее изолированных друг от друга локаль- ных популяций. Непрерывный процесс перекрестного опыления привел к появлению спонтанных мутаций и дал начало некото- рым новым комбинациям генов*. 1~ак известно, мутации возни- кают с определенной частотой в каждом поколении в зависи- мости от того, в какой степени им благоприятствуют или противодействуют природные факторы. Наличие малых локаль- ных популяций при соприкосновении и скрещивании позволяет возникающим компонентам расширить наследственную инфор- мацию. Благодаря этому у растений в горах возникают удачные комбинации новообразований, способных к дальнейшему рас- пространению. Разнообразные природные условия горных районов воздей- ствуют на различные новообразования, а также мутации. Бес- порядочно возникающие мутации подвергаются селективному действию естественного отбора, в результате чего в популяциях устанавливается равновесие. Однако, как показывают много- численные экспериментальные данные генетики популяций, про- цесс возникновения равновесия достаточно эффективен лишь в более постоянных условиях, чем в горах, и при наличии боль- ших, менее изолированных популяций. Поэтому у расте- ний в горах в связи с малыми размерами популяций
Глава 3. Структурные особенности, эводвция и фидогения 72 Е4:Э? CО z Е У б а ° ( Ц ~ в Е z Е ее а ax М Е Е 3 3 Е < Я ее в~ е ~ щ~ ф~ Е Е < О ф L е о ЗО 8 в � Яо< МЕЕ З D K 33 кй и в о > w ее hc Е 9 е~ кх ~? >м Ко «~ ма ~ЙЦ З Е Е < U о lo а ~ аф а«~ам ? а О SECT 1ЧCHLOROME ЕЕ 3 SECT. ЧG YMNOME ЕЕ 5 ЗЕШ Vl MALUS Рис. 19. Схема филогенетических отношений в роде Malus и разнообразием природных условий преобладают случайные комбинации признаков, мало контролируемых отбором. Это при- водит к возникновению новых качеств и быстрым темпам их эволюции. Видимо, это, наряду с описанным выше комплексным воздействием, явилось основной причиной возникновения в гор- ных районах Средней Азии огромного разнообразия форм и полиморфизма яблонь. В горах на предельных высотах и в аридных условиях про- исходит задержка развития некоторых тканей и органов растений. Организмы завершают свой годичный, а следова- тельно, и жизненный цикл в короткие сроки. Начальные пери- оды развития сокращаются, приобретают дефинитивные особен- ности, а поздние периоды как бы задерживаются и даже выпадают. Это приводит к изменению всей структуры растений. Они уменьшаются в росте, начинают рано плодоносить, стано-
Некоторые сведения о филогении рода Malus 73 вятся скороспелыми, и когда эти изменения получают наслед- ственный характер, возникают новообразования форм неотени- ческим путем. Нам представляется, что значительная часть многочисленных разнообразных признаков яблонь возникла именно неотенически. По существу, все яблони ксероморфного облика, обитающие в аридных условиях, во многих случаях яв- ляются результатом неотении. У неотенических яблонь встречаются случаи прогрессивного упрощения структуры, так как «всякая наследственная неотения представляет собой некоторую деспециализацию организма, что несомненно расширяет возможность эволюции в новых направ- лениях» .[Тахтаджян, 19б4, с. 18]. Различные неотенические формы в настоящее время можно наблюдать в горах Средней Азии (особенно в районе Гиссарского хребта), Кавказа, а также среди других яблонь. Анализ литературных данных и изучение обширного мате- риала из различных частей ареала рода Malus, сопоставление их морфологических, а также биологических особенностей и гео- графического распространения дают основание выявить основ- ные закономерности эволюционного развития рода. Начавшаяся еще в третичном периоде, а возможно и не- сколько раньше, дифференциация первичных форм яблони по- служила основой для обособления трех наиболее примитивных ветвей филогении рода Malus (рис. 19): пушистолопастных (секция Eriolobus), доциниевидных (секция Docyniopsis) и ря- биновидных яблонь (секция Sorbomalus) . Пушистололастные яблони (секция Eriolobus) представлены единственным, в высшей степени своеобразным и обособленным реликтовым видом — М. trilobata. Дизъюнкция ареала, а также структурные особенности цветка и листьев (пестик образован пятью не полностью сросшимися плодолистиками, верхние концы которых свободные, оттянутые, вдаются в эпигинную трубку чашечки, рыльца двурасщепленные, тычинок много (30 — 50), листья трехраздельные) дают основание рассматри- вать Eriolobus в качестве наиболее архаичной и близкой к тре- тичной яблоне формы, сохранившейся до нашего времени, од- нако не получившей дальнейшего развития. Доциниевидные яблони (секция Docyniopsis), несомненно, со- ставляют одну из древнейших ветвей рода Malus, о чем свиде- тельствуют их география (субтропическое распространение), а также ряд структурных особенностей. Из последних особо сле- дует подчеркнуть неполное срастание плодолистиков. Этот при- знак, равно как двурасщепленные рыльца и большое число тычинок, сближает доциниевидные яблони с Eriolobus и указы- вает на происхождение их от общих предков, имеющих пред- положительно в третичное время общее широкое распростране-
74 Глава 3. Структурные особенности, эволюция и филогения ние, по крайней мере от Юго-Восточной Азии до Средиземного моря. Доциниевидные яблони в настоящее время представлены четырьмя реликтовыми видами, из которых наиболее прими- тивными являются обитатели центра секционного ареала М. laosensis и М. formosana, а наиболее продвинутыми — пери- ферические виды M. tschonoskii (Япония) и М. sikkimensis (Ги- малаи) . Оба последних вида определенным образом связаны (действие закона гомологических рядов Н. И. Вавилова) с дру- гими ветвями эволюции яблонь: М. tschonoskii с секцией Chlo- romeles, а М. sikkimensis с видами секций Gymnomeles и Malus. Рябиновидные яблони (секция Sorbomalus), так же как до- циниевидные и пушистолопастные, имеют солидный возраст. Они возникли еще в третичном периоде и распространились по ог- рампой территории Центральной и Восточной Азии, Северной Америки и на запад, включая страны Средиземноморья, в рай- онах, которые в четвертичном периоде не подвергались оледе- нению. Таким образом, в ходе эволюции возникло большое разнообразие рябиновидных яблонь — от самых примитивных форм мезофильных деревьев с хорошо выраженными стволами до ксероморфных кустообразных деревьев, деревец или кустар- ников. У рябиновидных яблонь довольно отчетливо можно на- блюдать постепенную эволюцию признаков: сложные. листья с различной степенью лопастности, переходящие в упрощенные цельные, а также мелкие цветки, собранные в многоцветковые щитковидные, а затем малоцветковые зонтиковидные соцветия. Эволюция плодов привела к значительному уменьшению их размеров у примитивных видов и еще более .сильному умень- и~ению у видов, эволюционно продвинутых. Вслед за уменьше- нием размеров плода изменилась грубая (с каменистыми клет- ками) мякоть примитивных видов и возникла мелкозернистая и мучнистая мякоть, лишенная каменистых клеток, у видов продвинутых. Все современные рябиновидные яблони филогенетически можно разделить на четыре группы. Ю н ь н а н ь с к и е я бл он и (серия Yunnanenses). Деревья с хорошо выраженными стволами, крупными цельными или сла- болопастными листьями, цветками в щитковидных соцветиях, плодами с грубой мякотью, содержащей каменистые клетки, и остающимися при плодах чашелистиками. Виды юньнаньских яблонь: M. ombrophila, М. yunnanensis и М. honanens s — имеют ряд примитивных признаков. Однако встречаются признаки, свидетельствующие 06 эволюционной продвинутости. Ганьсу йские яблони (серия Kansuenses). Небольшие кустообразные деревья или деревца с 3 — 5- или многолопаст- ными листьями средней величины, цветками в многоцветковых соцветиях", мелкими, с нежной мякотью и без каменистых кле-
Некоторые сведения о филогении рода Malccs ток (за исключением М. kansuensis) плодами и опадающей при плодах чашечкой. Ганьсуйские яблони: М. kansuensis, М. torin- goi des, М. transitoria, М. centralasiatica и М. komarovii достигли высокой степени эволюционного развития. Это невысо- кие деревца (3 — 5 м) ксероморфного облика, обитают на высо- когорьях (2000 — 4000 м н. у. м.). Их филогенетические связи с юньнаньскими яблонями подтверждаются наличием общих при- знаков: присутствие каменистых клеток в мякоти плода, ха- рактерное для юньнаньских яблонь и свойственное М. kansuen- sLs. У других видов ганьсуйских яблонь мякоть плода мелко- зернистая. Ганьсуйские яблони — самая разнообразная и полиморфная группа рябиновидных яблонь — представлены видами, наиболее эволюционно продвинутыми. Я б л о н и т о р и н г о (серия Toringonae). Кустообразные деревца или кустарники с гетерофильными 3 — 5-лопастными листьями на коротких побегах, цельными — на длинных; цветки в плоских зонтиковидных соцветиях; плоды мелкие, с мучнистой мякотью и опадающей чашечкой. Эти яблони морфологически и географически обособлены— М. toringo и var. sargentii обитают в Восточной Азии, горных лесах Корейского п-ова и на островах Японии. В эволюционном отношении эти яблони в настоящее время не имеют потенци- ала для дальнейшего развития, о чем свидетельствуют их виды, состоящие из больших и прочных популяций. По- видимому, Toringonae представляет короткую боковую, т. е. наименее продвинутую в филогенетическом отношении ветвь эволюции рябиновидных яблонь, не получившую дальнейшего развития. Б у р ы е я б л о н и (серия Fuscae) с эндемичным видом М. (изса образовались в период прежней связи Азии и Север- ной Америки от общего ядра Sorbomalus. Однако кубаревидная форма ее плодов, равно как и опушенность вегетативных час- тей, подтверждают некоторую, видимо конвергентную, близость М. fusca к доциниевидным яблоням. Именно здесь сказалось действие закона гомологических рядов Н. И. Вавилова. В дальыейшем в филогении рода 18alus намечается значи- тельный прогресс: освоение новых территорий и приспособление к новым, более суровым и аридным условиям, а в связи с этим — возникновение приспособительных изменений (значи- тельная редукция и целесообразные упрощения) структур. Следующие ветви филогении рода 18alus имеют более высо- кий эволюционный уровень. Их образование в основном прохо- дило в четвертичном периоде, за исключением ягодных яблонь (секция бутпоте!еь), которые, видимо, начали развиваться несколько раньше, еще в третичное время.
Глава 3. Структурные особенности, эволюция и филогения Особая ветвь эволюции яблони наметилась в Северной Аме- рике и имела широкое распространение. Это крупноплодные яблони секции Chloromeles, приспособившиеся к распростране- нию семян с помощью наземных животных. Если бурые яблони в пределах секции Sorbomalus эволюционировали медленно, то крупноплодные североамериканские- яблони развивались чрез- вычайно быстро. Это подтверждается наличием множества мел- ких популяций, возникших на основе случайных мутаций. В про- цессе эволюции крупноплодных яблонь в Северной Америке создалась весьма пестрая смесь различных мелких популяций яблони, что послужило причиной для необоснованного описания многих локальных «видов» в Северной Америке, число которых уже достигает семи. Крупный очаг распространения третичных яблонь находился в Гималаях. Остатком его является современный М. sikkimen- sis. В результате его изучения высказано предположение, что еще в третичном периоде в Гималаях образовалась своеобраз- ная ветвь мелкоплодных яблонь — секция Gymnomeles, кото- рая имела широкое распространение на северо-восток и юго- восток. Примитивные ягодные яблони уже имели большие цель- ные листья и довольно крупные плоды (около 2 см в диаметре) с опадающей на них чашечкой, как у современной гималайской яблони, встречающейся в западной части Гималаев, от Инда на восток до Китая. Оледенение четвертичного периода способствовало отступ- лению третичных ягодных яблонь на восток Азии, севернее ареала рябиновидных яблонь, где оледенения не было. В ре- зультате дальнейшей эволюции, в основном по признакам из- мельчения плодов и наличия периода глубокого зимнего покоя, выделились современные ягодные яблони — М. baccata, М. mandshurica и М. sachalinensis, достигшие пределов холо- достойкости рода (М. baccata выдерживает морозы до — 50— — 55'С). В Гималаях обособился другой вид ягодных яблонь— М. himalaica, который не подвергался сильным изменениям. Для всех этих яблонь характерны цельные эллиптические или яйцевидные, зеленые при распускании листья, обладающие округленным основанием и зеленым черешком. другая ветвь эволюции ягодных яблонь имела распростра- нение в центральной части Азии. Это хубейские яблони (М. hu- pehensis), обитающие на высотах от 1000 до 2000 м н. у. м. в 1.~ентральном и Юго-Западном Китае, откуда они заходят в Бирму и Индию [Assami, 1927]. другой вид — М. halliana— изучен лишь в условиях культуры в Китае и Японии, в связи с чем при отсутствии сведений об ареале данные Ассами [As- sami, 1927] и Коидзуми о нахождении дикорастущих форм этого вида в Японии требуют проверки. Хубейские яблони оби- тают на высокогорьях, что способствует их обособлению. Хубей-
Некоторые сведения о филогении рода Malus 77 ские яблони отличаются от других ягодных более продолгова- тыми, довольно узкими, изредка едва заметно лопастными, пурпуровыми при распускании листьями с клиновидным или не- равносторонним основанием, красно-пурпуровыми черешками и главной жилкой листовой пластинки. Однако их родственная связь подтверждается целым рядом общих признаков. Это прежде всего строение цветков, а также форма и размеры плодов. А. М. Скибинская [1966] считает, что ягодные яблони явля- ются результатом эволюции рябиновидных. г~ействительно, некоторые общие признаки рябиновидных и ягодных яблонь на- водят на такую мысль. Однако, по нашему убеждению, некото- рое сближение видов Gymnomeles и Sorbomalus может рас- сматриваться в свете закона гомологических рядов Н. И. Вави- лова, а не на основе родства. Если это не так, тогда как объяснить произрастание в Гималаях М. himalaica (с первич- ными признаками ягодных яблонь), в известной степени связы- вающей М. baccata с М. hupehensis? Ведь в Гималаях рябино- видных яблонь никогда не было. муроме того, у ягодных яблонь листья в почкосложении всегда свернутые, что является характерным признаком яблонь гималайского очага. Свернутые в почкосложении листья имеют и крупноплодные настоящие яб- лони из секции 1Halus, которые происходят также из Гималаев. Восточно-азиатские рябиновидные яблони, от которых могли эволюционировать ягодные, имеют в почкосложении не сверну- тые, а сложенные листья. Последнее характерно для североаме- риканских видов, что подтверждает их связь с восточно-азиат- скими яблонями. К, концу третичного периода на огромных пространствах горных поднятий Гималаев, на Памире и в западной части Тянь-Шаня в результате дифференциации третичных (типа до- циниевидных) яблонь появилось новое направление в эволюции рода Маарив. Это крупноплодные яблони, из которых образова- лась самая крупная и полиморфная группа яблонь — секция настоящих яблонь — Маарив (по старой таксономии — Eumalus 2 abel) . Центром возникновения настоящих яблонь мы считаем За- падные и Северо-Западные Гималаи. Здесь в результате дифференциации от общих предков различных популяций тре- тичных яблонь, видимо еще в третичном периоде, образовались две ветви дальнейшей эволюции рода Маарив. Яблони одной из них распространились на восток и северо-восток и эволюциони- ровали по главному признаку — мелкоплодности, т. е. посте- пенно приспособились к распространению семян при помощи птиц — орнитохории. Это секция Gymnomeles. Яблони другой— в основном распространились на запад и северо-запад, а также на восток и юго-восток. Эта линия сформировалась по признаку крупноплодности с распространением семян посредством назем-
78 Глава 3. Структуриые особеииости, эволюция и филогеиия ных животных — териохорией. Так появились крупноплодные, настоящие яблони (секция Malus) Происхождение ягодных и крупноплодных яблонь от общих предков подтверждается наличием у обеих групп цельных (по. происхождению общих) свернутых в почкосложении листьев, а также способностью скрещиваться между собою, в результате чего образовались многочисленные гибриды (группа гибридов РгипчоИае = Malus )< Ggmnomeles) . Видовой состав настоящих яблонь весьма запутан, так как секция Malus представлена, по-видимому, весьма развитыми ви- дами, находящимися во многих случаях на начальных этапах дифференциации. Этим объясняется полиморфизм настоящих яблонь. В секции Malus сосредоточены наиболее ценные в прак- тическом отношении виды яблонь с крупными плодами, от кото- рых в результате длительной селекционной деятельности чело- века выведено множество культурных сортов яблони. Наиболее примитивные настоящие яблони еще встречаются в Средней Азии, Китае и на Кавказе. Все изученные яблони из Гималаев и соседних территорий, в том числе Передней Азии, несут на себе печать ксероморфизма. Это объясняется засушли- BbIM климатом данных территорий, вследствие чего потомки первичных яблонь мезофильного облика трансформировались и приобрели новые приспособительные признаки. Однако не ис- ключается вероятность прежнего нахождения в Западных Ги- малаях первичных форм настоящих яблонь, от которых еще в третичном и особенно в четвертичном периоде стала возможной эволюция этих яблонь в иных условиях. Из ныне известных настоящих яблонь наиболее древней и близкой к исходному третичному предку следует считать яб- лоню киргизов (M. kirg hisorum) . Это подтверждается ее мезо- фильностью (приуроченность к северным склонам гор и днищам речных долин) и совместным распространением с остатками реликтовых ореховых лесов. Большое внутривидовое разнообра- зие яблони киргизов свидетельствует о широких процессах ее дифференциации и позволяет считать, что в далеком прошлом данная яблоня имела более широкое распространение. Резкое повышение сухости климата способствовало образо- ванию ксерофильных форм яблони. Один из наиболее древних центров ксерофильной флоры располагается в Передней Азии Здесь мезофильные яблони оказались замененными ксерофиль- ными формами. Так, в Иране, Афганистане и Пакистане под влиянием крайне засушливого климата сформировались наибо- лее характерные формы ксерофильной яблони в результате про- цессов девиации (отклонение от прежнего хода развития) и терминальной аббревиации (сокращение продолжительности развития за счет выпадения поздних стадий) . Таким образом появились неотенические, засухоустойчивые, скороплодные и ско-
Некоторые сведения о филогении рода Ма!из 79 роспелые яблони низкого роста (3 — 5 м), крайне ксероморфного облика. Типичными представителями являются subsp. turkmeno- rum на Западном Копетдаге, имеющий связь с Кавказом, а также М. orientalis чаг. montana на Кавказе и новоописанные И. Т. Васильченко М. chitralensis — для Пакистана (пров. Читрал) и М. nuristanica — для Афганистана (пров. Нури- стан). Процесс ксероморфогенеза проходил и в других районах ареала настоящих яблонь. В Средней Азии рядом с мезофиль- ной яблоней киргизов встречается другая, более ксероморфная яблоня Сиверса (М. sieversii), произрастающая в кленовых ле- сах или кустарниковых зарослях, преимущественно на сухих солнечных склонах, на высотах до 2300 — 2500 м н. у. м. Зако- номерности распространения, а также ряд морфологических признаков и биологических свойств яблони Сиверса не позво- ляют сомневаться в близком родстве ее с яблоней киргизов. Яблоня гиссарская, обитающая в районе Гиссарского хребта в Средней Азии, явно примыкает с одной стороны к яблоне киргизов, с другой — к яблоне Сиверса. Вместе они являются полиморфными видами с общим происхождением. Относительно древний вид настоящих яблонь встречается на Кавказе. Это кавказская яблоня (М. orientalis), которая еще сохраняет третичные черты. Кавказские яблони, по-видимому, эволюционировали от общих с яблоней киргизов предков, не- когда имевших общий ареал. В настоящее время кавказская яблоня занимает большой ареал и встречается в горных широ- колиственных лесах Кавказа, откуда заходит в Малую Азию и произрастает в североанатолийских лесах Турции (на высоте 1000 — 1400 м н, у. м.) и в ее восточных районах (на высоте до 2000 м н. у. м.). Следует, однако, отметить, что действие ксероморфогенеза сказалось и на М. orientalis, особенно в южных частях ее ареала и на предельных высотах (1600 — 2000 м н. у. м.). Здесь встречаются небольшие ксероморфные популяции кавказской яблони, которая по своему облику напоминает М. turkmenorum. Мы ее называем М. orientalis var. montana. По существу она является результатом сложных явлений в образовании неотени- ческих форм, вызванных процессом ксероморфогенеза. С этим мы связываем существующее разнообразие яблонь в Малой и Передней Азии. Следует еще подчеркнуть родственную связь М. orientalis и М. orientalis чаг. montana c subsp. turkmenorum. В четвертичном периоде после отступления ледника из части северных ветвей ареала М. orientalis сформировался наиболее холодостойкий вид настоящих яблонь — М. sylvestris. Первич- ный центр этого вида располагался в Восточной Европе (бас- сейны Дона и Днепра). С отступлением ледника лесная яблоня распространилась на север и достигла пределов ареала рода
80 Глава 3. Структурные особенности,.эволюция и филогения Malus (Карельский перешеек, Скандинавия). В дальнейшем лесная яблоня образовала более широкий ареал, который охва- тил всю Восточную, Среднюю и Западную Европу (от Урала до Атлантического океана). С повышением сухости климата и образованием степного пояса в южных частях ареала М. sglvestris проходили процессы ее ксерофитизации. Особенно сильно они проявились в южной части Восточной Европы, лесостепных районах бассейнов Волги, Дона и Днепра и на Балканах. Образовалась ксерофильная раса лесной яблони — более низкого роста, скороплодная и скороспелая — яблоня ранняя (М. praecox). С Балкан ранняя яблоня имела возможность проникнуть в Малую Азию — в Тур- цию, а с Дона — на Северный Кавказ. Это привело к смешению яблонь ранней и кавказской, в результате чего на данных территориях, особенно в Крыму, появилась гибридная смесь с неопределенными признаками. Таким образом, следует еще обратить внимание на совре- менное разнообразие и распространение этих яблонь. По-види- мому, раньше настоящие яблони (секция Malus) имели общий обширный ареал. В результате описанных выше причин со временем произошла дизъюнкция общего ареала и обособление отдельных его частей, из которых к настоящему времени обра- зовались более или менее изолированные участки ареала секции Malus: китайский — с видом М. asiatica; гималайский — с М. caspiriensis, М. chitralensis и М. nuristanica; среднеазиат- ский — с М. sieversii. с разновидностями hissarica и kirghiso- гит; кавказский — с М. orientalis и его subsp. montana и turkmenorum и европейский — М. sglvestris и praecox. На Балканах (Древняя Греция) около 3000 лет назад распо- лагался крупный центр окультуривания яблонь. Здесь в резуль- тате деятельности человека были сосредоточены лучшие образцы крупноплодных яблонь, откуда они распространились в Древний Рим и далее — в Восточную и Западную Европу. Благодаря склонности к одичанию в районах широкого культивирования появились одичавшие яблони. Среди одичавших яблонь встречаются как мезофильные, так и ксерофильные популяции. Из ксерофильных популяций ан- глийский садовод Ф. Миллер [Miller, 1768] описал вид М. pu- mila, а мезофильные были сведены немецким ботаником М. Боркгаузеном '[Borkhausen, 1803] в вид М. dasgphglla. М. ритйа и М. dasgphglla — не определенные виды, а лишь популяции одичавших яблонь с весьма неясным происхожде- нием. Так как диагноз М. pumila содержит основные элементы, характерные для ксероморфных, низких яблонь, то не случайно эпитет М. pumila из-за недостаточной изученности других низ- ких яблонь получил неоправданно широкое применение. Осо- бенно часто им пользуются садоводы [Скибинская, 1966], не-
Некоторые сведения о филогении рода Malus 8! редко называя все низкорослые яблони Европы, Кавказа, Пе- редней и Средней Азии и даже Китая М. pumila. То же можно отнести и к М. dasyphylla. В ходе длительной интродукционной деятельности человека в садах выращивались различные виды яблонь. Это способст- вовало появлению некоторых межвидовых и даже межселекци- онных гибридов. Благодаря общему происхождению сравни- тельно легко скрещиваются крупноплодные — настоящие яб- лони (секция Malus) и ягодные яблони (секция Gymnomeles). В результате подобных скрещиваний образовался ряд гибрид- ных видов, получивших название Prunifoliae, с наиболее харак- терной для этой группы сливолистной яблоней, или китайкой (М. prunifolia) . Сливолистные и подобные ей яблони до сего времени явля- ются предметом дискуссий. Некоторые авторы, в частности А, M. Скибинская и А. Н. Ярагавцев, ссылаясь на китайского ботаника Чоу [Chow, 1934], считают сливолистную яблоню ди-- кой. Однако в диком состоянии эта яблоня не обнаружена.
Глава 4 ГЕОГРАФИЯИ ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИЯБЛОНИ РОДОВЫЕ АРЕАЛЫMALUS, PYRUS, SORBUS И CRATAEGUS Род Malus распространен преимущественно в лесных облас- тях умеренной зоны северного полушария (рис. 20). Виды, захо- дящие в субтропические и изредка в тропические широты, встречаются в основном в горах, где приурочены к умеренному или субтропическому типам климата Юго-Восточной Азии. По совокупности своих признаков они являются наименее продви- нутыми в эволюционном смысле. Общий родовой ареал Malus протянулся довольно широкой, но неравномерной полосой в пределах Евразии, от берегов Ат- лантики (Ирландия, Англия и Северная Испания) до Тихого океана (о-ва Хонсю и Хоккайдо Японского архипелага) и далее через Северную Америку, располагаясь к северу и югу от 45-й параллели. По обе стороны от этой линии область распростра- нения яблони то сужается, то расширяется в зависимости от климатических условий и особенностей земной поверхности. Наибольшего расширения ареал достигает к востоку от Гринвичского меридиана и охватывает страны Западной и Вос- точной Европы. Здесь его северная граница проходит от север- ной части Британских островов в Скандинавию и достигает 64' с. ш. (в Финляндии). Далее северная граница ареала про- ходит по линии Карельский перешеек — Вологда — Киров — Пермь, резко опускается от Перми к югу и проходит приблизительно по линии Уфа — Уральск — Астрахань, огибая с запада и юга Каспийское море (прикаспийская часть Ирана, горы Эльбруса). Далее граница ареала проходит по Туркмении (бассейн р. Ат- рек) и продвигается до 37' с. ш. Отсюда она идет по краю Копетдага, северо-восточной части Ирана и северной части Аф- ганистана. После небольшого перерыва в горах Памиро-Алая и Тянь-Шаня следует новый подъем границы к северу (до 55' с. ш.), т. е. примерно по линии Алма-Ата — Красноярск, от- куда она тянется на восток к Сахалину, Алеутским о-вам и на Аляску и тихоокеанское побережье Северной Америки (почти 60' с. ш.). На американском континенте граница распростране- ния яблонь резко поворачивает на юг (до 50' с. ш.), проходит
Родовые ареалы Malus, Ругиз, Sorbus и Crataegus 83 Рис. 20. Общий ареал рода Malus по линии Ванкувер — Виннипег — Мусони — Монреаль и достигает восточного побережья Северной Америки. Южная граница родового ареала Malus начинается от бе- регов Бискайского залива (в Северной Испании, приблизительно 43' с. ш.), проходит через Южную Францию (Марсель), Италию и, опускаясь до 42' с. ш., идет к Албании и Северной Греции (до 40' с. ш.). Огибая с юга Турцию и Иран, она опускается на юг до 35' с. ш. Далее тянется до Гималаев (Индия, пров. Каш- мир), идет на юг и, пройдя через индийский штат Сикким и китайскую пров. Юньнань, достигает Северного Лаоса, Вьет- нама, а также китайских пров. Гуаньдун, Фуцзянь и о-ва Тай- вань. Отсюда граница вновь уходит на север, захватывая Япо- нию. В пределах Северной Америки южная граница ареала Malus пересекает Калифорнию (у тихоокеанского побережья) и идет на восток по линии Даллас — Атланта, т. е. примерно по 32 — 33' с. ш. Сравнивая общий родовой ареал Malus с ареалами таких близких родов, как Pyrus (рис. 21), Зогоия (рис. 22) и Cratae- gus (рис. 23), можно видеть, что ареал Pyrus по своему геогра- фическому положению приурочен к умеренным широтам Евра- зии и совпадает в основном с горными лесами этого материка, где простирается в пределах Китая, Гималаев, Кавказа и средиземноморских стран. Наиболее продвинутые на север крылья родового ареала Pyrus располагаются в Западной и Восточной Европе (достигая 55' с. ш.) и на Дальнем Востоке (бассейн Амура, до 50' с. ш.). Южные пределы в Юго-Восточ- ной Азии (Китай) опускаются к югу до тропика Рака и даже южнее. Род Pyrus автохтонно совершенно отсутствует в преде- лах Америки.
$'ëàâà 4. ~'еография и экологические особенности яблони Рис. 21. Общий ареал рода Ругиь L. Рис. 22. Общий ареал рода Sorbus L. Рис. 28. Общий ареал рода Crataegus L.
Родовые ареалы Malus, Pyrus, Sorbus и Cretaegus Род Crataegus обитает как в Евразии, так и в Северной Америке. Его ареал имеет вид более или менее широкой полосы, северные пределы которой располагаются в Восточной Сибири (до 64' с. ш.), где они совпадают с хребтом Сунтар-Хаята и южными окраинами Верхоянского хребта. В Европе (Южная Скандинавия) северная граница ареала Crataegus достигает 62' с. ш., а в Северной Америке (на тихоокеанском побере- жье) — 53' с. ш. Южная граница ареала Crataegus располага- ется к северу и югу от 30-й параллели и лишь в Северной Америке (Калифорния) едва переходит линию тропической зоны. В Юго-Восточной Азии, у Восточно-Китайского моря и в Восточных Гималаях (долина Брахмапутры и прилегающие равнины) область распространения Crataegus едва переходит 30-ю параллель. Родовой ареал Sorbus расположен севернее, чем ареал рода Crataegus, и занимает как Евразию, так и Северную Америку. .На азиатском материке северная граница ареала переходит ли- нию Северного полярного круга, а в Европе, обогнув Сканди- навский п-ов, уходит к Северной Америке, также достигая Северного полярного круга. Южная граница ареала совпадает примерно с 40-й параллелью. Больше всего она продвигается к югу в Северной Африке, где проходит по горам Атласа. Затем она подходит к северным берегам Средиземного моря, тянется по территории Кавказа и северной части Малой Азии (Турция, Иран) и по горам Эльбруса достигает Копетдага. Здесь она прерывается и снова возникает в горах Памиро-Алая и Тянь- Шаня. После небольшого сужения ареала (пустыня Гоби) его южные пределы снова опускаются к югу, включая Корею. Сравнение родовых ареалов Ругия, Malus, Crataegus и Sor- bus позволяет усмотреть некоторые общие закономерности в географии этих близких между собой адов. В отличие от аре- ала рода Pterus общий ареал рода alus занимает более об- ширную территорию и простирается гораздо севернее. Род Pterus, как было сказано ранее, распространен исключительно в Старом Свете, тогда как Malus обитает и на материке Нового Света. Родственные Malus роды Crataegus и Sorbus имеют еще более северные и более широкие ареалы. Самые южные пределы ареалов Pterus, Malus, Crataegus и .Sorbus располагаются преимущественно в Юго-Восточной Азии, где достигают (Crataegus, Sorbus) или переходят (Pterus, Ma- lus) линию тропика Рака. Любопытен факт наличия в этих границах примитивных представителей родов и их реликтовых видов, которые довольно хорошо представлены в родах Pterus и Malus. Древние участки ареалов указанных родов находятся на об- ширной территории Восточной Азии (например, в бассейне Амура), где отсутствовало оледенение. Отсюда, вероятно, их
Глава 4. География и экологические особенности яблони предки распространились на север и заняли площади, освобо- дившиеся из-под ледникового покрова (Северо-Восточная Азия). Постепенное расширение северных границ ареалов Pyrus, Malus, Crataegus и Sorbus к северу Западной и Восточной Ев- ропы (особенно к Скандинавии) свидетельствует о том, что здесь сформировались виды более молодые, так как, например, в Скандинавии за время двукратного оледенения почти вся флора была уничтожена и восстановилась в послеледниковый период. Поэтому таксоны, обитающие ныне на данном участке ареала, представлены видами, возникшими сравнительно не- давно. Отсутствие на Скандинавском п-ове Pyrus (автохтонно) свидетельствует об относительной древности основного ядра этого рода. Любопытно также, что характер ареалов сравни- ваемых родов, распространенных в Передней Азии, соответст- вует ксероморфной флоре. Род Sorbus здесь почти отсутствует, Crataegus представлен несколько шире, но Malus и особенно Pyrus образуют ряд видов, несущих следы явного ксероморфо- генеза. Однако ксероморфные виды Pyrus и Malus, встречаю- щиеся в Передней Азии и других южных пределах своих ареа- лов, имеют древние связи, но не могут считаться примитивными в эволюционном смысле. Наличие тесных связей географического распространения родов Pyrus, Malus, Crataegus и Sorbus позволяет предполагать, что здесь когда-то существовали их общие предки, занимавшие более обширную территорию Евразии. Из них постепенно диф- ференцировались современные виды. РАСПРОСТРАНЕНИЕ РОДА MALUS Наиболее старые по происхождению яблони, ныне представ- ленные реликтовыми видами, приурочены к древнейшим участ- кам суши. Это прежде всего представители секций Erlolobus и Docynl opsЬ. Единственный вид секции Erl olobus — М. trl lobata имеет два довольно далеко отстоящих друг от друга участка ареала (рис. 24). Первый из них расположен на скалистых склонах среднегорной полосы Ливанскх гор на высоте от 1000 до 1800 м н. у. м., другой — на высоте от 800 до 1200 м в горных лесах Балканского п-ова, преимущественно в северной части Греции и местами — в Болгарии. Ареал доциниевидных яблонь (секция Docynlopsls) пред- ставлен четырьмя изолированными участками. Они занимают довольно обширную территорию от Восточных Гималаев до се- верной части современных Лаоса и Вьетнама, а также пров. Юньнань и южную часть пров. Фуцзянь в Китае, Тай~а~~ и о-ва Кюсю, Хонсю и Хоккайдо. В состав данной секции входят четыре
Распространение рода hfalus Рис. 24. Ареал М. trilobata (секция Eriolobus) вида: М. laosensis, М. fortnosana, М. tschonoskii и М. sikkimensis (рис. 25). М. laosensis распространена в горных субтропических лесах северной части современного Лаоса и Вьетнама и на юге Китая [Chevalier, 1920]. Ее ареал лежит в пределах отрогов Юнь- наньского нагорья (на высоте 1200 — 2000 м н. у. м.), которые на западе постепенно повышаются и граничат с высочайшими снежными хребтами Бирмы. На севере распространение этой яблони ограничено водораздельными горными цепями верховий рек Меконг, Черная, Юаньцзян (Красная), достигающими вы- сот 5500 — 6000 м н. у. м. Бассейны названных рек составляют основную часть ареала М, laosensis. Климат здесь субтропичес- кий, с явными признаками тропического. Средняя температура самого холодного месяца (январь) колеблется в пределах от 5 до 8'С, а самого теплого (июль) — достигает 22 — 30'С. Осадки выпадают в количестве свыше 1000 мм в год, причем около 70 — 80% их приходится на летний сезон. М. formosana — другой вид секции Docyniopsis, распростра- ненный в горных влажных субтропических лесах на высоте от 1600 до 2600 м н. у. м. и растущий в южной части пров. Фуц-
88 Глава 4. География и экологические особенности яблони Рис. 25. Ареал секции Dogyniopsis:/ — М. tschonoskii; 2 — f ormosana; 3 — М. laosensis; 4 — М. sikkimenst s зянь, а также на Тайване. Границы ареала М. formosana явля- ются наиболее южными пределами распространения рода Malus и заходят во влажно-тропическую зону. Средняя годовая темпе- ратура колеблется здесь от 20 до 24'С, в июле она достигает 28 — 30'С, а в январе — 15 — 20'С. Температурный максимум здесь составляет 38 — 39'С, абсолютный минимум обычно не ниже 6 — 9'С, лишь изредка, когда сюда проникают холодные воздушные массы из Сибири, наблюдаются резкие понижения температуры, приближающиеся к 0'С [Павлов, 1965]. Количе- ство осадков — 1800 мм в год (иногда больше), 90о/о которых приходится на летний сезон. М. tschonoskii — третий вид секции доциниевидных яблонь, распространен в Японии в составе широколиственных предгор- ных лесов, произрастающих на высоте от 400 до 500 м н. у. м. Здесь эта яблоня произрастает вместе с дубами (Quercus gros- seserrata, Q. glandulifera, Q. dentata), а выше 1600 — 1800 м- в области буковых, грабовых и кленовых лесов. Климат здесь мягкий, влажный муссонный, обусловленный теплым морским течением. Однако в умеренной зоне (о-в Хок-
Распространение рода Malus кайдо и северная часть о-ва Хонсю) наблюдаются значительные колебания температуры. Так, в наиболее холодные месяцы (ян- варь и февраль) на севере о-ва Хонсю средняя температура колеблется от — 12 до — 16'С (абсолютный минимум — 24'С). Зима длится 5 — 6 мес. Летняя температура — около 30'С [Павлов, 1965]. В пределах ареала М. tschonoskii, по данным Н. В. Павлова [1965], выпадает в среднем 1000 — 1500 мм осад- ков в год, а иногда и больше (на о-ве Кюсю — 2000 — 2500 мм). Сильное влияние на климат и распределение осадков на Япон- ских о-вах оказывают муссоны. Под воздействием летнего влаж- ного муссона на о-ве Кюсю, а также восточных и южных бере- гах других островов за четыре летних месяца (с июня по сен- тябрь) выпадает осадков гораздо больше, чем за остальное время года. На о-вах Хонсю и Хоккайдо, благодаря защитному действию горных хребтов, осадков летом выпадает мало. Зато западные части о-вов Хоккайдо и Хонсю наибольшее количе- ство осадков получают зимой в виде снега. Благодаря зимним сухим муссонам, дующим с северо-запада Азии и проходящим над Японским морем, воздух насыщается влагой и приносит сюда обильные осадки. М. sikkimensis обитает в Восточных Гималаях и горных ле- сах Сиккима на высоте 2000 — 3000 м н. у. м., откуда заходит в прилегающие районы Китая. Эта территория характеризуется умеренным довольно континентальным климатом. Средняя тем- пература самого теплого месяца (июнь) достигает 17'С (мак- симальная — 28 — 33'С, а минимальная составляет — 14'С). Безморозный период равен 140 — 150 дням. Среднее количество осадков — около 1600 мм в год (и больше), максимум прихо- дится н а лето [П авлов, 1965] . Значительную часть ареала рода Afalus занимают виды сек- ции Sorbomalus, распространенные на обширной территории Е~ентральной и Восточной Азии, а также вдоль тихоокеанского побережья Северной Америки (М. fusca) Центральная Азия — замкнутое плоскогорье — не имеет стока рек в море. Восточная Азия по своему рельефу пред- ставляет собой пологую равнину, постепенно опускающуюся к Тихому океану. Здесь протекают мощные реки, берущие начало в горных массивах на высоте от 3500 до 5000 м н. у. м. На пути к океану эти реки образовали глубокие русла и долины. Здесь так же как в горах экологические условия весьма разно- образны. Из числа видов секции Sorbomalus наиболее близкими к доциниевидным яблоням в Е~ентральной Азии являются М. yun- nanensis, М. ombrophila, М. prattii и М. honanensis (серия Yunnanenses) (рис. 26). Они не произрастают на почвах низ- менных территорий и обитают преимущественно в горах. Их ареалы приурочены к Юго-Западному, Центральному и час-
Глава 4. География и экологические особенности яблони 90 Рис. 2б. Ареал серии Yunnanenses (секция Яогбота1из1:1 — М. уиппапепзи; 2 — М. prattii; 3 — М. ombrophila; 4 — М. honanensis тично Восточному 1~итаю (пров. Сычуань, Ганьсу, Шэньси, Шаньси, Хэнань, Хубэй, Гуйчжоу) . Л1. yunnanensis распространена в 1~итае (пров. Юньнань, Гуйчжоу, Хубэй и восточная часть пров, Сычуань), где встреча- ется в горных лесах на высоте от 1600 до 2500 — 2700 м н. у. м. М. ombrophila также растет в пров. Юньнань и Сычуань на высоте 2250 — 2650 м н. у. м. и вместе с М. honanensis образует северные пределы ареала этой группы видов. М. honanensis занимает обширные пространства в пределах пров. Сычуань, Шэньси, Шаньси, Хэнань, Хубэй и южной части пров. Ганьсу на высоте от 1500 — 1700 до 2200 м н. у. м. М. prattii распростра- нена в пров. Сычуань и произрастает на высоте от 2000 — 3000 до 3200 м н. у. м. Основными факторами, определяющими экологические усло- вия в пределах ареалов рассматриваемых видов, являются сравнительно большое количество осадков (более 1000 мм в год, из которых до 65 приходится на летний сезон) [Павлов, 1965] и своеобразный температурный режим Центрального 1<итая (пров. Сычуань, Юньнань, Гуйчжоу и западная часть пров. Ху-
Распространение рода MaIus 91 бэй), который характеризуется следующими средними темпера- турами: года — около 19'С, самого холодного месяца (ян- варь) — около 8'С (минимумы ниже нуля бывают редко) и самого теплого месяца (июль) — 29 — 30'С. В следующую обширную группу видов секции Sorbomalus входят М. kansuensis, М. toringoides, М. transitoria, М. setok, М. centralasiatica и M. komarovii (серия E(ansuenses) (рис. 27). Ареал видов серии E(ansuenses в основном расположен в центральных и восточных провинциях Китая. В отличие от пре- дыдущей группы (серия Yunnanenses) ареалы видов Капвиеп- ses располагаются восточнее, а некоторые виды (М. komarovii) обитают исключительно в Восточной Азии (Корея) . В серию E(ansuenses входят полиморфные виды яблонь — обитатели вы- сокогорных лесов, достигающие крайних высотных пределов рас- пространения рода Malus — от 3000 — 3600 до 4000 м н. у. м. Вместе с тем эти яблони являются самыми устойчивыми к не- благоприятным условиям среды. Генетическая связь Kansuenses c Yunnanenses хорошо про- слеживается на М. kansuensis. Этот вид по некоторым призна- кам (частично опадающая чашечка, наличие каменистых клеток в плодах и др.) тесно примыкает к Yunnanenses, что вместе с хориологией (наложение ареалов друг на друга) подтверждает условность этих серий Редера. Основной областью распространения М. kansuensis stagsrexcsr Центральный Китай (пров. Ганьсу, Шэньси, Шаньси, Хубэй и восточная часть пров. Сычуань) . Яблоня ганьсуйская обитает в горных лесах на высоте от 2000 — 2200 до 3600 м н. у. м. М. toringoides имеет более западный и узкий ареал в пров. Ганьсу и Сычуань, вплоть до западной их части, ограниченной высокими горами. Основное местообитание ганьсуйской яб- лони — горные леса (в поясе гор до 3000 — 3600 м н. у. м.). Ареал М. transitoria приурочен к центральной части Китая (пров. Шэньси, Ганьсу и Цинхай). Из северной части пров. Ганьсу эта яблоня заходит во внутреннюю Монголию, о чем свидетельствует просмотренный нами экземпляр из гербария (цит. запись на этикетке: «Внутр. Монголия, Баян-Хото, в 30 км к востоку от города монастырей Байсы, хр. Алашань, елово-соси. лес, 2500 м. 6. VII 1957. М. П. Петров» — Le). Другой гербарный экземпляр, полученный нами из Вены, ука- зывает на нахождение М. transitoria в более южных районах Китая (пров. Юньнань) и, следовательно, на разрывы ареала этого вида (цит. запись на этикетке: «China, Prov. Yunnan: Prope vicum Sanyingpan ad septentr. urbis Yunnanfu, 26' lat., in regione calide temperata in declivibus siccis, substr. arenaceo, alt. 2400 т, 14 III 1914, n' 620, Handel-Mazzetti» — W). Из-за недостатка гербарного материала из Китая нам не удалось оп- ределить точные границы ареала М. transitoria в Китае.
92 Глава 4. География и экологические особенности яблони Рис. 27. Ареал серии Капвиепвев (секция Sorbomalus): 1 — М. kansuensi;s: 2 — М. 1огядоМез: g — Я. transitoria. 4 — М. seto'; 5 М. centralasiatica; б — Щ. komarovii М. transitoria обитает в горных лиственных лесах, по доли- нам рек и вокруг озер, например у оз. Кукунор (пров. UHHxBH) и доходит до 4000 м н. у. м., т. е. до высот, на которых не растет ни один из других видов яблони [Васильченко, 1963] В пределах ареала М. transitoria встречаются близкие к ней яблони: М. centralasiatica, распространенная в низких горах, и М. setok, вероятно, эндемичный вид, отличающийся от М. tran- sitoria более крупными размерами и обитающий в высокогор- ных долинах р. Кама (в Китае) на высоте 3000 — 3600 м н. у. м. М. komarovii значительно обособлена от других представи- телей данной группы как географически, так и морфологически. Эта яблоня распространена в лесах Кореи, откуда, возможно, заходит в прилегающие районы Китая [Васильченко, 1963]. Обитает в долинах рек, в лесах и горах Корейского хребта на высоте от 1000 до 2000 м н. у. м. «Растет на небольших про- талинах, на пологих склонах гор и на каменистой почве, как правило, одиночно или небольшими группами» [Комаров, 1950, с. 474].
Распространение рода Malus Климатические условия южной части ареала Kansuenses почти совпадают с таковыми ареала Yunnanenses. Однако в северной части ареала Капяиепяея господствует континенталь- ный засушливый климат с холодной и продолжительной зимой и жарким летом. Средняя температура самого холодного месяца колеблется от — 15 до — 19'С, а самого теплого — составляет 25'С (в приморских районах даже 28'С). Осадки здесь выпа- дают в количестве до 500 — 700 мм в год, причем до 70% их приходится на летний сезон [Павлов, 1965]. 1~онтинентально- стью и относительной сухостью климата обусловлен ксероморф- ный облик ганьсуйских яблонь, особенно М. transitoria. Небольшой участок ареала секции Sorbomalus занимает М. toringo, которая обитает на островах Японского архипелага, откуда заходит в Kopero и прилегающие провинции 1~итая. Здесь замыкаются восточные границы ареала Sorbomalus (рис. 28) . Самые северные пределы ареала секции Sorbomalus распо- лагаются в западной части Северной Америки. Здесь произрас- тает М. fusca на сравнительно узкой и растянутой полосе вдоль тихоокеанского побережья от Алеутских о-вов и Аляски до ка- лифорнии (рис. 29). Северная граница М. fusca достигает 60' с. ш., а южная — пересекает калифорнию примерно по 32' с. ш. Наличие выдвинутого крыла ареала М. fusca на Алеутских о-вах, по-видимому, свидетельствует о прежней связи североаме- риканских яблонь с другими рябиновидными и некоторыми до- циниевидными яблонями азиатского материка, обитающими в настоящее время на островах Японского архипелага, что под- тверждается и общностью некоторых признаков М. sieboldii, М. tschonoskii и М. fusca. Ареал секции Chloromeles расположен в основном в цен- тральных и восточных штатах США, а также частично в Южной канаде, главным образом в районе озер Верхнее, Гурон, Эри и Онтарио (рис. 30). Типичный вид секции Chloromeles — М. co- ronaria — наиболее известный и характерный для Северной Америки. Он занимает наибольший по территории ареал: берега озер Верхнее, Гурон, Мичиган, Эри и Онтарио, от Онтарио через штаты Нью-Йорк и Пенсильванию, вдоль Аппалачей до Ала- бамы, Миссури и Монтаны. Будучи растением мезофильного облика, М. coronaria тяго- теет к речным долинам, берегам и впадинам озер, где растет одиночно либо небольшими группировками, реже образует сплошные заросли. Произрастает в основном на богатых, хоро- шо увлажненных почвах. Встречается также в лесах, на опуш- юх и прогалинах, однако в лесах нередко затеняется и вытес- няется другими лесными породами. Ареал М. ioensis находится в центральных штатах США. Самые крупные местообитания яблони айовской встречаются в
Глава 4. География и экологические особеммости ябломи 94 Рис. 28. Ареал серии Toringonae (секция Sorbomalus):1 — М. tori ego; 2 — М. )< sar gentii бассейнах Миссисипи и Миссури, где она представлена большим разнообразием форм. В местах соприкосновения ареалов легко скрещивается с М. coronaria, образуя многочисленные естествен- ные гибриды. Многие формы и естественные гибриды яблони айовской были описаны в качестве новых видов без достаточных на то оснований [Henning, 1947]. М. angustifolia распространена в приатлантических штатах США — от Пенсильвании до Флориды и Луизианы и далее на север до Теннесси. Встречается в долинах рек, на лесных про- галинах. Секция Gymnomeles распространена на азиатском материке и занимает два обширных участка. Первый охватывает южные и юго-восточные районы Сибири, дальний Восток (к востоку от Енисея до Сахалина включительно), Монголию, северные про- винции 1~итая, откуда заходит в некоторые юго-западные рай- оны 1~итая, достигая Западных Гималаев. Основной вид секции М. baccata — яблоня ягодная имеет обширный ареал (рис. 31). В пределах СССР встречается в южных и юго-восточных районах Сибири (Предбайкалье, За-
Распространение рода Malus Рис. 29. Ареал М. fusca (секция Sorbomalus, серия Fyscae) байкалье), на Дальнем Востоке (Приморский край, южная часть Хабаровского края, Амурская область). Е~ентром ареала M. baccata в настоящее время является Забайкалье, где, по данным И. Т. Васильченко [1963], выделяются три обособлен- ных очага яблони сибирской: 1) Нерчинская Даурия, бассейны Шилки, Ингоды и Онона, откуда эта яблоня в 1767 г. впервые была описана К. Линнеем; 2) Селенгинская Даурия, бассейн Селенги, где имеются крупнейшие заросли этой яблони, занимающие сотни гектаров, нередко вместе с gibes discantha; 3) степная зона Забайкалья — Базинский, Быркинский, Александровский (южная часть) районы. На севере этого реги- она яблоня доходит до Ангары, Кудо и Иркута, где она часто растет вместе с черемухой. Приуроченность М. baccata к северным и восточным склонам гор и средней части пояса горной лесостепи Юго-Западного За- байкалья на высоте 1000 — 1100 м н. у. м. объясняется тем, что северные склоны отличаются более благоприятными климати- ческими условиями для развития лесной растительности, в том числе яблони. По условиям к северным склонам приближаются
Глава 4. География и экологические особенности яблони Рис. 80. Ареал секции СЯоiomeles: 1 — М. согопаг~а; 2 — М. ioenst',s; 3 — М. апдияЯо1~а восточные. Западные и южные — теплее и суше. Наиболее теп- лыми и сухими являются южные склоны, где яблоня не про- израстает из-за недостатка влаги. За пределами СССР М. baccata известна в Монголии, Китае (пров. Хэйлунцзян, Гирин, Ляонин (откуда заходит в Корею), Хэбэй, Шаньдун, Шаньси, Шэньси, Ганьсу (центральная часть) ), западнее — в Тибете, Гималаях, Непале и Индии (до северо- западной ее части, почти до Инда, где замещается М. hima- laica). Сведения о произрастании М. baccata в пров. Сычуань и 1Оньнань (Китай) кажутся сомнительными, хотя экземпляры, собранные Хандель-Маццетти (n'n' 5491, 7774, 10029, все W) и М.Делавеем (n'3767), несомненно, принадлежат к этому виду. Можно допустить, что сборы сделаны с культурных образцов в садах или с деревьев в одичавшем состоянии, но это маловеро- ятно, поскольку их авторы — известные и авторитетные исследо- ватели китайской флоры. Мы не смогли окончательно решить вопрос об истинном ареале М. baccata в Китае, поскольку имели весьма небогатый материал из этой страны. Однако наличие в
Распространение рода Ма1ия Рис. 81. Ареал М. baccata (секция Gymnomeles) Китае ягодной яблони свидетельствует о том, что здесь, по- видимому, сохранились остатки прежнего ареала М. baccata. Ягодная яблоня обитает одиночно или небольшими зарос- лями в долинах рек, на островах и в горах, также в лесах, среди кустарников, на лесных опушках (где развивается плохо). Весьма неприхотливая в отношении почв, «она часто растет на песчаных островах и песчаных же берегах рек, а иногда и на галечниках, имеющих лишь небольшое количество бурого суг- линка» [Васильченко, 1963, с. 42]. Небольшой, сравнительно узкий ареал в Западных Гималаях имеет М. himalaica (рис. 32). Этот эндемичный для Гималаев вид обитает в лесной полосе гор на высоте 1500 — 3500 м н. у. м. преимущественно в районе Инда, откуда некоторые местона- хождения выдвинуты на восток и более или менее отстоят от основного ареала в прилегающих районах Юго-Западного Ки- тая. Здесь встречаются популяции, которые по совокупности признаков отличаются от типичной М. himalaica, что, по-види- мому, связано с контактом ее с М: baccata и, вероятно, взаим- ной гибридизацией этих яблонь.
Глава 4. География и экологические особенности яблони 98 Рис. 32. Ареал М. himalaica (секция Gymnomeles) В восточных пределах ареала секции Gymnomeles произ- растает М. mandshurica (рис. 33) (яблоня маньчжурская). Она растет в Восточной Азии на территории, которая не подверга- лась оледенению, и представлена крупными деревьями мезо- фильного облика, достигающими 15 (30) м в высоту. Это позволяет предполагать древнее происхождение вида. Область распространения М. mandshurica в пределах СССР простира- ется в юго-восточной части Дальнего Востока, на юге При- морья — в бассейнах рек Лянчин, Суйфун, Майхэ, Супутинка, Раковка и др., на севере — в бассейнах рек Даубихэ, Улахэ, Иман и Бикин. По данным И. Т. Васильченко [1963], она произрастает вместе с М. baccata, с которой образует естествен- ные гибриды. Яблоня маньчжурская известна и в Китае, в северной части Маньчжурии и других, преимущественно северо-восточных про- винциях, а также в Корее и Японии (о-ва Хоккайдо, Хонсю), где встречается главным образом в долинах рек и на нижних склонах гор на высоте 1000 — 3000 м н. у. м. По данным Д. П. Воробьева, М. mandshurica обитает в дуб- няках из Quercus mongolica с участием березы, осины, клена и
Распространение рода Malus Рис. 33. Ареал М. mandshurica (секция Gymnomeles) подлеском из Lespedeza bicolor и Corylus heterophylla, в дуб- няках с подлеском из лещины разнолистной и в лиственных лесах с преобладанием лип Tillis amurensis и Т. mandshurica и кленов. В долинах рек она произрастает обычно на более ста- рых речных отложениях. Яблоня маньчжурская особенно ин- тенсивно разрастается во вторичных лесных группировках, воз- никающих после рубок или пожаров [Васильченко, 1963]. На Сахалине находится ареал М. sachalinensis (рис. 34). Она раопространена по всему Сахалину до 52' с. ш., но глав- ным образом на юге, откуда заходит на некоторые южные ос- трова Курильской гряды (Итуруп). Яблоня сахалинская входит в состав широколиственных лесов Сахалина и обитает преиму- щественно в долинах рек. Природные условия в пределах ареалов М, baccata, M. mand- shurica, subsp. sachalinensis характеризуются резко континен- тальным, суровым климатом с умеренно теплым летом, морозной и мало- или почти бесснежной зимой. Здесь встречаются клима- тические условия сибирской тайги, монгольской степи, а также муссонный климат умеренных широт дальнего Востока.
Глава 4. География и экологвческие особенкости яблони Рис. 84. Ареал subsp. sachalinensis (секция Gymnomeles) Другой участок ареала секции Оутиоте1ев располагается в Центральном и Юго-Западном Китае, Бирме и Индии [Аззат], 1927 и занят видами М. hupehensis (рис. 35) и М. halliana. . ЬиреЬеиыз довольно широко распространена в централь- ных и юго-западных провинциях Китая: Хубэй, Хунань, Фуц- зянь, Шаньси, Шэньси, Сычуань, юг Ганьсу и др. По данным Ассами [Assami, 1927], эта яблоня из юго-западных провинций Китая заходит в Бирму и Индию. В более восточных провинциях Китая (Шэньси и др.) оби- тает М. halliana. По сведениям Коидзуми, var. spontanea этого вида в диком состоянии встречается в Японии. Однако' досто- верно М. halliana известна лишь в культуре как декоративное растение. Кроме того, культивируются некоторые гибридные яб- лони, по своим признакам явно примыкающие к М. hupehensis. Наиболее известные из них М.)(hartwigii, М.)(atrosanguinea и др: Типичная секция рода — настоящие яблони (Malus) крупнейшая как по разнообразию, так и по занимаемой терри- тории группа яблонь, отличающаяся значительным полиморфиз-
Распространение рода Malus I0f Рис. 85. Ареал M hupehensis (секция Gymnomeles) мом. Автохтонные виды этой секции представлены исключи- тельно в Евразии, ареал которых в настоящее время составляют пять более или менее географически обособленных и изолиро- ванных друг от друга участков: китайский, гималайский, сред- неазиатский, ирано-кавказский и европейский (рис. 36). Первый участок соответствует ареалу вида М. asiatica*, который охватывает следующие провинции Китая: Юньнань, Гуйчжоу, Хунань, Хубэй, Хэнань, Шаньдун (частично), Шаньси, Шэньси, Ганьсу, Сычуань [Yii Те-tsun, Yen Chan-lung, 1956]. Во множестве разновидностей этот вид культивируется также в Корее и Японии. Ареал М. asiatica в основном располагается в пределах Индийско-Тихоокеанской климатической области суб- тропической зоны Китая. Лишь северные его пределы в провин- циях Шаньдун, Шаньси, Шэньси и Ганьсу незначительно захо- дят в Тихоокеанскую область умеренной зоны. Юго-западное крыло ареала охватывает северную часть Юньнаньского клима- ~ В номенклатуре китайских и восточно-азиатских настоящих яблонь вообще до сих пор существует некоторое разногласие, так как отдельные авторы употребляют ничем не обоснованное (по крайней мере для китайских яблонь) название Malus pumila.
Глава 4. География и экологические особенности яблони Рис. Зб. Участки ареала секции Afalus: — китайский; 2 — гималайский; 3 — среднеазиатский. 4 — ирано-; 5 европейский тического района (Индийско-Тихоокеанская область). На тер- ритории ареала М. asiatica встречаются самые разнообразные орографические, природные, а также климатические условия, определяющие весьма пеструю среду обитания. Второй участок ареала секции Afalus расположен в Гима- лаях, преимущественно в западной части. В своей южной части он отграничен от ареала М. айаПса высокими горными хреб- тами Бирмы и тянется через Непал, вдоль хребта Кайлас (Ти- бетский автономный район Китая). В Северной Индии (штат Кашмир, верхнее течение Инда) Afalus достигает кульминации. Здесь обитают слабо изученные в систематическом отношении яблони с архаическим названием Pyrus malus, которые мы пред- лагаем именовать Afalus caspiriensis — nom. nova. (рис. 37). На гималайском участке яблоня обитает в лесном поясе гор на высоте от 1600 до 2800 — 3220 м н. у. м. Совокупность при- родных условий и климатические особенности данного участка определяются в первую очередь сложной и мощной системой самих Гималаев, спускающихся в сторону Северной Индии и Непала гигантскими ступенями. Нижняя, южная ступень имеет
Распространение рода Malus 103 Рис. З7. Ареал Л1. casptriensis среднюю высоту 700 — 900 м н. у. м., и ее расчлененный край называется Сиваликскими горами. Вторая ступень — Малые Гималаи — достигает в высоту 3500 — 4000 м, а центральная ось хребта, называемая Главными, или Большими Гималаями, сло- жена горными массивами со средней высотой 5500 — 6000 м. 1~ северу от Инда начинается другой горный хребет — 1~арако- рум. Его южные, обращенные к Индии склоны сильно изрезаны верховьями рек, в ущельях и долинах которых произрастает яблоня. Высокогорьем гималайский участок отделен от соседних— китайского и среднеазиатского. Менее отграничены западные пределы данного участка, где горы переходят в крайне засуш- ливые территории Афганистана и Ирана, в так называемой Афгано-Иранской флористической области. На этой территории встречаются некоторые слабо изученные яблони ксероморфного, иногда карликового облика, принадлежащие скорее всего к со- седнему — ирано-кавказскому участку. И. Т. Васильченко опи- сал два новых вида этих яблонь: М. chitralensis из пров. Читрал (Пакистан) и М. nuristanica из пров. Нуристан (Афганистан).
404 Глава 4. География и экологические особенности яблони 1~лиматические условия гималайского участка разнообразны. В основном это муссонный, засушливый, резко континентальный климат (особенно в пределах Западных Гималаев), со значи- тельными колебаниями суточных температур и тремя выражен- ными сезонами: сухая и прохладная зима (с ноября по февраль}, сухая и жаркая весна (с марта по май} и жаркое и относи- тельно влажное лето (с июня по октябрь). Средняя температура января в Западных Гималаях около — 3'С. Ночи в это время года морозные — до — 15 и даже — 20'С. Летом жарко, даже ночные температуры держатся на уровне 24 — 27 С. Осадков мало — до 345 мм в год (в горах больше), причем они распре- делены неравномерно. В Восточных Гималаях средняя темпе- ратура января — 0,3'С. 1~оличество осадков превышает 1000 мм в год при более равномерном распределении по сезонам (до 75 их приходится на летний сезон}. Умеренная лесная полоса и особенно северные склоны гор и ущелья, где произрастает яблоня, достаточно увлажнены и, кроме того, обеспечиваются влагой за счет таяния горных снегов. Здесь господствуют хвойные породы. В пределах Западных Гималаев на высоте от 1500 — 3200 до 3500 м н. у. м. встречается M. caspiriensis. Она обитает преиму- щественно на северных, более влажных склонах, во втором ярусе вместе с вечнозелеными дубами (Quercus semecarpifolis, Q. dilatata), листопадными породами (Juglans kamaonia, Aescu- lus indica, Platanus orientalis, Padus asiatica, Ulmus nvallichi- ana, Carpinus faginera и др.), а в ущельях — с Acer pentapon- Бсшп, Quercus ilex, Fraxinus xanthoxyloides и др. [Павлов, 1965], Прослеживая взаимосвязь китайского, гималайского и кав- казского участков, следует отметить, что на востоке Гималаев происходит некоторое смешение признаков гималайских яблонь с китайскими, в то время как в Западных Гималаях у яблонь сочетаются признаки гималайского и кавказского участков. При- чиной этого, по-видимому, послужили климатические различия Восточных и Западных Гималаев, а также другие факторы, спо- собствующие определенной общности некоторых элементов флор Западных Гималаев и 1~авказа и даже Средиземноморья и Ев- ропы. Хотя географически М. caspiriensis обособлена на Западных Гималаях, по совокупности признаков она систематически при- мыкает к ирано-кавказским яблоням. Таким образом, гималай- ский участок ареала секции Malus сочетается с кавказским, от которого в настоящее время отделен крайне засушливыми тер- ритопиями Передней Азии. На третьем, среднеазиатском участке ареала секции Malus, расположенном в пределах мощных горных систем Тянь-Шаня и Памиро-Алая, сосредоточена основная часть настоящих яб-
Распространение рода Malus 105 Рис. 88. Ареал var. kirghisorum лонь. Среди них выделяют три вида: М. sieuersii (яблоня Си- верса), яблоня киргизов и гиссарская яблоня. Эти виды гео- графически, а также по своей морфологии (в меньшей мере по экологии) слабо обособлены. древняя яблоня этого участка — мезофильная яблоня кир- гизов — хорошо представлена на Тянь-Шане (Ферганский, Чат- кальский и Таласский хребты, Присырдарьинский Каратау. Заилийский и джунгарский Алатау) (рис. 38). Она обычно встречается в составе ореховых лесов или на их опушках, а также растет совместно с осиной и кленом. По Ал. А. Федорову и Ан. А. Федорову [1949], эта яблоня обитает преимущественно на северных склонах rop, на террасах и в речных долинах, на мощных, богатых и хорошо увлажненных почвах лесного пояса, кроме ярко освещенных, сухих, каменистых склонов южной экс- позиции. Встречается на высоте от 900 — 1000 до 2000 м н. у. м. Наиболее характерным местом произрастания яблони киргизов. является полоса гор в пределах от 1400 до 1800 м н. у. м. На Ферганском хребте яблоня киргизов широко распростра- нена и образует сплошные леса. В Южной Киргизии располо- жен центр ее ареала, откуда она и описана. Здесь яблоня кирги--
106 Глава 4. География и экологические особенности яблони зов постоянно сочетается с грецким орехом (Juglans regia). На высоте 1200 — 1400 м она экологически замещает грецкий орех, отличаясь от него способностью обитать в долинах рек и на склонах северной экспозиции. Так, на высоте 900 — 1500 м она преобладает, образуя сплошные яблоневые леса. На Чаткальском хребте яблоня киргизов встречается преи- мущественно на склонах северной экспозиции в нижней части гор в пределах 600 — 700 м и от 900 до 1500 м н. у. м., а также на северо-восточных склонах и в речных долинах, где растет на глубоких суглинистых почвах с высоким содержанием гу- муса. Древостой естественных насаждений данной яблони здесь имеет неодинаковую плотность. Местами ее мало, а иногда она образует довольно обширные массивы с примесью грецкого ореха, клена, дикой вишни, алычи и других пород, Природные условия в ущельях рек Чаткал, Пскем, Ак-Булак, Угам и Кок- Су весьма благоприятны, о чем свидетельствует наличие боль- шого разнообразия форм яблони, среди которых многие отли- чаются высоким качеством плодов. Основная масса этих форм принадлежит яблоне киргизов. На более сухих, мелкоземистых склонах и склонах южной экспозиции яблоню киргизов эколо- гически замещает яблоня Сиверса. На стыке Таласского Алатау и Присырдарьинского Каратау яблоня киргизов произрастает на территории Тюлькубасского и Боролдайского лесхозов, откуда заходит на северо-западные отроги Угамского хребта. Сплошные естественные насаждения яблони киргизов располагаются на склонах Заилийского Алатау ~~Ал. А. Федоров, Ан. А. Федоров, 1949]. Яблоневые леса хорошо сохранились в долинах Малой и Большой Алма-Атинки, близ Алма-Аты, а также в районах Талгара, Иссыка и Тургеня. Здесь яблоня киргизов сочетается с осиной. Второй ярус, как и на Ферганском хребте, здесь составляют различные кустар- ники, чаще всего виды рода Crataegus, а также спиреи, шипов- ники, жимолости, барбарисы и др. На Джунгарском Алатау яблоня киргизов сосредоточена в Алма-Атинской области и входит в состав яблоневых лесов. Здесь в полосе шириной от 3 до 4 км и длиной 80 — 100 км с юго-запада на северо-восток (80'45" — 81'45' в. д. и 45'02'— 45'44' с. ш.), на высоте от 300 до 1500 м н. у. м. яблоня кир- гизов встречается на мощных черноземах пологих склонов, где она не образует чистые сомкнутые насаждения. Постоянным ее спутником на Джунгарском, так же как на Заилийском Алатау, является осина, а на более увлажненных почвах — и черемуха. На крутых каменистых склонах долин яблоня киргизов встреча- ется реже. Здесь она растет в сочетании с сибирской пихтой и елью Шренка [Васильченко, 1963]. На более солнечных и сухих склонах на Джунгарском Ала- тау преобладает М. ясегегяй (яблоня Сиверса) (рис, 39). Она
Распространение рода Malus l07 Рис. 39. Ареал М. sieversii распространена также в пределах Тянь-Шаня и Памиро-Алая. Ареал данной яблони охватывает обширные районы Тарбага- тайского и Джунгарского хребтов, Заилийского Алатау, а также склоны Киргизского, Таласского и Чаткальского Алатау, Кара- тау и Ферганский хребет до Памиро-Алая включительно. За пределами СССР M. sieversii заходит в прилегающие районы Китая, особенно в Джунгарию, районы Кульджи и Кашгара. По сравнению с яблоней киргизов M. sieversii более ксеро- морфна, в связи с чем встречается на более сухих, солнечных и менее увлажненных склонах (преимущественно южной экспо- зиции), где растет на менее мощных почвах. На северных скло- нах и в глубоких речных долинах она, как сказано выше, усту- пает место яблоне киргизов. Эта закономерность наблюдается на всем протяжении обширного ареала M. sieversii, может быть, только за исключением Памиро-Алая. На Тарбагатайском хребте яблоня Сиверса изучена крайне недостаточно, хотя именно оттуда была описана Ледебуром под названием Ругив sieversiana. На Джунгарском и Заилийском Алатау яблоня Сиверса — обычное растение. Местами естест-
108 Глава 4. География и экологические особенности яблони венные насаждения яблони Сиверса здесь настолько значи- тельны, что образуют настоящие леса, которые сосредоточены на территории Саркандского, Андреевского, Чилийского, Энбек- ши-1~азахского и, 1~аскеленского районов. Однако чистые ябло- невые леса здесь встречаются редко. Чаще всего это смешанные насаждения с преобладанием яблони Сиверса и участием Abies sibirica и Picea schrenkiana (на высоте от 1800 до 2600 м н. у. м.) или осины (Populus tremula), тополя (P. laurifolia) и кустар- ников, главным образом, Amigdalus ledebouriana, Rosa spinosi- sima, Rhamnus catharica, Spiraea hypericifolia, Lonicera latarica и других, распространенных на высоте от 1000 до 1800 м н. у. м. В пределах Таласского и Чаткальского хребтов, а также в каратау и особенно на Ферганском хребте яблоня Сиверса обитает в среднегорном поясе, на высоте от 1000 до 2300 м н. у. м., где образует смешанные леса, но без характерного для Заилийского и Джунгарского хребтов спутника яблони — осины, которую здесь заменяют клен (Acer semenovii, А. turkestanicum) и орех (Juglans regia). Из кустарников, сопровождающих яб- лоню, наиболее распространены Crataegus sougorica, С. tur- kestanica, различные виды шиповника, алыча, вишня и др. Если в горных районах Тянь-Шаня яблоня Сиверса образует более или менее сплошные яблоневые леса, то на Памиро-Алае присутствуют только их фрагменты. Наиболее крупные яблоне- вые насаждения Памиро-Алая, образованные М. sieversii, со- средоточены в районах центральной части Таджикистана, где яблоня Сиверса обитает на высоте от 900 до 2300 (2600) м н. у. м. По данным В. И. Запрягаевой [1964], яблоня Сиверса в Таджикистане лучше развивается на северных склонах гор с типичными для них хорошо выраженными коричневыми и ко- ричневыми карбонатными почвами на высоте 1400 — 1500 м. Наиболее частыми спутниками яблони здесь являются орех (Juglans regia) и клен (Acer turkestanicum), реже — Cotonesster insignis, Crataegus turkestanica, Rosa corimbifera, Padus maha- 1eb, Prunus sogdiana, Berberis heterobotrys, Exochorda alberti, Lonicera nummulariifolia и др. Наличие одиночных деревьев и групповых насаждений яб- лони Сиверса в Таджикистане среди пашен или по берегам рек и ручьев, а также в труднодоступных для человека местаХ, по мнению В. И. Запрягаевой ~[1964], является следствием порубок, раскорчевки и выжигания лесов под пашню. Заслуживают вни- мания выводы В. И. Запрягаевой ~[1964] о том, что яблоневые насаждения в Таджикистане возникли на месте ореховых и кленовых лесов в результате уничтожения в них деревьев пер- вого яруса, так как яблоня слагает второй ярус. В бассейне Кашкадарьи, особенно в верховьях ее притока- Танхаса (Юго-Западный Гиссар), обитает эндемик этого рай- она — яблоня гиссарская. Эта яблоня произрастает между аре-
i09 Распространение рода Afalus ~ В летнее время высота солнца над горизонтом в широтах Средней Азии достигает 78', а в декабре она не ниже 25'. В этих широтах годовой объем одной лишь прямой солнечной радиаиии составляет 100 — 120 ккал сМ2, т. е. по сравнению с Закавказьем — в полтора раза больше [Борисов, 1967]. алами яблонь киргизов и Сиверса и занимает небольшой по площади ареал. По данным И. Т. Васильченко [1963], наиболее типичная гиссарская яблоня встречается в долинах рек Юго-Западного Гиссара, где растет единично или небольшими группами. При продвижении на восток (бассейны Сангардака и Тупаланга) она, выходя на мелкоземистые и щебнистые склоны, встреча- ется заметно чаще, причем здесь резко возрастает разнообра- зие ее форм. Последнее, по-видимому, связано с контактом ее с М. sieversii, с которой яблоня гиссарская иногда скрещива- ется, образуя (по общей совокупности признаков) множество отклоняющихся от типичной (шахрисабзской) форм., Типичная яблоня гиссарская в. пределах Юго-Западного Гис- сара обитает преимущественно на северных склонах речных долин и достигает высоты 1200 — 2500 м н. у. м., тогда как оптимальной для ее обитания является, высота от 1700 — 1800 до 2200 м н. у..м. Чистых яблоневых лесов эта яблоня не обра- зует, но иногда растет в виде небольших рощиц, где ей сопутст- вуют клен (Acer turkestanicum), арча (Juniperus seravschanica), боярышники (Crataegus turkestanica, С. pontica), миндаль (Amigdalus bucharica) и отчасти орех (Juglans regia), алыча (Prunus sogdiana) и др. Ареалы распространения среднеазиатских яблонь лежат в умеренно теплой зоне, которая зимой находится под влиянием воздушных масс, поступающих из умеренных широт, а летом подвергается вторжению теплого воздуха с юга. В пределах рас- пространения указанных видов яблони общими чертами климата являются интенсивная инсоляция~, большие суточные и годовые перепады температур, общая сухость воздуха при относительно обильных осадках (особенно в горных частях территории), хо- лодная зима и жаркое лето. Следует подчеркнуть, что к поясу от 1200 до 2500 м н. у. м., где располагаются крупнейшие массивы упомянутых выше ви- дов яблони, приурочен максимальныйуровеньконденсацни влаги и наибольшее количество выпадающих осадков. Средняя про- должительность безморозного периода в Средней Азии состав- ляет 236 дней (Наманган), а вегетационного — 202 дня (Прже- вальск). Высокие летние температуры воздуха, интенсивная инсоля- ция, продолжительный вегетационный период, достаточная увлажненность почвы (в горах) создают благоприятные условия для обитания названных выше яблонь. Различные комбинации этих факторов благоприятствуют возникновению видового раз-
Глава 4. География g экологические особенности яблони 110 Рис. 40. Ареал М. orientatis нообразия яблонь, а также многих других плодовых. Н. И. Ва- вилов [1931б] полагал, что горные районы с их разнообраз- ными условиями являются той природной лабораторией, где интенсивно идет процесс видообразования, в котором имеет место и спонтанная гибридизация. Такие гибриды нередко воз- никают, например, в результате скрещивания яблони киргизов с типичной М. ясeversi а также М. ясeversi и яблони гиссар- ской. Существование промежуточных форм с преобладанием признаков типичной M. sieversr видимо, послужило причиной отрицания В. И. Запрягаевой [1964] существования гиссарской яблони вообще. Наоборот, промежуточные формы яблонь, не укладывающиеся полностью в диагнозы уже описанных видов, были описаны различными авторами как новые виды. Ирано-кавказский — четвертый участок секции Malus находится в горах Кавказа, откуда, распространяясь на запад вдоль берегов Черного моря, достигает горной части Крыма, а на юге, в горах, занимает преимущественно северную часть Ма- лой Азии, а также прилегающие к Закавказью леса Эльбруса (в Северном Иране). Отсюда по верховьям Горгана и Атрека охватывает западную и центральную части Копетдага.
Распространение рода Malus Рис. 41. Ареал М. orientalis чаг. montana Этот обширный участок с востока и юго-востока отграничен от соседних (среднеазиатского и гималайского) пустынями Средней Азии и крайне засушливыми горными областями Пе- редней Азии. На севере и северо-западе яблоня встречается в долинах Кубани, Егорлыка, Еи, Дона и других рек, а также в горной части Крыма. Здесь находится зона перехода от кав- казского к наиболее молодому — европейскому участку ареала секции Ма1ия. Основной вид яблони, распространенной на Кавказе, — яб- лоня восточная (М.. orientalis) (рис. 40) . Это мезофильного облика яблоня, распространенная по долинам рек, на предгор- ных равнинах и в предгорьях Кавказа, где она обитает на высоте от 300 — 500 до 1600 м н. у. м., главным образом в пре- делах Краснодарского края, в северной части Черноморского побережья, в Карачаево-Черкесской АО, Кабардино-Балкарской, Северо-Осетинской, Чечено-Ингушской и Дагестанской АССР, а также на южных склонах Главного Кавказского хребта— в Абхазии, Юго-Осетинской АО и Закавказье — Грузии, Азер- байджане и Армении.
112 Глава 4. География и экологические особенности яблони Характерной особенностью распространения восточной яб- лони является ее приуроченность к достаточно увлажненным участкам, которые типичны для речных долин, и вообще пони- женным элементам рельефа. «Если иногда она и выходит на южные склоны, то, по-видимому, лишь в условиях хорошего под- почвенного увлажнения» [Васильченко, 1963, с. 28]. На сухих склонах и склонах южной экспозиции М. orientalis уступает место своей ксероморфной расе — var. montana (яблоня гор- ная) (рис. 41), а в крайних условиях аридного климата карликовой разновидности последней. Наилучшего развития восточная яблоня достигает на наносных почвах речных долин. Она растет также и на подзолистых серых суглинках, но на карбонатных, а также закисленных и песчаных почвах М. orien- talis, как правило, произрастает редко. Чистые естественные насаждения М. orientalis образует не- часто. Обычно она растет вместе с другими широколиственными лесными породами, преимущественно в дубово-грабовых и бу- ковых лесах речных долин и по горным склонам. Если в Сред- ней Азии характерными сопутствующими породами яблони являются грецкий орех (luglans regia), клен (Acer turkestani- cum) и осина (Populus tromula), то на Кавказе яблоне чаще всего сопутствуют дуб (Quercus sp.), граб (Carpinus orientalis, С. caucasica), бук (Fagus orientalis) и груша (Pyrus caucasica). Здесь яблоня нередко затеняется, находится в угнетенном со- стоянии и даже вытесняется совершенно. В Средней Азии подобное явление наблюдается редко. В северо-западной части Кавказа, по данным И. Т. Василь- ченко [1963], например в бассейне Кубани, восточная яблоня растет на предгорных равнинах и в горах (до 1300 — 1500 м н. у. м.), однако полоса массового ее распространения лежит в пределах от 800 до 1000 м. В северной части черноморского побережья восточная яблоня, образуя чистые насаждения плот- ностью до 10 экз./га, обитает в полосе гор до 1400 м н. у. м. В Кабардино-Балкарской АССР данная яблоня распростра- няется до 500 — 1600 м н. у. м., но чаще присутствует в горных районах, на высоте 400 — 600 м. В дагестане она чаще всего встречается на Верхне-Гунибском плато и по верхнему течению р. Аварское Койсу. Здесь она доходит до 1800 м н. у. м. В Аб- хазии, на горных хребтах Герзеул, Ацангвара, Губбис и в уро- чищах Псху, Псырцха, Азанта и других расположены крупней- шие массивы яблоневых лесов. В основном они произрастают в полосе гор от 400 до 800 м н. у. м., хотя иногда поднимаются до 1800 м. В !Ого-Осетинской АО яблоня встречается реже, чем груша (Pyrus caucasica), но продвигается в горы почти до 1800 м н. у. м., выше !300 м она слабо плодоносит. В Закавказье восточная яблоня выше 1600 м не поднимается. В Восточной
Распространение рода Afalus Грузии она представлена большим разнообразием форм, разли- чающихся внешним видом, величиной, окраской и вкусом пло- дов. Некоторые яблони напоминают местные культурные сорта, от которых отличаются лишь более мелкими плодами. В Азер- байджане М. orientalis встречается в дубовых, дубова-грабовых и смешанных лесах ущелий и речных долин. Лучше она разви- вается на открытых местах (полях и лесных опушках). В засушливых условиях, а также на высоте более 1200 м н. у. м. восточную яблоню замещает ее более ксероморфная раса — var. montana, которая наиболее часто встречается в горных районах Закавказья. Этот подвид не представлен во влажно-субтропических районах Западного Закавказья (в Абха- зии и Аджарии) на побережье Черного моря. Горный подвид восточной яблони широко распространен в Шемахинском и Исмайлинском районах Азербайджана, откуда он был описан. Здесь, в горных лесах южных склонов Главного Кавказского хребта (между Баскалом, Мюдзи, Бигуром и Ис- маиллы) он растет на высоте около 1900 — 2000 м. В Грузии горный подвид восточной яблони произрастает в ущельях р. Ба- курианис-LIxaau в урочище Оманети, окрестностях Бакуриани и в других местах. В Армении между Варденским и Зангезур- ским хребтами он распространен главным образом в районе Азизбекова и на Тегамском хребте (по р. Веди и на высоте от 1200 до 1400 м н. у. м.). Кроме того, на Кавказе, преимущественно в Закавказье, встречаются своеобразные яблони, называемые местным насе- лением по-разному. Так, в Азербайджане, особенно в Нахиче- ванской АССР, встречаются яблони, носящие названия Кёл алмасы, Тем алмасы, Яр алмасы. Азербайджанские названия носят также яблони в Дагестане (близ Дербента) — Дипчек алма или Дипчек алмасы, что свидетельствует о том, что эти яблони когда-то были завезены в Дагестан из Азербайджана. В Армении подобные яблони известны под названием Марга хндзор. Они встречаются главным образом в культуре (на при- усадебных участках Алавердского, Мегрийского, Аштаракского, Октемберянского и других районов, а также в садах Еревана). В Грузии, особенно в Кахетии, встречается яблоня Хомандули. Подобные яблони известны также в Иране. Все указанные выше культурные разновидности являются карликовой формой яблони и отличаются от типичного вида скороплодностью, а также обильным плодоношением. Эти кар- ликовые яблони, вероятно, происходят от диких яблонь, произ- растающих в аридных условиях Закавказья и особенно в об- ласти ксерофильной растительности Передней и Малой Азии (в частности, в Иране). В пользу предположений о местном (кавказском) происхождении этих яблонь свидетельствуют их
114 Глаиа 4. География и экологические особенности яблони самостоятельные азербайджанские, армянские и грузинские на- звания. На востоке кавказской части ареала секции Malus, а также на территории Туркмении (западный и восточный Копетдаг), равно как и в прилегающих районах Северного Ирана, обитает своеобразная, сильно ксероморфизированная яблоня — М. turk- тепогит (яблоня туркменов). Эта яблоня распространена на сухих склонах хребтов Хосар-Даг и Сюнт, где встречается в ущельях Иол-Дере, Гюен, Аксу и близ Койна-Косыра, откуда доходит на восток до с. Сулюкли. Она произрастает вместе с широколиственными породами, из которых наиболее характер- ными являются клен (Acer turkomanicum) и боярышник (Сга- 1аедия pontica) . Так же как и на Кавказе, на Копетдаге встречаются более или менее окультуренные карликовые яблони, широко известные под названием Баба-Араб. В отличие от кавказских, они по совокупности своих признаков, несомненно, принадлежат к ко- петдагской яблоне туркменов. Климат Кавказа характеризуется значительным разнообра- зием. Здесь можно найти районы с природными чертами, при- сущими едва ли не всем климатическим зонам СССР. По Н. Н. Иванову (цит. по [Борисов, 1967]), Кавказ может быть разделен на следующие основные климатические пояса: 1) субтропический, с температурой самого холодного ме- сяца в году не выше 6'С и абсолютным минимумом температур не ниже — 5'С. Он расположен на склонах гор до высоты 300— 350 м (Абхазия, каспийское побережье Азербайджана и Ирана); 2) умеренно-теплый, с мягкой зимой и температурой самого холодного месяца в году не ниже — 3 'С и абсолютным мини- мумом — 15'С. Это обширный пояс, занимающий весь бассейн Кубани, побережье Черного моря (от Таманского п-ова до Ту- апсе) до Сурамского хребта, долину Куры. В среднегорной и приморской частях северо-восточного склона Большого Кавказа расположен основной ареал М. orientalis; 3) умеренно-холодный со средней температурой самого хо- лодного месяца ниже — 3'С; охватывает Северный Кавказ до высоты 1800 — 2000 м на склонах Большого Кавказа; 4) холодный, с температурой самого холодного месяца — 38'С, расположенный на северных склонах Большого Кав- каза (на высоте от 1800 до 2600 м) и в Дагестане (от 1900 до 3000 м), а также на южных склонах Большого Кавказа (от 1600 до 2500 м); 5) пояс альпийских лугов; 6) область вечных снегов. М. orientalis распространена в основном в пределах первых трех климатических поясов. В четвертом — холодном — поясе
Распространение рода Malus Рис. 42. Ареал М. sylvest'I'i's она почти не встречается, а в пятом и шестом — полностью отсутствует. Последний — пятый участок ареала секции Malus располо- жен в Европе. !центр этого по возрасту несомненно наиболее молодого участка приурочен к территории Восточной Европы (европейская часть СССР). Это бассейн Дона, Воронежская, 1~урская и Харьковская области. Отсюда ареал яблони протя- нулся на восток до Волги, далее — на юго-восток (южнее Волгограда), потом на север почти до Уральска, Уфы и Перми, где северная его граница совпадает с линией Пермь — 1~иров- Вологда — 1~арельский перешеек. 1~роме того, яблоня растет в Скандинавии (Финляндия, Швеция, Норвегия), где достигает 64' с. ш. От Норвегии ареал вида секции Malus распространя- ется на юго-запад до 54' с. ш. по Гринвичскому меридиану (в Великобритании), включает Францию, далее протягивается на юг до Бискайского залива, захватывает Северную Испанию (43' с. ш.), Италию, Албанию, Северную Грецию и Малую Азию, В более северных районах преобладает европейский вид- лесная яблоня (М, sglvestris) (рис, 42), Характерными его осо-
116 Глава 4. География и экологические особенности яблони бенностями являются мезофильный облик, приуроченность к ле- сам увлажненных равнин и долин, где яблоня сопутствует дубу (Quercus гоЬиг), ясеню (Ргахьпиз excelsior), клену (Acer plata- noides и А. campestre), лещине (Corylus avelana) и другим ши- роколиственным породам. Больше естественные насаждения лесной яблони располага- ются в средней полосе европейской части СССР. Чистых ябло- невых лесов здесь нет. Яблоня обычно встречается вместе с перечисленными ранее лесными породами по долинам рек Сред- нерусской возвышенности. Здесь она растет в нижних частях склонов на темно- и светло-серых лесостепных, суглинистых и перегнойно-карбонатных, реже аллювиальных почвах. В отдель- ных случаях она заходит на более солнечные и сухие склоны плакоров, поднимаясь до 230 м н. у. м. На остальной территории РСФСР лесная яблоня рассеянно распространена в рамках указанного выше ареала. Она произ- растает здесь по долинам рек и берегам озер отдельными де- ревьями или небольшими рощами. Большие рощи лесной яблони встречаются в лесах Восточных Карпат (Закарпатская, Черно- вицкая и прилегающие к ним области) . Здесь М. splvestris приурочена к хорошо увлажненным лесным группировкам, хотя она довольно устойчива к засухе, затенению и холоду. Распро- странение ее в осветленных рубкой лесах, на лесных полянах, а также на лесных опушках свидетельствует о слабой способ- ности конкурировать с другими лесными породами. К югу от Карпат, будучи приуроченной к долинам Прута и Днестра, лесная яблоня заходит в северную часть Молдавии и прилегающие районы Румынии. Также довольно равномерно она распространена в северной части Украины, вплоть до вос- точной границы республики, где произрастает в светлых широ- колиственных и смешанных лесах, особенно в Донецкой области (бассейн Донца), в Харьковской и Луганской областях. В степ- ную зону лесная яблоня не заходит или встречается здесь из- редка и только по долинам рек. В Белоруссии лесная яблоня встречается сравнительно редко, растет преимущественно по долинам рек и в местах с более вы- раженным рельефом в основном на карбонатных почвах. В низ- ких заболоченных районах яблоня обычно не произрастает. Лесная яблоня растет также в Прибалтике, где распростра- няется довольно далеко к северу. В составе местных лесов она встречается сравнительно часто в условиях достаточного увлаж- нения и удовлетворительного освещения, на суглинистых, а также карбонатных почвах. Чаще всего яблоня растет в широколиственно-еловых ле- сах — сложных ельниках (Piccetum compositum), в которых наряду с елью (Picea abies) принимают участие липа (Tilia cordata), ясень (Fraxinus excelsior), дуб (Quercus гоЬиг), вяз
Распростраиеиие рода Malus 117 (Ulmus laevis), ильм (U. scabra), береза (Belula pendula), ольха серая (Ainus incana); реже (в более увлажненных мес- тах) — ольха черная (А. glulinosa), лещина (Corylus avelana), рябина (Sorbus aucuparis), боярышник (Cralaegus monogyna) и другие породы. Такие леса в Прибалтике произрастают на богатых суглинках, в связи с чем они были сильно вырублены для сельскохозяйственного использования их плодородных почв. Интересно отметить, что при расчистке лесных участков под пашню яблони, как правило, оставались нетронутыми, поскольку, согласно местным обычаям, люди считали яблоню неприкосно- венной. Таким образом, лесная яблоня распространялась на откры- тых местах, так как она плохо развивается под пологом леса и слабо плодоносит. Наиболее крупные рощи лесной яблони в Прибалтике встречаются в центральной и восточной частях Литвы, в юго-западных, западных и центральных районах Лат- вии. Местами, в частности в пределах южной части Земгаль- ской равнины (Бауский район, Латвия), яблоня растет на бо- гатых дерново-карбонатных почвах, где образует почти чистые яблоневые рощи, занимающие иногда несколько гектаров. В Эс- тонии яблоня встречается сравнительно редко, а в Калининград- ской области произрастает преимущественно на опушках и по долинам рек в незначительных количествах. За пределами СССР лесная яблоня встречается в Централь- нои и Западной Европе, главным образом в составе широко- лиственных лесов, развитых на выраженных элементах рельефа. В Румынии и Чехословакии, а также в прилегающих к Карпатам районах, в северной части Альп (Австрия, Швейцария) яблоня распространена довольно широко. Незначительные массивы М. sylveslrts произрастают также в северной части Аппенин (Италия) и у подножья Пиренеев (D' Espagne, Вагсе1опе: Ьа- папсо dU Rabassalet, Tibidabo, 20 Vll 1923, п' 4739, F. Seh- nen, W.). В Польше, Германии и Франции лесная яблоня распростра- нена неравномерно. В Великобритании М. sylveslris отмечена лишь в южной и средней частях. В скандинавских странах лес- ная яблоня распространена сравнительно редко. Следует еще отметить, что в пределах ареала М. sylveslris в различных условиях обитания лесная яблоня значительно варьирует и представлена большим числом форм. Среди них, характерных для аридных условий, встречаются карликовые. В южных, преимущественно лесостепных и степных районах, из яблонь секции Malus встречается ксероморфная и более засу- хоустойчивая и теплолюбивая разновидность — яблоня ранняя (var. praecox) (рис. 43). П. С. Паллас, впервые описывая раннюю яблоню, кроме ог- личительных ее признаков отметил, что она встречается «у нас
Глава 4. География и экологические особенности яблони Рис. 43. Ареал var. praecox в полуденных странах государства, наипаче по Волге и Яану в великом множестве~. Яблоня ранняя в основном распространена в южных и юго- восточных районах европейской части СССР (в бассейне Яона. в Нижнем и Среднем Поволжье), откуда ее ареал идет на за- пад, охватывает степную и лесостепную полосы Украины (в том числе Крым), а через Молдавию заходит в Румынию, Болгарию, Югославию, Австрию, Албанию, Италию, Грецию и северо-за- падную часть Турции. На севере и северо-западе ареал ранней яблони накладыва- ется на область распространения основного вида — М. sylues- iris, заходит в Венгрию, Чехословакию, а на севере достигает Прибалтики (57' с. ш.). В Центральной и Западной Европе, в частности в Германии, яблоня ранняя произрастает островками, далеко отстоящими ог основного ареал а. Яблоня ранняя связана с лесной множеством переходных форм. Климатические условия в пределах европейского участка ареала секции Malus характеризуются относительным однообра- зием в течение года, умеренно холодной зимой с частыми отте- пелями, сравнительно теплым летом с холодными периодами.
Глава 5СИСТЕМАТИЧЕСКИЙ ОБЗОРРОДА MALUS Genus (род) Malus Miller, 1752, Gard, Dict. ed. 6; ejusd. 1754, Gard. Dict. abridg. ed. 4; ejusd. 1768, Gard. Dict. ed. 8, descr. compl.; Tournefort, 1737, ex. Linn., Gen. ed. 1: 145, nom. nud.; Borkhausen, 1803, Theor.-prakt. Handb. Forstbot., 2: 1270; Decaisne, 1874, Nouv. Arch. Mus. Hist. Nat. Paris, 10: 153, t. 10, сит ic. 12 (Мет. Fam. Ратас.); goehne, 1890, Gatt. Ратас.; ejusd. 1893, Deutsche Dendr., 257; Britton et Brown, 1897, 111. Fl. N. U. S., 2: 234; Zabel, 1903. ех. Beissner et al., Handb. Laubh.-Berm., 185: С. К. Schneid., 1906, 111. Handb. Laubh., 1: 174; Rehder, 1915, in Sargent, Pl. Wilson, 2: 173; ejusd. 1940, in Man. Cult. Trees Shrubs, ed. 2, 390; Юзепчук, 1939, Фл. СССР, 9: 358; Федоров и Полетика, 1954, Дер. и куст. СССР, 3: 414.— Pyrus, Linnaeus, 1753, Sp. Pl. 459; ejusd. 1754, Gen. Р1. ed. 5, 214, N' 550, рр. Pyrus malus Weston, 1770, Bot. Univ., 1 '. 229; Ascherson et Graebner, 1906, Syn. Mitteleur. Fl., 6, 2: 74; Beiley, 1937, Stand. Сусlор. Hartic. — Apirophorum Necker, 1790, Elem, Hot., 2: 72, р. р. — Pyrenia Clairville, 1811, Man. Herb. Sisse, 161, р. р. — Pyrus sect. Malus De Candolle, 2: 635. — Pyrus subgen. Malus Focke, 1888, Nat, Pflanzenfam., III, 3: 24. Яблоня. Дерево средней высоты (6 — 12 м) или высокое деревце со стволом (2 — 6 м), иногда растущее кустообразно, реже мощное дерево высотой до 15 — 20 м (секция Docyniopsis). Многолетние ветви обычно коричневые, сероватые, реже красные, покрытые немногочисленными колючками или без них, с овальной, угловатой (в поперечном разрезе) сердцевиной, чаще, так же как и древесина, зеленовато-белого цвета. Годичные по- беги опушенные (по крайней мере в молодости), слегка опу- шенные или совершенно голые. Почки на годичных побегах очередные, прижатые или парал- лельные, длиной 3 — 8 мм (у культурных форм иногда больше), покрытые 2 — 6 (чаще 3) наружными, спирально расположен- ными, обычно у верхушки ясно трехзазубренными чешуями. Лис- товой рубец (на безлистных побегах) узкий, с тремя следами, листовая подушка ясно выделяется на поверхности побегов.
120 Глава 5. Систематический обзор рода Malus Листья с черешками и удлиненными прилистниками, более или менее опушенные, в почкосложении свернутые или сложен- ные, цельные или лопастные. Листорасположение очередное, прилистники иногда рано опадающие. Цветки с приятным ароматом, в соцветиях в виде ложных зонтиков или щитков по 5 — 8 (2 — 10), обоеполые; чашелисти- ков 5, треугольные или ланцетные, остающиеся при плодах или опадающие, свободные или частично сросшиеся, опушенные или голые; лепестков обычно 5 (у некоторых махровых и полумахро- вых форм до 12 — 20), округлые, эллиптические или обратно- яйцевидные с выраженным ноготком у основания, белые или розовые, реже красные или пурпурово-красные, слабоопушенные, железистые; тычинок много (18 — 50), с желтыми, реже фиолето- выми пыльниками (у североамериканских видов); стилодии (5, реже меньше), целиком или у самого основания срастающи- еся в столбик; рыльце чаще головчатое, реже — продолговатое, расширенное или двурасщепленное (секции Eriolobus и Docy- niopsis), расположенное немного выше уровня тычинок, наравне с ними, реже — несколько ниже. Завязь сросшаяся (нижняя), 5-гнездная (в каждом гнезде по 2 — 4 семяпочки), гнезда за- крытые, полузакрытые или открытые в осевую полость плода Плоды различной формы, обычно шаровидные, вытянутые или сплюснутые в направлении чашечки и плодоножки, мякоть сочная, обычно без каменистых клеток, редко — с небольшим их числом. 2n=34 (51,68) [Хромосомные числа цветковых рас- тений, 1969, с. 624]. Тип рода: Malus sylvestris (L.) Mill. Род Malus представлен 35 автохтонными видами, из которых почти каждый характеризуется значительным внутривидовым разнообразием различного таксономического ранга. Встречается множество гибридов самого различного происхождения. Почти все виды яблони так или иначе встречаются в культуре: одни— в качестве широко распространенных промышленных культурных сортов, число которых достигает 10000 [Жуковский, 1964], дру- гие — как декоративные растения с множеством различных садовых форм и сортов, а также в качестве подвоя. Ключ для определения секций 1. Мощные, слаборазветвленные деревья до 15 — 20 м в вы- соту, с хорошо выраженными, толстыми стволами; оби- татели Юго-Восточной Азии . Sect. П Docyniopsis С. К. Schneid. — С. 123 — Менее мощные, хорошо разветвленные деревья, от 8— 10 до 12 м в высоту, небольшие деревца или кустарники с ветвистыми стволами 2. Кустарники и небольшие деревца с глубоко трехраздель- ными, кожистыми, сверху голыми, снизу волосистыми
Глава 5. Систематический обзор рода Malus ]2] листьями и густо войлочно-опушенными цветоножками, гипантиями, чашечками; обитатели Передней Азии Sect. 1 Eriolobus С. К. Schneid. — С. 121 — Деревья средней высоты, небольшие деревья или кус- товидные деревца с цельными или слаболопастными, реже глубоко 3 — 5-лопастными (до раздельных) листьями; цветоножки, гипантии, чашечки, а также листья голые, слабоволосистые или слегка опушенные . 3. Листья в почкосложении свернутые, все цельные .. 4 — Листья в почкосложении сложенные, более или менее ло- пастные, реже цельные 4. Крупноплодные настоящие яблони, обитающие авто- хтонно в Евразии . Sect. VI Malus — С. 164 — Мелкоплодные, так называемые ягодные яблони, авто- хтонно обитающие в Азии Sect. V Gymnomeles Poehne — С. 154 5. Крупноплодные яблони Северной Америки. Плоды на толстых плодоножках . Sect. IV Chloromeles (Респе) Rehd. — С. 148 — Мелкоплодные яблони Восточной Азии. Плоды на тонких плодоножках . Sect. Ill Sorbomalus Zabel — С. 129 SECTIO (секция) I. ERIOLOBUS (С. К. Schneid.) Langenf.; С. К. Schneider, 1906, in Repert. Sp Nov. Reg. Veg., 3: 179; Rehder, 1920, in Journ. Arnold АгЬ., 2: 49. — Pyrus sect. Erio- lobus Seringe, 1825, in DeCandolle, Prodr., 2: 636. — Eriolobus Roemer, 1847, Fam. Nat. Reg. Veg. Syn., 3: 216. — Cormus sect. Eriolobus Decaisne, 1874, in Nouv. Arch. Mus. Hist. Nat. Paris, 10: 157. — Eriolobus sect. Eueriolobus С. К. Schneid., 1906. Ill. Нandb. Laubh., 1: 725, р. р. Fructiceta silve arbores non magnae. Folia circumscriptione rotundo-ovoidea ve1 vix rotunda, pro- funde tripartita sive 4 — 6 lobata serrato-dentatis triangula ris- busque сит lobis. Flores in corumbiformibus vel umbellulatis inflorescentiis, co- lore sunt albi, tomentosis pedicellis, hypanthium et calyx (extra) densitomentose pubescens. Fructus sphaerici че1 late ovoidei deciduo sive persistenti curn ca1yce. Typus sectionis: Malus trilobata (Labill.) С. К. Schneid. Кустарники или небольшие деревья. Листья в очертании округло- или широкояйцевидные, глубоко трехраздельные. Цветки в зонтиковидных соцветиях, гипантии и чашечка (сна- ружи) густо войлочно-опушенные, пестик из 5 в верхней части не полностью сросшихся остроконечных, наверху оттянутых
)2i Глава 5. Систематический обзор рода Ма1из Рис. 44. Манив trilobutu плодолистиков, которые вдаются в эпигинную часть трубки ча- шечки, где срастаются с беловойлочным столбиком; рыльца двурасщепленные. Плоды овальные или почти шаровидные. красные, диаметром 1 — 2 см, с каменистыми клетками и оста- ющейся чашечкой. Монотипичная секция Eriolobus представлена единственным весьма оригинальным и в высшей степени обособленным, свое- образным и примитивным видом — M. trilobata, обладающим узким дизъюнктивным ареалом в Ливане, отчасти в Греции и Болгарии, редко в Турции и Израиле. 1. Malus trilobata (Labill.) С. К. Schneid., 1906, in Repert. Sp. Nov. Reg. Veg., 3: 179. — Crataegus trilobata Labillardiere, 1812, 1соп. Pl. Svr., 4: 15, t. 10. — Pgrus trilobata de Candolle, 1812, Prodr. 2: 636. — Sorbus trilobata Heynhold, 1872, Nomencl. Bot., 1: 773; Boissier 1872, Fl. Or., 2: 65. — Cormus trilobata Decisne, 1874, in Nouv. Arch. Mus. Hist. Nat. Paris, 10: 157.— Eriolobus trilobatus Roemer, 1847, Fam. Nat. Reg. Veg. Syn., 3: 216. — Eriolobus trilobata С. К. Schneid., 1906. — 111. Handb. 1 аиЬп., 1: 726, сит ic. 407 а-d, 403 а-с. — К. Borwitz, 1969, in Arbor. gornickie, 14: 5 — 21. — Яблоня трехлопастная. Пряморастущий кустарник или небольшое деревце. Листья кожистые, голые, снизу волосистые, с обеих сторон светло-зеленые, осенью ярко-красные, пластинка трехраздел~-
Глава 5. Систематический обзор рода Ма!а~ 323 ная: боковые доли отстоящие, двухлопастные или двураздель- ные, верхушечная доля трехлопастная, трехраздельная или цель- ная; по краю пильчатые, в общем очертании округлые или ок- ругло-яйцевидные с сердцевидным основанием, длиной 3 — 8 см, шириной 5 — 9 см, на черешках длиной около 2,5 см, без при- Л ИСТНИ КОВ. Цветки по 6 — 8 в зонтиковидных соцветиях, венчик белый, диаметром 3,5 см, цветоножки, гипантии и чашечка густо вой- лочно-опушенные, тычинок много (30 — 50); пестик из 5 в верх- ней части не полностью сросшихся остроконечных, наверху от- тянутых плодолистиков, которые вдаются в эпигинную часть трубки чашечки, где срастаются с беловойлочным столбиком; рыльца двурасщепленные. Плоды овальные или почти шаровидные (шаровидно-яйце- видные), красные, диаметром 1 — 2 см, на тонких плодоножках длиной 2,5 см (рис. 44). Цветет V, плоды созревают IX, Х. Описана французским ботаником и путешественником Ла- биллардье из Ливанских гор (Ливан) под названием Crataegus frilobata. В 1906 г. Шнейдером этот вид перенесен в род Afalus. Тип неизвестен. Lectotypus: Liban, Silvan bei Lasa im Lfal, nahr Ibrahin, 1000 т, V 1899, fl., Juvernalk, in Herb. Inst. Bot пот. V. L. Komarovii Acad. Sci. URSS in Leningrad conservatur. SECTIO (секция) II. DOCYNIOPSIS (С. К. Schneid.) Lan- genf. С. К. Schneid., 1906, in Repert., Sp. Nov. Veg., 3: 179, exclud. М. docynioides Rehder, 1920, in Journ. Arnold. Arb., 2: 49. — Eriolobus Roemer, emend, Rehder, 1903, in Sargent, Trees and Trees and Shrubs, 1: 74, р. р. — Macromeles Koi. dzumi, 1930, Fl. Symb. Or.-As., 53. Arbores robustae longitudine usque ad 15 — 20 т bene сит expresso trunco crasso. Folia magna, ovalia, oblongata vel oblongato-ovoidea, rarius ob1ongato-lanceata, integres sive leviter lobata, interdum lucida, сопасеа. Flores magni usque ad 3,0 — 3,5 ст in diam., рейсе111з tomen- tosis. Fructus sphaerici 2 — 3 vel 4 — 5 ст in diam., sepalis сит persistentibus e solida crepaci ри!ра, cellulas lapideas conti- nenti. Туриз sectionis: Afalus tschonoskii (Maxim.) С. K,. Schneid. Мощные деревья высотой до 15 — 20 м, с хорошо выражен- ными толстыми стволами. Листья крупные, овальные, продолго- ватые или продолговато-яйцевидные, цельные или слегка ло- пастные, опушенные, иногда блестящие, кожистые. Цветки крупные, диаметром до 3,0 — 3,5 см, на войлочных цветоножках. Плоды шаровидные, диаметром 2 — 3 или 4 — 5 см, с остающи-
Гяава 5. Систематический обзор рода Afalus 124 Рис. 45. Виды секции 0осупй'орый: 1 — М, tschonoskii; 2 — М. f oã~иояапа; 3 — М. Laosensis; 4 М. si kki mensis мися при плодах чашелистиками и плотной хрустящей мякотью, содержащей каменистые клетки (рис. 45). Наличие ряда примитивных признаков (строение габитуса, листьев, со- цветий, цветков и плодов}, а также закономерности распространения (дизь- юнкция ареалов} реликтовых видов секции Docyniopsis позволяет предпо- .еа гать:
Глава 5. Систематический обзор рода Malus 325 1) доциниевидные яблони вместе с М. tritobata представляют наиболее древнюю ветвь рода Маарив; 2) доциниевидные яблони еще в третичном периоде имели общий обшир- ный ареал, приуроченный к Евразии; 3) по неизвестным причинам (вероятно, в связи с резким изменением климатических условий и неравномерным распределением природных фак- торов) образовались обособленные ареалы. где представлены современные реликты втих яблонь (описанные ниже 4 вида) (рис. 46). Ключ для определения видов 1. Плоды кубаревидные, 1,5 — 2,0 см в диаметре 5. М. sikkimensis (Wenz.) Koehne — Плоды шаровидные, 2 — 3 до 5 см в диаметре... 2 2. Листья толстые, кожистые, блестящие с обеих сторон . 3. М. formosana Kaw. et Koidz. — Листья опушенные с обеих сторон, особенно в молодости 3 3. Кора на стволах и многолетних ветвях светлая, свет- ло-серая 2. М. tschonoskii (Maxim.) С. К. Schneid. — Кора на стволах и многолетних ветвях коричнево-серая с красноватым оттенком . 4. М. laosensis А. Cheval. 2. Malus tschonoskii (Maxim.) С. К. Schneid., 1906, in Re- pert. Sp. Nov. Reg. Veg., 3: 179. — Pyrus tschonoskii Maxim., 1893, in Bull. Acad. Sci. St. Peter sb., 19: 169 (in -Mel. Biol., 9: 165). — Eriolobus tschonoskii Rehder, 1903, in Sargent, Tree and Shrub, 2: 73, t. 37; С. К. Schneid., 1906, 111. Handb. Laubh., 1: 727, сит ic., 403 h — i, 404 f. — Cormus tschonoskii Koidzumi, 1909, in Bot. Mag. Tokyo, 23: 171. — Macromeles tschonoskii, Koidzumi, 1930, Fl. Symb. Or-As., 53. — Docyniopsis tschonoskii, Koidzumi, 1934, in Acta Phytotax. Geobot., 3: 162, 196. Яблоня чоносски. Мощное дерево высотой 10 — 15 м, со стволом толщиной до 2 м, светлой корой и пирамидальной кроной (в ювенильном со- стоянии шаровидная). Многолетние ветви светло-серые, годич- ные побеги беловойлочные. Почки красные. Листья овальные или продолговато-яйцевидные, длиной 7— 12 см, шириной 5 — 8 см, при основании закругленные или серд. цевидные, на верхушке заостренные, по краю неравномерно пильчатые, иногда слегка лопастные, в молодости с обеих сто- рон беловойлочные, впоследствии сверху лоснящиеся, снизу тонко беловойлочные, на черешках длиной 2 — 4 см. 11ветки по 2 — 5 в щитовидных соцветиях, на опушенных цве- тоножках, длиной около 3 — 4 см, гипантии и чашечка густо беловойлочные; венчик белый, диаметром около 3 см, стилодии (5), сросшиеся у основания в мохнато-опушенный столбик, рыльце приподнято до уровня тычинок.
12Ь Глааа 5. Систеиатический обзор рода Маlиз Плоды почти шаровидные, диаметром 2 — 3 см, с остающейся чашечкой, желтовато-зеленые с красным бочком и плотной мя- котью, содержащей в небольшом количестве каменистые клетки. Описана из Японии, с подножья горы Фудзияма, где это растение было собрано К. И. Максимовичем в ноябре 1873 г. с плодами, но без листьев. Тип не указан. Lectotypus: Japan: Loc. Honshu Prei. Куопио, near Ebidani, Hiyoshicho Funaigun, alt. Ноот, 3 V 1963, fl. n' 18042, G. Mu- rata, in Herb. Inst. Bot. nom. V. 1. Komarovii Acad. Sci. USRR in Leningrad conservatur. Распространена в Японии на о-ве Хонсю (Хонда), где растет в предгорных лесах. 3. Malus formosana gawak. et Koidz., 1911, in Bot. Nag. Tokyo, 25, 146; Е. Н. Wilson, 1920, in Arn. Arb. Exped. Е.-Asia, 1917 — 1918, t. N 423, Hab. — Яблоня формозская. Мощное дерево высотой до 20 м, с выраженным стволом, пирамидальной кроной, темно-коричневой корой на многолетних ветвях. Листья овальные, продолговатые до продолговато-ланцет- ных, с заостренной верхушкой, закругленным или короткокли- новидным основанием, пильчато-городчатые, длиной 8 — 13 см, шириной 3,0 — 5,5 см, цельные, изредка едва трехлопастные, тол- стые, кожистые, блестящие с обеих сторон, с толстыми, на ниж- ней стороне пластинки выпуклыми жилками, на черешках дли- ной 2 — 3 см. Цветки в малоцветковых щитках по 3 — 5, на беловойлочных цветоножках длиной 1,5 — 3,0 см; диаметром 2,5 см, лепестки почти белые с желтоватым оттенком, стилодии (4 — 5), при ос- новании сросшиеся в шерстистые столбики. Плоды шаровидные, несколько продолговатые, варьирующие по размерам, диаметром в среднем 2 см, длиной 2,5 см, зелено- желтые с красным оттенком, твердой мякотью, содержащей ка- менистые клетки. Чашечка при плодах остающаяся, несколько приподнятая, у основания сросшаяся в мясистую трубку. Пло- доножки длиной 3 см. Вид описан японскими ботаниками Каваками и Коидзуми в 1911 г. из горных лесов о-ва Тайвань. Тип не указан. Неотип: Китай, пров. Фуцзянь у Чунань, Синцунь — Хуан- чжуао, 8. VIII 1952, пл. n' 106, Чжоу Чжэнь-ин, хранится в гербарии Ботанического ин-та им. В. Л. Комарова АН СССР в Ленинграде. Распространена в Китае — в горных лесах о-ва Тайвань и пров. Фуцзянь на высоте 1400 — 2000 м н. у. м. 4. Malus laosensis А. Cheval, 1920, in Compt. Rend., 170: 1129. — hfalus йоитегi А. Cheval, 1920, in op. cit. — hfalus
Глава 5. Систематический обзор рода hfalus 127 melliana Rehder, 1934, in Journ, Arnold Arb., 20: 313, — Яблоня лаосская. Крупное дерево со стволом высотой 15 — 20 м. Листья про- долговатые, продолговато-овальные до овальных, крупные, дли- ной 10 — 12 см, шириной около 4 — 5 см, почти цельнокрайные или едва заметно городчатые, изредка со слабовыраженной ло- пастностью, плотные, слегка кожистые, сверху блестящие, снизу вместе с черешком интенсивно войлочно-опушенные или опу- шенные с обеих сторон. Цветки в малоцветковых щитках, венчик почти белый с желтоватым оттенком, цветоножка, гипантии и чашечка сильно- опушенные. Плоды шаровидные, крупные, диаметром 3 — 5 см, желтые, несколько краснеющие или с зеленоватым оттенком, плотной грубоватой мякотью, содержащей каменистые клетки, сладкого вкуса, напоминающего вкус груши. Чашечка при плодах оста- ющаяся или опадающая (у более мелкоплодных форм). Тип не указан. NeotypUs: China Hoond Abisan prov., alt. 1166 — 2666 т, 29.Х 1928, fr., n' 10900; Е. Н. Wilson, in НегЬаг1пт Poyal Botanic Gardens in Kew, Gr. Britain conservatUr. Распространена в горных субтропических лесах северной части современного Лаоса (в ущелье Транид и центральной части Тонкина), а также в Северном Вьетнаме, откуда заходит в прилегающие провинции Китая (Юньнань, Гуанси, Гуандун). Ареал М. 1аозепиь располагается в пределах отрогов Юиьнаньского нагорья (на высоте от 800 — 1200 до 2000 м н. у. м.), которое на западе постепенно повышается и граничит с высочайшими хребтами Вирмы. На севере ареал лаосской яблони ограничен водораздельными хребтами верхо- вий рек Меконг, Черная и Юньцзян (Красная), которые достигают 5500— 6000 м н. у. м. Вассейны этих рек служат местом обитания М. 1aosensis, где существуют благоприятиые условия для почти непрерывного роста яб- лони в течение всего года. Описание М. laosensis сделано нами на основа- нии далеко не полных сведений по: А. Chevalier, 1920, Comptes Pendus des Seances, 170: 1129, Paris, Gauthier-Villars et Qie Imprimeurs Libraires, а также по фрагментарным сборам Э. Вильсона и P. Хинга, полученным нами из гербария Королевского ботанического сада в Кью (The Herbarium Royal Botanic Gardens, Кем, Gr. Britain). От близкого вида М. formosono лаосские яблони отличаются крупными листьями, более крупными плодами и неопределенностью признака — оста- ющейся при плодах или частично опадающей чашечкой; от другого соседнего восточно-гималайского вида М. sikkimensis — общим габитусом, строением листьев, цветков и плодов. У М. sikkimensis крона более густа я, ветви с большим числом колючек, листья более округлые — яйцевидные, цветки в зонтиках, плоды мелкие с остающимися при них чашелистиками. Мелкоплодность некоторых форм М. laosensis и опадающая чашечка позволяют предполагать, что эти третичные реликтовые яблони и их исход- ные примитивные формы были вероятными предками современных мелко- плодных рябиновидных яблонь (секция Sorbomolus), имеющих достаточно широкое современное распространение в центральных и восточных районах Азии. Это подтверждается более или менее ясными связями М. 1аояепи'.с
Глава 5. Систематический обзор рода Malus 128 Рис. 4б. Схема филогенетических отно-шений в секции Docyniopsis ПРЕДКОВЫЕ ФОРМЫ ТИПА ООСУИ!А с примитивными видами рябиновидных яблонь, в частности с М. ombrophila и М. уцппапепяз. Несомненно, крупноплодные формы М. laosensis имеют связь с крупно- плодными настоящими яблонями из секции Malus и, прежде всего, с вос- точно-азиатскими крупноплодными яблонями (серия Asiaiicae). а также с крупноплодными яблонями Северо-Западных Гималаев (Кашмир, Индия). 5. Malus sikkimensis (Wenzig) goehne, 1890, Gatt. Ратас. 27; С. K. Schneid., 1906, I11. Handb. Laubh., 1: 719, сит ic. 398 п, 404а. — Malus pashia p sikkimensis Wenzig, 1874, in Linnaea, 38: 49. — Pyrus sikkimensis Hooker, 1878, Fl. Brit. Ind. 2: 373.— Яблоня сиккимская. зверева высотой 5 — 8 м, с низко расположенной густой кро- ной и большим числом крепких колючек на стволе и ветвях. Го- дичные побеги опушены. Листья яйцевидные или продолговато-яйцевидные длиной 5 — 11 см, шириной 3 — 5 см, с длиннооттянутой верхушкой и за- кругленным основанием, по краю остропильчатые, сверху слегка опушенные, снизу густо войлочно-опушенные, на черешках дли- ной 1,5 — 3,5 см. Цветки по 4 — 9 в зонтиковидных соцветиях, на тонких опу- шенных цветоножках длиной 3,5 — 4,5 см, гипантии и чашечка густо войлочно-опушенные, венчик диаметром 2,5 см, распуска- ется одновременно с листьями, в бутонах — розовый, позже бе- лый или снаружи розоватый, лепестки округлые с едва замет- ным ноготком; тычинок много (30 — 50); пестик образован
Глава 5. Систематический обзор рода Malus 129 Рис. 47. Схема филогеиети- ческих отношений в секции Sorbomalus < CJ < Ф C р 4 IK lC й ~и 3 Я и М М z м~ О 1i Й 3 е е z p м le З g р М М з 5 44 ев М Ы й ф 5 плодолистиками, на верхушке свободными, вдающимися в зпи- гинную трубковидную часть чашечки; рыльца на верхушке дву- надреза нные; стилодии (5) гол ые. Плоды кубаревидные, несколько грушеобразные, диаметром около 1,5 — 2,0 см, желто-красные со светлыми точками, в моло- дости сильноопушенные, чашечка при плодах остающаяся, пло- доножки сравнительно тонкие длиной 3 — 4 см. Цветет V, плоды созревают IX. Описана из Восточных Гималаев (Сикким). Тип не указан. Neotypus: Sikkim, Himalaya, Lachen 8300 т, V 1885, fl., G. King' s in Central National Herbarium Botanical Survey of India in Calkutta conservatur. Распространена в Восточных Гималаях (Сикким), откуда заходит в прилегающие районы Китая, а также в Непал и не- которые штаты Индии. Обитает в высокогорных лесах на высоте от 2000 до 3000 м н. у. м., а иногда и выше — до 3900 м. Типичная М. sikkimensis характеризуется остающимися при плодах чаше- листиками. Указапие в литературе о наличии у М. sikkirnensis опадающей восле цветения чашечки относится только к окультуренным особям, однако это явилось причиной отнесения М. sikkimensis к мелкоплодным яблоням типа Baccatae. SECTIO (секция) Ш. SORBOMALUS Zabel in Beissner et a1., Handb. Laubh.-Ben., 189, nomen subnud. С. К. Schneid., 1906, Ill. Handb. Laubh., 1: 721. — Malus sect. GIImnomeles, Koehne, 1893, Deutsche Dendr., 257, р. р. — Рябиновидные яблони. Мелкоплодные яблони с опадающей, реже остающейся ча- шечкой.
Глава 5. Систеыатический обзор рода Ма1из Листья цельные, более или менее лопастные, иногда с вы- раженной гетероф илией, пильчатые, реже — зубчатые или цельнокрайные. Цветки в щитковидных или зонтиковидных соцветиях. Пыль- ники желтые. Стилодиев 3 — 5. Плоды мелкие, на тонких коротких ллодоножках, длина которых в 2 — 3 раза превышает размеры плода. Семенные гнезда на верхушке тупые. Тик секции: Malus kansuensis (Batal.) С. К. Schneid. Sorbomalus — наиболее обширная, разнообразная и одновре- менно примитивная секция рода Malus объединяет около 12 ви- дов, обитающих в Центральной и Восточной Азии, а также на тихоокеанском побережье Северной Америки (рис. 47). Виды этой секции принадлежат к различным эволюционным уровням и имеют в этом отношении некоторый параллелизм с секцией Docyniopsis. В пределах секции Sorbomalus различаются неко- торые внутрисекционные подразделения. Ключ для определения видов 1. Чашечка при плодах остающаяся (ser. Yunnanenses; Rehd ).............. 2 — Чашечка при плодах опадающая....... 4 2. Листья при основании косоусеченные, острозубчатые и двоякоострозубчатые . 7. М. отЬгорЫ!а Hand.-wazz. — Листья с округлым основанием, неравномерно дважды- пильчатые . 8. М. ргаШ! (Hemsel) С. К. Schneid 3. Листья продолговатые, при основании закругленные, слабосердцевидные, почти цельнокрайные 6. М. yunnanensis (Franch.) С. К. Schneid.. — Листья яйцевидные с закругленным или усеченным ос- нованием, мелкопильчатые . 9. М. honanensis Rehd. 4. Листья глубоко 3 — 5- или многолопастные, до раздель- ных (ser. gansuenses Rehd.)....,... 5 — Листья гетерофильные: на коротких побегах цельные, на длинных — 3-, реже 5-лопастные (ser. Toringonae (Rehd.) Langenf.) 16. M. toringo Siebold' 5. Листья З-лопастные, с острыми и широкими, по краю остропильчатыми лопастями 10. М. kansuensis (Batal.) С. К. Schneid.. — Листья 5-лопастные, с пильчатыми лопастями 11. М. toringoides (Rehd.) Hughes'
Глава 5. Систематический обзор рода Malus 6. Листья 3(5)-лопастные, раздельные, до рассеченных . 12. М. transitoria (Batal.) С. К. Schneid. — Листья перисто-раздельные, до трехрассеченных с лан- цетными или продолговато-яйцевидными, неравномерно пильчато-зубчатыми лопастями 13. М. setok Vass. 7. Листья 3 — 5-лопастные с клиновидным основанием . 14. М. centralasiatica Vass. — Листья 3-лопастные с сердцевидным основанием . 15. М. komarovi i (S ar g.) Rehd. Series (ряд) 1. Yиnnапеnses Rehder, 1940, Мап. Cult. Trees Shrubs, ed. 2: 390. — Malus sect. Sorbomalus subsect. Yunnanenses Rehd., 1920, in Jour. Агп. Arb., 2: 48. — Docyniop- sis зепзи Poidzumi, 1934, in Act. Phytotax. et Ceobot., 3: 162, р. р. — Malus sect. Docyniopsis С. Х. Schneid., 1906, in Repert. Sp. Nov. Veg., 3: 176. р. р. Большие деревья мезофильного облика, с мощными ство- лами, реже — небольшие деревья или кустарники. Листья крупные, длиной до 15 см, шириной 8 см, цельные или слаболопастные. Цветки в многоцветковых щитковидных соцветиях. Стилодии (3 — 5), сросшиеся до 1/3 длины в совершенно голый столбик. Плоды диаметром 1,0 — 1,5 см, с грубой мякотью, содержа- щей каменистые клетки, и с остающимися при плодах чашелис- тиками (рис. 48) . Тин серии: Malus yunnanensis (Franch.) С. К. Schneid. 6. Маlив yunnanensis (Franch.) С. К. Schneid., 1906, in Re- pert. Sp. Nov. Reg. Veg., 3: 179; Rehder, 1915, in Sargent, Pl. Wilson, 2: 281. — Pyrus yunnanensis Franchet, 1890, Pl. De- lavay, 228. — Eriolobus yunnanensis, С. К. Schneid., 1906, 111. Handb. Laubh., 1: 727, fig. 403 е-g, 404 Ь-с. — Cormus диппа- nensis Poidzumi, 1913, in Journ. Coll. Sci, Tokyo,34,2: 75 (Consp. Rosac. Jap.). — Docyniopsis yunnanensis Koidzumi, 1934, in Act. Phytotax. Geobot.: 196. — Яблоня юньнаньская. Мезофильное дерево высотой 8 — 13 м, с мощным стволом толщиной от 30 до 100 см и узкой компактной кроной. Молодые побеги войлочные. Листья продолговато-яйцевидные, длиной 6 — 12 см, при ос- новании закругленные, иногда слабосердцевидные, почти цель- нокрайные, на верхушке заостренные, по краю остро удвоенно- пильчатые или слаболопастные, снизу беловойлочно-опушенные. Цветки в многоцветковых (около 12) щитковидных соцве- тиях, цветоножки, гипантии и чашечка с обеих сторон беловой- лочно-опушенные, лепестки чисто-белые, с продолговатым ноготком; стилодии (5), сросшиеся у основания в голый столбик.
И2 Глава 5. Систематический обзор рода Malus Рис, 48, Виды серии Yunnanenses:1 — М, отЬгорЬйа, 2 — М. уиппапетЫ; 3 — М. prgttii; 4 — М. ho-nanensis (по 7И Te-tsun and Yen Cheng-lung [19563, с дополнением) Плоды шаровидные, диаметром 1,О — 1,5 см, красные, бело-точечные, с грубой мякотью, содержащей каменистые клетки Чашечка при плодах остающаяся.
Глава 5. Систематический обзор рода Malus Описана в 1890 г. французским ботаником и путешествен- ником А. Франше из Китая, пров. Юньнань. Тип неизвестен. Neotypus: China, Ргоч. Уип-пап Ьог. оссЫ., inter fluvios Landsang-djiang et 1.и-djiang, са 28, in pluvii silvio mixtis temperalis in valle а jugo si-la ad vicum Tseku descendente, sustr. granitico et schistaceo, alt. 3200 т, 30 IX 1915, 1г. n' 842, Handel-Mazzetti, in Herbario Naturhistorisches Museum in Wien (Austria) conservatur. Распространена в горных районах Китая на высоте 2500— 3500 (1600 — 3800) м н. у. м., в пров. Юньнань, восточных час- тях пров. Сычуань и Хубэй. 7. Malus ombrophila Hand.-Макк., 1926, in Anz. Acad. Wiss. Wien, Math. Nat. 62, 8, China (Yunnan). — Яблоня дожделю- бивая. дерево мезофильного облика, высотой 10 — 15 м, с выражен- ным стволом и весьма укороченными годичными побегами. Листья крупные, длиной до 13 см, шириной 7 см, цельные, яйцевидные, с заостренной или острой верхушкой, закругленным или косоусеченным основанием, по краю острозубчатые или двоякоострозубчатые, голые, на черешках длиной 1,5 — 3,0 см. Цветки по 3 — 5 в щитковидных или зонтиковидных соцве- тиях, белые, в бутонах едва розовые; пыльники желтые, распо- лагающиеся примерно на одном уровне с рыльцем. Стилодии (3), сросшиеся в нижней части в совершенно голый столбик. Плоды почти шаровидные, несколько приплюснутые, диамет- ром около 1 см, длиной 1,2 см, с остающимися при плодах ча- шелистиками, на тонких плодоножках, длиной около 2,5 — 3,0 см. Мякоть плода грубоватая. Описана в 1926 г. австрийским ботаником Х. Хандель-Мац- цетти из смешанных лесов Юго-Западного Китая (пров. Юнь- нань), в районе водораздельного хребта (на высоте 2250 — 2650м н. у. м.), между реками Лу-депанг (Салуин) и Яеноу-депанг (Иравади) . Typus: China, Proc, Yunnan Ьог.-occid.: Ргоре fines Tibeto- Birmanicas, inter fluvios 1.и-djiang (Salween) et Djion-djiang (Irrawadi orient. super.), in pluvii silvis calide temperatis vallis Tjiontson — lumba infra Tschamutong са 2250 — 2650 т, 28 VI 1916, n' 1916, n' 9119, Dr. Heinr. Frh. v. Handel-Mazzetti. In Herb. Mus. Nat. Hist. in Wien conservatur. Распространена во влажных горных районах Юго-Западного Китая, пров. Юньнань. 8. Malus prattii (Hemsel.) С. К. Schneid., 1906, V, Ill. Handb. Laubh., 1: 719, fig. 397 р., 398 k-m; С. К. Schneid., 1906, XII, in Repert. Sp. Nov. Reg. Veg., 3: 178; Rehder, 1915, in Sargent, Pl. Wilson, 2: 281. — Pyrus РгаШ~ Hemsley, 1895, in Кеж.
Глава 5. Систематический обзор рода Malus Bull. 1895: 16. — Docyniopsis Prattii, goidzumi, 1934, in Act. Phytotax. — Geobot., 3: 196. — Яблоня Пратта. Мезофильное дерево высотой 8 — 10 м, с мощным стволом до 80 — 100 см в обхвате. Молодые побеги белоопушенные, позже— голые. Листья яйцевидные, эллиптические (до продолговато-яйце- видных), крупные, длиной до 15 см, шириной 8 см, с округленным основанием, острой или заостренной верхушкой, цельные, не- равномерно дваждыпильчатые, голые или слабоопушенные лишь в молодости (по жилкам снизу). Цветки в многоцветковых (5 — 8) зонтиковидных, изредка щитковидных соцветиях, на тонких, голых или слабоопушенных цветоножках длиной около 2 (1,5 — 3,0) см. Венчик белый, диа- метром около 2 см. Гипантии и чашелистики голые (или чаше- листики слабошелковисто-опушенные изнутри). Пыльники жел- тые. Стилодии (5), сросшиеся у основания в голый столбик. Плоды шаровидно-сплюснутые, несколько яйцевидные, диа- метром около 1,0 — 1,5 см, желтые или красно-желтые, со свет- лыми точками и с остающимися чашелистиками. Описана в 1895 г. английским ботаником В. Хемсли из За- падного 1~итая (пров. Сычуань, близ Туэн-хуан-сиэн), где най- дена А. Праттом во влажных лесах. Тип неизвестен. Neotypus: China, Prov. Szechuan, Tiehchuan-hsien, alt. 2850 т, 29 V 1936, Н., n' 2658, К. L. Chu, in Herb. Naturhistori- sches Museum in Wien (Austria) consevatur. Яблоня Пратта распространена в Центральном и Юго-За- падном китае, чаще в пров. Сычуань, на высоте от 2000 до 3000 м н. у. м. 9. Malus honanensis gehder, 1920, in Jour. Arnold АгЬ., 2: 51. — Sinomalus honanensis Poidzumi, 1934, in Act. Phytotax. Geobot., 3: 196. — Яблоня хэнаньская. Небольшое дерево или кустарник ксероморфного облика вы- сотой до 6 м. Листья широкояйцевидные, реже удлиненно-яйцевидные, длиной 6 — 8 см, шириной 3 — 6 см, при основании закругленные или усеченные, с остропильчатым краем, на верхушке с 2 — 5 па- рами широкояйцевидных лопастей, по краю мелкопильчатые, снизу опушенные, осенью краснеющие, на черешках длиной 1,5 — 2,0 см. Цветки по 5 — 10 в щитковидных соцветиях. Венчик белый (в бутонах розовый), диаметр около 1,5 — 2,0 см. Стилодии (3 — 4), сросшиеся лишь у основания в голый столбик. Цвето- ножки слабоволосистые, чашечка снаружи опушенная. Плоды шаровидные, диаметром 1 см, желто-зеленые, со свет- лыми точками, грубоватой мякотью и остающимися чашелисти- ками.
Глава 5. Систематический обзор рода Malus l3S Описана в 1920 г. А. Редером из Китая, пров. Хэнань. Тип неизвестен. Neotypus: China, Prov. Honan, Sungshan-tà Yeh-mao-cha, 31 VII 1923, n' 195, J. Hers, in Herb. Royal Botanic Gardens in 3(ew (Great Britain) conservatur. Распространена в центральных районах Китая — пров. Хэ- нань и Шэньси. Series (ряд) 2. К а п s и en s e s (Rehd.) Rehder, 1940, Ìàn. Cult. Trees Shrubs, ed. 2: 390. — Afalus sect. Sorbomalus sub- sect. Kansuenses Rehd., 1920, in Jour. Arnold АгЬ., 2: 48. Eriolobus sensu С. К. Schneider, 1906, Ш. Handb. Laubh., 1: 725, р. р.: non Roemer, 1847. — Sinomalus 3(oidzumi, 1932, in Act. Phytotax. Geobot., 1: 11. Небольшие деревья, иногда кустарники ксероморфного об- лика. Листья средней величины, длиной до 3 см, шириной 4 см, глубоко 3 — 5- или многолопастные, до раздельных. Цветки в многоцветковых зонтиковидных соцветиях. Стило- дии (5) голые, сросшиеся у основания в столбик. Плоды мелкие, диаметром 0,5 — 1,0 см, с нежной мякотью, без каменистых клеток (за исключением M. kansuensis) и опа- дающей чашечкой (рис. 49). Тип серии: Malus kansuensis (Batal.) С. К. Schneider. Серия E(ansuenses — самый обширный и разнообразный ряд яблонь, представленный 6 видами, достигшими значительного полиморфизма. Филогенетические связи с юньнаньскими ябло- нями заключаются в наличии некоторых общих признаков. Можно предполагать, что между M. 'попапепзт и M. kansuensis имеет место близкое родство. 10. Malus kansuensis (Batal.) С. К. Schneid., 1906, in Re- pert. Sp. Nov. Reg. Veg., 3: 178; С. К. Schneid., 1912, Ill. Handb. Laubh., 2: 1001; Rehder, 1915, in Sarg. Pl. Wilson, 2: 286; Яей- der, 1920, in Journ. Агп. АгЬ., 2: 51; Henning, 1947, in Zuchter, 10 — 12: 296; Scaly R., 1955, in Curt. Bot. Mag. 1970, t. 3.— Pterus kansuensis Batal., 1893, в Тр. Петерб. Бот. сада, 13: (1)94. — Eriolobus kansuensis С. К. Schneid., 1906, 111. Hanb. Laubh., 1: 726; КоЫкипи', 1934, in Act. Phytotax et Geobot., 3: 196. — Яблоня ганьсуйская. Деревце или кустарник высотой 3 — 8 м. Молодые побеги во- лосистые, позже становятся голыми, красно-коричневыми. Листья яйцевидные, длиной 4 — 8 см, шириной 2 — 6 см, 3 — 5- лопастные, с острыми, широкими, по краю остропильчатыми ло- пастями, заостренными верхушками и закругленным, реже ши- рококлиновидным основанием, сверху темно-зеленые, снизу светло-зеленые, слегка волосистые, особенно по жилкам, на че- решках длиной 1,5 — 4,0 см.
Глава 5. Систематический обзор рода Malus Рис. 49. Виды серии Pansuenses". 1 — М. kansuensi' 2 — М. setok. 3 — М. centralasiatica; 4 M. transitorily; 5 — М. toringoides Цветки по 4 — 10 (12) в зонтиковидных соцветиях, венчик белый, в бутонах розоватый, диаметр 1,5 см; чашечки густо- опушенные; цветоножки тонкие, длинноволосистые.
Глава 5. Систематический обзор рода Ма!из 137 Плоды овально-эллиптические, длиной 1,0 — 1,5 см, диамет- ром 0,8 — 1,0 см, желтые (до красно-пурпуровых), с рассеянными светлыми точками; чашечка при плодах опадающая; мякоть плодов грубая, хрящеватая, содержащая каменистые клетки; плодоножки тонкие, длиной 2,5 см. Описана в 1893 г. русским ботаником А. Ф. Баталиным по сборам Г. H. Потанина из северной части пров. Сычуань (Ки- тай) под названием Ругиз kansuensis. В 1906 г. Шнейдером отнесена к роду Malus. Typus: China borealis: prov. Szetschuan septentrionalis, in valle fl. Honton, inter Ksernzo et Тsinyuan, 12 aug. 1885, S. N. Potanin, in Herb. Bot. Inst. Acad. Sci. URSS in Leningrad соп- яегvatur. Распространена в лесах горных областей Центрального и Юго-Западного Китая (пров. Ганьсу и Сычуань), где растет на высоте от 1800 — 2500 до 3500 м н. у. м. Отличается значитель- ной выносливостью к неблагоприятным условиям, в связи с чем заслуживает внимания селекционеров. Испытывается в ряде стран. 11. Malus toringoides (Rehd.) Hughes, 1920, in Кем. Bull., 1920: 205, сит. ic.; Stapf О., 1923, in Curt. Bot. Mag. 148 t. 8948, сит ic.; Henning, 1947, in Ziichter 10 — 12: 296. — Malus fransiforia var. foringoides Rehd., 1915, in Sarg. 01. Wilson, 2: 286; Bailey, 1916, in Rhodora, 18: 155; Osborn, 1919, in Gard. Chron., 1: 522. — Pyrus foringoides Osborn 1923, in Gard, Chron., ser. 3, 73: 89 сит ic., 42. — Sinomalus foringoides goidz. 1932, in Act. Phytotax. et Geobot., 1: 11. — Яблоня торннго- видная. ~зверева или высокий кустарник высотой 3 — 10 м. Молодые побеги слегка опушенные, позже — голые. Листья в очертании яйцевидные, удлиненно-яйцевидные или овальные (до продолговато-эллиптических), длиной 3 — 8 см, шириной 1,5 — 4,0 см, обычно 5-лопастные, по краю пильчатые или городчато-пильчатые, голые или лишь снизу по жилкам слегка опушенные, на черешках длиной 2 — 3 см. Цветки по 3 — 6 в зонтиковидных соцветиях, на слабоопу- шенных или голых цветоножках длиной 2 — 3 см; венчик белый с кремовым оттенком или розоватый, диаметром 2,0 (25) см; гипантий слабоопушенный; чашелистики опушенные, преимуще- ственно с внутренней стороны; стилодии (4 — 5) голые, сросши- еся у основания в слабоволосистый столбик. Плоды обратно-яйцевидные, несколько грушевидные или ша- ровидные, суженные к основанию, на верхушке вдавленные, дли- ной 1,5 см, диаметром 1,0 (1,2) см, желтые с красным бочком и опадающей чашечкой. Очень обильно плодоносит; плоды долго не осыпаются.
138 Глава 5. Систематический обзор рода Malus Описана в 1915 г. Редером из Китая (пров. Сычуань, запад- ные районы) по сборам Э. Вилсона в 1904 г. под названием М. 1гапзМог~а, var. toringoides. В 1920 г. английским ботаником Хугсом признана самостоятельным видом. Typus: China: West-Szechuan, west of Tachien-1и, 3300 т, october 1904, n' 1285, Wilson, in herb. Inst. Bot. Ac. Sc. URSS in Leningrad conservatur. Распространена в Юго-Западном Китае (западная часть пров. Сычуань) . Имеет узкий ареал в пределах от 3000 до 3600 м н. у. м. Одна из самых декоративных яблонь, особенно во время плодоношения. Как высокогорное растение представляет интерес для селекции. В 1913 г. была введена в культуру в Англии (Кью), где с 1919 г. обильно цветет и плодоносит. Зарекомендо- вала себя как превосходное декоративное растение. 12. Malus transitoria (Batal.) С. К. Schneid., 1906. in Repert. Sp. Nov. Reg. Veg., 3: 178; С. К. Schneid., 1906, lll. Handb. Laubh. 1: 726; Rehder, 1920, in Journ. Агп. АгЬ., 2: 721; Hug- hes, 1920, in pew. Виll. 215 сит ic.; Henning, 1947, in Zuchter, 10 — 12: 296. — Pyrus transitoria Batal., 1893, Тр. Петерб. Бот. сада, 13: 95. — Pyrus Malus transitoria Anon., 1922, in Garden, 86: 485. — Sinomalus transitoria Poidz., 1934, in Act. Phytotax. et Geobot., 3: 196. — Яблоня переходная («уасы» или «васы» согласно записи Г. Н. Потанина, in herb.). деревце или кустарник высотой 3 — 5 м; молодые побеги опушенные, ветви с красно-коричневой корой. Листья в очертании широкояйцевидные, 3 — 5-лопастные, раз- дельные (до рассеченных), длиной 2 — 3 см, шириной 1,5— 2,5 см, при основании неравносторонние, клиновидные, реже- слабосердцевидные; лопасти тупозаостренные, по краю город- чато-пильчатые, более или менее опушенные, на опушенных че- решках длиной 1,5 — 2,0 см. Цветки по 3 — 8 в зонтиковидных соцветиях, на тонких слабоволосистых цветоножках; гипантии и .особенно чашечка опушенные; венчик белый, диаметром 2 см; лепестки широко продолговатые. Плоды мелкие, шаровидные, диаметром 0,5 — 0,7 см, светло красные или желтоватые, с нежной мякотью, на точких глодо ножках длиной 1,2 — 1,5 см, с опадающими при плодах чаше- листиками. Цветет V — VI, плоды созревают IX. Описана русским ботаником А. Ф. Баталиным в 1893 г. по сборам Н. М. Пржевальского из восточной части пров. Ганьсу (Китай) . Typus: China: Pansu orientale jugum Sodis-kruxum apud И. Tetung in sylvis frondosis et abruptis regionis in ferioris humo-
Глава 5. Систематический обзор рода Malus 139 sis parce 16 (28) V 1873 fl. et 20 VIII (1 IX) 1873 fr., Przewal- sky, in herb. Bot. Inst. Ac. Sc. UPSS in Leningrad conservatur. Распространена в некоторых районах Центральной Азии (в Китае (пров. Ганьсу, Цинхай, Нинся) и Монголии), где оби- тает в горных лиственных лесах, в долинах рек и по берегам озер (оз. Кукунор и др.) . Доходит до предельных высотных границ распространения видов рода hfalus (4000 м н. у. м.). И. Т. Васильченко [1963, с. 123] предполагает, что «данное растение, вместе с другими, близкими к нему видами яблони, является самым выносливым к совокупности неблагоприятных условий внешней среды видом яблони на земном шаре». 13. Malus setok Vass., 1959, Бот. матр. Герб. Бот. ин-та АН СССР, 1: 199. — Яблоня сеток. Дерево высотой до 12 м, с темно-пурпуровой корой на моло- дых побегах и серо-бурой — на взрослых ветвях. Листья в очертании продолговато-яйцевидные или треуголь- ные, длиной 3 — 5 см, шириной 2,5 — 4,0 см, перисто-раздельные (до трехрассеченных), с ланцетными или продолговато-яйцевид- ными, по краю неравномерно пильчато-зубчатыми лопастями, сверху слабо рассеянно-волосистые, снизу мелкоопушенные; при основании короткоклиновидные или закругленные, на утолщен- ных, волосистых черешках длиной 2,0 — 2,5 см. Цветки по 4 — 6 в зонтиковидных соцветиях, на мелковоло- систых цветоножках длиной 1,5 — 2,0 см; венчик белый, диамет- ром около 1,5 — 2,0 см; чашелистики ланцетные, войлочно-опу- шенные. Плоды шаровидно-овальные, диаметром 0,8 — 1,0 см, рассеян- но-волосистые, темно-пурпуровые, с поздно опадающей чашеч- кой, на тонких плодоножках длиной около 2 см. Описана И. Т. Васильченко в 1959 г. по сборам В. Ладыгина из Камского нагорья в Китае (бассейны рек Меконг, Бар-Чю, Рэ-Чю, Ному-Чю, Цза-Чю и Цзэ-Чю). Тип: Китай, Тибет, Кам, бассейн р. Меконга, р. Бар-Чю, выс. 3150 — 3300 м, 2 XI 1900, пл., Ладыгин. Хранится в Герб. Бот. инст. АН СССР (Ленинград) . Распространена в Китае на Камском нагорье, на высоте 3000 — 3500 м. От близкого вида M. transitoria отличается более крупными размерами (высотой до 12 м), более крупными листь- ями, цветками, плодами и меньшей степенью опушенности. 14. Malus centralasiatica Vass., 1959, Бот. Матр. Герб. Бот. инст. АН СССР, 19: 202. — Яблоня центральноазиатская. Дерево высотой 7 — 8 м, иногда растет кустообразно. Листья мелкие, 3 — 5-лопастные или цельные, широколанцет- ные, по краю городчатые или почти цельнокрайные, голые или рассеянно-опушенные по жилкам. Цветки по 3 — 6 в зонтиковидных соцветиях, венчик белый, в бутонах розовый, диаметром 1,5 см.
140 Глава 5. Систематический обзор рода Afalus Плоды овальные, длиной 1,0 см, диаметром 0,5 — 0,8 см, с опадающей чашечкой. Описана в 1959 г. И. T. Васильченко по сборам Гинга из пров. Цинхай (Китай). Typus: China, prov. Zichai: Lao-Che-Tze-Shan mountins, зо- uth of Sining, alt. 3350 — 3900 т, 3 VII I 1923, 1г. n' 724, R. С. Ching, in herb. Bot. Inst. Ас. Ас. Sc. URSS, in Leningrad conservatur. Распространена в тех же районах, что и близкая к ней М. transitoria, но в более низкой полосе гор. Центральноазиат- ская яблоня замещает яблоню переходную в более южных (вы- сокогорных) районах Центральной Азии. 15. Malus komarovii (Sarg.) Rehd., 1920, in Journ. Arnold Arbor., 2: 51. — Crataegus 1епиЦо1т Кот., 1901, в Тр. Бот. сада, 17: 435, non Britt. 1904, Фл. Маньчж., 2: 470; С. К. Schneid., 1906, 111. Handb. Laubh., 1: 771; Nakai, 1909, in Journ. Coll. Sci. Tokyo, 24: 180. — Сга1аедия komarovii Sarg., 1912, Pl. Wilson, 1: 183; С. К. Schneid., 1. s., 2: 1005; Nakai, 1916, Fl. sylv. Когеапа, 4: 62. — Sinomalus 1епичо1т (Котаг.) 3(oidz., 1934, in Act. Phytotax. et Geobot., 3: 4,12. — Яблоня Комарова. Деревце высотой 3 — 5 м или кустарник высотой 1 — 2 м, с гонкими, извилистыми ветвями. Листья широкояйцевидные, глубоко З-лопастные, с продол- говато-яйцевидными, несколько заостренными, пильчато-зубча- тыми лопастями, при основании сердцевидные, голые или едва рассеянно-волосистые, на коротких черешках длиной 1,5 см. Цветки по 6 — 8 (12) в щитковидных соцветиях, диаметром около 1 см, на совершенно голых, тонких цветоножках длиной 1,5 см; венчик белый, в бутонах бледно-розоватый; гипантий голый; чашелистики с внутренней стороны мелкоопушенные. Плоды шаровидные, мелкие, диаметром 0,8 см, красные, с опадающими чашелистиками. Описана в 1901 г. В. Л. Комаровым по собственным сборам из лесной полосы Корейского хребта (на высоте 1000 — 2000 м н. у. м.) под названием Crataegus 1епш~ойа. В 1920 г. Редером перенесена в род Malus и названа яблоней Комарова. Тип: Корея: долина Сегельсу-Корани, басс. истоков р. Ту- мин-Чана, путь от г. Мусанга к р. Ялу, 16 — 18 VI 1897, цв., Комаров. Хранится в Герб. Бот. инст. АН СССР (Ленинград). Распространена в лесах Корейского хребта на высоте от 1000 до 2000 м н. у. м., откуда заходит в прилегающие районы Китая. Series (ряд) 3. Т о г i n g о п а е (Rehd.) Langenf. Rehder, 1940, Man. Cult. Trees Shrubs, ed. 2: 390. — Malus sect. Sorbo- malus subsect. Sieboldinae Rehd., 1920, in Journ. Агп. АгЬ., 2: 48.
Глава 6. Систематический обзор рода Malus Рис. 50. Malus toringo Деревья средней высоты или низкие кустарники. Листья мелкие (до средних), длиной 2,5 — 7,0 см, шириной 3,0 — 5,0 см, различной формы, на коротких побегах — цельные, на длинных — 3-, реже 5-лопастные. Цветки в плоских зонтиковидных соцветиях. Стилодии (3 — 4), сросшиеся до половины в мохнато-опушенный столбик. Плоды шаровидные, диаметром 0,8 — 1,5 см, с мучнистой мя- котью и опадающей чашечкой.
]42 Глава 5. Систематический обзор рода Malus Тип серии: Malus sieboldii (Regel) Rehd. Серия Toringonae представлена одним полиморфным ви- дом — М. toringo (рис. 50), который включает 2 экологически и морфологически обособленных разновидности. Различные формы этой яблони давно известны в культуре. Известны также некоторые гибриды, из числа которых выведены декоративные сорта. 16. Malus toringo Siebold, ех de Vriese, 1856, in Tiunbouw.— f1. Niederl., 3: 368 t. 17. 1) var. toringo Siebold 1856, Cat. Raison 4 sine descr.; Ca- riere, 1871, in Rev. Hort., 1870: 451, сит ic. 63; С. К. Scheid, 1906 Ill. Handb. Laubh. 1: 723, сит ic. 399 с, 400 g-h. — Malus зсеЬоЫсс (Regel) Rehd., 1915, in Sar gent, Pl. Wilson, 2: 293; Rehd., 1932, in Journ. Агп. АгЬ., 13: 308; Asami, 1927, СгаЬаррl. Nectar. Jap., 38, сит ic. 26 — 33; 35, 36; Suringar, 1928, in Me- ded., Rijks. Leid., 56; 43. — Pyrus riuularis sensu Gray, 1857, in Mem. Am. Асад. Arts Sci., n. ser. 6: 388, non Hocker, 1934.— Pyrus sieboldii Regel, 1859, in Ind. Sem. Hort. Petrop., 1858: 51; Regel, 1859, in Gartenf1., 8: 82. — Sorbus toringo K. Koch, 1864, in Ann. Mus. Bot. Lugd. — Bat., 1: 249. — Pyrus toringo Miquel, 1867, in Ann. Mus. Bot. Ludg. — Bat., 3: 41 (Ргоl. Fl. Jap. 229). — Pyrus mengo Siebold, 1869, ех K. Koch, Dendr., 1: 213, pro syn. — Pyrus toringo а. typica Maxim., 1873, ех Franchet et Savatier, Enum. Pl. Jap., 1: 139; Maxim., 1878? ех ор. cit., 2: 350. — Pyrus toringo Ь. incisa Franchet et Savatier, 1973, 1878, 1. s. — Pyrus riuularis Ь. toringo Wenzig, 1874, in Linnaea, 38: 39. — Malus microcarpa toringo Cariere, 1883, Et. Pomm. Microcarp., 61, сит ic. 11. — Malus baccata subsp. toringo Koidzumi, 1911, in Bot. Mag. Tokyo, 25 — 76. — Malus sieboldii var. Aoringo c. incisa (Fr. et Sav.) Koidzumi, 1916, in Bot. Mag. Tokyo, 30: 331. — Яблоня торинго. зверева или деревце, иногда имеющее вид куста, высотой 4 — 6 или 6 — 8 м, с широкой кроной, несколько колючими и пла- кучими ветвями, с пурпурово-коричневой корой. Молодые побеги опушенные. Почки мелкие, обычно яйцевидные. Листья различные по форме, яйцевидные или эллиптические, с клиновидным или закругленным основанием, на верхушке за- остренные, длиной 4 — 8 (2,5) см, шириной 2 — 4 (1,5) см; на коротких побегах — цельные, остропильчатые, на длинных (рос- товых) — 3-, реже 5-лопастные, по краю крупнопильчатые, зуб- чатые, в молодости опушенные с обеих сторон, а во взрослом состоянии — почти голые, на опушенных черешках длиной 1— 2 см. Цветки по 3 — 7 в зонтиковидных соцветиях, бледно-розовые или почти белые, в бутонах (до их распускания) ярко-розовые; венчик диаметром около 2 см; гипантий и чашелистики вой- лочно-опушенные; стилодии (3 — 4), у основания сросшиеся в
Глава 5. Систематический обзор рода Natus 143 мохнато-опушенный столбик; цветоножки тонкие, опушенные, длиной около 2,5 см. Плоды шаровидные, диаметром 0,8 — 1,0 см, желтовато-ко- ричневые или краснеющие, кислые, с мучнистой мякотью, на тонких плодоножках длиной 2,5 — 3,0 см. Описана немецким ботаником-японологом П. Зибольдом (Р. F. Siebold, 1796 — 1866) по экземплярам, интродуцирован- ным в 1856 г. из Японии. Тип не указан. Neotypus: Japan, Loc. Honshu. Pref. Nagano: en route from Senjongahara to Mt. Mikuni-yama, Azusayama, Pawakami-тига, Minamisaku-gun, alt. 1700 т, 25 V 1963, n' 12195, М. Hotta, in herb. Bot. Inst. Ac. Sc. URSS in Leningrad conservatur. Распространена в Японии, на Корейском п-ове, а также в восточной части Китая (пров. Хунань, Квейхоу, Куангси) (по гербарным образцам: сборы Хандель-Маццетти, 1917, 1918 и китайцев Vang-Te-Hui, 1919; Та-Но-Уеп, 1931; Fang-Ching-Ghan, 1931 и др.) . Эта разновидность распространена преимущественно по рав- нинам и долинам рек, на высоте от 600 до 1000 м н. у. м., реже встречается в горных лесах (до 1500 — 1700 м н. у. м.). Различные формы этой яблони давно известны в культуре не только в Японии, Корее, Китае, но также в Европе и Северной Америке. В Европу интродуцирована в 1856 г. М. loringo var. <oringo представляет собой довольно ксероморфное рас- тение. Имеет непосредственные родственные связи с древовидной разновид- ностью этой яблони. От последней отличается более низким, кустообразным ростом, лопастными листьями, интенсивно опушенными побегами, листьями и частями цветка, более мелкими цветками, собранными в мкогоцветковые зонтиковидные соцветия. Мнение некоторых авторов, что к var. toringo относятся только куль- турные формы яблони торинго, недостаточно обосновано. Яанная разновид- ность имеет широкое распространение в диком состоянии. Тем не менее ареал ее в Китае (особенно в крайних для яблони условиях — в субтропи- ческой зоне, пров. Хунань) необходимо тщательно исследовать. 2) Malus toringo var. arborescens Rehd., 1915, in Sargent, Pl. Wilson, 2: 294. — ? Pyrus Ьасса1а Thunnerg, 1784, Fl. Jap. 207. — Crataegus alnifolia sensu Regel, 1871, in Act. Hort. Pet- rop., 1: 125, non Sieboldii et Zuccarini, 1846. — Crataegus cava- /еперь Leveille et Vaniot, 1908, in Виll. $ос. Bot. France, 55: 58, 1915, Fl. gong.-Tcheon, 346, exclud. n' 1303. — Pyrus subcrata- egifolia Leveille, 1909, in Repert. Sp. Nov. Reg. Veg., 7: 199.— Photinia rubro-lutea Levei1le, 1911, in op. cit., 9: 460. — Malus toringo f. typiica Matsumura, 1912, Ind. Pl. Jap., 2, 2: 205; Koi- dzumi, 1913, in Jour. Coll. Sci. Tokyo, 34, 2: 81, non Pyrus toringo а. typica Franch. et Lav., 1873. — Crataegus taquetii Leveille, 1912, in Repert. Sp. Nov. Reg. Veg., 10: 377. — Pyrus esquirolii Leveille, 1913, in op. cit. 12: 189. — Malus toringo
Глава 5. Систематический обзор рода Ма1из 144 Я;ревовидная разновидность яблони торинго сохраняет родственные связи с некоторыми видами юньнаньских яблонь (М. yunnanensis, М. prattii). Сходство состоит главным образом в общности габитуса и структуры листьев. Тем ие менее, var. arborescens в значительной мере отличается от своих вероятных предков по ряду признаков и, прежде всего, по строению соцветий, цветков и плодов. В пределах этой разновидности существует ряд переходных форм, смыкающихся с типичной яблоней торинго. Г и б р и д ы, по совокупности признаков примыкающие к се- рии (Species hybridogena ad series Toringonae accedentes): Ма1икЯ1огйипйа (М. baccata)(M. sieboldii) Siebold, 1856, Cat. Raison, 1856. 5. — Яблоня обильноцветущая. Malus)(arnoldiana (М.)(floribunda)(M. baccata) Rehd., 1920 in Jour. Arnold Arb. 2: 37. — Яблоня Арнольда. Malus)(zumi (М. mandshurica)(M. sieboldii) (Matsum.) Reh der, 1905, in Sar gent, Trees et Shrubs, t. 91, 1: 191. Яблоня Цуми (рис. 51) . Malus)(sublobata (М.)(prunifolia)(M. sieboldii) Dipp. Rehd., 1920, in Jour. Arnold. Arb., 2: 56. — Яблоня лопастная. В культуре известны и другие сложные гибридные комбина- ции этой яблони, иногда описанные под бинарными названиями (например, М. floribunda)(M. prunifolia=M. scheideckeri и др.). >akai, 1916, Fl. Sylv. Ког., 6: 35, t. 10, чох Siebold, 1856, et vix Cariere, 1871. — Ма1ик sieboldii var. koringo b. vulgaris I(oidzumi, 1916, in Bot. Mag. Tokyo, 30: 331. — Ма1ик sieboldii Rehd., 1932, in Jour. Arn. Arb., 13: 308, р. р. jIepeao мезофильного облика, высотой до 10 м. Отличается от предыдущей разновидности более крупными, менее лопастными и в меньшей мере опушенными листьями чисто-белыми цветками (диаметром до 3 см), шаровидными плодами (диаметром 1 см) желто-красного цвета, удерживаю- щимися на дереве до декабря. Neotypus: Japan — Honshu: Abovethe I(ongo-fall, on the foot Mt. Imalkuma, Itsukaichi, Prov. Musashi, 27 V 1956, fl., n' 2129, Toshio Hamaina, in herb. Inst. Sylvabot. Univers. Tokyoensis in Tokyo conservatur. древовидная разновидность яблони Торинго является лесной ее расой. Встречается в горных лесах Японии и Корейского п-ова на высоте 1200 — 1700 м н. у. м. в зоне летне-зеленых широколиственных лесов, где растет вместе с широколиствен- ными древесными породами и кустарниками, из которых наибо- лее характерными являются Quercus dentata, Q. glandulifera, Асег ginnala, А. japonica, Juglans sieboldii, Fagus sieboldii, Aes- culus turbinata, Ulmus japonica, Fraxinus longicuspis, Prunus serrulata, Lapix leptolepis, а также некоторые лианы (Нуйгап- gea petiolaris, Hedera japonica) и множество кустарников из родов Syringa, Enonymus, Viburnum, Spireae и др.
Глава 5, Систематический обзор рода Malus 145 СМ Рис. 51. MalusXzumi Из этих гибридов выведены некоторые сорта декоративных яб- ЛОНЬ. Malus)(sargentii (Pehd.) Langenf. Malus sargentii Pehd., 1903, in Sargent, Trees Shrubs, 1: 71, t. 36; Bean., 1918, in Bot. Mag., 144: t. 8757. — Pyrus sargentii Bean, 1914, Trees Shrubs Brit. Isl., 2: 293. — Pyrus malus sargentii Anon, 1915, in Jour. Hort. Soc. Lond., 41: 96. — Malus sieboldii, var. koriingo а. sargentii Poidzumi, 1916, in Bot. Mag. Tokyo, 30: 331. — Malus
Глава 5. Систематический обзор рода Malus 146 sieboldii var. sargentii, f. adscendens Asami, 1927, СгаЬарр1. Nectar. Jap., 40, t. 21, 42 с. — Яблоня Саржента. Кустообразное деревце или кустарник высотой 1,5 — 2,5 м, с горизонтально расположенными ветвями и укороченными по- бегами, которые иногда заканчиваются колючкой. Кора ветвей темно-серая или почти черная, на годичных побегах — темно- серая с коричнево-пурпуровым оттенком. Листья яйцевидные или эллиптические, с закругленным осно- ванием и заостренной верхушкой; на укороченных побегах— обычно цельные, почти цельнокрайные или остропильчатые; на длинных — острозубчатые, обычно З-лопастные, длиной 5— 8 см, шириной 3 — 6 см. Цветки в зонтиковидных или пучковидных соцветиях по 5 — 7, белые, диаметром около 2,5 см, с округленным венчиком, на голых, пурпуровых цветоножках длиной 2 — 3 см; стилодии (4), срастающиеся в нижней половине в мохнато-опушенный столбик. Плоды шаровидные, диаметром около 1 см, темно-красные или оранжевые, с мучнистой мякотью. Описана по неизвестным образцам, вероятно, взятым из культуры, или особям, интродуцированным из Японии. От близкой разновидности M. toringo чаг. toringo яблоня Саржента отличается низким ростом, кустообразной формой роста и более ксеро- морфным обликом, а также горизонтально расположенными ветвями и на- личием множества укороченных колючих побегов, более широкими, круп- ными, яйцевидными листьями, с закрученным основанием, более интенсивным опушением вегетативных органов и частей цветка, обильным цветением и длодо ношением. Отсутствие образцов в просмотренных гербариях (Le, TIF, G, W, St, Р и др.) позволяет заключить, что М. sargentii едва ли может считаться самостоятельным видом, так как нет точных данных о распространении этой яблони в природе. Кроме того, при семенном размножении яблоня Саржента не сохраняет характерных признаков, а у ее потомства проявля- ется целый ряд форм гибридного характера. Все это послужило основанием для признания яблони Саржента яблоней гибридного происхождения. Series (ряд) 4. Fuscae Eseltine, 1933, in Agr. Exp. Stat. Bull., 208. Цветки в многоцветковых (6 — 12) щитковидных соцветиях; стилодии 3 — 4 (2), сросшиеся до половины в голый столбик. Плоды мелкие, диаметром около 1,5 см, овальные, с опадаю- щей чашечкой, желтые с красным румянцем. Цветки и плоды без аромата. Тип серии: Afalus fusca (Raf.) С. К. Schneid. 17. Malus fusca (Raf.) С. К. Schneid., 1906, 1:723, сит ic. 399 d, 400 1; Федоров и Полетика, 1954, ~ер. куст. СССР, 3:454; Васильченко, 1963, Новые для культ. виды ябл., 131.— Pyrus fusca Rafinisque, 1830, Med. Fl. 2: 254, nom. subnud.— Pyrus diversifolia Bongard, 1833, in Мет. Асад. Sci. St. Petersb.,
Глава 5. Систеыатический обзор рода Malus 147 Рис. 52. Malus musca вег. 6, 2: 133. — Pyrus rivularis Douglas, 1834, ех Hoocker, Fl. Вог.-Am., 1: 203, t. 68. — Sorbus rivularis Heynhold, 1840, No- тепс1. Bot. 1: 773. — Malus rivularis M. J. Roemer, 1847, Fam. Nat. Reg. Veg. Syn., 3: 215. — Malus diversifolia M. J. Roemer, 1847, 1. s. — Malus rivularis var. diversifolia goehne, 1893, Deu- tsche Dendr. 262. — Pyrus fusca var. diversifolia Bailey, 1916, Stand. Сус1. Hort 5: 2876. — Яблоня бурая, или приречная. Дерево высотой до 10 — 12 м, с выраженным стволом диамет- ром до 30 — 45 см. Иногда растет кустообразно (особенно в усло- виях культуры). Ветви темно- или каштаново-коричневые. По- беги ярко-красные, голые или слабоволосистые. Почки мелкие, тупые, каштаново-коричневые, длиной 1,5 мм. Листья яйцевидные, продолговато-яйцевидные или продолго- вато-ланцетные, с короткоклиновидным или несколько сердце-
Глава 5. Систематический обзор рода Malus 448 Ключ для определения видов 1. Листья продолговато-ланцетные 19. М. angustifolia (Ait.) Michx. — Листья яйцевидные, эллиптические, овальные или про- долговато-яйцевидные.......... 2 видным основанием, острой верхушкой, прижато-пильчатые, цельные или реже (на длинных побегах) слабо З-лопастные, сверху голые, снизу слегка опушенные, длиной 2,5 — 8,0 (10) см, шириной 1,2 — 3,0 (4) см, на черешках длиной 1,5 — 2,0 см. Цветки в многоцветковых (6 — 10) щитковидных соцветиях, белые, реже — розоватые, диаметром 2,0 см, на толстоватых слабоволосистых цветоножках длиной 2,5 — 3,0 см; стилодии (3 — 4 (2)), сросшиеся до половины в голый столбик. Плоды овальные, длиной 1,5 — 2,0 см, диаметром 0,5 — 0,8 см, зеленовато-желтые, с красным румянцем (рис. 52). Описана американским ботаником Рафинеском под назва- нием Ругив fusca с тихоокеанского побережья Северной Америки. Neotypus: Alaska: extra opidum Junean in latere alpis sylva- tica, lat. 58'30', long. 135'5', 5 IX 1913, S. J. Enander, in Her- barium Riksmuseum in Stockholm conservatur. Распространена на тихоокеанском побережье Северной Аме- рики (от Аляски и Алеутских о-вов до Калифорнии). Г и б р и д, по совокупности признаков примыкающий к се- рии Fuscae: Malus>(daavsoniana (М. fusca>(M. ритйа sensu Mill., 1768) Rehd., 1907, in Sargent, Тгеез Shrubs, 2: 23, t. 3. SECT[0 (секция) [Ч. CHOROMELES (Decne.) Rehd., 1920, in Jour. Arnold АгЬ., 2: 48; nom mutatum т. — Malus subgen. Cloromeles Decanisne, 1874, in Nouv. Arch. Mus. Hist. Nat., Pa- ris, 10: 155. — Chloromeles, Decanisne, 1883, in Fl. des Serr., 23: 156. — Malus sect. Calycomeles goehne, 1883, Deutsche Dendr., р. р. Malus subsect. Coronariae Rehd., 1913, in Sargent, Тгеез Shrubs, 2: 142. — Зеленоплодние яблони. Яблони Северной Америки. В основном деревья с хорошо выраженными стволами. Листья в почкосложении сложенные, более или менее ло- пастные (по крайней мере на длинных побегах), продолговато- яйцевидные, Зубчато-пильчатые, городчатые или почти цельно- крайные. Цветки в щитковидных соцветиях, на тонких цветоножках; венчик шаровидный, полуоткрытый; пыльники фиолетовые; сти- лодии (5), сросшиеся у основания в столбик. Плоды крупные (диаметром до 5 см), шаровидные, с оста- ющейся чашечкой (рис. 53). Тип секции: Malus coronaria (L.) Mill.
Глава 5. Систематический обзор рода Malus Рис. 53. Виды секции Chloronieles:1 — М. coronaria; 2 — М. angustigolia; 3 — М. ioensis 2 Плоды, сжатые с сплюснутые полюсов, плоско-шаровидные или 18. М. согопапа (L.) Mill до эллипсоидальных, несколько вы- и у плодоножки . . 20. М. ioensis (Wood) ВгШ. — Плоды шаровидные тянутые у чашечки Series (ряд) 5. Co ron ar lac (Rehd.) Langenf. сот. nova. Malus аиЬвес1. Coronariae Rehd., 1913, in Sargent, Trees Shrubs, 2: 142. Flores in paucifloris (4 — 6) corymboformibus inflorescentiis maxime fragrantes. Stylodia 5, concreta usque ad 1/3 longitudi- nis in stylum albopubescens.
Глава 5. Систематический обзор рода Malus Fructus magni, са 2,5 — 3,0 (1,5 — 3,5) usque ad 5 ст in diam., sphaerici, viridi (maturi соlоге sunt flavidi), maxime aromatici сит sepalis persistentibus. Typus series: Malus coronaria (L.) Mill. Цветки в немногоцветковых (4 — 6) щитковидных соцветиях, очень душистые. Стилодии (5), сросшиеся до 1/3 длины в бело- опушенный столбик. Плоды крупные, диаметром от 2,5 — 3,0 (1,5 — 3,5) до 5 см, шаровидные, зеленые (зрелые — желтоватые), очень аромат- ные, с остающимися чашелистиками. 18. Malus coronaria (L.) Mill., 1768, Gard. Dict., ed. 8: n' 2. Britton and Brown, 1897, 111. Fl. N. S. U., 2: 235, сит. ic. 1979; Rehder, 1915, im Mitt. Deutsch. Dendr. Ges. 1914 (23): 260; Федоров и Полетико, 1954, jIep. куст. СССР, 3: 456; Васильченко, 1963, Новые для культ. виды ябл., 133. Pterus coronaria Linnaeus, 1753, Sp. Рl. 480, р. р.: 0и Rof., 1772, Harbk. Baumz., 2: 729; Sargent, 1892, Silva N. Amer., 4: 71, 167. — Pterus malus coronaria Weston, 1770, Bot. Univ., 1: 230. — Pterus microcarpa coronaria Carriere, 1883, Et. Ротт. Microcarp. 133, сит ic. 17. — Malus glaucescens sensu Britton and Brown, 1913, 111. Fl. N. U. S. ed. 2, 2: 280, сит ic. 2323, р. р.; non Rehder, 1911. — Malus jragrans Rehd., 1913, in Sar- gent, Trees Shrubs, 2: 278. Яблоня венечная. Дерево высотой 5 — 10 м с хорошо выраженным стволом диа- метром 30 — 35 см. Кора на стволе растрескивается в продоль- ном направлении и слущивается узкими кусками коричневого или красно-коричневого цвета. Немногочисленные (3 — 5) расто- пыренные и колючие ветви, отходящие от ствола на высоте 2 — 3 см, обычно с яркой коричнево-красной корой, образуют широкую, прозрачную крону. Годичные побеги голые или слабо- опушенные, преимущественно на концах. Почки мелкие, длиной около 1,5 мм, тупые, с ярко-красными, реснитчатыми по краю чешуями. Листья тонкие, при распускании бронзово-красные, волосис- тые, позже — зеленые, почти голые, осенью — шарлахово-крас- ные, оранжевые, желтеющие, яйцевидные, продолговато-яйце- видные или овальные, длиной 5 — 10 см, шириной 3 — 7 см, с закругленным, едва сердцевидным или ширококлиновидным основанием и острой верхушкой, по краю неравномерно зубча- тые, зубчато-пильчатые или пильчатые, на длинных побегах, иногда слегка лопастные, на тонких черешках длиной 3 — 4 см, с парными, заостренными прилистниками. Цветки очень душистые, по 4 — 6 (3) в щитковидных соцве- тиях, на тонких, совершенно голых или слабоволосистых крас- неющих цветоножках длиной 2,5 — 3,0 см; венчик белый, светло- или ярко-розовый, диаметром до 3 см (в культуре — до 4,0— 4,5 см); лепестки неправильной формы или обратнояйцевидные,
Глава 5. Систематический обзор рода Malus 151 обычно на верхушке реснитчатые, слегка городчатые или зуб- чатые; стилодии (5), сросшиеся более чем на 1/3 длины в бело- войлочный столбик; рыльца двурасщепленные; чашелистики треугольные, снаружи голые, изнутри беловойлочные; гипантий обычно голый или слабоволосистый. Плоды сжатые с полюсов, плоскошаровидные или сплюсну- тые, зеленые, зрелые — желто-зеленые, слегка просвечивающи- еся, покрытые восковым налетом, диаметром 2,5 — 3,5 см (реже крупнее), очень душистые, с твердой, приятно-кислой, реже терпкой или горькой мякотью, долго удерживающиеся на де- реве. 11ветет Ч — VI (на родине — в начале VI). Плоды созревают Х — XI. Описана Линнеем в 1753 г. под названием Pyrus coronaria по образцам из Северной Америки (sine loco). Neotypus: Canada: Ontario, St. Thomas, 26 Ч 1902, fl., 9 Х 1904, fr., G. Fischer. 1п Herb. Conservatoire et Jardin botaniques, in Geneve, Switzerland, conservatur. Распространена в Северной Америке (от Онтарио через Нью- Лорк и Пенсильванию вдоль Аппалачей до Алабамы, Миссури и Монтаны). Обитает на лесных опушках, реже — в лесах. Растет на богатых и сильно увлажненных почвах, образуя сплошные за- росли. Реже встречается на глинистых склонах гор. Интродуцирована в Европу в 1724 г. [Ал. А. Федоров, По- летика, 1954]. 19. Malus angustifolia (Ait.) Michx., 1803, Fl. Вог. Ат., 1: 292; Ругиз согопаг~а Linnaeus, 1753, Sp. Pl. 480, р. р. Pyrus malus sempervirens Weston, 1770, Bot. Univ., 1: 230, nom. — Ругиз angustifolia Aiton, 1789, Hort. Kew, 2: 176; Sar- gent, 1892, Silva N. Атег., 4: 75, t. 169. — Malus sempervirens (Desfontatnes, 1807, Hist. АгЬ. Abriss., 2: 141. — Pyrus еетрего~- rens Willdenow, 1813, Епит. Hort. Вегаl. Suppl. 35. — Pyrus coro- naria чаг. angustitolia Wenzig, 1874, in Linnaea, 38: 41. — Ma- lus microcarpa sempervirens Carriere, 1883, Et. Ротт. Micro- carp., 136, сит ic. 18. — Pyrus coronaria sensu Rehder, 1913, in Sar gent, Tree Shrubs, 2: 229: non Miller, 1769. Яблоня узколистиая. Дерево с коротким стволом диаметром 25 — 30 см или кус- тарник высотой 5 — 7 (10) м. На сухих почвах растет кустами высотой 2 — 3 м, с серо-коричневой корой на стволах, коричне- вой — на многолетних ветвях; побеги голые, лишь в молодости слегка опушенные, иногда полувечнозеленые. Почки небольшие, длиной около 1,5 мм, каштановые или каштаново-коричневые, слегка опушенные. Листья кожистые, вытянутые (от продолговато-ланцетных до узкопродолговато-обратнояйцевидных), длиной 3 — 7 см, ши-
]52 Гяава 5. Систематический обзор рода Ма1из риной 1,5 — 2,5 см, с ширококлиновидным основанием и заос- тренной верхушкой, городчато-пильчатые (до почти цельнокрай- ных), на ростовых побегах грубозубчатые (до слаболопастных), в начале слабоопушенные, позже — почти голые, на черешках длиной 1,5 — 2,0 см. Цветки в немногоцветковых {3 — 5) щитковидных соцветиях, на голых цветоножках длиной 2 — 3 см, очень душистые; венчик бело-розовый, диаметром около 2,5 см; лепестки продолговато- овальные, с закругленной цельнокрайной верхушкой; стилодии (5), сросшиеся до 1/3 длины в опушенный столбик; гипантий белый, реже слабоопушенный; чашелистики треугольные, из- нутри беловойлочные. Плоды шаровидные, сжатые или слегка сжатые с боков, грушевидные, диаметром 1,5 — 2,5 см, на обоих концах углуб- ленные, светло-желто-зеленые, зрелые — очень душистые, с твердой кислой мякотью. Цветет VI, плоды созревают Х — XI. Описана английским ботаником Айтоном под названием Pterus angustifolia, по-видимому по образцам, интродуцирован- ным из Северной Америки. Neotypus: In quercetis, ad lac. Jamony, Florida, III 1845, Rugel. In Herb. Conservatoire et Jardin botaniques in Geneve, Swietzerland et Herb. Naturhistorisches Museum in Wien, Austria conservatUr. Распространена в Северной Америке (от Пенсильвании, вдоль приатлантических штатов, до Флориды, Луизианы и на север — до Теннесси) . Обитает преимущественно в долинах рек, на лесных прогалинах. В Европу интродуцирована в 1750 г. [Ал. А. Федоров, По- летико, 1954]. 20. Malus ioensis (Wood) Britton, 1897, in Britt. and Brown, Ill. Fl. N. U. S., 2: 235. Britton, 1901, Man. Fl. N. Stat. Can., 516; S ar gent, 1905, Man. Trees N. Amer., 354, сит ic. 278; Rehder, 1913, in Sargent, Тгеез Shrubs, 2: 230; Федоров и Полетика, 1954, Дер. куст. СССР, 3: 457; Васильченко, 1963, Новые для культ. виды ябл.: 35. — Pyrus coronaria var. ioensis Wood, 1860, Class-В. Bot. new ed., 353; Sargent, 1892, Silva N. Amer., 4: 72, t. 168. — Pyrus ioensis Bailey, 1891, in Am. Gard. 12: 473, сит ic. 7, 8; Bailey, 1892, in Bot. Gard. 17: 60. Malus coronaria var. ioensis С. К. Schneid., 1906, Ill. Handb. Laubh. 1: 724, сит ic. 40 q-г. — Malus coronaria var. typica Rehder, 1915, in Mitt. Deutsch. Ges. 1914 (23): 262. Яблоня айовская. зверева высотой 6 — 8 (10) м, со стволом диаметром 30— 45 см, иногда растет кустообразно, высотой 4 — 7 м. Немного- численные колючие ветви с ярко-красной корой образуют про-
Глава 5. Систематический обзор рода Malus l53 зрачную крону неправильной формы. Молодые побеги густо- серовойлочные, позднее — красно-коричневые, почти голые. Листья яйцевидные, эллиптические, продолговато-яйцевидные (до продолговатых}, длиной 5 — 10 см, шириной 3 — 6 см, с ок- ругленным ил и ширококли нов идным основ анием и острой, реже закругленной верхушкой, неравномерно пильчатые или нередко слаболопастные, с неравномерно остро- или грубопильчатыми лопастями, сверху темно-зеленые, блестящие, голые, снизу опу- шенные (до войлочных), реже голые, бледно-желтовато-зеленые, осенью темно-красные или желтые, на черешках длиной 1,5— 3,0 см. Цветки диаметром 2 — 3 см, на тонких, слабоопушенных или голых цветоножках длиной 2 — 3 см; лепестки белые, розовые (до ярко-розовых); стилодии (5), сросшиеся примерно до 1/3 длины в опушенный столбик; чашечка и гипантий опушенные; чашелистики удлиненно-треугольные, изнутри беловойлочные. Плоды шаровидные (до эллипсоидальных), несколько вытя- нутые у чашечки и плодоножки, сначала опушенные, позже— покрытые восковым налетом, диаметром 2 — 3 см, зеленые, очень душистые. Цветет (1V) V — Ч1, плоды созревают 1Х — Х. Описана английским ботаником Вудом в 1860 г. под назва- нием Pyrus coronaria var. ioensis. Neotypus: Атег1сае borealis: Missouri, thickets, limestone uplands, near Pattonsburg, Daviess County, 14 Vl 1924, Е1., n' 25466, Е. J. Ра1тег. ln Herb. Naturhistoriska Riksmuseum, in Stockholm, Sweden, conservatur. Распространена в Северной Америке (бассейны Миссисипи и Миссури}. Интродуцирована в Европу в 1885 г. [Ал. А. Федоров, По- летика, 1954]. С о м н и т е л ь н ы е в и д ы (species non satis notae) . Четыре североамериканских вида яблонь являются сомнительными и вряд ли могут быть описаны в качестве видов. Мы присоединя- емся к мнению В. Хеннинга [Henning, 1947] и Ф. Д. Лихоноса [1964] о том, что число североамериканских видов яблонь сле- дует сократить. Сюда относятся описанные ниже условные виды, так как точные данные об их ареалах отсутствуют, а признаки не позволяют отличать эти «виды» от других. Malus bracteata Pehd., 1913, in Sargent, Trees Shrubs, 2: 230. Malus glaucescens Rehd., 1911, in Sargent, Trees Shrubs, 2: 139, t. 187. Malus glabrata Rehd., 1913, in Sargent, Trees Shrubs, 2: 225, t. 188. Malus lancifolia Рейд., 1911, in Sar gent, Trees Shrubs, 2: 141, t. 139.
Глава 5. Систематический обзор рода Malus 164 М Я C ы о z Ь q T < ц < О З + u C 3 се Е ф о м z~ w L Р с( z % Ы 3 IS Рис. 54. Схема филогеиетических отио-шений в секции Gymnomeles Г и б р и д ы, по совокупности признаков примыкающие к серии Coronariae: Malus)(platусагра (М. coronaria)(M. pumila sensu Mill., 1768) Rehd., 1913, in Sargent, Trees Shrubs, 2: 227, t. 189. Malus)(heterophylla (М. coronaria)(M. pumila sensu Mill., 1768) Spach, 1832, Hist. Nat. Veg. Phsm., 2: 238. Malus)(soulardii (M. ioensis)(M. pumila sensu Mill., 1766) (Bailey) Britt, 1901, Man. Fl. N. Stat. Malus)(denboerii (М. ioensis)((?) М.',~ригригеа) gruessm., 1960, Handb. Laubgeh., 2: 113. SECTlO (секция) У. Gymnomeles Poehne 1893, Deutsche Dendr. 257. — Malus sect. Eumalus subsect. Baccatae Rehder, 1920, in Jour. Arnold Arbor., 2: 47. — Malus sect. Eumalus ser. Baccatae Rehder, 1940, Man. Cult. Trees Shruzs, ed. 2, 390, р. р. — Ягодные яблони. Мелкоплодные яблони, с опадающей при плодах чашечкой. Листья в почкосложении свернутые, цельные. Е~ветки в зонти- ках. Плоды мелкие, на длинных тонких плодоножках. Семенные гнезда на верхушке вытянутые, острые (рис. 54). Тип секции: М. baccata (L.) Borkh. Ключ для определения видов 1. Листья при распускании зеленые, овальные, яйцевидные или эллиптические ser. Baccatae Rehd. пурпуровые, продолговатые . ser. Hupehenses Langenf. — Листья при распускании
Глава 5. Систематический обзор рода Malus 155 Рис. 55. Виды серии Васса1ае:1 — М. Ьасса1а; 2 — М. rnandshurica; 3 — subsp. sachalinensts 2. Черешки у взрослых листьев голые . — Черешки у взрослых листьев опушенные..... 4 3. Листья крупнопильчатые 23. M. himalaica (Maxim.) Vass. — Листья тупогородчато-пильчатые . 21. M. baccata (1 .) Borkh. 4. Листья в нижней части или по всей длине цельнокрай- ные, на верхушке тупые или с коротким остроконечием 22. М. mandshurica (Maxim.) Кот. — Листья по всей длине остропильчатые с длиннооттяну- тым остроконечием . M. mandshurica subsp. sachalinensis (Кот.) Likh.
156 Глава 5. Систематический обзор рода Malus 5. Листья продолговато-яйцевидные, мелкоостропильчатые . 24. М. hupehensis (Ратр.) Rehd. — Листья продолговато-узкояйцевидные, зубчатые . 25. М. halliana (Anon.) КоеЬпе Series (ряд) 6. В а с с a t a е (Rehd.) Rehd., 1940, Man. Cult. Trees Shrubs, ed. 2, 390. — Malus sect. Eumalus subsect. Bacca- tae Rehd., 1920, in Jour. Arnold Arb., 2: 47. Листья при распускании зеленые, эллиптические, яйцевид- ные, заостренные, по краю пильчато-зубчатые, с округлым осно- ванием и зеленым черешком (рис. 55). Яблони сибирского (Восточная Сибирь) и дальневосточного очагов (Дальний Восток, о-в Сахалин, Маньчжурия, Северный Китай) . Тип серии: Malus baccata (L.) Borkh. 21. Malus baccata (L.) Borkh., 1803, Theor.-prakt. Handb. Forstbot., 2: 1280. 1) f. baccata Rehder, 1915, in Sargent, Р. Wilson, 2: 191; Юзепчук, 1939, Фл. СССР, 9: 370; Федоров, 1954, Рер. куст. СССР, 3: 442. — Pyrus baccata Linnaeus, 1767, Mant. Pl. 75; Maximowicz, 1875, in Bull. Acad. Sci., St. Petersb., 19: 70 (in Mel. Biol., 9: 166); Ascherson et Graebner, 1906, $уп. Mitteleur Fl. 6, 2: 70, — Malus rossica, Medicus, 1793, Gesch. Bot., 78.— Malus sibirica Borkh., 1798, in Arch. fur Bot. (Roemer), 1, 3: 89; Pomarow et К1оЬиКоча-Alisova, 1932, Кеу Pl. Far.-East. Reg., 2: 638. — Malus baccata var. leiostyla Ruprecht et Maximowicz, 1857, in Виll. Acad. Sci., St. Petersb., 15: 132. — Pyrus baccata а genuina Regel, 1862, in Gartenf1., 11: 207. — Ругила microcarpa Wendland, 1869, ех К. КосЬ, Dendr., 1: 211, pro. syn.; non De Candolle, 1825. — Pyrus baccata sibirica Maximowisz, 1875, in Виll. Acad. Sci., St. Petersb., 19: 170 (in Mel. Biol., 9: 166).— Malus microcarpa baccata Carriere, 1883, Et. Ротт. Microcarp., 68. — Malus baccata var. sibirica С. К. Schneid., 1906, 111. Handb., 1: 720. — Malus pallasiana Juz., 1939, Фл. СССР, 9: 370, t. 22, сит ic. 4; Федоров, 1954, Дер. куст. СССР, 3: 444, сит ic. 68. — Яблоня ягодная (сибирская). Небольшое дерево высотой 4 — 8 м, с низкой, округло-шаро- видной кроной и кривым стволом диаметром около 15 см, или крупный кустарник высотой 3 — 5 (8) м. В культуре иногда достигает 10 м. Кора ствола серая, трещиноватая. Ветви прутье- видные, распростертые, восходящие, с красно-коричневой, пур- пурово-бурой или серо-коричневой корой; побеги голые. Листья овальные, яйцевидные или короткоэллиптические, с округленно-клиновидным основанием, острой или заостренной верхушкой, длиной 3 — 8 (2 — 5) см, шириной 2,5 (1,5) см, по краю тупогородчато-зубчатые, обычно голые или слегка опу- шенные лишь в молодости и по главной жилке. Черешки тонкие
Глава 5. Систематический обзор рода Malus и сравнительно длинные — 2 — 4 (6) см, голые, с голыми, рано зеленеющими прилистниками. Цветки по 4 — 8 (10) в зонтиковидных соцветиях, на голых, тонких и длинных (2 — 6 см) цветоножках; венчик чисто-белый, диаметром около 3 (2,0 — 3,5) см, лепестки продолговато- или обратнояйцевидные с коротким ноготком, железистые; чаше- листики продолговато-ланцетные, распростертые или отверну- тые, вдвое длиннее гипантия, войлочно-волосистые, снаружи, как и гипантии, голые. Чашечка после цветения опадает и в обра- зовании плода не участвует. Стилодии (5 (3) ) едва длиннее тычинок, сросшиеся у основания в голый или слабомохнатый столбик. Плоды шаровидные со слабым углублением у основания и на верхушке, мелкие, диаметром около 1 см (у некоторых куль- турных форм и гибридов крупнее), желтые, на солнце красне- ющие, с горечью, терпкие, на тонких и длинных плодоножках, остающиеся на деревьях в течение зимы; охотно поедаются птицами (орнитохория). Семена коричневые до темно-коричневых, мелкие, вес 1000 семян 5,0 — 7,5 кг. Описана Линнеем из Забайкалья (Читинская обл., Нерчин- ский р-н), с берегов р. Шилка, под названием Pyrus baccata. Лектотип (топотип): Забайк. обл., Нерчинский окр., заросли по бер. р. Шилки, у поселка Куларки, n' 2063, 3 VI 1909, цв., Кузнецов; хранится в гербарии Вотан. инст. им. В. Л. Кома- рова АН СССР (Ленинград). Распространена в юго-восточных районах Сибири, к востоку от Енисея (Предбайкалье, Забайкалье), на Дальнем Востоке (Амурская обл.: вниз по Амуру до Благовещенска, вверх — по р. Зея до г. Зея и вверх по р. Селемдже до с. Февральского). За пределами СССР: в Монголии, Китае (северо-восточные пров. Хэйлунцзян, Гирин и Ляонин), откуда заходит в Корею; далее по Китаю (пров. Хэбэй, Шаньдун, Шаньси, Шэньси, Ганьсу (центральная часть) ) и в Тибет, через Гималаи в Непал и Индию (до северо-западной ее части). 2) f. chamardabanica (V. Vartapetjan et L. Solovjeva) Po- nom. 1988, Бот. журн. 73,1: 83. — Malus chamardabanica V. Vartapetjan et L. Solovjeva, 1980, Бюлл. Моск. общ. исп. прир. отд. биол. Т. 85,5: 81 — 82. Эта яблоня, которая, по данным В. В. Пономаренко [1988], оказалась прежде окультуренной, а теперь одичавшей, была найдена в Забайкалье — вблизи оз. паучьего Гусиноозерского р-на Бурятской АССР в подлеске соснового леса, на площади около 1 га. Ы 5accafa имеет непосредственные родственные связи с М. himalaica, от которой, по-видимому, возникла и распространилась на восток по юго- западной, центральной и восточной частям Китая. Ее формы, имеющие
158 Глава S. Систематический обзор рода Malus период глубокого зимнего покоя и надежную защиту почек, со временем распространились в более суровые условия Северо-Восточной Азии {Мон- голия, Сибирь, Дальний Восток) и достигли северных пределов восточного крыла ареала рода Malus {выдерживают морозы до — 50 — 55 'С [Василь- ченко, 1963]) .. Здесь проходил процесс дифференциации, в результате которого обосо- бились еще 2 близких к М. haccata вида: 1) совокупность более древних форм — высоких {до 10 м) деревьев, со стволом диаметром до 45 — 50 см и более, крупными, продолговато-эллипсоидальными плодами длиной 1,5 см, диаметром 1,0 см, сосредоточенных в Китае {Северный Китай), СССР {Дальний Восток, Приморский край), Корее и Японии {о-в Хоккайдо, север- ная часть о-ва Хонсю) — М. mandshurica; 2) совокупность менее древних форм, характеризующихся меньшим ростом или имеющих вид кустарников, с длиннозаостренными листьями, почти по всему краю остропильчато-зубча- тыми; встречающихся на о-ве Сахалин и некоторых южных островах Куриль- ской гряды — М. mandshurica subsp. sachali'nensis. 22. Malus mandshurica (Maxim.) Котагоч, 1917, Тр. Почв.-бот. эксп. исслед. Азиатск. России, 2, 1913, вып. 2:93. 1) subsp. mandshurica а) var. mandshurica Юзепчук, 1939, Фл. СССР, 9:371, t. 22, с рис. 5; Федоров и Полетика, 1954, pep. куст. СССР, 3: 446, с рис. 62: 2. — Pyrus baccata sensu Watson, 1825, Dendr. Brit., 2: t. 51, non Linnaeus, 1767. — Pyrus prunifolia sensu Maximo- wicz, 1859, in Мет. Div. Sav. Acad. Sci. St. Petersb., 9: 471 (Prim. Fl. Атиг.), non Willdenow, 1794. — Pyrus Ьассаlа var. mandshurica Maximowicz, 1873, in Bull. Acad. Sci. St. Petersb., 19: 170 (in Mel. Biol. 9: 166). — Malus baccata var. mand- shurica (Maxim.) С. К. Schneid, 1906, 111. Handb. Laubhk., 1: 721, сит ic. 397 n, «mandschurica»; А. Rehder (1927, 1940) 1958, Man. Cult. Trees Shrubs, ed. 3: 393. — Malus baccata var. mandshurica f. latifolia Matsumura, 1912, Ind. Pl. Jap., 2, 2: 202, nom.; Poidzumi, 1913, in Jour. Coll. Sci. Tokyo, 34, 2: 84 (Consp. Rosac. Jap.) . .— Malus mandshurica var. genuina Skvortzov, 1925, in Bull. Jard. Bot. Rep. Russe, 24: 146; 1928, in Linguan. Sci. Jour., 5: 345, t. 27. — Яблоня маньчжурская. Мезофильное дерево, высотой 10 — 15 м, с хорошо выражен- ным стволом диаметром 30 — 60 см, с темно-бурой или почти черной корой. Многолетние ветви темно-серые, годичные красновато-бурые с желтым оттенком, молодые побеги голые. Листья крупные, длиной до 8 (10) см, шириной 5 (6) см, широкоовальные, яйцевидные до широкояйцевидных или обрат- нояйцевидных, с округлым или несколько суженным основанием и туповатой, внезапно или короткозаостренной верхушкой, по краю почти цельнокрайные или в верхней половине мелкогород- чато-зубчатые, опушенные (особенно в молодости) и по нижней стороне пластинки, взрослые — сверху голые, снизу слабово- лосистые, главным образом вдоль жилок. Черешки длиной 2,0— 3,5 см, слабоопушенные с опушенными прилистниками. Цветки душистые, по 3 — 8 в зонтиковидных соцветиях, на различных по длине (от 3 — 4 до 5 — 6 см), голых или слабо-
Глава 5. Систематический обзор рода Malus 159 волосистых цветоножках; венчик белый, в бутонах розовый, диаметром 3 — 5 см; стилодии (5, реже 3) одинаковой длины с тычинками, при основании сросшиеся в мохнатый столбик; ча- шелистики продолговатые, ланцетные с обеих сторон, так же как гипантии, волосистые или войлочные, иногда голые; чашечка после цветения опадающая. Плоды продолговато-овальные или эллипсоидальные, а также грушевидные, реже шаровидные, длиной 0,8 — 1,5 см, шириной 0,8 — 1,0 (1,2) см, с притупленной верхушкой, несколько оттяну- тым основанием, красноватые (различных оттенков) или желтые (желтоватые) . Описана в 1873 г. русским ботаником К. И. Максимовичем под эпитетом Ругив baccata var. rnandshurica. Тип: Бухта Новика на Русском острове, вблизи Владивос- тока; цв. 25. V 1860, пл. 28. Vill 1860, Максимович. Хранится в Гербарии Вотан. ин-та им. В. Л. Комарова АН СССР в Ле- нинграде. Распространена на юге Приморья, в бассейнах рек Лянчин, Суйфун, Майхэ, Супутинка, Раковка и др., на севере — в бас- сейнах рек Даубихэ, Улахэ, Иман, Бикин; встречается вместе с M. baccata, с которой образует естественные гибриды [Ва- сильченко, 1963]; в Китае — преимущественно в северо-восточ- ных провинциях (Северная Маньчжурия), в долинах рек и на нижних склонах гор, на высоте 1000 — 3000 м н. у. м., в Корее и Японии (о-ва Хоккайдо, Хонсю). Ь) var. ШийооМу~ (Ponom.) Ропот. 1977, in Вотан. журн. 62,7: 829. subsp. Шийоозйу~ Ponomarenko, 1972, in Ви11. Арр. Bot. Genet. Pl. Breed. 46,2: 65 — 69. — Яблоня )Куковского. Сильноветвистое низкорослое деревце-карлик высотой 0,2— 1,5 м, с плотными кожистыми листьями. Плоды по 5 — 6, темно- красные, округло-сплюснутые (длиной 0,7 — 1,1 см, шириной 0,8 — 1,2 см), терпкого, кислого вкуса. Распространена в северо-восточных и северных частях о-вов Лавров, Клыков, )Келтухин по берегу моря в 15 — 30 м от по- лосы прибоя. 2) subsp. sachalinensis (Кот.) Likh. 1974, in Ви11. Арр. Bot. Genet., Pl. Breed. 52,3:28. — Malus sachalinensis (Кот.) Juz. 1939, Фл. СССР, 9: 372, 493; Федоров, Полетика, 1954, Дер. куст. СССР, 3: 447. — Яблоня сахалинская. gIepeBo или деревце высотой 6 — 8 (4) м, с темно-серой или почти черной корой. Многолетние ветви темные, темно-серые или с темно-коричневым и грязно-красновато-желтым оттенком; годичные побеги коричнево-красноватые или красновато- желтые. Листья длиннозаостренные, овальные или яйцевидные, с клиновидным основанием и обычно длиннооттянутой верхушкой,
36( Глава 5. Систематический обзор рода Malus остро-пильчатым краем, кверху обычно крупно- или острозуб- чатые, на слабоопушенных черешках. Цветки в зонтиковидных соцветиях, белые или розоватые; гипантии, как и чашечка, голые или слегка волосистые, реже войлочные. Плоды продолговатые или округлые, диаметром около 0,8— 1,0 см; твердые и кислые, долго удерживаются на дереве (не опадают даже зимой), при полном созревании мягкие, иногда прозрачные. Чашечка при плодах опадающая. Описана С. В. Юзепчуком в 1939 г. из Южного Сахалина. Тип: 1ns. Sachalin, Dr. Augustinowicz, in Herbario 1nst. Bot. nom. V. L. Котагочй Ас. Sc. URSS in Leningrad conservatur. Распространена на о-ве Сахалин и некоторых южных остро- вах Курильской гряды (о-в Итуруп — сборы Д. П. Воробьева и Е. Егорова). По данным Г. И. Никитина, subsp. sachalinensis распространена по всему Сахалину до 52' с. ш., но особенно на юге острова. 23. Malus himalaica (Maxim.) Vass., 1963. Новые для культ. виды ябл., 114. — Pyrus baccata var. himalaica Maximowicz, 1873, in Bull. Acad. Sci. St. Petersb., 19: 170 (Diagn. Pl. Nov. Jap., Mandsh. Dec., 15, in Mel. Biol., 9: 167). — Malus baccata var. himalaica С. К. Schneid., 1906, 111. Handb. Laubh., 1: 721, exclud. pl. chin. et сит ic. — Pyrus baccata sensu Brandis, 1874, Forest. Fl. 1nd., 205; Hoocker, И., 1878, Fl. Brit. 1nd., 2: 373; Non Linnaeus, 1753. — Malus pallasiana var. himalaica Al. Theod., 1954, Дер. куст. СССР, 3: 446. — Яблоня гималай- ская. Дерево высотой 4 — 8 (10) м, с хорошо выраженным стволом диаметром 20 — 30 (50) см и пирамидальной кроной. Листья крупные, шириной 2,5 — 3,5 см, длиной 4,5 — 7,0 см, овальные или широкоэллиптические, по краю крупно-, реже ос- тропильчатые, голые или слегка рассеянно-опушенные, преи- мущественно в молодости и по жилкам, главным образом с нижней стороны пластинки, на черешках длиной 3 0 — 3 5 см. Цветки по 3 — 5 в зонтиковидных соцветиях; венчик белый, в бутонах розоватый, диаметром около 3 см, с совершенно го- лым гипантием и голой или рассеянно-мохнатой внутри чашеч- кой; цветоножки тонкие, длиной около 3 см, голые. Плоды шаровидные диаметром 1,0 — 1,5 см, зеленовато-жел- тые с краснеющим бочком и опадающей при плодах чашечкой; плодоножки тонкие, длиной около 3,5 см. Описана в 1873 г. русским ботаником К. И. Максимовичем по образцам из западной части Гималаев под эпитетом Pyrus baccata var. himalaica. Neotypus: North-Western 1ndia: Tihri-Garhwal, alt. 8 — 9000 ft., n' 1064, 6 Vll 1883, J. Renstke, in Herb. Bot. Inst. As. Sc. URSS in Leningrad conservatur.
Глава 5. Систематический обзор рода Malus Распространена в Западных Гималаях от Инда до Кумаона и далее на восток, где заходит в прилегающие районы Юго- Западного Китая. Встречается на высоте 1500 — 3500 м н. у. м. М. Шта1а!са произошла в Гималаях в результате дифференциации тре- тичных доциниевидных яблонь типа M. si4kimensis, о чем свидетельствует сохраняющаяся до нашего времени общность некоторых нризнаков назван- ных яблонь: габитус растений, форма и строение листьев, цветков и отчасти плодов. По признакам плодов M.himalaica отличается от сиккимской яблони мелкоплодностью. При этом форма плодов М. sikkimensis кубаревидная, на верхушке вытянутая, с остающейся чашечкой и грубоватой мякостью, а для М. himalaica свойственны шаровидные плоды с несколько углубленной вер- хушкой, с опадающими после цветения чашелистиками и нежной мякотью. Сравнивая архаические особенности строения M. si44imensis с более продвинутыми признаками M. himalaica, можно представить облик древних исходных предков этих яблонь. Однако М. sjkkimensis в настоящее время носит ксероморфный облик, тогда как у М. Й!та1а~са эти черты выражены слабее. От M. himalaica, имеющей в настоящее время сравнительно узкий ареал, возникли наиболее эволюционно продвинутые виды рода Malus, от которых произошла секция Gymnomeles. М. himalaica принадлежит к этой секции и входит в состав серии викарирующих видов Baccatae. Г и б р и д ы, по совокупности признаков примыкающие к серии Baccatae (Species hybridogena ad series Baccatae accedentes): Malus)(robusta (M. baccata)<M)(prunifolia) (Carr.) Rehd., 1920, in Jour. Arnold Arb., 2: 54. Malus+cerasi fera (M.)(prunifolia+M. baccata) Spach 1834, Hist. Nat. Veget. Phan., 2: 152, t. 10, сит ic. (рис. 56). Series (ряд) 7. Н U p e h e n s e s Langenf., ser. nova. Folia in frondescentia plus minusve ригригеа, oblongato-ovoi- dea vel elliptica„ interdum sat. angusta, margine acuto-serrata vel crenato-serrata сит cuneata vel obliqua basi et pe tiolis rubris. Typus series: Malus hupehensis (Ратр.) Rehd. Листья при распускании более или менее пурпуровые, про- долговато-яйцевидные или эллиптические, иногда довольно уз- кие, по кр аю остропильчатые или городчато-зубчатые с клино- видным или неравносторонним основанием и красными череш- ками. 24. Malus hopehensis (Ратр.) Rehd., 1933, in Journ. Arnold Arbor., 14: 206; Scaly, 1946, in Bot. Mag., 146: 9667, сит ic. (р: 3); Федоров и Полетико, 1954, pep. куст. СССР, 3: 448.— Pyrus baccata sensu Hemsley, 1886. in Journ. Linn. Soc. Lond. Bot., 23: 255, quoad. pl. е Chekiang; Diels, 1900, in Bot. Jahrb. 29: 387, non Linnaeus, 1753. — Pyrus spectabilis sensu Hemsley, 1886, in op. cit. 258, quoad pl. e Piangsi et Hupeh.; Diels, 1900, 1. с.; non Aiton, 1789. — Malus baccata var. himalaica С. К. Schneid., 1906, 111. Handb. Laubh., 1: 721, сит ic. 397 s. quoad pl. chin. et fig.; non Pyrus baccata var. himalaica Ma- ximowicz, 1873. — Pyrus communis sensu Pavolini, 1908, in
Глава 5. Систематический обзор рода Malus 162 Рис. бб, Malus X cerasi fera Nuov. Giorn. Bot. Ital. n. ser., 15: 415; non Linnaeus, 1753.— Pterus hupehensis Pampanini, 1910, in Nuov. Giorn. Bot, Ital. n. ser., 17: 291; Rehder, 1915, in Sargent, Pl. Wilson, 2: 265, 300, «spec. dub.» — Malus theifera Rehder, 1915, in Sargent, Pl. Wi- son, 2: 283; Rehder, 1927, Man. Cult, Trees Shrubs, 295; Ни СЬип, 1927, leon. Pl. Sin., 1: 32, t. — Pterus theifera Bailey, 1916, in Rhodora, 18: 155. — Яблоня хубейская. Дерево, иногда растущее в виде куста, высотой до 8 м, с колючими, прямыми, растопыренными ветвями. Листья яйцевидные или продолговато-яйцевидные, на вер- хушке заостренные, с клиновидным или неравносторонним ос-
Глава 5. Систематический обзор рода hfalus 163 Рис. 57. Afalus hupehensi;s нованием, мелко остропильчатые, длиной 6 — 8 см, шириной 3— 6 см, голые или опушенные снизу по жилкам, при распускании пурпуровые. Цветки душистые, по 3 — 7 в зонтиковидных соцветиях, на тонких, голых или слабоопушенных цветоножках длиной 3— 4 см; венчик белый, розоватый или розовый, диаметром 34— 4,0 см; гипантии и чашелистики пурпуровые, голые; стило- диев 3 (4) . Плоды шаровидные, на верхушке несколько вдавленные, диаметром 0,8 — 1,2 см, зеленовато-желтые с красным бочком или красные; чашелистики при плоде полностью опадающие (рис. 57). Описана итальянским ботаником Пампанини в 1910 г. под названием Pyrus hupehensis. Лектотип: Kmail, пров. Фуцзянь, у Чунань, Саньган-Тинму- гуань, 11 Vill 1952, пл., n' 239, Чжоу-Чжэнь-ин; хранится в гер- барии Ботан. ин-та им. В. Л. Комарова АН СССР (Ленинград). Распространена в Центральном и 1Ого-Западном /mac, от- куда заходит в Бирму и Индию [Assami, 1927]. Обитает на высоте от 1000 до 2000 м н. у. м. 25. Malus halliana (Anon.) Koehne, 1890, Gatt. Ратас., 27; Koehne, 1893, Deutsche Dendr., 261; С. K. Schneid., 1906, П1. Handb. 1.aubh., 1: 719, curn ic. 397, 398 е-f; Rehder, 1915, in
Глава 5. Систематический обзор рода Ма1из 164 SECTIO (секция) Vl. MALUS. Malus sect. Calycomeles Ko- ehne 1893, Deutsche Dendr., 257. — Malus sect. Eumalus Zabel, Sargent, Рl. Wilson, 2: 285; Федоров и Полетика, 1954, Дер. куст. СССР, 3: 448, рис. 68,4. — Malus halllana Anon., 1882, in Garden, 22: 162, nom. — Pyrus halllana hort. ех Sargent, 1882, in Gard. For., 1: 152, pro syn. — Pyrus halllana Voss, Vilmor, 1894, Blumengart., 1: 277. — Pyrus spectabllls sensu Tanaka, Useful, 1895, PI. Jap., 156, сит ic. 634; non Aiton, 1789. Malus flortbunda var. parkmanntt goidzumi, 1911, in Bot. Mag. Tokyo, 25: 76; Poidzumi, 1913, in Jor, Coll Sci. Tokyo, 34, 2: 83 (Consp. Rosaae Jap.). — Яблоня Холла. Кустарник высотой 2 — 4 м или дерево высотой 3 — 5 м, с широкой, раскидистой кроной и красно-коричневыми ветвями. Молодые побеги сначала слабоопушенные, потом голые. Листья продолговато-яйцевидные или яйцевидные, заострен- ные к верхушке, суженные к основанию, иногда довольно узкие, длиной 4 — 8 см, шириной 2 — 3 см, по краю зубчатые или про- долговато-зубчатые, на молодых побегах красновато-пурпуро- вые, едва опушенные, позже темно-зеленые, совершенно голые, кожистые, сверху блестящие, главная жилка по нижней сто- роне пластинки, так же как черешки, красно-пурпуровая, слабо- волосистая, черешки длиной 0,5 — 2,0 см. Цветки по 4 — 7 в зонтиковидных соцветиях на тонких и длинных (4 — 5 см) поникающих, лишенных опущения цветонож- ках пурпурового оттенка; венчик диаметром 3,0 — 3,5 см, ярко- или темно-розовый (в бутонах красный), иногда махровый (число лепестков обычно больше 5), стилодиев 4 — 5; чашелис- тики треугольно-яйцевидные, пурпуровые, короче гипантия, ги- пантий голый, пурпуровый. Плоды шаровидные или несколько грушевидные, диаметром 6 — 8 мм, красно-пурпуровые или красно-коричневые, очень поздно созревающие, декоративные, с относительно крупными семенами. Описана немецким дендрологом Кене в 1890 г. Neotypus: China, Western Szechuan, South of Ching-chuan- growing, along rocky river bank, alt. 4000 ft., II1 1925, fl., n' 12026, J. Е. Rock, in Herb. Bot. Inst. Ac. Sci. URSS in Leningrad conservatur. Известна только в культуре в Китае и Японии. Интродуци- рована в Европу в середине XIX в. Г и б р и д ы, по совокупности признаков примыкающие к серии Hupehenses: Malus)(atrosangutnea (М. halltana)(M. иеЬоЫЙ) (Spath) С. К. Schneid., 1906, 111. Handb. Laubh., 1: 720. Malus)(hartw ща' (М. baccata)(M. halliana) КоеЬпе, 1907, in Mitt. Deutsch. Dendr. Ges., 1906 (15): 60.
Глава 5. Систематический обзор рода MaIus 1BS 1903, in Beissner et al. НалдЬ. Laubh.-Berm., 185, nom. subnud., С. К. Schneid., 1906, 111. Hand. Laubh., 1: 174; Юзепчук, 1939, Фл. СССР, 9: 358; Rehder, 1940, in Мал. Cult. Trees Shrubs, ed. 2, 390; ejusd. 1945, Bibliogr. Cult. Trees Shrubs, 267; Федоров и Полетика, 1954, julep. и куст. СССР, 3: 417. — На- стоящие яблони. Крупноплодные настоящие яблони с остающимися при пло- дах чашелистиками. Листья в почкосложении свернутые, все цельные, мелкозубчатые, пильчатые, городчатые, реже цельно- крайные. Цветки в зонтиковидных соцветиях; пыльники желтые, сти- лодиев 3 — 5. Плоды крупные, одиночные, имеющие характерную форму яблока, съедобные, округлые, шаровидные, сплюснутые или сдавленные с полюсов, иногда слегка ребристые, диаметром 2 — 3 или 3 — 8 см (у культурных форм крупнее). Семенные гнезда широкие. Тип секции: тип рода. Обширная секция настоящих яблонь в диком виде представ- лена исключительно лишь в Старом Свете (рис. 58). Клайч для определения видов 27. М. sieversii Ledeb. — Кора ветвей иного цвета 5. Кора стволов взрослых деревьев светло-серая, многолет- ние ветви серо-желтые или темно-коричневые (ser. Orientales Langenf.) 1. Чашелистики при плодах свободные, плоды с углубле- нием при основании.......... 2 — Чашелистики при плодах сросшиеся в короткую трубку, плоды без углубления у основания или едва вдавленные Gr.PPrunifoliae (Juz.) Langenf. 2. ~дикорастущие или окультуренные яблони, взятые в культуру из природы 3 — Культурные (сортовые) или одичавшие яблони . Gr. Domesticae Langenf. 3. Кора стволов взрослых деревьев серо-коричневая, мно- голетние ветви коричневые или красноватые (ser. An- atieae Langenf.) 26. М. asiatica Nakai — Кора стволов и ветвей иного цвета...... 4 4. Кора стволов и взрослых деревьев буровато-серая, мор- щинистая, многолетние ветви красноватые или красно- бурые (ser. Sieversinae Langenf.) .
Глава 5. Систематический обзор рода Malus F66 an C an ~ .,~ z ~ мю > ао IK о 3 д an ~+ z е~ gg + а О а 3 3 3 в w мю 2 3й ° ~ K IK Ф 5 3 Е Р IK 8 3 ? к к мг � an а, R c4 щ М ~ w 3 3 3 an IK > an > > е а. TICAE Рис.58. Схема филогенетических отношений в секции Malus — Кора стволов взрослых деревьев серая, многолетние ветви серые или серовато-бурые (ser. Afalus)... 9 6. деревья 8 — 10 (12 — 15) м в высоту, с широкой кроной 7 — деревца 3 — 6 м в высоту, с компактной кроной... 8 7. Листья крупные, яйцевидные с округлым основанием 29. М. caspiriensis Langenf. — Листья средней величины, продолговатые, с клиновид- ным основанием 28. М. orientalis (Uglitz.) Juz. 8. Ветви иногда с колючками М. orientalis var. montana (Uglitz.) Langenf. — Ветви без колючек М. orientalis subsp. turkmenorum (Juz. etМ. Рор.) Langenf. 9. Листья голые, ветви с колючками 31. М. sylvestris (1 .) Mill. — Листья опушенные ветви без колючек . М. sylvestris var. praecox (Pall.) Ропот. Series (ряд) 8. As i at i c a e Langenf. nomen provisorium. Mali area Asiae Orientalis (China, Когеа, Japonia). Cortex truncorum агЬогит adultorum griseo-cinnamomeus rami perennes brunneoli vel rubelli. Folia oblogato-ovalia sive ovalia, orbiculata vel laticuneata сит basi, apice acuta, serrulata. Flores in umbellis 5 — 7, 4 — 5 ст in diam., villosis pedicellis 2 — 3 ст lg.-sepala late triangularia.
Глава 5. Систематический обзор рода Malus 167 Typus series: Malus asiatica Nakai. Географически обособленные яблони восточно-азиатского очага (Китай, Корея, Япония). Эти яблони почти не изучены, и их видовая принадлежность трактуется по-разному, чаще всего «Pyrus malus» или «Malus pumila». Следует подчеркнуть, что эти неопределенные наименования (особенно М. pumila), упо- требляемые обычно для обозначения слабоизученных, иногда культурных, полукультурных и даже одичавших яблонь, стали постоянным источником ошибок. Поэтому, согласно рекоменда- циям Международного кодекса ботанической номенклатуры (1980, ст. 63 и 65), мы предлагаем отказаться от таких эпите- тов, как «М. ритйа», и подобных. 26. М. asiatica Nakai, 1915, in Matsumura, Icon. gaisik, 3, 155; idem, 1916, Fl. Sylv. Ког., 6: 40, t. 14; sp. non satis notae. 1) var. asiatica Pyrus malus Bunge, 1835, in Mem. Div. Sav. Acad. Sci. St. Petersb., 2: 101 (Enum., 1833, Pl. Chin. Bor. 27), non Linnaeus, 1753; Siebold. 1856, in Verh. Baataav. Genot Punst. Wetensch., 21, 1: 66 (Syn. Pl. Oecon.), nom. — Pyrus malus p tomentosa sensu Maximowicz, 1873, in Ви11. Acad. Sci. St. Petersb., 19: 170 (in Mel. Biol., 9: 165), поп КосЬ, 1837.— Pyrus praecox sensu Miquel, 1867, in Ann. Mus. Bot. Ludg.-Bat., 3: 40, non Pallas, 1784. — Malus matsumurae goidzumi, 1909, in Bot. Mag. Tokyo, 23: 172; Poidzumi, 1934, in Acta Phytotax Geobot. 3: 192. — Pyrus matsumurae Gordot, 1918, in Ви11. Mus. Hist. Nat. Paris, 24: 67. — Malus dulcissima var. asiatica Koi- dzumi, 1934, in Acta Phytotax. Geobot., 3: 190. — Malus prunifo- lia auct. рlиг. saltem р. р. Rehder. — Malus pumila auct. рlиг. saltem р. р. Jii Fet-sun and Jen-Chen-lung, 1956. — Яблоня азиатская. деревья высотой до 10 м и со стволом диаметром до 20 см. Листья овальные или продолговато-овальные, длиной 5— 10 см, шириной 3 — 5 см или более крупные, при основании за- кругленные, в молодости опушенные с обеих сторон, впоследст- вии сверху гладкие, темно-зеленые, снизу опушенные особенно по жилкам, по краю мелкопильчатые с прямыми зубцами, на черешках длиной около 2 — 3 см, с прилистниками. Цветки по 5 — 7 в зонтиковидных соцветиях; цветоножки, гипантии и чашечка особенно снаружи густоопушенные, венчик диаметром 3 — 5 см, бледно-розовый или белый, в бутонах розо- вый; стилодии (5), сросшиеся у основания опушенного стол- бика, рыльца приподняты едва выше тычинок. Плоды сжато-шаровидные, несколько сдавленные со стороны чашечки и плодоножки, длиной 2,5 и 3 — 4 см, диаметром 3,0 и 3,5 — 4,5 см, иногда более крупные, диаметром до 5 — 6 см, жел- тые, с розовым бочком и с рассеянными белыми точками, на
168 Глава 5. Систеыатический обзор рода A1alus коротких, длиной около 2 см, толстоватых плодоножках и с остающимися при плоде чашелистиками. Описана из Кореи. Тип неизвестен. В гербариях не обнаружена! Распространена в центральных районах Китая и в Корее, а также в Японии, где, по-видимому, широко культивируется. В просмотренных нами гербариях отсутствует. Точных данных об ареале этой яблони не имеется. По всей вероятности, М. asiatica является представителем малоизученных настоящих яблонь, встречающихся на террито- рии Восточной Азии или даже, возможно, представляет собой одичавшие особи культурной яблони [Лангенфельд, 1970]. 2) var. rinki Asami 1927, Crabapp. Neet. Jap., 6, сит ic. 3, 4, 7, t. 3, 37. — Malus ringo Siebold, 1856, Cat. Raison, 1856: 5, пот.; Carriere, 1874, in Rev. Hort., 1874: 170; Koidzumi, 1934, in Acta Phytotax. Geobot. 3: 191. — Ругила ringo Wenzig, 1874, in Linnaea, 38: 37; Stapf, 1909, in Bot. Mag., 135, t. 8265.— Malus prunifolia var. rinki Rehder, 1915, in Sargent, Pl. Wilson, 2:279; Bailey, 1949, Man. Cult. Pl. Федоров и Полетика, 1954, Яер. и куст. СССР, 3: 440. Деревце высотой 3 — 5 м или дерево высотой до 8 м, с широ- кой кроной и опушенными в молодости побегами. Листья продолговато-яйцевидные, молодые — опушенные с обеих сторон, взрослые — сверху голые, снизу опушенные. Цветки в зонтиковидных соцветиях, розовые или почти бе- лые, цветоножки короткие, длиной около 2 см, так же как гипантии и чашечка, опушенные. Плоды шаровидные, диаметром 3,5 см, зеленоватые или зе- леновато-желтые, сочные с горьковато-сладким привкусом, при созревании становятся мучнистыми. Это разновидность обнаружена Вильсоном в Юго-Западном Китае и описана П. Зибольдом в качестве отдельного вида— Malus ringo. Распространена в Юго-Западном Китае, пров. Юньнань, Сы- чуань, Шэньси и некоторых других. Series (ряд). 9. S i e v e r s i n а е Langenf. Mali areae Asiae Centralis. Cortex truncorum arborum adultorum fumigatus, rugosus, rami perennes rubelli vel rubuginosi. Folia oblongata vel oblongato-ovoidea cuneata sive laticuneata basi. apice acuta sive obtusa, denticulata. Flores in umbellis 3 — 5, 3,5 — 4,0 ст in diam., pedicellis levi- ter villosis, 1,5 — 2,0 ст Ig., sepala basi lata triangulari сит асшп~пе. Турия series: Malus sieversii (Ledeb.) М. J. Roem.
Глава 5. Систематический обзор рода Malus 169 Географически обособленные яблони среднеазиатского очага. Кора стволов взрослых деревьев буровато-серая, морщинис- тая, многолетние ветви красноватые или красно-бурые. Листья продолговатые или продолговато-яйцевидные, с кли- новидным или ширококлиновидным основанием, острой или за- тупленной верхушкой, мелкозубчатые. Цветки по 3 — 5 в зонтиках, диаметром 3,5 — 4,0 см, на слабо- мохнатых цветоножках длиной 1,5 — 2,0 см, чашелистики при основании широкие с треугольным остроконечием. Яля Средней Азии местные авторы различают до 12 видов яблони. Однако описания большинства из них сделаны на основании неполных ма- териалов, и в некоторых случаях они необоснованы, поскольку представляют лишь отдельные разновидности или даже формы локального значения, ли- шенные обособленных ареалов. В первую очередь это относится к ~видам», описанным Г. П. Сумневичем и П. П. Поляковым. Таким образом, коли- чество среднеазиатских видов подлежит значительному сокращению. Наиболее обычными и экологически обособленными среднеазиатскими видами принято считать М. А~~дШогит и М. яеоегзй. Первый является более древним видом мезофильного облика, обитает преимущественно на мощных и богатых почвах лесного пояса северных склонов и по днищам речных долин на высоте от 900 до 1800 (2000) м н. у. м. Второй, ксероморф- ный, также крайне разнообразный, замещает первый на более сухих солнеч- ных склонах в аридных условиях, где поднимается в горы до 2300 (2500) м н. у. м. Промежуточные, явно примыкающие, с одной стороны, к М. I<irghisorum, с другой — к M. sieoersii, яблони, обособленные в долинах рек на Юго- Западном Гиссаре с центром в Южном Узбекистане (бассейн р. Кашка- дарья, ее притока р. Танхас в районе Шахрисабза), можно отождествить q М. hissart',ca. Однако ареалы этих мнимых видов необособлены и полностью покры- ваются ареалом яблони Сиверса. Поэтому их следует признать экологичес- кими разновидностями основного вида — М. sieoersii. 27. Malus sieversii (Ledeb.) M. J. Roem., 1830, Syn. Rosif1. 21 1) var. sieversii: а) f. sieversii Юзепчук, 1939, Фл. СССР, 9:362; Федоров и Полетика, 1954, Рер. и куст. СССР, 3: 423; Кудряшев, 1955, Фл. Уз. ССР, 3: 280; Васильченко, 1963, Новые для культ. виды ябл. 67; Быков, 1961, Фл. Казахст., 4:402; Запрягаева, 1964, Яикораст. плод. Таджикист., 390. Pgrus зЕеаегпЕ Ledeb., Pyrus malus auct. fl. As. Med. — Afalus pumila auct. fl. As. Med., non Mill. — Яблоня Сиверса. Ксероморфное дерево или небольшое деревце высотой 2 — 6 (8) м с шатровидной, иногда узкой или округлой кроной. Кора на стволах взрослых деревьев слабоморщинистая, буровато-се- рая. Многолетние ветви колючие, особенно на порослевых побе- гах, отрастающих от корней при объедании растения живот- ными, бурые или красноватые. Годичные побеги толстоватые, иногда красные, в молодости обычно опушенные. Почки небольшие, длиной около 3 мм, рубиново-красные.
Глава 5. Систематический обзор рода Malus 170 Рис. 59. Malus si'eversi'i Листья кожистые, продолговатые или обратнояйцевндные, ширина 6 — 8 см, снизу сильноопушенные, по жилкам с красно- ватой пигментацией, округлым или ширококлиновидным осно- ванием, заостренной или тупой вершиной, по краю мелкозубча- тые, на густо сероопушенных черешках в 2 — 3 раза короче пластинки. 1Яветки по 3 5 в зонтиковидных соцветиях, розоватые или розовые, диаметром 3 — 4 см, чашечка густо сероопушенная, гипантии и цветоножки опушенные.
Глава 5. Смстематический обзор рода Malus 171 Плоды одиночные, шаровидные или сплюснутые с полюсов, диаметром 2 — 5 см, ярко-красные, малиновые, реже желтые, ароматные, горько-сладкие; длиной 1,5 — 2,0 см, в молодости опушенные. Чашечка при плодах открытая. Семена клиновид- ные или овальные, в сухом состоянии сероватые или буровато- коричневые, тусклые, гладкие, длиной 5 — 6 мм, шириной 3 — 4 мм (рис. 59). цветет IV — V, плоды созревают VIII — Х. Описана русским ботаником К. Ф. Ледебуром в 1844 г. из Тар багатая. Typus: Tarbagatai, prope Ust. Kamenogorsk (sine anno) leg. Dr. Sieverss, in Herb. Inst. Bot. nom. V. L. Komarovii Ac. Sci. in Leningrad conservatur. Распространена в Средней Азии (Тарбагатай, джунгарский и Заилийский Алатау, Ферганский, Чаткальский хребты, Кара- тау, Памиро-Алай). Будучи ксероморфным растением, произрас- тает на более сухих солнечных склонах южной экспозиции, где обитает преимущественно вблизи горных ручьев (на высоте бо- лее 1600 м н. у. м.). На северных склонах и в более глубоких речных долинах вытесняется и замещается яблоней киргизов. Встречается в лесной зоне на высоте 1000 — 1600 м н. у. м. на опушках кленовых или сосновых лесов, на почвах средней мощ- ности, вместе с боярышником, спиреей, шиповником, жимо- лостью, малиной и др., а также на участках с более бедными почвами, на каменистых склонах и скалах на высоте до 2200 (2500) м н. у. м. Размножается вегетативно-корневыми отпрыс- ками, а также семенами. Весьма оригинальная яблоня Недзвецкого, распространенная в пределах ареала М. sieversii, рассматривается нами в качестве разновидности М. sie- versii. При обзоре М. sieversii, представленной на территории Казахской ССР, мы используем внутривидовую классификацию Б. А. Быкова. Новые виды, описанные П. П. Поляковым, М. schischkinii и M. jarmolenkovii из долины р. Тургень (Заилийский Алатау) и М. linczevskii из заповедника Аксу-Дже- боглы, отнесены нами к основному политипичному виду — M. sieversii. Новоописанные Г. П. Сумневичем виды M. antsophyOa и M. kudrjaschevii из бассейна р. Ангрен (Узбекистан) рассматриваются нами как недоста- точно обоснованные. Они отнесены к основному виду М. sieversii в качестве разновидностей. М. persivifolia $итт. (М. pumila var. persifolia М. Pop.), описанная также из бассейна р. Ангрен, представляет собой окультуренную или одичавшую яблоню, довольно широко встречающуюся в культуре в не- которых районах Узбекистана. Ь) f. niedzrrvetzkyana (Dieck.) Langenf. (Malus niiedznvetzky- ana Dieck., 1891, Neucheiten-ОН. des Nat.Arb. Zochen,1891: 16). В культуре, в одичавшем и диком состоянии в районах Куль- джи, Кашгара, Алма-Аты, Каратау, Ферганского хребта и др. Нахождение оригинальных краснопигментированных яблонь в пределах указанной территории неоспоримо, однако наличие одной лишь красной пиг- ментации вегетативных частей, цветков и плодов растения не позволяет
172 Глава 5. Систематический обзор рода A1alus рассматривать эти яблони в качестве самостоятельного вида. Ведь в той или иной степени краснопигментированные формы встречаются и в пределах других видов. Основным критерием отрицания видовой самостоятельности М. niedzupetzkyana следует считать численность особей, так как в природе встречаются лишь отдельные краснопигментированные особи, не являющиеся надиндивидуальными образованиями. Следовательно, эти яблони не имеют определенного ареала, характерного для истинного вида. с) f. sphenocarpa (В. Bykov) Langenf. — Заилийский Алатау; d) f. longicarpa (В. Bykov) Langenf. — Заилийский и Та- ласский Алатау; е) f. linczevskii (Р. Ро1.) Langenf. — Западный Таласский Алатау; f) f. anisophylla (Sumn.) Langenf. — Западный Тянь-Шянь (р. Ангрен); g) f. kudrjaschevii (Sumn.) Langenf. — бассейн р. Ангрен. 2) var. hissarica (S. Kudr.) Langenf. Malus hissarica S. Кис1г., 1950, Плод. Шахрисябса, 1: 158; Федоров и Полетика, 1954, Дер. куст. СССР, 3: 424; Кудряшев, 1955, Фл. Узб. ССР, 3: 278; Васильченко, 1963, Новые для культ. виды ябл., 95. — Яблоня гиссарская. Дерево средней высоты (5 — 8 (8 — 12) м), со стволом диа- метром 35 — 50 см и серо- или темно-коричневой морщинистой корой. Крона широкая, раскидистая, шатровидная с толстыми ветвями с гладкой красновато-бурой корой, ветви обычно ли- шены колючек. Годичные побеги оливково-зеленые или красно- ватые, толстые, обычно опушенные. Листья продолговато-яйцевидные, обратнояйцевидные или эллиптические с заостренной верхушкой, с округленным, реже клиновидным основанием, по краю пильчато-зубчатые, сверху голые или слабоволосистые по жилкам, снизу опушенные, дли- ной 6 — 8 (4 — 10) см, шириной 3 — 4 (5 — 6) см, на слегка опу- шенных или голых черешках в 2 — 3 раза короче пластинки. Цветки в немногоцветковых (3 — 5) зонтиковидных соцве- тиях, чашечка и гипантии опушенные. Плоды слегка удлиненные или округлые, длиной 2,5— 4 (6) см, диаметром 3 — 4 (6) см, желто-зеленые с румянцем, реже — малиново-красные, у верхушки слегка ребристые, кис- лые, кисло-сладкие, сочные, но вяжущие. Плодоножка бурая, длиной 2,0 — 2,5 см. Чашечка при плодах полуоткрытая (рис. 60). Цветет V, плоды созревают VIII — IX. Описана С. Н. Кудряшевым (1950) из бассейна р. Кашка- Дарья, с верховий ее притока — р. Танхас, в районе Шахри- сабза (Южный Узбекистан) . Тип: Сев. склоны Гиссарского хребта, басс. р. Кашка-Дарья. Северное течение р. Танхас, выше к. Чопух, 1 VIII 1937, n' 1330, С. Н. Кудряшев; хранится в гербарии Института ботаники АН Узб. ССР (Ташкент).
Глава 5. Систематический обзор рода hfalus 173 Рис. 60. Malus sieoersii var. hissarica Распространена в Средней Азии (Памиро-Алай), эндемик преимущественно ущелий и речных долин Гиссарского и Зарав- шанского хребтов, реже на Каратечинском и Яарвазском хреб- тах. В более типичном виде встречается по долинам рек на Юго-Западном (Южный Узбекистан) и Южном (Западный Таджикистан) Гиссаре, где произрастает вместе с кленом и грец- ким орехом, одиночно или группами, образуя естественные на- саждения в виде небольших рощиц. Вообще обитает как на мощных почвах речных долин, так и на маломощных почвах склонов и скалистых участков. Однако приурочена к более ув- лажненным склонам в полосе от 1200 — 1800 до 2200 м н. у. м.
174 Глава 5. Систематический обзор рода Ма/из На более сухих и остепненных склонах и на высоте более 2200 м уступает место яблоне Сиверса, с которой нередко гиб- ридизирует, причем в потомстве иногда преобладают признаки М. sieversii. 3) var. kirghisorum (Al. et An. Theod.) Langenf. Afalus Иг- ghisorum Al. et An. Theod., 1949, Плод. лес. южн. Киргиз., 1:220; Федоров и Полетика, 1954, Дер. куст. СССР, 3:421; Васильченко, 1963. Нов. для культ. виды ябл., 84. — Afalus pu- mila auct. fl. As. Med. non Mill. — Яблоня киргизов. зверева мезофильного облика, высотой 10 — 12 (14) м, со стволом диаметром до 50 (65) см, широкой, шатровидной кро- ной; многолетние ветви сильные, неколючие, с темно-серой шероховатой корой, на стволах старых деревьев кора морщи- нистая, отпадающая продолговатыми кусками; годичные побеги также неколючие, длинные и тонкие, оливково-зеленые или красноватые, слабоопушенные в молодости, позже — почти го- лые или голые. Почки небольшие, длиной до 4 мм, так же как побеги олив- ково-зеленые или ярко-красные. Листья тонкие, бумажистые, продолговатые или широкооб- ратнояйцевидные, крупные — длиной до 8 (10) см, шириной 4 — 6 см, с клиновидным, постепенно оттянутым основанием, за- остренной или тупой верхушкой, по краю мелкозубчатые, почти голые или снизу слегка опушенные по жилкам, на слабоволо- систых или опушенных черешках в 2 — 3 раза короче пластинки. Цветки в немногоцветковых зонтиках, белые или розоватые, диаметром около 4 см, чашечка серовато-опушенная. Плоды одиночные, шаровидные, конические, или несколько цилиндрические, или сплюснутые с полюсов, у основания и вер- шины вдавленные, иногда несколько ребристые, диаметром 3— 8 см, длиной 3 — 8 или 5 — 10 см, зеленые или желтоватые с ру- мянцем, или полосатые, терпко-кислые, кислые или кисло-слад- кие; плодоножка длиной до 3 см, красновато-бурая, в молодости опушенная; чашечка при плодах открытая. Семена овально-кли- новидные, в сухом состоянии буроватые, серые или серовато- бурые, тусклые, гладкие, длиной 6 — 8 мм, шириной 3 — 5 мм (рис. 61). Цветет IV — Ч, плодоносит VII — X. Описана из Южной Киргизии и Западного Тянь-Шаня Ал. А. Федоровым и Ан. А. Федоровым в 1949 г. Typus: Pirghisia australis in districtu Dzhalalabad, in loco То taipan dicto. Ad ripam dextram fluminis Aktasch in silva jug1andina, 25 IX 1944, fr., Al. Theodorov, in Herb. Inst. Bot. nom. V. L. gomarovii Ac. Sci UgSS in Leningrad conservatur. Распространена в Средней Азии — Тянь-Шань (Ферганский, Таласский и Чаткальский хребты), Заилийский Алатау, джун- гарский Алатау.
Глава 5. Систематический обзор рода Malus 176 Рис. 61. Ма1из sieversii чаг. kirghisorum Встречается в лесной зоне на склонах преимущественно се- верных и северо-западных экспозиций, тяготеет к террасам и днищам речных долин. Приурочена к богатым, мощным, хорошо увлажненным почвам, входит в состав ореховых, кленовых или осиновых лесов и является сравнительно теневыносливым рас- тением мезофильного облика. Часто обитает на опушках, не- редко образует естественные насаждения с небольшой примесью других пород. Размножается вегетативно-корневыми отпрыс-
Глава 5. Систематический обзор рода Malus f76 ками, отводками и укоренением ветвей, реже пневой пророслью, что является характерной способностью этой яблони. Наблюда-ется и размножение семенным путем. Series (ряд) 10. О г i e n t а l e s Langenf. Mali агеае Caucasicae, Asiae Anterioris et Minoris. Cortex truncorum adultorum palido-griseus est, fragmentis separatis disruptus, rami annosi соlоге griso-flavidi vel atro- brunnei. Folia ovato-lanceata vel oblongo-elliptica sunt, basi cuneate angustata vel obtuse rotundata, apice obtuse acuminata, serrato- dentata. Flores 4 — 6 umbellis, 4,0 — 4,5 ст in diem., brevibus alboto- mentosis pedicellis, sepala angustotriangularia. Typus series: Ма1из orientalis (Uglitz.) Juz. Географически обособленные яблони кавказского, передне- и малоазиатского очагов. Кора стволов взрослых деревьев светло-серая, слущивающа- яся отдельными кусками, многолетние ветви серо-желтые или тем но-коричневые. Листья яйцевидно-ланцетные или продолговато-эллиптичес- кие с клиновидно-суженным или тупоокругленным основанием, тупозаостренной верхушкой, пильчато-зубчатые. Uaema по 4 — 6 в зонтиках, диаметром 4,0 — 4,5 см, на корот- ких беловойлочных цветоножках длиной 1,5 см, чашелистики узкотреугольн ые. Яблони данной серии распространились на обширной терри- тории, часть которой является наиболее древним центром ксеро- морфной флоры. В состав серии входят более или менее ксеро- морфные яблони Северной Индии (Западные Гималаи), Север- ного Пакистана (М. caspiriensis, М. chitralensis), ксероморфные яблони афганского и советского Туркестана (M nuristanica, subsp. turkmenorum) и Северо-Восточного Ирана (subsp. turk- menorum), а также более или менее ксероморфные яблони Кав- каза, Ирана и Турции (М. orientalis и его ксероморфная раса чаг. montana) (рис. 62). 28. Malus orientalis (Uglitz.) Juz. 1939, in Фл. СССР, 9: 362. 1) subsp. orientalis а) var. orientalis Гроссгейм, 1952, Фл. Кавказа, 5: 28; Фе- доров и Полетико, 1954, Дер. куст. СССР, 3: 419; Прилипко, 1954, Фл. Азербайджана, 5: 50; Федоров, 1958, Фл. Армении, 3: 248; Васильченко, 1963, Новые для культ. виды ябл., 27.— Pgrus malus auct. fl. cauc. Pyrus malus tomentosa Ldb. 1844, Fl. Ross., 2: 96, р. р. — Ма1из pumila Grossh. ег auct. fl. cauc., non Mill. — Яблоня восточная (кавказская). дерево высотой 8 — 10 (12 — 15) м, с широкой шатровидной, иногда неправильной кроной. Кора на старых стволах морщи-
Глава 5. Систематический обзор рода Malus 177 Рис. б2. Разновидности Malus orientali:s:1 — var. orientalis; 2 — var. montana: 3 — var. montana f. pygmea нистая, слущивающаяся небольшими кусками, серого цвета. Многолетние ветви обычно сильно развитые, гладкие, лишенные колючек, с темно-серой или серовато-коричневой корой. Годич- ные побеги темно-коричневые, слабовойлочные или волосистые. Листья продолговато-яйцевидные, продолговато-эллиптичес- кие, яйцевидно-ланцетные, на укороченных побегах, почти ок-
178 Глава 5. Систематический обзор рода Malus руглые, длиной 5 — 8 (2 — 10) см, шириной 3 — 4 (1,5 — 7,0) см, снизу густоопушенные или беловойлочные, сверху рассеянно-во- лосистые (во взрослом состоянии менее опущенные), по краю пильчато-зубчатые или городчато-зубчатые, при основании почти цельнокрайные, на верхушке тупые или заостренные, с тупоок- ругленным или клиновидно-суженным, реже неравносторонним основанием, на толстоватых войлочно-волосистых черешках в 2 — 4 раза короче пластинки. IIBeTKH по 4 — 6 в зонтиковидных соцветиях, на коротких (1,5 см) мохнатых или войлочных цветоножках; чашечка бело- войлочная; венчик белый, снаружи розоватый, диаметром 4,0— 4,5 см; стилодии (5), сросшиеся у основания в нижней части сильноопушенного столбика. Плоды шаровидные, округлые, конические, цилиндрические, с широкой воронковидной выемкой у основания, зеленовато- желтые, кремовые, белые с румянцем и полосатостью, розовые до красных, кислые, кисло-сладкие или терпкие, сочные. Пло- доножки толстоватые, длиной 1 — 2 (3) см, в молодости войлоч- ные или волосистые. Цветет IV — V; плоды созревают VIII — Х. Typus (paratypus): CoIchis, Guria- prope Maran, 28 V 1860, n' 876, Ruprecht, in Herb. Inst. Bot. nom. V. L. Котагоч11 Ас. Sci. URSS in Leningrad conservatur. Распространена на Кавказе, в Крыму, за пределами СССР— в Малой Азии (Иран). Наиболее типичная форма встречается на Кавказе, на предгорных равнинах и предгорьях, на высоте 200 — 500 м н. у. м. Входит в состав горных широколиственных (дубова-грабовых, буковых) лесов, встречается в долинах и по берегам рек, на лесных опушках, по горным склонам в виде примеси к другим лесным породам. Чистых естественных на- саждений не образует. Хорошо развивается на опушках и в рединах, в понижениях. В сгущенных лесах как порода свето- любивая затеняется, угнетается и вытесняется другими поро- дами. По всей территории Кавказа яблоня восточная представлена в большом разнообразии. На Северном Кавказе достигает круп- ных размеров — высота 15 — 20 м, окружность ствола 0,5 — 1,0 м, продолжительность жизни — 120 — 150 лет. Здесь можно выде- лить целый ряд форм, отличающихся главным образом внеш- ним видом плодов. Крупноплодные формы (диаметр 4 — 6 см) нередко встречаются на Северном Кавказе; по-видимому, это остатки прежней культуры. В Краснодарском и Ставропольском краях М. orientalis постепенно замещается соседним европейским видом М. sylves- tris и его ксероморфной разновидностью var. praecox. Контакт обоих видов яблони способствовал взаимной их гибридизации.
Глава 5. Систематический обзор рода Afalus 179 Ь) var. montana (Uglitz.) Langenf. сотЬ nova, Malus mon- tana Uglitz. 1932, Элем. мет. морфол. распозн. диких ябл.— Я бл он я горн ая. Arbuscula non magna, 2 — 3 т alt., interdum frutex cacumine compacto. Cortex trunci et stolonum annosorum griseolobrunneus, stolones interdum spinosi. $1о1опез annosi valde pebescentes, co- lore brunnei lenticel is сит distinctis, spinosi. Folia oblongata 4 — 6 ст lg, 2 — 3 ст lt, stolonibus abbrevia- tis fere rotunda, 2 — 3 ст lg, 1,8 — 2,5 ст lt, distincteinsuper levi- ter, subtus, 2 — 3 ст lg, 1,8 — 2,5 ст lt, distincteinsuper leviter, subtus valde-pubescentia sive albotomentosa pubescentibus 1,5— 2,0 lt petiolis. Flores in florescentiis umbelliformibus 3 — 5, colore albi че1 roseoli са 3 ст in diam., pedicellis pubescentibus 1,0 — 1,5 ст lg; calyx et hypanthium albotomentosum. Fructus solitarii vel bini, rarius terni, sphaerici 1,5 (1,8) ст in diam., sive раиlит polis compressi — 1,5 ст lg, 1,8 ст in diam., ЬгеиЬиз pedicellis 0,3 — 1,2 ст lg, rarius раиlит ovales- usque ad 2,0 ст lg, 1,8 ст in diam., pedicellis 1,8 ст lg (forma non atypice); colore sunt fIavo-virentes leviter rubescentes; acido- acerbi (icon). Area geographica Transcaucasia 1000 — 2200 (800 — 2500) т alt., unde evidenter transit fines ЫЯ$$ et pervenit in Turciam. Ira- пит, Iracum (coll. А Florenscii, 1916, ех Armenia Turcica; coll. А. В. Shelkovnikovii et В. Shinginskii, 1916, ех КЬигйз1апо Turcico et Persia Borea1i; etiam Handel-Mazzetti, 1907). Typus: Transcaucasia, Azerbajdzhan, distr. Ismailly, inter pa- gos Baskae et Njudzhi, in silva declivitalis orientalis, са 1200 т alt., 22 VIII 1940, fr., L. Prilipko et G. Gurvitsh. In Herb. Inst. Bot. Ac. Sc. Azerb. RSS in ВаКи conservatur. Affinitas: А specie principali М. orientatis (Uglitz.) Juz. subsp. montana differt habitu suo xeromorpho, crescentia humili- ore, aliis magis pubescentibus et frutibus sphaericis minutioribus (1,5 in diam.) atque habitatione sua а1Иийпе 1000 — 2200 (2800) т, aridis conditionibus. Небольшое деревце высотой 3 — 6 м, иногда растущее кус- том, с компактной кроной. Кора на стволах и многолетних побегах серовато-коричневая или коричневая. Побеги иногда колючие. Годичные побеги сильноопушенные, коричневые с за- метными чечевичками, колючие. Листья продолговатые, длиной 3 — 7 см, шириной 2 — 3 см, на укороченных побегах, почти округлые, заметно опушенные: сверху слегка, снизу сильно, или беловойлочные, на опушенных черешках длиной 1,5 — 2,0 см. 1Яветки по 3 — 5 в зонтиковидных соцветиях, белые или розо- ватые, диаметром около 3 см, на сильноопушенных цветонож- ках длиной 1,0 — 1,5 см; чашечка и гипантии беловойлочные.
180 Глава 5. Систематический обзор рода Malus Плоды одиночные или по 2, реже по 3, шаровидные, диа- метром 1,5 — 1,8 см, или едва сдавленные с полюсов, с корот- кими (0,3 — 1,2 (1,5) см) плодоножками, реже несколько оваль- ные — длиной до 2,0 см, диаметром 1,8 см, на плодоножках длиной 1,8 см (нетипичная форма!), желто-зеленоватые с лег- ким румянцем, кисло-терпкие. Географически обособлена в Закавказье на высоте 1000— 2200 (800 — 2500) м н. у. м., откуда, по-видимому, заходит в Турцию, Иран, Ирак (сборы А. Флоренского, 1916, из Турец- кой Армении; А. Б. Шелковникова и В. Шипчинского, 1916, из Турецкого Курдистана и Северной Персии, а также сборы Хан- дель-Маццетти, 1907). 2) subsp. turkmenorum (Juz. et М. Рор.) Langenf. Malus turkmenorum Juz. et М. Рор., 1939, in Фл. СССР, 9: 364, 493; Блиновский и Пояркова, 1949, Фл. Туркм., 4: 34; Федоров и Полетика, 1954, Дер. и куст. СССР, 3: 427; Васильченко, 1963, Новые для культ. виды ябл., 107. — Яблоня туркменов. Деревце высотой 3 (5) м, или более крупное дерево высотой до 6 — 8 м, с компактной или раскидистой кроной, нередко рас- тет кустообразно, образуя обильную проросль. Стволы диамет- ром 20 — 30 (40) см, с буровато-серой корой. Многолетние ветви красноватые или коричнево-бурые, годичные побеги иногда пиг- ментированные антоцианом и опушенные, в молодости густо белово йлоч ные. Листья эллиптические, обратнояйцевидные или овальные, длиной 6 — 8 см, шириной 3 — 4 см, с оттянутой верхушкой, кли- новидным или округленным основанием, хорошо выраженными жилками, сверху слабо-, снизу сильноопушенные или пушистые, на черешках в 2 — 3 раза короче пластинки (в среднем длиной 2,0 — 2,5 см). Цветки белые, в бутонах розоватые, диаметром 2,5 — 3,0 см, собраны по 3 — 5 в зонтиковидные соцветия, на коротких (1 см), сильноопушенных цветоножках, гипантии и чашечки сильноопу- шенные, чашелистики треугольно-ланцетные, с обеих сторон густо беловойлочные, тычинки на одном уровне с рыльцем или приподняты чуть выше, стилодии (5), сросшиеся на 1/3 длины в мохнатый столбик. Плоды диаметром 2 5 — 3,0 (3,0 — 5 0) см, шаровидные или овальные, сильно вдавленные с полюсов, иногда углубленные у чашечки и плодоножки, слегка ребристые, желтые, желтовато- зеленые с румянцем, плодоножка по длине равна плоду или немного короче (около 3 см), чашечка закрытая. Плоды пресно- сладкие, более крупные — с приятным ароматом. Цветет IV, плоды созревают (VI), VII — Ъ'П1 (IX). Описана из Копетдага (район горы Сют-Даг). Typus: Коре1-dagh, montes Sjunt-dagh; paratypus in fauc.
Глава 5. Систематический обзор рода Ма!из Guen, in Herb. Inst. Bot. nom. V. L. 3(omarovii Ас. Sci. UPSS in Leningrad conservatur. Распространена в Центральном и Западном Копетдаге (гора Сют-Даг, ущелье Иол-Даре (подъем на гору Сют-Даг); Хосар- Даг, урочище Чох-Агач (вершина Хасар-Дага), ущелья Гюен, Аксу, Теамиль, ущелья близ Койна-Косыра, по дороге Койна- Косыр — Нухур, Каргиле — Нухур). Восточная граница ареала проходит до Сулюкли, западная — по долинам рек Аврат и Артек до Кизыл-Артек и южнее, почти до северо-восточных пределов Эльбруса в Северном Иране, где постепенно замеща- ется родственным видом — М. опеп1а1сз, северо-восточное крыло ареала которого заходит в леса Эльбруса в Северном Иране. За пределами СССР яблоня туркменов встречается в Иране. Растет по сухим склонам, преимущественно северной экспо- зиции, среди зарослей клена туркменского, боярышника понтий- ского и других древесных пород. Низкорастущие формы размножаются главным образом кор- невыми отпрысками, образуя характерные особи порослевого происхождения. Семенами размножаются преимущественно бо- лее мощные деревья, однако обильному плодоношению нередко препятствуют неблагоприятные для цветения и опыления погод- ные условия (засуха). Крупноплодные формы яблонь с шаровидными плодами диаметром 3,5— 5,0 см, встречающиеся в ущельях Хосар-para и Иол-Даре, а также в неко- торых других ущельях, главным образом в пределах Западного Копетдага (бассейн р. Артек), имеют облик M. orientalis и представляют собой, по-ви- димому, менее ксероморфные популяции, переходные от M. orientalis к subsp. turkmenorum. Лучшие формы и клоны яблони туркменов с давних пор окультурены местным населением. В прикопетдагской полосе часто разводят кустарнико- вую форму яблони, известную под местным названием 'Баба-арабская'. По совокупности признаков она имеет прямое отношение к subsp. turkmenorum. Следует подчеркнуть, что естественные насаждения яблони туркменов в Копетдаге сильно сокращаются вследствие беспощадной вырубки, а также выпаса скота. На это указывает И. Т. Васильченко [1963, с. 109], который пишет: «Многочисленные названия местных урочищ и ущелий, как Алмалы, Алмалы-Даг, Алмаджик и т. п. свидетельствуют о б илом широком рас- пространении яблони туркменов в Копет-Дагер. 29. Malus caspiriensis. Langenf. Pyrus malus auct. р1иг. Fl. Ind. (Hocker, 1879, Fl. Brit. Ind.; Collet, 1902, Fl. Siml.), vix autem L. — Malus pumila auct. р1иг. saltem р. р. vix Mill.— Яблоня кашмирская. Arbores elatae, 8 — 10 т attingentes altitudinem. Folia magna usque ad 9 ст lg et 5 ст lt, ovoidea, breviter acuminata, margine crenata, pubescentia, subtus valde alboto- mentosa, insuper leviter pilosa, petiolis 2,5 — 3,5 ст lg. Flores albi sive roseo1i, 3,5 — 5,0 cm in diam., in alabastris гозе1.
Глава 5. Систематическик обзор рода Malus 182 Рис. бЗ. Malus caspiriensis, Typus~ Fructus sphaerici, sive plani (Typus.) utrisque polis compressi 3 ст lg, 4 ст in diam., lata et rotunda сит medulla са 2 ст in diam.; pediculi sunt breves 1,0 — 1,2 ст lg icon. Typus: N. India, Kashmir, locality Sunagai, 9 VIII 1891, fr., G. А. Gammi, in НегЬаг1ит Forest Research Inst, in Dehra Dun (India) conservatur. Affinitas: А territorialiter finitimis М. chitraiensis Vass., etiam а М. sieversii (Ledeb.) М. J. Roem. et aliis speciebus auto- chtonis subsectionis Malus species nostra cresscentia robustiore et foliis maioribus etiam floribus fructibusque та1ог!Ьиа differt. Дерево высотой 8 — 10 м.
Глава 5. Систематический обзор рода Malus 183 Листья крупные, длиной до 9 см, шириной 5 см, яйцевидные, короткозаостренные, по краю городчатые, опушенные, снизу сильно беловойлочные, сверху слегка волосистые, на черешках длиной 2,5 — 3,5 см. Цветки белые или розоватые, диаметром 3,5 — 5,0 см, в бу- тонах розовые. Плоды шаровидные или плоские (тип!), с обоих концов вдав- ленные, длиной в среднем 3 см, диаметром 4 см, с широкой, округлой сердцевиной диаметром около 2 см, плодоножки ко- роткие, толстоватые, длиной 1,0 — 1,2 см (рис. 63). Распространена в Западных Гималаях, в районе Кашмира и Джамму, в полосе от 1500 до 3500 м н. у. м., откуда, по- видимому, заходит в прилегающие районы Пакистана и неко- торые восточные районы Афганистана. На востоке эта яблоня отграничена хребтом Каракорум высотой до 8611 м (г. Чогоры), на севере — хребтом Гиндукуш высотой до 7690 м (г. Тигич- Мир). 30. Malus chitralensis Vass. 1969, in Fl. Jranica, 1: 38, сит. icon. tab. 12. — Яблоня читральская. Дерево средней высоты (?). Листья на плодоносящих побе- гах эллиптические, округлые, с закругленным основанием, на верхушке притупленные, по краю зазубренные, снизу опушен- ные, сверху едва волосистые, длиной 2,5 — 4,0 см, шириной 2— 3 см. Цветки в зонтиках; чашелистики опушенные, линейно-лан- цетные, отвернутые; лепестки эллиптические, бледно-розовые, длиной 15 — 16 мм, шириной 8 — 9 мм; цветоножка длиной 15 — 20 мм. Плоды не описаны. Турив: Pakistania, Chitral: Rumbour, 35'46' N, 71' 40' Е, 18 Ч 1958, n' 679, alt. 7500', S. А. Bowes 1.yon in Herb. Natur- historisches Museum in Wien, Austria conservatur. Распространена в Пакистане (пров. Читрал), Нуристане (Афганистан) . Series (ряд) 11. Ма 1 ив. Malus sect. Eumalus ser. Silveslres Juz., 1939, Фл. СССР. 9: 358. — Malus sect. Eumalus вег. Pu- milae Rehd., 1958, Man. Cult. Trees Shrubs 390, р. р. Malus areae Europeanae. Cortex truncorum adultorum агЬогит griseus, dehiscens; rami annosi sunt соlоге grisei vel griseolo-brunnei. Folia late ovata, late elliptica, infima parte гатогит vix rotunda, basi orbiculata vel emarginata et apice breviter acumi- nata, serrata. Flores paucifloris in umbellis, 2,5 — 3,5 ст in diam., nudis vel leviter pilosis pedicellis са 2 ст lg.; sepala triangularia sunt. Typus series: Typus generis. Географически обособленные яблони европейского очага.
Глава 5. Систематический обзор рода Malus Кора стволов взрослых деревьев серая, растрескивающаяся, многолетние ветви серые или серовато-бурые. Листья широкояйцевидные, широкоэллиптические, в нижней части побегов почти округлые с закругленным или выемчатым основанием и коротким остроконечием, пильчатые. Цветки в немногоцветковых зонтиках диаметром 2,5 — 3,5 см, на голых или слабоволосистых цветоножках длиной около 2 см, чашелистики треугольные. Тип серии: тип рода. 31. Malus sylvestris (L.) Mill., 1768, Gard. Dict. ed. 8, M. по 1. 1) var. sylvestris С. К. Schneid., 1906, Ill, Handb. Laubh., 1: 715, сит ic. 397 а-Ь; Юзепчук, 1939, Фл. СССР, 9: 358; Mans- feld, 1940, in Repert. Spec. Nov. Reg. Veg., 49: 45; Федоров и Полетика, 1954, Дер. куст. СССР, 3: 418; Васильченко, 1963, Новые для культ. виды ябл., 10. — Яблоня лесная. Небольшое дерево или кустарник высотой 6 — 10 (4 — 6, до 13) м. У отдельно растущих особей бывает широкая, раскидис- тая,шаровидная крона; в сомкнутых посадках крона вытянутая, узкая. Кора на старых стволах растрескивающаяся кусками 1,5)(3,0 см, темно-серого, мышино-серого или оливково-серого цвета. Многолетние ветви табачно-бурые, буроватые до буро- вато-желтых, образуют обычно небольшие горизонтальные от- ветвления, на концах которых располагаются мелкие почки или колючки. Двух-четырехлетние ветви имеют редкие мелкие чече- вички серовато-коричневого цвета. Годичные побеги оливково- зеленые или темно-коричневые, совершенно голые, редко слабоволосистые у самых верхушек, сравнительно тонкие. Почки небольшие, зеленовато-коричневые, оливковые или серо-фиолетовые, голые или слегка волосистые с тремя наруж- ными чешуя ми. Листья на однолетних побегах в большинстве широко ок- руглояйцевидные или почти округлые, реже эллиптические, на многолетних ветвях почти округлые или округло-яйцевидные, при основании слегка округленные, тупоокругленные, реже с ширококлиновидным, иногда выемчатым основанием плас- тинки и внезапно заостренной или короткоострой верхушкой, реже — без нее; край пластинки городчато-пильчатый или пиль- чато-зубчатый, реже — с двоякозазубренными или слегка заос- тренными зубцами, в молодости курчаво-войлочные (особенно по жилкам на нижней стороне пластинки), впоследствии почти голые, сверху ярко-зеленые, несколько лоснящиеся, снизу блед- но-зеленые, матовые, длиной 2 — 9 см, шириной 2 — 5 см, на черешках в 2 — 5 раз короче пластинки (1 — 2 (4) см), рыхло- войлочно-волосистых или впоследствии почти голых. Цветки белые или розоватые, в бутонах до распускания ро- зовые, реже — красные или розовато-фиолетовые, расположены
Глава 5. Систематический обзор рода Malus 185 Рис. б4. Malus sylvestri's в малоцветковых зонтиках на концах укороченных побегов; вен- чик диаметром 3 см, цветоножки длиной 1,8 (1,0 — 2,5) см, го- лые или слабоволосистые, особенно у гипантия; чашелистики длиной около 7 (5 — 9) мм, треугольные, длиннозаостренные, снаружи голые, внутри войлочно-мохнатые, при плодах остаю- щиеся, до низу свободные; лепестки длиной 1,3 — 2,0 см, шири- ной 1,0 — 1,2 см, округло-яйцевидные, внезапно суженные в ко- роткий, длиной 1 — 2 мм, ноготок; тычинки длиной 9 — 11 мм, желтые, голые или к основанию рассеянно-опушенные, стило- дии длиной 9 — 3 мм, в начале приблизительно одной длины, позднее — несколько длиннее тычинок, голые, сросшиеся у ос- нования в столбик, рыльца головчатые. Плоды варьируют по форме, окраске, срокам созревания и вкусу; в среднем небольшие, диаметром 2,0 — 3,5 см, 2,5 — 3,8 см
Глава 5. Систематический обзор рода Malus по вертикальной оси, по форме округло-яйцевидные, округлые или шаровидные, вдавленные при основании, желто-зеленые, со слабым зеленым или желтым оттенком и розоватым румянцем, часто слегка краснеющие на обращенной к солнцу стороне, кис- лые, горькие, терпкие; плодоножка короткая, приблизительно в 2 раза короче плода или равная ей; семенные камеры большие, шириной 1 — 2 см. В 1 кг — 50 — 65 плодов. Семена продолговатые, 6)(4)(2 мм, буроватые, бурые в све- жем виде и сероватые — в сухом состоянии (рис. 64). Цветет V — VI, плоды созревают Х — XI. Тип не указан. Neotypus: Latvia — destrictus Nadonensis, 4,5 km а4 viam Cesvaine — Vecpieba1ga silvae mixtag frondosus, ргоре villula зe- parata Grasi, 2 VI 1963 fl., 28 IX 1963 fr., n' 105, V. Langenfelds, in Herb. Universit. Latviensis in Riga (Rig) conservatur. Распространена в европейской части СССР (Карельский перешеек — Вологда — Пермь — Уфа — Уральск — Волгоград — Ма- риуполь — Кишинев), откуда заходит в Северную и Центральную Европу (Скандинавия, Польша, Чехословакия, Австрия, Герма- ния, Швейцария, Северная Италия, Британские о-ва, Франция и северо-восток Испании). Южную границу ареала точно про- следить трудно, так как по мере распространения на юг встре- чается множество промежуточных форм, которые частично но- сят характер лесной яблони или относятся к ксероморфной расе — ранней яблоне. В диком виде встречается рассеянно по всей указанной тер- ритории, главным образом на лесных опушках (реже в лесах), в поймах рек и на южных склонах возвышенностей. Приурочена к лесным, в особенности — к широколиственным и смешанным группировкам. Под пологом леса развивается плохо и встреча- ется сравнительно редко. Плотность естественных насаждений лесной яблони весьма неоднородна: в сплошных лесах — всего несколько деревьев на 1 га, на опушках лесов — 50 — 100, иногда (в благоприятных условиях) — 100 — 300 деревьев на 1 га. Предпочитает глинистую, карбонатную почву; избегает заболоченных, песчаных, закисленных и бедных почв. 2) var. praecox (Pall.) Ропот., 1977. Бот. журн. 62,6:824. Malus praecox (Pall.) Borkh., 1803, Нап4Ь. Forstbot., 2: 1271, saltem quo-ad nomen; Юзепчук, 1939, Фл. СССР, 9:361; Цит- терман, 1950, Фл. БССР, 3: 38; Федоров и Полетика, 1954, Дер. Куст. СССР, 3: 419; Яоброчаева, 1954, Фл. УССР, 4: 35 — 36.— Pyrus praecox Pallas, 1784, Fl. Ross, 1: 22, excl. cyn.; Ledeb., 1844, Fl. Ross, 2: 97. — Pyrus malus auct. р1иг. Fl. Ross saltem р. р. — Pyrus malus p tomentosa Ledeb., 1844, Ls. 96 р. p.— Malus var. rossica et var. woj ej kowii Litw., 1917, in Nta- евский. Фл. Ср. Росс., изд. 5: 198. — Malus ритila аuct.
Глава 5. Систематический обзор рода Malus 187 Рис. Я. Яц1из sylvestris var. р~аесох plur. saltem р. р., vix Mill. — Ма1из communis Woronow, 1931, 1п Acta Hort. Bot. Sc. URSS, t. 43, fasc. 49. Ic. Woronow, 1931, 1. s. f. 419 et 420. — Яблоня ранняя. деревце высотой 2 — 4 (6) м, с округлой компактной кроной (иногда растущее кустообразно), серой или серовато-коричне- вой корой на стволах и многолетних ветвях; годичные побеги коричневые, красновато-коричневые, в молодости слегка опу- шенные. Листья широкояйцевидные до округлых, эллиптические или обратнояйцевидные, длиной 3 — 10 см, шириной 1 — 6 см, у осно- вания округленные, реже клиновидные (на укороченных побе- гах), на верхушке тупые с короткими остроконечиями, по краю пильчатые, удвоенно-пильчатые или зубчатые, слегка опушен- ные, особенно в молодости и по нижней стороне пластинки, впоследствии почти голые или слабоволосистые по жилкам. Че- решки значительно, в 2 — 5 раз, короче пластинки, иногда слегка волосистые, слабо-красноватого оттенка. Цветки по нескольку в зонтиковидных соцветиях, белые, в бутонах розовые, розоватые, диаметром около 4 см; стилодии чуть длиннее тычинок, сросшиеся на 1/3 длины в войлочно-опу- шенный столбик; чашелистики треугольно-ланцетные, отверну- тые и вместе с гипантием беловойлочные. Плоды диаметром 2,0 — 2,5 (3) см, шаровидные или не- сколько удлиненные, желтые с румянцем, приятно кисло-слад-
Глава 5. Систематический обзор рода Malus ]88 Рис. бб. Одичавшая форма домашней яблони кие, сладкие, рано созревающие; плодоножки по длине равны плоду или едва короче, войлочно-опушенные (рис. 65). Цветет V — VI, плоды созревают VIII — IX. Описана из бассейна р. Яон. Лектотип: Ростовская обл. Маныгский лесхоз, близ хут. «Верхний Хамитец», Веселовского р-на, 28 VII 1949, пл., 108, Васильченко, Голубкова, Пояркова; хранится в гербарии Ботан. ин-та им. В. Л. Комарова АН СССР (Ленинград). Распространена в лесостепных районах европейской части СССР (бассейны Волги, Яана, Днепра, Днестра — Поволжье,
Глава 5. Систематический обзор рода Malus СМ Рис. б7. Безлепестная яблоня — М. domestica 'Apetala' (цветки без венчика и тычинок, плоды оез семян) центральные районы РСФСР, Северный 1~авказ, 1~рым, Укра- ина, Молдавия, а также в зоне широколиственных лесов Мол- давии, Украины, РСФСР, Белоруссии и Прибалтики). За пре- делы СССР выходит в Венгрию, Румынию, Болгарию, Югосла- вию, Албанию, Австрию, Италию, Грецию и Турцию. Встречается по речным долинам, на лесных опушках, преи- мущественно на открытых местах, реже — в широколиственных и смешанных лесах и кустарниках. Gr. cult. D о m e s t i с а е Langenf. nomen почит, grex culti- genus, ad sectionem Malus accedens. Malus sect. Eumalus, subsect. Pumi:lac, Rehder,1920, in Journ, Arnold Arb., 2: 47, р. р. — Malus sect. Ритйае, Юзепчук, 1939, Фл. СССР, 9: 358, р. р. — Malus sect. Eumalus, ser. Pumilae,
Глава 5. Систематический обзор рода Malus 190 Рис. 68. Malus X prunifoliu Rehder, 1940, Man. Cult. Trees Shrubs, ed. 2,390, р. p., sensu str.; Rehder, 1958, in op. cit. 391, p. p. Mali сиНае, semicultae sive quasi spontaneae. Folia magna, plerumque ruminata, utraque parte pubescentia, supra interdum nitida, atrovirentia, infra albotomentosa, crassis, brevibus pubescentibusque сит petiolis. Flores magni (4 — 5 ст in diam.) brevibus et albotomentosis pedicellis; sepala et hypanthium sunt dense albotomentosa.
Глава 5. Систематический обзор рода Ма1из СМ Рис. 69. Afa1us~spectabi1is Fructus magni ab 4 — 5 ст usque ad 8 ст in diam. (formac cultae (гис1из etiam maiores habent), brevibus pediculis, dulca- cidi, esculenti. Культурные, полукультурные или одичавшие яблони (рис.66, 67) . Листья крупные, обычно морщинистые, опушенные с обеих сторон, сверху иногда лоснящиеся, темно-зеленые, снизу бело- войлочные, на толстых, коротких и опушенных черешках. 1Яветки крупные, диаметром 4 — 5 см, на коротких беловой- лочных цветоножках; чашелистики и гипантии густо беловой- лочные. Плоды крупные, диаметром от 4 — 5 до 8 см (у культурных форм еще крупнее); на коротких ножках, кисло-сладкие. Malus domeslica Borkh. зепзи lato — культигенный комплекс.
Глава 5. Систематический обзор рода Malus 192 Рис. 70. Malus)(purpurea Некоторые ти пы культурной яблони. Gr.)(P r и n i f о11 а е (Juz.) 1.angenf. comb. nova, grex hybridus ad. sect. Malus accedens. Malus sect. Са1ycomeles)<sect. Gymnomeles goehne, 1893, Deutsche Dendr. р. р. — Malus sect. Eumalus)(sect. Sorbomalus, Zabel, 1903, in Beissner et al., Handb. 1.aubh.-Berm. — Malus sect. Eumalus ser. Prunifoliae Juz., 1939, Фл. СССР, 9: 336.— Malus sect. Calycomeles ser. Prunomeles 1.ikh., 1954, Сортовед. ябл. Hybridae interspecificae sunt et во1ит in cultu reperiuntur. Sunt in чаг11в combinationibus hybridis sectionis МаlивХОугг~- nomeles, rarius Malus)(Sorbomalus, гаго hybridae compositae (Malus)(Gymnomeles) ~ Sorbomalus. Folia longis tenuibus petio1is bis usque ter brevioribus lamina.
Глава 5. Систематический обзор рода Malus Рис. 7I. MalusXpurpurea 'Eleyi' Flores longis (ca 3 ст), tenuibus pedicellis, sepala lanceata, longiora hypanthio, basi tubulose sive torulose concreta, rarius calyx deciduus. Fructus connati in paucis 3 — 6, rarius solitarii, sine lugis fere сит basi sive plus minusve profunda, sed angusta areola ad pediculum. Pediculi tenues fructu longiores sunt. Combinatio characteristica totius gregis: Malus)(prunifolia (Willd.) Borkh. (М.domestica)(M. baccata).
Глава 5. Систематический обзор рода Malus Рис. 72. Malus)(purpurea 'Lemoinei' Межвидовые гибриды, встречающиеся лишь в кул ь тур е. Представляют собой различные гибридные комбинации сек- ций Malus)(Gymnomeles, реже Malus)(Sorbomalus, изредка сложные гибриды (Malus)(Gymnomeles)(Sorbomalus: М.'~p~u- nifolia (Willd.) Borkh., М.)<spectabilis (Ait.) Borkh., М.)(micro- malus Makino, М.)<magdeburgensis Hartw., М.><adstringens Zab., M.)<purpurea (Barbier) Rehd., М.>(scheideckeri Spath, М.)<gioriosa 1.emoine и др. (рис. 68 — 73).
Глава 5. Систематический обэор рода Malus Рис. 73. Malus Xsheideckeri
Глава 6КУЛЬТУРНЫЕ СОРТА ЯБЛОНИ Карельская АССР, Архангель- ская, Кировская, Вологодская, Пермская и Свердловская об- ласти, Удмуртская АССР, юж- ная часть Приморского края Грушовка московская, Китай-ка золотая, Боровинка и дру-гие местные сорта и сеянцы Культурные яблони (Gr. Domesticae) рассматриваются нами как сложный культигенный комплекс, образование которого в значительной мере связано с деятельностью человека. Вместе с тем им присущи и некоторые признаки крупноплодных яб- лонь. Так, среднерусские сорта по совокупности своих признаков стоят близко к лесной яблоне (М. sglvestris), местные кавказ- ские сорта напоминают М. orientalis, оригинальные среднеази- атские — М. я~еоегш и т. д. Однако близость некоторых куль- турных сортов к определенным автохтонным видам является весьма относительной и не может быть использована при клас- сификации. Большинство сортов культурной яблони создавались при скрещивании лучших, т. е. значительно отличающихся от типа образцов, прежде отобранных и окультуренных человеком. Та- ким образом, современные сорта культурной яблони далеко отошли от своих диких предков как по генетической основе, так и по экологии, географии, совокупности морфологических и других признаков. По существу, большинство наших культурных сортов являются искусственными, иногда случайными, слож- ными образованиями. Это подтверждается исчезновением мно- гих ценных свойств и признаков сортов при нормальном их половом размножении. Поэтому применение каких-либо извест- ных методов систематической ботаники для классификации сор- тов культурной яблони оказывается малоэффективным. Общий перечень основного сортимента и главнейших райо- нов промышленного выращивания культурной яблони в СССР по Ф. Д. Лихоносу [1959] приведен ниже:
Глава 6. Культурные сорта яблони !97 Ленинградская, Ярославская, Ивановская и Горьковская об- ласти, Башкирская АССР Псковская, Новгородская, Ка- лининская, Смоленская, Мос- ковская, Рязанская, Тульская и Куйбышевская области, Чу- вашская АССР, северная часть Белорусской ССР, Татарская и Мордовская АССР Тамбовская, Пензенская, Во- ронежская, Курская и Орлов- ская области, северная и се- веро-восточная части Украин- ской ССР Саратовская, Волгоградская области и северная часть Ас- траханской области Южная часть Белорусской ССР, Эстонская, Латвийская и Литовская ССР, )Китомир- ская, Черниговская, Харьков- ская и Ворошиловградская об- ласти Винницкая, Киевская, Кирово- градская, Днепропетровская, Донецкая и Ростовская обла- сти Пап ировка, Грушовка москов-ская, Китайка золотая, Корич-ное полосатое, Антоновка, Бо-ровинка, Осеннее полосатое Грушовка московская, Папи- ровка, Китайка золотая, Анто- новка, Анис, Бабушкино, Осен- нее полосатое, Славянка, Скри- жапель, Бельфлер-китайка Пепи н шафранный, Кальвиль анисовый, Антоновка, Варгуль, Диана, Пепин Черненко, Ба- бушкино, Коричное полосатое, Папировка, Грушовка москов- ская, Боровинка, Пепин литов- ский, Уэлси Анис, Антоновка, Малы бога- евский, Пепин литовский, Гру- шовка московская, Папировка, Коричное полосатое, Боровин- ка, Астраханское белое, Сла- вянка, Снежное, Бельфлер-ки- тайка, Пепин шафранный, Шафран-китайка, Пепин-ки- тайка Пепин шафранный, Бойкен, Ренет ландсбергский, Папи- ровка, Уэлси, Антоновка, Пе- пин литовский, Боровинка, Бельфлер-китайка, Бабушкино, Осеннее полосатое, Коричное полосатое, Диана, Пепин Чер- ненко, Буцкое, Кальвиль снеж- ный Ренет Симиренко, Кальвиль королевский, Ренет ландсберг- ский, Кальвиль снежный, Бой- кен
Глава 6. Культурные сорта яблони 198 Южная часть Астраханской области, Ставропольский и Краснодарский края, Калинин- градская область Южная часть Запорожской и Донецкой областей Узбекская ССР, Келесский район Казахской ССР, Крым- ская область, Таджикская и Туркменская ССР Черноморское побережье Крас- нодарского края, Кабардино- Балкарская и Северо-Осетин- ская АССР, Адыгейская АО Грузинская и Армянская ССР, Аджарская и Абхазская АССР Азербайджанская ССР, Дагес-танская АССР Папировка, Астраханское крас-ное, Ренет Баумана, Яндыков-ское, Кандиль-китайка, Пении литовский, Джонатан Пармен зимний золотой, Ренет Симиренко, Ренет бумажный, Кальвиль снежный, Кальвиль королевский, Ренет канадский, Розмарин белый, Бойкен, Ре- нет ландсбергский, Вагнер, Па- пировка, Пепин литовский Ренет Симиренко, Пармен зим- ний золотой, Золотое, Кандиль синан, Сары синан, Ренет бу- мажный, Розмарин белый, Ре- нет орлеанский, Кальвиль ко- ролевский, Наполеон, Вагнер, Бойкен, Бельфлер, Папировка Ренет Симиренко, Пармен зим- ний золотой, Ренет бумажный, Кальвиль королевский, Ренет ландсбергский, Ренет Баумана, Бельфлер, Пении шафранный, Шафран-китайка, Пении-ки- тайка, Диана, Пении Чернен- ко, Папировка, местные чер- кесские сорта Ренет шампанский, Пармен зимний золотой, Кальвиль ко- ролевский, Ренет канадский, Ренет серый, Банановое зим- нее, Бельфлер, Наполеон, мест- ные сорта Ренет бумажный, Кальвиль королевский, Ренет Симиренко, Ренет орлеанский, Ренет ка- надский, Розмарин белый, Пар- мен зимний золотой, Сары си- нан, Джир Гаджи и другие местные сорта
199 Глава 6. Культурные сорта яблони Апорт, Ренет орлеанский, Ре- нет Симиренко, Пармен зим- ний золотой, Розмарин белый, Бельфлер, Вагнер, Ренет ландс- бергский, Бойкен, Кандиль си- и ап, Кандиль-китайка, Суй- слепское, местные сорта Алма-Атинский и Джамбуль- ский районы Казахской ССР, Киргизская ССР Сорта открытой культуры типа китайской яблони: Непобеди- мое, Грелля, К,арагасс, Тунгус, Анисик, Таежное, Ермак и другие; сорта стелющейся культуры: Пепин шафранный, Китайка золотая, Еафран-ки- тайка, сорта яблони централь- ных областей европейской ча- сти СССР Челябинская, Томская, Ом- ская, Новосибирская и Кеме- ровская области, Краснояр- ский и Алтайский края В мировом сортименте известно до 10 000 культурных сортов яблони [Заец, 1960; Жуковский, 1964]. Разумеется, при таком обилии сортов уже давно возникла необходимость в их систе- матизации, чтобы облегчить распознавание, изучение, использо- вание и т. д. Наиболее полной и популярной в свое время была класси- фикация сортов, предложенная немецким помологом Дилем, видоизмененная и дополненная Лукасом. Согласно ей, культур- ные сорта яблони разделяются на 15 классов: 1) кальвили; 2) трещотки; 3) златки; 4) розовые (розовки); 5) голубки; 6) рамбуры; 7) рамбурные ренеты; 8) боксдорфские ренеты; 9) одноцветные ренеты; 10); 11) золотые ре- неты; 12) серые ренеты; 13) полосатые; 14) острые яблоки и 15) плоские яблоки. Система Диля — Лукаса является полностью искусственной, поскольку сорта классифицируются только по признакам плода, а не всего растения. В основу деления сортов на классы поло- жено несколько признаков. Однако многие из них, служащие критериями для образования классов Диля — Лукаса, являются столь неопределенными, что один и тот же сорт в различных географических зонах совершенно не соответствует определен- ному классу. Поэтому данная система в настоящее время не ислользуется. Американский помолог Хедрик считает невозможным созда- ние универсальной системы для всех культурных сортов яблони. По его мнению, можно классифицировать лишь сорта на срав- нительно небольшой территории, где деревья и плоды мало варьируют.
200 Глава 6. Культурные сорта яблони Общепризнанной в СССР является система классификации, предложенная советским помологом Ф.,Я. Лихоносом [1963, 1964]. В ее основу положены биологические особенности сортов, морфологическое строение цветков, сроки созревания плодов. Главнейший критерий системы Ф. Д. Лихоноса — эколого-гео- графический принцип с учетом зимостойкости сортов. Сорта яблони разделены на три секции и девять групп. Система пред- ставлена в следующем виде: 1 секция. Северные Группа 1. Вишнеплодные Группа 2. Сливолистные (китайки) II секция. Центральные Группа 3. Русские и типы русских, зимостойкие Группа 4. Русские и типы русских, менее зимостойкие Группа 5. Пепины (гибридные ренеты) Ш секция. Южные Группа 6. Южноевропейские (ренеты) Группа 7. Крымские Группа 8. Кавказские Группа 9. Среднеазиатские Элементы классификации сортов Ф.,Я. Лихоноса уже ис- пользуются на практике. ,Яанная система послужила основой для разработки некото- рых вариантов подразделения сортов яблони в СССР. Ниже приводится система В. 1<. Заеца [1960], в которой более по- дробно разработаны группы сортов в соответствии с важней- шими зонами и районами садоводства СССР. Однако ее недо- статком является то, что здесь отсутствуют высококачественные западно-европейские и североамериканские сорта яблони, до- вольно широко распространенные в отдельных районах СССР и имеющие промышленное значение. В помологической системе В. К. Заеца, как и у Ф. Д. Лихо- носа, сорта яблонь подразделяются на три отдела. I. Сорта теплого климата (южная зона садоводства). Г р у п п а 1. Закавказские: грузинские сорта и сеянцы типа,Язудзи-вашла, типа Турашаули, типа Китра, типа Аби- лаури, Кехура и др.; армянские — Марга хндзор, Кальбаджа- фар и др.; азербайджанские — Сары турш,,Яжир Гаджи, Кзыл Ахмед, Шихар джаны и др. Г р у п п а 2. Северокавказские: Розмарин черкесский, Го- лотлинское, Мигинц, Омаровское, Ренет ахтынский, Кахари г зимний, Раджабовское, Араканское красное, Оротинское желтое, Анис кубанский, Поляковское, Столбовка, Тавлинка и др. Г р у п п а 3. Туркменские. Г р у п п а 4. Таджикские: Касым саркары, Себи терамохи„ Аштарханы, Пангази, Себи шойн и др.
Глава 6. Культурные сорта яблони 20! Г р у п п а 5. Узбекские: Самаркандское раннее, Первенец Самарканда, Ренет ташкентский, Яжизакское, Боровинка таш- кентская; новые сорта — Афросияби, Зеравшанская розовка, Регистанли, Саратони и др. Г р у п п а 6. киргизские: Бель зимняя, Тянь-Шаньское, киргизское осеннее и др. Г р у п п а 7. Южно-казахстанские: Грушовка алма-атин- ская, Пеструшка, различные кульджинки; новые сорта — Заи- лийское, Алма-атинское зимнее, Бельфлер сипап, Бельфлер алма-атинский, Румянка алма-атинская и др. Г р у п п а 8. 1~рымские: Сары синап, кандиль синап, Су- дакский синап, Рара синап, Саблы синап, Челеби, Гульпембе и др. Г р у п п а 9. Молдавские: Лужанка, Нестрец, Голубок обыкновенный (Тиролька молдавская), Голубок днестровский, Яомнешты, Шафран серый, Цыганка молдавская и др. Г р у п п а 10. Украинские: Кальвиль снежный, Ренет Сими- ренко, Бутское, Зори, 1~ныш, Шлопак, Мухрик, Путивка осенняя, Репка меджибожская, Баковка, Зеленяк, Сивак, Гусак, Вин- ничка; новые сорта — Пепин Черненко, Яиана, Ренет украин- ский, Первомайское, Ренетное млеевское, Млеевская красавица, 1~альвиль млеевский, Слава победителям и др. II. Сорта умеренного климата (средняя зона садоводства). Г р у п п а 11. Белорусские: старинные народные и новые селекционные сорта (Ломицкое, Несравненное озимое и др.). Г р у п п а 12. Прибалтийские: Белый налив (Папировка), Пепин литовский, Осеннее полосатое, Серинка, Земляничное Ничнера, Рижское молочное, Борсдорфское луковичное, Видзем- ский золотой ренет, Сеянец Требу, Суйслепское, Целминю жел- тый, Анис бержаникайский, Монтвиховка, Литовское сахарное, Пайдаское, Грушовка ревельская и др. Г р у п п а 13. Среднерусские: Антоновка обыкновенная, Бо- ровинка, Грушовка московская, коричное полосатое, Бабуш- кино, Скрижапель, Варгуль воронежский, Аркад желтый, Ок- ское, Склянка курская, 1~урский золотой ренет, Коробовка и др.; сорта И. В. Мичурина и других советских селекционеров, выведенные в центральных областях РСФСР. Г р у п п а 14. Поволжские: Анис полосатый, Анис розово- полосатый, Анис алый, Анис винный, Малы богаевский, Малы крестовый, Малы украинский, Скрут белый, Скрут розовый, Хорошавка алая, Хорошавка сенгилеевская, Бель пучковая и др.; нижневолжские — Яндыковское, Мамутовское сладкое, Ма- мутовское красное, Ивановское, 1~алинчинское, китайка волж- ская и другие формы китайки, возникшие в этой зоне; новые сорта, выведенные в Поволжье. III. Сорта холодного климата (северо-восточная зона садо- водства).
202 Глава 6. Культурные сорта яблони Г р у п п а 15. Северорусские: сорта В. В. Спирина — Бес- семянка никольская, Пепинка Спирина, Коричное китайка, Ан- тоновка-китайка; сорта А. Н. Рудницкого — Кировское позднее, Янтарное, Синап непобедимый, Северная зорька, Пионер Севера и другие сорта, выведенные с 1917 г. в северных и северо-вос- точных областях европейской части РСФСР. Г р у п п а 16. Уральские: переходные от крупноплодных европейских к мелкоплодным сибирским сортам — Боровинка превосходная, Красавица Зигулева, Уральское наливное, Ураль- ское большое, Высокое, Призовое, Малютка, Янтарь, щедрая и др Г р у п п а 17. Сибирские: полукультурные, приближающи- еся к китайкам, — Анисик омский, Виновка желтая, Хорошавка фермская, Горно-алтайское, Алтайский голубок, Пепинка ал- тайская и др.; ранетки — Ранетка пурпуровая, Лалетино, Баг- рянка, Сеянец Пудовщины, Непобедимая Грелля, Янтарка ал- тайская и др. Г р у п п а 18. Дальневосточные: местные народные и новые селекционные сорта (Шураповское Ка2, Ефремовское Ка 1, Тон- коножка, Вкусное, Синап амурский, Хабаровское, Августовское дальневосточное и др.). Попытки классификации культурных яблонь не всегда были удачными, поскольку эта сложная группа создана человеком, а потому чрезвычайно многообразна. Здесь объединено множе- ство различных культурных сортов, декоративных садовых форм,при создании которых (в результате гибридизации, мута- ционного и модификационного процессов, явлений полиплоидии и т. п.) были использованы многие дикие виды яблонь, а также всевозможные гибриды и гибридные помеси, встречающиеся как в культуре, так и автохтонно. К этои группе примыкают также полукультурные и некоторые одичавшие яблони, являющиеся более или менее характерными для различных условий. Незам- кнутость ареала и искусственность распространения этих яблонь в природе почти всегда имеют антропогенный характер. Наши исследования (1957 — 1988) комплекса Domesticae прежде всего подтверждают мнение М. Г. Попова о том, что «основным путем формообразования, создания новых форм из старых всегда было отдаленное скрещивание между генетически различными видами» [Попов, 1963, с. 26]. Основываясь на этом, можно сделать следующие выводы. 1. На формирование разнообразия комплекса Domesticae в местах культивирования яблони и соприкосновения ее ареалов с некоторыми дикими видами большое влияние оказали как процесс гибридизации, так и различные экологические условия. 2. В роде Malus происходит процесс ксероморфогенеза, в результате которого образовались сравнительно молодые виды ксероморфного облика, отличающиеся (в своей генетической ос-
Глава 6. Культурнме сорта яблони 203 нове) от более древних мезофильных видов. В местах сопри- косновения этих генетически различных (мезо- и ксероморфных) яблонь при эффективном их скрещивании возникли новые, не- редко гетерозисные и полиплоидные формы, которые характе- ризуются хозяйственно важными качествами и в ряде случаев служат материалом для селекции культурных сортов яблони. В результате долгой селекционной деятельности человека из та- ких форм были отобраны лучшие по качествам крупноплодные яблони, из числа которых в процессе окультуривания и повтор- ной гибридизации было создано множество культурных сортов. 3. Благодаря склонности скрещиваться и дичать культурные яблони образовали сложное гибридное и весьма полиморфное потомство, где почти каждая особь обладает разнообразными признаками и свойствами. Среди форм яблони, окультуренных и одичавших, следует различать: а) низкие яблони ксероморфного облика, обитатели сухих, открытых мест. высотой до б — 8 м, с невыраженными стволами, растущие иногда кустообразно, отличающиеся от исходных ви- дов более толстыми опушенными побегами, крупными, войлочно- опушенными листьями, крупными кисло-сладкими плодами; распространенные иногда вблизи жилья или в окрестностях мест прежней культуры; б) высокие, опушенные (пушистые) яблони более мезофиль- ного облика, приуроченные к более влажным местам обитания, высотой 10 — 15 м, с хорошо выраженными, иногда мощными стволами (в обхвате от 1,0 до 1,5 м), отличающиеся от М. sgl- vestris более интенсивным опущением вегетативных органов (молодых побегов, листьев), а также частей цветка, более круп- ными и хозяйственно ценными плодами; распространенные в местах современного или прежнего культивирования яблони. Неясное систематическое положение, иногда гибридное про- исхождение, а также отсутствие обособленного ареала этих яб- лонь не позволяет отнести их к определенному виду. Однако во избежание дальнейшей путаницы можно предложить рассмат- ривать эти яблони в составе сборного культигенного ком- плекса — Malus domestica Borkh, sensu lato — вместе с круп- ноплодными культурными сортами. Такое объединение различ- ных сортов (и даже одичавших яблонь) в один сборный вид нельзя признать вполне удачным. Тем не менее мы полагаем, что при дальнейшем их изучении с помощью более совершенных методов исследования появится возможность различать и вы- делять ряд определенных типов, разных по происхождению и распространению в культуре.
Глава 7 ПЕРСПЕКТИВЫ ИСПОЛЬЗОВАНИЯ ЯБЛОНИ В НАРОДНОМ ХОЗЯЙСТВЕ Яблоня ценится прежде всего как хозяйственно-полезное плодовое дерево. По территории СССР дикая яблоня распро- странена сравнительно широко, однако наиболее крупные ес- тественные ее насаждения расположены в горных областях Средней Азии, на Кавказе, а также в европейской части СССР. В Юго-Восточной Сибири и на Дальнем Востоке произрастают мелкоплодные яблони. Общая площадь естественных насаждений яблони на терри- тории СССР составляет не менее 500 000 га, из которых более 250000 га приходится на Среднюю Азию Ал. А. Федоров, Ан. А. Федоров, 1949; Васильченко, 1963 и др., около 100000— на Кавказ, а в европейской части СССР яблоня занимает около 85000 га [Васильченко, 1963; Лангенфел~ьд, 1960б, 1961 и др.]. Площадь насаждений культурной яблони в СССР, по данным П. М. Жуковского [1964], составляет около 500 000 га (около 60 млн. деревьев) при среднем валовом сборе 2,5 — 3,0 млн. т свежих плодов в год. Ежегодная урожайность естественных насаждений диких яб- лонь по самым скромным подсчетам (при урожае 20 кг яблок с дерева и плотности насаждений 100 деревьев на 1 га) состав- ляет около 1 млн. т свежих плодов. При переработке этой про- дукции ежегодно можно получать не менее 150 000 т сушеных плодов, или 850 млн. л высококачественного сока, или 650000 т варенья, или не менее 700000 т повидла. Поскольку урожай- ность диких яблонь по годам и отдельным областям их рас- пространения значительно превышает в среднем 20 кг с дерева, общая их продуктивность иногда может достигать 1,5 — 2,0млн. т яблок в год. Однако эти дары природы используются крайне неудовлет- ворительно. В лучшем случае часть плодов диких яблонь соби- рает местное население, нередко повреждая при этом деревья. Много яблок остаются неубранными. Большой ущерб наносит естественным насаждениям и выпас скота. Следует централизовать работу по заготовке диких плодов, организовать их переработку, а также улучшить охрану естест-
Глава 7. Перспективы использования яблони 205 венных насаждений плодовых, более строго наказывать тех, кто нарушает установленный порядок охраны заповедных лесов и 30Н ПЛОДОВЫХ. Кроме того, дикие яблони в известной степени могут быть использованы для совершенствования культуры яблонь. Здесь можно вести селекционную работу по улучшению сортимента и созданию новых сортов яблони, а также по созданию новых подвоев, необходимых для прививки культурных сортов. Успех селекционной работы во многом зависит от правильного под- бора исходных форм. При создании новых сортов наилучшие результаты получают при скрещивании культурных сортов между собой. Однако определенная роль здесь принадлежит и некоторым диким видам, особенно крупноплодным и их формам с довольно высоким качеством плодов, а также местным фор- мам, которые приспособились к данным условиям. Следует об- ратить внимание на местные высококачественные дикие, а также полукультурные формы одичавших яблонь. Во многих районах СССР такие яблони очень разнообразны. У некоторых из них уже в естественных условиях проявляются высокие вкусовые качества плодов и значительная зимостой- кость. Такое разнообразие форм открывает широкие возмож- ности при подборе исходных форм для гибридизации. Полу- культурные формы одичавших яблонь, а также местные формы с высококачественными плодами, ценными биологическими и хозяйственными свойствами могут быть использованы в каче- стве материнских растений. Тем самым будет соблюдено поло- жение И. В. Мичурина об использовании в работах по гибри- дизации корнесобственных растений. Поскольку из родительской пары материнское растение полнее передает свои свойства по- томству, иногда следует воздерживаться от использования низ- кокачественных форм диких яблонь в качестве материнского растения. Из лучших форм известных видов следует обратить внимание на среднеазиатские яблони, произрастающие в пределах Фер- ганского хребта и Каратау, а также Заилийского Алатау. На Кавказе встречается немало интересных карликовых форм местной яблони, часть которых уже более или менее окульту- рена. Прежде всего, это яблони, известные под местными назва- ниями: в Дагестане — Дипчек-алма, в Азербайджане — Кёл- алмасы, Яр-алмасы, в Кахетии — Хомандули, в Армении Марга хндзор. В европейской части СССР перспективные ис- ходные формы для создания новых сортов следует искать среди ксероморфных М. praecox и лучше всего — среди одичавших полукультурных яблонь. Все дикие виды яблонь, особенно мел- коплодные сибирские, а также мелкоплодные азиатские менее перспективны в качестве исходного материала для выведения новых сортов в связи с низким качеством их плодов.
206 Глава 7. Перспективы использования яблони Особый интерес для выведения высококачественных, поздне- зимних сортов представляют некоторые североамериканские яблони, плоды которых характеризуются ароматом, мелкозер- нистой мякотью и хорошей лежкостью. 1~ультурные сорта обычно размножают путем прививки. Судьба будущего сортового дерева в значительной мере зави- сит от физиологической совместимости привоя и подвоя. По- скольку эти взаимоотношения могут быть весьма сложными и трудно обнаруживаемыми, то часто при подборе подвоя допус- каются серьезные ошибки. Производственные и опытные данные показали, что подвой оказывает большое влияние на привой. От него зависят продолжительность жизни привитого дерева, его размеры, время вступления в пору плодоношения, урожай- ность, периодичность плодоношения, качество плодов, сроки прохождения фенологических фаз развития, зимостойкость, ус- тойчивость к вредителям и болезням, реакция на почвенно-кли- матические условия и т. д. Одним из важнейших требований, предъявляемых к подвоям в культуре, является выносливость их в данных экологических условиях. В этой связи возникает настоятельная необходимость использования на практике прежде всего местного подвоя. Вторым, не менее важным требованием, является высокая степень физиологической совместимости привоя и подвоя. Иде- альное решение этого вопроса состоит в выделении определен- ного подвоя почти для каждого сорта или каждой группы род- ственных сортов. Но пока об этом не может быть и речи, хотя бы потому, что при семенном размножении яблоня дает весьма неоднородное потомство и мы пока еще не имеем таких опреде- ленных подвоев. Поэтому, решая вопрос о подвоях для яблони, следует изыс- кивать такие виды, разновидности, формы, популяции, эколо- гические и биологические расы и отдельные особи, которые по своим свойствам являлись бы более близкими к культурной яблоне, а их семенное потомство давало бы наиболее однород- ный материал. Этим требованиям больше всего отвечают сеянцы старых местных сортов. Очевидные перспективы имеют встре- чающиеся в диком состоянии формы одичавших яблонь, а для некоторых районов, в частности Средней Азии, — также высоко- качественные формы автохтонных видов, которые приспособи- лись к местным условиям, а их полезные признаки и свойства закрепились в потомстве. Менее перспективными в качестве подвоя являются прочие дикие виды яблони в связи с неоднородностью их сеянцев в условиях культуры. Из других яблонь в качестве генеративно размножаемых подвоев некоторые садоводы иногда рекомен- дуют сливолистную яблоню (М. >< prunifolia) . Однако следует отметить, что она, будучи гибридом, крайне неоднородна в за-
Глава 7. Перспективы использования яблони 207 висимости от происхождения отдельных ее популяций, а также условий культуры. Нередко почти все особи отличаются друг от друга по ряду признаков и свойств. Правда, в некоторых районах (особенно в Поволжье и др.) имеются перспективные клоны сливолистной яблони, дающие довольно однородный ма- териал. Однако на основании данных об отдельных клонах или популяциях нельзя сделать вывода о пригодности сливолистной яблони в качестве подвоя для всех условий. муроме того, ряд местных, особенно западно-европейских сортов, проявляет пло- хую совместимость со сливолистной яблоней. Мелкоплодные восточно-азиатские виды (секции Sorbomalus и Gymnomeles), в том числе сибирские ягодные яблони, в каче- стве подвоев для крупноплодных культурных сортов неперспек- тивны. То же относится и к их гибридным формам. Весьма перспективными являются вегетативно размножае- мые типы подвоев со строго определенными свойствами. В ка- честве таковых могут использоваться формы среднеазиатских яблонь, а также формы полукультурных низких яблонь При- балтики [Лангенфельд, 1961], которые даже в естественных условиях проявляют склонность к вегетативному размножению. Исходным материалом для выведения вегетативно размножае- мых подвоев могут служить и другие виды яблони. Особый ин- терес представляют некоторые формы и клоны кавказской яб- лони, особенно М. orientalis, а также М. praecox. По-видимому, эти две яблони являются самыми вероятными предками многих типов селекции подвоев яблони. Первую попытку классифицировать существующие формы подвоев сделало еще в 1872 г. Английское общество садовод- ства. Позже эта работа в более широком объеме проводилась в Англии Веллингтоном, а после него — Хеттоном на Ист- Моллингской опытной станции по садоводству, в Германии- Шиндлером, в Голландии — Шпренгером, а также в Бельгии, Франции и других европейских странах. За последнее время значительно увеличилось использование клоновых подвоев в Швеции и Норвегии, где наряду с ист-моллингскими подвоями стали применять подвой собственной селекции, например под- вой Иогансона А1 и А>. В СССР работа по инвентаризации су- ществующих и выведению новых подвоев яблони, а также их классификации проводится в Плодоовощном институте им. И. В. Мичурина под руководством проф. В. И. Будаговского, а также на многих опытных станциях страны и во Всесоюзном институте растениеводства. Среди подвоев яблони ист-моллингские типы II, III, V, VIII и IX имеют признаки культурных сортов. Видимо, они произо- шли не прямо от диких форм яблони. Большую часть ныне из- вестных парадизок и дусенов до введения их в культуру в качестве подвоев широко выращивали, размножая простейшими
209 Глава 7. Перспективы использования яблони способами — отпрысками, отводками, реже черенками. И для того, чтобы выработать присущие им теперь качества, эти формы должны были пройти длительный путь отбора. При изучении подвоев не так трудно установить их связь с некоторыми дикими яблонями. Так, почти все клановые низко- рослые, карликовые подвои, известные под общим названием парадизок, имеют схожие основные признаки с некоторыми по- пуляциями М. domestica. Более высокие, полукарликовые кла- новые подвои, так называемые дусены, по совокупности при- знаков во многих случаях примыкают к М. praecox. Следует, однако, отметить, что систематика существующих типов клановых подвоев, как и культурных сортов яблони, весьма запутана. Так, основную работу по типизации подвоев яблони в Англии проводили по давно установившейся традиции, преследуя лишь чисто практические цели, в связи с чем обе группы подвоев — парадизок и дусенов — нередко объединяли под одним названием — парадизки. Так были смешаны две ес- тественные и различные по происхождению группы подвоев. И наоборот, нередко один и тот же подвой распространялся под несколькими названиями. Кроме групп парадизок и дусе- нов, принадлежность клоновых типов к которым еще нечетко разработана, попыток применения других принципов система- тики при классификации подвоев яблони не было. Такого рода систематику еще следует разработать. Яблоня ценится не только как хозяйственно полезное де- рево. Она имеет большое декоративное и эстетическое значе- ние. Яблоня может быть одновременно и декоративным, и плодовым деревом. Наиболее пригодны для такого рода поса- док некоторые формы мелкоплодных яблонь из секции Ggmno- meles и особенно крупноплодные гибриды этой группы, напри- мер, типа М.>(prunifola, М.>(cerasifera и т. д. Эти яблони по своей природе являются весьма обильноцветущими, богато пло- доносят, в связи с чем они весьма декоративны. Их вишневид- ные плоды, небольшие, но сочные, дают высококачественный десертный сок, используются в консервировании и кулинарии. Мелкоплодные яблони и яблони с лопастными листьями, бе- лыми, розовыми или красно-пурпуровыми цветками и ярко окра- шенными плодами следует использовать исключительно для скверов. Низкорослые, а также и высокоствольные яблони, куль- турные и дикие, пригодны для аллейных и придорожных на- саждений. Все яблони, а особенно местные формы, с успехом могут быть использованы при создании прекрасных ландшаф- тов, садов и парков. В некоторых районах нашей страны яблоню давно уже ис- пользуют для придорожных насаждений. При этом высаживают главным образом привитые культурные сорта, тогда как для такой цели лучше использовать сеянцы (непривитые подвои} и
Глава 7. Перспективы использования яблони 209 местные дикие яблони. Придорожные насаждения, как пока- зывает опыт, не могут быть обеспечены хорошим уходом. По- этому для их создания рациональнее использовать местные ди- кие яблони. Так, в европейской части СССР вдоль дорог произ- растают различные формы одичавших и лесной яблонь: на %(авказе — М. orientalis, в Средней Азии — М. sieoersii и их подвиды. По габитусу наиболее пригодны для дорожных насаждений высокоствольные яблони. Менее пригодными являются естест- венные кустообразные формы, которые затрудняют обозрение дорог и окрестностей, задерживают ветер и тем самым замед- ляют просушку дорог, что важно для более северных районов европейской части СССР в пределах ареала М. sglvestris. Из интродуцированных яблонь для аллейных насаждений следует рекомендовать обильноцветущие и богато плодонося- щие сибирские и дальневосточные яблони — М. baccata и М. mandhsurica, а также некоторые гибридные яблони, такие, как М.)(prunifolia, М.)(cerasifera и др. Для создания неболь- ших аллей и особенно для оформления входов в парки, можно использовать и другие, низкие и кустообразные яблони. Однако при этом следует избегать применения интродуцированных, ори- гинальных видов, которые иногда слишком усложняют и нару- шают естественность насаждений, создают пестроту и не гармо- нируют с окружающим ландшафтом. Из яблонь можно создавать и живые изгороди, так как они поддаются стрижке (обрезке). Однако в подстриженных живых изгородях яблоня не будет обильно цвести и потеряет большую часть своей декоративной ценности. Поэтому яблони следует сажать в изгородях, не подвергающихся стрижке. В скверах яблони сажают как отдельными деревьями, так и небольшими группами, а в парках — небольшими группами. В крупнейших парках, где создаются целые ландшафты, яблони имеют боль- шое значение как элемент местной флоры. Так как яблоня принадлежит к группе светолюбивых расте- ний, то наибольший декоративный эффект получается при по- садке деревьев отдельно. Посаженные крупными группами, яб- лони образуют густые заросли, в то время как отдельно расту- щая яблоня всегда привлекает внимание своим внешним видом, цветами и плодами. ~(рупные группы яблонь можно сажать при входах в парки и оформлять их в виде аллей. Следует включать яблоню в пар- ковые насаждения, где она будет привлекать внимание и тем самым оживлять однородную группировку. В совместных посад- ках яблоня хорошо сочетается с шиповником, рябиной, грушей, боярышником и другими видами лиственных пород. Следует воздерживаться от совместных посадок яблони и хвойных пород.
Гла~а 7. Перспективы использования яблони 2]0 Таблица Рекомендации к использованию яблони I. Eriolobus М. trilobgta П. Docgniopsis M. tschonoskii М. formosana M. laosensis M. sikkimensis III. Sorbomalus М. ombrophila M. prat tii M. yunnanensis M. honanensis М. Aansaensis (f. kansuensis, f. с/а- га) M. toringoides M. transitoria M. setok M. centralasiatica M. Oomarovii М. 1огт~~о var. 1огс'про 'Moerl an dsii' 'Liset' 'Profusion' 'Win ter gold' чаг. arborescens чаг. sgrpenl'И К, Дстд, Дссл К, Дсел, (ф) К К К Дотд Дсел 6 ? 8 9 I0 6 7 8 9 10 К, Дсел, Д К, Д Дсел К> Д, Дсел К ° Д, Дсел К~ Д~ Дсел 1 — 2 1I 12 I3 14 ~б 16 К~ Д~ Дсел К, Д, Дсел К, Д К, Д К, Д!, Дсел Д~, Дсел II I2 I3 I4 I5 16 I7 I8 I9 20 2I 22 К, Д К, Д К, Д Е. Д К, Д а, Дд, Дгр, Н<ер К, Д 'Магу Potter' 23 В небольших скверах, которые создаются в строго регуляр- ном стиле и где предусмотрено сажать формовые деревья (плакучие, шаровидные и пр.) и цветы, создавать различные геометрические фигуры, рекомендуется высаживать интродуци- рованные виды и формы яблонь с оригинальными габитусом (плакучие и другие формы), листьями (пурпуровыми, лопаст- ными и т. д.) и др. Особое внимание следует уделять при этом декоративным сортам. В таких скверах яблони сажают от- дельно. Таким образом, возможности применения дикорастущих яб- лонь в народном хозяйстве достаточно широки (табл.). Следует- лишь полнее и рациональнее использовать их.
2!! Продолжение таблицы Ql Дотд1р Дсел1 К, Д1 К, Д1 Д}» Доел К,Д К, Д Д- Дгр, Дал, Дсел К, Д К, Д К, Д К, Д 24 25 26 27 28 29 30 10 ll 12 13 31 32 33 34 35 36 37 38 К К Д, Дгр, Доел К 17 39 18 14 15 19 20 16 17 18 9 10 19 20 21 22 23 67 21 К Далр Дсел (Фколо) К, Доел К. Дсел К Дотд К 68 69 70 71 'Соl цтпаг is' '8olgo' 'Gracilis' 'Lady ХогЬ Ьс1Ше' 24 25 26 27 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 Глава 7. Перспективы использования яблони М. y ftoribunda 'ОгтЫоп Roy' 'Peachblow' М.)< arnoldiana 'Henrieta бгозЬу' 'Henry' F. 0u- pont' М. X zumi 'Bob White' 'Са1осагра' 'Golden Hornet' 'Professor Spren- ger' М. >(sublobata М. y florentina М. fusca М Xdavsoniana Ж. Chloromeles М. coronaria 'Cha rlottae' чаг. dasycalyx чаг. elongata 'Nie Uwlandiana' М. angustifotia М. ioensis чаг. bushii var. creniserrala 'Fimbriata' var. раlтеп 'Рlena' 'Nova' чаг. spinosa var. texana М. bracteata М. glaucescens М. glabrata М. Iancifotia М. Х platycarра М. )(heteroрйуйа 'Hillieri !!' М. )С denboerii 'Evelyn' 'Lisa' М. Х soulardi'i 'Red f lech' 'Wynema' V. Gymnomeles М. baccata Ф, Д, (Поел}, (Плол) К, Л К,Д, (Ф} К,Д К, Д Д, К Д', К (@) Д, < Д,К К Д,К К К к, Д К,Д К К К К К, Псел К, Псел К К К К К К К
212 Продолжение та блицы Э [ 4 К, Дал Дсел, (Фкон с ) К, Дсел М. mandshurica 72 22 73 23 subsp. sachalinen- Ш М. himalaica М. Х cerasi~eãà 24 74 75 'Aurantica' 'Qen uina' 28 76 77 29 78 79 80 'Oblongata' >Масгосагра' M. Х rob usta 30 31 12 81 82 83 84 85 86 87 88 89 90 91 32 33 34 'Beauty' 'Fairy' 'Joan' М. hupehensis 'Ro sea' M. halliana 'Parkmanii' var. spontanea М. Х atrosang ut',nea М. Xhartuvig~~ Кайег1пе VI. Malus M. asiatica 25 35 26 36 13 14 37 92 27 93 28 M. sieversii 94 niedznuetzky- ana 95 38 'Aldenham Purple' 96 39 >Cowichan' 97 40 'gingsmere' 'Makamik' 'Oporto' 'КейИеЫ' 98 41 99 42 100 43 101 10с 44 45 'Rosybloom' 'Scugok' 10.' 46 'Wisley Grab' Глава 7. Перспективы использования яблони К, Д, Лсел К, Дал Дсел, Фконс Д, Дсел, (Фконс ) К. Л.~g. Л.р. Дал Д, Доел Д, Дсел Фсок Д, Дал, Дсел (ФKOAG) К, Д К. Д К. Д Д1, Дсел К, Дотд|, Дсел~ Д1, Дотд1, Лсел К, Доел, Дотд К. Д Дотд!, Доел!, Д К, Дотд Дсел К К, Ф> Д, Псел, П~под Псел, Ппод, Д, Дсел Д Дс ел> Дотд|, Фконс, + сок, ('р) К, Дотд1, Доел, (Ф), Фсок, Фконс К, Дотд!, Доел, Фс он, Фконс, (Ф) К, Дотд1, Доел, Фконс К Дотд>, Дс ел, Фконс, (Фсок) К, Дотд, Доел, Фконс К, Дотд, Дсел, Фконс К, Дотд1, Доел К» Дотд, Дсел Фкон с, Ф сок К, Дотд, Доел, Фсок~
213 Продолжение таблицы 104 105 106 107 108 109 10 110 12 112 13 113 12 114 29 13 115 М. orientalis subsp, orientalis var. montana 116 30 Ф Д) Пдод (Поел�} Ппод1 Д 117 sUbsp. turkme- norum М. caspiriensis 118 31 119 32 120 33 121 34 К, Псел, Паол , Ф к, л, п..., (Ф} Фсок1, Фсущ» Q, Лал, Лгр М. chitralensis hf. sylvestris var sylvestris f, sylvestris f. ucutitoliu 15 14 15 122 var. ruralis f. jrutescens f. папа f, macrocarpa <, atrocarðà var, praecox 123 16 16 124 17 125 18 126 19 127 128 35 17 Gr. Domes ticae hf. domestica Д, Ф, Псел, sensu la to Ппод 129 Глава 7. Перспективы использования яблони f. spherocarpa f. jarmolenkovii f. schichkinii f. pavlovii f, [ongicarра f. poljakovii f. dragavzevii f. linczevskii f. anisophylla f. ЙиШг!азсйеой var. kirghisorum var. hissarica Д Псел Паод Ф» Д Псел Пдод Ф» Д» Пс ел» Паод Д Псел Пдод Д Псел Ппод Д, Псел Пдод Д Псел, Пдод Д Псел Ппод Дал Ф, Д» Псел» Ппод Ф, Д, Псел Ппод Псел» Пдод» Д Дсел Д, Псел Пиод Л (Псел) Плод, Фсок, (+) Фсок Д, Дал, Дгр Д, Лал, Лгр, Фсок Д, (Лал }, Лгр, Ф сок е Фсуш Л Лгр Дотд, Пп,д Л, Лал, Лгр, Фсок Фсуш Д, Дал, Дгр» Дотд, Фсок Д, Ф, Проел, Ппод1
214 П родол жение та блицы 3 4 5 6 Д, Ф Псел П'...1 Д, Ф, Псел, (П.о.1 18 var. pumila 130 19 131 var. Шазур- hyle Gr. )( Ргипс) о- Йае М. >(pi ani~îéà Д Дал, Ф сок Фконс, Дс ел К, (~Реек}, (флокс), Лсел К, Дсел 132 15 133 47 'Сешеа Гз Grimson' 'Сешеа Гз Golden Gem' 'Hyslop' М. >(spectabilis 134 48 К, Дсел Фсок Д, Дотд, Дсел1 Фконс, Фсок К Дсел 135 136 49 16 137 50 138 К, Дсел 51 139 140 141 142 52 53 54 17 143 18 144 19 145 146 147 148 149 150 55 56 57 58 59 60 151 152 61 К, Дсел~ Дотд Д, Дотд1, Дсел1, Фконс ~, Фс ок1 К, Дотд Дс ел 20 153 'Al denha-mensis 'Amisk' 'Сг1п1зоп Brilliant' 'Eleyi' 154 155 63 64 К> Дотд, Дсел К, Лоg 156 65 Дотд-> Дсел тт Фконс, Фс ок1 К, Дотд Дс ел Д К Д сел~ Дотд 157 'Eleyi com- pacta' 'Рг derek Chopin' 'Helen' 66 158 67 159 68 Ло~л~ Л. Дсел Дотд!, Л, Дсел К> Дсел1 Дотд~ 160 69 'Hoser' 'Irene' 161 70 Глава 7. Перспективы использования яблони 'Blanche Ames' 'Frau LUise Dittmann' 'Plena' 'Я1чегзГ 'Van Eseltine' М. )(пгссгота- lus М. х mo,gde- Ьигцепаз М. )(adstri n- gens 'Almey' 'Нора' 'Os man' 'Patriciа' 'Robin' 'Transcen- dent' 'Wabiskaw' М. > ригригеа К, Дотд1 К, Дотд Дсел К, Дсел Д, Фконс, Фсон Дсел Д Дотд1, Фсок Дсел Д~ Дсел Фконс Фс ок К, Дотд, Дс ел К, Дотд К, Д К, До.дд, Д Д, Фсок. Фконс К, Дсел
215 Окончание таблицы 'Jadwiga' 71 162 'Jay Darling' 'Justina' 72 73 163 164 'Кооеп Ига' 'Lemoinei' 74 75 165 1б6 76 77 78 167 168 169 79 80 81 82 К, Дотд К, Дотд, Д К Д Лотд К, Дотд К> Лотд, Дсел Фконс К Д, Дотд!, Дс ед 170 171 172 173 174 21 175 176 83 84 177 178 179 85 К Д Дсел, К К, Дс ел Дотд 22 Обозначения: Д — в декоративном садоводстве для любых поса- док; Д~, — в декоративном садоводстве для аллейных насаждений; Дрр в декоративном садоводстве для групповых насаждений; Дотд — в декора- тивном садоводстве для посадки отдельными деревьями; Дсел — в декора- тивном садоводстве для селекции новык декоративных сортов; К — в кол- лекции для дальнейшего исследования; Псел — в плодоводстве для селекции новых сортов; П„,д — в плодоводстве для селекции новых подвоев; Ф для использования в свежем виде и различных видов переработки; Фсок- для переработки в сок; Фсу — для сушки; Фко — для консервирования. Восклицательным знаком отмечены те возможности использования яблони, которым следует отдавать предпочтение; в скобках — дополнительные воз- можностии. Глава 7. Перспективм использования яблони 'Мр1Ып g' 'Pink 61апС 'Purple Ъ'аwe' 'Red Silver' 'Sissipuk' 'Sophia' 'Ъ'ierdak' М. >(schei;- decker. '0огойеа' 'Exe1 benz ТЫеГ 'Hi 1 lieri' М. )( g tori'osa 'Oekonoin c- rat Echter- in eyer' К, Д, Дсел Фконс К, Д, Дсел К, Д, Дотд, Дсел К Д Дсел Д Дотд|, Дсел~ Фконс, Фсок К, Д, Doo-g! К, Д, Дотд К, Д, Дотд
ЗАКЛ lO× ЕН И Е В результате проведенного исследования рода Malus Mill. можно сделать следующие выводы. 1~аждая из существующих систем классификации видов рода Ма1из по-своему несовершенна. Нередко они противоречивы и не выдержаны во всех частях. В них недостаточно четко отра- жены родственные взаимоотношения видов, так как не всегда учитываются все признаки, выявляющие эволюционный уровень таксона. Наблюдаются коренные разногласия как в отношении качества видов, так и их объема: некоторые автохтонные виды, преимущественно наиболее примитивные, обитающие в Юго- Восточной Азии, пропущены, а некоторые явные гибриды или безареальные морфологические варианты нередко описаны в качестве автохтонных видов. В связи с этим не всегда соблю- дается географическая обособленность и экологическая опреде- ленность таксонов. Таким образом, систематика рода Malus до сих пор разра- ботана недостаточно, существующие классификации нуждаются в критическом пересмотре; необходимо разработать новую сис- тематику рода. На основании изучения структурных особенностей различ- ных видов яблони из разных географических зон земного шара нами сделана попытка выявления морфологической эволюции органов яблони, что послужило основой для определения эво- люционного уровня и установления родственных взаимоотно- шений видов в пределах всего рода. По эволюционному уровню и развитию морфологических признаков виды рода Malus раз- делены нами на три категории. Первично простые формы — мощные, слаборазветвленные (неопределенного моно- и симподиального ветвления) деревья с хорошо выраженными стволами. Эта категория характеризу- ется следующими признаками: листья первично простые, раз- дельные или лопастные; цветки собраны в многоцветковые со- цветия неопределенного типа (ложные щитки, лопастные зон- тики); плоды мелкие, с грубоватой мякотью, содержащей каменистые клетки, и остающеися чашечкой.
Заключение 217 Усложненные формы — хорошо разветвленные с симподи- альным ветвлением деревья средней высоты, со слабовыражен- HbIM одним или несколькими стволами, усложненными, иногда лопастными до раздельных листьями; цветки в многоцветковых щитковидных соцветиях; плоды различной формы и величины, чашечка при плодах остающаяся, опадающая или частично опа- дающая; намечается развитие крупно- и мелкоплодных форм. Вторично упрощенные формы — сильноразветвленные без- стволые деревца или кустарники с симподиальным ветвлением; листья вторично упрощенные, цельные, по краю зубчатые, пиль- чатые или городчатые; цветки в малоцветковых, редуцирован- ных зонтиковидных соцветиях; плоды либо крупные с остаю- щейся чашечкой (линия крупноплодности), либо мелкие с опа- дающей чашечкой (линия мелкоплодности). Данные о морфологической эволюции органов различных видов яблонь явились крайне важным критерием при разра- ботке систематики рода Malus. Анализ обширного материала по географии рода Malus, а также сопоставление родовых ареалов Pyrus, Malus, Sorbus и Crataegus, между видами которых существуют тесные связи, позволяют предполагать, что наиболее древние участки указан- ных ареалов находятся на обширной территории Юго-Восточной Азии и некоторых средиземноморских стран. Именно в Юго- Восточной Азии когда-то проходили процессы дифференциации общих предков упомянутых родов, из которых впоследствии вы- делились и обособились самостоятельные Pyrus, Sorbus, Malus и Crataegus, постепенно занимавшие все более обширную тер- ритории Евразии, а позже (за исключением Pyrus) и Северной Америки. Дизъюнкция ареала, свидетельствующая о древнем проис- хождении и более широком распространении в прошлом, наблю- дается в секции Eriolobus. Единственный вид этой секции (М. trilobata) встречается в среднегорной полосе Ливанских гор и в горных лесах Балканского п-ова. Примитивные яблони, ныне представленные реликтовыми ви- дами, встречаются в четырех изолированных участках обширной территории, простирающейся от Восточных Гималаев до север- ных районов современных Лаоса и Вьетнама, а также в китай- ской пров. Юньнань, южной части пров. Фуцзянь, на о-ве Тай- вань и островах Японии. Это доциниевидные яблони (секция Docyniopsis) . Значительную часть родового ареала Malus занимают виды секции Sorbomalus, распространенные на обширной территории Центральной и Восточной Азии, Северной Америки и стран Сре- диземного моря. Виды секции Chloromeles, имеющей связь с доциниевидными яблонями из Юго-Восточной Азии, произрас- тают в Северной Америке.
218 Заключение На азиатском материке располагается ареал более холодо- стойких яблонь — секция Gymnomeles, занимающая два обшир- ных участка. Первый охватывает южные и юго-восточные рай- оны Сибири, Дальний Восток, Монголию и северные провинции Китая, доходя до Западных Гималаев. Второй участок ареала располагается в Центральном и Юго-Западном Китае, захваты- вает Бирму и Индию. Наиболее обширная секция рода — настоящие яблони (сек- ция Malus) — в автохтонных видах представлена исключи- тельно в Евразии, где занимает пять более или менее изолиро- ванных друг от друга участков: китайский, гималайский, сред- неазиатский, ирано-кавказский и европейский. Географическое распространение рода Malus в значительной степени зависит от экологических условий и природных факто- ров местообитания и характеризуется наличием в роде двух основных экологических групп видов — мезофильных и ксеро- морфных. На основании изучения связей в морфологическом строении и закономерностей географического распространения, а также экологических особенностей различных видов яблонь можно вы- сказать следующие соображения о филогенезе рода Malus. Первичные формы яблонь появились еще в меловом пе- риоде от общих предков родов Docynia, Pyrus и Cydonia. Од- нако в основном яблони сформировались в третичном периоде, когда на обширной территории Евразии распространились раз- личные формы третичных яблонь. Из них до настоящего вре- мени сохранились некоторые, теперь уже сильно дифференциро- ванные, реликтовые виды (секции Eriolobus и Docyniopsis). В результате дифференциации первичных, по-видимому типа доциниевидных, яблонь в третичном периоде в Юго-Западном Китае возникла еще одна ветвь эволюции рода — рябиновид- ные яблони (секция Sorbomalus). Они распространились пер- воначально в Центральной и Восточной Азии, а потом проникли в Северную Америку и страны Средиземного моря. На основании исследования филогенетических связей наибо- лее примитивных североамериканских яблонь (М. fusca, распро- страненная ныне в узкой полосе вдоль тихоокеанского побе- режья) с некоторыми доциниевидными яблонями азиатского материка (М. tschonoskii, обитающая на японском о-ве Хонда) можно предположить, что еще в то время, когда Азия и Север- ная Америка составляли единый материк, в результате диффе- ренциации третичных яблонь в Северной Америке эволюциони- ровала самостоятельная ветвь рода Malus — секция Chloro- metes. Дифференциация третичных яблонь в Гималаях привела к образованию еще двух, наиболее продвинутых ветвей эволюции рода Malus. Одна из них — секция Gymnomeles — эволюцио-
Заключение 219 нировала по линии мелкоплодности и распространялась в Се- веро-Восточной Азии; из другой — крупноплодной — образова- лась секция настоящих яблонь — Malus, расселившихся в Евразии. Из-за полиморфизма различного происхождения даже у ди- корастущих видов, не говоря уже о гибридном, в том числе интрагрессивном смешении их с культурной и прежде окуль- туренной, т. е. одичавшей яблоней, в рамках рода Malus появились некоторые весьма трудноопределимые совокупности особей неясного систематического положения, крайне затруд- няющие всестороннее и подробное описание рода. Изучение этих комплексов является задачей будущего исследования с исполь- зованием более совершенных, комплексных (в том числе гене- тических, математических и т. п.) методов. Такие очаги полиморфизма и смешения различных как по происхождению, так и по генетической основе яблонь создались, главным образом, в результате длительной селекционной дея- тельности человека и окультуривания различных видов яблони. Очаги наибольшего разнообразия крупноплодных яблонь (секция Malus) оказались в Старом Свете, где приурочены главным образом к горным районам: Гималаи, Тянь-Шань, Па- мир, Кавказ и др. Они стали первичными центрами происхож- дения культурных сортов яблони [Вавилов, 1931а,в], из кото- рых, после интродукции лучших образцов в Европу и Северную Америку, были созданы вторичные центры происхождения куль- турной яблони. Согласно нашей систематике, род Malus содержит 35 авто- хтонных видов, распространенных в широтах умеренного кли- мата Старого и Нового Света, исключительно в северном полу- шарии. Кроме того, описан один культигенный, комплексный вид — М. domestica Borkh, sensu lato, объединяющий крупно- плодные окультуренные и одичавшие яблони. Гибридные виды, гибриды, безареальные морфологические варианты, а также прочие, по нашему убеждению, необоснованно описанные как виды, исключены. Они описаны как культивары, формы, разно- видности или гибридные виды, не входящие в число автохтон- ных видов. Особую группу в нашей системе составляют гибрид- ные виды, образовавшиеся при скрещивании некоторых окуль- туренных мелкоплодных яблонь с крупноплодными окультурен- ными, чаще всего М. baccataX>. domestica, или повторные, многократные гибриды подобных комбинаций. Они отнесены к группе гибридных видов Pruni joliae. Остальные гибридные виды описаны как примыкающие по совокупности своих признаков к соответствующей секции или автохтонному виду. В результате критической оценки видового состава в роде Malus выявлено 58 видов, в том числе 35 автохтонных (из них 9 полиморфных, содержащих несколько внутривидовых подраз-
220 Заключение делений), 1 культигенный и 22 гибридных. Среди описанных внутривидовых подразделений — 22 разновидности, 13 форм и 86 культиваров (декоративных сортов). Всего в роде описано 180 внутривидовых таксонов. Однако 4 вида яблонь из Север- ной Америки — М. bracteata, М. glabrata, М. glaucescens и М. lancifolia — остаются неясными и сомнительными. Описаны они А. Редером по крайне скудным материалам (типы не ука- заны), которые нам не удалось видеть, так как типового и во- обще фактического материала по этим видам в США не ока- залось. Однако если судить только по «голому» описанию, эти виды как по морфологии, так и по географии и экологии ка- жутся сомнительными. Принятая нами система рода hfalus в некоторых ее частях является оригинальной. Однако в ней сохранены удачные, на наш взгляд, подразделения, предложенные другими авторами. Такими, прежде всего, являются секции, из которых две Docyniopsis и Eriolobus восстановлены и трактуются в новом смысле. Окультуренные и гибридные виды подчинены группам Gr. Cult. Domesticae и Gr.><Prunifoliae. Первичным центром происхождения рода hfalus или, скорее всего, местом резервации наиболее примитивных видов рода в настоящее время мы считаем Юго-Восточную Азию, северо-вос- точные и восточные страны Средиземного моря. Наиболее крупные центры полиморфизма рода hfalus в СССР в настоящее время расположены главным образом в гор- ных областях Азии: в Казахстане — Тарбагатай, Джунгарский и Заилийский Алатау, Таласский Алатау, Каратау; Киргизии— Ферганский хребет, Чаткал; Узбекистане — Пскем, Угам, Кок- Су, Ангрен, Оби-Заранг, Шахрисабз; Таджикистане — Гиссар- ский хребет (особенно в Кондаре), Такоб, Вахшинский и Дар- вазский хребты и хребет Хозретуши; Туркмении — Копетдаг; на Кавказе, в Крыму, а также в средней полосе европейской части СССР, в Забайкалье и на Дальнем Востоке. Названные, а также многие другие естественные насажде- ния диких яблонь в СССР представляют собой ценнейшее на- родное достояние. Учитывая все большее уничтожение дикорас- тущей яблони, следует принять самые решительные меры для охраны диких яблоневых насаждений. Таким образом, содержание нашей работы — критический обзор рода hfalus на видовом уровне. В обзоре видового со- става яблонь мы учитывали также внутривидовую изменчивость полиморфных видов и пытались охватить их полиморфизм. При этом особое внимание было уделено сравнению амплитуд из- менчивости структурных особенностей различных таксонов в связи с экологией и географическим распространением видов, особенно при их продвижении с юга на север и в более аридных условиях. Сделана попытка выявления закономерностей и путей
Заключение 221 * Флора СССР. — М., 1934. — Т. 1. — С. 7. эволюции отдельных признаков и определения эволюционных уровней различных таксонов рода. Это и послужило основой выяснения филогении и создания новой классификации рода Malus. Мы убедились, что у яблони слишком ясна внутривидо- вая дифференциация, чтобы признать выводы В. Л. Комарова" о монотипичности вида (по крайней мере, в пределах рода Ma- lus). Поэтому в нашей работе полиморфизм и классификация видов рода Malus изложены на основе взглядов Н. И. Вавилова [19316] о виде как о системе внутривидовых таксонов, т. е. популяции.
СПИСОК ЛИТЕРАТУРЫ Борисов А. А. Климаты СССР. — М., 1967. — 295 с. Будаговский В. O. Карликовые подвои для яблони. — М., 1959. Вавилов H. И. дикие родичи плодовых деревьев Азиатской части СССР и Кавказа и проблема происхождения плодовых деревьев // Тр. по приклад- ной ботанике, генетике и селекции. — Л., 1931а. — Т. 26, вып. 3. Вавилов Н. O. Линнеевский вид как система // Тр. по прикладной бо- танике, генетике и селекции. — Л., 19316. — Т. 26, вып. 3. — С. 109 — 134. Вавилов H. O. Роль Центральной Азии в происхождении культурных растений // Тр. по прикладной ботанике, генетике и селекции. — Л., 1931в. — Т. 26, вып. 3. Вавилов Н. O. Пять континентов. — М., 1962. — 254 с. Васильченко O. Т. К познанию восточно-азиатских видов яблони // Бо- тан. материалы гербария Ботан. ин-та АН СССР. — М.— Л., 1959. Т. 19. Васильченко O. Т. Новые для культуры виды яблони. — М.— Л. 1963.— 152 с. Васильченко O. Т. Неотенические изменения у растений. — М.— Л., 1965.— 8 4 с. Ген В. Культурные растения и домашние животные при их переходе из Азии в Грецию и Италию ( Пер. с нем. — СПб., 1872. — 379 с. Горбунов М. Г. Об остатках плодов яблони (Ма1иэ/ из третичных отло- жений Западной Сибири // @AH СССР. — 1959. — Т. 128 № 3. С. 607 — 610. Д агавцев А. П. Яблоня гор~ых обитаний. — М.— Л., 1956. — 254 с. уковский П. М. Культурные растения и их сородичи. — Л., 1964.— С. 471 — 485. Забинкова Н. Н., Кирпичников М. Э. Справочное пособие по система- тике высших растений. — М.— Л., 1957. — Вып. 11. — 334 с. Заец В. К. Селекция и сортоведение яблони // Селекция и сортоведение. плодовых и ягодных культур. — М., 1960. — С. 198 — 243. Запряааева В. O. Дикорастущие плодовые Таджикистана. — М.— Л., 1964. — 695 с. Кейн А. Вид и его эволюция. — М., 1958. — 243 с. Кобель Ф. Плодоводства на физиологической основе. — М., 1957. 375 с. Колесников В. А., Реэниченко А. Г., Кузнецов М. Я'., Ефимов В. А. Пло- доводство. — М., 1966. — 431 с. Комаров В. Л. Избр. соч. — М.— Л. 1950. — Т. 4. — 374 с. Косых В. М. дикорастущие плодовые породы Крыма. — Симферополь, 1967. — 171 с. Лангенфельд В. Т. Яблони Латвийской ССР и их естественные гибриды /f Новые формы плодовых растений. — Рига, 1959. — С. 20 — 43. Лангенфельд В. Т. Йспользование различных видов яблони в зеленом строительстве // Интродукция растений и зеленое строительство. — Рига,
Список литературы 223 1960а. — С. 113 — 121. Лангенфельд В. Т. Перспективные для озеленения виды и формы яб- лони // Биологическая наука — сельскому и лесному хозяйству. — Рига, 19606. — С. 79 — 81. Лангенфельд В. Т. Виды яблони Латвийской ССР. — Рига, 1961. 107 с. Лангенфельд В. Т. География рода Malus Mill. // Растения лесов и парков. — Рига, 1970а. — С. 89 — 101. Лангенфельд В. Т. О филогении рода Malus Mill. // Растения лесов и парков. — Рига, 19706. — С. 102 — 110. Лангенфельд В. Т. Схема классификации рода Malus Mill. // Растения лесов и парков. — Рига, 1970в. — С. 111 — 138. Лангенфельд В. Т. Род Malus Mill. (Биология, география, систематика и филогения): Автореф. дис.... д-ра биол. наук. — Тарту, 1970. — 48 с. Лихонос Ф. Д. Яблоня. — М.— Л., 1959. — 192 с. Лихонос Ф. 4f. Некоторые данные по систематике яблони // Бюл. Гл. оотан. сада АН СССР. — М., 1963. — С. 59 — 67. Лихонос Ф. gf. К вопросу о систематике рода Malus Mill. — Яблоня // Тр. по прикладной ботанике, генетике и селекции. — Л., 1964. — Т. 36, вып. 3. — С. 5 — 16. Маевский П. Ф. Флора средней полосы Европейской части СССР. .М.— Л., 1964. — 880 с. Павлов Н. В. Ботаническая география зарубежных стран. — М. 1965.— Ч. 1 и 2. — 310 с. Пашкевич В. В. Яблоня // Природа. — 1938. — Ks 5. — С. 36 — 54. Пономаренко В.. В. Новый подвид маньчжурской яблони // Тр. по при- кладной ботанике, генетике и селекции. — Л., 1972a. — Т. 46, вып. 2. С. 65 — 69. Пономаренко В. В. Сибирская ягодная яблоня в Забайкалье // Раститель- ные есурсы. — Л., 19726. — Т. 8, — С. 21 — 28. ономаренко В. В. Ботанико-географические и таксономические данные о Malus sachalinensis (Кот.) Juz. // Ботан. журн. — 1973а. — Т. 58. С. 1629 — 1641 ° Пономаренко В. В. Дикорастущие яблони Восточной Сибири и Даль- него Востока СССР // Тр. по прикладной ботанике, генетике и селекции.— Л., 19736. — Т. 49, вып. 1. — С. 85 — 94. Пономаренко В. В. К интродукции Malus sieboldii (Regel) Rehd. на Сахалине // Бюл. Гл. ботан. сада. — М. 1974. — Вып. 93. — С. 24 — 29. Пономаренко В. В. Видовой состав дикорастущих яблонь СССР и центры их генетического разнообразия // Ботан. журн. — 1975а. — Т. 62, № 6. — С. 820 — 831. Пономаренко В. В. Материалы к познанию яблонь Кавказа // Ботан. журн. — 19756. — Т. 60, № 1. — С. 53 — 68. Пономаренко В. В. )то такое Malus pumila Mill.? // Ботан. журн. 1975в. — Т. 60 № 11. — С. 1574 — 1586. Пономаренко В. В. Существует ли вид Malus hissarica Кинг.? // Ботан. журн. — 1976. — Т. 61, № 7. — С. 993 — 998. Пономаренко В. В. Генетические центры разнообразия рода Ма~из Mill. // Сб. науч. тр. ВИР. — Л. 1988. — С. 11 — 21. Попов М. Г. Дикие плодовые деревья и кустарники Средней Азии // Тр. по прикладной ботанике, генетике и селекции. — 1929. — Т. 22, вып.3.— С. 429 — 451. Попов М. Г. Основы флорогенетики. — М. 1963. — 135 с. Серебряков И. Г. Экологическая морфология растений. — М., 1962. 378 с. Скибинская А. М. Историческая география рода Malus // Бюл. Гл. бо- тан. сада АН СССР. — М., 1966. — Вып. 61. — С. 52 — 59. Тарасенко Г. Г. Китайка (М. prunifolia Borak.) и ее происхождение //
224 Список литературы Вестн. сов. растениеводства. — 1940. — № 3. — С. 18 — 25. Тарасенко Г. Г. Яблоня. — М., 1941. — 171 с. Тактаджян А. Л. Происхождение покрытосеменных растений. — M., 1961. — 146 с. Тактаджян А. Л. Основы эволюционной морфологии покрытосеменных.— М.— Л., 1964. — 263 с. Федоров А. А. Тератогенез и его значение для форма- и видообразова- ния у растений // Проблемы вида в ботанике. — М.— Л., 1958. — Т. 1.— С. 213 — 293. Федоров Ал. А., Полетико О. М. Род Ма1из Mill. // деревья и кустар- ники СССР. — М.— Л., 1954. — Т. 3. — С. 414 — 458. Федоров Ан. А., Кирпичников М. 9. Справочное пособие по систематике высших растений. — M.— Л., 1954. — Вып, 1. — 109 с. Федоров Ал. А., Федоров Ан. А. Яблоня Южной Киргизии // Плодовые леса южной Киргизии. — Л., 1949. — С. 218 — 254. Хромосомные числа цветковых растений / Под ред. Ан. А. Федорова.— Л., 1969. — 624 с. Черепанов С. К. Сосудистые растения СССР. — Л., 1981. — 437 с. бритт П. Г. Учение о росте и развитии плодовых и ягодных растений.— М., 1958. — 447 с. Юзепчук С. В. Род Malus Mill. // Флора СССР. — М.— Л,, 1939. Т. 9. — С. 357 — 413. Юзепчук С. В. Комаровская концепция вида // Проблема вида в бота- нике. — М.— Л.. 1958. — С. 130 — 205. Азиат~ У. The crab-apples and nectarines of Japan. — Tokyo, 1927.— Р. 1 — 55: 78 — 86. Borkhausen М. В. Theoretisch-praktisches Handbuch der Forstbotanik und'. Forsttechnologie, 11. — Giessen; Darmstadt, 1803. — S. 1271 — 1280. Bronvicz К. Distribution of gosaceae in W. Asia. 111. Eriolobus trilobatus (Poir.) Roem. // Arboretum Kornickie. — 1969. — Т. 14. — S. 5 — 21. Bronviez К. Ма1из florentina — its histor~, systematic position and geo- graphical distribution // Fragm. Floristica Geobot. — 1970. — Ann. 16, pars 1. — Р. 61 — 83. Chevalier А. Ма1из laosensis А. Cheval // С. R. Acad. Sci., Paris. 1920. — Т. 170. — 1129 р. Choau Н. F. The familiar trees of hopey. — Peking, 1934. — 374 Р. Eieheeald К. Rosaeeae // Eesti NSV flora. — Та111пп, 1956. — Lk. 229— 238. Eseltine G., van. Notes on the species of apples // Ви11. New York Agr. Experimental Station. — 1933. — N 208. — Р. 214. Gutmani К. 1.atvijas PSR augJu biokfmiskais sastavs. — Riga: Zinatne, 1961. — 40.— 52. lрр. Oenning W'. Morphologisch-systematische und genetische Untersuchungen an Arten und Artbastarden der Gattung /Malus // Zuchter. — 1947. Н. 10/12. — S. 289 — 349. goehne Е. Deutsche Dendrologic. — Stuttgart, 1898. — S. 257 — 262. Koidzumi G. А synopsis of the genus Malus // Acta phytotax et geobot. Tokyo. — 1934. — Vol. 4. К~йтте1 Н. Die vegetativ vermehrbaren Unterlagen des Kern- und Stein- obstes. — Berlin, 1956. — S. 128. Кгйззтапп 6. Hanbuch der Laubgeholze, 2. — Berlin; Hamburg, 1960.— S. 110 — 125. I(uggler H. Blutenokologie. — Jena, 1970. — S. 59; 215. Langenfeld W. Die Evolution der Gattung Malus Mill. // Wissensch. 2tschr. Univ. Qostock. — 1971. — И. 1. — S. 49 — 51. Lauskis V. Auglu koku з~1гпез. — Riga, 1937. — 3 — 6.; 11.— 117. lрр. Linn,acus С. Species plantarum. — Stockholm, 1753. — N 1. — Р. 1200. Linnaeus С. Mantissae plantarum. — Stockholm 1767. — Vol. 1.— Р.75.
Список литературы 226 Linnaeus С. Sistema, Genera, Species plantarum. — Stockholm, 1835.— P. 487. Mansfeld R. Vorlaufiges Verzeichnis landwirtschaftlich oder gartnerisch ku1tivierter Pf1anzenarten. — Berlin 1962. — Beih. 2. — 659 s. Mildenberger Q. Studien zur Taxonomic der Gattung Malus. 1. Morpho- logisch-genetische Untersuchungen // Arch. Gartenbau. — 1963а. — Bd 11, Н. 3. Mildenberger Q. Studien zur Taxonomic der Gattung Malus. 11 Embryo- logische Untersuchungen // Arch. Gartenbau. — 1963b. — Bd 11, Н. 7. MilleI Ph. The gardener's dictionary. Abridged ed. 4. — London 1754. Miller Ph. The gardener's dictionary. Ed. 8. — London, 1768. Miller Ph. The gardener's and botanist's dictionary. — London, 1797. ЯеЫег А. Manual of cultivated trees and shrubs hardy in North-Ame- rica. — New York, 1958. — 996 р. Schneider С. К. 111ustriertes Handbuch der Laubholzkunde, 1. — Jena, 1906а. — 654 $. SIhneider С. К. Species varietatesque Ротасеагит novae, 1V // Feldus Pep. — 1960Ь. — Т. 3. — $. 177 — 183. Snarskis P. Obelis-Malus М111. // 1.ietuvos TSR flora. — Vilnius, 1971.— Т. 4. — P. 232 — 245. Тегро А. Malus Mill. // Flora Еигораеа. — Liverpool, 1968. — Vol. 2.— P. 66 — 67. Тоигпе/ог/ 1. P. 1пвШи11опев rei herbariae. — Рапв 1700. — Р. 634. Troll 5'. Praktische Einfuhrung in die Рflanzenmorphologie. — Jena, 1957. — $. 269 — 273. Vassilczenko I. Malus Mill. // Flora 1гап!са Graz. — 1969. — Vol.бб/30 N 4. — P. 36 — 39. Yii Te-tsun, Yen Chen'-lung. Study of the Chinese species of genus Malus // Acta Phytotaxonomica Sinica. — 1966. — Vol. 6 N 2. — Р. 1 — 110. Yii Te-csun, Ки Tsne-chih. Malus Mill. // Flora Рориlапв Sinicae. 1974. — Т. 36. — P. 372 — 402.
АЛФАВИТНЫЙ УКАЗАТЕЛЬЛАТИНСКИХ НАЗВАНИЙ* I. Род Ма(из и роды, имеющие непосредственную связь с ним Apirophorum Neck. 11 Crataegus L. 13, 52, 82, 86 Cydonia Mill. 10, 26, 69 Docynia Decne 26, 53, 68 — 69 Malus Mill. 5 — 13, 18, 25 — 26, 33, 36 — 39, 42 — 47, 52, 57, 68 — 69, 72 — 73, 77, 82 — 86, IIS — !20 Pyrenia C1airv. 11 Pyrus L. 10 — 11, 13, 26, 38, 52, 68 — 69, 82 — 86 Sorbus L. 13, 82 — 86 II. Надвидовые таксоны в пределах рода Ма/из Baccatae Rehd. (вег.) 17 — 19, 22, 42, 154, 156 СЫоготе1ев (Decne) Rehd. 15, 22, 39, 41, 43, 47, 54, 74,93, 121,148,211 Docyniopsis С. К. Schneid. 14, 16 — 17, 26, 39, 44 — 45, 52 — 53, 68 — 69, 71, 73, 88аг, 120, I23, 210 Domesticae Langenf. (gr. cult.) 189, 202, 213 Erio!obus С. К. Schneid. 14, 16 — 17, 26, 39, 40, 44, 52 — 53, 73, 86,!21,210 Eumalus Zabel 12 — 13, 15, 18 — 19, 22, 164 Fuscae Eselt. 16 — 17, 75, I46 Gymnomeles Koehne (sect.) 12, 17, 22, 39, 42 — 44, 47, 54, 74 — 77, 94, 121, I54, 211 Капвиепвев Rehd. (вег.) 15 — 17, 22, 55, 74, 135 Malus (sect. et вег.) Sl, 164, 183 Prunifoliae Juz. (вег.) 19 XPrunifoiiae (Juz.) Langenf. (gr. hybr.) 19, 78, Sl, 192, 214 Pumilae Rehd. (вег.) 15, 18, 189 Sieboldinae Rehd. (вег.) 15 — 16, 140 ' Светлым прямым шрифтом набраны основные названия родов, надвидовых таксонов и принятых нами видов, курсивом — производные (синонимы, гибриды и пр.). Полужирным шрифтом набраны страницы, на которых приведены основные данные о таксонах; «ic» — рисунок, «аг» — ареал.
Алфавитный указатель латинских названий 228 111. Виды, основные синонимы и гибридные виды Malus)(adstringens Zabel 194, 214 — angustifolia (Ait.) Michx. 14, 16, 18, 41, 94, 96аг, 149ic, 151 — anisophylla Sumn. 20, 23, 172, 213 — )(arnoldiana Rehd. 144, 211 — asiatica Nakai 101 — 102, 167, 212 — — rinki Asami 168 — astracanica Dum-Cours. 14 — X atrosanguinea (Spach.) С. К. Schneid. 164 — baccata (L.) Borkh. 11, 14 — 15, 17 — 20, 23, 34, 42, 51, 76 — 77, 97ar, 155ic — 156, 209, 211 — — var. himalaica (Maxim.) С. К. Schneid. 14, 160 — — var. sibirica С. К. Schneid. 17, 156 — bracteata Rehd. 211 — caspiriensis Langenf. 102, 103ar — 104, 181 — 182ic, 213 — centralasiatica Vass. 22, 91, 92ar, 136ic, 139, 210 — Xcerasifera Spach 20, 161, 162ic, 209, 212 — chitralensis Vass. 35, 79 — 80, 103, 182 — 183, 213 — coronaria (L.) Mill. 10, 14, 16 — 18, 41, 93, 96аг, 149ic — 150, 21 — dasyphylla Borkh. 11, 18, 80 — 81, 214 — )(denboerii Кгиьып. 154 — domestica Borkh. 11, 19, 188ic — 189, 208, 213 — Xflorentina (Zucc.) С. К. Schneid. 13 — 15, 24, 41, 211 — Xfloribunda Sieb. 15 — 16, 18, 144, 211 — formosana Kaw. et Koidz. 16, 18, 22, 34, 41, 45, 50, 69 — 70, 74, 124ic, 210 — fusca (Raf.) С. К. Schneid. 13 — 15, 18, 41, 69, 75, 93, 95аг, 147ic, 211 — glabrata Pehd. 153, 211 — glaucescens Rehd. 15, 18, 153, 211 — halliana (Anon.) Koehne 14, 18, 42, 76, 100, 163 — 164, 212 — )(ЬагШд~~ КоеЬпе 100, 164, 212 — )(heterophylla Spach 154, 211 — himalaica (Maxim.) Vass. 42, 76, 96, 97 — 98аг, 160, 212 — hissarica S. Kudr. 23, 80, 172 — 173ic, 213 — honanensis Rehd. 41, 74, 89 — 90ar, 132ic, 134, 210 — hortalana Mi11. 10 — hupehensis (Pamp.) Rehd. 42, 77, 101аг, 161, 163ic, 212 — ioensis (Vood) Вг111. 16, 18, 41, 93, 96аг, 149ic, 162, 211 — jarmolenkovii Р. Ро!. 21, 23, 213 211 94— 88ar, 146— Silvestres Juz. (зег.) 19, 183 $огЬота1из Zabel (sect.) 12, 13, 15 — 18, 39, 41, 43 — 44, 47, 52, 54 — 55, 73 — 74, 89, 92 — 93, 121, 129, 210 Yunnanenses Rehd. (зег.) 15 — 17, 22, 41, 55, 74, 131
Алфавитный указатель латинскик названий 229 juzepczukii Vass. 21, 23 kansuensis (Batal.) С. К. Schneid. 13 — 15, 91 — 92аг, 135 — 136ic, 210 kirghisorum Аl. et An. Theod. 20, 23, 42, 78, 80, 105аг, 169, 174— 175ic, 213 komarovii (Sarg.) Rehd. 40, 91, 92аг, 140, 210 Иийгjaschevii Sumn. 20, 23, 172, 213 lancifolia Rehd. 16, 153, 211 laosensis А. Cheval. 16, 34, 36, 41, 50, 52, 54, 69 — 70, 74, 87 — 88аг, 124ic, 126, 210 linczevskii P. Pol. 21, 23, 172, 213 mandshurica (Maxim.) Кот. 19, 23, 34, 42, 76, 98 — 99аг, 155ic, 158, 212 micromalus Pehd. 15 — 16, 194, 214 montana Uglitz. 179 niedzuuetzkgana Dieck. 19 — 20, 23, 212 nuristanica Vass. 35, 79 — 80, 103 obensis Gorb. 69 ombrophila Hand.— Ìàãã. 41, 74, 89 — 90аг, 132ic — 133, 210 orientalis (Uglitz.} Juz. 19, 34, 79, 110аг — 111, 113 — 114, 176 — 177ic, 196, 207, 209, 213 — var. montana (Uglitz.} Langenf. 35, 42, 79, lllаг, 177ic, 179, 213 pallasiana Лиг. 19, 156 persicifolia (М. Pop.) $итп. 20, 23 praecox (Pall.) Borkh. 11, 19, 23, 35, 42, 80, 117 — 118аг, 186 — 187ic, 207 — 208, 21~ рга11й (Hemsl.) С. К. Schneid. 13 — 15, 22, 41, 89 — 90аг, 132ic, 133— 134, 210 Xprurufolia (Willd.) Borkh. 11, 14 — 17, 19, 23, 190ic, 193, 194, 208— 209, 214 pumila Ni11. 10 — 11, 14 — 16, 18, 22, 80, 101, 167, 181, 186, 214 — var. domesticg С. К. Schneid. 14 — var. niedzssetzkyana (Dieck.} С. К. Schneid. 14 — var. paradisiaca С. К. Schneid. 14 — — f. apetala (Munch.} С. К. Schneid. 14 — — f. pendula (R. Miiller) С. К. Schneid. 14 Xpurpurea (ВагЫег) Rehd. 192ic — 194, 214 Xrobusta (Carr.) Rehd. 161, 212 sacha1inensis (Кот.) Juz. 19, 23, 76, 99 — 100аг, 155ic, 159, 212 Xsargentii pehd. 14 — 16, 18, 94аг, 146 — 147, 210 gscheideckeri Spaeth 144, 194 — 195ic, 215 schischkinii P. Po!. 21, 23, 213 setok Vass. 91 — 92аг, 136ic, 139, 210 sibirica Borkh. 156 sieboldii (Regel) Rehd. 15 — 16, 18, 23, 142 sieversii (Ledeb.} М. Roem. 19, 21, 23, 34, 105, 107аг — 110, 169, 170ic, 171, 209
230 Алфавитный указатель латинских названий Ру sikkimensis (Wenz.) goehne 16, 18, 34, 41 — 42, 54, 70, 74, 76, 87— 88ar, 124ic, 128, 210 )(яои1агЖ1 (Bailey) Britt. 211 X s pectabi lis (Ait.) Borkh. 11, 14 — 16, 18 — 191ic, 194, 214 sylvestris (L.) Mill. 10, 16, 18, 23, 63 — 64, 67, 79, 115ar, 184 — 185ic, 196, 209, 213 — var. hortulana С. К. Schneid. 13 — 14 tianschanica Sumn. 20, 23 toringo Siebold 14, 34, 41, 75, 93 — 94ar, 141ic — 142, 210 toringoides (Rehd.) Hughes 15, 40, 91 — 92ar, 136ic — 137, 210 transitoria (Batal.) С. К. Schneid. 40, 55, 91 — 92ar, 136ic, 138, 210 trilobata (Labill.) С. К. Schneid. 13 — 14, 16, 22, 24, 40, 69, 73, 86— 87ar, 122ic tschonoskii (Maxim ) С. К. Schneid. 13 — 14, 16, 22, 36, 50, 70, 74, 87 — 88ar, 124ic — 125, 210 turkmenorum Juz. et M. Pop. 19, 35, 180, 213 yunnanensis (Franch.) С. К. Schneid. 13 — 15, 41, 74, 89 — 90ar, 131— 132ic, 210 Хгит~ Rehd. 14 — 16, 144 — 145ic, 211 rus angustifolia Ait. 151 — 152 baccata L. 10, 19, 156 — 157 — var. mandshurica Maxim. 158 coronaria L. 150 — 151 — var. ioensis Vood 152 fusca Raf. 146 hupehensis Pamp. 162 — 163 kansuensis Batal. 135 Malus L. 10, 17, 167, 176, 181, 186 — var. paradisiaca L. 10 — var. sylvestris L. 10 ясеооЫсс Regel 142 sieversii Ledeb. 169 toringoides Osborn 137 transitoria Batal. 138 trilobata DC. 122 tschonoskii Maxim. 125 yunnanensis Franch. 131
V. 1.angenfelds APPLE-TREES:MORPHOLOGICAL EVOLUTION, PHYLOGENY,GEOGRAPHY, SYSTEMATICS Rfga «Zinatne» Publishers, 1991Summary The existing classification systems of the genus Afalus are incomplete. They are often contradictory, not balanced in all their parts. Therefore, it was necessary to revise the old classi- fication of the whole genus Afalus and to make а new classifi- cation zystern. On the basis of the comparison of the morphological structure (structure of plant parts) о apple species of different geographi- cal zones of the Earth we have made conclusions about the тог- phological evolution of apples, determining their evolutionary evels and mutual relationship (phylogenesis). The first apple populations appeared already in the Creta- ceoua period in South-East Asia about 70 million уеага ago. However, а large-scale development and differentiation of apples took place in the Palaeoneogene, i. е. 50 — 20 million years ago. Today only а few of them have survived and are greatly diffe- rentiated relict apple species (sect. Eriolobus and sect. Pocy- niopsis) . The docynio-like apples (sect. Docyniopsis) flourished in South-West China, then differentiated and gave rise to sorbo-like apples (sect. Sorbomalus) which spread in Central and East Asia. The green-fruit apples (sect. Chloromeles) evolved from the соттоп initial ancestors when they migrated to North America during the time when the Asian and North American continents were not separated yet. The differentiation of the original apples in the Himalayas produced two most highly developed branches of the genus Afalus, i. e. berry-like apples (sect. Gymnomeles) adapted to harsh con- ditions and evolved along the ornithophilous line tiny, berry-like fruit adapted to seed dispersal by means of birds) and true fruit apples (sect. Afalus) spread in Euroasia. Younger species, particularly true apples, are characterized by great diversity of forms of different origin, polymorphism, tending to mutual hybridization. Many cultivated apples have been bred from them by long selective breeding. Cultivated ap-
232 $шппчагу ples, in their turn, tend to grow wild. As а result of these two complicated, double-sided processes we can find in nature (espe- cially in areas of old culture) complexes with an abscure syste- matic status. Several sections of оиг classification are original. However, it retans sections of previous authors, two of them — Eriolobus and Docyniopsis паче been revised and interpreted in а пеъ way (emend. sensu novo). Cultivated and hybrid species are inclosed in the groups: Gr. cult. Domesticae and Gr.ßÐãèïÖî1éàå. As а result of critical inventory of the genus Malus we have found 58 species, including 35 autochthon (9 polymorphous with several taxons in one species), 1 cultigen and 22 hybrid species.
ОГЛАВЛ ЕН И Е Предисловие Глава 1. Материал и ыетоды исследоваиий Глава 2. Краткий обзор истории изучеиия рода Malus . Глава 3. Структуриые особеииости, эволюция и филогеиия яблоии . Кориевая систеыа и побег . Габитус и ветвлеиие Л Ист Соцветие ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Ы веток ° ° ° ° П лод Особеиности иидивидуального развития яблони . Некоторые сведеиия о филогеиии рода Malus Глава 4. География и экологические особеииости яблоии Родовые ареалы Malus, Pyrus, Sorbus и Crataegus . Распростраиеиие рода Malus Глава 5. Систеыатический обзор рода Malus . Глава 6. Культуриые сорта яблоии Глава 7. Перспективы использоваиия яблоии в иародиоы хозяйстве Заключеиие Список литературы Алфавитиый указатель латииских иазваиий Apple-trees: Morphological Evolution, Phylogeny, Geography, Systematics < Summary) 5 7 9 25 27 34 37 42 44 50 55 67 82 82 86 II9 196 204 2I6 222 227 23I
CONTENTS Preface Chapter 1. Research materials and methods Chapter 2. Short history of the investigations of the genus Malus . Chapter 3. Peculiarities of the structure of apple-trees, their evolution and phylogenesis Hoot system and sprout Habitus and branching . Leaves Floscule Blossom F ° ~ ГШ~ ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Peculiarities of apple tree individual development Briefy about phylogenesis of the genus Malus Chapter 4. Ecological peculiarities and geography of apple trees . Comparison of the areas of the genera Malus, Ругия, Sorbus and Crataegus Distribution of the genus Malus Chapter 5. Survey of systematics of the genus Malus Chapter 6. Apple tråå cultivars . Chapter 7. Prospects of the usage of apple trees ° Conclusion Bibliography . Alphabetical index of Latin names ° Apple trees: Morphological Evolution, Phylogeny, Geography, Systematics (Summary) 5 7 9 25 27 34 37 42 44 50 55 67 82 82 86 119 196 204 216 222 227 231
ВАЛДЕМАР ТЕОДОРОВИЧJIAH ГЕНФЕЛ ЬД ЯБЛОНЯМОРФОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ,ФИЛОГЕНИЯ, ГЕОГРДФИЯ,CHCTEMATH KA Редактор М. Про вороваХудожник В. КовалевХудожественный редактор И. ЕгереТехнический редактор Г. СлепковаКорректор Н. Ивлева ИБ № 3056 Сдано в набор 07.08.90. Подписано в оеч ать 14.01.91. Формат 60X90/16. Бумага типогр. № 1. Гар нитур а литер атурн а я. Печать высокая. 15 фиэ. печ. л.; 15 усл. печ. л.; 15 усл. кр.-отт.; 14,97 уч.-иэд. л. Гираж 1180 экэ. 3акаэ № 727, цех № 1. Цена 3 р. 30 к. Иэдательство «Зинатне», 226530 ГСП Рига, ул. Тур ге нева, 19. От- печатано в типографии «Рота», 226011 Рига, ул. Блауманя, 38/40.